Область техники, к которой относится изобретение
Настоящее изобретение относится к области органической химии, биохимии и молекулярной биологии, а именно, к способу получения комбинаторных модифицированных одноцепочечных ДНК-библиотек, предназначенных для специфического связывания с молекулярными мишенями. Сущность изобретения заключается в том, что для получения комбинаторной модифицированной одноцепочечной ДНК-библиотеки используется комбинаторная одноцепочечная ДНК-библиотека из природных нуклеотидов и комплементарная модифицированная ДНК синтезируется в ходе матрично-зависимой высокоспецифичной ферментативной реакции удлинения праймера (primer extension) при полной замене одного из природных нуклеотидов на модифицированный аналог, и последующем отщеплении синтезированных модифицированных цепей. Иммобилизованные праймеры содержат в своей структуре специальную группу, которая расщепляется фотохимическим или ферментативным методами. Предлагаемое изобретение может быть использовано для получения модифицированных фрагментов ДНК, модифицированных аптамеров, функциональных аналогов моноклональных антител.
Уровень техники, к которой относится изобретение
Наиболее перспективными областями применения изобретения является получение модифицированных аптамеров, которые по назначению могут использоваться для анализа белковых, пептидных и низкомолекулярных биологически-активных молекулярных мишеней, в том числе в датчиках и сенсорных устройствах, а также в качестве терапевтических средств. Модифицированные аптамеры, в отличие от антител, устойчивы к протеазам, что открывает новые возможности их применения, недостижимые для антител.
Известно, что для получения модифицированных фрагментов ДНК с функциональными группами на нуклеиновых основаниях применяют твердофазный химический метод синтеза с использованием фосфоамидитов модифицированных нуклеозидов. Недостатками такого метода является: сложность получения фосфоамидитов нуклеозидов с необходимыми модификациями по нуклеиновым основаниям; сложность твердофазного химического синтеза ДНК с использованием фосфоамидитов, несущих модифицирующие группы на нуклеиновых основаниях, условия химического синтеза достаточно жесткие и модифицирующие группы могут разрушаться; твердофазный фосфоамидитный метод синтеза имеет ограничения по длине синтезируемых молекул ДНК, с каждым последующим циклом наращивания цепи олигонуклеотида уменьшается его выход.
Авторы патента (Phosphinoamidite carboxylates and analogs thereof in the synthesis of oligonucleotide having reduced internucleotide charge, US patent, 8846898, - 2014, Lievre Cornu LLC, США) предлагают синтезировать модифицированные фрагменты ДНК с использованием модифицированных фосфоамидитов нуклеозидов, содержащих различные функциональные группы.
Известны методы получения одноцепочечных фрагментов немодифицированных ДНК с использованием твердофазной амплификации. В частности, известен ряд способов проведения твердофазной ПЦР, например, с использованием одного иммобилизованного праймера и другого праймера в растворе (Solid-phase nucleic acid amplification method and reagent kit therefor, US patent, 6274351, - 2001, GENSET SA, США; Single molecule nucleic acid sequence analysis processes and compositions, US patent, 9034580, - 2015, Sequenom INC, США; Multiplex branched-chain DNA assays, US patent, 8986931, - 2015, Asymetrix INC, США). В результате амплификации может быть получен ДНК-фрагмент, одна цепь которого связана с твердым носителем, тогда как другая находится в растворе. Метод не распространяется на получение ДНК, имеющих модификации по нуклеиновым основаниям.
Авторами патента (Method of producing heterogeneous set of single-chain DNA fragments for multiplex genetic analysis, Ru patent, 2604198, - 2016, Fed Gosudarstvennoe Bjudzhetnoe Uchrezhdenie Nauki Inst Molekuljarnoj Biologii Im V A Engelgardta Ro, Россия) предложен способ получения одноцепочечных фрагментов немодифицированных ДНК методом твердофазной ПЦР на агарозных микрочастицах с последующим фотохимическим или ферментативным отщеплением от носителя. Метод не распространяется на получение ДНК, имеющих модификации по нуклеиновым основаниям.
Заявляемое изобретение отличается от вышеописанных аналогов тем, что для получения модифицированных одноцепочечных молекул ДНК используется ДНК матрица из природных нуклеотидов, включающая вырожденную (комбинаторную) область, и комплементарная модифицированная ДНК синтезируется в ходе матрично-зависимой высокоспецифичной ферментативной реакции удлинения праймера (primer extension) при полной замене одного из природных нуклеотидов на модифицированный аналог. Для реакции удлинения праймера предлагаются Deep Vent(exo-) и KOD XL ДНК-полимеразы и модифицированные трифосфаты дезоксиуридина, которые полностью взаимосовместимы. При синтезе модифицированных ДНК используется праймер с фиксированной последовательностью, закрепленный на поверхности полиэтилентерефталатных микрочастиц, что позволяет количественно отделить одноцепочечные модифицированные ДНК, которые ковалентно связаны с ПЭТ-микрочастицами, от используемой ДНК матрицы и компонентов реакции. В состав праймера входят группы, позволяющие селективно отщепить модифицированную ДНК от носителя, не затрагивая ее функциональную часть, фотохимическим или ферментативным методом, что позволяет получить модифицированные ДНК-библиотеки в чистом виде.
Раскрытие изобретения
При получении модифицированных одноцепочечных фрагментов ДНК с модифицированными нуклеотидами необходимо обеспечить получение полноразмерного продукта и эффективную очистку от использованных реагентов.
Матрично-зависимый ферментативный синтез нуклеиновых кислот позволяет синтезировать комплементарные копии ДНК из природных нуклеотидов. Методология твердофазного синтеза биополимеров с использованием расщепляемых линкеров позволяет сначала удалить все используемые реагенты, а затем отщепить целевой биополимер и выделить биополимеры в чистом виде.
Целью данного изобретения является совмещение возможностей матрично-зависимого ферментативного и твердофазного синтеза ДНК для получения библиотеки одноцепочечных фрагментов ДНК с полной заменой природного нуклеотида на модифицированный аналог. Неполная замена природного нуклеотида на модифицированный приведет к получению нуклеотидных последовательностей, содержащих со статистической вероятностью в каждой позиции природный нуклеотид и модифицированный аналог. В данном изобретении реализована реакция удлинения праймера, при этом праймер ковалентно связан 5`-концом с поверхностью полиэтилентерефталатных (ПЭТ) микрочастиц.
