I. Уровень техники
[0001] Глипикан-3 (GPC3) представляет собой онкоэмбриональный антиген, который относится к семейству глипиканов гликозил-фосфатидилинозитол-заякоренных гепарансульфат-протеогликанов. GPC3 экспрессируется в эмбриональной печени и плаценте в ходе развития и является подавленным или выключенным в нормальных зрелых тканях. Мутации в гене GPC3 и его истощение обуславливают синдром Симпсона-Голаби-Бемеля или дисморфии Симпсона у людей. GPC3 экспрессируется при различных формах рака и, в частности, при гепатоцеллюлярной карциноме («HCC»), меланоме, карциноме из клеток Меркеля, опухоли Вильма и при гепатобластоме. (He, H. et al., Applied Immunohistochem Mol Morphol. 17:40-6 (2009); Jakubovic and Jothy; Ex. MoI. Path. 82:184-189 (2007); Nakatsura and Nishimura, Biodrugs 19(2):71-77 (2005).). HCC является третей ведущей причиной смертей, связанных с раком, во всем мире. Ежегодно на долю HCC приходится приблизительно 1 миллион смертей. (Nakatsura and Nishimura, Biodrugs 19(2):71-77 (2005)).
[0002] Для эффективного лечения видов рака, экспрессирующих GPC3, таких как HCC, требуются терапевтические соединения, которые целенаправленно воздействуют на GPC3, а также обеспечивают противоопухолевые эффекты.
[0003] CD137 является костимулирующим иммунным рецептором и представителем надсемейства рецепторов фактора некроза опухоли (TNFR). Он в основном экспрессируется на активированных CD4+ и CD8+ T-клетках, активированных B-клетках и клетках естественных киллерах (NK), но его также можно обнаружить на находящихся в покое моноцитах и дендритных клетках (Li, S. Y. et al., Clin Pharmacol 2013 5(Suppl 1):47-53) или эндотелиальных клетках (Snell, L. M. et al., Immunol Rev 2011 Nov; 244(1):197-217). CD137 играет важную роль в регуляции иммунных ответов, и таким образом является мишенью для иммунотерапии рака. Лиганд CD137 (CD137L) является единственным известным естественным лигандом CD137 и постоянно экспрессируется на некоторых типах APC, таких как активированные B-клетки, моноциты и дендритные клетки в селезенке, и его можно индуцировать на T-лимфоцитах.
[0004] CD137L представляет собой тримерный белок, который существует в виде мембраносвязанной формы и в виде растворимого варианта. Однако способность растворимого CD137L активировать CD137, например на экспрессирующих CD137 лимфоцитах, ограничена, и требуются большие концентрации для получения эффекта (Wyzgol, A. et al., J Immunol 2009 Aug 1; 183(3):1851-1861). Естественный путь активации CD137 представляет собой активацию посредством связывания CD137-положительной клетки с CD137L-положительной клеткой. Затем активация CD137, как считают, индуцируется путем образования скоплений с помощью CD137L на противоположной клетке, что приводит к передаче сигнала через TRAF1, 2 и 3 (Snell, L. M. et al., Immunol Rev 2011 Nov; 244(1):197-217, Yao, S. et al., Nat Rev Drug Disc 2013 Feb; 12(2):130-146) и далее сопутствующим нисходящим эффектам у CD137-положительной T-клетки. В случае T-клеток, активированных путем узнавания их соответствующих родственных мишеней, эффекты, вызванные путем костимуляции CD137, представляют собой дополнительную усиленную активацию, улучшенное выживание и повышенную пролиферацию, продуцирование провоспалительных цитокинов и улучшенную способность к уничтожению.
[0005] Преимущество костимуляции CD137 для устранения раковых клеток было продемонстрировано в ряде преклинических in-vivo моделей. Форсированная экспрессия CD137L на клетках опухоли, например, приводит к отторжению опухоли (Melero, I. et al., Eur J Immunol 1998 Mar; 28(3):1116-1121). Аналогичным образом форсированная экспрессия scFv к CD137 на клетках опухоли приводит к зависимому от CD4+ T-клеток и NK-клеток устранению опухоли (Ye, Z. et al., Nat Med 2002 Apr; 8(4):343-348, Zhang, H. et al., Mol Canc Ther 2006 Jan; 5(1):149-155, Yang, Y. et al., Canc Res 2007 Mar 1; 67(5):2339-2344). Также было продемонстрировано, что системно вводимое антитело к CD137 приводит к замедлению роста опухоли (Martinet, O. et al., Gene Ther 2002 Jun; 9(12):786-792).
[0006] Было показано, что CD137 является превосходным маркером для встречающихся в природе реагирующих на опухоль T-клеток в опухолях человека (Ye, Q. et al., Clin Canc Res: 2014 Jan 1; 20(1):44-55), и что антитела к CD137 можно использовать для усиления распространения и активности инфильтрирующих меланому CD8+ лимфоцитов для применения в адаптивной T-клеточной терапии (Chacon, J. A. et al., PloS One 2013 8(4):e60031).
[0007] Доклиническое обнаружение потенциального терапевтического преимущества костимуляции CD137 ускорило разработку терапевтических антител, целенаправленно воздействующих на CD137, BMS-663513 (Jure-Kunkel, M. et al., патент США 7288638) и PF-05082566 (Fisher, T. S. et al., Canc Immunol Immunother 2012 Oct; 61(10):1721-1733); оба в настоящее время проходят ранние фазы клинических исследований.
[0008] Однако только недавно выяснилось, что двухвалентной связывающей CD137 молекулы, подобной антителу, самой по себе может быть недостаточно для образования скоплений CD137 на T-клетках или NK-клетках и обеспечения эффективной активации, по аналогии с недостатком активности трехвалентного растворимого CD137L. В недавних публикациях, в которых использовали доклинические мышиные модели, были представлены in-vivo экспериментальные данные относительно того, что механизм действия других антител к TNFR фактически требует взаимодействия антитела через свою Fc-часть с Fc-гамма рецепторами на клетках, экспрессирующих Fc-гамма рецепторы (Bulliard, Y. et al., J Exp Med 2013 Aug 26; 210(9):1685-1693, Bulliard, Y. et al., Immunol Cell Biol 2014 Jul; 92(6):475-480). Вследствие этого механизм действия антител, находящихся в настоящее время на стадии клинических испытаний, может быть преобладающим за счет нецелевого образования скоплений через Fc-гамма рецепторы, которые могут практически произвольно зависеть от присутствия экспрессирующих Fc-α клеток в окружающем пространстве опухоли.
[0009] Таким образом, существует неудовлетворенная потребность в создании терапевтических средств, которые образуют скопления и активируют CD137 при помощи специфичного целенаправленно воздействующего на опухоль механизма действия.
[0010] Для удовлетворения этой неудовлетворенной потребности в настоящей заявке представлен новый подход одновременного связывания CD137 и опухолевого антигена GPC3 посредством слитого полипептида со следующими свойствами:
(a) со специфичностью связывания в отношении CD137 и
(b) специфичностью связывания в отношении GPC3.
[0011] Этот слитый полипептид сконструирован для обеспечения зависящей от мишени-опухоли активации CD137 на лимфоцитах при помощи GPC3, экспрессирующегося на опухолевых клетках. Ожидается, что такая молекула дополнительно активирует T-клетки и/или NK-клетки, которые расположены в окружающем пространстве GPC3-положительной опухоли. Такое биспецифическое средство может проявлять усиленные терапевтические эффекты по сравнению либо с антителами к GPC3, либо с антителами к CD137.
II. Определения
[0012] В приведенном ниже перечне определены термины, фразы и сокращения, используемые по всему данному описанию. Предполагается, что все термины, перечисленные и определенные в данном документе, охватывают все грамматические формы.
[0013] Как используется в данном документе, если не указано иное, «CD137» означает CD137 человека и включает варианты, изоформы и видовые гомологи CD137 человека. CD137 также известен как «4-1BB», или «представитель 9 надсемейства рецепторов фактора некроза опухоли (TNFRSF9)», или «индуцированный активацией лимфоцитов (ILA)». CD137 человека означает полноразмерный белок, обозначенный согласно UniProt как Q07011, его фрагмент или его вариант.
[0014] Как используется в данном документе, если не указано иное, «GPC3» означает GPC3 человека и включает варианты, изоформы и видовые гомологи GPC3 человека. GPC3 также известен как «глипикан-3», «протеогликан глипикан 3», «GPC3», «OTTHUMP00000062492», «GTR2-2», «SGB», «DGSX», «SDYS», «SGBS», «OCI-5» и «SGBSl», которые используются взаимозаменяемо. GPC3 человека означает полноразмерный белок, обозначенный согласно UniProt как P51654, его фрагмент или его вариант. Как используется в данном документе, «выявляемая аффинность» означает способность связываться с выбранной мишенью с константой аффинности, составляющей, как правило, по меньшей мере приблизительно 10-5 M или ниже. Более низкие значения аффинности, как правило, уже не поддаются измерению с помощью cтандартных способов, таких как ELISA, и, следовательно, имеют второстепенное значение.
[0015] Как используется в данном документе, «аффинность связывания» белка по настоящему раскрытию (например, мутеина липокалина) или его слитого полипептида с выбранной мишенью (в данном случае CD137 и/или GPC3), можно измерять (и тем самым определять значения KD для комплекса мутеин-лиганд) с помощью большого числа способов, известных специалисту в данной области. Такие способы включают без ограничения флуоресцентное титрование, конкурентный ELISA, калориметрические способы, такие как изотермическая титрационная калориметрия (ITC) и поверхностный плазмонный резонанс (BIAcore). Такие способы хорошо известны из уровня техники и их примеры также подробно описаны ниже.
[0016] Следует также отметить, что на образование комплекса между соответствующей связывающей молекулой и ее лигандом влияет много различных факторов, таких как концентрации соответствующих партнеров по связыванию, наличие конкурентов, показатель pH и ионная сила используемой буферной системы и экспериментальный способ, используемый для определения константы диссоциации, KD (например, флуоресцентное титрование, конкурентный ELISA или поверхностный плазмонный резонанс, при этом упомянуты лишь некоторые из них), или даже математический алгоритм, который используют для оценки экспериментальных данных.
[0017] Следовательно, специалисту в данной области также очевидно, что значения KD (константы диссоциации комплекса, образованного между соответствующей связывающей молекулой и ее мишенью/лигандом) могут варьироваться в пределах определенного экспериментального диапазона в зависимости от способа и экспериментальной установки, которую используют для определения аффинности конкретного мутеина липокалина в отношении данного лиганда. Это значит, что может иметь место незначительное отклонение в измеренных значениях KD или диапазоне допусков, зависящее, например, от того, было ли значение KD определено с помощью поверхностного плазмонного резонанса (Biacore), конкурентного ELISA или «прямого ELISA».
[0018] Как используется в данном документе, «мутеин», «подвергнутый мутации» целостный объект (или белок, или нуклеиновая кислота) или «мутант» относится к обмену, делеции или вставке одного или более из нуклеотидов или аминокислот по сравнению со встречающимся в природе (дикого типа) «эталонным» остовом нуклеиновой кислоты или белка. Указанный термин также включает фрагменты мутеина и вариантов, описанных в данном документе. Мутеины липокалина по настоящему изобретению, их фрагменты или варианты предпочтительно сохраняют функцию связывания с CD137 и/или GPC3, описанными в данном документе.
[0019] Термин «фрагмент», используемый в данном документе применительно к мутеинам по настоящему раскрытию, относится к белкам или пептидам, полученным из полноразмерного зрелого липокалина слезы человека или липокалина 2 человека, который укорочен с N-конца и/или C-конца, т. е. у него отсутствует по меньшей мере одна из N-концевых и/или C-концевых аминокислот. Такие фрагменты могут включать по меньшей мере 10 или больше, а именно 20 или 30 или больше, последовательных аминокислот первичной последовательности зрелого липокалина и обычно поддаются выявлению в иммуноанализе зрелого липокалина. Как правило, термин «фрагмент», используемый в данном документе в отношении соответствующего белкового лиганда CD137 и/или GPC3 для мутеина липокалина по настоящему раскрытию, или для комбинации согласно настоящему раскрытию, или для белка слияния, описанного в данном документе, относится к укороченным с N-конца и/или C-конца белковым или пептидным лигандам, которые сохраняют способность полноразмерного лиганда быть распознанными и/или связанными мутеином согласно настоящему раскрытию.
[0020] Термин «мутагенез», используемый в данном документе, означает, что экспериментальные условия выбраны таким образом, что аминокислота, встречающаяся в природе в данном положении в последовательности зрелого липокалина, может быть заменена по меньшей мере одной аминокислотой, которая не присутствует в этом конкретном положении в соответствующей природной полипептидной последовательности. Термин «мутагенез» также включает (дополнительную) модификацию длины сегментов последовательности путем делеции или вставки одной или более аминокислот. Таким образом, в пределы объема настоящего раскрытия подпадает, что, например, одну аминокислоту в выбранном положении в последовательности заменяют отрезком из трех случайных мутаций, следствием чего является вставка двух аминокислотных остатков по сравнению с длиной соответствующего сегмента белка дикого типа. Такая вставка или делеция могут быть введены независимо друг от друга в любой из сегментов пептидов, которые могут быть подвергнуты мутагенезу в настоящем раскрытии. Согласно одному иллюстративному варианту осуществления настоящего раскрытия вставка из нескольких мутаций может быть введена в петлю AB остова выбранного липокалина (см. международную патентную заявку WO 2005/019256, которая включена в данный документ в полном объеме посредством ссылки).
[0021] Термин «случайный мутагенез» означает, что в конкретном положении в последовательности отсутствует предопределенная отдельная аминокислота (мутация), однако по меньшей мере две аминокислоты могут быть включены с определенной вероятностью в заранее определенное положение в последовательности в ходе мутагенеза.
[0022] «Идентичность» является свойством последовательностей, с помощью которого определяют их сходство или родство. Термин «идентичность последовательностей» или «идентичность», используемый в настоящем раскрытии, означает процентную долю попарно идентичных остатков - последующее (гомологичное) выравнивание последовательности полипептида по настоящему раскрытию с последовательностью запроса - относительно количества остатков в более длинной из этих двух последовательностей. Идентичность последовательностей определяют путем деления количества идентичных аминокислотных остатков на общее количество остатков и умножения полученного результата на 100.
[0023] Термин «гомология» используют в данном документе в его общепринятом значении, и он включает идентичные аминокислоты, а также аминокислоты, которые рассматриваются как консервативные замены (например, обмен остатка глутамата на остаток аспартата) в аналогичных положениях в линейной аминокислотной последовательности полипептида по настоящему раскрытию (например, любого мутеина липокалина по настоящему раскрытию).
[0024] Процентную долю гомологии последовательностей или идентичности последовательностей, например, можно определять в данном документе с использованием программы BLASTP, версии blastp 2.2.5 (от 16 ноября 2002 года; см. Altschul, S. F. et al. (1997) Nucl. Acids Res. 25, 3389-3402). Согласно данному варианту осуществления процентная доля гомологии основана на выравнивании целых полипептидных последовательностей (матрица: BLOSUM 62; штрафы за введения гэпа: 11.1; значение порога отсечения устанавливали как 10-3), в том числе последовательностей пропептидов, предпочтительно с использованием остова белка дикого типа в качестве эталона в сравнительном анализе пар. Ее рассчитывают в виде процентной доли количеств «положительных» (гомологичных аминокислот), указанных в качестве итога, в результате расчета с помощью программы BLASTP, разделенных на общее количество аминокислот, выбранных программой для выравнивания.
[0025] В частности, для того, чтобы определить, соответствует ли аминокислотный остаток аминокислотной последовательности липокалина (мутеина), отличного от липокалина дикого типа, определенному положению в аминокислотной последовательности липокалина дикого типа, специалист в данной области может применять средства и способы, хорошо известные в данной области, например, выравнивания либо вручную, либо с использованием компьютерных программ, таких как BLAST2.0, которая означает средство поиска основного локального выравнивания, или ClustalW, или любой другой подходящей программы, которая подходит для получения выравниваний последовательностей. Соответственно, липокалин дикого типа может служить в качестве «последовательности для сравнения» или «эталонной последовательности», тогда как аминокислотная последовательность липокалина, отличного от липокалина дикого типа, описанного в данном документе, служит в качестве «искомой последовательности». Термины «эталонная последовательность» и «последовательность дикого типа» используют в данном документе взаимозаменяемо. Предпочтительная последовательность липокалина дикого типа показана под SEQ ID NO: 1 (Tlc) или SEQ ID NO: 2 (NGAL) соответственно. В зависимости от того, является ли основой для мутеина липокалина по настоящему изобретению соответственно Tlc или NGAL, соответствующий липокалин дикого типа можно использовать в качестве эталонной последовательности или последовательности дикого типа.
[0026] «Гэпы» представляют собой пространства в выравнивании, которые являются результатом добавлений или делеций аминокислот. Таким образом, две копии точно такой же последовательности характеризуются 100% идентичностью, но последовательности, которые в меньшей степени высоко консервативны и имеют делеции, добавления или замещения, могут характеризоваться более низкой степенью идентичности последовательностей. Специалисту в данной области будет понятно, что несколько компьютерных программ доступны для определения идентичности последовательностей с использованием стандартных параметров, например Blast (Altschul, et al. (1997) Nucleic Acids Res. 25, 3389-3402), Blast2 (Altschul, et al. (1990) J. Mol. Biol. 215, 403-410) и Smith-Waterman (Smith, et al. (1981) J. Mol. Biol. 147, 195-197).
[0027] Термин «вариант», используемый в настоящем раскрытии, относится к производным белка или пептида, которые предусматривают модификации аминокислотной последовательности, например путем замены, делеции, вставки или химической модификации. Согласно некоторым вариантам осуществления такие модификации не снижают функциональность белка или пептида. Такие варианты включают белки, в которых одна или более аминокислот были замещены их соответствующими D-стереоизомерами или аминокислотами, отличными от встречающихся в природе 20 аминокислот, такими как, например, орнитин, гидроксипролин, цитруллин, гомосерин, гидроксилизин, норвалин. Однако такие замены также могут быть консервативными, т. е. аминокислотный остаток замещают химически ему подобным аминокислотным остатком. Примеры консервативных замен представляют собой замещения из числа представителей следующих групп: 1) аланин, серин и треонин; 2) аспарагиновая кислота и глутаминовая кислота; 3) аспарагин и глутамин; 4) аргинин и лизин; 5) изолейцин, лейцин, метионин и валин и 6) фенилаланин, тирозин и триптофан. Термин «вариант», используемый в данном документе в отношении соответствующего белкового лиганда CD137 и/или GPC3 для мутеина липокалина по настоящему раскрытию, или для комбинации согласно настоящему раскрытию, или для белка слияния, описанного в данном документе, относится к CD137 или его фрагменту соответственно, который имеет одну или более, как например: 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 40, 50, 60, 70, 80 или больше, аминокислотных замен, делеций и/или вставок по сравнению с белком CD137 или GPC3 дикого типа соответственно, как, например, эталонным белком CD137 или GPC3, депонированным с идентификатором UniProt, описанным в данном документе. Соответственно,вариант CD137 предпочтительно характеризуется идентичностью аминокислотной последовательности, составляющей по меньшей мере 50%, 60%, 70%, 80%, 85%, 90% или 95%, с CD137 или GPC3 дикого типа человека, как, например, эталонным белком CD137 или GPC3, депонированным с идентификатором UniProt, описанным в данном документе.
[0028] Под «нативной последовательностью» липокалина подразумевают липокалин, который характеризуется такой же аминокислотной последовательностью, как и соответствующий полипептид, полученный из естественного окружения. Таким образом, нативная последовательность липокалина может характеризоваться аминокислотной последовательностью соответствующего встречающегося в природе липокалина из любого организма, в частности млекопитающего. Такая нативная последовательность полипептида может быть выделена из естественного окружения или может быть получена с помощью рекомбинантных способов или способов синтеза. Термин «нативная последовательность» полипептида, как правило, охватывает встречающиеся в природе усеченные или секретированные формы липокалина, встречающиеся в природе вариантные формы, такие как, в качестве альтернативы, сплайсированные формы, и встречающиеся в природе аллельные варианты липокалина. «Вариант» полипептида означает биологически активный полипептид, характеризующийся по меньшей мере приблизительно 50%, 60%, 70%, 80% или по меньшей мере приблизительно 85% идентичностью аминокислотной последовательности с нативной последовательностью полипептида. Такие варианты включают, к примеру, полипептиды, в которых один или более аминокислотных остатков добавлены или удалены на N- или C-конце полипептида. Как правило, вариант характеризуется по меньшей мере приблизительно 70%, в том числе по меньшей мере приблизительно 80%, как, например, по меньшей мере приблизительно 85% идентичностью аминокислотной последовательности, в том числе по меньшей мере приблизительно 90% идентичностью аминокислотной последовательности или по меньшей мере приблизительно 95% идентичностью аминокислотной последовательности с нативной последовательностью полипептида. В качестве иллюстративного примера первые 4 N-концевых аминокислотных остатка (His-His-Leu-Leu) и последние 2 C-концевых аминокислотных остатка (Ser-Asp) могут быть удалены в мутеине липокалина слезы (Tlc) по настоящему раскрытию без воздействия на биологическую функцию белка. Кроме того, в качестве другого иллюстративного примера определенные аминокислотные остатки могут быть удалены в мутеине липокалина 2 (NGAL) по настоящему раскрытию без воздействия на биологическую функцию белка, например (Lys-Asp-Pro, положения 46-48).
[0029] Термин «положение» при использовании в соответствии с настоящим раскрытием означает либо положение аминокислоты в аминокислотной последовательности, описанной в данном документе, либо положение нуклеотида в последовательности нуклеиновой кислоты, описанной в данном документе. Для понимания термина «соответствует» или «соответствующий», используемого в данном документе в отношении положений в аминокислотной последовательности одного или более мутеинов липокалина, соответствующее положение определено не только номером предыдущих нуклеотидов/аминокислот. Следовательно, положение указанной аминокислоты в соответствии с настоящим раскрытием, которая может быть замещена, может варьироваться за счет делеции или добавления аминокислот в любом месте в (мутантном или дикого типа) липокалине. Аналогично положение указанного нуклеотида в соответствии с настоящим раскрытием, который может быть замещен, может варьироваться за счет делеций или дополнительных нуклеотидов в любом месте в 5’-нетранслируемом участке (UTR) мутеина липокалина или липокалина дикого типа, в том числе промоторе, и/или любых других регуляторных последовательностях или гене (в том числе экзонах и интронах).
[0030] Таким образом, в отношении соответствующего положения в соответствии с настоящим раскрытием предпочтительно понимать, что положения нуклеотидов/аминокислот могут отличаться по обозначенным номерам от подобных соседних нуклеотидов/аминокислот, но указанные соседние нуклеотиды/аминокислоты, которые могут быть обменены, удалены или добавлены, также предусматриваются в одном или более соответствующих положениях.
[0031] Кроме того, в отношении соответствующего положения в мутеине липокалина на основе эталонного остова в соответствии с настоящим раскрытием предпочтительно понимать, что положения нуклеотидов/аминокислот являются структурно соответствующими положениям в любом месте в (мутантном или дикого типа) липокалине, даже если они могут отличаться по обозначенным номерам, что будет понятно специалисту в данной области, с учетом высококонсервативного общего паттерна фолдинга среди липокалинов.
[0032] Слово «выявлять», «выявление», «выявляемый» или «выявляющий», используемое в данном документе, понимается как на количественном, так и на качественном уровне, равно как и их комбинация. Таким образом, это включает количественные, полуколичественные и качественные измерения представляющей интерес молекулы.
[0033] «Субъект» представляет собой позвоночное, предпочтительно млекопитающее, более предпочтительно человека. Термин «млекопитающее» используется в данном документе в отношении любого животного, классифицированного как млекопитающее, в том числе без ограничения людей, домашних и сельскохозяйственных животных, а также животных в зоопарках, для занятий спортом или животных-питомцев, как например: овец, собак, лошадей, кошек, коров, крыс, свиней, высших приматов, таких как яванские макаки, и т. д., при этом упомянуты лишь некоторые иллюстративные примеры. Предпочтительно млекопитающим в данном документе является человек.
[0034] «Эффективное количество» представляет собой количество, достаточное для обеспечения полезных или необходимых результатов. Эффективное количество можно вводить за одно или более введений.
[0035] «Образец» определяют как биологический образец, взятый у любого субъекта. Биологические образцы включают без ограничения кровь, сыворотку крови, мочу, экскременты, семенную жидкость или ткань.
[0036] «Субъединица» слитого полипептида, раскрытого в данном документе, определяется как отрезок из аминокислот полипептида, при этом отрезок обозначает уникальную функциональную единицу указанного полипептида, как, например, предусматривает мотив связывания относительно мишени.
[0037] «Слитый полипептид», описанный в данном документе, содержит две или более субъединиц, при этом по меньшей мере одна из этих субъединиц связывается с GPC3, а дополнительная субъединица связывается с CD137. В слитом полипептиде эти субъединицы могут быть связаны ковалентной или не ковалентной связью. Предпочтительно слитый полипептид представляет собой трансляционный продукт слияния между двумя или более субъединицами. Трансляционный продукт слияния может быть получен при помощи генетически сконструированной кодирующей последовательности для одной субъединицы в рамке с кодирующей последовательностью дополнительной субъединицы. Между обеими субъединицами может быть вставлена нуклеотидная последовательность, кодирующая линкер. Однако субъединицы слитого полипептида по настоящему раскрытию также могут быть связаны при помощи химического линкера.
[0038] «Линкер», который может содержать слитый полипептид по настоящему раскрытию, связывает две или более субъединицы слитого полипептида, описанного в данном документе. Связь может быть ковалентной или не ковалентной. Предпочтительная ковалентная связь осуществляется с использованием пептидной связи, такой как пептидная связь между аминокислотами. Соответственно, в предпочтительном варианте осуществления указанный линкер содержит одну или более аминокислот, как, например: 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 или более аминокислот. Предпочтительные линкеры описаны в данном документе. Другие предпочтительне линкеры представляют собой химические линкеры.
iii. ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0039] На фигуре 1 представлен общий вид строения слитых полипептидов, описанных в данной заявке, которые являются биспецифическими в отношении мишеней, GPC3 и CD137. Использовали три разных подхода: на фигуре 1(A) представлен первый набор слитых полипептидов, которые основаны на специфичности антитела в отношении CD137 (например, антитела с SEQ ID NO: 34 и 35) и специфичности мутеина липокалина в отношении GPC3 (например, мутеина липокалина с SEQ ID NO: 10). Полученные полипептиды представляют собой отдельные слияния мутеина липокалина с любым из четырех концов антитела. Все слияния соединены с помощью линкера, такого как подвижный линкер (G4S)3 (например, линкер с SEQ ID NO: 49); на фигуре 1(B) представлен второй набор слитых полипептидов, которые основаны на двух мутеинах липокалина (например, специфичном к GPC3 мутеине липокалина с SEQ ID NO: 10 и специфичном к CD137 мутеине липокалина с SEQ ID NO: 26), слитых со сконструированным Fc-фрагментом IgG4 (SEQ ID NO: 73); и на фигуре 1(C) представлен третий набор белков слияния, которые основаны на двух мутеинах липокалина (например, SEQ ID NO: 10 и SEQ ID NO: 26), соединенных с помощью одного или более линкеров, таких как линкеры (G4S)2 (например, линкеры с SEQ ID NO: 48), при этом специфичный к GPC3 мутеин липокалина слит со специфичным к CD137 мутеином липокалина (например, в SEQ ID NO: 46) или специфичный к GPC3 мутеин липокалина и два специфичных к CD137 мутеина липокалина слиты вместе (например, в SEQ ID NO: 47).
[0040] На фигуре 2 представлен репрезентативный эксперимент, в котором определяли специфичность слитых полипептидов с SEQ ID NO: 36 и 37, SEQ ID NO: 38 и 39, SEQ ID NO: 40 и 41 и SEQ ID NO: 42 и 43 и мутеина липокалина с SEQ ID NO: 10 в отношении мишени GPC3. GPC3 наносили на микротитровальный планшет и оттитровывали тестируемые молекулы. Связанные молекулы выявляли с помощью меченного HRP специфичного к NGAL человека антитела, как описано в примере 2. Данные аппроксимировали с моделью связывания 1:1 со значением EC50 и наибольшим сигналом в качестве незаданных параметров и наклоном, который фиксировали для единства. Полученные в результате значения EC50 представлены в таблице 1.
[0041] На фигуре 3 представлен репрезентативный эксперимент, в котором определяли специфичность слитых полипептидов с SEQ ID NO: 36 и 37, SEQ ID NO: 38 и 39, SEQ ID NO: 40 и 41 и антитела с SEQ ID NO: 34 и 35 в отношении мишени CD137. Слитый CD137 человека с Fc наносили на микротитровальный планшет и оттитровывали тестируемые молекулы. Связанные молекулы выявляли с помощью меченного HRP антитела к Fc IgG человека, как описано в примере 3. Данные аппроксимировали с моделью связывания 1:1 со значением EC50 и наибольшим сигналом в качестве незаданных параметров и наклоном, который фиксировали для единства. Полученные в результате значения EC50 представлены в таблице 2.
[0042] На фигуре 4 представлен репрезентативный эксперимент, в котором определяли способность слитых полипептидов с SEQ ID NO: 36 и 37, SEQ ID NO: 38 и 39, SEQ ID NO: 40 и 41 одновременно связывать обе мишени, GPC3 и CD137. Белок слияния рекомбинантный CD137-Fc наносили на микротитровальный планшет с последующим титрованием белка слияния. Впоследствии добавляли одинаковую концентрацию биотинилированного GPC3 человека, который выявляли при помощи меченного HRP экстравидина, как описано в примере 4. Данные аппроксимировали с моделью связывания 1:1 со значением EC50 и наибольшим сигналом в качестве незаданных параметров и наклоном, который фиксировали для единства. Полученные в результате значения EC50 представлены в таблице 3.
[0043] На фигуре 5 представлен репрезентативный эксперимент, в котором определяли аффинность слитых полипептидов с SEQ ID NO: 36 и 37, SEQ ID NO: 38 и 39, SEQ ID NO: 40 и 41 и мутеина липокалина с SEQ ID NO: 10 относительно мишени GPC3 с помощью поверхностного плазмонного резонанса (SPR). Биотинилированный GPC3 иммобилизировали на сенсорном чипе и анализировали связывание слитых полипептидов и мутеина липокалина при разных концентрациях, как описано в примере 5. Полученные в результате значения KD представлены в таблице 4.
[0044] На фигуре 6 представлен репрезентативный эксперимент, в котором определяли аффинность слитых полипептидов с SEQ ID NO: 36 и 37, SEQ ID NO: 38 и 39, SEQ ID NO: 40 и 41 и антитела с SEQ ID NO: 34 и 35 относительно биотинилированного слияния CD137-Fc с помощью поверхностного плазмонного резонанса (SPR). Биотинилированный CD137-Fc иммобилизировали на сенсорном чипе и анализировали связывание белков слияния при разных концентрациях, как описано в примере 6. Полученные в результате значения KD представлены в таблице 5.
[0045] На фигуре 7 представлен репрезентативный эксперимент, в котором определяли специфичность слитых полипептидов мутеин липокалина-Fc с SEQ ID NO: 44 и SEQ ID NO: 45 и мутеина липокалина с SEQ ID NO: 10 в отношении мишени GPC3. GPC3 наносили на микротитровальный планшет и оттитровывали тестируемые молекулы. Связанные молекулы выявляли с помощью меченного HRP специфичного к NGAL человека антитела, как описано в примере 7. Данные аппроксимировали с моделью связывания 1:1 со значением EC50 и наибольшим сигналом в качестве незаданных параметров и наклоном, который фиксировали для единства. Полученные в результате значения EC50 представлены в таблице 6.
[0046] На фигуре 8 представлен репрезентативный эксперимент, в котором определяли специфичность слитых полипептидов мутеин липокалина-Fc с SEQ ID NO: 44 и SEQ ID NO: 45 и мутеина липокалина с SEQ ID NO: 26 в отношении CD137. Слитый CD137 человека с Fc наносили на микротитровальный планшет и оттитровывали тестируемые молекулы. Связанные молекулы выявляли с помощью меченного HRP антитела к Fc IgG человека, как описано в примере 8. Данные аппроксимировали с моделью связывания 1:1 со значением EC50 и наибольшим сигналом в качестве незаданных параметров и наклоном, который фиксировали для единства. Полученные в результате значения EC50 представлены в таблице 7.
[0047] На фигуре 9 представлен репрезентативный эксперимент, в котором определяли способность слитых полипептидов мутеин липокалина-Fc с SEQ ID NO: 44 и SEQ ID NO: 45 одновременно связывать мишени, GPC3 и CD137. Белок слияния рекомбинантный CD137-Fc наносили на микротитровальный планшет с последующим титрованием слитых полипептидов мутеин липокалина-Fc. Впоследствии добавляли одинаковую концентрацию биотинилированного GPC3 человека, который выявляли при помощи меченного HRP экстравидина, как описано в примере 9. Данные аппроксимировали с моделью связывания 1:1 со значением EC50 и наибольшим сигналом в качестве незаданных параметров и наклоном, который фиксировали для единства. Полученные в результате значения EC50 представлены в таблице 8.
[0048] На фигуре 10 представлен репрезентативный эксперимент, в котором определяли аффинность слитых полипептидов мутеин липокалина-Fc с SEQ ID NO: 44 и SEQ ID NO: 45 и мутеина липокалина с SEQ ID NO: 10 относительно мишени GPC3 с помощью поверхностного плазмонного резонанса (SPR). Биотинилированный GPC3 иммобилизировали на сенсорном чипе и анализировали связывание слитых полипептидов и мутеина липокалина при разных концентрациях. Полученные в результате значения KD представлены в таблице 9.
[0049] На фигуре 11 представлен репрезентативный эксперимент, в котором определяли аффинность слитых полипептидов мутеин липокалина-Fc с SEQ ID NO: 44 и SEQ ID NO: 45 и мутеина липокалина с SEQ ID NO: 26 относительно биотинилированного CD137-Fc с помощью поверхностного плазмонного резонанса (SPR). Биотинилированный CD137-Fc иммобилизировали на сенсорном чипе и анализировали связывание слитых полипептидов и мутеина липокалина при разных концентрациях. Полученные в результате значения KD представлены в таблице 10.
[0050] На фигуре 12 представлен репрезентативный эксперимент, в котором определяли специфичность слитого полипептида с SEQ ID NO: 53 и 54 и мутеина липокалина с SEQ ID NO: 10 в отношении мишени GPC3. GPC3 наносили на микротитровальный планшет и оттитровывали тестируемые молекулы. Связанные молекулы выявляли с помощью меченного HRP специфичного к NGAL человека антитела, как описано в примере 12. Данные аппроксимировали с моделью связывания 1:1 со значением EC50 и наибольшим сигналом в качестве незаданных параметров и наклоном, который фиксировали для единства. Полученные в результате значения EC50 представлены в таблице 11.
[0051] На фигуре 13 представлен репрезентативный эксперимент, в котором определяли способность слитого полипептида с SEQ ID NO: 53 и 54 одновременно связываться с обеими мишенями, GPC3 и CD137. Белок слияния рекомбинантный CD137-Fc наносили на микротитровальный планшет с последующим титрованием белка слияния. Впоследствии добавляли одинаковую концентрацию биотинилированного GPC3 человека, который выявляли при помощи меченного HRP экстравидина, как описано в примере 13. Данные аппроксимировали с моделью связывания 1:1 со значением EC50 и наибольшим сигналом в качестве незаданных параметров и наклоном, который фиксировали для единства.
[0052] На фигуре 14 представлен репрезентативный эксперимент, в котором определяли специфичность двух биспецифических слитых полипептидов с SEQ ID NO: 46 и SEQ ID NO: 47 и мутеина липокалина с SEQ ID NO: 8 в отношении мишени GPC3. GPC3 наносили на микротитровальный планшет и оттитровывали тестируемые молекулы. Связанные молекулы выявляли с помощью меченного HRP специфичного к NGAL человека антитела, как описано в примере 14. Данные аппроксимировали с моделью связывания 1:1 со значением EC50 и наибольшим сигналом в качестве незаданных параметров и наклоном, который фиксировали для единства. Полученные в результате значения EC50 представлены в таблице 12.
[0053] На фигуре 15 представлен репрезентативный эксперимент, в котором определяли специфичность двух биспецифических слитых полипептидов с SEQ ID NO: 46 и SEQ ID NO: 47 и мутеина липокалина с SEQ ID NO: 26 в отношении мишени CD137. Слитый CD137 человека с Fc наносили на микротитровальный планшет и оттитровывали тестируемые молекулы. Связанные молекулы выявляли с помощью меченного HRP антитела к Fc IgG человека, как описано в примере 15. Данные аппроксимировали с моделью связывания 1:1 со значением EC50 и наибольшим сигналом в качестве незаданных параметров и наклоном, который фиксировали для единства. Полученные в результате значения EC50 представлены в таблице 13.
[0054] На фигуре 16 представлен репрезентативный эксперимент, в котором определяли способность двух биспецифических слитых полипептидов с SEQ ID NO: 46 и SEQ ID NO: 47 одновременно связывать мишени, GPC3 и CD137. Белок слияния рекомбинантный CD137-Fc наносили на микротитровальный планшет с последующим титрованием белка слияния. Впоследствии добавляли одинаковую концентрацию биотинилированного GPC3 человека, который выявляли при помощи меченного HRP экстравидина, как описано в примере 16. Данные аппроксимировали с моделью связывания 1:1 со значением EC50 и наибольшим сигналом в качестве незаданных параметров и наклоном, который фиксировали для единства. Полученные в результате значения EC50 представлены в таблице 14.
[0055] На фигуре 17 представлен репрезентативный эксперимент, в котором определяли аффинность двух биспецифических слитых полипептидов с SEQ ID NO: 46 и SEQ ID NO: 47 и мутеина липокалина с SEQ ID NO: 8 относительно мишени GPC3 с помощью поверхностного плазмонного резонанса (SPR). Биотинилированный GPC3 иммобилизировали на сенсорном чипе и анализировали связывание слитых полипептидов при разных концентрациях. Полученные в результате значения KD представлены в таблице 15.
[0056] На фигуре 18 представлен репрезентативный эксперимент, в котором определяли аффинность двух биспецифических слитых полипептидов с SEQ ID NO: 46 и SEQ ID NO: 47 и мутеина липокалина с SEQ ID NO: 26 относительно CD137-Fc с помощью поверхностного плазмонного резонанса (SPR). CD137 человека-Fc иммобилизировали на сенсорном чипе и анализировали связывание белков слияния при разных концентрациях. Полученные в результате значения KD представлены в таблице 16.
[0057] На фигуре 19 представлен репрезентативный эксперимент, в котором изучали способность слитых полипептидов с SEQ ID NO: 36 и 37, SEQ ID NO: 38 и 39, SEQ ID NO: 40 и 41 и SEQ ID NO: 42 и 43 костимулировать ответы T-клеток при нанесении на пластиковую чашку для культивирования. Слитые полипептиды при разных концентрациях наносили на пластиковую чашку совместно с антителом к CD3 человека, а затем очищенные T-клетки инкубировали на покрытой поверхности в присутствии растворимого антитела к CD28 человека. Уровни интерлейкина 2 (IL-2) в супернатанте измеряли с помощью электрохемилюминесцентного (ELC) анализа, как описано в примере 19. В качестве отрицательного контроля использовали контроль, представляющий собой изотип IgG4 человека.
[0058] На фигуре 20 представлен репрезентативный эксперимент, в котором изучали способность слитых полипептидов с SEQ ID NO: 36 и 37, SEQ ID NO: 44 и SEQ ID NO: 45 костимулировать активацию T-клеток зависимым от мишени GPC3 образом. В качестве контроля авторы настоящего изобретения использовали моноспецифическое связывающее CD137 антитело с SEQ ID NO: 34 и 35. В эксперименте антитело к CD3 человека (+) или контроль, представляющий собой изотип (-), наносили на пластиковую чашку для культивирования, а затем GPC3-положительные клетки HepG2 культивировали на чашке в течение ночи. На следующий день очищенные T-клетки инкубировали на покрытой поверхности в присутствии 1 мкг/мл биспецифических слитых полипептидов с SEQ ID NO: 36 и 37, SEQ ID NO: 44, SEQ ID NO: 45 или контрольного антитела с SEQ ID NO: 34 и 35. Уровни интерлейкина 2 (IL-2) в супернатанте измеряли с помощью электрохемилюминесцентного (ELC) анализа, как описано в примере 20.
[0059] На фигуре 21 представлен репрезентативный эксперимент, в котором изучали способность слитых полипептидов с SEQ ID NO: 44 и SEQ ID NO: 45 костимулировать активацию T-клеток зависимым от мишени GPC3 образом. В эксперименте антитело к CD3 человека наносили на пластиковую чашку для культивирования, а затем GPC3-положительные клетки Hep3B культивировали на чашке в течение ночи. На следующий день очищенные T-клетки инкубировали на покрытой поверхности в присутствии разных концентраций биспецифических слитых полипептидов с SEQ ID NO: 44 (A) и SEQ ID NO: 45 (C). Уровень интерлейкина 2 (IL-2) в супернатанте определяли с помощью ELISA. Для блокирования связывания биспецифических слитых полипептидов с GPC3 эксперимент также проводили в присутствии избытка SEQ ID NO: 10, как для SEQ ID NO: 44 (B), так и для SEQ ID NO: 45 (D). Данные аппроксимировали с моделью связывания 1:1.
[0060] На фигуре 22 представлен репрезентативный эксперимент, в котором изучали способность тестируемых продуктов костимулировать активацию T-клеток с использованием разных клеточных линий. Используемыми клеточными линиями были GPC3-положительные HepG2 и GPC3-отрицательные SKBR-3 и MCF7. В эксперименте антитело к CD3 человека наносили на пластиковую чашку для культивирования, а затем исследуемую клеточную линию культивировали на чашке в течение ночи. На следующий день очищенные T-клетки инкубировали на покрытой поверхности в течение трех дней в присутствии разных концентраций биспецифических слитых полипептидов, указанных ниже: (A) SEQ ID NO: 44 (кружки), SEQ ID NO: 45 (квадраты) или контрольного антитела трастузумаба (треугольники). (B) антитело к CD137 с SEQ ID NO: 74 и 75. Уровни интерлейкина 2 в супернатанте определяли при помощи анализа на основе электрохемилюминесценции. Нанесенный на график относительный ответ IL-2 соответствует соотношению ответов, полученных в присутствии и в отсутствии («фоновое значение») тестируемых продуктов.
[0061] На фигуре 23 представлен результат in vitro оценки иммуногенности в отношении T-клеток биспецифических слитых полипептидов, контрольного антитела трастузумаба и положительного контроля, представляющего собой гемоцианин лимфы улитки (KLH). Анализ проводили с использованием основанного на PBMC формата, который описан в примере 23, с 32 донорами и аллотипами антигена лейкоцитов человека (HLA), отражающими распределение в мировой популяции. (A) Индекс стимуляции (пролиферация в присутствии тестируемого продукта в сравнении с отсутствием тестируемого продукта). Средние ответы показаны в виде линий. Пороговый уровень, который определяет прореагировавшего донора (индекс стимуляции > 2), показан в виде пунктирной линии. (B) Количество респондеров.
