Область техники, к которой относится изобретение
Изобретение относится к молекулярной биологии и обеспечивает определение субстратной пригодности модифицированных трифосфатов дезоксинуклеозидов в применении к селекции модифицированных аптамеров.
Уровень техники
Аптамеры - нуклеиновые аналоги моноклональных антител, обладающие рядом важных преимуществ. В настоящий момент аптамеры применяются как самостоятельные диагностические инструменты для выявления заболеваний различной этиологии, так и в качестве средств терапии. Для увеличения сродства аптамеров к их мишеням в последовательность аптамеров вводятся модификации. Отбор модифицированных аптамеров к белковым мишеням (mod-SELEX) - сложный и трудоемкий процесс (Rohloff J.C., Gelinas A.D., Jarvis Т.С., Ochsner U.A., Schneider D.J., Gold L., Janjic N. (2014) Nucleic Acid Ligands With Protein-like Side Chains: Modified Aptamers and Their Use as Diagnostic and Therapeutic Agents. Mol. Ther. Nucleic Acids. 3, e201; Лапа C.A., Павлов A.C., Кузнецова B.E., Шершов В.Е., Спицын М.А., Гусейнов Т.О., Радько С.Л., Заседателев А.С., Лисица А.В., Чудинов А.В. (2019) Ферментативное получение модифицированных ДНК: изучение кинетики ПЦР в режиме реального времени. Молекуляр. биология. 53, 513-523). Из-за низкой совместимости модифицированных дезоксинуклеозидтрифосфатов часто либо ингибируется ПЦР, либо происходит конкурентная амплификация с обогащением исходной ДНК-библиотеки не аптамерами с высоким сродством к мишени, а легко амплифицируемыми последовательностями (конкурентная амплификация) (Tolle F., Wilke J., Wengel J., Mayer G. (2014) By-product formation in repetitive PCR amplification of DNA libraries during SELEX. PLoS One. 9, el 14693).
В настоящее время для контроля обогащения библиотек применяется ряд методических подходов: секвенирование методом NGS комбинаторных библиотек, начиная с определенного раунда селекции ( Т., Wilhelm В., Greiner N., Braun Н., Peter F.,
Т., Wilhelm В., Greiner N., Braun Н., Peter F.,  , Erdmann V.A., Lehrach H., Konthur Z., Menger M, Arndt P.F.,
, Erdmann V.A., Lehrach H., Konthur Z., Menger M, Arndt P.F.,  (2011) Probing the SELEX process with next-generation sequencing. PLoS One. 6, e29604, Kolm C, Cervenka I., Aschl U.J., Baumann N., Jakwerth S., Krska R., Mach R.L., Sommer R., DeRosa M.C., Kirschner A.K.T., Farnleitner A.H., Reischer G.H. (2020) DNA aptamers against bacterial cells can be efficiently selected by a SELEX process using state-of-the art qPCR and ultra-deep sequencing. Sci. Rep.10, 20917), анализ кривых плавления комплексов библиотека-лиганд (Luo Z., Не L., Wang J., Fang X., Zhang L. (2017) Developing a combined strategy for monitoring the progress of aptamer selection. Analyst. 142, 3136-3139), ЯМР спектроскопия (Sanchez J.C., Zhang L., Evoli S., Schnicker N.J., Nunez-Hernandez M., Yu L., Wereszczynski J., Pufall M.A., Musselman C.A. (2020) The molecular basis of selective DNA binding by the BRG1 AT-hook and bromodomain. Biochim. Biophys. Acta Gene Regul. Mech. 1863, 194566; Amano R., Aoki K., Miyakawa S., Nakamura Y., Kozu Т., Kawai G., Sakamoto T. (2017) NMR monitoring of the SELEX process to confirm enrichment of structured RNA. Sci. Rep.7, 283), методы ВЭЖХ (Miiller J., El-Maarri O., Oldenburg J.,
