ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к клетке, которая содержит больше чем один химерных антигенный рецептор (CAR). Клетка может быть способна специфично распознавать клетку-мишень благодаря отличительному паттерну экспрессии (или неэкспрессии) двух или более антигенов клеткой-мишенью.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Описано множество иммунотерапевтических средств для использования в лечении злокачественных опухолей, в том числе терапевтические моноклональные антитела (mAb), иммуноконъюгированные mAb, радиоконъюгированные mAb и биспецифичные средства, вовлекающие T-клетки (Bi-specific T-cell Engager®).
Типично эти иммунотерапевтические средства направлены на один антиген: например, ритуксимаб направлен на CD20; миелотарг направлен на CD33; и алемтузумаб направлен на CD52.
Однако, относительно редко присутствия (или отсутствия) одного антигена достаточно для того, чтобы описать злокачественную опухоль, что может вести к недостатку специфичности.
Большинство злокачественных опухолей нельзя дифференцировать от нормальных тканей на основании одного антигена. Таким образом, при терапии возникает существенная токсичность «в мишени вне опухоли», посредством которой происходит повреждение нормальных тканей. Например, в то время, как направленно воздействуют на CD20 для того, чтобы лечить B-клеточные лимфомы ритуксимабом, происходит истощение компартмента нормальных B-клеток, в то время, как направленно воздействуют на CD52 для того, чтобы лечить хронический лимфоцитарный лейкоз, происходит истощение всего лимфоидного компартмента, в то время, как направленно воздействуют на CD33 для того, чтобы лечить острый миелолейкоз, происходит повреждение всего миелоидного компартмента, и т.д.
Предсказанная проблема токсичности «в мишени вне опухоли» обнаружена в клинических исследованиях. Например, подход, в котором направленно воздействуют на ERBB2, вызвал смерть у пациента со злокачественной опухолью ободочной кишки, метастазировавшей в легкие и печень. У некоторых пациентов со злокачественной опухолью ободочной кишки повышена экспрессия ERBB2, но также он экспрессирован в некоторых нормальных тканях, включая сердце и нормальное сосудистое русло.
Для некоторых злокачественных опухолей направленное воздействие на присутствие двух антигенов злокачественной опухоли может быть более избирательным и, следовательно, эффективным, чем направленное воздействие на один. Например, хронический B-лимфоцитарный лейкоз (B-CLL) представляет собой обыкновенный лейкоз, который в настоящее время лечат посредством направленного воздействия на CD19. Это лечит лимфому, но также истощает весь компартмент B-клеток, так что лечение имеет существенный токсический эффект. B-CLL имеет необычный фенотип в том отношении, что совместно экспрессируются CD5 и CD19. Посредством направленного воздействия только на клетки, которые экспрессируют CD5 и CD19, будет возможно значительно снижать токсичность в мишени вне опухоли.
Таким образом, существует потребность в иммунотерапевтических средствах, которые способны к дополнительному направленному воздействию, чтобы отражать комплексный паттерн экспрессии маркеров, который связан со многими злокачественными опухолями.
Химерные антигенные рецепторы (CAR)
Химерные антигенные рецепторы представляют собой белки, которые прививают специфичность моноклонального антитела (mAb) эффекторной функции T-клетки. Их обычная форма относится к белку с трансмембранным доменом I типа с антигенраспознающим амино-концом, спейсером, трансмембранным доменом, все соединены в составной эндодомен, который передает сигналы выживаемости и активации T-клеток (см. фиг. 1A).
Наиболее распространенной формой этих молекул являются слитные конструкции из одноцепочечных вариабельных фрагментов (scFv), полученных из моноклональных антител, распознающих антиген-мишень, которые слиты через спейсер и трансмембранный домен с сигнальным эндодоменом. Такие молекулы вызывают активацию T-клетки в ответ на распознавание с помощью scFv его мишени. Когда T-клетки экспрессируют такой CAR, они распознают и уничтожают клетки-мишени, которые экспрессируют антиген-мишень. Разработано несколько CAR против опухоль-ассоциированных антигенов, и в настоящее время клинические исследования проходят подходы адоптивного переноса с использованием таких CAR-экспрессирующих T-клеток для лечения различных злокачественных опухолей.
Однако использование CAR-экспрессирующих T-клеток также связано с токсичностью в мишени вне опухоли. Например, основанный на CAR подход, направленно воздействующий на карбоксиангидразу-IX (CAIX) для того, чтобы лечить почечноклеточную карциному, приводил к печеночной токсичности, которая предположительно обусловлена специфичной атакой на эпителиальные клетки желчного протока (Lamers et al (2013) Mol. Ther. 21:904-912.
Подходы двойного направленного воздействия с использованием CAR
Для того чтобы решить проблему токсичности «в мишени вне опухоли», разработаны CAR T-клетки с двойной антигенной специфичностью. В подходе «двойного направленного воздействия», два комплементарных CAR совместно экспрессируют в одной и той же популяции T-клеток, каждый направлен на отделенную опухолевую миешнь и сконструирован для того, чтобы предоставлять комплементарные сигналы.
Wlikie et al (2012 J Clin Immunol 32:1059-1070) описывают подход двойного направленного воздействия, в котором совместно экспрессируют ErbB2- и MUC1-специфичные CAR. ErbB2-специфичный CAR предоставлял только CD3ξ сигнал и MUC1-специфичный CAR предоставлял только CD28 костимуляторный сигнал. Обнаружено, что комплементарная передача сигналов возникала в присутствии обоих антигенов, что приводило к продукции IL-2. Однако продукция IL-2 была умеренной по сравнению с контрольными T-клетками со сконструированными CAR, в которых передачу сигналов обеспечивает слитный CD28+CD3ξ эндодомен.
Схожий подход описан у Kloss et al (2013 Nature Biotechnol. 31:71-75), в котором использовали CD-19-специфичный CAR, который предоставляет CD3ξ-опосредованный активационный сигнал в комбинации с химерным костимуляторным рецептором, специфичным к PSMA. С использованием этой конструкции «совместных CAR» CAR T-клетка получает активационный, сигнал когда она встречает клетку-мишень с одним антигеном, и костимуляторный сигнал, когда она встречает клетку-мишень с другим антигеном, и получает и активирующий и костимуляторный сигналы только когда встречает клетки-мишени, несущие оба антигена.
Это является ранней попыткой ограничения CAR активности только клеткой-мишенью, несущей два антигена. Однако этот подход ограничен: несмотря на то, что активность CAR T-клетки будет наибольшей в отношении мишеней, экспрессирующих оба антигена, CAR T-клетка все же будет уничтожать мишени, экспрессирующие только антиген, распознаваемый с помощью активирующего CAR; кроме того, костимуляция ведет к пролонгированным эффектам, оказываемым на T-клетки, которые длятся после ухода клетки-мишени. Таким образом, активность против положительных по одному антигену T-клеток, равная активности против двойных положительных, может быть возможна, например, в ситуации, когда одинарные положительные ткани находятся смежно с двойной положительной опухолью или на миграционном пути от нее.
Таким образом, существует потребность в усоверешенствованных терапевтических подходах на основе CAR со сниженной токсичностью в мишени вне опухоли, где активация T-клеток полностью ограничена клетками-мишенями, которые экспрессируют оба антигена.
ОПИСАНИЕ ФИГУР
Фиг. 1: (a) обобщенная архитектура CAR: связывающий домен распознает антиген; спейсер поднимает связывающий домен с клеточной поверхности; трансмембранный домен заякоривает белок в мембране и эндодомен передает сигналы. С (b) до (d): различные поколения и перестановки эндодоменов CAR: (b) начальные конструкции передавали ITAM сигналы только через эндодомен FcεR1-γ или CD3ξ, тогда как более поздние конструкции передавали дополнительный (c) один или (d) два костимуляторных сигнала в цис.
Фиг. 2: схематическое изображение, которое иллюстрирует изобретение
Изобретение относится к конструированию T-клеток, чтобы реагировать на логические правила антигенной экспрессии клетки-мишени. Это лучше всего проиллюстрировано с помощью воображаемой диаграммы рассеяния при FACS. Популяции клеток-мишеней экспрессируют оба, любой один или ни одного из антигенов «A» и «B». Различные популяции-мишени (помечены красным) подвержены уничтожению T-клетками, трансдуцированными парой CAR, соединенных с помощью различных логических элементов. При использовании рецепторов с логическим элементом ИЛИ будут уничтожены как одинарные положительные, так и двойные положительные клетки. При использовании рецепторов с логическим элементом И погибают только двойные положительные клетки-мишени. При использовании логического элемента И НЕ двойные положительные мишени сохранены, тогда как одинарные положительные мишени
Фиг. 3: создание популяций клеток-мишеней
Клетки SupT1 использовали в качестве клеток-мишеней. Эти клетки трансдуцировали для того, чтобы экспрессировать любое из CD19 и CD33 или оба CD19 и CD33. Клетки-мишени окрашивали подходящими антителами и анализировали с помощью проточной цитометрии.
Фиг. 4: конструкция кассеты для логического элемента ИЛИ
Одна открытая рамка считывания обеспечивает оба CAR с последовательностью FMD-2A с сохранением рамки, что ведет к двум белкам. Сигнал1 представляет собой сигнальный пептид, полученный из IgG1 (но может представлять собой любой эффективный сигнальный пептид). scFv1 представляет собой одноцепочечный вариабельный сегмент, который распознает CD19 (но может представлять собой scFv или пептидную петлю или лиганд или фактически любой домен, который распознает любую желаемую произвольную мишень). STK представляет собой «стебель» CD8, но может представлять собой любой подходящий внеклеточный домен. CD28tm представляет собой трансмембранный домен CD28, но может представлять собой любой стабильный трансмембранный домен белка I типа и CD3Z представляет собой дзета-эндодомен CD3, но может представлять собой любой эндодомен, который содержит ITAM. Сигнал2 представляет собой сигнальный пептид, полученный из CD8, но может представлять собой любой эффективный сигнальный пептид, который отличается по последовательности ДНК от сигнала1. scFv распознает CD33, но как и для scFv1, является произвольным. HC2CH3 представляет собой шарнир-CH2-CH3 из IgG1 человека, но может представлять собой любой внеклеточный домен, который не образует перекрестную пару со спейсером, используемым в первом CAR. CD28tm' и CD3Z' кодируют ту же белковую последовательность, что и CD28tm и CD3Z, но в них внесена неоднозначность кодонов для того, чтобы предотвращать гомологичную рекомбинацию.
Фиг. 5: схематическое представление химерных антигенных рецепторов (CAR) для логического элемента ИЛИ
Конструировали стимулирующие CAR, содержащие любой из N-концевой A) scFV-домен против CD19, за которым следует внеклеточная шарнирная область CD8 человека или B) scFV-домен против CD33, за которым следует область внеклеточного шарнира, CH2 и CH3 (содержащая мутацию pvaa для того, чтобы снижать связывание FcR) из IgG1 человека. Оба рецептора содержат трансмембранный домен CD28 человека и внутриклеточный дзета-домен CD3 человека (CD247). «S» обозначает присутствие дисульфидных связей.
Фиг. 6: экспрессионные данные, показывающие коэкспрессию обоих CAR на поверхности одной T-клетки.
Фиг. 7: функциональный анализ логического элемента ИЛИ
Эффекторные клетки (5×104 клеток), экспрессирующие конструкцию логического элемента ИЛИ, совместно инкубировали с различным числом клеток-мишеней и анализировали IL-2 после 16 часов с помощью ELISA. На графике представлен усредненная максимальная секреция IL-2 от химической стимуляции (PMA и иономицин) только эффекторных клеток и усредненный фоновый IL-2 от эффекторных клеток без какого-либо стимула из трех повторений.
Фиг. 8: схематическое представление, показывающее обе версии кассеты, используемой для экспрессии обоих логических элементов И
Активирующие и ингибирующие CAR совместно экспрессировали еще раз с использованием последовательности FMD-2A. Сигнал1 представляет собой сигнальный пептид, полученный из IgG1 (но может представлять собой любой эффективный сигнальный пептид). scFv1 представляет собой одноцепочечный вариабельный сегмент, который распознает CD19 (но может представлять собой scFv или пептидную петлю или лиганд или фактически любой домен, который распознает какую-либо желаемую произвольную мишень). STK представляет собой «стебель» CD8, но может представлять собой любой некрупный внеклеточный домен. CD28tm представляет собой трансмембранный домен CD28, но может представлять собой любой стабильный трансмембранный домен белка I типа и CD3Z представляет собой дзета-эндодомен CD3, но может представлять собой любой эндодомен, который содержит ITAM. Сигнал2 представляет собой сигнальный пептид, полученный из CD8, но может представлять собой любой эффективный сигнальный пептид, который отличается последовательностью ДНК от сигнала1. scFv распознает CD33, но как и для scFv1, является произвольным. HC2CH3 представляет собой шарнир-CH2-CH3 из IgG1 человека, но может представлять собой любой крупный внеклеточный домен. CD45 и CD148 представляют собой трансмембраные и эндодомены CD45 и CD148, соответственно, но могут быть получены из любого белка этого класса.
Фиг. 9: схематическое представление структуры белков химерных антигенных рецепторов (CAR) для логических элементов И
Стимулирующий CAR состоит из N-концевого scFV-домена против CD19, за которым следует область внеклеточного «стебля» из CD8 человека, трансмембранный домен CD28 человека и внутриклеточного дзета-домена CD3 человека (CD247). Тестировали два ингибирующих CAR. Они состоят из N-концевого scFV-домена против CD33, за которым следует область внеклеточного шарнира, CH2 и CH3 (содержащая мутацию pvaa для того, чтобы снижать связывание FcR) из IgG1 человека, за которой следует трансмембраный и внутриклеточный домен любого из CD148 или CD45 человека. «S» обозначает присутствие дисульфидных связей.
Фиг. 10: коэкспрессия активирующих и ингибирующих CAR
В качестве эффекторных клеток использовали клетки BW5147, которые трансдуцировали для того, чтобы экспрессировать как активирующий CAR против CD19, так и один из ингибирующих CAR против CD33. Эффекторные клетки окрашивали с использованием Fc мыши к CD19 и Fc кролика к CD33 и подходящих вторичных антител и анализировали с помощью проточной цитометрии.
Фиг. 11: функциональный анализ логических элементов И
Эффекторные клетки (5×104 клеток), экспрессирующие активирующий CAR против CD19 и ингибирующий CAR против CD33 с внутриклеточным доменом A) CD148 или B) CXD45, совместно инкубировали с различным числом клеток-мишеней и анализировали IL-2 после 16 часов с помощью ELISA. На графике представлена максимальная секреция IL-2 от химической стимуляции (PMA и иономицин) только эффекторных клеток и фоновый IL-2 от эффекторных клеток без какого-либо стимула из трех повторений.
Фиг. 12: схематическое представление трех версий кассеты, используемой для того, чтобы генерировать логический элемент И НЕ
Активирующие и ингибирующие CAR совместно экспрессировали еще раз с использованием последовательности FMD-2A. Сигнал1 представляет собой сигнальный пептид, полученный из IgG1 (но может представлять собой любой эффективный сигнальный пептид). scFv1 представляет собой одноцепочечный вариабельный сегмент, который распознает CD19 (но может представлять собой scFv или пептидную петлю или лиганд или фактически любой домен, который распознает любую желаемую произвольную мишень). STK представляет собой «стебель» CD8 человека, но может представлять собой любой некрупный внеклеточный домен. CD28tm представляет собой трансмембранный домен CD28, но может представлять собой любой стабильный трансмембранный домен белка I типа, и CD3Z представляет собой дзета-эндодомен CD3, но может представлять собой любой эндодомен, который содержит ITAM. Сигнал2 представляет собой сигнальный пептид, полученный из CD8, но может представлять собой любой эффективный сигнальный пептид, который отличается последовательностью ДНК от сигнала1. scFv распознает CD33, но как и для scFv1, является произвольным. muSTK представляет собой «стебель» CD8 мыши, но может представлять собой любой спейсер, который совместно локализован, но не образует перекрестные пары с таковым из активирующего CAR. dPTPN6 представляет собой фосфатазный домен из PTPN6. LAIR1 представляет собой трансмембраный и эндодомен из LAIR1. 2Aw представляет собой версию последовательности FMD-2A с неоднозначностью кодонов. SH2-CD148 представляет собой домен SH2 из PTPN6, слитый с фосфатазным доменом из CD148.
Фиг. 13: схематическое представление химерных антигенных рецепторов (CAR) для логических элементов НЕ И
A) стимулирующий CAR, состоящий из N-концевого scFV-домена против CD19, за которым следует область «стебля» из CD8 человека, трансмембранный домен CD28 человека и внутриклеточный домен CD247 человека. B) ингибирующий CAR, состоящий из N-концевого scFV-домена против CD33, за которым следует область «стебля» из CD8 мыши, трансмембранная область из CD8 мыши и фосфатазный домен из PTPN6. C) ингибирующий CAR, состоящий из N-концевого scFV-домена против CD33, за которым следует область «стебля» из CD8 мыши и трансмембраный и внутриклеточный сегменты из LAIR1. D) ингибирующий CAR, идентичный предыдущему CAR, за исключением того, что его коэкспрессируют со слитным белком из домена SH2 PTPN6 и фосфатазного домена CD148.
Фиг. 14: функциональный анализ логического элемента НЕ И
Эффекторные клетки (5×104 клеток), экспрессирующие A) полноразмерный SHP-1 или B) усеченную форму SHP-1, совместно инкубировали с различным числом клеток-мишеней и анализировали IL-2 после 16 часов с помощью ELISA. На графике представлена усредненная максимальная секреция IL-2 от химической стимуляции (PMA и иономицин) только эффекторных клеток и усредненный фоновый IL-2 от эффекторных клеток без какого-либо стимула из трех повторений.
Фиг. 15: аминокислотная последовательность логического элемента ИЛИ
Фиг. 16: аминокислотная последовательность логического элемента И на основе CD148 и CD145
Фиг. 17: аминокислотная последовательность двух логических элементов И НЕ
Фиг. 18: разбор функции логического элемента И
A. Прототип логического элемента И проиллюстрирован справа и его функция в ответ на одинарные CD19, CD33 и двойные CD19, CD33 положительные мишени представлена слева. B. ScFv меняют местами так, что активирующий эндодомен запускают с помощью CD33, а ингибирующий эндодомен активируют с помощью CD19. Этот логический элемент И остается функциональным, несмотря на то, что эти scFv меняют местами. C. «Стебель» CD8 мыши заменял Fc в спейсере ингибирующего CAR. При этой модификации логический элемент не способен реагировать на любую из одинарной CD19 положительной или двойной CD19, CD33 положительной мишеней.
Фиг. 19: экспрессия антигенов-мишеней на искусственных клетках-мишенях
A. Представлены диаграммы рассеяния при проточной цитометрии CD19 в сравнении с CD33 из исходного набора искусственных клеток-мишеней, полученных из клеток SupT1. Слева направо: двойные негативные клетки SupT1, клетки SupT1, положительные по CD19, положительные по CD33 и положительные по CD19 и CD33. B. Представлены диаграммы рассеяния при проточной цитометрии CD19 в сравнении с GD2 из искусственных клеток-мишеней, созданных для того, чтобы тестировать логический элемент CD19 И GD2. Слева направо: негативные клетки SupT1, клетки SupT1, экспрессирующие CD19, клетки SupT1, трансдуцированные векторами синтазы GD2 и GM3, которые становаятся GD2 положительными, и клетки SupT1, трансдуцированные с использованием CD19, а также синтазы GD2 и GM3, которые положительны по обоим GD2 и CD19. C. Представлены диаграммы рассеяния при проточной цитометрии для CD19 в сравнении с EGFRvIII из искусственных мишеней, созданных для того, чтобы тестировать логический элемент CD19 И EGFRvIII. Слева направо: негативные клетки SupT1, клетки SupT1, экспрессирующие CD19, клетки SupT1, трансдуцированные с использованием EGFRvIII, и клетки SupT1, трансдуцированные с использованием как CD19, так и EGFRvIII. D. Представлены диаграммы рассеяния при проточной цитометрии для CD19 в сравнении с CD5 из искусственных мишеней, созданных для того, чтобы тестировать логический элемент CD19 И CD5. Слева направо: негативные 293T клетки, 293T клетки, трансдуцированные с использованием CD19, 293T клетки, трансдуцированные с использованием CD5, 293T клетки, трансдуцированные с использованием обоих векторов CD5 и CD19.
Фиг. 20: Генерализуемость логического элемента И
A. Схематическое изображение логического элемента И, модифицированного так, что специфичность второго CAR изменена с исходной специфичности CD33, чтобы генерировать 3 новых CAR: CD19 И GD2, CD19 И EGFRvIII, CD19 И CD5. B. Логический элемент CD19 И GD2 И: слева: экспрессия логического элемента И показана окрашиванием рекомбинантным CD19-Fc (ось x) для CD19 CAR, в зависимости от окрашивания против Fc человека (ось y) для GD2 CAR. Справа: функция в ответ на одинарные положительные и двойные положительные мишени. C. Логический элемент CD19 И EGFRvIII И: слева: экспрессия логического элемента И показана окрашиванием рекомбинантным CD19-Fc (ось x) для CD19 CAR, в зависимости от окрашивания против Fc человека (ось y) для EGFRvIII CAR. Справа: функция в ответ на одинарные положительные и двойные положительные мишени. D. Логический элемент CD19 И CD5 И: слева: экспрессия логического элемента И показана окрашиванием рекомбинантного CD19-Fc (ось x) для CD19 CAR, в зависимости от окрашивания против Fc человека (ось y) для CD5 CAR. Справа: функция в ответ на одинарные положительные и двойные положительные мишени.
Фиг. 21: функция логических элементов И НЕ
Представлена функция трех реализаций логического элемента И НЕ. Схематическое представление протестированных логических элементов приведено справа, а функция в ответ на одинарные положительные и двойные положительные мишени приведена слева. A. Логический элемент И НЕ на основе PTPN6, посредством которого первый CAR распознает CD19, имеет спейсер «стебля» CD8 человека и ITAM-содержащий активирующий эндодомен; коэкспрессируют со вторым CAR, который распознает CD33, имеет спейсер «стебля» CD8 мыши и имеет эндодомен, который содержит фосфатазный домен PTPN6. B. логический элемент И НЕ на основе ITIM идентичен логическому элементу PTPN6, за исключением того, что эндодомен заменяют на эндодомен из LAIR1. C. Усиленный CD148 логический элемент И НЕ идентичен логическому элементу на основе ITIM, за исключением того, что экспрессируют дополнительное слияние между PTPN6 SH2 и эндодоменом из CD148. Все три логических элемента работают в соответствии с ожиданиями с активацией в ответ на CD19, но не в ответ на CD19 и CD33 вместе.
Фиг. 22: Разбор функции логического элемента И НЕ на основе PTPN6
Исходный логический элемент И НЕ на основе PTPN6 сравнивают с несколькими контролями для того, чтобы демонстрировать модель. Схематическое представление протестированных логических элементов приведено справа, а функция в ответ на одинарные положительные и двойные положительные мишени приведена слева. A. Исходный логический элемент И НЕ, с помощью которого первый CAR распознает CD19, имеет спейсер «стебля» CD8 человека и ITAM-содержащий активирующий эндодомен; коэкспрессируют со вторым CAR, который распознает CD33, имеет спейсер «стебля» CD8 мыши и имеет эндодомен, состоящий из фосфатазного домена PTPN6. B. Логический элемент И НЕ, модифицированный так, что спейсер «стебля» CD8 мыши заменен на Fc спейсер. C. Логический элемент И НЕ, модифицированный с тем, чтобы заменить фосфатазный домен PTPN6 на эндодомен из CD148. Исходный логический элемент И НЕ (A.) функционирует в соответствии с ожиданиями, запускаясь в ответ на CD19, но не в ответ на оба CD19 и CD33. Логический элемент в B. запускается как в ответ только на CD19 или вместе на CD19 и CD33. Логический элемент в C. не запускается в ответ на одну или обе мишени.
Фиг. 23: разбор логического элемента И НЕ на основе LAIR1
Показана функциональная активность против CD19 положительных, CD33 положительных и CD19, CD33 двойных положительных мишеней. A. Структура и активность исходного логического элемента И НЕ на основе ITIM. Этот логический элемент состоит из двух CAR: первый распознает CD19, имеет спейсер «стебля» CD8 человека и ITAM-содержащий эндодомен; второй CAR распознает CD33, имеет спейсер «стебля» CD8 мыши и ITIM-содержащий эндодомен. B. Структура и активность контрольного логического элемента на основе ITIM, где спейсер «стебля» CD8 мыши заменен на Fc домен. Этот логический элемент состоит из двух CAR: первый распознает CD19, имеет спейсер «стебля» CD8 человека и ITAM-содержащий эндодомен; второй CAR распознает CD33, имеет Fc спейсер и ITIM-содержащий эндодомен. Оба логических элемента отвечают на одинарные CD19 положительные мишени, тогда как только исходный логический элемент неактивен в ответ на двойные CD19 и CD33 положительные мишени.
Фиг. 24: Модель кинетической сегрегации логических элементов CAR
Модель кинетической сегрегации и поведения логического элемента И, логического элемента НЕ И и контролей. CAR распознают или CD19 или CD33. Иммунологический синапс можно представить между голубой линией, которая представляет мембрану клетки-мишени, и красной линией, которая представляет T-клеточную мембрану. «45» представляет нативный белок CD45, присутствующий на T-клетках. «H8» представляет эктодомен CAR со «стеблем» CD8 человека в качестве спейсера. «Fc» представляет эктодомен CAR с HCH2CH3 человека в качестве спейсера. «M8» представляет собой эктодомен CAR с мышиным «стеблем» CD8 в качестве спейсера. «19» представляет CD19 на поверхности клетки-мишени. «33» представляет CD33 на поверхности клетки-мишени. Символ «⊕» представляет активирующий эндодомен, содержащий ITAM. Символ « « представляет фосфатазу с медленной кинетикой - «включаемый лигированием» эндодомен, такой как тот, что содержит каталитический домен из PTPN6 или ITIM. Символ «∅« представляет фосфатазу с быстрой кинетикой - «выключаемый лигированием» эндодомен, такой как эндодомен из CD45 или CD148. Этот символ увеличен на фиг., чтобы подчеркнуть его высокую активность.
« представляет фосфатазу с медленной кинетикой - «включаемый лигированием» эндодомен, такой как тот, что содержит каталитический домен из PTPN6 или ITIM. Символ «∅« представляет фосфатазу с быстрой кинетикой - «выключаемый лигированием» эндодомен, такой как эндодомен из CD45 или CD148. Этот символ увеличен на фиг., чтобы подчеркнуть его высокую активность.
(a) Представлено предполагаемое поведение функционального логического элемента И, который содержит пару CAR, посредством чего первый CAR распознает CD19, имеет спейсер «стебля» CD8 человека и активирующий эндодомен; и второй CAR распознает CD33, имеет Fc спейсер и эндодомен CD148;
(b) Представлено предполагаемое поведение контрольного логического элемента И. Здесь первый CAR распознает CD19, имеет спейсер «стебля» CD8 человека и активирующий эндодомен; и второй CAR распознает CD33, но имеет спейсер «стебля» CD8 мыши и эндодомен CD148;
(c) Представлено поведение функционального логического элемента И НЕ, который содержит пару CAR, посредством чего первый CAR распознает CD19, имеет спейсер «стебля» CD8 человека и активирующий эндодомен; и второй CAR распознает CD33, имеет спейсер «стебля» CD8 мыши и эндодомен PTPN6;
(d) Представлено предполагаемое поведение контрольного логического элемента И НЕ, который содерж*т пару CAR, посредством чего первый CAR распознает CD19, имеет спейсер «стебля» CD8 человека и активирующий эндодомен; и второй CAR распознает CD33, но имеет Fc спейсер и эндодомен PTPN6;
В первой колонке клетки-мишени являются негативными как по CD19, так и по CD33. Во второй колонке мишени являются CD19 негативными и CD33 положительными. В третьей колонке клетки-мишени являются CD19 положительными и CD33 негативными. В четвертой колонке клетки-мишени являются положительными как по CD19, так и по CD33.
Фиг. 25: Конструкция CAR на основе APRIL.
Конструкцию CAR модифицировали с тем, чтобы заменять scFv на модифицированную форму индуцирующего пролиферацию лиганда (APRIL), который взаимодействует с BCMA, TACI и протеогликанами, чтобы действовать в качестве антигенсвязывающего домена: APRIL усечен с тем, чтобы отсутствовал амино-конец, связывающий протеогликаны. После этого сигнальный пептид прикрепляли к усеченному амино-концу APRIL, чтобы направлять белок на клеточную поверхность. Эти CAR генерировали с использованием этого связывающего домена на основе APRIL: A. В первом CAR домен «стебля» CD8 человека использовали в качестве спейсерного домена. B. Во втором CAR шарнир из IgG1 использовали в качестве спейсерного домена. C. В третьем CAR домены шарнира, CH2 и CH3 из IgG1 человека, модифицированные мутациями pva/a, описанными у Hombach et al (2010 Gene Ther. 17:1206-1213) для того, чтобы снижать связывание Fc рецептора, использовали в качестве спейсера (в дальнейшем обозначают как Fc-pvaa). Во всех CAR эти спейсеры соединяли с трансмембранным доменом CD28 и затем с тройным эндодоменом, содержащим слияние CD28, OX40 и дзета-эндодомена CD3 (Pule et al, Molecular Therapy, 2005: том 12; выпуск 5; стр. 933-41).
Фиг. 26: аннотированная аминокислотная последовательность трех вышеприведенных APRIL-CAR
A: представлена аннотированная аминокислотная последовательность CAR со «стеблем» CD8 и APRIL; B: представлена аннотированная аминокислотная последовательность CAR на основе шарнира IgG1 и APRIL; C: представлена аннотированная аминокислотная последовательность CAR на основе APRIL и Fc-pvaa.
Фиг. 27: экспрессия и связывание лиганда различными CAR на основе APRIL
A. Рецепторы совместно экспрессировали с маркерным геном усеченным CD34 в векторе с ретровирусными генами. Экспрессия маркерного гена на трансдуцированных клетках делает возможным подтверждение трансдукции. B. T-клетки трансдуцировали с использованием CAR на основе APRIL и спейсера из «стебля» CD8, шарнира IgG1 или Fc спейсера. Для того чтобы тестировать возможность стабильной экспрессии этих рецепторов на клеточной поверхности, затем T-клетки окрашивали с APC против APRIL-биотина/стрептавидина и против CD34. Осуществляли анализ проточной цитометрии. APRIL в равной мере обнаруживали на клеточной поверхности в трех CAR, что наводит на мысль об их стабильной экспрессии в равной мере. C. Затем определяли способность CAR распознавать TACI и BCMA. Трансдуцированные T-клетки окрашивали с использованием или рекомбинантного BCMA или TACI, слитого со слитной конструкцией IgG2a мыши Fc, наряду со вторичным против мыши и против CD34. Все три формата рецепторов демонстрировали связывание как с BCMA, так и с TACI. К удивлению, обнаруживали, что связывание BCMA казалось большим, чем с TACI. Кроме того, к удивлению обнаруживали, что несмотря на то, что все три CAR были экспрессированы в равной мере, CAR со «стеблем» CD8 и шарниром IgG1 выглядели лучше при распознавании BCMA и TACI, чем тот, что с Fc спейсером.
Фиг. 28: функция различных конструкций CAR.
Осуществляли функциональный анализ с использованием трех различных CAR на основе APRIL. T-клетки периферической крови нормального донора или не трансдуцировали (NT) или трансдуцировали для того, чтобы экспрессировать различные CAR. Трансдукцию осуществляли с использованием супернатантов с равными титрами. Затем эти T-клетки обедняли по CD56 для того, чтобы удалять неспецифичную NK активность и использовали в качестве эффекторов. Клетки SupT1, которые или не трансдуцировали (NT) или трансдуцировали для того, чтобы экспрессировать BCMA или TACI, использовали в качестве мишеней. Представленные данные представляют собой среднее и стандартное отклонение из 5 независимых экспериментов. A. Специфичное уничтожение T-клеток, экспрессирующих BCMA и TACI, определяли с использованием высвобождения хрома. B. Также определяли высвобождение интерферона-μ. Мишени и эффекторы совместно культивировали в соотношении 1:1. После 24 часов интерферон-μ в супернатанте анализировали с помощью ELISA. C. Пролиферация/выживаемость T-клеток с CAR также определяли посредством подсчета числа T-клеток с CAR в той же совместной культуре, которую инкубировали в течение дополнительных 6 суток. Все 3 CAR направляют ответ против мишеней, экспрессирующих BCMA и TACI. Ответы на BCMA были больше, чем для TACI.
Фиг. 29: функциональность логического элемента И в первичных клетках
PBMC выделяли из крови и стимулировали с использованием PHA и IL-2. Двумя сутками позже клетки трансдуцировали на покрытых ретронектином планшетах ретровирусом, содержащим конструкцию CD19:CD33 логического элемента И. На сутки 5 уровень экспрессии двух CAR, транслируемых с помощью конструкции логического элемента И, оценивали через проточную цитометрию, и клетки обедняли по CD56+ клеткам (преимущественно естественные киллерные клетки). В сутки 6 PBMC помещали в совместную культуру с клетками-мишенями с соотношением эффектора к клетке-мишени 1:2. В сутки 8 супернатант собирали и анализировали на секрецию IFN-γ с помощью ELISA
Фиг. 30: показан отбор/иерархия возможных спейсерных доменов увеличивающегося размера. Эктодомен CD3-дзета предложен в качестве самого короткого возможного спейсера, за которым следует (b) шарнир IgG1. (c) «стебель» CD8 мыши или человека, и эктодомены CD28 считают промежуточными по размеру и совместно сегрегирующимися. (d) домен шарнира, CH2 и CH3 из IgG1 больше и объемнее, и (e) шарнир, CH2, CH3 и CH4 домен из IgM еще больше. Учитывая свойства молекул-мишеней и эпитопа связывающих доменов на указанных молекулах-мишенях, возможно использовать эту иерархию спейсеров для того, чтобы создавать CAR систему передачи сигналов, которая или совместно сегрегируется или сегрегируется раздельно при формировании синапса.
Фиг. 31: правила конструирования для создания T-клеток с CAR с логическими элементами. CAR с логическими элементами ИЛИ, И НЕ и И представлены в схематическом формате с клеткой-мишенью сверху и T-клеткой внизу и с синапсом в середине. Клетки-мишени экспрессируют произвольные антигены-мишени A и B.
T-клетки экспрессируют два CAR, которые содержат распознающие домены против A и против B, спейсеры и эндодомены. Для логического элемента ИЛИ необходимы (1) спейсеры, которые просто делают возможным антигеное распознавание и активацию CAR, и (2) оба CAR, имеющие активирующие эндодомены; дл логического элемента И НЕ необходимы (1) спейсеры, которые ведут к совместной сегрегации обоих CAR при распознавании обоих антигенов и (2) один CAR с активирующим эндодоменом, и другой, эндодомен которого содержит или вовлекает слабую фосфатазу; для логического элемента И необходимы (1) спейсеры, которые ведут к сегрегации обоих CAR в различные части иммунологического синапса при распознавании обоих антигенов и (2) один CAR с активирующим эндодоменом, и другой, эндодомен которого содержит сильную фосфатазу.
КРАТКОЕ ИЗЛОЖЕНИЕ АСПЕКТОВ ИЗОБРЕТЕНИЯ
Авторы настоящего изобретения разработали панель пар химерных антигенных рецепторов «с логическими элементами», которые при экспрессии клеткой, такой как T-клетка, способны обнаруживать конкретный паттерн экспрессии по меньшей мере из двух антигенов-мишеней. Если по меньшей мере два антигена-мишени произвольно обозначены, как антиген A и антиген B, три возможных варианта представляют собой следующее:
«ЛОГИЧЕСКИЙ ЭЛЕМЕНТ ИЛИ» - T-клетка запускается, когда антиген A или антиген B присутствует на клетке-мишени
«ЛОГИЧЕСКИЙ ЭЛЕМЕНТ И» - T-клетка запускается только когда оба антигена A и B присутствуют на клетке-мишени
«ЛОГИЧЕСКИЙ ЭЛЕМЕНТ И НЕ» - T-клетка запускается, если только антиген A присутствует на клетке-мишени, но не если оба антигена A и B присутствуют на клетке-мишени
Сконструированные T-клетки, экспрессирующие эти комбинации CAR, можно адаптировать так, чтобы они обладали узкой специфичностью к клеткам злокачественной опухоли на основании их конкретной экспрессии (или отсутствия экспрессии) двух или более маркеров.
Таким образом, в первом аспекте настоящее изобретение предусматривает клетку, которая совместно экспрессирует первый химерный антигенный рецептор (CAR) и второй CAR на клеточной поверхности, каждый CAR содержит:
(i) антигенсвязывающий домен;
(ii) спейсер
(iii) трансмембранный домен; и
(iv) внутриклеточный домен T-клеточной сигнализации (эндодомен)
в которых антигенсвязывающие домены первого и второго CAR связывются с различными антигенами и в которых спейсер первого CAR отличается от спейсера второго CAR, так что первый и второй CAR не образуют гетеродимеры, и в которых
один из первого или второго CAR представляет собой активирующий CAR, который содержит активирующий внутриклеточный домен T-клеточной сигнализации, и другой CAR представляет собой ингибирующий CAR, который содержит «выключаемый лигированием» (как определено в настоящем документе) ингибирующий внутриклеточный домен T-клеточной сигнализации.
Клетка может представлять собой иммунную эффекторную клетку, такую как T-клетка или естественная киллерная клетка (NK). Признаки, отмеченные в настоящем документе применительно к T-клетке, применимы в равной мере к другим иммунным эффекторным клеткам, таким как естественные киллерные клетки.
Спейсер первого CAR может иметь длину и/или заряд и/или геометрическую форму и/или конфигурацию и/или гликозилирование, отличающиеся от спейсера второго CAR, так что когда первый CAR и второй CAR связывают соответствующие им антигены-мишени, первый CAR и второй CAR становятся пространственно разделенными на T-клетке. Лигирование первого и второго CAR с соответствующими им антигенами вызывает их компартментализацию вместе или отдельно в иммунологическом синапсе, что ведет к управлению активацией. Это можно понять, рассмотрев модель кинетического разделения активации T-клеток (см. далее).
Первый спейсер или второй спейсер может содержать «стебель» CD8 и другой спейсер может содержать домен шарнира, CH2 и CH3 из IgG1.
В настоящем изобретении, которое относется к логическому элементу «И», один из первого или второго CAR представляет собой активирующий CAR, который содержит активирующий эндодомен, и другой CAR представляет собой «выключаемый лигированием» ингибирующий CAR, который содержит ингибирующий эндодомен. Выключаемый лигированием ингибирующий CAR ингибирует активацию T-клеток с помощью активирующего CAR в отсутствие лигирования ингибирующего CAR, но не значительно ингибирует активацию T-клеток с помощью активирующего CAR, когда ингибирующий CAR лигирован. Поскольку спейсер первого CAR имеет длину и/или заряд и/или геометрическую форму и/или конфигурацию и/или гликозилирование, отличающееся от спейсера второго CAR, когда оба CAR лигированы, они сегрегируются. Это вызывает пространственное отделение ингибирующего CAR от активирующего CAR с тем, чтобы могла происходить активация T-клетки. Следовательно, активация T-клеток происходит только в ответ на клетку-мишень, несущую оба когнатных антигена.
Ингибирующий эндодомен может содержать весь или часть эндодомена из рецептороподобной тирозиновой фосфатазы, такой как CD148 или CD45.
Антигенсвязывающий домен первого CAR может связывать CD5 и антигенсвязывающий домен второго CAR может связывать CD19. Это имеет применение при направленном воздействии на хронический лимфоцитарный лейкоз (CLL). Это заболевание можно лечить посредством направленного воздействия только на CD19, но за счет истощения всего B-клеточного компартмента. CLL клетки отличаются тем, что они совместно экспрессируют CD5 и CD19. Направленное воздействие на эту пару антигенов с использованием логического элемента И будет увеличивать специфичность и снижать токсичность.
Во втором аспекте настоящее изобретение предусматривает последовательность нуклеиновой кислоты, которая кодирует как первый, так и второй химерные антигенные рецепторы (CAR), как определено в первом аспекте изобретения.
Соответственно, последовательность нуклеиновой кислоты может иметь следующую структуру: AgB1-спейсер1-TM1-эндо1-коэкспр-AgB2-спейсер2-TM2-эндо2,
в которой
AgB1 представляет собой последовательность нуклеиновой кислоты, которая кодирует антигенсвязывающий домен первого CAR;
спейсер1 представляет собой последовательность нуклеиновой кислоты, которая кодирует спейсер первого CAR;
TM1 представляет собой последовательность нуклеиновой кислоты, которая кодирует трансмембранный домен первого CAR;
эндо1 представляет собой последовательность нуклеиновой кислоты, которая кодирует эндодомен первого CAR;
коэкспр представляет собой последовательность нуклеиновой кислоты, которая допускает коэкспрессию двух CAR (например, сайт расщепления);
AgB2 представляет собой последовательность нуклеиновой кислоты, которая кодирует антигенсвязывающий домен второго CAR;
спейсер2 представляет собой последовательность нуклеиновой кислоты, которая кодирует спейсер второго CAR;
TM2 представляет собой последовательность нуклеиновой кислоты, которая кодирует трансмембранный домен второго CAR;
эндо2 представляет собой последовательность нуклеиновой кислоты, которая кодирует эндодомен второго CAR;
эта последовательность нуклеиновой кислоты, при экспрессии в T-клетке, кодирует полипептид, который расщепляют в сайте расщепления так, что первый и второй CAR совместно экспрессируют на поверхности T-клеток.
Последовательность нуклеиновой кислоты, допускающая коэкспрессию двух CAR, может кодировать саморасщепляющийся пептид или последовательность, которая допускает альтернативное средство коэкспрессирующих двух CAR, таких как внутренняя последовательность связывания рибосомы или второй промотор или другое такое средство, посредством которого специалист в данной области может экспрессировать два белка с одного и того же вектора.
Альтернативные кодоны можно использовать в областях последовательности, кодирующих одинаковые или схожие аминокислотные последовательности, во избежание гомологичной рекомбинации.
В третьем аспекте настоящее изобретение предусматривает набор, который содержит
(i) первую последовательность нуклеиновой кислоты, которая кодирует первый химерный антигенный рецептор (CAR), как определено в первом аспекте изобретения, эта последовательность нуклеиновой кислоты имеет следующую структуру:
AgB1-спейсер1-TM1-эндо1
в которой
AgB1 представляет собой последовательность нуклеиновой кислоты, которая кодирует антигенсвязывающий домен первого CAR;
спейсер1 представляет собой последовательность нуклеиновой кислоты, которая кодирует спейсер первого CAR;
TM1 представляет собой последовательность нуклеиновой кислоты, которая кодирует трансмембранный домен первого CAR;
эндо1 представляет собой последовательность нуклеиновой кислоты, которая кодирует эндодомен первого CAR; и
(ii) вторую последовательность нуклеиновой кислоты, которая кодирует второй химерный антигенный рецептор (CAR), как определено в первом аспекте изобретения, эта последовательность нуклеиновой кислоты имеет следующую структуру:
AgB2-спейсер2-TM2-эндо2
AgB2 представляет собой последовательность нуклеиновой кислоты, которая кодирует антигенсвязывающий домен второго CAR;
спейсер2 представляет собой последовательность нуклеиновой кислоты, которая кодирует спейсер второго CAR;
TM2 представляет собой последовательность нуклеиновой кислоты, которая кодирует трансмембранный домен второго CAR;
эндо2 представляет собой последовательность нуклеиновой кислоты, которая кодирует эндодомен второго CAR.
В четвертом аспекте настоящее изобретение предусматривает набор, который содержит: первый вектор, который содержит первую последовательность нуклеиновой кислоты, как определено выше; и второй вектор, который содержит первую последовательность нуклеиновой кислоты, как определено выше.
Векторы могут представлять собой плазмидные векторы, ретровирусные векторы или транспозонные векторы. Векторы могут представлять собой лентивирусные векторы.
В пятом аспекте настоящее изобретение предусматривает вектор, который содержит последовательность нуклеиновой кислоты согласно второму аспекту изобретения. Вектор может представлять собой лентивирусный вектор.
Вектор может представлять собой плазмидный вектор, ретровирусный вектор или транспозонный вектор.
В шестом аспекте настоящее изобретение включает коэкспрессию больше чем двух CAR таким образом, что на клетке-мишени можно распознавать комплексный паттерн больше чем из двух антигенов.
В седьмом аспекте настоящее изобретение предусматривает способ получения T-клетки согласно первому аспекту изобретения, который включает стадию введения одной или нескольких последовательностей нуклеиновой кислоты, кодирующих первый и второй CAR; или одного или нескольких векторов, как определено выше, в T-клетку.
T-клетка может быть из образца, выделенного у пациента, родственного или неродственного донора гематопоэтического трансплантата, полностью несвязанного донора, из пуповинной крови, дифференцирована из эмбриональной клеточной линии, дифференцирована из индуцибельной линии клеток-предшественников или получена из трансформированной линии Т-клеток.
В восьмом аспекте настоящее изобретение предусматривает фармацевтическую композицию, которая содержит множество T-клеток в соответствии с первым аспектом изобретения.
В девятом аспекте настоящее изобретение предусматривает способ лечения и/или предотвращения заболевания, который включает стадию введения субъекту фармацевтической композиции в соответствии с восьмым аспектом изобретения.
Способ может включать следующие стадии:
(i) выделение T-клетки, как перечислено выше.
(ii) трансдукция или трансфекция T-клеток одной или несколькими последовательностями нуклеиновой кислоты, которые кодируют первый и второй CAR, или одним или несколькими векторами, которые содержат такую последовательность(последовательности) нуклеиновой кислоты; и
(iii) введение T-клеток из (ii) субъекту.
Заболевание может представлять собой злокачественную опухоль.
В десятом аспекте настоящее изобретение предусматривает фармацевтическую композицию в соответствии с восьмым аспектом изобретения для использования в лечении и/или предотвращении заболевания.
Заболевание может представлять собой злокачественную опухоль.
В одиннадцатом аспекте настоящее изобретение предусматривает использование T-клетки в соответствии с первым аспектом изобретения при изготовлении лекарственного средства для лечения и/или предотвращения заболевания.
Заболевание может представлять собой злокачественную опухоль.
Настоящее изобретение также относится к последовательности нуклеиновой кислоты, которая содержит:
a) первую нуклеотидную последовательность, которая кодирует первый химерный антигенный рецептор (CAR);
b) вторую нуклеотидную последовательность, которая кодирует второй CAR;
c) последовательность, которая кодирует саморасщепляющийся пептид расположенный между первой и второй нуклеотидными последовательностями, так что два CAR экспрессируют в виде отдельных объектов.
Альтернативные кодоны можно использовать в одной или нескольких частях первой и второй нуклеотидных последовательностях в областях, которые кодируют одинаковые или схожие аминокислотные последовательности.
Настоящее изобретение также относится к вектору и клетке, которые содержат такую нуклеиновую кислоту.
Логический элемент И на основе кинетической сегрегации по настоящему изобретению дает значительное техническое преимущество относительно ранее описанных «совместных CAR», т. е. подход двойного направленного воздействия, в котором два антигена распознаются двумя CAR, которые подают или активирующий или костимулирующий сигнал T-клетке.
При использовании подхода с совместными CAR, несмотря на то, что наибольшую активность можно ожидать в отношении клеток-мишеней, несущих оба антигена, можно ожидать существенную активность в отношении тканей, несущих только антиген, распознаваемый с помощью активирующего CAR. Можно ожидать, что эта активность должна быть по меньшей мере таковой CAR первого поколения. CAR первого поколения приводили к существенной токсичности: например, билиарную токсичность наблюдали при клиническом тестировании CAR первого поколения, распознающего карбоангидразу IX, которая неожиданно была экспрессирована на билиарном эпителии (ссылка на Rotterdam). Стоит отметить, что терминально дифференцированные эффекторы не требуют или не отвечают на костимуляторные сигналы, так что любые терминально дифференцированные CAR T-клетки будут действовать максимально, несмотря на отсутствие костимуляторного CAR сигнала.
Кроме того, костимуляторные сигналы ведут к длительным эффектам, оказываемым на популяцию T-клеток. Эти эффекты длятся дольше, чем взаимодействие в синапсе T-клетка/мишень. Следовательно, CAR T-клетки, которые становятся полностью активированными в опухоли и мигрируют, могут иметь максимально высокую активность в отношении нормальных тканей, несущих один антиген. Этот эффект «выхода за пределы» может быть наиболее выражен в тканях внутри или вблизи опухоли или в которые дренируется опухоль. Фактически, предложены и тестированы стратегии на основании концепции активности CAR первого поколения, усиливающейся костимуляторными сигналами, не связаны с CAR активацией, но опосредованы отдельным рецептором (Rossig, Blood. 2002 Mar 15;99(6):2009-16.).
Таким образом, можно ожидать, что подход с совместными CAR даст в лучшем случае снижение, но не устранение токсичности в отношении нормальной ткани, экспрессирующей одиночный антиген. В настоящем изобретении используют кинетическую сегрегацию в иммунологическом синапсе, образованном между T-клеткой и клеткой-мишенью, чтобы регулировать запуск самой T-клетки. Следовательно, достигают абсолютно строгого контроля запуска в отсутствие второго антигена. Таким образом, вся активация T-клеток ограничена клетками-мишенями, экспрессирующими оба антигена, логический элемент И должен функционировать независимо от типа эффекторной клетки или состояния дифференциации и невозможен эффект выходалогического элемента И за пределы активации T-клеток.
ДОПОЛНИТЕЛЬНЫЕ АСПЕКТЫ ИЗОБРЕТЕНИЯ
Настоящее изобретение также относится к аспектам, перечисленным в следующих номерных абзацах:
1. T-клетка, которая совместно экспрессирует первый химерный антигенный рецептор (CAR) и второй CAR на клеточной поверхности, каждый CAR содержит:
(i) антигенсвязывающий домен;
(ii) спейсер;
(iii) трансмембранный домен; и
(iv) эндодомен
в которых антигенсвязывающие домены первого и второго CAR связываются с различными антигенами, в которых спейсер первого CAR отличается от спейсера второго CAR и в которых один из первого или второго CAR представляет собой активирующий CAR, который содержит активирующий эндодомен, и другой CAR представляет собой или активирующий CAR, который содержит активирующий эндодомен, или ингибирующий CAR, который содержит включаемый лигированием или выключаемый лигированием ингибирующий эндодомен.
2. T-клетка в соответствии с п. 1, в которой спейсер первого CAR имеет длину и/или заряд и/или размер и/или конфигурацию и/или гликозилирование, отличающиеся от спейсера второго CAR, так что когда первый CAR и второй CAR связывают соответствующие им антигены-мишени, первый CAR и второй CAR становятся пространственно разделенными на мембране T-клетки.
3. T-клетка в соответствии с п. 2, в которой или первый спейсер или второй спейсер содержит «стебель» CD8 и другой спейсер содержит домен шарнира, CH2 и CH3 из IgG1.
4. T-клетка в соответствии с п. 1, в которой как первый, так и второй CAR представляют собой активирующие CAR.
5. T-клетка в соответствии с п. 4, в которой один CAR связывает CD19 и другой CAR связывает CD20.
6. T-клетка в соответствии с п. 2 или 3, в которой один из первого или второго CAR представляет собой активирующий CAR, который содержит активирующий эндодомен, и другой CAR представляет собой ингибирующий CAR, который содержит выключаемый лигированием ингибирующий эндодомен, этот ингибирующий CAR ингибирует активацию T-клеток с помощью активирующего CAR в отсутствие лигирования ингибирующего CAR, но не значительно ингибирует активацию T-клеток с помощью активирующего CAR, когда ингибирующий CAR лигирован.
7. T-клетка в соответствии с п. 6, в которой ингибирующий эндодомен содержит весь эндодомен из CD148 или CD45 или его часть.
8. T-клетка в соответствии с п. 6 или 7, в которой антигенсвязывающий домен первого CAR связывает CD5 и антигенсвязывающий домен второго CAR связывает CD19.
9. T-клетка в соответствии с п. 1, в которой первый и второй спейсеры достаточно различны с тем, чтобы предотвращать образование перекрестных пар первого и второго CAR, но достаточно схожи, чтобы это приводило к совместной локализации первого и второго CAR после лигирования.
10. T-клетка в соответствии с п. 9, в которой один из первого или второго CAR представляет собой активирующий CAR, который содержит активирующий эндодомен, и другой CAR представляет собой ингибирующий CAR, который содержит включаемый лигированием ингибирующий эндодомен, этот ингибирующий CAR не значительно ингибирует активацию T-клеток с помощью активирующего CAR в отсутствие лигирования ингибирующего CAR, но ингибирует активацию T-клеток с помощью активирующего CAR, когда ингибирующий CAR лигирован.
11. T-клетка в соответствии с п. 10, в которой включаемый лигированием ингибирующий эндодомен содержит по меньшей мере часть фосфатазы.
12. T-клетка в соответствии с п. 11, в которой включаемый лигированием ингибирующий эндодомен содержит весь PTPN6 или его часть.
13. T-клетка в соответствии с п. 10, в которой включаемый лигированием ингибирующий эндодомен содержит по меньшей мере один ITIM-домен.
14. T-клетка в соответствии с п. 13, в которой активность включаемого лигированием ингибирующего эндодомена усиливают посредством коэкспрессии слитного белка PTPN6-CD45 или -CD148.
15. T-клетка в соответствии с любым из пп. с 10 до 14, в которой CAR, который содержит активирующий эндодомен, содержит антигенсвязывающий домен, который связывает CD33, и CAR, который содержит включаемый лигированием ингибирующий эндодомен, содержит антигенсвязывающий домен, который связывает CD34.
16. T-клетка, которая содержит больше чем два CAR, как определено в предшествующих пп., так что ее специфично стимулирует клетка, такая как T-клетка, несущая отличающий паттерн больше чем из двух антигенов.
17. Последовательность нуклеиновой кислоты, которая кодирует как первый, так и второй химерные антигенные рецепторы (CAR), как определено в любом из пп. с 1 до 16.
18. Последовательность нуклеиновой кислоты в соответствии с п. 17, которая имеет следующую структуру:
AgB1-спейсер1-TM1-эндо1-коэкспр-AbB2-спейсер2-TM2-эндо2
в которой
AgB1 представляет собой последовательность нуклеиновой кислоты, которая кодирует антигенсвязывающий домен первого CAR;
спейсер1 представляет собой последовательность нуклеиновой кислоты, которая кодирует спейсер первого CAR;
TM1 представляет собой последовательность нуклеиновой кислоты, которая кодирует трансмембранный домен первого CAR;
эндо1 представляет собой последовательность нуклеиновой кислоты, которая кодирует эндодомен первого CAR;
коэкспр представляет собой последовательность нуклеиновой кислоты, которая делает возможной коэкспрессию обоих CAR
AgB2 представляет собой последовательность нуклеиновой кислоты, которая кодирует антигенсвязывающий домен второго CAR;
спейсер2 представляет собой последовательность нуклеиновой кислоты, которая кодирует спейсер второго CAR;
TM2 представляет собой последовательность нуклеиновой кислоты, которая кодирует трансмембранный домен второго CAR;
эндо2 представляет собой последовательность нуклеиновой кислоты, которая кодирует эндодомен второго CAR;
эта последовательность нуклеиновой кислоты, когда ее экспрессируют в T-клетке, кодирует полипептид, который расщепляют в сайте расщепления так, что на поверхности T-клеток совместно экспрессируют первый и второй CAR.
19. Последовательность нуклеиновой кислоты в соответствии с п. 18, в которой коэкспр кодирует последовательность, которая содержит саморасщепляющийся пептид.
20. Последовательность нуклеиновой кислоты в соответствии с п. 18 или 19, в которой альтернативные кодоны используют в областях последовательности, которая кодирует одинаковые или схожие аминокислотные последовательности, во избежание гомологичной рекомбинации.
21. Набор, который содержит
(i) первую последовательность нуклеиновой кислоты, которая кодирует первый химерный антигенный рецептор (CAR), как определено в любом из пп. с 1 до 16, эта последовательность нуклеиновой кислоты имеет следующую структуру:
AgB1-спейсер1-TM1-эндо1
в которой
AgB1 представляет собой последовательность нуклеиновой кислоты, которая кодирует антигенсвязывающий домен первого CAR;
спейсер1 представляет собой последовательность нуклеиновой кислоты, которая кодирует спейсер первого CAR;
TM1 представляет собой последовательность нуклеиновой кислоты, которая кодирует трансмембранный домен первого CAR;
эндо1 представляет собой последовательность нуклеиновой кислоты, которая кодирует эндодомен первого CAR; и
(ii) вторую последовательность нуклеиновой кислоты, которая кодирует второй химерный антигенный рецептор (CAR), как определено в любом из пп. с 1 до 16, эта последовательность нуклеиновой кислоты имеет следующую структуру:
AgB2-спейсер2-TM2-эндо2
AgB2 представляет собой последовательность нуклеиновой кислоты, которая кодирует антигенсвязывающий домен второго CAR;
спейсер2 представляет собой последовательность нуклеиновой кислоты, которая кодирует спейсер второго CAR;
TM2 представляет собой последовательность нуклеиновой кислоты, которая кодирует трансмембранный домен второго CAR;
эндо2 представляет собой последовательность нуклеиновой кислоты, которая кодирует эндодомен второго CAR.
22. Набор, который содержит: первый вектор, который содержит первую последовательность нуклеиновой кислоты, как определено в п. 21; и второй вектор, который содержит первую последовательность нуклеиновой кислоты, как определено в п. 21.
23. Набор в соответствии с п. 22, в котором векторы представляют собой интегрирующие вирусные векторы или транспозоны.
24. Вектор, который содержит последовательность нуклеиновой кислоты в соответствии с любым из пп. с 17 до 20.
25. Ретровирусный вектор или лентивирусный вектор или транспозон в соответствии с п. 24.
26. Способ получения T-клетки в соответствии с любым из пп. с 1 до 16, который включает стадию введения: последовательности нуклеиновой кислоты в соответствии с любым из пп. с 17 до 20; первой последовательности нуклеиновой кислоты и второй последовательности нуклеиновой кислоты, как определено в п. 21; и/или первого вектора и второго вектора, как определено в п. 22, или вектора в соответствии с п. 24 или 25 в T-клетку.
27. Способ в соответствии с п. 24, в котором T-клетка происходит из образца, выделенного у субъекта.
28. Фармацевтическая композиция, которая содержит множество T-клеток в соответствии с любым из пп. с 1 до 16.
29. Способ лечения и/или предотвращения заболевания, который включает стадию введения фармацевтической композиции в соответствии с п. 28 субъекту.
30. Способ в соответствии с п. 29, который включает следующие стадии:
(i) выделение образца, содержащего T-клетки, у субъекта;
(ii) трансдукция или трансфекция T-клеток с использованием: последовательности нуклеиновой кислоты в соответствии с любым из пп. с 17 до 20; первой последовательности нуклеиновой кислоты и второй последовательности нуклеиновой кислоты, как определено в п. 21; первого вектора и второго вектора, как определено в п. 22 или 23, или вектора в соответствии с п. 24 или 25; и
(iii) введение T-клеток из (ii) субъекту.
31. Способ в соответствии с п. 29 или 30, в котором заболевание представляет собой злокачественную опухоль.
32. Фармацевтическая композиция в соответствии с п. 28 для использования в лечении и/или предотвращении заболевания.
33. Использование T-клетки в соответствии с любым из пп. с 1 до 16 при изготовлении лекарственного средства для лечения и/или предотвращения заболевания.
ПОДРОБНОЕ ОПИСАНИЕ
ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ (CAR)
CAR, которые схематически представлены на фиг. 1, представляют собой химерные трансмембранные белки I типа, которые соединяют внеклеточный антигенраспознающий домен (связывающее средство) с внутриклеточным сигнальным доменом (эндодомен). Связывающее средство типично представляет собой одноцепочечный вариабельный фрагмент (scFv), полученный из моноклонального антитела (mAb), но оно может быть основано на других форматах, которые содержат антителоподобный антигенсвязывающий участок. Спейсерный домен обычно необходим для того, чтобы отделять связывающее средство от мембраны и сделать возможной его подходящую ориентацию. Обыкновенный используемый спейсерный домен представляет собой Fc из IgG1. Может быть достаточно более компактного спейсера, например, «стебля» из CD8α и даже просто только шарнира IgG1, в зависимости от антигена. Трансмембранный домен заякоривает белок в клеточной мембране и соединяет спейсер с эндодоменом.
Ранние конструкции CAR содержали эндодомены, полученные из внутриклеточных частей или γ цепи из FcεR1 или CD3ξ. Следовательно, эти рецепторы первого поколения передавали иммунологический сигнал 1, которого было достаточно для того, чтобы запускать уничтожение T-клетками когнатных клеток-мишеней, но не достаточно для того, чтобы полностью активировать T-клетку для пролиферации и выживания. Чтобы преодолеть это ограничение, сконструированы составные эндодомены: слияние внутриклеточной части из костимуляторной молекулы T-клеткы с таковой из CD3ξ дает рецепторы второго поколения, которые могут передавать активирующий и костимуляторный сигнал одновременно после распознавания антигена. Наиболее широко используют костимуляторный домен из CD28. Это обеспечивает наиболее сильный костимуляторный сигнал, а именно, иммунологический сигнал 2, который запускает T-клеточную пролиферацию. Также описаны некоторые рецепторы, которые содержат эндодомены семейства рецепторов TNF, такие как близкородственные OX40 и 41BB, которые передают сигналы выживания. Сейчас даже описаны еще более сильные CAR третьего поколения, которые имеют эндодомены, способные передавать сигналы активации, пролиферации и выживания.
Кодирующие CAR нуклеиновые кислоты можно переносить в T-клетки с использованием, например, ретровирусных векторов. Можно использовать лентивирусные векторы. Таким образом, большое число T-клеток со специфичностью к злокачественной опухоли можно создавать для адоптивного переноса клеток. Когда CAR связывает антиген-мишень, это ведет к передаче активирующего сигнала в T-клетку, на которой он экспрессирован. Таким образом CAR определяет специфичность и цитотоксичность T-клетки в направлении опухолевых клеток, которые экспрессируют антиген-мишень.
Первый аспект изобретения относится к T-клетке, которая совместно экспрессирует первый CAR и второй CAR так, что T-клетка может распознавать желаемый паттерн экспрессии на клетках-мишенях наподобие логического элемента, как подробно изложено в таблицах истинности: таблица 1, 2 и 3.
И первый и второй (и необязательно последующие) CAR содержат:
(i) антигенсвязывающий домен;
(ii) спейсер;
(iii) трансмембранный домен; и
(iii) внутриклеточный домен.
Таблица истинности для CAR логического элемента ИЛИ
Таблица истинности для CAR логического элемента И
Таблица истинности для CAR логического элемента И НЕ
Первый и второй CAR T-клетки по настоящему изобретению можно получать в виде полипептида, который содержит оба CAR, вместе с сайтом расщепления.
В SEQ ID № с 1 до 5 приведены примеры таких полипептидов, каждый из которых содержит два CAR. Следовательно, CAR может содержать одну или другую часть следующих аминокислотных последовательностей, которые соответствуют одному CAR.
SEQ ID № 1 представляет собой CAR логический элемент ИЛИ, который распознает CD19 ИЛИ CD33
SEQ ID № 2 представляет собой CAR логический элемент И, который распознает CD19 И CD33 с использованием фосфатазы CD148
SEQ ID № 3 представляет собой альтернативную реализацию CAR логического элемента И, который распознает CD19 И CD33, который использует фосфатазу CD45
SEQ ID № 4 представляет собой CAR логический элемент И НЕ, который распознает CD19 И НЕ CD33 на основании фосфатазы PTPN6
SEQ ID № 5 представляет собой альтернативную реализацию CAR логического элемента И НЕ, который распознает CD19 И НЕ CD33 и основан на ITIM-содержащем эндодомене из LAIR1
SEQ ID № 6 представляет собой дополнительную альтернативную реализацию CAR логического элемента И НЕ, который распознает CD19 И НЕ CD33 и рекрутирует слитный белок PTPN6-CD148 в эндодомен, содержащий ITIM.
