Область техники
Настоящее изобретение относится к анти-CD137 антигенсвязывающим молекулам и способам их применения.
Предшествующий уровень техники
Рак является смертельно опасным заболеванием, которое трудно полностью вылечить, за исключением некоторых случаев. Результаты основного метода лечения, заключающегося в применении химиотерапевтических средств, нельзя признать удовлетворительными. Было высказано предположение, что не только гетерогенность самих раковых клеток, но и микроокружение опухоли играют важную роль в качестве фактора, затрудняющего лечение рака (NPL 1). Недавно было показано, что неоперабельная злокачественная меланома и другие подобные заболевания потенциально излечимы анти-CTLA-4 антителом, которое подавляет иммуносупрессивную функцию CTLA-4 и тем самым способствует активации Т-клеток (NPL 2). В 2011 г. моноклональное антитело против CTLA-4 человека (ипилимумаб) было одобрено Управлением по санитарному надзору за качеством пищевых продуктов и медикаментов США (FDA) как первое в мире иммуноактивирующее антитело. Кроме того, сообщалось, что ингибирующие антитела против PD-1 и PD-L1, других отличных от CTLA-4 молекул иммунных контрольных точек, обладают терапевтическим действием (NPL 3) и одобрены FDA.
Известно, что Т-клетки, которые играют важную роль в опухолевом иммунитете, активируются двумя сигналами: 1) связыванием рецептора Т-клеток (TCR) с антигенным пептидом, представленным молекулами класса I главного комплекса гистосовместимости (МНС), и активацией TCR; и 2) связыванием костимулирующей молекулы на поверхности Т-клеток с ее лигандами на антигенпрезентирующих клетках и активацией костимулирующей молекулы. Кроме того, активацию костимулирующих молекул, принадлежащих к надсемейству рецепторов фактора некроза опухоли (TNFRSF, tumor necrosis factor receptor superfamily), включая CD137 (4-1BB), на поверхности Т-клеток, оценивают как важную функцию для активации Т-клеток (NPL 4).
К надсемейству TNFRSF относят CD137, CD40, ОХ40, RANK, GITR и подобные молекулы. Установлено, что CD137 экспрессируется не только на поверхности Т-клеток, но также и на поверхности других иммунных клеток, таких как дендритные клетки (ДК), В-клетки, клетки-киллеры (NK), макрофаги и нейтрофилы (NPL 5).
Агонистические антитела против CD137 уже продемонстрировали противоопухолевый эффект на модели мышей, и было показано, что это является результатом активации CDS-положительных Т-клеток и клеток NK в экспериментах на модели мышей (NPL 6). Однако побочные эффекты, обусловленные неспецифической гепатотоксичностью агонистического антитела против CD137, стали клиническими и неклиническими проблемами, препятствуя требуемому прогрессу при разработке лекарств (NPL 7, NPL 8). Предполагают, что побочные эффекты вызваны в основном активацией иммунных клеток в неопухолевых, неиммунных тканях, таких как печень, что включает связывание антитела с рецептором Fcγ через константную область антитела (NPL 9). С другой стороны, показано, что для того, чтобы агонистические антитела, входящие в надсемейство антител против TNF рецепторов, проявляли агонистическую активность in vivo, антитело должно быть перекрестно связано клетками, экспрессирующими рецептор Fcγ (клетками, экспрессирующими FcγRII) (NPL 10). То есть связывание агонистического антитела против CD137 с рецептором Fcγ вовлечено как в лекарственную эффективность противоопухолевого действия антитела, так и в его побочные эффекты, такие как гепатотоксичность. Таким образом, увеличение связывания между антителом и рецептором Fcγ предположительно повысит эффективность лекарственного средства, но также может усилить гепатотоксические побочные эффекты, а уменьшение связывания между антителом и рецептором Fcγ может уменьшить побочные эффекты, но также понизить эффективность лекарственного средства. До сих пор не было сообщений об агонистических антителах против CD137, эффективность и побочные эффекты которых были бы разделены. Более того, противоопухолевый эффект агонистического антитела против CD137 никоим образом не является сильным клиническим средством, и желательно дальнейшее повышение эффективности лекарственного средства наряду с предотвращением токсичности. Соответственно, желательно разработать новое лекарственное средство, которое способно вызывать противоопухолевые иммунные ответы при одновременном уменьшении побочных эффектов.
Когда терапевтическое антитело вводят в живой организм, желательно, чтобы его антиген-мишень экспрессировался специфически только в месте поражения. Однако во многих случаях тот же антиген экспрессируется также в непораженных участках, то есть в нормальных тканях, и это может быть причиной побочных эффектов, нежелательных с точки зрения лечения. Например, хотя антитела против опухолевых антигенов могут проявлять цитотоксическую активность в отношении опухолевых клеток посредством антителозависимой клеточной цитотоксичности (ADCC) и т.д., они также могут повреждать нормальные клетки, если тот же антиген экспрессируется в нормальных клетках. Чтобы решить вышеупомянутые проблемы, основное внимание было уделено явлению, при котором определенные соединения в изобилии присутствуют в тканях-мишенях (например, в опухолевых тканях), и разработана технология поиска антигенсвязывающих молекул с различной антигенсвязывающей активностью в зависимости от концентрации таких соединений (например, PTL 1).
Цитируемые источники
Патентная литература
[PTL 1] WO 2013/180200
Непатентная литература
[NPL 1] Hanahan, Cell, 144, 2011, 646-674
[NPL 2] Prieto, Clin Cancer Res. 18, 2012, 2039-2047
[NPL 3] Hamid, Expert Opin. Biol. Ther., 6, 2013, 847-861
[NPL 4] Summers, Nat Rev Immunol, 12, 2012, 339-351
[NPL 5] Vinay, Cellular & Molecular Immunology, 8, 2011, 281-284
[NPL 6] Houot, Blood, 114, 2009, 3431-3438
[NPL 7] Ascierto, Semin Oncol, 37, 2010, 508-516
[NPL 8] Dubrot, Cancer Immunol Immunother, 59, 2010, 1223-1233
[NPL 9] Schabowsky, Vaccine, 28, 2009, 512-522
[NPL 10] Li, Proc Natl Acad Sci USA, 110(48), 2013, 19501-19506
Краткое описание изобретения
Техническая проблема
Настоящее изобретение относится к анти-CD137 антигенсвязывающим молекулам и способам их применения.
