ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННУЮ ЗАЯВКУ
В настоящей заявке на изобретение испрашивается приоритет заявки на патент Китая №202011128926.0, поданной 21 октября 2020 года, включенной в данное описание путем ссылки.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение в общем относится к области генетической инженерии и лекарственному средству на основе антитела. В частности, настоящее изобретение относится к антителам, направленным против столбнячного токсина, и их применению.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Столбняк представляет собой острое и смертельное заболевание, вызываемое нейротоксинами, секретируемыми Clostridium tetani, и поражает людей и животных. Наиболее восприимчивые животные представляют собой лошадей, мулов или ослов из отряда Perssodactyla. Люди также восприимчивы к нейротоксинам, секретируемым Clostridium tetani. Это заболевание может быть выявлено в любом возрасте. Столбняк новорожденных является одной из основных причин смертности новорожденных в экономически слаборазвитых районах. Инкубационный период столбняка обычно составляет 3-21 суток, в основном приблизительно 10 суток, но он также может составлять от одних суток до нескольких месяцев в зависимости от характеристик, степени и расположения раны. Типичными клиническими проявлениями столбняка являются сжатие зубов, усиление стресса двигательного нервного центра и локальный или системный мышечный спазмолитический паралич. Пациенты часто умирают от асфиксии и системной органной недостаточности. Даже в современных реанимациях уровень смертности при этом заболевании очень высок. Смертность от столбняка может составлять от 19% до 31% особенно после серьезных стихийных бедствий. Существует три клинических типа столбняка, а именно системный столбняк, местный столбняк и столбняк головы, которые составляют во всех случаях, соответственно, приблизительно 88%, приблизительно 12% и приблизительно 1%.
Clostridium tetani представляет собой строгую анаэробную грамположительную бактерию. У нее есть жгутик, но нет капсулы. Ее споры широко встречаются в удобренной почве, уличной пыли, протухшей грязи, в кишечниках животных и загрязненных поверхностях объектов. Clostridium tetani проникает в организм человека преимущественно через рану. Если рана узкая и глубокая или если рана одновременно заражена аэробными пиогенными бактериями, то Clostridium tetani будет размножаться в большом количестве в анаэробных условиях и продуцировать три экзотоксина, среди которых столбнячный токсин вызывает характерные симптомы столбняка и приводит к продукции защитных антител [1]. Тем не менее, приблизительно 20% пациентов не имеют явных инвазивных ран, что позволяет предположить, что Clostridium tetani также может проникать в организм через небольшие царапины. Clostridium tetani могут быть разделены на десять серотипов в зависимости от реакций агглютинации флагеллярного антигена, но все серотипы продуцируют один и тот же нейротоксин. Токсины, продуцируемые отдельными типами штаммов, могут быть нейтрализованы любым типом антитоксина. Столбнячный токсин является высокотоксичным, вторым только по отношению к ботулиническому токсину. Его летальная доза для мышей составляет всего 2-6 нг/кг.
Столбнячный токсин, также известный как столбнячный нейротоксин, является одноцепочечным белком, который имеет относительную молекулярную массу приблизительно 150 кДа, состоит из 1315 аминокислотных остатков и может быть расщеплен на легкую цепь (фрагмент А, 50 кДа) и тяжелую цепь (НС, 100 кДа), связанные через дисульфидную связь, тем самым образуя его активную форму [2]. Фрагмент А представляет собой металлопротеиназу цинка, которая при попадании в цитоплазму расщепляет ассоциированный с везикулами мембранный белок-2 (VAMP-2) и блокирует высвобождение ингибирующих нейромедиаторов, таких как глицин и гшилш-аминомасляная кислота (ГАМК), приводя в результате к мышечному спазмолитическому параличу [3]. Тяжелая цепь содержит два функциональных домена. С-концевой домен (фрагмент С) связывается с поверхностями нервных клеток таким образом, что молекула токсина подвергается эндоцитозу в везикулы. N-концевой домен (фрагмент В) проходит через везикульную мембрану и транспортирует фрагмент А в цитоплазму нейрона [4]. Фрагмент С подразделяется на С-концевой субдомен (НСС) и N-концевой субдомен (HCN) [5]. Процесс, с помощью которого столбнячный токсин попадает в нервные клетки, не до конца понятен. В настоящее время общепринят механизм двойного рецептора, и он включает ганглиозидный рецептор, в частности GTlb и GDlb, и белковый рецептор [6].
Столбняк является предотвратимым заболеванием. Субъекты могут получать активную и пассивную иммунизационную обработку после травмы. Гуморальный иммунитет оказывает защитное действие против столбняка. Нейтрализующие антитела связываются с токсином и препятствуют взаимодействию токсина с рецепторами на клетках-мишенях и последующей интернализации токсина в клетку. Клинически пассивная иммунизация рекомендована для травмированных пациентов, которые имеют неясную историю иммунизации или не иммунизированы, не получили полной первичной иммунизации или получили полную первичную иммунизацию, но с последней бустерной иммунизацией, полученной более 5 лет назад. В настоящее время существует два вида препаратов пассивной иммуннизации для предупреждения и лечения столбняка в клинике. Один из них представляет собой столбнячный антитоксин (ТАТ), который получают из лошадиной плазмы крови, иммунизированной столбнячным анатоксином, путем ферментативного расщепления и обессоливания. Перед использованием ТАТ необходимо провести кожный тест. ТАТ часто вызывает аллергические реакции с частотой возникновения 5-30%. Иногда возникает анафилактический шок. Другой вид препаратов пассивной иммуннизации представляет собой человеческий иммуноглобулин против столбняка (HIGT), который получают путем отбора плазмы или сыворотки крови с высоким титром столбнячного антитела от доноров крови, последовательно иммунизированных вакциной против гепатита В и столбнячным анатоксином, и обработки плазмы или сыворотки крови с использованием низкотемпературного способа с этанолом. HIGT могут быть инъецированы непосредственно без кожного теста. Тем не менее, поскольку HIGT относится к препаратам крови, существует потенциальный риск инфекционных заболеваний, таких как гепатит С и СПИД. Кроме того, HIGT ограничен источниками и имеет низкий выход, высокую стоимость и нестабильное качество между партиями [7].
Лекарственные средства на основе антитела могут преодолеть некоторые вышеупомянутые недостатки и имеют определенные преимущества, такие как продуцирование путем стабильной экспрессии экспрессирующимися системами на основе клеток млекопитающего и определенный состав.
Из-за клинических потребностей большое медицинское значение заключается именно в разработке антител, которые могут нейтрализовать столбнячный токсин и быть использованы в предупреждении и лечении столбняка.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В первом аспекте настоящего изобретения предложено биспецифическое антитело, содержащее первый антигенсвязывающий фрагмент и второй антигенсвязывающий фрагмент, который связывается с разными эпитопами столбнячного токсина, где биспецифическое антитело обладает активностью, нейтрализующей столбнячный токсин.
В некоторых воплощениях первого аспекта указанный первый антигенсвязывающий фрагмент содержит HCDR1, имеющую аминокислотную последовательность SYWIY (SEQ ID NO: 1), HCDR2, имеющую аминокислотную последовательность EINPTNGFANYNEKFKT (SEQ ID NO: 2), или EINPTAGFANYNEKFKT (SEQ ID NO: 3), или EINPTNAFANYNEKFKT (SEQ ID NO: 4), HCDR3, имеющую аминокислотную последовательность HFRFPY (SEQ ID NO: 5), LCDR1, имеющую аминокислотную последовательность RASQDIGSSLT (SEQ ID NO: 6), LCDR2, имеющую аминокислотную последовательность ATSSLDS (SEQ ID NO: 7), и LCDR3, имеющую аминокислотную последовательность LQYASSPYT (SEQ ID NO: 8); где аминокислотные последовательности HCDR и LCDR определены согласно Kabat.
В некоторых воплощениях первого аспекта указанный второй антигенсвязывающий фрагмент содержит HCDR1, имеющую аминокислотную последовательность DYGVN (SEQ ID NO: 9), HCDR2, имеющую аминокислотную последовательность MIWSDGTTDYSSALKS (SEQ ID NO: 10), HCDR3, имеющую аминокислотную последовательность VDGYSHYYAMDY (SEQ ID NO: 11), LCDR1, имеющую аминокислотную последовательность RASENIYSYLA (SEQ ID NO: 12), LCDR2, имеющую аминокислотную последовательность NAKTLAE (SEQ ID NO: 13), и LCDR3, имеющую аминокислотную последовательность QHHYGLPFT (SEQ ID NO: 14); где аминокислотные последовательности HCDR и LCDR определены согласно Kabat.
В некоторых воплощениях первого аспекта аминокислотная последовательность вариабельной области тяжелой цепи первого антигенсвязывающего фрагмента является такой, как представлено в SEQ ID NO: 15, и аминокислотная последовательность вариабельной области легкой цепи первого антигенсвязывающего фрагмента является такой, как представлено в SEQ ID NO: 18; или аминокислотная последовательность вариабельной области тяжелой цепи первого антигенсвязывающего фрагмента является такой, как представлено в SEQ ID NO: 16, и аминокислотная последовательность вариабельной области легкой цепи первого антигенсвязывающего фрагмента является такой, как представлено в SEQ ID NO: 18; или аминокислотная последовательность вариабельной области тяжелой цепи первого антигенсвязывающего фрагмента является такой, как представлено в SEQ ID NO: 17, и аминокислотная последовательность вариабельной области легкой цепи первого антигенсвязывающего фрагмента является такой, как представлено в SEQ ID NO: 18.
В некоторых воплощениях первого аспекта, аминокислотная последовательность вариабельной области тяжелой цепи второго антигенсвязывающего фрагмента является такой, как представлено в SEQ ID NO: 19, и аминокислотная последовательность вариабельной области легкой цепи второго антигенсвязывающего фрагмента является такой, как представлено в SEQ ID NO: 20.
В некоторых воплощениях первого аспекта формы первого антигенсвязывающего фрагмента и второго антигенсвязывающего фрагмента независимо выбраны из одноцепочечного вариабельного фрагмента (scFv) или фрагмента Fab.
Во втором аспекте изобретения предложено моноклональное антитело, которое связывается со столбнячным токсином, содержащее:
HCDR1, имеющую аминокислотную последовательность SYWIY (SEQ ID NO: 1), HCDR2, имеющую аминокислотную последовательность EINPTNGFANYNEKFKT (SEQ ID NO: 2), или EINPTAGFANYNEKFKT (SEQ ID NO: 3), или EINPTNAFANYNEKFKT (SEQ ID NO: 4), HCDR3, имеющую аминокислотную последовательность HFRFPY (SEQ ID NO: 5), LCDR1, имеющую аминокислотную последовательность RASQDIGSSLT (SEQ ID NO: 6), LCDR2, имеющую аминокислотную последовательность ATSSLDS (SEQ ID NO: 7), и LCDR3, имеющую аминокислотную последовательность LQYASSPYT (SEQ ID NO: 8); где аминокислотные последовательности HCDR и LCDR определены согласно Kabat.
В третьем аспекте изобретения предложено моноклональное антитело, которое связывается со столбнячным токсином, содержащее:
HCDR1, имеющую аминокислотную последовательность DYGVN (SEQ ID NO: 9), HCDR2, имеющую аминокислотную последовательность MIWSDGTTDYSSALKS (SEQ ID NO: 10), HCDR3, имеющую аминокислотную последовательность VDGYSHYYAMDY (SEQ ID NO: 11), LCDR1, имеющую аминокислотную последовательность RASENIYSYLA (SEQ ID NO: 12), LCDR2, имеющую аминокислотную последовательность NAKTLAE (SEQ ID NO: 13), и LCDR3, имеющую аминокислотную последовательность QHHYGLPFT (SEQ ID NO: 14); где аминокислотные последовательности HCDR и LCDR определены согласно Kabat.
В четвертом аспекте изобретения предложена фармацевтическая композиция, содержащая биспецифическое антитело в соответствии с первым аспектом или моноклональное антитело в соответствии со вторым или третьим аспектом и фармацевтически приемлемый эксципиент, разбавитель или носитель.
В пятом аспекте изобретения предложено применение биспецифического антитела в соответствии с первым аспектом, моноклонального антитела в соответствии со вторым или третьим аспектом или фармацевтической композиции в соответствии с четвертым аспектом в изготовлении лекарственного средства для предупреждения или лечения столбняка.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг. 1 демонстрирует результаты анализа ИФА (иммуноферментный анализ), включающие оценку того, каким образом моноклональные антитела против ТТ-Нс S22E2-mIgG2a и S24B8-mIgG2a ингибируют связывание очищенного фага S22E2 с ТТ-Нс.
Фиг. 2 демонстрирует результаты анализа ИФА, включающие оценку того, каким образом моноклональные антитела против ТТ-Нс S22E2-mIgG2a и S24B8-mIgG2a ингибируют связывание очищенного фага S24B8 с ТТ-Нс.
Фиг. 3 демонстрирует результаты анализа ИФА, включающие оценку того, каким образом моноклональные антитела против ТТ-Нс S22E2-mIgG2a и S24B8-mIgG2a ингибируют связывание ТТ-Нс с ганглиозидом GTlb.
Фиг. 4 демонстрирует нейтрализующие эффекты моноклональных антител против столбнячного токсина в отношении столбнячного токсина.
Фиг. 5 демонстрирует защитное действие комбинации моноклональных антител против столбнячного токсина против летального заражения столбнячным токсином у мышей Balb/c.
Фиг. 6 демонстрирует, что биспецифическое антитело S22E2hl-scFv-FcHl+S24B8h3-IgG1K ингибирует связывание ТТ-Нс с ганглиозидом GM1.
Фиг. 7 демонстрирует, что биспецифическое антитело S22E2hl-scFv-FcHl+S24B8h3-IgG1K ингибирует связывание ТТ-Нс с ганглиозидом GD3.
Фиг. 8 демонстрирует, что биспецифическое антитело S22E2hl-scFv-FcHl+S24B8h3-IgG1K ингибирует связывание ТТ-Нс с ганглиозидом GT1b.