SEQ ID № 1
MSLPVTALLLPLALLLHAARPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITKAGGGGSGGGGSGGGGSGGGGSEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSDPTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIFWVLVVVGGVLACYSLLVTVAFIIFWVRRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRRAEGRGSLLTCGDVEENPGPMAVPTQVLGLLLLWLTDARCDIQMTQSPSSLSASVGDRVTITCRASEDIYFNLVWYQQKPGKAPKLLIYDTNRLADGVPSRFSGSGSGTQYTLTISSLQPEDFATYYCQHYKNYPLTFGQGTKLEIKRSGGGGSGGGGSGGGGSGGGGSRSEVQLVESGGGLVQPGGSLRLSCAASGFTLSNYGMHWIRQAPGKGLEWVSSISLNGGSTYYRDSVKGRFTISRDNAKSTLYLQMNSLRAEDTAVYYCAAQDAYTGGYFDYWGQGTLVTVSSMDPAEPKSPDKTHTCPPCPAPPVAGPSVFLFPPKPKDTLMIARTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGKKDPKFWVLVVVGGVLACYSLLVTVAFIIFWVRSRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR
SEQ ID № 2
MSLPVTALLLPLALLLHAARPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITKAGGGGSGGGGSGGGGSGGGGSEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSDPTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIFWVLVVVGGVLACYSLLVTVAFIIFWVRRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRRAEGRGSLLTCGDVEENPGPMAVPTQVLGLLLLWLTDARCDIQMTQSPSSLSASVGDRVTITCRASEDIYFNLVWYQQKPGKAPKLLIYDTNRLADGVPSRFSGSGSGTQYTLTISSLQPEDFATYYCQHYKNYPLTFGQGTKLEIKRSGGGGSGGGGSGGGGSGGGGSRSEVQLVESGGGLVQPGGSLRLSCAASGFTLSNYGMHWIRQAPGKGLEWVSSISLNGGSTYYRDSVKGRFTISRDNAKSTLYLQMNSLRAEDTAVYYCAAQDAYTGGYFDYWGQGTLVTVSSMDPAEPKSPDKTHTCPPCPAPPVAGPSVFLFPPKPKDTLMIARTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGKKDPKAVFGCIFGALVIVTVGGFIFWRKKRKDAKNNEVSFSQIKPKKSKLIRVENFEAYFKKQQADSNCGFAEEYEDLKLVGISQPKYAAELAENRGKNRYNNVLPYDISRVKLSVQTHSTDDYINANYMPGYHSKKDFIATQGPLPNTLKDFWRMVWEKNVYAIIMLTKCVEQGRTKCEEYWPSKQAQDYGDITVAMTSEIVLPEWTIRDFTVKNIQTSESHPLRQFHFTSWPDHGVPDTTDLLINFRYLVRDYMKQSPPESPILVHCSAGVGRTGTFIAIDRLIYQIENENTVDVYGIVYDLRMHRPLMVQTEDQYVFLNQCVLDIVRSQKDSKVDLIYQNTTAMTIYENLAPVTTFGKTNGYIA
SEQ ID № 3
MSLPVTALLLPLALLLHAARPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITKAGGGGSGGGGSGGGGSGGGGSEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSDPTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIFWVLVVVGGVLACYSLLVTVAFIIFWVRRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRRAEGRGSLLTCGDVEENPGPMAVPTQVLGLLLLWLTDARCDIQMTQSPSSLSASVGDRVTITCRASEDIYFNLVWYQQKPGKAPKLLIYDTNRLADGVPSRFSGSGSGTQYTLTISSLQPEDFATYYCQHYKNYPLTFGQGTKLEIKRSGGGGSGGGGSGGGGSGGGGSRSEVQLVESGGGLVQPGGSLRLSCAASGFTLSNYGMHWIRQAPGKGLEWVSSISLNGGSTYYRDSVKGRFTISRDNAKSTLYLQMNSLRAEDTAVYYCAAQDAYTGGYFDYWGQGTLVTVSSMDPAEPKSPDKTHTCPPCPAPPVAGPSVFLFPPKPKDTLMIARTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGKKDPKALIAFLAFLIIVTSIALLVVLYKIYDLHKKRSCNLDEQQELVERDDEKQLMNVEPIHADILLETYKRKIADEGRLFLAEFQSIPRVFSKFPIKEARKPFNQNKNRYVDILPYDYNRVELSEINGDAGSNYINASYIDGFKEPRKYIAAQGPRDETVDDFWRMIWEQKATVIVMVTRCEEGNRNKCAEYWPSMEEGTRAFGDVVVKINQHKRCPDYIIQKLNIVNKKEKATGREVTHIQFTSWPDHGVPEDPHLLLKLRRRVNAFSNFFSGPIVVHCSAGVGRTGTYIGIDAMLEGLEAENKVDVYGYVVKLRRQRCLMVQVEAQYILIHQALVEYNQFGETEVNLSELHPYLHNMKKRDPPSEPSPLEAEFQRLPSYRSWRTQHIGNQEENKSKNRNSNVIPYDYNRVPLKHELEMSKESEHDSDESSDDDSDSEEPSKYINASFIMSYWKPEVMIAAQGPLKETIGDFWQMIFQRKVKVIVMLTELKHGDQEICAQYWGEGKQTYGDIEVDLKDTDKSSTYTLRVFELRHSKRKDSRTVYQYQYTNWSVEQLPAEPKELISMIQVVKQKLPQKNSSEGNKHHKSTPLLIHCRDGSQQTGIFCALLNLLESAETEEVVDIFQVVKALRKARPGMVSTFEQYQFLYDVIASTYPAQNGQVKKNNHQEDKIEFDNEVDKVKQDANCVNPLGAPEKLPEAKEQAEGSEPTSGTEGPEHSVNGPASPALNQGS
SEQ ID № 4
MSLPVTALLLPLALLLHAARPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITKAGGGGSGGGGSGGGGSGGGGSEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSDPTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIFWVLVVVGGVLACYSLLVTVAFIIFWVRRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRRAEGRGSLLTCGDVEENPGPMAVPTQVLGLLLLWLTDARCDIQMTQSPSSLSASVGDRVTITCRASEDIYFNLVWYQQKPGKAPKLLIYDTNRLADGVPSRFSGSGSGTQYTLTISSLQPEDFATYYCQHYKNYPLTFGQGTKLEIKRSGGGGSGGGGSGGGGSGGGGSRSEVQLVESGGGLVQPGGSLRLSCAASGFTLSNYGMHWIRQAPGKGLEWVSSISLNGGSTYYRDSVKGRFTISRDNAKSTLYLQMNSLRAEDTAVYYCAAQDAYTGGYFDYWGQGTLVTVSSMDPATTTKPVLRTPSPVHPTGTSQPQRPEDCRPRGSVKGTGLDFACDIYWAPLAGICVALLLSLIITLICYHRSRKRVCKSGGGSFWEEFESLQKQEVKNLHQRLEGQRPENKGKNRYKNILPFDHSRVILQGRDSNIPGSDYINANYIKNQLLGPDENAKTYIASQGCLEATVNDFWQMAWQENSRVIVMTTREVEKGRNKCVPYWPEVGMQRAYGPYSVTNCGEHDTTEYKLRTLQVSPLDNGDLIREIWHYQYLSWPDHGVPSEPGGVLSFLDQINQRQESLPHAGPIIVHCSAGIGRTGTIIVIDMLMENISTKGLDCDIDIQKTIQMVRAQRSGMVQTEAQYKFIYVAIAQFIETTKKKL
SEQ ID № 5
MSLPVTALLLPLALLLHAARPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITKAGGGGSGGGGSGGGGSGGGGSEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSDPTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIFWVLVVVGGVLACYSLLVTVAFIIFWVRRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRRAEGRGSLLTCGDVEENPGPMAVPTQVLGLLLLWLTDARCDIQMTQSPSSLSASVGDRVTITCRASEDIYFNLVWYQQKPGKAPKLLIYDTNRLADGVPSRFSGSGSGTQYTLTISSLQPEDFATYYCQHYKNYPLTFGQGTKLEIKRSGGGGSGGGGSGGGGSGGGGSRSEVQLVESGGGLVQPGGSLRLSCAASGFTLSNYGMHWIRQAPGKGLEWVSSISLNGGSTYYRDSVKGRFTISRDNAKSTLYLQMNSLRAEDTAVYYCAAQDAYTGGYFDYWGQGTLVTVSSMDPATTTKPVLRTPSPVHPTGTSQPQRPEDCRPRGSVKGTGLDFACDILIGVSVVFLFCLLLLVLFCLHRQNQIKQGPPRSKDEEQKPQQRPDLAVDVLERTADKATVNGLPEKDRETDTSALAAGSSQEVTYAQLDHWALTQRTARAVSPQSTKPMAESITYAAVARH
SEQ ID № 6
MSLPVTALLLPLALLLHAARPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITKAGGGGSGGGGSGGGGSGGGGSEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSDPTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIFWVLVVVGGVLACYSLLVTVAFIIFWVRRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRRAEGRGSLLTCGDVEENPGPMAVPTQVLGLLLLWLTDARCDIQMTQSPSSLSASVGDRVTITCRASEDIYFNLVWYQQKPGKAPKLLIYDTNRLADGVPSRFSGSGSGTQYTLTISSLQPEDFATYYCQHYKNYPLTFGQGTKLEIKRSGGGGSGGGGSGGGGSGGGGSRSEVQLVESGGGLVQPGGSLRLSCAASGFTLSNYGMHWIRQAPGKGLEWVSSISLNGGSTYYRDSVKGRFTISRDNAKSTLYLQMNSLRAEDTAVYYCAAQDAYTGGYFDYWGQGTLVTVSSMDPATTTKPVLRTPSPVHPTGTSQPQRPEDCRPRGSVKGTGLDFACDILIGVSVVFLFCLLLLVLFCLHRQNQIKQGPPRSKDEEQKPQQRPDLAVDVLERTADKATVNGLPEKDRETDTSALAAGSSQEVTYAQLDHWALTQRTARAVSPQSTKPMAESITYAAVARHRAEGRGSLLTCGDVEENPGPWYHGHMSGGQAETLLQAKGEPWTFLVRESLSQPGDFVLSVLSDQPKAGPGSPLRVTHIKVMCEGGRYTVGGLETFDSLTDLVEHFKKTGIEEASGAFVYLRQPYSGGGGSFEAYFKKQQADSNCGFAEEYEDLKLVGISQPKYAAELAENRGKNRYNNVLPYDISRVKLSVQTHSTDDYINANYMPGYHSKKDFIATQGPLPNTLKDFWRMVWEKNVYAIIMLTKCVEQGRTKCEEYWPSKQAQDYGDITVAMTSEIVLPEWTIRDFTVKNIQTSESHPLRQFHFTSWPDHGVPDTTDLLINFRYLVRDYMKQSPPESPILVHCSAGVGRTGTFIAIDRLIYQIENENTVDVYGIVYDLRMHRPLMVQTEDQYVFLNQCVLDIVRSQKDSKVDLIYQNTTAMTIYENLAPVTTFGKTNGYIASGS
CAR может содержать вариант CAR-кодирующей части последовательности, представленной в качестве SEQ ID № 1, 2, 3, 4, 5 или 6, которая имеет по меньшей мере 80, 85, 90, 95, 98 или 99% идентичность последовательностей, при условии, что вариант последовательности представляет собой CAR, который обладает необходимыми свойствами.
Способы выравнивания последовательностей хорошо известны в данной области, и их реализуют с использованием подходящих программ для выравнивания. Процент идентичности последовательностей относится к процентной доле остатков аминокислот или нуклеотид, которые идентичны в двую последовательностях, когда они выровнены оптимально. Гомологию или идентичность нуклеотидных и белковых последовательностей можно определять с использованием стандартных алгоритмов, таких как программа BLAST (Basic Local Alignment Search Tool в National Center for Biotechnology Information) с параметрами по умолчанию, которая общедоступна на http://blast.ncbi.nlm.nih.gov. Другие алгоритмы для определения идентичности или гомологии последовательностей включают: LALIGN (http://www.ebi.ac.uk/Tools/psa/lalign/ и http://www.ebi.ac.uk/Tools/psa/lalign/nucleotide.html), AMAS (Analysis of Multiply Aligned Sequences, http://www.compbio.dundee.ac.uk/Software/Amas/amas.html), FASTA (http://www.ebi.ac.uk/Tools/sss/fasta/), Clustal Omega (http://www.ebi.ac.uk/Tools/msa/clustalo/), SIM (http://web.expasy.org/sim/) и EMBOSS Needle (http://www.ebi.ac.uk/Tools/psa/emboss_needle/nucleotide.html).
CAR ЛОГИЧЕСКИЙ ЭЛЕМЕНТ ИЛИ
В этом варианте осуществления антигенсвязывающие домены первого и второго CAR по настоящему изобретению связываются с различными антигенами, и оба CAR содержат активирующий эндодомен. Оба CAR имеют различные спейсерные домены для того, чтобы предотвращать образование перекрестных пар из двух различных рецепторов. Таким образом, можно конструировать T-клетки для того, чтобы активация происходила при распознавании любого или обоих антигенов. Это можно использовать в области онкологии, как показан в гипотизе Goldie-Coldman: одиночное направленное воздействие на один антиген может вести к ускользанию опухоли посредством модуляции указанного антигена из-за высокой частоты мутаций, свойственной большинству злокачественных опухолей. Посредством одновременного направленного воздействия на два антигена вероятность такого ускользания снижается экспоненциально.
Известны различные опухоль-ассоциированные антигены, как показано в следующей таблице 4. Для данного заболевания первый CAR и второй CAR может связываться с двумя различными TAA, связанными с этим заболеванием. Таким образом, предотвращено ускользание опухоли посредством модуляции одного антигена, поскольку на второй антиген также воздействуют направленно. Например, при направленном воздействии на B-клеточное злокачественное новообразование, можно одновременно направленно воздействовать на CD19 и CD20. В этом варианте осуществления важно, что два CAR не образуют гетеродимеров.
МОДЕЛЬ КИНЕТИЧЕСКОЙ СЕГРЕГАЦИИ
Последующее образование пар CAR для создания логического элемента И и логического элемента И НЕ основано на модели кинетической сегрегации (KS) активации T-клеток. Она представляет собой функциональную модель, подтвержденную экспериментальными данными, которая объясняет, как распознавание антигена T-клеточным рецептором превращается в нисходящие сигналы активации. В кратком изложении: в основном состоянии сигнальные компоненты на мембране T-клетки находятся в динамическом гомеостазе, за счет чего дефосфорилированные ITAM предпочтительнее фосфорилированных ITAM. Это обусловлено более высокой активностью трансмембранного CD45/фосфатазы CD148, чем у киназ, связанных с мембраной, таких как lck. Когда T-клетка связывается с клеткой-мишенью через распознавание когнатного антигена T-клеточным рецептором (или CAR), образуются плотные иммунологические синапсы. Такое тесное расположение T-клетки и мембран-мишеней исключает CD45/CD148 из-за их больших эктодоменов, которые не могут поместиться в синапсе. Сегрегация высокой концентрации ассоциированных с T-клеточным рецептором ITAM и киназ в синапсе, в отсутствие фосфатазы, ведет к состоянию, в котором предпочтительны фосфорилированные ITAM. ZAP70 распознает порог фосфорилированных ITAM и распространяет сигнал активации T-клеток. Это глубокое понимание активации T-клеток используют в настоящем изобретении. В частности, изобретение основано на этом понимании того, как эктодомены с различной длиной и/или объемом и/или зарядом и/или конфигурацией и/или гликозилированием ведут к дифференциальной сегрегации при формировании синапса.
CAR ЛОГИЧЕСКИЙ ЭЛЕМЕНТ И
В этом варианте осуществления один CAR содержит активирующий эндодомен и один CAR содержит ингибирующий эндодомен, посредством чего ингибирующий CAR конститутивно ингибирует первый активирующий CAR, но при распознавании своего когнатного антигена, снимает свое ингибирование активирующего CAR. Таким образом, можно конструировать T-клетку для запуска, только если клетка-мишень экспрессирует оба когнатных антигена. Этого поведения достигают с помощью активирующего CAR, который содержит активирующий эндодомен, содержащий ITAM-домены, например эндодомен CD3 дзета, и ингибирующего CAR, который содержит эндодомен из фосфатазы, способный дефосфорилировать ITAM (например, CD45 или CD148). Самое главное, спейсерные домены обоих CAR значительно отличаются по размеру и/или геометрической форме и/или заряду и т.д. Когда лигирован только активирующий CAR, ингибирующий CAR находится в растворе на поверхности T-клетки и может диффундировать в и из синапса, ингибируя активирующий CAR. Когда лигированы оба CAR, из-за различий в свойствах спейсера, активирующий и ингибирующий CAR дифференциально сегрегированны, что позволяет активирующему CAR запускать активацию T-клеток без пространственного затруднения со стороны ингибирующего CAR.
Это обладает существенной полезностью в области терапии злокачественных опухолей. В настоящее время иммунотерапия типично направлена на один антиген. Большинство злокачественных опухолей нельзя отличить от нормальных тканей на основании одного антигена. Таким образом, имеет место существенная токсичность «в мишени вне опухоли», за счет которой при терапии происходит повреждение нормальных тканей. Например, при направленном воздействии на CD20 для того, чтобы лечить B-клеточные лимфомы ритуксимабом, происходит истощение всего компартмента нормальных B-клеток. Например, при направленном воздействии на CD52 для того, чтобы лечить хронический лимфоцитарный лейкоз, происходит истощение всего лимфоидного компартмента. Например, при направленном воздействии на CD33 для того, чтобы лечить острый миелолейкоз, происходит истощение всего миелоидного компартмента и т.д. С помощью ограничения активности парой антигенов можно разработать значительно более точное направленное воздействие и, таким образом, менее токсичную терапию. Практическим примером является направленное воздействие на CLL, который экспрессирует и CD5 и CD19. Только небольшая доля нормальных B-клеток экспрессирует оба антигена, так что токсичность вне мишени при направленном воздействии на оба антигена с использованием логического элемента И по существу меньше, чем индивидуальное направленное воздействие на каждый антиген.
Конструкция по настоящему изобретению представляет собой существенное усовершенствование предыдущей реализации, как описано у Wilkie et al. ((2012). J. Clin. Immunol. 32, 1059-1070) и затем протестировано in vivo (Kloss et al (2013) Nat. Biotechnol. 31, 71-75). В этой реализации первый CAR содержит активирующий эндодомен, а второй - костимуляторный домен. Таким образом, T-клетка получает только активирующий и костимуляторный сигнал, когда присутствуют оба антигена. Однако T-клетка все же будет активировать в присутствии только первого антигена, что ведет к возможной токсичности вне мишени. Кроме того, реализация по настоящему изобретению допускает множественные составные сцепленные логические элементы, посредством которых клетка может интерпретировать комплексный паттерн антигенов.
CAR ЛОГИЧЕСКИЙ ЭЛЕМЕНТ И НЕ
В этом варианте осуществления один CAR содержит активирующий эндодомен и один CAR содержит ингибирующий эндодомен так, что этот ингибирующий CAR активен только когда он распознает свой когнатный антиген. Таким образом, T-клетка, сконструированная таким образом, активируется в ответ на присутствие только первого антигена, но не активируется, когда присутствуют оба антигена. Это изобретение реализовано с помощью ингибирующих CAR со спейсером, который может быть локализован совместно с первым CAR, но или фосфатазная активность ингибирующего CAR не должна быть настолько высокой, чтобы он ингибировал в растворе, или ингибирующий эндодомен фактически рекрутирует фосфатазу только когда ингибирующий CAR распознает свою когнатную мишень. В настоящем документе такие эндодомены называют «включаемыми лигированием» или полуингибирующими.
Это изобретение полезно для уточнения направленного воздействия, когда опухоль можно отличать от нормальной ткани по присутствию опухоль-ассоциированного антигена и утрате антигена, экспрессированного на нормальной ткани. Логический элемент И НЕ обладает существенной полезностью в области онкологии, поскольку он делает возможным направленное воздействие на антиген, который экспрессирует нормальная клетка, эта нормальная клетка также экспрессирует антиген, распознаваемый с помощью CAR, который содержит активирующий эндодомен. Примером такго антигена является CD33, который экспрессируют нормальные стволовые клетки и клетки острого миелолейкоза (AML). CD34 экспрессируют стволовые клетки, но типично не экспрессируют клетки AML. T-клетка, распознающая CD33 И НЕ CD34, будет вести к разрушению клеток лейкоза, но беречь нормальные стволовые клетки.
Возможные пары антигенов для использования с логическими элементами И НЕ приведены в таблице 6.
СОСТАВНЫЕ ЛОГИЧЕСКИЕ ЭЛЕМЕНТЫ
Модель кинетической сегрегации с вышеуказанными компонентами допускает создание составных логических элементов, например, T-клетку, которая запускается в ответ на паттерны из больше чем двух антигенов-мишеней. Например, возможно создать T-клетку, которая запускается только когда присутствуют три антигена (A И B И C). Здесь клетка экспрессирует три CAR, каждый распознает антигены A, B и C. Один CAR является возбуждающим, а два являются ингибирующими, где каждый CAR имеет спейсерные домены, которые ведут к дифференциальной сегрегации. Только когда все три лигированы, T-клетка будет активна. Дополнительный пример: (A ИЛИ B) И C: здесь CAR, распознающие антигены A и B, являются активирующими и имеют спейсеры которые могут быть совместно локализованы, тогда как CAR, распознающий антиген C, является ингибирующим и имеет спейсер, результатом чего является другая совместная сегрегация. Дополнительный пример (A И НЕ B) И C: здесь CAR против антигена A имеет активирующий эндодомен и совместно локализован с CAR против антигена B, который имеет эндодомен, ингибирующий при определенных условиях. CAR против антигена C имеет спейсер, который сегрегируется отлично от A или B, и является ингибирующим. Фактически, с использованием этих простых компонентов по изобретению и любого числа CAR и спейсеров можно программировать даже более сложную булеву логика.
СИГНАЛЬНЫЙ ПЕПТИД
CAR T-клетки по настоящему изобретению могут содержать сигнальный пептид с тем, чтобы когда CAR экспрессируют внутри клетки, такой как T-клетка, направлять появляющийся белок на эндоплазматический ретикулум и впоследствии на клеточную поверхность, где он будет экспрессирован.
Сердцевина сигнального пептида может содержать длинный участок гидрофобных аминокислот, который имеет склонность к формированию одной альфа-спирали. Сигнальный пептид может начинаться коротким положительно заряженным участком аминокислот, который помогает укреплять надлежащую топологию полипептида во время транслокации. На конце сигнального пептида типично имеет место участок аминокислот, которые распознает и отщепляет сигнальная пептидаза. Сигнальная пептидаза может осуществлять отщепление или во время транслокации или по ее завершении, чтобы создавать свободный сигнальный пептид и зрелый белок. После этого специфичные протеазы расщепляют свободные сигнальные пептиды.
Сигнальный пептид может находиться на амино-конце молекулы.
Сигнальный пептид может содержать SEQ ID № 7, 8 или 9 или их вариант, который имеет 5, 4, 3, 2 или 1 аминокислотную мутацию (инсерцию, замену или вставку) при условии, что сигнальный пептид все еще функционирует, обеспечивая экспрессию CAR на клеточной поверхности.
SEQ ID № 7: MGTSLLCWMALCLLGADHADG
Сигнальный пептид SEQ ID № 7 является компактным и высоко эффективным. Предсказано, что он дает расщепление приблизительно 95% после концевого глицина, давая эффективное удаление сигнальной пептидазой.
SEQ ID № 8: MSLPVTALLLPLALLLHAARP
Сигнальный пептид SEQ ID № 8 получают из IgG1.
SEQ ID № 9: MAVPTQVLGLLLLWLTDARC
Сигнальный пептид SEQ ID № 9 получают из CD8.
Сигнальный пептид для первого CAR может иметь последовательность, отличную от сигнального пептида из второго CAR (и от третьего CAR и четвертого CAR и т. п.).
АНТИГЕНСВЯЗЫВАЮЩИЙ ДОМЕН
Антигенсвязывающий домен представляет собой часть CAR, которая распознает антиген. Многие антигенсвязывающие домены известны в данной области, в том числе те, которые основаны на антигенсвязывающем участке антитела, миметике антитела и T-клеточных рецепторах. Например, антигенсвязывающий домен может содержать: одноцепочечный вариабельный фрагмент (scFv), полученный из моноклонального антитела; природный лиганд антигена-мишени; пептид с достаточной аффинностью к мишени; однодоменное антитело; искусственное одиночное связывающее средство, такое как Darpin (сконструированный белок с анкириновым повтором); или одна цепь, полученная из T-клеточного рецептора.
Антигенсвязывающий домен может содержать домен, который не основан на антигенсвязывающем участке антитела. Например, антигенсвязывающий домен может содержать домен на основании белка/пептида, который представляет собой растворимый лиганд для рецептора клеточной поверхности опухоли (например, растворимый пептид, такой как цитокин или хемокин); или внеклеточный домен лиганда или рецептора, заякоренного в мембране, для которого вторая часть пары связывания экспрессирована на опухолевой клетке.
Примеры с 11 до 13 относятся к CAR, который связывает BCMA, в котором антигенсвязывающий домен содержит APRIL, лиганд для BCMA.
Антигенсвязывающий домен может быть основан на природном лиганде антигена.
Антигенсвязывающий домен может содержать аффинный пептид из комбинаторной библиотеки или сконструированный de novo аффинный белок/пептид.
СПЕЙСЕРНЫЙ ДОМЕН
CAR содержит спейсерную последовательность для соединения антигенсвязывающего домена с трансмембранным доменом и пространственного отделения антигенсвязывающего домен от эндодомена. Гибкий спейсер позволяет антигенсвязывающему домену ориентироваться в различных направлениях для того, чтобы содействовать связыванию.
В T-клетке по настоящему изобретению первый и второй CAR содержат различные спейсерные молекулы. Например, спейсерная последовательность, например, может содержать Fc-область IgG1, шарнир IgG1 или «стебель» CD8 человека или «стебель» CD8 мыши. Спейсер альтернативно может содержать альтернативную линкерную последовательность, которая имеет схожую длину и/или свойства разнесения доменов, как у Fc-области IgG1, шарнира IgG1 или «стебля» CD8. Спейсер IgG1 человека можно изменять для того, чтобы удалять мотивы связывания Fc.
Примеры аминокислотных последовательностей для этих спейсеров приведены ниже:
SEQ ID № 10 (шарнир-CH2CH3 из IgG1 человека)
AEPKSPDKTHTCPPCPAPPVAGPSVFLFPPKPKDTLMIARTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGKKD
SEQ ID № 11 («стебель» CD8 человека):
TTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDI
SEQ ID № 12 (шарнир IgG1 человека):
AEPKSPDKTHTCPPCPKDPK
SEQ ID № 13 (эктодомен CD2)
KEITNALETWGALGQDINLDIPSFQMSDDIDDIKWEKTSDKKKIAQFRKEKETFKEKDTYKLFKNGTLKIKHLKTDDQDIYKVSIYDTKGKNVLEKIFDLKIQERVSKPKISWTCINTTLTCEVMNGTDPELNLYQDGKHLKLSQRVITHKWTTSLSAKFKCTAGNKVSKESSVEPVSCPEKGLD
SEQ ID № 14 (эктодомен CD34)
SLDNNGTATPELPTQGTFSNVSTNVSYQETTTPSTLGSTSLHPVSQHGNEATTNITETTVKFTSTSVITSVYGNTNSSVQSQTSVISTVFTTPANVSTPETTLKPSLSPGNVSDLSTTSTSLATSPTKPYTSSSPILSDIKAEIKCSGIREVKLTQGICLEQNKTSSCAEFKKDRGEGLARVLCGEEQADADAGAQVCSLLLAQSEVRPQCLLLVLANRTEISSKLQLMKKHQSDLKKLGILDFTEQDVASHQSYSQKT
Поскольку CAR типично представляют собой гомодимеры (см. фиг. 1a), перекрестное образование пар может вести к гетеродимерному химерному антигенному рецептору. Это нежелательно по различным причинам, например: (1) эпитоп может не быть на том же «уровне» на клетке-мишени, так что перекрестно-спареный CAR может быть способен только связываться с одним антигеном; (2) VH и VL из двух различных scFv могут меняться местами и оказываться неспособными распознавать мишень или, хуже того, распознавать неожиданный и непредсказуемый антиген. Для логического элемента «ИЛИ» и логического элемента «И НЕ» спейсер первого CAR достаточно отличен от спейсера второго CAR во избежание образования перекрестных пар. Аминокислотная последовательность первого спейсера может иметь меньше чем 50%, 40%, 30% или 20% идентичности со вторым спейсером на аминокислотном уровне.
В других аспектах изобретения (например, логический элемент И) важно, что спейсер первого CAR имеет отличающуюся длину и/или заряд и/или геометрическую форму и/или конфигурацию и/или гликозилирование, так что когда первый и второй CAR связывают свой антиген-мишень, различия в заряде или размерах спейсера ведет к пространственному разделению CAR двух типов на различные части мембраны, чтобы приводить к активации, как предсказано с помощью модели кинетического разделения. В этих аспектах отличающуюся длину, геометрическую форму и/или конфигурацию спейсеров осторожно выбирают, принимая во внимание размер и связывающий эпитоп на антигене-мишени для того, чтобы сделать возможным дифференциальную сегрегацию при распознавании когнатной мишени. Например, предположительно шарнир IgG1, «стебель» CD8, Fc из IgG1, эктодомен CD34, эктодомен CD45 способны к дифференциальной сегрегации.
Примеры пар спейсеров, которые способны к дифференциальной сегрегации и, следовательно, пригодны для использования с логическим элементом И, представлены в следующей таблице:
В других аспектах изобретения (например, логический элемент И НЕ) важно, чтобы спейсер достаточно отличался для того, чтобы предотвращать образование перекрестных пар, но был достаточно схожим для совместной локализации. Можно использовать пары ортологичных спейсерных последовательностей. Примерами являются «стебли» CD8 мыши и человека или альтернативно спейсерные домены, которые являются мономерными, например, эктодомен CD2.
Примеры пар спейсеров, которые совместно локализуются и, следовательно, пригодны для использования с логическим элементом И НЕ, представлены в следующей таблице:
Все указанные выше спейсерные домены формируют гомодимеры. Однако механизм не ограничен использованием гомодимерных рецепторов и будет работать с мономерными рецепторами до тех пор, пока спейсер является достаточно жестким. Примером такого спейсера является CD2 или усеченный CD22.
ТРАНСМЕМБРАННЫЙ ДОМЕН
Трансмембранный домен представляет собой последовательность CAR, которая пронизывает мембрану.
Трансмембранный домен может представлять собой любую белковую структуру, которая термодинамически стабильна в мембране. Типично она представляет собой альфа-спираль, которая содержит несколько гидрофобных остатков. Трансмембранный домен любого трансмембранного белка можно использовать для предоставления трансмембранной части по изобретению. Присутствие и размер трансмембранного домена белка могут определять специалисты в данной области с использованием алгоритма TMHMM (http://www.cbs.dtu.dk/services/TMHMM-2.0/). Кроме того, учитывая, что трансмембранный домен белка представляет собой относительно простую структуру, т. е. полипептидная последовательность, согласно прогнозу, может формировать гидрофобную альфа-спираль достаточной длины, чтобы пронизывать мембрану, также можно использовать искусственно сконструированный TM домен (в US 7052906 B1 описаны синтетические трансмембранные компоненты).
Трансмембранный домен можно получать из CD28, который дает хорошую стабильность рецепторов.
АКТИВИРУЮЩИЙ ЭНДОДОМЕН
Эндодомен представляет собой передающую сигналы часть CAR. После распознавания антигена, рецепторы образуют кластер, нативные CD45 и CD148 исключены из синапса и сигнал передается в клетку. Наиболее широко используемый эндодоменным компонентом является таковой из CD3-дзета, который содержит 3 ITAM. Он передает активационный сигнал в T-клетку после связывания антигена. CD3-дзета может не предоставлять полностью достаточный активационный сигнал и может быть необходима передача дополнительных костимуляторных сигналов. Например, химерный CD28 и OX40 можно использовать с CD3-дзета для того, чтобы передавать пролиферативный сигнал/сигнал выживания, или можно использовать вместе все три.
Когда T-клетка по настоящему изобретению содержит CAR с активирующим эндодоменом, он может содержать только эндодомен CD3-дзета, эндодомен CD3-дзета с таковым из любого из CD28 или OX40 или эндодомен CD28 и OX40 и эндодомен CD3-дзета.
Любой эндодомен, который содержит мотив ITAM, в этом изобретении может действовать в качестве активационного эндодомена. Известно, что некоторые белки содержат эндодомены с одним или несколькими мотивами ITAM. Примеры таких белков, среди прочего, включают эпсилон-цепь CD3, гамма-цепь CD3 и дельта-цепь CD3. Мотив ITAM можно легко распознавать как тирозин, отделенный от лейцина или изолейцина какими-либо двумя другими аминокислотами, дающими сигнатуру YxxL/I. Типично, но не всегда, два из этих мотивов разделены 6-8 аминокислотами в хвосте молекулы (YxxL/Ix(6-8)YxxL/I). Таким образом, специалист в данной области может легко найти существующие белки, которые содержат один или несколько ITAM, чтобы передавать активационный сигнал. Кроме того, данный мотив является простым, и сложная вторичная структура не требуется, специалист в данной области может конструировать полипептиды, содержащие искусственные ITAM для того, чтобы передавать активационн* сигнал (см. WO 2000063372, которая относится к синтетическим сигнальным молекулам).
T-клеточный трансмембраный и внутриклеточный сигнальный домен (эндодомен) в CAR с активирующим эндодоменом может содержать последовательность, представленную как SEQ ID № 15, 16 или 17 или ее вариант, который имеет по меньшей мере 80% идентичности последовательностей.
SEQ ID № 15, содержащая трансмембранный домен CD28 и эндодомен CD3 Z
FWVLVVVGGVLACYSLLVTVAFIIFWVRRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR
SEQ ID № 16, содержащая трансмембранный домен CD28 и эндодомены CD28 и CD3-дзета
FWVLVVVGGVLACYSLLVTVAFIIFWVRSKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRSRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR
SEQ ID № 17, содержащая трансмембранный домен CD28 и эндодомены CD28, OX40 и CD3-дзета.
FWVLVVVGGVLACYSLLVTVAFIIFWVRSKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRSRDQRLPPDAHKPPGGGSFRTPIQEEQADAHSTLAKIRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR
Вариант последовательности может иметь по меньшей мере 80%, 85%, 90%, 95%, 98% или 99% идентичности последовательностей с SEQ ID № 15, 16 или 17, при условии, что последовательность предусматривает эффективный трансмембранный домен и эффективный внутриклеточный домен T-клеточной сигнализации.
«ВЫКЛЮЧАЕМЫЙ ЛИГИРОВАНИЕМ» ИНГИБИРУЮЩИЙ ЭНДОДОМЕН
В варианте осуществления, указанном выше в качестве логического элемента И, один из CAR содержит ингибирующий эндодомен так, что ингибирующий CAR ингибирует активацию T-клеток с помощью активирующего CAR в отсутствие лигирования ингибирующего CAR, но не значительно ингибирует активацию T-клеток с помощью активирующего CAR, когда ингибирующий CAR лигирован. Это называют «выключаемым лигированием» ингибирующим эндодоменом.
В этом случае, спейсер ингибирующего CAR имеет длину, заряд, геометрическую форму и/или конфигурацию и/или гликозилирование, отличающиеся от спейсера активирующего CAR, так что когда оба рецептора лигированы, различия в размерах спейсеров ведут к выделению активирующих CAR и ингибирующих CAR в различные мембранные компартменты иммунологического синапса с тем, чтобы снять ингибирование активирующего эндодомена ингибирующим эндодоменом.
Следовательно, ингибирующие эндодомены для использования в выключаемом лигированием ингибирующем CAR могут содержать какую-либо последовательность, которая ингибирует T-клеточную сигнализацию с помощью активирующего CAR, когда она находится в том же мембранном компартменте (т. е. в отсутствие антигена для ингибирующего CAR), но которая не значительно ингибирует T-клеточную сигнализацию, когда она изолирована в отдельной части мембраны от ингибирующего CAR.
Выключаемый лигированием ингибирующий эндодомен может представлять собой или содержать тирозиновую фосфатазу, такую как рецептороподобная тирозиновая фосфатаза. Ингибирующий эндодомен может представлять собой или содержать любую тирозиновую фосфатазу, которая способна ингибировать передачу сигнала TCR, когда лигирован только стимулирующий рецептор. Ингибирующий эндодомен может представлять собой или содержать любую тирозиновую фосфатазу с достаточно высокой скоростью катализа для фосфорилированных ITAM, которая способна ингибировать передачу сигнала TCR, когда лигирован только стимулирующий рецептор.
Например, ингибирующий эндодомен логического элемента И может содержать эндодомен из CD148 или CD45. Показано, что CD148 и CD45 действует естественным путем на фосфорилированные тирозины выше по каскаду реакций от передачи сигнала TCR.
CD148 представляет собой рецептороподобную белковую тирозиновую фосфатазу, которая отрицательно регулирует передачу сигнала TCR посредством препятствования фосфорилированию и функционированию PLCγ1 и LAT.
CD45 присутствует на всех гематопоэтических клетках, представляет собой белковую тирозиновую фосфатазу, которая способна регулировать сигнальную трансдукцию и функциональные реации, также посредством фосфорилирования PLCγ1.
Ингибирующий эндодомен может содержать всю рецептороподобную тирозиновую фосфатазу или ее часть. Фосфатаза может препятствовать фосфорилированию и/или функционированию элементов, вовлеченных в передачу T-клеточных сигналов, таких как PLCγ1 и/или LAT.
Трансмембраный и эндодомен из CD45 и CD148 представлены как SEQ ID № 18 и № 19, соответственно.
SEQ ID 18 - последовательность трансмембранного и эндодомена CD45
ALIAFLAFLIIVTSIALLVVLYKIYDLHKKRSCNLDEQQELVERDDEKQLMNVEPIHADILLETYKRKIADEGRLFLAEFQSIPRVFSKFPIKEARKPFNQNKNRYVDILPYDYNRVELSEINGDAGSNYINASYIDGFKEPRKYIAAQGPRDETVDDFWRMIWEQKATVIVMVTRCEEGNRNKCAEYWPSMEEGTRAFGDVVVKINQHKRCPDYIIQKLNIVNKKEKATGREVTHIQFTSWPDHGVPEDPHLLLKLRRRVNAFSNFFSGPIVVHCSAGVGRTGTYIGIDAMLEGLEAENKVDVYGYVVKLRRQRCLMVQVEAQYILIHQALVEYNQFGETEVNLSELHPYLHNMKKRDPPSEPSPLEAEFQRLPSYRSWRTQHIGNQEENKSKNRNSNVIPYDYNRVPLKHELEMSKESEHDSDESSDDDSDSEEPSKYINASFIMSYWKPEVMIAAQGPLKETIGDFWQMIFQRKVKVIVMLTELKHGDQEICAQYWGEGKQTYGDIEVDLKDTDKSSTYTLRVFELRHSKRKDSRTVYQYQYTNWSVEQLPAEPKELISMIQVVKQKLPQKNSSEGNKHHKSTPLLIHCRDGSQQTGIFCALLNLLESAETEEVVDIFQVVKALRKARPGMVSTFEQYQFLYDVIASTYPAQNGQVKKNNHQEDKIEFDNEVDKVKQDANCVNPLGAPEKLPEAKEQAEGSEPTSGTEGPEHSVNGPASPALNQGS
SEQ ID 19 - последовательность трансмембраного и эндодомена CD148
AVFGCIFGALVIVTVGGFIFWRKKRKDAKNNEVSFSQIKPKKSKLIRVENFEAYFKKQQADSNCGFAEEYEDLKLVGISQPKYAAELAENRGKNRYNNVLPYDISRVKLSVQTHSTDDYINANYMPGYHSKKDFIATQGPLPNTLKDFWRMVWEKNVYAIIMLTKCVEQGRTKCEEYWPSKQAQDYGDITVAMTSEIVLPEWTIRDFTVKNIQTSESHPLRQFHFTSWPDHGVPDTTDLLINFRYLVRDYMKQSPPESPILVHCSAGVGRTGTFIAIDRLIYQIENENTVDVYGIVYDLRMHRPLMVQTEDQYVFLNQCVLDIVRSQKDSKVDLIYQNTTAMTIYENLAPVTTFGKTNGYIA
Ингибирующий CAR может содержать всю SEQ ID № 18 или 19 или ее часть (например, он может содержать фосфатазную функцию эндодомена). Он может содержать вариант последовательности или ее части, имеющий по меньшей мере 80% идентичности последовательностей до тех пор, пока вариант сохраняет способность базово ингибировать передачу T-клеточных сигналов с помощью активирующего CAR.
Другие спейсеры и эндодомены можно тестировать, например, с использованием модельной системы, проиллюстрированной в настоящем документе. Популяции клеток-мишеней можно создавать посредством трансдукции подходящей клеточной линии, такой как клеточная линия SupT1, или однократно или двукратно для того, чтобы создавать клетки, негативные по обоим антигенам (дикого типа), положительные по любому и положительные по обоим (например CD19-CD33-, CD19+CD33-, CD19-CD33+ и CD19+CD33+). T-клетки, такие как линия Т-клеток мыши BW5147, которые высвобождают IL-2 при активации, можно трансдуцировать парами CAR, и измерять их способность функционировать в логическом элементе через измерение высвобождения IL-2 (например, с помощью ELISA). Например, в примере 4 показано, что оба эндодомена CD148 и CD45 могут выполнять функцию ингибирующих CAR в комбинации с активирующим CAR, содержащим дзета-эндодомен CD3. Эти CAR основаны на коротком/некрупном спейсере из «стебля» CD8 на одном CAR и крупном Fc спейсере на другом CAR, чтобы получать логическую функцию И. Когда оба рецептора лигированы, различие в размерах спейсеров ведет к выделению различных рецепторов в различные мембранные компартменты, что снимает ингибирование рецептора CD3-дзета эндодоменами CD148 или CD45. Таким образом, активация возникает только когда активируют оба рецептора. Можно легко видеть, что эту модульную систему можно использовать для того, чтобы тестировать альтернативные пары спейсеров и ингибирующих эндодоменов. Если спейсеры не достигают изоляции после лигирования обоих рецепторов, ингибирование не будет устранено и поэтому не произойдет активация. Если тестируемый ингибирующий эндодомен не эффективен, следует ожидать активации в присутствии лигирования активирующего CAR независимо от состояния лигирования ингибирующего CAR.
«ВКЛЮЧАЕМЫЙ ЛИГИРОВАНИЕМ» ЭНДОДОМЕН
В варианте осуществления, упомянутом выше в качестве логического элемента И НЕ, один из CAR содержит «включаемый лигированием» ингибирующий эндодомен так, что ингибирующий CAR не значительно ингибирует активацию T-клеток с помощью активирующего CAR в отсутствие лигирования ингибирующего CAR, но ингибирует активацию T-клеток с помощью активирующего CAR, когда ингибирующий CAR лигирован.
«Включаемый лигированием» ингибирующий эндодомен может представлять собой или содержать тирозиновую фосфатазу, которая не способна ингибировать передачу сигнала TCR, когда лигирован только стимулирующий рецептор.
«Включаемый лигированием» ингибирующий эндодомен может представлять собой или содержать тирозиновую фосфатазу с достаточно низкой скоростью катализа для фосфорилированных ITAM, которая не способна ингибировать передачу сигнала TCR, когда лигирован только стимулирующий рецептор, но она способна ингибировать ответ на передачу сигнала TCR, когда сконцентрирована в синапсе. Концентрация в синапс достигается через лигирование ингибирующего рецептора.
Если тирозиновая фосфатаза имеет скорость катализа, которая слишком велика для «включаемого лигированием» ингибирующего эндодомена, то возможно снижать скорость катализа фосфатазы через модификацию, такую как точечные мутации и короткие линкеры (которые вызывают стерические затруднения), чтобы делать ее подходящей для «включаемого лигированием» ингибирующего эндодомена.
В этом первом варианте осуществления эндодомен может представлять собой или содержать фосфатазу, которая значительно менее активна, чем CD45 или CD148, так что значимое дефосфорилирование ITAM возникает только когда активирующие и ингибирующие эндодомены совместно локализованы. В данной области известны многие подходящие последовательности. Например, ингибирующий эндодомен логического элемента НЕ И может содержать всю белковую тирозиновую фосфатазу, такую как PTPN6, или ее часть.
Белковые тирозиновые фосфатазы (PTP) представляют собой сигнальные молекулы, которые регулируют различные клеточные процессы, в том числе клеточный рост, дифференциацию, митотический цикл и онкогенная трансформация. N-концевая часть этой PTP содержит два тандемных гомологичных (SH2) домена Src, которые действуют в качестве белковых фосфотирозин-связывающих доменов и опосредуют взаимодействие этой PTP с ее субстратом. Эту PTP экспрессируют в первую очередь гематопоэтические клетки, и она функционирует в качестве важного регулятора множества путей передачи сигналов в гематопоэтических клетках.
Ингибирующий домен может содержать всю PTPN6 (SEQ ID № 20) или только фосфатазный домен (SEQ ID № 21).
SEQ ID 20 - последовательность PTPN6
MVRWFHRDLSGLDAETLLKGRGVHGSFLARPSRKNQGDFSLSVRVGDQVTHIRIQNSGDFYDLYGGEKFATLTELVEYYTQQQGVLQDRDGTIIHLKYPLNCSDPTSERWYHGHMSGGQAETLLQAKGEPWTFLVRESLSQPGDFVLSVLSDQPKAGPGSPLRVTHIKVMCEGGRYTVGGLETFDSLTDLVEHFKKTGIEEASGAFVYLRQPYYATRVNAADIENRVLELNKKQESEDTAKAGFWEEFESLQKQEVKNLHQRLEGQRPENKGKNRYKNILPFDHSRVILQGRDSNIPGSDYINANYIKNQLLGPDENAKTYIASQGCLEATVNDFWQMAWQENSRVIVMTTREVEKGRNKCVPYWPEVGMQRAYGPYSVTNCGEHDTTEYKLRTLQVSPLDNGDLIREIWHYQYLSWPDHGVPSEPGGVLSFLDQINQRQESLPHAGPIIVHCSAGIGRTGTIIVIDMLMENISTKGLDCDIDIQKTIQMVRAQRSGMVQTEAQYKFIYVAIAQFIETTKKKLEVLQSQKGQESEYGNITYPPAMKNAHAKASRTSSKHKEDVYENLHTKNKREEKVKKQRSADKEKSKGSLKRK
SEQ ID 21 - последовательность фосфатазного домена из PTPN6
FWEEFESLQKQEVKNLHQRLEGQRPENKGKNRYKNILPFDHSRVILQGRDSNIPGSDYINANYIKNQLLGPDENAKTYIASQGCLEATVNDFWQMAWQENSRVIVMTTREVEKGRNKCVPYWPEVGMQRAYGPYSVTNCGEHDTTEYKLRTLQVSPLDNGDLIREIWHYQYLSWPDHGVPSEPGGVLSFLDQINQRQESLPHAGPIIVHCSAGIGRTGTIIVIDMLMENISTKGLDCDIDIQKTIQMVRAQRSGMVQTEAQYKFIYVAIAQF
Второй вариант осуществления включаемого лигированием ингибирующего эндодомена представляет собой ITIM (иммунорецепторный тирозиновый ингибирующий мотив)-содержащий эндодомен, такой как таковой из CD22, LAIR-1, семейства рецепторов подавления цитотоксичности (KIR), LILRB1, CTLA4, PD-1, BTLA и т.д. Когда фосфорилированы, ITIM рекрутируют эндогенную PTPN6 через свой домен SH2. При совместной локализации с ITAM-содержащим эндодоменом, происходит дефосфорилирование и ингибирование активирующего CAR.
ITIM представляет собой консервативную последовательность аминокислот (S/I/V/LxYxxI/V/L), которую находят в цитоплазматических хвостах многих ингибирующих рецепторов иммунной системы. Специалист в данной области может легко найти белковые домены, содержащие ITIM. Список вероятных ITIM-содержащих белков человека создан посредством сканирования всего протеома (Staub, et al (2004) Cell. Signal. 16, 435-456). Кроме того, поскольку хорошо известна консенсусная последовательность и, по-видимому, необходима небольшая вторичная структура, специалист в данной области может создавать искусственные ITIM.
ITIM-эндодомены из PDCD1, BTLA4, LILRB1, LAIR1, CTLA4, KIR2DL1, KIR2DL4, KIR2DL5, KIR3DL1 и KIR3DL3 представлены в SEQ ID с 22 до 31, соответственно.
SEQ ID 22 эндодомен PDCD1
CSRAARGTIGARRTGQPLKEDPSAVPVFSVDYGELDFQWREKTPEPPVPCVPEQTEYATIVFPSGMGTSSPARRGSADGPRSAQPLRPEDGHCSWPL
SEQ ID 23 BTLA4
KLQRRWKRTQSQQGLQENSSGQSFFVRNKKVRRAPLSEGPHSLGCYNPMMEDGISYTTLRFPEMNIPRTGDAESSEMQRPPPDCDDTVTYSALHKRQVGDYENVIPDFPEDEGIHYSELIQFGVGERPQAQENVDYVILKH
SEQ ID 24 LILRB1
LRHRRQGKHWTSTQRKADFQHPAGAVGPEPTDRGLQWRSSPAADAQEENLYAAVKHTQPEDGVEMDTRSPHDEDPQAVTYAEVKHSRPRREMASPPSPLSGEFLDTKDRQAEEDRQMDTEAAASEAPQDVTYAQLHSLTLRREATEPPPSQEGPSPAVPSIYATLAIH
SEQ ID 25 LAIR1
HRQNQIKQGPPRSKDEEQKPQQRPDLAVDVLERTADKATVNGLPEKDRETDTSALAAGSSQEVTYAQLDHWALTQRTARAVSPQSTKPMAESITYAAVARH
SEQ ID 26 CTLA4
FLLWILAAVSSGLFFYSFLLTAVSLSKMLKKRSPLTTGVYVKMPPTEPECEKQFQPYFIPIN
SEQ ID 27 KIR2DL1
GNSRHLHVLIGTSVVIIPFAILLFFLLHRWCANKKNAVVMDQEPAGNRTVNREDSDEQDPQEVTYTQLNHCVFTQRKITRPSQRPKTPPTDIIVYTELPNAESRSKVVSCP
SEQ ID 28 KIR2DL4
GIARHLHAVIRYSVAIILFTILPFFLLHRWCSKKKENAAVMNQEPAGHRTVNREDSDEQDPQEVTYAQLDHCIFTQRKITGPSQRSKRPSTDTSVCIELPNAEPRALSPAHEHHSQALMGSSRETTALSQTQLASSNVPAAGI
SEQ ID 29 KIR2DL5
TGIRRHLHILIGTSVAIILFIILFFFLLHCCCSNKKNAAVMDQEPAGDRTVNREDSDDQDPQEVTYAQLDHCVFTQTKITSPSQRPKTPPTDTTMYMELPNAKPRSLSPAHKHHSQALRGSSRETTALSQNRVASSHVPAAGI
SEQ ID 30 KIR3DL1
KDPRHLHILIGTSVVIILFILLLFFLLHLWCSNKKNAAVMDQEPAGNRTANSEDSDEQDPEEVTYAQLDHCVFTQRKITRPSQRPKTPPTDTILYTELPNAKPRSKVVSCP
SEQ ID 31 KIR3DL3
KDPGNSRHLHVLIGTSVVIIPFAILLFFLLHRWCANKKNAVVMDQEPAGNRTVNREDSDEQDPQEVTYAQLNHCVFTQRKITRPSQRPKTPPTDTSV
Третий вариант осуществления включаемого лигированием ингибирующего эндодомена представляет собой ITIM-содержащий эндодомен, совместно экспрессируемый со слитным белком. Слитный белок может содержать по меньшей мере часть белковой тирозиновой фосфатазы и по меньшей мере часть рецептороподобной тирозиновой фосфатазы. Слитная конструкция может содержать один или несколько доменов SH2 из белковой тирозиновой фосфатазы. Например, слитная конструкция может быть между доменом SH2 PTPN6 и эндодоменом CD45 или между доменом SH2 PTPN6 и эндодоменом CD148. Когда фосфорилированы, ITIM-домены рекрутируют слитный белок, приводя высоко активную фосфатазу CD45 или CD148 в непосредственную близость с активирующим эндодоменом, блокируя активацию.
ПОСЛЕДОВАТЕЛЬНОСТИ слитных белков перечислены 32 и 33
SEQ ID 32 слитный белок PTPN6-CD45
WYHGHMSGGQAETLLQAKGEPWTFLVRESLSQPGDFVLSVLSDQPKAGPGSPLRVTHIKVMCEGGRYTVGGLETFDSLTDLVEHFKKTGIEEASGAFVYLRQPYKIYDLHKKRSCNLDEQQELVERDDEKQLMNVEPIHADILLETYKRKIADEGRLFLAEFQSIPRVFSKFPIKEARKPFNQNKNRYVDILPYDYNRVELSEINGDAGSNYINASYIDGFKEPRKYIAAQGPRDETVDDFWRMIWEQKATVIVMVTRCEEGNRNKCAEYWPSMEEGTRAFGDVVVKINQHKRCPDYIIQKLNIVNKKEKATGREVTHIQFTSWPDHGVPEDPHLLLKLRRRVNAFSNFFSGPIVVHCSAGVGRTGTYIGIDAMLEGLEAENKVDVYGYVVKLRRQRCLMVQVEAQYILIHQALVEYNQFGETEVNLSELHPYLHNMKKRDPPSEPSPLEAEFQRLPSYRSWRTQHIGNQEENKSKNRNSNVIPYDYNRVLKHELEMSKESEHDSDESSDDDSDSEEPSKYINASFIMSYWKPEVMIAAQGPLKETIGDFMIQRKVKVIVMLTELKHGDQEICAQYWGEGKQTYGDIEVDLKDTDKSSTYTLRVFELRHSKRKDSRTVYQYQYTNWSVEQLPAEPKELISMIQVVKQKLPQKNSSEGNKHHKSTPLLIHCRDGSQQTGIFCALLNLLESAETEEVVDIFQVVKALRKARPGMVSTFEQYQFLYDVIASTYPAQNGQVKKNNHQEDKIEFDNEVDKVKQDANCVNPLGAPEKLPEAKEQAEGSEPTSGTEGPEHSVNGPASPALNQGS
SEQ ID 33 слитный белок PTPN6-CD148
ETLLQAKGEPWTFLVRESLSQPGDFVLSVLSDQPKAGPGSPLRVTHIKVMCEGGRYTVGGLETFDSLTDLVEHFKKTGIEEASGAFVYLRQPYRKKRKDAKNNEVSFSQIKPKKSKLIRVENFEAYFKKQQADSNCGFAEEYEDLKLVGISQPKYAAELAENRGKNRYNNVLPYDISRVKLSVQTHSTDDYINANYMPGYHSKKDFIATQGPLPNTLKDFWRMVWEKNVYAIIMLTKCVEQGRTKCEEYWPSKQAQDYGDITVAMTSEIVLPEWTIRDFTVKNIQTSESHPLRQFHFTSWPDHGVPDTTDLLINFRYLVRDYMKQSPPESPILVHCSAGVGRTGTFIAIDRLIYQIENENTVDVYGIVYDLRMHRPLMVQTEDQYVFLNQCVLDIVRSQKDSKVDLIYQNTTAMTIYENLAPVTTFGKTNGYIA
Включаемый лигированием ингибирующий CAR может содержать всю SEQ ID № 20 или 21 или ее часть. Он может содержать всю SEQ ID с 22 до 31 или ее часть. Он может содержать всю SEQ ID с 22 до 31 или ее часть, совместно экспрессированную с любой SEQ ID 32 или 33. Он может содержать вариант последовательности или его часть, который имеет по меньшей мере 80% идентичности последовательностей, до тех пор, пока вариант сохраняет способность ингибировать T-клеточную сигнализацию с помощью активирующего CAR при лигировании ингибирующего CAR.
Как указано выше, альтернативные спейсеры и эндодомены можно тестировать, например, с использованием модельной системы, приведенной в качестве примера в настоящем документе. В примере 5 показано, что эндодомен PTPN6 может выполнять функцию полуингибирующего CAR в комбинации с активирующим CAR, который содержит дзета-эндодомен CD3. Эти CAR основаны на спейсере из «стебля» CD8 человека на одном CAR и спейсере из «стебля» CD8 мыши на другом CAR. Ортологичные последовательности препятствуют образованию перекрестных пар. Однако, когда оба рецептора лигирвоаны, сходство между спейсерами ведет к совместной сегрегации различных рецепторов в одних и тех же мембранных компартментах. Это ведет к ингибированию CD3 дзета рецептора эндодоменом PTPN6. Если лигирован только активирующий CAR, эндодомен PTPN6 не достаточно активен для того, чтобы предотвращать активацию T-клетки. Таким образом, активация происходит, если лигирован только активирующий CAR, а ингибирующий CAR не лигирован (логическая функция И НЕ). Можно легко видеть, что модульную систему можно использовать для того, чтобы тестировать альтернативные пары спейсеров и ингибирующие домены. Если спейсеры не достигают совместной сегрегации после лигирования обоих рецепторов, ингибирование не будет эффективным и, таким образом, будет происходить активация. Если тестируемый полуингибирующий эндодомен неэффективен, следует ожидать активации в присутствии лигирования активирующего CAR, независимо от состояния лигирования полуингибирующего CAR.
МЕСТО КОЭКСПРЕССИИ
Второй аспект изобретения относится к нуклеиновой кислоте, которая кодирует первый и второй CAR.
Нуклеиновая кислота может давать полипептид, который содержит две молекулы CAR, соединенные через сайт расщепления. Сайт расщепления может быть саморасщепляющимся, так что при получении полипептида, он незамедлительно расщепляется на первый и второй CAR без потребности в какой-либо внешней расщепляющей активности.
Известны различные саморасщепляющиеся сайты, в том числе саморасщепляющийся пептид 2a вируса ящура (FMDV), который имеет последовательность, представленную в качестве SEQ ID № 34:
SEQ ID № 34
RAEGRGSLLTCGDVEENPGP
Коэкспрессирующая последовательность может представлять собой внутреннюю последовательность связывания рибосомы (IRES). Коэкспрессирующая последовательность может представлять собой внутренний промотор.
КЛЕТКА
Первый аспект изобретения относится к клетке, которая совместно экспрессирует первый CAR и второй CAR на клеточной поверхности.
Клетка может представлять собой какую-либо эукариотическую клетка, способную экспрессировать CAR на поверхности клетки, такой как иммунная клетка.
В частности, клетка может представлять собой иммунную эффекторную клетку, такую как T-клетка или естественная киллерная (NK) клетка.
T-клетки или T-лимфоциты представляют собой тип лимфоцитов, который играет центральную роль в клеточно-опосредованном иммунитете. Их можно отличать от других лимфоцитов, таких как B-клетки и естественные киллерные клетки, с помощью присутствия T-клеточного рецептора (TCR) на клеточной поверхности. Существуют различные типы T-клетки, как кратко изложено далее.
T-хелперные клетки (TH клетки) помогают другим белым клеткам крови в иммунологических процессах, в том числе в созревании B-клеток в плазматические клетки и B-клетки памяти и активации цитотоксических T-клеток и макрофагов. На своей поверхности TH клетки экспрессируют CD4. Активация TH клеток происходит, когда молекулы MHC II класса на поверхности антигенпредставляющих клеток (APC) презентируют им пептидные антигены. Эти клетки можно отнести к одному из нескольких подтипов, в том числе TH1, TH2, TH3, TH17, Th9 или TFH, которые секретируют различные цитокины для того, чтобы содействовать различными типам иммунного ответа.
Цитотоксические T-клетки (TC клетки или CTL) уничтожают инфицированные вирусами клетки и опухолевые клетки, и также участвуют в отторжении трансплантата. CTL экспрессируют CD8 на своей поверхности. Эти клетки распознают свои мишени посредством связывания с антигеном, связанным с MHC I класса, который присутствует на поверхности всех ядросодержащих клеток. Через IL-10, аденозин и другие молекулы, секретируемые регуляторными T-клетками, CD8+ клетки можно инактивировать до анергического состояния, которое препятствует аутоиммунным заболеваниям, таким как экспериментальный аутоиммунный энцефаломиелит.
T-клетки памяти представляют собой подмножество антигенспецифичных T-клеток, которые персистируют длительное время после разрешения инфекции. Они быстро размножаютя до большого числа эффекторных T-клеток при повторном воздействии их когнатного антигена, таким образом обеспечивая «память» иммунной системы о прошлых инфекциях. T-клетки памяти содержат три подтипа: центральные T-клетки памяти (TCM клетки) и два типа эффекторных T-клеток памяти (TEM клетки и TEMRA клетки). Клетки памяти могут быть или CD4+ или CD8+. T-клетки памяти типично экспрессируют белок клеточной поверхности CD45RO.
Регуляторные T-клетки (клетки Treg), ранее известные как супрессорные T-клетки, критически важны для поддержания иммунологической толерантности. Их основная роль состоит в том, чтобы переключать T-клеточно-опосредованный иммунитет в направлении окончания иммунной реакции и продавлять аутореактивные T-клетки, которые избежали процесса негативной селекции в тимусе.
Описано два основных класса CD4+ клеток Treg - встречающиеся в природе клетки Treg и адаптивные клетки Treg.
Встречающиеся в природе клетки Treg (также известные как CD4+CD25+FoxP3+ клетки Treg) образуются в тимусе и связаны с взаимодействиями развивающихся T-клеток как с миелоидными (CD11c+), так и с плазмоцитоидными (CD123+) дендритными клетками, которые активированы с использованием TSLP. Встречающиеся в природе клетки Treg можно отличать от других T-клеток по присутствию внутриклеточной молекулы, называемой FoxP3. Мутации гена FOXP3 могут препятствовать развитию регуляторных T-клеток, вызывая смертельное аутоиммунное заболевание IPEX.
Адаптивные клетки Treg (также известные как клетки Tr1 или клеткми Th3) могут появляться во время нормального иммунного ответа.
T-клетка по изобретению может относиться к любому из типов T-клеток, которые указаны выше, в частности, к CTL.
Естественные киллерные (NK) клетки относятся к типу цитолитических клеток, которые образуют часть врожденной иммунной системы. Естественные киллерные клетки обеспечивают быстрый ответ на врожденный сигнал от инфицированных вирусами клеток независимым от MHC образом.
Естественные киллерные клетки (относящиеся к группе врожденных лимфоидных клеток) определяют как большие гранулярные лимфоциты (LGL), который составляют третий тип клеток, дифференцированный от общего лимфоидного предшественника, образующего B- и T-лимфоциты. Известно, что естественные киллерные клетки дифференцируются и созревают в костном мозге, лимфатических узлах, селезенке, миндалинах и тимусе, где после этого они выходят в циркуляцию.
CAR клетки по изобретению могут относиться к любому указанному выше типу клеток.
CAR-экспрессирующие клетки, такие как CAR-экспрессирующие T-клетки или естественные киллерные клетки, можно или создавать ex vivo или брать из собственной периферической крови пациента (1-я сторона), или в условиях трансплантата гематопоэтических стволовых клеток из донорской периферической крови (2-я сторона) или периферической крови от неродственного донора (3-я сторона).
Настоящее изобретение также предусматривает клеточную композицию, которая содержит CAR-экспрессирующие T-клетки и/или CAR-экспрессирующие естественные киллерные клетки в соответствии с настоящим изобретением. Клеточную композицию можно создавать посредством трансдукции или трансфекции образца крови ex vivo с использованием нуклеиновой кислоты в соответствии с настоящим изобретением.
Альтернативно, CAR-экспрессирующие клетки можно получать путем дифференциации ex vivo индуцибельных клеток-предшественников или эмбриональных клетлк-предшественников в клетки релевантного типа, такие как T-клетки. Альтернативно, можно использовать бессмертную клеточную линию, такую как линия Т-клеток, которая сохраняет свою литическую функцию и может действовать в качестве терапевтического средства.
Во всех этих вариантах осуществления CAR клетки получают посредством введения ДНК или РНК, кодирующих CAR, с помощью одного из множества средств, включая трансдукцию с использованием вирусного вектора, трансфекцию с использованием ДНК или РНК.
CAR T-клетка по изобретению может представлять собой T-клетку ex vivo от субъекта. T-клетка может быть из образца мононуклеарных клеток периферической крови (PBMC). T-клетки можно активирвоать и/или размножать перед трансдукцией кодирующей CAR нуклеиновой кислотой, например, посредством обработки моноклональным антителом против CD3.
CAR T-клетку по изобретению можно создавать путем:
(i) выделения содержащего T-клетки образца у субъекта или из других источников, перечисленных выше; и
(ii) трансдукции или трансфекции T-клеток одной или несколькими последовательностями нуклеиновых кислот, которые кодируют первый и второй CAR.
Затем T-клетки можно очищать, например, отбирать на основании коэкспрессии первого и второго CAR.
ПОСЛЕДОВАТЕЛЬНОСТИ НУКЛЕИНОВОЙ КИСЛОТЫ
Второй аспект изобретения относится к одной или нескольким последовательностям нуклеиновых кислот, которые кодируют первый CAR и второй CAR, как определено в первом аспекте изобретения.