Решение проблемы
Чтобы получить анти-CD137 антигенсвязывающие молекулы, которые активируют иммуноциты, обладают цитотоксической или противоопухолевой активностью и при этом проявляют пониженное действие на неопухолевые ткани, такие как нормальные ткани, и имеют меньше побочных эффектов, а также, чтобы разработать способы их применения, в настоящем изобретении получают анти-CD137 антигенсвязывающие молекулы, характеризующиеся тем, что их активность связывания с CD137 варьирует в зависимости от различных соединений (например, низкомолекулярных соединений) в тканях-мишенях (например, в опухолевых тканях), а также разрабатывают способы их применения, фармацевтические составы и тому подобное. В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающие молекулы по настоящему изобретению обладают низкими побочными эффектами, и, таким образом, дозировка может быть увеличена без опасений по поводу побочных эффектов, в результате они могут проявлять более высокую лекарственную эффективность (цитотоксическую активность или противоопухолевую активность).
Конкретно в настоящем изобретении предусматривают анти-CD137 антигенсвязывающие молекулы, способы их применения, фармацевтические составы и тому подобное, что описано в ниже приведенных примерах.
[1]
Анти-CD137 антигенсвязывающая молекула, которая обладает CD137-связывающей активностью в зависимости от низкомолекулярного соединения.
[2]
Анти-CD137 антигенсвязывающая молекула по п. [1], причем связывающая активность с CD137 в присутствии 10 мкМ, 50 мкМ, 100 мкМ, 150 мкМ, 200 мкМ или 250 мкМ низкомолекулярного соединения в два или более раз выше по сравнению со связывающей активностью с CD137 при отсутствии низкомолекулярного соединения.
[2.1]
Анти-CD137 антигенсвязывающая молекула по п.п. [1] или [2], причем связывающая активность с CD137 в присутствии 10 мкМ или более низкомолекулярного соединения в два раза или более раз более чем связывающая активность с CD137 при отсутствии низкомолекулярного соединения.
[2.2]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [1] - [2.1], причем величина KD в отношении CD137 в присутствии 10 мкМ или более низкомолекулярного соединения составляет 5×10-7 М или менее.
[2.3]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [1] - [2.2], причем величина KD в отношении CD137 в отсутствие низкомолекулярного соединения составляет 5×10-6 М или более.
[2.4]
Анти-CD137 антигенсвязывающая молекула по п. [1], причем величина KD в отношении CD137 в растворе, который получают таким образом, что концентрация низкомолекулярного соединения составляет 10 мкМ или более, составляет 5×10-7 М или менее, и величина KD для CD137 в растворе, в который не добавляют низкомолекулярное соединение, составляет 1×10-6 М или более.
[2.5]
Анти-CD137 антигенсвязывающая молекула по п. [1], причем величину KD в отношении CD137 в растворе, который получают таким образом, что концентрация низкомолекулярного соединения составляет 10 мкМ или более, и величину KD в отношении CD137 в растворе, в который низкомолекулярное соединение не вносят, каждую измеряют методом Biacore в течение 24 ч после контакта CD137 и антигенсвязывающей молекулы анти-CD137 в растворе.
[2.6]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [1] - [2.5], которая образует тримолекулярный комплекс с низкомолекулярным соединением и CD137.
[2.7]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [1] - [2.6], которая связывается с молекулой CD137, происходящей от человека или обезьяны.
[2.8]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [1] - [2.7], в которых низкомолекулярным соединением является аденозин-содержащее. [2.9]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [1] - [2.8], в которых низкомолекулярным соединением является АТФ. [3]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [1] - [2.9], которая включает любую комбинацию гипервариабельных участков HVR-H1, HVR-H2 и HVR-H3, выбранных из представленных ниже п.п. (а) - (л):
(а) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 8, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 17;
(б) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 9, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 17;
(в) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 10, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 17;
(г) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 11, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 18;
(д) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 8, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 18;
(е) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 12, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 18;
(ж) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 13, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 18;
(з) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 14, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 19;
(и) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 15, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 20;
(к) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 16, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 20; и
(л) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 14, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 17.
[3.1]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [1] - [3], которая включает любую комбинацию гипервариабельных участков HVR-L1, HVR-L2 и HVR-L3, выбранную из приведенных ниже п.п. (а) - (ж):
(а) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 21, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 27;
(б) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 22, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 27;
(в) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 21, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 28;
(г) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 21, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 29;
(д) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 23, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 27;
(е) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 24, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 27; и
(ж) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 25, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 27.
[4]
Анти-CD137 антигенсвязывающая молекула, которая включает любую комбинацию гипервариабельных участков HVR-H1, HVR-H2, HVR-H3, HVR-L1, HVR-L2 и HVR-L3, выбранных из представленных ниже п.п. (а) - (н):
(а) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 8, HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 17, HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 21, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 27;
(б) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 9, HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 17, HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 22, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 27;
(в) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 10, HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 17, HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 22, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 27;
(г) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 11, HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 18, HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 21, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 27;
(д) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 8, HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 18, HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 21, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 27;
(е) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 12, HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 18, HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 21, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 28;
(ж) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 13, HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 18, HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 21, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 29;
(з) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 14, HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 19, HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 23, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 27;
(и) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 15, HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 20, HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 24, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 27;
(к) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 15, HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 20, HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 25, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 27;
(л) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 16, HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 20, HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 25, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 27;
(м) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 14, HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 19, HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 24, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 27; и
(н) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 14, HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 17, HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 21, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 27.
[5]
Анти-CD137 антигенсвязывающая молекула, включающая:
(а) вариабельную область тяжелой цепи (VH), последовательность которой по меньшей мере на 95% идентична любой аминокислотной последовательности SEQ ID NO: 43 - 53; или
(б) вариабельную область легкой цепи (VL), последовательность которой по меньшей мере на 95% идентична любой аминокислотной последовательности SEQ ID NO: 54 - 60.