Фиг. 9 демонстрирует нейтрализующую активность биспецифического антитела против столбнячного токсина S22E2h1-scFv-FcH1+S24B8h3-IgG1K.
ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
SEQ ID NO: 1 демонстрирует аминокислотную последовательность HCDR1 мутантов гуманизированной вариабельной области тяжелой цепи S24B8VH-h1, S24B8VH-h2 и S24B8VH-h3.
SEQ ID NO: 2 демонстрирует аминокислотную последовательность HCDR2 мутанта гуманизированной вариабельной области тяжелой цепи S24B8VH-h1.
SEQ ID NO: 3 демонстрирует аминокислотную последовательность HCDR2 мутанта гуманизированной вариабельной области тяжелой цепи S24B8VH-h2.
SEQ ID NO: 4 демонстрирует аминокислотную последовательность HCDR2 мутанта гуманизированной вариабельной области тяжелой цепи S24B8VH-h3.
SEQ ID NO: 5 демонстрирует аминокислотную последовательность HCDR3 мутантов гуманизированной вариабельной области тяжелой цепи S24B8VH-h1, S24B8VH-h2 и S24B8VH-h3.
SEQ ID NO: 6-8 демонстрируют соответственно аминокислотные последовательности LCDR1, LCDR2 и LCDR3 мутанта гуманизированной вариабельной области легкой цепи S24B8VK-h1.
SEQ ID NO: 9-11 демонстрируют соответственно аминокислотные последовательности HCDR1, HCDR2 и HCDR3 мутанта гуманизированной вариабельной области тяжелой цепи S22E2VH-h1.
SEQ ID NO: 12-14 демонстрируют соответственно аминокислотные последовательности LCDR1, LCDR2 и LCDR3 мутанта гуманизированной вариабельной области легкой цепи S22E2VK-h1.
SEQ ID NO: 15 демонстрирует аминокислотную последовательность мутанта гуманизированной вариабельной области тяжелой цепи S24B8VH-h1.
SEQ ID NO: 16 демонстрирует аминокислотную последовательность мутанта гуманизированной вариабельной области тяжелой цепи S24B8VH-h2.
SEQ ID NO: 17 демонстрирует аминокислотную последовательность мутанта гуманизированной вариабельной области тяжелой цепи S24B8VH-h3.
SEQ ID NO: 18 демонстрирует аминокислотную последовательность мутанта гуманизированной вариабельной области легкой цепи S24B8VK-h1.
SEQ ID NO: 19 демонстрирует аминокислотную последовательность мутанта гуманизированной вариабельной области тяжелой цепи S22E2VH-h1.
SEQ ID NO: 20 демонстрирует аминокислотную последовательность мутанта гуманизированной вариабельной области легкой цепи S22E2VK-h1.
SEQ ID NO: 21 демонстрирует аминокислотную последовательность S24B8VH-h1+CH-IgG1K.
SEQ ID NO: 22 демонстрирует аминокислотную последовательность S24B8VH-h2+CH-IgG1K.
SEQ ID NO: 23 демонстрирует аминокислотную последовательность S24B8VH-h3+CH-IgG1K.
SEQ ID NO: 24 демонстрирует аминокислотную последовательность S24B8VK-h1+CK.
SEQ ID NO: 25 демонстрирует аминокислотную последовательность S22E2-h1-scFv-FcHl.
SEQ ID NO: 26 демонстрирует аминокислотную последовательность рекомбинантного белка С-концевого домена (ТТ-Нс) тяжелой цепи столбнячного токсина.
SEQ ID NO: 27 демонстрирует аминокислотную последовательность His концевой последовательности.
SEQ ID NO: 28 демонстрирует аминокислотную последовательность константной области тяжелой цепи человеческого (homo sapiens) подтипа IgG1.
SEQ ID NO: 29 демонстрирует аминокислотную последовательность константной области тяжелой цепи мышиного (mus musculus) подтипа IgG2a.
SEQ ID NO: 30 демонстрирует аминокислотную последовательность мутанта IgG1H человеческого подтипа IgG1.
SEQ ID NO: 31 демонстрирует аминокислотную последовательность мутанта IgG1K человеческого подтипа IgG1.
SEQ ID NO: 32 демонстрирует аминокислотную последовательность мутанта mIgG2a-H мышиного подтипа IgG2a.
SEQ ID NO: 33 демонстрирует аминокислотную последовательность мутанта mIgG2aK мышиного подтипа IgG2a.
SEQ ID NO: 34 демонстрирует аминокислотную последовательность константной области легкой цепи человеческого (homo sapiens) подтипа каппа.
SEQ ID NO: 35 демонстрирует аминокислотную последовательность константной области легкой цепи человеческого (homo sapiens) подтипа лямбда.
SEQ ID NO: 36 демонстрирует аминокислотную последовательность константной области легкой цепи мышиного (mus musculus) подтипа лимбда.
SEQ ID NO: 37 демонстрирует аминокислотную последовательность константной области легкой цепи мышиного (mus musculus) подтипа лямбда.
SEQ ID NO: 38 демонстрирует нуклеотидную последовательность праймера PmCGR.
SEQ ID NO: 39 демонстрирует нуклеотидную последовательность праймера PmCKR.
SEQ ID NO: 40 демонстрирует аминокислотную последовательность сегмента Fc, содержащего мутацию Knob (FcK).
SEQ ID NO: 41 демонстрирует аминокислотную последовательность сегмента Fc, содержащего мутацию Hole (FcHl).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Авторы настоящего изобретения провели обширное исследование и разработку лекарственных средств на основе антител против столбняка и получили новые биспецифические антитела и моноклональные антитела против столбнячного токсина с использованием технологии инжиниринга антител. Столбнячный токсин представляет собой макромолекулярный белок, который состоит из нескольких доменов. Теоретически, он содержит множество эпитопов, которые могут вызывать гуморальный иммунитет, и антитела, которые связываются с разными эпитопами, могут быть получены у иммунизированных субъектов. Фрагмент С столбнячного токсина представляет собой рецептор-связывающий домен, и вполне вероятно, что антитела против фрагмента С обладают нейтрализующими активностями, предотвращая связывание токсина с ганглиозидами GT1b, GD1b и GM1, и достигая нейтрализующих эффектов в отношении токсина.
В различных аспектах настоящего изобретения предложены новые биспецифические антитела и моноклональные антитела против столбнячного токсина, полинуклеотиды, кодирующие такие биспецифические антитела или моноклональные антитела, векторы, содержащие такие полинуклеотиды, клетки-хозяева, содержащие такие полинуклеотиды или векторы, способы получения и очистки таких биспецифических антител или моноклональных антител и медицинское и биологическое применение таких биспецифических антител или моноклональных антител. На основе предложенных здесь последовательностей вариабельных областей биспецифических антител или моноклональных антител молекулы полноразмерного биспецифического антитела или молекулы моноклонального антитела могут быть сконструированы для клинического применения в качестве лекарственного средства для предупреждения или лечения столбняка.
Если не указано иное, изобретение в соответствии с настоящей заявкой может быть реализовано практически с использованием обычной молекулярной биологии, микробиологии, клеточной биологии, биохимии и иммунологических способов в данной области техники.
Если не указано иное, термины, используемые в настоящем описании изобретения, имеют значения, обычно понимаемые специалистом в данной области техники.
ОПРЕДЕЛЕНИЯ
Использованный здесь термин "антитело" относится к молекуле иммуноглобулина, которая способна специфически связываться с мишенью при помощи по меньшей мере одного сайта распознавания антигена, расположенного в вариабельной области молекулы иммуноглобулина. Мишени включают без ограничения углеводы, полинуклеотиды, липиды и полипептиды. Использованное здесь "антитело" включает не только интактное (т.е. полноразмерное) антитело, но также и его антигенсвязывающий фрагмент (например, Fab, Fab', F(ab')2, Fv), его вариант, слитой белок, содержащий фрагменты антитела, гуманизированное антитело, химерное антитело, диатело, линейное антитело, одноцепочечное антитело, мультиспецифическое антитело (например биспецифическое антитело) и любые другие модифицированные форматы молекулы иммуноглобулина, содержащие желаемый специфический сайт распознавания антигена, включая гликозилированный вариант антитела, вариант аминокислотной последовательности и ковалентно модифицированное антитело.
Как правило, интактное или полноразмерное антитело содержит две тяжелые цепи и две легкие цепи. Каждая тяжелая цепь содержит вариабельную область тяжелой цепи (VH) и первую, вторую и третью константные области (CH1, СН2 и СН3). Каждая легкая цепь содержит вариабельную область (VL) и константную область (CL) легкой цепи. Полноразмерное антитело может представлять собой антитело любого типа, такое как антитело IgD, IgE, IgG, IgA или IgM (или их подтипы), но не обязательно принадлежит к какому-либо конкретному типу. Иммуноглобулины могут быть отнесены к разным типам в зависимости от их аминокислотных последовательностей константных доменов тяжелой цепи. Как правило, у иммуноглобулинов существует пять основных типов, т.е. IgA, IgD, IgE, IgG и IgM, и некоторые из этих типов могут быть дополнительно классифицированы на подтипы (изотипы), такие как IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелой цепи, соответствующие отдельным типам иммуноглобулина, упомянуты, соответственно, как α δ, ε, γ, и μ. Структуры субъединиц и трехмерные структуры различных типов иммуноглобулинов хорошо известны.
Использованный здесь термин "биспецифическое антитело" представляет собой антитело, обладающее способностью связываться с двумя разными антигенами. Биспецифическое антитело может обладать различными структурными конфигурациями. Например, биспецифическое антитело может состоять из двух фрагментов Fc и двух антигенсвязывающих фрагментов, слитых с ними, соответственно (подобно природному антителу, за исключением того, что эти два плеча связываются с разными антигенными мишенями или эпитопами). Антигенсвязывающие фрагменты могут быть в форме scfv или фрагментов Fab. Когда приведены два антигена, тогда каждый из двух отличающихся антигенсвязывающих фрагментов биспецифического антитела связывается с N-концом одного фрагмента Fc. Конфигурация антигенсвязывающих фрагментов двух плечей может иметь четыре комбинации, т.е. scfv+фрагмент Fab, фрагмент Fab+scfv, scfv+scfv и фрагмент Fab+фрагмент Fab. Фрагменты Fc могут содержать мутации, которые могут обеспечить гетеромеризацию тяжелой цепи. Способ KIH (выступ-во-впадине) является одной из стратегий, относящихся к гетеромеризации тяжелой цепи. Как правило, способ KIH относится к формированию структуры, которая облегчает спаривание гетерогенных полутел путем инженерии аминокислотной последовательности области СН3. Последнее позволяет максимально поддерживать структуру нормального антитела при формировании биспецифического антитела. В отношении руководства по способу KIH см., например, "An efficient route to human bispecific IgG", A. Margaret Merchant et al., Nature Biotechnology, Volume 16, 1998, которое включено в данное описание путем ссылки в полном объеме. Кроме того, биспецифическое антитело может быть конфигурировано таким образом, что у антитела, которое связывается с первым антигеном (например, в форме природного антитела), имеется антигенсвязывающий фрагмент, который может связываться со вторым антигеном, простирающимся от С-конца области СН3 (например, посредством гибкого линкера).
Использованные здесь термины "антигенсвязывающая часть" или "антигенсвязывающий фрагмент" могут быть использованы взаимозаменяемо, и относятся к части или области молекулы интактного антитела, ответственной за связывание с антигеном. Антигенсвязывающий домен может содержать вариабельную область тяжелой цепи (VH), вариабельную область легкой цепи (VL) или обе. Каждая из VH и VL как правило содержит три области, определяющие комплементарность, т.е. CDR1, CDR2 и CDR3.
Специалисту в данной области техники хорошо известно, что области, определяющие комплементарность (CDR, обычно включающие CDR1 CDR2 и CDR3), представляют собой части вариабельной области, которые в основном влияют на аффинность и специфичность антитела. Последовательности CDR VH или VL имеют два общих определения, то есть определение Kabat и определение Chothia (см., например, Kabat, "Sequences of Proteins of Immunological Interest", National Institutes of Health, Bethesda, Md.(1991); Al-Lazikani et al, J. Mol. Biol. 273: 927-948 (1997); и Martin et al, Proc. Natl. Acad. Sci. USA 86: 9268-9272 (1989)). Для последовательностей вариабельной области данного антитела последовательности областей CDR в VH и VL могут быть определены в соответствии с определением Kabat или определением Chothia. В воплощении настоящего изобретения последовательности CDR определены согласно Kabat.
Для последовательностей вариабельной области данного антитела последовательности областей CDR в последовательностях вариабельной области могут быть проанализированы различными путями, например, с использованием онлайн программного обеспечения Abysis (http://www.abysis.org/).
Для обычных антител примеры антигенсвязывающего фрагмента включают без ограничения (1) фрагмент Fab, который может представлять собой одновалентный фрагмент, имеющий цепь VL-CL и цепь VH-CH1; (2) фрагмент F(ab')2, который может представлять собой двухвалентный фрагмент, имеющий два фрагмента Fab', связанные дисульфидной мостиковой связью шарнирной области (т.е. димер Fab'); (3) фрагмент Fv, имеющий домены VL и VH в одном плече антитела; (4) одноцепочечный Fv (scFv), который может представлять собой единичную полипептидную цепь, состоящую из домена VH и домена VL, связанные полипептидным линкером; и (5) (scFv)2, который может содержать два домена VH, связанные пептидным линкером, и два домена VL, которые комбинированы с двумя доменами VH через дисульфидную мостиковую связь.
При конструировании биспецифического антитела "антигенсвязывающий фрагмент" включает, без ограничения, фрагмент Fab или одноцепочечный вариабельный фрагмент (scFv).