Последовательность нуклеиновой кислоты может содержать одну из следующих последовательностей или ее вариант
SEQ ID 35 логический элемент ИЛИ
SEQ ID 36 логический элемент И с использованием CD45
SEQ ID 37 логический элемент И с использованием CD148
SEQ ID 38 логический элемент И НЕ с использованием PTPN6 в качестве эндодомена
SEQ ID 39 логический элемент И НЕ с использованием эндодомена LAIR1
SEQ ID 40 логический элемент И НЕ с использованием слитой конструкции LAIR1 и PTPN6 SH2 с фосфатазой CD148
SEQ ID № 35:
>MP13974.SFG.aCD19fmc63-CD8STK-CD28tmZ-2A-aCD33glx-HCH2CH3pvaa-CD28tmZw
ATGAGCCTGCCCGTGACCGCCCTGCTGCTGCCCCTGGCCCTGCTGCTGCACGCCGCCAGACCAGACATCCAGATGACCCAGACCACCAGCAGCCTGAGCGCCAGCCTGGGCGACCGGGTGACCATCAGCTGCAGAGCCAGCCAGGACATCAGCAAGTACCTGAACTGGTACCAGCAGAAGCCCGACGGCACCGTGAAGCTGCTGATCTACCACACCAGCCGGCTGCACAGCGGCGTGCCCAGCCGGTTCAGCGGCAGCGGCAGCGGCACCGACTACAGCCTGACCATCAGCAACCTGGAGCAGGAGGACATCGCCACCTACTTCTGCCAGCAGGGCAACACCCTGCCCTACACCTTCGGAGGCGGCACCAAGCTGGAGATCACCAAGGCCGGAGGCGGAGGCTCTGGCGGAGGCGGCTCTGGCGGAGGCGGCTCTGGCGGAGGCGGCAGCGAGGTGAAGCTGCAGGAGTCTGGCCCAGGCCTGGTGGCCCCAAGCCAGAGCCTGAGCGTGACCTGCACCGTGAGCGGCGTGAGCCTGCCCGACTACGGCGTGAGCTGGATCAGGCAGCCCCCACGGAAGGGCCTGGAGTGGCTGGGCGTGATCTGGGGCAGCGAGACCACCTACTACAACAGCGCCCTGAAGAGCCGGCTGACCATCATCAAGGACAACAGCAAGAGCCAGGTGTTCCTGAAGATGAACAGCCTGCAGACCGACGACACCGCCATCTACTACTGCGCCAAGCACTACTACTATGGCGGCAGCTACGCTATGGACTACTGGGGCCAGGGCACCAGCGTGACCGTGAGCTCAGATCCCACCACGACGCCAGCGCCGCGACCACCAACACCGGCGCCCACCATCGCGTCGCAGCCCCTGTCCCTGCGCCCAGAGGCGTGCCGGCCAGCGGCGGGGGGCGCAGTGCACACGAGGGGGCTGGACTTCGCCTGTGATATCTTTTGGGTGCTGGTGGTGGTTGGTGGAGTCCTGGCTTGCTATAGCTTGCTAGTAACAGTGGCCTTTATTATTTTCTGGGTGAGGAGAGTGAAGTTCAGCAGGAGCGCAGACGCCCCCGCGTACCAGCAGGGCCAGAACCAGCTCTATAACGAGCTCAATCTAGGACGAAGAGAGGAGTACGATGTTTTGGACAAGAGACGTGGCCGGGACCCTGAGATGGGGGGAAAGCCGAGAAGGAAGAACCCTCAGGAAGGCCTGTACAATGAACTGCAGAAAGATAAGATGGCGGAGGCCTACAGTGAGATTGGGATGAAAGGCGAGCGCCGGAGGGGCAAGGGGCACGATGGCCTTTACCAGGGTCTCAGTACAGCCACCAAGGACACCTACGACGCCCTTCACATGCAGGCCCTGCCTCCTCGCAGAGCCGAGGGCAGGGGAAGTCTTCTAACATGCGGGGACGTGGAGGAAAATCCCGGGCCCATGGCCGTGCCCACTCAGGTCCTGGGGTTGTTGCTACTGTGGCTTACAGATGCCAGATGTGACATCCAGATGACACAGTCTCCATCTTCCCTGTCTGCATCTGTCGGAGATCGCGTCACCATCACCTGTCGAGCAAGTGAGGACATTTATTTTAATTTAGTGTGGTATCAGCAGAAACCAGGAAAGGCCCCTAAGCTCCTGATCTATGATACAAATCGCTTGGCAGATGGGGTCCCATCACGGTTCAGTGGCTCTGGATCTGGCACACAGTATACTCTAACCATAAGTAGCCTGCAACCCGAAGATTTCGCAACCTATTATTGTCAACACTATAAGAATTATCCGCTCACGTTCGGTCAGGGGACCAAGCTGGAAATCAAAAGATCTGGTGGCGGAGGGTCAGGAGGCGGAGGCAGCGGAGGCGGTGGCTCGGGAGGCGGAGGCTCGAGATCTGAGGTGCAGTTGGTGGAGTCTGGGGGCGGCTTGGTGCAGCCTGGAGGGTCCCTGAGGCTCTCCTGTGCAGCCTCAGGATTCACTCTCAGTAATTATGGCATGCACTGGATCAGGCAGGCTCCAGGGAAGGGTCTGGAGTGGGTCTCGTCTATTAGTCTTAATGGTGGTAGCACTTACTATCGAGACTCCGTGAAGGGCCGATTCACTATCTCCAGGGACAATGCAAAAAGCACCCTCTACCTTCAAATGAATAGTCTGAGGGCCGAGGACACGGCCGTCTATTACTGTGCAGCACAGGACGCTTATACGGGAGGTTACTTTGATTACTGGGGCCAAGGAACGCTGGTCACAGTCTCGTCTATGGATCCCGCCGAGCCCAAATCTCCTGACAAAACTCACACATGCCCACCGTGCCCAGCACCTCCCGTGGCCGGCCCGTCAGTCTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCGCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAACCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCCCTGCACAATCACTATACCCAGAAATCTCTGAGTCTGAGCCCAGGCAAGAAGGACCCCAAGTTCTGGGTCCTGGTGGTGGTGGGAGGCGTGCTGGCCTGTTACTCTCTCCTGGTGACCGTGGCCTTCATCATCTTTTGGGTGCGCTCCCGGGTGAAGTTTTCTCGCTCTGCCGATGCCCCAGCCTATCAGCAGGGCCAGAATCAGCTGTACAATGAACTGAACCTGGGCAGGCGGGAGGAGTACGACGTGCTGGATAAGCGGAGAGGCAGAGACCCCGAGATGGGCGGCAAACCACGGCGCAAAAATCCCCAGGAGGGACTCTATAACGAGCTGCAGAAGGACAAAATGGCCGAGGCCTATTCCGAGATCGGCATGAAGGGAGAGAGAAGACGCGGAAAGGGCCACGACGGCCTGTATCAGGGATTGTCCACCGCTACAAAAGATACATATGATGCCCTGCACATGCAGGCCCTGCCACCCAGATGA
SEQ ID № 36
>MP14802.SFG.aCD19fmc63_clean-CD8STK-CD28tmZ-2A-aCD33glx-HCH2CH3pvaa-dCD45
ATGAGCCTGCCCGTGACCGCCCTGCTGCTGCCCCTGGCCCTGCTGCTGCACGCCGCCAGACCAGACATCCAGATGACCCAGACCACCAGCAGCCTGAGCGCCAGCCTGGGCGACCGGGTGACCATCAGCTGCAGAGCCAGCCAGGACATCAGCAAGTACCTGAACTGGTACCAGCAGAAGCCCGACGGCACCGTGAAGCTGCTGATCTACCACACCAGCCGGCTGCACAGCGGCGTGCCCAGCCGGTTCAGCGGCAGCGGCAGCGGCACCGACTACAGCCTGACCATCAGCAACCTGGAGCAGGAGGACATCGCCACCTACTTCTGCCAGCAGGGCAACACCCTGCCCTACACCTTCGGAGGCGGCACCAAGCTGGAGATCACCAAGGCCGGAGGCGGAGGCTCTGGCGGAGGCGGCTCTGGCGGAGGCGGCTCTGGCGGAGGCGGCAGCGAGGTGAAGCTGCAGGAGTCTGGCCCAGGCCTGGTGGCCCCAAGCCAGAGCCTGAGCGTGACCTGCACCGTGAGCGGCGTGAGCCTGCCCGACTACGGCGTGAGCTGGATCAGGCAGCCCCCACGGAAGGGCCTGGAGTGGCTGGGCGTGATCTGGGGCAGCGAGACCACCTACTACAACAGCGCCCTGAAGAGCCGGCTGACCATCATCAAGGACAACAGCAAGAGCCAGGTGTTCCTGAAGATGAACAGCCTGCAGACCGACGACACCGCCATCTACTACTGCGCCAAGCACTACTACTATGGCGGCAGCTACGCTATGGACTACTGGGGCCAGGGCACCAGCGTGACCGTGAGCTCAGATCCCACCACGACGCCAGCGCCGCGACCACCAACACCGGCGCCCACCATCGCGTCGCAGCCCCTGTCCCTGCGCCCAGAGGCGTGCCGGCCAGCGGCGGGGGGCGCAGTGCACACGAGGGGGCTGGACTTCGCCTGTGATATCTTTTGGGTGCTGGTGGTGGTTGGTGGAGTCCTGGCTTGCTATAGCTTGCTAGTAACAGTGGCCTTTATTATTTTCTGGGTGAGGAGAGTGAAGTTCAGCAGGAGCGCAGACGCCCCCGCGTACCAGCAGGGCCAGAACCAGCTCTATAACGAGCTCAATCTAGGACGAAGAGAGGAGTACGATGTTTTGGACAAGAGACGTGGCCGGGACCCTGAGATGGGGGGAAAGCCGAGAAGGAAGAACCCTCAGGAAGGCCTGTACAATGAACTGCAGAAAGATAAGATGGCGGAGGCCTACAGTGAGATTGGGATGAAAGGCGAGCGCCGGAGGGGCAAGGGGCACGATGGCCTTTACCAGGGTCTCAGTACAGCCACCAAGGACACCTACGACGCCCTTCACATGCAGGCCCTGCCTCCTCGCAGAGCCGAGGGCAGGGGAAGTCTTCTAACATGCGGGGACGTGGAGGAAAATCCCGGGCCCATGGCCGTGCCCACTCAGGTCCTGGGGTTGTTGCTACTGTGGCTTACAGATGCCAGATGTGACATCCAGATGACACAGTCTCCATCTTCCCTGTCTGCATCTGTCGGAGATCGCGTCACCATCACCTGTCGAGCAAGTGAGGACATTTATTTTAATTTAGTGTGGTATCAGCAGAAACCAGGAAAGGCCCCTAAGCTCCTGATCTATGATACAAATCGCTTGGCAGATGGGGTCCCATCACGGTTCAGTGGCTCTGGATCTGGCACACAGTATACTCTAACCATAAGTAGCCTGCAACCCGAAGATTTCGCAACCTATTATTGTCAACACTATAAGAATTATCCGCTCACGTTCGGTCAGGGGACCAAGCTGGAAATCAAAAGATCTGGTGGCGGAGGGTCAGGAGGCGGAGGCAGCGGAGGCGGTGGCTCGGGAGGCGGAGGCTCGAGATCTGAGGTGCAGTTGGTGGAGTCTGGGGGCGGCTTGGTGCAGCCTGGAGGGTCCCTGAGGCTCTCCTGTGCAGCCTCAGGATTCACTCTCAGTAATTATGGCATGCACTGGATCAGGCAGGCTCCAGGGAAGGGTCTGGAGTGGGTCTCGTCTATTAGTCTTAATGGTGGTAGCACTTACTATCGAGACTCCGTGAAGGGCCGATTCACTATCTCCAGGGACAATGCAAAAAGCACCCTCTACCTTCAAATGAATAGTCTGAGGGCCGAGGACACGGCCGTCTATTACTGTGCAGCACAGGACGCTTATACGGGAGGTTACTTTGATTACTGGGGCCAAGGAACGCTGGTCACAGTCTCGTCTATGGATCCCGCCGAGCCCAAATCTCCTGACAAAACTCACACATGCCCACCGTGCCCAGCACCTCCCGTGGCCGGCCCGTCAGTCTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCGCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAACCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCCCTGCACAATCACTATACCCAGAAATCTCTGAGTCTGAGCCCAGGCAAGAAGGACCCCAAGGCACTGATAGCATTTCTGGCATTTCTGATTATTGTGACATCAATAGCCCTGCTTGTTGTTCTCTACAAAATCTATGATCTACATAAGAAAAGATCCTGCAATTTAGATGAACAGCAGGAGCTTGTTGAAAGGGATGATGAAAAACAACTGATGAATGTGGAGCCAATCCATGCAGATATTTTGTTGGAAACTTATAAGAGGAAGATTGCTGATGAAGGAAGACTTTTTCTGGCTGAATTTCAGAGCATCCCGCGGGTGTTCAGCAAGTTTCCTATAAAGGAAGCTCGAAAGCCCTTTAACCAGAATAAAAACCGTTATGTTGACATTCTTCCTTATGATTATAACCGTGTTGAACTCTCTGAGATAAACGGAGATGCAGGGTCAAACTACATAAATGCCAGCTATATTGATGGTTTCAAAGAACCCAGGAAATACATTGCTGCACAAGGTCCCAGGGATGAAACTGTTGATGATTTCTGGAGGATGATTTGGGAACAGAAAGCCACAGTTATTGTCATGGTCACTCGATGTGAAGAAGGAAACAGGAACAAGTGTGCAGAATACTGGCCGTCAATGGAAGAGGGCACTCGGGCTTTTGGAGATGTTGTTGTAAAGATCAACCAGCACAAAAGATGTCCAGATTACATCATTCAGAAATTGAACATTGTAAATAAAAAAGAAAAAGCAACTGGAAGAGAGGTGACTCACATTCAGTTCACCAGCTGGCCAGACCACGGGGTGCCTGAGGATCCTCACTTGCTCCTCAAACTGAGAAGGAGAGTGAATGCCTTCAGCAATTTCTTCAGTGGTCCCATTGTGGTGCACTGCAGTGCTGGTGTTGGGCGCACAGGAACCTATATCGGAATTGATGCCATGCTAGAAGGCCTGGAAGCCGAGAACAAAGTGGATGTTTATGGTTATGTTGTCAAGCTAAGGCGACAGAGATGCCTGATGGTTCAAGTAGAGGCCCAGTACATCTTGATCCATCAGGCTTTGGTGGAATACAATCAGTTTGGAGAAACAGAAGTGAATTTGTCTGAATTACATCCATATCTACATAACATGAAGAAAAGGGATCCACCCAGTGAGCCGTCTCCACTAGAGGCTGAATTCCAGAGACTTCCTTCATATAGGAGCTGGAGGACACAGCACATTGGAAATCAAGAAGAAAATAAAAGTAAAAACAGGAATTCTAATGTCATCCCATATGACTATAACAGAGTGCCACTTAAACATGAGCTGGAAATGAGTAAAGAGAGTGAGCATGATTCAGATGAATCCTCTGATGATGACAGTGATTCAGAGGAACCAAGCAAATACATCAATGCATCTTTTATAATGAGCTACTGGAAACCTGAAGTGATGATTGCTGCTCAGGGACCACTGAAGGAGACCATTGGTGACTTTTGGCAGATGATCTTCCAAAGAAAAGTCAAAGTTATTGTTATGCTGACAGAACTGAAACATGGAGACCAGGAAATCTGTGCTCAGTACTGGGGAGAAGGAAAGCAAACATATGGAGATATTGAAGTTGACCTGAAAGACACAGACAAATCTTCAACTTATACCCTTCGTGTCTTTGAACTGAGACATTCCAAGAGGAAAGACTCTCGAACTGTGTACCAGTACCAATATACAAACTGGAGTGTGGAGCAGCTTCCTGCAGAACCCAAGGAATTAATCTCTATGATTCAGGTCGTCAAACAAAAACTTCCCCAGAAGAATTCCTCTGAAGGGAACAAGCATCACAAGAGTACACCTCTACTCATTCACTGCAGGGATGGATCTCAGCAAACGGGAATATTTTGTGCTTTGTTAAATCTCTTAGAAAGTGCGGAAACAGAAGAGGTAGTGGATATTTTTCAAGTGGTAAAAGCTCTACGCAAAGCTAGGCCAGGCATGGTTTCCACATTCGAGCAATATCAATTCCTATATGACGTCATTGCCAGCACCTACCCTGCTCAGAATGGACAAGTAAAGAAAAACAACCATCAAGAAGATAAAATTGAATTTGATAATGAAGTGGACAAAGTAAAGCAGGATGCTAATTGTGTTAATCCACTTGGTGCCCCAGAAAAGCTCCCTGAAGCAAAGGAACAGGCTGAAGGTTCTGAACCCACGAGTGGCACTGAGGGGCCAGAACATTCTGTCAATGGTCCTGCAAGTCCAGCTTTAAATCAAGGTTCATAG
SEQ ID № 37:
>MP14801.SFG.aCD19fmc63_clean-CD8STK-CD28tmZ-2A-aCD33glx-HCH2CH3pvaa-dCD148
ATGAGCCTGCCCGTGACCGCCCTGCTGCTGCCCCTGGCCCTGCTGCTGCACGCCGCCAGACCAGACATCCAGATGACCCAGACCACCAGCAGCCTGAGCGCCAGCCTGGGCGACCGGGTGACCATCAGCTGCAGAGCCAGCCAGGACATCAGCAAGTACCTGAACTGGTACCAGCAGAAGCCCGACGGCACCGTGAAGCTGCTGATCTACCACACCAGCCGGCTGCACAGCGGCGTGCCCAGCCGGTTCAGCGGCAGCGGCAGCGGCACCGACTACAGCCTGACCATCAGCAACCTGGAGCAGGAGGACATCGCCACCTACTTCTGCCAGCAGGGCAACACCCTGCCCTACACCTTCGGAGGCGGCACCAAGCTGGAGATCACCAAGGCCGGAGGCGGAGGCTCTGGCGGAGGCGGCTCTGGCGGAGGCGGCTCTGGCGGAGGCGGCAGCGAGGTGAAGCTGCAGGAGTCTGGCCCAGGCCTGGTGGCCCCAAGCCAGAGCCTGAGCGTGACCTGCACCGTGAGCGGCGTGAGCCTGCCCGACTACGGCGTGAGCTGGATCAGGCAGCCCCCACGGAAGGGCCTGGAGTGGCTGGGCGTGATCTGGGGCAGCGAGACCACCTACTACAACAGCGCCCTGAAGAGCCGGCTGACCATCATCAAGGACAACAGCAAGAGCCAGGTGTTCCTGAAGATGAACAGCCTGCAGACCGACGACACCGCCATCTACTACTGCGCCAAGCACTACTACTATGGCGGCAGCTACGCTATGGACTACTGGGGCCAGGGCACCAGCGTGACCGTGAGCTCAGATCCCACCACGACGCCAGCGCCGCGACCACCAACACCGGCGCCCACCATCGCGTCGCAGCCCCTGTCCCTGCGCCCAGAGGCGTGCCGGCCAGCGGCGGGGGGCGCAGTGCACACGAGGGGGCTGGACTTCGCCTGTGATATCTTTTGGGTGCTGGTGGTGGTTGGTGGAGTCCTGGCTTGCTATAGCTTGCTAGTAACAGTGGCCTTTATTATTTTCTGGGTGAGGAGAGTGAAGTTCAGCAGGAGCGCAGACGCCCCCGCGTACCAGCAGGGCCAGAACCAGCTCTATAACGAGCTCAATCTAGGACGAAGAGAGGAGTACGATGTTTTGGACAAGAGACGTGGCCGGGACCCTGAGATGGGGGGAAAGCCGAGAAGGAAGAACCCTCAGGAAGGCCTGTACAATGAACTGCAGAAAGATAAGATGGCGGAGGCCTACAGTGAGATTGGGATGAAAGGCGAGCGCCGGAGGGGCAAGGGGCACGATGGCCTTTACCAGGGTCTCAGTACAGCCACCAAGGACACCTACGACGCCCTTCACATGCAGGCCCTGCCTCCTCGCAGAGCCGAGGGCAGGGGAAGTCTTCTAACATGCGGGGACGTGGAGGAAAATCCCGGGCCCATGGCCGTGCCCACTCAGGTCCTGGGGTTGTTGCTACTGTGGCTTACAGATGCCAGATGTGACATCCAGATGACACAGTCTCCATCTTCCCTGTCTGCATCTGTCGGAGATCGCGTCACCATCACCTGTCGAGCAAGTGAGGACATTTATTTTAATTTAGTGTGGTATCAGCAGAAACCAGGAAAGGCCCCTAAGCTCCTGATCTATGATACAAATCGCTTGGCAGATGGGGTCCCATCACGGTTCAGTGGCTCTGGATCTGGCACACAGTATACTCTAACCATAAGTAGCCTGCAACCCGAAGATTTCGCAACCTATTATTGTCAACACTATAAGAATTATCCGCTCACGTTCGGTCAGGGGACCAAGCTGGAAATCAAAAGATCTGGTGGCGGAGGGTCAGGAGGCGGAGGCAGCGGAGGCGGTGGCTCGGGAGGCGGAGGCTCGAGATCTGAGGTGCAGTTGGTGGAGTCTGGGGGCGGCTTGGTGCAGCCTGGAGGGTCCCTGAGGCTCTCCTGTGCAGCCTCAGGATTCACTCTCAGTAATTATGGCATGCACTGGATCAGGCAGGCTCCAGGGAAGGGTCTGGAGTGGGTCTCGTCTATTAGTCTTAATGGTGGTAGCACTTACTATCGAGACTCCGTGAAGGGCCGATTCACTATCTCCAGGGACAATGCAAAAAGCACCCTCTACCTTCAAATGAATAGTCTGAGGGCCGAGGACACGGCCGTCTATTACTGTGCAGCACAGGACGCTTATACGGGAGGTTACTTTGATTACTGGGGCCAAGGAACGCTGGTCACAGTCTCGTCTATGGATCCCGCCGAGCCCAAATCTCCTGACAAAACTCACACATGCCCACCGTGCCCAGCACCTCCCGTGGCCGGCCCGTCAGTCTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCGCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAACCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCCCTGCACAATCACTATACCCAGAAATCTCTGAGTCTGAGCCCAGGCAAGAAGGACCCCAAGGCGGTTTTTGGCTGTATCTTTGGTGCCCTGGTTATTGTGACTGTGGGAGGCTTCATCTTCTGGAGAAAGAAGAGGAAAGATGCAAAGAATAATGAAGTGTCCTTTTCTCAAATTAAACCTAAAAAATCTAAGTTAATCAGAGTGGAGAATTTTGAGGCCTACTTCAAGAAGCAGCAAGCTGACTCCAACTGTGGGTTCGCAGAGGAATACGAAGATCTGAAGCTTGTTGGAATTAGTCAACCTAAATATGCAGCAGAACTGGCTGAGAATAGAGGAAAGAATCGCTATAATAATGTTCTGCCCTATGATATTTCCCGTGTCAAACTTTCGGTCCAGACCCATTCAACGGATGACTACATCAATGCCAACTACATGCCTGGCTACCACTCCAAGAAAGATTTTATTGCCACACAAGGACCTTTACCGAACACTTTGAAAGATTTTTGGCGTATGGTTTGGGAGAAAAATGTATATGCCATCATTATGTTGACTAAATGTGTTGAACAGGGAAGAACCAAATGTGAGGAGTATTGGCCCTCCAAGCAGGCTCAGGACTATGGAGACATAACTGTGGCAATGACATCAGAAATTGTTCTTCCGGAATGGACCATCAGAGATTTCACAGTGAAAAATATCCAGACAAGTGAGAGTCACCCTCTGAGACAGTTCCATTTCACCTCCTGGCCAGACCACGGTGTTCCCGACACCACTGACCTGCTCATCAACTTCCGGTACCTCGTTCGTGACTACATGAAGCAGAGTCCTCCCGAATCGCCGATTCTGGTGCATTGCAGTGCTGGGGTCGGAAGGACGGGCACTTTCATTGCCATTGATCGTCTCATCTACCAGATAGAGAATGAGAACACCGTGGATGTGTATGGGATTGTGTATGACCTTCGAATGCATAGGCCTTTAATGGTGCAGACAGAGGACCAGTATGTTTTCCTCAATCAGTGTGTTTTGGATATTGTCAGATCCCAGAAAGACTCAAAAGTAGATCTTATCTACCAGAACACAACTGCAATGACAATCTATGAAAACCTTGCGCCCGTGACCACATTTGGAAAGACCAATGGTTACATCGCCTAA
SEQ ID № 38
>16076.SFG.aCD19fmc63-CD8STK-CD28tmZ-2A-aCD33glx-muCD8STK-tm-dPTPN6
ATGAGCCTGCCCGTGACCGCCCTGCTGCTGCCCCTGGCCCTGCTGCTGCACGCCGCCAGACCAGACATCCAGATGACCCAGACCACCAGCAGCCTGAGCGCCAGCCTGGGCGACCGGGTGACCATCAGCTGCAGAGCCAGCCAGGACATCAGCAAGTACCTGAACTGGTACCAGCAGAAGCCCGACGGCACCGTGAAGCTGCTGATCTACCACACCAGCCGGCTGCACAGCGGCGTGCCCAGCCGGTTCAGCGGCAGCGGCAGCGGCACCGACTACAGCCTGACCATCAGCAACCTGGAGCAGGAGGACATCGCCACCTACTTCTGCCAGCAGGGCAACACCCTGCCCTACACCTTCGGAGGCGGCACCAAGCTGGAGATCACCAAGGCCGGAGGCGGAGGCTCTGGCGGAGGCGGCTCTGGCGGAGGCGGCTCTGGCGGAGGCGGCAGCGAGGTGAAGCTGCAGGAGTCTGGCCCAGGCCTGGTGGCCCCAAGCCAGAGCCTGAGCGTGACCTGCACCGTGAGCGGCGTGAGCCTGCCCGACTACGGCGTGAGCTGGATCAGGCAGCCCCCACGGAAGGGCCTGGAGTGGCTGGGCGTGATCTGGGGCAGCGAGACCACCTACTACAACAGCGCCCTGAAGAGCCGGCTGACCATCATCAAGGACAACAGCAAGAGCCAGGTGTTCCTGAAGATGAACAGCCTGCAGACCGACGACACCGCCATCTACTACTGCGCCAAGCACTACTACTATGGCGGCAGCTACGCTATGGACTACTGGGGCCAGGGCACCAGCGTGACCGTGAGCTCAGATCCCACCACGACGCCAGCGCCGCGACCACCAACACCGGCGCCCACCATCGCGTCGCAGCCCCTGTCCCTGCGCCCAGAGGCGTGCCGGCCAGCGGCGGGGGGCGCAGTGCACACGAGGGGGCTGGACTTCGCCTGTGATATCTTTTGGGTGCTGGTGGTGGTTGGTGGAGTCCTGGCTTGCTATAGCTTGCTAGTAACAGTGGCCTTTATTATTTTCTGGGTGAGGAGAGTGAAGTTCAGCAGGAGCGCAGACGCCCCCGCGTACCAGCAGGGCCAGAACCAGCTCTATAACGAGCTCAATCTAGGACGAAGAGAGGAGTACGATGTTTTGGACAAGAGACGTGGCCGGGACCCTGAGATGGGGGGAAAGCCGAGAAGGAAGAACCCTCAGGAAGGCCTGTACAATGAACTGCAGAAAGATAAGATGGCGGAGGCCTACAGTGAGATTGGGATGAAAGGCGAGCGCCGGAGGGGCAAGGGGCACGATGGCCTTTACCAGGGTCTCAGTACAGCCACCAAGGACACCTACGACGCCCTTCACATGCAGGCCCTGCCTCCTCGCAGAGCCGAGGGCAGGGGAAGTCTTCTAACATGCGGGGACGTGGAGGAAAATCCCGGGCCCATGGCCGTGCCCACTCAGGTCCTGGGGTTGTTGCTACTGTGGCTTACAGATGCCAGATGTGACATCCAGATGACACAGTCTCCATCTTCCCTGTCTGCATCTGTCGGAGATCGCGTCACCATCACCTGTCGAGCAAGTGAGGACATTTATTTTAATTTAGTGTGGTATCAGCAGAAACCAGGAAAGGCCCCTAAGCTCCTGATCTATGATACAAATCGCTTGGCAGATGGGGTCCCATCACGGTTCAGTGGCTCTGGATCTGGCACACAGTATACTCTAACCATAAGTAGCCTGCAACCCGAAGATTTCGCAACCTATTATTGTCAACACTATAAGAATTATCCGCTCACGTTCGGTCAGGGGACCAAGCTGGAAATCAAAAGATCTGGTGGCGGAGGGTCAGGAGGCGGAGGCAGCGGAGGCGGTGGCTCGGGAGGCGGAGGCTCGAGATCTGAGGTGCAGTTGGTGGAGTCTGGGGGCGGCTTGGTGCAGCCTGGAGGGTCCCTGAGGCTCTCCTGTGCAGCCTCAGGATTCACTCTCAGTAATTATGGCATGCACTGGATCAGGCAGGCTCCAGGGAAGGGTCTGGAGTGGGTCTCGTCTATTAGTCTTAATGGTGGTAGCACTTACTATCGAGACTCCGTGAAGGGCCGATTCACTATCTCCAGGGACAATGCAAAAAGCACCCTCTACCTTCAAATGAATAGTCTGAGGGCCGAGGACACGGCCGTCTATTACTGTGCAGCACAGGACGCTTATACGGGAGGTTACTTTGATTACTGGGGCCAAGGAACGCTGGTCACAGTCTCGTCTATGGATCCCGCCACCACAACCAAGCCCGTGCTGCGGACCCCAAGCCCTGTGCACCCTACCGGCACCAGCCAGCCTCAGAGACCCGAGGACTGCCGGCCTCGGGGCAGCGTGAAGGGCACCGGCCTGGACTTCGCCTGCGACATCTACTGGGCACCTCTGGCCGGAATATGCGTGGCACTGCTGCTGAGCCTCATCATCACCCTGATCTGTTATCACCGAAGCCGCAAGCGGGTGTGTAAAAGTGGAGGCGGAAGCTTCTGGGAGGAGTTTGAGAGTTTGCAGAAGCAGGAGGTGAAGAACTTGCACCAGCGTCTGGAAGGGCAGCGGCCAGAGAACAAGGGCAAGAACCGCTACAAGAACATTCTCCCCTTTGACCACAGCCGAGTGATCCTGCAGGGACGGGACAGTAACATCCCCGGGTCCGACTACATCAATGCCAACTACATCAAGAACCAGCTGCTAGGCCCTGATGAGAACGCTAAGACCTACATCGCCAGCCAGGGCTGTCTGGAGGCCACGGTCAATGACTTCTGGCAGATGGCGTGGCAGGAGAACAGCCGTGTCATCGTCATGACCACCCGAGAGGTGGAGAAAGGCCGGAACAAATGCGTCCCATACTGGCCCGAGGTGGGCATGCAGCGTGCTTATGGGCCCTACTCTGTGACCAACTGCGGGGAGCATGACACAACCGAATACAAACTCCGTACCTTACAGGTCTCCCCGCTGGACAATGGAGACCTGATTCGGGAGATCTGGCATTACCAGTACCTGAGCTGGCCCGACCACGGGGTCCCCAGTGAGCCTGGGGGTGTCCTCAGCTTCCTGGACCAGATCAACCAGCGGCAGGAAAGTCTGCCTCACGCAGGGCCCATCATCGTGCACTGCAGCGCCGGCATCGGCCGCACAGGCACCATCATTGTCATCGACATGCTCATGGAGAACATCTCCACCAAGGGCCTGGACTGTGACATTGACATCCAGAAGACCATCCAGATGGTGCGGGCGCAGCGCTCGGGCATGGTGCAGACGGAGGCGCAGTACAAGTTCATCTACGTGGCCATCGCCCAGTTCATTGAAACCACTAAGAAGAAGCTGTGA
SEQ ID № 39
>MP16091.SFG.aCD19fmc63-CD8STK-CD28tmZ-2A-aCD33glx-muCD8STK-LAIR1tm-endo
ATGAGCCTGCCCGTGACCGCCCTGCTGCTGCCCCTGGCCCTGCTGCTGCACGCCGCCAGACCAGACATCCAGATGACCCAGACCACCAGCAGCCTGAGCGCCAGCCTGGGCGACCGGGTGACCATCAGCTGCAGAGCCAGCCAGGACATCAGCAAGTACCTGAACTGGTACCAGCAGAAGCCCGACGGCACCGTGAAGCTGCTGATCTACCACACCAGCCGGCTGCACAGCGGCGTGCCCAGCCGGTTCAGCGGCAGCGGCAGCGGCACCGACTACAGCCTGACCATCAGCAACCTGGAGCAGGAGGACATCGCCACCTACTTCTGCCAGCAGGGCAACACCCTGCCCTACACCTTCGGAGGCGGCACCAAGCTGGAGATCACCAAGGCCGGAGGCGGAGGCTCTGGCGGAGGCGGCTCTGGCGGAGGCGGCTCTGGCGGAGGCGGCAGCGAGGTGAAGCTGCAGGAGTCTGGCCCAGGCCTGGTGGCCCCAAGCCAGAGCCTGAGCGTGACCTGCACCGTGAGCGGCGTGAGCCTGCCCGACTACGGCGTGAGCTGGATCAGGCAGCCCCCACGGAAGGGCCTGGAGTGGCTGGGCGTGATCTGGGGCAGCGAGACCACCTACTACAACAGCGCCCTGAAGAGCCGGCTGACCATCATCAAGGACAACAGCAAGAGCCAGGTGTTCCTGAAGATGAACAGCCTGCAGACCGACGACACCGCCATCTACTACTGCGCCAAGCACTACTACTATGGCGGCAGCTACGCTATGGACTACTGGGGCCAGGGCACCAGCGTGACCGTGAGCTCAGATCCCACCACGACGCCAGCGCCGCGACCACCAACACCGGCGCCCACCATCGCGTCGCAGCCCCTGTCCCTGCGCCCAGAGGCGTGCCGGCCAGCGGCGGGGGGCGCAGTGCACACGAGGGGGCTGGACTTCGCCTGTGATATCTTTTGGGTGCTGGTGGTGGTTGGTGGAGTCCTGGCTTGCTATAGCTTGCTAGTAACAGTGGCCTTTATTATTTTCTGGGTGAGGAGAGTGAAGTTCAGCAGGAGCGCAGACGCCCCCGCGTACCAGCAGGGCCAGAACCAGCTCTATAACGAGCTCAATCTAGGACGAAGAGAGGAGTACGATGTTTTGGACAAGAGACGTGGCCGGGACCCTGAGATGGGGGGAAAGCCGAGAAGGAAGAACCCTCAGGAAGGCCTGTACAATGAACTGCAGAAAGATAAGATGGCGGAGGCCTACAGTGAGATTGGGATGAAAGGCGAGCGCCGGAGGGGCAAGGGGCACGATGGCCTTTACCAGGGTCTCAGTACAGCCACCAAGGACACCTACGACGCCCTTCACATGCAGGCCCTGCCTCCTCGCAGAGCCGAGGGCAGGGGAAGTCTTCTAACATGCGGGGACGTGGAGGAAAATCCCGGGCCCATGGCCGTGCCCACTCAGGTCCTGGGGTTGTTGCTACTGTGGCTTACAGATGCCAGATGTGACATCCAGATGACACAGTCTCCATCTTCCCTGTCTGCATCTGTCGGAGATCGCGTCACCATCACCTGTCGAGCAAGTGAGGACATTTATTTTAATTTAGTGTGGTATCAGCAGAAACCAGGAAAGGCCCCTAAGCTCCTGATCTATGATACAAATCGCTTGGCAGATGGGGTCCCATCACGGTTCAGTGGCTCTGGATCTGGCACACAGTATACTCTAACCATAAGTAGCCTGCAACCCGAAGATTTCGCAACCTATTATTGTCAACACTATAAGAATTATCCGCTCACGTTCGGTCAGGGGACCAAGCTGGAAATCAAAAGATCTGGTGGCGGAGGGTCAGGAGGCGGAGGCAGCGGAGGCGGTGGCTCGGGAGGCGGAGGCTCGAGATCTGAGGTGCAGTTGGTGGAGTCTGGGGGCGGCTTGGTGCAGCCTGGAGGGTCCCTGAGGCTCTCCTGTGCAGCCTCAGGATTCACTCTCAGTAATTATGGCATGCACTGGATCAGGCAGGCTCCAGGGAAGGGTCTGGAGTGGGTCTCGTCTATTAGTCTTAATGGTGGTAGCACTTACTATCGAGACTCCGTGAAGGGCCGATTCACTATCTCCAGGGACAATGCAAAAAGCACCCTCTACCTTCAAATGAATAGTCTGAGGGCCGAGGACACGGCCGTCTATTACTGTGCAGCACAGGACGCTTATACGGGAGGTTACTTTGATTACTGGGGCCAAGGAACGCTGGTCACAGTCTCGTCTATGGATCCCGCCACCACAACCAAGCCCGTGCTGCGGACCCCAAGCCCTGTGCACCCTACCGGCACCAGCCAGCCTCAGAGACCCGAGGACTGCCGGCCTCGGGGCAGCGTGAAGGGCACCGGCCTGGACTTCGCCTGCGACATTCTCATCGGGGTCTCAGTGGTCTTCCTCTTCTGTCTCCTCCTCCTGGTCCTCTTCTGCCTCCATCGCCAGAATCAGATAAAGCAGGGGCCCCCCAGAAGCAAGGACGAGGAGCAGAAGCCACAGCAGAGGCCTGACCTGGCTGTTGATGTTCTAGAGAGGACAGCAGACAAGGCCACAGTCAATGGACTTCCTGAGAAGGACCGGGAGACCGACACCAGCGCCCTGGCTGCAGGGAGTTCCCAGGAGGTGACGTATGCTCAGCTGGACCACTGGGCCCTCACACAGAGGACAGCCCGGGCTGTGTCCCCACAGTCCACAAAGCCCATGGCCGAGTCCATCACGTATGCAGCCGTTGCCAGACACTGA
SEQ ID № 40
>MP16092.SFG.aCD19fmc63-CD8STK-CD28tmZ-2A-aCD33glx-muCD8STK-LAIR1tm-endo-2A-PTPN6_SH2-dCD148
ATGAGCCTGCCCGTGACCGCCCTGCTGCTGCCCCTGGCCCTGCTGCTGCACGCCGCCAGACCAGACATCCAGATGACCCAGACCACCAGCAGCCTGAGCGCCAGCCTGGGCGACCGGGTGACCATCAGCTGCAGAGCCAGCCAGGACATCAGCAAGTACCTGAACTGGTACCAGCAGAAGCCCGACGGCACCGTGAAGCTGCTGATCTACCACACCAGCCGGCTGCACAGCGGCGTGCCCAGCCGGTTCAGCGGCAGCGGCAGCGGCACCGACTACAGCCTGACCATCAGCAACCTGGAGCAGGAGGACATCGCCACCTACTTCTGCCAGCAGGGCAACACCCTGCCCTACACCTTCGGAGGCGGCACCAAGCTGGAGATCACCAAGGCCGGAGGCGGAGGCTCTGGCGGAGGCGGCTCTGGCGGAGGCGGCTCTGGCGGAGGCGGCAGCGAGGTGAAGCTGCAGGAGTCTGGCCCAGGCCTGGTGGCCCCAAGCCAGAGCCTGAGCGTGACCTGCACCGTGAGCGGCGTGAGCCTGCCCGACTACGGCGTGAGCTGGATCAGGCAGCCCCCACGGAAGGGCCTGGAGTGGCTGGGCGTGATCTGGGGCAGCGAGACCACCTACTACAACAGCGCCCTGAAGAGCCGGCTGACCATCATCAAGGACAACAGCAAGAGCCAGGTGTTCCTGAAGATGAACAGCCTGCAGACCGACGACACCGCCATCTACTACTGCGCCAAGCACTACTACTATGGCGGCAGCTACGCTATGGACTACTGGGGCCAGGGCACCAGCGTGACCGTGAGCTCAGATCCCACCACGACGCCAGCGCCGCGACCACCAACACCGGCGCCCACCATCGCGTCGCAGCCCCTGTCCCTGCGCCCAGAGGCGTGCCGGCCAGCGGCGGGGGGCGCAGTGCACACGAGGGGGCTGGACTTCGCCTGTGATATCTTTTGGGTGCTGGTGGTGGTTGGTGGAGTCCTGGCTTGCTATAGCTTGCTAGTAACAGTGGCCTTTATTATTTTCTGGGTGAGGAGAGTGAAGTTCAGCAGGAGCGCAGACGCCCCCGCGTACCAGCAGGGCCAGAACCAGCTCTATAACGAGCTCAATCTAGGACGAAGAGAGGAGTACGATGTTTTGGACAAGAGACGTGGCCGGGACCCTGAGATGGGGGGAAAGCCGAGAAGGAAGAACCCTCAGGAAGGCCTGTACAATGAACTGCAGAAAGATAAGATGGCGGAGGCCTACAGTGAGATTGGGATGAAAGGCGAGCGCCGGAGGGGCAAGGGGCACGATGGCCTTTACCAGGGTCTCAGTACAGCCACCAAGGACACCTACGACGCCCTTCACATGCAGGCCCTGCCTCCTCGCAGAGCCGAGGGCAGGGGAAGTCTTCTAACATGCGGGGACGTGGAGGAAAATCCCGGGCCCATGGCCGTGCCCACTCAGGTCCTGGGGTTGTTGCTACTGTGGCTTACAGATGCCAGATGTGACATCCAGATGACACAGTCTCCATCTTCCCTGTCTGCATCTGTCGGAGATCGCGTCACCATCACCTGTCGAGCAAGTGAGGACATTTATTTTAATTTAGTGTGGTATCAGCAGAAACCAGGAAAGGCCCCTAAGCTCCTGATCTATGATACAAATCGCTTGGCAGATGGGGTCCCATCACGGTTCAGTGGCTCTGGATCTGGCACACAGTATACTCTAACCATAAGTAGCCTGCAACCCGAAGATTTCGCAACCTATTATTGTCAACACTATAAGAATTATCCGCTCACGTTCGGTCAGGGGACCAAGCTGGAAATCAAAAGATCTGGTGGCGGAGGGTCAGGAGGCGGAGGCAGCGGAGGCGGTGGCTCGGGAGGCGGAGGCTCGAGATCTGAGGTGCAGTTGGTGGAGTCTGGGGGCGGCTTGGTGCAGCCTGGAGGGTCCCTGAGGCTCTCCTGTGCAGCCTCAGGATTCACTCTCAGTAATTATGGCATGCACTGGATCAGGCAGGCTCCAGGGAAGGGTCTGGAGTGGGTCTCGTCTATTAGTCTTAATGGTGGTAGCACTTACTATCGAGACTCCGTGAAGGGCCGATTCACTATCTCCAGGGACAATGCAAAAAGCACCCTCTACCTTCAAATGAATAGTCTGAGGGCCGAGGACACGGCCGTCTATTACTGTGCAGCACAGGACGCTTATACGGGAGGTTACTTTGATTACTGGGGCCAAGGAACGCTGGTCACAGTCTCGTCTATGGATCCCGCCACCACAACCAAGCCCGTGCTGCGGACCCCAAGCCCTGTGCACCCTACCGGCACCAGCCAGCCTCAGAGACCCGAGGACTGCCGGCCTCGGGGCAGCGTGAAGGGCACCGGCCTGGACTTCGCCTGCGACATTCTCATCGGGGTCTCAGTGGTCTTCCTCTTCTGTCTCCTCCTCCTGGTCCTCTTCTGCCTCCATCGCCAGAATCAGATAAAGCAGGGGCCCCCCAGAAGCAAGGACGAGGAGCAGAAGCCACAGCAGAGGCCTGACCTGGCTGTTGATGTTCTAGAGAGGACAGCAGACAAGGCCACAGTCAATGGACTTCCTGAGAAGGACCGGGAGACCGACACCAGCGCCCTGGCTGCAGGGAGTTCCCAGGAGGTGACGTATGCTCAGCTGGACCACTGGGCCCTCACACAGAGGACAGCCCGGGCTGTGTCCCCACAGTCCACAAAGCCCATGGCCGAGTCCATCACGTATGCAGCCGTTGCCAGACACAGGGCAGAAGGAAGAGGTAGCCTGCTGACTTGCGGGGACGTGGAAGAGAACCCAGGGCCATGGTATCATGGCCACATGTCTGGCGGGCAGGCAGAGACGCTGCTGCAGGCCAAGGGCGAGCCCTGGACGTTTCTTGTGCGTGAGAGCCTCAGCCAGCCTGGAGACTTCGTGCTTTCTGTGCTCAGTGACCAGCCCAAGGCTGGCCCAGGCTCCCCGCTCAGGGTCACCCACATCAAGGTCATGTGCGAGGGTGGACGCTACACAGTGGGTGGTTTGGAGACCTTCGACAGCCTCACGGACCTGGTGGAGCATTTCAAGAAGACGGGGATTGAGGAGGCCTCAGGCGCCTTTGTCTACCTGCGGCAGCCGTACAGCGGTGGCGGTGGCAGCTTTGAGGCCTACTTCAAGAAGCAGCAAGCTGACTCCAACTGTGGGTTCGCAGAGGAATACGAAGATCTGAAGCTTGTTGGAATTAGTCAACCTAAATATGCAGCAGAACTGGCTGAGAATAGAGGAAAGAATCGCTATAATAATGTTCTGCCCTATGATATTTCCCGTGTCAAACTTTCGGTCCAGACCCATTCAACGGATGACTACATCAATGCCAACTACATGCCTGGCTACCACTCCAAGAAAGATTTTATTGCCACACAAGGACCTTTACCGAACACTTTGAAAGATTTTTGGCGTATGGTTTGGGAGAAAAATGTATATGCCATCATTATGTTGACTAAATGTGTTGAACAGGGAAGAACCAAATGTGAGGAGTATTGGCCCTCCAAGCAGGCTCAGGACTATGGAGACATAACTGTGGCAATGACATCAGAAATTGTTCTTCCGGAATGGACCATCAGAGATTTCACAGTGAAAAATATCCAGACAAGTGAGAGTCACCCTCTGAGACAGTTCCATTTCACCTCCTGGCCAGACCACGGTGTTCCCGACACCACTGACCTGCTCATCAACTTCCGGTACCTCGTTCGTGACTACATGAAGCAGAGTCCTCCCGAATCGCCGATTCTGGTGCATTGCAGTGCTGGGGTCGGAAGGACGGGCACTTTCATTGCCATTGATCGTCTCATCTACCAGATAGAGAATGAGAACACCGTGGATGTGTATGGGATTGTGTATGACCTTCGAATGCATAGGCCTTTAATGGTGCAGACAGAGGACCAGTATGTTTTCCTCAATCAGTGTGTTTTGGATATTGTCAGATCCCAGAAAGACTCAAAAGTAGATCTTATCTACCAGAACACAACTGCAATGACAATCTATGAAAACCTTGCGCCCGTGACCACATTTGGAAAGACCAATGGTTACATCGCCAGCGGTAGCTAA
Последовательность нуклеиновой кислоты может кодировать ту же аминокислотную последовательность, как та, которую кодирует SEQ ID № 35, 36, 37, 38, 39 или 40, но может иметь отличающуюся последовательность нуклеиновой кислоты из-за вырожденности генетического кода. Последовательность нуклеиновой кислоты может иметь по меньшей мере 80, 85, 90, 95, 98 или 99% идентичности с последовательностью, представленной в качестве SEQ ID № 35, 36, 37, 38, 39 или 40, при условии, что она кодирует первый CAR и второй CAR, как определено в первом аспекте изобретения.
ВЕКТОР
Настоящее изобретение также относится к вектору или набору векторов, которые содержат одну или несколько CAR-кодирующих последовательностей нуклеиновых кислот. Такой вектор можно использовать для того, чтобы вводить последовательность (последовательности) нуклеиновой кислоты в клетку-хозяина с тем, чтобы она экспрессировала первый и второй CAR.
Вектор может представлять собой, например, плазмидный или вирусный вектор, такой как ретровирусный вектор или лентивирусный вектор, или вектор на основе транспозона или синтетическую мРНК.
Вектор может быть способен к трансфекции или трансдукции T-клетки.
ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ
Настоящее изобретение также относится к фармацевтической композиции, которая содержит множество CAR-экспрессирующих клеток, таких как T-клетки или естественные киллерные клетки, в соответствии с первым аспектом изобретения. Фармацевтическая композиция дополнительно может содержать фармацевтически приемлемый носитель, разбавитель или эксципиент. Фармацевтическая композиция необязательно может содержать один или несколько дополнительных фармацевтически активных полипептидов и/или соединений. Такой состав, например, может быть в форме, подходящей для внутривенной инфузии.
СПОСОБ ЛЕЧЕНИЯ
T-клетки по настоящему изобретению могут быть способны уничтожать клетки-мишени, такие как клетки злокачественной опухоли. Клетка-мишень может быть распознана с помощью определенного паттерна антигенной экспрессии, например, экспрессии антигена A И антигена B; экспрессии антигена A ИЛИ антигена B; или экспрессии антигена A И НЕ антигена B или комплексных итераций этих логических элементов.
T-клетки по настоящему изобретению можно использовать для лечения инфекции, такой как вирусная инфекция.
T-клетки по изобретению также можно использовать для управления патогенными иммунными ответами, например, при аутоиммунных заболеваниях, аллергиях и реакциях трансплантат против хозяина.
T-клетки по изобретению можно использовать для лечения злокачественных заболеваний, таких как злокачественная опухоль мочевого пузыря, злокачественная опухоль молочной железы, злокачественная опухоль ободочной кишки, злокачественная опухоль эндометрия, злокачественная опухоль почки (клеток почки), лейкоз, злокачественная опухоль легких, меланома, неходжкинская лимфома, злокачественная опухоль поджелудочной железы, злокачественная опухоль предстательной железы и злокачественная опухоль щитовидной железы.
В частности, это подходит для лечения солидных опухолей, когда доступность хороших избирательных одинарных мишеней ограничена.
T-клетки по изобретению можно использовать для того, чтобы лечить: злокачественные опухоли полости рта и глотки, которые включают злокачественную опухоль языка, рта и глотки; злокачественные опухоли пищеварительной системы, которые включают злокачественные опухоли пищевода, желудка и толстой кишки; злокачественные опухоли печени и желчных протоков, которые включают гепатоцеллюлярные карциномы и холангиокарциномы; злокачественные опухоли дыхательной системы, которые включают бронхогенные злокачественные опухоли и злокачественные опухоли гортани; злокачественные опухоли костей и суставов, которые включают остеосаркому; злокачественные опухоли кожи, которые включают меланому; злокачественную опухоль молочной железы; злокачественные опухоли полового тракта, которые включают злокачественные опухоли матки, яичников и шейки матки у женщин, злокачественные опухоли предстательной железы и яичек у мужчин; злокачественные опухоли мочевыделительной системы, которые включают почечноклеточную карциному и переходно-клеточные карциномы уретры или мочевого пузыря; злокачественные опухоли головного мозга, включая глиомы, мультиформную глиобластому и медуллобластомы; злокачественные опухоли эндокринной системы, включая злокачественную опухоль щитовидной железы, карциному надпочечника, и злокачественные опухоли, связанные с синдромами множественных эндокринных новообразований; лимфомы, включая лимфому Ходжкина и неходжкинскую лимфому; множественную миелому и плазмоцитомы; лейкозы, как острые, так и хронические, миелоидные или лимфоидные; и злокачественные опухоли других и неустановленных мест, включая нейробластому.
Лечение с использованием T-клеток по изобретению может помогать предотвращать ускользание или освобождение опухолевых клеток, которые часто возникают при стандартных подходах.
Далее изобретение дополнительно описано в виде примеров, которые должны служить в качестве помощи специалисту в данной области при выполнении изобретения и не предназначены каким-либо образом ограничивать объем изобретения.
ПРИМЕРЫ
Пример 1 - создание популяций клеток-мишеней
С целью подтверждения принципа изобретения произвольно выбирали рецепторы на основании средств против CD19 и против CD33. Используя ретровирусные векторы, клонировали CD19 и CD33. Эти белки усекали с тем, чтобы они не передавали сигнал и могли быть стабильно экспрессированы в течение длительных периодов. Затем эти векторы использовали для трансдукции клеточной линии SupT1, или однократно или двукратно, чтобы создавать клетки, негативные по обоим антигенам (дикого типа), положительные по любому и положительные по обоим. Экспрессионные данные представлены на фиг. 3.
Пример 2 - конструкция и функция логического элемента ИЛИ
Для того чтобы конструировать логический элемент ИЛИ, совместно экспрессировали пару рецепторов, распознающих CD19 и CD33. Различные спейсеры использовали для того, чтобы предотвращать образование перекрестных пар. Оба рецептора имели трансмембранный домен, полученный из CD28, чтобы повышать поверхностную стабильность, и эндодомен, полученный из такового в CD3 дзета, чтобы предоставлять простой активирующий сигнал. Таким образом, совместно экспрессировали пару независимых CAR первого поколения. В ретровирусной векторной кассете, используемой для того, чтобы совместно экспрессировать последовательности, используют саморасщепляющийся пептид 2A вируса ящура, чтобы сделать возможным коэкспрессию обоих рецепторов 1:1. Конструкция кассеты показана на фиг. 4, а структуры белков на фиг. 5. В нуклеотидную последовательность гомологичных областей вносили неоднозначность кодонов для того, чтобы предотвращать рекомбинацию во время обратной транскрипции ретровирусного вектора.
Пример 3 - тестирование логического элемента ИЛИ
Экспрессию обоих CAR тестировали на поверхности T-клеток посредством окрашивания когнатным антигеном, слитым с Fc. Используя Fc домены различных видов (мышиные для CD19 и кроличьи для CD33), коэкспрессию обоих CAR определяли на клеточной поверхности посредством окрашивания различными вторичными антителами, конъюгированными с различными флуорофорами. Это показано на фиг. 6.
Затем осуществляли функциональное тестирование с использованием линии Т-клеток мыши BW5147. Эта клеточная линия секретирует IL-2 при активации, что делает возможным простое количественное считывание. Эти T-клетки совместно культивировали с возрастающими количествами искусственных клеток-мишеней, описанных выше. T-клетки реагировали на клетки-мишени, экспрессирующие их антиген, как показано с помощью секреции IL-2, измеряемой посредством ELISA. Показана экспрессия обоих CAR на клеточных поверхностях и показано, что T-клетки реагировали на любой или оба антигена. Эти данные представлены на фиг. 7.
Пример 4 - конструкция и функция логического элемента И
Логический элемент И объединяет простой активирующий рецептор с рецептором, который осуществляет базовое ингибирование активности, но у которого ингибирование выключается, когда рецептор лигирован. Этого достигали посредством объединения стандартного CAR первого поколения с коротким/некрупным спейсером из «стебля» CD8 и дзета-эндодоменом CD3 со вторым рецептором с крупным Fc спейсером, эндодомен которого содержал или эндодомен CD148 или эндодомен CD45. Когда оба рецептора лигированы, различия в размерах спейсеров ведут к выделению различных рецепторов в различные мембранные компартменты, устранению ингибирования CD3 дзета рецептора с помощью эндодоменов CD148 или CD45. Таким образом, активация происходит только когда активируют оба рецептора. Для этого выбрали CD148 и CD45, поскольку они функционируют так нативным образом: например, очень крупный CD45 эктодомен исключает весь рецептор из иммунологического синапса. Экспрессирующая кассета изображена на фиг. 8 и последующие белки на фиг. 9.
Окрашивание поверхности по другой специфичности демонстрировало, что обе пары рецепторов могут быть эффективно экспрессированы на клеточной поверхности, представленной на фиг. 10. Функционирование в BW5147 показывает, что активация T-клеток происходит только в присутствии обоих антигенов (фиг. 11).
Пример 5: демонстрация генерализуемости логического элемента И
Чтобы гарантировать, что наблюдения не представляли собой манифестацию некоторой конкретной характеристики CD19/CD33 и использованных их связывающих средств, два направленных scFv меняли местами так, что после этого передача активирующего (ITAM) сигнала происходила при распознавании CD33 вместо CD19; а передача ингибирующего (CD148) сигнала происходила при распознавании CD19 вместо CD33. Поскольку эндодомены CD45 и CD148 считают функционально схожими, эксперименты ограничивали логическими элементами И с эндодоменом CD148. Это все еще должно давать функциональный логический элемент И. T-клетки, экспрессирующие новый логический элемент, стимулировали мишенями, несущими отдельно CD19 или CD33 или их оба. T-клетки отвечали на мишени, экспрессирующие как CD19, так и CD33, но не на мишени, экспрессирующие только один или ни одного из этих антигенов. Это демонстрирует, что логический элемент И все еще функционален в этом формате (фиг. 18B).
На тех же линиях предполагали установить, насколько генерализуемым является логический элемент И: логический элемент И должен быть генерализуемым среди различных мишеней. Хотя может иметь место меньшая или большая точность логического элемента с учетом относительной антигенной плотности, кинетики когнатного связывания scFv и конкретного расстояния до связывающего эпитопа scFv, следует ожидать увидеть некоторые манифестации логического элемента И с широким набором мишеней и связывающих средств. Для того чтобы тестировать это, создавали три дополнительных логических элемента И. Еще раз, эксперименты ограничивали версией логического элемента И с CD148. Второй scFv из исходного логического элемента И с CD148 заменяли на scFv huK666 против GD2 (SEQ ID 41 и SEQ ID 42) или на scFv против CD5 (SEQ ID 43 и SEQ ID 44) или scFv MR1.1 против EGFRvIII (SEQ ID 45 И SEQ ID 46) для того, чтобы создавать следующие CAR логические элементы И: CD19 И GD2; CD19 И CD5; CD19 И EGFRvIII. Также создавали следующие искусственные экспрессирующие антиген клеточные линии: посредством трансдукции SupT1 и SupT1.CD19 по изобретению синтазами GM3 и GD2 создавали SupT1.GD2 и SupT1.CD19.GD2. Посредством трансдукции SupT1 и SupT1.CD19 ретровирусным вектором, кодирующим EGFRvIII, создавали SupT1.EGFRvIII и SupT1.CD19.EGFRvIII. Поскольку CD5 экспрессирован на клетках SupT1, использовали другую клеточную линию для того, чтобы создавать клетки-мишени: создавали 293T клетки, которые экспрессируют только CD19, только CD5 и вместе CD5 и CD19. Экспрессию подтверждали с помощью проточной цитометрии (фиг. 19). T-клетки, экспрессирующие три новых CAR логических элемента И, стимулировали с использованием SupT1.CD19 и соответствующих когнатных двойных положительных и одинарных положительных клеток-мишеней. Все три логических элемента И демонстрировали пониженную активацию двойными положительными клеточными линиями в сравнении с одинарными положительными мишенями (фиг. 20). Это демонстрирует генерализуемость конструкции логического элемента И на произвольные мишени и когнатные связывающие средства.
Пример 6: экспериментальное подтверждение модели кинетической сегрегации CAR логического элемента И
Цель заключалась в подтверждении моделированием того, что дифференциальная сегрегация, обусловленная различными спейсерами, представляет собой центральный механизм, лежаший в основе способности генерировать эти CAR логические элементы. Модель представляет собой то, что если лигирован только активирующий CAR, сильный ингибирующий «выключаемый лигированием» CAR находится в растворе в мембране и может ингибировать активирующий CAR. Когда лигированы оба CAR, если оба CAR спейсера достаточно различны, они будут сегрегироваться в синапсе и не локализоваться совместно. Таким образом, ключевое требование состоит в том, чтобы спейсеры было достаточно различными. Если модель верна, если оба спейсера достаточно схожи, что они были совместно локализованы, когда оба рецептора лигированы, логический элемент не сможет функционировать. Для того чтобы тестировать это, «крупный» Fc спейсер в исходом CAR заменяли на мышиный CD8 спейсер. Предсказано, что он имеет схожую длину, объем и заряд с CD8 человека, но при этом не образует перекрестных пар с ним. Таким образом, новый логический элемент имел первый CAR, который распознает CD19, спейсер «стебля» CD8 человека и активирующий эндодомен; тогда как второй CAR распознает CD33, имеет спейсер «стебля» CD8 мыши и эндодомен CD148 (фиг. 18C). T-клетки трансдуцировали для того, чтобы экспрессировать этот новый CAR логический элемент. Затем эти T-клетки стимулировали клетками SupT1, экспрессирующими только CD19, только CD33 или оба CD19 и CD33 вместе. T-клетки не отвечали на клетки SupT1, экспрессирующие любой антиген отдельно, как и исходный логический элемент И. Однако CAR T-клетки не могли отвечать на клетки SupT1, экспрессирующие оба антигена, тем самым подтверждая модель (фиг. 18C). Для функционального логического элемента И необходимо, чтобы оба CAR имели спейсеры, достаточно различные, чтобы они не локализовались совместно в иммунологическом синапсе (фиг. 23A и B).
Пример 7 - Конструкция и функция логического элемента И НЕ
Фосфатазы, такие как CD45 и CD148, достаточно активны, чтобы даже малое количество, входящее в иммунологический синапс, могло ингибировать ITAM активацию. Это является основной ингибирования логического элемента И. Другие классы фосфатаз не настолько активны, например, PTPN6 и родственные фосфатазы. Предсказано, что небольшое количество PTPN6, попадающее в синапс посредством диффузии, не будет ингибировать активацию. Кроме того, предсказано, что если ингибирующий CAR имеет достаточно схожи спейсер с активирующим CAR, они могут локализоваться совместно в синапсе, если оба CAR лигированы. В этом случае большие количества ингибирующего эндодомена будут достаточны для того, чтобы останавливать ITAM активацию, когда оба антигена присутствуют. Таким образом, можно создавать логический элемент И НЕ.
Для логического элемента НЕ И второй сигнал должен запрещать активацию. Это выполняют посредством привнесения ингибирующего сигнала в иммунологический синапс, например, посредством привнесения фосфатазы фермента, такого как PTPN6. Таким образом, авторы изобретения создавали начальный логический элемент И НЕ следующим образом: совместно экспрессировали два CAR, где первый распознает CD19, имеет спейсер «стебля» CD8 человека и активирующий эндодомен; и CAR против CD33 со спейсером «стебля» CD8 мыши и эндодоменом, который содержит каталитический домен из PTPN6 (SEQ ID 38, фиг. 13A и B). На фиг. 12 представлена подходящая кассета, а на фиг. 14 представлены предварительные функциональные данные.
Кроме того, для создания логического элемента И НЕ разрабатывали альтернативную стратегию. Иммунорецепторные тирозиновые ингибирующие мотивы (ITIM) активировали подобно ITAM в том отношении, что они становятся фосфорилированными с помощью lck при кластеризации и исключении фосфатаз. Взамен запуска активации посредством связывания ZAP70, фосфорилированные ITIM привлекают фосфатазы, такие как PTPN6, через их когнатные домены SH2. ITIM может выполнять функцию ингибирующего эндодомена до тех пор, пока спейсеры на активирующих и ингибирующих CAR могут локализоваться совместно. Для того чтобы создать эту конструкцию, логический элемент И НЕ создавали следующим образом: совместно экспрессировали два CAR - первый распознает CD19, имеет спейсер «стебля» CD8 человека и активирующий эндодомен; и CAR против CD33 со спейсером «стебля» CD8 мыши и ITIM-содержащим эндодоменом, полученным из такового из LAIR1 (SEQ ID 39, фиг. 13A и C).
Также разрабатывали более сложный логический элемент И НЕ также, посредством чего ITIM усиливали с помощью присутствия дополнительного химерного белка: внутриклеточное слияние домена SH2 из PTPN6 и эндодомена из CD148. В этой конструкции экспрессируют три белка - первый распознает CD19, имеет спейсер «стебля» CD8 человека и активирующий эндодомен; совместно экспрессировали с CAR против CD33 со спейсером «стебля» CD8 мыши и ITIM-содержащим эндодоменом, полученным из такового из LAIR1. Дополнительный пептид 2A делает возможной коэкспрессию слитного белка PTPN6-CD148 (SEQ ID 40, фиг. 13A и D). Предсказано, что эти логические элементы И НЕ будут иметь различный диапазон ингибирования: PTPN6-CD148 > PTPN6 >> ITIM.
T-клетки трансдуцировали с использованием этих логических элементов и стимулировали мишенями, экспрессирующими отдельно CD19 или CD33 или оба CD19 и CD33 вместе. Все три логических элемента отвечали на мишени, экспрессирующие только CD19, но не мишени, экспрессирующие оба CD19 и CD33 вместе (фиг. 21), что подтверждало, что все три логических элемента И НЕ были функциональны.
Пример 8: экспериментальное подтверждение модели кинетической сегрегации логического элемента И НЕ на основе PTPN6.
Модель логического элемента И НЕ основана на том факте, что свойства спейсеров, используемых в обоих CAR, являются ключевыми для правильного функционирования логического элемента. В функциональном логическом элементе И НЕ с использованием PTPN6, оба спейсера CAR достаточно схожи, чтобы, когда оба CAR лигированы, оба совместно локализовались в синапсе, так что высокая концентрация даже слабой PTPN6 является достаточной для того, чтобы ингибировать активацию. Если спейсеры различны, сегрегация в синапсе будет изолировать PTPN6 от ITAM, что делает возможной активацию, нарушающую логический элемент И НЕ. Для того чтобы тестировать это, создавали контроль, в котором заменяли мышиный спейсер из «стебля» CD8 на таковой из Fc. В этом случае, тестовый логический элемент состоял из двух CAR, первый распознает CD19, имеет спейсер «стебля» CD8 человека и ITAM эндодомен; тогда как второй CAR распознает CD33, имеет Fc спейсер и эндодомен, состоящий из фосфатазы из PTPN6. Этот логический элемент активирует в ответ на CD19, но также активирует в ответ на CD19 и CD33 вместе (фиг. 22B, где функционирование этого логического элемента сравнивают с таковой у исходного И НЕ и контрольного варианта логического элемента И, описанного в примере 6). Эти экспериментальные данные подтверждают модель, в которой для функционального логического элемента И НЕ с использованием PTPN6 необходимы совместно локализующие спейсеры.
Пример 9: экспериментальное подтверждение модели кинетической сегрегации логического элемента И НЕ на основе ITIM.
