Область техники
Настоящее изобретение относится к области биотехнологии, а именно к двухвалентным биспецифическим химерным антителам, которые включают гетеродимер на основе околомембранных доменов МНС (главный комплекс гистосовместимости, ГКГС) или МНС-подобных белков (CD1 (кластер дифференцировки 1) или HFE (белок гемохроматоза)), а также к технологии получения данных биспецифических антител. Изобретение также относится к нуклеиновой кислоте, кодирующей данное биспецифическое антитело, вектору экспрессии, клетке-хозяину для получения данного двухвалентного химерного биспецифического антитела и способу получения данной клетки.
Предшествующий уровень техники
Моноклональные антитела в виде химерных, гуманизированных или полностью человеческих молекул доказали свою ценность в качестве эффективных лекарств для лечения ряда нарушений и заболеваний.
Природные молекулы антител человека состоят из двух гомодимеров тяжелых цепей, каждый из которых образует гетеродимер в партнерстве с двумя идентичными молекулами легкой цепи. Обычные моноклональные антитела в виде целых молекул состоят из двухвалентных («двухруких») гетеродимеров тяжелых и легких цепей.
Часто заболевания возникают в результате нескольких патологий и сопровождаются множеством сопутствующих заболеваний. Биспецифичные антитела обладают способностью связывать и тем самым нейтрализовать два или более отличных друг от друга антигена на молекулу антитела. Потенциал значительного улучшения терапевтических свойств (и ценности) лекарственных средств по сравнению с моноклональными антителами сделал биспецифические антитела активной областью исследований. За последние двадцать лет в научной литературе было описано множество решений в отношении сконструированных версий биспецифических антител, как описано в Brinfcmann, U and RE Kontermann, 2017, The Making of Bispecific Antibodies, MAbs; 209 Feb/Mar; 9(2): 182-212, doi: 10.1080/19420862.2016.1268307.
Как было указано выше, существует много путей создания молекул со смешанными антигенсвязывающими доменами, то есть с антигенсвязывающими доменами, отличными друг от друга. Но каждый из указанных методов характеризуется своими недостатками.
Перекрестное сшивание химическими методами является трудоемким процессом, так как соответствующие части необходимо очистить от гомодимеров и других нежелательных побочных продуктов. Кроме того, стадии химической модификации могут изменить целостность белков, что ухудшает их устойчивость. Таким образом, указанный метод часто оказывается неэффективным и может привести к утрате активности антитела.
Метод, основанный на слияние клеток (например, получение гибридом), представляет собой произвольную сборку двух тяжелых и двух легких цепей, в результате чего образуется 10 комбинаций антител. Целевые гетеромультимерные антитела составляют небольшую часть полученных таким образом антител. Выделение целевых гетеромультимерных белков существенно уменьшает выход продукта и увеличивает производственные затраты.
Методы рекомбинантных ДНК используют для создания разных гетеромультимерных антител, например, одноцепочечных Fv фрагментов, диател и т.д., не содержащих Fc фрагмент.Основным недостатком молекулы антитела такого типа является отсутствие Fc домена и, следовательно, неспособность такого антитела запускать эффекторную функцию (такую как, например, активация комплемента, связывание с Fc-рецептором и т.д.). Таким образом, необходимо биспецифическое антитело, содержащее функциональный Fc домен.
Методы рекомбинантных ДНК также используют для создания биспецифических антител, соединяемых по типу «выступ во впадину» (Knob-into-Holes). См. международные заявки WO 9627011 и WO 9850431, а также статью Merchant AM ЕТ ALL., An efficient route to human bispecific IgG, Nat Biotechnol. 1998 Jul; 16(7):677-81. Одним фактором, ограничивающим применение данного метода, является то, что легкие цепи двух исходных антител должны быть идентичными для предотвращения ошибочного спаривания и образования нежелательных и/или неактивных молекул при экспрессии в одной клетке.
Чистота продукта биспецифических антител зависит от двух факторов:
a) гетеродимерной сборки двух разных тяжелых цепей, коэкспрессируемых в клетке, и
b) корректного спаривания двух разных легких цепей с соответствующими тяжелыми цепями.
Использование технологии «выступ во впадину» (Knob-into-Holes) для создания биспецифических антител позволяет решить проблему правильной гетеродимерной сборки двух разных тяжелых цепей, коэкспрессируемых в клетке. Однако использование только Knob-into-Holes для создания биспецифических антител позволяет достигнуть выхода правильно собранного биспецифика только около 25%, так как остается не решенной проблема правильного спаривания двух разных легких цепей с соответствующими тяжелыми цепями.
Проблема правильного спаривания двух разных легких цепей с соответствующими тяжелыми цепями решается различными способами:
1. Использование одинаковой легкой цепи в первой и второй антигенсвязывающих частях антитела (Van Blarcom Т ЕТ AL., Productive common light chain libraries yield diverse panels of high affinity bispecific antibodies, MAbs. 2018 Feb/Mar; 10(2):256-268. doi: 10.1080/19420862.2017.1406570).
Недостатком данного решения является его не универсальность, так как не всегда удается подобрать подходящую для обеих валентностей легкую цепь. В дополнении к этому, при замене аминокислот в легкой цепи для оптимизации свойств антигенсвязывающего фрагмента, замены затронут обе валентности. При этом связывание антитела ко второму антигену может быть нарушено.
2. Использование single-chain (одноцепочечных) форматов, то есть форматов, где легкая и тяжелая цепи антигенсвязывающего фрагмента, специфичного к первому антигену, соединены между собой через линкер из нескольких аминокислот.
У данного формата есть технологические недостатки, так как в нем используются линкеры либо для слияния основной структуры антитела (IgA, IgD, IgE, IgG или IgM) с дополнительным связывающим белком (например, scFv или scFab), либо для слияния, например, легкого и тяжелого вариабельных доменов (VH и VL) в составе scFv или легкой цепи (VL-CK(или CL)) с VH-CH1 в составе scFab. Линкеры могут представлять проблемы терапевтического плана. Фактически эти чужеродные пептиды могут вызывать иммунный ответ против самого линкера или области стыка между белком и линкером. Кроме того, гибкая природа этих пептидов и их подвижность делает их более чувствительными к протеолитическому расщеплению, что может приводить к плохой стабильности, агрегации и повышению иммуногенности антитела.
3. Модификация СН1-CK доменов в биспецифическом антителе, что позволит в технологиях экспрессии биспецифических антител изменять интерфейс взаимодействия таким образом, чтобы исключить неправильную ассоциацию легких цепей. Например, в международной заявке WO 2017059551 указываются различные аминокислотные замены в СН1 и/или CK, которые способствуют предпочтительному спариванию нужной тяжелой цепи с нужной легкой цепью.
Несмотря на вышеуказанные различные технологии экспрессии биспецифических антител, в данной области техники, по-прежнему, существует потребность в улучшенной чистоте продукта биспецифических антител, а также в масштабируемом производственном решении для получения правильно собранных биспецифических антител.
Краткое описание изобретения
Разработанный авторами данного изобретения новый формат биспецифических химерных антител, которые включают гетеродимер на основе околомембранных доменов МНС или МНС-подобных белков, например, CD1 (cluster of differentiation 1, кластер дифференцировки 1) или HFE (Human homeostatic iron regulator protein), а также технология получения данных биспецифических химерных антител, неожиданно позволяют получить с высоким выходом продукт с правильной гетеродимерной сборкой двух разных тяжелых цепей, коэкспрессируемых в клетке, и правильным спариванием двух разных легких цепей с соответствующими тяжелыми цепями.
Разработанный авторами данного изобретения новый формат биспецифических химерных антител, которые включают гетеродимер на основе околомембранных доменов МНС или МНС-подобных белков, а также технология получения данных биспецифических антител, неожиданно позволяют получить правильно собранный продукт биспецифических химерных антител с высокой чистотой.
Вышеуказанные результаты, как следствие, уменьшают производственные затраты и приводят к масштабируемому производственному решению для получения правильно собранных биспецифических антител.
В одном из аспектов настоящее изобретение относится к двухвалентному биспецифическому химерному антителу, где указанное антитело содержит:
а) первую легкую цепь и первую тяжелую цепь антитела, специфически связывающегося с первым антигеном;
где первая легкая цепь содержит вариабельный домен легкой цепи и константный домен легкой цепи;
где первая тяжелая цепь содержит вариабельный домен тяжелой цепи и константные домены тяжелой цепи антитела, включающие первый (СН1) константный домен тяжелой цепи и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи;
б) вторую легкую цепь и вторую тяжелую цепь антитела, специфически связывающегося со вторым антигеном,
где вторая легкая цепь содержит вариабельный домен легкой цепи и константный домен, который выбирают из группы:
первого околомембранного домена МНС (главный комплекс гистосовместимости) или
первого околомембранного домена МНС-подобного белка;
где вторая тяжелая цепь, содержит вариабельный домен тяжелой цепи, константный домен, который выбирают из группы:
второго околомембранного домена МНС (главный комплекс гистосовместимости) или
второго околомембранного домена МНС-подобного белка;
и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи;
при этом первый околомембранный домен МНС или МНС-подобного белка и второй околомембранный домен МНС или МНС-подобного белка образуют между собой гетеродимер, который стабилизирован дисульфидной связью;
где СН3-домен одной тяжелой цепи и СН3-домен другой тяжелой цепи соприкасаются друг с другом поверхностями, которые изменены для формирования двухвалентного биспецифического химерного антитела, причем данные изменения в СН3 доменах тяжелых цепей представляют собой замены для обеспечения гетеродимеризации.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС или МНС-подобного белка и второй околомембранный домен МНС или МНС-подобного белка, которые образуют между собой гетеродимер, который стабилизирован дисульфидной связью за счет мутации или мутаций в первом и/или втором константном домене для образования S-S связи (дисульфидной связи, цистеинового мостика) между первым и вторым околомембранными доменами МНС или МНС-подобного белка.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС или МНС-подобного белка и второй околомембранный домен МНС или МНС-подобного белка, которые образуют между собой гетеродимер, который стабилизирован дисульфидной связью за счет удлинения первого околомембранного домена МНС или МНС-подобного белка на одну или несколько (от 1 до 10) аминокислот на С-конце и с терминальным Cys на С-конце для образования S-S связи (дисульфидной связи, цистеинового мостика) между первым околомембранным доменом МНС или МНС-подобного белка и шарниром (hinge).
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает удлинение первого околомембранного домена МНС или МНС-подобного белка, которое представляет собой последовательность из трех аминокислот GSC.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает:
первую легкую цепь, которая содержит вариабельный домен легкой цепи (VL1) и константный домен легкой цепи;
первую тяжелую цепь, которая содержит вариабельный домен тяжелой цепи (VH1) и константные домены тяжелой цепи антитела, включающие первый (СН1) константный домен тяжелой цепи и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи;
вторую легкую цепь, которая содержит вариабельный домен легкой цепи (VL2) и первый околомембранный домен МНС или первый околомембранный домен МНС-подобного белка;
вторую тяжелую цепь, которая содержит вариабельный домен тяжелой цепи (VH2), второй околомембранный домен МНС или второй околомембранный домен МНС-подобного белка и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи;
при этом первый околомембранный домен МНС или МНС-подобного белка и второй околомембранный домен МНС или МНС-подобного белка образуют между собой гетеродимер, который стабилизирован дисульфидной связью за счет мутации или мутаций в первом и/или втором константном домене для образования S-S связи (дисульфидной связи, цистеинового мостика) между первым и вторым околомембранными доменами МНС или МНС-подобного белка;
где между первым вариантом Fc и вторым вариантом Fc в СН3 домене образована структура «выступ (Knob) во впадину (Hole)».
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает:
первую легкую цепь, которая содержит вариабельный домен легкой цепи (VL1) и константный домен легкой цепи;
первую тяжелую цепь, которая содержит вариабельный домен тяжелой цепи (VH1) и константные домены тяжелой цепи антитела, включающие первый (СН1) константный домен тяжелой цепи и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи;
вторую легкую цепь, которая содержит вариабельный домен легкой цепи (VL2) и первый околомембранный домен МНС или первый околомембранный домен МНС-подобного белка;
вторую тяжелую цепь, которая содержит вариабельный домен тяжелой цепи (VH2), второй околомембранный домен МНС или второй околомембранный домен МНС-подобного белка и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи;
где между первым вариантом Fc и вторым вариантом Fc в СН3 домене образована структура «выступ (Knob) во впадину (Hole)»
где первый околомембранный домен МНС или первый околомембранный домен МНС-подобного белка дополнительно содержит удлинение первого околомембранного домена МНС или МНС-подобного белка на одну или несколько (от 1 до 10) аминокислот на С-конце с терминальным Cys на С-конце для образования S-S связи (дисульфидной связи, цистеинового мостика) между первым околомембранным доменом МНС или МНС-подобного белка и шарниром (hinge).
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает:
первую легкую цепь, которая содержит вариабельный домен легкой цепи (VL1) и константный домен легкой цепи;
первую тяжелую цепь, которая содержит вариабельный домен тяжелой цепи (VH1) и константные домены тяжелой цепи антитела, включающие первый (СН1) константный домен тяжелой цепи и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи;
вторую легкую цепь, которая содержит вариабельный домен легкой цепи (VL2) и первый околомембранный домен МНС или первый околомембранный домен МНС-подобного белка;
вторую тяжелую цепь, которая содержит вариабельный домен тяжелой цепи (VH2), второй околомембранный домен МНС или второй околомембранный домен МНС-подобного белка и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи;
где между первым вариантом Fc и вторым вариантом Fc в СН3 домене образована структура «выступ (Knob) во впадину (Hole)»
где первый околомембранный домен МНС или первый околомембранный домен МНС-подобного белка дополнительно содержит удлинение первого околомембранного домена МНС или МНС-подобного белка на последовательность аминокислот GSC для образования S-S связи (дисульфидной связи, цистеинового мостика) между первым околомембранным доменом МНС или МНС-подобного белка и шарниром (hinge).
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС, который может быть выбран из группы, включающей:
первый околомембранный домен МНС класса I (главный комплекс гистосовместимости класса I),
первый околомембранный домен МНС класса II (главный комплекс гистосовместимости класса II),
модифицированный вариант первого околомембранного домена МНС класса I или
модифицированный вариант первого околомембранного домена МНС класса II.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает второй околомембранный домен МНС, который может быть выбран из группы, включающей:
второй околомембранный домен МНС класса I (главный комплекс гистосовместимости класса I),
второй околомембранный домен МНС класса II (главный комплекс гистосовместимости класса II),
модифицированный вариант второго околомембранного домена МНС класса I или
модифицированный вариант второго околомембранного домена МНС класса II.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает околомембранный домен МНС класса I человека.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает околомембранный домен МНС класса II человека.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает околомембранный домен МНС класса II, который выбирают из группы: HLA-DM, HLA-DO, HLA-DP, HLA-DQ илиНЬА-DR.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает околомембранный домен МНС класса I, который выбирают из группы: HLA-A, HLA-B, HLA-C, HLA-E, HLA-F или HLA-G.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает околомембранный домен МНС-подобного белка человека.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС-подобного белка, который может быть выбран из группы, включающей:
первый околомембранный домен CD1 (кластер дифференцировки 1),
первый околомембранный домена HFE (белок гемохроматоза),
модифицированный вариант первого околомембранного домена CD1 или
модифицированный вариант первого околомембранного домена HFE.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает второй околомембранный домен МНС-подобного белка, который может быть выбран из группы, включающей:
второй околомембранный домен CD1 (кластер дифференцировки 1),
второй околомембранный домена HFE (белок гемохроматоза),
модифицированный вариант второго околомембранного домена CD1 или
модифицированный вариант второго околомембранного домена HFE.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает околомембранный домен CD1, который выбирают из группы: CD1a, CD1b, CD1c, CD1d или CD1e.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает вариабельный фрагмент второй легкой цепи (VL), который отделен от первого околомембранного домена МНС или МНС-подобного белка линкером длиной от 1 до 25 аминокислот, и/или вариабельный фрагмент второй тяжелой цепи (VH), который отделен от второго околомембранного домена МНС или МНС-подобного белка линкером длиной от 1 до 25 аминокислот.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает:
а) СН3-домен одной тяжелой цепи, который изменен так, что на поверхности СН3-домена одной тяжелой цепи, которая соприкасается с поверхностью СН3-домена второй тяжелой цепи в двухвалентном биспецифическом антителе, аминокислотный остаток заменен на аминокислотный остаток, который имеет большую по объему боковую цепь, что приводит к созданию выпуклости на поверхности СН3-домена одной тяжелой цепи, которая может помещаться в полость на поверхности СН3-домена другой тяжелой цепи, и
б) СН3-домен другой тяжелой цепи, который изменен так, что на поверхности СН3-домена второй тяжелой цепи, которая соприкасается с поверхностью СН3-домена первой тяжелой в двухвалентном биспепифическом антителе, аминокислотный остаток заменен на аминокислотный остаток, который имеет меньшую по объему боковую цепь, что приводит к созданию полости на поверхности СН3-домена второй тяжелой цепи, в которую может помещаться выпуклость на поверхности раздела СН3-домена первой тяжелой цепи;
где указанный аминокислотный остаток, который имеет большую по объему боковую цепь, выбран из группы, включающей аргинин (R), фенилаланин (F), тирозин (Y), триптофан (W),
и где указанный аминокислотный остаток, который имеет меньшую по объему боковую цепь, выбран из группы, включающей аланин (А), серии (S), треонин (Т), валин (V).
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает константный домен первой легкой цепи антитела, который выбирают из CK или CL.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает СН3-домены антитела, которые дополнительно изменены путем интродукции цистеина (С) в качестве аминокислоты в соответствующие положения каждого СН3-домена, так что между СН3-доменами может образоваться дисульфидный мостик.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает СН3-домен одной тяжелой цепи, который изменен с образованием Knob, и СН3-домен другой тяжелой цепи, который изменен с образованием Hole, или наоборот.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает СН3-домен одной тяжелой цепи, который имеет аминокислотные замены S354C/T366W, и СН3-домен другой тяжелой цепи, который имеет аминокислотные замены Y349C/T366S/L368A/Y407V.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает СН3-домен одной тяжелой цепи, который имеет аминокислотные замены Y349C/T366S/L368A/Y407, и СН3-домен другой тяжелой цепи, который имеет аминокислотные замены S354C/T366W.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС и второй околомембранный домен МНС, которые, соответственно, представляют собой α2 домен МНС II и β2 домен МНС II, и образуют между собой гетеродимер.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС и второй околомембранный домен МНС, которые, соответственно, представляют собой β2 домен МНС II и α2 домен МНС II, и образуют между собой гетеродимер.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает α2 домен МНС II, который имеет аминокислотную последовательность, которую выбирают из группы: SEQ ID NO: 30, SEQ ID NO: 32, SEQ ID NO: 34, SEQ ID NO: 36 или SEQ ID NO: 38, и p2 домен МНС II, который имеет аминокислотную последовательность, которую выбирают из группы: SEQ ID NO: 31, SEQ ID NO: 33, SEQ ID NO: 35, SEQ ID NO: 37 или SEQ ID NO: 39.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС и второй околомембранный домен МНС, которые, соответственно, представляют собой модифицированный вариант α2 домена МНС II и модифицированный вариант β2 домена МНС II, и образуют между собой гетеродимер.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС и второй околомембранный домен МНС, которые, соответственно, представляют собой модифицированный вариант β2 домена МНС II и модифицированный вариант α2 домена МНС II, и образуют между собой гетеродимер.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС и второй околомембранный домен МНС, которые, соответственно, представляют собой α3 домен МНС I и β2 микроглобулин (β2М), и образуют между собой гетеродимер.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС и второй околомембранный домен МНС, которые, соответственно, представляют собой β2 микроглобулин (β2М) и α3 домен МНС I, и образуют между собой гетеродимер.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает α3 домен МНС I, который имеет аминокислотную последовательность, которую выбирают из группы SEQ ID NO: 1-29, и β2 микроглобулин (β2М), который имеет аминокислотную последовательность SEQ ID NO: 46.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС и второй околомембранный домен МНС, которые, соответственно, представляют собой модифицированный вариант α3 домена МНС I и модифицированный вариант β2 микроглобулина (β2М), и образуют между собой гетеродимер.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС и второй околомембранный домен МНС, которые, соответственно, представляют собой модифицированный вариант β2 микроглобулина (β2М) и модифицированный вариант α3 домена МНС I, и образуют между собой гетеродимер.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает модифицированный вариант β2 микроглобулина (β2М), который имеет аминокислотную последовательность, которую выбирают из SEQ ID NO: 47 или SEQ ID NO: 48.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС-подобного белка и второй околомембранный домен МНС-подобного белка, которые, соответственно, представляют собой α3 домен CD1 и β2 микроглобулин (β2М), и образуют между собой гетеродимер.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС-подобного белка и второй околомембранный домен МНС-подобного белка, которые, соответственно, представляют собой β2 микроглобулин (β2М) и α3 домен CD1, и образуют между собой гетеродимер.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает α3 домен CD1, который имеет аминокислотную последовательность, которую выбирают из группы SEQ ID NO: 40-44, и β2 микроглобулин (β2М), который имеет аминокислотную последовательность SEQ ID NO: 46.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС-подобного белка и второй околомембранный домен МНС-подобного белка, которые, соответственно, представляют собой модифицированный вариант α3 домена CD1 и модифицированный вариант β2 микроглобулина (β2М), и образуют между собой гетеродимер.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС-подобного белка и второй околомембранный домен МНС-подобного белка, которые, соответственно, представляют собой модифицированный вариант β2 микроглобулина (β2М) и модифицированный вариант α3 домена CD1, и образуют между собой гетеродимер.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает модифицированный вариант α3 домена CD1, который имеет аминокислотную последовательность, которую выбирают из группы SEQ ID NO: 49-56 и или SEQ ID NO: 109, и модифицированный вариант β2 микроглобулина (β2М), который имеет аминокислотную последовательность, которую выбирают из SEQ ID NO: 47 или SEQ ID NO: 48.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС-подобного белка и второй околомембранный домен МНС-подобного белка, которые, соответственно, представляют собой α3 домен HFE и β2 микроглобулин (β2М), и образуют между собой гетеродимер.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС-подобного белка и второй околомембранный домен МНС-подобного белка, которые, соответственно, представляют собой β2 микроглобулин (β2М) и α3 домен HFE, и образуют между собой гетеродимер.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает α3 домен HFE, который имеет аминокислотную последовательность SEQ ID NO: 45, и β2 микроглобулин (β2М), который имеет аминокислотную последовательность SEQ ID NO: 46.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС-подобного белка и второй околомембранный домен МНС-подобного белка, которые, соответственно, представляют собой модифицированный вариант α3 домена HFE и модифицированный вариант β2 микроглобулина (β2М), и образуют между собой гетеродимер.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС-подобного белка и второй околомембранный домен МНС-подобного белка, которые, соответственно, представляют собой модифицированный вариант β2 микроглобулина (β2М) и модифицированный вариант α3 домена HFE, и образуют между собой гетеродимер.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает модифицированный вариант β2 микроглобулина (β2М), который имеет аминокислотную последовательность, которую выбирают из SEQ ID NO: 47 или SEQ ID NO: 48.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает модифицированный вариант МНС или МНС-подобного белка, где под модифицированным вариантом подразумевается вариант, включающий замены на цистеин (С) для образования дисульфидного мостика между цепями гетеродимера, полученного из первого и второго околомембранных доменов МНС или МНС-подобного белка.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает модифицированный вариант МНС или МНС-подобного белка, где под модифицированным вариантом подразумевается вариант, включающий одну или более замен в различных позициях околомембранных доменов МНС или МНС-подобного белка, приводящие к увеличению термодинамической стабильности Tm более чем на 1 градус Цельсия в сравнении с дикой формой околомембранных доменов МНС или МНС-подобного белка, соответственно.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает модифицированный вариант МНС или МНС-подобного белка, где под модифицированным вариантом подразумевается вариант, включающий одну или более замен в различных позициях околомембранных доменов МНС или МНС-подобного белка, приводящие к уменьшению количества агрегатов на более чем 5% при концентрации выше 10 мг/ мл в сравнении с дикой формой околомембранных доменов МНС или МНС-подобного белка, соответственно.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает модифицированный вариант МНС или МНС-подобного белка, где под модифицированным вариантом подразумевается вариант, включающий одну или более замен в различных позициях околомембранных доменов МНС или МНС-подобного белка, приводящие к удалению сайтов гликозилирования в сравнении с дикой формой околомембранных доменов МНС или МНС-подобного белка, соответственно.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает вариабельный домен первой легкой цепи и вариабельный домен второй легкой цепи, которые идентичны.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает Fc-фрагмент, который относится к IgG.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает Fc-фрагмент, который выбирают из группы: IgG1, IgG2, или IgG4 человека.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает мономер Fc-фрагмента, где дополнительно вводят замены, приводящие к отсутствию ADCC, CDC и/или ADCP свойств у двухвалентного биспецифического антитела.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает мономер Fc-фрагмента, где дополнительно вводят замены LALA (L234A и L235A).
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает мономер Fc-фрагмента, где дополнительно вводят замены, приводящие к пролонгированному действию антитела.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает мономер Fc-фрагмента, где дополнительно вводят замены YTE (M252Y, S254T и Т256Е).
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает мономер Fc-фрагмента, где дополнительно вводят замены, приводящие к усилению ADCC, CDC и/или ADCP свойств у двухвалентного биспецифического антитела.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает мономер Fc-фрагмента, где дополнительно вводят замену E345R.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело специфически связывается с CD20 и CD3.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело специфически связывается с ВСМА и CD3.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело специфически связывается с PD-L1 и CD47.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело специфически связывается с фактором свертывания крови 9 (FIX) и фактором свертывания крови 10 (FX).
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело специфически связывается с GD2 и CD3.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело специфически связывается с AXL и CD3.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело специфически связывается с PD-L1 и TGF beta.
В одном из аспектов настоящее изобретение относится к выделенной нуклеиновой кислоте, которая кодирует любое из вышеуказанных двухвалентных биспецифических химерных антител.
В некоторых вариантах осуществления изобретения нуклеиновая кислота представляет собой ДНК.
В одном из аспектов настоящее изобретение относится к экспрессионному вектору, содержащему любую из вышеуказанных нуклеиновых кислот.
В одном из аспектов настоящее изобретение относится к способу получения клетки-хозяина для получения любого из вышеуказанных двухвалентных биспецифических химерных антител и включает трансформирование клетки вышеуказанным экспрессионным вектором.
В одном из аспектов настоящее изобретение относится к клетке-хозяину для получения любого из вышеуказанных двухвалентных биспецифических химерных антител, которая содержит любую из вышеуказанных нуклеиновых кислот.
В одном из аспектов настоящее изобретение относится к способу получения любого из вышеуказанных двухвалентных биспецифических химерных антител, который включает следующие этапы:
а) трансформация клетки-хозяина
- экспрессионными векторами, которые содержат молекулы нуклеиновых кислот, кодирующие первую легкую цепь и первую тяжелую цепь биспецифического химерного антитела,
- экспрессионными векторами, которые содержат молекулы нуклеиновых кислот, кодирующие вторую легкую цепь и вторую тяжелую цепь биспецифического химерного антитела,
б) культивирование клетки-хозяина в условиях, которые позволяют синтезировать указанное двухвалентное биспецифическое химерное антитело; и
в) выделение указанного двухвалентного биспецифического антитела из культуры клеток.
Краткое описание чертежей
Фигура 1 представляет собой схематическое изображение формата двухвалентного биспецифического химерного антитела.
А - «прямая» ориентация димеризующего блока, Б - «обратная» ориентация димеризующего блока
VH1, VL1 - вариабельные домены тяжелой и легкой цепи, соответственно, отвечающие за связывание с антигеном 1;
VH2, VL2 - вариабельные домены, отвечающие за связывание с антигеном 2.
CH1, СК - константные домены тяжелой и легкой цепи, соответственно;
b2M - β2-микроглобулин,
CD1b - α3 домен белка CD1b.
Knob, hole - мутации в СН3-домене антитела, обеспечивающие гетеродимеризацию тяжелых цепей.
Фигура 2 представляет собой схематичное изображение комплекса тяжелой и легкой цепей химерного антитела с заменой доменов СН1 и CK на β2-микроглобулин и околомембранный домен α3 белка CD1b.
А - «прямая» ориентация доменов димеризующего блока;
Б - «обратная» ориентация димеризующих блоков.
VH, VL - вариабельные домены антитела,
b2M - β2-микроглобулин,
CD1b - околомембранный α3 домен белка CD1b,
hinge - шарнирная область антитела, первые 5 аминокислот показаны в явном виде,
Fc - Fc-фрагмент антитела;
аминокислоты с С-конца доменов b2M, α3 CD1b и с N-конца шарнирного региона показаны в явном виде;
SS-связь показана пунктиром.
Фигура 3 представляет собой электрофореграмму химерных антител, содержащих и не содержащих дополнительные дисульфидные связи между тяжелой и легкой цепями, 7,5% ПААГ, нередуцирующие условия.
Дорожки:
М - стандартный маркер Precision Plus Protein™ Dual Color Standards (BIO-RAD), молекулярные массы соответствующих полос указаны в кДа в столбце слева от геля;
1 - Контроль Prolgolimab - IgG1;
2 - Моноспецифическое химерное антитело на основе вариабельных доменов Prolgolimab и доменов МНС-подобных белков, заменяющих СН1-CK, с дополнительным дисульфидным мостиком;
3 - Моноспецифическое химерное антитело на основе вариабельных доменов Prolgolimab и доменов МНС-подобных белков, заменяющих СН1-CK, без дополнительного дисульфидного мостика;
4 - Моноспецифическое химерное антитело на основе вариабельных доменов Prolgolimab и доменов МНС-подобных белков, заменяющих СН1-CK, в «обратной» ориентации с дополнительным дисульфидным мостиком.
Фигура 4 представляет собой схему эксперимента по проверке влияния МНС-димеризующего блока на некорректное спариванию тяжелых и легких цепей. Овалами обозначены домены, заменяющие СН1 и СК, взятые из МНС-подобных белков.
Группа 1: образцы 01-001 - 01-003 - антитела с «корректным» сочетанием VH и VL и «некорректной» парой константных доменов.
Группа 2: образцы 01-004 - 01-007 - антитела с «некорректным» сочетанием VH и VL и «корректной» парой константных доменов.
Группа 3: образцы 01-008 - 01-010 - антитела с «некорректным» сочетанием VH и VL и «некорректной» парой константных доменов.
Группа 4: образцы 01-011 и 01-012- контрольные антитела, представляющие собой химерное антитело, в котором константные домены заменены на МНС-димеризующие блоки, и классическое антитело формата IgG1, соответственно.
Под «корректным» сочетанием VH и VL подразумевается пара VH и VF, взятая из одного антитела. Под «некорректным» сочетанием VH и VL подразумеваются VH и VF, взятые из разных антител.
Под «корректной» парой константных доменов подразумевается либо пара СН1-CK либо пара доменов из МНС-подобных белков (в данном случае β2-микроглобулин и α3-домен белка CD1b). Под «некорректной» парой константных доменов подразумеваются пары CH1-CD1b или β2-микроглобулин-CK.
Фигура 5 представляет собой электрофореграмму образцов, полученных в эксперименте по неправильному спариванию цепей. 7,5% ПААГ, нередуцирующие условия.
А - Дорожки:
М - стандартный маркер Precision Plus Protein™ Dual Color Standards (BIO-RAD), молекулярные массы соответствующих полос указаны в кДа в столбце слева от геля;
1 - 01-001;
2 - 01-002;
3 - 01-007;
4 - 01-008;
5 - 01-009;
6 - 01-012;
Б - Дорожки:
М - стандартный маркер Precision Plus Protein™ Dual Color Standards (BIO-RAD);
1 - 01-003;
2 - 01-004;
3 - 01-005;
4 - 01-006;
5 - 01-007,
6 - 01-012;
В - Дорожки:
М - стандартный маркер Precision Plus Protein™ Dual Color Standards (BIO-RAD);
1 - 01-010;
2 - 01-011.
Фигура 6 представляет собой электрофореграмму биспецифичеких химерных антител.
А - 7,5% ПААГ, нередуцирующие условия,
Б - 12.5% ПААГ, редуцирующие условия.
Дорожки:
М - стандартный маркер Precision Plus Protein™ Dual Color Standards (BIO-RAD), молекулярные массы соответствующих полос указаны в кДа в столбце слева от геля;
1 - 02-004,
2 - 02-005,
3 - 02-006,
4 - 02-007,
5 - 02-008,
6 - 02-009.
Фигура 7 представляет собой сенсограмму эксперимента по одновременному взаимодействию антител 02-006 и 02-007 с двумя различными антигенами (hPDlex-H6F и hCSF1R_His). Этапы (шаги эксперимента) пронумерованы в верхней части изображения и разделены вертикальными линиями. Для каждого антитела приведены 2 сенсограммы - взаимодействие с hCSF1R_His на 8 шаге (наблюдается увеличение уровня сигнала) и референсный сигнал в буфере 1хКБ без hCSF1R_His на 8 шаге (нет увеличения уровня сигнала).
Фигура 8 представляет собой деконволюированный масс-спектр хроматограммы полного ионного тока для образца 02-004.
А - пик 4,
Б - пик 5,
В - пик 6.
Фигура 9 представляет собой электрофореграмму SDS-гель электрофореза биспецифичеких химерных антител просле протеолиза протеазой GingisKHAN. 7,5% ПААГ, нередуцирующие условия.
А - дорожки:
М - стандартный маркер Precision Plus Protein™ Dual Color Standards (BIO-RAD), молекулярные массы соответствующих полос указаны в кДа в столбце слева от геля;
1 - 02-004 без обработки GingisKHAN,
2 - 02-009 без обработки GingisKHAN,
3 - Ocrelizumab без обработки GingisKHAN,
4 - 01-011 без обработки GingisKHAN,
5 - Prolgolimab без обработки GingisKHAN,
6 - 02-004 без обработки GingisKHAN в буфере 100 мМ бикарбонат аммония рН 7.2;
Б - дорожки:
М - стандартный маркер Precision Plus Protein™ Dual Color Standards (BIO-RAD);
1 - 02-004 после протеолиза GingisKHAN,
2 - 02-009 после протеолиза GingisKHAN,
3 - Ocrelizumab после протеолиза GingisKHAN,
4 - 01-011 после протеолиза GingisKHAN,
5 - Prolgolimab после протеолиза GingisKHAN,
6 - 02-004 после протеолиза GingisKHAN в буфере 100 мМ бикарбонат аммония рН 7.2,
В - дорожки:
М - стандартный маркер Precision Plus Protein™ Dual Color Standards (BIO-RAD),
1 - 02-005 без обработки GingisKHAN,
2 - 02-005 после протеолиза GingisKHAN,
3 - 02-006 без обработки GingisKHAN,
4 - 02-006 после протеолиза GingisKHAN,
5 - 02-007 без обработки GingisKHAN,
6 - 02-007 после протеолиза GingisKHAN,
Г - дорожки:
М - стандартный маркер Precision Plus Protein™ Dual Color Standards (BIO-RAD),
1 - 02-008 без обработки GingisKHAN,
2 - 02-008 после протеолиза GingisKHAN,
3 - 02-004 без обработки GingisKHAN,
4 - 02-004 после протеолиза GingisKHAN.
Фигура 10 представляет собой электрофореграмму образцов с модификациями, внесенными в димеризующий блок на основе околомембранных доменов МНС-подобного белка. 7,5% ПААГ, нередуцирующие условия.
А - Дорожки:
М - стандартный маркер Precision Plus Protein™ Dual Color Standards (BIO-RAD), молекулярные массы соответствующих полос указаны в кДа в столбце слева от геля;
1 - Prolgolimab;
2 - 01-011;
3 - 03-003;
4 - 03-004;
5 - 03-005;
6 - 03-006;
7 - 03-007;
8 - 03-008.
Б - Дорожки:
М - стандартный маркер Precision Plus Protein™ Dual Color Standards (BIO-RAD);
1 - Prolgolimab;
2 - 01-011;
3 - 03-001;
4 - 03-002.
Описание изобретения
Общие определения и общие методы
Если иное не определено в настоящем документе, научные и технические термины, используемые в связи с настоящим изобретением, будут иметь значения, которые обычно понятны специалистам в данной области.
Кроме того, если по контексту не требуется иное, термины в единственном числе включают в себя термины во множественном числе, и термины во множественном числе включают в себя термины в единственном числе. Как правило, используемая классификация и методы культивирования клеток, молекулярной биологии, иммунологии, микробиологии, генетики, аналитической химии, химии органического синтеза, медицинской и фармацевтической химии, а также гибридизации и химии белка и нуклеиновых кислот, описанные в настоящем документе, хорошо известны специалистам и широко применяются в данной области. Ферментативные реакции и способы очистки осуществляют в соответствии с инструкциями производителя, как это обычно осуществляется в данной области, или как описано в настоящем документе.
Термин «KD» в данном описании относится к константе аффинности (или константе равновесия), которую получают из отношения Kd к Ka (т.е. Kd/Ka), и ее выражают в виде молярной концентрации (М).
«Аффинность связывания» обычно относится к силе совокупных нековалентных взаимодействий между единичным сайтом связывания молекулы (например, антитела) и ее партнером по связыванию (например, антигеном). Если не указано иначе, «аффинность связывания» относится к внутренней (характерной, истинной) аффинности связывания, которая отражает 1:1 взаимодействие между членами пары связывания (например, антителом и антигеном). Аффинность молекулы X к своему партнеру Y обычно можно представить константой аффинности (KD). Желательно, чтобы величина KD составляла, примерно, 200 нМ, 150 нМ, 100 нМ, 60 нМ, 50 нМ, 40 нМ, 30 нМ, 20 нМ, 10 нМ, 8 нМ, 6 нМ, 4 нМ, 2 нМ, 1 нМ или менее. Аффинность можно измерять обычными методами, известными в уровне техники, включая методы по данному описанию. Низкоаффинные антитела обычно медленно связываются с антигеном и имеют тенденцию легко диссоциировать, тогда как высокоаффинные антитела обычно быстрее связывают антиген и имеют тенденцию дольше оставаться в связанном состоянии. В уровне техники известны различные методы измерения аффинности связывания, любой из этих методов можно использовать для целей настоящего изобретения.
Термин «Kd», «Koff» или «kdis» относится к константе скорости диссоциации конкретного взаимодействия связывающей молекулы и антигена. Константу скорости диссоциации koff можно измерить посредством биослойной интерферометрии, например, с помощью системы Octet™.
Термин «Ka», «kon» или «on-rate» относится к константе скорости ассоциации.
Термин «R2» относится к коэффициенту детерминации.
Термин «Response» относится к сигналу связывания антитела с антигеном.
В настоящем описании и в последующей формуле изобретения, если контекстом не предусмотрено иное, слова «включать» и «содержать» или их вариации, такие как «включает», «включающий», «содержит» или «содержащий», следует понимать как включение указанного целого или группы целых, но не исключение любого другого целого или группы целых.
Подробное описание изобретения
Двухвалентное биспецифическое химерное антитело
Настоящее изобретение относится к двухвалентному биспецифическому химерному антителу.
Антитело по изобретению является моноклональным антителом.
Термин «моноклональное антитело» или «nAb» относится к антителу, которое синтезировано и выделено отдельной клональной популяцией клеток.
Антитело по изобретению является рекомбинантным антителом.
Термин «рекомбинантное антитело» означает антитело, которое экспрессируется в клетке или клеточной линии, содержащей нуклеотидную последовательность (нуклеотидные последовательности), которая кодирует антитело, при этом указанная нуклеотидная последовательность (нуклеотидные последовательности) не ассоциирована с клеткой в природе.
Двухвалентные биспецифические химерные антитела по изобретению является выделенным антителом.
Определение «выделенный» («изолированный»), применяемое для описания различных антител по данному описанию, означает антитело, идентифицированное и выделенное и/или регенерированное из клетки или клеточной культуры, в которой оно экспрессируется. Примеси (загрязняющие компоненты) из природной среды представляют собой материалы, которые, как правило, мешают диагностическому или терапевтическому применению полипептида, и могут включать ферменты, гормоны и другие белковые или небелковые растворенные вещества. Обычно выделенный полипептид получают в результате по меньшей мере одной стадии очистки.
В одном из аспектов настоящее изобретение относится к двухвалентному биспецифическому химерному антителу, где указанное антитело содержит:
а) первую легкую цепь и первую тяжелую цепь антитела, специфически связывающегося с первым антигеном;
где первая легкая цепь содержит вариабельный домен легкой цепи и константный домен легкой цепи;
где первая тяжелая цепь содержит вариабельный домен тяжелой цепи и константные домены тяжелой цепи антитела, включающие первый (СН1) константный домен тяжелой цепи и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи;
б) вторую легкую цепь и вторую тяжелую цепь антитела, специфически связывающегося со вторым антигеном,
где вторая легкая цепь содержит вариабельный домен легкой цепи и константный домен, который выбирают из группы:
первого околомембранного домена МНС (главный комплекс гистосовместимости) или
первого околомембранного домена МНС-подобного белка;
где вторая тяжелая цепь, содержит вариабельный домен тяжелой цепи, константный домен, который выбирают из группы:
второго околомембранного домена МНС (главный комплекс гистосовместимости) или
второго околомембранного домена МНС-подобного белка;
и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи;
при этом первый околомембранный домен МНС или МНС-подобного белка и второй околомембранный домен МНС или МНС-подобного белка образуют между собой гетеродимер, который стабилизирован дисульфидной связью;
где СН3-домен одной тяжелой цепи и СН3-домен другой тяжелой цепи соприкасаются друг с другом поверхностями, которые изменены для формирования двухвалентного биспецифического химерного антитела, причем данные изменения в СН3 доменах тяжелых цепей представляют собой замены для обеспечения гетеродимеризации.
Под гетеродимером, которой образован первым околомембранным доменом МНС или МНС-подобного белка и вторым околомембранным доменом МНС или МНС-подобного белка, и стабилизирован дисульфидной связью понимается:
1) гетеродимер, который включает мутацию или мутации в первом и/или втором околомембранном домене МНС или МНС-подобного белка для образования S-S связи (дисульфидной связи, цистеинового мостика) между первым и вторым околомембранным доменом МНС или МНС-подобного белка; или
2) удлинение первого околомембранного домена МНС или МНС-подобного белка на одну или несколько (от 1 до 10) аминокислот на С-конце и с терминальным Cys на С-конце для образования S-S связи (дисульфидной связи, цистеинового мостика) между первым околомембранным доменом МНС или МНС-подобного белка и шарниром (hinge).
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС или МНС-подобного белка и второй околомембранный домен МНС или МНС-подобного белка, которые образуют между собой гетеродимер, который стабилизирован дисульфидной связью за счет мутации или мутаций в первом и/или втором околомембранном домене МНС или МНС-подобного белка для образования S-S связи (дисульфидной связи, цистеинового мостика) между первым и вторым околомембранным доменом МНС или МНС-подобного белка.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС или МНС-подобного белка и второй околомембранный домен МНС или МНС-подобного белка, которые образуют между собой гетеродимер, который стабилизирован дисульфидной связью за счет удлинения первого околомембранного домена МНС или МНС-подобного белка на одну или несколько (от 1 до 10) аминокислот на С-конце и с терминальным Cys на С-конце для образования S-S связи (дисульфидной связи, цистеинового мостика) между первым околомембранным доменом МНС или МНС-подобного белка и шарниром (hinge).
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает удлинение первого околомембранного домена МНС или МНС-подобного белка, которое представляет собой представляет собой последовательность из трех аминокислот GSC.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает:
первую легкую цепь, которая содержит вариабельный домен легкой цепи (VL1) и константный домен легкой цепи;
первую тяжелую цепь, которая содержит вариабельный домен тяжелой цепи (VH1) и константные домены тяжелой цепи антитела, включающие первый (СН1) константный домен тяжелой цепи и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи;
вторую легкую цепь, которая содержит вариабельный домен легкой цепи (VL2) и первый околомембранный домен МНС или первый околомембранный домен МНС-подобного белка;
вторую тяжелую цепь, которая содержит вариабельный домен тяжелой цепи (VH2), второй околомембранный домен МНС или второй околомембранный домен МНС-подобного белка и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи;
при этом первый околомембранный домен МНС или МНС-подобного белка и второй околомембранный домен МНС или МНС-подобного белка образуют между собой гетеродимер, который стабилизирован дисульфидной связью за счет мутации или мутаций в первом и/или втором околомембранном домене МНС или МНС-подобного белка для образования S-S связи (дисульфидной связи, цистеинового мостика) между первым и вторым околомембранным доменом МНС или МНС-подобного белка;
где между первым вариантом Fc и вторым вариантом Fc в СН3 домене образована структура «выступ (Knob) во впадину (Hole)».
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает:
первую легкую цепь, которая содержит вариабельный домен легкой цепи (VL1) и константный домен легкой цепи;
первую тяжелую цепь, которая содержит вариабельный домен тяжелой цепи (VH1) и константные домены тяжелой цепи антитела, включающие первый (СН1) константный домен тяжелой цепи и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи;
вторую легкую цепь, которая содержит вариабельный домен легкой цепи (VL2) и первый околомембранный домен МНС или первый околомембранный домен МНС-подобного белка;
вторую тяжелую цепь, которая содержит вариабельный домен тяжелой цепи (VH2), второй околомембранный домен МНС или второй околомембранный домен МНС-подобного белка и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи;
где между первым вариантом Fc и вторым вариантом Fc в СН3 домене образована структура «выступ (Knob) во впадину (Hole)»,
где первый околомембранный домен МНС или первый околомембранный домен МНС-подобного белка дополнительно содержит удлинение первого околомембранного домена МНС или МНС-подобного белка на одну или несколько (от 1 до 10) аминокислот на С-конце и с терминальным Cys на С-конце для образования S-S связи (дисульфидной связи, цистеинового мостика) между первым околомембранным доменом МНС или МНС-подобного белка и шарниром (hinge).
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает:
первую легкую цепь, которая содержит вариабельный домен легкой цепи (VL1) и константный домен легкой цепи;
первую тяжелую цепь, которая содержит вариабельный домен тяжелой цепи (VH1) и константные домены тяжелой цепи антитела, включающие первый (СН1) константный домен тяжелой цепи и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи;
вторую легкую цепь, которая содержит вариабельный домен легкой цепи (VL2) и первый околомембранный домен МНС или первый околомембранный домен МНС-подобного белка;
вторую тяжелую цепь, которая содержит вариабельный домен тяжелой цепи (VH2), второй околомембранный домен МНС или второй околомембранный домен МНС-подобного белка и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи;
где между первым вариантом Fc и вторым вариантом Fc в СН3 домене образована структура «выступ (Knob) во впадину (Hole)»
где первый околомембранный домен МНС или первый околомембранный домен МНС-подобного белка дополнительно содержит удлинение первого околомембранного домена МНС или МНС-подобного белка на GSC для образования S-S связи (дисульфидной связи, цистеинового мостика) между первым околомембранным доменом МНС или МНС-подобного белка и шарниром (hinge).
В вышеуказанном двухвалентном биспецифическом химерном антителе вариабельные и константные домены расположены в тяжелых и легких цепях в следующей последовательности:
первая легкая цепь:
1) вариабельный домен легкой цепи,
2) константный домен легкой цепи; первая тяжелая цепь антитела:
1) вариабельный домен тяжелой цепи,
2) первый (СН1) константный домен тяжелой цепи,
3) второй (СН2) константный домен тяжелой цепи и
4) третий (СН3) константный домен тяжелой цепи;
вторая легкая цепь:
1) вариабельный домен легкой цепи,
2) первый околомембранный домен МНС или первый околомембранный домен МНС-подобного белка;
вторая тяжелая цепь антитела:
1) вариабельный домен тяжелой цепи,
2) второй околомембранный домен МНС или второй околомембранный домен МНС-подобного белка,
3) второй (СН2) константный домен тяжелой цепи и
4) третий (СН3) константный домен тяжелой цепи.
Некоторые варианты формата вышеуказанного двухвалентного биспецифического химерного антитела представлены на фигуре 1.
Термин «антитело» или «иммуноглобулин» (Ig), как использовано в данном описании, включает целые антитела. Термин «антитело» относится к гликопротеину, содержащему по меньшей мере две тяжелые (Н) цепи и две легкие (L) цепи, взаимосвязанные дисульфидными связями, или его антигенсвязывающей части. Каждая тяжелая цепь содержит вариабельную область тяжелой цепи (сокращенно называемую в данном описании как VH) и константную область тяжелой цепи. Известно пять типов тяжелых цепей антител млекопитающих, которые обозначают греческими буквами: α, δ, ε, γ и μ. (Janeway С.A., Jr. и др., Immunobiology, 5-е изд., изд-во Garland Publishing, 2001). Присутствующий тип тяжелой цепи определяет класс антитела; указанные цепи обнаружены в антителах типа IgA, IgD, IgE, IgG и IgM соответственно (Rhoades R.A., Pflanzer R.G., Human Physiology, 4-е изд., изд-во Thomson Learning, 2002). Различные тяжелые цепи отличаются по размеру и составу; α и γ содержат примерно 450 аминокислот, а μ и ε состоят примерно из 550 аминокислот. Константная область является идентичной во всех антителах одного и того же изотипа, но отличается в антителах различного изотипа. Тяжелые цепи γ, α и δ содержат константную область, которая состоит из трех константных доменов CH1, СН2 и СН3 (выстроены в ряд) и шарнирной области, которая придает гибкость (Woof J., Burton D., Nat Rev Immunol 4, 2004, cc. 89-99); тяжелые цепи μ и ε содержат константную область, которая состоит из четырех константных доменов CH1, СН2, СН3 и СН4 (Janeway С.A., Jr. и др., Immunobiology, 5-е изд., изд-во Garland Publishing, 2001). У млекопитающих известно только два типа легких цепей, которые обозначают как лямбда (λ) и каппа (κ). Каждая легкая цепь состоит из вариабельной области легкой цепи (сокращенно называемой в данном описании как VL) и константной области легкой цепи. Примерная длина легкой цепи составляет 211-217 аминокислот. Предпочтительно легкая цепь представляет собой легкую каппа (κ)-цепь, а константный домен CL предпочтительно представляет собой С-каппа (κ).
«Антитела» согласно изобретению могут представлять собой антитела любого класса (например, IgA, IgD, IgE, IgG и IgM, предпочтительно IgG) или подкласса (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2, предпочтительно IgG1).
Области VH и VL могут быть дополнительно подразделены на области гипервариабельности, называемые определяющими комплементарность областями (CDR), расположенные между областями, которые являются более консервативными, называемыми каркасными областями (FR). Каждая VH и VL состоит из трех CDR и четырех FR, расположенных от амино-конца к карбокси-концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Вариабельные области тяжелой и легкой цепей содержат домен связывания, который взаимодействует с антигеном. Константные области антител могут опосредовать связывание иммуноглобулина с тканями хозяина или факторами, включая различные клетки иммунной системы (например, эффекторными клетками), и первый компонент (C1q) классической системы комплемента.
Термин «антигенсвязывающая часть» антитела или «антигенсвязывающий фрагмент» (или просто «часть антитела» или «фрагмент антитела»), как использовано в данном описании, относится к одному или нескольким фрагментам антитела, которые сохраняют способность специфически связываться с антигеном. Пример связывающего фрагмента, включенного в термин «антигенсвязывающая часть» антитела включает Fab-фрагмент, то есть одновалентный фрагмент, состоящий из доменов VL, VH, CL и СН1 или Fab-подобный фрагмент, то есть одновалентный фрагмент, состоящий из доменов VL, VH, первого околомембранного домена МНС или МНС-подобного белка и второго околомембранного домена МНС или МНС-подобного белка.
Предпочтительно CDR антигенсвязывающего участка или весь антигенсвязывающий участок антител по изобретению имеет происхождение из мыши, ламы или донорской человеческой библиотеки или по существу человеческое происхождение с определенными аминокислотными остатками, измененными, например, замещенными разными аминокислотными остатками с тем, чтобы оптимизировать конкретные свойства антитела, например KD, koff, IC50, ЕС50, ED50. Предпочтительно каркасные участки антитела по изобретению имеют человеческое происхождение или по существу человеческое происхождение (по крайней мере на 80, 85, 90, 95, 96, 97, 98 или 99% человеческое происхождение).
В других вариантах осуществления антигенсвязывающий участок антитела по изобретению может происходить из других нечеловеческих видов, включая мышь, ламу, кролика, крысу или хомяка, но не ограничиваясь ими. Альтернативно, антигенсвязывающий участок может происходить из человеческих видов.
Термин «вариабельный» относится к тому факту, что определенные сегменты вариабельных доменов широко отличаются в последовательности среди антител. Домен V опосредует связывание антигена и определяет специфичность конкретного антитела к его конкретному антигену. Однако вариабельность неравномерно распределяется на участке вариабельных доменов из ПО аминокислот. Напротив, V области состоят из инвариантных фрагментов, называемых каркасными областями (FR) из 15-30 аминокислот, разделенных более короткими участками чрезвычайной вариабельности, называемых «гипервариабельными областями» или CDR. Каждый вариабельный домен нативных тяжелых и легких цепей содержит четыре FR, в основном принимающих конфигурацию бета-листов, связанных тремя гипервариабельными областями, которые образуют петли, связывающие, и в некоторых случаях являющиеся частью бета-складчатой структуры. Гипервариабельные области в каждой цепи удерживаются вместе в тесной близости с помощью FR и с гипервариабельными областями другой цепи вносят вклад в образование антигенсвязывающего сайта антител (см. Kabat et al., Sequences of Proteins of Immunological Interest. 5 th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991)). Константные домены не принимают непосредственного участия в связывании антитела с антигеном, но проявляют различные эффекторные функции, такие как участие антитела в антителозависимой клеточной цитотоксичности (АЗКЦ, ADCC).
Термин «гипервариабельная область» по данному описанию относится к аминокислотным остаткам антитела, которые отвечают за связывание антигена. Обычно гипервариабельная область содержит аминокислотные остатки из «области, определяющей комплементарность» или «CDR», и/или такие остатки из «гипервариабельной петли».
«Kabat номенклатура» или «номенклатура по Kabat» применяются в данной заявке к системе нумерации аминокислотных остатков, которые являются более вариабельными (т.е. гипервариабельными), чем остальные аминокислотные остатки в вариабельных участках тяжелой и легкой цепи антитела (Kabat et al. Arm. N.Y. Acad. Sci., 190:382-93 (1971); Kabat etal. Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242(1991)).
Антитело по данному изобретению, «которое связывает» целевой антиген, представляет собой антитело, которое связывает антиген с достаточной аффинностью так, что антитело можно применять в качестве диагностического и/или терапевтического агента при нацеливании на белок или клетку или ткань, экспрессирующую антиген, и в незначительной степени перекрестно реагирует с другими белками. По данным аналитических методов: сортинга флуоресцентно-активированных клеток (FACS), радиоиммунопреципитации (RIA) или ИФА (ELISA), в таких вариантах изобретения степень связывания антитела с белком, не являющимся «мишенью» (с «нецелевым белком»), составляет менее 10% от связывания антитела с конкретным белком-мишенью. По отношению к связыванию антитела с молекулой-мишенью термин «специфическое связывание» или выражения «специфически связывается с» или «специфический к» конкретному полипептиду или эпитопу на конкретном полипептиде-мишени означает связывание, которое заметно (измеримо) отличается от неспецифического взаимодействия.
Специфическое связывание можно определять количественно, например, определяя связывание молекулы по сравнению со связыванием контрольной молекулы. Например, специфическое связывание можно определять конкурентной реакцией с другой молекулой, аналогичной мишени, например, с избытком немеченой мишени. В этом случае специфическое связывание указывается, если связывание меченой мишени с зондом конкурентно ингибируется избытком немеченой мишени. В данном описании термин «специфическое связывание» или выражения «специфически связывается с» или «специфический к» конкретному полипептиду или эпитопу на конкретном полипептиде-мишени можно характеризовать на примере молекулы, имеющей KD к мишени по меньшей мере около 200 нМ, или же по меньшей мере около 150 нМ, или же по меньшей мере около 100 нМ, или же по меньшей мере около 60 нМ, или же по меньшей мере около 50 нМ, или же по меньшей мере около 40 нМ, или же по меньшей мере около 30 нМ, или же по меньшей мере около 20 нМ, или же по меньшей мере около 10 нМ, или же по меньшей мере около 8 нМ, или же по меньшей мере около 6 нМ, или же по меньшей мере около 4 нМ, или же по меньшей мере около 2 нМ, или же по меньшей мере около 1 нМ или выше. В одном варианте изобретения термин «специфическое связывание» относится к связыванию, при котором молекула связывается с конкретным полипептидом или эпитопом на конкретном полипептиде, практически не связываясь с каким-либо другим полипептидом или эпитопом на полипептиде.
Термин «биспецифическое антитело» означает антитело, содержащее антигенсвязывающие домены, которые способны к специфическому связыванию с двумя различными эпитопами на одной биологической молекуле или способны к специфическому связыванию с эпитопами на двух различных биологических молекулах. Биспецифичное антитело также упоминается в настоящем документе, как обладающее «двойной специфичностью» или как являющееся антителом с «двойной специфичностью».
Кристаллизующийся фрагмент иммуноглобулина (англ. fragment crystallizable region, Fc region, Fc) - это концевая часть молекулы иммуноглобулина, которая взаимодействует с Fc-рецептором на поверхности клетки и с некоторыми белками системы комплемента. Данное свойство позволяет антителам активировать иммунную систему. Fc-участок IgG, IgA и IgD изотипов состоит из двух одинаковых белковых фрагментов, соответственно, второго и третьего константных доменов двух тяжелых цепей; в случае изотипов IgM и IgE Fc содержит три константных домена тяжелых цепей (домены СН2, СН3 и СН4) в каждой полипептидной цепочке.
Под «мономером Fc-фрагмента» подразумевается Fc-участок из второго и третьего константных доменов любой одной из двух тяжелых цепей (для IgG, IgA и IgD изотипов); в случае изотипов IgM и IgE мономер Fc содержит три константных домена одной из двух тяжелых цепей (домены СН2, СН3 и СН4).
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает:
первую легкую цепь, которая содержит вариабельный домен легкой цепи (VL1) и константный домен легкой цепи;
первую тяжелую цепь, которая содержит вариабельный домен тяжелой цепи (VH1) и константные домены тяжелой цепи антитела, включающие первый (СН1) константный домен тяжелой цепи и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи;
вторую легкую цепь, которая содержит вариабельный домен легкой цепи (VL2) и первый околомембранный домен МНС или первый околомембранный домен МНС-подобного белка;
вторую тяжелую цепь, которая содержит вариабельный домен тяжелой цепи (VH2), второй околомембранный домен МНС или второй околомембранный домен МНС-подобного белка и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи;
и где между первым вариантом Fc и вторым вариантом Fc в СН3 домене образована структура «выступ (Knob) во впадину (Hole)»
и между вторым околомембранным доменом МНС и/или вторым околомембранным доменом МНС-подобного белка и шарниром (hinge) вставили дополнительный пептидный линкер.
Термин «пептидный линкер» в настоящем документе означает любой пептид с возможностью соединения доменов с длиной в зависимости от доменов, которые он связывает между собой, содержащий любую аминокислотную последовательность. Предпочтительно пептидный линкер имеет длину 4 аминокислоты и более и состоит из любого набора аминокислот, выбранного из G, A, S, Р, Е, Т, D, K.
Под МНС (major histocompatibility complex, главный комплекс гистосовместимости, ГКГС) подразумевается молекула главного комплекса гистосовместимости, которая представляет собой центральный компонент иммунной системы позвоночных, находящийся на поверхности всех ядросодержащих клеток. МНС находится в двух основных формах, а именно МНС класса I и II.
Под МНС-подобными белками подразумеваются белки, которые имеют структурное сходство с экстраклеточной частью молекул МНС класса I или молекул МНС класса II. В частности, под МНС-подобными белками подразумеваются, например, белок CD1 (cluster of differentiation 1, кластер дифференцировки 1) или белок HFE (Hereditary hemochromatosis protein, белок гемохроматоза).
Общее структурное сходство между молекулами МНС I, II, CD1 и HFE очевидно. Это - однотипность пространственной организации всей молекулы, количество доменов (четыре), структурное сходство околомембранных доменов МНС и МНС-подобных белков, а также антигенсвязывающего участка, близкие мол. веса.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС, который может быть выбран из группы, включающей:
первый околомембранный домен МНС класса I (главный комплекс гистосовместимости класса I),
первый околомембранный домен МНС класса II (главный комплекс гистосовместимости класса II),
модифицированный вариант первого околомембранного домена МНС класса I или
модифицированный вариант первого околомембранного домена МНС класса II.
У людей классические гены МНС I называются HLA-A, HLA-B, HLA-C, HLA-E, HLA-F или HLA-G. Молекулы класса I состоят из двух полипептидных цепей: полиморфной а цепи (в некоторых случаях имеющей название тяжелая цепь) и меньшей цепи, которая называется β2 микроглобулин (также известная как легкая цепь), которая в основном не является полиморфной. Эти две цепи образуют нековалентный гетеродимер на клеточной поверхности, а цепь содержит три домена (α1, α2 и α3). Экзон 1 гена α цепи кодирует лидерную последовательность, экзоны 2 и 3 кодируют α1 и α2 домены, экзон 4 кодирует α3 домен, экзон 5 кодирует трансмембранный домен, и экзоны 6 и 7 кодируют цитоплазматический хвост. α цепь образует антигенсвязывающую область, включающую в себя α1 и α2 домены.
За α2 доменом следует α3 домен, который расположен с С-конца экстраклеточной части а цепи МНС I и образует с β2 микроглобулином гетеродимерный нековалентный комплекс. Данный гетеродимерный нековалентный комплекс, состоящий из α3 домена МНС I и β2 микроглобулина, в описании данного изобретения называется гетеродимер на основе околомембранных доменов МНС I.
Молекулы МНС класса I экспрессируются на всех ядросодержащих клетках, включающих в себя опухолевые клетки. Они экспрессируются специфически на Т- и В-лимфоцитах, макрофагах, дендритных клетках и нейтрофилах, среди прочих клеток, и функционируют для представления пептидных фрагментов (как правило 8-10 аминокислот в длину) на поверхности CD8+ цитотоксическим Т-лимфоцитам (CTL).
У людей классические гены МНС II называются HLA-DM, HLA-DO, HLA-DP, HLA-DQ или HLA-DR. Молекулы МНС класса II являются гетеродимерами, построенными из нековалентно сцепленных альфа- и бета-цепей. Внеклеточная часть каждой из цепей состоит из двух доменов (α1 (альфа1), α2 (альфа2) и β1 (бета1), β2 (бета2), соответственно) и соединена коротким пептидом с трансмембранным сегментом (длиной примерно 30 аминокислотных остатков). Трансмембранный сегмент переходит в цитоплазматический домен, содержащий примерно 10-15 остатков.
Антигенсвязывающая область молекул МНС класса II формируется альфа-спиральными участками взаимодействующих цепей подобно молекулам класса I, но при одном существенном отличии: антигенсвязывающая полость молекул МНС класса II формируется не двумя доменами одной альфа-цепи, а двумя доменами разных цепей - доменами α1 и β1.
За α1 доменом следует α2 домен, который образует с β2 доменом гетеродимерный нековалентный комплекс. Данный гетеродимерный нековалентный комплекс, состоящий из α2 домена МНС II и β2 домена МНС II, в описании данного изобретения называется гетеродимер на основе околомембранных доменов МНС II.
В структуре молекул МНС класса II антигенсвязывающая полость открыта больше, чем у молекул МНС класса I, поэтому в ней могут поместиться более длинные пептиды.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает второй околомембранный домен МНС, который может быть выбран из группы, включающей:
второй околомембранный домен МНС класса I (главный комплекс гистосовместимости класса I),
второй околомембранный домен МНС класса II (главный комплекс гистосовместимости класса II),
модифицированный вариант второго околомембранного домена МНС класса I или
модифицированный вариант второго околомембранного домена МНС класса II.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает околомембранный домен МНС класса I человека.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает околомембранный домен МНС класса II человека.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС, который может быть выбран из группы, включающей:
первый околомембранный домен МНС класса I (главный комплекс гистосовместимости класса I),
первый околомембранный домен МНС класса II (главный комплекс гистосовместимости класса II),
модифицированный вариант первого околомембранного домена МНС класса I или
модифицированный вариант первого околомембранного домена МНС класса II,
и
второй околомембранный домен МНС, который может быть выбран из группы, включающей:
второй околомембранный домен МНС класса I (главный комплекс гистосовместимости класса I),
второй околомембранный домен МНС класса II (главный комплекс гистосовместимости класса II),
модифицированный вариант второго околомембранного домена МНС класса I или
модифицированный вариант второго околомембранного домена МНС класса II.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает:
первый околомембранный домен МНС, который представляет собой первый околомембранный домен МНС класса I (главный комплекс гистосовместимости класса I),
и
второй околомембранный домен МНС, который представляет собой второй околомембранный домен МНС класса I (главный комплекс гистосовместимости класса I).
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает:
первый околомембранный домен МНС, который представляет собой первый околомембранный домен МНС класса II (главный комплекс гистосовместимости класса II),
и
второй околомембранный домен МНС, который представляет собой второй околомембранный домен МНС класса II (главный комплекс гистосовместимости класса II).
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает:
первый околомембранный домен МНС, который представляет собой модифицированный вариант первого околомембранного домена МНС класса I,
и
второй околомембранный домен МНС, который представляет собой модифицированный вариант второго околомембранного домена МНС класса I.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает:
первый околомембранный домен МНС, который представляет собой модифицированный вариант первого околомембранного домена МНС класса II,
и
второй околомембранный домен МНС, который представляет собой модифицированный вариант второго околомембранного домена МНС класса II.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает:
первый околомембранный домен МНС, который представляет собой первый околомембранный домен МНС класса I (главный комплекс гистосовместимости класса I),
и
второй околомембранный домен МНС, который представляет собой модифицированный вариант второго околомембранного домена МНС класса I.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает:
первый околомембранный домен МНС, который представляет собой модифицированный вариант первого околомембранного домена МНС класса I,
и
второй околомембранный домен МНС, который представляет собой второй околомембранный домен МНС класса I (главный комплекс гистосовместимости класса I).
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает:
первый околомембранный домен МНС, который представляет собой первый околомембранный домен МНС класса II (главный комплекс гистосовместимости класса II),
и
второй околомембранный домен МНС, который представляет собой модифицированный вариант второго околомембранного домена МНС класса II.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает:
первый околомембранный домен МНС, который представляет собой модифицированный вариант первого околомембранного домена МНС класса II,
и
второй околомембранный домен МНС, который представляет собой второй околомембранный домен МНС класса II (главный комплекс гистосовместимости класса II).
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает околомембранный домен МНС класса II, котрый выбирают из группы: HLA-DM, HLA-DO, HLA-DP, HLA-DQ или HLA-DR.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает околомембранный домен МНС класса I, который выбирают из группы: HLA-A, HLA-B, HLA-C, HLA-E, HLA-F или HLA-G.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает околомембранный домен МНС-подобного белка человека.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС-подобного белка, который может быть выбран из группы, включающей:
первый околомембранный домен CD1 (кластер дифференцировки 1),
первый околомембранный домена HFE (белок гемохроматоза),
модифицированный вариант первого околомембранного домена CD1 или
модифицированный вариант первого околомембранного домена HFE.
Под CD1 (cluster of differentiation 1, кластер дифференцировки 1) подразумевается молекула кластера дифференцировки 1, которая является компонентом иммунной системы, находящейся на поверхности различных антиген-презентирующих клеток, таких как дендритные, клетки, макрофаги и прочие клетки. Так же, как и МНС классов I, II, CD1 презентируют антигены для распознавания Т-клетками через взаимодействие с Т-клеточным рецептором. В отличие от МНС классов I, II, белки CD1 презентируют липиды и их производные, а не пептиды.
Множество вариантов CD1, встречающихся у людей подразделяют на 5 групп: CD1a, CD1b, CD1c, CD1d, CD1e, которые отличаются структурой антигенсвязывающего фрагмента и, как следствие, специфичностью по отношению к липидам разной структуры. Белки группы CD1e, в отличие от белков остальных групп, не экспрессируются на поверхности клеток, а являются растворимыми и отвечают за транспорт липидов (Kaczmarek, R., Pasciak, М., Szymczak-Kulus, K., & Czerwinski, М. (2017). CD1: A Singed Cat of the Three Antigen Presentation Systems. Archivum Immunologiae et Therapiae Experimentalise 65(3), 201-214).
Молекулы CD1 схожи по структуре с МНС класса I. По аналогии, одна молекула CD1 является нековалентным комплексом двух полипептидных цепей: полиморфной α цепи (в некоторых случаях имеющей название тяжелая цепь) и меньшей цепи, которая называется β2 микроглобулин (также известная как легкая цепь), которая в основном не является полиморфной.
α цепь образует антигенсвязывающую область, включающую в себя α1 и α2 домены. За α2 доменом следует α3 домен, который расположен с С-конца экстраклеточной части а цепи CD1 и образует с β2 микроглобулином гетеродимерный нековалентный комплекс. Данный гетеродимерный нековалентный комплекс, состоящий из α3 домена CD1 и β2 микроглобулина, в описании данного изобретения называется гетеродимер на основе околомембранных доменов МНС-подобных белков.
Под HFE (Hereditary hemochromatosis protein, белок гемохроматоза) подразумевается молекула белка гемохроматоза. Локус гена HFE человека расположен в 6й хромосоме и состоит из 5 экзонов и 4 интронов. Белок гемохроматоза - это мембранный белок, ассоциированный с β2 микроглобулином, отвечает за регуляцию абсорбции железа через регуляцию взаимодействия трансферрина с рецептором трансферрина. Мутации в гене HFE приводят к гемохроматозу - рецессивной патологии обмена железа, связанной с его накоплением в различных органах. Молекула HFE по структуре аналогична с МНС I. Одна молекула HFE является нековалентным комплексом двух полипептидных цепей: полиморфной α цепи (в некоторых случаях имеющей название тяжелая цепь) и меньшей цепи, которая называется β2 микроглобулин (также известная как легкая цепь), которая в основном не является полиморфной.
α цепь включает в себя α1, α2 и α3 домены. α3 домен следует за α2 домен и располагается с С-конца экстраклеточной части α цепи HFE и образует с β2 микроглобулином гетеродимерный нековалентный комплекс. Данный гетеродимерный нековалентный комплекс, состоящий из α3 домена HFE и β2 микроглобулина, в описании данного изобретения называется гетеродимер на основе околомембранных доменов МНС-подобных белков.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает второй околомембранный домен МНС-подобного белка, который может быть выбран из группы, включающей:
второй околомембранный домен CD1 (кластер дифференцировки 1),
второй околомембранный домена HFE (белок гемохроматоза),
модифицированный вариант второго околомембранного домена CD1 или
модифицированный вариант второго околомембранного домена HFE.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает:
первый околомембранный домен МНС-подобного белка, который может быть выбран из группы, включающей:
первый околомембранный домен CD1 (кластер дифференцировки 1),
первый околомембранный домена HFE (белок гемохроматоза),
модифицированный вариант первого околомембранного домена CD1 или
модифицированный вариант первого околомембранного домена HFE,
и
второй околомембранный домен МНС-подобного белка, который может быть выбран из группы, включающей:
второй околомембранный домен CD1 (кластер дифференцировки 1),
второй околомембранный домен HFE (белок гемохроматоза),
модифицированный вариант второго околомембранного домена CD1 или
модифицированный вариант второго околомембранного домена HFE.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает:
первый околомембранный домен МНС-подобного белка, который представляет собой первый околомембранный домен CD1,
и
второй околомембранный домен МНС-подобного белка, который представляет собой второй околомембранный домен CD1.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает:
первый околомембранный домен МНС-подобного белка, который представляет собой первый околомембранный домена HFE,
и
второй околомембранный домен МНС-подобного белка, который представляет собой второй околомембранный домена HFE,
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает:
первый околомембранный домен МНС-подобного белка, который представляет собой модифицированный вариант первого околомембранного домена CD1,
и
второй околомембранный домен МНС-подобного белка, который представляет собой модифицированный вариант второго околомембранного домена CD1.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает:
первый околомембранный домен МНС-подобного белка, который представляет собой модифицированный вариант первого околомембранного домена HFE,
и
второй околомембранный домен МНС-подобного белка, который представляет собой модифицированный вариант второго околомембранного домена HFE.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает:
первый околомембранный домен МНС-подобного белка, который представляет собой первый околомембранный домен CD1,
и
второй околомембранный домен МНС-подобного белка, который представляет собой модифицированный вариант второго околомембранного домена CD1.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает:
первый околомембранный домен МНС-подобного белка, который представляет собой модифицированный вариант первого околомембранного домена CD1,
и
второй околомембранный домен МНС-подобного белка, который представляет собой второй околомембранный домен CD1.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает:
первый околомембранный домен МНС-подобного белка, который представляет собой первый околомембранный домен HFE,
и
второй околомембранный домен МНС-подобного белка, который представляет собой модифицированный вариант второго околомембранного домена HFE.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает:
первый околомембранный домен МНС-подобного белка, который представляет собой модифицированный вариант первого околомембранного домена HFE,
и
второй околомембранный домен МНС-подобного белка, который представляет собой второй околомембранный домен HFE.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает околомембранный домен CD1, который выбирают из группы: CD1a, CD1b, CD1c, CD1d или CD1e.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает вариабельный фрагмент второй легкой цепи (VL), который отделен от первого околомембранного домена МНС или МНС-подобного белка линкером длиной от 1 до 25 аминокислот, и/или вариабельный фрагмент второй тяжелой цепи (VH), который отделен от второго околомембранного домена МНС или МНС-подобного белка линкером длиной от 1 до 25 аминокислот.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает:
а) СН3-домен одной тяжелой цепи, который изменен так, что на поверхности СН3-домена одной тяжелой цепи, которая соприкасается с поверхностью СН3-домена второй тяжелой цепи в двухвалентном биспецифическом антителе, аминокислотный остаток заменен на аминокислотный остаток, который имеет большую по объему боковую цепь, что приводит к созданию выпуклости на поверхности СН3-домена одной тяжелой цепи, которая может помещаться в полость на поверхности СН3-домена другой тяжелой цепи,
и
б) СН3-домен другой тяжелой цепи, который изменен так, что на поверхности СН3-домена второй тяжелой цепи, которая соприкасается с поверхностью СН3-домена первой тяжелой в двухвалентном биспецифическом антителе, аминокислотный остаток заменен на аминокислотный остаток, который имеет меньшую по объему боковую цепь, что приводит к созданию полости на поверхности СН3-домена второй тяжелой цепи, в которую может помещаться выпуклость на поверхности раздела СН3-домена первой тяжелой цепи;
где указанный аминокислотный остаток, который имеет большую по объему боковую цепь, выбран из группы, включающей аргинин (R), фенилаланин (F), тирозин (Y), триптофан (W),
и где указанный аминокислотный остаток, который имеет меньшую по объему боковую цепь, выбран из группы, включающей аланин (А), серии (S), треонин (Т), валин (V).
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает константный домен первой легкой цепи антитела, который выбирают из CK или CL.
У млекопитающих известно только два типа легких цепей, которые обозначают как лямбда (λ) и каппа (κ). Константной домен лямбда легкой цепи обозначается как CL, а каппа легкой цепи обозначается как CK.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает СН3-домены антитела, которые дополнительно изменены путем интродукции цистеина (С) в качестве аминокислоты в соответствующие положения каждого СН3-домена, так что дисульфидный мостик может образовываться между обоими СН3-доменами.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает СН3-домен одной тяжелой цепи, который изменен с образованием Knob, и СН3-домен другой тяжелой цепи, который изменен с образованием Hole, или наоборот.
«knobs -into-holes» (взаимодействие по типу «выступ-впадина») это подход, с помощью которого можно обойти проблему, связанную с имеющими ошибочные спаривания побочными продуктами. Данный подход направлен на усиление спаривания двух различных тяжелых цепей антитела путем интродукции мутаций в СН3-домены для модификации поверхности раздела в области контакта. На одной цепи имеющие большие размеры аминокислоты заменяли на аминокислоты с короткими боковыми цепями для создания «впадины». И, наоборот, аминокислоты с более крупными боковыми цепями интродуцировали в другой СН3-домен, создавая «выступ». Путем совместной экспрессии этих двух тяжелых цепей получали высокий выход гетеродимерной формации («выступ-впадина») относительно гомодимерной формации («впадина-впадина» или «выступ-выступ») (WO 9627011 и WO9850431, а также статью Merchant AM ЕТ ALL., An efficient route to human bispecific IgG, Nat Biotechnol. 1998 Jul; 16(7):677-81).
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает СН3-домен одной тяжелой цепи, который имеет аминокислотные замены S354C/T366W, и СН3-домен другой тяжелой цепи, который имеет аминокислотные замены Y349C/T366S/L368A/Y407V.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает СН3-домен одной тяжелой цепи, который имеет аминокислотные замены Y349C/T366S/L368A/Y407, и СН3-домен другой тяжелой цепи, который имеет аминокислотные замены S354C/T366W.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС и второй околомембранный домен МНС, которые, соответственно, представляют собой α2 домен МНС II и β2 домен МНС II, и образуют между собой гетеродимер.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС и второй околомембранный домен МНС, которые, соответственно, представляют собой β2 домен МНС II и α2 домен МНС II, и образуют между собой гетеродимер.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает α2 домен МНС II, который имеет аминокислотную последовательность, которую выбирают из группы: SEQ ID NO: 30, SEQ ID NO: 32, SEQ ID NO: 34, SEQ ID NO: 36 или SEQ ID NO: 38, и p2 домен МНС II, который имеет аминокислотную последовательность, которую выбирают из группы: SEQ ID NO: 31, SEQ ID NO: 33, SEQ ID NO: 35, SEQ ID NO: 37 или SEQ ID NO: 39.
SEQ ID NO: 30 представляет собой аминокислотную последовательность α2 околомембранного домена МНС класса II HLA-DM (HLA-DMA*01:01:01:01).
SEQ ID NO: 31 представляет собой аминокислотную последовательность последовательность β2 околомембранного домена МНС класса II HLA-DM (HLA-DMB*01:01:01:01).
SEQ ID NO: 32 представляет собой аминокислотную последовательность последовательность α2 околомембранного домена МНС класса II HLA-DO (HLA-DOA*01:01:01).
SEQ ID NO: 33 представляет собой аминокислотную последовательность последовательность β2 околомембранного домена МНС класса II HLA-DO (HLA-DOB*01:01:01:01).
SEQ ID NO: 34 представляет собой аминокислотную последовательность последовательность α2 околомембранного домена МНС класса II HLA-DP (HLA-DPA1*01:03:01:01).
SEQ ID NO: 35 представляет собой аминокислотную последовательность β2 околомембранного домена МНС класса II HLA-DP (HLA-DPB1*01:01:01:01).
SEQ ID NO: 36 представляет собой аминокислотную последовательность α2 околомембранного домена МНС класса II HLA-DQ (HLA-DQA1*01:01:01:01).
SEQ ID NO: 37 представляет собой аминокислотную последовательность β2 околомембранного домена МНС класса II HLA-DQ (HLA-DQB1*05:01:01:01).
SEQ ID NO: 38 представляет собой аминокислотную последовательность α2 околомембранного домена МНС 2 класса II HLA-DR (HLA-DRA*01:01:01:01).
SEQ ID NO: 39 представляет собой аминокислотную последовательность β2 околомембранного домена МНС класса HLA-DR (HLA-DRB1*01:01:01:01).
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС и второй околомембранный домен МНС, которые, соответственно, представляют собой модифицированный вариант α2 домена МНС II и модифицированный варианта β2 домена МНС II, и образуют между собой гетеродимер.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС и второй околомембранный домен МНС, которые, соответственно, представляют собой модифицированный вариант β2 домена МНС II и модифицированный вариант α2 домена МНС II, и образуют между собой гетеродимер.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС и второй околомембранный домен МНС, которые, соответственно, представляют собой α3 домен МНС I и β2 микроглобулин (β2М), и образуют между собой гетеродимер.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС и второй околомембранный домен МНС, которые, соответственно, представляют собой β2 микроглобулин (β2М) и α3 домен МНС I, и образуют между собой гетеродимер.
β2 микроглобулин представляет собой негликозилированный белок массой 12 кДа; одной из его функций является стабилизация а цепи МНС класса I, а также белков CD1 и HFE. В составе вышеуказанных белков β2 микроглобулин человека имеет практически идентичную структуру и поэтому в описании данного изобретения β2 микроглобулин человека упоминается без указания белка, в состав которого он входит.
В отличие от α цепи β2 микроглобулин не пересекает мембрану. Локус β2 микроглобулина человека находится на 15 хромосоме. Ген β2 микроглобулина состоит из 4 экзонов и 3 нитронов.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает α3 домен МНС I, который имеет аминокислотную последовательность, которую выбирают из группы SEQ ID NO: 1-29, и β2 микроглобулин (β2М), который имеет аминокислотную последовательность SEQ ID NO: 46.
SEQ ID NO: 1 представляет собой аминокислотную последовательность α3 околомембранного домена МНС класса I HLA-A (аллель А*01:01:01:01).
SEQ ID NO: 2 представляет собой аминокислотную последовательность α3 околомембранного домена МНС класса I HLA-A (А*02:01:01:01).
SEQ ID NO: 3 представляет собой аминокислотную последовательность α3 околомембранного домена МНС класса I HLA-A (А*03:01:01:01).
SEQ ID NO: 4 представляет собой аминокислотную последовательность α3 околомембранного домена МНС класса I HLA-A (А*11:01:01:01).
SEQ ID NO: 5 представляет собой аминокислотную последовательность α3 околомембранного домена МНС класса I HLA-A (А*23:01:01:01).
SEQ ID NO: 6 представляет собой аминокислотную последовательность α3 околомембранного домена МНС класса I HLA-B (В*07:02:01:01).
SEQ ID NO: 7 представляет собой аминокислотную последовательность α3 околомембранного домена МНС класса I HLA-B (В*08:01:01:01).
SEQ ID NO: 8 представляет собой аминокислотную последовательность α3 околомембранного домена МНС класса1 HLA-B (В*13:01:01:01).
SEQ ID NO: 9 представляет собой аминокислотную последовательность α3 околомембранного домена МНС класса I HLA-B (В*14:01:01:01).
SEQ ID NO: 10 представляет собой аминокислотную последовательность α3 околомембранного домена МНС класса I HLA-B (В*15:01:01:01).
SEQ ID NO: 11 представляет собой аминокислотную последовательность α3 околомембранного домена МНС класса I HLA-C (С*01:02:01:01).
SEQ ID NO: 12 представляет собой аминокислотную последовательность α3 околомембранного домена МНС класса I HLA-C (С*03:02:01).
SEQ ID NO: 13 представляет собой аминокислотную последовательность α3 околомембранного домена МНС класса I HLA-C (С*04:01:01:01).
SEQ ID NO: 14 представляет собой аминокислотную последовательность α3 околомембранного домена МНС класса I HLA-C (С*05:01:01:01).
SEQ ID NO: 15 представляет собой аминокислотную последовательность α3 околомембранного домена МНС класса I HLA-C (С*06:02:01:01).
SEQ ID NO: 16 представляет собой аминокислотную последовательность α3 околомембранного домена МНС класса I HLA-E (Е*01:01:01:01).
SEQ ID NO: 17 представляет собой аминокислотную последовательность α3 околомембранного домена МНС класса I HLA-E (Е*01:03:01:01).
SEQ ID NO: 18 представляет собой аминокислотную последовательность α3 околомембранного домена МНС класса I HLA-E (Е*01:05).
SEQ ID NO: 19 представляет собой аминокислотную последовательность α3 околомембранного домена МНС класса I HLA-E (Е*01:06).
SEQ ID NO: 20 представляет собой аминокислотную последовательность α3 околомембранного домена МНС класса I HLA-E (Е*01:09).
SEQ ID NO: 21 представляет собой аминокислотную последовательность α3 околомембранного домена МНС класса I HLA-F (F*01:01:01:01).
SEQ ID NO: 22 представляет собой аминокислотную последовательность α3 околомембранного домена МНС класса I HLA-F (F*01:02).
SEQ ID NO: 23 представляет собой аминокислотную последовательность α3 околомембранного домена МНС класса I HLA-F (F*01:03:01:01).
SEQ ID NO: 24 представляет собой аминокислотную последовательность α3 околомембранного домена МНС класса I HLA-F (F*01:04:01:01).
SEQ ID NO: 25 представляет собой аминокислотную последовательность α3 околомембранного домена МНС класса I HLA-F (F*01:05).
SEQ ID NO: 26 представляет собой аминокислотную последовательность α3 околомембранного домена МНС класса I HLA-G (G*01:01:01:01).
SEQ ID NO: 27 представляет собой аминокислотную последовательность α3 околомембранного домена МНС класса I HLA-G (G*01:03:01:01).
SEQ ID NO: 28 представляет собой аминокислотную последовательность α3 околомембранного домена МНС класса I HLA-G (G*01:04:01:01).
SEQ ID NO: 29 представляет собой аминокислотную последовательность α3 околомембранного домена МНС класса I HLA-G (G*01:06).
SEQ ID NO: 46 представляет собой аминокислотную последовательность универсального β2 микроглобулина (β2М) МНС класса I или МНС-подобных белков (CD1, HFE).
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС и второй околомембранный домен МНС, которые, соответственно, представляют собой модифицированный вариант α3 домена МНС I и модифицированный вариант β2 микроглобулина (β2М), и образуют между собой гетеродимер.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС и второй околомембранный домен МНС, которые, соответственно, представляют собой модифицированный вариант β2 микроглобулина (β2М) и модифицированный вариант α3 домена МНС I, и образуют между собой гетеродимер.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает модифицированный вариант β2 микроглобулина (β2М), который имеет аминокислотную последовательность, которую выбирают из SEQ ID NO: 47 или SEQ ID NO: 48.
SEQ ID NO: 47 представляет собой аминокислотную последовательность универсального β2 микроглобулина (β2М) МНС класса I или МНС-подобных белков (CD1, HFE) с мутацией R12C (SS мостик внутри блока).
SEQ ID NO: 48 представляет собой аминокислотную последовательность универсального β2 микроглобулина (β2М) МНС класса I или МНС-подобных белков (CD1, HFE) с мутацией R12C (SS мостик внутри блока) + шарнир (hinge) с мутацией С220А (SS мостик убран с С-конца блока и перенесен внутрь блока).
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС-подобного белка и второй околомембранный домен МНС-подобного белка, которые, соответственно, представляют собой α3 домен CD1 и β2 микроглобулин (β2М), и образуют между собой гетеродимер.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС-подобного белка и второй околомембранный домен МНС-подобного белка, которые, соответственно, представляют собой β2 микроглобулин (β2М) и α3 домен CD1, и образуют между собой гетеродимер.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает α3 домен CD1, который имеет аминокислотную последовательность, которую выбирают из группы SEQ ID NO: 40-44, и β2 микроглобулин (β2М), который имеет аминокислотную последовательность SEQ ID NO: 46.
SEQ ID NO: 40 представляет собой аминокислотную последовательность α3 околомембранного домена CD1a.
SEQ ID NO: 41 представляет собой аминокислотную последовательность α3 околомембранного домена CD1b.
SEQ ID NO: 42 представляет собой аминокислотную последовательность α3 околомембранного домена CD1e.
SEQ ID NO: 43 представляет собой аминокислотную последовательность α3 околомембранного домена CD1d.
SEQ ID NO: 44 представляет собой аминокислотную последовательность α3 околомембранного домена CD1e.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС-подобного белка и второй околомембранный домен МНС-подобного белка, которые, соответственно, представляют собой модифицированный вариант α3 домена CD1 и модифицированный вариант β2 микроглобулина (β2М), и образуют между собой гетеродимер.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС-подобного белка и второй околомембранный домен МНС-подобного белка, которые, соответственно, представляют собой модифицированный вариант β2 микроглобулина (β2М) и модифицированный вариант α3 домена CD1, и образуют между собой гетеродимер.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает модифицированный вариант α3 домена CD1, который имеет аминокислотную последовательность, которую выбирают из группы SEQ ID NO: 49-56 или SEQ ID NO: 109, и модифицированный вариант β2 микроглобулина (β2М), который имеет аминокислотную последовательность, которую выбирают из SEQ ID NO: 47 или SEQ ID NO: 48.
SEQ ID NO: 49 представляет собой аминокислотную последовательность α3 околомембранного домена CD1b с мутацией N57C (дополнительный SS мостик внутри блока) и удлинением на 3 аминокислоты GSC на С-конце.
SEQ ID NO: 50 представляет собой аминокислотную последовательность α3 околомембранного домена CD1b с мутацией N57C (SS мостик внутри блока) и удлинением на 2 аминокислоты GS на С-конце (SS мостик убран с С-конца блока и перенесен внутрь блока).
SEQ ID NO: 51 представляет собой аминокислотную последовательность α3 околомембранного домена CD1b с мутацией N59A (Замена в предсказанном сайте N-гликозилирования) и удлинением на 3 аминокислоты GSC на С-конце.
SEQ ID NO: 52 представляет собой аминокислотную последовательность α3 околомембранного домена CD1b с мутацией N59D (Замена в предсказанном сайте N-гликозилирования) и удлинением на 3 аминокислоты GSC на С-конце.
SEQ ID NO: 53 представляет собой аминокислотную последовательность α3 околомембранного домена CD1b с мутациями N57C, N59A и удлинением на 3 аминокислоты GSC на С-конце (дополнительный SS-мостик внутри блока + убран предсказанный сайт N-гликозилирования).
SEQ ID NO: 54 представляет собой аминокислотную последовательность α3 околомембранного домена CD1b с мутациями N57C, N59D и удлинением на 3 аминокислоты GSC на С-конце (дополнительный SS-мостик внутри блока + убран предсказанный сайт N-гликозилирования).
SEQ ID NO: 55 представляет собой аминокислотную последовательность α3 околомембранного домена CD1b с мутациями N57C, N59A и удлинением на 2 аминокислоты GS на С-конце (SS мостик перенесен внутрь блока + убран предсказанный сайт N-гликозилирования).
SEQ ID NO: 56 представляет собой аминокислотную последовательность α3 околомембранного домена CD1b с мутациями N57C, N59D и удлинением на 2 аминокислоты GS на С-конце (SS мостик перенесен внутрь блока + убран предсказанный сайт N-гликозилирования).
SEQ ID NO: 109 представляет собой аминокислотную последовательность α3 околомембранного домена CD1b с удлинением на 3 аминокислоты GSC на С-конце.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС-подобного белка и второй околомембранный домен МНС-подобного белка, которые, соответственно, представляют собой α3 домен HFE и β2 микроглобулин (β2М), и образуют между собой гетеродимер.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС-подобного белка и второй околомембранный домен МНС-подобного белка, которые, соответственно, представляют собой β2 микроглобулин (β2М) и α3 домен HFE, и образуют между собой гетеродимер.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает α3 домен HFE, который имеет аминокислотную последовательность SEQ ID NO: 45, и β2 микроглобулин (β2М), который имеет аминокислотную последовательность SEQ ID NO: 46.
SEQ ID NO: 45 представляет собой аминокислотную последовательность α3 околомембранного домена HFE.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС-подобного белка и второй околомембранный домен МНС-подобного белка, которые, соответственно, представляют собой модифицированный вариант α3 домена HFE и модифицированный вариант β2 микроглобулина (β2М), и образуют между собой гетеродимер.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает первый околомембранный домен МНС-подобного белка и второй околомембранный домен МНС-подобного белка, которые, соответственно, представляют собой модифицированный вариант β2 микроглобулина (β2М) и модифицированный вариант α3 домена HFE, и образуют между собой гетеродимер.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает модифицированный вариант β2 микроглобулина (β2М), который имеет аминокислотную последовательность, которую выбирают из SEQ ID NO: 47 или SEQ ID NO: 48.
Известны более 1000 охарактеризованных структур МНС, CD1 и HFE. Любая из вышеуказанных структур может быть использована в биспецифическом антителе по изобретению.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает модифицированный вариант МНС или МНС-подобного белка, где под модифицированным вариантом подразумевается вариант, включающий замены на цистеин (С) для образования дисульфидного мостика между цепями гетеродимера, полученного из первого и второго околомембранных доменов МНС или МНС-подобного белка.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает модифицированный вариант МНС или МНС-подобного белка, где под модифицированным вариантом подразумевается вариант, включающий одну или более замен в различных позициях околомембранных доменов МНС или МНС-подобного белка, приводящие к увеличению термодинамической стабильности Tm более чем на 1 градус Цельсия в сравнении с дикой формой околомембранных доменов МНС или МНС-подобного белка, соответственно.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает модифицированный вариант МНС или МНС-подобного белка, где под модифицированным вариантом подразумевается вариант, включающий одну или более замен в различных позициях околомембранных доменов МНС или МНС-подобного белка, приводящие к уменьшению количества агрегатов на более чем 5% при концентрации выше 10 мг/мл в сравнении с дикой формой околомембранных доменов МНС или МНС-подобного белка, соответственно.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает модифицированный вариант МНС или МНС-подобного белка, где под модифицированным вариантом подразумевается вариант, включающий одну или более замен в различных позициях околомембранных доменов МНС или МНС-подобного белка, приводящие удалению сайтов гликозилирования в сравнении с дикой формой околомембранных доменов МНС или МНС-подобного белка, соответственно.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает вариабельный домен первой легкой цепи и вариабельный домен второй легкой цепи, которые идентичны.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает Fc-фрагмент, который относится к IgG.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает Fc-фрагмент, который выбирают из группы: IgG1, IgG2, или IgG4 человека.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает мономер Fc-фрагмента, где дополнительно вводят замены, приводящие к отсутствию ADCC, CDC и/или ADCP свойств у двухвалентного биспецифического антитела.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает мономер Fc-фрагмента, где дополнительно вводят замены LALA (L234A и L235A).
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает мономер Fc-фрагмента, где дополнительно вводят замены, приводящие к пролонгированному действия антитела.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает мономер Fc-фрагмента, где дополнительно вводят замены YTE (M252Y, S254T и Т256Е).
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает мономер Fc-фрагмента, где дополнительно вводят замены, приводящие к усилению ADCC, CDC и/или ADCP свойств у двухвалентного биспецифического антитела.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело включает мономер Fc-фрагмента, где дополнительно вводят замену E345R.
Вышеуказанные мутации в Fc фрагменте пронумерованы согласно нумерации аминокислот цепей антител EU (Edelman G.M. и др., Proc. Natl. Acad. Sci. USA 63,1969, ее. 78-85; Kabat E.A. и др., Sequences of Proteins of Immunological Interest, 5-ое изд., изд-во Public Health Service, National Institutes of Health, Bethesda, MD, 1991)».
Под мутациями в Fc-фрагменте предлагается модификация(и) аминокислотных последовательностей антител, описанных в настоящей заявке. Варианты аминокислотной последовательности антитела получают введением соответствующих изменений нуклеотидов в нуклеиновую кислоту антитела или пептидным синтезом. Такие модификации включают, например, делеции, и/или инсерции, и/или замены остатков в аминокислотных последовательностях антитела. Осуществляют любое сочетание делеции, инсерции и замены, чтобы получить конечную конструкцию, при условии, что конечная конструкция обладает требуемыми характеристиками.
Вариант модификации аминокислотных последовательностей антител с помощью аминокислотных замен представляет собой замену, по меньшей мере, одного аминокислотного остатка в молекуле антитела на другой остаток.
Консервативные замены показаны в таблице А под заголовком «предпочтительные замены».
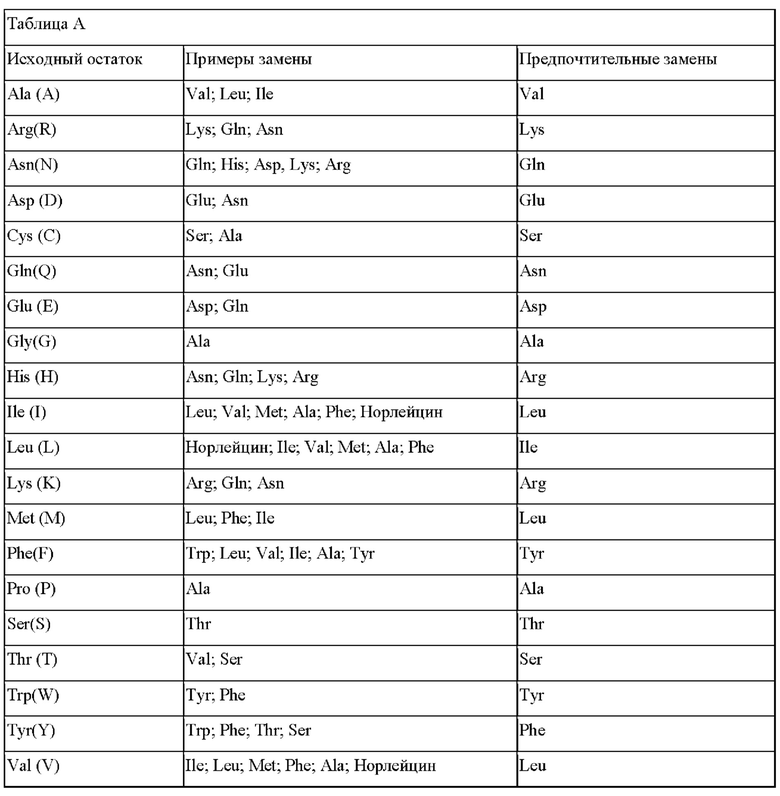
Понятие «эффекторная функция» антитела относится к видам биологической активности, связанным с Fc-областью (нативной последовательностью Fc-области или с вариантами аминокислотной последовательности Fc-области) антитела, и варьируют в зависимости от изотипа антитела. Примерами эффекторных функций антитела являются: Clq- связывание; комплементзависимая цитотоксичность; связывание Fc-рецептора; антитело-зависимая клеточно-опосредованная цитотоксичность (ADCC); фагоцитоз; понижающая регуляция рецепторов клеточной поверхности (например, В-клеточного рецептора, BCR) и В-клеточная активация.
«Антитело-зависимая клеточная цитотоксичность» или «ADCC» относится к опосредованному клетками ответу, при котором неспецифичные цитотоксические клетки, которые экспрессируют рецепторы Fc (FcR) (например, природные клетки-киллеры (NK), нейтрофилы и макрофаги), узнают связанное антитело на клетке-мишени и затем вызывают лизис или фагоцитоз клетки-мишени. Первичные клетки для опосредования ADCC, NK-клетки, экспрессируют только FcγRIII, тогда как моноциты экспрессируют FcγRI, FcγRII и FcγRIII. Экспрессия FcR на гематопоэтических клетках суммирована в таблице 3 на странице 464 в публикации Ravetch and Kinet, Annu. Rev. Immunol 9: 457-92 (1991). Чтобы оценить активность в ADCC представляющей интерес молекулы можно осуществить анализы ADCC in vitro, такие как анализы, описанные в патентах США №5500362 или 5821337. Применимые эффекторные клетки для таких анализов включают мононуклеарные клетки периферической крови (РВМС) и природные клетки-киллеры (NK). Альтернативно или дополнительно ADCC-активность представляющей интерес молекулы можно оценить in vivo, например, в животной модели, такой как модель, описанная в Clynes et al. PNAS (USA) 95: 652-656 (1998).
«Эффекторными клетками человека» являются лейкоциты, которые экспрессируют один или несколько FcR и осуществляют эффекторные функции. Предпочтительно клетки экспрессируют, по меньшей мере, FcγRIII и осуществляют ADCC-эффекторную функцию. Примеры лейкоцитов человека, которые опосредуют ADCC, включают мононуклеарные клетки периферической крови (РВМС), природные клетки-киллеры (NK), моноциты, цитотоксические Т-клетки и нейтрофилы; при этом предпочтительны РВМС и NK-клетки. Эффекторные клетки могут быть выделены из их природного источника, например из крови или РВМС, как описано в настоящей публикации.
«Комплемент-зависимая цитотоксичность» и «CDC» относятся к способности молекулы лизировать мишень в присутствии комплемента. Путь активации комплемента инициируется связыванием первого компонента системы комплемента (C1q) с молекулой (например, антителом) в комплексе со своим антигеном. Чтобы оценить активацию комплемента можно осуществить анализ CDC, например, как описано в Gazzano-Santoro et al., J. Immunol. Methods 202: 163 (1996).
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело специфически связывается с CD20 и CD3.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело специфически связывается с ВСМА и CD3.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело специфически связывается с PD-L1 и CD47.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело специфически связывается с фактором свертывания крови 9 (FIX) и фактором свертывания крови 10 (FX).
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело специфически связывается с GD2 и CD3.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело специфически связывается с AXL и CD3.
В некоторых вариантах осуществления изобретения двухвалентное биспецифическое химерное антитело специфически связывается с PD-L1 и TGF beta.
В материалах заявки приведены следующие антитела и антитело-подобные молекулы: 01-001, 01-002, 01-003, 01-004, 01-005, 01-006, 01-007, 01-008, 01-009, 01-010, 01-011, 01-012, 02-004, 02-005, 02-006, 02-007, 02-008, 02-009, 03-001, 03-002, 03-003, 03-004, 03-005, 03-006, 03-007, 03-008.
01-001 представляет собой модельную моноспецифическую антитело-подобную молекулу, которая включает тяжелую цепь на основе VH и СН1, а также легкую цепь на основе VL и CD1b, где VH представляет собой аминокислотную последовательность вариабельного домена тяжелой цепи антитела Пролголимаб (Prolgolimab_VH) с SEQ ID NO: 103, VL представляет собой аминокислотную последовательность вариабельного домена легкой цепи антитела антитела Пролголимаб (Prolgolimab_VL) с SEQ ID NO: 104, a CD1b представляет собой α3 домен CD1b, который имеет аминокислотную последовательность SEQ ID NO: 109.
01-002 представляет собой модельную моноспецифическую антитело-подобную молекулу, которая включает тяжелую цепь на основе VH и b2M, а также легкую цепь на основе VL и CK, где VH представляет собой аминокислотную последовательность вариабельного домена тяжелой цепи антитела Пролголимаб (Prolgolimab_VH) с SEQ ID NO: 103, b2M представляет собой β2 микроглобулин, который имеет аминокислотную последовательность SEQ ID NO: 46, a VL представляет собой аминокислотную последовательность вариабельного домена легкой цепи антитела Пролголимаб (Prolgolimab_VL) с SEQ ID NO: 104.
01-003 представляет собой модельную моноспецифическую антитело-подобную молекулу, которая включает тяжелую цепь на основе VH и СН1, а также легкую цепь на основе VL и CD1b, где VH представляет собой аминокислотную последовательность вариабельного домена тяжелой цепи антитела антитела Окрелизумаб (Ocrelizumab_VH) с SEQ ID NO: 105, VL представляет собой аминокислотную последовательность вариабельного домена легкой цепи антитела антитела Окрелизумаб (Ocrelizumab_VL) с SEQ ID NO: 106, a CD1b представляет собой α3 домен CD1b, который имеет аминокислотную последовательность SEQ ID NO: 109.
01-004 представляет собой модельную моноспецифическую антитело-подобную молекулу, которая включает тяжелую цепь на основе VH и СН1, а также легкую цепь на основе VL и CK, где VH представляет собой аминокислотную последовательность вариабельного домена тяжелой цепи антитела Пролголимаб (Prolgolimab_VH) с SEQ ID NO: 103, a VL представляет собой аминокислотную последовательность вариабельного домена легкой цепи антитела Окрелизумаб (Ocrelizumab_VL) с SEQ ID NO: 106.
01-005 представляет собой модельную моноспецифическую антитело-подобную молекулу, которая включает тяжелую цепь на основе VH и b2M, а также легкую цепь на основе VL и CD1b, где VH представляет собой аминокислотную последовательность вариабельного домена тяжелой цепи антитела Пролголимаб (Prolgolimab_VH) с SEQ ID NO: 103, b2M представляет собой β2 микроглобулин, который имеет аминокислотную последовательность SEQ ID NO: 46, VL представляет собой аминокислотную последовательность вариабельного домена легкой цепи антитела антитела Окрелизумаб (Ocrelizumab_VL) с SEQ ID NO: 106, a CD1b представляет собой α3 домен CD1b, который имеет аминокислотную последовательность SEQ ID NO: 109.
01-006 представляет собой модельную моноспецифическую антитело-подобную молекулу, которая включает тяжелую цепь на основе VH и СН1, а также легкую цепь на основе VL и CK, где VH представляет собой аминокислотную последовательность вариабельного домена тяжелой цепи антитела Окрелизумаб (Ocrelizimiab_VH) с SEQ ID NO: 105, a VL представляет собой аминокислотную последовательность вариабельного домена легкой цепи антитела Пролголимаб (Prolgolimab_VL) с SEQ ID NO: 104.
01-007 представляет собой модельную моноспецифическую антитело-подобную молекулу, которая включает тяжелую цепь на основе VH и b2M, а также легкую цепь на основе VL и CD1b, где VH представляет собой аминокислотную последовательность вариабельного домена тяжелой цепи антитела Окрелизумаб (Ocrelizimiab_VH) с SEQ ID NO: 105, b2M представляет собой β2 микроглобулин, который имеет аминокислотную последовательность SEQ ID NO: 46, VL представляет собой аминокислотную последовательность вариабельного домена легкой цепи антитела антитела Пролголимаб (Prolgolimab_VL) с SEQ ID NO: 104, a CD1b представляет собой α3 домен CD1b, который имеет аминокислотную последовательность SEQ ID NO: 109.
01-008 представляет собой модельную моноспецифическую антитело-подобную молекулу, которая включает тяжелую цепь на основе VH и СН1, а также легкую цепь на основе VL и CD1b, где VH представляет собой аминокислотную последовательность вариабельного домена тяжелой цепи антитела антитела Пролголимаб (Prolgolimab_VH) с SEQ ID NO: 103, VL представляет собой аминокислотную последовательность вариабельного домена легкой цепи антитела антитела Окрелизумаб (Ocrelizumab_VL) с SEQ ID NO: 106, a CD1b представляет собой α3 домен CD1b, который имеет аминокислотную последовательность SEQ ID NO: 109.
01-009 представляет собой модельную моноспецифическую антитело-подобную молекулу, которая включает тяжелую цепь на основе VH и b2M, а также легкую цепь на основе VL и CK, где VH представляет собой аминокислотную последовательность вариабельного домена тяжелой цепи антитела Пролголимаб (Prolgolimab_VH) с SEQ ID NO: 103, b2M представляет собой β2 микроглобулин, который имеет аминокислотную последовательность SEQ ID NO: 46, a VL представляет собой аминокислотную последовательность вариабельного домена легкой цепи антитела Окрелизумаб (Ocrelizumab_VL) с SEQ ID NO: 106.
01-010 представляет собой модельную моноспецифическую антитело-подобную молекулу, которая включает тяжелую цепь на основе VH и СН1, а также легкую цепь на основе VL и CD1b, где VH представляет собой аминокислотную последовательность вариабельного домена тяжелой цепи антитела Окрелизумаб (Ocrelizimiab_VH) с SEQ ID NO: 105, VL представляет собой аминокислотную последовательность вариабельного домена легкой цепи антитела антитела Пролголимаб (Prolgolimab_VL) с SEQ ID NO: 104, a CD1b представляет собой α3 домен CD1b, который имеет аминокислотную последовательность SEQ ID NO: 109.
01-011 представляет собой модельную моноспецифическую антитело-подобную молекулу, которая включает тяжелую цепь на основе VH и b2M, а также легкую цепь на основе VL и CD1b, где VH представляет собой аминокислотную последовательность вариабельного домена тяжелой цепи антитела Пролголимаб (Prolgolimab_VH) с SEQ ID NO: 103, b2M представляет собой β2 микроглобулин, который имеет аминокислотную последовательность SEQ ID NO: 46, VL представляет собой аминокислотную последовательность вариабельного домена легкой цепи антитела антитела Пролголимаб (Prolgolimab_VL) с SEQ ID NO: 104, a CD1b представляет собой α3 домен CD1b, который имеет аминокислотную последовательность SEQ ID NO: 109.
01-012 представляет собой модельную моноспецифическую антитело-подобную молекулу, которая включает тяжелую цепь на основе VH и СН1, а также легкую цепь на основе VL и CK, где VH представляет собой аминокислотную последовательность вариабельного домена тяжелой цепи антитела Пролголимаб (Prolgolimab_VH) с SEQ ID NO: 103, a VL представляет собой аминокислотную последовательность вариабельного домена легкой цепи антитела Пролголимаб (Prolgolimab_VL) с SEQ ID NO: 104.
02-004 представляет собой двухвалентное биспецифическое химерное антитело, которое специфически связывается с PD1 и CD20, и включает:
а) первую легкую цепь и первую тяжелую цепь антитела, специфически связывающегося с PD1;
где первая легкая цепь содержит вариабельный домен легкой цепи антитела Пролголимаб (Prolgolimab_VL) с SEQ ID NO: 104 и константный домен легкой цепи;
где первая тяжелая цепь содержит вариабельный домен тяжелой цепи антитела Пролголимаб (Prolgolimab_VH) с SEQ ID NO: 103 и константные домены тяжелой цепи антитела, включающие первый (СН1) константный домен тяжелой цепи и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи, при этом СН3 включает аминокислотные замены для образования Hole;
б) вторую легкую цепь и вторую тяжелую цепь антитела, специфически связывающегося с CD20,
где вторая легкая цепь содержит вариабельный домен легкой цепи антитела Окрелизумаб (Ocrelizumab_VL) с SEQ ID NO: 106 и околомембранный α3 домен CD1b, который имеет аминокислотную последовательность SEQ ID NO: 109;
где вторая тяжелая цепь, содержит вариабельный домен тяжелой цепи антитела Окрелизумаб (Ocrelizumab_VH) с SEQ ID NO: 105, околомембранный домен β2 микроглобулина, который имеет аминокислотную последовательность SEQ ID NO: 46, и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи, при этом СН3 включает аминокислотные замены для образования Knob.
02-004 представляет собой двухвалентное биспецифическое химерное антитело, которое специфически связывается с PD1 и CD20, и включает:
а) первую легкую цепь и первую тяжелую цепь антитела, специфически связывающегося с PD1;
где первая легкая цепь содержит аминокислотную последовательность SEQ ID NO: 58 (Prolgolimab_VL_CK);
где первая тяжелая цепь содержит аминокислотную последовательность SEQ ID NO: 57 (Prolgolimab_VH_HC_hole);
б) вторую легкую цепь и вторую тяжелую цепь антитела, специфически связывающегося с CD20,
где вторая легкая цепь содержит аминокислотную последовательность SEQ ID NO: 60 (Ocrelizumab_VL_CD1b);
где вторая тяжелая цепь содержит аминокислотную последовательность SEQ ID NO: 59 (Ocrelizumab_VH_b2m_Fc_knob).
02-005 представляет собой двухвалентное биспецифическое химерное антитело, которое специфически связывается с PD1 и CD20, и включает:
а) первую легкую цепь и первую тяжелую цепь антитела, специфически связывающегося с CD20;
где первая легкая цепь содержит вариабельный домен легкой цепи антитела Окрелизумаб (Ocrelizumab_VL) с SEQ ID NO: 106 и константный домен легкой цепи;
где первая тяжелая цепь содержит вариабельный домен тяжелой цепи антитела Окрелизумаб (Ocrelizumab_VH) с SEQ ID NO: 105 и константные домены тяжелой цепи антитела, включающие первый (СН1) константный домен тяжелой цепи и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи, при этом СН3 включает аминокислотные замены для образования Hole;
б) вторую легкую цепь и вторую тяжелую цепь антитела, специфически связывающегося с PD1,
где вторая легкая цепь содержит вариабельный домен легкой цепи антитела Пролголимаб (Prolgolimab_VL) с SEQ ID NO: 104 и околомембранный α3 домен CD1b, который имеет аминокислотную последовательность SEQ ID NO: 109;
где вторая тяжелая цепь, содержит вариабельный домен тяжелой цепи антитела Пролголимаб (Prolgolimab_VH) с SEQ ID NO: 103, околомембранный домен β2 микроглобулина, который имеет аминокислотную последовательность SEQ ID NO: 46, и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи, при этом СН3 включает аминокислотные замены для образования Knob.
02-005 представляет собой двухвалентное биспецифическое химерное антитело, которое специфически связывается с PD1 и CD20, и включает:
а) первую легкую цепь и первую тяжелую цепь антитела, специфически связывающегося с CD20;
где первая легкая цепь содержит аминокислотную последовательность SEQ ID NO: 62 (Ocrelizumab_VL_CK);
где первая тяжелая цепь содержит аминокислотную последовательность SEQ ID NO: 61 (Ocrelizumab_VH_ HC_hole);
б) вторую легкую цепь и вторую тяжелую цепь антитела, специфически связывающегося с PD1,
где вторая легкая цепь содержит аминокислотную последовательность SEQ ID NO: 64 (Prolgolimab_VL_CD1b);
где вторая тяжелая цепь содержит аминокислотную последовательность SEQ ID NO: 63 (Prolgolimab_VH_b2m_Fc_knob).
02-006 представляет собой двухвалентное биспецифическое химерное антитело, которое специфически связывается с PD1 и CSF1R, и включает:
а) первую легкую цепь и первую тяжелую цепь антитела, специфически связывающегося с PD1;
где первая легкая цепь содержит вариабельный домен легкой цепи антитела Пролголимаб (Prolgolimab_VL) с SEQ ID NO: 104 и константный домен легкой цепи;
где первая тяжелая цепь содержит вариабельный домен тяжелой цепи антитела Пролголимаб (Prolgolimab_VH) с SEQ ID NO: 103 и константные домены тяжелой цепи антитела, включающие первый (СН1) константный домен тяжелой цепи и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи, при этом СН3 включает аминокислотные замены для образования Hole;
б) вторую легкую цепь и вторую тяжелую цепь антитела, специфически связывающегося с CSF1R,
где вторая легкая цепь содержит вариабельный домен легкой цепи антитела к CSF1R с SEQ ID NO: 108 и околомембранный α3 домен CD1b, который имеет аминокислотную последовательность SEQ ID NO: 109;
где вторая тяжелая цепь, содержит вариабельный домен тяжелой цепи антитела к CSF1R с SEQ ID NO: 107, околомембранный домен β2 микроглобулина, который имеет аминокислотную последовательность SEQ ID NO: 46 и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи, при этом СН3 включает аминокислотные замены для образования Knob.
02-006 представляет собой двухвалентное биспецифическое химерное антитело, которое специфически связывается с PD1 и CSF1R, и включает:
а) первую легкую цепь и первую тяжелую цепь антитела, специфически связывающегося с PD1;
где первая легкая цепь содержит аминокислотную последовательность SEQ ID NO: 58 (Prolgolimab_VL_CK);
где первая тяжелая цепь содержит аминокислотную последовательность SEQ ID NO: 57 (Prolgolimab_VH_HC_hole);
б) вторую легкую цепь и вторую тяжелую цепь антитела, специфически связывающегося с CSF1R,
где вторая легкая цепь содержит аминокислотную последовательность SEQ ID NO: 66 (Анти-CSF1R_VL_CD1b);
где вторая тяжелая цепь содержит аминокислотную последовательность SEQ ID NO: 65 (Анти-CSF1R_VH_b2m_Fc_knob).
02-007 представляет собой двухвалентное биспецифическое химерное антитело, которое специфически связывается с PD1 и CSF1R, и включает:
а) первую легкую цепь и первую тяжелую цепь антитела, специфически связывающегося с CSF1R;
где первая легкая цепь содержит вариабельный домен легкой цепи антитела к CSF1R с SEQ ID NO: 108 и константный домен легкой цепи;
где первая тяжелая цепь содержит вариабельный домен тяжелой цепи антитела к CSF1R с SEQ ID NO: 107 и константные домены тяжелой цепи антитела, включающие первый (СН1) константный домен тяжелой цепи и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи, при этом СН3 включает аминокислотные замены для образования Hole;
б) вторую легкую цепь и вторую тяжелую цепь антитела, специфически связывающегося с PD1,
где вторая легкая цепь содержит вариабельный домен легкой цепи антитела Пролголимаб (Prolgolimab_VL) с SEQ ID NO: 104 и околомембранный α3 домен CD1b, который имеет аминокислотную последовательность SEQ ID NO: 109;
где вторая тяжелая цепь, содержит вариабельный домен тяжелой цепи антитела Пролголимаб (Prolgolimab_VH) с SEQ ID NO: 103, околомембранный домен β2 микроглобулина, который имеет аминокислотную последовательность SEQ ID NO: 46, и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи, при этом СН3 включает аминокислотные замены для образования Knob.
02-007 представляет собой двухвалентное биспецифическое химерное антитело, которое специфически связывается с PD1 и CSF1R, и включает:
а) первую легкую цепь и первую тяжелую цепь антитела, специфически связывающегося с CSF1R;
где первая легкая цепь содержит аминокислотную последовательность SEQ ID NO: 68 (Анти-CSF1R_VL_CK);
где первая тяжелая цепь содержит аминокислотную последовательность SEQ ID NO: 67 (Анти-CSF1R_VH_HC_hole);
б) вторую легкую цепь и вторую тяжелую цепь антитела, специфически связывающегося с PD1,
где вторая легкая цепь содержит аминокислотную последовательность SEQ ID NO: 64 (Prolgolimab_VL_CD1b);
где вторая тяжелая цепь содержит аминокислотную последовательность SEQ ID NO: 63 (Prolgolimab_VH_b2m_Fc_knob).
02-008 представляет собой двухвалентное биспецифическое химерное антитело, которое специфически связывается с CD20 и CSF1R, и включает:
а) первую легкую цепь и первую тяжелую цепь антитела, специфически связывающегося с CD20;
где первая легкая цепь содержит вариабельный домен легкой цепи антитела Окрелизумаб (Ocrelizumab_VL) с SEQ ID NO: 106 и константный домен легкой цепи;
где первая тяжелая цепь содержит вариабельный домен тяжелой цепи антитела Окрелизумаб (Ocrelizumab_VH) с SEQ ID NO: 105 и константные домены тяжелой цепи антитела, включающие первый (СН1) константный домен тяжелой цепи и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи, при этом СН3 включает аминокислотные замены для образования Hole;
б) вторую легкую цепь и вторую тяжелую цепь антитела, специфически связывающегося с CSF1R,
где вторая легкая цепь содержит вариабельный домен легкой цепи антитела к CSF1R с SEQ ID NO: 108 и околомембранный α3 домен CD1b, который имеет аминокислотную последовательность SEQ ID NO: 109;
где вторая тяжелая цепь, содержит вариабельный домен тяжелой цепи антитела к CSF1R с SEQ ID NO: 107, околомембранный домен β2 микроглобулина, который имеет аминокислотную последовательность SEQ ID NO: 46, и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи, при этом СН3 включает аминокислотные замены для образования Knob.
02-008 представляет собой двухвалентное биспецифическое химерное антитело, которое специфически связывается с CD20 и CSF1R, и включает:
а) первую легкую цепь и первую тяжелую цепь антитела, специфически связывающегося с CD20;
где первая легкая цепь содержит аминокислотную последовательность SEQ ID NO: 62 (Ocrelizumab_VL_CK);
где первая тяжелая цепь содержит аминокислотную последовательность SEQ ID NO: 61 (Ocrelizumab_VH_ HC_hole);
б) вторую легкую цепь и вторую тяжелую цепь антитела, специфически связывающегося с CSF1R,
где вторая легкая цепь содержит аминокислотную последовательность SEQ ID NO: 66 (Анти-CSF1R_VL_CD1b);
где вторая тяжелая цепь содержит аминокислотную последовательность SEQ ID NO: 65 (Анти-CSF1R_VH_b2m_Fc_knob).
02-009 представляет собой двухвалентное биспецифическое химерное антитело, которое специфически связывается с CD20 и CSF1R, и включает:
а) первую легкую цепь и первую тяжелую цепь антитела, специфически связывающегося с CSF1R;
где первая легкая цепь содержит вариабельный домен легкой цепи антитела к CSF1R с SEQ ID NO: 108 и константный домен легкой цепи;
где первая тяжелая цепь содержит вариабельный домен тяжелой цепи антитела к CSF1R с SEQ ID NO: 107 и константные домены тяжелой цепи антитела, включающие первый (СН1) константный домен тяжелой цепи и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи, при этом СН3 включает аминокислотные замены для образования Hole;
б) вторую легкую цепь и вторую тяжелую цепь антитела, специфически связывающегося с CD20,
где вторая легкая цепь содержит вариабельный домен легкой цепи антитела Окрелизумаб (Ocrelizumab_VL) с SEQ ID NO: 106 и околомембранный α3 домен CD1b, который имеет аминокислотную последовательность SEQ ID NO: 109;
где вторая тяжелая цепь, содержит вариабельный домен тяжелой цепи антитела Окрелизумаб (Ocrelizumab_VH) с SEQ ID NO: 105, околомембранный домен β2 микроглобулина, который имеет аминокислотную последовательность SEQ ID NO: 46, и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи, при этом СН3 включает аминокислотные замены для образования Knob.
02-009 представляет собой двухвалентное биспецифическое химерное антитело, которое специфически связывается с CD20 и CSF1R, и включает:
а) первую легкую цепь и первую тяжелую цепь антитела, специфически связывающегося с CSF1R;
где первая легкая цепь содержит аминокислотную последовательность SEQ ID NO: 68 (Анти-CSF1R_VL_CK);
где первая тяжелая цепь содержит аминокислотную последовательность SEQ ID NO: 67 (Анти-CSF1R_VH_HC_hole);
б) вторую легкую цепь и вторую тяжелую цепь антитела, специфически связывающегося с CD20,
где вторая легкая цепь содержит аминокислотную последовательность SEQ ID NO: 60 (Ocrelizimiab_VL_CD1b);
где вторая тяжелая цепь содержит аминокислотную последовательность SEQ ID NO: 59 (Ocrelizimiab_VH_b2m_Fc_knob).
03-001 представляет собой двухвалентное моноспецифическое химерное антитело, которое специфически связывается с PD1, и включает:
легкую цепь, которая содержит вариабельный домен легкой цепи антитела Пролголимаб (Prolgolimab_VL) с SEQ ID NO: 104 и α3 околомембранный домен CD1b с мутацией N57C (дополнительный SS мостик внутри блока) и удлинением на 3 аминокислоты GSC на С-конце, который имеет аминокислотную последовательность SEQ ID NO: 49;
тяжелую цепь, которая содержит вариабельный домен тяжелой цепи антитела Пролголимаб (Prolgolimab_VH) с SEQ ID NO: 103, околомембранный домен β2 микроглобулина с мутацией R12C, который имеет аминокислотную последовательность SEQ ID NO: 47, и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи.
03-001 представляет собой двухвалентное моноспецифическое химерное антитело, которое специфически связывается с PD1, и включает:
легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 72 и
тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 69.
03-002 представляет собой двухвалентное моноспецифическое химерное антитело, которое специфически связывается с PD1, и включает:
легкую цепь, которая содержит вариабельный домен легкой цепи антитела Пролголимаб (Prolgolimab_VL) с SEQ ID NO: 104 и α3 околомембранный домен CD1b с мутацией N57C (SS мостик внутри блока) и удлинением на 2 аминокислоты GS на С-конце (SS мостик убран с С-конца блока и перенесен внутрь блока), который имеет аминокислотную последовательность SEQ ID NO: 50;
тяжелую цепь, которая содержит вариабельный домен тяжелой цепи антитела Пролголимаб (Prolgolimab_VH) с SEQ ID NO: 103, околомембранный домен β2 микроглобулина с мутацией R12C и шарниром (hinge) с мутацией С220А, которые имеют аминокислотную последовательность SEQ ID NO: 48 и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи.
03-002 представляет собой двухвалентное моноспецифическое химерное антитело, которое специфически связывается с PD1, и включает:
легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 73 и
тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 71.
03-003 представляет собой двухвалентное моноспецифическое химерное антитело, которое специфически связывается с PD1, и включает:
легкую цепь, которая содержит вариабельный домен легкой цепи антитела Пролголимаб (Prolgolimab_VL) с SEQ ID NO: 104 и α3 околомембранный домен CD1b с мутацией N59A (Замена в предсказанном сайте N-гликозилирования на легкой цепи) и удлинением на 3 аминокислоты GSC на С-конце, который имеет аминокислотную последовательность SEQ ID NO: 51;
тяжелую цепь, которая содержит вариабельный домен тяжелой цепи антитела Пролголимаб (Prolgolimab_VH) с SEQ ID NO: 103, околомембранный домен β2 микроглобулина, который имеет аминокислотную последовательность SEQ ID NO: 46, и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи.
03-003 представляет собой двухвалентное моноспецифическое химерное антитело, которое специфически связывается с PD1, и включает:
легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 74 и
тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 70.
03-004 представляет собой двухвалентное моноспецифическое химерное антитело, которое специфически связывается с PD1, и включает:
легкую цепь, которая содержит вариабельный домен легкой цепи антитела Пролголимаб (Prolgolimab_VL) с SEQ ID NO: 104 и α3 околомембранный домен CD1b с мутацией N59D (Замена в предсказанном сайте N-гликозилирования на легкой цепи) и удлинением на 3 аминокислоты GSC на С-конце, который имеет аминокислотную последовательность SEQ ID NO: 52;
тяжелую цепь, которая содержит вариабельный домен тяжелой цепи антитела Пролголимаб (Prolgolimab_VH) с SEQ ID NO: 103, околомембранный домен β2 микроглобулина, который имеет аминокислотную последовательность SEQ ID NO: 46, и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи.
03-004 представляет собой двухвалентное моноспецифическое химерное антитело, которое специфически связывается с PD1, и включает:
легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 75 и
тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 70.
03-005 представляет собой двухвалентное моноспецифическое химерное антитело, которое специфически связывается с PD1, и включает:
легкую цепь, которая содержит вариабельный домен легкой цепи антитела Пролголимаб (Prolgolimab_VL) с SEQ ID NO: 104 и α3 околомембранный домен CD1b с мутациями N57C, N59A и удлинением на 3 аминокислоты GSC на С-конце (дополнительный SS-мостик внутри блока + убран предсказанный сайт N-гликозилирования), который имеет аминокислотную последовательность SEQ ID NO: 53;
тяжелую цепь, которая содержит вариабельный домен тяжелой цепи антитела Пролголимаб (Prolgolimab_VH) с SEQ ID NO: 103, околомембранный домен β2 микроглобулина с мутацией R12C, который имеет аминокислотную последовательность SEQ ID NO: 47, и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи.
03-005 представляет собой двухвалентное моноспецифическое химерное антитело, которое специфически связывается с PD1, и включает:
легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 76 и
тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 69.
03-006 представляет собой двухвалентное моноспецифическое химерное антитело, которое специфически связывается с PD1, и включает:
легкую цепь, которая содержит вариабельный домен легкой цепи антитела Пролголимаб (Prolgolimab_VL) с SEQ ID NO: 104 и α3 околомембранный домен CD1b с мутациями N57C, N59D и удлинением на 3 аминокислоты GSC на С-конце (дополнительный SS-мостик внутри блока + убран предсказанный сайт N-гликозилирования), который имеет аминокислотную последовательность SEQ ID NO: 54;
тяжелую цепь, которая содержит вариабельный домен тяжелой цепи антитела Пролголимаб (Prolgolimab_VH) с SEQ ID NO: 103, околомембранный домен β2 микроглобулина с мутацией R12C, который имеет аминокислотную последовательность SEQ ID NO: 47, и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи.
03-006 представляет собой двухвалентное моноспецифическое химерное антитело, которое специфически связывается с PD1, и включает:
легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 77 и
тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 69.
03-007 представляет собой двухвалентное моноспецифическое химерное антитело, которое специфически связывается с PD1, и включает:
легкую цепь, которая содержит вариабельный домен легкой цепи антитела Пролголимаб (Prolgolimab_VL) с SEQ ID NO: 104 и α3 околомембранный домен CD1b с мутациями N57C, N59A и удлинением на 2 аминокислоты GS на С-конце (SS мостик перенесен внутрь блока + убран предсказанный сайт N-гликозилирования), который имеет аминокислотную последовательность SEQ ID NO: 55;
тяжелую цепь, которая содержит вариабельный домен тяжелой цепи антитела Пролголимаб (Prolgolimab_VH) с SEQ ID NO: 103, околомембранный домен β2 микроглобулина с мутацией R12C и шарниром (hinge) с мутацией С220А, которые имеют аминокислотную последовательность SEQ ID NO: 48, и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи.
03-007 представляет собой двухвалентное моноспецифическое химерное антитело, которое специфически связывается с PD1, и включает:
легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 78 и
тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 71.
03-008 представляет собой двухвалентное моноспецифическое химерное антитело, которое специфически связывается с PD1, и включает:
легкую цепь, которая содержит вариабельный домен легкой цепи антитела Пролголимаб (Prolgolimab_VL) с SEQ ID NO: 104 и α3 околомембранный домен CD1b с мутациями N57C, N59D и удлинением на 2 аминокислоты GS на С-конце (SS мостик перенесен внутрь блока + убран предсказанный сайт N-гликозилирования), который имеет аминокислотную последовательность SEQ ID NO: 56;
тяжелую цепь, которая содержит вариабельный домен тяжелой цепи антитела Пролголимаб (Prolgolimab_VH) с SEQ ID NO: 103, околомембранный домен β2 микроглобулин с мутацией R12C и шарниром (hinge) с мутацией С220А, которые имеют аминокислотную последовательность SEQ ID NO: 48 и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи.
03-008 представляет собой двухвалентное моноспецифическое химерное антитело, которое специфически связывается с PD1, и включает:
легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 79 и
тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 71.
Данные антитела приведены в иллюстративных целях для подтвеждения работоспособности формата двухвалентного биспецифического химерного антитела по изобретения, а также неожиданных его свойств. Данные антитела не должны рассматриваться как каким-то образом ограничивающие двухвалентное биспецифическое химерное антитело по изобретению.
Параметры выхода продукта с правильной гетеродимерной сборкой двух разных тяжелых цепей и правильным спариванием двух разных легких цепей с соответствующими тяжелыми цепями не зависят от вариабельных фрагментов тяжелой и легкой цепей биспецифического химерного антитела и их специфичности к антигенам.
Двухвалентные биспецифические химерные антитела по изобретению могут применяться для лечения различных заболеваний, в частности, онкологических заболеваний, аутоиммунных заболеваний или заболеваний, которые связаны с нарушением свертывания крови (коагуляции).
Нуклеиновая кислота
В одном из аспектов настоящее изобретение относится к выделенной нуклеиновой кислоте, которая кодирует любое из вышеуказанных двухвалентных биспецифических химерных антител или ее фрагментов.
Термины «нуклеиновая кислота», «нуклеиновая последовательность» или «нуклеиновокислотная последовательность», «полинуклеотид», «олигонуклеотид», «полинуклеотидная последовательность» и «нуклеотидная последовательность», которые используются равнозначно в данном описании, обозначают четкую последовательность нуклеотидов, модифицированных или не модифицированных, определяющую фрагмент или участок нуклеиновой кислоты, содержащую или не содержащую неприродные нуклеотиды и являющуюся либо двухцепочечной ДНК или РНК, либо одноцепочечной ДНК или РНК, либо продуктами транскрипции указанных ДНК.
Здесь также следует упомянуть, что данное изобретение не относится к нуклеотидным последовательностям в их природной хромосомной среде, т.е. в природном состоянии. Последовательности данного изобретения были выделены и/или очищены, т.е. были взяты прямо или косвенно, например, путем копирования, при этом их среда была по меньшей мере частично модифицирована. Таким образом, также здесь следует подразумевать изолированные нуклеиновые кислоты, полученные путем генетической рекомбинации, например, с помощью принимающих клеток (клеток-хозяев), или полученные путем химического синтеза.
Ссылка на нуклеотидную последовательность охватывает его комплимент, если не указано иное. Таким образом, ссылка на нуклеиновую кислоту, имеющую определенную последовательность следует понимать как охватывающие ее комплементарную цепь с ее комплементарной последовательностью.
В любом из указанных выше вариантов осуществления изобретения молекулы нуклеиновых кислот могут быть выделенными.
«Выделенная» молекула нуклеиновой кислоты представляет собой молекулу нуклеиновой кислоты, которая идентифицирована и отделена от по меньшей мере одной молекулы нуклеиновой кислоты-примеси, с которой она обычно связана в естественном источнике нуклеиновой кислоты антитела. Выделенная молекула нуклеиновой кислоты отличается от той формы или набора, в которых она находится в естественных условиях. Таким образом, выделенная молекула нуклеиновой кислоты отличается от молекулы нуклеиновой кислоты, существующей в клетках в естественных условиях.
В одном из аспектов настоящее изобретение относится к молекуле нуклеиновой кислоты, содержащей нуклеотидную последовательность, которая кодирует аминокислотную последовательность, выбранную из SEQ ID NO: 1-79 или SEQ ID NO: 109. Молекула нуклеиновой кислоты может также содержать любую комбинацию указанных нуклеотидных последовательностей.
В некоторых вариантах нуклеиновая кислота представляет собой ДНК.
В одном из вариантов настоящее изобретение относится к молекуле нуклеиновой кислоты, содержащей нуклеотидную последовательность, которая кодирует аминокислотную последовательность легкой или тяжелой цепи вышеуказанного биспецифического антитела по изобретению, выбранную из:
аминокислотной последовательности первой легкой цепи, которая содержит вариабельный домен легкой цепи и константный домен легкой цепи;
аминокислотной последовательности первой тяжелой цепи, которая содержит вариабельный домен тяжелой цепи и константные домены тяжелой цепи антитела, включающие первый (СН1) константный домен тяжелой цепи и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи;
аминокислотной последовательности второй легкой цепи, которая содержит вариабельный домен легкой цепи и константный домен, который выбирают из группы:
первого околомембранного домена МНС (главный комплекс гистосовместимости) или
первого околомембранного домена МНС-подобного белка;
аминокислотной последовательности второй тяжелой цепи, которая содержит вариабельный домен тяжелой цепи, константный домен, который выбирают из группы:
второго околомембранного домена МНС (главный комплекс гистосовместимости) или
второго околомембранного домена МНС-подобного белка,
и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи.
Молекула нуклеиновой кислоты может также содержать любую комбинацию указанных выше нуклеотидных последовательностей.
Специалисту в данной области будет очевидно, что пептид с аминокислотной последовательностью легкой или тяжелой цепи вышеуказанного биспецифического антитела по изобретению, выбранной из:
аминокислотной последовательности первой легкой цепи, которая содержит вариабельный домен легкой цепи и константный домен легкой цепи;
аминокислотной последовательности первой тяжелой цепи, которая содержит вариабельный домен тяжелой цепи и константные домены тяжелой цепи антитела, включающие первый (СН1) константный домен тяжелой цепи и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи;
аминокислотной последовательности второй легкой цепи, которая содержит вариабельный домен легкой цепи и константный домен, который выбирают из группы:
первого околомембранного домена МНС (главный комплекс гистосовместимости) или
первого околомембранного домена МНС-подобного белка;
аминокислотной последовательности второй тяжелой цепи, которая содержит вариабельный домен тяжелой цепи, константный домен, который выбирают из группы:
второго околомембранного домена МНС (главный комплекс гистосовместимости) или
второго околомембранного домена МНС-подобного белка,
и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи,
может быть кодирован широким рядом различных ДНК-последовательностей с учетом вырожденности генетического кода. Специалистам в данной области хорошо известно получение таких альтернативных ДНК-последовательностей, кодирующих одни и те же аминокислотные последовательности. Такие вариантные ДНК-последовательности находятся в объеме настоящего изобретения.
Молекула нуклеиновой кислоты по данному изобретению может быть выделена из любого источника, который продуцирует биспецифическое антитело по изобретению. В определенных вариантах осуществления изобретения молекула нуклеиновой кислоты по данному изобретению может быть синтезирована, а не выделена.
В некоторых вариантах осуществления изобретения нуклеиновая кислота представляет собой нуклеиновую кислоту, которая кодирует аминокислотную последовательность первой тяжелой цепи кандидатов 02-004 и 02-006 (Prolgolimab_VH_HC_hole), и включает нуклеотидную последовательность с SEQ ID NO: 80.
В некоторых вариантах осуществления изобретения нуклеиновая кислота представляет собой нуклеиновую кислоту, которая кодирует аминокислотную последовательность первой легкой цепи кандидатов 02-004 и 02-006 (Prolgolimab_VL_CK), и включает нуклеотидную последовательность с SEQ ID NO: 81.
В некоторых вариантах осуществления изобретения нуклеиновая кислота представляет собой нуклеиновую кислоту, которая кодирует аминокислотную последовательность второй тяжелой цепи кандидатов 02-004 и 02-009 (Ocrelizimiab_VH_b2m_Fc_knob), и включает нуклеотидную последовательность с SEQ ID NO: 82.
В некоторых вариантах осуществления изобретения нуклеиновая кислота представляет собой нуклеиновую кислоту, которая кодирует аминокислотную последовательность второй легкой цепи кандидатов 02-004 и 02-009 (Ocrelizimiab_VL_CD1b), и включает нуклеотидную последовательность с SEQ ID NO: 83.
В некоторых вариантах осуществления изобретения нуклеиновая кислота представляет собой нуклеиновую кислоту, которая кодирует аминокислотную последовательность первой тяжелой цепи кандидатов 02-005 и 02-008 (Ocrelizimiab_VH_ HC_hole), и включает нуклеотидную последовательность с SEQ ID NO: 84.
В некоторых вариантах осуществления изобретения нуклеиновая кислота представляет собой нуклеиновую кислоту, которая кодирует аминокислотную последовательность первой легкой цепи кандидатов 02-005 и 02-008 (Ocrelizimiab_VL_CK), и включает нуклеотидную последовательность с SEQ ID NO: 85.
В некоторых вариантах осуществления изобретения нуклеиновая кислота представляет собой нуклеиновую кислоту, которая кодирует аминокислотную последовательность второй тяжелой цепи кандидатов 02-005 и 02-007 (Prolgolimab_VH_b2m_Fc_knob), и включает нуклеотидную последовательность с SEQ ID NO: 86.
В некоторых вариантах осуществления изобретения нуклеиновая кислота представляет собой нуклеиновую кислоту, которая кодирует аминокислотную последовательность второй легкой цепи кандидатов 02-005 и 02-007 (Prolgolimab_VL_CD1b), и включает нуклеотидную последовательность с SEQ ID NO: 87.
В некоторых вариантах осуществления изобретения нуклеиновая кислота представляет собой нуклеиновую кислоту, которая кодирует аминокислотную последовательность второй тяжелой цепи кандидатов 02-006 и 02-008 (Анти-CSFlR_VH_b2m_Fc_knob), и включает нуклеотидную последовательность с SEQ ID NO: 88.
В некоторых вариантах осуществления изобретения нуклеиновая кислота представляет собой нуклеиновую кислоту, которая кодирует аминокислотную последовательность второй легкой цепи кандидатов 02-006 и 02-008 (Анти-CSF1R_VL_CD1b), и включает нуклеотидную последовательность с SEQ ID NO: 89.
В некоторых вариантах осуществления изобретения нуклеиновая кислота представляет собой нуклеиновую кислоту, которая кодирует аминокислотную последовательность первой тяжелой цепи кандидатов 02-007 и 02-009 (Анти-CSF1R_VH_HC_hole), и включает нуклеотидную последовательность с SEQ ID NO: 90.
В некоторых вариантах осуществления изобретения нуклеиновая кислота представляет собой нуклеиновую кислоту, которая кодирует аминокислотную последовательность первой легкой цепи кандидатов 02-007 и 02-009 (Анти-CSF1R_VL_CK), и включает нуклеотидную последовательность с SEQ ID NO: 91.
В некоторых вариантах осуществления изобретения нуклеиновая кислота представляет собой нуклеиновую кислоту, которая кодирует аминокислотную последовательность тяжелой цепи кандидатов 03-001 и 03-005, которая содержит универсальный β2 микроглобулина (β2М) с мутацией R12C, и включает нуклеотидную последовательность с SEQ ID NO: 92.
В некоторых вариантах осуществления изобретения нуклеиновая кислота представляет собой нуклеиновую кислоту, которая кодирует аминокислотную последовательность тяжелой цепи кандидатов 03-003 и 03-004, которая содержит универсальный β2 микроглобулина (β2М), и включает нуклеотидную последовательность с SEQ ID NO: 93.
В некоторых вариантах осуществления изобретения нуклеиновая кислота представляет собой нуклеиновую кислоту, которая кодирует аминокислотную последовательность тяжелой цепи кандидатов 03-002, 03-007 и 03-008, которая содержит универсальный β2 микроглобулина (β2М) с мутацией R12C и шарнир (hinge) с мутацией С220А, и включает нуклеотидную последовательность с SEQ ID NO: 94.
В некоторых вариантах осуществления изобретения нуклеиновая кислота представляет собой нуклеиновую кислоту, которая кодирует аминокислотную последовательность легкой цепи кандидата 03-001, которая содержит α3 околомембранный домен CD1b с мутацией N57C и удлинением на 3 аминокислоты GSC на С-конце, и включает нуклеотидную последовательность с SEQ ID NO: 95.
В некоторых вариантах осуществления изобретения нуклеиновая кислота представляет собой нуклеиновую кислоту, которая кодирует аминокислотную последовательность легкой цепи кандидата 03-002, которая содержит α3 околомембранный домен CD1b с мутацией N57C и удлинением на 2 аминокислоты GS на С-конце, и включает нуклеотидную последовательность с SEQ ID NO: 96.
В некоторых вариантах осуществления изобретения нуклеиновая кислота представляет собой нуклеиновую кислоту, которая кодирует аминокислотную последовательность легкой цепи кандидата 03-003, которая содержит α3 околомембранный домен CD1b с мутацией N59A и удлинением на 3 аминокислоты GSC на С-конце, и включает нуклеотидную последовательность с SEQ ID NO: 97.
В некоторых вариантах осуществления изобретения нуклеиновая кислота представляет собой нуклеиновую кислоту, которая кодирует аминокислотную последовательность легкой цепи кандидата 03-004, которая содержит α3 околомембранный домен CD1b с мутацией N59D и удлинением на 3 аминокислоты GSC на С-конце, и включает нуклеотидную последовательность с SEQ ID NO: 98.
В некоторых вариантах осуществления изобретения нуклеиновая кислота представляет собой нуклеиновую кислоту, которая кодирует аминокислотную последовательность легкой цепи кандидата 03-005, которая содержит α3 околомембранный домен CD1b с мутациями N57C, N59A и удлинением на 3 аминокислоты GSC на С-конце, и включает нуклеотидную последовательность с SEQ ID NO: 99.
В некоторых вариантах осуществления изобретения нуклеиновая кислота представляет собой нуклеиновую кислоту, которая кодирует аминокислотную последовательность легкой цепи кандидата 03-006, которая содержит α3 околомембранный домен CD1b с мутациями N57C, N59D и удлинением на 3 аминокислоты GSC на С-конце, и включает нуклеотидную последовательность с SEQ ID NO: 100.
В некоторых вариантах осуществления изобретения нуклеиновая кислота представляет собой нуклеиновую кислоту, которая кодирует аминокислотную последовательность легкой цепи кандидата 03-007, которая содержит α3 околомембранный домен CD1b с мутациями N57C, N59A и удлинением на 2 аминокислоты GS на С-конце, и включает нуклеотидную последовательность с SEQ ID NO: 101.
В некоторых вариантах осуществления изобретения нуклеиновая кислота представляет собой нуклеиновую кислоту, которая кодирует аминокислотную последовательность легкой цепи кандидата 03-008, которая содержит α3 околомембранный домен CD1b с мутациями N57C, N59D и удлинением на 2 аминокислоты GS на С-конце, и включает нуклеотидную последовательность с SEQ ID NO: 102.
Молекулы нуклеиновой кислоты могут использоваться для экспрессии двухвалентных биспецифических химерных антител по изобретению.
Экспрессионной вектор
В одном из аспектов настоящее изобретение относится к экспрессионному вектору, содержащему вышеуказанную выделенную нуклеиновую кислоту. Настоящее изобретение относится к вектору, подходящему для экспрессии любой из нуклеотидных последовательностей, описанных в настоящем документе.
Термин «вектор» при использовании в настоящем документе означает молекулу нуклеиновой кислоты, способную транспортировать другую нуклеиновую кислоту, с которой она соединена. В некоторых вариантах осуществления изобретения вектор представляет собой плазмиду, т.е. кольцевую двуцепочечную часть ДНК, в которую могут быть лигированы дополнительные сегменты ДНК. В некоторых вариантах осуществления изобретения вектор представляет собой вирусный вектор, в котором дополнительные сегменты ДНК могут быть лигированы в вирусный геном. В некоторых вариантах осуществления изобретения векторы способны к автономной репликации в клетке-хозяине, в которую они введены (например, бактериальные векторы, имеющие бактериальный сайт инициации репликации и эписомные векторы млекопитающих). В других вариантах осуществления изобретения векторы (например, неэписомальные векторы млекопитающих) могут быть интегрированы в геном клетки-хозяина при введении в клетку-хозяина, и таким образом реплицируются вместе с геном хозяина. Более того, некоторые векторы способны направлять экспрессию генов, с которыми они функционально соединены. Такие векторы упоминаются в данном документе как «рекомбинантные экспрессирующие векторы» (или просто «экспрессирующие векторы» («вектор экспрессии» или «экспрессионный вектор»)).
Настоящее изобретение относится к векторам, содержащим вышеуказанные молекулы нуклеиновых кислот, которые кодируют любое из вышеуказанных двухвалентных биспецифических антител или его конструктивных частей, выбранных из:
аминокислотной последовательности первой легкой цепи, которая содержит вариабельный домен легкой цепи и константный домен легкой цепи;
аминокислотной последовательности первой тяжелой цепи, которая содержит вариабельный домен тяжелой цепи и константные домены тяжелой цепи антитела, включающие первый (СН1) константный домен тяжелой цепи и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи;
аминокислотной последовательности второй легкой цепи, которая содержит вариабельный домен легкой цепи и константный домен, который выбирают из группы:
первого околомембранного домена МНС (главный комплекс гистосовместимости) или
первого околомембранного домена МНС-подобного белка;
аминокислотной последовательности второй тяжелой цепи, которая содержит вариабельный домен тяжелой цепи, константный домен, который выбирают из группы:
второго околомембранного домена МНС (главный комплекс гистосовместимости) или
второго околомембранного домена МНС-подобного белка,
и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи,
как описано в настоящем документе.
Экспрессионные векторы включают плазмиды, ретровирусы, аденовирусы, аденоассоциированные вирусы (AAV), вирусы растений, такие как вирус мозаики цветной капусты, вирусы табачной мозаики, космиды, YAC, EBV полученные эписомы и тому подобное. Молекулы ДНК могут быть лигированы в вектор таким образом, что последовательности, контролирующие транскрипцию и трансляцию в векторе, выполняют предусмотренную функцию регуляции транскрипции и трансляции ДНК. Экспрессионный вектор и последовательности контроля экспрессии могут быть выбраны таким образом, чтобы быть совместимыми с используемой экспрессирующей клеткой-хозяином. Молекулы ДНК, кодирующие частично или по всей длине последовательности первого и второго связывающих доменов (например, последовательности тяжелой и легкой цепи, где связывающий домен содержит последовательность тяжелой и легкой цепи) могут быть введены в отдельные векторы. В одном варианте осуществления изобретения любая комбинация указанных выше молекул ДНК вводится в тот же экспрессионный вектор. Молекулы ДНК могут быть введены в экспрессионный вектор стандартными способами (например, лигированием комплементарных сайтов рестрикции на фрагменте гена антитела и вектора или лигированием тупых концов, если сайты рестрикции отсутствуют).
В некоторых вариантах осуществления изобретения подходящим вектором является тот, который включает места рестрикции так, что любая последовательность VH или VL может быть легко включена и экспрессирована, как описано выше. Прекращение полиаденилирования и транскрипции может произойти вниз по ходу сайта нативной хромосомы кодируемых участков. Рекомбинантный экспрессионный вектор также может кодировать сигнальный пептид, который облегчает выработку цепочки антитела клеткой-хозяином. Ген цепочки антитела может быть клонирован в вектор таким образом, что сигнальный пептид соединен с рамкой считывания аминоконца цепи иммуноглобулина. Сигнальным пептидом может быть сигнальный пептид иммуноглобулина или гетерологичный сигнальный пептид (то есть, сигнальный пептид белка не иммуноглобулиновой природы).
В некоторых вариантах осуществления изобретения помимо цепочки генов антител, рекомбинантная экспрессия векторов по данному изобретению может нести регулирующие последовательности, которые контролируют экспрессию генов цепи антител в клетке-хозяине. Специалистам в этой области будет понятно, что дизайн экспрессионного вектора, включая выбор регулирующих последовательностей, может зависеть от таких факторов, как селекция клетки-хозяина для трансформации, уровень экспрессии желаемого белка, и т.д. Предпочтительные регулирующие последовательности для экспрессирующей клетки-хозяина млекопитающих включают вирусные элементы обеспечивающие высокий уровень экспрессии белков в клетках млекопитаю1 ггих, таких как промоторы и/или энхансеры, полученные из ретровирусной LTR, цитомегаловируса (CMV) (например, CMV промотора/энхансера), обезьяньего вируса 40 (SV40) (например, SV40 промотора/энхансера), аденовируса, (например, большого позднего промотора аденовируса (AdMLP)), вирус полиомы, а также сильных промоторов млекопитающих, таких как промотор нативных иммуноглобулинов или промотор актина. Методы экспрессии полипептидов в бактериальных клетках или клетках грибов, например, дрожжевых клетках, также хорошо известны в данной области техники.
В некоторых вариантах осуществления изобретения в дополнение к генам цепи антитела и регулирующим последовательностям, рекомбинантные векторы экспрессии изобретения могут нести дополнительные последовательности, такие как последовательности, которые регулируют репликацию вектора в клетках-хозяевах (например, точки начала репликации) и гены селектируемого маркера. Ген селектируемого маркера облегчает селекцию клеток-хозяев, в которые был введен вектор.
В некоторых вариантах осуществления изобретения вектор может включать последовательность контроля экспрессии. Термин «последовательность контроля экспрессии», используемый в данном описании, означает полинуклеотидные последовательности, которые необходимы для воздействия на экспрессию и процессинг кодирующих последовательностей, к которым они лигированы. Контролирующие экспрессию последовательности включают соответствующие последовательности инициации транскрипции, терминации, промотора и энхансера; эффективные сигналы процессинга РНК, такие как сплайсинг и сигналы полиаденилирования; последовательности, которые стабилизируют цитоплазматическую мРНК; последовательности, которые повышают эффективность трансляции (т.е. консенсусная последовательность Козака); последовательности, которые повышают стабильность белка; и, при желании, последовательности, которые усиливают секрецию белка. Характер таких контролирующих последовательностей различается в зависимости от организма-хозяина; в прокариотах такие контролирующие последовательности, как правило, включают промотор, сайт связывания рибосомы, а также последовательности терминации транскрипции; в эукариотах, как правило, такие контролирующие последовательности включают промоторы и последовательности терминации транскрипции. Термин «контролирующие последовательности» включает, как минимум, все компоненты, наличие которых имеет важное значение для экспрессии и процессинга, и может также включать дополнительные компоненты, чье присутствие является полезным, например, лидирующие последовательности и последовательности слившихся клеток.
Клетки-хозяева и способ их получения
В одном из аспектов настоящее изобретение относится к способу получения клетки-хозяина для получения любого из вышеуказанных двухвалентных биспецифических химерных антител и включает трансформирование клетки вышеуказанным экспрессионным вектором.
В одном из аспектов настоящее изобретение относится к клетке-хозяину для получения любого из вышеуказанных двухвалентных биспецифических химерных антител, которая содержит любую из вышеуказанных нуклеиновых кислот.
Термин «рекомбинантная клетка-хозяин» (или просто «клетка-хозяин») при использовании в данном документе означает клетку, в которую введен рекомбинантный экспрессионный вектор. Настоящее изобретение относится к клеткам-хозяевам, которые могут включать, например, вектор в соответствии с настоящим изобретением, описанным выше.
Настоящее изобретение относится также к клеткам-хозяевам, которые включают, например, нуклеотидную последовательность, кодирующую первую тяжелую цепь двухвалентного биспецифического химерного антитела по изобретению, нуклеотидную последовательность, кодирующую первую легкую цепь двухвалентного биспецифического химерного антитела по изобретению, нуклеотидную последовательность, кодирующую вторую тяжелую цепь двухвалентного биспецифического химерного антитела по изобретению или нуклеотидную последовательность, кодирующую вторую легкую цепь двухвалентного биспецифического химерного антитела по изобретению или все четыре вышеуказанные последовательности. Следует понимать, что «рекомбинантная клетка-хозяин» и «клетка-хозяин» означают не только конкретную заявленную клетку, но также и потомство такой клетки. Поскольку модификации могут проходить в последующих поколениях вследствие мутации или воздействий окружающей среды, такое потомство не может, на самом деле, быть идентичным родительской клетке, но такие клетки по-прежнему включены в объем термина «клетка-хозяин» при использовании в настоящем документе.
Молекулы нуклеиновой кислоты, содержащей нуклеотидную последовательность, которая кодирует аминокислотную последовательность легкой или тяжелой цепи вышеуказанного биспецифического антитела, выбранную из:
аминокислотной последовательности первой легкой цепи, которая содержит вариабельный домен легкой цепи и константный домен легкой цепи;
аминокислотной последовательности первой тяжелой цепи, которая содержит вариабельный домен тяжелой цепи и константные домены тяжелой цепи антитела, включающие первый (СН1) константный домен тяжелой цепи и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи;
аминокислотной последовательности второй легкой цепи, которая содержит вариабельный домен легкой цепи и константный домен, который выбирают из группы:
первого околомембранного домена МНС (главный комплекс гистосовместимости) или
первого околомембранного домена МНС-подобного белка;
аминокислотной последовательности второй тяжелой цепи, которая содержит вариабельный домен тяжелой цепи, константный домен, который выбирают из группы:
второго околомембранного домена МНС (главный комплекс гистосовместимости) или
второго околомембранного домена МНС-подобного белка;
и мономер Fc-фрагмента, содержащий второй (СН2) и третий (СН3) константные домены тяжелой цепи,
и векторы, содержащие эти молекулы нуклеиновой кислоты, могут быть использованы для трансфекции подходящего млекопитающего или его клетки, растения или его клетки, бактериальной или дрожжевой клетки-хозяина. Преобразование может происходить любым известным способом для введения полинуклеотидов в клетку хозяина. Способы введения гетерологичных полинуклеотидов в клетки млекопитающих хорошо известны в данной области и включают декстран опосредованную трансфекцию, трансфекцию комплексом нуклеиновой кислоты и позитивно заряженного полимера, трансфекцию преципитатом нуклеиновой кислоты и фосфата кальция, полибрен опосредованную трансфекцию, слияние протопластов, трансфекцию инкапсулированными в липосомы полинуклеотидами и прямую микроинъекцию ДНК в ядра. В дополнение молекулы нуклеиновых кислот могут быть введены в клетки млекопитающих вирусными векторами.
Клеточные линии млекопитающих, используемые в качестве хозяев для трансформации, хорошо известны в данной области и включают множество иммортализованных доступных клеточных линий. К ним относятся, например, клетки яичников китайского хомячка (СНО), NS0 клетки, клетки SP2, HEK-293Т клетки, 293 Фристайл клетки (Invitrogen), NIH-3T3 клетки, клетки HeLa, клетки почек хомячка (BHK), клетки почек африканских зеленых мартышек (COS), клетки гепатоцеллюлярной карциномы человека (например, Hep G2), А549 клетки и ряд других клеточных линий. Клеточные линии выбираются путем определения, какие клеточные линии имеют высокие уровни экспрессии и обеспечивают необходимые характеристики продуцируемого белка. Другими клеточными линиями, которые могут быть использованы, являются клеточные линии насекомых, такие как Sf9 или Sf21 клетки. Когда векторы рекомбинантной экспрессии, кодирующие вышеуказанное биспецифическое антитело или его часть, вводятся в клетки-хозяева млекопитающих, вышеуказанное биспецифическое антитело продуцируются путем культивирования клеток-хозяев в течение времени, достаточного для экспрессии вышеуказанного биспецифического антитела по изобретению в клетках-хозяевах или, предпочтительнее, выделения вышеуказанного биспецифического антитела в питательную среду, в которой выращиваются клетки-хозяева. Вышеуказанное биспецифическое антитело может быть выделено из питательной среды с использованием стандартных методов очистки белка. Клетки-хозяева растений, например, включают Nicotiana, Arabidopsis, ряску, кукурузу, пшеницу, картофель и т.д. Клетки бактерий хозяина включают виды Escherichia и Streptomyces. Дрожжевые клетки-хозяева включают Schizosaccharomyces pombe, Saccharomyces cerevisiae и Pichia pastoris.
Кроме того, уровень продукции биспецифического антитела по данному изобретению из продуцирующей клеточной линии можно усилить с помощью ряда известных методов. Например, система экспрессии гена глутамин синтетазы (система GS) является достаточно распространенной для усиления экспрессии при определенных условиях. Система GS обсуждается в целом или частично в связи с патентами ЕР 0216846, 0256055, 0323997 и 0338841.
Вполне вероятно, что биспецифическое химерное антитело по изобретению в различных клеточных линиях или клетках-хозяевах будут отличаться друг от друга профилем гликозилирования. Однако биспецифическое антитело, описанное в данном документе, является частью данного изобретения, независимо от состояния гликозилирования связывающих молекул и в целом, независимо от наличия или отсутствия посттрансляционных модификаций.
Вышеуказанная клетка-хозяин не относится к клетке-хозяину, полученной с использованием человеческих эмбрионов.
Вышеуказанная клетка-хозяин не относится к клетке-хозяину, полученной с модификации генетической целостности клеток зародышевой линии человека.
Способ получения антитела
В одном из аспектов настоящее изобретение относится к способу получения любого из вышеуказанных двухвалентных биспецифических химерных антител, который включает следующие этапы:
а) трансформация клетки-хозяина
- экспрессионными векторами, которые содержат молекулы нуклеиновых кислот, кодирующие первую легкую цепь и первую тяжелую цепь двухвалентного биспецифического химерного антитела по изобретению,
- экспрессионными векторами, которые содержат молекулы нуклеиновых кислот, кодирующие вторую легкую цепь и вторую тяжелую цепь двухвалентного биспецифического химерного антитела по изобретению,
б) культивирование клетки-хозяина в условиях, которые позволяют синтезировать указанное двухвалентное биспецифическое антитело; и
в) выделение указанного двухвалентного биспецифического антитела из культуры клеток.
Настоящего изобретения относится к способам получения двухвалентных биспецифических химерных антител по данному изобретению. Один вариант осуществления изобретения относится к способу получения двухвалентных биспецифических химерных антител как определено в настоящем документе, содержащему получение рекомбинантной клетки-хозяина, способной экспрессировать двухвалентное биспецифическое химерное антитело, культивированию указанной клетки-хозяина в условиях, подходящих для экспрессии двухвалентных биспецифических химерных антител, и выделение полученных двухвалентных биспецифических химерных антител. Двухвалентное биспецифическое химерное антитело, полученное такой экспрессией в таких рекомбинантных клетках-хозяевах упоминается в данном документе как «двухвалентное биспецифическое химерное антитела».
Примеры
Для наилучшего понимания изобретения приводятся следующие примеры. Эти примеры приведены только в иллюстративных целях и не должны толковаться как ограничивающие сферу применения изобретения в любой форме.
Все публикации, патенты и патентные заявки, указанные в этой спецификации включены в данный документ путем отсылки. Хотя вышеупомянутое изобретение было довольно подробно описано путем иллюстрации и примера в целях исключения двусмысленного толкования, специалистам в данной области на основе идей, раскрытых в данном изобретении, будет вполне понятно, что могут быть внесены определенные изменения и модификации без отклонения от сущности и объема прилагаемых вариантов осуществления изобретения.
Материалы и общие методы
Методы рекомбинантной ДНК
Для манипуляций с ДНК использовали стандартные методы, описанные у Sambrook J. и др., Molecular cloning: A laboratory manual; Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1989. Реагенты для молекулярной биологии использовали согласно инструкциям производителей.
Синтез генов
Требуемые сегменты генов получали из олигонуклеотидов, созданных путем химического синтеза. Генные сегменты длиной от 300 до 4000 п.н., которые фланкированы уникальными сайтами рестрикции, собирали путем отжига и лигирования олигонуклеотидов, включая ПЦР-амплификацию и последующее клонирование через указанные сайты рестрикции. Последовательности ДНК субклонированных генных фрагментов подтверждали путем секвенирования ДНК.
Определение последовательностей ДНК
Последовательности ДНК определяли путем секвенирования по Сэнгеру.
Анализ последовательностей ДНК и белков и обработка данных о последовательностях
Применяли пакет программ Ylab2 (Биокад) для создания, картирования, анализа, аннотирования и иллюстрации последовательностей.
Экспрессионные векторы
Для кратковременной экспрессии описанных антител, антитело-подобных белков и антигенов в клетках эукариот (например, в клетках СНО) применяли экспрессионные варианты плазмид. Помимо кассеты экспрессии целевого белка плазмиды (векторы) содержали: сайт инициации репликации, обеспечивающий репликацию указанной плазмиды в E. coli, гены, придающие устойчивость в E. coli к различным антибиотикам (например, к ампициллину и канамицину).
Слияние генов, содержащих описанные цепи антитела, как указано выше, осуществляли с помощью ПЦР и/или синтеза и сборки генов с использованием известных методов и процедур путем соединения соответствующих сегментов нуклеиновых кислот, например, с использованием уникальных сайтов рестрикции в соответствующих векторах. Субклонированные нуклеотидные последовательности подтверждали секвенированием ДНК. Необходимые количества плазмид для транзиентной трансфекции нарабатывали в культурах клеток Е. coli и выделяли с помощью известных методик.
Наработка и очистка рекомбинантных антигенов в суспензионной культуре клеток млекопитающих
Рекомбинантные белки продуцировали в клетках постоянной клеточной линии, полученной из клеток яичника китайского хомячка (линия СНО). Суспензионное культивирование осуществляли в колбах на орбитальном шейкере-инкубаторе, с использованием бессывороточных сред с добавлением 8 мМ L-глутамина и 1 г/л плюроника 68. Для транзиентной экспрессии клетки в концентрации 2-2,2×106 кл/мл трансфецировали с помощью линейного полиэтиленимина. Через 9 дней после трансфекции культуральную жидкость отделяли от клеток фильтрацией через глубинный фильтр с размером пор 0,5/0,22 мкм.
Белки с гистидиновым тагом очищали путем металл-хелатной хроматографии. Очищенные белки фильтровали через 0.22 мкм и хранили при -70°С.
Чистоту полученного раствора белка оценивали с помощью SDS-гель-электрофореза, электрофорез проводили в денатурирующем 12% ПААГ с добавлением меркаптоэтанола и нативном 8% ПААГ.
Продукция антител и антитело-подобных белков в суспензионной культуре клеток млекопитающих
Контрольные антитела, биспецифические химерные антитела по изобретению и антитело-подобные белки продуцировали в клетках постоянной клеточной линии, полученной из клеток яичника китайского хомячка (линия СНО). Суспензионное культивирование осуществляли в колбах на орбитальном шейкере-инкубаторе, с использованием бессывороточных сред с добавлением 8 мМ L-глутамина и 1 г/л плюроника 68. Для транзиентной экспрессии клетки в концентрации 2-2,2×106 кл/мл трансфецировали с помощью линейного полиэтиленимина. Соотношение ДНК/ПЭИ составляло 1:3/1:10. Через 9 дней после трансфекции культуральную жидкость отделяли от клеток фильтрацией через глубинный фильтр с размером пор 0,5/0,22 мкм, а затем проводили измерение титра белка на ForteBio по стандартной методике. Осветленную культуральную жидкость пропускали через колонку с аффинным сорбентом Protein А в нагрузке 10-20 мг на мл сорбента, уравновешенную фосфатно-солевым буфером (ФСБ, рН 7,4). Затем колонку промывали 5 колоночными объемами ФСБ, чтобы удалить неспецифически связывающиеся компоненты. Связанный белок элюировали, используя 0,1 М глициновый буфер рН 3. Главный протеиновый пик элюции собирали и доводили его рН до значений 6,8-7,0 с помощью 1 М буфера Tris-HCl (рН 8). Все стадии проводили при скорости потока ПО см/ч. Далее белок переводили в ФСБ (рН 7,4) с помощью диализа, фильтровали (0,22 мкм), переносили в пробирки и хранили при -70°С.
Чистоту полученного раствора белка оценивали с помощью SDS-гель-электрофореза в редуцирующих и нередуцирующих условиях, а также с помощью эксклюзионной высокоэффективной жидкостной хроматографии (эВЭЖХ).
ЭВЭЖХ проводили на колонке TSK-Gel G3000SWXL, 7.8×300 мм, размер частиц 5 мкм, размер пор 250А и предколонке TSK-Gel Guard SWx1.
Пример 1. Выбор замены для СН1/CK доменов в биспецифическом антителе
Для корректной гетеродимеризации тяжелых и легких цепей в биспецифическом антителе одна из двух пар СН1/CK доменов заменялась на структурно схожие домены из других белков (фиг. 1). Выбор подходящих для замены доменов включал в себя три этапа:
1) предподготовка трехмерной модели доменов СН1/CK;
2) Структурное выравнивание трехмерных моделей СН1/CK доменов со всеми объектами базы данных Protein Data Bank (www.rcsb.org) за исключением антител и Т-клеточных рецепторов;
3) Выбор подходящей замены по результатам структурного выравнивания на основе метрики RMSD и числа аминокислот, образующих идентичные элементы вторичной структуры
Первый этап - предподготовка трехмерной модели доменов СН1/CK - представляет собой достраивание недостающих атомов в СН1/CK доменах из структуры PDBid 4nyl с помощью утилиты Prepwizard из пакета программ Schrodinger Suite. На втором этапе полученную модель поочередно выравнивали на все структуры из Protein Data Bank (за исключением антител и Т-клеточных рецепторов) с помощью утилиты Protein Structure Alignment из пакета программ Schrodinger Suite. Степень сходства произвольной структуры с моделью СН1/CK доменов оценивали по двум метрикам: 1) RMSD, отражающей среднеквадратичное отклонение атомов одной структуры от другой (расчет проводился по атомам остова полипептидной цепи), 2) количество аминокислот, образующих по итогам выравнивания идентичные элементы вторичной структуры белка. Лучшие результаты выравнивания по метрике RMSD представлены в таблице 1.