Использованный здесь термин "одноцепочечный вариабельный фрагмент (scFv)" относится к антителу с одноцепочечной структурой, обычно сконструированному с использованием способов генной инженерии, содержащему полипептидную цепь, содержащую вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL). Гибкий линкер, как правило, сконструирован между вариабельной областью тяжелой цепи и вариабельной областью легкой цепи таким образом, чтобы вариабельная область тяжелой цепи и вариабельная область легкой цепи могли складываться в правильную конформацию для связывания с антигеном.
Использованный здесь термин "Fab (антигенсвязывающий) фрагмент", "фрагмент Fab" и т.п. относится к фрагменту антитела, который получают после обработки интактного антитела папаином и который способен связываться с антигеном, включающему интактную легкую цепь (VL-CL), вариабельную область тяжелой цепи и фрагмент CHI (VH-CH1).
Использованный здесь термин "моноклональное антитело" относится к антителу из по существу гомогенной популяции антител, означая что антитела, составляющие популяцию, являются одинаковыми, за исключением природных мутаций, которые могут возникнуть в небольшом количестве индивидуальных антител. Описанные здесь моноклональные антитела, в частности, включают "химерные" антитела, в которых фрагмент тяжелой и/или легкой цепи идентичен или гомологичен соответствующей последовательности в антителе, полученной из конкретных видов или относящейся к конкретному типу или подтипу антитела, тогда как оставшаяся часть тяжелой и/или легкой цепи идентична или гомологична соответствующей последовательности в антителе, происходящей от других видов или относящейся к другому типу или подтипу антитела, и также включают фрагменты таких антител, при условии что они демонстрируют желаемую биологическую активность (см. патент США №4816567; и Morrison et al, Proc. Natl. Acad. Sci. USA 81: 6851-6855 (1984)).
Использованный здесь термин "специфическое связывание" относится к реакции непроизвольного связывания между двумя молекулами, например, связывание антитела с эпитопом антигена.
В первом аспекте настоящего изобретения предложено биспецифическое антитело, содержащее первый антигенсвязывающий фрагмент и второй антигенсвязывающий фрагмент, который связывается с разными эпитопами столбнячного токсина, где биспецифическое антитело обладает активностью, нейтрализующей столбнячный токсин.
В некоторых воплощениях первого аспекта указанный первый антигенсвязывающий фрагмент содержит HCDR1, имеющую аминокислотную последовательность SYWIY (SEQ ID NO: 1), HCDR2, имеющую аминокислотную последовательность EINPTNGFANYNEKFKT (SEQ ID NO: 2), или EINPTAGFANYNEKFKT (SEQ ID NO: 3), или EINPTNAFANYNEKFKT (SEQ ID NO: 4), HCDR3, имеющую аминокислотную последовательность HFRFPY (SEQ ID NO: 5), LCDR1, имеющую аминокислотную последовательность RASQDIGSSLT (SEQ ID NO: 6), LCDR2, имеющую аминокислотную последовательность ATSSLDS (SEQ ID NO: 7), и LCDR3, имеющую аминокислотную последовательность LQYASSPYT (SEQ ID NO: 8); где аминокислотные последовательности HCDR и LCDR определены согласно Kabat.
В некоторых воплощениях первого аспекта указанный второй антигенсвязывающий фрагмент содержит HCDR1, имеющую аминокислотную последовательность DYGVN (SEQ ID NO: 9), HCDR2, имеющую аминокислотную последовательность MIWSDGTTDYSSALKS (SEQ ID NO: 10), HCDR3, имеющую аминокислотную последовательность VDGYSHYYAMDY (SEQ ID NO: 11), LCDR1, имеющую аминокислотную последовательность RASENIYSYLA (SEQ ID NO: 12), LCDR2, имеющую аминокислотную последовательность NAKTLAE (SEQ ID NO: 13), и LCDR3, имеющую аминокислотную последовательность QHHYGLPFT (SEQ ID NO: 14); где аминокислотные последовательности HCDR и LCDR определены согласно Kabat.
В некоторых воплощениях первого аспекта аминокислотная последовательность вариабельной области тяжелой цепи первого антигенсвязывающего фрагмента является такой, как представлено в SEQ ID NO: 15, и аминокислотная последовательность вариабельной области легкой цепи является такой, как представлено в SEQ ID NO: 18; или аминокислотная последовательность вариабельной области тяжелой цепи первого антигенсвязывающего фрагмента является такой, как представлено в SEQ ID NO: 16, и аминокислотная последовательность вариабельной области легкой цепи первого антигенсвязывающего фрагмента является такой, как представлено в SEQ ID NO: 18; или аминокислотная последовательность вариабельной области тяжелой цепи первого антигенсвязывающего фрагмента является такой, как представлено в SEQ ID NO: 17, и аминокислотная последовательность вариабельной области легкой цепи первого антигенсвязывающего фрагмента является такой, как представлено в SEQ ID NO: 18.
В некоторых воплощениях первого аспекта аминокислотная последовательность вариабельной области тяжелой цепи второго антигенсвязывающего фрагмента является такой, как представлено в SEQ ID NO: 19, и аминокислотная последовательность вариабельной области легкой цепи второго антигенсвязывающего фрагмента является такой, как представлено в SEQ ID NO: 20.
В некоторых воплощениях первого аспекта формы первого антигенсвязывающего фрагмента и второго антигенсвязывающего фрагмента независимо выбраны из одноцепочечного вариабельного фрагмента (scFv) или фрагмента Fab.
В некоторых воплощениях первого аспекта указанный первый антигенсвязывающий фрагмент представляет собой фрагмент Fab, а указанный второй антигенсвязывающий фрагмент представляет собой одноцепочечный вариабельный фрагмент (scFv).
В некоторых воплощениях первого аспекта биспецифическое антитело содержит аминокислотную последовательность, представленную в SEQ ID NO: 21, 22 и 23, и аминокислотную последовательность, представленную в SEQ ID NO: 24.
В некоторых воплощениях первого аспекта биспецифическое антитело содержит аминокислотную последовательность, представленную в SEQ ID NO: 25.
Во втором аспекте изобретения предложено моноклональное антитело, которое связывается со столбнячным токсином, содержащее:
HCDR1, имеющую аминокислотную последовательность SYWIY (SEQ ID NO: 1), HCDR2, имеющую аминокислотную последовательность EINPTNGFANYNEKFKT (SEQ ID NO: 2), или EINPTAGFANYNEKFKT (SEQ ID NO: 3), или EINPTNAFANYNEKFKT (SEQ ID NO: 4), HCDR3, имеющую аминокислотную последовательность HFRFPY (SEQ ID NO: 5), LCDR1, имеющую аминокислотную последовательность RASQDIGSSLT (SEQ ID NO: 6), LCDR2, имеющую аминокислотную последовательность ATSSLDS (SEQ ID NO: 7), и LCDR3, имеющую аминокислотную последовательность LQYASSPYT (SEQ ID NO: 8); где аминокислотные последовательности HCDR и LCDR определены согласно Kabat.
В некоторых воплощениях второго аспекта моноклональное антитело содержит аминокислотную последовательность вариабельной области тяжелой цепи, представленную в SEQ ID NO: 15, и аминокислотную последовательность вариабельной области легкой цепи, представленную в SEQ ID NO: 18; или аминокислотную последовательность вариабельной области тяжелой цепи, представленную в SEQ ID NO: 16, и аминокислотную последовательность вариабельной области легкой цепи, представленную в SEQ ID NO: 18; или аминокислотную последовательность вариабельной области тяжелой цепи, представленную в SEQ ID NO: 17, и аминокислотную последовательность вариабельной области легкой цепи, представленную в SEQ ID NO: 18.
В третьем аспекте изобретения предложено моноклональное антитело, которое связывается со столбнячным токсином, содержащее:
HCDR1, имеющую аминокислотную последовательность DYGVN (SEQ ID NO: 9), HCDR2, имеющую аминокислотную последовательность MIWSDGTTDYSSALKS (SEQ ID NO: 10), HCDR3, имеющую аминокислотную последовательность VDGYSHYYAMDY (SEQ ID NO: 11), LCDR1, имеющую аминокислотную последовательность RASENIYSYLA (SEQ ID NO: 12), LCDR2, имеющую аминокислотную последовательность NAKTLAE (SEQ ID NO: 13), и LCDR3, имеющую аминокислотную последовательность QHHYGLPFT (SEQ ID NO: 14); где аминокислотные последовательности HCDR и LCDR определены согласно Kabat.
В некоторых воплощениях третьего аспекта моноклональное антитело содержит аминокислотную последовательность вариабельной области тяжелой цепи, представленную в SEQ ID NO: 19, и аминокислотную последовательность вариабельной области легкой цепи, представленную в SEQ ID NO: 20.
В четвертом аспекте изобретения предложена фармацевтическая композиция, содержащая биспецифическое антитело в соответствии с первым аспектом или моноклональное антитело в соответствии со вторым или третьим аспектом и фармацевтически приемлемый эксципиент, разбавитель или носитель.
В некоторых воплощениях четвертого аспекта фармацевтическая композиция предназначена для предупреждения или лечения столбняка.
В некоторых воплощениях четвертого аспекта фармацевтическая композиция может дополнительно содержать один или более чем один из смазывающего вещества, такого как тальк, стеарат магния и минеральное масло; смачивающего агента; эмульгатора; суспендирующего агента; консерванта, такого как бензойная кислота, сорбиновая кислота и пропионат кальция; подсластителя и/или корригента.
В некоторых воплощениях четвертого аспекта фармацевтическая композиция здесь может быть приготовлена в виде таблетки, пилюли, порошка, лепешки, эликсира, суспензии, эмульсии, раствора, сиропа, суппозитория или капсулы.
В некоторых воплощениях четвертого аспекта фармацевтическая композиция в соответствии с настоящим изобретением может быть доставлена с использованием любого физиологически приемлемого пути введения, включающего, без ограничения, пероральное введение, парентеральное введение, назальное введение, ректальное введение, внутрибрюшинное введение, внутрисосудистую инъекцию, подкожное введение, трансдермальное введение или ингаляционное введение.
В некоторых воплощениях четвертого аспекта фармацевтическая композиция для терапевтического применения может быть приготовлена для хранения в лиофилизированной композиции или в форме водного раствора путем смешивания агента, обладающего желаемой чистотой, с фармацевтически приемлемым носителем или эксципиентом, что является подходящим.
В пятом аспекте изобретения предложено применение биспецифического антитела в соответствии с первым аспектом, моноклонального антитела в соответствии со вторым или третьим аспектом или фармацевтической композиции в соответствии с четвертым аспектом в изготовлении лекарственного средства для предупреждения или лечения столбняка.
В шестом аспекте изобретения предложен способ предупреждения или лечения столбняка, включающий введение субъекту, нуждающемуся в этом, биспецифического антитела в соответствии с первым аспектом, моноклонального антитела в соответствии с вторым или третьим аспектом или фармацевтической композиции в соответствии с четвертым аспектом.
В других аспектах изобретения предложена молекула нуклеиновой кислоты, кодирующая биспецифическое антитело в соответствии с первым аспектом или моноклональное антитело в соответствии со вторым или третьим аспектом. В некоторых воплощениях молекула нуклеиновой кислоты функционально связана с регуляторной последовательностью, которая может распознаваться клеткой-хозяином, трансформированной вектором.
В изобретении также предложен вектор, содержащий выделенную молекулу нуклеиновой кислоты, кодирующую биспецифическое антитело в соответствии с первым аспектом или моноклональное антитело в соответствии со вторым или третьим аспектом, и клетка-хозяин, содержащая молекулу нуклеиновой кислоты или вектор.
В других аспектах изобретения предложен способ получения биспецифического антитела в соответствии с первым аспектом или моноклонального антитела в соответствии со вторым или третьим аспектом. В некоторых воплощениях способ получения биспецифического антитела в соответствии с первым аспектом или моноклонального антитела в соответствии со вторым или третьим аспектом включает культивирование клетки-хозяина таким образом, чтобы облегчить экспрессию нуклеиновой кислоты. В некоторых воплощениях способ получения биспецифического антитела в соответствии с первым аспектом или моноклонального антитела в соответствии со вторым или третьим аспектом дополнительно включает выделение биспецифического антитела или моноклонального антитела из среды для культивирования клетки-хозяина.
Следует понимать, что вышеприведенное подробное описание предназначено только для того, чтобы обеспечить специалисту в данной области техники возможность лучшего понимания настоящего изобретения, и не предполагается, что оно каким-либо путем приводит к ограничению. Различные модификации и вариации могут быть осуществлены с описанными воплощениями специалистом в данной области техники.
Следующие примеры приведены исключительно для иллюстративных задач и не предполагается, что они ограничивают объем настоящего изобретения.
Примеры
Пример 1: Получение рекомбинантного антигена столбнячного токсина и рекомбинантных антител
1.1 Получение рекомбинантного антигена столбнячного токсина Рекомбинантный белок С-концевого домена тяжелой цепи столбнячного токсина (ТТ-Нс, SEQ ID NO: 26) использовали в получении моноклональных антител против столбнячного токсина. Этот белок был получен из Clostridium tetani и был лишен любой посттрансляционной модификации, и поэтому он может экспрессироваться с использованием системы экспрессии Е. coli. Кроме того, к С-концу рекомбинантного белка была добавлена His концевая последовательность (His, SEQ ID NO: 27), чтобы облегчить очистку рекомбинантного белка и идентификацию функции моноклонального антитела.