[5.1]
Анти-CD137 антигенсвязывающая молекула, включающая любую комбинацию вариабельных областей VH и VL, выбранных из представленных ниже п.п. (а) - (н):
(а) VH, имеющей по меньшей мере 95% идентичности с аминокислотной последовательностью SEQ ID NO: 43, и VL, имеющей по меньшей мере 95% идентичности с аминокислотной последовательностью SEQ ID NO: 54;
(б) VH, имеющей по меньшей мере 95% идентичности с аминокислотной последовательностью SEQ ID NO: 44, и VL, имеющей по меньшей мере 95% идентичности с аминокислотной последовательностью SEQ ID NO: 55;
(в) VH, имеющей по меньшей мере 95% идентичности с аминокислотной последовательностью SEQ ID NO: 45, и VL, имеющей по меньшей мере 95% идентичности с аминокислотной последовательностью SEQ ID NO: 55;
(г) VH, имеющей по меньшей мере 95% идентичности с аминокислотной последовательностью SEQ ID NO: 46, и VL, имеющей по меньшей мере 95% идентичности с аминокислотной последовательностью SEQ ID NO: 54;
(д) VH, имеющей по меньшей мере 95% идентичности с аминокислотной последовательностью SEQ ID NO: 47, и VL, имеющей по меньшей мере 95% идентичности с аминокислотной последовательностью SEQ ID NO: 54;
(е) VH, имеющей по меньшей мере 95% идентичности с аминокислотной последовательностью SEQ ID NO: 48, и VL, имеющей по меньшей мере 95% идентичности с аминокислотной последовательностью SEQ ID NO: 56;
(ж) VH, имеющей по меньшей мере 95% идентичности с аминокислотной последовательностью SEQ ID NO: 449, и VL, имеющей по меньшей мере 95% идентичности с аминокислотной последовательностью SEQ ID NO: 57;
(з) VH, имеющей по меньшей мере 95% идентичности с аминокислотной последовательностью SEQ ID NO: 50, и VL, имеющей по меньшей мере 95% идентичности с аминокислотной последовательностью SEQ ID NO: 58;
(и) VH, имеющей по меньшей мере 95% идентичности с аминокислотной последовательностью SEQ ID NO: 51, и VL, имеющей по меньшей мере 95% идентичности с аминокислотной последовательностью SEQ ID NO: 59;
(к) VH, имеющей по меньшей мере 95% идентичности с аминокислотной последовательностью SEQ ID NO: 51, и VL, имеющей по меньшей мере 95% идентичности с аминокислотной последовательностью SEQ ID NO: 60;
(л) VH, имеющей по меньшей мере 95% идентичности с аминокислотной последовательностью SEQ ID NO: 52, и VL, имеющей по меньшей мере 95% идентичности с аминокислотной последовательностью SEQ ID NO: 60;
(м) VH, имеющей по меньшей мере 95% идентичности с аминокислотной последовательностью SEQ ID NO: 50, и VL, имеющей по меньшей мере 95% идентичности с аминокислотной последовательностью SEQ ID NO: 59; и
(н) VH, имеющей по меньшей мере 95% идентичности с аминокислотной последовательностью SEQ ID NO: 53, и VL, имеющей по меньшей мере 95% идентичности с аминокислотной последовательностью SEQ ID NO: 54.
[5.2]
Анти-CD137 антигенсвязывающая молекула, включающая любую комбинацию вариабельных областей VH и VL, выбранных из представленных ниже п.п. (а) - (н):
(а) VH, содержащая аминокислотную последовательность SEQ ID NO: 43, и VL, содержащий аминокислотную последовательность SEQ ID NO: 54;
(б) VH, содержащая аминокислотную последовательность SEQ ID NO: 44, и VL, содержащий аминокислотную последовательность SEQ ID NO: 55;
(в) VH, содержащая аминокислотную последовательность SEQ ID NO: 45, и VL, содержащий аминокислотную последовательность SEQ ID NO: 55;
(г) VH, содержащая аминокислотную последовательность SEQ ID NO: 46, и VL, содержащая аминокислотную последовательность SEQ ID NO: 54;
(д) VH, содержащая аминокислотную последовательность SEQ ID NO: 47, и VL, содержащая аминокислотную последовательность SEQ ID NO: 54;
(е) VH, содержащая аминокислотную последовательность SEQ ID NO: 48, и VL, содержащая аминокислотную последовательность SEQ ID NO: 56;
(ж) VH, содержащая аминокислотную последовательность SEQ ID NO: 49, и VL, содержащая аминокислотную последовательность SEQ ID NO: 57;
(з) VH, содержащая аминокислотную последовательность SEQ ID NO: 50, и VL, содержащая аминокислотную последовательность SEQ ID NO: 58;
(и) VH, содержащая аминокислотную последовательность SEQ ID NO: 51, и VL, содержащая аминокислотную последовательность SEQ ID NO: 59;
(к) VH, содержащая аминокислотную последовательность SEQ ID NO: 51, и VL, содержащая аминокислотную последовательность SEQ ID NO: 60;
(л) VH, содержащая аминокислотную последовательность SEQ ID NO: 52, и VL, содержащая аминокислотную последовательность SEQ ID NO: 60;
(м) VH, содержащая аминокислотную последовательность SEQ ID NO: 50, и VL, содержащая аминокислотную последовательность SEQ ID NO: 59; и
(н) VH, содержащая аминокислотную последовательность SEQ ID NO: 53, и VL, содержащая аминокислотную последовательность SEQ ID NO: 54. [5.3]
Анти-CD137 антигенсвязывающая молекула, значение которой [связывающая активность (связывающееся количество) с CD137 в присутствии 10 мкМ или более низкомолекулярного соединения]/[связывающая активность (связывающееся количество) с CD137 в отсутствие низкомолекулярного соединения] равно или больше, чем значение контрольной антигенсвязывающей молекулы, где контрольная антигенсвязывающая молекула представляет собой анти-CD137 антигенсвязывающую молекулу, содержащую комбинацию гипервариабельного участка HVR-H1, содержащего аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащего аминокислотную последовательность SEQ ID NO: 8, HVR-H3, содержащего аминокислотную последовательность SEQ ID NO: 17, HVR-L1, содержащего аминокислотную последовательность SEQ ID NO: 21, HVR- L2, содержащего аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащего аминокислотную последовательность SEQ ID NO: 27.
