I. Предпосылки создания изобретения
[0001] Глипикан-3 (GPC3, также называемый DGSX, GTR2-2, MXR7, OCI-5, SDYS, SGB, SGBS и SGBS1) представляет собой онкофетальный антиген, принадлежащий к глипикановому семейству заякоренных посредством гликозил-фосфатидилинозитола гепаринсульфатных протеогликанов. GPC3 в ходе развития экспрессируется в различных тканях, играя важные роли в морфогенезе и росте, например, через сигнальный путь с участием фактора роста фибробластов (FGF), неканонического Wnt или инсулиноподобного фактора роста (Cheng et al., Carcinogenesis, 2008, Song et al., J Biol Chem, 2005, Song et al., J Biol Chem, 1997). Тем не менее, в большинстве здоровых тканей взрослого человека экспрессия GPC3 подавлена или подвергнута сайленсингу. Кроме того, GPC3 может как положительно, так и отрицательно регулировать клеточный рост в зависимости от типа клеток. Мутации GPC3 с потерей его функции ответственны за развитие синдрома Симпсона-Голаби-Бемеля или дизморфического синдрома Симпсона (SGBS), редкого сцепленного с X-хромосомой нарушения, проявляющегося в виде избыточного роста, а пациенты с SGBS имеют повышенный риск развития эмбриональных опухолей (Pilia et al., Nat Genet, 1996), в том числе опухолей Вильмса.
[0002] GPC3 экспрессируется при различных типах рака, в том числе при карциноме печени, желудка, меланоме, уротелиальной карциноме высокой степени злокачественности, раке яичка и некоторых формах рака матки и влагалища (Aydin et al., Diagn Pathol, 2015, Ushiku et al., Cancer Sci, 2009, Gailey and Bellizzi, Am J Clin Pathol, 2013, Yamanaka et al., Oncology, 2007, Nakatsura et al., Clin Cancer Res, 2004, Zynger et al., Am J Surg Pathol, 2006, Montalbano et al., Int J Oncol, 2016, Midorikawa et al., Int J Cancer, 2003). В частности, GPC3 имеет высокий уровень экспрессии при гепатоклеточной карциноме (HCC) (Capurro et al., Gastroenterology, 2003, Nakatsura et al., Biochem Biophys Res Commun, 2003, Sung et al., Cancer Sci, 2003, Zhu et al., Gut, 2001), основной форме рака печени, на долю которой приходится 90% всех случаев рака печени и которая приводит по меньшей мере к 500000 смертей в год (Jelic et al., Ann Oncol, 2010). Было проведено обширное исследование по изучению GPC3 в качестве диагностического биомаркера и терапевтической мишени в случае HCC и других типов рака. Было создано несколько антител, нацеленных на GPC3, в том числе гуманизированных мышиных антител YP7 и GC33 и человеческих антител HN3 и MDX-1414, однако не была продемонстрирована их способность ингибировать пролиферацию клеток HCC или индуцировать апоптоз (Feng and Ho, FEBS Lett, 2014). В порядке исключения следует отметить, что GC33 в настоящее время проходит оценку в клинических испытаниях в отношении HCC. Механизм функции GC33 включает антителозависимую клеточноопосредованную цитотоксичность (ADCC), но GC33 напрямую не ингибирует пролиферацию GPC3-положительных опухолевых клеток (Takai et al., Cancer Biol Ther, 2009, Nakano et al., Biochem Biophys Res Commun, 2009, Ishiguro et al., Cancer Res, 2008). Тем не менее хотя и было показано, что GC33 оказывает ингибирующее действие на рост опухоли in vivo, он не продемонстрировал удовлетворительной клинической эффективности. Между тем результаты экспериментов по нокдауну и использованию siRNA свидетельствуют, что GPC3 не является летальным геном для клеток HCC, что делает неопределенным, могут ли антитела к GPC3 вызывать эффективную регрессию опухоли.
[0003] Кластер дифференцировки 137 или CD137 (также известный как 4-1BB или TNFRS9) является костимулирующим иммунным рецептором и представителем суперсемейства рецепторов фактора некроза опухоли (TNFR). Он преимущественно экспрессируется на активированных CD4+ и CD8+ T-клетках, активированных B-клетках и клетках-натуральных киллерах (NK), но также может встречаться на покоящихся моноцитах и дендритных клетках (Li and Liu, Clin Pharmacol, 2013) или эндотелиальных клетках (Snell et al., Immunol Rev, 2011). CD137 играет важную роль в регуляции иммунного ответа и поэтому является мишенью при противораковой иммунотерапии. Лиганд CD137 (CD137L) является единственным известным природным лигандом CD137 и конститутивно экспрессируется на нескольких типах антигенпрезентирующих клеток, таких как активированные В-клетки, моноциты и дендритные клетки селезенки, и может быть индуцирован на Т-лимфоцитах.
[0004] CD137L представляет собой тримерный белок, который существует в мембраносвязанной форме и в виде растворимого варианта. Тем не менее способность растворимого CD137L активировать CD137, например на CD137-экспрессирующих лимфоцитах, ограничена, и для достижения эффекта необходимы большие концентрации (Wyzgol et al., J Immunol, 2009). Природный путь активации CD137 заключается в воздействии CD137-положительной клетки на CD137L-положительную клетку. Считается, что затем активация CD137 индуцируется кластеризацией с участием CD137L на противоположной клетке, что приводит к передаче сигналов посредством TRAF1, 2 и 3 (Yao et al., Nat Rev Drug Discov, 2013, Snell et al., Immunol Rev, 2011) и дополнительным сопутствующим последующим эффектам у CD137-положительной Т-клетки. В случае Т-клеток, активируемых путем распознавания их соответствующих родственных мишеней, эффекты, вызываемые костимуляцией CD137, заключаются в дополнительной усиленной активации, усиленной выживаемости и пролиферации, продуцировании провоспалительных цитокинов и улучшенной способности к уничтожению.
[0005] Польза костимуляции CD137 в отношении устранения раковых клеток была продемонстрирована на ряде моделей in vivo. Принудительная экспрессия CD137L в опухоли, например, приводит к отторжению опухоли (Melero et al., Eur J Immunol, 1998). Аналогичным образом, принудительная экспрессия scFv к CD137 на опухоли приводит к зависимому от CD4+ Т-лимфоцитов и NK-клеток устранению опухоли (Yang et al., Cancer Res, 2007, Zhang et al., Mol Cancer Ther, 2006, Ye et al., Nat Med, 2002). Также было продемонстрировано, что системно вводимое антитело к CD137 приводит к замедлению роста опухоли (Martinet et al., Gene Ther, 2002).
[0006] Было показано, что CD137 является отличным маркером встречающихся в природе реактивных в отношении опухоли Т-клеток в опухолях человека (Ye et al., Clin Cancer Res, 2014), и что можно использовать антитела к CD137 для улучшения размножения и активности CD8+ эффекторных лимфоцитов против меланомы для применения в адоптивной Т-клеточной терапии (Chacon et al., PLoS One, 2013).
[0007] Доклиническая демонстрация потенциальной терапевтической полезности костимуляции CD137 подстегнула разработку терапевтических антител, нацеленных на CD137, в том числе BMS-663513 (описанного в патенте США №7288638) и PF-05082566 (Fisher et al., Cancer Immunol Immunother, 2012).
[0008] В настоящем изобретении предусмотрены, среди прочего, новые подходы по одновременному задействованию CD137 и GPC3 с помощью одного или более слитых белков, обладающих свойствами специфичности связывания в отношении CD137 и специфичности связывания в отношении GPC3.
II. ОПРЕДЕЛЕНИЯ
[0009] В представленном далее перечне определены термины, фразы и сокращения, используемые в настоящем описании. Подразумевается, что все термины, перечисленные и определенные в данном документе, охватывают все грамматические формы.
[0010] В контексте данного документа, если не указано иное, «CD137» означает CD137 человека (huCD137). CD137 человека означает полноразмерный белок, определенный в UniProt Q07011, его фрагмент или вариант. CD137 также известен как 4-1BB, представитель суперсемейства рецепторов фактора некроза опухоли 9 (TNFRSF9), и индуцируется при активации лимфоцитов (ILA). В некоторых конкретных вариантах осуществления применяют CD137 от отличных от человека видов, например CD137 яванского макака и CD137 мыши.
[0011] В контексте данного документа, если не указано иное, «глипикан-3» или «GPC3» означает GPC3 человека (huGPC3). GPC3 человека означает полноразмерный белок, определенный в UniProt P51654, его фрагмент или вариант. GPC3 человека кодируется геном GPC3. GPC3 также известен как DGSX, GTR2-2, MXR7, OCI-5, SDYS, SGB, SGBS или SGBS1. В некоторых конкретных вариантах осуществления применяют GPC3 от отличных от человека видов, например GPC3 яванского макака и GPC3 мыши.
[0012] В контексте данного документа термин «аффинность связывания» описывает способность биомолекулы (например, полипептида или белка) по настоящему изобретению (например, мутеина липокалина, антитела, слитого белка или любого другого пептида или белка) связывать избранную мишень и образовывать с ней комплекс. Аффинность связывания измеряют с помощью ряда способов, известных специалистам в данной области, в том числе без ограничения, с помощью флуоресцентного титрования, твердофазного иммуноферментного анализа (ELISA), включая прямой и конкурентный ELISA, калориметрических способов, таких как калориметрия изотермического титрования (ITC), и поверхностного плазмонного резонанса (SPR). Данные способы хорошо известны из уровня техники, и в данном документе дополнительно описаны некоторые примеры таких способов. При этом аффинность связывания регистрируют как величину константы диссоциации (KD), полумаксимальной эффективной концентрации (EC50) или полумаксимальной ингибирующей концентрации (IC50), измеряемые с помощью данных способов. Более низкое значение KD, EC50 или IC50 отражает лучшую (более высокую) связывающую способность (аффинность). Соответственно, можно измерить и сравнить значения аффинности связывания двух биомолекул в отношении выбранной мишени. При сравнении значений аффинности связывания двух биомолекул с выбранной мишенью термин «приблизительно такая же», «практически такая же» или «практически схожая» означает, что одна биомолекула имеет аффинность связывания, выраженную в виде значения KD, EC50 или IC50, которое идентично или похоже на таковое другой молекулы в пределах колебаний показаний измерения аффинности связывания от эксперимента к эксперименту. Колебание показаний измерения аффинности связывания от эксперимента к эксперименту зависит от конкретного применяемого способа и известны специалистам в данной области.
[0013] В контексте данного документа термин «практически» может также относиться к качественному условию проявления полной или почти полной меры или степени представляющей интерес характеристики или представляющего интерес свойства. Специалист в области биологии поймет, что биологические и химические явления редко, если вообще когда-либо, доходят до завершенности и/или проходят до завершенности или достигают или не достигают абсолютного результата. Таким образом, термин «практически» в данном документе применяют для обозначения потенциального отсутствия завершенности, присущей многим биологическим и химическим явлениям.
[0014] В контексте данного документа термин «выявлять», «выявление», «поддающийся выявлению» или «выявляющий» понимают как на количественном, так и на качественном уровне, а также в их комбинации. Таким образом, он включает количественные, полуколичественные и качественные измерения, производимые на биомолекуле по настоящему изобретению.
[0015] В контексте данного документа термин “поддающаяся выявлению аффинность” обычно означает способность связывания биомолекулы с ее мишенью, регистрируемую при помощи значения KD, EC50 или IC50, максимум приблизительно 10-5 M или ниже. Аффинность связывания, регистрируемая при помощи значения KD, EC50 или IC50, которая превышает 10-5 M, обычно более не поддается измерению с помощью традиционных способов, таких как ELISA и SPR, и поэтому имеет второстепенное значение.
[0016] Следует отметить, что на образование комплекса между биомолекулой по настоящему изобретению и ее мишенью оказывает влияние множество различных факторов, таких как концентрации соответствующей мишени, присутствие конкурентов, pH и ионная сила применяемой буферной системы, способ проведения эксперимента, применяемый для определения аффинности связывания (например, флуоресцентное титрование, конкурентный ELISA (также называемый ELISA в условиях конкурентности) и поверхностный плазмонный резонанс), и даже математический алгоритм, применяемый для оценки экспериментальных данных. Следовательно, специалисту в данной области будет понятно, что аффинность связывания, регистрируемая с помощью значения KD, EC50 или IC50, может варьироваться в пределах определенного экспериментального диапазона, в зависимости от способа и схемы эксперимента. Это означает, что может иметь место незначительное отклонение в измеренных значениях KD, EC50 или IC50 или диапазоне допустимых значений в зависимости, например, от того, были ли такие значения определены с помощью ELISA (в том числе прямого или конкурентного ELISA), SPR или другого способа.
[0017] В контексте данного документа «специфический в отношении», «специфическое связывание», «специфически связываются» или «специфичностью связывания» относятся к способности биомолекулы отличать требуемую мишень (например, CD137 и GPC3) от одной или более контрольных мишеней (например, клеточного рецептора для липокалина, ассоциированного с желатиназой нейтрофилов). Понятно, что такая специфичность является не абсолютным, а относительным свойством, ее можно определить, например, согласно результатам SPR, вестерн-блоттинга, ELISA, сортировки флуоресцентно-активированных клеток (FACS), радиоиммуноанализа (RIA), электрохемилюминесцентного (ECL), иммунорадиометрического анализа (IRMA), иммуногистохимии (IHC) и пептидного сканирования.
[0018] При использовании в данном документе в контексте слитого белка по настоящему изобретению, который связывается с CD137 и GPC3, термин «специфический в отношении», «специфическое связывание», «специфически связывается» или «специфичность связывания» означает, что слитый белок связывается, вступает в реакцию или направлен против CD137 и GPC3, как описано в данном документе, но фактически не связывает другой белок. Термин «другой белок» включает любые белки, которые не являются CD137 или GPC3, или белки, которые являются близкородственными CD137 или GPC3 или гомологичными с ними. Тем не менее, термином «другой белок» не исключаются CD137 или GPC3, полученные от видов, отличных от человека, и фрагменты и/или варианты CD137 или GPC3. Термин «фактически не связывает» означает, что слитые белки по настоящему изобретению связывают другой белок с более низкой аффинностью связывания, чем CD137 и/или GPC3, т.е. демонстрируют перекрестную реактивность, составляющую менее 30%, предпочтительно 20%, более предпочтительно 10%, особенно предпочтительно менее 9, 8, 7, 6 или 5%. Способен ли слитый белок специфически вступать в реакцию так, как определено в данном документе выше, можно легко протестировать, среди прочего, путем сравнения реакции слитого белка по настоящему изобретению с CD137 и/или GPC3 и реакции указанного слитого белка с другим (другими) белком (белками).
[0019] В контексте данного документа термин «липокалин» относится к мономерному белку весом примерно 18-20 кДа, имеющему область со сверхвторичной структурой из цилиндрического β-складчатого листа, содержащую множество β-цепей (предпочтительно восемь β-цепей, обозначенных от A до H), попарно соединенных посредством множества (предпочтительно четырех) петель на одном конце так, чтобы они в результате составляли лиганд-связывающий карман и определяли вход в лиганд-связывающий карман. Предпочтительно, петли, составляющие лиганд-связывающий карман, в контексте настоящего изобретения, представляют собой петли, соединяющие открытые концы β-цепей A и B, C и D, E и F и G и H, и являются петлями с обозначениями AB, CD, EF и GH. Хорошо известно, что разнообразие указанных петель в остальном жестком каркасе липокалина приводит к возникновению множества различных вариантов связывания среди представителей семейства липокалинов, каждый из которых способен приспособиться к мишеням разного размера, формы и химического характера (рассмотрено, например, в Skerra, Biochim Biophys Acta, 2000, Flower et al., Biochim Biophys Acta, 2000, Flower, Biochem J, 1996). Понятно, что липокалиновое семейство белков естественным образом эволюционировало для связывания широкого спектра лигандов, обладающих необычно низкими уровнями общей консервативности последовательностей (зачастую со значениями идентичности последовательностей менее 20%), но сохранив при этом высококонсервативный общий паттерн фолдинга. Соответствие между положениями в различных липокалинах также хорошо известно специалистам в данной области (см., например, патент США №7250297). Белки, подпадающие под определение «липокалин», в контексте данного документа включают без ограничения липокалины человека, в том числе липокалин слезы (Tlc, Lcn1), липокалин-2 (Lcn2) или липокалин, ассоциированный с желатиназой нейтрофилов (NGAL), аполипопротеин D (ApoD), аполипопротеин M, α1-кислый гликопротеин 1, α1-кислый гликопротеин 2, α1-микроглобулин, компонент 8γ системы комплемента, ретинол-связывающий белок (RBP), эпидидимальный белок, связывающий ретиноевую кислоту, гликоделин, одоранто-связывающий белок IIa, одоранто-связывающий белок IIb, липокалин-15 (Lcn15) и простагландин-D-синтазу.
[0020] В контексте данного документа, если не указано иное, «липокалин слезы» относится к липокалину слезы человека (hTlc) и дополнительно относится к зрелому липокалину слезы человека. Термин «зрелый» при его использовании для определения характеристики белка означает белок, фактически не содержащий сигнальный пептид. «Зрелый hTlc» по настоящему изобретению относится к зрелой форме липокалина слезы человека, не содержащей сигнальный пептид. Зрелый hTlc описывается остатками 19-176 последовательности, депонированной в банке данных SWISS-PROT под регистрационным номером P31025, и его аминокислотная последовательность приведена под SEQ ID NO: 1.
[0021] В контексте данного документа «липокалин-2» или «липокалин, ассоциированный с желатиназой нейтрофилов» относится к человеческому липокалину-2 (hLcn2) или человеческому липокалину, ассоциированному с желатиназой нейтрофилов (hNGAL), а также дополнительно относится к зрелому человеческому липокалину-2 или зрелому человеческому липокалину, ассоциированному с желатиназой нейтрофилов. Термин «зрелый» при его использовании для определения характеристики белка означает белок, фактически не содержащий сигнальный пептид. «Зрелый hNGAL» по настоящему изобретению относится к зрелой форме человеческого липокалина, ассоциированного с желатиназой нейтрофилов, не содержащей сигнальный пептид. Зрелый hNGAL описывается остатками 21-198 последовательности, депонированной в банке данных SWISS-PROT под регистрационным номером P80188, и его аминокислотная последовательность приведена под SEQ ID NO: 2.
[0022] В контексте данного документа термин «нативная последовательность» относится к белку или полипептиду, имеющему последовательность, которая встречается в природе, или имеющему последовательность дикого типа, независимо от способа ее получения. Белок или полипептид с такой нативной последовательностью можно выделять из природных источников или можно получать другими способами, такими как рекомбинантные или синтетические способы.
[0023] «Липокалин с нативной последовательностью» относится к липокалину, имеющему такую же аминокислотную последовательность, что и соответствующий полипептид, полученный из природных источников. Таким образом, липокалин с нативной последовательностью может иметь аминокислотную последовательность соответствующего встречающегося в природе липокалина (дикого типа) из любого организма, в частности из млекопитающего. Термин «нативная последовательность» при его использовании в контексте липокалина, в частности, охватывает встречающиеся в природе укороченные или секретируемые формы липокалина, встречающиеся в природе вариантные формы, такие как полученные в результате альтернативного сплайсинга формы и встречающиеся в природе аллельные варианты липокалина. Термины «липокалин с нативной последовательностью» и «липокалин дикого типа» в данном документе используют взаимозаменяемо.
[0024] В контексте данного документа «мутеин», «мутантная» молекула (будь то белок или нуклеиновая кислота) или «мутант» относится к изменению, делеции или вставке одной или более аминокислот или одного или более нуклеотидов в сравнении с встречающимся в природе белком (дикого типа) или встречающейся в природе нуклеиновой кислотой (дикого типа). Указанный термин также включает фрагменты описываемого в данном документе мутеина. Настоящим изобретением явно охвачены описываемые в данном документе мутеины липокалина, имеющие область со сверхвторичной структурой из цилиндрического β-складчатого листа, содержащую восемь β-цепей, попарно соединенных посредством четырех петель на одном конце так, чтобы они в результате составляли лиганд-связывающий карман и определяли вход в лиганд-связывающий карман, при этом по меньшей мере одна аминокислота каждой из по меньшей мере трех из указанных четырех петель была подвергнута мутации в сравнении с липокалином с нативной последовательностью. Мутеины липокалина по настоящему изобретению предпочтительно обладают функцией связывания CD137, как описано в данном документе.
[0025] В контексте данного документа термин «фрагмент» в связи с мутеинами липокалина по настоящему изобретению относится к белкам или полипептидам, полученным из полноразмерных зрелых hTlc или hNGAL или мутеинов липокалина, которые укорочены на N-конце и/или C-конце, т.е. у которых отсутствует по меньшей мере одна из N-концевых и/или C-концевых аминокислот. Такие фрагменты могут включать по меньшей мере 10 или более, например 20 или 30 или более последовательных аминокислот первичной последовательности зрелого hTlc или hNGAL или мутеина липокалина, из которого он получен, и обычно они поддаются выявлению в ходе иммуноанализа зрелого hTlc или hNGAL. В таком фрагменте могут отсутствовать до 2, до 3, до 4, до 5, до 10, до 15, до 20, до 25 или до 30 (включая все числа между ними) N-концевых и/или C-концевых аминокислот. В качестве иллюстративного примера, в таком фрагменте могут отсутствовать одна, две, три или четыре N-концевые (His-His-Leu-Leu) и/или одна или две C-концевые аминокислоты (Ser-Asp) зрелого hTlc. Понятно, что фрагмент предпочтительно является функциональным фрагментом зрелого hTlc или hNGAL или мутеина липокалина, из которого он получен, что означает, что он предпочтительно сохраняет специфичность связывания, предпочтительно с CD137, зрелого hTlc/hNGAL или мутеина липокалина, из которого он получен. В качестве иллюстративного примера, такой функциональный фрагмент может содержать по меньшей мере аминокислоты в положениях 5-153, 5-150, 9-148, 12-140, 20-135 или 26-133, соответствующих линейной полипептидной последовательности зрелого hTlc. В качестве другого иллюстративного примера, такой функциональный фрагмент может содержать по меньшей мере аминокислоты в положениях 13-157, 15-150, 18-141, 20-134, 25-134 или 28-134, соответствующих линейной полипептидной последовательности зрелого hNGAL.
[0026] «Фрагмент» по отношению к соответствующему целевому CD137 или GPC3 слитого белка по настоящему изобретению относится к укороченному на N-конце и/или C-конце CD137 или GPC3 или к белковым доменам CD137 или GPC3. Описываемые в данном документе фрагменты CD137 или фрагменты GPC3 сохраняют способность полноразмерного CD137 или GPC3 распознаваться и/или связываться слитым белком по настоящему изобретению. В качестве иллюстративного примера, фрагмент может представлять собой внеклеточный домен CD137 или GPC3. В качестве иллюстративного примера, такой внеклеточный домен может содержать аминокислоты внеклеточных субдоменов CD137, такие как отдельные или взятые в сочетании аминокислотные последовательности домена 1 (остатки 24-45 из UniProt Q07011), домена 2 (остатки 46-86), домена 3 (87-118) и домена 4 (остатки 119-159).
[0027] В контексте данного документа термин «вариант» относится к производным белка или полипептида, которые включают мутации, например путем замен, делеций, вставок и/или химических модификаций, аминокислотной последовательности или нуклеотидной последовательности. В некоторых вариантах осуществления такие мутации и/или химические модификации не уменьшают функциональность белка или пептида. Такие замены могут быть консервативными, т.е. аминокислотный остаток заменяют химически схожим аминокислотным остатком. Примерами консервативных замен являются замены среди членов следующих групп: 1) аланин, серин, треонин и валин; 2) аспарагиновая кислота, глутаминовая кислота, глутамин и аспарагин, а также гистидин; 3) аргинин, лизин, глутамин, аспарагин и гистидин; 4) изолейцин, лейцин, метионин, валин, аланин, фенилаланин, треонин и пролин и 5) изолейцин, лейцин, метионин, фенилаланин, тирозин и триптофан. К таким вариантам относятся белки или полипептиды, в которых одна или более аминокислот были заменены их соответствующими D-стереоизомерами или аминокислотами, отличными от встречающихся в природе 20 аминокислот, такими как, например, орнитин, гидроксипролин, цитруллин, гомосерин, гидроксилизин, норвалин. К таким вариантам также относятся, например, белки или полипептиды, у которых на N- и/или C-конце добавлены или удалены один или более аминокислотных остатков. Как правило, вариант имеет по меньшей мере приблизительно 50%, 60%, 70%, 75%, 80%, 85%, 90%, 92%, 95% или по меньшей мере около 98% идентичности аминокислотной последовательности с белком или полипептидом с нативной последовательностью. Вариант предпочтительно сохраняет биологическую активность, например связывание с той же мишенью, белка или полипептида, из которого он получен.
[0028] Термин «вариант», используемый в данном документе в отношении соответствующего белкового лиганда CD137 или GPC3 слитого белка по настоящему изобретению, относится к CD137 или GPC3 или его фрагменту соответственно, который имеет одну или более, как, например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 40, 50, 60, 70, 80 или более аминокислотных замен, делеций и/или вставок по сравнению с нативной последовательностью CD137 или GPC3 (CD137 или GPC3 дикого типа), такой как CD137, депонированный под номером UniProt Q07011, или GPC3, депонированный под номером UniProt P51654, как описано в данном документе. Вариант CD137 или вариант GPC3 соответственно предпочтительно имеет аминокислотную идентичность по меньшей мере 50%, 60%, 70%, 80%, 85%, 90% или 95% с CD137 или GPC3 дикого типа. Описываемый в данном документе вариант CD137 или вариант GPC3 сохраняет способность связывать слитые белки, специфические в отношении CD137 и GPC3, которые раскрыты в настоящем изобретении.
[0029] Термин «вариант», используемый в данном документе по отношению к мутеину липокалина, относится к мутеину липокалина или его фрагменту по настоящему изобретению, при этом последовательность имеет мутации, в том числе замены, делеции и вставки, и/или химические модификации. Описываемый в данном документе вариант мутеина липокалина сохраняет биологическую активность, например связывание с CD137, мутеина липокалина, из которого он получен. Как правило, вариант мутеина липокалина имеет по меньшей мере приблизительно 50%, 60%, 70%, 75%, 80%, 85%, 90%, 92%, 95%, 98% идентичности аминокислотной последовательности с мутеином липокалина, из которого он получен.
[0030] В контексте данного документа термин «мутагенез» относится к введению мутаций в полинуклеотидную или аминокислотную последовательность. Мутации предпочтительно вводят в таких экспериментальных условиях, чтобы могла быть изменена аминокислота, встречающаяся в природе в заданном положении последовательности белка или полипептида, например, заменена по меньшей мере одной аминокислотой. Термин «мутагенез» также включает (дополнительную) модификацию длины сегментов последовательности путем делеции или вставки одной или более аминокислот. Таким образом, в объеме настоящего изобретения, например, одну аминокислоту в выбранном положении последовательности заменяют участком из трех аминокислот, что приводит к добавлению двух аминокислотных остатков по сравнению с длиной соответствующего сегмента нативной аминокислотной последовательности белка или полипептида. Такую вставку или делецию можно ввести независимо друг от друга в любой из сегментов последовательности, которые в настоящем изобретении могут быть подвергнуты мутагенезу. В одном иллюстративном варианте осуществления настоящего изобретения вставку можно ввести в сегмент аминокислотной последовательности, соответствующий петле AB липокалина с нативной последовательностью (см. международную патентную публикацию № WO 2005/019256, которая в полном объеме включена в данный документ посредством ссылки).
[0031] В контексте данного документа термин «случайный мутагенез» означает, что никакой заданной мутации (изменения аминокислоты) не присутствует в определенном положении последовательности, но что в ходе мутагенеза в заранее определенное положение последовательности с определенной вероятностью могут быть включены по меньшей мере две аминокислоты.
[0032] В контексте данного документа термин «идентичность последовательностей» или «идентичность» обозначает свойство последовательностей, которое позволяет измерить их сходство или родственную связь. Термин «идентичность последовательности» или «идентичность», используемый в данном документе, означает полученный после (гомологичного) выравнивания последовательности белка или полипептида по настоящему изобретению с рассматриваемой последовательностью процент попарно идентичных остатков относительно количества остатков в более длинной из этих двух последовательностей. Идентичность последовательностей измеряют путем деления количества идентичных аминокислотных остатков на общее количество остатков и умножения произведения на 100.
[0033] В контексте данного документа термин «гомология последовательности» или «гомология» имеет свое обычное значение, а к гомологичной аминокислоте относятся идентичные аминокислоты, а также аминокислоты, которые считаются консервативными заменами в эквивалентных положениях в линейной аминокислотной последовательности белка, или полипептид по настоящему изобретению (например, любые слитые белки или мутеины липокалина по настоящему изобретению).
[0034] Специалисту будут известны доступные компьютерные программы, например BLAST (Altschul et al., Nucleic Acids Res, 1997), BLAST2 (Altschul et al., J Mol Biol, 1990), а также Smith-Waterman (Smith and Waterman, J Mol Biol, 1981), для определения гомологии последовательностей или идентичности последовательностей с применением стандартных параметров. Процент гомологии последовательностей или идентичности последовательностей в данном случае можно определить, например, с помощью программы BLASTP, версии 2.2.5 от 16 ноября 2002 года (Altschul et al., Nucleic Acids Res, 1997). В данном варианте осуществления процент гомологии основан на результатах выравнивания всех белковых или полипептидных последовательностей (матрица: BLOSUM 62; штрафы за открытие гэпа: 11.1; предельное значение, выставленное на 10-3), в том числе пропептидных последовательностей, предпочтительно с использованием каркаса белка дикого типа в качестве эталона при попарном сравнении. Его рассчитывают как процентное значение количества «положительных» (гомологичных аминокислот), указанных в результате вывода программы BLASTP, деленного на общее количество аминокислот, выбранных программой для выравнивания.
[0035] В частности, для определения того, отличается ли аминокислотный остаток из аминокислотной последовательности мутеина липокалина от такового у липокалина дикого типа, соответствующего определенному положению в аминокислотной последовательности липокалина дикого типа, специалист в данной области может использовать средства и способы, хорошо известные из уровня техники, например различные способы выравнивания вручную или с использованием компьютерных программ, таких как BLAST 2.0, что означает средство поиска основного локального выравнивания, или ClustalW, или любую другую подходящую программу, которая подходит для получения результатов выравнивания последовательностей. Соответственно, последовательность липокалина дикого типа может служить «рассматриваемой последовательностью» или «эталонной последовательностью», тогда как аминокислотная последовательность мутеина липокалина, отличного от описанного в данном документе липокалина дикого типа, служит «последовательностью запроса». Термины «последовательность дикого типа», «эталонная последовательность» и «рассматриваемая последовательность» в данном документе используют взаимозаменяемо. Предпочтительной последовательностью липокалина дикого типа является последовательность hTLc, представленная под SEQ ID NO: 1, или hNGAL, представленная под SEQ ID NO: 2.
[0036] «Гэпы» представляют собой пробелы в результатах выравнивания, которые являются следствием добавления или удаления аминокислот. Так, две копии одной и той же последовательности имеют 100% идентичности, но последовательности, которые менее консервативны и имеют делеции, добавления или замены, могут иметь более низкую степень идентичности.
[0037] В контексте данного документа термин «положение» означает положение либо аминокислоты в аминокислотной последовательности, изображенной в данном документе, либо положение нуклеотида в последовательности нуклеиновой кислоты, изображенной в данном документе. Следует понимать, что при использовании в данном документе термина «соответствует» или «соответствующий» в контексте положений в аминокислотной последовательности одного или более мутеинов липокалина, соответствующее положение определяется не только количеством предшествующих нуклеотидов или аминокислот. Следовательно, абсолютное положение заданной аминокислоты в соответствии с настоящим изобретением может отличаться от соответствующего положения из-за делеции или добавления аминокислот в другом месте липокалина (мутантного или дикого типа). Аналогичным образом, абсолютное положение заданного нуклеотида в соответствии с настоящим изобретением может отличаться от соответствующего положения из-за делеций или дополнительных нуклеотидов в другом месте в 5'-нетранслируемой области (UTR) мутеина липокалина или липокалина дикого типа, в том числе в промоторе и/или любых других регуляторных последовательностях или в гене (включающем экзоны и интроны).
[0038] «Соответствующее положение» в соответствии с настоящим изобретением может быть положением в последовательности, которое совмещается с положением в последовательности, которому оно соответствует при попарном или множественном выравнивании последовательностей согласно настоящему изобретению. Предпочтительно следует понимать, что для «соответствующего положения» в соответствии с настоящим изобретением абсолютные положения нуклеотидов или аминокислот могут отличаться относительно соседних нуклеотидов или аминокислот, но указанные соседние нуклеотиды или аминокислоты, которые могли быть заменены, удалены или добавлены, могут содержаться в том же одном или более «соответствующих положениях».
[0039] Кроме того, для соответствующего положения в мутеине липокалина, исходя из эталонной последовательности в соответствии с настоящим изобретением, предпочтительно следует понимать, что положения нуклеотидов или аминокислот в мутеине липокалина могут структурно соответствовать положениям в другом месте эталонного липокалина (липокалин дикого типа) или другого мутеина липокалина, даже если они могут отличаться по абсолютным номерам положений, что будет понятно специалистами в свете высококонсервативного общего паттерна фолдинга среди липокалинов.
[0040] Взаимозаменяемо используемые в данном документе термины «конъюгировать», «конъюгация», «сливать», «слияние» или «соединенный» относятся к объединению друг с другом двух или более субъединиц с помощью всех форм ковалентной или нековалентной связи посредством без ограничения генетического слияния, химической конъюгации, связывания посредством линкера или сшивающего средства и нековалентной связи.
[0041] В контексте данного документа термин «слитый полипептид» или «слитый белок» относится к полипептиду или белку, содержащему две или более субъединиц. В некоторых вариантах осуществления описываемый в данном документе слитый белок содержит две или более субъединиц, при этом по меньшей мере одна из этих субъединиц способна специфически связываться с CD137, а другая субъединица способна специфически связываться с GPC3. В слитом белке данные субъединицы могут быть соединены при помощи ковалентной или нековалентной связи. Предпочтительно, слитый белок представляет собой результат трансляционного слияния двух или более субъединиц. Трансляционное слияние можно осуществить с помощью генетической инженерии кодирующей последовательности для одной субъединицы в рамке считывания с кодирующей последовательностью другой субъединицы. Обе субъединицы могут разделяться нуклеотидной последовательностью, кодирующей линкер. Тем не менее, субъединицы слитого белка по настоящему изобретению также могут быть соединены при помощи химической конъюгации. Субъединицы, формирующие слитый белок, обычно соединены друг с другом, С-конец одной субъединицы с N-концом другой субъединицы, или С-конец одной субъединицы с С-концом другой субъединицы, или N-конец одной субъединицы с N-концом другой субъединицы, или N-конец одной субъединицы с С-концом другой субъединицы. Субъединицы слитого белка могут быть соединены в любом порядке и могут включать более одной из составляющих субъединиц. Если одна или более субъединиц являются частью белка (комплекса), который состоит из более чем одной полипептидной цепи, то термин «слитый белок» может также относиться к белку, содержащему слитые последовательности и все остальные полипептидные цепи белка (комплекса). В качестве иллюстративного примера, если полноразмерный иммуноглобулин слит с мутеином липокалина через тяжелую или легкую цепь иммуноглобулина, то термин «слитый белок» может относиться к единой полипептидной цепи, содержащей мутеин липокалина и тяжелую или легкую цепь иммуноглобулина. Термин «слитый белок» может также относиться ко всему иммуноглобулину (как легкой, так и тяжелой цепей) и мутеину липокалина, слитому с одной из или обеими его тяжелой и/или легкой цепями.
[0042] В контексте данного документа термин «субъединица» раскрываемого в данном документе слитого белка относится к одному белку или отдельной полипептидной цепи, которые сами по себе могут образовывать стабильную сложенную структуру и определять уникальную функцию формирования связывающего мотива к мишени. В некоторых вариантах осуществления предпочтительной субъединицей по настоящему изобретению является мутеин липокалина. В некоторых других вариантах осуществления предпочтительной субъединицей по настоящему изобретению является полноразмерный иммуноглобулин или его антигенсвязывающий домен.
[0043] «Линкер», который может входить в состав слитого белка по настоящему изобретению, объединяет вместе две или более субъединиц описываемого в данном документе слитого белка. Связь может быть ковалентной или нековалентной. Предпочтительная ковалентная связь образована посредством пептидной связи, такой как пептидная связь между аминокислотами. Предпочтительным линкером является пептидный линкер. Соответственно, в предпочтительном варианте осуществления указанный линкер содержит одну или более аминокислот, например 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 или более аминокислот. В данном документе описаны предпочтительные пептидные линкеры, в том числе глицин-сериновые (GS) линкеры, гликозилированные GS-линкеры и линкеры на основе пролин-аланин-серинового полимера (PAS). В некоторых предпочтительных вариантах осуществления GS-линкер представляет собой (G4S)3, который описан под SEQ ID NO: 13, и его применяют для соединения субъединиц слитого белка. К другим предпочтительным линкерам относятся химические линкеры.
[0044] В контексте данного документа термин «альбумин» включает все альбумины млекопитающих, такие как человеческий сывороточный альбумин, или бычий сывороточный альбумин, или крысиный сывороточный альбумин.
[0045] В контексте данного документа термин «органическая молекула» или «малая органическая молекула» обозначает органическую молекулу, содержащую по меньшей мере два атома углерода, но предпочтительно не более 7 или 12 вращающихся углеродных связей, имеющую молекулярный вес в диапазоне от 100 до 2000 дальтон, предпочтительно от 100 до 1000 дальтон, и необязательно включающую один или два атома металла.
[0046] «Образец» определяется как биологический образец, взятый у любого субъекта. К биологическим образцам относятся без ограничения кровь, сыворотка, моча, кал, сперма или ткань, в том числе опухолевая ткань.
[0047] «Субъект» представляет собой позвоночное животное, предпочтительно млекопитающее, более предпочтительно человека. Термин «млекопитающее» используют в данном документе для отсылки к любому животному, классифицируемому как млекопитающее, в том числе без ограничения к людям, ручным и сельскохозяйственным животным и к животным из зоопарков, спортивным животным или домашним животным, таким как овцы, собаки, лошади, кошки, коровы, крысы, свиньи, человекообразные обезьяны, такие как яванский макак, и это лишь несколько из иллюстративных примеров. Предпочтительно, в контексте данного документа «млекопитающим» является человек.
[0048] «Эффективное количество» представляет собой количество, достаточное для получения полезных или требуемых результатов. Эффективное количество можно вводить за одно или более отдельных введений или доз.
[0049] В контексте данного документа к «антителу» относятся цельные антитела или любой его антигенсвязывающий фрагмент (т.е. «антигенсвязывающий участок») или отдельная цепь. Под цельным антителом понимают гликопротеин, содержащий по меньшей мере две тяжелые цепи (HC) и две легкие цепи (LC), соединенные друг с другом дисульфидными связями. Каждая тяжелая цепь состоит из вариабельного домена тяжелой цепи (VH или HCVR) и константной области тяжелой цепи (CH). Константная область тяжелой цепи состоит из трех доменов: CH1, CH2 и CH3. Каждая легкая цепь состоит из вариабельного домена легкой цепи (VL или LCVR) и константной области легкой цепи (CL). Константная область легкой цепи состоит из одного домена CL. Области VH и VL можно дополнительно подразделить на области гипервариабельности, называемые определяющими комплементарность областями (CDR), со вставками областей, которые являются более консервативными и называются каркасными областями (FR). Каждая из VH и VL состоит из трех CDR и четырех FR, расположенных в следующем порядке от аминоконца к карбокси-концу: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Вариабельные области тяжелой и легкой цепей содержат связывающий домен, который взаимодействует с антигеном (например, GPC3). Константные области антител необязательно могут опосредовать связывание иммуноглобулина с тканями или факторами хозяина, в том числе различными клетками иммунной системы (например, эффекторными клетками) и первым компонентом (C1q) классической системы комплемента.
[0050] В контексте данного документа «антигенсвязывающий фрагмент» антитела относится к одному или более фрагментам антитела, которые сохраняют способность специфически связываться с антигеном (например, GPC3). Было продемонстрировано, что антигенсвязывающая функция антитела может выполняться фрагментами полноразмерного антитела. Примеры связывающих фрагментов, охватываемых термином «антигенсвязывающий фрагмент» антитела включают (i) Fab-фрагмент, состоящий из доменов VH, VL, CL и CH1; (ii) F(ab')2-фрагмент, содержащий два Fab-фрагмента, связанные дисульфидным мостиком в шарнирной области; (iii) Fab'-фрагмент, состоящий из доменов VH, VL, CL и CH1 и области между доменами CH1 и CH2; (iv) Fd-фрагмент, состоящий из доменов VH и CH1; (v) одноцепочечный Fv-фрагмент, состоящий из доменов VH и VL одного плеча антитела, (vi) dAb-фрагмент (Ward et al., Nature, 1989), состоящий из домена VH; и (vii) выделенную определяющую комплементарность область (CDR) или комбинацию двух или более выделенных CDR, которые необязательно могут быть соединены синтетическим линкером; (viii) a «диатело», содержащее VH и VL, соединенные в одну полипептидную цепь с помощью короткого линкера (см., например, патентные документы EP 404097; WO 93/11161; и Holliger et al., Proc Natl Acad Sci U S A, 1993); (ix) «доменный фрагмент антитела», содержащий только VH или VL, при этом в некоторых случаях две или более областей VH соединены ковалентной связью.
[0051] Антитела могут быть поликлональными или моноклональными; ксеногенными, аллогенными или сингенными или их модифицированными формами (например, гуманизированными, химерными или мультиспецифическими). Антитела также могут быть полностью человеческими.
[0052] В контексте данного документа «каркас» или «FR» относится к остаткам вариабельного домена, отличным от остатков гипервариабельной области (CDR).
[0053] «Кристаллизующийся фрагмент иммуноглобулина» или «Fc-область» относится к C-концевой области тяжелой цепи иммуноглобулина, в том числе к Fc-областям с нативной последовательностью и вариантным Fc-областям. Хотя границы Fc-области тяжелой цепи иммуноглобулина могут варьировать, Fc-область тяжелой цепи человеческого IgG обычно определяют как проходящую от аминокислотного остатка в положении Cys226 или от Pro230 до его карбоксильного конца, нумерация которого соответствует EU-индексу по Kabat (Johnson and Wu, Nucleic Acids Res, 2000). С-концевой лизин (остаток 447 согласно EU-индексу по Kabat) Fc-области можно удалить, например, во время получения или очистки антитела или путем рекомбинантной инженерии нуклеиновой кислоты, кодирующей тяжелую цепь антитела. Соответственно, композиция с интактными антителами может содержать популяции антител с удаленными остатками K447, популяции антител без удаленных остатков K447 и популяции антител, содержащие смесь антител с остатком K447 и без него. К подходящим Fc-областям с нативной последовательностью для применения в антителах по настоящему изобретению относятся человеческий IgG1, IgG2 (IgG2A, IgG2B), IgG3 и IgG4.
[0054] «Fc-рецептор» или «FcR» относится к рецептору, который связывается с Fc-областью антитела.
[0055] В контексте данного документа «выделенное антитело» относится к антителу, которое практически не содержит своего природного окружения. Например, выделенное антитело практически не содержит клеточного материала и других белков из клеточного или тканевого источника, из которого оно получено. «Выделенное антитело» дополнительно относится к антителу, которое практически не содержит других антител, имеющих иные антигенные специфичности. В данном случае выделенное антитело, которое специфически связывает GPC3, практически не содержит антитела, которые специфически связывают антигены, отличные от GPC3. Тем не менее, выделенное антитело, которое специфически связывает GPC3, может обладать перекрестной реактивностью с другими антигенами, такими как молекулы GPC3 от других видов.
[0056] В контексте данного документа «моноклональное антитело» относится к препарату молекул антитела с единой молекулярной композицией. Композиция с моноклональными антителами характеризуется единой специфичностью связывания и аффинностью связывания в отношении конкретного эпитопа.
[0057] В контексте данного документа «гуманизированное антитело» относится к антителу, которое состоит из CDR антител, полученных от млекопитающих, отличных от человека, а также FR-области и константной области человеческого антитела или антитела, полученного из человеческого антитела. В некоторых вариантах осуществления гуманизированное антитело содержит вариабельный домен, который имеет аминокислотную последовательность вариабельной области, которая, при анализе в целом, ближе к человеческой, чем к другим видам, по результатам оценки с использованием инструментария DomainGapAlign базы данных Immunogenetics Information System (IMGT), как описано у Ehrenmann et al. (2010). В некоторых вариантах осуществления гуманизированное антитело может быть применимым в качестве эффективного компонента терапевтического средства из-за его пониженной антигенности. В контексте данного документа термин «терапевтическое средство» или «терапевтически активное средство» относится к средству, которое является терапевтически применимым. Терапевтическое средство может представлять собой любое средство для предупреждения, уменьшения интенсивности или лечения заболеваний, физиологического состояния, симптома или для их оценки или диагностики.
[0058] В контексте данного документа к «человеческому антителу» относятся антитела, имеющие вариабельные области, у которых как каркасная область, так и CDR-области получены из последовательностей человеческого иммуноглобулина эмбрионального типа. Более того, если антитело содержит константную область, то константная область также получена из последовательностей человеческого иммуноглобулина эмбрионального типа. Человеческие антитела по настоящему изобретению могут включать аминокислотные остатки, не кодируемые последовательностями человеческого иммуноглобулина эмбрионального типа (например, мутации, введенные в результате случайного или сайт-специфического мутагенеза in vitro или в результате соматической мутации in vivo). Тем не менее, в контексте данного документа термин «человеческое антитело» не подразумевается как включающий антитела, в которых последовательности CDR, полученные из зародышевой линии другого вида млекопитающих, такого как мышь, были привиты к человеческим каркасным последовательностям.
III. Описание графических материалов
[0059] На фигуре 1 представлен обзор конструкции типичных слитых белков, описываемых в настоящей заявке, которые являются биспецифическими в отношении мишеней CD137 и GPC3. Типичные слитые белки получали на основании антитела, специфического в отношении GPC3 (например, антитела, тяжелая цепь которого представлена под SEQ ID NO: 81, или содержит вариабельный домен тяжелой цепи под SEQ ID NO: 78, или содержит последовательности CDR, представляющие собой GYTFTDYE (HCDR1, SEQ ID NO: 72), LDPKTGDT (HCDR2, SEQ ID NO: 73) и TRFYSYTY (HCDR3; SEQ ID NO: 74), а легкие цепи представлены под SEQ ID NO: 82, или содержат вариабельный домен легкой цепи под SEQ ID NO: 79, или содержат последовательности CDR, представляющие собой QSLVHSNRNTY (LCDR1, SEQ ID NO: 75), KVS (LCDR2, SEQ ID NO: 76) и SQNTHVPPT (LCDR3; SEQ ID NO: 77), и один или более мутеинов липокалина, специфических в отношении CD137 (например, мутеинов липокалина с последовательностью под SEQ ID NO: 40 или мутеин липокалина, обладающий 97% идентичности последовательности с последовательностью под SEQ ID NO: 49 и обозначаемый CD137Ac1). Один или более мутеинов липокалина генетическими способами сливали с C- и/или N-концом либо тяжелой цепи, либо легкой цепи специфического в отношении GPC3 антитела, как изображено на фигуре 1A-1I, что приводило к получению слитых белков, например SEQ ID NO: 87 и 82, SEQ ID NO: 88 и 82, SEQ ID NO: 81 и 89, SEQ ID NO: 81 и 90, а также CD137Ac1-слитый белок 1, CD137Ac1-слитый белок 2, CD137Ac1-слитый белок 3, CD137Ac1-слитый белок 4, CD137Ac1-слитый белок 5, CD137Ac1-слитый белок 6 и CD137Ac1-слитый белок 7 (слитые белки, обладающие 97% идентичности последовательности с последовательностями под SEQ ID NO: 91 и 82, SEQ ID NO: 92 и 82, SEQ ID NO: 81 и 93, SEQ ID NO: 81 и 94, SEQ ID NO: 95 и 82 или SEQ ID NO: 96 и 82). Кроме того, созданные слитые белки могут быть двухвалентными по отношению к CD137 (например, как изображено на фигуре 1A-1D) или четырехвалентеными по отношению к CD137 (например, как изображено на фигуре 1E-1H) или имеют еще более высокую валентность по отношению к CD137 (например, как изображено на фигуре 1I). Дополнительные моноспецифические слитые белки получали путем слияния одного или более мутеинов липокалина, специфических в отношении GPC-3, или мутеинов липокалина, специфических в отношении CD137, с С-концом Fc-области предусмотренного антитела так, как описано в данном документе, через пептидный линкер (например, как изображено на фигуре 1J-1K). Полученные моноспецифические слитые белки представлены, например, под SEQ ID NO: 97 и SEQ ID NO: 98.
[0060] На фигуре 2 показаны результаты экспериментов с применением ELISA, в которых связывание с типичными слитыми белками с человеческим CPC3 (фигура 2A), GPC3 яванского макака (фигура 2B) и человеческим CD137 (фигура 2C) определяли так, как описано в примере 3. GPC3 или C-концевой His-меченный CD137 наносили слоем на микротитровальный планшет и тестируемые средства титровали, начиная с наивысшей концентрации, составлявшей 100 нМ. Исследуемые связанные средства выявляли с помощью антител к человеческому IgG Fc, конъюгированных с пероксидазой хрена (HRP), или антител к NGAL, конъюгированных с HRP, соответственно. Данные согласовывали с моделью связывания 1:1 со значением EC50 и максимальным сигналом в качестве свободных параметров, а также угловым коэффициентом, который был зафиксирован на уровне единицы. Полученные значения EC50 представлены в таблице 4.
[0061] На фигуре 3 проиллюстрированы результаты иллюстративных экспериментов с применением ELISA, в которых способность типичных слитых белков одновременно связывать обе мишени, GPC3 и CD137, определяли так, как описано в примере 4. Рекомбинантный huGPC3 (фигура 3A) или huCD137-His (фигура 3B) наносили слоем на микротитровальный планшет с последующим титрованием слитых белков. Затем вносили биотинилированный huCD137-His (фигура 3A) или биотинилированный huGPC3 в постоянной концентрации (фигура 3B) соответственно, который выявляли с помощью комплекса авидина и пероксидазы ExtrAvidin-Peroxidase. Данные согласовывали с моделью связывания 1:1 со значением EC50 и максимальным сигналом в качестве свободных параметров, а также угловым коэффициентом, который был зафиксирован на едином уровне. Полученные значения EC50 представлены в таблице 3.
[0062] На фигуре 4 показаны результаты оценки связывания с мишенями слитых белков, полученные с помощью проточной цитометрии с использованием клеток СНО, экспрессирующих человеческий CD137 (фигура 4A), а также клеток SK-Hep1, экспрессирующих человеческий GPC3 (фигура 4B), как описано в примере 6. При использовании ложнотрансфицированных клеток связывания не наблюдали. Для расчета значений EC50, которые представлены в таблице 6, использовали средние геометрические значения интенсивности флуоресценции.
[0063] На фигуре 5 показано связывание слитых белков с GPC3-положительными опухолевыми клетками, оцениваемое с помощью проточной цитометрии. Линии опухолевых клеток с разными уровнями экспрессии GPC3 (от высокой до умеренной экспрессии: HepG2 (фигура 5A) > Hep3B (фигура 5B) > MKN-45 (фигура 5C)) и линию GPC3-отрицательных клеток NCI-N87 (фигура 5D) инкубировали с различными слитыми белками и контролями, как описано в примере 7, а соответствующие значения аффинности связывания (значения EC50) подытожены в таблице 7.
[0064] На фигуре 6 показан потенциал типичных слитых белков совместно стимулировать активацию Т-клеток зависимым от GPC3-мишени образом, оцениваемый с помощью биоанализа CD137. NFκB-luc2/CD137 клетки Jurkat совместно культивировали с линиями GPC3-экспрессирующих опухолевых клеток (от высокой до умеренной экспрессии: HepG2 > Hep3B > MKN-45, отрицательный по GPC3 контроль: NCI-N87, фигура 6A, 6B, 6C и 6D соответственно) в присутствии различных концентраций слитых белков или контролей. Через 4 часа добавляли реагент для люциферазного анализа и измеряли люминесцентные сигналы. Для расчета значений EC50 (см. таблицу 7) проводили четырехпараметрический анализ логистической кривой. Слитые белки совместно стимулируют активацию Т-клеток только в избытке GPC3 (фигура 6A и 6B), но не в условиях недостаточного количества или отсутствия GPC3 (фигура 6C и 6D). В отличие от этого, эталонное mAb к CD137 (SEQ ID NO: 26 и 27) проявляет схожую активацию независимо от уровня экспрессии GPC3, в отсутствие каких-либо GPC3-экспрессирующих клеток и в отсутствие каких-либо опухолевых клеток.
[0065] На фигуре 7 продемонстрирована способность типичного слитого белка совместно стимулировать активацию Т-клеток в присутствии GPC3. Антитело к GPC3, которое входит в состав слитого белка (SEQ ID NO: 81 и 82), GPC3-специфический мутеин липокалина (продукт слияния с Fc) (SEQ ID NO: 97), уже известный как биспецифический в отношении CD137/GPC3 слитый белок (SEQ ID NO: 83), и изотипический контроль (SEQ ID NO: 24 и 25) тестировали в параллели. Клетки SK-Hep1, либо трансфицированные человеческим GPC3 (фигура 7A), либо ложнотрансфицированные (отрицательные по человеческому GPC3, фигура 7B), либо линию GPC3-экспрессирующих (фигура 7C) опухолевых клеток Hep-G2 высевали в планшеты со слоем антител к человеческому CD3. Добавляли все T-клетки, а также различные концентрации тестируемых молекул и инкубировали в течение трех дней. Уровни секретируемого IL-2 в супернатанте определяли с помощью электрохемолюминесцентного анализа, как описано в примере 9. Слитый белок с SEQ ID NO: 87 и 82 индуцирует сильное дозозависимое увеличение секреции IL-2 только в присутствии GPC3, сильнее, чем уже известный биспецифический в отношении CD137/GPC3 слитый белок с SEQ ID NO: 83.
[0066] На фигуре 8 показана способность типичных слитых белков совместно стимулировать активацию Т-клеток зависимым от GPC3-мишени образом. Различные линии опухолевых клеток, экспрессирующих разные уровни GPC3 (от высокой до умеренной экспрессии: HepG2 > Hep3B > MKN-45, фигура 8A, 8B и 8C соответственно; отрицательный по GPC3 контроль: NCI-N87, фигура 8D) высевали в планшеты со слоем антител к человеческому CD3. Добавляли все T-клетки и различные концентрации слитых белков и отдельных строительных блоков и инкубировали в течение 3 дней. Уровни секретируемого IL-2 определяли с помощью электрохемолюминесцентного анализа, как описано в примере 10. Слитый белок способен увеличивать секрецию IL-2 зависимым от GPC3 образом. В отличие от этого, секреция IL-2, индуцированная параллельно протестированным эталонным антителом к CD137 (SEQ ID NO: 26 и 27), не имеет зависимости от GPC3.
[0067] На фигуре 9 показана способность типичных слитых белков активировать костимулирующий сигнальный путь с участием CD137 и индуцировать опосредованный Т-клетками цитолиз GPC3-экспрессирующих опухолевых клеток, что продемонстрировано в импедансном анализе Т-клеточного уничтожения. GPC3-экспрессирующие клетки HepG2 или GPC3-отрицательные клетки NCI-N87 высевали и давали им прикрепиться к электронному микротитровальному планшету с последующим добавлением антитела к CD3, тестируемой молекулы и неадгезивных CD8+ Т-клеток. Импеданс измерялся как безразмерный параметр «клеточный коэффициент (CI)» каждые 15 минут в течение трех дней, как описано в примере 11. Слитый белок с SEQ ID NO: 87 и 82 способен активировать путь с участием CD137 зависимым от GPC3 образом и индуцировать зависимый от дозы, опосредованный Т-клетками лизис GPC3-экспрессирующих опухолевых клеток на более высоком уровне, чем в случае уже известного биспецифического к CD137/GPC3 слитого белка с SEQ ID NO: 83.
[0068] На фигуре 10 показан объем опухоли с течением времени после обработки посредством 0,5, 5 и 20 мг/кг иллюстративного слитого белка, эквимолярными концентрациями (0,39 и 3,9 мг/кг) антитела к GPC3, которое включено в слитый белок, эквимолярной концентрацией (3,9 мг/кг) эталонного антитела к CD137 или контрольной среды-носителя (PBS) в гуманизированной мышиной модели опухоли, как описано в примере 12. Мышам NOG подкожно подсаживали опухоли HepG2, которым давали возможность вырасти до размера 80-100 мм3. Затем мышей рандомизировали в группы обработки (или контрольные группы) и вводили им внутривенно 5×106 свежих человеческих PBMC и внутрибрюшинные инъекции тестируемых молекул в указанных дозах в день 1, день 8 и день 15. Рост опухоли регистрировали каждые 3-4 дня. Данные представлены в виде медианных значений с планками погрешностей межквартильного размаха. Пунктирной (пунктирными) линией (линиями) показано уменьшение количества животных в группе по сравнению с предыдущими днями. Слитый белок полностью ингибировал рост опухоли в ходе исследования при более высоких уровнях доз (5 и 20 мг/кг), при этом при более низких концентрациях (0,5 или 0,39 мг/кг соответственно) он оказывал дозозависимое, но незначительное влияние на рост опухоли.
