I. УРОВЕНЬ ТЕХНИКИ
[1] Ангиогенез или образование новых кровеносных сосудов из существующих важен для многих физиологических и патологических процессов. Обычно ангиогенез жестко регулируется про- и антиангиогенными факторами, но в случае заболеваний, таких как рак, неоваскулярные заболевания глаз, артрит и псориаз, это процесс может пойти неправильно. Folkman, J., Nat. Med., 1:27-31 (1995). Известно большое количество заболеваний, связанных с дерегулированным или нежелательным ангиогенезом. Такие заболевания включают, но не ограничиваются этим, неоваскуляризацию глаз, такую как ретинопатии (включая диабетическую ретинопатию), возрастную макулярную дегенерацию, псориаз, гемангиобластому, гемангиому, артериосклероз, воспалительные заболевания, такие как ревматоидные или ревматические воспалительные заболевания, в особенности артрит (включая ревматоидный артрит), или другие хронические воспалительные нарушения, такие как хроническая астма, артериальный или посттрансплантационный атеросклероз, эндометриоз, и неопластические заболевания, например, так называемые солидных опухоли и жидкие (или гемопоэтические) опухоли (такие как лейкозы и лимфомы). Другие заболевания, связанные с нежелательным ангиогенезом, очевидны для специалистов в данной области техники.
[2] Хотя в регуляцию ангиогенеза вовлечено много систем сигнальной трансдукции, одна из наиболее изученных и наиболее избирательных в отношении эндотелиальных клеток систем включает рецепторную тирозинкиназу Tie-2, которая избирательно экспрессируется в сосудистом эндотелии (и называется «Tie-2» или «Tie-2R» (также называется «ORK»); мышиная Tie-2 также называется «tek»), и ее лиганды, ангиопоэтины (Yancopoulos, G. D., et al., Nature 407 [2000] 242-48; Gale, N. W. and Yancopoulos, G. D., Genes Dev. 13:1055- 1066 [1999]).
[3] Известно 4 ангиопоэтина; от ангиопоэтина-1 («Ang-1», альтернативная аббревиатура ANGPT1 или Ang1) до ангиопоэтина-4 («Ang-4»). Эти ангиопоэтины также называются «лигандами Tie-2» (Davis, S., et al., Cell, §7:1161-1169 [1996]; Grosios, K., et al., Cytogenet Cell Genet, §4:118-120 [1999]; Holash, J., et al., Investigative Ophthalmology & Visual Science, 42:1611-1625 [1999]; Koblizek, T. I., et al., Current Biology, S:529-532 [1998]; Lin, P., et al., Proc Natl Acad Sci USA, 95:8829-8834 [1998]; Maisonpierre, P. C, et al., Science, 277:55- 60 [1997]; Papapetropoulos, A., et al., Lab Invest, 79:213-223 [1999]; Sato, T. N., et al., Nature, 375:70-74 [1998]; Shyu, K. G., et al., Circulation, 95:2081-2087 [1998]; Suri, C, et al., Cell, <37:1171-1180 [1996]; Suri, C, et al., Science, 252:468-471[1998]; Valenzuela, D. M., et al., Proceedings of the National Academy of Sciences of the USA, 96:1904-1909 [1999]; Witzenbichler, B., et al., J Biol Chem, 273:18514-18521 [1998]).
[4] Как Ang-1, так и -2 связываются с Tie-2 с аффинностью 3 нM (Kд) (Maisonpierre, P. C., et al., Science 277 (1997) 55-60). В то время как связывание Ang-1 с Tie-2 стимулирует фосфорилирование рецептора в культивируемых эндотелиальных клетках, Ang-2, согласно наблюдениям, как агонизирует, так и антагонизирует фосфорилирование рецептора Tie-2 (Davis, S., et al., [1996], выше; Maisonpierre, P.C., et al., [1997], выше; Kim, I, J.H. Kim, et al., Oncogene 19(39): 4549-4552 (2000); Teichert-Kuliszewska, K., P.C. Maisonpierre, et al., Cardiovascular Research 49(3): 659-70 (2001)). Фенотипы мышиных нокаутов Tie-2 и Ang-1 сходны между собой, что дает основание предположить, что Ang-1-стимулированное фосфорилирование Tie-2 опосредует ремоделирование и стабилизацию внутриутробного развития сосудов посредством поддержания адгезии опорных эндотелиальных клеток (Dumont, D. J., et al., Genes & Development, 8:1897-1909 [1994]; Sato, T. N., et al., Nature, 376:10-14 [1995]; Suri, C, et al., [1996], выше). Считается, что роль Ang-1 в стабилизации сосудов сохраняется во взрослом организме, где он широко и постоянно экспрессируется (Hanahan, D., Science, 277:48-50 [1997]; Zagzag, D., et al., Experimental Neurology, 59:391-400 [1999]). В противоположность этому, экспрессия Ang-2 ограничена в основном участками ремоделирования сосудов, где он, как считается, блокирует функцию Ang-1, тем самым индуцируя состояние пластичности сосудов, способствующее ангиогенезу (Hanahan, D., [1997], выше; Holash, J., et al., Science, 284:1994-1998 [1999]; Maisonpierre, P. C, et al., [1997], выше).
[5] Человеческий ангиопоэтин-2 (Ang-2) (альтернативная аббревиатура ANGPT2 или Ang2) описан в Maisonpierre, P. C., et al., Science 277 (1997) 55-60 и Cheung, A. H., et al., Genomics 48 (1998) 389-91. В большом количестве опубликованных исследований было предположительно продемонстрировано наличие избирательной в отношении сосудов экспрессии Ang-2 при болезненных состояниях, связанных с дерегуляцией ангиогенеза (Bunone, G., et al., American Journal of Pathology, 155:1961-1916; Etoh, T., et al., Cancer Research, 67:2145-2153 [2001]; Hangai, M., et al., Investigative Ophthalmology & Visual Science, 42:1611-1625 [2001]; Holash, J., et al., [1999] выше; Kuroda, K., et al., Journal of Investigative Dermatology, 116:113-120 [2001]; Otani1A., et al., Investigative Ophthalmology & Visual Science, 40:1912-1920 [1999]; Stratmann, A., et al., American Journal of Pathology, 153: 1459-1466 [1998]; Tanaka, S., et al., J Clin Invest, 203:34-345 [1999]; Yoshida, Y., et al., International Journal of Oncology, 25:1221-1225 [1999]; Yuan, K., et al., Journal of Periodontal Research, 35:165-171 [2000]; Zagzag, D., et al., [1999] выше). Эффективная анти-Ang-2 терапия будет полезной для огромной популяции пациентов со связанными с ангиогенезом заболеваниями, такими как рак, ретинопатии, артрит и псориаз.
[6] Следовательно, существует большая необходимость в выявлении новых соединений, которые специфически распознают и связывают Ang-2. Такие соединения будут полезны для проведения диагностического исследования и терапевтического вмешательства при болезненных состояниях, которые связаны с активностью Ang-2. Соответственно, целью настоящего изобретения является обеспечение специфических связывающих Ang-2 соединений для модуляции активности Ang-2. Такие соединения, раскрытые в данном документе, представлены в виде мутеинов, полученных из человеческого липокалина 2 (также известного как липокалин, ассоциированный с желатиназой нейтрофилов, «hNGAL»).
II. ОПРЕДЕЛЕНИЯ
[7] В нижеследующем перечне приведены определения терминов, выражений и сокращений, употребляемых в тексте настоящей заявки. Подразумевается, что все перечисленные и определенные в данном документе термины включают все грамматические формы.
[8] В контексте данного документа «Ang-1», если не указано его происхождение от отличных от человека видов (например, «мышиный Ang-1», «обезьяний Ang-1» и т.д.), означает человеческий Ang-1, полноразмерный белок согласно определению Swiss Prot Q15389 или его биологически активный фрагмент (например, фрагмент белка Ang-1, который способен индуцировать ангиогенез in vitro или in vivo).
[9] В контексте данного документа «Ang-2», если не указано его происхождение от отличных от человека видов (например, «мышиный Ang-2», «обезьяний Ang-2» и т.д.), означает человеческий Ang-2, полноразмерный белок согласно определению Swiss Prot 015123, (также смотрите Фиг. 6 патента США № 6166185; в полном объеме включенного в данный документ посредством ссылки) или его биологически активный фрагмент (например, фрагмент белка Ang-2, который способен индуцировать ангиогенез in vitro или in vivo).
[10] Термин «Tie-2» (также называемая в данной области техники «tek»), если не указано ее происхождение от отличных от человека видов (например, «мышиная Tie-2», «обезьянья Tie-2» и т.д.), относится к человеческой Tie-2 или ее биологически активному фрагменту. Человеческая Tie-2 имеет аминокислотную последовательность, приведенную в базе данных белковых последовательностей NCBI под номером доступа № AAA61130.
[11] В контексте данного документа «выявляемая аффинность» означает способность связываться с выбранной мишенью с константой аффинности, составляющей по меньшей мере около 10-5 M или меньше. Более низкие значения аффинности в общем случае невозможно измерить обычными способами, такими как ELISA, и, следовательно, они представляют собой второстепенную важность.
[12] В контексте данного документа «аффинность связывания» белка согласно изобретению (например, мутеина человеческого липокалина 2) с выбранной мишенью (в настоящем случае Ang-1 или Ang-2) можно измерять (и таким образом определять значения KД комплекса мутеин-лиганд) множеством способов, известных специалистам в данной области техники. Такие способы включают, но не ограничиваются этим, флуоресцентное титрование, прямой ELISA-анализ, конкурентный ELISA-анализ, калориметрические способы, такие как изотермическая титрационная калориметрия (ИТК), и поверхностный плазмонный резонанс (BIAcore). Такие способы являются общепринятыми в данной области техники и также подробно описаны ниже.
[13] Также следует отметить, что на образование комплекса между соответствующей связывающей молекулой и ее лигандом влияет много разных факторов, таких как концентрации соответствующих партнеров по связыванию, наличие конкурирующих молекул, рН и ионная сила применяемой буферной системы, а также применяемый экспериментальный способ для определения константы диссоциации KД (например, помимо прочего, флуоресцентное титрование, прямой ELISA-анализ, конкурентный ELISA-анализ или поверхностный плазмонный резонанс) или даже математический алгоритм, который применяют для оценки экспериментальных данных.
[14] Следовательно, специалисту также понятно, что значения KД (константы диссоциации комплекса, образуемого между соответствующей связывающей молекулой и ее мишенью/лигандом) могут варьироваться в пределах некоторого экспериментального диапазона в зависимости от способа и экспериментальной установки, применяемой для определения аффинности конкретного мутеина в отношении данного лиганда. Это означает, что для измеряемых значений KД может существовать небольшое отклонение или допустимый диапазон значений в зависимости, например, от того, измеряли значение KД с помощью поверхностного плазмонного резонанса (Biacore), конкурентного ELISA-анализа или «прямого ELISA-анализа».
[15] В контексте данного документа соединение, такое как мутеин согласно изобретению, «специфически связывает» мишень (например, Ang-1 или Ang-2) или обладает «специфичностью связывания» в отношении мишени, если оно способно к распознаванию этой мишени среди одной или более стандартных мишеней, так как специфичность связывания является не абсолютным, а относительным свойством. «Специфическое связывание» можно определить, например, в соответствии с методами вестерн-блоттинга, анализом ELISA, РИА, ЭХЛ, ИРМА, ИГХ и сканированием пептидов.
[16] В контексте данного документа термин «человеческий липокаин 2» или «человеческий Lcn 2», или «человеческий NGAL», или «hNGAL» относится к зрелому человеческому липокалину, ассоциированному с желатиназой нейтрофилов (NGAL), имеющему номер доступа P80188 в банке данных SWISS-PROT/UniProt. Мутеин человеческого липокаина 2 согласно изобретению также может называться в данном документе «мутеином hNGAL». Аминокислотную последовательность, приведенную под номером доступа P80188 в банке данных SWISS-PROT/UniProt, можно использовать в качестве предпочтительной «стандартной последовательности», более предпочтительно в качестве стандартной последовательности применяют аминокислотную последовательность, приведенную в SEQ ID NO: 16.
[17] В контексте данного документа «мутеин», «мутированный» компонент (белок или нуклеиновая кислота) или «мутант» относится к замене, удалению или вставке одного или более нуклеотидов или одной или более аминокислот по сравнению со «стандартной» структурой нуклеиновой кислоты или белка природного происхождения (дикого типа). В контексте данного документа термин «мутеин» также включает его функциональные фрагменты или варианты. Фрагменты или варианты конкретных мутеинов, описанных в настоящем изобретении, предпочтительно сохраняют функцию связывания с Ang-1 или Ang-2, например, с выявляемой или даже более высокой аффинностью, а такие фрагменты или варианты представляют собой «функциональные фрагменты или варианты» раскрытых в данном документе стандартных мутеинов.
[18] В контексте данного документа термин «фрагмент», применяемый в связи с мутеинами согласно изобретению, относится к белкам или пептидам, полученным из полноразмерного зрелого человеческого липокалина 2, которые были укорочены в N-конце и/или С-конце, т.е. в которых отсутствует по меньшей мере одна из N-концевых и/или С-концевых аминокислот. Такие фрагменты могут содержать по меньшей мере 10, чаще 20 или 30, или более последовательных аминокислот первичной последовательности зрелого человеческого липокалина 2 и обычно выявляются при иммуноанализе зрелого человеческого липокалина 2.
[19] В общем случае термин «фрагмент», употребляемый в отношении соответствующего белкового лиганда мутеина согласно изобретению или комбинации в соответствии с изобретением, относится к укороченным в N-конце и/или С-конце белкам или пептидным лигандам, которые сохраняют способность полноразмерного лиганда распознаваться и/или связываться мутеином согласно изобретению.
[20] В контексте данного документа термин «мутагенез» означает, что экспериментальные условия выбраны так, что аминокислота, в природном состоянии находящаяся в заданном положении последовательности зрелого человеческого липокалина 2, может быть замещена по меньшей мере одной аминокислотой, которая не присутствует в этом конкретном положении в соответствующей природной полипептидной последовательности. Термин «мутагенез» также включает (дополнительную) модификацию длины сегментов последовательности путем удаления или вставки одной или более аминокислот. Таким образом, в объем настоящего изобретения входит, например, то, что одна аминокислота в выбранном положении последовательности замещена участком из трех случайных мутаций, что приводит к вставке двух аминокислотных остатков по сравнению с длиной соответствующего сегмента белка дикого типа. Такие вставки или удаления можно вносить независимо друг от друга в любой из пептидных сегментов, которые можно подвергать мутагенезу в данном изобретении.
[21] Термин «случайный мутагенез» означает, что в определенном положении последовательности присутствует не одна предопределенная аминокислота (мутация), а что по меньшей мере две аминокислоты могут быть с некоторой вероятностью внесены в заданное положение последовательности во время мутагенеза.
[22] «Идентичность» – это свойство последовательностей, которое определяет их сходство или взаимосвязь. В контексте данного изобретения термин «идентичность последовательностей» или «идентичность» означает процентное содержание попарно сравниваемых идентичных остатков, после (гомологичного) выравнивания последовательности полипептида согласно изобретению с рассматриваемой последовательностью, по отношению к числу остатков в более длинной из этих двух последовательностей. Идентичность последовательностей определяют, деля число идентичных аминокислотных остатков на общее число остатков и умножая результат на 100.
[23] Термин «гомология» употребляется в данном документе в своем обычном значении и включает идентичные аминокислоты, а также аминокислоты, которые считаются консервативными заменами (например, замена остатка глутамата остатком аспартата) в эквивалентных положениях в линейной аминокислотной последовательности полипептида согласно изобретению (например, любого мутеина согласно изобретению).
[24] Согласно данному документу гомологию последовательностей или идентичность последовательностей можно определить, например, используя программу BLASTP, версии blastp 2.2.5 (16 ноября 2002 г.; cf. Altschul, S. F. et al. (1997) Nucl. Acids Res. 25, 3389-3402). В этом варианте реализации изобретения процент гомологии основан на выравнивании целых полипептидных последовательностей (матрица: BLOSUM 62; штраф за гэп: 11,1; предельное значение установлено на 10-3), включая пропептидные последовательности, предпочтительно с использованием для попарного сравнения белковой структуры дикого типа. Его рассчитывают как процентное содержание количества «положительных» (гомологичных) аминокислот, получаемого на выходе с помощью программы BLASTP, деленное на общее количество аминокислот, выбранных программой для выравнивания.
[25] В частности, чтобы определить, соответствует ли аминокислотный остаток аминокислотной последовательности мутеина, отличного от человеческого липокалина 2 дикого типа, определенному положению в аминокислотной последовательности человеческого липокалина 2 дикого типа, специалист в данной области техники использует средства и способы, хорошо известные в данной области, например, выравнивание, как вручную, так и с помощью компьютерных программ, таких как BLAST2.0, что расшифровывается как Basic Local Alignment Search Tool (основное средство поиска локального выравнивания), или ClustalW или любой другой подходящей программы, которая подходит для выполнения выравнивания последовательностей. Соответственно, человеческий липокалин 2 дикого типа может служить в качестве «эталонной последовательности» или «стандартной последовательности», тогда как аминокислотная последовательность мутеина, отличного от описанного в данном документе человеческого липокалина 2 дикого типа, служит в качестве «запрашиваемой последовательности». Термины «стандартная последовательность» и «последовательность дикого типа» взаимозаменяемо употребляются в данном документе.
[26] «Гэпы» представляют собой промежутки в выравнивании, которые являются результатом добавлений или удалений аминокислот. Таким образом, две копии одной единственной последовательности имеют 100% идентичности, но последовательности, являющиеся не настолько высококонсервативными и содержащие удаления, добавления или замещения, могут иметь меньшую степень идентичности последовательностей. Специалистам в данной области техники понятно, что для определения идентичности последовательностей доступно несколько компьютерных программ, использующих стандартные параметры, например, Blast (Altschul, et al. (1997) Nucleic Acids Res. 25, 3389-3402), Blast2 (Altschul, et al. (1990) J. Mol. Biol. 215, 403-410), и Smith-Waterman (Smith, et al. (1981) J. Mol. Biol. 147, 195-197).
[27] В контексте данного изобретения термин «вариант» относится к производным белка или пептида, которые содержат модификации аминокислотной последовательности, например, путем замены, удаления, вставки или химической модификации. Такие модификации, в некоторых вариантах реализации изобретения, не снижают функциональность белка или пептида. Такие варианты включают белки, в которых одна или более аминокислот были замещены соответствующими им D-стериоизомерами или аминокислотами, отличными от 20 аминокислот природного происхождения, такими как, например, орнитин, гидроксипролин, цитруллин, гомосерин, гидроксилизин, норвалин. При этом такие замены также могут быть консервативными, т.е. когда аминокислотный остаток замещен химически сходным аминокислотным остатком. Примерами консервативных замен являются замещения среди представителей следующих групп: 1) аланин, серин и треонин; 2) аспарагиновая кислота и глутаминовая кислота; 3) аспарагин и глутамин; 4) аргинин и лизин; 5) изолейцин, лейцин, метионин и валин; и 6) фенилаланин, тирозин и триптофан.
[28] Под «нативной последовательностью» человеческого липокалина 2 подразумевается человеческий липокалин 2, который имеет такую же аминокислотную последовательность, что и соответствующий полипептид, полученный из природного источника. Следовательно, нативная последовательность человеческого липокалина 2 может иметь аминокислотную последовательность соответствующего человеческого липокалина 2 природного происхождения. Такой полипептид с нативной последовательностью можно выделять из природного источника или получать рекомбинантными или синтетическими способами. Термин полипептид с «нативной последовательностью» в частности включает усеченные или секретируемые формы человеческого липокалина 2 природного происхождения, вариантные формы природного происхождения, такие как формы с альтернативным сплайсингом, и аллельные варианты человеческого липокалина 2 природного происхождения. Полипептидный «вариант» означает биологически активный полипептид, имеющий по меньшей мере около 50%, 60%, 70%, 80% или по меньшей мере около 85% идентичности аминокислотной последовательности с последовательностью нативного полипептида. Такие варианты включают, например, полипептиды, в которых были добавлены или удалены один или более аминокислотных остатков в N- или C-конце полипептида. В общем случае вариант имеет по меньшей мере около 70%, включая по меньшей мере около 80%, например, по меньшей мере около 85% идентичности аминокислотной последовательности, включая по меньшей мере около 90% идентичности аминокислотной последовательности или по меньшей мере около 95% идентичности аминокислотной последовательности с последовательностью нативного полипептида.
[29] Термин «положение», употребляемый в соответствии с изобретением, означает положение любой аминокислоты в аминокислотной последовательности, проиллюстрированной в данном документе, или положение нуклеотида в нуклеотидной последовательности, проиллюстрированной в данном документе. Для понимания, в случае термина «соответствовать» или «соответствующий», употребляемого в данном документе в контексте положений аминокислотной последовательности одного или более мутеинов, соответствующее положение определяется не только числом предшествующих нуклеотидов/аминокислот. Соответственно, положение заданной аминокислоты согласно изобретению, которая может быть заменена, может варьироваться вследствие добавления или удаления аминокислот в любом месте человеческого липокалина 2 (мутантного или дикого типа). Аналогично, положение заданного нуклеотида согласно изобретению, который может быть заменен, может варьироваться вследствие добавления или удаления нуклеотидов в любом месте 5’-нетранслируемой области (НТО), включая промотор и/или любые другие регуляторные последовательности или гены (включая экзоны и интроны) человеческого липокалина 2, мутеина или дикого типа.
[30] Таким образом, в случае соответствующего положения согласно изобретению предпочтительно понимать, что положение нуклеотидов/аминокислот могут отличаться по указанному номеру от аналогичных соседних нуклеотидов/аминокислот, но указанные соседние нуклеотиды/аминокислоты, которые могут быть заменены, удалены или добавлены, также включены в одно или более соответствующих положений.
[31] Кроме того, в случае соответствующего положения в мутеине на основании стандартной структуры согласно изобретению предпочтительно понимать, что положения нуклеотидов/аминокислот структурно соответствуют положениям в любом месте человеческого липокалина 2, мутеина или дикого типа, даже если они отличаются по указанному номеру.
[32] Термин «органическая молекула» или «небольшая органическая молекула», употребляемый в данном документе в отношении к неприродной мишени, означает органическую молекулу, содержащую по меньшей мере два атома углерода, но предпочтительно не более 7 или 12 вращающихся углеродных связей, имеющую молекулярную массу в диапазоне от 100 до 2000 Дальтон, предпочтительно от 100 до 1000 Дальтон, и необязательно содержащую один или два атома металла.
[33] В контексте данного документа слово «выявлять», «выявление» или «выявляемый» следует понимать как на количественном, так и на качественном уровне, а также в смысле их комбинации. Следовательно, это включает количественные, полуколичественные и качественные измерения, проводимые для представляющей интерес молекулы.
[34] «Субъект» представляет собой позвоночное, предпочтительно млекопитающее, более предпочтительно человека. Термин «млекопитающее» употребляется в данном документе для обозначения любого животного, классифицируемого как млекопитающее, включая, без ограничений, людей, домашних и сельскохозяйственных животных, животных из зоопарков, животных для спорта или домашних любимцев, таких как овцы, собаки, лошади, кошки, коровы, крысы, свиньи, обезьяны, такие как яванские макаки и т.д., если называть только несколько иллюстративных примеров. Предпочтительно млекопитающее согласно данному документу представляет собой человека.
[35] «Эффективное количество» представляет количество, достаточное для достижения благоприятных или желаемых результатов. Эффективное количество можно вводить за один или более раз.
[36] «Образец» определяется как биологический образец, полученный от любого субъекта. Биологические образцы включают, но не ограничиваются этим, кровь, сыворотку крови, мочу, кал, сперму или ткань.
[37] Термин «метастаз» в соответствии с изобретением относится к переносу раковых клеток от первичной опухоли в один или более участков в другом месте организма субъекта, где развивается вторичная опухоль. Средства для определения того, метастазировал ли рак, известны в данной области техники и включают остеосцинтиграфию, рентген грудной клетки, компьютерную аксиальную томографию (КАТ), магнито-резонансную томографию (МРТ) и тесты на опухолевые маркеры. Термин «предотвращение метастазов» означает предотвращение, замедление или уменьшение количества метастазов первичной опухоли или рака и, следовательно, предотвращение, замедление или уменьшение развития вторичных опухолей. Предпочтительно происходит предотвращение или уменьшение количества метастазов, т.е. вторичных опухолей, легкого, что означает, что происходит предотвращение или уменьшение метастатического переноса раковых клеток из первичной опухоли в легкое.
[38] В контексте данного документа термин «рак» относится к пролиферативным заболеваниям, таким как лимфомы, лимфоцитарные лейкозы, рак легкого, немелкоклеточный рак легкого (НМКРЛ), рак легкого из бронхоальвеолярных клеток, рак костей, рак поджелудочной железы, рак кожи, рак головы и шеи, кожная или интраокулярная меланома, рак матки, рак яичника, рак прямой кишки, рак анальной области, рак желудка, рак желудочно-кишечного тракта, рак толстой кишки, рак молочной железы, рак матки, карцинома фаллопиевых труб, карцинома эндометрия, карцинома шейки матки, карцинома вагины, карцинома вульвы, болезнь Ходжкина, рак пищевода, рак тонкого кишечника, рак эндокринной системы, рак щитовидной железы, рак паращитовидной железы, рак надпочечника, саркома мягких тканей, рак уретры, рак пениса, рак простаты, рак мочевого пузыря, рак почки или уретры, почечно-клеточная карцинома, карцинома почечной лоханки, мезотелиома, гепатоцеллюлярный рак, рак желчных протоков, новообразования центральной нервной системы (ЦНС), опухоли оси позвоночника, глиома из клеток ствола головного мозга, мультиформная глиобластома, астроцитомы, шванномы, эпендимомы, медуллобластомы, менингиомы, плоскоклеточные карциномы, аденома гипофиза и саркома Юинга, включая рефрактерные версии любого из вышеприведенных видов рака или комбинации одного или более вышеприведенных видов рака.
[39] Термин «сосудистые заболевания» включает рак, воспалительные заболевания, атеросклероз, ишемию, травму, сепсис, ХОБЛ, астму, диабет, ВМД, ретинопатию, инсульт, ожирение, острое повреждение легких, кровоизлияние, пропотевание сосудов, например, индуцированное цитокинами, аллергию, болезнь Грейвcа, аутоиммунный тиреоидит Хашимото, идиопатическую тромбоцитопеническую пурпуру, гигантоклеточный артериит, ревматоидный артрит, системную красную волчанку (СКВ), волчаночный нефрит, болезнь Крона, множественный склероз, язвенный колит, в особенности в отношении солидных опухолей, интраокулярные неоваскулярные синдромы (такие как пролиферативные ретинопатии или возрастная макулярная дегенерация (ВМД)), ревматоидный артрит и псориаз (Folkman, J., et al., J. Biol. Chem. 267 (1992) 10931- 10934; Klagsbrun, M., et al., Annu. Rev. Physiol. 53 (1991) 217-239; и Garner, A., Vascular diseases, In: Pathobiology of ocular disease, A dynamic approach, Garner, A., and Klintworth, G. K. (eds.), 2nd edition, Marcel Dekker, New York (1994), pp. 1625-1710).
[40] Подразумевается, что в контексте данного документа термин «антитело» относится к молекулам иммуноглобулина, содержащим четыре полипептидные цепи, две тяжелые (Н) цепи и две легкие (L) цепи, связанные между собой дисульфидными связями, а также их мультимеры (например, IgM). Каждая тяжелая цепь содержит вариабельную область тяжелой цепи (в данном документе сокращаемую до HCVR или VH) и константную область тяжелой цепи. Константная область тяжелой цепи содержит три домена, CH1, CH2 и Ch3. Каждая легкая цепь содержит вариабельную область легкой цепи (в данном документе сокращаемую до LCVR или VL) и константную область легкой цепи. Константная область легкой цепи содержит один домен (CL1). Области Vh и Vl могут быть дополнительно подразделены на области гипервариабельности, называемые определяющими комплементарность областями (CDR), разделенные областями, являющимися более консервативными и называющимися каркасными областями (FR). Каждая Vh и Vl состоит из трех CDR и четырех FR, расположенных от амино-конца до карбокси-конца в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. В разных вариантах реализации изобретения FR антитела к Ang-2 (или его антигенсвязывающего фрагмента) могут быть идентичными с последовательностями человеческой зародышевой линии, или могут быть модифицированы природным или искусственным путем. Аминокислотную консенсусную последовательность можно определить на основании параллельного анализа одной или более CDR.