Структуры, обладающие максимальным структурным сходством с СН1/CK доменами по метрике RMSD относятся к белкам главного комплекса гистосовместимости I-го и II-го классов (МНС), а также к их структурным аналогам: CD1 и HFE. Исходя из сравнимых значений степени структурного сходства (RMSD и количества аминокислот из идентичных вторичных структур) для замены могут быть выбраны любые из вышеперечисленных белков. Для дальнейших работ по замене СН1/CK доменов в иллюстративных целях был выбран CD1b, а именно его околомембранные домены.
Пример 2. Выбор расположения дисульфидной связи между тяжелой и легкой цепью в молекуле антитела с заменой СН1/CK на околомембранные домены МНС или МНС-подобных белков.
Молекула антитела человека изотипа IgG состоит из двух легких и двух тяжелых цепей. Каждая из двух легких цепей антитела связана с тяжелой цепью через SS-мостик - ковалентную связь между атомами серы в остатках цистеина. Эта связь образована (в случае изотипа IgG1) С-концевым остатком цистеина легкой цепи и с ближайшим к N-конпу шарнирной области (т.н. хиндж-регион (hinge)) остатком цистеина тяжелой цепи.
Околомембранные домены МНС и МНС-подобных белков связаны только нековалентными связями и не образуют SS-мостиков между собой, поэтому замена СН1/CK доменов на околомембранные домены МНС потенциально снижает термостабильность молекулы антитела и выход целевого продукта при наработке. Во избежание этого в легкой цепи химерной молекулы антитела потребовалась точечная замена на цистеин в позиции, пригодной для образования SS-мостика с остатком цистеина, расположенного ближе к N-концу хиндж-региона.
Выбор позиции для введения остатка цистеина в околомембранный домен МНС или МНС-подобного белка основан на структурном выравнивании белка МНС или МНС-подобного белка и молекулы полноразмерного антитела. По итогам выравнивания остаток цистеина вводится в околомембранный домен МНС или МНС-подобного белка, расположенный на месте исходного домена CL (CK) антитела, в позицию, ближайшую к остатку цистеина хиндж-региона, который образует SS-мостик с легкой цепью в нативной молекуле антитела изотипа IgG1. По аналогии с легкой цепью нативного антитела введенный остаток цистеина является терминальным. Для того, чтобы расстояние между остатками цистеинов было достаточным для образования дисульфидной связи легкую цепь удлинили на две аминокислоты GS. На фиг.2 схематично представлено расположение SS-мостика между тяжелой и легкой цепью в химерном антителе с заменой СН1-CK доменов на околомембранные домены МНС-подобного белка CD1b.
Пример 3
Получение генетических конструкций для наработки двухвалентных биспецифических химерных антител
Для получения конструкций, кодирующих последовательности первой легкой и первой тяжелой цепей антитела, специфически связывающегося с первым антигеном, нарабатывали ПЦР-продукты, содержащие гены вариабельных доменов тяжелой и легкой цепей антител, с праймерами содержащими сайты рестрикции. Вариабельный домен тяжелой цепи клонировали в вектор pSXn-HChole-NR_VH1 по сайтам рестрикции SalI/XbaI. Вариабельный домен легкой цепи клонировали в вектор pSXn-CL-BR_VL1 по сайтам рестрикции SalI/XbaI.
Для получения конструкций, кодирующих последовательности второй легкой и второй тяжелой цепей антитела, специфически связывающегося со вторым антигеном, были синтезированы конструкции, содержащие гены вариабельных доменов тяжелой и легкой цепей антител и околомембранные домены последовательности CD1 (https://www.rcsb.org/structure/5wl1) человека с различными модификациями или без таких модификаций.
Последовательность была синтезирована из олигонуклеотидов посредством ПЦР с праймерами содержащими сайты рестрикции. Вариабельный домен тяжелой цепи с первым околомембранным доменом последовательности CD1 человека с модификациями или без таких модификаций клонировали в вектор pSX-FCknob-PR по сайтам рестрикции SalI/XbaI. Вариабельный домен легкой цепи со вторым околомембранным доменом последовательности CD1 человека с модификациями или без таких модификаций клонировали в вектор pSX-HR по сайтам рестрикции SalI/XbaI.
Вышеуказанные четыре вектора были скомбинированы на стадии трансфекции для получения двухвалентных биспецифических химерных антител по изобретению.
Все указанные выше плазмиды нарабатывали в необходимых количествах в клетках Е. coli и очищали при помощи набора Maxiprep Qiagen.
Пример 4. Влияние введенной дисульфидной связи на сборку полноразмерного химерного антитела.
Для проверки работоспособности введенной дисульфидной связи были наработаны модельные моноспецифические химерные антитела. Данный формат представляет собой антитело, состоящее из двух тяжелых и двух легких цепей, в которых СН1 тяжелой цепи и СК домен легкой цепи заменены на β2-микроглобулин и α3-домен белка CD1b, соответственно.
Последовательности вариабельных доменов VH (SEQ ID NO: 103) и VL (SEQ ID NO: 104) взяты из антитела Prolgolimab (ЗАО Биокад). В первом варианте образец не содержал дисульфидной связи между тяжелой и легкой цепью. Во втором варианте на С-конец легкой цепи был добавлен цистеин (как описано в примере 2), который, согласно расчетам, должен образовывать дисульфидную связь с цистеином верхнего хиндж-региона, тем самым стабилизируя гетеродимер тяжелой и легкой цепи.
На фиг.3 представлен результат SDS-гель-электрофореза наработанных образцов в нередуцирующих условиях. В качестве контроля на дорожке 3 нанесен образец Prolgolimab (классический IgG1). Из представленных результатов видно, что полноразмерная молекула собралась только в случае наличия дополнительной дисульфидной связи (дорожка 2), в случае же ее отсутствия наблюдается только фрагмент, соответствующий по подвижности димеру тяжелых цепей (дорожка 3).
Также был наработан химерное антитело с обратной ориентацией димеризующих блоков относительно тяжелых и легких цепей (СН1-домен тяжелой цепи и СК-домен легкой цепи заменены на α3-домен белка CD1b и β2-микроглобулин, соответственно, см. фиг 1Б). На фиг. 3 (дорожка 4) представлен результат SDS-гель-электрофореза данного образца в нередуцирующих условиях, и как видно из представленных результатов, данная ориентация также позволяет собирать химерные антитела.
Пример 5. Проверка влияния димеризующего блока на основе МНС или МНС-подобного белка на корректность спаривания тяжелых и легких цепей.
Для доказательства того, что димеризующий блок на основе МНС или МНС-подобного белка выполняет свою функцию, а именно, препятствует некорректному спариванию легких цепей с несоответствующими тяжелыми цепями, был проведен следующий эксперимент на моноспецифических молекулах. Были наработаны 4 группы молекул:
1. Молекулы с «корректным» сочетанием VH и VL и «некорректной» парой константных доменов.
2. Молекулы с «некорректным» сочетанием VH и VL и «корректной» парой константных доменов.
3. Молекулы с «некорректным» сочетанием VH и VL и «некорректной» парой константных доменов.
4. Контрольные молекулы, представляющие собой классическое антитело формата IgG1 и химерное антитело, в котором константные домены заменены на околомембранные домены из МНС или МНС-подобных белков (в данном случае β2-микроглобулин и α3-домен белка CD1b).
Под «корректным» сочетанием VH и VL подразумевается пара VH и VL, взятая из одного антитела. Под «некорректным» сочетанием VH и VL подразумеваются VH и VL, взятые из разных антител.
Под «корректной» парой константных доменов подразумевается либо пара СН1-CK либо пара взаимодействующих доменов из МНС-подобных белков (в данном случае β2-микроглобулин и α3-домен белка CD1b). Под «некорректной» парой константных доменов подразумеваются пары CH1-CD1b или β2-микроглобулин-CK.
Общая схема эксперимента представлена на фиг. 4. В качестве вариабельных доменов VH1 и VL1 были взяты последовательности вариабельных доменов антитела Prolgolimab, в качестве вариабельных доменов VH2 и VL2 были взяты последовательности соответствующих вариабельных доменов антитела Ocrelizumab (Genentech).
Данные по продуктивности белков представлены в таблице 2. Из представленных результатов видно, что кандидаты, у которых в тяжелой цепи был константный домен СН1, а в легкой цепи околомембранный домен белка CD1b продемонстрировали низкую продуктивность (антитела 01-001, 01-003,01-008, 01-010). Это согласуется с общеизвестным фактом, что для корректного фолдинга и экспорта из эндоплазматического ретикулума (ЭПР) необходимо взаимодействие домена СН1 с доменом CL (Feige MJ, Groscurth S, Marcinowski M, Shimizu Y, Kessler H, Hendershot LM, Buchner J. An unfolded CH1 domain controls the assembly and secretion of IgG antibodies. Mol Cell. 2009 Jun 12; 34(5):569-79. doi: 10.1016/j.molcel.2009.04.028. PMLD: 19524537; PMCID: PMC2908990). Антитела 01-002 и 01-009 не проходили такой контроль качества в ЭПР и продемонстрировали продуктивность выше 200 мг/л, однако несмотря на высокие значения продуктивности, их подвижности в электрофорезе в ПААГ не соответствовали подвижности полноразмерной молекулы (фиг.5А, дорожки 2, 5).