[0069] На фигуре 11 представлены результаты фармакокинетических анализов у мышей иллюстративного слитого белка и антитела к GPC3, которое включено в слитый белок, по сравнению с двумя уже известными биспецифическими в отношении CD137/GPC3 слитыми белками (SEQ ID NO: 83 и SEQ ID NO: 84), как описано в примере 13. Самцам CD-1 мышей (2 мышам на каждый момент времени) внутривенно вводили слитые белки в дозе 2 мг/кг или 10 мг/кг. Уровни лекарственного средства выявляли с помощью сэндвич-ELISA, позволяющего выявить полную молекулу с помощью GPC3-мишени и антитела к человеческому Fc или антитела к NGAL. Слитый белок по настоящему изобретению (SEQ ID NO: 87 и 82) демонстрирует сильно улучшенные фармакокинетические свойства и увеличенные периоды полужизни по сравнению с ранее известными биспецифическими в отношении CD137/GPC3 слитыми белками (SEQ ID NO: 83 и SEQ ID NO: 84).
IV. Подробное описание изобретения
[0070] Как описано в данном документе, настоящим изобретением охвачено признание того, что двухвалентной CD137-связывающей конструкции, такой как антитело, самой по себе может быть недостаточно для кластеризации CD137 на Т-клетках или NK-клетках и достижения эффективной активации. Кроме того, недавняя работа по изучению представителей семейства TNFR иллюстрирует механизмы действия антител к TNFR, посредством которых антитела взаимодействуют через свои Fc-области с Fc-гамма-рецепторами, задействуя активирующие иммунные клетки, экспрессирующие Fc-гамма-рецептор, и способствуют последующей их противоопухолевой активности (Bulliard et al., Immunol Cell Biol, 2014, Bulliard et al., J Exp Med, 2013). Следовательно, это позволяет предположить, что антитело к CD137 может запускать кластеризацию CD137 в зависимости от избытка положительных по Fc-гамма-рецептору клеток, которые не локализованы избирательно в опухоли, а распределены по всему организму. Соответственно, эффективность и целевая специфичность монотерапии антителом к CD137 могут представлять определенный интерес. Известно, что, некоторые проходящие клинические исследования терапевтические средства на основе антител к CD137, такие как урелумаб и утомилумаб, демонстрируют неутешительные результаты в отношении эффективности в низкой дозе и/или проявляют токсичность в высокой дозе или в эффективной дозе (Bulliard et al., Immunol Cell Biol, 2014, Bulliard et al., J Exp Med, 2013). Таким образом, существует неудовлетворенная потребность в нацеленных на CD137 терапевтических средствах, которые были бы как эффективными, так и безопасными. Идеальное нацеленное на CD137 средство должно приводить к кластеризации CD137 и должно делать это локализованным в месторасположении опухоли способом на противоопухолевых эффекторных лимфоцитах. Как описано в данном документе, для получения такого нацеленного на CD137 средства можно разработать биспецифическое средства для нацеливания на CD137 на одном конце и на дифференциально экспрессируемую опухолевую мишень на другом конце.
[0071] Настоящим изобретением охвачено признание того, что нацеленное на GPC3 моноспецифическое средство может быть недостаточно эффективным для радикального лечения опухолей, таких как HCC, и что может иметь значение противоопухолевая активность средства на основе антитела к GPC3, которое способно рекрутировать иммунные клетки к местоположению опухоли и/или оказывать прямое ингибирующее действие на пролиферацию и/или выживаемость раковых клеток. Следовательно, существует неудовлетворенная потребность в нацеленных на GPC3 терапевтических средствах с дополнительными противоопухолевыми эффектами, и которые обладают способностью обеспечивать усиленную противоопухолевую активность по сравнению с отдельным моноспецифическим средством к GPC3-мишени.
[0072] Настоящим изобретением предусмотрены, среди прочего, новые подходы по одновременному задействованию CD137 и GPC3 с помощью слитых белков, обладающих специфичностью связывания к CD137 и специфичностью связывания к GPC3. Например, в WO2016/184882 были описаны слитые белки из мутеина липокалина, специфического в отношении CD137, и антитела к GPC3 для задействования CD137 и GPC3. Тем не менее, все еще требуется, чтобы слитые белки индуцировали GPC3-зависимые Т-клеточные уничтожение/иммунный ответ для подавления с высокой эффективностью и селективностью роста GPC3-экспрессирующей опухоли. Различными аспектами настоящего изобретения предусмотрены слитые белки, которые стимулируют кластеризацию CD137 путем связывания мостиком CD137-положительных T-клеток с GPC3, экспрессируемым в опухолевом микроокружении, вследствие чего сочетают CD137-индуцированную активацию и размножение T-клеток с опосредованным антителами к GPC3 цитотоксичным уничтожением опухолевых клеток. Настоящим изобретением также предусмотрены слитые белки, которые обеспечивают возможности комбинаторной терапии в одной молекуле и в то же время делают возможной локализованную индукцию антиген-специфических Т-клеток в опухолевом микроокружении, потенциально снижая периферическую токсичность. В данном отношении слитые белки по настоящему изобретению могут обеспечивать усиленные продуцирование IL-2, иммунные ответы, опосредованный Т-клетками цитолиз и/или противоопухолевые эффекты по сравнению с монотерапией в отношении CD137 или в отношении GPC3.
[0073] Кроме того, поскольку яванские макаки широко используются в фармакокинетических исследованиях или исследованиях безопасности лекарственных средств при разработке новых средств терапии, в том числе новых биологических средств, и дополнительно такие исследования могут быть необходимыми предварительными обязательными условиями для получения одобрения регулирующих органов, желательно получить слитые белки с GPC3-связывающим фрагментом, который вступает в перекрестную реакцию как с человеческим GPC3 человека, так и с GPC3 яванского макака. Такие слитые белки с GPC3-связывающим фрагментом, обладающие такими характеристиками перекрестной реактивности, ранее описаны не были.
[0074] Более того, поскольку биспецифические или мультиспецифические молекулы могут отличаться от своих структурных элементов по определенным свойствам, в том числе по аффинности и специфичности к мишени, стабильности, фармакокинетике в крови и механизму действия (Spiess et al., Mol Immunol, 2015, Sedykh et al., Drug Des Devel Ther., 2018), как предусмотрено в данном документе, в некоторых вариантах осуществления для получения качественных клинических кандидатов предусмотрены тщательный выбор структурных элементов и оптимизация производства и других практических способов. Например, биспецифические или мультиспецифические молекулы могут частично или полностью утратить аффинность и специфичность связывания с мишенью структурных элементов, что приводит к нежелательному нецелевому связыванию. Нецелевое связывание может дополнительно повлиять на фармакокинетику, распределение в тканях, эффективность и токсичность таких биспецифических или мультиспецифических молекул для вариантов терапевтического применения. В частности, плохие фармакокинетические профили могут привести к трудностям в достижении профиля распределения доз, необходимого для терапевтической эффективности, и к несоблюдению пациентом режима лечения (Alavijeh and Palmer, IDrugs, 2004), а благоприятная фармакокинетика позволяет снизить дозу, позволяет разработать подкожные составы и снизить значения COG, а также продлить интервал между приемами доз, например, до одного раза в неделю, раза в 2 недели, раза в 3 недели или раза в 4 недели (Ryman and Meibohm, CPT Pharmacometrics Syst Pharmacol, 2017). На фармакокинетические свойства также оказывает влияние изоэлектрическая точка (pI). Повышение pI может привести к увеличению клиренса из крови и увеличению удержания в тканях с более коротким периодом полужизни, тогда как более низкая pI может снизить поглощение тканями и привести к более длительному периоду полужизни. Хотя результаты наблюдений могут быть противоречивыми в том, что касается корреляции между клиренсом белка и pI, терапевтические антитела зачастую имеют значения pI немного выше физиологического pH 7,4, так как большинство клеточных поверхностей имеют отрицательный заряд. В данном отношении различными аспектами настоящего изобретения предусмотрены слитые белки, которые являются биспецифическими в отношении CD137 и GPC3, с высокой аффинностью и специфичностью, повышенной стабильностью, подходящей pI и благоприятными фармакокинетическими свойствами.
[0075] Такие слитые белки, обладающие такими признаками, пригодными для вариантов применения, предусмотренных настоящим изобретением, ранее описаны не были. В отличие от предусмотренных в данном документе слитых белков, уже известные слитые белки, нацеленные как на CD137, так и на GPC3, характеризовались одним или более из плохой PK, неприемлемой степени нецелевого связывания, неприемлемой степени неспецифической (например, независимой от GPC3) активации, например, иммунной системы и/или сниженной или иным образом нарушенной способности опосредовать активацию, пролиферацию и/или инфильтрацию Т-клеток.
A. Иллюстративные слитые белки, специфические в отношении CD137 и GPC3, по настоящему изобретению
[0076] В некоторых вариантах осуществления предусмотренный слитый белок содержит по меньшей мере две субъединицы в любом порядке: (1) первую субъединицу, которая предусматривает полноразмерный иммуноглобулин или его антигенсвязывающий домен, специфической в отношении GPC3, и (2) вторую субъединицу, которая предусматривает мутеин липокалина, специфической в отношении CD137.
[0077] В некоторых вариантах осуществления предусмотренный слитый белок содержит по меньшей мере две субъединицы, где первая субъединица предусматривает полноразмерный иммуноглобулин или его антигенсвязывающий домен и является специфической в отношении GPC3, и где вторая субъединица предусматривает мутеин липокалина и является специфической в отношении CD137, причем вторая субъединица соединена на N-конце с C-концом каждой тяжелой цепи (HC) первой субъединицы, необязательно посредством линкера.
[0078] В некоторых вариантах осуществления предусмотренный слитый белок содержит по меньшей мере две субъединицы, где первая субъединица предусматривает полноразмерный иммуноглобулин или его антигенсвязывающий домен и является специфической в отношении GPC3, и где вторая субъединица предусматривает мутеин липокалина и является специфической в отношении CD137, причем вторая субъединица соединена на С-конце с N-концом каждой НС первой субъединицы, необязательно посредством линкера.
[0079] В некоторых вариантах осуществления предусмотренный слитый белок содержит по меньшей мере две субъединицы, где первая субъединица предусматривает полноразмерный иммуноглобулин или его антигенсвязывающий домен и является специфической в отношении GPC3, и где вторая субъединица предусматривает мутеин липокалина и является специфической в отношении CD137, причем вторая субъединица соединена на N-конце с C-концом каждой легкой цепи (LC) первой субъединицы, необязательно посредством линкера.
[0080] В некоторых вариантах осуществления предусмотренный слитый белок содержит по меньшей мере две субъединицы, где первая субъединица предусматривает полноразмерный иммуноглобулин или его антигенсвязывающий домен и является специфической в отношении GPC3, и где вторая субъединица предусматривает мутеин липокалина и является специфической в отношении CD137, причем вторая субъединица соединена на С-конце с N-концом каждой LC первой субъединицы, необязательно посредством линкера.
[0081] В некоторых вариантах осуществления предусмотренный слитый белок содержит по меньшей мере две субъединицы, где первая субъединица предусматривает полноразмерный иммуноглобулин или его антигенсвязывающий домен и является специфической в отношении GPC3, и где вторая субъединица предусматривает мутеин липокалина и является специфической в отношении CD137, причем вторая субъединица соединена на N-конце с C-концом константной области каждой тяжелой цепи (CH) первой субъединицы, необязательно посредством линкера.
[0082] В некоторых вариантах осуществления предусмотренный слитый белок содержит по меньшей мере две субъединицы, где первая субъединица предусматривает полноразмерный иммуноглобулин или его антигенсвязывающий домен и является специфической в отношении GPC3, и где вторая субъединица предусматривает мутеин липокалина и является специфической в отношении CD137, причем вторая субъединица соединена на N-конце с C-концом константной области каждой легкой цепи (CL) первой субъединицы, необязательно посредством линкера.
[0083] В некоторых вариантах осуществления предусмотренный слитый белок также может содержать по меньшей мере одну дополнительную субъединицу, например третью субъединицу. Например, слитый белок может содержать третью субъединицу, специфическую в отношении CD137. В некоторых вариантах осуществления третья субъединица может представлять собой или предусматривать мутеин липокалина, специфический в отношении CD137. Например, два мутеина липокалина могут быть слиты с первой субъединицей иммуноглобулина, один на С-конце и один на N-конце иммуноглобулина. В некоторых вариантах осуществления мутеины липокалина могут быть слиты с тяжелой цепью или легкой цепью иммуноглобулина.
[0084] В некоторых вариантах осуществления предусмотренные слитые белки могут содержать одну или более дополнительных субъединиц (например, четвертую, пятую или шестую субъединицу).
[0085] В некоторых вариантах осуществления по меньшей мере одна субъединица может быть слита своим N-концом и/или своим C-концом с другой субъединицей.
[0086] В некоторых вариантах осуществления по меньшей мере одна субъединица может быть соединена с другой субъединицей посредством линкера. В некоторых дополнительных вариантах осуществления линкер представляет собой пептидный линкер, например неструктурированный глицин-сериновый (GS) линкер, гликозилированный GS-линкер или линкер на основе пролин-аланин-серинового полимера (PAS). В некоторых вариантах осуществления GS-линкер представляет собой (Gly4Ser)3-линкер ((G4S)3), который представлен под SEQ ID NO: 13. Другие иллюстративные линкеры представлены под SEQ ID NO: 14-23. В некоторых вариантах осуществления пептидный линкер может иметь от 1 до 50 аминокислот, как, например, 1, 2, 3, 4, 5, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 25, 30, 35, 40, 45 или 50 аминокислот. Например, если первая субъединица предусматривает полноразмерный иммуноглобулин, то вторая субъединица может быть присоединена посредством пептидного линкера между N-концом второй субъединицы и C-концом константной области тяжелой цепи (СН) указанного иммуноглобулина. В некоторых дополнительных вариантах осуществления третья субъединица может быть присоединена посредством пептидного линкера между N-концом третьей субъединицы и C-концом константной области легкой цепи (CL) указанного иммуноглобулина.
[0087] В некоторых вариантах осуществления одну субъединицу можно соединить с другой субъединицей так, как фактически описано на фигуре 1. В целом, одна субъединица может быть слита своим N-концом и/или своим C-концом с другой субъединицей. Например, в некоторых вариантах осуществления субъединица, представляющая собой мутеин липокалина, может быть слита своим N-концом и/или своим C-концом с субъединицей, представляющей собой иммуноглобулин, предпочтительно посредством пептидной связи. В качестве дополнительных примеров субъединица, представляющая собой мутеин липокалина, может быть соединена своим N-концом с C-концом домена тяжелой цепи (HC) субъединицы, представляющей собой иммуноглобулин (фигура 1A), субъединица, представляющая собой мутеин липокалина, может быть соединена своим C-концом с N-концом HC субъединицы, представляющей собой иммуноглобулин (фигура 1С), субъединица, представляющая собой мутеин липокалина, может быть соединена своим N-концом с C-концом легкой цепи (LC) субъединицы, представляющей собой иммуноглобулин (фигура 1B), и/или субъединица, представляющая собой мутеин липокалина, может быть соединена своим C-концом с N-концом LC субъединицы, представляющей собой иммуноглобулин (фигура 1D).
[0088] В некоторых вариантах осуществления субъединица, представляющая собой мутеин липокалина, может быть слита своим N-концом и/или своим C-концом с фрагментом, представляющим собой иммуноглобулин. Например, в некоторых вариантах осуществления субъединица, представляющая собой мутеин липокалина, может быть соединена, предпочтительно посредством пептидного линкера, своим N-концом с C-концом константной области тяжелой цепи (СН) субъединицы, представляющей собой иммуноглобулин, или субъединица, представляющая собой мутеин липокалина, может быть соединена, предпочтительно посредством пептидного линкера, своим N-концом с C-концом константной области легкой цепи (CL) субъединицы, представляющей собой иммуноглобулин.
[0089] В некоторых вариантах осуществления, если одна субъединица предусматривает полноразмерный иммуноглобулин, то вторая субъединица может быть присоединена между N-концом второй субъединицы и C-концом константной области тяжелой цепи (СН) указанного иммуноглобулина.
[0090] В некоторых вариантах осуществления третья субъединица может быть присоединена между N-концом третьей субъединицы и C-концом константной области легкой цепи (CL) указанного иммуноглобулина.
[0091] В некоторых вариантах осуществления, что касается слитого белка по настоящему изобретению, в котором по меньшей мере одна субъединица может представлять собой или предусматривать полноразмерный иммуноглобулин, то Fc-функцию Fc-области полноразмерного иммуноглобулина по отношению к положительной по Fc-рецептору клетке можно при этом сохранить, в то же время слитый белок одновременно будет воздействовать на CD137 и GPC3.
[0092] В некоторых вариантах осуществления, где по меньшей мере одна субъединица предусмотренного слитого белка может представлять собой или предусматривать полноразмерный иммуноглобулин, Fc-функцию Fc-области полноразмерного иммуноглобулина по отношению к положительной по Fc-рецептору клетке можно снизить или полностью подавить посредством белковой инженерии, в то же время слитый белок одновременно будет воздействовать на CD137 и GPC3. В некоторых вариантах осуществления этого можно достичь, например, путем замены основной цепи IgG1 на IgG4, поскольку известно, что IgG4 проявляет сниженные взаимодействия Fc-гамма-рецептора по сравнению с IgG1. В некоторых вариантах осуществления для дополнительного снижения остаточного связывания с Fc-гамма-рецепторами в остов IgG4 можно вводить мутации, такие как F234A и L235A. В некоторых вариантах осуществления для минимизации замены полуантитела IgG4 также в остов IgG4 можно водить мутацию S228P (Silva et al., J Biol Chem, 2015). В некоторых вариантах осуществления можно водить мутации F234A и L235A для снижения ADCC и ADCP (Glaesner et al., Diabetes Metab Res Rev, 2010), и/или мутации M428L и N434S, или мутации M252Y, S254T и T256E для продления периода полужизни в сыворотке (Dall'Acqua et al., J Biol Chem, 2006, Zalevsky et al., Nat Biotechnol, 2010). В некоторых вариантах осуществления для удаления природного мотива гликозилирования в тяжелой цепи иммуноглобулина в слитом белке может присутствовать дополнительная мутация N297A.
[0093] В некоторых вариантах осуществления Fc-часть иммуноглобулина, включенная в слитый белок по настоящему изобретению, может способствовать поддержанию сывороточных уровней слитого белка. Например, при связывании Fc-части с Fc-рецепторами на эндотелиальных клетках и фагоцитах слитый белок может интернализироваться и возвращаться обратно в кровоток, что увеличивает его период полужизни в организме.
[0094] В одном аспекте слитые белки по настоящему изобретению с высокой аффинностью связывают CD137. В другом аспекте предусмотренные слитые белки с высокой аффинностью связывают GPC3. В некоторых предпочтительных вариантах осуществления предусмотренные слитые белки одновременно связывают CD137 и GPC3. В некоторых вариантах осуществления одновременное связывание с CD137 и GPC3 позволяет предусмотренным слитым белкам проявлять стойкий противоопухолевый или противоинфекционный ответ.
[0095] В некоторых вариантах осуществления слитый белок по настоящему изобретению может иметь изоэлектрическую точку (pI) приблизительно 6,5 или выше, как, например, приблизительно 6,8 или выше, приблизительно 7,1 или выше, приблизительно 7,4 или выше, приблизительно 7,5 или выше, приблизительно 7,7 или выше, приблизительно 8,0 или выше, приблизительно 8,5 или выше или приблизительно 9,0 или еще выше. В некоторых вариантах осуществления слитый белок по настоящему изобретению может иметь pI выше, чем у уже известного слитого белка, который связывает как CD137, так и GPC3 и содержит аминокислотную последовательность SEQ ID NO: 83.
[0096] В некоторых вариантах осуществления слитый белок по настоящему изобретению может быть способен связывать GPC3 со значением KD, которое составляет максимум приблизительно 2 нМ или еще ниже, как, например, приблизительно 1,5 нМ или ниже, приблизительно 1 нМ или ниже, приблизительно 0,8 нМ или ниже или приблизительно 0,7 нМ или ниже. В некоторых вариантах осуществления слитый белок по настоящему изобретению может быть способен связывать GPC3 со значением KD, сравнимым со значением KD или ниже него у специфического в отношении GPC3 иммуноглобулина, включенного в такой слитый белок, такого как антитело с тяжелой и легкой цепями, представленными под SEQ ID NO: 81 и 82. Значения KD предусмотренных слитых белков можно измерить, например, в анализе поверхностного плазмонного резонанса (SPR), таком как SPR-анализ, который фактически описан в примере 2.
[0097] В некоторых вариантах осуществления слитый белок по настоящему изобретению может быть способен связывать GPC3 со значением EC50, которое составляет максимум приблизительно 1 нМ или еще ниже, такое как приблизительно 0,5 нМ или ниже, приблизительно 0,3 нМ или ниже, приблизительно 0,2 нМ или ниже, приблизительно 0,15 нМ или ниже или приблизительно 0,1 нМ или ниже. В некоторых вариантах осуществления слитый белок по настоящему изобретению может быть способен связывать GPC3 со значением EC50, сравнимым со значением EC50 или ниже него у специфического в отношении GPC3 иммуноглобулина, включенного в конкретный слитый белок, такого как антитело с тяжелой и легкой цепями, представленными под SEQ ID NO: 81 и 82. Значения EC50 предусмотренных слитых белков можно измерить, например, в твердофазном иммуноферментном анализе (ELISA) , таком как ELISA-анализ, который фактически описан в примере 3.
[0098] В некоторых вариантах осуществления слитый белок по настоящему изобретению может быть способен связывать CD137 со значением EC50, которое составляет максимум приблизительно 5 нМ или еще ниже, такое как приблизительно 3 нМ или ниже, приблизительно 2 нМ или ниже, приблизительно 1 нМ или ниже, приблизительно 0,5 нМ или ниже, приблизительно 0,2 нМ или ниже или приблизительно 0,1 нМ или ниже. Значения EC50 предусмотренных слитых белков можно измерить, например, в ELISA-анализе, таком как ELISA-анализ, который фактически описан в примере 3.
[0099] В некоторых вариантах осуществления слитый белок по настоящему изобретению может вступать в перекрестную реакцию с GPC3 яванского макака. В некоторых вариантах осуществления предусмотренный слитый белок может быть способен связывать GPC3 яванского макака со значением KD, которое составляет максимум приблизительно 2 нМ или еще ниже, такое как приблизительно 1,5 нМ или ниже, приблизительно 1 нМ или ниже, приблизительно 0,8 нМ или ниже или приблизительно 0,7 нМ или ниже. В некоторых вариантах осуществления слитый белок по настоящему изобретению может быть способен связывать GPC3 яванского макака со значением KD, сравнимым со значением KD или ниже него у специфического в отношении GPC3 иммуноглобулина, включенного в такой слитый белок, такого как антитело с тяжелой и легкой цепями, представленными под SEQ ID NO: 81 и 82. Значения KD предусмотренных слитых белков можно измерить, например, в SPR-анализе, таком как SPR-анализ, который фактически описан в примере 2. В некоторых вариантах осуществления предусмотренный слитый белок может быть способен связывать GPC3 яванского макака со значением EC50, которое составляет максимум приблизительно 1 нМ или еще ниже, как, например, приблизительно 0,5 нМ или ниже, как, например, приблизительно 0,2 нМ или ниже, приблизительно 0,1 нМ или ниже. Значения EC50 предусмотренных слитых белков можно измерить, например измерить в ELISA-анализе, таком как ELISA-анализ, который фактически описан в примере 3.
[00100] В некоторых вариантах осуществления слитый белок по настоящему изобретению может быть способен одновременно связывать CD137 и GPC3. В некоторых вариантах осуществления предусмотренный слитый белок может быть способен одновременно связывать CD137 и GPC3 со значением EC50, которое составляет максимум приблизительно 10 нМ или еще ниже, как, например, 5 нм или ниже, 3 нМ или ниже, 2 нМ или ниже, 1 нМ или ниже или 0,5 нМ или ниже. В некоторых других вариантах осуществления предусмотренный слитый белок может быть способен одновременно связывать CD137 и GPC3 со значением EC50, которое составляет максимум приблизительно 10 нМ или еще ниже, как, например, 8 нМ или ниже, 5 нМ или ниже, 3 нМ или ниже или 2 нМ или ниже. Одновременное связывание можно определить, например, в ELISA-анализе, таком как ELISA-анализ, который фактически описан в примере 4.
[00101] В некоторых вариантах осуществления слитый белок по настоящему изобретению может быть способен связывать CD137, экспрессируемый на клетке, со значением EC50, которое составляет максимум приблизительно 50 нМ или еще ниже, как, например, приблизительно 30 нМ или еще ниже, приблизительно 25 нМ или еще ниже, приблизительно 20 нМ или ниже, приблизительно 15 нМ или ниже, приблизительно 10 нМ или ниже, приблизительно 5 нМ или ниже, приблизительно 3 нМ или ниже или приблизительно 1 нМ или еще ниже. Значение EC50 предусмотренного слитого белка можно измерить, например, в анализе способом проточной цитометрии, который фактически описан в примере 6. Клетка, экспрессирующая CD137, может быть, например, клеткой СНО, трансфицированной CD137 человека.
[00102] В некоторых вариантах осуществления слитый белок по настоящему изобретению может быть способен связывать GPC3, экспрессируемый на клетке, со значением EC50, которое составляет максимум приблизительно 50 нМ или еще ниже, как, например, приблизительно 30 нМ или ниже, приблизительно 25 нМ или еще ниже, приблизительно 20 нМ или ниже, приблизительно 15 нМ или ниже, приблизительно 10 нМ или ниже, приблизительно 5 нМ или ниже, приблизительно 3 нМ или ниже или приблизительно 1 или еще ниже. Значение EC50 предусмотренного слитого белка можно измерить, например, в анализе способом проточной цитометрии, который фактически описан в примере 6. Клетка, экспрессирующая GPC3, может быть, например, клеткой SK-Hep1, трансфицированной GPC3 человека.
[00103] В некоторых вариантах осуществления слитые белки по настоящему изобретению могут быть способны связывать GPC3, экспрессируемый на опухолевых клетках. В некоторых вариантах осуществления предусмотренный слитый белок может быть способен связывать GPC3, экспрессируемый на опухолевой клетке, со значением EC50, которое составляет максимум приблизительно 50 нМ или еще ниже, как, например, приблизительно 30 нМ или ниже, приблизительно 25 нМ или еще ниже, приблизительно 20 нМ или ниже, приблизительно 15 нМ или ниже, приблизительно 10 нМ или ниже, приблизительно 5 нМ или ниже, приблизительно 3 нМ или ниже или приблизительно 1 или еще ниже. Значение EC50 слитого белка для связывания экспрессирующих GPC3 опухолевых клеток можно измерить, например, в анализе способом проточной цитометрии, который фактически описан в примере 7. Экспрессирующие GPC3 опухолевые клетки могут быть, например, клетками HepG2, Hep3B и MKN-45.
[00104] В некоторых вариантах осуществления слитые белки по настоящему изобретению практически не связывают мишени, отличные от CD137 или GPC3. В некоторых вариантах осуществления предусмотренный слитый белок имеет пониженное нецелевое связывание с мишенями, отличными от CD137 или GPC3, по сравнению с таковым у уже известного слитого белка, который связывает как CD137, так и GPC3 и содержит аминокислотную последовательность SEQ ID NO: 83. Такую оценку целевого связывания можно провести с помощью ELISA-анализа, как описано в примере 5.
[00105] В некоторых вариантах осуществления слитые белки по настоящему изобретению могут быть способны совместно стимулировать Т-клеточные ответы. В некоторых вариантах осуществления предусмотренные слитые белки приводят к сравнимой или более сильной активации Т-клеток по сравнению с антителом к GPC3, включенным в слитый белок, имеющим тяжелую и легкую цепи, представленные под SEQ ID NO: 81 и 82, эталонным антителом к CD137, имеющим тяжелую и легкую цепи, представленные под SEQ ID NO: 26 и 27, или уже известным слитым белком, который связывает как CD137, так и GPC3, содержащим аминокислотную последовательность под SEQ ID NO: 83. В некоторых вариантах осуществления предусмотренные слитые белки приводят к активации Т-клеток с сопоставимой или лучшей эффективностью по сравнению с антителом к GPC3, включенным в слитый белок, имеющим тяжелую и легкую цепи, представленные под SEQ ID NO: 81 и 82, эталонным антителом к CD137, имеющим тяжелую и легкую цепи, представленные под SEQ ID NO: 26 и 27, или уже известным слитым белком, который связывает как CD137, так и GPC3, содержащим аминокислотную последовательность под SEQ ID NO: 83. Стимулируемые Т-клеточный ответ или активация Т-клеток можно измерить, например, в биоанализе CD137, который фактически описан в примере 8, или в функциональном анализе активации Т-клеток, который фактически описан в примере 9, примере 10 и примере 11.
[00106] В некоторых вариантах осуществления слитые белки по настоящему изобретению могут быть способны индуцировать повышенную секрецию IL-2. В некоторых предпочтительных вариантах осуществления предусмотренные слитые белки могут индуцировать зависимую от концентрации секрецию IL-2 и/или при более высоких концентрациях характеризуются тенденцией к индукции усиленной секреции IL-2. В некоторых вариантах осуществления предусмотренные слитые белки могут приводить к повышенной секреции IL-2 с сопоставимой или лучшей эффективностью по сравнению с антителом к GPC3, входящим в состав слитого белка, имеющим тяжелую и легкую цепи, представленные под SEQ ID NO: 81 и 82, или уже известным слитым белком, который связывает как CD137, так и GPC3, содержащим аминокислотную последовательность под SEQ ID NO: 83. Секрецию IL-2 можно измерить, например, в функциональном анализе активации Т-клеток, который фактически описан в примере 9 и примере 10.
[00107] В некоторых вариантах осуществления слитые белки по настоящему изобретению могут быть способны индуцировать опосредованную Т-клетками цитотоксичность. В некоторых вариантах осуществления предусмотренные слитые белки могут приводить к дозозависимому опосредованному Т-клетками цитолизу. В некоторых вариантах осуществления предусмотренные слитые белки могут индуцировать активацию цитотоксических Т-клеток с сопоставимой или лучшей эффективностью по сравнению с антителом к GPC3, включенным в предусмотренные слитые белки, имеющим тяжелую и легкую цепи, представленные под SEQ ID NO: 81 и 82. В некоторых вариантах осуществления предусмотренные слитые белки могут индуцировать активацию цитотоксических Т-клеток с сопоставимой или лучшей эффективностью по сравнению с антителом к CD137, имеющим тяжелую и легкую цепи, представленные под SEQ ID NO: 26 и 27, эталонным антителом к CD137, имеющим тяжелую и легкую цепи, представленные под SEQ ID NO: 26 и 27, или уже известным слитым белком, который связывает как CD137, так и GPC3, содержащим аминокислотную последовательность под SEQ ID NO: 83. Опосредованную Т-клетками цитотоксичность можно определить, например, с помощью импедансного анализа Т-клеточного уничтожения, который фактически описан в примере 11.
[00108] В некоторых вариантах осуществления слитые белки по настоящему изобретению могут быть способны совместно стимулировать Т-клеточные ответы зависимым от GPC3 образом. В некоторых вариантах осуществления предусмотренные слитые белки могут приводить к локальной индукции продуцирования IL-2 Т-клетками поблизости от GPC3-положительных клеток, таких как клетки, трансфицированные посредством GPC3, или опухолевые клетки, экспрессирующие GPC3. «Поблизости от GPC3-положительных клеток» в контексте данного документа относится к Т-клетке и GPC3-положительной клетке, которые приведены в близкое друг относительно друга местоположение посредством предусмотренного слитого белка, который одновременно связывает CD137 и GPC3. В некоторых вариантах осуществления предусмотренные слитые белки могут приводить к локализованному опосредованному Т-клетками уничтожению опухолевых клеток, экспрессирующих GPC3. GPC3-зависимую активацию Т-клеток предусмотренными слитыми белками можно измерить, например, в биоанализе CD137, который фактически описан в примере 8, или в функциональном анализе активации Т-клеток, который фактически описан в примере 9, примере 10 и примере 11.
[00109] В некоторых вариантах осуществления предусмотренные слитые белки не способны совместно стимулировать Т-клеточные ответы в отсутствие GPC3. В некоторых вариантах осуществления предусмотренные слитые белки не способны совместно стимулировать Т-клеточные ответы в отсутствие клеток, экспрессирующих GPC3. В некоторых вариантах осуществления предусмотренный слитый белок может распознавать присутствие GPC3 и приводить к соответствующей активации Т-клеток лучше, чем антитело к CD137, имеющее тяжелую и легкую цепи, представленные под SEQ ID NO: 26 и 27. GPC3-зависимое действие слитых белков можно измерить, например, в биоанализе CD137, который фактически описан в примере 8, или в функциональном анализе активации Т-клеток, который фактически описан в примере 9, примере 10 и примере 11.
[00110] В некоторых вариантах осуществления предусмотренные слитые белки могут приводить к формированию разновидностей противоопухолевой активности. В некоторых вариантах осуществления предусмотренные слитые белки могут ингибировать опухолевый рост клеток HepG2 гепатоклеточной карциномы. В некоторых вариантах осуществления предусмотренные слитые белки могут ингибировать опухолевый рост клеток HepG2 гепатоклеточной карциномы с сопоставимой или лучшей эффективностью по сравнению с антителом к GPC3, включенным в слитые белки, имеющим тяжелую и легкую цепи, представленные под SEQ ID NO: 81 и 82. Такие разновидности противоопухолевой активности можно определить, например, с помощью ксенотрансплантатной модели HepG2, как фактически описано в примере 12.
[00111] В некоторых вариантах осуществления предусмотренные слитые белки могут повышать уровни противоопухолевых эффекторных лимфоцитов. В некоторых вариантах осуществления предусмотренные слитые белки могут приводить к повышению внутриопухолевой инфильтрации CD3, CD4 или CD8 T-клеток по сравнению с антителом к GPC3, включенным в слитые белки, имеющие тяжелую и легкую цепи, представленные под SEQ ID NO: 81 и 82. Противоопухолевые эффекторные лимфоциты можно проанализировать, например, так, как описано в примере 13.
[00112] В некоторых вариантах осуществления предусмотренные слитые белки характеризуются благоприятными профилями стабильности и фармакокинетики. В некоторых вариантах осуществления предусмотренный слитый белок характеризуется профилем фармакокинетики, сопоставимым в сравнении с таковым у антитела к GPC3, включенного в слитый белок, имеющего тяжелую и легкую цепи, представленные под SEQ ID NO: 81 и 82. В некоторых вариантах осуществления предусмотренный слитый белок имеет фармакокинетику, подобную таковой у антитела. В некоторых вариантах осуществления предусмотренный слитый белок имеет конечный период полужизни приблизительно 50 часов или дольше, 75 часов или дольше, 100 часов или дольше, 125 часов или дольше, приблизительно 150 часов или дольше, приблизительно 175 часов или дольше, приблизительно 200 часов или дольше, приблизительно 250 часов или дольше или еще дольше. В некоторых вариантах осуществления предусмотренный слитый белок имеет конечный период полужизни у мышей приблизительно 50 часов или дольше, 75 часов или дольше, 100 часов или дольше, 125 часов или дольше, примерно 150 часов или дольше, приблизительно 175 часов или дольше, приблизительно 200 часов или дольше, приблизительно 250 часов или дольше или еще дольше. Профили фармакокинетики предусмотренных слитых белков можно анализировать так, как описано в примере 13.
[00113] В некоторых вариантах осуществления предусмотренный слитый белок содержит аминокислотную последовательность, представленную под любым из SEQ ID NO: 87-96.
[00114] В некоторых вариантах осуществления предусмотренный слитый белок содержит аминокислотную последовательность, обладающую по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 92%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 98% или еще более высокой идентичностью последовательности с аминокислотными последовательностями, представленными под любым из SEQ ID NO: 87-96.
[00115] В некоторых вариантах осуществления предусмотренный слитый белок содержит аминокислотные последовательности, представленные под SEQ ID NO: 87 и 82, SEQ ID NO: 88 и 82, SEQ ID NO: 81 и 89, SEQ ID NO: 81 и 90, SEQ ID NO: 91 и 82, SEQ ID NO: 92 и 82, SEQ ID NO: 81 и 93, SEQ ID NO: 81 и 94, SEQ ID NO: 95 и 82 или SEQ ID NO: 96 и 82.
[00116] В некоторых вариантах осуществления предусмотренный слитый белок содержит аминокислотные последовательности, обладающие по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 92%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 98% или еще более высокой идентичностью последовательности с аминокислотными последовательностями, представленными под SEQ ID NO: 87 и 82, SEQ ID NO: 88 и 82, SEQ ID NO: 81 и 89, SEQ ID NO: 81 и 90, SEQ ID NO: 91 и 82, SEQ ID NO: 92 и 82, SEQ ID NO: 81 и 93, SEQ ID NO: 81 и 94, SEQ ID NO: 95 и 82 или SEQ ID NO: 96 и 82.
B. Иллюстративные иммуноглобулины, включенные в слитые белки
[00117] В некоторых вариантах осуществления, что касается предусмотренного слитого белка, первая субъединица может представлять собой или предусматривать полноразмерный иммуноглобулин или его антигенсвязывающий домен, специфический в отношении GPC3. В некоторых вариантах осуществления иммуноглобулин, например, может представлять собой IgG1, IgG2 или IgG4. В некоторых вариантах осуществления иммуноглобулин представляет собой или предусматривает IgG4. В некоторых вариантах осуществления иммуноглобулин представляет собой моноклональное антитело к GPC3.
[00118] Иллюстративные примеры GPC3-связывающих антител по настоящему изобретению могут включать антигенсвязывающую область, которая перекрестно блокирует или связывается с тем же эпитопом, что и GPC3-связывающее антитело, содержащее области вариабельного домена тяжелой цепи (VH) и вариабельного домена легкой цепи (VL) такого известного антитела, как кодритузумаб (также известное как GC33 или RO5137382), YP7 (в том числе гуманизированное YP7), HN3 и HS20. В некоторых вариантах осуществления GPC3-связывающее антитело по настоящему изобретению может содержать антигенсвязывающую область, такую как любую из трех определяющих комплементарность областей тяжелой цепи (CDR) (HCDR1, HCDR2 и HCDR3) и трех CDR легкой цепи (LCDR1, LCDR2 и LCDR3) из антитела, выбранного из группы, состоящей из кодритузумаба, YP7, HN3 и HS20.
[00119] В некоторых вариантах осуществления предусмотренное антитело к GPC3 или его антигенсвязывающий домен может иметь вариабельную область тяжелой цепи (HCVR), выбранную из группы, состоящей из SEQ ID NO: 78, 114, 119, 126 и 129, и/или вариабельную область легкой цепи (LCVR), выбранную из группы, состоящей из SEQ ID NO: 79, 115 и 127.
[00120] В некоторых вариантах осуществления пара тяжелой цепи и легкой цепи предусмотренного антитела к GPC3 или его антигенсвязывающего домена представляют собой или содержат соответственно следующие HCVR и LCVR: SEQ ID NO: 78 и 79, SEQ ID NO: 129 и 79, SEQ ID NO: 114 и 115 или SEQ ID NO: 126 и 127.
[00121] В некоторых вариантах осуществления пара тяжелой цепи и легкой цепи предусмотренного антитела к GPC3 или его антигенсвязывающего домена представляет собой или содержит соответственно HCVR и LCVR, которые имеют последовательность, имеющую по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 92%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 98% или более высокую идентичность последовательности с аминокислотными последовательностями, представленными под SEQ ID NO: 78 и 79, SEQ ID NO: 129 и 79, SEQ ID NO: 114 и 115 или SEQ ID NO: 126 и 127.
[00122] В некоторых вариантах осуществления предусмотренное антитело к GPC3 или его антигенсвязывающий домен может иметь тяжелую цепь, которая представлена под любым из SEQ ID NO: 80 и 81, и/или легкую цепь, которая представлена под SEQ ID NO: 82.
[00123] В некоторых вариантах осуществления пара тяжелой цепи и легкой цепи предусмотренного антитела к GPC3 представляет собой или содержит аминокислотные последовательности, представленные под SEQ ID NO: 80 и 82 или SEQ ID NO: 81 и 82.
[00124] В некоторых вариантах осуществления пара тяжелой цепи и легкой цепи предусмотренного антитела к GPC3 представляет собой или содержит тяжелую цепь и легкую цепь, которые имеют последовательность, имеющую по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 92%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 98% или более высокую идентичность последовательности с аминокислотными последовательностями, представленными под SEQ ID NO: 80 и 82 или SEQ ID NO: 81 и 82.
[00125] В некоторых вариантах осуществления предусмотренное антитело к GPC3 или его антигенсвязывающий домен может иметь HCVR с по меньшей мере 70%, с по меньшей мере 75%, с по меньшей мере 80%, с по меньшей мере 85%, с по меньшей мере 90%, с по меньшей мере 92%, с по меньшей мере 95%, с по меньшей мере 97%, с по меньшей мере 98% или еще более высокой идентичностью последовательности с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 78, 114, 119, 126 и 129, и/или LCVR с по меньшей мере 70%, с по меньшей мере 75%, с по меньшей мере 80%, с по меньшей мере 85%, с по меньшей мере 90%, с по меньшей мере 92%, с по меньшей мере 95%, с по меньшей мере 97%, с по меньшей мере 98% или еще более высокой идентичностью последовательности с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 79, 115 и 127. В других вариантах осуществления предусмотренное антитело к GPC3 или его антигенсвязывающий домен может иметь тяжелую цепь с по меньшей мере 70%, с по меньшей мере 75%, с по меньшей мере 80%, с по меньшей мере 85%, с по меньшей мере 90%, с по меньшей мере 92%, с по меньшей мере 95%, с по меньшей мере 97%, с по меньшей мере 98% или еще более высокой идентичностью последовательности с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 80 и 81, и/или легкую цепь с по меньшей мере 70%, с по меньшей мере 75%, с по меньшей мере 80%, с по меньшей мере 85%, с по меньшей мере 90%, с по меньшей мере 92%, с по меньшей мере 95%, с по меньшей мере 97%, с по меньшей мере 98% или еще более высокой идентичностью последовательности с аминокислотной последовательностью под SEQ ID NO: 82.
[00126] В некоторых вариантах осуществления вариабельная область тяжелой цепи предусмотренного антитела к GPC3 или его антигенсвязывающего домена может иметь три CDR, имеющие следующие последовательности: GYTFTDYE (HCDR1, SEQ ID NO: 72), LDPKTGDT (HCDR2, SEQ ID NO: 73), TRFYSYTY (HCDR3; SEQ ID NO: 74). В некоторых вариантах осуществления вариабельная область тяжелой цепи предусмотренного антитела к GPC3 или его антигенсвязывающего домена может иметь три CDR, имеющие следующие последовательности: GFTFNKNA (HCDR1, SEQ ID NO: 108), IRNKTNNYAT (HCDR2, SEQ ID NO: 109), VAGNSFAY (HCDR3; SEQ ID NO: 110). В некоторых вариантах осуществления вариабельная область тяжелой цепи предусмотренного антитела к GPC3 или его антигенсвязывающего домена может иметь три CDR, имеющие следующие последовательности: YFDFDSYE (HCDR1, SEQ ID NO: 116), IYHSGST (HCDR2, SEQ ID NO: 117), ARVNMDRFDY (HCDR3; SEQ ID NO: 118). В некоторых вариантах осуществления вариабельная область тяжелой цепи предусмотренного антитела к GPC3 или его антигенсвязывающего домена может иметь три CDR, имеющие следующие последовательности: GFTFSSYA (HCDR1, SEQ ID NO: 120), IQKQGLPT (HCDR2, SEQ ID NO: 121), AKNRAKFDY (HCDR3; SEQ ID NO: 122).
[00127] В некоторых вариантах осуществления вариабельная область легкой цепи предусмотренного антитела к GPC3 или его антигенсвязывающего домена может иметь три CDR, имеющие следующие последовательности: QSLVHSNRNTY (LCDR1, SEQ ID NO: 75), KVS (LCDR2), SQNTHVPPT (LCDR3; SEQ ID NO: 77). В некоторых вариантах осуществления вариабельная область легкой цепи предусмотренного антитела к GPC3 или его антигенсвязывающего домена может иметь три CDR, имеющие следующие последовательности: QSLLYSSNQKNY (LCDR1, SEQ ID NO: 111), WAS (LCDR2), QQYYNYPLT (LCDR3; SEQ ID NO: 113). В некоторых вариантах осуществления вариабельная область легкой цепи предусмотренного антитела к GPC3 или его антигенсвязывающего домена может иметь три CDR, имеющие следующие последовательности: QSISSY (LCDR1, SEQ ID NO: 123), NAS (LCDR2), QQNRGFPLT (LCDR3; SEQ ID NO: 125).
[00128] В некоторых вариантах осуществления предусмотренное антитело к GPC3 или его антигенсвязывающий домен содержит вариабельную область тяжелой цепи, которая имеет три CDR, имеющие следующие последовательности: GYTFTDYE (HCDR1, SEQ ID NO: 72), LDPKTGDT (HCDR2, SEQ ID NO: 73), TRFYSYTY (HCDR3; SEQ ID NO: 74), и вариабельную область легкой цепи, которая имеет три CDR, имеющие следующие последовательности: QSLVHSNRNTY (LCDR1, SEQ ID NO: 75), KVS (LCDR2), SQNTHVPPT (LCDR3; SEQ ID NO: 77). В некоторых вариантах осуществления предусмотренное антитело к GPC3 или его антигенсвязывающий домен содержит вариабельную область тяжелой цепи, которая имеет три CDR, имеющие следующие последовательности: GFTFNKNA (HCDR1, SEQ ID NO: 108), IRNKTNNYAT (HCDR2, SEQ ID NO: 109), VAGNSFAY (HCDR3; SEQ ID NO: 110), и вариабельную область легкой цепи, которая имеет три CDR, имеющие следующие последовательности: QSLLYSSNQKNY (LCDR1, SEQ ID NO: 111), WAS (LCDR2), QQYYNYPLT (LCDR3; SEQ ID NO: 113). В некоторых вариантах осуществления предусмотренное антитело к GPC3 или его антигенсвязывающий домен содержит вариабельную область тяжелой цепи, которая имеет три CDR, имеющие следующие последовательности: GFTFSSYA (HCDR1, SEQ ID NO: 120), IQKQGLPT (HCDR2, SEQ ID NO: 121), AKNRAKFDY (HCDR3; SEQ ID NO: 122), и вариабельную область легкой цепи, которая имеет три CDR, имеющие следующие последовательности: QSISSY (LCDR1, SEQ ID NO: 123), NAS (LCDR2), QQNRGFPLT (LCDR3; SEQ ID NO: 125).
[00129] Если не указано иное, то все раскрываемые в данном документе последовательности CDR определены в соответствии со способом IMGT, который описан в Lefranc, M.-P., The Immunologist, 7, 132-136 (1999). CDR1 состоит из положений 27-38, CDR2 состоит из положений 56-65, CDR3 для генов V зародышевой линии состоит из положений 105-116, CDR3 для перестроенных V-J-генов или V-D-J-генов состоит из положений 105-117 (положение, предшествующее J-PHE или J-TRP в положении 118) с промежутками в верхней части петли для перестроенного CDR3-IMGT с менее чем 13 аминокислотами или с дополнительными положениями 112.1, 111.1, 112.2, 111.2 и т.д. для перестроенного CDR3-IMGT с более чем 13 аминокислотами. Положения, приведенные в данном абзаце, соответствуют нумерации IMGT, описанной в Lefranc, M.-P., The Immunologist, 7, 132-136 (1999).
[00130] Антитела, специфически связывающиеся с GPC3, включенные в слитые белки по настоящему изобретению, могут содержать Fc-часть, которая позволяет увеличивать in vivo период полужизни биспецифической связывающей молекулы по настоящему изобретению. В некоторых вариантах осуществления такая Fc-часть предпочтительно имеет человеческое происхождение, более предпочтительно представляет собой человеческую Fc-часть антитела IgG1 или lgG4, еще более предпочтительно сконструированную человеческую Fc-часть IgG1 или lgG4 с активацией или сайленсингом эффекторных функций. В некоторых вариантах осуществления сайленсинг эффекторных функций может быть предпочтительнее активации эффекторных функций. В некоторых вариантах осуществления такая Fc-часть модифицирована способами инженерии для сайленсинга эффекторных функций посредством мутации(мутаций) в положениях 234 и/или 235, нумерация которых соответствует EU-индексу по Kabat (Johnson and Wu, Nucleic Acids Res, 2000). В некоторых вариантах осуществления мутации для сайленсинга эффекторных функций можно ввести в положениях F234 и L235 предусмотренного антитела к GPC3. В других вариантах осуществления мутации для сайленсинга эффекторной функции можно ввести в положениях D265 и P329 предусмотренного антитела к GPC3. Нумерация обоих наборов данных потенциальных мутаций соответствует EU-индексу по Kabat (Shields et al., J Biol Chem, 2001).
[00131] Различные методики получения антител и их фрагментов хорошо известны из уровня техники и описаны, например, в Altshuler et al. (2010). Так, например, поликлональные антитела можно получать из крови животного после иммунизации антигеном в смеси с добавками и адъювантами, а моноклональные антитела можно получать с помощью любой методики, которая позволяет получить антитела, продуцируемые непрерывными культурами клеточных линий. Примеры таких методик описаны, например, в Harlow and Lane (1999), (1988), и включают гибридомную методику, первоначально описанную Köhler и Milstein, 1975, триомную методику, гибридомную методику с В-клетками человека (см., например, Li et al., Proc Natl Acad Sci U S A, 2006, Kozbor and Roder, Immunol Today, 1983) и EBV-гибридомную методику получения человеческих моноклональных антител (Cole et al., Cancer Res, 1984). Более того, из моноклональных антител можно получить рекомбинантные антитела или их можно получить de novo с помощью различных способов дисплея, таких как фаговый, рибосомальный, mRNA- или клеточный дисплей. В некоторых вариантах осуществления подходящую систему для экспрессии рекомбинантных (гуманизированных) антител или их фрагментов можно выбрать, например, из линий клеток бактерий, дрожжей, насекомых, млекопитающих или трансгенных животных или растений (см., например, патент США № 6080560; Holliger and Hudson, Nat Biotechnol, 2005). Дополнительно, описанные способы получения одноцепочечных антител (см., в частности, патент США №4946778) можно адаптировать для получения одноцепочечных антител, специфических в отношении мишени по настоящему изобретению. Для увеличения эффективности фаговых антител можно применять поверхностный плазмонный резонанс, используемый в системе BIAcore.
C. Иллюстративные мутеины липокалина по настоящему изобретению
[00132] Липокалины представляют собой белковые связывающие молекулы, которые естественным образом в ходе эволюции развили в себе способность связывать лиганды. Липокалины встречаются во многих организмах, в том числе у позвоночных, насекомых, растений и бактерий. Представители семейства липокалиновых белков (Pervaiz and Brew, FASEB J, 1987), как правило, представляют собой небольшие секретируемые белки и имеют одну полипептидную цепь. Они характеризуются спектром различных свойств молекулярного распознавания: их связыванием с различными, преимущественно гидрофобными, небольшими молекулами (такими как ретиноиды, жирные кислоты, холестерины, простагландины, биливердины, феромоны, тастанты и одоранты) и их связыванием с конкретными рецепторам на клеточной поверхности, а также образованием с ними макромолекулярных комплексов. Хотя в прошлом их классифицировали преимущественно как транспортные белки, теперь стало ясно, что липокалины выполняют ряд физиологических функций. Сюда относится роль в транспорте ретинола, обонянии, передаче сигналов от феромонов и синтезе простагландинов. Липокалины также задействованы в регуляции иммунного ответа и опосредовании клеточного гомеостаза (рассмотрено, например, в Flower et al., Biochim Biophys Acta, 2000, Flower, Biochem J, 1996).
[00133] Липокалины обладают необычно низкими уровнями общей консервативности последовательности, зачастую со значениями идентичности последовательностей менее 20%. В противоположность этому, их общий паттерн фолдинга имеет высокую степень консервативности. Центральная часть структуры липокалина состоит из одного восьмицепочечного антипараллельного β-слоя, замкнутого на себя с образованием непрерывно связанного водородными связями β-бочонка. Данный β-бочонок образует центральную полость. Один конец бочонка стерически заблокирован N-концевым пептидным сегментом, который проходит через его дно, а также тремя пептидными петлями, соединяющими β-цепи. Другой конец β-бочонка открыт для растворителя и охватывает связывающийся с мишенью сайт, который образован четырьмя гибкими пептидными петлями (AB, CD, EF и GH). Именно разнообразие петель в оставшейся части жесткого каркаса липокалина приводит к возникновению множества различных вариантов связывания, каждый из которых способен приспособиться к мишеням разного размера, формы и химического характера (рассмотрено, например, в Skerra, Biochim Biophys Acta, 2000, Flower et al., Biochim Biophys Acta, 2000, Flower, Biochem J, 1996).
[00134] Мутеин липокалина согласно настоящему изобретению может быть мутеином любого липокалина. Примеры подходящих липокалинов (также иногда обозначаемых как «эталонный липокалин», «липокалин дикого типа», «эталонные белковые каркасы» или просто «каркасы»), из которых можно использовать мутеин, включают без ограничения липокалин слезы (липокалин-1, Tlc или белок железы фон Эбнера), ретинол-связывающий белок, простагландин-D-синтаза липокалинового типа нейтрофилов, β-лактоглобулин, билин-связывающий белок (BBP), аполипопротеин D (APOD), липокалин, ассоциированный с желатиназой нейтрофилов (NGAL), белок, родственный α2-микроглобулину (A2m), 24p3/утерокалин (24p3), белок 1 железы фон Эбнера (VEGP 1), белок 2 железы фон Эбнера (VEGP 2) и основной аллерген Can f 1 (ALL-1). В сопутствующих вариантах осуществления мутеин липокалина получен из группы липокалинов, состоящей из липокалина слезы человека (hTlc), липокалина, ассоциированного с желатиназой нейтрофилов человека (hNGAL), аполипопротеина D человека (hAPOD) и билин-связывающего белка из Pieris brassicae.
[00135] Аминокислотная последовательность мутеина липокалина согласно настоящему изобретению может обладать высокой идентичностью последовательности в сравнении с эталонным липокалином (или липокалином дикого типа), из которого он получен, например hTlc или hNGAL, по сравнению со значениями идентичности последовательности с другим липокалином (см. также выше). В данном общем контексте аминокислотная последовательность мутеина липокалина согласно настоящему изобретению по меньшей мере практически схожа с аминокислотной последовательностью соответствующего эталонного липокалина (липокалина дикого типа), при условии, что могут иметь место гэпы (определение которым дано в данном документе) при выравнивании, которые являются следствием добавления или удаления аминокислот. Соответствующая последовательность мутеина липокалина по настоящему изобретению, будучи практически схожей с последовательностями соответствующего эталонного липокалина (липокалина дикого типа), в некоторых вариантах осуществления обладает по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 82%, по меньшей мере 85%, по меньшей мере 87%, по меньшей мере 90% идентичности, в том числе по меньшей мере 95% идентичности с последовательностью соответствующего липокалина. В данном отношении мутеин липокалина по настоящему изобретению, конечно, может содержать описанные в данном документе замены, которые делают мутеин липокалина способным связываться с CD137.
[00136] Как правило, мутеин липокалина содержит один или более мутантных аминокислотных остатков, относительно аминокислотной последовательности липокалина дикого типа или эталонного липокалина, например hTlc и hNGAL, в четырех петлях на открытом конце, которые составляют лиганд-связывающий карман и определяют вход в лиганд-связывающий карман (см. выше). Как объяснено выше, данные области необходимы для определения специфичности связывания мутеина липокалина с требуемой мишенью. В некоторых вариантах осуществления мутеин липокалина по настоящему изобретению также может содержать области мутантных аминокислотных остатков за пределами четырех петель. В некоторых вариантах осуществления мутеин липокалина по настоящему изобретению может содержать один или более мутантных аминокислотных остатков в одной или более из трех пептидных петель (обозначаемых BC, DE и FG), соединяющих β-цепи на закрытом конце липокалина. В некоторых вариантах осуществления мутеин, полученный из липокалина слезы, липокалина NGAL или его гомолога, может иметь 1, 2, 3, 4 или более мутантных аминокислотных остатков в любом положении последовательности в N-концевой области и/или в трех пептидных петлях BC, DE и FG, расположенные на конце структуры β-бочонка, которая расположена напротив природного связывающего кармана липокалина. В некоторых вариантах осуществления мутеин, полученный из липокалина слезы, липокалина NGAL или его гомолога, может не иметь мутантных аминокислотных остатков в пептидной петле DE, расположенной на конце структуры β-бочонка, по сравнению с последовательностью дикого типа липокалина слезы.