[41] В контексте данного документа термин «антитело» также включает антигенсвязывающие фрагменты полноразмерных молекул антител. В контексте данного документа термины «антигенсвязывающая часть» антитела, «антигенсвязывающий фрагмент» антитела и т.д. включают любой природный, получаемый ферментативно, синтетический или генетически сконструированный полипептид или гликопротеин, который специфически связывается с антигеном, образуя комплекс. Антигенсвязывающие фрагменты антитела можно получить, например, из полноразмерных молекул антител, используя любые подходящие стандартные методы, такие как протеолитическое расщепление или технологии рекомбинантного генетического конструирования, включающие обработку и экспрессию ДНК, кодирующей вариабельные и, необязательно, константные домены антитела. Такая ДНК известна и/или легко доступна, например, из коммерческих источников, библиотек ДНК (включая, например, фаговые библиотеки антител), или может быть синтезирована. Можно проводить секвенирование или химическую обработку ДНК или использовать методы молекулярной биологии, например, для упорядочения одного или более вариабельных и/или константных доменов в подходящую конфигурацию или для внесения кодонов, создания цистеиновых остатков, модификации, добавления или удаления аминокислот и т.д. Неограничивающие примеры антигенсвязывающих фрагментов включают: (i) фрагменты Fab; (ii) фрагменты F(ab')2; (iii) фрагменты Fd; (iv) фрагменты Fv; (v) одноцепочечные молекулы Fv (scFv); (vi) фрагменты dAb; и (vii) минимальные распознающие единицы, состоящие из аминокислотных остатков, которые имитируют гипервариабельную область антитела (например, выделенную определяющую комплементарность область (CDR)). Другие сконструированные молекулы, такие как диатела, триатела, тетратела и минитела, также включены в выражение «антигенсвязывающий фрагмент» в контексте данного документа. Антигенсвязывающий фрагмент антитела, как правило, содержит по меньшей мере один вариабельный домен. Вариабельный домен может иметь любой размер или аминокислотный состав и в общем случае содержит по меньшей мере одну CDR, которая соседствует или находится в рамке с одной или более каркасными последовательностями. В антигенсвязывающих фрагментах, содержащих домен Vh, связанный с доменом Vl, домены Vh и Vl могут быть расположены по отношению друг к другу в любом подходящем упорядочении. Например, вариабельная область может быть димерной и содержать димеры Vh-Vh, Vh-Vl или Vl-Vl. В альтернативном варианте антигенсвязывающий фрагмент антитела может содержать мономерный домен Vh или Vl.
III. ОПИСАНИЕ ФИГУР
[42] Фиг. 1: демонстрирует, что мутеины липокалина (SEQ ID NO: 1, 3, 7, 8, 9 и 11) способны блокировать взаимодействие между человеческим Ang-2 и его рецептором Tie-2 с IC50 в субнаномолярном диапазоне. Человеческий Ang-2 преинкубировали с разными концентрациями мутеинов липокалина, и проводили количественную оценку не нейтрализованного человеческого Ang-2 на планшете ELISA с иммобилизованным растворимым человеческим Tie-2Fc. Эталонное антитело (SEQ ID NO: 19/20) использовали в качестве положительного контроля. Отрицательный контроль (SEQ ID NO: 16) не обладал конкурентным действием. Данные аппроксимировали моделью связывания в одном участке.
[43] Фиг. 2: иллюстрирует профиль перекрестной реактивности и специфичности мутеинов липокалина (SEQ ID NO: 7 и 8) согласно данным, полученным в конкурентном формате ELISA. Мутеины липокалина перекрестно реагируют с человеческим Ang-1, человеческим Ang-2 и мышиным Ang-2. Данные аппроксимировали с помощью модели связывания в одном участке.
[44] Фиг. 3: демонстрирует, что мутеины липокалина (SEQ ID NO: 1, 3, 6-14) способны блокировать взаимодействие между человеческим Ang-2 и его человеческим рецептором Tie-2, сверхэкспрессируемым в клетках HEK. Человеческий Ang-2 преинкубировали с разными концентрациями SEQ ID NO: 1,3, 6-14, не нейтрализованный Аng-2 выявляли с помощью антитела против HIS-метки. Эталонное антитело (SEQ ID NO: 19/20) использовали в качестве положительного контроля. Данные аппроксимировали моделью связывания в одном участке.
[45] Фиг. 4: демонстрирует, что мутеины липокалина (SEQ ID NO: 1, 3, 6-14) способны блокировать взаимодействие между мышиным Ang-2 и человеческим рецептором Tie-2, сверхэкспрессируемым в клетках HEK. Мышиный Ang-2 преинкубировали с разными концентрациями SEQ ID NO: 1, 3, 6-14, а не нейтрализованный мышиный Ang-2 выявляли с помощью антитела против HIS-метки. Эталонное антитело (SEQ ID NO: 19/20) использовали в качестве положительного контроля. Данные аппроксимировали с помощью модели связывания в одном участке.
[46] Фиг. 5: демонстрирует, что мутеины липокалина SEQ ID NO: 7, 8, 9 и 11 способны блокировать биологическую активность hAng-2 в анализе пролиферации клеток. В данном анализе SEQ ID NO: 7, 8, 9, 11 и 16, изотипический отрицательный контрольный IgG и два эталонных антитела добавляли к обедненным HLEC. Эксперимент показывает, что пролиферация LEC блокируется SEQ ID NO: 7, 8, 9, 11 и эталонными антителами 1 и 2 (эталонное антитело 1: SEQ ID NO: 17/18; эталонное антитело 2: SEQ ID NO: 19/20) со сходным диапазоном значений IC50. Изотипический контрольный IgG и отрицательный контроль SEQ ID NO: 16 не оказывали влияние на пролиферацию клеток. Данные аппроксимировали с помощью сигмоидальной модели доза-ответ.
IV. ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[47] В настоящем изобретении предложен полипептид, обладающий специфичностью связывания в отношении Ang-2, причем полипептид содержит мутеин hNGAL, который связывает Ang-2 с выявляемой аффинностью.
[48] В некоторых вариантах реализации изобретения мутеин hNGAL, связывающий Ang-2 с выявляемой аффинностью, может содержать по меньшей мере одну аминокислотную замену нативного цистеинового остатка другой аминокислотой, например, остатком серина. В некоторых других вариантах реализации изобретения мутеин, связывающий Ang-2 с выявляемой аффинностью, может содержать один или более ненативных остатков цистеина, замещающих одну или более аминокислот hNGAL дикого типа. В дополнительном конкретном варианте реализации изобретения мутеин hNGAL в соответствии с настоящим изобретением содержит по меньшей мере две аминокислотные замены нативной аминокислоты цистеиновым остатком для образования таким образом одного или более цистеиновых мостиков. В некоторых вариантах реализации изобретения указанный цистеиновый мостик может соединять по меньшей мере две петлевые области. Определение этих областей используется в данном документе в соответствии с Flower (Flower, 1996, выше, Flower, et al., 2000, выше) и Breustedt et al. (2005, выше).
[49] Мутеин или его композиция, обладающие специфичностью в отношении раскрытого в данном документе Ang-2, могут обладать антагонизирующей или нейтрализующей, или блокирующей активностью в отношении по меньшей мере одного вида биологической активности Ang-2.
[50] В одном аспекте настоящее изобретение включает различные мутеины hNGAL, которые связывают Ang-2 с по меньшей мере выявляемой аффинностью. В этом смысле Ang-2 считается неприродным лигандом стандартного hNGAL дикого типа, где «неприродный лиганд» относится к соединению, которое не связывается с человеческим липокалином 2 дикого типа в физиологических условиях. Конструируя hNGAL дикого типа с одной или более мутациями в определенных положениях последовательности, авторы настоящего изобретения продемонстрировали, что возможна высокая аффинность и высокая специфичность в отношении неприродного лиганда, Ang-2. В некоторых вариантах реализации изобретения при 1,2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 или даже более нуклеотидных триплетах, кодирующих определенные положения последовательности человеческого липокалина 2 дикого типа, можно проводить случайный мутагенез путем проведения замещения в этих положениях подгруппой нуклеотидных триплетов.
[51] Кроме того, мутеины согласно изобретению могут содержать мутированный аминокислотный остаток в любой одной или более, в том числе по меньшей мере в одном, двух, трех, четырех, пяти, шести, семи, восьми, девяти, десяти, одиннадцати или двенадцати положениях последовательности, соответствующих определенным положениям последовательности линейной полипептидной последовательности hNGAL, таких как положения последовательности 28, 36, 40, 41,49, 52, 65, 68, 70, 72-74, 77, 79, 81, 87, 96, 100, 103, 106, 116, 125, 126, 127, 129, 132 и 134 линейной полипептидной последовательности человеческого NGAL (SEQ ID NO: 16).
[52] Мутеин согласно изобретению может содержать аминокислотную последовательность дикого типа (природную) «родительской» белковой структуры (такой как hNGAL) за пределами мутированных положений аминокислотной последовательности. В некоторых вариантах реализации изобретения мутеин hNGAL в соответствии с изобретением также может нести одну или больше аминокислотных мутаций в положении/положениях последовательности до тех пор, пока такая мутация не затрудняет или не мешает, по меньшей мере существенно, связывающей активности и сворачиванию мутеина. Такие мутации очень легко можно осуществить на уровне ДНК, используя общепринятые стандартные способы (Sambrook, J. et al. (2001) Molecular Cloning: A Laboratory Manual, 3rd Ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY). Иллюстративными примерами изменений аминокислотной последовательности являются вставки и удаления, а также аминокислотные замены. Такие замены могут быть консервативными, т.е. когда аминокислотный остаток замещен аминокислотным остатком с химически сходными свойствами, в частности, в отношении полярности, а также размера. Примерами консервативных замен являются замещения среди представителей следующих групп: 1) аланин, серин и треонин; 2) аспарагиновая кислота и глутаминовая кислота; 3) аспарагин и глутамин; 4) аргинин и лизин; 5) изолейцин, лейцин, метионин и валин; и 6) фенилаланин, тирозин и триптофан. С другой стороны, также возможно вносить в аминокислотную последовательность неконсервативные изменения. Кроме того, вместо замещения одиночных аминокислотных остатков также возможно вставлять или удалять одну или более смежных аминокислот первичной структуры человеческого липокалина 2, до тех пор, пока эти удаления или вставки приводят к получению стабильного свернутого/функционального мутеина (например, мутеинов hNGAL с усеченным N- и C-концом). В таком мутеине в N- или C-конце полипептида, например, добавлены или удалены один или более аминокислотных остатков. В общем случае такой мутеин может иметь по меньшей мере около 70%, включая по меньшей мере около 80%, например, по меньшей мере около 85% идентичности аминокислотной последовательности с аминокислотной последовательностью зрелого hNGAL.
[53] Аминокислотная последовательность раскрытого в данном документе мутеина hNGAL обладает высокой идентичностью последовательностей со зрелым hNGAL (SEQ ID NO: 16) по сравнению с идентичностью последовательностей с другими липокалинами. В этом общем контексте аминокислотная последовательность мутеина согласно изобретению является по меньшей мере в значительной степени сходной с аминокислотной последовательностью природного hNGAL дикого типа, при условии возможности наличия в выравнивании гэпов (согласно определению ниже), которые являются результатом добавления или удаления аминокислот. Соответствующая последовательность мутеина согласно изобретению, являясь в значительной степени сходной с последовательностями зрелого hNGAL, имеет, в некоторых вариантах реализации изобретения, по меньшей мере 70% идентичности или гомологии последовательности, по меньшей мере 75% идентичности или гомологии последовательности, по меньшей мере 80% идентичности или гомологии последовательности, по меньшей мере 82% идентичности или гомологии последовательности, по меньшей мере 85% идентичности или гомологии последовательности, по меньшей мере 87% идентичности или гомологии последовательности или по меньшей мере 90% идентичности или гомологии последовательности, включая по меньшей мере 95% идентичности или гомологии последовательности с последовательностью зрелого hNGAL, при условии сохранения измененного положения или последовательности и возможности наличия одного или более гэпов.
[54] В контексте данного документа мутеин согласно изобретению «специфически связывает» мишень (например, Ang-2), если он способен к распознаванию между этой мишенью и одной или более стандартными мишенями, так как специфичность связывания является не абсолютным, а относительным свойством. «Специфическое связывание» можно определить, например, в соответствии с методами вестерн-блоттинга, анализом ELISA, РИА, ЭХЛ, ИРМА, ИГХ и сканированием пептидов.
[55] В одном варианте реализации изобретения мутеины согласно изобретению слиты по N-концу и/или C-концу с партнером по слиянию, который, в некоторых вариантах реализации изобретения, представляет собой белок или белковый домен, или пептид. В некоторых вариантах реализации изобретения белковый домен может продлевать период полужизни мутеина в сыворотке крови. В дополнительных конкретных вариантах реализации изобретения белковый домен представляет собой Fс-часть иммуноглобулина, CH3-домен иммуноглобулина, CH4-домен иммуноглобулина, альбуминсвязывающий пептид или альбуминсвязывающий белок.
[56] В другом варианте реализации изобретения мутеины согласно изобретению конъюгированы с соединением, которое продлевает период полужизни мутеина в сыворотке крови. Более предпочтительно мутеин конъюгирован с соединением, выбранным из группы, состоящей из молекулы полиалкиленгликоля, гидроэтилкрахмала, Fс-части иммуноглобулина, CH3-домена иммуноглобулина, CH4-домена иммуноглобулина, альбуминсвязывающего пептида или альбуминсвязывающего белка.
[57] В другом варианте реализации настоящее изобретение относится к молекуле нуклеиновой кислоты, содержащей нуклеотидную последовательность, кодирующую раскрытый в данном документе мутеин. Изобретение включает клетку-хозяина, содержащую указанную молекулу нуклеиновой кислоты.
A. Типовые мутеины, специфические в отношении Ang-2
[58] В одном аспекте настоящее изобретение относится к новым, характеризующимся специфическим связыванием мутеинам человеческого липокалина 2 (человеческого Lcn2 или hNGAL), специфическим в отношении Ang-2.
[59] Один вариант реализации настоящего изобретения относится к мутеину, который способен связывать Ang-2 с выявляемой аффинностью, такой как аффинность, определяемая Kд около 200 нМ или меньше, например, около 150 нМ или меньше.
[60] В одном аспекте в настоящем изобретении предложен мутеин hNGAL, который способен связывать Ang-2 с Kд около 5 нМ или меньше согласно измерениям на приборе Biacore T200 методом поверхностного плазмонного резонанса (ППР), по сути, описанным в Примере 6.
[61] В некоторых дополнительных вариантах реализации изобретения один или более мутеинов hNGAL согласно данному изобретению способны связывать Ang-2 с аффинностью, измеренной посредством определения значения EC50, составляющего около 5 нM или меньше, согласно измерениям в ELISA-анализе, по сути, описанном в Примере 4.
[62] В некоторых других вариантах реализации изобретения один или более мутеинов hNGAL согласно данному изобретению способны связывать Ang-2 с аффинностью, измеренной посредством определения значения EC50, составляющего около 5 нM или меньше, согласно измерениям в конкурентном ELISA-анализе, по сути, описанном в Примере 5.
[63] В некоторых других вариантах реализации изобретения один или более мутеинов hNGAL согласно данному изобретению способны ингибировать или снижать пролиферацию эндотелиальных клеток лимфатических капилляров, опосредованную Ang-2, со значением IC50 около 5 нМ или меньше согласно анализу пролиферации клеток, по сути, описанному в Примере 9.
[64] В некоторых конкретных вариантах реализации изобретения Ang-2-связывающий мутеин hNGAL способен связываться как с Ang-2, так и с Ang-1 с выявляемой аффинностью, такой как аффинность, определяемая Kд около 200 нМ или меньше, например, около 150 нМ или меньше. В некоторых вариантах реализации изобретения один или более мутеинов hNGAL согласно данному изобретению перекрестно реагируют с человеческим Ang-1 и с человеческим Ang-2.
[65] В некоторых дополнительных вариантах реализации изобретения мутеин способен связывать Ang-1 с аффинностью, измеренной посредством определения значения EC50, составляющего около 150 нM или меньше, согласно измерениям в ELISA-анализе, по сути, описанном в Примере 7.
[66] В некоторых других вариантах реализации изобретения один или более мутеинов hNGAL согласно данному изобретению перекрестно реагируют с человеческим Ang-2 и мышиным Ang-2. В некоторых вариантах реализации изобретения один или более таких мутеинов способны связывать как человеческий Ang-2, так и мышиный Ang-2 с выявляемой аффинностью, такой как аффинность, определяемая Kд около 200 нМ или меньше, например, около 150 нМ или меньше.
[67] В некоторых дополнительных вариантах реализации изобретения один или более таких мутеинов способны связывать мышиный Ang-2 с аффинностью, измеренной посредством определения значения EC50, составляющего около 5 нM или меньше, согласно измерениям в ELISA-анализе, по сути, описанном в Примере 7.
[68] В некоторых дополнительных вариантах реализации изобретения один или более таких мутеинов способны блокировать связывание человеческого Ang-2 с hTie-2 и мышиного Ang-2 с hTie-2 со значением IC50 около 25 нМ или меньше, соответственно, в конкурентном формате клеточного анализа ЭХЛ, по сути, описанного в Примере 8.
[69] В некоторых вариантах реализации изобретения один или более мутеинов hNGAL согласно данному изобретению не обладают перекрестной реактивностью с человеческим Ang-4. В некоторых вариантах реализации изобретения один или более мутеинов hNGAL согласно данному изобретению не обладают перекрестной реактивностью с мышиным Ang-3. В некоторых вариантах реализации изобретения один или более мутеинов hNGAL согласно данному изобретению не обладают перекрестной реактивностью с человеческим VEGF-A.
[70] В этом отношении изобретение относится к полипептиду, причем указанный полипептид содержит мутеин hNGAL, а указанный hNGAL, по сравнению с линейной полипептидной последовательностью зрелого hNGAL, содержит по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21 или даже больше мутированных аминокислотных остатков в положениях последовательности 28, 36, 40, 41, 49, 52, 65, 68, 70, 72-74, 77, 79, 81, 87, 96, 100, 103, 106, 116, 125, 126, 127, 129, 132 и 134, и при этом указанный полипептид связывает Ang-2 с выявляемой аффинностью.
[71] В некоторых вариантах реализации изобретения Ang-2-связывающий мутеин hNGAL согласно изобретению содержит в любом одном или более положениях последовательности 36, 40, 41, 49, 52, 68, 70, 72-73, 77, 79, 81, 96, 100, 103, 106, 125, 127, 132 и 134 линейной полипептидной последовательности зрелого hNGAL (SEQ ID NO: 16) один или более из следующих мутированных аминокислотных остатков: Leu 36 → Gln, Glu, His, Val, Met или Phe; Ala 40 → Val, Tyr, His или Trp; Ile 41 → His, Tyr, Trp или Val; Gln 49 → Gly, Ile, Val, Glu или Val; Tyr 52 → Trp, His, Thr или Ser; Ser 68 → Gly, Asp, Gln, Glu или Ile; Leu 70 → Ser, Thr, Gly, Arg, Tyr или Ala; Arg 72 → Gly, Ala, Trp, Thr или Glu; Lys 73 → Pro, Phe, Leu, Arg, Ala или Gln; Asp 77 → Asn, Lys, Ser или Val; Trp 79 → Thr, Arg, Ser или Asn; Arg 81 → Trp, His или Tyr; Asn 96 → Gly, Ala, Pro, Gln или Asp; Tyr 100 → Pro, Trp, Gly, Ser, Leu или Asp; Leu 103 → Gly, Glu, Aso, Met или Gln; Tyr 106 → Thr, Leu или Phe; Lys 125 → His, Thr или Gly; Ser 127 → Leu или Met; Tyr 132 → Phe, Trp или Val; и Lys 134 → Ala, Glu или Trp. В некоторых вариантах реализации изобретения мутеин hNGAL согласно изобретению содержит два или более, например, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16 или даже более, или все мутированные аминокислотные остатки в этих положениях последовательности зрелого hNGAL.
[72] Кроме того, Ang-2-связывающий мутеин hNGAL согласно изобретению также может содержать следующую замену по сравнению с линейной полипептидной последовательностью зрелого hNGAL: Gln 28 → His; Asn 65 → Asp; Lys 74 → Glu; Cys 87 → Ser; Asn 116 → Asp; Val 126 → Met и Asn 129 → Asp.
[73] В некоторых дополнительных вариантах реализации изобретения мутеин hNGAL согласно изобретению, который связывается с Ang-2, содержит следующие аминокислотные замены по сравнению с линейной полипептидной последовательностью зрелого hNGAL:
(a) Gln 28 → His; Leu 36 → Gln; Ala 40 → Tyr; Gln 49 → Gly; Tyr 52 → Trp; Ser 68 → Gly; Leu 70 → Ser; Arg 72 → Gly; Lys 73 → Pro; Asp 77 → Asn; Trp 79 → Thr; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Gly; Tyr 100 → Pro; Leu 103 → Gly; Tyr 106 → Thr; Lys 125 → His; Ser 127 → Leu; Tyr 132 → Phe; Lys 134 → Glu;
(b) Gln 28 → His; Leu 36 → Phe; Ala 40 His; Ile 41 → Arg; Gln 49 → Gly; Tyr 52 → His; Ser 68 → Asp; Leu 70 → Thr; Arg 72 → Ala; Lys 73 → Phe; Asp 77→ Asn; Trp 79 → Arg; Arg 81 → His; Cys 87 → Ser; Tyr 100 → Trp; Leu 103 → Glu; Tyr 106 → Thr; Lys 125 → Thr; Ser 127 → Met; Tyr 132 → Trp; Lys 134 → Trp;
(c) Gln 28 → His; Leu 36 → Val; Ala 40 → Trp; Ile 41 → Tyr; Gln 49 → Ile; Tyr 52 → Thr; Ser 68 → Gln; Leu 70 → Gly; Arg 72 → Glu; Lys 73 → Gln; Asp 77 → Lys; Trp 79 → Ser; Arg 81 → His; Cys 87 → Ser; Tyr 100 → Trp; Leu 103 → Asp; Tyr 106 → Leu; Lys 125 → Gly; Ser 127 → Met; Tyr 132 → Val; Lys 134 Ala;
(d) Gln 28 → His; Leu 36 → Glu, Ala 40 →Val; Ile 41 → Glu; Gln 49 → Val; Tyr 52 → Thr Ser 68 → Glu; Leu 70 → Arg; Arg 72 → Trp; Lys 73 → Leu; Asp 77 → Lys; Trp 79 → Asn; Arg 81 → His; Cys 87 → Ser; Asn 96 → Ala; Tyr 100 → Gly; Leu 103 → Met; Tyr 106 → Thr; Lys 125 → Thr; Ser 127 → Met; Tyr 132 → Trp; Lys 134 → Trp;
(e) Gln 28 → His; Leu 36 → Gln; Ala 40 → Tyr; Ile 41 → Trp; Gln 49 → Ile; Tyr 52 → Ser; Ser 68 → Ile; Leu 70 → Tyr; Arg 72 → Thr; Lys 73 → Arg; Asp 77 → Ser; Trp 79 → Arg; Arg 81 → Tyr; Cys 87 → Ser; Asn 96 → Pro; Leu 103 → Asp; Tyr 106 → Thr; Lys 125 → His; Ser 127 → Tyr; Tyr 132 → Trp; Lys 134 → Glu;
(f) Gln 28 → His; Leu 36 → Gln; Ala 40 → Tyr; Gln 49 → Glu; Tyr 52 → Trp; Asn 65 → Asp; Ser 68 → Gly; Leu 70 → Ser; Arg 72 → Gly; Lys 73 → Pro; Asp 77 → Asn; Trp 79 → Arg; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Gly; Tyr 100 → Ser; Leu 103 → Gln; Tyr 106 → Thr; Lys 125 → His; Ser 127 → Leu; Tyr 132 → Phe; Lys 134 → Glu;
(g) Gln 28 → His; Leu 36 → His; Ala 40 → Tyr; Gln 49 → Glu; Tyr 52 → Trp; Asn 65 → Asp; Ser 68 → Glu; Leu 70 → Ser; Arg 72 → Gly; Lys 73 → Pro; Asp 77 → Asn; Trp 79 → Arg; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Gly; Tyr 100 → Pro; Leu 103 → Asp; Tyr 106 → Thr; Lys 125 → His; Ser 127 → Leu; Tyr 132 → Phe; Lys 134 → Glu;
(h) Gln 28 → His; Leu 36 → Gln; Ala 40 → Tyr; Gln 49 → Gly; Tyr 52 → Trp; Asn 65 → Asp; Ser 68 → Glu; Leu 70 → Ser; Arg 72 → Gly; Lys 73 → Ala; Asp 77 → Asn; Trp 79 → Arg; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Gly; Tyr 100 → Asp; Leu 103 → Gly; Tyr 106 → Thr; Lys 125 → His; Ser 127 → Leu; Tyr 132 → Phe; Lys 134 → Glu;
(i) Gln 28 → His; Leu 36 → His; Ala 40 →Tyr; Gln 49 → Gly; Tyr 52 → Trp; Asn 65 → Asp; Ser 68 → Glu; Leu 70 → Ser; Arg 72 → Gly; Lys 73 → Pro; Asp 77 → Asn; Trp 79 → Arg; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Gly; Tyr 100 → Pro; Leu 103 → Gly; Tyr 106 → Thr; Lys 125 → His; Ser 127 → Leu; Tyr 132 → Phe; Lys 134 → Glu;
(j) Gln 28 → His; Leu 36 → Gln; Ala 40 → Tyr; Gln 49 → Gly; Tyr 52 → Trp; Asn 65 → Asp; Ser 68 → Gly; Leu 70 → Ser; Arg 72 → Gly; Lys 73 → Ala; Asp 77 → Val; Trp 79 → Arg; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Gly; Tyr 100 → Pro; Leu 103 → Gly; Tyr 106 → Thr; Lys 125 → His; Ser 127 → Leu; Tyr 132 → Phe; Lys 134 → Glu;
(k) Gln 28 → His; Leu 36 → Gln; Ala 40 → Tyr; Gln 49 → Val; Tyr 52 → Trp; Asn 65 → Asp; Ser 68 → Glu; Leu 70 → Ser; Arg 72 → Gly; Lys 73 → Pro; Asp 77 → Asn; Trp 79 → Arg; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Gly; Tyr 100 → Leu; Leu 103 → Gly; Tyr 106 → Thr; Lys 125 → His; Ser 127 → Leu; Tyr 132 → Phe; Lys 134 → Glu;
(l) Gln 28 → His; Leu 36 → Val; Ala 40 → Tyr; Ile 41 → Tyr; Gln 49 → Ile; Tyr 52 → Thr; Asn 65 → Asp; Ser 68 → Gln; Leu 70 → Gly; Arg 72 → Glu; Lys 73 → Gln; Lys 74 → Glu; Asp 77 → Lys; Trp 79 → Ser; Arg 81 → His; Cys 87 → Ser; Tyr 100 → Trp; Leu 103 → Asp; Tyr 106 → Pro; Asn 116 → Asp; Lys 125 → Gly; Ser 127 → Met; Asn 129 → Asp; Tyr 132 → Val; Lys 134 Ala;
(m) Gln 28 → His; Leu 36 → Val; Ala 40 → Tyr; Ile 41 → Tyr; Gln 49 → Ile; Tyr 52 → Thr; Asn 65 → Asp; Ser 68 → Gln; Leu 70 → Gly; Arg 72 → Glu; Lys 73 → Gln; Lys 74 → Glu; Asp 77 → Lys; Trp 79 → Ser; Arg 81 → His; Cys 87 → Ser; Asn 96 → Asp; Tyr 100 → Trp; Leu 103 → Asp; Tyr 106 → Pro; Lys 125 → Gly; Val 126 → Met; Ser 127 → Met; Asn 129 → Asp; Tyr 132 → Val; Lys 134 → Ala; или
(n) Gln 28 → His; Leu 36 → Met; Ala 40 →Tyr; Ile 41 → Asp; Gln 49 → Ile; Tyr 52 → Thr; Asn 65 → Asp; Ser 68 → Gln; Leu 70 → Gly; Arg 72 → Glu; Lys 73 → Gln; Asp 77 → Lys; Trp 79 → Ser; Arg 81 → His; Cys 87 → Ser; Asn 96 → Gln; Tyr 100 → Trp; Leu 103 → Asp; Tyr 106 → Pro; Lys 125 → Gly; Ser 127 → Met; Tyr 132 → Val; Lys 134 →Ala.
[74] В оставшейся области, т.е. области, отличной от положений последовательности 28, 36, 40, 41,49, 52, 65, 68, 70, 72-74, 77, 79, 81, 87, 96, 100, 103, 106, 116, 125, 126, 127, 129, 132 и 134, мутеин hNGAL согласно изобретению может содержать аминокислотную последовательность дикого типа (природную) за пределами мутированных положений аминокислотной последовательности.
[75] В дополнительных конкретных вариантах реализации изобретения мутеин согласно настоящему изобретению содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-14 или их функционального фрагмента или варианта. В некоторых вариантах реализации изобретения такой фрагмент или вариант является структурным гомологом мутеина, определяемого любой из SEQ ID NO: 1-14.
[76] Аминокислотная последовательность Ang-2-связывающего мутеина hNGAL согласно изобретению может иметь высокую идентичность последовательности, такую как по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 82%, по меньшей мере 85%, по меньшей мере 87%, по меньшей мере 90% идентичности, включая по меньшей мере 95% идентичности, с последовательностью, выбранной из группы, состоящей из SEQ ID NO: 1-14.
[77] В некоторых других вариантах реализации изобретения мутеин hNGAL в соответствии с изобретением, перекрестно реагирующий или связывающийся с человеческим Ang-1, человеческим Ang-2 и/или мышиным Ang-2, содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-14 и их функциональных фрагментов или вариантов.
[78] Также изобретение включает структурные гомологи мутеина hNGAL, имеющие аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-14, при этом структурные гомологи обладают гомологией аминокислотной последовательности или идентичностью последовательности, составляющей более чем около 60%, предпочтительно более чем 65%, более чем 70%, более чем 75%, более чем 80%, более чем 85%, более чем 90%, более чем 92% и наиболее предпочтительно более чем 95%, по отношению к указанному мутеину hNGAL.