Подобно логическому элементу И НЕ на основе PTPN6, для логического элемента на основе ITIM также необходима совместная локализация в иммунологическом синапсе, чтобы функционировать в качестве логического элемента И НЕ. Чтобы подтвердить эту гипотезу, создавали контрольный логический элемент на основе ITIM следующим образом: совместно экспрессировали два CAR - первый распознает CD19, имеет спейсер «стебля» CD8 человека и активирующий эндодомен; второй CAR против CD33 с Fc спейсером и ITIM-содержащим эндодоменом, полученным из такового из LAIR1. Активность этого логического элемента сравнивали с таковой у исходного логического элемента И НЕ на основе ITIM. В этом случае активация модифицированного логического элемента происходила в ответ на мишени, экспрессирующие CD19, но также происходила в ответ на клетки, экспрессирующие оба CD19 и CD33. Эти данные показывают, что логические элементы И НЕ на основе ITIM придерживаются модели, основанной на кинетической сегрегации, и правильный спейсер следует выбирать для того, чтобы создавать функциональный логический элемент (фиг. 23B).
Пример 10: краткое описание модели логических элементов CAR, созданной посредством кинетической сегрегации
На основании текущего понимания модели кинетической сегрегации и экспериментальных данных, описанных в настоящем документе, краткое описание модели для логического элемента из двух CAR представлено на фиг. 24. На фиг. Показана клетка, которая экспрессирует два CAR, каждый распознает отличающийся антиген. Когда любой один или оба CAR распознают антиген-мишень на клетке, происходит формирование синапса и исключение нативных CD45 и CD148 из синапса из-за массивности их эктодомена. Это задает основу для активации T-клеток. В том случае, когда клетка-мишень несет только один когнатных антиген, когнатный CAR лигирован и когнатный CAR сегрегируется в синапс. Нелигированный CAR остается в растворе на мембране T-клетки и может диффундировать в синапс и из него с тем, чтобы происходило формирование области высокой локальной концентрации лигированного CAR с низкой концентрацией нелигированного CAR. В этом случае, если лигированный CAR имеет ITAM и нелигированный CAR имеет ингибирующий эндодомен «выключаемого лигированием», типа такой как таковой из CD148, количество нелигированного CAR является достаточным для того, чтобы ингибировать активацию и выключать логический элемент. В отличие от этого, в том случае, если лигированный CAR имеет ITAM и нелигированный CAR имеет ингибирующий эндодомен «включаемого лигированием» типа, такой как PTPN6, количество нелигированного CAR является недостаточным для того, чтобы осуществлять ингибирование, и логический элемент включен. Когда стимулируют клеткой-мишенью, несущей оба когнатных антигена, оба когнатных CAR лигированы и образуют часть иммунологического синапса. Важно, если CAR спейсеры достаточно схожи, CAR совместно локализованы в синапсе, но если CAR спейсере достаточно различны, CAR сегрегируются внутри синапса. В этом последнем случае, происходит формирование областей мембраны, в которых присутствуют высокие концентрации одного CAR, но другой CAR отсутствует. В этом случае, поскольку сегрегация выполнена, даже если ингибирующий эндодомен относится к «выключаемому лигированием» типу, логический элемент включен. В первом случае, происходит формирование областей мембраны с высокими концентрациями обоих CAR, смешанных вместе. В этом случае, поскольку оба эндодомена концентрированы, даже если ингибирующий эндодомен относится ко «включаемому лигированием» типу, логический элемент выключен. Посредством отбора в CAR T-клетке можно программировать правильную логическую комбинацию спейсера и эндодомена.
На основании вышеописанной работы авторов изобретения, они установили ряд правил разработки для того, чтобы сделать возможным поколение CAR для логических элементов (проиллюстрировано на фиг. 31). Для того чтобы создавать T-клетку с CAR логическим элементом «антиген A ИЛИ антиген B», следует создавать CAR против A и против B так, что (1) каждый CAR имеет спейсер, который делает возможным легкий доступ к антигену и формирование синапса так, что CAR функционирует, и (2) каждый CAR имеет активирующий эндодомен; для того чтобы генерировать T-клетку с CAR логическим элементом «антиген A И НЕ B», CAR против A и против B следует создавать так, что (1) оба CAR имеют спейсеры, которые не образуют перекрестных пар, но которые допускают совместную сегрегацию CAR при распознавании обоих когнатных антигенов на клетке-мишени, (2) и один CAR имеет активирующий эндодомен, тогда как другой CAR имеет эндодомен, который содержит или рекрутирует слабую фосфатазу (например, PTPN6); (3) для того чтобы генерировать T-клетку с CAR логическим элементов «антиген A И антиген B», следует создавать CAR против A и против B так, что (1) один CAR имеет спейсер, достаточно отличающийся от другого CAR так, что оба CAR не будет совместно сегрегироваться при распознавании обоих когнатных антигенов на клетке-мишени, (2) один CAR имеет активирующий эндодомен, тогда как другой CAR имеет эндодомен, который состоит из сильной фосфатазы (например, таковой из CD45 или CD148). Правильные спейсеры для достижения желаемого эффекта можно выбирать из набора спейсеров с известными размером/геометрической формой и т.п., а также принимая во внимание размер/геометрическую форму и т.п. антигена-мишени (например, см. фиг. 30) и местоположение когнатного эпитопа на антигене-мишени.
SEQ ID № 41: SFG.aCD19-CD8STK-CD28tmZ-2A-aGD2-HCH2CH3pvaa-dCD148
MSLPVTALLLPLALLLHAARPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITKAGGGGSGGGGSGGGGSGGGGSEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSDPTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIFWVLVVVGGVLACYSLLVTVAFIIFWVRRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRRAEGRGSLLTCGDVEENPGPMETDTLLLWVLLLWVPGSTGQVQLQESGPGLVKPSQTLSITCTVSGFSLASYNIHWVRQPPGKGLEWLGVIWAGGSTNYNSALMSRLTISKDNSKNQVFLKMSSLTAADTAVYYCAKRSDDYSWFAYWGQGTLVTVSSGGGGSGGGGSGGGGSENQMTQSPSSLSASVGDRVTMTCRASSSVSSSYLHWYQQKSGKAPKVWIYSTSNLASGVPSRFSGSGSGTDYTLTISSLQPEDFATYYCQQYSGYPITFGQGTKVEIKRSDPAEPKSPDKTHTCPPCPAPPVAGPSVFLFPPKPKDTLMIARTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGKKDPKAVFGCIFGALVIVTVGGFIFWRKKRKDAKNNEVSFSQIKPKKSKLIRVENFEAYFKKQQADSNCGFAEEYEDLKLVGISQPKYAAELAENRGKNRYNNVLPYDISRVKLSVQTHSTDDYINANYMPGYHSKKDFIATQGPLPNTLKDFWRMVWEKNVYAIIMLTKCVEQGRTKCEEYWPSKQAQDYGDITVAMTSEIVLPEWTIRDFTVKNIQTSESHPLRQFHFTSWPDHGVPDTTDLLINFRYLVRDYMKQSPPESPILVHCSAGVGRTGTFIAIDRLIYQIENENTVDVYGIVYDLRMHRPLMVQTEDQYVFLNQCVLDIVRSQKDSKVDLIYQNTTAMTIYENLAPVTTFGKTNGYIA
SEQ ID № 42: SFG.aCD19-CD8STK-CD28tmZ-2A-aGD2-HCH2CH3pvaa-dCD148
ATGAGCCTGCCCGTGACCGCCCTGCTGCTGCCCCTGGCCCTGCTGCTGCACGCCGCCAGACCAGACATCCAGATGACCCAGACCACCAGCAGCCTGAGCGCCAGCCTGGGCGACCGGGTGACCATCAGCTGCAGAGCCAGCCAGGACATCAGCAAGTACCTGAACTGGTACCAGCAGAAGCCCGACGGCACCGTGAAGCTGCTGATCTACCACACCAGCCGGCTGCACAGCGGCGTGCCCAGCCGGTTCAGCGGCAGCGGCAGCGGCACCGACTACAGCCTGACCATCAGCAACCTGGAGCAGGAGGACATCGCCACCTACTTCTGCCAGCAGGGCAACACCCTGCCCTACACCTTCGGAGGCGGCACCAAGCTGGAGATCACCAAGGCCGGAGGCGGAGGCTCTGGCGGAGGCGGCTCTGGCGGAGGCGGCTCTGGCGGAGGCGGCAGCGAGGTGAAGCTGCAGGAGTCTGGCCCAGGCCTGGTGGCCCCAAGCCAGAGCCTGAGCGTGACCTGCACCGTGAGCGGCGTGAGCCTGCCCGACTACGGCGTGAGCTGGATCAGGCAGCCCCCACGGAAGGGCCTGGAGTGGCTGGGCGTGATCTGGGGCAGCGAGACCACCTACTACAACAGCGCCCTGAAGAGCCGGCTGACCATCATCAAGGACAACAGCAAGAGCCAGGTGTTCCTGAAGATGAACAGCCTGCAGACCGACGACACCGCCATCTACTACTGCGCCAAGCACTACTACTATGGCGGCAGCTACGCTATGGACTACTGGGGCCAGGGCACCAGCGTGACCGTGAGCTCAGATCCCACCACGACGCCAGCGCCGCGACCACCAACACCGGCGCCCACCATCGCGTCGCAGCCCCTGTCCCTGCGCCCAGAGGCGTGCCGGCCAGCGGCGGGGGGCGCAGTGCACACGAGGGGGCTGGACTTCGCCTGTGATATCTTTTGGGTGCTGGTGGTGGTTGGTGGAGTCCTGGCTTGCTATAGCTTGCTAGTAACAGTGGCCTTTATTATTTTCTGGGTGAGGAGAGTGAAGTTCAGCAGGAGCGCAGACGCCCCCGCGTACCAGCAGGGCCAGAACCAGCTCTATAACGAGCTCAATCTAGGACGAAGAGAGGAGTACGATGTTTTGGACAAGAGACGTGGCCGGGACCCTGAGATGGGGGGAAAGCCGAGAAGGAAGAACCCTCAGGAAGGCCTGTACAATGAACTGCAGAAAGATAAGATGGCGGAGGCCTACAGTGAGATTGGGATGAAAGGCGAGCGCCGGAGGGGCAAGGGGCACGATGGCCTTTACCAGGGTCTCAGTACAGCCACCAAGGACACCTACGACGCCCTTCACATGCAGGCCCTGCCTCCTCGCAGAGCCGAGGGCAGGGGAAGTCTTCTAACATGCGGGGACGTGGAGGAAAATCCCGGGCCCATGGAGACCGACACCCTGCTGCTGTGGGTGCTGCTGCTGTGGGTGCCAGGCAGCACCGGCCAGGTGCAGCTGCAGGAGTCTGGCCCAGGCCTGGTGAAGCCCAGCCAGACCCTGAGCATCACCTGCACCGTGAGCGGCTTCAGCCTGGCCAGCTACAACATCCACTGGGTGCGGCAGCCCCCAGGCAAGGGCCTGGAGTGGCTGGGCGTGATCTGGGCTGGCGGCAGCACCAACTACAACAGCGCCCTGATGAGCCGGCTGACCATCAGCAAGGACAACAGCAAGAACCAGGTGTTCCTGAAGATGAGCAGCCTGACAGCCGCCGACACCGCCGTGTACTACTGCGCCAAGCGGAGCGACGACTACAGCTGGTTCGCCTACTGGGGCCAGGGCACCCTGGTGACCGTGAGCTCTGGCGGAGGCGGCTCTGGCGGAGGCGGCTCTGGCGGAGGCGGCAGCGAGAACCAGATGACCCAGAGCCCCAGCAGCTTGAGCGCCAGCGTGGGCGACCGGGTGACCATGACCTGCAGAGCCAGCAGCAGCGTGAGCAGCAGCTACCTGCACTGGTACCAGCAGAAGAGCGGCAAGGCCCCAAAGGTGTGGATCTACAGCACCAGCAACCTGGCCAGCGGCGTGCCCAGCCGGTTCAGCGGCAGCGGCAGCGGCACCGACTACACCCTGACCATCAGCAGCCTGCAGCCCGAGGACTTCGCCACCTACTACTGCCAGCAGTACAGCGGCTACCCCATCACCTTCGGCCAGGGCACCAAGGTGGAGATCAAGCGGTCGGATCCCGCCGAGCCCAAATCTCCTGACAAAACTCACACATGCCCACCGTGCCCAGCACCTCCCGTGGCCGGCCCGTCAGTCTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCGCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAACCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCCCTGCACAATCACTATACCCAGAAATCTCTGAGTCTGAGCCCAGGCAAGAAGGACCCCAAGGCGGTTTTTGGCTGTATCTTTGGTGCCCTGGTTATTGTGACTGTGGGAGGCTTCATCTTCTGGAGAAAGAAGAGGAAAGATGCAAAGAATAATGAAGTGTCCTTTTCTCAAATTAAACCTAAAAAATCTAAGTTAATCAGAGTGGAGAATTTTGAGGCCTACTTCAAGAAGCAGCAAGCTGACTCCAACTGTGGGTTCGCAGAGGAATACGAAGATCTGAAGCTTGTTGGAATTAGTCAACCTAAATATGCAGCAGAACTGGCTGAGAATAGAGGAAAGAATCGCTATAATAATGTTCTGCCCTATGATATTTCCCGTGTCAAACTTTCGGTCCAGACCCATTCAACGGATGACTACATCAATGCCAACTACATGCCTGGCTACCACTCCAAGAAAGATTTTATTGCCACACAAGGACCTTTACCGAACACTTTGAAAGATTTTTGGCGTATGGTTTGGGAGAAAAATGTATATGCCATCATTATGTTGACTAAATGTGTTGAACAGGGAAGAACCAAATGTGAGGAGTATTGGCCCTCCAAGCAGGCTCAGGACTATGGAGACATAACTGTGGCAATGACATCAGAAATTGTTCTTCCGGAATGGACCATCAGAGATTTCACAGTGAAAAATATCCAGACAAGTGAGAGTCACCCTCTGAGACAGTTCCATTTCACCTCCTGGCCAGACCACGGTGTTCCCGACACCACTGACCTGCTCATCAACTTCCGGTACCTCGTTCGTGACTACATGAAGCAGAGTCCTCCCGAATCGCCGATTCTGGTGCATTGCAGTGCTGGGGTCGGAAGGACGGGCACTTTCATTGCCATTGATCGTCTCATCTACCAGATAGAGAATGAGAACACCGTGGATGTGTATGGGATTGTGTATGACCTTCGAATGCATAGGCCTTTAATGGTGCAGACAGAGGACCAGTATGTTTTCCTCAATCAGTGTGTTTTGGATATTGTCAGATCCCAGAAAGACTCAAAAGTAGATCTTATCTACCAGAACACAACTGCAATGACAATCTATGAAAACCTTGCGCCCGTGACCACATTTGGAAAGACCAATGGTTACATCGCCTAA
SEQ ID № 43: SFG.aCD19-CD8STK-CD28tmZ-2A-aCD5-HCH2CH3pvaa-dCD148
MSLPVTALLLPLALLLHAARPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITKAGGGGSGGGGSGGGGSGGGGSEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSDPTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIFWVLVVVGGVLACYSLLVTVAFIIFWVRRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRRAEGRGSLLTCGDVEENPGPMETDTLLLWVLLLWVPGSTGQVTLKESGPGILKPSQTLSLTCSFSGFSLSTSGMGVGWIRQPSGKGLEWLAHIWWDDDVYYNPSLKNQLTISKDASRDQVFLKITNLDTADTATYYCVRRRATGTGFDYWGQGTTLTVSSGGGGSGGGGSGGGGSNIVMTQSHKFMSTSVGDRVSIACKASQDVGTAVAWYQQKPGQSPKLLIYWTSTRHTGVPDRFTGSGSGTDFTLTITNVQSEDLADYFCHQYNSYNTFGSGTRLELKRSDPAEPKSPDKTHTCPPCPAPPVAGPSVFLFPPKPKDTLMIARTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGKKDPKAVFGCIFGALVIVTVGGFIFWRKKRKDAKNNEVSFSQIKPKKSKLIRVENFEAYFKKQQADSNCGFAEEYEDLKLVGISQPKYAAELAENRGKNRYNNVLPYDISRVKLSVQTHSTDDYINANYMPGYHSKKDFIATQGPLPNTLKDFWRMVWEKNVYAIIMLTKCVEQGRTKCEEYWPSKQAQDYGDITVAMTSEIVLPEWTIRDFTVKNIQTSESHPLRQFHFTSWPDHGVPDTTDLLINFRYLVRDYMKQSPPESPILVHCSAGVGRTGTFIAIDRLIYQIENENTVDVYGIVYDLRMHRPLMVQTEDQYVFLNQCVLDIVRSQKDSKVDLIYQNTTAMTIYENLAPVTTFGKTNGYIA
SEQ ID № 44: SFG.aCD19-CD8STK-CD28tmZ-2A-aCD5-HCH2CH3pvaa-dCD148
ATGAGCCTGCCCGTGACCGCCCTGCTGCTGCCCCTGGCCCTGCTGCTGCACGCCGCCAGACCAGACATCCAGATGACCCAGACCACCAGCAGCCTGAGCGCCAGCCTGGGCGACCGGGTGACCATCAGCTGCAGAGCCAGCCAGGACATCAGCAAGTACCTGAACTGGTACCAGCAGAAGCCCGACGGCACCGTGAAGCTGCTGATCTACCACACCAGCCGGCTGCACAGCGGCGTGCCCAGCCGGTTCAGCGGCAGCGGCAGCGGCACCGACTACAGCCTGACCATCAGCAACCTGGAGCAGGAGGACATCGCCACCTACTTCTGCCAGCAGGGCAACACCCTGCCCTACACCTTCGGAGGCGGCACCAAGCTGGAGATCACCAAGGCCGGAGGCGGAGGCTCTGGCGGAGGCGGCTCTGGCGGAGGCGGCTCTGGCGGAGGCGGCAGCGAGGTGAAGCTGCAGGAGTCTGGCCCAGGCCTGGTGGCCCCAAGCCAGAGCCTGAGCGTGACCTGCACCGTGAGCGGCGTGAGCCTGCCCGACTACGGCGTGAGCTGGATCAGGCAGCCCCCACGGAAGGGCCTGGAGTGGCTGGGCGTGATCTGGGGCAGCGAGACCACCTACTACAACAGCGCCCTGAAGAGCCGGCTGACCATCATCAAGGACAACAGCAAGAGCCAGGTGTTCCTGAAGATGAACAGCCTGCAGACCGACGACACCGCCATCTACTACTGCGCCAAGCACTACTACTATGGCGGCAGCTACGCTATGGACTACTGGGGCCAGGGCACCAGCGTGACCGTGAGCTCAGATCCCACCACGACGCCAGCGCCGCGACCACCAACACCGGCGCCCACCATCGCGTCGCAGCCCCTGTCCCTGCGCCCAGAGGCGTGCCGGCCAGCGGCGGGGGGCGCAGTGCACACGAGGGGGCTGGACTTCGCCTGTGATATCTTTTGGGTGCTGGTGGTGGTTGGTGGAGTCCTGGCTTGCTATAGCTTGCTAGTAACAGTGGCCTTTATTATTTTCTGGGTGAGGAGAGTGAAGTTCAGCAGGAGCGCAGACGCCCCCGCGTACCAGCAGGGCCAGAACCAGCTCTATAACGAGCTCAATCTAGGACGAAGAGAGGAGTACGATGTTTTGGACAAGAGACGTGGCCGGGACCCTGAGATGGGGGGAAAGCCGAGAAGGAAGAACCCTCAGGAAGGCCTGTACAATGAACTGCAGAAAGATAAGATGGCGGAGGCCTACAGTGAGATTGGGATGAAAGGCGAGCGCCGGAGGGGCAAGGGGCACGATGGCCTTTACCAGGGTCTCAGTACAGCCACCAAGGACACCTACGACGCCCTTCACATGCAGGCCCTGCCTCCTCGCAGAGCCGAGGGCAGGGGAAGTCTTCTAACATGCGGGGACGTGGAGGAAAATCCCGGGCCCATGGAGACCGACACCCTGCTGCTGTGGGTGCTGCTGCTGTGGGTGCCCGGCAGCACCGGCCAGGTGACCCTGAAGGAGAGCGGTCCCGGCATCCTGAAGCCCAGCCAGACCCTGAGCCTGACCTGCAGCTTCAGCGGCTTCAGCCTGAGCACCAGCGGCATGGGCGTGGGCTGGATTCGGCAGCCCAGCGGCAAGGGCCTGGAGTGGCTGGCCCACATCTGGTGGGACGACGACGTGTACTACAACCCCAGCCTGAAGAACCAGCTGACCATCAGCAAGGACGCCAGCCGGGACCAGGTGTTCCTGAAGATCACCAACCTGGACACCGCCGACACCGCCACCTACTACTGCGTGCGGCGCCGGGCCACCGGCACCGGCTTCGACTACTGGGGCCAGGGCACCACCCTGACCGTGAGCAGCGGTGGCGGTGGCAGCGGCGGCGGCGGAAGCGGAGGTGGTGGCAGCAACATCGTGATGACCCAGAGCCACAAGTTCATGAGCACCAGCGTGGGCGACCGGGTGAGCATCGCCTGCAAGGCCAGCCAGGACGTGGGCACCGCCGTGGCCTGGTACCAGCAGAAGCCTGGCCAGAGCCCCAAGCTGCTGATCTACTGGACCAGCACCCGGCACACCGGCGTGCCCGACCGGTTCACCGGCAGCGGCAGCGGCACCGACTTCACCCTGACCATCACCAACGTGCAGAGCGAGGACCTGGCCGACTACTTCTGCCACCAGTACAACAGCTACAACACCTTCGGCAGCGGCACCCGGCTGGAGCTGAAGCGGTCGGATCCCGCCGAGCCCAAATCTCCTGACAAAACTCACACATGCCCACCGTGCCCAGCACCTCCCGTGGCCGGCCCGTCAGTCTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCGCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAACCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCCCTGCACAATCACTATACCCAGAAATCTCTGAGTCTGAGCCCAGGCAAGAAGGACCCCAAGGCGGTTTTTGGCTGTATCTTTGGTGCCCTGGTTATTGTGACTGTGGGAGGCTTCATCTTCTGGAGAAAGAAGAGGAAAGATGCAAAGAATAATGAAGTGTCCTTTTCTCAAATTAAACCTAAAAAATCTAAGTTAATCAGAGTGGAGAATTTTGAGGCCTACTTCAAGAAGCAGCAAGCTGACTCCAACTGTGGGTTCGCAGAGGAATACGAAGATCTGAAGCTTGTTGGAATTAGTCAACCTAAATATGCAGCAGAACTGGCTGAGAATAGAGGAAAGAATCGCTATAATAATGTTCTGCCCTATGATATTTCCCGTGTCAAACTTTCGGTCCAGACCCATTCAACGGATGACTACATCAATGCCAACTACATGCCTGGCTACCACTCCAAGAAAGATTTTATTGCCACACAAGGACCTTTACCGAACACTTTGAAAGATTTTTGGCGTATGGTTTGGGAGAAAAATGTATATGCCATCATTATGTTGACTAAATGTGTTGAACAGGGAAGAACCAAATGTGAGGAGTATTGGCCCTCCAAGCAGGCTCAGGACTATGGAGACATAACTGTGGCAATGACATCAGAAATTGTTCTTCCGGAATGGACCATCAGAGATTTCACAGTGAAAAATATCCAGACAAGTGAGAGTCACCCTCTGAGACAGTTCCATTTCACCTCCTGGCCAGACCACGGTGTTCCCGACACCACTGACCTGCTCATCAACTTCCGGTACCTCGTTCGTGACTACATGAAGCAGAGTCCTCCCGAATCGCCGATTCTGGTGCATTGCAGTGCTGGGGTCGGAAGGACGGGCACTTTCATTGCCATTGATCGTCTCATCTACCAGATAGAGAATGAGAACACCGTGGATGTGTATGGGATTGTGTATGACCTTCGAATGCATAGGCCTTTAATGGTGCAGACAGAGGACCAGTATGTTTTCCTCAATCAGTGTGTTTTGGATATTGTCAGATCCCAGAAAGACTCAAAAGTAGATCTTATCTACCAGAACACAACTGCAATGACAATCTATGAAAACCTTGCGCCCGTGACCACATTTGGAAAGACCAATGGTTACATCGCCTAA
SEQ ID № 45: SFG.aCD19-CD8STK-CD28tmZ-2A-aEGFRvIII-HCH2CH3pvaa-dCD148
MSLPVTALLLPLALLLHAARPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITKAGGGGSGGGGSGGGGSGGGGSEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSDPTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIFWVLVVVGGVLACYSLLVTVAFIIFWVRRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRRAEGRGSLLTCGDVEENPGPMETDTLLLWVLLLWVPGSTGQVKLQQSGGGLVKPGASLKLSCVTSGFTFRKFGMSWVRQTSDKRLEWVASISTGGYNTYYSDNVKGRFTISRENAKNTLYLQMSSLKSEDTALYYCTRGYSSTSYAMDYWGQGTTVTVSSGGGGSGGGGSGGGGSDIELTQSPASLSVATGEKVTIRCMTSTDIDDDMNWYQQKPGEPPKFLISEGNTLRPGVPSRFSSSGTGTDFVFTIENTLSEDVGDYYCLQSFNVPLTFGDGTKLEIKRSDPAEPKSPDKTHTCPPCPAPPVAGPSVFLFPPKPKDTLMIARTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGKKDPKAVFGCIFGALVIVTVGGFIFWRKKRKDAKNNEVSFSQIKPKKSKLIRVENFEAYFKKQQADSNCGFAEEYEDLKLVGISQPKYAAELAENRGKNRYNNVLPYDISRVKLSVQTHSTDDYINANYMPGYHSKKDFIATQGPLPNTLKDFWRMVWEKNVYAIIMLTKCVEQGRTKCEEYWPSKQAQDYGDITVAMTSEIVLPEWTIRDFTVKNIQTSESHPLRQFHFTSWPDHGVPDTTDLLINFRYLVRDYMKQSPPESPILVHCSAGVGRTGTFIAIDRLIYQIENENTVDVYGIVYDLRMHRPLMVQTEDQYVFLNQCVLDIVRSQKDSKVDLIYQNTTAMTIYENLAPVTTFGKTNGYIA
SEQ ID № 46: SFG.aCD19-CD8STK-CD28tmZ-2A-aEGFRvIII-HCH2CH3pvaa-dCD148
ATGAGCCTGCCCGTGACCGCCCTGCTGCTGCCCCTGGCCCTGCTGCTGCACGCCGCCAGACCAGACATCCAGATGACCCAGACCACCAGCAGCCTGAGCGCCAGCCTGGGCGACCGGGTGACCATCAGCTGCAGAGCCAGCCAGGACATCAGCAAGTACCTGAACTGGTACCAGCAGAAGCCCGACGGCACCGTGAAGCTGCTGATCTACCACACCAGCCGGCTGCACAGCGGCGTGCCCAGCCGGTTCAGCGGCAGCGGCAGCGGCACCGACTACAGCCTGACCATCAGCAACCTGGAGCAGGAGGACATCGCCACCTACTTCTGCCAGCAGGGCAACACCCTGCCCTACACCTTCGGAGGCGGCACCAAGCTGGAGATCACCAAGGCCGGAGGCGGAGGCTCTGGCGGAGGCGGCTCTGGCGGAGGCGGCTCTGGCGGAGGCGGCAGCGAGGTGAAGCTGCAGGAGTCTGGCCCAGGCCTGGTGGCCCCAAGCCAGAGCCTGAGCGTGACCTGCACCGTGAGCGGCGTGAGCCTGCCCGACTACGGCGTGAGCTGGATCAGGCAGCCCCCACGGAAGGGCCTGGAGTGGCTGGGCGTGATCTGGGGCAGCGAGACCACCTACTACAACAGCGCCCTGAAGAGCCGGCTGACCATCATCAAGGACAACAGCAAGAGCCAGGTGTTCCTGAAGATGAACAGCCTGCAGACCGACGACACCGCCATCTACTACTGCGCCAAGCACTACTACTATGGCGGCAGCTACGCTATGGACTACTGGGGCCAGGGCACCAGCGTGACCGTGAGCTCAGATCCCACCACGACGCCAGCGCCGCGACCACCAACACCGGCGCCCACCATCGCGTCGCAGCCCCTGTCCCTGCGCCCAGAGGCGTGCCGGCCAGCGGCGGGGGGCGCAGTGCACACGAGGGGGCTGGACTTCGCCTGTGATATCTTTTGGGTGCTGGTGGTGGTTGGTGGAGTCCTGGCTTGCTATAGCTTGCTAGTAACAGTGGCCTTTATTATTTTCTGGGTGAGGAGAGTGAAGTTCAGCAGGAGCGCAGACGCCCCCGCGTACCAGCAGGGCCAGAACCAGCTCTATAACGAGCTCAATCTAGGACGAAGAGAGGAGTACGATGTTTTGGACAAGAGACGTGGCCGGGACCCTGAGATGGGGGGAAAGCCGAGAAGGAAGAACCCTCAGGAAGGCCTGTACAATGAACTGCAGAAAGATAAGATGGCGGAGGCCTACAGTGAGATTGGGATGAAAGGCGAGCGCCGGAGGGGCAAGGGGCACGATGGCCTTTACCAGGGTCTCAGTACAGCCACCAAGGACACCTACGACGCCCTTCACATGCAGGCCCTGCCTCCTCGCAGAGCCGAGGGCAGGGGAAGTCTTCTAACATGCGGGGACGTGGAGGAAAATCCCGGGCCCATGGAGACCGACACCCTGCTGCTGTGGGTGCTGCTGCTGTGGGTGCCCGGCAGCACCGGCCAGGTGAAGCTGCAGCAGAGCGGCGGAGGCCTGGTGAAGCCCGGCGCCAGCCTGAAGCTGAGCTGCGTGACCAGCGGCTTCACCTTCCGGAAGTTCGGCATGAGCTGGGTGCGGCAGACCAGCGACAAGCGGCTGGAGTGGGTGGCCAGCATCAGCACCGGCGGCTACAACACCTACTACAGCGACAACGTGAAGGGCCGGTTCACCATCAGCCGGGAGAACGCCAAGAACACCCTGTACCTGCAGATGAGCAGCCTGAAGAGCGAGGACACCGCCCTGTACTACTGCACCCGGGGCTACAGCAGCACCAGCTACGCTATGGACTACTGGGGCCAGGGCACCACCGTGACAGTGAGCAGCGGCGGAGGAGGCAGTGGTGGGGGTGGATCTGGCGGAGGTGGCAGCGACATCGAGCTGACCCAGAGCCCCGCCAGCCTGAGCGTGGCCACCGGCGAGAAGGTGACCATCCGGTGCATGACCAGCACCGACATCGACGACGACATGAACTGGTACCAGCAGAAGCCCGGCGAGCCCCCAAAGTTCCTGATCAGCGAGGGCAACACCCTGCGGCCCGGCGTGCCCAGCCGGTTCAGCAGCAGCGGCACCGGCACCGACTTCGTGTTCACCATCGAGAACACCCTGAGCGAGGACGTGGGCGACTACTACTGCCTGCAGAGCTTCAACGTGCCCCTGACCTTCGGCGACGGCACCAAGCTGGAGATCAAGCGGTCGGATCCCGCCGAGCCCAAATCTCCTGACAAAACTCACACATGCCCACCGTGCCCAGCACCTCCCGTGGCCGGCCCGTCAGTCTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCGCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAACCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCCCTGCACAATCACTATACCCAGAAATCTCTGAGTCTGAGCCCAGGCAAGAAGGACCCCAAGGCGGTTTTTGGCTGTATCTTTGGTGCCCTGGTTATTGTGACTGTGGGAGGCTTCATCTTCTGGAGAAAGAAGAGGAAAGATGCAAAGAATAATGAAGTGTCCTTTTCTCAAATTAAACCTAAAAAATCTAAGTTAATCAGAGTGGAGAATTTTGAGGCCTACTTCAAGAAGCAGCAAGCTGACTCCAACTGTGGGTTCGCAGAGGAATACGAAGATCTGAAGCTTGTTGGAATTAGTCAACCTAAATATGCAGCAGAACTGGCTGAGAATAGAGGAAAGAATCGCTATAATAATGTTCTGCCCTATGATATTTCCCGTGTCAAACTTTCGGTCCAGACCCATTCAACGGATGACTACATCAATGCCAACTACATGCCTGGCTACCACTCCAAGAAAGATTTTATTGCCACACAAGGACCTTTACCGAACACTTTGAAAGATTTTTGGCGTATGGTTTGGGAGAAAAATGTATATGCCATCATTATGTTGACTAAATGTGTTGAACAGGGAAGAACCAAATGTGAGGAGTATTGGCCCTCCAAGCAGGCTCAGGACTATGGAGACATAACTGTGGCAATGACATCAGAAATTGTTCTTCCGGAATGGACCATCAGAGATTTCACAGTGAAAAATATCCAGACAAGTGAGAGTCACCCTCTGAGACAGTTCCATTTCACCTCCTGGCCAGACCACGGTGTTCCCGACACCACTGACCTGCTCATCAACTTCCGGTACCTCGTTCGTGACTACATGAAGCAGAGTCCTCCCGAATCGCCGATTCTGGTGCATTGCAGTGCTGGGGTCGGAAGGACGGGCACTTTCATTGCCATTGATCGTCTCATCTACCAGATAGAGAATGAGAACACCGTGGATGTGTATGGGATTGTGTATGACCTTCGAATGCATAGGCCTTTAATGGTGCAGACAGAGGACCAGTATGTTTTCCTCAATCAGTGTGTTTTGGATATTGTCAGATCCCAGAAAGACTCAAAAGTAGATCTTATCTACCAGAACACAACTGCAATGACAATCTATGAAAACCTTGCGCCCGTGACCACATTTGGAAAGACCAATGGTTACATCGCCTAA
Пример 11: разработка и конструкция CAR на основе APRIL.
APRIL в его природной форме представляет собой секретируемый белок II типа. Использование APRIL в качестве BCMA связывающего домена для CAR требует превращения этого секретируемого белка II типа в связанный с мембраной белок I типа, и для этого белок должен быть стабильным и сохранять связывание с BCMA в этой форме. Для того чтобы создавать возможно подходящие молекулы, дальний амино-конец APRIL удаляли для того, чтобы удалять связывание с протеогликанами. Затем добавляли сигнальный пептид для того, чтобы направлять появляющийся белок в эндоплазматический ретикулум, и таким образом, на клеточную поверхность. Также, поскольку свойства используемого спейсера могут менять функционирование CAR, тестировали три различных спейсерных доменах: создавали CAR на основе APRIL, содержащий (i) спейсер IgG1 человека, измененный для того, чтобы удалять Fc связывающие мотивы; (ii) «стебель» CD8; и (iii) отдельно шарнир IgG1 (рисунок на фиг. 25 и аминокислотные последовательности на фиг. 26). Эти CAR экспрессировали в бицистронном ретровирусном векторе (фиг. 27A) с тем, чтобы маркерный белок - усеченный CD34 можно было совместно экспрессировать в качестве удобного маркерного гена.
Пример 12: экспрессия и функционирование CAR на основе APRIL.
Цель этого исследования состояла в том, чтобы тестировать экспрессию сконструированных CAR на основе APRIL на клеточной поверхности и формирует ли укладка APRIL нативный белок. T-клетки трансдуцировали этими конструкциями с различными CAR и окрашивали с использованием коммерчески доступного mAb против APRIL наряду с окрашиванием по маркерному гену и анализировали с помощью проточной цитометрии. Результаты этого эксперимента представлены на фиг. 27B, где на график нанесено связывание APRIL в зависимости от флуоресценции маркерного гена. Эти данные показывают, что в этом формате CAR на основе APRIL экспрессированы на клеточной поверхности и APRIL укладывается достаточно, чтобы быть распознанным с помощью mAb против APRIL.
Затем определяли, может ли APRIL в этом формате распознавать BCMA и TACI. Рекомбинантные BCMA и TACI создавали в виде слитных белков с IgG2a-Fc мыши. Эти рекомбинантные белки инкубировали с трансдуцированными T-клетками. После этого клетки промывали и окрашивали антимышиным антителом, конъюгированным с флуорофором, и антителом для обнаружения маркерного гена, конъюгированным с другим флуорофором. Клетки анализировали с помощью проточной цитометрии и результаты представлены на фиг. 27C. Различные CAR способны связывать как BCMA, так и TACI. К удивлению, CAR способны связывать BCMA лучше, чем TACI. Также, к удивлению, CAR со «стеблем» CD8 или шарнирным IgG1 спейсером способны лучше связывать BCMA и TACI, чем CAR с Fc спейсером.
Пример 13: химерные антигенные рецепторы на основе APRIL активны против экспрессирующих BCMA клеток
T-клетки от нормальных доноров трансдуцировали различными APRIL CAR и тестировали на клетках SupT1 или дикого типа или сконструированных для того, чтобы экспрессировать BCMA и TACI. Использовали несколько различных анализов для того, чтобы определять функцию. Осуществляли классический анализ на высвобождение хрома. Здесь клетки-мишени (клетки SupT1) метили с использованием 51Cr и смешивали с эффекторами (трансдуцированные T-клетки) в различных соотношениях. Лизис клеток-мишеней определяли посредством подсчета 51Cr в супернатанте совместной культуры (на фиг. 28A представлены кумулятивные данные).
Кроме того, супернатант от T-клеток, которые культивировали 1:1 с клетками SupT1, анализировали с помощью ELISA на интерферон-γ (на фиг. 28B представлены кумулятивные данные). Также осуществляли измерение размножения T-клеток после одной недели совместного культивирования с клетками SupT1 (фиг. 28C). T-клетки считали с помощью проточного цитометра, который калибровали с использованием счетных гранул. Эти экспериментальные данные показывают, что CAR на основе APRIL могут уничтожать мишени, экспрессирующие BCMA. Кроме того, эти данные показывают, что CAR, основанные на «стебле» CD8 или шарнире IgG1, работали лучше, чем CAR на основе Fc-pvaa.
Пример 14: функциональный анализ логического элемента И в первичных клетках
PBMC выделяли из крови и стимулировали с использованием PHA и IL-2. Двумя сутками позже клетки трансдуцировали в покрытых ретронектином планшетах с использованием ретровируса, содержащего конструкцию логического элемента И CD19:CD33. В сутки 5 уровень экспрессии двух CAR, транслируемых конструкцией логического элемента И, оценивали через проточную цитометрию, и клетки обедняли по CD56+ клеткам (преимущественно естественные киллерные клетки). В сутки 6 PBMC помещали в совместную культуру с клетками-мишенями в соотношении эффекторы к клеткам-мишеням 1:2. В сутки 8 супернатант собирали и анализировали на секрецию IFN-γ с помощью ELISA (фиг. 29).
Эти данные демонстрируют, что логический элемент И функционирует в первичных клетках.
Все публикации, указанные в приведенном выше описании, включены в настоящий документ посредством ссылки. Различные модификации и вариации описанных способов и систем по изобретению будут очевидны специалисты в данной области, не отступая от объема и сущности изобретения. Несмотря на то, что изобретение описан применительно к конкретным предпочтительным вариантам осуществления, следует понимать, что изобретение, как заявлено, не должно быть чрезмерно ограничено такими конкретными вариантами осуществления. В действительности, предусмотрено, что различные модификации описанных способов осуществления изобретения, которые очевидны специалистам в молекулярной биологии, клеточной биологии или связанных областях, включены в объем следующей формулы изобретения.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> UCL Business PLC
<120> T-клетка
<130> P103294PCT1
<150> GB 1410934.2
<151> 2014-06-19
<160> 53
<170> PatentIn version 3.5
<210> 1
<211> 1129
<212> Белок
<213> Искусственная последовательность
<220>
<223> Химерный антигенный рецептор (CAR)
<400> 1
Met Ser Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Asp Ile Gln Met Thr Gln Thr Thr Ser Ser Leu
20 25 30
Ser Ala Ser Leu Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln
35 40 45
Asp Ile Ser Lys Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr
50 55 60
Val Lys Leu Leu Ile Tyr His Thr Ser Arg Leu His Ser Gly Val Pro
65 70 75 80
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile
85 90 95
Ser Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly
100 105 110
Asn Thr Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Thr
115 120 125
Lys Ala Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Gly Gly Gly Gly Ser Glu Val Lys Leu Gln Glu Ser Gly Pro Gly
145 150 155 160
Leu Val Ala Pro Ser Gln Ser Leu Ser Val Thr Cys Thr Val Ser Gly
165 170 175
Val Ser Leu Pro Asp Tyr Gly Val Ser Trp Ile Arg Gln Pro Pro Arg
180 185 190
Lys Gly Leu Glu Trp Leu Gly Val Ile Trp Gly Ser Glu Thr Thr Tyr
195 200 205
Tyr Asn Ser Ala Leu Lys Ser Arg Leu Thr Ile Ile Lys Asp Asn Ser
210 215 220
Lys Ser Gln Val Phe Leu Lys Met Asn Ser Leu Gln Thr Asp Asp Thr
225 230 235 240
Ala Ile Tyr Tyr Cys Ala Lys His Tyr Tyr Tyr Gly Gly Ser Tyr Ala
245 250 255
Met Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Asp Pro
260 265 270
Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala
275 280 285
Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly
290 295 300
Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp Ile Phe Trp
305 310 315 320
Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val
325 330 335
Thr Val Ala Phe Ile Ile Phe Trp Val Arg Arg Val Lys Phe Ser Arg
340 345 350
Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn
355 360 365
Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg
370 375 380
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro
385 390 395 400
Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala
405 410 415
Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His
420 425 430
Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp
435 440 445
Ala Leu His Met Gln Ala Leu Pro Pro Arg Arg Ala Glu Gly Arg Gly
450 455 460
Ser Leu Leu Thr Cys Gly Asp Val Glu Glu Asn Pro Gly Pro Met Ala
465 470 475 480
Val Pro Thr Gln Val Leu Gly Leu Leu Leu Leu Trp Leu Thr Asp Ala
485 490 495
Arg Cys Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser
500 505 510
Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Asp Ile Tyr
515 520 525
Phe Asn Leu Val Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu
530 535 540
Leu Ile Tyr Asp Thr Asn Arg Leu Ala Asp Gly Val Pro Ser Arg Phe
545 550 555 560
Ser Gly Ser Gly Ser Gly Thr Gln Tyr Thr Leu Thr Ile Ser Ser Leu
565 570 575
Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln His Tyr Lys Asn Tyr
580 585 590
Pro Leu Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Ser Gly
595 600 605
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
610 615 620
Gly Gly Ser Arg Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu
625 630 635 640
Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe
645 650 655
Thr Leu Ser Asn Tyr Gly Met His Trp Ile Arg Gln Ala Pro Gly Lys
660 665 670
Gly Leu Glu Trp Val Ser Ser Ile Ser Leu Asn Gly Gly Ser Thr Tyr
675 680 685
Tyr Arg Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala
690 695 700
Lys Ser Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr
705 710 715 720
Ala Val Tyr Tyr Cys Ala Ala Gln Asp Ala Tyr Thr Gly Gly Tyr Phe
725 730 735
Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Met Asp Pro
740 745 750
Ala Glu Pro Lys Ser Pro Asp Lys Thr His Thr Cys Pro Pro Cys Pro
755 760 765
Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
770 775 780
Lys Asp Thr Leu Met Ile Ala Arg Thr Pro Glu Val Thr Cys Val Val
785 790 795 800
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
805 810 815
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
820 825 830
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
835 840 845
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
850 855 860
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
865 870 875 880
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
885 890 895
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
900 905 910
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
915 920 925
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
930 935 940
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
945 950 955 960
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
965 970 975
Ser Leu Ser Leu Ser Pro Gly Lys Lys Asp Pro Lys Phe Trp Val Leu
980 985 990
Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val Thr Val
995 1000 1005
Ala Phe Ile Ile Phe Trp Val Arg Ser Arg Val Lys Phe Ser Arg
1010 1015 1020
Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr
1025 1030 1035
Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp
1040 1045 1050
Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg
1055 1060 1065
Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
1070 1075 1080
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg
1085 1090 1095
Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala
1100 1105 1110
Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro
1115 1120 1125
Arg
<210> 2
<211> 1350
<212> Белок
<213> Искусственная последовательность
<220>
<223> Химерный антигенный рецептор (CAR)
<400> 2
Met Ser Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Asp Ile Gln Met Thr Gln Thr Thr Ser Ser Leu
20 25 30
Ser Ala Ser Leu Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln
35 40 45
Asp Ile Ser Lys Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr
50 55 60
Val Lys Leu Leu Ile Tyr His Thr Ser Arg Leu His Ser Gly Val Pro
65 70 75 80
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile
85 90 95
Ser Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly
100 105 110
Asn Thr Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Thr
115 120 125
Lys Ala Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Gly Gly Gly Gly Ser Glu Val Lys Leu Gln Glu Ser Gly Pro Gly
145 150 155 160
Leu Val Ala Pro Ser Gln Ser Leu Ser Val Thr Cys Thr Val Ser Gly
165 170 175
Val Ser Leu Pro Asp Tyr Gly Val Ser Trp Ile Arg Gln Pro Pro Arg
180 185 190
Lys Gly Leu Glu Trp Leu Gly Val Ile Trp Gly Ser Glu Thr Thr Tyr
195 200 205
Tyr Asn Ser Ala Leu Lys Ser Arg Leu Thr Ile Ile Lys Asp Asn Ser
210 215 220
Lys Ser Gln Val Phe Leu Lys Met Asn Ser Leu Gln Thr Asp Asp Thr
225 230 235 240
Ala Ile Tyr Tyr Cys Ala Lys His Tyr Tyr Tyr Gly Gly Ser Tyr Ala
245 250 255
Met Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Asp Pro
260 265 270
Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala
275 280 285
Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly
290 295 300
Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp Ile Phe Trp
305 310 315 320
Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val
325 330 335
Thr Val Ala Phe Ile Ile Phe Trp Val Arg Arg Val Lys Phe Ser Arg
340 345 350
Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn
355 360 365
Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg
370 375 380
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro
385 390 395 400
Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala
405 410 415
Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His
420 425 430
Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp
435 440 445
Ala Leu His Met Gln Ala Leu Pro Pro Arg Arg Ala Glu Gly Arg Gly
450 455 460
Ser Leu Leu Thr Cys Gly Asp Val Glu Glu Asn Pro Gly Pro Met Ala
465 470 475 480
Val Pro Thr Gln Val Leu Gly Leu Leu Leu Leu Trp Leu Thr Asp Ala
485 490 495
Arg Cys Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser
500 505 510
Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Asp Ile Tyr
515 520 525
Phe Asn Leu Val Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu
530 535 540
Leu Ile Tyr Asp Thr Asn Arg Leu Ala Asp Gly Val Pro Ser Arg Phe
545 550 555 560
Ser Gly Ser Gly Ser Gly Thr Gln Tyr Thr Leu Thr Ile Ser Ser Leu
565 570 575
Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln His Tyr Lys Asn Tyr
580 585 590
Pro Leu Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Ser Gly
595 600 605
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
610 615 620
Gly Gly Ser Arg Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu
625 630 635 640
Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe
645 650 655
Thr Leu Ser Asn Tyr Gly Met His Trp Ile Arg Gln Ala Pro Gly Lys
660 665 670
Gly Leu Glu Trp Val Ser Ser Ile Ser Leu Asn Gly Gly Ser Thr Tyr
675 680 685
Tyr Arg Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala
690 695 700
Lys Ser Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr
705 710 715 720
Ala Val Tyr Tyr Cys Ala Ala Gln Asp Ala Tyr Thr Gly Gly Tyr Phe
725 730 735
Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Met Asp Pro
740 745 750
Ala Glu Pro Lys Ser Pro Asp Lys Thr His Thr Cys Pro Pro Cys Pro
755 760 765
Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
770 775 780
Lys Asp Thr Leu Met Ile Ala Arg Thr Pro Glu Val Thr Cys Val Val
785 790 795 800
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
805 810 815
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
820 825 830
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
835 840 845
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
850 855 860
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
865 870 875 880
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
885 890 895
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
900 905 910
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
915 920 925
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
930 935 940
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
945 950 955 960
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
965 970 975
Ser Leu Ser Leu Ser Pro Gly Lys Lys Asp Pro Lys Ala Val Phe Gly
980 985 990
Cys Ile Phe Gly Ala Leu Val Ile Val Thr Val Gly Gly Phe Ile Phe
995 1000 1005
Trp Arg Lys Lys Arg Lys Asp Ala Lys Asn Asn Glu Val Ser Phe
1010 1015 1020
Ser Gln Ile Lys Pro Lys Lys Ser Lys Leu Ile Arg Val Glu Asn
1025 1030 1035
Phe Glu Ala Tyr Phe Lys Lys Gln Gln Ala Asp Ser Asn Cys Gly
1040 1045 1050
Phe Ala Glu Glu Tyr Glu Asp Leu Lys Leu Val Gly Ile Ser Gln
1055 1060 1065
Pro Lys Tyr Ala Ala Glu Leu Ala Glu Asn Arg Gly Lys Asn Arg
1070 1075 1080
Tyr Asn Asn Val Leu Pro Tyr Asp Ile Ser Arg Val Lys Leu Ser
1085 1090 1095
Val Gln Thr His Ser Thr Asp Asp Tyr Ile Asn Ala Asn Tyr Met
1100 1105 1110
Pro Gly Tyr His Ser Lys Lys Asp Phe Ile Ala Thr Gln Gly Pro
1115 1120 1125
Leu Pro Asn Thr Leu Lys Asp Phe Trp Arg Met Val Trp Glu Lys
1130 1135 1140
Asn Val Tyr Ala Ile Ile Met Leu Thr Lys Cys Val Glu Gln Gly
1145 1150 1155
Arg Thr Lys Cys Glu Glu Tyr Trp Pro Ser Lys Gln Ala Gln Asp
1160 1165 1170
Tyr Gly Asp Ile Thr Val Ala Met Thr Ser Glu Ile Val Leu Pro
1175 1180 1185
Glu Trp Thr Ile Arg Asp Phe Thr Val Lys Asn Ile Gln Thr Ser
1190 1195 1200
Glu Ser His Pro Leu Arg Gln Phe His Phe Thr Ser Trp Pro Asp
1205 1210 1215
His Gly Val Pro Asp Thr Thr Asp Leu Leu Ile Asn Phe Arg Tyr
1220 1225 1230
Leu Val Arg Asp Tyr Met Lys Gln Ser Pro Pro Glu Ser Pro Ile
1235 1240 1245
Leu Val His Cys Ser Ala Gly Val Gly Arg Thr Gly Thr Phe Ile
1250 1255 1260
Ala Ile Asp Arg Leu Ile Tyr Gln Ile Glu Asn Glu Asn Thr Val
1265 1270 1275
Asp Val Tyr Gly Ile Val Tyr Asp Leu Arg Met His Arg Pro Leu
1280 1285 1290
Met Val Gln Thr Glu Asp Gln Tyr Val Phe Leu Asn Gln Cys Val
1295 1300 1305
Leu Asp Ile Val Arg Ser Gln Lys Asp Ser Lys Val Asp Leu Ile
1310 1315 1320
Tyr Gln Asn Thr Thr Ala Met Thr Ile Tyr Glu Asn Leu Ala Pro
1325 1330 1335
Val Thr Thr Phe Gly Lys Thr Asn Gly Tyr Ile Ala
1340 1345 1350
<210> 3
<211> 1717
<212> Белок
<213> Искусственная последовательность
<220>
<223> Химерный антигенный рецептор (CAR)
<400> 3
Met Ser Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Asp Ile Gln Met Thr Gln Thr Thr Ser Ser Leu
20 25 30
Ser Ala Ser Leu Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln
35 40 45
Asp Ile Ser Lys Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr
50 55 60
Val Lys Leu Leu Ile Tyr His Thr Ser Arg Leu His Ser Gly Val Pro
65 70 75 80
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile
85 90 95
Ser Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly
100 105 110
Asn Thr Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Thr
115 120 125
Lys Ala Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Gly Gly Gly Gly Ser Glu Val Lys Leu Gln Glu Ser Gly Pro Gly
145 150 155 160
Leu Val Ala Pro Ser Gln Ser Leu Ser Val Thr Cys Thr Val Ser Gly
165 170 175
Val Ser Leu Pro Asp Tyr Gly Val Ser Trp Ile Arg Gln Pro Pro Arg
180 185 190
Lys Gly Leu Glu Trp Leu Gly Val Ile Trp Gly Ser Glu Thr Thr Tyr
195 200 205
Tyr Asn Ser Ala Leu Lys Ser Arg Leu Thr Ile Ile Lys Asp Asn Ser
210 215 220
Lys Ser Gln Val Phe Leu Lys Met Asn Ser Leu Gln Thr Asp Asp Thr
225 230 235 240
Ala Ile Tyr Tyr Cys Ala Lys His Tyr Tyr Tyr Gly Gly Ser Tyr Ala
245 250 255
Met Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Asp Pro
260 265 270
Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala
275 280 285
Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly
290 295 300
Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp Ile Phe Trp
305 310 315 320
Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val
325 330 335
Thr Val Ala Phe Ile Ile Phe Trp Val Arg Arg Val Lys Phe Ser Arg
340 345 350
Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn
355 360 365
Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg
370 375 380
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro
385 390 395 400
Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala
405 410 415
Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His
420 425 430
Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp
435 440 445
Ala Leu His Met Gln Ala Leu Pro Pro Arg Arg Ala Glu Gly Arg Gly
450 455 460
