I. Уровень техники
[0001] CD137 представляет собой молекулу надсемейства рецепторов фактора некроза опухоли (TNFR), активность которой может быть вовлечена во многие иммуноопосредованные аутоиммунные и воспалительные заболевания. Она также является мишенью для иммунотерапии рака.
[0002] Как было показано, передача сигнала CD137 является первоначальной для поддержания и развития иммунного ответа на антигены, а также для образования T-клеток памяти. Многочисленные исследования T-клеток мышей и человека указывают на то, что CD137 обеспечивает усиление клеточной пролиферации, выживание и продуцирование цитокинов (Croft, 2009, Nat Rev Immunol 9:271-285). Исследования показали, что некоторые mAb-агонисты CD137 повышают экспрессию костимулирующих молекул и значительно усиливают ответы цитолитических T-лимфоцитов, обусловливая противоопухолевое действие в различных моделях. Была показана эффективность mAb-агонистов CD137 в профилактических и терапевтических мероприятиях. К тому же по результатам монотерапии и комбинированной терапии на моделях рака в отношении CD137 установили наличие продолжительных противоопухолевых защитных ответов T-клеток памяти (Lynch, 2008, Immunol Rev. 22: 277-286). Также было показано, что агонисты CD137 подавляют аутоиммунные реакции в ряде принятых в данной области аутоиммунных моделей (Vinay, 2006, J Mol Med 84:726-736). Эта двойная активность CD137 создает возможность обеспечивать противоопухолевую активность, ослабляя в то же время аутоиммунные побочные эффекты, которые могут быть ассоциированы с иммунотерапевтическими подходами, которые нарушают иммунную толерантность.
[0003] Следовательно, с учетом ролей CD137 в модуляции иммунного ответа, существует давно назревшая неудовлетворенная потребность в соединениях, которые связывают CD137 человека, усиливают CD137-опосредованный ответ и тем самым обеспечивают перспективное терапевтическое средство для лечения или предупреждения различных заболеваний и состояний, таких как рак, инфекционные заболевания и аутоиммунные заболевания.
[0004] Соответственно, объектом настоящего изобретения является обеспечение таких соединений, которые представляют собой мутеины, полученные из липокалинов. Мутеины различных липокалинов представляют собой стремительно увеличивающийся класс терапевтических средств и они могут быть сконструированы посредством весьма сложной искусственной разработки с проявлением высокой аффинности и специфичности в отношении мишени, которая отличается от естественного лиганда липокалинов дикого типа (см., например, WO 99/16873, WO 00/75308, WO 03/029463, WO 03/029471 и WO 05/19256).
II. Определения
[0005] В приведенном ниже перечне определены термины, фразы и сокращения, используемые по всему данному описанию. Предполагается, что все термины, перечисленные и определенные в данном документе, охватывают все грамматические формы.
[0006] Как используется в данном документе, если не указано иное, «CD137» означает CD137 человека. CD137 также известен как «4-1BB» или «представитель 9 надсемейства рецепторов фактора некроза опухоли (TNFRSF9)» или «индуцированный активацией лимфоцитов (ILA)». CD137 человека означает полноразмерный белок, обозначенный согласно UniProt как Q07011, его фрагмент или его вариант.
[0007] Как используется в данном документе, «выявляемая аффинность» означает способность связываться с выбранной мишенью с константой аффинности, составляющей, как правило, по меньшей мере приблизительно 10-5 M или ниже. Более низкие значения аффинности, как правило, уже не поддаются измерению с помощью стандартных способов, таких как ELISA, и, следовательно, имеют второстепенное значение.
[0008] Как используется в данном документе, «аффинность связывания» белка по настоящему раскрытию (например, мутеина липокалина) или его полипептида слияния с выбранной мишенью (в данном случае CD137) можно измерять (и таким образом определять значения KD для комплекса мутеин-лиганд) с помощью большого числа способов, известных специалисту в данной области. Такие способы включают без ограничения флуоресцентное титрование, конкурентный ELISA, калориметрические способы, такие как изотермическая титрационная калориметрия (ITC) и поверхностный плазмонный резонанс (BIAcore). Такие способы хорошо известны в уровне техники и их примеры также подробно описаны ниже.
[0009] Следует также отметить, что на образование комплекса между соответствующей связывающей молекулой и ее лигандом влияет много различных факторов, таких как концентрации соответствующих партнеров по связыванию, наличие конкурентов, показатель pH и ионная сила используемой буферной системы и экспериментальный способ, используемый для определения константы диссоциации, KD (например, флуоресцентное титрование, конкурентный ELISA или поверхностный плазмонный резонанс, при этом упомянуто лишь несколько), или даже математический алгоритм, который используют для оценки экспериментальных данных.
[0010] Следовательно, специалисту в данной области также очевидно, что значения KD (константы диссоциации комплекса, образованного между соответствующей связывающей молекулой и ее мишенью/лигандом) могут варьироваться в пределах определенного экспериментального диапазона в зависимости от способа и экспериментальной установки, которую используют для определения аффинности конкретного мутеина липокалина в отношении данного лиганда. Это значит, что может иметь место незначительное отклонение в измеренных значениях KD или диапазоне допусков, зависящее, например, от того, было ли значение KD определено с помощью поверхностного плазмонного резонанса (Biacore), конкурентного ELISA или «прямого ELISA».
[0011] Как используется в данном документе, «мутеин», «подвергнутый мутации» целостный объект (или белок, или нуклеиновая кислота) или «мутант» относится к обмену, делеции или вставке одного или более из нуклеотидов или аминокислот по сравнению со встречающимся в природе (дикого типа) «эталонным» остовом нуклеиновой кислоты или белка. Указанный термин также включает фрагменты мутеина и вариантов, описанных в данном документе. Мутеины липокалина по настоящему изобретению, их фрагменты или варианты предпочтительно сохраняют функцию связывания с CD137, описанным в данном документе.
[0012] Термин «фрагмент», используемый в данном документе применительно к мутеинам по настоящему раскрытию, относится к белкам или пептидам, полученным из полноразмерного зрелого липокалина слезы человека, который укорочен с N-конца и/или C-конца, т.е. у него отсутствует по меньшей мере одна из N-концевых и/или C-концевых аминокислот. Такие фрагменты могут включать по меньшей мере 10 или больше, а именно 20 или 30 или больше, последовательных аминокислот первичной последовательности зрелого липокалина и обычно поддаются выявлению в иммуноанализе зрелого липокалина. Как правило, термин «фрагмент», используемый в данном документе в отношении соответствующего белкового лиганда CD137 для мутеина липокалина по настоящему раскрытию, или для комбинации согласно настоящему раскрытию, или для белка слияния, описанного в данном документе, относится к укороченным с N-конца и/или C-конца белковым или пептидным лигандам, которые сохраняют способность полноразмерного лиганда быть распознанными и/или связанными мутеином согласно настоящему раскрытию.
[0013] Термин «мутагенез», используемый в данном документе, означает, что экспериментальные условия выбраны таким образом, что аминокислота, встречающаяся в природе в данном положении в последовательности зрелого липокалина, может быть заменена по меньшей мере одной аминокислотой, которая не присутствует в этом конкретном положении в соответствующей природной полипептидной последовательности. Термин «мутагенез» также включает (дополнительную) модификацию длины сегментов последовательности путем делеции или вставки одной или более аминокислот. Таким образом, в пределы объема настоящего раскрытия подпадает, что, например, одну аминокислоту в выбранном положении в последовательности заменяют отрезком из трех случайных мутаций, следствием чего является вставка двух аминокислотных остатков по сравнению с длиной соответствующего сегмента белка дикого типа. Такая вставка или делеция могут быть введены независимо друг от друга в любой из сегментов пептидов, которые могут быть подвергнуты мутагенезу в настоящем раскрытии. Согласно одному иллюстративному варианту осуществления настоящего раскрытия вставка из нескольких мутаций может быть введена в петлю AB остова выбранного липокалина (см. международную патентную заявку WO 2005/019256, которая включена в данный документ в полном объеме посредством ссылки).
[0014] Термин «случайный мутагенез» означает, что в конкретном положении в последовательности отсутствует предопределенная отдельная аминокислота (мутация), однако по меньшей мере две аминокислоты могут быть включены с определенной вероятностью в заранее определенное положение в последовательности во время мутагенеза.
[0015] «Идентичность» является свойством последовательностей, с помощью которого определяют их сходство или родство. Термин «идентичность последовательностей» или «идентичность», используемый в настоящем раскрытии, означает процентную долю попарно идентичных остатков - последующее (гомологичное) выравнивание последовательности полипептида по настоящему раскрытию с последовательностью запроса - относительно количества остатков в более длинной из этих двух последовательностей. Идентичность последовательностей определяют путем деления количества идентичных аминокислотных остатков на общее количество остатков и умножения полученного результата на 100.
[0016] Термин «гомология» используют в данном документе в его общепринятом значении, и он включает идентичные аминокислоты, а также аминокислоты, которые рассматриваются как консервативные замены (например, обмен остатка глутамата на остаток аспартата) в аналогичных положениях в линейной аминокислотной последовательности полипептида по настоящему раскрытию (например, любого мутеина липокалина по настоящему раскрытию).
[0017] Процентную долю гомологии последовательностей или идентичности последовательностей, например, можно определять в данном документе с использованием программы BLASTP, версии blastp 2.2.5 (от 16 ноября 2002 года; см. Altschul, S.F. et al. (1997) Nucl. Acids Res. 25, 3389-3402). Согласно данному варианту осуществления процентная доля гомологии основана на выравнивании целых полипептидных последовательностей (матрица: BLOSUM 62; штрафы за введения гэпа: 11.1; значение порога отсечения устанавливали как 10-3), в том числе последовательностей пропептидов, предпочтительно с использованием остова белка дикого типа в качестве эталона в сравнительном анализе пар. Ее рассчитывают в виде процентной доли количеств «положительных» (гомологичных аминокислот), указанных в качестве итога в результате расчета с помощью программы BLASTP, разделенных на общее количество аминокислот, выбранных программой для выравнивания.
[0018] В частности, для того, чтобы определить, соответствует ли аминокислотный остаток аминокислотной последовательности липокалина (мутеина), отличного от липокалина дикого типа, определенному положению в аминокислотной последовательности липокалина дикого типа, специалист в данной области может применять средства и способы, хорошо известные в данной области, например, выравнивания либо вручную, либо с использованием компьютерных программ, таких как BLAST2.0, которая означает средство поиска основного локального выравнивания, или ClustalW, или любой другой подходящей программы, которая подходит для получения выравниваний последовательностей. Соответственно, липокалин дикого типа может служить в качестве «последовательности для сравнения» или «эталонной последовательности», тогда как аминокислотная последовательность липокалина, отличного от липокалина дикого типа, описанного в данном документе, служит в качестве «искомой последовательности». Термины «эталонная последовательность» и «последовательность дикого типа» используют в данном документе взаимозаменяемо. Предпочтительная последовательность липокалина дикого типа показана под SEQ ID NO: 1 (Tlc) или SEQ ID NO: 2 (NGAL) соответственно. В зависимости от того, является ли основой для мутеина липокалина по настоящему изобретению соответственно Tlc или NGAL, соответствующий липокалин дикого типа можно использовать в качестве эталонной последовательности или последовательности дикого типа.
[0019] «Гэпы» представляют собой пространства в выравнивании, которые являются результатом добавлений или делеций аминокислот. Таким образом, две копии точно такой же последовательности характеризуются 100% идентичностью, но последовательности, которые в меньшей степени высоко консервативны и имеют делеции, добавления или замещения, могут характеризоваться более низкой степенью идентичности последовательностей. Специалисту в данной области будет понятно, что несколько компьютерных программ доступны для определения идентичности последовательностей с использованием стандартных параметров, например Blast (Altschul, et al. (1997) Nucleic Acids Res. 25, 3389-3402), Blast2 (Altschul, et al. (1990) J. Mol. Biol. 215, 403-410) и Smith-Waterman (Smith, et al. (1981) J. Mol. Biol. 147, 195-197).
[0020] Термин «вариант», используемый в настоящем раскрытии, относится к производным белка или пептида, которые предусматривают модификации аминокислотной последовательности, например путем замены, делеции, вставки или химической модификации. Согласно некоторым вариантам осуществления такие модификации не снижают функциональность белка или пептида. Такие варианты включают белки, в которых одна или более аминокислот были замещены их соответствующими D-стереоизомерами или аминокислотами, отличными от встречающихся в природе 20 аминокислот, такими как, например, орнитин, гидроксипролин, цитруллин, гомосерин, гидроксилизин, норвалин. Однако такие замены также могут быть консервативными, т.е. аминокислотный остаток замещают химически ему подобным аминокислотным остатком. Примеры консервативных замен представляют собой замещения из числа представителей следующих групп: 1) аланин, серин и треонин; 2) аспарагиновая кислота и глутаминовая кислота; 3) аспарагин и глутамин; 4) аргинин и лизин; 5) изолейцин, лейцин, метионин и валин и 6) фенилаланин, тирозин и триптофан. Термин «вариант», используемый в данном документе в отношении соответствующего белкового лиганда CD137 для мутеина липокалина по настоящему раскрытию, или для комбинации согласно настоящему раскрытию, или для белка слияния, описанного в данном документе, относится к CD137 или его фрагменту соответственно, который имеет одну или более, а именно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 40, 50, 60, 70, 80 или больше, аминокислотных замен, делеций и/или вставок по сравнению с белком CD137 дикого типа соответственно, как, например, эталонным белком CD137, депонированным с идентификатором SwissProt, описанным в данном документе. Вариант CD137, соответственно, предпочтительно характеризуется аминокислотной идентичностью, составляющей по меньшей мере 50%, 60%, 70%, 80%, 85%, 90% или 95%, с CD137 дикого типа человека, как, например, эталонным белком CD137, депонированным с идентификатором SwissProt, описанным в данном документе.
[0021] Под «нативной последовательностью» липокалина подразумевают липокалин, который характеризуется такой же аминокислотной последовательностью, что и соответствующий полипептид, полученный из естественного окружения. Таким образом, нативная последовательность липокалина может характеризоваться аминокислотной последовательностью соответствующего встречающегося в природе липокалина из любого организма, в частности млекопитающего. Такая нативная последовательность полипептида может быть выделена из естественного окружения или может быть получена с помощью рекомбинантных способов или способов синтеза. Термин «нативная последовательность» полипептида, как правило, охватывает встречающиеся в природе усеченные или секретированные формы липокалина, встречающиеся в природе вариантные формы, такие как, в качестве альтернативы, сплайсированные формы, и встречающиеся в природе аллельные варианты липокалина. «Вариант» полипептида означает биологически активный полипептид, характеризующийся по меньшей мере приблизительно 50%, 60%, 70%, 80% или по меньшей мере приблизительно 85% идентичностью аминокислотной последовательности с нативной последовательностью полипептида. Такие варианты включают, к примеру, полипептиды, в которых один или более аминокислотных остатков добавлены или удалены на N- или C-конце полипептида. Как правило, вариант характеризуется по меньшей мере приблизительно 70%, в том числе по меньшей мере приблизительно 80%, как, например, по меньшей мере приблизительно 85% идентичностью аминокислотной последовательности, в том числе по меньшей мере приблизительно 90% идентичностью аминокислотной последовательности или по меньшей мере приблизительно 95% идентичностью аминокислотной последовательности с нативной последовательностью полипептида. В качестве иллюстративного примера первые 4 N-концевых аминокислотных остатка (His-His-Leu-Leu) и последние 2 C-концевых аминокислотных остатка (Ser-Asp) могут быть удалены в мутеине липокалина слезы (Tlc) по настоящему раскрытию без воздействия на биологическую функцию белка, например SEQ ID NO: 5-11. Кроме того, в качестве другого иллюстративного примера, определенные аминокислотные остатки могут быть удалены в мутеине липокалина 2 (NGAL) по настоящему раскрытию без воздействия на биологическую функцию белка, например (Lys-Asp-Pro, положения 46-48) относительно SEQ ID NO: 16.
[0022] Термин «положение» при использовании в соответствии с настоящим раскрытием означает либо положение аминокислоты в аминокислотной последовательности, описанной в данном документе, либо положение нуклеотида в последовательности нуклеиновой кислоты, описанной в данном документе. Для понимания термина «соответствует» или «соответствующий», используемого в данном документе в отношении положений в аминокислотной последовательности одного или более мутеинов липокалина, соответствующее положение определено не только номером предыдущих нуклеотидов/аминокислот. Следовательно, положение указанной аминокислоты в соответствии с настоящим раскрытием, которая может быть замещена, может варьироваться за счет делеции или добавления аминокислот в любом месте в (мутантном или дикого типа) липокалине. Аналогично положение указанного нуклеотида в соответствии с настоящим раскрытием, который может быть замещен, может варьироваться за счет делеции или дополнительных нуклеотидов в любом месте в 5'-нетранслируемом участке (UTR) мутеина липокалина или липокалина дикого типа, в том числе промоторе, и/или любых других регуляторных последовательностях или гене (в том числе экзонах и интронах).
[0023] Таким образом, в отношении соответствующего положения в соответствии с настоящим раскрытием предпочтительно понимать, что положения нуклеотидов/аминокислот могут отличаться по обозначенным номерам от подобных соседних нуклеотидов/аминокислот, но указанные соседние нуклеотиды/аминокислоты, которые могут быть обменены, удалены или добавлены, также предусматриваются в одном или более соответствующих положениях.
[0024] Кроме того, в отношении соответствующего положения в мутеине липокалина на основе эталонного остова в соответствии с настоящим раскрытием предпочтительно понимать, что положения нуклеотидов/аминокислот являются структурно соответствующими положениям в любом месте в (мутантном или дикого типа) липокалине, даже если они могут отличаться по обозначенным номерам, что будет понятно специалисту в данной области, с учетом высококонсервативного общего паттерна фолдинга среди липокалинов.
[0025] Термин «альбумин» включает альбумины всех млекопитающих, как например: сывороточный альбумин человека, или бычий сывороточный альбумин, или сывороточный альбумин крысы.
[0026] Термин «органическая молекула» или «малая органическая молекула», используемый в данном документе в отношении отличной от естественной мишени, означает органическую молекулу, содержащую по меньшей мере два атома углерода, но предпочтительно не более 7 или 12 вращающихся углеродных связей с молекулярной массой в диапазоне от 100 до 2000 дальтон, предпочтительно от 100 до 1000 дальтон, и необязательно включающую один или два атома металла.
[0027] Слово «выявлять», «выявление», «выявляемый» или «выявляющий», используемое в данном документе, понимается как на количественном, так и на качественном уровне, а также их комбинация. Таким образом, это включает количественные, полуколичественные и качественные измерения представляющей интерес молекулы.
[0028] «Субъект» представляет собой позвоночное, предпочтительно млекопитающее, более предпочтительно человека. Термин «млекопитающее» используется в данном документе в отношении любого животного, классифицированного как млекопитающее, в том числе без ограничения людей, домашних и сельскохозяйственных животных, а также животных в зоопарках, для занятий спортом или животных-питомцев, как например: овец, собак, лошадей, кошек, коров, крыс, свиней, высших приматов, таких как яванские макаки, и т.д., при этом упомянуты лишь некоторые иллюстративные примеры. Предпочтительно, млекопитающим в данном документе является человек.
[0029] «Эффективное количество» представляет собой количество, достаточное для обеспечения полезных или необходимых результатов. Эффективное количество можно вводить за одно или более введений.
[0030] «Образец» определяют как биологический образец, взятый у любого субъекта. Биологические образцы включают без ограничения кровь, сыворотку крови, мочу, экскременты, семенную жидкость или ткань.
[0031] На фигуре 1 представлены характерные измерения скорости ассоциации и скорости диссоциации, полученные с помощью поверхностного плазмонного резонанса (SPR), в отношении взаимодействия разных репрезентативных мутеинов липокалина (SEQ ID NO показаны на графике) с CD137 человека (слияние с Fc) в качестве мишени. Мишени иммобилизировали с использованием антитела к Fc IgG человека, которое было, в свою очередь, иммобилизировано на сенсорном чипе с использованием общепринятой химии сшивания аминов. Мутеины липокалина использовали в качестве растворимого аналита, который проходил при разных концентрациях по поверхности чипа. Имеют место отчетливые сигналы связывания, обнаруженные с помощью SPR, относительно мишени у человека, белка слияния CD137 человека-Fc (huCD137-Fc), для всех тестируемых мутеинов, тогда как отрицательные контроли, представляющие собой SEQ ID NO: 3 и SEQ ID NO: 4, не проявляют связывание. Константы диссоциации, полученные в результате выравнивания (модель связывания 1:1) показанных данных для всех SEQ ID NO, представлены в таблице 1.
[0032] На фигуре 2 представлены репрезентативные примеры основанного на SPR эксперимента, предназначенного для изучения того, нарушают ли мутеины с SEQ ID NO: 5, SEQ ID NO: 12 и SEQ ID NO: 13 связывание лиганда CD137 (CD137L) с CD137. Это изучали путем создания комплекса CD137 и CD137L на сенсорном чипе для SPR и проверки того, могут ли тестируемые мутеины липокалина связываться с этим комплексом или нет. В качестве эталона CD137 в отсутствие CD137L инкубировали с мутеинами липокалина. На фигурах представлены только релевантные сегменты сенсограмм. Кривая, полученная с помощью SPR, в отношении связывания соответствующего мутеина липокалина только с huCD137-Fc обозначена стрелкой с непрерывной линией. Кривая, полученная с помощью SPR, в отношении связывания соответствующего мутеина липокалина с huCD137-Fc, который был насыщен CD137L, обозначена стрелкой с прерывистой линией. На фигуре 2(A) показано, что SEQ ID NO: 5 не может связываться с huCD137-Fc в присутствии CD137L. На фигуре 2(B) и на фигуре 2(C) показано, что SEQ ID NO: 12 и SEQ ID NO: 13 связываются с huCD137-Fc с очень похожим ответом как в отсутствие, так и в присутствии CD137L, демонстрируя, что конкуренция в связывании между двумя мутеинами липокалина и CD137L отсутствует.
[0033] На фигуре 3 показаны репрезентативные примеры исследований с использованием сортировки флуоресцентно-активированных клеток (FACS), осуществленных для того, чтобы оценить специфичное связывание репрезентативных мутеинов липокалина (SEQ ID NO показаны на графике) с CD137 человека, экспрессированным на клетках млекопитающих. Ложно-трансфицированные клетки служили в качестве отрицательного контроля.
[0034] На фигуре 4 показаны результаты анализа активации T-клеток, который осуществляли для того, чтобы оценить способность набора репрезентативных связывающих CD137 мутеинов липокалина (SEQ ID NO: 13, SEQ ID NO: 14 и SEQ ID NO: 15) к костимуляции ответов T-клеток, при нанесении на пластиковую чашку, путем индуцирования образования скоплений и сопутствующей нисходящей передачи сигнала, связанного с CD137. Кроме того, активацию T-клеток путем инкубации с растворимыми мутеинами липокалина тестировали с целью изучения того, проявляют ли соответствующие связывающие молекулы агонистическую активность в отсутствии скоплений. На фигуре 4(A) мутеины липокалина наносили на пластиковую чашку совместно с антителом к CD3 человека, а затем очищенные T-клетки инкубировали на покрытой поверхности в присутствии растворимого антитела к CD28 человека. На фигуре 4(B) антитело к CD3 человека наносили на пластиковую чашку, а затем очищенные T-клетки инкубировали на покрытой поверхности в присутствии растворимого антитела к CD28 человека и мутеинов липокалина в растворе. В обоих случаях уровни интрелейкина 2 (IL-2) в супернатанте служили в качестве показателей для считывания. В качестве отрицательного контроля использовали SEQ ID NO: 4. В эксперименте с фигуры 4(A), имеет место отчетливо повышенная концентрация IL-2 в супернатанте вследствие активации T-клеток в ответ на мутеины липокалина с SEQ ID NO: 13, SEQ ID NO: 14 и SEQ ID NO: 15 по сравнению с отрицательным контролем, представляющим собой SEQ ID NO: 4. В отношении эксперимента, в котором используют мутеины липокалина в растворе, на фигуре 4(B) отсутствует значимое повышение концентрации IL-2 в супернатанте в ответ на любой из тестируемых мутеинов липокалина по сравнению с отрицательным контролем, представляющим собой SEQ ID NO: 4. В совокупности на фигурах 4(A) и 4(B) показано, что тестируемые мутеины липокалина проявляют необходимое поведение: образование скоплений CD137 на поверхности T-клетки с помощью нанесенных на пластик мутеинов к CD137 приводит к необходимой костимуляции T-клеток, тогда как соответствующий мутеин в растворе, при связывании с CD137, которое показано в примере 4 и на фигуре 3, не индуцирует какую-либо костимуляцию T-клеток.
[0035] На фигуре 5 представлены результаты эксперимента активации T-клеток, в котором использовали связывающий CD137 мутеин липокалина с SEQ ID NO: 13 в качестве тестовой молекулы. SEQ ID NO: 4 использовали в качестве отрицательного контроля. Эксперимент проводили с использованием дополнительной стимуляции на субоптимальном уровне антителом к CD3 и антителом к CD28 T-клеток с регистрируемыми показателями, представленными на фигуре 5(A) в виде длительной пролиферации T-клеток после трех дней инкубации с использованием импульсного мечения BrdU в течение 4 ч, на фигуре 5(C) в виде концентрации IL-2 в супернатанте и на фигуре 5(E) в виде уровней IFN-g в супернатанте. В качестве альтернативы использовали только субоптимальную концентрацию антитела к CD3 с регистрируемыми показателями, представленными на фигуре 5(B) в виде длительной пролиферации, на фигуре 5(D) в виде концентрации IL-2 в супернатанте и на фигуре 5(F) в виде уровней IFN-g в супернатанте. В эксперименте показаны зависимые от дозы SEQ ID NO: 13 повышения уровней пролиферации, IL-2 и IFN-гамма с использованием как стимуляции антителом к CD3/антителом к CD28, так и стимуляции только антителом к CD3.
IV. Подробное описание раскрытия
[0036] Как используется в данном документе, «липокалин» определен как мономерный белок весом примерно 18-20 кДа, имеющий участок с супервторичной структурой, представляющей собой цилиндрический β-складчатый лист, содержащий множество (предпочтительно восемь) β-нитей, соединенных попарно множеством петель (предпочтительно четырьмя) на одном конце с образованием таким образом кармана связывания. Именно разнообразие петель в остальной части жесткого остова липокалина приводит к появлению разнообразия различных видов связывания среди представителей семейства липокалинов, при этом каждый способен приспосабливаться к мишеням различных размера, формы и с различными химическими особенностями (рассмотрены, например, в Flower, D.R. (1996), ранее; Flower, D.R. et al. (2000), ранее, или Skerra, A. (2000) Biochim. Biophys. Acta 1482, 337-350). В действительности липокалиновое семейство белков естественным образом развивалось для связывания широкого спектра лигандов, при этом они характеризуются необычайно низкими уровнями общей консервативности последовательностей между собой (часто с идентичностями последовательностей менее 20%), сохраняя по-прежнему высококонсервативный общий паттерн фолдинга. Соответствие между положениями в разных липокалинах хорошо известно специалисту в данной области. См., например, патент США № 7250297.