[00137] В некоторых вариантах осуществления мутеин липокалина согласно настоящему изобретению может включать один или более, как, например, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 или даже больше мутантных аминокислотных остатков по сравнению с аминокислотной последовательностью соответствующего эталонного липокалина (липокалина дикого типа) при условии, что такой мутеин липокалина будет способен связываться с CD137. В некоторых вариантах осуществления мутеин липокалина по настоящему изобретению включает по меньшей мере два, в том числе 2, 3, 4, 5 или еще больше, мутантных аминокислотных остатка, где нативный аминокислотный остаток соответствующего эталонного липокалина (липокалина дикого типа) замещен на остаток аргинина.
[00138] Предусмотрены любые типы и количества мутаций, в том числе замен, делеций и вставок, при условии, что предусмотренный мутеин липокалина сохраняет свою способность связывать CD137, и/или он обладает идентичностью последовательности, составляющей по меньшей мере 60%, как, например, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, или более высокой идентичностью с аминокислотной последовательностью эталонного липокалина (липокалина дикого типа), например зрелого hTlc или зрелого hNGAL.
[00139] В некоторых вариантах осуществления замена представляет собой консервативную замену. В некоторых вариантах осуществления замена представляет собой неконсервативную замену или одну или более из приставленных ниже иллюстративных замен.
[00140] В частности, для определения того, отличается ли аминокислотный остаток из аминокислотной последовательности мутеина липокалина от такового у эталонного липокалина (липокалина дикого типа), соответствующего определенному положению в аминокислотной последовательности эталонного липокалина (липокалина дикого типа), специалист в данной области может использовать средства и способы, хорошо известные в уровне техники, например различные способы выравнивания вручную или с использованием компьютерных программ, таких как BLAST 2.0, что означает средство поиска основного локального выравнивания, или ClustalW, или любую другую подходящую программу, которая подходит для получения результатов выравнивания последовательностей. Соответственно, аминокислотная последовательность эталонного липокалина (липокалина дикого типа) может служить «рассматриваемой последовательностью» или «эталонной последовательностью», тогда как аминокислотная последовательность мутеина липокалина служит «последовательностью запроса» (см. также выше).
[00141] Консервативные замены обычно представляют собой следующие замены, перечисленные в соответствии с подлежащей мутированию аминокислотой, за каждой из которых следует одна или более замен, которые можно считать консервативными: Ala → Ser, Thr или Val; Arg → Lys, Gln, Asn или His; Asn → Gln, Glu, Asp или His; Asp → Glu, Gln, Asn или His; Gln → Asn, Asp, Glu или His; Glu → Asp, Asn, Gln или His; His → Arg, Lys, Asn, Gln, Asp или Glu; Ile → Thr, Leu, Met, Phe, Val, Trp, Tyr, Ala или Pro; Leu → Thr, Ile, Val, Met, Ala, Phe, Pro, Tyr или Trp; Lys → Arg, His, Gln или Asn; Met → Thr, Leu, Tyr, Ile, Phe, Val, Ala, Pro или Trp; Phe → Thr, Met, Leu, Tyr, Ile, Pro, Trp, Val или Ala; Ser → Thr, Ala или Val; Thr → Ser, Ala, Val, Ile, Met, Val, Phe, Pro или Leu; Trp → Tyr, Phe, Met, Ile или Leu; Tyr → Trp, Phe, Ile, Leu или Met; Val → Thr, Ile, Leu, Met, Phe, Ala, Ser или Pro. Также допустимы и другие замены, и их можно определить опытным путем или согласно другим известным консервативным или неконсервативным заменам. В качестве дополнительной ориентации каждая из следующих групп содержит аминокислоты, которые обычно можно взять для определения консервативных замен друг друга:
(a) аланин (Ala), серин (Ser), треонин (Thr), валин (Val);
(b) аспарагиновая кислота (Asp), глутаминовая кислота (Glu), глутамин (Gln), аспарагин (Asn), гистидин (His);
(c) аргинин (Arg), лизин (Lys), глутамин (Gln), аспарагин (Asn), гистидин (His);
(d) изолейцин (Ile), лейцин (Leu), метионин (Met), валин (Val), аланин (Ala), фенилаланин (Phe), треонин (Thr), пролин (Pro);
(e) изолейцин (Ile), лейцин (Leu), метионин (Met), фенилаланин (Phe), тирозин (Tyr), триптофан (Trp).
[00142] Если такие консервативные замены приводят к изменению биологической активности, то можно внести более существенные изменения, такие как представленные далее или дополнительно описанные ниже в отношении классов аминокислот, а продукты можно подвергнуть скринингу на предмет требуемой характеристики. Примерами таких более существенных изменений являются: Ala → Leu или Phe; Arg → Glu; Asn → Ile, Val или Trp; Asp → Met; Cys → Pro; Gln → Phe; Glu → Arg; His → Gly; Ile → Lys, Glu или Gln; Leu → Lys или Ser; Lys → Tyr; Met → Glu; Phe → Glu, Gln или Asp; Trp → Cys; Tyr → Glu или Asp; Val → Lys, Arg, His.
[00143] В некоторых вариантах осуществления существенные модификации физических и биологических свойств (мутеина) липокалина осуществляют путем выбора замен, которые значительно различаются по своему влиянию на поддержание (а) структуры полипептидного остова в области замены, например, в виде листовой или спиральной конформации, (b) заряда или гидрофобности молекулы в целевом сайте или (c) объемных характеристик боковой цепи.
[00144] Встречающиеся в природе остатки подразделяют на группы, исходя из общих свойств боковой цепи: (1) гидрофобные: метионин, аланин, валин, лейцин, изолейцин; (2) нейтральные гидрофильные: цистеин, серин, треонин, аспарагин, глутамин; (3) кислые: аспарагиновая кислота, глутаминовая кислота; (4) основные: гистидин, лизин, аргинин; (5) остатки, которые влияют на ориентацию цепи: глицин, пролин; и (6) ароматические: триптофан, тирозин, фенилаланин. В некоторых вариантах осуществления замены могут повлечь за собой замену представителя одного из данных классов на представителя другого класса.
[00145] Любой остаток цистеина, не участвующий в поддержании надлежащей конформации соответствующего липокалина, также можно заменить, обычно на серин, для улучшения окислительной стабильности молекулы и предупреждения ошибочного сшивания. И наоборот, в липокалин можно внести цистеиновую связь(цистеиновые связи) для улучшения его стабильности.
D. Иллюстративные CD137-специфические мутеины липокалина по настоящему изобретению
[00146] Как отмечалось выше, липокалин представляет собой полипептид, определяемый его сверхвторичной структурой, а именно областью со сверхвторичной структурой из цилиндрического β-складчатого листа, содержащей восемь β-цепей, попарно соединенных четырьмя петлями на одном конце, определяя таким образом связывающий карман. Настоящее изобретение не ограничено конкретно раскрытыми в данном документе мутеинами липокалина. В данном отношении настоящее изобретение относится к мутеину липокалина, имеющему область со сверхвторичной структурой из цилиндрического β-складчатого листа, содержащую восемь β-цепей, попарно соединенных посредством четырех петель на одном конце так, чтобы они в результате определяли связывающий карман, при этом по меньшей мере одна аминокислота каждой из по меньшей мере трех из указанных четырех петель была подвергнута мутации, и при этом указанный липокалин с поддающейся выявлению аффинностью эффективен в связывании с CD137.
[00147] В некоторых вариантах осуществления раскрытые в данном документе мутеины липокалина могут представлять собой или предусматривать мутеин зрелого липокалина слезы человека (hTlc). Мутеин зрелого hTlc в данном документе может быть обозначен как «мутеин hTlc». В некоторых других вариантах осуществления раскрываемый в данном документе мутеин липокалина представляет собой мутеин зрелого липокалина, ассоциированного с желатиназой нейтрофилов человека (hNGAL). Мутеин зрелого hNGAL в данном документе может быть обозначен как «мутеин hNGAL».
[00148] В одном аспекте настоящим изобретением предусмотрено любое количество мутеинов липокалина, полученных из эталонного липокалина (липокалина дикого типа), предпочтительно полученного из зрелого hTlc или зрелого hNGAL, которые с поддающейся выявлению аффинностью связывают CD137. В сопутствующем аспекте настоящим изобретением предусмотрены различные мутеины липокалина, которые способны активировать последующие сигнальные пути CD137 путем связывания с CD137. В этом смысле CD137 можно рассматривать в качестве неприродной мишени эталонного липокалина (липокалина дикого типа), предпочтительно hTlc или hNGAL, где «неприродная мишень» относится к веществу, которое не связывается с эталонными липокалинами (липокалинами дикого типа) в физиологических условиях. Посредством модификации способами инженерии эталонных липокалинов (липокалинов дикого типа) с внесением одной или более мутаций в определенных положениях последовательности авторы настоящего изобретения продемонстрировали, что возможна высокая аффинность и высокая специфичность в отношении неприродной мишени, CD137. В некоторых вариантах осуществления в 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 или даже большем количестве нуклеотидных триплетов, кодирующих определенные положения в последовательностях липокалинов дикого типа, можно подвергнуть случайному мутагенезу путем замены в данных положениях на подгруппу нуклеотидных триплетов с целью получения мутеина липокалина, способного связывать CD137.
[00149] В некоторых вариантах осуществления мутеины липокалина по настоящему изобретению могут иметь мутантный(-ые), в том числе замененный(-ые), удаленный(-ые) и вставленный(-ые) аминокислотный(-ые) остаток(остатки) в одном или более положениях последовательности, соответствующей линейной полипептидной последовательности эталонного липокалина, предпочтительно hTlc или hNGAL. В некоторых вариантах осуществления количество аминокислотных остатков мутеина липокалина по настоящему изобретению, которые являются мутантными в сравнении с аминокислотной последовательностью эталонного липокалина, предпочтительно hTlc или hNGAL, составляет 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 или более, как, например, 25, 30, 35, 40, 45 или 50, при этом предпочтительными являются 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или 11, а еще более предпочтительными являются 9, 10 или 11. Однако предпочтительно, чтобы мутеин липокалина по настоящему изобретению все еще был способен связывать CD137.
[00150] В некоторых вариантах осуществления мутеин липокалина по настоящему изобретению может не иметь 1, 2, 3, 4 или более аминокислот на своем N-конце и/или 1, 2 или более аминокислот на своем C-конце в сравнении с соответствующим эталонным липокалином (липокалином дикого типа); например, SEQ ID NO: 32-38. В некоторых вариантах осуществления настоящим изобретением охвачены мутеины hTlc, которые определены выше, у которых были удалены первые четыре, один, два, три N-концевых аминокислотных остатка последовательности зрелого hTlc (His-His-Leu-Leu; положения 1-4) и/или последние один или два C-концевых аминокислотных остатка (Ser-Asp; положения 157-158) линейной полипептидной последовательности зрелого hTlc (например, SEQ ID NO: 32-38). В некоторых вариантах осуществления настоящим изобретением охвачены мутеины hNGAL, которые определены выше, у которых удалены аминокислотные остатки (Lys-Asp-Pro, положения 46-48) линейной полипептидной последовательности зрелого hNGAL (SEQ ID NO: 43). Дополнительно, мутеин липокалина по настоящему изобретению может включать аминокислотную последовательность дикого типа (природную аминокислотную последовательность) эталонного липокалина (липокалина дикого типа), предпочтительно hTlc или hNGAL, вне мутантных положений аминокислотной последовательности.
[00151] В некоторых вариантах осуществления один или более мутантных аминокислотных остатков, включенных в мутеин липокалина по настоящему изобретению, практически не затрудняют или не препятствуют активности связывания с обозначенной мишенью и фолдингу мутеина. Такие мутации, в том числе замену, делецию и вставку, можно осуществить на уровне ДНК с помощью общепризнанных стандартных способов (Sambrook and Russell, 2001, Molecular cloning: a laboratory manual). В некоторых вариантах осуществления мутантный(-ые) аминокислотный(-ые) остаток (остатки) в одном или более положениях последовательности, соответствующих линейной полипептидной последовательности эталонного липокалина (липокалина дикого типа), предпочтительно hTlc или hNGAL, вводят посредством случайного мутагенеза путем замены нуклеотидного(-ых) триплета(-ов), кодирующего(-их) соответствующие положения последовательности эталонного липокалина, на подгруппу нуклеотидных триплетов.
[00152] В некоторых вариантах осуществления предусмотренный мутеин липокалина, который с поддающейся выявлению аффинностью связывает CD137, может включать по меньшей мере одну аминокислотную замену нативного остатка цистеина на другую аминокислоту, например остаток серина. В некоторых вариантах осуществления мутеин липокалина, который с поддающейся выявлению аффинностью связывает CD137, может включать один или более ненативных остатков цистеина, заменяющих одну или более аминокислот эталонного липокалина (липокалина дикого типа), предпочтительно hTlc или hNGAL. В некоторых вариантах осуществления мутеин липокалина согласно настоящему изобретению включает по меньшей мере две аминокислотные замены нативной аминокислоты на остаток цистеина с образованием в результате одного или более цистеиновых мостиков. В некоторых вариантах осуществления указанный цистеиновый мостик может соединять по меньшей мере две петлевые области. В данном документе используют определение данных областей согласно (Biochim Biophys Acta, 2000), Flower (1996) и Breustedt et al. (2005).
[00153] Как правило, мутеин липокалина по настоящему изобретению может обладать по меньшей мере 70%, в том числе по меньшей мере приблизительно 80%, как, например, по меньшей мере приблизительно 85%, идентичности аминокислотной последовательности с аминокислотной последовательностью зрелого hTlc (SEQ ID NO: 1) или зрелого hNGAL (SEQ ID NO: 2).
[00154] В некоторых аспектах настоящим изобретением предусмотрены связывающие CD137 мутеины hTlc. В данном отношении настоящим изобретением предусмотрен один или более мутеинов hTlc, которые способны связывать CD137 с аффинностью, измеряемой по KD, составляющей приблизительно 300 нМ, 200 нМ, 150 нМ, 100 нМ или ниже. В некоторых вариантах осуществления предусмотренные мутеины hTlc способны связывать CD137 со значением EC50, составляющим приблизительно 250 нМ, 150 нМ, 100 нМ, 50 нМ, 20 нМ или еще ниже. В некоторых других вариантах осуществления связывающие CD137 мутеины hTlc могут быть перекрестно реактивными с CD137 яванского макака (cyCD137).
[00155] В некоторых вариантах осуществления мутеин hTlc по настоящему изобретению может препятствовать связыванию CD137L с CD137.
[00156] В некоторых вариантах осуществления предусмотренные мутеины hTlc могут содержать мутантный аминокислотный остаток в одном или более положениях, соответствующих положениям 5, 26-31, 33-34, 42, 46, 52, 56, 58, 60-61, 65, 71, 85, 94, 101, 104-106, 108, 111, 114, 121, 133, 148, 150 и 153 линейной полипептидной последовательности зрелого hTlc (SEQ ID NO: 1).
[00157] В некоторых вариантах осуществления предусмотренные мутеины hTlc могут содержать мутантный аминокислотный остаток в одном или более положениях, соответствующих положениям 26-34, 55-58, 60-61, 65, 104-106 и 108 линейной полипептидной последовательности зрелого hTlc. (SEQ ID NO: 1).
[00158] В некоторых вариантах осуществления предусмотренные мутеины hTlc могут дополнительно содержать мутантный аминокислотный остаток в одном или более положениях, соответствующих положениям 101, 111, 114 и 153 линейной полипептидной последовательности зрелого hTlc (SEQ ID NO: 1).
[00159] В некоторых вариантах осуществления предусмотренные мутеины hTlc могут содержать мутантный аминокислотный остаток в 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26 или еще большем количестве положений, соответствующих положениям 5, 26-31, 33-34, 42, 46, 52, 56, 58, 60-61, 65, 71, 85, 94, 101, 104-106, 108, 111, 114, 121, 133, 148, 150 и 153 линейной полипептидной последовательности зрелого hTlc (SEQ ID NO: 1). В некоторых предпочтительных вариантах осуществления предусмотренные мутеины hTlc способны связывать CD137, в частности CD137 человека.
[00160] В некоторых вариантах осуществления предусмотренные мутеины hTlc могут содержать мутантный аминокислотный остаток в 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 или еще большем количестве положений, соответствующих положениям 26-34, 55-58, 60-61, 65, 104-106 и 108 линейной полипептидной последовательности зрелого hTlc. (SEQ ID NO: 1). В некоторых предпочтительных вариантах осуществления предусмотренные мутеины hTlc способны связывать CD137, в частности CD137 человека.
[00161] В некоторых вариантах осуществления мутеин липокалина согласно настоящему изобретению может включать по меньшей мере одну аминокислотную замену нативного остатка цистеина, например, на остаток серина. В некоторых вариантах осуществления мутеин hTlc согласно настоящему изобретению включает аминокислотную замену нативного остатка цистеина в положениях, соответствующих положениям 61 и/или 153 линейной полипептидной последовательности зрелого hTlc (SEQ ID NO:1), на другую аминокислоту, такую как остаток серина. В данном контексте следует отметить, что было обнаружено, что удаление структурной дисульфидной связи (на уровне соответствующей библиотеки интактных нуклеиновых кислот) hTlc дикого типа, которая образована остатками цистеина 61 и 153 (см. Breustedt et al., J Biol Chem, 2005), может привести к получению мутеинов hTlc, которые не только характеризуются стабильным фолдингом, но также способны с высокой аффинностью связывать заданную неприродную мишень. В некоторых вариантах осуществления устранение структурной дисульфидной связи может дать дополнительное преимущество, делающее возможным создание или преднамеренное введение в мутеины по настоящему изобретению неприродных дисульфидных связей, тем самым повышая стабильность мутеинов. Тем не менее, частью настоящего изобретения также являются мутеины hTlc, которые связывают CD137 и которые имеют дисульфидный мостик, образованный между Cys 61 и Cys 153.
[00162] В некоторых конкретных вариантах осуществления мутеин hTlc по настоящему изобретению может включать одну или более аминокислотных замен Cys 61 → Ala, Phe, Lys, Arg, Thr, Asn, Gly, Gln, Asp, Asn, Leu, Tyr, Met, Ser, Pro или Trp и/или Cys 153 → Ser или Ala в положениях, соответствующих положениям 61 и/или 153 линейной полипептидной последовательности зрелого hTlc (SEQ ID NO:1).
[00163] В некоторых вариантах осуществления два или все три кодона цистеина в положениях, соответствующих положениям 61, 101 и 153 линейной полипептидной последовательности зрелого hTlc (SEQ ID NO:1), заменены кодоном другой аминокислоты. Дополнительно, в некоторых вариантах осуществления мутеин hTlc согласно настоящему изобретению включает аминокислотную замену нативного остатка цистеина в положении, соответствующем положению 101 линейной полипептидной последовательности зрелого hTlc (SEQ ID NO:1), на остаток серина или остаток гистидина.
[00164] В некоторых вариантах осуществления мутеин согласно настоящему изобретению содержит аминокислотную замену нативной аминокислоты на остаток цистеина в положениях, соответствующих положениям 28 или 105 линейной полипептидной последовательности зрелого hTlc (SEQ ID NO: 1). Дополнительно, в некоторых вариантах осуществления мутеин согласно настоящему изобретению содержит аминокислотную замену нативного остатка аргинина в положении, соответствующем положению 111 линейной полипептидной последовательности зрелого hTlc (SEQ ID NO:1), на остаток пролина. Дополнительно, в некоторых вариантах осуществления мутеин согласно настоящему изобретению содержит аминокислотную замену нативного остатка лизина в положении, соответствующем положению 114 линейной полипептидной последовательности зрелого hTlc (SEQ ID NO:1), на остаток триптофана или глутаминовую кислоту.
[00165] В некоторых вариантах осуществления предусмотренные связывающие CD137 мутеины hTlc могут содержать в одном или более положениях, соответствующих положениям 5, 26-31, 33-34, 42, 46, 52, 56, 58, 60-61, 65, 71, 85, 94, 101, 104-106, 108, 111, 114, 121, 133, 148, 150 и 153 линейной полипептидной последовательности зрелого hTlc (SEQ ID NO: 1), один или более из следующих мутантных аминокислотных остатков: Ala 5 → Val или Thr; Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Thr 42 → Ser; Gly 46 → Asp; Lys 52 → Glu; Leu 56 →Ala; Ser 58 → Asp; Arg 60 →Pro; Cys 61 → Ala; Lys 65 → Arg или Asn; Thr 71 → Ala; Val 85 → Asp; Lys 94 → Arg или Glu; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Lys 121 → Glu; Ala 133 → Thr; Arg 148 → Ser; Ser 150 → Ile и Cys 153 → Ser. В некоторых вариантах осуществления мутеин hTlc по настоящему изобретению содержит два или более, как, например, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26 или более или даже все мутантные аминокислотные остатки в данных положениях последовательности зрелого hTlc (SEQ ID NO: 1).
[00166] В некоторых вариантах осуществления предусмотренные связывающие CD137 мутеины hTlc могут содержать один из следующих наборов мутантных аминокислотных остатков в сравнении с линейной полипептидной последовательностью зрелого hTlc (SEQ ID NO: 1):
(a) Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp и Cys 153 → Ser;
(b) Ala 5 → Thr; Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Lys 65 → Arg; Val 85 → Asp; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Lys 121 →Glu; Ala 133 → Thr и Cys 153 → Ser;
(c) Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Lys 65 → Asn; Lys 94 → Arg; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Lys 121 → Glu; Ala 133 → Thr и Cys 153 → Ser;
(d) Ala 5 → Val; Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Lys 65 → Arg; Lys 94 → Glu; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Lys 121 → Glu; Ala 133 → Thr и Cys 153 → Ser;
(e) Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 →Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Thr 42 → Ser; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Ser 150 → Ile и Cys 153 → Ser;
(f) Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Lys 52 → Glu; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Thr 71 → Ala; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Ala 133 → Thr; Arg 148 → Ser; Ser 150 → Ile и Cys 153 → Ser и
(g) Ala 5 → Thr; Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Gly 46 → Asp; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Thr 71 → Ala; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Ser 150 → Ile и Cys 153 → Ser.
[00167] В некоторых вариантах осуществления остальная область, т.е. область, отличающаяся от положений, соответствующих положениям 5, 26-31, 33-34, 42, 46, 52, 56, 58, 60-61, 65, 71, 85, 94, 101, 104-106, 108, 111, 114, 121, 133, 148, 150 и 153 линейной полипептидной последовательности зрелого hTlc (SEQ ID NO: 1) мутеина hTlc по настоящему изобретению, может содержать (природную аминокислотную последовательность) аминокислотную последовательность дикого типа линейной полипептидной последовательности зрелого hTlc вне положений мутантной аминокислотной последовательности.
[00168] В некоторых вариантах осуществления мутеин hTlc по настоящему изобретению обладает по меньшей мере 70% идентичности последовательности или по меньшей мере 70% гомологии последовательности с последовательностью зрелого hTlc (SEQ ID NO: 1). В качестве иллюстративного примера мутеин SEQ ID NO: 32 обладает идентичностью аминокислотной последовательности или гомологией последовательности, составляющей примерно 84%, с аминокислотной последовательностью зрелого hTlc.
[00169] В некоторых вариантах осуществления мутеин hTlc по настоящему изобретению содержит аминокислотную последовательность, изложенную под любым из SEQ ID NO: 32-38, или ее фрагмент или вариант.
[00170] В некоторых вариантах осуществления мутеин hTlc по настоящему изобретению обладает по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или более высокой идентичностью последовательности с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 32-38.
[00171] Настоящим изобретением также предусмотрены структурные гомологи мутеина hTlc, имеющего аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 32-38, при этом структурные гомологи обладают гомологией аминокислотных последовательностей или идентичностью последовательностей, составляющей более чем приблизительно 60%, предпочтительно более 65%, более 70%, более 75%, более 80%, более 85%, более 90%, более 92% и наиболее предпочтительно более 95% по отношению к указанному мутеину hTlc.
[00172] В некоторых аспектах настоящим изобретением предусмотрены CD137-связывающие hNGAL. В данном отношении настоящим изобретением предусмотрен один или более мутеинов hNGAL которые способны связывать CD137 с аффинностью, измеряемой по KD, составляющей приблизительно 800 нМ, 700 нМ, 200 нМ, 140 нМ, 100 нМ или ниже, предпочтительно приблизительно 70 нМ, 50 нМ, 30 нМ, 10 нМ, 5 нМ, 2 нМ или еще ниже. В некоторых вариантах осуществления предусмотренные мутеины hNGAL способны связывать CD137 со значением EC50, составляющим приблизительно 1000 нМ, 500 нМ, 100 нМ, 80 нМ, 50 нМ, 25 нМ, 18 нМ, 15 нМ, 10 нМ, 5 нМ или ниже.
[00173] В некоторых вариантах осуществления предусмотренные связывающие CD137 мутеины hNGAL могут быть перекрестно реактивными с CD137 яванского макака. В некоторых вариантах осуществления предусмотренные мутеины hNGAL способны связывать CD137 яванского макака с аффинностью, измеряемой с помощью KD, составляющей приблизительно 50 нМ, 20 нМ, 10 нМ, 5 нМ, 2 нМ или еще ниже. В некоторых вариантах осуществления предусмотренные мутеины hNGAL способны связывать CD137 яванского макака со значением EC50, составляющим приблизительно 100 нМ, 80 нМ, 50 нМ, 30 нМ или еще ниже.
[00174] В некоторых вариантах осуществления мутеин hNGAL по настоящему изобретению может препятствовать связыванию или конкурировать за связывание CD137L с CD137. В некоторых других вариантах осуществления мутеин hNGAL по настоящему изобретению может быть способен связывать CD137 в присутствии CD137L и/или связывать комплекс CD137/CD137L.
[00175] В некоторых вариантах осуществления предусмотренные мутеины hNGAL могут содержать мутантный аминокислотный остаток в одном или более положениях, соответствующих положениям 28, 36, 40-41, 49, 52, 65, 68, 70, 72-73, 77, 79, 81, 83, 87, 94, 96, 100, 103, 106, 125, 127, 132 и 134 линейной полипептидной последовательности зрелого hNGAL (SEQ ID NO: 2).
[00176] В некоторых вариантах осуществления предусмотренные мутеины hNGAL могут содержать мутантный аминокислотный остаток в 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21 или еще большем количестве положений, соответствующих положению 28, 36, 40-41, 49, 52, 65, 68, 70, 72-73, 77, 79, 81, 83, 87, 94, 96, 100, 103, 106, 125, 127, 132 и 134 линейной полипептидной последовательности зрелого hNGAL (SEQ ID NO: 2). В некоторых предпочтительных вариантах осуществления предусмотренные мутеины hNGAL способны связывать CD137, в частности CD137 человека.
[00177] В некоторых вариантах осуществления предусмотренные мутеины hNGAL могут содержать мутантный аминокислотный остаток в одном или более положениях, соответствующих положениям 28, 36, 40-41, 49, 52, 65, 68, 70, 72-73, 77, 79, 81, 87, 96, 100, 103, 106, 125, 127, 132 и 134 линейной полипептидной последовательности зрелого hNGAL (SEQ ID NO: 2). В некоторых предпочтительных вариантах осуществления предусмотренные мутеины hNGAL способны связывать CD137, в частности CD137 человека.
[00178] В некоторых вариантах осуществления предусмотренные мутеины hNGAL могут содержать мутантный аминокислотный остаток в одном или более положениях, соответствующих положениям 36, 87 и 96 линейной полипептидной последовательности зрелого hNGAL (SEQ ID NO: 2), и в одном или более положениях, соответствующих положениям 28, 40-41, 49, 52, 65, 68, 70, 72-73, 77, 79, 81, 83, 94, 100, 103, 106, 125, 127, 132 и 134 линейной полипептидной последовательности зрелого hNGAL (SEQ ID NO: 2).
[00179] В некоторых других вариантах осуществления предусмотренные мутеины hNGAL могут содержать мутантный аминокислотный остаток в одном или более положениях, соответствующих положениям 20, 25, 28, 33, 36, 40-41, 44, 49, 52, 59, 68, 70-73, 77-82, 87, 92, 96, 98, 100, 101, 103, 122, 125, 127, 132 и 134 линейной полипептидной последовательности зрелого hNGAL (SEQ ID NO: 2).
[00180] В других вариантах осуществления предусмотренные мутеины hNGAL могут содержать мутантный аминокислотный остаток в одном или более положениях, соответствующих положениям 36, 40, 41, 49, 52, 68, 70, 72, 73, 77, 79, 81, 96, 100, 103, 125, 127, 132 и 134 линейной полипептидной последовательности зрелого hNGAL (SEQ ID NO: 2), и в одном или более положениях, соответствующих положениям 20, 25, 33, 44, 59, 71, 78, 80, 82, 87, 92, 98, 101 и 122 линейной полипептидной последовательности зрелого hNGAL (SEQ ID NO: 2).
[00181] В некоторых вариантах осуществления мутеин липокалина согласно настоящему изобретению может содержать по меньшей мере одну аминокислотную замену нативного остатка цистеина, например, на остаток серина. В некоторых вариантах осуществления мутеин hNGAL согласно настоящему изобретению может содержать аминокислотную замену нативного остатка цистеина в положениях, соответствующих положениям 76 и/или 175 линейной полипептидной последовательности hNGAL (SEQ ID NO: 2), на другую аминокислоту, такую как остаток серина. В данном контексте следует отметить, что было обнаружено, что удаление структурной дисульфидной связи (на уровне соответствующей библиотеки интактных нуклеиновых кислот) hNGAL дикого типа, которая образована остатками цистеина 76 и 175 (см. Breustedt et al., J Biol Chem, 2005), может привести к получению мутеинов hNGAL, которые не только характеризуются стабильным фолдингом, но также способны с высокой аффинностью связывать заданную неприродную мишень. В некоторых вариантах осуществления устранение структурной дисульфидной связи может дать дополнительное преимущество, делающее возможным создание или преднамеренное введение в мутеины по настоящему изобретению неприродных дисульфидных связей, тем самым повышая стабильность мутеинов. Тем не менее, частью настоящего изобретения также являются мутеины hNGAL, которые связывают CD137 и которые имеют дисульфидный мостик, образованный между Cys 76 и Cys 175.
[00182] В некоторых вариантах осуществления предусмотренные связывающие CD137 мутеины hNGAL могут содержать в одном или более положениях, соответствующих положениям 28, 36, 40-41, 49, 52, 65, 68, 70, 72-73, 77, 79, 81, 83, 87, 94, 96, 100, 103, 106, 125, 127, 132 и 134 линейной полипептидной последовательности зрелого hNGAL (SEQ ID NO: 2), один или более из следующих мутантных аминокислотных остатков: Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Arg или Lys; Gln 49 → Val, Ile, His, Ser или Asn; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Met, Ala или Gly; Leu 70 → Ala, Lys, Ser или Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Met, Arg, Thr или Asn; Trp 79 → Ala или Asp; Arg 81 → Met, Trp или Ser; Phe 83 → Leu; Cys 87 → Ser; Leu 94 → Phe; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu и Lys 134 → Tyr. В некоторых вариантах осуществления мутеин hNGAL по настоящему изобретению содержит два или более, как, например, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, еще более, как, например, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или все мутантные аминокислотные остатки в данных положениях последовательности зрелого hNGAL (SEQ ID NO: 2).
[00183] В некоторых вариантах осуществления предусмотренные связывающие CD137 мутеины hNGAL могут содержать в одном или более положениях, соответствующих положениям 20, 25, 28, 33, 36, 40-41, 44, 49, 52, 59, 68, 70-73, 77-82, 87, 92, 96, 98, 100, 101, 103, 122, 125, 127, 132 и 134 линейной полипептидной последовательности зрелого hNGAL (SEQ ID NO: 2), один или более из следующих мутантных аминокислотных остатков: Gln 20 → Arg; Asn 25 → Tyr или Asp; Gln 28 → His; Val 33 → Ile; Leu 36 → Met; Ala 40 → Asn; Ile 41 → Leu; Glu 44 → Val или Asp; Gln 49 → His; Tyr 52 → Ser или Gly; Lys 59 → Asn; Ser 68 → Asp; Leu 70 → Met; Phe 71 → Leu; Arg 72 → Leu; Lys 73 → Asp; Asp 77 → Gln или His; Tyr 78 → His; Trp 79 → Ile; Ile 80 → Asn; Arg 81 → Trp или Gln; Thr 82 → Pro; Cys 87 → Ser; Phe 92 → Leu или Ser; Asn 96 → Phe; Lys 98 → Arg; Tyr 100 → Asp; Pro 101 → Leu; Leu 103 → His или Pro; Phe 122 → Tyr; Lys 125 → Ser; Ser 127 → Ile; Tyr 132 → Trp и Lys 134 → Gly. В некоторых вариантах осуществления мутеин hNGAL по настоящему изобретению содержит два или более, как, например, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33 или 34, мутантных аминокислотных остатков в данных положениях последовательности зрелого hNGAL (SEQ ID NO: 2).
[00184] В некоторых вариантах осуществления предусмотренные связывающие CD137 мутеины hNGAL могут содержать в одном или более, как, например, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, положениях, соответствующих положениям 36, 40, 41, 49, 52, 68, 70, 72, 73, 77, 79, 81, 96, 100, 103, 125, 127, 132 и 134 линейной полипептидной последовательности зрелого hNGAL (SEQ ID NO: 2), один или более из следующих мутантных аминокислотных остатков: Leu 36 → Met; Ala 40 → Asn; Ile 41 → Leu; Gln 49 → His; Tyr 52 → Ser или Gly; Ser 68 → Asp; Leu 70 → Met; Arg 72 → Leu; Lys 73 → Asp; Asp 77 → Gln или His; Trp 79 → Ile; Arg 81 → Trp или Gln; Asn 96 → Phe; Tyr 100 → Asp; Leu 103 → His или Pro; Lys 125 → Ser; Ser 127 → Ile; Tyr 132 → Trp и Lys 134 → Gly. В некоторых вариантах осуществления предусмотренные связывающие CD137 мутеины hNGAL могут дополнительно содержать в одном или более, как, например, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, положениях, соответствующих положениям 20, 25, 33, 44, 59, 71, 78, 80, 82, 87, 92, 98, 101 и 122 линейной полипептидной последовательности зрелого hNGAL (SEQ ID NO: 2), один или более из следующих мутантных аминокислотных остатков: Gln 20 → Arg; Asn 25 → Tyr или Asp; Val 33 → Ile; Glu 44 → Val или Asp; Lys 59 → Asn; Phe 71 → Leu; Tyr 78 → His; Ile 80 → Asn; Thr 82 → Pro; Phe 92 → Leu или Ser; Lys 98 → Arg; Pro 101 → Leu и Phe 122 → Tyr.
[00185] В некоторых вариантах осуществления предусмотренные связывающие CD137 мутеины hNGAL могут содержать один из следующих наборов мутантных аминокислотных остатков в сравнении с линейной полипептидной последовательностью зрелого hNGAL (SEQ ID NO: 2):
(a) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Lys; Gln 49 → Asn; Tyr 52 → Met; Ser 68 → Gly; Leu 70 → Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Ala; Arg 81 → Ser; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu и Lys 134 → Tyr;
(b) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Arg; Gln 49 → Ile; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Met; Leu 70 → Lys; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Met; Trp 79 → Asp; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu и Lys 134 → Tyr;
(c) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Arg; Gln 49 → Asn; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Ala; Leu 70 → Ala; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Asp; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu и Lys 134 → Tyr;
(d) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Lys; Gln 49 → Asn; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Ala; Leu 70 → Ala; Arg 72 →Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Asp; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 →Phe; Tyr 132 → Glu и Lys 134 → Tyr;
(e) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Lys; Gln 49 → Ser; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Gly; Leu 70 → Ser; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Ala; Arg 81 → Met; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 →Phe; Tyr 132 → Glu и Lys 134 → Tyr;
(f) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Lys; Gln 49 →Val; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Gly; Leu 70 → Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Arg; Trp 79 → Asp; Arg 81 → Ser; Cys 87 → Ser; Leu 94 → Phe; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu и Lys 134 → Tyr;
(g) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Arg; Gln 49 → His; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Gly; Leu 70 → Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Ala; Arg 81 → Ser; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu и Lys 134 → Tyr;
(h) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Lys; Gln 49 → Asn; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Gly; Leu 70 → Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Ala; Arg 81 → Ser; Phe 83 → Leu; Cys 87 → Ser; Leu 94 → Phe; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu и Lys 134 → Tyr или
(i) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Arg; Gln 49 → Ser; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Ala; Leu 70 → Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Asn; Trp 79 → Ala; Arg 81 → Ser; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu и Lys 134 → Tyr.
[00186] В некоторых дополнительных вариантах осуществления в остальной области, т.е. области, отличающейся от положений 28, 36, 40-41, 49, 52, 65, 68, 70, 72-73, 77, 79, 81, 83, 87, 94, 96, 100, 103, 106, 125, 127, 132 и 134 линейной полипептидной последовательности hNGAL (SEQ ID NO: 2) мутеина hNGAL по настоящему изобретению, может включать (природную аминокислотную последовательность) аминокислотную последовательность дикого типа зрелого hNGAL вне положений мутантной аминокислотной последовательности.
[00187] В некоторых других вариантах осуществления предусмотренные связывающие CD137 мутеины hNGAL могут содержать один из следующих наборов мутантных аминокислотных остатков в сравнении с линейной полипептидной последовательностью зрелого hNGAL (SEQ ID NO: 2):
(a) Leu 36 → Met; Ala 40 → Asn; Ile 41 → Leu; Gln 49 → His; Tyr 52 → Ser; Ser 68 → Asp; Leu 70 → Met; Arg 72 → Leu; Lys 73 → Asp; Asp 77 → Gln; Trp 79 → Ile; Arg 81 → Trp; Asn 96 → Phe; Tyr 100 → Asp; Leu 103 → His; Lys 125 → Ser; Ser 127 → Ile; Tyr 132 → Trp и Lys 134 → Gly;
(b) Leu 36 → Met; Ala 40 → Asn; Ile 41 → Leu; Gln 49 → His; Tyr 52 → Ser; Ser 68 → Asp; Leu 70 → Met; Arg 72 → Leu; Lys 73 → Asp; Asp 77 → Gln; Trp 79 → Ile; Arg 81 → Trp; Phe 92 → Leu; Asn 96 → Phe; Lys 98 → Arg; Tyr 100 → Asp; Pro 101 → Leu; Leu 103 → His; Lys 125 → Ser; Ser 127 → Ile; Tyr 132 → Trp и Lys 134 → Gly;
(c) Asn 25 → Tyr; Leu 36 → Met; Ala 40 → Asn; Ile 41 → Leu; Gln 49 → His; Tyr 52 → Gly; Ser 68 → Asp; Leu 70 → Met; Phe 71 → Leu; Arg 72 → Leu; Lys 73 → Asp; Asp 77 → Gln; Trp 79 → Ile; Arg 81 → Gln; Phe 92 → Ser; Asn 96 → Phe; Tyr 100 → Asp; Leu 103 → His; Lys 125 → Ser; Ser 127 → Ile; Tyr 132 → Trp и Lys 134 → Gly;
(d) Leu 36 → Met; Ala 40 → Asn; Ile 41 → Leu; Gln 49 → His; Tyr 52 → Gly; Ser 68 → Asp; Leu 70 → Met; Arg 72 → Leu; Lys 73 →Asp; Asp 77 → Gln; Tyr 78 → His; Trp 79 → Ile; Arg 81 → Trp; Phe 92 → Leu; Asn 96 → Phe; Tyr 100 → Asp; Leu 103 → His; Lys 125 → Ser; Ser 127 → Ile; Tyr 132 → Trp и Lys 134 → Gly;
(e) Asn 25 → Asp; Leu 36 → Met; Ala 40 → Asn; Ile 41 → Leu; Gln 49 → His; Tyr 52 → Gly; Ser 68 → Asp; Leu 70 → Met; Arg 72 → Leu; Lys 73 → Asp; Asp 77 → Gln; Trp 79 → Ile; Arg 81 → Trp; Phe 92 → Leu; Asn 96 → Phe; Tyr 100 → Asp; Leu 103 → His; Lys 125 → Ser; Ser 127 → Ile; Tyr 132 → Trp и Lys 134 → Gly;
(f) Val 33 → Ile; Leu 36 → Met; Ala 40 → Asn; Ile 41 → Leu; Gln 49 → His; Tyr 52 → Gly; Ser 68 → Asp; Leu 70 → Met; Arg 72 → Leu; Lys 73 → Asp; Asp 77 → Gln; Trp 79 → Ile; Arg 81 → Trp; Phe 92 → Leu; Asn 96 → Phe; Tyr 100 →Asp; Leu 103 → His; Lys 125 → Ser; Ser 127 → Ile; Tyr 132 → Trp и Lys 134 → Gly;
(g) Gln 20 → Arg; Leu 36 → Met; Ala 40 → Asn; Ile 41 → Leu; Glu 44 → Val; Gln 49 → His; Tyr 52 → Gly; Ser 68 → Asp; Leu 70 → Met; Arg 72 → Leu; Lys 73 → Asp; Asp 77 → Gln; Trp 79 → Ile; Arg 81 → Trp; Phe 92 → Leu; Asn 96 → Phe; Tyr 100 → Asp; Leu 103 → His; Phe 122 → Tyr; Lys 125 → Ser; Ser 127 → Ile; Tyr 132 → Trp и Lys 134 → Gly;
(h) Leu 36 → Met; Ala 40 → Asn; Ile 41 → Leu; Gln 49 → His; Tyr 52 → Ser; Ser 68 → Asp; Leu 70 → Met; Arg 72 → Leu; Lys 73 → Asp; Asp 77 → Gln; Trp 79 → Ile; Ile 80 → Asn; Arg 81 → Trp; Thr 82 → Pro; Asn 96 → Phe; Tyr 100 → Asp; Pro 101 → Leu; Leu 103 → Pro; Lys 125 → Ser; Ser 127 → Ile; Tyr 132 → Trp и Lys 134 → Gly;
(i) Leu 36 → Met; Ala 40 → Asn; Ile 41 → Leu; Gln 49 → His; Tyr 52 → Gly; Lys 59 → Asn; Ser 68 → Asp; Leu 70 → Met; Arg 72 → Leu; Lys 73 → Asp; Asp 77 → Gln; Trp 79 → Ile; Arg 81 → Trp; Phe 92 → Leu; Asn 96 → Phe; Tyr 100 → Asp; Leu 103 → His; Lys 125 → Ser; Ser 127 → Ile; Tyr 132 → Trp и Lys 134 → Gly и
(j) Leu 36 → Met; Ala 40 → Asn; Ile 41 → Leu; Glu 44 → Asp; Gln 49 → His; Tyr 52 → Ser; Ser 68 → Asp; Leu 70 → Met; Phe 71 → Leu; Arg 72 → Leu; Lys 73 → Asp; Asp 77 → His; Trp 79 → Ile; Arg 81 → Trp; Phe 92 → Leu; Asn 96 → Phe; Tyr 100 → Asp; Leu 103 → His; Lys 125 → Ser; Ser 127 → Ile; Tyr 132 → Trp и Lys 134 → Gly.
[00188] В некоторых вариантах осуществления предусмотренный связывающий CD137 мутеин hNGAL может содержать следующий набор мутантных аминокислотных остатков в сравнении с линейной полипептидной последовательностью зрелого hNGAL (SEQ ID NO: 2) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Arg; Gln 49 → Ile; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Met; Leu 70 → Lys; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Met; Trp 79 → Asp; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu и Lys 134 → Tyr, и/или предусмотренный мутеин может обладать по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или более высокой идентичностью последовательности с аминокислотной последовательностью под SEQ ID NO: 40.
[00189] В некоторых вариантах осуществления в остальной области, т.е. области, отличающейся от положений 20, 25, 28, 33, 36, 40-41, 44, 49, 52, 59, 68, 70-73, 77-82, 87, 92, 96, 98, 100, 101, 103, 122, 125, 127, 132 и 134 линейной полипептидной последовательности hNGAL (SEQ ID NO: 2) мутеина hNGAL по настоящему изобретению, может включать (природную аминокислотную последовательность) аминокислотную последовательность дикого типа зрелого hNGAL вне положений мутантной аминокислотной последовательности.
[00190] В некоторых вариантах осуществления мутеин hNGAL по настоящему изобретению обладает по меньшей мере 70% идентичности последовательности или по меньшей мере 70% гомологии последовательности с последовательностью зрелого hNGAL (SEQ ID NO: 2). В качестве иллюстративного примера мутеин SEQ ID NO: 40 обладает идентичностью аминокислотной последовательности или гомологией последовательности, составляющей примерно 87%, с аминокислотной последовательностью зрелого hNGAL.
[00191] В некоторых вариантах осуществления мутеин hNGAL по настоящему изобретению содержит аминокислотную последовательность, изложенную под любым из SEQ ID NO: 39-57, или ее фрагмент или вариант.
[00192] В некоторых вариантах осуществления мутеин hNGAL по настоящему изобретению обладает по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или более высокой идентичностью последовательности с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 39-57.
[00193] Настоящим изобретением также предусмотрены структурные гомологи мутеина hNGAL, имеющего аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 39-57, при этом структурные гомологи обладают гомологией аминокислотных последовательностей или идентичностью последовательностей, составляющей более чем приблизительно 60%, предпочтительно более 65%, более 70%, более 75%, более 80%, более 85%, более 90%, более 92% и наиболее предпочтительно более 95% по отношению к указанному мутеину hNGAL.
[00194] В некоторых вариантах осуществления настоящим изобретением предусмотрен мутеин липокалина, который связывает CD137 с аффинностью, измеряемой с помощью KD, составляющей приблизительно 5 нМ или ниже, при этом мутеин липокалина обладает по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или более высокой идентичностью последовательности с аминокислотной последовательностью под SEQ ID NO: 40.
[00195] В некоторых вариантах осуществления настоящим изобретением предусмотрен мутеин липокалина, который связывает CD137 с аффинностью, измеряемой с помощью KD, составляющей приблизительно 5 нМ или ниже, при этом мутеин липокалина обладает по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или более высокой идентичностью последовательности с аминокислотной последовательностью под SEQ ID NO: 49.
[00196] В некоторых вариантах осуществления мутеин липокалина по настоящему изобретению может содержать гетерологичную аминокислотную последовательность на своем N- или C-конце, предпочтительно на C-конце, такую как стрептавидиновая метка Strep II (SEQ ID NO: 12) или последовательность сайта расщепления для определенных рестрикционных ферментов, которая не влияет на биологическую активность (связывание со своей мишенью, например CD137) мутеина липокалина.
[00197] В некоторых вариантах осуществления можно ввести дополнительные модификации мутеина липокалина для модуляции определенных характеристик мутеина, таких как повышение стабильности фолдинга, стабильности в сыворотке, устойчивости к другим белкам или растворимость в воде, или для уменьшения тенденции к агрегации, или для внесения в мутеин новых характеристик. В некоторых вариантах осуществления модификация(модификации) может(могут) привести к модуляции двух или более (например, 2, 3, 4, 5, 6, 7, 8, 9 или 10) характеристик предусмотренного мутеина.
[00198] Например, можно изменить одно или более положений в аминокислотной последовательности мутеина липокалина с введением новых реакционно-способных групп, например, для конъюгации с другими соединениями, такими как полиэтиленгликоль (PEG), гидроксиэтилкрахмал (HES), биотин, пептиды или белки, или для образования не встречающихся в природе дисульфидных связей. Конъюгированные соединения, например PEG и HES, в некоторых случаях могут увеличивать период полужизни в сыворотке соответствующего мутеина липокалина.
[00199] В некоторых вариантах осуществления реакционно-способная группа у мутеина липокалина может встречаться в природе в его аминокислотной последовательности, как, например, во встречающихся в природе остатках цистеина в указанной аминокислотной последовательности. В других вариантах осуществления такую реакционно-способную группу можно ввести посредством мутагенеза. В случае введения реакционно-способной группы посредством мутагенеза одной возможностью является мутация аминокислоты в соответствующем положении на остаток цистеина. К иллюстративным вариантам такой мутации для введения остатка цистеина в аминокислотную последовательность мутеина hTlc относятся замены Thr 40 → Cys, Glu 73 → Cys, Arg 90 → Cys, Asp 95 → Cys и Glu 131 → Cys последовательности дикого типа hTlc (SEQ ID NO: 1). К иллюстративным вариантам такой мутации для введения остатка цистеина в аминокислотную последовательность мутеина hNGAL относится введение остатка цистеина в одно или более положений последовательности, которые соответствуют положениям последовательности 14, 21, 60, 84, 88, 116, 141, 145, 143, 146 или 158 последовательности дикого типа hNGAL (SEQ ID NO: 2). Полученный тиоловый фрагмент можно применять для пегилирования или гэкилирования мутеина, например, для увеличения периода полужизни соответствующего мутеина липокалина в сыворотке.
[00200] В некоторых вариантах осуществления с целью получения подходящих боковых цепей аминокислот в качестве новых реакционно-способных групп для конъюгирования одного из вышеупомянутых соединений с мутеином липокалина в аминокислотную последовательность мутеина липокалина можно ввести искусственные аминокислоты. Обычно, такие искусственные аминокислоты предназначены для большей реакционной способности, а значит для облегчения конъюгации с требуемым соединением. Такие искусственные аминокислоты можно ввести путем мутагенеза, например, с применением искусственной tRNA, которая представляет собой пара-ацетил-фенилаланин.
[00201] В некоторых вариантах осуществления мутеин липокалина по настоящему изобретению слит на своем N-конце или своем C-конце с белком, белковым доменом или пептидом, например антителом, сигнальной последовательностью и/или аффинной меткой. В некоторых других вариантах осуществления мутеин липокалина по настоящему изобретению конъюгирован на своем N-конце или на своем C-конце с партнером, который представляет собой белок, белковый домен или пептид; например антитело, сигнальную последовательность и/или аффинную метку.
[00202] Примерами подходящих партнеров по слиянию являются аффинные метки, такие как стрептавидиновые метки Strep-метка или Strep-метка II (Schmidt et al., J Mol Biol, 1996), c-myc-метка, FLAG-метка, His-метка или HA-метка, или белки, такие как глутатион-S-трансфераза, которые обеспечивают легкое выявление и/или очистку рекомбинантных белков. Подходящими партнерами по слиянию для мутеинов липокалина по настоящему изобретению также являются белки с хромогенными или флуоресцентными свойствами, такие как зеленый флуоресцентный белок (GFP) или желтый флуоресцентный белок (YFP). В целом, мутеины липокалина по настоящему изобретению можно пометить любым подходящим химическим веществом или ферментом, которые прямо или косвенно дают поддающееся выявлению соединение или сигнал в химической, физической, оптической или ферментативной реакции. Например, с мутеином липокалина можно конъюгировать флуоресцентную или радиоактивную метку с получением флуоресценции или рентгеновских лучей в качестве поддающегося выявлению сигнала. Примерами ферментных меток (и в то же время оптических меток), которые катализируют образование хромогенных продуктов реакции, являются щелочная фосфатаза, пероксидаза хрена и β-галактозидаза. В общем, для конъюгации с мутеинами липокалина по настоящему изобретению также можно применять все метки, обычно применяемые в случае антител (за исключением меток, исключительно применяемых с сахарным фрагментом в Fc-части иммуноглобулинов).
[00203] В некоторых вариантах осуществления мутеин липокалина по настоящему изобретению может быть слит или конъюгирован с фрагментом, который продлевает период полужизни мутеина в сыворотке (по этому поводу см. также международную патентную публикацию № WO 2006/056464, где такие стратегии описаны с привязкой к мутеинам липокалина, ассоциированного с желатиназой нейтрофилов человека (hNGAL), с аффинностью связывания с CTLA-4). Фрагмент, который продлевает период полужизни в сыворотке, может представлять собой молекулу PEG, молекулу HES, молекулу жирной кислоты, такой как пальмитиновая кислота (Vajo and Duckworth, Pharmacol Rev, 2000), Fc-часть иммуноглобулина, CH3-домен иммуноглобулина, CH4-домен иммуноглобулина, альбумин-связывающий пептид, альбумин-связывающий белок или трансферрин, и это названы лишь некоторые из них.
[00204] В некоторых вариантах осуществления при применении PEG в качестве партнера по конъюгации молекула PEG может быть замещенной, незамещенной, линейной или разветвленной. Она также может представлять собой активированное производное полиэтилена. Примерами подходящих соединений являются молекулы PEG, которые описаны в международной патентной публикации № WO 1999/64016, в патенте США №6177074 или в патенте США №6403564 в отношении интерферона или которые описаны для других белков, таких как PEG-модифицированная аспарагиназа, PEG-аденозиндезаминаза (PEG-ADA) или PEG-супероксиддисмутаза (Fuertges and Abuchowski, Journal of Controlled Release, 1990). Молекулярный вес такого полимера, такого как полиэтиленгликоль, может находиться в диапазоне от приблизительно 300 до приблизительно 70000 дальтон, в том числе, например, полиэтиленгликоль с молекулярным весом приблизительно 10000, приблизительно 20000, приблизительно 30000 или приблизительно 40000 дальтон. Более того, как, например, описано в патентах США №6500930 или №6620413, с целью продления периода полужизни в сыворотке с мутеином по настоящему изобретению можно конъюгировать углеводные олигомеры и полимеры, такие как HES.
[00205] В некоторых вариантах осуществления при применении Fc-части иммуноглобулина с целью продления периода полужизни мутеинов липокалина по настоящему изобретению в сыворотке можно применять технологию SynFusion™, которая коммерчески доступна от Syntonix Pharmaceuticals, Inc. (штат Массачусетс, США). Применение данной технологии слияния с Fc позволяет создавать биофармацевтические средства длительного действия, а полученный с помощью такой технологии продукт может, например, состоять из двух копий мутеина, связанного с Fc-областью антитела, для улучшения фармакокинетики, растворимости и эффективности продуцирования.
[00206] Примерами альбумин-связывающих пептидов, которые можно применять для увеличения периода полужизни мутеина липокалина в сыворотке, являются, например, пептиды, имеющие консенсусную последовательность Cys-Xaa1-Xaa2-Xaa3-Xaa4-Cys, где Xaa1 представляет собой Asp, Asn, Ser, Thr или Trp; Xaa2 представляет собой Asn, Gln, His, Ile, Leu или Lys; Xaa3 представляет собой Ala, Asp, Phe, Trp или Tyr; и Xaa4 представляет собой Asp, Gly, Leu, Phe, Ser или Thr, как описано в патентной публикации США № 20030069395 или Dennis et al. (2002). Альбумин-связывающий белок, слитый или конъюгированный с мутеином липокалина для увеличения периода полужизни в сыворотке, может представлять собой бактериальный альбумин-связывающий белок, антитело, фрагмент антитела, включающий доменные антитела (см., например, патент США 6696245), или мутеин липокалина со связывающей активностью к альбумину. Примеры бактериальных альбумин-связывающих белков включают стрептококковый белок G (Konig and Skerra, J Immunol Methods, 1998).
[00207] В некоторых вариантах осуществления, если альбумин-связывающий белок представляет собой фрагмент антитела, он может быть доменным антителом. Доменные антитела (dAb) сконструированы для обеспечения точного контроля за биофизическими свойствами и in vivo периодом полужизни для создания оптимального профиля безопасности и эффективности продукта. Доменные антитела, например, коммерчески доступны от Domantis Ltd. (Кембридж, Великобритания и штат Массачусетс, США).
[00208] В некоторых вариантах осуществления в качестве партнера мутеина липокалина по настоящему изобретению для увеличения периода полужизни в сыворотке можно применять и сам по себе альбумин (Osborn et al., J Pharmacol Exp Ther, 2002) или биологически активный фрагмент альбумина. Термин «альбумин» включает все альбумины млекопитающих, такие как человеческий сывороточный альбумин, или бычий сывороточный альбумин, или крысиный альбумин. Альбумин или его фрагмент можно получить рекомбинантными методиками, как описано в патенте США №5728553 или в европейских патентных публикациях № EP0330451 и № EP0361991. Соответственно, с мутеином липокалина по настоящему изобретению можно конъюгировать или сливать рекомбинантный человеческий альбумин (например, Recombumin® от Novozymes Delta Ltd., Ноттингем, Великобритания).
[00209] В некоторых вариантах осуществления при применении трансферрина в качестве партнера для увеличения периода полужизни мутеинов липокалина по настоящему изобретению в сыворотке мутеины можно генетическими способами слить с N- или C-концом, или как с первым, так и со вторым, негликозилированного трансферрина. Негликозилированный трансферрин имеет период полужизни 14-17 дней, а слитый с трансферрином белок аналогичным образом будет иметь увеличенный период полужизни. Трансферриновый носитель также обеспечивает высокую биодоступность, биораспределение и стабильность циркуляции в кровотоке. Данная технология коммерчески доступна от BioRexis (BioRexis Pharmaceutical Corporation, штат Пенсильвания, США). Рекомбинантный человеческий трансферрин (DeltaFerrin™) для применения в качестве стабилизатора белка /партнера для увеличения периода полужизни также коммерчески доступен от Novozymes Delta Ltd. (Ноттингем, Великобритания).
[00210] Еще одна альтернатива для увеличения периода полужизни мутеинов липокалина по настоящему изобретению заключается в слиянии с N- или С-концом мутеина длинных, неструктурированных, гибких богатых глицином последовательностей (например, полиглицина с приблизительно 20-80 последовательными остатками глицина). Данный подход, раскрытый, например, в международной патентной публикации № WO2007/038619, также называется «rPEG» (рекомбинантный PEG).