ПЭТ, представляющий собой продукт линейной поликонденсации терефталевой кислоты и этиленгликоля, содержит сложноэфирные группы, которые в результате щелочного гидролиза могут быть трансформированы в реакционно-способные карбоксильные группы. Карбоксильные группы могут быть использованы для ковалентного связывания с биомолекулами, имеющими первичные аминогруппы (Р.А. Мифтахов и др., Получение активных карбоксильных групп на поверхности полиэтилентерефталатной пленки и количественный анализ этих групп с помощью цифровой люминесцентной микроскопии, Биофизика. 2018, т. 63, №4, с. 661-668). В данной публикации предложен щелочной гидролиз поверхности ПЭТ-микрочастиц с образованием поверхностных карбоксильных групп. Для контроля удельной концентрации карбоксильных групп их активировали бис-(4-нитрофенил)карбонатом и связывали с аминосодержащим индодикарбоцианиновым красителем (Фиг. 1).
Праймер, используемый в заявляемом изобретении, является отщепляемым и состоит из функциональной части, специфичной к участку матричной ДНК, со свободным 3`-концом. Последовательность функциональной части праймера представлена ниже (Фиг. 2А-Б, Олиг 2):
5’-CTG TCA GCT CCA ТАС TGG TAG СС-3’
По 5`-концу праймера находится расщепляемая группа, линкерный олигонуклеотид (Фиг. 2А-Б, Олиг 1) с последовательностью (Т)5, и затем С6-аминолинкер с аминогруппой для ковалентной иммобилизации на ПЭТ носителе (Фиг. 2А-Б).
Метод отщепления праймера может быть фотохимическим или ферментативным. Для синтеза фоторасщепляемого линкера предлагается использовать фосфоамидит, содержащий 2-нитробензильную группу (Фиг. 2А). В качестве расщепляемой последовательности предлагается использовать участки с последовательностью рибонуклеотидов, которые расщепляются РНК-азами (Фиг. 2Б). Могут быть использованы участки ДНК, которые узнаются эндонуклеазами рестрикции.
Для контроля удельной концентрации праймеров, иммобилизованных на ПЭТ-микрочастицах, проводили их отщепление УФ-светом (Фиг. 3) и с помощью РНК-азы (Фиг. 4).
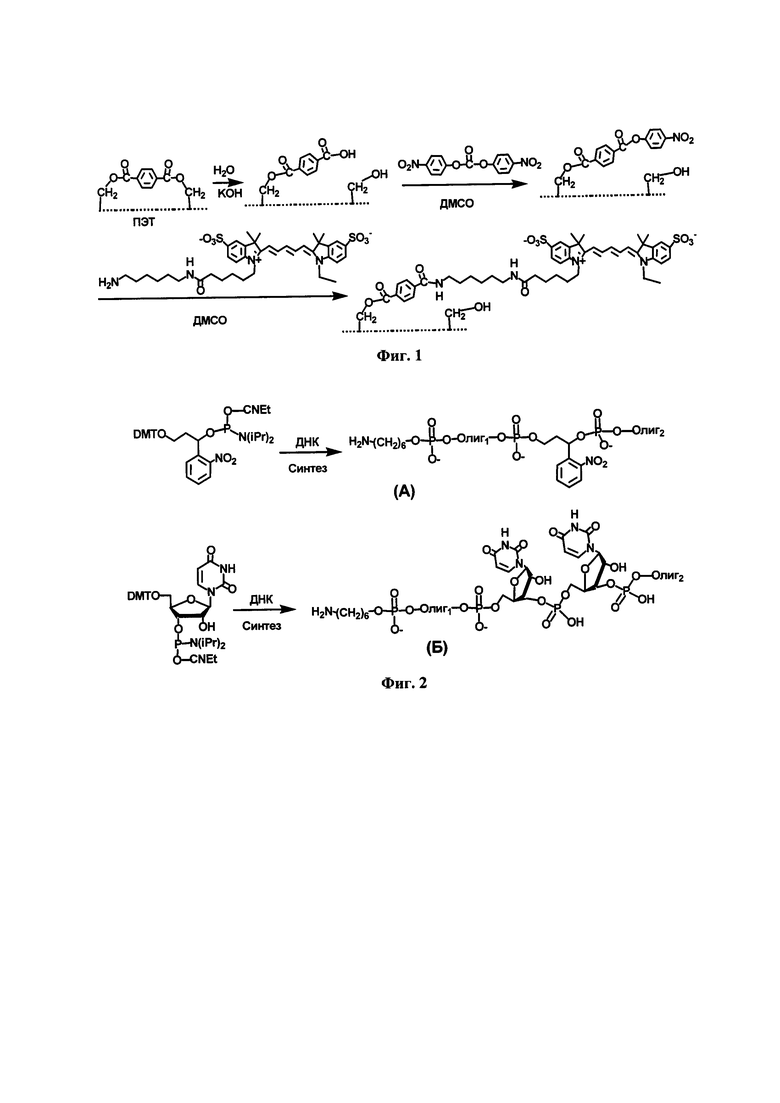
Модифицированные 5`-трифосфаты-2`-дезоксиуридина, связанные по С5-положению уридинового основания с фрагментами аминокислот фенилаланина (Фиг. 5, dUTP4-6), триптофана (Фиг. 5, dUTP1-3), валина и лейцина (Фиг. 5, dUTP7-8), и аланина (Фиг. 5, dUTP9), получены химическим путем. Соединения dUTP1dUTP6 и dUTP7-dUTP9 получены в соответствии с методикой, представленной в публикации (С.А. Лапа и др., Получение модифицированных комбинаторных ДНК-библиотек методом ПЦР в обратной эмульсии с последующим разделением цепей. Молекулярная биология, 2018, т. 52, №6, с. 984-996).
В заявляемом изобретении для получения модифицированных одноцепочечных молекул ДНК использовали синтетическую универсальную ДНК библиотеку с комплементарной к праймеру фланкирующей частью и вырожденной частью (N)40, где N смесь всех четырех нуклеотидов А, Т, G, С.