Результаты SDS-гель-электрофореза в нередуцирующих условиях представлены на фиг.5. Как видно из представленных результатов, электрофоретическая подвижность антител из группы 1 (Фиг. 5А дорожки 1, 2, Фиг. 5Б дорожка 1) и группы 3 (Фиг. 5А дорожки 4,5, Фиг. 5В дорожка 1) оказалась выше, чем у полноразмерной молекулы (Фиг. 5В дорожка 2), что говорит о том, что молекулы не собрались. Для антител из группы 2, несмотря на то что последовательности VH и VL были взяты из разных антител, их электрофоретическая подвижность (Фиг. 5Б дорожки 2, 3, 4, 5 (то же, что и фиг 5А дорожка 3)) соответствует подвижности полноразмерной молекулы (Фиг. 5В дорожка 2). Антитела из контрольной группы собрались, как и ожидалось (фиг. 5В дорожка 2, фиг. 5А дорожка 6 (то же, что и фиг. 5Б дорожка 6)
Полученные данные указывают на то, что димеризующий блок на основе околомембранных доменов МНС или МНС-подобных белков препятствует образованию неправильных пар между тяжелой и легкой цепью, если эти цепи содержат константные домены различной природы.
Пример 6. Наработка двухвалентных биспецифических химерных антител, содержащих МНС-подобный димеризующий блок.
Для проверки универсальности предложенного подхода как платформенного решения сборки биспецифических молекул с использованием любой пары антиген-связьгвающих фрагментов (далее вариабельные фрагменты легкой и тяжелой цепей) были выбраны три случайных пары вариабельных фрагментов легкой и тяжелой цепей из известных антител (см. таблицу 3.) и на основе их были наработаны 6 биспецифических химерных антител. Результаты продуктивности полученных белков представлены в таблице 3. Результаты SDS-гель-электрофореза наработанных образцов в нередуцирующих и редуцирую1пих условиях представлены на фиг 6. Для всех шести образцов присутствует мажорная полоса в области 150 кДа, соответствующая полноразмерной молекуле. Результаты чистоты образцов по эВЭЖХ также представлены в таблице 3.