Ser Leu Leu Thr Cys Gly Asp Val Glu Glu Asn Pro Gly Pro Met Ala
465 470 475 480
Val Pro Thr Gln Val Leu Gly Leu Leu Leu Leu Trp Leu Thr Asp Ala
485 490 495
Arg Cys Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser
500 505 510
Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Asp Ile Tyr
515 520 525
Phe Asn Leu Val Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu
530 535 540
Leu Ile Tyr Asp Thr Asn Arg Leu Ala Asp Gly Val Pro Ser Arg Phe
545 550 555 560
Ser Gly Ser Gly Ser Gly Thr Gln Tyr Thr Leu Thr Ile Ser Ser Leu
565 570 575
Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln His Tyr Lys Asn Tyr
580 585 590
Pro Leu Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Ser Gly
595 600 605
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
610 615 620
Gly Gly Ser Arg Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu
625 630 635 640
Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe
645 650 655
Thr Leu Ser Asn Tyr Gly Met His Trp Ile Arg Gln Ala Pro Gly Lys
660 665 670
Gly Leu Glu Trp Val Ser Ser Ile Ser Leu Asn Gly Gly Ser Thr Tyr
675 680 685
Tyr Arg Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala
690 695 700
Lys Ser Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr
705 710 715 720
Ala Val Tyr Tyr Cys Ala Ala Gln Asp Ala Tyr Thr Gly Gly Tyr Phe
725 730 735
Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Met Asp Pro
740 745 750
Ala Glu Pro Lys Ser Pro Asp Lys Thr His Thr Cys Pro Pro Cys Pro
755 760 765
Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
770 775 780
Lys Asp Thr Leu Met Ile Ala Arg Thr Pro Glu Val Thr Cys Val Val
785 790 795 800
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
805 810 815
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
820 825 830
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
835 840 845
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
850 855 860
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
865 870 875 880
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
885 890 895
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
900 905 910
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
915 920 925
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
930 935 940
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
945 950 955 960
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
965 970 975
Ser Leu Ser Leu Ser Pro Gly Lys Lys Asp Pro Lys Ala Leu Ile Ala
980 985 990
Phe Leu Ala Phe Leu Ile Ile Val Thr Ser Ile Ala Leu Leu Val Val
995 1000 1005
Leu Tyr Lys Ile Tyr Asp Leu His Lys Lys Arg Ser Cys Asn Leu
1010 1015 1020
Asp Glu Gln Gln Glu Leu Val Glu Arg Asp Asp Glu Lys Gln Leu
1025 1030 1035
Met Asn Val Glu Pro Ile His Ala Asp Ile Leu Leu Glu Thr Tyr
1040 1045 1050
Lys Arg Lys Ile Ala Asp Glu Gly Arg Leu Phe Leu Ala Glu Phe
1055 1060 1065
Gln Ser Ile Pro Arg Val Phe Ser Lys Phe Pro Ile Lys Glu Ala
1070 1075 1080
Arg Lys Pro Phe Asn Gln Asn Lys Asn Arg Tyr Val Asp Ile Leu
1085 1090 1095
Pro Tyr Asp Tyr Asn Arg Val Glu Leu Ser Glu Ile Asn Gly Asp
1100 1105 1110
Ala Gly Ser Asn Tyr Ile Asn Ala Ser Tyr Ile Asp Gly Phe Lys
1115 1120 1125
Glu Pro Arg Lys Tyr Ile Ala Ala Gln Gly Pro Arg Asp Glu Thr
1130 1135 1140
Val Asp Asp Phe Trp Arg Met Ile Trp Glu Gln Lys Ala Thr Val
1145 1150 1155
Ile Val Met Val Thr Arg Cys Glu Glu Gly Asn Arg Asn Lys Cys
1160 1165 1170
Ala Glu Tyr Trp Pro Ser Met Glu Glu Gly Thr Arg Ala Phe Gly
1175 1180 1185
Asp Val Val Val Lys Ile Asn Gln His Lys Arg Cys Pro Asp Tyr
1190 1195 1200
Ile Ile Gln Lys Leu Asn Ile Val Asn Lys Lys Glu Lys Ala Thr
1205 1210 1215
Gly Arg Glu Val Thr His Ile Gln Phe Thr Ser Trp Pro Asp His
1220 1225 1230
Gly Val Pro Glu Asp Pro His Leu Leu Leu Lys Leu Arg Arg Arg
1235 1240 1245
Val Asn Ala Phe Ser Asn Phe Phe Ser Gly Pro Ile Val Val His
1250 1255 1260
Cys Ser Ala Gly Val Gly Arg Thr Gly Thr Tyr Ile Gly Ile Asp
1265 1270 1275
Ala Met Leu Glu Gly Leu Glu Ala Glu Asn Lys Val Asp Val Tyr
1280 1285 1290
Gly Tyr Val Val Lys Leu Arg Arg Gln Arg Cys Leu Met Val Gln
1295 1300 1305
Val Glu Ala Gln Tyr Ile Leu Ile His Gln Ala Leu Val Glu Tyr
1310 1315 1320
Asn Gln Phe Gly Glu Thr Glu Val Asn Leu Ser Glu Leu His Pro
1325 1330 1335
Tyr Leu His Asn Met Lys Lys Arg Asp Pro Pro Ser Glu Pro Ser
1340 1345 1350
Pro Leu Glu Ala Glu Phe Gln Arg Leu Pro Ser Tyr Arg Ser Trp
1355 1360 1365
Arg Thr Gln His Ile Gly Asn Gln Glu Glu Asn Lys Ser Lys Asn
1370 1375 1380
Arg Asn Ser Asn Val Ile Pro Tyr Asp Tyr Asn Arg Val Pro Leu
1385 1390 1395
Lys His Glu Leu Glu Met Ser Lys Glu Ser Glu His Asp Ser Asp
1400 1405 1410
Glu Ser Ser Asp Asp Asp Ser Asp Ser Glu Glu Pro Ser Lys Tyr
1415 1420 1425
Ile Asn Ala Ser Phe Ile Met Ser Tyr Trp Lys Pro Glu Val Met
1430 1435 1440
Ile Ala Ala Gln Gly Pro Leu Lys Glu Thr Ile Gly Asp Phe Trp
1445 1450 1455
Gln Met Ile Phe Gln Arg Lys Val Lys Val Ile Val Met Leu Thr
1460 1465 1470
Glu Leu Lys His Gly Asp Gln Glu Ile Cys Ala Gln Tyr Trp Gly
1475 1480 1485
Glu Gly Lys Gln Thr Tyr Gly Asp Ile Glu Val Asp Leu Lys Asp
1490 1495 1500
Thr Asp Lys Ser Ser Thr Tyr Thr Leu Arg Val Phe Glu Leu Arg
1505 1510 1515
His Ser Lys Arg Lys Asp Ser Arg Thr Val Tyr Gln Tyr Gln Tyr
1520 1525 1530
Thr Asn Trp Ser Val Glu Gln Leu Pro Ala Glu Pro Lys Glu Leu
1535 1540 1545
Ile Ser Met Ile Gln Val Val Lys Gln Lys Leu Pro Gln Lys Asn
1550 1555 1560
Ser Ser Glu Gly Asn Lys His His Lys Ser Thr Pro Leu Leu Ile
1565 1570 1575
His Cys Arg Asp Gly Ser Gln Gln Thr Gly Ile Phe Cys Ala Leu
1580 1585 1590
Leu Asn Leu Leu Glu Ser Ala Glu Thr Glu Glu Val Val Asp Ile
1595 1600 1605
Phe Gln Val Val Lys Ala Leu Arg Lys Ala Arg Pro Gly Met Val
1610 1615 1620
Ser Thr Phe Glu Gln Tyr Gln Phe Leu Tyr Asp Val Ile Ala Ser
1625 1630 1635
Thr Tyr Pro Ala Gln Asn Gly Gln Val Lys Lys Asn Asn His Gln
1640 1645 1650
Glu Asp Lys Ile Glu Phe Asp Asn Glu Val Asp Lys Val Lys Gln
1655 1660 1665
Asp Ala Asn Cys Val Asn Pro Leu Gly Ala Pro Glu Lys Leu Pro
1670 1675 1680
Glu Ala Lys Glu Gln Ala Glu Gly Ser Glu Pro Thr Ser Gly Thr
1685 1690 1695
Glu Gly Pro Glu His Ser Val Asn Gly Pro Ala Ser Pro Ala Leu
1700 1705 1710
Asn Gln Gly Ser
1715
<210> 4
<211> 1114
<212> Белок
<213> Искусственная последовательность
<220>
<223> Химерный антигенный рецептор (CAR)
<400> 4
Met Ser Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Asp Ile Gln Met Thr Gln Thr Thr Ser Ser Leu
20 25 30
Ser Ala Ser Leu Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln
35 40 45
Asp Ile Ser Lys Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr
50 55 60
Val Lys Leu Leu Ile Tyr His Thr Ser Arg Leu His Ser Gly Val Pro
65 70 75 80
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile
85 90 95
Ser Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly
100 105 110
Asn Thr Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Thr
115 120 125
Lys Ala Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Gly Gly Gly Gly Ser Glu Val Lys Leu Gln Glu Ser Gly Pro Gly
145 150 155 160
Leu Val Ala Pro Ser Gln Ser Leu Ser Val Thr Cys Thr Val Ser Gly
165 170 175
Val Ser Leu Pro Asp Tyr Gly Val Ser Trp Ile Arg Gln Pro Pro Arg
180 185 190
Lys Gly Leu Glu Trp Leu Gly Val Ile Trp Gly Ser Glu Thr Thr Tyr
195 200 205
Tyr Asn Ser Ala Leu Lys Ser Arg Leu Thr Ile Ile Lys Asp Asn Ser
210 215 220
Lys Ser Gln Val Phe Leu Lys Met Asn Ser Leu Gln Thr Asp Asp Thr
225 230 235 240
Ala Ile Tyr Tyr Cys Ala Lys His Tyr Tyr Tyr Gly Gly Ser Tyr Ala
245 250 255
Met Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Asp Pro
260 265 270
Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala
275 280 285
Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly
290 295 300
Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp Ile Phe Trp
305 310 315 320
Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val
325 330 335
Thr Val Ala Phe Ile Ile Phe Trp Val Arg Arg Val Lys Phe Ser Arg
340 345 350
Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn
355 360 365
Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg
370 375 380
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro
385 390 395 400
Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala
405 410 415
Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His
420 425 430
Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp
435 440 445
Ala Leu His Met Gln Ala Leu Pro Pro Arg Arg Ala Glu Gly Arg Gly
450 455 460
Ser Leu Leu Thr Cys Gly Asp Val Glu Glu Asn Pro Gly Pro Met Ala
465 470 475 480
Val Pro Thr Gln Val Leu Gly Leu Leu Leu Leu Trp Leu Thr Asp Ala
485 490 495
Arg Cys Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser
500 505 510
Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Asp Ile Tyr
515 520 525
Phe Asn Leu Val Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu
530 535 540
Leu Ile Tyr Asp Thr Asn Arg Leu Ala Asp Gly Val Pro Ser Arg Phe
545 550 555 560
Ser Gly Ser Gly Ser Gly Thr Gln Tyr Thr Leu Thr Ile Ser Ser Leu
565 570 575
Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln His Tyr Lys Asn Tyr
580 585 590
Pro Leu Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Ser Gly
595 600 605
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
610 615 620
Gly Gly Ser Arg Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu
625 630 635 640
Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe
645 650 655
Thr Leu Ser Asn Tyr Gly Met His Trp Ile Arg Gln Ala Pro Gly Lys
660 665 670
Gly Leu Glu Trp Val Ser Ser Ile Ser Leu Asn Gly Gly Ser Thr Tyr
675 680 685
Tyr Arg Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala
690 695 700
Lys Ser Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr
705 710 715 720
Ala Val Tyr Tyr Cys Ala Ala Gln Asp Ala Tyr Thr Gly Gly Tyr Phe
725 730 735
Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Met Asp Pro
740 745 750
Ala Thr Thr Thr Lys Pro Val Leu Arg Thr Pro Ser Pro Val His Pro
755 760 765
Thr Gly Thr Ser Gln Pro Gln Arg Pro Glu Asp Cys Arg Pro Arg Gly
770 775 780
Ser Val Lys Gly Thr Gly Leu Asp Phe Ala Cys Asp Ile Tyr Trp Ala
785 790 795 800
Pro Leu Ala Gly Ile Cys Val Ala Leu Leu Leu Ser Leu Ile Ile Thr
805 810 815
Leu Ile Cys Tyr His Arg Ser Arg Lys Arg Val Cys Lys Ser Gly Gly
820 825 830
Gly Ser Phe Trp Glu Glu Phe Glu Ser Leu Gln Lys Gln Glu Val Lys
835 840 845
Asn Leu His Gln Arg Leu Glu Gly Gln Arg Pro Glu Asn Lys Gly Lys
850 855 860
Asn Arg Tyr Lys Asn Ile Leu Pro Phe Asp His Ser Arg Val Ile Leu
865 870 875 880
Gln Gly Arg Asp Ser Asn Ile Pro Gly Ser Asp Tyr Ile Asn Ala Asn
885 890 895
Tyr Ile Lys Asn Gln Leu Leu Gly Pro Asp Glu Asn Ala Lys Thr Tyr
900 905 910
Ile Ala Ser Gln Gly Cys Leu Glu Ala Thr Val Asn Asp Phe Trp Gln
915 920 925
Met Ala Trp Gln Glu Asn Ser Arg Val Ile Val Met Thr Thr Arg Glu
930 935 940
Val Glu Lys Gly Arg Asn Lys Cys Val Pro Tyr Trp Pro Glu Val Gly
945 950 955 960
Met Gln Arg Ala Tyr Gly Pro Tyr Ser Val Thr Asn Cys Gly Glu His
965 970 975
Asp Thr Thr Glu Tyr Lys Leu Arg Thr Leu Gln Val Ser Pro Leu Asp
980 985 990
Asn Gly Asp Leu Ile Arg Glu Ile Trp His Tyr Gln Tyr Leu Ser Trp
995 1000 1005
Pro Asp His Gly Val Pro Ser Glu Pro Gly Gly Val Leu Ser Phe
1010 1015 1020
Leu Asp Gln Ile Asn Gln Arg Gln Glu Ser Leu Pro His Ala Gly
1025 1030 1035
Pro Ile Ile Val His Cys Ser Ala Gly Ile Gly Arg Thr Gly Thr
1040 1045 1050
Ile Ile Val Ile Asp Met Leu Met Glu Asn Ile Ser Thr Lys Gly
1055 1060 1065
Leu Asp Cys Asp Ile Asp Ile Gln Lys Thr Ile Gln Met Val Arg
1070 1075 1080
Ala Gln Arg Ser Gly Met Val Gln Thr Glu Ala Gln Tyr Lys Phe
1085 1090 1095
Ile Tyr Val Ala Ile Ala Gln Phe Ile Glu Thr Thr Lys Lys Lys
1100 1105 1110
Leu
<210> 5
<211> 918
<212> Белок
<213> Искусственная последовательность
<220>
<223> Химерный антигенный рецептор (CAR)
<400> 5
Met Ser Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Asp Ile Gln Met Thr Gln Thr Thr Ser Ser Leu
20 25 30
Ser Ala Ser Leu Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln
35 40 45
Asp Ile Ser Lys Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr
50 55 60
Val Lys Leu Leu Ile Tyr His Thr Ser Arg Leu His Ser Gly Val Pro
65 70 75 80
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile
85 90 95
Ser Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly
100 105 110
Asn Thr Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Thr
115 120 125
Lys Ala Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Gly Gly Gly Gly Ser Glu Val Lys Leu Gln Glu Ser Gly Pro Gly
145 150 155 160
Leu Val Ala Pro Ser Gln Ser Leu Ser Val Thr Cys Thr Val Ser Gly
165 170 175
Val Ser Leu Pro Asp Tyr Gly Val Ser Trp Ile Arg Gln Pro Pro Arg
180 185 190
Lys Gly Leu Glu Trp Leu Gly Val Ile Trp Gly Ser Glu Thr Thr Tyr
195 200 205
Tyr Asn Ser Ala Leu Lys Ser Arg Leu Thr Ile Ile Lys Asp Asn Ser
210 215 220
Lys Ser Gln Val Phe Leu Lys Met Asn Ser Leu Gln Thr Asp Asp Thr
225 230 235 240
Ala Ile Tyr Tyr Cys Ala Lys His Tyr Tyr Tyr Gly Gly Ser Tyr Ala
245 250 255
Met Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Asp Pro
260 265 270
Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala
275 280 285
Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly
290 295 300
Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp Ile Phe Trp
305 310 315 320
Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val
325 330 335
Thr Val Ala Phe Ile Ile Phe Trp Val Arg Arg Val Lys Phe Ser Arg
340 345 350
Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn
355 360 365
Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg
370 375 380
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro
385 390 395 400
Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala
405 410 415
Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His
420 425 430
Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp
435 440 445
Ala Leu His Met Gln Ala Leu Pro Pro Arg Arg Ala Glu Gly Arg Gly
450 455 460
Ser Leu Leu Thr Cys Gly Asp Val Glu Glu Asn Pro Gly Pro Met Ala
465 470 475 480
Val Pro Thr Gln Val Leu Gly Leu Leu Leu Leu Trp Leu Thr Asp Ala
485 490 495
Arg Cys Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser
500 505 510
Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Asp Ile Tyr
515 520 525
Phe Asn Leu Val Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu
530 535 540
Leu Ile Tyr Asp Thr Asn Arg Leu Ala Asp Gly Val Pro Ser Arg Phe
545 550 555 560
Ser Gly Ser Gly Ser Gly Thr Gln Tyr Thr Leu Thr Ile Ser Ser Leu
565 570 575
Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln His Tyr Lys Asn Tyr
580 585 590
Pro Leu Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Ser Gly
595 600 605
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
610 615 620
Gly Gly Ser Arg Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu
625 630 635 640
Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe
645 650 655
Thr Leu Ser Asn Tyr Gly Met His Trp Ile Arg Gln Ala Pro Gly Lys
660 665 670
Gly Leu Glu Trp Val Ser Ser Ile Ser Leu Asn Gly Gly Ser Thr Tyr
675 680 685
Tyr Arg Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala
690 695 700
Lys Ser Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr
705 710 715 720
Ala Val Tyr Tyr Cys Ala Ala Gln Asp Ala Tyr Thr Gly Gly Tyr Phe
725 730 735
Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Met Asp Pro
740 745 750
Ala Thr Thr Thr Lys Pro Val Leu Arg Thr Pro Ser Pro Val His Pro
755 760 765
Thr Gly Thr Ser Gln Pro Gln Arg Pro Glu Asp Cys Arg Pro Arg Gly
770 775 780
Ser Val Lys Gly Thr Gly Leu Asp Phe Ala Cys Asp Ile Leu Ile Gly
785 790 795 800
Val Ser Val Val Phe Leu Phe Cys Leu Leu Leu Leu Val Leu Phe Cys
805 810 815
Leu His Arg Gln Asn Gln Ile Lys Gln Gly Pro Pro Arg Ser Lys Asp
820 825 830
Glu Glu Gln Lys Pro Gln Gln Arg Pro Asp Leu Ala Val Asp Val Leu
835 840 845
Glu Arg Thr Ala Asp Lys Ala Thr Val Asn Gly Leu Pro Glu Lys Asp
850 855 860
Arg Glu Thr Asp Thr Ser Ala Leu Ala Ala Gly Ser Ser Gln Glu Val
865 870 875 880
Thr Tyr Ala Gln Leu Asp His Trp Ala Leu Thr Gln Arg Thr Ala Arg
885 890 895
Ala Val Ser Pro Gln Ser Thr Lys Pro Met Ala Glu Ser Ile Thr Tyr
900 905 910
Ala Ala Val Ala Arg His
915
<210> 6
<211> 1363
<212> Белок
<213> Искусственная последовательность
<220>
<223> Химерный антигенный рецептор (CAR)
<400> 6
Met Ser Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Asp Ile Gln Met Thr Gln Thr Thr Ser Ser Leu
20 25 30
Ser Ala Ser Leu Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln
35 40 45
Asp Ile Ser Lys Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr
50 55 60
Val Lys Leu Leu Ile Tyr His Thr Ser Arg Leu His Ser Gly Val Pro
65 70 75 80
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile
85 90 95
Ser Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly
100 105 110
Asn Thr Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Thr
115 120 125
Lys Ala Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Gly Gly Gly Gly Ser Glu Val Lys Leu Gln Glu Ser Gly Pro Gly
145 150 155 160
Leu Val Ala Pro Ser Gln Ser Leu Ser Val Thr Cys Thr Val Ser Gly
165 170 175
Val Ser Leu Pro Asp Tyr Gly Val Ser Trp Ile Arg Gln Pro Pro Arg
180 185 190
Lys Gly Leu Glu Trp Leu Gly Val Ile Trp Gly Ser Glu Thr Thr Tyr
195 200 205
Tyr Asn Ser Ala Leu Lys Ser Arg Leu Thr Ile Ile Lys Asp Asn Ser
210 215 220
Lys Ser Gln Val Phe Leu Lys Met Asn Ser Leu Gln Thr Asp Asp Thr
225 230 235 240
Ala Ile Tyr Tyr Cys Ala Lys His Tyr Tyr Tyr Gly Gly Ser Tyr Ala
245 250 255
Met Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Asp Pro
260 265 270
Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala
275 280 285
Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly
290 295 300
Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp Ile Phe Trp
305 310 315 320
Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val
325 330 335
Thr Val Ala Phe Ile Ile Phe Trp Val Arg Arg Val Lys Phe Ser Arg
340 345 350
Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn
355 360 365
Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg
370 375 380
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro
385 390 395 400
Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala
405 410 415
Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His
420 425 430
Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp
435 440 445
Ala Leu His Met Gln Ala Leu Pro Pro Arg Arg Ala Glu Gly Arg Gly
450 455 460
Ser Leu Leu Thr Cys Gly Asp Val Glu Glu Asn Pro Gly Pro Met Ala
465 470 475 480
Val Pro Thr Gln Val Leu Gly Leu Leu Leu Leu Trp Leu Thr Asp Ala
485 490 495
Arg Cys Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser
500 505 510
Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Asp Ile Tyr
515 520 525
Phe Asn Leu Val Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu
530 535 540
Leu Ile Tyr Asp Thr Asn Arg Leu Ala Asp Gly Val Pro Ser Arg Phe
545 550 555 560
Ser Gly Ser Gly Ser Gly Thr Gln Tyr Thr Leu Thr Ile Ser Ser Leu
565 570 575
Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln His Tyr Lys Asn Tyr
580 585 590
Pro Leu Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Ser Gly
595 600 605
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
610 615 620
Gly Gly Ser Arg Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu
625 630 635 640
Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe
645 650 655
Thr Leu Ser Asn Tyr Gly Met His Trp Ile Arg Gln Ala Pro Gly Lys
660 665 670
Gly Leu Glu Trp Val Ser Ser Ile Ser Leu Asn Gly Gly Ser Thr Tyr
675 680 685
Tyr Arg Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala
690 695 700
Lys Ser Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr
705 710 715 720
Ala Val Tyr Tyr Cys Ala Ala Gln Asp Ala Tyr Thr Gly Gly Tyr Phe
725 730 735
Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Met Asp Pro
740 745 750
Ala Thr Thr Thr Lys Pro Val Leu Arg Thr Pro Ser Pro Val His Pro
755 760 765
Thr Gly Thr Ser Gln Pro Gln Arg Pro Glu Asp Cys Arg Pro Arg Gly
770 775 780
Ser Val Lys Gly Thr Gly Leu Asp Phe Ala Cys Asp Ile Leu Ile Gly
785 790 795 800
Val Ser Val Val Phe Leu Phe Cys Leu Leu Leu Leu Val Leu Phe Cys
805 810 815
Leu His Arg Gln Asn Gln Ile Lys Gln Gly Pro Pro Arg Ser Lys Asp
820 825 830
Glu Glu Gln Lys Pro Gln Gln Arg Pro Asp Leu Ala Val Asp Val Leu
835 840 845
Glu Arg Thr Ala Asp Lys Ala Thr Val Asn Gly Leu Pro Glu Lys Asp
850 855 860
Arg Glu Thr Asp Thr Ser Ala Leu Ala Ala Gly Ser Ser Gln Glu Val
865 870 875 880
Thr Tyr Ala Gln Leu Asp His Trp Ala Leu Thr Gln Arg Thr Ala Arg
885 890 895
Ala Val Ser Pro Gln Ser Thr Lys Pro Met Ala Glu Ser Ile Thr Tyr
900 905 910
Ala Ala Val Ala Arg His Arg Ala Glu Gly Arg Gly Ser Leu Leu Thr
915 920 925
Cys Gly Asp Val Glu Glu Asn Pro Gly Pro Trp Tyr His Gly His Met
930 935 940
Ser Gly Gly Gln Ala Glu Thr Leu Leu Gln Ala Lys Gly Glu Pro Trp
945 950 955 960
Thr Phe Leu Val Arg Glu Ser Leu Ser Gln Pro Gly Asp Phe Val Leu
965 970 975
Ser Val Leu Ser Asp Gln Pro Lys Ala Gly Pro Gly Ser Pro Leu Arg
980 985 990
Val Thr His Ile Lys Val Met Cys Glu Gly Gly Arg Tyr Thr Val Gly
995 1000 1005
Gly Leu Glu Thr Phe Asp Ser Leu Thr Asp Leu Val Glu His Phe
1010 1015 1020
Lys Lys Thr Gly Ile Glu Glu Ala Ser Gly Ala Phe Val Tyr Leu
1025 1030 1035
Arg Gln Pro Tyr Ser Gly Gly Gly Gly Ser Phe Glu Ala Tyr Phe
1040 1045 1050
Lys Lys Gln Gln Ala Asp Ser Asn Cys Gly Phe Ala Glu Glu Tyr
1055 1060 1065
Glu Asp Leu Lys Leu Val Gly Ile Ser Gln Pro Lys Tyr Ala Ala
1070 1075 1080
Glu Leu Ala Glu Asn Arg Gly Lys Asn Arg Tyr Asn Asn Val Leu
1085 1090 1095
Pro Tyr Asp Ile Ser Arg Val Lys Leu Ser Val Gln Thr His Ser
1100 1105 1110
Thr Asp Asp Tyr Ile Asn Ala Asn Tyr Met Pro Gly Tyr His Ser
1115 1120 1125
Lys Lys Asp Phe Ile Ala Thr Gln Gly Pro Leu Pro Asn Thr Leu
1130 1135 1140
Lys Asp Phe Trp Arg Met Val Trp Glu Lys Asn Val Tyr Ala Ile
1145 1150 1155
Ile Met Leu Thr Lys Cys Val Glu Gln Gly Arg Thr Lys Cys Glu
1160 1165 1170
Glu Tyr Trp Pro Ser Lys Gln Ala Gln Asp Tyr Gly Asp Ile Thr
1175 1180 1185
Val Ala Met Thr Ser Glu Ile Val Leu Pro Glu Trp Thr Ile Arg
1190 1195 1200
Asp Phe Thr Val Lys Asn Ile Gln Thr Ser Glu Ser His Pro Leu
1205 1210 1215
Arg Gln Phe His Phe Thr Ser Trp Pro Asp His Gly Val Pro Asp
1220 1225 1230
Thr Thr Asp Leu Leu Ile Asn Phe Arg Tyr Leu Val Arg Asp Tyr
1235 1240 1245
Met Lys Gln Ser Pro Pro Glu Ser Pro Ile Leu Val His Cys Ser
1250 1255 1260
Ala Gly Val Gly Arg Thr Gly Thr Phe Ile Ala Ile Asp Arg Leu
1265 1270 1275
Ile Tyr Gln Ile Glu Asn Glu Asn Thr Val Asp Val Tyr Gly Ile
1280 1285 1290
Val Tyr Asp Leu Arg Met His Arg Pro Leu Met Val Gln Thr Glu
1295 1300 1305
Asp Gln Tyr Val Phe Leu Asn Gln Cys Val Leu Asp Ile Val Arg
1310 1315 1320
Ser Gln Lys Asp Ser Lys Val Asp Leu Ile Tyr Gln Asn Thr Thr
1325 1330 1335
Ala Met Thr Ile Tyr Glu Asn Leu Ala Pro Val Thr Thr Phe Gly
1340 1345 1350
Lys Thr Asn Gly Tyr Ile Ala Ser Gly Ser
1355 1360
<210> 7
<211> 21
<212> Белок
<213> Искусственная последовательность
<220>
<223> Сигнальный пептид
<400> 7
Met Gly Thr Ser Leu Leu Cys Trp Met Ala Leu Cys Leu Leu Gly Ala
1 5 10 15
Asp His Ala Asp Gly
20
<210> 8
<211> 21
<212> Белок
<213> Искусственная последовательность
<220>
<223> Сигнальный пептид
<400> 8
Met Ser Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro
20
<210> 9
<211> 20
<212> Белок
<213> Искусственная последовательность
<220>
<223> Сигнальный пептид
<400> 9
Met Ala Val Pro Thr Gln Val Leu Gly Leu Leu Leu Leu Trp Leu Thr
1 5 10 15
Asp Ala Arg Cys
20
<210> 10
<211> 234
<212> Белок
<213> Искусственная последовательность
<220>
<223> Спейсер (шарнир-CH2CH3 из IgG1 человека)
<400> 10
Ala Glu Pro Lys Ser Pro Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
20 25 30
Lys Asp Thr Leu Met Ile Ala Arg Thr Pro Glu Val Thr Cys Val Val
35 40 45
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
50 55 60
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
65 70 75 80
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
85 90 95
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
100 105 110
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
115 120 125
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
130 135 140
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
145 150 155 160
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
165 170 175
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
180 185 190
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
195 200 205
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
210 215 220
Ser Leu Ser Leu Ser Pro Gly Lys Lys Asp
225 230
<210> 11
<211> 46
<212> Белок
<213> Искусственная последовательность
<220>
<223> Спейсер (стебель CD8 человека)
<400> 11
Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala
1 5 10 15
Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly
20 25 30
Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp Ile
35 40 45
<210> 12
<211> 20
<212> Белок
<213> Искусственная последовательность
<220>
<223> Спейсер (шарнир IgG1 человека)
<400> 12
Ala Glu Pro Lys Ser Pro Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
Lys Asp Pro Lys
20
<210> 13
<211> 185
<212> Белок
<213> Искусственная последовательность
<220>
<223> Спейсер (эктодомен CD2)
<400> 13
Lys Glu Ile Thr Asn Ala Leu Glu Thr Trp Gly Ala Leu Gly Gln Asp
1 5 10 15
Ile Asn Leu Asp Ile Pro Ser Phe Gln Met Ser Asp Asp Ile Asp Asp
20 25 30
Ile Lys Trp Glu Lys Thr Ser Asp Lys Lys Lys Ile Ala Gln Phe Arg
35 40 45
Lys Glu Lys Glu Thr Phe Lys Glu Lys Asp Thr Tyr Lys Leu Phe Lys
50 55 60
Asn Gly Thr Leu Lys Ile Lys His Leu Lys Thr Asp Asp Gln Asp Ile
65 70 75 80
Tyr Lys Val Ser Ile Tyr Asp Thr Lys Gly Lys Asn Val Leu Glu Lys
85 90 95
Ile Phe Asp Leu Lys Ile Gln Glu Arg Val Ser Lys Pro Lys Ile Ser
100 105 110
Trp Thr Cys Ile Asn Thr Thr Leu Thr Cys Glu Val Met Asn Gly Thr
115 120 125
Asp Pro Glu Leu Asn Leu Tyr Gln Asp Gly Lys His Leu Lys Leu Ser
130 135 140
Gln Arg Val Ile Thr His Lys Trp Thr Thr Ser Leu Ser Ala Lys Phe
145 150 155 160
Lys Cys Thr Ala Gly Asn Lys Val Ser Lys Glu Ser Ser Val Glu Pro
165 170 175
Val Ser Cys Pro Glu Lys Gly Leu Asp
180 185
<210> 14
<211> 259
<212> Белок
<213> Искусственная последовательность
<220>
<223> Спейсер (эктодомен CD34)
<400> 14
Ser Leu Asp Asn Asn Gly Thr Ala Thr Pro Glu Leu Pro Thr Gln Gly
1 5 10 15
Thr Phe Ser Asn Val Ser Thr Asn Val Ser Tyr Gln Glu Thr Thr Thr
20 25 30
Pro Ser Thr Leu Gly Ser Thr Ser Leu His Pro Val Ser Gln His Gly
35 40 45
Asn Glu Ala Thr Thr Asn Ile Thr Glu Thr Thr Val Lys Phe Thr Ser
50 55 60
Thr Ser Val Ile Thr Ser Val Tyr Gly Asn Thr Asn Ser Ser Val Gln
65 70 75 80
Ser Gln Thr Ser Val Ile Ser Thr Val Phe Thr Thr Pro Ala Asn Val
85 90 95
Ser Thr Pro Glu Thr Thr Leu Lys Pro Ser Leu Ser Pro Gly Asn Val
100 105 110
Ser Asp Leu Ser Thr Thr Ser Thr Ser Leu Ala Thr Ser Pro Thr Lys
115 120 125
Pro Tyr Thr Ser Ser Ser Pro Ile Leu Ser Asp Ile Lys Ala Glu Ile
130 135 140
Lys Cys Ser Gly Ile Arg Glu Val Lys Leu Thr Gln Gly Ile Cys Leu
145 150 155 160
Glu Gln Asn Lys Thr Ser Ser Cys Ala Glu Phe Lys Lys Asp Arg Gly
165 170 175
Glu Gly Leu Ala Arg Val Leu Cys Gly Glu Glu Gln Ala Asp Ala Asp
180 185 190
Ala Gly Ala Gln Val Cys Ser Leu Leu Leu Ala Gln Ser Glu Val Arg
195 200 205
Pro Gln Cys Leu Leu Leu Val Leu Ala Asn Arg Thr Glu Ile Ser Ser
210 215 220
Lys Leu Gln Leu Met Lys Lys His Gln Ser Asp Leu Lys Lys Leu Gly
225 230 235 240
Ile Leu Asp Phe Thr Glu Gln Asp Val Ala Ser His Gln Ser Tyr Ser
245 250 255
Gln Lys Thr
<210> 15
<211> 140
<212> Белок
<213> Искусственная последовательность
<220>
<223> Трансмембранный домен CD28 и эндодомены CD3 Z
<400> 15
Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu
1 5 10 15
Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val Arg Arg Val Lys Phe
20 25 30
Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu
35 40 45
Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp
50 55 60
Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys
65 70 75 80
Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala
85 90 95
Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys
100 105 110
Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr
115 120 125
Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
130 135 140
<210> 16
<211> 180
<212> Белок
<213> Искусственная последовательность
<220>
<223> Трансмембранный домен CD28 и эндодомены CD28 и CD3 дзета
<400> 16
Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu
1 5 10 15
Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val Arg Ser Lys Arg Ser
20 25 30
Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr Pro Arg Arg Pro Gly
35 40 45
Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro Arg Asp Phe Ala
50 55 60
Ala Tyr Arg Ser Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala
65 70 75 80
Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg
85 90 95
Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu
100 105 110
Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn
115 120 125
Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met
130 135 140
Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly
145 150 155 160
Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala
165 170 175
Leu Pro Pro Arg
180
<210> 17
<211> 216
<212> Белок
<213> Искусственная последовательность
<220>
<223> Трансмембранный домен CD28 и эндодомены CD28, OX40 и CD3 дзета
<400> 17
Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu
1 5 10 15
Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val Arg Ser Lys Arg Ser
20 25 30
Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr Pro Arg Arg Pro Gly
35 40 45
Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro Arg Asp Phe Ala
50 55 60
Ala Tyr Arg Ser Arg Asp Gln Arg Leu Pro Pro Asp Ala His Lys Pro
65 70 75 80
Pro Gly Gly Gly Ser Phe Arg Thr Pro Ile Gln Glu Glu Gln Ala Asp
85 90 95
Ala His Ser Thr Leu Ala Lys Ile Arg Val Lys Phe Ser Arg Ser Ala
100 105 110
Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu
115 120 125
Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly
130 135 140
Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu
145 150 155 160
Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser
165 170 175
Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly
180 185 190
Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu
195 200 205
His Met Gln Ala Leu Pro Pro Arg
210 215
<210> 18
<211> 729
<212> Белок
<213> Искусственная последовательность
<220>
<223> Трансмембранный и эндодомен CD45
<400> 18
Ala Leu Ile Ala Phe Leu Ala Phe Leu Ile Ile Val Thr Ser Ile Ala
1 5 10 15
Leu Leu Val Val Leu Tyr Lys Ile Tyr Asp Leu His Lys Lys Arg Ser
20 25 30
Cys Asn Leu Asp Glu Gln Gln Glu Leu Val Glu Arg Asp Asp Glu Lys
35 40 45
Gln Leu Met Asn Val Glu Pro Ile His Ala Asp Ile Leu Leu Glu Thr
50 55 60
Tyr Lys Arg Lys Ile Ala Asp Glu Gly Arg Leu Phe Leu Ala Glu Phe
65 70 75 80
Gln Ser Ile Pro Arg Val Phe Ser Lys Phe Pro Ile Lys Glu Ala Arg
85 90 95
Lys Pro Phe Asn Gln Asn Lys Asn Arg Tyr Val Asp Ile Leu Pro Tyr
100 105 110
Asp Tyr Asn Arg Val Glu Leu Ser Glu Ile Asn Gly Asp Ala Gly Ser
115 120 125
Asn Tyr Ile Asn Ala Ser Tyr Ile Asp Gly Phe Lys Glu Pro Arg Lys
130 135 140
Tyr Ile Ala Ala Gln Gly Pro Arg Asp Glu Thr Val Asp Asp Phe Trp
145 150 155 160
Arg Met Ile Trp Glu Gln Lys Ala Thr Val Ile Val Met Val Thr Arg
165 170 175
Cys Glu Glu Gly Asn Arg Asn Lys Cys Ala Glu Tyr Trp Pro Ser Met
180 185 190
Glu Glu Gly Thr Arg Ala Phe Gly Asp Val Val Val Lys Ile Asn Gln
195 200 205
His Lys Arg Cys Pro Asp Tyr Ile Ile Gln Lys Leu Asn Ile Val Asn
210 215 220
Lys Lys Glu Lys Ala Thr Gly Arg Glu Val Thr His Ile Gln Phe Thr
225 230 235 240
Ser Trp Pro Asp His Gly Val Pro Glu Asp Pro His Leu Leu Leu Lys
245 250 255
Leu Arg Arg Arg Val Asn Ala Phe Ser Asn Phe Phe Ser Gly Pro Ile
260 265 270
Val Val His Cys Ser Ala Gly Val Gly Arg Thr Gly Thr Tyr Ile Gly
275 280 285
Ile Asp Ala Met Leu Glu Gly Leu Glu Ala Glu Asn Lys Val Asp Val
290 295 300
Tyr Gly Tyr Val Val Lys Leu Arg Arg Gln Arg Cys Leu Met Val Gln
305 310 315 320
Val Glu Ala Gln Tyr Ile Leu Ile His Gln Ala Leu Val Glu Tyr Asn
325 330 335
Gln Phe Gly Glu Thr Glu Val Asn Leu Ser Glu Leu His Pro Tyr Leu
340 345 350
His Asn Met Lys Lys Arg Asp Pro Pro Ser Glu Pro Ser Pro Leu Glu
355 360 365
Ala Glu Phe Gln Arg Leu Pro Ser Tyr Arg Ser Trp Arg Thr Gln His
370 375 380
Ile Gly Asn Gln Glu Glu Asn Lys Ser Lys Asn Arg Asn Ser Asn Val
385 390 395 400
Ile Pro Tyr Asp Tyr Asn Arg Val Pro Leu Lys His Glu Leu Glu Met
405 410 415
Ser Lys Glu Ser Glu His Asp Ser Asp Glu Ser Ser Asp Asp Asp Ser
420 425 430
Asp Ser Glu Glu Pro Ser Lys Tyr Ile Asn Ala Ser Phe Ile Met Ser
435 440 445
Tyr Trp Lys Pro Glu Val Met Ile Ala Ala Gln Gly Pro Leu Lys Glu
450 455 460
Thr Ile Gly Asp Phe Trp Gln Met Ile Phe Gln Arg Lys Val Lys Val
465 470 475 480
Ile Val Met Leu Thr Glu Leu Lys His Gly Asp Gln Glu Ile Cys Ala
485 490 495
Gln Tyr Trp Gly Glu Gly Lys Gln Thr Tyr Gly Asp Ile Glu Val Asp
500 505 510
Leu Lys Asp Thr Asp Lys Ser Ser Thr Tyr Thr Leu Arg Val Phe Glu
515 520 525
Leu Arg His Ser Lys Arg Lys Asp Ser Arg Thr Val Tyr Gln Tyr Gln
530 535 540
Tyr Thr Asn Trp Ser Val Glu Gln Leu Pro Ala Glu Pro Lys Glu Leu
545 550 555 560
Ile Ser Met Ile Gln Val Val Lys Gln Lys Leu Pro Gln Lys Asn Ser
565 570 575
Ser Glu Gly Asn Lys His His Lys Ser Thr Pro Leu Leu Ile His Cys
580 585 590
Arg Asp Gly Ser Gln Gln Thr Gly Ile Phe Cys Ala Leu Leu Asn Leu
595 600 605
Leu Glu Ser Ala Glu Thr Glu Glu Val Val Asp Ile Phe Gln Val Val
610 615 620
Lys Ala Leu Arg Lys Ala Arg Pro Gly Met Val Ser Thr Phe Glu Gln
625 630 635 640
Tyr Gln Phe Leu Tyr Asp Val Ile Ala Ser Thr Tyr Pro Ala Gln Asn
645 650 655
Gly Gln Val Lys Lys Asn Asn His Gln Glu Asp Lys Ile Glu Phe Asp
660 665 670
Asn Glu Val Asp Lys Val Lys Gln Asp Ala Asn Cys Val Asn Pro Leu
675 680 685
Gly Ala Pro Glu Lys Leu Pro Glu Ala Lys Glu Gln Ala Glu Gly Ser
690 695 700
Glu Pro Thr Ser Gly Thr Glu Gly Pro Glu His Ser Val Asn Gly Pro
705 710 715 720
Ala Ser Pro Ala Leu Asn Gln Gly Ser
725
<210> 19
<211> 362
<212> Белок
<213> Искусственная последовательность
<220>
<223> Трансмембранный и эндодомен CD148
<400> 19
Ala Val Phe Gly Cys Ile Phe Gly Ala Leu Val Ile Val Thr Val Gly
1 5 10 15
Gly Phe Ile Phe Trp Arg Lys Lys Arg Lys Asp Ala Lys Asn Asn Glu
20 25 30
Val Ser Phe Ser Gln Ile Lys Pro Lys Lys Ser Lys Leu Ile Arg Val
35 40 45
Glu Asn Phe Glu Ala Tyr Phe Lys Lys Gln Gln Ala Asp Ser Asn Cys
50 55 60
Gly Phe Ala Glu Glu Tyr Glu Asp Leu Lys Leu Val Gly Ile Ser Gln
65 70 75 80
Pro Lys Tyr Ala Ala Glu Leu Ala Glu Asn Arg Gly Lys Asn Arg Tyr
85 90 95
Asn Asn Val Leu Pro Tyr Asp Ile Ser Arg Val Lys Leu Ser Val Gln
100 105 110
Thr His Ser Thr Asp Asp Tyr Ile Asn Ala Asn Tyr Met Pro Gly Tyr
115 120 125
His Ser Lys Lys Asp Phe Ile Ala Thr Gln Gly Pro Leu Pro Asn Thr
130 135 140
Leu Lys Asp Phe Trp Arg Met Val Trp Glu Lys Asn Val Tyr Ala Ile
145 150 155 160
Ile Met Leu Thr Lys Cys Val Glu Gln Gly Arg Thr Lys Cys Glu Glu
165 170 175
Tyr Trp Pro Ser Lys Gln Ala Gln Asp Tyr Gly Asp Ile Thr Val Ala
180 185 190
Met Thr Ser Glu Ile Val Leu Pro Glu Trp Thr Ile Arg Asp Phe Thr
195 200 205
Val Lys Asn Ile Gln Thr Ser Glu Ser His Pro Leu Arg Gln Phe His
210 215 220
Phe Thr Ser Trp Pro Asp His Gly Val Pro Asp Thr Thr Asp Leu Leu
225 230 235 240
Ile Asn Phe Arg Tyr Leu Val Arg Asp Tyr Met Lys Gln Ser Pro Pro
245 250 255
Glu Ser Pro Ile Leu Val His Cys Ser Ala Gly Val Gly Arg Thr Gly
260 265 270
Thr Phe Ile Ala Ile Asp Arg Leu Ile Tyr Gln Ile Glu Asn Glu Asn
275 280 285
Thr Val Asp Val Tyr Gly Ile Val Tyr Asp Leu Arg Met His Arg Pro
290 295 300
Leu Met Val Gln Thr Glu Asp Gln Tyr Val Phe Leu Asn Gln Cys Val
305 310 315 320
Leu Asp Ile Val Arg Ser Gln Lys Asp Ser Lys Val Asp Leu Ile Tyr
325 330 335
Gln Asn Thr Thr Ala Met Thr Ile Tyr Glu Asn Leu Ala Pro Val Thr
340 345 350
Thr Phe Gly Lys Thr Asn Gly Tyr Ile Ala
355 360
<210> 20
<211> 595
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность PTPN6
<400> 20
Met Val Arg Trp Phe His Arg Asp Leu Ser Gly Leu Asp Ala Glu Thr
1 5 10 15
Leu Leu Lys Gly Arg Gly Val His Gly Ser Phe Leu Ala Arg Pro Ser
20 25 30
Arg Lys Asn Gln Gly Asp Phe Ser Leu Ser Val Arg Val Gly Asp Gln
35 40 45
Val Thr His Ile Arg Ile Gln Asn Ser Gly Asp Phe Tyr Asp Leu Tyr
50 55 60
Gly Gly Glu Lys Phe Ala Thr Leu Thr Glu Leu Val Glu Tyr Tyr Thr
65 70 75 80
Gln Gln Gln Gly Val Leu Gln Asp Arg Asp Gly Thr Ile Ile His Leu
85 90 95
Lys Tyr Pro Leu Asn Cys Ser Asp Pro Thr Ser Glu Arg Trp Tyr His
100 105 110
Gly His Met Ser Gly Gly Gln Ala Glu Thr Leu Leu Gln Ala Lys Gly
115 120 125
Glu Pro Trp Thr Phe Leu Val Arg Glu Ser Leu Ser Gln Pro Gly Asp
130 135 140
Phe Val Leu Ser Val Leu Ser Asp Gln Pro Lys Ala Gly Pro Gly Ser
145 150 155 160
Pro Leu Arg Val Thr His Ile Lys Val Met Cys Glu Gly Gly Arg Tyr
165 170 175
Thr Val Gly Gly Leu Glu Thr Phe Asp Ser Leu Thr Asp Leu Val Glu
180 185 190
His Phe Lys Lys Thr Gly Ile Glu Glu Ala Ser Gly Ala Phe Val Tyr
195 200 205
Leu Arg Gln Pro Tyr Tyr Ala Thr Arg Val Asn Ala Ala Asp Ile Glu
210 215 220
Asn Arg Val Leu Glu Leu Asn Lys Lys Gln Glu Ser Glu Asp Thr Ala
225 230 235 240
Lys Ala Gly Phe Trp Glu Glu Phe Glu Ser Leu Gln Lys Gln Glu Val
245 250 255
Lys Asn Leu His Gln Arg Leu Glu Gly Gln Arg Pro Glu Asn Lys Gly
260 265 270
Lys Asn Arg Tyr Lys Asn Ile Leu Pro Phe Asp His Ser Arg Val Ile
275 280 285
Leu Gln Gly Arg Asp Ser Asn Ile Pro Gly Ser Asp Tyr Ile Asn Ala
290 295 300
Asn Tyr Ile Lys Asn Gln Leu Leu Gly Pro Asp Glu Asn Ala Lys Thr
305 310 315 320
Tyr Ile Ala Ser Gln Gly Cys Leu Glu Ala Thr Val Asn Asp Phe Trp
325 330 335
Gln Met Ala Trp Gln Glu Asn Ser Arg Val Ile Val Met Thr Thr Arg
340 345 350
Glu Val Glu Lys Gly Arg Asn Lys Cys Val Pro Tyr Trp Pro Glu Val
355 360 365
Gly Met Gln Arg Ala Tyr Gly Pro Tyr Ser Val Thr Asn Cys Gly Glu
370 375 380
His Asp Thr Thr Glu Tyr Lys Leu Arg Thr Leu Gln Val Ser Pro Leu
385 390 395 400
Asp Asn Gly Asp Leu Ile Arg Glu Ile Trp His Tyr Gln Tyr Leu Ser
405 410 415
Trp Pro Asp His Gly Val Pro Ser Glu Pro Gly Gly Val Leu Ser Phe
420 425 430
Leu Asp Gln Ile Asn Gln Arg Gln Glu Ser Leu Pro His Ala Gly Pro
435 440 445
Ile Ile Val His Cys Ser Ala Gly Ile Gly Arg Thr Gly Thr Ile Ile
450 455 460
Val Ile Asp Met Leu Met Glu Asn Ile Ser Thr Lys Gly Leu Asp Cys
465 470 475 480
Asp Ile Asp Ile Gln Lys Thr Ile Gln Met Val Arg Ala Gln Arg Ser
485 490 495
Gly Met Val Gln Thr Glu Ala Gln Tyr Lys Phe Ile Tyr Val Ala Ile
500 505 510
Ala Gln Phe Ile Glu Thr Thr Lys Lys Lys Leu Glu Val Leu Gln Ser
515 520 525
Gln Lys Gly Gln Glu Ser Glu Tyr Gly Asn Ile Thr Tyr Pro Pro Ala
530 535 540
Met Lys Asn Ala His Ala Lys Ala Ser Arg Thr Ser Ser Lys His Lys
545 550 555 560
Glu Asp Val Tyr Glu Asn Leu His Thr Lys Asn Lys Arg Glu Glu Lys
565 570 575
Val Lys Lys Gln Arg Ser Ala Asp Lys Glu Lys Ser Lys Gly Ser Leu
580 585 590
Lys Arg Lys
595
<210> 21
<211> 272
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность фосфатазного домена из PTPN6
<400> 21
Phe Trp Glu Glu Phe Glu Ser Leu Gln Lys Gln Glu Val Lys Asn Leu
1 5 10 15
His Gln Arg Leu Glu Gly Gln Arg Pro Glu Asn Lys Gly Lys Asn Arg
20 25 30
Tyr Lys Asn Ile Leu Pro Phe Asp His Ser Arg Val Ile Leu Gln Gly
35 40 45
Arg Asp Ser Asn Ile Pro Gly Ser Asp Tyr Ile Asn Ala Asn Tyr Ile
50 55 60
Lys Asn Gln Leu Leu Gly Pro Asp Glu Asn Ala Lys Thr Tyr Ile Ala
65 70 75 80
Ser Gln Gly Cys Leu Glu Ala Thr Val Asn Asp Phe Trp Gln Met Ala
85 90 95
Trp Gln Glu Asn Ser Arg Val Ile Val Met Thr Thr Arg Glu Val Glu
100 105 110
Lys Gly Arg Asn Lys Cys Val Pro Tyr Trp Pro Glu Val Gly Met Gln
115 120 125
Arg Ala Tyr Gly Pro Tyr Ser Val Thr Asn Cys Gly Glu His Asp Thr
130 135 140
Thr Glu Tyr Lys Leu Arg Thr Leu Gln Val Ser Pro Leu Asp Asn Gly
145 150 155 160
Asp Leu Ile Arg Glu Ile Trp His Tyr Gln Tyr Leu Ser Trp Pro Asp
165 170 175
His Gly Val Pro Ser Glu Pro Gly Gly Val Leu Ser Phe Leu Asp Gln
180 185 190
Ile Asn Gln Arg Gln Glu Ser Leu Pro His Ala Gly Pro Ile Ile Val
195 200 205
His Cys Ser Ala Gly Ile Gly Arg Thr Gly Thr Ile Ile Val Ile Asp
210 215 220
Met Leu Met Glu Asn Ile Ser Thr Lys Gly Leu Asp Cys Asp Ile Asp
225 230 235 240
Ile Gln Lys Thr Ile Gln Met Val Arg Ala Gln Arg Ser Gly Met Val
245 250 255
Gln Thr Glu Ala Gln Tyr Lys Phe Ile Tyr Val Ala Ile Ala Gln Phe
260 265 270
<210> 22
<211> 97
<212> Белок
<213> Искусственная последовательность
<220>