[0037] Как отмечалось выше, липокалин представляет собой полипептид, определяемый его супервторичной структурой, а именно участком с супервторичной структурой, представляющей собой цилиндрический β-складчатый лист, содержащим восемь β-нитей, соединенных попарно четырьмя петлями на одном конце с образованием таким образом кармана связывания. Настоящее раскрытие не ограничено мутеинами липокалина, конкретно раскрытыми в данном документе. В этом отношении настоящее раскрытие относится к мутеину липокалина, имеющему участок с супервторичной структурой, представляющей собой цилиндрический β-складчатый лист, содержащий восемь β-нитей, соединенных попарно четырьмя петлями на одном конце с образованием таким образом кармана связывания, где по меньшей мере одна аминокислота из каждой по меньшей мере из трех из указанных четырех петель была подвергнута мутации и где указанный липокалин является эффективным в связывании CD137 с выявляемой аффинностью.
[0038] Согласно одному конкретному варианту осуществления мутеин липокалина, раскрытый в данном документе, представляет собой мутеин липокалина слезы человека (TLPC или Tlc), также называемый липокалином-1, преальбумином слезы или белком железы фон Эбнера. Термин «липокалин слезы человека», или «Tlc», или «липокалин-1», используемый в данном документе, относится к зрелому липокалину слезы человека с номером доступа в базе данных SWISS-PROT/UniProt P31025 (изоформа 1). Аминокислотную последовательность, показанную под номером доступа в базе данных SWISS-PROT/UniProt P31025, можно использовать в качестве предпочтительной «эталонной последовательности», более предпочтительно аминокислотную последовательность, показанную под SEQ ID NO: 1, используют в качестве эталонной последовательности.
[0039] Согласно другому конкретному варианту осуществления мутеин липокалина, раскрытый в данном документе, представляет собой мутеин липокалина 2 человека. Термин «липокалин 2 человека», или «Lcn 2 человека», или «NGAL человека», используемый в данном документе, относится к зрелому человеческому липокалину, ассоциированному с желатиназой нейтрофилов (NGAL), с номером доступа в базе данных SWISS-PROT/UniProt P80188. Мутеин липокалина 2 человека по настоящему раскрытию также можно назвать в данном документе «мутеином hNGAL». Аминокислотную последовательность, показанную под номером доступа в базе данных SWISS-PROT/UniProt P80188, можно использовать в качестве предпочтительной «эталонной последовательности», более предпочтительно аминокислотную последовательность, показанную под SEQ ID NO: 2, используют в качестве эталонной последовательности.
[0040] Согласно некоторым вариантам осуществления, связывающий CD137 мутеин липокалина с выявляемой аффинностью может включать по меньшей мере одну аминокислотную замену нативного остатка цистеина другой аминокислотой, например остатком серина. Согласно некоторым другим вариантам осуществления, связывающий CD137 мутеин липокалина с выявляемой аффинностью может включать один или более ненативных остатков цистеина, замещающих одну или более аминокислот липокалина дикого типа. Согласно дополнительному конкретному варианту осуществления мутеин липокалина согласно настоящему раскрытию включает по меньшей мере две аминокислотные замены нативной аминокислоты остатком цистеина с образованием тем самым одного или более цистеиновых мостиков. Согласно некоторым вариантам осуществления, указанный цистеиновый мостик может соединять по меньшей мере два участка петли. Определение этих участков используют в данном документе в соответствии с Flower (Flower, 1996, ранее, Flower, et al., 2000, ранее) и Breustedt et al. (2005, ранее). В родственном варианте осуществления в раскрытии сообщается об одном или более мутеинах липокалина, которые способны к активации нисходящих сигнальных путей, связанных с CD137, путем связывания с CD137.
[0041] Белки по настоящему раскрытию, которые направлены против или специфичны в отношении CD137, включают любое количество мутеинов-белков со специфическим связыванием, в которых за основу взят остов определенного белка. Предпочтительно количество нуклеотидов или аминокислот соответственно, которое обменивают, подвергают делеции или вставляют, составляет 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 или больше, а именно 25, 30, 35, 40, 45 или 50, при этом предпочтительными являются 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или 11, и даже более предпочтительными являются 9, 10 или 11. Однако предпочтительно, чтобы мутеин липокалина по настоящему раскрытию по-прежнему был способен к связыванию CD137.
[0042] Согласно одному аспекту настоящее раскрытие включает разные мутеины липокалина, которые связывают CD137 по меньшей мере с выявляемой аффинностью. В этом смысле CD137 может быть рассмотрен как отличный от естественного лиганд эталонного липокалина дикого типа, где «отличный от естественного лиганд» относится к соединению, которое не связывается с липокалинами дикого типа в физиологических условиях. Путем внесения в липокалины дикого типа одной или более мутаций в определенные положения в последовательности авторы настоящего изобретения показали, что возможны высокая аффинность и высокая специфичность в отношении отличного от естественного лиганда, CD137. Согласно некоторым вариантам осуществления в 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 или даже большем количестве нуклеотидных триплетов, кодирующих определенные положения в последовательности в липокалинах дикого типа, случайный мутагенез можно осуществлять посредством замены в этих положениях подмножеством нуклеотидных триплетов.
[0043] К тому же мутеины липокалина по настоящему раскрытию могут иметь подвергнутый мутации аминокислотный остаток в любом одном или более, в том числе по меньшей мере в любом одном, двух, трех, четырех, пяти, шести, семи, восьми, девяти, десяти, одиннадцати или двенадцати, из положений в последовательности, соответствующих определенным положениям в последовательности линейной полипептидной последовательности эталонного липокалина.
[0044] Белок по настоящему раскрытию может включать дикого типа (природную) аминокислотную последовательность «исходного» остова белка (такого как липокалин) вне подвергнутых мутации положений в аминокислотной последовательности. Согласно некоторым вариантам осуществления мутеин липокалина согласно настоящему раскрытию также может нести одну или более аминокислотных мутаций в положении/положениях в последовательности, если только такая мутация по меньшей мере в значительной степени не препятствует связывающей активности и фолдингу мутеина или не нарушает их. Такие мутации могут быть очень легко выполнены на уровне ДНК с использованием разработанных стандартных способов (Sambrook, J. et al. (2001) Molecular Cloning: A Laboratory Manual, 3rd Ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY). Иллюстративные примеры перестроек аминокислотной последовательности представляют собой вставки или делеции, а также аминокислотные замены. Такие замены могут быть консервативными, т. е. аминокислотный остаток замещается аминокислотным остатком с химически подобными свойствами, в частности в отношении полярности, а также размера. Примеры консервативных замен представляют собой замещения из числа представителей следующих групп: 1) аланин, серин и треонин; 2) аспарагиновая кислота и глутаминовая кислота; 3) аспарагин и глутамин; 4) аргинин и лизин; 5) изолейцин, лейцин, метионин и валин и 6) фенилаланин, тирозин и триптофан. С другой стороны, существует также возможность введения неконсервативных перестроек в аминокислотную последовательность. К тому же вместо замещения отдельных аминокислотных остатков существует также возможность либо вставлять, либо подвергать делеции одну или более последовательных аминокислот первичной структуры липокалина слезы человека, если только эти делеции или вставка приводят в результате к стабильно свернутому/функциональному мутеину (например, мутеинам Tlc с усеченными N- и C-концами). В таком мутеине, к примеру, один или более аминокислотных остатков добавлены или подвергнуты делеции на N- или C-конце полипептида. В целом такой мутеин может характеризоваться приблизительно по меньшей мере 70%, в том числе по меньшей мере приблизительно 80%, а именно по меньшей мере приблизительно 85% идентичностью аминокислотной последовательности с аминокислотной последовательностью зрелого липокалина слезы человека. В качестве иллюстративного примера, настоящее раскрытие также охватывает мутеины Tlc, определенные выше, в которых первых четыре N-концевых аминокислотных остатка последовательности зрелого липокалина слезы человека (His-His-Leu-Leu; положения 1-4) и/или последних два C-концевых аминокислотных остатка (Ser-Asp; положения 157-158) линейной полипептидной последовательности зрелого липокалина слезы человека были подвергнуты делеции (SEQ ID NO: 5-11). Кроме того, в качестве другого иллюстративного примера настоящее раскрытие также охватывает мутеины NGAL, определенные выше, в которых были подвергнуты делеции аминокислотные остатки (Lys-Asp-Pro, положения 46-48) линейной полипептидной последовательности зрелого липокалина 2 человека (hNGAL) (SEQ ID NO: 16).
[0045] Аминокислотная последовательность мутеина липокалина, раскрытого в данном документе, характеризуется высокой идентичностью последовательности с эталонным липокалином по сравнению с идентичностями последовательностей с другими липокалинами. В этом общем смысле аминокислотная последовательность мутеина липокалина по настоящему раскрытию по меньшей мере в значительной степени подобна аминокислотной последовательности эталонного липокалина, при условии, что в ней возможны гэпы (как определено ниже) при выравнивании, которые являются результатом добавлений или делеций аминокислот. Соответствующая последовательность мутеина липокалина по настоящему раскрытию, которая в значительной степени подобна последовательностям эталонного липокалина, согласно некоторым вариантам осуществления характеризуется по меньшей мере 70% идентичностью или гомологией последовательности, по меньшей мере 75% идентичностью или гомологией последовательности, по меньшей мере 80% идентичностью или гомологией последовательности, по меньшей мере 82% идентичностью или гомологией последовательности, по меньшей мере 85% идентичностью или гомологией последовательности, по меньшей мере 87% идентичностью или гомологией последовательности или по меньшей мере 90% идентичностью или гомологией последовательности, в том числе по меньшей мере 95% идентичностью или гомологией последовательности с последовательностью эталонного липокалина, при условии, что сохраняются измененные положение или последовательность и что возможны один или более гэпов.
[0046] Как используется в данном документе, мутеин липокалина по настоящему раскрытию «специфически связывает» мишень (например, CD137), если он способен отличать ту мишень от одной или более эталонных мишеней, поскольку специфичность связывания является не абсолютным, а относительным свойством. «Специфичное связывание» можно определять, например, согласно результатам вестерн-блоттингов, анализов ELISA, RIA, ECL, IRMA, с использованием FACS, IHC и пептидных сканирований.
[0047] Согласно одному варианту осуществления мутеины липокалина по настоящему раскрытию слиты на своем N-конце и/или своем C-конце с партнером по слиянию, который представляет собой белковый домен, который удлиняет время полужизни мутеина в сыворотке крови. Согласно дополнительным конкретным вариантам осуществления белковый домен представляет собой Fc-часть иммуноглобулина, домен CH3 иммуноглобулина, домен CH4 иммуноглобулина, альбумин-связывающий пептид или альбумин-связывающий белок.
[0048] Согласно другому варианту осуществления мутеины липокалина по настоящему раскрытию конъюгированы с соединением, которое удлиняет время полужизни мутеина в сыворотке крови. Более предпочтительно мутеин конъюгирован с соединением, выбранным из группы, состоящей из молекулы полиалкиленгликоля, гидроксиэтилкрахмала, Fc-части иммуноглобулина, домена CH3 иммуноглобулина, домена CH4 иммуноглобулина, альбумин-связывающего пептида и альбумин-связывающего белка.
[0049] Согласно еще одному варианту осуществления настоящее раскрытие относится к молекуле нуклеиновой кислоты, содержащей нуклеотидную последовательность, кодирующую мутеин липокалина, раскрытый в данном документе. Настоящее раскрытие охватывает клетку-хозяина, содержащую указанную молекулу нуклеиновой кислоты.
A. Мутеины липокалина, специфичные в отношении CD137
[0050] Согласно одному аспекту настоящее раскрытие предусматривает мутеины липокалина человека, которые связывают CD137, и их полезные пути применения. Настоящее раскрытие также предусматривает способы получения белков, связывающих CD137, описанных в данном документе, а также композиций, содержащих такие белки. Белки, связывающие CD137, по настоящему раскрытию, а также их композиции можно применять в способах выявления CD137 в образцах или в способах связывания CD137 в субъекте. Ранее не были описаны такие мутеины липокалина человека с такими качествами, сопутствующими путям применения, предусмотренным настоящим раскрытием.
1. Иллюстративные мутеины липокалина, специфичные в отношении CD137
[0051] Один вариант осуществления настоящего раскрытия относится к мутеину липокалина, который способен к связыванию CD137 с аффинностью, измеренной по KD, составляющей приблизительно 300 нМ, 100 нМ, 75 нМ, 50 нМ, 25 нМ, 10 нМ или даже ниже, а именно 2 нМ, например, как определено с помощью анализа с использованием поверхностного плазмонного резонанса (SPR), описанного, по сути, в примере 4.
[0052] Согласно другому варианту осуществления мутеин липокалина способен к связыванию CD137 со значением EC50, составляющим приблизительно 250 нМ или ниже, приблизительно 100 нМ или ниже, приблизительно 50 нМ или ниже, приблизительно 18 нМ или ниже, например, как определено с помощью анализа с использованием FACS, описанного, по сути, в примере 6.
[0053] Другой вариант осуществления настоящего раскрытия предусматривает мутеин липокалина, который способен к активации нисходящих сигнальных путей, связанных с CD137, путем связывания с CD137.
[0054] Согласно некоторым вариантам осуществления по сравнению с отрицательным контролем, представляющим собой SEQ ID NO: 4, мутеин липокалина по настоящему раскрытию способен индуцировать более высокую концентрацию IL-2, например, при измерении в функциональном анализе активации T-клеток, описанном, по сути, в примере 7.
[0055] Согласно некоторым другим вариантам осуществления по сравнению с отрицательным контролем, представляющим собой SEQ ID NO: 4, мутеин липокалина по настоящему раскрытию не приводит к более высокой концентрации IL-2, например, при измерении в функциональном анализе активации T-клеток, описанном, по сути, в примере 8.
[0056] Согласно некоторым вариантам осуществления по сравнению с отрицательным контролем, представляющим собой SEQ ID NO: 4, мутеин липокалина по настоящему раскрытию способен индуцировать пролиферацию с обеспечением более высоких уровней IL-2 и IFN-γ, например, при измерении в функциональном анализе активации T-клеток, описанном, по сути, в примере 9.
[0057] Согласно одному аспекту настоящее раскрытие предусматривает связывающие CD137 мутеины липокалина слезы человека.
[0058] В связи с этим настоящее раскрытие предусматривает один или более мутеинов Tlc, которые способны к связыванию CD137 с аффинностью, измеренной по KD, составляющей приблизительно 300 нМ или ниже и даже приблизительно 100 нМ или ниже.
[0059] Согласно некоторым вариантам осуществления такой мутеин Tlc содержит подвергнутый мутации аминокислотный остаток в одном или более положениях, соответствующих положениям 5, 26-31, 33-34, 42, 46, 52, 56, 58, 60-61, 65, 71, 85, 94, 101, 104-106, 108, 111, 114, 121, 133, 148, 150 и 153 в линейной полипептидной последовательности зрелого липокалина слезы человека (SEQ ID NO: 1).
[0060] Согласно некоторым конкретным вариантам осуществления такой мутеин Tlc может содержать подвергнутый мутации аминокислотный остаток в одном или более положениях, соответствующих положениям 26-34, 55-58, 60-61, 65, 104-106 и 108 в линейной полипептидной последовательности зрелого липокалина слезы человека.
[0061] Согласно дополнительным конкретным вариантам осуществления такой мутеин Tlc может дополнительно включать подвергнутый мутации аминокислотный остаток в одном или более положениях, соответствующих положениям 101, 111, 114 и 153 в линейной полипептидной последовательности зрелого липокалина слезы человека.
[0062] Согласно другим конкретным вариантам осуществления Tlc может содержать подвергнутый мутации аминокислотный остаток в одном или более положениях, соответствующих положениям 5, 26-31, 33-34, 42, 46, 52, 56, 58, 60-61, 65, 71, 85, 94, 101, 104-106, 108, 111, 114, 121, 133, 148, 150 и 153 в линейной полипептидной последовательности зрелого липокалина слезы человека.
[0063] Согласно некоторым дополнительным вариантам осуществления мутеин Tlc может содержать по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26 или даже больше подвергнутых мутации аминокислотных остатков в одном или более положениях в последовательности, соответствующих положениям 5, 26-31, 33-34, 42, 46, 52, 56, 58, 60-61, 65, 71, 85, 94, 101, 104-106, 108, 111, 114, 121, 133, 148, 150 и 153 последовательности в линейной полипептидной последовательности зрелого липокалина слезы человека, и где указанный полипептид связывает CD137, в частности CD137 человека.
[0064] Согласно еще некоторым дополнительным вариантам осуществления настоящее раскрытие относится к полипептиду, где указанный полипептид представляет собой мутеин Tlc, по сравнению с линейной полипептидной последовательностью зрелого липокалина слезы человека, содержащий по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 или даже больше подвергнутых мутации аминокислотных остатков в положениях 526-34, 55-58, 60-61, 65, 104-106 и 108 в последовательности, и где указанный полипептид связывает CD137, в частности CD137 человека.
[0065] Согласно некоторым вариантам осуществления мутеин липокалина согласно настоящему раскрытию может включать по меньшей мере одну аминокислотную замену нативного остатка цистеина, например, остатком серина. Согласно некоторым вариантам осуществления мутеин Tlc согласно настоящему раскрытию включает аминокислотную замену нативного остатка цистеина в положениях 61 и/или 153 другой аминокислотой, такой как остаток серина. В данном случае отмечается, что было обнаружено, что удаление дисульфидной связи в структуре (на уровне соответствующей библиотеки не подвергнутых мутации нуклеиновых кислот) липокалина дикого типа слезы, которая образована остатками цистеина 61 и 153 (см. Breustedt, et al., 2005, ранее), может обеспечить мутеины липокалина слезы, которые не только стабильно сворачиваются, но и также способны связывать указанный отличный от естественного лиганд с высокой аффинностью. Согласно некоторым конкретным вариантам осуществления мутеин Tlc согласно настоящему раскрытию включает аминокислотные замены Cys 61 → Ala, Phe, Lys, Arg, Thr, Asn, Gly, Gln, Asp, Asn, Leu, Tyr, Met, Ser, Pro или Trp и Cys 153 → Ser или Ala. Как было доказано, такая замена удобна для предупреждения образования встречающегося в природе дисульфидного мостика, соединяющего Cys 61 и Cys 153, и облегчения тем самым манипуляции с мутеином. Однако мутеины липокалина слезы, которые связывают CD137 и которые имеют дисульфидный мостик, образованный между Cys 61 и Cys 153, также являются частью настоящего раскрытия.
[0066] Согласно некоторым вариантам осуществления устранение дисульфидной связи в структуре может обеспечивать дополнительное преимущество предоставления возможности (самопроизвольного) образования или преднамеренного введения отличных от естественных искусственных дисульфидных связей в мутеины по настоящему раскрытию, повышая тем самым стабильность мутеинов. Например, согласно некоторым вариантам осуществления или два, или все три из кодонов цистеина в положениях 61, 101 и 153 заменены кодоном другой аминокислоты. Кроме того, согласно некоторым вариантам осуществления мутеин Tlc согласно настоящему раскрытию включает аминокислотную замену нативного остатка цистеина в положении 101 остатком серина или остатком гистидина.
[0067] Согласно некоторым вариантам осуществления мутеин согласно настоящему раскрытию включает аминокислотную замену нативной аминокислоты остатком цистеина в положениях 28 или 105 в отношении аминокислотной последовательности зрелого липокалина слезы человека.
[0068] Кроме того, согласно некоторым вариантам осуществления мутеин согласно настоящему раскрытию включает аминокислотную замену нативного остатка аргинина в положении 111 остатком пролина. Кроме того, согласно некоторым вариантам осуществления мутеин согласно настоящему раскрытию включает аминокислотную замену нативного остатка лизина в положении 114 остатком триптофана или глутаминовой кислотой.
[0069] Согласно некоторым вариантам осуществления связывающий CD137 мутеин Tlc согласно настоящему раскрытию включает в одном или более положениях, соответствующих положениям 5, 26-31, 33-34, 42, 46, 52, 56, 58, 60-61, 65, 71, 85, 94, 101, 104-106, 108, 111, 114, 121, 133, 148, 150 и 153 в линейной полипептидной последовательности зрелого липокалина слезы человека (SEQ ID NO: 1), один или более из следующих подвергнутых мутации аминокислотных остатков: Ala 5 → Val или Thr; Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Thr 42 → Ser; Gly 46 → Asp; Lys 52 → Glu; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Lys 65 → Arg или Asn; Thr 71 → Ala; Val 85 → Asp; Lys 94 → Arg или Glu; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Lys 121 → Glu; Ala 133 → Thr; Arg 148 → Ser; Ser 150 → Ile и Cys 153 → Ser. Согласно некоторым вариантам осуществления мутеин Tlc согласно настоящему раскрытию включает два или более, а именно 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, более того, как например: 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26 или все подвергнутые мутации аминокислотные остатки в этих положениях в последовательности зрелого липокалина слезы человека.
[0070] Согласно некоторым дополнительным вариантам осуществления связывающий CD137 мутеин Tlc предусматривает один из следующих наборов аминокислотных замен по сравнению с линейной полипептидной последовательностью зрелого липокалина слезы человека:
1. Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Cys 153 → Ser;
2. Ala 5 → Thr; Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Lys 65 → Arg; Val 85 → Asp; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Lys 121 → Glu; Ala 133 → Thr; Cys 153 → Ser; 157 → Pro;
3. Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Lys 65 → Asn; Lys 94 → Arg; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Lys 121 → Glu; Ala 133 → Thr; Cys 153 → Ser;
4. Ala 5 → Val; Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Lys 65 → Arg; Lys 94 → Glu; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Lys 121 → Glu; Ala 133 → Thr; Cys 153 → Ser; 157 → Pro;
5. Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Thr 42 → Ser; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Ser 150 → Ile; Cys 153 → Ser; 157 → Pro;
6. Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Lys 52 → Glu; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Thr 71 → Ala; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Ala 133 → Thr; Arg 148 → Ser; Ser 150 → Ile; Cys 153 → Ser; 157 → Pro или
7. Ala 5 → Thr; Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Gly 46 → Asp; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Thr 71 → Ala; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Ser 150 → Ile; Cys 153 → Ser; 157 → Pro.
[0071] В остальном участке, т. е. участке, отличающемся от положений 5, 26-31, 33-34, 42, 46, 52, 56, 58, 60-61, 65, 71, 85, 94, 101, 104-106, 108, 111, 114, 121, 133, 148, 150 и 153 в последовательности, мутеин Tlc по настоящему раскрытию может включать аминокислотную последовательность дикого типа (природную) вне подвергнутых мутации положений аминокислотной последовательности.
[0072] В еще дополнительных вариантах осуществления мутеин Tlc согласно настоящему раскрытию характеризуется по меньшей мере 70% идентичностью последовательности или по меньшей мере 70% гомологией последовательности с последовательностью зрелого липокалина слезы человека (SEQ ID NO: 1). В качестве иллюстративного примера мутеин с SEQ ID NO: 7 характеризуется идентичностью аминокислотной последовательности или гомологией последовательности, составляющей примерно 81%, с аминокислотной последовательностью зрелого липокалина слезы человека.
[0073] Согласно дополнительным конкретным вариантам осуществления мутеин Tlc по настоящему раскрытию содержит аминокислотную последовательность, изложенную под любой из SEQ ID NO: 5-11, или ее фрагмент или вариант.
[0074] Согласно дополнительным конкретным вариантам осуществления мутеин Tlc по настоящему раскрытию характеризуется по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85% или более высокой идентичностью последовательности с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 5-11.
[0075] Настоящее раскрытие также включает структурные гомологи мутеина Tlc с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 5-11, структурные гомологи которой характеризуются гомологией аминокислотной последовательности или идентичностью последовательностей, составляющей более чем приблизительно 60%, предпочтительно более чем 65%, более чем 70%, более чем 75%, более чем 80%, более чем 85%, более чем 90%, более чем 92% и наиболее предпочтительно более чем 95% относительно указанного мутеина Tlc.
[0076] Мутеин Tlc согласно настоящему раскрытию можно получать с помощью мутагенеза встречающейся в природе формы липокалина слезы человека. Согласно некоторым вариантам осуществления мутагенеза замена (или замещение) представляет собой консервативную замену. Тем не менее, любая замена, в том числе неконсервативная замена или одна или более из приведенных ниже иллюстративных замен, предусматривается, если только мутеин липокалина сохраняет свою способность связываться с CD137, и/или он характеризуется идентичностью последовательности с впоследствии замещенной последовательностью в том смысле, что имеет место по меньшей мере 60%, а именно по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85% или более идентичность последовательности с аминокислотной последовательностью зрелого липокалина слезы человека (номер доступа в базе данных SWISS-PROT - P31025).
[0077] Согласно некоторым дополнительным вариантам осуществления мутеин hNGAL по настоящему раскрытию способен нарушать связывание CD137L с CD137, например, как измерено в анализе с использованием поверхностного плазмонного резонанса (SPR), описанном, по сути, в примере 5.
[0078] Согласно некоторым конкретным вариантам осуществления настоящее раскрытие предусматривает мутеин липокалина, который связывает CD137 с аффинностью, измеренной по KD, составляющей приблизительно 200 нМ или ниже, при этом мутеин липокалина характеризуется по меньшей мере 90% или более высокой, как, например, 95% идентичностью с аминокислотной последовательностью под SEQ ID NO: 5.
[0079] Согласно другому аспекту настоящее раскрытие относится к новым мутеинам липокалина 2 человека (Lcn2 человека или hNGAL) со специфичностью связывания, направленным против или специфичным в отношении CD137.
[0080] В связи с этим настоящее раскрытие предусматривает один или более мутеинов hNGAL, которые способны к связыванию CD137 с аффинностью, измеренной по KD, составляющей 200 нМ или ниже, приблизительно 140 нМ или ниже, приблизительно 50 нМ или ниже и даже приблизительно 10 нМ или ниже. Более предпочтительно мутеины hNGAL могут характеризоваться аффинностью, измеренной по KD, составляющей приблизительно 5 нМ или ниже.
[0081] Согласно некоторым вариантам осуществления мутеин hNGAL по настоящему раскрытию включает в одном или более положениях, соответствующих положениям 28, 36, 40-41, 49, 52, 65, 68, 70, 72-73, 77, 79, 81, 83, 87, 94, 96, 100, 103, 106, 125, 127, 132 и 134 в линейной полипептидной последовательности зрелого hNGAL (SEQ ID NO: 2), замену.