5’-(N)40-GG СТА CCA GTA TGG AGC TGA CAG-3`
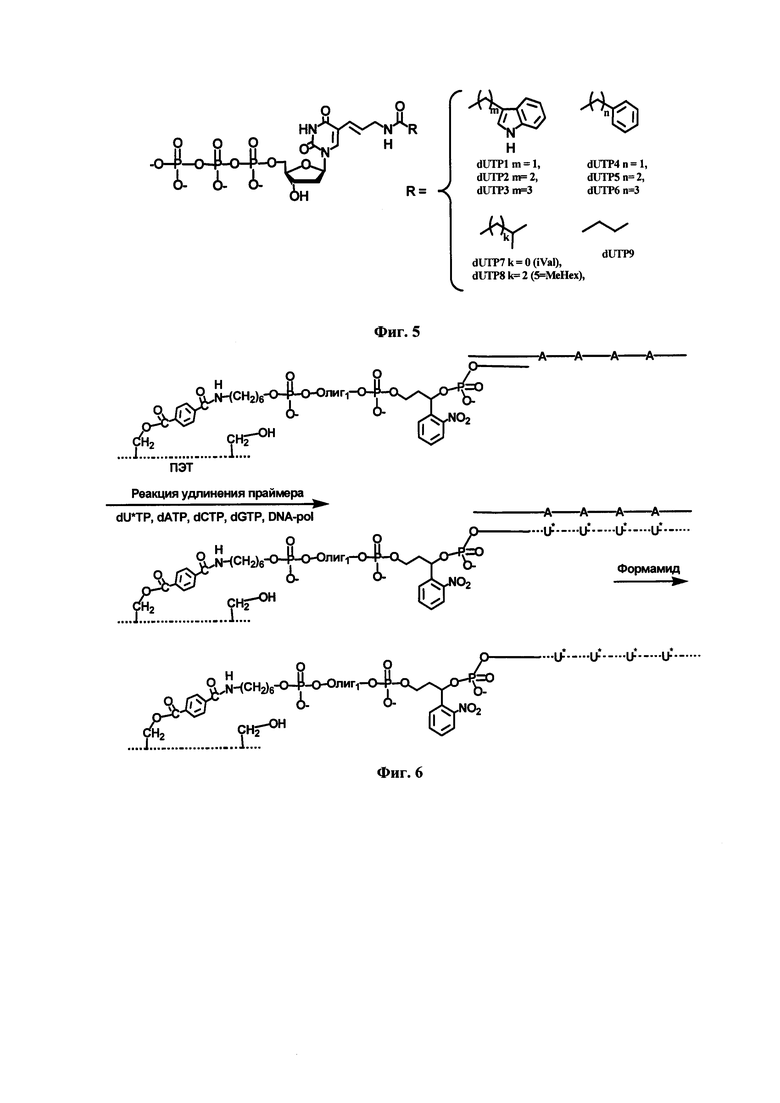
Комплементарные модифицированные ДНК получали в ходе матрично-зависимой высокоспецифичной ферментативной реакции удлинения праймера (primer extension, РЕХ) на твердой фазе при полной замене одного из природных нуклеотидов на соответствующий модифицированный. Для реакции удлинения праймера предлагаются матрично-зависимые Deep Vent(exo-) и KOD XL ДНК-полимеразы и модифицированные трифосфаты дезоксиуридина, которые полностью взаимосовместимы. При синтезе модифицированных ДНК используется праймер, закрепленный на поверхности твердых ПЭТ-микрочастиц, что позволяет отделить одноцепочечные модифицированные ДНК от используемой ДНК матрицы и компонентов реакции (Фиг. 6).
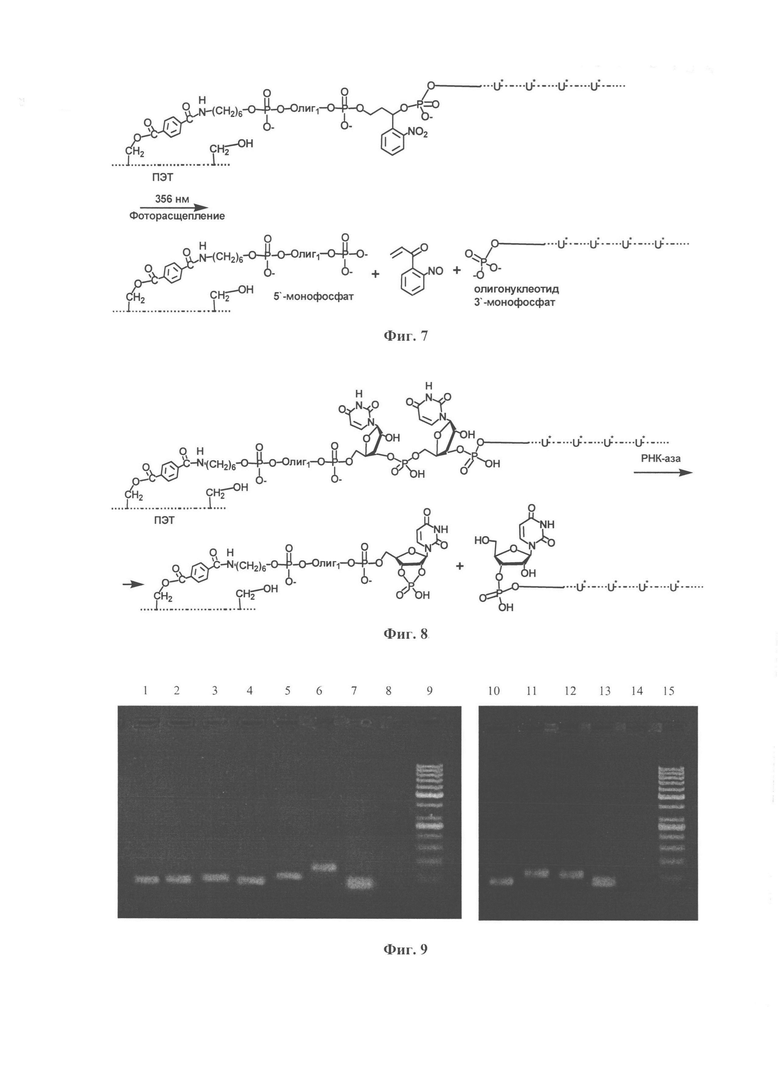
В состав праймеров входят группы, позволяющие селективно отщепить модифицированную ДНК от носителя, не затрагивая ее функциональную часть, фотохимическим (Фиг. 7) или ферментативным методом (Фиг. 8), что позволяет получить одноцепочечные модифицированные ДНК в чистом виде. Удельный выход модифицированного одноцепочечного продукта твердофазной реакции удлинения праймера определяли спектрофотометрически.