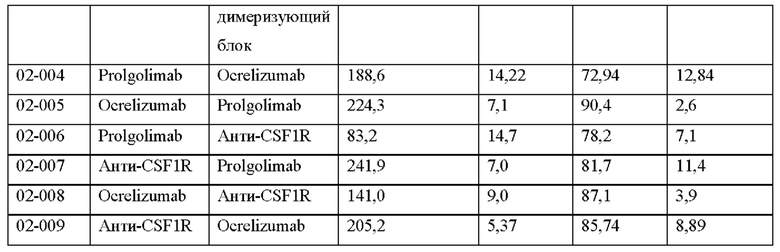
Пример 7 Определение аффинности полноразмерных биспецифических химерных антител на Forte Bio Octert RED 384
Чтобы удостовериться, что наработанные биспецифические химерные антитела не утратили свою антиген-связьшаюгдую способность был проведен анализ аффинности связывания на приборе Forte Bio Octert RED 384. Для антител 02-004 и 02-005 измеряли аффинность к экстраклеточному домену белка PD-1 человека (hPD1ex-H6F) и к биотинилированному пептиду [NH2]CEPANPSEKNSPSTQYCYSIQS[CH2CH2]biotm, содержащему в аминокислотной последовательности фрагмент последовательности CD20 человека (далее обозначен CD20-пептид), для антител 02-006 и 02-007 измерялась аффинность к экстраклеточному домену белка PD-1 человека (hPD1ex-H6F) и к экстраклеточному домену белка CSF1R человека (hCSF1R_His), для антител 02-008 и 02-009 измерялась аффинность к экстраклеточному домену белка CSF1R человека (hCSF1R_His) и к CD20-пептиду.
В качестве антигена hCSF1R была взята последовательность 20-512 ак белка CSF1R человека с С-концевыми His и FLAG-тагами, молекулярная масса 57,4 кДа. В качестве антигена hPD-lex-H6F была взята последовательность 21-170 ак белка PD-1 человека с С-концевыми His и FLAG-тагами, молекулярная масса 20,6 кДа (см. таблицу 6).
Генетические последовательности, кодирующие антигены, были синтезированы de novo, клонированы в экспрессионный вектор, наработаны в клетках СНО и очищены с помощью аффинной хроматографии, как описано общих примерах.
Эксперименты проводились с применением кинетического буферного раствора (далее в тексте 1xКБ) состава: 4,3 мМ Na2HPO4; 136,9 мМ NaCl, 1,5 мМ KH2PO4; 2,7 мМ KCl; объемная доля добавленного Tween 20 составляла 0,1%; массовая доля добавленного БСА (бычий сывороточный альбумин) составляла 0,1%; рН 7,4. Перед измерением сенсоры ProA регенерировались раствором 50 мМ глицина и соляной кислоты с рН 1.8 (5 с в регенерирующем растворе, 5 секунд в 1×КБ, три повтора).
В случае антигенов hPD1ex-H6F и hCSF1R_His сенсоры ProA (Protein А (ProA) biosensors, ForteBio) погружались в раствор с антителами в концентрации 10 мкг/мл на 60 с для их иммобилизации. Запись базовой линии проводилась в 1хКБ в течение 60 с. Затем сенсоры с загруженным антителом погружались в лунки с раствором целевого антигена (аналит) в кинетическом буфере на 90 с. Измерения проводились для растворов с концентрациями аналита hPD1ex-H6F 2,50 мкг/мл (121,4 нМ), 1,25 мкг/мл (60,7 нМ), 0,625 мкг/мл (30,35 нМ). Измерения проводились для растворов с концентрациями аналита hCSF1R_His 2,50 мкг/мл (43,6 нМ), 1,25 мкг/мл (21,8 нМ), 0,625 мкг/мл (10,9 нМ). Затем детектировалась диссоциация комплекса в 1хКБ в течение 210 с для hPD1ex-H6F, 300 с для hCSF1R_His.
Референсные сенсоры проходили все этапы аналогично сенсорам, применяемым для записи сенсограмм аналита, за исключением этапа ассоциации -на этапе ассоциации данные сенсоры погружались в 1хКБ раствор без аналита (измерение сигнала референсных сенсоров осуществлялось параллельно с записью основных сенсограмм). Референсный сигнал вычитался из сигнала, полученного на сенсорах, взаимодействующих с аналитом при обработке сенсограмм.
Для проверки неспецифического взаимодействия аналита с сенсорами применялся сенсор, не загруженный антителом (на этапе загрузки сенсор погружался в 1хКБ раствор, все остальные этапы аналогичны загруженному антителом сенсору).
В случае биотинилированного CD20-пептида данный пептид в концентрации 5 мкг/мл иммобилизовали на поверхности сенсоров SAX (High Precision Streptavidin (SAX) biosensors, ForteBio) в течение 120 c. Запись базовой линии проводилась в 1хКБ растворе в течение 120 с. Затем сенсоры с загруженным пептидом погружались в лунки с раствором антитела (аналит) в 1хКБ на 15 с. Измерения проводились для растворов с концентрациями аналита (антитела в случае CD20-пептида) 150 мкг/мл, 75 мкг/мл, 37,5 мкг/мл. Запись диссоциации комплекса проводилась в 1хКБ в течение 120 с.
Все измерения проводились при 30°С, скорость орбитального перемешивания составляла 1000 оборотов в минуту.
Для получения численных значений кинетических констант (kon - константа скорости ассоциации, kdis - константа скорости диссоциации, KD - равновесная константа диссоциации или константа аффинности) полученные сенсограммы обрабатывались по модели взаимодействия 1:1 («один к одному») при помощи метода Global Fit (подбор одного набора констант kon, kdis, KD для анализа нескольких сенсограмм разных концентраций) программного обеспечения ForteBio Octet Data Analysis 9.0. Результаты представлены в таблице 4.

