ССЫЛКА НА РОДСТВЕННЫЕ ИЗОБРЕТЕНИЯ
В настоящей заявке на изобретение заявлен приоритет Китайской заявки на патент №201910532734.7, поданной 19 июня 2019 года, включенной здесь посредством ссылки во всей полноте.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящая заявка на изобретение в общем относится к области лекарственных средств на основе антитела. В частности, настоящая заявка на изобретение относится к биспецифическому антителу, содержащему антигенсвязывающий фрагмент против человеческого CD3E (кластер дифференцировки 3Е) и/или антигенсвязывающий фрагмент против человеческого ВСМА (антиген созревания В-клеток), и его медицинским и биологическим применениям.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Биспецифическое антитело (BsAb) представляет собой тип искусственного антитела, которое содержит два различных антигенсвязывающих сайта. Биспецифические антитела широко используют в области биомедицины, особенно иммунотерапии опухолей. Одно плечо биспецифических антител, нацеленное против CD3, может связываться с субъединицей CD3E в комплексе рецептора TCR на поверхности Т-клеток, обеспечивая первый сигнал для активации Т-клеток, а другое плечо нацелено против опухолевого антигена. Биспецифические антитела могут сближать вместе опухолевые клетки и Т-клетки и непосредственно уничтожать опухолевые клетки при активации Т-клеток.
Существуют многочисленные платформы для биспецифических антител, и их структуры являются сложными. С точки зрения структур антитела биспецифические антитела можно разделить на две категории: антитела с сегментами Fc и без сегментов Fc. Биспецифические антитела без сегментов Fc состоят из VH и VL-областей или Fab-фрагментов от двух антител. Основными представителями таких биспецифических антител являются BiTE, DART, TandAb, би-нанотело и т.п. Преимущество таких биспецифических антител заключается в отсутствии несоответствия между тяжелой и легкой цепями, а недостатком является то, что период полувыведения является коротким, и клиническое применение является неудобным. Биспецифические антитела с сегментами Fc сохраняют структуры обычных моноклональных антител и могут опосредовать биологическую функцию сегментов Fc. Представителями таких биспецифических антител являются KIH IgG crossmab, DVD-Ig, Triomab и т.п., которые имеют длительный период полувыведения in vivo и могут обладать активностями ADCC (антителозависимая клеточная цитотоксичность) и CDC (комплементзависимая цитотоксичность) (Hongyan Liu, Abhishek Saxena, Sachdev S. Sidhu, et al.. Fc engineering for Developing Therapeutic Bispecifc Antibodies and Novel Scaffolds. Front. Immunol. 2017; 8: 38).
Таким образом, ввиду широкой применимости биспецифических антител существует необходимость в разработке новых биспецифических антител в данной области техники.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В первом аспекте в настоящей заявке на изобретение предложено биспецифическое антитело, содержащее антигенсвязывающий фрагмент против человеческого CD3E, где антигенсвязывающий фрагмент против человеческого CD3E содержит:
HCDR1 (CDR1 тяжелой цепи), как представлено в SEQ ID NO: 1, HCDR2 (CDR2 тяжелой цепи), как представлено в SEQ ID NO: 2, HCDR3 (CDR3 тяжелой цепи), как представлено в SEQ ID NO: 3, LCDR1 (CDR1 легкой цепи), как представлено в SEQ ID NO: 4, LCDR2 (CDR2 легкой цепи), как представлено в SEQ ID NO: 5 и LCDR3 (CDR3 легкой цепи), как представлено в SEQ ID NO: 6; где HCDR и LCDR определены в соответствии с системой нумерации Кабата. Во втором аспекте в настоящей заявке на изобретение предложено биспецифическое антитело, содержащее антигенсвязывающий фрагмент против человеческого ВСМА, где антигенсвязывающий фрагмент против человеческого ВСМА содержит:
HCDR1 (CDR1 тяжелой цепи), как представлено в SEQ ID NO: 7, HCDR2 (CDR2 тяжелой цепи), как представлено в SEQ ID NO: 8, HCDR3 (CDR3 тяжелой цепи), как представлено в SEQ ID NO: 9, LCDR1 (CDR1 легкой цепи), как представлено в SEQ ID NO: 4, LCDR2 (CDR2 легкой цепи), как представлено в SEQ ID NO: 5 и LCDR3 (CDR3 легкой цепи), как представлено в SEQ ID NO: 6;
где HCDR и LCDR определены в соответствии с системой нумерации Кабата.
В третьем аспекте в настоящей заявке на изобретение предложено биспецифическое антитело, содержащее антигенсвязывающий фрагмент против человеческого CD3E и антигенсвязывающий фрагмент против человеческого ВСМА.
В некоторых воплощениях третьего аспекта антигенсвязывающий фрагмент против человеческого CD3E содержит:
HCDR1, как представлено в SEQ ID NO: 1,
HCDR2, как представлено в SEQ ID NO: 2,
HCDR3, как представлено в SEQ ID NO: 3,
LCDR1, как представлено в SEQ ID NO: 4,
LCDR2, как представлено в SEQ ID NO: 5, и
LCDR3, как представлено в SEQ ID NO: 6;
где HCDR и LCDR определены в соответствии с системой нумерации Кабата. В некоторых воплощениях третьего аспекта антигенсвязывающий фрагмент против человеческого ВСМА содержит:
HCDR1, как представлено в SEQ ID NO: 7,
HCDR2, как представлено в SEQ ID NO: 8,
HCDR3, как представлено в SEQ ID NO: 9,
LCDR1, как представлено в SEQ ID NO: 4,
LCDR2, как представлено в SEQ ID NO: 5, и
LCDR3, как представлено в SEQ ID NO: 6;
где HCDR и LCDR определены в соответствии с системой нумерации Кабата.
В некоторых воплощениях третьего аспекта антигенсвязывающий фрагмент против человеческого CD3E и антигенсвязывающий фрагмент против человеческого ВСМА содержат одну и ту же вариабельную область легкой цепи.
В некоторых воплощениях третьего аспекта биспецифическое антитело представляет собой антитело IgG1, содержащее две константные области тяжелой цепи, имеющие одинаковую шарнирную область, и аминокислотная последовательность шарнирной области представлена в SEQ ID NO: 15.
В некоторых воплощениях третьего аспекта биспецифическое антитело представляет собой антитело IgG1, содержащее первую константную область тяжелой цепи и вторую константную область тяжелой цепи, где аминокислоты в положениях 354 и 366 первой константной области тяжелой цепи представляют собой С и W, соответственно, и аминокислоты в положениях 349, 366, 368 и 407 второй константной области тяжелой цепи представляют собой С, S, А и V, соответственно; положение аминокислот константной области антитело определено в соответствии с нумерацией EU (Евросоюза).
В некоторых воплощениях третьего аспекта биспецифическое антитело представляет собой антитело IgGl, содержащее первую константную область тяжелой цепи и вторую константную область тяжелой цепи, где аминокислоты в положениях 234, 235 и 331 первой и второй константных областей тяжелой цепи представляют собой F, Е, и S, соответственно; положение аминокислот константной области антитела определено в соответствии с нумерацией EU.
В некоторых воплощениях первого и третьего аспектов антигенсвязывающий фрагмент против человеческого CD3E содержит вариабельную область тяжелой цепи, как представлено в SEQ ID NO: 12, и вариабельную область легкой цепи, как представлено в SEQ ID NO: 14.
В некоторых воплощениях второго и третьего аспектов антигенсвязывающий фрагмент против человеческого ВСМА содержит вариабельную область тяжелой цепи, как представлено в SEQ ID NO: 10, и вариабельную область легкой цепи, как представлено в SEQ ID NO: 14.
В некоторых воплощениях любого из вышеприведенных аспектов антигенсвязывающий фрагмент против человеческого CD3E содержит одноцепочечное антитело (scfv) или Fab-фрагмент.
В некоторых воплощениях любого из вышеприведенных аспектов антигенсвязывающий фрагмент против человеческого ВСМА содержит одноцепочечное антитело (scfv) или Fab-фрагмент.
В некоторых воплощениях третьего аспекта антитело имеет первое плечо и второе плечо, где первое плечо содержит антигенсвязывающий фрагмент против человеческого CD3E и второе плечо содержит антигенсвязывающий фрагмент против человеческого ВСМА:
первое плечо содержит аминокислотную последовательность вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 12, аминокислотную последовательность константной области тяжелой цепи, как представлено в SEQ ID NO: 19, аминокислотную последовательность вариабельной области легкой цепи, как представлено в SEQ ID NO: 14, и аминокислотную последовательность константной области легкой цепи, как представлено в SEQ ID N0:20;
второе плечо содержит аминокислотную последовательность вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 10, аминокислотную последовательность константной области тяжелой цепи, как представлено в SEQ ID NO: 18, аминокислотную последовательность вариабельной области легкой цепи, как представлено в SEQ ID NO: 14, и аминокислотную последовательность константной области легкой цепи, как представлено в SEQ ID NO: 20.
В четвертом аспекте в настоящей заявке на изобретение предложена фармацевтическая композиция, содержащая биспецифическое антитело по любому из первого-третьего аспектов.
В пятом аспекте в настоящей заявке на изобретение предложено применение биспецифического антитела по любому из первого-третьего аспектов или фармацевтической композиции в соответствии с четвертым аспектом при приготовлении лекарственного средства для предупреждения или лечения множественной миеломы.
В шестом аспекте в настоящей заявке на изобретение предложен способ предупреждения или лечения множественной миеломы, включающий введение биспецифического антитела по любому из первого-третьего аспектов или фармацевтической композиции в соответствии с четвертым аспектом субъекту, нуждающемуся в этом.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг. 1 демонстрирует результаты обнаружения одновременного связывания биспецифического антитела CD3ExBCMA с обоими антигенами CD3E и ВСМА посредством метода ИФА (иммуноферментного анализа).
Фиг. 2 демонстрирует результаты анализа связывания биспецифического антитела CD3ExBCMA с CD3E на поверхности клетки Jurkat человека при остром Т-лимфоцитарном лейкозе посредством проточной цитометрии.
Фиг. 3 демонстрирует результаты анализа связывания биспецифического антитела CD3ExBCMA с ВСМА на поверхности клетки человека при плазмоклеточном лейкозе NCI-H929 посредством проточной цитометрии.
Фиг. 4 демонстрирует специфическую активацию jurkat-двойных клеток ВСМА-положительными опухолевыми клетками, опосредованную биспецифическим антителом CD3ExBCMA.
Фиг. 5 демонстрирует уничтожающее действие РВМС (мононуклеарные клетки периферической крови) в отношении ВСМА-положительных опухолевых клеток, опосредованное биспецифическим антителом CD3ExBCMA.
Фиг. 6 демонстрирует результаты стимулирования биспецифическим антителом CD3ExBCMA экспрессии CD69 на поверхности Т-клеток в присутствии ВСМА-положительных опухолевых клеток.
Фиг. 7 демонстрирует изменения объема опухоли в РВМС гуманизированной мышиной модели с человеческой миеломой NCI-H929, обработанной биспецифическим антителом CD3ExBCMA.
Фиг. 8 демонстрирует изменения массы тела в РВМС гуманизированной мышиной модели с человеческой миеломой NCI-H929, обработанной биспецифическим антителом CD3ExBCMA.
Фиг. 9 демонстрирует изменения массы тела в hCD34+ гуманизированной мышиной модели с человеческой миеломой RPMI-8226, обработанной биспецифическим антителом CD3ExBCMA.
Фиг. 10 демонстрирует изменения объема опухоли в hCD34+ гуманизированной мышиной модели с человеческой миеломой RPMI-8226, обработанной биспецифическим антителом CD3ExBCMA.
ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТИ
SEQ ID NO: 1 демонстрирует аминокислотную последовательность вариабельной области HCDR1 тяжелой цепи Н10В7 моноклонального антитела H10B7+L1G10 против человеческого CD3E.
SEQ ID NO: 2 демонстрирует аминокислотную последовательность вариабельной области HCDR2 тяжелой цепи Н10В7 моноклонального антитела H10B7+L1G10 против человеческого CD3E.
SEQ ID NO: 3 демонстрирует аминокислотную последовательность вариабельной области HCDR3 тяжелой цепи Н10В7 моноклонального антитела H10B7+L1G10 против человеческого CD3E.
SEQ ID NO: 4 демонстрирует аминокислотную последовательность вариабельной области LCDR1 легкой цепи L9B9.
SEQ ID NO: 5 демонстрирует аминокислотную последовательность вариабельной области LCDR2 легкой цепи L9B9.
SEQ ID NO: 6 демонстрирует аминокислотную последовательность вариабельной области LCDR3 легкой цепи L9B9.
SEQ ID NO: 7 демонстрирует аминокислотную последовательность вариабельной области HCDR1 тяжелой цепи мутанта H13F1 моноклонального антитела С4 против человеческого ВСМА.
SEQ ID NO: 8 демонстрирует аминокислотную последовательность вариабельной области HCDR2 тяжелой цепи мутанта H13F1 моноклонального антитела С4 против человеческого ВСМА.
SEQ ID NO: 9 демонстрирует аминокислотную последовательность вариабельной области HCDR3 тяжелой цепи мутанта H13F1 моноклонального антитела С4 против человеческого ВСМА.
SEQ ID NO: 10 демонстрирует аминокислотную последовательность вариабельной области тяжелой цепи мутанта H13F1 моноклонального антитела С4 против человеческого ВСМА.
SEQ ID NO: 11 демонстрирует аминокислотную последовательность вариабельной области тяжелой цепи C4VH моноклонального антитела С4 против человеческого ВСМА.
SEQ ID NO: 12 демонстрирует аминокислотную последовательность вариабельной области тяжелой цепи Н10В7 моноклонального антитела H10B7+L1G10 против человеческого CD3E.
SEQ ID NO: 13 демонстрирует аминокислотную последовательность вариабельной области легкой цепи L1G10 моноклонального антитела H10B7+L1G10 против человеческого CD3E.
SEQ ID NO: 14 демонстрирует аминокислотную последовательность вариабельной области легкой цепи L9B9.
SEQ ID NO: 15 демонстрирует аминокислотную последовательность шарнирной области.