[0062] На фигуре 24 представлен репрезентативный эксперимент на аффинность полипептидов к FcgRI, FcgRIII и FcRn, как описано в примерах 24 и 25.
[0063] На фигуре 25 представлен результат фармакокинетического анализа биспецифических слитых полипептидов с SEQ ID NO: 44 и SEQ ID NO: 45 на мышах. Самцам мышей линии CD-1 (3 мыши на момент времени) вводили внутривенно слитые полипептиды в дозе, составляющей 10 мг/кг. Уровни лекарственного средства выявляли при помощи ELISA сэндвич-типа, при котором выявляли целую биспецифическую конструкцию посредством мишеней GPC3 и CD137. Данные аппроксимировали с использованием двухкамерной модели.
[0064] На фигуре 26 представлен результат фармакокинетического анализа биспецифических слитых полипептидов с SEQ ID NO: 44 и SEQ ID NO: 45 на макаках крабоедах. Самцы макаков-крабоедов получали тестируемые продукты в виде внутривенного вливания продолжительностью, составляющей 60 минут, в дозе 3 мг/кг. Уровни лекарственного средства выявляли при помощи ELISA сэндвич-типа, при котором выявляли целую биспецифическую конструкцию посредством мишеней GPC3 и CD137. Данные аппроксимировали с использованием двухкамерной модели.
IV. ПОДРОБНОЕ ОПИСАНИЕ РАСКРЫТИЯ
[0065] Согласно некоторым вариантам осуществления слитый полипептид содержит по меньшей мере две субъединицы в любом порядке: первую субъединицу, которая предусматривает полноразмерный иммуноглобулин, его антиген-связывающий домен или мутеин липокалина, специфичный в отношении GPC3, и вторую субъединицу, которая предусматривает полноразмерный иммуноглобулин, его антиген-связывающий домен или мутеин липокалина, специфичный в отношении CD137.
[0066] Согласно некоторым вариантам осуществления слитый полипептид также может содержать третью субъединицу. К примеру, полипептид может содержать субъединицу, специфичную в отношении CD137. Согласно некоторым вариантам осуществления указанная третья субъединица предусматривает мутеин липокалина, специфичный в отношении CD137.
[0067] Согласно некоторым вариантам осуществления одна субъединица может быть присоединена к другой субъединице, как, по сути, показано на фигуре 1.
[0068] Например, один мутеин липокалина может быть присоединен посредством пептидной связи к C-концу домена тяжелой цепи (VH) иммуноглобулина, N-концу VH, C-концу легкой цепи (VL) иммуноглобулина и/или N-концу VL, как изображено на фигуре 1А. Согласно некоторым конкретным вариантам осуществления субъединица, представляющая собой мутеин липокалина, может быть слита на своем N-конце и/или своем C-конце с субъединицей, представляющей собой иммуноглобулин. Например, мутеин липокалина может быть присоединен посредством пептидной связи к C-концу константного участка тяжелой цепи (CH) или C-концу константного участка легкой цепи (CL) иммуноглобулина. Согласно еще некоторым дополнительным вариантам осуществления пептидная связь может представлять собой линкер, в частности неструктурированный линкер (G4S)3, например, который показан под SEQ ID NO: 49.
[0069] В качестве другого иллюстративного примера один мутеин липокалина может быть присоединен посредством пептидной связи к C-концу или N-концу Fc-фрагмента иммуноглобулина, как показано на фигуре 1B.
[0070] В качестве дополнительного примера один мутеин липокалина может быть присоединен посредством пептидной связи к одному или более другим мутеинам липокалина, как показано на фигуре 1C.
[0071] В связи с этим одна субъединица может быть слита на своем N-конце и/или своем C-конце с другой субъединицей. Например, в тех случаях, когда одна субъединица предусматривает полноразмерный иммуноглобулин, другая субъединица может быть присоединена посредством пептидной связи к N-концу второй субъединицы и C-концу константного участка тяжелой цепи (CH) указанного иммуноглобулина. Согласно некоторым дополнительным вариантам осуществления третья субъединица может быть присоединена посредством пептидной связи к N-концу третьего связывающего домена и C-концу константного участка легкой цепи (CL) указанного иммуноглобулина. Согласно еще некоторым дополнительным вариантам осуществления пептидная связь может представлять собой линкер, в частности неструктурированный линкер (G4S)3, например, который показан под SEQ ID NO: 49, или может представлять собой неструктурированный линкер (G4S)2, например, который показан под SEQ ID NO: 48.
[0072] огласно некоторым вариантам осуществления третья субъединица присоединена к первой субъединице посредством пептидной связи к N-концу мутеина липокалина третьей субъединицы и C-концу константного участка легкой цепи (CL) иммуноглобулина первой субъединицы.
[0073] Согласно некоторым вариантам осуществления относительно слитого полипептида по настоящему раскрытию, одна из субъединиц которого предусматривает полноразмерный иммуноглобулин, в то время как полипептид одновременно связывает GPC3 и CD137, одновременно может быть сохранена Fc-функция Fc-участка полноразмерного иммуноглобулина в отношении положительной по Fc-рецептору клетки.
[0074] Согласно некоторым другим вариантам осуществления относительно слитого полипептида по настоящему раскрытию, одна из субъединиц которого предусматривает полноразмерный иммуноглобулин, в то время как полипептид одновременно связывает GPC3 и CD137, Fc-функцию Fc-участка полноразмерного иммуноглобулина, т. е. связывание с положительными по рецептору Fc-гамма или FcRn клетками, можно ослаблять или полностью подавлять при помощи белковой инженерии. Этого можно добиваться, например, путем использования каркаса, который проявляет низкую степень взаимодействий с рецепторами Fc-гамма или FcRn, такого как IgG2 или IgG4. Для того, чтобы ослабить остаточное связывание с Fc-гамма рецепторами, в каркас IgG можно вводить мутации, такие как мутация F234A и/или мутация L235A. Кроме того, что касается каркаса IgG4, то мутацию S228P можно вводить для сведения к минимуму обмена в половине антитела IgG4. Согласно еще некоторым дополнительным вариантам осуществления дополнительная мутация N297A может присутствовать в тяжелой цепи иммуноглобулина слитого полипептида для удаления естественного мотива гликозилирования.
[0075] Согласно некоторым вариантам осуществления в результате одновременного связывания с GPC3 на опухолевых клетках и CD137 на поверхности эффекторных клеток иммунной системы, таких как T-клетки или NK-клетки, слитые полипептиды по настоящему раскрытию могут проявлять зависимую от GPC3 активацию эффекторных клеток, тем самым эффекторная клетка иммунной системы активно лизирует экспрессирующую GPC3 опухолевую клетку.
[0076] Согласно некоторым дополнительным вариантам осуществления слитый полипептид способен проявлять сопоставимый или более высокий уровень зависимой от GPC3 активации CD137, как и иммуноглобулин, включенный в такой слитый полипептид, например, при измерении в анализе, в котором показано зависимое от мишени распространение инфильтрирующих опухоль лимфоцитов ex-vivo, по сути, описанном в Chacon, J. A. et al., PloS one 2013 8(4):e60031. Согласно некоторым дополнительным вариантам осуществления слитый полипептид способен проявлять сопоставимый или более высокий уровень зависимой от GPC3 активации CD137, как и иммуноглобулин, включенный в такой слитый полипептид, например, при измерении в in-vivo полученной при помощи ксенотрансплантата модели гепатоцеллюлярной карциномы («HCC»), меланомы, карциномы из клеток Меркеля, опухоли Вильма и гепатобластомы человека, по аналогии с тем, что, по сути, описано в Kohrt, H. et al, J Clin Invest. 2012 Mar;122(3):1066-75).
[0077] Согласно некоторым вариантам осуществления Fc-часть иммуноглобулина, включенного в слитый полипептид по настоящему раскрытию, может способствовать поддержанию уровней слитого полипептида в сыворотке крови, имеющих решающее значение для его стабильности и нахождения в организме. Например, в тех случаях, когда Fc-часть связывается с Fc-рецепторами на эндотелиальных клетках и на фагоцитах, слитый полипептид может становиться интернализированным и возвращаться обратно в кровоток, что увеличивает время его полужизни в организме.
[0078] Согласно некоторым вариантам осуществления CD137-специфичная субъединица, включенная в слитый полипептид по настоящему раскрытию, может представлять собой мутеин липокалина, который специфичен в отношении CD137, такой как мутеин липокалина с SEQ ID NO: 26. Согласно некоторым вариантам осуществления CD137-специфичная субъединица, включенная в слитый полипептид по настоящему раскрытию, может представлять собой полноразмерный иммуноглобулин или его антиген-связывающий домен, который специфичен в отношении CD137, такой как моноклональное антитело (например, антитело с SEQ ID NO: 34 и 35 или антитело с SEQ ID NO: 51 и 52).
[0079] Согласно некоторым вариантам осуществления GPC3-специфичная субъединица, включенная в слитый полипептид по настоящему раскрытию, может представлять собой мутеин липокалина, который специфичен в отношении GPC3, такой как мутеин липокалина с SEQ ID NO: 8 или мутеин липокалина с SEQ ID NO: 10. Согласно некоторым вариантам осуществления CD137-специфичная субъединица, включенная в слитый полипептид по настоящему раскрытию, может представлять собой полноразмерный иммуноглобулин или его антиген-связывающий домен, который специфичен в отношении GPC3.
[0080] Согласно некоторым вариантам осуществления в слитом полипептиде по настоящему раскрытию CD137-специфичная субъединица слита с GPC3-специфичной субъединицей.
[0081] Согласно некоторым более конкретным вариантам осуществления специфичная к GPC3 субъединица предусматривает мутеин липокалина, а специфичная к CD137 субъединица предусматривает моноклональное антитело.
[0082] Согласно некоторым дополнительным вариантам осуществления слитый полипептид по настоящему раскрытию содержит две специфичные к GPC3 субъединицы и одну специфичную к CD137 субъединицу. Согласно некоторым более конкретным вариантам осуществления каждая из специфичной к GPC3 субъединицы предусматривает мутеин липокалина, а каждая из специфичной к CD137 субъединицы предусматривает моноклональное антитело. Согласно некоторым дополнительным вариантам осуществления две специфичные к GPC3 субъединицы являются идентичными. Согласно еще некоторым дополнительным вариантам осуществления три субъединицы слиты друг с другом, как структурно показано на фигуре 1A. Согласно некоторым вариантам осуществления слитый полипептид предусматривает аминокислотные последовательности, выбранные из группы, состоящей из SEQ ID NO: 36 и 37, 38 и 39, 40 и 41 или 42 и 43.
[0083] Согласно некоторым другим определенным вариантам осуществления специфичная к GPC3 субъединица предусматривает мутеин липокалина, а специфичная к CD137 субъединица предусматривает мутеин липокалина. Согласно некоторым дополнительным вариантам осуществления две субъединицы слиты друг с другом, как структурно показано на фигуре 1C. Согласно некоторым вариантам осуществления слитый полипептид предусматривает аминокислотную последовательность с SEQ ID NO: 46.
[0084] Согласно некоторым дополнительным конкретным вариантам осуществления слитый полипептид по настоящему раскрытию содержит две специфичные к CD137 субъединицы и одну специфичную к GPC3 субъединицу. Согласно некоторым более конкретным вариантам осуществления специфичная к GPC3 субъединица предусматривает мутеин липокалина, и каждая из специфичной к CD137 субъединицы предусматривает мутеин липокалина. Согласно некоторым дополнительным вариантам осуществления две специфичные к CD137 субъединицы являются идентичными. Согласно некоторым дополнительным вариантам осуществления три субъединицы слиты друг с другом, как структурно показано на фигуре 1C. Согласно некоторым вариантам осуществления слитый полипептид предусматривает аминокислотную последовательность с SEQ ID NO: 47.
[0085] Согласно некоторым дополнительным вариантам осуществления в слитом полипептиде по настоящему раскрытию специфичная к GPC3 субъединица предусматривает мутеин липокалина, и специфичная к CD137 субъединица предусматривает мутеин липокалина, и две субъединицы слиты с Fc-фрагментом иммуноглобулина. Согласно некоторым дополнительным вариантам осуществления две субъединицы слиты с Fc-фрагментом иммуноглобулина, как структурно показано на фигуре 1B. Согласно некоторым конкретным вариантам осуществления Fc-фрагмент иммуноглобулина представляет собой Fc-фрагмент IgG4. Согласно некоторым дополнительным вариантам осуществления Fc-фрагмент IgG4 сконструирован с тем, чтобы в нем была мутация S228P и чтобы сводить к минимуму обмен в половине антитела IgG4 in-vitro и in-vivo. Согласно некоторым вариантам осуществления Fc-фрагмент IgG4 характеризуется аминокислотной последовательностью с SEQ ID NO: 73. Согласно некоторым вариантам осуществления слитый полипептид предусматривает аминокислотную последовательность с SEQ ID NO: 44 или с SEQ ID NO: 45.
[0086] Согласно некоторым вариантам осуществления иммуноглобулин, включенный в слитый полипептид по настоящему раскрытию, содержит каркас IgG2 или IgG4. Согласно некоторым дополнительным вариантам осуществления в каркасе IgG4 имеется любая из следующих мутаций, выбранных из группы, состоящей из S228P, N297A, F234A и L235A. Согласно некоторым дополнительным вариантам осуществления в каркасе IgG2 имеется любая из следующих мутаций, выбранных из группы, состоящей из N297A, F234A и L235A.
[0087] Согласно некоторым вариантам осуществления слитый полипептид может быть способен связывать CD137 со значением EC50, составляющим по меньшей мере приблизительно 5 нМ или даже ниже, как например: приблизительно 1 нМ или ниже, приблизительно 0,6 нМ или ниже, приблизительно 0,5 нМ или ниже, приблизительно 0,4 нМ или ниже или приблизительно 0,3 нМ или ниже, например, при оценке полипептида в анализе ELISA, по сути, описанном в примере 3, примере 8 или примере 15.
[0088] Согласно некоторым вариантам осуществления слитый полипептид по настоящему раскрытию может быть способен связывать CD137 со значением EC50, по меньшей мере таким же как или превосходящим значение EC50 для мутеина липокалина, специфичного в отношении CD137, который включен в такой слитый полипептид, такого как мутеин липокалина с SEQ ID NO: 26, или антитела, специфичного в отношении CD137, которые включены в такой слитый полипептид, такого как антитело с SEQ ID NO: 34 и 35 или антитело с SEQ ID NO: 51 и 52, например, при оценке указанного мутеина липокалина или антитела и полипептида в анализе ELISA, по сути, описанном в примере 8 или примере 15.
[0089] Согласно некоторым вариантам осуществления слитый полипептид может быть способен связывать CD137 с аффинностью, характеризующейся значением KD, составляющим по меньшей мере приблизительно 5 нМ или даже ниже, как например: приблизительно 1 нМ или ниже, приблизительно 0,6 нМ или ниже, приблизительно 0,5 нМ или ниже, приблизительно 0,3 нМ или ниже, приблизительно 200 пМ или ниже, приблизительно 150 пМ или ниже, приблизительно 100 пМ или ниже, или приблизительно 70 пМ или ниже, или приблизительно 2 пМ или ниже, например, при измерении в анализе с применением поверхностного плазмонного резонанса (SPR), по сути, описанном в примере 6, примере 11 или примере 18.
[0090] Согласно другому аспекту слитый полипептид может быть способен связывать GPC3 со значением EC50, составляющим по меньшей мере приблизительно 5 нМ или даже ниже, как например: приблизительно 1 нМ или ниже, приблизительно 0,6 нМ или ниже, приблизительно 0,5 нМ или ниже, приблизительно 0,4 нМ или ниже, приблизительно 0,3 нМ или ниже или приблизительно 0,2 нМ или ниже, например, при оценке полипептида в анализе ELISA, по сути, описанном в примере 2, примере 7, примере 12 или примере 14.
[0091] Согласно некоторым вариантам осуществления слитый полипептид по настоящему раскрытию может быть способен связывать GPC3 со значением EC50, сопоставимым со значением EC50 для мутеина липокалина, специфичного в отношении GPC3, который включен в такой слитый полипептид, такого как мутеин липокалина с SEQ ID NO: 8 или мутеин липокалина с SEQ ID NO: 10, например, при оценке указанного мутеина липокалина и слитого полипептида в анализе ELISA, по сути, описанном в примере 7, примере 12 или примере 14.
[0092] Согласно некоторым вариантам осуществления слитый полипептид может быть способен связывать GPC3 с аффинностью, характеризующейся значением KD, составляющим по меньшей мере приблизительно 5 нМ или даже ниже, как например: приблизительно 1 нМ, приблизительно 0,3 нМ, приблизительно 100 пМ, приблизительно 50 пМ или ниже, приблизительно 20 пМ или ниже или приблизительно 10 пМ или ниже, например, при измерении в анализе с применением поверхностного плазмонного резонанса (SPR), по сути, описанном в примере 5, примере 10 или примере 17.
[0093] Согласно некоторым вариантам осуществления слитые полипептиды по настоящему раскрытию, специфичные в отношении как CD137, так и GPC3, могут быть способны одновременно связывать CD137 и GPC3, например, при оценке указанного слитого полипептида в анализе ELISA, по сути, описанном в примере 4, примере 9, примере 13 или примере 16.
[0094] Согласно некоторым вариантам осуществления слитые полипептиды по настоящему раскрытию, специфичные в отношении как CD137, так и GPC3, могут быть способны одновременно связывать CD137 и GPC3 со значением EC50, составляющим по меньшей мере приблизительно 10 нМ или даже ниже, как например: приблизительно 8 нМ или ниже, приблизительно 5 нМ или ниже, приблизительно 2,5 нМ или ниже, приблизительно 2 нМ или ниже или приблизительно 1,5 нМ или ниже, например, при оценке указанного слитого полипептида в анализе ELISA, по сути, описанном в примере 4, примере 9, примере 13 или примере 16.
[0095] Согласно некоторым вариантам осуществления слитые полипептиды по настоящему раскрытию, специфичные в отношении как CD137, так и GPC3, могут быть способны костимулировать ответы T-клеток в функциональном анализе активации T-клеток, по сути, описанном в примере 19. Согласно некоторым вариантам осуществления слитые полипептиды по настоящему раскрытию могут быть способны индуцировать продуцирование IL-2 в присутствии стимуляции T-клеток в функциональном анализе активации T-клеток, по сути, описанном в примере 19, и даже могут проявлять тенденцию к более сильной индукции продуцирования IL-2 при более высоких концентрациях в покрывающем слое. Согласно некоторым вариантам осуществления слитые полипептиды по настоящему раскрытию не индуцируют продуцирование IL-2 в отсутствие стимуляции антителом к CD3 T-клеток в функциональном анализе активации T-клеток, по сути, описанном в примере 19. Согласно некоторым дополнительным вариантам осуществления слитые полипептиды по настоящему раскрытию, специфичные в отношении как CD137, так и GPC3, могут быть способны костимулировать активацию T-клеток, стимулированных антителами к CD3 и к CD28 при субоптимальных концентрациях в функциональном анализе активации T-клеток, по сути, описанном в примере 19.
[0096] Согласно некоторым вариантам осуществления слитые полипептиды по настоящему раскрытию, специфичные в отношении как CD137, так и GPC3, могут быть способны костимулировать ответы T-клеток в функциональном анализе активации T-клеток, по сути, описанном в примере 20. Согласно некоторым вариантам осуществления слитые полипептиды по настоящему раскрытию могут быть способны индуцировать продуцирование IL-2 в функциональном анализе активации T-клеток, по сути, описанном в примере 20. Согласно некоторым вариантам осуществления слитые полипептиды по настоящему раскрытию могут быть способны костимулировать активацию T-клеток зависимым от мишени, экспрессирующей GPC3, образом в функциональном анализе активации T-клеток, по сути, описанном в примере 20.
A. Примерные иммуноглобулины, которые включены в слитые полипептиды
[0097] Согласно некоторым вариантам осуществления относительно слитого полипептида первый связывающий домен предусматривает полноразмерный иммуноглобулин или его антиген-связывающий домен, специфичный в отношении GPC3 или CD137. Иммуноглобулин, например, может представлять собой IgG1, IgG2 или IgG4. Согласно дополнительным вариантам осуществления иммуноглобулин представляет собой моноклональное антитело к GPC3 или CD137. Иллюстративным примером связывающего GPC3 иммуноглобулина является GC33 (Cancer Sci. 2014 Apr;105(4):455-62.). Иллюстративными примерами связывающих CD137 антител являются BMS-663513 (Jure-Kunkel, M. et al., патент США 7288638) и PF-05082566 (Fisher, T. S. et al., Canc Immunol Immunother 2012 Oct; 61(10):1721-1733).
B. Примерные специфичные к GPC3 мутеины липокалина, которые включены в слитые полипептиды
[0098] В одном аспекте настоящего раскрытия предусмотрен мутеин липокалина, который способен связывать глипикан-3 (GPC3) человека с аффинностью, измеренной по KD, составляющей приблизительно 1 нМ или ниже. Более предпочтительно мутеин может характеризоваться аффинностью, измеренной по KD, составляющей приблизительно 1 нМ или 0,2 нМ или ниже.
[0099] В другом варианте осуществления настоящее раскрытие относится к мутеину липокалина, где указанный мутеин содержит в одном или более положениях, соответствующих положению 36, 40, 41, 49, 52, 65, 68, 70, 72, 73, 77, 79, 81, 87, 96, 100, 103, 105, 106, 125, 127, 132, 134, 136 и/или 175 в линейной полипептидной последовательности hNGAL (SEQ ID NO: 2), замену, предпочтительно замену, описанную в данном документе.
[00100] Согласно конкретным вариантам осуществления мутеин по настоящему раскрытию содержит по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 или даже больше, как например: 21, 22, 23, 24, 25 и 26, замен в положении последовательности, соответствующем положению 36, 40, 41, 49, 52, 65, 68, 70, 72, 73, 77, 79, 81, 87, 96, 100, 103, 105, 106, 125, 127, 132, 134, 136 и/или 175 последовательности в линейной полипептидной последовательности зрелого hNGAL (SEQ ID NO: 2).
[00101] Согласно дополнительным конкретным вариантам осуществления мутеин липокалина согласно настоящему раскрытию содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4-17. В другом варианте осуществления мутеин характеризуется по меньшей мере 70% идентичностью с последовательностью зрелого hNGAL (SEQ ID NO: 2). Предпочтительно указанный мутеин содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 или даже больше, как например: 21, 22, 23, 24, 25 и 26, подвергнутых мутации аминокислотных остатков в положении 36, 40, 41, 49, 52, 65, 68, 70, 72, 73, 77, 79, 81, 87, 96, 100, 103, 105, 106, 125, 127, 132, 134, 136 и/или 175 последовательности в линейной полипептидной последовательности зрелого hNGAL (SEQ ID NO: 2).
[00102] В некоторых дополнительных вариантах осуществления для облегчения экспрессии в эукариотических клетках природный сайт Asn N-гликозилирования в положении 65 линейной полипептидной последовательности зрелого hNGAL (SEQ ID NO: 2) удаляют в соответствующем положении в последовательности мутеина липокалина согласно настоящему раскрытию, например, посредством мутации с заменой Asn на Asp в положении 65. Кроме того, предпочтительно, чтобы сайты N-гликозилирования (Asn-X-Ser/Thr) не существовали на мутеине липокалина согласно настоящему раскрытию.
[00103] В некоторых других вариантах осуществления мутеин липокалина согласно настоящему раскрытию не содержит мутацию в положении последовательности, соответствующем положению 28 последовательности в линейной полипептидной последовательности зрелого hNGAL (SEQ ID NO: 2), что, например, позволяет дополнительно оптимизировать стабильность.
[00104] В другом варианте осуществления мутеин по настоящему раскрытию является антагонистом GPC3.
[00105] Как используется в данном документе, мутеин липокалина по настоящему раскрытию «специфически связывает» мишень (в данном документе GPC3), если он способен отличать ту мишень от одной или более эталонных мишеней, поскольку специфичность связывания является не абсолютным, а относительным свойством. «Специфичное связывание» можно определять, например, согласно результатам вестерн-блоттингов, анализов ELISA, RIA, ECL, IRMA, с использованием FACS, IHC и пептидных сканирований.
[00106] Аналогично этому в другом аспекте настоящее раскрытие относится к мутеину hNGAL, где указанный мутеин содержит в одном или более положениях, соответствующих положению 36, 40, 41, 49, 52, 68, 70, 72, 73, 77, 79, 81, 96, 100, 103, 106, 125, 127, 132 и/или 134 в линейной полипептидной последовательности зрелого hNGAL (SEQ ID NO: 2), замену, предпочтительно замену, описанную в данном документе.
[00107] Согласно альтернативному аспекту настоящее раскрытие относится к полипептиду, предусматривающему мутеин hNGAL, где мутеин hNGAL содержит в 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20, или даже большем количестве, как например: 21, 22, 23, 24, 25 или 26, положений аминокислот, соответствующих положениям 36, 40, 41, 49, 52, 65, 68, 70, 72, 73, 77, 79, 81, 87, 96, 100, 103, 105, 106, 125, 127, 132, 134, 136 и/или 175 в линейной полипептидной последовательности зрелого hNGAL (SEQ ID NO: 2), замену, предпочтительно замену, описанную в данном документе.
[00108] Подобным образом настоящее раскрытие относится к мутеину липокалина, полученному из hNGAL, с участком с супервторичной структурой, представляющей собой цилиндрический β-складчатый лист, содержащим восемь
-нитей, соединенных попарно четырьмя петлями на одном конце с образованием таким образом кармана связывания, где по меньшей мере одна аминокислота из каждой по меньшей мере из трех из указанных четырех петель была подвергнута мутации и где указанный липокалин является эффективным в связывании GPC3 в качестве заданной отличной от естественной мишени с выявляемой аффинностью. Преимущественно мутеин липокалина содержит в 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 аминокислотных положениях, соответствующих аминокислоте в положении 36, 40, 41, 49, 52, 65, 68, 70, 72, 73, 77, 79, 81, 87, 96, 100, 103, 105, 106, 125, 127, 132, 134, 136 и/или 175 в линейной полипептидной последовательности hNGAL (SEQ ID NO: 1), замену, предпочтительно замену, описанную в данном документе. Настоящее раскрытие также относится к нуклеиновым кислотам, кодирующим эти белки.
[00109] Таким образом, с учетом вышеизложенного специалист в данной области может легко определить, какое аминокислотное положение, подвергнутое мутации в hNGAL, описанном в данном документе, соответствует аминокислоте остова, отличного от hNGAL. В частности, специалист в данной области может выровнять аминокислотную последовательность мутеина, описанного в данном документе, в частности мутеина hNGAL по настоящему раскрытию с аминокислотной последовательностью отличного мутеина для определения того, какая(какие) аминокислота(аминокислоты) указанного мутеина соответствует(соответствуют) соответствующей(соответствующим) аминокислоте(аминокислотам) аминокислотной последовательности указанного отличного липокалина. Говоря более конкретно, специалист в данной области может таким образом определить, какая аминокислота в аминокислотной последовательности указанного отличного липокалина соответствует аминокислоте в положении(положениях) 36, 40, 41, 49, 52, 65, 68, 70, 72, 73, 77, 79, 81, 87, 96, 100, 103, 105, 106, 125, 127, 132, 134, 136 и/или 175 в линейной полипептидной последовательности hNGAL (SEQ ID NO: 2).
[00110] Белки по настоящему раскрытию, которые направлены против или являются специфическими в отношении GPC3, включают любое количество специфически связывающих белков-мутеинов, в которых за основу взят остов определенного белка. Как используется в данном документе, «мутеин», «подвергнутый мутации» объект (либо белок, либо нуклеиновая кислота) или «мутант» относится к обмену, делеции или вставке соответственно одного или более из нуклеотидов или аминокислот по сравнению со встречающимся в природе (дикого типа) «эталонным» остовом нуклеиновой кислоты или белка. Предпочтительно количество нуклеотидов или аминокислот соответственно, которое обменивают, подвергают делеции или вставляют, составляет 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 или даже больше, например, 21, 22, 23, 24, 25 и 26. Однако предпочтительно, чтобы мутеин по настоящему раскрытию по-прежнему был способен к связыванию GPC3.
[00111] В некоторых предпочтительных вариантах осуществления мутеин согласно настоящему раскрытию связывает GPC3 человека или мыши с KD, составляющей приблизительно 1 нМ или меньше, в том числе 0,5 нМ или меньше, 0,3 нМ или меньше и или 0,2 нМ или меньше. Мутеин по настоящему раскрытию может специфически связывать один или более смежных, несмежных или конформационных эпитопов зрелой свернутой биологически активной формы GPC3.
[00112] Аффинность связывания белка по настоящему раскрытию (например, мутеина липокалина) с выбранной мишенью (в данном случае GPC3) можно измерять (и таким образом определять значения KD для комплекса мутеин-лиганд) с помощью большого количества способов, известных специалисту в данной области. Такие способы включают без ограничения флуоресцентное титрование, конкурентный ELISA, калориметрические способы, такие как изотермическая титрационная калориметрия (ITC) и поверхностный плазмонный резонанс (BIAcore). Такие способы хорошо известны из уровня техники и их примеры также подробно описаны ниже.
[00113] Аминокислотная последовательность мутеина по настоящему раскрытию может характеризоваться высокой идентичностью последовательности со зрелым липокалином 2 человека. В данном случае белок по настоящему раскрытию может характеризоваться по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 82%, по меньшей мере 85%, по меньшей мере 87%, по меньшей мере 90% идентичностью, в том числе по меньшей мере 95% идентичностью с белком, выбранным из группы, состоящей из последовательности под SEQ ID NO: 2, причем такой мутеин с аминокислотной последовательностью выбран из группы, состоящей из SEQ ID NO: 4-17.
[00114] Настоящее раскрытие также включает структурные гомологи белков, выбранных из группы, состоящей из последовательностей под SEQ ID NO: 4-17, которые характеризуются гомологией аминокислотной последовательности или идентичностью последовательностей, составляющей более чем приблизительно 60%, предпочтительно более 65%, более 70%, более 75%, более 80%, более 85%, более 90%, более 92 % и наиболее предпочтительно более 95% относительно них.
[00115] В соответствии с вышеизложенным мутеин по настоящему раскрытию предпочтительно действует как антагонист GPC3. Согласно некоторым вариантам осуществления мутеин по настоящему раскрытию может действовать как антагонист GPC3 посредством ингибирования способности молекулы GPC3 связываться или иным образом взаимодействовать с ее когнатным лигандом.
[00116] Согласно еще одному аспекту настоящее раскрытие включает мутеины липокалина 2 человека, которые специфически связывают GPC3. В этом смысле GPC3 может быть рассмотрен как отличный от естественного лиганд дикого типа липокалина 2 человека, где «отличный от естественного лиганд» относится к соединению, которое не связывается с липокалином 2 человека в физиологических условиях. Путем внесения в липокалины дикого типа, такие как липокалин 2 человека, мутаций в определенные положения авторы настоящего изобретения показали, что возможны высокая аффинность и высокая специфичность в отношении отличного от естественного лиганда. Согласно одному аспекту по меньшей мере в 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 и/или 20 нуклеотидных триплетах, кодирующих любые положения 36, 40, 41, 49, 52, 65, 68, 70, 72, 73, 77, 79, 81, 87, 96, 100, 103, 105, 106, 125, 127, 132, 134, 136 и/или 175 последовательности в линейной полипептидной последовательности зрелого липокалина 2 человека (SEQ ID NO: 2), случайный мутагенез можно осуществлять путем обеспечения замены в этих положениях подмножеством нуклеотидных триплетов.
[00117] К тому же липокалины можно применять для создания мутеинов, которые имеют подвергнутый мутации аминокислотный остаток в любом одном или более, в том числе по меньшей мере в любых двух, трех, четырех, пяти, шести, семи, восьми, девяти, десяти, одиннадцати, двенадцати, тринадцати, четырнадцати, пятнадцати, шестнадцати, семнадцати, восемнадцати, девятнадцати или двадцати положениях в последовательности, соответствующих положениям 36, 40, 41, 49, 52, 65, 68, 70, 72, 73, 77, 79, 81, 87, 96, 100, 103, 105, 106, 125, 127, 132, 134, 136 и/или 175 последовательности в линейной полипептидной последовательности зрелого липокалина 2 человека (SEQ ID NO: 2).
[00118] Замена в положении 36 в последовательности может, например, представлять собой замену Leu 36 → Val или Arg. Замена в положении 40 в последовательности может, например, представлять собой замену Ala 40 → Leu, Val или Gly. Замена в положении 41 в последовательности может, например, представлять собой замену Ile 41 → Leu, Arg, Met, Gly или Ala. Замена в положении 49 в последовательности может, например, представлять собой замену Gln 49 → Pro или Leu. Замена в положении 52 в последовательности может, например, представлять собой замену Tyr 52 → Arg или Trp. Замена в положении 68 в последовательности может, например, представлять собой замену Asn 65 → Asp. Замена в положении 68 в последовательности может, например, представлять собой замену Ser 68 → Val, Gly, Asn или Ala. Замена в положении 70 в последовательности может, например, представлять собой замену Leu 70 → Arg, Ser, Ala или Val. Замена в положении 72 в последовательности может, например, представлять собой замену Arg 72 → Asp, Trp, Ala или Gly. Замена в положении 73 в последовательности может, например, представлять собой замену Lys 73 → Gly, Arg, Asn, Glu или Ser. Замена в положении 76 в последовательности может, например, представлять собой замену Cys 76 → Val или Ile. Замена в положении 77 в последовательности может, например, представлять собой замену Asp 77 → His, Met, Val, Leu, Thr или Lys. Замена в положении 79 в последовательности может, например, представлять собой замену Trp 79 → Lys, Ser или Thr. Замена в положении 81 в последовательности может, например, представлять собой замену Arg 81 → Gly. Замена в положении 81 в последовательности может, например, представлять собой замену Cys 87 → Ser. Замена в положении 96 в последовательности может, например, представлять собой замену Asn 96 → Arg, Asp, Gln или Pro. Замена в положении 100 в последовательности может, например, представлять собой замену Tyr 100 → Gly, Glu, Pro или Gln. Замена в положении 103 в последовательности может, например, представлять собой замену Leu 103 → Glu, Gln, Asn, Gly, Ser или Tyr. Замена в положении 106 в последовательности может, например, представлять собой замену Ser 105 → Ala. Замена в положении 106 в последовательности может, например, представлять собой замену Tyr 106 → Asn, Ser или Thr. Замена в положении 125 в последовательности может, например, представлять собой замену Lys 125 → Glu. Замена в положении 127 в последовательности может, например, представлять собой замену Ser 127 → Arg или Tyr. Замена в положении 132 в последовательности может, например, представлять собой замену Tyr 132 → Trp или Ile. Замена в положении 134 в последовательности может, например, представлять собой замену Lys 134 → Ala или Phe. Замена в положении 134 в последовательности может, например, представлять собой замену Thr 136 → Ile. Замена в положении 175 в последовательности может, например, представлять собой замену Cys 175 → Ala. Следует отметить, что любые из аминокислот, которые замещают соответствующую аминокислоту в эталонной последовательности, можно заменить соответствующей консервативной аминокислотой. В частности, консервативные замещения представляют собой замещения из числа представителей следующих групп: 1) аланин, серин и треонин; 2) аспарагиновая кислота и глутаминовая кислота; 3) аспарагин и глутамин; 4) аргинин и лизин; 5) изолейцин, лейцин, метионин и валин и 6) фенилаланин, тирозин и триптофан.
[00119] В одном варианте осуществления мутеин по настоящему раскрытию, который связывается с GPC3, включает следующие аминокислотные замещения:
(a) Leu 36 → Val; Ile 41 → Leu; Gln 49 → Leu; Tyr 52 → Arg; Asn 65 → Asp; Ser 68 → Val; Leu 70 → Ser; Arg 72 → Trp; Lys 73 → Arg; Asp 77 → His; Trp 79 → Lys; Arg 81 → Gly; Cys 87 → Ser; Asn 96 → Asp; Tyr 100 → Gly; Leu 103 → Gln; Tyr 106 → Asn; Lys 125 → Glu; Ser 127 → Arg; Tyr 132 → Trp; Lys 134 → Ala;
(b) Leu 36 → Val; Ala 40 → Val; Ile 41 → Arg; Gln 49 → Pro; Tyr 52 → Arg; Asn 65 → Asp; Ser 68 → Gly; Leu 70 → Ser; Lys 73 → Gly; Asp 77 → His; Trp 79 → Lys; Arg 81 → Gly; Cys 87 → Ser; Asn 96 → Asp; Tyr 100 → Gly; Leu 103 → Glu; Tyr 106 → Asn; Lys 125 → Glu; Ser 127 → Arg; Tyr 132 → Trp; Lys 134 → Phe;
(c) Leu 36 → Val; Ala 40 → Gly; Ile 41 → Met; Gln 49 → Leu; Tyr 52 → Arg; Asn 65 → Asp; Leu 70 → Ala; Lys 73 → Asn; Asp 77 → His; Trp 79 → Lys; Arg 81 → Gly; Cys 87 → Ser; Asn 96 → Gln; Tyr 100 → Gly; Leu 103 → Glu; Tyr 106 → Asn; Lys 125 → Glu; Ser 127 → Arg; Tyr 132 → Trp; Lys 134 → Phe;
(d) Leu 36 → Arg; Ala 40 → Val; Ile 41→ Gly; Gln 49 → Pro; Tyr 52 → Trp; Asn 65 → Asp; Ser 68 → Asn; Leu 70 → Arg; Arg 72 → Ala; Lys 73 → Arg; Asp 77 → Leu; Trp 79 → Ser; Arg 81 → Gly; Cys 87 → Ser; Asn 96 → Gln; Tyr 100 → Glu; Leu 103 → Asn; Ser 105 → Ala; Tyr 106 → Asn; Lys 125 → Glu; Ser 127 → Tyr; Tyr 132 → Ile; Lys 134 → Phe; Thr 136 → Ile;
(e) Leu 36 → Arg; Ala 40 → Val; Ile 41→ Gly; Gln 49 → Pro; Tyr 52 → Trp; Asn 65 → Asp; Ser 68 → Asn; Leu 70 → Arg; Arg 72 → Ala; Lys 73 → Arg; Asp 77 → Thr; Trp 79 → Ser; Arg 81 → Gly; Cys 87 → Ser; Asn 96 → Gln; Tyr 100 → Glu; Leu 103 → Gly; Ser 105 → Ala; Tyr 106 → Asn; Lys 125 → Glu; Ser 127 → Tyr; Tyr 132 → Ile; Lys 134 → Phe; Thr 136 → Ile;
(f) Leu 36 → Arg; Ala 40 → Gly; Ile 41→ Ala; Gln 49 → Pro; Tyr 52 → Trp; Asn 65 → Asp; Ser 68 → Asn; Leu 70 → Arg; Arg 72 → Ala; Lys 73 → Arg; Asp 77 → Val; Trp 79 → Ser; Arg 81 → Gly; Cys 87 → Ser; Asn 96 → Pro; Tyr 100 → Glu; Leu 103 → Asn; Ser 105 → Ala; Tyr 106 → Ser; Lys 125 → Glu; Ser 127 → Tyr; Tyr 132 → Ile; Lys 134 → Phe; Thr 136 → Ile;
(g) Leu 36 → Arg; Ala 40 → Val; Ile 41→ Ala; Gln 49 → Pro; Tyr 52 → Arg; Asn 65 → Asp; Ser 68 → Ala; Leu 70 → Arg; Arg 72 → Ala; Lys 73 → Arg; Asp 77 → Leu; Trp 79 → Ser; Arg 81 → Gly; Cys 87 → Ser; Asn 96 → Arg; Tyr 100 → Glu; Leu 103 → Tyr; Ser 105 → Ala; Tyr 106 → Asn; Lys 125 → Glu; Ser 127 → Tyr; Tyr 132 → Ile; Lys 134 → Phe; Thr 136 → Ile;
(h) Leu 36 → Arg; Ala 40 → Val; Ile 41→ Ala; Gln 49 → Pro; Tyr 52 → Arg; Asn 65 → Asp; Ser 68 → Asn; Leu 70 → Val; Arg 72 → Ala; Lys 73 → Gly; Asp 77 → Lys; Trp 79 → Ser; Arg 81 → Gly; Cys 87 → Ser; Asn 96 → Arg; Tyr 100 → Pro; Leu 103 → Asn; Ser 105 → Ala; Tyr 106 → Asn; Lys 125 → Glu; Ser 127 → Tyr; Tyr 132 → Ile; Lys 134 → Phe; Thr 136 → Ile;
(i) Leu 36 → Arg; Ala 40 → Leu; Ile 41→ Gly; Gln 49 → Pro; Tyr 52 → Trp; Asn 65 → Asp; Ser 68 → Asn; Leu 70 → Arg; Arg 72 → Ala; Lys 73 → Arg; Asp 77 → Met; Trp 79 → Ser; Arg 81 → Gly; Cys 87 → Ser; Asn 96 → Gln; Tyr 100 → Glu; Leu 103 → Ser; Ser 105 → Ala; Tyr 106 → Asn; Lys 125 → Glu; Ser 127 → Tyr; Tyr 132 → Ile; Lys 134 → Phe;
(j) Leu 36 → Arg; Ala 40 → Val; Ile 41→ Gly; Gln 49 → Pro; Tyr 52 → Trp; Asn 65 → Asp; Ser 68 → Asn; Leu 70 → Arg; Arg 72 → Ala; Lys 73 → Gly; Cys 76 → Val; Asp 77 → Lys; Trp 79 → Thr; Arg 81 → Gly; Cys 87 → Ser; Asn 96 → Gln; Tyr 100 → Glu; Leu 103 → Asn; Ser 105 → Ala; Tyr 106 → Thr; Lys 125 → Glu; Ser 127 → Tyr; Tyr 132 → Ile; Lys 134 → Phe; Cys 175 → Ala;
(k) Leu 36 → Arg; Ala 40 → Val; Ile 41→ Gly; Gln 49 → Pro; Tyr 52 → Arg; Asn 65 → Asp; Ser 68 → Gly; Leu 70 → Arg; Arg 72 → Gly; Lys 73 → Glu; Cys 76 → Ile; Asp 77 → Lys; Trp 79 → Ser; Arg 81 → Gly; Cys 87 → Ser; Asn 96 → Gln; Tyr 100 → Gln; Leu 103 → Asp; Ser 105 → Ala; Tyr 106 → Thr; Lys 125 → Glu; Ser 127 → Tyr; Tyr 132 → Ile; Lys 134 → Phe; Thr 136 → Ile; Cys 175 → Ala или
(l) Leu 36 → Arg; Ala 40 → Val; Ile 41→ Gly; Gln 49 → Pro; Tyr 52 → Arg; Asn 65 → Asp; Ser 68 → Gly; Leu 70 → Arg; Arg 72 → Asp; Lys 73 → Ser; Cys 76 → Val; Asp 77 → Thr; Trp 79 → Ser; Arg 81 → Gly; Cys 87 → Ser; Asn 96 → Gln; Tyr 100 → Glu; Leu 103 → Asn; Ser 105 → Ala; Tyr 106 → Thr; Lys 125 → Glu; Ser 127 → Tyr; Tyr 132 → Ile; Lys 134 → Phe; Thr 136 → Ile; Cys 175 → Ala.