(2011) Probing the SELEX process with next-generation sequencing. PLoS One. 6, e29604, Kolm C, Cervenka I., Aschl U.J., Baumann N., Jakwerth S., Krska R., Mach R.L., Sommer R., DeRosa M.C., Kirschner A.K.T., Farnleitner A.H., Reischer G.H. (2020) DNA aptamers against bacterial cells can be efficiently selected by a SELEX process using state-of-the art qPCR and ultra-deep sequencing. Sci. Rep.10, 20917), анализ кривых плавления комплексов библиотека-лиганд (Luo Z., Не L., Wang J., Fang X., Zhang L. (2017) Developing a combined strategy for monitoring the progress of aptamer selection. Analyst. 142, 3136-3139), ЯМР спектроскопия (Sanchez J.C., Zhang L., Evoli S., Schnicker N.J., Nunez-Hernandez M., Yu L., Wereszczynski J., Pufall M.A., Musselman C.A. (2020) The molecular basis of selective DNA binding by the BRG1 AT-hook and bromodomain. Biochim. Biophys. Acta Gene Regul. Mech. 1863, 194566; Amano R., Aoki K., Miyakawa S., Nakamura Y., Kozu Т., Kawai G., Sakamoto T. (2017) NMR monitoring of the SELEX process to confirm enrichment of structured RNA. Sci. Rep.7, 283), методы ВЭЖХ (Miiller J., El-Maarri O., Oldenburg J., , Mayer G. (2008) Monitoring the progression of the in vitro selection of nucleic acid aptamers by denaturing high-performance liquid chromatography. Anal Bioanal. Chem. 390, 1033-1037) и другие (Thevendran R., Citartan M. (2022) Assays to Estimate the Binding Affinity of Aptamers. Talanta. 238, 122971). Для cell-SELEX предложено секвенирование после каждого шага селекции (Gooch J., Tungsirisurp S., Costanzo H., Napier R., Frascione N. (2021) Generating aptamers towards human sperm cells using massively parallel sequencing. Anal. Bioanal. Chem. 413, 5821-5834), что является весьма трудозатратным и дорогим подходом.
, Mayer G. (2008) Monitoring the progression of the in vitro selection of nucleic acid aptamers by denaturing high-performance liquid chromatography. Anal Bioanal. Chem. 390, 1033-1037) и другие (Thevendran R., Citartan M. (2022) Assays to Estimate the Binding Affinity of Aptamers. Talanta. 238, 122971). Для cell-SELEX предложено секвенирование после каждого шага селекции (Gooch J., Tungsirisurp S., Costanzo H., Napier R., Frascione N. (2021) Generating aptamers towards human sperm cells using massively parallel sequencing. Anal. Bioanal. Chem. 413, 5821-5834), что является весьма трудозатратным и дорогим подходом.
Все описанные методы основаны на контроле библиотек в процессе проведения SELEX. Однако, в случае использования модификаций более рациональным подходом является предварительная проверка субстратной применимости модифицированных субстратов (dNTPs) и оценка «нецелевого» обогащения модифицированной библиотеки, обусловленного конкурентной амплификацией, до проведения селекции аптамеров. Несмотря на ряд работ по субстратной совместимости модифицированных dNTPs с ДНК-полимеразами (Rohloff J.C., Gelinas A.D., Jarvis Т.С., Ochsner U.A., Schneider D.J., Gold L., Janjic N. (2014) Nucleic Acid Ligands With Protein-like Side Chains: Modified Aptamers and Their Use as Diagnostic and Therapeutic Agents. Mol. Ther. Nucleic Acids. 3, e201; Лапа C.A., Павлов A.C., Кузнецова B.E., Шершов В.Е., Спицын М.А., Гусейнов Т.О., Радько С.П., Заседателев А.С., Лисица А.В., Чудинов А.В. (2019) Ферментативное получение модифицированных ДНК: изучение кинетики ПЦР в режиме реального времени. Молекуляр. биология. 53, 513-523; Чудинов А.В., Шершов В.Е., Павлов А.С., Волкова О.С, Кузнецова В.Е., Заседателев А.С., Лапа С.А. (2020) Одновременное встраивание модифицированных производных dU и dC в растущую цепь ДНК в реакции удлинения праймера и ПЦР. Биоорг.химия. 