Ген, кодирующий ТТ-Нс (содержащий His концевую последовательность), разрабатывали и синтезировали в соответствии с аминокислотными последовательностями рекомбинантных белков ТТ-Нс в базе данных Uniprot. Синтезированный ген клонировали в подходящий вектор для эукариотической экспрессии (такой как pET-22b, mvitrogen, Inc.) с использованием обычных способов молекулярной биологии. Экспрессионные плазмиды трансформировали в компетентные клетки Е. coli (такие как BL21 (DE3), Solarbio, Inc.). Е. coli, экспрессирующие рекомбинантный белок TT-Hc-His, индуцировали для экспрессии с помощью IPTG (изопропил-бета-0-1-тиогалактопиранозид) (такой как 18070-lg, Solarbio, Inc.). Затем бактерии собирали путем центрифугирования и разрушали с помощью ультразвукового разрушителя для клеток (такой как VCX130, Sonics, Inc.). Супернатанты собирали путем центрифугирования. Рекомбинантный белок в супернатантах дополнительно очищали на колонке для аффинной хроматографии с хелатированным металлом (такой как HisTrap FF, GE, Inc.). Буфер для консервирования рекомбинантного белка затем заменяли на PBS (физиологический раствор, забуференный фосфатом) (рН 7,0) или другие подходящие буферы при помощи колонки для обессоливания (такой как Hitrap desaulting, GE, Inc.). При необходимости образец может быть стерилизован путем фильтрования и затем храниться в аликвотах при -20°С.
1.2. Получение рекомбинантных антител
Нуклеотидные последовательности, кодирующие вариабельную область тяжелой цепи и вариабельную область легкой цепи антитела клонировали в эукариотические экспрессионные векторы (такие как pcDNA3.1, Invitrogen, Inc.), несущие, соответственно, нуклеотидные последовательности, кодирующие константную область тяжелой цепи и константную область легкой цепи, с использованием обычных молекулярно-биологических способов. Интактные антитела экспрессировали в комбинации. Константные области тяжелой цепи антител могут иметь человеческий подтип IgG1 (SEQ ID NO: 28) или мышиный подтип IgG2a (SEQ ID NO: 29), или могут представлять собой мутант IgG1H человеческого подтипа IgG1 (SEQ ID NO: 30) и IgG1K (SEQ ID NO: 31) на основе технологии KIH (выступ-во-впадине), или могут представлять собой мутант mIgG2a-H мышиного подтипа IgG2a (SEQ ID NO: 32) и mIgG2aK (SEQ ID NO: 33). Константные области легкой цепи могут иметь человеческий подтип каппа (SEQ ID NO: 34), человеческий подтип лямбда (SEQ ID NO: 35), мышиный подтип каппа (SEQ ID NO: 36) или мышиный подтип лямбда (SEQ ID NO: 37).
Полученные плазмиды для экспрессиии рекомбинантного антитела трансфицировали в клетки НЕK293 (такие как HEK293F, Invitrogen, Inc.) с использованием липосом (таких как 293fectin, Invitrogen, Inc.) или других реагентов для трансфекции (таких как PEI). Клетки культивировали в суспензии в бессывороточных средах в течение 3-5 суток. Затем культуральные супернатанты собирали путем центрифугирования. Рекомбинантный белок в супернатанте далее очищали на колонке для аффинной хроматографии ProteinA/G (такой как Mabselect SURE, GE, Inc.). Буфер для консервирования рекомбинантного белка затем заменяли на PBS (рН 7,0) или другие подходящие буферы при помощи колонки для обессоливания (такой как Hitrap desaulting, GE, Inc.). При необходимости, образец антитела может быть стерилизован путем фильтрования и затем храниться в аликвотах при -20°С.
Пример 2: Иммунизация мышей и конструирование иммунной библиотеки Мышей BALB/c в возрасте от 6 до 8 недель иммунизировали рекомбинантным белком TT-Hc-His, полученным в Примере 1, в качестве антигена при дозе иммунизации 50 мкг на мышь. Бустерную иммунизацию проводили каждые 14 суток. Мышей умерщвляли через 8 недель после первичной иммунизации и отбирали клетки селезенки. Мышиные лимфоциты селезенки выделяли с использованием раствора для выделения мышиных лимфоцитов (Dakewe Biotech Co., Ltd., CAT # DKW33-R0100). Общую РНК экстрагировали из выделенных лимфоцитов с использованием набора для клеточной экстракции общей РНК (Tiangen Biotech (Beijing) Co., Ltd., CAT # DP430). Используя выделенную общую РНК в качестве матрицы, синтезировали вариабельную область тяжелой цепи и вариабельную область легкой цепи антитела, используя набор для синтеза первой цепи кДНК (Thermo scientific, САТ#K1621). Используемые праймеры для обратной транскрипции представляли собой генспецифические праймеры, и парные для праймеров области были расположены, соответственно, в константной области тяжелой цепи и константной области легкой цепи антитела. Конкретные последовательности представляли собой, соответственно, PmCGR: TGCATTTGAACTCCTGCC (SEQ ID NO: 38), и PmCKR: CCATCAATCTCCACTGAC (SEQ ID NO: 39). Синтезированную кДНК немедленно сохраняли при -70°С для последующего использования. Затем, используя кДНК, полученную путем обратной транскрипции в качестве матрицы, праймеры синтезировали в соответствии с Krebber A, Bornhauser S, Burmester J, et al. Reliable cloning of functional antibody variable domains from hybridomas and spleen cell repertoires employing a reengineered phage display system. J Immunol Methods. 1997; 201(1):35-55, полное содержание которой включено в данное описание путем ссылки, и нуклеотидные последовательности, кодирующие VH и VK мышиных антител, соответственно амплифицировали с помощью ПЦР. Затем гены одноцепочечного вариабельного фрагмента (scFv) конструировали с помощью технологии ПЦР с перекрытием с расширением. Наконец, нуклеотидные последовательности полученных мышиных одноцепочечных антител клонировали в вектор pADSCFV-S (см. пример 1 в заявке на Китайский патент No 201510097117.0 в отношении подробных экспериментальных протоколов, содержание которых включено в данное описание путем ссылки) для конструирования библиотеки scFv. Библиотека антител имеет емкость 5 × 10Е8 и точность 50%.
Пример 3: Скрининг и предварительная идентификация мышиной иммунной библиотеки
3.1. Скрининг мышиной иммунной библиотеки
Ссылаясь на заявку на патент Китая №201510097117.0 (полное содержание которой включено в описание настоящего изобретения путем ссылки), библиотеку фагов, демонстрирующую мышиные одно цепочечные антитела, сконструированную в Примере 2, подвергали скринингу с использованием стратегии твердофазного скрининга (в отношении экспериментального протокола см. Phage Display: A Practical Approach, Tim Clackson and Henry B. Lowman (ed.), translated by Lan Ma et al., Chemical Industry Press, May 2008) с использованием рекомбинантного человеческого TT-Hc-His, полученного в Примере 1, в качестве антигена. Три раунда скрининга проводили путем связывания, элюирования, нейтрализации, инфицирования и амплификации. Наконец, получили два одноцепочечных антитела S22E2 и S24B8, специфически связывающихся с человеческим TT-Hc-His.
Нуклеотидные последовательности, кодирующие тяжелую цепь и легкие цепи S22E2 и S24B8, соответственно, клонировали в эукариотические экспрессионные векторы, несущие нуклеотидные последовательности, кодирующие константную область тяжелой цепи мышиного mIgG2a и константную область легкой цепи мышиного каппа, путем использования обычных молекулярно-биологических способов для получения рекомбинантных мышиных антител.
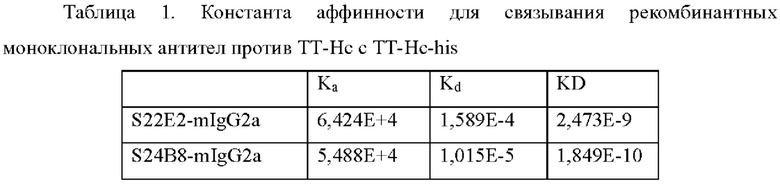
3.2. Анализ аффинности рекомбинантных моноклональных антител против ТТ-Нс Аффинности антител против ТТ-Нс определяли методом поверхностного плазмонного резонанса с использованием Biacore Х100. Реактивы и расходные материалы, такие как набор для аминосвязывания (BR-1000-50), набор для иммобилизации мышиного антитела (BR-1008-38), чип СМ5 (BR100012) и 10xHBS-EP, рН 7,4 (BR100669), были приобретены в GE Healthcare. Карбоксилированную СМ5 поверхность чипа активировали гидрохлоридом 1-этил-3-(3-диметиламинопропил)карбодиимида (EDC) и N-гидроксисукцинимидом (NHS) в соответствии с инструкциями к набору. Антитело против мышиного IgG (Fc) (иммобилизованное антитело) разбавляли до 25 мкг/мл с использованием 10 мМ ацетата натрия (рН 5,0) и затем инъецировали при скорости потока 10 мкл/мин для того, чтобы достичь величины связывания приблизительно вплоть до 10000 единиц ответа (RU). После инъекции иммобилизованного антитела инъецировали 1М этаноламин для блокирования непрореагировавших групп.Для измерения кинетики антитело против ТТ-Нс разбавляли до 0,5-1 мкг/мл и инъецировали при 10 мкл/мин, чтобы гарантировать то, что приблизительно 50 RU антитела захватывается антителом против мышиного Fc. TT-Hc-His затем подвергали сериям градиентов концентрации (например, 6,17 нм, 18,5 нм, 55,6 нм, 167 нм и 500 нм) и инъецировали при 25°С от более низкой к более высоким концентрациям при 30 мкл/мин со временем ассоциации 120 с и временем диссоциации 3600 с. 10 мМ раствор глицина-HCl (рН 2,0) инъецировали при 10 мкл/мин в течение 30 с для регенерации поверхности чипа. Скорость ассоциации (Ка) и скорость диссоциации (Kd) рассчитывали путем подгонки сенсограмм связывания и диссоциации с моделью связывания 1:1 с использованием версии 2.0.1 программного обеспечения для оценки Biacore Х100. Константу равновесия диссоциации (KD) рассчитывали как отношение Kd/Ka. Результаты подгонки представлены в Таблице 1.
Пример 4: Анализ эпитопов моноклональных антител против столбнячного токсина 96-луночные планшеты для ИФА покрывали рекомбинантный белком TT-Hc-His (3 мкг/мл, 100 мкл/лунку) в течение ночи в холодильнике при 4°С и затем блокировали блокирующим раствором (PBS-0,1% Tween20-3% молоко) при 37°С в течение 1 часа. Мышиные антитела S22E2-mIgG2a и S24B8-mIgG2a градиентно разбавляли очищенным фагом S22E2 (фаг S22E2) с фиксированной концентрацией (1*10п КОЕ/мл). Исходная концентрация антитела составляла 50 мкг/мл, и 3-кратное разбавление осуществляли в общей сложности для 11 градиентов концентрации. Разведения антитела добавляли в блокированные 96-луночные планшеты для ИФА в количестве 100 мкл/лунку и планшеты инкубировали при 37°С в течение 1 часа. Затем планшеты для ИФА промывали PBS-0,1% Tween20, а затем добавляли вторичное антитело анти-М13 с пероксидазой хрена (HRP) (Sino Biological Inc., 11973-ММ05Т-Н) и планшеты инкубировали при 37°С в течение 1 часа. Планшеты для ИФА промывали PBS-0,1% Tween20 и добавляли раствор для окрашивания с субстратом OPD. После инкубации в течение 5-10 минут окрашивание прекращали при помощи 1М H2SO4. Значения оптической плотности определяли с использованием микропланшетного ридера при двойной длине волны 492 нм/630 нм. Результаты анализа ИФА (Фиг. 1) продемонстрировали, что S22E2-mIgG2a полностью ингибировало сигнал связывания между фагом S22E2 и рекомбинантным белком ТТ-Нс, но S24B8-mIgG2a не оказывало никакого действия на связывание фага S22E2 с ТТ-Нс.
Аналогично, S22E2-mIgG2a и S24B8-mIgG2a подвергали градиентному разведению с очищенным фагом S24B8 (фаг S24B8) в фиксированной концентрации. Сигнал связывания между фагом S24B8 и ТТ-Нс обнаруживали при помощи вторичного антитела анти-М13 с HRP. Результаты ИФА (Фиг. 2) продемонстрировали, что S24B8-mIgG2a полностью ингибировало сигнал связывания между фагом S24B8 и рекомбинантным белком ТТ-Нс, но S22E2-mIgG2a оказывало минимальное действие на связывание фага S24B8 с ТТ-Нс.
Таким образом, моноклональные антитела против ТТ-Нс S22E2-mIgG2a и S24B8-mIgG2a связываются с разными эпитопами ТТ-Нс.
Пример 5: Ингибирование связывания ТТ-Нс с ганглиозидами при помощи моноклональных антител против столбнячного токсина
Столбнячный токсин может связываться со специфическими рецепторами на поверхностях нервных клеток через домен Нc. В настоящее время общепринятым механизмом является механизм двойного рецептора, включающий ганглиозидный рецептор и белковый рецептор. 96-луночные планшеты для ИФА покрывали ганглиозидами GT1b (100 мкл/лунку), которые разбавляли до 10 мкг/мл метанолом, и планшеты оставляли на ночь при комнатной температуре для обеспечения испарения метанола. Затем планшеты блокировали блокирующим раствором (PBS-0,1% Tween20-1% BSA (бычий сывороточный альбумин)) при 37°С в течение 1 часа. ТТ-Нс подвергали градиентному разведению. Исходная концентрация ТТ-Нс составляла 100 мкг/мл, и 3-кратное разведение проводили в общей сложности для 7 градиентов концентрации. Мышиные моноклональные антитела против ТТ-Нс разбавляли до 40 мкг/мл при помощи PBS и смешивали с разведениями ТТ-Нс в равных объемах, и затем смешанные растворы добавляли к блокированным 96-луночным планшетам для ИФА в количестве 100 мкл/лунку. Была установлена контрольная группа ТТ-Нс без антитела. Планшеты инкубировали при 37°С в течение 1 часа. Планшеты для ИФА промывали PBS-0,1% Tween20 и добавляли поликлональное антитело против ТТ с HRP (приготовленное в лаборатории) в количестве 100 мкл/лунку. Планшеты инкубировали при 37°С в течение 1 часа. Затем планшеты для ИФА промывали PBS-0,1% Tween20 и добавляли раствор для окрашивания с субстратом OPD. После инкубации в течение 5-10 минут окрашивание прекращали при помощи 1 М раствора H2SO4, и значения оптической плотности определяли с использованием микропланшетного ридера при двойной длине волны 492 нм/630 нм. Результаты анализа ИФА (Фиг. 3) показали, что по отношению к кривой связывания ТТ-Нс в качестве основы, кривая связывания значительно смещалась вправо после добавления S22E2-mIgG2a, указывая на то, что S22E2-mIgG2a эффективно ингибирует связывание ТТ-Нс с ганглиозидом GT1b. Напротив, S24B8-mIgG2a не ингибировало связывание ТТ-Нс с ганглиозидом, и вместо этого проявляло в некоторой степени стимулирующее действие.