Е. Иллюстративные варианты применения и области применения новых слитых белков, специфических в отношении CD137 и GPC3
[00211] В некоторых вариантах осуществления слитые белки по настоящему изобретению могут производить синергический эффект посредством двойного нацеливания на CD137 и GPC3. В некоторых вариантах осуществления слитые белки по настоящему изобретению могут производить локализованный противоопухолевый эффект посредством двойного нацеливания на CD137 и GPC3. Следовательно, многочисленные возможные области применения слитых белков по настоящему изобретению существуют в медицине.
[00212] В некоторых вариантах осуществления настоящим изобретением охвачено применение одного или более раскрываемых в данном документе слитых белков или одной или более композиций, содержащих такие слитые белки, для одновременного связывания CD137 и GPC3.
[00213] Настоящим изобретением также предусмотрено применение одного или более описываемых слитых белков для образования комплекса с CD137 и/или GPC3.
[00214] Следовательно, в одном аспекте настоящего изобретения предусмотренные слитые белки можно применять для выявления CD137 и GPC3. Такое применение может предусматривать стадии приведения в контакт одного или более указанных слитых белков в подходящих условиях с образцом, предположительно содержащим CD137 и/или GPC3, тем самым обеспечивая возможность образования комплекса между слитыми белками и CD137 и/или GPC3, и выявление комплекса с помощью подходящего сигнала. Поддающийся выявлению сигнал может быть обусловлен меткой, как поясняется выше, или изменением физических свойств из-за связывания, т.е. образования собственно комплекса. Одним примером является поверхностный плазмонный резонанс, значение которого изменяется во время связывания партнеров по связыванию, один из которых иммобилизован на поверхности, такой как золотая фольга.
[00215] Слитые белки по настоящему изобретению также можно применять для разделения CD137 и/или GPC3. Такое применение может предусматривать стадии приведения в контакт одного или более указанных слитых белков в подходящих условиях с образцом, который предположительно содержит CD137 и/или GPC3, тем самым обеспечивая возможность образования комплекса между слитыми белками и CD137 и/или GPC3, и отделение комплекса от образца.
[00216] В некоторых вариантах осуществления настоящим изобретением предусмотрены диагностические и/или аналитические наборы, содержащие один или более слитых белков согласно настоящему изобретению.
[00217] Помимо их применения в диагностике, в еще одном аспекте настоящим изобретением предусмотрены фармацевтические композиции, содержащие один или более слитых белков по настоящему изобретению и фармацевтически приемлемое вспомогательное вещество.
[00218] Более того, в некоторых вариантах осуществления настоящим изобретением предусмотрены слитые белки, которые одновременно связывают CD137 и/или GPC3, для применения таких белков в качестве противоопухолевых и/или противоинфекционных средств, а также иммуномодуляторов. В некоторых вариантах осуществления слитые белки по настоящему изобретению предусмотрены для применения в способе предупреждения, уменьшения интенсивности или лечения заболеваний человека, таких как различные формы рака, в том числе GPC3-положительные формы рака, такие как гепатоклеточная карцинома (HCC), меланома, карцинома из клеток Меркеля, опухоль Вильма и гепатобластома. Соответственно, также предусмотрены способы предупреждения, уменьшения интенсивности или лечения заболеваний человека, таких как различные формы рака, в том числе GPC3-положительный рак, такой как гепатоклеточная карцинома (HCC), меланома, карцинома из клеток Меркеля, у нуждающегося в этом субъекта, предусматривающие введение указанному субъекту терапевтически эффективного количества одного или более слитых белков по настоящему изобретению.
[00219] Другие примеры форм рака, которые можно лечить с применением слитых белков по настоящему изобретению, включают рак печени, рак костей, рак поджелудочной железы, рак кожи, рак головы и шеи, рак молочной железы, рак легких, кожную или внутриглазную злокачественную меланому, рак почек, рак матки, рак яичников, колоректальный рак, рак толстой кишки, рак прямой кишки, рак анальной области, рак желудка, рак яичек, рак матки, карциному фаллопиевых труб, карциному эндометрия, карциному шейки матки, карциному влагалища, карциному вульвы, неходжкинскую лимфому, рак пищевода, рак тонкой кишки, рак эндокринной системы, рак щитовидной железы, рак паращитовидной железы, рак надпочечников, саркому мягких тканей, рак уретры, рак полового члена, солидные опухоли в детском возрасте, лимфоцитарную лимфому, рак мочевого пузыря, рак почки или мочеточника, рак почечной лоханки, новообразование в центральной нервной системе (CNS), первичную лимфому CNS, опухолевый ангиогенез, опухоль в оси позвоночника, глиому ствола головного мозга, аденому гипофиза, саркому Капоши, эпидермоидный рак, плоскоклеточный рак, индуцированные внешними факторами формы рака, в том числе индуцированные асбестом, гематологические злокачественные новообразования, в том числе, например, множественную миелому, В-клеточную лимфому, лимфому Ходжкина/первичную В-клеточную лимфому средостения, неходжкинские лимфомы, острую миелоидную лимфому, хронический миелогенный лейкоз, хронический лимфоидный лейкоз, фолликулярную лимфому, диффузную крупноклеточную B-клеточную лимфому, лимфому Беркита, иммунобластную крупноклеточную лимфому, В-лимфобластную лимфому из клеток-предшественников, лимфому из клеток мантийной зоны, острый лимфобластный лейкоз, фунгоидный микоз, анапластическую крупноклеточную лимфому, Т-клеточную лимфому и Т-лимфобластную лимфому из клеток-предшественников и любые комбинации указанных форм рака. В некоторых вариантах осуществления настоящее изобретение также применимо для лечения метастатических форм рака.
[00220] В некоторых вариантах осуществления слитые белки по настоящему изобретению могут быть одновременно нацелены на опухолевые клетки, у которых экспрессируется GPC3, такие как клетки HCC, меланомы, карциномы из клеток Меркеля, опухоли Вильма и гепатобластомы, и активировать лимфоциты иммунной системы хозяина, находящиеся вблизи таких опухолевых клеток. В некоторых вариантах осуществления слитые белки по настоящему изобретению могут увеличивать целенаправленную противоопухолевую активность Т-клеток, повышать противоопухолевый иммунитет, индуцировать опосредованный Т-клетками цитолиз и/или оказывать прямое ингибирующее действие на рост опухоли, тем самым давая противоопухолевые результаты. В некоторых вариантах осуществления слитые белки по настоящему изобретению могут активировать иммунные ответы в опухолевом микроокружении. В некоторых вариантах осуществления слитые белки по настоящему изобретению могут уменьшать побочные эффекты от эффекторных лимфоцитов по отношению к здоровым клеткам, т.е. нецелевую токсичность, например, посредством локального ингибирования активности онкогена и индукции опосредованной клетками цитотоксичности, реализуемой NK-клетками и/или Т-клетками.
[00221] В некоторых вариантах осуществления настоящим изобретением охвачено применение слитого белка по настоящему изобретению или композиции, содержащей предусмотренный слитый белок, для индукции локализованного лимфоцитарного ответа в непосредственной близости от GPC3-положительных опухолевых клеток, таких как клетки HCC, меланомы, карциномы из клеток Меркеля, опухоли Вильма и гепатобластомы. Соответственно, в некоторых вариантах осуществления настоящим изобретением предусмотрены способы индукции локализованного лимфоцитарного ответа в непосредственной близости от GPC3-положительных опухолевых клеток, таких как клетки HCC, меланомы, карциномы из клеток Меркеля, опухоли Вильмса и гепатобластомы, предусматривающие применение одного или более слитых белков по настоящему изобретению или одной или более композиций, содержащих такие слитые белки. «Локализованный» означает, что при одновременном связывании Т-клеток через CD137 и воздействии на GPC3-положительные опухолевые клетки Т-клетки продуцируют цитокины, в частности IL-2 и/или IFN-гамма, в непосредственной близости от GPC3-положительных клеток. Такие цитокины отражают активацию Т-клеток, которые затем могут быть способны уничтожать GPC3-положительные клетки либо прямо, либо опосредованно путем активации других клеток-киллеров, таких как Т-клетки или NK-клетки.
[00222] В некоторых вариантах осуществления настоящим изобретением охвачено применение слитого белка по настоящему изобретению или композиции, содержащей такой слитый белок, для совместной стимуляции T-клеток и/или активации последующих сигнальных путей CD137. Предпочтительно, предусмотренный слитый белок обеспечивает совместную стимулцию Т-клеток и/или активирует последующие сигнальные пути CD137 при воздействии на опухолевые клетки, у которых экспрессируется GPC3. Соответственно, настоящим изобретением предусмотрены способы индукции пролиферации Т-лимфоцитов и/или активации последующих сигнальных путей CD137, предпочтительно при воздействии на опухолевые клетки, у которых экспрессируется GPC3, такие как клетки HCC, меланомы, карциномы из клеток Меркеля, опухоли Вильмса и гепатобластомы, предусматривающие применение одного или более слитых белков по настоящему изобретению и/или одной или более композиций, содержащих такие слитые белки.
[00223] В некоторых вариантах осуществления настоящим изобретением охвачено применение слитого белка по настоящему изобретению или композиции, содержащей такой слитый белок, для индукции кластеризации и активации CD137 на Т-клетках и направления таких Т-клеток к опухолевым клеткам, у которых экспрессируется GPC3, таким как клетки HCC, меланомы, карциномы из клеток Меркеля, опухоли Вильма и гепатобластомы.
[00224] Дополнительные цели, преимущества и признаки настоящего изобретения станут очевидны для специалистов в данной области после изучения представленных далее примеров и прилагаемых к ним графических материалов, которые не подразумеваются как ограничивающие. Таким образом, следует понимать, что хотя настоящее изобретение, в частности, раскрыто с помощью иллюстративных вариантов осуществления и дополнительных признаков, специалисты в данной области могут прибегнуть к модификациям и вариациям представленных вариантов осуществления настоящего изобретения, и что такие модификации и вариации считаются подпадающими под объем настоящего изобретения.
F. Получение иллюстративных предусмотренных новых слитых белков, специфических в отношении CD137 и GPC3
[00225] В некоторых вариантах осуществления настоящим изобретением предусмотрены молекулы нуклеиновой кислоты (ДНК и РНК), которые включают нуклеотидные последовательности, кодирующие предусмотренные слитые белки. В некоторых вариантах осуществления настоящим изобретением охвачена клетка-хозяин, содержащая предусмотренную молекулу нуклеиновой кислоты. Поскольку вырожденность генетического кода допускает замены определенных кодонов на другие кодоны, определяющими ту же аминокислоту, настоящее изобретение не ограничено конкретной молекулой нуклеиновой кислоты, кодирующей описываемый в данном документе слитый белок, скорее, охватывая все молекулы нуклеиновой кислоты, которые включают нуклеотидные последовательности, кодирующие функциональный слитый белок. В данном отношении настоящее изобретение также относится к нуклеотидным последовательностям, кодирующим предусмотренные слитые белки.
[00226] Молекулу нуклеиновой кислоты, такую как ДНК, называют «способной экспрессировать молекулу нуклеиновой кислоты» или «способной сделать возможной экспрессию нуклеотидной последовательности», если она включает элементы последовательности, которые содержат информацию, касающуюся регуляции транскрипции и/или трансляции, и такие последовательности «функционально связаны» с нуклеотидной последовательностью, кодирующей белок. Функциональная связь представляет собой связь, в которой регуляторные элементы последовательности и подлежащая экспрессии последовательность связаны таким образом, чтобы обеспечивать экспрессию гена. Точная природа регуляторных областей, необходимых для экспрессии генов, может различаться у разных видов, но в целом к этим областям относятся промотор, который у прокариот содержит как сам промотор, т.е. элементы ДНК, управляющие запуском транскрипции, так и элементы ДНК, которые при транскрибировании в РНК будут сигнализировать о запуске трансляции. Такие промоторные области обычно включают 5'-некодирующие последовательности, участвующие в запуске транскрипции и трансляции, такие как -35/-10 боксы и элемент Шайна-Дальгарно у прокариот или ТАТА-бокс, СААТ-последовательности и 5'-кэппирующие элементы у эукариот. К этим областям также могут относиться энхансерные или репрессорные элементы, а также транслируемые сигнальные и лидерные последовательности для нацеливания нативного белка в конкретный компартмент клетки-хозяина.
[00227] Кроме того, 3'-некодирующие последовательности могут содержать регуляторные элементы, участвующие в окончании транскрипции, полиаденилировании и др. Тем не менее, если эти последовательности окончания транскрипции неудовлетворительно функционируют в конкретной клетке-хозяине, то они могут быть заменены сигналами, являющимися функциональными в такой клетке.
[00228] Следовательно, молекула нуклеиновой кислоты по настоящему изобретению может быть «функционально связана» с одной или более регуляторными последовательностями, такими как промоторная последовательность, для того, чтобы сделать возможной экспрессию данной молекулы нуклеиновой кислоты. В некоторых вариантах осуществления молекула нуклеиновой кислоты по настоящему изобретению включает промоторную последовательность и последовательность окончания транскрипции. Подходящими прокариотическими промоторами являются, например, промотор tet, промотор lacUV5 или промотор T7. Примерами промоторов, применимых для экспрессии в эукариотических клетках, являются промотор SV40 или промотор CMV.
[00229] В некоторых вариантах осуществления молекула нуклеиновой кислоты, кодирующая раскрываемый в настоящей заявке мутеин липокалина, может быть «функционально связана» с другой молекулой нуклеиновой кислоты, кодирующей иммуноглобулин по настоящему изобретению, для того, чтобы сделать возможной экспрессию раскрываемого в данном документе слитого белка.
[00230] В некоторых вариантах осуществления предусмотренные способы могут предусматривать подвергание по меньшей мере одной молекулы нуклеиновой кислоты, кодирующей зрелый hTlc, мутагенезу в одном или более триплетах, кодирующих одно или более положений, соответствующих положениям 5, 26-31, 33-34, 42, 46, 52, 56, 58, 60-61, 65, 71, 85, 94, 101, 104-106, 108, 111, 114, 121, 133, 148, 150 и 153 линейной полипептидной последовательности hTlc (SEQ ID NO: 1), с получением мутеинов липокалина, которые включены в предусмотренные слитые белки. В некоторых вариантах осуществления предусмотренные способы могут предусматривать подвергание по меньшей мере одной молекулы нуклеиновой кислоты, кодирующей зрелый hNGAL, мутагенезу в одном или более триплетах, кодирующих одно или более положений, соответствующих положениям 28, 36, 40-41, 49, 52, 65, 68, 70, 72-73, 77, 79, 81, 83, 87, 94, 96, 100, 103, 106, 125, 127, 132 и 134 линейной полипептидной последовательности hNGAL (SEQ ID NO: 2), с получением мутеинов липокалина, которые включены в предусмотренные слитые белки. В некоторых вариантах осуществления предусмотренный способ может предусматривать подвергание по меньшей мере одной молекулы нуклеиновой кислоты, кодирующей зрелый hNGAL, мутагенезу в одном или более триплетах, кодирующих одно или более положений, соответствующих положениям 20, 25, 28, 33, 36, 40-41, 44, 49, 52, 59, 68, 70-73, 77-82, 87, 92, 96, 98, 100, 101, 103, 122, 125, 127, 132 и 134 линейной полипептидной последовательности hNGAL (SEQ ID NO: 2), с получением мутеинов липокалина, которые включены в предусмотренные слитые белки.
[00231] Кроме того, что касается мутеинов hTlc или мутеинов hNGAL по настоящему изобретению, которые входят в состав слитых белков, в некоторых вариантах осуществления может быть удалена встречающаяся в природе дисульфидная связь между Cys 61 и Cys 153 или Cys 76 и Cys 175 соответственно. Соответственно, такие мутеины могут продуцироваться в компартменте клетки, имеющем восстанавливающую окислительно-восстановительную среду, например, в цитоплазме грамотрицательных бактерий.
[00232] Что касается следующих предусмотренных мутеинов hTlc или мутеинов hNGAL по настоящему изобретению, которые включены в слитые белки, настоящее изобретение также включает молекулы нуклеиновой кислоты, кодирующие такие мутеины, которые в некоторых вариантах осуществления могут включать одну или более дополнительных мутаций вне указанных положений последовательности экспериментального мутагенеза. Такие мутации часто допускаются или даже могут оказаться преимущественными, например, если они вносят вклад в улучшенную эффективность фолдинга, стабильность в сыворотке, термостабильность или аффинность связывания лиганда у мутеинов липокалина и/или слитых белков.
[00233] В некоторых вариантах осуществления предусмотренные молекулы нуклеиновой кислоты также могут быть частью вектора или любого другого типа носителя для клонирования, такого как плазмида, фагмида, фаг, бакуловирус, космида или искусственная хромосома.
[00234] В некоторых вариантах осуществления предусмотренная молекула нуклеиновой кислоты может быть включена в фагмиду. В данном контексте фагмидный вектор обозначает вектор, кодирующий межгенную область умеренного фага, такого как M13 или f1, или его функциональную часть, слитую с представляющей интерес cDNA. Например, в некоторых вариантах осуществления после заражения посредством суперинфекции бактериальных клеток-хозяев таким предусмотренным фагмидным вектором и подходящим хелперным фагом (например, M13K07, VCS-M13 или R408) получают интактные фаговые частицы, тем самым обеспечивая физическое связывание закодированной гетерологичной cDNA с его соответствующим полипептидом, экспонированном на поверхности фага (Lowman, Annu Rev Biophys Biomol Struct, 1997, Rodi and Makowski, Curr Opin Biotechnol, 1999).
[00235] В соответствии с различными вариантами осуществления носители для клонирования могут включать, помимо описанных выше регуляторных последовательностей и последовательности нуклеиновой кислоты, кодирующей описываемый в данном документе слитый белок, репликационные и управляющие последовательности, полученные от видов, совместимых с клеткой-хозяином, которую применяют для экспрессии, а также маркеры отбора, придающие трансформированным или трансфицированным клеткам селектируемый фенотип. Из уровня техники известны и коммерчески доступны многочисленные подходящие векторы для клонирования.
[00236] Настоящее изобретение в некоторых вариантах осуществления также относится к способам получения слитых белков по настоящему изобретению, исходя из нуклеиновой кислоты, кодирующей слитый белок или любые субъединицы в нем, с применением способов генной инженерии. В некоторых вариантах осуществления представленный способ можно осуществлять in vivo, при этом предусмотренный слитый белок можно, например, получить в бактериальном или эукариотическом организме-хозяине, а затем выделить из этого организма-хозяина или его культуры. Слитый белок по настоящему изобретению также можно получить in vitro, например, с применением системы трансляции in vitro.
[00237] При получении слитого белка in vivo нуклеиновую кислоту, кодирующую такой слитый белок, можно ввести в подходящий бактериальный или эукариотический организм-хозяин с применением технологии рекомбинантной ДНК, которая хорошо известна из уровня техники. В некоторых вариантах осуществления молекулу ДНК, кодирующую описываемый в данном документе слитый белок, и, в частности, вектор для клонирования, содержащий последовательность, кодирующую такой слитый белок, можно ввести путем трансформации в клетку-хозяин, способную экспрессировать данный ген. Трансформацию можно осуществить с применением стандартных методик. Таким образом, настоящее изобретение также относится к клеткам-хозяевам, содержащим раскрываемую в данном документе молекулу нуклеиновой кислоты.
[00238] В некоторых вариантах осуществления трансформированные клетки-хозяева можно культивировать в условиях, подходящих для экспрессии нуклеотидной последовательности, кодирующей слитый белок по настоящему изобретению. В некоторых вариантах осуществления клетки-хозяева могут быть прокариотическими, такими как Escherichia coli (E. coli) или Bacillus subtilis, или эукариотическими, такими как Saccharomyces cerevisiae, Pichia pastoris, SF9 или клетки насекомых High5, иммортализованными линиями клеток млекопитающих (например, клетками HeLa или клетками CHO) или первичными клетками млекопитающих.
[00239] В некоторых вариантах осуществления, если мутеин липокалина по настоящем изобретению, в том числе содержащийся в описываемом в данном документе слитом белке, включает внутримолекулярные дисульфидные связи, предпочтительным может быть направить формирующийся белок, с помощью соответствующей сигнальной последовательности, в компартмент клетки, имеющий окислительно-восстановительную среду. Такая окислительная среда может обеспечиваться периплазмой грамотрицательных бактерий, таких как E. coli, во внеклеточной среде грамположительных бактерий или в просвете эндоплазматического ретикулума эукариотических клеток и обычно способствует образованию структурных дисульфидных связей.
[00240] В некоторых вариантах осуществления также возможно получить слитый белок по настоящему изобретению в цитозоле клетки-хозяина, предпочтительно E. coli. В этом случае предусмотренный слитый белок можно либо непосредственно получить в растворимом и подвергнутом фолдингу состоянии, либо выделить в форме телец включения с последующей ренатурацией in vitro. Дополнительным вариантом является применение конкретных штаммов-хозяев, имеющих окислительную внутриклеточную среду, которая, таким образом, может обеспечить образование дисульфидных связей в цитозоле (Venturi et al., J Mol Biol, 2002).
[00241] В некоторых вариантах осуществления слитый белок по настоящему изобретению, описанному в данном документе, не всегда можно получить или продуцировать полностью или частично с применением генной инженерии. Скорее, такой белок также можно получить с помощью любой из многих традиционных и хорошо известных методик, таких как стандартные стратегии органического синтеза, методики твердофазного синтеза, коммерчески доступные автоматизированные синтезаторы или с помощью транскрипции и трансляции in vitro. Например, возможно, чтобы перспективные слитые белки или мутеины липокалина, включенные в такие слитые белки, были идентифицированы с помощью молекулярного моделирования, синтезированы in vitro, и исследованы в отношении активности связывания с представляющей(-ими) интерес мишенью(-ями). Способы твердофазного и/или жидкофазного синтеза белков хорошо известны в уровне техники (см., например, Bruckdorfer et al., Curr Pharm Biotechnol, 2004).
[00242] В некоторых вариантах осуществления слитый белок по настоящему изобретению можно получить с помощью in vitro транскрипции/трансляции с использованием общеизвестных способов, известных специалистам в данной области.
[00243] В некоторых дополнительных вариантах осуществления описываемые в данном документе слитые белки также можно получить с помощью традиционных рекомбинантных методик отдельно или в комбинации с традиционными методиками синтеза.
[00244] Более того, в некоторых вариантах осуществления слитый белок согласно настоящему изобретению можно получить путем конъюгирования вместе отдельных субъединиц, например иммуноглобулинов и мутеинов, включенных в слитый белок. Такую конъюгацию можно осуществить, например, посредством всех форм ковалентного или нековалентного связывания с применением традиционных способов.
[00245] Специалисту в данной области будут известны способы, применимые для получения слитых белков, предусмотренных настоящим изобретением, но белковые или нуклеиновые последовательности которых не раскрыты явно в данном документе. В качестве обзора к таким модификациям аминокислотной последовательности относятся, например, направленный мутагенез отдельных аминокислотных положений для упрощения субклонирования гена белка или его частей путем включения сайтов расщепления для определенных рестрикционных ферментов. Также данные мутации можно включить для дополнительного улучшения аффинности слитого белка в отношении его мишеней (например, CD137 и GPC3). Более того, при необходимости, можно ввести мутации для модуляции одной или более характеристик белка, таких как повышение стабильности фолдинга, стабильности в сыворотке, устойчивости к другим белкам или растворимость в воде, или для уменьшения тенденции к агрегации.
[00246] Настоящее изобретение можно дополнительно охарактеризовать следующими пунктами.
[00247] Пункт 1. Слитый белок, способный связывать как CD137, так и GPC3, где слитый белок содержит по меньшей мере две субъединицы в любом порядке, где первая субъединица предусматривает полноразмерный иммуноглобулин или его антигенсвязывающий домен и является специфической в отношении GPC3, и где вторая субъединица предусматривает мутеин липокалина и является специфической в отношении CD137.
[00248] Пункт 2. Слитый белок, где слитый белок содержит по меньшей мере две субъединицы, где первая субъединица предусматривает полноразмерный иммуноглобулин или его антигенсвязывающий домен и является специфической в отношении GPC3, и где вторая субъединица предусматривает мутеин липокалина и является специфической в отношении CD137, причем вторая субъединица соединена на N-конце с C-концом каждой тяжелой цепи первой субъединицы, необязательно посредством линкера.
[00249] Пункт 3. Слитый белок, где слитый белок содержит по меньшей мере две субъединицы, где первая субъединица предусматривает полноразмерный иммуноглобулин или его антигенсвязывающий домен и является специфической в отношении GPC3, и где вторая субъединица предусматривает мутеин липокалина и является специфической в отношении CD137, причем вторая субъединица соединена на С-конце с N-концом каждой тяжелой цепи первой субъединицы, необязательно посредством линкера.
[00250] Пункт 4. Слитый белок, где слитый белок содержит по меньшей мере две субъединицы, где первая субъединица предусматривает полноразмерный иммуноглобулин или его антигенсвязывающий домен и является специфической в отношении GPC3, и где вторая субъединица предусматривает мутеин липокалина и является специфической в отношении CD137, причем вторая субъединица соединена на N-конце с C-концом каждой легкой цепи первой субъединицы, необязательно посредством линкера.
[00251] Пункт 5. Слитый белок, где слитый белок содержит по меньшей мере две субъединицы, где первая субъединица предусматривает полноразмерный иммуноглобулин или его антигенсвязывающий домен и является специфической в отношении GPC3, и где вторая субъединица предусматривает мутеин липокалина и является специфической в отношении CD137, причем вторая субъединица соединена на С-конце с N-концом каждой легкой цепи первой субъединицы, необязательно посредством линкера.
[00252] Пункт 6. Слитый белок по любому из пунктов 1-5, где слитый белок способен связывать GPC3 со значением KD, составляющим максимум приблизительно 1 нМ или сравнимым или ниже значения KD иммуноглобулина или его антигенсвязывающего домена, который включен исключительно в первую субъединицу.
[00253] Пункт 7. Слитый белок по пункту 6, где значение KD определено с помощью анализа поверхностного плазмонного резонанса (SPR).
[00254] Пункт 8. Слитый белок по любому из пунктов 1-7, где слитый белок способен связывать GPC3 со значением EC50, составляющим максимум приблизительно 0,5 нМ или сравнимым или ниже значения EC50 у иммуноглобулина или его антигенсвязывающего домена, который включен исключительно в первую субъединицу.
[00255] Пункт 9. Слитый белок по любому из пунктов 1-8, где слитый белок способен связывать CD137 со значением EC50, составляющим максимум приблизительно 3 нМ или сравнимым или ниже значения EC50 мутеина липокалина, специфического в отношении CD137, который включен исключительно во вторую субъединицу.
[00256] Пункт 10. Слитый белок по любому из пунктов 8-9, где значение EC50 определено с помощью анализа, представляющего собой твердофазный иммуноферментный анализ (ELISA).
[00257] Пункт 11. Слитый белок по любому из пунктов 1-10, где слитый белок вступает в перекрестную реакцию с GPC3 яванского макака.
[00258] Пункт 12. Слитый белок по любому из пунктов 1-11, где слитый белок способен одновременно связывать CD137 и GPC3 со значениями EC50, составляющими максимум приблизительно 10 нМ, при измерении указанного слитого белка в ELISA-анализе.
[00259] Пункт 13. Слитый белок по любому из пунктов 1-12, где слитый белок способен связывать CD137, экспрессируемый на клетке, со значениями EC50, составляющими максимум приблизительно 30 нМ, при измерении указанного слитого белка в анализе способом проточной цитометрии.
[00260] Пункт 14. Слитый белок по любому из пунктов 1-13, где слитый белок способен связывать GPC3, экспрессируемый на клетке, со значениями EC50, составляющими максимум приблизительно 30 нМ, при измерении указанного слитого белка в анализе способом проточной цитометрии.
[00261] Пункт 15. Слитый белок по любому из пунктов 1-14, где слитый белок способен связывать GPC3-экспрессирующие опухолевые клетки.
[00262] Пункт 16. Слитый белок по любому из пунктов 1-15, где слитый белок способен стимулировать T-клеточные ответы.
[00263] Пункт 17. Слитый белок по любому из пунктов 1-16, где слитый белок способен индуцировать повышенную секрецию IL-2.
[00264] Пункт 18. Слитый белок по любому из пунктов 1-17, где слитый белок способен индуцировать повышенную секрецию IL-2 до более высокого уровня, чем SEQ ID NO: 83, и/или с лучшей эффективностью в сравнении с SEQ ID NO: 83.
[00265] Пункт 19. Слитый белок по любому из пунктов 1-18, где слитый белок способен индуцировать опосредуемую лимфоцитами цитотоксичность.
[00266] Пункт 20. Слитый белок по любому из пунктов 1-19, где слитый белок способен индуцировать усиленное уничтожение GPC3-экспрессирующих опухолевых клеток, опосредованное Т-клетками в сравнении с SEQ ID NO: 83 и/или индуцировать активацию цитотоксических Т-клеток с лучшей эффективностью в сравнении с SEQ ID NO: 83.
[00267] Пункт 21. Слитый белок по любому из пунктов 1-20, где слитый белок способен совместно стимулировать Т-клеточные ответы зависимым от GPC3 образом.
[00268] Пункт 22. Слитый белок по любому из пунктов 1-21, где слитый белок способен совместно стимулировать Т-клеточные ответы в опухолевом микроокружении.
[00269] Пункт 23. Слитый белок по любому из пунктов 1-22, где слитый белок не производит совместной стимуляции Т-клеточных ответов в отсутствие GPC3.
[00270] Пункт 24. Слитый белок по любому из пунктов 1-23, где слитый белок обладает фармакокинетическим профилем, подобным таковому у антитела.
[00271] Пункт 25. Слитый белок по любому из пунктов 1-24, где слитый белок обладает периодом полужизни у мышей, составляющим по меньшей мере 50 часов, по меньшей мере 75 часов, по меньшей мере 100 часов, по меньшей мере 125 часов, по меньшей мере 150 часов, по меньшей мере 175 часов, по меньшей мере 200 часов, по меньшей мере 250 часов или еще дольше, и/или где слитый белок обладает периодом полужизни у мышей, который длиннее, чем у SEQ ID NO: 83.
[00272] Пункт 26. Слитый белок по любому из пунктов 1-25, где слитый белок характеризуется изоэлектрической точкой, составляющей по меньшей мере 6,5, по меньшей мере 6,8, по меньшей мере 7,1, по меньшей мере 7,4, по меньшей мере 7,5, по меньшей мере 7,7 или еще выше, и/или где слитый белок характеризуется изоэлектрической точкой, которая выше чем у SEQ ID NO: 83.
[00273] Пункт 27. Слитый белок по любому из пунктов 1-26, где мутеин липокалина содержит один или более мутантных аминокислотных остатков в положениях, соответствующих положениям 5, 26-31, 33-34, 42, 46, 52, 56, 58, 60-61, 65, 71, 85, 94, 101, 104-106, 108, 111, 114, 121, 133, 148, 150 и 153 линейной полипептидной последовательности зрелого липокалина слезы человека (SEQ ID NO: 1).
[00274] Пункт 28. Слитый белок по любому из пунктов 1-27, где аминокислотная последовательность мутеина липокалина содержит в одном или более положениях, соответствующих положениям 5, 26-31, 33-34, 42, 46, 52, 56, 58, 60-61, 65, 71, 85, 94, 101, 104-106, 108, 111, 114, 121, 133, 148, 150 и 153 линейной полипептидной последовательности зрелого hTlc (SEQ ID NO: 1), один или более из следующих мутантных аминокислотных остатков: Ala 5 → Val или Thr; Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Thr 42 → Ser; Gly 46 → Asp; Lys 52 → Glu; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Lys 65 → Arg или Asn; Thr 71 → Ala; Val 85 → Asp; Lys 94 → Arg или Glu; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Lys 121 → Glu; Ala 133 → Thr; Arg 148 → Ser; Ser 150 → Ile и Cys 153 → Ser.
[00275] Пункт 29. Слитый белок по любому из пунктов 1-28, где аминокислотная последовательность мутеина липокалина содержит один из следующих наборов мутантных аминокислотных остатков в сравнении с линейной полипептидной последовательностью зрелого липокалина слезы человека (SEQ ID NO: 1):
(a) Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp и Cys 153 → Ser;
(b) Ala 5 → Thr; Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Lys 65 → Arg; Val 85 → Asp; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Lys 121 → Glu; Ala 133 → Thr и Cys 153 → Ser;
(c) Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Lys 65 → Asn; Lys 94 → Arg; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Lys 121 → Glu; Ala 133 → Thr и Cys 153 → Ser;
(d) Ala 5 → Val; Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Lys 65 → Arg; Lys 94 → Glu; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Lys 121 → Glu; Ala 133 → Thr и Cys 153 → Ser;
(e) Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Thr 42 → Ser; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Ser 150 → Ile и Cys 153 → Ser;
(f) Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Lys 52 → Glu; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Thr 71 → Ala; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Ala 133 → Thr; Arg 148 → Ser; Ser 150 → Ile и Cys 153 → Ser и
(g) Ala 5 → Thr; Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Gly 46 → Asp; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Thr 71 → Ala; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Ser 150 → Ile и Cys 153 → Ser.
[00276] Пункт 30. Слитый белок по любому из пунктов 1-29, где аминокислотная последовательность мутеина липокалина содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 32-38, или ее фрагмент или вариант.
[00277] Пункт 31. Слитый белок по любому из пунктов 1-30, где аминокислотная последовательность мутеина липокалина обладает по меньшей мере 85% идентичностью последовательности с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 32-38.
[00278] Пункт 32. Слитый белок по любому из пунктов 1-26, где мутеин липокалина содержит один или более мутантных аминокислотных остатков в положениях, соответствующих положениям 28, 36, 40-41, 49, 52, 65, 68, 70, 72-73, 77, 79, 81, 83, 87, 94, 96, 100, 103, 106, 125, 127, 132 и 134 линейной полипептидной последовательности зрелого липокалина, ассоциированного с желатиназой нейтрофилов человека (hNGAL) (SEQ ID NO: 2).
[00279] Пункт 33. Слитый белок по любому из пунктов 1-26 и 32, где аминокислотная последовательность мутеина липокалина содержит в положениях, соответствующих положениям 28, 36, 40-41, 49, 52, 65, 68, 70, 72-73, 77, 79, 81, 83, 87, 94, 96, 100, 103, 106, 125, 127, 132 и 134 линейной полипептидной последовательности зрелого липокалина, ассоциированного с желатиназой нейтрофилов человека (hNGAL) (SEQ ID NO: 2), один или более из следующих мутантных аминокислотных остатков: Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Arg или Lys; Gln 49 → Val, Ile, His, Ser или Asn; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Met, Ala или Gly; Leu 70 → Ala, Lys, Ser или Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Met, Arg, Thr или Asn; Trp 79 → Ala или Asp; Arg 81 → Met, Trp или Ser; Phe 83 → Leu; Cys 87 → Ser; Leu 94 → Phe; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu и Lys 134 → Tyr.
[00280] Пункт 34. Слитый белок по любому из пунктов 1-26 и 33, где мутеин липокалина содержит один или более мутантных аминокислотных остатков в положениях, соответствующих положениям 20, 25, 28, 33, 36, 40-41, 44, 49, 52, 59, 68, 70-73, 77-82, 87, 92, 96, 98, 100, 101, 103, 122, 125, 127, 132 и 134 линейной полипептидной последовательности зрелого липокалина, ассоциированного с желатиназой нейтрофилов человека (hNGAL) (SEQ ID NO: 2).
[00281] Пункт 35. Слитый белок по любому из пунктов 1-26 и 34, где аминокислотная последовательность мутеина липокалина содержит в положениях, соответствующих положениям 20, 25, 28, 33, 36, 40-41, 44, 49, 52, 59, 68, 70-73, 77-82, 87, 92, 96, 98, 100, 101, 103, 122, 125, 127, 132 и 134 линейной полипептидной последовательности зрелого hNGAL (SEQ ID NO: 2), один или более из следующих мутантных аминокислотных остатков: Gln 20 → Arg; Asn 25 → Tyr или Asp; Gln 28 → His; Val 33 → Ile; Leu 36 → Met; Ala 40 → Asn; Ile 41 → Leu; Glu 44 → Val или Asp; Gln 49 → His; Tyr 52 → Ser или Gly; Lys 59 → Asn; Ser 68 → Asp; Leu 70 → Met; Phe 71 → Leu; Arg 72 → Leu; Lys 73 → Asp; Asp 77 → Gln или His; Tyr 78 → His; Trp 79 → Ile; Ile 80 → Asn; Arg 81 → Trp или Gln; Thr 82 → Pro; Cys 87 → Ser; Phe 92 → Leu или Ser; Asn 96 → Phe; Lys 98 → Arg; Tyr 100 → Asp; Pro 101 → Leu; Leu 103 → His или Pro; Phe 122 → Tyr; Lys 125 → Ser; Ser 127 → Ile; Tyr 132 → Trp и Lys 134 → Gly.
[00282] Пункт 36. Слитый белок по любому из пунктов 1-26 и 32-35, где аминокислотная последовательность мутеина липокалина содержит один из следующих наборов мутантных аминокислотных остатков в сравнении с линейной полипептидной последовательностью зрелого hNGAL (SEQ ID NO: 2):
(a) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Lys; Gln 49 → Asn; Tyr 52 → Met; Ser 68 → Gly; Leu 70 → Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Ala; Arg 81 → Ser; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu и Lys 134 → Tyr;
(b) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Arg; Gln 49 → Ile; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Met; Leu 70 → Lys; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Met; Trp 79 → Asp; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu и Lys 134 → Tyr;
(c) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Arg; Gln 49 → Asn; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Ala; Leu 70 → Ala; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Asp; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu и Lys 134 → Tyr;
(d) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Lys; Gln 49 → Asn; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Ala; Leu 70 → Ala; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Asp; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu и Lys 134 → Tyr;
(e) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Lys; Gln 49 → Ser; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Gly; Leu 70 → Ser; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Ala; Arg 81 → Met; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu и Lys 134 → Tyr;
(f) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Lys; Gln 49 → Val; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Gly; Leu 70 → Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Arg; Trp 79 → Asp; Arg 81 → Ser; Cys 87 → Ser; Leu 94 → Phe; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu и Lys 134 → Tyr;
(g) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Arg; Gln 49 → His; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Gly; Leu 70 → Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Ala; Arg 81 → Ser; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu и Lys 134 → Tyr;
(h) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Lys; Gln 49 → Asn; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Gly; Leu 70 → Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Ala; Arg 81 → Ser; Phe 83 → Leu; Cys 87 → Ser; Leu 94 → Phe; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu и Lys 134 → Tyr;
(i) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Arg; Gln 49 → Ser; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Ala; Leu 70 → Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Asn; Trp 79 → Ala; Arg 81 → Ser; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu и Lys 134 → Tyr;
(j) Leu 36 → Met; Ala 40 → Asn; Ile 41 → Leu; Gln 49 → His; Tyr 52 → Ser; Ser 68 → Asp; Leu 70 → Met; Arg 72 → Leu; Lys 73 → Asp; Asp 77 → Gln; Trp 79 → Ile; Arg 81 → Trp; Asn 96 → Phe; Tyr 100 → Asp; Leu 103 → His; Lys 125 → Ser; Ser 127 → Ile; Tyr 132 → Trp и Lys 134 → Gly;
(k) Leu 36 → Met; Ala 40 → Asn; Ile 41 → Leu; Gln 49 → His; Tyr 52 → Ser; Ser 68 → Asp; Leu 70 → Met; Arg 72 → Leu; Lys 73 → Asp; Asp 77 → Gln; Trp 79 → Ile; Arg 81 → Trp; Phe 92 → Leu; Asn 96 → Phe; Lys 98 → Arg; Tyr 100 → Asp; Pro 101 → Leu; Leu 103 → His; Lys 125 → Ser; Ser 127 → Ile; Tyr 132 → Trp и Lys 134 → Gly;
(l) Asn 25 → Tyr; Leu 36 → Met; Ala 40 → Asn; Ile 41 → Leu; Gln 49 → His; Tyr 52 → Gly; Ser 68 → Asp; Leu 70 → Met; Phe 71 → Leu; Arg 72 → Leu; Lys 73 → Asp; Asp 77 → Gln; Trp 79 → Ile; Arg 81 → Gln; Phe 92 → Ser; Asn 96 → Phe; Tyr 100 → Asp; Leu 103 → His; Lys 125 → Ser; Ser 127 → Ile; Tyr 132 → Trp и Lys 134 → Gly;
(m) Leu 36 → Met; Ala 40 → Asn; Ile 41 → Leu; Gln 49 → His; Tyr 52 → Gly; Ser 68 → Asp; Leu 70 → Met; Arg 72 → Leu; Lys 73 → Asp; Asp 77 → Gln; Tyr 78 → His; Trp 79 → Ile; Arg 81 → Trp; Phe 92 → Leu; Asn 96 → Phe; Tyr 100 → Asp; Leu 103 → His; Lys 125 → Ser; Ser 127 → Ile; Tyr 132 → Trp и Lys 134 → Gly;
(n) Asn 25 → Asp; Leu 36 → Met; Ala 40 → Asn; Ile 41 → Leu; Gln 49 → His; Tyr 52 → Gly; Ser 68 → Asp; Leu 70 → Met; Arg 72 → Leu; Lys 73 → Asp; Asp 77 → Gln; Trp 79 → Ile; Arg 81 → Trp; Phe 92 → Leu; Asn 96 → Phe; Tyr 100 → Asp; Leu 103 → His; Lys 125 → Ser; Ser 127 → Ile; Tyr 132 → Trp и Lys 134 → Gly;
(o) Val 33 → Ile; Leu 36 → Met; Ala 40 → Asn; Ile 41 → Leu; Gln 49 → His; Tyr 52 → Gly; Ser 68 → Asp; Leu 70 → Met; Arg 72 → Leu; Lys 73 → Asp; Asp 77 → Gln; Trp 79 → Ile; Arg 81 → Trp; Phe 92 → Leu; Asn 96 → Phe; Tyr 100 → Asp; Leu 103 → His; Lys 125 → Ser; Ser 127 → Ile; Tyr 132 →Trp и Lys 134 → Gly;
(p) Gln 20 → Arg; Leu 36 → Met; Ala 40 → Asn; Ile 41 → Leu; Glu 44 → Val; Gln 49 → His; Tyr 52 → Gly; Ser 68 → Asp; Leu 70 → Met; Arg 72 → Leu; Lys 73 → Asp; Asp 77 → Gln; Trp 79 → Ile; Arg 81 → Trp; Phe 92 → Leu; Asn 96 → Phe; Tyr 100 → Asp; Leu 103 → His; Phe 122 → Tyr; Lys 125 → Ser; Ser 127 → Ile; Tyr 132 → Trp и Lys 134 → Gly;
(q) Leu 36 →Met; Ala 40 → Asn; Ile 41 → Leu; Gln 49 → His; Tyr 52 → Ser; Ser 68 → Asp; Leu 70 → Met; Arg 72 → Leu; Lys 73 → Asp; Asp 77 → Gln; Trp 79 → Ile; Ile 80 → Asn; Arg 81 → Trp; Thr 82 → Pro; Asn 96 → Phe; Tyr 100 → Asp; Pro 101 → Leu; Leu 103 → Pro; Lys 125 → Ser; Ser 127 → Ile; Tyr 132 → Trp и Lys 134 → Gly;
(r) Leu 36 → Met; Ala 40 → Asn; Ile 41 → Leu; Gln 49 → His; Tyr 52 → Gly; Lys 59 → Asn; Ser 68 → Asp; Leu 70 → Met; Arg 72 → Leu; Lys 73 → Asp; Asp 77 → Gln; Trp 79 → Ile; Arg 81 → Trp; Phe 92 → Leu; Asn 96 → Phe; Tyr 100 → Asp; Leu 103 → His; Lys 125 → Ser; Ser 127 → Ile; Tyr 132 → Trp и Lys 134 → Gly и
(s) Leu 36 → Met; Ala 40 → Asn; Ile 41 → Leu; Glu 44 → Asp; Gln 49 → His; Tyr 52 → Ser; Ser 68 → Asp; Leu 70 → Met; Phe 71 → Leu; Arg 72 → Leu; Lys 73 → Asp; Asp 77 → His; Trp 79 → Ile; Arg 81 → Trp; Phe 92 → Leu; Asn 96 → Phe; Tyr 100 → Asp; Leu 103 → His; Lys 125 → Ser; Ser 127 →Ile; Tyr 132 → Trp и Lys 134 → Gly.
[00283] Пункт 37. Слитый белок по любому из пунктов 1-26 и 32-36, где аминокислотная последовательность мутеина липокалина содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 39-57, или ее фрагмент или вариант.
[00284] Пункт 38. Слитый белок по любому из пунктов 1-26 и 32-36, где аминокислотная последовательность мутеина липокалина обладает по меньшей мере 85% идентичностью последовательности с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 39-57.
[00285] Пункт 39. Слитый белок по любому из пунктов 1-26 и 32-38 где аминокислотная последовательность мутеина липокалина содержит следующий набор мутантных аминокислотных остатков в сравнении с линейной полипептидной последовательностью зрелого hNGAL (SEQ ID NO: 2) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Arg; Gln 49 → Ile; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Met; Leu 70 → Lys; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Met; Trp 79 → Asp; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu и Lys 134 → Tyr, и/или где мутеин липокалина обладает по меньшей мере 85% идентичностью последовательности с аминокислотной последовательностью под SEQ ID NO: 40.
[00286] Пункт 40. Слитый белок по любому из пунктов 1-39, где одна субъединица присоединена к другой субъединицы посредством линкера.
[00287] Пункт 41. Слитый белок по любому из пунктов 1-40, где вторая субъединица соединена на N-конце посредством линкера с N- или C-концом константной области каждой тяжелой цепи (СН) первой субъединицы или с N- или C-концом константной области каждой легкой цепи (CL) первой субъединицы.
[00288] Пункт 42. Слитый белок по любому из пунктов 1-41, где третья субъединица соединена на N-конце посредством линкера с N- или C-концом константной области каждой тяжелой цепи (СН) первой субъединицы, N- или C-концом константной области каждой легкой цепи (CL) первой субъединицы или C-концом каждой второй субъединицы.
[00289] Пункт 43. Слитый белок по любому из пунктов 40-42, где линкер представляет собой неструктурированный (Gly-Gly-Gly-Gly-Ser)3 линкер (SEQ ID NO: 13).
[00290] Пункт 44. Слитый белок по любому из пунктов 40-43, где линкер представляет собой неструктурированный глицин-сериновый линкер, полипролиновый линкер, пролин-аланин-сериновый полимер или линкер, выбранный из группы, состоящей из SEQ ID NO: 13-23.
[00291] Пункт 45. Слитый белок по любому из пунктов 1-44, где первая субъединица представляет собой антитело.
[00292] Пункт 46. Слитый белок по любому из пунктов 1-45, где вариабельная область тяжелой цепи антитела выбрана из группы, состоящей из SEQ ID NO: 78, 114, 119, 126 и 129 или последовательности, обладающей по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 92%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 98% или более высокой идентичностью последовательности с аминокислотными последовательностями, представленными под SEQ ID NO: 78, 114, 119, 126 и 129, и где вариабельная область легкой цепи антитела выбрана из группы, состоящей из SEQ ID NO: 79, 115 и 127 или последовательности, обладающей по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 92%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 98% или более высокой идентичностью последовательности с аминокислотными последовательностями, представленными под SEQ ID NO: 79, 115 и 127.
[00293] Пункт 47. Слитый белок по любому из пунктов 1-46, где антитело содержит тяжелую цепь, которая представляет собой любую из SEQ ID NO: 80 и 81 или последовательность, обладающую по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 92%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 98% или более высокой идентичностью последовательности с аминокислотными последовательностями, представленными под SEQ ID NO: 80 или 81, и легкую цепь SEQ ID NO: 82 или последовательность, обладающую по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 92%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 98% или более высокой идентичностью последовательности с аминокислотными последовательностями, представленными под SEQ ID NO: 82.
[00294] Пункт 48. Слитый белок по любому из пунктов 1-47, причем антитело содержит соответственно следующую вариабельную область тяжелой цепи и следующую вариабельную область легкой цепи: SEQ ID NO: 78 и 79, SEQ ID NO: 129 и 79, SEQ ID NO: 114 и 115 или SEQ ID NO: 126 и 127, или вариабельную область тяжелой цепи и вариабельную область легкой цепи, которые имеют последовательность, обладающую по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 92%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 98% или более высокой идентичностью последовательности с аминокислотными последовательностями, представленными под SEQ ID NO: 78 и 79, SEQ ID NO: 129 и 79, SEQ ID NO: 114 и 115 или SEQ ID NO: 126 и 127.
[00295] Пункт 49. Слитый белок по любому из пунктов 1-48, причем антитело содержит соответственно следующую тяжелую цепь и следующую легкую цепь: SEQ ID NO: 80 и 82 или SEQ ID NO: 81 и 82, или тяжелую цепь и легкую цепь, которые имеют последовательность, обладающую по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 92%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 98% или более высокой идентичностью последовательности с аминокислотными последовательностями, представленными под SEQ ID NO: 80 и 82 или SEQ ID NO: 81 и 82.
[00296] Пункт 50. Слитый белок по пункту 36, причем тяжелая цепь антитела содержит один из следующих наборов последовательностей CDR:
(a) GYTFTDYE (HCDR1, SEQ ID NO: 72), LDPKTGDT (HCDR2, SEQ ID NO: 73), TRFYSYTY (HCDR3; SEQ ID NO: 74);
(b) GFTFNKNA (HCDR1, SEQ ID NO: 108), IRNKTNNYAT (HCDR2, SEQ ID NO: 109), VAGNSFAY (HCDR3, SEQ ID NO: 110);
(c) YFDFDSYE (HCDR1, SEQ ID NO: 116), IYHSGST (HCDR2, SEQ ID NO: 117), ARVNMDRFDY (HCDR3, SEQ ID NO: 108) или
(d) GFTFSSYA (HCDR1, SEQ ID NO: 120), IQKQGLPT (HCDR2, SEQ ID NO: 121), AKNRAKFDY (HCDR3, SEQ ID NO: 122).
[00297] Пункт 51. Слитый белок по любому из пунктов 1-50, причем легкая цепь антитела содержит один из следующих наборов последовательностей CDR:
(a) QSLVHSNRNTY (LCDR1, SEQ ID NO: 75), KVS (LCDR2), SQNTHVPPT (LCDR3; SEQ ID NO: 77);
(b) QSLLYSSNQKNY (LCDR1, SEQ ID NO: 111), WAS (LCDR2), QQYYNYPLT (LCDR3, SEQ ID NO: 113) или
(c) QSISSY (LCDR1, SEQ ID NO: 123), NAS (LCDR2), QQNRGFPLT (LCDR3, SEQ ID NO: 125).
[00298] Пункт 52. Слитый белок по любому из пунктов 1-51, причем тяжелая цепь антитела содержит следующий набор последовательностей CDR: GYTFTDYE (HCDR1, SEQ ID NO: 72), LDPKTGDT (HCDR2, SEQ ID NO: 73), TRFYSYTY (HCDR3; SEQ ID NO: 74), а легкая цепь антитела содержит следующий набор последовательностей CDR: QSLVHSNRNTY (LCDR1, SEQ ID NO: 75), KVS (LCDR2), SQNTHVPPT (LCDR3; SEQ ID NO: 77).
[00299] Пункт 53. Слитый белок по любому из пунктов 1-51, причем антитело содержит следующий набор последовательностей CDR:
(a) GFTFNKNA (HCDR1, SEQ ID NO: 108), IRNKTNNYAT (HCDR2, SEQ ID NO: 109), VAGNSFAY (HCDR3, SEQ ID NO: 110), QSLLYSSNQKNY (LCDR1, SEQ ID NO: 111), WAS (LCDR2), QQYYNYPLT (LCDR3, SEQ ID NO: 113) или
(b) GFTFSSYA (HCDR1, SEQ ID NO: 120), IQKQGLPT (HCDR2, SEQ ID NO: 121), AKNRAKFDY (HCDR3, SEQ ID NO: 122), QSISSY (LCDR1, SEQ ID NO: 123), NAS (LCDR2), QQNRGFPLT (LCDR3, SEQ ID NO: 125).
[00300] Пункт 54. Слитый белок по любому из пунктов 1-53, где антитело имеет остов IgG4.
[00301] Пункт 55. Слитый белок по пункту 54, где остов IgG4 имеет одну или более из следующих мутаций: S228P, N297A, F234A, L235A, M428L, N434S, M252Y, S254T и T256E.
[00302] Пункт 56. Слитый белок по любому из пунктов 1-55, где слитый белок содержит аминокислотную последовательность, представленную под любым из SEQ ID NO: 87-96, или где слитый белок содержит аминокислотную последовательность, обладающую по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 92%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 98% или более высокой идентичностью последовательности с аминокислотными последовательностями, представленными под любым из SEQ ID NO: 87-96.
[00303] Пункт 57. Слитый белок по любому из пунктов 1-56, где слитый белок содержит аминокислоты, представленные под SEQ ID NO: 87 и 82, аминокислоты, представленные под SEQ ID NO: 88 и 82, аминокислоты, представленные под SEQ ID NO: 81 и 89, аминокислоты, представленные под SEQ ID NO: 81 и 90, аминокислоты, представленные под SEQ ID NO: 91 и 82, аминокислоты, представленные под SEQ ID NO: 92 и 82, аминокислоты, представленные под SEQ ID NO: 81 и 93, аминокислоты, представленные под SEQ ID NO: 81 и 94, аминокислоты, представленные под SEQ ID NO: 95 и 82, или аминокислоты, представленные под SEQ ID NO: 96 и 82.
[00304] Пункт 58. Слитый белок по любому из пунктов 1-57, где слитый белок содержит аминокислотные последовательности, обладающие по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 92%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 98% или более высокой идентичностью последовательности с аминокислотными последовательностями, представленными под SEQ ID NO: 87 и 82, SEQ ID NO: 88 и 82, SEQ ID NO: 81 и 89, SEQ ID NO: 81 и 90, SEQ ID NO: 91 и 82, SEQ ID NO: 92 и 82, SEQ ID NO: 81 и 93, SEQ ID NO: 81 и 94, SEQ ID NO: 95 и 82 или SEQ ID NO: 96 и 82.
[00305] Пункт 59. Молекула нуклеиновой кислоты, содержащая нуклеотидную последовательность, кодирующую слитый белок по любому из пунктов 1-58.
[00306] Пункт 60. Молекула нуклеиновой кислоты по пункту 59, где молекула нуклеиновой кислоты функционально связана с регуляторной последовательностью для того, чтобы сделать возможной экспрессию указанной молекулы нуклеиновой кислоты.
[00307] Пункт 61. Молекула нуклеиновой кислоты по пункту 59 или пункту 60, где молекула нуклеиновой кислоты содержится в векторе или в фагмидном векторе.
[00308] Пункт 62. Клетка-хозяин, содержащая молекулу нуклеиновой кислоты по любому из пунктов 59-61.
[00309] Пункт 63. Способ получения слитого белка по любому из пунктов 1-58, где слитый белок получают, исходя из нуклеиновой кислоты, кодирующей слитый белок.
[00310] Пункт 64. Способ по пункту 63, где слитый белок получают в бактериальном или эукариотическом организме-хозяине и выделяют из данного организма-хозяина или его культуры.
[00311] Пункт 65. Применение слитого белка по любому из пунктов 1-58 или композиции, содержащей такой слитый белок, для одновременной активации последующих сигнальных путей CD137 и воздействия на GPC3-положительные опухолевые клетки.
[00312] Пункт 66. Способ одновременной активации последующих сигнальных путей CD137 и воздействия на GPC3-положительные опухолевые клетки, предусматривающий применение одного или более слитых белков по любому из пунктов 1-58 или одной или более композиций, содержащих такой слитый белок, по отношению к ткани, содержащей опухоль.
[00313] Пункт 67. Способ одновременной совместной стимуляции Т-клеток и воздействия на GPC3-положительные опухолевые клетки, предусматривающий применение одного или более слитых белков по любому из пунктов 1-58 или одной или более композиций, содержащих такой слитый белок, по отношению к ткани, содержащей опухоль.
[00314] Пункт 68. Способ одновременной индукции активности лимфоцитов и воздействия на GPC3-положительные опухолевые клетки, предусматривающий применением одного или более слитых белков по любому из пунктов 1-58 или одной или более композиций, содержащих такой слитый белок, по отношению к ткани, содержащей опухоль.
[00315] Пункт 69. Способ индукции кластеризации и активации CD137 на T-клетках и направления указанных T-клеток к GPC3-положительным опухолевым клеткам, предусматривающий применение одного или более слитых белков по любому из пунктов 1-58 или одной или более композиций, содержащих такой слитый белок, по отношению к ткани, содержащей опухоль.
[00316] Пункт 70. Способ индукции локализованного лимфоцитарного ответа в непосредственной близости от GPC3-положительных опухолевых клеток, предусматривающий применение одного или более слитых белков по любому из пунктов 1-58 или одной или более композиций, содержащих такой слитый белок, по отношению к ткани, содержащей опухоль.
[00317] Пункт 71. Способ индукции повышенной секреции IL-2 Т-клетками в непосредственной близости от GPC3-положительных опухолевых клеток, предусматривающий применение одного или более слитых белков по любому из пунктов 1-58 или одной или более композиций, содержащих такой слитый белок, по отношению к ткани, содержащей опухоль.
[00318] Пункт 72. Способ индукции повышенного опосредованного лимфоцитами цитолиза GPC3-положительных опухолевых клеток, предусматривающий применение одного или более слитых белков по любому из пунктов 1-58 или одной или более композиций, содержащих такой слитый белок, по отношению к ткани, содержащей опухоль.