SEQ ID NO: 16 демонстрирует аминокислотную последовательность вариабельной области легкой цепи C4VK моноклонального антитела С4 против человеческого ВСМА.
SEQ ID NO: 17 демонстрирует аминокислотную последовательность константной области тяжелой цепи мутантного IgG1K подтипа IgG1 человеческого антитела.
SEQ ID NO: 18 демонстрирует аминокислотную последовательность константной области тяжелой цепи мутантного IgG1m3-H подтипа IgG1 человеческого антитела.
SEQ ID NO: 19 демонстрирует аминокислотную последовательность константной области тяжелой цепи мутантного IgG1m3-K подтипа IgG1 человеческого антитела.
SEQ ID NO: 20 демонстрирует аминокислотную последовательность константной области легкой цепи подтипа каппа (κ) человека.
SEQ ID NO: 21 демонстрирует аминокислотную последовательность константной области легкой цепи подтипа лямбда (λ) человека.
SEQ ID NO: 22 демонстрирует аминокислотную последовательность внеклеточной области человеческого CD3E.
SEQ ID NO: 23 демонстрирует аминокислотную последовательность внеклеточной области человеческого CD3D.
SEQ ID NO: 24 демонстрирует аминокислотную последовательность внеклеточной области обезьяньего CD3E.
SEQ ID NO: 25 демонстрирует аминокислотную последовательность внеклеточной области обезьяньего CD3D.
SEQ ID NO: 26 демонстрирует аминокислотную последовательность внеклеточной области мышиного CD3E.
SEQ ID NO: 27 демонстрирует аминокислотную последовательность внеклеточной области мышиного CD3D.
SEQ ID NO: 28 демонстрирует аминокислотную последовательность внеклеточной области человеческого ВСМА.
SEQ ID NO: 29 демонстрирует аминокислотную последовательность внеклеточной области обезьяньего ВСМА.
SEQ ID NO: 30 демонстрирует аминокислотную последовательность внеклеточной области мышиного ВСМА.
SEQ ID NO: 31 демонстрирует аминокислотную последовательность метки His.
SEQ ID NO: 32 демонстрирует аминокислотную последовательность сегмента Fc мышиного антитела IgG2a (mFc).
SEQ ID NO: 33 демонстрирует аминокислотную последовательность Fc мутантного FcK подтипа человеческого IgG1 гетеродимера.
SEQ ID NO: 34 демонстрирует аминокислотную последовательность Fc мутантного FcH подтипа человеческого IgG1 гетеродимера.
SEQ ID NO: 35 демонстрирует аминокислотную последовательность константной области тяжелой цепи человеческого антитела подтипа IgG1.
SEQ ID NO: 36 демонстрирует аминокислотную последовательность константной области тяжелой цепи мутантного IgG1H подтипа IgG1 человеческого антитела.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
ОПРЕДЕЛЕНИЯ
Следующие определения и методы приведены для лучшего определения настоящей заявки на изобретение и направления специалиста в данной области техники к практическому применению настоящей заявки на изобретение. Если указано иное, то термины, использованные в настоящей заявке на изобретение, имеют значения, обычно понимаемые специалистом в данной области техники. Все патентные документы, научные работы и другие публикации, процитированные здесь, включены здесь путем ссылки во всей полноте.
Описывая здесь структуру антитела, делают ссылку на определение нумерации EU человеческого антитела IgG1 в отношении описания нумерации положений аминокислот, которое хорошо известно и легко доступно специалистам в данной области техники. Кроме того, когда здесь описывают мутацию в связи с положением нумерации EU, она относится к мутации, полученной относительно нативной последовательности антитела.
Используемый здесь термин «фрагмент Fc», «домен Fc», «участок Fc» и т.п.относится к участку константной области тяжелой цепи антитела, включающему шарнирную область, сегмент СН2 и сегмент СН3 константной области. Со ссылкой на определение нумерации EU человеческого антитела IgG1, фрагмент Fc относится к аминокислотной последовательности в позициях 216-447 в константной области антитела.
Используемый здесь термин «Fab-фрагмент (антигенсвязывающий фрагмент)», «участок Fab» и т.п.относится к фрагменту антитела, способному связываться с антигеном, который образуется после обработки интактного антитела папаином, включая интактную легкую цепь (VL-CL), вариабельную область тяжелой цепи и фрагмент CH1 (VH-CH1).
Используемый здесь термин «одноцепочечный вариабельный фрагмент (scFv)» относится к антителу одноцепочечной структуры, содержащему полипептидную цепь, содержащую вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), которое обычно конструируют с использованием способов генной инженерии. Гибкий линкер обычно конструируют между вариабельной областью тяжелой цепи и вариабельной областью легкой цепи, таким образом, что вариабельная область тяжелой цепи и вариабельная область легкой цепи могут складываться в правильную конформацию, способную связываться с антигеном.
Используемый здесь термин «антигенсвязывающий фрагмент» относится к части структуры антитела, которая определяет антигенсвязывающую способность. Специалистам в данной области техники должно быть понятно, что основными частями структуры антитела, которая определяет антигенсвязывающую способность, являются CDR, поэтому CDR также являются основными компонентами антигенсвязывающего фрагмента. При конструировании биспецифического антитела примеры «антигенсвязывающего фрагмента» включают без ограничения одноцепочечное антитело (scfv) или Fab-фрагмент.
Используемый здесь термин «биспецифическое антитело» относится к антителу, обладающему способностью связаться с двумя различными антигенами, которые могут состоять из двух фрагментов Fc и слитых с ними соответственно двух антигенсвязывающих фрагментов.
В некоторых воплощениях используемое здесь «биспецифическое антитело» относится к биспецифическому антителу на основе человеческого антитела IgG1, и в дополнение к измененным структурам, описанным здесь, оно обладает основными характеристиками и функцией человеческого антитела IgG1. Специалисту в данной области техники известно, что используемое здесь «биспецифическое антитело» может также представлять собой биспецифическое антитело на основе другого подтипа иммуноглобулина, такого как человеческое антитело IgG2.
Специалистам в данной области техники хорошо известно, что области, определяющие комплементарность (CDR, обычно включающие CDR1, CDR2 и CDR3), являются областями вариабельной области, которые в основном влияют на аффинность и специфичность антитела. Последовательности CDR VH или VL имеют два общих определения, то есть определение в соответствии с нумерацией Кабата и определение в соответствии с нумерацией Чотиа (смотри, например, Kabat, "Sequences of Proteins of Immunological Interest", National Institutes of Health, Bethesda, Md. (1991); A1-Lazikani et al., J. Mol. Biol. 273:927-948 (1997); и Martin et al., Proc. Natl. Acad. Sci. USA, 86:9268-9272 (1989)). Для последовательностей вариабельных областей данного антитела последовательности областей CDR в VH и VL можно определить в соответствии с определением нумерации Кабата или определением нумерации Чотиа. В некоторых воплощениях в соответствии с настоящей заявкой на изобретение последовательности CDR определены в соответствии с определением нумерации Кабата.
Для последовательностей вариабельных областей данного антитела последовательности областей CDR в последовательностях вариабельной области можно анализировать различными способами, например, с использованием программного обеспечения Abysis онлайн (http://www.abysis.org/).
Используемый здесь термин «специфическое связывание» относится к неслучайной реакции связывания между двумя молекулами, например связывания антитела с эпитопом антигена.
Молекула CD3 является важным дифференцировочным антигеном на мембране Т-клеток и также является характерным маркером зрелых Т-клеток. Молекула CD3 состоит из четырех цепей γ, δ, ε и ζ, или пяти цепей γ, δ, ε, ζ, и η (ζ и η представляют собой гомологичные изомеры), состоит из трех димеров CD3γε, CD3δε и CD3ζζ (или CD3ζη) и экспрессируется на Т-клеточной мембране. Три цепи CD3γ, δ и λ содержат высококонсервативные кислые аминокислотные остатки (глутаминовая кислота в γ цепи, и аспарагиновая кислота в цепях δ и ε), которые могут быть нековалентно связаны с основными аминокислотными остатками в α и β-цепях Т-клеточного рецептора (TCR) при помощи солевой мостиковой связи с образованием стабильной структуры комплекса TCR-CD3. Комплекс может передавать сигналы активации Т-клеток и стабилизирует структуру TCR. Внутриклеточный домен каждой цепи CD3 содержит структуру ITAM (иммунорецепторный тирозиновый активирующий мотив), которая является основой для внутриклеточной передачи сигналов, опосредованной молекулой CD3. Когда TCR специфически распознает антиген и связывается с ним (антигенный пептид, представленный молекулой МНС (главного комплекса гистосовместимости)), тогда тирозиновые протеинкиназы в Т-клетках фосфорилируют остатки тирозина на ITAM и рекрутируют тирозиновые протеинкиназы, содержащие домены SH2 (ZAP-70). Сигнал передается в цитоплазму Т-клеток для инициирования механизма активации внутри клеток. Таким образом, CD3 выполняет функцию передачи сигнала активации, генерируемого после того, как TCR распознает антиген, и сигнал является первым сигналом для индукции активации Т-клеток.
Антиген созревания В-клеток (ВСМА) является 17-м членом суперсемейства рецепторов TNF. В качестве негликозилированного трансмембранного белкового рецептора III типа ВСМА состоит из 184 аминокислотных остатков с 80 аминокислотными остатками во внутриклеточной области и только одного домена распознавания углеводов во внеклеточной области. ВСМА участвует в созревании и дифференцировке В-клеток в качестве специфического антигена на поверхности плазматических клеток. ВСМА также участвует в долгосрочной выживаемости плазматических клеток в качестве основного вещества. ВСМА, TACI и BAFFR соответственно связываются с двумя лигандами, то есть индуцирующим пролиферацию лигандом (APRIL) и фактором активации В-клеток (BAFF), и участвуют в активации молекул сигнальной трансдукции, таких как р38, Elk и c-Jun через путь NFκB, таким образом, влияя на созревание, рост и выживание В-клеток. Тем не менее, ВСМА не является критическим для выживания В-клеток. Было показано, что после нокаута ВСМА не оказывается влияние на продукцию краткосрочного иммуноглобулина, ранний гуморальный иммунный ответ и развитие В-лимфоцитов в мышиных плазматических клетках (Christine М. Coquery Loren D. Erickson. Regulatory Roles of the Tumor Necrosis Factor Receptor BCMA. Crit Rev Immunol. 2012; 32(4): 287-305). Экспрессия ВСМА является избирательной. Он не экспрессируется в наивных В-клетках, В-клетках памяти, гематопоэтических клетках CD34+ и других нормальных тканях, и селективно индуцируется для экспрессии при дифференцировке плазматических клеток, и в основном экспрессируется на плазмоцитоидных дендритных клетках и плазматических клетках костного мозга. Множественная миелома представляет собой В-клеточную злокачественную опухоль, вызванную злокачественной пролиферацией и злокачественным перерождением клеток. Оно в основном проявляется неконтролируемым разрастанием плазматических клеток в костном мозге и выработкой большого количества моноклональных иммуноглобулинов, которые в результате приводят к ряду симптомов, таких как разрушение кости, повышенный уровень кальция в крови, анемия, поражение почек, снижение иммунитета и т.п. Уровень экспрессии ВСМА в клетках миеломы значительно выше, чем в плазматических клетках и плазмабластах. ВСМА экспрессируется на высоком уровне и в значительной степени на всем протяжении злокачественных заболеваний плазматических клеток, от моноклональной гаммопатии до вялотекущей миеломы, и далее до множественной миеломы (Shih-Feng Cho, Kenneth С. Anderson, Yu-Tzu Tai. Targeting В Cell Maturation Antigen (BCMA) in Multiple Myeloma: Potential Uses of BCMA-Based immunotherapy. Front. Immunol, 2018; 9:1821).
В первом аспекте в настоящей заявке на изобретение предложено биспецифическое антитело, содержащее антигенсвязывающий фрагмент против человеческого CD3E, где антигенсвязывающий фрагмент против человеческого CD3E содержит:
HCDR1, как представлено в SEQ ID NO: 1,
HCDR2, как представлено в SEQ ID NO: 2,
HCDR3, как представлено в SEQ ID NO: 3,
LCDR1, как представлено в SEQ ID NO: 4,
LCDR2, как представлено в SEQ ID NO: 5, и
LCDR3, как представлено в SEQ ID NO: 6;
где HCDR и LCDR определены в соответствии с системой нумерации Кабата.
Во втором аспекте в настоящей заявке на изобретение предложено биспецифическое антитело, содержащее антигенсвязывающий фрагмент против человеческого ВСМА, где антигенсвязывающий фрагмент против человеческого ВСМА содержит:
HCDR1, как представлено в SEQ ID NO: 7,
HCDR2, как представлено в SEQ ID NO: 8,
HCDR3, как представлено в SEQ ID NO: 9,
LCDR1, как представлено в SEQ ID NO: 4,
LCDR2, как представлено в SEQ ID NO: 5, и
LCDR3, как представлено в SEQ ID NO: 6;
где HCDR и LCDR определены в соответствии с системой нумерации Кабата.
В третьем аспекте в настоящей заявке на изобретение предложено биспецифическое антитело, содержащее антигенсвязывающий фрагмент против человеческого CD3E и антигенсвязывающий фрагмент против человеческого ВСМА.
В некоторых воплощениях третьего аспекта антигенсвязывающий фрагмент против человеческого CD3E содержит:
HCDR1, как представлено в SEQ ID NO: 1,
HCDR2, как представлено в SEQ ID NO: 2,
HCDR3, как представлено в SEQ ID NO: 3,
LCDR1, как представлено в SEQ ID NO: 4,
LCDR2, как представлено в SEQ ID NO: 5, и
LCDR3, как представлено в SEQ ID NO: 6;
где HCDR и LCDR определены в соответствии с системой нумерации Кабата. В некоторых воплощениях третьего аспекта антигенсвязывающий фрагмент против человеческого ВСМА содержит:
HCDR1, как представлено в SEQ ID NO: 7,
HCDR2, как представлено в SEQ ID NO: 8,
HCDR3, как представлено в SEQ ID NO: 9,
LCDR1, как представлено в SEQ ID NO: 4,
LCDR2, как представлено в SEQ ID NO: 5, и
LCDR3, как представлено в SEQ ID NO: 6;
где HCDR и LCDR определены в соответствии с системой нумерации Кабата.