Результаты проверки неспецифического взаимодействия аналита с незагруженными сенсорами представлены в таблице 5. Не выявлено неспецифического взаимодействия аналита с незагруженными сенсорами.
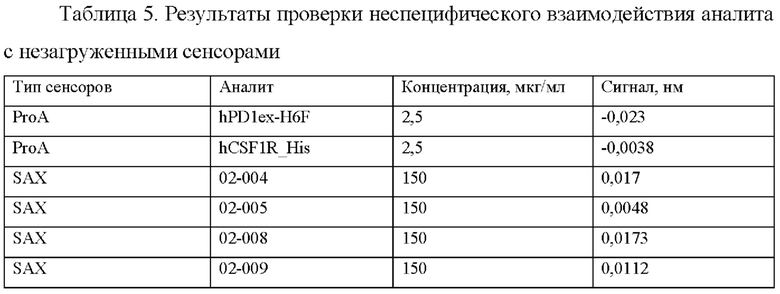
Из представленных результатов следует, что все биспецифические химерные антитела 02-004-02-009 демонстрируют специфичное связывание с целевыми антигенами, причем в независимости от положения соответсвующих антиген-связывающих фрагментов (в составе Fab-фрагмента или в составе Fab-подобного фрагмента, содержащего β2-микроглобулин и α3-домен белка CD1b вместо CK- и СН1-доменов) численные значения KD имеют один и тот же порядок величины.
Пример 8. Анализ неспецифического связывания биспецифических химерных антител с нецелевыми антигенами
Эксперименты по исследованию неспецифического связывания биспецифических химерных антител с нецелевыми антигенами проводились на приборе Forte Bio Octert RED 384.
Сенсоры ProA (Protein А (РгоА) biosensors, ForteBio) погружали в раствор с антителами в концентрации 10 мкг/мл на 150 секунд для их иммобилизации. Запись базовой линии проводилась в 1хКБ растворе в течение 30 с. Затем сенсоры с загруженным антителом погружали в лунки с растворами нецелевых антигенов в концентрации 30 мкг/мл на 150 с. Затем записывалась диссоциацию комплекса в течение 30 секунд.
Референсные сенсоры проходили все этапы аналогично сенсорам, применяемым для записи сенсограмм аналита, за исключением этапа ассоциации - на этапе ассоциации данные сенсоры погружались в кинетический буферный раствор без аналита (измерение сигнала референсных сенсоров осуществлялось параллельно с записью основных сенсограмм). Референсный сигнал вычитался из сигнала, полученного на сенсорах, взаимодействующих с аналитом при обработке сенсограмм.
В панель антигенов входили: hAng2_H6F, hPD1ex-H6F, hCSF1R_His, IL5R-His, hIL4R-His, hCD38-Avi, C-Avi-His-TEVs-HSA_hBCMA, Her3-H6F, hisOX40, Macaca CSF1R_C-His (см. таблицу 6) Для каждого исследуемого антитела в данной панели антигенов есть специфичный (целевой) антиген, результаты взаимодействия с целевым антигеном приведены в качестве положительного контроля.
Обработка сенсограмм проводилась с применение программного обеспечения ForteBio Octet Data Analysis 9.0. Для заключения об отсутствии неспецифического взаимодействия проверялся регистрируемый уровень сигнала в конце этапа ассоциации (параметр response). Результаты проверки связывания биспецифических химерных антител представлены в таблице 7. В результате эксперимента не выявлено взаимодействия антител 02-004 - 02-009 с нецелевыми антигенами.


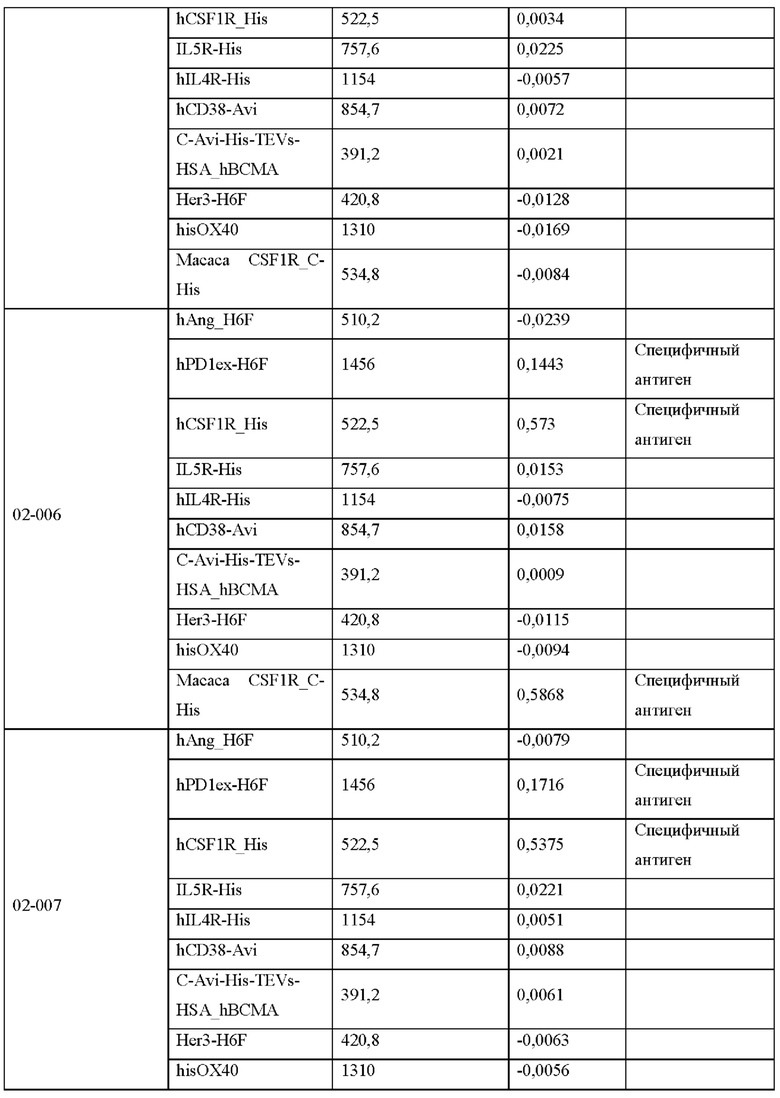


Пример 9 Анализ одновременного связывания биспецифических химерных антител по изобретению с двумя антигенами.
Для антител 02-006 и 02-007 был проведен анализ одновременного связывания с двумя различными целевыми антигенами (hPD1ex-H6F и hCSF1R_His) на приборе Forte Bio Octet RED 384. Эксперимент проводился на сенсорах AR2G (Amine Reactive Second-Generation (AR2G) biosensors, ForteBio). Этапы эксперимента представлены в таблице 8.
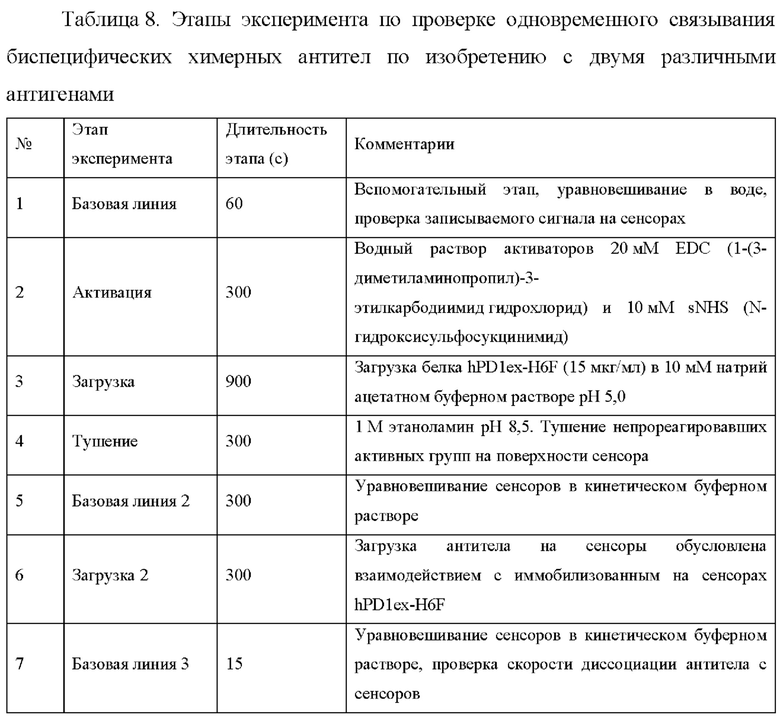

Активация сенсоров осуществлялась в водном растворе, содержащем 20 мМ EDC (1-(3-диметиламинопропил)-3-этилкарбодиимид гидрохлорид) и 10 мМ sNHS (N-гидроксисульфосукцинимид) в течении 300 с. Загрузка антигена (hPDlex-H6F) на поверхность биосенсоров осуществлялась в 10 мМ натрий-ацетатном буфере с рН 5,0 в течении 300 с. Концентрация белка для загрузки составляла 15 мкг/мл. Тушение непрореагировавших активных центров на поверхности сенсоров осуществлялось в 1М водном растворе этаноламина с рН 8,5 в течение 300 с. Базовая линия на пятом этапе эксперимента и все последующие шаги проводились в кинетическом буферном растворе (1хКБ) состава: 4,3 мМ Na2HPO4; 136,9 мМ NaCl, 1,5 мМ KH2PO4; 2,7 мМ KCl; объемная доля добавленного Tween 20 составляла 0,1%; массовая доля добавленного БСА составляла 0,1%; рН 7,4. После записи базовой линии (этап 5) проходила загрузка антител на сенсоры, концентрация загружаемого антитела 30 мкг/мл (этап 6). Далее проводилась запись третьей базовой линии (этап 7). Данный этап демонстрирует отсутствие быстрого спада сигнала, что указывает на специфичное взаимодействие загруженного антител с hPDlex-H6F иммобилизированным ранее на сенсоры. На следующем этапе ассоциации (этап 8) сенсоры с иммобилизированным hPD1ex-H6F и связавшимися антителами погружаются в раствор антигена hCSF1R_His с концентрацией 100 мкг/мл. Рост сигнала на данном этапе обусловлен наличием у антител, загруженных на сенсоры, FAB фрагмента к антигену hCSF1R_His.
Анализа сенсограмм проводился с применением программного обеспечения ForteBio Octet Data Analysis 9.0. Основные выводы представлены в таблице 9. Сенсограммы шагов (этапов) эксперимента представлены на фиг.7. Из представленных результатов следует, что образцы 02-006 и 02-007 демонстрируют одновременное связывание с антигенами hPDlex-H6F и hCSF1R_His.

Из представленных результатов видно, что антитела 02-006 и 02-007 демонстрируют одновременное связывание с антигенами hPD1ex-H6F и hCSF1R_His.
Пример 10. Анализ молекулярной массы биспецифических химерных антител по изобретению с помощью обращенно-фазовой ультравысокоэффективной жидкостной хроматографии (ОФ УВЭЖХ) с масс-спектрометрическим детектированием
Для подтверждения правильности сборки молекул был проведен анализ молекулярных масс полноразмерных биспецифических химерных антител по изобретению. Данный метод позволяет выявить полноразмерные биспецифические химерные антитела по изобретению, состоягцие из 4х различных цепей и отличить их от побочных продуктов, образованных из другого набора цепей. Эти данные в совокупности с данными, полученными в примере 5, говорят о том, что технологическое решение, основанное на замене пары константных доменов CHICK на пару околомембранных доменов из МНС или МНС-подобных белков, обеспечивает правильную сборку биспецифических химерных антител по изобретению.
Анализ проводился с помощью хромато-масс-спектрометрического комплекса Agilent 1290 Infinity II UPLC - Agilent 6530 Q-Tof колонке BioResolve Polypheny 1 RP (2,1×50 мм, диаметр частиц 2,7 мкм; также использовали предколонку BioResolve Polyphenyl RP 2,1×5 мм, диаметр частиц 2,7 мкм). Перед анализом все молекулы были обработаны ферментом PNGase F (Promega) для удаления N-гликанов. Для этого фермент разводили водой до концентрации 1 ед активности в мкл. Образец с содержанием белка 120 мкг смешивали с буферным раствором (100 мМ бикарбонат аммония рН 7,2), добавляли раствор PNGase F в соотношении фермент:белок - 1ед:50 мкг, и инкубировали в термостате в течение 18 часов при (37,0×0,1)°С.
Для анализа отбирали 10 мкг испытуемого раствора в соответствии с измеренными концентрациями белка и доводили концентрацию подвижной фазой А до 0,2 мг/мл. Объем ввода образца составлял 7 мкл.
Перед началом анализа хроматографическую систему уравновешивали подвижной фазой при исходном соотношении подвижных фаз А: 95%, В: 5% (фаза А содержит 0,1%) раствор муравьиной кислоты, 0,02% раствор трифторуксусной кислоты в воде, фаза В - 0,1%) раствор муравьиной кислоты, 30% ацетонитрил в изопропиловом спирте) в течение не менее 30 мин до достижения стабильного давления. Проводили калибровку масс-спектрометра с помощью стандарта для калибровки в соответствии с рекомендациями производителя.
Разделение образца проводили при скорости потока 0,5 мл/мин в градиенте концентрации подвижной фазы В (с 5% до 27% с 5 до 6 мин после ввода образца, далее с 27% до 37%) с 6 по 30 мин) при температуре колонки (60±1)°С, снимали хроматограмму при длине волны 280 нм.
Обработка данных проводилась в программном обеспечении (ПО) PMi Intact.
Названия цепей, составляющих биспецифические химерные антитела, представлены в таблице 10.

Результаты анализа масс для антител 02-004, 02-006, 02-009 (как наиболее репрезентативные) представлены в таблице 11. Также на фиг. 8 представлены изображения деконволюированного масс-спектра пиков 4-6 хроматограммы полного ионного тока для антитела 02-004, для остальных антител пики выглядели сходным образом.
Для каждого антитела в таблице представлена масса вещества в соответствующих пиках хроматограммы, также проведена аннотация пиков, в результате чего были идентифицированы цепи, из которых состоят фрагменты определенной массы. Для всех образцов наблюдалось схожее распределение масс: основной пик в районе 146 кДа, а также минорные пики масс порядка 23 кДа (соответствует массе одиночной легкой цепи) и масс порядка 74 кДа (соответствуют гетеродимеру тяжелой и легкой цепи). Основные пики были проаннотированы, исходя из теоретических значений средних масс.
Пик 4 антитела 02-004 не был проаннотирован ПО, так как настройки включали допущение, что все N-концевые глутамины находятся в пироформе (соответственно, масса пироглутамина на 18 Да меньше массы глутамина). Таким образом массы, полученные в пике 4 соответвуют массам молекул, аннотированных в пике 5, в которых один из N-концевых глутаминов находится в непироформе.
Пики 5 и 6 антитела 02-004 несут в себе по 3 массы, соответсвующие полноразмерной молекуле и двум безлизиновым формам полноразмерной молекулы (с одним отсутствующим С-концевым цистеином и с двумя отсутствующими С-концевыми лизинами). Для антител 02-006 и 02-009 характерно аналогичное распределение масс.

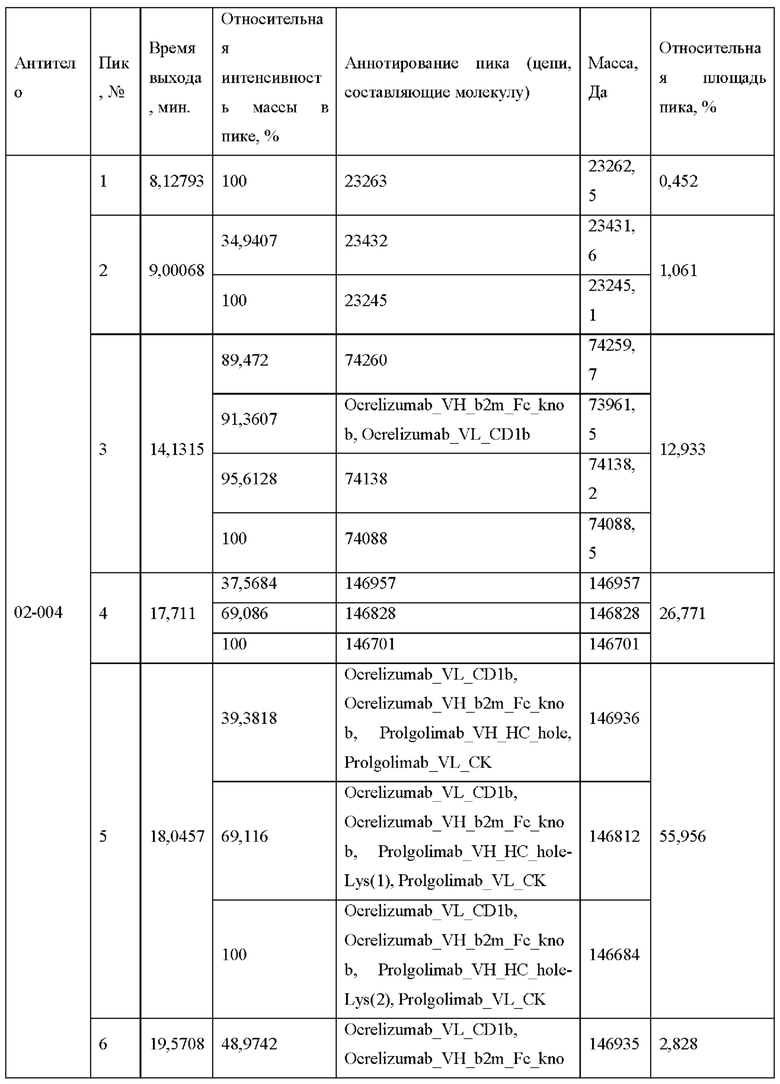



Суммируя все вышесказанное, результаты демонстрируют, что масса полноразмерных биспецифических химерных антител соответствует массе молекулы, состоящей из четырех различных цепей. Также в препаратах присутствуют низкомолекулярные фрагменты, но в минорном количестве.
Пример 11. Подтверждение правильности сборки биспецифических химерных антител по изобретению с помощью протеолиза и анализа масс полученных фрагментов
Для прямой проверки корректного спаривания легких и тяжелых цепей проводили расщепление биспецифических химерных антител по изобретению протеазой GingisKHAN (Genovis), сайт узнавания которой находится в хиндж-регионе антитела (…KSCDK/THTCPPCP…). После такого протеолиза полноразмерное антитело распадается на Fc-фрагмент, Fab-фрагмент и Fab-подобный фрагмент, содержащий β2-микроглобулин и α3-домен белка CD1b вместо CK- и СН1-доменов. Фрагменты биспецифической молекулы после протеолиза анализировали при помощи вертикального электрофореза в нерецуцирующих условиях, а также проводили масс-спектрометрический анализ полученных фрагментов.
Реакционная смесь для протеолиза содержала 40 мкг биспецифического антитела и 40 единиц фермента GingisKHAN (соотношение 1 единица фермента на 1 мкг белка) в буфере состава 100 мМ Трис-HCl, рН 8,0, 1 мМ цистеин в объеме 60 мкл. Реакцию проводили в термостате при (37,0±0,1)°С в течение 1 ч и останавливали добавлением йодацетамида до концентрации 10 мМ. Для проведения электрофореза к полученным образцам добавляли буфер, содержащий SDS (до концентрации 1% SDS), и проводили SDS-гель-электрофорез в нередуцирующих условиях.
Результаты SDS-гель-электрофореза представлены на фиг.9. После обработки биспецифических химерных антител (02-004 - 02-009) протеазой GingisKHAN образуется 3 мажорных фрагмента, наблюдаемых на электрофореграмме в области 40-50 кДа и обладающих разной подвижностью в ПААГ. Моноспецифические молекулы, содержащие (01-011) и не содержащие димеризующий блок на основе околомембранных доменов МНС-подобного белка CD1b (Prolgolimab, Ocrelizumab), были также обработаны протеазой GingisKHAN (фиг. 9Б, дорожки 3, 4, 5). В результате сопоставления электрофоретических подвижностей, образующихся в результате протеолиза фрагментов, был сделан вывод, что фрагмент с наименьшей подвижностью соответствует Fc-фрагменту, со средней подвижностью - Fab-фрагменту, а фрагмент с наивысшей подвижностью - Fab-подобному фрагменту, содержащему β2-микроглобулин и α3-домен белка CD1b вместо CK- и СН1-доменов. В случае антител 02-006 - 02-009 на электрофореграмме наблюдался дополнительный фрагмент, образующийся в результате неспецифического разрезания вариабельного домена анти-CSF1R фрагмента. Чтобы установить точные массы образующихся в результате протеолиза фрагментов был проведен масс-спектрометрический анализ.
Перед масс-спектрометрическим анализом образцы сразу после добавления йодоацетамида переводили в 50 мМ бикарбонат аммония, рН (7,6±0,2), на колонках Zeba (MWCO 7 kDa), согласно рекомендациям производителя, после чего к образцам добавляли PNGase F в соотношении 1 ед. фермента на 50 мкг белка и инкубировали в термостате в течение 18 часов при (37,0±0,1)°С.
Анализ масс полученных фрагментов проводился по методике, описанной в примере 10.
Результаты ОФ УВЭЖХ с масс-спектрометрическим детектированием представлены в таблице 12.
Для антител 02-006 и 02-008 наблюдались фрагменты массой 11424 Да и 37035 Да, которые в сумме дают 48459 Да, что соответствует массе Fab-подобному фрагменту, содержащему β2-микроглобулин и α3-домен белка CD1b вместо CK- и СН1-доменов и вариабельным фрагментам легкой и тяжелой цепей к CSF1R. Для антител 02-007 и 02-009 наблюдались фрагменты массой 11424 Да и 36062 Да, которые в сумме дают 47486 Да, что соответствует массе Fab фрагмента с вариабельными фрагментами легкой и тяжелой цепей к CSF1R, что согласуется с данными, полученными после SDS-гель-электрофореза данных образцов и подтверждает предположение о дополнительном сайте протеолиза в вариабельном домене к CSF1R (фиг. 9).




Массы 97811,8 Да и 97693 Да в случае образца 02-009 соответствуют молекуле с одним отрезанным Fab-подобным фрагментом.
Из представленных результатов видно, что все биспецифические химерные антитела по изобретению после протеолиза разделяются на фрагменты, соответствующие по массе Fc-фрагменту, Fab-фрагменту и Fab-подобному фрагменту, содержащему β2-микроглобулин и α3-домен белка CD1b вместо CK- и СН1-доменов, что говорит о том, что технологическое решение, основанное на замене пары константных доменов СН1-CK на пару околомембранных доменов из МНС или МНС-подобных белков, обеспечивает правильную сборку биспецифических химерных антител по изобретению.
Пример 12. Проверка влияния аминокислотных замен в димеризующем блоке на основе около мембранных доменов МНС-подобного белка на термостабильность химерных антител.
Также были протестированы различные замены в димеризующем блоке и их влияние на чистоту и стабильность получаемых химерных антител. Тестирование проводилось в моноспецифическом формате, представляющем собой антитело, состоящее из двух тяжелых и двух легких цепей, в которых СН1 тяжелой цепи и СК домен легкой цепи заменены на β2-микроглобулин и а3-домен белка CD1b, соответственно («прямая» ориентация димеризующего блока). Последовательности вариабельных доменов VH и VL взяты из антитела Prolgolimab (ЗАО Биокад). В последовательности β2-микроглобулина и α3-домена белка CD1b были введены мутации, указанные в таблице 13. Последовательности α3-домена белка CD1b также содержали С-концевые удлинения, указанные в таблице 13. В таблице 13 нумерация позиций, в которых вводились мутации, представлена от начала димеризующего блока, а также приведено соответствие позиций, пронумерованных согласно нумерации аминокислот цепей антител EU (Edelman G.M. и др., Proc. Natl. Acad. Sci. USA 63, 1969, ее. 78-85; Kabat E.A. и др., Sequences of Proteins of Immunological Interest, 5-ое изд., изд-во Public Health Service, National Institutes of Health, Bethesda, MD, 1991)». Далее в тексте приведена нумерация позиций, в которых вводились замены, от начала димеризующего блока, положение замен в хиндж-регионе указано согласно нумерации EU.
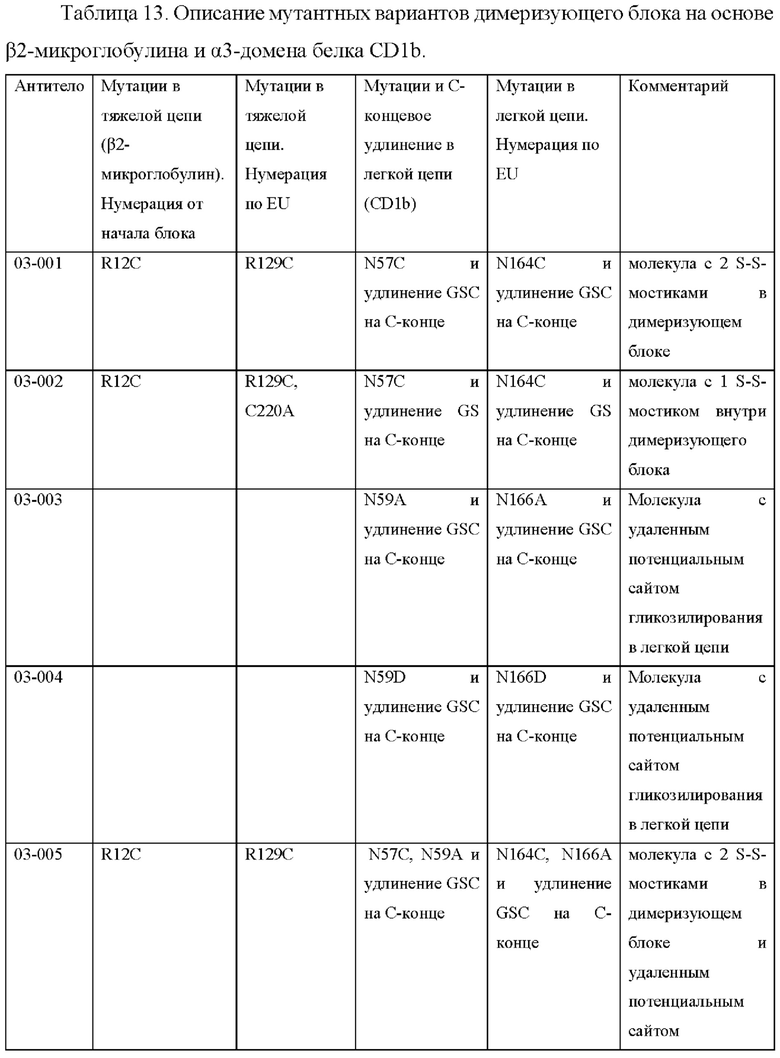

На фиг.10 представлен результат SDS-гель-электрофореза наработанных антител в нередуцирующих условиях. В качестве контролен на дорожках 1 и 2 (фиг. 10А и 10Б) нанесены образцы Prolgolimab (классический IgG1) и 01-011 (образец с исходным димеризующим блоком на основе околомембранных доменов МНС-подобного белка, см. пример 5), соответственно. Из представленных результатов видно, что подвижность мажорного фрагмента всех модифицированных образцов соответствует полноразмерной молекуле (см. фиг.10 А дорожки 3-8, фиг. 10Б дорожки 3-4), что говорит о том, что внесенные модификации не влияют на сборку полноразмерной молекулы. По данным эВЭЖХ (таблица 14) видно, что некоторые модификации приводят к уменьшению содержания мономера и повышению количества низкомолекулярных фрагментов (антитела 03-001, 03-003, 03-005, 03-006, 03-007), однако среди них есть и те, кто продемонстрировал увеличение процентного содержания мономера (03-002, 03-004, 03-008). Также был проведен анализ температуры агрегации методом динамического светорассеяния полученных антител, результат которого также представлен в таблице 14.