[0082] Согласно конкретным вариантам осуществления мутеин липокалина по настоящему раскрытию содержит по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21 или даже больше замен в положении последовательности, соответствующем положению 28, 36, 40-41, 49, 52, 65, 68, 70, 72-73, 77, 79, 81, 83, 87, 94, 96, 100, 103, 106, 125, 127, 132 и 134 последовательности в линейной полипептидной последовательности зрелого hNGAL (номер доступа базы данных SWISS-PROT P80188; SEQ ID NO: 2). Предпочтительно предусматривают, что настоящее раскрытие относится к мутеину липокалина, который содержит, в дополнение к одной или более заменам в положениях, соответствующих положениям 36, 87 и/или 96 в линейной полипептидной последовательности зрелого NGAL человека, в одном или более положениях, соответствующих положениям 28, 40-41, 49, 52, 65, 68, 70, 72-73, 77, 79, 81, 83, 94, 100, 103, 106, 125, 127, 132 и 134 в линейной полипептидной последовательности зрелого hNGAL, замену.
[0083] Согласно еще некоторым дополнительным вариантам осуществления настоящее раскрытие относится к полипептиду, где указанный полипептид представляет собой мутеин hNGAL, который по сравнению с линейной полипептидной последовательностью зрелого hNGAL (номер доступа базы данных SWISS-PROT P80188; SEQ ID NO: 2) содержит по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21 или даже больше подвергнутых мутации аминокислотных остатков в положениях 28, 36, 40-41, 49, 52, 65, 68, 70, 72-73, 77, 79, 81, 87, 96, 100, 103, 106, 125, 127, 132 и 134 последовательности, и где указанный полипептид связывает CD137, в частности CD137 человека.
[0084] Согласно некоторым вариантам осуществления связывающий CD137 мутеин hNGAL по настоящему раскрытию включает в любом одном или более из положений 28, 36, 40-41, 49, 52, 65, 68, 70, 72-73, 77, 79, 81, 83, 87, 94, 96, 100, 103, 106, 125, 127, 132 и 134 последовательности в линейной полипептидной последовательности зрелого hNGAL (SEQ ID NO: 2) один или более из следующих подвергнутых мутации аминокислотных остатков: Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Arg или Lys; Gln 49 → Val, Ile, His, Ser или Asn; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Met, Ala или Gly; Leu 70 → Ala, Lys, Ser или Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Met, Arg, Thr или Asn; Trp 79 → Ala или Asp; Arg 81 → Met, Trp или Ser; Phe 83 → Leu; Cys 87 → Ser; Leu 94 → Phe; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu и Lys 134 → Tyr.
[0085] Согласно некоторым вариантам осуществления мутеин hNGAL по настоящему раскрытию включает два или более, а именно 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, более того, как например: 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или все подвергнутые мутации аминокислотные остатки в этих положениях в последовательности зрелого hNGAL.
[0086] Согласно некоторым дополнительным вариантам осуществления мутеин hNGAL по настоящему раскрытию, который связывается с CD137, включает следующие аминокислотные замещения по сравнению с линейной полипептидной последовательностью зрелого hNGAL:
(a) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Lys; Gln 49 → Asn; Tyr 52 → Met; Ser 68 → Gly; Leu 70 → Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Ala; Arg 81 → Ser; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; Lys 134 → Tyr;
(b) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Arg; Gln 49 → Ile; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Met; Leu 70 → Lys; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Met; Trp 79 → Asp; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; Lys 134 → Tyr;
(c) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Arg; Gln 49 → Asn; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Ala; Leu 70 → Ala; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Asp; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; Lys 134 → Tyr;
(d) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Lys; Gln 49 → Asn; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Ala; Leu 70 → Ala; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Asp; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; Lys 134 → Tyr;
(e) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Lys; Gln 49 → Ser; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Gly; Leu 70 → Ser; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Ala; Arg 81 → Met; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; Lys 134 → Tyr;
(f) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Lys; Gln 49 → Val; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Gly; Leu 70 → Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Arg; Trp 79 → Asp; Arg 81 → Ser; Cys 87 → Ser; Leu 94 → Phe; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; Lys 134 → Tyr;
(g) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Arg; Gln 49 → His; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Gly; Leu 70 → Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Ala; Arg 81 → Ser; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; Lys 134 → Tyr;
(h) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Lys; Gln 49 → Asn; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Gly; Leu 70 → Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Ala; Arg 81 → Ser; Phe 83 → Leu; Cys 87 → Ser; Leu 94 → Phe; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; Lys 134 → Tyr или
(i) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Arg; Gln 49 → Ser; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Ala; Leu 70 → Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Asn; Trp 79 → Ala; Arg 81 → Ser; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; Lys 134 → Tyr.
[0087] В остальном участке, т. е. участке, отличающемся от положений 28, 36, 40-41, 49, 52, 65, 68, 70, 72-73, 77, 79, 81, 83, 87, 94, 96, 100, 103, 106, 125, 127, 132 и 134 в последовательности, мутеин hNGAL по настоящему раскрытию может включать аминокислотную последовательность дикого типа (природную) вне подвергнутых мутации положений аминокислотной последовательности.
[0088] Согласно другому варианту осуществления мутеин hNGAL характеризуется по меньшей мере 70% или даже большей идентичностью последовательности с аминокислотной последовательностью зрелого липокалина 2 человека (номер доступа в базе данных SWISS-PROT P80188). В качестве иллюстративного примера мутеин с SEQ ID NO: 17 характеризуется идентичностью аминокислотной последовательности или гомологией последовательности, составляющей примерно 86,5%, с аминокислотной последовательностью зрелого hNGAL.
[0089] Согласно дополнительным конкретным вариантам осуществления мутеин липокалина согласно настоящему раскрытию содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 12-20 или их фрагмента или варианта.
[0090] Аминокислотная последовательность связывающего CD137 мутеина hNGAL по настоящему раскрытию может характеризоваться высокой идентичностью последовательности, а именно по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 82%, по меньшей мере 85%, по меньшей мере 87%, по меньшей мере 90% идентичностью, в том числе по меньшей мере 95% идентичностью с последовательностью, выбранной из группы, состоящей из SEQ ID NO: 12-20.
[0091] Настоящее раскрытие также включает структурные гомологи мутеина hNGAL с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 12-20, структурные гомологи которой характеризуются гомологией аминокислотной последовательности или идентичностью последовательностей, составляющей более чем приблизительно 60%, предпочтительно более чем 65%, более чем 70%, более чем 75%, более чем 80%, более чем 85%, более чем 90%, более чем 92% и наиболее предпочтительно более чем 95% относительно указанного мутеина hNGAL.
[0092] Мутеин hNGAL согласно настоящему раскрытию можно получать с помощью мутагенеза встречающейся в природе формы липокалина 2 человека. Согласно некоторым вариантам осуществления мутагенеза замена (или замещение) представляет собой консервативную замену. Тем не менее любая замена, в том числе неконсервативная замена или одна или более из приведенных ниже иллюстративных замен, предусматривается в том случае, если только мутеин липокалина сохраняет свою способность связываться с CD137, и/или он характеризуется идентичностью с впоследствии замещенной последовательностью в том смысле, что имеет место по меньшей мере 60%, а именно по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85% или больше идентичность с аминокислотной последовательностью зрелого липокалина 2 человека (номер доступа базы данных SWISS-PROT P80188).
[0093] Согласно некоторым дополнительным вариантам осуществления мутеин Tlc по настоящему раскрытию не нарушает связывание CD137L с CD137, например, как измерено в анализе с использованием поверхностного плазмонного резонанса (SPR), описанном, по сути, в примере 5.
[0094] Согласно некоторым конкретным вариантам осуществления настоящее раскрытие предусматривает мутеин липокалина, который связывает CD137 с аффинностью, измеренной по KD, составляющей приблизительно 5 нМ или ниже, при этом мутеин липокалина характеризуется по меньшей мере 90% или более высокой, а именно 95% идентичностью с аминокислотной последовательностью под SEQ ID NO: 13.
2. Пути применения мутеинов липокалина, специфичных в отношении CD137
[0095] CD137 представляет собой T-клеточный костимулирующий рецептор, индуцируемый при активации T-клеточного рецептора (TCR) (Nam et al., Curr. Cancer Drug Targets, 5:357-363 (2005); Watts et al., Annu. Rev. Immunol., 23:23-68 (2005)). Помимо его экспрессии на активированных CD4+ и CD8+ T-клетках, CD137 также экспрессируется на CD4+CD25+ регуляторных T-клетках, клетках естественных киллерах (NK) и NK-T-клетках, моноцитах, нейтрофилах и дендритных клетках. Его естественный лиганд, CD137L, был описан для антиген-презентирующих клеток, в том числе B-клеток, моноцита/макрофага и дендритных клеток (Watts et al., Annu. Rev. Immunol., 23:23-68 (2005)). При взаимодействии со своим лигандом CD137 вызывает повышенную индуцированную TCR пролиферацию T-клеток, продукцию цитокинов, функциональное созревание и пролонгированное выживание CD8+ T-клеток (Nam et al., Curr. Cancer Drug Targets, 5:357-363 (2005), Watts et al., Annu. Rev. Immunol., 23:23-68 (2005)).
[0096] Взаимодействие CD137/CD137L вовлечено в разные аспекты иммунного ответа. Оно, по-видимому, важно в подавлении индуцированной активацией клеточной смерти у T-клеток (Hurtado et al., J. Immunol. 158:2600, 1997), но аннулирует антиапоптические эффекты других цитокинов у нейтрофилов (Heinisch et al., Eur. J. Immunol. 30:3441, 2001). Таким образом, CD137 может играть некоторую роль в гомеостазе, относящемся к иммунной функции (Ebata et al., Eur. J. Immunol. 31:1210, 2001), и может представлять целевую костимулирующую систему, на которую можно целенаправленно воздействовать при лечении рака или воспалительного ответа (Blazar et al., J. Immunol. 166:174, 2001; Takahashi et al., Immunol. Lett. 76:183, 2001; Kim and Broxmeyer, J. Hematother. Stem Cell Res. 10:441, 2001; Kim et al., Cancer Res. 61:2031, 2001).
[0097] Ввиду этого в медицине существует множество возможных путей применения связывающих CD137 мутеинов липокалина по настоящему раскрытию. Согласно одному дополнительному аспекту настоящее раскрытие относится к применению связывающего CD137 мутеина липокалина, раскрытого в данном документе, для выявления CD137 в образце, а к также соответствующему способу диагностирования.
[0098] Настоящее раскрытие также включает применение одного или более связывающих CD137 мутеинов липокалина, которые описаны, для образования комплекса с CD137.
[0099] Ввиду этого в другом аспекте настоящего раскрытия раскрытые мутеины липокалина применяют для выявления CD137. Такое применение может включать стадии приведения одного или более указанных мутеинов, при соответствующих условиях, в контакт с образцом, который, как подозревают, содержит CD137, с обеспечением таким образом возможности образования комплекса между мутеинами и CD137 и выявления комплекса по соответствующему сигналу.
[00100] Выявляемый сигнал может быть обусловлен меткой, как разъяснено выше, или изменением физических свойств в результате связывания, т. е. образования комплекса как такового. Одним примером является поверхностный плазмонный резонанс, значение которого изменяется в ходе связывания партнеров по связыванию, из которых один иммобилизован на поверхности, такой как золотая фольга.
[00101] Связывающие CD137 мутеины липокалина, раскрытые в данном документе, также можно применять для отделения CD137. Такое применение может включать стадии приведения одного или более указанных мутеинов, при соответствующих условиях, в контакт с образцом, который, как полагают, содержит CD137, с обеспечением таким образом возможности образования комплекса между мутеинами и CD137 и отделения комплекса от образца.
[00102] При применении раскрытых мутеинов для выявления CD137, а также отделения CD137, мутеины и/или CD137 или их домен или фрагмент могут быть иммобилизованы на подходящей твердой фазе.
[00103] Согласно еще одному аспекту в настоящем раскрытии описан диагностический или аналитический набор, содержащий связывающий CD137 мутеин липокалина согласно настоящему раскрытию.
[00104] Помимо их применения в диагностике, согласно еще другому аспекту настоящее раскрытие предполагает фармацевтическую композицию, содержащую мутеин по настоящему раскрытию и фармацевтически приемлемый наполнитель.
[00105] Кроме того, настоящее раскрытие предусматривает мутеины липокалина человека, которые связывают CD137, для применения в качестве противораковых средств и/или иммуномодуляторов. В связи с этим мутеины липокалина по настоящему раскрытию, которые связывают CD137, предусматриваются как подлежащие применению в способе лечения или предупреждения заболеваний человека, таких как рак, инфекционные заболевания и аутоиммунные заболевания. Соответственно, также представлены способы лечения или предупреждения заболеваний человека, таких как рак, инфекционные заболевания и аутоиммунные заболевания, у нуждающегося в этом субъекта, включающие введение указанному субъекту терапевтически эффективного количества мутеина липокалина по настоящему изобретению, который связывает CD137.
[00106] В T-клетках опосредованная CD137 передача сигнала приводит к накоплению представителей семейства TRAF и активации некоторых киназ, в том числе ASK-1, MKK, MAPK3/MAPK4, p38 и JNK/SARK. Активация киназ затем сопровождается активаций и транслокацией в ядро некоторых факторов транскрипции, в том числе ATF-2, Jun и NF-κB. Помимо усиления индуцированной TCR пролиферации на субоптимальном уровне, опосредованная CD137 передача сигнала защищает T-клетки, и в частности CD8+T-клетки, от индуцированной активацией клеточной смерти (AICD).
[00107] Настоящее раскрытие охватывает применение связывающего CD137 мутеина липокалина по настоящему раскрытию или композиции, содержащей такой мутеин липокалина, для связывания CD137, костимуляции T-клеток и/или активации нисходящих сигнальных путей, связанных с CD137, путем связывания с CD137, в том числе усиления секреции IL-2 и продуцирования интерферона IFN-γ.
[00108] Настоящее раскрытие также описывает способ связывания CD137 или костимуляции T-клеток, включающий применение одного или более связывающих CD137 мутеинов липокалина по настоящему раскрытию или одной или более композиций, содержащих такие мутеины липокалина.
[00109] Кроме того, настоящее раскрытие предусматривает способ активации нисходящих сигнальных путей, связанных с CD137, в том числе усиления секреции IL-2 и продуцирования интерферона IFN-γ, включающий применение одного или более связывающих CD137 мутеинов липокалина по настоящему раскрытию или одной или более композиций, содержащих такие мутеины липокалина.
[00110] Настоящее раскрытие также предполагает способ индуцирования пролиферации T-лимфоцитов, включающий применение одного или более связывающих CD137 мутеинов липокалина по настоящему раскрытию или одной или более композиций, содержащих такие мутеины липокалина.
[00111] Более того, отсутствие взаимодействий CD137/CD137L предупреждает развитие определенных аутоиммунных заболеваний (Seo et al., 2003, 2004). Взаимодействия CD137/CD137L вовлечены в сеть гематопоэтических и отличных от гематопоэтических клеток, помимо хорошо изученных взаимодействий антиген-презентирующая клетка - T-клетка. Передача сигнала через CD137L играет ключевую роль в дифференцировке миелоидных клеток и их видах клеточной активности, позволяя предположить, что сигналы CD137L запускают и поддерживают воспаление. [Immune Network 2009;9(3):84-89].
[00112] Настоящее раскрытие охватывает применение связывающего CD137 мутеина липокалина по настоящему раскрытию, который способен к связыванию с CD137 на конкурентной основе с CD137L, или композицию, содержащую такой мутеин липокалина, для нарушения связывания CD137L с CD137 и/или естественной передачи сигнала CD137L.
В настоящем раскрытии также описан способ нарушения связывания CD137L с CD137, включающий применение одного или более конкурентно связывающих CD137 мутеинов липокалина по настоящему раскрытию или одной или более композиций, содержащих такие мутеины липокалина.
[00114] Кроме того, настоящее раскрытие включает способ нарушения естественной передачи сигнала CD137L, включающий применение одного или более конкурентно связывающих CD137 мутеинов липокалина по настоящему раскрытию или одной или более композиций, содержащих такие мутеины липокалина.
[00115] Настоящее раскрытие также предполагает способ снижения уровня продуцирования провоспалительных цитокинов и хемокинов, включающий применение одного или более конкурентно связывающих CD137 мутеинов липокалина по настоящему раскрытию или одной или более композиций, содержащих такие мутеины липокалина.
B. Мутеины липокалина по настоящему раскрытию
[00116] Липокалины являются белок-подобными связывающими молекулами, которые естественным образом развивались для связывания лигандов. Липокалины встречаются у многих организмов, в том числе у позвоночных, насекомых, растений и бактерий. Представители семейства белков липокалина (Pervaiz, S., & Brew, K. (1987) FASEB J. 1, 209-214), как правило, представляют собой малые секретируемые белки и имеют одиночную полипептидную цепь. Они характеризуются целым рядом различных свойств молекулярного распознавания: своей способностью связывать разные, в основном, гидрофобные молекулы (такие как ретиноиды, жирные кислоты, холестерины, простагландины, биливердины, феромоны, вкусовые добавки и отдушки), своим связыванием со специфичными рецепторами клеточной поверхности и своим образованием макромолекулярных комплексов. Хотя в прошлом их первоначально классифицировали как транспортные белки, теперь ясно, что липокалины выполняют множество физиологических функций. Они включают некоторые роли в транспорте ретинола, обонянии, передаче сигнала с помощью феромонов и синтезе простагландинов. Липокалины также вовлечены в регуляцию иммунного ответа и опосредование гомеостаза клеток (рассмотрено, например, в Flower, D.R. (1996) Biochem. J. 318, 1-14 и Flower, D.R. et al. (2000) Biochim. Biophys. Acta 1482, 9-24).
[00117] Липокалины характеризуются необычайно низкими уровнями общей консервативности последовательностей, часто с идентичностями последовательностей менее 20%. В то же время их общий паттерн фолдинга является высококонсервативным. Центральная часть структуры липокалина образована отдельным встречно-параллельным β-листом с восемью нитями, замкнутым самом на себе, с образованием β-бочонка с непрерывной сеткой водородных связей. Такой β-бочонок образует центральную полость. Один конец бочонка стерически блокирован N-концевым пептидным сегментом, который пересекает его дно, а также тремя пептидными петлями, соединяющими β-нити. Другой конец β-бочонка открыт для растворителя и содержит сайт связывания мишени, который образован четырьмя гибкими пептидными петлями. Именно это разнообразие петель в остальной части жесткого остова липокалина приводит к образованию разнообразия различных видов связывания, при этом каждый способен приспосабливаться к мишеням различных размера, формы и химических особенностей (рассмотрены, например, в Flower, D.R. (1996), ранее; Flower, D.R. et al. (2000), ранее, или Skerra, A. (2000) Biochim. Biophys. Acta 1482, 337-350).
[00118] Мутеин липокалина согласно настоящему раскрытию может быть мутеином любого выбранного липокалина. Примеры подходящих липокалина (также иногда называемых «белковыми 'эталонными' остовами» или просто «остовами»), мутеины которых можно использовать, включают без ограничения липокалин слезы (липокалин-1, белок железы фон Эбнера), ретинол-связывающий белок, нейтрофил, простагландин-D-синтазу липокалинового типа, β-лактоглобулин, билин-связывающий белок (BBP), аполипопротеин D (APO D), липокалин, ассоциированный с желатиназой нейтрофилов (NGAL), липокалин слезы (Tlc), родственный α2-микроглобулину белок (A2m), 24p3/утерокалин (24p3), белок 1 железы фон Эбнера (VEGP 1), белок 2 железы фон Эбнера (VEGP 2) и предшественник главного аллергена Can f1 (ALL-1). Согласно родственным вариантам осуществления мутеин липокалина выбран из группы, состоящей из человеческого липокалина, ассоциированного с желатиназой нейтрофилов (NGAL), липокалина слезы (Tlc) человека, аполипопротеина D (APO D) человека и билин-связывающего белка Pieris brassicae.
[00119] При использовании в данном документе в контексте мутеинов липокалина по настоящему раскрытию, которые связываются с CD137, термин «специфичный в отношении» включает то, что мутеин липокалина направлен соответственно против CD137, связывается или вступает в реакцию с ним. Таким образом, состояние «направленный на», «связывающийся с» или «вступающий в реакцию с» включает то, что мутеин липокалина соответственно специфически связывается с CD137. Термин «специфически» в данном случае означает, что мутеин липокалина вступает в реакцию с белком CD137, который описан в данном документе, но по существу не с другим белком. Термин «другой белок» включает любой отличный от CD137 белок, соответственно, в том числе белки близкородственные или являющиеся гомологичными CD137, против которых направлены липокалины, раскрытые в данном документе. Однако белки, фрагменты и/или варианты CD137 от видов, отличных от человека, как, например, тех, которые описаны в контексте определения «субъект», не исключены посредством термина «другой белок». Термин «фактически не связывает» означает, что мутеин липокалина по настоящему раскрытию не связывает другой белок, т. е. демонстрирует перекрестную реактивность менее 30%, предпочтительно 20%, более предпочтительно 10%, в частности предпочтительно менее 9, 8, 7, 6 или 5%. Вступает ли липокалин в реакцию специфически, как определено в данном документе выше, можно легко протестировать, в частности, путем сравнения реакции мутеина липокалина по настоящему раскрытию с CD137 и реакции указанного липокалина с другим(другими) белком(белками). «Специфичное связывание» также можно определять, например, согласно результатам вестерн-блоттингов, анализов ELISA, RIA, ECL, IRMA, с использованием FACS, IHC и пептидных сканирований.
[00120] Аминокислотная последовательность мутеина липокалина согласно настоящему раскрытию характеризуется высокой идентичностью последовательностей с соответствующим липокалином по сравнению с идентичностями последовательностей с другим липокалином (также см. выше). В таком общем смысле аминокислотная последовательность мутеина липокалина комбинации согласно настоящему раскрытию по меньшей мере в значительной степени подобна аминокислотной последовательности соответствующего липокалина (липокалина дикого типа или эталонного липокалина). Соответствующая последовательность мутеина липокалина комбинации согласно настоящему раскрытию, будучи в значительной степени подобной последовательностям соответствующего липокалина, имеет в некоторой части относительно липокалина дикого типа (или эталонного липокалина) один или более аминокислотных вариантов по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 82%, по меньшей мере 85%, по меньшей мере 87%, по меньшей мере 90% идентичности, в том числе по меньшей мере 95% идентичности, с последовательностью соответствующего липокалина. В связи с этим мутеин липокалина по настоящему раскрытию, разумеется, может содержать, в сравнении, замены, описанные в данном документе, которые делают мутеин липокалина способным к связыванию с CD137 соответственно. Как правило, мутеин липокалина включает одну или более мутаций - по отношению к нативной последовательности липокалина - аминокислот в четырех петлях на открытом конце сайта связывания лиганда липокалина (см. выше). Как разъяснено выше, эти участки являются важными в определении специфичности связывания мутеина липокалина в отношении необходимой мишени. В качестве иллюстративного примера мутеин, полученный из полипептида липокалина слезы, липокалина NGAL или их гомологов, может иметь один, два, три, четыре или более подвергнутых мутации аминокислотных остатков в любом положении последовательности в N-концевом участке и/или в трех пептидных петлях BC, DE и FG, расположенных на конце структуры в виде β-бочонка, которая расположена напротив естественного кармана связывания липокалина. В качестве дополнительного иллюстративного примера мутеин, полученный из полипептида липокалина слезы или его гомолога, может не иметь подвергнутых мутации аминокислотных остатков в пептидной петле DE, расположенной на конце структуры в виде β-бочонка, по сравнению с последовательностью дикого типа липокалина слезы.
[00121] Мутеин липокалина согласно настоящему раскрытию включает одну или более, как, например: две, три, четыре, пять, шесть, семь, восемь, девять, десять, одиннадцать, двенадцать, тринадцать, четырнадцать, пятнадцать, шестнадцать, семнадцать, восемнадцать, девятнадцать или даже двадцать замен по сравнению с соответствующим нативным липокалином при условии, что такой мутеин липокалина должен быть способным к связыванию с CD137 соответственно. Например, мутеин липокалина может иметь замену в положении, соответствующем отличному положению (т.е. в соответствующем положении) липокалина дикого типа с последовательностью дикого типа, например, липокалина слезы, липокалина NGAL или любого другого липокалина, описанного в данном документе. Согласно некоторым вариантам осуществления мутеин липокалина комбинации согласно настоящему раскрытию включает по меньшей мере две аминокислотные замены, в том числе 2, 3, 4, 5 или даже больше аминокислотных замен, нативной аминокислоты остатком аргинина. Соответственно, нуклеиновую кислоту белкового 'эталонного' остова, который описан в данном документе, подвергают мутагенезу с целью создания мутеина липокалина, который способен к связыванию с CD137 соответственно.
[00122] Также мутеин липокалина по настоящему раскрытию может содержать гетерологичную аминокислотную последовательность на своем N- или C-конце, предпочтительно C-конце, такой как с SEQ ID NO: 23, например, Strep-метку II, без воздействия на биологическую активность (связывание со своей мишенью, например, CD137 соответственно) мутеина липокалина.
[00123] Аналогично у мутеина липокалина по настоящему раскрытию может отсутствовать 1, 2, 3, 4 или более аминокислот на его N-конце и/или 1, 2 или более аминокислот на его C-конце по сравнению с соответствующим липокалином дикого типа; например с SEQ ID NO: 5-11 и 16.
[00124] В частности, для того, чтобы определить, соответствует ли аминокислотный остаток аминокислотной последовательности мутеина липокалина, отличного от липокалина дикого типа, определенному положению в аминокислотной последовательности липокалина дикого типа, специалист в данной области может применять средства и способы, хорошо известные в данной области, например, выравнивания либо вручную, либо с использованием компьютерных программ, таких как BLAST2.0, которая означает средство поиска основного локального выравнивания, или ClustalW, или любой другой подходящей программы, которая подходит для получения выравниваний последовательностей. Соответственно, липокалин дикого типа может служить в качестве «последовательности для сравнения» или «эталонной последовательности», тогда как аминокислотная последовательность липокалина, отличного от липокалина дикого типа, описанного в данном документе, служит в качестве «искомой последовательности». Термины «эталонная последовательность» и «последовательность дикого типа» используют в данном документе взаимозаменяемо.
[00125] Согласно некоторым вариантам осуществления замена (или замещение) представляет собой консервативную замену. Тем не менее, любая замена, в том числе неконсервативная замена или одна или более из перечисленных ниже иллюстративных замен, рассматривается как возможная лишь в том случае, если только мутеин липокалина сохраняет свою способность связываться с CD137 соответственно, и/или он характеризуется идентичностью с впоследствии замещенной последовательностью в том смысле, что имеет место по меньшей мере 60%, как, например, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85% или более высокая идентичность с «исходной» последовательностью.