46, 546-549), до настоящего времени не был разработан экспресс-метод оценки применимости различных модифицированных dNTPs для селекции аптамеров без проведения раундов SELEX. Субстратная несовместимость в случае комбинаторных библиотек может не детектироваться методом ПЦР в режиме реального времени, поскольку быстрая кинетика накопления сигнала при амплификации может быть обусловлена конкурентной амплификацией «в обход» сложных последовательностей. Так происходит накопление сигнала за счет «нецелевого» обогащения библиотеки (конкурентная амплификация). Это приводит к необходимости использования сложных процедур проверки, осуществляемых не до, а уже в процессе mod-SELEX. Учитывая многостадийность и трудозатратность процедуры селекции с модифицированными dNTPs, особенно в случае cell-SELEX (Rahimizadeh K., AlShamaileh Н., Fratini М., Chakravarthy М., Stephen М., Shigdar S., Veedu R.N. (2017) Development of Cell-Specific Aptamers: Recent Advances and Insight into the Selection Procedures. Molecules. 22, 2070; Sefah K., Shangguan D., Xiong X., O'Donoghue M.B., Tan W. (2010) Development of DNA aptamers using Cell-SELEX. Nat. Protoc. 5, 1169-1185; Gooch J., Tungsirisurp S., Costanzo H., Napier R., Frascione N. (2021) Generating aptamers towards human sperm cells using massively parallel sequencing. Anal Bioanal. Chem. 413, 5821-5834), где требуется проведение трудоемких дополнительных процедур (наращивание культур чистых клеточных линий, мягкие методы создания комплексов для предотвращения разрушения клеточных стенок, повышенные требования к специфичности амплификации библиотек из-за присутствия нецелевых ДНК и РНК), разработка такого способа весьма актуальна.
Изобретение описывает новый способ определения субстратной эффективности модифицированных дезоксинуклеозидов в применении к селекции аптамеров, осуществляемый до начала процедуры SELEX.
Раскрытие сущности изобретения
В настоящем изобретении предлагается новый способ оценки субстратной эффективности дезоксинуклеозидтрифосфатов, содержащих функциональные группы, для селекции модифицированных аптамеров (mod-SELEX). Способ предполагает проведение трех последовательных раундов ПЦР с комбинаторной библиотекой и модифицированным dNTP - кандидатом для проведения SELEX. Вывод о применимости конкретного производного dNTP осуществляется по характеру изменения кривой накопления сигнала в течение трех раундов ПЦР в режиме реального времени и не требует проведения раундов селекции аптамеров. Если в процессе амплификации библиотека вырождается (становится менее представительной), значит, конкретная модификация dNTP не может быть использована с выбранной полимеразой и другими выбранными условиями амплификации библиотеки, поскольку приводит к конкурентной амплификации. В случае, когда характер кривой накопления сигнала не меняется, делается вывод, что используемый модифицированный трифосфат дезоксинуклеозида не влияет на распределение олигонуклеотидов с различными последовательностями в составе библиотеки, то есть не приводит к изменению ее состава с точки зрения применяемого метода детекции. Именно такие производные могут быть применимы в выбранных условиях проведения селекции аптамеров.
Способ применим для быстрой оценки субстратной пригодности модифицированных dNTPs, для проведения mod-SELEX и будет полезен при отборе аптамеров для клинической диагностики, медицины и научных исследований.
Краткое описание фигур и таблиц Фигура 1. Кривые амплификации с модифицированными дезоксиуридинтрифосфатами и ДНК-полимеразами: a) Taq и б) Vent (ехо-). Использованная библиотека и праймеры для ее амплификации - как в (С.А. Лапа, К.С. Ромашова, М.А. Спицын, В.Е. Шершов, В.Е. Кузнецова, Т.О. Гусейнов, О.А. Заседателева, С.П. Радько, Э.Н. Тимофеев, А.В. Лисица, А.В. Чудинов. Получение модифицированных комбинаторных ДНК-библиотек методом ПЦР в обратной эмульсии с последующим разделением цепей. Молекулярная биология. 2018, 52, 6, 984-996).