[00120] Нумерация предпочтительно связана с линейной полипептидной последовательностью зрелого hNGAL (SEQ ID NO: 2). Соответственно, с учетом идеи настоящего раскрытия специалист в данной области может легко определить, какие аминокислоты в предпочтительной эталонной последовательности зрелого hNGAL (SEQ ID NO: 2) соответствуют описанным выше в (a) - (l); таким образом, чтобы подвергнуть мутации указанные аминокислоты в эталонной последовательности.
C. Примерные специфичные к CD137 мутеины липокалина, которые включены в слитые полипептиды
[00121] Согласно одному аспекту настоящее раскрытие предусматривает мутеины липокалина человека, которые связывают CD137, и их полезные пути применения. Настоящее раскрытие также предусматривает способы получения белков, связывающих CD137, описанных в данном документе, а также композиций, содержащих такие белки. Белки, связывающие CD137, по настоящему раскрытию, а также их композиции можно применять в способах выявления CD137 в образцах или в способах связывания CD137 в субъекте. Ранее не были описаны такие мутеины липокалина человека с такими характеристиками, сопутствующими путям применения, предусмотренным настоящим раскрытием.
[00122] Другой вариант осуществления настоящего раскрытия предусматривает мутеин липокалина, который способен к активации нисходящих сигнальных путей, связанных с CD137, путем связывания с CD137.
[00123] Согласно одному варианту осуществления настоящее раскрытие предусматривает связывающие CD137 мутеины липокалина слезы человека.
[00124] В связи с этим настоящее раскрытие предусматривает один или более мутеинов Tlc, которые способны к связыванию CD137 с аффинностью, измеренной по KD, составляющей приблизительно 300 нМ или ниже и даже приблизительно 100 нМ или ниже.
[00125] Согласно некоторым вариантам осуществления такой мутеин Tlc содержит подвергнутый мутации аминокислотный остаток в одном или более положениях, соответствующих положениям 5, 26-31, 33-34, 42, 46, 52, 56, 58, 60-61, 65, 71, 85, 94, 101, 104-106, 108, 111, 114, 121, 133, 148, 150 и 153 в линейной полипептидной последовательности зрелого липокалина слезы человека (SEQ ID NO: 1).
[00126] Согласно некоторым конкретным вариантам осуществления такой мутеин Tlc может содержать подвергнутый мутации аминокислотный остаток в одном или более положениях, соответствующих положениям 26-34, 55-58, 60-61, 65, 104-106 и 108 в линейной полипептидной последовательности зрелого липокалина слезы человека.
[00127] Согласно дополнительным конкретным вариантам осуществления такой мутеин Tlc может дополнительно включать подвергнутый мутации аминокислотный остаток в одном или более положениях, соответствующих положениям 101, 111, 114 и 153 в линейной полипептидной последовательности зрелого липокалина слезы человека.
[00128] Согласно другим конкретным вариантам осуществления Tlc может содержать подвергнутый мутации аминокислотный остаток в одном или более положениях, соответствующих положениям 5, 26-31, 33-34, 42, 46, 52, 56, 58, 60-61, 65, 71, 85, 94, 101, 104-106, 108, 111, 114, 121, 133, 148, 150 и 153 в линейной полипептидной последовательности зрелого липокалина слезы человека.
[00129] Согласно некоторым дополнительным вариантам осуществления мутеин Tlc может содержать по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26 или даже больше подвергнутых мутации аминокислотных остатков в одном или более положениях в последовательности, соответствующих положениям 5, 26-31, 33-34, 42, 46, 52, 56, 58, 60-61, 65, 71, 85, 94, 101, 104-106, 108, 111, 114, 121, 133, 148, 150 и 153 последовательности в линейной полипептидной последовательности зрелого липокалина слезы человека, и где указанный полипептид связывает CD137, в частности CD137 человека.
[00130] Согласно еще некоторым дополнительным вариантам осуществления настоящее раскрытие относится к полипептиду, где указанный полипептид представляет собой мутеин Tlc, по сравнению с линейной полипептидной последовательностью зрелого липокалина слезы человека, содержащий по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 или даже больше подвергнутых мутации аминокислотных остатков в положениях 526-34, 55-58, 60-61, 65, 104-106 и 108 в последовательности, и где указанный полипептид связывает CD137, в частности CD137 человека.
[00131] Согласно некоторым вариантам осуществления мутеин липокалина согласно настоящему раскрытию может включать по меньшей мере одну аминокислотную замену нативного остатка цистеина, например, остатком серина. Согласно некоторым вариантам осуществления мутеин Tlc согласно настоящему раскрытию включает аминокислотную замену нативного остатка цистеина в положениях 61 и/или 153 другой аминокислотой, такой как остаток серина. В данном случае отмечается, что было обнаружено, что удаление дисульфидной связи в структуре (на уровне соответствующей библиотеки не подвергнутых мутации нуклеиновых кислот) липокалина дикого типа слезы, которая образована остатками цистеина 61 и 153 (см. Breustedt, et al., 2005, ранее), может обеспечить мутеины липокалина слезы, которые не только стабильно сворачиваются, но и также способны связывать указанный отличный от естественного лиганд с высокой аффинностью. Согласно некоторым конкретным вариантам осуществления мутеин Tlc согласно настоящему раскрытию включает аминокислотные замены Cys 61 → Ala, Phe, Lys, Arg, Thr, Asn, Gly, Gln, Asp, Asn, Leu, Tyr, Met, Ser, Pro или Trp и Cys 153 → Ser или Ala. Было доказано, что такая замена удобна для предупреждения образования встречающегося в природе дисульфидного мостика, соединяющего Cys 61 и Cys 153, и облегчения тем самым манипуляции с мутеином. Однако мутеины липокалина слезы, которые связывают CD137 и которые имеют дисульфидный мостик, образованный между Cys 61 и Cys 153, также являются частью настоящего раскрытия.
[00132] Согласно некоторым вариантам осуществления устранение дисульфидной связи в структуре может обеспечивать дополнительное преимущество предоставления возможности (самопроизвольного) образования или преднамеренного введения отличных от естественных искусственных дисульфидных связей в мутеины по настоящему раскрытию, повышая тем самым стабильность мутеинов. Например, согласно некоторым вариантам осуществления или два, или все три из кодонов цистеина в положениях 61, 101 и 153 заменены кодоном другой аминокислоты. Кроме того, согласно некоторым вариантам осуществления мутеин Tlc согласно настоящему раскрытию включает аминокислотную замену нативного остатка цистеина в положении 101 остатком серина или остатком гистидина.
[00133] Согласно некоторым вариантам осуществления мутеин согласно настоящему раскрытию включает аминокислотную замену нативной аминокислоты остатком цистеина в положениях 28 или 105 в отношении аминокислотной последовательности зрелого липокалина слезы человека.
[00134] Кроме того, согласно некоторым вариантам осуществления мутеин согласно настоящему раскрытию включает аминокислотную замену нативного остатка аргинина в положении 111 остатком пролина. Кроме того, согласно некоторым вариантам осуществления мутеин согласно настоящему раскрытию включает аминокислотную замену нативного остатка лизина в положении 114 остатком триптофана или глутаминовой кислотой.
[00135] Согласно некоторым вариантам осуществления связывающий CD137 мутеин Tlc согласно настоящему раскрытию включает в одном или более положениях, соответствующих положениям 5, 26-31, 33-34, 42, 46, 52, 56, 58, 60-61, 65, 71, 85, 94, 101, 104-106, 108, 111, 114, 121, 133, 148, 150 и 153 в линейной полипептидной последовательности зрелого липокалина слезы человека (SEQ ID NO: 1) один или более из следующих подвергнутых мутации аминокислотных остатков: Ala 5 → Val или Thr; Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Thr 42 → Ser; Gly 46 → Asp; Lys 52 → Glu; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Lys 65 → Arg или Asn; Thr 71 → Ala; Val 85 → Asp; Lys 94 → Arg или Glu; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Lys 121 → Glu; Ala 133 → Thr; Arg 148 → Ser; Ser 150 → Ile и Cys 153 → Ser. Согласно некоторым вариантам осуществления мутеин Tlc согласно настоящему раскрытию включает два или более, а именно 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, более того, как например: 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26 или все подвергнутые мутации аминокислотные остатки в этих положениях в последовательности зрелого липокалина слезы человека.
[00136] Согласно некоторым дополнительным вариантам осуществления связывающий CD137 мутеин Tlc предусматривает один из следующих наборов аминокислотных замен по сравнению с линейной полипептидной последовательностью зрелого липокалина слезы человека:
1. Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Cys 153 → Ser;
2. Ala 5 → Thr; Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Lys 65 → Arg; Val 85 → Asp; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Lys 121 → Glu; Ala 133 → Thr; Cys 153 → Ser; 157 → Pro;
3. Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Lys 65 → Asn; Lys 94 → Arg; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Lys 121 → Glu; Ala 133 → Thr; Cys 153 → Ser;
4. Ala 5 → Val; Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Lys 65 → Arg; Lys 94 → Glu; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Lys 121 → Glu; Ala 133 → Thr; Cys 153 → Ser; 157 → Pro;
5. Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Thr 42 → Ser; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Ser 150 → Ile; Cys 153 → Ser; 157 → Pro;
6. Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Lys 52 → Glu; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Thr 71 → Ala; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Ala 133 → Thr; Arg 148 → Ser; Ser 150 → Ile; Cys 153 → Ser; 157 → Pro или
7. Ala 5 → Thr; Arg 26 → Glu; Glu 27 → Gly; Phe 28 → Cys; Pro 29 → Arg; Glu 30 → Pro; Met 31 → Trp; Leu 33 → Ile; Glu 34 → Phe; Gly 46 → Asp; Leu 56 → Ala; Ser 58 → Asp; Arg 60 → Pro; Cys 61 → Ala; Thr 71 → Ala; Cys 101 → Ser; Glu 104 → Val; Leu 105 → Cys; His 106 → Asp; Lys 108 → Ser; Arg 111 → Pro; Lys 114 → Trp; Ser 150 → Ile; Cys 153 → Ser; 157 → Pro.
[00137] В остальном участке, т. е. участке, отличающемся от положений 5, 26-31, 33-34, 42, 46, 52, 56, 58, 60-61, 65, 71, 85, 94, 101, 104-106, 108, 111, 114, 121, 133, 148, 150 и 153 в последовательности, мутеин Tlc по настоящему раскрытию может включать аминокислотную последовательность дикого типа (природную) вне подвергнутых мутации положений в аминокислотной последовательности.
[00138] В еще дополнительных вариантах осуществления мутеин Tlc согласно настоящему раскрытию характеризуется по меньшей мере 70% идентичностью последовательности или по меньшей мере 70% гомологией последовательности с последовательностью зрелого липокалина слезы человека (SEQ ID NO: 1).
[00139] Мутеин Tlc согласно настоящему раскрытию можно получать с помощью мутагенеза встречающейся в природе формы липокалина слезы человека. Согласно некоторым вариантам осуществления мутагенеза замена (или замещение) представляет собой консервативную замену. Тем не менее, любая замена, в том числе неконсервативная замена или одна или более из приведенных ниже иллюстративных замен, предусматривается, если только мутеин липокалина сохраняет свою способность связываться с CD137, и/или он характеризуется идентичностью последовательности с последовательностью, с впоследствии содержащей замену, в том смысле, что имеет место по меньшей мере 60%, а именно по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85% или большая идентичность последовательности с аминокислотной последовательностью зрелого липокалина слезы человека (номер доступа в базе данных SWISS-PROT - P31025).
[00140] Согласно другому аспекту настоящее раскрытие относится к новым мутеинам hNGAL со специфичностью связывания, направленным против или специфичным в отношении CD137.
[00141] В связи с этим настоящее раскрытие предусматривает один или более мутеинов hNGAL, которые способны к связыванию CD137 с аффинностью, измеренной по KD, составляющей 200 нМ или ниже, приблизительно 140 нМ или ниже, приблизительно 50 нМ или ниже и даже приблизительно 10 нМ или ниже. Более предпочтительно мутеины hNGAL могут характеризоваться аффинностью, измеренной по KD, составляющей приблизительно 5 нМ или ниже.
[00142] Согласно некоторым вариантам осуществления мутеин hNGAL по настоящему раскрытию включает в одном или более положениях, соответствующих положениям 28, 36, 40-41, 49, 52, 65, 68, 70, 72-73, 77, 79, 81, 83, 87, 94, 96, 100, 103, 106, 125, 127, 132 и 134 в линейной полипептидной последовательности зрелого hNGAL (SEQ ID NO: 2), замену.
[00143] Согласно конкретным вариантам осуществления мутеин липокалина по настоящему раскрытию содержит по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21 или даже больше замен в положении последовательности, соответствующем положению 28, 36, 40-41, 49, 52, 65, 68, 70, 72-73, 77, 79, 81, 83, 87, 94, 96, 100, 103, 106, 125, 127, 132 и 134 последовательности в линейной полипептидной последовательности зрелого hNGAL (номер доступа базы данных SWISS-PROT - P80188; SEQ ID NO: 2). Предпочтительно предусматривают, что настоящее раскрытие относится к мутеину липокалина, который содержит, в дополнение к одной или более заменам в положениях, соответствующих положениям 36, 87 и/или 96 в линейной полипептидной последовательности зрелого NGAL человека, в одном или более положениях, соответствующих положениям 28, 40-41, 49, 52, 65, 68, 70, 72-73, 77, 79, 81, 83, 94, 100, 103, 106, 125, 127, 132 и 134 в линейной полипептидной последовательности зрелого hNGAL, замену.
[00144] Согласно еще некоторым дополнительным вариантам осуществления настоящее раскрытие относится к полипептиду, где указанный полипептид представляет собой мутеин hNGAL, который по сравнению с линейной полипептидной последовательностью зрелого hNGAL (номер доступа базы данных SWISS-PROT - P80188; SEQ ID NO: 2) содержит по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21 или даже больше подвергнутых мутации аминокислотных остатков в положениях 28, 36, 40-41, 49, 52, 65, 68, 70, 72-73, 77, 79, 81, 87, 96, 100, 103, 106, 125, 127, 132 и 134 в последовательности, и где указанный полипептид связывает CD137, в частности CD137 человека.
[00145] Согласно некоторым вариантам осуществления связывающий CD137 мутеин hNGAL по настоящему раскрытию включает в любом одном или более из положений 28, 36, 40-41, 49, 52, 65, 68, 70, 72-73, 77, 79, 81, 83, 87, 94, 96, 100, 103, 106, 125, 127, 132 и 134 последовательности в линейной полипептидной последовательности зрелого hNGAL (SEQ ID NO: 2) один или более из следующих подвергнутых мутации аминокислотных остатков: Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Arg или Lys; Gln 49 → Val, Ile, His, Ser или Asn; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Met, Ala или Gly; Leu 70 → Ala, Lys, Ser или Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Met, Arg, Thr или Asn; Trp 79 → Ala или Asp; Arg 81 → Met, Trp или Ser; Phe 83 → Leu; Cys 87 → Ser; Leu 94 → Phe; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu и Lys 134 → Tyr.
[00146] Согласно некоторым вариантам осуществления мутеин hNGAL по настоящему раскрытию включает два или более, а именно 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, более того, как например: 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или все подвергнутые мутации аминокислотные остатки в этих положениях в последовательности зрелого hNGAL.
[00147] Согласно некоторым дополнительным вариантам осуществления мутеин hNGAL по настоящему раскрытию, который связывается с CD137, включает следующие аминокислотные замещения по сравнению с линейной полипептидной последовательностью зрелого hNGAL:
(a) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Lys; Gln 49 → Asn; Tyr 52 → Met; Ser 68 → Gly; Leu 70 → Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Ala; Arg 81 → Ser; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; Lys 134 → Tyr;
(b) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Arg; Gln 49 → Ile; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Met; Leu 70 → Lys; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Met; Trp 79 → Asp; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; Lys 134 → Tyr;
(c) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Arg; Gln 49 → Asn; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Ala; Leu 70 → Ala; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Asp; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; Lys 134 → Tyr;
(d) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Lys; Gln 49 → Asn; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Ala; Leu 70 → Ala; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Asp; Arg 81 → Trp; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; Lys 134 → Tyr;
(e) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Lys; Gln 49 → Ser; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Gly; Leu 70 → Ser; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Ala; Arg 81 → Met; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; Lys 134 → Tyr;
(f) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Lys; Gln 49 → Val; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Gly; Leu 70 → Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Arg; Trp 79 → Asp; Arg 81 → Ser; Cys 87 → Ser; Leu 94 → Phe; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; Lys 134 → Tyr;
(g) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Arg; Gln 49 → His; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Gly; Leu 70 → Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Ala; Arg 81 → Ser; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; Lys 134 → Tyr;
(h) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Lys; Gln 49 → Asn; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Gly; Leu 70 → Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Thr; Trp 79 → Ala; Arg 81 → Ser; Phe 83 → Leu; Cys 87 → Ser; Leu 94 → Phe; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; Lys 134 → Tyr или
(i) Gln 28 → His; Leu 36 → Gln; Ala 40 → Ile; Ile 41 → Arg; Gln 49 → Ser; Tyr 52 → Met; Asn 65 → Asp; Ser 68 → Ala; Leu 70 → Thr; Arg 72 → Asp; Lys 73 → Asp; Asp 77 → Asn; Trp 79 → Ala; Arg 81 → Ser; Cys 87 → Ser; Asn 96 → Lys; Tyr 100 → Phe; Leu 103 → His; Tyr 106 → Ser; Lys 125 → Phe; Ser 127 → Phe; Tyr 132 → Glu; Lys 134 → Tyr.
[00148] В остальном участке, т. е. участке, отличающемся от положений 28, 36, 40-41, 49, 52, 65, 68, 70, 72-73, 77, 79, 81, 83, 87, 94, 96, 100, 103, 106, 125, 127, 132 и 134 в последовательности, мутеин hNGAL по настоящему раскрытию может включать аминокислотную последовательность дикого типа (природную) вне подвергнутых мутации положений в аминокислотной последовательности.
[00149] Согласно другому варианту осуществления мутеин hNGAL характеризуется по меньшей мере 70% или даже большей идентичностью последовательности с аминокислотной последовательностью зрелого липокалина 2 человека (номер доступа в базе данных SWISS-PROT - P80188).
[00150] Согласно дополнительным конкретным вариантам осуществления связывающий CD137 мутеин липокалина согласно настоящему раскрытию содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 18-33 или ее фрагмента или варианта.
[00151] Аминокислотная последовательность связывающего CD137 мутеина липокалина по настоящему раскрытию может характеризоваться высокой идентичностью последовательности, а именно по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 82%, по меньшей мере 85%, по меньшей мере 87%, по меньшей мере 90% идентичностью, в том числе по меньшей мере 95% идентичностью, с последовательностью, выбранной из группы, состоящей из SEQ ID NO: 18-33.
[00152] Настоящее раскрытие также включает структурные гомологи мутеина липокалина с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 18-33, структурные гомологи которой характеризуются гомологией аминокислотной последовательности или идентичностью последовательностей, составляющей более чем приблизительно 60%, предпочтительно более чем 65%, более чем 70%, более чем 75%, более чем 80%, более чем 85%, более чем 90%, более чем 92% и наиболее предпочтительно более чем 95% относительно указанного мутеина.
D. Примерные пути применения, применения и получение слитых полипептидов
[00153] Согласно некоторым вариантам осуществления слитые полипептиды по настоящему раскрытию могут обеспечивать синергический эффект посредством двойного целенаправленного воздействия на CD137 и GPC3.
[00154] Вследствие этого в медицине существует множество возможных применений слитых полипептидов по настоящему раскрытию.
[00155] Согласно одному аспекту настоящее раскрытие относится к применению слитых полипептидов, раскрытых в данном документе, для выявления CD137 и GPC3 в образце, а также к соответствующему способу диагностирования.
[00156] Согласно другому аспекту в настоящем раскрытии описано применение одного или более слитых полипептидов, раскрытых в данном документе, или одной или более композиций, содержащих такие полипептиды, для одновременного связывания CD137 и GPC3.
[00157] Настоящее раскрытие также включает применение описанных одного или более слитых полипептидов для образования комплекса с CD137 и GPC3.
[00158] Таким образом, согласно еще дополнительному аспекту настоящего раскрытия раскрытые один или более слитых полипептидов используют для выявления уровня CD137 и GPC3. Такое применение может включать стадии приведения одного или более указанных слитых полипептидов, при соответствующих условиях, в контакт с образцом, который, как полагают, содержит CD137 и GPC3, с обеспечением таким образом возможности образования комплекса между слитыми полипептидами и CD137 и GPC3 и выявления комплекса по соответствующему сигналу. Выявляемый сигнал может быть обусловлен меткой, как разъяснено выше, или изменением физических свойств в результате связывания, т. е. собственно образования комплекса. Одним примером является поверхностный плазмонный резонанс, значение которого изменяется в ходе связывания партнеров по связыванию, из которых один иммобилизован на поверхности, такой как золотая фольга.
[00159] Слитые полипептиды, раскрытые в данном документе, также можно применять для отделения CD137 и GPC3. Такое применение может включать стадии приведения одного или более указанных слитых полипептидов, при соответствующих условиях, в контакт с образцом, который, как полагают, содержит CD137 и GPC3, с обеспечением таким образом возможности образования комплекса между слитыми полипептидами и CD137 и GPC3 и отделения комплекса от образца.
[00160] Согласно еще одному аспекту в настоящем раскрытии описан диагностический или аналитический набор, содержащий слитый полипептид согласно настоящему раскрытию.
[00161] Помимо их применения в диагностике, согласно еще одному аспекту настоящее раскрытие предполагает фармацевтическую композицию, содержащую слитый полипептид по настоящему раскрытию и фармацевтически приемлемый наполнитель.
[00162] Кроме того, настоящее раскрытие предусматривает слитые полипептиды, которые одновременно связывают CD137 и GPC3, для применения в качестве противораковых средств и/или иммуномодуляторов. В силу этого слитые полипептиды по настоящему раскрытию предусматриваются для применения в способе лечения или предупреждения заболеваний человека, а именно ряда опухолей, в том числе гепатоцеллюлярной карциномы («HCC»), меланомы, карциномы из клеток Меркеля, опухоли Вильма и гепатобластомы. Соответственно, также представлены способы лечения или предупреждения заболеваний человека, а именно ряда опухолей, в том числе гепатоцеллюлярной карциномы («HCC»), меланомы, карциномы из клеток Меркеля, опухоли Вильма и гепатобластомы, у нуждающегося в этом субъекта, включающие введение указанному субъекту терапевтически эффективного количества одного или более слитых полипептидов по настоящему раскрытию.
[00163] Путем одновременно целенаправленного воздействия на опухолевые клетки, в которых экспрессируется GPC3, как например: при гепатоцеллюлярной карциноме («HCC»), меланоме, карциноме из клеток Меркеля, опухоли Вильма и гепатобластоме, и активации клеток естественных киллеров (NK) врожденной иммунной системы хозяина, находящихся в зоне таких опухолевых клеток, или T-клеток адаптивной иммунной системы, слитый полипептид по настоящему раскрытию может повышать целенаправленную активность клеток-лимфоцитов против опухоли, усиливать противоопухолевый иммунитет и в то же время обладать непосредственным ингибирующим действием на рост опухоли, обеспечивая таким образом синергичные противоопухолевые ответы. Кроме того, путем локального ингибирования активности онкогена и индуцирования клеточно-опосредованной цитотоксичности с помощью NK-клеток и/или T-клеток, слитый полипептид по настоящему раскрытию может уменьшать побочные эффекты эффекторных лимфоцитов в отношении здоровых клеток, т. е. нецелевую токсичность.
[00164] В T-клетках опосредованная CD137 передача сигнала приводит к накоплению представителей семейства TRAF и активации некоторых киназ, в том числе ASK-1, MKK, MAPK3/MAPK4, p38 и JNK/SAPK. Активация киназ затем сопровождается активацией и транслокацией в ядро некоторых факторов транскрипции, в том числе ATF-2, Jun и NF-κB. Помимо усиления индуцированной TCR пролиферации на субоптимальном уровне, опосредованная CD137 передача сигнала защищает T-клетки, и в частности CD8+T-клетки, от индуцированной активацией клеточной смерти (AICD).
[00165] Настоящее раскрытие охватывает применение слитого полипептида по настоящему раскрытию или композиции, содержащей такой слитый полипептид, для костимуляции T-клеток и/или активации нисходящих сигнальных путей, связанных с CD137, при воздействии на опухолевые клетки, в которых экспрессируется GPC3, как например: при гепатоцеллюлярной карциноме («HCC»), меланоме, карциноме из клеток Меркеля, опухоли Вильма и гепатобластоме.
[00166] В настоящем раскрытии также описан способ костимуляции T-клеток и/или активации нисходящих сигнальных путей, связанных с CD137, при воздействии на опухолевые клетки, в которых экспрессируется GPC3, как например: при гепатоцеллюлярной карциноме («HCC»), меланоме, карциноме из клеток Меркеля, опухоли Вильма и гепатобластоме, включающий применение одного или более слитых полипептидов по настоящему раскрытию или одной или более композиций, содержащих такие слитые полипептиды.
[00167] К тому же в настоящее раскрытие включен способ активации нисходящих сигнальных путей, связанных с CD137, при воздействии на опухолевые клетки, в которых экспрессируется GPC3, при гепатоцеллюлярной карциноме («HCC»), меланоме, карциноме из клеток Меркеля, опухоли Вильма и гепатобластоме, включающий применение одного или более слитых полипептидов по настоящему раскрытию или одной или более композиций, содержащих такие слитые полипептиды.
[00168] Настоящее раскрытие также предполагает способ индуцирования пролиферации T-лимфоцитов при воздействии на опухолевые клетки, в которых экспрессируется GPC3, при гепатоцеллюлярной карциноме («HCC»), меланоме, карциноме из клеток Меркеля, опухоли Вильма и гепатобластоме, включающий применение одного или более слитых полипептидов по настоящему раскрытию или одной или более композиций, содержащих такие слитые полипептиды.
[00169] Настоящее раскрытие охватывает применение слитого полипептида по настоящему раскрытию или композиции, содержащей такой слитый полипептид, для направления образования скоплений CD137 и активации на T-клетках по отношению к опухолевым клеткам, в которых экспрессируется GPC3, как, например, при гепатоцеллюлярной карциноме («HCC»), меланоме, карциноме из клеток Меркеля, опухоли Вильма и гепатобластоме.
[00170] Согласно другому варианту осуществления настоящее раскрытие также относится к молекулам нуклеиновых кислот (ДНК и РНК), которые включают нуклеотидные последовательности, кодирующие слитые полипептиды, раскрытые в данном документе. Согласно еще одному варианту осуществления настоящее раскрытие охватывает клетку-хозяина, содержащую указанную молекулу нуклеиновой кислоты. Поскольку вырожденность генетического кода допускает замены определенных кодонов на другие кодоны, обозначающие ту же самую аминокислоту, настоящее раскрытие не ограничено конкретной молекулой нуклеиновой кислоты, кодирующей слитый полипептид, описанный в данном документе, а охватывает все молекулы нуклеиновой кислоты, которые включают нуклеотидные последовательности, кодирующие функциональный полипептид. В связи с этим настоящее раскрытие также относится к нуклеотидным последовательностям, кодирующим слитые полипептиды по настоящему раскрытию.
[00171] Согласно некоторым вариантам осуществления молекула нуклеиновой кислоты, кодирующая мутеин липокалина, раскрытый в данной заявке, такая как ДНК, может быть «функционально связана» с другой молекулой нуклеиновой кислоты, кодирующей иммуноглобулин по настоящему раскрытию, для обеспечения экспрессии слитого полипептида, раскрытого в данном документе. В связи с этим функциональная связь представляет собой связь, при которой элементы последовательности одной молекулы нуклеиновой кислоты и элементы последовательности другой молекулы нуклеиновой кислоты соединены таким образом, который обеспечивает экспрессию слитого полипептида в виде единого полипептида.
[00172] Настоящее раскрытие также относится к способу получения слитого полипептида, при этом слитый полипептид по настоящему раскрытию получают, исходя из нуклеиновой кислоты, кодирующей полипептид или любую его субъединицу, при помощи способов генной инженерии. Согласно некоторым вариантам осуществления способ можно осуществлять in vivo, при этом полипептид можно, например, получать в бактериальном или эукариотическом организме-хозяине, а затем выделять из этого организма-хозяина или его культуры. Существует также возможность получения слитого полипептида по настоящему раскрытию in vitro, например путем использования системы трансляции in vitro.
[00173] При получении слитого полипептида in vivo нуклеиновую кислоту, кодирующую такой полипептид, вводят в подходящий организм-хозяина бактериального или эукариотического происхождения с помощью технологии рекомбинантной ДНК (как уже подчеркивалось выше). С этой целью клетку-хозяина сперва трансформируют вектором для клонирования, который включает молекулу нуклеиновой кислоты, кодирующую слитый полипептид, описанный в данном документе, с применением разработанных стандартных способов. Клетку-хозяина затем культивируют в условиях, которые обеспечивают экспрессию гетерологичной ДНК и таким образом синтез соответствующего полипептида. Впоследствии полипептид выделяют либо из клетки, либо из среды для культивирования.
[00174] Согласно одному варианту осуществления настоящего раскрытия способ включает воздействие по меньшей мере на одну молекулу нуклеиновой кислоты, кодирующей hNGAL, мутагенезом в нуклеотидных триплетах, кодирующих по меньшей мере одно, иногда даже больше из положений в последовательности, соответствующее положениям 28, 40-52, 60, 68, 65, 70, 71-81, 87, 89, 96, 98, 100-106, 114, 118, 120, 125-137 и 145 последовательности в линейной полипептидной последовательности hNGAL (SEQ ID NO: 2).
[00175] К тому же согласно некоторым вариантам осуществления встречающаяся в природе дисульфидная связь между Cys 76 и Cys 175 может быть удалена в мутеинах hNGAL по настоящему раскрытию. Соответственно, такие мутеины можно получать в компартменте клетки со средой с восстановительным окислительно-восстановительным потенциалом, например в цитоплазме грамотрицательных бактерий.
[00176] Настоящее раскрытие также включает молекулы нуклеиновой кислоты, кодирующие мутеины липокалина по настоящему раскрытию, которые включают дополнительные мутации вне указанных положений в последовательности с экспериментальным мутагенезом. Такие мутации часто допустимы или могут даже оказаться преимущественными, например, если они способствуют улучшенной эффективности сворачивания, стабильности в сыворотке крови, термостойкости или аффинности связывания с лигандом мутеинов липокалина.
[00177] Молекула нуклеиновой кислоты, раскрытая в данной заявке, может быть «функционально связана» с регуляторной последовательностью (или регуляторными последовательностями) для обеспечения экспрессии этой молекулы нуклеиновой кислоты.
[00178] Молекулу нуклеиновой кислоты, как, например, ДНК, называют «способной к экспрессии молекулой нуклеиновой кислоты» или способной «обеспечивать экспрессию нуклеотидной последовательности», если она включает элементы последовательности, которые содержат информацию относительно транскрипционной и/или трансляционной регуляции, и такие последовательности «функционально связаны» с нуклеотидной последовательностью, кодирующей полипептид. Функциональная связь представляет собой связь, при которой элементы регуляторной последовательности и последовательность, подлежащая экспрессии, соединены таким образом, который обеспечивает экспрессию гена. Определенные особенности регуляторных участков, необходимых для экспрессии гена, могут варьировать между видами, но в общем эти участки включают промотор, который у прокариот содержит как промотор per se, т. е. элементы ДНК, направляющие инициацию транскрипции, так и элементы ДНК, которые при транскрибировании в РНК, будут запускать сигнал об инициации трансляции. Такие промоторные участки, как правило, включают 5'-некодирующие последовательности, вовлеченные в инициацию транскрипции и трансляции, такие как боксы -35/-10 и элемент Шайн-Дальгарно у прокариот или TATA-бокс, последовательности CAAT и 5'-кэппирующие элементы у эукариот. Такие участки также могут включать энхансерные или репрессорные элементы, а также сигнальные последовательности трансляции и лидерные последовательности для нацеливания нативного полипептида на конкретный компартмент клетки-хозяина.
[00179] К тому же 3'-некодирующие последовательности могут содержать регуляторные элементы, вовлеченные в терминацию транскрипции, полиаденилирование или им подобные. Однако, если такие последовательности терминации не удовлетворительны с точки зрения функциональности в конкретной клетке-хозяине, тогда их можно заменить сигналами, функциональными в такой клетке.
[00180] Ввиду этого молекула нуклеиновой кислоты по настоящему раскрытию может включать регуляторную последовательность, как, например, промоторную последовательность. Согласно некоторым вариантам осуществления молекула нуклеиновой кислоты по настоящему раскрытию включает промоторную последовательность и последовательность терминации транскрипции. Подходящими прокариотическими промоторами являются, например, промотор tet, промотор lacUV5 или промотор T7. Примерами промоторов, применимых для экспрессии в эукариотических клетках, являются промотор SV40 или промотор CMV.
[00181] Молекулы нуклеиновой кислоты по настоящему раскрытию также могут быть частью вектора или любого другого типа средства для клонирования, а именно плазмиды, фагмиды, фага, бакуловируса, космиды или искусственной хромосомы.
[00182] Согласно одному варианту осуществления молекула нуклеиновой кислоты включена в фазмиду. Фазмидный вектор означает вектор, кодирующий межгенный участок умеренного фага, как, например, M13 или f1, или его функциональную часть, слитую с представляющей интерес кДНК. После суперинфекции бактериальных клеток-хозяев таким фагмидным вектором и соответствующим хелперным фагом (например, M13K07, VCS-M13 или R408) продуцируются интактные фаговые частицы, обеспечивая таким образом физическое сопряжение закодированной гетерологичной кДНК с ее соответствующим полипептидом, выведенным на поверхность фага (см., например, Lowman, H.B. (1997) Annu. Rev. Biophys. Biomol. Struct. 26, 401-424 или Rodi, D.J., and Makowski, L. (1999) Curr. Opin. Biotechnol. 10, 87-93).
[00183] Такие средства для клонирования могут включать, помимо регуляторных последовательностей, описанных выше, и последовательности нуклеиновой кислоты, кодирующей слитый полипептид, описанный в данном документе, последовательности для репликации и контрольные последовательности, полученные от видов, совместимых с клеткой-хозяином, которую используют для экспрессии, а также маркеры отбора, придающие селектируемый фенотип трансформированным или трансфицированным клеткам. В данной области известно большое число подходящих векторов для клонирования, и они являются коммерчески доступными.
[00184] Молекулой ДНК, кодирующей слитый полипептид, который описан в данном документе (например, SEQ ID NO: 20 и 31), и в частности вектором для клонирования, содержащим кодирующую последовательность такого полипептида, можно трансформировать клетку-хозяина, способную экспрессировать ген. Трансформацию можно осуществлять с использованием стандартных методик. Таким образом, настоящее раскрытие также направлено на клетку-хозяина, содержащую молекулу нуклеиновой кислоты, которая раскрыта в данном документе.
[00185] Трансформированные клетки-хозяева культивируют в условиях, подходящих для экспрессии нуклеотидной последовательности, кодирующей слитый полипептид по настоящему раскрытию. Подходящие клетки-хозяева могут быть прокариотическими, такими как Escherichia coli (E. coli) или Bacillus subtilis, или эукариотическими, такими как Saccharomyces cerevisiae, Pichia pastoris, клетками насекомых SF9 или High5, иммортализованными клеточными линиями млекопитающих (например, клетками HeLa или клетками CHO) или первичными клетками млекопитающих.
[00186] Согласно некоторым вариантам осуществления в том случае, если мутеин липокалина по настоящему раскрытию, в том числе который содержится в слитом полипептиде, раскрытом в данном документе, включает внутримолекулярные дисульфидные связи, может быть предпочтительным направлять образующийся полипептид в компартмент клетки со средой с окисляющим окислительно-восстановительным потенциалом с помощью соответствующей сигнальной последовательности. Такая окисляющая среда может обеспечиваться периплазмой грамотрицательных бактерий, таких как E. coli, во внеклеточной среде грамположительных бактерий или в просвете эндоплазматической сети эукариотических клеток и, как правило, она способствует образованию дисульфидных связей в структуре.
[00187] Согласно некоторым вариантам осуществления также возможно получать слитый полипептид по настоящему раскрытию в цитозоле клетки-хозяина, предпочтительно E. coli. В данном случае полипептид может быть либо непосредственно получен в растворимом и свернутом состоянии, либо выделен в форме телец включения с последующей ренатурацией in vitro. Дополнительной возможностью является применение конкретных штаммов-хозяев с окисляющей внутриклеточной средой, которая таким образом может обеспечивать образование дисульфидных связей в цитозоле (Venturi et al. (2002) J. Mol. Biol. 315, 1-8.).
[00188] Согласно некоторым вариантам осуществления слитый полипептид по настоящему раскрытию, который описан в данном документе, можно создавать или получать необязательно только с применением генной инженерии. Предпочтительнее такой полипептид можно также получать путем химического синтеза, такого как твердофазный синтез полипептидов Меррифилда, или путем транскрипции и трансляции in vitro. Например, возможно, что перспективные мутации определяют с помощью молекулярного моделирования и затем синтезируют необходимый (сконструированный) мутеин или полипептид in vitro и исследуют активность связывания в отношении представляющей интерес мишени. Способы твердофазного синтеза и/или синтеза в жидкой фазе белков хорошо известны в данной области (см., например, Bruckdorfer, T. et al. (2004) Curr. Pharm. Biotechnol. 5, 29-43).
[00189] Согласно другому варианту осуществления слитый полипептид по настоящему раскрытию можно получать путем транскрипции/трансляции in vitro, используя стандартные способы, известные специалистам в данной области.
[00190] Специалисту в данной области будут понятны способы, применимые для получения слитых полипептидов, предполагаемых настоящим раскрытием, но чьи последовательности белков или нуклеиновых кислот конкретно не раскрыты в данном документе. В качестве обзора такие модификации аминокислотной последовательности включают, например, направленный мутагенез отдельных положений аминокислот для упрощения субклонирования гена полипептида или его частей путем введения сайтов расщепления для определенных рестрикционных ферментов. К тому же такие мутации можно также вводить для дополнительного улучшения аффинности слитого полипептида в отношении его мишеней (например, CD137 и GPC3). Кроме того, мутации можно вводить для модулирования определенных характеристик полипептида, а именно для улучшения стабильности фолдинга, стабильности в сыворотке крови, устойчивости белка или растворимости в воде или для снижения склонности к агрегированию, если в этом существует необходимость. Например, встречающиеся в природе остатки цистеина можно подвергать мутации в другие аминокислоты для предотвращения образования дисульфидного мостика.
[00191] Дополнительные объекты, преимущества и особенности настоящего раскрытия станут очевидными специалисту в данной области при изучении следующих примеров и прилагаемых фигур, включающих их, которые не предназначены для ограничения. Таким образом, следует понимать, что хотя настоящее раскрытие конкретно раскрыто с помощью примерных вариантов осуществления и необязательных особенностей, к модификации и варианту раскрываемой информации, приведенной в них, раскрытой в данном документе, может прибегнуть специалист в данной области, и что такие модификации и варианты рассматриваются как часть объема настоящего раскрытия.
V. ПРИМЕРЫ
[00192] Пример 1. Экспрессия и анализ слитых полипептидов антитело-мутеин липокалина
[00193] Авторы настоящего изобретения применяли три подхода для создания биспецифических конструкций, которые могут связывать мишени, GPC3 и CD137, одновременно.
[00194] При первом подходе авторы настоящего изобретения создавали слитые полипептиды антитело-мутеин липокалина на основе специфичного к CD137 антитела, например, у которого есть тяжелая и легкая цепи, представленные под SEQ ID NO: 34 и 35, и мутеина липокалина к GPC3, например, под SEQ ID NO: 10. Неструктурированный нечувствительный к протеазе линкер (G4S)3 (SEQ ID NO: 49) использовали для слияния белков друг с другом во всех случаях. Различные форматы, которые были сконструированы, показаны на фигуре 1A. Созданные варианты представляют собой слияния мутеина липокалина с любым из четырех концов антитела, которое содержит каркас IgG4, подвергнутый мутации для сведения к минимуму обмена в половине антитела (мутация S228P, см. SEQ ID NO: 34): SEQ ID NO: 36 и 37, SEQ ID NO: 38 и 39, SEQ ID NO: 40 и 41, SEQ ID NO: 42 и 43.
[00195] При втором подходе авторы настоящего изобретения создавали слияния двух мутеинов липокалина (SEQ ID NO: 10, связывающего GPC3, и SEQ ID NO: 26, связывающего CD137) со сконструированным Fc-фрагментом IgG4 (SEQ ID NO: 73), который содержал мутацию S228P для сведения к минимуму обмена в половине антитела IgG4 in-vitro и in-vivo (см. Silva 2015), а также мутации F234A и L235A для ослабления взаимодействий с Fc-гамма рецептором (Alegre 1992). Полученные в результате слитые полипептиды (SEQ ID NO: 44 и SEQ ID NO: 45) структурно показаны на фигуре 1B.
[00196] Конструкции из первого и второго подходов получали путем синтеза генов и клонировали в вектор экспрессии для млекопитающих. Затем их транзиентно экспрессировали в клетках CHO. Концентрацию слитых полипептидов антитело-мутеин липокалина и слитых полипептидов IgG4Fc-мутеин липокалина в среде с клеточной культурой измеряли при помощи сенсорного устройства с белком A от ForteBio (Pall Corp.) и количественно оценивали при помощи стандарта, представляющего собой IgG1 человека (данные не показаны).
[00197] При третьем подходе авторы настоящего изобретения создавали слияния двух мутеинов липокалина (SEQ ID NO: 10 и SEQ ID NO: 26), соединенных одним или большим количеством линкеров (G4S)2 (SEQ ID NO: 48), и с применением двух различных структурных вариантов, показанных на фигуре 1C. В первом структурном варианте SEQ ID NO: 26 был слит на C-конце с SEQ ID NO: 10, что давало в результате слитый полипептид с SEQ ID NO: 46; во втором структурном варианте две копии SEQ ID NO: 26 были слиты на C-конце с SEQ ID NO: 10, что давало в результате слитый полипептид с SEQ ID NO: 47. Конструкции содержали Strep-метку (SEQ ID NO: 50) для очистки с помощью аффинной хроматографии. Конструкции клонировали с помощью стандартных способов и экспрессировали в E. coli с задействованием секреции в периплазму.