<223> Эндодомен PDCD1
<400> 22
Cys Ser Arg Ala Ala Arg Gly Thr Ile Gly Ala Arg Arg Thr Gly Gln
1 5 10 15
Pro Leu Lys Glu Asp Pro Ser Ala Val Pro Val Phe Ser Val Asp Tyr
20 25 30
Gly Glu Leu Asp Phe Gln Trp Arg Glu Lys Thr Pro Glu Pro Pro Val
35 40 45
Pro Cys Val Pro Glu Gln Thr Glu Tyr Ala Thr Ile Val Phe Pro Ser
50 55 60
Gly Met Gly Thr Ser Ser Pro Ala Arg Arg Gly Ser Ala Asp Gly Pro
65 70 75 80
Arg Ser Ala Gln Pro Leu Arg Pro Glu Asp Gly His Cys Ser Trp Pro
85 90 95
Leu
<210> 23
<211> 141
<212> Белок
<213> Искусственная последовательность
<220>
<223> Эндодомен BTLA4
<400> 23
Lys Leu Gln Arg Arg Trp Lys Arg Thr Gln Ser Gln Gln Gly Leu Gln
1 5 10 15
Glu Asn Ser Ser Gly Gln Ser Phe Phe Val Arg Asn Lys Lys Val Arg
20 25 30
Arg Ala Pro Leu Ser Glu Gly Pro His Ser Leu Gly Cys Tyr Asn Pro
35 40 45
Met Met Glu Asp Gly Ile Ser Tyr Thr Thr Leu Arg Phe Pro Glu Met
50 55 60
Asn Ile Pro Arg Thr Gly Asp Ala Glu Ser Ser Glu Met Gln Arg Pro
65 70 75 80
Pro Pro Asp Cys Asp Asp Thr Val Thr Tyr Ser Ala Leu His Lys Arg
85 90 95
Gln Val Gly Asp Tyr Glu Asn Val Ile Pro Asp Phe Pro Glu Asp Glu
100 105 110
Gly Ile His Tyr Ser Glu Leu Ile Gln Phe Gly Val Gly Glu Arg Pro
115 120 125
Gln Ala Gln Glu Asn Val Asp Tyr Val Ile Leu Lys His
130 135 140
<210> 24
<211> 168
<212> Белок
<213> Искусственная последовательность
<220>
<223> Эндодомен LILRB1
<400> 24
Leu Arg His Arg Arg Gln Gly Lys His Trp Thr Ser Thr Gln Arg Lys
1 5 10 15
Ala Asp Phe Gln His Pro Ala Gly Ala Val Gly Pro Glu Pro Thr Asp
20 25 30
Arg Gly Leu Gln Trp Arg Ser Ser Pro Ala Ala Asp Ala Gln Glu Glu
35 40 45
Asn Leu Tyr Ala Ala Val Lys His Thr Gln Pro Glu Asp Gly Val Glu
50 55 60
Met Asp Thr Arg Ser Pro His Asp Glu Asp Pro Gln Ala Val Thr Tyr
65 70 75 80
Ala Glu Val Lys His Ser Arg Pro Arg Arg Glu Met Ala Ser Pro Pro
85 90 95
Ser Pro Leu Ser Gly Glu Phe Leu Asp Thr Lys Asp Arg Gln Ala Glu
100 105 110
Glu Asp Arg Gln Met Asp Thr Glu Ala Ala Ala Ser Glu Ala Pro Gln
115 120 125
Asp Val Thr Tyr Ala Gln Leu His Ser Leu Thr Leu Arg Arg Glu Ala
130 135 140
Thr Glu Pro Pro Pro Ser Gln Glu Gly Pro Ser Pro Ala Val Pro Ser
145 150 155 160
Ile Tyr Ala Thr Leu Ala Ile His
165
<210> 25
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> Эндодомен LAIR1
<400> 25
His Arg Gln Asn Gln Ile Lys Gln Gly Pro Pro Arg Ser Lys Asp Glu
1 5 10 15
Glu Gln Lys Pro Gln Gln Arg Pro Asp Leu Ala Val Asp Val Leu Glu
20 25 30
Arg Thr Ala Asp Lys Ala Thr Val Asn Gly Leu Pro Glu Lys Asp Arg
35 40 45
Glu Thr Asp Thr Ser Ala Leu Ala Ala Gly Ser Ser Gln Glu Val Thr
50 55 60
Tyr Ala Gln Leu Asp His Trp Ala Leu Thr Gln Arg Thr Ala Arg Ala
65 70 75 80
Val Ser Pro Gln Ser Thr Lys Pro Met Ala Glu Ser Ile Thr Tyr Ala
85 90 95
Ala Val Ala Arg His
100
<210> 26
<211> 62
<212> Белок
<213> Искусственная последовательность
<220>
<223> Эндодомен CTLA4
<400> 26
Phe Leu Leu Trp Ile Leu Ala Ala Val Ser Ser Gly Leu Phe Phe Tyr
1 5 10 15
Ser Phe Leu Leu Thr Ala Val Ser Leu Ser Lys Met Leu Lys Lys Arg
20 25 30
Ser Pro Leu Thr Thr Gly Val Tyr Val Lys Met Pro Pro Thr Glu Pro
35 40 45
Glu Cys Glu Lys Gln Phe Gln Pro Tyr Phe Ile Pro Ile Asn
50 55 60
<210> 27
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Эндодомен KIR2DL1
<400> 27
Gly Asn Ser Arg His Leu His Val Leu Ile Gly Thr Ser Val Val Ile
1 5 10 15
Ile Pro Phe Ala Ile Leu Leu Phe Phe Leu Leu His Arg Trp Cys Ala
20 25 30
Asn Lys Lys Asn Ala Val Val Met Asp Gln Glu Pro Ala Gly Asn Arg
35 40 45
Thr Val Asn Arg Glu Asp Ser Asp Glu Gln Asp Pro Gln Glu Val Thr
50 55 60
Tyr Thr Gln Leu Asn His Cys Val Phe Thr Gln Arg Lys Ile Thr Arg
65 70 75 80
Pro Ser Gln Arg Pro Lys Thr Pro Pro Thr Asp Ile Ile Val Tyr Thr
85 90 95
Glu Leu Pro Asn Ala Glu Ser Arg Ser Lys Val Val Ser Cys Pro
100 105 110
<210> 28
<211> 143
<212> Белок
<213> Искусственная последовательность
<220>
<223> Эндодомен KIR2DL4
<400> 28
Gly Ile Ala Arg His Leu His Ala Val Ile Arg Tyr Ser Val Ala Ile
1 5 10 15
Ile Leu Phe Thr Ile Leu Pro Phe Phe Leu Leu His Arg Trp Cys Ser
20 25 30
Lys Lys Lys Glu Asn Ala Ala Val Met Asn Gln Glu Pro Ala Gly His
35 40 45
Arg Thr Val Asn Arg Glu Asp Ser Asp Glu Gln Asp Pro Gln Glu Val
50 55 60
Thr Tyr Ala Gln Leu Asp His Cys Ile Phe Thr Gln Arg Lys Ile Thr
65 70 75 80
Gly Pro Ser Gln Arg Ser Lys Arg Pro Ser Thr Asp Thr Ser Val Cys
85 90 95
Ile Glu Leu Pro Asn Ala Glu Pro Arg Ala Leu Ser Pro Ala His Glu
100 105 110
His His Ser Gln Ala Leu Met Gly Ser Ser Arg Glu Thr Thr Ala Leu
115 120 125
Ser Gln Thr Gln Leu Ala Ser Ser Asn Val Pro Ala Ala Gly Ile
130 135 140
<210> 29
<211> 143
<212> Белок
<213> Искусственная последовательность
<220>
<223> Эндодомен KIR2DL5
<400> 29
Thr Gly Ile Arg Arg His Leu His Ile Leu Ile Gly Thr Ser Val Ala
1 5 10 15
Ile Ile Leu Phe Ile Ile Leu Phe Phe Phe Leu Leu His Cys Cys Cys
20 25 30
Ser Asn Lys Lys Asn Ala Ala Val Met Asp Gln Glu Pro Ala Gly Asp
35 40 45
Arg Thr Val Asn Arg Glu Asp Ser Asp Asp Gln Asp Pro Gln Glu Val
50 55 60
Thr Tyr Ala Gln Leu Asp His Cys Val Phe Thr Gln Thr Lys Ile Thr
65 70 75 80
Ser Pro Ser Gln Arg Pro Lys Thr Pro Pro Thr Asp Thr Thr Met Tyr
85 90 95
Met Glu Leu Pro Asn Ala Lys Pro Arg Ser Leu Ser Pro Ala His Lys
100 105 110
His His Ser Gln Ala Leu Arg Gly Ser Ser Arg Glu Thr Thr Ala Leu
115 120 125
Ser Gln Asn Arg Val Ala Ser Ser His Val Pro Ala Ala Gly Ile
130 135 140
<210> 30
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Эндодомен KIR3DL1
<400> 30
Lys Asp Pro Arg His Leu His Ile Leu Ile Gly Thr Ser Val Val Ile
1 5 10 15
Ile Leu Phe Ile Leu Leu Leu Phe Phe Leu Leu His Leu Trp Cys Ser
20 25 30
Asn Lys Lys Asn Ala Ala Val Met Asp Gln Glu Pro Ala Gly Asn Arg
35 40 45
Thr Ala Asn Ser Glu Asp Ser Asp Glu Gln Asp Pro Glu Glu Val Thr
50 55 60
Tyr Ala Gln Leu Asp His Cys Val Phe Thr Gln Arg Lys Ile Thr Arg
65 70 75 80
Pro Ser Gln Arg Pro Lys Thr Pro Pro Thr Asp Thr Ile Leu Tyr Thr
85 90 95
Glu Leu Pro Asn Ala Lys Pro Arg Ser Lys Val Val Ser Cys Pro
100 105 110
<210> 31
<211> 97
<212> Белок
<213> Искусственная последовательность
<220>
<223> Эндодомен KIR3DL3
<400> 31
Lys Asp Pro Gly Asn Ser Arg His Leu His Val Leu Ile Gly Thr Ser
1 5 10 15
Val Val Ile Ile Pro Phe Ala Ile Leu Leu Phe Phe Leu Leu His Arg
20 25 30
Trp Cys Ala Asn Lys Lys Asn Ala Val Val Met Asp Gln Glu Pro Ala
35 40 45
Gly Asn Arg Thr Val Asn Arg Glu Asp Ser Asp Glu Gln Asp Pro Gln
50 55 60
Glu Val Thr Tyr Ala Gln Leu Asn His Cys Val Phe Thr Gln Arg Lys
65 70 75 80
Ile Thr Arg Pro Ser Gln Arg Pro Lys Thr Pro Pro Thr Asp Thr Ser
85 90 95
Val
<210> 32
<211> 807
<212> Белок
<213> Искусственная последовательность
<220>
<223> Слитный белок PTPN6-CD45
<400> 32
Trp Tyr His Gly His Met Ser Gly Gly Gln Ala Glu Thr Leu Leu Gln
1 5 10 15
Ala Lys Gly Glu Pro Trp Thr Phe Leu Val Arg Glu Ser Leu Ser Gln
20 25 30
Pro Gly Asp Phe Val Leu Ser Val Leu Ser Asp Gln Pro Lys Ala Gly
35 40 45
Pro Gly Ser Pro Leu Arg Val Thr His Ile Lys Val Met Cys Glu Gly
50 55 60
Gly Arg Tyr Thr Val Gly Gly Leu Glu Thr Phe Asp Ser Leu Thr Asp
65 70 75 80
Leu Val Glu His Phe Lys Lys Thr Gly Ile Glu Glu Ala Ser Gly Ala
85 90 95
Phe Val Tyr Leu Arg Gln Pro Tyr Lys Ile Tyr Asp Leu His Lys Lys
100 105 110
Arg Ser Cys Asn Leu Asp Glu Gln Gln Glu Leu Val Glu Arg Asp Asp
115 120 125
Glu Lys Gln Leu Met Asn Val Glu Pro Ile His Ala Asp Ile Leu Leu
130 135 140
Glu Thr Tyr Lys Arg Lys Ile Ala Asp Glu Gly Arg Leu Phe Leu Ala
145 150 155 160
Glu Phe Gln Ser Ile Pro Arg Val Phe Ser Lys Phe Pro Ile Lys Glu
165 170 175
Ala Arg Lys Pro Phe Asn Gln Asn Lys Asn Arg Tyr Val Asp Ile Leu
180 185 190
Pro Tyr Asp Tyr Asn Arg Val Glu Leu Ser Glu Ile Asn Gly Asp Ala
195 200 205
Gly Ser Asn Tyr Ile Asn Ala Ser Tyr Ile Asp Gly Phe Lys Glu Pro
210 215 220
Arg Lys Tyr Ile Ala Ala Gln Gly Pro Arg Asp Glu Thr Val Asp Asp
225 230 235 240
Phe Trp Arg Met Ile Trp Glu Gln Lys Ala Thr Val Ile Val Met Val
245 250 255
Thr Arg Cys Glu Glu Gly Asn Arg Asn Lys Cys Ala Glu Tyr Trp Pro
260 265 270
Ser Met Glu Glu Gly Thr Arg Ala Phe Gly Asp Val Val Val Lys Ile
275 280 285
Asn Gln His Lys Arg Cys Pro Asp Tyr Ile Ile Gln Lys Leu Asn Ile
290 295 300
Val Asn Lys Lys Glu Lys Ala Thr Gly Arg Glu Val Thr His Ile Gln
305 310 315 320
Phe Thr Ser Trp Pro Asp His Gly Val Pro Glu Asp Pro His Leu Leu
325 330 335
Leu Lys Leu Arg Arg Arg Val Asn Ala Phe Ser Asn Phe Phe Ser Gly
340 345 350
Pro Ile Val Val His Cys Ser Ala Gly Val Gly Arg Thr Gly Thr Tyr
355 360 365
Ile Gly Ile Asp Ala Met Leu Glu Gly Leu Glu Ala Glu Asn Lys Val
370 375 380
Asp Val Tyr Gly Tyr Val Val Lys Leu Arg Arg Gln Arg Cys Leu Met
385 390 395 400
Val Gln Val Glu Ala Gln Tyr Ile Leu Ile His Gln Ala Leu Val Glu
405 410 415
Tyr Asn Gln Phe Gly Glu Thr Glu Val Asn Leu Ser Glu Leu His Pro
420 425 430
Tyr Leu His Asn Met Lys Lys Arg Asp Pro Pro Ser Glu Pro Ser Pro
435 440 445
Leu Glu Ala Glu Phe Gln Arg Leu Pro Ser Tyr Arg Ser Trp Arg Thr
450 455 460
Gln His Ile Gly Asn Gln Glu Glu Asn Lys Ser Lys Asn Arg Asn Ser
465 470 475 480
Asn Val Ile Pro Tyr Asp Tyr Asn Arg Val Leu Lys His Glu Leu Glu
485 490 495
Met Ser Lys Glu Ser Glu His Asp Ser Asp Glu Ser Ser Asp Asp Asp
500 505 510
Ser Asp Ser Glu Glu Pro Ser Lys Tyr Ile Asn Ala Ser Phe Ile Met
515 520 525
Ser Tyr Trp Lys Pro Glu Val Met Ile Ala Ala Gln Gly Pro Leu Lys
530 535 540
Glu Thr Ile Gly Asp Phe Met Ile Gln Arg Lys Val Lys Val Ile Val
545 550 555 560
Met Leu Thr Glu Leu Lys His Gly Asp Gln Glu Ile Cys Ala Gln Tyr
565 570 575
Trp Gly Glu Gly Lys Gln Thr Tyr Gly Asp Ile Glu Val Asp Leu Lys
580 585 590
Asp Thr Asp Lys Ser Ser Thr Tyr Thr Leu Arg Val Phe Glu Leu Arg
595 600 605
His Ser Lys Arg Lys Asp Ser Arg Thr Val Tyr Gln Tyr Gln Tyr Thr
610 615 620
Asn Trp Ser Val Glu Gln Leu Pro Ala Glu Pro Lys Glu Leu Ile Ser
625 630 635 640
Met Ile Gln Val Val Lys Gln Lys Leu Pro Gln Lys Asn Ser Ser Glu
645 650 655
Gly Asn Lys His His Lys Ser Thr Pro Leu Leu Ile His Cys Arg Asp
660 665 670
Gly Ser Gln Gln Thr Gly Ile Phe Cys Ala Leu Leu Asn Leu Leu Glu
675 680 685
Ser Ala Glu Thr Glu Glu Val Val Asp Ile Phe Gln Val Val Lys Ala
690 695 700
Leu Arg Lys Ala Arg Pro Gly Met Val Ser Thr Phe Glu Gln Tyr Gln
705 710 715 720
Phe Leu Tyr Asp Val Ile Ala Ser Thr Tyr Pro Ala Gln Asn Gly Gln
725 730 735
Val Lys Lys Asn Asn His Gln Glu Asp Lys Ile Glu Phe Asp Asn Glu
740 745 750
Val Asp Lys Val Lys Gln Asp Ala Asn Cys Val Asn Pro Leu Gly Ala
755 760 765
Pro Glu Lys Leu Pro Glu Ala Lys Glu Gln Ala Glu Gly Ser Glu Pro
770 775 780
Thr Ser Gly Thr Glu Gly Pro Glu His Ser Val Asn Gly Pro Ala Ser
785 790 795 800
Pro Ala Leu Asn Gln Gly Ser
805
<210> 33
<211> 434
<212> Белок
<213> Искусственная последовательность
<220>
<223> Слитный белок PTPN6-CD148
<400> 33
Glu Thr Leu Leu Gln Ala Lys Gly Glu Pro Trp Thr Phe Leu Val Arg
1 5 10 15
Glu Ser Leu Ser Gln Pro Gly Asp Phe Val Leu Ser Val Leu Ser Asp
20 25 30
Gln Pro Lys Ala Gly Pro Gly Ser Pro Leu Arg Val Thr His Ile Lys
35 40 45
Val Met Cys Glu Gly Gly Arg Tyr Thr Val Gly Gly Leu Glu Thr Phe
50 55 60
Asp Ser Leu Thr Asp Leu Val Glu His Phe Lys Lys Thr Gly Ile Glu
65 70 75 80
Glu Ala Ser Gly Ala Phe Val Tyr Leu Arg Gln Pro Tyr Arg Lys Lys
85 90 95
Arg Lys Asp Ala Lys Asn Asn Glu Val Ser Phe Ser Gln Ile Lys Pro
100 105 110
Lys Lys Ser Lys Leu Ile Arg Val Glu Asn Phe Glu Ala Tyr Phe Lys
115 120 125
Lys Gln Gln Ala Asp Ser Asn Cys Gly Phe Ala Glu Glu Tyr Glu Asp
130 135 140
Leu Lys Leu Val Gly Ile Ser Gln Pro Lys Tyr Ala Ala Glu Leu Ala
145 150 155 160
Glu Asn Arg Gly Lys Asn Arg Tyr Asn Asn Val Leu Pro Tyr Asp Ile
165 170 175
Ser Arg Val Lys Leu Ser Val Gln Thr His Ser Thr Asp Asp Tyr Ile
180 185 190
Asn Ala Asn Tyr Met Pro Gly Tyr His Ser Lys Lys Asp Phe Ile Ala
195 200 205
Thr Gln Gly Pro Leu Pro Asn Thr Leu Lys Asp Phe Trp Arg Met Val
210 215 220
Trp Glu Lys Asn Val Tyr Ala Ile Ile Met Leu Thr Lys Cys Val Glu
225 230 235 240
Gln Gly Arg Thr Lys Cys Glu Glu Tyr Trp Pro Ser Lys Gln Ala Gln
245 250 255
Asp Tyr Gly Asp Ile Thr Val Ala Met Thr Ser Glu Ile Val Leu Pro
260 265 270
Glu Trp Thr Ile Arg Asp Phe Thr Val Lys Asn Ile Gln Thr Ser Glu
275 280 285
Ser His Pro Leu Arg Gln Phe His Phe Thr Ser Trp Pro Asp His Gly
290 295 300
Val Pro Asp Thr Thr Asp Leu Leu Ile Asn Phe Arg Tyr Leu Val Arg
305 310 315 320
Asp Tyr Met Lys Gln Ser Pro Pro Glu Ser Pro Ile Leu Val His Cys
325 330 335
Ser Ala Gly Val Gly Arg Thr Gly Thr Phe Ile Ala Ile Asp Arg Leu
340 345 350
Ile Tyr Gln Ile Glu Asn Glu Asn Thr Val Asp Val Tyr Gly Ile Val
355 360 365
Tyr Asp Leu Arg Met His Arg Pro Leu Met Val Gln Thr Glu Asp Gln
370 375 380
Tyr Val Phe Leu Asn Gln Cys Val Leu Asp Ile Val Arg Ser Gln Lys
385 390 395 400
Asp Ser Lys Val Asp Leu Ile Tyr Gln Asn Thr Thr Ala Met Thr Ile
405 410 415
Tyr Glu Asn Leu Ala Pro Val Thr Thr Phe Gly Lys Thr Asn Gly Tyr
420 425 430
Ile Ala
<210> 34
<211> 20
<212> Белок
<213> Вирус ящура
<400> 34
Arg Ala Glu Gly Arg Gly Ser Leu Leu Thr Cys Gly Asp Val Glu Glu
1 5 10 15
Asn Pro Gly Pro
20
<210> 35
<211> 3390
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательности нуклеиновой кислоты, кодирующие CAR
(MP13974.SFG.aCD19fmc63-CD8STK-CD28tmZ-2A-aCD33glx-HCH2CH3pvaa-CD
28tmZw)
<400> 35
atgagcctgc ccgtgaccgc cctgctgctg cccctggccc tgctgctgca cgccgccaga 60
ccagacatcc agatgaccca gaccaccagc agcctgagcg ccagcctggg cgaccgggtg 120
accatcagct gcagagccag ccaggacatc agcaagtacc tgaactggta ccagcagaag 180
cccgacggca ccgtgaagct gctgatctac cacaccagcc ggctgcacag cggcgtgccc 240
agccggttca gcggcagcgg cagcggcacc gactacagcc tgaccatcag caacctggag 300
caggaggaca tcgccaccta cttctgccag cagggcaaca ccctgcccta caccttcgga 360
ggcggcacca agctggagat caccaaggcc ggaggcggag gctctggcgg aggcggctct 420
ggcggaggcg gctctggcgg aggcggcagc gaggtgaagc tgcaggagtc tggcccaggc 480
ctggtggccc caagccagag cctgagcgtg acctgcaccg tgagcggcgt gagcctgccc 540
gactacggcg tgagctggat caggcagccc ccacggaagg gcctggagtg gctgggcgtg 600
atctggggca gcgagaccac ctactacaac agcgccctga agagccggct gaccatcatc 660
aaggacaaca gcaagagcca ggtgttcctg aagatgaaca gcctgcagac cgacgacacc 720
gccatctact actgcgccaa gcactactac tatggcggca gctacgctat ggactactgg 780
ggccagggca ccagcgtgac cgtgagctca gatcccacca cgacgccagc gccgcgacca 840
ccaacaccgg cgcccaccat cgcgtcgcag cccctgtccc tgcgcccaga ggcgtgccgg 900
ccagcggcgg ggggcgcagt gcacacgagg gggctggact tcgcctgtga tatcttttgg 960
gtgctggtgg tggttggtgg agtcctggct tgctatagct tgctagtaac agtggccttt 1020
attattttct gggtgaggag agtgaagttc agcaggagcg cagacgcccc cgcgtaccag 1080
cagggccaga accagctcta taacgagctc aatctaggac gaagagagga gtacgatgtt 1140
ttggacaaga gacgtggccg ggaccctgag atggggggaa agccgagaag gaagaaccct 1200
caggaaggcc tgtacaatga actgcagaaa gataagatgg cggaggccta cagtgagatt 1260
gggatgaaag gcgagcgccg gaggggcaag gggcacgatg gcctttacca gggtctcagt 1320
acagccacca aggacaccta cgacgccctt cacatgcagg ccctgcctcc tcgcagagcc 1380
gagggcaggg gaagtcttct aacatgcggg gacgtggagg aaaatcccgg gcccatggcc 1440
gtgcccactc aggtcctggg gttgttgcta ctgtggctta cagatgccag atgtgacatc 1500
cagatgacac agtctccatc ttccctgtct gcatctgtcg gagatcgcgt caccatcacc 1560
tgtcgagcaa gtgaggacat ttattttaat ttagtgtggt atcagcagaa accaggaaag 1620
gcccctaagc tcctgatcta tgatacaaat cgcttggcag atggggtccc atcacggttc 1680
agtggctctg gatctggcac acagtatact ctaaccataa gtagcctgca acccgaagat 1740
ttcgcaacct attattgtca acactataag aattatccgc tcacgttcgg tcaggggacc 1800
aagctggaaa tcaaaagatc tggtggcgga gggtcaggag gcggaggcag cggaggcggt 1860
ggctcgggag gcggaggctc gagatctgag gtgcagttgg tggagtctgg gggcggcttg 1920
gtgcagcctg gagggtccct gaggctctcc tgtgcagcct caggattcac tctcagtaat 1980
tatggcatgc actggatcag gcaggctcca gggaagggtc tggagtgggt ctcgtctatt 2040
agtcttaatg gtggtagcac ttactatcga gactccgtga agggccgatt cactatctcc 2100
agggacaatg caaaaagcac cctctacctt caaatgaata gtctgagggc cgaggacacg 2160
gccgtctatt actgtgcagc acaggacgct tatacgggag gttactttga ttactggggc 2220
caaggaacgc tggtcacagt ctcgtctatg gatcccgccg agcccaaatc tcctgacaaa 2280
actcacacat gcccaccgtg cccagcacct cccgtggccg gcccgtcagt cttcctcttc 2340
cccccaaaac ccaaggacac cctcatgatc gcccggaccc ctgaggtcac atgcgtggtg 2400
gtggacgtga gccacgaaga ccctgaggtc aagttcaact ggtacgtgga cggcgtggag 2460
gtgcataatg ccaagacaaa gccgcgggag gagcagtaca acagcacgta ccgtgtggtc 2520
agcgtcctca ccgtcctgca ccaggactgg ctgaatggca aggagtacaa gtgcaaggtc 2580
tccaacaaag ccctcccagc ccccatcgag aaaaccatct ccaaagccaa agggcagccc 2640
cgagaaccac aggtgtacac cctgccccca tcccgggatg agctgaccaa gaaccaggtc 2700
agcctgacct gcctggtcaa aggcttctat cccagcgaca tcgccgtgga gtgggagagc 2760
aatgggcaac cggagaacaa ctacaagacc acgcctcccg tgctggactc cgacggctcc 2820
ttcttcctct acagcaagct caccgtggac aagagcaggt ggcagcaggg gaacgtcttc 2880
tcatgctccg tgatgcatga ggccctgcac aatcactata cccagaaatc tctgagtctg 2940
agcccaggca agaaggaccc caagttctgg gtcctggtgg tggtgggagg cgtgctggcc 3000
tgttactctc tcctggtgac cgtggccttc atcatctttt gggtgcgctc ccgggtgaag 3060
ttttctcgct ctgccgatgc cccagcctat cagcagggcc agaatcagct gtacaatgaa 3120
ctgaacctgg gcaggcggga ggagtacgac gtgctggata agcggagagg cagagacccc 3180
gagatgggcg gcaaaccacg gcgcaaaaat ccccaggagg gactctataa cgagctgcag 3240
aaggacaaaa tggccgaggc ctattccgag atcggcatga agggagagag aagacgcgga 3300
aagggccacg acggcctgta tcagggattg tccaccgcta caaaagatac atatgatgcc 3360
ctgcacatgc aggccctgcc acccagatga 3390
<210> 36
<211> 5154
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательности нуклеиновой кислоты, кодирующие CAR
(MP14802.SFG.aCD19fmc63_clean-CD8STK-CD28tmZ-2A-aCD33glx-HCH2CH3p
vaa-dCD45)
<400> 36
atgagcctgc ccgtgaccgc cctgctgctg cccctggccc tgctgctgca cgccgccaga 60
ccagacatcc agatgaccca gaccaccagc agcctgagcg ccagcctggg cgaccgggtg 120
accatcagct gcagagccag ccaggacatc agcaagtacc tgaactggta ccagcagaag 180
cccgacggca ccgtgaagct gctgatctac cacaccagcc ggctgcacag cggcgtgccc 240
agccggttca gcggcagcgg cagcggcacc gactacagcc tgaccatcag caacctggag 300
caggaggaca tcgccaccta cttctgccag cagggcaaca ccctgcccta caccttcgga 360
ggcggcacca agctggagat caccaaggcc ggaggcggag gctctggcgg aggcggctct 420
ggcggaggcg gctctggcgg aggcggcagc gaggtgaagc tgcaggagtc tggcccaggc 480
ctggtggccc caagccagag cctgagcgtg acctgcaccg tgagcggcgt gagcctgccc 540
gactacggcg tgagctggat caggcagccc ccacggaagg gcctggagtg gctgggcgtg 600
atctggggca gcgagaccac ctactacaac agcgccctga agagccggct gaccatcatc 660
aaggacaaca gcaagagcca ggtgttcctg aagatgaaca gcctgcagac cgacgacacc 720
gccatctact actgcgccaa gcactactac tatggcggca gctacgctat ggactactgg 780
ggccagggca ccagcgtgac cgtgagctca gatcccacca cgacgccagc gccgcgacca 840
ccaacaccgg cgcccaccat cgcgtcgcag cccctgtccc tgcgcccaga ggcgtgccgg 900
ccagcggcgg ggggcgcagt gcacacgagg gggctggact tcgcctgtga tatcttttgg 960
gtgctggtgg tggttggtgg agtcctggct tgctatagct tgctagtaac agtggccttt 1020
attattttct gggtgaggag agtgaagttc agcaggagcg cagacgcccc cgcgtaccag 1080
cagggccaga accagctcta taacgagctc aatctaggac gaagagagga gtacgatgtt 1140
ttggacaaga gacgtggccg ggaccctgag atggggggaa agccgagaag gaagaaccct 1200
caggaaggcc tgtacaatga actgcagaaa gataagatgg cggaggccta cagtgagatt 1260
gggatgaaag gcgagcgccg gaggggcaag gggcacgatg gcctttacca gggtctcagt 1320
acagccacca aggacaccta cgacgccctt cacatgcagg ccctgcctcc tcgcagagcc 1380
gagggcaggg gaagtcttct aacatgcggg gacgtggagg aaaatcccgg gcccatggcc 1440
gtgcccactc aggtcctggg gttgttgcta ctgtggctta cagatgccag atgtgacatc 1500
cagatgacac agtctccatc ttccctgtct gcatctgtcg gagatcgcgt caccatcacc 1560
tgtcgagcaa gtgaggacat ttattttaat ttagtgtggt atcagcagaa accaggaaag 1620
gcccctaagc tcctgatcta tgatacaaat cgcttggcag atggggtccc atcacggttc 1680
agtggctctg gatctggcac acagtatact ctaaccataa gtagcctgca acccgaagat 1740
ttcgcaacct attattgtca acactataag aattatccgc tcacgttcgg tcaggggacc 1800
aagctggaaa tcaaaagatc tggtggcgga gggtcaggag gcggaggcag cggaggcggt 1860
ggctcgggag gcggaggctc gagatctgag gtgcagttgg tggagtctgg gggcggcttg 1920
gtgcagcctg gagggtccct gaggctctcc tgtgcagcct caggattcac tctcagtaat 1980
tatggcatgc actggatcag gcaggctcca gggaagggtc tggagtgggt ctcgtctatt 2040
agtcttaatg gtggtagcac ttactatcga gactccgtga agggccgatt cactatctcc 2100
agggacaatg caaaaagcac cctctacctt caaatgaata gtctgagggc cgaggacacg 2160
gccgtctatt actgtgcagc acaggacgct tatacgggag gttactttga ttactggggc 2220
caaggaacgc tggtcacagt ctcgtctatg gatcccgccg agcccaaatc tcctgacaaa 2280
actcacacat gcccaccgtg cccagcacct cccgtggccg gcccgtcagt cttcctcttc 2340
cccccaaaac ccaaggacac cctcatgatc gcccggaccc ctgaggtcac atgcgtggtg 2400
gtggacgtga gccacgaaga ccctgaggtc aagttcaact ggtacgtgga cggcgtggag 2460
gtgcataatg ccaagacaaa gccgcgggag gagcagtaca acagcacgta ccgtgtggtc 2520
agcgtcctca ccgtcctgca ccaggactgg ctgaatggca aggagtacaa gtgcaaggtc 2580
tccaacaaag ccctcccagc ccccatcgag aaaaccatct ccaaagccaa agggcagccc 2640
cgagaaccac aggtgtacac cctgccccca tcccgggatg agctgaccaa gaaccaggtc 2700
agcctgacct gcctggtcaa aggcttctat cccagcgaca tcgccgtgga gtgggagagc 2760
aatgggcaac cggagaacaa ctacaagacc acgcctcccg tgctggactc cgacggctcc 2820
ttcttcctct acagcaagct caccgtggac aagagcaggt ggcagcaggg gaacgtcttc 2880
tcatgctccg tgatgcatga ggccctgcac aatcactata cccagaaatc tctgagtctg 2940
agcccaggca agaaggaccc caaggcactg atagcatttc tggcatttct gattattgtg 3000
acatcaatag ccctgcttgt tgttctctac aaaatctatg atctacataa gaaaagatcc 3060
tgcaatttag atgaacagca ggagcttgtt gaaagggatg atgaaaaaca actgatgaat 3120
gtggagccaa tccatgcaga tattttgttg gaaacttata agaggaagat tgctgatgaa 3180
ggaagacttt ttctggctga atttcagagc atcccgcggg tgttcagcaa gtttcctata 3240
aaggaagctc gaaagccctt taaccagaat aaaaaccgtt atgttgacat tcttccttat 3300
gattataacc gtgttgaact ctctgagata aacggagatg cagggtcaaa ctacataaat 3360
gccagctata ttgatggttt caaagaaccc aggaaataca ttgctgcaca aggtcccagg 3420
gatgaaactg ttgatgattt ctggaggatg atttgggaac agaaagccac agttattgtc 3480
atggtcactc gatgtgaaga aggaaacagg aacaagtgtg cagaatactg gccgtcaatg 3540
gaagagggca ctcgggcttt tggagatgtt gttgtaaaga tcaaccagca caaaagatgt 3600
ccagattaca tcattcagaa attgaacatt gtaaataaaa aagaaaaagc aactggaaga 3660
gaggtgactc acattcagtt caccagctgg ccagaccacg gggtgcctga ggatcctcac 3720
ttgctcctca aactgagaag gagagtgaat gccttcagca atttcttcag tggtcccatt 3780
gtggtgcact gcagtgctgg tgttgggcgc acaggaacct atatcggaat tgatgccatg 3840
ctagaaggcc tggaagccga gaacaaagtg gatgtttatg gttatgttgt caagctaagg 3900
cgacagagat gcctgatggt tcaagtagag gcccagtaca tcttgatcca tcaggctttg 3960
gtggaataca atcagtttgg agaaacagaa gtgaatttgt ctgaattaca tccatatcta 4020
cataacatga agaaaaggga tccacccagt gagccgtctc cactagaggc tgaattccag 4080
agacttcctt catataggag ctggaggaca cagcacattg gaaatcaaga agaaaataaa 4140
agtaaaaaca ggaattctaa tgtcatccca tatgactata acagagtgcc acttaaacat 4200
gagctggaaa tgagtaaaga gagtgagcat gattcagatg aatcctctga tgatgacagt 4260
gattcagagg aaccaagcaa atacatcaat gcatctttta taatgagcta ctggaaacct 4320
gaagtgatga ttgctgctca gggaccactg aaggagacca ttggtgactt ttggcagatg 4380
atcttccaaa gaaaagtcaa agttattgtt atgctgacag aactgaaaca tggagaccag 4440
gaaatctgtg ctcagtactg gggagaagga aagcaaacat atggagatat tgaagttgac 4500
ctgaaagaca cagacaaatc ttcaacttat acccttcgtg tctttgaact gagacattcc 4560
aagaggaaag actctcgaac tgtgtaccag taccaatata caaactggag tgtggagcag 4620
cttcctgcag aacccaagga attaatctct atgattcagg tcgtcaaaca aaaacttccc 4680
cagaagaatt cctctgaagg gaacaagcat cacaagagta cacctctact cattcactgc 4740
agggatggat ctcagcaaac gggaatattt tgtgctttgt taaatctctt agaaagtgcg 4800
gaaacagaag aggtagtgga tatttttcaa gtggtaaaag ctctacgcaa agctaggcca 4860
ggcatggttt ccacattcga gcaatatcaa ttcctatatg acgtcattgc cagcacctac 4920
cctgctcaga atggacaagt aaagaaaaac aaccatcaag aagataaaat tgaatttgat 4980
aatgaagtgg acaaagtaaa gcaggatgct aattgtgtta atccacttgg tgccccagaa 5040
aagctccctg aagcaaagga acaggctgaa ggttctgaac ccacgagtgg cactgagggg 5100
ccagaacatt ctgtcaatgg tcctgcaagt ccagctttaa atcaaggttc atag 5154
<210> 37
<211> 4053
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательности нуклеиновой кислоты, кодирующие CAR
(MP14801.SFG.aCD19fmc63_clean-CD8STK-CD28tmZ-2A-aCD33glx-HCH2CH3p
vaa-dCD148)
<400> 37
atgagcctgc ccgtgaccgc cctgctgctg cccctggccc tgctgctgca cgccgccaga 60
ccagacatcc agatgaccca gaccaccagc agcctgagcg ccagcctggg cgaccgggtg 120
accatcagct gcagagccag ccaggacatc agcaagtacc tgaactggta ccagcagaag 180
cccgacggca ccgtgaagct gctgatctac cacaccagcc ggctgcacag cggcgtgccc 240
agccggttca gcggcagcgg cagcggcacc gactacagcc tgaccatcag caacctggag 300
caggaggaca tcgccaccta cttctgccag cagggcaaca ccctgcccta caccttcgga 360
ggcggcacca agctggagat caccaaggcc ggaggcggag gctctggcgg aggcggctct 420
ggcggaggcg gctctggcgg aggcggcagc gaggtgaagc tgcaggagtc tggcccaggc 480
ctggtggccc caagccagag cctgagcgtg acctgcaccg tgagcggcgt gagcctgccc 540
gactacggcg tgagctggat caggcagccc ccacggaagg gcctggagtg gctgggcgtg 600
atctggggca gcgagaccac ctactacaac agcgccctga agagccggct gaccatcatc 660
aaggacaaca gcaagagcca ggtgttcctg aagatgaaca gcctgcagac cgacgacacc 720
gccatctact actgcgccaa gcactactac tatggcggca gctacgctat ggactactgg 780
ggccagggca ccagcgtgac cgtgagctca gatcccacca cgacgccagc gccgcgacca 840
ccaacaccgg cgcccaccat cgcgtcgcag cccctgtccc tgcgcccaga ggcgtgccgg 900
ccagcggcgg ggggcgcagt gcacacgagg gggctggact tcgcctgtga tatcttttgg 960
gtgctggtgg tggttggtgg agtcctggct tgctatagct tgctagtaac agtggccttt 1020
attattttct gggtgaggag agtgaagttc agcaggagcg cagacgcccc cgcgtaccag 1080
cagggccaga accagctcta taacgagctc aatctaggac gaagagagga gtacgatgtt 1140
ttggacaaga gacgtggccg ggaccctgag atggggggaa agccgagaag gaagaaccct 1200
caggaaggcc tgtacaatga actgcagaaa gataagatgg cggaggccta cagtgagatt 1260
gggatgaaag gcgagcgccg gaggggcaag gggcacgatg gcctttacca gggtctcagt 1320
acagccacca aggacaccta cgacgccctt cacatgcagg ccctgcctcc tcgcagagcc 1380
gagggcaggg gaagtcttct aacatgcggg gacgtggagg aaaatcccgg gcccatggcc 1440
gtgcccactc aggtcctggg gttgttgcta ctgtggctta cagatgccag atgtgacatc 1500
cagatgacac agtctccatc ttccctgtct gcatctgtcg gagatcgcgt caccatcacc 1560
tgtcgagcaa gtgaggacat ttattttaat ttagtgtggt atcagcagaa accaggaaag 1620
gcccctaagc tcctgatcta tgatacaaat cgcttggcag atggggtccc atcacggttc 1680
agtggctctg gatctggcac acagtatact ctaaccataa gtagcctgca acccgaagat 1740
ttcgcaacct attattgtca acactataag aattatccgc tcacgttcgg tcaggggacc 1800
aagctggaaa tcaaaagatc tggtggcgga gggtcaggag gcggaggcag cggaggcggt 1860
ggctcgggag gcggaggctc gagatctgag gtgcagttgg tggagtctgg gggcggcttg 1920
gtgcagcctg gagggtccct gaggctctcc tgtgcagcct caggattcac tctcagtaat 1980
tatggcatgc actggatcag gcaggctcca gggaagggtc tggagtgggt ctcgtctatt 2040
agtcttaatg gtggtagcac ttactatcga gactccgtga agggccgatt cactatctcc 2100
agggacaatg caaaaagcac cctctacctt caaatgaata gtctgagggc cgaggacacg 2160
gccgtctatt actgtgcagc acaggacgct tatacgggag gttactttga ttactggggc 2220
caaggaacgc tggtcacagt ctcgtctatg gatcccgccg agcccaaatc tcctgacaaa 2280
actcacacat gcccaccgtg cccagcacct cccgtggccg gcccgtcagt cttcctcttc 2340
cccccaaaac ccaaggacac cctcatgatc gcccggaccc ctgaggtcac atgcgtggtg 2400
gtggacgtga gccacgaaga ccctgaggtc aagttcaact ggtacgtgga cggcgtggag 2460
gtgcataatg ccaagacaaa gccgcgggag gagcagtaca acagcacgta ccgtgtggtc 2520
agcgtcctca ccgtcctgca ccaggactgg ctgaatggca aggagtacaa gtgcaaggtc 2580
tccaacaaag ccctcccagc ccccatcgag aaaaccatct ccaaagccaa agggcagccc 2640
cgagaaccac aggtgtacac cctgccccca tcccgggatg agctgaccaa gaaccaggtc 2700
agcctgacct gcctggtcaa aggcttctat cccagcgaca tcgccgtgga gtgggagagc 2760
aatgggcaac cggagaacaa ctacaagacc acgcctcccg tgctggactc cgacggctcc 2820
ttcttcctct acagcaagct caccgtggac aagagcaggt ggcagcaggg gaacgtcttc 2880
tcatgctccg tgatgcatga ggccctgcac aatcactata cccagaaatc tctgagtctg 2940
agcccaggca agaaggaccc caaggcggtt tttggctgta tctttggtgc cctggttatt 3000
gtgactgtgg gaggcttcat cttctggaga aagaagagga aagatgcaaa gaataatgaa 3060
gtgtcctttt ctcaaattaa acctaaaaaa tctaagttaa tcagagtgga gaattttgag 3120
gcctacttca agaagcagca agctgactcc aactgtgggt tcgcagagga atacgaagat 3180
ctgaagcttg ttggaattag tcaacctaaa tatgcagcag aactggctga gaatagagga 3240
aagaatcgct ataataatgt tctgccctat gatatttccc gtgtcaaact ttcggtccag 3300
acccattcaa cggatgacta catcaatgcc aactacatgc ctggctacca ctccaagaaa 3360
gattttattg ccacacaagg acctttaccg aacactttga aagatttttg gcgtatggtt 3420
tgggagaaaa atgtatatgc catcattatg ttgactaaat gtgttgaaca gggaagaacc 3480
aaatgtgagg agtattggcc ctccaagcag gctcaggact atggagacat aactgtggca 3540
atgacatcag aaattgttct tccggaatgg accatcagag atttcacagt gaaaaatatc 3600
cagacaagtg agagtcaccc tctgagacag ttccatttca cctcctggcc agaccacggt 3660
gttcccgaca ccactgacct gctcatcaac ttccggtacc tcgttcgtga ctacatgaag 3720
cagagtcctc ccgaatcgcc gattctggtg cattgcagtg ctggggtcgg aaggacgggc 3780
actttcattg ccattgatcg tctcatctac cagatagaga atgagaacac cgtggatgtg 3840
tatgggattg tgtatgacct tcgaatgcat aggcctttaa tggtgcagac agaggaccag 3900
tatgttttcc tcaatcagtg tgttttggat attgtcagat cccagaaaga ctcaaaagta 3960
gatcttatct accagaacac aactgcaatg acaatctatg aaaaccttgc gcccgtgacc 4020
acatttggaa agaccaatgg ttacatcgcc taa 4053
<210> 38
<211> 3345
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательности нуклеиновой кислоты, кодирующие CAR
(16076.SFG.aCD19fmc63-CD8STK-CD28tmZ-2A-aCD33glx-muCD8STK-tm-dPTP
N6)
<400> 38
atgagcctgc ccgtgaccgc cctgctgctg cccctggccc tgctgctgca cgccgccaga 60
ccagacatcc agatgaccca gaccaccagc agcctgagcg ccagcctggg cgaccgggtg 120
accatcagct gcagagccag ccaggacatc agcaagtacc tgaactggta ccagcagaag 180
cccgacggca ccgtgaagct gctgatctac cacaccagcc ggctgcacag cggcgtgccc 240
agccggttca gcggcagcgg cagcggcacc gactacagcc tgaccatcag caacctggag 300
caggaggaca tcgccaccta cttctgccag cagggcaaca ccctgcccta caccttcgga 360
ggcggcacca agctggagat caccaaggcc ggaggcggag gctctggcgg aggcggctct 420
ggcggaggcg gctctggcgg aggcggcagc gaggtgaagc tgcaggagtc tggcccaggc 480
ctggtggccc caagccagag cctgagcgtg acctgcaccg tgagcggcgt gagcctgccc 540
gactacggcg tgagctggat caggcagccc ccacggaagg gcctggagtg gctgggcgtg 600
atctggggca gcgagaccac ctactacaac agcgccctga agagccggct gaccatcatc 660
aaggacaaca gcaagagcca ggtgttcctg aagatgaaca gcctgcagac cgacgacacc 720
gccatctact actgcgccaa gcactactac tatggcggca gctacgctat ggactactgg 780
ggccagggca ccagcgtgac cgtgagctca gatcccacca cgacgccagc gccgcgacca 840
ccaacaccgg cgcccaccat cgcgtcgcag cccctgtccc tgcgcccaga ggcgtgccgg 900
ccagcggcgg ggggcgcagt gcacacgagg gggctggact tcgcctgtga tatcttttgg 960
gtgctggtgg tggttggtgg agtcctggct tgctatagct tgctagtaac agtggccttt 1020
attattttct gggtgaggag agtgaagttc agcaggagcg cagacgcccc cgcgtaccag 1080
cagggccaga accagctcta taacgagctc aatctaggac gaagagagga gtacgatgtt 1140
ttggacaaga gacgtggccg ggaccctgag atggggggaa agccgagaag gaagaaccct 1200
caggaaggcc tgtacaatga actgcagaaa gataagatgg cggaggccta cagtgagatt 1260
gggatgaaag gcgagcgccg gaggggcaag gggcacgatg gcctttacca gggtctcagt 1320
acagccacca aggacaccta cgacgccctt cacatgcagg ccctgcctcc tcgcagagcc 1380
gagggcaggg gaagtcttct aacatgcggg gacgtggagg aaaatcccgg gcccatggcc 1440
gtgcccactc aggtcctggg gttgttgcta ctgtggctta cagatgccag atgtgacatc 1500
cagatgacac agtctccatc ttccctgtct gcatctgtcg gagatcgcgt caccatcacc 1560
tgtcgagcaa gtgaggacat ttattttaat ttagtgtggt atcagcagaa accaggaaag 1620
gcccctaagc tcctgatcta tgatacaaat cgcttggcag atggggtccc atcacggttc 1680
agtggctctg gatctggcac acagtatact ctaaccataa gtagcctgca acccgaagat 1740
ttcgcaacct attattgtca acactataag aattatccgc tcacgttcgg tcaggggacc 1800
aagctggaaa tcaaaagatc tggtggcgga gggtcaggag gcggaggcag cggaggcggt 1860
ggctcgggag gcggaggctc gagatctgag gtgcagttgg tggagtctgg gggcggcttg 1920
gtgcagcctg gagggtccct gaggctctcc tgtgcagcct caggattcac tctcagtaat 1980
tatggcatgc actggatcag gcaggctcca gggaagggtc tggagtgggt ctcgtctatt 2040
agtcttaatg gtggtagcac ttactatcga gactccgtga agggccgatt cactatctcc 2100
agggacaatg caaaaagcac cctctacctt caaatgaata gtctgagggc cgaggacacg 2160
gccgtctatt actgtgcagc acaggacgct tatacgggag gttactttga ttactggggc 2220
caaggaacgc tggtcacagt ctcgtctatg gatcccgcca ccacaaccaa gcccgtgctg 2280
cggaccccaa gccctgtgca ccctaccggc accagccagc ctcagagacc cgaggactgc 2340
cggcctcggg gcagcgtgaa gggcaccggc ctggacttcg cctgcgacat ctactgggca 2400
cctctggccg gaatatgcgt ggcactgctg ctgagcctca tcatcaccct gatctgttat 2460
caccgaagcc gcaagcgggt gtgtaaaagt ggaggcggaa gcttctggga ggagtttgag 2520
agtttgcaga agcaggaggt gaagaacttg caccagcgtc tggaagggca gcggccagag 2580
aacaagggca agaaccgcta caagaacatt ctcccctttg accacagccg agtgatcctg 2640
cagggacggg acagtaacat ccccgggtcc gactacatca atgccaacta catcaagaac 2700
cagctgctag gccctgatga gaacgctaag acctacatcg ccagccaggg ctgtctggag 2760
gccacggtca atgacttctg gcagatggcg tggcaggaga acagccgtgt catcgtcatg 2820
accacccgag aggtggagaa aggccggaac aaatgcgtcc catactggcc cgaggtgggc 2880
atgcagcgtg cttatgggcc ctactctgtg accaactgcg gggagcatga cacaaccgaa 2940
tacaaactcc gtaccttaca ggtctccccg ctggacaatg gagacctgat tcgggagatc 3000
tggcattacc agtacctgag ctggcccgac cacggggtcc ccagtgagcc tgggggtgtc 3060
ctcagcttcc tggaccagat caaccagcgg caggaaagtc tgcctcacgc agggcccatc 3120
atcgtgcact gcagcgccgg catcggccgc acaggcacca tcattgtcat cgacatgctc 3180
atggagaaca tctccaccaa gggcctggac tgtgacattg acatccagaa gaccatccag 3240
atggtgcggg cgcagcgctc gggcatggtg cagacggagg cgcagtacaa gttcatctac 3300
gtggccatcg cccagttcat tgaaaccact aagaagaagc tgtga 3345
<210> 39
<211> 2757
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательности нуклеиновой кислоты, кодирующие CAR
(MP16091.SFG.aCD19fmc63-CD8STK-CD28tmZ-2A-aCD33glx-muCD8STK-LAIR1
tm-endo)
<400> 39
atgagcctgc ccgtgaccgc cctgctgctg cccctggccc tgctgctgca cgccgccaga 60
ccagacatcc agatgaccca gaccaccagc agcctgagcg ccagcctggg cgaccgggtg 120
accatcagct gcagagccag ccaggacatc agcaagtacc tgaactggta ccagcagaag 180
cccgacggca ccgtgaagct gctgatctac cacaccagcc ggctgcacag cggcgtgccc 240
agccggttca gcggcagcgg cagcggcacc gactacagcc tgaccatcag caacctggag 300
caggaggaca tcgccaccta cttctgccag cagggcaaca ccctgcccta caccttcgga 360
ggcggcacca agctggagat caccaaggcc ggaggcggag gctctggcgg aggcggctct 420
ggcggaggcg gctctggcgg aggcggcagc gaggtgaagc tgcaggagtc tggcccaggc 480
ctggtggccc caagccagag cctgagcgtg acctgcaccg tgagcggcgt gagcctgccc 540
gactacggcg tgagctggat caggcagccc ccacggaagg gcctggagtg gctgggcgtg 600
atctggggca gcgagaccac ctactacaac agcgccctga agagccggct gaccatcatc 660
aaggacaaca gcaagagcca ggtgttcctg aagatgaaca gcctgcagac cgacgacacc 720
gccatctact actgcgccaa gcactactac tatggcggca gctacgctat ggactactgg 780
ggccagggca ccagcgtgac cgtgagctca gatcccacca cgacgccagc gccgcgacca 840
ccaacaccgg cgcccaccat cgcgtcgcag cccctgtccc tgcgcccaga ggcgtgccgg 900
ccagcggcgg ggggcgcagt gcacacgagg gggctggact tcgcctgtga tatcttttgg 960
gtgctggtgg tggttggtgg agtcctggct tgctatagct tgctagtaac agtggccttt 1020
attattttct gggtgaggag agtgaagttc agcaggagcg cagacgcccc cgcgtaccag 1080
cagggccaga accagctcta taacgagctc aatctaggac gaagagagga gtacgatgtt 1140
ttggacaaga gacgtggccg ggaccctgag atggggggaa agccgagaag gaagaaccct 1200
caggaaggcc tgtacaatga actgcagaaa gataagatgg cggaggccta cagtgagatt 1260
gggatgaaag gcgagcgccg gaggggcaag gggcacgatg gcctttacca gggtctcagt 1320
acagccacca aggacaccta cgacgccctt cacatgcagg ccctgcctcc tcgcagagcc 1380
gagggcaggg gaagtcttct aacatgcggg gacgtggagg aaaatcccgg gcccatggcc 1440
gtgcccactc aggtcctggg gttgttgcta ctgtggctta cagatgccag atgtgacatc 1500
cagatgacac agtctccatc ttccctgtct gcatctgtcg gagatcgcgt caccatcacc 1560
tgtcgagcaa gtgaggacat ttattttaat ttagtgtggt atcagcagaa accaggaaag 1620
gcccctaagc tcctgatcta tgatacaaat cgcttggcag atggggtccc atcacggttc 1680
agtggctctg gatctggcac acagtatact ctaaccataa gtagcctgca acccgaagat 1740
ttcgcaacct attattgtca acactataag aattatccgc tcacgttcgg tcaggggacc 1800
aagctggaaa tcaaaagatc tggtggcgga gggtcaggag gcggaggcag cggaggcggt 1860
ggctcgggag gcggaggctc gagatctgag gtgcagttgg tggagtctgg gggcggcttg 1920
gtgcagcctg gagggtccct gaggctctcc tgtgcagcct caggattcac tctcagtaat 1980
tatggcatgc actggatcag gcaggctcca gggaagggtc tggagtgggt ctcgtctatt 2040
agtcttaatg gtggtagcac ttactatcga gactccgtga agggccgatt cactatctcc 2100
agggacaatg caaaaagcac cctctacctt caaatgaata gtctgagggc cgaggacacg 2160
gccgtctatt actgtgcagc acaggacgct tatacgggag gttactttga ttactggggc 2220
caaggaacgc tggtcacagt ctcgtctatg gatcccgcca ccacaaccaa gcccgtgctg 2280
cggaccccaa gccctgtgca ccctaccggc accagccagc ctcagagacc cgaggactgc 2340
cggcctcggg gcagcgtgaa gggcaccggc ctggacttcg cctgcgacat tctcatcggg 2400
gtctcagtgg tcttcctctt ctgtctcctc ctcctggtcc tcttctgcct ccatcgccag 2460
aatcagataa agcaggggcc ccccagaagc aaggacgagg agcagaagcc acagcagagg 2520
cctgacctgg ctgttgatgt tctagagagg acagcagaca aggccacagt caatggactt 2580
cctgagaagg accgggagac cgacaccagc gccctggctg cagggagttc ccaggaggtg 2640
acgtatgctc agctggacca ctgggccctc acacagagga cagcccgggc tgtgtcccca 2700
cagtccacaa agcccatggc cgagtccatc acgtatgcag ccgttgccag acactga 2757
<210> 40
<211> 4092
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательности нуклеиновой кислоты, кодирующие CAR
(MP16092.SFG.aCD19fmc63-CD8STK-CD28tmZ-2A-aCD33glx-muCD8STK-LAIR1
tm-endo-2A-PTPN6_SH2-dCD148)
<400> 40
atgagcctgc ccgtgaccgc cctgctgctg cccctggccc tgctgctgca cgccgccaga 60
ccagacatcc agatgaccca gaccaccagc agcctgagcg ccagcctggg cgaccgggtg 120
accatcagct gcagagccag ccaggacatc agcaagtacc tgaactggta ccagcagaag 180
cccgacggca ccgtgaagct gctgatctac cacaccagcc ggctgcacag cggcgtgccc 240
agccggttca gcggcagcgg cagcggcacc gactacagcc tgaccatcag caacctggag 300
caggaggaca tcgccaccta cttctgccag cagggcaaca ccctgcccta caccttcgga 360
ggcggcacca agctggagat caccaaggcc ggaggcggag gctctggcgg aggcggctct 420
ggcggaggcg gctctggcgg aggcggcagc gaggtgaagc tgcaggagtc tggcccaggc 480
ctggtggccc caagccagag cctgagcgtg acctgcaccg tgagcggcgt gagcctgccc 540
gactacggcg tgagctggat caggcagccc ccacggaagg gcctggagtg gctgggcgtg 600
atctggggca gcgagaccac ctactacaac agcgccctga agagccggct gaccatcatc 660
aaggacaaca gcaagagcca ggtgttcctg aagatgaaca gcctgcagac cgacgacacc 720
gccatctact actgcgccaa gcactactac tatggcggca gctacgctat ggactactgg 780
ggccagggca ccagcgtgac cgtgagctca gatcccacca cgacgccagc gccgcgacca 840
ccaacaccgg cgcccaccat cgcgtcgcag cccctgtccc tgcgcccaga ggcgtgccgg 900
ccagcggcgg ggggcgcagt gcacacgagg gggctggact tcgcctgtga tatcttttgg 960
gtgctggtgg tggttggtgg agtcctggct tgctatagct tgctagtaac agtggccttt 1020
attattttct gggtgaggag agtgaagttc agcaggagcg cagacgcccc cgcgtaccag 1080
cagggccaga accagctcta taacgagctc aatctaggac gaagagagga gtacgatgtt 1140
ttggacaaga gacgtggccg ggaccctgag atggggggaa agccgagaag gaagaaccct 1200
caggaaggcc tgtacaatga actgcagaaa gataagatgg cggaggccta cagtgagatt 1260
gggatgaaag gcgagcgccg gaggggcaag gggcacgatg gcctttacca gggtctcagt 1320
acagccacca aggacaccta cgacgccctt cacatgcagg ccctgcctcc tcgcagagcc 1380
gagggcaggg gaagtcttct aacatgcggg gacgtggagg aaaatcccgg gcccatggcc 1440
gtgcccactc aggtcctggg gttgttgcta ctgtggctta cagatgccag atgtgacatc 1500
cagatgacac agtctccatc ttccctgtct gcatctgtcg gagatcgcgt caccatcacc 1560
tgtcgagcaa gtgaggacat ttattttaat ttagtgtggt atcagcagaa accaggaaag 1620
gcccctaagc tcctgatcta tgatacaaat cgcttggcag atggggtccc atcacggttc 1680
agtggctctg gatctggcac acagtatact ctaaccataa gtagcctgca acccgaagat 1740
ttcgcaacct attattgtca acactataag aattatccgc tcacgttcgg tcaggggacc 1800
aagctggaaa tcaaaagatc tggtggcgga gggtcaggag gcggaggcag cggaggcggt 1860
ggctcgggag gcggaggctc gagatctgag gtgcagttgg tggagtctgg gggcggcttg 1920
gtgcagcctg gagggtccct gaggctctcc tgtgcagcct caggattcac tctcagtaat 1980
tatggcatgc actggatcag gcaggctcca gggaagggtc tggagtgggt ctcgtctatt 2040
agtcttaatg gtggtagcac ttactatcga gactccgtga agggccgatt cactatctcc 2100
agggacaatg caaaaagcac cctctacctt caaatgaata gtctgagggc cgaggacacg 2160
gccgtctatt actgtgcagc acaggacgct tatacgggag gttactttga ttactggggc 2220
caaggaacgc tggtcacagt ctcgtctatg gatcccgcca ccacaaccaa gcccgtgctg 2280
cggaccccaa gccctgtgca ccctaccggc accagccagc ctcagagacc cgaggactgc 2340
cggcctcggg gcagcgtgaa gggcaccggc ctggacttcg cctgcgacat tctcatcggg 2400
gtctcagtgg tcttcctctt ctgtctcctc ctcctggtcc tcttctgcct ccatcgccag 2460
aatcagataa agcaggggcc ccccagaagc aaggacgagg agcagaagcc acagcagagg 2520
cctgacctgg ctgttgatgt tctagagagg acagcagaca aggccacagt caatggactt 2580
cctgagaagg accgggagac cgacaccagc gccctggctg cagggagttc ccaggaggtg 2640
acgtatgctc agctggacca ctgggccctc acacagagga cagcccgggc tgtgtcccca 2700
cagtccacaa agcccatggc cgagtccatc acgtatgcag ccgttgccag acacagggca 2760
gaaggaagag gtagcctgct gacttgcggg gacgtggaag agaacccagg gccatggtat 2820
catggccaca tgtctggcgg gcaggcagag acgctgctgc aggccaaggg cgagccctgg 2880
acgtttcttg tgcgtgagag cctcagccag cctggagact tcgtgctttc tgtgctcagt 2940
gaccagccca aggctggccc aggctccccg ctcagggtca cccacatcaa ggtcatgtgc 3000
gagggtggac gctacacagt gggtggtttg gagaccttcg acagcctcac ggacctggtg 3060
gagcatttca agaagacggg gattgaggag gcctcaggcg cctttgtcta cctgcggcag 3120