[5.4]
Анти-CD137 антигенсвязывающая молекула по п. [5.3], причем контрольная антигенсвязывающая молекула представляет собой анти-CD137 антигенсвязывающую молекулу, содержащую комбинацию VH, содержащую аминокислотную последовательность SEQ ID NO: 43, и VL, содержащую аминокислотную последовательность SEQ ID NO: 54.
[5.5]
Анти-CD137 антигенсвязывающая молекула, значение которой [связывающая активность (KD) с CD137 в присутствии 1 мкМ низкомолекулярного соединения]/[связывающая активность (KD) с CD137 в присутствии 10 мкМ или более низкомолекулярное соединения] равно или больше значения контрольной антигенсвязывающей молекулы, где контрольная антигенсвязывающая молекула представляет собой анти-CD137 антигенсвязывающую молекулу, содержащую комбинацию HVR-H1, содержащего аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащего аминокислотную последовательность SEQ ID NO: 8, HVR-H3, содержащего аминокислотную последовательность SEQ ID NO: 17, HVR-L1, содержащего аминокислотную последовательность SEQ ID NO: 21, HVR-L2, содержащего аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащего аминокислотная последовательность SEQ ID NO: 27. [5.6]
Анти-CD137 антигенсвязывающая молекула по п. [5.5], причем контрольная антигенсвязывающая молекула представляет собой анти-CD137 антигенсвязывающую молекулу, содержащую комбинацию VH, содержащую аминокислотную последовательность SEQ ID NO: 43, и VL, содержащую аминокислотную последовательность SEQ ID NO: 54.
[5.7]
Анти-CD137 антигенсвязывающая молекула, которая обладает CD137-антигенсвязывающей активностью, зависящей от низкомолекулярного соединения, причем анти-CD137 антигенсвязывающая молекула конкурирует с антигенсвязывающей молекулой по любому из п.п. [3] - [5.2] за связывание с CD137 в присутствии 10 мкМ или более, 50 мкМ или более, 100 мкМ или более, 150 мкМ или более, 200 мкМ или более или 250 мкМ или более низкомолекулярного соединения.
[5.8]
Анти-CD137 антигенсвязывающая молекула, которая обладает CD137-антигенсвязывающей активностью, зависящей от низкомолекулярного соединения, причем анти-CD137 антигенсвязывающая молекула связывается с тем же эпитопом CD137, связанным с антигенсвязывающей молекулой по любому из п.п. [3] - [5.2] в присутствии 10 мкМ или более, 50 мкМ или более, 100 мкМ или более, 150 мкМ или более, 200 мкМ или более или 250 мкМ или более низкомолекулярного соединения.
[5.8А]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [5.3] - [5.8], где низкомолекулярное соединение представляет собой аденозинсодержащее соединение.
[5.8Б]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [5.3] - [5.8А], где низкомолекулярное соединение представляет собой АТФ. [5.9]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [1] - [5.8Б], которая является моноклональным антителом или его антигенсвязывающим фрагментом.
[5.10]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [1] - [5.9], которая является антителом человека, гуманизированным антителом или химерным антителом, или их антигенсвязывающий фрагментом.
[5.11]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [1] - [5.10], которая является антителом IgG1 полной длины. [5.12]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [1] - [5.11], которая содержит измененную область Fc, в которой изменена по меньшей мере одна аминокислота, причем измененная область Fc имеет повышенную связывающую активность с FcγRIIb по сравнению с исходной областью Fc, которая не содержит изменений аминокислот.
[5.13]
Анти-CD137 антигенсвязывающая молекула по п. [5.12], в которой связывающая активность измененной Fc с FcγRIIb равна или выше, чем у контрольной области Fc, причем контрольная область Fc является областью Fc IgG1 человека, содержащей комбинацию аминокислотных замен G236N/H268D/A330K согласно нумерации EU.
[5.14]
Анти-CD137 антигенсвязывающая молекула по п. [5.12] или [5.13], причем контрольная область Fc содержит аминокислотную последовательность SEQ ID NO: 153.
[5.15]
Анти-CD137 антигенсвязывающая молекула по п. [5.12], в которой по меньшей мере одно аминокислотное изменение представляет собой по меньшей мере одну аминокислотную замену, выбранную из группы, состоящей из G236N, H268D и АЗЗОК в соответствии с нумерацией EU.
[5.16]
Анти-CD137 антигенсвязывающая молекула по п.п. [5.12] или [5.15], в которой по меньшей мере одно аминокислотное изменение представляет собой комбинацию аминокислотных замен G236N/H268D/A330K в соответствии с нумерацией EU. [5.17]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [5.12] - [5.16], в которой исходная область Fc является производной от области Fc IgG1 человека.
[5.18]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [1] - [5.17], которая содержит измененную область Fc, в которой изменена по меньшей мере одна аминокислота, причем анти-CD137 антигенсвязывающая молекула имеет повышенную изоэлектрическую точку (pi) по сравнению с изоэлектрической точкой исходной анти-CD137 антигенсвязывающей молекулы, содержащей исходную область Fc, которая не содержит изменения аминокислот.
[5.19]
Анти-CD137 антигенсвязывающая молекула по п. [5.18], в которой по меньшей мере одно аминокислотное изменение представляет собой изменение аминокислотного остатка, который может быть экспонирован на поверхности исходной области Fc.