[00198] Слитые полипептиды антитело-мутеин липокалина и слитые полипептиды IgG4Fc-фрагмент-мутеин липокалина очищали с помощью хроматографии с белком A, после которой проводили эксклюзионную хроматографию (SEC) в 10 мМ гистидина, pH 5,5, 150 мМ NaCl или PBS, pH7,4. После очистки при помощи SEC фракции, содержащие мономерный белок, объединяли и снова анализировали при помощи аналитической SEC. Согласно этому анализу слитые полипептиды были полностью мономерными без выявляемых мультимерных молекул или агрегатов (данные не показаны).
[00199] Пример 2. Специфичность слитых полипептидов относительно GPC3
[00200] Авторы настоящего изобретения использовали анализ ELISA для определения специфичности слитых полипептидов с SEQ ID NO: 36 и 37, SEQ ID NO: 38 и 39, SEQ ID NO: 40 и 41 и SEQ ID NO: 42 и 43 в отношении рекомбинантного GPC3 человека (R&D Systems № 2119-GP-050/CF). Мишень растворяли в PBS (1 мкг/мл) и наносили на микротитровальные планшеты на ночь при 4°C. Планшет промывали после каждой стадии инкубации при помощи 100 мкл PBS, дополненного 0,1% (объем/объем) Tween 20 (PBS-T), пять раз. Планшеты блокировали при помощи 2% BSA (вес/объем) в PBS-T в течение 1 ч при комнатной температуре и затем промывали. В лунки добавляли различные концентрации мутеина липокалина с SEQ ID NO: 10) или слитых полипептидов и инкубировали в течение 1 ч при комнатной температуре с последующей стадией промывания. Связанный белок слияния или мутеин липокалина выявляли после инкубации с разведенным 1:1000 антителом к NGAL человека, конъюгированным с HRP, в PBS-T, дополненном 2% (вес/объем) BSA (PBS-TB). После дополнительной стадии промывания в каждую лунку добавляли флуорогенный субстрат с HRP (QuantaBlu, Thermo) и интенсивность флуоресценции выявляли при помощи считывающего уровень флуоресценции устройства для микропланшетов.
[00201] Результат эксперимента изображен на фигуре 2 совместно с аппроксимированными кривыми, полученными из сигмоидальной функции связывания 1:1, где значение EC50 и наибольший сигнал были незаданными параметрами, и наклон фиксировали для единства. Полученные в результате значения EC50 представлены в таблице 1, в том числе погрешности сигмоидальной функции данных, которая является примером для всех данных, представленных в таблицах в данном документе. Все наблюдаемые значения EC50, которые находятся в аналогичном диапазоне для всех слитых полипептидов антитело-мутеин липокалина (0,25–0,28 нМ), несколько лучше, чем для мутеина липокалина (SEQ ID NO: 10), значение для которого составляло 0,55 нМ Из эксперимента видно, что при включении в слитые полипептиды, описанные выше, мутеин липокалина может быть слит с любым из четырех концов антитела без потери активности относительно GPC3.
[00202] Таблица 1. Данные ELISA в отношении связывания GPC3
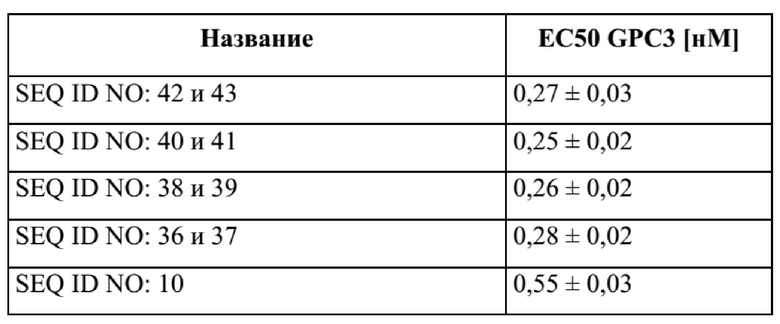
[00203] Пример 3. Специфичность слитых полипептидов относительно CD137
[00204] Авторы настоящего изобретения использовали анализ ELISA для определения специфичности слитых полипептидов с SEQ ID NO: 36 и 37, SEQ ID NO: 38 и 39, SEQ ID NO: 40 и 41 и SEQ ID NO: 42 и 43 в отношении белка слияния рекомбинантный CD137-Fc (№ 838-4B-100, R&D Systems). Антитело с SEQ ID NO: 34 и 35 служило в качестве положительного контроля. Мишень растворяли в PBS (1 мкг/мл) и наносили на микротитровальные планшеты на ночь при 4°C. Планшет промывали после каждой стадии инкубации при помощи 100 мкл PBS-T пять раз. Планшеты блокировали при помощи 2% BSA (вес/объем) в PBS-T в течение 1 ч при комнатной температуре и затем промывали. В лунки добавляли различные концентрации специфичного к CD137 антитела или слитых полипептидов и инкубировали в течение 1 ч при комнатной температуре с последующей стадией промывания. Связанный белок слияния выявляли после инкубации в течение 1 ч при комнатной температуре с разведенным 1:5000 антителом мыши к Fab IgG человека, конъюгированным с HRP (Jackson Laboratories), в PBS-TB. После дополнительной стадии промывания в каждую лунку добавляли флуорогенный субстрат с HRP (QuantaBlu, Thermo) и интенсивность флуоресценции выявляли при помощи считывающего уровень флуоресценции устройства для микропланшетов.
[00205] Результат эксперимента изображен на фигуре 3 совместно с аппроксимированными кривыми, полученными из сигмоидальной функции связывания 1:1, где значение EC50 и наибольший сигнал были незаданными параметрами, и наклон фиксировали для единства. Полученные в результате значения EC50 представлены в таблице 2. Наблюдаемые значения EC50 для всех тестируемых молекул были очень сходными и варьировались от 1,5 нМ до 2,3 нМ. Из эксперимента видно, что при включении в описанные слитые полипептиды антитело может быть слито с мутеином липокалина на любом из четырех концов антитела без потери активности относительно CD137.
[00206] Таблица 2. Данные ELISA в отношении связывания CD137

[00207] Пример 4. Демонстрация одновременного связывания мишени слитых полипептидов в условиях на основе ELISA
[00208] Для того, чтобы продемонстрировать одновременное связывание слитых полипептидов с SEQ ID NO: 36 и 37, SEQ ID NO: 38 и 39, SEQ ID NO: 40 и 41 и SEQ ID NO: 42 и 43 как с GPC3, так и с CD137 использовали формат ELISA с двойным связыванием. Белком слияния рекомбинантный CD137 человека-Fc (R&D Systems) в PBS (1 мкг/мл) покрывали микротитровальные планшеты и оставляли на ночь при 4°C. Планшет промывали пять раз после каждой стадии инкубации при помощи 100 мкл PBS-T. Планшеты блокировали при помощи 2% BSA (вес/объем) в PBS-T в течение 1 ч при комнатной температуре и затем опять промывали. В лунки добавляли различные концентрации слитых полипептидов и инкубировали в течение 1 ч при комнатной температуре с последующей стадией промывания. Затем добавляли биотинилированный GPC3 человека в постоянной концентрации, составляющей 1 мкг/мл, в PBS-TB на 1 ч. После промывания в лунки на 1 ч добавляли экстравидин-HRP (Sigma-Adrich, 1:5000 в PBS-TB). После дополнительной стадии промывания в каждую лунку добавляли флуорогенный субстрат с HRP (QuantaBlu, Thermo) и интенсивность флуоресценции выявляли при помощи планшет-ридера, считывающего флуоресцентный сигнал.
[00209] Результат эксперимента изображен на фигуре 4 совместно с аппроксимированными кривыми, полученными из сигмоидальной функции связывания 1:1, где значение EC50 и наибольший сигнал были незаданными параметрами, и наклон фиксировали для единства. Полученные в результате значения EC50 представлены в таблице 3. Все слитые полипептиды характеризовались отчетливыми сигналами связывания со значениями EC50, находящимися в диапазоне от 1,7 до 2,1 нМ, демонстрируя, что слитые полипептиды способны одновременно связывать GPC3 и CD137.
[00210] Таблица 3. Данные ELISA в отношении одновременного связывания мишени
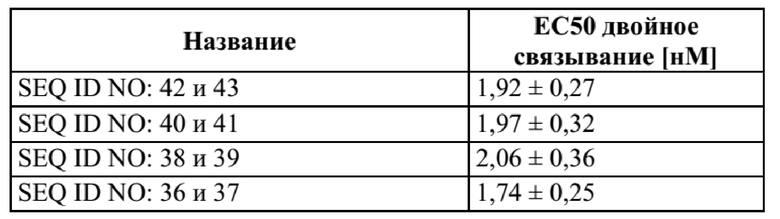
[00211] Пример 5. Аффинность антитела и слитых полипептидов в отношении GPC3 человека
[00212] Аффинности связывания мутеина липокалина с SEQ ID NO: 10 и слитых полипептидов с SEQ ID NO: 36 и 37, SEQ ID NO: 38 и 39, SEQ ID NO: 40 и 41, а также SEQ ID NO: 42 и 43 в отношении рекомбинантного GPC3 человека (R&D Systems № 2119-GP-050/CF) определяли с помощью поверхностного плазмонного резонанса (SPR) с использованием прибора Biacore T200 (GE Healthcare). В анализе аффинности с помощью SPR биотинилированный GPC3 захватывали на сенсорном чипе («чип CAP») с использованием набора для захвата биотина Biotin CAPture Kit (GE Healthcare): на сенсорном чипе CAP предварительно иммобилизировали олигонуклеотид ssDNA. Неразбавленный реагент набора для захвата биотина Biotin CAPture Reagent (стрептавидин, конъюгированный с комплементарным олигонуклеотидом ss-DNA) наносили при скорости потока, составляющей 2 мкл/мин, в течение 300 с. Для анализа мутеина липокалина использовали биотинилированный GPC3 в концентрации, составляющей 1 мкг/мл, и 0,25 мкг/мл биотинилированного GPC3 для белков слияний. Биотнилирванный GPC3 использовали в течение 300 с при скорости потока, составляющей 5 мкл/мин. GPC3 биотинилировали путем инкубации с NHS-PEG4-биотином EZ-Link® (5-кратный молярный избыток (Thermo Scientific)) на протяжении двух часов при комнатной температуре. Избыток непрореагировавшего реагента с биотином удаляли путем загрузки реакционной смеси в планшет для обессоливания для применения в формате с вращением ZebaTM Spin Desalting Plate (Thermo Scientific). В эталонный канал загружали только реагент набора для захвата биотина Biotin CAPture Reagent.
[00213] Для определения аффинности GCP3 иммобилизировали на поверхности чипа и готовили четыре различные концентрации (11,1, 3,7, 1,2 и 0,4 нМ) каждого тестированного средства (слитых полипептидов или мутеина липокалина) в подвижном буфере (10 мМ HEPES, 150 мМ NaCl, 0,05% об./об. поверхностно-активного вещества P20, 3 мМ EDTA, pH 7,4 (GE Healthcare)) и наносили на поверхность чипа. При нанесении со скоростью потока, составляющей 30 мкл/мин, время контакта образца составляло 180 с, а время диссоциации составляло 1200 с. Все измерения проводили при 25°C. Восстановление поверхности сенсорного чипа CAP выполняли при помощи введения 6 M гуанидиния-HCl с 0,25 M NaOH с последующими дополнительным промыванием подвижным буфером и периодом стабилизации, составляющим 120 с. Перед измерениями уровня белка проводили три цикла восстановления с целью приведения к требуемым условиям. Данные оценивали с помощью программного обеспечения Biacore T200 Evaluation (V 2.0). Использовали двойное сравнение с образцом и для аппроксимирования первичных данных использовали модель связывания 1:1.
[00214] Данные показаны на фигуре 5, и аппроксимированные результаты представлены в таблице 4. Из этих данных можно сделать вывод, что слитые полипептиды связывают GPC3 с аффинностями, которые очень подобны таковым для мутеина липокалина с SEQ ID NO: 10. Выявляемые аффинности связывания находились в диапазоне от 17 до 30 пМ для слитых полипептидов и выявляемая аффинность связывания составляла 12 пМ для мутеина липокалина с SEQ ID NO: 10.
[00215] Таблица 4. Аффинности связывания для GPC3

[00216] Пример 6. Аффинность антитела и слитых полипептидов в отношении CD137 человека
[00217] Аффинности связывания антитела с SEQ ID NO: 34 и 35 и слитых полипептидов с SEQ ID NO: 36 и 37, SEQ ID NO: 38 и 39, SEQ ID NO: 40 и 41 и SEQ ID NO: 42 и 43 в отношении белка слияния рекомбинантный CD137 человека-Fc (№ 838-4B-100, R&D Systems) определяли с помощью поверхностного плазмонного резонанса (SPR) по аналогии с примером 5. Вкратце, биотинилированный CD137-Fc захватывали на сенсорном чипе CAP и готовили четыре разведения (20, 5, 1,3 и 0,3 нМ) каждого тестированного средства (белок слияния или SEQ ID NO: 34 и 35) в подвижном буфере и наносили на поверхность чипа. При нанесении со скоростью потока, составляющей 30 мкл/мин, время контакта образца составляло 180 с, и время диссоциации составляло 600 с. Все измерения проводили иным способом и анализировали как описано в примере 5.
[00218] Результаты представлены в таблице 5. Из данных видно, что слитые полипептиды связывают CD137 с аффинностями, которые очень подобны таковым для антитела. Выявляемые аффинности связывания находились в диапазоне от 71 до 179 пМ для белков слияния и выявляемая аффинность связывания составляла 92 пМ для антитела 20H4.9 (SEQ ID NO: 34 и 35).
[00219] Таблица 5. Аффинности связывания для CD137

[00220] Пример 7. Специфичность слитых полипептидов мутеин липокалина-Fc относительно GPC3
[00221] Авторы настоящего изобретения использовали анализ ELISA, как описано в примере 2, для определения специфичности слитых полипептидов, SEQ ID NO: 44 и SEQ ID NO: 45, в отношении рекомбинантного GPC3 человека.
[00222] Результат эксперимента изображен на фигуре 7 совместно с аппроксимированными кривыми, полученными из сигмоидальной функции связывания 1:1, где значение EC50 и наибольший сигнал были незаданными параметрами, и наклон фиксировали для единства. Полученные в результате значения EC50 представлены в таблице 6. Наблюдаемые значения EC50 для обоих слитых полипептидов мутеин липокалина-Fc были лучше, чем такое значение для связывающего GPC3 мутеина липокалина (SEQ ID NO: 10).
[00223] Таблица 6. Данные ELISA в отношении связывания GPC3

[00224] Пример 8. Специфичность слитых полипептидов мутеин липокалина-Fc относительно CD137 человека
[00225] Авторы настоящего изобретения использовали анализ ELISA для определения специфичности слитых полипептидов мутеин липокалина-Fc с SEQ ID NO: 44 и SEQ ID NO: 45 в отношении слитого полипептида рекомбинантный CD137-Fc, как описано в примере 3.
[00226] Результат эксперимента изображен на фигуре 8 совместно с подогнанными кривыми, полученными из сигмоидальной функции связывания 1:1, где значение EC50 и наибольший сигнал были незаданными параметрами, и наклон фиксировали для единства. Полученные в результате значения EC50 приведены в таблице 7. Наблюдаемые значения EC50 для обоих слитых полипептидов мутеин липокалина-Fc были лучше, чем наблюдаемое значение EC50 для связывающего CD137 мутеина липокалина (SEQ ID NO: 26).
[00227] Таблица 7. Данные ELISA в отношении связывания CD137

[00228] Пример 9. Демонстрация одновременного связывания мишени слитых полипептидов мутеин липокалина-Fc в условиях на основе ELISA
[00229] Для того, чтобы продемонстрировать одновременное связывание слитых полипептидов с SEQ ID NO: 44 и SEQ ID NO: 45 в отношении GPC3 и CD137 использовали формат ELISA с двойным связыванием по аналогии с примером 4.
[00230] Результат эксперимента изображен на фигуре 9 совместно с аппроксимированными кривыми, полученными из сигмоидальной функции связывания 1:1, где значение EC50 и наибольший сигнал были незаданными параметрами, и наклон фиксировали для единства. Полученные в результате значения EC50 представлены в таблице 8. Оба слитых полипептида характеризовались отчетливыми сигналами связывания со значениями EC50, близкими к 1,7 нМ, демонстрируя, что слитые полипептиды способны одновременно связывать GPC3 и CD137.
[00231] Таблица 8. Данные ELISA в отношении одновременного связывания мишени

[00232] Пример 10. Аффинность антитела и слитых полипептидов в отношении GPC3 человека
[00233] Аффинности связывания связывающего GPC3 мутеина липокалина с SEQ ID NO: 10 и слитых полипептидов с SEQ ID NO: 44 и SEQ ID NO: 45 в отношении рекомбинантного GPC3 человека определяли с помощью поверхностного плазмонного резонанса, как описано в примере 5.
[00234] Данные показаны на фигуре 10, и аппроксимированные значения KD представлены в таблице 9. Из данных видно, что слитые полипептиды связывают GPC3 с аффинностями, которые очень подобны таковым для мутеина липокалина. Выявляемые аффинности связывания составляют 23 пМ и 29 пМ для слитых полипептидов соответственно по сравнению с выявляемой аффинностью связывания, составляющей 33 пМ для мутеина липокалина.
[00235] Таблица 9. Аффинности связывания для GPC3

[00236] Пример 11. Аффинность антитела и слитых полипептидов в отношении CD137 человека
[00237] Аффинности связывания связывающего CD137 мутеина липокалина с SEQ ID NO: 26 и слитых полипептидов с SEQ ID NO: 44 и SEQ ID NO: 45 в отношении белка слияния рекомбинантный CD137 человека-Fc определяли по аналогии с примером 6.
[00238] Данные показаны на фигуре 11 для слитых полипептидов с SEQ ID NO: 44 и SEQ ID NO: 45, и аппроксимированные значения KD для всех тестированных молекул приведены в таблице 10. Из данных видно, что слитые полипептиды связывают CD137 с аффинностями, составляющими 1 нМ или 1,1 нМ соответственно, превосходящими значение KD для мутеина липокалина, который характеризуется значением, составляющим 2,3 нМ.
[00239] Таблица 10. Аффинности связывания для CD137

[00240] Пример 12. Специфичность слитого полипептида относительно GPC3
[00241] Авторы настоящего изобретения создавали дополнительный слитый полипептид на основе связывающего CD137 антитела с SEQ ID NO: 51 и 52 и связывающего GPC3 мутеина липокалина с SEQ ID NO: 10. Мутеин липокалина был слит на C-конце с тяжелой цепью с помощью линкера (G4S)3 с получением слитого полипептида с SEQ ID NO: 53 и 54.
[00242] Авторы настоящего изобретения использовали анализ ELISA, как описано в примере 2, для определения специфичности слитого полипептида с SEQ ID NO: 53 и 54 в отношении рекомбинантного GPC3 человека.
[00243] Результат эксперимента изображен на фигуре 12 совместно с аппроксимированными кривыми, полученными из сигмоидальной функции связывания 1:1, где значение EC50 и наибольший сигнал были незаданными параметрами, и наклон фиксировали для единства. Полученные в результате значения EC50 представлены в таблице 11. Значение EC50 относительно GPC3 сопоставимо для слитого полипептида и мутеина липокалина. Из данных видно, что при включении в слитый полипептид мутеин липокалина может быть слит с антителом без потери активности относительно GPC3.
[00244] Таблица 11. Данные ELISA в отношении связывания GPC3

[00245] Пример 13. Демонстрация одновременного связывания мишени слитого полипептида в условиях на основе ELISA
[00246] Для того, чтобы продемонстрировать одновременное связывание слитых полипептидов с SEQ ID NO: 53 и 54 как с GPC3, так и с CD137 использовали формат ELISA с двойным связыванием по аналогии с примером 4.
[00247] Результат эксперимента изображен на фигуре 13 совместно с аппроксимированными кривыми, полученными из сигмоидальной функции связывания 1:1, где значение EC50 и наибольший сигнал были незаданными параметрами, и наклон фиксировали для единства. Слитый полипептид характеризовался отчетливыми сигналами связывания со значением EC50, составляющим 4,66±0,65 нМ, демонстрируя, что полипептид способен одновременно связывать GPC3 и CD137.
[00248] Пример 14. Специфичность слитых полипептидов относительно GPC3
[00249] Авторы настоящего изобретения использовали анализ ELISA, как описано в примере 2, для определения специфичности биспецифических слитых полипептидов с SEQ ID NO: 46 и SEQ ID NO: 47, а также мутеина липокалина с SEQ ID NO: 8 в отношении рекомбинантного GPC3 человека.
[00250] Результат эксперимента изображен на фигуре 14 совместно с аппроксимированными кривыми, полученными из сигмоидальной функции связывания 1:1, где значение EC50 и наибольший сигнал были незаданными параметрами, и наклон фиксировали для единства. Полученные в результате значения EC50 представлены в таблице 12. Значения EC50 для слитых полипептидов по меньшей мере такие же как или даже превосходящие значение EC50 для мутеина липокалина. Из данных видно, что при включении в два слитых полипептида мутеин липокалина может быть слит с антителом без потери активности относительно GPC3.
[00251] Таблица 12. Данные ELISA в отношении связывания GPC3
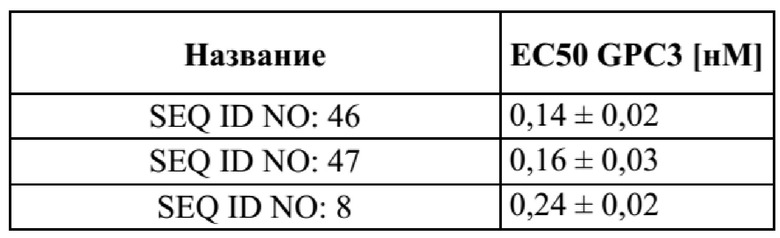
[00252] Пример 15. Специфичность слитых полипептидов относительно CD137
[00253] Авторы настоящего изобретения использовали анализ ELISA для определения специфичности биспецифических полипептидов с SEQ ID NO: 46 и SEQ ID NO: 47, а также мутеина липокалина с SEQ ID NO: 26 в отношении белка слияния рекомбинантный CD137-Fc, как описано в примере 3.
[00254] Результат эксперимента наносили на график на фигуре 15 совместно с кривыми зависимости, полученными из сигмоидальной функции связывания 1:1, где значение EC50 и наибольший сигнал были незаданными параметрами, и наклон фиксировали для единства. Полученные в результате значения EC50 представлены в таблице 13. Значения EC50 для слитых полипептидов по меньшей мере такие же как или даже превосходящие значение EC50 для мутеина липокалина. Из данных видно, что при включении в два слитых полипептида антитело может быть слито с мутеином липокалина без потери активности относительно CD137.
[00255] Таблица 13. Данные ELISA в отношении связывания CD137

[00256] Пример 16. Демонстрация одновременного связывания мишени слитых полипептидов в условиях на основе ELISA
[00257] Для того, чтобы продемонстрировать одновременное связывание биспецифических полипептидов с SEQ ID NO: 46 и SEQ ID NO: 47 в отношении GPC3 и CD137 использовали формат ELISA с двойным связыванием по аналогии с примером 4.
[00258] Результат эксперимента изображен на фигуре 16 совместно с аппроксимированными кривыми, полученными из сигмоидальной функции связывания 1:1, где значение EC50 и наибольший сигнал были незаданными параметрами, и наклон фиксировали для единства. Оба слитых полипептида характеризовались отчетливыми сигналами связывания со значениями EC50, составляющими от 7,3 до 7,5 нМ, демонстрируя, что оба слитых полипептида способны одновременно связывать GPC3 и CD137.
[00259] Таблица 14. Данные ELISA в отношении одновременного связывания мишени

[00260] Пример 17. Аффинность слитых полипептидов в отношении GPC3 человека
[00261] Аффинности связывания связывающего GPC3 мутеина липокалина и биспецифических полипептидов SEQ ID NO: 46 и SEQ ID NO: 47 в отношении рекомбинантного GPC3 человека и рекомбинантного CD137 человека определяли с помощью поверхностного плазмонного резонанса на приборе Biacore T200 (GE Healthcare) с использованием HBS-EP+ (1x; BR-1006-69; GE Healthcare) в качестве подвижного буфера по аналогии с процедурой, описанной в примере 5.
[00262] Набор для захвата биотина Biotin CAPture (GE Healthcare) использовали для иммобилизации биотинилированных биспецифических полипептидов на поверхности чипа. Биспецифические полипептиды биотинилировали с помощью стандартной химии NHS. Неразведенный реагент из набора для захвата биотина Biotin CAPture (стрептавидин, конъюгированный с олигонуклеотидом ss-DNA) фиксировали на сенсорном чипе CAP с предварительно иммобилизированным комплементарным олигонуклеотидом ss-DNA. Затем биотинилированные мутеины в концентрации, составляющей 1 мкг/мл, использовали в течение 300 с при скорости потока, составляющей 5 мкл/мин.
[00263] GPC3 наносили в четырех концентрациях (300 нМ, 100 нМ, 33 нМ и 11 нМ) при скорости потока, составляющей 30 мкл/мин. GPC3 вводили в течение 180 с, а затем время диссоциации устанавливали как 1200 с. Восстановление поверхности чипа выполняли путем введения 6 M гуанидиния-HCl + 0,25 M NaOH (120 с) со скоростью потока, составляющей 10 мкл/мин. После введения восстанавливающих растворов следовала дополнительная стадия промывания с помощью подвижного буфера HBS-EP+ (1x; BR-1006-69; GE Healthcare) и период стабилизации, составляющий 120 с.
[00264] Данные сравнивали с таковыми для двух контрольных образцов посредством вычитания соответствующих сигналов, измеренных для контрольного канала (загруженный только реагентом для захвата биотина Biotin CAPture) и посредством вычитания введений буфера из ответов связывания. Константу скорости ассоциации ka и константу скорости диссоциации kd для реакции связывания определяли с использованием программного обеспечения для оценки V2.0 для Biacore T200 для обработки данных и аппроксимации кинетических измерений.
[00265] Соответствующие сенсограммы показаны на фигуре 17. Результаты представлены в таблице 15. Из данных видно, что биспецифические полипептиды связывают GPC3 с аффинностями, составляющими 4,3 нМ (SEQ ID NO: 46) и 3,5 нМ (SEQ ID NO: 47) соответственно.
[00266] Таблица 15. Аффинности связывания для GPC3

[00267] Пример 18. Аффинность слитых полипептидов в отношении CD137 человека
[00268] Аффинности связывания связывающего CD137 мутеина липокалина и биспецифических полипептидов SEQ ID NO: 46 и SEQ ID NO: 47 в отношении белка слияния рекомбинантный CD137 человека-Fc (№ 838-4B-100, R&D Systems) определяли с помощью поверхностного плазмонного резонанса с использованием прибора Biacore T200 (GE Healthcare) по аналогии с примером 6. Перед анализом аффинности с помощью SPR сенсорный чип CM5 получали с помощью антитела к Fc человека с помощью набора для захвата антитела человека Human Antibody Capture Kit (GE Healthcare № BR-1008-39) согласно инструкциям изготовителя.
[00269] Для определения аффинности белок слияния CD137 человека-Fc иммобилизировали на чипе в концентрации, составляющей 0,25 мг/мл, при скорости потока, составляющей 10 мкл/мин, и времени контакта, составляющем 180. Готовили четыре различные концентрации (1000 нМ, 200 нМ, 40 нМ и 8 нМ) биспецифических полипептидов в подвижном буфере (10 мМ HEPES, 150 мМ NaCl, 0,05% об./об. поверхностно-активного вещества P20, 3 мМ EDTA, pH 7,4 (GE Healthcare)) и наносили на поверхность чипа. При нанесении со скоростью потока, составляющей 30 мкл/мин, время контакта с образцом составляло 180 с, а время диссоциации составляло 600 с. Все измерения проводили при 25°C. Восстановление поверхности сенсорного чипа выполняли при помощи введения 10 мМ глицина, pH 1,7, с последующими дополнительным промыванием подвижным буфером и периодом стабилизации, составляющим 120 с. Перед измерениями уровня белка проводили три цикла восстановления с целью приведения к требуемым условиям. Данные оценивали с помощью программного обеспечения Biacore T200 Evaluation (V 2.0). Использовали двойное сравнение с образцом и для аппроксимирования первичных данных использовали модель связывания 1:1.
[00270] Результаты показаны на фигуре 18 и представлены в таблице 16. Из данных видно, что биспецифические полипептиды связывают CD137 с аффинностями, которые по меньшей мере такие же как аффинность мутеина липокалина относительно CD137.
[00271] Таблица 16. Аффинности связывания для CD137

[00272] Пример 19. Функциональный анализ активации T-клеток с использованием нанесенных слитых полипептидов
[00273] Авторы настоящего изобретения использовали анализ активации T-клеток для оценки способности слитых полипептидов с SEQ ID NO: 36 и 37, SEQ ID NO: 38 и 39, SEQ ID NO: 40 и 41 и SEQ ID NO: 42 и 43 костимулировать ответы T-клеток. Для этой цели слитые полипептиды при разных концентрациях наносили на пластиковую чашку совместно с антителом к CD3 человека (OKT3, eBioscience) и очищенные T-клетки затем инкубировали на покрытых поверхностях в присутствии растворимого антитела к CD28 человека (клон 28.2; eBioscience). В качестве показателя для считывания авторы настоящего изобретения измеряли уровни интерлейкина 2 (IL-2) в супернатанте. В качестве отрицательного контроля использовали контроль, представляющий собой изотип IgG4 человека, в качестве отрицательного контроля. Ниже авторы настоящего изобретения приводят подробное описание эксперимента.
[00274] Мононуклеарные клетки периферической крови человека (PBMC) от здоровых доноров-добровольцев выделяли из лейкоцитарных пленок путем центрифугирования в градиенте плотности полисахарозы (Biocoll 1,077 г/мл от Biochrom), следуя протоколу компании Biochrom. T-лимфоциты выделяли из полученных в результате PBMC c использованием набора для очистки Pan T-cell purification Kit (Miltenyi Biotec GmbH) и протоколов изготовителя. Очищенные T-клетки повторно суспендировали в буфере, состоящем из 90% FCS и 10% DMSO, немедленно замораживали с помощью жидкого азота и хранили в жидком азоте до последующего использования. Для анализа T-клетки оттаивали в течение 16 ч, и их культивировали в культуральной среде (RPMI 1640, Life Technologies), дополненной 10% FCS и 1% смеси пенициллин-стрептомицин (Life Technologies).
[00275] Следующую процедуру проводили с использованием трех повторов для каждого экспериментального условия. На плоскодонные планшеты для культур тканей наносили на ночь при 4°C 200 мкл смеси из 0,5 мкг/мл антитела к CD3 и серии разведений слитых полипептидов с SEQ ID NO: 36 и 37, SEQ ID NO: 38 и 39, SEQ ID NO: 40 и 41 и SEQ ID NO: 42 и 43 (25 мкг/мл, 2,5 мкг/мл и 0,25 мкг/мл) или отрицательного контроля, представляющего собой изотип IgG4 (25 мкг/мл). В другой постановке при таком же экспериментальном условии слитые полипептиды наносили вместе с изотипом IgG1 (в качестве дополнительного отрицательного контроля) вместо антитела к CD3. На следующий день лунки промывали дважды с использованием PBS и в каждую лунку добавляли 100 мкл суспензии T-клеток (соответствующей 5×104 T-клеток) в культуральной среде, дополненной 2 мкг/мл антитела к hCD28. Планшеты накрывали газопроницаемой уплотняющей пленкой (4titude) и инкубировали при 37°C в увлажненной атмосфере с 5% CO2 в течение 3 дней. Далее оценивали концентрацию IL-2 в супернатанте, а также клеточную пролиферацию.
[00276] Уровни IL-2 человека в объединенных супернатантах клеточных культур количественно измеряли с помощью набора DuoSet для IL-2 от R&D Systems. Процедуру выполняли как описано ниже. На первой стадии на 384-луночный планшет наносили при комнатной температуре на 2 ч 1 мкг/мл «антитела для захвата IL-2 человека» (R&D System), разведенного в PBS. Затем лунки промывали 5 раз с помощью 80 мкл PBS-T (PBS, содержащего 0,05% Tween20) с использованием установки для избирательного промывания планшетов с глубокими лунками Biotek EL405 select CW (Biotek). После блокирования в течение 1 ч в PBS-T, дополнительно содержащем 1% казеина (вес/вес), объединенный супернатант и серию концентраций стандарта IL-2, разведенного в культуральной среде, инкубировали в 384-луночном планшете в течение ночи при 4°C. Для обеспечения возможности выявления и количественного измерения захваченного IL-2 смесь из 100 нг/мл биотинилированного антитела козы к hIL-2-Bio для выявления (R&D System) и 1 мкг/мл меченного сульфо-меткой стрептавидина (Mesoscale Discovery) добавляли в PBS-T, содержащий 0,5% казеина, и инкубировали при комнатной температуре в течение 1 ч. После промывания 25 мкл буфера для считывания добавляли в каждую лунку и электрохемилюминесцентный (ECL) сигнал каждой лунки считывали с помощью планшет-ридера Mesoscale Discovery. Анализ и количественное измерение проводили с помощью программного обеспечения от Mesoscale Discovery.
[00277] Результат эксперимента показан на фигуре 19. Для всех четырех слитых полипептидов (SEQ ID NO: 36 и 37, SEQ ID NO: 38 и 39, SEQ ID NO: 40 и 41 и SEQ ID NO: 42 и 43) имела место отчетливая индукция продуцирования IL-2 с помощью используемых T-клеток по сравнению с отрицательным контролем, представляющим собой изотип IgG4. Из данных дополнительно видна склонность к более сильной индукции продуцирования IL-2 при более высоких концентрациях в покрывающем слое слияний полипептидов. В отсутствие стимуляции антителом к CD3 T-клеток слитые полипептиды не индуцируют продуцирование IL-2 T-клетками. Из этого видно, что слитые полипептиды способны костимулировать активацию T-клеток, стимулированных антителом к CD3 и антителом к CD28 при субоптимальных концентрациях.
[00278] Пример 20. Функциональный анализ активации T-клеток с использованием связанных с опухолевыми клетками слитых полипептидов
[00279] Авторы настоящего изобретения использовали анализ зависимой от клетки-мишени активации T-клеток для оценки способности слитых полипептидов с SEQ ID NO: 36 и 37, SEQ ID NO: 44 и SEQ ID NO: 45, способных к связыванию CD137 и GPC3 в одно и то же время, костимулировать ответы T-клеток при иммобилизации на GPC3-положительной клеточной линии. В качестве отрицательного контроля авторы настоящего изобретения использовали моноспецифическое связывающее CD137 антитело с SEQ ID NO: 34 и 35. В эксперименте антитело к CD3 человека (OKT3, eBioscience) наносили на пластиковую чашку для культивирования, а затем GPC3-положительные клетки HepG2 культивировали на чашке в течение ночи. На следующий день очищенные T-клетки инкубировали на покрытой поверхности в присутствии 1 мкг/мл слитых полипептидов с SEQ ID NO: 36 и 37, SEQ ID NO: 44 и SEQ ID NO: 45 или контрольного антитела с SEQ ID NO: 34 и 35. В качестве показателя для считывания авторы настоящего изобретения измеряли уровни интерлейкина 2 (IL-2) в супернатанте. Более подробно эксперимент описан ниже.
[00280] Мононуклеарные клетки периферической крови человека (PBMC) от здоровых доноров-добровольцев выделяли из лейкоцитарных пленок путем центрифугирования в градиенте плотности полисахарозы (Biocoll 1,077 г/мл от Biochrom), следуя протоколу компании Biochrom. T-лимфоциты выделяли из полученных в результате PBMC c использованием набора для очистки Pan T-cell purification Kit (Miltenyi Biotec GmbH) и протоколов изготовителя. Очищенные T-клетки повторно суспендировали в буфере, состоящем из 90% FCS и 10% DMSO, немедленно замораживали с помощью жидкого азота и хранили в жидком азоте до последующего использования. Для анализа T-клетки оттаивали в течение 16 ч, и их культивировали в культуральной среде (RPMI 1640, Life Technologies), дополненной 10% FCS и 1% смеси пенициллин-стрептомицин (Life Technologies).
[00281] Следующую процедуру проводили с использованием трех повторов для каждого экспериментального условия. На плоскодонные планшеты для культур тканей предварительно наносили на 1 ч при 37°C 200 мкл 0,25 мкг/мл антитела к CD3 или не наносили его. Затем лунки дважды промывали с использованием PBS. Высевали 1,25×104 опухолевых клеток HepG2 на лунку и обеспечивали возможность прилипания в течение ночи при 37°C в увлажненной атмосфере с 5% CO2. Клетки HepG2 предварительно выращивали в культуре при стандартных условиях, отделяли при помощи Accutase и повторно суспендировали в культуральной среде.
[00282] В последующие дни опухолевые клетки обрабатывали 2 часа при 37°C митомицином C (Sigma Aldrich) в концентрации, составляющей 10 мкг/мл, для блокирования их пролиферации. Планшеты дважды промывали с использованием PBS и в каждую лунку добавляли 100 мкл суспензии T-клеток (соответствующей 5×104 T-клеток) и слитые полипептиды или отрицательный контроль в концентрации, составляющей 1 мкг/мл. Планшеты накрывали газопроницаемой уплотняющей пленкой (4titude) и инкубировали при 37°C в увлажненной атмосфере с 5% CO2 в течение 3 дней. Затем оценивали концентрацию IL-2 в супернатанте, как описано ниже.
[00283] Уровни IL-2 человека в супернатантах клеточных культур количественно измеряли с помощью набора DuoSet для IL-2 от R&D Systems. Процедуру осуществляли так, как она описана ниже. На первой стадии на 384-луночный планшет наносили при комнатной температуре на 2 ч 1 мкг/мл «антитела для захвата IL-2 человека» (R&D System), разведенного в PBS. Затем лунки промывали 5 раз с помощью 80 мкл PBS-T (PBS, содержащего 0,05% Tween20) с использованием установки для избирательного промывания планшетов с глубокими лунками Biotek EL405 select CW (Biotek). После блокирования в течение 1 ч в PBS-T, дополнительно содержащем 1% казеина (вес/вес), объединенный супернатант и серию концентраций стандарта IL-2, разведенного в культуральной среде, инкубировали в 384-луночном планшете в течение ночи при 4°C. Для обеспечения возможности выявления и количественного измерения захваченного IL-2 смесь из 100 нг/мл биотинилированного антитела козы к hIL-2-Bio для выявления (R&D System) и 1 мкг/мл меченного сульфо-меткой стрептавидина (Mesoscale Discovery) добавляли в PBS-T, содержащий 0,5% казеина, и инкубировали при комнатной температуре в течение 1 ч. После промывания 25 мкл буфера для считывания добавляли в каждую лунку и электрохемилюминесцентный (ECL) сигнал каждой лунки считывали с помощью планшет-ридера Mesoscale Discovery. Анализ и количественное измерение проводили с помощью программного обеспечения от Mesoscale Discovery.
[00284] Результат эксперимента показан на фигуре 20. Для трех слитых полипептидов с SEQ ID NO: 36 и 37, SEQ ID NO: 44 и SEQ ID NO: 45 имела место отчетливая индукция продуцирования IL-2 с помощью используемых T-клеток по сравнению с контрольным антителом с SEQ ID NO: 34 и 35. Из этого видно, что слитые полипептиды по настоящему раскрытию способны костимулировать активацию T-клеток зависимым от мишени образом, и это подтверждается тем, что связывающие GPC3 слитые полипептиды проявляют более высокие уровни продуцирования IL-2, чем контрольное антитело.
[00285] Пример 21. Функциональный анализ активации T-клеток с использованием связанных с опухолевыми клетками слитых полипептидов с блокированием биспецифического связывания и без него
[00286] Авторы настоящего изобретения использовали анализ зависимой от клетки-мишени активации T-клеток, подобно эксперименту, описанному в примере 20, для оценки способности слитых полипептидов с SEQ ID NO: 44 и SEQ ID NO: 45, способных к одновременному связыванию CD137 и GPC3, костимулировать ответы T-клеток при связывании с GPC3-положительной клеточной линией. В качестве контроля эксперимент проводили в присутствии избытка моноспецифического связывающего GPC3 антикалина с SEQ ID NO: 10 для вытеснения биспецифических конструкций с SEQ ID NO: 44 или SEQ ID NO: 45 из связывания с GPC3-положительными клетками. В эксперименте антитело к CD3 человека (OKT3, eBioscience) наносили на пластиковую чашку для культивирования, а затем GPC3-положительные клетки Hep3B культивировали на чашке в течение ночи. На следующий день очищенные T-клетки инкубировали на покрытой поверхности в присутствии четырех концентраций слитых полипептидов с SEQ ID NO: 44 и SEQ ID NO: 45 (1 мкг/мл, 0,1 мкг/мл, 0,01 мкг/мл, 0,001 мкг/мл). Параллельно эксперимент проводили с добавлением избытка SEQ ID NO: 10 (1 мг/мл). В качестве показателя для считывания авторы настоящего изобретения измеряли уровни интерлейкина 2 (IL-2) в супернатанте. Более подробно эксперимент описан ниже.
[00287] Мононуклеарные клетки периферической крови человека (PBMC) от здоровых доноров-добровольцев выделяли из лейкоцитарных пленок путем центрифугирования в градиенте плотности полисахарозы (Biocoll 1,077 г/мл от Biochrom), следуя протоколу компании Biochrom. T-лимфоциты выделяли из полученных в результате PBMC c использованием набора для очистки Pan T-cell purification Kit (Miltenyi Biotec GmbH) и протоколов изготовителя. Очищенные T-клетки культивировали в культуральной среде (RPMI 1640, Life Technologies), дополненной 10% FCS и 1% смеси пенициллин-стрептомицин (Life Technologies).
[00288] Следующую процедуру проводили с использованием трех повторов для каждого экспериментального условия. На плоскодонные планшеты для культур тканей предварительно наносили на 1 ч при 37°C 200 мкл 0,25 мкг/мл антитела к CD3. Затем лунки дважды промывали с использованием PBS. Высевали 1,25×104 опухолевых клеток Hep3B на лунку и обеспечивали возможность прилипания в течение ночи при 37°C в увлажненной атмосфере с 5% CO2. Клетки Hep3B предварительно выращивали в культуре при стандартных условиях, отделяли при помощи Accutase и повторно суспендировали в культуральной среде.
[00289] На следующий день опухолевые клетки обрабатывали 2 часа при 37°C митомицином C (Sigma Aldrich) в концентрации, составляющей 10 мкг/мл, для блокирования их пролиферации. Планшеты дважды промывали с использованием PBS и добавляли 100 мкл суспензии T-клеток (соответствующей 5×104 T-клеток) и слитые полипептиды с SEQ ID NO: 44 и SEQ ID NO: 45 в концентрации, составляющей 1 мкг/мл, 0,1 мкг/мл, 0,01 мкг/мл, 0,001 мкг/мл, в присутствии избытка SEQ ID NO: 10 (1 мг/мл) или при его отсутствии. Планшеты накрывали газопроницаемой уплотняющей пленкой (4titude) и инкубировали при 37°C в увлажненной атмосфере с 5% CO2 в течение 3 дней. Затем концентрацию IL-2 в супернатанте определяли с помощью ELISA с использованием набора для ELISA для IL-2 человека от BD Bioscience согласно инструкциям изготовителя.