[79] Ang-2-связывающий мутеин hNGAL в соответствии с настоящим изобретением можно получить посредством мутагенеза формы природного происхождения человеческого липокалина 2. В некоторых вариантах реализации мутагенеза замена (или замещение) представляет собой консервативную замену. Не смотря на это, предусмотрена любая замена, включая неконсервативную замену или одну или более из типовых замен, приведенных ниже, до тех пор, пока мутеин сохраняет свою способность связываться с Ang-2 и/или он имеет идентичность с замещенной последовательностью, в которой по меньшей мере 60%, например, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85% или более идентичности с аминокислотной последовательностью зрелого человеческого липокалина 2 (номер доступа в банке данных SWISS-PROT P80188).
[80] Настоящее изобретение также относится к фармацевтической композиции, которая содержит по меньшей мере один раскрытый в данном документе Ang-2-связывающий мутеин hNGAL или его конъюгат, или продукт слияния, как описано в данном документе, и, необязательно, фармацевтически приемлемое вспомогательное вещество.
[81] Соответственно, Ang-2-связывающие мутеины hNGAL согласно изобретению можно получать в виде композиций, используя фармацевтически приемлемые ингредиенты, а также общепринятые способы приготовления (Gennaro and Gennaro (2000) Remington: The Science and Practice of Pharmacy, 20th Ed., Lippincott Williams & Wilkins, Philadelphia, PA). Для приготовления фармацевтических композиций можно использовать фармацевтически инертные неорганические или органические вспомогательные вещества.
B. Применения мутеинов, специфических в отношении Ang-2
[82] Недавно, используя мышиную модель с нокаутом ANG-2, группа Yancopoulos сообщила, что ANG-2 необходим для постнатального ангиогенеза (Gale, N. W., et al., Dev. Cell 3 (2002) 411-23). Они показали, что связанная с развитием запрограммированная регрессия гиалоидной сосудистой системы в глазу не происходит у мышей с нокаутом ANG-2, а кровеносные сосуды сетчатки не вырастают из центральной ретинальной артерии (Gale, N. W., et al., Dev. Cell 3 (2002) 411-23). Также они обнаружили, что удаление ANG-2 приводит к сильным дефектам формирования и функции лимфатической сосудистой системы (Gale, N. W., et al., Dev. Cell 3 (2002) 411-23). Кроме того, считается, что эффективная анти-Ang-2 терапия будет полезной для лечения заболеваний, таких как рак, прогрессирование которых зависит от аберрантного ангиогенеза, когда блокирование такого процесса может привести в предотвращению усугубления заболевания (Folkman, J., Nature Medicine. 1, (1995) 27-31). Более того, было показано, что экспрессия Ang-2 коррелирует с тяжестью различных воспалительных и/или инфекционных заболеваний (смотрите, например, Siner et al., 2009, Shock 31 :348-353; Yeo et al., 2008, Proc. Natl. Acad. Sci. (USA): 105: 17097-17102).
[83] Следовательно, в медицине, например, в офтальмологии и онкологии, существует множество возможных применений мутеинов, обладающих аффинностью связывания в отношении Ang-2 согласно изобретению. В одном дополнительном аспекте изобретение относится к применению такого раскрытого в данном документе мутеина для выявления Ang-2 в образце, а также к соответствующему способу диагностики.
[84] Настоящее изобретение также включает применение одного или более мутеинов, обладающих аффинностью связывания в отношении описанного Ang-2, для образования комплекса с Ang-2.
[85] Следовательно, в другом аспекте изобретения раскрытые мутеины применяют для выявления Ang-2. Такое применение может включать этапы приведения в подходящих условиях одного или более указанных мутеинов в контакт с образцом, в котором подозревается наличие Ang-2, тем самым обеспечивая возможность образования комплекса между мутеинами и Ang-2, и выявление комплекса по подходящему сигналу.
[86] Выявляемым сигналом может быть метка, как описано выше, или изменение физических свойств вследствие связывания, т.е. само по себе образование комплекса. Одним из примеров является поверхностный плазмонный резонанс, значение которого меняется во время связывания партнеров по связыванию, один из которых иммобилизован на поверхности, такой как золотая фольга.
[87] Раскрытые в данном документе мутеины также можно применять для отделения Ang-2. Такое применение может включать этапы приведения в подходящих условиях одного или более указанных мутеинов в контакт с образцом, в котором предполагается наличие Ang-2, тем самым обеспечивая возможность образования комплекса между мутеинами и Ang-2, и отделение комплекса от образца.
[88] При применении раскрытых мутеинов для выявления Ang-2, а также для отделения Ang-2, мутеины и/или Ang-2, или их домен, или фрагмент могут быть иммобилизованы на подходящей твердой подложке.
[89] Соответственно, можно определить наличие или отсутствие молекулы, такой как Ang-2, например, в образце, а также ее концентрацию или уровень.
[90] Следовательно, мутеины согласно настоящему изобретению можно применять для выявления и/или измерения Ang-2 в образце, например, в диагностических целях. Например, мутеины можно применять для диагностики патологического состояния или заболевания, характеризуемого аберрантной экспрессией (например, сверхэкспрессией, недостаточной экспрессией или отсутствием экспрессии и т.д.) Ang-2. Типовые диагностические методы анализа Ang-2 могут включать, например, приведение образца, полученного от пациента, в контакт с мутеинами, причем мутеины помечены выявляемой меткой или репортерной молекулой. В альтернативном варианте в диагностических применениях можно использовать немеченые мутеины в комбинации с вторичной молекулой, которая сама по себе выявляемо помечена. Выявляемая метка или репортерная молекула может представлять собой радиоактивный изотоп, такой как 3H, 14C, 32P, 35S или 125I; флуоресцентный или хемилюминесцентный компонент, такой как флуоресцеинизотиоцианат или родамин; или фермент, такой как щелочная фосфатаза, β-галактозидаза, пероксидаза хрена или люцифераза. Конкретные типовые методы анализа, которые можно применять для выявления или измерения Ang-2 в образце, включают ферментный иммуносорбентный анализ (ELISA), радиоиммуноанализ (РИА) и сортировку флуоресцентно-активированных клеток (FACS).
[91] В другом аспекте настоящее изобретение относится к диагностическому или аналитическому набору, содержащему мутеин, обладающий аффинностью связывания в отношении Ang-2 в соответствии с изобретением.
[92] В дополнительном аспекте изобретение также включает применение раскрытых Ang-2-связывающих мутеинов hNGAL или комбинаций, содержащих такие описанные в данном документе мутеины, для производства фармацевтической композиции. Полученная таким образом фармацевтическая композиция может быть пригодна для лечения, предотвращения и/или уменьшения выраженности заболеваний или нарушений, связанных с дерегулированным ангиогенезом, таких как рак, неоваскулярные заболевания глаз (такие как ретинопатии), артрит и псориаз. Фармацевтическую композицию можно применять в качестве монотерапии или комбинированной терапии. Соответственно, в изобретении также предложены Ang-2-связывающие мутеины hNGAL для лечения заболевания или нарушения, связанного с дерегулированным ангиогенезом, такого как рак, неоваскулярные заболевания глаз (такие как ретинопатии), артрит и псориаз.
[93] Кроме применения в диагностике, в другом аспекте изобретение включает применение такого мутеина согласно изобретению или композиции или комбинации, содержащей такой мутеин, для связывания Ang-2 в организме субъекта и/или ингибирования, или снижения интенсивности ангиогенеза в организме субъекта. В некоторых вариантах реализации изобретения такой субъект может страдать от заболеваний или нарушений, связанных с дерегулированным ангиогенезом, таких как рак, неоваскулярные заболевания глаз (такие как ретинопатии), артрит и псориаз.
[94] В другом аспекте настоящее изобретение относится к способу связывания Ang-2 в организме субъекта, включающему введение указанному субъекту эффективного количества одного или более мутеинов, обладающих аффинностью связывания в отношении Ang-2 согласно изобретению, или одной или более композиций или комбинаций, содержащих такой мутеин.
[95] В другом аспекте настоящее изобретение включает способ ингибирования или снижения интенсивности ангиогенеза в организме субъекта, включающий введение указанному субъекту эффективного количества одного или более мутеинов, обладающих аффинностью связывания в отношении Ang-2 согласно изобретению, или одной или более композиций или комбинаций, содержащих такой мутеин. В некоторых вариантах реализации изобретения такой субъект может страдать от заболеваний или нарушений, связанных с дерегулированным ангиогенезом, таких как рак, неоваскулярные заболевания глаз (такие как ретинопатии), артрит и псориаз.
[96] Мутеины согласно изобретению или композиции, или комбинации, содержащие такие мутеины, применимы, помимо прочего, для лечения, предотвращения и/или уменьшения выраженности любого заболевания или нарушения, связанного с активностью Ang-2, включая заболевания или нарушения, связанные с дерегулированным ангиогенезом.
[97] Например, мутеины или композиции или комбинации, содержащие такие мутеины согласно изобретению, можно применять для ингибирования или снижения роста опухоли, характеризуемого нежелательным ангиогенезом, в организме субъекта, например, в отношении первичных и/или метастатических опухолей, растущих в головном мозге и оболочке головного мозга, ротовой части глотки, легких и бронхиальном дереве, желудочно-кишечном тракте, мужском и женском репродуктивном тракте, мышцах, костях, коже и придатках, соединительной ткани, селезенке, иммунной системе, кроветворных клетках и костном мозге, печени и мочевыводящем тракте, и специальных сенсорных органах, таких как глаз. В определенных вариантах реализации антитела или антигенсвязывающие фрагменты согласно изобретению применяют для лечения одного или более из следующих видов рака: почечно-клеточной карциномы, карциномы поджелудочной железы, рака молочной железы, рака простаты, злокачественных глиом, остеосаркомы, колоректального рака, злокачественной мезотелиомы, множественной миеломы, рака яичника, мелкоклеточного рака легкого, немелкоклеточного рака легкого, синовиальной саркомы, рака щитовидной железы или меланомы.
[98] В некоторых других вариантах реализации изобретения мутеины или композиции или комбинации, содержащие такие мутеины согласно настоящему изобретению, также можно применять для лечения, предотвращения и/или уменьшения выраженности одного или более глазных нарушений, таких как ретинопатия. Типовые глазные нарушения, которые можно лечить, предотвращать и/или уменьшать выраженность с помощью или посредством мутеинов или композиций, содержащих такие мутеины, включают, например, возрастную макулярную дегенерацию, диабетическую ретинопатию и другие глазные нарушения, связанные с хориоидальной неоваскуляризацией, пропотеванием сосудов, отеком сетчатки и воспалением. Кроме того, мутеины можно вводить в качестве вспомогательного средства вместе с хирургией глаукомы для предотвращения раннего гем- и лимфангиогенеза и накопления макрофагов в фильтрационной подушке после хирургии глаукомы и улучшения клинического результата.
[99] В некоторых дополнительных вариантах реализации изобретения мутеины или композиции или комбинации, содержащие такие мутеины, применяют для лечения, предотвращения или уменьшения выраженности сосудистых заболеваний, таких как гипертензия, диабет (включая инсулинонезависимый сахарный диабет), псориаз, артрит (включая ревматоидный артрит), астма, сепсис, болезнь почек и отек. В некоторых дополнительных вариантах реализации изобретения эти заболевания или нарушения связаны с повреждением, инсультом или опухолью.
[100] Кроме того, мутеины или композиции, или комбинации, содержащие такие мутеины, раскрытые в данном документе, можно применять для лечения, предотвращения или уменьшения выраженности одного или более воспалительных или инфекционных заболеваний. Типовые инфекционные заболевания, которые можно лечить, предотвращать и/или уменьшать выраженность посредством введения анти-Ang-2 мутеинов согласно изобретению или композиций, содержащих такие мутеины, включают, но не ограничиваются этим, малярию (инфекцию Plasmodium falciparum), вирусные геморрагические лихорадки (например, лихорадку денге), риккетсиоз, токсический шок, сепсис, гепатит C, инфекцию Bartonella bacilliformis, лейшманиоз, микобактериальную инфекцию и инфекцию вирусом Эпштейна-Барр.
[101] Настоящее изобретение также предполагает применение раскрытых в данном документе мутеинов или композиций или комбинаций, содержащих такие мутеины, для приготовления медикамента для предотвращения появления метастазов или лечения рака или сосудистых заболеваний.
[102] Ang-2-связывающие мутеины hNGAL в соответствии с изобретением или композиции или комбинации, содержащие такие мутеины, можно вводить любым парентеральным или непарентеральным (например, энтеральным) путем, являющимся терапевтически эффективным. Терапевтически эффективный путь обеспечивает доставку агента в необходимый компартмент, систему или участок. Например, терапевтически эффективный путь – это такой путь, которым агент можно вводить так, чтобы обеспечить в желаемом участке действия количество агента, достаточное для достижения благоприятного или желаемого клинического результата.
C. Комбинирование Ang-2-связывающего мутеина с одним или более антиангиогенными агентами
[103] Для ангиогенеза требуется связывание сигнальных молекул, таких как фактор роста эндотелия сосудов (VEGF), с рецепторами на поверхности нормальных эндотелиальных клеток. Когда VEGF и другие эндотелиальные факторы роста связываются со своими рецепторами на эндотелиальных клетках, в этих клетках инициируются сигналы, которые стимулируют рост и выживаемость новых кровеносных сосудов. Одним из подходов к разработке эффективных антиангиогенных методов лечения является комбинирование агентов, которые действуют на разные мишени, вовлеченные в ангиогенез, предпочтительно мишени, которые действуют на хорошо изолированные сигнальные пути.
[104] Следовательно, в настоящее изобретение включено применение мутеина hNGAL, специфического в отношении Ang-2, в комбинации с одним или более антиангиогенными агентами. В контексте данного документа «антиангиогенный агент» означает любое вещество, способное ингибировать или препятствовать связыванию одной из таких сигнальных молекул со своим рецептором. В некоторых вариантах реализации изобретения антиангиогенный агент способен блокировать или принимает участие в блокировании одного из сигналов, который стимулирует рост и выживаемость новых кровеносных сосудов.
[105] В некоторых конкретных вариантах реализации изобретения антиангиогенные агенты включают (i) антагонисты Ang-1, Ang-2, Ang-3, Ang-4 и/или Tie-2; (ii) антагонисты Fltl, KDR, Flt4, VEGF-A, VEGF-B, VEGF-C, VEGF-D, VEGF-E, PlGF антагонисты EG-VEGF; (iii) антагонисты дельта-подобного лиганда 4 (DLL4, сосудисто-специфический лиганд Notch), (iv) антагонисты рецептора эпидермального фактора роста (EGFR) и (v) ингибиторы цитокинов.
[106] В некоторых дополнительных вариантах реализации изобретения антагонист Ang-2 может представлять собой мутеин hNGAL согласно изобретению, антитело к Ang-2 (смотрите патент США № 8133979; в полном объеме включенный в данный документ посредством ссылки), пептитело (смотрите патент США № 8129331; в полном объеме включенный в данный документ посредством ссылки) или CovX-тело (такое как CVX-060, смотрите патент США № 7521425; в полном объеме включенный в данный документ посредством ссылки). В некоторых дополнительных вариантах реализации изобретения антагонист DLL4 может представлять собой антитело к DLL4 (например, антитело к DLL4, раскрытое в заявке на патент США № 2009/0142354, такое как REGN421 и т.д.). В некоторых дополнительных вариантах реализации изобретения антагонист EGFR может представлять собой антитело к EGFR или низкомолекулярный ингибитор активности EGFR. Другие антиангиогенные агенты, которые могут обеспечивать преимущество при введении в комбинации с анти-Ang-2 мутеинами hNGAL согласно изобретению, включают ингибиторы цитокинов, включая низкомолекулярные ингибиторы цитокинов, и антитела, которые связываются с цитокинами, таким как IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-8, IL-9, IL-11, IL-12, IL-13, IL-17, IL-18 и/или их соответствующими рецепторами.
[107] В связи с этим, настоящее изобретение также включает терапевтические комбинации, содержащие любой из анти-Ang-2 мутеинов hNGAL, упоминаемых в данном документе, и антиангиогенный агент, такой как антагонист одного или более из VEGF, DLL4, EGFR или любого из вышеуказанных цитокинов, при этом антагонист может быть аптамером, антисмысловой молекулой, рибозимом, миРНК, пептителом, нанотелом, антителом, фрагментом антитела (например, фрагментом Fab; фрагментом F(ab')2; фрагментом Fd; фрагментом Fv; scFv; фрагментом dAb; сконструированной молекулой (такой как диатело, триатело, тетратело, минитело и минимальная распознающая единица); противовирусным средством, антибиотиком, анальгетиком, кортикостероидом и/или нестероидным противовоспалительным лекарственным препаратом (NSAID).
[108] В некоторых вариантах реализации изобретения указанная сконструированная молекула может представлять собой EGF-подобный домен, крингл-домен, домен фибронектина типа I, домен фибронектина типа II, домен фибронектина типа III, домен PAN, домен G1a, домен SRCR, домен ингибитора трипсина Кунитца/трипсина поджелудочной железы быка, тендамистат, домен ингибитора сериновых протеаз типа Kazal, домен трилистника (P-типа), домен фактора Виллебранда типа С, анафилотоксин-подобный домен, домен CUB, тироглобулиновый повтор типа I, домен рецептора ЛПНП класса А, домен Sushi, домен Link, домен тромбоспондина типа I, домен иммуноглобулина или иммуноглобулин-подобный домен (например, доменные антитела или верблюжьи антитела, содержащие только тяжелые цепи), домен лектина С-типа, домен МАМ, домен фактора Виллебранда типа А, домен соматомедина В, коровый домен с четырьмя дисульфидными связями WAP-типа, домен F5/8 типа С, домен гемопексина, домен SH2, домен SH3, EGF-подобный домен типа ламинина, домен C2, «каппатело» (III. et al. “Design and construction of a hybrid immunoglobulin domain with properties of both heavy and light chain variable regions” Protein Eng 10:949-57 (1997)), «минитело» (Martin et al. “The affinity-selection of a minibody polypeptide inhibitor of human interleukin-6” EMBO J 13:5303-9 (1994)), «диатело» (Holliger et al. “‘Diabodies’: small bivalent and bispecific antibody fragments” PNAS USA 90:6444-6448 (1993)), «янусин» (Traunecker et al. “Bispecific single chain molecules (Janusins) target cytotoxic lymphocytes on HIV infected cells” EMBO J 10:3655-3659 (1991) и Traunecker et al. “Janusin: new molecular design for bispecific reagents” Int J Cancer Suppl 7:51-52 (1992)), нанотело, аднектин, тетранектин, микротело, аффилин, аффитело, анкирин, кристаллин, ноттин, убиквитин, цинк-пальцевый белок, автофлуоресцирующий белок, анкирин или белок с анкириновым повтором или белок с богатыми лейцином повторами, или авимер (Silverman, Lu Q, Bakker A, To W, Duguay A, Alba BM, Smith R, Rivas A, Li P, Le H, Whitehorn E, Moore KW, Swimmer C, Perlroth V, Vogt M, Kolkman J, Stemmer WP 2005, Nat Biotech, Dec;23(12):1556-61, E-Publication in Nat Biotech. 2005 Nov 20 edition).
[109] При комбинации с одним или более дополнительными агентами согласно изобретению анти-Ang-2 мутеины hNGAL можно вводить до, одновременно (например, в одном составе или в разных составах) или после введения другого(их) агента(ов). Мутеин hNGAL и антиангиогенный агент можно вводить в комбинации, включая одновременное, параллельное или последовательное введение. В некоторых вариантах реализации изобретения комбинации согласно изобретению, мутеины hNGAL и антиангиогенные агенты могут быть включены в одну композицию, предназначенную для введения. Композиция может содержать эффективное количество мутеина hNGAL и антиангиогенного агента в качестве активных ингредиентов в сочетании по меньшей мере с одним фармацевтически приемлемым вспомогательным средством, разбавителем или носителем. В связи с этим, комбинации согласно изобретению можно получать в виде композиций, используя фармацевтически приемлемые ингредиенты, а также общепринятые способы приготовления (Gennaro and Gennaro (2000) Remington: The Science and Practice of Pharmacy, 20th Ed., Lippincott Williams & Wilkins, Philadelphia, PA). Для приготовления фармацевтических композиций можно использовать фармацевтически инертные неорганические или органические вспомогательные вещества.
[110] Мутеин hNGAL и антиангиогенный агент также можно вводить независимо друг от друга, в том числе через индивидуальные интервалы времени в независимые моменты времени. Комбинации мутеина hNGAL и антиангиогенного агента можно обеспечивать в различных формах и в любой ориентации.
[111] Анти-Ang-2 мутеины hNGAL согласно изобретению и их комбинации также можно вводить в качестве части схемы лечения, которая также включает лучевую терапию и/или традиционную химиотерапию.
[112] В некоторых конкретных вариантах реализации изобретения антиангиогенный агент представляет собой антагонист любого компонента рецепторных систем VEGF/VEGF и рецепторной системы ангиопоэтин/Tie-2; то есть, одного из Fltl, KDR, Flt4, VEGF-A, VEGF-B, VEGF-C, VEGF-D, VEGF-E, PlGF, EG-VEGF, Ang-1, Ang-2, Ang-3, Ang-4 или Tie-2. Путь VEGF-VEGFR и путь Tie-2 следует рассматривать как два независимых медиатора, существенных для процесса in vivo ангиогенеза (Siemeister, G., et al., Cancer Res. 59:3 (1999) 3185-91; Jendreyko, N., et al., Journal of Biological Chemistry, 278:47812-47819 (2003); Jendreyko, N., et al., PNAS, 102:8293-8298 (2005)).
[113] В некоторых дополнительных вариантах реализации изобретения антиангиогенный агент представляет собой ингибитор VEGF. Семейство VEGF включает VEGF-A, VEGF-B, VEGF-C, VEGF-D, VEGF-E, плацентарный фактор роста (PlGF) и VEGF эндокринной железы (EG-VEGF). Известно, что представители семейства VEGF связываются с разной аффинностью с тремя родственными рецепторными тирозинкиназами; VEGFR1 (рецептор fms-подобной тирозинкиназы, Fltl), VEGFR2 (рецептор, имеющий в составе домен, содержащий киназу, KDR) и VEGFR3 (рецептор другой fms-подобной тирозинкиназы, Flt4).
[114] В контексте данного документа «ингибитор VEGF» означает любое вещество, которое снижает сигнализацию через путь VEGF-VEGFR. Следовательно, ингибитор VEGF может ингибировать или создавать помехи для любого представителя семейства VEGF и/или любой из рецепторных тирозинкиназ. Ингибиторы VEGF могут представлять собой, если называть только некоторые примеры, небольшие молекулы, пептиды, полипептиды, белки, в частности, включая антитела, в том числе антитела к VEGF, антитела к VEGFR, интратела, макситела, минитела, диатела, Fc-слитые белки, такие как пептитела, рецептитела, растворимые белки и фрагменты рецептора VEGF и много других. Многие ингибиторы VEGF действуют путем связывания с VEGF или с рецептором VEGF. Другие действуют косвенным путем, связываясь с факторами, которые связываются с VEGF или с рецептором VEGF, или с другими компонентами сигнального пути VEGF. Другие ингибиторы VEGF действуют путем изменения регуляторных посттрансляционных модификаций, которые модулируют сигнализацию пути VEGF-VEGFR. Ингибиторы VEGF в соответствии с изобретением также могут действовать посредством непрямых механизмов. Вне зависимости от механизма, в контексте данного документа ингибитор VEGF снижает эффективную активность сигнального пути VEGF-VEGFR в заданных условиях, какие были бы в отсутствие ингибитора.
[115] В некоторых конкретных вариантах реализации изобретения ингибитор VEGF может представлять собой VEGF-Trap (смотрите, например, патент США № 7087411; в полном объеме включенный в данный документ посредством ссылки), антитело к VEGF (например, бевацизумаб), низкомолекулярный киназный ингибитор рецептора VEGF (например, сунитиниб, сорафениб или пазопаниб), анти-VEGF мутеин липокалина слезной жидкости человека (смотрите, например, заявку согласно PCT PCT/EP2007/057971, в полном объеме включенную в данный документ посредством ссылки) или анти-VEGFR мутеин липокалина слезной жидкости человека (смотрите, например, заявку согласно PCT PCT/EP2007/057971, в полном объеме включенную в данный документ посредством ссылки).
[116] В некоторых вариантах реализации изобретения ингибитор VEGF в качестве антиангиогенного агента выбран из группы, состоящей из VEGF-Trap (смотрите, например, патент США № 7087411; в полном объеме включенный в данный документ посредством ссылки), бевацизумаб (Avastin®), сорафениб (Nexavar®), сунитиниб (Sutent®) и пазопаниб (Votrient®).
[117] В некоторых дополнительных вариантах реализации изобретения ингибитор VEGF может представлять собой антагонист VEGF-A, такой как антитело к VEGF-A (например, бевацизумаб). В некоторых других вариантах реализации изобретения антагонист VEGF-A может представлять собой мутеин липокалина, который обладает специфичностью связывания в отношении VEGF-A (смотрите, например, заявку согласно PCT PCT/EP2007/057971; в полном объеме включенную в данный документ посредством ссылки).
[118] В некоторых дополнительных вариантах реализации изобретения ингибитор VEGF может представлять собой антагонист VEGF-C, такой как антитело к VEGF-C (смотрите, например, патенты США № 6403088 и 8486397; в полном объеме включенные в данный документ посредством ссылки). В некоторых других вариантах реализации изобретения антагонист VEGF-C может представлять собой мутеин липокалина, который обладает специфичностью связывания в отношении VEGF-C.
[119] В некоторых вариантах реализации изобретения мутеин hNGAL согласно изобретению, специфический в отношении Ang-2, представлен в комбинации с более чем одним антиангиогенным агентом, например, двумя, причем один агент представляет собой антагонист VEGF-A, упоминаемый в данном документе, а второй агент представляет собой антагонист VEGF-C, упоминаемый в данном документе.
[120] В некоторых вариантах реализации настоящее изобретение включает применение (i) мутеина hNGAL согласно изобретению, специфического в отношении Ang-2, и (ii) одного или более антиангиогенных агентов для ингибирования дерегулированного ангиогенеза у субъекта. Такое применение включает этап введения субъекту эффективного количества (i) мутеина hNGAL согласно изобретению, специфического в отношении Ang-2, и (ii) одного или более антиангиогенных агентов.
[121] Аналогично, настоящее изобретение включает применение (i) мутеина hNGAL согласно изобретению, специфического в отношении Ang-2, и (ii) одного или более антиангиогенных агентов для лечения, предотвращения или уменьшения выраженности заболеваний или нарушений, связанных с дерегулированным ангиогенезом, у субъекта. В некоторых дополнительных вариантах реализации изобретения заболевания или нарушения, связанные с дерегулированным ангиогенезом, включают рак, неоваскулярные заболевания глаз (такие как ретинопатии), артрит и псориаз. В некоторых вариантах реализации изобретения один антиангиогенный агент представляет собой антагонист VEGF-A, упоминаемый в данном документе, а второй антиангиогенный агент представляет собой антагонист VEGF-C, упоминаемый в данном документе.
[122] Дополнительные подробности относительно мутеинов hNGAL с выявляемой аффинностью в отношении Ang-2 можно найти в Разделе А настоящего описания.
[123] В предпочтительном варианте реализации изобретения мутеин hNGAL, специфический в отношении Ang-2, выбран из группы, состоящей из SEQ ID NO: 1-14 и их функциональных фрагментов или вариантов. В некоторых вариантах реализации изобретения такие фрагменты или варианты являются структурными гомологами мутеина, определяемого любой из SEQ ID NO: 1-14.
[124] Настоящее изобретение также относится к фармацевтической композиции, содержащей по меньшей мере один из следующих компонентов: (i) мутеин hNGAL согласно изобретению, специфический в отношении Ang-2, и (ii) один или более антиангиогенных агентов, при этом композицию можно применять для ингибирования дерегулированного ангиогенеза. В некоторых вариантах реализации изобретения один антиангиогенный агент представляет собой антагонист VEGF-A, упоминаемый в данном документе, а второй антиангиогенный агент представляет собой антагонист VEGF-C, упоминаемый в данном документе.
[125] В связи с этим, комбинации согласно изобретению можно получать в виде композиций, используя фармацевтически приемлемые ингредиенты, а также общепринятые способы приготовления (Gennaro and Gennaro (2000) Remington: The Science and Practice of Pharmacy, 20th Ed., Lippincott Williams & Wilkins, Philadelphia, PA). Для приготовления фармацевтических композиций можно использовать фармацевтически инертные неорганические или органические вспомогательные вещества.
[126] В другом аспекте настоящее изобретение относится к способу лечения, предотвращения или уменьшения выраженности заболеваний или нарушений, связанных с дерегулированным ангиогенезом, у субъекта, включающему введение указанному субъекту эффективного количества композиции, которая содержит по меньшей мере следующее: (i) мутеин hNGAL, который обладает выявляемой аффинностью в отношении Ang-2, и (ii) один или более антиангиогенных агентов. В некоторых дополнительных вариантах реализации изобретения заболевания или нарушения, связанные с дерегулированным ангиогенезом, включают рак, неоваскулярные заболевания глаз (такие как ретинопатии), артрит и псориаз. В некоторых вариантах реализации изобретения один антиангиогенный агент представляет собой антагонист VEGF-A, упоминаемый в данном документе, а второй антиангиогенный агент представляет собой антагонист VEGF-C, упоминаемый в данном документе.