В некоторых воплощениях третьего аспекта антигенсвязывающий фрагмент против человеческого CD3E и антигенсвязывающий фрагмент против человеческого ВСМА содержат одну и ту же вариабельную область легкой цепи.
В некоторых специфических воплощениях третьего аспекта антигенсвязывающий фрагмент против человеческого CD3E и антигенсвязывающий фрагмент против человеческого ВСМА содержат одинаковую легкую цепь. Это воплощение облегчает надлежащую сборку легкой и тяжелой цепей, и также представляет собой предпочтительное воплощение.
В некоторых воплощениях третьего аспекта биспецифическое антитело представляет собой антитело IgG1, содержащее две константные области тяжелой цепи, имеющие одинаковую шарнирную область, и аминокислотная последовательность шарнирной области представлена в SEQ ID NO: 15, которая заменяет последовательности в положениях 216-230 константной области природного человеческого антитела IgG1; положение аминокислот константной области антитела определено в соответствии с нумерацией EU.
В некоторых воплощениях третьего аспекта биспецифическое антитело представляет собой антитело IgG1, содержащее первую константную область тяжелой цепи и вторую константную область тяжелой цепи, где аминокислоты в положениях 354 и 366 первой константной области тяжелой цепи представляют собой С и W, соответственно, и аминокислоты в положениях 349, 366, 368 и 407 второй константной области тяжелой цепи представляют собой С, S, А и V, соответственно; положение аминокислот константной области антитела определено в соответствии с нумерацией EU.
При конструировании биспецифического антитела с сохранением домена Fc, структура биспецифического антитела может быть оптимизирована посредством следующих двух перспектив: гетеромеризация тяжелой цепи и надлежащая сборка легкой и тяжелой цепей. В некоторых воплощениях два фрагмента Fc содержат мутации, которые могут обеспечить гетеромеризацию тяжелой цепи. Методика KIH (выступ во впадину) представляет собой стратегию, которая обеспечивает гетеромеризацию тяжелой цепи. Как правило, методика KIH относится к образованию структуры, которая облегчает спаривание друг с другом гетерологических половин путем модифицирования аминокислотной последовательности области СН3, которая может максимально возможно поддерживать структуру нормального антитела при конструировании биспецифического антитела. В некоторых воплощениях используемая методика KIH включает то, что аминокислоты в положениях 354 и 366 одного фрагмента Fc представляют собой, соответственно, С и W, а аминокислоты в положениях 349, 366, 368 и 407 другого фрагмента Fc представляют собой, соответственно, С, S, А и V. Руководство по проведению методики KIH можно найти, например, в "An efficient route to human bispecific IgG", A. Margaret Merchant et al., Nature Biotechnology, Volume 16, 1998", включенному здесь посредством ссылки во всей полноте.
В некоторых воплощениях третьего аспекта биспецифическое антитело представляет собой антитело IgG1, содержащее первую константную область тяжелой цепи и вторую константную область тяжелой цепи, где аминокислоты в положениях 234, 235 и 331 первой и второй константных областей тяжелой цепи представляют собой F, Е и S, соответственно; положение аминокислот константной области антитела определено в соответствии с нумерацией EU.
В некоторых воплощениях третьего аспекта аминокислоты в положениях 234, 235 и 331 фрагмента СН2 двух константных областей тяжелой цепи представляют собой, соответственно, F, Е и S, что может уменьшить антителозависимую цитотоксичность (ADCC), опосредованную фрагментом Fc антитела, таким образом, потенциально уменьшая побочные действия, вызванные биспецифическим антителом in vivo. В отношении руководства по вышеуказанным мутациям смотри, например, "The binding affinity of human IgG for its high affinity Fc receptor is determined by multiple amino acids in the CH2 domain and is modulated by the hinge region", Stephen M. Canfield et al., J. Exp.Med. Volume 173, 1991, включенной здесь посредством ссылки во всей полноте.
В некоторых воплощениях первого и третьего аспектов антигенсвязывающий фрагмент против человеческого CD3E содержит вариабельную область тяжелой цепи, как представлено в SEQ ID NO: 12 (содержащее HCDR1, как представлено в SEQ ID NO: 1, HCDR2, как представлено в SEQ ID NO: 2, и HCDR3, как представлено в SEQ ID NO: 3) и вариабельную область легкой цепи, как представлено в SEQ ID NO: 14 (содержащее LCDR1, как представлено в SEQ ID NO: 4, LCDR2, как представлено в SEQ ID NO: 5, и LCDR3, как представлено в SEQ ID NO: 6).
В некоторых воплощениях второго и третьего аспектов антигенсвязывающий фрагмент против человеческого ВСМА содержит вариабельную область тяжелой цепи, как представлено в SEQ ID NO: 10 (содержащее HCDR1, как представлено в SEQ ID NO: 7, HCDR2, как представлено в SEQ ID NO: 8, и HCDR3, как представлено в SEQ ID NO: 9) и вариабельную область легкой цепи, как представлено в SEQ ID NO: 14 (содержащее LCDR1, как представлено в SEQ ID NO: 4, LCDR2, как представлено в SEQ ID NO: 5, и LCDR3, как представлено в SEQ ID NO: 6).
В некоторых воплощениях любого из вышеприведенных аспектов антигенсвязывающий фрагмент против человеческого CD3E содержит одноцепочечное антитело (scfv) или Fab-фрагмент.
В некоторых воплощениях любого из вышеприведенных аспектов антигенсвязывающий фрагмент против человеческого ВСМА содержит одноцепочечное антитело (scfv) или Fab-фрагмент.
Биспецифическое антитело имеет два отличающихся антигенсвязывающих фрагмента против двух отличающихся антигенов, и антигенсвязывающие фрагменты могут содержать две формы одноцепочечного антитела (scfv) или Fab-фрагмента. Конфигурация антигенсвязывающего фрагмента биспецифического антитела может иметь четыре комбинации для двух заданных антигенов: Fab+Fab, Fab+scfv, scfv+Fab и scfv+scfv.
В некоторых конкретных воплощениях любого из вышеприведенных аспектов антигенсвязывающий фрагмент против человеческого CD3E содержит Fab-фрагмент и антигенсвязывающий фрагмент против человеческого ВСМА содержит Fab-фрагмент.
В некоторых конкретных воплощениях любого из вышеприведенных аспектов антигенсвязывающий фрагмент против человеческого CD3E содержит Fab-фрагмент и антигенсвязывающий фрагмент против человеческого ВСМА содержит одноцепочечное антитело (scfv).
В некоторых конкретных воплощениях любого из вышеприведенных аспектов антигенсвязывающий фрагмент против человеческого CD3E содержит одноцепочечное антитело (scfv) и антигенсвязывающий фрагмент против человеческого ВСМА содержит Fab-фрагмент.
В некоторых конкретных воплощениях любого из вышеприведенных аспектов антигенсвязывающий фрагмент против человеческого CD3E содержит одноцепочечное антитело (scfv) и антигенсвязывающий фрагмент против человеческого ВСМА содержит одноцепочечное антитело (scfv).
Биспецифическое антитело также описано здесь, как имеющее два «плеча». Биспецифическое антитело можно разделить на два плеча, связанные центральной осью. Плечи биспецифического антитела могут состоять из фрагмента Fc и антигенсвязывающего фрагмента (Fab-фрагмент или одноцепочечное антитело). Для плеча, состоящего из фрагмента Fc и фрагмента Fab, его структура похожа на структуру обычного антитела, содержащего интактные тяжелые и легкие цепи, и, таким образом, структура такого плеча может быть представлена как Fc+Fab, или может быть представлена как тяжелая цепь (Fc+ вариабельная область тяжелой цепи Fab и фрагмент СН1)+ легкая цепь (фрагмент легкой цепи Fab). Когда оба плеча содержат антигенсвязывающие фрагменты в форме Fab-фрагмента, тогда структура образующегося таким образом биспецифического антитела похожа на структуру нативного антитела и представляет собой предпочтительное воплощение.
В некоторых воплощениях третьего аспекта антитело имеет первое плечо и второе плечо, где первое плечо содержит антигенсвязывающий фрагмент против человеческого CD3E и второе плечо содержит антигенсвязывающий фрагмент против человеческого ВСМА:
первое плечо содержит аминокислотную последовательность вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 12, аминокислотную последовательность константной области тяжелой цепи, как представлено в SEQ ID NO: 19, аминокислотную последовательность вариабельной области легкой цепи, как представлено в SEQ ID NO: 14, и аминокислотную последовательность константной области легкой цепи, как представлено в SEQ ID NO: 20;
второе плечо содержит аминокислотную последовательность вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 10, аминокислотную последовательность константной области тяжелой цепи, как представлено в SEQ ID NO: 18, аминокислотную последовательность вариабельной области легкой цепи, как представлено в SEQ ID NO: 14, и аминокислотную последовательность константной области легкой цепи, как представлено в SEQ ID NO: 20.
В некоторых воплощениях любого из вышеприведенных аспектов константная область тяжелой цепи биспецифического антитела представляет собой подтип человеческого IgG1 или различные мутанты выбранного подтипа человеческого IgG1, такие как IgG1H, IgG1K, IgG1m3-H или IgG1m3-K.
В некоторых воплощениях любого из вышеприведенных аспектов константная область легкой цепи биспецифического антитела представляет собой подтип к человека или подтип λ человека, предпочтительно подтип κ человека.
В четвертом аспекте в настоящей заявке на изобретение предложена фармацевтическая композиция, содержащая биспецифическое антитело по любому из первого-третьего аспектов.
В некоторых воплощениях фармацевтическая композиция дополнительно содержит фармацевтически приемлемый носитель, эксципиент, разбавитель и т.п.
В некоторых воплощениях фармацевтическую композицию используют для предупреждения или лечения множественной миеломы.
В некоторых воплощениях фармацевтическая композиция может дополнительно содержать смазывающее вещество, такое как тальк, стеарат магния и минеральное масло; увлажнитель; эмульгатор; суспендирующий агент; консервант, такой как бензойная кислота, сорбиновая кислота и пропионат кальция; подсластитель и/или корригент.
В некоторых воплощениях, фармацевтическая композиция в соответствии с настоящей заявкой на изобретение может быть приготовлена в виде таблетки, пилюли, порошка, лепешки, эликсира, суспензии, эмульсии, раствора, сиропа, суппозитория или капсулы.
В некоторых воплощениях фармацевтическая композиция в соответствии с настоящей заявкой на изобретение может быть доставлена с использованием любого физиологически приемлемого пути введения, включающего без ограничения пероральное введение, парентеральное введение, назальное введение, ректальное введение, внутрибрюшинное введение, внутрисосудистую инъекцию, подкожное введение, трансдермальное введение, введение путем ингаляции и т.п.
В некоторых воплощениях фармацевтическая композиция для терапевтического применения может быть приготовлена для хранения в форме лиофилизированной композиции или в форме водного раствора путем смешивания агента, имеющего желаемую чистоту, с подходящим фармацевтически приемлемым носителем или эксципиентом.
В пятом аспекте в настоящей заявке на изобретение предложено применение биспецифического антитела по любому из первого-третьего аспектов или фармацевтической композиции в соответствии с четвертым аспектом для изготовления лекарственного средства для предупреждения или лечения множественной миеломы.
В шестом аспекте в настоящей заявке на изобретение предложен способ предупреждения или лечения множественной миеломы, включающий введение биспецифического антитела по любому из первого-третьего аспектов или фармацевтической композиции в соответствии с четвертым аспектом субъекту, нуждающемуся в этом.
Понятно, что вышеприведенное подробное описание предназначено только для того, чтобы представить специалисту в данной области техники более хорошее понимание настоящей заявки на изобретение и не предполагается, что оно каким-либо образом ограничивает объем заявки на изобретение. Различные модификации и вариации могут быть осуществлены специалистом в данной области техники в отношении описанных воплощений.
Следующие примеры приведены исключительно для иллюстрации и не предполагается, что они ограничивают объем настоящей заявки на изобретение.
ПРИМЕРЫ
Пример 1: Получение рекомбинантных белков
Множество различных рекомбинантных белков требовалось при получении и идентификации биспецифического антитела CD3ExBCMA, включающего внеклеточную область человеческого CD3E (hCD3E, SEQ ID NO: 22), внеклеточную область человеческого CD3D (hCD3D, SEQ ID NO: 23), внеклеточную область обезьяньего CD3E (mfCD3E, SEQ ID NO: 24), внеклеточную область обезьяньего CD3D (mfCD3D, SEQ ID NO: 25), внеклеточную область мышиного CD3E (mCD3E, SEQ ID NO: 26), внеклеточную область мышиного CD3D (mCD3D, SEQ ID NO: 27) и внеклеточную область человеческого ВСМА (hBCMA, SEQ ID NO: 28), внеклеточную область обезьяньего ВСМА (mfBCMA, SEQ ID NO: 29) и внеклеточную область мышиного ВСМА (mBCMA, SEQ ID NO: 30). Все эти рекомбинантные белки имеют большое количество посттрансляционных модификаций (например, гликозилирование или дисульфидные связи и т.п.), и, таким образом, применение экспрессионной системы на основе клеток млекопитающих может быть более благоприятным при поддержании структур и функций рекомбинантных белков. Кроме того, для облегчения очистки добавляли к С-концу рекомбинантных белков, отличающихся от антитела, His-метки (SEQ ID NO: 31) или фрагменты Fc мышиного антитела IgG2a (mFc, SEQ ID NO: 32), или мутанты Fc (FcK, SEQ ID NO: 33 или FcH, SEQ ID NO: 34) гетеродимера подтипа IgG1 человека получали на основе способа KIH (Knob-Into-Hole, выступ во впадину). При получении рекомбинантного антитела константная область тяжелой цепи антитела может представлять собой подтип IgG1 человека (SEQ ID NO: 35) или различные мутанты выбранного подтипа IgG1 человека, такие как IgG1H (SEQ ID NO: 36), IgG1K (SEQ ID NO: 17), IgG1m3-H (SEQ ID NO: 18) или IgG1m3-K (SEQ ID NO: 19), и константная область легкой цепи может представлять собой подтип к человека (SEQ ID NO: 20) или подтип λ человека (SEQ ID NO: 21).