[00319] Пункт 73. Фармацевтическая композиция, содержащая один или более слитых белков по любому из пунктов 1-58.
[00320] Пункт 74. Способ предупреждения, уменьшения интенсивности или лечения GPC3-положительных форм рака, предусматривающий применение слитого белка по любому из пунктов 1-58 или одной или более композиций, содержащих такой слитый белок, по отношению к ткани, содержащей опухоль.
[00321] Пункт 75. Способ предупреждения, уменьшения интенсивности или лечения гепатоклеточной карциномы, предусматривающий применение слитого белка по любому из пунктов 1-58 или одной или более композиций, содержащих такой слитый белок, по отношению к ткани, содержащей опухоль.
[00322] Пункт 76. Слитый белок по любому из пунктов 1-58 для применения в терапии.
[00323] Пункт 77. Слитый белок для применения по пункту 76, где применение производят при лечении рака.
[00324] Пункт 78. Применение слитого белка по любому из пунктов 1-58 для производства лекарственного препарата.
[00325] Пункт 79. Применение по пункту 78, где лекарственный препарат предназначен для лечения рака.
V. ПРИМЕРЫ
[00326] Пример 1. Экспрессия и анализ репрезентативных слитых белков
[00327] В данном примере репрезентативные слитые белки из антитела и мутеина липокалина создавали путем слияния друг с другом GPC3-специфического антитела, имеющего тяжелую цепь, представленную под SEQ ID NO: 81, или содержащую вариабельный домен тяжелой цепи под SEQ ID NO: 78, или содержащую CDR GYTFTDYE (HCDR1, SEQ ID NO: 72), LDPKTGDT (HCDR2, SEQ ID NO: 73) и TRFYSYTY (HCDR3; SEQ ID NO: 74), и легкие цепи, представленные под SEQ ID NO: 82, или содержащие вариабельный домен легкой цепи под SEQ ID NO: 79, или содержащие CDR QSLVHSNRNTY (LCDR1, SEQ ID NO: 75), KVS (LCDR2, SEQ ID NO: 76) и SQNTHVPPT (LCDR3; SEQ ID NO: 77), и CD137-специфического мутеина липокалина с SEQ ID NO: 40 или CD137-специфического липокалина, обладающего 97% идентичности последовательности с SEQ ID NO: 49 и обозначенного CD137Ac1, посредством линкера, такого как неструктурированный (G4S)3 линкер с SEQ ID NO: 13, для одновременного воздействия на GPC3 и CD137. Различные форматы, которые были созданы, изображены на фигуре 1. Например, такие слитые белки, например, SEQ ID NO: 87 и 82, SEQ ID NO: 88 и 82, SEQ ID NO: 81 и 89, SEQ ID NO: 81 и 90, а также CD137Ac1-слитый белок 1, CD137Ac1-слитый белок 2, CD137Ac1-слитый белок 3, CD137Ac1-слитый белок 4, CD137Ac1-слитый белок 5, CD137Ac1-слитый белок 6 и CD137Ac1-слитый белок 7 (слитые белки, обладающие 97% идентичности последовательности с последовательностями под SEQ ID NO: 91 и 82, SEQ ID NO: 92 и 82, SEQ ID NO: 81 и 93, SEQ ID NO: 81 и 94, SEQ ID NO: 95 и 82 или SEQ ID NO: 96 и 82), создавали путем слияния одного или более мутеинов липокалина с SEQ ID NO: 40 или обладающих 97% идентичности последовательности с SEQ ID NO: 49, с одним или более из четырех концов антитела, содержащего тяжелую цепь, представленную тяжелой цепью, представленной под SEQ ID NO: 81, или содержащей вариабельный домен тяжелой цепи под SEQ ID NO: 78, или содержащей CDR GYTFTDYE (HCDR1, SEQ ID NO: 72), LDPKTGDT (HCDR2, SEQ ID NO: 73) и TRFYSYTY (HCDR3; SEQ ID NO: 74), и легкие цепи, представленные под SEQ ID NO: 82, или содержащие вариабельный домен легкой цепи под SEQ ID NO: 79, или содержащие CDR QSLVHSNRNTY (LCDR1, SEQ ID NO: 75), KVS (LCDR2, SEQ ID NO: 76) и SQNTHVPPT (LCDR3; SEQ ID NO: 77). Созданные слитые белки могут быть двухвалентными по отношению к CD137 (например, как изображено на фигуре 1A-1D) или четырехвалентеныим по отношению к CD137 (например, как изображено на фигуре 1E-1H) или имеют еще более высокую валентность по отношению к CD137 (например, как изображено на фигуре 1I).
[00328] GPC3-специфические антитела, а также все слитые белки из антитела и мутеина липокаина, описанные в данном примере, имели сконструированный остов IgG4, который содержал мутацию S228P для минимизации обмена полуантитела IgG4 в условиях in-vitro и in-vivo (Silva et al., J Biol Chem, 2015). Во всех описываемых в данном документе антителах и слитых белках могут также существовать дополнительные мутации в остове IgG4, в том числе любая одна или более мутаций F234A, L235A, M428L, N434S, M252Y, S254T и T256E. Можно ввести мутации F234A и L235A для снижения ADCC и ADCP (Glaesner et al., Diabetes Metab Res Rev, 2010). Мутации M428L и N434S или мутации M252Y, S254T и T256E можно ввести для увеличения периода полужизни в сыворотке (Dall'Acqua et al., J Biol Chem, 2006, Zalevsky et al., Nat Biotechnol, 2010). Во избежание гетерогенности все антитела были экспрессированы без лизина на карбокси-конце.
[00329] Кроме того, моноспецифические слитые белки из мутеина липокалина с Fc создавали путем слияния одного или более CD137-специфического мутеина липокалина с SEQ ID NO: 40 или GPC3-специфического липокалина с SEQ ID NO: 64 посредством линкера, например неструктурированного (G4S)3 линкера с SEQ ID NO: 13, с C-концом Fc-области антитела, представленного под SEQ ID NO: 28, как изображено на фигуре 1J-1K. Полученные конструкции представлены под SEQ ID NO: 98.
[00330] Настоящее изобретение также предусматривает асимметричные форматы слияния антитела с мутеином липокалина, где, например, одна легкая цепь антитела может быть слита с мутеином липокалина, а другая - нет.
[00331] Конструкции слитых белков создавали посредством генного синтеза и клонировали в вектор экспрессии у млекопитающих. Затем осуществляли их временную экспрессию в клетках Expi293FTM (Life Technologies). Концентрацию слитых белков в среде для культивирования клеток измеряли с помощью HPLC (Agilent Technologies) с использованием аффинной колонки POROS® с белком A (Applied Biosystems). Титры слитых белков подытожены в таблице 1.
[00332] Слитые белки очищали с помощью хроматографии с белком А с последующей эксклюзионной хроматографией (SEC) в фосфатно-солевом буферном растворе (PBS). После очистки с помощью SEC фракции, содержащие мономерный белок, объединяли в пул и снова анализировали с использованием аналитической SEC и процентного содержания мономеров.
[00333] Содержание мономерных белков в иллюстративных слитых белках после очистки с помощью SEC подытожено в таблице 1, где также приведены значения изоэлектрической точки (pI). Значения pI предусмотренных слитых белков (SEQ ID NO: 87 и 82) были повышены по сравнению с определенным уже известным биспецифическим в отношении CD137/GPC3 слитым белком SEQ ID NO: 83.
[00334] Таблица 1. Титры в ходе временной экспрессии
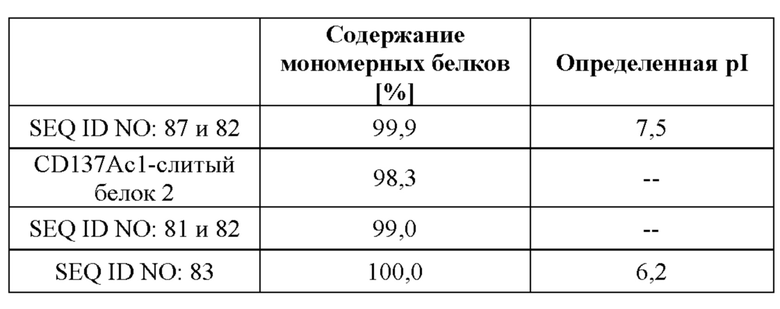
[00335] Пример 2. Связывание слитых белков с GPC3, определенное с помощью поверхностного плазмонного резонанса (SPR)
[00336] Кинетику и аффинность связывания у иллюстративных слитых белков с рекомбинантным GPC3 человека (huGPC3) и GPC3 яванского макака (cyGPC3) (R&D Systems) определяли с помощью поверхностного плазмонного резонанса (SPR) с использованием прибора Biacore T200 (GE Healthcare).
[00337] Антитело к Fc IgG человека (GE Healthcare) иммобилизировали на сенсорном чипе CM5 с помощью стандартной химии аминов: карбоксильные группы на чипе активировали с использованием 1-этил-3-(3-диметиламинопропил)-карбодиимида (EDC) и N-гидроксисукцинимида (NHS). После этого раствор антител к Fc IgG человека (GE Healthcare) в концентрации 25 мкг/мл в 10 мМ ацетате натрия (pH 5,0) вносили со скоростью потока 5 мкл/мин. до достижения уровня иммобилизации 6000-10000 резонансных единиц (RU). Оставшиеся непрореагировавшие сложноэфирные группы NHS блокировали путем пропускания по поверхности 1 М раствора этаноламина. Аналогичным образом обрабатывали сравнительный канал. После этого тестируемые слитые белки (SEQ ID NO: 87 и 82 и CD137Ac1-слитый белок 1) или антитело к GPC3, включенное в слитый белок (SEQ ID NO: 81 и 82), в концентрации 0,2 мкг/мл в HBS-EP+ буфере захватывали посредством антитела к Fc IgG человека на поверхности чипа в течение 180 секунд при скорости потока 10 мкл/мин. Иглу промывали после каждой стадии захвата.
[00338] Для определения аффинности готовили разведения (100 нМ, 25 нМ, 6,25 нМ и 1,56 нМ) huGPC3 или cyGPC3 в HBS-EP+ буфере (GE Healthcare) и вносили их на подготовленную поверхность чипа. Анализ связывания проводили с временем контакта 180 с, временем диссоциации 900 с и скоростью потока 30 мкл/мин. После каждой инъекции иглу очищали посредством 40 мМ NaOH + 20% изопропанола. Все измерения проводили при 25°C. Регенерацию поверхности чипа осуществляли путем введения 3 М MgCl2 на 120 с. Перед измерениями белка производили три пусковых цикла с целью кондиционирования. Данные оценивали с помощью программного обеспечения Biacore T200 Evaluation (вер. 2.0). Для аппроксимации необработанных данных использовали двойное сравнение и модель связывания 1:1.
[00339] Значения, определенные для kon, koff и полученной равновесной константы диссоциации (KD), для иллюстративных слитых белков, подытожены в таблице 2. Протестированные слитые белки (SEQ ID NO: 87 и 82 и CD137Ac1-слитый белок 1) связывали huGPC3, а также cyGPC3 с субнаномолярными значениями аффинности, сравнимыми с таковыми у антитела GPC3, входящего в состав слитых белков, и определенного уже известного биспецифического в отношении CD137/GPC3 слитого белка (SEQ ID NO: 83).
[00340] Таблица 2. Кинетические константы и значения аффинности у слитых белков, определенные с помощью SPR
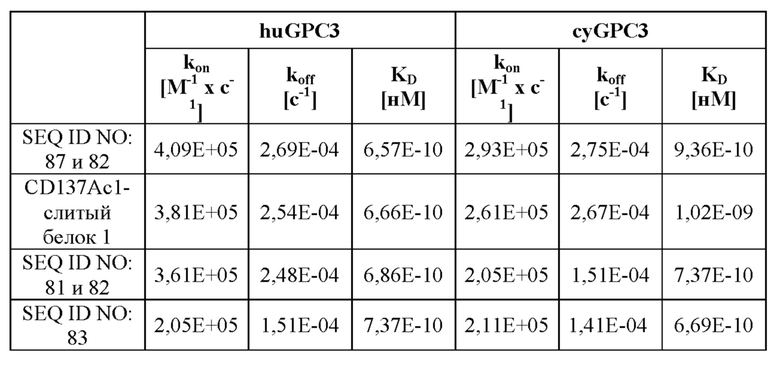
[00341] Пример 3. Связывание слитых белков с GPC3 или CD137 в твердофазном иммуноферментном анализе (ELISA)
[00342] Твердофазный иммуноферментный анализ (ELISA) использовали для определения эффективности связывания иллюстративных слитых белков с GPC3 и CD137.
[00343] Рекомбинантный huGPC3 (R&D Systems) в концентрации 1 мкг/мл в PBS наносили в течение ночи на микротитровальные планшеты при 4°C. После пяти промывок по 100 мкл PBS-0,05%T (PBS с дополнением 0,05% (об./об.) Tween 20) планшеты блокировали посредством 2% BSA (вес/об.) в PBS-0,1%T (PBS-0,1%Т-2%BSA) в течение 1 ч. при комнатной температуре. После пяти промывок по 100 мкл PBS-0,05%T (PBS с дополнением 0,05% (об./об.) Tween 20) иллюстративные слитые белки (SEQ ID NO: 87 и 92 и CD137Ac1-слитый белок 1-7) или антитело к GPC3, входящее в состав слитых белков (SEQ ID NO: 81 и 82), в различных концентрациях вносили в лунки и инкубировали в течение 1 ч. при комнатной температуре с последующей еще одной стадией промывки. Исследуемые связанные молекулы выявляли путем инкубации с разведенным 1:5000 конъюгата HRP с Fc к IgG человека (Jackson Laboratory) в PBS-0,1%T-2%BSA. После дополнительной стадии промывки в каждую лунку вносили флуорогенный субстрат для HRP (QuantaBlu, Thermo) и выявляли интенсивность флуоресценции с помощью считывающего флуоресценцию устройства для микропланшетов.
[00344] Такие же условия ELISA также применяли для определения действенности связывания слитых белков с GPC3 яванского макака и huCD137, при этом вместо этого на микротитровальный планшет наносили рекомбинантный cyGPC3 (R&D Systems) или huCD137-His (рекомбинантный человеческий CD137 с С-концевой полигистидиновой меткой, R&D Systems). Тестируемые средства имели аналогичные титры разведения, а связанные средства выявляли с помощью конъюгатов HRP с антителом к NGAL.
[00345] Иллюстративные результаты изображены на фигуре 2A-2D вместе с аппроксимирующими кривыми, полученными в результате сигмоидальной аппроксимации связывания 1:1, при этом значение EC50 и максимальный сигнал были свободными параметрами, а угловой коэффициент был зафиксирован равным единице. Полученные значения EC50 представлены в таблице 3.
[00346] Наблюдаемые значения EC50 к GPC3 человека у представленных слитых белков (SEQ ID NO: 87 и 82 и CD137Ac1-слитого белка 1-7) были очень схожими или сравнимыми со значениями у уже известного биспецифического в отношении CD137/GPC3 слитого белка (SEQ ID NO: 83) и/или антитела к GPC3, включенного в слитые белки (SEQ ID NO: 81 и 82). Некоторые протестированные слитые белки (SEQ ID NO: 87 и 82, CD137Ac1-слитый белок 3 и CD137Ac1-слитый белок 4) сохраняли сильную аффинность связывания с CD137 человека. Кроме того, у определенного протестированного слитого белка (SEQ ID NO: 87 и 82) также наблюдали перекрестную реактивность с GPC3 яванского макака на уровне, сравнимым с таковым для GPC3 человека, т.е. он связывал GPC3 яванского макака со значениями EC50 в том же диапазоне, что и соответствующие значения EC50 для GPC3 человека.
[00347] Таблица 3. Данные ELISA для связывания GPC3 или CD137
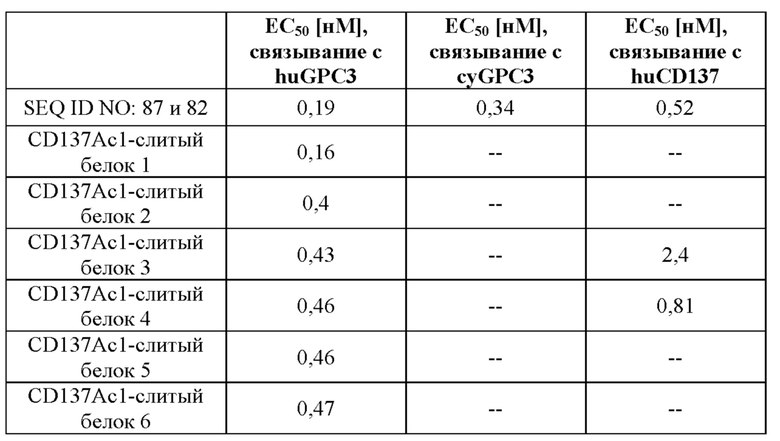
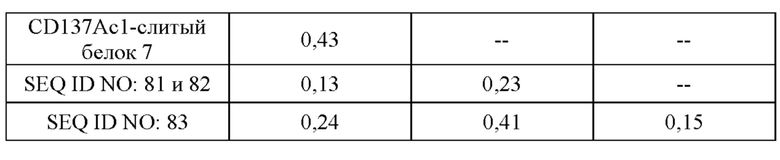
[00348] Пример 4. Одновременное связывание слитых белков с GPC3 и с CD137 в ELISA
[00349] Для демонстрации одновременного связывания иллюстративных слитых белков с GPC3 и CD137 использовали формат ELISA с двойным связыванием.
[00350] Рекомбинантный huCD137-His (R&D Systems) в PBS (1 мкг/мл) наносили покрытием в течение ночи на микротитровальные планшеты при 4°C. После каждой стадии инкубации планшеты пять раз промывали 100 мкл PBS-0,05%T. Планшеты блокировали посредством PBS-0,1%T-2%BSA в течение 1 ч. при комнатной температуре, а затем снова промывали. В лунки вносили различные концентрации тестируемых слитых белков и инкубировали в течение 1 ч. при комнатной температуре с последующей стадией промывки. Затем добавляли биотинилированный huGPC3 в постоянной концентрации 1 мкг/мл в PBS-0,1%T-2%BSA на 1 ч. После промывки в лунки вносили разведенный 1:5000 раствор коньюгата ExtrAvidin-HRP (Sigma-Aldrich) в PBS-0,1%T-2%BSA и инкубировали в течение 1 ч. После дополнительной стадии промывки в каждую лунку вносили флуорогенный субстрат HRP (QuantaBlu, Thermo) и выявляли интенсивность флуоресценции с помощью считывающего флуоресценцию устройства для микропланшетов.
[00351] Двойное связывание слитых белков также тестировали с обратной схемой, когда рекомбинантный huGPC3 (R&D Systems) наносили на микротитровальные планшеты в концентрации 1 мкг/мл, а связанные слитые белки выявляли посредством внесения 5 мкг/мл биотинилированного huCD137-His.
[00352] Иллюстративные данные по двойному связыванию результаты представлены на фигуре 3 вместе с аппроксимирующими кривыми, полученными в результате сигмоидальной аппроксимации связывания 1:1, при этом значение EC50 и максимальный сигнал были свободными параметрами, а угловой коэффициент был зафиксирован равным единице. Значения EC50 подытожены в таблице 4. У слитых белков (SEQ ID NO:87 и 82 и CD137Ac1-слитые белки 1-7) наблюдали четкие сигналы связывания, что свидетельствует, что они могут одновременно воздействовать на GPC3 и CD137. Большинство таких слитых белков способны одновременно воздействовать на GPC3 и CD137 на уровне, сравнимом с уже известным биспецифическим в отношении CD137/GPC3 слитым белком SEQ ID NO: 83.
[00353] Таблица 4. Данные ELISA по одновременному целевому связыванию как с GPC3, так и с CD37
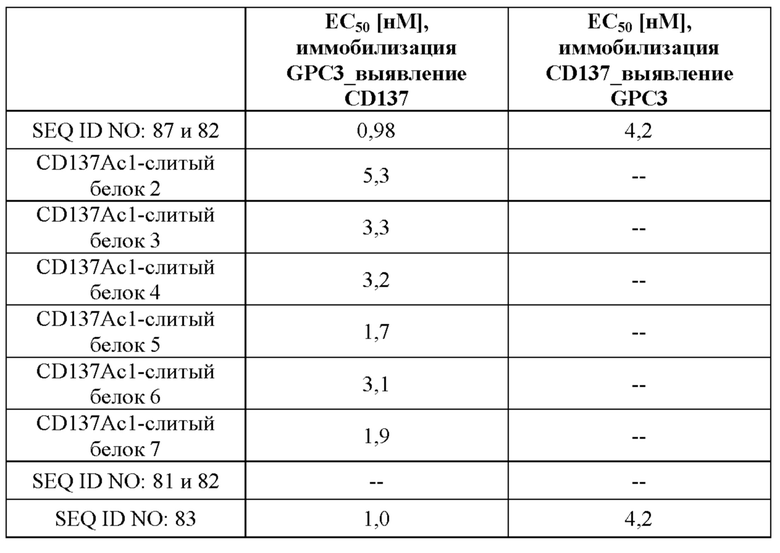
[00354] Пример 5. Нецелевое связывание слитых белков по результатам анализа с помощью ELISA
[00355] Анализ на основе ELISA применяли для оценки нецелевого связывания слитых белков с 32 различными мишенями, в том числе с GPC3, GPC5 и другими рецепторными белками TNF (Frese et al., MAbs, 2013). Мишени в концентрации 5 мкг/мл в PBS наносили в течение ночи на микротитровальные планшеты при 4°C. После промывки посредством PBS-0,05%T планшеты блокировали посредством PBS-0,1%Т-2%BSA течение 1 ч. при комнатной температуре. После пяти циклов промывки по 100 мкл PBS-0,05%T в лунки вносили тестируемые слитые белки в концентрации 100 нМ или 10 нМ и инкубировали в течение 1 ч. при комнатной температуре с последующей еще одной стадией промывки. Исследуемые связанные антитела выявляли путем инкубации с разведенным 1:5000 конъюгата HRP с Fc козы к IgG человека (Jackson Laboratory) в PBS-0,1%T-2%BSA. После дополнительной стадии промывки в каждую лунку вносили флуорогенный субстрат для HRP (QuantaBlu, Thermo) и инкубировали в течение 1 ч. Интенсивность флуоресценции выявляли с помощью считывающего флуоресценцию устройства для микропланшетов и нормализовали к сигналу контрольного антитела (SEQ ID NO: 106 и 107). Нормализованные значения сигнала связывания каждой тестируемой молекулы с 32 мишенями суммировали с получением кумулятивного коэффициента связывания для антитела при заданной концентрации, т. е. 100 нМ или 10 нМ. Кумулятивные коэффициенты связывания при 100 нМ и 10 нМ для каждой тестируемой молекулы дополнительно суммировали с получением суммы кумулятивных коэффициентов связывания. Например, контрольное антитело (SEQ ID NO: 106 и 107), к которому нормализовали интенсивность флуоресценции по результатам связывания с каждой из 32 мишеней, имеет кумулятивный коэффициент связывания, равный 32, при тестировании в концентрации либо 100 нМ, либо 10 нМ и сумму кумулятивных коэффициентов связывания, равную 64. Результаты представлены в таблице 5. Более высокий кумулятивный коэффициент связывания коррелирует с более сильным нецелевым связыванием.
[00356] Предусмотренный слитый белок с SEQ ID NO: 87 и 82 демонстрировал отсутствие или незначительное нецелевое связывание (сумма кумулятивных коэффициентов связывания <150), равно как и антитело к GPC3 и мутеин липокалина, специфический в отношении CD137 (продукт слияния с Fc), включенные в слиты белок (SEQ ID NO: 81 и 82 и SEQ ID NO: 98 соответственно), тогда как определенный уже известный биспецифический в отношении CD137/GPC3 слитый белок (SEQ ID NO: 83) демонстрировал нежелательное нецелевое связывание (сумма кумулятивных коэффициентов связывания >250).
[00357] Таблица 5. Нецелевое связывание слитых белков
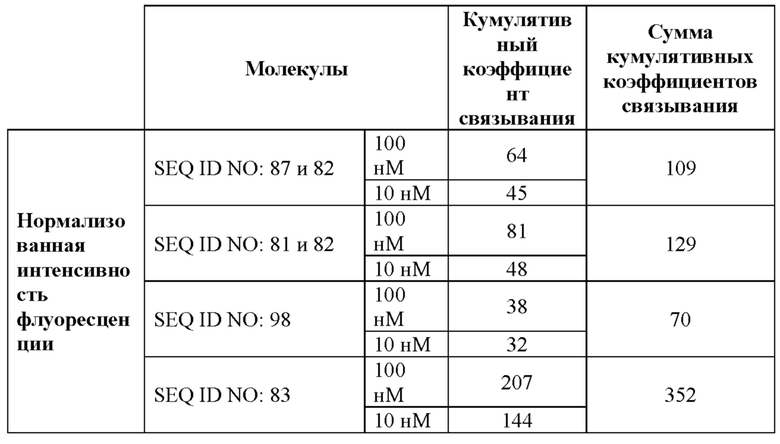
[00358] Пример 6. Анализ способом проточной цитометрии связывания слитых белков с клетками, экспрессирующими CD137 или GPC3
[00359] Целевое специфическое связывание иллюстративных слитых белков с клетками, экспрессирующими человеческий CD137, и клетками, экспрессирующими человеческий GPC3, оценивали с помощью проточной цитометрии.
[00360] В клетки СНО с помощью стабильной трансфекции вводили человеческий CD137 или холостой контроль с использованием системы Flp-In (Life technologies) в соответствии с инструкциями производителя. В клетки SK-Hep1 с помощью стабильной трансфекции вводили человеческий GPC3 или холостой контроль с использованием Lipofectamine2000 (Invitrogen) в соответствии с инструкциями производителя. Трансфицированные клетки СНО поддерживали в среде F12 Хэма (Gibco), дополненной 10% эмбриональной телячьей сывороткой (Sigma-Aldrich) и 500 мкг/мл гигромицина B (Roth). Трансфицированные клетки SK-Hep1 культивировали в среде RPMI 1640 + GlutaMAX (Gibco), дополненной 20% эмбриональной телячьей сывороткой (Sigma-Aldrich) и 500 мкг/мл G418 (Gibco). Клетки культивировали в колбах для культивирования клеток в соответствии с инструкциями производителя (37°C, атмосфера с 5% CO2).
[00361] Для анализа способом проточной цитометрии соответствующие клеточные линии инкубировали с иллюстративным слитым белком (SEQ ID NO: 87 и 82), антителом к GPC3 или мутеином липокалина, специфическим в отношении CD137 (продукт слияния с Fc), включенным в слитый белок (SEQ ID NO: 81 и 82 и SEQ ID NO: 98 соответственно), эталонным антителом к CD137 с SEQ ID NO: 26 и 27 или изотипическим контролем (SEQ ID NO: 24 и 25) и выявляли с помощью меченного флуоресцентной меткой антитела к человеческому IgG в FACS-анализе так, как описано ниже:
[00362] 5×104 клеток на лунку инкубировали в течение 1 ч. в ледяном PBS, содержащем 5% эмбриональной телячьей сыворотки (PBS-FCS). К клеткам добавляли серию разведений тестируемой молекулы и инкубировали в течение 1 ч. на льду. Клетки дважды промывали посредством PBS, а затем инкубировали с меченным посредством Alexa647 козьим антителом к hIgG или меченным посредством Alexa488 козьим антителом к hIgG в течение 30 минут на льду. Затем клетки промывали и анализировали с применением цитометра iQue Flow (Intellicyte Screener). Средние геометрические значения флуоресцентных сигналов представляли в виде графика и аппроксимировали с помощью программного обеспечения Graphpad. Для расчета значений EC50 с помощью четырехпараметрической логистической регрессии, при этом наиболее низкое значение принимали за фоновое, использовали средние геометрические значения интенсивности флуоресценции.
[00363] Способность тестируемых молекул связывать GPC3 и CD137 человека изображена на фигуре 4. Значения аффинности связывания (EC50) у слитого белка SEQ ID NO: 87 и 82 с клетками, экспрессирующими человеческий GPC3 и человеческий CD137, были в нижнем наномолярном диапазоне, сравнимым с таковым для антитела к GPC3 (SEQ ID NO: 81 и 82) и для мутеина липокалина, специфического в отношении CD137 (SEQ ID NO: 98) или для антитела (SEQ ID NO: 26 и 27) соответственно (подытожены в таблице 6). Определенный уже известный биспецифический в отношении CD137/GPC3 слитый белок (SEQ ID NO: 83), тем не менее, демонстрировал ограниченное дозозависимое связывание с клетками, экспрессирующими GPC3, поскольку кривая связывания не достигала плато при концентрациях до 100 нМ (фигура 4B). Ни одна из протестированных молекул не связывалась с трансфицированными холостым контролем клетками (данные не показаны).
[00364] Таблица 6. Значения аффинности связывания слитых белков с клетками, экспрессирующими GPC3 или CD137
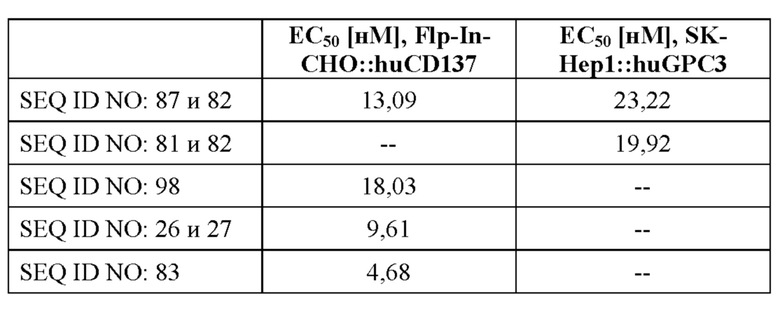
[00365] Пример 7. Аффинность связывания слитых белков с GPC3-положительными опухолевыми клетками
[00366] Связывание слитых белков с опухолевыми клетками, экспрессирующими различные уровни GPC3 - HepG2, Hep3B, MKN-45 и NCI-N87 - оценивали с помощью проточной цитометрии.
[00367] Клеточную линию HepG2 культивировали в модифицированной по Дульбекко среде Игла (DMSO, Pan Biotech), дополненной 10% эмбриональной телячьей сывороткой (Sigma-Aldrich). Клеточную линию Hep3B культивировали в минимальной необходимой среде (с солями Эрла) (Gibco), дополненной 10% эмбриональной телячьей сывороткой (Sigma-Aldrich) и 2 мМ L-глутамина (Gibco). Клеточные линии MKN-45 и NCI-N87 культивировали в среде RPMI-1640 + GlutaMAX (Gibco), дополненной 20% или 10% эмбриональной телячьей сывороткой (Sigma-Aldrich) соответственно. Все линии опухолевых клеток культивировали в колбах для культивирования клеток в соответствии с инструкциями производителя (37°C, атмосфера с 5% CO2).
[00368] Для анализа способом проточной цитометрии линии опухолевых клеток с разными уровнями экспрессии GPC3 (от высокой до умеренной экспрессии: HepG2 > Hep3B > MKN-45) и GPC3-отрицательную клеточную линию NCI-N87 инкубировали с иллюстративным слитым белком SEQ ID NO: 87 и 82, антителом к GPC3 или специфическим в отношении CD137 мутеином липокалина (продуктом слияния с Fc), включенными в слитый белок (SEQ ID NO: 81 и 82 и SEQ ID NO: 98 соответственно), эталонным антителом к CD137 с SEQ ID NO: 26 и 27 или изотипическим контролем (SEQ ID NO: 24 и 25) и выявляли с помощью меченного флуоресцентной меткой антитела к человеческому IgG так, как описано в примере 6.
[00369] Способность слитого белка SEQ ID NO: 87 и 82 связывать GPC3-положительные опухолевые клетки изображена на фигуре 5, а соответствующие значения аффинности связывания (EC50) подытожены в таблице 7. Значения аффинности связывания слитого белка с опухолевыми клетками, экспрессирующими GPC3, лежали в нижнем наномолярном диапазоне, сравнимом с таковым у антитела к GPC3, включенном в слитые белки (SEQ ID NO: 81 и 82). Результаты дополнительно свидетельствовали, что слитый белок не связывается с опухолевыми клетками, отрицательными по GPC3.
[00370] Таблица 7. Аффинность связывания слитых белков с GPC3-положительными опухолевыми клетками
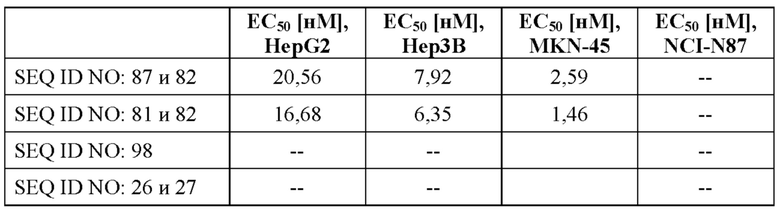
[00371] Пример 8. Зависимая от GPC3 совместная стимуляция Т-клеток с применением биоанализа CD137
[00372] Потенциал у выбранных слитых белков индуцировать активацию сигнального пути CD137 зависимым от GPC3 образом оценивали с применением коммерчески доступной клеточной линии Jurkat с двойной стабильной трансфекцией, экспрессирующей CD137 и ген luc2 (гуманизированная версия люциферазы светлячка), при этом экспрессия luc2 находится под управлением NFκB-чувствительного элемента. В данном биоанализе воздействие на CD137 приводит к внутриклеточной передаче сигналов CD137, что приводит к NFκB-опосредованной люминесценции.
[00373] Линии клеток гепатоклеточного рака HepG2 и Hep3B, которые имеют высокий уровень экспрессии GPC3, и линии клеток рака желудка MKN-45, которые имеют умеренный уровень экспрессии GPC3, и NCI-N87, которая является GPC3-отрицательной, культивировали так, как описано в примере 7. За день до анализа соответствующие опухолевые клетки высевали в количестве 6,25 x 103 клеток на лунку и оставляли их для прикрепления на ночь при 37°C в увлажненной атмосфере с 5% CO2.
[00374] На следующий день в каждую лунку вносили 3,75×104 NF-kB-Luc2/CD137 клеток Jurkat с последующим добавлением различных концентраций, как правило в диапазоне от 0,00488 нМ до 10 нМ, иллюстративного слитого белка (SEQ ID NO: 87 и 82), антитела к GPC3 или мутеина липокалина, специфического в отношении CD137 (продукта слияния с Fc), включенных в слитый белок (SEQ ID NO: 81 и 82 и SEQ ID NO: 98 соответственно), эталонного антитела к CD137 с SEQ ID NO: 26 и 27 или изотипического контроля (SEQ ID NO: 24 и 25). Планшеты накрывали газопроницаемой крышкой и инкубировали при 37°C в увлаженной атмосфере с 5% CO2. Спустя 4 ч. в каждую лунку вносили 30 мкл реагента Bio-Glo™ и количественно определяли биолюминесцентный сигнал с помощью люминометра (PHERAstar). Для расчета значений EC50 (с общим нижним пределом), которые подытожены в таблице 8, проводили четырехпараметрический анализ логистической кривой с помощью GraphPad Prism®. Для демонстрации зависимости от GPC3 при воздействии слитых белков на CD137 параллельно проводили тот же эксперимент в отсутствие опухолевых клеток с использованием наиболее высокой концентрации тестируемых молекул. Анализ проводили в трех повторностях.
[00375] Результаты репрезентативного эксперимента изображены на фигуре 6. Данные, представленные на фигуре 6A и 6B, свидетельствуют, что иллюстративный слитый белок SEQ ID NO: 87 и 82 индуцировал сильную CD137-опосредованную совместную стимуляцию T-клеток в присутствии линии опухолевых клеток с высоким уровнем экспрессии GPC3. На фигуре 6C-6E показано, что активация CD137 слитым белком зависит от GPC3, поскольку активация NF-kB-Luc2/CD137 клеток Jurkat не была выявлена в присутствии опухолевых клеток с умеренной или низкой экспрессией GPC3 или в отсутствие GPC3-экспрессирующих опухолевых клеток. В отличие от этого, эталонное mAb к CD137 (SEQ ID NO: 26 и 27) демонстрировало CD137-опосредованную совместную стимуляцию Т-клеток независимо от уровня экспрессии GPC3 и в отсутствие клеток-мишеней.
[00376] Таблица 8. Результаты оценки активации Т-клеток с помощью биоанализа CD137
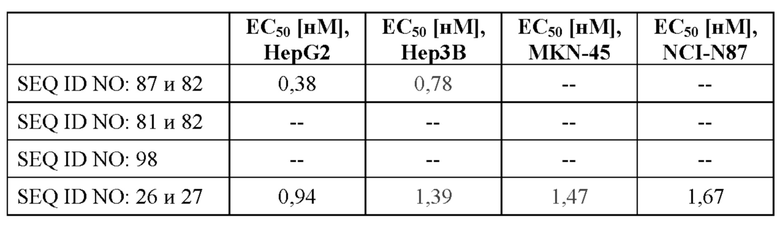
[00377] Пример 9. Оценка зависимой от GPC3 активации Т-клеток, индуцированной слитыми белками
[00378] Зависимую от GPC3-мишени совместную стимуляцию Т-клеток слитыми белками анализировали с помощью анализа активации Т-клеток. Слитые белки вносили в различных концентрациях к простимулированным антителами к CD3 Т-клеткам, совместно культивированным с клетками SK-Hep1, трансфицированными человеческими GPC3 или трансфицированными холостым контролем, или с линией HepG2 GPC3-положительных опухолевых клеток. Уровни секреции IL-2 измеряли в супернатантах.
[00379] Из лейкоцитарных пленок от здоровых добровольцев-доноров выделяли PBMC путем центрифугирования в градиенте плотности полисахарозы (Biocoll, 1,077 г/мл, Biochrom) согласно протоколам Biochrom. Т-Лимфоциты дополнительно очищали от PBMC с помощью магнитной сортировки клеток с применением набора для очистки всех T-клеток (Miltenyi Biotec GmbH) согласно инструкциям производителя. Очищенные все T-клетки ресуспендировали в буфере, состоящем из 90% FCS и 10% DMSO, сразу подвергали заморозке и хранили в жидком азоте до дальнейшего использования. Для анализа Т-клетки оттаивали и помещали в культуральную среду (среду RPMI-1640 + GlutaMAX, Gibco) с добавлением 10% эмбриональной телячьей сыворотки (Sigma-Aldrich) и 1% смеси пенициллина и стрептомицина (Gibco) на ночь при 37°C в увлажненную атмосферу с 5% CO2. Клетки SK-Hep1 и HepG2 культивировали так, как описано соответственно в примере 6 и примере 7.
[00380] С помощью трех повторностей для каждого экспериментального условия проводили следующую процедуру: на плоскодонные планшеты для культивирования тканей предварительно наносили 0,25 мкг/мл антитела к CD3 в течение 2 ч. при 37°C, а затем дважды промывали PBS. Клетки SK-Hep1, трансфицированные человеческим GPC3 или трансфицированные холостым контролем, и GPC3-положительные опухолевые клетки HepG2 обрабатывали в течение 30 минут посредством 30 мкг/мл митомицина C (Sigma Aldrich) для блокировки пролиферации. Обработанные митомицином клетки затем дважды промывали посредством PBS и высевали в количестве 1,0×104 клеток на лунку в культуральную среду, оставляя их для прикрепления на ночь при 37°C в увлажненной атмосфере с 5% CO2. Экспрессирующие мишень клетки были вырощены в стандартных условиях, отделены с помощью аккутазы (PAA Laboratories) и ресуспендированы в культуральной среде.
[00381] В последующие дни после двукратной промывки планшетов посредством PBS в каждую лунку вносили 2,5×104 T-клеток. Серию разведений слитых белков (SEQ ID NO:87 и 82), антитела к GPC3, включенного в слитый белок (SEQ ID NO: 81 и 82), уже известного как биспецифический в отношении CD137/GPC3 слитого белка (SEQ ID NO: 83), специфического в отношении GPC3 мутеина липокалина (продукта слияния с Fc) (SEQ ID NO: 97), эталонного антитела к CD137 (SEQ ID NO: 26 и 27) или изотипического контроля (SEQ ID NO: 24 и 25), как правило, в диапазоне от 0,003 нМ до 10 нМ вносили в соответствующие лунки. Планшеты накрывали газопроницаемой крышкой и инкубировали при 37°C в увлаженной атмосфере с 5% CO2 в течение 3 дней. Уровни IL-2 в супернатанте оценивали с помощью набора Human IL-2 DuoSet (R&D Systems), как описано в указанных далее процедурах.
[00382] В 384-луночных планшетах наносили в течение 2 ч. при комнатной температуре 1 мкг/мл «антитела для захвата человеческого IL-2» в PBS. После этого лунки 5 раз промывали посредством 80 мкл PBS-0,05%T. После 1 ч. блокировки в PBS-0,05%Т, содержащем 1% казеина (вес/вес), аналитические супернатанты и серию концентраций стандарта IL-2, разведенного в культуральной среде, переносили в соответствующие лунки и инкубировали в течение ночи при 4°C. На следующий день вносили смесь 100 нг/мл выявляющего козьего антитела к hIL-2-Bio (R&D Systems) и 1 мкг/мл меченного сульфометком стрептавидина (Mesoscale Discovery) в PBS-0,05%T, содержащем 0,5% казеина, и инкубировали при комнатной температуре в течение 1 ч. После промывки в каждую лунку вносили 25 мкл буфера для считывания (Mesoscale Discovery) и образующийся электрохемилюминесцентный сигнал (ECL) выявляли с помощью считывающего устройства Mesoscale Discovery. Анализ и количественную оценку производили с использованием программного обеспечения Mesoscale Discovery.
[00383] Также схожим образом применяли анализ активации Т-клеток для оценки способности дополнительных уже известных биспецифических в отношении CD137/GPC3 слитых белков SEQ ID NO: 99 и 27, SEQ ID NO: 83 и SEQ ID NO: 84 совместно стимулировать Т-клеточные ответы. В эксперименте в планшеты с культурами тканей предварительно наносили в течение 1 ч. при 37°C 200 мкл 0,25 мкг/мл антитела к CD3 и дважды промывали посредством PBS. Высевали 1,25×104 опухолевых клеток HepG2 на лунку, оставляли их на ночь для прикрепления при 37°C в увлажненной атмосфере с 5% CO2 и обрабатывали в течение 2 часов при 37°C с помощью 10 мкг/мл митомицина C. Планшеты дважды промывали посредством PBS и в каждую лунку вносили 5×104 T-клеток и тестовых молекул в концентрации 1 мкг/мл. Планшеты накрывали газопроницаемой крышкой и инкубировали при 37°C в увлаженной атмосфере с 5% CO2 в течение 3 дней. После этого оценивали концентрацию IL-2 в супернатанте.
[00384] Иллюстративные данные представлены на фигуре 7. Совместное культивирование всех Т-клеток с клетками SK-Hep1, трансфицированными человеческим GPC3 (фигура 7B), или GPC-3-экспрессирующими опухолевыми клетками HepG2 (фигура 7C) в присутствии слитого белка (SEQ ID NO: 87 и 82) приводило к сильной дозозависимой секреции IL-2 по сравнению с изотипическим контролем и намного более сильным, чем в случае антитела к GPC3 (SEQ ID NO: 81 и 82, мутеина липокалина, специфического в отношении GPC3 (SEQ ID NO: 129) или уже известного как биспецифический в отношении CD137/GPC3 слитый белок (SEQ ID NO: 83), где не наблюдали увеличение IL-2 или незначительного увеличения секреции IL-2 при высоких уровнях доз. При совместном культивировании с трансфицированными холостым контролем клетками SK-Hep1 (отрицательными по GPC3) ни одна тестируемая молекула не вызывала дозозависимое увеличение секреции IL-2. Из результатов видно, что активация Т-клеток представленными слитыми белками имела зависимый от GPC3 характер.
[00385] Пример 10. Оценка активации Т-клеток в присутствии опухолевых клеток, экспрессирующих разный уровень GPC3
[00386] Для оценки способности типичных слитых белков совместно стимулировать активацию Т-клеток зависимым от GPC3-мишени образом применяли дополнительный анализ Т-клеток. Слитые белки вносили в различных концентрациях к Т-клеткам, простимулированным антителами к CD3, в присутствии линий опухолевых клеток с разными уровнями экспрессии GPC3. К тестируемым линиям опухолевых клеток относились HepG2, Hep3B, MKN-45 и NCI-N87 (от высокого до среднего уровня экспрессии: HepG2 > Hep3B > MKN-45, отрицательный по GPC3 контроль: NCI-N87). Уровни секреции IL-2 измеряли в супернатантах.
[00387] РВМС от здоровых добровольцев-доноров выделяли из лейкоцитарных пленок, а лимфоциты очищали из РВМС так, как описано в примере 9.
[00388] Для анализа Т-клетки оттаивали и помещали в культуральную среду (среду RPMI-1640 + GlutaMAX, Gibco) с добавлением 10% эмбриональной телячьей сыворотки (Sigma-Aldrich) и 1% смеси пенициллина и стрептомицина (Gibco) на 16 ч. при 37°C в увлажненную атмосферу с 5% CO2.
[00389] С помощью трех повторностей для каждого экспериментального условия проводили следующую процедуру: на плоскодонные планшеты для культивирования тканей предварительно наносили 0,25 мкг/мл антитела к CD3 в течение 2 ч. при 37°C, а затем дважды промывали PBS. Для блокировки пролиферации линию опухолевых клеток HepG2, Hep3B, MKN-45 или NIC-N87 обрабатывали в течение 30 мин. посредством 30 мкг/мл митомицина C (Sigma Aldrich). Обработанные митомицином опухолевые клетки затем дважды промывали посредством PBS и высевали в количестве 8,3×103 клеток на лунку в культуральную среду, оставляя их для прикрепления на ночь при 37°C в увлажненной атмосфере с 5% CO2. Целевые клетки были вырощены в стандартных условиях, отделены с помощью аккутазы (PAA Laboratories) и ресуспендированы в культуральной среде.
[00390] В последующие дни после двукратной промывки планшетов посредством PBS в каждую лунку к опухолевым клеткам вносили 2,5×104 T-клеток. Серию разведений иллюстративного слитого белка (SEQ ID NO: 87 и 82), антитела к GPC3 или мутеина липокалина, специфического в отношении CD137 (продукта слияния с Fc), включенных в слитый белок (SEQ ID NO: 81 и 82 и SEQ ID NO: 98 соответственно), эталонного антитела к CD137 (SEQ ID NO: 26 и 27) или изотипического контроля (SEQ ID NO: 24 и 25), как правило, в диапазоне от 0,26 нМ до 10 нМ вносили в соответствующие лунки. Планшеты накрывали газопроницаемой крышкой и инкубировали при 37°C в увлаженной атмосфере с 5% CO2 в течение 3 дней.
[00391] Через 3 дня совместного культивирования уровень IL-2 в супернатанте оценивали так, как описано в примере 9.
[00392] Иллюстративные данные представлены на фигуре 8. Совместное культивирование всех T-клеток с клетками HepG2 и Hep3B, имеющими высокие уровни экспрессии GPC3, в присутствии слитого белка SEQ ID NO: 87 и 82 приводило к явному увеличению секреции IL-2 по сравнению с изотипическим контролем hIgG4. В дополнение, совместное культивирование с MKN-45 (умеренный уровень GPC3) или NIC-N87 (отрицательная по GPC3) не увеличивало уровни секреции IL-2 со слитым белком. Из данных видно, что функциональная активность слитых белков по результатам измерения их способности активировать Т-клетки или увеличивать секрецию IL-2 имела зависимый от GPC3 характер. В отличие от этого, активация Т-клеток или секреция IL-2, индуцированная эталонным антителом к CD137 (SEQ ID NO: 26 и 27), не всегда имела зависимый от GPC3 характер и была трудно прогнозируемой.
[00393] Пример 11. Оценка опосредованного Т-лимфоцитами цитолиза GPC3-экспрессирующих опухолевых клеток, индуцированного слитыми белками
[00394] Для оценки способности слитых белков активировать костимулирующий сигнальный путь с участием CD137 и индуцировать опосредованный Т-клетками цитолиз GPC3-экспрессирующих опухолевых клеток использовали импедансный анализ Т-клеточного уничтожения. С этой целью адгезивные опухолевые клетки высевали в лунки электронного микротитровального планшета (Э-планшета). Адгезия клеток на золотых микроэлектродах препятствует прохождению электрического тока между электродами. Данный импеданс измеряют как безразмерный параметр «клеточный коэффициент». Клеточный коэффициент (CI) увеличивается по мере того, как клетки прикрепляются, а затем пролиферируют с течение времени. К клеткам HepG2 добавляли неадгезивные CD8+ Т-клетки, вырабатываемые которыми антитела к CD3 независимо активировали Т-клеточные антигены, и тестируемые молекулы в различных концентрациях. Непосредственно добавление не вызывало изменений импеданса. Если тестируемая молекула оказывает костимулирующее действие на цитотоксические Т-клетки путем активации сигнального пути CD137, опосредуемый Т-клетками цитолиз клеток-мишеней будет увеличиваться, а разрушение адгезивных опухолевых клеток эффекторными клетками можно будет выявить в виде уменьшения CI, что делает возможным построение кривой уничтожения в реальном времени.
[00395] PBMC от здоровых добровольцев-доноров выделяли из лейкоцитарных пленок. Из данных PBMC выделяли CD8+ Т-клетки и хранили их в жидком азоте до дальнейшего применения. Для анализа CD8+ Т-клетки размораживали и помещали в среду для анализа, состоявшую из среды RPMI 1640, дополненной 10% FBS и 1% смеси пенициллина и стрептомицина, на 24 ч. при 37°C в увлажненной атмосфере с 5% CO2.
[00396] Описанную далее процедуру выполняли с использованием трех повторностей для каждого условия эксперимента. Линии опухолевых клеток HepG2 (линию GPC3-экспрессирующих клеток гепатоклеточной карциномы) или NCI-N87 (линию GPC3-отрицательных клеток карциномы желудка) высевали в Э-планшет в аналитическую среду и инкубировали в течение 24 ч. при 37°C в увлажненной атмосфере с 5% CO2 для обеспечения адгезии и пролиферации клеток.
[00397] На следующий день к опухолевым клеткам добавляли антитело к CD3 с последующей серией разведений иллюстративного слитого белка (SEQ ID NO: 87 и 82), антитела к GPC3 или мутеина липокалина, специфического в отношении CD137 (продукта слияния с Fc), включенных в слитый белок (SEQ ID NO: 81 и 82 и SEQ ID NO: 98 соответственно), эталонного антитела к CD137 (SEQ ID NO: 26 и 27), определенного уже известного как биспецифический в отношении CD137/GPC3 слитого белка (SEQ ID NO: 83), специфического в отношении GPC3 мутеина липокалина (продукта слияния с Fc) (SEQ ID NO: 97) или изотипического контроля (SEQ ID NO: 24 и 25). Затем к опухолевым клеткам добавляли находящиеся в состоянии покоя CD8+ Т-клетки в соотношении 5:1. Планшеты накрывали газопроницаемой крышкой и инкубировали в течение 3 дней, в течение которых периодически измеряли импеданс.
[00398] Значения CI отображали на графике в виде зависимости от времени с использованием программного обеспечения RTCA HT Software V 1.0.1 (ACEA Biosciences). Нормализованный CI рассчитывали как CI в заданный момент времени (20 ч., 30 ч., 40 ч., 50 ч., 60 ч., 70 ч. после добавления эффекторных клеток), деленный на CI в момент времени нормализации (первое измерение после добавления эффекторных клеток). В случае каждых трех повторностей для расчета конкретного значения уничтожения использовали среднее нормализованное значение CI и соответствующее значение SD по следующей формуле: 100 - (нормализованный средний CI тестируемой молекулы/нормализованный средний CI клеток-мишеней и эффекторных клеток) x 100. Конкретные значения уничтожения экспортировали в GraphPad Prism v7 и представляли в виде графика с соответствующими значениями SD в зависимости от представляющих интерес интервалов времени на графике с осями X и Y.
[00399] Результаты иллюстративных экспериментов представлены на фигуре 9. Из данных видно, что слитый белок SEQ ID NO: 87 и 82 индуцировал дозозависимый опосредованный Т-клетками лизис GPC3-экспрессирующих клеток HepG2 (фигура 9A-9C), тогда как специфического лизиса целевых отрицательных клеток NCI-N87 не наблюдали (фигура 9D), что свидетельствовало о том, что индуцированная слитым белком активация пути с участием CD137, ведущая к опосредованному Т-клетками уничтожению, имела зависимый от GPC3 характер. Предусмотренный слитый белок SEQ ID NO: 87 и 82 индуцировал гораздо более высокий уровень уничтожения GPC3-положительных клеток-мишеней цитотоксическими Т-клетками по сравнению с эквимолярным количеством уже известного биспецифического в отношении CD137/GPC3 слитого белка (SEQ ID NO: 83), антитела к GPC3 (SEQ ID NO: 81 и 82) или специфического в отношении GPC3 мутеина липокалина (продукта слияния с Fc) (SEQ ID NO: 97) (фигура 9C). Кроме того, специфический в отношении CD137 мутеин липокалина (продукт слияния с Fc) (SEQ ID NO: 98), эталонное антитело к CD137 (SEQ ID NO: 26 и 27) и изотипический контроль hIgG4 (SEQ ID NO: 24 и 25) не индуцировали опосредованное CD8+ T-клетками уничтожение (фигура 9C).
[00400] Проводили дополнительный эксперимент для демонстрации того, что сильный опосредованный Т-лимфоцитами лизис клеток HepG2 биспецифическим в отношении CD137/GPC3 слитым белком (SEQ ID NO: 87 и 82) имел зависимый от строения биспецифического белка характер, а не только от комбинации антитела к GPC3 и антитела к CD137. Эксперимент проводили в целом так же, как описано выше, но все конструкции добавляли в концентрации 10 нМ и сравнивали со смесью антител к GPC3 (SEQ ID NO: 81 и 82) и эталонного антитела к CD137 (SEQ ID NO: 26 и 27), а также смесью антитела к GPC3 (SEQ ID NO: 81 и 82) и специфического в отношении CD137 мутеина липокалина (продукта слияния с Fc) (SEQ ID NO: 98). Хотя и в этот раз был выявлен сильный опосредованный Т-клетками лизис в случае слитого белка SEQ ID NO: 87 и 82, специфический лизис, опосредованный обеими смесями, был лишь умеренным, что свидетельствовало о преимуществе данного биспецифического формата для достижения опосредованного Т-клетками лизиса.
[00401] Пример 12. Оценка функциональной in vivo активности у ксенотрансплантатной мышиной модели, которой были подсажены человеческие PBMC
[00402] Для исследования in vivo активности представленных слитых белков использовали иммунодефицитных мышей NOG с подсаженными человеческими опухолевыми клетками HepG2 и человеческими РВМС.
[00403] Мышам NOG возрастом 4-6 недель подкожно (s.c.) вводили 5×106 клеток HepG2 в растворе матригеля/PBS (1:1). Опухолям давали дорасти до размеров 80-100 мм3, момент времени, который был определен как день 0 эксперимента. В день 0 мышей рандомизировали на группы обработки (или контрольную группу) в соответствии с размером опухоли и весом животного. Мышам посредством инъекции в хвостовую вену вводили 5×106 свежих человеческих PBMC внутривенно (i.v.). Мышам вводили средство обработки или контроль (PBS) посредством внутрибрюшинной (i.p.) инъекции после i.v. инъекции РВМС в день 1 и снова вводили средство обработки или контроль в день 8 и день 15. К исследуемым молекулами относились иллюстративный слитый белок SEQ ID NO: 87 и 82 (0,5, 5 или 20 мг/кг), антитело к GPC3, включенное в слитый белок (SEQ ID NO: 81 и 82) (0,39 или 3,9 мг/кг, концентрация, эквимолярная для 0,5 или 5 мг/кг обработки слитым белком), и эталонное антитело к CD137 (SEQ ID NO: 26 и 27) (3,9 мг/кг, концентрация, эквимолярная для 5 мг/кг обработки слитым белком). Рост опухоли регистрировали каждые 3-4 дня. Животные, которых не обрабатывали лекарственным средством, были исключены из анализа данных.
[00404] В конце исследования (день 16) мышей умерщвляли, опухоли фиксировали формалином и заливали парафином (FFPE). Гистологический и иммуногистохимический анализ опухолей проводили в лаборатории BioSiteHisto. Делали срезы опухолевой ткани ксенотрансплантата FFPE и окрашивали с помощью H&E или в отношении Т-клеточных маркеров CD3, CD4 или CD8. Окрашенные препараты фотографировали и оцифровывали с использованием инструментария 3D Histech Pannoramic MIDI с объективом с 20 x увеличением. Для микроскопического изучения цифровых снимков использовали CaseViewer 2.2, а для анализа площади опухоли использовали Image J. Рассчитывали процент противоопухолевых эффекторных лимфоцитов (TIL) на площадь опухоли за вычетом некротической площади.