[127] В другом аспекте настоящее изобретение включает способ ингибирования или снижения дерегулированного ангиогенеза, у субъекта, включающий введение указанному субъекту эффективного количества композиции, которая содержит по меньшей мере следующее: (i) мутеин hNGAL, который обладает выявляемой аффинностью в отношении Ang-2, и (ii) один или более антиангиогенных агентов. В некоторых вариантах реализации изобретения один антиангиогенный агент представляет собой антагонист VEGF-A, упоминаемый в данном документе, а второй антиангиогенный агент представляет собой антагонист VEGF-C, упоминаемый в данном документе.
D. Мутеины согласно изобретению
[128] При употреблении в контексте данного документа мутеинов согласно настоящему изобретению, которые связываются с Ang-2, термин «специфический в отношении» включает то, что мутеин направлен против, связывается с или реагирует с Ang-2. Таким образом, направленность на, связывание с или вступление в реакцию с включает то, что мутеин специфически связывается с Ang-2. В этом контексте термин «специфически» означает, что мутеин вступает в реакцию с Ang-2, как описано в данном документе, но преимущественно не с другой мишенью. Вступает ли мутеин специфическим образом в реакцию по вышеприведенному определению данного документа, можно легко проверить, помимо прочего, путем сравнения реакции мутеина hNGAL согласно настоящему изобретению с Ang-2 и реакции указанного мутеина с другой(ими) мишенью(ями). «Специфическое связывание» также можно определить, например, в соответствии с методами вестерн-блоттинга, анализом ELISA, РИА, ЭХЛ, ИРМА, FACS, ИГХ и сканированием пептидов.
[129] Аминокислотная последовательность мутеина в соответствии с изобретением имеет высокую идентичность последовательности с человеческим липокалином 2 по сравнению с идентичностью последовательности с другим липокалином (также смотрите выше). В этом общем контексте аминокислотная последовательность мутеина в комбинации в соответствии с изобретением является по меньшей мере в значительной степени сходной с аминокислотной последовательностью соответствующего липокалина (hNGAL дикого типа). Соответствующая последовательность мутеина в комбинации в соответствии с изобретением является в значительной степени сходной с последовательностью зрелого hNGAL, например, имеет по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 82%, по меньшей мере 85%, по меньшей мере 87%, по меньшей мере 90% идентичности, включая по меньшей мере 95% идентичности с последовательностью зрелого hNGAL. В связи с этим, мутеин согласно изобретению, конечно, может содержать описанные в данном документе замены, которые придают мутеину способность связываться с Ang-2. Как правило, мутеин hNGAL содержит по сравнению с нативной последовательностью hNGAL одну или более мутаций в четырех петлях в открытом конце лигандсвязывающего участка hNGAL. Как объяснялось выше, эти области важны для определения специфичности связывания мутеина в отношении Ang-2. Полученный из мутеина hNGAL или его гомолог может содержать один, два, три, четыре или более мутированных аминокислотных остатков в любом положении последовательности в N-концевой области и/или в трех пептидных петлях BC, DE и FG, расположенных в конце структуры β-бочки, которая находится напротив природного связывающего кармана.
[130] Мутеин в соответствии с изобретением содержит одну или более, например, две, три, четыре, пять, шесть, семь, восемь, девять, десять, одиннадцать, двенадцать, тринадцать, четырнадцать, пятнадцать, шестнадцать, семнадцать, восемнадцать, девятнадцать или даже двадцать замен по сравнению с соответствующим нативным липокалином hNGAL, при условии, что такой мутеин способен связываться с Ang-2. Например, мутеин может содержать замену в положении, соответствующем отдельному положению (т.е. в соответствующем положении) hNGAL. В некоторых вариантах реализации изобретения мутеин в комбинации в соответствии с изобретением содержит по меньшей мере две аминокислотные замены, включая 2, 3, 4, 5 или даже более аминокислотных замен нативной аминокислоты остатком аргинина. Соответственно, нуклеиновая кислота «стандартной» белковой структуры, описанной в данном документе, является объектом мутагенеза с целью создания мутеина, который способен связываться с Ang-2.
[131] Также мутеин согласно настоящему изобретению может содержать гетерологичную аминокислотную последовательность в N- или C-конце, предпочтительно С-конце, такую как Strep-метка, например, Strep II-метка, без влияния на биологичскую активность (связывание со своей мишенью, например, Ang-2) мутеина.
[132] В частности, чтобы определить, соответствует ли аминокислотный остаток аминокислотной последовательности мутеина, отличного от hNGAL дикого типа, определенному положению в аминокислотной последовательности hNGAL дикого типа, специалист может использовать средства и методы, хорошо известные в данной области техники, например, выравнивание, проводимое как вручную, так и с помощью компьютерных программ, таких как BLAST2.0, что расшифровывается как Basic Local Alignment Search Tool (основное средство поиска локального выравнивания), или ClustalW или любой другой подходящей программы, которая подходит для создания выравнивания последовательностей. Соответственно, hNGAL дикого типа может служить в качестве «эталонной последовательности» или «стандартной последовательности», тогда как аминокислотная последовательность мутеина, отличного от описанного в данном документе hNGAL дикого типа, служит в качестве «запрашиваемой последовательности». Термины «стандартная последовательность» и «последовательность дикого типа» взаимозаменяемо употребляются в данном документе.
[133] В некоторых вариантах реализации изобретения замена (или замещение) представляет собой консервативную замену. Не смотря на это, предусмотрена любая замена, включая неконсервативную замену или одну или более из типовых замен, приведенных ниже, до тех пор, пока мутеин сохраняет свою способность связываться с Ang-2 и/или он имеет идентичность с замещенной последовательностью, в которой по меньшей мере 60%, например, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85% или более идентичности с «исходной» последовательностью.
[134] В общем случае консервативные замены представляют собой следующие замены, приведенные в соответствии с предназначенной для мутации аминокислотой, за которой следует одно или более замещений, которые могут считаться консервативными: Ala → Gly, Ser, Val; Arg Lys; Asn Gln, His; Asp → Glu; Cys → Ser; Gln → Asn; Glu → Asp; Gly → Ala; His → Arg, Asn, Gln; Ile → Leu, Val; Leu → Ile, Val; Lys → Arg, Gln, Glu; Met → Leu, Tyr, Ile; Phe → Met, Leu, Tyr; Ser → Thr; Thr → Ser; Trp → Tyr; Tyr → Trp, Phe; Val → Ile, Leu. Разрешены также другие замены, которые можно определить эмпирически или в соответствии с другими известными консервативными или неконсервативными заменами. В качестве дополнительного руководства приведены следующие восемь групп, каждая из которых содержит аминокислоты, которые, как правило, можно рассматривать как консервативные замены по отношению друг к другу:
a. аланин (Ala), глицин (Gly);
b. аспарагиновая кислота (Asp), глутаминовая кислота (Glu);
c. аспарагин (Asn), глутамин (Gln);
d. аргинин (Arg), лизин (Lys);
e. изолейцин (Ile), лейцин (Leu), метионин (Met), валин (Val);
f. фенилаланин (Phe), тирозин (Tyr), триптофан (Trp);
g. серин (Ser), треонин (Thr); и
h. цистеин (Cys), метионин (Met)
[135] Если такие замены приводят к изменению биологической активности, тогда можно вносить более существенные замены, такие как приведены ниже или описаны ниже в отношении классов аминокислот, и проводить скрининг продуктов в отношении желаемых характеристик. Примерами таких более существенных замен являются: Ala → Leu, Ile; Arg Gln; Asn → Asp, Lys, Arg, His; Asp → Asn; Cys → Ala; Gln → Glu; Glu → Gln; His → Lys; Ile → Met, Ala, Phe; Leu → Ala, Met, норлейцин; Lys → Asn; Met → Phe; Phe → Val, Ne, Ala; Trp → Phe; Tyr → Thr, Ser; Val → Met, Phe, Ala.
[136] Существенные модификации биологических свойств hNGAL осуществляют, выбирая замены, которые значительно отличаются по своему действию на поддержание (a) структуры полипептидного скелета в области замены, например, в виде складчатой или спиральной конформации, (b) заряда или гидрофобности молекулы в целевом участке или (c) объема боковой цепи. Остатки природного происхождения делятся на группы на основании общих свойств боковой цепи: (1) гидрофобные: норлейцин, метионин, аланин, валин, лейцин, изолейцин; (2) нейтральные гидрофильные: цистеин, серин, треонин; (3) кислые: аспарагиновая кислота, глутаминовая кислота; (4) основные: аспарагин, глутамин, гистидин, лизин, аргинин; (5) остатки, которые влияют на ориентацию цепи: глицин, пролин; и (6) ароматические: триптофан, тирозин, фенилаланин.
[137] Неконсервативные замены включают замещение представителя одного из этих классов другим классом. Также может быть замещен любой цистеиновый остаток, не вовлеченный в поддержание правильной конформации hNGAL, в общем случае серином, для улучшения устойчивости к окислению молекулы и предотвращения аберрантного перекрестного связывания. И наоборот, цистеиновые связи можно добавлять для улучшения стабильности.
[138] Любую мутацию, включая обсуждаемую выше вставку, можно очень легко провести в нуклеиновой кислоте, например, на уровне ДНК, используя общепринятые стандартные способы. Иллюстративными примерами изменений аминокислотной последовательности являются вставки или удаления, а также аминокислотные замены. Такие замены могут быть консервативными, т.е. аминокислотный остаток замещают аминокислотным остатком со сходными химическими свойствами, в частности, в отношении полярности, а также размера. Примерами консервативных замен являются замещения между представителями следующих групп: 1) аланин, серин и треонин; 2) аспарагиновая кислота и глутаминовая кислота; 3) аспарагин и глутамин; 4) аргинин и лизин; 5) изолейцин, лейцин, метионин и валин; и 6) фенилаланин, тирозин и триптофан. С другой стороны также возможно вносить неконсервативные изменения в аминокислотную последовательность. Кроме того, вместо замещения одиночных аминокислотных остатков также возможно вставлять или удалять одну или более последовательных аминокислот первичной структуры hNGAL до тех пор, пока эти удаления или вставки приводят к получению стабильного свернутого/функционального мутеина.
[139] Модификации аминокислотной последовательности включают направленный мутагенез единичных аминокислотных положений с целью упрощения субклонирования мутированного гена hNGAL или его частей путем включения участков расщепления для определенных рестрикционных ферментов. Кроме того, эти мутации также можно вносить для дополнительного улучшения аффинности мутеина в отношении заданной мишени, такой как Ang-2. Помимо этого, мутации можно вводить с целью модуляции определенных характеристик мутеина, такой как улучшение стабильности сворачивания, сывороточной стабильности, устойчивости белка или растворимости в воде, или для снижения тенденции к агрегации в случае необходимости. Например, цистеиновые остатки природного происхождения можно мутировать на другие аминокислоты для предотвращения образования дисульфидных мостиков. Также возможно преднамеренно мутировать другие положения аминокислотной последовательности на цистеин с целью внесения новых реактивных групп, например, для конъюгации с другими соединениями, такими как полиэтиленгликоль (ПЭГ), гидроэтилкрахмал (ГЭК), биотин, пептиды или белки, или для образования дисульфидных связей, не существующих в природе. Созданный тиоловый компонент можно использовать для ПЭГилирования или ГЕКилирования мутеина, например, с целью повышения периода полужизни соответствующего мутеина в сыворотке крови.
[140] Также возможно преднамеренно мутировать другие положения аминокислотной последовательности на цистеин с целью внесения новых реактивных групп, например, для конъюгации с другими соединениями, такими как полиэтиленгликоль (ПЭГ), гидроэтилкрахмал (ГЭК), биотин, пептиды или белки, или для образования дисульфидных связей, не существующих в природе.
[141] В некоторых вариантах реализации изобретения, если один из вышеуказанных компонентов конъюгирован с мутеином согласно изобретению, преимущество может обеспечить конъюгация с боковой цепью аминокислоты. Подходящие боковые цепи аминокислот в аминокислотной последовательности hNGAL могут иметь природное происхождение или могут быть внесены с помощью мутагенеза. В случае внесения подходящего связывающего участка с помощью мутагенеза, одной из возможностей является замещение аминокислоты в соответствующем положении цистеиновым остатком.
[142] В отношении мутеина человеческого липокалина 2 типовой возможностью такой мутации является внесение цистеинового остатка в аминокислотную последовательность липокалина, включая мутеин человеческого липокалина 2, для внесения цистеинового остатка (Cys) по меньшей мере в одно из положений последовательности, которые соответствуют положениям последовательности 14, 21, 60, 84, 88, 116, 141, 145, 143, 146 или 158 последовательности человеческого NGAL дикого типа. В некоторых вариантах реализации изобретения, в которых мутеин человеческого липокалина 2 согласно изобретению имеет последовательность, в которой, по сравнению с последовательностью с номером доступа в банке данных SWISS-PROT/UniProt P80188, цистеин был замещен другим аминокислотным остатком, соответствующий цистеин может быть повторно внесен в последовательность. В качестве иллюстративного примера цистеиновый остаток в аминокислотном положении 87 можно вносить в таком случае путем возвращения цистеина, исходно присутствующего в последовательности с номером доступа SWISS-PROT № P80188. Созданный тиоловый компонент в участке любом из аминокислотных положений 14, 21,60, 84, 88, 116, 141, 145, 143, 146 и/или 158 можно использовать для ПЭГилирования или ГЕКилирования мутеина, например, с целью повышения периода полужизни соответствующего мутеина человеческого липокалина 2 в сыворотке крови.
[143] В другом варианте реализации изобретения с целью обеспечения подходящих аминокислотных боковых цепей для конъюгации одного из вышеуказанных соединений с мутеином согласно настоящему изобретению можно вносить искусственные аминокислоты с помощью мутагенеза. В общем случае такие искусственные аминокислоты конструируют так, чтобы они были более реактивными и, следовательно, облегчали конъюгацию с желаемым компонентом. Одним примером такой искусственной аминокислоты, которую можно вносить с помощью искусственной тРНК, является параацетилфенилаланин.
[144] В некоторых вариантах реализации изобретения мутеин согласно изобретению может быть слит по N-концу или С-концу с белком, белковым доменом или пептидом, например, сигнальной последовательностью и/или аффинной меткой.
[145] Аффинные метки, такие как Strep-метка® или Strep-метка® II (Schmidt, T.G.M. et al. (1996) J. Mol. Biol. 255, 753-766), myc-метка, FLAG-метка, Hisg-метка или HA-метка, или белки, такие как глутатион-S-трансфераза, также обеспечивают возможность легкого выявления и/или очистки рекомбинантных белков и являются дополнительными примерами партнеров по слиянию. И наконец, белки с хромогенными или флуоресцентными свойствами, такие как зеленый флуоресцентный белок (ЗФБ) или желтый флуоресцентный белок (ЖФБ), также являются подходящими партнерами для слияния с мутеинами согласно изобретению.
[146] В общем случае возможно метить мутеины согласно изобретению любым подходящим химическим веществом или ферментом, который прямым или непрямым образом генерирует выявляемое соединение или сигнал в химической, физической, оптической или ферментативной реакции. Примером физической реакции и одновременно оптической реакции/оптического маркера является флуоресценция после облучения или рентгеновское излучение в случае применения радиоактивной метки. Щелочная фосфатаза, пероксидаза хрена и β-галактозидаза являются примерами ферментных меток (и в то же время оптических меток), которые катализируют образование продуктов хромогенной реакции. В общем случае все метки, обычно используемые для антител (за исключением тех, которые используются исключительно с сахарным компонентом или Fс-частью иммуноглобулинов), также можно использовать для конъюгации с мутеинами согласно изобретению. Мутеины согласно изобретению также можно конъюгировать с любым подходящим терапевтически активным агентом, например, для направленной доставки таких агентов в заданную клетку, ткань или орган или для избирательного нацеливания на клетки, например, опухолевые клетки, не влияя на окружающие их нормальные клетки. Примеры таких терапевтически активных агентов включают радионуклиды, токсины, небольшие органические молекулы и терапевтические пептиды (такие как пептиды, действующие в качестве агонистов/антагонистов рецепторов клеточной поверхности, или пептиды, конкурирующие за участок связывания белка на заданной клеточной мишени). При этом мутеины согласно изобретению также можно конъюгировать с терапевтически активными молекулами нуклеиновых кислот, такими как антисмысловые молекулы нуклеиновых кислот, малые инферферирующие РНК, микроРНК или рибозимы. Такие конъюгаты можно получать способами, хорошо известными в данной области техники.
[147] Как указано выше, мутеин согласно изобретению может в некоторых вариантах реализации изобретения быть конъюгирован с компонентом, который продлевает период полужизни мутеина в сыворотке крови (по данной теме смотрите также публикацию согласно РСТ WO 2006/56464, в которой такие стратегии конъюгации описаны в отношении мутеинов человеческого липокалина, ассоциированного с желатиназой нейтрофилов, обладающего аффинностью связывания в отношении CTLA-4). Компонентом, который продлевает период полужизни в сыворотке крови, может быть молекула полиалкиленгликоля, гидроксиэтиловый крохмал, молекулы жирных кислот, таких как пальмитиновая кислота (Vajo & Duckworth 2000, Pharmacol. Rev. 52, 1-9), Fс-часть иммуноглобулина, CH3-домен иммуноглобулина, CH4-домен иммуноглобулина, альбуминсвязывающий пептид или альбуминсвязывающий белок, трансферрин, если называть только некоторые. Альбуминсвязывающий белок может представлять собой бактериальный альбуминсвязывающий белок, антитело, фрагмент антитела, включая доменные антитела (см. патент США 6696245, например), или мутеин, проявляющий активность связывания в отношении альбумина. Соответственно, подходящие партнеры по конъюгации для продления времени полужизни мутеина согласно изобретению включают альбуминсвязывающий белок, например, бактериальный альбуминсвязывающий белок, такой как представитель стрептококковых протеинов G (Konig, T, & Skerra, A. (1998) J. Immunol. Methods 218, 73-83). Другими примерами альбуминсвязывающих пептидов, которые можно использовать в качестве партнеров по конъюгации, являются, например, пептиды, имеющие консенсусную последовательность Cys-Xaa1-Xaa2-Xaa3-Xaa4-Cys, где Xaai представляет собой Asp, Asn, Ser, Thr или Trp; Xaa2 представляет собой Asn, Gln, His, Ile, Leu или Lys; Xaa3 представляет собой Ala, Asp, Phe, Trp или Tyr; а Xaa4 представляет собой Asp, Gly, Leu, Phe, Ser или Thr, как описано в заявке на патент США 2003/0069395 (в полном объеме включенной в данный документ посредством ссылки) или в Dennis et al. (Dennis, M. S., Zhang, M., Meng, Y. G., Kadkhodayan, M., Kirchhofer, D., Combs, D. & Damico, L. A. (2002) J Biol Chem 277, 35035-35043).
[148] В других вариантах реализации изобретения сам альбумин (Osborn, B.L. et al., 2002, J. Pharmacol. Exp. Then 303, 540-548) или его биологический фрагмент можно использовать в качестве партнера по конъюгации мутеина согласно изобретению. Термин «альбумин» включает все альбумины млекопитающих, такие как человеческий сывороточный альбумин или бычий сывороточный альбумин, или крысиный альбумин. Альбумин или его фрагмент может быть получен рекомбинантным способом, как описано в патенте США № 5728553 или заявках на Европейский патент EP 0 330 451 и EP 0 361 991 (в полном объеме включенных в данный документ посредством ссылки). Рекомбинантный человеческий альбумин (Рекомбумин®) от Novozymes Delta Ltd. (Nottingham, UK) можно конъюгировать или сливать с мутеином согласно изобретению с целью продления времени полужизни мутеина.
[149] Если альбуминсвязывающий белок представляет собой фрагмент антитела, он может представлять собой доменное антитело. Доменные антитела (dAb) конструируют так, чтобы обеспечить точный контроль биофизических свойств и in vivo времени полужизни для создания профиля продукта с оптимальной безопасностью и эффективностью. Доменные антитела доступны, например, от Domantis Ltd. (Cambridge, UK и MA, USA).
[150] При использовании трансферрина для продления времени полужизни мутеинов согласно изобретению мутеины могут, в общем случае, быть слиты с N- или С-концом или обеими концами негликозилированного трансферрина. Время полужизни негликозилированного трансферрина составляет 14-17 дней, и слитый белок трансферрина также будет иметь аналогично продленное время полужизни. Трансферриновый носитель также обеспечивает повышение биодоступности, биораспределения и циркуляционной стабильности. Эта технология доступна на коммерческой основе от BioRexis (BioRexis Pharmaceutical Corporation, PA, USA). Рекомбинантный человеческий трансферрин (DeltaFerrinТМ) для применения в качестве партнера для стабилизации/продления времени полужизни также доступен на коммерческой основе от Novozymes Delta Ltd. (Nottingham, UK).
[151] Если в целях продления времени полужизни мутеина согласно изобретению используют Fс-часть иммуноглобулина, можно применять технологию SynFusionТМ, доступную на коммерческой основе от Syntonix Pharmaceuticals, Inc (MA, USA). Применение этой технологии Fc-слияния позволяет создавать биофармацевтические препараты более продолжительного действия и может, например, включать использование двух копий мутеина, связанных с Fс-областью антитела, для улучшения фармакокинетики, растворимости и эффективности.
[152] Другой альтернативой продления времени полужизни мутеинов согласно изобретению является слияние с N- или C-концом мутеинов длинных, неструктурированных, гибких, богатых глицином последовательностей (например, полиглицина, содержащего от около 20 до 80 последовательных остатков глицина). Этот подход раскрыт, например, в W02007/038619 и также называется «rPEG» (рекомбинатный PEG).
[153] Если в качестве партнера по конъюгации используется полиалкиленгликоль, полиалкиленгликоль может быть замещенным, незамещенным, линейным или разветвленным. Также он может представлять собой активированное производное полиалкилена. Примерами подходящих соединений являются молекулы полиэтиленгликоля (ПЭГ), описанные в WO 99/64016, в патенте США 6177074 или в патенте США 6403564 в отношении интерферона, или описанные для других белков, таких как ПЭГ-модифицированная аспарагиназа, ПЭГ-аденозиндеаминаза (ПЭГ-АДА) или ПЭГ-скпероксиддисмутаза (смотрите, например, Fuertges et al. (1990) The Clinical Efficacy of Poly(Ethylene Glycol)-Modified Proteins J. Control. Release 11, 139-148). Молекулярная масса такого полимера, например, полиэтиленгликоля, может находиться в диапазоне от около 300 до около 70000 дальтон, включая, например, полиэтиленгликоль с молекулярной массой около 10000, около 20000, около 30000 или около 40000 дальтон. Кроме того, например, как описано в патентах США 6500930 или 6620413, в целях продления периода полужизни в сыворотке крови к мутеину согласно изобретению можно конъюгировать углеводные олиго- и полимеры, такие как крахмал или гидроксиэтиловый крахмал (ГЭК).
[154] Кроме того, раскрытый в данном документе мутеин можно сливать с компонентом, который может придавать мутеинам согласно изобретению новые характеристики, такие как ферментативная активность или аффинность связывания в отношении других молекул. Примерами подходящих партнеров по слиянию являются щелочная фосфатаза, пероксидаза хрена, глутатион-S-трансфераза, альбуминсвязывающий домен протеина G, протеина A, фрагменты антител, олигомеризационные домены или токсины.
[155] В частности, возможно слить раскрытый в данном документе мутеин с отдельным активным участком фермента так, чтобы оба «компонента» полученного в результате слитого белка вместе действовали на заданную терапевтическую мишень. Связывающий домен мутеина присоединяется к вызывающей заболевание мишени, позволяя ферментному домену аннулировать биологическую функцию мишени.
[156] Настоящее изобретение также относится к молекулам нуклеиновых кислот (ДНК и РНК), которые содержат нуклеотидные последовательности, кодирующие мутеины согласно изобретению. Так как вырожденность генетического кода позволяет проводить замещения определенных кодонов другими кодонами, отвечающими за такую же аминокислоту, изобретение не ограничено конкретной молекулой нуклеиновой кислоты, кодирующей описанный в данном документе мутеин, но включает все молекулы нуклеиновых кислот, которые содержат нуклеотидные последовательности, кодирующие функциональный мутеин. В связи с этим, в настоящем изобретении предложены нуклеотидные последовательности, приведенные в SEQ ID NO: 21-34, кодирующие некоторые мутеины согласно изобретению.
[157] В одном варианте реализации изобретения способ включает подвержение молекулы нуклеиновой кислоты мутагенезу в нуклеотидных триплетах, кодирующих по меньшей мере одно или даже более положений последовательности, соответствующих положениям последовательности 28, 36, 40, 41, 49, 52, 65, 68, 70, 72-74, 77, 79, 81, 87, 96, 100, 103, 106, 116, 125, 126, 127, 129, 132 и 134 линейной полипептидной последовательности человеческого NGAL (SEQ ID NO: 16).
[158] Изобретение также включает молекулы нуклеиновых кислот, кодирующие мутеины согласно изобретению, которые содержат дополнительные мутации за пределами указанных положений последовательности экспериментального мутагенеза. Такие мутации часто хорошо переносятся и даже могут обеспечивать преимущество, например, если они улучшают эффективность сворачивания, сывороточную стабильность, температурную стабильность или лигандсвязывающую аффинность мутеинов.
[159] Раскрытая в данной заявке молекула нуклеиновой кислоты может быть «функционально связана» с регуляторной последовательностью (или регуляторными последовательностями), обеспечивающей экспрессию этой молекулы нуклеиновой кислоты.
[160] Говорят, что молекула нуклеиновой кислоты, такая как ДНК, «способна экспрессировать молекулу нуклеиновой кислоты» или способна «обеспечивать экспрессию молекулы нуклеиновой кислоты», если она содержит элементы, содержащие информацию относительно транскрипционной и/или трансляционной регуляции, и такие последовательности «функционально связаны» с нуклеотидной последовательностью, кодирующей полипептид. Функциональная связь представляет собой связь, в которой элементы регуляторной последовательности и предназначенная для экспрессии последовательность связаны способом, обеспечивающим генную экспрессию. Точная природа регуляторных областей, необходимых для генной экспрессии, может варьироваться для разных видов, но в общем случае эти области включают промотор, который у прокариотов содержит как промотор per se, т.е. элементы ДНК, управляющие инициацией транскрипции, так и элементы ДНК, которые при транскрибировании в РНК будут сигнализировать об инициации трансляции. Такие области промотора обычно содержат 5' некодирующие последовательности, вовлеченные в инициацию транскрипции и трансляции, такие как -35/-10 боксы и элемент Шайна-Дальгарно у прокариотов или TATA-бокс, последовательности CAAT и 5'-кэпирующие элементы у эукариотов. Эти области также могут содержать энхансерные или репрессорные элементы, а также транслируемые сигнальные и лидерные последовательности для нацеливания нативного полипептида на специфический компартмент в клетке-хозяине.
[161] Кроме того, 3' некодирующие последовательности могут содержать регуляторные элементы, вовлеченные в терминацию транскрипции, полиаденилирование и т.д. Однако, если эти терминационные последовательности недостаточно функциональны в конкретной клетке-хозяине, они могут быть замещены сигналами, функциональными в этой клетке.
[162] Следовательно, молекула нуклеиновой кислоты согласно изобретению может содержать регуляторную последовательность, такую как промоторная последовательность. В некоторых вариантах реализации изобретения молекула нуклеиновой кислоты согласно изобретению содержит промоторную последовательность и последовательность терминации транскрипции. Подходящими прокариотическими промоторами являются, например, промотор tet, промотор lacUV5 или промотор T7. Примерами промоторов, применяемых в эукариотических клетках, являются промотор SV40 или промотор ЦМВ.
[163] Молекулы нуклеиновых кислот согласно изобретению также могут быть частью вектора или любого другого средства клонирования, такого как плазмида, фагмида, фаг, бакуловирус, космида или искусственная хромосома.
[164] В одном варианте реализации изобретения молекула нуклеиновой кислоты включена в фазмиду. Фазмидный вектор означает вектор, кодирующий межгенную область умеренного фага, такого как M13 или f1, или ее функциональную область, слитую с представляющей интерес кДНК. После суперинфекции бактериальных клеток-хозяев таким фагмидным вектором и соответствующим хелперным фагом (например, M13K07, VCS-M13 или R408) вырабатываются интактные фаговые частицы, обеспечивая физическое сопряжение кодируемой гетерологической кДНК с соответствующим полипептидом, представленным на поверхности фага (смотрите, например, Lowman, H.B. (1997) Annu. Rev. Biophys. Biomol. Struct. 26, 401-424, или Rodi, D.J., and Makowski, L. (1999) Curr. Opin. Biotechnol. 10, 87-93).
[165] Такие средства клонирования могут включать, помимо описанных выше регуляторных последовательностей и нуклеотидной последовательности, кодирующей описанный в данном документе мутеин, репликационные и контрольные последовательности, полученные от видов, совместимых с клеткой-хозяином, применяемой для экспрессии, а также селективные маркеры, придающие трансформированным или трансфицированным клеткам выбранный фенотип. В данной области техники известно и доступно на коммерческой основе большое количество подходящих клонирующих векторов.