[00126] Консервативные замены, как правило, представляют собой следующие замены, перечисленные с учетом аминокислоты, подвергаемой мутации, за каждой следует одно или более замещений, которые могут быть приняты за консервативные: Ala → Gly, Ser, Val; Arg → Lys; Asn → Gln, His; Asp → Glu; Cys → Ser; Gln → Asn; Glu → Asp; Gly → Ala; His → Arg, Asn, Gln; Ile → Leu, Val; Leu → Ile, Val; Lys → Arg, Gln, Glu; Met → Leu, Tyr, Ile; Phe → Met, Leu, Tyr; Ser → Thr; Thr → Ser; Trp → Tyr; Tyr → Trp, Phe; Val → Ile, Leu. Другие замены также возможны и они могут быть определены эмпирически или в соответствии с другими известными консервативными или неконсервативными заменами. В качестве дополнительной ориентировки, каждая из следующих восьми групп содержит аминокислоты, которые, как правило, могут быть взяты для определения консервативных замен в отношении друг друга:
a. аланин (Ala), глицин (Gly);
b. аспарагиновая кислота (Asp), глутаминовая кислота (Glu);
c. аспарагин (Asn), глутамин (Gln);
d. аргинин (Arg), лизин (Lys);
e. изолейцин (Ile), лейцин (Leu), метионин (Met), валин (Val);
f. фенилаланин (Phe), тирозин (Tyr), триптофан (Trp);
g. серин (Ser), треонин (Thr) и
h. цистеин (Cys), метионин (Met).
[00127] Если такие замены приводят в результате к изменению биологической активности, тогда может быть введено больше существенных изменений, таких как следующие, или которые дополнительно описаны ниже относительно классов аминокислот, и продукты подвергают скринингу в отношении требуемой характеристики. Примерами таких более существенных изменений являются Ala → Leu, Ile; Arg → Gln; Asn → Asp, Lys, Arg, His; Asp → Asn; Cys → Ala; Gln → Glu; Glu → Gln; His → Lys; Ile → Met, Ala, Phe; Leu → Ala, Met, норлейцин; Lys → Asn; Met → Phe; Phe → Val, Ile, Ala; Trp → Phe; Tyr → Thr, Ser; Val → Met, Phe, Ala.
[00128] Существенные модификации биологических свойств липокалина выполняют путем выбора замен, которые существенно отличаются по их влиянию на поддержание (a) структуры полипептидного скелета в области замены, например, в виде листовой или спиральной конформации, (b) заряда или гидрофобности молекулы в целевом сайте или (c) большей части боковой цепи. Встречающиеся в природе остатки разделены на группы на основе общих свойств боковых цепей: (1) гидрофобные: норлейцин, метионин, аланин, валин, лейцин, изолейцин; (2) нейтральные гидрофильные: цистеин, серин, треонин; (3) кислые: аспарагиновая кислота, глутаминовая кислота; (4) основные: аспарагин, глутамин, гистидин, лизин, аргинин; (5) остатки, которые влияют на ориентацию цепей: глицин, пролин и (6) ароматические: триптофан, тирозин, фенилаланин.
[00129] Неконсервативные замены будут включать в себя обмен представителя одного из этих классов на другой класс. Любой остаток цистеина, не вовлеченный в поддержание правильной конформации соответствующего липокалина, также может быть заменен, как правило, серином, для улучшения стойкости молекулы к окислению и предупреждения неправильного образования поперечных связей. В свою очередь, цистеиновая(цистеиновые) связь(связи) может(могут) быть добавлена(добавлены) к липокалину для улучшения его стабильности.
[00130] Любая мутация, в том числе вставка, которая рассматривалась выше, может быть легко выполнена на уровне нуклеиновой кислоты, например, ДНК, с использованием разработанных стандартных способов. Иллюстративные примеры перестроек аминокислотной последовательности представляют собой вставки или делеции, а также аминокислотные замены. Такие замены могут быть консервативными, т. е. аминокислотный остаток замещается аминокислотным остатком с химически подобными свойствами, в частности в отношении полярности, а также размера. Примеры консервативных замен представляют собой замещения из числа представителей следующих групп: 1) аланин, серин и треонин; 2) аспарагиновая кислота и глутаминовая кислота; 3) аспарагин и глутамин; 4) аргинин и лизин; 5) изолейцин, лейцин, метионин и валин и 6) фенилаланин, тирозин и триптофан. С другой стороны, существует также возможность введения неконсервативных перестроек в аминокислотную последовательность. К тому же вместо замещения отдельных аминокислотных остатков, существует также возможность либо вставлять, либо подвергать делеции одну или более последовательных аминокислот первичной структуры липокалина слезы, если только эти делеции или вставка приводят в результате к стабильно свернутому/функциональному мутеину.
[00131] Модификации аминокислотной последовательности включают направленный мутагенез отдельных аминокислотных положений для упрощения субклонирования подвергнутого мутации гена липокалина или его частей путем введения сайтов расщепления для определенных рестрикционных ферментов. Дополнительно эти мутации также можно вводить для дополнительного улучшения аффинности мутеина липокалина в отношении указанной мишени, такой как CD137. Кроме того, мутации можно вводить для модулирования определенных характеристик мутеина, как, например: для улучшения стабильности фолдинга, стабильности в сыворотке крови, устойчивости белка или растворимости в воде или для снижения склонности к агрегации, если в этом существует необходимость. Например, встречающиеся в природе остатки цистеина можно подвергать мутации в другие аминокислоты для предупреждения образования дисульфидного мостика. Существует также возможность подвергнуть преднамеренной мутации другие аминокислотные положения последовательности в цистеин для введения новых реакционноспособных групп, например, для конъюгации с другими соединениями, такими как полиэтиленгликоль (ПЭГ), гидроксиэтилкрахмал (ГЭК), биотин, пептиды или белки, или для образования не встречающихся в природе дисульфидных связей. Полученный тиоловый фрагмент можно использовать для ПЭГилирования или ГЭКилирования мутеина, например, для увеличения времени полужизни соответствующего мутеина липокалина в сыворотке крови. Иллюстративные варианты такой мутации для введения остатка цистеина в аминокислотную последовательность мутеина Tlc включают замены Thr 40→ Cys, Glu 73→ Cys, Arg 90→ Cys, Asp 95→ Cys и Glu 131→ Cys. Полученный тиоловый фрагмент с боковой стороны любого из аминокислотных положений 40, 73, 90, 95 и/или 131 можно использовать для ПЭГилирования или ГЭКилирования мутеина, например, для увеличения времени полужизни соответствующего мутеина Tlc в сыворотке крови.
[00132] Существует также возможность подвергнуть мутации другие аминокислотные положения последовательности в цистеин для введения новых реакционноспособных групп, например, для конъюгации с другими соединениями, такими как полиэтиленгликоль (ПЭГ), гидроксиэтилкрахмал (ГЭК), биотин, пептиды или белки, или для образования не встречающихся в природе дисульфидных связей.
[00133] Согласно некоторым вариантам осуществления, если один из приведенных выше фрагментов конъюгируется с мутеином липокалина по настоящему раскрытию, то конъюгация с боковой цепью аминокислоты может быть преимущественной. Подходящие боковые цепи аминокислот могут встречаться в природе в аминокислотной последовательности липокалина человека, или их можно вводить с помощью мутагенеза. В том случае, если подходящий сайт связывания вводят посредством мутагенеза, то один вариант представляет собой замещение аминокислоты в соответствующем положении остатком цистеина. Например, такая мутация включает по меньшей мере одну из замен Thr 40→ Cys, Glu 73→ Cys, Arg 90→ Cys, Asp 95→ Cys или Glu 131→ Cys в последовательности дикого типа липокалина слезы человека. Вновь созданный остаток цистеина в любом из этих положений в дальнейшем можно использовать для конъюгации мутеина с фрагментом, пролонгирующим время полужизни мутеина в сыворотке крови, таким как ПЭГ или его активированное производное.
[00134] В отношении мутеина липокалина 2 человека иллюстративными вариантами такой мутации являются введение остатка цистеина в аминокислотную последовательность липокалина, в том числе мутеина липокалина 2 человека, с включением введения остатка цистеина (Cys) по меньшей мере в одно из положений в последовательности, которое соответствует положениям 14, 21, 60, 84, 88, 116, 141, 145, 143, 146 или 158 последовательности в последовательности дикого типа NGAL человека. Согласно некоторым вариантам осуществления, в которых мутеин липокалина 2 человека по настоящему раскрытию характеризуется последовательностью, в которой по сравнению с последовательностью с номером доступа из базы данных SWISS-PROT/UniProt P80188 цистеин был замещен другим аминокислотным остатком, соответствующий цистеин может быть повторно введен в последовательность. В качестве иллюстративного примера, остаток цистеина в аминокислотном положении 87 может быть введен в таком случае путем возвращения цистеина, который первоначально присутствует в последовательности под номером доступа SWISS-PROT P80188. Полученный тиоловый фрагмент с боковой стороны любого из аминокислотных положений 14, 21, 60, 84, 88, 116, 141, 145, 143, 146 и/или 158 можно использовать для ПЭГилирования или ГЭКилирования мутеина, например, для увеличения времени полужизни соответствующего мутеина липокалина 2 человека в сыворотке крови.
[00135] Согласно другому варианту осуществления путем мутагенеза можно вводить искусственные аминокислоты с обеспечением подходящих боковых цепей аминокислот для конъюгирования одного из приведенных выше соединений с мутеином липокалина согласно настоящему раскрытию. Как правило, такие искусственные аминокислоты конструируют таким образом, чтобы они были более реакционноспособными и таким образом облегчали конъюгацию с необходимым соединением. Одним примером такой искусственной аминокислоты, которую можно вводить посредством искусственной тРНК, является пара-ацетил-фенилаланин.
[00136] Для некоторых путей применения мутеинов, раскрытых в данном документе, может быть преимущественным использование их в форме белков слияния. Согласно некоторым вариантам осуществления мутеин липокалина по настоящему раскрытию слит на своем N-конце или своем C-конце с белком, белковым доменом или пептидом, к примеру, сигнальной последовательностью и/или аффинной меткой.
[00137] Аффинные метки, как, например: Strep-tag® или Strep-метка II (Schmidt, T.G.M. et al. (1996) J. Mol. Biol. 255, 753-766), myc-метка, FLAG-метка, His6-метка или HA-метка, или белки, как, например, глутатион-S-трансфераза, также обеспечивающие легкое выявление и/или легкую очистку рекомбинантных белков, являются дополнительными примерами подходящих партнеров по слиянию. И наконец, белки с хромогенными или флуоресцентными свойствами, как, например, зеленый флуоресцентный белок (GFP) или желтый флуоресцентный белок (YFP), также являются подходящими партнерами по слиянию для мутеинов липокалина по настоящему раскрытию.
[00138] В целом существует возможность мечения мутеинов липокалина по настоящему раскрытию любым соответствующим химическим веществом или ферментом, которые непосредственно или опосредованно образуют выявляемые соединение или сигнал при химической, физической, оптической или ферментативной реакции. Примером физической реакции и в то же время оптической реакции/маркера является испускание флуоресценции при облучении или испускание X-лучей при использовании радиоактивной метки. Щелочная фосфатаза, пероксидаза хрена и β-галактозидаза являются примерами ферментных меток (и в то же время оптических меток), которые катализируют образование продуктов хромогенной реакции. В целом все метки, широко используемые для антител (за исключением тех, которые используют только с сахарными фрагментами в Fc-части иммуноглобулинов), также можно использовать для конъюгации с мутеинами липокалина по настоящему раскрытию. Мутеины липокалина по настоящему раскрытию также можно конъюгировать с любым подходящим терапевтически активным средством, например, для целенаправленной доставки таких средств к указанным клетке, ткани или органу или для селективного целенаправленного воздействия на клетки, например, клетки опухоли, без воздействия на окружающие нормальные клетки. Примеры таких терапевтически активных средств включают радионуклиды, токсины, малые органические молекулы и терапевтические пептиды (такие как пептиды, действующие в качестве агонистов/антагонистов рецептора клеточной поверхности, или пептиды, конкурирующие за сайт связывания белка на данной мишени, представляющей собой клетку). Однако мутеины липокалина по настоящему раскрытию также можно конъюгировать с терапевтически активными нуклеиновыми кислотами, как, например: антисмысловыми молекулами нуклеиновых кислот, малыми интерферирующими РНК, микро-РНК или рибозимами. Такие конъюгаты можно получать посредством способов, хорошо известных в данной области.
[00139] Как указано выше, мутеин липокалина по настоящему раскрытию согласно некоторым вариантам осуществления можно конъюгировать с фрагментом, который удлиняет время полужизни мутеина в сыворотке крови (в связи с этим также см. публикацию заявки согласно PCT WO 2006/56464, в которой такие стратегии конъюгации описаны в отношении мутеинов человеческого липокалина, ассоциированного с желатиназой нейтрофилов, с аффинностью связывания в отношении CTLA-4). Фрагментом, который удлиняет время полужизни в сыворотке крови, может быть молекула полиалкиленгликоля, гидроксиэтилкрахмал, молекулы жирных кислот, как, например, пальмитиновой кислоты (Vajo & Duckworth 2000, Pharmacol. Rev. 52, 1-9), Fc-часть иммуноглобулина, домен CH3 иммуноглобулина, домен CH4 иммуноглобулина, альбумин-связывающий пептид или альбумин-связывающий белок, трансферрин, при этом упомянуты лишь некоторые. Альбумин-связывающий белок может быть бактериальным альбумин-связывающим белком, антителом, фрагментом антитела, в том числе доменными антителами (см., например, патент США 6696245), или мутеином липокалина с активностью связывания в отношении альбумина. Соответственно, подходящие партнеры по конъюгации для удлинения времени полужизни мутеина липокалина по настоящему раскрытию включают альбумин-связывающий белок, например, бактериальный альбумин-связывающий домен, такой, например, как белка G стрептококка (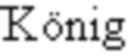 , T., & Skerra, A. (1998) J. Immunol. Methods 218, 73-83). Другими примерами альбумин-связывающих пептидов, которые можно использовать в качестве партнера по конъюгации, к примеру, являются таковыми с консенсусной последовательностью Cys-Xaa1-Xaa2-Xaa3-Xaa4-Cys, где Xaa1 представляет собой Asp, Asn, Ser, Thr или Trp; Xaa2 представляет собой Asn, Gln, His, Ile, Leu или Lys; Xaa3 представляет собой Ala, Asp, Phe, Trp или Tyr; и Xaa4 представляет собой Asp, Gly, Leu, Phe, Ser или Thr, которая описана в патентной заявке США 2003/0069395 или в Dennis et al. (Dennis, M. S., Zhang, M., Meng, Y. G., Kadkhodayan, M., Kirchhofer, D., Combs, D. & Damico, L. A. (2002) J Biol Chem 277, 35035-35043).
, T., & Skerra, A. (1998) J. Immunol. Methods 218, 73-83). Другими примерами альбумин-связывающих пептидов, которые можно использовать в качестве партнера по конъюгации, к примеру, являются таковыми с консенсусной последовательностью Cys-Xaa1-Xaa2-Xaa3-Xaa4-Cys, где Xaa1 представляет собой Asp, Asn, Ser, Thr или Trp; Xaa2 представляет собой Asn, Gln, His, Ile, Leu или Lys; Xaa3 представляет собой Ala, Asp, Phe, Trp или Tyr; и Xaa4 представляет собой Asp, Gly, Leu, Phe, Ser или Thr, которая описана в патентной заявке США 2003/0069395 или в Dennis et al. (Dennis, M. S., Zhang, M., Meng, Y. G., Kadkhodayan, M., Kirchhofer, D., Combs, D. & Damico, L. A. (2002) J Biol Chem 277, 35035-35043).
[00140] Согласно другим вариантам осуществления собственно альбумин (Osborn, B.L. et al., 2002, J. Pharmacol. Exp. Ther. 303, 540-548) или биологический активный фрагмент альбумина можно использовать в качестве партнера по конъюгации мутеина липокалина по настоящему раскрытию. Термин «альбумин» включает альбумины всех млекопитающих, как, например: сывороточный альбумин человека, или бычий сывороточный альбумин, или альбумин крысы. Альбумин или его фрагмент можно получать рекомбинантными способами, как описано в патенте США 5728553 или европейской патентной заявке EP 0330451 и EP 0361991. Рекомбинантный альбумин человека (Recombumin®) от Novozymes Delta Ltd. (Ноттингем, Великобритания) можно конъюгировать или сливать с мутеином липокалина по настоящему раскрытию для удлинения времени полужизни мутеина.
[00141] Если альбумин-связывающий белок представляет собой фрагмент антитела, то он может быть доменным антителом. Доменные антитела (dAb) сконструированы для обеспечения точного контроля биофизических свойств и времени полужизни in vivo с обеспечением оптимальной безопасности и эффективности профиля продукта. Доменные антитела, например, являются коммерчески доступными от Domantis Ltd. (Кембридж, Великобритания и Массачусетс, США).
[00142] При использовании трансферрина в качестве фрагмента для удлинения времени полужизни мутеинов липокалина по настоящему раскрытию в сыворотке крови, мутеины могут быть генетически слиты с N- или C-концом, или с ними обоими, негликозилированного трансферрина. Негликозилированный трансферрин характеризуется временем полужизни, составляющим 14-17 суток, и белок слияния с трансферрином будет точно так же характеризоваться удлиненным временем полужизни. Носитель, представляющий собой трансферрин, также обеспечивает высокие показатели биодоступности, биораспределения и стабильности в кровотоке. Такая технология является коммерчески доступной от BioRexis (BioRexis Pharmaceutical Corporation, Пенсильвания, США). Рекомбинантный трансферрин человека (DeltaFerrin™) для использования в качестве стабилизатора белка/партнера для удлинения времени полужизни также является коммерчески доступным от Novozymes Delta Ltd. (Ноттингем, Великобритания).
[00143] Если Fc-часть иммуноглобулина используют с целью пролонгирования времени полужизни мутеинов липокалина по настоящему раскрытию в сыворотке крови, то можно использовать технологию SynFusion™, коммерчески доступную от Syntonix Pharmaceuticals, Inc (Массачусетс, США). Применение этой технологии слияния с Fc предоставляет возможность создания длительно действующих биофармацевтических средств и, например, может предусматривать две копии мутеина, соединенного с Fc-участком антитела, для улучшения фармакокинетики, растворимости и эффективности продуцирования.
[00144] Еще одной альтернативой для пролонгирования времени полужизни мутеинов липокалина по настоящему раскрытию является слияние с N- или C-концом мутеинов длинных неструктурированных гибких богатых глицином последовательностей (например, полиглицина с приблизительно 20-80 последовательными остатками глицина). Такой подход, раскрытый в WO2007/038619, например, также был назван «рПЭГ» (рекомбинантный ПЭГ).
[00145] Если используют полиалкиленгликоль в качестве партнера по конъюгации, то полиалкиленгликоль может быть замещенным, незамещенным, линейным или разветвленным. Он также может быть активированным производным полиалкилена. Примерами подходящих соединений являются молекулы полиэтиленгликоля (ПЭГ), которые описаны в WO 99/64016, в патенте США 6177074 или в патенте США 6403564 касательно интерферона, или которые описаны в отношении других белков, как, например: модифицированная ПЭГ аспарагиназа, ПЭГ-аденозиндезаминаза (PEG-ADA) или ПЭГ-супероксиддисмутаза (см., например, Fuertges et al. (1990) The Clinical Efficacy of Poly(Ethylene Glycol)-Modified Proteins J. Control. Release 11, 139-148). Молекулярный вес такого полимера, как, например, полиэтиленгликоля, может варьировать от приблизительно 300 до приблизительно 70000 дальтон, в том числе, например, полиэтиленгликоль с молекулярным весом, составляющим приблизительно 10000, приблизительно 20000, приблизительно 30000 или приблизительно 40000 дальтон. Более того, например, как описано в патентах США 6500930 или 6620413, углеводные олиго- и полимеры, как, например, крахмал или гидроксиэтилкрахмал (ГЭК), могут быть конъюгированы с мутеином по настоящему раскрытию с целью удлинения времени полужизни в сыворотке крови.
[00146] К тому же мутеин липокалина, раскрытый в данном документе, можно сливать с фрагментом, который может придавать новые характеристики мутеинам липокалина по настоящему раскрытию, как, например, ферментативную активность или аффинность связывания в отношении других молекул. Примерами подходящих партнеров по слиянию являются щелочная фосфатаза, пероксидаза хрена, глутатион-S-трансфераза, альбумин-связывающий домен белка G, белок A, фрагменты антител, домены олигомеризации или токсины.
[00147] В частности, существует возможность слияния мутеина липокалина, раскрытого в данном документе, с активным сайтом отдельного фермента, как, например, оба «компонента» полученного в результате белка слияния действуют совместно на указанную терапевтическую мишень. Связывающий домен мутеина липокалина присоединяется к обусловливающей заболевание мишени, предоставляя домену фермента возможность нарушать биологическую функцию мишени.
[00148] Настоящее раскрытие также относится к молекулам нуклеиновых кислот (ДНК и РНК), которые включают нуклеотидные последовательности, кодирующие мутеины липокалина по настоящему раскрытию. Поскольку вырожденность генетического кода допускает замены определенных кодонов на другие кодоны, обозначающие ту же самую аминокислоту, настоящее раскрытие не ограничено конкретной молекулой нуклеиновой кислоты, кодирующей мутеин липокалина, который описан в данном документе, а охватывает все молекулы нуклеиновой кислоты, которые включают нуклеотидные последовательности, кодирующие функциональный мутеин. В связи с этим настоящее раскрытие предусматривает нуклеотидные последовательности, кодирующие некоторые мутеины липокалина по настоящему раскрытию, последовательности которых показаны под SEQ ID NO: 24-39.
[00149] Согласно одному варианту осуществления настоящего раскрытия способ включает воздействие на молекулы нуклеиновой кислоты мутагенезом в нуклеотидных триплетах, кодирующих по меньшей мере одно или даже больше из положений в последовательности, соответствующих положениям 28, 36, 40-41, 49, 52, 65, 68, 70, 72-73, 77, 79, 81, 83, 87, 94, 96, 100, 103, 106, 125, 127, 132 и 134 последовательности в линейной полипептидной последовательности NGAL человека (SEQ ID NO: 2).
[00150] Согласно другому варианту осуществления способа согласно настоящему раскрытию молекулу нуклеиновой кислоты, кодирующую липокалин слезы человека, сперва подвергают мутагенезу в одном или более из положений 5, 26-31, 33-34, 42, 46, 52, 56, 58, 60-61, 65, 71, 85, 94, 101, 104-106, 108, 111, 114, 121, 133, 148, 150 и 153 аминокислотной последовательности в линейной полипептидной последовательности липокалина слезы человека (SEQ ID NO: 1). После этого молекулу нуклеиновой кислоты, кодирующую липокалин слезы человека, также подвергают мутагенезу в одном или более из положений 101, 111, 114 и 153 аминокислотной последовательности в линейной полипептидной последовательности зрелого липокалина слезы человека.
[00151] Настоящее раскрытие также включает молекулы нуклеиновых кислот, кодирующие мутеины липокалина по настоящему раскрытию, которые включают дополнительные мутации вне указанных положений в последовательности экспериментального мутагенеза. Такие мутации часто допустимы или могут даже оказаться преимущественными, например, если они способствуют улучшенной эффективности сворачивания, стабильности в сыворотке крови, термостойкости или аффинности связывания с лигандом мутеинов.
[00152] Молекула нуклеиновой кислоты, раскрытая в данной заявке, может быть «функционально соединена» с регуляторной последовательностью (или регуляторными последовательностями) для обеспечения экспрессии этой молекулы нуклеиновой кислоты.
[00153] Молекулу нуклеиновой кислоты, как, например, ДНК, называют «способной к экспрессии молекулой нуклеиновой кислоты» или способной «обеспечивать экспрессию нуклеотидной последовательности», если она включает элементы последовательности, которые содержат информацию относительно транскрипционной и/или трансляционной регуляции, и такие последовательности «функционально связаны» с нуклеотидной последовательностью, кодирующей полипептид. Функциональная связь представляет собой связь, при которой элементы регуляторной последовательности и последовательность, подлежащая экспрессии, соединены таким образом, который обеспечивает экспрессию гена. Определенные особенности регуляторных участков, необходимых для экспрессии гена, могут варьировать между видами, но в общем эти участки включают промотор, который у прокариотов содержит как промотор per se, т.е. элементы ДНК, управляющие инициацией транскрипции, так и элементы ДНК, которые при транскрибировании в РНК, будут запускать сигнал об инициации трансляции. Такие промоторные участки, как правило, включают 5'-некодирующие последовательности, вовлеченные в инициацию транскрипции и трансляции, такие как боксы -35/-10 и элемент Шайн-Дальгарно у прокариот или TATA-бокс, последовательности CAAT и 5'-кэппирующие элементы у эукариот. Такие участки также могут включать энхансерные или репрессорные элементы, а также сигнальные последовательности трансляции и лидерные последовательности для нацеливания нативного полипептида на конкретный компартмент клетки-хозяина.
[00154] К тому же 3'-некодирующие последовательности могут содержать регуляторные элементы, вовлеченные в терминацию транскрипции, полиаденилирование или им подобные. Однако, если такие последовательности терминации не удовлетворительны с точки зрения функциональности в конкретной клетке-хозяине, тогда их можно заменить сигналами, функциональными в такой клетке.
[00155] Ввиду этого молекула нуклеиновой кислоты по настоящему раскрытию может включать регуляторную последовательность, как, например, промоторную последовательность. Согласно некоторым вариантам осуществления молекула нуклеиновой кислоты по настоящему раскрытию включает промоторную последовательность и последовательность терминации транскрипции. Подходящими прокариотическими промоторами являются, например, промотор tet, промотор lacUV5 или промотор T7. Примерами промоторов, применимых для экспрессии в эукариотических клетках, являются промотор SV40 или промотор CMV.
[00156] Молекулы нуклеиновой кислоты по настоящему раскрытию также могут быть частью вектора или любого другого типа средства для клонирования, а именно плазмиды, фагмиды, фага, бакуловируса, космиды или искусственной хромосомы.
[00157] Согласно одному варианту осуществления молекула нуклеиновой кислоты включена в фазмиду. Фазмидный вектор означает вектор, кодирующий межгенный участок умеренного фага, как, например, M13 или f1, или его функциональную часть, слитую с представляющей интерес кДНК. После суперинфекции бактериальных клеток-хозяев таким фагмидным вектором и соответствующим хелперным фагом (например, M13K07, VCS-M13 или R408) продуцируются интактные фаговые частицы, обеспечивая таким образом физическое сопряжение закодированной гетерологичной кДНК с ее соответствующим полипептидом, выведенным на поверхность фага (см., например, Lowman, H.B. (1997) Annu. Rev. Biophys. Biomol. Struct. 26, 401-424 или Rodi, D.J., and Makowski, L. (1999) Curr. Opin. Biotechnol. 10, 87-93).