Контроль продуктов отщепления ДНК от ПЭТ-микрочастиц проводили методом электрофореза в 4%-ном агарозном геле (Фиг. 9). В результате получены полноразмерные модифицированная комбинаторные ДНК-библиотеки олигонуклеотидов с вырожденной частью.
Краткое описание схем и фигур
Фиг. 1. Схема химической модификации поверхности ПЭТ микрочастиц и связывания с аминосодержащим красителем.
Фиг. 2. Схема строения праймера фоторасщепляемого (А) и расщепляемого РНК-азой (Б).
Фиг. 3. Схема фоторасщепления праймеров, иммобилизованных на поверхности ПЭТ микрочастиц.
Фиг. 4. Схема расщепления РНК-азой А праймеров, иммобилизованых на поверхности ПЭТ микрочастиц.
Фиг. 5. Схема строения модифицированных по С5- положению 2`-дезоксиуридин-5`-трифосфатов.
Фиг. 6. Схема синтеза модифицированных одноцепочечных фрагментов ДНК методом твердофазной реакции удлинения праймера на ПЭТ микрочастицах. U*-модифицированный дезоксииридинфосфат, соответствующий дезоксиуридинтрифосфатам dUTP1-9.
Фиг. 7. Схема фотоотщепления модифицированого одноцепочечного продукта реакции удлинения праймера от ПЭТ микрочастиц. U*-модифицированный дезоксииридинфосфат, соответствующий дезоксиуридинтрифосфатам dUTP1-9.
Фиг. 8. Схема отщепления рРНК-азой А модифицированного одноцепочечного продукта реакции удлинения праймера от ПЭТ микрочастиц. U*-модифицированный дезоксииридинфосфат, соответствующий дезоксиуридинтрифосфатам dUTP1-9.
Фиг. 9. Анализ продуктов отщепления ДНК от ПЭТ-микрочастиц в 4%-ном агарозном геле, прокрашенных этидиум бромидом: 1 - dUTP1, 2 - dUTP2, 3 - dUTP3, 4 - dUTP4, 5 - dUTP5, 6 - dUTP6, 7 - dTTP природный - положительный контроль, 8 - отрицательный контроль, 9 - маркер длин двухцепочечных ДНК GeneRuler 50 bp (ThermoScientific), 10 - dUTP7, 11 - dUTP8, 12 - dUTP9, 13 - dTTP природный - положительный контроль, 14 - отрицательный контроль, 15 - маркер длин двухцепочечных ДНК GeneRuler 50 bp (ThermoScientific).
Осуществление изобретения
Для осуществления изобретения коммерчески доступный порошкообразный полиэтилентерефталат подготавливали специальным образом. С помощью сит высевали фракцию ПЭТ микрочастиц диаметром 40-63 мкм. Методом седиментации в воде отделяли мелкие пылевидные частицы. Микрочастицы ПЭТ промывали ацетоном с центрифугированием и высушивали в вакуум-эксикаторе. Получали мелкодисперсный полиэтилентерефталат с размером частиц 40-63 мкм, свободный от мелких пылевидных частиц.
Карбоксилированные ПЭТ-микрочастицы получали действием спиртового раствора гидроксида калия (Фиг. 1). При деструкции полиэтилентерефталата происходит разрыв сложноэфирных связей макромолекул с образованием на поверхности полимера карбоксильных и гидроксильных групп. Последующую активацию карбоксильных групп на поверхности ПЭТ-микрочастиц проводили в ДМСО при действии бис-(4-нитрофенил)карбоната в присутствии диизопропилэтиламина (DIPEA). Реакцию активации проводили в течение 16 ч при комнатной температуре.
Для определения концентрации 4-нитрофенилоксикарбонильных групп на поверхности ПЭТ-микрочастиц использовали флуоресцентное маркирование при помощи аминосодержащего индодикарбоцианинового красителя, флуоресцирующего в ближней ИК-области спектра. Реакцию конденсации активных 4-нитрофенилоксикарбонильных групп ПЭТ-микрочастиц и цианинового красителя осуществляли в ДМСО в присутствии DIPEA. Спектрофотометрически определяли количество красителя, связавшегося с карбоксильными группами ПЭТ-микрочастиц, путем определения концентрации раствора до и после проведения реакции. Удельная концентрация 4-нитрофенилоксикарбонильных групп составила 0.60 наномоль на 1 мг ПЭТ-микрочастиц.
Зная ориентировочную концентрацию активных групп на поверхности ПЭТ-микрочастиц, проводили иммобилизацию олигонуклеотидных праймеров с 5`-концевой аминогруппой. Реакцию проводили в смеси ДМСО / карбонатбикарбонатный буферный раствор при +4°С в течение 16 ч. Проведение реакции при охлаждении позволило сместить равновесие в сторону образования пептидной связи.
Определение концентрации олигонуклеотидных праймеров, иммобилизованных на поверхности ПЭТ-микрочастиц, осуществляли по принципу, представленному для определения концентрации индодикарбоцианинового красителя, измеряя поглощение раствора в воде на длине волны 260 нм. Для этого необходимо отщепить олигонуклеотидные праймеры с поверхности ПЭТ-микрочастиц. Метод отщепления праймера может быть фотохимическим (облучение УФ-светом) (Фиг. 3) или ферментативным (с помощью РНК-азы) (Фиг. 4), поскольку по 5`-концу праймера находится расщепляемая группа (Фиг. 2А-Б, Олиг 1) с последовательностью (Т)5, и затем С6-аминолинкер с аминогруппой для ковалентной иммобилизации на ПЭТ носителе (Фиг. 2А-Б). Удельная концентрация олигонуклеотидов составила 0.54 наномоль на 1 мг ПЭТ-микрочастиц.
В заявляемом изобретении для получения модифицированных одноцепочечных молекул ДНК использовали синтетическую универсальную ДНК библиотеку с комплементарной к праймеру фланкирующей частью и вырожденной частью (N)40, где N - смесь всех четырех нуклеотидов А, Т, G, С.