Пример 6: Нейтрализующие активности моноклональных антител против столбнячного токсина
Нейтрализующую активность антитела определяли путем смешивания антитела и летальной дозы столбнячного токсина in vitro и затем инъекции данной смеси мышам Balb/c. В частности, столбнячный токсин смешивали с моноклональным антителом против столбнячного токсина или антитоксином и эту смесь оставляли при 37°С на 1 час, и затем инъецировали в мышцу правых задних лап мышей Balb/c. Инъецируемая доза столбнячного токсина составляла 20×LD50 (средняя летальная доза)/мышь. Инъецируемая доза имеющегося в продаже лошадиного столбнячного иммуноглобулина (F(ab')2) (меченого как ТАТ, Shanghai Serum Bio-technology Co., LTD) составляла 1 МЕ/мышь. Инъецируемая доза моноклонального антитела S22E2 или S24B8 составляла 10 мкг/мышь. Доза S24B8 и S22E2 в комбинации составляла 1,25 мкг/мышь для каждого антитела. Мышей Balb/c, которым вводили 20×LD50 столбнячного токсина, использовали в качестве контрольной группы. В общей сложности было 5 групп, в каждой из которых было по 8 мышей. Столбнячный токсин разбавляли до 800×LD50/мл буфером на основе борной кислоты (т.е. 20×LD50 на 50 мкл инъекции после смешивания с антителом в равных объемах). Лошадиный столбнячный иммуноглобулин разбавляли до 40 МЕ/мл буфером на основе борной кислоты (т.е. 1 ME на 50 мкл инъекции после смешивания со столбнячным токсином в равных объемах). Антитела S22E2 и S24B8 разбавляли до 0,4 мг/мл (или 0,1 мг/мл) буфером на основе борной кислоты (то есть для обработки S22E2 или S24B8, 10 мкг антитела на 50 мкл инъекции после смешивания со столбнячным токсином в равных объемах; и для обработки S24B8 и S22E2 в комбинации 1,25 мкг каждого антитела на 50 мкл инъекции после смешивания смеси двух антител (смешанных в равных объемах) со столбнячным токсином в равных объемах). Каждому животному инъецировали 50 мкл смеси столбнячного токсина и антитела. Экспериментальные результаты представлены на Фиг. 4. Все мыши Balb/c в контрольной группе умерли от столбняка в течение 3 суток после введения. В экспериментальных условиях моноклональное антитело S24B8 или S22E2 частично нейтрализовало токсин и продлило выживание мышей. Напротив, комбинация S24B8 и S22E2 полностью нейтрализовала токсин, и все мыши доживали до конца экспериментального периода, так же как и нейтрализующую активность обнаруживали для лошадиного столбнячного иммуноглобулина ТАТ.
Пример 7: Защита от комбинирования моноклональных антител против столбнячного токсина у мышей Balb/c, получивших летальное воздействие столбнячным токсином
Мышей Balb/c подвергали иммунизации перед воздействием иммуноглобулином, и затем мышей, которым вводили антитело, вводили летальную дозу столбнячного токсина для определения защитных эффектов антитела. Инъецируемая доза моноклональных антител S22E2 или S24B8 составляла 10 мкг/мышь. Инъецируемая доза S24B8 и S22E2 в комбинации составляла 1,25 мкг/мышь для каждого антитела. Мышей Balb/c, которым вводили 20×LD50 столбнячного токсина, использовали в качестве контрольной группы. Всего было 4 группы, каждая из которых включала 8 мышей.
Столбнячный токсин разбавляли до 400×LD50/мл буфером на основе борной кислоты (т.е. содержащим 20×LD50 на 50 мкл инъекции). Антитела S22E2 и S24B8 разбавляли до 0,2 мг/мл (или 0,05 мг/мл) буфером на основе борной кислоты (то есть для обработки S22E2 или S24B8, 10 мкг антитела на 50 мкл инъекции; и для обработки S24B8 и S22E2 в комбинации 1,25 мкг каждого антитела на 50 мкл инъекции после смешивания двух антител в равных объемах). 50 мкл антител против столбнячного токсина вводили мышам Balb/c, и через 24 часа мышам вводили столбнячный токсин в мышцы правых задних лап.Каждому животному инъецировали 50 мкл столбнячного токсина. Экспериментальные результаты представлены на Фиг. 5. Все мыши Balb/c в контрольной группе умерли от столбняка в течение 4 суток после введения. В данных экспериментальных условиях комбинация S24B8 и S22E2 полностью защищала мышей Balb/c, все из которых выживали к концу экспериментального периода.
Пример 8: Гуманизация и мутация сайта дезаминирования моноклональных антител против столбнячного токсина
8.1. Гуманизация мышиного моноклонального антитела S24B8 Мышиное моноклональное антитело S24B8 гуманизировали для снижения его иммуногенности. Схема гуманизации была адаптирована к классической рамочной стратегии трансплантации (см. Tan Р, Mitchell DA, Buss TN, Holmes MA, Anasetti C, Foote J. "Superhumanized" antibodies: reduction of immunogenic potential by complementarity-determining region grafting with human germline sequences: application to an anti-CD28. J Immunol. 2002;169(2):1119-1125, полное содержание которой включено в данное описание путем ссылки). Гены, кодирующие вариабельные области тяжелой и легкой цепей S24B8, соответственно сравнивали с терминальными генетическими последовательностями человеческого антитела в базе данных IMGT. Подходящие терминальные генетические последовательности отбирали для получения каркасной области 1-3 антитела (FR1+FR2+FR3), и подходящую генетическую последовательность области J отбирали для того, чтобы получить каркасную область 4 (FR4). Эта матрица может быть выбрана в зависимости от множества факторов, таких как относительная общая длина антитела, размеры CDR, аминокислотные остатки в точке соединения каркасной области (FR) и гипервариабельной области (CDR) антитела, и гомология всей последовательности. Выбранная матрица может представлять собой смесь множества последовательностей или общую матрицу для того, чтобы с масксимальной возможностью поддерживать подходящую конформацию родительской области, определяющей комплементарность (CDR). В то же время, для того чтобы избежать гетерогенности белка, возможно вызванной дезаминированием сайта NG в гипервариабельной области (CDR) антитела, вариабельную область тяжелой цепи гуманизированного антитела конструировали для мутаций. В итоге получили три гуманизированных мутанта вариабельной области тяжелой цепи S24B8VH-h1 (SEQ ID NO: 15), S24B8VH-h2 (SEQ ID NO: 16), S24B8VH-h3 (SEQ ID NO: 17) и один гуманизированный мутант вариабельной области легкой цепи S24B8VK-h1 (SEQ ID NO: 18).
8.2. Гуманизация мышиного моноклонального антитела S22E2 Гуманизация мышиного моноклонального антитела S22E2 адаптировала классическую рамочную стратегию трансплантации. Со ссылкой на Пример 8.1, пересадка CDR вариабельной области легкой и тяжелой цепей S22E2 привела к гуманизированному мутанту вариабельной области тяжелой цепи S22E2VH-h1 (SEQ ID NO: 19) и гуманизированному мутанту вариабельной области легкой цепи S22E2VK-h1 (SEQ ID NO: 20).
Гены вариабельной области антитела конструировали и синтезировали в соответствии с аминокислотными последовательностями гуманизированных антител S24B8 и S22E2, и клонировали в подходящие экспрессионные векторы для эукариотического гена. Интактные антитела получали в соответствии с Примером 1. Три гуманизированных варианта S24B8 получили название S24B8-h1, S24B8-h2 и S24B8-h3. Гуманизированный вариант S22E2 получил название S22E2-h1.
8.3. Анализ аффинности гуманизированных S24B8 и S22E2
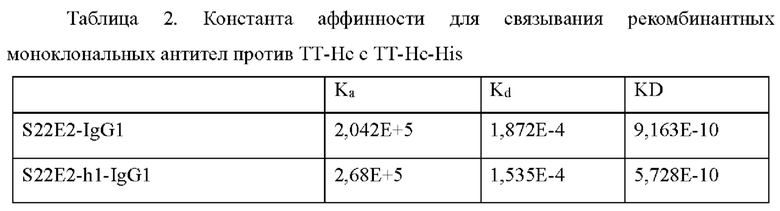
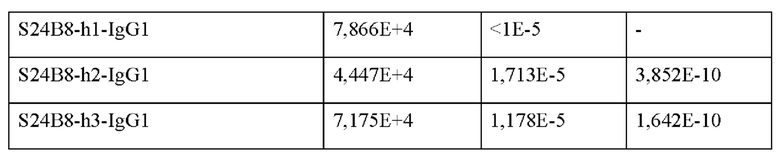
Со ссылкой на Пример 3.2, связывающую аффинность между гуманизированным антителом против ТТ-Нс и ТТ-Нс определяли при помощи Biacore XI00 с использованием набора для иммобилизации человеческого антитела (BR-1008-39). Результаты подгонки представлены в Таблице 2. Все константные области тяжелой цепи индивидуальных гуманизированных антител имели подтип человеческого IgG1.
Пример 9: Получение биспецифических антител
На основе превосходящей нейтрализующей активности комбинации моноклональных антител против столбнячного токсина S24B8 и S22E2 по сравнению с обработками отдельно каждым из них, антигенсвязывающие фрагменты, связывающиеся с разными эпитопами белка Не столбнячного токсина из S24B8-h3 и S22E2-h1, конструировали в виде Fab и scFv, соответственно, для конструирования гетеродимера человеческого IgG1, основанного на технологии KIH (выступ-во-впадину). То есть, происхоящий из S24B8-h3 антигенсвязывающий фрагмент в форме Fab подвергали слиянию с N-концом Fc-сегмента, содержащего мутацию Knob (FcK, SEQ ID NO: 40), a происходящий из S22E2-h1 антигенсвязывающий фрагмент в форме scFv подвергали слиянию с N-концом сегмента Fc, содержащего мутацию Hole (FcH, SEQ ID NO: 41), таким образом конструируя белок биспецифического антитела против столбнячного токсина (Нc).
Три сконструированных эукариотических экспрессионных вектора, экспрессирующих S22E2-h1-scFv-FcHl, S24B8VH-h3+CH-IgG1K и S24B8VK-h1+CK, соответственно, котрансфицировали в клетки HEK293F, используя липосомы, и клетки культивировали в суспензии в бессывороточной среде в течение 3-5 суток. Культуральный супернатант собирали путем центрифугирования. Биспецифические антитела в кулыуральном супернатанте очищали с использованием колонки для аффинной хроматографии Protein A/G (например, Mabselect SURE, GE Inc.). Затем буфер для консервирования рекомбинантного антитела заменяли на буфер PBS (рН 7,0) или другие подходящие буферы, используя колонку для обессоливания (например, Hitrap desaulting, GE Inc.). Обессоленный белковый раствор очищали при помощи гель-фильтрационной хроматографии (SEC), используя Superdex 200 (GE), тем самым получая биспецифическое антитело S22E2h1-scFv-FcH1+S24B8h3-IgG1K. При необходимости образцы антитела могут быть стерилизованы путем фильтрования и затем храниться в аликвотах при -20°С.
Пример 10: Функциональная валидация биспецифических антител
10.1. Ингибирование связывания ТТ-Нс с ганглиозидами
Со ссылкой на Пример 5, определяли ингибирующие способности антител против ТТ-Нс в отношении связывания ТТ-Нс с тремя ганглиозидами GD3, GT1b и GM1a. Результаты ИФА (Фиг. 6, 7 и 8) продемонстрировали, что моноклональное антитело S22E2-h1-scFv-FcHl и биспецифическое антитело S22E2h1-scFv-FcH1+S24B8h3-IgG1K обладали значительными ингибирующими действиями на связывание ТТ-Нс с этими тремя ганглиозидами, и моноклональное антитело S24B8-h3-IgG1 не влияло на связывание ТТ-Нс с этими тремя ганглиозидами.
10.2. Нейтрализующая активность
Со ссылкой на Пример 6, инъецируемая доза столбнячного токсина составляла 20×LD50/мышь. Инъецируемая доза моноклонального антитела S24B8-h3 или S22E2-h1, при их использовании по отдельности, составляла 10 мкг/мышь. Инъецируемая доза моноклонального антитела S24B8-h3 и S22E2-h1 в комбинации составляла 1,2 мкг/мышь для каждого антитела. Инъецируемая доза биспецифического антитела S22E2h1-scFv-FcH1+S24B8h3-IgG1K составляла 2,4 мкг/мышь. Мышей Balb/c, которым инъецировали 20×LD50 столбнячного токсина, использовали в качестве контрольной группы. В общей сложности было 5 групп с 8 мышами на группу. Столбнячный токсин разбавляли до 800×LD50/мл буфером на основе борной кислоты (т.е. 20×LD50 на 50 мкл инъекции после смешивания с антителом в равных объемах). Антитела S22E2 и S24B8 разбавляли до 0,4 мг/мл (или 0,096 мг/мл) буфером на основе борной кислоты (то есть, для обработки S22E2 или S24B8, 10 мкг антитела на 50 мкл инъекции после смешивания со столбнячным токсином в равных объемах; и для обработки S24B8 и S22E2 в комбинации 1,2 мкг каждого антитела на 50 мкл инъекции после смешивания смеси двух антител (смешанных в равных объемах) со столбнячным токсином в равных объемах). Биспецифическое антитело S22E2h1-scFv-FcH1+S24B8h3-IgG1K разбавляли до 0,096 мг/мл буфером на основе борной кислоты (т.е. 2,4 мкг биспецифического антитела на 50 мкл инъекции после смешивания со столбнячным токсином в равных объемах). Каждому животному инъецировали 50 мкл смеси столбнячного токсина и антитела. Экспериментальные результаты представлены на Фиг. 9. Все мыши Balb/c в контрольной группе умерли от столбняка в течение 3 суток после введения, а все мыши Balb/c в группе введения комбинации S24B8-h3 и S22E2-h1 и мыши Balb/c в группе введения биспецифического антитела S22E2h1-scFv-FcH1+S24B8h3-IgG1K выжили к концу экспериментального периода. Нейтрализующая активность биспецифического антитела S22E2h1-scFv-FcH1+S24B8h3-IgG1K превосходила нейтрализующую активность моноклональных антител.