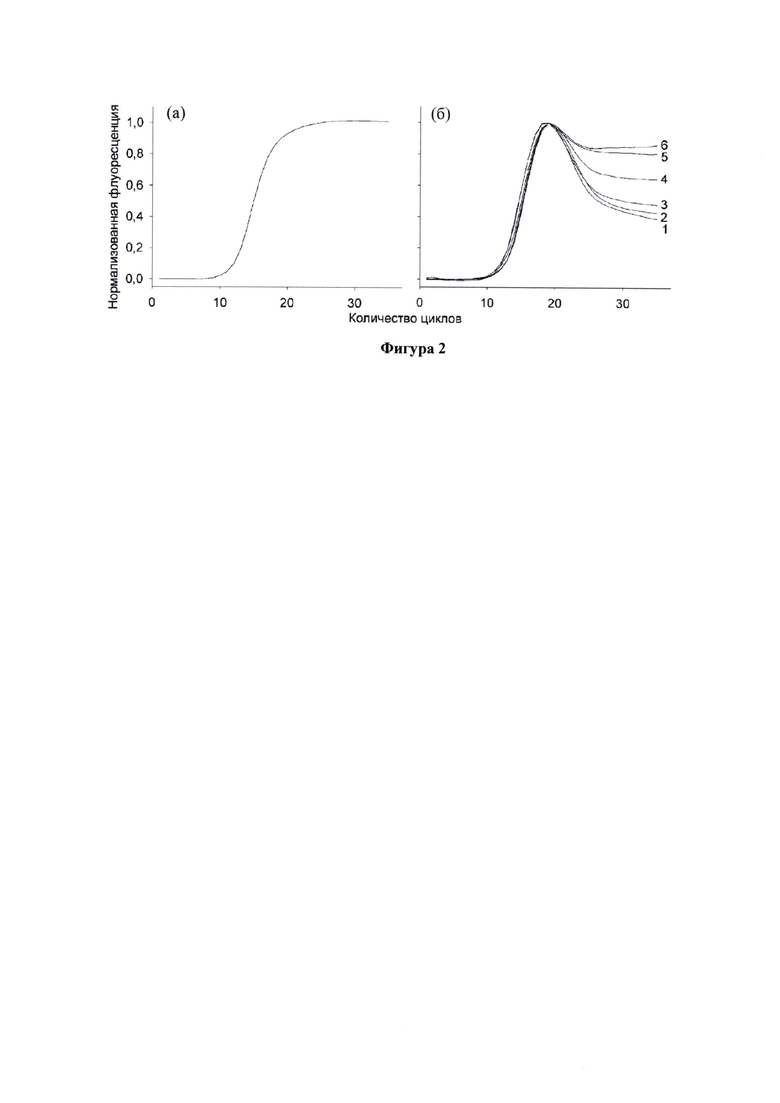
Фигура 2. Кривые накопления сигнала при амплификации различных матриц, а) матрица с фиксированной последовательностью, кривая имеет канонический вид сигмоиды, б) вырожденная комбинаторная библиотека (1-6 - стадии обогащения библиотеки, в процессе которых кривая накопления сигнала постепенно приобретает канонический сигмоидный вид). По данным (Радько С.П., Лапа С.А., Чудинов А.В., Хмелева С.А., Маннанова М.М., Курбатов Л.К., Киселева Я.Ю., Заседателев А.С., Лисица А.В. (2019) Оценка разнообразия комбинаторных ДНК-библиотек на основе анализа формы амплификационных кривых для мониторинга эффективности селекции аптамеров. Биомед. хим. 65, 477-84).
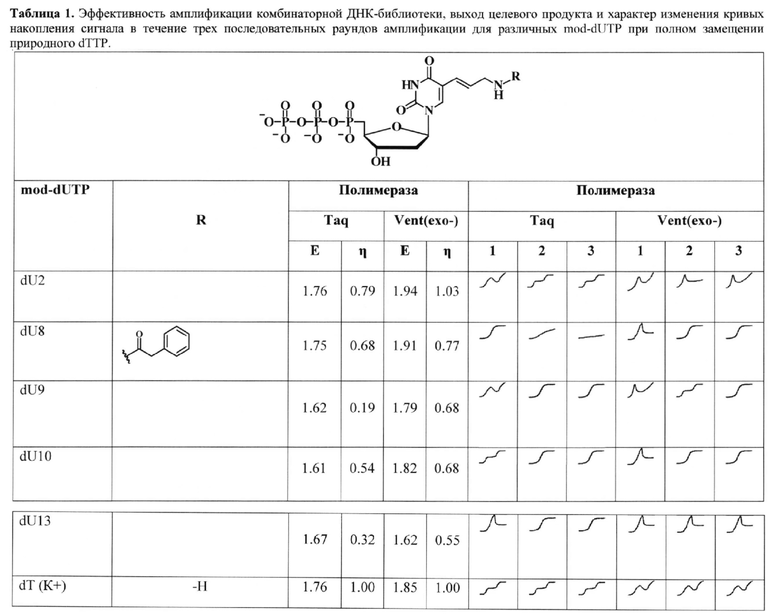
Таблица 1. Эффективность амплификации комбинаторной ДНК-библиотеки, выход целевого продукта и характер изменения кривых накопления сигнала в течение трех последовательных раундов амплификации для различных mod-dUTP при полном замещении природного dTTP.