[00158] Такие средства для клонирования могут включать, помимо регуляторных последовательностей, описанных выше, и последовательность нуклеиновой кислоты, кодирующей мутеин липокалина, который описан в данном документе, последовательности для репликации и контрольные последовательности, полученные от видов, совместимых с клеткой-хозяином, которую используют для экспрессии, а также маркеры отбора, придающие селектируемый фенотип трансформированным или трансфицированным клеткам. В данной области известно большое число подходящих векторов для клонирования, и они являются коммерчески доступными.
[00159] Молекулой ДНК, кодирующей мутеин липокалина, который описан в данном документе, и в частности вектором клонирования, содержащим кодирующую последовательность такого мутеина, можно трансформировать клетку-хозяина, способную экспрессировать ген. Трансформацию можно осуществлять с использованием стандартных методик. Таким образом, настоящее раскрытие также направлено на клетку-хозяина, содержащую молекулу нуклеиновой кислоты, которая раскрыта в данном документе.
[00160] Трансформированные клетки-хозяева культивируют в условиях, подходящих для экспрессии нуклеотидной последовательности, кодирующей белок слияния по настоящему раскрытию. Подходящие клетки-хозяева могут быть прокариотическими, такими как Escherichia coli (E. coli) или Bacillus subtilis, или эукариотическими, такими как Saccharomyces cerevisiae, Pichia pastoris, клетками насекомых SF9 или High5, иммортализованными клеточными линиями млекопитающих (например, клетками HeLa или клетками CHO) или первичными клетками млекопитающих.
[00161] Настоящее раскрытие также относится к способу получения мутеина липокалина, который описан в данном документе, где мутеин, фрагмент мутеина или белок слияния мутеина и другого полипептида (например, другого мутеина липокалина) получают, исходя из нуклеиновой кислоты, кодирующей мутеин, с помощью способов генной инженерии. Способ можно осуществлять in vivo, при этом мутеин липокалина, например, можно получать в бактериальном или эукариотическом организме-хозяине, а затем выделять из этого организма-хозяина или его культуры. Существует также возможность получения белка in vitro, например, путем использования системы трансляции in vitro.
[00162] При получении мутеина липокалина in vivo нуклеиновую кислоту, кодирующую такой мутеин, вводят в подходящий бактериальный или эукариотический организм-хозяин с помощью технологии рекомбинантной ДНК (как уже подчеркивалось выше). С этой целью клетку-хозяина сперва трансформируют вектором для клонирования, который включает молекулу нуклеиновой кислоты, кодирующую мутеин липокалина, который описан в данном документе, с применением разработанных стандартных способов. Клетку-хозяина затем культивируют в условиях, которые обеспечивают экспрессию гетерологичной ДНК и таким образом синтез соответствующего полипептида. Впоследствии полипептид выделяют либо из клетки, либо из среды для культивирования.
[00163] Согласно некоторым вариантам осуществления молекула нуклеиновой кислоты, такая как ДНК, раскрытая в данной заявке, может быть «функционально связана» с другой молекулой нуклеиновой кислоты по настоящему раскрытию для обеспечения экспрессии белка слияния по настоящему раскрытию. В связи с этим функциональная связь представляет собой связь, при которой элементы последовательности первой молекулы нуклеиновой кислоты и элементы последовательности второй молекулы нуклеиновой кислоты соединены таким образом, который обеспечивает экспрессию белка слияния в виде единого полипептида.
[00164] К тому же согласно некоторым вариантам осуществления встречающаяся в природе дисульфидная связь между Cys 76 и Cys 175 может быть удалена в мутеинах NGAL по настоящему раскрытию. Согласно некоторым вариантам осуществления для мутеинов Tlc по настоящему раскрытию встречающаяся в природе дисульфидная связь между Cys 61 и Cys 153 может быть удалена. Соответственно, такие мутеины можно получать в компартменте клетки со средой с восстановительным окислительно-восстановительным потенциалом, например, в цитоплазме грамотрицательных бактерий.
[00165] В том случае, если мутеин липокалина по настоящему раскрытию включает внутримолекулярные дисульфидные связи, может быть предпочтительным направлять образующийся полипептид в компартмент клетки со средой с окисляющим окислительно-восстановительным потенциалом с помощью соответствующей сигнальной последовательности. Такая окисляющая среда может обеспечиваться периплазмой грамотрицательных бактерий, таких как E. coli, во внеклеточной среде грамположительных бактерий или в просвете эндоплазматической сети эукариотических клеток и, как правило, она способствует образованию дисульфидных связей в структуре.
[00166] Однако также существует возможность получения мутеина по настоящему раскрытию в цитозоле клетки-хозяина, предпочтительно E. coli. В данном случае полипептид может быть либо непосредственно получен в растворимом и свернутом состоянии, либо выделен в форме телец включения с последующей ренатурацией in vitro. Дополнительной возможностью является применение конкретных штаммов-хозяев с окисляющей внутриклеточной средой, которая таким образом может обеспечивать образование дисульфидных связей в цитозоле (Venturi et al. (2002) J. Mol. Biol. 315, 1-8.).
[00167] Однако мутеин липокалина, который описан в данном документе, не обязательно можно создавать или получать только путем применения генной инженерии. Предпочтительнее такой мутеин можно также получать путем химического синтеза, такого как твердофазный синтез полипептидов Меррифилда, или путем транскрипции и трансляции in vitro. Например, возможно, что представляющие интерес мутации определяют с помощью молекулярного моделирования, а затем синтезируют необходимый (разработанный) полипептид in vitro и изучают активность связывания в отношении CD137. Способы твердофазного синтеза и/или синтеза в жидкой фазе белков хорошо известны в данной области (см., например, Bruckdorfer, T. et al. (2004) Curr. Pharm. Biotechnol. 5, 29-43).
[00168] Согласно другому варианту осуществления мутеины липокалина по настоящему раскрытию можно получать путем транскрипции/трансляции in vitro, используя стандартные способы, известные специалистам в данной области.
[00169] Специалисту в данной области будут понятны способы, применимые для получения мутеинов липокалина, предполагаемые настоящим раскрытием, но чьи последовательности белков или нуклеиновых кислот явным образом не раскрыты в данном документе. В качестве обзора такие модификации аминокислотной последовательности включают, например, направленный мутагенез отдельных положений аминокислот для упрощения субклонирования подвергнутого мутации гена липокалина или его частей путем введения сайтов расщепления для определенных рестрикционных ферментов. К тому же такие мутации можно также вводить для дополнительного улучшения аффинности мутеина липокалина в отношении его мишени (например, CD137 соответственно). Кроме того, мутации можно вводить для модулирования определенных характеристик мутеина, а именно для улучшения стабильности фолдинга, стабильности в сыворотке крови, устойчивости белка или растворимости в воде или для снижения склонности к агрегации, если в этом существует необходимость. Например, встречающиеся в природе остатки цистеина можно подвергать мутации в другие аминокислоты для предупреждения образования дисульфидного мостика.
[00170] Мутеины липокалина, раскрытые в данном документе, и их производные можно применять во многих областях, как и в случае их антител или фрагментов. Например, мутеины липокалина можно использовать для мечения ферментом, антителом, радиоактивным веществом или любой другой группой с биохимической активностью или определенными характеристиками связывания. При помощи такого подхода их соответствующие мишени, или конъюгаты, или белки слияния можно выявлять или приводить в контакт с ними. К тому же мутеины липокалина по настоящему раскрытию могут служить для выявления химических структур с помощью разработанных аналитических способов (например, ELISA или вестерн-блоттинга) или с помощью микроскопического исследования или иммуносенсорных анализов. В связи с этим сигнал для выявления может быть получен либо непосредственно путем применения подходящего конъюгата мутеина или белка слияния, либо опосредованно путем иммунохимического выявления связанного мутеина посредством антитела.
[00171] Дополнительные объекты, преимущества и особенности настоящего раскрытия станут очевидными специалисту в данной области при изучении следующих примеров и прилагаемых фигур, включающих их, которые не предназначены для ограничения. Таким образом, следует понимать, что хотя настоящее раскрытие конкретно раскрыто с помощью примерных вариантов осуществления и необязательных особенностей, к модификации и варианту раскрываемой информации, приведенной в них, раскрытой в данном документе, может прибегнуть специалист в данной области, и что такие модификации и варианты рассматриваются как часть объема настоящего раскрытия.
V. Примеры
[00172] Пример 1. Выбор и оптимизация мутеинов, специфически связывающихся с CD137
[00173] Репрезентативные специфичные в отношении CD137 мутеины липокалина, раскрытие в данной заявке, выбирали из библиотек мутантов, полученных с помощью ненаправленного случайного мутагенеза, основанных на NGAL человека и TLc человека. Разные стратегии и мишени использовали для получения связывающих CD137 мутеинов. Используемые рекомбинантные мишени представляли собой коммерчески доступное слияние с Fc, представляющие собой полный внеклеточный домен CD137 человека (huCD137-Fc, R&D Systems 838-4B) и отдельные субдомены CD137 человека, при этом все получены в виде элементов для слияний с Fc-фрагментом человека. В качестве альтернативы не слитой с Fc мишени авторы настоящего изобретения использовали меченный His внеклеточный домен CD137 человека (Invitrogen, 10041-H08H-250). В качестве альтернативы, использовали пэннинг с клетками с использованием клеток CHO, трансфицированных полной кДНК CD137 человека. Типы пэннинга с использованием белков и клеток осуществляли с помощью стандартных процедур. Клоны, полученные после отбора, подвергали процессу скрининга, который описан в примере 2.
[00174] Пример 2. Идентификация мутеинов, специфически связывающихся с CD137, с использованием высокопроизводительного скрининга с ELISA
[00175] Отдельные мутеины липокалина, слитые с со Strep-меткой на C-конце (SEQ ID NO: 23, см. пример 3), использовали для инокуляции среды 2xYT/Amp и культивировали в течение ночи (14-18 ч) до стационарной фазы. Затем проводили посев 50 мкл 2xYT/Amp из культур в стационарной фазе, и инкубировали в течение 3 ч при 37°C, а затем снижали до 22°C, пока не достигали OD595, составляющую 0,6-0,8. Продуцирование мутеинов индуцировали добавлением 10 мкл 2xYT/Amp, дополненной 1,2 мкг/мл ангидротетрациклина. Культуры инкубировали при 22°C до следующего дня. После добавления 40 мкл 5% (вес/об.) BSA в PBS/T и инкубации в течение 1 ч при 25°C культуры были готовы для использования в скрининговых анализах.
[00176] Связывание выделенных мутеинов с CD137 человека тестировали путем нанесения на микротитровальные планшеты huCD137-Fc (5 мкг/мл в PBS) релевантных видов на ночь при 4°C. После блокирования планшета с помощью PBST, содержащего 2% BSA, 20 мкл блокированных с помощью BSA культур добавляли в микротитровальные планшеты и инкубировали в течение 1 ч при 25°C. Связанные мутеины выявляли с помощью антитела к StrepTag, конъюгированного с пероксидазой хрена (1 ч инкубация; IBA, Геттинген). Для количественного измерения добавляли 20 мкл флуорогенного субстрата для пероксидазы QuantaBlu и полученную в результате флуоресценцию определяли при длине волны возбуждения, составляющей 330 нМ, и длине волны излучения, составляющей 420 нМ.
[00177] Для отбора в отношении мутеинов с повышенной термостойкостью блокированные с помощью BSA культуры инкубировали в течение 1 ч при 60°C, а затем обеспечивали охлаждение до комнатной температуры перед их добавлением в микротитровальные планшеты с нанесенным CD137 и блокированных с помощью BSA, которые описаны в предыдущем абзаце. Мутеины далее обрабатывали как описано в предыдущем абзаце и отбирали для бактериальной экспрессии, очистки и дополнительного определения характеристик.
[00178] Пример 3. Экспрессия мутеинов
[00179] Уникальные мутеины экспрессировали с C-концевой меткой, представляющей собой SAWSHPQFEK (SEQ ID NO: 21) или PSAWSHPQFEK (SEQ ID NO: 22); в том числе линкером SA или PSA и Strep-tag® II, WSHPQFEK (SEQ ID NO: 23) в E. coli в среде 2YT-Amp с очисткой мутеинов после экспрессии с использованием аффинной хроматографии Streptactin и препаративной эксклюзионной хроматографии. И в завершении, мутеины липокалина подвергали стадии очистки от эндотоксина с использованием колонок Mustang E. Очищенные мутеины липокалина затем характеризовали как подробно описано во всех изложенных ниже примерах.
[00180] Пример 4. Аффинность мутеинов, связывающихся с белком слияния CD137 человека-Fc, определенная с помощью поверхностного плазмонного резонанса (SPR)
[00181] Поверхностный плазмонный резонанс (SPR) использовали для измерения кинетики и аффинности связывания репрезентативных мутеинов липокалина, раскрытых в данном документе.
[00182] Анализ с использованием SPR связывания репрезентативных мутеинов с белком слияния CD137 человека-Fc (huCD137-Fc) проводили при 37°C на приборе Biacore T200 (GE Healthcare) с использованием HBS-EP+ (1x; BR-1006-69; GE Healthcare) в качестве подвижного буфера.
[00183] Перед измерениями уровня белка проводили три цикла восстановления с целью приведения к требуемым условиям. Восстановления полученной поверхности чипа достигали путем нанесения 3 М MgCl2 на период 60 с, после которого шел 10 мМ глицин, pH 1,7, в течение 180 с. Антитело к IgG человека-Fc использовали для иммобилизации huCD137-Fc на следующей стадии, и его брали из набора для захвата антител человека (GE Healthcare, BR-1008-39). Его иммобилизировали на сенсорном чипе CM5 с использованием общепринятой химии сшивания аминов и буфера для иммобилизации, включенного в набор (10 мМ ацетата натрия, pH 5,0), с достижением в результате плотности лиганда, составляющей приблизительно 13000 резонансных единиц (RU). Эталонный канал обрабатывали соответственно.
[00184] HuCD137-Fc в концентрации, составляющей 0,5 мкг/мл, фиксировали на этой поверхности в течение 180 с при скорости потока, составляющей 10 мкл/мин в буфере HBS-EP+. Целевой белок не применяли для эталонного канала. Далее, мутеины липокалина использовали в соответствующей серии разведений в буфере HBS-EP+ при скорости потока, составляющей 30 мкл/мин. Восстановления полученной поверхности чипа достигали, как описано выше. Данные оценивали с помощью программного обеспечения Biacore T200 Evaluation (V 2.0). Использовали двойное сравнение с образцом и для аппроксимирования первичных данных использовали модель связывания 1:1.
[00185] На фигуре 1 показаны кривые, полученные с помощью SPR, и аппроксимированные кривые, определенные для тестируемых мутеинов липокалина, с соответствующими SEQ ID NO, представленными на графиках. Данные показаны в отношении связывания с huCD137-Fc. Имеют место отчетливые сигналы связывания, обнаруженные с помощью SPR, относительно мишени у человека, тогда как отрицательные контроли, представляющие собой SEQ ID NO: 3 и SEQ ID NO: 4, не проявляли связывание. Значения аффинности, полученные в результате аппроксимации этих данных, представлены в таблице 1 ниже.
[00186] Таблица 1.

[00187] Пример 5. Анализ с использованием поверхностного плазмонного резонанса (SPR) для определения конкуренции между CD137L человека и мутеинами в связывании с белком слияния CD137 человека-Fc
[00188] В отношении мутеина липокалина, описанного в данной заявке, который связывает CD137, возможны, как правило, два механизма связывания: в первом случае сайт связывания мутеина перекрывается с сайтом связывания лиганда CD137 человека (CD137L) с CD137. В тех случаях, когда такой мутеин липокалина связывается с CD137, это нарушает связывание CD137L с CD137 и одновременно приводит к вмешательству в естественную передачу сигнала CD137L («конкурентное связывание»); во втором случае сайт связывания мутеина не перекрывается с сайтом связывания CD137L, и такой мутеин липокалина может связываться с CD137 без нарушения связывания CD137L и естественной передачи сигнала CD137L («неконкурентное связывание»).
[00189] Образование скопления CD137 через его лиганд активирует нисходящие сигнальные пути, связанные с CD137. В случае T-клеток активация CD137 приводит к костимуляции активаторных ответов T-клеток, как, например, пролиферации, и к продуцированию провоспалительных цитокинов.
[00190] Другой путь индуцирования образования скоплений CD137 представляет собой применение связывающих CD137 иммобилизованных средств. В случае нанесения на планшет (например, на пластиковую чашку для культивирования и инкубирования T-клеток в чашке) как конкурентные, так и неконкурентные связывающие CD137 молекулы обеспечивают образование скопления CD137 и таким образом активируют нисходящую передачу сигнала.
[00191] Следовательно, с одной стороны, как конкурентные, так и неконкурентные связывающие CD137 молекулы, при применении как описано выше, могут активировать нисходящие сигнальные пути, связанные с CD137.
[00192] Молекулу, конкурентно связывающую CD137, с другой стороны, можно применять для подавления естественного взаимодействия CD137/CD137L и таким образом супрессировать естественную передачу сигнала, индуцированную встречей CD137-положительных клеток с клетками, экспрессирующими CD137L, например, антиген-презентирующими клетками. Такой механизм действия является необходимым в тех случаях, когда необходимо супрессировать недопустимо сильную воспалительную или аутоиммунную реакцию.
[00193] Для того, чтобы показать, что данная заявка предусматривает как мутеины конкурентного типа, так и мутеины неконкурентного типа, авторы настоящего изобретения прибегали к эксперименту с использованием поверхностного плазмонного резонанса (SPR). Авторы настоящего изобретения использовали его для изучения конкуренции между CD137L и тремя репрезентативными мутеинами липокалина, раскрытыми в данном документе, в связывании с белком слияния CD137 человека-Fc (huCD137-Fc). В этом анализе изучали, может ли мутеин липокалина связываться с предварительно образованным комплексом CD137 и CD137L; если это не так, то это является доказательством того, что связывающий мутеин липокалина эпитоп на CD137 перекрывается с эпитопом, связывающим CD137L, на CD137. Соответствующий мутеин таким образом связывается с CD137 на конкурентной основе в отношении взаимодействия CD137/CD137L. Если как CD137L, так и мутеин липокалина могут связываться в одно и то же время, то связывание является неконкурентным в отношении взаимодействия CD137/CD137L.
[00194] Конкурентный анализ проводили при 37°C на приборе Biacore T200 (GE Healthcare) с использованием HBS-EP+ (1x; BR-1006-69; GE Healthcare) в качестве подвижного буфера. Набор для захвата биотина (GE Healthcare) использовали для иммобилизации биотинилированного huCD137-Fc на поверхности чипа. Белки CD137-Fc подвергали биотинилированию с использованием общепринятой химии NHS. Неразведенный реагент для захвата биотина Biotin CAPture Kit (стрептавидин, конъюгированный с олигонуклеотидом ss-DNA) фиксировали на сенсорном чипе CAP с предварительно иммобилизованным комплементарным олигонуклеотидом ss-DNA. Затем биотинилированный белок CD137-Fc в концентрации, составляющей 2 мкг/мл, использовали в течение 300 с при скорости потока, составляющей 5 мкл/мин. Восстановления поверхности чипа достигали путем использования 6 М гуанидиний-HCl в 250 мМ NaOH в течение 120 с при скорости потока, составляющей 10 мкл/мин.
[00195] В первых двух циклах измерения определяли успешное связывание CD137L с CD137 в экспериментальных условиях и получали эталонный уровень для отдельного связывания тестируемого мутеина липокалина в отсутствие лиганда. В третьем цикле CD137 насыщали CD137L перед добавлением мутеина липокалина, как подробно описано ниже.
[00196] Лиганд CD137 человека - Fc-лиганд (R&D Systems 2295-4L-025/CF) использовали в отношении иммобилизованного белка CD137-Fc в концентрации, составляющей 500 нМ, и при скорости потока, составляющей 30 мкл/мин, в течение 30 с. После восстановления мутеины липокалина наносили в концентрации, составляющей 5 мкМ, при скорости потока, составляющей 30 мкл/мин, в течение 30 с. И в завершении, после дополнительного цикла восстановления лиганд CD137 человека-Fc применяли в отношении иммобилизованных белков CD137-Fc в концентрации, составляющей 500 нМ, в течение 30 с, после которого непосредственно шли мутеины в концентрации, составляющей 5 мкМ, в течение 30 с, оба при скорости потока, составляющей 30 мкл/мин. Восстановления поверхности чипа достигали путем использования 6 М гуанидиний-HCl в 250 мМ NaOH в течение 120 с при скорости потока, составляющей 10 мкл/мин. Полученные в результате сенсограммы визуально анализировали и определяли, оказывает ли связанный лиганд CD137-Fc влияние на взаимодействие мутеинов с иммобилизованными белками CD137-Fc. Сенсограммы циклов представляли собой взаимодействие лиганда CD137-Fc, или мутеины, наносимые по отдельности, служили в качестве контролей.
[00197] Репрезентативные примеры для релевантного сегмента полученных в результате сенсограмм представлены на фигуре 2 для мутеинов с SEQ ID NO: 5, SEQ ID NO: 12 и SEQ ID NO: 13. Кривая, полученная с помощью SPR, в отношении связывания соответствующего мутеина липокалина только с huCD137-Fc обозначена стрелкой с непрерывной линией. Кривая, полученная с помощью SPR, в отношении связывания мутеина липокалина с huCD137-Fc, который был насыщен CD137L, обозначена стрелкой с прерывистым стержнем. Из данных видно, что мутеин с SEQ ID NO: 5, например, не может связываться с huCD137-Fc в присутствии CD137L (фигура 2A). В отличие от этого, как мутеин с SEQ ID NO: 12, так и мутеин с SEQ ID NO: 13 связываются с huCD137-Fc с очень похожим ответом как в отсутствие, так и в присутствии CD137L, что показывает то, что конкуренция в связывании между мутеинами липокалина и CD137L отсутствует. Эти данные подытожены в таблице 2.
Таблица 2

[00198] Пример 6. Анализ с использованием FACS мутеинов липокалина, связывающихся с клетками, экспрессирующими CD137 человека
[00199] Авторы настоящего изобретения прибегали к исследованиям с использованием FACS для того, чтобы оценить специфичное связывание мутеинов липокалина и отрицательных контролей с клетками яичника китайского хомячка (CHO), стабильно трансфицированными с помощью CD137 человека (CHO-huCD137). Клеточную линию получали с помощью системы Flp-In (Invitrogen) согласно инструкциям изготовителя. Ложно трансфицированные клетки CHO Flp-In служили в качестве отрицательного контроля.
[00200] Трансфицированные клетки CHO поддерживали в среде F12 Хэма (Invitrogen), дополненной 10% фетальной телячьей сыворотки (FCS, Biochrom) и 500 мкг/мл гигромицина B (Roth). Клетки культивировали в сосудах для клеточных культур в стандартных условиях согласно инструкциям изготовителя (37°C, атмосфера с 5% CO2). Для того, чтобы отделить адгезивные клетки для субкультивирования или экспериментов с использованием FACS, использовали Accutase (PAA) согласно инструкциям изготовителя.
[00201] Для проведения эксперимента CD137-положительные и -отрицательные клетки CHO Flp-In инкубировали с мутеинами липокалина и связанный мутеин метили с помощью первичных антител к липокалину и флуоресцентно меченных вторичных антител, которые выявляли с помощью анализа с использованием FACS, который описан ниже.
[00202] 1×105 клеток на лунку предварительно инкубировали в течение 1 ч в ледяном PBS, содержащем 5% фетальной телячьей сыворотки (PBS-FCS). Затем серию разведений мутеинов липокалина и отрицательных контролей, как правило, находящихся в диапазоне от 10 мкМ до 1 нМ, добавляли к клеткам и инкубацию продолжали на льду в течение 1 ч. Клетки дважды промывали ледяным PBS с использованием центрифугирования при 300 g, а затем инкубировали с первичным антителом кролика к липокалину (Pieris, (поликлональное антитело кролика к hNGAL и антитело кролика к hTLC; Pieris) в течение 30 мин на льду. Клетки дважды промывали ледяным PBS, повторно суспендировали в PBS-FCS и инкубировали 30 мин на льду с вторичным антителом к антителу кролика, меченным флуоресцентным красителем Alexa488 (Life Technologies). Клетки впоследствии промывали и анализировали с использованием проточного цитометра Guava easyCyte HT. Как правило, гейтирование производили для исключения нежизнеспособных клеток и регистрировали 5000 объектов. Численные результаты выражали в виде среднего геометрического значений интенсивности флуоресценции.
[00203] Гистограммы, полученные с помощью FACS, для всех тестируемых клонов представлены на фигуре 3. На соответствующих графических изображениях показаны SEQ ID NO соответствующих мутеинов липокалина. Наряду с данными, полученными с помощью SPR (фигура 1, таблица 1), все мутеины показывают отчетливое связывание с CD137, экспрессируемым клетками. Значение EC50, полученное в результате аппроксимации этих данных, представлено в таблице 3 ниже.
[00204] Таблица 3

[00205] Пример 7. Функциональный анализ активации T-клеток с использованием нанесенных мутеинов липокалина
[00206] Авторы настоящего изобретения прибегали к анализу активации T-клеток для оценки способности набора репрезентативных связывающих CD137 мутеинов липокалина костимулировать ответы T-клеток. У тестируемых мутеинов (SEQ ID NO: 13, SEQ ID NO: 14 и SEQ ID NO: 15) перекрывается значение аффинности, полученное с помощью SPR, находящееся в диапазоне от 2 нМ до >23 нМ, в примере 4 (см. таблицу 1). Как обсуждалось в примере 5, существует несколько путей индуцирования образования скоплений CD137, и в этом эксперименте авторы настоящего изобретения использовали связывающие CD137 иммобилизованные средства. В этом эксперименте мутеины липокалина наносили на пластиковую чашку совместно с антителом к CD3 человека (мурономаб, Janssen-Cilag) и очищенные T-клетки затем инкубировали на покрытых поверхностях в присутствии растворимого антитела к CD28 человека (клон 28.2; eBioscience). Антитела к CD3 и CD28 использовали для обеспечения стимула ниже порогового в отношении T-клеток, которые можно было костимулировать путем костимуляции с помощью CD137. В качестве показателя для считывания авторы настоящего изобретения измеряли уровни интерлейкина 2 (IL-2) в супернатанте. Усиленное продуцирование IL-2 представляет собой один из признаков активации T-клеток, и повышение уровней IL-2 путем костимуляции антителом к CD137 было описано в литературе (Fisher T.S. et al., Cancer Immunol Immunother (2012) 61:1721-1733). В качестве отрицательного контроля использовали SEQ ID NO: 4. Ниже авторы настоящего изобретения приводят подробное описание эксперимента.