[5.20]
Анти-CD137 антигенсвязывающая молекула по п.п. [5.18] или [5.19], в которой по меньшей мере одно аминокислотное изменение представляет собой:
(i) замену по меньшей мере одного аминокислотного остатка, имеющего отрицательный заряд в боковой цепи в исходной области Fc, на аминокислотный остаток, не имеющий заряда в боковой цепи,
(ii) замену по меньшей мере одного аминокислотного остатка, не имеющего заряда в боковой цепи в исходной области Fc, на аминокислотный остаток, имеющий положительный заряд в боковой цепи, и/или
(iii) замену по меньшей мере одного аминокислотного остатка, имеющего отрицательный заряд в боковой цепи в исходной области Fc, на аминокислотный остаток, имеющий положительный заряд в боковой цепи,
[5.21]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [5.18] - [5.20], в которой по меньшей мере одно аминокислотное изменение представляет собой комбинацию аминокислотных замен, и в которой аминокислотные замены расположены в положениях, конформационно близко расположенных друг к другу.
[5.22]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [5.18] - [5.21], в которой связывающая активность измененной области Fc с рецептором Fcγ (FcγR) существенно не снижена по сравнению со связывающей активностью у исходной области Fc.
[5.23]
Анти-CD137 антигенсвязывающая молекула по п. [5.22], в которой рецептором Fcγ (FcγR) является FcγRIIb. [5.24]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [5.18] - [5.23], в которой по меньшей мере одно аминокислотное изменение представляет собой по меньшей мере одну аминокислотную замену, выбранную из группы, состоящей из Q311R, P343R и D413K в соответствии с нумерацией EU.
[5.25]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [5.18] - [5.24], в которой по меньшей мере одно аминокислотное изменение представляет собой (i) аминокислотную замену P343R, (ii) комбинацию аминокислотных замен Q311R/P343R или (iii) комбинацию аминокислотных замен Q311R/D413K, согласно нумерации EU.
[6]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [1] - [5.25], которая содержит измененную область Fc, причем измененная область Fc содержит любую комбинацию аминокислотных изменений, выбранных из следующих:
L235W/G236N/H268D/Q295L/K326T/A330K/P343R/D413K;
K214R/L235W/G236N/H268D/Q295L/K326T/A330K/P343R/D413K;
L234Y/P238D/T250V/V264I/T307P/A330K/P343R/D413K;
L234Y/P238D/V264I/A330K/P343R/D413K;
L234Y/G237D/P238D/T250V/T307P/A330K/P343R/D413K;
L234Y/G237D/P238D/A330K/P343R/D413K;
L235W/G236N/H268D/Q295L/K326T/A330K/Q311R/P343R;
L234Y/P238D/T250V/V264I/T307P/A330K/Q311R/P343R;
L234Y/P238D/V264I/A330K/Q311R/P343R;
L234Y/G237D/P238D/T250V/T307P/A330K/Q311R/P343R;
L234Y/G237D/P238D/A330K/Q311R/P343R;
L235W/G236N/H268D/Q295L/K326T/A330K/P343R;
K214R/L235W/G236N/H268D/Q295L/K326T/A330K/P343R;
L235W/G236N/H268D/Q295L/K326T/A330K/D413K;
K214R/G236N/H268D/A330K/P343R;
K214R/L235W/G236N/H268D/A330K/P343R;
K214R/G236N/H268D/A330K/D413K;
K214R/G236N/H268D/A330K/P343R/D413K;
K214R/L235W/G236N/H268D/A330K/P343R/D413K;
K214R/G236N/H268D/A330K/Q311R;
K214R/L235W/G236N/H268D/A330K/Q311R;
K214R/G236N/H268D/A330K/Q311R/P343R;
K214R/L235W/G236N/H268D/A330K/Q311R/P343R;
K214R/G236N/H268D/A330K/Q311R/D413K;
K214R/L235W/G236N/H268D/A330K/Q311R/D413K; и
K214R/L235W/G236N/H268D/Q295L/K326T/A330K/Q311R,
Нумерация по EU.
[6.1]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [1] - [6], в которой измененная область Fc является производной от области Fc IgG1 человека.
[6.2]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [1] - [6.1], в которой измененная область Fc дополнительно включает делеции в положениях 446 и 447 в соответствии с нумерацией EU.
[7]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [1] - [6.2], которая включает константную область тяжелой цепи, содержащую любую из аминокислотных последовательностей SEQ ID NO: 64-85.
[7.1]
Анти-CD137 антигенсвязывающая молекула, содержащая любую комбинацию областей VH, VL, СН и CL, выбранных из приведенных ниже п.п. (i) - (xxxviii):
(i) VH, содержащей аминокислотную последовательность SEQ ID NO: 43, СН, содержащей аминокислотную последовательность SEQ ID NO: 64, VL, содержащей аминокислотную последовательность SEQ ID NO: 54, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(ii) VH, содержащей аминокислотную последовательность SEQ ID NO: 43, СН, содержащей аминокислотную последовательность SEQ ID NO: 66, VL, содержащей аминокислотную последовательность SEQ ID NO: 54, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(iii) VH, содержащей аминокислотную последовательность SEQ ID NO: 43, СН, содержащей аминокислотную последовательность SEQ ID NO: 67, VL, содержащей аминокислотную последовательность SEQ ID NO: 54, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(iv) VH, содержащей аминокислотную последовательность SEQ ID NO: 43, СН, содержащей аминокислотную последовательность SEQ ID NO: 68, VL, содержащей аминокислотную последовательность SEQ ID NO: 54, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(v) VH, содержащей аминокислотную последовательность SEQ ID NO: 43, СН, содержащей аминокислотную последовательность SEQ ID NO: 69, VL, содержащей аминокислотную последовательность SEQ ID NO: 54, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(vi) VH, содержащей аминокислотную последовательность SEQ ID NO: 43, СН, содержащей аминокислотную последовательность SEQ ID NO: 70, VL, содержащей аминокислотную последовательность SEQ ID NO: 54, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(vii) VH, содержащей аминокислотную последовательность SEQ ID NO: 43, СН, содержащей аминокислотную последовательность SEQ ID NO: 71, VL, содержащей аминокислотную последовательность SEQ ID NO: 54, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(viii) VH, содержащей аминокислотную последовательность SEQ ID NO: 43, СН, содержащей аминокислотную последовательность SEQ ID NO: 73, VL, содержащей аминокислотную последовательность SEQ ID NO: 54, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(ix) VH, содержащей аминокислотную последовательность SEQ ID NO: 43, CH, содержащей аминокислотную последовательность SEQ ID NO: 75, VL, содержащей аминокислотную последовательность SEQ ID NO: 54, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(x) VH, содержащей аминокислотную последовательность SEQ ID NO: 43, СН, содержащей аминокислотную последовательность SEQ ID NO: 78, VL, содержащей аминокислотную последовательность SEQ ID NO: 54, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(xi) VH, содержащей аминокислотную последовательность SEQ ID NO: 43, CH, содержащей аминокислотную последовательность SEQ ID NO: 80, VL, содержащей аминокислотную последовательность SEQ ID NO: 54, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(xii) VH, содержащей аминокислотную