Эффективность амплификации и выход продукта (левая часть таблицы). В качестве матрицы использована вырожденная составная ДНК-библиотека (длина 86 нт). Е -эффективность амплификации (Е=10tgα, где Е - эффективность амплификации, tgα -тангенс угла наклона прямого участка сигмоидной кривой накопления флуоресцентного сигнала, представленной в логарифмическом масштабе (Ramakers С., Ruijter J.M., Deprez R.H., Moorman A.F. (2003) Assumption-free analysis of quantitative realtime polymerase chain reaction (PCR) data. Neurosci. Lett. 339, 62-66), η - выход продукта, нормированный на выход немодифицированной ДНК (ПЦР с использованием природного dTTP). К+- положительный контроль (природный dTTP). Характер кривых накопления сигнала (правая часть таблицы). 1, 2, 3 - последовательные раунды ПЦР. Приведены схематичные кривые накопления сигнала амплификации для каждого раунда ПЦР.
Осуществление изобретения
Для оценки влияния субстрата (модифицированного дезоксинуклеозидтрифосфата) на поведение кривой амплификации комбинаторной ДНК-библиотеки проводили три последовательных раунда ПЦР в режиме реального времени (без проведения стадий селекции аптамеров). В качестве матрицы для каждого следующего этапа брали 1 мкл реакционной смеси из пробирки предыдущего этапа.
Наблюдали несколько типов кривых накопления сигнала, форма которых зависела от степени влияния конкретного модифицированного dNTP на процесс амплификации библиотеки. На Фигуре 1 приведены результаты проведения трех раундов ПЦР с различными производными дезоксиуридинтрифосфата с полимеразами Taq и Vent (ехо-). Из Фигуры 1 видно, что кривая амплификации может сохранять свою форму на протяжении всех трех раундов ПЦР, может постепенно приходить к каноническому виду сигмоиды, характерной для фиксированных нуклеотидных последовательностей матрицы либо вырожденных (обогащенных) комбинаторных библиотек, также может наблюдаться ингибирование ПЦР с постепенным затуханием амплификации. В качестве контроля использовали образец, не содержащий модифицированных дезоксинуклеозидов.
В Таблице 1 приведены структуры использованных в работе дезоксиуридинтрифосфатов с различными функциональными группами-заместителями и показатели их субстратной эффективности. В правой части таблицы схематично показаны основные характеристические формы кривых амплификации, наблюдаемые в зависимости от использованного модифицированного дезоксиуридинтрифосфата и ДНК-полимеразы.
Важным аспектом успешного отбора модифицированных аптамеров (SELEX) является субстратная совместимость модифицированных dNTPs с выбранной полимеразой. При их недостаточной совместимости может происходить конкурентная амплификация последовательностей исходной ДНК-библиотеки, содержащих меньшее количество нуклеотидов или их повторов, комплементарных модифицированному dNTP. Часто низкая эффективность встраивания модификаций остается незамеченной в процессе проведения SELEX. В результате библиотека будет обеднена модифицированными аптамерами.