[00290] Результат эксперимента показан на фигуре 21. Для двух слитых полипептидов с SEQ ID NO: 44 (фигура 21A) и SEQ ID NO: 45 (фигура 21C) имела место отчетливая индукция продуцирования IL-2 с помощью использованных T-клеток, которая возрастала с повышением концентрации. В отличие от этого индукция продуцирования IL-2 исчезала в присутствии избытка SEQ ID NO: 10, который подавляет связывание биспецифических средств с SEQ ID NO: 44 и SEQ ID NO: 45 с клетками Hep3B. Из этого видно, что слитые полипептиды по настоящему раскрытию способны костимулировать активацию T-клеток зависимым от мишени образом.
[00291] Примечательно, что уровень индуцированого IL-2 выше для SEQ ID NO: 44 по сравнению с SEQ ID NO: 45, что указывает на то, что геометрические параметры биспецифического слияния GPC3/CD137 играют важную роль в определении интенсивности активации T-клеток.
[00292] Пример 22. Функциональный анализ активации T-клеток с использованием опухолевых клеток с высоким и низким уровнями GPC3
[00293] Авторы настоящего изобретения использовали анализ зависимой от клетки-мишени активации T-клеток, подобно эксперименту, описанному в примере 20, для оценки способности слитых полипептидов с SEQ ID NO: 44 и SEQ ID NO: 45 костимулировать ответы T-клеток в зависимости от уровня GPC3 в используемой клеточной линии. В качестве отрицательного контроля авторы настоящего изобретения использовали антитело, связывающее HER2, трастузумаб. Для сравнения авторы настоящего изобретения исследовали поведение эталонного моноклонального антитела к CD137 с SEQ ID NO: 74 и 75. В эксперименте антитело к CD3 человека (OKT3, eBioscience) наносили на пластиковые чашки для культивирования, а затем клетки HepG2, SKBR3 или MCF7 отдельно культивировали на чашках в течение ночи. На следующий день очищенные T-клетки инкубировали на покрытой поверхности в присутствии разных концентраций слитого полипептида с SEQ ID NO: 44, SEQ ID NO: 45, эталонным антителом с SEQ ID NO: 74 и 75 и отрицательных контролей, представляющих собой трастузумаб и среду (т.е. без добавления тестируемого продукта). В качестве показателя для считывания авторы настоящего изобретения измеряли уровни интерлейкина 2 (IL-2) в супернатанте. Более подробно эксперимент описан ниже.
[00294] Мононуклеарные клетки периферической крови человека (PBMC) от здоровых доноров-добровольцев выделяли из лейкоцитарных пленок путем центрифугирования в градиенте плотности полисахарозы (Biocoll 1,077 г/мл от Biochrom), следуя протоколу компании Biochrom. T-лимфоциты выделяли из полученных в результате PBMC c использованием набора для очистки Pan T-cell purification Kit (Miltenyi Biotec GmbH) и протоколов изготовителя. Очищенные T-клетки повторно суспендировали в буфере, состоящем из 90% FCS и 10% DMSO, немедленно замораживали с помощью жидкого азота и хранили в жидком азоте до последующего использования. Для анализа T-клетки оттаивали в течение 16 ч, и их культивировали в культуральной среде (RPMI 1640, Life Technologies), дополненной 10% FCS и 1% смеси пенициллин-стрептомицин (Life Technologies).
[00295] Следующую процедуру проводили с использованием трех повторов для каждого экспериментального условия. На плоскодонные планшеты для культур тканей предварительно наносили на 1 ч при 37°C 200 мкл 0,25 мкг/мл антитела к CD3. Затем планшеты дважды промывали с использованием PBS. Высевали 5×104 целевых опухолевых клеток на лунку и обеспечивали возможность прилипания в течение ночи при 37°C в увлажненной атмосфере с 5% CO2. Целевые клетки предварительно выращивали в культуре при стандартных условиях, отделяли при помощи Accutase и повторно суспендировали в культуральной среде.
[00296] На следующий день опухолевые клетки обрабатывали 2 часа при 37°C митомицином C (Sigma Aldrich) в концентрации, составляющей 30 мкг/мл, для блокирования их пролиферации. Планшеты дважды промывали с использованием PBS и в каждую лунку добавляли 100 мкл суспензии T-клеток (соответствующей 5×104 T-клеток) совместно с тестируемыми продуктами с SEQ ID NO: 44, SEQ ID NO: 45, эталонным антителом с SEQ ID NO: 74 и 75 и отрицательным контролем трастузумабом в концентрации, находящейся в диапазоне от 0,05 нМ до 5 нМ. Планшеты накрывали газопроницаемой уплотняющей пленкой (4titude) и инкубировали при 37°C в увлажненной атмосфере с 5% CO2 в течение 3 дней. Затем оценивали концентрацию IL-2 в супернатанте, как описано ниже.
[00297] Уровни IL-2 человека в супернатантах клеточных культур количественно измеряли с помощью набора DuoSet для IL-2 от R&D Systems. Процедуру осуществляли так, как она описана ниже. На первой стадии на 384-луночный планшет наносили при комнатной температуре на 2 ч 1 мкг/мл «антитела для захвата IL-2 человека» (R&D System), разведенного в PBS. Затем лунки промывали 5 раз с помощью 80 мкл PBS-T (PBS, содержащего 0,05% Tween20) с использованием установки для избирательного промывания планшетов с глубокими лунками Biotek EL405 select CW (Biotek). После блокирования в течение 1 ч в PBS-T, дополнительно содержащем 1% казеина (вес/вес), объединенный супернатант и серию концентраций стандарта IL-2, разведенного в культуральной среде, инкубировали в 384-луночном планшете в течение ночи при 4°C. Для обеспечения возможности выявления и количественного измерения захваченного IL-2 смесь из 100 нг/мл биотинилированного антитела козы к hIL-2-Bio для выявления (R&D System) и 1 мкг/мл меченного сульфо-меткой стрептавидина (Mesoscale Discovery) добавляли в PBS-T, содержащий 0,5% казеина, и инкубировали при комнатной температуре в течение 1 ч. После промывания 25 мкл буфера для считывания добавляли в каждую лунку и электрохемилюминесцентный (ECL) сигнал каждой лунки считывали с помощью планшет-ридера Mesoscale Discovery. Анализ и количественное измерение проводили с помощью программного обеспечения от Mesoscale Discovery.
[00298] Результат репрезентативного эксперимента показан на фигуре 22. На этой фигуре значения наносили на график относительно фонового уровня продуцирования IL-2 при отсутствии тестируемого соединения, и таким образом они представляют кратное изменение по сравнению с фоном. Хотя отрицательный контроль трастузумаб (фигура 22A, треугольники) не приводил к индукции IL-2 на T-клетках с любой из трех клеточных линий, повышающиеся концентрации биспецифических слитых полипептидов SEQ ID NO: 44 (фигура 22A, кружки) и SEQ ID NO: 45 (фигура 22A, квадраты) индуцируют продуцирование IL-2 T-клетками в присутствии экспрессирующих GPC3 клеток HepG2. Однако какого-либо увеличения уровня IL-2 в результате воздействия SEQ ID NO: 44 и SEQ ID NO: 45 не наблюдали для GPC3-отрицательных клеток SKBR3 и MCF7 (фигура 22). Такое поведение заметно отличается от антитела к CD137 с SEQ ID NO: 74 и 75, которое индуцирует IL-2 на T-клетках в присутствии всех трех клеточных линий (фигура 22B).
[00299] В эксперименте отчетливо показано, что SEQ ID NO: 44 и SEQ ID NO: 45 активируют T-клетки таким образом, что это зависит от присутствия GPC3 на целевых клетках. Хотя GPC3-положительная клеточная линия HepG2 проявляет отчетливую активацию T-клеток, которая измерена по продуцированию IL-2, такого эффекта не наблюдали с клетками SKBR3 и MCF7, которые не экспрессируют выявляемые уровни GPC3. То, что этот эффект объясняется наличием GPC3, а не исследуемыми GPC3-отрицательными клеточными линиями, которые потенциально делают передачу сигнала CD137 неэффективной, становится очевидным вследствие того факта, что антитело к CD137 с SEQ ID NO: 74 и 75 способно активировать T-клетки посредством передачи сигнала CD137 во всех трех типах клеток.
[00300] Пример 23. Ex vivo оценка иммуногенности слитых полипептидов в отношении T-клеток
Для изучения риска образования антител к лекарственному средству у человека проводили in vitro оценку иммуногенности в отношении T-клеток биспецифических слитых полипептидов с SEQ ID NO: 44 и SEQ ID NO: 45, контрольного антитела, представляющего собой трастузумаб, и положительного контроля, представляющего собой гемоцианин лимфы улитки (KLH). Для выполнения эксперимента PBMC от 32 доноров, выбранных для охвата аллотипов HLA, отражающих распределение в мировой популяции, размораживали, промывали и высевали на 96-луночные планшеты с плотностью, составляющей 3×105 клеток на лунку. Тестируемые продукты, разведенные в средах для анализа, добавляли к клеткам в концентрации, составляющей 30 мкг/мл. Среду для анализа использовали отдельно в качестве холостой пробы и гемоцианин лимфы улитки (KLH) использовали в качестве не подвергнутого воздействию положительного контроля. PBMC инкубировали в течение 7 дней в увлажненной атмосфере при 37°C и 5% CO2. В день 7 PBMC метили в отношении поверхностных фенотипических маркеров CD3+ и CD4+ и в отношении включенного в ДНК EdU (5-этинил-2’-дезоксиуридина), используемого в качестве маркера клеточной пролиферации. Процентную долю пролиферирующих CD3+CD4+EdU+ клеток измеряли с помощью проточного цитометра Guava easyCyte 8HT и анализировали с помощью программного обеспечения GuavaSoft InCyte.
На фигуре 23 представлены результаты этого анализа для всех 32 доноров и всех тестируемых исследуемых молекул. На фигуре 23A на график наносили индекс стимуляции, который получали по соотношению пролиферации в присутствии тестируемого продукта в сравнении с пролиферацией в отсутствие тестируемого продукта. Пороговый уровень, который определяет прореагировавшего донора (индекс стимуляции > 2), показан в виде пунктирной линии. На фигуре 23B на график наносили количество прореагировавших доноров, определенное по этому пороговому уровню. Очевидно, что количество доноров, прореагировавших на эталонный препарат, представляющий собой трастузумаб, остается на уровне одного и в связи с этим является незначительным, тогда как все 32 донора прореагировали на положительный контроль, представляющий собой KLH, сильной пролиферацией выше порогового уровня. Для биспецифических слитых полипептидов SEQ ID NO: 44 и SEQ ID NO: 45, количество прореагировавших доноров также остается на уровне одного в обоих случаях.
[00301] Из эксперимента таким образом видно, что биспецифические слитые полипептиды индуцируют незначительный ответ при in vitro оценке иммуногенности в отношении T-клеток, что указывает на то, что риск индукции иммуногенных ответов низок.
[00302] Пример 24. Аффинность в отношении Fc-гамма рецепторов hFcγ RI/CD64 и hFcγ RIIIA/CD16a
[00303] Для измерения аффинностей связывания слияний полипептидов со сконструированным каркасом на основе IgG4 (SEQ ID NO: 44 и SEQ ID NO: 45) в отношении Fc-гамма рецепторов hFcγ RI/CD64 (R&D Systems) и hFcγ RIIIA/CD16a (R&D Systems) использовали анализ на основе поверхностного плазмонного резонанса (SPR). Трастузумаб служил в качестве контроля, представляющего собой моноспецифическое антитело с каркасом IgG1. В анализе аффинности с помощью SPR слияния полипептидов биотинилировали и захватывали на сенсорном чипе CAP с помощью набора для захвата биотина Biotin CAPture Kit (GE Healthcare). Сенсорный чип CAP был предварительно иммобилизирован с использованием олигонуклеотида ssDNA. Неразведенный реагент для захвата биотина Biotin CAPture Reagent (стрептавидин, конъюгированный с комплементарным олигонуклеотидом ss-DNA) наносили при скорости потока, составляющей 2 мкл/мин, в течение 300 с. Затем 10 мкг/мл биотинилированного слияния полипептидов наносили в течение 300 с при скорости потока, составляющей 5 мкл/мин. Трастузумаб и слияния полипептидов биотинилировали путем инкубации с NHS-PEG4-биотином EZ-Link® (Thermo Scientific) в течение двух часов при комнатной температуре. Избыток непрореагировавшего реагента с биотином удаляли путем загрузки реакционной смеси в планшет для обессоливания для применения в формате с вращением ZebaTM Spin Desalting Plate (Thermo Scientific). В эталонный канал загружали только реагент набора для захвата биотина Biotin CAPture Reagent.
[00304] Для определения аффинности готовили четыре разведения hFcγ RI/CD64 (в концентрации 100, 25, 6,25 и 1,6) или от четыре до пяти разведений hFcγ RIIIA/CD16a (в концентрации 1000, 333, 111, 37 и 12 нМ) в подвижном буфере (10 мМ HEPES, 150 мМ NaCl, 0,05% об./об. поверхностно-активного вещества P20, 3 мМ EDTA, pH 7,4 (GE Healthcare)) и наносили на поверхность чипа. При нанесении со скоростью потока, составляющей 30 мкл/мин, время контакта образца составляло 180 с, и время диссоциации составляло 1800/2700 с для hFcγ RI/CD64 или 300 с для hFcγ RIIIA/CD16a. Все измерения проводили при 25°C. Восстановление поверхности сенсорного чипа CAP выполняли при помощи введения 6 M Gua-HCl с 0,25 M NaOH с последующими дополнительным промыванием подвижным буфером и периодом стабилизации, составляющим 120 с. Перед измерениями уровня белка проводили три цикла восстановления с целью приведения к требуемым условиям. Данные оценивали с помощью программного обеспечения Biacore T200 Evaluation (V 2.0). Применяли двойное сравнение с образцом. Для hFcγ RI/CD64 использовали модель связывания 1:1 для аппроксимирования первичных данных. Для hFcγ RIIIA/CD16a использовали модель аффинности в равновесном состоянии для аппроксимирования первичных данных.
[00305] В таблице 17 показаны результаты аппроксимирования данных для hFcγ RI/CD64. Тестируемый продукт на основе IgG1, представляющий собой трастузумаб, проявляет аффинность, составляющую 0,3 нМ. Слияния полипептидов с SEQ ID NO: 44 и SEQ ID NO: 45 не проявили существенного связывания с hFcγ RI/CD64. Из этих данных видно, что связывание с hFcγ RI/CD64 может быть снижено до несущественных уровней путем смены изотипа с IgG1 на сконструированный IgG4.
[00306] Таблица 17
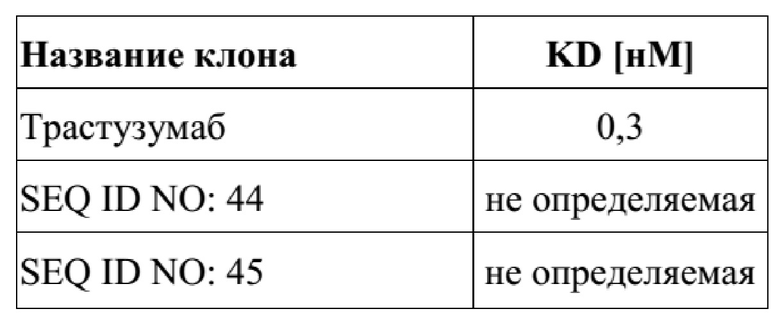
[00307] В таблице 18 показаны результаты приведения в соответствие данных для hFcγ RIIIA/CD16a. Полученная в результате аффинность связывания в отношении hFcγ RIIIA/CD16a тестируемых продуктов на основе IgG1, представляющих собой трастузумаб, составляла около 350 нМ, тогда как слияния полипептидов с SEQ ID NO: 44 и SEQ ID NO: 45 не проявили существенного связывания с hFcγ RIIIA/CD16a. Из этих данных видно, что связывание с hFcγ RI/CD64 может быть снижено до несущественных уровней путем смены изотипа с IgG1 на сконструированный IgG4.
[00308] Таблица 18
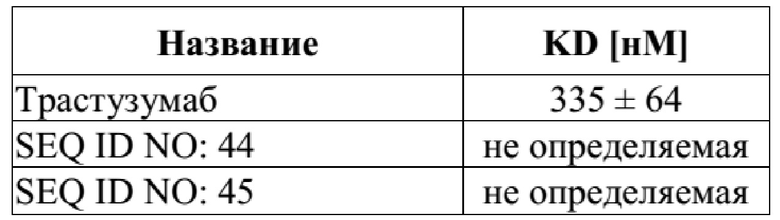
[00309] Пример 25. Аффинность в отношении неонатального Fc-рецептора
[00310] Для измерения аффинностей связывания слияний полипептидов со сконструированным каркасом на основе IgG4 (SEQ ID NO: 44 и SEQ ID NO: 45) в отношении неонатального Fc-рецептора (FcRn, Sino Biologicals, № CT009-H08H) использовали анализ на основе поверхностного плазмонного резонанса (SPR). Трастузумаб служил в качестве контроля, представляющего собой моноспецифическое антитело с каркасом IgG1. В анализе аффинности с помощью SPR FcRn ковалентно иммобилизировали на сенсорном чипе CM5 (GE Healthcare) согласно инструкциям изготовителя. Вкратце, после активации карбоксильных групп декстрановой матрицы 1-этил-3-(3-диметиламинпропил)-карбодиимидом (EDC) и N-гидроксисукцинимидом (NHS) первичным аминам белка FcRn позволяли вступать в реакцию со сложным эфиром NHS на поверхности до достижения сигнала, составляющего ~200 RU. В конечном итоге непрореагировавшие сложные эфиры NHS блокировали путем пускания раствора 1 M этаноламина по поверхности. Скорость потока на протяжении всей процедуры иммобилизации составляла 10 мкл/мин.
[00311] Для определения их аффинности готовили шесть разведений (1000 нМ, 333 нМ, 111 нМ, 37 нМ, 12 нМ и 4 нМ) всех конструкций в подвижном буфере (10 мМ HEPES, 150 мМ NaCl, 0,05% об./об. поверхностно-активного вещества P20, 3 мМ EDTA, pH 6,0) и наносили на поверхность чипа. При нанесении со скоростью потока, составляющей 30 мкл/мин, время контакта образца составляло 180 с, и время диссоциации составляло 30 с. Все измерения проводили при 25°C. Восстановление поверхности сенсорного чипа CAP выполняли при помощи введения 10 мМ глицина, pH 3,0. Перед измерениями уровня белка проводили три цикла восстановления с целью приведения к требуемым условиям. Данные оценивали с помощью программного обеспечения Biacore T200 Evaluation (V 2.0) с двойным сравнением. Для аппроксимирования первичных данных использовали модель аффинности в равновесном состоянии.
[00312] Отраженные в таблице 19, полученные в результате аффинности связывания для всех слияний полипептидов с FcRn составляли около 2 мкМ, что указывает на то, что смена изотипа с IgG1 на сконструированный каркас IgG4 не оказывает выявляемого влияния на связывание FcRn.
[00313] Таблица 19

[00314] Пример 26. Фармакокинетика слитых полипептидов у мышей
[00315] Анализ фармакокинетики слитых полипептидов, обозначенных как SEQ ID NO: 44 и SEQ ID NO: 45, проводили на мышах. Самцам мышей линии CD-1 возрастом примерно 5 недель (3 мыши на момент времени; Charles River Laboratories, Research Models and Services, Germany GmbH) вводили в хвостовую вену слитый полипептид в дозе 10 мг/кг. Тестируемые продукты вводили в виде однократной дозы в объеме 5 мл/кг. Образцы плазмы крови от мышей получали в моменты времени, соответствующие 5 мин, 1 ч, 2 ч, 4 ч, 8 ч, 24 ч, 48 ч, 4 дням, 8 дням и 14 дням. Достаточное количество цельной крови, взятое под анестезией с использованием изофлурана, собирали для получения по меньшей мере 100 мкл плазмы крови в пробирке Li-Heparin на животное и временную точку. Уровни лекарственного средства выявляли при помощи ELISA сэндвич-типа, при котором выявляли целую биспецифическую конструкцию посредством мишеней GPC3 и CD137. Данные аппроксимировали с использованием двухкомпартментной модели с использованием программного обеспечения Prism GraphPad 5.
[00316] На фигуре 25 показаны графики зависимости концентрации в плазме от времени для конструкций с SEQ ID NO: 44 и SEQ ID NO: 45 со вставной секцией, на которой показаны те же данные на графике в полулогарифмическом масштабе. Фармакокинетика выглядит аналогичной в обоих случаях. Начиная с концентрации в плазме крови, составляющей около 150 мкг/мл, уровни в плазме крови падали до фоновых уровней в пределах около 100 часов. Биэкспоненциальный распад при двухкомпартментной модели успешно применяли для точного описания данных, и аппроксимация данных (фигура 25) с использованием этой модели давала в результате периоды полувыведения в конечной фазе, составляющие 13,7 ч для SEQ ID NO: 44 и 10,0 ч. Для SEQ ID NO: 45.
[00317] Из данных видно, что биспецифические слияния характеризуются периодами полувыведения, которые находятся в промежуточном диапазоне того, что можно ожидать для белков слияния с Fc.
[00318] Пример 27. Фармакокинетика слитых полипептидов у яванского макака
[00319] Анализ фармакокинетики слитых полипептидов, обозначенных как SEQ ID NO: 44 и SEQ ID NO: 45, проводили на яванских макаках. Самцы яванских макаков получали внутривенное вливание в течение 60 минут с дозой, составляющей 3 мг/кг тестируемого продукта. Образцы плазмы крови от яванских макаков получали в моменты времени, соответствующие 15 мин, 2 ч, 4 ч, 8 ч, 24 ч, 48 ч, 3 дням, 4 дням, 5 дням, 6 дням, 7 дням, 9 дням, 11 дням, 14 дням, 18 дням и 24 дням. Уровни лекарственного средства выявляли при помощи ELISA сэндвич-типа, при котором выявляли целую биспецифическую конструкцию посредством мишеней HER2 и CD137. Уровни трастузумаба в плазме крови определяли при помощи ELISA сэндвич-типа с мишенями HER2 и Fc человека. Данные аппроксимировали с использованием двухкомпартментной модели с использованием программного обеспечения Prism GraphPad 5.
На фигуре 26 показаны графики зависимости концентрации в плазме от времени для конструкций с SEQ ID NO: 44 и SEQ ID NO: 45 со вставной секцией, на которой показаны те же данные на графике в полулогарифмическом масштабе. Фармакокинетика выглядит аналогичной в обоих случаях, при этом SEQ ID NO: 44 проявляет явно более продолжительный период полувыведения. Начиная с концентрации в плазме крови, составляющей около 70 мкг/мл, уровни в плазме крови падали до уровней, близких к нулю, на протяжении времени, составляющего 200 ч. Биэкспоненциальный распад при двухкомпартментной модели успешно применяли для точного описания данных, и аппроксимация данных (фигура 26) с использованием этой модели давала в результате периоды полувыведения в конечной фазе, составляющие 39 ч (SEQ ID NO: 44) и 24,1 ч (SEQ ID NO: 45) соответственно.
[00320] Из данных таким образом видно, что биспецифические слияния характеризуются периодами полувыведения в конечной фазе у яванских макаков, которые повышены по сравнению с периодом полувыведения у мышей, и находятся в приемлемом диапазоне для терапевтического применения биологического препарата.
[00321] Варианты осуществления, иллюстративно описанные в данном документе, надлежащим образом можно осуществлять на практике в отсутствие любых элемента или элементов, ограничения или ограничений, не раскрытых конкретно в данном документе. Таким образом, например, термины «содержащий», «включающий», «состоящий из» и т. д. следует читать как открытые и неограничивающие. Дополнительно термины и выражения, используемые в данном документе, использовали в качестве терминов описания, а не ограничения, и в использовании таких терминов и выражений отсутствует намерение исключить любые эквиваленты показанных и описанных характеристик или их частей, но следует понимать, что разные модификации возможны в пределах объема заявленного изобретения. Таким образом, следует понимать, что несмотря на то, что настоящие варианты осуществления были конкретно раскрыты с помощью предпочтительных вариантов осуществления и необязательных характеристик, к их модификации и вариациям может прибегнуть специалист в данной области, и что такие модификации и вариации рассматриваются как часть объема настоящего изобретения. Все патенты, патентные заявки, руководства и прошедшие рецензию публикации, описанные в данном документе, в данный документ включены с помощью ссылки в полном их объеме. Кроме того, в тех случаях, когда определение или использование термина в ссылочном материале, который включен посредством ссылки в данный документ, не согласуется или противоречит определению такого термина, представленному в данном документе, тогда используют определение термина, представленное в данном документе, а определение термина из ссылки не используют. Каждое из более узких видов и субродовых группировок, находящихся в пределах родового раскрытия, также образует часть настоящего изобретения. Это включает описание настоящего изобретения в обобщенном виде с оговоркой или отрицательными признаками, удаляющими любой объект из рода, независимо от того, исключенный материал конкретно изложен в данном документе или нет. К тому же в тех случаях, когда особенности описаны в контексте групп Маркуша, специалисту в данной области будет понятно, что настоящее раскрытие также таким образом раскрыто в контексте любого отдельного представителя или подгруппы представителей группы Маркуша. Дополнительные варианты осуществления будут очевидны из прилагаемой формулы изобретения.
Эквиваленты: специалистам в данной области будут понятны, или они будут способны установить с помощью всего лишь проведения обычных экспериментов, многие эквиваленты конкретных вариантов осуществления настоящего изобретения, описанного в данном документе. Как предполагается, такие эквиваленты охватываются прилагаемой формулой изобретения. Все публикации, патенты и патентные заявки, упомянутые в этом описании, таким образом включены с помощью ссылки в описание в такой же степени, как если бы каждая отдельная публикация, отдельный патент или отдельная патентная заявка были конкретно и отдельно указаны как включенные в данном документе с помощью ссылки.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Пиерис АГ
<120> Слитый полипептид с противораковой активностью
<130> PIE15362PCT
<150> EP15167927.1
<151> 2015-05-18
<150> EP16150508.6
<151> 2016-01-08
<160> 75
<170> PatentIn версия 3.5
<210> 1
<211> 156
<212> Белок
<213> искусственный
<220>
<223> Tlc дикого типа
<400> 1
His His Leu Leu Ala Ser Asp Glu Glu Ile Gln Asp Val Ser Gly Thr
1 5 10 15
Trp Tyr Leu Lys Ala Met Thr Val Asp Arg Glu Phe Pro Glu Met Asn
20 25 30
Leu Glu Ser Val Thr Pro Met Thr Leu Thr Thr Leu Glu Gly Gly Asn
35 40 45
Leu Glu Ala Lys Val Thr Met Leu Ile Ser Gly Arg Cys Gln Glu Val
50 55 60
Lys Ala Val Leu Glu Lys Thr Asp Glu Pro Gly Lys Tyr Thr Ala Asp
65 70 75 80
Gly Gly Lys His Val Ala Tyr Ile Ile Arg Ser His Val Lys Asp His
85 90 95
Tyr Ile Phe Tyr Cys Glu Gly Glu Leu His Gly Lys Pro Val Arg Gly
100 105 110
Val Lys Leu Val Gly Arg Asp Pro Lys Asn Asn Leu Glu Ala Leu Glu
115 120 125
Asp Phe Glu Lys Ala Ala Gly Ala Arg Gly Leu Ser Thr Glu Ser Ile
130 135 140
Leu Ile Pro Arg Gln Ser Glu Thr Cys Ser Pro Gly
145 150 155
<210> 2
<211> 178
<212> Белок
<213> искусственный
<220>
<223> NGAL дикого типа
<400> 2
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Leu Ala Gly Asn Ala Ile Leu Arg Glu Asp Lys Asp Pro
35 40 45
Gln Lys Met Tyr Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asn Val Thr Ser Val Leu Phe Arg Lys Lys Lys Cys Asp Tyr Trp Ile
65 70 75 80
Arg Thr Phe Val Pro Gly Cys Gln Pro Gly Glu Phe Thr Leu Gly Asn
85 90 95
Ile Lys Ser Tyr Pro Gly Leu Thr Ser Tyr Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Lys Val Ser Gln
115 120 125
Asn Arg Glu Tyr Phe Lys Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 3
<211> 178
<212> Белок
<213> искусственный
<220>
<223> контроль, представляющий собой NGAL98
<400> 3
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Leu Ala Gly Asn Ala Ile Leu Arg Glu Asp Lys Asp Pro
35 40 45
Gln Lys Met Tyr Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asn Val Thr Ser Val Leu Phe Arg Lys Lys Lys Cys Asp Tyr Trp Ile
65 70 75 80
Arg Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Asn
85 90 95
Ile Lys Ser Tyr Pro Gly Leu Thr Ser Tyr Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Lys Val Ser Gln
115 120 125
Asn Arg Glu Tyr Phe Lys Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 4
<211> 178
<212> Белок
<213> искусственный
<220>
<223> Мутеин липокалина
<400> 4
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Val Ala Gly Asn Ala Met Leu Arg Glu Asp Lys Asp Pro
35 40 45
Leu Lys Met Arg Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asn Val Thr Gly Val Ser Phe Trp Arg Lys Lys Cys His Tyr Lys Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Asp
85 90 95
Ile Lys Ser Gly Pro Gly Gln Thr Ser Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Arg Gln
115 120 125
Asn Arg Glu Trp Phe Ala Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 5
<211> 178
<212> Белок
<213> искусственный
<220>
<223> Мутеин липокалина
<400> 5
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Arg Ala Gly Asn Val Ala Leu Arg Glu Asp Lys Asp Pro
35 40 45
Pro Lys Met Arg Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asn Val Thr Asn Val Arg Phe Ala Met Lys Lys Cys Met Tyr Ser Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln
85 90 95
Ile Lys Ser Glu Pro Gly Asn Thr Ser Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Tyr Gln
115 120 125
Asn Arg Glu Ile Phe Phe Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 6
<211> 178
<212> Белок
<213> искусственный
<220>
<223> Мутеин липокалина
<400> 6
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Val Ala Gly Asn Ala Leu Leu Arg Glu Asp Lys Asp Pro
35 40 45
Leu Lys Met Arg Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Val Val Ser Phe Trp Arg Lys Lys Cys His Tyr Lys Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Asp
85 90 95
Ile Lys Ser Gly Pro Gly Gln Thr Ser Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Arg Gln
115 120 125
Asn Arg Glu Trp Phe Ala Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 7
<211> 178
<212> Белок
<213> искусственный
<220>
<223> Мутеин липокалина
<400> 7
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Val Ala Gly Asn Val Arg Leu Arg Glu Asp Lys Asp Pro
35 40 45