[00405] На фигуре 10 изображено изменение объема опухоли по результатам измерения на день 2, день 6, день 9, день 13 и день 16 исследования. Ингибирование роста опухоли осуществляли слитым белком SEQ ID NO: 87 и 82 и антителом к GPC3, включенным в слитый белок SEQ ID NO: 81 и 82. Как слитый белок, так и антитело к GPC3 (в концентрации 5 или 3,9 мг/кг соответственно) полностью ингибировали рост опухоли в ходе исследования. Воздействие слитого белка и антитела к GPC3 имело дозозависимый характер, поскольку более низкие концентрации (0,5 или 0,39 мг/кг) оказывали незначительное воздействие на рост опухоли. Было достигнуто лишь ограниченное воздействие на рост опухоли при обработке эталонным антителом к CD137 (SEQ ID NO: 26 и 27).
[00406] В таблице 9 подытожены данные, полученные в ходе гистологического и иммуногистохимического анализа противоопухолевых эффекторных лимфоцитов. Внутриопухолевая инфильтрация CD3, CD4 или CD8 Т-клеток представлена как % TIL на (площадь опухоли - некротическая площадь). Обработка слитым белком приводила к инфильтрации Т-клеток до 10% (площадь опухоли - некротическая площадь) (все группы обработки). Внутриопухолевая инфильтрации Т-клеток не индуцировалась обработкой антителом к GPC3 или эталонным антителом к CD137 в сравнении с контрольной средой.
[00407] Таблица 9. Процент противоопухолевых эффекторных лимфоцитов (TIL) на площадь опухоли за вычетом некротической площади
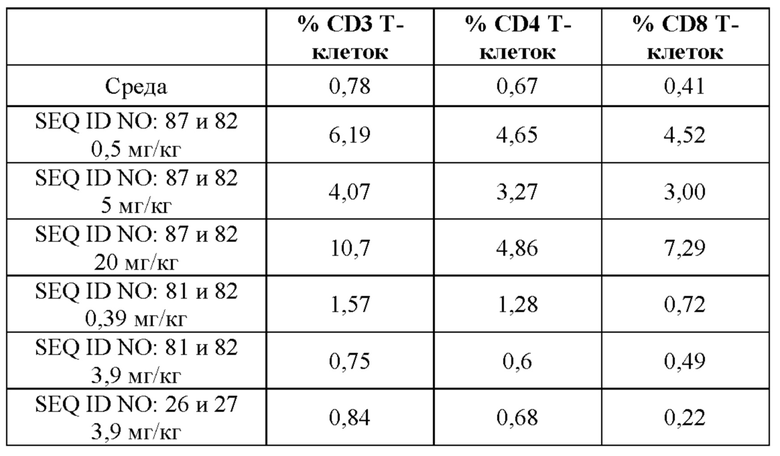
[00408] Пример 13. Фармакокинетика слитых белков у мышей
[00409] Анализы фармакокинетики репрезентативного слитого белка (SEQ ID NO: 87 и 82) и антитела к GPC3, включенного в слитый белок (SEQ ID NO: 81 и 82), проводили на мышах и сравнивали результатами анализа двух уже известных биспецифических в отношении CD137/GPC3 слитых белков (SEQ ID NO: 83 и SEQ ID NO: 84). Самцам мышей CD-1 в возрасте примерно 5 недель (2 мыши на момент времени; Charles River Laboratories) в хвостовую вену вводили соответствующую конструкцию в дозе 2 мг/кг (SEQ ID NO: 87 и 82, SEQ ID NO: 81 и 82) или 10 мг/кг (SEQ ID NO: 83, SEQ ID NO: 84). Образцы плазмы мышей отбирали в моменты времени 5 мин, 24 ч, 168 ч и 336 ч в случае конструкций на основе антител и 5 мин, 2 ч, 4 ч, 8 ч, 24 ч, 2 дня, 3 дня, 4 дня, 7 дней, 9 дней, 11 дней, 14 дней и 21 день. Производили забор достаточного количества цельной крови, которую брали под изофлурановой анестезией, чтобы получить по меньшей мере 30-50 мкл плазмы с добавлением Li и гепарина на животное и момент времени. Затем уровни лекарственного средства в плазме анализировали с помощью ELISA.
[00410] В случае SEQ ID NO: 87 и 82 или SEQ ID NO: 81 и 82 использовали следующий протокол: человеческий GPC3 растворяли в PBS (1 мкг/мл) и наносили покрытием в течение ночи на микротитровальные планшеты при 4°C. После каждой стадии инкубации планшеты пять раз промывали 80 мкл PBS-0,05%T. Планшеты блокировали посредством PBS-0,1%T-2%BSA в течение 1 ч. при комнатной температуре, а затем промывали. Образцы плазмы разводили в PBS-0,1%T-2%BSA до концентрации 20% в плазме, вносили в лунки и инкубировали в течение 1 ч при комнатной температуре. После этого шла еще одна стадия промывания. Исследуемые связанные средства выявляли после 1 ч инкубации с меченным сульфометкой антителом к человеческим антителам (Mesoscale Discovery) или поликлональной антисывороткой, очищенной на колонке для аффинной хроматографии к NGAL, в концентрации 1 мкг/мл, разведенной в PBS-0,1%T-2%BSA. После дополнительной стадии промывания в каждую лунку вносили 25 мкл буфера для считывания и считывали электрохемилюминесцентный сигнал (ECL) из каждой лунки с помощью считывающего устройства Mesoscale Discovery.
[00411] В случае SEQ ID NO: 83 или SEQ ID NO: 84 использовали следующий протокол: человеческий CD137 растворяли в PBS (1 мкг/мл) и наносили покрытием в течение ночи на микротитровальные планшеты при 4°C. После каждой стадии инкубации планшеты промывали 80 мкл PBS-0,05%T. Планшеты блокировали посредством PBS-0,1%T-2%BSA в течение 1 ч. при комнатной температуре, а затем промывали. Образцы плазмы разводили в PBS-0,1%T-2%BSA до концентрации 20% в плазме, вносили в лунки и инкубировали в течение 1 ч. при комнатной температуре. После этого шла еще одна стадия промывания. Исследуемые связанные средства выявляли после 1 ч. инкубации с меченным сульфометкой человеческим Glypican-bio и стрептавидином в количестве 1 мкг/мл, каждый из которых был разведен в PBS-0,1%T-2%BSA. После дополнительной стадии промывания в каждую лунку вносили 25 мкл буфера для считывания и считывали ECL-сигнал из каждой лунки с помощью считывающего устройства Mesoscale Discovery.
[00412] Для анализа и количественной оценки данных также строили калибровочную кривую с разведениями стандартных белков. Концентрация в плазме с течением времени для тестируемых молекул в иллюстративном эксперименте отображена на графике на фигуре 12. Данные подвергали анализу без учета компартментов с использованием Phoenix WinNonlin версии 8.1, а результаты представлены в таблице 10.
[00413] Из данных видно, что предусмотренный слитый белок SEQ ID NO: 87 и 82 характеризовался типичной фармакокинетикой антител, тогда как уже известный биспецифический в отношении CD137/GPC3 слитый белок SEQ ID NO: 83 и SEQ ID NO: 84 имел значительно худшие фармакокинетические профили.
[00414] Таблица 10. Фармакокинетика у мышей
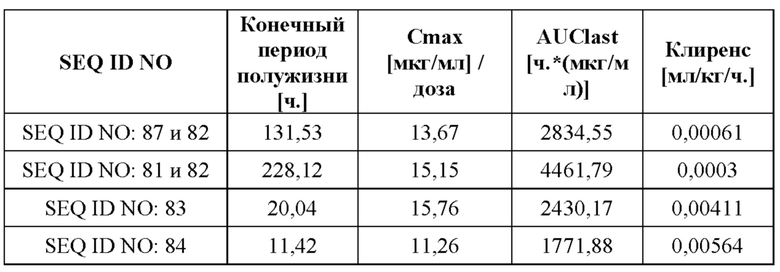
[00415] Пример 14. Оценка термостабильности слитых белков
[00416] Для определения температур плавления (Tm) слитых белков, которая является общим показателем общей стабильности, тестируемые молекулы с концентрацией белка 1 мг/мл в PBS (Gibco) подвергали сканированию (25-100°C) с шагом 1°C/мин с помощью капиллярного прибора nanoDSC (CSC 6300, TA Instruments). Значения Tm рассчитывали, исходя из получаемых термограмм, с использованием встроенного программного обеспечения Nano Analyze.
[00417] Полученные максимальные температуры плавления, а также температура начала плавления для иллюстративных слитых белков представлены в приведенной ниже таблице 11. Слитые белки SEQ ID NO: 72 и 82, а также CD137Ac1-слитый белок 2 обладали улучшенной термостабильностью в сравнении с определенным уже известными биспецифическими в отношении CD137/GPC3 слитыми белками SEQ ID NO: 83, SEQ ID NO: 84 и SEQ ID NO: 26 и 102.
[00418] Таблица 11. Tm и температура начала плавления, определенные с помощью nanoDSC
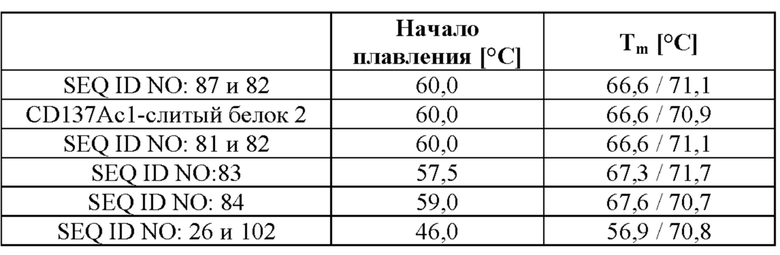
[00419] Варианты осуществления, описанные в данном документе в качестве иллюстрации, в зависимости от ситуации можно реализовать на практике при отсутствии какого-либо элемента или элементов, ограничения или ограничений, которые конкретно не раскрыты в данном документе. Так, например, термины «входящий в состав», «включающий», «содержащий» и т.д. следует читать в наиболее широком смысле и без ограничения. Кроме того, используемые в данном документе термины и выражения были использованы в качестве терминов описания, а не ограничения, и при применении данных терминов и выражений не подразумевается исключение каких-либо эквивалентов показанных и описанных признаков или их частей, а вместо этого считается, что в объеме заявляемого изобретения возможны различные модификации. Таким образом, следует понимать, что хотя настоящие варианты осуществления, в частности, были раскрыты с помощью предпочтительных вариантов осуществления и дополнительных признаков, специалисты в данной области могут прибегнуть к их модификациям и вариациям, и что такие модификации и вариации считаются подпадающими под объем настоящего изобретения. Все патенты, патентные заявки, научные пособия и рецензируемые публикации, описанные в данном документе, включены в данный документ посредством ссылки в полном их объеме. Кроме того, в случае, если определение или применения термина в литературном источнике, который включен в данный документ посредством ссылки, несовместимо или противоречит определению данного термина, приведенному в данном документе, применимо определение данного термина, приведенное в данном документе, а определение такого термина из литературного источника не применимо. Каждый из более узких видов и каждая из более узких субродовых групп, подпадающих под общее раскрытие, также составляют часть настоящего изобретения. Сюда относится общее описание настоящего изобретения с условием или отрицательным ограничением, исключающим любой предмет из рода, независимо от того, упомянут ли конкретно в данном документе вырезанный материал. Кроме того, если признаки описаны по принципу групп Маркуша, специалисты в данной области поймут, что настоящее изобретение также таким образом описано в категориях любого отдельного члена или подгруппы членов группы Маркуша. Дополнительные варианты осуществления станут очевидны из представленной далее формулы изобретения.
[00420] Эквиваленты: специалисты в данной области поймут или с помощью постановки простейшего эксперимента смогут установить многие эквиваленты конкретных вариантов осуществления настоящего изобретения, которые описаны в данном документе. Подразумевается, что такие эквиваленты охватываются представленной далее формулой изобретения. Все публикации, патенты и патентные заявки, упомянутые в данном описании, настоящим включены в данное описание посредством ссылки в той же степени, как если бы каждая отдельная публикация, патент или патентная заявка были специально и отдельно указаны как включенные в данный документ посредством ссылки.
VI. НЕПАТЕНТНЫЕ ЛИТЕРАТУРНЫЕ ИСТОЧНИКИ
1. CHENG, W., TSENG, C. J., LIN, T. T., CHENG, I., PAN, H. W., HSU, H. C. & LEE, Y. M. 2008. Glypican-3-mediated oncogenesis involves the Insulin-like growth factor-signaling pathway. Carcinogenesis, 29, 1319-26.
2. SONG, H. H., SHI, W., XIANG, Y. Y. & FILMUS, J. 2005. The loss of glypican-3 induces alterations in Wnt signaling. J Biol Chem, 280, 2116-25.
3. SONG, H. H., SHI, W. & FILMUS, J. 1997. OCI-5/rat glypican-3 binds to fibroblast growth factor-2 but not to insulin-like growth factor-2. J Biol Chem, 272, 7574-7.
4. PILIA, G., HUGHES-BENZIE, R. M., MACKENZIE, A., BAYBAYAN, P., CHEN, E. Y., HUBER, R., NERI, G., CAO, A., FORABOSCO, A. & SCHLESSINGER, D. 1996. Mutations in GPC3, a glypican gene, cause the Simpson-Golabi-Behmel overgrowth syndrome. Nat Genet, 12, 241-7.
5. AYDIN, O., YILDIZ, L., BARIS, S., DUNDAR, C. & KARAGOZ, F. 2015. Expression of Glypican 3 in low and high grade urothelial carcinomas. Diagn Pathol, 10, 34.
6. USHIKU, T., UOZAKI, H., SHINOZAKI, A., OTA, S., MATSUZAKA, K., NOMURA, S., KAMINISHI, M., ABURATANI, H., KODAMA, T. & FUKAYAMA, M. 2009. Glypican 3-expressing gastric carcinoma: distinct subgroup unifying hepatoid, clear-cell, and alpha-fetoprotein-producing gastric carcinomas. Cancer Sci, 100, 626-32.
7. GAILEY, M. P. & BELLIZZI, A. M. 2013. Immunohistochemistry for the novel markers glypican 3, PAX8, and p40 (DeltaNp63) in squamous cell and urothelial carcinoma. Am J Clin Pathol, 140, 872-80.
8. YAMANAKA, K., ITO, Y., OKUYAMA, N., NODA, K., MATSUMOTO, H., YOSHIDA, H., MIYAUCHI, A., CAPURRO, M., FILMUS, J. & MIYOSHI, E. 2007. Immunohistochemical study of glypican 3 in thyroid cancer. Oncology, 73, 389-94.
9. NAKATSURA, T., KAGESHITA, T., ITO, S., WAKAMATSU, K., MONJI, M., IKUTA, Y., SENJU, S., ONO, T. & NISHIMURA, Y. 2004. Identification of glypican-3 as a novel tumor marker for melanoma. Clin Cancer Res, 10, 6612-21.
10. ZYNGER, D. L., DIMOV, N. D., LUAN, C., TEH, B. T. & YANG, X. J. 2006. Glypican 3: a novel marker in testicular germ cell tumors. Am J Surg Pathol, 30, 1570-5.
11. MONTALBANO, M., RASTELLINI, C., WANG, X., CORSELLO, T., ELTORKY, M. A., VENTO, R. & CICALESE, L. 2016. Transformation of primary human hepatocytes in hepatocellular carcinoma. Int J Oncol, 48, 1205-17.
12. MIDORIKAWA, Y., ISHIKAWA, S., IWANARI, H., IMAMURA, T., SAKAMOTO, H., MIYAZONO, K., KODAMA, T., MAKUUCHI, M. & ABURATANI, H. 2003. Glypican-3, overexpressed in hepatocellular carcinoma, modulates FGF2 and BMP-7 signaling. Int J Cancer, 103, 455-65.
13. CAPURRO, M., WANLESS, I. R., SHERMAN, M., DEBOER, G., SHI, W., MIYOSHI, E. & FILMUS, J. 2003. Glypican-3: a novel serum and histochemical marker for hepatocellular carcinoma. Gastroenterology, 125, 89-97.
14. NAKATSURA, T., YOSHITAKE, Y., SENJU, S., MONJI, M., KOMORI, H., MOTOMURA, Y., HOSAKA, S., BEPPU, T., ISHIKO, T., KAMOHARA, H., ASHIHARA, H., KATAGIRI, T., FURUKAWA, Y., FUJIYAMA, S., OGAWA, M., NAKAMURA, Y. & NISHIMURA, Y. 2003. Glypican-3, overexpressed specifically in human hepatocellular carcinoma, is a novel tumor marker. Biochem Biophys Res Commun, 306, 16-25.
15. SUNG, Y. K., HWANG, S. Y., PARK, M. K., FAROOQ, M., HAN, I. S., BAE, H. I., KIM, J. C. & KIM, M. 2003. Glypican-3 is overexpressed in human hepatocellular carcinoma. Cancer Sci, 94, 259-62.
16. ZHU, Z. W., FRIESS, H., WANG, L., ABOU-SHADY, M., ZIMMERMANN, A., LANDER, A. D., KORC, M., KLEEFF, J. & BUCHLER, M. W. 2001. Enhanced glypican-3 expression differentiates the majority of hepatocellular carcinomas from benign hepatic disorders. Gut, 48, 558-64.
17. JELIC, S., SOTIROPOULOS, G. C. & GROUP, E. G. W. 2010. Hepatocellular carcinoma: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann Oncol, 21 Suppl 5, v59-64.
18. FENG, M. & HO, M. 2014. Glypican-3 antibodies: a new therapeutic target for liver cancer. FEBS Lett, 588, 377-82.
19. TAKAI, H., KATO, A., KINOSHITA, Y., ISHIGURO, T., TAKAI, Y., OHTANI, Y., SUGIMOTO, M. & SUZUKI, M. 2009. Histopathological analyses of the antitumor activity of anti-glypican-3 antibody (GC33) in human liver cancer xenograft models: The contribution of macrophages. Cancer Biol Ther, 8, 930-8.
20. NAKANO, K., ORITA, T., NEZU, J., YOSHINO, T., OHIZUMI, I., SUGIMOTO, M., FURUGAKI, K., KINOSHITA, Y., ISHIGURO, T., HAMAKUBO, T., KODAMA, T., ABURATANI, H., YAMADA-OKABE, H. & TSUCHIYA, M. 2009. Anti-glypican 3 antibodies cause ADCC against human hepatocellular carcinoma cells. Biochem Biophys Res Commun, 378, 279-84.
21. ISHIGURO, T., SUGIMOTO, M., KINOSHITA, Y., MIYAZAKI, Y., NAKANO, K., TSUNODA, H., SUGO, I., OHIZUMI, I., ABURATANI, H., HAMAKUBO, T., KODAMA, T., TSUCHIYA, M. & YAMADA-OKABE, H. 2008. Anti-glypican 3 antibody as a potential antitumor agent for human liver cancer. Cancer Res, 68, 9832-8.
22. LI, S. Y. & LIU, Y. 2013. Immunotherapy of melanoma with the immune costimulatory monoclonal antibodies targeting CD137. Clin Pharmacol, 5, 47-53.
23. SNELL, L. M., LIN, G. H., MCPHERSON, A. J., MORAES, T. J. & WATTS, T. H. 2011. T-cell intrinsic effects of GITR and 4-1BB during viral infection and cancer immunotherapy. Immunol Rev, 244, 197-217.
24. WYZGOL, A., MULLER, N., FICK, A., MUNKEL, S., GRIGOLEIT, G. U., PFIZENMAIER, K. & WAJANT, H. 2009. Trimer stabilization, oligomerization, and antibody-mediated cell surface immobilization improve the activity of soluble trimers of CD27L, CD40L, 41BBL, and glucocorticoid-induced TNF receptor ligand. J Immunol, 183, 1851-61.
25. YAO, S., ZHU, Y. & CHEN, L. 2013. Advances in targeting cell surface signalling molecules for immune modulation. Nat Rev Drug Discov, 12, 130-46.
26. MELERO, I., BACH, N., HELLSTROM, K. E., ARUFFO, A., MITTLER, R. S. & CHEN, L. 1998. Amplification of tumor immunity by gene transfer of the co-stimulatory 4-1BB ligand: synergy with the CD28 co-stimulatory pathway. Eur J Immunol, 28, 1116-21.
27. YANG, Y., YANG, S., YE, Z., JAFFAR, J., ZHOU, Y., CUTTER, E., LIEBER, A., HELLSTROM, I. & HELLSTROM, K. E. 2007. Tumor cells expressing anti-CD137 scFv induce a tumor-destructive environment. Cancer Res, 67, 2339-44.
28. ZHANG, H., KNUTSON, K. L., HELLSTROM, K. E., DISIS, M. L. & HELLSTROM, I. 2006. Antitumor efficacy of CD137 ligation is maximized by the use of a CD137 single-chain Fv-expressing whole-cell tumor vaccine compared with CD137-specific monoclonal antibody infusion. Mol Cancer Ther, 5, 149-55.
29. YE, Z., HELLSTROM, I., HAYDEN-LEDBETTER, M., DAHLIN, A., LEDBETTER, J. A. & HELLSTROM, K. E. 2002. Gene therapy for cancer using single-chain Fv fragments specific for 4-1BB. Nat Med, 8, 343-8.
30. MARTINET, O., DIVINO, C. M., ZANG, Y., GAN, Y., MANDELI, J., THUNG, S., PAN, P. Y. & CHEN, S. H. 2002. T cell activation with systemic agonistic antibody versus local 4-1BB ligand gene delivery combined with interleukin-12 eradicate liver metastases of breast cancer. Gene Ther, 9, 786-92.
31. YE, Q., SONG, D. G., POUSSIN, M., YAMAMOTO, T., BEST, A., LI, C., COUKOS, G. & POWELL, D. J., JR. 2014. CD137 accurately identifies and enriches for naturally occurring tumor-reactive T cells in tumor. Clin Cancer Res, 20, 44-55.
32. CHACON, J. A., WU, R. C., SUKHUMALCHANDRA, P., MOLLDREM, J. J., SARNAIK, A., PILON-THOMAS, S., WEBER, J., HWU, P. & RADVANYI, L. 2013. Co-stimulation through 4-1BB/CD137 improves the expansion and function of CD8(+) melanoma tumor-infiltrating lymphocytes for adoptive T-cell therapy. PLoS One, 8, e60031.
33. FISHER, T. S., KAMPERSCHROER, C., OLIPHANT, T., LOVE, V. A., LIRA, P. D., DOYONNAS, R., BERGQVIST, S., BAXI, S. M., ROHNER, A., SHEN, A. C., HUANG, C., SOKOLOWSKI, S. A. & SHARP, L. L. 2012. Targeting of 4-1BB by monoclonal antibody PF-05082566 enhances T-cell function and promotes anti-tumor activity. Cancer Immunol Immunother, 61, 1721-33.
34. SKERRA, A. 2000. Lipocalins as a scaffold. Biochim Biophys Acta, 1482, 337-50.
35. FLOWER, D. R., NORTH, A. C. & SANSOM, C. E. 2000. The lipocalin protein family: structural and sequence overview. Biochim Biophys Acta, 1482, 9-24.
36. FLOWER, D. R. 1996. The lipocalin protein family: structure and function. Biochem J, 318 ( Pt 1), 1-14.
37. ALTSCHUL, S. F., MADDEN, T. L., SCHAFFER, A. A., ZHANG, J., ZHANG, Z., MILLER, W. & LIPMAN, D. J. 1997. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res, 25, 3389-402.
38. ALTSCHUL, S. F., GISH, W., MILLER, W., MYERS, E. W. & LIPMAN, D. J. 1990. Basic local alignment search tool. J Mol Biol, 215, 403-10.
39. SMITH, T. F. & WATERMAN, M. S. 1981. Identification of common molecular subsequences. J Mol Biol, 147, 195-7.
40. WARD, E. S., GUSSOW, D., GRIFFITHS, A. D., JONES, P. T. & WINTER, G. 1989. Binding activities of a repertoire of single immunoglobulin variable domains secreted from Escherichia coli. Nature, 341, 544-6.
41. HOLLIGER, P., PROSPERO, T. & WINTER, G. 1993. "Diabodies": small bivalent and bispecific antibody fragments. Proc Natl Acad Sci U S A, 90, 6444-8.
42. JOHNSON, G. & WU, T. T. 2000. Kabat database and its applications: 30 years after the first variability plot. Nucleic Acids Res, 28, 214-8.
43. EHRENMANN, F., KAAS, Q. & LEFRANC, M. P. 2010. IMGT/3Dstructure-DB and IMGT/DomainGapAlign: a database and a tool for immunoglobulins or antibodies, T cell receptors, MHC, IgSF and MhcSF. Nucleic Acids Res, 38, D301-7.
44. BULLIARD, Y., JOLICOEUR, R., ZHANG, J., DRANOFF, G., WILSON, N. S. & BROGDON, J. L. 2014. OX40 engagement depletes intratumoral Tregs via activating FcgammaRs, leading to antitumor efficacy. Immunol Cell Biol, 92, 475-80.
45. BULLIARD, Y., JOLICOEUR, R., WINDMAN, M., RUE, S. M., ETTENBERG, S., KNEE, D. A., WILSON, N. S., DRANOFF, G. & BROGDON, J. L. 2013. Activating Fc gamma receptors contribute to the antitumor activities of immunoregulatory receptor-targeting antibodies. J Exp Med, 210, 1685-93.
46. SPIESS, C., ZHAI, Q. & CARTER, P. J. 2015. Alternative molecular formats and therapeutic applications for bispecific antibodies. Mol Immunol, 67, 95-106.
47. SEDYKH, S. E., PRINZ, V. V., BUNEVA, V. N. & NEVINSKY, G. A. 2018. Bispecific antibodies: design, therapy, perspectives. Drug Des Devel Ther, 12, 195-208.
48. ALAVIJEH, M. S. & PALMER, A. M. 2004. The pivotal role of drug metabolism and pharmacokinetics in the discovery and development of new medicines. IDrugs, 7, 755-63.
49. RYMAN, J. T. & MEIBOHM, B. 2017. Pharmacokinetics of Monoclonal Antibodies. CPT Pharmacometrics Syst Pharmacol, 6, 576-588.
50. SILVA, J. P., VETTERLEIN, O., JOSE, J., PETERS, S. & KIRBY, H. 2015. The S228P mutation prevents in vivo and in vitro IgG4 Fab-arm exchange as demonstrated using a combination of novel quantitative immunoassays and physiological matrix preparation. J Biol Chem, 290, 5462-9.
51. GLAESNER, W., VICK, A. M., MILLICAN, R., ELLIS, B., TSCHANG, S. H., TIAN, Y., BOKVIST, K., BRENNER, M., KOESTER, A., PORKSEN, N., ETGEN, G. & BUMOL, T. 2010. Engineering and characterization of the long-acting glucagon-like peptide-1 analogue LY2189265, an Fc fusion protein. Diabetes Metab Res Rev, 26, 287-96.
52. DALL'ACQUA, W. F., KIENER, P. A. & WU, H. 2006. Properties of human IgG1s engineered for enhanced binding to the neonatal Fc receptor (FcRn). J Biol Chem, 281, 23514-24.
53. ZALEVSKY, J., CHAMBERLAIN, A. K., HORTON, H. M., KARKI, S., LEUNG, I. W., SPROULE, T. J., LAZAR, G. A., ROOPENIAN, D. C. & DESJARLAIS, J. R. 2010. Enhanced antibody half-life improves in vivo activity. Nat Biotechnol, 28, 157-9.
54. SHIELDS, R. L., NAMENUK, A. K., HONG, K., MENG, Y. G., RAE, J., BRIGGS, J., XIE, D., LAI, J., STADLEN, A., LI, B., FOX, J. A. & PRESTA, L. G. 2001. High resolution mapping of the binding site on human IgG1 for Fc gamma RI, Fc gamma RII, Fc gamma RIII, and FcRn and design of IgG1 variants with improved binding to the Fc gamma R. J Biol Chem, 276, 6591-604.
55. ALTSHULER, E. P., SEREBRYANAYA, D. V. & KATRUKHA, A. G. 2010. Generation of recombinant antibodies and means for increasing their affinity. Biochemistry (Mosc), 75, 1584-605.
56. HARLOW, E. & LANE, D. 1999. Using antibodies : a laboratory manual, Cold Spring Harbor, N.Y., Cold Spring Harbor Laboratory Press.
57. HARLOW, E. & LANE, D. 1988. Antibodies : a laboratory manual, Cold Spring Harbor, NY, Cold Spring Harbor Laboratory.
58. LI, J., SAI, T., BERGER, M., CHAO, Q., DAVIDSON, D., DESHMUKH, G., DROZDOWSKI, B., EBEL, W., HARLEY, S., HENRY, M., JACOB, S., KLINE, B., LAZO, E., ROTELLA, F., ROUTHIER, E., RUDOLPH, K., SAGE, J., SIMON, P., YAO, J., ZHOU, Y., KAVURU, M., BONFIELD, T., THOMASSEN, M. J., SASS, P. M., NICOLAIDES, N. C. & GRASSO, L. 2006. Human antibodies for immunotherapy development generated via a human B cell hybridoma technology. Proc Natl Acad Sci U S A, 103, 3557-62.
59. KOZBOR, D. & RODER, J. C. 1983. The production of monoclonal antibodies from human lymphocytes. Immunol Today, 4, 72-9.
60. COLE, S. P., CAMPLING, B. G., LOUWMAN, I. H., KOZBOR, D. & RODER, J. C. 1984. A strategy for the production of human monoclonal antibodies reactive with lung tumor cell lines. Cancer Res, 44, 2750-3.
61. HOLLIGER, P. & HUDSON, P. J. 2005. Engineered antibody fragments and the rise of single domains. Nat Biotechnol, 23, 1126-36.
62. PERVAIZ, S. & BREW, K. 1987. Homology and structure-function correlations between alpha 1-acid glycoprotein and serum retinol-binding protein and its relatives. FASEB J, 1, 209-14.
63. SAMBROOK, J. & RUSSELL, D. W. 2001. Molecular cloning: a laboratory manual, Cold Spring Harbor, N.Y., Cold Spring Harbor Laboratory Press.
64. FLOWER, D. R. 2000. Beyond the superfamily: the lipocalin receptors. Biochim Biophys Acta, 1482, 327-36.
65. BREUSTEDT, D. A., KORNDORFER, I. P., REDL, B. & SKERRA, A. 2005. The 1.8-A crystal structure of human tear lipocalin reveals an extended branched cavity with capacity for multiple ligands. J Biol Chem, 280, 484-93.
66. SCHMIDT, T. G., KOEPKE, J., FRANK, R. & SKERRA, A. 1996. Molecular interaction between the Strep-tag affinity peptide and its cognate target, streptavidin. J Mol Biol, 255, 753-66.
67. VAJO, Z. & DUCKWORTH, W. C. 2000. Genetically engineered insulin analogs: diabetes in the new millenium. Pharmacol Rev, 52, 1-9.
68. FUERTGES, F. & ABUCHOWSKI, A. 1990. The clinical efficacy of poly(ethylene glycol)-modified proteins. Journal of Controlled Release, 11, 139-148.
69. DENNIS, M. S., ZHANG, M., MENG, Y. G., KADKHODAYAN, M., KIRCHHOFER, D., COMBS, D. & DAMICO, L. A. 2002. Albumin binding as a general strategy for improving the pharmacokinetics of proteins. J Biol Chem, 277, 35035-43.
70. KONIG, T. & SKERRA, A. 1998. Use of an albumin-binding domain for the selective immobilisation of recombinant capture antibody fragments on ELISA plates. J Immunol Methods, 218, 73-83.
71. OSBORN, B. L., OLSEN, H. S., NARDELLI, B., MURRAY, J. H., ZHOU, J. X., GARCIA, A., MOODY, G., ZARITSKAYA, L. S. & SUNG, C. 2002. Pharmacokinetic and pharmacodynamic studies of a human serum albumin-interferon-alpha fusion protein in cynomolgus monkeys. J Pharmacol Exp Ther, 303, 540-8.
72. LOWMAN, H. B. 1997. Bacteriophage display and discovery of peptide leads for drug development. Annu Rev Biophys Biomol Struct, 26, 401-24.
73. RODI, D. J. & MAKOWSKI, L. 1999. Phage-display technology--finding a needle in a vast molecular haystack. Curr Opin Biotechnol, 10, 87-93.
74. VENTURI, M., SEIFERT, C. & HUNTE, C. 2002. High level production of functional antibody Fab fragments in an oxidizing bacterial cytoplasm. J Mol Biol, 315, 1-8.
75. BRUCKDORFER, T., MARDER, O. & ALBERICIO, F. 2004. From production of peptides in milligram amounts for research to multi-tons quantities for drugs of the future. Curr Pharm Biotechnol, 5, 29-43.
76. FRESE, K., EISENMANN, M., OSTENDORP, R., BROCKS, B. & PABST, S. 2013. An automated immunoassay for early specificity profiling of antibodies. MAbs, 5, 279-87.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Пиерис Фармасьютикалс ГмбХ
<120> Новые слитые белки, специфические в отношении CD137
и GPC3
<130> PIE16678PCT
<150> EP 19000100.8
<151> 2019-02-26
<160> 129
<170> PatentIn версия 3.5
<210> 1
<211> 158
<212> Белок
<213> человек
<400> 1
His His Leu Leu Ala Ser Asp Glu Glu Ile Gln Asp Val Ser Gly Thr
1 5 10 15
Trp Tyr Leu Lys Ala Met Thr Val Asp Arg Glu Phe Pro Glu Met Asn
20 25 30
Leu Glu Ser Val Thr Pro Met Thr Leu Thr Thr Leu Glu Gly Gly Asn
35 40 45
Leu Glu Ala Lys Val Thr Met Leu Ile Ser Gly Arg Cys Gln Glu Val
50 55 60
Lys Ala Val Leu Glu Lys Thr Asp Glu Pro Gly Lys Tyr Thr Ala Asp
65 70 75 80
Gly Gly Lys His Val Ala Tyr Ile Ile Arg Ser His Val Lys Asp His
85 90 95
Tyr Ile Phe Tyr Cys Glu Gly Glu Leu His Gly Lys Pro Val Arg Gly
100 105 110
Val Lys Leu Val Gly Arg Asp Pro Lys Asn Asn Leu Glu Ala Leu Glu
115 120 125
Asp Phe Glu Lys Ala Ala Gly Ala Arg Gly Leu Ser Thr Glu Ser Ile
130 135 140
Leu Ile Pro Arg Gln Ser Glu Thr Cys Ser Pro Gly Ser Asp
145 150 155
<210> 2
<211> 178
<212> Белок
<213> человек
<400> 2
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Leu Ala Gly Asn Ala Ile Leu Arg Glu Asp Lys Asp Pro
35 40 45
Gln Lys Met Tyr Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asn Val Thr Ser Val Leu Phe Arg Lys Lys Lys Cys Asp Tyr Trp Ile
65 70 75 80
Arg Thr Phe Val Pro Gly Cys Gln Pro Gly Glu Phe Thr Leu Gly Asn
85 90 95
Ile Lys Ser Tyr Pro Gly Leu Thr Ser Tyr Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Lys Val Ser Gln
115 120 125
Asn Arg Glu Tyr Phe Lys Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 3
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 3
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Leu Ala Gly Asn Ala Ile Leu Arg Glu Asp Lys Asp Pro
35 40 45
Gln Lys Met Tyr Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asn Val Thr Ser Val Leu Phe Arg Lys Lys Lys Cys Asp Tyr Trp Ile
65 70 75 80
Arg Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Asn
85 90 95
Ile Lys Ser Tyr Pro Gly Leu Thr Ser Tyr Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Lys Val Ser Gln
115 120 125
Asn Arg Glu Tyr Phe Lys Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 4
<211> 152
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 4
Ala Ser Asp Glu Glu Ile Gln Asp Val Ser Gly Thr Trp Tyr Leu Lys
1 5 10 15
Ala Met Thr Val Asp Arg Glu Cys Pro Glu Met Asn Leu Glu Ser Val
20 25 30
Thr Pro Met Thr Leu Thr Thr Leu Glu Gly Gly Asn Leu Glu Ala Lys
35 40 45
Val Thr Met Leu Ile Ser Gly Arg Ser Gln Glu Val Lys Ala Val Leu
50 55 60
Glu Lys Thr Asp Glu Pro Gly Lys Tyr Thr Ala Asp Gly Gly Lys His
65 70 75 80
Val Ala Tyr Ile Ile Arg Ser His Val Lys Asp His Tyr Ile Phe Tyr
85 90 95
Ser Glu Gly Glu Cys His Gly Lys Pro Val Pro Gly Val Trp Leu Val
100 105 110
Gly Arg Asp Pro Lys Asn Asn Leu Glu Ala Leu Glu Asp Phe Glu Lys
115 120 125
Ala Ala Gly Ala Arg Gly Leu Ser Thr Glu Ser Ile Leu Ile Pro Arg
130 135 140
Gln Ser Glu Thr Ser Ser Pro Gly
145 150
<210> 5
<211> 4
<212> Белок
<213> человек
<400> 5
His His Leu Leu
1
<210> 6
<211> 535
<212> Белок
<213> человек
<400> 6
Gln Pro Pro Pro Pro Pro Pro Asp Ala Thr Cys His Gln Val Arg Ser
1 5 10 15
Phe Phe Gln Arg Leu Gln Pro Gly Leu Lys Trp Val Pro Glu Thr Pro
20 25 30
Val Pro Gly Ser Asp Leu Gln Val Cys Leu Pro Lys Gly Pro Thr Cys
35 40 45
Cys Ser Arg Lys Met Glu Glu Lys Tyr Gln Leu Thr Ala Arg Leu Asn
50 55 60
Met Glu Gln Leu Leu Gln Ser Ala Ser Met Glu Leu Lys Phe Leu Ile
65 70 75 80
Ile Gln Asn Ala Ala Val Phe Gln Glu Ala Phe Glu Ile Val Val Arg
85 90 95
His Ala Lys Asn Tyr Thr Asn Ala Met Phe Lys Asn Asn Tyr Pro Ser
100 105 110
Leu Thr Pro Gln Ala Phe Glu Phe Val Gly Glu Phe Phe Thr Asp Val
115 120 125
Ser Leu Tyr Ile Leu Gly Ser Asp Ile Asn Val Asp Asp Met Val Asn
130 135 140
Glu Leu Phe Asp Ser Leu Phe Pro Val Ile Tyr Thr Gln Leu Met Asn
145 150 155 160
Pro Gly Leu Pro Asp Ser Ala Leu Asp Ile Asn Glu Cys Leu Arg Gly
165 170 175
Ala Arg Arg Asp Leu Lys Val Phe Gly Asn Phe Pro Lys Leu Ile Met
180 185 190
Thr Gln Val Ser Lys Ser Leu Gln Val Thr Arg Ile Phe Leu Gln Ala
195 200 205
Leu Asn Leu Gly Ile Glu Val Ile Asn Thr Thr Asp His Leu Lys Phe
210 215 220
Ser Lys Asp Cys Gly Arg Met Leu Thr Arg Met Trp Tyr Cys Ser Tyr
225 230 235 240
Cys Gln Gly Leu Met Met Val Lys Pro Cys Gly Gly Tyr Cys Asn Val
245 250 255
Val Met Gln Gly Cys Met Ala Gly Val Val Glu Ile Asp Lys Tyr Trp
260 265 270
Arg Glu Tyr Ile Leu Ser Leu Glu Glu Leu Val Asn Gly Met Tyr Arg
275 280 285
Ile Tyr Asp Met Glu Asn Val Leu Leu Gly Leu Phe Ser Thr Ile His
290 295 300
Asp Ser Ile Gln Tyr Val Gln Lys Asn Ala Gly Lys Leu Thr Thr Thr
305 310 315 320
Ile Gly Lys Leu Cys Ala His Ser Gln Gln Arg Gln Tyr Arg Ser Ala
325 330 335
Tyr Tyr Pro Glu Asp Leu Phe Ile Asp Lys Lys Val Leu Lys Val Ala
340 345 350
His Val Glu His Glu Glu Thr Leu Ser Ser Arg Arg Arg Glu Leu Ile
355 360 365
Gln Lys Leu Lys Ser Phe Ile Ser Phe Tyr Ser Ala Leu Pro Gly Tyr
370 375 380
Ile Cys Ser His Ser Pro Val Ala Glu Asn Asp Thr Leu Cys Trp Asn
385 390 395 400
Gly Gln Glu Leu Val Glu Arg Tyr Ser Gln Lys Ala Ala Arg Asn Gly
405 410 415
Met Lys Asn Gln Phe Asn Leu His Glu Leu Lys Met Lys Gly Pro Glu
420 425 430
Pro Val Val Ser Gln Ile Ile Asp Lys Leu Lys His Ile Asn Gln Leu
435 440 445
Leu Arg Thr Met Ser Met Pro Lys Gly Arg Val Leu Asp Lys Asn Leu
450 455 460
Asp Glu Glu Gly Phe Glu Ser Gly Asp Cys Gly Asp Asp Glu Asp Glu
465 470 475 480
Cys Ile Gly Gly Ser Gly Asp Gly Met Ile Lys Val Lys Asn Gln Leu
485 490 495
Arg Phe Leu Ala Glu Leu Ala Tyr Asp Leu Asp Val Asp Asp Ala Pro
500 505 510
Gly Asn Ser Gln Gln Ala Thr Pro Lys Asp Asn Glu Ile Ser Thr Phe
515 520 525
His Asn Leu Gly Asn Val His
530 535
<210> 7
<211> 232
<212> Белок
<213> человек
<400> 7
Leu Gln Asp Pro Cys Ser Asn Cys Pro Ala Gly Thr Phe Cys Asp Asn
1 5 10 15
Asn Arg Asn Gln Ile Cys Ser Pro Cys Pro Pro Asn Ser Phe Ser Ser
20 25 30
Ala Gly Gly Gln Arg Thr Cys Asp Ile Cys Arg Gln Cys Lys Gly Val
35 40 45
Phe Arg Thr Arg Lys Glu Cys Ser Ser Thr Ser Asn Ala Glu Cys Asp
50 55 60
Cys Thr Pro Gly Phe His Cys Leu Gly Ala Gly Cys Ser Met Cys Glu
65 70 75 80
Gln Asp Cys Lys Gln Gly Gln Glu Leu Thr Lys Lys Gly Cys Lys Asp
85 90 95
Cys Cys Phe Gly Thr Phe Asn Asp Gln Lys Arg Gly Ile Cys Arg Pro
100 105 110
Trp Thr Asn Cys Ser Leu Asp Gly Lys Ser Val Leu Val Asn Gly Thr
115 120 125
Lys Glu Arg Asp Val Val Cys Gly Pro Ser Pro Ala Asp Leu Ser Pro
130 135 140
Gly Ala Ser Ser Val Thr Pro Pro Ala Pro Ala Arg Glu Pro Gly His
145 150 155 160
Ser Pro Gln Ile Ile Ser Phe Phe Leu Ala Leu Thr Ser Thr Ala Leu
165 170 175
Leu Phe Leu Leu Phe Phe Leu Thr Leu Arg Phe Ser Val Val Lys Arg
180 185 190
Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met Arg Pro
195 200 205
Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe Pro Glu
210 215 220
Glu Glu Glu Gly Gly Cys Glu Leu
225 230
<210> 8
<211> 535
<212> Белок
<213> Macaca fascicularis
<400> 8
Gln Pro Pro Pro Pro Pro Pro Asp Ala Thr Cys His Gln Val Arg Ser
1 5 10 15
Phe Phe Gln Arg Leu Gln Pro Gly Leu Lys Trp Val Pro Glu Thr Pro
20 25 30
Val Pro Gly Ser Asp Leu Gln Val Cys Leu Pro Lys Gly Pro Thr Cys
35 40 45
Cys Ser Arg Lys Met Glu Glu Lys Tyr Gln Leu Thr Ala Arg Leu Asn
50 55 60
Met Glu Gln Leu Leu Gln Ser Ala Ser Met Glu Leu Lys Phe Leu Ile
65 70 75 80
Ile Gln Asn Ala Ala Val Phe Gln Glu Ala Phe Glu Ile Val Val Arg
85 90 95
His Ala Lys Asn Tyr Thr Asn Ala Met Phe Lys Asn Asn Tyr Pro Ser
100 105 110
Leu Thr Pro Gln Ala Phe Glu Phe Val Gly Glu Phe Phe Thr Asp Val
115 120 125
Ser Leu Tyr Ile Leu Gly Ser Asp Ile Asn Val Asp Asp Met Val Asn
130 135 140
Glu Leu Phe Asp Ser Leu Phe Pro Val Ile Tyr Thr Gln Leu Met Asn
145 150 155 160
Pro Gly Leu Pro Asp Ser Ala Leu Asp Ile Asn Glu Cys Leu Arg Gly
165 170 175
Ala Arg Arg Asp Leu Lys Val Phe Gly Asn Phe Pro Lys Leu Ile Met
180 185 190
Thr Gln Val Ser Lys Ser Leu Gln Val Thr Arg Ile Phe Leu Gln Ala
195 200 205
Leu Asn Leu Gly Ile Glu Val Ile Asn Thr Thr Asp His Leu Lys Phe
210 215 220
Ser Lys Asp Cys Gly Arg Met Leu Thr Arg Met Trp Tyr Cys Ser Tyr
225 230 235 240
Cys Gln Gly Leu Met Met Val Lys Pro Cys Gly Gly Tyr Cys Asn Val
245 250 255
Val Met Gln Gly Cys Met Ala Gly Val Val Glu Ile Asp Lys Tyr Trp
260 265 270
Arg Glu Tyr Ile Leu Ser Leu Glu Glu Leu Val Asn Gly Met Tyr Arg
275 280 285
Ile Tyr Asp Met Glu Asn Val Leu Leu Gly Leu Phe Ser Thr Ile His
290 295 300
Asp Ser Ile Gln Tyr Val Gln Lys Asn Ala Gly Lys Leu Thr Thr Thr
305 310 315 320
Ile Gly Lys Leu Cys Ala His Ser Gln Gln Arg Gln Tyr Arg Ser Ala
325 330 335
Tyr Tyr Pro Glu Asp Leu Phe Ile Asp Lys Lys Val Leu Lys Val Ala
340 345 350
His Val Glu His Glu Glu Thr Leu Ser Ser Arg Arg Arg Glu Leu Ile
355 360 365
Gln Lys Leu Lys Ser Phe Ile Ser Phe Tyr Ser Ala Leu Pro Gly Tyr
370 375 380
Ile Cys Ser His Ser Pro Val Ala Glu Asn Asp Thr Leu Cys Trp Asn
385 390 395 400
Gly Gln Glu Leu Val Glu Arg Tyr Ser Gln Lys Ala Ala Arg Asn Gly
405 410 415
Met Lys Asn Gln Phe Asn Leu His Glu Leu Lys Met Lys Gly Pro Glu
420 425 430
Pro Val Val Ser Gln Ile Ile Asp Lys Leu Lys His Ile Asn Gln Leu
435 440 445
Leu Arg Thr Met Ser Val Pro Lys Gly Arg Val Leu Asp Lys Asn Leu
450 455 460
Asp Glu Glu Gly Phe Glu Ser Gly Asp Cys Gly Asp Asp Glu Asp Glu
465 470 475 480
Cys Ile Gly Gly Ser Gly Asp Gly Met Met Lys Val Lys Asn Gln Leu
485 490 495
Arg Phe Leu Ala Glu Leu Ala Tyr Asp Leu Asp Val Asp Asp Val Pro
500 505 510
Gly Asn Asn Gln Gln Ala Thr Pro Lys Asp Asn Glu Ile Ser Thr Phe
515 520 525
His Asn Leu Gly Asn Val His
530 535
<210> 9
<211> 231
<212> Белок
<213> Macaca fascicularis
<400> 9
Leu Gln Asp Leu Cys Ser Asn Cys Pro Ala Gly Thr Phe Cys Asp Asn
1 5 10 15
Asn Arg Ser Gln Ile Cys Ser Pro Cys Pro Pro Asn Ser Phe Ser Ser
20 25 30
Ala Gly Gly Gln Arg Thr Cys Asp Ile Cys Arg Gln Cys Lys Gly Val
35 40 45
Phe Lys Thr Arg Lys Glu Cys Ser Ser Thr Ser Asn Ala Glu Cys Asp
50 55 60
Cys Ile Ser Gly Tyr His Cys Leu Gly Ala Glu Cys Ser Met Cys Glu
65 70 75 80
Gln Asp Cys Lys Gln Gly Gln Glu Leu Thr Lys Lys Gly Cys Lys Asp
85 90 95
Cys Cys Phe Gly Thr Phe Asn Asp Gln Lys Arg Gly Ile Cys Arg Pro
100 105 110
Trp Thr Asn Cys Ser Leu Asp Gly Lys Ser Val Leu Val Asn Gly Thr
115 120 125
Lys Glu Arg Asp Val Val Cys Gly Pro Ser Pro Ala Asp Leu Ser Pro
130 135 140
Gly Ala Ser Ser Ala Thr Pro Pro Ala Pro Ala Arg Glu Pro Gly His
145 150 155 160
Ser Pro Gln Ile Ile Phe Phe Leu Ala Leu Thr Ser Thr Val Val Leu
165 170 175
Phe Leu Leu Phe Phe Leu Val Leu Arg Phe Ser Val Val Lys Arg Ser
180 185 190
Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met Arg Pro Val
195 200 205
Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe Pro Glu Glu
210 215 220
Glu Glu Gly Gly Cys Glu Leu
225 230
<210> 10
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> сайт расщепления
<400> 10
Ile Glu Gly Arg
1
<210> 11
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> слитый пептид
<400> 11
Ser Ala Trp Ser His Pro Gln Phe Glu Lys
1 5 10
<210> 12
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> пептид со стрептовидиновой меткой II
<400> 12
Trp Ser His Pro Gln Phe Glu Lys
1 5
<210> 13
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> линкер
<400> 13
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 14
<211> 18
<212> Белок
<213> Искусственная последовательность
<220>
<223> линкер
<400> 14
Pro Ser Thr Pro Pro Thr Asn Ser Ser Ser Thr Pro Pro Thr Pro Ser
1 5 10 15
Pro Ser
<210> 15
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> линкер
<400> 15
Gly Gly Ser Gly Asn Ser Ser Gly Ser Gly Gly Ser Pro Val
1 5 10
<210> 16
<211> 20
<212> Белок
<213> Искусственная последовательность
<220>
<223> линкер
<400> 16
Ala Ser Pro Ala Ala Pro Ala Pro Ala Ser Pro Ala Ala Pro Ala Pro
1 5 10 15
Ser Ala Pro Ala
20
<210> 17
<211> 66
<212> Белок
<213> Искусственная последовательность
<220>
<223> линкер
<400> 17
Ala Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Pro Val Pro Ser
1 5 10 15
Thr Pro Pro Thr Pro Ser Pro Ser Thr Pro Pro Thr Pro Ser Pro Ser
20 25 30
Gly Gly Ser Gly Asn Ser Ser Gly Ser Gly Gly Ser Pro Val Pro Ser
35 40 45
Thr Pro Pro Thr Pro Ser Pro Ser Thr Pro Pro Thr Pro Ser Pro Ser
50 55 60
Ala Ser
65
<210> 18
<211> 32
<212> Белок
<213> Искусственная последовательность
<220>
<223> линкер
<400> 18
Pro Ser Thr Pro Pro Thr Pro Ser Pro Ser Thr Pro Pro Thr Pro Ser
1 5 10 15
Pro Ser Gly Gly Ser Gly Asn Ser Ser Gly Ser Gly Gly Ser Pro Val
20 25 30
<210> 19
<211> 74
<212> Белок
<213> Искусственная последовательность
<220>
<223> линкер
<400> 19
Ala Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Pro Val Pro Ser
1 5 10 15
Thr Pro Pro Thr Asn Ser Ser Ser Thr Pro Pro Thr Pro Ser Pro Ser
20 25 30
Pro Val Pro Ser Thr Pro Pro Thr Asn Ser Ser Ser Thr Pro Pro Thr
35 40 45
Pro Ser Pro Ser Pro Val Pro Ser Thr Pro Pro Thr Asn Ser Ser Ser
50 55 60
Thr Pro Pro Thr Pro Ser Pro Ser Ala Ser
65 70
<210> 20
<211> 40
<212> Белок
<213> Искусственная последовательность
<220>
<223> линкер
<400> 20
Ala Ser Pro Ala Ala Pro Ala Pro Ala Ser Pro Ala Ala Pro Ala Pro
1 5 10 15
Ser Ala Pro Ala Ala Ser Pro Ala Ala Pro Ala Pro Ala Ser Pro Ala
20 25 30
Ala Pro Ala Pro Ser Ala Pro Ala
35 40
<210> 21
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> линкер
<400> 21
Val Asp Asp Ile Glu Gly Arg Met Asp Glu
1 5 10
<210> 22
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> линкер
<400> 22
Glu Asn Leu Tyr Phe Gln Gly Arg Met Asp Glu
1 5 10
<210> 23
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> линкер
<400> 23
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210> 24
<211> 327
<212> Белок
<213> Искусственная последовательность
<220>
<223> тяжелая цепь антитела
<400> 24
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 25
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> легкая цепь антитела
<400> 25
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Ser Ser Asn Trp Pro Arg
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 26
<211> 448
<212> Белок
<213> Искусственная последовательность
<220>
<223> тяжелая цепь антитела
<400> 26
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn His Gly Gly Tyr Val Thr Tyr Asn Pro Ser Leu Glu
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Tyr Gly Pro Gly Asn Tyr Asp Trp Tyr Phe Asp Leu Trp Gly
100 105 110
Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly
210 215 220
Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro
260 265 270
Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
<210> 27
<211> 216
<212> Белок
<213> Искусственная последовательность
<220>
<223> легкая цепь антитела
<400> 27
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Pro
85 90 95
Ala Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val
100 105 110
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
115 120 125
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
130 135 140
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
145 150 155 160
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
165 170 175
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
180 185 190
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr
195 200 205
Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 28
<211> 228
<212> Белок
<213> Искусственная последовательность
<220>
<223> Fc-область антитела
<400> 28
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala
1 5 10 15
Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
20 25 30
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
35 40 45
Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val
50 55 60
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser
65 70 75 80
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
85 90 95
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser
100 105 110
Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
115 120 125
Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln
130 135 140
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
145 150 155 160
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
165 170 175
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu
180 185 190
Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser
195 200 205
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
210 215 220
Leu Ser Leu Gly
225
<210> 29
<211> 228
<212> Белок
<213> Искусственная последовательность
<220>
<223> Fc-область антитела
<400> 29
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe
1 5 10 15
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
20 25 30
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
35 40 45
Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val
50 55 60
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser
65 70 75 80
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
85 90 95
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser
100 105 110
Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
115 120 125
Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln
130 135 140
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
145 150 155 160
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
165 170 175
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu
180 185 190
Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser
195 200 205
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
210 215 220
Leu Ser Leu Gly
225
<210> 30
<211> 228
<212> Белок
<213> Искусственная последовательность
<220>
<223> Fc-область антитела
<400> 30
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe
1 5 10 15
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
20 25 30
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
35 40 45
Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val
50 55 60
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser
65 70 75 80
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
85 90 95
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser
100 105 110
Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
115 120 125
Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln
130 135 140
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