Кривые амплификации исходных комбинаторных библиотек отличаются от кривых, характерных для амплификации матриц с фиксированными последовательностями. Различие заключается в том, что накопление сигнала при амплификации фиксированной последовательности имеет сигмоидную форму, в случае комбинаторной библиотеки - проходит через один или более максимумов. По всей видимости, это связано с тем, что после определенного цикла ПЦР и распределения амплифицированных продуктов по реакционному объему образуются локальные очаги, в которых велика концентрация образующихся дуплексов с некомплементарной внутренней частью. Это приводит к временному замедлению амплификации и снижению наблюдаемого накопления сигнала. Однако, по мере образования новых комплементарных дуплексов - продуктов амплификации - кривая проходит через максимум. По-видимому, этот процесс может приводить более чем к одному максимуму, что и наблюдается при аплификации комбинаторных библиотек (Фигура 1).
Ранее было показано, что в процессе обогащении ДНК-библиотеки кривая при ее амплификации в режиме реального времени с каждым следующим раундом приближается по форме к канонической сигмоидной кривой, характерной для фиксированной последовательности ДНК (Luo Z., Не L., Wang J., Fang X., Zhang L. (2017) Developing a combined strategy for monitoring the progress of aptamer selection. Analyst. 142, 3136-3139; Радько С.П., Лапа C.A., Чудинов А.В., Хмелева С.А., Маннанова М.М., Курбатов Л.К., Киселева Я.Ю., Заседателев А.С., Лисица А.В. (2019) Оценка разнообразия комбинаторных ДНК-библиотек на основе анализа формы амплификационных кривых для мониторинга эффективности селекции аптамеров. Биомед. хим. 65, 477-484). Это является показателем обогащения библиотеки специфичными к мишени олигонуклеотидами (Фигура 2.). То есть, в процессе обогащения постепенно ослабевают факторы, указанные выше (образование дуплексов с некомплементарными внутренними частями олигонуклеотидов библиотеки).
Однако, библиотека может обогащаться не специфичными, а наиболее легко амплифицируемыми последовательностями (Tolle F., Wilke J., Wengel J., Mayer G. (2014) By-product formation in repetitive PCR amplification of DNA libraries during SELEX. PLoS One. 9, el 14693). Наиболее актуальной эта проблема становится при использовании модифицированных dNTP, которые являются, как правило, «плохими» субстратами для полимераз, характеризуются медленной кинетикой встраивания и/или в той или иной степени ингибируют ПЦР.
Из Таблицы 1 видно, что характер кривых накопления сигнала зависит как от использованного модифицированного dNTP, так и от полимеразы.
Taq-полимераза оказалась способна амплифицировать библиотеки с применением модифицированного dUTP с линейным алкановым заместителем (dU2). Это подтверждается как показателями эффективности ПЦР «Е» и значением выхода целевого продукта «л», так и относительно неизменным характером кривой амплификации. При этом гетероциклические производные оказываются для нее сложными субстратами. Это приводит к изменению формы кривой амплификации - за три цикла ПЦР она либо приходит к сигмоиде, либо ингибируется (как в случае dU8). При этом может наблюдаться как быстрая кинетика накопления сигнала, так и выход полноразмерного продукта, то есть эти показатели не подходят для оценки влияния mod-dNTP на поведение комбинаторных библиотек в процессе SELEX.
Для производных с бензильным заместителем, вне зависимости от длины используемого линкера, наблюдали вырождение библиотек и сигмоидной кривой амплификаци после третьего раунда ПЦР. Причем, это было характерно также и в случае применения Vent (ехо-) полимеразы.
Интересно, но производное dU13 с конденсированным гетероциклом оказывало незначительное влияние на характер кривой амплификации в случае применения Vent (ехо-) полимеразы, хотя показатели эффективности амплификации и выхода были низкими.
В случае изменения характера кривой амплификации делали вывод о неприменимости конкретного модифицированного dNTP в выбранных условиях амплификации, которые могут содержать несколько значимых факторов (используемая полимераза, выбранные условия амплификации, комбинаторная библиотека и праймеры для ее амплификации и др.).
В случае, когда характер кривой амплификации не менялся на протяжении трех раундов ПЦР, делали вывод о том, что влияние трифосфата на распределение последовательностей внутри комбинаторной библиотеки минимально (либо отсутствует), то есть, такое производное не приводит к вырождению библиотеки при амплификации и может быть применено для проведения mod-SELEX.
Способ позволяет за три загрузки амплификатора проверить весь необходимый набор трифосфатов - кандидатов для планируемого mod-SELEX.