Pro Lys Met Arg Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Gly Val Ser Phe Arg Gly Lys Lys Cys His Tyr Lys Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Asp
85 90 95
Ile Lys Ser Gly Pro Gly Glu Thr Ser Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Arg Gln
115 120 125
Asn Arg Glu Trp Phe Phe Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 8
<211> 178
<212> Белок
<213> искусственный
<220>
<223> Мутеин липокалина
<400> 8
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Val Ala Gly Asn Gly Met Leu Arg Glu Asp Lys Asp Pro
35 40 45
Leu Lys Met Arg Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Ser Val Ala Phe Arg Asn Lys Lys Cys His Tyr Lys Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln
85 90 95
Ile Lys Ser Gly Pro Gly Glu Thr Ser Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Arg Gln
115 120 125
Asn Arg Glu Trp Phe Phe Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 9
<211> 178
<212> Белок
<213> искусственный
<220>
<223> Мутеин липокалина
<400> 9
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Arg Ala Gly Asn Val Gly Leu Arg Glu Asp Lys Asp Pro
35 40 45
Pro Lys Met Trp Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Asn Val Arg Phe Ala Arg Lys Lys Cys Leu Tyr Ser Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln
85 90 95
Ile Lys Ser Glu Pro Gly Asn Thr Ala Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Tyr Gln
115 120 125
Asn Arg Glu Ile Phe Phe Ile Ile Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 10
<211> 178
<212> Белок
<213> искусственный
<220>
<223> Мутеин липокалина
<400> 10
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Arg Ala Gly Asn Val Gly Leu Arg Glu Asp Lys Asp Pro
35 40 45
Pro Lys Met Trp Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Asn Val Arg Phe Ala Arg Lys Lys Cys Thr Tyr Ser Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln
85 90 95
Ile Lys Ser Glu Pro Gly Gly Thr Ala Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Tyr Gln
115 120 125
Asn Arg Glu Ile Phe Phe Ile Ile Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 11
<211> 178
<212> Белок
<213> искусственный
<220>
<223> Мутеин липокалина
<400> 11
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Arg Ala Gly Asn Gly Ala Leu Arg Glu Asp Lys Asp Pro
35 40 45
Pro Lys Met Trp Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Asn Val Arg Phe Ala Arg Lys Lys Cys Val Tyr Ser Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Pro
85 90 95
Ile Lys Ser Glu Pro Gly Asn Thr Ala Ser Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Tyr Gln
115 120 125
Asn Arg Glu Ile Phe Phe Ile Ile Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 12
<211> 178
<212> Белок
<213> искусственный
<220>
<223> Мутеин липокалина
<400> 12
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Arg Ala Gly Asn Val Ala Leu Arg Glu Asp Lys Asp Pro
35 40 45
Pro Lys Met Arg Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Ala Val Arg Phe Ala Arg Lys Lys Cys Leu Tyr Ser Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Arg
85 90 95
Ile Lys Ser Glu Pro Gly Tyr Thr Ala Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Tyr Gln
115 120 125
Asn Arg Glu Ile Phe Phe Ile Ile Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 13
<211> 178
<212> Белок
<213> искусственный
<220>
<223> Мутеин липокалина
<400> 13
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Arg Ala Gly Asn Val Ala Leu Arg Glu Asp Lys Asp Pro
35 40 45
Pro Lys Met Arg Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Asn Val Val Phe Ala Gly Lys Lys Cys Lys Tyr Ser Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Arg
85 90 95
Ile Lys Ser Pro Pro Gly Asn Thr Ala Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Tyr Gln
115 120 125
Asn Arg Glu Ile Phe Phe Ile Ile Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 14
<211> 178
<212> Белок
<213> искусственный
<220>
<223> Мутеин липокалина
<400> 14
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Arg Ala Gly Asn Leu Gly Leu Arg Glu Asp Lys Asp Pro
35 40 45
Pro Lys Met Trp Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Asn Val Arg Phe Ala Arg Lys Lys Cys Met Tyr Ser Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln
85 90 95
Ile Lys Ser Glu Pro Gly Ser Thr Ala Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Tyr Gln
115 120 125
Asn Arg Glu Ile Phe Phe Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 15
<211> 178
<212> Белок
<213> искусственный
<220>
<223> Мутеин липокалина
<400> 15
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Arg Ala Gly Asn Val Gly Leu Arg Glu Asp Lys Asp Pro
35 40 45
Pro Lys Met Trp Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Asn Val Arg Phe Ala Gly Lys Lys Val Lys Tyr Thr Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln
85 90 95
Ile Lys Ser Glu Pro Gly Asn Thr Ala Thr Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Tyr Gln
115 120 125
Asn Arg Glu Ile Phe Phe Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Ala Ile
165 170 175
Asp Gly
<210> 16
<211> 178
<212> Белок
<213> искусственный
<220>
<223> Мутеин липокалина
<400> 16
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Arg Ala Gly Asn Val Gly Leu Arg Glu Asp Lys Asp Pro
35 40 45
Pro Lys Met Arg Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Gly Val Arg Phe Gly Glu Lys Lys Ile Lys Tyr Ser Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln
85 90 95
Ile Lys Ser Gln Pro Gly Asp Thr Ala Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Tyr Gln
115 120 125
Asn Arg Glu Ile Phe Phe Ile Ile Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Ala Ile
165 170 175
Asp Gly
<210> 17
<211> 178
<212> Белок
<213> искусственный
<220>
<223> Мутеин липокалина
<400> 17
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Arg Ala Gly Asn Val Gly Leu Arg Glu Asp Lys Asp Pro
35 40 45
Pro Lys Met Arg Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Gly Val Arg Phe Asp Ser Lys Lys Val Thr Tyr Ser Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln
85 90 95
Ile Lys Ser Glu Pro Gly Asn Thr Ala Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Tyr Gln
115 120 125
Asn Arg Glu Ile Phe Phe Ile Ile Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Ala Ile
165 170 175
Asp Gly
[
<210> 18
<211> 152
<212> Белок
<213> искусственный
<220>
<223> Мутеин липокалина
<400> 18
Ala Ser Asp Glu Glu Ile Gln Asp Val Ser Gly Thr Trp Tyr Leu Lys
1 5 10 15
Ala Met Thr Val Asp Glu Gly Cys Arg Pro Trp Asn Ile Phe Ser Val
20 25 30
Thr Pro Met Thr Leu Thr Thr Leu Glu Gly Gly Asn Leu Glu Ala Lys
35 40 45
Val Thr Met Ala Ile Asp Gly Pro Ala Gln Glu Val Lys Ala Val Leu
50 55 60
Glu Lys Thr Asp Glu Pro Gly Lys Tyr Thr Ala Asp Gly Gly Lys His
65 70 75 80
Val Ala Tyr Ile Ile Arg Ser His Val Lys Asp His Tyr Ile Phe Tyr
85 90 95
Ser Glu Gly Val Cys Asp Gly Ser Pro Val Pro Gly Val Trp Leu Val
100 105 110
Gly Arg Asp Pro Lys Asn Asn Leu Glu Ala Leu Glu Asp Phe Glu Lys
115 120 125
Ala Ala Gly Ala Arg Gly Leu Ser Thr Glu Ser Ile Leu Ile Pro Arg
130 135 140
Gln Ser Glu Thr Ser Ser Pro Gly
145 150
<210> 19
<211> 152
<212> Белок
<213> искусственный
<220>
<223> Мутеин липокалина
<400> 19
Thr Ser Asp Glu Glu Ile Gln Asp Val Ser Gly Thr Trp Tyr Leu Lys
1 5 10 15
Ala Met Thr Val Asp Glu Gly Cys Arg Pro Trp Asn Ile Phe Ser Val
20 25 30
Thr Pro Met Thr Leu Thr Thr Leu Glu Gly Gly Asn Leu Glu Ala Lys
35 40 45
Val Thr Met Ala Ile Asp Gly Pro Ala Gln Glu Val Arg Ala Val Leu
50 55 60
Glu Lys Thr Asp Glu Pro Gly Lys Tyr Thr Ala Asp Gly Gly Lys His
65 70 75 80
Asp Ala Tyr Ile Ile Arg Ser His Val Lys Asp His Tyr Ile Phe Tyr
85 90 95
Ser Glu Gly Val Cys Asp Gly Ser Pro Val Pro Gly Val Trp Leu Val
100 105 110
Gly Arg Asp Pro Glu Asn Asn Leu Glu Ala Leu Glu Asp Phe Glu Lys
115 120 125
Thr Ala Gly Ala Arg Gly Leu Ser Thr Glu Ser Ile Leu Ile Pro Arg
130 135 140
Gln Ser Glu Thr Ser Ser Pro Gly
145 150
<210> 20
<211> 152
<212> Белок
<213> искусственный
<220>
<223> Мутеин липокалина
<400> 20
Ala Ser Asp Glu Glu Ile Gln Asp Val Ser Gly Thr Trp Tyr Leu Lys
1 5 10 15
Ala Met Thr Val Asp Glu Gly Cys Arg Pro Trp Asn Ile Phe Ser Val
20 25 30
Thr Pro Met Thr Leu Thr Thr Leu Glu Gly Gly Asn Leu Glu Ala Lys
35 40 45
Val Thr Met Ala Ile Asp Gly Pro Ala Gln Glu Val Asn Ala Val Leu
50 55 60
Glu Lys Thr Asp Glu Pro Gly Lys Tyr Thr Ala Asp Gly Gly Lys His
65 70 75 80
Val Ala Tyr Ile Ile Arg Ser His Val Arg Asp His Tyr Ile Phe Tyr
85 90 95
Ser Glu Gly Val Cys Asp Gly Ser Pro Val Pro Gly Val Trp Leu Val
100 105 110
Gly Arg Asp Pro Glu Asn Asn Leu Glu Ala Leu Glu Asp Phe Glu Lys
115 120 125
Thr Ala Gly Ala Arg Gly Leu Ser Thr Glu Ser Ile Leu Ile Pro Arg
130 135 140
Gln Ser Glu Thr Ser Ser Pro Gly
145 150
<210> 21
<211> 152
<212> Белок
<213> искусственный
<220>
<223> Мутеин липокалина
<400> 21
Val Ser Asp Glu Glu Ile Gln Asp Val Ser Gly Thr Trp Tyr Leu Lys
1 5 10 15
Ala Met Thr Val Asp Glu Gly Cys Arg Pro Trp Asn Ile Phe Ser Val
20 25 30
Thr Pro Met Thr Leu Thr Thr Leu Glu Gly Gly Asn Leu Glu Ala Lys
35 40 45
Val Thr Met Ala Ile Asp Gly Pro Ala Gln Glu Val Arg Ala Val Leu
50 55 60
Glu Lys Thr Asp Glu Pro Gly Lys Tyr Thr Ala Asp Gly Gly Lys His
65 70 75 80
Val Ala Tyr Ile Ile Arg Ser His Val Glu Asp His Tyr Ile Phe Tyr
85 90 95
Ser Glu Gly Val Cys Asp Gly Ser Pro Val Pro Gly Val Trp Leu Val
100 105 110
Gly Arg Asp Pro Glu Asn Asn Leu Glu Ala Leu Glu Asp Phe Glu Lys
115 120 125
Thr Ala Gly Ala Arg Gly Leu Ser Thr Glu Ser Ile Leu Ile Pro Arg
130 135 140
Gln Ser Glu Thr Ser Ser Pro Gly
145 150
<210> 22
<211> 152
<212> Белок
<213> искусственный
<220>
<223> Мутеин липокалина
<400> 22
Ala Ser Asp Glu Glu Ile Gln Asp Val Ser Gly Thr Trp Tyr Leu Lys
1 5 10 15
Ala Met Thr Val Asp Glu Gly Cys Arg Pro Trp Asn Ile Phe Ser Val
20 25 30
Thr Pro Met Thr Leu Ser Thr Leu Glu Gly Gly Asn Leu Glu Ala Lys
35 40 45
Val Thr Met Ala Ile Asp Gly Pro Ala Gln Glu Val Lys Ala Val Leu
50 55 60
Glu Lys Thr Asp Glu Pro Gly Lys Tyr Thr Ala Asp Gly Gly Lys His
65 70 75 80
Val Ala Tyr Ile Ile Arg Ser His Val Lys Asp His Tyr Ile Phe Tyr
85 90 95
Ser Glu Gly Val Cys Asp Gly Ser Pro Val Pro Gly Val Trp Leu Val
100 105 110
Gly Arg Asp Pro Lys Asn Asn Leu Glu Ala Leu Glu Asp Phe Glu Lys
115 120 125
Ala Ala Gly Ala Arg Gly Leu Ser Thr Glu Ser Ile Leu Ile Pro Arg
130 135 140
Gln Ile Glu Thr Ser Ser Pro Gly
145 150
<210> 23
<211> 152
<212> Белок
<213> искусственный
<220>
<223> Мутеин липокалина
<400> 23
Ala Ser Asp Glu Glu Ile Gln Asp Val Ser Gly Thr Trp Tyr Leu Lys
1 5 10 15
Ala Met Thr Val Asp Glu Gly Cys Arg Pro Trp Asn Ile Phe Ser Val
20 25 30
Thr Pro Met Thr Leu Thr Thr Leu Glu Gly Gly Asn Leu Glu Ala Glu
35 40 45
Val Thr Met Ala Ile Asp Gly Pro Ala Gln Glu Val Lys Ala Val Leu
50 55 60
Glu Lys Ala Asp Glu Pro Gly Lys Tyr Thr Ala Asp Gly Gly Lys His
65 70 75 80
Val Ala Tyr Ile Ile Arg Ser His Val Lys Asp His Tyr Ile Phe Tyr
85 90 95
Ser Glu Gly Val Cys Asp Gly Ser Pro Val Pro Gly Val Trp Leu Val
100 105 110
Gly Arg Asp Pro Lys Asn Asn Leu Glu Ala Leu Glu Asp Phe Glu Lys
115 120 125
Thr Ala Gly Ala Arg Gly Leu Ser Thr Glu Ser Ile Leu Ile Pro Ser
130 135 140
Gln Ile Glu Thr Ser Ser Pro Gly
145 150
<210> 24
<211> 152
<212> Белок
<213> искусственный
<220>
<223> Мутеин липокалина
<400> 24
Thr Ser Asp Glu Glu Ile Gln Asp Val Ser Gly Thr Trp Tyr Leu Lys
1 5 10 15
Ala Met Thr Val Asp Glu Gly Cys Arg Pro Trp Asn Ile Phe Ser Val
20 25 30
Thr Pro Met Thr Leu Thr Thr Leu Glu Asp Gly Asn Leu Glu Ala Lys
35 40 45
Val Thr Met Ala Ile Asp Gly Pro Ala Gln Glu Val Lys Ala Val Leu
50 55 60
Glu Lys Ala Asp Glu Pro Gly Lys Tyr Thr Ala Asp Gly Gly Lys His
65 70 75 80
Val Ala Tyr Ile Ile Arg Ser His Val Lys Asp His Tyr Ile Phe Tyr
85 90 95
Ser Glu Gly Val Cys Asp Gly Ser Pro Val Pro Gly Val Trp Leu Val
100 105 110
Gly Arg Asp Pro Lys Asn Asn Leu Glu Ala Leu Glu Asp Phe Glu Lys
115 120 125
Ala Ala Gly Ala Arg Gly Leu Ser Thr Glu Ser Ile Leu Ile Pro Arg
130 135 140
Gln Ile Glu Thr Ser Ser Pro Gly
145 150
<210> 25
<211> 178
<212> Белок
<213> искусственный
<220>
<223> Мутеин липокалина
<400> 25
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Ile Lys Leu Arg Glu Asp Lys Asp Pro
35 40 45
Asn Lys Met Met Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asn Val Thr Gly Val Thr Phe Asp Asp Lys Lys Cys Thr Tyr Ala Ile
65 70 75 80
Ser Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Lys
85 90 95
Ile Lys Ser Phe Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln
115 120 125
Asn Arg Glu Glu Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 26
<211> 178
<212> Белок
<213> искусственный
<220>
<223> Мутеин липокалина
<400> 26
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Ile Arg Leu Arg Glu Asp Lys Asp Pro
35 40 45
Ile Lys Met Met Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Met Val Lys Phe Asp Asp Lys Lys Cys Met Tyr Asp Ile
65 70 75 80
Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Lys
85 90 95
Ile Lys Ser Phe Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln
115 120 125
Asn Arg Glu Glu Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 27
<211> 178
<212> Белок
<213> искусственный
<220>
<223> Мутеин липокалина
<400> 27
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Ile Arg Leu Arg Glu Asp Lys Asp Pro
35 40 45
Asn Lys Met Met Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Ala Val Ala Phe Asp Asp Lys Lys Cys Thr Tyr Asp Ile
65 70 75 80
Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Lys
85 90 95
Ile Lys Ser Phe Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln
115 120 125
Asn Arg Glu Glu Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 28
<211> 178
<212> Белок
<213> искусственный
<220>
<223> Мутеин липокалина
<400> 28
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Ile Lys Leu Arg Glu Asp Lys Asp Pro
35 40 45
Asn Lys Met Met Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Ala Val Ala Phe Asp Asp Lys Lys Cys Thr Tyr Asp Ile
65 70 75 80
Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Lys
85 90 95
Ile Lys Ser Phe Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln
115 120 125
Asn Arg Glu Glu Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 29
<211> 175
<212> Белок
<213> искусственный
<220>
<223> Мутеин липокалина
<400> 29
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Ile Lys Leu Arg Glu Asp Ser Lys Met
35 40 45
Met Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr Asp Val Thr
50 55 60
Gly Val Ser Phe Asp Asp Lys Lys Cys Thr Tyr Ala Ile Met Thr Phe
65 70 75 80
Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Lys Ile Lys Ser
85 90 95
Phe Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser Thr Asn Tyr
100 105 110
Asn Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln Asn Arg Glu
115 120 125
Glu Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu Thr Ser Glu
130 135 140
Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly Leu Pro Glu
145 150 155 160
Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile Asp Gly
165 170 175
<210> 30
<211> 178
<212> Белок
<213> искусственный
<220>
<223> Мутеин липокалина
<400> 30
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Ile Lys Leu Arg Glu Asp Lys Asp Pro
35 40 45
Val Lys Met Met Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Gly Val Thr Phe Asp Asp Lys Lys Cys Arg Tyr Asp Ile
65 70 75 80
Ser Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Phe Gly Lys
85 90 95
Ile Lys Ser Phe Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln
115 120 125
Asn Arg Glu Glu Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 31
<211> 178
<212> Белок
<213> искусственный
<220>
<223> Мутеин липокалина
<400> 31
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Ile Arg Leu Arg Glu Asp Lys Asp Pro
35 40 45
His Lys Met Met Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Gly Val Thr Phe Asp Asp Lys Lys Cys Thr Tyr Ala Ile
65 70 75 80
Ser Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Lys
85 90 95
Ile Lys Ser Phe Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln
115 120 125
Asn Arg Glu Glu Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 32
<211> 178
<212> Белок
<213> искусственный
<220>
<223> Мутеин липокалина
<400> 32
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Ile Lys Leu Arg Glu Asp Lys Asp Pro
35 40 45
Asn Lys Met Met Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Gly Val Thr Phe Asp Asp Lys Lys Cys Thr Tyr Ala Ile
65 70 75 80
Ser Thr Leu Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Phe Gly Lys
85 90 95
Ile Lys Ser Phe Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln
115 120 125
Asn Arg Glu Glu Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 33
<211> 178
<212> Белок
<213> искусственный
<220>
<223> Мутеин липокалина
<400> 33
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr
20 25 30
Val Val Gly Gln Ala Gly Asn Ile Arg Leu Arg Glu Asp Lys Asp Pro
35 40 45
Ser Lys Met Met Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Ala Val Thr Phe Asp Asp Lys Lys Cys Asn Tyr Ala Ile
65 70 75 80
Ser Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Lys
85 90 95
Ile Lys Ser Phe Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln
115 120 125
Asn Arg Glu Glu Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly
<210> 34
<211> 448
<212> Белок
<213> искусственный
<220>
<223> тяжелая цепь антитела
<400> 34
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn His Gly Gly Tyr Val Thr Tyr Asn Pro Ser Leu Glu
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Tyr Gly Pro Gly Asn Tyr Asp Trp Tyr Phe Asp Leu Trp Gly
100 105 110
Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly
210 215 220
Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro
260 265 270
Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
<210> 35
<211> 216
<212> Белок
<213> искусственный
<220>
<223> полипептидная цепь антитела
<400> 35
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Pro
85 90 95
Ala Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val
100 105 110
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
115 120 125
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
130 135 140
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
145 150 155 160
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
165 170 175
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
180 185 190
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr
195 200 205
Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 36
<211> 641
<212> Белок
<213> искусственный
<220>
<223> тяжелая цепь слитого полипептида
<400> 36
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn His Gly Gly Tyr Val Thr Tyr Asn Pro Ser Leu Glu
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Tyr Gly Pro Gly Asn Tyr Asp Trp Tyr Phe Asp Leu Trp Gly
100 105 110
Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly
210 215 220
Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro
260 265 270
Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln
450 455 460
Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val Pro
465 470 475 480
Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr Val
485 490 495
Val Gly Arg Ala Gly Asn Val Gly Leu Arg Glu Asp Lys Asp Pro Pro
500 505 510
Lys Met Trp Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr Asp
515 520 525
Val Thr Asn Val Arg Phe Ala Arg Lys Lys Cys Thr Tyr Ser Ile Gly
530 535 540
Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln Ile
545 550 555 560
Lys Ser Glu Pro Gly Gly Thr Ala Asn Leu Val Arg Val Val Ser Thr
565 570 575
Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Tyr Gln Asn
580 585 590
Arg Glu Ile Phe Phe Ile Ile Leu Tyr Gly Arg Thr Lys Glu Leu Thr
595 600 605
Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly Leu
610 615 620
Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile Asp
625 630 635 640
Gly
<210> 37
<211> 216
<212> Белок
<213> искусственный
<220>
<223> легкая цепь слитого полипептида
<400> 37
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Pro
85 90 95
Ala Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val
100 105 110
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
115 120 125
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
130 135 140
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
145 150 155 160
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
165 170 175
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
180 185 190
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr
195 200 205
Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 38
<211> 641
<212> Белок
<213> искусственный
<220>
<223> тяжелая цепь слитого полипептида
<400> 38
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Arg Ala Gly Asn Val Gly Leu Arg Glu Asp Lys Asp Pro
35 40 45
Pro Lys Met Trp Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Asn Val Arg Phe Ala Arg Lys Lys Cys Thr Tyr Ser Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln
85 90 95
Ile Lys Ser Glu Pro Gly Gly Thr Ala Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Tyr Gln
115 120 125
Asn Arg Glu Ile Phe Phe Ile Ile Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
180 185 190
Ser Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser
195 200 205
Glu Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly
210 215 220
Tyr Tyr Trp Ser Trp Ile Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp
225 230 235 240
Ile Gly Glu Ile Asn His Gly Gly Tyr Val Thr Tyr Asn Pro Ser Leu
245 250 255
Glu Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser
260 265 270
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
275 280 285
Ala Arg Asp Tyr Gly Pro Gly Asn Tyr Asp Trp Tyr Phe Asp Leu Trp
290 295 300
Gly Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
305 310 315 320
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
325 330 335
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
340 345 350
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
355 360 365
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
370 375 380
Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp
385 390 395 400
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
405 410 415
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro
420 425 430
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
435 440 445
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
450 455 460
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
465 470 475 480
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
485 490 495
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
500 505 510
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
515 520 525
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
530 535 540
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
545 550 555 560
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
565 570 575
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
580 585 590
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
595 600 605
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
610 615 620
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
625 630 635 640
Lys
<210> 39
<211> 216
<212> Белок
<213> искусственный
<220>
<223> легкая цепь слитого полипептида
<400> 39
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Pro
85 90 95
Ala Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val
100 105 110
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
115 120 125
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
130 135 140
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
145 150 155 160
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
165 170 175
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
180 185 190
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr
195 200 205
Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 40
<211> 448
<212> Белок
<213> искусственный
<220>
<223> тяжелая цепь слитого полипептида
<400> 40
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn His Gly Gly Tyr Val Thr Tyr Asn Pro Ser Leu Glu
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Tyr Gly Pro Gly Asn Tyr Asp Trp Tyr Phe Asp Leu Trp Gly
100 105 110
Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly
210 215 220
Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro
260 265 270
Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
<210> 41
<211> 409
<212> Белок
<213> искусственный
<220>
<223> легкая цепь слитого полипептида
<400> 41
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Arg Ala Gly Asn Val Gly Leu Arg Glu Asp Lys Asp Pro
35 40 45
Pro Lys Met Trp Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Asn Val Arg Phe Ala Arg Lys Lys Cys Thr Tyr Ser Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln
85 90 95
Ile Lys Ser Glu Pro Gly Gly Thr Ala Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Tyr Gln
115 120 125
Asn Arg Glu Ile Phe Phe Ile Ile Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
180 185 190
Ser Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro
195 200 205
Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser
210 215 220
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
225 230 235 240
Ile Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser
245 250 255
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu
260 265 270
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro
275 280 285
Pro Ala Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr
290 295 300
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
305 310 315 320
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
325 330 335
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
340 345 350
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
355 360 365
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
370 375 380
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
385 390 395 400
Thr Lys Ser Phe Asn Arg Gly Glu Cys
405
<210> 42
<211> 448
<212> Белок
<213> искусственный
<220>
<223> тяжелая цепь слитого полипептида
<400> 42
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn His Gly Gly Tyr Val Thr Tyr Asn Pro Ser Leu Glu
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Tyr Gly Pro Gly Asn Tyr Asp Trp Tyr Phe Asp Leu Trp Gly
100 105 110
Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly
210 215 220
Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro
260 265 270
Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
<210> 43
<211> 408
<212> Белок
<213> искусственный
<220>
<223> легкая цепь слитого полипептида
<400> 43
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Pro
85 90 95
Ala Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val
100 105 110
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
115 120 125
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
130 135 140
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
145 150 155 160
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
165 170 175
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
180 185 190
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr
195 200 205
Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser Gly Gly Gly
210 215 220
Gly Ser Gly Gly Gly Gly Ser Gln Asp Ser Thr Ser Asp Leu Ile Pro
225 230 235 240
Ala Pro Pro Leu Ser Lys Val Pro Leu Gln Gln Asn Phe Gln Asp Asn
245 250 255
Gln Phe Gln Gly Lys Trp Tyr Val Val Gly Arg Ala Gly Asn Val Gly
260 265 270
Leu Arg Glu Asp Lys Asp Pro Pro Lys Met Trp Ala Thr Ile Tyr Glu
275 280 285
Leu Lys Glu Asp Lys Ser Tyr Asp Val Thr Asn Val Arg Phe Ala Arg
290 295 300
Lys Lys Cys Thr Tyr Ser Ile Gly Thr Phe Val Pro Gly Ser Gln Pro
305 310 315 320
Gly Glu Phe Thr Leu Gly Gln Ile Lys Ser Glu Pro Gly Gly Thr Ala
325 330 335
Asn Leu Val Arg Val Val Ser Thr Asn Tyr Asn Gln His Ala Met Val
340 345 350
Phe Phe Lys Glu Val Tyr Gln Asn Arg Glu Ile Phe Phe Ile Ile Leu
355 360 365
Tyr Gly Arg Thr Lys Glu Leu Thr Ser Glu Leu Lys Glu Asn Phe Ile
370 375 380
Arg Phe Ser Lys Ser Leu Gly Leu Pro Glu Asn His Ile Val Phe Pro
385 390 395 400
Val Pro Ile Asp Gln Cys Ile Asp
405
<210> 44
<211> 615
<212> Белок
<213> искусственный
<220>
<223> слитый полипептид