На основе аминокислотных последовательностей различных интересующих рекомбинантных белков, записанных в базе данных Uniprot, разрабатывали и синтезировали гены (содержащие His-метку, ген, кодирующий mFc или Fc) вышеописанных рекомбинантных белков. При помощи обычных способов молекулярной биологии клонировали в подходящих эукариотических экспрессирующихся векторах (например pcDNA3.1 от Invitrogen Inc.) синтезированные гены, кодирующие рекомбинантные белки. Затем липосомы (например 293 фектин от Invitrogen Inc.) или другие агенты трансфекции (например PEI (полиэтиленимин)) использовали для трансфекции плазмид, экспрессирующих рекомбинантный белок, в клетки HEK293 (например HEK293F от Invitrogen Inc.). Клетки инкубировали в суспензии в бессывороточных условиях в течение 3-5 суток. Затем культуральный супернатант собирали центрифугированием.
Для рекомбинантных белков, слитых с His-метками, рекомбинантные белки в супернатанте дополнительно очищали с использованием аффинных хроматографических колонок на основе хелата металла (например, HisTrap FF от GE Inc.). Рекомбинантные белки и антитела, слитые с mFc, дополнительно очищали с использованием колонки для аффинной хроматографии Protein A/G (например, Mabselect SURE от GE Inc.). Затем буфер для консервирования рекомбинантного белка заменяли на буфер PBS (физиологический раствор, забуференный фосфатом) (рН 7,0) или другие подходящие буферы с использованием колонки для обессоливания (например, Hitrap desaulting, GE Inc.). При необходимости образцы антитела могут быть стерилизованы путем фильтрования, и затем их можно хранить в аликвотах при -20°С для использования позже.
Пример 2: Скрининг и идентификации общих легких цепей
2.1 Скрининг общих легких цепей
H10B7+L1G10 представляет собой моноклональное антитело, которое связывается с человеческим CD3E, полученным посредством методики библиотек человеческого антитела. Аминокислотная последовательность вариабельной области тяжелой цепи Н10 В7 из H10B7+L1G10 представлена в SEQ ID NO: 12, а аминокислотная последовательность вариабельной области легкой цепи L1G10 представлена в SEQ ID NO: 13 (смотри аминокислотные последовательности, представленные в SEQ ID NO: 19 и SEQ ID NO: 20 в Китайской заявке на патент №201910372193.6).
С4 представляет собой моноклональное антитело, нацеленное на опухолевый антиген ВСМА, аминокислотная последовательность вариабельной области тяжелой цепи C4VH представлена в SEQ ID NO: 11, а аминокислотная последовательность вариабельной области легкой цепи C4VK представлена в SEQ ID NO: 16 (смотри последовательность моноклонального антитела CA8-J7M0 в US 9273141 В2).
Функции и свойства моноклональных антител С4 и H10B7+L1G10 были подтверждены экспериментально.
На основе признанной двухвекторной системы для фагового дисплея исходную библиотеку легкой цепи подвергали двум раундам скрининга и обогащения на основе вариабельной области тяжелой цепи Н10В7 моноклонального антитела H10B7+L1G10 против CD3E с CD3E/CD3D в качестве скринингового антигена путем использования стратегии замещения легкой цепи (смотри пример 4 в Китайской заявке на патент №201510097117.0 в отношении подробных экспериментальных протоколов). Затем библиотеку легкой цепи, обогащенную CD3E/CD3D, подвергали двум раундам скрининга и обогащения на основе тяжелой цепи моноклонального антитела С4 против ВСМА с ВСМА в качестве антигена. Наконец, полученную в результате легкую цепь идентифицировали с получением общей вариабельной области легко цепи L9B9 (SEQ ID NO: 14), которая может одновременно поддерживать активность антитела против CD3E и антитела против ВСМА.
Вариабельную область тяжелой цепи Н10В7, вариабельную область тяжелой цепи C4VH в С4 и вариабельную область легкой цепи L9B9, соответственно, клонировали в эукариотические экспрессирующиеся векторы, слитые с константной областью тяжелой цепи IgGl человека и константной областью легкой цепи к путем использования обычных средств молекулярной биологии, таким образом, чтобы в комбинации экспрессировать полноразмерные антитела H10B7+L9B9 и C4VH+L9B9.
2.2 Определение аффинности связывания антитела C4VH+L9B9 против ВСМА с общей легкой цепью с человеческим ВСМА
Аффинность антител против ВСМА (С4 и C4VH+L9B9) определяли при помощи метода поверхностного плазмонного резонанса с использованием Biacore Х100. Реактивы и расходные материалы, такие как набор для связывания по N-концу (BR-1000-50), набор для присоединения человеческого антитела (BR-1008-39), чип СМ5 (BR100012) и 10×HBS-EP, рН 7,4 (BR100669) приобретали в GE healthcare. Поверхность карбоксилированного чипа СМ5 активировали при помощи 1-этил-3-(3-диметиламинопропил)карбодиимида хлоргидрата (EDC) и N-гидроксисукцинимида (NHS) в соответствии с инструкциями к набору. Антитело против человеческого IgG (Fc) (захватывающее антитело) разбавляли до 25 мкг/мл при помощи 10 мМ раствора ацетата натрия (рН 5,0) с последующей инжекцией при скорости потока 10 мкл/мин для достижения связывающего количества приблизительно до 10000 единиц ответа (RU). После инжекции захватывающего антитела 1М этаноламин инжектировали для блокирования непрореагировавших групп.Для измерения кинетики антитело против ВСМА разбавляли до 0,5-1 мкг/мл, а затем инжектировали при скорости потока 10 мкл/мин для обеспечения того, что приблизительно 400 RU антитела захватывались антителом против человеческого Fc. Затем готовили серии градиентов концентрации для hBCMA-his (например, 0,617 нМ, 1,85 нМ, 5,56 нМ, 16,7 нМ и 50 нМ), и инжектировали от низкой концентрации до высокой концентрации при 30 мкл/мин при 25°С. Время ассоциации составило 120 с, а время диссоциации составило 1800 с. Поверхность чипа регенерировали путем инжекции 3М раствора MgCl2 при 10 мкл/мин в течение 30 с. Скорость ассоциации (Kon) и скорость диссоциации (Koff) рассчитывали путем подгонки сенсограмм ассоциации и диссоциации под модель ассоциации 1:1 с использованием программного обеспечения для оценки Biacore Х100, версия 2.0.1. Равновесную константу диссоциации (KD) рассчитывали как отношение Koff/Kon. Результаты подгонки представлены в Таблице 1.
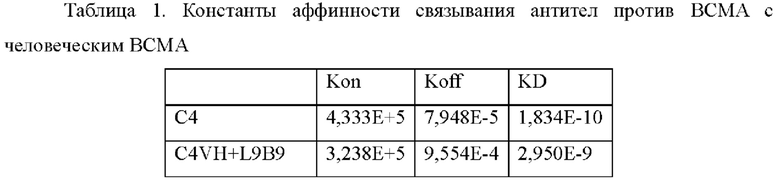
Пример 3: Созревание аффинности антитела против ВСМА
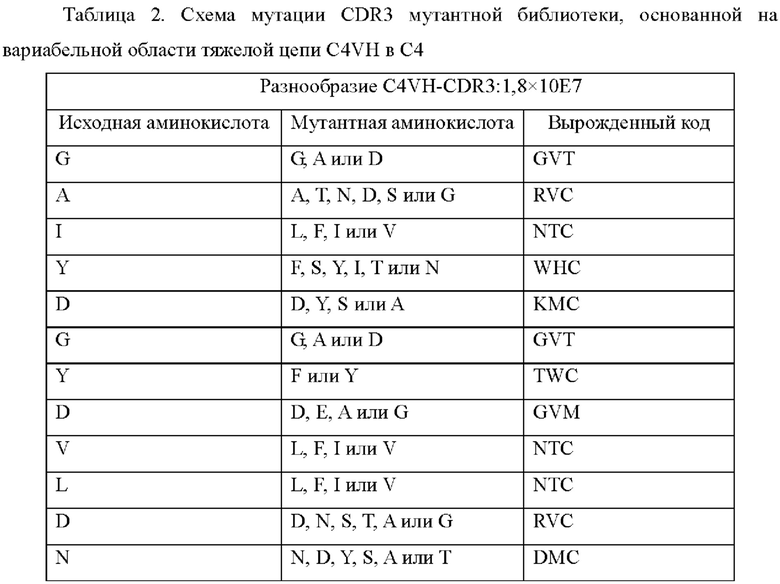
3.1 Скрининг библиотеки созревания тяжелой цепи С4
Мутантную библиотеку CDR3, основанную на вариабельной области тяжелой цепи C4VH, конструировали путем введения мутации в область CDR3 вариабельной области тяжелой цепи C4VH с использованием обычных средств молекулярной биологии. Разработанная схема мутации представлена в Таблице 2, и 1,2×10Е8 емкости библиотеки достигали с точностью 86,7%.
На основе двухвекторной системы для фагового дисплея (смотри пример 5 в Китайской заявке на патент №201510097117.0) сконструированную мутантную библиотеку C4VH-CDR3 подвергали трем раундам скрининга и обогащения антигеном hBCMA-His при помощи способа твердофазного скрининга. Наконец, получали вариабельную область тяжелой цепи мутантного H13F1 (SEQ ID NO: 10) с увеличенной аффинностью.
3.2 Анализ аффинности мутанта тяжелой цепи С4
Нуклеотидные последовательности, кодирующие вариабельную область тяжелой цепи мутантного H13F1 в С4 и вариабельную область легкой цепи L9B9, соответственно, клонировали в эукариотические экспрессирующиеся векторы, слитые с нуклеотидными последовательностями, кодирующими человеческую константную область тяжелой цепи и константную область легкой цепи путем использования обычных средств молекулярной биологии, таким образом, чтобы экспрессировать полноразмерные антитела в комбинации. Со ссылкой на пример 2.2 мутант С4 (подтип IgG1) подвергали анализу аффинности путем использования Biacore Х100, и результат представлен в Таблице 3.
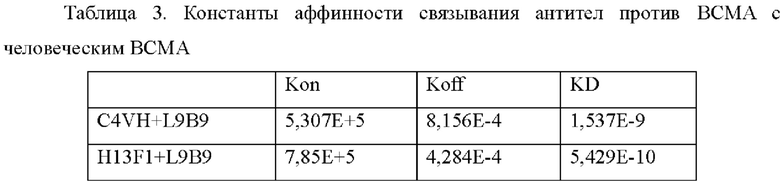
Пример 4: Получение биспецифических антител
Нуклеотидные последовательности, кодирующие вариабельную область тяжелой цепи Н10В7 моноклонального антитела против CD3E, и вариабельную область тяжелой цепи H13F1 моноклонального антитела против ВСМА, соответственно, клонировали в подходящие эукариотические экспрессирующиеся векторы для конструирования гетеродимеров, основанных на общей легкой цепи. То есть, нуклеотидную последовательность, кодирующую вариабельную область тяжелой цепи антитела против CD3E, клонировали в эукариотические экспрессирующиеся векторы, слитые с нуклеотидной последовательностью, кодирующей константную область IgG1 с мутацией Knob IgG1m3-K, нуклеотидную последовательность, кодирующую вариабельную область тяжелой цепи антитела против ВСМА, клонировали в эукариотические экспрессирующиеся векторы, содержащие нуклеотидную последовательность, кодирующую константную область IgG1 с мутацией Hole IgG1m3-Н, и нуклеотидную последовательность, кодирующую вариабельную область VK общей легкой цепи L9B9 клонировали в эукариотические экспрессирующиеся векторы, слитые с нуклеотидной последовательностью, кодирующей человеческую константную область легкой цепи CK.
Три сконструированных эукариотических экспрессионных вектора, экспрессирующих H10B7-IgG1m3-K, H13F1-IgG1m3-H и L9B9VK-CK, совместно трансфицировали в клетки HEK293F с использованием липосом, и клетки выращивали в суспензии в бессывороточной среде в течение 3-5 суток. Культуральный супернатант собирали путем центрифугирования. Биспецифические антитела в культуральном супернатанте очищали с использованием аффинной хроматографической колонки Protein A/G (например Mabselect SURE, GE Inc.). Буфер для консервации рекомбинантного белка затем заменяли на буфер PBS (рН 7,0) или другие подходящие буферы с использованием колонки для обессоливания (например Hitrap desaulting, GE Inc.). Обессоленный белковый раствор очищали при помощи гель-фильтрационной хроматографии (SEC) с использованием Superdex 200 (GE), таким образом получая интересующий белок. При необходимости образцы антитела могут быть стерилизованы путем фильтрования, и затем их можно хранить в аликвотах при -20°С для более использования позже.
Пример 5: Анализ аффинности биспецифического антитела
Ссылаясь на пример 2.2, анализы аффинности провели в отношении моноклонального антитела H13F1+L9B9 против ВСМА, моноклонального антитела H10B7+L9B9 против CD3E и биспецифического антитела CD3ExBCMA при помощи способа поверхностного плазмонного резонанса с использованием Biacore X100.
При определении аффинности моноклонального антитела против ВСМА и биспецифического антитела CD3ExBCMA к антигену ВСМА антитело против человеческого IgG (Fc) конъюгировали с поверхностью чипа СМ5. Белок антитела разбавляли до 0,5-1 мкг/мл и инжектировали со скоростью 10 мкл/мин. Обеспечивали захват приблизительно 400 RU моноклонального антитела ВСМА антителом против человеческого Fc, и обеспечивали захват приблизительно 800 RU биспецифического антитела CD3ExBCMA антителом против человеческого Fc. Затем готовили серии градиентов концентрации для BCMA-his (например, 0,617 нМ, 1,85 нМ, 5,56 нМ, 16,7 нМ и 50 нМ), и инжектировали от низкой концентрации до высокой концентрации со скоростью 30 мкл/мин при 25°С. Время ассоциации составляло 120 с, и время диссоциации составило 1800 с. Поверхность чипа регенерировали путем инжекции 3М раствора MgCl2 со скоростью 10 мкл/мин в течение 30 с. Результаты подгонки аффинности представлены в Таблице 4 и Таблице 5.