Следует заметить, что предлагаемый подход не исключает контроля обогащения библиотек в процессе селекции, но в случае mod-SELEX позволяет предсказать влияние модификаций на обогащение библиотек и избежать неудач в последующем процессе отбора модифицированных аптамеров.
Пример 1. ПЦР в режиме реального времени.
1. Реакционную смесь готовят с содержанием природных dATP, dCTP и dGTP в концентрации 0.2 мМ каждого, а также различных модифицированных дезоксинуклеозидтрифосфатов (в отдельных пробирках), структуры которых указаны в Таблице 1 (при полной замене природного dTTP на один из модифицированных аналогов), также в концентрации 0.2 мМ.
2. В реакционную смесь вносят комбинаторную библиотеку (выбирается в зависимости от целей исследования) в начальной концентрации 105 копий на реакционный объем (20 мкл) и праймеры для ее амплификации.
3. В реакционную пробирку добавляют ДНК-полимеразу (например, 1.5U Taq-или 0.5U Vent (ехо-) ДНК-полимеразы, или другие) в реакционном буфере, соответствующем применяемой полимеразе.
4. Для визуализации процесса в реакционную смесь добавляют 0,5 мкл красителя EvaGreen (Biotium, Россия).
5. Проводят ПЦР в режиме реального времени на ДНК-амплификаторе IQ5 (Bio-Rad, США) по следующей программе: предварительная денатурация при 95°С в течение 3 мин, за которой следовало 50 циклов: 95°С в течение 30 с, 65°С в течение 30 с, 72°С в течение 40 с и далее завершающая инкубация при 72°С в течение 5 мин.
6. После первого раунда амплификации отбирают 1 мкл и используют его в качестве матрицы для проведения второго раунда ПЦР; аналогично, после второго раунда отбирают 1 мкл реакционного объема для проведения третьего раунда ПЦР.
Пример 2. Контроль продуктов реакции.
1. Для контроля продуктов реакции осуществляют ПЦР указанного в Примере 1 состава и по тому же температурному-временному профилю, за исключением того, что количество циклов ПЦР сокращают до 32 во избежание появления артефактов и побочных продуктов ферментативного разложения ДНК; реакцию проводят на ДНК-амплификаторе MiniCycler (MJ Research, США).
2. Проводят электрофорез ПЦР-продуктов в денатурирующих условиях, для чего образцы прогревают 1 мин при 90°С в присутствии 75%-ного формамида и наносят на полиакриламидный гель (Т=15%, С=7%), содержащий 20 М формамида.
3. Разделение продуктов проводят в ТВЕ-буфере, после чего гели окрашивают красителем SYBR Green.
4. Осуществляют визуальный контроль наличия полноразмерных целевых продуктов по электрофореграмме.
Примечание: этапы, описанные в Примере 2, необходимы только в процессе разработки способа для контроля образования побочных продуктов реакции при оптимизации способа.
Пример 3. Интерпретация результатов ПЦР в режиме реального времени.
1. Проводят визуальный анализ полученных кривых амплификации.
2. Визуально разделяют несколько типов характеристических кривых накопления сигнала: классическая сигмоида, кривая с одним максимумом, кривая с двумя максимумами, кривая «затухающей» амплификации (характеризуется низким выходом продукта и невыраженной формой, стремящейся к прямой линии с низкими значениями tg угла наклона); характеристические типы кривых схематично представлены в Таблице 1.