<400> 44
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Arg Ala Gly Asn Val Gly Leu Arg Glu Asp Lys Asp Pro
35 40 45
Pro Lys Met Trp Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Asn Val Arg Phe Ala Arg Lys Lys Cys Thr Tyr Ser Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln
85 90 95
Ile Lys Ser Glu Pro Gly Gly Thr Ala Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Tyr Gln
115 120 125
Asn Arg Glu Ile Phe Phe Ile Ile Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
180 185 190
Ser Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu
195 200 205
Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
210 215 220
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
225 230 235 240
Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
245 250 255
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
260 265 270
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
275 280 285
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
290 295 300
Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
305 310 315 320
Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn
325 330 335
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
340 345 350
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
355 360 365
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg
370 375 380
Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys
385 390 395 400
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
405 410 415
Ser Leu Ser Leu Gly Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
420 425 430
Gly Gly Gly Gly Ser Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro
435 440 445
Pro Leu Ser Lys Val Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe
450 455 460
His Gly Lys Trp Tyr Val Val Gly Gln Ala Gly Asn Ile Arg Leu Arg
465 470 475 480
Glu Asp Lys Asp Pro Ile Lys Met Met Ala Thr Ile Tyr Glu Leu Lys
485 490 495
Glu Asp Lys Ser Tyr Asp Val Thr Met Val Lys Phe Asp Asp Lys Lys
500 505 510
Cys Met Tyr Asp Ile Trp Thr Phe Val Pro Gly Ser Gln Pro Gly Glu
515 520 525
Phe Thr Leu Gly Lys Ile Lys Ser Phe Pro Gly His Thr Ser Ser Leu
530 535 540
Val Arg Val Val Ser Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe
545 550 555 560
Lys Phe Val Phe Gln Asn Arg Glu Glu Phe Tyr Ile Thr Leu Tyr Gly
565 570 575
Arg Thr Lys Glu Leu Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe
580 585 590
Ser Lys Ser Leu Gly Leu Pro Glu Asn His Ile Val Phe Pro Val Pro
595 600 605
Ile Asp Gln Cys Ile Asp Gly
610 615
<210> 45
<211> 610
<212> Белок
<213> искусственный
<220>
<223> слитый полипептид
<400> 45
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Arg Ala Gly Asn Val Gly Leu Arg Glu Asp Lys Asp Pro
35 40 45
Pro Lys Met Trp Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Asn Val Arg Phe Ala Arg Lys Lys Cys Thr Tyr Ser Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln
85 90 95
Ile Lys Ser Glu Pro Gly Gly Thr Ala Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Tyr Gln
115 120 125
Asn Arg Glu Ile Phe Phe Ile Ile Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Asp Ser Thr
180 185 190
Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val Pro Leu Gln Gln
195 200 205
Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr Val Val Gly Gln
210 215 220
Ala Gly Asn Ile Arg Leu Arg Glu Asp Lys Asp Pro Ile Lys Met Met
225 230 235 240
Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr Asp Val Thr Met
245 250 255
Val Lys Phe Asp Asp Lys Lys Cys Met Tyr Asp Ile Trp Thr Phe Val
260 265 270
Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Lys Ile Lys Ser Phe
275 280 285
Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser Thr Asn Tyr Asn
290 295 300
Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln Asn Arg Glu Glu
305 310 315 320
Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu Thr Ser Glu Leu
325 330 335
Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly Leu Pro Glu Asn
340 345 350
His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile Asp Gly Gly Gly
355 360 365
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Ser Lys
370 375 380
Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
385 390 395 400
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
405 410 415
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
420 425 430
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
435 440 445
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg
450 455 460
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
465 470 475 480
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
485 490 495
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
500 505 510
Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu
515 520 525
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
530 535 540
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
545 550 555 560
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp
565 570 575
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His
580 585 590
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu
595 600 605
Gly Lys
610
<210> 46
<211> 366
<212> Белок
<213> искусственный
<220>
<223> слитый полипептид
<400> 46
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Val Ala Gly Asn Gly Met Leu Arg Glu Asp Lys Asp Pro
35 40 45
Leu Lys Met Arg Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Ser Val Ala Phe Arg Asn Lys Lys Cys His Tyr Lys Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln
85 90 95
Ile Lys Ser Gly Pro Gly Glu Thr Ser Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Arg Gln
115 120 125
Asn Arg Glu Trp Phe Phe Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Asp Ser Thr
180 185 190
Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val Pro Leu Gln Gln
195 200 205
Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr Val Val Gly Gln
210 215 220
Ala Gly Asn Ile Arg Leu Arg Glu Asp Lys Asp Pro Ile Lys Met Met
225 230 235 240
Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr Asp Val Thr Met
245 250 255
Val Lys Phe Asp Asp Lys Lys Cys Met Tyr Asp Ile Trp Thr Phe Val
260 265 270
Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Lys Ile Lys Ser Phe
275 280 285
Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser Thr Asn Tyr Asn
290 295 300
Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln Asn Arg Glu Glu
305 310 315 320
Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu Thr Ser Glu Leu
325 330 335
Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly Leu Pro Glu Asn
340 345 350
His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile Asp Gly
355 360 365
<210> 47
<211> 554
<212> Белок
<213> искусственный
<220>
<223> слитый полипептид
<400> 47
Gln Asp Ser Thr Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val
1 5 10 15
Pro Leu Gln Gln Asn Phe Gln Asp Asn Gln Phe Gln Gly Lys Trp Tyr
20 25 30
Val Val Gly Val Ala Gly Asn Gly Met Leu Arg Glu Asp Lys Asp Pro
35 40 45
Leu Lys Met Arg Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr
50 55 60
Asp Val Thr Ser Val Ala Phe Arg Asn Lys Lys Cys His Tyr Lys Ile
65 70 75 80
Gly Thr Phe Val Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Gln
85 90 95
Ile Lys Ser Gly Pro Gly Glu Thr Ser Asn Leu Val Arg Val Val Ser
100 105 110
Thr Asn Tyr Asn Gln His Ala Met Val Phe Phe Lys Glu Val Arg Gln
115 120 125
Asn Arg Glu Trp Phe Phe Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu
130 135 140
Thr Ser Glu Leu Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly
145 150 155 160
Leu Pro Glu Asn His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile
165 170 175
Asp Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Asp Ser Thr
180 185 190
Ser Asp Leu Ile Pro Ala Pro Pro Leu Ser Lys Val Pro Leu Gln Gln
195 200 205
Asn Phe Gln Asp Asn Gln Phe His Gly Lys Trp Tyr Val Val Gly Gln
210 215 220
Ala Gly Asn Ile Arg Leu Arg Glu Asp Lys Asp Pro Ile Lys Met Met
225 230 235 240
Ala Thr Ile Tyr Glu Leu Lys Glu Asp Lys Ser Tyr Asp Val Thr Met
245 250 255
Val Lys Phe Asp Asp Lys Lys Cys Met Tyr Asp Ile Trp Thr Phe Val
260 265 270
Pro Gly Ser Gln Pro Gly Glu Phe Thr Leu Gly Lys Ile Lys Ser Phe
275 280 285
Pro Gly His Thr Ser Ser Leu Val Arg Val Val Ser Thr Asn Tyr Asn
290 295 300
Gln His Ala Met Val Phe Phe Lys Phe Val Phe Gln Asn Arg Glu Glu
305 310 315 320
Phe Tyr Ile Thr Leu Tyr Gly Arg Thr Lys Glu Leu Thr Ser Glu Leu
325 330 335
Lys Glu Asn Phe Ile Arg Phe Ser Lys Ser Leu Gly Leu Pro Glu Asn
340 345 350
His Ile Val Phe Pro Val Pro Ile Asp Gln Cys Ile Asp Gly Gly Gly
355 360 365
Gly Gly Ser Gly Gly Gly Gly Ser Gln Asp Ser Thr Ser Asp Leu Ile
370 375 380
Pro Ala Pro Pro Leu Ser Lys Val Pro Leu Gln Gln Asn Phe Gln Asp
385 390 395 400
Asn Gln Phe His Gly Lys Trp Tyr Val Val Gly Gln Ala Gly Asn Ile
405 410 415
Arg Leu Arg Glu Asp Lys Asp Pro Ile Lys Met Met Ala Thr Ile Tyr
420 425 430
Glu Leu Lys Glu Asp Lys Ser Tyr Asp Val Thr Met Val Lys Phe Asp
435 440 445
Asp Lys Lys Cys Met Tyr Asp Ile Trp Thr Phe Val Pro Gly Ser Gln
450 455 460
Pro Gly Glu Phe Thr Leu Gly Lys Ile Lys Ser Phe Pro Gly His Thr
465 470 475 480
Ser Ser Leu Val Arg Val Val Ser Thr Asn Tyr Asn Gln His Ala Met
485 490 495
Val Phe Phe Lys Phe Val Phe Gln Asn Arg Glu Glu Phe Tyr Ile Thr
500 505 510
Leu Tyr Gly Arg Thr Lys Glu Leu Thr Ser Glu Leu Lys Glu Asn Phe
515 520 525
Ile Arg Phe Ser Lys Ser Leu Gly Leu Pro Glu Asn His Ile Val Phe
530 535 540
Pro Val Pro Ile Asp Gln Cys Ile Asp Gly
545 550
<210> 48
<211> 11
<212> Белок
<213> искусственный
<220>
<223> пептидный линкер
<400> 48
Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210> 49
<211> 15
<212> Белок
<213> искусственный
<220>
<223> пептидный линкер
<400> 49
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 50
<211> 10
<212> Белок
<213> искусственный
<220>
<223> Strep-метка
<400> 50
Ser Ala Trp Ser His Pro Gln Phe Glu Lys
1 5 10
<210> 51
<211> 442
<212> Белок
<213> искусственный
<220>
<223> тяжелая цепь антитела
<400> 51
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Arg Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Ser Thr Tyr
20 25 30
Trp Ile Ser Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Lys Ile Tyr Pro Gly Asp Ser Tyr Thr Asn Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Gly Tyr Gly Ile Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe
180 185 190
Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro
210 215 220
Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro
225 230 235 240
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
245 250 255
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp
260 265 270
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
275 280 285
Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val
290 295 300
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
305 310 315 320
Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly
325 330 335
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
340 345 350
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
355 360 365
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
370 375 380
Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe
385 390 395 400
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
405 410 415
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
420 425 430
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 52
<211> 214
<212> Белок
<213> искусственный
<220>
<223> легкая цепь антитела
<400> 52
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly Asp Asn Ile Gly Asp Gln Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Lys Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Tyr Thr Gly Phe Gly Ser Leu
85 90 95
Ala Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys
100 105 110
Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln
115 120 125
Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly
130 135 140
Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly
145 150 155 160
Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala
165 170 175
Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser
180 185 190
Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val
195 200 205
Ala Pro Thr Glu Cys Ser
210
<210> 53
<211> 635
<212> Белок
<213> искусственный
<220>
<223> тяжелая цепь слитого полипептида
<400> 53
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Arg Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Ser Thr Tyr
20 25 30
Trp Ile Ser Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Lys Ile Tyr Pro Gly Asp Ser Tyr Thr Asn Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Gly Tyr Gly Ile Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe
180 185 190
Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro
210 215 220
Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro
225 230 235 240
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
245 250 255
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp
260 265 270
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
275 280 285
Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val
290 295 300
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
305 310 315 320
Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly
325 330 335
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
340 345 350
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
355 360 365
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
370 375 380
Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe
385 390 395 400
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
405 410 415
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
420 425 430
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly
435 440 445
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Asp Ser Thr Ser Asp Leu
450 455 460
Ile Pro Ala Pro Pro Leu Ser Lys Val Pro Leu Gln Gln Asn Phe Gln
465 470 475 480
Asp Asn Gln Phe Gln Gly Lys Trp Tyr Val Val Gly Arg Ala Gly Asn
485 490 495
Val Gly Leu Arg Glu Asp Lys Asp Pro Pro Lys Met Trp Ala Thr Ile
500 505 510
Tyr Glu Leu Lys Glu Asp Lys Ser Tyr Asp Val Thr Asn Val Arg Phe
515 520 525
Ala Arg Lys Lys Cys Thr Tyr Ser Ile Gly Thr Phe Val Pro Gly Ser
530 535 540
Gln Pro Gly Glu Phe Thr Leu Gly Gln Ile Lys Ser Glu Pro Gly Gly
545 550 555 560
Thr Ala Asn Leu Val Arg Val Val Ser Thr Asn Tyr Asn Gln His Ala
565 570 575
Met Val Phe Phe Lys Glu Val Tyr Gln Asn Arg Glu Ile Phe Phe Ile
580 585 590
Ile Leu Tyr Gly Arg Thr Lys Glu Leu Thr Ser Glu Leu Lys Glu Asn
595 600 605
Phe Ile Arg Phe Ser Lys Ser Leu Gly Leu Pro Glu Asn His Ile Val
610 615 620
Phe Pro Val Pro Ile Asp Gln Cys Ile Asp Gly
625 630 635
<210> 54
<211> 214
<212> Белок
<213> искусственный
<220>
<223> легкая цепь слитого полипептида
<400> 54
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly Asp Asn Ile Gly Asp Gln Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Lys Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Tyr Thr Gly Phe Gly Ser Leu
85 90 95
Ala Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys
100 105 110
Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln
115 120 125
Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly
130 135 140
Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly
145 150 155 160
Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala
165 170 175
Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser
180 185 190
Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val
195 200 205
Ala Pro Thr Glu Cys Ser
210
<210> 55
<211> 1344
<212> ДНК
<213> искусственная
<220>
<223> тяжелая цепь антитела
<400> 55
caggtccagc tgcagcagtg gggggctggt ctgctgaagc caagtgaaac tctgtcactg 60
acctgcgctg tgtacggcgg atcattctcc ggctactatt ggtcttggat cagacagagt 120
cccgagaaag gcctggaatg gatcggagag attaaccacg ggggttacgt gacctataat 180
cctagcctgg agtctagggt gaccatttcc gtcgacacaa gcaagaacca gttctctctg 240
aaactgtcca gcgtgacagc cgctgacact gcagtctact attgtgccag ggattatggc 300
cccggaaatt acgactggta ttttgatctg tgggggcggg gtaccctggt gacagtctcg 360
agcgctagca ccaagggccc ctccgtgttc cccctggccc cttgctcccg gtccacctcc 420
gagtctaccg ccgctctggg ctgcctggtg aaagactact tccccgagcc tgtgaccgtg 480
agctggaact ctggcgccct gacctccggc gtgcacacct tccctgccgt gctgcaatcc 540
tccggcctgt actccctgtc ctccgtggtg acagtgccct cctccagcct gggcaccaag 600
acctacacct gtaacgtgga ccacaagccc tccaacacca aggtggacaa gcgggtggaa 660
tctaaatacg gccctccctg ccccccctgc cctgcccctg aatttctggg cggaccttcc 720
gtgtttctgt tccccccaaa gcccaaggac accctgatga tctcccggac ccccgaagtg 780
acctgcgtgg tggtggacgt gtcccaggaa gatccagagg tgcagttcaa ctggtatgtt 840
gacggcgtgg aagtgcacaa cgccaagacc aagcccagag aggaacagtt caactccacc 900
taccgggtgg tgtccgtgct gaccgtgctg caccaggact ggctgaacgg caaagagtac 960
aagtgcaagg tgtccaacaa gggcctgccc tccagcatcg aaaagaccat ctccaaggcc 1020
aagggccagc cccgcgagcc ccaggtgtac accctgcccc ctagccagga agagatgacc 1080
aagaaccagg tgtccctgac ctgtctggtg aaaggcttct acccctccga cattgccgtg 1140
gaatgggagt ccaacggcca gcccgagaac aactacaaga ccaccccccc tgtgctggac 1200
tccgacggct ccttcttcct gtactctcgg ctgacagtgg ataagtcccg gtggcaggaa 1260
ggcaacgtgt tctcctgcag cgtgatgcac gaggccctgc acaaccacta tacccagaag 1320
tccctgtccc tgagcctggg caag 1344
<210> 56
<211> 648
<212> ДНК
<213> искусственная
<220>
<223> легкая цепь антитела
<400> 56
gagattgtcc tgacccagtc ccccgcaaca ctgtcactgt cccccggcga gagggctact 60
ctgagttgcc gggcaagcca gtctgtgtcc agctacctgg cctggtatca gcagaagcca 120
gggcaggctc ccagactgct gatctacgac gcatctaaca gagccaccgg aattcctgcc 180
cgcttctcgg gttcaggctc cggaacagac tttaccctga caatctctag tctggagcca 240
gaagatttcg cagtctacta ttgtcagcag cgaagcaatt ggccccctgc tctgactttt 300
ggcggaggga ccaaggtgga gatcaagcgt acggtcgcgg cgccttctgt gttcattttc 360
cccccatctg atgaacagct gaaatctggc actgcttctg tggtctgtct gctgaacaac 420
ttctacccta gagaggccaa agtccagtgg aaagtggaca atgctctgca gagtgggaat 480
tcccaggaat ctgtcactga gcaggactct aaggatagca catactccct gtcctctact 540
ctgacactga gcaaggctga ttacgagaaa cacaaagtgt acgcctgtga agtcacacat 600
caggggctgt ctagtcctgt gaccaaatcc ttcaataggg gagagtgc 648
<210> 57
<211> 1923
<212> ДНК
<213> искусственная
<220>
<223> тяжелая цепь слитого полипептида
<400> 57
caggtccagc tgcagcagtg gggggctggt ctgctgaagc caagtgaaac tctgtcactg 60
acctgcgctg tgtacggcgg atcattctcc ggctactatt ggtcttggat cagacagagt 120
cccgagaaag gcctggaatg gatcggagag attaaccacg ggggttacgt gacctataat 180
cctagcctgg agtctagggt gaccatttcc gtcgacacaa gcaagaacca gttctctctg 240
aaactgtcca gcgtgacagc cgctgacact gcagtctact attgtgccag ggattatggc 300
cccggaaatt acgactggta ttttgatctg tgggggcggg gtaccctggt gacagtctcg 360
agcgctagca ccaagggccc ctccgtgttc cccctggccc cttgctcccg gtccacctcc 420
gagtctaccg ccgctctggg ctgcctggtg aaagactact tccccgagcc tgtgaccgtg 480
agctggaact ctggcgccct gacctccggc gtgcacacct tccctgccgt gctgcaatcc 540
tccggcctgt actccctgtc ctccgtggtg acagtgccct cctccagcct gggcaccaag 600
acctacacct gtaacgtgga ccacaagccc tccaacacca aggtggacaa gcgggtggaa 660
tctaaatacg gccctccctg ccccccctgc cctgcccctg aatttctggg cggaccttcc 720
gtgtttctgt tccccccaaa gcccaaggac accctgatga tctcccggac ccccgaagtg 780
acctgcgtgg tggtggacgt gtcccaggaa gatccagagg tgcagttcaa ctggtatgtt 840
gacggcgtgg aagtgcacaa cgccaagacc aagcccagag aggaacagtt caactccacc 900
taccgggtgg tgtccgtgct gaccgtgctg caccaggact ggctgaacgg caaagagtac 960
aagtgcaagg tgtccaacaa gggcctgccc tccagcatcg aaaagaccat ctccaaggcc 1020
aagggccagc cccgcgagcc ccaggtgtac accctgcccc ctagccagga agagatgacc 1080
aagaaccagg tgtccctgac ctgtctggtg aaaggcttct acccctccga cattgccgtg 1140
gaatgggagt ccaacggcca gcccgagaac aactacaaga ccaccccccc tgtgctggac 1200
tccgacggct ccttcttcct gtactctcgg ctgacagtgg ataagtcccg gtggcaggaa 1260
ggcaacgtgt tctcctgcag cgtgatgcac gaggccctgc acaaccacta tacccagaag 1320
tccctgtccc tgagcctggg caagggcggt ggaggatccg ggggtggggg aagcggcgga 1380
ggaggtagcc aggactccac tagcgatctg atcccggctc cccctctgag taaggtgccc 1440
ctgcaacaaa acttccaaga caatcagttt cagggcaaat ggtacgtggt cgggagagct 1500
ggtaacgtgg gactgcgaga ggacaaggac ccccccaaaa tgtgggccac catctacgag 1560
ctgaaggaag acaaaagcta tgatgtgaca aatgtcaggt tcgcacggaa gaaatgcact 1620
tactcaatcg gcaccttcgt gcccggctcc cagcctgggg agtttacact gggccagatt 1680
aagagcgaac ctggcggaac agccaacctg gtgcgggtgg tctctactaa ctataatcag 1740
cacgctatgg tgttctttaa agaggtctac cagaaccgag aaatcttctt tatcatcctg 1800
tacggccgta ccaaggagct gacatccgag ctgaaagaaa acttcatccg cttttctaag 1860
agtctgggac tgccagaaaa tcatattgtg tttcctgtcc caatcgacca gtgtattgat 1920
ggg 1923
<210> 58
<211> 648
<212> ДНК
<213> искусственная
<220>
<223> легкая цепь слитого полипептида
<400> 58
gagattgtcc tgacccagtc ccccgcaaca ctgtcactgt cccccggcga gagggctact 60
ctgagttgcc gggcaagcca gtctgtgtcc agctacctgg cctggtatca gcagaagcca 120
gggcaggctc ccagactgct gatctacgac gcatctaaca gagccaccgg aattcctgcc 180
cgcttctcgg gttcaggctc cggaacagac tttaccctga caatctctag tctggagcca 240
gaagatttcg cagtctacta ttgtcagcag cgaagcaatt ggccccctgc tctgactttt 300
ggcggaggga ccaaggtgga gatcaagcgt acggtcgcgg cgccttctgt gttcattttc 360
cccccatctg atgaacagct gaaatctggc actgcttctg tggtctgtct gctgaacaac 420
ttctacccta gagaggccaa agtccagtgg aaagtggaca atgctctgca gagtgggaat 480
tcccaggaat ctgtcactga gcaggactct aaggatagca catactccct gtcctctact 540
ctgacactga gcaaggctga ttacgagaaa cacaaagtgt acgcctgtga agtcacacat 600
caggggctgt ctagtcctgt gaccaaatcc ttcaataggg gagagtgc 648
<210> 59
<211> 1923
<212> ДНК
<213> искусственная
<220>
<223> тяжелая цепь слитого полипептида
<400> 59
caggactcca ctagcgatct gatcccggct ccccctctga gtaaggtgcc cctgcaacaa 60
aacttccaag acaatcagtt tcagggcaaa tggtacgtgg tcgggagagc tggtaacgtg 120
ggactgcgag aggacaagga cccccccaaa atgtgggcca ccatctacga gctgaaggaa 180
gacaaaagct atgatgtgac aaatgtcagg ttcgcacgga agaaatgcac ttactcaatc 240
ggcaccttcg tgcccggctc ccagcctggg gagtttacac tgggccagat taagagcgaa 300
cctggcggaa cagccaacct ggtgcgggtg gtctctacta actataatca gcacgctatg 360
gtgttcttta aagaggtcta ccagaaccga gaaatcttct ttatcatcct gtacggccgt 420
accaaggagc tgacatccga gctgaaagaa aacttcatcc gcttttctaa gagtctggga 480
ctgccagaaa atcatattgt gtttcctgtc ccaatcgacc agtgtattga tgggggcggt 540
ggaggatccg ggggtggggg aagcggcgga ggaggtagcc aggtgcaact gcaacaatgg 600
ggcgcaggct tgttgaagcc ctctgagaca ctgagtctta cctgtgctgt gtacggtggc 660
agcttttccg gatactattg gtcctggatt cggcagtctc cagagaaagg actggaatgg 720
atcggggaga tcaatcatgg aggctacgtt acttacaatc cctccctgga gagtcgagtg 780
acgatctctg tggatacatc taaaaaccag ttttctctca agctgagttc tgttacagcg 840
gctgacaccg ccgtatacta ttgtgcgcgg gactatggcc ctggcaacta cgactggtac 900
tttgacctgt gggggagagg cacgctggtg acagtctcga gcgctagcac caagggcccc 960
tccgtgttcc ccctggcccc ttgctcccgg tccacctccg agtctaccgc cgctctgggc 1020
tgcctggtga aagactactt ccccgagcct gtgaccgtga gctggaactc tggcgccctg 1080
ctgcaatcct ccggcctgta ctccctgtcc 1140
acctccggcg tgcacacctt ccctgccgtg
tccgtggtga cagtgccctc ctccagcctg ggcaccaaga cctacacctg taacgtggac 1200
cacaagccct ccaacaccaa ggtggacaag cgggtggaat ctaaatacgg ccctccctgc 1260
cccccctgcc ctgcccctga atttctgggc ggaccttccg tgtttctgtt ccccccaaag 1320
cccaaggaca ccctgatgat ctcccggacc cccgaagtga cctgcgtggt ggtggacgtg 1380
tcccaggaag atccagaggt gcagttcaac tggtatgttg acggcgtgga agtgcacaac 1440
gccaagacca agcccagaga ggaacagttc aactccacct accgggtggt gtccgtgctg 1500
accgtgctgc accaggactg gctgaacggc aaagagtaca agtgcaaggt gtccaacaag 1560
ggcctgccct ccagcatcga aaagaccatc tccaaggcca agggccagcc ccgcgagccc 1620
caggtgtaca ccctgccccc tagccaggaa gagatgacca agaaccaggt gtccctgacc 1680
tgtctggtga aaggcttcta cccctccgac attgccgtgg aatgggagtc caacggccag 1740
cccgagaaca actacaagac caccccccct gtgctggact ccgacggctc cttcttcctg 1800
tactctcggc tgacagtgga taagtcccgg tggcaggaag gcaacgtgtt ctcctgcagc 1860
gtgatgcacg aggccctgca caaccactat acccagaagt ccctgtccct gagcctgggc 1920
aag 1923
<210> 60
<211> 648
<212> ДНК
<213> искусственная
<220>
<223> легкая цепь слитого полипептида
<400> 60
gagattgtcc tgacccagtc ccccgcaaca ctgtcactgt cccccggcga gagggctact 60
ctgagttgcc gggcaagcca gtctgtgtcc agctacctgg cctggtatca gcagaagcca 120
gggcaggctc ccagactgct gatctacgac gcatctaaca gagccaccgg aattcctgcc 180
cgcttctcgg gttcaggctc cggaacagac tttaccctga caatctctag tctggagcca 240
gaagatttcg cagtctacta ttgtcagcag cgaagcaatt ggccccctgc tctgactttt 300
ggcggaggga ccaaggtgga gatcaagcgt acggtcgcgg cgccttctgt gttcattttc 360
cccccatctg atgaacagct gaaatctggc actgcttctg tggtctgtct gctgaacaac 420
ttctacccta gagaggccaa agtccagtgg aaagtggaca atgctctgca gagtgggaat 480
tcccaggaat ctgtcactga gcaggactct aaggatagca catactccct gtcctctact 540
ctgacactga gcaaggctga ttacgagaaa cacaaagtgt acgcctgtga agtcacacat 600
caggggctgt ctagtcctgt gaccaaatcc ttcaataggg gagagtgc 648
<210> 61
<211> 1344
<212> ДНК
<213> искусственная
<220>
<223> тяжелая цепь слитого полипептида
<400> 61
caggtccagc tgcagcagtg gggggctggt ctgctgaagc caagtgaaac tctgtcactg 60
acctgcgctg tgtacggcgg atcattctcc ggctactatt ggtcttggat cagacagagt 120
cccgagaaag gcctggaatg gatcggagag attaaccacg ggggttacgt gacctataat 180
cctagcctgg agtctagggt gaccatttcc gtcgacacaa gcaagaacca gttctctctg 240
aaactgtcca gcgtgacagc cgctgacact gcagtctact attgtgccag ggattatggc 300
cccggaaatt acgactggta ttttgatctg tgggggcggg gtaccctggt gacagtctcg 360
agcgctagca ccaagggccc ctccgtgttc cccctggccc cttgctcccg gtccacctcc 420
gagtctaccg ccgctctggg ctgcctggtg aaagactact tccccgagcc tgtgaccgtg 480
agctggaact ctggcgccct gacctccggc gtgcacacct tccctgccgt gctgcaatcc 540
tccggcctgt actccctgtc ctccgtggtg acagtgccct cctccagcct gggcaccaag 600
acctacacct gtaacgtgga ccacaagccc tccaacacca aggtggacaa gcgggtggaa 660
tctaaatacg gccctccctg ccccccctgc cctgcccctg aatttctggg cggaccttcc 720
gtgtttctgt tccccccaaa gcccaaggac accctgatga tctcccggac ccccgaagtg 780
acctgcgtgg tggtggacgt gtcccaggaa gatccagagg tgcagttcaa ctggtatgtt 840
gacggcgtgg aagtgcacaa cgccaagacc aagcccagag aggaacagtt caactccacc 900
taccgggtgg tgtccgtgct gaccgtgctg caccaggact ggctgaacgg caaagagtac 960
aagtgcaagg tgtccaacaa gggcctgccc tccagcatcg aaaagaccat ctccaaggcc 1020
aagggccagc cccgcgagcc ccaggtgtac accctgcccc ctagccagga agagatgacc 1080
aagaaccagg tgtccctgac ctgtctggtg aaaggcttct acccctccga cattgccgtg 1140
gaatgggagt ccaacggcca gcccgagaac aactacaaga ccaccccccc tgtgctggac 1200
tccgacggct ccttcttcct gtactctcgg ctgacagtgg ataagtcccg gtggcaggaa 1260
ggcaacgtgt tctcctgcag cgtgatgcac gaggccctgc acaaccacta tacccagaag 1320
tccctgtccc tgagcctggg caag 1344
<210> 62
<211> 1227
<212> ДНК
<213> искусственная
<220>
<223> легкая цепь слитого полипептида
<400> 62
caggactcca ctagcgatct gatcccggct ccccctctga gtaaggtgcc cctgcaacaa 60
aacttccaag acaatcagtt tcagggcaaa tggtacgtgg tcgggagagc tggtaacgtg 120
ggactgcgag aggacaagga cccccccaaa atgtgggcca ccatctacga gctgaaggaa 180
gacaaaagct atgatgtgac aaatgtcagg ttcgcacgga agaaatgcac ttactcaatc 240
ggcaccttcg tgcccggctc ccagcctggg gagtttacac tgggccagat taagagcgaa 300
cctggcggaa cagccaacct ggtgcgggtg gtctctacta actataatca gcacgctatg 360
gtgttcttta aagaggtcta ccagaaccga gaaatcttct ttatcatcct gtacggccgt 420
accaaggagc tgacatccga gctgaaagaa aacttcatcc gcttttctaa gagtctggga 480
ctgccagaaa atcatattgt gtttcctgtc ccaatcgacc agtgtattga tgggggcggt 540
ggaggatccg ggggtggggg aagcggcgga ggaggtagcg agattgtcct tacacagtca 600
ccagccaccc tgtccttgtc acccggtgag cgcgccacac tgtcctgccg ggcatcacaa 660
agcgttagct cctacttggc atggtaccag cagaagcctg gacaggcccc aaggctgctg 720
atctatgatg ctagcaacag ggccaccggc attcccgccc gtttctctgg tagtgggagc 780
ggcactgact ttacattgac aatctcttca ttggagcccg aggactttgc tgtgtactac 840
tgtcagcagc ggagcaactg gcctcccgcc ctgaccttcg gcgggggcac aaaggtggag 900
attaagcgta cggtcgcggc gccttctgtg ttcattttcc ccccatctga tgaacagctg 960
aaatctggca ctgcttctgt ggtctgtctg ctgaacaact tctaccctag agaggccaaa 1020
gtccagtgga aagtggacaa tgctctgcag agtgggaatt cccaggaatc tgtcactgag 1080
caggactcta aggatagcac atactccctg tcctctactc tgacactgag caaggctgat 1140
tacgagaaac acaaagtgta cgcctgtgaa gtcacacatc aggggctgtc tagtcctgtg 1200
accaaatcct tcaatagggg agagtgc 1227
<210> 63
<211> 1029
<212> ДНК
<213> искусственная
<220>
<223> тяжелая цепь слитого полипептида
<400> 63
tggtattttg atctgtgggg gcggggtacc ctggtgacag tctcgagcgc tagcaccaag 60
ggcccctccg tgttccccct ggccccttgc tcccggtcca cctccgagtc taccgccgct 120