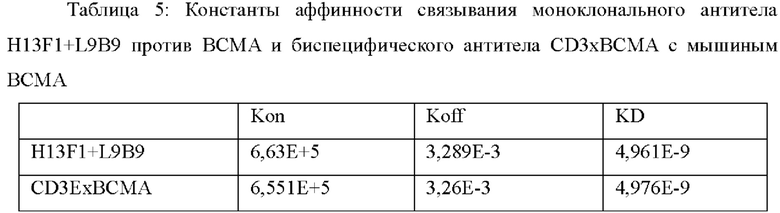
При определении аффинности моноклонального антитела против CD3E и биспецифического антитела CD3ExBCMA в отношении антигена CD3E антитело против человеческого Fab (Human Fab Capture Kit, GE, 28-9583-25) конъюгировали с поверхностью чипа СМ5. Белок антитела разбавляли до 0,5-1 мкг/мл и инжектировали со скоростью 10 мкл/мин. Обеспечивали захват приблизительно 70 RU моноклонального антитела CD3E антителом против человеческого Fab, и обеспечивали захват приблизительно 150 RU биспецифического антитела CD3ExBCMA антителом против человеческого Fab. Затем готовили серии градиентов концентрации для человеческого гетеродимера CD3E CD3E-FcK/CD3D-FcH (например, 12,5 нМ, 25 нМ, 50 нМ, 100 нМ и 200 нМ), и инжектировали от низкой концентрации до высокой концентрации со скорость 30 мкл/мин при 25°С. Время ассоциации составляло 120 с, и время диссоциации составило 600 с. Поверхность чипа регенерировали путем инжекции 10 мМ глицин-HCl (рН 2,1) со скоростью 10 мкл/мин в течение 60 с. Результаты подгонки аффинности представлены в Таблице 6.
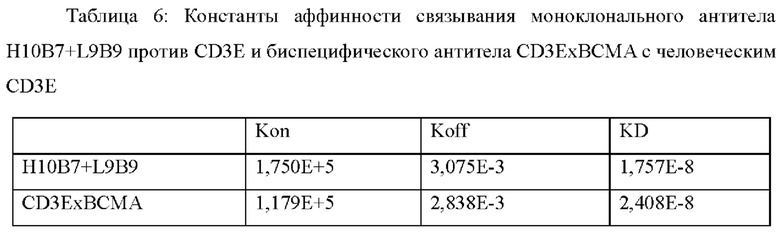
Пример 6: Идентификация способности биспецифического антитела одновременно распознавать антигены CD3E и ВСМА
Способность биспецифического антитела CD3ExBCMA (CD3ExBCMA BsAb) одновременно связываться с антигенами CD3E и ВСМА определяли с использованием обычных способов ELISA (иммуноферментного анализа).
96-луночный планшет для ELIS А покрывали антигеном CD3E-FcK/CD3D-FcH (3 мкг/мл, 100 мкл/лунку) и оставляли на ночь в холодильнике при 4°С. После блокирования блокирующим раствором PBS-0,1% Tween 20-3% молоко при 37°С в течение 1 часа моноклональное антитело H13F1+L9B9 против ВСМА, моноклональное антитело H10B7+L9B9 против CD3E и биспецифическое антитело CD3ExBCMA (10 мкг/мл, 100 мкл/лунку) соответственно добавляли в планшет в двух параллелях и инкубировали при 37°С в течение 1 часа. Планшет для ELISA промывали PBS-0,1% Tween 20, а затем добавляли антиген BCMA-His (1 мкг/мл, 100 мкл/лунку) и инкубировали при 37°С в течение 1 часа. Планшет ELISA промывали PBS-0,1% Tween 20, а затем добавляли конъюгированный с HRP (пероксидаза хрена) мышиный IgG против his (Beijing ComWin Biotech Co., Ltd., cw0285M) и инкубировали при 37°C в течение 1 часа. Планшет для ELISA промывали PBS-0,1% Tween 20, и добавляли раствор для развития окрашивания на основе субстрата OPD. Развитие окрашивания останавливали при помощи 1М H2SO4 через 5-10 минут. Измеряли величину оптической плотности при двух длинах волн 492 нм/630 нм с использованием микропланшетного ридера. Результат анализа ELISA представлен на Фиг. 1. Биспецифическое антитело CD3ExBCMA может одновременно распознавать антигены CD3E и ВСМА.
Пример 7: Идентификация способности биспецифического антитела распознавать CD3E и ВСМА на клеточных поверхностях
Человеческие клетки острого Т-лимфоцитарного лейкоза Jurkat (Cell Resource Center, Institute of Basic Medicine, Chinese Academy of Medical Sciences) собирали в логарифмической фазе роста, центрифугировали и ресуспендировали в буфере PBS, содержащем 1% BSA (бычий сывороточный альбумин), до 2×106 клеток/мл, и высевали в количестве 100 мкл/лунку в 96-лун очные планшеты с V-образным дном. Моноклональное антитело H10B7+L9B9 против CD3E и биспецифическое антитело CD3ExBCMA готовили для градиентного разведения. Моноклональное антитело против CD3E имело исходную концентрацию 3 мкг/мл, и его разводили в 3-кратном градиенте в общей сложности с 8 точками концентраций. Биспецифическое антитело имело исходную концентрацию 6 мкг/мл, и его разводили в 3-кратном градиенте в общей сложности с 8 точками концентраций. 100 мкл моноклонального антитела против CD3E или 100 мкл биспецифического антитела добавляли в лунки, содержащие клетки, и инкубировали при 4°С в течение 1 часа. Затем клетки трижды промывали 200 мкл раствора PBS и инкубировали с козьим антителом против человеческого IgG-FITC (флуоресцеин изотиоцианат) (Beijing Zhongshan Golden Bridge Biotechnology Co., Ltd., ZF-0308) (100 мкл/лунку) при 4°C в течение 30 минут в темноте. Затем клетки трижды промывали 200 мкл раствора PBS и суспендировали в 100 мкл раствора PBS. Затем канал FITC детектировали при помощи проточного питометра (ACEA, Novocyte). Результаты продемонстрировали, что биспецифическое антитело CD3ExBCMA может хорошо связываться с CD3-положительными клетками Jurkat (Фиг. 2). Величина KD для биспецифического антитела CD3ExBCMA составляла 16,79 нМ, и величина KD для моноклонального антитела против CD3E составляла 1,43 нМ.
Человеческие клетки плазмоклеточного лейкоза NCI-H929 (Cell Resource Center, Institute of Basic Medicine, Chinese Academy of Medical Sciences) собирали в фазе логарифмического роста, центрифугировали и ресуспендировали в буфере PBS, содержащем 1% BSA, до 2×106 клеток/мл, и высевали в количестве 100 мкл/лунку в 96-луночные планшеты с V-образным дном. Моноклональное антитело H13F1+L9B9 против ВСМА и биспецифическое антитело CD3ExBCMA готовили для градиентного разведения. Моноклональное антитело против ВСМА имело исходную концентрацию 10 мкг/мл, и его разводили в 3-кратном градиенте в общей сложности с 8 точками концентраций. Биспецифическое антитело имело исходную концентрацию 20 мкг/мл, и его разводили в 3-кратном градиенте в общей сложности с 8 точками концентраций. 100 мкл моноклонального антитела против ВСМА или 100 мкл биспецифического антитела добавляли в лунки, содержащие клетки, и инкубировали при 4°С в течение 1 часа. Затем клетки трижды промывали 200 мкл раствора PBS и инкубировали с козьим антителом против человеческого IgG-FITC (Beijing Zhongshan Golden Bridge Biotechnology Co., Ltd., ZF-0308) (100 мкл/лунку) при 4°C в течение 30 минут в темноте. Затем клетки трижды промывали 200 мкл раствора PBS и суспендировали в 100 мкл раствора PBS. Затем канал FITC детектировали при помощи проточного цитометра (ACEA, Novocyte). Результаты продемонстрировали, что биспецифическое антитело CD3ExBCMA может хорошо связываться с ВСМА-положительными клетками NCI-H929 (Фиг. 3). Величина KD для биспецифического антитела CD3ExBCMA составляла 8,27 нМ, и величина KD для моноклонального антитела против ВСМА составляла 8,47 нМ.
Пример 8: Биспецифическое антитело опосредует специфическую активацию Jurkat-двойных клеток ВСМА-положительными опухолевыми клетками
Человеческие клетки плазмоклеточного лейкоза NCI-H929 (высокая степень экспрессии ВСМА, приобретены в Cell Resource Center, Institute of Basic Medicine, Chinese Academy of Medical Sciences) собирали в логарифмической фазе роста. После центрифугирования клетки ресуспендировали в среде 1640 до 4×105 клеток/мл и высевали в клеточный планшет в количестве 50 мкл/лунку. Jurkat-двойные клетки (приобретены в Invivogen) собирали в логарифмической фазе роста, центрифугировали и ресуспендировали в среде 1640 до 1×106 клеток/мл, и добавляли в клеточный планшет в количестве 100 мкл/лунку с получением окончательного отношения Е:Т 5:1. Затем в клеточный планшет добавляли биспецифическое антитело CD3ExBCMA (50 мкл/лунку), моноклональное антитело H10B7+L9B9 против CD3E (50 мкл/лунку) или комбинацию моноклонального антитела H10B7+L9B9 против CD3E и моноклонального антитела H13F1+L9B9 против ВСМА, где биспецифическое антитело имело исходную концентрацию 10 мкг/мл, и его разводили в 3-кратном градиенте в общей сложности с 10 точками концентрации; моноклональное антитело H10B7+L9B9 против CD3E имело исходную концентрацию 5 мкг/мл, и его разводили в 3-кратном градиенте в общей сложности с 10 точками концентрации; и для комбинации моноклонального антитела H10B7+L9B9 против CD3E и моноклонального антитела H13F1+L9B9 против ВСМА каждое из них имело исходную концентрацию 5 мкг/мл, и ее разводили в 3-кратном градиенте в общей сложности с 10 точками концентрации. После 20 часов инкубации супернатант отбирали, и обнаруживали и анализировали специфическую активацию Jurkat-двойных клеток ВСМА-положительными опухолевыми клетками, опосредованную биспецифическим антителом CD3ExBCMA, моноклональным антителом против CD3E и комбинацией моноклонального антитела против CD3E и моноклонального антитела против ВСМА, в соответствии с указаниями QUANTI-Luc™ (QUANTI-Luc™, Invivogen, rep-qlc2). Результаты демонстрируют, что только биспецифическое антитело CD3ExBCMA может опосредовать активацию Jurkat-двойных клеток ВСМА-положительными опухолевыми клетками, и ни само по себе моноклональное антитело против CD3E, ни комбинация моноклонального антитела против CD3E и моноклонального антитела против ВСМА не могут опосредовать активацию Jurkat-двойных клеток ВСМА-положительными опухолевыми клетками (Фиг. 4).
Пример 9: Биспецифическое антитело опосредует уничтожение ВСМА-положительных опухолевых клеток Т-клетками
9.1 Выделение человеческих мононуклеарных клеток периферической крови (РВМС)
Пробы крови (50 мл каждой) отбирали у нормальных добровольцев. Пробы крови отбирали у авторов изобретения и их коллег в качестве добровольцев, все из которых подписывали информированное согласие. Критерии включения для добровольцев были следующими:
1. Возраст больше 18 лет;
2. Отсутствие инфекции HIV (вирус иммунодефицита человека) и HBV (вирус гепатита В);
3. Нормальный общий анализ крови;
4. Не беременные или не кормящие женщины.
РВМС выделяли из цельной крови добровольцев с использованием центрифугирования в градиенте плотности фиколла и выращивали в среде 1640.
9.2 Обнаружение уничтожения клетками РВМС ВСМА-положительных опухолевых клеток, опосредованного биспецифическим антителом
Человеческие клетки плазмоклеточного лейкоза NCI-H929 (высокий уровень экспрессии ВСМА), человеческие клетки множественной миеломы RPMI-8226 (умеренный уровень экспрессии ВСМА) и человеческие клетки острого промиелоцитарного лейкоза HL60 (ВСМА отрицательные) приобретали в Cell Resource Center, Institute of Basic Medicine, Chinese Academy of Medical Sciences. Клетки в логарифмической фазе роста собирали, центрифугировали и ресуспендировали в среде 1640 до 4×105 клеток/мл, и высевали в клеточные планшеты в количестве 50 мкл/лунку. Затем в клеточный планшет добавляли биспецифическое антитело CD3ExBCMA (50 мкл/лунку), которое имело исходную концентрацию 1 мкг/мл и разводили в 4-кратном градиенте в общей сложности с 10 точками концентраций. Наконец, добавляли 100 мкл/лунку РВМС (эффекторы) с получением конечного отношения Е:Т 5:1. В то же самое время, готовили контроль исключительно клеток-мишеней (клетки NCI-H929, клетки RPMI-8226 или клетки HL60), контроль исключительно эффекторных клеток (РВМС) и контроль исключительно среды и их объемы доводили средой до 200 мкл. После 20 часов инкубации супернатант отбирали. Скорость уничтожения Т-клетками опухолевых клеток, опосредованного биспецифическим антителом, детектировали и анализировали в соответствии с инструкциями cytoTox96® Non-Radioactive Cytotoxicity Assay (Promega, G1780).
Результаты продемонстрировали, что в присутствии биспецифического антитела CD3ExBCMA эффекторные клетки обладали существенным уничтожающим действием в отношении высоко экспрессирующихся клеток NCI-H929 и умеренно экспрессирующихся клеток RPMI-8226, но не оказывали уничтожающего действия в отношении отрицательных HL60 (Фиг. 5), свидетельствуя о том, что биспецифическое антитело CD3ExBCMA может эффективно опосредовать уничтожение Т-клетками клеток с различными положительными уровнями экспрессии ВСМА, и не может опосредовать уничтожение ВСМА-отрицательных клеток.