Из представленных результатов видно, что варианты антител, продемонстрировавшие увеличение относительного содержания мономера по эВЭЖХ, также имеют и повышенную по сравнению с немодифицированным вариантом димеризующего блока на основе МНС-подобных белков температуру агрегации.
--->
Перечень последовательностей
<110> ЗАО "БИОКАД"
<120> Двухвалентное биспецифическое химерное антитело, включающее гетеродимер на
основе MHC или MHC-подобных белков
<160> 109
<170> BiSSAP 1.3.6
<210> 1
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена MHC
класса I HLA-A (аллель A*01:01:01:01)
<400> 1
Gln Arg Thr Asp Pro Pro Lys Thr His Met Thr His His Pro Ile Ser
1 5 10 15
Asp His Glu Ala Thr Leu Arg Cys Trp Ala Leu Gly Phe Tyr Pro Ala
20 25 30
Glu Ile Thr Leu Thr Trp Gln Arg Asp Gly Glu Asp Gln Thr Gln Asp
35 40 45
Thr Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Gly Thr Phe Gln Lys
50 55 60
Trp Ala Ala Val Val Val Pro Ser Gly Glu Glu Gln Arg Tyr Thr Cys
65 70 75 80
His Val Gln His Glu Gly Leu Pro Lys Pro Leu Thr Leu Arg Trp Glu
85 90 95
Leu Ser Ser
<210> 2
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена MHC
класса I HLA-A (A*02:01:01:01)
<400> 2
Gln Arg Thr Asp Ala Pro Lys Thr His Met Thr His His Ala Val Ser
1 5 10 15
Asp His Glu Ala Thr Leu Arg Cys Trp Ala Leu Ser Phe Tyr Pro Ala
20 25 30
Glu Ile Thr Leu Thr Trp Gln Arg Asp Gly Glu Asp Gln Thr Gln Asp
35 40 45
Thr Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Gly Thr Phe Gln Lys
50 55 60
Trp Ala Ala Val Val Val Pro Ser Gly Gln Glu Gln Arg Tyr Thr Cys
65 70 75 80
His Val Gln His Glu Gly Leu Pro Lys Pro Leu Thr Leu Arg Trp Glu
85 90 95
Pro Ser Ser
<210> 3
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена MHC
класса I HLA-A (A*03:01:01:01)
<400> 3
Gln Arg Thr Asp Pro Pro Lys Thr His Met Thr His His Pro Ile Ser
1 5 10 15
Asp His Glu Ala Thr Leu Arg Cys Trp Ala Leu Gly Phe Tyr Pro Ala
20 25 30
Glu Ile Thr Leu Thr Trp Gln Arg Asp Gly Glu Asp Gln Thr Gln Asp
35 40 45
Thr Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Gly Thr Phe Gln Lys
50 55 60
Trp Ala Ala Val Val Val Pro Ser Gly Glu Glu Gln Arg Tyr Thr Cys
65 70 75 80
His Val Gln His Glu Gly Leu Pro Lys Pro Leu Thr Leu Arg Trp Glu
85 90 95
Leu Ser Ser
<210> 4
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена MHC
класса I HLA-A (A*11:01:01:01)
<400> 4
Gln Arg Thr Asp Pro Pro Lys Thr His Met Thr His His Pro Ile Ser
1 5 10 15
Asp His Glu Ala Thr Leu Arg Cys Trp Ala Leu Gly Phe Tyr Pro Ala
20 25 30
Glu Ile Thr Leu Thr Trp Gln Arg Asp Gly Glu Asp Gln Thr Gln Asp
35 40 45
Thr Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Gly Thr Phe Gln Lys
50 55 60
Trp Ala Ala Val Val Val Pro Ser Gly Glu Glu Gln Arg Tyr Thr Cys
65 70 75 80
His Val Gln His Glu Gly Leu Pro Lys Pro Leu Thr Leu Arg Trp Glu
85 90 95
Leu Ser Ser
<210> 5
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена MHC
класса I HLA-A (A*23:01:01:01)
<400> 5
Gln Arg Thr Asp Pro Pro Lys Thr His Met Thr His His Pro Ile Ser
1 5 10 15
Asp His Glu Ala Thr Leu Arg Cys Trp Ala Leu Gly Phe Tyr Pro Ala
20 25 30
Glu Ile Thr Leu Thr Trp Gln Arg Asp Gly Glu Asp Gln Thr Gln Asp
35 40 45
Thr Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Gly Thr Phe Gln Lys
50 55 60
Trp Ala Ala Val Val Val Pro Ser Gly Glu Glu Gln Arg Tyr Thr Cys
65 70 75 80
His Val Gln His Glu Gly Leu Pro Lys Pro Leu Thr Leu Arg Trp Glu
85 90 95
Pro Ser Ser
<210> 6
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена MHC
класса I HLA-B (B*07:02:01:01)
<400> 6
Glu Arg Ala Asp Pro Pro Lys Thr His Val Thr His His Pro Ile Ser
1 5 10 15
Asp His Glu Ala Thr Leu Arg Cys Trp Ala Leu Gly Phe Tyr Pro Ala
20 25 30
Glu Ile Thr Leu Thr Trp Gln Arg Asp Gly Glu Asp Gln Thr Gln Asp
35 40 45
Thr Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Arg Thr Phe Gln Lys
50 55 60
Trp Ala Ala Val Val Val Pro Ser Gly Glu Glu Gln Arg Tyr Thr Cys
65 70 75 80
His Val Gln His Glu Gly Leu Pro Lys Pro Leu Thr Leu Arg Trp Glu
85 90 95
Pro Ser Ser
<210> 7
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена MHC
класса I HLA-B (B*08:01:01:01)
<400> 7
Glu Arg Ala Asp Pro Pro Lys Thr His Val Thr His His Pro Ile Ser
1 5 10 15
Asp His Glu Ala Thr Leu Arg Cys Trp Ala Leu Gly Phe Tyr Pro Ala
20 25 30
Glu Ile Thr Leu Thr Trp Gln Arg Asp Gly Glu Asp Gln Thr Gln Asp
35 40 45
Thr Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Arg Thr Phe Gln Lys
50 55 60
Trp Ala Ala Val Val Val Pro Ser Gly Glu Glu Gln Arg Tyr Thr Cys
65 70 75 80
His Val Gln His Glu Gly Leu Pro Lys Pro Leu Thr Leu Arg Trp Glu
85 90 95
Pro Ser Ser
<210> 8
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена MHC
класса I HLA-B (B*13:01:01:01)
<400> 8
Gln Arg Ala Asp Pro Pro Lys Thr His Val Thr His His Pro Ile Ser
1 5 10 15
Asp His Glu Ala Thr Leu Arg Cys Trp Ala Leu Gly Phe Tyr Pro Ala
20 25 30
Glu Ile Thr Leu Thr Trp Gln Arg Asp Gly Glu Asp Gln Thr Gln Asp
35 40 45
Thr Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Arg Thr Phe Gln Lys
50 55 60
Trp Ala Ala Val Val Val Pro Ser Gly Glu Glu Gln Arg Tyr Thr Cys
65 70 75 80
His Val Gln His Glu Gly Leu Pro Lys Pro Leu Thr Leu Arg Trp Glu
85 90 95
Pro Ser Ser
<210> 9
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена MHC
класса I HLA-B (B*14:01:01:01)
<400> 9
Gln Arg Ala Asp Pro Pro Lys Thr His Val Thr His His Pro Ile Ser
1 5 10 15
Asp His Glu Ala Thr Leu Arg Cys Trp Ala Leu Gly Phe Tyr Pro Ala
20 25 30
Glu Ile Thr Leu Thr Trp Gln Arg Asp Gly Glu Asp Gln Thr Gln Asp
35 40 45
Thr Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Arg Thr Phe Gln Lys
50 55 60
Trp Ala Ala Val Val Val Pro Ser Gly Glu Glu Gln Arg Tyr Thr Cys
65 70 75 80
His Val Gln His Glu Gly Leu Pro Lys Pro Leu Thr Leu Arg Trp Glu
85 90 95
Pro Ser Ser
<210> 10
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена MHC
класса I HLA-B (B*15:01:01:01)
<400> 10
Gln Arg Ala Asp Pro Pro Lys Thr His Val Thr His His Pro Ile Ser
1 5 10 15
Asp His Glu Ala Thr Leu Arg Cys Trp Ala Leu Gly Phe Tyr Pro Ala
20 25 30
Glu Ile Thr Leu Thr Trp Gln Arg Asp Gly Glu Asp Gln Thr Gln Asp
35 40 45
Thr Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Arg Thr Phe Gln Lys
50 55 60
Trp Ala Ala Val Val Val Pro Ser Gly Glu Glu Gln Arg Tyr Thr Cys
65 70 75 80
His Val Gln His Glu Gly Leu Pro Lys Pro Leu Thr Leu Arg Trp Glu
85 90 95
Pro Ser Ser
<210> 11
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена MHC
класса I HLA-C (C*01:02:01:01)
<400> 11
Gln Arg Ala Glu His Pro Lys Thr His Val Thr His His Pro Val Ser
1 5 10 15
Asp His Glu Ala Thr Leu Arg Cys Trp Ala Leu Gly Phe Tyr Pro Thr
20 25 30
Glu Ile Thr Leu Thr Trp Gln Arg Asp Gly Glu Asp Gln Thr Gln Asp
35 40 45
Thr Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Gly Thr Phe Gln Lys
50 55 60
Trp Ala Ala Val Val Val Pro Ser Gly Glu Glu Gln Arg Tyr Thr Cys
65 70 75 80
His Val Gln His Glu Gly Leu Pro Glu Pro Leu Thr Leu Arg Trp Glu
85 90 95
Pro Ser Ser
<210> 12
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена MHC
класса I HLA-C (C*03:02:01)
<400> 12
Gln Arg Ala Glu His Pro Lys Thr His Val Thr His His Pro Val Ser
1 5 10 15
Asp His Glu Ala Thr Leu Arg Cys Trp Ala Leu Gly Phe Tyr Pro Ala
20 25 30
Glu Ile Thr Leu Thr Trp Gln Trp Asp Gly Glu Asp Gln Thr Gln Asp
35 40 45
Thr Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Gly Thr Phe Gln Lys
50 55 60
Trp Ala Ala Val Val Val Pro Ser Gly Glu Glu Gln Arg Tyr Thr Cys
65 70 75 80
His Val Gln His Glu Gly Leu Pro Glu Pro Leu Thr Leu Arg Trp Glu
85 90 95
Pro Ser Ser
<210> 13
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена MHC
класса I HLA-C (C*04:01:01:01)
<400> 13
Gln Arg Ala Glu His Pro Lys Thr His Val Thr His His Pro Val Ser
1 5 10 15
Asp His Glu Ala Thr Leu Arg Cys Trp Ala Leu Gly Phe Tyr Pro Ala
20 25 30
Glu Ile Thr Leu Thr Trp Gln Trp Asp Gly Glu Asp Gln Thr Gln Asp
35 40 45
Thr Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Gly Thr Phe Gln Lys
50 55 60
Trp Ala Ala Val Val Val Pro Ser Gly Glu Glu Gln Arg Tyr Thr Cys
65 70 75 80
His Val Gln His Glu Gly Leu Pro Glu Pro Leu Thr Leu Arg Trp Lys
85 90 95
Pro Ser Ser
<210> 14
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена MHC
класса I HLA-C (C*05:01:01:01)
<400> 14
Gln Arg Ala Glu His Pro Lys Thr His Val Thr His His Pro Val Ser
1 5 10 15
Asp His Glu Ala Thr Leu Arg Cys Trp Ala Leu Gly Phe Tyr Pro Ala
20 25 30
Glu Ile Thr Leu Thr Trp Gln Arg Asp Gly Glu Asp Gln Thr Gln Asp
35 40 45
Thr Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Gly Thr Phe Gln Lys
50 55 60
Trp Ala Ala Val Val Val Pro Ser Gly Glu Glu Gln Arg Tyr Thr Cys
65 70 75 80
His Val Gln His Glu Gly Leu Pro Glu Pro Leu Thr Leu Arg Trp Gly
85 90 95
Pro Ser Ser
<210> 15
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена MHC
класса I HLA-C (C*06:02:01:01)
<400> 15
Gln Arg Ala Glu His Pro Lys Thr His Val Thr His His Pro Val Ser
1 5 10 15
Asp His Glu Ala Thr Leu Arg Cys Trp Ala Leu Gly Phe Tyr Pro Ala
20 25 30
Glu Ile Thr Leu Thr Trp Gln Arg Asp Gly Glu Asp Gln Thr Gln Asp
35 40 45
Thr Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Gly Thr Phe Gln Lys
50 55 60
Trp Ala Ala Val Val Val Pro Ser Gly Glu Glu Gln Arg Tyr Thr Cys
65 70 75 80
His Val Gln His Glu Gly Leu Pro Glu Pro Leu Thr Leu Arg Trp Glu
85 90 95
Pro Ser Ser
<210> 16
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена MHC
класса I HLA-E (E*01:01:01:01)
<400> 16
Leu His Leu Glu Pro Pro Lys Thr His Val Thr His His Pro Ile Ser
1 5 10 15
Asp His Glu Ala Thr Leu Arg Cys Trp Ala Leu Gly Phe Tyr Pro Ala
20 25 30
Glu Ile Thr Leu Thr Trp Gln Gln Asp Gly Glu Gly His Thr Gln Asp
35 40 45
Thr Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Gly Thr Phe Gln Lys
50 55 60
Trp Ala Ala Val Val Val Pro Ser Gly Glu Glu Gln Arg Tyr Thr Cys
65 70 75 80
His Val Gln His Glu Gly Leu Pro Glu Pro Val Thr Leu Arg Trp Lys
85 90 95
Pro Ala Ser
<210> 17
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена MHC
класса I HLA-E (E*01:03:01:01)
<400> 17
Leu His Leu Glu Pro Pro Lys Thr His Val Thr His His Pro Ile Ser
1 5 10 15
Asp His Glu Ala Thr Leu Arg Cys Trp Ala Leu Gly Phe Tyr Pro Ala
20 25 30
Glu Ile Thr Leu Thr Trp Gln Gln Asp Gly Glu Gly His Thr Gln Asp
35 40 45
Thr Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Gly Thr Phe Gln Lys
50 55 60
Trp Ala Ala Val Val Val Pro Ser Gly Glu Glu Gln Arg Tyr Thr Cys
65 70 75 80
His Val Gln His Glu Gly Leu Pro Glu Pro Val Thr Leu Arg Trp Lys
85 90 95
Pro Ala Ser
<210> 18
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена MHC
класса I HLA-E (E*01:05)
<400> 18
Leu His Leu Glu Pro Pro Lys Thr His Val Thr His His Pro Ile Ser
1 5 10 15
Asp His Glu Ala Thr Leu Arg Cys Trp Ala Leu Gly Phe Tyr Pro Ala
20 25 30
Glu Ile Thr Leu Thr Trp Gln Gln Asp Gly Glu Gly His Thr Gln Asp
35 40 45
Thr Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Gly Thr Phe Gln Lys
50 55 60
Trp Ala Ala Val Val Val Pro Ser Gly Glu Glu Gln Arg Tyr Thr Cys
65 70 75 80
His Val Gln His Glu Gly Leu Pro Glu Pro Val Thr Leu Arg Trp Lys
85 90 95
Pro Ala Ser
<210> 19
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена MHC
класса I HLA-E (E*01:06)
<400> 19
Leu His Leu Glu Pro Pro Lys Thr His Val Thr His His Pro Ile Ser
1 5 10 15
Asp His Glu Ala Thr Leu Arg Cys Trp Ala Leu Gly Phe Tyr Pro Ala
20 25 30
Glu Ile Thr Leu Thr Trp Gln Gln Asp Gly Glu Gly His Thr Gln Asp
35 40 45
Thr Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Gly Thr Phe Gln Lys
50 55 60
Trp Ala Ala Val Val Val Pro Ser Gly Glu Glu Gln Arg Tyr Thr Cys
65 70 75 80
His Val Gln His Glu Gly Leu Ser Glu Pro Val Thr Leu Arg Trp Lys
85 90 95
Pro Ala Ser
<210> 20
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена MHC
класса I HLA-E (E*01:09)
<400> 20
Leu His Leu Glu Pro Pro Lys Thr His Val Thr His His Pro Ile Ser
1 5 10 15
Asp His Glu Ala Thr Leu Arg Cys Trp Ala Leu Gly Phe Tyr Pro Ala
20 25 30
Glu Ile Thr Leu Thr Trp Gln Gln Asp Gly Glu Gly His Thr Gln Asp
35 40 45
Thr Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Gly Thr Phe Gln Lys
50 55 60
Trp Ala Ala Val Val Val Pro Ser Gly Glu Glu Pro Arg Tyr Thr Cys
65 70 75 80
His Val Gln His Glu Gly Leu Pro Glu Pro Val Thr Leu Arg Trp Lys
85 90 95
Pro Ala Ser
<210> 21
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена MHC
класса I HLA-F (F*01:01:01:01)
<400> 21
Gln Arg Ala Asp Pro Pro Lys Ala His Val Ala His His Pro Ile Ser
1 5 10 15
Asp His Glu Ala Thr Leu Arg Cys Trp Ala Leu Gly Phe Tyr Pro Ala
20 25 30
Glu Ile Thr Leu Thr Trp Gln Arg Asp Gly Glu Glu Gln Thr Gln Asp
35 40 45
Thr Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Gly Thr Phe Gln Lys
50 55 60
Trp Ala Ala Val Val Val Pro Ser Gly Glu Glu Gln Arg Tyr Thr Cys
65 70 75 80
His Val Gln His Glu Gly Leu Pro Gln Pro Leu Ile Leu Arg Trp Glu
85 90 95
Gln Ser Pro
<210> 22
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена MHC
класса I HLA-F (F*01:02)
<400> 22
Gln Arg Ala Asp Pro Pro Lys Ala His Val Ala His His Pro Ile Ser
1 5 10 15
Asp His Glu Ala Thr Leu Arg Cys Trp Ala Leu Gly Phe Tyr Pro Ala
20 25 30
Glu Ile Thr Leu Thr Trp Gln Arg Asp Gly Glu Glu Gln Thr Gln Asp
35 40 45
Thr Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Gly Thr Phe Gln Lys
50 55 60
Trp Ala Ala Val Val Val Pro Ser Gly Glu Glu Gln Arg Tyr Thr Cys
65 70 75 80
His Val Gln His Glu Gly Leu Pro Gln Pro Leu Ile Leu Arg Trp Glu
85 90 95
Gln Ser Pro
<210> 23
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена MHC
класса I HLA-F (F*01:03:01:01)
<400> 23
Gln Arg Ala Asp Pro Pro Lys Ala His Val Ala His His Pro Ile Ser
1 5 10 15
Asp His Glu Ala Thr Leu Arg Cys Trp Ala Leu Gly Phe Tyr Pro Ala
20 25 30
Glu Ile Thr Leu Thr Trp Gln Arg Asp Gly Glu Glu Gln Thr Gln Asp
35 40 45
Thr Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Gly Thr Phe Gln Lys
50 55 60
Trp Ala Ala Val Val Val Pro Pro Gly Glu Glu Gln Arg Tyr Thr Cys
65 70 75 80
His Val Gln His Glu Gly Leu Pro Gln Pro Leu Ile Leu Arg Trp Glu
85 90 95
Gln Ser Pro
<210> 24
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена MHC
класса I HLA-F (F*01:04:01:01)
<400> 24
Gln Arg Ala Asp Pro Pro Lys Ala His Val Ala His His Pro Ile Ser
1 5 10 15
Asp His Glu Ala Thr Leu Arg Cys Trp Ala Leu Gly Phe Tyr Pro Ala
20 25 30
Glu Ile Thr Leu Thr Trp Gln Arg Asp Gly Glu Glu Gln Thr Gln Asp
35 40 45
Thr Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Gly Thr Phe Gln Lys
50 55 60
Trp Ala Ala Val Val Val Pro Ser Gly Glu Glu Gln Arg Tyr Thr Cys
65 70 75 80
His Val Gln His Glu Gly Leu Pro Gln Pro Leu Ile Leu Arg Trp Glu
85 90 95
Gln Ser Pro
<210> 25
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена MHC
класса I HLA-F (F*01:05)
<400> 25
Gln Arg Ala Asp Pro Pro Lys Ala His Val Ala His His Pro Ile Ser
1 5 10 15
Asp His Glu Ala Thr Leu Arg Cys Trp Ala Leu Gly Phe Tyr Pro Ala
20 25 30
Glu Ile Thr Leu Thr Trp Gln Arg Asp Gly Glu Glu Gln Thr Gln Asp
35 40 45
Thr Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Gly Thr Phe Gln Lys
50 55 60
Trp Ala Ala Val Val Val Pro Ser Gly Glu Glu Gln Arg Tyr Thr Cys
65 70 75 80
His Val Gln His Glu Gly Leu Pro Gln Pro Leu Ile Leu Arg Trp Glu
85 90 95
Gln Ser Leu
<210> 26
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена MHC
класса I HLA-G (G*01:01:01:01)
<400> 26
Gln Arg Ala Asp Pro Pro Lys Thr His Val Thr His His Pro Val Phe
1 5 10 15
Asp Tyr Glu Ala Thr Leu Arg Cys Trp Ala Leu Gly Phe Tyr Pro Ala
20 25 30
Glu Ile Ile Leu Thr Trp Gln Arg Asp Gly Glu Asp Gln Thr Gln Asp
35 40 45
Val Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Gly Thr Phe Gln Lys
50 55 60
Trp Ala Ala Val Val Val Pro Ser Gly Glu Glu Gln Arg Tyr Thr Cys
65 70 75 80
His Val Gln His Glu Gly Leu Pro Glu Pro Leu Met Leu Arg Trp Lys
85 90 95
Gln Ser Ser
<210> 27
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена MHC
класса I HLA-G (G*01:03:01:01)
<400> 27
Gln Arg Ala Asp Pro Pro Lys Thr His Val Thr His His Pro Val Phe
1 5 10 15
Asp Tyr Glu Ala Thr Leu Arg Cys Trp Ala Leu Gly Phe Tyr Pro Ala
20 25 30
Glu Ile Ile Leu Thr Trp Gln Arg Asp Gly Glu Asp Gln Thr Gln Asp
35 40 45
Val Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Gly Thr Phe Gln Lys
50 55 60
Trp Ala Ala Val Val Val Pro Ser Gly Glu Glu Gln Arg Tyr Thr Cys
65 70 75 80
His Val Gln His Glu Gly Leu Pro Glu Pro Leu Met Leu Arg Trp Lys
85 90 95
Gln Ser Ser
<210> 28
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена MHC
класса I HLA-G (G*01:04:01:01)
<400> 28
Gln Arg Ala Asp Pro Pro Lys Thr His Val Thr His His Pro Val Phe
1 5 10 15
Asp Tyr Glu Ala Thr Leu Arg Cys Trp Ala Leu Gly Phe Tyr Pro Ala
20 25 30
Glu Ile Ile Leu Thr Trp Gln Arg Asp Gly Glu Asp Gln Thr Gln Asp
35 40 45
Val Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Gly Thr Phe Gln Lys
50 55 60
Trp Ala Ala Val Val Val Pro Ser Gly Glu Glu Gln Arg Tyr Thr Cys
65 70 75 80
His Val Gln His Glu Gly Leu Pro Glu Pro Leu Met Leu Arg Trp Lys
85 90 95
Gln Ser Ser
<210> 29
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена MHC
класса I HLA-G (G*01:06)
<400> 29
Gln Arg Ala Asp Pro Pro Lys Thr His Val Thr His His Pro Val Phe
1 5 10 15
Asp Tyr Glu Ala Thr Leu Arg Cys Trp Ala Leu Gly Phe Tyr Pro Ala
20 25 30
Glu Ile Ile Leu Thr Trp Gln Arg Asp Gly Glu Asp Gln Thr Gln Asp
35 40 45
Val Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Gly Thr Phe Gln Lys
50 55 60
Trp Ala Ala Val Val Val Pro Ser Gly Glu Glu Gln Arg Tyr Met Cys
65 70 75 80
His Val Gln His Glu Gly Leu Pro Glu Pro Leu Met Leu Arg Trp Lys
85 90 95
Gln Ser Ser
<210> 30
<211> 98
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α2 околомембранного домена MHC
класса II HLA-DM (HLA-DMA*01:01:01:01)
<400> 30
Ser Arg Gly Phe Pro Ile Ala Glu Val Phe Thr Leu Lys Pro Leu Glu
1 5 10 15
Phe Gly Lys Pro Asn Thr Leu Val Cys Phe Val Ser Asn Leu Phe Pro
20 25 30
Pro Met Leu Thr Val Asn Trp Gln His His Ser Val Pro Val Glu Gly
35 40 45
Phe Gly Pro Thr Phe Val Ser Ala Val Asp Gly Leu Ser Phe Gln Ala
50 55 60
Phe Ser Tyr Leu Asn Phe Thr Pro Glu Pro Ser Asp Ile Phe Ser Cys
65 70 75 80
Ile Val Thr His Glu Ile Asp Arg Tyr Thr Ala Ile Ala Tyr Trp Val
85 90 95
Pro Arg
<210> 31
<211> 100
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность β2 околомембранного домена MHC
класса II HLA-DM (HLA-DMB*01:01:01:01)
<400> 31
Arg Thr Arg Pro Pro Ser Val Gln Val Ala Lys Thr Thr Pro Phe Asn
1 5 10 15
Thr Arg Glu Pro Val Met Leu Ala Cys Tyr Val Trp Gly Phe Tyr Pro
20 25 30
Ala Glu Val Thr Ile Thr Trp Arg Lys Asn Gly Lys Leu Val Met Pro
35 40 45
His Ser Ser Ala His Lys Thr Ala Gln Pro Asn Gly Asp Trp Thr Tyr
50 55 60
Gln Thr Leu Ser His Leu Ala Leu Thr Pro Ser Tyr Gly Asp Thr Tyr
65 70 75 80
Thr Cys Val Val Glu His Ile Gly Ala Pro Glu Pro Ile Leu Arg Asp
85 90 95
Trp Thr Pro Gly
100
<210> 32
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α2 околомембранного домена MHC
класса II HLA-DO (HLA-DOA*01:01:01)
<400> 32
Ile Asn Val Pro Pro Arg Val Thr Val Leu Pro Lys Ser Arg Val Glu
1 5 10 15
Leu Gly Gln Pro Asn Ile Leu Ile Cys Ile Val Asp Asn Ile Phe Pro
20 25 30
Pro Val Ile Asn Ile Thr Trp Leu Arg Asn Gly Gln Thr Val Thr Glu
35 40 45
Gly Val Ala Gln Thr Ser Phe Tyr Ser Gln Pro Asp His Leu Phe Arg
50 55 60
Lys Phe His Tyr Leu Pro Phe Val Pro Ser Ala Glu Asp Val Tyr Asp
65 70 75 80
Cys Gln Val Glu His Trp Gly Leu Asp Ala Pro Leu Leu Arg His Trp
85 90 95
Glu Leu Gln
<210> 33
<211> 100
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность β2 околомембранного домена MHC
класса II HLA-DO (HLA-DOB*01:01:01:01)
<400> 33
Arg Lys Val Gln Pro Glu Val Thr Val Tyr Pro Glu Arg Thr Pro Leu
1 5 10 15
Leu His Gln His Asn Leu Leu His Cys Ser Val Thr Gly Phe Tyr Pro
20 25 30
Gly Asp Ile Lys Ile Lys Trp Phe Leu Asn Gly Gln Glu Glu Arg Ala
35 40 45
Gly Val Met Ser Thr Gly Pro Ile Arg Asn Gly Asp Trp Thr Phe Gln
50 55 60
Thr Val Val Met Leu Glu Met Thr Pro Glu Leu Gly His Val Tyr Thr
65 70 75 80
Cys Leu Val Asp His Ser Ser Leu Leu Ser Pro Val Ser Val Glu Trp
85 90 95
Arg Ala Gln Ser
100
<210> 34
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α2 околомембранного домена MHC
класса II HLA-DP (HLA-DPA1*01:03:01:01)
<400> 34
Thr Asn Asp Pro Pro Glu Val Thr Val Phe Pro Lys Glu Pro Val Glu
1 5 10 15
Leu Gly Gln Pro Asn Thr Leu Ile Cys His Ile Asp Lys Phe Phe Pro
20 25 30
Pro Val Leu Asn Val Thr Trp Leu Cys Asn Gly Glu Leu Val Thr Glu
35 40 45
Gly Val Ala Glu Ser Leu Phe Leu Pro Arg Thr Asp Tyr Ser Phe His
50 55 60
Lys Phe His Tyr Leu Thr Phe Val Pro Ser Ala Glu Asp Phe Tyr Asp
65 70 75 80
Cys Arg Val Glu His Trp Gly Leu Asp Gln Pro Leu Leu Lys His Trp
85 90 95
Glu Ala Gln
<210> 35
<211> 100
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность β2 околомембранного домена MHC
класса II HLA-DP (HLA-DPB1*01:01:01:01)
<400> 35
Arg Arg Val Gln Pro Lys Val Asn Val Ser Pro Ser Lys Lys Gly Pro
1 5 10 15
Leu Gln His His Asn Leu Leu Val Cys His Val Thr Asp Phe Tyr Pro
20 25 30
Gly Ser Ile Gln Val Arg Trp Phe Leu Asn Gly Gln Glu Glu Thr Ala
35 40 45
Gly Val Val Ser Thr Asn Leu Ile Arg Asn Gly Asp Trp Thr Phe Gln
50 55 60
Ile Leu Val Met Leu Glu Met Thr Pro Gln Gln Gly Asp Val Tyr Ile
65 70 75 80
Cys Gln Val Glu His Thr Ser Leu Asp Ser Pro Val Thr Val Glu Trp
85 90 95
Lys Ala Gln Ser
100
<210> 36
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α2 околомембранного домена MHC
класса II HLA-DQ (HLA-DQA1*01:01:01:01)
<400> 36
Thr Asn Glu Val Pro Glu Val Thr Val Phe Ser Lys Ser Pro Val Thr
1 5 10 15
Leu Gly Gln Pro Asn Thr Leu Ile Cys Leu Val Asp Asn Ile Phe Pro
20 25 30
Pro Val Val Asn Ile Thr Trp Leu Ser Asn Gly Gln Ser Val Thr Glu
35 40 45
Gly Val Ser Glu Thr Ser Phe Leu Ser Lys Ser Asp His Ser Phe Phe
50 55 60
Lys Ile Ser Tyr Leu Thr Phe Leu Pro Ser Ala Asp Glu Ile Tyr Asp
65 70 75 80
Cys Lys Val Glu His Trp Gly Leu Asp Gln Pro Leu Leu Lys His Trp
85 90 95
Glu Pro Glu
<210> 37
<211> 100
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность β2 околомембранного домена MHC
класса II HLA-DQ (HLA-DQB1*05:01:01:01)
<400> 37
Arg Arg Val Glu Pro Thr Val Thr Ile Ser Pro Ser Arg Thr Glu Ala
1 5 10 15
Leu Asn His His Asn Leu Leu Ile Cys Ser Val Thr Asp Phe Tyr Pro
20 25 30
Ser Gln Ile Lys Val Arg Trp Phe Arg Asn Asp Gln Glu Glu Thr Ala
35 40 45
Gly Val Val Ser Thr Pro Leu Ile Arg Asn Gly Asp Trp Thr Phe Gln
50 55 60
Ile Leu Val Met Leu Glu Met Thr Pro Gln Arg Gly Asp Val Tyr Thr
65 70 75 80
Cys His Val Glu His Pro Ser Leu Gln Ser Pro Ile Thr Val Glu Trp
85 90 95
Arg Ala Gln Ser
100
<210> 38
<211> 99
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α2 околомембранного домена MHC
класса II HLA-DR (HLA-DRA*01:01:01:01)
<400> 38
Thr Asn Val Pro Pro Glu Val Thr Val Leu Thr Asn Ser Pro Val Glu
1 5 10 15
Leu Arg Glu Pro Asn Val Leu Ile Cys Phe Ile Asp Lys Phe Thr Pro
20 25 30
Pro Val Val Asn Val Thr Trp Leu Arg Asn Gly Lys Pro Val Thr Thr
35 40 45
Gly Val Ser Glu Thr Val Phe Leu Pro Arg Glu Asp His Leu Phe Arg
50 55 60
Lys Phe His Tyr Leu Pro Phe Leu Pro Ser Thr Glu Asp Val Tyr Asp
65 70 75 80
Cys Arg Val Glu His Trp Gly Leu Asp Glu Pro Leu Leu Lys His Trp
85 90 95
Glu Phe Asp
<210> 39
<211> 100
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность β2 околомембранного домена MHC
класса II HLA-DR (HLA-DRB1*01:01:01:01)
<400> 39
Arg Arg Val Glu Pro Lys Val Thr Val Tyr Pro Ser Lys Thr Gln Pro
1 5 10 15
Leu Gln His His Asn Leu Leu Val Cys Ser Val Ser Gly Phe Tyr Pro
20 25 30
Gly Ser Ile Glu Val Arg Trp Phe Arg Asn Gly Gln Glu Glu Lys Ala
35 40 45
Gly Val Val Ser Thr Gly Leu Ile Gln Asn Gly Asp Trp Thr Phe Gln
50 55 60
Thr Leu Val Met Leu Glu Thr Val Pro Arg Ser Gly Glu Val Tyr Thr
65 70 75 80
Cys Gln Val Glu His Pro Ser Val Thr Ser Pro Leu Thr Val Glu Trp
85 90 95
Arg Ala Arg Ser
100
<210> 40
<211> 100
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена CD1a
<400> 40
Gln Arg Gln Val Lys Pro Glu Ala Trp Leu Ser His Gly Pro Ser Pro
1 5 10 15
Gly Pro Gly His Leu Gln Leu Val Cys His Val Ser Gly Phe Tyr Pro
20 25 30
Lys Pro Val Trp Val Met Trp Met Arg Gly Glu Gln Glu Gln Gln Gly
35 40 45
Thr Gln Arg Gly Asp Ile Leu Pro Ser Ala Asp Gly Thr Trp Tyr Leu
50 55 60
Arg Ala Thr Leu Glu Val Ala Ala Gly Glu Ala Ala Asp Leu Ser Cys
65 70 75 80
Arg Val Lys His Ser Ser Leu Glu Gly Gln Asp Ile Val Leu Tyr Trp
85 90 95
Glu His His Ser
100
<210> 41
<211> 97
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена CD1b
<400> 41
Gln Arg Gln Val Lys Pro Glu Ala Trp Leu Ser Ser Gly Pro Ser Pro
1 5 10 15
Gly Pro Gly Arg Leu Gln Leu Val Cys His Val Ser Gly Phe Tyr Pro
20 25 30
Lys Pro Val Trp Val Met Trp Met Arg Gly Glu Gln Glu Gln Gln Gly
35 40 45
Thr Gln Leu Gly Asp Ile Leu Pro Asn Ala Asn Trp Thr Trp Tyr Leu
50 55 60
Arg Ala Thr Leu Asp Val Ala Asp Gly Glu Ala Ala Gly Leu Ser Cys
65 70 75 80
Arg Val Lys His Ser Ser Leu Glu Gly Gln Asp Ile Ile Leu Tyr Trp
85 90 95
Arg
<210> 42
<211> 100
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена CD1c
<400> 42
His Arg Gln Val Arg Pro Glu Ala Trp Leu Ser Ser Arg Pro Ser Leu
1 5 10 15
Gly Ser Gly Gln Leu Leu Leu Val Cys His Ala Ser Gly Phe Tyr Pro
20 25 30
Lys Pro Val Trp Val Thr Trp Met Arg Asn Glu Gln Glu Gln Leu Gly
35 40 45
Thr Lys His Gly Asp Ile Leu Pro Asn Ala Asp Gly Thr Trp Tyr Leu
50 55 60
Gln Val Ile Leu Glu Val Ala Ser Glu Glu Pro Ala Gly Leu Ser Cys
65 70 75 80
Arg Val Arg His Ser Ser Leu Gly Gly Gln Asp Ile Ile Leu Tyr Trp
85 90 95
Gly His His Phe
100
<210> 43
<211> 100
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена CD1d
<400> 43
Lys Lys Gln Val Lys Pro Lys Ala Trp Leu Ser Arg Gly Pro Ser Pro
1 5 10 15
Gly Pro Gly Arg Leu Leu Leu Val Cys His Val Ser Gly Phe Tyr Pro
20 25 30
Lys Pro Val Trp Val Lys Trp Met Arg Gly Glu Gln Glu Gln Gln Gly
35 40 45
Thr Gln Pro Gly Asp Ile Leu Pro Asn Ala Asp Glu Thr Trp Tyr Leu
50 55 60
Arg Ala Thr Leu Asp Val Val Ala Gly Glu Ala Ala Gly Leu Ser Cys
65 70 75 80
Arg Val Lys His Ser Ser Leu Glu Gly Gln Asp Ile Val Leu Tyr Trp
85 90 95
Gly Gly Ser Tyr
100
<210> 44
<211> 100
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена CD1e
<400> 44
Lys Arg Lys Val Lys Pro Glu Ala Trp Leu Ser Cys Gly Pro Ser Pro
1 5 10 15
Gly Pro Gly Arg Leu Gln Leu Val Cys His Val Ser Gly Phe Tyr Pro
20 25 30
Lys Pro Val Trp Val Met Trp Met Arg Gly Glu Gln Glu Gln Arg Gly
35 40 45
Thr Gln Arg Gly Asp Val Leu Pro Asn Ala Asp Glu Thr Trp Tyr Leu
50 55 60
Arg Ala Thr Leu Asp Val Ala Ala Gly Glu Ala Ala Gly Leu Ser Cys
65 70 75 80
Arg Val Lys His Ser Ser Leu Gly Gly His Asp Leu Ile Ile His Trp
85 90 95
Gly Gly Tyr Ser
100
<210> 45
<211> 101
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена HFE
<400> 45
Asp Gln Gln Val Pro Pro Leu Val Lys Val Thr His His Val Thr Ser
1 5 10 15
Ser Val Thr Thr Leu Arg Cys Arg Ala Leu Asn Tyr Tyr Pro Gln Asn
20 25 30
Ile Thr Met Lys Trp Leu Lys Asp Lys Gln Pro Met Asp Ala Lys Glu
35 40 45
Phe Glu Pro Lys Asp Val Leu Pro Asn Gly Asp Gly Thr Tyr Gln Gly
50 55 60
Trp Ile Thr Leu Ala Val Pro Pro Gly Glu Glu Gln Arg Tyr Thr Cys
65 70 75 80
Gln Val Glu His Pro Gly Leu Asp Gln Pro Leu Ile Val Ile Trp Glu
85 90 95
Pro Ser Pro Ser Gly
100
<210> 46
<211> 98
<212> PRT
<213> Природная последовательность
<220>
<223> аминокислотная последовательность универсального β2
микроглобулина (β2Μ) МНС класса I или МНС-подобных белков (CD1,
HFE)
<400> 46
Ile Gln Arg Thr Pro Lys Ile Gln Val Tyr Ser Arg His Pro Ala Glu
1 5 10 15
Asn Gly Lys Ser Asn Phe Leu Asn Cys Tyr Val Ser Gly Phe His Pro
20 25 30
Ser Asp Ile Glu Val Asp Leu Leu Lys Asn Gly Glu Arg Ile Glu Lys
35 40 45
Val Glu His Ser Asp Leu Ser Phe Ser Lys Asp Trp Ser Phe Tyr Leu
50 55 60
Leu Tyr Tyr Thr Glu Phe Thr Pro Thr Glu Lys Asp Glu Tyr Ala Cys
65 70 75 80
Arg Val Asn His Val Thr Leu Ser Gln Pro Lys Ile Val Lys Trp Asp
85 90 95
Arg Asp
<210> 47
<211> 98
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность универсального β2
микроглобулина (β2Μ) МНС класса I или МНС-подобных белков (CD1,
HFE) с мутацией R12C
<400> 47
Ile Gln Arg Thr Pro Lys Ile Gln Val Tyr Ser Cys His Pro Ala Glu
1 5 10 15
Asn Gly Lys Ser Asn Phe Leu Asn Cys Tyr Val Ser Gly Phe His Pro
20 25 30
Ser Asp Ile Glu Val Asp Leu Leu Lys Asn Gly Glu Arg Ile Glu Lys
35 40 45
Val Glu His Ser Asp Leu Ser Phe Ser Lys Asp Trp Ser Phe Tyr Leu
50 55 60
Leu Tyr Tyr Thr Glu Phe Thr Pro Thr Glu Lys Asp Glu Tyr Ala Cys
65 70 75 80
Arg Val Asn His Val Thr Leu Ser Gln Pro Lys Ile Val Lys Trp Asp
85 90 95
Arg Asp
<210> 48
<211> 110
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность универсального β2
микроглобулина (β2Μ) МНС класса I или МНС-подобных белков (CD1,
HFE) с мутацией R12C + шарнир (hinge) с мутацией C220A
<400> 48
Ile Gln Arg Thr Pro Lys Ile Gln Val Tyr Ser Cys His Pro Ala Glu
1 5 10 15
Asn Gly Lys Ser Asn Phe Leu Asn Cys Tyr Val Ser Gly Phe His Pro
20 25 30
Ser Asp Ile Glu Val Asp Leu Leu Lys Asn Gly Glu Arg Ile Glu Lys
35 40 45
Val Glu His Ser Asp Leu Ser Phe Ser Lys Asp Trp Ser Phe Tyr Leu
50 55 60
Leu Tyr Tyr Thr Glu Phe Thr Pro Thr Glu Lys Asp Glu Tyr Ala Cys
65 70 75 80
Arg Val Asn His Val Thr Leu Ser Gln Pro Lys Ile Val Lys Trp Asp
85 90 95
Arg Asp Glu Pro Lys Ser Ala Asp Lys Thr His Thr Cys Pro
100 105 110
<210> 49
<211> 100
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена CD1b
с мутацией N57C и удлинением на 3 аминокислоты GSС на С-конце
<400> 49
Gln Arg Gln Val Lys Pro Glu Ala Trp Leu Ser Ser Gly Pro Ser Pro
1 5 10 15
Gly Pro Gly Arg Leu Gln Leu Val Cys His Val Ser Gly Phe Tyr Pro
20 25 30
Lys Pro Val Trp Val Met Trp Met Arg Gly Glu Gln Glu Gln Gln Gly
35 40 45
Thr Gln Leu Gly Asp Ile Leu Pro Cys Ala Asn Trp Thr Trp Tyr Leu
50 55 60
Arg Ala Thr Leu Asp Val Ala Asp Gly Glu Ala Ala Gly Leu Ser Cys
65 70 75 80
Arg Val Lys His Ser Ser Leu Glu Gly Gln Asp Ile Ile Leu Tyr Trp
85 90 95
Arg Gly Ser Cys
100
<210> 50
<211> 99
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена CD1b
с мутацией N57C и удлинением на 2 аминокислоты GS на С-конце
<400> 50
Gln Arg Gln Val Lys Pro Glu Ala Trp Leu Ser Ser Gly Pro Ser Pro
1 5 10 15
Gly Pro Gly Arg Leu Gln Leu Val Cys His Val Ser Gly Phe Tyr Pro
20 25 30
Lys Pro Val Trp Val Met Trp Met Arg Gly Glu Gln Glu Gln Gln Gly
35 40 45
Thr Gln Leu Gly Asp Ile Leu Pro Cys Ala Asn Trp Thr Trp Tyr Leu
50 55 60
Arg Ala Thr Leu Asp Val Ala Asp Gly Glu Ala Ala Gly Leu Ser Cys
65 70 75 80
Arg Val Lys His Ser Ser Leu Glu Gly Gln Asp Ile Ile Leu Tyr Trp
85 90 95
Arg Gly Ser
<210> 51
<211> 100
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена CD1b
с мутацией N59A и удлинением на 3 аминокислоты GSС на С-конце
<400> 51
Gln Arg Gln Val Lys Pro Glu Ala Trp Leu Ser Ser Gly Pro Ser Pro
1 5 10 15
Gly Pro Gly Arg Leu Gln Leu Val Cys His Val Ser Gly Phe Tyr Pro
20 25 30
Lys Pro Val Trp Val Met Trp Met Arg Gly Glu Gln Glu Gln Gln Gly
35 40 45
Thr Gln Leu Gly Asp Ile Leu Pro Asn Ala Ala Trp Thr Trp Tyr Leu
50 55 60
Arg Ala Thr Leu Asp Val Ala Asp Gly Glu Ala Ala Gly Leu Ser Cys
65 70 75 80
Arg Val Lys His Ser Ser Leu Glu Gly Gln Asp Ile Ile Leu Tyr Trp
85 90 95
Arg Gly Ser Cys
100
<210> 52
<211> 100
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена CD1b
с мутацией N59D и удлинением на 3 аминокислоты GSС на С-конце
<400> 52
Gln Arg Gln Val Lys Pro Glu Ala Trp Leu Ser Ser Gly Pro Ser Pro
1 5 10 15
Gly Pro Gly Arg Leu Gln Leu Val Cys His Val Ser Gly Phe Tyr Pro
20 25 30
Lys Pro Val Trp Val Met Trp Met Arg Gly Glu Gln Glu Gln Gln Gly
35 40 45
Thr Gln Leu Gly Asp Ile Leu Pro Asn Ala Asp Trp Thr Trp Tyr Leu
50 55 60
Arg Ala Thr Leu Asp Val Ala Asp Gly Glu Ala Ala Gly Leu Ser Cys
65 70 75 80
Arg Val Lys His Ser Ser Leu Glu Gly Gln Asp Ile Ile Leu Tyr Trp
85 90 95
Arg Gly Ser Cys
100
<210> 53
<211> 100
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена CD1b
с мутациями N57C, N59A и удлинением на 3 аминокислоты GSС на
С-конце
<400> 53
Gln Arg Gln Val Lys Pro Glu Ala Trp Leu Ser Ser Gly Pro Ser Pro
1 5 10 15
Gly Pro Gly Arg Leu Gln Leu Val Cys His Val Ser Gly Phe Tyr Pro
20 25 30
Lys Pro Val Trp Val Met Trp Met Arg Gly Glu Gln Glu Gln Gln Gly
35 40 45
Thr Gln Leu Gly Asp Ile Leu Pro Cys Ala Ala Trp Thr Trp Tyr Leu
50 55 60
Arg Ala Thr Leu Asp Val Ala Asp Gly Glu Ala Ala Gly Leu Ser Cys
65 70 75 80
Arg Val Lys His Ser Ser Leu Glu Gly Gln Asp Ile Ile Leu Tyr Trp
85 90 95
Arg Gly Ser Cys
100
<210> 54
<211> 100
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена CD1b
с мутациями N57C, N59D и удлинением на 3 аминокислоты GSС на
С-конце
<400> 54
Gln Arg Gln Val Lys Pro Glu Ala Trp Leu Ser Ser Gly Pro Ser Pro
1 5 10 15
Gly Pro Gly Arg Leu Gln Leu Val Cys His Val Ser Gly Phe Tyr Pro
20 25 30
Lys Pro Val Trp Val Met Trp Met Arg Gly Glu Gln Glu Gln Gln Gly
35 40 45
Thr Gln Leu Gly Asp Ile Leu Pro Cys Ala Asp Trp Thr Trp Tyr Leu
50 55 60
Arg Ala Thr Leu Asp Val Ala Asp Gly Glu Ala Ala Gly Leu Ser Cys
65 70 75 80
Arg Val Lys His Ser Ser Leu Glu Gly Gln Asp Ile Ile Leu Tyr Trp
85 90 95
Arg Gly Ser Cys
100
<210> 55
<211> 99
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена CD1b
с мутациями N57C, N59A и удлинением на 2 аминокислоты GS на
С-конце
<400> 55
Gln Arg Gln Val Lys Pro Glu Ala Trp Leu Ser Ser Gly Pro Ser Pro
1 5 10 15
Gly Pro Gly Arg Leu Gln Leu Val Cys His Val Ser Gly Phe Tyr Pro
20 25 30
Lys Pro Val Trp Val Met Trp Met Arg Gly Glu Gln Glu Gln Gln Gly
35 40 45
Thr Gln Leu Gly Asp Ile Leu Pro Cys Ala Ala Trp Thr Trp Tyr Leu
50 55 60
Arg Ala Thr Leu Asp Val Ala Asp Gly Glu Ala Ala Gly Leu Ser Cys
65 70 75 80
Arg Val Lys His Ser Ser Leu Glu Gly Gln Asp Ile Ile Leu Tyr Trp
85 90 95
Arg Gly Ser
<210> 56
<211> 99
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена CD1b
с мутациями N57C, N59D и удлинением на 2 аминокислоты GS на
С-конце
<400> 56
Gln Arg Gln Val Lys Pro Glu Ala Trp Leu Ser Ser Gly Pro Ser Pro
1 5 10 15
Gly Pro Gly Arg Leu Gln Leu Val Cys His Val Ser Gly Phe Tyr Pro
20 25 30
Lys Pro Val Trp Val Met Trp Met Arg Gly Glu Gln Glu Gln Gln Gly
35 40 45
Thr Gln Leu Gly Asp Ile Leu Pro Cys Ala Asp Trp Thr Trp Tyr Leu
50 55 60
Arg Ala Thr Leu Asp Val Ala Asp Gly Glu Ala Ala Gly Leu Ser Cys
65 70 75 80
Arg Val Lys His Ser Ser Leu Glu Gly Gln Asp Ile Ile Leu Tyr Trp
85 90 95
Arg Gly Ser
<210> 57
<211> 459
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность первой тяжелой цепи кандидатов
02-004 и 02-006 (Prolgolimab_VH_HC_hole)
<400> 57
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met Tyr Trp Val Arg Gln Val Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Asp Thr Gly Gly Gly Arg Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Ala Ile Ser Arg Val Asn Ala Lys Asn Thr Met Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Glu Gly Gly Gly Thr Gly Trp Gly Val Leu Lys Asp Trp
100 105 110
Pro Tyr Gly Leu Asp Ala Trp Gly Gln Gly Thr Leu Val Thr Val Ser
115 120 125
Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser
130 135 140
Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp
145 150 155 160
Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr
165 170 175
Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr
180 185 190
Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln
195 200 205
Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp
210 215 220
Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro
225 230 235 240
Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro
245 250 255
Pro Lys Pro Lys Asp Thr Leu Tyr Ile Thr Arg Glu Pro Glu Val Thr
260 265 270
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
275 280 285
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
290 295 300
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
305 310 315 320
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
325 330 335
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
340 345 350
Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Asp
355 360 365
Glu Leu Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe
370 375 380
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
385 390 395 400
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
405 410 415
Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
420 425 430
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
435 440 445
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
450 455
<210> 58
<211> 214
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность первой легкой цепи кандидатов
02-004 и 02-006 (Prolgolimab_VL_CK)
<400> 58
Gln Pro Val Leu Thr Gln Pro Leu Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Asn Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Arg Asp Ser Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Thr Ala Val
85 90 95
Phe Gly Thr Gly Thr Lys Leu Thr Val Leu Gln Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 59
<211> 452
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность второй тяжелой цепи кандидатов
02-004 и 02-009 (Ocrelizumab_VH_b2m_Fc_knob)
<400> 59
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Asn Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ala Ile Tyr Pro Gly Asn Gly Asp Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Lys Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Val Val Tyr Tyr Ser Asn Ser Tyr Trp Tyr Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ile Gln Arg Thr Pro Lys
115 120 125
Ile Gln Val Tyr Ser Arg His Pro Ala Glu Asn Gly Lys Ser Asn Phe
130 135 140
Leu Asn Cys Tyr Val Ser Gly Phe His Pro Ser Asp Ile Glu Val Asp
145 150 155 160
Leu Leu Lys Asn Gly Glu Arg Ile Glu Lys Val Glu His Ser Asp Leu
165 170 175
Ser Phe Ser Lys Asp Trp Ser Phe Tyr Leu Leu Tyr Tyr Thr Glu Phe
180 185 190
Thr Pro Thr Glu Lys Asp Glu Tyr Ala Cys Arg Val Asn His Val Thr
195 200 205
Leu Ser Gln Pro Lys Ile Val Lys Trp Asp Arg Asp Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Tyr Ile Thr Arg Glu Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 60
<211> 206
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность второй легкой цепи кандидатов
02-004 и 02-009 (Ocrelizumab_VL_CD1b)
<400> 60
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Pro Leu Ile Tyr
35 40 45
Ala Pro Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Phe Asn Pro Pro Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Gln Arg Gln Val Lys Pro
100 105 110
Glu Ala Trp Leu Ser Ser Gly Pro Ser Pro Gly Pro Gly Arg Leu Gln
115 120 125
Leu Val Cys His Val Ser Gly Phe Tyr Pro Lys Pro Val Trp Val Met
130 135 140
Trp Met Arg Gly Glu Gln Glu Gln Gln Gly Thr Gln Leu Gly Asp Ile
145 150 155 160
Leu Pro Asn Ala Asn Trp Thr Trp Tyr Leu Arg Ala Thr Leu Asp Val
165 170 175
Ala Asp Gly Glu Ala Ala Gly Leu Ser Cys Arg Val Lys His Ser Ser
180 185 190
Leu Glu Gly Gln Asp Ile Ile Leu Tyr Trp Arg Gly Ser Cys
195 200 205
<210> 61
<211> 452
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность первой тяжелой цепи кандидатов
02-005 и 02-008 (Ocrelizumab_VH_ HC_hole)
<400> 61
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Asn Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ala Ile Tyr Pro Gly Asn Gly Asp Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Lys Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Val Val Tyr Tyr Ser Asn Ser Tyr Trp Tyr Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Tyr Ile Thr Arg Glu Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 62
<211> 213
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность первой легкой цепи кандидатов
02-005 и 02-008 (Ocrelizumab_VL_CK)
<400> 62
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Pro Leu Ile Tyr
35 40 45
Ala Pro Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Phe Asn Pro Pro Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 63
<211> 459
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность второй тяжелой цепи кандидатов
02-005 и 02-007 (Prolgolimab_VH_b2m_Fc_knob)
<400> 63
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met Tyr Trp Val Arg Gln Val Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Asp Thr Gly Gly Gly Arg Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Ala Ile Ser Arg Val Asn Ala Lys Asn Thr Met Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Glu Gly Gly Gly Thr Gly Trp Gly Val Leu Lys Asp Trp
100 105 110
Pro Tyr Gly Leu Asp Ala Trp Gly Gln Gly Thr Leu Val Thr Val Ser
115 120 125
Ser Ile Gln Arg Thr Pro Lys Ile Gln Val Tyr Ser Arg His Pro Ala
130 135 140
Glu Asn Gly Lys Ser Asn Phe Leu Asn Cys Tyr Val Ser Gly Phe His
145 150 155 160
Pro Ser Asp Ile Glu Val Asp Leu Leu Lys Asn Gly Glu Arg Ile Glu
165 170 175
Lys Val Glu His Ser Asp Leu Ser Phe Ser Lys Asp Trp Ser Phe Tyr
180 185 190
Leu Leu Tyr Tyr Thr Glu Phe Thr Pro Thr Glu Lys Asp Glu Tyr Ala
195 200 205
Cys Arg Val Asn His Val Thr Leu Ser Gln Pro Lys Ile Val Lys Trp
210 215 220
Asp Arg Asp Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro
225 230 235 240
Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro
245 250 255
Pro Lys Pro Lys Asp Thr Leu Tyr Ile Thr Arg Glu Pro Glu Val Thr
260 265 270
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
275 280 285
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
290 295 300
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
305 310 315 320
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
325 330 335
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
340 345 350
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Asp
355 360 365
Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe
370 375 380
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
385 390 395 400
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
405 410 415
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
420 425 430
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
435 440 445
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
450 455
<210> 64
<211> 207
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность второй легкой цепи кандидатов
02-005 и 02-007 (Prolgolimab_VL_CD1b)
<400> 64
Gln Pro Val Leu Thr Gln Pro Leu Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Asn Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Arg Asp Ser Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Thr Ala Val
85 90 95
Phe Gly Thr Gly Thr Lys Leu Thr Val Leu Gln Gln Arg Gln Val Lys
100 105 110
Pro Glu Ala Trp Leu Ser Ser Gly Pro Ser Pro Gly Pro Gly Arg Leu
115 120 125
Gln Leu Val Cys His Val Ser Gly Phe Tyr Pro Lys Pro Val Trp Val
130 135 140
Met Trp Met Arg Gly Glu Gln Glu Gln Gln Gly Thr Gln Leu Gly Asp
145 150 155 160
Ile Leu Pro Asn Ala Asn Trp Thr Trp Tyr Leu Arg Ala Thr Leu Asp
165 170 175
Val Ala Asp Gly Glu Ala Ala Gly Leu Ser Cys Arg Val Lys His Ser
180 185 190
Ser Leu Glu Gly Gln Asp Ile Ile Leu Tyr Trp Arg Gly Ser Cys
195 200 205
<210> 65
<211> 454
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность второй тяжелой цепи кандидатов
02-006 и 02-008 (Анти-CSF1R_VH_b2m_Fc_knob)
<400> 65
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Arg Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Glu Phe Thr Phe Asp Asp Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Trp Asn Gly Gly Ser Thr Asn Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Ser Thr Val Glu Val Ala His Arg Arg Leu Tyr Lys Tyr Leu Glu
100 105 110
Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ile Gln Arg Thr
115 120 125
Pro Lys Ile Gln Val Tyr Ser Arg His Pro Ala Glu Asn Gly Lys Ser
130 135 140
Asn Phe Leu Asn Cys Tyr Val Ser Gly Phe His Pro Ser Asp Ile Glu
145 150 155 160
Val Asp Leu Leu Lys Asn Gly Glu Arg Ile Glu Lys Val Glu His Ser
165 170 175
Asp Leu Ser Phe Ser Lys Asp Trp Ser Phe Tyr Leu Leu Tyr Tyr Thr
180 185 190
Glu Phe Thr Pro Thr Glu Lys Asp Glu Tyr Ala Cys Arg Val Asn His
195 200 205
Val Thr Leu Ser Gln Pro Lys Ile Val Lys Trp Asp Arg Asp Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Tyr Ile Thr Arg Glu Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn
355 360 365
Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly Lys
450
<210> 66
<211> 208
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность второй легкой цепи кандидатов
02-006 и 02-008 (Анти-CSF1R_VL_CD1b)
<400> 66
Gln Ala Gly Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Val Tyr
35 40 45
Gly Asp Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ile Ser Ser Asp His
85 90 95
Val Val Phe Gly Gly Gly Thr Gln Leu Thr Val Leu Gln Arg Gln Val
100 105 110
Lys Pro Glu Ala Trp Leu Ser Ser Gly Pro Ser Pro Gly Pro Gly Arg
115 120 125
Leu Gln Leu Val Cys His Val Ser Gly Phe Tyr Pro Lys Pro Val Trp
130 135 140
Val Met Trp Met Arg Gly Glu Gln Glu Gln Gln Gly Thr Gln Leu Gly
145 150 155 160
Asp Ile Leu Pro Asn Ala Asn Trp Thr Trp Tyr Leu Arg Ala Thr Leu
165 170 175
Asp Val Ala Asp Gly Glu Ala Ala Gly Leu Ser Cys Arg Val Lys His
180 185 190
Ser Ser Leu Glu Gly Gln Asp Ile Ile Leu Tyr Trp Arg Gly Ser Cys
195 200 205
<210> 67
<211> 454
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность первой тяжелой цепи кандидатов
02-007 и 02-009 (Анти-CSF1R_VH_HC_hole)
<400> 67
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Arg Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Glu Phe Thr Phe Asp Asp Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Trp Asn Gly Gly Ser Thr Asn Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Ser Thr Val Glu Val Ala His Arg Arg Leu Tyr Lys Tyr Leu Glu
100 105 110
Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Tyr Ile Thr Arg Glu Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
355 360 365
Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly Lys
450
<210> 68
<211> 215
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность первой легкой цепи кандидатов
02-007 и 02-009 (Анти-CSF1R_VL_CK)
<400> 68
Gln Ala Gly Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Val Tyr
35 40 45
Gly Asp Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ile Ser Ser Asp His
85 90 95
Val Val Phe Gly Gly Gly Thr Gln Leu Thr Val Leu Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 69
<211> 459
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность тяжелой цепи кандидатов 03-001,
03-005 и 03-006, которая содержит универсальный β2 микроглобулина
(β2Μ) с мутацией R12C
<400> 69
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met Tyr Trp Val Arg Gln Val Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Asp Thr Gly Gly Gly Arg Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Ala Ile Ser Arg Val Asn Ala Lys Asn Thr Met Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Glu Gly Gly Gly Thr Gly Trp Gly Val Leu Lys Asp Trp
100 105 110
Pro Tyr Gly Leu Asp Ala Trp Gly Gln Gly Thr Leu Val Thr Val Ser
115 120 125
Ser Ile Gln Arg Thr Pro Lys Ile Gln Val Tyr Ser Cys His Pro Ala
130 135 140
Glu Asn Gly Lys Ser Asn Phe Leu Asn Cys Tyr Val Ser Gly Phe His
145 150 155 160
Pro Ser Asp Ile Glu Val Asp Leu Leu Lys Asn Gly Glu Arg Ile Glu
165 170 175
Lys Val Glu His Ser Asp Leu Ser Phe Ser Lys Asp Trp Ser Phe Tyr
180 185 190
Leu Leu Tyr Tyr Thr Glu Phe Thr Pro Thr Glu Lys Asp Glu Tyr Ala
195 200 205
Cys Arg Val Asn His Val Thr Leu Ser Gln Pro Lys Ile Val Lys Trp
210 215 220
Asp Arg Asp Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro
225 230 235 240
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
245 250 255
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
260 265 270
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
275 280 285
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
290 295 300
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
305 310 315 320
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
325 330 335
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
340 345 350
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
355 360 365
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
370 375 380
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
385 390 395 400
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
405 410 415
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
420 425 430
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
435 440 445
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
450 455
<210> 70
<211> 459
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность тяжелой цепи кандидатов 03-003
и 03-004, которая содержит универсальный β2 микроглобулина (β2Μ)
<400> 70
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met Tyr Trp Val Arg Gln Val Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Asp Thr Gly Gly Gly Arg Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Ala Ile Ser Arg Val Asn Ala Lys Asn Thr Met Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Glu Gly Gly Gly Thr Gly Trp Gly Val Leu Lys Asp Trp
100 105 110
Pro Tyr Gly Leu Asp Ala Trp Gly Gln Gly Thr Leu Val Thr Val Ser
115 120 125
Ser Ile Gln Arg Thr Pro Lys Ile Gln Val Tyr Ser Arg His Pro Ala
130 135 140
Glu Asn Gly Lys Ser Asn Phe Leu Asn Cys Tyr Val Ser Gly Phe His
145 150 155 160
Pro Ser Asp Ile Glu Val Asp Leu Leu Lys Asn Gly Glu Arg Ile Glu
165 170 175
Lys Val Glu His Ser Asp Leu Ser Phe Ser Lys Asp Trp Ser Phe Tyr
180 185 190
Leu Leu Tyr Tyr Thr Glu Phe Thr Pro Thr Glu Lys Asp Glu Tyr Ala
195 200 205
Cys Arg Val Asn His Val Thr Leu Ser Gln Pro Lys Ile Val Lys Trp
210 215 220
Asp Arg Asp Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro
225 230 235 240
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
245 250 255
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
260 265 270
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
275 280 285
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
290 295 300
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
305 310 315 320
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
325 330 335
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
340 345 350
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
355 360 365
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
370 375 380
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
385 390 395 400
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
405 410 415
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
420 425 430
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
435 440 445
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
450 455
<210> 71
<211> 459
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность тяжелой цепи кандидатов 03-002,
03-007 и 03-008, которая содержит универсальный β2 микроглобулина
(β2Μ) с мутацией R12C и шарнир (hinge) с мутацией C220A
<400> 71
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met Tyr Trp Val Arg Gln Val Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Asp Thr Gly Gly Gly Arg Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Ala Ile Ser Arg Val Asn Ala Lys Asn Thr Met Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Glu Gly Gly Gly Thr Gly Trp Gly Val Leu Lys Asp Trp
100 105 110
Pro Tyr Gly Leu Asp Ala Trp Gly Gln Gly Thr Leu Val Thr Val Ser
115 120 125
Ser Ile Gln Arg Thr Pro Lys Ile Gln Val Tyr Ser Cys His Pro Ala
130 135 140
Glu Asn Gly Lys Ser Asn Phe Leu Asn Cys Tyr Val Ser Gly Phe His
145 150 155 160
Pro Ser Asp Ile Glu Val Asp Leu Leu Lys Asn Gly Glu Arg Ile Glu
165 170 175
Lys Val Glu His Ser Asp Leu Ser Phe Ser Lys Asp Trp Ser Phe Tyr
180 185 190
Leu Leu Tyr Tyr Thr Glu Phe Thr Pro Thr Glu Lys Asp Glu Tyr Ala
195 200 205
Cys Arg Val Asn His Val Thr Leu Ser Gln Pro Lys Ile Val Lys Trp
210 215 220
Asp Arg Asp Glu Pro Lys Ser Ala Asp Lys Thr His Thr Cys Pro Pro
225 230 235 240
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
245 250 255
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
260 265 270
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
275 280 285
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
290 295 300
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
305 310 315 320
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
325 330 335
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
340 345 350
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
355 360 365
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
370 375 380
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
385 390 395 400
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
405 410 415
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
420 425 430
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
435 440 445
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
450 455
<210> 72
<211> 207
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность легкой цепи кандидата 03-001,
которая содержит α3 околомембранный домен CD1b с мутацией N57C
<400> 72
Gln Pro Val Leu Thr Gln Pro Leu Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Asn Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Arg Asp Ser Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Thr Ala Val
85 90 95
Phe Gly Thr Gly Thr Lys Leu Thr Val Leu Gln Gln Arg Gln Val Lys
100 105 110
Pro Glu Ala Trp Leu Ser Ser Gly Pro Ser Pro Gly Pro Gly Arg Leu
115 120 125
Gln Leu Val Cys His Val Ser Gly Phe Tyr Pro Lys Pro Val Trp Val
130 135 140
Met Trp Met Arg Gly Glu Gln Glu Gln Gln Gly Thr Gln Leu Gly Asp
145 150 155 160
Ile Leu Pro Cys Ala Asn Trp Thr Trp Tyr Leu Arg Ala Thr Leu Asp
165 170 175
Val Ala Asp Gly Glu Ala Ala Gly Leu Ser Cys Arg Val Lys His Ser
180 185 190
Ser Leu Glu Gly Gln Asp Ile Ile Leu Tyr Trp Arg Gly Ser Cys
195 200 205
<210> 73
<211> 206
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность легкой цепи кандидата 03-002,
которая содержит α3 околомембранный домен CD1b с мутацией N57C
<400> 73
Gln Pro Val Leu Thr Gln Pro Leu Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Asn Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Arg Asp Ser Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Thr Ala Val
85 90 95
Phe Gly Thr Gly Thr Lys Leu Thr Val Leu Gln Gln Arg Gln Val Lys
100 105 110
Pro Glu Ala Trp Leu Ser Ser Gly Pro Ser Pro Gly Pro Gly Arg Leu
115 120 125
Gln Leu Val Cys His Val Ser Gly Phe Tyr Pro Lys Pro Val Trp Val
130 135 140
Met Trp Met Arg Gly Glu Gln Glu Gln Gln Gly Thr Gln Leu Gly Asp
145 150 155 160
Ile Leu Pro Cys Ala Asn Trp Thr Trp Tyr Leu Arg Ala Thr Leu Asp
165 170 175
Val Ala Asp Gly Glu Ala Ala Gly Leu Ser Cys Arg Val Lys His Ser
180 185 190
Ser Leu Glu Gly Gln Asp Ile Ile Leu Tyr Trp Arg Gly Ser
195 200 205
<210> 74
<211> 207
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность легкой цепи кандидата 03-003,
которая содержит α3 околомембранный домен CD1b с мутацией N59A
<400> 74
Gln Pro Val Leu Thr Gln Pro Leu Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Asn Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Arg Asp Ser Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Thr Ala Val
85 90 95
Phe Gly Thr Gly Thr Lys Leu Thr Val Leu Gln Gln Arg Gln Val Lys
100 105 110
Pro Glu Ala Trp Leu Ser Ser Gly Pro Ser Pro Gly Pro Gly Arg Leu
115 120 125
Gln Leu Val Cys His Val Ser Gly Phe Tyr Pro Lys Pro Val Trp Val
130 135 140
Met Trp Met Arg Gly Glu Gln Glu Gln Gln Gly Thr Gln Leu Gly Asp
145 150 155 160
Ile Leu Pro Asn Ala Ala Trp Thr Trp Tyr Leu Arg Ala Thr Leu Asp
165 170 175
Val Ala Asp Gly Glu Ala Ala Gly Leu Ser Cys Arg Val Lys His Ser
180 185 190
Ser Leu Glu Gly Gln Asp Ile Ile Leu Tyr Trp Arg Gly Ser Cys
195 200 205
<210> 75
<211> 207
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность легкой цепи кандидата 03-004,
которая содержит α3 околомембранный домен CD1b с мутацией N59D
<400> 75
Gln Pro Val Leu Thr Gln Pro Leu Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Asn Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Arg Asp Ser Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Thr Ala Val
85 90 95
Phe Gly Thr Gly Thr Lys Leu Thr Val Leu Gln Gln Arg Gln Val Lys
100 105 110
Pro Glu Ala Trp Leu Ser Ser Gly Pro Ser Pro Gly Pro Gly Arg Leu
115 120 125
Gln Leu Val Cys His Val Ser Gly Phe Tyr Pro Lys Pro Val Trp Val
130 135 140
Met Trp Met Arg Gly Glu Gln Glu Gln Gln Gly Thr Gln Leu Gly Asp
145 150 155 160
Ile Leu Pro Asn Ala Asp Trp Thr Trp Tyr Leu Arg Ala Thr Leu Asp
165 170 175
Val Ala Asp Gly Glu Ala Ala Gly Leu Ser Cys Arg Val Lys His Ser
180 185 190
Ser Leu Glu Gly Gln Asp Ile Ile Leu Tyr Trp Arg Gly Ser Cys
195 200 205
<210> 76
<211> 207
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность легкой цепи кандидата 03-005,
которая содержит α3 околомембранный домен CD1b с мутациями N57C,
N59A
<400> 76
Gln Pro Val Leu Thr Gln Pro Leu Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Asn Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Arg Asp Ser Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Thr Ala Val
85 90 95
Phe Gly Thr Gly Thr Lys Leu Thr Val Leu Gln Gln Arg Gln Val Lys
100 105 110
Pro Glu Ala Trp Leu Ser Ser Gly Pro Ser Pro Gly Pro Gly Arg Leu
115 120 125
Gln Leu Val Cys His Val Ser Gly Phe Tyr Pro Lys Pro Val Trp Val
130 135 140
Met Trp Met Arg Gly Glu Gln Glu Gln Gln Gly Thr Gln Leu Gly Asp
145 150 155 160
Ile Leu Pro Cys Ala Ala Trp Thr Trp Tyr Leu Arg Ala Thr Leu Asp
165 170 175
Val Ala Asp Gly Glu Ala Ala Gly Leu Ser Cys Arg Val Lys His Ser
180 185 190
Ser Leu Glu Gly Gln Asp Ile Ile Leu Tyr Trp Arg Gly Ser Cys
195 200 205
<210> 77
<211> 207
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность легкой цепи кандидата 03-006,
которая содержит α3 околомембранный домен CD1b с мутациями N57C,
N59D
<400> 77
Gln Pro Val Leu Thr Gln Pro Leu Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Asn Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Arg Asp Ser Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Thr Ala Val
85 90 95
Phe Gly Thr Gly Thr Lys Leu Thr Val Leu Gln Gln Arg Gln Val Lys
100 105 110
Pro Glu Ala Trp Leu Ser Ser Gly Pro Ser Pro Gly Pro Gly Arg Leu
115 120 125
Gln Leu Val Cys His Val Ser Gly Phe Tyr Pro Lys Pro Val Trp Val
130 135 140
Met Trp Met Arg Gly Glu Gln Glu Gln Gln Gly Thr Gln Leu Gly Asp
145 150 155 160
Ile Leu Pro Cys Ala Asp Trp Thr Trp Tyr Leu Arg Ala Thr Leu Asp
165 170 175
Val Ala Asp Gly Glu Ala Ala Gly Leu Ser Cys Arg Val Lys His Ser
180 185 190
Ser Leu Glu Gly Gln Asp Ile Ile Leu Tyr Trp Arg Gly Ser Cys
195 200 205
<210> 78
<211> 206
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность легкой цепи кандидата 03-007,
которая содержит α3 околомембранный домен CD1b с мутациями N57C,
N59A
<400> 78
Gln Pro Val Leu Thr Gln Pro Leu Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Asn Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Arg Asp Ser Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Thr Ala Val
85 90 95
Phe Gly Thr Gly Thr Lys Leu Thr Val Leu Gln Gln Arg Gln Val Lys
100 105 110
Pro Glu Ala Trp Leu Ser Ser Gly Pro Ser Pro Gly Pro Gly Arg Leu
115 120 125
Gln Leu Val Cys His Val Ser Gly Phe Tyr Pro Lys Pro Val Trp Val
130 135 140
Met Trp Met Arg Gly Glu Gln Glu Gln Gln Gly Thr Gln Leu Gly Asp
145 150 155 160
Ile Leu Pro Cys Ala Ala Trp Thr Trp Tyr Leu Arg Ala Thr Leu Asp
165 170 175
Val Ala Asp Gly Glu Ala Ala Gly Leu Ser Cys Arg Val Lys His Ser
180 185 190
Ser Leu Glu Gly Gln Asp Ile Ile Leu Tyr Trp Arg Gly Ser
195 200 205
<210> 79
<211> 206
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность легкой цепи кандидата 03-008,
которая содержит α3 околомембранный домен CD1b с мутациями N57C,
N59D
<400> 79
Gln Pro Val Leu Thr Gln Pro Leu Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Asn Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Arg Asp Ser Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Thr Ala Val
85 90 95
Phe Gly Thr Gly Thr Lys Leu Thr Val Leu Gln Gln Arg Gln Val Lys
100 105 110
Pro Glu Ala Trp Leu Ser Ser Gly Pro Ser Pro Gly Pro Gly Arg Leu
115 120 125
Gln Leu Val Cys His Val Ser Gly Phe Tyr Pro Lys Pro Val Trp Val
130 135 140
Met Trp Met Arg Gly Glu Gln Glu Gln Gln Gly Thr Gln Leu Gly Asp
145 150 155 160
Ile Leu Pro Cys Ala Asp Trp Thr Trp Tyr Leu Arg Ala Thr Leu Asp
165 170 175
Val Ala Asp Gly Glu Ala Ala Gly Leu Ser Cys Arg Val Lys His Ser
180 185 190
Ser Leu Glu Gly Gln Asp Ile Ile Leu Tyr Trp Arg Gly Ser
195 200 205
<210> 80
<211> 1377
<212> DNA
<213> Искусственная последовательность
<220>
<223> Нуклеиновая кислота, которая кодирует аминокислотную
последовательность первой тяжелой цепи кандидатов 02-004 и 02-006
(Prolgolimab_VH_HC_hole)
<400> 80
caggttcagc tggtgcagtc tgggggaggc ttggtgcagc ctggggggtc tctgagactc 60
tcctgtgcag cctctggatt caccttcagt agctactgga tgtattgggt ccgccaggtt 120
ccagggaagg ggctcgagtg ggtctcagct attgatactg gtggtggtag gacatactat 180
gcagactccg tgaagggccg gttcgccatc tccagagtca acgccaagaa cacaatgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagagatgaa 300
ggcgggggta cgggttgggg tgttttgaag gattggccct acggcttgga cgcatggggc 360
caggggaccc tggtcaccgt ctcctcagct agcaccaagg gcccatcggt cttccccctg 420
gcaccctcct ccaagagcac ctctgggggc acagcggccc tgggctgcct ggtcaaggac 480
tacttccccg aaccggtgac ggtgtcgtgg aactcaggcg ccctgaccag cggcgtgcac 540
accttcccgg ctgtcctaca gtcctcagga ctctactccc tcagcagcgt ggtgaccgtg 600
ccctccagca gcttgggcac ccagacctac atctgcaacg tgaatcacaa gcccagcaac 660
accaaggtgg acaagagagt tgagcccaaa tcttgtgaca aaactcacac atgcccaccg 720
tgcccagcac ctgaagccgc agggggaccg tcagtcttcc tcttcccccc aaaacccaag 780
gacaccctct acatcacccg ggagcctgag gtcacatgcg tggtggtgga cgtgagccac 840
gaagaccctg aggtcaagtt caactggtac gtggacggcg tggaggtgca taatgccaag 900
acaaagccgc gggaggagca gtacaacagc acgtaccgtg tggtcagcgt cctcaccgtc 960
ctgcaccagg actggctgaa tggcaaggag tacaagtgca aggtctccaa caaagccctc 1020
ccagccccca tcgagaaaac catctccaaa gccaaagggc agccccgaga accacaggtg 1080
tgcaccctgc ccccatcccg ggatgagctg accaagaacc aggtcagcct gtcctgcgcc 1140
gtcaaaggct tctatcccag cgacatcgcc gtggagtggg agagcaatgg gcagccggag 1200
aacaactaca agaccacgcc tcccgtgctg gactccgacg gctccttctt cctcgtcagc 1260
aagctcaccg tggacaagag caggtggcag caggggaacg tcttctcatg ctccgtgatg 1320
catgaggctc tgcacaacca ctacacgcag aagagcctct ccctgtcccc gggtaaa 1377
<210> 81
<211> 642
<212> DNA
<213> Искусственная последовательность
<220>
<223> Нуклеиновая кислота, которая кодирует аминокислотную
последовательность первой легкой цепи кандидатов 02-004 и 02-006
(Prolgolimab_VL_CK)
<400> 81
cagcctgtgc tgactcagcc actctcagtg tcagtggccc tgggacagac ggccaggatt 60
acctgtgggg gaaacaacat tggaagtaaa aatgtgcact ggtaccagca gaagccaggc 120
caggcccctg tgctggtcat ctatagggat agcaaccggc cctctgggat ccctgagcga 180
ttctctggct ccaactcggg gaacacggcc accctgacca tcagcagagc ccaagccggg 240
gatgaggctg actattactg tcaggtgtgg gacagcagca ctgctgtctt cggaactggg 300
accaagctga ccgtcctaca gcgtacggtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gt 642
<210> 82
<211> 1356
<212> DNA
<213> Искусственная последовательность
<220>
<223> Нуклеиновая кислота, которая кодирует аминокислотную
последовательность второй тяжелой цепи кандидатов 02-004 и 02-009
(Ocrelizumab_VH_b2m_Fc_knob)
<400> 82
gaagtccaat tagtagaatc aggcggcggc ttagtgcaac ccggtggctc actgcgtttg 60
agctgtgctg ctagtggcta tacctttacg tcctacaaca tgcattgggt gagacaagca 120
cctggcaagg gtctcgaatg ggtaggcgct atctatccag gaaatggtga cacctcttac 180
aaccagaagt ttaagggtcg ctttaccatc tcggtcgata agagcaagaa caccctgtac 240
ctacagatga attcgttacg ggcagaggac acagccgtct attactgcgc tcgggtggta 300
tattactcca actcttactg gtacttcgac gtttggggcc agggaactct tgtgaccgtg 360
agctctatcc aacgcacacc aaagatccag gtctattcgc ggcatccggc agaaaacgga 420
aagagtaact tcctgaactg ctatgtatcg ggctttcatc caagtgatat cgaggtggac 480
ctgctgaaga acggagagag aatcgagaag gtggaacact cggacctgag cttctccaag 540
gactggagtt tctacctgct gtactatacc gagttcaccc cgacagaaaa ggacgaatac 600
gcatgtcggg tcaatcatgt caccctgagt cagccgaaaa tcgtaaagtg ggatcgggac 660
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gcccagcacc tgaagccgct 720
gggggaccgt cagtcttcct cttcccccca aaacccaagg acaccctcta catcacccgg 780
gaacctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 840
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 900
tacaacagca cgtaccgtgt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 960
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccat cgagaaaacc 1020
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatgccgg 1080
gatgagctga ccaagaacca ggtcagcctg tggtgcctgg tcaaaggctt ctatcccagc 1140
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 1200
cccgtgctgg actccgacgg ctccttcttc ctctatagca agctcaccgt ggacaagagc 1260
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1320
tacacgcaga agagcctctc cctgtccccg ggtaaa 1356
<210> 83
<211> 618
<212> DNA
<213> Искусственная последовательность
<220>
<223> Нуклеиновая кислота, которая кодирует аминокислотную
последовательность второй легкой цепи кандидатов 02-004 и 02-009
(Ocrelizumab_VL_CD1b)
<400> 83
gacattcaaa tgacccaatc gccatcttcg ctttcagctt cggtaggaga tcgcgtaacc 60
attacatgcc gggcctcgtc ttcggtctcg tacatgcatt ggtatcagca aaaaccagga 120
aaagctccca aacccctgat ctacgcaccc tctaacctgg cttctggcgt cccgtcgagg 180
ttttcgggtt ctggctctgg cacagacttt acacttacca tctcgtcgct gcagccagag 240
gattttgcca cctactactg tcagcaatgg tcttttaatc caccaacatt cggccagggg 300
accaaagtag aaatcaaaca gagacaggtt aaaccagagg catggctaag ttctggacct 360
tcacctggac ctggtagatt acaattagta tgtcatgtgt cgggatttta tcctaaacct 420
gtttgggtca tgtggatgag gggcgagcaa gagcagcaag gaacacagtt aggcgacatc 480
ctgccgaatg caaactggac atggtactta agagcaaccc tggatgtagc ggacggcgaa 540
gccgctggat tatcgtgccg ggtaaagcat tcgtcgctgg agggccaaga cataatactc 600
tattggcgcg gtagttgc 618
<210> 84
<211> 1356
<212> DNA
<213> Искусственная последовательность
<220>
<223> Нуклеиновая кислота, которая кодирует аминокислотную
последовательность первой тяжелой цепи кандидатов 02-005 и 02-008
(Ocrelizumab_VH_ HC_hole)
<400> 84
gaagtccaat tagtagaatc aggcggcggc ttagtgcaac ccggtggctc actgcgtttg 60
agctgtgctg ctagtggcta tacctttacg tcctacaaca tgcattgggt gagacaagca 120
cctggcaagg gtctcgaatg ggtaggcgct atctatccag gaaatggtga cacctcttac 180
aaccagaagt ttaagggtcg ctttaccatc tcggtcgata agagcaagaa caccctgtac 240
ctacagatga attcgttacg ggcagaggac acagccgtct attactgcgc tcgggtggta 300
tattactcca actcttactg gtacttcgac gtttggggcc agggaactct tgtgaccgtg 360
agctctgcta gcaccaaggg cccatcggtc ttccccctgg caccctcctc caagagcacc 420
tctgggggca cagcggccct gggctgcctg gtcaaggact acttccccga accggtgacg 480
gtgtcgtgga actcaggcgc cctgaccagc ggcgtgcaca ccttcccggc tgtcctacag 540
tcctcaggac tctactccct cagcagcgtg gtgaccgtgc cctccagcag cttgggcacc 600
cagacctaca tctgcaacgt gaatcacaag cccagcaaca ccaaggtgga caagagagtt 660
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gcccagcacc tgaagccgca 720
gggggaccgt cagtcttcct cttcccccca aaacccaagg acaccctcta catcacccgg 780
gagcctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 840
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 900
tacaacagca cgtaccgtgt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 960
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccat cgagaaaacc 1020
atctccaaag ccaaagggca gccccgagaa ccacaggtgt gcaccctgcc cccatcccgg 1080
gatgagctga ccaagaacca ggtcagcctg tcctgcgccg tcaaaggctt ctatcccagc 1140
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 1200
cccgtgctgg actccgacgg ctccttcttc ctcgtcagca agctcaccgt ggacaagagc 1260
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1320
tacacgcaga agagcctctc cctgtccccg ggtaaa 1356
<210> 85
<211> 639
<212> DNA
<213> Искусственная последовательность
<220>
<223> Нуклеиновая кислота, которая кодирует аминокислотную
последовательность первой легкой цепи кандидатов 02-005 и 02-008
(Ocrelizumab_VL_CK)
<400> 85
gacattcaaa tgacccaatc gccatcttcg ctttcagctt cggtaggaga tcgcgtaacc 60
attacatgcc gggcctcgtc ttcggtctcg tacatgcatt ggtatcagca aaaaccagga 120
aaagctccca aacccctgat ctacgcaccc tctaacctgg cttctggcgt cccgtcgagg 180
ttttcgggtt ctggctctgg cacagacttt acacttacca tctcgtcgct gcagccagag 240
gattttgcca cctactactg tcagcaatgg tcttttaatc caccaacatt cggccagggg 300
accaaagtag aaatcaaacg tacggtggct gcaccatctg tcttcatctt cccgccatct 360
gatgagcagt tgaaatctgg aactgcctct gttgtgtgcc tgctgaataa cttctatccc 420
agagaggcca aagtacagtg gaaggtggat aacgccctcc aatcgggtaa ctcccaggag 480
agtgtcacag agcaggacag caaggacagc acctacagcc tcagcagcac cctgacgctg 540
agcaaagcag actacgagaa acacaaagtc tacgcctgcg aagtcaccca tcagggcctg 600
agctcgcccg tcacaaagag cttcaacagg ggagagtgt 639
<210> 86
<211> 1377
<212> DNA
<213> Искусственная последовательность
<220>
<223> Нуклеиновая кислота, которая кодирует аминокислотную
последовательность второй тяжелой цепи кандидатов 02-005 и 02-007
(Prolgolimab_VH_b2m_Fc_knob)
<400> 86
caggttcagc tggtgcagtc tgggggaggc ttggtgcagc ctggggggtc tctgagactc 60
tcctgtgcag cctctggatt caccttcagt agctactgga tgtattgggt ccgccaggtt 120
ccagggaagg ggctcgagtg ggtctcagct attgatactg gtggtggtag gacatactat 180
gcagactccg tgaagggccg gttcgccatc tccagagtca acgccaagaa cacaatgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagagatgaa 300
ggcgggggta cgggttgggg tgttttgaag gattggccct acggcttgga cgcatggggc 360
caggggaccc tggtcaccgt ctcctcaatc caacgcacac caaagatcca ggtctattcg 420
cggcatccgg cagaaaacgg aaagagtaac ttcctgaact gctatgtatc gggctttcat 480
ccaagtgata tcgaggtgga cctgctgaag aacggagaga gaatcgagaa ggtggaacac 540
tcggacctga gcttctccaa ggactggagt ttctacctgc tgtactatac cgagttcacc 600
ccgacagaaa aggacgaata cgcatgtcgg gtcaatcatg tcaccctgag tcagccgaaa 660
atcgtaaagt gggatcggga cgagcccaaa tcttgtgaca aaactcacac atgcccaccg 720
tgcccagcac ctgaagccgc tgggggaccg tcagtcttcc tcttcccccc aaaacccaag 780
gacaccctct acatcacccg ggaacctgag gtcacatgcg tggtggtgga cgtgagccac 840
gaagaccctg aggtcaagtt caactggtac gtggacggcg tggaggtgca taatgccaag 900
acaaagccgc gggaggagca gtacaacagc acgtaccgtg tggtcagcgt cctcaccgtc 960
ctgcaccagg actggctgaa tggcaaggag tacaagtgca aggtctccaa caaagccctc 1020
ccagccccca tcgagaaaac catctccaaa gccaaagggc agccccgaga accacaggtg 1080
tacaccctgc ccccatgccg ggatgagctg accaagaacc aggtcagcct gtggtgcctg 1140
gtcaaaggct tctatcccag cgacatcgcc gtggagtggg agagcaatgg gcagccggag 1200
aacaactaca agaccacgcc tcccgtgctg gactccgacg gctccttctt cctctatagc 1260
aagctcaccg tggacaagag caggtggcag caggggaacg tcttctcatg ctccgtgatg 1320
catgaggctc tgcacaacca ctacacgcag aagagcctct ccctgtcccc gggtaaa 1377
<210> 87
<211> 621
<212> DNA
<213> Искусственная последовательность
<220>
<223> Нуклеиновая кислота, которая кодирует аминокислотную
последовательность второй легкой цепи кандидатов 02-005 и 02-007
(Prolgolimab_VL_CD1b)
<400> 87
cagcctgtgc tgactcagcc actctcagtg tcagtggccc tgggacagac ggccaggatt 60
acctgtgggg gaaacaacat tggaagtaaa aatgtgcact ggtaccagca gaagccaggc 120
caggcccctg tgctggtcat ctatagggat agcaaccggc cctctgggat ccctgagcga 180
ttctctggct ccaactcggg gaacacggcc accctgacca tcagcagagc ccaagccggg 240
gatgaggctg actattactg tcaggtgtgg gacagcagca ctgctgtctt cggaactggg 300
accaagctga ccgtcctaca gcagagacag gttaaaccag aggcatggct aagttctgga 360
ccttcacctg gacctggtag attacaatta gtatgtcatg tgtcgggatt ttatcctaaa 420
cctgtttggg tcatgtggat gaggggcgag caagagcagc aaggaacaca gttaggcgac 480
atcctgccga atgcaaactg gacatggtac ttaagagcaa ccctggatgt agcggacggc 540
gaagccgctg gattatcgtg ccgggtaaag cattcgtcgc tggagggcca agacataata 600
ctctattggc gcggtagttg c 621
<210> 88
<211> 1362
<212> DNA
<213> Искусственная последовательность
<220>
<223> Нуклеиновая кислота, которая кодирует аминокислотную
последовательность второй тяжелой цепи кандидатов 02-006 и 02-008
(Анти-CSF1R_VH_b2m_Fc_knob)
<400> 88
caagtccagt tggtagaaag cggcggaggt ttggttaggc ccggaggcag cctgagatta 60
tcctgtgcag caagcgagtt cacttttgat gactatgcga tgagttgggt gcgacaagct 120
cctggaaagg gattggaatg ggtgtcagcg atctcatgga atggcggatc aactaactac 180
gcagattctg tcaagggcag gtttaccatc tcccgggata acgcaaagaa tagtctgtac 240
cttcaaatga acagtctgcg cgctgaggac actgccttat attattgcgc aagcactgtg 300
gaagtagccc accgacggtt gtataagtac ctcgaagtat ggggtcaggg aacactggtc 360
acagtaagct caatccaacg cacaccaaag atccaggtct attcgcggca tccggcagaa 420
aacggaaaga gtaacttcct gaactgctat gtatcgggct ttcatccaag tgatatcgag 480
gtggacctgc tgaagaacgg agagagaatc gagaaggtgg aacactcgga cctgagcttc 540
tccaaggact ggagtttcta cctgctgtac tataccgagt tcaccccgac agaaaaggac 600
gaatacgcat gtcgggtcaa tcatgtcacc ctgagtcagc cgaaaatcgt aaagtgggat 660
cgggacgagc ccaaatcttg tgacaaaact cacacatgcc caccgtgccc agcacctgaa 720
gccgctgggg gaccgtcagt cttcctcttc cccccaaaac ccaaggacac cctctacatc 780
acccgggaac ctgaggtcac atgcgtggtg gtggacgtga gccacgaaga ccctgaggtc 840
aagttcaact ggtacgtgga cggcgtggag gtgcataatg ccaagacaaa gccgcgggag 900
gagcagtaca acagcacgta ccgtgtggtc agcgtcctca ccgtcctgca ccaggactgg 960
ctgaatggca aggagtacaa gtgcaaggtc tccaacaaag ccctcccagc ccccatcgag 1020
aaaaccatct ccaaagccaa agggcagccc cgagaaccac aggtgtacac cctgccccca 1080
tgccgggatg agctgaccaa gaaccaggtc agcctgtggt gcctggtcaa aggcttctat 1140
cccagcgaca tcgccgtgga gtgggagagc aatgggcagc cggagaacaa ctacaagacc 1200
acgcctcccg tgctggactc cgacggctcc ttcttcctct atagcaagct caccgtggac 1260
aagagcaggt ggcagcaggg gaacgtcttc tcatgctccg tgatgcatga ggctctgcac 1320
aaccactaca cgcagaagag cctctccctg tccccgggta aa 1362
<210> 89
<211> 624
<212> DNA
<213> Искусственная последовательность
<220>
<223> Нуклеиновая кислота, которая кодирует аминокислотную
последовательность второй легкой цепи кандидатов 02-006 и 02-008
(Анти-CSF1R_VL_CD1b)
<400> 89
caggcagggc tgactcagcc accctcggtg tcagtggccc caggacagac ggccaggatt 60
acctgtgggg gaaacaacat tggaagtaaa agtgtgcact ggtaccagca gaagccaggc 120
caggcccctg tgctggtcgt ctatggtgat agcgaccggc cctcagggat ccctgagcga 180
ttctctggct ccaactctgg gaacacggcc accctgacca tcagcagggt cgaagccggg 240
gatgaggccg actattactg tcaggtgtgg gatattagta gtgatcatgt ggtattcggc 300
ggagggaccc agctcaccgt tttacagaga caggttaaac cagaggcatg gctaagttct 360
ggaccttcac ctggacctgg tagattacaa ttagtatgtc atgtgtcggg attttatcct 420
aaacctgttt gggtcatgtg gatgaggggc gagcaagagc agcaaggaac acagttaggc 480
gacatcctgc cgaatgcaaa ctggacatgg tacttaagag caaccctgga tgtagcggac 540
ggcgaagccg ctggattatc gtgccgggta aagcattcgt cgctggaggg ccaagacata 600
atactctatt ggcgcggtag ttgc 624
<210> 90
<211> 1362
<212> DNA
<213> Искусственная последовательность
<220>
<223> Нуклеиновая кислота, которая кодирует аминокислотную
последовательность первой тяжелой цепи кандидатов 02-007 и 02-009
(Анти-CSF1R_VH_HC_hole)
<400> 90
caagtccagt tggtagaaag cggcggaggt ttggttaggc ccggaggcag cctgagatta 60
tcctgtgcag caagcgagtt cacttttgat gactatgcga tgagttgggt gcgacaagct 120
cctggaaagg gattggaatg ggtgtcagcg atctcatgga atggcggatc aactaactac 180
gcagattctg tcaagggcag gtttaccatc tcccgggata acgcaaagaa tagtctgtac 240
cttcaaatga acagtctgcg cgctgaggac actgccttat attattgcgc aagcactgtg 300
gaagtagccc accgacggtt gtataagtac ctcgaagtat ggggtcaggg aacactggtc 360
acagtaagct cagctagcac caagggccca tcggtcttcc ccctggcacc ctcctccaag 420
agcacctctg ggggcacagc ggccctgggc tgcctggtca aggactactt ccccgaaccg 480
gtgacggtgt cgtggaactc aggcgccctg accagcggcg tgcacacctt cccggctgtc 540
ctacagtcct caggactcta ctccctcagc agcgtggtga ccgtgccctc cagcagcttg 600
ggcacccaga cctacatctg caacgtgaat cacaagccca gcaacaccaa ggtggacaag 660
agagttgagc ccaaatcttg tgacaaaact cacacatgcc caccgtgccc agcacctgaa 720
gccgcagggg gaccgtcagt cttcctcttc cccccaaaac ccaaggacac cctctacatc 780
acccgggagc ctgaggtcac atgcgtggtg gtggacgtga gccacgaaga ccctgaggtc 840
aagttcaact ggtacgtgga cggcgtggag gtgcataatg ccaagacaaa gccgcgggag 900
gagcagtaca acagcacgta ccgtgtggtc agcgtcctca ccgtcctgca ccaggactgg 960
ctgaatggca aggagtacaa gtgcaaggtc tccaacaaag ccctcccagc ccccatcgag 1020
aaaaccatct ccaaagccaa agggcagccc cgagaaccac aggtgtgcac cctgccccca 1080
tcccgggatg agctgaccaa gaaccaggtc agcctgtcct gcgccgtcaa aggcttctat 1140
cccagcgaca tcgccgtgga gtgggagagc aatgggcagc cggagaacaa ctacaagacc 1200
acgcctcccg tgctggactc cgacggctcc ttcttcctcg tcagcaagct caccgtggac 1260
aagagcaggt ggcagcaggg gaacgtcttc tcatgctccg tgatgcatga ggctctgcac 1320
aaccactaca cgcagaagag cctctccctg tccccgggta aa 1362
<210> 91
<211> 645
<212> DNA
<213> Искусственная последовательность
<220>
<223> Нуклеиновая кислота, которая кодирует аминокислотную
последовательность первой легкой цепи кандидатов 02-007 и 02-009
(Анти-CSF1R_VL_CK)
<400> 91
caggcagggc tgactcagcc accctcggtg tcagtggccc caggacagac ggccaggatt 60
acctgtgggg gaaacaacat tggaagtaaa agtgtgcact ggtaccagca gaagccaggc 120
caggcccctg tgctggtcgt ctatggtgat agcgaccggc cctcagggat ccctgagcga 180
ttctctggct ccaactctgg gaacacggcc accctgacca tcagcagggt cgaagccggg 240
gatgaggccg actattactg tcaggtgtgg gatattagta gtgatcatgt ggtattcggc 300
ggagggaccc agctcaccgt tttacgtacg gtggctgcac catctgtctt catcttcccg 360
ccatctgatg agcagttgaa atctggaact gcctctgttg tgtgcctgct gaataacttc 420
tatcccagag aggccaaagt acagtggaag gtggataacg ccctccaatc gggtaactcc 480
caggagagtg tcacagagca ggacagcaag gacagcacct acagcctcag cagcaccctg 540
acgctgagca aagcagacta cgagaaacac aaagtctacg cctgcgaagt cacccatcag 600
ggcctgagct cgcccgtcac aaagagcttc aacaggggag agtgt 645
<210> 92
<211> 1377
<212> DNA
<213> Искусственная последовательность
<220>
<223> Нуклеиновая кислота, которая кодирует аминокислотную
последовательность тяжелой цепи кандидатов 03-001 и 03-005,
которая содержит универсальный β2 микроглобулина (β2Μ) с мутацией
R12C
<400> 92
caggttcagc tggtgcagtc tgggggaggc ttggtgcagc ctggggggtc tctgagactc 60
tcctgtgcag cctctggatt caccttcagt agctactgga tgtattgggt ccgccaggtt 120
ccagggaagg ggctcgagtg ggtctcagct attgatactg gtggtggtag gacatactat 180
gcagactccg tgaagggccg gttcgccatc tccagagtca acgccaagaa cacaatgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagagatgaa 300
ggcgggggta cgggttgggg tgttttgaag gattggccct acggcttgga cgcatggggc 360
caggggaccc tggtcaccgt ctcctcaatc caacgcacac caaagatcca ggtctattcg 420
tgccatccgg cagaaaacgg aaagagtaac ttcctgaact gctatgtatc gggctttcat 480
ccaagtgata tcgaggtgga cctgctgaag aacggagaga gaatcgagaa ggtggaacac 540
tcggacctga gcttctccaa ggactggagt ttctacctgc tgtactatac cgagttcacc 600
ccgacagaaa aggacgaata cgcatgtcgg gtcaatcatg tcaccctgag tcagccgaaa 660
atcgtaaagt gggatcggga cgagcccaaa tcttgtgaca aaactcacac atgcccaccg 720
tgcccagcac ctgaactcct ggggggaccg tcagtcttcc tcttcccccc aaaacccaag 780
gacaccctca tgatctcccg gacccctgag gtcacatgcg tggtggtgga cgtgagccac 840
gaagaccctg aggtcaagtt caactggtac gtggacggcg tggaggtgca taatgccaag 900
acaaagccgc gggaggagca gtacaacagc acgtaccgtg tggtcagcgt cctcaccgtc 960
ctgcaccagg actggctgaa tggcaaggag tacaagtgca aggtctccaa caaagccctc 1020
ccagccccca tcgagaaaac catctccaaa gccaaagggc agccccgaga accacaggtg 1080
tacaccctgc ccccatcccg ggaggagatg accaagaacc aggtcagcct gacctgcctg 1140
gtcaaaggct tctatcccag cgacatcgcc gtggagtggg agagcaatgg gcagccggag 1200
aacaactaca agaccacgcc tcccgtgctg gactccgacg gctccttctt cctctatagc 1260
aagctcaccg tggacaagag caggtggcag caggggaacg tcttctcatg ctccgtgatg 1320
catgaggctc tgcacaacca ctacacgcag aaaagcctct ccctgtcccc gggtaaa 1377
<210> 93
<211> 1377
<212> DNA
<213> Искусственная последовательность
<220>
<223> Нуклеиновая кислота, которая кодирует аминокислотную
последовательность тяжелой цепи кандидатов 03-003 и 03-004,
которая содержит универсальный β2 микроглобулина (β2Μ)
<400> 93
caggttcagc tggtgcagtc tgggggaggc ttggtgcagc ctggggggtc tctgagactc 60
tcctgtgcag cctctggatt caccttcagt agctactgga tgtattgggt ccgccaggtt 120
ccagggaagg ggctcgagtg ggtctcagct attgatactg gtggtggtag gacatactat 180
gcagactccg tgaagggccg gttcgccatc tccagagtca acgccaagaa cacaatgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagagatgaa 300
ggcgggggta cgggttgggg tgttttgaag gattggccct acggcttgga cgcatggggc 360
caggggaccc tggtcaccgt ctcctcaatc caacgcacac caaagatcca ggtctattcg 420
cggcatccgg cagaaaacgg aaagagtaac ttcctgaact gctatgtatc gggctttcat 480
ccaagtgata tcgaggtgga cctgctgaag aacggagaga gaatcgagaa ggtggaacac 540
tcggacctga gcttctccaa ggactggagt ttctacctgc tgtactatac cgagttcacc 600
ccgacagaaa aggacgaata cgcatgtcgg gtcaatcatg tcaccctgag tcagccgaaa 660
atcgtaaagt gggatcggga cgagcccaaa tcttgtgaca aaactcacac atgcccaccg 720
tgcccagcac ctgaactcct ggggggaccg tcagtcttcc tcttcccccc aaaacccaag 780
gacaccctca tgatctcccg gacccctgag gtcacatgcg tggtggtgga cgtgagccac 840
gaagaccctg aggtcaagtt caactggtac gtggacggcg tggaggtgca taatgccaag 900
acaaagccgc gggaggagca gtacaacagc acgtaccgtg tggtcagcgt cctcaccgtc 960
ctgcaccagg actggctgaa tggcaaggag tacaagtgca aggtctccaa caaagccctc 1020
ccagccccca tcgagaaaac catctccaaa gccaaagggc agccccgaga accacaggtg 1080
tacaccctgc ccccatcccg ggaggagatg accaagaacc aggtcagcct gacctgcctg 1140
gtcaaaggct tctatcccag cgacatcgcc gtggagtggg agagcaatgg gcagccggag 1200
aacaactaca agaccacgcc tcccgtgctg gactccgacg gctccttctt cctctatagc 1260
aagctcaccg tggacaagag caggtggcag caggggaacg tcttctcatg ctccgtgatg 1320
catgaggctc tgcacaacca ctacacgcag aaaagcctct ccctgtcccc gggtaaa 1377
<210> 94
<211> 1377
<212> DNA
<213> Искусственная последовательность
<220>
<223> Нуклеиновая кислота, которая кодирует аминокислотную
последовательность тяжелой цепи кандидатов 03-002, 03-007 и
03-008, которая содержит универсальный β2 микроглобулина (β2Μ) с
мутацией R12C и шарнир (hinge) с мутацией C220A
<400> 94
caggttcagc tggtgcagtc tgggggaggc ttggtgcagc ctggggggtc tctgagactc 60
tcctgtgcag cctctggatt caccttcagt agctactgga tgtattgggt ccgccaggtt 120
ccagggaagg ggctcgagtg ggtctcagct attgatactg gtggtggtag gacatactat 180
gcagactccg tgaagggccg gttcgccatc tccagagtca acgccaagaa cacaatgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagagatgaa 300
ggcgggggta cgggttgggg tgttttgaag gattggccct acggcttgga cgcatggggc 360
caggggaccc tggtcaccgt ctcctcaatc caacgcacac caaagatcca ggtctattcg 420
tgccatccgg cagaaaacgg aaagagtaac ttcctgaact gctatgtatc gggctttcat 480
ccaagtgata tcgaggtgga cctgctgaag aacggagaga gaatcgagaa ggtggaacac 540
tcggacctga gcttctccaa ggactggagt ttctacctgc tgtactatac cgagttcacc 600
ccgacagaaa aggacgaata cgcatgtcgg gtcaatcatg tcaccctgag tcagccgaaa 660
atcgtaaagt gggatcggga cgagcccaaa tctgccgaca aaactcacac atgcccaccg 720
tgcccagcac ctgaactcct ggggggaccg tcagtcttcc tcttcccccc aaaacccaag 780
gacaccctca tgatctcccg gacccctgag gtcacatgcg tggtggtgga cgtgagccac 840
gaagaccctg aggtcaagtt caactggtac gtggacggcg tggaggtgca taatgccaag 900
acaaagccgc gggaggagca gtacaacagc acgtaccgtg tggtcagcgt cctcaccgtc 960
ctgcaccagg actggctgaa tggcaaggag tacaagtgca aggtctccaa caaagccctc 1020
ccagccccca tcgagaaaac catctccaaa gccaaagggc agccccgaga accacaggtg 1080
tacaccctgc ccccatcccg ggaggagatg accaagaacc aggtcagcct gacctgcctg 1140
gtcaaaggct tctatcccag cgacatcgcc gtggagtggg agagcaatgg gcagccggag 1200
aacaactaca agaccacgcc tcccgtgctg gactccgacg gctccttctt cctctatagc 1260
aagctcaccg tggacaagag caggtggcag caggggaacg tcttctcatg ctccgtgatg 1320
catgaggctc tgcacaacca ctacacgcag aaaagcctct ccctgtcccc gggtaaa 1377
<210> 95
<211> 621
<212> DNA
<213> Искусственная последовательность
<220>
<223> Нуклеиновая кислота, которая кодирует аминокислотную
последовательность легкой цепи кандидата 03-001, которая содержит
α3 околомембранный домен CD1b с мутацией N57C
<400> 95
cagcctgtgc tgactcagcc actctcagtg tcagtggccc tgggacagac ggccaggatt 60
acctgtgggg gaaacaacat tggaagtaaa aatgtgcact ggtaccagca gaagccaggc 120
caggcccctg tgctggtcat ctatagggat agcaaccggc cctctgggat ccctgagcga 180
ttctctggct ccaactcggg gaacacggcc accctgacca tcagcagagc ccaagccggg 240
gatgaggctg actattactg tcaggtgtgg gacagcagca ctgctgtctt cggaactggg 300
accaagctga ccgtcctaca gcagagacag gttaaaccag aggcatggct aagttctgga 360
ccttcacctg gacctggtag attacaatta gtatgtcatg tgtcgggatt ttatcctaaa 420
cctgtttggg tcatgtggat gaggggcgag caagagcagc aaggaacaca gttaggcgac 480
atcctgccgt gcgcaaactg gacatggtac ttaagagcaa ccctggatgt agcggacggc 540
gaagccgctg gattatcgtg ccgggtaaag cattcgtcgc tggagggcca agacataata 600
ctctattggc gcggtagttg c 621
<210> 96
<211> 618
<212> DNA
<213> Искусственная последовательность
<220>
<223> Нуклеиновая кислота, которая кодирует аминокислотную
последовательность легкой цепи кандидата 03-002, которая содержит
α3 околомембранный домен CD1b с мутацией N57C
<400> 96
cagcctgtgc tgactcagcc actctcagtg tcagtggccc tgggacagac ggccaggatt 60
acctgtgggg gaaacaacat tggaagtaaa aatgtgcact ggtaccagca gaagccaggc 120
caggcccctg tgctggtcat ctatagggat agcaaccggc cctctgggat ccctgagcga 180
ttctctggct ccaactcggg gaacacggcc accctgacca tcagcagagc ccaagccggg 240
gatgaggctg actattactg tcaggtgtgg gacagcagca ctgctgtctt cggaactggg 300
accaagctga ccgtcctaca gcagagacag gttaaaccag aggcatggct aagttctgga 360
ccttcacctg gacctggtag attacaatta gtatgtcatg tgtcgggatt ttatcctaaa 420
cctgtttggg tcatgtggat gaggggcgag caagagcagc aaggaacaca gttaggcgac 480
atcctgccgt gcgcaaactg gacatggtac ttaagagcaa ccctggatgt agcggacggc 540
gaagccgctg gattatcgtg ccgggtaaag cattcgtcgc tggagggcca agacataata 600
ctctattggc gcggtagt 618
<210> 97
<211> 621
<212> DNA
<213> Искусственная последовательность
<220>
<223> Нуклеиновая кислота, которая кодирует аминокислотную
последовательность легкой цепи кандидата 03-003, которая содержит
α3 околомембранный домен CD1b с мутацией N59A
<400> 97
cagcctgtgc tgactcagcc actctcagtg tcagtggccc tgggacagac ggccaggatt 60
acctgtgggg gaaacaacat tggaagtaaa aatgtgcact ggtaccagca gaagccaggc 120
caggcccctg tgctggtcat ctatagggat agcaaccggc cctctgggat ccctgagcga 180
ttctctggct ccaactcggg gaacacggcc accctgacca tcagcagagc ccaagccggg 240
gatgaggctg actattactg tcaggtgtgg gacagcagca ctgctgtctt cggaactggg 300
accaagctga ccgtcctaca gcagagacag gttaaaccag aggcatggct aagttctgga 360
ccttcacctg gacctggtag attacaatta gtatgtcatg tgtcgggatt ttatcctaaa 420
cctgtttggg tcatgtggat gaggggcgag caagagcagc aaggaacaca gttaggcgac 480
atcctgccga atgcagcctg gacatggtac ttaagagcaa ccctggatgt agcggacggc 540
gaagccgctg gattatcgtg ccgggtaaag cattcgtcgc tggagggcca agacataata 600
ctctattggc gcggtagttg c 621
<210> 98
<211> 621
<212> DNA
<213> Искусственная последовательность
<220>
<223> Нуклеиновая кислота, которая кодирует аминокислотную
последовательность легкой цепи кандидата 03-004, которая содержит
α3 околомембранный домен CD1b с мутацией N59D
<400> 98
cagcctgtgc tgactcagcc actctcagtg tcagtggccc tgggacagac ggccaggatt 60
acctgtgggg gaaacaacat tggaagtaaa aatgtgcact ggtaccagca gaagccaggc 120
caggcccctg tgctggtcat ctatagggat agcaaccggc cctctgggat ccctgagcga 180
ttctctggct ccaactcggg gaacacggcc accctgacca tcagcagagc ccaagccggg 240
gatgaggctg actattactg tcaggtgtgg gacagcagca ctgctgtctt cggaactggg 300
accaagctga ccgtcctaca gcagagacag gttaaaccag aggcatggct aagttctgga 360
ccttcacctg gacctggtag attacaatta gtatgtcatg tgtcgggatt ttatcctaaa 420
cctgtttggg tcatgtggat gaggggcgag caagagcagc aaggaacaca gttaggcgac 480
atcctgccga atgcagactg gacatggtac ttaagagcaa ccctggatgt agcggacggc 540
gaagccgctg gattatcgtg ccgggtaaag cattcgtcgc tggagggcca agacataata 600
ctctattggc gcggtagttg c 621
<210> 99
<211> 621
<212> DNA
<213> Искусственная последовательность
<220>
<223> Нуклеиновая кислота, которая кодирует аминокислотную я
последовательность легкой цепи кандидата 03-005, которая содержит
α3 околомембранный домен CD1b с мутациями N57C, N59A
<400> 99
cagcctgtgc tgactcagcc actctcagtg tcagtggccc tgggacagac ggccaggatt 60
acctgtgggg gaaacaacat tggaagtaaa aatgtgcact ggtaccagca gaagccaggc 120
caggcccctg tgctggtcat ctatagggat agcaaccggc cctctgggat ccctgagcga 180
ttctctggct ccaactcggg gaacacggcc accctgacca tcagcagagc ccaagccggg 240
gatgaggctg actattactg tcaggtgtgg gacagcagca ctgctgtctt cggaactggg 300
accaagctga ccgtcctaca gcagagacag gttaaaccag aggcatggct aagttctgga 360
ccttcacctg gacctggtag attacaatta gtatgtcatg tgtcgggatt ttatcctaaa 420
cctgtttggg tcatgtggat gaggggcgag caagagcagc aaggaacaca gttaggcgac 480
atcctgccgt gcgcagcctg gacatggtac ttaagagcaa ccctggatgt agcggacggc 540
gaagccgctg gattatcgtg ccgggtaaag cattcgtcgc tggagggcca agacataata 600
ctctattggc gcggtagttg c 621
<210> 100
<211> 621
<212> DNA
<213> Искусственная последовательность
<220>
<223> Нуклеиновая кислота, которая кодирует аминокислотную
последовательность легкой цепи кандидата 03-006, которая содержит
α3 околомембранный домен CD1b с мутациями N57C, N59D
<400> 100
cagcctgtgc tgactcagcc actctcagtg tcagtggccc tgggacagac ggccaggatt 60
acctgtgggg gaaacaacat tggaagtaaa aatgtgcact ggtaccagca gaagccaggc 120
caggcccctg tgctggtcat ctatagggat agcaaccggc cctctgggat ccctgagcga 180
ttctctggct ccaactcggg gaacacggcc accctgacca tcagcagagc ccaagccggg 240
gatgaggctg actattactg tcaggtgtgg gacagcagca ctgctgtctt cggaactggg 300
accaagctga ccgtcctaca gcagagacag gttaaaccag aggcatggct aagttctgga 360
ccttcacctg gacctggtag attacaatta gtatgtcatg tgtcgggatt ttatcctaaa 420
cctgtttggg tcatgtggat gaggggcgag caagagcagc aaggaacaca gttaggcgac 480
atcctgccgt gcgcagactg gacatggtac ttaagagcaa ccctggatgt agcggacggc 540
gaagccgctg gattatcgtg ccgggtaaag cattcgtcgc tggagggcca agacataata 600
ctctattggc gcggtagttg c 621
<210> 101
<211> 618
<212> DNA
<213> Искусственная последовательность
<220>
<223> Нуклеиновая кислота, которая кодирует аминокислотную
последовательность легкой цепи кандидата 03-007, которая содержит
α3 околомембранный домен CD1b с мутациями N57C, N59A
<400> 101
cagcctgtgc tgactcagcc actctcagtg tcagtggccc tgggacagac ggccaggatt 60
acctgtgggg gaaacaacat tggaagtaaa aatgtgcact ggtaccagca gaagccaggc 120
caggcccctg tgctggtcat ctatagggat agcaaccggc cctctgggat ccctgagcga 180
ttctctggct ccaactcggg gaacacggcc accctgacca tcagcagagc ccaagccggg 240
gatgaggctg actattactg tcaggtgtgg gacagcagca ctgctgtctt cggaactggg 300
accaagctga ccgtcctaca gcagagacag gttaaaccag aggcatggct aagttctgga 360
ccttcacctg gacctggtag attacaatta gtatgtcatg tgtcgggatt ttatcctaaa 420
cctgtttggg tcatgtggat gaggggcgag caagagcagc aaggaacaca gttaggcgac 480
atcctgccgt gcgcagcctg gacatggtac ttaagagcaa ccctggatgt agcggacggc 540
gaagccgctg gattatcgtg ccgggtaaag cattcgtcgc tggagggcca agacataata 600
ctctattggc gcggtagt 618
<210> 102
<211> 618
<212> DNA
<213> Искусственная последовательность
<220>
<223> Нуклеиновая кислота, которая кодирует аминокислотную
последовательность легкой цепи кандидата 03-008, которая содержит
α3 околомембранный домен CD1b с мутациями N57C, N59D
<400> 102
cagcctgtgc tgactcagcc actctcagtg tcagtggccc tgggacagac ggccaggatt 60
acctgtgggg gaaacaacat tggaagtaaa aatgtgcact ggtaccagca gaagccaggc 120
caggcccctg tgctggtcat ctatagggat agcaaccggc cctctgggat ccctgagcga 180
ttctctggct ccaactcggg gaacacggcc accctgacca tcagcagagc ccaagccggg 240
gatgaggctg actattactg tcaggtgtgg gacagcagca ctgctgtctt cggaactggg 300
accaagctga ccgtcctaca gcagagacag gttaaaccag aggcatggct aagttctgga 360
ccttcacctg gacctggtag attacaatta gtatgtcatg tgtcgggatt ttatcctaaa 420
cctgtttggg tcatgtggat gaggggcgag caagagcagc aaggaacaca gttaggcgac 480
atcctgccgt gcgcagactg gacatggtac ttaagagcaa ccctggatgt agcggacggc 540
gaagccgctg gattatcgtg ccgggtaaag cattcgtcgc tggagggcca agacataata 600
ctctattggc gcggtagt 618
<210> 103
<211> 129
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность вариабельного домена тяжелой
цепи антитела Пролголимаб (Prolgolimab_VH)
<400> 103
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met Tyr Trp Val Arg Gln Val Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Asp Thr Gly Gly Gly Arg Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Ala Ile Ser Arg Val Asn Ala Lys Asn Thr Met Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Glu Gly Gly Gly Thr Gly Trp Gly Val Leu Lys Asp Trp
100 105 110
Pro Tyr Gly Leu Asp Ala Trp Gly Gln Gly Thr Leu Val Thr Val Ser
115 120 125
Ser
<210> 104
<211> 107
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность вариабельного домена легкой
цепи антитела Пролголимаб (Prolgolimab_VL)
<400> 104
Gln Pro Val Leu Thr Gln Pro Leu Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Asn Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Arg Asp Ser Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Thr Ala Val
85 90 95
Phe Gly Thr Gly Thr Lys Leu Thr Val Leu Gln
100 105
<210> 105
<211> 122
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность вариабельного домена тяжелой
цепи антитела Окрелизумаб (Ocrelizumab_VH)
<400> 105
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Asn Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ala Ile Tyr Pro Gly Asn Gly Asp Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Lys Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Val Val Tyr Tyr Ser Asn Ser Tyr Trp Tyr Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 106
<211> 106
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность вариабельного домена легкой
цепи антитела Окрелизумаб (Ocrelizumab_VL)
<400> 106
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Pro Leu Ile Tyr
35 40 45
Ala Pro Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Phe Asn Pro Pro Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 107
<211> 124
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность вариабельного домена тяжелой
цепи антитела к CSF1R (Анти-CSF1R_VH)
<400> 107
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Arg Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Glu Phe Thr Phe Asp Asp Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Trp Asn Gly Gly Ser Thr Asn Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Ser Thr Val Glu Val Ala His Arg Arg Leu Tyr Lys Tyr Leu Glu
100 105 110
Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 108
<211> 108
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность вариабельного домена легкой
цепи антитела к CSF1R (Анти-CSF1R_VL)
<400> 108
Gln Ala Gly Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Val Tyr
35 40 45
Gly Asp Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ile Ser Ser Asp His
85 90 95
Val Val Phe Gly Gly Gly Thr Gln Leu Thr Val Leu
100 105
<210> 109
<211> 100
<212> PRT
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность α3 околомембранного домена CD1b
с удлинением GSC на С-конце
<400> 109
Gln Arg Gln Val Lys Pro Glu Ala Trp Leu Ser Ser Gly Pro Ser Pro
1 5 10 15
Gly Pro Gly Arg Leu Gln Leu Val Cys His Val Ser Gly Phe Tyr Pro
20 25 30
Lys Pro Val Trp Val Met Trp Met Arg Gly Glu Gln Glu Gln Gln Gly
35 40 45
Thr Gln Leu Gly Asp Ile Leu Pro Asn Ala Asn Trp Thr Trp Tyr Leu
50 55 60
Arg Ala Thr Leu Asp Val Ala Asp Gly Glu Ala Ala Gly Leu Ser Cys
65 70 75 80
Arg Val Lys His Ser Ser Leu Glu Gly Gln Asp Ile Ile Leu Tyr Trp
85 90 95
Arg Gly Ser Cys
100
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО ПРОТИВ ВИРУСА БЕШЕНСТВА И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2764740C1 |
| МУЛЬТИСПЕЦИФИЧЕСКАЯ АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА, ОБЛАДАЮЩАЯ ЗАМЕЩАЮЩЕЙ ФУНКЦИОНАЛЬНОЙ АКТИВНОСТЬЮ КОФАКТОРА КОАГУЛИРУЮЩЕГО ФАКТОРА КРОВИ VIII, И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ УКАЗАННУЮ МОЛЕКУЛУ В КАЧЕСТВЕ АКТИВНОГО ИНГРЕДИЕНТА | 2018 |
|
RU2812909C2 |
| ИММУНОЦИТОКИН, ВКЛЮЧАЮЩИЙ ГЕТЕРОДИМЕРНЫЙ БЕЛКОВЫЙ КОМПЛЕКС НА ОСНОВЕ IL-15/IL-15Rα, И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2753282C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, СПЕЦИФИЧЕСКИ СВЯЗЫВАЮЩИЕСЯ С PD1 И LAG3 | 2018 |
|
RU2778805C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С TFR | 2019 |
|
RU2810756C2 |
| БИСПЕЦИФИЧЕСКИЙ БЕЛОК | 2019 |
|
RU2784486C1 |
| АНТИТЕЛА, СПЕЦИФИЧНЫЕ К CD47 и PD-L1 | 2020 |
|
RU2779652C2 |
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2837821C1 |
| Выделенный альтернативный внутриклеточный сигнальный домен химерного антигенного рецептора и включающий его химерный антигенный рецептор | 2019 |
|
RU2742000C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СОДЕРЖАЩИЕ АНТИ-FAP КЛОН 212 | 2019 |
|
RU2799429C2 |