5’-(N)40-GG СТА CCA GTA TGG AGC TGA CAG-3`
Комплементарные модифицированные одноцепочечные фрагменты ДНК получали в ходе реакции удлинения праймера на ПЭТ-микрочастицах при полной замене одного из природных нуклеотидов на соответствующий модифицированный. Для проведения реакции удлинения праймера использованы матрично-зависимые Deep Vent(exo-) и KOD XL ДНК-полимеразы и модифицированные трифосфаты дезоксиуридина, которые содержат в своей структуре фрагменты аминокислот. При синтезе модифицированных ДНК используется праймер, закрепленный на поверхности ПЭТ-микрочастиц, что позволяет отделить одноцепочечные модифицированные ДНК от используемой ДНК матрицы и компонентов реакции (Фиг. 6).
Наличие в составе праймеров групп, позволяющих отщепить фотохимическим (Фиг. 7) или ферментативным (Фиг. 8) методами модифицированную ДНК от носителя, позволяет получить одноцепочечные модифицированные ДНК в чистом виде. Количество ДНК определяют спектрофотометрически, измеряя поглощение раствора в воде на длине волны 260 нм. Контроль продуктов отщепления ДНК от ПЭТ-микрочастиц проводили методом электрофореза (Фиг. 9), по результатам которого получены полноразмерные модифицированные комбинаторные ДНК-библиотеки олигонуклеотидов с вырожденной частью.
Примеры соединений и их применение
Использовали полиэтилентерефталат порошкообразный производства Goodfellow
Cambridge Ltd, Enghland марки ES306030, каталожный номер 264-007-76 (www.goodfellow.com). Материал натурального цвета с плотностью 1.39 г/см3, полидисперсный с максимальным размером микрочастиц 300 микрон.
Аминосодержащий индодикарбоцианиновый краситель синтезировали по схеме, представленной в публикации (Р.А. Мифтахов и др., Получение активных карбоксильных групп на поверхности полиэтилентерефталатной пленки и количественный анализ этих групп с помощью цифровой люминесцентной микроскопии, Биофизика. 2018, т. 63, №4, с. 661-668).
Для синтеза фоторасщепляемого линкера используется фосфоамидит, содержащий 2-нитробензильную группу (Gene Link (www.genelink.com) PC Linker (photocleavable) Cattalog No 26-6888-10).
Модифицированные 5`-трифосфаты-2`-дезоксиуридина, связанные по С5-положению уридинового основания с фрагментами аминокислот фенилаланина, тирозина, триптофана, валина, лейцина и изолейцина, получены химическим путем (Фиг. 5).
Реакцию удлинения праймера (primer extension, РЕХ) проводили на амплификаторе DNA Engine Tetrad 2 ("Bio-Rad", США). Температурно-временной профиль реакции: 5 мин при 95°С, 30 сек при 60°С и 30 мин при 72°С.
Количество ДНК определяли на спектрофотометре NanoDrop 1000 ("Thermo Scientific", США).
Электрофоретический анализ продуктов реакции удлинения праймера осуществляли 4%-ном агарозном геле. Гели окрашивали красителем этидий бромидом.
В работе использованы полимеразы Deep Vent ("BioLabs", Великобритания) и KOD XL ("Novagen", США) с буферными растворами соответствующих производителей; трифосфаты дезоксинуклеозидов производства "Fermentas" (США);
Остальные реактивы и растворители марок "ОСЧ", "ХЧ" или "ЧДА" приобретены в фирмах Aldrich, Alfa Aesar, Fluka, Химмед. Воду для приготовления растворов получали на установке Milli-Q (EMD Millipore, США).
Олигонуклеотиды для реакции удлинения праймера РЕХ синтезировали на ДНК/РНК-синтезаторе ASM-800 ("Биоссет", Россия).
Пример 1. Подготовка ПЭТ-микрочастиц.
С помощью сит высевают фракцию ПЭТ-микрочастиц диаметром 40-63 мкм. Методом седиментации в воде отделяют мелкие пылевидные частицы. Микрочастицы ПЭТ промывают ацетоном с центрифугированием, а супернатант удаляют. Высушивают в вакуум-эксикаторе над P2O5. Получают мелкодисперсный полиэтилентерефталат с размером частиц 40-63 мкм, свободный от мелких пылевидных частиц.
Пример 2. Получение карбоксилированных ПЭТ-микрочастиц.
К навеске ПЭТ-микрочастиц (100 мг) добавляют раствор КОН в 96%-ном этиловом спирте (0.5 М, 1.5 мл) и перемешивают встряхиванием на шейкере (Вортекс) в течение 60 мин. Затем промывают водой, 0.05М HCl, водой. Высушивают и хранят в вакуум-эксикаторе над Р2О5.
Пример 3. Активация карбоксильных групп на поверхности ПЭТ-микрочастиц.
К карбоксилированным ПЭТ-микрочастицам (100 мг) добавляют 1.5 мл раствора бис-(4-нитрофенил)карбоната (30 нмоль/мкл) и диизопропилэтиламина (60 нмоль/мкл) в ДМСО. Смесь перемешивают встряхиванием на шейкере (Вортекс) при комнатной температуре в течение 16 ч. Осадок отделяют центрифугированием и промывают безводным ацетоном (3×1 мл) с центрифугированием. Высушивают и хранят в вакуум-эксикаторе над Р2О5.
Пример 4. Определение концентрации 4-нитрофенилоксикарбонильных групп на поверхности ПЭТ микрочастиц.
К навеске ПЭТ-микрочастиц с активными 4-нитрофенилоксикарбонильными группами (100 мг) добавляют 1 мл раствора аминосодержащего красителя (0.1 ммоль/мкл) и диизопропилэтиламина (0.25 ммоль/мкл) в ДМСО. Смесь перемешивают встряхиванием на шейкере (Вортекс) при комнатной температуре в течение 4 ч. Смесь центрифугируют, супернатант отделяют и осадок промывают 50%-ным ацетонитрилом в 50 мМ триэтиламмоний гидрокарбонатном буферном растворе (ТЕАН) (5×100 мкл) с перемешиванием на шейкере (Вортекс) и центрифугированием. Супернатант и промывные жидкости объединяют, и добавляют 50%-ный раствор ацетонитрила в 50 мМ ТЕАН до объема 1 мл. Спектрофотометрически определяют концентрацию красителя. Для этого 60 мкл раствора добавляют к 3 мл 50%-ного ацетонитрила в 50 мМ ТЕАН. Разбавление в 51 раз. Коэффициент молярной экстинкции аминосодержащего красителя на длине волны 647 нм равен 2.5*105 л*М-1*см-1. Измеряют оптическую плотность раствора на длине волны 647 нм. Измеряемая оптическая плотность при длине светового пути 1 см равна 0.196, что соответствует концентрации 0.196*51*1/2.5*105=3.998*10-5 М/л=0.3998*10-4 М/л=0.04 мМ.