Пример 10: Биспецифические антитела могут специфически стимулировать экспрессию активирующей молекулы на поверхности Т-клеток
Человеческие клетки плазмоклеточного лейкоза NCI-H929 (высокий уровень экспрессии ВСМА), человеческие клетки множественной миеломы RPMI-8226 (умеренный уровень экспрессии ВСМА) и человеческие клетки острого промиелоцитарного лейкоза HL60 (ВСМА отрицательные) собирали в логарифмической фазе роста, центрифугировали и ресуспендировали в среде 1640 до 4×105 клеток/мл, и высевали в клеточные планшеты в количестве 50 мкл/лунку. Затем в клеточный планшет добавляли биспецифическое антитело CD3ExBCMA (50 мкл/лунку), которое имело исходную концентрацию 1 мкг/мл и разводили в 4-кратном градиенте в общей сложности с 10 точками концентраций. Наконец, добавляли 100 мкл/лунку РВМС (эффекторы) с получением конечного отношения Е:Т 5:1. После 20 часов инкубации клетки центрифугировали при 350g в течение 5 минут, однократно промывали PBS и инкубировали с антителами для проточной цитометрии, т.е. антителом против человеческого CD3 (Ebioscience, 17-0037-42) и антителом против человеческого CD69 (Ebioscience, 11-0069-42) при 4°С в течение 30 минут в темноте. Затем клетки дважды промывали 200 мкл раствора PBS, ресуспендировали в 100 мкл раствора PBS и обнаруживали при помощи проточного питометра (ACEA, Novocyte) для сравнения различия в экспрессии маркера активации CD69 популяции CD3-положительных клеток после обработки биспецифическим антителом CD3ExBCMA.
Результаты продемонстрировали, что биспецифическое антитело CD3ExBCMA может специфически активировать Т-клетки в присутствии NCI-H929 с высокой степенью экспрессии ВСМА или RPMI-8226 с умеренной степенью экспрессии ВСМА, и биспецифическое антитело CD3ExBCMA не может активировать Т-клетки в присутствии HL60 с отрицательным ВСМА (Фиг. 6), свидетельствуя о том, что биспецифическое антитело CD3ExBCMA может эффективно активировать Т-клетки в присутствии клеток с различными положительными уровнями экспрессии ВСМА, и не могут активировать Т-клетки в присутствии ВСМА-отрицательных клеток.
Пример 11: Противоопухолевая активность биспецифического антитела в мышиной модели восстановления иммунитета РВМС
Пробы крови (50 мл каждой) отбирали у нормальных добровольцев. Пробы крови отбирали у авторов изобретения и их коллег в качестве добровольцев, все из которых подписывали информированное согласие. Критерии включения для добровольцев были следующими:
1. Возраст больше 18 лет;
2. Отсутствие инфекции HIV (вирус иммунодефицита человека) и HBV (вирус гепатита В);
3. Нормальный общий анализ крови;
4. Не беременные или не кормящие женщины.
Человеческие мононуклеарные клетки периферической крови (РВМС) выделяли из периферической крови здоровых людей с использованием центрифугирования в градиенте плотности фиколла. Отобрали двадцать пять самок мышей NPG в возрасте 7-8 недель (Beijing Viktor Biotechnology Co., Ltd.). 5×106 клеток NCI-H929 подкожно инокулировали в правый бок мышей NPG, и сутки инокуляции были определены как 0 сутки. Через два часа после инокуляции опухолевых клеток каждой мыши внутрибрюшинно инокулировали 5×106 отдельных РВМС. Когда средний объем опухоли достигал 95 мм3, тогда мышей случайным образом разделяли на три группы в соответствии с объемом опухоли. Тестируемые группы могут быть разделены на 3 группы: группа 2, биспецифическое антитело CD3ExBCMA, 0,1 мг/кг; группа 3, биспецифическое антитело CD3ExBCMA, 0,02 мг/кг; и группа 1, контрольная группа, которой вводили растворитель. Каждая группа состояла из шести мышей. Биспецифическое антитело вводили однократно путем инъекции в хвостовую вену. Через четверо суток после первого введения дозу в группе 2 изменяли на 0,5 мг/кг, дозу в группе 3 изменяли на 0,1 мг/кг, и биспецифическое антитело вводили еще три раза с частотой два раза в неделю. Терапевтические эффекты оценивали в соответствии с величиной относительного ингибирования опухолевого роста (TGI), и безопасность оценивали в соответствии с изменением массы тела и смертью животных.
Результаты продемонстрировали, что биспецифическое антитело CD3ExBCMA значительно ингибировало опухолевый рост в дозе 0,1/0,5 мг/кг с TGI (%) 95%, р менее 0,001 (Фиг. 7). В то же самое время, в дозах 0,02/0,1 мг/кг и 0,1/0,5 мг/кг биспецифического антитела CD3ExBCMA не обнаружено смерти животных и никакой очевидной токсичности лекарственного средства в каждой обрабатываемой группе, и животные хорошо переносили лечение (Фиг. 8).
Пример 12: Противоопухолевая активность биспецифического антитела в модели гуманизированных мышей hCD34+
Отобрали двадцать самок гуманизированных мышей hCD34+ в возрасте 20-24 недель (приобретенных в Pengli Biomedical Technology (Shanghai) Co., Ltd). 100 мкл 1×107 клеток RPMI8226 и 100 мкл Матригель хорошо перемешивали и затем смесь инокулировали в правую часть спины мышей посредством подкожной инъекции. Мышей анестезировали при помощи 3-4% изофлурана перед инокуляцией. Когда средний объем опухоли достигал приблизительно 50-80 мм3, тогда 16 мышей, несущих опухоль, случайным образом разделяли на 2 группы в соответствии с отношением hCD34+ в периферической крови и объемами опухоли по 8 мышей в каждой группе. Сутки разделения на группы и введения были определены как 0 сутки. Тестируемые группы могут быть разделены на 2 группы: группа, которой осуществляли введение биспецифического антитела CD3ExBCMA, 0,01 мг/кг, и отрицательная контрольная группа, IgGlm3, 0,01 мг/кг. Каждая группа состояла из восьми мышей. Биспецифическое антитело вводили путем инъекции в хвостовую вену в общей сложности 4 раза с частотой один раз в неделю. Терапевтические эффекты оценивали в соответствии с величиной относительного ингибирования опухолевого роста (TGI), и безопасность оценивали в соответствии с изменением массы тела и смертью животных.
Во время эксперимента животные в общем находились в хорошей умственном состоянии. К концу эксперимента in vivo (34 сутки) отсутствовало значимое различие в массе тела (Р более 0,05) в группе, которой осуществлялось введение (группа G2), по сравнению с отрицательной контрольной группой (IgG1m3, в/в, 0,01 мг/кг, группа G1). Тенденция к изменению массы тела в каждый момент времени в каждой группе представлена на Фиг. 9. Биспецифическое антитело CD3ExBCMA значительно ингибировало опухолевый рост в дозе 0,01 мг/кг с TGI (%) 61,17%. Опухолевый рост представлен на Фиг. 10.
Хотя настоящая заявка на изобретение подробно описана со ссылкой на общее описание и конкретные воплощения, специалистам в данной области техники понятно, что изменения или улучшения могут быть осуществлены в отношении настоящего изобретения на основе настоящей заявки на изобретение. Соответственно, все эти изменения или улучшения, осуществляемые без отступления от сущности в соответствии с настоящей заявкой на изобретение, оказываются в заявленном объеме изобретения.
--->
Sequence Listing
<110> Beijing Wisdomab Biotechnology Co., Ltd
<120> ANTI-CD3E/BCMA BISPECIFIC ANTIBODY AND USE THEREOF
<150> CN 201910532734.7
<151> 2019-06-19
<160> 36
<170> CNIPASequenceListing 1.0
<210> 1
<211> 5
<212> PRT
<213> Artificial Sequence
<400> 1
Gly Tyr Gly Met His
1 5
<210> 2
<211> 17
<212> PRT
<213> Artificial Sequence
<400> 2
Val Ile Trp Phe Asp Gly Ser Arg Lys Tyr Tyr Val Asp Ser Val Lys
1 5 10 15
Gly
<210> 3
<211> 9
<212> PRT
<213> Artificial Sequence
<400> 3
Gln Met Gly Tyr Trp His Phe Gly Leu
1 5
<210> 4
<211> 11
<212> PRT
<213> Artificial Sequence
<400> 4
Arg Ala Ser Gln Ser Ile Ser Asn Tyr Leu Thr
1 5 10
<210> 5
<211> 7
<212> PRT
<213> Artificial Sequence
<400> 5
Glu Ala Ser Ser Arg Pro Ser
1 5
<210> 6
<211> 9
<212> PRT
<213> Artificial Sequence
<400> 6
Gln Gln Trp Ser Arg Leu Pro Val Thr
1 5
<210> 7
<211> 5
<212> PRT
<213> Artificial Sequence
<400> 7
Asn Tyr Trp Met His
1 5
<210> 8
<211> 17
<212> PRT
<213> Artificial Sequence
<400> 8
Ala Thr Tyr Arg Gly His Ser Asp Thr Tyr Tyr Asn Gln Lys Phe Lys
1 5 10 15
Gly
<210> 9
<211> 12
<212> PRT
<213> Artificial Sequence
<400> 9
Gly Ala Val Tyr Ala Gly Tyr Asp Val Leu Asp Tyr
1 5 10
<210> 10
<211> 121
<212> PRT
<213> Artificial Sequence
<400> 10
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ala Thr Tyr Arg Gly His Ser Asp Thr Tyr Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Gly Ala Val Tyr Ala Gly Tyr Asp Val Leu Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 11
<211> 121
<212> PRT
<213> Artificial Sequence
<400> 11