3. По характеру кривой амплификации делают вывод о применимости конкретного модифицированного дезоксинуклеозидтрифосфата для отбора модифицированных аптамеров в процедуре SELEX.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ОПРЕДЕЛЕНИЯ СУБСТРАТНОЙ СОВМЕСТИМОСТИ МОДИФИЦИРОВАННЫХ ТРИФОСФАТОВ ДЕЗОКСИУРИДИНА И ДНК-ПОЛИМЕРАЗ СОЧЕТАНИЕМ МЕТОДОВ КОЛИЧЕСТВЕННОЙ ПЦР И ЭЛЕКТРОФОРЕЗА | 2019 |
|
RU2736969C1 |
| Способ определения субстратной эффективности производных трифосфатов дезоксиуридина для ДНК-полимераз методом реакции достраивания праймера | 2019 |
|
RU2731739C1 |
| Способ ферментативного получения модифицированных ДНК для создания реагентов, специфично связывающихся с гидрофобными участками высокомолекулярных органических соединений | 2017 |
|
RU2699522C2 |
| Способ термической диссоциации для проведения селекции ДНК-аптамеров | 2019 |
|
RU2723373C1 |
| СПОСОБ СОЗДАНИЯ И СЕЛЕКЦИИ БИБЛИОТЕКИ МОДИФИЦИРОВАННЫХ АПТАМЕРОВ | 2016 |
|
RU2652952C1 |
| Способ селекции одноцепочечных олигонуклеотидов с повышенной аффинностью к молекулярной мишени | 2019 |
|
RU2736789C1 |
| СПОСОБ СЕЛЕКЦИИ АПТАМЕРОВ К ЗАДАННЫМ БЕЛКОВЫМ МИШЕНЯМ НА ПОВЕРХНОСТИ КЛЕТОК | 2012 |
|
RU2518368C1 |
| Аптамер, проходящий через гематоэнцефалический барьер головного мозга мыши | 2016 |
|
RU2711912C2 |
| Способ направленного истощения олигонуклеотидных библиотек для снижения неспецифической адсорбции при твердофазной селекции аптамеров на основе нуклеиновых кислот | 2015 |
|
RU2618872C1 |
| ДНК аптамеры, связывающие сердечный тропонин I человека | 2019 |
|
RU2716409C1 |

Изобретение относится к биотехнологии, в частности способ предполагает проведение трех последовательных раундов ПЦР с комбинаторной библиотекой и модифицированным dNTP - кандидатом для проведения SELEX. Вывод о применимости конкретного производного dNTP осуществляется по характеру изменения кривой накопления сигнала в течение трех раундов ПЦР в режиме реального времени и не требует проведения раундов селекции аптамеров. Если в процессе амплификации библиотека вырождается (становится менее представительной), значит конкретная модификация dNTP не может быть использована с выбранной полимеразой и другими выбранными условиями амплификации библиотеки, поскольку приводит к конкурентной амплификации. В случае когда характер кривой накопления сигнала не меняется, делается вывод, что используемый модифицированный трифосфат дезоксинуклеозида не влияет на распределение олигонуклеотидов с различными последовательностями в составе библиотеки, то есть не приводит к изменению ее состава с точки зрения применяемого метода детекции. Способ применим для быстрой оценки субстратной пригодности модифицированных dNTPs для проведения mod-SELEX и будет полезен при отборе аптамеров для клинической диагностики, медицины и научных исследований. 2 ил., 1 табл., 3 пр.
Способ оценки влияния модифицированных трифосфатов дезоксинуклеозидов на олигонуклеотидный состав комбинаторных ДНК-библиотек для проведения селекции модифицированных аптамеров, включающий:
а - амплификацию комбинаторных библиотек с исследуемыми модифицированными трифосфатами в заданных экспериментом условиях в течение трех раундов ПЦР в режиме реального времени, используя в качестве матрицы для каждого последующего раунда реакционную смесь из предыдущего раунда;
б - построение кривых накопления сигнала амплификации для каждого из трех раундов ПЦР;
в - визуальный анализ кривых амплификации и формирование заключения о применимости конкретного трифосфата дезоксинуклеозида на основании анализа изменения характеристической кривой амплификации в течение трех раундов.
| Лапа C.A | |||
| и др., Ферментативное получение модифицированных ДНК: изучение кинетики ПЦР в режиме реального времени | |||
| Молекуляр | |||
| биология, 2019, 53, стр | |||
| СПОСОБ ПОЛУЧЕНИЯ ПРОДУКТОВ КОНДЕНСАЦИИ ФЕНОЛОВ С ФОРМАЛЬДЕГИДОМ ИЛИ ЕГО ПОЛИМЕРАМИ | 1925 |
|
SU513A1 |
| Способ ферментативного получения модифицированных ДНК для создания реагентов, специфично связывающихся с гидрофобными участками высокомолекулярных органических соединений | 2017 |
|
RU2699522C2 |
| US 7138565 B2, 21.11.2006 | |||
| US 20080213870 A1, 04.09.2008060. | |||