последовательность SEQ ID NO: 43, CH, содержащей аминокислотную последовательность SEQ ID NO: 82, VL, содержащей аминокислотную последовательность SEQ ID NO: 54, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(xiii) VH, содержащей аминокислотную последовательность SEQ ID NO: 43, CH, содержащей аминокислотную последовательность SEQ ID NO: 84, VL, содержащей аминокислотную последовательность SEQ ID NO: 54, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(xiv) VH, содержащей аминокислотную последовательность SEQ ID NO: 43, CH, содержащей аминокислотную последовательность SEQ ID NO: 85, VL, содержащей аминокислотную последовательность SEQ ID NO: 54, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(xv) VH, содержащей аминокислотную последовательность SEQ ID NO: 51, CH, содержащей аминокислотную последовательность SEQ ID NO: 65, VL, содержащей аминокислотную последовательность SEQ ID NO: 59, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(xvi) VH, содержащей аминокислотную последовательность SEQ ID NO: 51, CH, содержащей аминокислотную последовательность SEQ ID NO: 72, VL, содержащей аминокислотную последовательность SEQ ID NO: 59, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(xvii) VH, содержащей аминокислотную последовательность SEQ ID NO: 51, CH, содержащей аминокислотную последовательность SEQ ID NO: 74, VL, содержащей аминокислотную последовательность SEQ ID NO: 59, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(xviii) VH, содержащей аминокислотную последовательность SEQ ID NO: 51, CH, содержащей аминокислотную последовательность SEQ ID NO: 75, VL, содержащей аминокислотную последовательность SEQ ID NO: 59, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(xix) VH, содержащей аминокислотную последовательность SEQ ID NO: 51, CH, содержащей аминокислотную последовательность SEQ ID NO: 77, VL, содержащей аминокислотную последовательность SEQ ID NO: 59, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(xx) VH, содержащей аминокислотную последовательность SEQ ID NO: 51, CH, содержащей аминокислотную последовательность SEQ ID NO: 78, VL, содержащей аминокислотную последовательность SEQ ID NO: 59, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(xxi) VH, содержащей аминокислотную последовательность SEQ ID NO: 51, CH, содержащей аминокислотную последовательность SEQ ID NO: 79, VL, содержащей аминокислотную последовательность SEQ ID NO: 59, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(xxii) VH, содержащей аминокислотную последовательность SEQ ID NO: 51, CH, содержащей аминокислотную последовательность SEQ ID NO: 80, VL, содержащей аминокислотную последовательность SEQ ID NO: 59, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(xxiii) VH, содержащей аминокислотную последовательность SEQ ID NO: 51, CH, содержащей аминокислотную последовательность SEQ ID NO: 81, VL, содержащей аминокислотную последовательность SEQ ID NO: 59, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(xxiv) VH, содержащей аминокислотную последовательность SEQ ID NO: 51, CH, содержащей аминокислотную последовательность SEQ ID NO: 82, VL, содержащей аминокислотную последовательность SEQ ID NO: 59, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(xxv) VH, содержащей аминокислотную последовательность SEQ ID NO: 51, CH, содержащей аминокислотную последовательность SEQ ID NO: 83, VL, содержащей аминокислотную последовательность SEQ ID NO: 59, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(xxvi) VH, содержащей аминокислотную последовательность SEQ ID NO: 51, СН, содержащей аминокислотную последовательность SEQ ID NO: 84, VL, содержащей аминокислотную последовательность SEQ ID NO: 59, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(xxvii) VH, содержащей аминокислотную последовательность SEQ ID NO: 51, СН, содержащей аминокислотную последовательность SEQ ID NO: 72, VL, содержащей аминокислотную последовательность SEQ ID NO: 60, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(xxviii) VH, содержащей аминокислотную последовательность SEQ ID NO: 51, СН, содержащей аминокислотную последовательность SEQ ID NO: 74, VL, содержащей аминокислотную последовательность SEQ ID NO: 60, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(xxix) VH, содержащей аминокислотную последовательность SEQ ID NO: 51, СН, содержащей аминокислотную последовательность SEQ ID NO: 75, VL, содержащей аминокислотную последовательность SEQ ID NO: 60, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(xxx) VH, содержащей аминокислотную последовательность SEQ ID NO: 51, СН, содержащей аминокислотную последовательность SEQ ID NO: 77, VL, содержащей аминокислотную последовательность SEQ ID NO: 60, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(xxxi) VH, содержащей аминокислотную последовательность SEQ ID NO: 51, СН, содержащей аминокислотную последовательность SEQ ID NO: 78, VL, содержащей аминокислотную последовательность SEQ ID NO: 60, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(xxxii) VH, содержащей аминокислотную последовательность SEQ ID NO: 51, СН, содержащей аминокислотную последовательность SEQ ID NO: 79, VL, содержащей аминокислотную последовательность SEQ ID NO: 60, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(xxxiii) VH, содержащей аминокислотную последовательность SEQ ID NO: 51, СН, содержащей аминокислотную последовательность SEQ ID NO: 80, VL, содержащей аминокислотную последовательность SEQ ID NO: 60, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(xxxiv) VH, содержащей аминокислотную последовательность SEQ ID NO: 51, CH, содержащей аминокислотную последовательность SEQ ID NO: 81, VL, содержащей аминокислотную последовательность SEQ ID NO: 60, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(xxxv) VH, содержащей аминокислотную последовательность SEQ ID NO: 51, CH, содержащей аминокислотную последовательность SEQ ID NO: 82, VL, содержащей аминокислотную последовательность SEQ ID NO: 60, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(xxxvi) VH, содержащей аминокислотную последовательность SEQ ID NO: 51, CH, содержащей аминокислотную последовательность SEQ ID NO: 83, VL, содержащей аминокислотную последовательность SEQ ID NO: 60, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63;
(xxxvii) VH, содержащей аминокислотную последовательность SEQ ID NO: 51, CH, содержащей аминокислотную последовательность SEQ ID NO: 84, VL, содержащей аминокислотную последовательность SEQ ID NO: 60, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63; и
(xxxviii) VH, содержащей аминокислотную последовательность SEQ ID NO: 51, CH, содержащей аминокислотную последовательность SEQ ID NO: 85, VL, содержащей аминокислотную последовательность SEQ ID NO: 60, и CL, содержащей аминокислотную последовательность SEQ ID NO: 63.