При измерении исходной концентрации красителя в ДМСО отбирают пробу 30 мкл и добавляют к 3 мл 50%-ного ацетонитрила в 50 мм ТЕАН. Разбавление 101 раз. Коэффициент молярной экстинкции аминосодержащего красителя на длине волны 647 нм равен 2.5*105 л*М-1*см-1. Измеряемая оптическая плотность при длине светового пути 1 см, равна 0.248, что соответствует концентрации 0.248*101*1/2.5*105=10*10-5 М/л=1*10-4 М=0.1 мМ.
Количество красителя, связавшегося с ПЭТ-микрочастицами, вычисляют по разности красителя, взятого в реакцию и не связавшегося с ПЭТ-микрочастицами в объеме 1 мл (0.1-0.04)мМ/л*10-3 л=6*10-8 м=60 наномоль. Удельная концентрация 4-нитрофенилоксикарбонильных групп 60/100=0.60 наномоль на 1 мг ПЭТ-микрочастиц.
Пример 5. Иммобилизация олигонуклеотидных праймеров на активированных ПЭТ-микрочастицах.
К навеске ПЭТ-микрочастиц (20 мг) с 4-нитрофенилоксикарбонильными группами (0.6 наномоль/мг) добавляют 25 мкл раствора, содержащего 20 наномоль олигонуклеотидного праймера с 5`-концевой аминогруппой в 50%-ном ДМСО в 0.2 М NaHCO3/Na2CO3 (рН 9.5). Смесь перемешивают на шейкере при +4°С в течение 16 ч. ПЭТ-микрочастицы с иммобилизованными праймерами отделяют центрифугированием, промывают 50%-ным ДМСО в дистиллированной воде (3×150 мкл), водой (3×150 мкл), ацетоном (3×150 мкл) при температуре 25°С. Высушивают в вакуум-эксикаторе над Р2О5. Хранят при -18°С.
Пример 6. Определение концентрации олигонуклеотидных праймеров, иммобилизованных на поверхности ПЭТ микрочастиц.
Навеску ПЭТ-микрочастиц с иммобилизованными олигонуклеотидами (20 мг), содержащими в своем составе фоторасщепляемый линкер (0.6 наномоль/мг), суспендируют в 60 мкл воды milliQ. Смесь облучают 10 мин под УФ-лампами (четыре лампы 15W Sylvania F15T8/350BL, 356 nm). Затем добавляют 40 мкл формамида и нагревают при 95°С в течение 1 мин. ПЭТ-микрочастицы отделяют фильтрованием и промывают на фильтре 40%-ным формамидом (2×20 мкл). Фильтраты объединяют. ДНК выделяют из объединенных фильтратов осаждением смесью (1:2) этилового спирта и 2% перхлората лития в ацетоне.
При отщеплении с помощью РНК-азы используют праймер, содержащий в своем составе два уридиновых нуклеотида. Навеску ПЭТ-микрочастиц с иммобилизованными олигонуклеотидами (20 мг), содержащие в своем составе два уридиновых нуклеотида, (примерно 0.6 наномоль/мг) промывают водой и суспендируют в 30 мкл 0.1 М ацетата натрия. Расщепление проводят при 60°С в течение 1 ч в присутствии РНК-азы А. Затем добавляют 20 мкл формамида и нагревают при 95°С в течение 1 мин. ПЭТ-микрочастицы отделяют фильтрованием и промывают на фильтре 40%-ным формамидом (2×20 мкл). Фильтраты объединяют. ДНК выделяют из объединенных фильтратов осаждением смесью (1:2) этилового спирта и 2% перхлората лития в ацетоне.
Количество ДНК определяют спектрофотометрически, измеряя поглощение раствора в воде на длине волны 260 нм. Коэффициент экстинкции рассчитывают, исходя из длины и соотношения азотистых оснований. Выход около 10.8 наномоль, что составляет 90% от емкости ПЭТ-микрочастиц. Удельная концентрация олигонуклеотидов 0.54 наномоль на 1 мг ПЭТ-микрочастиц.
Пример 7. Получение модифицированных одноцепочечных фрагментов ДНК методом твердофазной реакции удлинения праймера на ПЭТ-микрочастицах.
Реакцию удлинения праймера (primer extension, РЕХ) проводят с использованием ПЭТ-микрочастиц с иммобилизованным праймером (20 мг, ~0,54 наномоль/мг) и 50 мкл смеси, содержащей 0.2 мМ каждого dCTP, dATP, dGTP и модифицированного dU*TP, 12 наномоль матричной ДНК, 1.5 мМ MgCl2, 5 ед. ДНК полимеразы (Deep Vent(exo-) или KOD XL) и 5% ДМСО в 1×ПЦР буфере (рН 8.5).
Проводят реакцию удлинения праймера (РЕХ) по следующей программе: 5 мин при 95°С, 30 сек при 60°С и 30 мин при 72°С. Охлаждают до комнатной температуры. Реакцию останавливают добавлением Na-EDTA (рН 8) в конечной концентрации 20 мМ. Затем добавляют 35 мкл формамида и нагревают при 95°С в течение 1 мин. ПЭТ-микрочастицы отделяют фильтрованием и промывают на фильтре 40%-ным формамидом, водой, ацетоном порциями по 150 мкл по 3 раза при температуре 25°С. Высушивают в вакуум-эксикаторе над Р2О5. Хранят при -18°С.
Пример 8. Отщепление модифицированного одноцепочечного продукта реакции удлинения праймера от ПЭТ-микрочастиц.