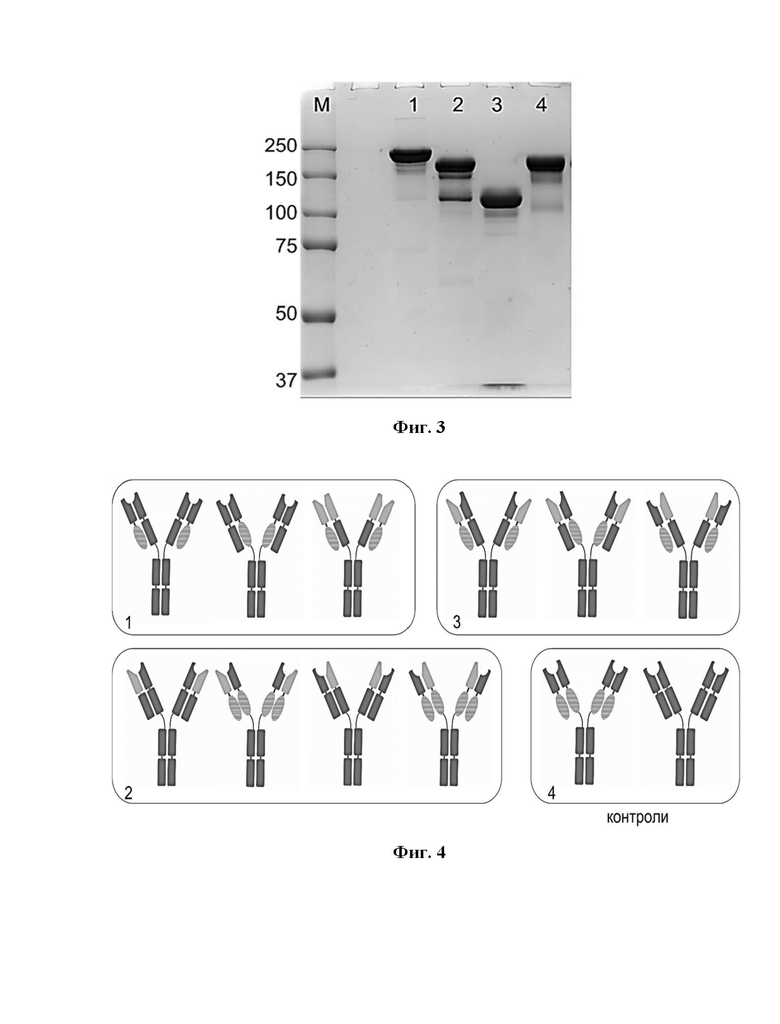
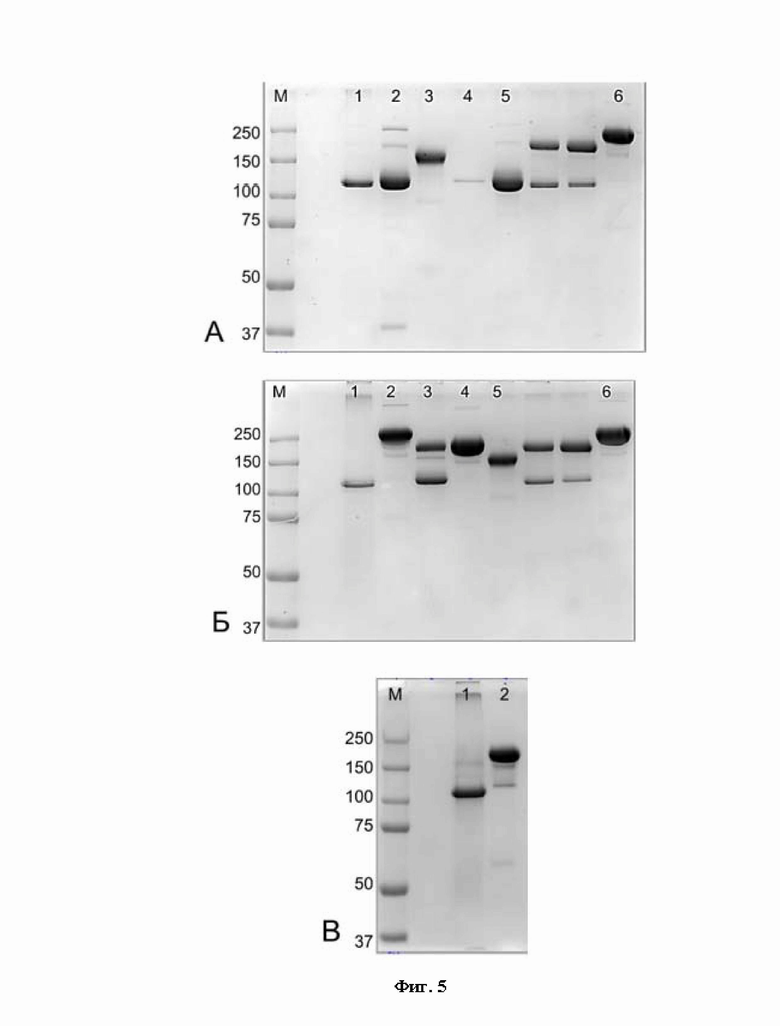
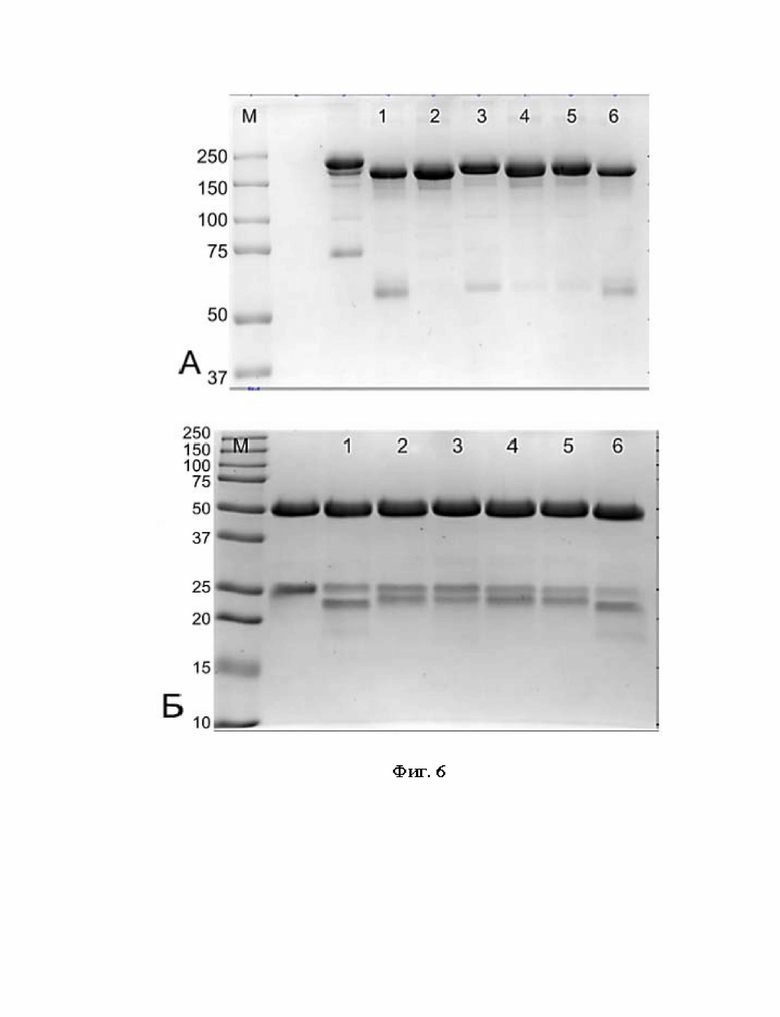