145 150 155 160
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
165 170 175
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu
180 185 190
Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser
195 200 205
Val Leu His Glu Ala Leu His Ser His Tyr Thr Gln Lys Ser Leu Ser
210 215 220
Leu Ser Leu Gly
225
<210> 31
<211> 228
<212> Белок
<213> Искусственная последовательность
<220>
<223> Fc-область антитела
<400> 31
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe
1 5 10 15
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
20 25 30
Leu Tyr Ile Thr Arg Glu Pro Glu Val Thr Cys Val Val Val Asp Val
35 40 45
Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val
50 55 60
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser
65 70 75 80
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
85 90 95
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser
100 105 110
Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
115 120 125
Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln
130 135 140
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
145 150 155 160
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
165 170 175
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu
180 185 190
Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser
195 200 205
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
210 215 220
Leu Ser Leu Gly
225
<210> 32
<211> 152
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 32
Ala Ser Asp Glu Glu Ile Gln Asp Val Ser Gly Thr Trp Tyr Leu Lys
1 5 10 15
Ala Met Thr Val Asp Glu Gly Cys Arg Pro Trp Asn Ile Phe Ser Val
20 25 30
Thr Pro Met Thr Leu Thr Thr Leu Glu Gly Gly Asn Leu Glu Ala Lys
35 40 45
Val Thr Met Ala Ile Asp Gly Pro Ala Gln Glu Val Lys Ala Val Leu
50 55 60
Glu Lys Thr Asp Glu Pro Gly Lys Tyr Thr Ala Asp Gly Gly Lys His
65 70 75 80
Val Ala Tyr Ile Ile Arg Ser His Val Lys Asp His Tyr Ile Phe Tyr
85 90 95
Ser Glu Gly Val Cys Asp Gly Ser Pro Val Pro Gly Val Trp Leu Val
100 105 110
Gly Arg Asp Pro Lys Asn Asn Leu Glu Ala Leu Glu Asp Phe Glu Lys
115 120 125
Ala Ala Gly Ala Arg Gly Leu Ser Thr Glu Ser Ile Leu Ile Pro Arg
130 135 140
Gln Ser Glu Thr Ser Ser Pro Gly
145 150
<210> 33
<211> 152
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 33
Thr Ser Asp Glu Glu Ile Gln Asp Val Ser Gly Thr Trp Tyr Leu Lys
1 5 10 15
Ala Met Thr Val Asp Glu Gly Cys Arg Pro Trp Asn Ile Phe Ser Val
20 25 30
Thr Pro Met Thr Leu Thr Thr Leu Glu Gly Gly Asn Leu Glu Ala Lys
35 40 45
Val Thr Met Ala Ile Asp Gly Pro Ala Gln Glu Val Arg Ala Val Leu
50 55 60
Glu Lys Thr Asp Glu Pro Gly Lys Tyr Thr Ala Asp Gly Gly Lys His
65 70 75 80
Asp Ala Tyr Ile Ile Arg Ser His Val Lys Asp His Tyr Ile Phe Tyr
85 90 95
Ser Glu Gly Val Cys Asp Gly Ser Pro Val Pro Gly Val Trp Leu Val
100 105 110
Gly Arg Asp Pro Glu Asn Asn Leu Glu Ala Leu Glu Asp Phe Glu Lys
115 120 125
Thr Ala Gly Ala Arg Gly Leu Ser Thr Glu Ser Ile Leu Ile Pro Arg
130 135 140
Gln Ser Glu Thr Ser Ser Pro Gly
145 150
<210> 34
<211> 152
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 34
Ala Ser Asp Glu Glu Ile Gln Asp Val Ser Gly Thr Trp Tyr Leu Lys
1 5 10 15
Ala Met Thr Val Asp Glu Gly Cys Arg Pro Trp Asn Ile Phe Ser Val
20 25 30
Thr Pro Met Thr Leu Thr Thr Leu Glu Gly Gly Asn Leu Glu Ala Lys
35 40 45
Val Thr Met Ala Ile Asp Gly Pro Ala Gln Glu Val Asn Ala Val Leu
50 55 60
Glu Lys Thr Asp Glu Pro Gly Lys Tyr Thr Ala Asp Gly Gly Lys His
65 70 75 80
Val Ala Tyr Ile Ile Arg Ser His Val Arg Asp His Tyr Ile Phe Tyr
85 90 95
Ser Glu Gly Val Cys Asp Gly Ser Pro Val Pro Gly Val Trp Leu Val
100 105 110
Gly Arg Asp Pro Glu Asn Asn Leu Glu Ala Leu Glu Asp Phe Glu Lys
115 120 125
Thr Ala Gly Ala Arg Gly Leu Ser Thr Glu Ser Ile Leu Ile Pro Arg
130 135 140
Gln Ser Glu Thr Ser Ser Pro Gly
145 150
<210> 35
<211> 152
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 35
Val Ser Asp Glu Glu Ile Gln Asp Val Ser Gly Thr Trp Tyr Leu Lys
1 5 10 15
Ala Met Thr Val Asp Glu Gly Cys Arg Pro Trp Asn Ile Phe Ser Val
20 25 30
Thr Pro Met Thr Leu Thr Thr Leu Glu Gly Gly Asn Leu Glu Ala Lys
35 40 45
Val Thr Met Ala Ile Asp Gly Pro Ala Gln Glu Val Arg Ala Val Leu
50 55 60
Glu Lys Thr Asp Glu Pro Gly Lys Tyr Thr Ala Asp Gly Gly Lys His
65 70 75 80
Val Ala Tyr Ile Ile Arg Ser His Val Glu Asp His Tyr Ile Phe Tyr
85 90 95
Ser Glu Gly Val Cys Asp Gly Ser Pro Val Pro Gly Val Trp Leu Val
100 105 110
Gly Arg Asp Pro Glu Asn Asn Leu Glu Ala Leu Glu Asp Phe Glu Lys
115 120 125
Thr Ala Gly Ala Arg Gly Leu Ser Thr Glu Ser Ile Leu Ile Pro Arg
130 135 140
Gln Ser Glu Thr Ser Ser Pro Gly
145 150
<210> 36
<211> 152
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 36
Ala Ser Asp Glu Glu Ile Gln Asp Val Ser Gly Thr Trp Tyr Leu Lys
1 5 10 15
Ala Met Thr Val Asp Glu Gly Cys Arg Pro Trp Asn Ile Phe Ser Val
20 25 30
Thr Pro Met Thr Leu Ser Thr Leu Glu Gly Gly Asn Leu Glu Ala Lys
35 40 45
Val Thr Met Ala Ile Asp Gly Pro Ala Gln Glu Val Lys Ala Val Leu
50 55 60
Glu Lys Thr Asp Glu Pro Gly Lys Tyr Thr Ala Asp Gly Gly Lys His
65 70 75 80
Val Ala Tyr Ile Ile Arg Ser His Val Lys Asp His Tyr Ile Phe Tyr
85 90 95
Ser Glu Gly Val Cys Asp Gly Ser Pro Val Pro Gly Val Trp Leu Val
100 105 110
Gly Arg Asp Pro Lys Asn Asn Leu Glu Ala Leu Glu Asp Phe Glu Lys
115 120 125
Ala Ala Gly Ala Arg Gly Leu Ser Thr Glu Ser Ile Leu Ile Pro Arg
130 135 140
Gln Ile Glu Thr Ser Ser Pro Gly
145 150
<210> 37
<211> 152
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 37
Ala Ser Asp Glu Glu Ile Gln Asp Val Ser Gly Thr Trp Tyr Leu Lys
1 5 10 15
Ala Met Thr Val Asp Glu Gly Cys Arg Pro Trp Asn Ile Phe Ser Val
20 25 30
Thr Pro Met Thr Leu Thr Thr Leu Glu Gly Gly Asn Leu Glu Ala Glu
35 40 45
Val Thr Met Ala Ile Asp Gly Pro Ala Gln Glu Val Lys Ala Val Leu
50 55 60
Glu Lys Ala Asp Glu Pro Gly Lys Tyr Thr Ala Asp Gly Gly Lys His
65 70 75 80
Val Ala Tyr Ile Ile Arg Ser His Val Lys Asp His Tyr Ile Phe Tyr
85 90 95
Ser Glu Gly Val Cys Asp Gly Ser Pro Val Pro Gly Val Trp Leu Val
100 105 110
Gly Arg Asp Pro Lys Asn Asn Leu Glu Ala Leu Glu Asp Phe Glu Lys
115 120 125
Thr Ala Gly Ala Arg Gly Leu Ser Thr Glu Ser Ile Leu Ile Pro Ser
130 135 140
Gln Ile Glu Thr Ser Ser Pro Gly
145 150
<210> 38
<211> 152
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 38
Thr Ser Asp Glu Glu Ile Gln Asp Val Ser Gly Thr Trp Tyr Leu Lys
1 5 10 15
Ala Met Thr Val Asp Glu Gly Cys Arg Pro Trp Asn Ile Phe Ser Val
20 25 30
Thr Pro Met Thr Leu Thr Thr Leu Glu Asp Gly Asn Leu Glu Ala Lys
35 40 45
Val Thr Met Ala Ile Asp Gly Pro Ala Gln Glu Val Lys Ala Val Leu
50 55 60
Glu Lys Ala Asp Glu Pro Gly Lys Tyr Thr Ala Asp Gly Gly Lys His
65 70 75 80
Val Ala Tyr Ile Ile Arg Ser His Val Lys Asp His Tyr Ile Phe Tyr
85 90 95
Ser Glu Gly Val Cys Asp Gly Ser Pro Val Pro Gly Val Trp Leu Val
100 105 110
Gly Arg Asp Pro Lys Asn Asn Leu Glu Ala Leu Glu Asp Phe Glu Lys
115 120 125
Ala Ala Gly Ala Arg Gly Leu Ser Thr Glu Ser Ile Leu Ile Pro Arg
130 135 140
Gln Ile Glu Thr Ser Ser Pro Gly
145 150
<210> 39
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 39
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Ile Lys Leu Arg Glu Asp Lys Asp Pro
35 40 45
Asn Lys Met Met Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asn Val Thr Gly Val Thr Phe Asp Asp Lys Lys Cys Thr Tyr Ala Ile
65 70 75 80
Ser Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Lys
85 90 95
Ile Lys Ser Phe Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln
115 120 125
Asn Arg Glu Glu Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 40
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 40
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Ile Arg Leu Arg Glu Asp Lys Asp Pro
35 40 45
Ile Lys Met Met Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Met Val Lys Phe Asp Asp Lys Lys Cys Met Tyr Asp Ile
65 70 75 80
Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Lys
85 90 95
Ile Lys Ser Phe Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln
115 120 125
Asn Arg Glu Glu Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 41
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 41
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Ile Arg Leu Arg Glu Asp Lys Asp Pro
35 40 45
Asn Lys Met Met Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Ala Val Ala Phe Asp Asp Lys Lys Cys Thr Tyr Asp Ile
65 70 75 80
Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Lys
85 90 95
Ile Lys Ser Phe Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln
115 120 125
Asn Arg Glu Glu Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 42
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 42
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Ile Lys Leu Arg Glu Asp Lys Asp Pro
35 40 45
Asn Lys Met Met Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Ala Val Ala Phe Asp Asp Lys Lys Cys Thr Tyr Asp Ile
65 70 75 80
Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Lys
85 90 95
Ile Lys Ser Phe Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln
115 120 125
Asn Arg Glu Glu Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 43
<211> 175
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 43
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Ile Lys Leu Arg Glu Asp Ser Lys Met
35 40 45
Met Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr Asp Val Thr
50 55 60
Gly Val Ser Phe Asp Asp Lys Lys Cys Thr Tyr Ala Ile Met Thr Phe
65 70 75 80
Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Lys Ile Lys Ser
85 90 95
Phe Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser Thr Asn Tyr
100 105 110
Asn Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln Asn Arg Glu
115 120 125
Glu Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu Thr Ser Glu
130 135 140
Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly Leu Pro Glu
145 150 155 160
Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile Asp Gly
165 170 175
<210> 44
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 44
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Ile Lys Leu Arg Glu Asp Lys Asp Pro
35 40 45
Val Lys Met Met Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Gly Val Thr Phe Asp Asp Lys Lys Cys Arg Tyr Asp Ile
65 70 75 80
Ser Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Phe Gly Lys
85 90 95
Ile Lys Ser Phe Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln
115 120 125
Asn Arg Glu Glu Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 45
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 45
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Ile Arg Leu Arg Glu Asp Lys Asp Pro
35 40 45
His Lys Met Met Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Gly Val Thr Phe Asp Asp Lys Lys Cys Thr Tyr Ala Ile
65 70 75 80
Ser Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Lys
85 90 95
Ile Lys Ser Phe Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln
115 120 125
Asn Arg Glu Glu Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 46
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 46
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Ile Lys Leu Arg Glu Asp Lys Asp Pro
35 40 45
Asn Lys Met Met Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Gly Val Thr Phe Asp Asp Lys Lys Cys Thr Tyr Ala Ile
65 70 75 80
Ser Thr Leu Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Phe Gly Lys
85 90 95
Ile Lys Ser Phe Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln
115 120 125
Asn Arg Glu Glu Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 47
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 47
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Ile Arg Leu Arg Glu Asp Lys Asp Pro
35 40 45
Ser Lys Met Met Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Ala Val Thr Phe Asp Asp Lys Lys Cys Asn Tyr Ala Ile
65 70 75 80
Ser Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Lys
85 90 95
Ile Lys Ser Phe Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln
115 120 125
Asn Arg Glu Glu Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 48
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 48
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Met Ala Gly Asn Asn Leu Leu Arg Glu Asp Lys Asp Pro
35 40 45
His Lys Met Ser Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asn Val Thr Asp Val Met Phe Leu Asp Lys Lys Cys Gln Tyr Ile Ile
65 70 75 80
Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Phe
85 90 95
Ile Lys Ser Asp Pro Gly His Thr Ser Tyr Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Ser Val Ile Gln
115 120 125
Asn Arg Glu Trp Phe Gly Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 49
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 49
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Met Ala Gly Asn Asn Leu Leu Arg Glu Asp Lys Asp Pro
35 40 45
His Lys Met Ser Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asn Val Thr Asp Val Met Phe Leu Asp Lys Lys Cys Gln Tyr Ile Ile
65 70 75 80
Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Leu Thr Leu Gly Phe
85 90 95
Ile Arg Ser Asp Leu Gly His Thr Ser Tyr Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Ser Val Ile Gln
115 120 125
Asn Arg Glu Trp Phe Gly Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 50
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 50
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Tyr Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Met Ala Gly Asn Asn Leu Leu Arg Glu Asp Lys Asp Pro
35 40 45
His Lys Met Gly Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asn Val Thr Asp Val Met Leu Leu Asp Lys Lys Cys Gln Tyr Ile Ile
65 70 75 80
Gln Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Ser Thr Leu Gly Phe
85 90 95
Ile Lys Ser Asp Pro Gly His Thr Ser Tyr Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Ser Val Ile Gln
115 120 125
Asn Arg Glu Trp Phe Gly Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 51
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 51
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Met Ala Gly Asn Asn Leu Leu Arg Glu Asp Lys Asp Pro
35 40 45
His Lys Met Gly Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asn Val Thr Asp Val Met Phe Leu Asp Lys Lys Cys Gln His Ile Ile
65 70 75 80
Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Leu Thr Leu Gly Phe
85 90 95
Ile Lys Ser Asp Pro Gly His Thr Ser Tyr Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Ser Val Ile Gln
115 120 125
Asn Arg Glu Trp Phe Gly Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 52
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 52
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asp Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Met Ala Gly Asn Asn Leu Leu Arg Glu Asp Lys Asp Pro
35 40 45
His Lys Met Gly Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asn Val Thr Asp Val Met Phe Leu Asp Lys Lys Cys Gln Tyr Ile Ile
65 70 75 80
Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Leu Thr Leu Gly Phe
85 90 95
Ile Lys Ser Asp Pro Gly His Thr Ser Tyr Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Ser Val Ile Gln
115 120 125
Asn Arg Glu Trp Phe Gly Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 53
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 53
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Ile Val Gly Met Ala Gly Asn Asn Leu Leu Arg Glu Asp Lys Asp Pro
35 40 45
His Lys Met Gly Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asn Val Thr Asp Val Met Phe Leu Asp Lys Lys Cys Gln Tyr Ile Ile
65 70 75 80
Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Leu Thr Leu Gly Phe
85 90 95
Ile Lys Ser Asp Pro Gly His Thr Ser Tyr Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Ser Val Ile Gln
115 120 125
Asn Arg Glu Trp Phe Gly Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 54
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 54
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Arg Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Met Ala Gly Asn Asn Leu Leu Arg Val Asp Lys Asp Pro
35 40 45
His Lys Met Gly Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asn Val Thr Asp Val Met Phe Leu Asp Lys Lys Cys Gln Tyr Ile Ile
65 70 75 80
Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Leu Thr Leu Gly Phe
85 90 95
Ile Lys Ser Asp Pro Gly His Thr Ser Tyr Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Tyr Phe Lys Ser Val Ile Gln
115 120 125
Asn Arg Glu Trp Phe Gly Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 55
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 55
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Met Ala Gly Asn Asn Leu Leu Arg Glu Asp Lys Asp Pro
35 40 45
His Lys Met Ser Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asn Val Thr Asp Val Met Phe Leu Asp Lys Lys Cys Gln Tyr Ile Asn
65 70 75 80
Trp Pro Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Phe
85 90 95
Ile Lys Ser Asp Leu Gly Pro Thr Ser Tyr Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Ser Val Ile Gln
115 120 125
Asn Arg Glu Trp Phe Gly Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 56
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 56
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Met Ala Gly Asn Asn Leu Leu Arg Glu Asp Lys Asp Pro
35 40 45
His Lys Met Gly Ala Thr Ile Tyr Glu Leu Asn Glu Asp Lys Ser Tyr
50 55 60
Asn Val Thr Asp Val Met Phe Leu Asp Lys Lys Cys Gln Tyr Ile Ile
65 70 75 80
Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Leu Thr Leu Gly Phe
85 90 95
Ile Lys Ser Asp Pro Gly His Thr Ser Tyr Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Ser Val Ile Gln
115 120 125
Asn Arg Glu Trp Phe Gly Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 57
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 57
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Met Ala Gly Asn Asn Leu Leu Arg Asp Asp Lys Asp Pro
35 40 45
His Lys Met Ser Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asn Val Thr Asp Val Met Leu Leu Asp Lys Lys Cys His Tyr Ile Ile
65 70 75 80
Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Leu Thr Leu Gly Phe
85 90 95
Ile Lys Ser Asp Pro Gly His Thr Ser Tyr Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Ser Val Ile Gln
115 120 125
Asn Arg Glu Trp Phe Gly Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 58
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 58
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Val Ala Gly Asn Ala Met Leu Arg Glu Asp Lys Asp Pro
35 40 45
Leu Lys Met Arg Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asn Val Thr Gly Val Ser Phe Trp Arg Lys Lys Cys His Tyr Lys Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Asp
85 90 95
Ile Lys Ser Gly Pro Gly Gln Thr Ser Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Arg Gln
115 120 125
Asn Arg Glu Trp Phe Ala Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 59
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 59
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Arg Ala Gly Asn Val Ala Leu Arg Glu Asp Lys Asp Pro
35 40 45
Pro Lys Met Arg Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asn Val Thr Asn Val Arg Phe Ala Met Lys Lys Cys Met Tyr Ser Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln
85 90 95
Ile Lys Ser Glu Pro Gly Asn Thr Ser Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Tyr Gln
115 120 125
Asn Arg Glu Ile Phe Phe Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 60
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 60
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Val Ala Gly Asn Ala Leu Leu Arg Glu Asp Lys Asp Pro
35 40 45
Leu Lys Met Arg Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Val Val Ser Phe Trp Arg Lys Lys Cys His Tyr Lys Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Asp
85 90 95
Ile Lys Ser Gly Pro Gly Gln Thr Ser Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Arg Gln
115 120 125
Asn Arg Glu Trp Phe Ala Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 61
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 61
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Val Ala Gly Asn Val Arg Leu Arg Glu Asp Lys Asp Pro
35 40 45
Pro Lys Met Arg Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Gly Val Ser Phe Arg Gly Lys Lys Cys His Tyr Lys Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Asp
85 90 95
Ile Lys Ser Gly Pro Gly Glu Thr Ser Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Arg Gln
115 120 125
Asn Arg Glu Trp Phe Phe Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 62
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 62
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Val Ala Gly Asn Gly Met Leu Arg Glu Asp Lys Asp Pro
35 40 45
Leu Lys Met Arg Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Ser Val Ala Phe Arg Asn Lys Lys Cys His Tyr Lys Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln
85 90 95
Ile Lys Ser Gly Pro Gly Glu Thr Ser Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Arg Gln
115 120 125
Asn Arg Glu Trp Phe Phe Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 63
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 63
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Arg Ala Gly Asn Val Gly Leu Arg Glu Asp Lys Asp Pro
35 40 45
Pro Lys Met Trp Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Asn Val Arg Phe Ala Arg Lys Lys Cys Leu Tyr Ser Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln
85 90 95
Ile Lys Ser Glu Pro Gly Asn Thr Ala Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Tyr Gln
115 120 125
Asn Arg Glu Ile Phe Phe Ile Ile Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 64
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 64
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Arg Ala Gly Asn Val Gly Leu Arg Glu Asp Lys Asp Pro
35 40 45
Pro Lys Met Trp Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Asn Val Arg Phe Ala Arg Lys Lys Cys Thr Tyr Ser Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln
85 90 95
Ile Lys Ser Glu Pro Gly Gly Thr Ala Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Tyr Gln
115 120 125
Asn Arg Glu Ile Phe Phe Ile Ile Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 65
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 65
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Arg Ala Gly Asn Gly Ala Leu Arg Glu Asp Lys Asp Pro
35 40 45
Pro Lys Met Trp Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Asn Val Arg Phe Ala Arg Lys Lys Cys Val Tyr Ser Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Pro
85 90 95
Ile Lys Ser Glu Pro Gly Asn Thr Ala Ser Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Tyr Gln
115 120 125
Asn Arg Glu Ile Phe Phe Ile Ile Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 66
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 66
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Arg Ala Gly Asn Val Ala Leu Arg Glu Asp Lys Asp Pro
35 40 45
Pro Lys Met Arg Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Ala Val Arg Phe Ala Arg Lys Lys Cys Leu Tyr Ser Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Arg
85 90 95
Ile Lys Ser Glu Pro Gly Tyr Thr Ala Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Tyr Gln
115 120 125
Asn Arg Glu Ile Phe Phe Ile Ile Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 67
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 67
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Arg Ala Gly Asn Val Ala Leu Arg Glu Asp Lys Asp Pro
35 40 45
Pro Lys Met Arg Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Asn Val Val Phe Ala Gly Lys Lys Cys Lys Tyr Ser Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Arg
85 90 95
Ile Lys Ser Pro Pro Gly Asn Thr Ala Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Tyr Gln
115 120 125
Asn Arg Glu Ile Phe Phe Ile Ile Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 68
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 68
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Arg Ala Gly Asn Leu Gly Leu Arg Glu Asp Lys Asp Pro
35 40 45
Pro Lys Met Trp Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Asn Val Arg Phe Ala Arg Lys Lys Cys Met Tyr Ser Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln
85 90 95
Ile Lys Ser Glu Pro Gly Ser Thr Ala Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Tyr Gln
115 120 125
Asn Arg Glu Ile Phe Phe Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 69
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 69
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Arg Ala Gly Asn Val Gly Leu Arg Glu Asp Lys Asp Pro
35 40 45
Pro Lys Met Trp Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Asn Val Arg Phe Ala Gly Lys Lys Val Lys Tyr Thr Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln
85 90 95
Ile Lys Ser Glu Pro Gly Asn Thr Ala Thr Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Tyr Gln
115 120 125
Asn Arg Glu Ile Phe Phe Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Ala Ile
165 170 175
Asp Gly
<210> 70
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 70
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Arg Ala Gly Asn Val Gly Leu Arg Glu Asp Lys Asp Pro
35 40 45
Pro Lys Met Arg Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Gly Val Arg Phe Gly Glu Lys Lys Ile Lys Tyr Ser Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln
85 90 95
Ile Lys Ser Gln Pro Gly Asp Thr Ala Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Tyr Gln
115 120 125
Asn Arg Glu Ile Phe Phe Ile Ile Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Ala Ile
165 170 175
Asp Gly
<210> 71
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин липокалина
<400> 71
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Arg Ala Gly Asn Val Gly Leu Arg Glu Asp Lys Asp Pro
35 40 45
Pro Lys Met Arg Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Gly Val Arg Phe Asp Ser Lys Lys Val Thr Tyr Ser Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln
85 90 95
Ile Lys Ser Glu Pro Gly Asn Thr Ala Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Tyr Gln
115 120 125
Asn Arg Glu Ile Phe Phe Ile Ile Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Ala Ile
165 170 175
Asp Gly
<210> 72
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR1 тяжелой цепи антитела
<400> 72
Gly Tyr Thr Phe Thr Asp Tyr Glu
1 5
<210> 73
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR2 тяжелой цепи антитела
<400> 73
Leu Asp Pro Lys Thr Gly Asp Thr
1 5
<210> 74
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR3 тяжелой цепи антитела
<400> 74
Thr Arg Phe Tyr Ser Tyr Thr Tyr
1 5
<210> 75
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR1 легкой цепи антитела
<400> 75
Gln Ser Leu Val His Ser Asn Arg Asn Thr Tyr
1 5 10
<210> 76
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR1 легкой цепи антитела
<400> 76
Gln Ser Leu Val His Ser Asn Arg Asn Thr Tyr
1 5 10
<210> 77
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR3 легкой цепи антитела
<400> 77
Ser Gln Asn Thr His Val Pro Pro Thr
1 5
<210> 78
<211> 116
<212> Белок
<213> Искусственная последовательность
<220>
<223> вариабельный домен тяжелой цепи антитела
<400> 78
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Glu Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ala Leu Asp Pro Lys Thr Gly Asp Thr Ala Tyr Ser Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Tyr Ser Tyr Thr Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Leu Val Ser Ser
115
<210> 79
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> вариабельный домен легкой цепи антитела
<400> 79
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Arg Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ser Gln Asn
85 90 95
Thr His Val Pro Pro Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 80
<211> 445
<212> Белок
<213> Искусственная последовательность
<220>
<223> тяжелая цепь антитела
<400> 80
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Glu Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ala Leu Asp Pro Lys Thr Gly Asp Thr Ala Tyr Ser Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Tyr Ser Tyr Thr Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val
130 135 140
Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly
165 170 175
Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly
180 185 190
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
195 200 205
Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 81
<211> 442
<212> Белок
<213> Искусственная последовательность
<220>
<223> тяжелая цепь антитела
<400> 81
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Glu Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ala Leu Asp Pro Lys Thr Gly Asp Thr Ala Tyr Ser Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Tyr Ser Tyr Thr Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Leu Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro
210 215 220
Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440
<210> 82
<211> 219
<212> Белок
<213> Искусственная последовательность
<220>
<223> легкая цепь антитела
<400> 82
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Arg Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ser Gln Asn
85 90 95
Thr His Val Pro Pro Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 83
<211> 615
<212> Белок
<213> Искусственная последовательность
<220>
<223> слитый белок
<400> 83
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Arg Ala Gly Asn Val Gly Leu Arg Glu Asp Lys Asp Pro
35 40 45
Pro Lys Met Trp Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Asn Val Arg Phe Ala Arg Lys Lys Cys Thr Tyr Ser Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln
85 90 95
Ile Lys Ser Glu Pro Gly Gly Thr Ala Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Tyr Gln
115 120 125
Asn Arg Glu Ile Phe Phe Ile Ile Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
180 185 190
Ser Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu
195 200 205
Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
210 215 220
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
225 230 235 240
Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
245 250 255
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
260 265 270
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
275 280 285
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
290 295 300
Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
305 310 315 320
Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn
325 330 335
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
340 345 350
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
355 360 365
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg
370 375 380
Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys
385 390 395 400
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
405 410 415
Ser Leu Ser Leu Gly Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
420 425 430
Gly Gly Gly Gly Ser Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro
435 440 445
Pro Leu Ser Lys Val Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe
450 455 460
His Gly Lys Trp Tyr Val Val Gly Gln Ala Gly Asn Ile Arg Leu Arg
465 470 475 480
Glu Asp Lys Asp Pro Ile Lys Met Met Ala Thr Ile Tyr Glu Leu Lys
485 490 495
Glu Asp Lys Ser Tyr Asp Val Thr Met Val Lys Phe Asp Asp Lys Lys
500 505 510
Cys Met Tyr Asp Ile Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu
515 520 525
Phe Thr Leu Gly Lys Ile Lys Ser Phe Pro Gly His Thr Ser Ser Leu
530 535 540
Val Arg Val Val Ser Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe
545 550 555 560
Lys Phe Val Phe Gln Asn Arg Glu Glu Phe Tyr Ile Thr Leu Tyr Gly
565 570 575
Arg Thr Lys Glu Leu Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe
580 585 590
Ser Lys Ser Leu Gly Leu Pro Glu Asn His Ile Val Phe Pro Val Pro
595 600 605
Ile Asp Gln Cys Ile Asp Gly
610 615
<210> 84
<211> 610
<212> Белок
<213> Искусственная последовательность
<220>
<223> слитый белок
<400> 84
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Arg Ala Gly Asn Val Gly Leu Arg Glu Asp Lys Asp Pro
35 40 45
Pro Lys Met Trp Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Asn Val Arg Phe Ala Arg Lys Lys Cys Thr Tyr Ser Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln
85 90 95
Ile Lys Ser Glu Pro Gly Gly Thr Ala Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Tyr Gln
115 120 125
Asn Arg Glu Ile Phe Phe Ile Ile Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Asp Ser Thr
180 185 190
Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val Pro Leu Gln Gln
195 200 205
Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr Val Val Gly Gln
210 215 220
Ala Gly Asn Ile Arg Leu Arg Glu Asp Lys Asp Pro Ile Lys Met Met
225 230 235 240
Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr Asp Val Thr Met
245 250 255
Val Lys Phe Asp Asp Lys Lys Cys Met Tyr Asp Ile Trp Thr Phe Val
260 265 270
Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Lys Ile Lys Ser Phe
275 280 285
Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser Thr Asn Tyr Asn
290 295 300
Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln Asn Arg Glu Glu
305 310 315 320
Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu Thr Ser Glu Leu
325 330 335
Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly Leu Pro Glu Asn
340 345 350
His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile Asp Gly Gly Gly
355 360 365
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Ser Lys
370 375 380
Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
385 390 395 400
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
405 410 415
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
420 425 430
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
435 440 445
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg
450 455 460
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
465 470 475 480
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
485 490 495
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
500 505 510
Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu
515 520 525
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
530 535 540
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
545 550 555 560
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp
565 570 575
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His
580 585 590
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu
595 600 605
Gly Lys
610
<210> 85
<211> 366
<212> Белок
<213> Искусственная последовательность
<220>
<223> слитый белок
<400> 85
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Val Ala Gly Asn Gly Met Leu Arg Glu Asp Lys Asp Pro
35 40 45
Leu Lys Met Arg Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Ser Val Ala Phe Arg Asn Lys Lys Cys His Tyr Lys Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln
85 90 95
Ile Lys Ser Gly Pro Gly Glu Thr Ser Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Arg Gln
115 120 125
Asn Arg Glu Trp Phe Phe Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Asp Ser Thr
180 185 190
Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val Pro Leu Gln Gln
195 200 205
Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr Val Val Gly Gln
210 215 220
Ala Gly Asn Ile Arg Leu Arg Glu Asp Lys Asp Pro Ile Lys Met Met
225 230 235 240
Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr Asp Val Thr Met
245 250 255
Val Lys Phe Asp Asp Lys Lys Cys Met Tyr Asp Ile Trp Thr Phe Val
260 265 270
Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Lys Ile Lys Ser Phe
275 280 285
Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser Thr Asn Tyr Asn
290 295 300
Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln Asn Arg Glu Glu
305 310 315 320
Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu Thr Ser Glu Leu
325 330 335
Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly Leu Pro Glu Asn
340 345 350
His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile Asp Gly
355 360 365
<210> 86
<211> 554
<212> Белок
<213> Искусственная последовательность
<220>
<223> слитый белок
<400> 86
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Val Ala Gly Asn Gly Met Leu Arg Glu Asp Lys Asp Pro
35 40 45
Leu Lys Met Arg Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Ser Val Ala Phe Arg Asn Lys Lys Cys His Tyr Lys Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln
85 90 95
Ile Lys Ser Gly Pro Gly Glu Thr Ser Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Arg Gln
115 120 125
Asn Arg Glu Trp Phe Phe Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Asp Ser Thr
180 185 190
Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val Pro Leu Gln Gln
195 200 205
Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr Val Val Gly Gln
210 215 220
Ala Gly Asn Ile Arg Leu Arg Glu Asp Lys Asp Pro Ile Lys Met Met
225 230 235 240
Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr Asp Val Thr Met
245 250 255
Val Lys Phe Asp Asp Lys Lys Cys Met Tyr Asp Ile Trp Thr Phe Val
260 265 270
Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Lys Ile Lys Ser Phe
275 280 285
Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser Thr Asn Tyr Asn
290 295 300
Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln Asn Arg Glu Glu
305 310 315 320
Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu Thr Ser Glu Leu
325 330 335
Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly Leu Pro Glu Asn
340 345 350
His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile Asp Gly Gly Gly
355 360 365
Gly Gly Ser Gly Gly Gly Gly Ser Gln Asp Ser Thr Ser Asp Leu Ile
370 375 380
Pro Ala Pro Pro Leu Ser Lys Val Pro Leu Gln Gln Asn Phe Gln Asp
385 390 395 400
Asn Gln Phe His Gly Lys Trp Tyr Val Val Gly Gln Ala Gly Asn Ile
405 410 415
Arg Leu Arg Glu Asp Lys Asp Pro Ile Lys Met Met Ala Thr Ile Tyr
420 425 430
Glu Leu Lys Glu Asp Lys Ser Tyr Asp Val Thr Met Val Lys Phe Asp
435 440 445
Asp Lys Lys Cys Met Tyr Asp Ile Trp Thr Phe Val Pro Gly Ser Gln
450 455 460
Pro Gly Glu Phe Thr Leu Gly Lys Ile Lys Ser Phe Pro Gly His Thr
465 470 475 480
Ser Ser Leu Val Arg Val Val Ser Thr Asn Tyr Asn Gln His Ala Met
485 490 495
Val Phe Phe Lys Phe Val Phe Gln Asn Arg Glu Glu Phe Tyr Ile Thr
500 505 510
Leu Tyr Gly Arg Thr Lys Glu Leu Thr Ser Glu Leu Lys Glu Asn Phe
515 520 525
Ile Arg Phe Ser Lys Ser Leu Gly Leu Pro Glu Asn His Ile Val Phe
530 535 540
Pro Val Pro Ile Asp Gln Cys Ile Asp Gly
545 550
<210> 87
<211> 635
<212> Белок
<213> Искусственная последовательность
<220>
<223> слитый белок
<400> 87
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Glu Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ala Leu Asp Pro Lys Thr Gly Asp Thr Ala Tyr Ser Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Tyr Ser Tyr Thr Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Leu Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro
210 215 220
Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly Ser Gly
435 440 445
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Asp Ser Thr Ser Asp Leu
450 455 460
Ile Pro Ala Pro Pro Leu Ser Lys Val Pro Leu Gln Gln Asn Phe Gln
465 470 475 480
Asp Asn Gln Phe His Gly Lys Trp Tyr Val Val Gly Gln Ala Gly Asn
485 490 495
Ile Arg Leu Arg Glu Asp Lys Asp Pro Ile Lys Met Met Ala Thr Ile
500 505 510
Tyr Glu Leu Lys Glu Asp Lys Ser Tyr Asp Val Thr Met Val Lys Phe
515 520 525
Asp Asp Lys Lys Cys Met Tyr Asp Ile Trp Thr Phe Val Pro Gly Ser
530 535 540
Gln Pro Gly Glu Phe Thr Leu Gly Lys Ile Lys Ser Phe Pro Gly His
545 550 555 560
Thr Ser Ser Leu Val Arg Val Val Ser Thr Asn Tyr Asn Gln His Ala
565 570 575
Met Val Phe Phe Lys Phe Val Phe Gln Asn Arg Glu Glu Phe Tyr Ile
580 585 590
Thr Leu Tyr Gly Arg Thr Lys Glu Leu Thr Ser Glu Leu Lys Glu Asn
595 600 605
Phe Ile Arg Phe Ser Lys Ser Leu Gly Leu Pro Glu Asn His Ile Val
610 615 620
Phe Pro Val Pro Ile Asp Gln Cys Ile Asp Gly
625 630 635
<210> 88
<211> 828
<212> Белок
<213> Искусственная последовательность
<220>
<223> слитый белок
<400> 88
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Ile Arg Leu Arg Glu Asp Lys Asp Pro
35 40 45
Ile Lys Met Met Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Met Val Lys Phe Asp Asp Lys Lys Cys Met Tyr Asp Ile
65 70 75 80
Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Lys
85 90 95
Ile Lys Ser Phe Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln
115 120 125
Asn Arg Glu Glu Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
180 185 190
Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly
195 200 205
Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp
210 215 220
Tyr Glu Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp
225 230 235 240
Met Gly Ala Leu Asp Pro Lys Thr Gly Asp Thr Ala Tyr Ser Gln Lys
245 250 255
Phe Lys Gly Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala
260 265 270
Tyr Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr
275 280 285
Cys Thr Arg Phe Tyr Ser Tyr Thr Tyr Trp Gly Gln Gly Thr Leu Val
290 295 300
Thr Leu Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
305 310 315 320
Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys
325 330 335
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
340 345 350
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
355 360 365
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
370 375 380
Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn
385 390 395 400
Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro
405 410 415
Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe
420 425 430
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
435 440 445
Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe
450 455 460
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
465 470 475 480
Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
485 490 495
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
500 505 510
Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala
515 520 525
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln
530 535 540
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
545 550 555 560
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
565 570 575
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
580 585 590
Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu
595 600 605
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
610 615 620
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly Ser
625 630 635 640
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Asp Ser Thr Ser Asp
645 650 655
Leu Ile Pro Ala Pro Pro Leu Ser Lys Val Pro Leu Gln Gln Asn Phe
660 665 670
Gln Asp Asn Gln Phe His Gly Lys Trp Tyr Val Val Gly Gln Ala Gly
675 680 685
Asn Ile Arg Leu Arg Glu Asp Lys Asp Pro Ile Lys Met Met Ala Thr
690 695 700
Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr Asp Val Thr Met Val Lys
705 710 715 720
Phe Asp Asp Lys Lys Cys Met Tyr Asp Ile Trp Thr Phe Val Pro Gly
725 730 735
Ser Gln Pro Gly Glu Phe Thr Leu Gly Lys Ile Lys Ser Phe Pro Gly
740 745 750
His Thr Ser Ser Leu Val Arg Val Val Ser Thr Asn Tyr Asn Gln His
755 760 765
Ala Met Val Phe Phe Lys Phe Val Phe Gln Asn Arg Glu Glu Phe Tyr
770 775 780
Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu Thr Ser Glu Leu Lys Glu
785 790 795 800
Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly Leu Pro Glu Asn His Ile
805 810 815
Val Phe Pro Val Pro Ile Asp Gln Cys Ile Asp Gly
820 825
<210> 89
<211> 412
<212> Белок
<213> Искусственная последовательность
<220>
<223> слитый белок
<400> 89
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Arg Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ser Gln Asn
85 90 95