[00207] Мононуклеарные клетки периферической крови человека (PBMC) от здоровых доноров-добровольцев выделяли из лейкоцитарных пленок путем центрифугирования в градиенте плотности полисахарозы (Biocoll 1,077 г/мл от Biochrom), следуя протоколу компании Biochrom. T-лимфоциты выделяли из полученных в результате PBMC с использованием набора для очистки T-клеток Pan T-cell purification Kit (Miltenyi Biotec GmbH) и протоколов изготовителя. Очищенные T-клетки повторно суспендировали в буфере, состоящем из 90% FCS и 10% DMSO, немедленно замораживали с использованием жидкого азота и хранили в жидком азоте до последующего использования. Для анализа T-клетки оттаивали в течение 16 ч и культивировали в культуральной среде (RPMI 1640, Life Technologies), дополненной 10% FCS и 1% смеси пенициллина-стрептомицина (Life Technologies).
[00208] Следующую процедуру проводили с использованием трех повторов для каждого экспериментального условия. На плоскодонные планшеты для культур тканей наносили 200 мкл смеси из 0,5 мкг/мл антитела к CD3 и 25 мкг/мл антител кролика к липокалину (поликлональное антитело кролика к hNGAL, Pieris) и выдерживали в течение ночи при 4°C. Последнее использовали для обеспечения возможности иммобилизации мутеинов липокалина путем аффинного захвата. На следующий день лунки промывали дважды с использованием PBS и 50 мкл связывающих CD137 мутеинов липокалина с SEQ ID NO: 13, SEQ ID NO: 14 и SEQ ID NO: 15, все в концентрации, составляющей 25 мкг/мл, захватывали на предварительно покрытых планшетах в течение 1 ч при 37°C. SEQ ID NO: 4 использовали подобным образом, и она служила в качестве отрицательного контроля. После еще одного двухкратного промывания с использованием PBS 100 мкл суспензии T-клеток (соответствующей 5×104 T-клеток) в культуральной среде, дополненной 2 мкг/мл антитела к hCD28, добавляли в каждую лунку. Планшеты накрывали газопроницаемой уплотняющей пленкой (4titude) и инкубировали при 37°C в увлажненной атмосфере с 5% CO2 в течение 3 дней. Затем оценивали уровень IL-2 в супернатанте.
[00209] Уровни IL-2 человека в объединенных супернатантах клеточных культур количественно измеряли с помощью набора DuoSet для IL-2 от R&D Systems. На первой стадии на 384-луночный планшет наносили при комнатной температуре на 2 ч 1 мкг/мл «антитела для захвата IL-2 человека» (R&D System), разведенного в PBS. Затем лунки промывали 5 раз с использованием 80 мкл PBS-T (PBS, содержащего 0,05% Tween20) с использованием установки для избирательного промывания планшетов с глубокими лунками Biotek EL405 Select CW (Biotek). После блокирования в течение 1 ч в PBS-T, дополнительно содержащем 1% казеина (вес/вес), объединенный супернатант и серию концентраций стандарта IL-2, разведенного в культуральной среде, инкубировали в 384-луночном планшете в течение ночи при 4°C. Для обеспечения возможности выявления и количественного измерения захваченного IL-2 смесь из 100 нг/мл биотинилированного антитела козы к hIL-2-Bio для выявления (R&D System) и 1 мкг/мл меченного сульфо-меткой стрептавидина (Mesoscale Discovery) добавляли в PBS-T, содержащий 0,5% казеина, и инкубировали при комнатной температуре в течение 1 ч. После промывания 25 мкл буфера для считывания добавляли в каждую лунку и электрохемилюминесцентный (ECL) сигнал каждой лунки считывали с использованием планшет-ридера Mesoscale Discovery. Анализ и количественное измерение проводили с использованием программного обеспечения от Mesoscale Discovery.
[00210] Из полученных в результате данных строили графики, показанные на фигуре 4A. Имеет место отчетливо повышенная концентрация IL-2 в супернатанте вследствие активации T-клеток в ответ на мутеины липокалина с SEQ ID NO: 13, SEQ ID NO: 14 и SEQ ID NO: 15 по сравнению с отрицательным контролем, представляющим собой SEQ ID NO: 4. Из эксперимента видно, что все тестируемые мутеины способны костимулировать ответ T-клеток при нанесении на пластиковую чашку для культивирования.
[00211] Пример 8. Функциональный анализ активации T-клеток с использованием мутеинов липокалина в растворе
[00212] Для тестирования того, активируют ли репрезентативные мутеины липокалина CD137 также путем простого связывания без образования скоплений, анализ из примера 7 осуществляли аналогичным примеру 5 образом, но с использованием растворимых, а не захваченных мутеинов липокалина. В этом анализе плоскодонные планшеты для культур тканей покрывали, как описано выше, но с использованием только антитела к CD3. Планшеты обрабатывали, как описано выше, но только после стадии добавления T-клеток (включая 2 мкг/мл hCD28) добавляли 50 мкл мутеинов липокалина в растворе в концентрации, составляющей 25 мкг/мл.
[00213] Из полученных в результате данных строили графики, показанные на фигуре 4B. Не наблюдали значимого повышения концентрации IL-2 в супернатанте вследствие активации T-клеток в ответ на любой из тестируемых мутеинов липокалина в сравнении с отрицательным контролем, представляющим собой SEQ ID NO: 4. Данный эксперимент свидетельствует о том, что мономерные мутеины липокалина в растворе в концентрации, которой достаточно для насыщения всех рецепторов CD137, не обеспечивают костимуляцию T-клеток.
Пример 9. Функциональный анализ активации T-клеток с использованием нанесенных мутеинов липокалина
[00215] Для более подробного изучения способности мутеина с SEQ ID NO: 13 костимулировать ответы T-клеток, авторы настоящего изобретения прибегали к анализу активации T-клеток, как в примере 7. В качестве показателей для считывания авторы настоящего изобретения оценивали длительную пролиферацию T-клеток после трех дней инкубации с использованием импульсного мечения BrdU в течение 4 ч и измеряли уровни IL-2 и интерферона-гамма (IFN-g) в супернатанте. Помимо пролиферации и продуцирования IL-2 усиленное продуцирование IFN-g является дополнительным признаком активации T-клеток, а повышение уровней IFN-γ путем костимуляции антителом к CD137 было описано в литературе (Jure-Kunkel, M. et al., патент США 7288638).
[00216] В качестве отрицательного контроля использовали подобный дикому типу мутеин липокалина с SEQ ID NO: 4. В некоторых аспектах этот эксперимент осуществляли таким образом, который аналогичен эксперименту, описанному в примере 8. Ниже авторы настоящего изобретения приводят подробное описание эксперимента.
[00217] Мононуклеарные клетки периферической крови человека (PBMC) от здоровых доноров-добровольцев выделяли из лейкоцитарных пленок путем центрифугирования в градиенте плотности полисахарозы (Biocoll 1,077 г/мл от Biochrom), следуя протоколу компании Biochrom. T-лимфоциты выделяли из полученных в результате PBMC c использованием набора для очистки Pan T-cell purification Kit (Miltenyi Biotec GmbH) и протоколов изготовителя. Очищенные T-клетки повторно суспендировали в буфере, состоящем из 90% FCS и 10% DMSO, немедленно замораживали с помощью жидкого азота и хранили в жидком азоте до последующего использования. Для анализа T-клетки оттаивали в течение 16 ч и культивировали в культуральной среде (RPMI 1640, Life Technologies), дополненной 10% FCS и 1% смеси пенициллина-стрептомицина (Life Technologies).
[00218] Следующую процедуру проводили с использованием трех повторов для каждого экспериментального условия. На плоскодонные планшеты для культур тканей наносили 200 мкл смеси из 5 мкг/мл антитела к CD3 и 25 мкг/мл антитела кролика к остову липокалина (поликлональное антитело кролика к hNGAL, Pieris) и выдерживали в течение ночи при 4°C. Последнее использовали для обеспечения возможности иммобилизации SEQ ID NO: 13 путем аффинного захвата. В качестве отрицательного контроля наносили контроль, представляющий собой изотип IgG1, в концентрации, составляющей 5 мкг/мл, вместо антитела к CD3 наряду с 25 мкг/мл антитела кролика к остову липокалина. На следующий день лунки промывали дважды с использованием PBS и 50 мкл серии разведений SEQ ID NO: 13, находящейся в диапазоне от 50 мкг/мл до 0,8 мкг/мл, в семь стадий захватывали на предварительно покрытых планшетах в течение 1 ч при 37°C. В качестве отрицательного контроля SEQ ID NO: 4 захватывали в трех концентрациях (50 мкг/мл, 25 мкг/мл, 12,5 мкг/мл). В качестве дополнительного отрицательного контроля SEQ ID NO: 13 захватывали в концентрации, составляющей 50 мкг/мл, в лунках, которые были покрыты изотипом IgG1 и антителом к hNGAL для захвата (см. выше). После еще одного двухкратного промывания с использованием PBS 100 мкл суспензии T-клеток (соответствующей 5 ×104 T-клеток) в культуральной среде добавляли в лунки. Это проводили либо в присутствии, либо в отсутствие антитела к hCD28 в концентрации, составляющей 2 мкг/мл. Планшеты накрывали газопроницаемой уплотняющей пленкой (4 titude) и инкубировали при 37°C в увлажненной атмосфере с 5% CO2 в течение 3 дней. Затем оценивали концентрацию IL-2 и IFN-γ в супернатанте, а также клеточную пролиферацию.
[00219] Для того, чтобы количественно оценить T-клеточную пролиферацию, использовали набор для ELISA для определения хемилюминесцентного сигнала при клеточной пролиферации, основанный на включении BrdU (Roche), согласно инструкциям изготовителя. Вкратце, на 3 день 10 мкл раствора для мечения с BrdU добавляли в каждую лунку и пролиферации предоставляли возможность проходить в течение дополнительных 4 ч при 37°C в условиях увлажненной атмосферы с 5% CO2. Планшеты центрифугировали при 300 g в течение 10 мин, и супернатанты трех повторов объединяли и сразу помещали для хранения при -20°C для дальнейшего количественного измерения уровней IL-2 и IFN-γ. Затем планшеты сушили при 60°C в течение 1 часа. 200 мкл раствора «FixDenat» добавляли в каждую лунку и планшеты инкубировали при комнатной температуре в течение 30 мин. Включенный BRDU метили меченным пероксидазой антителом к BrdU путем 2 ч инкубации при комнатной температуре. Уровни BrdU оценивали путем количественного анализа катализируемой пероксидазой, испускающей хемилюминесцентный сигнал, реакции в планшет-ридере PheraStar FS.
[00220] Уровни IL-2 и IFN-γ человека в объединенных супернатантах клеточных культур количественно измеряли с помощью наборов DuoSet для IL-2 и DuoSet для IFN-γ от R&D Systems. Процедуру осуществляли аналогично для двух цитокинов и описывали ниже только для IL-2. На первой стадии на 384-луночный планшет наносили при комнатной температуре на 2 ч 1 мкг/мл «антитела для захвата IL-2 человека» (R&D System), разведенного в PBS. Затем лунки промывали 5 раз с использование 80 мкл PBS-T (PBS, содержащего 0,05% Tween20) с использованием установки для избирательного промывания Biotek EL405 Select CW (Biotek). После блокирования в течение 1 ч в PBS-T, дополнительно содержащем 1% казеина (вес/вес), объединенный супернатант и серию концентраций стандарта IL-2, разведенного в культуральной среде, инкубировали в 384-луночном планшете в течение ночи при 4°C. Для обеспечения возможности выявления и количественного измерения захваченного IL-2 смесь из 100 нг/мл биотинилированного антитела козы к hIL-2-Bio для выявления (R&D System) и 1 мкг/мл меченного сульфо-меткой стрептавидина (Mesoscale Discovery) добавляли в PBS-T, содержащий 0,5% казеина, и инкубировали при комнатной температуре в течение 1 ч. После промывания 25 мкл буфера для считывания добавляли в каждую лунку и электрохемилюминесцентный (ECL) сигнал каждой лунки считывали с использованием планшет-ридера Mesoscale Discovery. Анализ и количественное измерение проводили с использованием программного обеспечения от Mesoscale Discovery.
[00221] Результат эксперимента показан на фигуре 5. Показатели для считывания пролиферации, уровни IL-2 и IFN-γ в супернатанте для эксперимента с использованием как антител к CD3, так и антител к CD28 представлены на фигурах 5A, 5C и 5E соответственно. Те же показатели для считывания для эксперимента, выполненного исключительно с антителом к CD3, представлены на фигурах 5B, 5D и 5F.
[00222] В эксперименте, в котором применяли стимуляцию с помощью антител к CD3 и к CD28, имеет место отчетливое зависимое от дозы увеличение скорости пролиферации (фигура 5A), которое до 14-раз больше, чем для отрицательного контроля, представляющего собой SEQ ID NO: 4. Пролиферация в отсутствие mAb к CD3 (столбец, помеченный как «IgG1») несущественна. При считывании уровня продуцирования IL-2 (фигура 5C) также имеет место отчетливое зависимое от дозы повышение, при котором уровни при максимальном ответе при концентрации SEQ ID NO: 13 в нанесенном слое, составляющей 6,25 мкг/мл, и при более высоких концентрациях остаются неизменно на уровнях вплоть до около 6 раз выше по сравнению с отрицательным контролем. При считывании уровня продуцирования IFN-γ (фигура 5E) паттерн является весьма схожим, при этом максимальные уровни IFN-g достигают значений вплоть до 2,5 раза больше по сравнению с отрицательным контролем.
[00223] В эксперименте, в котором использовали стимуляцию с помощью только mAb к CD3, авторы настоящего изобретения снова обнаруживали отчетливое зависимое от дозы увеличение скорости пролиферации (фигура 5A), которое составляет вплоть до 4 раз больше, чем для отрицательного контроля, представляющего собой SEQ ID NO: 4. По-видимому, максимальный ответ будет при концентрации, составляющей 6,25 мкг/мл, в нанесенном слое SEQ ID NO: 13, который достигает значения в 15 раз больше по сравнению с отрицательным контролем. И как при более высокой, так и при более низкой концентрациях ответ является менее выраженным. При считывании уровня продуцирования IFN-γ имеет место зависимое от дозы повышение, при котором уровни при максимальном ответе при концентрации SEQ ID NO: 13 в нанесенном слое, составляющей 6,25 мкг/мл, и при более высоких концентрациях остаются неизменными на уровнях, составляющих вплоть до около 2,5 раза выше по сравнению с отрицательным контролем.
[00224] В целом из эксперимента, показанного в данном примере 9, отчетливо видно существенную костимуляцию ответа T-клеток с помощью мутеина с SEQ ID NO: 13 относительно пролиферации, продуцирования IL-2 и продуцирования IFN-γ как в присутствии, так и в отсутствие стимуляции CD28.
[00225] Варианты осуществления, иллюстративно описанные в данном документе, надлежащим образом можно осуществлять на практике в отсутствие любых элемента или элементов, ограничения или ограничений, не раскрытых конкретно в данном документе. Таким образом, например, термины «содержащий», «включающий», «состоящий из» и т. д. следует читать как открытые и неограничивающие. Дополнительно термины и выражения, используемые в данном документе, использовали в качестве терминов описания, а не ограничения, и в использовании таких терминов и выражений отсутствует намерение исключить любые эквиваленты показанных и описанных характеристик или их частей, но следует понимать, что разные модификации возможны в пределах объема заявленного изобретения. Таким образом, следует понимать, что несмотря на то, что настоящие варианты осуществления были конкретно раскрыты с помощью предпочтительных вариантов осуществления и необязательных характеристик, к их модификации и вариациям может прибегать специалист в данной области, и что такие модификации и вариации рассматриваются как часть объема настоящего изобретения. Все патенты, патентные заявки, руководства и прошедшие рецензию публикации, описанные в данном документе, в данный документ включены с помощью ссылки в полном объеме. Кроме того, в тех случаях, когда определение или использование термина в ссылочном материале, который включен посредством ссылки в данный документ, не согласуется или противоречит определению такого термина, представленному в данном документе, тогда используют определение термина, представленное в данном документе, а определение термина из ссылки не используют. Каждое из более узких видов и субродовых группировок, находящихся в пределах раскрытия информации в обобщенном виде, также образует часть настоящего изобретения. Это включает описание настоящего изобретения в обощенном виде с оговоркой или отрицательными признаками, удаляющими любой объект из рода, независимо от того, исключен материал, конкретно изложенный в данном документе, или нет. К тому же в тех случаях, когда особенности описаны в контексте группы Маркуша, тогда специалисту в данной области будет понятно, что настоящее раскрытие также таким образом раскрыто в контексте любого отдельного представителя или подгруппы представителей группы Маркуша. Дополнительные варианты осуществления будут очевидны из прилагаемой формулы изобретения.
[00226] Эквиваленты. Специалисту в данной области будут понятны, или он будет способен установить с помощью всего лишь проведения обычных экспериментов, многие эквиваленты конкретных вариантов осуществления настоящего изобретения, описанного в данном документе. Как предполагается, такие эквиваленты охватываются прилагаемой формулой изобретения. Все публикации, патенты и патентные заявки, упомянутые в этом описании, таким образом включены с помощью ссылки в описание в такой же степени, как если бы каждая отдельная публикация, отдельный патент или отдельная патентная заявка были конкретно и отдельно указаны как включенные в данном документе с помощью ссылки.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Пиерис АГ
<120> Новые белки, специфичные в отношении CD137
<130> PIE15347PCT
<150> EP15166184.0
<151> 2015-05-04
<160> 39
<170> PatentIn версия 3.5
<210> 1
<211> 156
<212> БЕЛОК
<213> искусственный
<220>
<223> Tlc дикого типа человека
<400> 1
His His Leu Leu Ala Ser Asp Glu Glu Ile Gln Asp Val Ser Gly Thr
1 5 10 15
Trp Tyr Leu Lys Ala Met Thr Val Asp Arg Glu Phe Pro Glu Met Asn
20 25 30
Leu Glu Ser Val Thr Pro Met Thr Leu Thr Thr Leu Glu Gly Gly Asn
35 40 45
Leu Glu Ala Lys Val Thr Met Leu Ile Ser Gly Arg Cys Gln Glu Val
50 55 60
Lys Ala Val Leu Glu Lys Thr Asp Glu Pro Gly Lys Tyr Thr Ala Asp
65 70 75 80
Gly Gly Lys His Val Ala Tyr Ile Ile Arg Ser His Val Lys Asp His
85 90 95
Tyr Ile Phe Tyr Cys Glu Gly Glu Leu His Gly Lys Pro Val Arg Gly
100 105 110
Val Lys Leu Val Gly Arg Asp Pro Lys Asn Asn Leu Glu Ala Leu Glu
115 120 125
Asp Phe Glu Lys Ala Ala Gly Ala Arg Gly Leu Ser Thr Glu Ser Ile
130 135 140
Leu Ile Pro Arg Gln Ser Glu Thr Cys Ser Pro Gly Ser Asp
145 150 155
<210> 2
<211> 178
<212> БЕЛОК
<213> искусственный
<220>
<223> hNGAL дикого типа
<400> 2
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Leu Ala Gly Asn Ala Ile Leu Arg Glu Asp Lys Asp Pro
35 40 45
Gln Lys Met Tyr Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asn Val Thr Ser Val Leu Phe Arg Lys Lys Lys Cys Asp Tyr Trp Ile
65 70 75 80
Arg Thr Phe Val Pro Gly Cys Gln Pro Gly Glu Phe Thr Leu Gly Asn
85 90 95
Ile Lys Ser Tyr Pro Gly Leu Thr Ser Tyr Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Lys Val Ser Gln
115 120 125
Asn Arg Glu Tyr Phe Lys Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 3
<211> 152
<212> БЕЛОК
<213> искусственный
<220>
<223> отрицательный контроль
<400> 3
Ala Ser Asp Glu Glu Ile Gln Asp Val Ser Gly Thr Trp Tyr Leu Lys
1 5 10 15
Ala Met Thr Val Asp Arg Glu Cys Pro Glu Met Asn Leu Glu Ser Val
20 25 30
Thr Pro Met Thr Leu Thr Thr Leu Glu Gly Gly Asn Leu Glu Ala Lys
35 40 45
Val Thr Met Leu Ile Ser Gly Arg Ser Gln Glu Val Lys Ala Val Leu
50 55 60
Glu Lys Thr Asp Glu Pro Gly Lys Tyr Thr Ala Asp Gly Gly Lys His
65 70 75 80
Val Ala Tyr Ile Ile Arg Ser His Val Lys Asp His Tyr Ile Phe Tyr
85 90 95
Ser Glu Gly Glu Cys His Gly Lys Pro Val Pro Gly Val Trp Leu Val
100 105 110
Gly Arg Asp Pro Lys Asn Asn Leu Glu Ala Leu Glu Asp Phe Glu Lys
115 120 125
Ala Ala Gly Ala Arg Gly Leu Ser Thr Glu Ser Ile Leu Ile Pro Arg
130 135 140
Gln Ser Glu Thr Ser Ser Pro Gly
145 150
<210> 4
<211> 178
<212> БЕЛОК
<213> искусственный
<220>
<223> отрицательный контроль
<400> 4
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Leu Ala Gly Asn Ala Ile Leu Arg Glu Asp Lys Asp Pro
35 40 45
Gln Lys Met Tyr Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asn Val Thr Ser Val Leu Phe Arg Lys Lys Lys Cys Asp Tyr Trp Ile
65 70 75 80
Arg Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Asn
85 90 95
Ile Lys Ser Tyr Pro Gly Leu Thr Ser Tyr Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Lys Val Ser Gln
115 120 125
Asn Arg Glu Tyr Phe Lys Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 5
<211> 152
<212> БЕЛОК
<213> искусственный
<220>
<223> мутеин липокалина