ctgggctgcc tggtgaaaga ctacttcccc gagcctgtga ccgtgagctg gaactctggc 180
gccctgacct ccggcgtgca caccttccct gccgtgctgc aatcctccgg cctgtactcc 240
ctgtcctccg tggtgacagt gccctcctcc agcctgggca ccaagaccta cacctgtaac 300
gtggaccaca agccctccaa caccaaggtg gacaagcggg tggaatctaa atacggccct 360
ccctgccccc cctgccctgc ccctgaattt ctgggcggac cttccgtgtt tctgttcccc 420
ccaaagccca aggacaccct gatgatctcc cggacccccg aagtgacctg cgtggtggtg 480
gacgtgtccc aggaagatcc agaggtgcag ttcaactggt atgttgacgg cgtggaagtg 540
cacaacgcca agaccaagcc cagagaggaa cagttcaact ccacctaccg ggtggtgtcc 600
gtgctgaccg tgctgcacca ggactggctg aacggcaaag agtacaagtg caaggtgtcc 660
aacaagggcc tgccctccag catcgaaaag accatctcca aggccaaggg ccagccccgc 720
gagccccagg tgtacaccct gccccctagc caggaagaga tgaccaagaa ccaggtgtcc 780
ctgacctgtc tggtgaaagg cttctacccc tccgacattg ccgtggaatg ggagtccaac 840
ggccagcccg agaacaacta caagaccacc ccccctgtgc tggactccga cggctccttc 900
ttcctgtact ctcggctgac agtggataag tcccggtggc aggaaggcaa cgtgttctcc 960
tgcagcgtga tgcacgaggc cctgcacaac cactataccc agaagtccct gtccctgagc 1020
ctgggcaag 1029
<210> 64
<211> 1224
<212> ДНК
<213> искусственная
<220>
<223> легкая цепь слитого полипептида
<400> 64
gagattgtcc tgacccagtc ccccgcaaca ctgtcactgt cccccggcga gagggctact 60
ctgagttgcc gggcaagcca gtctgtgtcc agctacctgg cctggtatca gcagaagcca 120
gggcaggctc ccagactgct gatctacgac gcatctaaca gagccaccgg aattcctgcc 180
cgcttctcgg gttcaggctc cggaacagac tttaccctga caatctctag tctggagcca 240
gaagatttcg cagtctacta ttgtcagcag cgaagcaatt ggccccctgc tctgactttt 300
ggcggaggga ccaaggtgga gatcaagcgt acggtcgcgg cgccttctgt gttcattttc 360
cccccatctg atgaacagct gaaatctggc actgcttctg tggtctgtct gctgaacaac 420
ttctacccta gagaggccaa agtccagtgg aaagtggaca atgctctgca gagtgggaat 480
tcccaggaat ctgtcactga gcaggactct aaggatagca catactccct gtcctctact 540
ctgacactga gcaaggctga ttacgagaaa cacaaagtgt acgcctgtga agtcacacat 600
caggggctgt ctagtcctgt gaccaaatcc ttcaataggg gagagtgcgg cggcggagga 660
tccgggggtg ggggaagcgg cggaggaggt agccaggact ccactagcga tctgatcccg 720
gctccccctc tgagtaaggt gcccctgcaa caaaacttcc aagacaatca gtttcagggc 780
aaatggtacg tggtcgggag agctggtaac gtgggactgc gagaggacaa ggaccccccc 840
aaaatgtggg ccaccatcta cgagctgaag gaagacaaaa gctatgatgt gacaaatgtc 900
aggttcgcac ggaagaaatg cacttactca atcggcacct tcgtgcccgg ctcccagcct 960
ggggagttta cactgggcca gattaagagc gaacctggcg gaacagccaa cctggtgcgg 1020
gtggtctcta ctaactataa tcagcacgct atggtgttct ttaaagaggt ctaccagaac 1080
cgagaaatct tctttatcat cctgtacggc cgtaccaagg agctgacatc cgagctgaaa 1140
gaaaacttca tccgcttttc taagagtctg ggactgccag aaaatcatat tgtgtttcct 1200
gtcccaatcg accagtgtat tgat 1224
<210> 65
<211> 1845
<212> ДНК
<213> искусственная
<220>
<223> слитый полипептид
<400> 65
caggactcca ctagcgatct gatcccggct ccccctctga gtaaggtgcc cctgcaacaa 60
aacttccaag acaatcagtt tcagggcaaa tggtacgtgg tcgggagagc tggtaacgtg 120
ggactgcgag aggacaagga cccccccaaa atgtgggcca ccatctacga gctgaaggaa 180
gacaaaagct atgatgtgac aaatgtcagg ttcgcacgga agaaatgcac ttactcaatc 240
ggcaccttcg tgcccggctc ccagcctggg gagtttacac tgggccagat taagagcgaa 300
cctggcggaa cagccaacct ggtgcgggtg gtctctacta actataatca gcacgctatg 360
gtgttcttta aagaggtcta ccagaaccga gaaatcttct ttatcatcct gtacggccgt 420
accaaggagc tgacatccga gctgaaagaa aacttcatcc gcttttctaa gagtctggga 480
ctgccagaaa atcatattgt gtttcctgtc ccaatcgacc agtgtattga tgggggcggt 540
ggaggatccg ggggtggggg aagcggcgga ggaggtagcg aatcgaaata cggccctccc 600
tgccccccct gccctgcccc tgaagctgcg ggcggacctt ccgtgtttct gttcccccca 660
aagcccaagg acaccctgat gatctcccgg acccccgaag tgacctgcgt ggtggtggac 720
gtgtcccagg aagatccaga ggtgcagttc aactggtatg ttgacggcgt ggaagtgcac 780
aacgccaaga ccaagcccag agaggaacag ttcaactcca cctaccgggt ggtgtccgtg 840
ctgaccgtgc tgcaccagga ctggctgaac ggcaaagagt acaagtgcaa ggtgtccaac 900
aagggcctgc cctccagcat cgaaaagacc atctccaagg ccaagggcca gccccgcgag 960
ccccaggtgt acaccctgcc ccctagccag gaagagatga ccaagaacca ggtgtccctg 1020
acctgtctgg tgaaaggctt ctacccctcc gacattgccg tggaatggga gtccaacggc 1080
cagcccgaga acaactacaa gaccaccccc cctgtgctgg actccgacgg ctccttcttc 1140
ctgtactctc ggctgacagt ggataagtcc cggtggcagg aaggcaatgt gttctcctgc 1200
agcgtgatgc acgaggccct gcacaaccac tatacccaga agtccctgtc cctgagcctg 1260
ggcaagggcg gtggaggatc cgggggtggg ggaagcggcg gaggaggtag ccaggactct 1320
actagtgatc tgatcccggc accgccactg tcaaaagtcc ctctgcaaca aaactttcaa 1380
gacaatcagt ttcacggcaa atggtatgtg gtcggccagg ccggaaacat taggctgcgg 1440
gaggacaagg accccatcaa aatgatggct accatctacg agctgaagga agacaaatct 1500
tatgatgtga caatggtcaa gttcgacgat aagaaatgca tgtacgacat ctggaccttc 1560
gtgcccggct cccagccggg agagttcacc ctgggcaaga tcaagtcctt ccccggccac 1620
acttccagcc tggtccgcgt ggtctcgacc aactataatc agcatgctat ggtgttcttc 1680
aagttcgtct ttcagaatag agaggagttc tacatcacac tgtatggacg caccaaggag 1740
ctgacaagcg agctgaaaga aaacttcatc aggttttcaa agtccctggg gctgcccgaa 1800
aatcatatcg tgttcccagt ccccatcgac cagtgtattg atggt 1845
<210> 66
<211> 1830
<212> ДНК
<213> искусственная
<220>
<223> слитый полипептид
<400> 66
caggactcta ctagtgatct gattccagcc cctccactga gtaaggtgcc cctgcagcag 60
aacttccagg acaatcagtt tcagggcaag tggtatgtcg tgggaagagc aggaaacgtg 120
ggactgcgtg aggacaagga ccctccaaaa atgtgggcca ctatctacga gctgaaggaa 180
gacaaatcct atgatgtgac caatgtcaga ttcgctcgca agaaatgtac atactccatc 240
ggaactttcg tgcctggaag ccagccaggg gagtttaccc tggggcagat taagtcagaa 300
ccaggcggaa ccgccaacct ggtgcgagtg gtctccacaa actataatca gcacgctatg 360
gtgttcttta aagaggtcta ccagaacagg gaaatcttct ttatcatcct gtacggccgg 420
accaaggagc tgacatccga gctgaaagaa aacttcatcc gtttttcaaa gtccctggga 480
ctgcccgaaa atcatatcgt gttccccgtc cctatcgacc agtgcattga tgggggtggc 540
ggagggtccg gtggcggagg gagccaggac agcacatcgg atctgatccc ggcaccgcca 600
ctgtcgaaag tcccactgca acaaaatttt caagacaatc agtttcacgg caagtggtac 660
gtggtcggtc aggctggcaa cattaggctg cgggaggaca aggaccccat caagatgatg 720
gcaacaatct acgagctgaa ggaggacaaa tcttatgatg tgactatggt caagttcgac 780
gataagaaat gtatgtacga catctggacc ttcgtgcccg gctcccagcc tggcgagttc 840
acactgggca agatcaagtc cttccccggc cacacttcca gcctagtcag agtggtcagc 900
accaactata atcagcatgc tatggtgttc tttaagttcg tctttcagaa tagagaggag 960
ttctacatca ccctgtatgg ccgcactaag gagctgacct ctgagctgaa agagaatttc 1020
atccggttta gtaagtcact gggcctgcct gagaatcata tcgtgttccc agtccccatt 1080
gatcagtgca ttgatggtgg cggaggggga tccgggggtg ggggaagcgg cggaggaggt 1140
agcgaatcga aatacggccc tccctgcccc ccctgccctg cccctgaagc tgcgggcgga 1200
ccttccgtgt ttctgttccc cccaaagccc aaggacaccc tgatgatctc ccggaccccc 1260
gaagtgacct gcgtggtggt ggacgtgtcc caggaagatc cagaggtgca gttcaactgg 1320
tatgttgacg gcgtggaagt gcacaacgcc aagaccaagc ccagagagga acagttcaac 1380
tccacctacc gggtggtgtc cgtgctgacc gtgctgcacc aggactggct gaacggcaaa 1440
gagtacaagt gcaaggtgtc caacaagggc ctgccctcca gcatcgaaaa gaccatctcc 1500
aaggccaagg gccagccccg cgagccccag gtgtacaccc tgccccctag ccaggaagag 1560
atgaccaaga accaggtgtc cctgacctgt ctggtgaaag gcttctaccc ctccgacatt 1620
gccgtggaat gggagtccaa cggccagccc gagaacaact acaagaccac cccccctgtg 1680
ctggactccg acggctcctt cttcctgtac tctcggctga cagtggataa gtcccggtgg 1740
caggaaggca atgtgttctc ctgcagcgtg atgcacgagg ccctgcacaa ccactatacc 1800
cagaagtccc tgtccctgag cctgggcaag 1830
<210> 67
<211> 1098
<212> ДНК
<213> искусственная
<220>
<223> слитый полипептид
<400> 67
caggactcca cctcagacct gatcccagcc ccacctctga gcaaggtccc tctgcagcag 60
aacttccagg acaaccaatt ccaagggaaa tggtatgtcg tgggcgttgc cggaaatggt 120
atgctgcgtg aggataagga tccgcttaaa atgagggcga ccatttacga gttgaaagaa 180
gataaatcat atgacgtcac cagtgtggct tttcggaata agaaatgcca ttacaagatt 240
gggacctttg tgccggggag ccagccgggc gagtttactt taggccagat taaaagtggt 300
ccgggcgaga catcaaattt ggtccgcgtc gtgagcacca actacaacca gcatgccatg 360
gtgttcttca aggaggtgag gcagaaccgc gagtggtttt ttatcacact gtacgggcgc 420
acgaaagaac tgacaagcga gctgaaggaa aattttatcc gcttttccaa atctctgggc 480
ctccctgaaa accacatcgt cttccctgtc ccaatcgacc agtgtatcga cggcggtggt 540
ggtggttctg gtggtggtgg atcgcaggac tccacctcag acctgatccc agccccacct 600
ctgagcaagg tccctctgca gcagaacttc caggacaacc aattccatgg gaaatggtac 660
gttgtcgggc aggccggaaa tattaggctg cgtgaggata aggatccgat taaaatgatg 720
gcgaccattt acgagttgaa agaagataaa tcatatgacg tcaccatggt gaagtttgat 780
gataagaaat gcatgtacga tatttggacc tttgtgccgg ggagccagcc gggcgagttt 840
actttaggca agattaaaag ttttccgggc catacatcat cgttggtccg cgtcgtgagc 900
accaactaca accagcatgc catggtgttc ttcaagtttg tgtttcagaa ccgcgaggag 960
ttttatatca cactgtacgg gcgcacgaaa gaactgacaa gcgagctgaa ggaaaatttt 1020
atccgctttt ccaaatctct gggcctccct gaaaaccaca tcgtcttccc tgtcccaatc 1080
gaccagtgta tcgacggc 1098
<210> 68
<211> 1662
<212> ДНК
<213> искусственная
<220>
<223> слитый полипептид
<400> 68
caggactcca cctcagacct gatcccagcc ccacctctga gcaaggtccc tctgcagcag 60
aacttccagg acaaccaatt ccaagggaaa tggtatgtcg tgggcgttgc cggaaatggt 120
atgctgcgtg aggataagga tccgcttaaa atgagggcga ccatttacga gttgaaagaa 180
gataaatcat atgacgtcac cagtgtggct tttcggaata agaaatgcca ttacaagatt 240
gggacctttg tgccggggag ccagccgggc gagtttactt taggccagat taaaagtggt 300
ccgggcgaga catcaaattt ggtccgcgtc gtgagcacca actacaacca gcatgccatg 360
gtgttcttca aggaggtgag gcagaaccgc gagtggtttt ttatcacact gtacgggcgc 420
acgaaagaac tgacaagcga gctgaaggaa aattttatcc gcttttccaa atctctgggc 480
ctccctgaaa accacatcgt cttccctgtc ccaatcgacc agtgtatcga cggcggtggt 540
ggtggttctg gtggtggtgg atcgcaggac tccacctcag acctgatccc agccccacct 600
ctgagcaagg tccctctgca gcagaacttc caggacaacc aattccatgg gaaatggtac 660
gttgtcgggc aggccggaaa tattaggctg cgtgaggata aggatccgat taaaatgatg 720
gcgaccattt acgagttgaa agaagataaa tcatatgacg tcaccatggt gaagtttgat 780
gataagaaat gcatgtacga tatttggacc tttgtgccgg ggagccagcc gggcgagttt 840
actttaggca agattaaaag ttttccgggc catacatcat cgttggtccg cgtcgtgagc 900
accaactaca accagcatgc catggtgttc ttcaagtttg tgtttcagaa ccgcgaggag 960
ttttatatca cactgtacgg gcgcacgaaa gaactgacaa gcgagctgaa ggaaaatttt 1020
atccgctttt ccaaatctct gggcctccct gaaaaccaca tcgtcttccc tgtcccaatc 1080
gaccagtgta tcgacggcgg cggaggtggc tcaggaggtg gcggatccca ggactccacc 1140
tcagacctga tcccagcccc acctctgagc aaggtccctc tgcagcagaa cttccaggac 1200
aaccaattcc atgggaaatg gtacgttgtc gggcaggccg gaaatattag gctgcgtgag 1260
gataaggatc cgattaaaat gatggcgacc atttacgagt tgaaagaaga taaatcatat 1320
gacgtcacca tggtgaagtt tgatgataag aaatgcatgt acgatatttg gacctttgtg 1380
ccggggagcc agccgggcga gtttacttta ggcaagatta aaagttttcc gggccataca 1440
tcatcgttgg tccgcgtcgt gagcaccaac tacaaccagc atgccatggt gttcttcaag 1500
tttgtgtttc agaaccgcga ggagttttat atcacactgt acgggcgcac gaaagaactg 1560
acaagcgagc tgaaggaaaa ttttatccgc ttttccaaat ctctgggcct ccctgaaaac 1620
cacatcgtct tccctgtccc aatcgaccag tgtatcgacg gc 1662
<210> 69
<211> 1326
<212> ДНК
<213> искусственная
<220>
<223> тяжелая цепь антитела
<400> 69
gaggtccagc tggtgcagag cggtgccgaa gtcaagaagc ctggcgagag cctgcggatt 60
tcctgcaaag gaagcggata ctcattcagc acatactgga tctcttgggt caggcagatg 120
ccaggcaagg gactggagtg gatggggaaa atctaccccg gagacagtta cactaactat 180
tctccgagtt tccaaggcca ggtcactatc agcgctgata agtcaatttc caccgcctac 240
ctgcaatggt ccagcctgaa agcctccgac accgctatgt actattgcgc tcgggggtac 300
ggtatctttg attattgggg ccagggaacc ctggtgacag tctcgagcgc tagcaccaag 360
ggcccctccg tgtttcctct ggccccttgc tccagatcca cctccgagtc taccgccgct 420
ctgggctgcc tcgtgaagga ctacttcccc gagcctgtga ccgtgtcctg gaactctggc 480
gctctgacct ctggcgtgca caccttccct gctgtgctgc agtctagcgg cctgtactcc 540
ctgtcctccg tcgtgaccgt gccctcctct aacttcggca cccagaccta cacctgtaac 600
gtggaccaca agccctccaa caccaaggtg gacaagaccg tggaacggaa gtgctgcgtg 660
gaatgccccc cttgtcctgc ccctcctgtg gctggcccta gcgtgttcct gttcccccca 720
aagcccaagg acaccctgat gatctcccgg acccccgaag tgacctgcgt ggtggtggat 780
gtgtcccacg aggaccccga ggtgcagttc aattggtacg tggacggcgt ggaagtgcac 840
aacgccaaga ccaagcccag agaggaacag ttcaactcca ccttccgggt ggtgtccgtg 900
ctgaccgtgg tgcatcagga ctggctgaac ggcaaagagt acaagtgcaa ggtgtccaac 960
aagggcctgc ctgcccccat cgaaaagacc atctctaaga ccaagggaca gccccgcgag 1020
ccccaggtgt acacactgcc tccatcacgg gaagagatga ccaagaacca ggtgtccctg 1080
acctgtctcg tgaaaggctt ctacccctcc gatatcgccg tggaatggga gtccaacggc 1140
cagcccgaga acaactacaa gaccaccccc cccatgctgg actccgacgg ctcattcttc 1200
ctgtactcca agctgacagt ggacaagtcc cggtggcagc agggcaacgt gttctcctgc 1260
tccgtgatgc acgaggccct gcacaaccac tacacccaga agtccctgtc cctgagcccc 1320
gggaaa 1326
<210> 70
<211> 642
<212> ДНК
<213> искусственная
<220>
<223> легкая цепь антитела
<400> 70
tcctatgaac tgacacagcc tccttccgtg agcgtgagcc ctggacagac tgcttctatt 60
acttgtagcg gggataacat cggggatcag tacgcccact ggtatcagca gaagcccgga 120
cagagtcctg tgctggtcat ctaccaggac aaaaacaggc catcaggcat tcccgagcgg 180
ttctccggaa gcaactctgg gaataccgct acactgacta tctccggaac acaggcaatg 240
gacgaagccg attactattg cgctacctat acaggtttcg gctctctggc agtgtttggc 300
ggagggacta agctgaccgt cctgggccag cctaaagcgg cgccatccgt caccctgttc 360
cctccctcat ccgaggaact gcaggccaat aaggctacac tggtctgtct gattagcgac 420
ttctaccctg gggccgtgac tgtggcttgg aaagccgatt cttctcccgt gaaagctgga 480
gtggaaacaa ccaccccctc taaacagagc aacaacaaat acgctgcctc ttcatacctg 540
tccctgaccc ctgaacagtg gaaatctcac cggtcttact catgccaggt gacacacgag 600
ggatcaactg tggagaaaac cgtggctcct accgaatgtt ca 642
<210> 71
<211> 1905
<212> ДНК
<213> искусственная
<220>
<223> тяжелая цепь слитого полипептида
<400> 71
gaggtccagc tggtgcagag cggtgccgaa gtcaagaagc ctggcgagag cctgcggatt 60
tcctgcaaag gaagcggata ctcattcagc acatactgga tctcttgggt caggcagatg 120
ccaggcaagg gactggagtg gatggggaaa atctaccccg gagacagtta cactaactat 180
tctccgagtt tccaaggcca ggtcactatc agcgctgata agtcaatttc caccgcctac 240
ctgcaatggt ccagcctgaa agcctccgac accgctatgt actattgcgc tcgggggtac 300
ggtatctttg attattgggg ccagggaacc ctggtgacag tctcgagcgc tagcaccaag 360
ggcccctccg tgtttcctct ggccccttgc tccagatcca cctccgagtc taccgccgct 420
ctgggctgcc tcgtgaagga ctacttcccc gagcctgtga ccgtgtcctg gaactctggc 480
gctctgacct ctggcgtgca caccttccct gctgtgctgc agtctagcgg cctgtactcc 540
ctgtcctccg tcgtgaccgt gccctcctct aacttcggca cccagaccta cacctgtaac 600
gtggaccaca agccctccaa caccaaggtg gacaagaccg tggaacggaa gtgctgcgtg 660
gaatgccccc cttgtcctgc ccctcctgtg gctggcccta gcgtgttcct gttcccccca 720
aagcccaagg acaccctgat gatctcccgg acccccgaag tgacctgcgt ggtggtggat 780
gtgtcccacg aggaccccga ggtgcagttc aattggtacg tggacggcgt ggaagtgcac 840
aacgccaaga ccaagcccag agaggaacag ttcaactcca ccttccgggt ggtgtccgtg 900
ctgaccgtgg tgcatcagga ctggctgaac ggcaaagagt acaagtgcaa ggtgtccaac 960
aagggcctgc ctgcccccat cgaaaagacc atctctaaga ccaagggaca gccccgcgag 1020
ccccaggtgt acacactgcc tccatcacgg gaagagatga ccaagaacca ggtgtccctg 1080
acctgtctcg tgaaaggctt ctacccctcc gatatcgccg tggaatggga gtccaacggc 1140
cagcccgaga acaactacaa gaccaccccc cccatgctgg actccgacgg ctcattcttc 1200
ctgtactcca agctgacagt ggacaagtcc cggtggcagc agggcaacgt gttctcctgc 1260
tccgtgatgc acgaggccct gcacaaccac tacacccaga agtccctgtc cctgagcccc 1320
gggaaaggcg gcggaggatc cgggggtggg ggaagcggcg gaggaggtag ccaggactcc 1380
actagcgatc tgatcccggc tccccctctg agtaaggtgc ccctgcaaca aaacttccaa 1440
gacaatcagt ttcagggcaa atggtacgtg gtcgggagag ctggtaacgt gggactgcga 1500
gaggacaagg acccccccaa aatgtgggcc accatctacg agctgaagga agacaaaagc 1560
tatgatgtga caaatgtcag gttcgcacgg aagaaatgca cttactcaat cggcaccttc 1620
gtgcccggct cccagcctgg ggagtttaca ctgggccaga ttaagagcga acctggcgga 1680
acagccaacc tggtgcgggt ggtctctact aactataatc agcacgctat ggtgttcttt 1740
aaagaggtct accagaaccg agaaatcttc tttatcatcc tgtacggccg taccaaggag 1800
ctgacatccg agctgaaaga aaacttcatc cgcttttcta agagtctggg actgccagaa 1860
aatcatattg tgtttcctgt cccaatcgac cagtgtattg atggg 1905
<210> 72
<211> 642
<212> ДНК
<213> искусственная
<220>
<223> легкая цепь слитого полипептида
<400> 72
tcctatgaac tgacacagcc tccttccgtg agcgtgagcc ctggacagac tgcttctatt 60
acttgtagcg gggataacat cggggatcag tacgcccact ggtatcagca gaagcccgga 120
cagagtcctg tgctggtcat ctaccaggac aaaaacaggc catcaggcat tcccgagcgg 180
ttctccggaa gcaactctgg gaataccgct acactgacta tctccggaac acaggcaatg 240
gacgaagccg attactattg cgctacctat acaggtttcg gctctctggc agtgtttggc 300
ggagggacta agctgaccgt cctgggccag cctaaagcgg cgccatccgt caccctgttc 360
cctccctcat ccgaggaact gcaggccaat aaggctacac tggtctgtct gattagcgac 420
ttctaccctg gggccgtgac tgtggcttgg aaagccgatt cttctcccgt gaaagctgga 480
gtggaaacaa ccaccccctc taaacagagc aacaacaaat acgctgcctc ttcatacctg 540
tccctgaccc ctgaacagtg gaaatctcac cggtcttact catgccaggt gacacacgag 600
ggatcaactg tggagaaaac cgtggctcct accgaatgtt ca 642
<210> 73
<211> 229
<212> Белок
<213> искусственный
<220>
<223> IgG4-Fc с мутациями
<400> 73
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala
1 5 10 15
Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
20 25 30
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
35 40 45
Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val
50 55 60
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser
65 70 75 80
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
85 90 95
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser
100 105 110
Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
115 120 125
Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln
130 135 140
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
145 150 155 160
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
165 170 175
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu
180 185 190
Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser
195 200 205
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
210 215 220
Leu Ser Leu Gly Lys
225
<210> 74
<211> 448
<212> Белок
<213> искусственный
<220>
<223> эталонное антитело
<400> 74
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn His Gly Gly Tyr Val Thr Tyr Asn Pro Ser Leu Glu
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Tyr Gly Pro Gly Asn Tyr Asp Trp Tyr Phe Asp Leu Trp Gly
100 105 110
Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly
210 215 220
Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro
260 265 270
Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
<210> 75
<211> 216
<212> Белок
<213> эталонное антитело
<400> 75
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Pro
85 90 95
Ala Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val
100 105 110
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
115 120 125
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
130 135 140
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
145 150 155 160
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
165 170 175
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
180 185 190
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr
195 200 205
Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| НОВЫЕ СЛИТЫЕ БЕЛКИ, СПЕЦИФИЧЕСКИЕ В ОТНОШЕНИИ CD137 И GPC3 | 2020 |
|
RU2814653C2 |
| БЕЛКИ, СПЕЦИФИЧНЫЕ В ОТНОШЕНИИ CD137 | 2016 |
|
RU2736312C2 |
| МУТЕИНЫ ЛИПОКАЛИНА 2 ЧЕЛОВЕКА С АФФИННОСТЬЮ В ОТНОШЕНИИ ГЛИПИКАНА-3 (GPC3) | 2016 |
|
RU2756318C2 |
| НОВЫЙ СЛИТЫЙ БЕЛОК, СПЕЦИФИЧНЫЙ ДЛЯ CD137 И PD-L1 | 2019 |
|
RU2818349C2 |
| НОВЫЕ ПОЛИПЕПТИДЫ СО СПЕЦИФИЧЕСКИМ СВЯЗЫВАНИЕМ И ПУТИ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2723034C2 |
| НОВЫЕ БЕЛКИ, СПЕЦИФИЧЕСКИЕ В ОТНОШЕНИИ АНГИОГЕНЕЗА | 2016 |
|
RU2720688C2 |
| ИММУНОАКТИВИРУЮЩАЯ АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА | 2015 |
|
RU2722788C2 |
| СЛИТЫЙ ПОЛИПЕПТИД С ПРОТИВОРАКОВОЙ АКТИВНОСТЬЮ | 2016 |
|
RU2727165C2 |
| МУТЕИНЫ ЛИПОКАЛИНА 2 ЧЕЛОВЕКА (Lcn2, hNGAL) С АФФИННОСТЬЮ ДЛЯ ОПРЕДЕЛЕННОЙ МИШЕНИ | 2019 |
|
RU2804336C2 |
| МУТЕИНЫ ЛИПОКАЛИНА 2 ЧЕЛОВЕКА (Lcn2, hNGAL) С АФФИННОСТЬЮ ДЛЯ ОПРЕДЕЛЕННОЙ МИШЕНИ | 2010 |
|
RU2707126C2 |
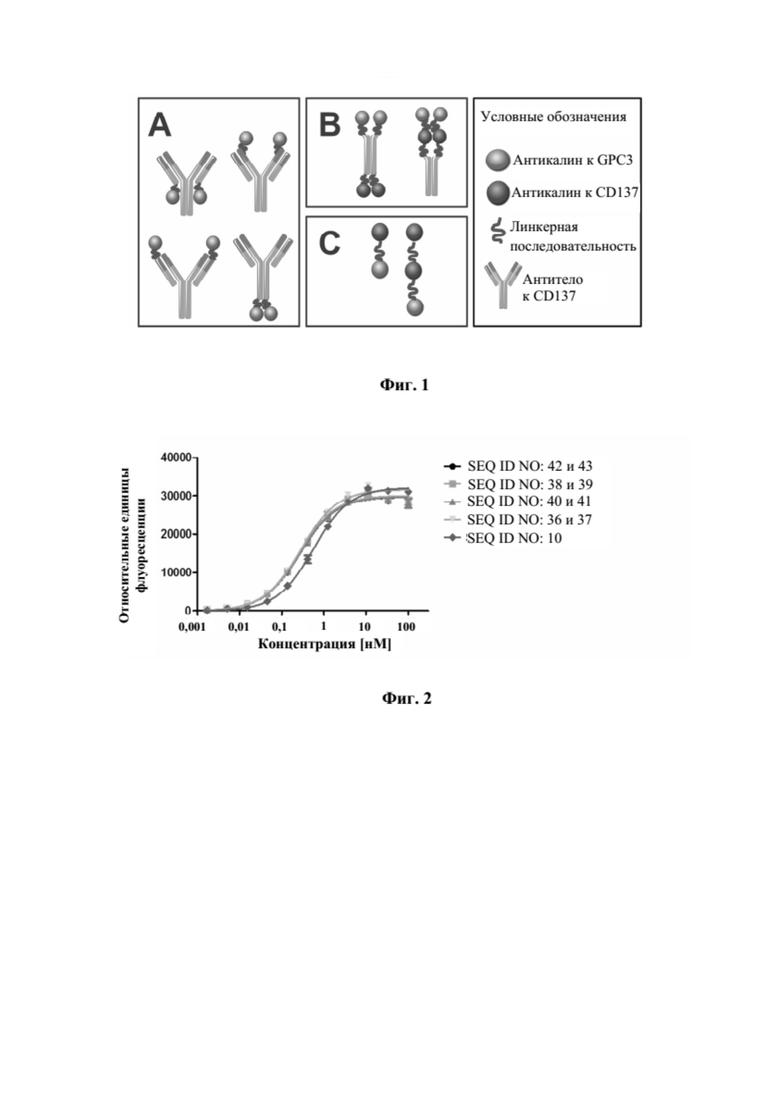
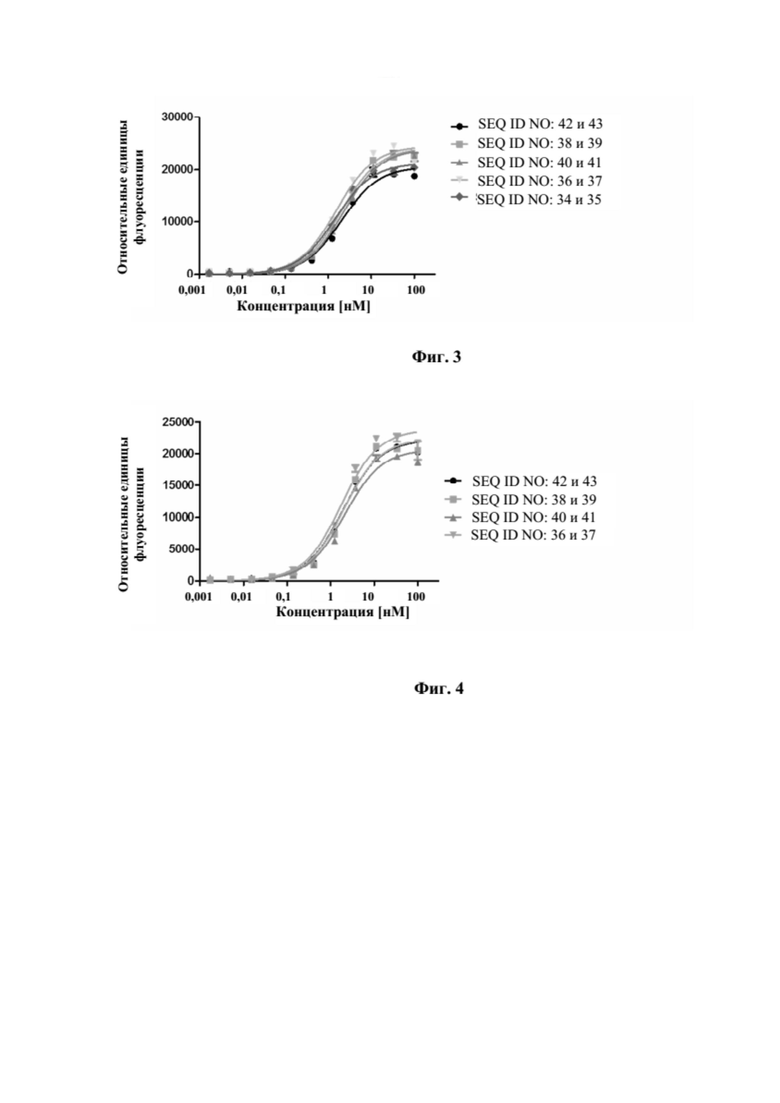
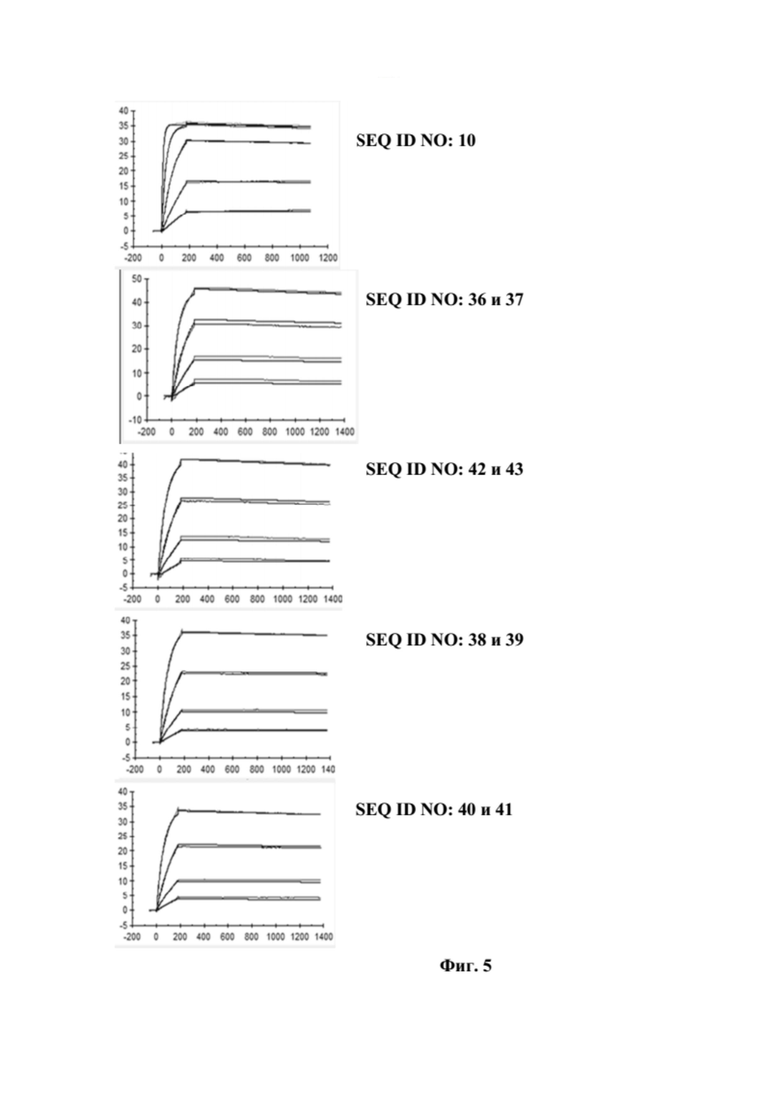
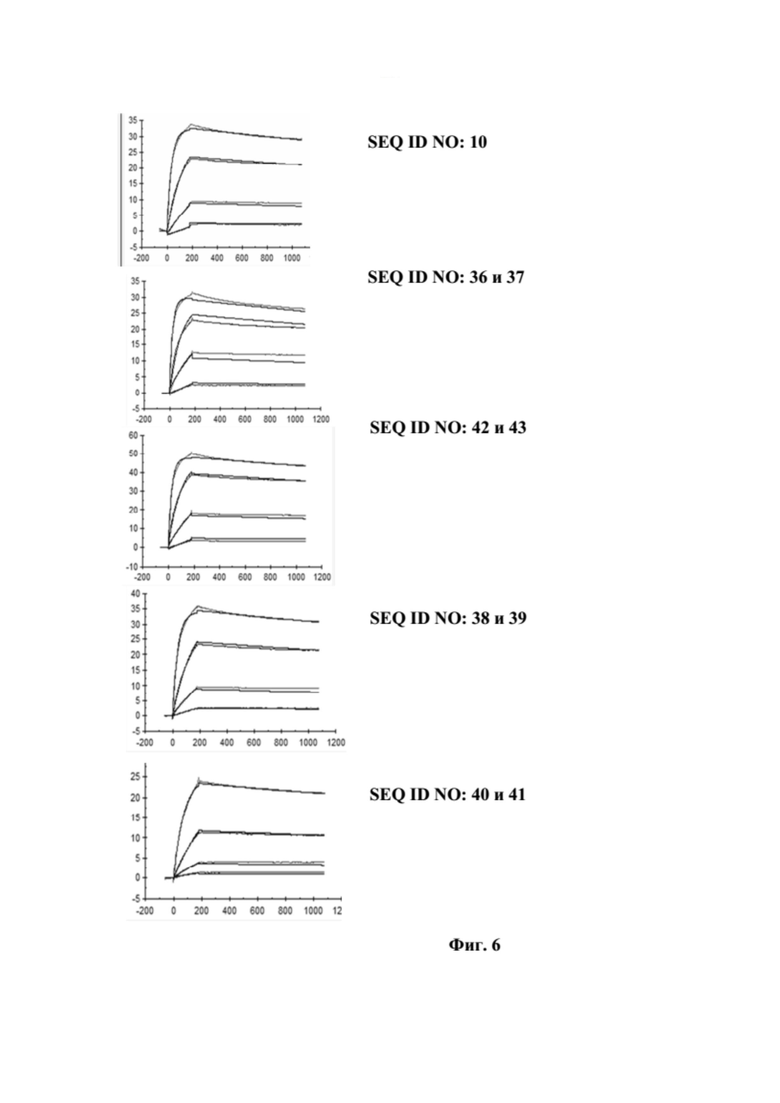
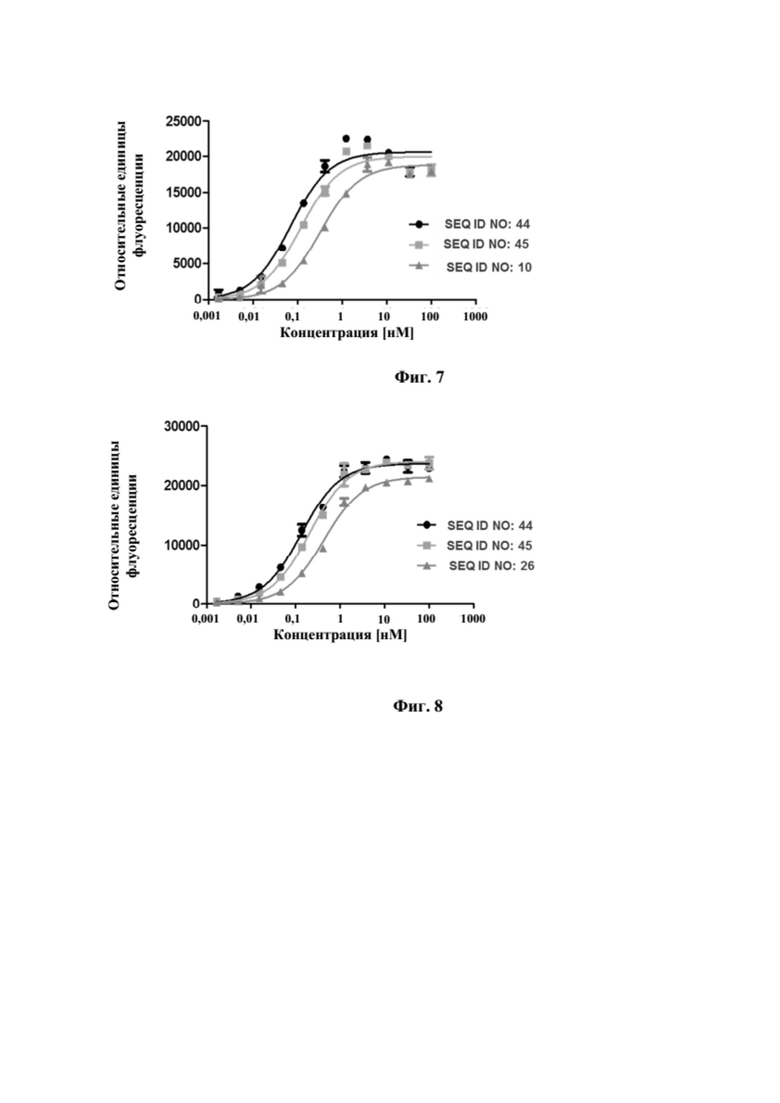
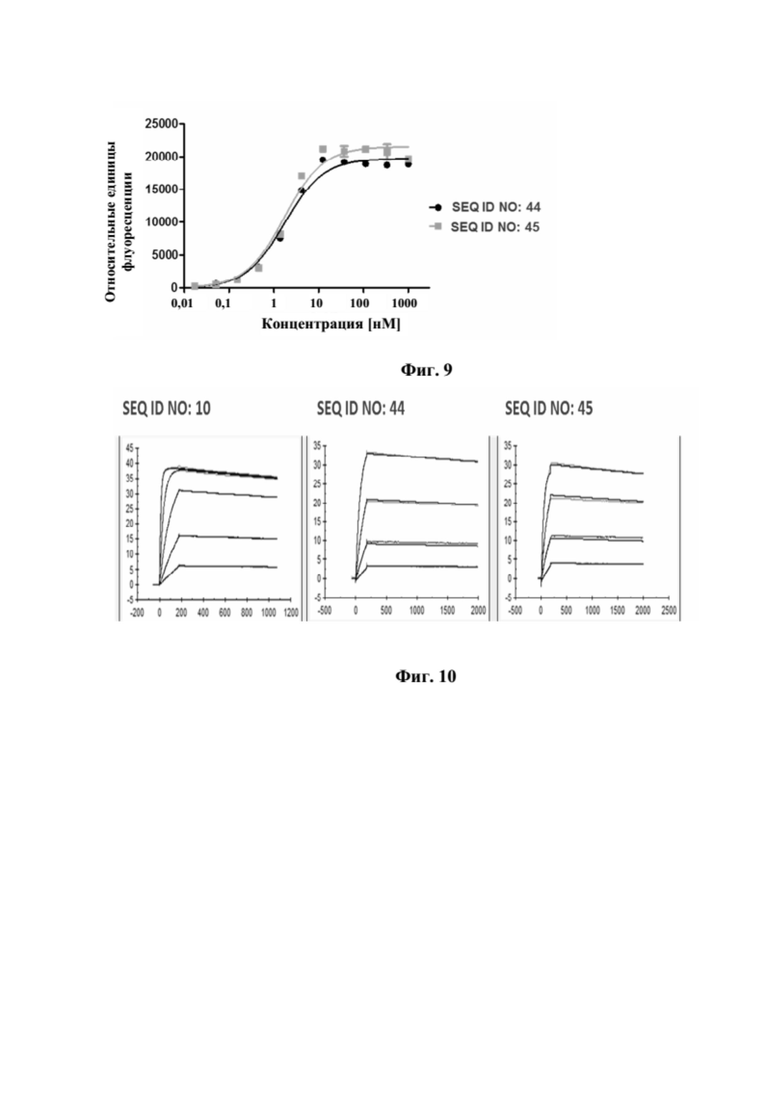
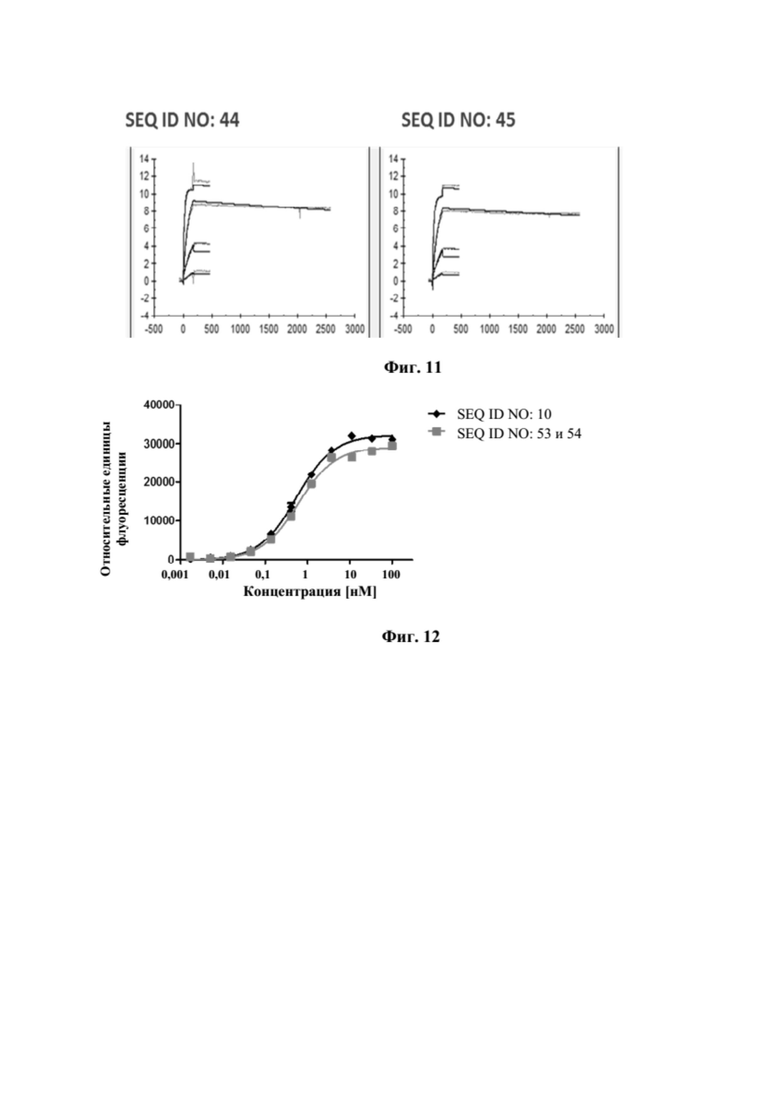
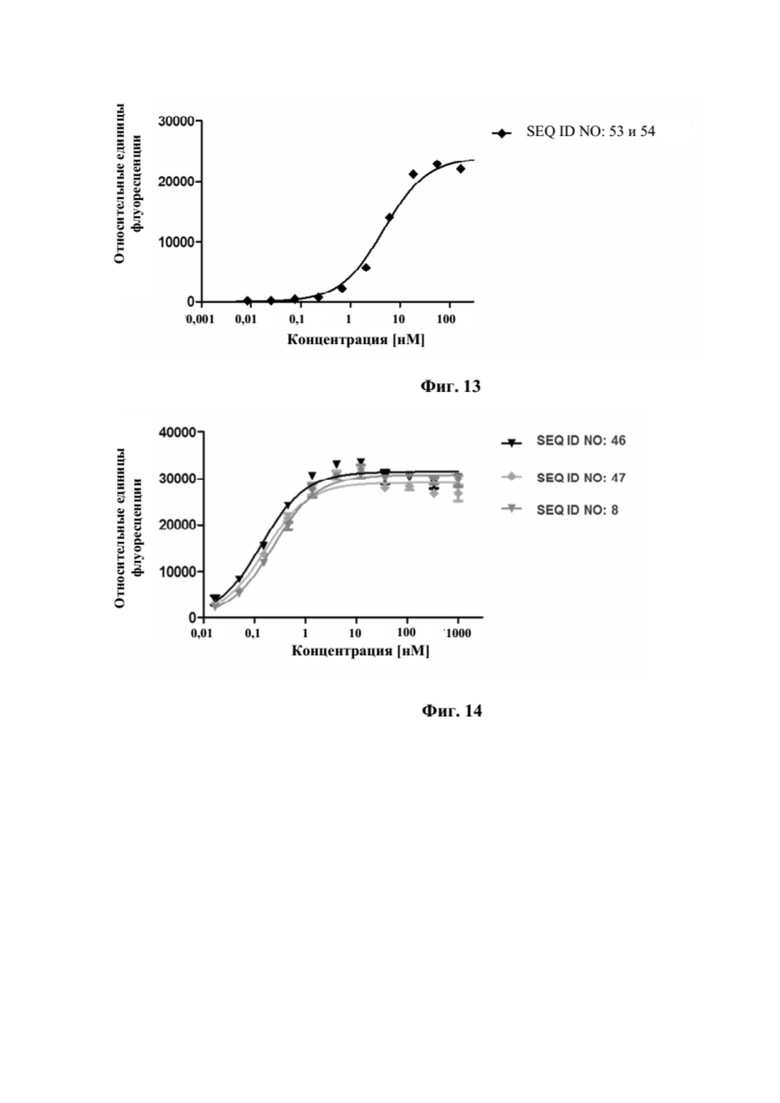
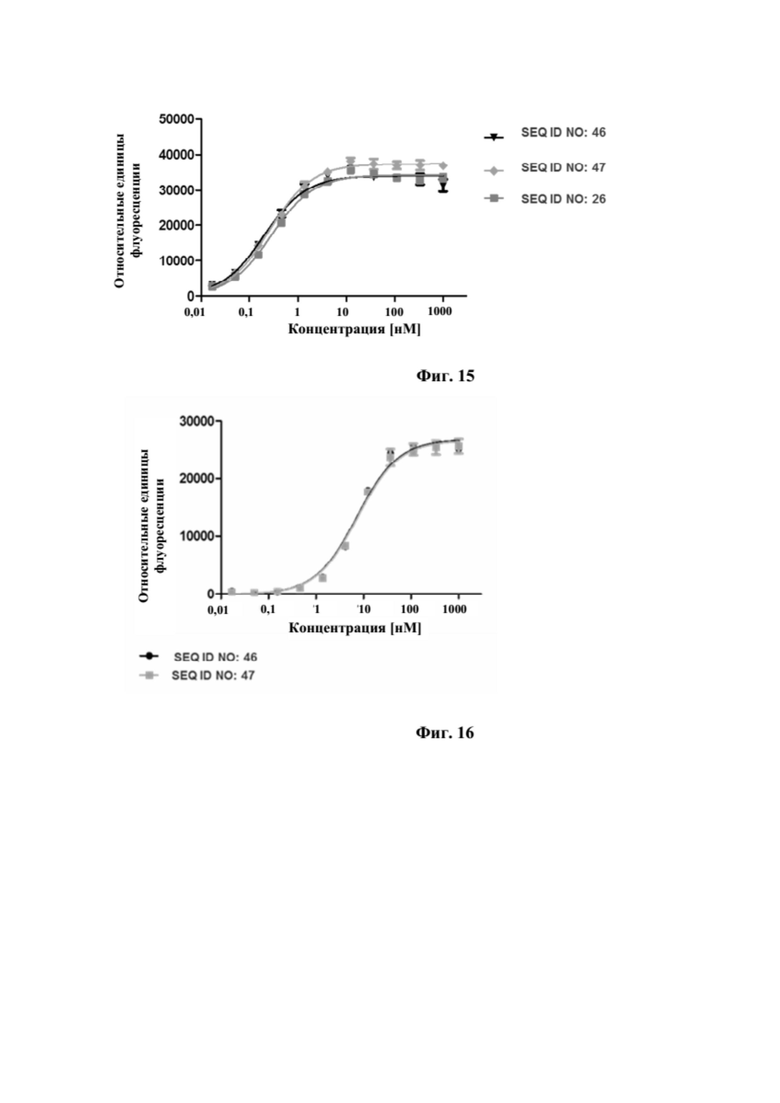
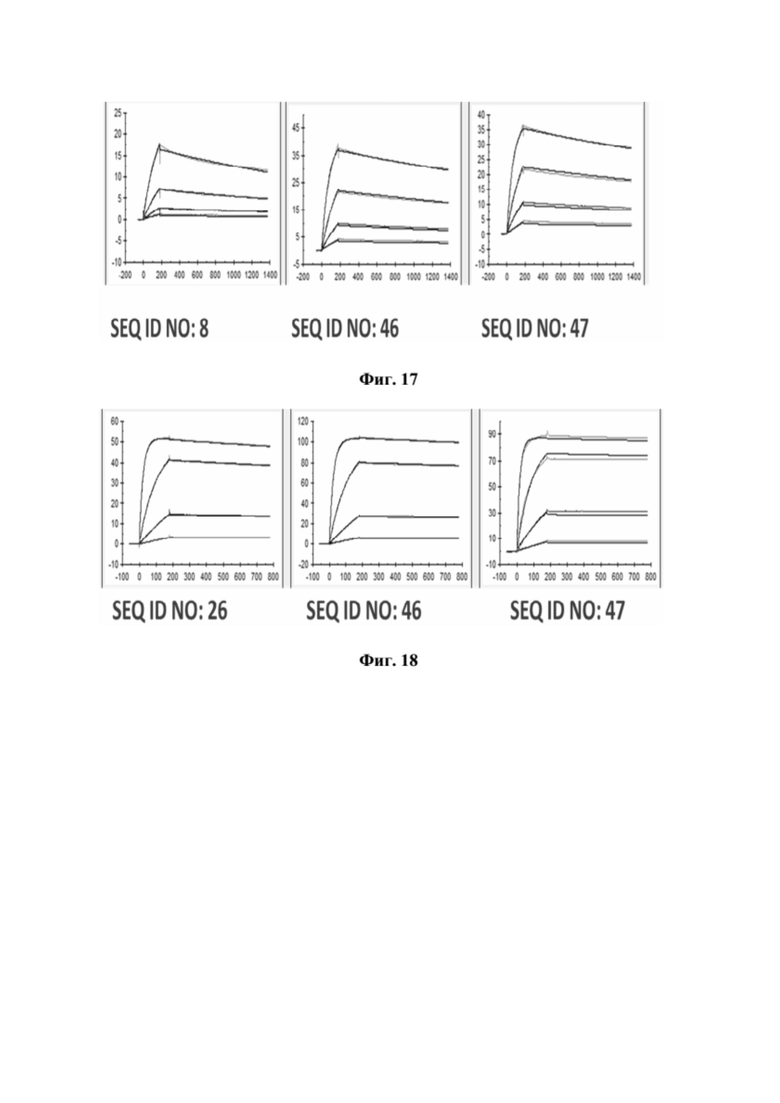
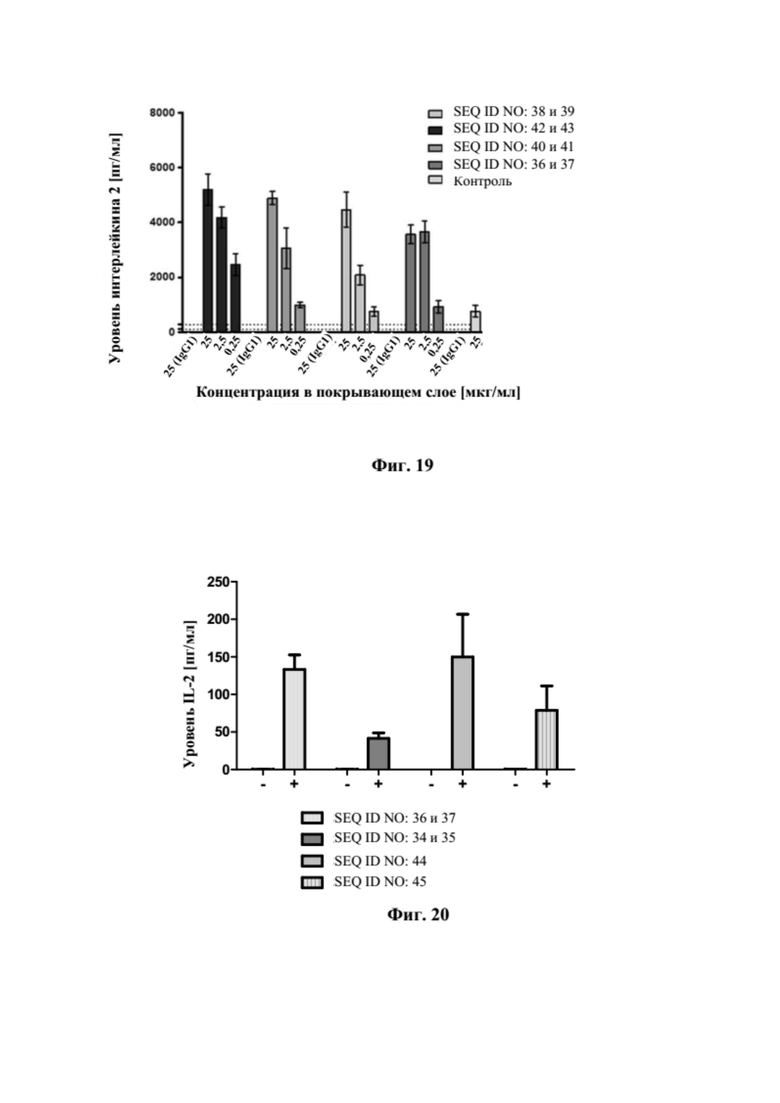
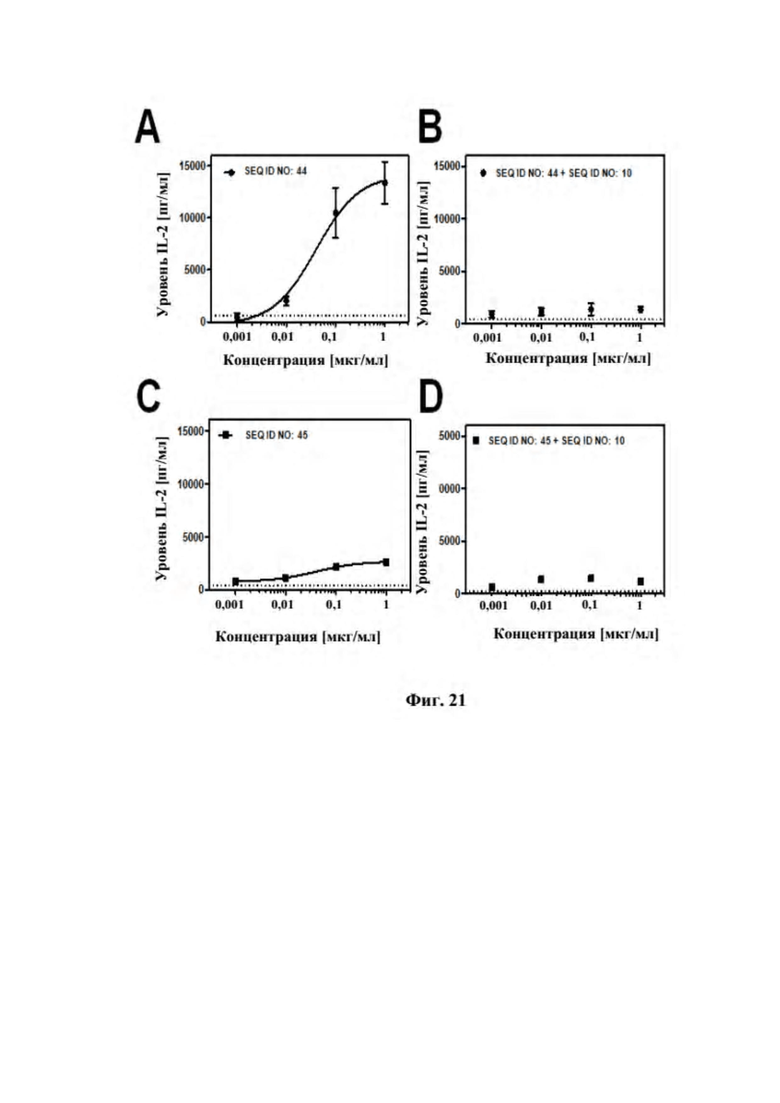
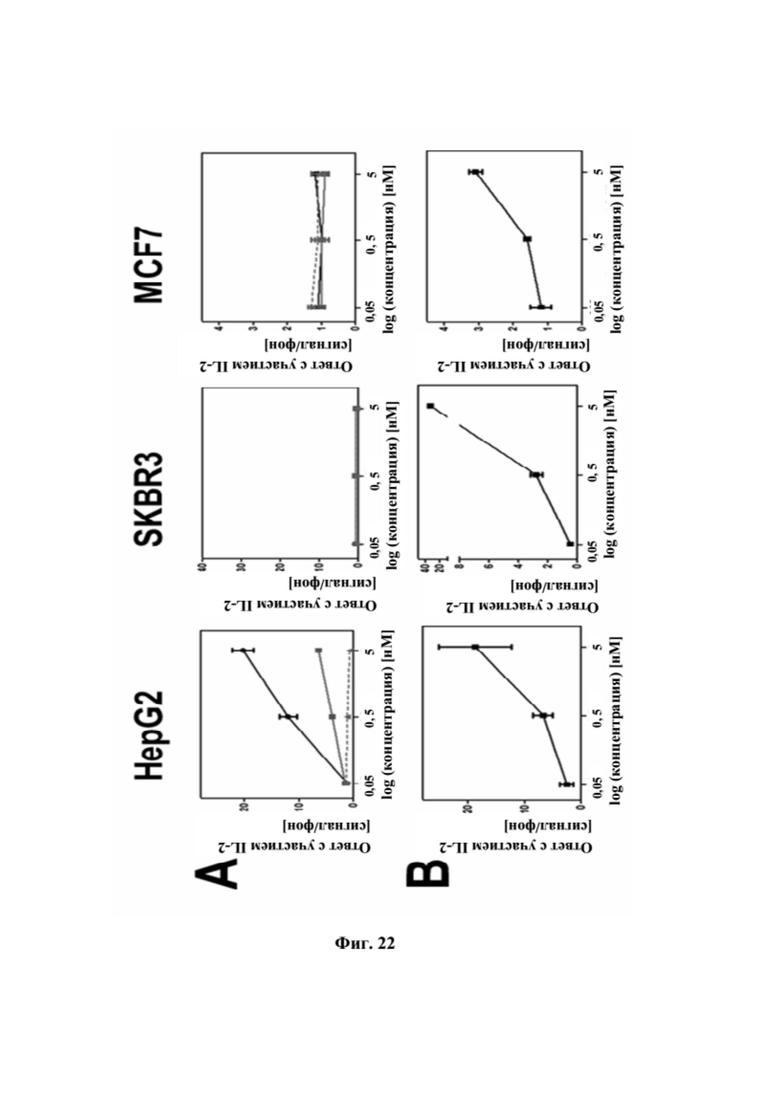
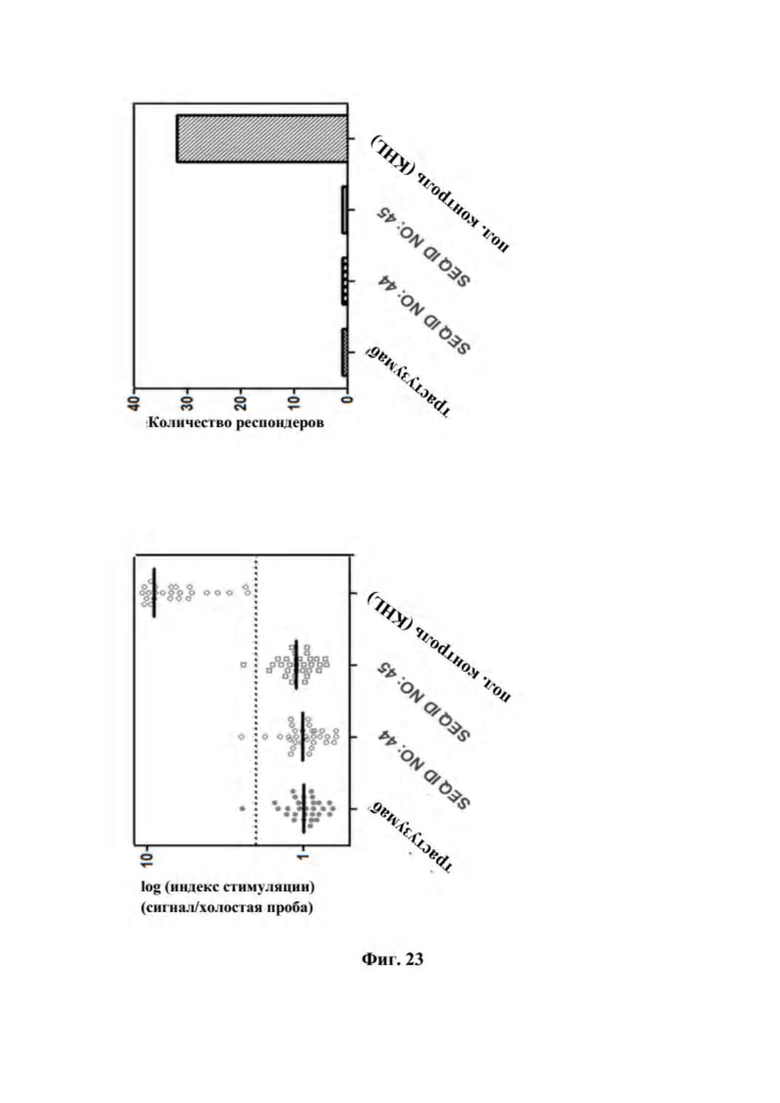
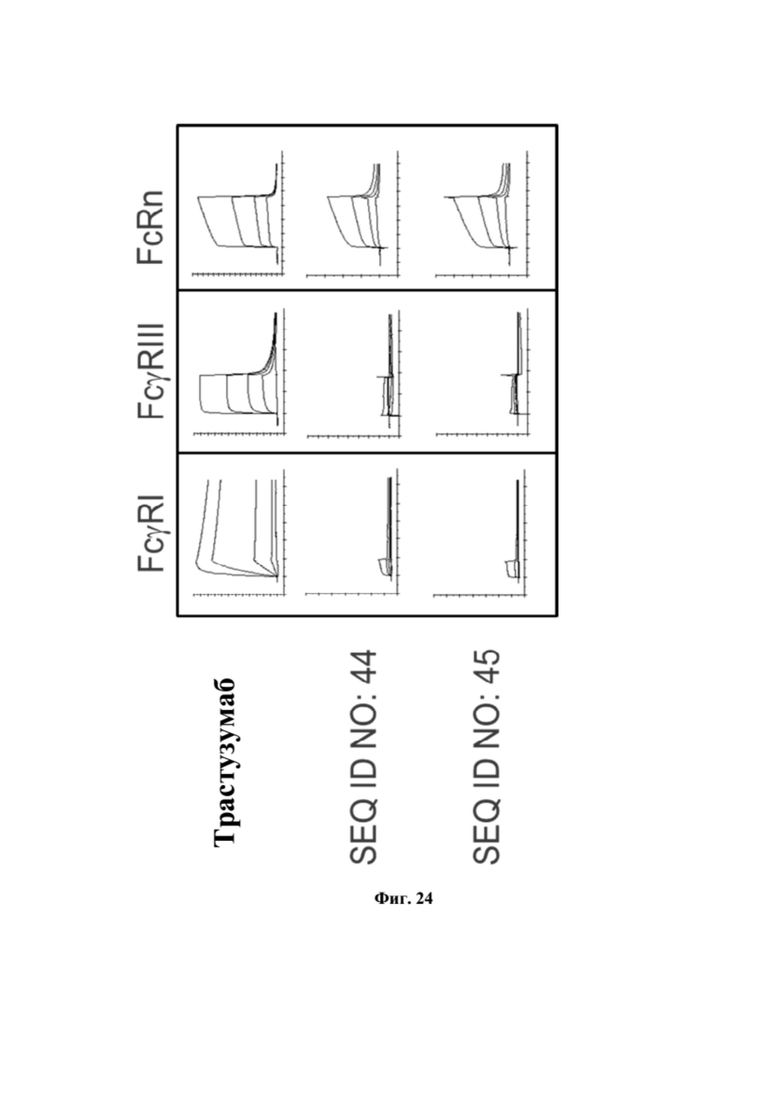
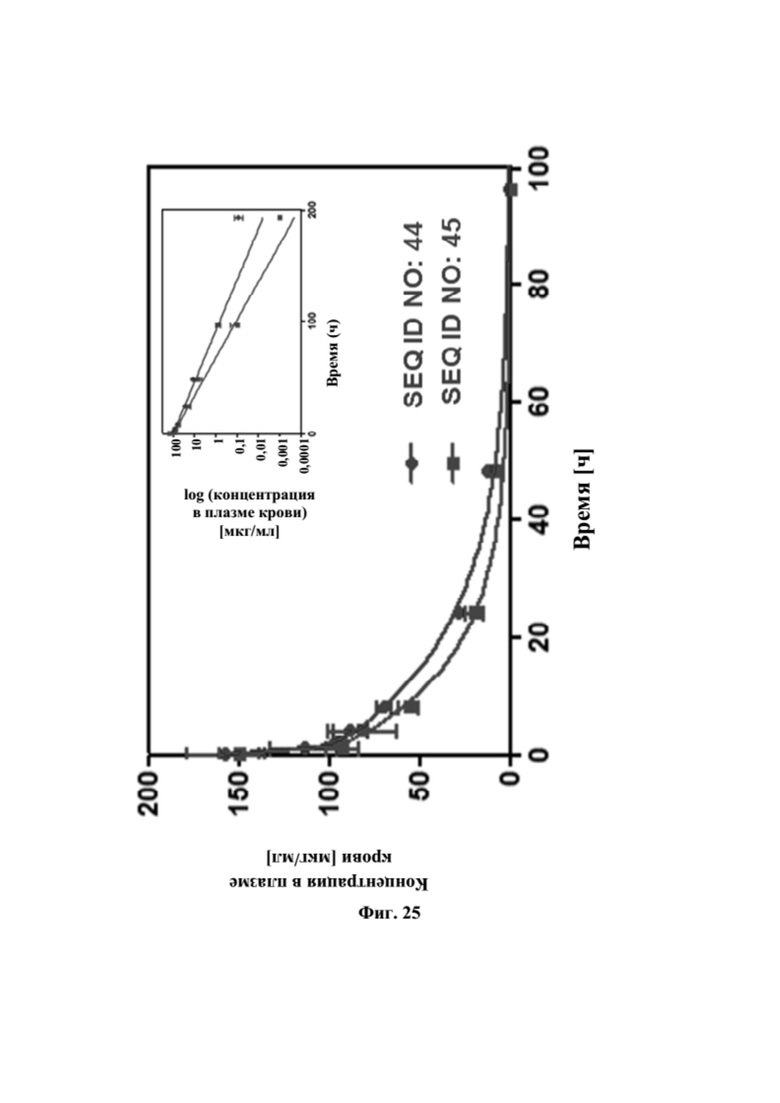
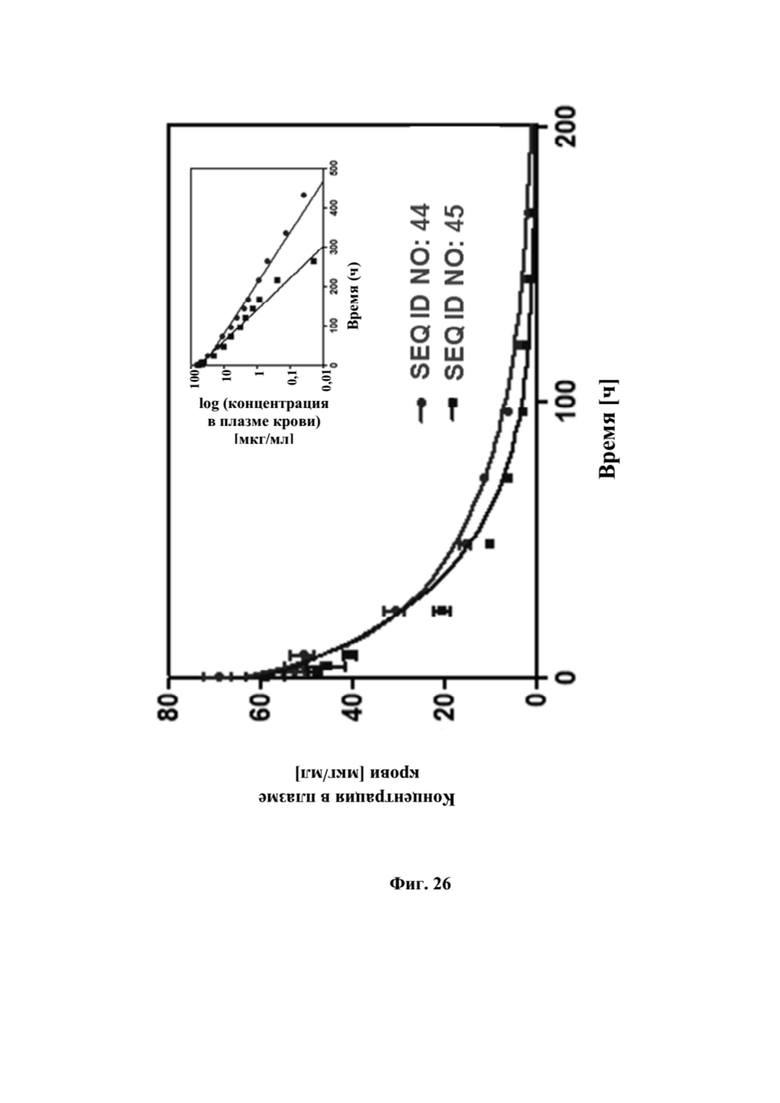
Настоящее изобретение относится к области иммунологии. Предложен слитый пептид, способный связываться как CD137, так и глипикан-3 (GPC3), молекула нуклеиновой кислоты, клетка-хозяин, способы получения слитого полипептида. Кроме того, рассмотрено применение слитого полипептида, способ одновременной активации нисходящих сигнальных путей, способ одновременной костимуляции Т-клеток и воздействия на GPC3-положительные опухолевые клетки. Также предложен способ направления образования скоплений и активации CD137 на Т-клетках в отношении GPC3-положительных опухолевых клеток, способ лечения рака и фармацевтическая композиция. Данное изобретение может найти дальнейшее применение в терапии рака, ассоциированного с CD137 и GPC3. 12 н. и 22 з.п. ф-лы, 26 ил., 19 табл., 27 пр.
1. Слитый полипептид, который способен связывать как CD137, так и глипикан-3 (GPC3), где слитый полипептид содержит по меньшей мере две субъединицы в любом порядке, при этом первая субъединица предусматривает мутеин липокалина со специфичностью связывания в отношении CD137 и с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 18-33, а вторая субъединица предусматривает мутеин липокалина со специфичностью связывания в отношении GPC3 и с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 4-17,
где слитый полипептид содержит линкер, причем линкер представляет собой:
- линкер (G4S)3, показанный под SEQ ID NO: 49;
- сконструированный Fc-фрагмент IgG4, показанный под SEQ ID NO: 73; и
- линкер (G4S)2, показанный под SEQ ID NO: 48.
2. Слитый полипептид по п. 1, где слитый полипептид дополнительно содержит Fc-фрагмент иммуноглобулина.
3. Слитый полипептид по п. 1, где слитый полипептид дополнительно содержит третью субъединицу, специфичную в отношении CD137.
4. Слитый полипептид по п. 3, где третья субъединица предусматривает мутеин липокалина со специфичностью связывания в отношении CD137.
5. Слитый полипептид по п. 1, где слитый полипептид способен связывать CD137 со значением EC50, составляющим по меньшей мере приблизительно 5 нМ.
6. Слитый полипептид по п. 1, где слитый полипептид способен связывать CD137 со значением EC50, по меньшей мере таким же как или превосходящим значение EC50 для мутеина липокалина или антитела, специфичного в отношении CD137, которые включены в такой слитый полипептид.
7. Слитый полипептид по п. 1, где слитый полипептид способен связывать CD137 с аффинностью, характеризующейся значением KD, составляющим по меньшей мере приблизительно 5 нМ.
8. Слитый полипептид по п. 1, где слитый полипептид способен связывать GPC3 со значением EC50, составляющим по меньшей мере приблизительно 5 нМ.
9. Слитый полипептид по п. 1, где слитый полипептид способен связывать GPC3 со значением EC50, сопоставимым со значением EC50 для мутеина липокалина, специфичного в отношении GPC3, который включен в такой слитый полипептид.
10. Слитый полипептид по п. 1, где слитый полипептид способен связывать GPC3 с аффинностью, характеризующейся значением KD, составляющим по меньшей мере приблизительно 5 нМ.
11. Слитый полипептид по п. 1, где слитый полипептид способен одновременно связывать CD137 и GPC3.
12. Слитый полипептид по п. 1, где слитый полипептид способен одновременно связывать CD137 и GPC3 со значением EC50, составляющим по меньшей мере приблизительно 10 нМ.
13. Слитый полипептид по п. 1, где слитый полипептид способен костимулировать ответы T-клеток.
14. Слитый полипептид по п. 1, где слитый полипептид способен индуцировать продуцирование IL-2 в присутствии стимуляции T-клеток.
15. Слитый полипептид по п. 1, где слитый полипептид не индуцирует продуцирование IL-2 в отсутствие стимуляции антителом к CD3 T-клеток.
16. Слитый полипептид по п. 1, где слитый полипептид способен костимулировать активацию T-клеток, стимулированных антителом к CD3 и антителом к CD28 при субоптимальных концентрациях.
17. Слитый полипептид по п. 1, где слитый полипептид способен костимулировать ответы T-клеток.
18. Слитый полипептид по п. 1, где слитый полипептид способен индуцировать продуцирование IL-2.
19. Слитый полипептид по п. 1, где слитый полипептид способен костимулировать активацию T-клеток зависимым от мишени, экспрессирующей GPC3, образом.
20. Слитый полипептид по любому из пп. 1-19, где слитый полипептид содержит аминокислоты, показанные в SEQ ID NO: 44, аминокислоты, показанные в SEQ ID NO: 45, аминокислоты, показанные в SEQ ID NO: 46, или аминокислоты, показанные в SEQ ID NO: 47.
21. Молекула нуклеиновой кислоты, которая имеет нуклеотидную последовательность, кодирующую полипептид по любому из пп. 1-20.
22. Молекула нуклеиновой кислоты по п. 21, где молекула нуклеиновой кислоты функционально связана с регуляторной последовательностью для обеспечения экспрессии указанной молекулы нуклеиновой кислоты.
23. Молекула нуклеиновой кислоты по п. 21 или 22, где молекула нуклеиновой кислоты входит в состав вектора или фагмидного вектора.
24. Клетка-хозяин для получения слитого полипептида, содержащая молекулу нуклеиновой кислоты по любому из пп. 21-23.
25. Способ получения слитого полипептида по любому из пп. 1-20, где способ включает культивирование клетки-хозяина по п. 24 с получением слитого полипептида исходя из нуклеиновой кислоты по любому из пп. 21-23, кодирующей мутеин, при помощи способов генной инженерии.
26. Способ по п. 25, где слитый полипептид получают в бактериальном или эукариотическом организме-хозяине и выделяют из этого организма-хозяина или его культуры.
27. Способ получения слитого полипептида по любому из пп. 1-20, где полипептид получают путем химического синтеза или путем транскрипции и трансляции in vitro.
28. Применение слитого полипептида по любому из пп. 1-20 или композиции, содержащей такой слитый полипептид, для одновременной активации нисходящих сигнальных путей, связанных с CD137, и воздействия на GPC3-положительные опухолевые клетки.
29. Способ одновременной активации нисходящих сигнальных путей, связанных с CD137, и воздействия на GPC3-положительные опухолевые клетки, включающий применение слитых полипептидов по любому из пп. 1-20 или композиции, содержащей такой слитый полипептид.
30. Способ одновременной костимуляции T-клеток и воздействия на GPC3-положительные опухолевые клетки, включающий применение слитых полипептидов по любому из пп. 1-20 или композиции, содержащей такой слитый полипептид.
31. Способ одновременной индукции пролиферации T-лимфоцитов и воздействия на GPC3-положительные опухолевые клетки, включающий применение слитых полипептидов по любому из пп. 1-20 или композиции, содержащей такой слитый полипептид.
32. Способ направления образования скоплений и активации CD137 на
T-клетках в отношении GPC3-положительных опухолевых клеток, включающий применение слитых полипептидов по любому из пп. 1-20 или композиции, содержащей такой слитый полипептид.
33. Способ лечения рака, ассоциированного с CD137 и GPC, где указанный способ включает введение эффективного количества слитого полипептида по любому из пп. 1-20 субъекту, нуждающемуся в этом.
34. Фармацевтическая композиция для лечения рака, ассоциированного с CD137 и GPC, содержащая эффективное количество слитого полипептида по любому из пп. 1-20 и фармацевтически приемлемый носитель.
| WYZGOL A et | |||
| al | |||
| "Trimer Stabilization, Oligomerization, and Antibody-Mediated Cell Surface Immobilization Improve the Activity of Soluble Trimers of CD27L, CD40L, 41BBL, and Glucocorticoid-Induced TNF Receptor Ligand", J Immunology, 2009, Vol | |||
| Переносная мусоросжигательная печь-снеготаялка | 1920 |
|
SU183A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Улей | 1923 |
|
SU1851A1 |
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| SKERRA A "‘Anticalins’: a new class of | |||