Пример 11: Определение титров антитела
Титр биспецифического антитела S22E2h1-scFv-FcH1+S24B8h3-IgG1K определяли на основе способа определения нейтрализующих активностей антител в Примере 6. Количество международных единиц (ME) антитела против столбнячного токсина, содержащихся в 1 мг тестируемого образца, рассчитывали путем сравнения тестируемого образца и стандартного образца. Разбавление столбнячного токсина включало разбавление столбнячного токсина буфером на основе борной кислоты до 40 тестируемых доз (L+1/10) на миллилитр (т.е. 1 тестируемая доза (L+1/10) на 50 мкл инъекции после смешивания с антителом в равных объемах). Разведение стандарта человеческого столбнячного иммуноглобулина включало разведение стандарта человеческого столбнячного иммуноглобулина буфером на основе борной кислоты до 4 МЕ/мл (т.е. 1/10 ME на 50 мкл инъекции после смешивания с токсином в равных объемах). Биспецифическое антитело S22E2h1-scFv-FcH1+S24B8h3-IgG1K последовательно разбавляли буфером на основе борной кислоты до пяти концентраций с 15% интервалами, т.е. 14,40 мкг/мл, 16,80 мкг/мл, 20 мкг/мл, 23,6 мкг/мл и 27,6 мкг/мл (т.е. 0,36 мкг, 0,42 мкг, 0,50 мкг, 0,59 мкг и 0,69 мкг биспецифических антител на 50 мкл инъекции после смешивания с токсином в равных объемах). Разбавленные стандартные растворы человеческого столбнячного иммуноглобулина и растворы биспецифического антитела в различных разбавлениях соответственно смешивали с равным объемом разбавленного столбнячного токсина. Немедленно после инкубирования в течение 1 часа при 37°С смеси инъецировали мышам с внутримышечно инъецируемым объемом 50 мкл на мышь. За мышами наблюдали один раз в сутки для регистрации заболевания или смерти. Результаты продемонстрировали, что все мыши Balb/c в контрольной группе погибли от столбняка в течение 72 часов после введения, и самая высокая доза биспецифического антитела в группе, при которой мыши погибали одновременно с мышами в контрольной группе, составляла 0,42 мкг, рассчитанный титр для которой составил 238 МЕ/мг.
Хотя настоящее изобретение подробно описано со ссылкой на общее описание и конкретные примеры, специалистам в данной области техники понятно, что в настоящее изобретение могут быть внесены некоторые модификации и улучшения. Соответственно, такие модификации и улучшения, осуществленные без отклонения от сущности изобретения, попадают под объем формулы изобретения.
ССЫЛКИ
1. Thwaites CL, Beeching NJ, Newton CR (2015) Maternal and neonatal tetanus. Lancet 385:362-370.
2. Yousefi M, Tahmasebi F, Younesi V, Razavi A, Khoshnoodi J, Bayat AA, Abbasi E, Rabbani H, Jeddi-tehrani M, Shokri F (2014a) Characterization of neutralizing monoclonal antibodies directed against tetanus toxin fragment C. J hnmunotoxicol 11:28 34.
3. Ashton AC, Li Y, Doussau F, WellerU, Dougan G, Poulain B, Dolly JO (1995)tetanus toxin inhibits neuroexocytosis even when its Zn21-dependent protease activity is removed. J Biol Chem 270:31386 31390.
4. Scott N, Qazi O, Wright MJ, Fairweather NF, Deonarain MP (2010) Characterisation of a panel of anti-tetanus toxin single-chain Fvs reveals cooperative binding. Mol Immunol 47:1931 1941.
5. Yousefi M, Khosravi-Eghbal R, Reza Mahmoudi A, Jeddi-Tehrani M, Rabbani H, Shokri F (2014b) Comparative in vitro and in vivo assessment of toxin neutralization by antitetanus toxin monoclonal antibodies. Hum Vaccin Immunother 10:344-351.
6. Petrusic V, Zivkovic I, Stojanovic M, Stojicevic I, Marinkovic E,Dimitrijevic L (2012) Production, characterization and applications of a tetanus toxin specific monoclonal antibody T-62. Acta Histochem 114:480-486.
7. Lang AB, Cryz SJ Jr, Schurch U, Ganss MT, Bruderer U (1993immunotherapy with human monoclonal antibodies. Fragment A specificity of polyclonal and monoclonal antibodies is crucial for full protection against tetanus toxin. J Immunol 151:466 472.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Beijing Wisdomab Biotechnology Co., Ltd
GENRIX (Shanghai) Biopharmaceutical Co.Ltd.
Chongqing Genrix Biopharmaceutical Co., Ltd.
<120> АНТИТЕЛО ПРОТИВ СТОЛБНЯЧНОГО ТОКСИНА И ЕГО ПРИМЕНЕНИЕ
<150> CN 202011128926.0
<151> 2020-10-21
<160> 41
<170> SIPOSequenceListing 1.0
<210> 1
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 1
Ser Tyr Trp Ile Tyr
1 5
<210> 2
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 2
Glu Ile Asn Pro Thr Asn Gly Phe Ala Asn Tyr Asn Glu Lys Phe Lys
1 5 10 15
Thr
<210> 3
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 3
Glu Ile Asn Pro Thr Ala Gly Phe Ala Asn Tyr Asn Glu Lys Phe Lys
1 5 10 15
Thr
<210> 4
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 4
Glu Ile Asn Pro Thr Asn Ala Phe Ala Asn Tyr Asn Glu Lys Phe Lys
1 5 10 15
Thr
<210> 5
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 5
His Phe Arg Phe Pro Tyr
1 5
<210> 6
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 6
Arg Ala Ser Gln Asp Ile Gly Ser Ser Leu Thr
1 5 10
<210> 7
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 7
Ala Thr Ser Ser Leu Asp Ser
1 5
<210> 8
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 8
Leu Gln Tyr Ala Ser Ser Pro Tyr Thr
1 5
<210> 9
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 9
Asp Tyr Gly Val Asn
1 5
<210> 10
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 10
Met Ile Trp Ser Asp Gly Thr Thr Asp Tyr Ser Ser Ala Leu Lys Ser
1 5 10 15
<210> 11
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 11
Val Asp Gly Tyr Ser His Tyr Tyr Ala Met Asp Tyr
1 5 10
<210> 12
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 12
Arg Ala Ser Glu Asn Ile Tyr Ser Tyr Leu Ala
1 5 10
<210> 13
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 13
Asn Ala Lys Thr Leu Ala Glu
1 5
<210> 14
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 14
Gln His His Tyr Gly Leu Pro Phe Thr
1 5
<210> 15
<211> 115
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 15
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Ile Tyr Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn Pro Thr Asn Gly Phe Ala Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Thr Lys Ala Thr Ile Thr Val Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ile His Phe Arg Phe Pro Tyr Trp Gly Gln Gly Thr Met Val Thr
100 105 110
Val Ser Ser
115
<210> 16
<211> 115
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 16
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Ile Tyr Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn Pro Thr Ala Gly Phe Ala Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Thr Lys Ala Thr Ile Thr Val Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ile His Phe Arg Phe Pro Tyr Trp Gly Gln Gly Thr Met Val Thr
100 105 110
Val Ser Ser
115
<210> 17
<211> 115
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 17
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Ile Tyr Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn Pro Thr Asn Ala Phe Ala Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Thr Lys Ala Thr Ile Thr Val Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ile His Phe Arg Phe Pro Tyr Trp Gly Gln Gly Thr Met Val Thr
100 105 110
Val Ser Ser
115
<210> 18
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 18
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Gly Ser Ser
20 25 30
Leu Thr Trp Leu Gln Gln Lys Pro Gly Lys Ala Pro Lys Arg Leu Ile
35 40 45
Tyr Ala Thr Ser Ser Leu Asp Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Tyr Ala Ser Ser Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 19
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 19
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Asp Tyr
20 25 30
Gly Val Asn Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu Trp Leu
35 40 45
Gly Met Ile Trp Ser Asp Gly Thr Thr Asp Tyr Ser Ser Ala Leu Lys
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Ser Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Val Asp Gly Tyr Ser His Tyr Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 20
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 20
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Asn Ile Tyr Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Gln Gly Lys Ala Pro Gln Leu Leu Ile
35 40 45
Tyr Asn Ala Lys Thr Leu Ala Glu Gly Val Ser Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln His His Tyr Gly Leu Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 21
<211> 445
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 21
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Ile Tyr Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn Pro Thr Asn Gly Phe Ala Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Thr Lys Ala Thr Ile Thr Val Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ile His Phe Arg Phe Pro Tyr Trp Gly Gln Gly Thr Met Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val
130 135 140
Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly
165 170 175
Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly
180 185 190
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
195 200 205
Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys
340 345 350
Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 22
<211> 445
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 22
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Ile Tyr Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn Pro Thr Ala Gly Phe Ala Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Thr Lys Ala Thr Ile Thr Val Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ile His Phe Arg Phe Pro Tyr Trp Gly Gln Gly Thr Met Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val
130 135 140
Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly