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ala Thr Tyr Arg Gly His Ser Asp Thr Tyr Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Gly Ala Ile Tyr Asp Gly Tyr Asp Val Leu Asp Asn Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 12
<211> 118
<212> PRT
<213> Artificial Sequence
<400> 12
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Lys Phe Ser Gly Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Trp Phe Asp Gly Ser Arg Lys Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Met Gly Tyr Trp His Phe Gly Leu Trp Gly Arg Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 13
<211> 107
<212> PRT
<213> Artificial Sequence
<400> 13
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Asn Asn Ser
20 25 30
Leu Thr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Asn Arg Glu Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Leu Lys Leu Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 14
<211> 107
<212> PRT
<213> Artificial Sequence
<400> 14
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Asn Tyr
20 25 30
Leu Thr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Glu Ala Ser Ser Arg Pro Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Arg Leu Pro Val
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 15
<211> 12
<212> PRT
<213> Artificial Sequence
<400> 15
Glu Arg Lys Ser Cys Val Glu Cys Pro Pro Cys Pro
1 5 10
<210> 16
<211> 107
<212> PRT
<213> Artificial Sequence
<400> 16
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Asn Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Arg Lys Leu Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 17
<211> 330
<212> PRT
<213> Artificial Sequence
<400> 17
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 18
<211> 330
<212> PRT
<213> Artificial Sequence
<400> 18
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Phe Glu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 19
<211> 330
<212> PRT
<213> Artificial Sequence
<400> 19
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Phe Glu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 20
<211> 107
<212> PRT
<213> Homo sapiens
<400> 20
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 21
<211> 106
<212> PRT
<213> Homo sapiens
<400> 21
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
35 40 45
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 22
<211> 105
<212> PRT
<213> Homo sapiens
<400> 22
Gln Asp Gly Asn Glu Glu Met Gly Gly Ile Thr Gln Thr Pro Tyr Lys
1 5 10 15
Val Ser Ile Ser Gly Thr Thr Val Ile Leu Thr Cys Pro Gln Tyr Pro
20 25 30
Gly Ser Glu Ile Leu Trp Gln His Asn Asp Lys Asn Ile Gly Gly Asp
35 40 45
Glu Asp Asp Lys Asn Ile Gly Ser Asp Glu Asp His Leu Ser Leu Lys
50 55 60
Glu Phe Ser Glu Leu Glu Gln Ser Gly Tyr Tyr Val Cys Tyr Pro Arg
65 70 75 80
Gly Ser Lys Pro Glu Asp Ala Asn Phe Tyr Leu Tyr Leu Arg Ala Arg
85 90 95
Val Cys Glu Asn Cys Met Glu Met Asp
100 105
<210> 23
<211> 84
<212> PRT
<213> Homo sapiens
<400> 23
Phe Lys Ile Pro Ile Glu Glu Leu Glu Asp Arg Val Phe Val Asn Cys
1 5 10 15
Asn Thr Ser Ile Thr Trp Val Glu Gly Thr Val Gly Thr Leu Leu Ser
20 25 30
Asp Ile Thr Arg Leu Asp Leu Gly Lys Arg Ile Leu Asp Pro Arg Gly
35 40 45
Ile Tyr Arg Cys Asn Gly Thr Asp Ile Tyr Lys Asp Lys Glu Ser Thr
50 55 60
Val Gln Val His Tyr Arg Met Cys Gln Ser Cys Val Glu Leu Asp Pro
65 70 75 80
Ala Thr Val Ala
<210> 24
<211> 96
<212> PRT
<213> Macaca fascicularis
<400> 24
Gln Asp Gly Asn Glu Glu Met Gly Ser Ile Thr Gln Thr Pro Tyr Gln
1 5 10 15
Val Ser Ile Ser Gly Thr Thr Val Ile Leu Thr Cys Ser Gln His Leu
20 25 30
Gly Ser Glu Ala Gln Trp Gln His Asn Gly Lys Asn Lys Glu Asp Ser
35 40 45
Gly Asp Arg Leu Phe Leu Pro Glu Phe Ser Glu Met Glu Gln Ser Gly
50 55 60
Tyr Tyr Val Cys Tyr Pro Arg Gly Ser Asn Pro Glu Asp Ala Ser His
65 70 75 80
His Leu Tyr Leu Lys Ala Arg Val Cys Glu Asn Cys Met Glu Met Asp
85 90 95
<210> 25
<211> 84
<212> PRT
<213> Macaca fascicularis
<400> 25
Phe Lys Ile Pro Val Glu Glu Leu Glu Asp Arg Val Phe Val Lys Cys
1 5 10 15
Asn Thr Ser Val Thr Trp Val Glu Gly Thr Val Gly Thr Leu Leu Thr
20 25 30
Asn Asn Thr Arg Leu Asp Leu Gly Lys Arg Ile Leu Asp Pro Arg Gly
35 40 45
Ile Tyr Arg Cys Asn Gly Thr Asp Ile Tyr Lys Asp Lys Glu Ser Ala
50 55 60
Val Gln Val His Tyr Arg Met Cys Gln Asn Cys Val Glu Leu Asp Pro
65 70 75 80
Ala Thr Leu Ala
<210> 26
<211> 87
<212> PRT
<213> Mus musculus
<400> 26
Asp Asp Ala Glu Asn Ile Glu Tyr Lys Val Ser Ile Ser Gly Thr Ser
1 5 10 15
Val Glu Leu Thr Cys Pro Leu Asp Ser Asp Glu Asn Leu Lys Trp Glu
20 25 30
Lys Asn Gly Gln Glu Leu Pro Gln Lys His Asp Lys His Leu Val Leu
35 40 45
Gln Asp Phe Ser Glu Val Glu Asp Ser Gly Tyr Tyr Val Cys Tyr Thr
50 55 60
Pro Ala Ser Asn Lys Asn Thr Tyr Leu Tyr Leu Lys Ala Arg Val Cys
65 70 75 80
Glu Tyr Cys Val Glu Val Asp
85
<210> 27
<211> 84
<212> PRT
<213> Mus musculus
<400> 27
Phe Lys Ile Gln Val Thr Glu Tyr Glu Asp Lys Val Phe Val Thr Cys
1 5 10 15
Asn Thr Ser Val Met His Leu Asp Gly Thr Val Glu Gly Trp Phe Ala
20 25 30
Lys Asn Lys Thr Leu Asn Leu Gly Lys Gly Val Leu Asp Pro Arg Gly
35 40 45
Ile Tyr Leu Cys Asn Gly Thr Glu Gln Leu Ala Lys Val Val Ser Ser
50 55 60
Val Gln Val His Tyr Arg Met Cys Gln Asn Cys Val Glu Leu Asp Ser
65 70 75 80
Gly Thr Met Ala
<210> 28
<211> 54
<212> PRT
<213> Homo sapiens
<400> 28
Met Leu Gln Met Ala Gly Gln Cys Ser Gln Asn Glu Tyr Phe Asp Ser
1 5 10 15
Leu Leu His Ala Cys Ile Pro Cys Gln Leu Arg Cys Ser Ser Asn Thr
20 25 30
Pro Pro Leu Thr Cys Gln Arg Tyr Cys Asn Ala Ser Val Thr Asn Ser
35 40 45
Val Lys Gly Thr Asn Ala
50
<210> 29
<211> 52
<212> PRT
<213> Macaca fascicularis
<400> 29
Met Leu Gln Met Ala Arg Gln Cys Ser Gln Asn Glu Tyr Phe Asp Ser
1 5 10 15
Leu Leu His Asp Cys Lys Pro Cys Gln Leu Arg Cys Ser Ser Thr Pro
20 25 30
Pro Leu Thr Cys Gln Arg Tyr Cys Asn Ala Ser Met Thr Asn Ser Val
35 40 45
Lys Gly Met Asn
50
<210> 30
<211> 49
<212> PRT
<213> Mus musculus
<400> 30
Met Ala Gln Gln Cys Phe His Ser Glu Tyr Phe Asp Ser Leu Leu His
1 5 10 15
Ala Cys Lys Pro Cys His Leu Arg Cys Ser Asn Pro Pro Ala Thr Cys
20 25 30
Gln Pro Tyr Cys Asp Pro Ser Val Thr Ser Ser Val Lys Gly Thr Tyr
35 40 45
Thr
<210> 31
<211> 6
<212> PRT
<213> Artificial Sequence
<400> 31
His His His His His His
1 5
<210> 32
<211> 232
<212> PRT
<213> Mus musculus
<400> 32
Pro Arg Gly Pro Thr Ile Lys Pro Cys Pro Pro Cys Lys Cys Pro Ala
1 5 10 15
Pro Asn Leu Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Ile
20 25 30
Lys Asp Val Leu Met Ile Ser Leu Ser Pro Ile Val Thr Cys Val Val
35 40 45
Val Asp Val Ser Glu Asp Asp Pro Asp Val Gln Ile Ser Trp Phe Val
50 55 60
Asn Asn Val Glu Val His Thr Ala Gln Thr Gln Thr His Arg Glu Asp
65 70 75 80
Tyr Asn Ser Thr Leu Arg Val Val Ser Ala Leu Pro Ile Gln His Gln
85 90 95
Asp Trp Met Ser Gly Lys Glu Phe Lys Cys Lys Val Asn Asn Lys Asp
100 105 110
Leu Pro Ala Pro Ile Glu Arg Thr Ile Ser Lys Pro Lys Gly Ser Val
115 120 125
Arg Ala Pro Gln Val Tyr Val Leu Pro Pro Pro Glu Glu Glu Met Thr
130 135 140
Lys Lys Gln Val Thr Leu Thr Cys Met Val Thr Asp Phe Met Pro Glu
145 150 155 160
Asp Ile Tyr Val Glu Trp Thr Asn Asn Gly Lys Thr Glu Leu Asn Tyr
165 170 175
Lys Asn Thr Glu Pro Val Leu Asp Ser Asp Gly Ser Tyr Phe Met Tyr
180 185 190
Ser Lys Leu Arg Val Glu Lys Lys Asn Trp Val Glu Arg Asn Ser Tyr
195 200 205
Ser Cys Ser Val Val His Glu Gly Leu His Asn His His Thr Thr Lys
210 215 220
Ser Phe Ser Arg Thr Pro Gly Lys
225 230
<210> 33
<211> 232
<212> PRT
<213> Artificial Sequence
<400> 33
Glu Pro Lys Ser Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
1 5 10 15
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
20 25 30
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
35 40 45
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
50 55 60
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
65 70 75 80
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
85 90 95
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
100 105 110
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
115 120 125
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met Thr
130 135 140
Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser
145 150 155 160
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
165 170 175
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
180 185 190
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
195 200 205
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
210 215 220
Ser Leu Ser Leu Ser Pro Gly Lys
225 230
<210> 34
<211> 232
<212> PRT
<213> Artificial Sequence
<400> 34
Glu Pro Lys Ser Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
1 5 10 15
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