При УФ-отщеплении используют ДНК с праймером, содержащим в своем составе фоторасщепляемый линкер. Навеску 20 мг ПЭТ микрочастиц с одноцепочечными продуктами реакции удлинения праймера (20 мг, ~0.54 наномоль/мг) суспендируют в 60 мкл воды MilliQ. Смесь облучают 10 мин под УФ-лампами (четыре лампы 15W Sylvania F15T8/350BL, 356 nm). Затем добавляют 40 мкл формамида и нагревают при 95°С в течение 1 мин. ПЭТ-микрочастицы отделяют фильтрованием и промывают на фильтре 40%-ным формамидом (2×20 мкл). Фильтраты объединяют. ДНК выделяют из объединенных фильтратов осаждением смесью (1:2) этилового спирта и 2% перхлората лития в ацетоне.
При отщеплении с помощью РНК-азы используют ДНК с праймером, содержащим в своем составе два уридиновых нуклеотида. Навеску 20 мг ПЭТ-микрочастиц с одноцепочечными продуктами реакции удлинения праймера (20 мг, ~0.54 наномоль/мг) промывают водой и суспендируют в 30 мкл 0.1 М ацетата натрия. Расщепление проводят при 60°С в течение 1 ч в присутствии РНК-азы А. Затем добавляют 20 мкл формамида и нагревают при 95°С в течение 1 мин. ПЭТ-микрочастицы отделяют фильтрованием и промывают на фильтре 40%-ным формамидом (2×20 мкл). Фильтраты объединяют. ДНК выделяют из объединенных фильтратов осаждением смесью (1:2) этилового спирта и 2% перхлората лития в ацетоне.
Количество ДНК определяют спектрофотометрически, измеряя поглощение раствора в воде на длине волны 260 нм. Коэффициент экстинкции рассчитывается, исходя из длины и соотношения азотистых оснований. Длину продуктов и их чистоту (полноразмерность, индивидуальность) оценивают методом электрофореза в 4%-ном агарозном геле. Модифицированные по основаниям нуклеотиды замедляют скорость движения ДНК в сравнении с природной ДНК равной длины.
Выход ~9.7 наномоль. Удельный выход модифицированного одноцепочечного продукта твердофазной реакции удлинения праймера 0.49 наномоль на 1 мг ПЭТ-микрочастиц, что составляет 80% от емкости ПЭТ-микрочастиц.
Пример 9. Электрофоретический анализ продуктов удлинения праймера.
Контроль продуктов отщепления ДНК от ПЭТ-микрочастиц проводят методом электрофореза в 4%-ном агарозном геле. Перед проведением анализа ДНК осаждают 2% перхлоратом лития в ацетоне, супернатант удаляют, дважды промывают ацетоном. Осадок высушивают и растворяют в минимальном объеме 10 мМ фосфатного буфера (PBS), рН 7.2. Смесь исследуемого образца (10 мкл) и специального загрузочного буфера (3 мкл), содержащего электроподвижные красители для контроля прохождения электрофоретического разделения, наносят в лунки агарозного геля. Проводят электрофорез при 10 В/см в течение 30 мин. Гель окрашивают бромистым этидием. Результаты визуализируют с помощью ультрафилетового трансиллюминатора, регистрируют системой гель-документации, либо цифровым фотоаппаратом и сохраняют в виде JPEG-файлов на электронных носителях информации.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ определения субстратной эффективности производных трифосфатов дезоксиуридина для ДНК-полимераз методом реакции достраивания праймера | 2019 |
|
RU2731739C1 |
| Способ ферментативного получения модифицированных ДНК для создания реагентов, специфично связывающихся с гидрофобными участками высокомолекулярных органических соединений | 2017 |
|
RU2699522C2 |
| Способ термической диссоциации для проведения селекции ДНК-аптамеров | 2019 |
|
RU2723373C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ СУБСТРАТНОЙ СОВМЕСТИМОСТИ МОДИФИЦИРОВАННЫХ ТРИФОСФАТОВ ДЕЗОКСИУРИДИНА И ДНК-ПОЛИМЕРАЗ СОЧЕТАНИЕМ МЕТОДОВ КОЛИЧЕСТВЕННОЙ ПЦР И ЭЛЕКТРОФОРЕЗА | 2019 |
|
RU2736969C1 |
| СПОСОБ СОЗДАНИЯ И СЕЛЕКЦИИ БИБЛИОТЕКИ МОДИФИЦИРОВАННЫХ АПТАМЕРОВ | 2016 |
|
RU2652952C1 |
| Способ оценки влияния модифицированных трифосфатов дезоксинуклеозидов на олигонуклеотидный состав комбинаторных ДНК-библиотек для проведения селекции модифицированных аптамеров | 2021 |
|
RU2779058C1 |
| СПОСОБ ПОЛУЧЕНИЯ ГЕТЕРОГЕННОГО НАБОРА ОДНОЦЕПОЧЕЧНЫХ ФРАГМЕНТОВ ДНК ДЛЯ МУЛЬТИПЛЕКСНОГО ГЕНЕТИЧЕСКОГО АНАЛИЗА | 2015 |
|
RU2604198C1 |
| КОМПОЗИЦИИ И СПОСОБЫ ХИМИЧЕСКОГО РАСЩЕПЛЕНИЯ И СНЯТИЯ ЗАЩИТЫ ДЛЯ СВЯЗАННЫХ С ПОВЕРХНОСТЬЮ ОЛИГОНУКЛЕОТИДОВ | 2019 |
|
RU2766688C2 |
| Способ получения альфа-фетопротеина, иммобилизованного на ПЭТ-микрочастицах | 2018 |
|
RU2731415C2 |
| Флуоресцентно-меченые дезоксиуридинтрифосфаты | 2016 |
|
RU2637310C1 |

Настоящее изобретение относится к области органической химии, биохимии и молекулярной биологии. Описан способ получения комбинаторных ДНК-библиотек, содержащих модифицированные нуклеотиды, предназначенные для специфического связывания с молекулярными мишенями. Сущность изобретение заключается в том, что для получения комбинаторной модифицированной одноцепочечной ДНК-библиотеки используется комбинаторная одноцепочечная ДНК-библиотека из природных нуклеотидов и комплементарная модифицированная ДНК синтезируется в ходе матрично-зависимой высокоспецифичной ферментативной реакции удлинения праймера (primer extension) при полной замене одного из природных нуклеотидов на модифицированный аналог. Для реакции удлинения праймера предлагаются Deep Vent(exo-) и KOD XL ДНК-полимеразы и модифицированные трифосфаты дезоксиуридина с заместителями по С5-положению уридинового основания, которые полностью взаимосовместимы. При синтезе модифицированных ДНК используется один тип праймеров, закрепленных на поверхности полиэтилентерефталатных микрочастиц, что позволяет отделить и отмыть микрочастицы носителя с полученными закрепленными одноцепочечными модифицированными ДНК от используемой ДНК матрицы и компонентов реакции. В состав праймеров входят группы, позволяющие селективно отщепить модифицированную ДНК от носителя, не затрагивая ее функциональную часть фотохимическим или ферментативным методами, что позволяет получить комбинаторную модифицированную одноцепочечную ДНК-библиотеку в чистом виде. Кроме того, представлена соответствующая библиотека. Изобретение расширяет способы получения ДНК-библиотек. 2 н. и 15 з.п. ф-лы, 9 ил., 9 пр.