[166] Молекула ДНК, кодирующая описанный в данном документе мутеин, и, в частности, клонирующий вектор, содержащий кодирующую последовательность такого мутеина, можно трансформировать в клетку-хозяина, способную экспрессировать ген. Трансформацию можно осуществлять, используя стандартные методы. Таким образом, изобретение также относится к клетке-хозяину, содержащей раскрытую в данном документе молекулу нуклеиновой кислоты.
[167] Трансформированные клетки-хозяев культивируют в условиях, подходящих для экспрессии нуклеотидной последовательности, кодирующей слитый белок согласно изобретению. Подходящие клетки-хозяева могут быть прокариотическими, такими как Escherichia coli (E. coli) или Bacillus subtilis, или эукариотическими, такими как Saccharomyces cerevisiae, Pichia pastoris, SF9 или High5 клетки насекомых, иммортализованные линии клеток млекопитающих (например, клеток HeLa или клеток CHO) или первичные клетки млекопитающих.
[168] Изобретение также относится к способу получения описанного в данном документе мутеина, причем мутеин или полипептид, фрагмент мутеина или слитый белок мутеина получают, начиная с нуклеиновой кислоты, кодирующей мутеин или полипептид, с помощью методов генетической инженерии. Способ можно осуществлять in vivo, например, мутеин или полипептид можно получать в бактериальном или эукариотическом организме-хозяине, а затем выделять из организма-хозяина или культуры. Также возможно получать белок in vitro, например, используя in vitro систему трансляции.
[169] При получении мутеина in vivo нуклеиновую кислоту, кодирующую такой мутеин или полипептид, вносят в подходящий бактериальный или эукариотический организм-хозяин посредством технологии рекомбинантных ДНК (как уже было отмечено выше). С этой целью клетку-хозяина сначала трансформируют клонирующим вектором, который содержит молекулу нуклеиновой кислоты, кодирующую описанный в данном документе мутеин, используя общепринятые стандартные методы. Затем клетку-хозяина культивируют в условиях, которые обеспечивают экспрессию гетерологичной ДНК и, следовательно, синтез соответствующего полипептида. Впоследствии полипептид выделяют из клетки-хозяина или культуральной среды.
[170] В некоторых вариантах реализации изобретения молекула нуклеиновой кислоты, такая как ДНК, раскрытая в данной заявке, может быть «функционально связана» с другой молекулой нуклеиновой кислоты согласно изобретению для обеспечения экспрессии слитого белка согласно изобретению. В связи с этим функциональная связь представляет собой такую связь, в которой элементы последовательности первой молекулы нуклеиновой кислоты и элементы последовательности второй молекулы нуклеиновой кислоты связаны таким образом, который обеспечивает экспрессию слитого белка в виде одного полипептида.
[171] Кроме того, в некоторых вариантах реализации в мутеинах hNGAL согласно изобретению можно удалять природную дисульфидную связь между Cys 76 и Cys 175. Соответственно, такие мутеины можно получать в клеточном компартменте, содержащем восстановительную среду, например, в цитоплазме грамотрицательных бактерий.
[172] В случае, когда мутеин согласно изобретению содержит внутримолекулярные дисульфидные связи, может быть предпочтительно направлять образующийся полипептид в клеточный компартмент, содержащий окислительную среду, используя соответствующую сигнальную последовательность. Такая окислительная среда может находиться в периплазме грамотрицательных бактерий, таких как E coli, во внеклеточной среде грамположительных бактерий или в полости эндоплазматического ретикулума эукариотических клеток и обычно способствует образованию структурных дисульфидных связей.
[173] Однако также возможно получать мутеин согласно изобретению в цитозоле клетки-хозяина, предпочтительно E. coli. В этом случае мутеин или полипептид можно получать как прямым образом в растворимом и свернутом состоянии, так и восстанавливать в форме телец включения с последующей ренатурацией in vitro. Дополнительной опцией является применение специфических штаммов хозяев, содержащих окислительную внутриклеточную среду, что может, таким образом, обеспечивать образование дисульфидных связей в цитозоле (Venturi et al. (2002) J. Mol. Biol. 315, 1-8).
[174] При этом описанный в данном документе мутеин или полипептид не обязательно должен быть создан или получен только с помощью генной инженерии. Наоборот, такой мутеин или полипептид также может быть получен с помощью химического синтеза, такого как твердофазный полипептидный синтез по Меррифилду, или с помощью in vitro транскрипции и трансляции. Например, возможна ситуация, когда перспективные мутации определяют с помощью молекулярного моделирования, а затем синтезируют желаемый (сконструированный) полипептид in vitro и исследуют активность связывания в отношении Ang-2. Способы твердофазного и/или жидкофазного синтеза белков хорошо известны в данной области техники (смотрите, например, Bruckdorfer, T. et al. (2004) Curr. Pharm. Biotechnol. 5, 29-43).
[175] В другом варианте реализации изобретения мутеин или полипептид согласно изобретению можно получать путем in vitro транскрипции/трансляции, применяя общепринятые методы, известные специалистам в данной области техники.
[176] Для специалиста очевидны методы, которые можно применять для получения мутеинов или их полипептидов, предусмотренных настоящим изобретением, но чьи белковые или нуклеотидные последовательности явным образом не раскрыты в данном документе. В качестве обзора, такие модификации аминокислотной последовательности включают, например, направленный мутагенез единичных аминокислотных положений с целью упрощения субклонирования мутированного гена hNGAL или его частей путем внесения участков расщепления для определенных рестрикционных ферментов. Кроме того, эти мутации также можно включать для дополнительного улучшения аффинности мутеина в отношении его мишени (например, Ang-2 или Ang-1 соответственно). Помимо этого, мутации можно вводить с целью модуляции определенных характеристик мутеина, таких как улучшение стабильности сворачивания, сывороточной стабильности, устойчивости белка или растворимости в воде, или для снижения тенденции к агрегации в случае необходимости. Например, цистеиновые остатки природного происхождения можно мутировать на другие аминокислоты для предотвращения образования дисульфидных мостиков.
[177] Раскрытые в данном документе мутеины или их полипептиды, а также их производные можно применять во многих областях, аналогично антителам или их фрагментам. Например, мутеины можно применять для мечения ферментом, антителом, радиоактивным веществом или любой другой группой, имеющей биохимическую активность или определенные характеристики связывания. Таким образом, возможно выявление или приведение в контакт их соответствующих мишеней или конъюгатов, или слитых белков. Кроме того, мутеины или их полипептиды согласно изобретению могут служить для выявления химических структур с помощью принятых аналитических методов (например, ELISA или вестерн-блоттинга) или с помощью микроскопии или иммуносенсорики. В связи с этим, выявляемый сигнал может генерироваться прямым образом, посредством применения подходящего конъюгата или слитого белка мутеина, или непрямым образом, посредством иммунохимического выявления связанного мутеина с помощью антитела.
[178] Дополнительные объекты, преимущества и признаки этого изобретения станут очевидны для специалистов в данной области техники после изучения нижеприведенных Примеров и присоединенных графических материалов, которые не являются ограничивающими. Таким образом, следует понимать, что хотя настоящее изобретение специально раскрыто с помощью типовых вариантов реализации и необязательных признаков, специалисты в данной области техники могут прибегнуть к модификации и вариации изобретения, раскрытого в данном документе, и что такие модификации и вариации считаются входящими в объем данного изобретения.
V. ПРИМЕРЫ
[179] Пример 1: Отбор мутеинов, специфически связывающихся с Ang-2
[180] Для отбора мутеинов, специфически связывающихся с Ang-2, использовали библиотеки, полученные путем случайного мутагенеза зрелого hNGAL.
[181] 2x1012 фагмид из этих библиотек инкубировали с 200 нM биотинилированного человеческого Ang-2 (R & D System). Парамагнитные гранулы, покрытые нейтравидином или стрептавидином использовали для захвата комплексов мишень/фагмида, которые впоследствии выделяли с помощью магнита. Несвязанные фагмиды удаляли, промывая гранулы ФСБТ или ФСБ. Связанные фагмиды сначала элюировали с помощью 300 мкл 70 мM триэтиламина с последующей незамедлительной нейтрализацией супернатанта с помощью 100 мкл 1 M Трис-Cl, pH 6,0. После одного промежуточного промывочного цикла оставшиеся фагмиды элюировали с помощью 100 мM глицина, pH 2,2, в течение 10 мин с последующей незамедлительной нейтрализацией с помощью 50 мкл 0,5 M Трис-основания. Обе элюированные фракции объединяли и использовали для инфицирования 4 мл культуры E. coli XL1-blue для повторной амплификации.
[182] Проводили четыре последовательных этапа отбора. Фагмидную ДНК получали из клеток E. coli, инфицированных продуктом четвертого этапа селекции, а кассету мутеина hNGAL выделяли путем расщепления. Кассету мутеина hNGAL вставляли в аналогично вырезанный вектор, который обеспечивает бактериальную выработку мутеинов hNGAL под управлением тетрациклинового промотора. CaCl2-компетентные клетки TG1-F’ трансформировали лигационной смесью и высевали на планшеты LB/Amp.
[183] Для оптимизации Ang-2-специфических мутеинов создавали дополнительные библиотеки на основании последовательностей мутеина SEQ ID NO: 1 и SEQ ID NO: 3. Библиотеки создавали, используя методы на основании содержащей ошибку рандомизации выбранных положений или полимеразной цепной реакции (ПЦР) с внесением ошибок. Отбор оптимизированных мутеинов проводили, как описано выше, но в условиях повышенной жесткости.
[184] Пример 2: Определение мутеинов, специфически связывающихся с Ang-2 с помощью высокопроизводительного скрининга ELISA
[185] Для инокуляции среды 2xYT/Amp использовали индивидуальные колонии и выращивали в течение ночи (14-18 ч) до стационарной фазы. После этого из культур стационарной фазы инокулировали 50 мкл 2xYT/Amp и инкубировали в течение 3 ч при 37°C, а затем температуру снижали до 22°C до достижения OП595 0,6-0,8. Получение мутеинов индуцировали добавлением 10 мкл 2xYT/Amp, дополненной 1,2 мкг/мл ангидротетрациклина. Культуры инкубировали при 22°C до следующего дня. После добавления 40 мкл 5% (масс./об.) БСА в ФСБ/Т и инкубации в течение 1 ч при 25°C культуры были готовы для применения в скрининговом анализе.
[186] Специфическое связывание выделенных мутеинов с человеческим Ang-2 исследовали, покрывая микротитровальные планшеты 1:1 смесью нейтравидина и стрептавидина (5 мкг/мл в ФСБ) в течение ночи при 4°C. После блокирования планшета в течение 1 ч с помощью 2% БСА в ФСБТ проводили захват используемой для отбора биотинилированной мишени на покрытых микротитровальных планшетах в концентрации 1 мкг/мл в ФСБ/Т. Планшеты, покрытые таким же образом, но без биотинилированной мишени, использовали в качестве отрицательной контрольной мишени при проведении скрининга. После этого добавляли 20 мкл заблокированных БСА культур в покрытый микротитровальный планшет, содержащий захваченную мишень или альдостерон, и инкубировали в течение 1 ч при 25°C. Связанные мутеины выявляли через 1 ч инкубации с антителом против стрептавидиновой метки, конъюгированным с пероксидазой хрена (IBA, Boettingen). Для проведения количественной оценки добавляли 20 мкл флуорогенного пероксидазного субстрата QuantaBlu и определяли флуоресценцию при длине волны возбуждения 320 нм и длине волны испускания 430 нм. Затем проводили секвенирование мутеинов, специфически связывающихся с Ang-2.
[187] Чтобы отобрать мутеины с повышенной аффинностью и стабильностью, проводили скрининг i) со сниженной концентрацией антигена и/или ii) с конкуренцией с Tie-2 Fс (Reliatech), и/или iii) инкубацией скринингового супернатанта при 65°C или 70°C перед добавлением в целевой планшет, и/или iv) с применением обратного формата скрининга, в котором мутеины захватывали с помощью стрептавидиновой метки на микротитровальных планшетах, покрытых антителом против стрептавидиновой метки (Qiagen), а разные концентрации мишени добавляли и выявляли с помощью HRP-меченого антитела против His-метки (Abeam).
[188] Пример 3: Экспрессия мутеинов
[189] Уникальные мутеины экспрессировали с С-концевой последовательностью (SEQ ID NO: 15), включая Strep-метку® II, в среде 2YT-Amp, чтобы провести очистку мутеинов после экспрессии методом Strep-Tactin аффинной хроматографии и препаративной эксклюзионной хроматографии.
[190] Пример 4: Аффинность мутеинов к Ang-2, определяемая в ELISA-анализе
[191] Связывание мутеинов исследовали методом сэндвич-анализа ELISA. В частности, 384-луночный планшет, подходящий для измерения флуоресценции (Greiner FLUOTRACТМ 600, черное плоское дно, высокая степень связывания) покрывали 20 мкл Ang-2 в концентрации 5 мкг/мл в ФСБ в течение ночи при 4 C. После промывки покрытые Ang-2 лунки блокировали 100 мкл блокирующего буфера, содержащего 0,1% Твин 20 и 2% БСА (ФСБ-T/БСА) в течение 1 ч при комнатной температуре.
[192] 20 мкл серийно разведенных мутеинов инкубировали в ФСБ-T/БСА в течение 1 ч при комнатной температуре (КТ).
[193] Оставшиеся супернатанты утилизировали и добавляли 20 мкл HRP-меченого антитела против стрептавидиновой метки в преопределенной оптимальной концентрации в ФСБ-T/БСА и инкубировали в течение 1 ч при КТ. После промывки в каждую лунку добавляли 20 мкл флуорогенного субстрата HRP (QuantaBlu, Thermo) и оставляли реакцию на период от 15 до 60 минут. Интенсивность флуоресценции каждой лунки на планшете считывали, используя флуоресцентное считывающее устройство для микропланшетов (Tecan или Molecular Devices). Аппроксимацию кривых проводили, используя программное обеспечение GraphPad Prism 4. Полученные в результате значения EC50 обобщены в Таблице 1, ниже.
[194] Таблица 1:
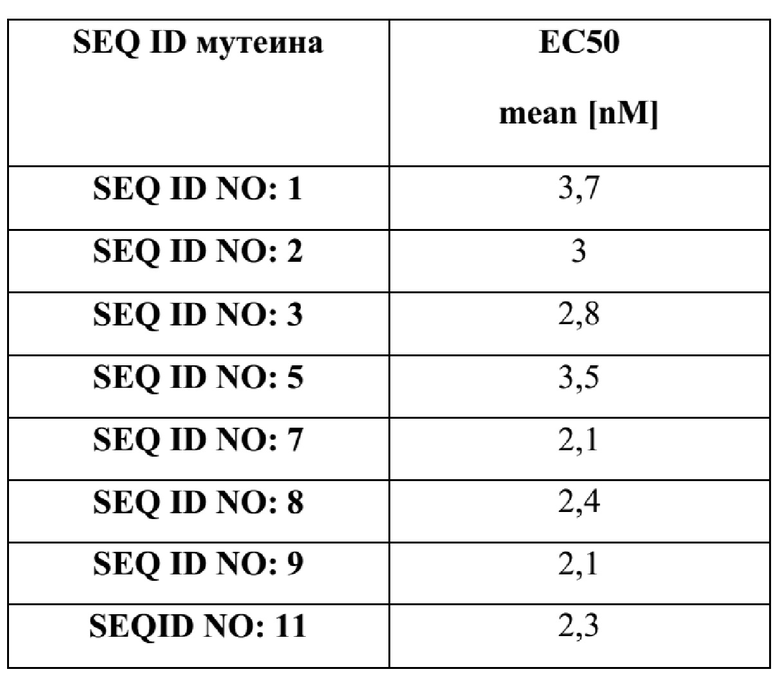
[195] Пример 5: Конкурентный режим действия мутеина липокалина, связывающегося с Ang-2
[196] Связываются ли отобранные мутеины с человеческим Ang-2 в конкурентном режиме, исследовали in vitro, используя конкурентный формат ELISA-анализа (смотрите типовые мутеины липокалина на Фиг. 1). В этом эксперименте неизменную концентрацию человеческого Ang-2 инкубировали с разными концентрациями мутеинов липокалина в течение 1 ч. После этой предварительной инкубации в растворе аликвоту смеси мутеина липокалина/Ang-2 переносили в планшет ELISA, покрытый рецептором человеческой Tie-2, чтобы определить концентрацию hAng-2, который не был заблокирован для связывания с hTie-2.
[197] Все этапы инкубации проводили со смешиванием при 300 об/мин, а планшет промывали после каждого этапа инкубации с помощью 80 мкл буфера ФСБ-T (ФСБ, 0,05 % Твин 20) пять раз, используя селективный промывочный буфер Biotek EL405 CW (Biotek). На первом этапе 384-луночный буфер прямым образом покрывали 20 мкл растворимого человеческого hTie-2 Fс (Reliatech) в концентрации 2 мкг/мл в ФСБ в течение ночи при 4 °C. После промывки лунки, покрытые hTie-2 Fс, блокировали с помощью 60 мкл ФСБ-T/БСА (2% БСА в ФСБ, содержащем 0,1% Твин 20) в течение 1 ч при комнатной температуре.
[198] Фиксированную концентрацию 0,5 нM человеческого Ang-2 инкубировали в растворе с разными концентрациями SEQ ID NO: 1-14 или с SEQ ID NO: 16 в качестве отрицательного контроля, и с эталонными антителами 1 и 2 (SEQ ID NO: 17/18 и 19/20 соответственно) в качестве положительных контролей, используя стабильную стартовую концентрацию, которая была серийно разведена в соотношении 1:3 до пикомолярного диапазона в буфере ФСБ-T/БСА. Через 1 ч инкубации при комнатной температуре 20 мкл реакционной смеси переносили в покрытый hTie-2 Fc планшет для захвата несвязанного (свободного) или неконкурентно связанного hAng-2 в течение 20 мин при КТ. Чтобы обеспечить пересчет результатов ELISA в абсолютную концентрацию свободного Ang-2 (ср. ниже), получали стандартную кривую, включающую разные концентрации hAng-2 в ФСБ-T/БСА, и инкубировали в течение 20 мин в том же планшете.
[199] Чтобы сделать возможным выявление и количественную оценку связанного hAng-2, удаляли оставшиеся супернатанты и добавляли 20 мкл HRP-меченого антитела против HIS-метки в концентрации в ФСБ-T/БСА и инкубировали в течение 1 ч при КТ. После промывки в каждую лунку добавляли 20 мкл quanta blue, а интенсивность флуоресценции каждой лунки считывали, используя флуоресцентное считывающее устройство для микропланшетов (Tecan или Molecular Devices).
[200] Оценку проводили следующим образом: рассчитывали концентрацию свободного hAng-2 к(Ang-2)своб. (по параллельно определяемой стандартной кривой) и наносили на график в сравнении с концентрацией SEQ ID NO: 1-14, 16 и эталонных антител 1 и 2 (SEQ ID NO: 17/18 и 19/20 соответственно), к(SEQ ID NO: 1-14, 16, 17/18 и 19/20). Чтобы получить концентрацию SEQ ID NO: 1-14, 16, 17/18 и 19/20, при которой образование комплекса hAng-2/hTie-2 Fc блокируется на 50% (IC50), кривые аппроксимировали нелинейной регрессионной моделью со связыванием в одном участке в соответствии с к(Ang-2)своб. =c(Ang-2)общ./(1+к(SEQ ID NO: 1-14, 16, 17/18 и 19/20)/1050)), с общей следовой концентрацией к(Ang-2)общ. и значением IC50 в качестве свободных параметров (как показано на Фиг. 1). Аппроксимацию кривых проводили, используя программное обеспечение GraphPad Prism 4. Полученные в результате значения IC50 обобщены в Таблице 2, ниже.
[201] Таблица 2:
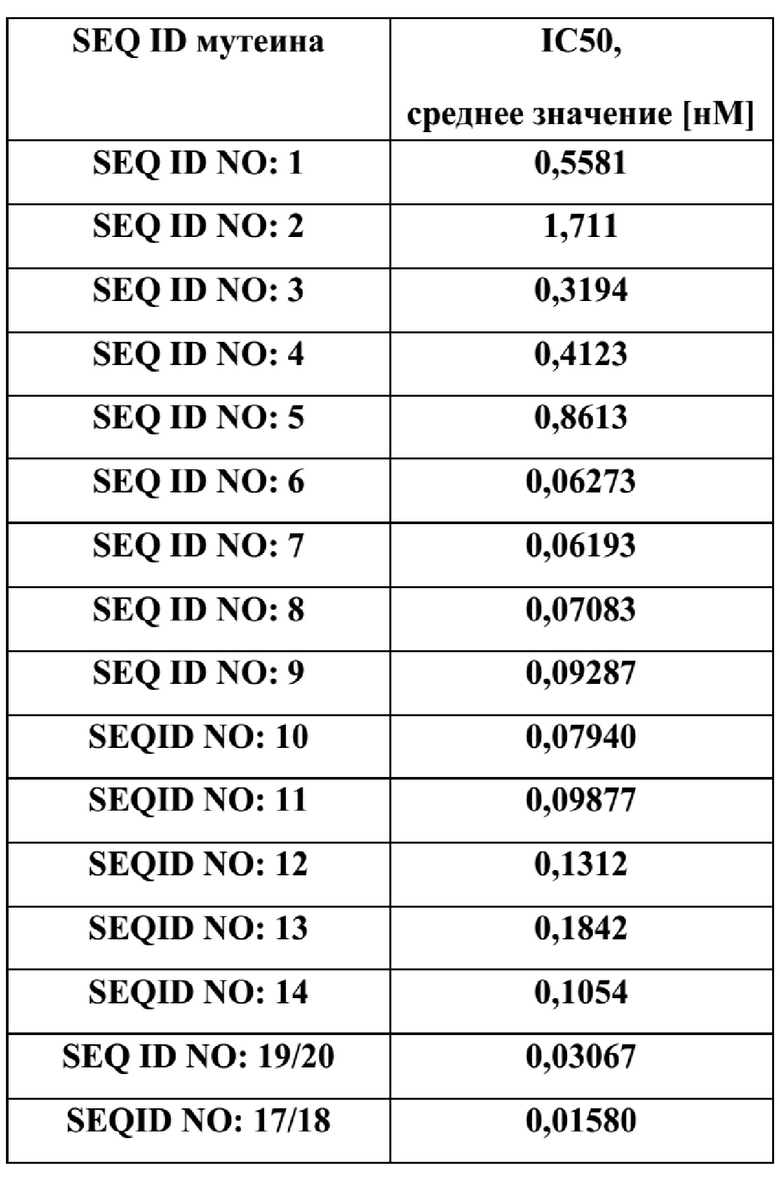
[202] Пример 6: Аффинность мутеинов, связывающихся с Ang-2, определенная с помощью Biacore
[203] В анализе методом поверхностного плазмонного резонанса (ППР) использовали прибор Biacore T200 (GE Healthcare) для измерения аффинности связывания мутеинов с hAng-2. Проводили биотинилирование мутеинов, отобранных для связывания с Ang-2, и отрицательного контроля (SEQ ID NO: 16) в течение 2 ч при комнатной температуре, применяя соответствующий избыток EZ-Link NHS-PEG4-Biotin (Thermo). Биотинилированные образцы очищали от непрореагировавшего биотина, используя Zeba Spin Desalting Plate (Thermo) в соответствии с инструкциями производителя.
[204] В ППР-анализе аффинности проводили захват биотинилированных мутеинов и отрицательного контроля на сенсорном чипе CAP, используя набор Biotin CAPture (GE Healthcare): сенсорный чип CAP предварительно иммобилизуют олиго оцДНК. Неразведенный реагент Biotin CAPture (стрептавидин, конъюгированный с комплементарным олиго оцДНК) применяли при скорости потока 2 мкл/мин в течение 300 с. После этого применяли 0,02 мкг/мл биотинилированных мутеинов или отрицательного контроля в течение 300 с при скорости потока 5 мкл/мин. Так как аналит Ang-2 является мультимерной молекулой, цель состояла в создании поверхности с минимальной плотностью лигандов для снижения бивалентных взаимодействий на поверхности чипа до минимума. Стандартный канал загружали только реагентом Biotin CAPture.
[205] Чтобы определить аффинность связывания, готовили четыре разведения человеческого Ang-2 с концентрациями в диапазоне 1-27 нM в буфере HBS-EP+ (GE Healthcare) и применяли к приготовленной поверхности чипа. При применении при скорости потока 30 мкл/мин использовали многоцикловый кинетический подход со временем контакта образца 180 с и временем диссоциации 4600 с. Все измерения проводили при 25°C. Восстановление поверхности сенсорного чипа CAP осуществляли с помощью инъекции 6 M Gua-HCl с 0,25 M NaOH с последующей дополнительной промывкой экспериментальным буфером и периодом стабилизации 120 с. Оценку данных проводили с помощью программного обеспечения Biacore T200 (V 1.0). Использовали двойное сравнение со стандартом. Для аппроксимации исходных данных использовали 1:1 модель связывания, даже если было известно, что аналит является мультимером.
[206] Полученные в результате кинетические константы для выборки мутеинов липокалина обобщены ниже в Таблице 3. Такие типовые мутеины липокалина (SEQ ID NO: 7, 8, 9 и 11) связывают человеческий Ang-2 с аффинностью в диапазоне 0,2-1,4 нM, тогда как для отрицательного контроля связывание не определяется (SEQ ID NO: 16). Следует отметить, что аппроксимированные величины были получены на основании предположения, что применение очень низкой плотности лигандов со значениями Rмакс ниже десяти снижает мультивалентные взаимодействия до пренебрежимо малого уровня.
[207] Таблица 3:
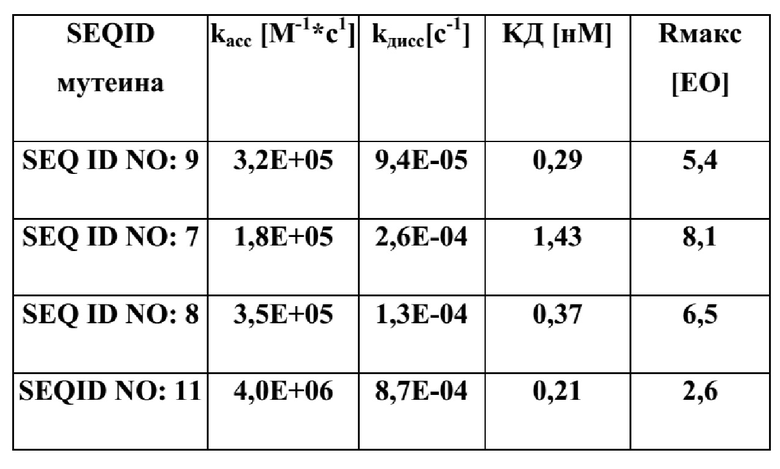
[208] Пример 7: Специфичность и межвидовая перекрестная реактивность мутеина липокалина в отношении Ang-1 и Ang-2
[209] Специфичность и межвидовую перекрестную реактивность мутеинов липокалина (SEQ ID NO: 1, 3, 7, 8, 9, 11) и эталонного антитела (SEQ ID NO: 17/18) исследовали методом «конкурентного ELISA-анализа в растворе» (смотрите типовые мутеины липокалина, Фиг. 2), принцип которого в следующем: постоянную концентрацию SEQ ID NO: 1, 3, 7, 8, 9, 11 и эталонного антитела (SEQ ID NO: 17/18) инкубировали с разными концентрациями лигандов (человеческий Ang-2, человеческий Ang-1, человеческий Ang-4, мышиный Ang2 и мышиный Ang 3, а также hVEGF-A в качестве отрицательного контроля (R&D system)) в течение 1 ч. После этой предварительной инкубации в растворе аликвоту смеси мутеин/лиганд переносили в планшет ELISA с иммобилизованным hAng-2 для определения концентрации оставшихся свободных мутеинов липокалина SEQ ID NO: 1,3, 7, 8, 9, 11 и свободного эталонного антитела. Концентрацию свободных (несвязанных) SEQ ID NO: 1, 3, 7, 8, 9, 11 и свободного эталонного антитела определяли с помощью установки для количественного ELISA-анализа. Следует обратить внимание на то, что этот анализ был основан на том, что все лиганды были нацелены на тот же самый участок связывания в SEQ ID NO: 1, 3, 7, 8, 9, 11 и 17/18, т.е. что лиганды, связанные с SEQ ID NO: 1,3,7,8,9,11 и 17/18, конкурируют друг с другом.
[210] В следующем подробном экспериментальном протоколе этапы инкубации и промывки проводили, как описано выше, согласно протоколу конкурентного ELISA-анализа (смотрите Пример 5). 384-луночный планшет, подходящий для измерения флуоресценции (Greiner FLUOTRAC™ 600, черное плоское дно, высокая степень связывания) покрывали 20 мкл человеческого ангиопоэтина 2 в концентрации 5 мкг/мл в ФСБ в течение ночи при 4 °C. После промывки покрытые Ang-2 лунки блокировали 100 мкл блокирующего буфера (ФСБ-T/БСА) в течение 1 ч при комнатной температуре.