[8]
Выделенная нуклеиновая кислота, кодирующая анти-CD137 антигенсвязывающую молекулу по любому из п.п. [1] - [7.1].
[9]
Вектор, содержащий нуклеиновую кислоту по п. [8].
[10]
Клетка-хозяин, содержащая нуклеиновую кислоту по п. [8] или вектор по п. [9].
[11]
Метод получения анти-CD137 антигенсвязывающей молекулы, который включает культивирование клетки-хозяина по п. [10] таким образом, чтобы получить анти-CD137 антигенсвязывающую молекулу.
[12]
Иммуноконъюгат, включающий анти-CD137 антигенсвязывающую молекулу по любому из п.п. [1] - [7.1] и цитотоксический агент.[13]
Фармацевтический состав, включающий анти-CD137 антигенсвязывающую молекулу по любому из п.п. [1] - [7.1] или иммуноконъюгат по п. [12]; и фармацевтически приемлемый носитель.
[14]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [1] - [7.1] или иммуноконъюгат по п. [12], которые предназначены для применения в качестве фармацевтического средства.
[14.1]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [1] - [7.1], иммуноконъюгат по п. [12] или фармацевтический состав по п. [13], которые предназначены для применения в лечении рака.
[14.2]
Анти-CD137 антигенсвязывающая молекула, иммуноконъюгат или фармацевтический состав по п. [14.1], в котором опухоль является солидной опухолью, инфильтрованной В-клетками, дендритными клетками, природными клетками-киллерами, макрофагами и/или CDS-положительными Т-клетками.
[14.3]
Анти-CD137 антигенсвязывающая молекула, иммуноконъюгат или фармацевтический состав по п. [14.1], в котором опухоль является солидной опухолью, инфильтрованной регуляторными Т-клетками (Treg).
[15]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [1] - [7.1], иммуноконъюгат по п. [12] или фармацевтический состав по п. [13], которые предназначены для применения в активации иммунных клеток.
[15.1]
Анти-CD137 антигенсвязывающая молекула, иммуноконъюгат или фармацевтический состав по п. [15], в котором иммунные клетки являются В-клетками, дендритными клетками, природными клетками-киллерами, макрофагами и/или Т-клетками.
[15.2]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [1] - [7.1] или фармацевтический состав по п. [13], которые предназначены для активации иммунных клеток в опухолевой ткани.
[15.3]
Анти-CD137 антигенсвязывающая молекула или фармацевтический состав по п. [15.2], в которых иммунными клетками являются В-клетки, дендритные клетки, природные клетки-киллеры, макрофаги и/или Т-клетки.
[15.4]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [1] - [7.1], иммуноконъюгат по п. [12] или фармацевтический состав по п. [13], которые предназначены для применения в разрушении клеток.
[16]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [1] - [7.1], иммуноконъюгат по п. [12] или фармацевтический состав по п. [13], чей уровень активации иммунитета в неопухолевой ткани ниже, чем уровень активации иммунитета у анти-CD137 антигенсвязывающей молекулы, которая не обладает СВ137-связывающей активностью, зависящей от низкомолекулярного соединения.
[16.1]
Анти-CD137 антигенсвязывающая молекула, иммуноконъюгат или фармацевтический состав по п. [16], в котором неопухолевой тканью являются лимфоузлы, селезенка и/или печень.
[16.2]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [1] - [7.1] или иммуноконъюгат по п. [12], которые существенным образом не связываются с CD137, экспрессируемым в неопухолевой ткани.
[16.3]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [1] - [7.1] или иммуноконъюгат по п. [12], которые имеют более длительный период полужизни в крови по сравнению с анти-CD137 антигенсвязывающей молекулой, которая не обладает СВ137-связывающей активностью, зависящей от низкомолекулярного соединения.
[17]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [1] - [7.1], иммуноконъюгат по п. [12] или фармацевтический состав по п. [13], которые имеют пониженный уровень побочных эффектов по сравнению с анти-CD137 антигенсвязывающей молекулой, которая не обладает CD137-связывающей активностью, зависящей от низкомолекулярного соединения.
[17.1]
Анти-CD137 антигенсвязывающая молекула, иммуноконъюгат или фармацевтический состав по п. [17], в котором побочные эффекты представлены повышенным AST, повышенным ALT, лихорадкой, тошнотой, острым гепатитом, гепатопатией, спленомегалией, энтеритом, гнойным воспалением кожи, уменьшением нейтрофилов, уменьшением лимфоцитов, уменьшением тромбоцитов, экспрессией трансаминазы и/или гипербилирубинемией.