165 170 175
Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly
180 185 190
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
195 200 205
Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys
340 345 350
Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 23
<211> 445
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 23
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Ile Tyr Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn Pro Thr Asn Ala Phe Ala Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Thr Lys Ala Thr Ile Thr Val Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ile His Phe Arg Phe Pro Tyr Trp Gly Gln Gly Thr Met Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val
130 135 140
Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly
165 170 175
Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly
180 185 190
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
195 200 205
Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys
340 345 350
Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 24
<211> 214
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 24
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Gly Ser Ser
20 25 30
Leu Thr Trp Leu Gln Gln Lys Pro Gly Lys Ala Pro Lys Arg Leu Ile
35 40 45
Tyr Ala Thr Ser Ser Leu Asp Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Tyr Ala Ser Ser Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 25
<211> 474
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 25
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Asp Tyr
20 25 30
Gly Val Asn Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu Trp Leu
35 40 45
Gly Met Ile Trp Ser Asp Gly Thr Thr Asp Tyr Ser Ser Ala Leu Lys
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Ser Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Val Asp Gly Tyr Ser His Tyr Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly
115 120 125
Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser Pro Ser
130 135 140
Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala
145 150 155 160
Ser Glu Asn Ile Tyr Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Gln Gly
165 170 175
Lys Ala Pro Gln Leu Leu Ile Tyr Asn Ala Lys Thr Leu Ala Glu Gly
180 185 190
Val Ser Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu
195 200 205
Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln
210 215 220
His His Tyr Gly Leu Pro Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu
225 230 235 240
Ile Lys Glu Pro Lys Ser Ser Asp Lys Thr His Thr Cys Pro Pro Cys
245 250 255
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
260 265 270
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
275 280 285
Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp
290 295 300
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
305 310 315 320
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
325 330 335
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
340 345 350
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
355 360 365
Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Gln Glu Glu
370 375 380
Met Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr
385 390 395 400
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
405 410 415
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
420 425 430
Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn
435 440 445
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
450 455 460
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
465 470
<210> 26
<211> 451
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 26
Lys Asn Leu Asp Ala Trp Val Asp Asn Glu Glu Asp Ile Asp Val Ile
1 5 10 15
Leu Lys Lys Ser Thr Ile Leu Asn Leu Asp Ile Asn Asn Asp Ile Ile
20 25 30
Ser Asp Ile Ser Gly Phe Asn Ser Ser Val Ile Thr Tyr Pro Asp Ala
35 40 45
Gln Leu Val Pro Gly Ile Asn Gly Lys Ala Ile His Leu Val Asn Asn
50 55 60
Glu Ser Ser Glu Val Ile Val His Lys Ala Met Asp Ile Glu Tyr Asn
65 70 75 80
Asp Met Phe Asn Asn Phe Thr Val Ser Phe Trp Leu Arg Val Pro Lys
85 90 95
Val Ser Ala Ser His Leu Glu Gln Tyr Gly Thr Asn Glu Tyr Ser Ile
100 105 110
Ile Ser Ser Met Lys Lys His Ser Leu Ser Ile Gly Ser Gly Trp Ser
115 120 125
Val Ser Leu Lys Gly Asn Asn Leu Ile Trp Thr Leu Lys Asp Ser Ala
130 135 140
Gly Glu Val Arg Gln Ile Thr Phe Arg Asp Leu Pro Asp Lys Phe Asn
145 150 155 160
Ala Tyr Leu Ala Asn Lys Trp Val Phe Ile Thr Ile Thr Asn Asp Arg
165 170 175
Leu Ser Ser Ala Asn Leu Tyr Ile Asn Gly Val Leu Met Gly Ser Ala
180 185 190
Glu Ile Thr Gly Leu Gly Ala Ile Arg Glu Asp Asn Asn Ile Thr Leu
195 200 205
Lys Leu Asp Arg Cys Asn Asn Asn Asn Gln Tyr Val Ser Ile Asp Lys
210 215 220
Phe Arg Ile Phe Cys Lys Ala Leu Asn Pro Lys Glu Ile Glu Lys Leu
225 230 235 240
Tyr Thr Ser Tyr Leu Ser Ile Thr Phe Leu Arg Asp Phe Trp Gly Asn
245 250 255
Pro Leu Arg Tyr Asp Thr Glu Tyr Tyr Leu Ile Pro Val Ala Ser Ser
260 265 270
Ser Lys Asp Val Gln Leu Lys Asn Ile Thr Asp Tyr Met Tyr Leu Thr
275 280 285
Asn Ala Pro Ser Tyr Thr Asn Gly Lys Leu Asn Ile Tyr Tyr Arg Arg
290 295 300
Leu Tyr Asn Gly Leu Lys Phe Ile Ile Lys Arg Tyr Thr Pro Asn Asn
305 310 315 320
Glu Ile Asp Ser Phe Val Lys Ser Gly Asp Phe Ile Lys Leu Tyr Val
325 330 335
Ser Tyr Asn Asn Asn Glu His Ile Val Gly Tyr Pro Lys Asp Gly Asn
340 345 350
Ala Phe Asn Asn Leu Asp Arg Ile Leu Arg Val Gly Tyr Asn Ala Pro
355 360 365
Gly Ile Pro Leu Tyr Lys Lys Met Glu Ala Val Lys Leu Arg Asp Leu
370 375 380
Lys Thr Tyr Ser Val Gln Leu Lys Leu Tyr Asp Asp Lys Asn Ala Ser
385 390 395 400
Leu Gly Leu Val Gly Thr His Asn Gly Gln Ile Gly Asn Asp Pro Asn
405 410 415
Arg Asp Ile Leu Ile Ala Ser Asn Trp Tyr Phe Asn His Leu Lys Asp
420 425 430
Lys Ile Leu Gly Cys Asp Trp Tyr Phe Val Pro Thr Asp Glu Gly Trp
435 440 445
Thr Asn Asp
450
<210> 27
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 27
His His His His His His
1 5
<210> 28
<211> 330
<212> PRT
<213> Homo sapiens
<400> 28
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 29
<211> 330
<212> PRT
<213> Mus musculus
<400> 29
Ala Lys Thr Thr Ala Pro Ser Val Tyr Pro Leu Ala Pro Val Cys Gly
1 5 10 15
Asp Thr Thr Gly Ser Ser Val Thr Leu Gly Cys Leu Val Lys Gly Tyr
20 25 30
Phe Pro Glu Pro Val Thr Leu Thr Trp Asn Ser Gly Ser Leu Ser Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Asp Leu Tyr Thr Leu
50 55 60
Ser Ser Ser Val Thr Val Thr Ser Ser Thr Trp Pro Ser Gln Ser Ile
65 70 75 80
Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys Lys
85 90 95
Ile Glu Pro Arg Gly Pro Thr Ile Lys Pro Cys Pro Pro Cys Lys Cys
100 105 110
Pro Ala Pro Asn Leu Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro
115 120 125
Lys Ile Lys Asp Val Leu Met Ile Ser Leu Ser Pro Ile Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Glu Asp Asp Pro Asp Val Gln Ile Ser Trp
145 150 155 160
Phe Val Asn Asn Val Glu Val His Thr Ala Gln Thr Gln Thr His Arg
165 170 175
Glu Asp Tyr Asn Ser Thr Leu Arg Val Val Ser Ala Leu Pro Ile Gln
180 185 190
His Gln Asp Trp Met Ser Gly Lys Glu Phe Lys Cys Lys Val Asn Asn
195 200 205
Lys Asp Leu Pro Ala Pro Ile Glu Arg Thr Ile Ser Lys Pro Lys Gly
210 215 220
Ser Val Arg Ala Pro Gln Val Tyr Val Leu Pro Pro Pro Glu Glu Glu
225 230 235 240
Met Thr Lys Lys Gln Val Thr Leu Thr Cys Met Val Thr Asp Phe Met
245 250 255
Pro Glu Asp Ile Tyr Val Glu Trp Thr Asn Asn Gly Lys Thr Glu Leu
260 265 270
Asn Tyr Lys Asn Thr Glu Pro Val Leu Asp Ser Asp Gly Ser Tyr Phe
275 280 285
Met Tyr Ser Lys Leu Arg Val Glu Lys Lys Asn Trp Val Glu Arg Asn
290 295 300
Ser Tyr Ser Cys Ser Val Val His Glu Gly Leu His Asn His His Thr
305 310 315 320
Thr Lys Ser Phe Ser Arg Thr Pro Gly Lys
325 330
<210> 30
<211> 330
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 30
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 31
<211> 330
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 31
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 32
<211> 330
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 32
Ala Lys Thr Thr Ala Pro Ser Val Tyr Pro Leu Ala Pro Val Cys Gly
1 5 10 15
Asp Thr Thr Gly Ser Ser Val Thr Leu Gly Cys Leu Val Lys Gly Tyr
20 25 30
Phe Pro Glu Pro Val Thr Leu Thr Trp Asn Ser Gly Ser Leu Ser Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Asp Leu Tyr Thr Leu
50 55 60
Ser Ser Ser Val Thr Val Thr Ser Ser Thr Trp Pro Ser Gln Ser Ile
65 70 75 80
Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys Lys
85 90 95
Ile Glu Pro Arg Gly Pro Thr Ile Lys Pro Cys Pro Pro Cys Lys Cys
100 105 110
Pro Ala Pro Asn Leu Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro
115 120 125
Lys Ile Lys Asp Val Leu Met Ile Ser Leu Ser Pro Ile Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Glu Asp Asp Pro Asp Val Gln Ile Ser Trp
145 150 155 160
Phe Val Asn Asn Val Glu Val His Thr Ala Gln Thr Gln Thr His Arg
165 170 175
Glu Asp Tyr Asn Ser Thr Leu Arg Val Val Ser Ala Leu Pro Ile Gln
180 185 190
His Gln Asp Trp Met Ser Gly Lys Glu Phe Lys Cys Lys Val Asn Asn
195 200 205
Lys Asp Leu Pro Ala Pro Ile Glu Arg Thr Ile Ser Lys Pro Lys Gly
210 215 220
Ser Val Arg Ala Pro Gln Val Cys Val Leu Pro Pro Pro Glu Glu Glu
225 230 235 240
Met Thr Lys Lys Gln Val Thr Leu Ser Cys Ala Val Thr Asp Phe Met
245 250 255
Pro Glu Asp Ile Tyr Val Glu Trp Thr Asn Asn Gly Lys Thr Glu Leu
260 265 270
Asn Tyr Lys Asn Thr Glu Pro Val Leu Asp Ser Asp Gly Ser Tyr Phe
275 280 285
Met Val Ser Lys Leu Arg Val Glu Lys Lys Asn Trp Val Glu Arg Asn
290 295 300
Ser Tyr Ser Cys Ser Val Val His Glu Gly Leu His Asn His His Thr
305 310 315 320
Thr Lys Ser Phe Ser Arg Thr Pro Gly Lys
325 330
<210> 33
<211> 330
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 33
Ala Lys Thr Thr Ala Pro Ser Val Tyr Pro Leu Ala Pro Val Cys Gly