[211] Фиксированную концентрацию 0,1 нM SEQ ID NO: 1, 3, 7, 8, 9, 11 и 17/18 инкубировали в растворе с разными концентрациями лигандов (hAng-2, mAng-2, hAn1, mAng-3 и hAng-4, и VEGF-A), используя подходящую стартовую концентрацию, которую серийно разводили в соотношении 1:3 до пикомолярного диапазона в ФСБ-T/БСА. Через 1 ч инкубации при комнатной температуре 20 мкл реакционной смеси переносили в 384-планшет с иммобилизованным hAng-2 для захвата несвязанных компонентов (свободных SEQ ID NO: 1, 3, 7, 8, 9, 11 и 17/18) в течение 20 мин при КТ. Чтобы обеспечить пересчет результатов ELISA в абсолютную концентрацию свободных SEQ ID NO: 1, 3, 7, 8, 9, 11 и 17/18 (ср. ниже), получали стандартную кривую, включающую разные концентрации SEQ ID NO: 1, 3, 7, 8, 9, 11 и 17/18 в ФСБ- T/БСА, и инкубировали в течение 20 мин в том же планшете ELISA.
[212] Удаляли остаточные супернатанты и добавляли 20 мкл HRP-меченого антитела к мутеину липокалина в предопределенной оптимальной концентрации в ФСБ-T/БСА и инкубировали в течение 1 ч при КT. Антитело против мутеина липокалина получали путем иммунизации кроликов смесью мутеинов липокалина и после этого связывали с HRP, используя набор (EZ-link Plus Activated Peroxidase, Thermo Scientific) в соответствии с инструкциями производителя, для получения конъюгата антитело-HRP. После промывки в каждую лунку добавляли 20 мкл флуорогенного субстрата HRP (Quantablue, Pierce) и оставляли реакцию на 60 минут. Интенсивность флуоресценции каждой лунки на планшете считывали, используя считывающее устройство для микропланшетов Genios Plus (Tecan). Чтобы провести оценку данных рассчитывали концентрацию свободных мутеинов к(мутеин)своб. на основании результатов стандартной кривой и наносили на график против концентрации лигандов, к(лиганд). Чтобы получить концентрацию лигандов, при которой образование комплекса hAng-2/мутеин блокируется на 50% (IC50), кривые аппроксимировали нелинейной регрессионной моделью со связыванием в одном участке в соответствии с к(мутеин)своб. = к(мутеин)общ./(1+к(лиганд)/IC50)), с общей следовой концентрацией к(мутеин)своб. и значениями IC50 в качестве свободных параметров. Аппроксимацию кривой проводили, используя программное обеспечение GraphPad Prism 4. Указанная аппроксимация кривой дала результаты, которые обобщены в нижеследующей Таблице 4.
[213] Таблица 4:
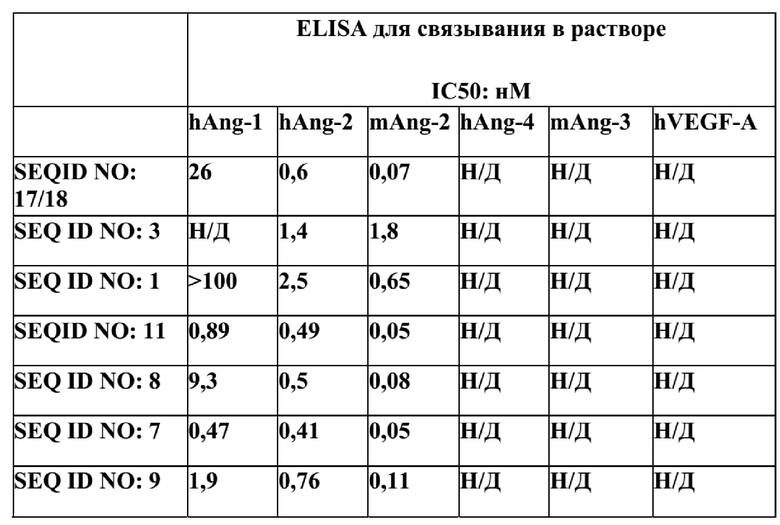
[214] Как проиллюстрировано на Фиг. 2, данные демонстрируют, что SEQ ID NO: 8 проявляла высокую аффинность в отношении человеческого и мышиного Ang-2, а SEQ ID NO: 7 проявляла высокую аффинность не только в отношении человеческого и мышиного Ang-2, но также в отношении человеческого Ang-1.
[215] Пример 8: Мутеины липокалина, блокирующие связывание человеческого и мышиного Ang-2 с экспрессирующими hTie-2 клетками
[216] Связываются ли мутеины липокалина с человеческим Ang-2 и мышиным Ang-2 конкурентным образом, исследовали на сверхэкспрессирующих hTie-2 клетках HEK, используя формат анализа конкурентной клеточной электрохемолюминесценции (ЭХЛ) (смотрите Фиг. 3 и 4). В этом эксперименте постоянную концентрацию человеческого Ang-2 и мышиного Ang-2 инкубировали с разными концентрациями мутеинов липокалина (SEQ ID NO: 1, 3, 6-14) и эталонным антителом (SEQ ID NO: 19/20) в течение 1 ч. После этой предварительной инкубации в растворе аликвоту смеси мутеина липокалина/Ang-2 переносили в планшет MSD, покрытый сверхэкспрессирующими hTie-2 клетками HEK, для определения концентрации hAng-2 или mAng-2, связывание которых с hTie-2 не было заблокировано.
[217] Все этапы инкубации проводили при комнатной температуре и два раза промывали планшеты после каждого этапа инкубации 80 мкл буфера ФСБ, используя промывочный буфер Biotek EL405 select CW (Biotek). На первом этапе 384-луночный планшет предварительно покрывали в течение 5 минут поли-D-лизином и дважды промывали ФСБ. Высевали 104 клеток HEK:hTie-2 на лунку и оставляли для прикрепления к поверхности лунок на ночь при 37°C. После промывки покрытые клетками лунки блокировали 60 мкл ФСБ/казеина (2% казеин в ФСБ) в течение 1 ч при комнатной температуре.
[218] Фиксированную концентрацию человеческого Ang-2 или мышиного Ang-2 инкубировали в растворе с разными концентрациями SEQ ID NO: 1, 3, 6-14 и эталонным антителом (SEQ ID NO: 19/20), используя подходящую стартовую концентрацию SEQ ID NO: 1,3, 6-14 и эталонного антитела, которые серийно разводили в соотношении 1:3 до пикомолярного диапазона в буфере ФСБ/казеин. Через 1 ч инкубации при комнатной температуре 20 мкл реакционной смеси переносили в покрытый HEK:hTie2 планшет для захвата не связанного конкурентно hAng-2 в течение 1 ч при КТ. Получали стандартную кривую, включающую разные концентрации hAng-2 или мышиного Ang-2 в ФСБ/казеине, и инкубировали в течение 1 ч в том же планшете.
[219] Чтобы обеспечить выявление и количественную оценку связанного hAng-2 и мышиного Ang-2, удаляли оставшиеся супернатанты и добавляли 20 мкл смеси антитела против HIS-метки (Abeam) и меченого сульфо-меткой антикозьего антитела (Mesoscale Discovery) в концентрации 1 мкг/мл в ФСБ/казеине и инкубировали в течение 1 ч при КТ. После промывки в каждую лунку добавляли 35 мкл не содержащего сурфактанта считывающего буфера, а сигнал ЭХЛ каждой лунки считывали с помощью считывающего устройства Mesoscale Discovery.
[220] Оценку и аппроксимацию кривой проводили, используя программное обеспечение GraphPad Prism 4, а полученные результаты обобщены в Таблице 5, ниже.
[221] Таблица 5:
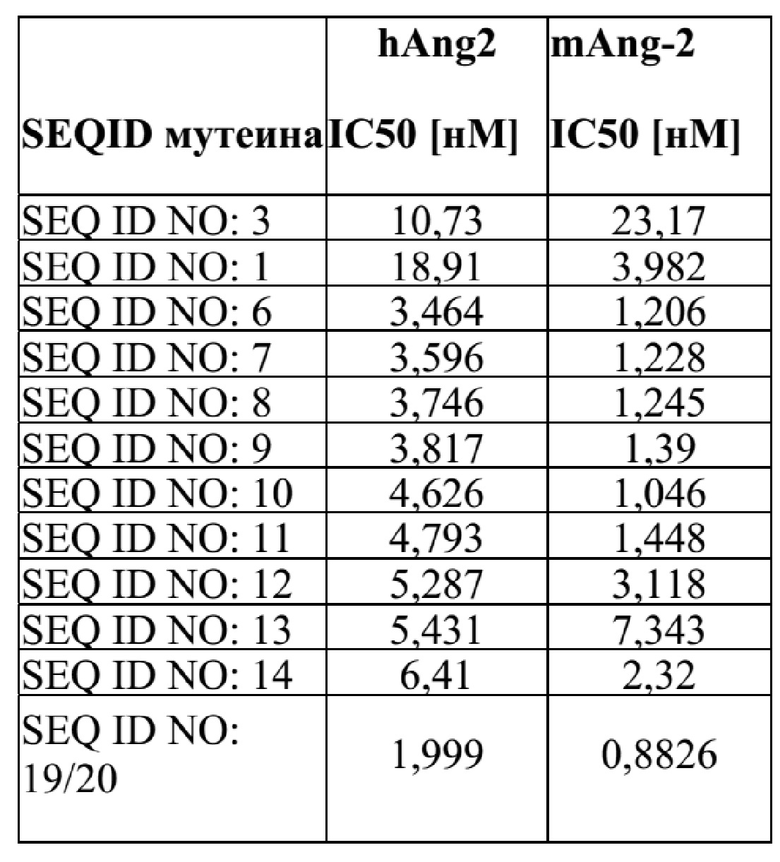
[222] Пример 9: Опосредованная мутеином липокалина блокировка Ang-2 в клеточном анализе пролиферации
[223] Способность мутеинов липокалина SEQ ID NO: 1, 3, 7-9 и 11 нейтрализовать биологическую активность hAng-2 оценивали, применяя краткосрочный биоанализ пролиферации, используя эндотелиальные клетки лимфатических микрососудов (LEC). Пролиферация LEC может ингибироваться агентами, имеющими hAng-2-нейтрализующее действие. В этом анализе SEQ ID NO: 1, 3, 7-9 и 11 добавляли в обедненную сыворотку клеток LEC в культуре. Через три дня в культуре оценивали степень пролиферации, путем количественной оценки числа жизнеспособных клеток. Это осуществляли с помощью анализа жизнеспособности клеток CellTiter-Glo Luminescent (Promega) чтобы определить уровни АТФ, которые коррелируют с числом метаболически активных клеток. Способность SEQ ID NO: 1, 3, 7-9 и 11 нейтрализовать hAng-2 оценивали по их значению IC50, т.е. концентрации мутеинов липокалина, которая приводит к полумаксимальному ингибированию опосредованной hAng-2 пролиферации.
[224] Подробная процедура проведения анализа описывается следующим образом. LEC поддерживали в EBM, 5% фетальной телячьей сыворотки и дополняющем наборе MV2.
Анализ проводили в 96-луночных белых планшетах с прозрачным дном (Greiner) в объеме 25 мкл на лунку.
[225] Клетки LEC культивировали в пробирках для клеточного культивирования в стандартных условиях в соответствии с инструкциями производителя (PAA Laboratories), 37°C, атмосфера 5% CO2).
[226] В 1 день эксперимента адгезивные клетки отделяли от их субстрата с помощью трипсина/ЭДТА в соответствии с инструкциями производителя. После этого клетки центрифугировали в течение 5 минут при 1000 об/мил, ресуспендировали в EBM и фильтровали через 100 мкм клеточный фильтр (Falcon), чтобы удалить клеточные агрегаты. Затем клетки высевали в 96-луночные планшеты для тканевого культивирования с плоским дном (Greiner) при плотности 3200 клеток на лунку, используя конечный объем 100 мкл. Их инкубировали 1 час в стандартных условиях.
[227] Через 1 час в лунки добавляли серийные разведения SEQ ID NO: 1, 3, 7-9, 11 отрицательных контролей (SEQ ID NO: 16 и человеческого IgG изотипического антитела (Dianova)) и эталонных антител 1 и 2 (SEQ ID NO: 17/18 и 19/20 соответственно). Все титрационные серии проводили при серийном разведении 1:3 в аналитической среде и при подходящей стартовой концентрации. После этого клетки оставляли для пролиферации в течение 72 часов при 37°C. Для проведения количественной оценки клеточной пролиферации через 72 часа к клеткам добавляли по 25 мкл реагентов CellTiter-Glo в каждую лунку и инкубировали в течение 2 минут на орбитальной качалке, чтобы индуцировать лизис клеток, а люминесценцию измеряли, используя считывающее устройство PheraStar FS.
[228] Значение IC50 определяли, используя программное обеспечение GraphPad Prism (GraphPad Software Inc.), путем нанесения на график стандартизированных сигналов против концентрации образца и нелинейной регрессии данных с сигмоидальной моделью доза-ответ.
[229] Результаты эксперимента показаны на Фиг. 5. Раскрытый выше анализ пролиферации проведен по трем независимым экспериментам. Результаты обобщены в Таблице 6, ниже.
[230] Таблица 6:
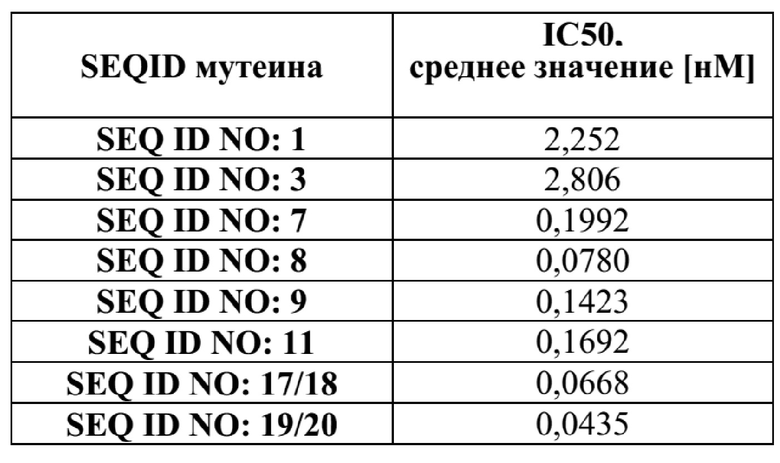
[231] SEQ ID NO: 8 демонстрировала среднее значение EC50 0,07 нМ, эталонное антитело 1 (SEQ ID NO: 17/18) демонстрировало значение EC50 0,06 нМ, а эталонное антитело 2 (SEQ ID NO: 19/20) демонстрировало значение EC50 0,04 нМ. Отрицательные контроли не оказывали влияние на пролиферацию. Следовательно, данные демонстрируют, что SEQ ID NO: 8 и эталонные антитела демонстрировали сравнимую эффективность в этом функциональном анализе.
[232] Пример 10: Оценка стабильности мутеинов
[233] Полученные в результате температуры плавления (Tm), а также начало плавления мутеинов липокалина (SEQ ID NO: 1,3, 7-9, 11) приведены в Таблице 7, ниже, по сравнению с эталонным антителом (SEQ ID NO: 17/18). Отобранные мутеины липокалина имели Tm в диапазоне от 65 до 77°C, что свидетельствует о хорошей стабильности молекул.
[234] Таблица 7: Tm и начало плавления согласно данным nanoDSC для Ang-2-специфических мутеинов липокалина и эталонного антитела
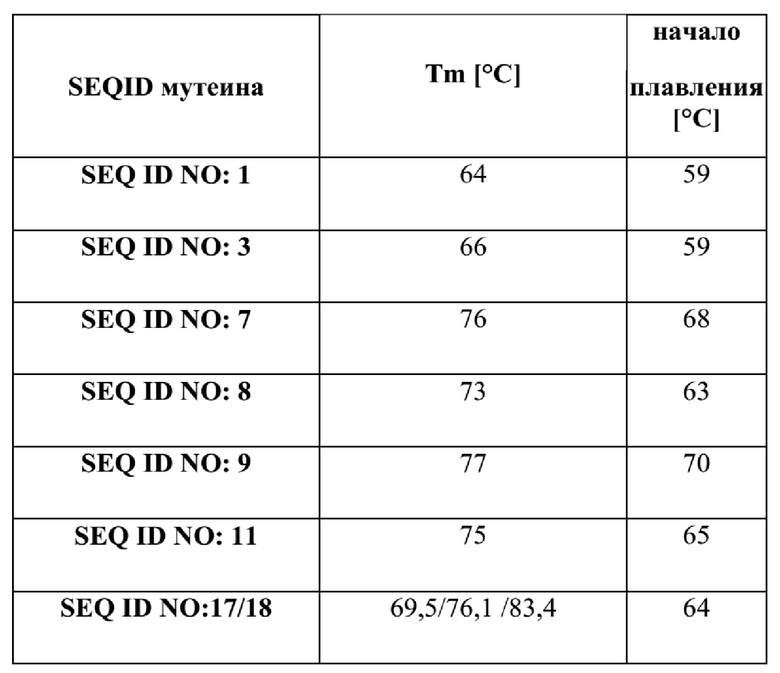
[235] Чтобы оценить стабильность при хранении, типовые мутеины инкубировали в пробирках в концентрации 1 мг/мл в течение 1 недели при 37°C. Определение активных мутеинов проводили методом количественного ELISA-анализа. Определение мономерного белка проводили методом аналитической эксклюзионной хроматографии. Полученные в результате данные для SEQ ID NO: 1, 3, 7-9 и 11 приведены в Таблице 8, ниже.
[236] Для исследования белковой активности применяли следующий ELISA-анализ: 384-луночный планшет, подходящий для измерения флуоресценции (Greiner FLUOTRACТМ 600, черное плоское дно, высокая степень связывания) покрывали 20 мкл Ang-2 (Thermo Scientific) в концентрации 5 мкг/мл в ФСБ в течение ночи при 4 °C. После промывки покрытые Ang-2 лунки блокировали 100 мкл блокирующего буфера (2% масс./об. БСА в ФСБ, содержащего 0,1% об./об. Твин-20) в течение 1 ч. Планшет промывали и переносили 20 мкл соответствующим образом разведенного белкового стандарта, не подвергавшегося стрессовым условиям стандартного образца или подвергавшегося стрессовым условиям образца в планшет ELISA и инкубировали. Чтобы провести количественную оценку связанного с планшетом белка, планшет ELISA промывали, удаляли оставшиеся супернатанты и добавляли 20 мкл HRP-меченого антитела к hNGAL в предопределенной оптимальной концентрации в блокирующем буфере и инкубировали. После промывки в каждую лунку добавляли 20 мкл флуорогенного субстрата HRP (QuantaBlu, Pierce) и оставляли реакцию на 20-30 минут. Интенсивность флуоресценции каждой лунки считывали, используя флуоресцентное считывающее устройство для микропланшетов (Tecan).
[237] Если не указано иное, все этапы инкубации проводили в течение 1 ч при комнатной температуре, а после каждого этапа инкубации планшет пять раз промывали 100 мкл буфера ФСБ-T (ФСБ, 0,05 % Твин 20), используя промывочный буфер Biotek ELx405 select CW.
[238] Для вышеописанного ELISA-анализа получали калибровочную кривую, включающую 11 разведений, как правило, в диапазоне 0,008-500 нг/мл, и для каждого образца готовили три разных, независимых разведения в линейном диапазоне калибровочной кривой. Для разведений использовали блокирующий буфер, необязательно дополненный 1% человеческой или мышиной плазмой.
[239] Аппроксимацию калибровочной кривой проводили, используя 4-параметрическую логистическую (4PL) нелинейно-регрессионную модель, и использовали для расчета концентрации активного белка для исследуемых образцов. Определенные концентрации активного белка сравнивали с не подвергавшимся стрессовым условиям образцом в той же концентрации и в той же матрице.
[240] Аналитическую эксклюзионную хроматографию проводили на ВЭЖХ-системе Agilent с двумя колонками Superdex 75 5/150 GL (GE Healthcare) в линии, используя ФСБ (Gibco) в качестве элюента при скорости потока 0,3 мл/мин.
[241] Чтобы оценить стабильность при хранении в плазме, типовые мутеины инкубировали в пробирках в концентрации 0,5 мг/мл в течение 1 недели при 37°C в человеческой и мышиной плазме. Определение активных мутеинов проводили методом количественного ELISA-анализа, как описано выше.
[242] Все исследуемые мутеины липокалина оказались стабильными во всех условиях исследования, а результаты исследований обобщены в Таблице 8, ниже.
[243] Таблица 8: Стабильность после 3 циклов заморозка/разморозка (З/Р); хранение в течение 1 недели в ФСБ при 37°C и хранение в течение 1 недели в человеческой (чел.) или мышиной (мыш.) плазме оценивали по восстановлению активности в анализе qELISA и мономерному содержанию в ЭХ: стабильны в случае qELISA = 100 +/- 15%; стабильны в случает аЭХ = 100 +/- 5% (восстановление площади пика мономеров по сравнению с не подвергавшимся стрессовым условиям образцом); для всех образцов, включая стандарты, было обнаружено содержание мономеров на 100 процентах площади.