<400> 5
Ala Ser Asp Glu Glu Ile Gln Asp Val Ser Gly Thr Trp Tyr Leu Lys
1 5 10 15
Ala Met Thr Val Asp Glu Gly Cys Arg Pro Trp Asn Ile Phe Ser Val
20 25 30
Thr Pro Met Thr Leu Thr Thr Leu Glu Gly Gly Asn Leu Glu Ala Lys
35 40 45
Val Thr Met Ala Ile Asp Gly Pro Ala Gln Glu Val Lys Ala Val Leu
50 55 60
Glu Lys Thr Asp Glu Pro Gly Lys Tyr Thr Ala Asp Gly Gly Lys His
65 70 75 80
Val Ala Tyr Ile Ile Arg Ser His Val Lys Asp His Tyr Ile Phe Tyr
85 90 95
Ser Glu Gly Val Cys Asp Gly Ser Pro Val Pro Gly Val Trp Leu Val
100 105 110
Gly Arg Asp Pro Lys Asn Asn Leu Glu Ala Leu Glu Asp Phe Glu Lys
115 120 125
Ala Ala Gly Ala Arg Gly Leu Ser Thr Glu Ser Ile Leu Ile Pro Arg
130 135 140
Gln Ser Glu Thr Ser Ser Pro Gly
145 150
<210> 6
<211> 152
<212> БЕЛОК
<213> искусственный
<220>
<223> мутеин липокалина
<400> 6
Thr Ser Asp Glu Glu Ile Gln Asp Val Ser Gly Thr Trp Tyr Leu Lys
1 5 10 15
Ala Met Thr Val Asp Glu Gly Cys Arg Pro Trp Asn Ile Phe Ser Val
20 25 30
Thr Pro Met Thr Leu Thr Thr Leu Glu Gly Gly Asn Leu Glu Ala Lys
35 40 45
Val Thr Met Ala Ile Asp Gly Pro Ala Gln Glu Val Arg Ala Val Leu
50 55 60
Glu Lys Thr Asp Glu Pro Gly Lys Tyr Thr Ala Asp Gly Gly Lys His
65 70 75 80
Asp Ala Tyr Ile Ile Arg Ser His Val Lys Asp His Tyr Ile Phe Tyr
85 90 95
Ser Glu Gly Val Cys Asp Gly Ser Pro Val Pro Gly Val Trp Leu Val
100 105 110
Gly Arg Asp Pro Glu Asn Asn Leu Glu Ala Leu Glu Asp Phe Glu Lys
115 120 125
Thr Ala Gly Ala Arg Gly Leu Ser Thr Glu Ser Ile Leu Ile Pro Arg
130 135 140
Gln Ser Glu Thr Ser Ser Pro Gly
145 150
<210> 7
<211> 152
<212> БЕЛОК
<213> искусственный
<220>
<223> мутеин липокалина
<400> 7
Ala Ser Asp Glu Glu Ile Gln Asp Val Ser Gly Thr Trp Tyr Leu Lys
1 5 10 15
Ala Met Thr Val Asp Glu Gly Cys Arg Pro Trp Asn Ile Phe Ser Val
20 25 30
Thr Pro Met Thr Leu Thr Thr Leu Glu Gly Gly Asn Leu Glu Ala Lys
35 40 45
Val Thr Met Ala Ile Asp Gly Pro Ala Gln Glu Val Asn Ala Val Leu
50 55 60
Glu Lys Thr Asp Glu Pro Gly Lys Tyr Thr Ala Asp Gly Gly Lys His
65 70 75 80
Val Ala Tyr Ile Ile Arg Ser His Val Arg Asp His Tyr Ile Phe Tyr
85 90 95
Ser Glu Gly Val Cys Asp Gly Ser Pro Val Pro Gly Val Trp Leu Val
100 105 110
Gly Arg Asp Pro Glu Asn Asn Leu Glu Ala Leu Glu Asp Phe Glu Lys
115 120 125
Thr Ala Gly Ala Arg Gly Leu Ser Thr Glu Ser Ile Leu Ile Pro Arg
130 135 140
Gln Ser Glu Thr Ser Ser Pro Gly
145 150
<210> 8
<211> 152
<212> БЕЛОК
<213> искусственный
<220>
<223> мутеин липокалина
<400> 8
Val Ser Asp Glu Glu Ile Gln Asp Val Ser Gly Thr Trp Tyr Leu Lys
1 5 10 15
Ala Met Thr Val Asp Glu Gly Cys Arg Pro Trp Asn Ile Phe Ser Val
20 25 30
Thr Pro Met Thr Leu Thr Thr Leu Glu Gly Gly Asn Leu Glu Ala Lys
35 40 45
Val Thr Met Ala Ile Asp Gly Pro Ala Gln Glu Val Arg Ala Val Leu
50 55 60
Glu Lys Thr Asp Glu Pro Gly Lys Tyr Thr Ala Asp Gly Gly Lys His
65 70 75 80
Val Ala Tyr Ile Ile Arg Ser His Val Glu Asp His Tyr Ile Phe Tyr
85 90 95
Ser Glu Gly Val Cys Asp Gly Ser Pro Val Pro Gly Val Trp Leu Val
100 105 110
Gly Arg Asp Pro Glu Asn Asn Leu Glu Ala Leu Glu Asp Phe Glu Lys
115 120 125
Thr Ala Gly Ala Arg Gly Leu Ser Thr Glu Ser Ile Leu Ile Pro Arg
130 135 140
Gln Ser Glu Thr Ser Ser Pro Gly
145 150
<210> 9
<211> 152
<212> БЕЛОК
<213> искусственный
<220>
<223> мутеин липокалина
<400> 9
Ala Ser Asp Glu Glu Ile Gln Asp Val Ser Gly Thr Trp Tyr Leu Lys
1 5 10 15
Ala Met Thr Val Asp Glu Gly Cys Arg Pro Trp Asn Ile Phe Ser Val
20 25 30
Thr Pro Met Thr Leu Ser Thr Leu Glu Gly Gly Asn Leu Glu Ala Lys
35 40 45
Val Thr Met Ala Ile Asp Gly Pro Ala Gln Glu Val Lys Ala Val Leu
50 55 60
Glu Lys Thr Asp Glu Pro Gly Lys Tyr Thr Ala Asp Gly Gly Lys His
65 70 75 80
Val Ala Tyr Ile Ile Arg Ser His Val Lys Asp His Tyr Ile Phe Tyr
85 90 95
Ser Glu Gly Val Cys Asp Gly Ser Pro Val Pro Gly Val Trp Leu Val
100 105 110
Gly Arg Asp Pro Lys Asn Asn Leu Glu Ala Leu Glu Asp Phe Glu Lys
115 120 125
Ala Ala Gly Ala Arg Gly Leu Ser Thr Glu Ser Ile Leu Ile Pro Arg
130 135 140
Gln Ile Glu Thr Ser Ser Pro Gly
145 150
<210> 10
<211> 152
<212> БЕЛОК
<213> искусственный
<220>
<223> мутеин липокалина
<400> 10
Ala Ser Asp Glu Glu Ile Gln Asp Val Ser Gly Thr Trp Tyr Leu Lys
1 5 10 15
Ala Met Thr Val Asp Glu Gly Cys Arg Pro Trp Asn Ile Phe Ser Val
20 25 30
Thr Pro Met Thr Leu Thr Thr Leu Glu Gly Gly Asn Leu Glu Ala Glu
35 40 45
Val Thr Met Ala Ile Asp Gly Pro Ala Gln Glu Val Lys Ala Val Leu
50 55 60
Glu Lys Ala Asp Glu Pro Gly Lys Tyr Thr Ala Asp Gly Gly Lys His
65 70 75 80
Val Ala Tyr Ile Ile Arg Ser His Val Lys Asp His Tyr Ile Phe Tyr
85 90 95
Ser Glu Gly Val Cys Asp Gly Ser Pro Val Pro Gly Val Trp Leu Val
100 105 110
Gly Arg Asp Pro Lys Asn Asn Leu Glu Ala Leu Glu Asp Phe Glu Lys
115 120 125
Thr Ala Gly Ala Arg Gly Leu Ser Thr Glu Ser Ile Leu Ile Pro Ser
130 135 140
Gln Ile Glu Thr Ser Ser Pro Gly
145 150
<210> 11
<211> 152
<212> БЕЛОК
<213> искусственный
<220>
<223> мутеин липокалина
<400> 11
Thr Ser Asp Glu Glu Ile Gln Asp Val Ser Gly Thr Trp Tyr Leu Lys
1 5 10 15
Ala Met Thr Val Asp Glu Gly Cys Arg Pro Trp Asn Ile Phe Ser Val
20 25 30
Thr Pro Met Thr Leu Thr Thr Leu Glu Asp Gly Asn Leu Glu Ala Lys
35 40 45
Val Thr Met Ala Ile Asp Gly Pro Ala Gln Glu Val Lys Ala Val Leu
50 55 60
Glu Lys Ala Asp Glu Pro Gly Lys Tyr Thr Ala Asp Gly Gly Lys His
65 70 75 80
Val Ala Tyr Ile Ile Arg Ser His Val Lys Asp His Tyr Ile Phe Tyr
85 90 95
Ser Glu Gly Val Cys Asp Gly Ser Pro Val Pro Gly Val Trp Leu Val
100 105 110
Gly Arg Asp Pro Lys Asn Asn Leu Glu Ala Leu Glu Asp Phe Glu Lys
115 120 125
Ala Ala Gly Ala Arg Gly Leu Ser Thr Glu Ser Ile Leu Ile Pro Arg
130 135 140
Gln Ile Glu Thr Ser Ser Pro Gly
145 150
<210> 12
<211> 178
<212> БЕЛОК
<213> искусственный
<220>
<223> мутеин липокалина
<400> 12
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Ile Lys Leu Arg Glu Asp Lys Asp Pro
35 40 45
Asn Lys Met Met Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asn Val Thr Gly Val Thr Phe Asp Asp Lys Lys Cys Thr Tyr Ala Ile
65 70 75 80
Ser Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Lys
85 90 95
Ile Lys Ser Phe Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln
115 120 125
Asn Arg Glu Glu Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 13
<211> 178
<212> БЕЛОК
<213> искусственный
<220>
<223> мутеин липокалина
<400> 13
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Ile Arg Leu Arg Glu Asp Lys Asp Pro
35 40 45
Ile Lys Met Met Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Met Val Lys Phe Asp Asp Lys Lys Cys Met Tyr Asp Ile
65 70 75 80
Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Lys
85 90 95
Ile Lys Ser Phe Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln
115 120 125
Asn Arg Glu Glu Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 14
<211> 178
<212> БЕЛОК
<213> искусственный
<220>
<223> мутеин липокалина
<400> 14
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Ile Arg Leu Arg Glu Asp Lys Asp Pro
35 40 45
Asn Lys Met Met Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Ala Val Ala Phe Asp Asp Lys Lys Cys Thr Tyr Asp Ile
65 70 75 80
Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Lys
85 90 95
Ile Lys Ser Phe Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln
115 120 125
Asn Arg Glu Glu Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 15
<211> 178
<212> БЕЛОК
<213> искусственный
<220>
<223> мутеин липокалина
<400> 15
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Ile Lys Leu Arg Glu Asp Lys Asp Pro
35 40 45
Asn Lys Met Met Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Ala Val Ala Phe Asp Asp Lys Lys Cys Thr Tyr Asp Ile
65 70 75 80
Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Lys
85 90 95
Ile Lys Ser Phe Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln
115 120 125
Asn Arg Glu Glu Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 16
<211> 175
<212> БЕЛОК
<213> искусственный
<220>
<223> мутеин липокалина
<400> 16
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Ile Lys Leu Arg Glu Asp Ser Lys Met
35 40 45
Met Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr Asp Val Thr
50 55 60
Gly Val Ser Phe Asp Asp Lys Lys Cys Thr Tyr Ala Ile Met Thr Phe
65 70 75 80
Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Lys Ile Lys Ser
85 90 95
Phe Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser Thr Asn Tyr
100 105 110
Asn Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln Asn Arg Glu
115 120 125
Glu Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu Thr Ser Glu
130 135 140
Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly Leu Pro Glu
145 150 155 160
Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile Asp Gly
165 170 175
<210> 17
<211> 178
<212> БЕЛОК
<213> искусственный
<220>
<223> мутеин липокалина
<400> 17
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Ile Lys Leu Arg Glu Asp Lys Asp Pro
35 40 45
Val Lys Met Met Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Gly Val Thr Phe Asp Asp Lys Lys Cys Arg Tyr Asp Ile
65 70 75 80
Ser Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Phe Gly Lys
85 90 95
Ile Lys Ser Phe Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln
115 120 125
Asn Arg Glu Glu Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 18
<211> 178
<212> БЕЛОК
<213> искусственный
<220>
<223> мутеин липокалина
<400> 18
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Ile Arg Leu Arg Glu Asp Lys Asp Pro
35 40 45
His Lys Met Met Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Gly Val Thr Phe Asp Asp Lys Lys Cys Thr Tyr Ala Ile
65 70 75 80
Ser Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Lys
85 90 95
Ile Lys Ser Phe Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln
115 120 125
Asn Arg Glu Glu Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 19
<211> 178
<212> БЕЛОК
<213> искусственный
<220>
<223> мутеин липокалина
<400> 19
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Ile Lys Leu Arg Glu Asp Lys Asp Pro
35 40 45
Asn Lys Met Met Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Gly Val Thr Phe Asp Asp Lys Lys Cys Thr Tyr Ala Ile
65 70 75 80
Ser Thr Leu Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Phe Gly Lys
85 90 95
Ile Lys Ser Phe Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln
115 120 125
Asn Arg Glu Glu Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 20
<211> 178
<212> БЕЛОК
<213> искусственный
<220>
<223> мутеин липокалина
<400> 20
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Ile Arg Leu Arg Glu Asp Lys Asp Pro
35 40 45
Ser Lys Met Met Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Ala Val Thr Phe Asp Asp Lys Lys Cys Asn Tyr Ala Ile
65 70 75 80
Ser Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Lys
85 90 95
Ile Lys Ser Phe Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln
115 120 125
Asn Arg Glu Glu Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 21
<211> 10
<212> БЕЛОК
<213> искусственный
<220>
<223> C-концевая метка 1
<400> 21
Ser Ala Trp Ser His Pro Gln Phe Glu Lys
1 5 10
<210> 22
<211> 11
<212> БЕЛОК
<213> искусственный
<220>
<223> C-концевая метка 2
<400> 22
Pro Ser Ala Trp Ser His Pro Gln Phe Glu Lys
1 5 10
<210> 23
<211> 8
<212> БЕЛОК
<213> искусственный
<220>
<223> Strep-метка
<400> 23
Trp Ser His Pro Gln Phe Glu Lys
1 5
<210> 24
<211> 456
<212> ДНК
<213> искусственная
<220>
<223> мутеин липокалина
<400> 24
gcctcagacg aggagattca ggatgtgtca gggacgtggt atctgaaggc catgacggtg 60
gatgaggggt gtcgtccttg gaatatattt tcagttacgc caatgactct gactaccctt 120
gaaggcggca atctggaggc taaggtcacc atggcaatag atgggccggc acaggaggtg 180
aaagcagtgt tagagaagac agatgaaccg ggtaaatata cggccgatgg cggtaaacat 240
gttgcctata tcattcgcag ccatgtgaaa gatcattaca tcttttatag cgagggcgtg 300
tgcgatgggt ctcctgttcc aggggtgtgg ctcgtgggca gagaccccaa gaacaacctg 360
gaagccttgg aggactttga gaaagccgca ggagcccgcg gactcagcac ggagagcatc 420
ctcatcccca ggcagagcga aaccagctct ccaggg 456
<210> 25
<211> 459
<212> ДНК
<213> искусственная
<220>
<223> мутеин липокалина
<400> 25
acctcagacg aggagattca ggatgtgtca gggacgtggt atctgaaggc gatgacggtg 60
gatgaggggt gtcgtccttg gaatatattt tcagttacgc caatgactct gactaccctt 120
gaaggcggca atctggaggc taaggtcacc atggcaatag atgggccggc acaggaggtg 180
agagcagtgt tagagaagac agatgaaccg ggtaaatata cggccgacgg cggtaaacat 240
gatgcctata tcattcgcag ccatgtgaaa gatcattaca tcttttatag cgagggcgtg 300
tgcgatgggt ctcctgttcc gggggtgtgg ctcgtgggca gagaccccga gaacaacctg 360
gaagccttgg aggactttga gaaaaccgca ggagcccgcg gactcagcac ggagagcatc 420
ctcatcccca ggcagagcga aaccagctct ccagggcca 459
<210> 26
<211> 456
<212> ДНК
<213> искусственная
<220>
<223> мутеин липокалина
<400> 26
gcctcagacg aggagattca ggatgtgtca gggacgtggt atctgaaggc gatgacggtg 60
gatgaggggt gtcgtccttg gaatatattt tcagttacgc caatgactct gactaccctt 120
gaaggcggca atctggaggc taaggtcacc atggcaatag atgggccggc acaggaggtg 180
aacgcagtgt tagagaagac agatgaaccg ggtaaatata cggccgatgg cggtaaacat 240
gttgcctata tcattcgcag ccatgtgaga gatcattaca tcttttatag cgagggcgtg 300
tgcgatgggt ctcctgttcc gggggtgtgg ctcgtgggca gggaccccga gaacaacctg 360
gaagccttgg aggactttga gaaaaccgca ggagcccgcg gactcagcac ggagagcatt 420
ctcattccca ggcagagcga aaccagctct ccaggg 456
<210> 27
<211> 459
<212> ДНК
<213> искусственная
<220>
<223> мутеин липокалина
<400> 27
gtctcagacg aggagattca ggatgtgtca gggacgtggt atctgaaggc gatgacggtg 60
gatgaggggt gtcgtccttg gaatatattt tcagttacgc caatgactct gactaccctt 120
gaaggcggca atctggaggc taaggtcacc atggcaatag atgggccggc acaggaggtg 180
agagcagtgt tagagaagac agatgaaccg ggtaaatata cggccgatgg cggtaaacat 240
gttgcctata tcattcgcag ccatgtggaa gatcattaca tcttttatag cgagggcgtg 300
tgcgatgggt ctcctgttcc gggggtgtgg ctcgtgggca gagaccccga gaacaacctg 360
gaagccttgg aggactttga gaaaaccgca ggagcccgcg gactcagcac ggagagcatc 420
ctcatcccca ggcagagcga aaccagctct ccagggcca 459
<210> 28
<211> 459
<212> ДНК
<213> искусственная
<220>
<223> мутеин липокалина
<400> 28
gcctcagacg aggagattca ggatgtgtca gggacgtggt atctgaaggc gatgacggtg 60
gatgaggggt gtcgtccttg gaatatattt tcagttacgc caatgactct gtctaccctt 120
gaaggcggca atctggaggc taaggtcacc atggcaatag atgggccggc acaggaggtg 180
aaagcagtgt tagagaagac agatgaaccg ggtaaatata cggccgatgg cggtaaacat 240
gttgcctata tcattcgcag ccatgtgaaa gatcattaca tcttttatag cgagggcgtg 300
tgcgatgggt ctcctgttcc gggggtgtgg ctcgtgggca gagaccccaa gaacaacctg 360
gaagccttgg aggactttga gaaagccgca ggagcccgcg gactcagcac ggagagcatc 420
ctcatcccca ggcagatcga aaccagctct ccagggcca 459
<210> 29
<211> 459
<212> ДНК
<213> искусственная
<220>
<223> мутеин липокалина
<400> 29
gcctcagacg aggagattca ggatgtgtca gggacgtggt atctgaaggc gatgacggtg 60
gatgaggggt gtcgtccttg gaatatattt tcagttacgc caatgactct gactaccctt 120
gaaggcggca atctggaggc tgaggtcacc atggcaatag atgggccggc acaggaggtg 180
aaagcagtgt tagagaaggc agatgaaccg ggtaaatata cggccgatgg cggtaaacat 240
gttgcctata tcattcgcag ccatgtgaaa gatcattaca tcttttatag cgagggcgtg 300
tgcgatgggt ctcctgttcc gggggtgtgg ctcgtgggca gagaccccaa gaacaacctg 360
gaagccttgg aggactttga gaaaaccgca ggagcccgcg gactcagcac ggagagcatc 420
ctcatcccca gtcagatcga aaccagctct ccagggcca 459
<210> 30
<211> 459
<212> ДНК
<213> искусственная
<220>
<223> мутеин липокалина
<400> 30
acctcagacg aggagattca ggatgtgtca gggacgtggt atctgaaggc gatgacggtg 60
gatgaggggt gtcgtccttg gaatatattt tcagttacgc caatgactct gactaccctt 120
gaagacggca atctggaggc taaggtcacc atggcaatag atgggccggc acaggaggtg 180
aaagcagtgt tagagaaggc agatgaaccg ggtaaatata cggccgatgg cggtaaacat 240
gttgcctata tcattcgcag ccatgtgaaa gatcattaca tcttttatag cgagggcgtg 300
tgcgatgggt ctcctgttcc gggggtgtgg ctcgtgggca gagaccccaa gaacaacctg 360
gaagccttgg aggactttga gaaagccgca ggagcccgcg gactcagcac ggagagcatc 420
ctcatcccca ggcagatcga aaccagctct ccagggcca 459
<210> 31
<211> 534
<212> ДНК
<213> искусственная
<220>
<223> мутеин липокалина
<400> 31
caggactcca cctcagacct gatcccagcc ccacctctga gcaaggtccc tctgcagcag 60
aacttccagg acaaccaatt ccatgggaag tggtatgtgg taggtcaggc agggaatatc 120
aaactcagag aagacaaaga cccgaacaag atgatggcca ccatctatga gctgaaagaa 180
gacaagagct acaatgtcac cggtgtcact tttgacgaca agaagtgtac ttacgctatc 240
tctacttttg ttccaggttc ccagccaggc gagttcacgc tgggcaaaat taagagtttc 300
cctggacata cgagttctct cgtccgagtg gtgagcacca actacaacca gcatgctatg 360
gtgttcttca agttcgtttt ccaaaacagg gaggaattct acatcaccct ctacgggaga 420
accaaggagc tgacttcgga actaaaggag aacttcatcc gcttctccaa atctctgggc 480
ctccctgaaa accacatcgt cttccctgtc ccaatcgacc agtgtatcga cggc 534
<210> 32
<211> 534
<212> ДНК
<213> искусственная
<220>
<223> мутеин липокалина
<400> 32
caggactcca cctcagacct gatcccagcc ccacctctga gcaaggtccc tctgcagcag 60
aacttccagg acaaccaatt ccatgggaaa tggtacgttg tcgggcaggc cggaaatatt 120
aggctgcgtg aggataagga tccgattaaa atgatggcga ccatttacga gttgaaagaa 180
gataaatcat atgacgtcac catggtgaag tttgatgata agaaatgcat gtacgatatt 240
tggacctttg tgccggggag ccagccgggc gagtttactt taggcaagat taaaagtttt 300
ccgggccata catcatcgtt ggtccgcgtc gtgagcacca actacaacca gcatgccatg 360
gtgttcttca agtttgtgtt tcagaaccgc gaggagtttt atatcacact gtacgggcgc 420
acgaaagaac tgacaagcga gctgaaggaa aattttatcc gcttttccaa atctctgggc 480
ctccctgaaa accacatcgt cttccctgtc ccaatcgacc agtgtatcga cggc 534
<210> 33
<211> 534
<212> ДНК
<213> искусственная
<220>
<223> мутеин липокалина
<400> 33
caggactcca cctcagacct gatcccagcc ccacctctga gcaaggtccc tctgcagcag 60
aacttccagg acaaccaatt ccatgggaaa tggtacgttg tcgggcaggc cggaaatatt 120
aggctgcgtg aggataagga tccgaataaa atgatggcga ccatttacga gttgaaagaa 180
gataaatcat atgacgtcac cgcggtggcg tttgatgata agaaatgcac gtacgatatt 240
tggacctttg tgccggggag ccagccgggc gagtttactt taggcaagat taaaagtttt 300
ccgggccata catcatcgtt ggtccgcgtc gtgagcacca actacaacca gcatgccatg 360
gtgttcttca agtttgtgtt tcagaaccgc gaggagtttt atatcacact gtacgggcgc 420
acgaaagaac tgacaagcga gctgaaggaa aattttatcc gcttttccaa atctctgggc 480
ctccctgaaa accacatcgt cttccctgtc ccaatcgacc agtgtatcga cggc 534
<210> 34
<211> 534
<212> ДНК
<213> искусственная
<220>
<223> мутеин липокалина
<400> 34
caggactcca cctcagacct gatcccagcc ccacctctga gcaaggtccc tctgcagcag 60
aacttccagg acaaccaatt ccatgggaaa tggtacgttg tcgggcaggc cggaaatatt 120
aagctgcgtg aggataagga tccgaataaa atgatggcga ccatttacga gttgaaagaa 180
gataaatcat atgacgtcac cgcggtggcg tttgatgata agaaatgcac gtacgatatt 240
tggacctttg tgccggggag ccagccgggc gagtttactt taggcaagat taaaagtttt 300
ccgggccata catcatcttt ggtccgcgtc gtgagcacca actacaacca gcatgccatg 360
gtgttcttca agtttgtgtt tcagaaccgc gaggagtttt atatcacact gtacgggcgc 420
acgaaagaac tgacaagcga gctgaaggaa aattttatcc gcttttccaa atctctgggc 480
ctccctgaaa accacatcgt cttccctgtc ccaatcgacc agtgtatcga cggc 534
<210> 35
<211> 525
<212> ДНК
<213> искусственная
<220>
<223> мутеин липокалина
<400> 35
caggactcca cctcagacct gatcccagcc ccacctctga gcaaggtccc tctgcagcag 60
aacttccagg acaaccaatt ccatgggaaa tggtacgttg tcgggcaggc cggaaatatt 120
aagctgcgtg aggatagtaa aatgatggcg accatttacg agttgaaaga agataaatca 180
tatgacgtca ccggtgtgag ttttgatgat aagaaatgca cgtacgctat tatgaccttt 240
gtgccgggga gccagccggg cgagtttact ttaggcaaga ttaaaagttt tccgggccat 300
acatcatcgt tggtccgcgt cgtgagcacc aactacaacc agcatgccat ggtgttcttc 360
aagtttgtgt ttcagaaccg cgaggagttt tatatcacac tgtacgggcg cacgaaagaa 420
ctgacaagcg agctgaagga aaattttatc cgcttttcca aatctctggg cctccctgaa 480
aaccacatcg tcttccctgt cccaatcgac cagtgtatcg acggc 525
<210> 36
<211> 534
<212> ДНК
<213> искусственная
<220>
<223> мутеин липокалина
<400> 36
caggactcca cctcagacct gatcccagcc ccacctctga gcaaggtccc tctgcagcag 60
aacttccagg acaaccaatt ccatgggaaa tggtacgttg tcgggcaggc cggaaatatt 120
aagctgcgtg aggataagga tccggttaaa atgatggcga ccatttacga gttgaaagaa 180
gataaatcat atgacgtcac cggggtgacg tttgatgata agaaatgcag gtacgatatt 240
tcgacctttg tgccggggag ccagccgggc gagtttactt ttggcaagat taaaagtttt 300
ccgggccata catcatcgtt ggtccgcgtc gtgagcacca actacaacca gcatgccatg 360
gtgttcttca agtttgtgtt tcagaaccgc gaggagtttt atatcacact gtacgggcgc 420
acgaaagaac tgacaagcga gctgaaggaa aattttatcc gcttttccaa atctctgggc 480
ctccctgaaa accacatcgt cttccctgtc ccaatcgacc agtgtatcga cggc 534
<210> 37
<211> 534
<212> ДНК
<213> искусственная
<220>
<223> мутеин липокалина
<400> 37
caggactcca cctcagacct gatcccagcc ccacctctga gcaaggtccc tctgcagcag 60
aacttccagg acaaccaatt ccatgggaaa tggtacgttg tcgggcaggc cggaaatatt 120
aggctgcgtg aggataagga tccgcataaa atgatggcga ccatttacga gttgaaagaa 180
gataaatcat atgacgtcac cggggtgact tttgatgata agaaatgcac gtacgctatt 240
tcgacctttg tgccggggag ccagccgggc gagtttactt taggcaagat taaaagtttt 300