Thr His Val Pro Pro Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser
210 215 220
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Asp Ser Thr Ser Asp
225 230 235 240
Leu Ile Pro Ala Pro Pro Leu Ser Lys Val Pro Leu Gln Gln Asn Phe
245 250 255
Gln Asp Asn Gln Phe His Gly Lys Trp Tyr Val Val Gly Gln Ala Gly
260 265 270
Asn Ile Arg Leu Arg Glu Asp Lys Asp Pro Ile Lys Met Met Ala Thr
275 280 285
Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr Asp Val Thr Met Val Lys
290 295 300
Phe Asp Asp Lys Lys Cys Met Tyr Asp Ile Trp Thr Phe Val Pro Gly
305 310 315 320
Ser Gln Pro Gly Glu Phe Thr Leu Gly Lys Ile Lys Ser Phe Pro Gly
325 330 335
His Thr Ser Ser Leu Val Arg Val Val Ser Thr Asn Tyr Asn Gln His
340 345 350
Ala Met Val Phe Phe Lys Phe Val Phe Gln Asn Arg Glu Glu Phe Tyr
355 360 365
Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu Thr Ser Glu Leu Lys Glu
370 375 380
Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly Leu Pro Glu Asn His Ile
385 390 395 400
Val Phe Pro Val Pro Ile Asp Gln Cys Ile Asp Gly
405 410
<210> 90
<211> 412
<212> Белок
<213> Искусственная последовательность
<220>
<223> слитый белок
<400> 90
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Ile Arg Leu Arg Glu Asp Lys Asp Pro
35 40 45
Ile Lys Met Met Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Met Val Lys Phe Asp Asp Lys Lys Cys Met Tyr Asp Ile
65 70 75 80
Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Lys
85 90 95
Ile Lys Ser Phe Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln
115 120 125
Asn Arg Glu Glu Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
180 185 190
Ser Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro
195 200 205
Gly Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His
210 215 220
Ser Asn Arg Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln
225 230 235 240
Ser Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val
245 250 255
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys
260 265 270
Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ser Gln
275 280 285
Asn Thr His Val Pro Pro Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile
290 295 300
Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
305 310 315 320
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
325 330 335
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu
340 345 350
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
355 360 365
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
370 375 380
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
385 390 395 400
Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
405 410
<210> 91
<211> 635
<212> Белок
<213> Искусственная последовательность
<220>
<223> слитый белок
<400> 91
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Glu Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ala Leu Asp Pro Lys Thr Gly Asp Thr Ala Tyr Ser Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Tyr Ser Tyr Thr Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Leu Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro
210 215 220
Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly Ser Gly
435 440 445
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Asp Ser Thr Ser Asp Leu
450 455 460
Ile Pro Ala Pro Pro Leu Ser Lys Val Pro Leu Gln Gln Asn Phe Gln
465 470 475 480
Asp Asn Gln Phe Gln Gly Lys Trp Tyr Val Val Gly Met Ala Gly Asn
485 490 495
Asn Leu Leu Arg Glu Asp Lys Asp Pro His Lys Met Ser Ala Thr Ile
500 505 510
Tyr Glu Leu Lys Glu Asp Lys Ser Tyr Asn Val Thr Asp Val Met Phe
515 520 525
Leu Asp Lys Lys Cys Gln Tyr Ile Ile Trp Thr Phe Val Pro Gly Ser
530 535 540
Gln Pro Gly Glu Leu Thr Leu Gly Phe Ile Arg Ser Asp Leu Gly His
545 550 555 560
Thr Ser Tyr Leu Val Arg Val Val Ser Thr Asn Tyr Asn Gln His Ala
565 570 575
Met Val Phe Phe Lys Ser Val Ile Gln Asn Arg Glu Trp Phe Gly Ile
580 585 590
Thr Leu Tyr Gly Arg Thr Lys Glu Leu Thr Ser Glu Leu Lys Glu Asn
595 600 605
Phe Ile Arg Phe Ser Lys Ser Leu Gly Leu Pro Glu Asn His Ile Val
610 615 620
Phe Pro Val Pro Ile Asp Gln Cys Ile Asp Gly
625 630 635
<210> 92
<211> 635
<212> Белок
<213> Искусственная последовательность
<220>
<223> слитый белок
<400> 92
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Met Ala Gly Asn Asn Leu Leu Arg Glu Asp Lys Asp Pro
35 40 45
His Lys Met Ser Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asn Val Thr Asp Val Met Phe Leu Asp Lys Lys Cys Gln Tyr Ile Ile
65 70 75 80
Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Leu Thr Leu Gly Phe
85 90 95
Ile Arg Ser Asp Leu Gly His Thr Ser Tyr Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Ser Val Ile Gln
115 120 125
Asn Arg Glu Trp Phe Gly Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
180 185 190
Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly
195 200 205
Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp
210 215 220
Tyr Glu Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp
225 230 235 240
Met Gly Ala Leu Asp Pro Lys Thr Gly Asp Thr Ala Tyr Ser Gln Lys
245 250 255
Phe Lys Gly Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala
260 265 270
Tyr Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr
275 280 285
Cys Thr Arg Phe Tyr Ser Tyr Thr Tyr Trp Gly Gln Gly Thr Leu Val
290 295 300
Thr Leu Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
305 310 315 320
Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys
325 330 335
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
340 345 350
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
355 360 365
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
370 375 380
Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn
385 390 395 400
Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro
405 410 415
Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe
420 425 430
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
435 440 445
Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe
450 455 460
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
465 470 475 480
Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
485 490 495
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
500 505 510
Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala
515 520 525
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln
530 535 540
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
545 550 555 560
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
565 570 575
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
580 585 590
Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu
595 600 605
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
610 615 620
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
625 630 635
<210> 93
<211> 412
<212> Белок
<213> Искусственная последовательность
<220>
<223> слитый белок
<400> 93
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Arg Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ser Gln Asn
85 90 95
Thr His Val Pro Pro Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser
210 215 220
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Asp Ser Thr Ser Asp
225 230 235 240
Leu Ile Pro Ala Pro Pro Leu Ser Lys Val Pro Leu Gln Gln Asn Phe
245 250 255
Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr Val Val Gly Met Ala Gly
260 265 270
Asn Asn Leu Leu Arg Glu Asp Lys Asp Pro His Lys Met Ser Ala Thr
275 280 285
Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr Asn Val Thr Asp Val Met
290 295 300
Phe Leu Asp Lys Lys Cys Gln Tyr Ile Ile Trp Thr Phe Val Pro Gly
305 310 315 320
Ser Gln Pro Gly Glu Leu Thr Leu Gly Phe Ile Arg Ser Asp Leu Gly
325 330 335
His Thr Ser Tyr Leu Val Arg Val Val Ser Thr Asn Tyr Asn Gln His
340 345 350
Ala Met Val Phe Phe Lys Ser Val Ile Gln Asn Arg Glu Trp Phe Gly
355 360 365
Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu Thr Ser Glu Leu Lys Glu
370 375 380
Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly Leu Pro Glu Asn His Ile
385 390 395 400
Val Phe Pro Val Pro Ile Asp Gln Cys Ile Asp Gly
405 410
<210> 94
<211> 412
<212> Белок
<213> Искусственная последовательность
<220>
<223> слитый белок
<400> 94
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Met Ala Gly Asn Asn Leu Leu Arg Glu Asp Lys Asp Pro
35 40 45
His Lys Met Ser Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asn Val Thr Asp Val Met Phe Leu Asp Lys Lys Cys Gln Tyr Ile Ile
65 70 75 80
Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Leu Thr Leu Gly Phe
85 90 95
Ile Arg Ser Asp Leu Gly His Thr Ser Tyr Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Ser Val Ile Gln
115 120 125
Asn Arg Glu Trp Phe Gly Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
180 185 190
Ser Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro
195 200 205
Gly Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His
210 215 220
Ser Asn Arg Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln
225 230 235 240
Ser Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val
245 250 255
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys
260 265 270
Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ser Gln
275 280 285
Asn Thr His Val Pro Pro Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile
290 295 300
Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
305 310 315 320
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
325 330 335
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu
340 345 350
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
355 360 365
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
370 375 380
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
385 390 395 400
Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
405 410
<210> 95
<211> 640
<212> Белок
<213> Искусственная последовательность
<220>
<223> слитый белок
<400> 95
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Glu Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ala Leu Asp Pro Lys Thr Gly Asp Thr Ala Tyr Ser Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Tyr Ser Tyr Thr Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Leu Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro
210 215 220
Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly Ser Gly
435 440 445
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Asp
450 455 460
Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val Pro Leu
465 470 475 480
Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr Val Val
485 490 495
Gly Met Ala Gly Asn Asn Leu Leu Arg Glu Asp Lys Asp Pro His Lys
500 505 510
Met Ser Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr Asn Val
515 520 525
Thr Asp Val Met Phe Leu Asp Lys Lys Cys Gln Tyr Ile Ile Trp Thr
530 535 540
Phe Val Pro Gly Ser Gln Pro Gly Glu Leu Thr Leu Gly Phe Ile Arg
545 550 555 560
Ser Asp Leu Gly His Thr Ser Tyr Leu Val Arg Val Val Ser Thr Asn
565 570 575
Tyr Asn Gln His Ala Met Val Phe Phe Lys Ser Val Ile Gln Asn Arg
580 585 590
Glu Trp Phe Gly Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu Thr Ser
595 600 605
Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly Leu Pro
610 615 620
Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile Asp Gly
625 630 635 640
<210> 96
<211> 645
<212> Белок
<213> Искусственная последовательность
<220>
<223> слитый белок
<400> 96
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Glu Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ala Leu Asp Pro Lys Thr Gly Asp Thr Ala Tyr Ser Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Tyr Ser Tyr Thr Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Leu Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro
210 215 220
Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly Ser Gly
435 440 445
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
450 455 460
Gly Gly Ser Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu
465 470 475 480
Ser Lys Val Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly
485 490 495
Lys Trp Tyr Val Val Gly Met Ala Gly Asn Asn Leu Leu Arg Glu Asp
500 505 510
Lys Asp Pro His Lys Met Ser Ala Thr Ile Tyr Glu Leu Lys Glu Asp
515 520 525
Lys Ser Tyr Asn Val Thr Asp Val Met Phe Leu Asp Lys Lys Cys Gln
530 535 540
Tyr Ile Ile Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Leu Thr
545 550 555 560
Leu Gly Phe Ile Arg Ser Asp Leu Gly His Thr Ser Tyr Leu Val Arg
565 570 575
Val Val Ser Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Ser
580 585 590
Val Ile Gln Asn Arg Glu Trp Phe Gly Ile Thr Leu Tyr Gly Arg Thr
595 600 605
Lys Glu Leu Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys
610 615 620
Ser Leu Gly Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp
625 630 635 640
Gln Cys Ile Asp Gly
645
<210> 97
<211> 421
<212> Белок
<213> Искусственная последовательность
<220>
<223> слитый белок
<400> 97
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Arg Ala Gly Asn Val Gly Leu Arg Glu Asp Lys Asp Pro
35 40 45
Pro Lys Met Trp Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Asn Val Arg Phe Ala Arg Lys Lys Cys Thr Tyr Ser Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln
85 90 95
Ile Lys Ser Glu Pro Gly Gly Thr Ala Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Tyr Gln
115 120 125
Asn Arg Glu Ile Phe Phe Ile Ile Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
180 185 190
Ser Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu
195 200 205
Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
210 215 220
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
225 230 235 240
Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
245 250 255
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
260 265 270
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
275 280 285
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
290 295 300
Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
305 310 315 320
Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn
325 330 335
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
340 345 350
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
355 360 365
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg
370 375 380
Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys
385 390 395 400
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
405 410 415
Ser Leu Ser Leu Gly
420
<210> 98
<211> 422
<212> Белок
<213> Искусственная последовательность
<220>
<223> слитый белок
<400> 98
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala
1 5 10 15
Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
20 25 30
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
35 40 45
Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val
50 55 60
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser
65 70 75 80
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
85 90 95
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser
100 105 110
Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
115 120 125
Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln
130 135 140
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
145 150 155 160
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
165 170 175
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu
180 185 190
Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser
195 200 205
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
210 215 220
Leu Ser Leu Gly Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
225 230 235 240
Gly Gly Gly Ser Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro
245 250 255
Leu Ser Lys Val Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His
260 265 270
Gly Lys Trp Tyr Val Val Gly Gln Ala Gly Asn Ile Arg Leu Arg Glu
275 280 285
Asp Lys Asp Pro Ile Lys Met Met Ala Thr Ile Tyr Glu Leu Lys Glu
290 295 300
Asp Lys Ser Tyr Asp Val Thr Met Val Lys Phe Asp Asp Lys Lys Cys
305 310 315 320
Met Tyr Asp Ile Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe
325 330 335
Thr Leu Gly Lys Ile Lys Ser Phe Pro Gly His Thr Ser Ser Leu Val
340 345 350
Arg Val Val Ser Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys
355 360 365
Phe Val Phe Gln Asn Arg Glu Glu Phe Tyr Ile Thr Leu Tyr Gly Arg
370 375 380
Thr Lys Glu Leu Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser
385 390 395 400
Lys Ser Leu Gly Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile
405 410 415
Asp Gln Cys Ile Asp Gly
420
<210> 99
<211> 641
<212> Белок
<213> Искусственная последовательность
<220>
<223> слитый белок
<400> 99
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn His Gly Gly Tyr Val Thr Tyr Asn Pro Ser Leu Glu
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Tyr Gly Pro Gly Asn Tyr Asp Trp Tyr Phe Asp Leu Trp Gly
100 105 110
Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly
210 215 220
Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro
260 265 270
Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln
450 455 460
Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val Pro
465 470 475 480
Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr Val
485 490 495
Val Gly Arg Ala Gly Asn Val Gly Leu Arg Glu Asp Lys Asp Pro Pro
500 505 510
Lys Met Trp Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr Asp
515 520 525
Val Thr Asn Val Arg Phe Ala Arg Lys Lys Cys Thr Tyr Ser Ile Gly
530 535 540
Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln Ile
545 550 555 560
Lys Ser Glu Pro Gly Gly Thr Ala Asn Leu Val Arg Val Val Ser Thr
565 570 575
Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Tyr Gln Asn
580 585 590
Arg Glu Ile Phe Phe Ile Ile Leu Tyr Gly Arg Thr Lys Glu Leu Thr
595 600 605
Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly Leu
610 615 620
Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile Asp
625 630 635 640
Gly
<210> 100
<211> 641
<212> Белок
<213> Искусственная последовательность
<220>
<223> слитый белок
<400> 100
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Arg Ala Gly Asn Val Gly Leu Arg Glu Asp Lys Asp Pro
35 40 45
Pro Lys Met Trp Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Asn Val Arg Phe Ala Arg Lys Lys Cys Thr Tyr Ser Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln
85 90 95
Ile Lys Ser Glu Pro Gly Gly Thr Ala Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Tyr Gln
115 120 125
Asn Arg Glu Ile Phe Phe Ile Ile Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
180 185 190
Ser Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser
195 200 205
Glu Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly
210 215 220
Tyr Tyr Trp Ser Trp Ile Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp
225 230 235 240
Ile Gly Glu Ile Asn His Gly Gly Tyr Val Thr Tyr Asn Pro Ser Leu
245 250 255
Glu Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser
260 265 270
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
275 280 285
Ala Arg Asp Tyr Gly Pro Gly Asn Tyr Asp Trp Tyr Phe Asp Leu Trp
290 295 300
Gly Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
305 310 315 320
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
325 330 335
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
340 345 350
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
355 360 365
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
370 375 380
Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp
385 390 395 400
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
405 410 415
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro
420 425 430
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
435 440 445
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
450 455 460
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
465 470 475 480
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
485 490 495
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
500 505 510
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
515 520 525
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
530 535 540
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
545 550 555 560
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
565 570 575
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
580 585 590
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
595 600 605
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
610 615 620
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
625 630 635 640
Lys
<210> 101
<211> 409
<212> Белок
<213> Искусственная последовательность
<220>
<223> слитый белок
<400> 101
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Arg Ala Gly Asn Val Gly Leu Arg Glu Asp Lys Asp Pro
35 40 45
Pro Lys Met Trp Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Asn Val Arg Phe Ala Arg Lys Lys Cys Thr Tyr Ser Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln
85 90 95
Ile Lys Ser Glu Pro Gly Gly Thr Ala Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Tyr Gln
115 120 125
Asn Arg Glu Ile Phe Phe Ile Ile Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
180 185 190
Ser Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro
195 200 205
Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser
210 215 220
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
225 230 235 240
Ile Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser
245 250 255
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu
260 265 270
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro
275 280 285
Pro Ala Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr
290 295 300
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
305 310 315 320
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
325 330 335
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
340 345 350
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
355 360 365
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
370 375 380
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
385 390 395 400
Thr Lys Ser Phe Asn Arg Gly Glu Cys
405
<210> 102
<211> 408
<212> Белок
<213> Искусственная последовательность
<220>
<223> слитый белок
<400> 102
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Pro
85 90 95
Ala Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val
100 105 110
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
115 120 125
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
130 135 140
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
145 150 155 160
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
165 170 175
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
180 185 190
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr
195 200 205
Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser Gly Gly Gly
210 215 220
Gly Ser Gly Gly Gly Gly Ser Gln Asp Ser Thr Ser Asp Leu Ile Pro
225 230 235 240
Ala Pro Pro Leu Ser Lys Val Pro Leu Gln Gln Asn Phe Gln Asp Asn
245 250 255
Gln Phe Gln Gly Lys Trp Tyr Val Val Gly Arg Ala Gly Asn Val Gly
260 265 270
Leu Arg Glu Asp Lys Asp Pro Pro Lys Met Trp Ala Thr Ile Tyr Glu
275 280 285
Leu Lys Glu Asp Lys Ser Tyr Asp Val Thr Asn Val Arg Phe Ala Arg
290 295 300
Lys Lys Cys Thr Tyr Ser Ile Gly Thr Phe Val Pro Gly Ser Gln Pro
305 310 315 320
Gly Glu Phe Thr Leu Gly Gln Ile Lys Ser Glu Pro Gly Gly Thr Ala
325 330 335
Asn Leu Val Arg Val Val Ser Thr Asn Tyr Asn Gln His Ala Met Val
340 345 350
Phe Phe Lys Glu Val Tyr Gln Asn Arg Glu Ile Phe Phe Ile Ile Leu
355 360 365
Tyr Gly Arg Thr Lys Glu Leu Thr Ser Glu Leu Lys Glu Asn Phe Ile
370 375 380
Arg Phe Ser Lys Ser Leu Gly Leu Pro Glu Asn His Ile Val Phe Pro
385 390 395 400
Val Pro Ile Asp Gln Cys Ile Asp
405
<210> 103
<211> 635
<212> Белок
<213> Искусственная последовательность
<220>
<223> слитый белок
<400> 103
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Arg Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Ser Thr Tyr
20 25 30
Trp Ile Ser Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Lys Ile Tyr Pro Gly Asp Ser Tyr Thr Asn Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Gly Tyr Gly Ile Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe
180 185 190
Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro
210 215 220
Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro
225 230 235 240
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
245 250 255
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp
260 265 270
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
275 280 285
Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val
290 295 300
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
305 310 315 320
Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly
325 330 335
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
340 345 350
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
355 360 365
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
370 375 380
Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe
385 390 395 400
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
405 410 415
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
420 425 430
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly
435 440 445
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Asp Ser Thr Ser Asp Leu
450 455 460
Ile Pro Ala Pro Pro Leu Ser Lys Val Pro Leu Gln Gln Asn Phe Gln
465 470 475 480
Asp Asn Gln Phe Gln Gly Lys Trp Tyr Val Val Gly Arg Ala Gly Asn
485 490 495
Val Gly Leu Arg Glu Asp Lys Asp Pro Pro Lys Met Trp Ala Thr Ile
500 505 510
Tyr Glu Leu Lys Glu Asp Lys Ser Tyr Asp Val Thr Asn Val Arg Phe
515 520 525
Ala Arg Lys Lys Cys Thr Tyr Ser Ile Gly Thr Phe Val Pro Gly Ser
530 535 540
Gln Pro Gly Glu Phe Thr Leu Gly Gln Ile Lys Ser Glu Pro Gly Gly
545 550 555 560
Thr Ala Asn Leu Val Arg Val Val Ser Thr Asn Tyr Asn Gln His Ala
565 570 575
Met Val Phe Phe Lys Glu Val Tyr Gln Asn Arg Glu Ile Phe Phe Ile
580 585 590
Ile Leu Tyr Gly Arg Thr Lys Glu Leu Thr Ser Glu Leu Lys Glu Asn
595 600 605
Phe Ile Arg Phe Ser Lys Ser Leu Gly Leu Pro Glu Asn His Ile Val
610 615 620
Phe Pro Val Pro Ile Asp Gln Cys Ile Asp Gly
625 630 635
<210> 104
<211> 442
<212> Белок
<213> Искусственная последовательность
<220>
<223> тяжелая цепь антитела
<400> 104
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Arg Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Ser Thr Tyr
20 25 30
Trp Ile Ser Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Lys Ile Tyr Pro Gly Asp Ser Tyr Thr Asn Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Gly Tyr Gly Ile Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe
180 185 190
Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro
210 215 220
Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro
225 230 235 240
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
245 250 255
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp
260 265 270
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
275 280 285
Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val
290 295 300
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
305 310 315 320
Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly
325 330 335
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
340 345 350
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
355 360 365
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
370 375 380
Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe
385 390 395 400
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
405 410 415
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
420 425 430
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 105
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> легкая цепь антитела
<400> 105
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly Asp Asn Ile Gly Asp Gln Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Lys Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Tyr Thr Gly Phe Gly Ser Leu
85 90 95
Ala Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys
100 105 110
Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln
115 120 125
Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly
130 135 140
Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly
145 150 155 160
Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala
165 170 175
Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser
180 185 190
Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val
195 200 205
Ala Pro Thr Glu Cys Ser
210
<210> 106
<211> 439
<212> Белок
<213> Искусственная последовательность
<220>
<223> тяжелая цепь антитела
<400> 106
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Asp Cys Lys Ala Ser Gly Ile Thr Phe Ser Asn Ser
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Trp Tyr Asp Gly Ser Lys Arg Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Asn Asp Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser
100 105 110
Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser
115 120 125
Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp
130 135 140
Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr
145 150 155 160
Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr
165 170 175
Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys
180 185 190
Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp
195 200 205
Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala
210 215 220
Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
225 230 235 240
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
245 250 255
Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val
260 265 270
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
275 280 285
Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
290 295 300
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly
305 310 315 320
Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
325 330 335
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr
340 345 350
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
355 360 365
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
370 375 380
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
385 390 395 400
Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe
405 410 415
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
420 425 430
Ser Leu Ser Leu Ser Leu Gly
435
<210> 107
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> легкая цепь антитела
<400> 107
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Ser Ser Asn Trp Pro Arg
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 108
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR1 тяжелой цепи антитела
<400> 108
Gly Phe Thr Phe Asn Lys Asn Ala
1 5
<210> 109
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR2 тяжелой цепи антитела
<400> 109
Ile Arg Asn Lys Thr Asn Asn Tyr Ala Thr
1 5 10
<210> 110
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR3 тяжелой цепи антитела
<400> 110
Val Ala Gly Asn Ser Phe Ala Tyr
1 5
<210> 111
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR1 легкой цепи антитела
<400> 111
Gln Ser Leu Leu Tyr Ser Ser Asn Gln Lys Asn Tyr
1 5 10
<210> 112
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR1 легкой цепи антитела
<400> 112
Gln Ser Leu Leu Tyr Ser Ser Asn Gln Lys Asn Tyr
1 5 10
<210> 113
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR легкой цепи антитела
<400> 113
Gln Gln Tyr Tyr Asn Tyr Pro Leu Thr
1 5
<210> 114
<211> 116
<212> Белок
<213> Искусственная последовательность
<220>
<223> вариабельный домен тяжелой цепи антитела
<400> 114
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Lys Asn
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Arg Asn Lys Thr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Ala Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Ala Gly Asn Ser Phe Ala Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser
115
<210> 115
<211> 113
<212> Белок
<213> Искусственная последовательность
<220>
<223> вариабельный домен легкой цепи антитела
<400> 115
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu Tyr Ser
20 25 30
Ser Asn Gln Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Ser Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Tyr Tyr Asn Tyr Pro Leu Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile
100 105 110
Lys
<210> 116
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR1 тяжелой цепи антитела
<400> 116
Tyr Phe Asp Phe Asp Ser Tyr Glu
1 5
<210> 117
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR2 тяжелой цепи антитела
<400> 117
Ile Tyr His Ser Gly Ser Thr
1 5
<210> 118
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR3 тяжелой цепи антитела
<400> 118
Ala Arg Val Asn Met Asp Arg Phe Asp Tyr
1 5 10
<210> 119
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> вариабельный домен тяжелой цепи антитела
<400> 119
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Tyr Phe Asp Phe Asp Ser Tyr
20 25 30
Glu Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ser Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Thr Leu Arg Ala Glu Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Val Asn Met Asp Arg Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ser
115
<210> 120
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR1 тяжелой цепи антитела
<400> 120
Gly Phe Thr Phe Ser Ser Tyr Ala
1 5
<210> 121
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR2 тяжелой цепи антитела
<400> 121
Ile Gln Lys Gln Gly Leu Pro Thr
1 5
<210> 122
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR3 тяжелой цепи антитела
<400> 122
Ala Lys Asn Arg Ala Lys Phe Asp Tyr
1 5
<210> 123
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR1 легкой цепи антитела
<400> 123
Gln Ser Ile Ser Ser Tyr
1 5
<210> 124
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR1 легкой цепи антитела
<400> 124
Gln Ser Ile Ser Ser Tyr
1 5
<210> 125
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR3 легкой цепи антитела
<400> 125
Gln Gln Asn Arg Gly Phe Pro Leu Thr
1 5
<210> 126
<211> 116
<212> Белок
<213> Искусственная последовательность
<220>
<223> вариабельный домен тяжелой цепи антитела
<400> 126
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Thr Ile Gln Lys Gln Gly Leu Pro Thr Gln Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asn Arg Ala Lys Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 127
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> вариабельный домен легкой цепи антитела
<400> 127
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asn Ala Ser Met Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Asn Arg Gly Phe Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 128
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> вариабельный домен легкой цепи антитела
<400> 128
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Met Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Asn Arg Gly Phe Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 129
<211> 115
<212> Белок
<213> Искусственная последовательность
<220>
<223> вариабельный домен тяжелой цепи антитела
<400> 129
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Glu Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ala Leu Asp Pro Lys Thr Gly Asp Thr Ala Tyr Ser Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Tyr Ser Tyr Thr Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser
115
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СЛИТЫЙ ПОЛИПЕПТИД С ПРОТИВОРАКОВОЙ АКТИВНОСТЬЮ | 2016 |
|
RU2754466C2 |
| МУТЕИНЫ ЛИПОКАЛИНА 2 ЧЕЛОВЕКА С АФФИННОСТЬЮ В ОТНОШЕНИИ ГЛИПИКАНА-3 (GPC3) | 2016 |
|
RU2756318C2 |
| НОВЫЙ СЛИТЫЙ БЕЛОК, СПЕЦИФИЧНЫЙ ДЛЯ CD137 И PD-L1 | 2019 |
|
RU2818349C2 |
| НОВЫЕ ПОЛИПЕПТИДЫ СО СПЕЦИФИЧЕСКИМ СВЯЗЫВАНИЕМ И ПУТИ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2723034C2 |
| БЕЛКИ, СПЕЦИФИЧНЫЕ В ОТНОШЕНИИ CD137 | 2016 |
|
RU2736312C2 |
| НОВЫЕ БЕЛКИ, СПЕЦИФИЧЕСКИЕ В ОТНОШЕНИИ АНГИОГЕНЕЗА | 2016 |
|
RU2720688C2 |
| МУТЕИНЫ ЛИПОКАЛИНА 2 ЧЕЛОВЕКА (Lcn2, hNGAL) С АФФИННОСТЬЮ ДЛЯ ОПРЕДЕЛЕННОЙ МИШЕНИ | 2019 |
|
RU2804336C2 |
| МУТЕИНЫ ЛИПОКАЛИНА 2 ЧЕЛОВЕКА (Lcn2, hNGAL) С АФФИННОСТЬЮ ДЛЯ ОПРЕДЕЛЕННОЙ МИШЕНИ | 2010 |
|
RU2707126C2 |
| ИНДУЦИРУЮЩИЙ ЦИТОТОКСИЧНОСТЬ ТЕРАПЕВТИЧЕСКИЙ АГЕНТ | 2022 |
|
RU2824285C2 |
| ИММУНОАКТИВИРУЮЩАЯ АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА | 2015 |
|
RU2722788C2 |
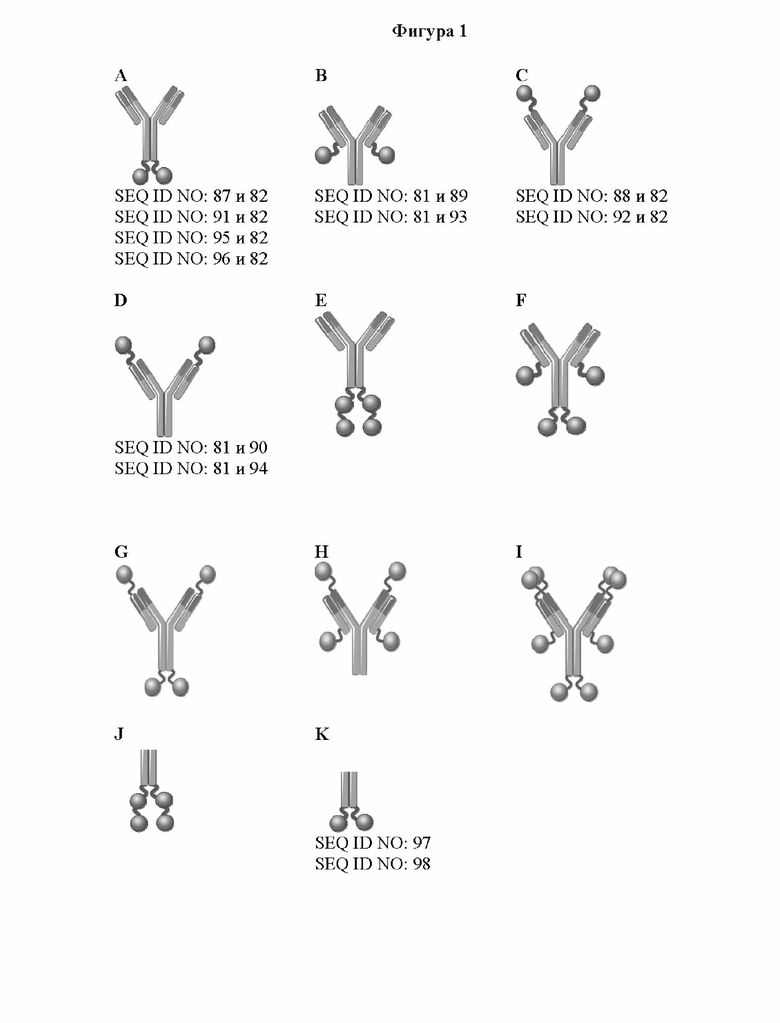
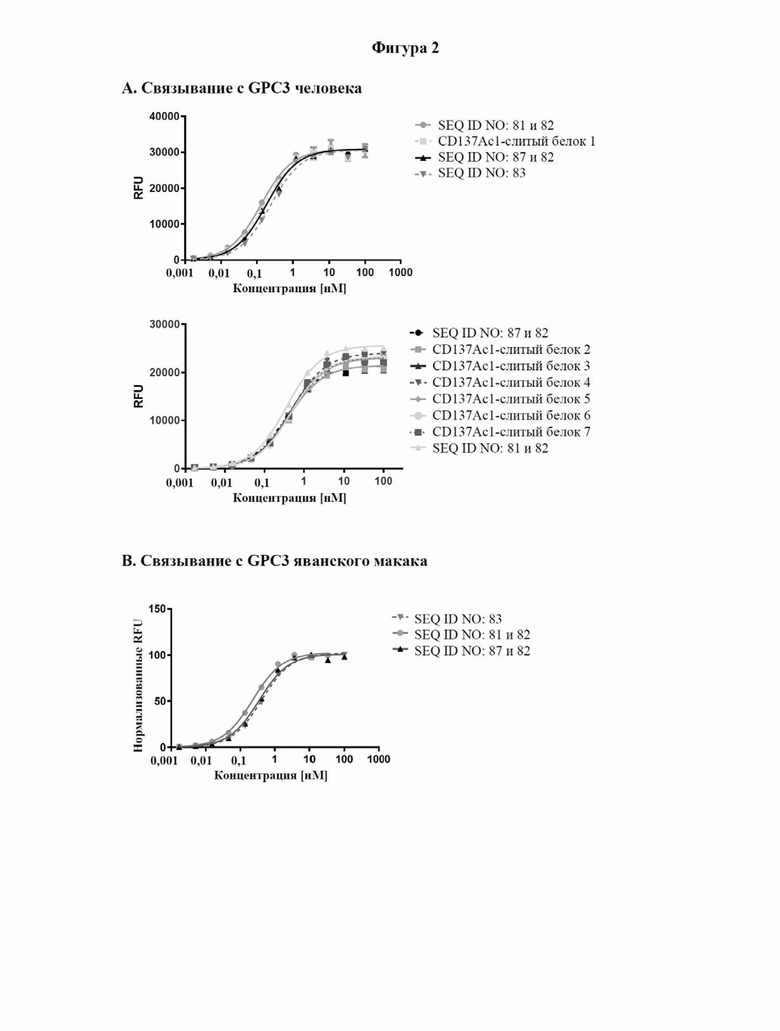
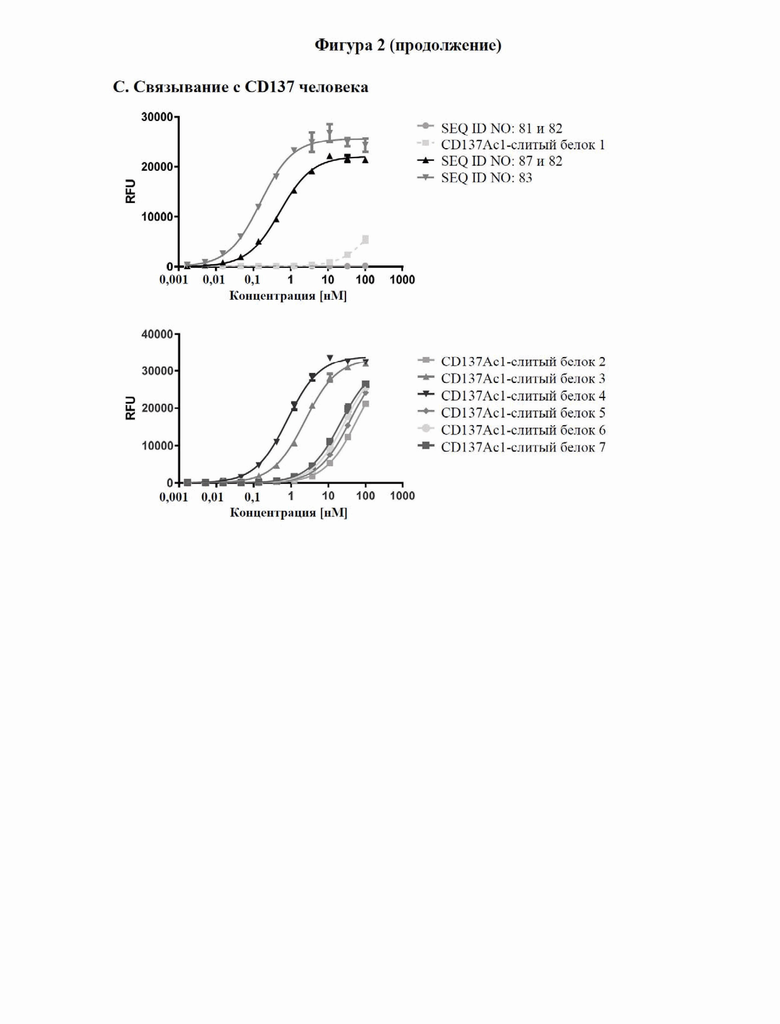
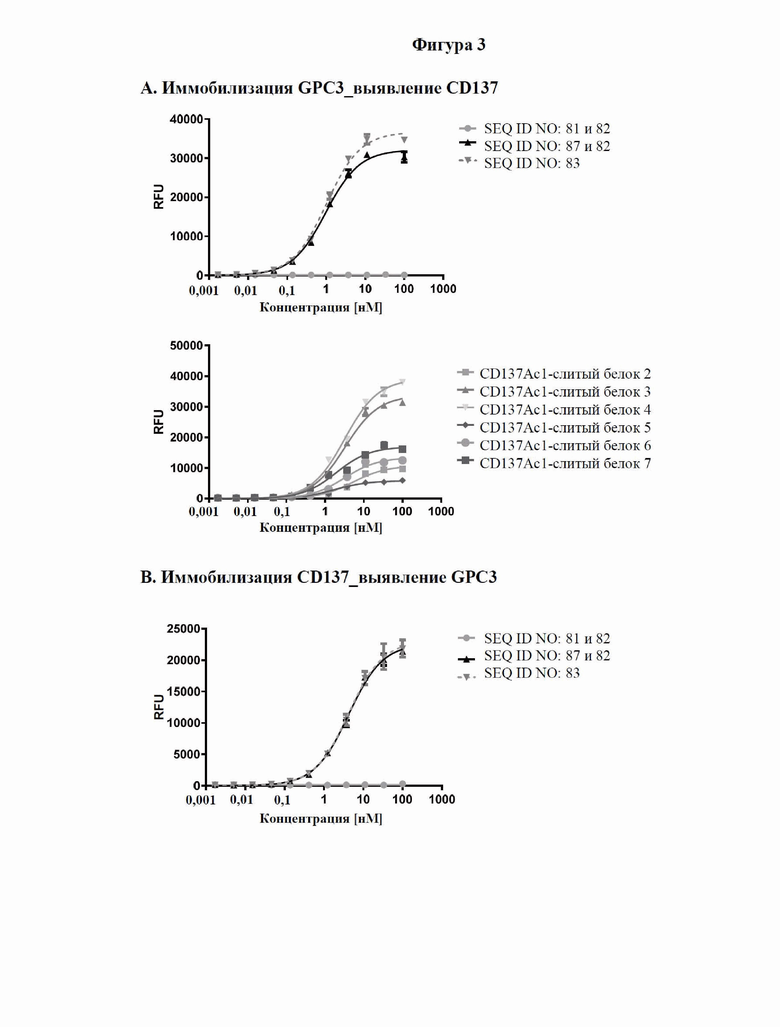
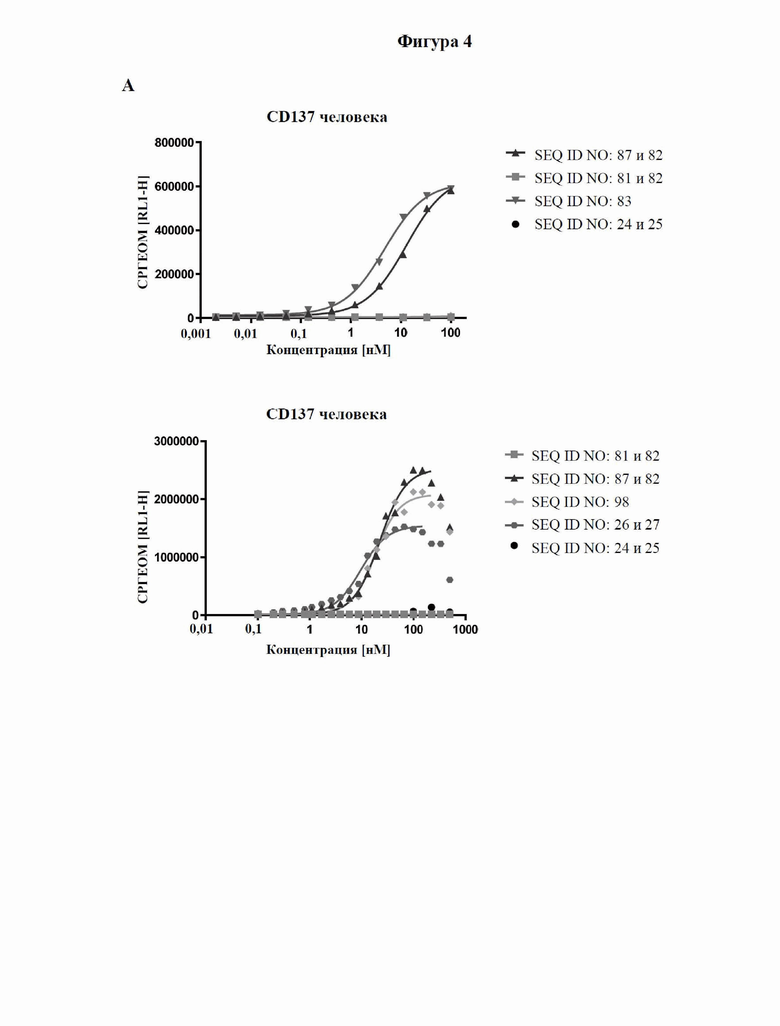
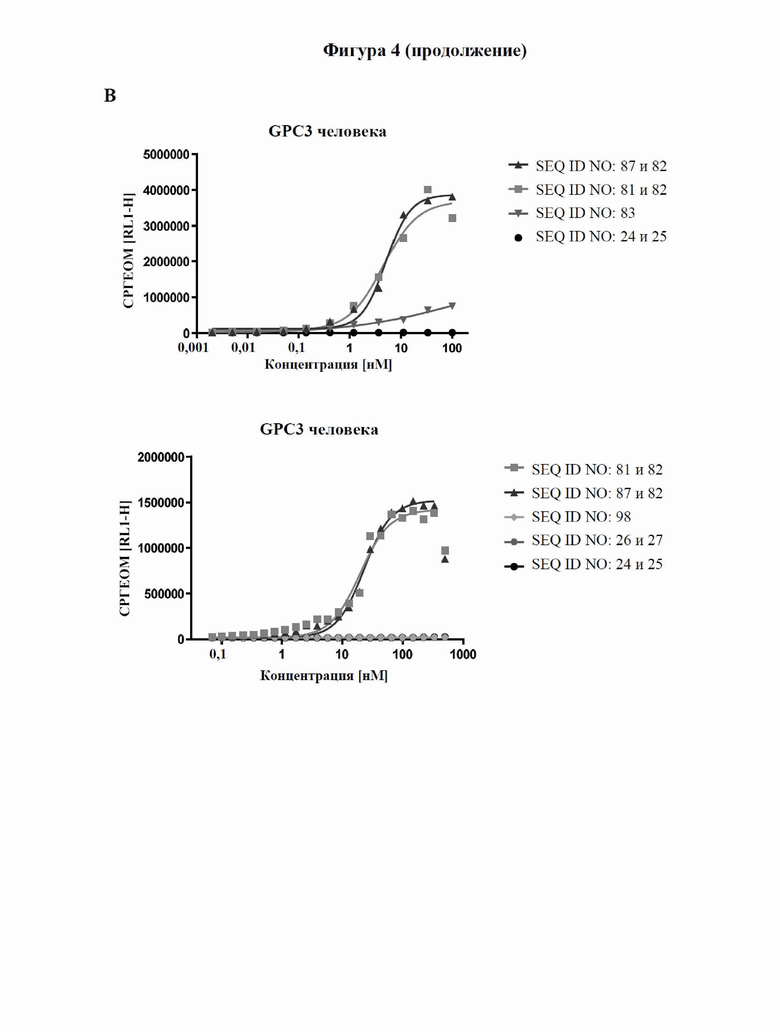
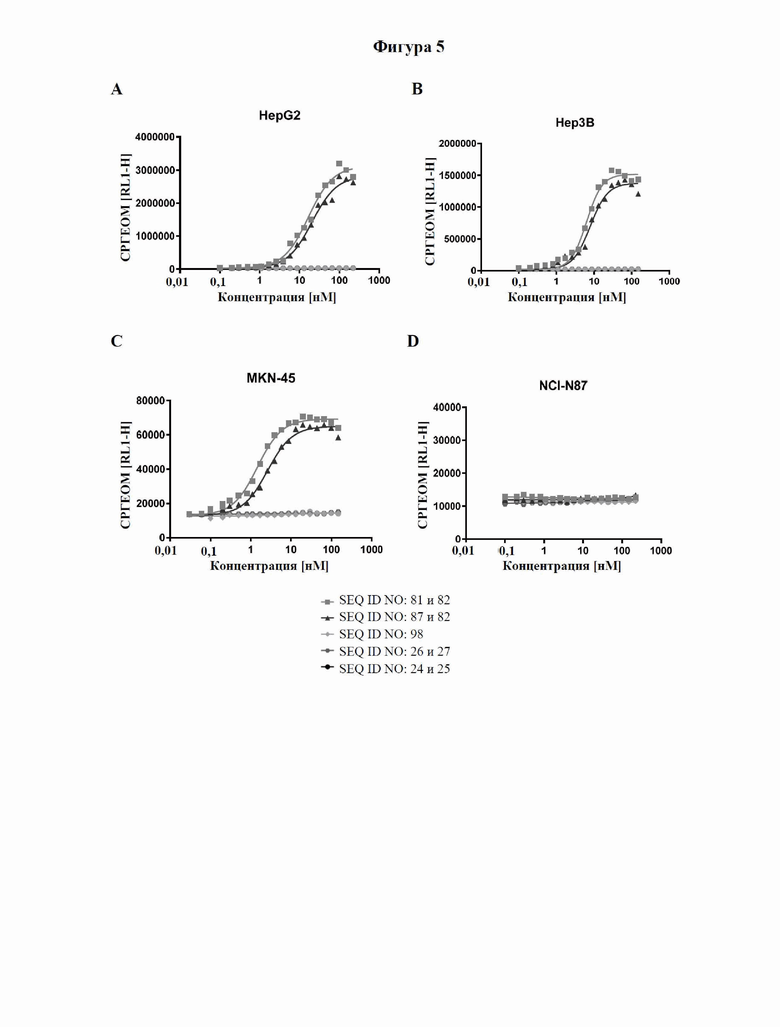
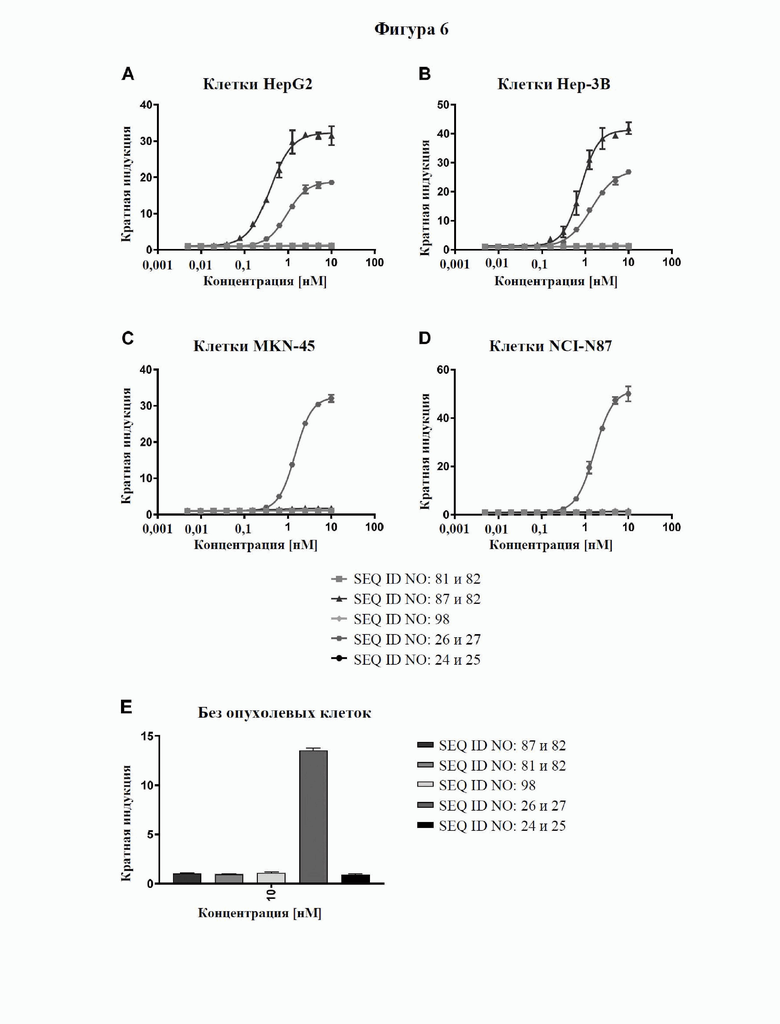
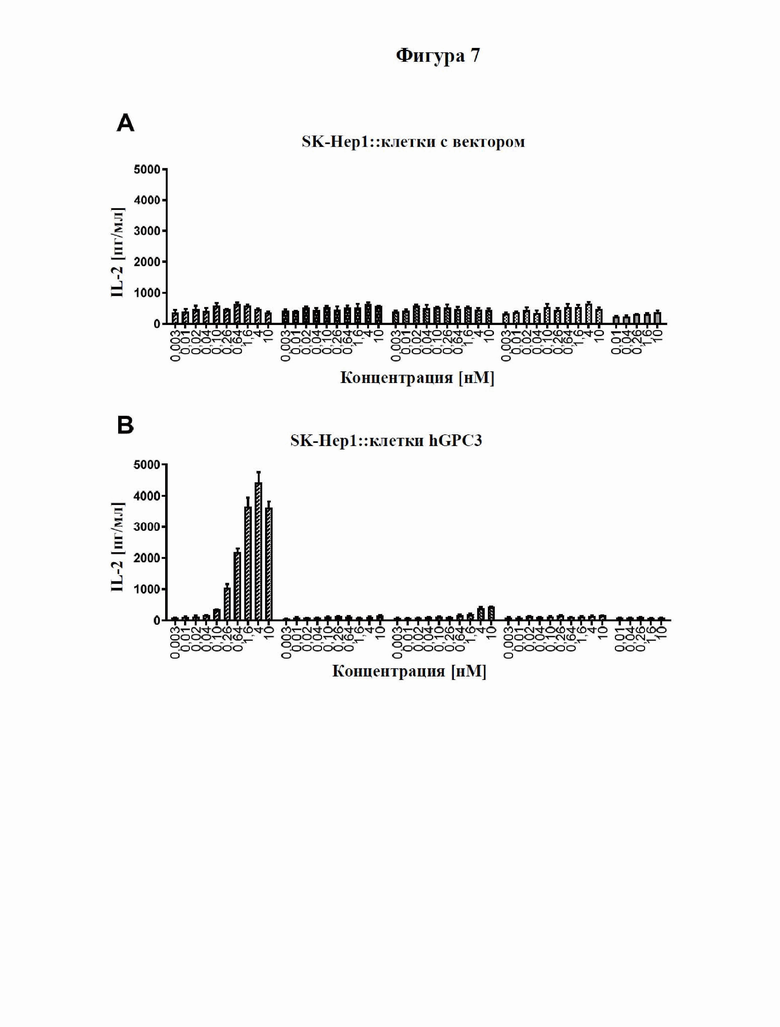
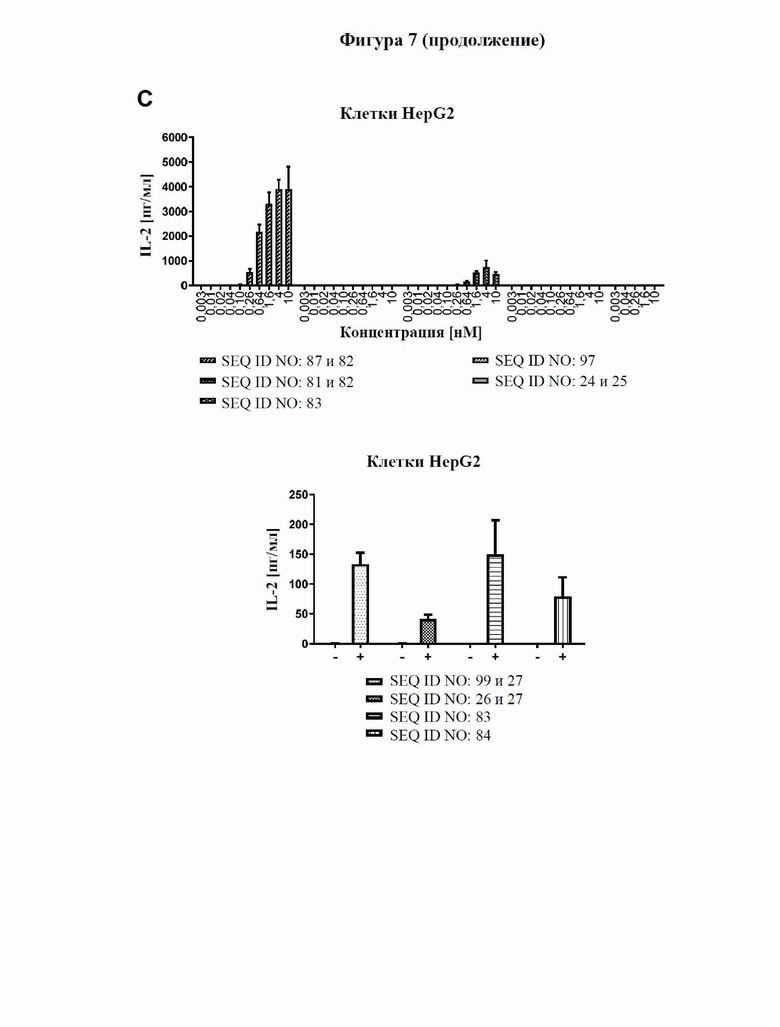
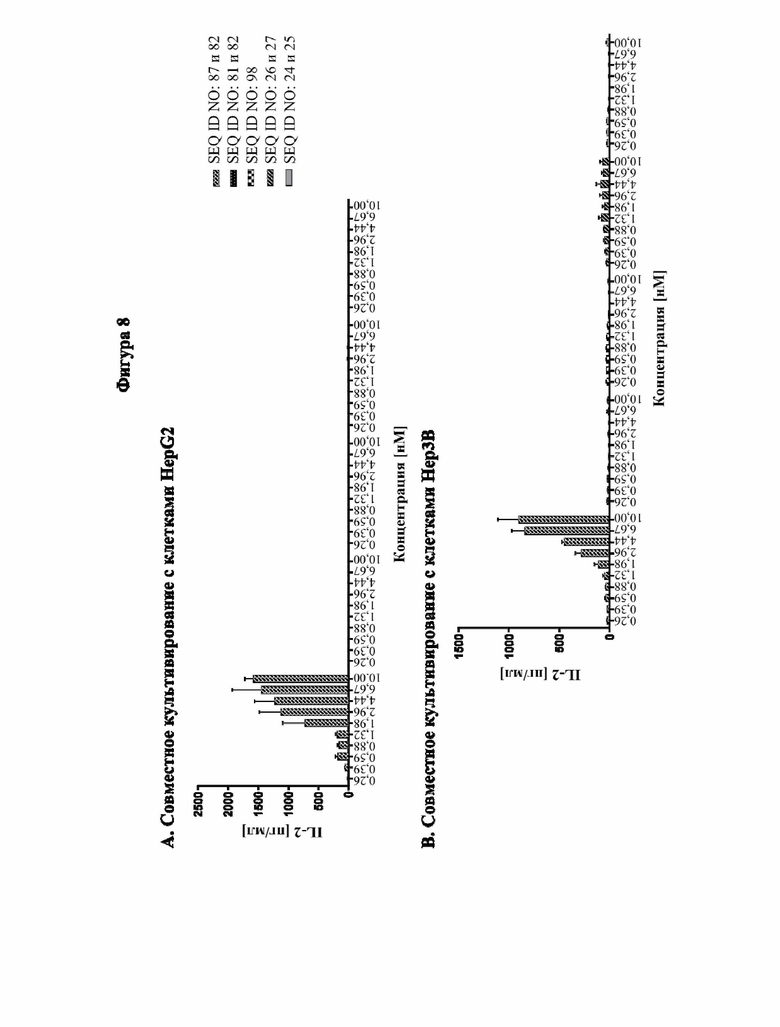
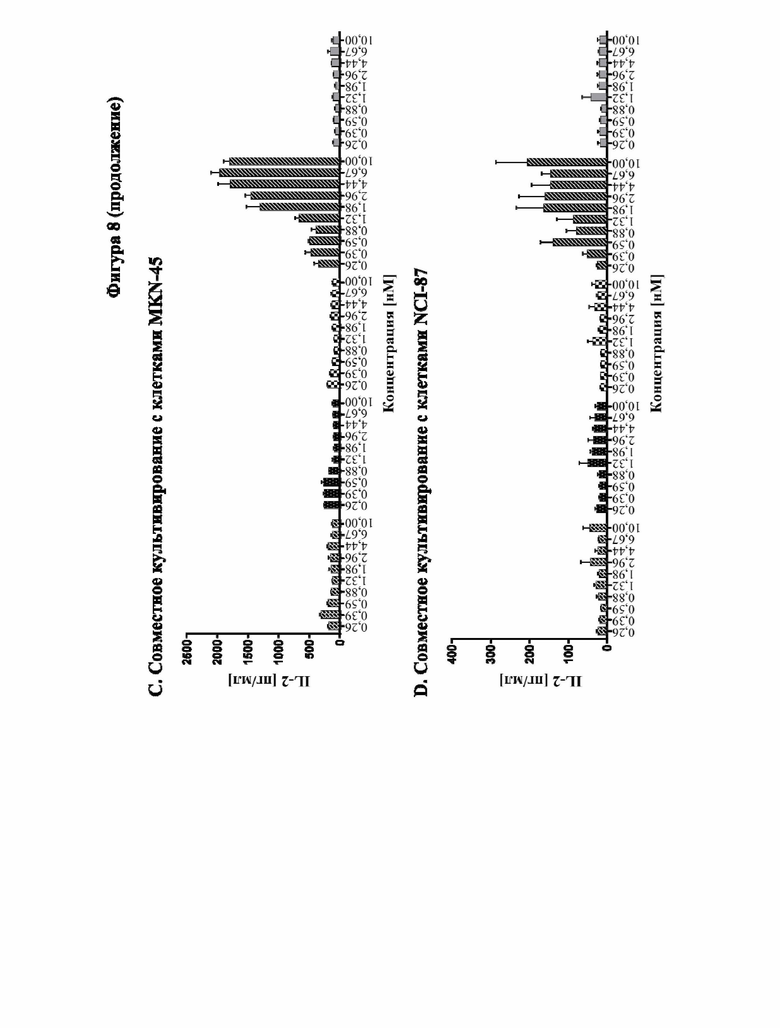
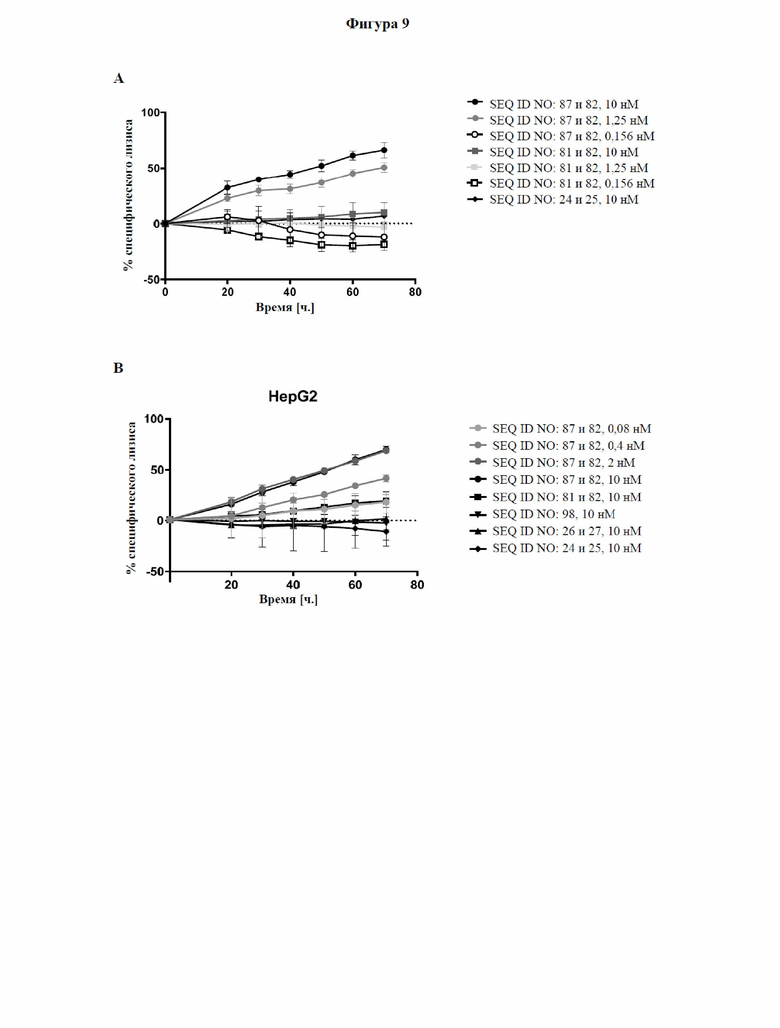
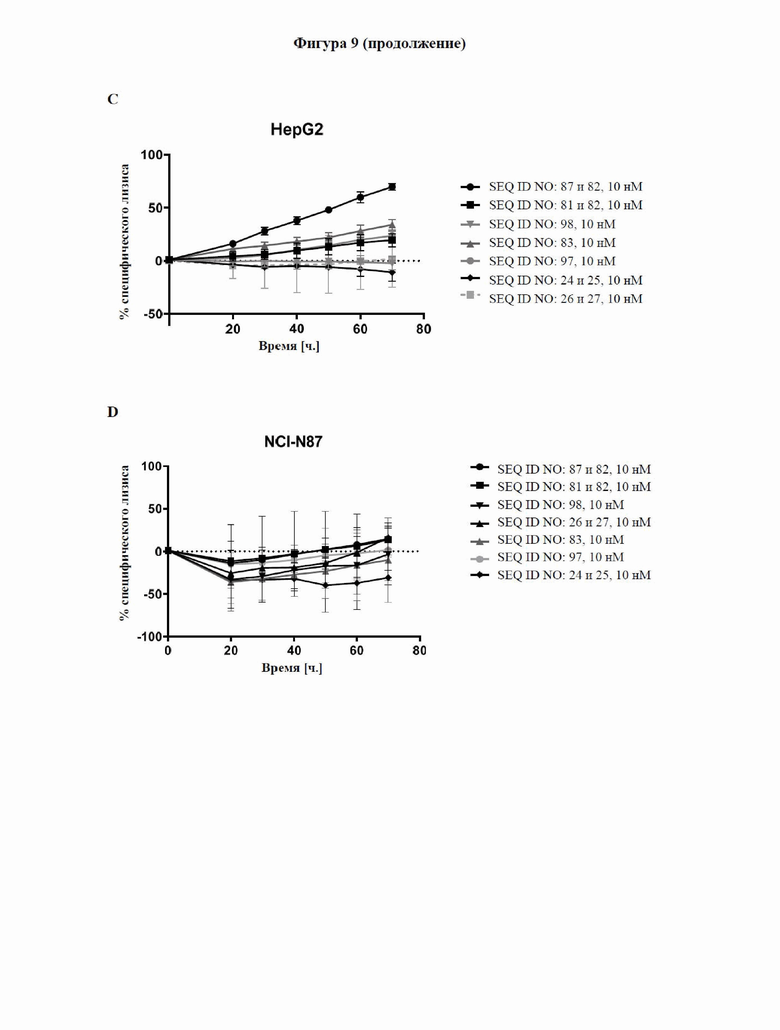
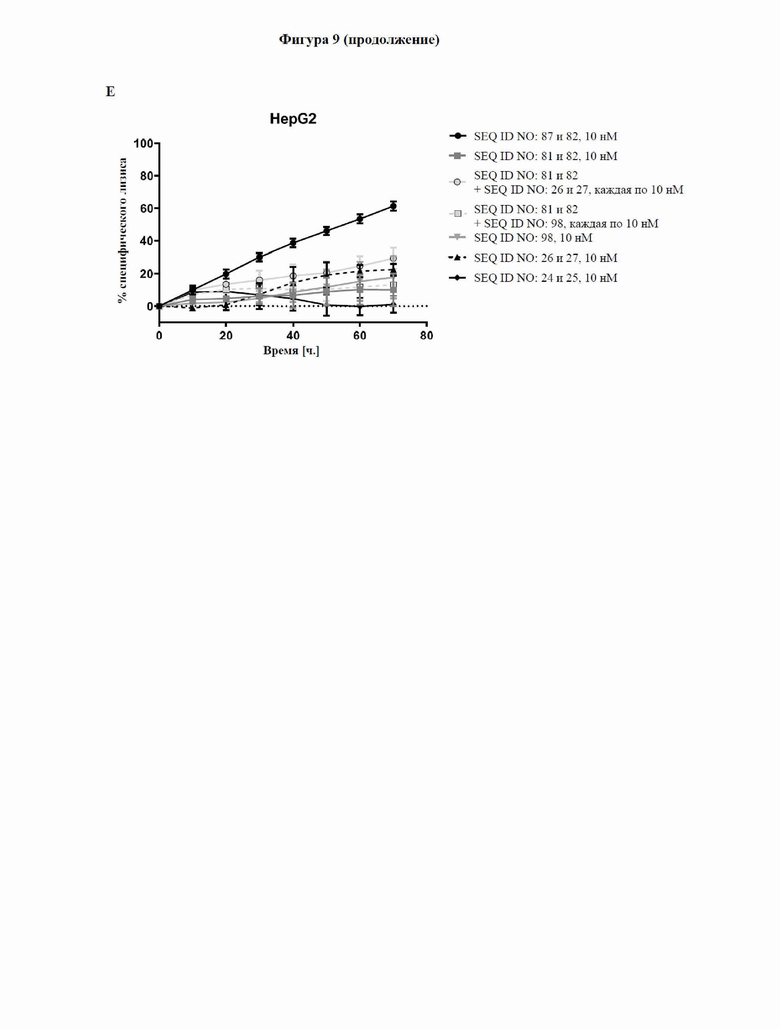
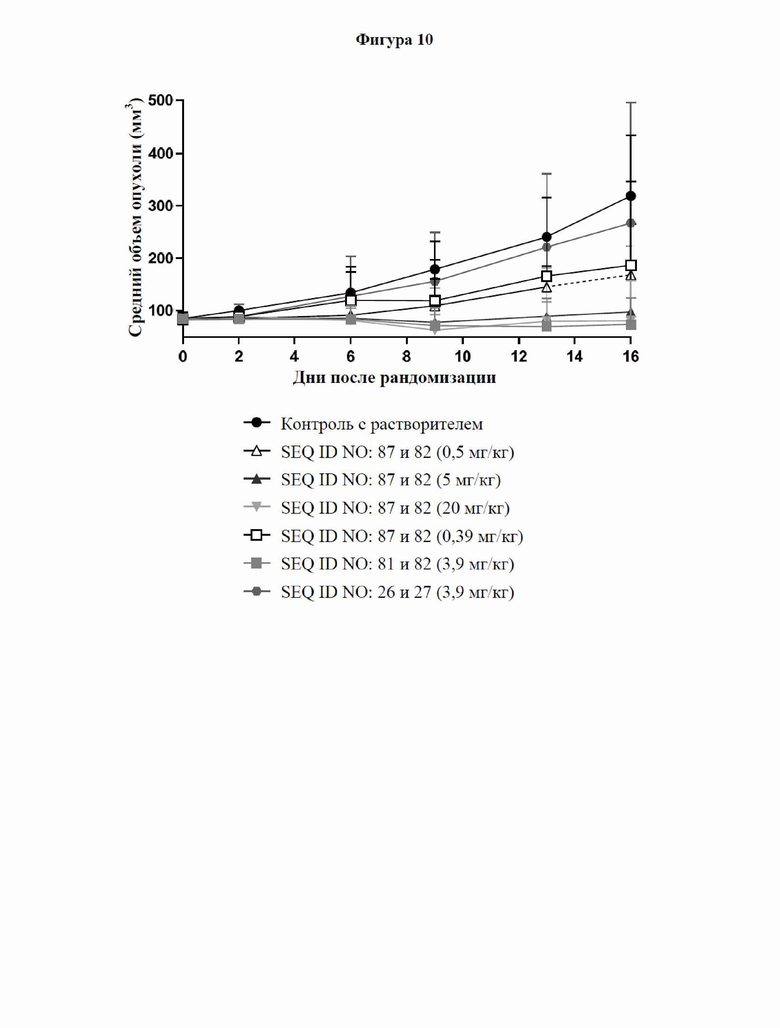
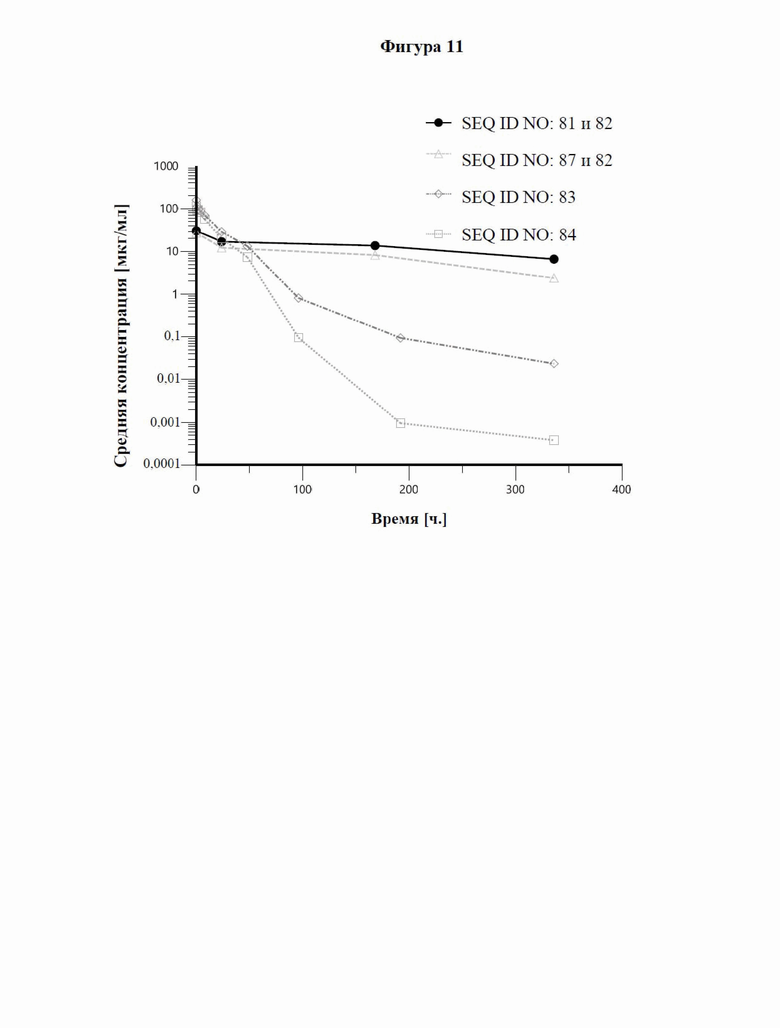
Изобретение относится к области биотехнологии, конкретно к рекомбинантному получению слитых белков, связывающих глипикан-3 (GPC3) и кластер дифференцировки 137 (CD137), и может быть использовано в медицине для терапии GPC3-положительных форм рака. Предложен слитый белок, который включает полноразмерный иммуноглобулин, который специфически связывается с GPC3, соединенный с C-конца посредством пептидного линкера с N-концом мутеина липокалина, который специфически связывается с CD137. Изобретение обеспечивает получение слитого белка, способного одновременно специфически связывать как CD137, так и GPC3. 9 н. и 6 з.п. ф-лы, 11 ил., 11 табл., 14 пр.
1. Слитый белок, специфически связывающий как кластер дифференцировки 137 (CD137), так и глипикан-3 (GPC3), где слитый белок включает две субъединицы, где первая субъединица представляет собой полноразмерный иммуноглобулин и специфически связывается с GPC3, и где вторая субъединица представляет собой мутеин липокалина и специфически связывается с CD137, причем вторая субъединица соединена на N-конце с С-концом первой субъединицы посредством пептидного линкера, и причем слитый белок содержит SEQ ID NO: 87 и 82.
2. Слитый белок по п. 1,
(a) где слитый белок способен связывать GPC3 со значением KD, составляющим максимум 1 нМ или сравнимым или ниже значения KD у иммуноглобулина, который включен исключительно в первую субъединицу;
(b) где слитый белок способен связывать CD137 со значением ЕС50, составляющим максимум 3 нМ или сравнимым или ниже значения ЕС50 у мутеина липокалина, специфического в отношении CD137, который включен исключительно во вторую субъединицу;
(c) где слитый белок вступает в перекрестную реакцию с GPC3 яванского макака;
(d) где слитый белок способен одновременно связывать CD137 и GPC3 со значениями ЕС50, составляющими максимум 10 нМ, при измерении указанного слитого белка в ELISA-анализе;
(e) где слитый белок способен связывать GРС3-экспрессирующие опухолевые клетки;
(f) где слитый белок способен индуцировать повышенную секрецию IL-2;
(g) где слитый белок способен индуцировать повышенную секрецию IL-2 до более высокого уровня, чем SEQ ID NO: 83, и/или с лучшей эффективностью в сравнении с SEQ ID NO: 83;
(h) где слитый белок способен индуцировать опосредуемую лимфоцитами цитотоксичность;
(i) где слитый белок способен индуцировать усиленное уничтожение GPC3-экспрессирующих опухолевых клеток, опосредованное Т-клетками, в сравнении с SEQ ID NO: 83 и/или индуцировать активацию цитотоксических Т-клеток с лучшей эффективностью в сравнении с SEQ ID NO: 83;
(j) где слитый белок способен совместно стимулировать Т-клеточные ответы зависимым от GPC3 образом; или
(k) где слитый белок способен совместно стимулировать Т-клеточные ответы в опухолевом микроокружении.
3. Слитый белок по п. 1 или 2, где слитый белок обладает периодом полужизни у мышей, составляющим по меньшей мере 50 часов, по меньшей мере 75 часов, по меньшей мере 100 часов, по меньшей мере 125 часов, по меньшей мере 150 часов, по меньшей мере 175 часов, по меньшей мере 200 часов, по меньшей мере 250 часов или еще дольше, и/или где слитый белок обладает периодом полужизни у мышей, который длиннее, чем у SEQ ID NO: 83.
4. Слитый белок по любому из пп. 1-3, где слитый белок характеризуется изоэлектрической точкой, составляющей по меньшей мере 6,5, по меньшей мере 6,8, по меньшей мере 7,1, по меньшей мере 7,4, по меньшей мере 7,5, по меньшей мере 7,7 или еще выше, и/или где слитый белок характеризуется изоэлектрической точкой, которая выше, чем у SEQ ID NO: 83.
5. Слитый белок по любому из пп. 1-4, где вторая субъединица соединена на N-конце посредством линкера с С-концом константной области тяжелой цепи (СН) первой субъединицы.
6. Слитый белок по любому из пп. 1-5, где первая субъединица содержит следующий набор последовательностей CDR: GYTFTDYE HCDR1, SEQ ID NO: 72, LDPKTGDT HCDR2, SEQ ID NO: 73, TRFYSYTY HCDR3; SEQ ID NO: 74, и следующий набор последовательностей CDR: QSLVHSNRNTY LCDR1, SEQ ID NO: 75, KVS LCDR2, SQNTHVPPT LCDR3; SEQ ID NO: 77.
7. Слитый белок по любому из пп. 1-6, где иммуноглобулин имеет остов IgG4.
8. Молекула нуклеиновой кислоты, предназначенная для экспрессии слитого белка, специфически связывающего как CD137, так и GPC3, содержащая нуклеотидную последовательность, кодирующую слитый белок по любому из пп. 1-7, при этом указанная последовательность функционально связана с регуляторной последовательностью.
9. Способ получения слитого белка по любому из пп. 1-7, где способ включает культивирование клетки-хозяина для получения слитого белка и трансформирование клетки-хозяина молекулой нуклеиновой кислоты по п. 8.
10. Фармацевтическая композиция для предупреждения, уменьшения интенсивности или лечения GРС3-положительных форм рака, содержащая эффективное количество слитого белка по любому из пп. 1-7 и фармацевтически приемлемого носителя.
11. Способ одновременной активации последующих сигнальных путей CD137 и воздействия на GPC3-положительные опухолевые клетки, предусматривающий применение по отношению к ткани, содержащей опухоль, слитого белка по любому из пп. 1-7 или фармацевтической композиции по п. 10.
12. Способ одновременной совместной стимуляции Т-клеток и воздействия на GРС3-положительные опухолевые клетки, предусматривающий применение по отношению к ткани, содержащей опухоль, слитого белка по любому из пп. 1-7 или фармацевтической композиции по п. 10.
13. Способ индукции локализованного лимфоцитарного ответа в непосредственной близости от GРС3-положительных опухолевых клеток, предусматривающий применение по отношению к ткани, содержащей опухоль, слитого белка по любому из пп. 1-7 или фармацевтической композиции по п. 10.
14. Способ индукции повышенного опосредованного лимфоцитами цитолиза GРС3-положительных опухолевых клеток, предусматривающий применение по отношению к ткани, содержащей опухоль, слитого белка по любому из пп. 1-7 или фармацевтической композиции по п. 10.
15. Способ предупреждения, уменьшения интенсивности или лечения GPC3-положительных форм рака, предусматривающий применение по отношению к ткани, содержащей опухоль, слитого белка по любому из пп. 1-7 или фармацевтической композиции по п. 10.
| US 2018148485 A1, 31.05.2018, WO 2016177802 А1, 10.11.2016, US 2017267783 А1, 21.09.2017, WO 2018087108 А1, 17.05.2018, WO 2018228442 А1, 20.12.2018, RU 2658504 С9, 21.08.2018, PAKULA A.A | |||
| et al., Genetic analysis of protein stability and function | |||
| Anna | |||
| Rev | |||
| Genet | |||
| Механизм для сообщения поршню рабочего цилиндра возвратно-поступательного движения | 1918 |
|
SU1989A1 |
| et al., Characterization of diphtheria | |||