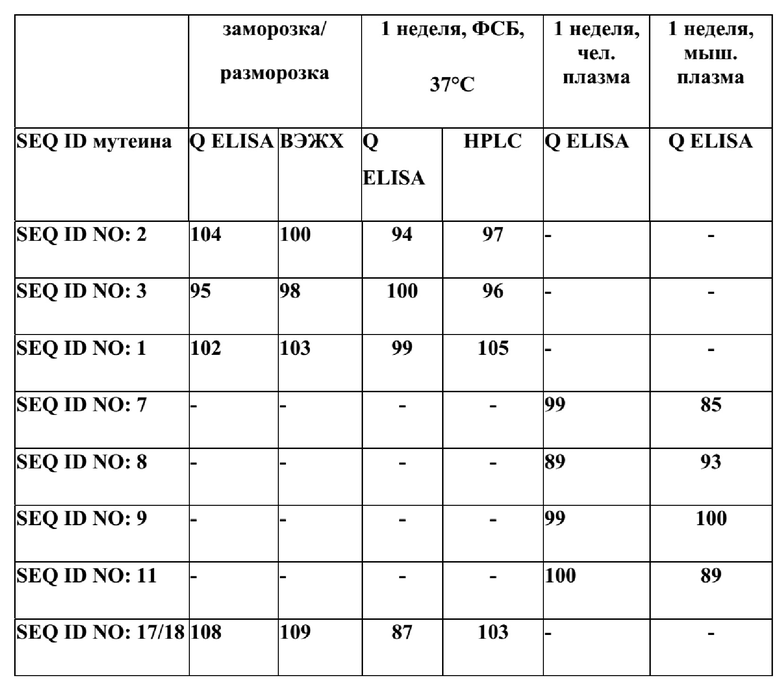
[244] Варианты реализации, иллюстративно описанные в данном документе, легко можно реализовать на практике в отсутствие любого элемента или элементов, ограничения или ограничений, специальным образом не раскрытых в данном документе. Таким образом, например, термины «содержащий», «включающий» и т.д. следует воспринимать в расширенном смысле и без ограничений. Кроме того, употребляемые в данном документе термины и выражения использовались как термины для описания, но не ограничения, а использование этих терминов и выражений не предусматривает исключение любых эквивалентов приведенных и описанных признаков или их частей, но предполагается, что возможны различные модификации, входящие в объем заявляемого изобретения. Таким образом, следует понимать, что хотя настоящие варианты реализации были специальным образом раскрыты с помощью предпочтительных вариантов реализации и необязательных признаков, специалисты в данной области техники могут прибегнуть к их модификациям и вариациям, а такие модификации и вариации считаются входящими в объем этого изобретения. Все описанные в данном документе патенты, патентные заявки, руководства и рецензируемые публикации в полном объеме включены в данный документ посредством ссылки. Кроме того, если определение или применение термина в ссылочном материале не согласуется или противоречит определению этого термина, приведенному в данном документе, приоритет имеет определение этого термина, приведенное в данном документе, а определение этого термина в ссылочном материале не имеет приоритета. Каждый из более узко определенных компонентов и субгенерических групп, попадающих в общее описание, также составляет часть этого изобретения. Это включает общее описание изобретения при условии или отрицательном ограничении, подразумевающем удаление из перечня любого предмета, вне зависимости от того, приведен ли исключаемый материал специальным образом в данном документе. Кроме того, если признаки описаны в терминах групп Маркуша, специалистам в данной области техники понятно, что изобретение также описано в терминах любого отдельного представителя или подгруппы представителей группы Маркуша. Дополнительные варианты реализации станут понятны из нижеприведенной формулы изобретения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Pieris Pharmaceuticals GmbH
<120> Новые белки, специфические в отношении ангиогенеза
<130> PIE15265PCT
<150> EP15152826.2
<151> 2015-01-28
<160> 34
<170> PatentIn версия 3.5
<210> 1
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин hNGAL
<400> 1
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Tyr Ile Leu Arg Glu Asp Lys Asp Pro
35 40 45
Gly Lys Met Trp Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asn Val Thr Gly Val Ser Phe Gly Pro Lys Lys Cys Asn Tyr Thr Ile
65 70 75 80
Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gly
85 90 95
Ile Lys Ser Pro Pro Gly Gly Thr Ser Thr Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys His Val Leu Gln
115 120 125
Asn Arg Glu Phe Phe Glu Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 2
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин hNGAL
<400> 2
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Phe Ala Gly Asn His Arg Leu Arg Glu Asp Lys Asp Pro
35 40 45
Gly Lys Met His Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asn Val Thr Asp Val Thr Phe Ala Phe Lys Lys Cys Asn Tyr Arg Ile
65 70 75 80
His Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Asn
85 90 95
Ile Lys Ser Trp Pro Gly Glu Thr Ser Thr Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Thr Val Met Gln
115 120 125
Asn Arg Glu Trp Phe Trp Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 3
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин hNGAL
<400> 3
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Val Ala Gly Asn Trp Tyr Leu Arg Glu Asp Lys Asp Pro
35 40 45
Ile Lys Met Thr Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asn Val Thr Gln Val Gly Phe Glu Gln Lys Lys Cys Lys Tyr Ser Ile
65 70 75 80
His Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Asp
85 90 95
Ile Lys Ser Trp Pro Gly Asp Thr Ser Leu Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Gly Val Met Gln
115 120 125
Asn Arg Glu Val Phe Ala Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 4
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин hNGAL
<400> 4
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Glu Ala Gly Asn Val Glu Leu Arg Glu Asp Lys Asp Pro
35 40 45
Val Lys Met Thr Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asn Val Thr Glu Val Arg Phe Trp Leu Lys Lys Cys Lys Tyr Asn Ile
65 70 75 80
His Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Ala
85 90 95
Ile Lys Ser Gly Pro Gly Met Thr Ser Thr Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Thr Val Met Gln
115 120 125
Asn Arg Glu Trp Phe Trp Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 5
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин hNGAL
<400> 5
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Tyr Trp Leu Arg Glu Asp Lys Asp Pro
35 40 45
Ile Lys Met Ser Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asn Val Thr Ile Val Tyr Phe Thr Arg Lys Lys Cys Ser Tyr Arg Ile
65 70 75 80
Tyr Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Pro
85 90 95
Ile Lys Ser Tyr Pro Gly Asp Thr Ser Thr Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys His Val Tyr Gln
115 120 125
Asn Arg Glu Trp Phe Glu Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 6
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин hNGAL
<400> 6
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Tyr Ile Leu Arg Glu Asp Lys Asp Pro
35 40 45
Glu Lys Met Trp Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Gly Val Ser Phe Gly Pro Lys Lys Cys Asn Tyr Arg Ile
65 70 75 80
Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gly
85 90 95
Ile Lys Ser Ser Pro Gly Gln Thr Ser Thr Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys His Val Leu Gln
115 120 125
Asn Arg Glu Phe Phe Glu Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 7
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин hNGAL
<400> 7
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly His Ala Gly Asn Tyr Ile Leu Arg Glu Asp Lys Asp Pro
35 40 45
Glu Lys Met Trp Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Glu Val Ser Phe Gly Pro Lys Lys Cys Asn Tyr Arg Ile
65 70 75 80
Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gly
85 90 95
Ile Lys Ser Pro Pro Gly Asp Thr Ser Thr Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys His Val Leu Gln
115 120 125
Asn Arg Glu Phe Phe Glu Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 8
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин hNGAL
<400> 8
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Tyr Ile Leu Arg Glu Asp Lys Asp Pro
35 40 45
Gly Lys Met Trp Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Glu Val Ser Phe Gly Ala Lys Lys Cys Asn Tyr Arg Ile
65 70 75 80
Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gly
85 90 95
Ile Lys Ser Asp Pro Gly Gly Thr Ser Thr Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys His Val Leu Gln
115 120 125
Asn Arg Glu Phe Phe Glu Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 9
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин hNGAL
<400> 9
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly His Ala Gly Asn Tyr Ile Leu Arg Glu Asp Lys Asp Pro
35 40 45
Gly Lys Met Trp Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Glu Val Ser Phe Gly Pro Lys Lys Cys Asn Tyr Arg Ile
65 70 75 80
Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gly
85 90 95
Ile Lys Ser Pro Pro Gly Gly Thr Ser Thr Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys His Val Leu Gln
115 120 125
Asn Arg Glu Phe Phe Glu Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 10
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин hNGAL
<400> 10
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Tyr Ile Leu Arg Glu Asp Lys Asp Pro
35 40 45
Gln Lys Met Trp Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Gly Val Ser Phe Gly Ala Lys Lys Cys Val Tyr Arg Ile
65 70 75 80
Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gly
85 90 95
Ile Lys Ser Pro Pro Gly Gly Thr Ser Thr Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys His Val Leu Gln
115 120 125
Asn Arg Glu Phe Phe Glu Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 11
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин hNGAL
<400> 11
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Tyr Ile Leu Arg Glu Asp Lys Asp Pro
35 40 45
Val Lys Met Trp Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Glu Val Ser Phe Gly Pro Lys Lys Cys Asn Tyr Arg Ile
65 70 75 80
Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gly
85 90 95
Ile Lys Ser Leu Pro Gly Gly Thr Ser Thr Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys His Val Leu Gln
115 120 125
Asn Arg Glu Phe Phe Glu Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 12
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин hNGAL
<400> 12
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Val Ala Gly Asn Trp Tyr Leu Arg Glu Asp Lys Asp Pro
35 40 45
Ile Lys Met Thr Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Gln Val Gly Phe Glu Gln Glu Lys Cys Lys Tyr Ser Ile
65 70 75 80
His Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Asn
85 90 95
Ile Lys Ser Trp Pro Gly Asp Thr Ser Pro Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asp Gln His Ala Met Val Phe Phe Lys Gly Val Met Gln
115 120 125
Asp Arg Glu Val Phe Ala Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 13
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин hNGAL
<400> 13
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Val Ala Gly Asn Trp Tyr Leu Arg Glu Asp Lys Asp Pro
35 40 45
Ile Lys Met Thr Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Gln Val Gly Phe Glu Gln Glu Lys Cys Lys Tyr Ser Ile
65 70 75 80
His Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Asp
85 90 95
Ile Lys Ser Trp Pro Gly Asp Thr Ser Pro Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Gly Met Met Gln
115 120 125
Asp Arg Glu Val Phe Ala Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 14
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутеин hNGAL
<400> 14
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Met Ala Gly Asn Trp Asp Leu Arg Glu Asp Lys Asp Pro
35 40 45
Ile Lys Met Thr Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Gln Val Ala Phe Glu Gln Lys Lys Cys Lys Tyr Ser Ile
65 70 75 80
His Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln
85 90 95
Ile Lys Ser Trp Pro Gly Asp Thr Ser Pro Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Gly Val Met Gln
115 120 125
Asn Arg Glu Val Phe Ala Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 15
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Стреп-метка
<400> 15
Ser Ala Trp Ser His Pro Gln Phe Glu Lys
1 5 10
<210> 16
<211> 178
<212> Белок
<213> Искусственная последовательность
<220>
<223> hNGAL дикого типа
<400> 16
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Leu Ala Gly Asn Ala Ile Leu Arg Glu Asp Lys Asp Pro
35 40 45
Gln Lys Met Tyr Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asn Val Thr Ser Val Leu Phe Arg Lys Lys Lys Cys Asp Tyr Trp Ile
65 70 75 80
Arg Thr Phe Val Pro Gly Cys Gln Pro Gly Glu Phe Thr Leu Gly Asn
85 90 95
Ile Lys Ser Tyr Pro Gly Leu Thr Ser Tyr Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Lys Val Ser Gln
115 120 125
Asn Arg Glu Tyr Phe Lys Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 17
<211> 459
<212> Белок
<213> Искусственная последовательность
<220>
<223> тяжелая цепь эталонного антитела 1
<400> 17
Gln Val Gln Leu Val Glu Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gly Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Pro Asn Ser Gly Gly Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Pro Asn Pro Tyr Tyr Tyr Asp Ser Ser Gly Tyr Tyr Tyr
100 105 110
Pro Gly Ala Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser
115 120 125
Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser
130 135 140
Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp
145 150 155 160
Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr
165 170 175
Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr
180 185 190
Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln
195 200 205
Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp
210 215 220
Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro
225 230 235 240
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
245 250 255
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
260 265 270
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
275 280 285
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
290 295 300
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
305 310 315 320
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
325 330 335
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
340 345 350
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
355 360 365
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
370 375 380
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
385 390 395 400
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
405 410 415
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
420 425 430
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
435 440 445
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
450 455
<210> 18
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> легкая цепь эталонного антитела 1
<400> 18
Gln Pro Gly Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Val Tyr
35 40 45
Asp Asp Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ser Asp His
85 90 95
Tyr Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu Gly Gln Pro Lys
100 105 110
Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln
115 120 125
Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly
130 135 140
Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly
145 150 155 160
Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala
165 170 175
Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser
180 185 190
Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val
195 200 205
Ala Pro Thr Glu Cys Ser
210
<210> 19
<211> 452
<212> Белок
<213> Искусственная последовательность
<220>
<223> тяжелая цепь эталонного антитела 2
<400> 19
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Ile His Trp Val Arg Gln Ala Thr Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Gly Pro Ala Gly Asp Thr Tyr Tyr Pro Gly Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Glu Asn Ala Lys Asn Ser Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Gly Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gly Leu Ile Thr Phe Gly Gly Leu Ile Ala Pro Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 20
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> легкая цепь эталонного антитела 2
<400> 20
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Thr
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln His Tyr Asp Asn Ser Gln
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 21
<211> 536
<212> ДНК
<213> Искусственная последовательность
<220>
<223> мутеин hNGAL
<400> 21
caggactcca cctcagacct gatcccagcc ccacctctga gcaaggtccc tctgcagcag 60
aacttccagg acaaccaatt ccatgggaaa tggtatgtcg tgggccaagc cggaaattac 120
atcctgcgtg aggataagga tccgggaaaa atgtgggcga ccatttacga gttgaaagaa 180
gataaatcat ataacgtcac cggagtgagc tttggaccta agaaatgcaa ttacaccatt 240
tggacctttg tgccggggag ccagccgggc gagtttactt taggcggaat taaaagtcct 300
ccgggcggaa catcaacctt ggtccgcgtc gtgagcacca actacaacca gcatgccatg 360
gtgttcttca agcacgtgct gcagaaccgc gagttctttg agatcacact gtacgggcgc 420
acgaaagaac tgacaagcga gctgaaggaa aattttatcc gcttttccaa atctctgggc 480
ctccctgaaa accacatcgt cttccctgtc ccaatcgacc agtgtatcga cggcag 536
<210> 22
<211> 536
<212> ДНК
<213> Искусственная последовательность
<220>
<223> мутеин hNGAL
<400> 22
caggactcca cctcagacct gatcccagcc ccacctctga gcaaggtccc tctgcagcag 60
aacttccagg acaaccaatt ccatgggaaa tggtatgtcg tgggcttcgc cggaaatcac 120
cgtctgcgtg aggataagga tccgggaaaa atgcacgcga ccatttacga gttgaaagaa 180
gataaatcat ataacgtcac cgacgtgacc tttgcattca agaaatgcaa ttaccgtatt 240
cacacctttg tgccggggag ccagccgggc gagtttactt taggcaatat taaaagttgg 300
ccgggcgaga catcaacctt ggtccgcgtc gtgagcacca actacaacca gcatgccatg 360
gtgttcttca agaccgtgat gcagaaccgc gagtggtttt ggatcacact gtacgggcgc 420
acgaaagaac tgacaagcga gctgaaggaa aattttatcc gcttttccaa atctctgggc 480
ctccctgaaa accacatcgt cttccctgtc ccaatcgacc agtgtatcga cggcag 536
<210> 23
<211> 536
<212> ДНК
<213> Искусственная последовательность
<220>
<223> мутеин hNGAL
<400> 23
caggactcca cctcagacct gatcccagcc ccacctctga gcaaggtccc tctgcagcag 60
aacttccagg acaaccaatt ccatgggaaa tggtatgtcg tgggcgttgc cggaaattgg 120
tacctgcgtg aggataagga tccgatcaaa atgaccgcga ccatttacga gttgaaagaa 180
gataaatcat ataacgtcac ccaagtggga tttgagcaaa agaaatgcaa atacagcatt 240
cacacctttg tgccggggag ccagccgggc gagtttactt taggcgacat taaaagttgg 300
ccgggcgaca catcactgtt ggtccgcgtc gtgagcacca actacaacca gcatgccatg 360
gtgttcttca agggagtgat gcagaaccgc gaggtttttg caatcacact gtacgggcgc 420
acgaaagaac tgacaagcga gctgaaggaa aattttatcc gcttttccaa atctctgggc 480
ctccctgaaa accacatcgt cttccctgtc ccaatcgacc agtgtatcga cggcag 536
<210> 24
<211> 536
<212> ДНК
<213> Искусственная последовательность
<220>
<223> мутеин hNGAL
<400> 24
caggactcca cctcagacct gatcccagcc ccacctctga gcaaggtccc tctgcagcag 60
aacttccagg acaaccaatt ccatgggaaa tggtatgtcg tgggcgaggc cggaaatgtt 120
gagctgcgtg aggataagga tccggttaaa atgaccgcga ccatttacga gttgaaagaa 180
gataaatcat ataacgtcac cgaggtgcgt ttttggctga agaaatgcaa atacaatatt 240
cacacctttg tgccggggag ccagccgggc gagtttactt taggcgcaat taaaagtgga 300
ccgggcatga catcaacctt ggtccgcgtc gtgagcacca actacaacca gcatgccatg 360
gtgttcttca agaccgtgat gcagaaccgc gagtggtttt ggatcacact gtacgggcgc 420
acgaaagaac tgacaagcga gctgaaggaa aattttatcc gcttttccaa atctctgggc 480
ctccctgaaa accacatcgt cttccctgtc ccaatcgacc agtgtatcga cggcag 536
<210> 25
<211> 536
<212> ДНК
<213> Искусственная последовательность
<220>
<223> мутеин hNGAL
<400> 25
caggactcca cctcagacct gatcccagcc ccacctctga gcaaggtccc tctgcagcag 60
aacttccagg acaaccaatt ccatgggaaa tggtatgtcg tgggccaagc cggaaattac 120
tggctgcgtg aggataagga tccgatcaaa atgtctgcga ccatttacga gttgaaagaa 180
gataaatcat ataacgtcac catcgtgtac tttacccgta agaaatgcag ctaccgtatt 240
tacacctttg tgccggggag ccagccgggc gagtttactt taggccctat taaaagttac 300
ccgggcgaca catcaacctt ggtccgcgtc gtgagcacca actacaacca gcatgccatg 360
gtgttcttca agcacgtgta ccagaaccgc gagtggtttg agatcacact gtacgggcgc 420
acgaaagaac tgacaagcga gctgaaggaa aattttatcc gcttttccaa atctctgggc 480
ctccctgaaa accacatcgt cttccctgtc ccaatcgacc agtgtatcga cggcag 536
<210> 26
<211> 535
<212> ДНК
<213> Искусственная последовательность
<220>
<223> мутеин hNGAL
<400> 26
caggactcca cctcagacct gatcccagcc ccacctctga gcaaggtccc tctgcagcag 60
aacttccagg acaaccaatt ccatgggaaa tggtacgtag ttgggcaggc cggaaattat 120
attctgcgtg aggataagga tccggagaaa atgtgggcga ccatttacga gttgaaagaa 180
gataaatcat atgacgtcac cggggtgtcg tttgggccta agaaatgcaa ttacaggatt 240
tggacctttg tgccggggag ccagccgggc gagtttactt taggcgggat taaaagttct 300
ccgggccaga catcaacgtt ggtccgcgtc gtgagcacca actacaacca gcatgccatg 360
gtgttcttca agcatgtgct tcagaaccgc gagttttttg agatcacact gtacgggcgc 420
acgaaagaac tgacaagcga gctgaaggaa aattttatcc gcttttccaa atctctgggc 480
ctccctgaaa accacatcgt cttccctgtc ccaatcgacc agtgtatcga cggca 535
<210> 27
<211> 536
<212> ДНК
<213> Искусственная последовательность
<220>
<223> мутеин hNGAL
<400> 27
caggactcca cctcagacct gatcccagcc ccacctctga gcaaggtccc tctgcagcag 60
aacttccagg acaaccaatt ccatgggaaa tggtacgtag ttgggcatgc cggaaattat 120
attctgcgtg aggataagga tccggagaaa atgtgggcga ccatttacga gttgaaagaa 180
gataaatcat atgacgtcac cgaggtgtct tttgggccga agaaatgcaa ttacaggatt 240
tggacctttg tgccggggag ccagccgggc gagtttactt taggcggtat taaaagtcct 300
ccgggcgata catcaacgtt ggtccgcgtc gtgagcacca actacaacca gcatgccatg 360
gtgttcttca agcatgtgct tcagaaccgc gagttttttg agatcacact gtacgggcgc 420
acgaaagaac tgacaagcga gctgaaggaa aattttatcc gcttttccaa atctctgggc 480
ctccctgaaa accacatcgt cttccctgtc ccaatcgacc agtgtatcga cggcag 536
<210> 28
<211> 536
<212> ДНК
<213> Искусственная последовательность
<220>
<223> мутеин hNGAL
<400> 28
caggactcca cctcagacct gatcccagcc ccacctctga gcaaggtccc tctgcagcag 60
aacttccagg acaaccaatt ccatgggaaa tggtacgtag ttgggcaggc cggaaattat 120
attctgcgtg aggataagga tccggggaaa atgtgggcga ccatttacga gttgaaagaa 180
gataaatcat atgacgtcac cgaggtgtcg tttggggcta agaaatgcaa ttacaggatt 240
tggacctttg tgccggggag ccagccgggc gagtttactt taggcggtat taaaagtgat 300
ccgggcggga catcaacgtt ggtccgcgtc gtgagcacca actacaacca gcatgccatg 360
gtgttcttca agcatgtgct tcagaaccgc gagttttttg agatcacact gtacgggcgc 420
acgaaagaac tgacaagcga gctgaaggaa aattttatcc gcttttccaa atctctgggc 480
ctccctgaaa accacatcgt cttccctgtc ccaatcgacc agtgtatcga cggcag 536
<210> 29
<211> 536
<212> ДНК
<213> Искусственная последовательность
<220>
<223> мутеин hNGAL
<400> 29
caggactcca cctcagacct gatcccagcc ccacctctga gcaaggtccc tctgcagcag 60
aacttccagg acaaccaatt ccatgggaaa tggtacgtag ttgggcatgc cggaaattat 120
attctgcgtg aggataagga tccggggaaa atgtgggcga ccatttacga gttgaaagaa 180
gataaatcat atgacgtcac cgaggtgtct tttgggccta agaaatgcaa ttaccgtatt 240
tggacctttg tgccggggag ccagccgggc gagtttactt taggcggtat taaaagtccg 300
ccgggcggga catcaacgtt ggtccgcgtc gtgagcacca actacaacca gcatgccatg 360
gtgttcttca agcatgtgct tcagaaccgc gagttttttg agatcacact gtacgggcgc 420
acgaaagaac tgacaagcga gctgaaggaa aattttatcc gcttttccaa atctctgggc 480
ctccctgaaa accacatcgt cttccctgtc ccaatcgacc agtgtatcga cggcag 536
<210> 30
<211> 535
<212> ДНК
<213> Искусственная последовательность
<220>
<223> мутеин hNGAL
<400> 30
caggactcca cctcagacct gatcccagcc ccacctctga gcaaggtccc tctgcagcag 60
aacttccagg acaaccaatt ccatgggaaa tggtacgtag ttgggcaggc cggaaattat 120
attctgcgtg aggataagga tccgcagaaa atgtgggcga ccatttacga gttgaaagaa 180
gataaatcat atgacgtcac cggtgtgtct tttggggcta agaaatgcgt ttacaggatt 240
tggacctttg tgccggggag ccagccgggc gagtttactt taggcggtat taaaagtccg 300
ccgggcggta catcaacgtt ggtccgcgtc gtgagcacca actacaacca gcatgccatg 360
gtgttcttca agcatgtgct tcagaaccgc gagttttttg agatcacact gtacgggcgc 420
acgaaagaac tgacaagcga gctgaaggaa aattttatcc gcttttccaa atctctgggc 480
ctccctgaaa accacatcgt cttccctgtc ccaatcgacc agtgtatcga cggca 535
<210> 31
<211> 536
<212> ДНК
<213> Искусственная последовательность
<220>
<223> мутеин hNGAL
<400> 31
caggactcca cctcagacct gatcccagcc ccacctctga gcaaggtccc tctgcagcag 60
aacttccagg acaaccaatt ccatgggaaa tggtacgtag ttgggcaggc cggaaattat 120
attctgcgtg aggataagga tccggttaaa atgtgggcga ccatttacga gttgaaagaa 180
gataaatcat atgacgtcac cgaggtgtct tttggtccta agaaatgcaa ttacaggatt 240
tggacctttg tgccggggag ccagccgggc gagtttactt taggcgggat taaaagtctt 300
ccgggcggta catcaacttt ggtccgcgtc gtgagcacca actacaacca gcatgccatg 360
gtgttcttca agcatgtgct tcagaaccgc gagttttttg agatcacact gtacgggcgc 420
acgaaagaac tgacaagcga gctgaaggaa aattttatcc gcttttccaa atctctgggc 480
ctccctgaaa accacatcgt cttccctgtc ccaatcgacc agtgtatcga cggcag 536
<210> 32
<211> 535
<212> ДНК
<213> Искусственная последовательность
<220>
<223> мутеин hNGAL
<400> 32
caggactcca cctcagacct gatcccagcc ccacctctga gcaaggtccc tctgcagcag 60
aacttccagg acaaccaatt ccatgggaaa tggtatgtcg tgggcgttgc cggaaattgg 120
tacctgcgtg aggataagga tccgatcaaa atgaccgcga ccatttacga gttgaaagaa 180
gataaatcat atgacgtcac ccaagtggga tttgagcaag agaaatgcaa atacagcatt 240
cacacctttg tgccggggag ccagccgggc gagtttactt taggcaacat taaaagttgg 300
ccgggcgaca catcaccgtt ggtccgcgtc gtgagcacca actacgacca gcatgccatg 360
gtgttcttca agggagtgat gcaggatcgc gaggtttttg caatcacact gtacgggcgc 420
acgaaagaac tgacaagcga gctgaaggaa aattttatcc gcttttccaa atctctgggc 480
ctccctgaaa accacatcgt cttccctgtc ccaatcgacc agtgtatcga cggca 535
<210> 33
<211> 535
<212> ДНК
<213> Искусственная последовательность
<220>
<223> мутеин hNGAL
<400> 33
caggactcca cctcagacct gatcccagcc ccacctctga gcaaggtccc tctgcagcag 60
aacttccagg acaaccaatt ccatgggaaa tggtatgtcg tgggcgttgc cggaaattgg 120
tacctgcgtg aggataagga tccgatcaaa atgaccgcga ccatttacga gttgaaagaa 180
gataaatcat atgacgtcac ccaagtggga tttgagcaag agaaatgcaa atacagcatt 240
cacacctttg tgccggggag ccagccgggc gagtttactt taggcgacat taagagttgg 300
ccgggcgaca catcaccgtt ggtccgcgtc gtgagcacca actacaacca gcatgccatg 360
gtgttcttca agggaatgat gcaggaccgc gaggtttttg caatcacact gtacgggcgc 420
acgaaagaac tgacaagcga gctgaaggaa aattttatcc gcttttccaa atctctgggc 480
ctccctgaaa accacatcgt cttccctgtc ccaatcgacc agtgtatcga cggca 535
<210> 34
<211> 535
<212> ДНК
<213> Искусственная последовательность
<220>
<223> мутеин hNGAL
<400> 34
caggactcca cctcagacct gatcccagcc ccacctctga gcaaggtccc tctgcagcag 60
aacttccagg acaaccaatt ccatgggaaa tggtacgtag taggtatggc cggaaattgg 120
gatctgcgtg aggataagga tccgattaaa atgacggcga ccatttacga gttgaaagaa 180
gataaatcat atgacgtcac ccaggtggcg tttgagcaga agaaatgcaa gtactcgatt 240
catacctttg tgccggggag ccagccgggc gagtttactt taggccagat taaaagttgg 300
ccgggcgata catcaccttt ggtccgcgtc gtgagcacca actacaacca gcatgccatg 360
gtgttcttca agggggtgat gcagaaccgc gaggtgtttg ctatcacact gtacgggcgc 420
acgaaagaac tgacaagcga gctgaaggaa aattttatcc gcttttccaa atctctgggc 480
ctccctgaaa accacatcgt cttccctgtc ccaatcgacc agtgtatcga cggca 535
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| НОВЫЕ СЛИТЫЕ БЕЛКИ, СПЕЦИФИЧЕСКИЕ В ОТНОШЕНИИ CD137 И GPC3 | 2020 |
|
RU2814653C2 |
| НОВЫЙ СЛИТЫЙ БЕЛОК, СПЕЦИФИЧНЫЙ ДЛЯ CD137 И PD-L1 | 2019 |
|
RU2818349C2 |
| БЕЛКИ, СПЕЦИФИЧНЫЕ В ОТНОШЕНИИ CD137 | 2016 |
|
RU2736312C2 |
| МУТЕИНЫ ЛИПОКАЛИНА 2 ЧЕЛОВЕКА С АФФИННОСТЬЮ В ОТНОШЕНИИ ГЛИПИКАНА-3 (GPC3) | 2016 |
|
RU2756318C2 |
| СЛИТЫЙ ПОЛИПЕПТИД С ПРОТИВОРАКОВОЙ АКТИВНОСТЬЮ | 2016 |
|
RU2754466C2 |
| НОВЫЕ ПОЛИПЕПТИДЫ СО СПЕЦИФИЧЕСКИМ СВЯЗЫВАНИЕМ И ПУТИ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2723034C2 |
| МУТЕИНЫ ЛИПОКАЛИНА 2 ЧЕЛОВЕКА (Lcn2, hNGAL) С АФФИННОСТЬЮ ДЛЯ ОПРЕДЕЛЕННОЙ МИШЕНИ | 2019 |
|
RU2804336C2 |
| МУТЕИНЫ ЛИПОКАЛИНА 2 ЧЕЛОВЕКА (Lcn2, hNGAL) С АФФИННОСТЬЮ ДЛЯ ОПРЕДЕЛЕННОЙ МИШЕНИ | 2010 |
|
RU2707126C2 |
| Способ оценки ранозаживляющего потенциала популяции мезенхимальных стволовых клеток и родственные способы отбора мезенхимальных стволовых клеток и идентификации ткани в качестве исходного материала для продуцирования популяции мезенхимальных стволовых клеток | 2020 |
|
RU2827447C1 |
| БИСПЕЦИФИЧНЫЙ СЛИТЫЙ БЕЛОК И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2801528C2 |
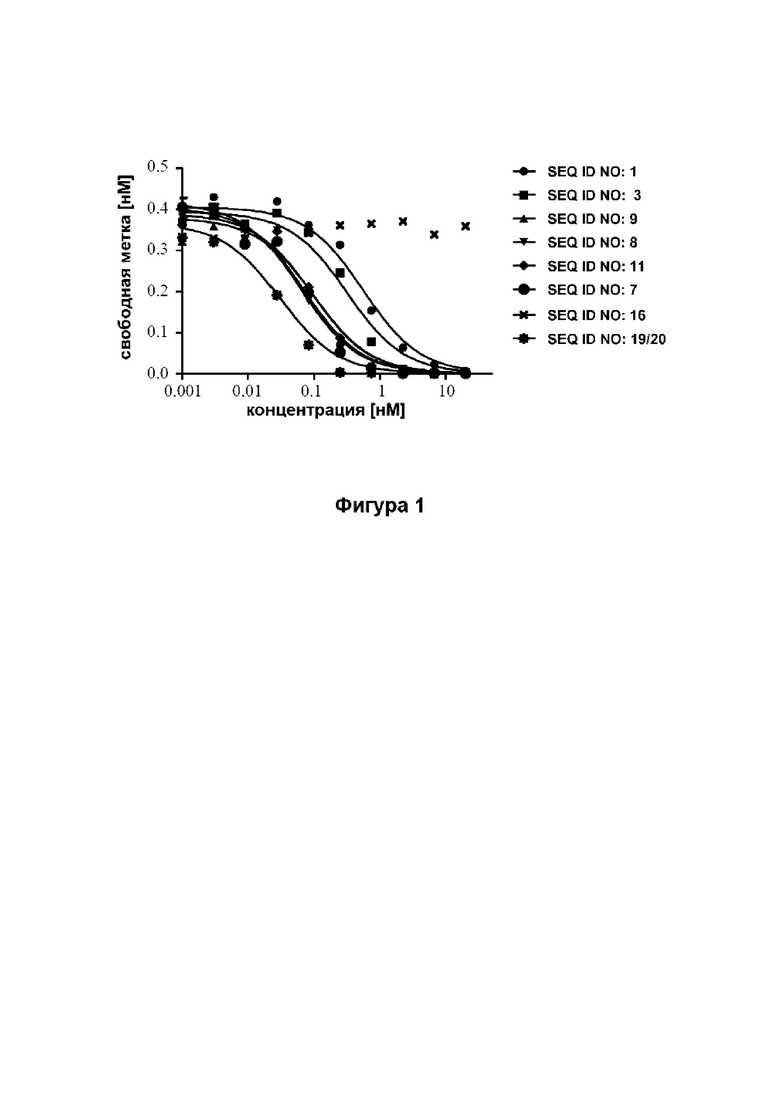
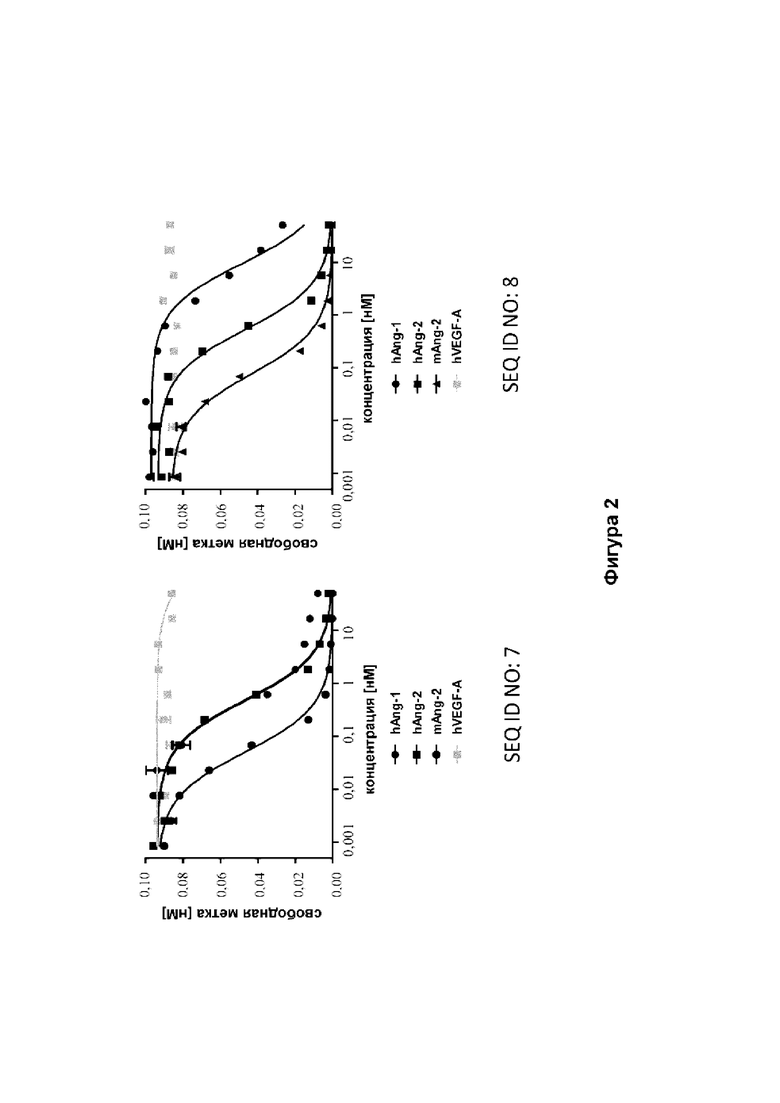
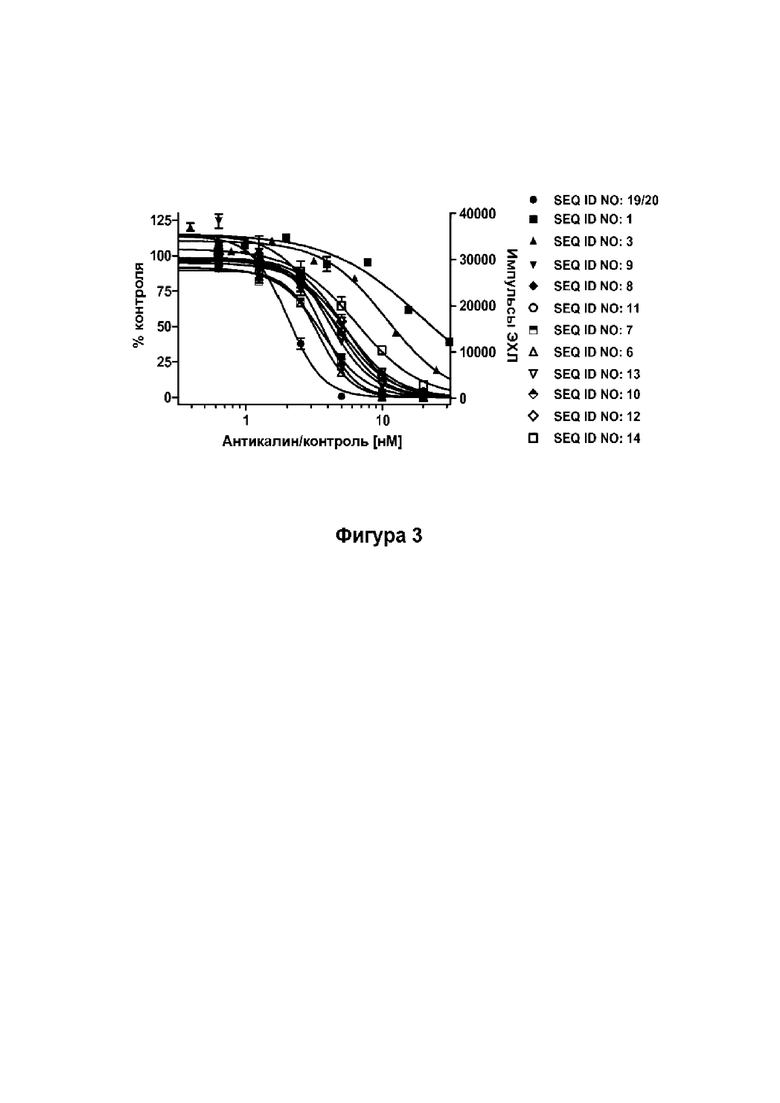
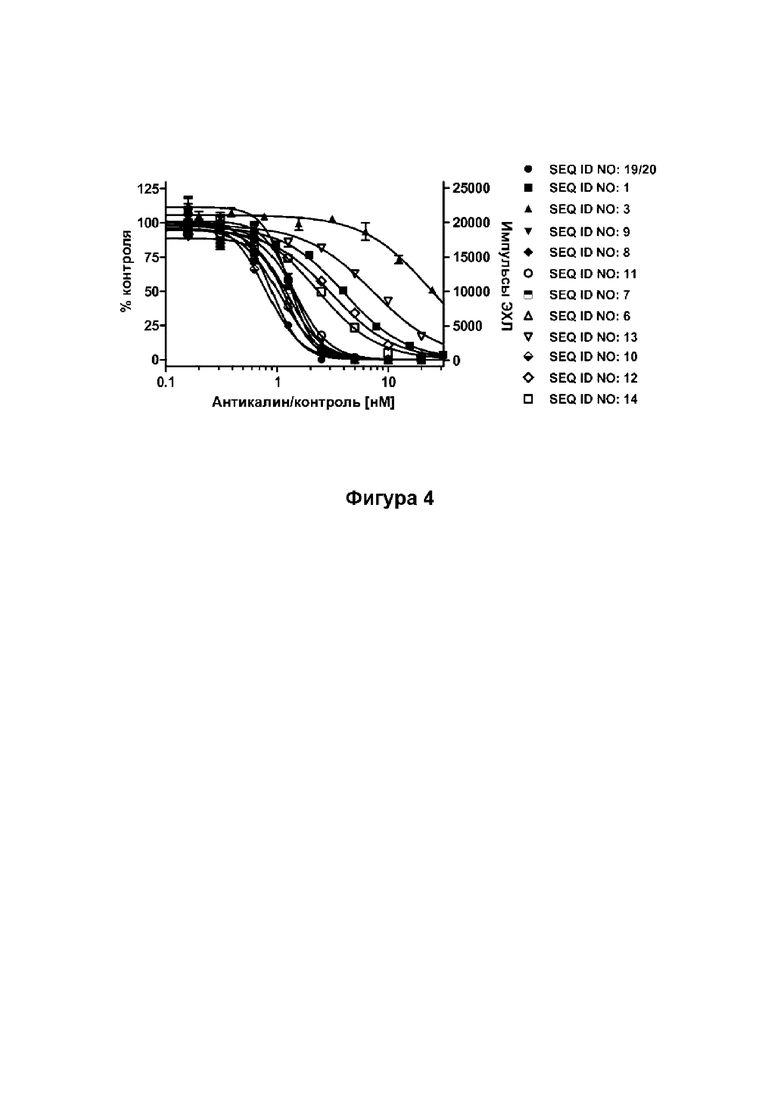
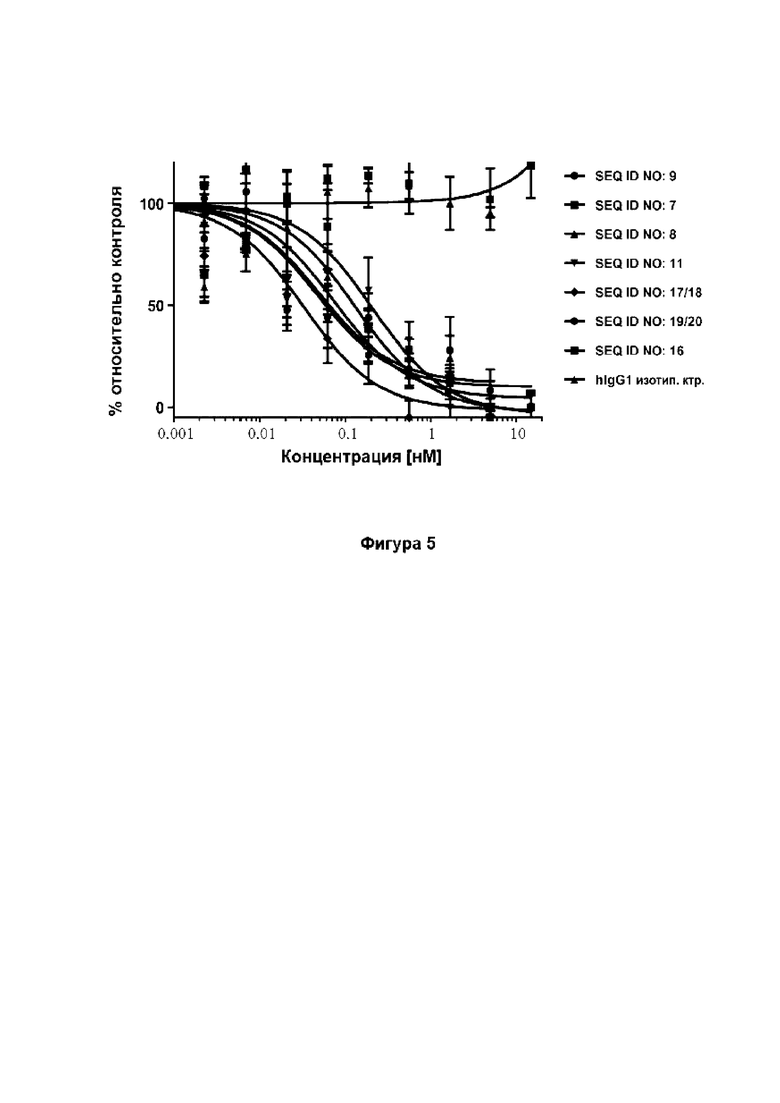
Группа изобретений относится к биотехнологии, а именно к новым соединениям, которые специфически распознают и связывают Ang-2. Предложен мутеин человеческого липокалина, ассоциированного с желатиназой нейтрофилов (hNGAL), способный связывать Ang-2. Аминокислотная последовательность мутеина hNGAL содержит по меньшей мере шестнадцать мутированных аминокислотных остатков в положениях последовательности 28, 36, 40, 41, 49, 52, 68, 70, 72, 73, 77, 79, 81, 96, 100, 103, 106, 125, 127, 132 и 134 по сравнению с линейной полипептидной последовательностью зрелого hNGAL (SEQ ID NO: 16) и обладает по меньшей мере 85% идентичностью по отношению к последовательности, выбранной из SEQ ID NO: 1-14. Также предложены способ получения hNGAL, слитый белок, композиция и комбинация, содержащие hNGAL, молекула нуклеиновой кислоты, кодирующая hNGAL, и клетка-хозяин для генной экспрессии hNGAL. Мутеин hNGAL может быть использован в различных применениях, включая фармацевтические применения для ингибирования или снижения интенсивности ангиогенеза. 23 н. и 32 з.п. ф-лы, 5 ил., 8 табл., 10 пр.