ccgtacagcg gtggcggtgg cagctttgag gcctacttca agaagcagca agctgactcc 3180
aactgtgggt tcgcagagga atacgaagat ctgaagcttg ttggaattag tcaacctaaa 3240
tatgcagcag aactggctga gaatagagga aagaatcgct ataataatgt tctgccctat 3300
gatatttccc gtgtcaaact ttcggtccag acccattcaa cggatgacta catcaatgcc 3360
aactacatgc ctggctacca ctccaagaaa gattttattg ccacacaagg acctttaccg 3420
aacactttga aagatttttg gcgtatggtt tgggagaaaa atgtatatgc catcattatg 3480
ttgactaaat gtgttgaaca gggaagaacc aaatgtgagg agtattggcc ctccaagcag 3540
gctcaggact atggagacat aactgtggca atgacatcag aaattgttct tccggaatgg 3600
accatcagag atttcacagt gaaaaatatc cagacaagtg agagtcaccc tctgagacag 3660
ttccatttca cctcctggcc agaccacggt gttcccgaca ccactgacct gctcatcaac 3720
ttccggtacc tcgttcgtga ctacatgaag cagagtcctc ccgaatcgcc gattctggtg 3780
cattgcagtg ctggggtcgg aaggacgggc actttcattg ccattgatcg tctcatctac 3840
cagatagaga atgagaacac cgtggatgtg tatgggattg tgtatgacct tcgaatgcat 3900
aggcctttaa tggtgcagac agaggaccag tatgttttcc tcaatcagtg tgttttggat 3960
attgtcagat cccagaaaga ctcaaaagta gatcttatct accagaacac aactgcaatg 4020
acaatctatg aaaaccttgc gcccgtgacc acatttggaa agaccaatgg ttacatcgcc 4080
agcggtagct aa 4092
<210> 41
<211> 1341
<212> Белок
<213> Искусственная последовательность
<220>
<223> Одноцепочечный вариабельный фрагмент (scFv)
SFG.aCD19-CD8STK-CD28tmZ-2A-aGD2-HCH2CH3pvaa-dCD148
<400> 41
Met Ser Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Asp Ile Gln Met Thr Gln Thr Thr Ser Ser Leu
20 25 30
Ser Ala Ser Leu Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln
35 40 45
Asp Ile Ser Lys Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr
50 55 60
Val Lys Leu Leu Ile Tyr His Thr Ser Arg Leu His Ser Gly Val Pro
65 70 75 80
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile
85 90 95
Ser Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly
100 105 110
Asn Thr Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Thr
115 120 125
Lys Ala Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Gly Gly Gly Gly Ser Glu Val Lys Leu Gln Glu Ser Gly Pro Gly
145 150 155 160
Leu Val Ala Pro Ser Gln Ser Leu Ser Val Thr Cys Thr Val Ser Gly
165 170 175
Val Ser Leu Pro Asp Tyr Gly Val Ser Trp Ile Arg Gln Pro Pro Arg
180 185 190
Lys Gly Leu Glu Trp Leu Gly Val Ile Trp Gly Ser Glu Thr Thr Tyr
195 200 205
Tyr Asn Ser Ala Leu Lys Ser Arg Leu Thr Ile Ile Lys Asp Asn Ser
210 215 220
Lys Ser Gln Val Phe Leu Lys Met Asn Ser Leu Gln Thr Asp Asp Thr
225 230 235 240
Ala Ile Tyr Tyr Cys Ala Lys His Tyr Tyr Tyr Gly Gly Ser Tyr Ala
245 250 255
Met Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Asp Pro
260 265 270
Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala
275 280 285
Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly
290 295 300
Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp Ile Phe Trp
305 310 315 320
Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val
325 330 335
Thr Val Ala Phe Ile Ile Phe Trp Val Arg Arg Val Lys Phe Ser Arg
340 345 350
Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn
355 360 365
Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg
370 375 380
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro
385 390 395 400
Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala
405 410 415
Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His
420 425 430
Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp
435 440 445
Ala Leu His Met Gln Ala Leu Pro Pro Arg Arg Ala Glu Gly Arg Gly
450 455 460
Ser Leu Leu Thr Cys Gly Asp Val Glu Glu Asn Pro Gly Pro Met Glu
465 470 475 480
Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro Gly Ser
485 490 495
Thr Gly Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro
500 505 510
Ser Gln Thr Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Ala
515 520 525
Ser Tyr Asn Ile His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu
530 535 540
Trp Leu Gly Val Ile Trp Ala Gly Gly Ser Thr Asn Tyr Asn Ser Ala
545 550 555 560
Leu Met Ser Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Val
565 570 575
Phe Leu Lys Met Ser Ser Leu Thr Ala Ala Asp Thr Ala Val Tyr Tyr
580 585 590
Cys Ala Lys Arg Ser Asp Asp Tyr Ser Trp Phe Ala Tyr Trp Gly Gln
595 600 605
Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly
610 615 620
Gly Ser Gly Gly Gly Gly Ser Glu Asn Gln Met Thr Gln Ser Pro Ser
625 630 635 640
Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Met Thr Cys Arg Ala
645 650 655
Ser Ser Ser Val Ser Ser Ser Tyr Leu His Trp Tyr Gln Gln Lys Ser
660 665 670
Gly Lys Ala Pro Lys Val Trp Ile Tyr Ser Thr Ser Asn Leu Ala Ser
675 680 685
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Thr
690 695 700
Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
705 710 715 720
Gln Gln Tyr Ser Gly Tyr Pro Ile Thr Phe Gly Gln Gly Thr Lys Val
725 730 735
Glu Ile Lys Arg Ser Asp Pro Ala Glu Pro Lys Ser Pro Asp Lys Thr
740 745 750
His Thr Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val
755 760 765
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ala Arg Thr
770 775 780
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
785 790 795 800
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
805 810 815
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
820 825 830
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
835 840 845
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
850 855 860
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
865 870 875 880
Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
885 890 895
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
900 905 910
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
915 920 925
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
930 935 940
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
945 950 955 960
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Lys
965 970 975
Asp Pro Lys Ala Val Phe Gly Cys Ile Phe Gly Ala Leu Val Ile Val
980 985 990
Thr Val Gly Gly Phe Ile Phe Trp Arg Lys Lys Arg Lys Asp Ala Lys
995 1000 1005
Asn Asn Glu Val Ser Phe Ser Gln Ile Lys Pro Lys Lys Ser Lys
1010 1015 1020
Leu Ile Arg Val Glu Asn Phe Glu Ala Tyr Phe Lys Lys Gln Gln
1025 1030 1035
Ala Asp Ser Asn Cys Gly Phe Ala Glu Glu Tyr Glu Asp Leu Lys
1040 1045 1050
Leu Val Gly Ile Ser Gln Pro Lys Tyr Ala Ala Glu Leu Ala Glu
1055 1060 1065
Asn Arg Gly Lys Asn Arg Tyr Asn Asn Val Leu Pro Tyr Asp Ile
1070 1075 1080
Ser Arg Val Lys Leu Ser Val Gln Thr His Ser Thr Asp Asp Tyr
1085 1090 1095
Ile Asn Ala Asn Tyr Met Pro Gly Tyr His Ser Lys Lys Asp Phe
1100 1105 1110
Ile Ala Thr Gln Gly Pro Leu Pro Asn Thr Leu Lys Asp Phe Trp
1115 1120 1125
Arg Met Val Trp Glu Lys Asn Val Tyr Ala Ile Ile Met Leu Thr
1130 1135 1140
Lys Cys Val Glu Gln Gly Arg Thr Lys Cys Glu Glu Tyr Trp Pro
1145 1150 1155
Ser Lys Gln Ala Gln Asp Tyr Gly Asp Ile Thr Val Ala Met Thr
1160 1165 1170
Ser Glu Ile Val Leu Pro Glu Trp Thr Ile Arg Asp Phe Thr Val
1175 1180 1185
Lys Asn Ile Gln Thr Ser Glu Ser His Pro Leu Arg Gln Phe His
1190 1195 1200
Phe Thr Ser Trp Pro Asp His Gly Val Pro Asp Thr Thr Asp Leu
1205 1210 1215
Leu Ile Asn Phe Arg Tyr Leu Val Arg Asp Tyr Met Lys Gln Ser
1220 1225 1230
Pro Pro Glu Ser Pro Ile Leu Val His Cys Ser Ala Gly Val Gly
1235 1240 1245
Arg Thr Gly Thr Phe Ile Ala Ile Asp Arg Leu Ile Tyr Gln Ile
1250 1255 1260
Glu Asn Glu Asn Thr Val Asp Val Tyr Gly Ile Val Tyr Asp Leu
1265 1270 1275
Arg Met His Arg Pro Leu Met Val Gln Thr Glu Asp Gln Tyr Val
1280 1285 1290
Phe Leu Asn Gln Cys Val Leu Asp Ile Val Arg Ser Gln Lys Asp
1295 1300 1305
Ser Lys Val Asp Leu Ile Tyr Gln Asn Thr Thr Ala Met Thr Ile
1310 1315 1320
Tyr Glu Asn Leu Ala Pro Val Thr Thr Phe Gly Lys Thr Asn Gly
1325 1330 1335
Tyr Ile Ala
1340
<210> 42
<211> 4026
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Одноцепочечный вариабельный фрагмент (scFv)
SFG.aCD19-CD8STK-CD28tmZ-2A-aGD2-HCH2CH3pvaa-dCD148
<400> 42
atgagcctgc ccgtgaccgc cctgctgctg cccctggccc tgctgctgca cgccgccaga 60
ccagacatcc agatgaccca gaccaccagc agcctgagcg ccagcctggg cgaccgggtg 120
accatcagct gcagagccag ccaggacatc agcaagtacc tgaactggta ccagcagaag 180
cccgacggca ccgtgaagct gctgatctac cacaccagcc ggctgcacag cggcgtgccc 240
agccggttca gcggcagcgg cagcggcacc gactacagcc tgaccatcag caacctggag 300
caggaggaca tcgccaccta cttctgccag cagggcaaca ccctgcccta caccttcgga 360
ggcggcacca agctggagat caccaaggcc ggaggcggag gctctggcgg aggcggctct 420
ggcggaggcg gctctggcgg aggcggcagc gaggtgaagc tgcaggagtc tggcccaggc 480
ctggtggccc caagccagag cctgagcgtg acctgcaccg tgagcggcgt gagcctgccc 540
gactacggcg tgagctggat caggcagccc ccacggaagg gcctggagtg gctgggcgtg 600
atctggggca gcgagaccac ctactacaac agcgccctga agagccggct gaccatcatc 660
aaggacaaca gcaagagcca ggtgttcctg aagatgaaca gcctgcagac cgacgacacc 720
gccatctact actgcgccaa gcactactac tatggcggca gctacgctat ggactactgg 780
ggccagggca ccagcgtgac cgtgagctca gatcccacca cgacgccagc gccgcgacca 840
ccaacaccgg cgcccaccat cgcgtcgcag cccctgtccc tgcgcccaga ggcgtgccgg 900
ccagcggcgg ggggcgcagt gcacacgagg gggctggact tcgcctgtga tatcttttgg 960
gtgctggtgg tggttggtgg agtcctggct tgctatagct tgctagtaac agtggccttt 1020
attattttct gggtgaggag agtgaagttc agcaggagcg cagacgcccc cgcgtaccag 1080
cagggccaga accagctcta taacgagctc aatctaggac gaagagagga gtacgatgtt 1140
ttggacaaga gacgtggccg ggaccctgag atggggggaa agccgagaag gaagaaccct 1200
caggaaggcc tgtacaatga actgcagaaa gataagatgg cggaggccta cagtgagatt 1260
gggatgaaag gcgagcgccg gaggggcaag gggcacgatg gcctttacca gggtctcagt 1320
acagccacca aggacaccta cgacgccctt cacatgcagg ccctgcctcc tcgcagagcc 1380
gagggcaggg gaagtcttct aacatgcggg gacgtggagg aaaatcccgg gcccatggag 1440
accgacaccc tgctgctgtg ggtgctgctg ctgtgggtgc caggcagcac cggccaggtg 1500
cagctgcagg agtctggccc aggcctggtg aagcccagcc agaccctgag catcacctgc 1560
accgtgagcg gcttcagcct ggccagctac aacatccact gggtgcggca gcccccaggc 1620
aagggcctgg agtggctggg cgtgatctgg gctggcggca gcaccaacta caacagcgcc 1680
ctgatgagcc ggctgaccat cagcaaggac aacagcaaga accaggtgtt cctgaagatg 1740
agcagcctga cagccgccga caccgccgtg tactactgcg ccaagcggag cgacgactac 1800
agctggttcg cctactgggg ccagggcacc ctggtgaccg tgagctctgg cggaggcggc 1860
tctggcggag gcggctctgg cggaggcggc agcgagaacc agatgaccca gagccccagc 1920
agcttgagcg ccagcgtggg cgaccgggtg accatgacct gcagagccag cagcagcgtg 1980
agcagcagct acctgcactg gtaccagcag aagagcggca aggccccaaa ggtgtggatc 2040
tacagcacca gcaacctggc cagcggcgtg cccagccggt tcagcggcag cggcagcggc 2100
accgactaca ccctgaccat cagcagcctg cagcccgagg acttcgccac ctactactgc 2160
cagcagtaca gcggctaccc catcaccttc ggccagggca ccaaggtgga gatcaagcgg 2220
tcggatcccg ccgagcccaa atctcctgac aaaactcaca catgcccacc gtgcccagca 2280
cctcccgtgg ccggcccgtc agtcttcctc ttccccccaa aacccaagga caccctcatg 2340
atcgcccgga cccctgaggt cacatgcgtg gtggtggacg tgagccacga agaccctgag 2400
gtcaagttca actggtacgt ggacggcgtg gaggtgcata atgccaagac aaagccgcgg 2460
gaggagcagt acaacagcac gtaccgtgtg gtcagcgtcc tcaccgtcct gcaccaggac 2520
tggctgaatg gcaaggagta caagtgcaag gtctccaaca aagccctccc agcccccatc 2580
gagaaaacca tctccaaagc caaagggcag ccccgagaac cacaggtgta caccctgccc 2640
ccatcccggg atgagctgac caagaaccag gtcagcctga cctgcctggt caaaggcttc 2700
tatcccagcg acatcgccgt ggagtgggag agcaatgggc aaccggagaa caactacaag 2760
accacgcctc ccgtgctgga ctccgacggc tccttcttcc tctacagcaa gctcaccgtg 2820
gacaagagca ggtggcagca ggggaacgtc ttctcatgct ccgtgatgca tgaggccctg 2880
cacaatcact atacccagaa atctctgagt ctgagcccag gcaagaagga ccccaaggcg 2940
gtttttggct gtatctttgg tgccctggtt attgtgactg tgggaggctt catcttctgg 3000
agaaagaaga ggaaagatgc aaagaataat gaagtgtcct tttctcaaat taaacctaaa 3060
aaatctaagt taatcagagt ggagaatttt gaggcctact tcaagaagca gcaagctgac 3120
tccaactgtg ggttcgcaga ggaatacgaa gatctgaagc ttgttggaat tagtcaacct 3180
aaatatgcag cagaactggc tgagaataga ggaaagaatc gctataataa tgttctgccc 3240
tatgatattt cccgtgtcaa actttcggtc cagacccatt caacggatga ctacatcaat 3300
gccaactaca tgcctggcta ccactccaag aaagatttta ttgccacaca aggaccttta 3360
ccgaacactt tgaaagattt ttggcgtatg gtttgggaga aaaatgtata tgccatcatt 3420
atgttgacta aatgtgttga acagggaaga accaaatgtg aggagtattg gccctccaag 3480
caggctcagg actatggaga cataactgtg gcaatgacat cagaaattgt tcttccggaa 3540
tggaccatca gagatttcac agtgaaaaat atccagacaa gtgagagtca ccctctgaga 3600
cagttccatt tcacctcctg gccagaccac ggtgttcccg acaccactga cctgctcatc 3660
aacttccggt acctcgttcg tgactacatg aagcagagtc ctcccgaatc gccgattctg 3720
gtgcattgca gtgctggggt cggaaggacg ggcactttca ttgccattga tcgtctcatc 3780
taccagatag agaatgagaa caccgtggat gtgtatggga ttgtgtatga ccttcgaatg 3840
cataggcctt taatggtgca gacagaggac cagtatgttt tcctcaatca gtgtgttttg 3900
gatattgtca gatcccagaa agactcaaaa gtagatctta tctaccagaa cacaactgca 3960
atgacaatct atgaaaacct tgcgcccgtg accacatttg gaaagaccaa tggttacatc 4020
gcctaa 4026
<210> 43
<211> 1341
<212> Белок
<213> Искусственная последовательность
<220>
<223> Одноцепочечный вариабельный фрагмент (scFv)
SFG.aCD19-CD8STK-CD28tmZ-2A-aCD5-HCH2CH3pvaa-dCD148
<400> 43
Met Ser Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Asp Ile Gln Met Thr Gln Thr Thr Ser Ser Leu
20 25 30
Ser Ala Ser Leu Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln
35 40 45
Asp Ile Ser Lys Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr
50 55 60
Val Lys Leu Leu Ile Tyr His Thr Ser Arg Leu His Ser Gly Val Pro
65 70 75 80
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile
85 90 95
Ser Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly
100 105 110
Asn Thr Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Thr
115 120 125
Lys Ala Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Gly Gly Gly Gly Ser Glu Val Lys Leu Gln Glu Ser Gly Pro Gly
145 150 155 160
Leu Val Ala Pro Ser Gln Ser Leu Ser Val Thr Cys Thr Val Ser Gly
165 170 175
Val Ser Leu Pro Asp Tyr Gly Val Ser Trp Ile Arg Gln Pro Pro Arg
180 185 190
Lys Gly Leu Glu Trp Leu Gly Val Ile Trp Gly Ser Glu Thr Thr Tyr
195 200 205
Tyr Asn Ser Ala Leu Lys Ser Arg Leu Thr Ile Ile Lys Asp Asn Ser
210 215 220
Lys Ser Gln Val Phe Leu Lys Met Asn Ser Leu Gln Thr Asp Asp Thr
225 230 235 240
Ala Ile Tyr Tyr Cys Ala Lys His Tyr Tyr Tyr Gly Gly Ser Tyr Ala
245 250 255
Met Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Asp Pro
260 265 270
Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala
275 280 285
Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly
290 295 300
Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp Ile Phe Trp
305 310 315 320
Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val
325 330 335
Thr Val Ala Phe Ile Ile Phe Trp Val Arg Arg Val Lys Phe Ser Arg
340 345 350
Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn
355 360 365
Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg
370 375 380
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro
385 390 395 400
Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala
405 410 415
Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His
420 425 430
Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp
435 440 445
Ala Leu His Met Gln Ala Leu Pro Pro Arg Arg Ala Glu Gly Arg Gly
450 455 460
Ser Leu Leu Thr Cys Gly Asp Val Glu Glu Asn Pro Gly Pro Met Glu
465 470 475 480
Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro Gly Ser
485 490 495
Thr Gly Gln Val Thr Leu Lys Glu Ser Gly Pro Gly Ile Leu Lys Pro
500 505 510
Ser Gln Thr Leu Ser Leu Thr Cys Ser Phe Ser Gly Phe Ser Leu Ser
515 520 525
Thr Ser Gly Met Gly Val Gly Trp Ile Arg Gln Pro Ser Gly Lys Gly
530 535 540
Leu Glu Trp Leu Ala His Ile Trp Trp Asp Asp Asp Val Tyr Tyr Asn
545 550 555 560
Pro Ser Leu Lys Asn Gln Leu Thr Ile Ser Lys Asp Ala Ser Arg Asp
565 570 575
Gln Val Phe Leu Lys Ile Thr Asn Leu Asp Thr Ala Asp Thr Ala Thr
580 585 590
Tyr Tyr Cys Val Arg Arg Arg Ala Thr Gly Thr Gly Phe Asp Tyr Trp
595 600 605
Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly
610 615 620
Gly Gly Gly Ser Gly Gly Gly Gly Ser Asn Ile Val Met Thr Gln Ser
625 630 635 640
His Lys Phe Met Ser Thr Ser Val Gly Asp Arg Val Ser Ile Ala Cys
645 650 655
Lys Ala Ser Gln Asp Val Gly Thr Ala Val Ala Trp Tyr Gln Gln Lys
660 665 670
Pro Gly Gln Ser Pro Lys Leu Leu Ile Tyr Trp Thr Ser Thr Arg His
675 680 685
Thr Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe
690 695 700
Thr Leu Thr Ile Thr Asn Val Gln Ser Glu Asp Leu Ala Asp Tyr Phe
705 710 715 720
Cys His Gln Tyr Asn Ser Tyr Asn Thr Phe Gly Ser Gly Thr Arg Leu
725 730 735
Glu Leu Lys Arg Ser Asp Pro Ala Glu Pro Lys Ser Pro Asp Lys Thr
740 745 750
His Thr Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val
755 760 765
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ala Arg Thr
770 775 780
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
785 790 795 800
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
805 810 815
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
820 825 830
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
835 840 845
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
850 855 860
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
865 870 875 880
Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
885 890 895
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
900 905 910
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
915 920 925
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
930 935 940
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
945 950 955 960
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Lys
965 970 975
Asp Pro Lys Ala Val Phe Gly Cys Ile Phe Gly Ala Leu Val Ile Val
980 985 990
Thr Val Gly Gly Phe Ile Phe Trp Arg Lys Lys Arg Lys Asp Ala Lys
995 1000 1005
Asn Asn Glu Val Ser Phe Ser Gln Ile Lys Pro Lys Lys Ser Lys
1010 1015 1020
Leu Ile Arg Val Glu Asn Phe Glu Ala Tyr Phe Lys Lys Gln Gln
1025 1030 1035
Ala Asp Ser Asn Cys Gly Phe Ala Glu Glu Tyr Glu Asp Leu Lys
1040 1045 1050
Leu Val Gly Ile Ser Gln Pro Lys Tyr Ala Ala Glu Leu Ala Glu
1055 1060 1065
Asn Arg Gly Lys Asn Arg Tyr Asn Asn Val Leu Pro Tyr Asp Ile
1070 1075 1080
Ser Arg Val Lys Leu Ser Val Gln Thr His Ser Thr Asp Asp Tyr
1085 1090 1095
Ile Asn Ala Asn Tyr Met Pro Gly Tyr His Ser Lys Lys Asp Phe
1100 1105 1110
Ile Ala Thr Gln Gly Pro Leu Pro Asn Thr Leu Lys Asp Phe Trp
1115 1120 1125
Arg Met Val Trp Glu Lys Asn Val Tyr Ala Ile Ile Met Leu Thr
1130 1135 1140
Lys Cys Val Glu Gln Gly Arg Thr Lys Cys Glu Glu Tyr Trp Pro
1145 1150 1155
Ser Lys Gln Ala Gln Asp Tyr Gly Asp Ile Thr Val Ala Met Thr
1160 1165 1170
Ser Glu Ile Val Leu Pro Glu Trp Thr Ile Arg Asp Phe Thr Val
1175 1180 1185
Lys Asn Ile Gln Thr Ser Glu Ser His Pro Leu Arg Gln Phe His
1190 1195 1200
Phe Thr Ser Trp Pro Asp His Gly Val Pro Asp Thr Thr Asp Leu
1205 1210 1215
Leu Ile Asn Phe Arg Tyr Leu Val Arg Asp Tyr Met Lys Gln Ser
1220 1225 1230
Pro Pro Glu Ser Pro Ile Leu Val His Cys Ser Ala Gly Val Gly
1235 1240 1245
Arg Thr Gly Thr Phe Ile Ala Ile Asp Arg Leu Ile Tyr Gln Ile
1250 1255 1260
Glu Asn Glu Asn Thr Val Asp Val Tyr Gly Ile Val Tyr Asp Leu
1265 1270 1275
Arg Met His Arg Pro Leu Met Val Gln Thr Glu Asp Gln Tyr Val
1280 1285 1290
Phe Leu Asn Gln Cys Val Leu Asp Ile Val Arg Ser Gln Lys Asp
1295 1300 1305
Ser Lys Val Asp Leu Ile Tyr Gln Asn Thr Thr Ala Met Thr Ile
1310 1315 1320
Tyr Glu Asn Leu Ala Pro Val Thr Thr Phe Gly Lys Thr Asn Gly
1325 1330 1335
Tyr Ile Ala
1340
<210> 44
<211> 4026
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Одноцепочечный вариабельный фрагмент (scFv)
SFG.aCD19-CD8STK-CD28tmZ-2A-aCD5-HCH2CH3pvaa-dCD148
<400> 44
atgagcctgc ccgtgaccgc cctgctgctg cccctggccc tgctgctgca cgccgccaga 60
ccagacatcc agatgaccca gaccaccagc agcctgagcg ccagcctggg cgaccgggtg 120
accatcagct gcagagccag ccaggacatc agcaagtacc tgaactggta ccagcagaag 180
cccgacggca ccgtgaagct gctgatctac cacaccagcc ggctgcacag cggcgtgccc 240
agccggttca gcggcagcgg cagcggcacc gactacagcc tgaccatcag caacctggag 300
caggaggaca tcgccaccta cttctgccag cagggcaaca ccctgcccta caccttcgga 360
ggcggcacca agctggagat caccaaggcc ggaggcggag gctctggcgg aggcggctct 420
ggcggaggcg gctctggcgg aggcggcagc gaggtgaagc tgcaggagtc tggcccaggc 480
ctggtggccc caagccagag cctgagcgtg acctgcaccg tgagcggcgt gagcctgccc 540
gactacggcg tgagctggat caggcagccc ccacggaagg gcctggagtg gctgggcgtg 600
atctggggca gcgagaccac ctactacaac agcgccctga agagccggct gaccatcatc 660
aaggacaaca gcaagagcca ggtgttcctg aagatgaaca gcctgcagac cgacgacacc 720
gccatctact actgcgccaa gcactactac tatggcggca gctacgctat ggactactgg 780
ggccagggca ccagcgtgac cgtgagctca gatcccacca cgacgccagc gccgcgacca 840
ccaacaccgg cgcccaccat cgcgtcgcag cccctgtccc tgcgcccaga ggcgtgccgg 900
ccagcggcgg ggggcgcagt gcacacgagg gggctggact tcgcctgtga tatcttttgg 960
gtgctggtgg tggttggtgg agtcctggct tgctatagct tgctagtaac agtggccttt 1020
attattttct gggtgaggag agtgaagttc agcaggagcg cagacgcccc cgcgtaccag 1080
cagggccaga accagctcta taacgagctc aatctaggac gaagagagga gtacgatgtt 1140
ttggacaaga gacgtggccg ggaccctgag atggggggaa agccgagaag gaagaaccct 1200
caggaaggcc tgtacaatga actgcagaaa gataagatgg cggaggccta cagtgagatt 1260
gggatgaaag gcgagcgccg gaggggcaag gggcacgatg gcctttacca gggtctcagt 1320
acagccacca aggacaccta cgacgccctt cacatgcagg ccctgcctcc tcgcagagcc 1380
gagggcaggg gaagtcttct aacatgcggg gacgtggagg aaaatcccgg gcccatggag 1440
accgacaccc tgctgctgtg ggtgctgctg ctgtgggtgc ccggcagcac cggccaggtg 1500
accctgaagg agagcggtcc cggcatcctg aagcccagcc agaccctgag cctgacctgc 1560
agcttcagcg gcttcagcct gagcaccagc ggcatgggcg tgggctggat tcggcagccc 1620
agcggcaagg gcctggagtg gctggcccac atctggtggg acgacgacgt gtactacaac 1680
cccagcctga agaaccagct gaccatcagc aaggacgcca gccgggacca ggtgttcctg 1740
aagatcacca acctggacac cgccgacacc gccacctact actgcgtgcg gcgccgggcc 1800
accggcaccg gcttcgacta ctggggccag ggcaccaccc tgaccgtgag cagcggtggc 1860
ggtggcagcg gcggcggcgg aagcggaggt ggtggcagca acatcgtgat gacccagagc 1920
cacaagttca tgagcaccag cgtgggcgac cgggtgagca tcgcctgcaa ggccagccag 1980
gacgtgggca ccgccgtggc ctggtaccag cagaagcctg gccagagccc caagctgctg 2040
atctactgga ccagcacccg gcacaccggc gtgcccgacc ggttcaccgg cagcggcagc 2100
ggcaccgact tcaccctgac catcaccaac gtgcagagcg aggacctggc cgactacttc 2160
tgccaccagt acaacagcta caacaccttc ggcagcggca cccggctgga gctgaagcgg 2220
tcggatcccg ccgagcccaa atctcctgac aaaactcaca catgcccacc gtgcccagca 2280
cctcccgtgg ccggcccgtc agtcttcctc ttccccccaa aacccaagga caccctcatg 2340
atcgcccgga cccctgaggt cacatgcgtg gtggtggacg tgagccacga agaccctgag 2400
gtcaagttca actggtacgt ggacggcgtg gaggtgcata atgccaagac aaagccgcgg 2460
gaggagcagt acaacagcac gtaccgtgtg gtcagcgtcc tcaccgtcct gcaccaggac 2520
tggctgaatg gcaaggagta caagtgcaag gtctccaaca aagccctccc agcccccatc 2580
gagaaaacca tctccaaagc caaagggcag ccccgagaac cacaggtgta caccctgccc 2640
ccatcccggg atgagctgac caagaaccag gtcagcctga cctgcctggt caaaggcttc 2700
tatcccagcg acatcgccgt ggagtgggag agcaatgggc aaccggagaa caactacaag 2760
accacgcctc ccgtgctgga ctccgacggc tccttcttcc tctacagcaa gctcaccgtg 2820
gacaagagca ggtggcagca ggggaacgtc ttctcatgct ccgtgatgca tgaggccctg 2880
cacaatcact atacccagaa atctctgagt ctgagcccag gcaagaagga ccccaaggcg 2940
gtttttggct gtatctttgg tgccctggtt attgtgactg tgggaggctt catcttctgg 3000
agaaagaaga ggaaagatgc aaagaataat gaagtgtcct tttctcaaat taaacctaaa 3060
aaatctaagt taatcagagt ggagaatttt gaggcctact tcaagaagca gcaagctgac 3120
tccaactgtg ggttcgcaga ggaatacgaa gatctgaagc ttgttggaat tagtcaacct 3180
aaatatgcag cagaactggc tgagaataga ggaaagaatc gctataataa tgttctgccc 3240
tatgatattt cccgtgtcaa actttcggtc cagacccatt caacggatga ctacatcaat 3300
gccaactaca tgcctggcta ccactccaag aaagatttta ttgccacaca aggaccttta 3360
ccgaacactt tgaaagattt ttggcgtatg gtttgggaga aaaatgtata tgccatcatt 3420
atgttgacta aatgtgttga acagggaaga accaaatgtg aggagtattg gccctccaag 3480
caggctcagg actatggaga cataactgtg gcaatgacat cagaaattgt tcttccggaa 3540
tggaccatca gagatttcac agtgaaaaat atccagacaa gtgagagtca ccctctgaga 3600
cagttccatt tcacctcctg gccagaccac ggtgttcccg acaccactga cctgctcatc 3660
aacttccggt acctcgttcg tgactacatg aagcagagtc ctcccgaatc gccgattctg 3720
gtgcattgca gtgctggggt cggaaggacg ggcactttca ttgccattga tcgtctcatc 3780
taccagatag agaatgagaa caccgtggat gtgtatggga ttgtgtatga ccttcgaatg 3840
cataggcctt taatggtgca gacagaggac cagtatgttt tcctcaatca gtgtgttttg 3900
gatattgtca gatcccagaa agactcaaaa gtagatctta tctaccagaa cacaactgca 3960
atgacaatct atgaaaacct tgcgcccgtg accacatttg gaaagaccaa tggttacatc 4020
gcctaa 4026
<210> 45
<211> 1342
<212> Белок
<213> Искусственная последовательность
<220>
<223> Одноцепочечный вариабельный фрагмент (scFv)
SFG.aCD19-CD8STK-CD28tmZ-2A-aEGFRvIII-HCH2CH3pvaa-dCD148
<400> 45
Met Ser Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Asp Ile Gln Met Thr Gln Thr Thr Ser Ser Leu
20 25 30
Ser Ala Ser Leu Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln
35 40 45
Asp Ile Ser Lys Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr
50 55 60
Val Lys Leu Leu Ile Tyr His Thr Ser Arg Leu His Ser Gly Val Pro
65 70 75 80
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile
85 90 95
Ser Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly
100 105 110
Asn Thr Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Thr
115 120 125
Lys Ala Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Gly Gly Gly Gly Ser Glu Val Lys Leu Gln Glu Ser Gly Pro Gly
145 150 155 160
Leu Val Ala Pro Ser Gln Ser Leu Ser Val Thr Cys Thr Val Ser Gly
165 170 175
Val Ser Leu Pro Asp Tyr Gly Val Ser Trp Ile Arg Gln Pro Pro Arg
180 185 190
Lys Gly Leu Glu Trp Leu Gly Val Ile Trp Gly Ser Glu Thr Thr Tyr
195 200 205
Tyr Asn Ser Ala Leu Lys Ser Arg Leu Thr Ile Ile Lys Asp Asn Ser
210 215 220
Lys Ser Gln Val Phe Leu Lys Met Asn Ser Leu Gln Thr Asp Asp Thr
225 230 235 240
Ala Ile Tyr Tyr Cys Ala Lys His Tyr Tyr Tyr Gly Gly Ser Tyr Ala
245 250 255
Met Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Asp Pro
260 265 270
Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala
275 280 285
Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly
290 295 300
Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp Ile Phe Trp
305 310 315 320
Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val
325 330 335
Thr Val Ala Phe Ile Ile Phe Trp Val Arg Arg Val Lys Phe Ser Arg
340 345 350
Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn
355 360 365
Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg
370 375 380
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro
385 390 395 400
Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala
405 410 415
Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His
420 425 430
Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp
435 440 445
Ala Leu His Met Gln Ala Leu Pro Pro Arg Arg Ala Glu Gly Arg Gly
450 455 460
Ser Leu Leu Thr Cys Gly Asp Val Glu Glu Asn Pro Gly Pro Met Glu
465 470 475 480
Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro Gly Ser
485 490 495
Thr Gly Gln Val Lys Leu Gln Gln Ser Gly Gly Gly Leu Val Lys Pro
500 505 510
Gly Ala Ser Leu Lys Leu Ser Cys Val Thr Ser Gly Phe Thr Phe Arg
515 520 525
Lys Phe Gly Met Ser Trp Val Arg Gln Thr Ser Asp Lys Arg Leu Glu
530 535 540
Trp Val Ala Ser Ile Ser Thr Gly Gly Tyr Asn Thr Tyr Tyr Ser Asp
545 550 555 560
Asn Val Lys Gly Arg Phe Thr Ile Ser Arg Glu Asn Ala Lys Asn Thr
565 570 575
Leu Tyr Leu Gln Met Ser Ser Leu Lys Ser Glu Asp Thr Ala Leu Tyr
580 585 590
Tyr Cys Thr Arg Gly Tyr Ser Ser Thr Ser Tyr Ala Met Asp Tyr Trp
595 600 605
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly
610 615 620
Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Glu Leu Thr Gln Ser
625 630 635 640
Pro Ala Ser Leu Ser Val Ala Thr Gly Glu Lys Val Thr Ile Arg Cys
645 650 655
Met Thr Ser Thr Asp Ile Asp Asp Asp Met Asn Trp Tyr Gln Gln Lys
660 665 670
Pro Gly Glu Pro Pro Lys Phe Leu Ile Ser Glu Gly Asn Thr Leu Arg
675 680 685
Pro Gly Val Pro Ser Arg Phe Ser Ser Ser Gly Thr Gly Thr Asp Phe
690 695 700
Val Phe Thr Ile Glu Asn Thr Leu Ser Glu Asp Val Gly Asp Tyr Tyr
705 710 715 720
Cys Leu Gln Ser Phe Asn Val Pro Leu Thr Phe Gly Asp Gly Thr Lys
725 730 735
Leu Glu Ile Lys Arg Ser Asp Pro Ala Glu Pro Lys Ser Pro Asp Lys
740 745 750
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser
755 760 765
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ala Arg
770 775 780
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
785 790 795 800
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
805 810 815
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
820 825 830
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
835 840 845
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
850 855 860
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
865 870 875 880
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
885 890 895
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
900 905 910
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
915 920 925
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
930 935 940
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
945 950 955 960
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
965 970 975
Lys Asp Pro Lys Ala Val Phe Gly Cys Ile Phe Gly Ala Leu Val Ile
980 985 990
Val Thr Val Gly Gly Phe Ile Phe Trp Arg Lys Lys Arg Lys Asp Ala
995 1000 1005
Lys Asn Asn Glu Val Ser Phe Ser Gln Ile Lys Pro Lys Lys Ser
1010 1015 1020
Lys Leu Ile Arg Val Glu Asn Phe Glu Ala Tyr Phe Lys Lys Gln
1025 1030 1035
Gln Ala Asp Ser Asn Cys Gly Phe Ala Glu Glu Tyr Glu Asp Leu
1040 1045 1050
Lys Leu Val Gly Ile Ser Gln Pro Lys Tyr Ala Ala Glu Leu Ala
1055 1060 1065
Glu Asn Arg Gly Lys Asn Arg Tyr Asn Asn Val Leu Pro Tyr Asp
1070 1075 1080
Ile Ser Arg Val Lys Leu Ser Val Gln Thr His Ser Thr Asp Asp
1085 1090 1095
Tyr Ile Asn Ala Asn Tyr Met Pro Gly Tyr His Ser Lys Lys Asp
1100 1105 1110
Phe Ile Ala Thr Gln Gly Pro Leu Pro Asn Thr Leu Lys Asp Phe
1115 1120 1125
Trp Arg Met Val Trp Glu Lys Asn Val Tyr Ala Ile Ile Met Leu
1130 1135 1140
Thr Lys Cys Val Glu Gln Gly Arg Thr Lys Cys Glu Glu Tyr Trp
1145 1150 1155
Pro Ser Lys Gln Ala Gln Asp Tyr Gly Asp Ile Thr Val Ala Met
1160 1165 1170
Thr Ser Glu Ile Val Leu Pro Glu Trp Thr Ile Arg Asp Phe Thr
1175 1180 1185
Val Lys Asn Ile Gln Thr Ser Glu Ser His Pro Leu Arg Gln Phe
1190 1195 1200
His Phe Thr Ser Trp Pro Asp His Gly Val Pro Asp Thr Thr Asp
1205 1210 1215
Leu Leu Ile Asn Phe Arg Tyr Leu Val Arg Asp Tyr Met Lys Gln
1220 1225 1230
Ser Pro Pro Glu Ser Pro Ile Leu Val His Cys Ser Ala Gly Val
1235 1240 1245
Gly Arg Thr Gly Thr Phe Ile Ala Ile Asp Arg Leu Ile Tyr Gln
1250 1255 1260
Ile Glu Asn Glu Asn Thr Val Asp Val Tyr Gly Ile Val Tyr Asp
1265 1270 1275
Leu Arg Met His Arg Pro Leu Met Val Gln Thr Glu Asp Gln Tyr
1280 1285 1290
Val Phe Leu Asn Gln Cys Val Leu Asp Ile Val Arg Ser Gln Lys
1295 1300 1305
Asp Ser Lys Val Asp Leu Ile Tyr Gln Asn Thr Thr Ala Met Thr
1310 1315 1320
Ile Tyr Glu Asn Leu Ala Pro Val Thr Thr Phe Gly Lys Thr Asn
1325 1330 1335
Gly Tyr Ile Ala
1340
<210> 46
<211> 4029
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Одноцепочечный вариабельный фрагмент (scFv)
SFG.aCD19-CD8STK-CD28tmZ-2A-aEGFRvIII-HCH2CH3pvaa-dCD148
<400> 46
atgagcctgc ccgtgaccgc cctgctgctg cccctggccc tgctgctgca cgccgccaga 60
ccagacatcc agatgaccca gaccaccagc agcctgagcg ccagcctggg cgaccgggtg 120
accatcagct gcagagccag ccaggacatc agcaagtacc tgaactggta ccagcagaag 180
cccgacggca ccgtgaagct gctgatctac cacaccagcc ggctgcacag cggcgtgccc 240
agccggttca gcggcagcgg cagcggcacc gactacagcc tgaccatcag caacctggag 300
caggaggaca tcgccaccta cttctgccag cagggcaaca ccctgcccta caccttcgga 360
ggcggcacca agctggagat caccaaggcc ggaggcggag gctctggcgg aggcggctct 420
ggcggaggcg gctctggcgg aggcggcagc gaggtgaagc tgcaggagtc tggcccaggc 480
ctggtggccc caagccagag cctgagcgtg acctgcaccg tgagcggcgt gagcctgccc 540
gactacggcg tgagctggat caggcagccc ccacggaagg gcctggagtg gctgggcgtg 600
atctggggca gcgagaccac ctactacaac agcgccctga agagccggct gaccatcatc 660
aaggacaaca gcaagagcca ggtgttcctg aagatgaaca gcctgcagac cgacgacacc 720
gccatctact actgcgccaa gcactactac tatggcggca gctacgctat ggactactgg 780
ggccagggca ccagcgtgac cgtgagctca gatcccacca cgacgccagc gccgcgacca 840
ccaacaccgg cgcccaccat cgcgtcgcag cccctgtccc tgcgcccaga ggcgtgccgg 900
ccagcggcgg ggggcgcagt gcacacgagg gggctggact tcgcctgtga tatcttttgg 960
gtgctggtgg tggttggtgg agtcctggct tgctatagct tgctagtaac agtggccttt 1020
attattttct gggtgaggag agtgaagttc agcaggagcg cagacgcccc cgcgtaccag 1080
cagggccaga accagctcta taacgagctc aatctaggac gaagagagga gtacgatgtt 1140
ttggacaaga gacgtggccg ggaccctgag atggggggaa agccgagaag gaagaaccct 1200
caggaaggcc tgtacaatga actgcagaaa gataagatgg cggaggccta cagtgagatt 1260
gggatgaaag gcgagcgccg gaggggcaag gggcacgatg gcctttacca gggtctcagt 1320
acagccacca aggacaccta cgacgccctt cacatgcagg ccctgcctcc tcgcagagcc 1380
gagggcaggg gaagtcttct aacatgcggg gacgtggagg aaaatcccgg gcccatggag 1440
accgacaccc tgctgctgtg ggtgctgctg ctgtgggtgc ccggcagcac cggccaggtg 1500
aagctgcagc agagcggcgg aggcctggtg aagcccggcg ccagcctgaa gctgagctgc 1560
gtgaccagcg gcttcacctt ccggaagttc ggcatgagct gggtgcggca gaccagcgac 1620
aagcggctgg agtgggtggc cagcatcagc accggcggct acaacaccta ctacagcgac 1680
aacgtgaagg gccggttcac catcagccgg gagaacgcca agaacaccct gtacctgcag 1740
atgagcagcc tgaagagcga ggacaccgcc ctgtactact gcacccgggg ctacagcagc 1800
accagctacg ctatggacta ctggggccag ggcaccaccg tgacagtgag cagcggcgga 1860
ggaggcagtg gtgggggtgg atctggcgga ggtggcagcg acatcgagct gacccagagc 1920
cccgccagcc tgagcgtggc caccggcgag aaggtgacca tccggtgcat gaccagcacc 1980
gacatcgacg acgacatgaa ctggtaccag cagaagcccg gcgagccccc aaagttcctg 2040
atcagcgagg gcaacaccct gcggcccggc gtgcccagcc ggttcagcag cagcggcacc 2100
ggcaccgact tcgtgttcac catcgagaac accctgagcg aggacgtggg cgactactac 2160
tgcctgcaga gcttcaacgt gcccctgacc ttcggcgacg gcaccaagct ggagatcaag 2220
cggtcggatc ccgccgagcc caaatctcct gacaaaactc acacatgccc accgtgccca 2280
gcacctcccg tggccggccc gtcagtcttc ctcttccccc caaaacccaa ggacaccctc 2340
atgatcgccc ggacccctga ggtcacatgc gtggtggtgg acgtgagcca cgaagaccct 2400
gaggtcaagt tcaactggta cgtggacggc gtggaggtgc ataatgccaa gacaaagccg 2460
cgggaggagc agtacaacag cacgtaccgt gtggtcagcg tcctcaccgt cctgcaccag 2520
gactggctga atggcaagga gtacaagtgc aaggtctcca acaaagccct cccagccccc 2580
atcgagaaaa ccatctccaa agccaaaggg cagccccgag aaccacaggt gtacaccctg 2640
cccccatccc gggatgagct gaccaagaac caggtcagcc tgacctgcct ggtcaaaggc 2700
ttctatccca gcgacatcgc cgtggagtgg gagagcaatg ggcaaccgga gaacaactac 2760
aagaccacgc ctcccgtgct ggactccgac ggctccttct tcctctacag caagctcacc 2820
gtggacaaga gcaggtggca gcaggggaac gtcttctcat gctccgtgat gcatgaggcc 2880
ctgcacaatc actataccca gaaatctctg agtctgagcc caggcaagaa ggaccccaag 2940
gcggtttttg gctgtatctt tggtgccctg gttattgtga ctgtgggagg cttcatcttc 3000
tggagaaaga agaggaaaga tgcaaagaat aatgaagtgt ccttttctca aattaaacct 3060
aaaaaatcta agttaatcag agtggagaat tttgaggcct acttcaagaa gcagcaagct 3120
gactccaact gtgggttcgc agaggaatac gaagatctga agcttgttgg aattagtcaa 3180
cctaaatatg cagcagaact ggctgagaat agaggaaaga atcgctataa taatgttctg 3240
ccctatgata tttcccgtgt caaactttcg gtccagaccc attcaacgga tgactacatc 3300
aatgccaact acatgcctgg ctaccactcc aagaaagatt ttattgccac acaaggacct 3360
ttaccgaaca ctttgaaaga tttttggcgt atggtttggg agaaaaatgt atatgccatc 3420
attatgttga ctaaatgtgt tgaacaggga agaaccaaat gtgaggagta ttggccctcc 3480
aagcaggctc aggactatgg agacataact gtggcaatga catcagaaat tgttcttccg 3540
gaatggacca tcagagattt cacagtgaaa aatatccaga caagtgagag tcaccctctg 3600
agacagttcc atttcacctc ctggccagac cacggtgttc ccgacaccac tgacctgctc 3660
atcaacttcc ggtacctcgt tcgtgactac atgaagcaga gtcctcccga atcgccgatt 3720
ctggtgcatt gcagtgctgg ggtcggaagg acgggcactt tcattgccat tgatcgtctc 3780
atctaccaga tagagaatga gaacaccgtg gatgtgtatg ggattgtgta tgaccttcga 3840
atgcataggc ctttaatggt gcagacagag gaccagtatg ttttcctcaa tcagtgtgtt 3900
ttggatattg tcagatccca gaaagactca aaagtagatc ttatctacca gaacacaact 3960
gcaatgacaa tctatgaaaa ccttgcgccc gtgaccacat ttggaaagac caatggttac 4020
atcgcctaa 4029
<210> 47
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Иммунорецепторный тирозиновый ингибирующий мотив (ITIM)
<220>
<221> смешанные свойства
<222> (1)..(1)
<223> Xaa может представлять собой Ser, Ile, Val или Leu
<220>
<221> смешанные свойства
<222> (2)..(2)
<223> Xaa может представлять собой любую встречаюущся в природе аминокислоту
<220>
<221> смешанные свойства
<222> (4)..(5)
<223> Xaa может представлять собой любую встречаюущся в природе аминокислоту
<220>
<221> смешанные свойства
<222> (6)..(6)
<223> Xaa может представлять собой Ile, Val или Leu
<400> 47
Xaa Xaa Tyr Xaa Xaa Xaa
1 5
<210> 48
<211> 1114
<212> Белок
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность логического элемента И НЕ
(16076.SFG.aCD19fmc63-CD8STK-CD28tmZ-2A-aCD33glx-muCD8STK-tm-dPTP
N6)
<400> 48
Met Ser Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Asp Ile Gln Met Thr Gln Thr Thr Ser Ser Leu
20 25 30
Ser Ala Ser Leu Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln
35 40 45
Asp Ile Ser Lys Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr
50 55 60
Val Lys Leu Leu Ile Tyr His Thr Ser Arg Leu His Ser Gly Val Pro
65 70 75 80
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile
85 90 95
Ser Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly
100 105 110
Asn Thr Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Thr
115 120 125
Lys Ala Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Gly Gly Gly Gly Ser Glu Val Lys Leu Gln Glu Ser Gly Pro Gly
145 150 155 160
Leu Val Ala Pro Ser Gln Ser Leu Ser Val Thr Cys Thr Val Ser Gly
165 170 175
Val Ser Leu Pro Asp Tyr Gly Val Ser Trp Ile Arg Gln Pro Pro Arg
180 185 190
Lys Gly Leu Glu Trp Leu Gly Val Ile Trp Gly Ser Glu Thr Thr Tyr
195 200 205
Tyr Asn Ser Ala Leu Lys Ser Arg Leu Thr Ile Ile Lys Asp Asn Ser
210 215 220
Lys Ser Gln Val Phe Leu Lys Met Asn Ser Leu Gln Thr Asp Asp Thr
225 230 235 240
Ala Ile Tyr Tyr Cys Ala Lys His Tyr Tyr Tyr Gly Gly Ser Tyr Ala
245 250 255
Met Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Asp Pro
260 265 270
Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala
275 280 285
Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly
290 295 300
Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp Ile Phe Trp
305 310 315 320
Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val
325 330 335
Thr Val Ala Phe Ile Ile Phe Trp Val Arg Arg Val Lys Phe Ser Arg
340 345 350
Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn
355 360 365
Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg
370 375 380
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro
385 390 395 400
Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala
405 410 415
Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His
420 425 430
Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp
435 440 445
Ala Leu His Met Gln Ala Leu Pro Pro Arg Arg Ala Glu Gly Arg Gly
450 455 460
Ser Leu Leu Thr Cys Gly Asp Val Glu Glu Asn Pro Gly Pro Met Ala
465 470 475 480
Val Pro Thr Gln Val Leu Gly Leu Leu Leu Leu Trp Leu Thr Asp Ala
485 490 495
Arg Cys Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser
500 505 510
Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Asp Ile Tyr
515 520 525
Phe Asn Leu Val Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu
530 535 540
Leu Ile Tyr Asp Thr Asn Arg Leu Ala Asp Gly Val Pro Ser Arg Phe
545 550 555 560
Ser Gly Ser Gly Ser Gly Thr Gln Tyr Thr Leu Thr Ile Ser Ser Leu
565 570 575
Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln His Tyr Lys Asn Tyr
580 585 590
Pro Leu Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Ser Gly
595 600 605
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
610 615 620
Gly Gly Ser Arg Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu
625 630 635 640
Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe
645 650 655
Thr Leu Ser Asn Tyr Gly Met His Trp Ile Arg Gln Ala Pro Gly Lys
660 665 670
Gly Leu Glu Trp Val Ser Ser Ile Ser Leu Asn Gly Gly Ser Thr Tyr
675 680 685
Tyr Arg Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala
690 695 700
Lys Ser Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr
705 710 715 720
Ala Val Tyr Tyr Cys Ala Ala Gln Asp Ala Tyr Thr Gly Gly Tyr Phe
725 730 735
Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Met Asp Pro
740 745 750
Ala Thr Thr Thr Lys Pro Val Leu Arg Thr Pro Ser Pro Val His Pro
755 760 765