1. Библиотеки модифицированных одноцепочечных фрагментов ДНК предназначены для получения аптамеров, функциональных аналогов моноклональных антител, включающие праймерную часть из природных нуклеотидов и комбинаторную часть, в которой природный дезокситимидин полностью заменен на модифицированный дезоксиуридин.
2. Способ получения библиотеки модифицированных одноцепочечных фрагментов ДНК, включающий проведение щелочного гидролиза поверхности ПЭТ-микрочастиц с образованием поверхностных карбоксильных групп, затем связывание поверхностных активированных карбоксильных групп с праймером, содержащим аминогруппу, фоторасщепляемую или ферментативно расщепляемую группу и функциональную часть, с образованием иммобилизованных на ПЭТ-микрочастицах праймеров, при этом не менее 0.5 наномоль праймеров иммобилизуется на 1 мг сухих микрочастиц полиэтилентерефталата, затем проведение реакции удлинения праймера (primer extension, РЕХ) с синтетической универсальной ДНК библиотекой с комплементарной к праймеру фланкирующей частью и вырожденной частью, с полной заменой природного дезокситимидинтрифосфата на модифицированный дезоксиуридинтрифосфат, с ДНК-полимеразой, затем отщепление синтезированной библиотеки модифицированных одноцепочечных фрагментов ДНК от ПЭТ-микрочастиц.
3. Способ по п. 2, отличающийся тем, что используются ПЭТ-микрочастицы размером 40-63 мкм, свободные от мелких пылевидных частиц.
4. Способ по п. 2, отличающийся тем, что для иммобилизации на ПЭТ-микрочастицах с последующим фотоотщеплением модифицированных одноцепочечных фрагментов ДНК используется праймер, содержащий аминогруппу, фоторасщепляемую группу и функциональную часть:
5'-NH2-(CH2)6-O-PO2-(T)6-PO2-O-(CH2)2-CH(2-NO2-C6H4)-O-PO2-CGTCGTTCA TCAATGCAACT-3'
5. Способ по п. 2, отличающийся тем, что для иммобилизации на ПЭТ-микрочастицах с последующим ферментативным отщеплением модифицированных одноцепочечных фрагментов ДНК используется праймер, содержащий аминогруппу, ферментативно расщепляемую группу и функциональную часть:
5'-NH2-(CH2)6-O-PO2-(T)6-PO2-(U)2-O-PO2-CGTCGTTCA TCAATGCAACT-3'
6. Способ по п. 2, отличающийся тем, что в качестве матрицы в реакции удлинения праймера (primer extension, РЕХ) используется синтетическая универсальная ДНК библиотека с комплементарной к праймеру фланкирующей частью 20 нуклеотидов и вырожденной частью (N)40, где N - смесь всех четырех нуклеотидов А, Т, G, С:
5'- AGT TGC ATT GAT GAC AGA CG -(N)40-3'
7. Способ по п. 2, отличающийся тем, что в качестве модифицированных дезоксиуридинтрифосфатов в реакции удлинения праймера (primer extension, РЕХ) используют 2'-дезоксиуридинтрифосфаты, модифицированные по С5-положению фрагментами аминокислот, а также линкер, связывающий их с С5-положением пиримидинового основания.
8. Способ по п. 2, отличающийся тем, что для модификации 2'-дезоксиуридинтрифосфата использован фрагмент аминокислоты фенилаланин.
9. Способ по п. 2, отличающийся тем, что для модификации 2-дезоксиуридинтрифосфата использован фрагмент аминокислоты триптофан.
10. Способ по п. 2, отличающийся тем, что для модификации 2'-дезоксиуридинтрифосфата использован фрагмент аминокислоты валин.
11. Способ по п. 2, отличающийся тем, что для модификации 2'-дезоксиуридинтрифосфата использован фрагмент аминокислоты лейцин.
12. Способ по п. 2, отличающийся тем, что для модификации 2'-дезоксиуридинтрифосфата использован фрагмент аминокислоты аланин.
13. Способ по п. 2, отличающийся тем, что использованы ДНК-полимеразы Deep Vent(exo-) или KOD XL.
14. Способ по п. 2, отличающийся тем, что для отщепления синтезированной библиотеки модифицированных одноцепочечных фрагментов ДНК от ПЭТ-микрочастиц используется ферментативный метод.
15. Способ по п. 2, отличающийся тем, что для ферментативного отщепления синтезированной библиотеки модифицированных одноцепочечных фрагментов ДНК от ПЭТ-микрочастиц используется фермент РНК-аза.
16. Способ по п. 2, отличающийся тем, что для отщепления синтезированной библиотеки модифицированных одноцепочечных фрагментов ДНК от ПЭТ-микрочастиц используется фотохимический метод.
17. Способ по п. 2, отличающийся тем, что для фотохимического отщепления синтезированной библиотеки модифицированных одноцепочечных фрагментов ДНК от ПЭТ-микрочастиц используется УФ-свет с длиной волны излучения с максимумом 356 нм, с диапазоном 350-400 нм.
| RU 2017141790 A, 30.11.2017 | |||
| СПОСОБ ПОЛУЧЕНИЯ ГЕТЕРОГЕННОГО НАБОРА ОДНОЦЕПОЧЕЧНЫХ ФРАГМЕНТОВ ДНК ДЛЯ МУЛЬТИПЛЕКСНОГО ГЕНЕТИЧЕСКОГО АНАЛИЗА | 2015 |
|
RU2604198C1 |
| ВОГСО.ЭСНАЯ jПАт:н;1й--;.;:;;:г:г^гц?|БЧБЛИОГЕКА | 0 |
|
SU285058A1 |