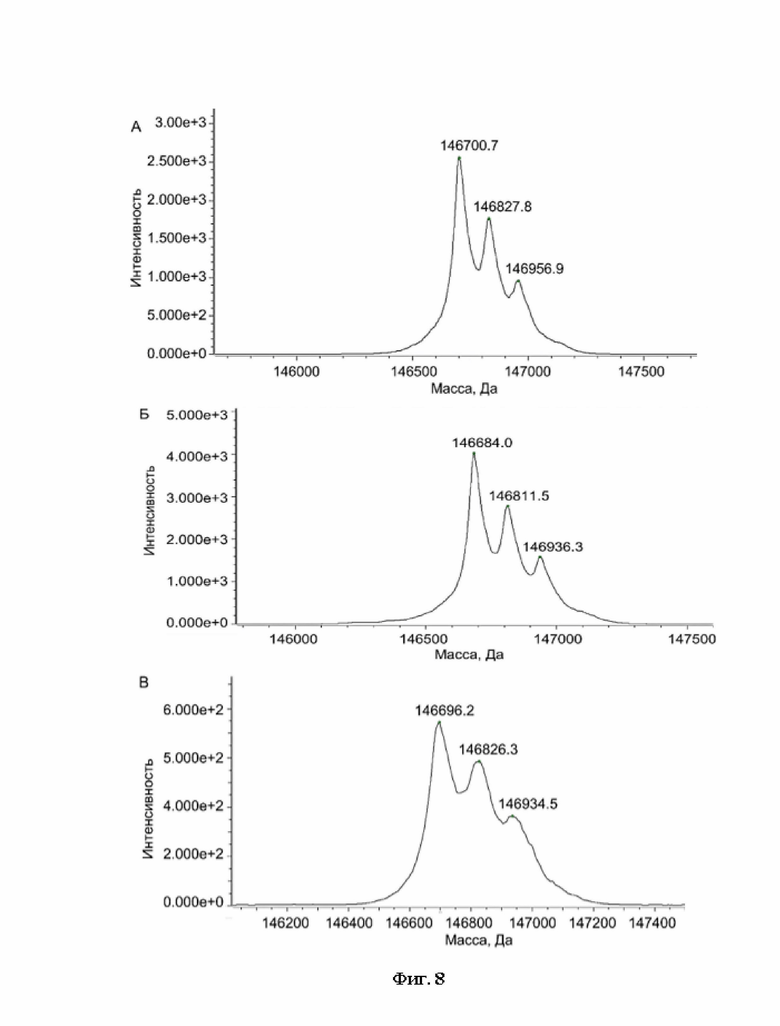
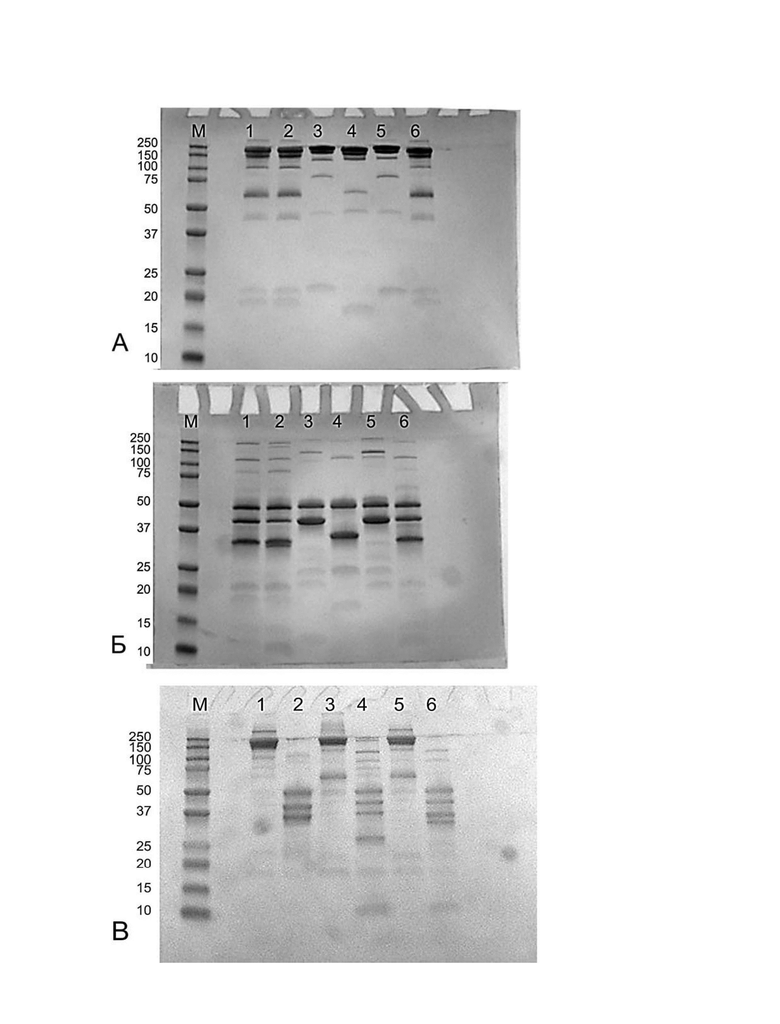

Изобретение относится к области биохимии, в частности к двухвалентному биспецифическому химерному антителу, которое включают гетеродимер на основе околомембранных доменов MHC. Также раскрыт способ получения указанного антитела. Изобретение позволяет эффективно получать биспецифические антитела. 2 н. и 58 з.п. ф-лы, 14 табл., 10 ил., 12 пр.
1. Двухвалентное биспецифическое химерное антитело, специфически связывающееся с первым антигеном и вторым антигеном, где указанное антитело содержит:
а) первую легкую цепь и первую тяжелую цепь антитела, специфически связывающегося с первым антигеном;
где первая легкая цепь содержит вариабельный домен легкой цепи и константный домен легкой цепи;
где первая тяжелая цепь содержит вариабельный домен тяжелой цепи и константные домены тяжелой цепи антитела, включающие первый константный домен тяжелой цепи и мономер Fc-фрагмента, содержащий второй и третий константные домены тяжелой цепи;
б) вторую легкую цепь и вторую тяжелую цепь антитела, специфически связывающегося со вторым антигеном,
где вторая легкая цепь содержит вариабельный домен легкой цепи и константный домен, который выбирают из группы:
первого околомембранного домена МНС или
первого околомембранного домена МНС-подобного белка;
где вторая тяжелая цепь, содержит вариабельный домен тяжелой цепи, константный домен, который выбирают из группы:
второго околомембранного домена МНС или
второго околомембранного домена МНС-подобного белка;
и мономер Fc-фрагмента, содержащий второй и третий константные домены тяжелой цепи;
при этом первый околомембранный домен МНС или МНС-подобного белка и второй околомембранный домен МНС или МНС-подобного белка образуют между собой гетеродимер, который стабилизирован дисульфидной связью;
где третий константный домен одной тяжелой цепи и третий константный домен другой тяжелой цепи соприкасаются друг с другом поверхностями, которые изменены для формирования двухвалентного биспецифического химерного антитела, причем данные изменения в третьих константных доменах тяжелых цепей представляют собой замены для обеспечения гетеродимеризации.
2. Двухвалентное биспецифическое химерное антитело по п. 1, где первый околомембранный домен МНС может быть выбран из группы, включающей:
первый околомембранный домен МНС класса I,
первый околомембранный домен МНС класса II,
модифицированный вариант первого околомембранного домена МНС класса I или
модифицированный вариант первого околомембранного домена МНС класса II.
3. Двухвалентное биспецифическое химерное антитело по п. 1, где второй околомембранный домен МНС может быть выбран из группы, включающей:
второй околомембранный домен МНС класса I,
второй околомембранный домен МНС класса II,
модифицированный вариант второго околомембранного домена МНС класса I или
модифицированный вариант второго околомембранного домена МНС класса II.
4. Двухвалентное биспецифическое химерное антитело по любому из пп. 2, 3, где МНС класса II выбирают из группы: HLA-DM, HLA-DO, HLA-DP, HLA-DQ или HLA-DR.
5. Двухвалентное биспецифическое химерное антитело по любому из пп. 2, 3, где МНС класса I выбирают из группы: HLA-A, HLA-B, HLA-C, HLA-E, HLA-F или HLA-G.
6. Двухвалентное биспецифическое химерное антитело по п. 1, где первый околомембранный домен МНС-подобного белка может быть выбран из группы, включающей:
первый околомембранный домен CD1,
первый околомембранный домен HFE,
модифицированный вариант первого околомембранного домена CD1 или
модифицированный вариант первого околомембранного домена HFE.
7. Двухвалентное биспецифическое химерное антитело по п. 1, где второй околомембранный домен МНС-подобного белка может быть выбран из группы, включающей:
второй околомембранный домен CD1,
второй околомембранный домен HFE,
модифицированный вариант второго околомембранного домена CD1 или
модифицированный вариант второго околомембранного домена HFE.
8. Двухвалентное биспецифическое химерное антитело по любому из пп. 6, 7, где CD1 выбирают из группы: CD1a, CD1b, CD1c, CD1d или CD1e.
9. Двухвалентное биспецифическое химерное антитело по п. 1, где вариабельный фрагмент второй легкой цепи отделен от первого околомембранного домена МНС или МНС-подобного белка линкером длиной от 1 до 25 аминокислот и/или вариабельный фрагмент второй тяжелой цепи отделен от второго околомембранного домена МНС или МНС-подобного белка линкером длиной от 1 до 25 аминокислот.
10. Двухвалентное биспецифическое химерное антитело по п. 1, где
а) СН3-домен одной тяжелой цепи изменен так, что на поверхности СН3-домена одной тяжелой цепи, которая соприкасается с поверхностью СН3-домена второй тяжелой цепи в двухвалентном биспецифическом антителе, аминокислотный остаток заменен на аминокислотный остаток, который имеет большую по объему боковую цепь, что приводит к созданию выпуклости на поверхности СН3-домена одной тяжелой цепи, которая может помещаться в полость на поверхности СН3-домена другой тяжелой цепи, и
б) СН3-домен другой тяжелой цепи изменен так, что на поверхности СН3-домена второй тяжелой цепи, которая соприкасается с поверхностью СН3-домена первой тяжелой в двухвалентном биспецифическом антителе, аминокислотный остаток заменен на аминокислотный остаток, который имеет меньшую по объему боковую цепь, что приводит к созданию полости на поверхности СН3-домена второй тяжелой цепи, в которую может помещаться выпуклость на поверхности раздела СН3-домена первой тяжелой цепи;
где указанный аминокислотный остаток, который имеет большую по объему боковую цепь, выбран из группы, включающей аргинин, фенилаланин, тирозин, триптофан, и
где указанный аминокислотный остаток, который имеет меньшую по объему боковую цепь, выбран из группы, включающей аланин, серин, треонин, валин.
11. Двухвалентное биспецифическое химерное антитело по п. 1, где константный домен первой легкой цепи антитела выбирают из CK или CL.
12. Двухвалентное биспецифическое химерное антитело по п. 1, где СН3-домены антитела дополнительно изменены путем интродукции цистеина в качестве аминокислоты в соответствующие положения каждого СН3-домена, так что дисульфидный мостик может образовываться между обоими СН3-доменами.
13. Двухвалентное биспецифическое химерное антитело по п. 10, где СН3-домен одной тяжелой цепи изменен с образованием Knob, а СН3-домен другой тяжелой цепи изменен с образованием Hole или наоборот.
14. Двухвалентное биспецифическое химерное антитело по п. 13, где СН3-домен одной тяжелой цепи имеет аминокислотные замены S354C/T366W, а СН3-домен другой тяжелой цепи имеет аминокислотные замены Y349C/T366S/L368A/Y407V.
15. Двухвалентное биспецифическое химерное антитело по п. 13, где СН3-домен одной тяжелой цепи имеет аминокислотные замены Y349C/T366S/L368A/Y407, а СН3-домен другой тяжелой цепи имеет аминокислотные замены S354C/T366W.
16. Двухвалентное биспецифическое химерное антитело по п. 1, где первый околомембранный домен МНС и второй околомембранный домен МНС соответственно представляют собой α2 домен МНС II и β2 домен МНС II, которые образуют между собой гетеродимер.
17. Двухвалентное биспецифическое химерное антитело по п. 16, где α2 домен МНС II имеет аминокислотную последовательность, которую выбирают из группы: SEQ ID NO: 30, SEQ ID NO: 32, SEQ ID NO: 34, SEQ ID NO: 36 или SEQ ID NO: 38, а β2 домен МНС II имеет аминокислотную последовательность, которую выбирают из группы: SEQ ID NO: 31, SEQ ID NO: 33, SEQ ID NO: 35, SEQ ID NO: 37 или SEQ ID NO: 39.
18. Двухвалентное биспецифическое химерное антитело по п. 1, где первый околомембранный домен МНС и второй околомембранный домен МНС соответственно представляют собой β2 домен МНС II и α2 домен МНС II, которые образуют между собой гетеродимер.
19. Двухвалентное биспецифическое химерное антитело по п. 18, где β2 домен МНС II имеет аминокислотную последовательность, которую выбирают из группы: SEQ ID NO: 31, SEQ ID NO: 33, SEQ ID NO: 35, SEQ ID NO: 37 или SEQ ID NO: 39, а α2 домен МНС II имеет аминокислотную последовательность, которую выбирают из группы: SEQ ID NO: 30, SEQ ID NO: 32, SEQ ID NO: 34, SEQ ID NO: 36 или SEQ ID NO: 38.
20. Двухвалентное биспецифическое химерное антитело по п. 1, где первый околомембранный домен МНС и второй околомембранный домен МНС соответственно представляют собой модифицированный вариант α2 домена МНС II и модифицированный вариант β2 домена МНС II, которые образуют между собой гетеродимер.
21. Двухвалентное биспецифическое химерное антитело по п. 1, где первый околомембранный домен МНС и второй околомембранный домен МНС соответственно представляют собой модифицированный вариант β2 домена МНС II и модифицированный вариант α2 домена МНС II, которые образуют между собой гетеродимер.
22. Двухвалентное биспецифическое химерное антитело по п. 1, где первый околомембранный домен МНС и второй околомембранный домен МНС соответственно представляют собой α3 домен МНС I и β2 микроглобулин, которые образуют между собой гетеродимер.
23. Двухвалентное биспецифическое химерное антитело по п. 22, где α3 домен МНС I имеет аминокислотную последовательность, которую выбирают из группы SEQ ID NO: 1-29, а β2 микроглобулин имеет аминокислотную последовательность SEQ ID NO: 46.
24. Двухвалентное биспецифическое химерное антитело по п. 1, где первый околомембранный домен МНС и второй околомембранный домен МНС соответственно представляют собой β2 микроглобулин и α3 домен МНС I, которые образуют между собой гетеродимер.
25. Двухвалентное биспецифическое химерное антитело по п. 24, где β2 микроглобулин имеет аминокислотную последовательность SEQ ID NO: 46, а α3 домен МНС I имеет аминокислотную последовательность, которую выбирают из группы SEQ ID NO: 1-29.
26. Двухвалентное биспецифическое химерное антитело по п. 1, где первый околомембранный домен МНС и второй околомембранный домен МНС соответственно представляют собой модифицированный вариант α3 домена МНС I и модифицированный вариант β2 микроглобулина, которые образуют между собой гетеродимер.
27. Двухвалентное биспецифическое химерное антитело по п. 1, где первый околомембранный домен МНС и второй околомембранный домен МНС соответственно представляют собой модифицированный вариант β2 микроглобулина и модифицированный вариант α3 домена МНС I, которые образуют между собой гетеродимер.
28. Двухвалентное биспецифическое химерное антитело по пп. 26, 27, где модифицированный вариант β2 микроглобулина имеет аминокислотную последовательность, которую выбирают из SEQ ID NO: 47 или SEQ ID NO: 48.
29. Двухвалентное биспецифическое химерное антитело по п. 1, где первый околомембранный домен МНС-подобного белка и второй околомембранный домен МНС-подобного белка соответственно представляют собой α3 домен CD1 и β2 микроглобулин, которые образуют между собой гетеродимер.
30. Двухвалентное биспецифическое химерное антитело по п. 29, где α3 домен CD1 имеет аминокислотную последовательность, которую выбирают из группы SEQ ID NO: 40-44, а β2 микроглобулин имеет аминокислотную последовательность SEQ ID NO: 46.
31. Двухвалентное биспецифическое химерное антитело по п. 1, где первый околомембранный домен МНС-подобного белка и второй околомембранный домен МНС-подобного белка соответственно представляют собой β2 микроглобулин и α3 домен CD1, которые образуют между собой гетеродимер.
32. Двухвалентное биспецифическое химерное антитело по п. 31, где β2 микроглобулин имеет аминокислотную последовательность SEQ ID NO: 46, а α3 домен CD1 имеет аминокислотную последовательность, которую выбирают из группы SEQ ID NO: 40-44.
33. Двухвалентное биспецифическое химерное антитело по п. 1, где первый околомембранный домен МНС-подобного белка и второй околомембранный домен МНС-подобного белка соответственно представляют собой модифицированный вариант α3 домена CD1 и модифицированный вариант β2 микроглобулина, которые образуют между собой гетеродимер.
34. Двухвалентное биспецифическое химерное антитело по п. 33, где модифицированный вариант α3 домена CD1 имеет аминокислотную последовательность, которую выбирают из группы SEQ ID NO: 49-56, а модифицированный вариант β2 микроглобулина имеет аминокислотную последовательность, которую выбирают из SEQ ID NO: 47 или SEQ ID NO: 48.
35. Двухвалентное биспецифическое химерное антитело по п. 1, где первый околомембранный домен МНС-подобного белка и второй околомембранный домен МНС-подобного белка соответственно представляют собой модифицированный вариант β2 микроглобулина и модифицированный вариант α3 домена CD1, которые образуют между собой гетеродимер.
36. Двухвалентное биспецифическое химерное антитело по п. 35, где модифицированный вариант β2 микроглобулина имеет аминокислотную последовательность, которую выбирают из SEQ ID NO: 47 или SEQ ID NO: 48, а модифицированный вариант α3 домена CD1 имеет аминокислотную последовательность, которую выбирают из группы SEQ ID NO: 49-56 или SEQ ID NO: 109.
37. Двухвалентное биспецифическое химерное антитело по п. 1, где первый околомембранный домен МНС-подобного белка и второй околомембранный домен МНС-подобного белка соответственно представляют собой α3 домен HFE и β2 микроглобулин, которые образуют между собой гетеродимер.
38. Двухвалентное биспецифическое химерное антитело по п. 37, где α3 домен HFE имеет аминокислотную последовательность SEQ ID NO: 45, а β2 микроглобулин имеет аминокислотную последовательность SEQ ID NO: 46.
39. Двухвалентное биспецифическое химерное антитело по п. 1, где первый околомембранный домен МНС-подобного белка и второй околомембранный домен МНС-подобного белка соответственно представляют собой β2 микроглобулин и α3 домен HFE, которые образуют между собой гетеродимер.
40. Двухвалентное биспецифическое химерное антитело по п. 39, где β2 микроглобулин имеет аминокислотную последовательность SEQ ID NO: 46, а α3 домен HFE имеет аминокислотную последовательность SEQ ID NO: 45.
41. Двухвалентное биспецифическое химерное антитело по п. 1, где первый околомембранный домен МНС-подобного белка и второй околомембранный домен МНС-подобного белка соответственно представляют собой модифицированный вариант α3 домена HFE и модифицированный вариант β2 микроглобулина, которые образуют между собой гетеродимер.
42. Двухвалентное биспецифическое химерное антитело по п. 1, где первый околомембранный домен МНС-подобного белка и второй околомембранный домен МНС-подобного белка соответственно представляют собой модифицированный вариант β2 микроглобулина и модифицированный вариант α3 домена HFE, которые образуют между собой гетеродимер.
43. Двухвалентное биспецифическое химерное антитело по пп. 41, 42, где модифицированный вариант β2 микроглобулина имеет аминокислотную последовательность, которую выбирают из SEQ ID NO: 47 или SEQ ID NO: 48.
44. Двухвалентное биспецифическое химерное антитело по любому из пп. 2, 3, 6, 7, 20, 21, 26, 27, 33, 35 и 41, 42, где под модифицированным вариантом подразумевается вариант, включающий замены на цистеин для образования дисульфидного мостика между цепями гетеродимера, полученного из первого и второго околомембранных доменов МНС или МНС-подобного белка.
45. Двухвалентное биспецифическое химерное антитело по любому из пп. 2, 3, 6, 7, 20, 21, 26, 27, 33, 35 и 41, 42, где под модифицированным вариантом подразумевается вариант, включающий одну или более замены в различных позициях околомембранных доменов МНС или МНС-подобного белка, приводящие к увеличению термодинамической стабильности Tm более чем на 1 градус Цельсия в сравнении с дикой формой околомембранных доменов МНС или МНС-подобного белка, соответственно.
46. Двухвалентное биспецифическое химерное антитело по любому из пп. 2, 3, 6, 7, 20, 21, 26, 27, 33, 35 и 41, 42, где под модифицированным вариантом подразумевается вариант, включающий одну или более замены в различных позициях околомембранных доменов МНС или МНС-подобного белка, приводящие к уменьшению количества агрегатов в сравнении с дикой формой околомембранных доменов МНС или МНС-подобного белка, соответственно.
47. Двухвалентное биспецифическое химерное антитело по любому из пп. 2, 3, 6, 7, 20, 21, 26, 27, 33, 35 и 41, 42, где под модифицированным вариантом подразумевается вариант, включающий одну или более замены в различных позициях околомембранных доменов МНС или МНС-подобного белка, приводящие к удалению сайтов гликозилирования в сравнении с дикой формой околомембранных доменов МНС или МНС-подобного белка, соответственно.
48. Двухвалентное биспецифическое химерное антитело по п. 1, где вариабельный домен первой легкой цепи и вариабельный домен второй легкой цепи идентичны.
49. Двухвалентное биспецифическое химерное антитело по п. 1, где Fc-фрагмент относится к IgG.
50. Двухвалентное биспецифическое химерное антитело по п. 49, где изотип Fc-фрагмента выбирают из группы: IgG1, IgG2 или IgG4 человека.
51. Двухвалентное биспецифическое химерное антитело по п. 1, где в мономере Fc-фрагмента дополнительно вводят замены, приводящие к отсутствию ADCC, CDC и/или ADCP свойств у двухвалентного биспецифического антитела.
52. Двухвалентное биспецифическое химерное антитело по п. 51, где в мономере Fc-фрагмента дополнительно вводят замены L234A и L235A.
53. Двухвалентное биспецифическое химерное антитело по п. 1, где в мономере Fc-фрагмента дополнительно вводят замены, приводящие к пролонгированному действию антитела.
54. Двухвалентное биспецифическое химерное антитело по п. 53, где в мономере Fc-фрагмента дополнительно вводят замены M252Y, S254T и Т256Е.
55. Двухвалентное биспецифическое химерное антитело по п. 1, где в мономере Fc-фрагмента дополнительно вводят замены, приводящие к усилению ADCC, CDC и/или ADCP свойств у двухвалентного биспецифического антитела.
56. Двухвалентное биспецифическое химерное антитело по п. 55, где в мономере Fc-фрагмента дополнительно вводят замену E345R.
57. Двухвалентное биспецифическое химерное антитело по п. 1, где первый околомембранный домен МНС или МНС-подобного белка и второй околомембранный домен МНС или МНС-подобного белка образуют между собой гетеродимер, который стабилизирован дисульфидной связью за счет мутации или мутаций в первом и/или втором околомембранном домене для образования цистеинового мостика между первым и вторым околомембранными доменами МНС или МНС-подобного белка.
58. Двухвалентное биспецифическое химерное антитело по п. 1, где первый околомембранный домен МНС или МНС-подобного белка и второй околомембранный домен МНС или МНС-подобного белка образуют между собой гетеродимер, который стабилизирован дисульфидной связью за счет удлинения первого околомембранного домена МНС или МНС-подобного белка на одну или несколько аминокислот на С-конце и с терминальным Cys на С-конце для образования цистеинового мостика между первым околомембранным доменом МНС или МНС-подобного белка и шарниром.
59. Двухвалентное биспецифическое химерное антитело по п. 58, где удлинение первого околомембранного домена МНС или МНС-подобного белка представляет собой GSC.
60. Способ получения двухвалентного биспецифического химерного антитела по пп. 1-59, где способ включает следующие этапы:
а) трансформация клетки-хозяина
- экспрессионными векторами, которые содержат молекулы нуклеиновых кислот, кодирующие первую легкую цепь и первую тяжелую цепь биспецифического химерного антитела,
- экспрессионными векторами, которые содержат молекулы нуклеиновых кислот, кодирующие вторую легкую цепь и вторую тяжелую цепь биспецифического химерного антитела,
б) культивирование клетки-хозяина в условиях, которые позволяют синтезировать указанное двухвалентное биспецифическое химерное антитело; и
в) выделение указанного двухвалентного биспецифического антитела из культуры клеток.
| M | |||
| SCHMITTNAEGEL, A New Class of Bifunctional Major Histocompatibility Class I Antibody Fusion Molecules to Redirect CD8 T Cells, Molecular Cancer Therapeutics, 2016, 15(9), pp.2130-2142 | |||
| WO 2016049641 A1, 31.03.2016 | |||
| WO 2018053069 A1, 22.03.2018 | |||
| ВАРИАНТЫ FC-ОБЛАСТИ С УЛУЧШЕННОЙ СПОСОБНОСТЬЮ СВЯЗЫВАТЬСЯ С БЕЛКОМ А | 2015 |
|
RU2698969C2 |
| АНТИГЕННЫЕ TAU-ПЕПТИДЫ И ИХ ПРИМЕНЕНИЯ | 2010 |
|
RU2518291C2 |