ccgggccata catcatcttt ggtccgcgtc gtgagcacca actacaacca gcatgccatg 360
gtgttcttca agtttgtgtt tcagaaccgc gaggagtttt atatcacact gtacgggcgc 420
acgaaagaac tgacaagcga gctgaaggaa aattttatcc gcttttccaa atctctgggc 480
ctccctgaaa accacatcgt cttccctgtc ccaatcgacc agtgtatcga cggc 534
<210> 38
<211> 534
<212> ДНК
<213> искусственная
<220>
<223> мутеин липокалина
<400> 38
caggactcca cctcagacct gatcccagcc ccacctctga gcaaggtccc tctgcagcag 60
aacttccagg acaaccaatt ccatgggaaa tggtacgttg tcgggcaggc cggaaatatt 120
aagctgcgtg aggataagga tccgaataaa atgatggcga ccatttacga gttgaaagaa 180
gataaatcat atgacgtcac cggggtgact tttgatgata agaaatgcac gtacgctatt 240
tctacccttg tgccggggag ccagccgggc gagtttactt ttggcaagat taaaagtttt 300
ccgggccata catcatcgtt ggtccgcgtc gtgagcacca actacaacca gcatgccatg 360
gtgttcttca agtttgtgtt tcagaaccgc gaggagtttt atatcacact gtacgggcgc 420
acgaaagaac tgacaagcga gctgaaggaa aattttatcc gcttttccaa atctctgggc 480
ctccctgaaa accacatcgt cttccctgtc ccaatcgacc agtgtatcga cggc 534
<210> 39
<211> 534
<212> ДНК
<213> искусственная
<220>
<223> мутеин липокалина
<400> 39
caggactcca cctcagacct gatcccagcc ccacctctga gcaaggtccc tctgcagcag 60
aacttccagg acaaccaatt ccatgggaaa tggtacgttg tcgggcaggc cggaaatatt 120
aggctgcgtg aggataagga tccgtctaaa atgatggcga ccatttacga gttgaaagaa 180
gataaatcat atgacgtcac cgctgtgacg tttgatgata agaaatgcaa ttacgctatt 240
tctacctttg tgccggggag ccagccgggc gagtttactt taggcaagat taaaagtttt 300
ccgggccata catcatcgtt ggtccgcgtc gtgagcacca actacaacca gcatgccatg 360
gtgttcttca agtttgtgtt tcagaaccgc gaggagtttt atatcacact gtacgggcgc 420
acgaaagaac tgacaagcga gctgaaggaa aattttatcc gcttttccaa atctctgggc 480
ctccctgaaa accacatcgt cttccctgtc ccaatcgacc agtgtatcga cggc 534
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СЛИТЫЙ ПОЛИПЕПТИД С ПРОТИВОРАКОВОЙ АКТИВНОСТЬЮ | 2016 |
|
RU2754466C2 |
| НОВЫЕ СЛИТЫЕ БЕЛКИ, СПЕЦИФИЧЕСКИЕ В ОТНОШЕНИИ CD137 И GPC3 | 2020 |
|
RU2814653C2 |
| НОВЫЙ СЛИТЫЙ БЕЛОК, СПЕЦИФИЧНЫЙ ДЛЯ CD137 И PD-L1 | 2019 |
|
RU2818349C2 |
| НОВЫЕ ПОЛИПЕПТИДЫ СО СПЕЦИФИЧЕСКИМ СВЯЗЫВАНИЕМ И ПУТИ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2723034C2 |
| МУТЕИНЫ ЛИПОКАЛИНА 2 ЧЕЛОВЕКА С АФФИННОСТЬЮ В ОТНОШЕНИИ ГЛИПИКАНА-3 (GPC3) | 2016 |
|
RU2756318C2 |
| НОВЫЕ БЕЛКИ, СПЕЦИФИЧЕСКИЕ В ОТНОШЕНИИ АНГИОГЕНЕЗА | 2016 |
|
RU2720688C2 |
| СПОСОБЫ КУЛЬТИВИРОВАНИЯ КЛЕТОК И НАБОРЫ И УСТРОЙСТВО ДЛЯ НИХ | 2016 |
|
RU2761557C2 |
| АНТИ-CD137 АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2829536C2 |
| МУТЕИНЫ ЛИПОКАЛИНА 2 ЧЕЛОВЕКА (Lcn2, hNGAL) С АФФИННОСТЬЮ ДЛЯ ОПРЕДЕЛЕННОЙ МИШЕНИ | 2010 |
|
RU2707126C2 |
| МУТЕИНЫ ЛИПОКАЛИНА 2 ЧЕЛОВЕКА (Lcn2, hNGAL) С АФФИННОСТЬЮ ДЛЯ ОПРЕДЕЛЕННОЙ МИШЕНИ | 2019 |
|
RU2804336C2 |
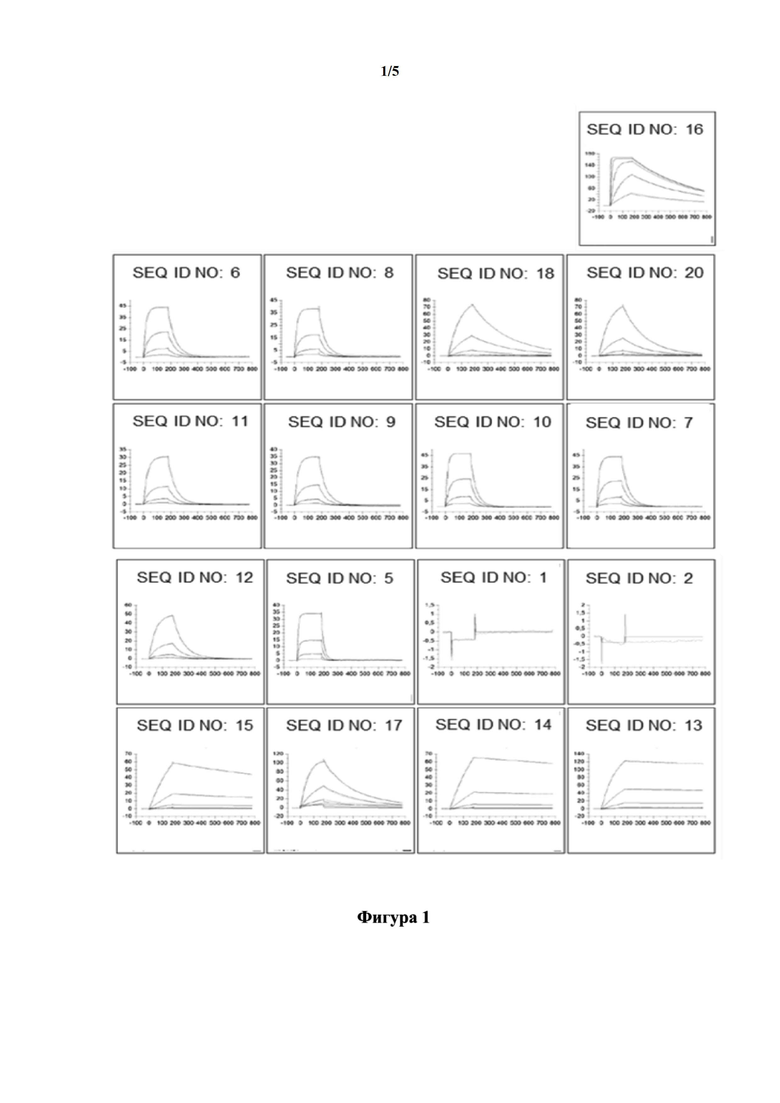
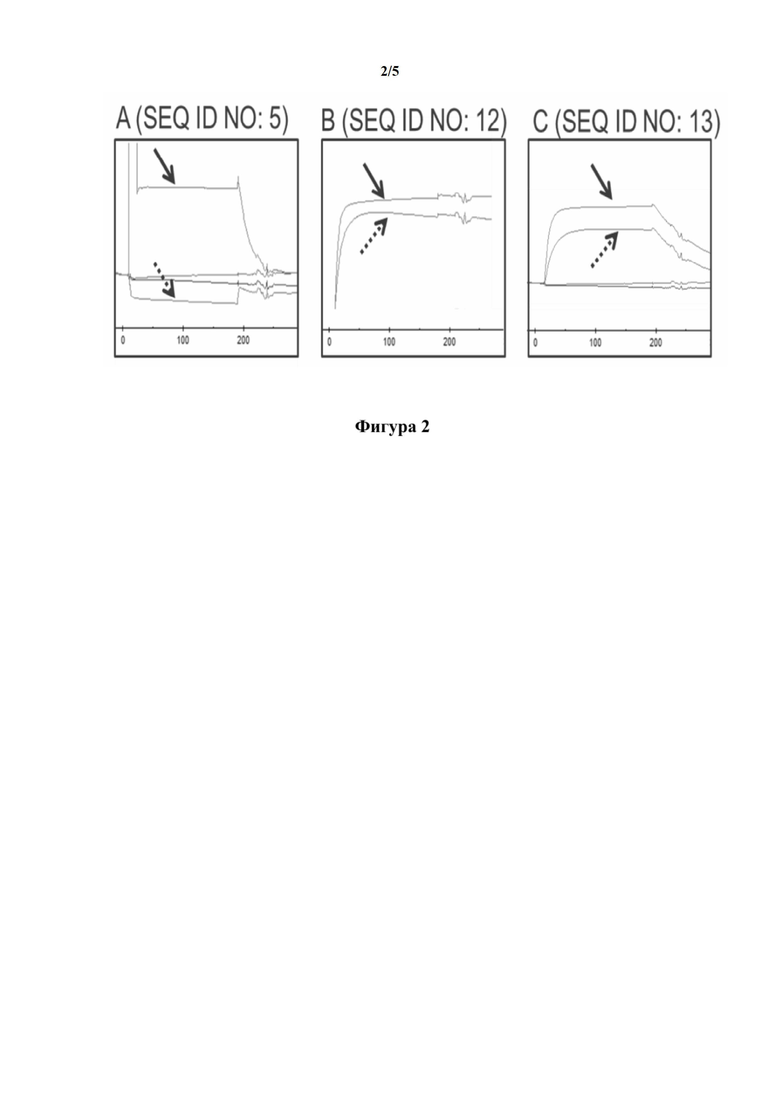
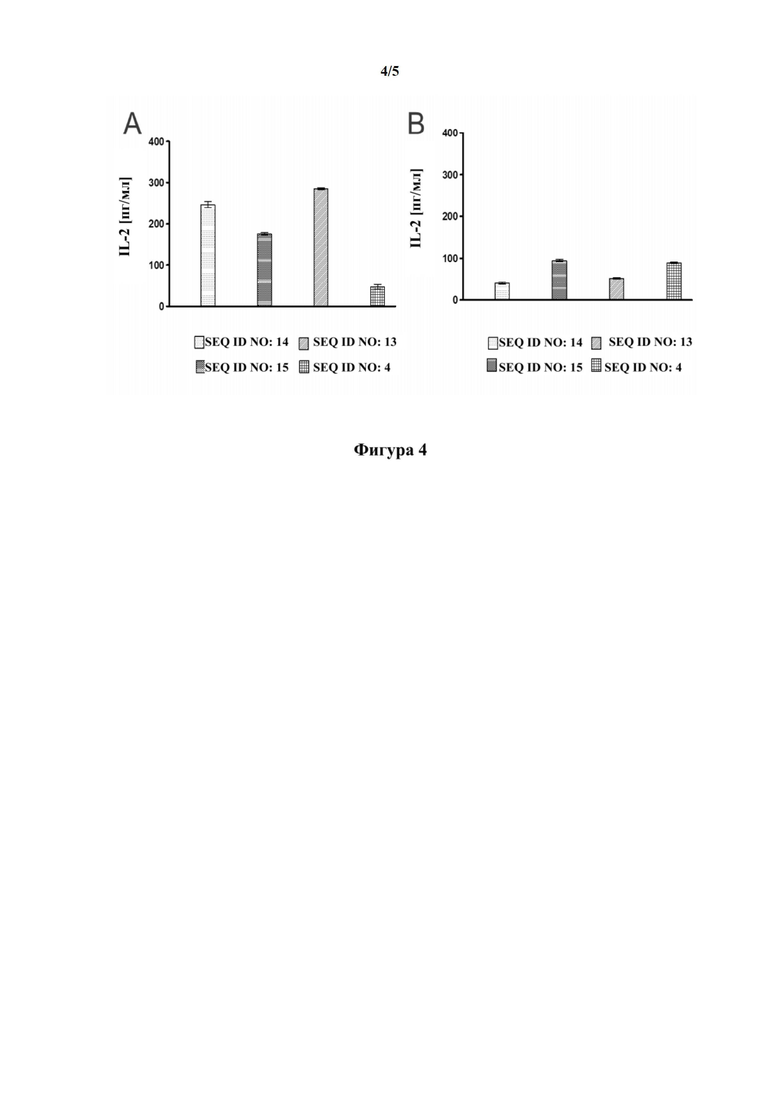
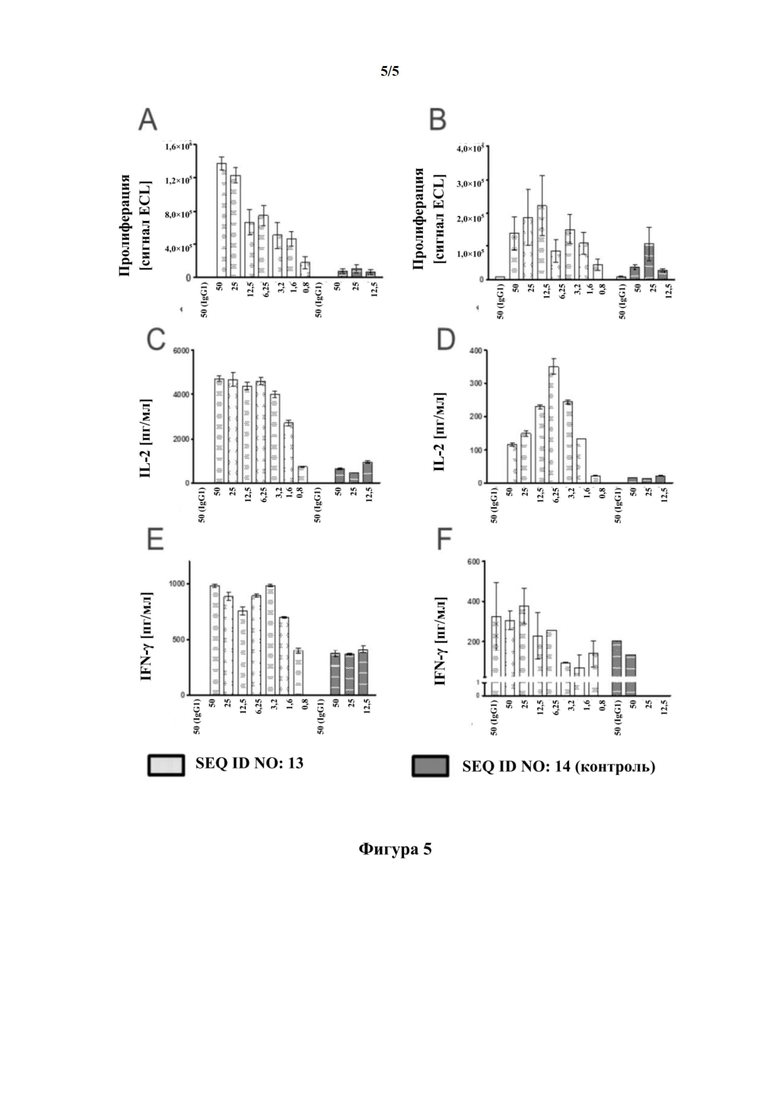
Настоящее изобретение относится к области биотехнологии, конкретно к получению мутеинов липокалина человека, которые связывают CD137, и может быть использовано в медицине для терапии рака, инфекционных и аутоиммунных заболеваний. Мутантный липокалин получают на основе зрелого липокалина слезы человека или зрелого липокалина 2 человека (hNGAL). Полученные мутеины используют также в составе конъюгатов и слитых белков с соединением, которое удлиняет время полужизни мутеина в сыворотке крови. Изобретение позволяет получить высокоаффинные к CD137 мутеины липокалина, способные нарушать связывание CD137L с CD137. 21 н. и 17 з.п. ф-лы, 3 табл., 9 пр., 5 ил.
1. Мутеин липокалина, связывающий CD137,
(i) причем мутеин представляет собой полипептид, характеризующийся аминокислотной последовательностью, которая обладает по меньшей мере 90% идентичностью по отношению к аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 5-11, и где аминокислотная последовательность мутеина предусматривает один из следующих наборов мутированных аминокислотных остатков по сравнению с линейной полипептидной последовательностью зрелого липокалина слезы человека (SEQ ID NO: 1):
(a) Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; и Cys 153 → Ser;
(b) Ala 5 → Thr; Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Lys 65 → Arg; Val 85 → Asp; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Lys 121 → Glu; Ala 133 → Thr; и Cys 153 → Ser;
(c) Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Lys 65 → Asn; Lys 94 → Arg; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Lys 121 → Glu; Ala 133 → Thr; и Cys 153 → Ser;
(d) Ala 5 → Val; Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Lys 65 → Arg; Lys 94 → Glu; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Lys 121 → Glu; Ala 133 → Thr; и Cys 153 → Ser;
(e) Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Thr 42 → Ser; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Ser 150 → Ile; и Cys 153 → Ser;
(f) Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Lys 52 → Glu; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Thr 71 → Ala; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Ala 133 → Thr; Arg 148 → Ser; Ser 150 → Ile; и Cys 153 → Ser; или
(g) Ala 5 → Thr; Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Gly 46 → Asp; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Thr 71 → Ala; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Ser 150 → Ile; и Cys 153 → Ser;
или
(ii) причем мутеин представляет собой полипептид, характеризующийся аминокислотной последовательностью, которая обладает по меньшей мере 90% идентичностью по отношению к аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 12-20, и где аминокислотная последовательность мутеина предусматривает один из следующих наборов мутированных аминокислотных остатков по сравнению с линейной полипептидной последовательностью зрелого липокалина 2 человека (hNGAL, SEQ ID NO: 2):
(a) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Lys; Gln 49 → Asn; Tyr 52 → Met; Ser 68 → Gly; Leu 70 → Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Ala; Arg 81 → Ser; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; и Lys 134 → Tyr;
(b) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Arg; Gln 49 → Ile; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Met; Leu 70 → Lys; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Met; Trp 79 → Asp; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; и Lys 134 → Tyr;
(c) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Arg; Gln 49 → Asn; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Ala; Leu 70 → Ala; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Asp; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; и Lys 134 → Tyr;
(d) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Lys; Gln 49 → Asn; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Ala; Leu 70 → Ala; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Asp; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; и Lys 134 → Tyr;
(e) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Lys; Gln 49 → Ser; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Gly; Leu 70 → Ser; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Ala; Arg 81 → Met; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; и Lys 134 → Tyr;
(f) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Lys; Gln 49 → Val; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Gly; Leu 70 → Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Arg; Trp 79 → Asp; Arg 81 → Ser; Cys 87 → Ser; Leu 94 → Phe; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; и Lys 134 → Tyr;
(g) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Arg; Gln 49 → His; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Gly; Leu 70 → Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Ala; Arg 81 → Ser; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; и Lys 134 → Tyr;
(h) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Lys; Gln 49 → Asn; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Gly; Leu 70 → Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Ala; Arg 81 → Ser; Phe 83 → Leu; Cys 87 → Ser; Leu 94 → Phe; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; и Lys 134 → Tyr; или
(i) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Arg; Gln 49 → Ser; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Ala; Leu 70 → Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Asn; Trp 79 → Ala; Arg 81 → Ser; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; и Lys 134 → Tyr.
2. Мутеин по п. 1, где мутеин представляет собой полипептид, характеризующийся аминокислотной последовательностью, которая обладает по меньшей мере 90% идентичностью по отношению к аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 5-11,
причем аминокислотная последовательность мутеина предусматривает один из следующих наборов мутированных аминокислотных остатков по сравнению с линейной полипептидной последовательностью зрелого липокалина слезы человека (SEQ ID NO: 1):
(a) Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; и Cys 153 → Ser;
(b) Ala 5 → Thr; Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Lys 65 → Arg; Val 85 → Asp; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Lys 121 → Glu; Ala 133 → Thr; и Cys 153 → Ser;
(c) Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Lys 65 → Asn; Lys 94 → Arg; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Lys 121 → Glu; Ala 133 → Thr; и Cys 153 → Ser;
(d) Ala 5 → Val; Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Lys 65 → Arg; Lys 94 → Glu; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Lys 121 → Glu; Ala 133 → Thr; и Cys 153 → Ser;
(e) Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Thr 42 → Ser; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Ser 150 → Ile; и Cys 153 → Ser;
(f) Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Lys 52 → Glu; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Thr 71 → Ala; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Ala 133 → Thr; Arg 148 → Ser; Ser 150 → Ile; и Cys 153 → Ser; или
(g) Ala 5 → Thr; Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Gly 46 → Asp; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Thr 71 → Ala; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Ser 150 → Ile; и Cys 153 → Ser; и
где мутеин связывает CD137 с аффинностью, характеризующейся значением KD, составляющим приблизительно 270 нМ или ниже.
3. Мутеин по п. 2, где мутеин связывает CD137 со значением EC50, составляющим приблизительно 250 нМ или ниже.
4. Мутеин по п. 2, где мутеин способен нарушать связывание CD137L с CD137.
5. Мутеин по п. 2, характеризующийся по меньшей мере 95% идентичностью последовательности с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 5-11.
6. Мутеин по п. 2, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 5-11.
7. Мутеин по п. 1,
где мутеин представляет собой полипептид, характеризующийся аминокислотной последовательностью, которая обладает по меньшей мере 90% идентичностью по отношению к аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 12-20, и где аминокислотная последовательность мутеина предусматривает один из следующих наборов мутированных аминокислотных остатков по сравнению с линейной полипептидной последовательностью зрелого липокалина 2 человека (hNGAL, SEQ ID NO: 2):
(а) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Lys; Gln 49 → Asn; Tyr 52 → Met; Ser 68 → Gly; Leu 70 → Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Ala; Arg 81 → Ser; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; и Lys 134 → Tyr;
(b) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Arg; Gln 49 → Ile; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Met; Leu 70 → Lys; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Met; Trp 79 → Asp; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; и Lys 134 → Tyr;
(c) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Arg; Gln 49 → Asn; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Ala; Leu 70 → Ala; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Asp; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; и Lys 134 → Tyr;
(d) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Lys; Gln 49 → Asn; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Ala; Leu 70 → Ala; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Asp; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; и Lys 134 → Tyr;
(e) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Lys; Gln 49 → Ser; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Gly; Leu 70 → Ser; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Ala; Arg 81 → Met; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; и Lys 134 → Tyr;
(f) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Lys; Gln 49 → Val; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Gly; Leu 70 → Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Arg; Trp 79 → Asp; Arg 81 → Ser; Cys 87 → Ser; Leu 94 → Phe; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; и Lys 134 → Tyr;
(g) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Arg; Gln 49 → His; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Gly; Leu 70 → Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Ala; Arg 81 → Ser; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; и Lys 134 → Tyr;
(h) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Lys; Gln 49 → Asn; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Gly; Leu 70 → Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Ala; Arg 81 → Ser; Phe 83 → Leu; Cys 87 → Ser; Leu 94 → Phe; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; и Lys 134 → Tyr; или
(i) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Arg; Gln 49 → Ser; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Ala; Leu 70 → Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Asn; Trp 79 → Ala; Arg 81 → Ser; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; и Lys 134 → Tyr; и
где мутеин связывает CD137 с аффинностью, характеризующейся значением KD, составляющим приблизительно 150 нМ или ниже.
8. Мутеин по п. 7, где мутеин связывает CD137 со значением EC50, составляющим приблизительно 18 нМ или ниже.
9. Мутеин по п. 7, где мутеин способен индуцировать более высокую концентрацию IL-2 по сравнению с отрицательным контролем, представляющим собой SEQ ID NO: 4.
10. Мутеин по п. 7, где мутеин не приводит к более высокой концентрации IL-2 по сравнению с отрицательным контролем, представляющим собой SEQ ID NO: 4.
11. Мутеин по п. 7, где мутеин способен индуцировать пролиферацию с обеспечением более высоких уровней IL-2 и IFN-γ по сравнению с отрицательным контролем, представляющим собой SEQ ID NO: 4.
12. Мутеин по п. 7, где мутеин не нарушает связывание CD137L с CD137.
13. Мутеин по п. 7, характеризующийся по меньшей мере 95% идентичностью последовательности с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 12-20.
14. Мутеин по п. 7, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 12-20.
15. Мутеин по п. 7, содержащий аминокислотную последовательность SEQ ID NO: 12.
16. Мутеин по п. 7, содержащий аминокислотную последовательность SEQ ID NO: 13.
17. Мутеин по п. 7, содержащий аминокислотную последовательность SEQ ID NO: 14.
18. Конъюгат для связывания CD137, содержащий мутеин липокалина по любому из пп. 1-17, где мутеин конъюгирован с соединением, которое удлиняет время полужизни мутеина в сыворотке крови, выбранным из группы, состоящей из молекулы полиалкиленгликоля, полиэтилена (ПЭГ) или его активированного производного, и гидроксиэтилкрахмала.
19. Слитый белок для связывания CD137, содержащий мутеин липокалина по любому из пп. 1-17, где мутеин слит на своем N-конце и/или своем C-конце с партнером по слиянию, который удлиняет время полужизни в сыворотке крови и представляет собой белок, белковый домен или пептид.
20. Слитый белок по п. 19, где белок, белковый домен или пептид выбран из группы, состоящей из Fc-части иммуноглобулина, домена CH3 иммуноглобулина, домена CH4 иммуноглобулина, альбумин-связывающего пептида и альбумин-связывающего белка.
21. Молекула нуклеиновой кислоты для экспрессии мутеина по любому из пп. 1-17, содержащая нуклеотидную последовательность, кодирующую мутеин.
22. Молекула нуклеиновой кислоты для экспрессии слитого белка по п. 19 или 20, содержащая нуклеотидную последовательность, кодирующую слитый белок.
23. Молекула нуклеиновой кислоты для клонирования мутеина по любому из пп. 1-17, содержащая нуклеотидную последовательность, кодирующую мутеин.
24. Молекула нуклеиновой кислоты для клонирования слитого белка по п. 19 или 20, содержащая нуклеотидную последовательность, кодирующую слитый белок.
25. Клетка-хозяин для экспрессии мутеина, кодируемого нуклеиновой кислотой, содержащая молекулу нуклеиновой кислоты по п. 21, причем клетка-хозяин не является клеткой эмбриона человека или клеткой зародышевой линии человека.
26. Клетка-хозяин для экспрессии слитого белка, кодируемого нуклеиновой кислотой, содержащая молекулу нуклеиновой кислоты по п. 22, причем клетка-хозяин не является клеткой эмбриона человека или клеткой зародышевой линии человека.
27. Клетка-хозяин для клонирования мутеина, кодируемого нуклеиновой кислотой, содержащая молекулу нуклеиновой кислоты по п. 23, причем клетка-хозяин не является клеткой эмбриона человека или клеткой зародышевой линии человека.
28. Клетка-хозяин для клонирования слитого белка, кодируемого нуклеиновой кислотой, содержащая молекулу нуклеиновой кислоты по п. 24, причем клетка-хозяин не является клеткой эмбриона человека или клеткой зародышевой линии человека.
29. Фармацевтическая композиция для связывания CD137, содержащая эффективное количество мутеина липокалина по любому из пп. 1-17, конъюгата по п. 18 или слитого белка по п. 19 или 20 и фармацевтически приемлемый наполнитель.
30. Способ получения мутеина по любому из пп. 1-17, где мутеин получают путем экспрессирования мутеина в клетке-хозяине по п. 25 или in vitro трансляции вектора экспрессии, содержащего молекулу нуклеиновой кислоты по п. 21.
31. Способ получения слитого белка по п. 19 или 20, где слитый белок получают путем экспрессирования слитого белка в клетке-хозяине по п. 26 или in vitro трансляции вектора экспрессии, содержащего молекулу нуклеиновой кислоты по п. 22.
32. Способ связывания CD137, включающий введение субъекту эффективного количества одного или более мутеинов липокалина по любому из пп. 1-17, конъюгата по п. 18, слитого белка по п. 19 или 20 или фармацевтической композиции по п. 29.
33. Способ активации нисходящих сигнальных путей, связанных с CD137, включающий введение субъекту эффективного количества одного или более мутеинов липокалина по любому из пп. 1-17, конъюгата по п. 18, слитого белка по п. 19 или 20 или фармацевтической композиции по п. 29.
34. Способ индуцирования пролиферации T-лимфоцитов, включающий введение субъекту эффективного количества одного или более мутеинов липокалина по любому из пп. 1-17, конъюгата по п. 18, слитого белка по п. 19 или 20 или фармацевтической композиции по п. 29.
35. Способ нарушения связывания CD137L с CD137, включающий введение субъекту эффективного количества одного или более мутеинов липокалина по п. 4 или одной или более фармацевтических композиций по п. 29, где фармацевтическая композиция содержит мутеин липокалина по п. 4.
36. Способ лечения рака, включающий введение субъекту эффективного количества мутеина липокалина по любому из пп. 1-17, конъюгата по п. 18, слитого белка по п. 19 или 20 или фармацевтической композиции по п. 29.
37. Способ лечения инфекционного заболевания, включающий введение субъекту эффективного количества мутеина липокалина по любому из пп. 1-17, конъюгата по п. 18, слитого белка по п. 19 или 20 или фармацевтической композиции по п. 29.
38. Способ лечения аутоиммунного заболевания, включающий введение субъекту эффективного количества мутеина липокалина по любому из пп. 1-17, конъюгата по п. 18, слитого белка по п. 19 или 20 или фармацевтической композиции по п. 29.
| US 2014288008 A1, 25.09.2014 | |||
| WO 2009156456 A1, 30.12.2009 | |||
| LYNCH D | |||
| H., The promise of 4-1BB (CD137)-mediated immunomodulation and the immunotherapy of cancer, IMMUNOLOGICAL REVIEWS, v.222, p.277 - 286 | |||
| SCHLEHUBER S | |||
| et al., Lipocalins in drug discovery: from natural ligand-binding proteins to "anticalins", Drug Discov Today, 2005, v.10, n.1, |