20 25 30
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
35 40 45
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
50 55 60
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
65 70 75 80
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
85 90 95
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
100 105 110
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
115 120 125
Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
130 135 140
Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser
145 150 155 160
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
165 170 175
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val
180 185 190
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
195 200 205
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
210 215 220
Ser Leu Ser Leu Ser Pro Gly Lys
225 230
<210> 35
<211> 330
<212> PRT
<213> Homo sapiens
<400> 35
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 36
<211> 330
<212> PRT
<213> Artificial Sequence
<400> 36
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО ПРОТИВ ВИРУСА БЕШЕНСТВА И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2764740C1 |
| АНТИТЕЛО ПРОТИВ СТОЛБНЯЧНОГО ТОКСИНА И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2815280C1 |
| Антитела к TSLP человека и их применение | 2021 |
|
RU2825460C1 |
| АНТИ-TSLP АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2834227C1 |
| Антитело к BCMA, его антигенсвязывающий фрагмент и их медицинское применение | 2020 |
|
RU2819660C2 |
| АНТИТЕЛО К CLDN-18.2 И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2829997C1 |
| АНТИТЕЛО К B7-H4, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2019 |
|
RU2792748C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ АНТИТЕЛА К PCSK-9, И ЕЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2782792C2 |
| Антитело против FXI/FXIA, его антигенсвязывающий фрагмент и фармацевтическое применение | 2021 |
|
RU2838408C1 |
| АНТИТЕЛО К CD3 И ЕГО ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2019 |
|
RU2802272C2 |
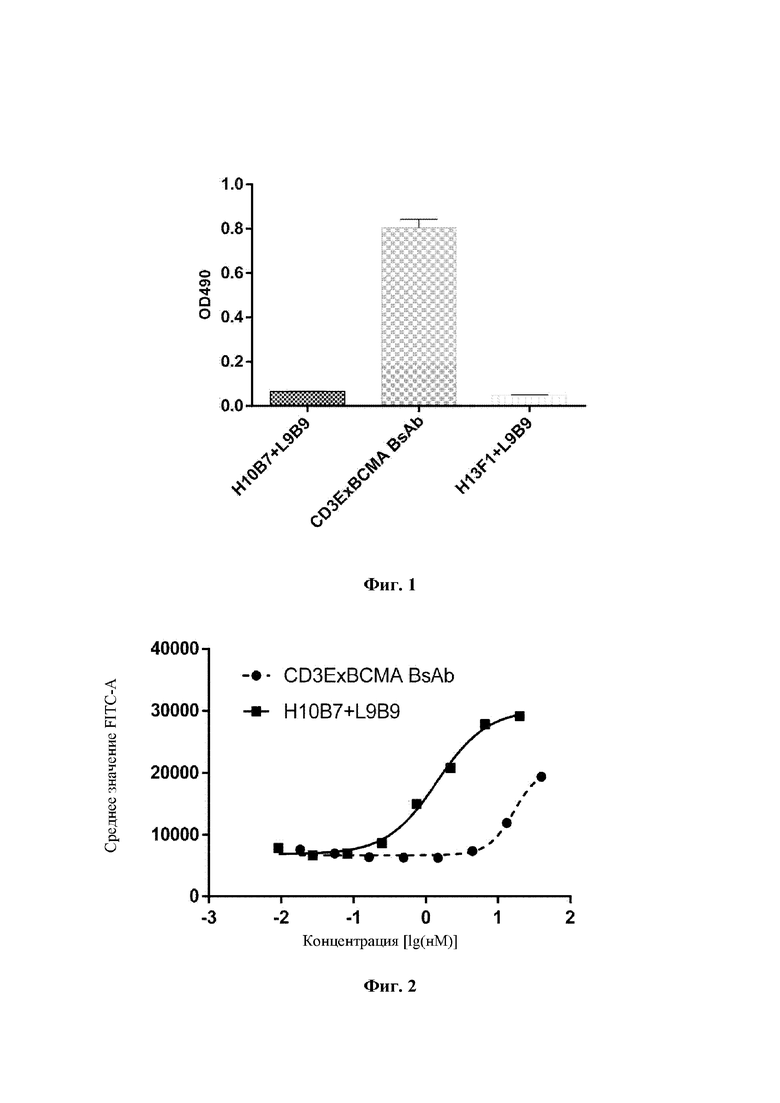
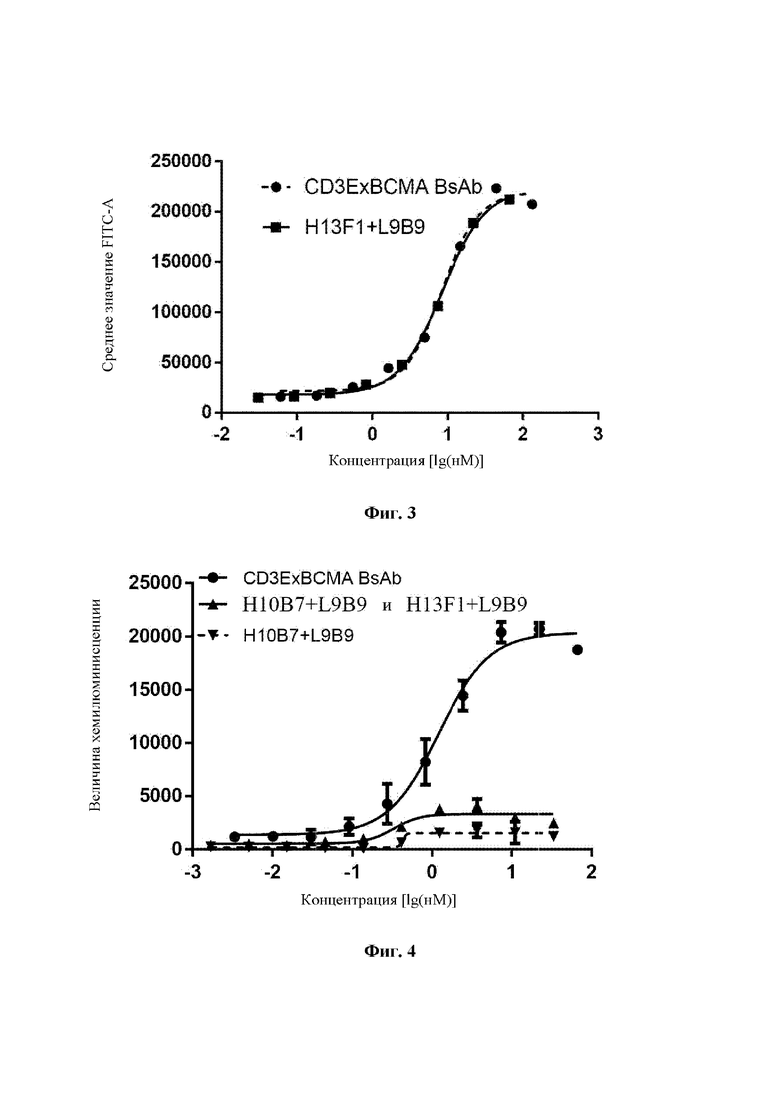
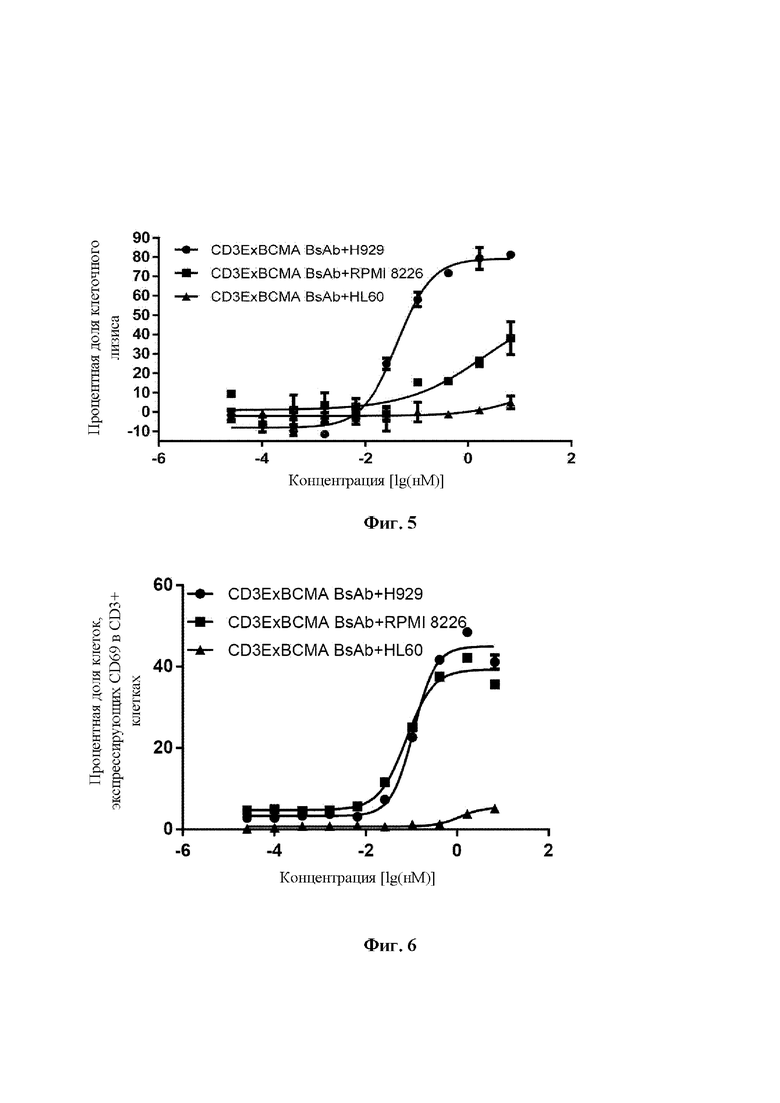
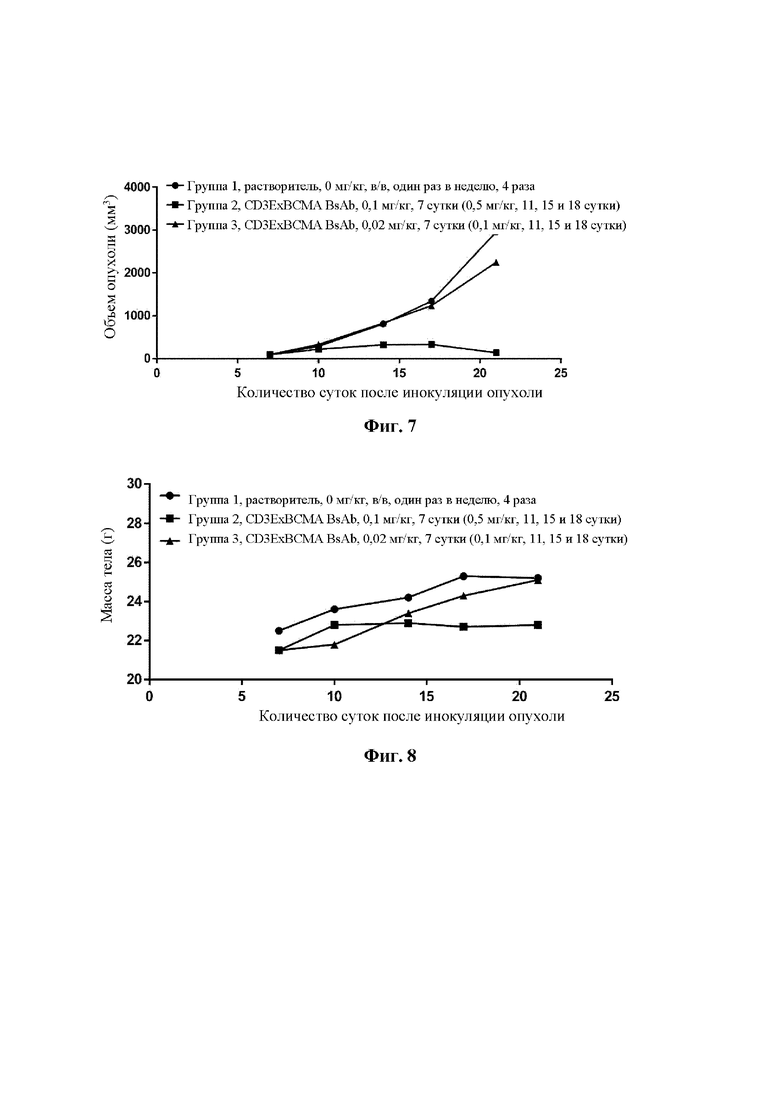
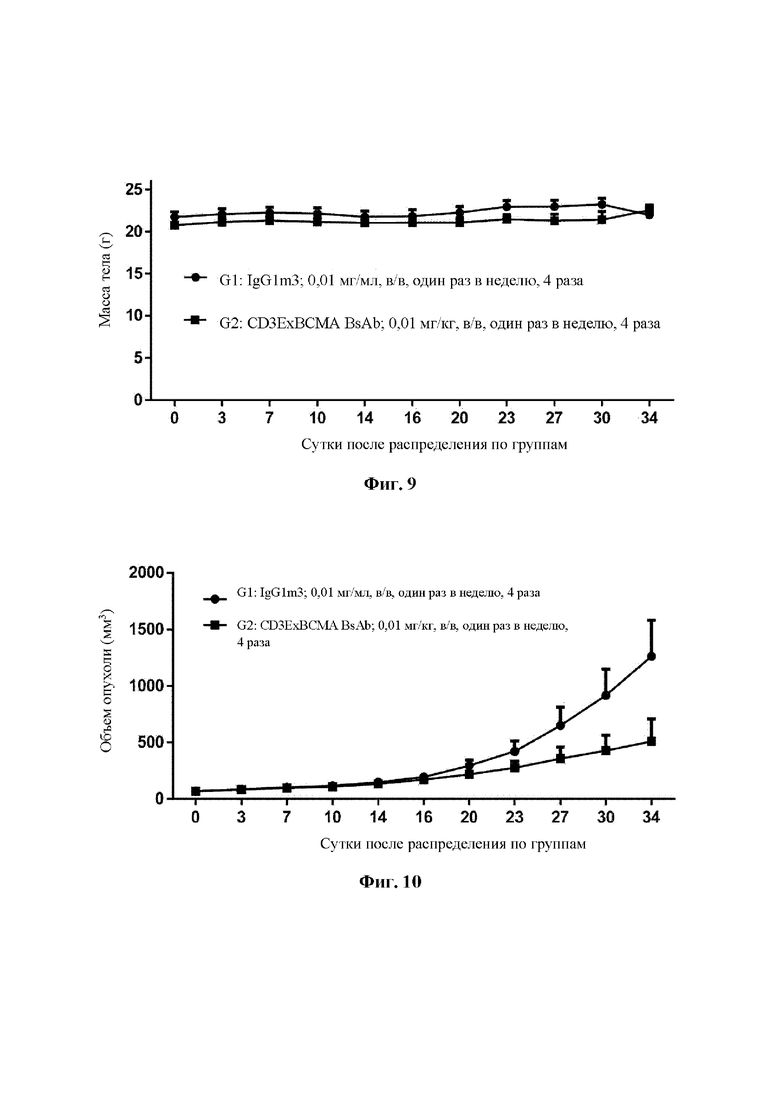
Изобретение относится к биотехнологии. Представлена фармацевтическая композиция, содержащая эффективное количество биспецифического антитела, содержащего антигенсвязывающий фрагмент против человеческого CD3E и антигенсвязывающий фрагмент против человеческого BCMA, и фармацевтически приемлемый носитель, эксципиент или разбавитель. Изобретение позволяет применять его для предупреждения или лечения множественной миеломы. 5 з.п. ф-лы, 10 ил., 6 табл., 12 пр.
1. Фармацевтическая композиция для использования в предупреждении или лечении множественной миеломы, содержащая эффективное количество биспецифического антитела, содержащего антигенсвязывающий фрагмент против человеческого CD3E и антигенсвязывающий фрагмент против человеческого ВСМА, и фармацевтически приемлемый носитель, эксципиент или разбавитель, где антигенсвязывающий фрагмент против человеческого CD3E содержит:
HCDR1, как представлено в SEQ ID NO: 1,
HCDR2, как представлено в SEQ ID NO: 2,
HCDR3, как представлено в SEQ ID NO: 3,
LCDR1, как представлено в SEQ ID NO: 4,
LCDR2, как представлено в SEQ ID NO: 5, и
LCDR3, как представлено в SEQ ID NO: 6; и
антигенсвязывающий фрагмент против человеческого ВСМА содержит:
HCDR1, как представлено в SEQ ID NO: 7,
HCDR2, как представлено в SEQ ID NO: 8,
HCDR3, как представлено в SEQ ID NO: 9,
LCDR1, как представлено в SEQ ID NO: 4,
LCDR2, как представлено в SEQ ID NO: 5, и
LCDR3 как представлено в SEQ ID NO: 6;
где HCDR и LCDR определены в соответствии с системой нумерации Кабата.
2. Фармацевтическая композиция по п. 1, где антигенсвязывающий фрагмент против человеческого CD3E и антигенсвязывающий фрагмент против человеческого ВСМА содержат одну и ту же вариабельную область легкой цепи, предпочтительно одинаковую легкую цепь; и/или
биспецифическое антитело представляет собой антитело IgG1, содержащее две константные области тяжелой цепи, имеющие одинаковую шарнирную область, и аминокислотная последовательность шарнирной области представлена в SEQ ID NO: 15.
3. Фармацевтическая композиция по п. 1, где биспецифическое антитело представляет собой антитело IgG1, содержащее первую константную область тяжелой цепи и вторую константную область тяжелой цепи, где
аминокислоты в положениях 354 и 366 первой константной области тяжелой цепи представляют собой соответственно С и W, а аминокислоты в положениях 349, 366, 368 и 407 второй константной области тяжелой цепи представляют собой соответственно С, S, А и V; и/или
аминокислоты в положениях 234, 235 и 331 первой и второй константных областей тяжелой цепи представляют собой соответственно F, Е и S;
где положение аминокислот константной области антитела определено в соответствии с нумерацией EU.
4. Фармацевтическая композиция по п. 1, где антигенсвязывающий фрагмент против человеческого CD3E содержит вариабельную область тяжелой цепи, как представлено в SEQ ID NO: 12, и вариабельную область легкой цепи, как представлено в SEQ ID NO: 14; и/или
антигенсвязывающий фрагмент против человеческого ВСМА содержит вариабельную область тяжелой цепи, как представлено в SEQ ID NO: 10, и вариабельную область легкой цепи, как представлено в SEQ ID NO: 14.
5. Фармацевтическая композиция по п. 1, где антигенсвязывающий фрагмент против человеческого CD3E и/или антигенсвязывающий фрагмент против человеческого ВСМА представляет собой одноцепочечное антитело (scfv) или Fab-фрагмент;
предпочтительно антигенсвязывающий фрагмент против человеческого CD3E представляет собой Fab-фрагмент и антигенсвязывающий фрагмент против человеческого ВСМА представляет собой Fab-фрагмент; или
антигенсвязывающий фрагмент против человеческого CD3E представляет собой Fab-фрагмент и антигенсвязывающий фрагмент против человеческого ВСМА представляет собой одноцепочечное антитело (scfv); или
антигенсвязывающий фрагмент против человеческого CD3E представляет собой одноцепочечное антитело (scfv) и антигенсвязывающий фрагмент против человеческого ВСМА представляет собой Fab-фрагмент; или
антигенсвязывающий фрагмент против человеческого CD3E представляет собой одноцепочечное антитело (scfv) и антигенсвязывающий фрагмент против человеческого ВСМА представляет собой одноцепочечное антитело (scfv).
6. Фармацевтическая композиция по п. 1, где антитело имеет первое плечо и второе плечо, где первое плечо содержит антигенсвязывающий фрагмент против человеческого CD3E, а второе плечо содержит антигенсвязывающий фрагмент против человеческого ВСМА:
первое плечо содержит аминокислотную последовательность вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 12, аминокислотную последовательность константной области тяжелой цепи, как представлено в SEQ ID NO: 19, аминокислотную последовательность вариабельной области легкой цепи, как представлено в SEQ ID NO: 14, и аминокислотную последовательность константной области легкой цепи, как представлено в SEQ ID NO: 20;
второе плечо содержит аминокислотную последовательность вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 10, аминокислотную последовательность константной области тяжелой цепи, как представлено в SEQ ID NO: 18, аминокислотную последовательность вариабельной области легкой цепи, как представлено в SEQ ID NO: 14, и аминокислотную последовательность константной области легкой цепи, как представлено в SEQ ID NO: 20.
| EP 2982692 A1, 10.02.2016 | |||
| Устройство для проволочной радиофикации домов | 1929 |
|
SU28162A1 |
| SECKINGER A | |||
| et al., Target Expression, Generation, Preclinical Activity, and Pharmacokinetics of the BCMA-T Cell Bispecific Antibody EM801 for Multiple Myeloma Treatment | |||
| Cancer Cell | |||
| Автомобиль-сани, движущиеся на полозьях посредством устанавливающихся по высоте колес с шинами | 1924 |
|
SU2017A1 |
| Vol | |||
| Способ очистки нефти и нефтяных продуктов и уничтожения их флюоресценции | 1921 |
|
SU31A1 |
| No | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Pp | |||
| Приспособление для подъема падающих гребней в машинах льнопрядильного, джутового и т.п. производств | 1913 |
|
SU396A1 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, СПЕЦИФИЧНЫЕ К АНТИГЕНАМ, АКТИВИРУЮЩИМ Т-КЛЕТКИ, И ОПУХОЛЕВОМУ АНТИГЕНУ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2605390C2 |