Thr Gly Thr Ser Gln Pro Gln Arg Pro Glu Asp Cys Arg Pro Arg Gly
770 775 780
Ser Val Lys Gly Thr Gly Leu Asp Phe Ala Cys Asp Ile Tyr Trp Ala
785 790 795 800
Pro Leu Ala Gly Ile Cys Val Ala Leu Leu Leu Ser Leu Ile Ile Thr
805 810 815
Leu Ile Cys Tyr His Arg Ser Arg Lys Arg Val Cys Lys Ser Gly Gly
820 825 830
Gly Ser Phe Trp Glu Glu Phe Glu Ser Leu Gln Lys Gln Glu Val Lys
835 840 845
Asn Leu His Gln Arg Leu Glu Gly Gln Arg Pro Glu Asn Lys Gly Lys
850 855 860
Asn Arg Tyr Lys Asn Ile Leu Pro Phe Asp His Ser Arg Val Ile Leu
865 870 875 880
Gln Gly Arg Asp Ser Asn Ile Pro Gly Ser Asp Tyr Ile Asn Ala Asn
885 890 895
Tyr Ile Lys Asn Gln Leu Leu Gly Pro Asp Glu Asn Ala Lys Thr Tyr
900 905 910
Ile Ala Ser Gln Gly Cys Leu Glu Ala Thr Val Asn Asp Phe Trp Gln
915 920 925
Met Ala Trp Gln Glu Asn Ser Arg Val Ile Val Met Thr Thr Arg Glu
930 935 940
Val Glu Lys Gly Arg Asn Lys Cys Val Pro Tyr Trp Pro Glu Val Gly
945 950 955 960
Met Gln Arg Ala Tyr Gly Pro Tyr Ser Val Thr Asn Cys Gly Glu His
965 970 975
Asp Thr Thr Glu Tyr Lys Leu Arg Thr Leu Gln Val Ser Pro Leu Asp
980 985 990
Asn Gly Asp Leu Ile Arg Glu Ile Trp His Tyr Gln Tyr Leu Ser Trp
995 1000 1005
Pro Asp His Gly Val Pro Ser Glu Pro Gly Gly Val Leu Ser Phe
1010 1015 1020
Leu Asp Gln Ile Asn Gln Arg Gln Glu Ser Leu Pro His Ala Gly
1025 1030 1035
Pro Ile Ile Val His Cys Ser Ala Gly Ile Gly Arg Thr Gly Thr
1040 1045 1050
Ile Ile Val Ile Asp Met Leu Met Glu Asn Ile Ser Thr Lys Gly
1055 1060 1065
Leu Asp Cys Asp Ile Asp Ile Gln Lys Thr Ile Gln Met Val Arg
1070 1075 1080
Ala Gln Arg Ser Gly Met Val Gln Thr Glu Ala Gln Tyr Lys Phe
1085 1090 1095
Ile Tyr Val Ala Ile Ala Gln Phe Ile Glu Thr Thr Lys Lys Lys
1100 1105 1110
Leu
<210> 49
<211> 918
<212> Белок
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность логического элемента И НЕ
(MP16091.SFG.aCD19fmc63-CD8STK-CD28tmZ-2A-aCD33glx-muCD8STK-LAIR1
tm-endo)
<400> 49
Met Ser Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Asp Ile Gln Met Thr Gln Thr Thr Ser Ser Leu
20 25 30
Ser Ala Ser Leu Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln
35 40 45
Asp Ile Ser Lys Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr
50 55 60
Val Lys Leu Leu Ile Tyr His Thr Ser Arg Leu His Ser Gly Val Pro
65 70 75 80
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile
85 90 95
Ser Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly
100 105 110
Asn Thr Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Thr
115 120 125
Lys Ala Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Gly Gly Gly Gly Ser Glu Val Lys Leu Gln Glu Ser Gly Pro Gly
145 150 155 160
Leu Val Ala Pro Ser Gln Ser Leu Ser Val Thr Cys Thr Val Ser Gly
165 170 175
Val Ser Leu Pro Asp Tyr Gly Val Ser Trp Ile Arg Gln Pro Pro Arg
180 185 190
Lys Gly Leu Glu Trp Leu Gly Val Ile Trp Gly Ser Glu Thr Thr Tyr
195 200 205
Tyr Asn Ser Ala Leu Lys Ser Arg Leu Thr Ile Ile Lys Asp Asn Ser
210 215 220
Lys Ser Gln Val Phe Leu Lys Met Asn Ser Leu Gln Thr Asp Asp Thr
225 230 235 240
Ala Ile Tyr Tyr Cys Ala Lys His Tyr Tyr Tyr Gly Gly Ser Tyr Ala
245 250 255
Met Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Asp Pro
260 265 270
Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala
275 280 285
Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly
290 295 300
Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp Ile Phe Trp
305 310 315 320
Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val
325 330 335
Thr Val Ala Phe Ile Ile Phe Trp Val Arg Arg Val Lys Phe Ser Arg
340 345 350
Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn
355 360 365
Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg
370 375 380
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro
385 390 395 400
Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala
405 410 415
Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His
420 425 430
Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp
435 440 445
Ala Leu His Met Gln Ala Leu Pro Pro Arg Arg Ala Glu Gly Arg Gly
450 455 460
Ser Leu Leu Thr Cys Gly Asp Val Glu Glu Asn Pro Gly Pro Met Ala
465 470 475 480
Val Pro Thr Gln Val Leu Gly Leu Leu Leu Leu Trp Leu Thr Asp Ala
485 490 495
Arg Cys Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser
500 505 510
Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Asp Ile Tyr
515 520 525
Phe Asn Leu Val Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu
530 535 540
Leu Ile Tyr Asp Thr Asn Arg Leu Ala Asp Gly Val Pro Ser Arg Phe
545 550 555 560
Ser Gly Ser Gly Ser Gly Thr Gln Tyr Thr Leu Thr Ile Ser Ser Leu
565 570 575
Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln His Tyr Lys Asn Tyr
580 585 590
Pro Leu Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Ser Gly
595 600 605
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
610 615 620
Gly Gly Ser Arg Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu
625 630 635 640
Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe
645 650 655
Thr Leu Ser Asn Tyr Gly Met His Trp Ile Arg Gln Ala Pro Gly Lys
660 665 670
Gly Leu Glu Trp Val Ser Ser Ile Ser Leu Asn Gly Gly Ser Thr Tyr
675 680 685
Tyr Arg Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala
690 695 700
Lys Ser Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr
705 710 715 720
Ala Val Tyr Tyr Cys Ala Ala Gln Asp Ala Tyr Thr Gly Gly Tyr Phe
725 730 735
Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Met Asp Pro
740 745 750
Ala Thr Thr Thr Lys Pro Val Leu Arg Thr Pro Ser Pro Val His Pro
755 760 765
Thr Gly Thr Ser Gln Pro Gln Arg Pro Glu Asp Cys Arg Pro Arg Gly
770 775 780
Ser Val Lys Gly Thr Gly Leu Asp Phe Ala Cys Asp Ile Leu Ile Gly
785 790 795 800
Val Ser Val Val Phe Leu Phe Cys Leu Leu Leu Leu Val Leu Phe Cys
805 810 815
Leu His Arg Gln Asn Gln Ile Lys Gln Gly Pro Pro Arg Ser Lys Asp
820 825 830
Glu Glu Gln Lys Pro Gln Gln Arg Pro Asp Leu Ala Val Asp Val Leu
835 840 845
Glu Arg Thr Ala Asp Lys Ala Thr Val Asn Gly Leu Pro Glu Lys Asp
850 855 860
Arg Glu Thr Asp Thr Ser Ala Leu Ala Ala Gly Ser Ser Gln Glu Val
865 870 875 880
Thr Tyr Ala Gln Leu Asp His Trp Ala Leu Thr Gln Arg Thr Ala Arg
885 890 895
Ala Val Ser Pro Gln Ser Thr Lys Pro Met Ala Glu Ser Ile Thr Tyr
900 905 910
Ala Ala Val Ala Arg His
915
<210> 50
<211> 1362
<212> Белок
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность логического элемента И НЕ
(MP16092.SFG.aCD19fmc63-CD8STK-CD28tmZ-2A-aCD33glx-muCD8STK-LAIR1
tm-endo-2A-PTPN6_SH2-dCD148)
<400> 50
Met Ser Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Asp Ile Gln Met Thr Gln Thr Thr Ser Ser Leu
20 25 30
Ser Ala Ser Leu Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln
35 40 45
Asp Ile Ser Lys Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr
50 55 60
Val Lys Leu Leu Ile Tyr His Thr Ser Arg Leu His Ser Gly Val Pro
65 70 75 80
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile
85 90 95
Ser Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly
100 105 110
Asn Thr Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Thr
115 120 125
Lys Ala Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Gly Gly Gly Gly Ser Glu Val Lys Leu Gln Glu Ser Gly Pro Gly
145 150 155 160
Leu Val Ala Pro Ser Gln Ser Leu Ser Val Thr Cys Thr Val Ser Gly
165 170 175
Val Ser Leu Pro Asp Tyr Gly Val Ser Trp Ile Arg Gln Pro Pro Arg
180 185 190
Lys Gly Leu Glu Trp Leu Gly Val Ile Trp Gly Ser Glu Thr Thr Tyr
195 200 205
Tyr Asn Ser Ala Leu Lys Ser Arg Leu Thr Ile Ile Lys Asp Asn Ser
210 215 220
Lys Ser Gln Val Phe Leu Lys Met Asn Ser Leu Gln Thr Asp Asp Thr
225 230 235 240
Ala Ile Tyr Tyr Cys Ala Lys His Tyr Tyr Tyr Gly Gly Ser Tyr Ala
245 250 255
Met Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Asp Pro
260 265 270
Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala
275 280 285
Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly
290 295 300
Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp Ile Phe Trp
305 310 315 320
Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val
325 330 335
Thr Val Ala Phe Ile Ile Phe Trp Val Arg Arg Val Lys Phe Ser Arg
340 345 350
Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn
355 360 365
Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg
370 375 380
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro
385 390 395 400
Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala
405 410 415
Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His
420 425 430
Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp
435 440 445
Ala Leu His Met Gln Ala Leu Pro Pro Arg Arg Ala Glu Gly Arg Gly
450 455 460
Ser Leu Leu Thr Cys Gly Asp Val Glu Glu Asn Pro Gly Pro Met Ala
465 470 475 480
Val Pro Thr Gln Val Leu Gly Leu Leu Leu Leu Trp Leu Thr Asp Ala
485 490 495
Arg Cys Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser
500 505 510
Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Asp Ile Tyr
515 520 525
Phe Asn Leu Val Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu
530 535 540
Leu Ile Tyr Asp Thr Asn Arg Leu Ala Asp Gly Val Pro Ser Arg Phe
545 550 555 560
Ser Gly Ser Gly Ser Gly Thr Gln Tyr Thr Leu Thr Ile Ser Ser Leu
565 570 575
Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln His Tyr Lys Asn Tyr
580 585 590
Pro Leu Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Ser Gly
595 600 605
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
610 615 620
Gly Gly Ser Arg Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu
625 630 635 640
Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe
645 650 655
Thr Leu Ser Asn Tyr Gly Met His Trp Ile Arg Gln Ala Pro Gly Lys
660 665 670
Gly Leu Glu Trp Val Ser Ser Ile Ser Leu Asn Gly Gly Ser Thr Tyr
675 680 685
Tyr Arg Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala
690 695 700
Lys Ser Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr
705 710 715 720
Ala Val Tyr Tyr Cys Ala Ala Gln Asp Ala Tyr Thr Gly Gly Tyr Phe
725 730 735
Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Met Asp Pro
740 745 750
Thr Thr Thr Lys Pro Val Leu Arg Thr Pro Ser Pro Val His Pro Thr
755 760 765
Gly Thr Ser Gln Pro Gln Arg Pro Glu Asp Cys Arg Pro Arg Gly Ser
770 775 780
Val Lys Gly Thr Gly Leu Asp Phe Ala Cys Asp Ile Leu Ile Gly Val
785 790 795 800
Ser Val Val Phe Leu Phe Cys Leu Leu Leu Leu Val Leu Phe Cys Leu
805 810 815
His Arg Gln Asn Gln Ile Lys Gln Gly Pro Pro Arg Ser Lys Asp Glu
820 825 830
Glu Gln Lys Pro Gln Gln Arg Pro Asp Leu Ala Val Asp Val Leu Glu
835 840 845
Arg Thr Ala Asp Lys Ala Thr Val Asn Gly Leu Pro Glu Lys Asp Arg
850 855 860
Glu Thr Asp Thr Ser Ala Leu Ala Ala Gly Ser Ser Gln Glu Val Thr
865 870 875 880
Tyr Ala Gln Leu Asp His Trp Ala Leu Thr Gln Arg Thr Ala Arg Ala
885 890 895
Val Ser Pro Gln Ser Thr Lys Pro Met Ala Glu Ser Ile Thr Tyr Ala
900 905 910
Ala Val Ala Arg His Arg Ala Glu Gly Arg Gly Ser Leu Leu Thr Cys
915 920 925
Gly Asp Val Glu Glu Asn Pro Gly Pro Trp Tyr His Gly His Met Ser
930 935 940
Gly Gly Gln Ala Glu Thr Leu Leu Gln Ala Lys Gly Glu Pro Trp Thr
945 950 955 960
Phe Leu Val Arg Glu Ser Leu Ser Gln Pro Gly Asp Phe Val Leu Ser
965 970 975
Val Leu Ser Asp Gln Pro Lys Ala Gly Pro Gly Ser Pro Leu Arg Val
980 985 990
Thr His Ile Lys Val Met Cys Glu Gly Gly Arg Tyr Thr Val Gly Gly
995 1000 1005
Leu Glu Thr Phe Asp Ser Leu Thr Asp Leu Val Glu His Phe Lys
1010 1015 1020
Lys Thr Gly Ile Glu Glu Ala Ser Gly Ala Phe Val Tyr Leu Arg
1025 1030 1035
Gln Pro Tyr Ser Gly Gly Gly Gly Ser Phe Glu Ala Tyr Phe Lys
1040 1045 1050
Lys Gln Gln Ala Asp Ser Asn Cys Gly Phe Ala Glu Glu Tyr Glu
1055 1060 1065
Asp Leu Lys Leu Val Gly Ile Ser Gln Pro Lys Tyr Ala Ala Glu
1070 1075 1080
Leu Ala Glu Asn Arg Gly Lys Asn Arg Tyr Asn Asn Val Leu Pro
1085 1090 1095
Tyr Asp Ile Ser Arg Val Lys Leu Ser Val Gln Thr His Ser Thr
1100 1105 1110
Asp Asp Tyr Ile Asn Ala Asn Tyr Met Pro Gly Tyr His Ser Lys
1115 1120 1125
Lys Asp Phe Ile Ala Thr Gln Gly Pro Leu Pro Asn Thr Leu Lys
1130 1135 1140
Asp Phe Trp Arg Met Val Trp Glu Lys Asn Val Tyr Ala Ile Ile
1145 1150 1155
Met Leu Thr Lys Cys Val Glu Gln Gly Arg Thr Lys Cys Glu Glu
1160 1165 1170
Tyr Trp Pro Ser Lys Gln Ala Gln Asp Tyr Gly Asp Ile Thr Val
1175 1180 1185
Ala Met Thr Ser Glu Ile Val Leu Pro Glu Trp Thr Ile Arg Asp
1190 1195 1200
Phe Thr Val Lys Asn Ile Gln Thr Ser Glu Ser His Pro Leu Arg
1205 1210 1215
Gln Phe His Phe Thr Ser Trp Pro Asp His Gly Val Pro Asp Thr
1220 1225 1230
Thr Asp Leu Leu Ile Asn Phe Arg Tyr Leu Val Arg Asp Tyr Met
1235 1240 1245
Lys Gln Ser Pro Pro Glu Ser Pro Ile Leu Val His Cys Ser Ala
1250 1255 1260
Gly Val Gly Arg Thr Gly Thr Phe Ile Ala Ile Asp Arg Leu Ile
1265 1270 1275
Tyr Gln Ile Glu Asn Glu Asn Thr Val Asp Val Tyr Gly Ile Val
1280 1285 1290
Tyr Asp Leu Arg Met His Arg Pro Leu Met Val Gln Thr Glu Asp
1295 1300 1305
Gln Tyr Val Phe Leu Asn Gln Cys Val Leu Asp Ile Val Arg Ser
1310 1315 1320
Gln Lys Asp Ser Lys Val Asp Leu Ile Tyr Gln Asn Thr Thr Ala
1325 1330 1335
Met Thr Ile Tyr Glu Asn Leu Ala Pro Val Thr Thr Phe Gly Lys
1340 1345 1350
Thr Asn Gly Tyr Ile Ala Ser Gly Ser
1355 1360
<210> 51
<211> 424
<212> Белок
<213> Искусственная последовательность
<220>
<223> CAR на основе APRIL (на основе индуцирующего пролиферацию лиганда),
APRIL CAR со стеблем CD8
<400> 51
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Ser Val Leu His Leu Val Pro Ile Asn Ala Thr Ser
20 25 30
Lys Asp Asp Ser Asp Val Thr Glu Val Met Trp Gln Pro Ala Leu Arg
35 40 45
Arg Gly Arg Gly Leu Gln Ala Gln Gly Tyr Gly Val Arg Ile Gln Asp
50 55 60
Ala Gly Val Tyr Leu Leu Tyr Ser Gln Val Leu Phe Gln Asp Val Thr
65 70 75 80
Phe Thr Met Gly Gln Val Val Ser Arg Glu Gly Gln Gly Arg Gln Glu
85 90 95
Thr Leu Phe Arg Cys Ile Arg Ser Met Pro Ser His Pro Asp Arg Ala
100 105 110
Tyr Asn Ser Cys Tyr Ser Ala Gly Val Phe His Leu His Gln Gly Asp
115 120 125
Ile Leu Ser Val Ile Ile Pro Arg Ala Arg Ala Lys Leu Asn Leu Ser
130 135 140
Pro His Gly Thr Phe Leu Gly Phe Val Lys Leu Ser Gly Gly Gly Ser
145 150 155 160
Asp Pro Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr
165 170 175
Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala
180 185 190
Ala Gly Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp Ile
195 200 205
Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu
210 215 220
Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val Arg Ser Lys Arg Ser
225 230 235 240
Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr Pro Arg Arg Pro Gly
245 250 255
Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro Arg Asp Phe Ala
260 265 270
Ala Tyr Arg Ser Arg Asp Gln Arg Leu Pro Pro Asp Ala His Lys Pro
275 280 285
Pro Gly Gly Gly Ser Phe Arg Thr Pro Ile Gln Glu Glu Gln Ala Asp
290 295 300
Ala His Ser Thr Leu Ala Lys Ile Arg Val Lys Phe Ser Arg Ser Ala
305 310 315 320
Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu
325 330 335
Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly
340 345 350
Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu
355 360 365
Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser
370 375 380
Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly
385 390 395 400
Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu
405 410 415
His Met Gln Ala Leu Pro Pro Arg
420
<210> 52
<211> 398
<212> Белок
<213> Искусственная последовательность
<220>
<223> CAR на основе APRIL (на основе индуцирующего пролиферацию лиганда),
APRIL CAR на основе шарнира IgG1
<400> 52
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Ser Val Leu His Leu Val Pro Ile Asn Ala Thr Ser
20 25 30
Lys Asp Asp Ser Asp Val Thr Glu Val Met Trp Gln Pro Ala Leu Arg
35 40 45
Arg Gly Arg Gly Leu Gln Ala Gln Gly Tyr Gly Val Arg Ile Gln Asp
50 55 60
Ala Gly Val Tyr Leu Leu Tyr Ser Gln Val Leu Phe Gln Asp Val Thr
65 70 75 80
Phe Thr Met Gly Gln Val Val Ser Arg Glu Gly Gln Gly Arg Gln Glu
85 90 95
Thr Leu Phe Arg Cys Ile Arg Ser Met Pro Ser His Pro Asp Arg Ala
100 105 110
Tyr Asn Ser Cys Tyr Ser Ala Gly Val Phe His Leu His Gln Gly Asp
115 120 125
Ile Leu Ser Val Ile Ile Pro Arg Ala Arg Ala Lys Leu Asn Leu Ser
130 135 140
Pro His Gly Thr Phe Leu Gly Phe Val Lys Leu Ser Gly Gly Gly Ser
145 150 155 160
Asp Pro Ala Glu Pro Lys Ser Pro Asp Lys Thr His Thr Cys Pro Pro
165 170 175
Cys Pro Lys Asp Pro Lys Phe Trp Val Leu Val Val Val Gly Gly Val
180 185 190
Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp
195 200 205
Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met
210 215 220
Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala
225 230 235 240
Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Arg Asp Gln Arg Leu Pro
245 250 255
Pro Asp Ala His Lys Pro Pro Gly Gly Gly Ser Phe Arg Thr Pro Ile
260 265 270
Gln Glu Glu Gln Ala Asp Ala His Ser Thr Leu Ala Lys Ile Arg Val
275 280 285
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn
290 295 300
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
305 310 315 320
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg
325 330 335
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
340 345 350
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg
355 360 365
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys
370 375 380
Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
385 390 395
<210> 53
<211> 614
<212> Белок
<213> Искусственная последовательность
<220>
<223> CAR на основе APRIL (на основе индуцирующего пролиферацию лиганда),
APRIL CAR на основе Fc-pvaa
<400> 53
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Ser Val Leu His Leu Val Pro Ile Asn Ala Thr Ser
20 25 30
Lys Asp Asp Ser Asp Val Thr Glu Val Met Trp Gln Pro Ala Leu Arg
35 40 45
Arg Gly Arg Gly Leu Gln Ala Gln Gly Tyr Gly Val Arg Ile Gln Asp
50 55 60
Ala Gly Val Tyr Leu Leu Tyr Ser Gln Val Leu Phe Gln Asp Val Thr
65 70 75 80
Phe Thr Met Gly Gln Val Val Ser Arg Glu Gly Gln Gly Arg Gln Glu
85 90 95
Thr Leu Phe Arg Cys Ile Arg Ser Met Pro Ser His Pro Asp Arg Ala
100 105 110
Tyr Asn Ser Cys Tyr Ser Ala Gly Val Phe His Leu His Gln Gly Asp
115 120 125
Ile Leu Ser Val Ile Ile Pro Arg Ala Arg Ala Lys Leu Asn Leu Ser
130 135 140
Pro His Gly Thr Phe Leu Gly Phe Val Lys Leu Ser Gly Gly Gly Ser
145 150 155 160
Asp Pro Ala Glu Pro Lys Ser Pro Asp Lys Thr His Thr Cys Pro Pro
165 170 175
Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro
180 185 190
Lys Pro Lys Asp Thr Leu Met Ile Ala Arg Thr Pro Glu Val Thr Cys
195 200 205
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
210 215 220
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
225 230 235 240
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
245 250 255
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
260 265 270
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
275 280 285
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
290 295 300
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
305 310 315 320
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
325 330 335
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
340 345 350
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
355 360 365
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
370 375 380
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Lys Asp Pro Lys Phe Trp
385 390 395 400
Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val
405 410 415
Thr Val Ala Phe Ile Ile Phe Trp Val Arg Ser Lys Arg Ser Arg Leu
420 425 430
Leu His Ser Asp Tyr Met Asn Met Thr Pro Arg Arg Pro Gly Pro Thr
435 440 445
Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro Arg Asp Phe Ala Ala Tyr
450 455 460
Arg Ser Arg Asp Gln Arg Leu Pro Pro Asp Ala His Lys Pro Pro Gly
465 470 475 480
Gly Gly Ser Phe Arg Thr Pro Ile Gln Glu Glu Gln Ala Asp Ala His
485 490 495
Ser Thr Leu Ala Lys Ile Arg Val Lys Phe Ser Arg Ser Ala Asp Ala
500 505 510
Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu
515 520 525
Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp
530 535 540
Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu
545 550 555 560
Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile
565 570 575
Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr
580 585 590
Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met
595 600 605
Gln Ala Leu Pro Pro Arg
610
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| КЛЕТКА | 2014 |
|
RU2732236C2 |
| КЛЕТКА | 2015 |
|
RU2768019C2 |
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР | 2016 |
|
RU2752880C2 |
| СИГНАЛЬНАЯ СИСТЕМА | 2015 |
|
RU2731638C2 |
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР | 2019 |
|
RU2810092C2 |
| ХИМЕРНЫЙ ЦИТОКИНОВЫЙ РЕЦЕПТОР | 2016 |
|
RU2826122C1 |
| СВЯЗЫВАЮЩИЙ ДОМЕН | 2019 |
|
RU2829934C2 |
| ХИМЕРНЫЕ РЕЦЕПТОРЫ АНТИГЕНОВ | 2015 |
|
RU2745705C2 |
| УЛУЧШЕННЫЙ СИНТЕТИЧЕСКИЙ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР И АНТИГЕННЫЙ РЕЦЕПТОР | 2021 |
|
RU2820497C1 |
| Т-КЛЕТКИ С КОСТИМУЛИРУЮЩИМ ХИМЕРНЫМ АНТИГЕННЫМ РЕЦЕПТОРОМ, НАЦЕЛЕННЫЕ НА IL13Rα2 | 2015 |
|
RU2749922C2 |
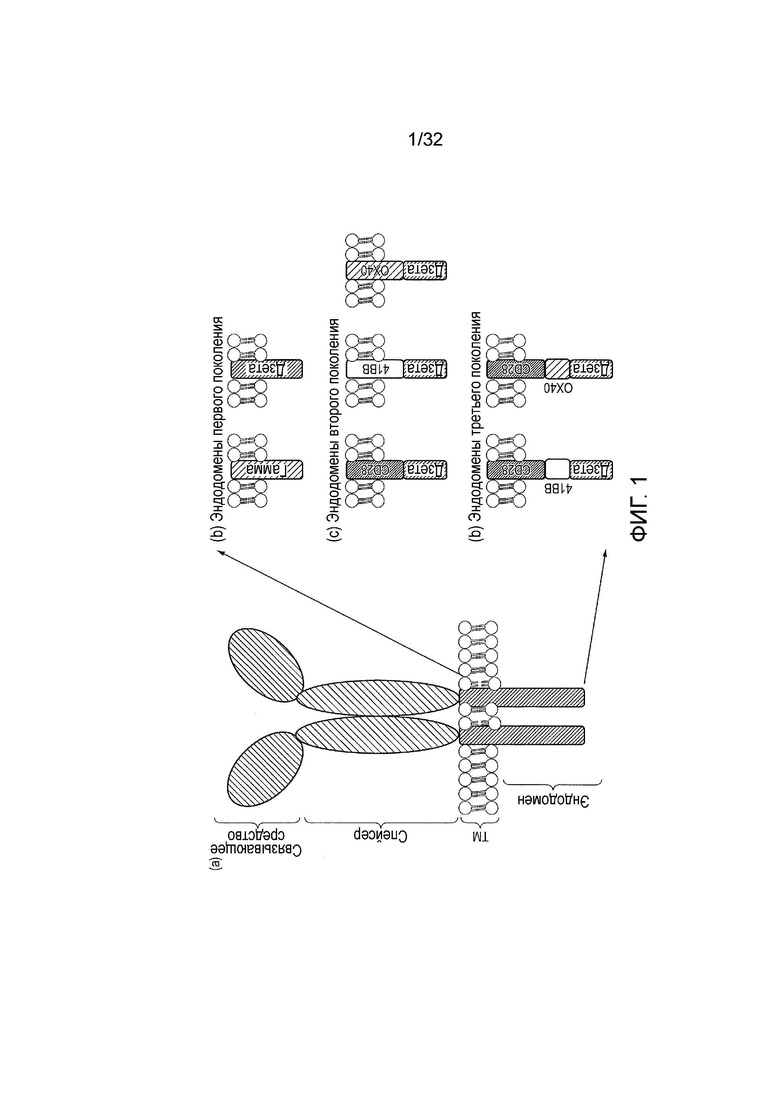
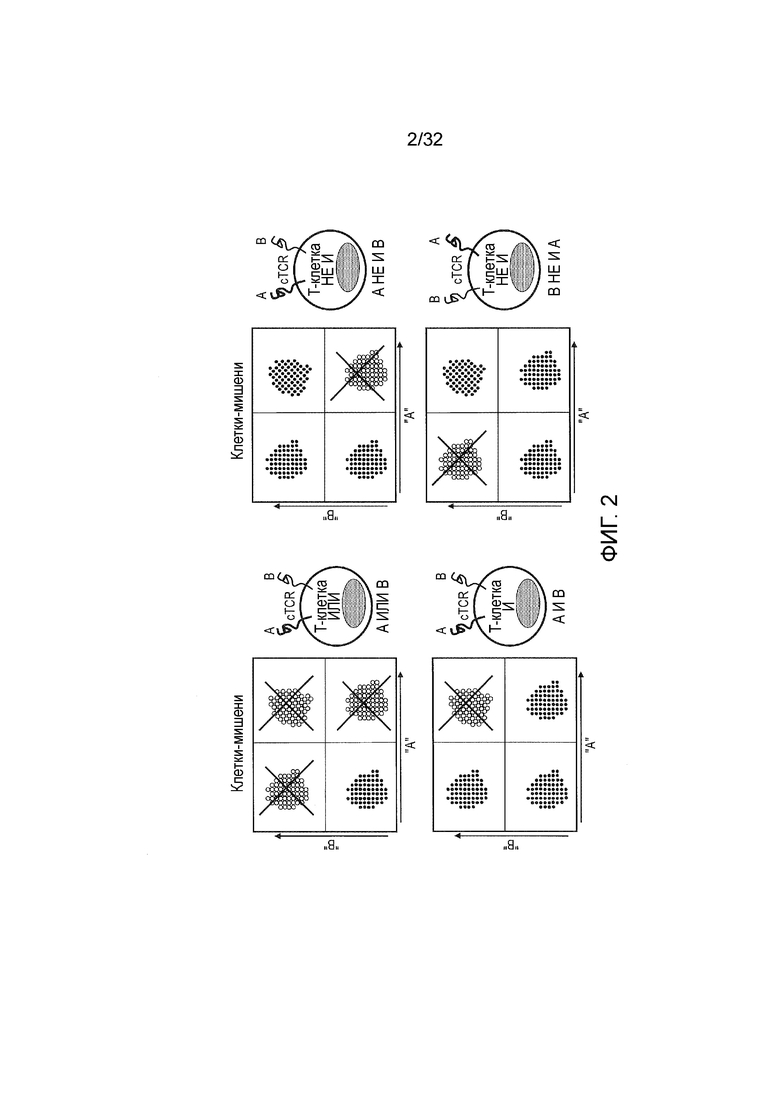
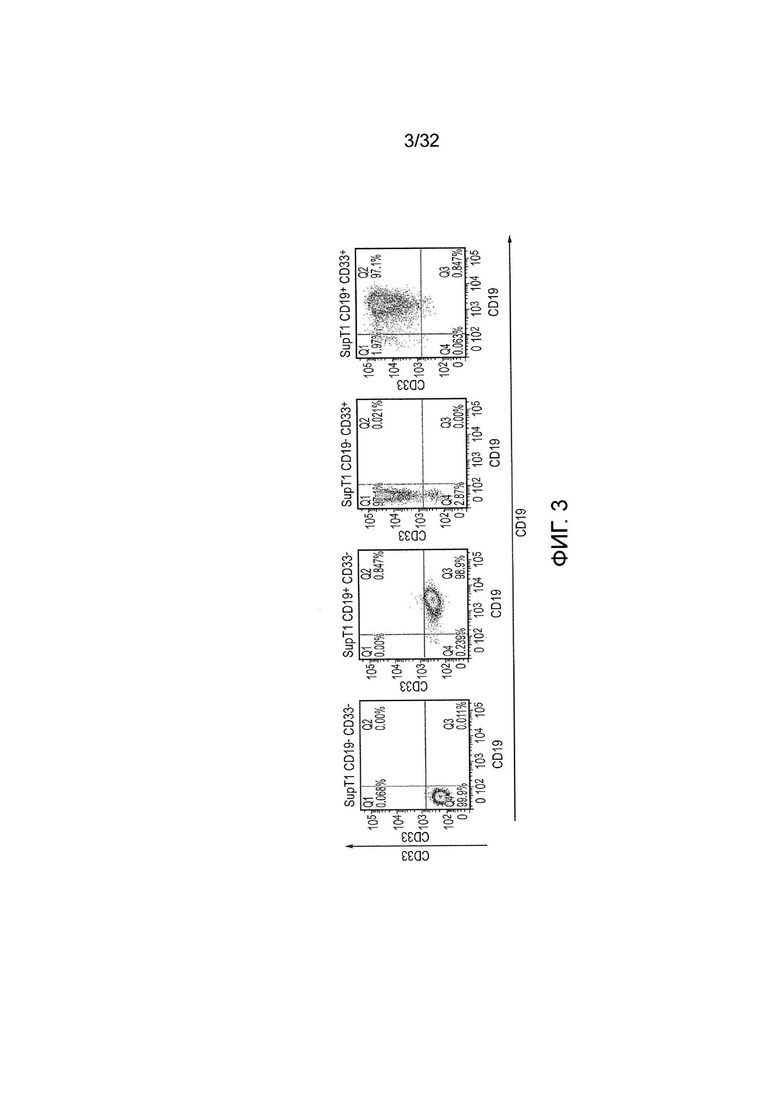

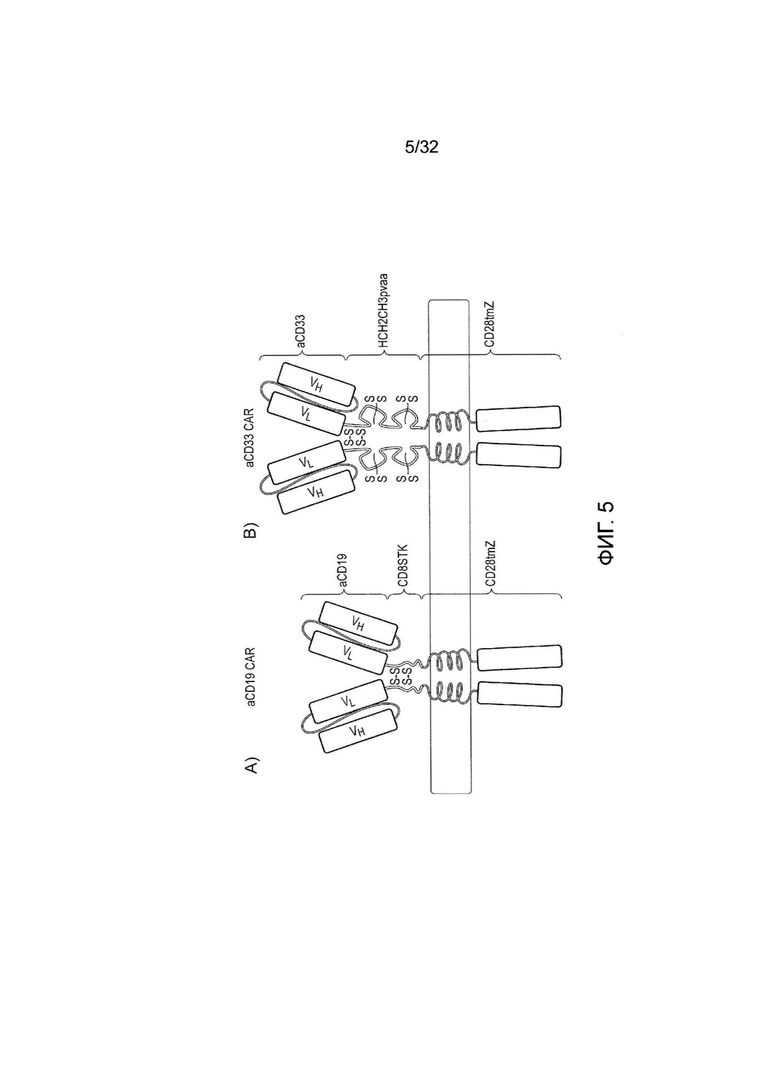

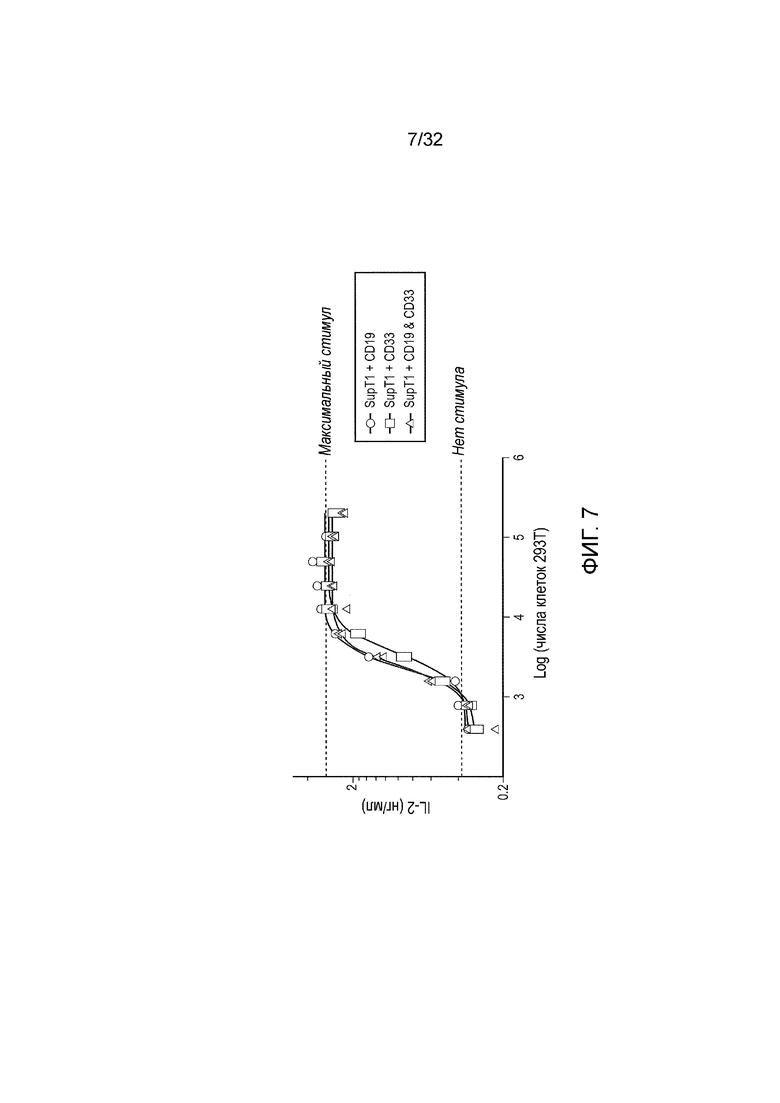

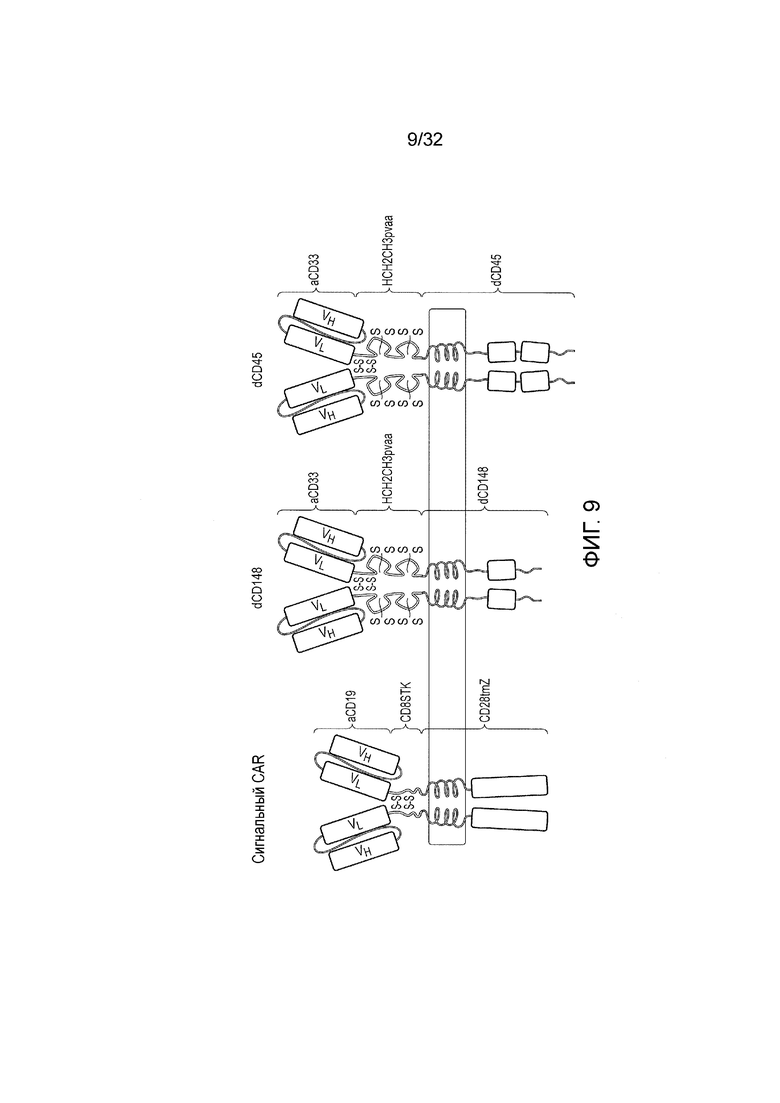
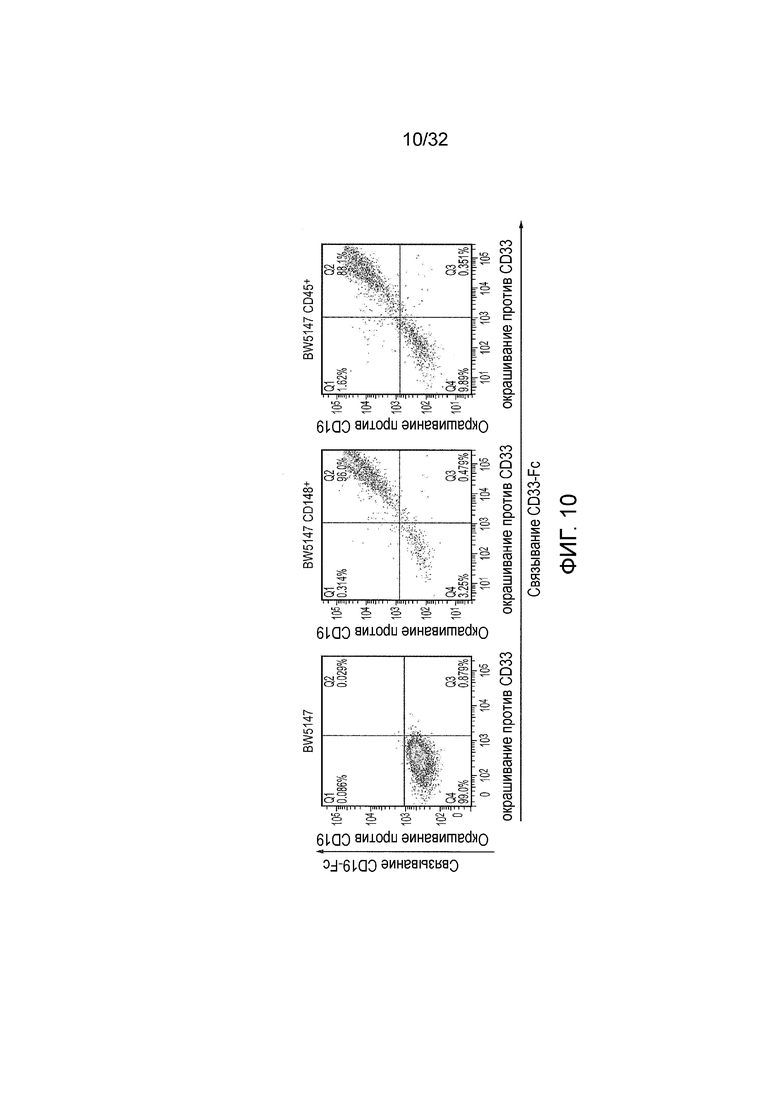
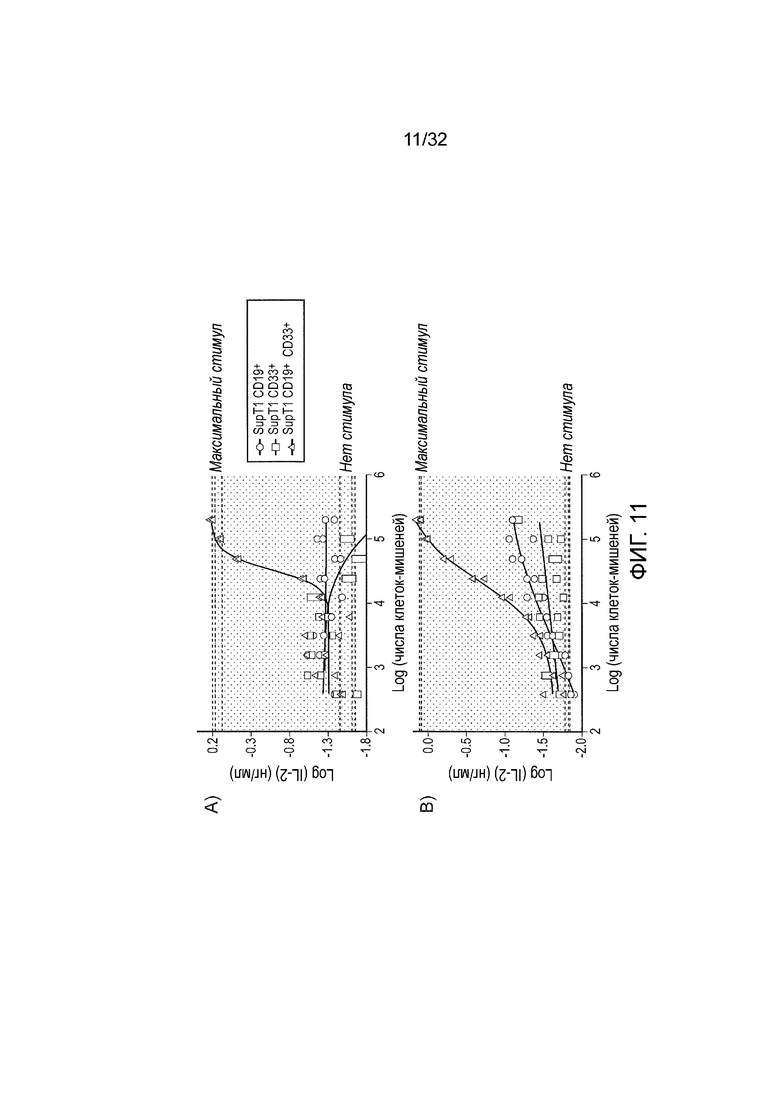

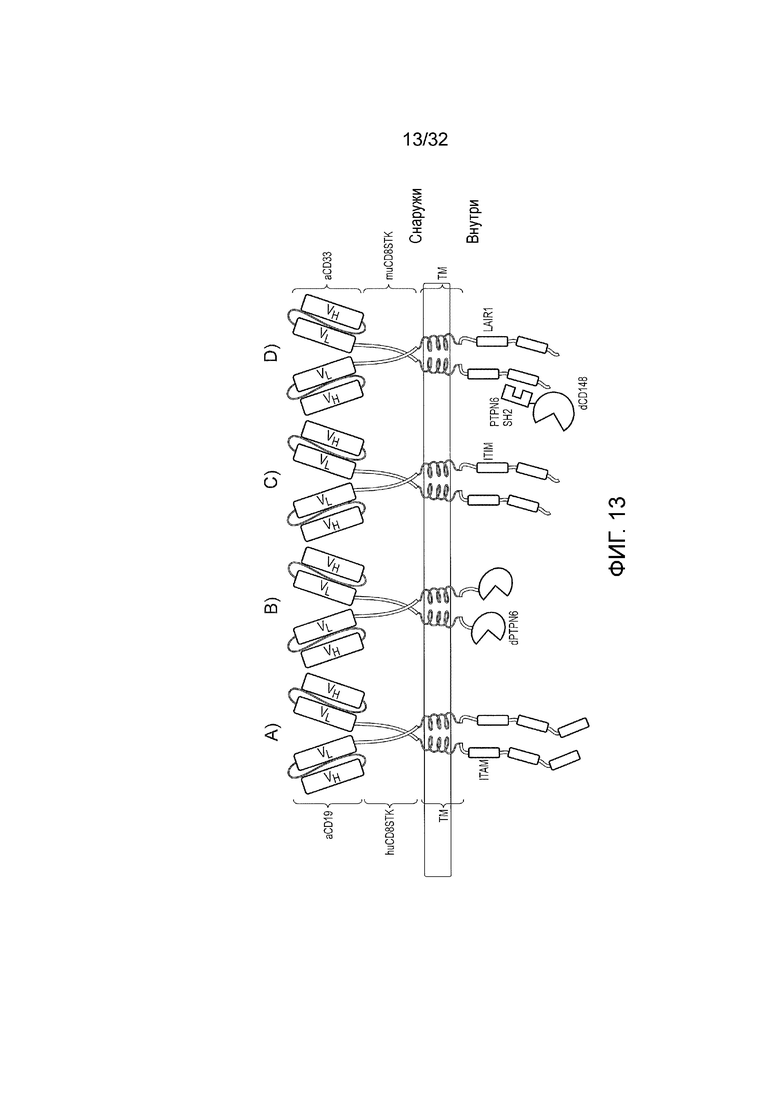
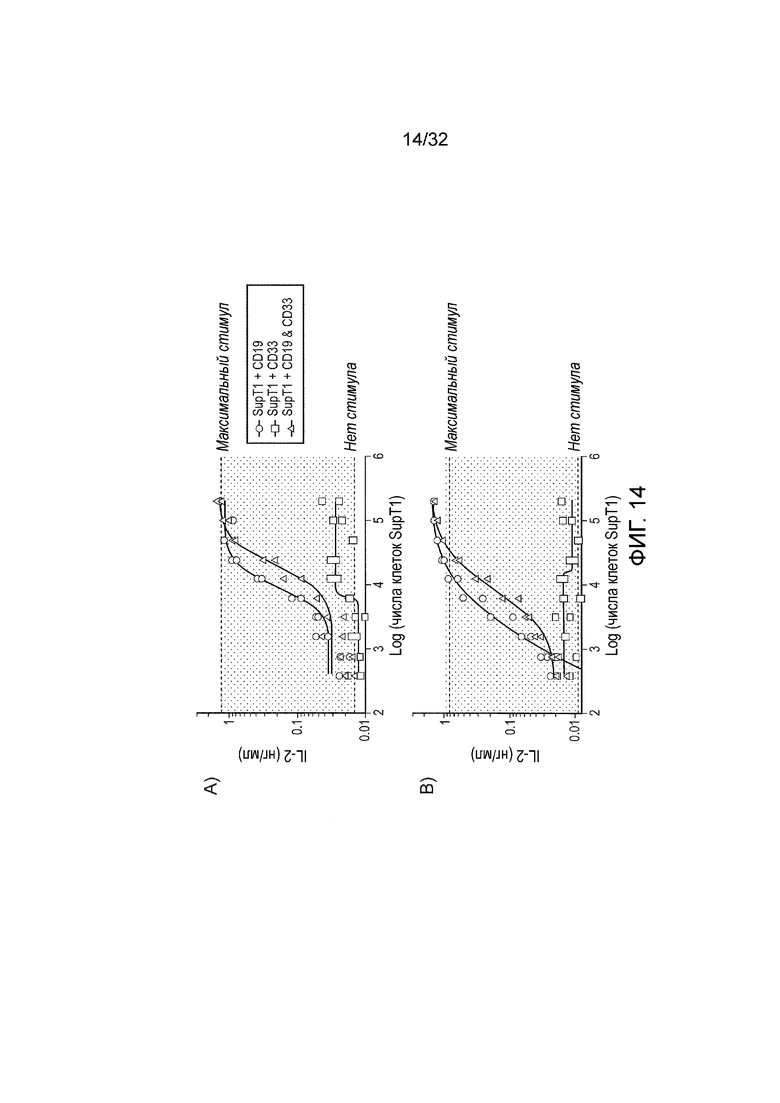



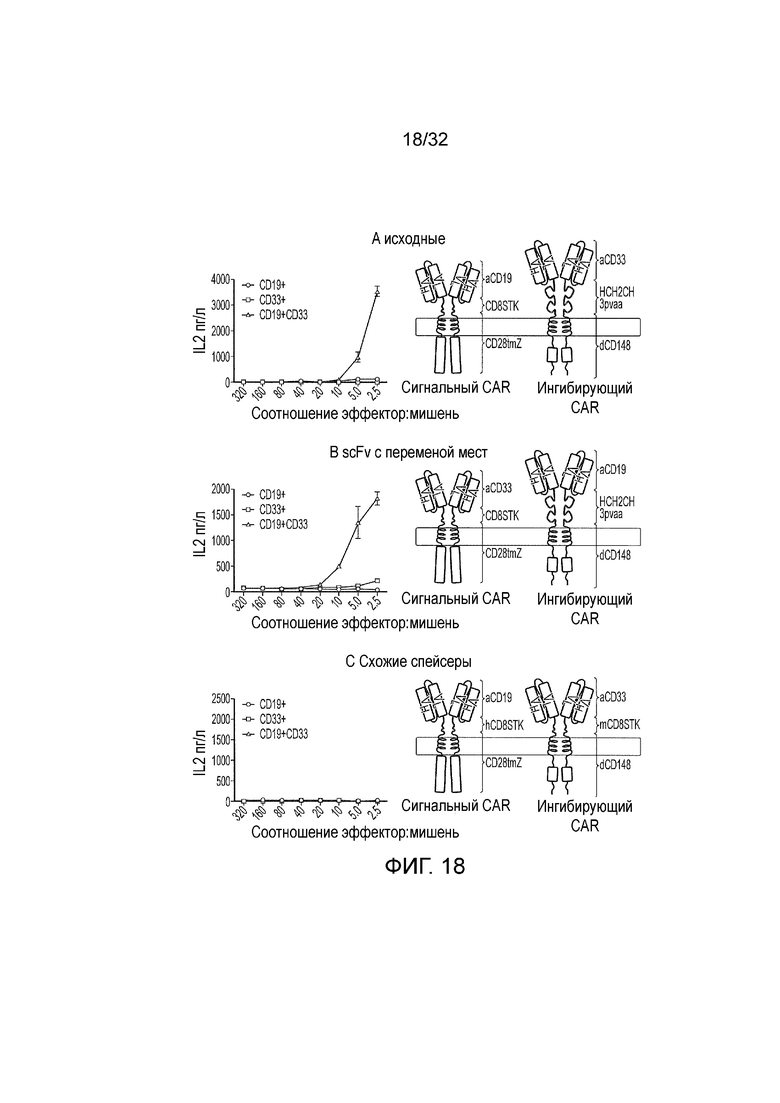
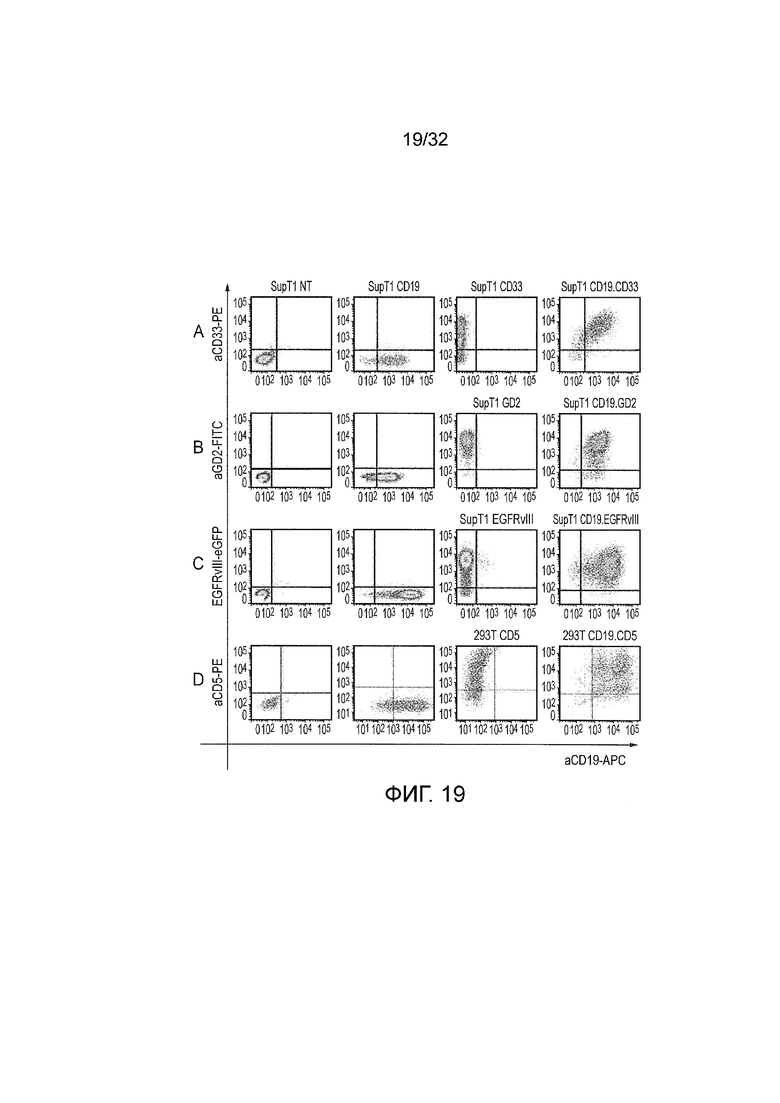
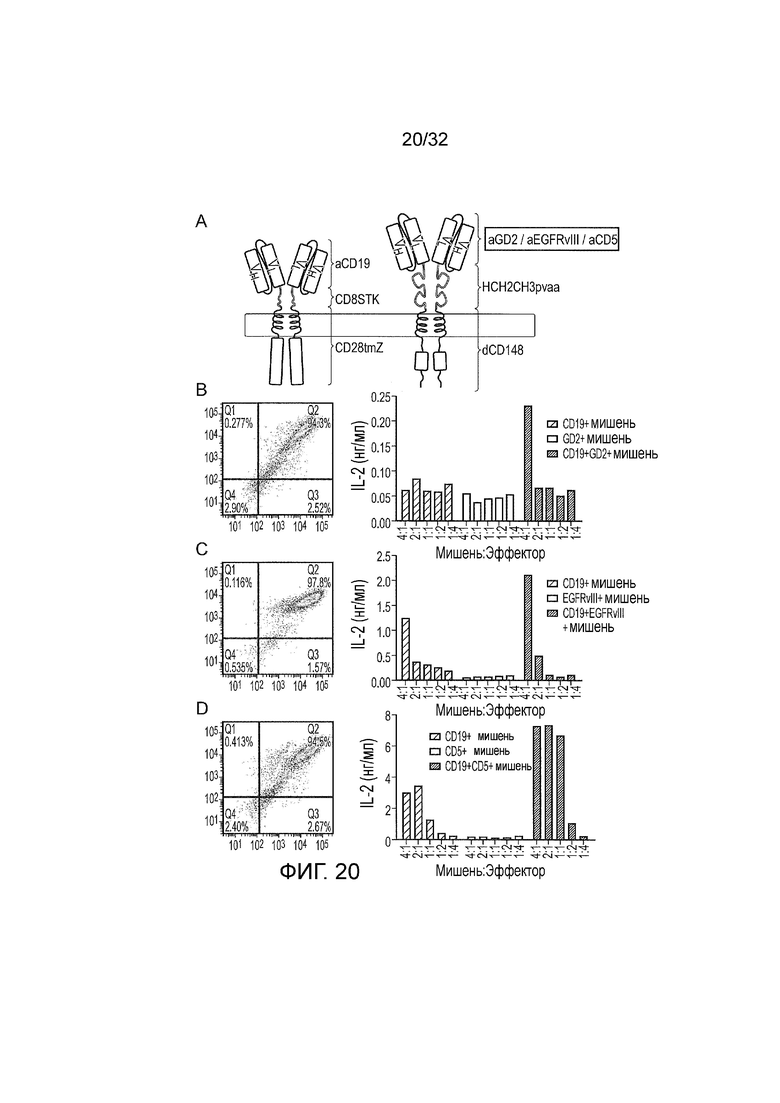
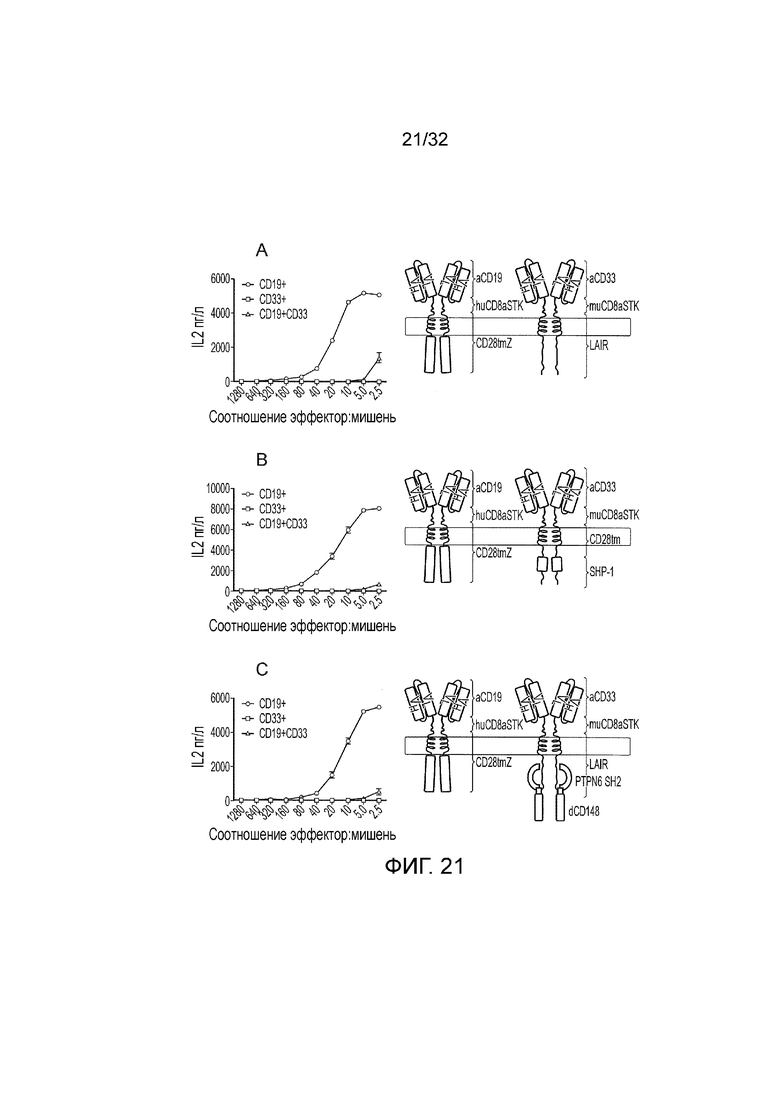
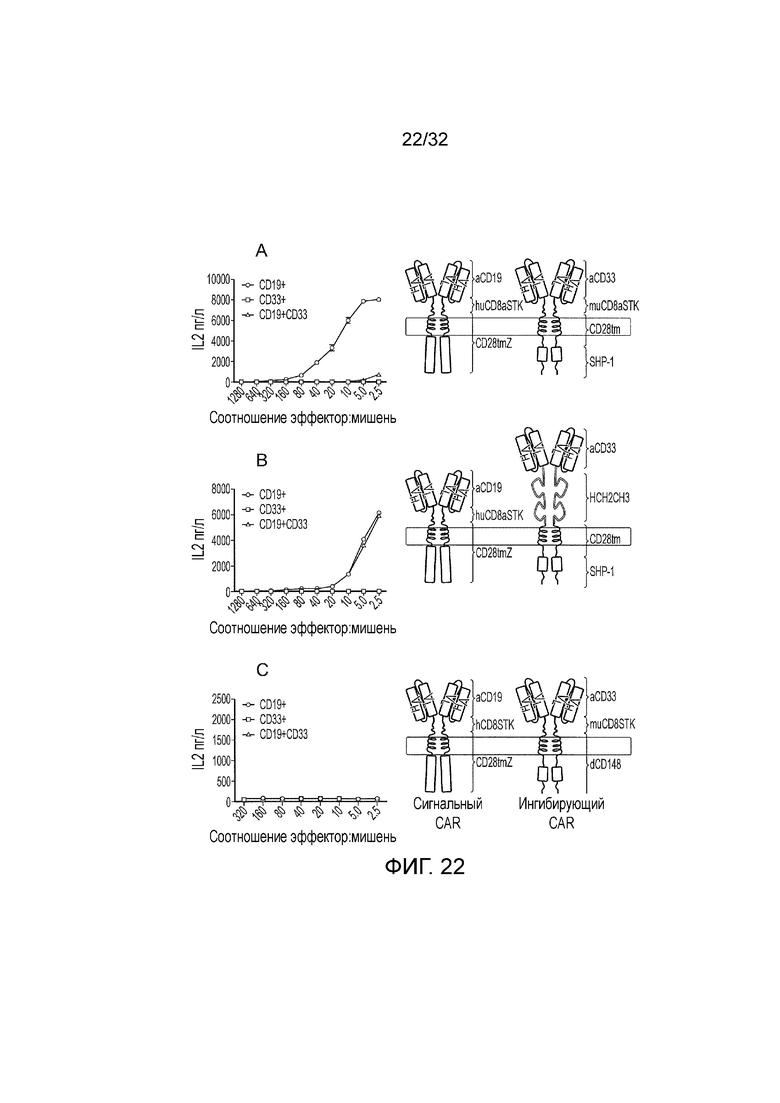
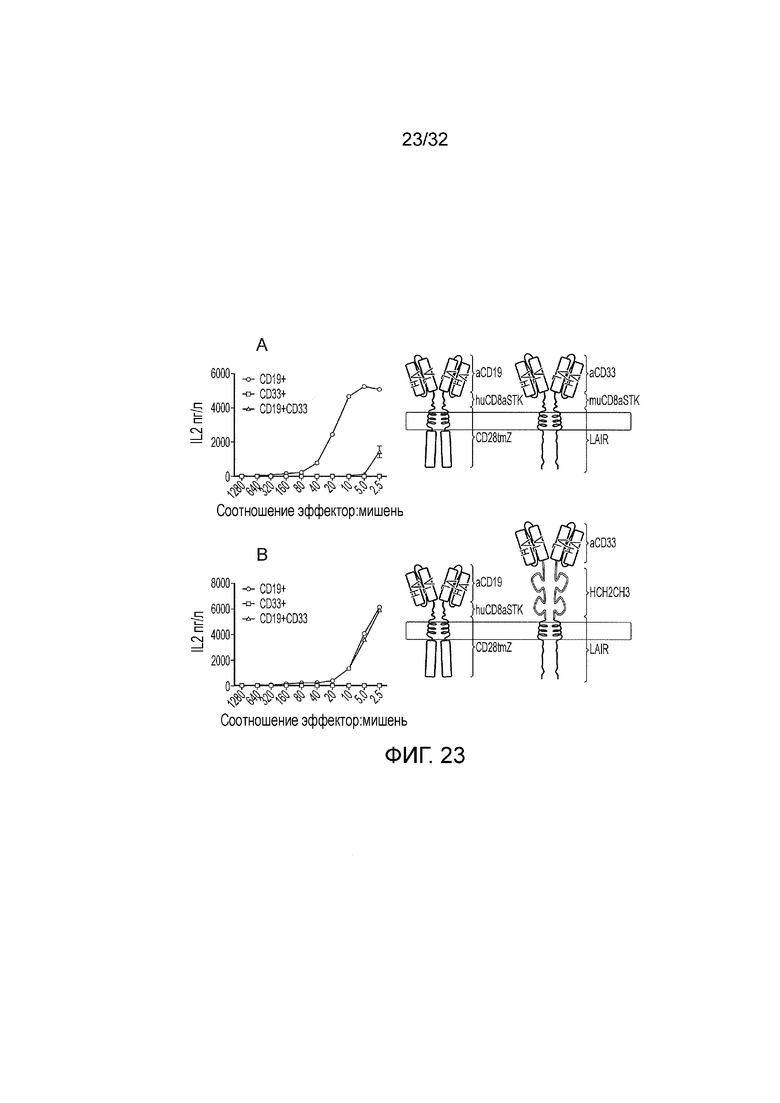
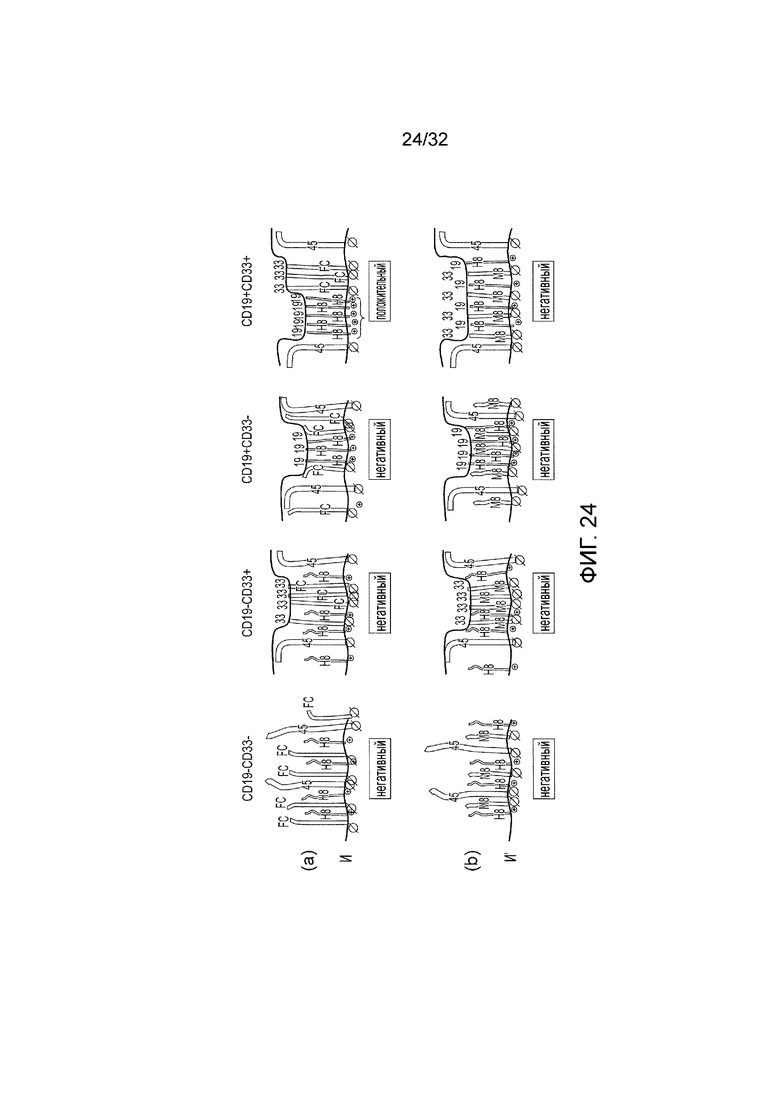
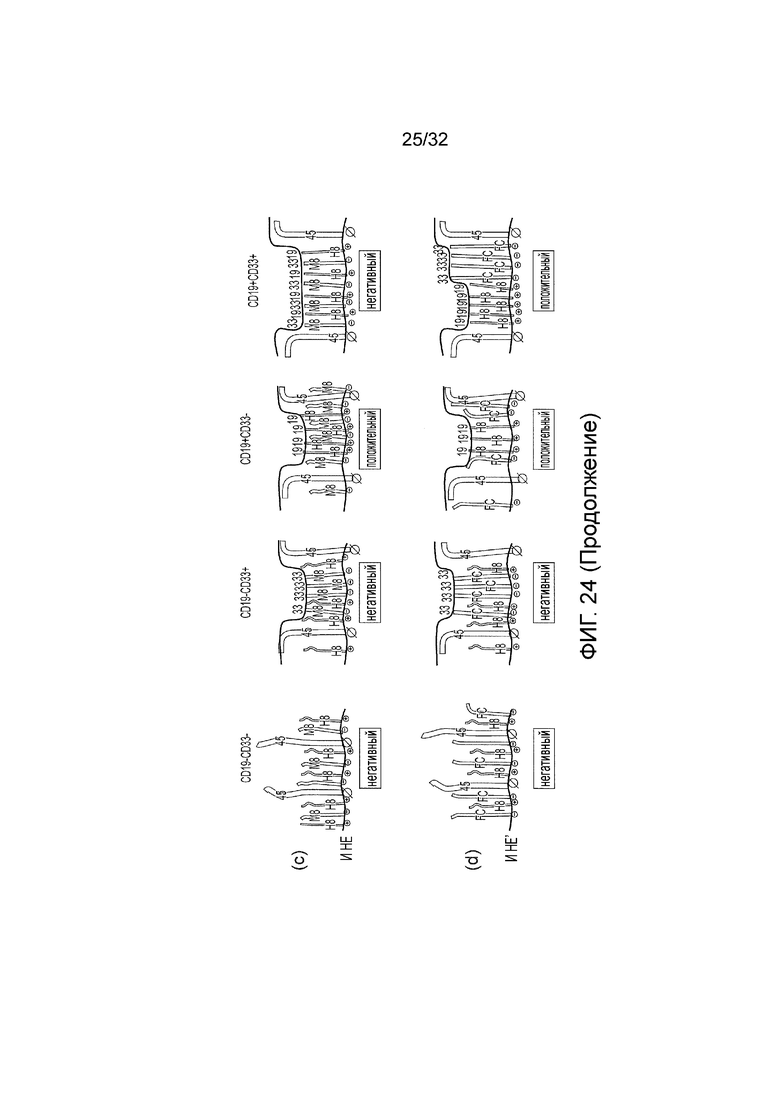
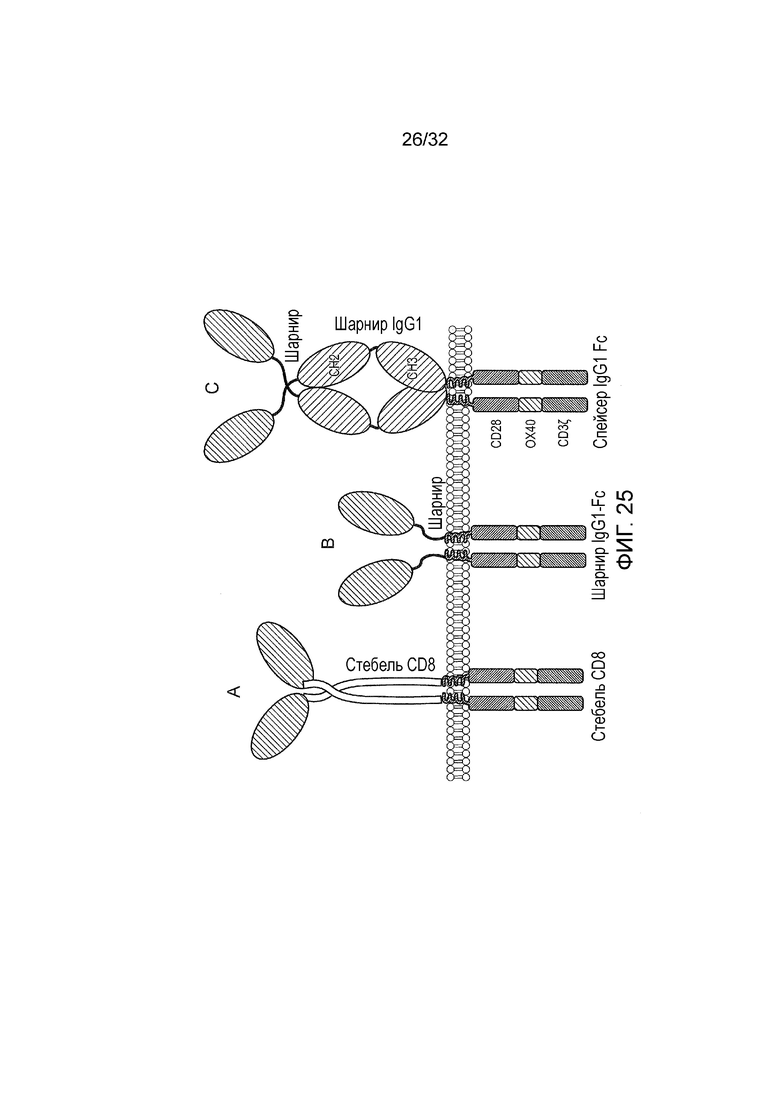

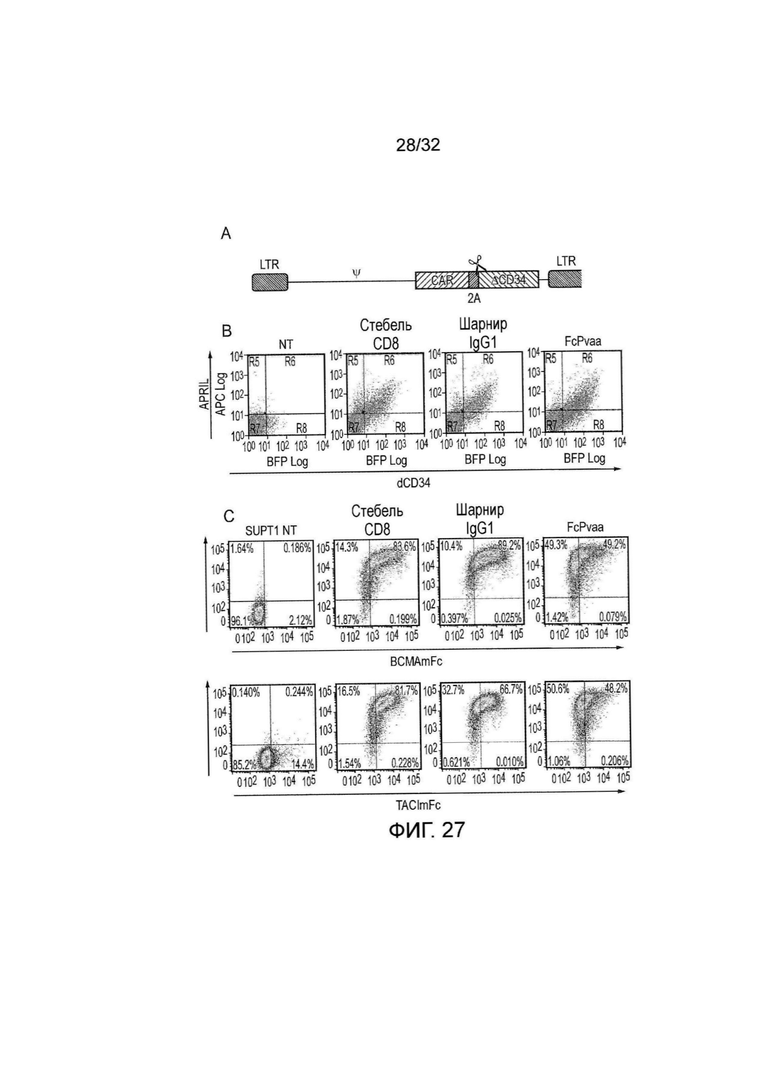
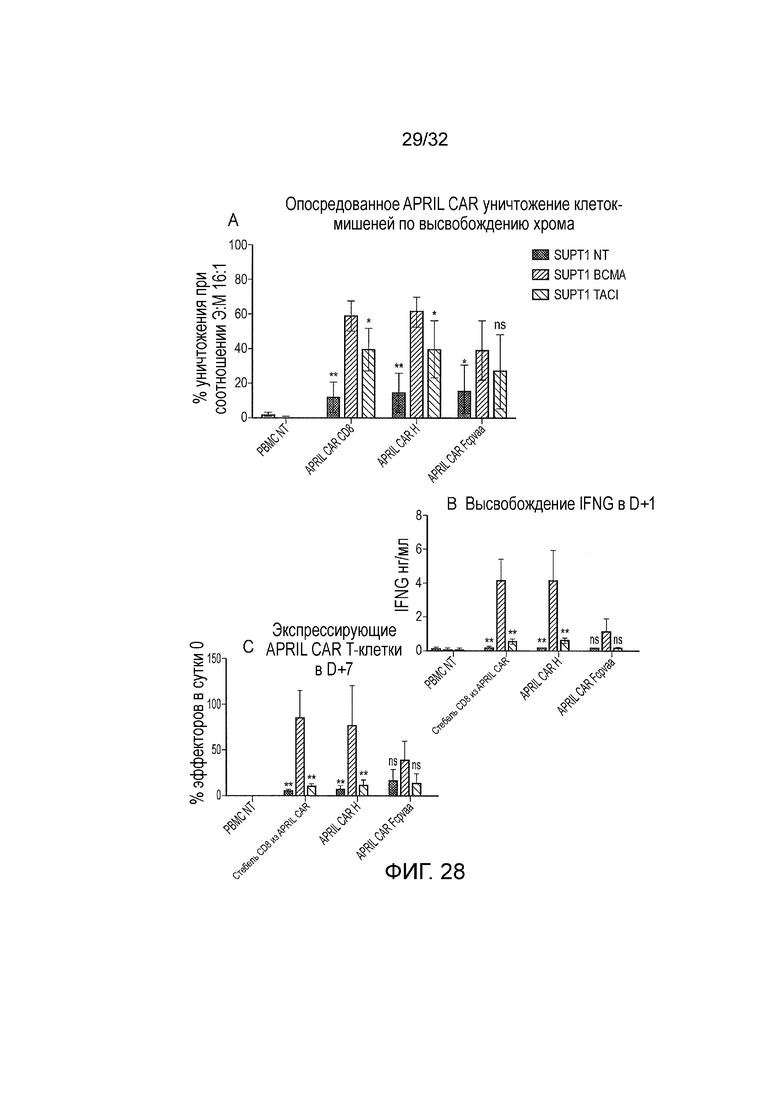
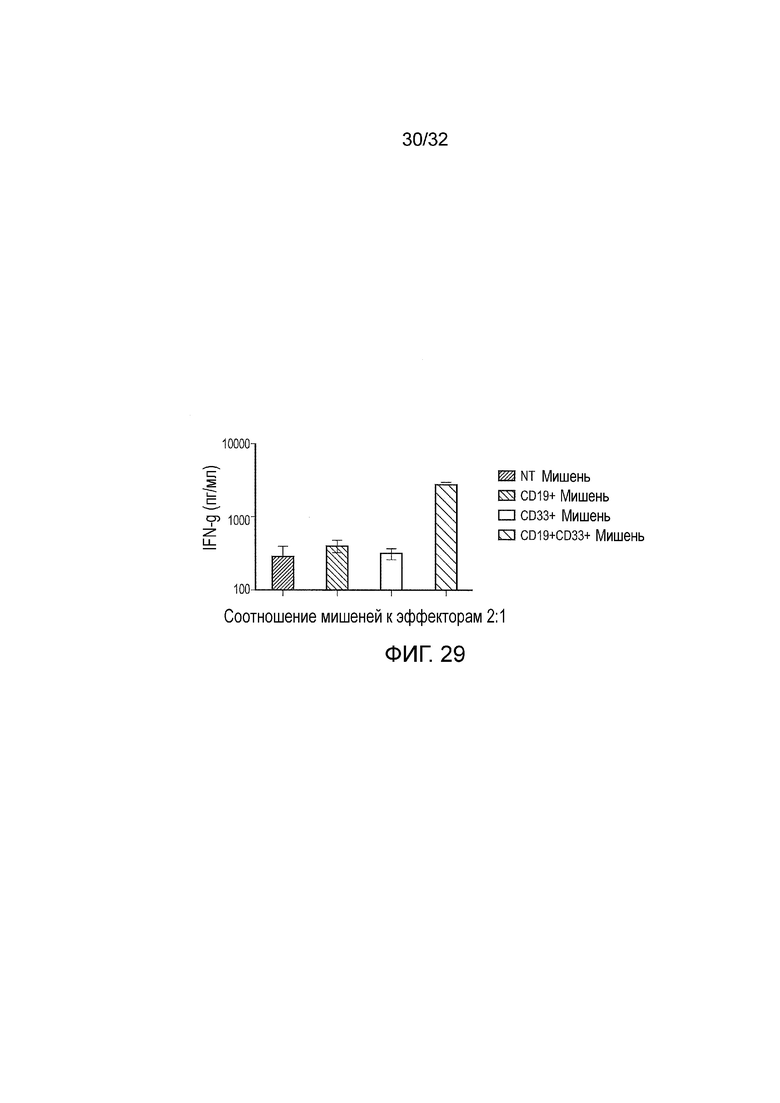
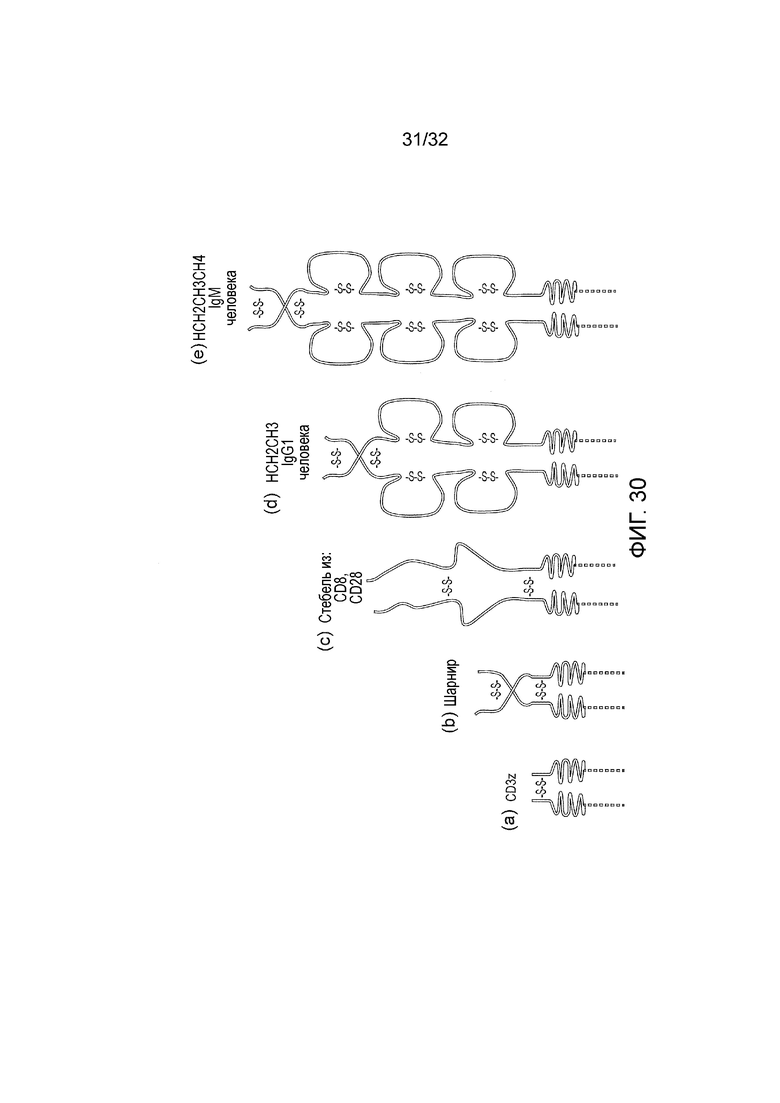
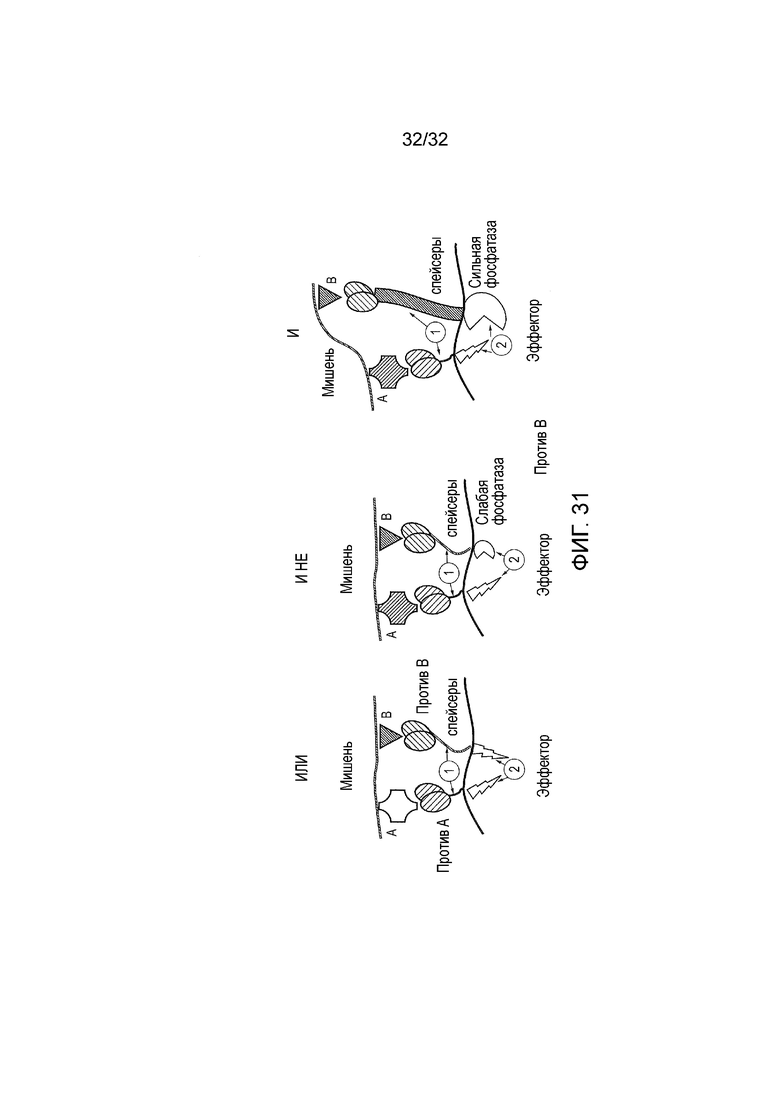
Настоящая группа изобретений относится к иммунологии и клеточной биологии. Предложены цитотоксическая Т-клетка и клетка естественного киллера (NK-клетка), содержащие по два химерных антигенных рецептора (CAR), один из которых содержит эндодомен CD3ξ, активирующий клетку в присутствии первого антигена, а второй - домен тирозиновой фосфатазы CD148 или CD45, ингибирующий клетку в отсутствии второго антигена. Такая система CAR позволяет клетке функционировать по принципу логического оператора "И", т.е. клетка активируется только в присутствии и первого, и второго антигенов. Дополнительно представлены: нуклеиновая кислота и наборы нуклеиновых кислот, кодирующих упомянутые CAR; вектор и способ получения цитотоксической T-клетки; а также фармацевтическая композиция и способ лечения и/или предотвращения заболевания путем уничтожения опухолевых клеток-мишеней, экспрессирующих первый антиген и второй антиген; применение фармацевтической композиции и T-клетки при изготовлении лекарственного средства и для лечения и/или предотвращения заболевания. 13 н. и 14 з.п. ф-лы, 31 ил., 6 табл., 14 пр.
1. Цитотоксическая T-клетка, способная уничтожить опухолевую клетку-мишень, экспрессирующую первый антиген и второй антиген, где Т-клетка совместно экспрессирует первый химерный антигенный рецептор (CAR) и второй CAR на клеточной поверхности, где каждый CAR содержит:
(i) антигенсвязывающий домен;
(ii) спейсер;
(iii) трансмембранный домен; и
(iv) эндодомен,
где спейсер первого CAR отличается от спейсера второго CAR, в которой первый CAR связывает первый антиген и представляет собой активирующий CAR, который содержит активирующий эндодомен, содержащий домен иммунорецепторного тирозинового активирующего мотива (ITAM), где активирующий эндодомен содержит эндодомен CD3ξ, и
где второй CAR связывает второй антиген и представляет собой ингибирующий CAR, который содержит ингибирующий эндодомен, где ингибирующий эндодомен содержит домен тирозиновой фосфатазы CD148 или CD45.
2. T-клетка по п.1, в которой спейсер первого CAR имеет длину, и/или заряд, и/или размер, и/или конфигурацию, и/или гликозилирование, отличающиеся от спейсера второго CAR, так что, когда первый CAR и второй CAR связывают соответствующие им антигены-мишени, первый CAR и второй CAR становятся пространственно разделенными на мембране T-клетки.
3. T-клетка по любому из пп. 1 или 2, в которой или первый спейсер, или второй спейсер содержит «стебель» CD8, и другой спейсер содержит шарнирную область, домен CH2 и домен CH3 из IgG1.
4. T-клетка по любому из пп. 1-3, в которой антигенсвязывающий домен первого CAR связывает CD5, и антигенсвязывающий домен второго CAR связывает CD19.
5. Нуклеиновая кислота, которая кодирует первый химерный антигенный рецептор (CAR) и второй CAR на клеточной поверхности, где каждый CAR содержит:
(i) антигенсвязывающий домен;
(ii) спейсер;
(iii) трансмембранный домен; и
(iv) эндодомен,
где спейсер первого CAR отличается от спейсера второго CAR, в которой первый CAR связывает первый антиген и представляет собой активирующий CAR, который содержит активирующий эндодомен, содержащий домен иммунорецепторного тирозинового активирующего мотива (ITAM), где активирующий эндодомен содержит эндодомен CD3ξ, и
где второй CAR связывает второй антиген и представляет собой ингибирующий CAR, который содержит ингибирующий эндодомен, где ингибирующий эндодомен содержит домен тирозиновой фосфатазы CD148 или CD45.
6. Нуклеиновая кислота по п. 5, в которой спейсер первого CAR имеет длину, и/или заряд, и/или размер, и/или конфигурацию, и/или гликозилирование, отличающиеся от спейсера второго CAR, так что, когда первый CAR и второй CAR связывают соответствующие им антигены-мишени, первый CAR и второй CAR становятся пространственно разделенными на мембране T-клетки.
7. Нуклеиновая кислота по любому из пп. 5 или 6, в которой или первый спейсер, или второй спейсер содержит «стебель» CD8, и другой спейсер содержит шарнирную область, CH2 домен и CH3 домен из IgG1.
8. Нуклеиновая кислота по любому из пп. 5-7, в которой антигенсвязывающий домен первого CAR связывает CD5, и антигенсвязывающий домен второго CAR связывает CD19.
9. Нуклеиновая кислота по любому из пп. 5-8, которая имеет следующую структуру:
AgB1-спейсер1-TM1-эндо1-коэкспр-AbB2-спейсер2-TM2-эндо2,
в которой
AgB1 представляет собой нуклеиновую кислоту, которая кодирует антигенсвязывающий домен первого CAR;
спейсер1 представляет собой нуклеиновую кислоту, которая кодирует спейсер первого CAR;
TM1 представляет собой нуклеиновую кислоту, которая кодирует трансмембранный домен первого CAR;
эндо1 представляет собой нуклеиновую кислоту, которая кодирует эндодомен первого CAR;
коэкспр представляет собой нуклеиновую кислоту, которая делает возможной коэкспрессию обоих CAR;
AgB2 представляет собой нуклеиновую кислоту, которая кодирует антигенсвязывающий домен второго CAR;
спейсер2 представляет собой нуклеиновую кислоту, которая кодирует спейсер второго CAR;
TM2 представляет собой нуклеиновую кислоту, которая кодирует трансмембранный домен второго CAR;
эндо2 представляет собой нуклеиновую кислоту, которая кодирует эндодомен второго CAR;
где нуклеиновая кислота, когда экспрессирована в T-клетке, кодирует полипептид, который расщепляется в сайте расщепления так, что первый и второй CAR совместно экспрессированы на поверхности T-клетки.
10. Нуклеиновая кислота по п. 9, в которой коэкспр кодирует последовательность, которая содержит саморасщепляющийся пептид.
11. Нуклеиновая кислота по любому из пп. 5-10, в которой в областях, кодирующих те же или схожие аминокислотные последовательности, во избежание гомологичной рекомбинации используют альтернативные кодоны.
12. Набор нуклеиновых кислот для получения клетки по любому из пп. 1-4, который содержит
(i) первую нуклеиновую кислоту, которая кодирует первый химерный антигенный рецептор (CAR), где нуклеиновая кислота имеет следующую структуру:
AgB1-спейсер1-TM1-эндо1, в которой
AgB1 представляет собой нуклеиновую кислоту, которая кодирует антигенсвязывающий домен первого CAR;
спейсер1 представляет собой нуклеиновую кислоту, которая кодирует спейсер первого CAR;
TM1 представляет собой нуклеиновую кислоту, которая кодирует трансмембранный домен первого CAR;
эндо1 представляет собой нуклеиновую кислоту, которая кодирует эндодомен первого CAR; и
(ii) вторую нуклеиновую кислоту, которая кодирует второй химерный антигенный рецептор (CAR), где указанная нуклеиновая кислота имеет следующую структуру:
AgB2-спейсер2-TM2-эндо2, где
AgB2 представляет собой нуклеиновую кислоту, которая кодирует антигенсвязывающий домен второго CAR;
спейсер2 представляет собой нуклеиновую кислоту, которая кодирует спейсер второго CAR;
TM2 представляет собой нуклеиновую кислоту, которая кодирует трансмембранный домен второго CAR;
эндо2 представляет собой нуклеиновую кислоту, которая кодирует эндодомен второго CAR, где спейсер первого CAR отличается от спейсера второго CAR, причём первый CAR связывает первый антиген и представляет собой активирующий CAR, который содержит активирующий эндодомен, содержащий домен иммунорецепторного тирозинового активирующего мотива (ITAM), где активирующий эндодомен содержит эндодомен CD3ξ, и
при этом второй CAR связывает второй антиген и представляет собой ингибирующий CAR, который содержит ингибирующий эндодомен, где ингибирующий эндодомен содержит домен тирозиновой фосфатазы CD148 или CD45.
13. Набор нуклеиновых кислот для получения клетки по любому из пп. 1-4, который содержит:
(i) первый вектор, который содержит первую нуклеиновую кислоту, кодирующую первый химерный антигенный рецептор (CAR), где нуклеиновая кислота имеет следующую структуру:
AgB1-спейсер1-TM1-эндо1, в которой
AgB1 представляет собой нуклеиновую кислоту, которая кодирует антигенсвязывающий домен первого CAR;
спейсер1 представляет собой нуклеиновую кислоту, которая кодирует спейсер первого CAR;
TM1 представляет собой нуклеиновую кислоту, которая кодирует трансмембранный домен первого CAR;
эндо1 представляет собой нуклеиновую кислоту, которая кодирует эндодомен первого CAR; и
(ii) второй вектор, который содержит вторую нуклеиновую кислоту, кодирующую второй химерный антигенный рецептор (CAR), где указанная нуклеиновая кислота имеет следующую структуру:
AgB2-спейсер2-TM2-эндо2, где
AgB2 представляет собой нуклеиновую кислоту, которая кодирует антигенсвязывающий домен второго CAR;
спейсер2 представляет собой нуклеиновую кислоту, которая кодирует спейсер второго CAR;
TM2 представляет собой нуклеиновую кислоту, которая кодирует трансмембранный домен второго CAR;
эндо2 представляет собой нуклеиновую кислоту, которая кодирует эндодомен второго CAR, где спейсер первого CAR отличается от спейсера второго CAR, причём первый CAR связывает первый антиген и представляет собой активирующий CAR, который содержит активирующий эндодомен, содержащий домен иммунорецепторного тирозинового активирующего мотива (ITAM), где активирующий эндодомен содержит эндодомен CD3ξ, и
при этом второй CAR связывает второй антиген и представляет собой ингибирующий CAR, который содержит ингибирующий эндодомен, где ингибирующий эндодомен содержит домен тирозиновой фосфатазы CD148 или CD45.
14. Набор по п. 13, в котором векторы представляют собой интегрирующие вирусные векторы или транспозоны.
15. Вектор для получения Т-клетки по любому из пп. 1-4, который содержит нуклеиновую кислоту по пп. 5-11.
16. Вектор для получения Т-клетки по п. 15, который представляет собой ретровирусный вектор или лентивирусный вектор или транспозон.
17. Способ получения цитотоксической T-клетки по любому из пп. 1-4, который включает стадию введения:
(а) первого вектора экспрессии и второго вектора экспрессии, где
(i) первый вектор экспрессии содержит первую нуклеиновую кислоту, кодирующую первый химерный антигенный рецептор (CAR), где нуклеиновая кислота имеет следующую структуру:
AgB1-спейсер1-TM1-эндо1, в которой
AgB1 представляет собой нуклеиновую кислоту, которая кодирует антигенсвязывающий домен первого CAR;
спейсер1 представляет собой нуклеиновую кислоту, которая кодирует спейсер первого CAR;
TM1 представляет собой нуклеиновую кислоту, которая кодирует трансмембранный домен первого CAR;
эндо1 представляет собой нуклеиновую кислоту, которая кодирует эндодомен первого CAR; и
(ii) второй вектор экспрессии содержит вторую нуклеиновую кислоту, кодирующую второй химерный антигенный рецептор (CAR), где указанная нуклеиновая кислота имеет следующую структуру:
AgB2-спейсер2-TM2-эндо2, где
AgB2 представляет собой нуклеиновую кислоту, которая кодирует антигенсвязывающий домен второго CAR;
спейсер2 представляет собой нуклеиновую кислоту, которая кодирует спейсер второго CAR;
TM2 представляет собой нуклеиновую кислоту, которая кодирует трансмембранный домен второго CAR;
эндо2 представляет собой нуклеиновую кислоту, которая кодирует эндодомен второго CAR, где спейсер первого CAR отличается от спейсера второго CAR, причём первый CAR связывает первый антиген и представляет собой активирующий CAR, который содержит активирующий эндодомен, содержащий домен иммунорецепторного тирозинового активирующего мотива (ITAM), где активирующий эндодомен содержит эндодомен CD3ξ, и
при этом второй CAR связывает второй антиген и представляет собой ингибирующий CAR, который содержит ингибирующий эндодомен, где ингибирующий эндодомен содержит домен тирозиновой фосфатазы CD148 или CD45; или
(b) вектора по любому из пп. 15 или 16 в цитотоксическую T-клетку.
18. Способ по п. 17, в котором указанная цитотоксическая T-клетка происходит из образца, выделенного у субъекта.
19. Фармацевтическая композиция для лечения заболевания путем уничтожения опухолевых клеток-мишеней, экспрессирующих первый антиген и второй антиген, которая содержит множество T-клеток по любому из пп. 1-4.
20. Способ лечения и/или предотвращения заболевания путем уничтожения опухолевых клеток-мишеней, экспрессирующих первый антиген и второй антиген, который включает стадию введения эффективного количества фармацевтической композиции по п.19 субъекту.
21. Способ по п.20, который включает следующие стадии:
(i) выделение содержащего T-клетки образца у субъекта;
(ii) трансдукция или трансфекция T-клеток с использованием:
(а) нуклеиновой кислоты по любому из пп. 5-11;
(b) первой нуклеиновой кислоты и второй нуклеиновой кислоты, где
i. первая нуклеиновая кислота кодирует первый химерный антигенный рецептор (CAR), где нуклеиновая кислота имеет следующую структуру:
AgB1-спейсер1-TM1-эндо1, в которой
AgB1 представляет собой нуклеиновую кислоту, которая кодирует антигенсвязывающий домен первого CAR;
спейсер1 представляет собой нуклеиновую кислоту, которая кодирует спейсер первого CAR;
TM1 представляет собой нуклеиновую кислоту, которая кодирует трансмембранный домен первого CAR;
эндо1 представляет собой нуклеиновую кислоту, которая кодирует эндодомен первого CAR; и
ii. вторая нуклеиновая кислота кодирует второй химерный антигенный рецептор (CAR), где указанная нуклеиновая кислота имеет следующую структуру:
AgB2-спейсер2-TM2-эндо2, где
AgB2 представляет собой нуклеиновую кислоту, которая кодирует антигенсвязывающий домен второго CAR;
спейсер2 представляет собой нуклеиновую кислоту, которая кодирует спейсер второго CAR;
TM2 представляет собой нуклеиновую кислоту, которая кодирует трансмембранный домен второго CAR;
эндо2 представляет собой нуклеиновую кислоту, которая кодирует эндодомен второго CAR, где спейсер первого CAR отличается от спейсера второго CAR, причём первый CAR связывает первый антиген и представляет собой активирующий CAR, который содержит активирующий эндодомен, содержащий домен иммунорецепторного тирозинового активирующего мотива (ITAM), где активирующий эндодомен содержит эндодомен CD3ξ, и
при этом второй CAR связывает второй антиген и представляет собой ингибирующий CAR, который содержит ингибирующий эндодомен, где ингибирующий эндодомен содержит домен тирозиновой фосфатазы CD148 или CD45;
(с) первого вектора и второго вектора, где
i. первый вектор содержит первую нуклеиновую кислоту, кодирующую первый химерный антигенный рецептор (CAR), где нуклеиновая кислота имеет следующую структуру:
AgB1-спейсер1-TM1-эндо1, в которой
AgB1 представляет собой нуклеиновую кислоту, которая кодирует антигенсвязывающий домен первого CAR;
спейсер1 представляет собой нуклеиновую кислоту, которая кодирует спейсер первого CAR;
TM1 представляет собой нуклеиновую кислоту, которая кодирует трансмембранный домен первого CAR;
эндо1 представляет собой нуклеиновую кислоту, которая кодирует эндодомен первого CAR; и
ii. второй вектор содержит вторую нуклеиновую кислоту, кодирующую второй химерный антигенный рецептор (CAR), где указанная нуклеиновая кислота имеет следующую структуру:
AgB2-спейсер2-TM2-эндо2, где
AgB2 представляет собой нуклеиновую кислоту, которая кодирует антигенсвязывающий домен второго CAR;
спейсер2 представляет собой нуклеиновую кислоту, которая кодирует спейсер второго CAR;
TM2 представляет собой нуклеиновую кислоту, которая кодирует трансмембранный домен второго CAR;
эндо2 представляет собой нуклеиновую кислоту, которая кодирует эндодомен второго CAR, где спейсер первого CAR отличается от спейсера второго CAR, причём первый CAR связывает первый антиген и представляет собой активирующий CAR, который содержит активирующий эндодомен, содержащий домен иммунорецепторного тирозинового активирующего мотива (ITAM), где активирующий эндодомен содержит эндодомен CD3ξ, и
при этом второй CAR связывает второй антиген и представляет собой ингибирующий CAR, который содержит ингибирующий эндодомен, где ингибирующий эндодомен содержит домен тирозиновой фосфатазы CD148 или CD45; или
(d) вектора по п.15 или 16; и
(iii) введение T-клеток из (ii) субъекту.
22. Способ по любому из пп. 20 или 21, в котором заболевание представляет собой злокачественную опухоль.
23. Применение фармацевтической композиции по п.19 для лечения и/или предотвращения заболевания путем уничтожения опухолевых клеток-мишеней, экспрессирующих первый антиген и второй антиген.
24. Применение T-клетки по любому из пп. 1-4 при изготовлении лекарственного средства для лечения и/или предотвращения заболевания путем уничтожения опухолевых клеток-мишеней, экспрессирующих первый антиген и второй антиген.
25. Применение T-клетки по любому из пп. 1-4 для лечения и/или предотвращения заболевания путем уничтожения опухолевых клеток-мишеней, экспрессирующих первый антиген и второй антиген.
26. Клетка - естественный киллер (NK-клетка), способная уничтожить опухолевую клетку-мишень, экспрессирующую первый антиген и второй антиген, где NK-клетка совместно экспрессирует первый CAR и второй CAR на клеточной поверхности, где каждый CAR содержит:
(i) антигенсвязывающий домен;
(ii) спейсер;
(iii) трансмембранный домен; и
(iv) эндодомен,
где спейсер первого CAR отличается от спейсера второго CAR, где первый CAR связывает первый антиген и представляет собой активирующий CAR, который содержит активирующий эндодомен, содержащий домен иммунорецепторного тирозинового активирующего мотива (ITAM), где активирующий эндодомен содержит эндодомен CD3ξ, и
где второй CAR связывает второй антиген и представляет собой ингибирующий CAR, который содержит ингибирующий эндодомен, где ингибирующий эндодомен содержит домен тирозиновой фосфатазы CD148 или CD45.
27. Композиция клеток для лечения заболевания путем уничтожения опухолевых клеток-мишеней, экспрессирующих первый антиген и второй антиген, которая содержит экспрессирующие CAR T-клетки по любому из пп. 1-4 и/или экспрессирующие CAR NK-клетки по п.26, полученные посредством трансдукции образца крови ex vivo с использованием нуклеиновой кислоты, кодирующей первый и второй CAR, по любому из пп. 5-11.
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| CHICAYBAM L ET AL, "P60: A conditional system for the activation of lymphocytes expressing activating and inhibitory CARs", HUMAN GENE THERAPY, 2010, 21(10): 1418 | |||
| FEDOROV V | |||
| et al., "Inhibitory chimeric antigen receptors (iCARs) limit undesirable side effects of T-cell therapies", Experimental Hematology | |||
| Устройство для усиления микрофонного тока с применением самоиндукции | 1920 |
|
SU42A1 |