1 5 10 15
Asp Thr Thr Gly Ser Ser Val Thr Leu Gly Cys Leu Val Lys Gly Tyr
20 25 30
Phe Pro Glu Pro Val Thr Leu Thr Trp Asn Ser Gly Ser Leu Ser Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Asp Leu Tyr Thr Leu
50 55 60
Ser Ser Ser Val Thr Val Thr Ser Ser Thr Trp Pro Ser Gln Ser Ile
65 70 75 80
Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys Lys
85 90 95
Ile Glu Pro Arg Gly Pro Thr Ile Lys Pro Cys Pro Pro Cys Lys Cys
100 105 110
Pro Ala Pro Asn Leu Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro
115 120 125
Lys Ile Lys Asp Val Leu Met Ile Ser Leu Ser Pro Ile Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Glu Asp Asp Pro Asp Val Gln Ile Ser Trp
145 150 155 160
Phe Val Asn Asn Val Glu Val His Thr Ala Gln Thr Gln Thr His Arg
165 170 175
Glu Asp Tyr Asn Ser Thr Leu Arg Val Val Ser Ala Leu Pro Ile Gln
180 185 190
His Gln Asp Trp Met Ser Gly Lys Glu Phe Lys Cys Lys Val Asn Asn
195 200 205
Lys Asp Leu Pro Ala Pro Ile Glu Arg Thr Ile Ser Lys Pro Lys Gly
210 215 220
Ser Val Arg Ala Pro Gln Val Tyr Val Leu Pro Pro Cys Glu Glu Glu
225 230 235 240
Met Thr Lys Lys Gln Val Thr Leu Trp Cys Met Val Thr Asp Phe Met
245 250 255
Pro Glu Asp Ile Tyr Val Glu Trp Thr Asn Asn Gly Lys Thr Glu Leu
260 265 270
Asn Tyr Lys Asn Thr Glu Pro Val Leu Asp Ser Asp Gly Ser Tyr Phe
275 280 285
Met Tyr Ser Lys Leu Arg Val Glu Lys Lys Asn Trp Val Glu Arg Asn
290 295 300
Ser Tyr Ser Cys Ser Val Val His Glu Gly Leu His Asn His His Thr
305 310 315 320
Thr Lys Ser Phe Ser Arg Thr Pro Gly Lys
325 330
<210> 34
<211> 107
<212> PRT
<213> Homo sapiens
<400> 34
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 35
<211> 106
<212> PRT
<213> Homo sapiens
<400> 35
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
35 40 45
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 36
<211> 107
<212> PRT
<213> Mus musculus
<400> 36
Arg Ala Asp Ala Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu
1 5 10 15
Gln Leu Thr Ser Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe
20 25 30
Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg
35 40 45
Gln Asn Gly Val Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Met Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu
65 70 75 80
Arg His Asn Ser Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser
85 90 95
Pro Ile Val Lys Ser Phe Asn Arg Asn Glu Cys
100 105
<210> 37
<211> 106
<212> PRT
<213> Mus musculus
<400> 37
Gly Gln Pro Lys Ser Ser Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Glu Thr Asn Lys Ala Thr Leu Val Cys Thr Ile Thr Asp
20 25 30
Phe Tyr Pro Gly Val Val Thr Val Asp Trp Lys Val Asp Gly Thr Pro
35 40 45
Val Thr Gln Gly Met Glu Thr Thr Gln Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Met Ala Ser Ser Tyr Leu Thr Leu Thr Ala Arg Ala Trp Glu
65 70 75 80
Arg His Ser Ser Tyr Ser Cys Gln Val Thr His Glu Gly His Thr Val
85 90 95
Glu Lys Ser Leu Ser Arg Ala Asp Cys Ser
100 105
<210> 38
<211> 19
<212> DNA
<213> Artificial Sequence
<220>
<223> Синтетический полинуклеотид
<400> 38
tgcatttgaa ctccttgcc
19
<210> 39
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Синтетический полинуклеотид
<400> 39
ccatcaatct tccacttgac
20
<210> 40
<211> 232
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 40
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
1 5 10 15
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
20 25 30
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
35 40 45
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
50 55 60
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
65 70 75 80
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
85 90 95
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
100 105 110
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
115 120 125
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met Thr
130 135 140
Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser
145 150 155 160
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
165 170 175
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
180 185 190
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
195 200 205
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
210 215 220
Ser Leu Ser Leu Ser Pro Gly Lys
225 230
<210> 41
<211> 232
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетический полипептид
<400> 41
Glu Pro Lys Ser Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
1 5 10 15
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
20 25 30
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
35 40 45
Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val
50 55 60
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
65 70 75 80
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
85 90 95
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
100 105 110
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
115 120 125
Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Gln Glu Glu Met Thr
130 135 140
Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser
145 150 155 160
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
165 170 175
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val
180 185 190
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe
195 200 205
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
210 215 220
Ser Leu Ser Leu Ser Pro Gly Lys
225 230
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО ПРОТИВ ВИРУСА БЕШЕНСТВА И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2764740C1 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО ПРОТИВ CD3E/BCMA И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2800164C2 |
| Антитела к TSLP человека и их применение | 2021 |
|
RU2825460C1 |
| АНТИ-TSLP АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2834227C1 |
| АНТИТЕЛО К CLDN-18.2 И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2829997C1 |
| АНТИТЕЛО К B7-H4, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2019 |
|
RU2792748C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ АНТИТЕЛА К PCSK-9, И ЕЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2782792C2 |
| Антитело против FXI/FXIA, его антигенсвязывающий фрагмент и фармацевтическое применение | 2021 |
|
RU2838408C1 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ АНТИТЕЛО К LAG-3, И ЕЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2771384C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ НА ОСНОВЕ АНТИТЕЛА К PD-L1 И ЕЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2766590C2 |
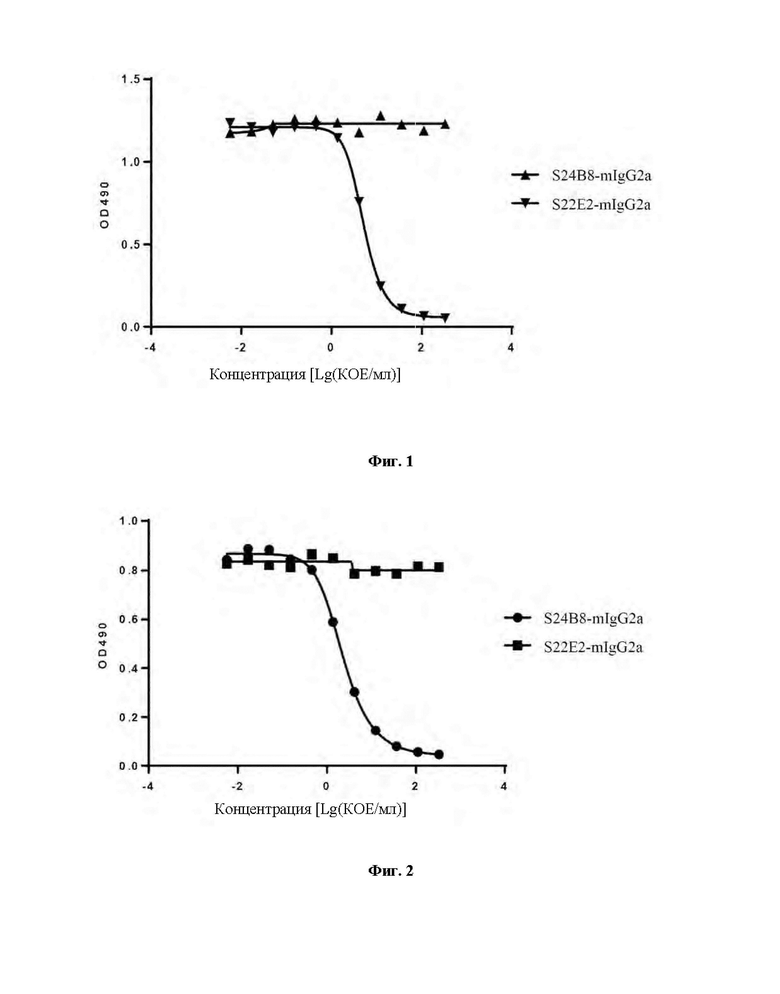
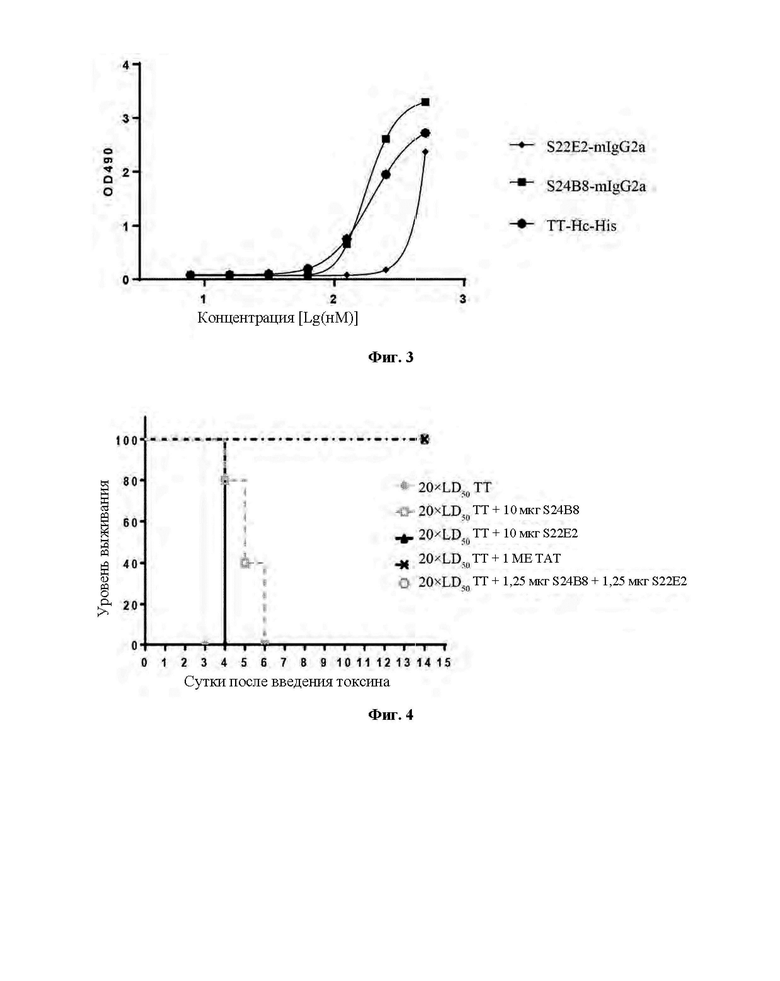
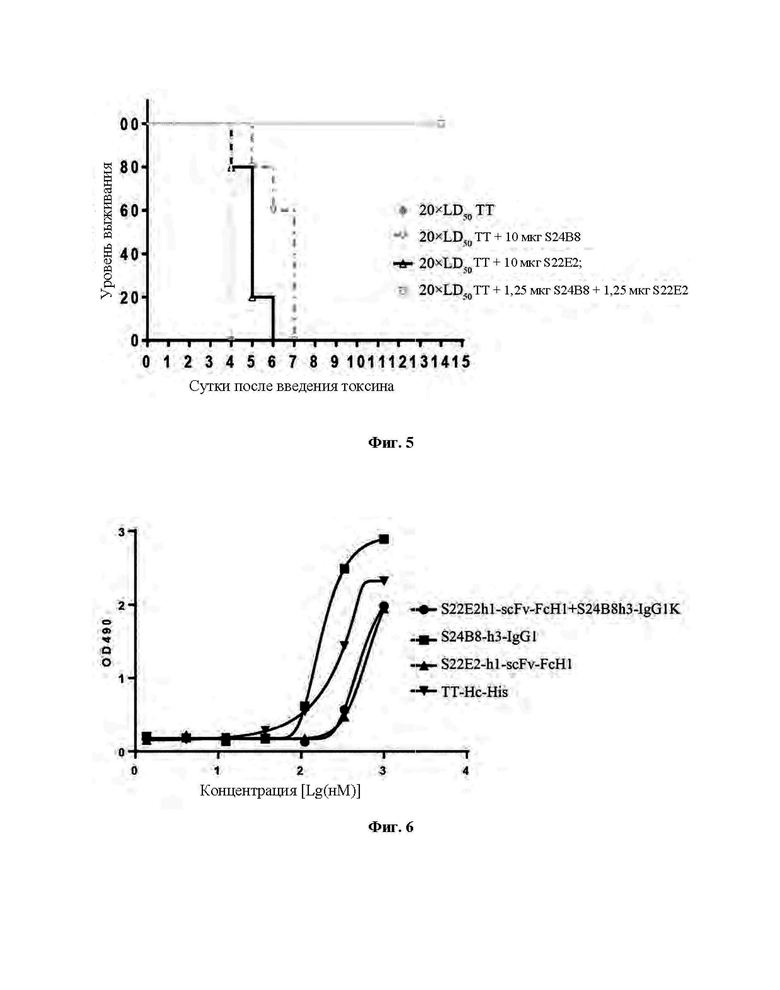
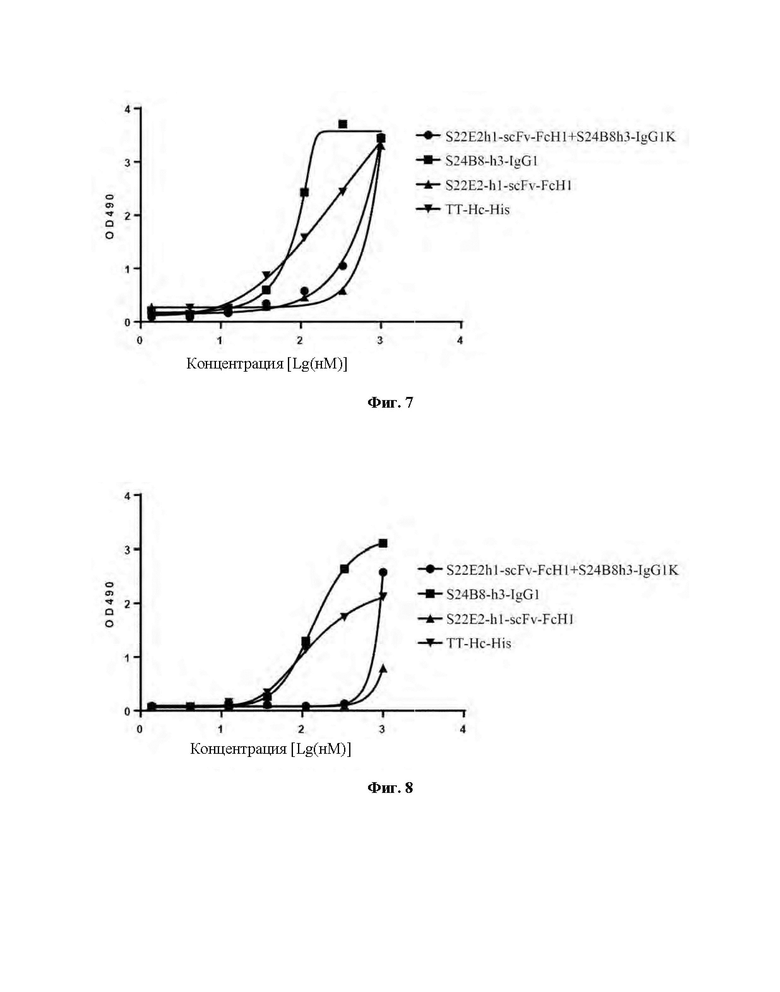
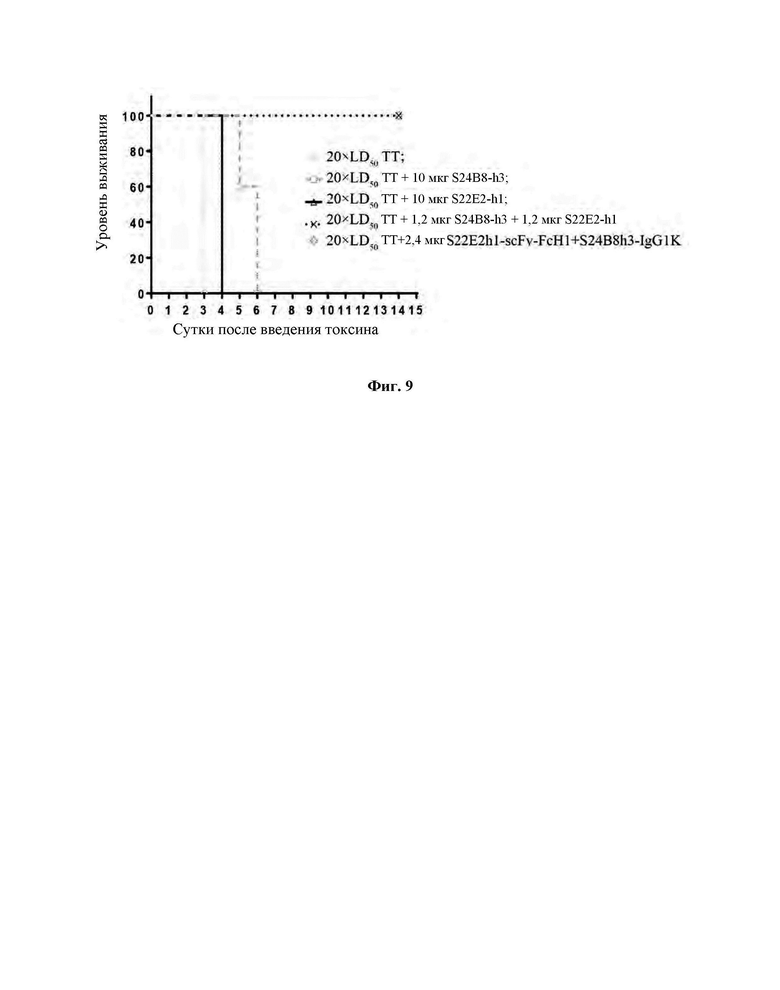
Изобретение относится к области биотехнологии. Описана группа изобретений, включающая моноклональное антитело, которое связывается со столбнячным токсином, применение моноклонального антитела в изготовлении лекарственного средства для предупреждения или лечения столбняка и способ предупреждения или лечения столбняка, включающий введение субъекту вышеуказанного моноклонального антитела. В одном из вариантов реализации моноклональное антитело содержит аминокислотную последовательность вариабельной области тяжелой цепи, выбранную из SEQ ID NO: 15-17, и аминокислотную последовательность вариабельной области легкой цепи SEQ ID NO: 18. Изобретение расширяет арсенал средств, связывающихся со столбнячным токсином. 3 н. и 1 з.п. ф-лы, 9 ил., 2 табл., 11 пр.
1. Моноклональное антитело, которое связывается со столбнячным токсином, содержащее:
HCDR1, имеющую аминокислотную последовательность SYWIY (SEQ ID NO: 1), HCDR2, имеющую аминокислотную последовательность EINPTNGFANYNEKFKT (SEQ ID NO: 2), или EINPTAGFANYNEKFKT (SEQ ID NO: 3), или EINPTNAFANYNEKFKT (SEQ ID NO: 4), HCDR3, имеющую аминокислотную последовательность HFRFPY (SEQ ID NO: 5), LCDR1, имеющую аминокислотную последовательность RASQDIGSSLT (SEQ ID NO: 6), LCDR2, имеющую аминокислотную последовательность ATSSLDS (SEQ ID NO: 7), и LCDR3, имеющую аминокислотную последовательность LQYASSPYT (SEQ ID NO: 8);
где аминокислотные последовательности HCDR и LCDR определены согласно Kabat.
2. Моноклональное антитело по п. 1, где указанное моноклональное антитело содержит:
аминокислотную последовательность вариабельной области тяжелой цепи, представленную в SEQ ID NO: 15, и аминокислотную последовательность вариабельной области легкой цепи, представленную в SEQ ID NO: 18; или
аминокислотную последовательность вариабельной области тяжелой цепи, представленную в SEQ ID NO: 16, и аминокислотную последовательность вариабельной области легкой цепи, представленную в SEQ ID NO: 18; или
аминокислотную последовательность вариабельной области тяжелой цепи, представленную в SEQ ID NO: 17, и аминокислотную последовательность вариабельной области легкой цепи, представленную в SEQ ID NO: 18.
3. Применение моноклонального антитела по п. 1 или 2 в изготовлении лекарственного средства для предупреждения или лечения столбняка.
4. Способ предупреждения или лечения столбняка, включающий введение субъекту, нуждающемуся в этом, моноклонального антитела по п. 1 или 2.
| WO 2019128121 A1, 04.07.2019 | |||
| WO 2019129214 A1, 04.07.2019 | |||
| CN 105153305 B, 01.03.2019 | |||
| Буркин М | |||
| А., Свиридов В | |||
| В., Перелыгина О | |||
| В | |||
| Иммуноферментное определение столбнячного токсина и анатоксина с использованием моноклональных антител // Прикладная биохимия и микробиология | |||
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| Двухколесный автомобиль для формовки кирпичей из разлитой по полю сушки торфяной массы | 1923 |
|
SU478A1 |