1. Мутеин человеческого липокалина, ассоциированного с желатиназой нейтрофилов (hNGAL), способный связывать Ang-2, где аминокислотная последовательность мутеина hNGAL содержит по меньшей мере шестнадцать из следующих мутированных аминокислотных остатков по сравнению с линейной полипептидной последовательностью зрелого hNGAL (SEQ ID NO: 16): Gln28 → His; Leu 36 → Gln, Glu, His, Val, Met или Phe; Ala 40 → Val, Tyr, His или Trp; Ile 41 → His, Tyr, Trp, Arg, Glu, Asp или Val; Gln 49 → Gly, Ile, Glu или Val; Tyr 52 → Trp, His, Thr или Ser; Ser 68 → Gly, Asp, Gln, Glu или Ile; Leu 70 → Ser, Thr, Gly, Arg, Tyr или Ala; Arg 72 → Gly, Ala, Trp, Thr или Glu; Lys 73 → Pro, Phe, Leu, Arg, Ala или Gln; Asp 77 → Asn, Lys, Ser или Val; Trp 79 → Thr, Arg, Ser или Asn; Arg 81 → Trp, His или Tyr; Asn 96 → Gly, Ala, Pro, Gln или Asp; Tyr 100 → Pro, Trp, Gly, Ser, Leu или Asp; Leu 103 → Gly, Glu, Asp, Met или Gln; Tyr 106 → Thr, Leu, Pro или Phe; Lys 125 → His, Thr или Gly; Ser 127 → Leu, Tyr или Met; Tyr 132 → Phe, Trp или Val; и Lys 134 → Ala, Glu или Trp, при этом мутеин hNGAL обладает идентичностью последовательности, составляющей по меньшей мере 85% по отношению к последовательности, выбранной из группы, состоящей из SEQ ID NO: 1-14.
2. Мутеин hNGAL по п. 1, где мутеин hNGAL способен связывать Ang-2 с Kд около 5 нM или меньше.
3. Мутеин hNGAL по п. 1, где мутеин hNGAL способен связывать Ang-2 с аффинностью, измеренной посредством определения значения EC50, составляющего около 5 нM или меньше, согласно измерениям в ELISA-анализе.
4. Мутеин hNGAL по п. 1, где мутеин hNGAL способен связывать Ang-2 с аффинностью, измеренной посредством определения значения IC50, составляющего около 5 нM или меньше, согласно измерениям в конкурентном ELISA-анализе.
5. Мутеин hNGAL по п. 1, где мутеин hNGAL способен ингибировать или снижать пролиферацию эндотелиальных клеток лимфатических микрососудов, опосредованную Ang-2, со значением IC50 около 5 нM или меньше согласно данным анализа клеточной пролиферации.
6. Мутеин hNGAL по любому из пп. 1-5, где аминокислотная последовательность мутеина hNGAL содержит один или более из следующих мутированных аминокислотных остатков по сравнению с линейной полипептидной последовательностью зрелого hNGAL (SEQ ID NO: 16): Asn 65 → Asp; Lys 74 → Glu; Cys 87 → Ser; Asn 116 →Asp; Val 126 → Met и Asn 129 →Asp.
7. Мутеин hNGAL по любому из пп. 1-5, где аминокислотная последовательность мутеина hNGAL содержит один из следующих наборов мутированных аминокислотных остатков по сравнению с линейной полипептидной последовательностью зрелого hNGAL (SEQ ID NO: 16):
(a) Gln 28 → His; Leu 36 → Gln; Ala 40 →Tyr; Gln 49 → Gly; Tyr 52 → Trp; Ser 68 → Gly; Leu 70 → Ser; Arg 72 → Gly; Lys 73 → Pro; Asp 77 → Asn; Trp 79 → Thr; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Gly; Tyr 100 → Pro; Leu 103 → Gly; Tyr 106 → Thr; Lys 125 → His; Ser 127 → Leu; Tyr 132 → Phe; Lys 134 → Glu;
(b) Gln 28 → His; Leu 36 → Phe; Ala 40 → His; Ile 41 → Arg; Gln 49 → Gly; Tyr 52 → His; Ser 68 → Asp; Leu 70 → Thr; Arg 72 → Ala; Lys 73 → Phe; Asp 77→ Asn; Trp 79 → Arg; Arg 81 → His; Cys 87 → Ser; Tyr 100 → Trp; Leu 103 → Glu; Tyr 106 → Thr; Lys 125 → Thr; Ser 127 → Met; Tyr 132 → Trp; Lys 134 → Trp;
(c) Gln 28 → His; Leu 36 → Val; Ala 40 → Trp; Ile 41 → Tyr; Gln 49 → Ile; Tyr 52 → Thr; Ser 68 → Gln; Leu 70 → Gly; Arg 72 → Glu; Lys 73 → Gln; Asp 77 → Lys; Trp 79 → Ser; Arg 81 → His; Cys 87 → Ser; Tyr 100 → Trp; Leu 103 → Asp; Tyr 106 → Leu; Lys 125→ Gly; Ser 127 → Met; Tyr 132 → Val; Lys 134 → Ala;
(d) Gln 28 → His; Leu 36 → Glu, Ala 40 →Val; Ile 41 → Glu; Gln 49 → Val; Tyr 52 → Thr; Ser 68 → Glu; Leu 70 → Arg; Arg 72 → Trp; Lys 73 → Leu; Asp 77 → Lys; Trp 79 → Asn; Arg 81 → His; Cys 87 → Ser; Asn 96 → Ala; Tyr 100 → Gly; Leu 103 → Met; Tyr 106 → Thr; Lys 125 → Thr; Ser 127 → Met; Tyr 132 → Trp; Lys 134 → Trp;
(e) Gln 28 → His; Leu 36 → Gln; Ala 40 → Tyr; Ile 41 → Trp; Gln 49 → Ile; Tyr 52 → Ser; Ser 68 → Ile; Leu 70 → Tyr; Arg 72 → Thr; Lys 73 → Arg; Asp 77 → Ser; Trp 79 → Arg; Arg 81 → Tyr; Cys 87 → Ser; Asn 96 → Pro; Leu 103 → Asp; Tyr 106 → Thr; Lys 125 → His; Ser 127 → Tyr; Tyr 132 → Trp; Lys 134 → Glu;
(f) Gln 28 → His; Leu 36 → Gln; Ala 40 → Tyr; Gln 49 → Glu; Tyr 52 → Trp; Asn 65 → Asp; Ser 68 → Gly; Leu 70 → Ser; Arg 72 → Gly; Lys 73 → Pro; Asp 77 → Asn; Trp 79 → Arg; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Gly; Tyr 100 → Ser; Leu 103 → Gln; Tyr 106 → Thr; Lys 125 → His; Ser 127 → Leu; Tyr 132 → Phe; Lys 134 → Glu;
(g) Gln 28 → His; Leu 36 → His; Ala 40 →Tyr; Gln 49 → Glu; Tyr 52 → Trp; Asn 65 → Asp; Ser 68 → Glu; Leu 70 → Ser; Arg 72 → Gly; Lys 73 → Pro; Asp 77 → Asn; Trp 79 → Arg; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Gly; Tyr 100 → Pro; Leu 103 → Asp; Tyr 106 → Thr; Lys 125 → His; Ser 127 → Leu; Tyr 132 → Phe; Lys 134 → Glu;
(h) Gln 28 → His; Leu 36 → Gln; Ala 40 →Tyr; Gln 49 → Gly; Tyr 52 → Trp; Asn 65 → Asp; Ser 68 → Glu; Leu 70 → Ser; Arg 72 → Gly; Lys 73 → Ala; Asp 77 → Asn; Trp 79 → Arg; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Gly; Tyr 100 → Asp; Leu 103 → Gly; Tyr 106 → Thr; Lys 125 → His; Ser 127 → Leu; Tyr 132 → Phe; Lys 134 → Glu;
(i) Gln 28 → His; Leu 36 → His; Ala 40 →Tyr; Gln 49 → Gly; Tyr 52 → Trp; Asn 65 → Asp; Ser 68 → Glu; Leu 70 → Ser; Arg 72 → Gly; Lys 73 → Pro; Asp 77 → Asn; Trp 79 → Arg; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Gly; Tyr 100 → Pro; Leu 103 → Gly; Tyr 106 → Thr; Lys 125 → His; Ser 127 → Leu; Tyr 132 → Phe; Lys 134 → Glu;
(j) Gln 28 → His; Leu 36 → Gln; Ala 40 → Tyr; Gln 49 → Gly; Tyr 52 → Trp; Asn 65 → Asp; Ser 68 → Gly; Leu 70 → Ser; Arg 72 → Gly; Lys 73 → Ala; Asp 77 → Val; Trp 79 → Arg; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Gly; Tyr 100 → Pro; Leu 103 → Gly; Tyr 106 → Thr; Lys 125 → His; Ser 127 → Leu; Tyr 132 → Phe; Lys 134 → Glu;
(k) Gln 28 → His; Leu 36 → Gln; Ala 40 → Tyr; Gln 49 → Val; Tyr 52 → Trp; Asn 65 → Asp; Ser 68 → Glu; Leu 70 → Ser; Arg 72 → Gly; Lys 73 → Pro; Asp 77 → Asn; Trp 79 → Arg; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Gly; Tyr 100 → Leu; Leu 103 → Gly; Tyr 106 → Thr; Lys 125 → His; Ser 127 → Leu; Tyr 132 → Phe; Lys 134 → Glu;
(l) Gln 28 → His; Leu 36 → Val; Ala 40 → Tyr; Ile 41 → Tyr; Gln 49 → Ile; Tyr 52 → Thr; Asn 65 → Asp; Ser 68 → Gln; Leu 70 → Gly; Arg 72 → Glu; Lys 73 → Gln; Lys 74 → Glu; Asp 77 → Lys; Trp 79 → Ser; Arg 81 → His; Cys 87 → Ser; Tyr 100 → Trp; Leu 103 → Asp; Tyr 106 → Pro; Asn 116 → Asp; Lys 125 → Gly; Ser 127 → Met; Asn 129 → Asp; Tyr 132 → Val; Lys 134 → Ala;
(m) Gln 28 → His; Leu 36 → Val; Ala 40 → Tyr; Ile 41 → Tyr; Gln 49 → Ile; Tyr 52 → Thr; Asn 65 → Asp; Ser 68 → Gln; Leu 70 → Gly; Arg 72 → Glu; Lys 73 → Gln; Lys 74 → Glu; Asp 77 → Lys; Trp 79 → Ser; Arg 81 → His; Cys 87 → Ser; Asn 96 → Asp; Tyr 100 → Trp; Leu 103 → Asp; Tyr 106 → Pro; Lys 125 → Gly; Val 126 → Met; Ser 127 → Met; Asn 129 → Asp; Tyr 132 → Val; Lys 134 → Ala; или
(n) Gln 28 → His; Leu 36 → Met; Ala 40 →Tyr; Ile 41 → Asp; Gln 49 → Ile; Tyr 52 → Thr; Asn 65 → Asp; Ser 68 → Gln; Leu 70 → Gly; Arg 72 → Glu; Lys 73 → Gln; Asp 77 → Lys; Trp 79 → Ser; Arg 81 → His; Cys 87 → Ser; Asn 96 → Gln; Tyr 100 → Trp; Leu 103 → Asp; Tyr 106 → Pro; Lys 125 → Gly; Ser 127 → Met; Tyr 132 → Val; Lys 134 → Ala.
8. Мутеин hNGAL по любому из пп. 1-7, где мутеин hNGAL способен связывать Ang-1 с аффинностью, измеренной посредством определения значения IC50, составляющего около 150 нM или меньше.
9. Мутеин hNGAL по любому из пп. 1-8, где мутеин hNGAL перекрестно реагирует как с человеческим Ang-2, так и с мышиным Ang-2.
10. Мутеин hNGAL по п. 9, где мутеин hNGAL способен связывать мышиный Ang-2 с аффинностью, измеренной посредством определения значения IC50, составляющего около 5 нM или меньше.
11. Мутеин hNGAL по любому из пп. 1-10, где мутеин hNGAL способен блокировать связывание человеческого Ang-2 с hTie-2 и мышиного Ang-2 с hTie-2 со значением IC50 около 25 нM или меньше, соответственно.
12. Мутеин hNGAL по любому из пп. 1-11, где мутеин hNGAL содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-14 и их фрагментов, где во фрагменте отсутствует по меньшей мере одна из N-концевых и/или С-концевых аминокислот мутеина hNGAL.
13. Мутеин hNGAL по любому из пп. 1-11, где мутеин hNGAL обладает идентичностью последовательности, составляющей по меньшей мере 90% по отношению к последовательности, выбранной из группы, состоящей из SEQ ID NO: 1-14.
14. Мутеин hNGAL по любому из пп. 1-13, где мутеин hNGAL содержит один или более ненативных цистеиновых остатков, замещающих одну или более аминокислот hNGAL в положениях последовательности, соответствующих положениям последовательности 14, 21, 60, 84, 88, 116, 141, 145, 143, 146 или 158 линейной полипептидной последовательности зрелого hNGAL (SEQ ID NO: 16).
15. Мутеин hNGAL по любому из пп. 1-13, где указанный мутеин hNGAL содержит по меньшей мере одну аминокислотную замену нативного цистеинового остатка другой аминокислотой, выбранной из лейцина, метионина, аланина, валина, изолейцина, серина, треонина, аспарагиновой кислоты, глутаминовой кислоты, аспарагина, глутамина, гистидина, лизина, аргинина, глицина, пролина, триптофана, тирозина, фенилаланина.
16. Мутеин hNGAL по п. 15, где указанная другая аминокислота представляет собой сериновый остаток.
17. Конъюгат, содержащий мутеин hNGAL по любому из пп. 1-13 для связывания Ang-2, где мутеин hNGAL конъюгирован с соединением, выбранным из группы, состоящей из органической молекулы, ферментной метки, радиоактивной метки, цветной метки, флуоресцентной метки, хромогенной метки, люминесцентной метки, гаптена, дигоксигенина, биотина, цитостатического агента, токсинов, комплекса металла, металла, коллоидного золота и соединения, которое обеспечивает продление периода полужизни мутеина hNGAL в сыворотке крови.
18. Слитый белок, содержащий мутеин hNGAL по любому из пп. 1-13 для связывания Ang-2, где мутеин hNGAL слит по его N-концу и/или его C-концу с партнером по слиянию, который представляет собой белок, белковый домен или пептид.
19. Слитый белок по п. 18, где соединение, которое обеспечивает продление периода полужизни мутеина hNGAL в сыворотке крови, выбрано из группы, состоящей из молекулы полиалкиленгликоля, молекулы полиэтиленгликоля (ПЭГ), гидроксиэтилкрахмала, Fс-части иммуноглобулина, CH3-домена иммуноглобулина, CH4-домена иммуноглобулина, альбуминсвязывающего пептида и альбуминсвязывающего белка.
20. Молекула нуклеиновой кислоты для обеспечения экспрессии мутеина hNGAL в клетке-хозяине, содержащая нуклеотидную последовательность, кодирующую мутеин hNGAL по любому из пп. 1-16.
21. Молекула нуклеиновой кислоты по п. 20, где молекула нуклеиновой кислоты содержится в векторе или фагмидном векторе.
22. Клетка-хозяин для генной экспрессии, содержащая молекулу нуклеиновой кислоты по п. 20 или 21.
23. Способ получения мутеина hNGAL по любому из пп. 1-16, где полипептид получают начиная с нуклеиновой кислоты, кодирующей мутеин hNGAL, где мутеин hNGAL получают в бактериальном или эукариотическом организме-хозяине и выделяют из этого организма-хозяина, или его культуры, или в in vitro системе трансляции.
24. Фармацевтическая композиция для лечения заболевания или нарушения, связанного с дерегулированным ангиогенезом, содержащая эффективное количество мутеина hNGAL по любому из пп. 1-16.
25. Композиция по п. 24, где указанная композиция дополнительно содержит по меньшей мере одно из фармацевтически приемлемого вспомогательного средства, разбавителя или носителя.
26. Применение мутеина hNGAL по любому из пп. 1-16 для выявления Ang-2 в образце.
27. Комбинация по меньшей мере (a) мутеина hNGAL по любому из пп. 1-16 и (b) антиангиогенного агента для антиангиогенного лечения.
28. Комбинация по п. 27, где мутеин hNGAL и антиангиогенный агент вводятся в комбинации, включая одновременное, параллельное или последовательное введение.
29. Комбинация по п. 27, где мутеин hNGAL и антиангиогенный агент вводятся независимо друг от друга, в том числе через индивидуальные интервалы времени в независимые моменты времени.
30. Комбинация по п. 27, где комбинация содержится в одной композиции.
31. Комбинация по любому из пп. 27-30, где антиангиогенный агент выбран из группы, состоящей из: (i) антагонистов Ang-1, Ang-2, Ang-3, Ang-4 и/или Tie-2; (ii) антагонистов Flt1, KDR, Flt4, VEGF-A, VEGF-B, VEGF-C, VEGF-D, VEGF-E, PlGF и/или EG-VEGF; (iii) антагонистов дельта-подобного лиганда 4 (DLL4, специфического в отношении сосудов лиганда Notch); (iv) антагонистов рецепторов эпидермального фактора роста (EGFR); и (v) ингибиторов цитокинов.
32. Комбинация по любому из пп. 27-30, где антиангиогенный агент представляет собой ингибитор VEGF.
33. Комбинация по п. 32, где ингибитор VEGF выбран из группы, состоящей из: VEGF-Trap, бевацизумаба (Avastin®), сорафениба (Nexavar®), сунитиниба (Sutent®) и пазопаниба (Votrient®).
34. Комбинация по п. 32 или 33, где ингибитор VEGF является антагонистом VEGF-A и/или антагонистом VEGF-C.
35. Комбинация по любому из пп. 27-34, где комбинация дополнительно содержит дополнительный антиангиогенный агент.
36. Комбинация по п. 35, где первый антиангиогенный агент является антагонистом VEGF-A, а второй антиангиогенный агент является антагонистом VEGF-C.
37. Комбинация по п. 34 или 36, где антагонист VEGF-A представляет собой антитело к VEGF-A или мутеин липокалина, который обладает специфичностью связывания в отношении VEGF-A.
38. Комбинация по п. 34 или 36, где антагонист VEGF-C представляет собой антитело к VEGF-C или мутеин липокалина, который обладает специфичностью связывания в отношении VEGF-C.
39. Применение мутеина hNGAL по любому из пп. 1-16 для производства фармацевтической композиции, подходящей для лечения заболевания или нарушения, связанного с дерегулированным ангиогенезом.
40. Применение комбинации по любому из пп. 27-38 для производства фармацевтической композиции, подходящей для лечения заболевания или нарушения, связанного с дерегулированным ангиогенезом.
41. Способ связывания Ang-2 у субъекта, включающий введение указанному субъекту эффективного количества мутеина hNGAL по любому из пп. 1-16, или композиции по п. 24 или 25, или комбинации по любому из пп. 27-38.
42. Способ ингибирования ангиогенеза у субъекта, включающий введение указанному субъекту эффективного количества мутеина hNGAL по любому из пп. 1-16, или композиции по п. 24 или 25, или комбинации по любому из пп. 27-38.
43. Способ лечения заболевания или нарушения, связанного с дерегулированным ангиогенезом, у субъекта, включающий введение указанному субъекту эффективного количества мутеина hNGAL по любому из пп. 1-16, или композиции по п. 24 или 25, или комбинации по любому из пп. 27-38.
44. Способ по п. 43, где заболевание или нарушение выбрано из группы, состоящей из: роста опухоли, глазных нарушений, сосудистых заболеваний, воспалительных или инфекционных заболеваний, рака, неоваскулярных заболеваний глаза, артрита и псориаза.
45. Применение мутеина hNGAL по любому из пп. 1-16 для связывания Ang-2 у субъекта.
46. Применение композиции по п. 24 или 25 для связывания Ang-2 у субъекта.
47. Применение комбинации по любому из пп. 27-38 для связывания Ang-2 у субъекта.
48. Применение мутеина hNGAL по любому из пп. 1-16 для ингибирования ангиогенеза у субъекта.
49. Применение композиции по п. 24 или 25 для ингибирования ангиогенеза у субъекта.
50. Применение комбинации по любому из пп. 27-38 для ингибирования ангиогенеза у субъекта.
51. Применение мутеина hNGAL по любому из пп. 1-16 для лечения заболевания или нарушения, связанного с дерегулированным ангиогенезом.
52. Применение композиции по п. 24 или 25 для лечения заболевания или нарушения, связанного с дерегулированным ангиогенезом.
53. Применение по п. 52, где заболевание или нарушение выбрано из группы, состоящей из роста опухоли, глазных нарушений, сосудистых заболеваний, воспалительных или инфекционных заболеваний, рака, неоваскулярных заболеваний глаз, артрита и псориаза.
54. Применение комбинации по любому из пп. 27-38 для лечения заболевания или нарушения, связанного с дерегулированным ангиогенезом.
55. Применение по п. 54, где заболевание или нарушение выбрано из группы, состоящей из роста опухоли, глазных нарушений, сосудистых заболеваний, воспалительных или инфекционных заболеваний, рака, неоваскулярных заболеваний глаз, артрита и псориаза.
| US 2013316962 A1, 28.11.2013 | |||
| WO 2014076321 A1, 22.05.2014 | |||
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| МУТЕИНЫ ЛИПОКАЛИНА СЛЕЗНОЙ ЖИДКОСТИ, ОБЛАДАЮЩИЕ АФФИННОСТЬЮ К С-МЕТ РЕЦЕПТОРНОЙ ТИРОЗИНКИНАЗЕ ЧЕЛОВЕКА, И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2009 |
|
RU2515063C9 |
| RU 2012128586 A, 20.01.2014. | |||