[18]
Анти-CD137 антигенсвязывающая молекула, которая обладает агонистической активностью, зависящей от низкомолекулярного соединения. [18.1]
Анти-CD137 антигенсвязывающая молекула по п. [18], в котором агонистическая активность против CD137 в присутствии 10 мкМ, 50 мкМ, 100 мкМ, 150 мкМ, 200 мкМ или 250 мкМ низкомолекулярного соединения в два или более раз больше по сравнению с агонистической активностью против CD137 при отсутствии низкомолекулярного соединения.
[18.2]
Анти-CD137 антигенсвязывающая молекула по п.п. [18] или [18.1], в которых агонистическая активность против CD137 в присутствии 10 мкМ или более низкомолекулярного соединения в два или более раз больше по сравнению с агонистической активностью против CD137 при отсутствии низкомолекулярного соединения.
[18.3]
Анти-CD137 антигенсвязывающая молекула по п.п. [18] или [18.1], в которых агонистическая активность против CD137 в присутствии 50 мкМ или более низкомолекулярного соединения в два или более раз больше по сравнению с агонистической активностью против CD137 при отсутствии низкомолекулярного соединения.
[18.4]
Анти-CD137 антигенсвязывающая молекула по п.п. [18] или [18.1], в которых агонистическая активность против CD137 в присутствии 250 мкМ или более низкомолекулярного соединения в два или более раз больше по сравнению с агонистической активностью против CD137 при отсутствии низкомолекулярного соединения.
[18.5]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [18] - [18.4], в которых агонистическую активность против CD137 оценивают по количеству IL-2 и/или IFN-y, вырабатываемых CD137-экспрессирующими клетками.
[18.6]
Анти-CD137 антигенсвязывающая молекула по п. [18.5], в котором CD137-экспрессирующими клетками являются выделенные мононуклеарные клетки периферической крови (МКПК) человека или Т-клетки, производные от МКПК человека.
[18.7]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [18] - [18.4], в которых агонистическую активность против CD137 оценивают по анализу репортерного гена.
[18.8]
Анти-CD137 антигенсвязывающая молекула по п. [18], которая проявляет агонистическую активность в отношении CD137 в растворе, который приготовлен таким образом, что конечная концентрация низкомолекулярного соединения составляет 50 мкМ или более, и по существу не проявляет агонистической активности в отношении CD137 в растворе, в который не добавлено низкомолекулярное соединение.
[18.9]
Анти-CD137 антигенсвязывающая молекула по п. [18.8], в котором агонистическую активность против CD137 в растворе, который приготовлен таким образом, что конечная концентрация низкомолекулярного соединения составляет 50 мкМ или более, и агонистическую активность против CD137 в растворе, в который не добавляют низкомолекулярное соединение, каждую оценивают с количеством IL-2, IFN-γ и/или IL-6 по измерениям в течение 72 ч после контакта клетки, экспрессирующей CD137, и анти-CD137 антигенсвязывающей молекулы в растворе.
[18.10]
Анти-CD137 антигенсвязывающая молекула по п. [18.8], в котором агонистическую активность против CD137 в растворе, который приготовлен таким образом, что конечная концентрация низкомолекулярного соединения составляет 50 мкМ или более, и агонистическую активность против CD137 в растворе, в который не добавляют низкомолекулярное соединение, каждую оценивают по сигналу люминесценции люциферазы, который измеряют в течение 6 ч после контакта Т-клеток, экспрессирующих NF-kappaB-люцифераза репортерную конструкцию и CD137 с анти-CD137 антигенсвязывающей молекулой.
[18.11]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [18] - [18.10], в которых низкомолекулярным соединением является соединение, содержащее аденозин.
[18.12]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [18] - [18.11], в которых низкомолекулярным соединением является АТФ. [19]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [1] - [7.1], которая обладает агонистической активностью против CD137, зависящей от низкомолекулярного соединения.
[19.1]
Анти-CD137 антигенсвязывающая молекула по п. [19], в котором агонистическая активность против CD137 в присутствии 10 мкМ, 50 мкМ, 100 мкМ, 150 мкМ, 200 мкМ или 250 мкМ низкомолекулярного соединения в два или более раз выше, чем агонистическая активность против CD137 при отсутствии низкомолекулярного соединения.
[19.2]
Анти-CD137 антигенсвязывающая молекула по п.п. [19] или [19.1], в которых агонистическая активность против CD137 в присутствии 10 мкМ или более низкомолекулярного соединения в два или более раз выше, чем агонистическая активность против CD137 при отсутствии низкомолекулярного соединения.
[19.3]
Анти-CD137 антигенсвязывающая молекула по п. [19] или [19.1], в которых агонистическая активность против CD137 в присутствии 50 мкМ или более низкомолекулярного соединения в два или более раз выше, чем агонистическая активность против CD137 при отсутствии низкомолекулярного соединения.
[19.4]
Анти-CD137 антигенсвязывающая молекула по п. [19] или [19.1], в которых агонистическая активность против CD137 в присутствии 250 мкМ или более низкомолекулярного соединения в два или более раз выше, чем агонистическая активность против CD137 при отсутствии низкомолекулярного соединения.
[19.5]
Анти-CD137 антигенсвязывающая молекула по п. [19] или [19.1], в которых агонистическую активность против CD137 оценивают по количеству IL-2 и/или IFN-y, продуцируемых клеткой, экспрессирующей CD137.
[19.6]
Анти-CD137 антигенсвязывающая молекула по п. [19.5], в котором CD137-экспрессирующая клетка представляет собой выделенную мононуклеарную клетку периферической крови (МКПК) человека или Т-клетку, происходящую из МКПК человека.
[19.7]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [19] или [19.4], в которых агонистическую активность CD137 оценивают с помощью анализа репортерного гена.
[19.8]
Анти-CD137 антигенсвязывающая молекула по п. [19], которая проявляет агонистическую активность в отношении CD137 в растворе, который приготовлен таким образом, что конечная концентрация низкомолекулярного соединения составляет 50 мкМ или более, и по существу не проявляет агонистической активности в отношении CD137 в растворе, в который не добавлено низкомолекулярное соединение.
[19.9]
Анти-CD137 антигенсвязывающая молекула по п. [19.8], в котором агонистическую активность против CD137 в растворе, который приготовлен таким образом, что конечная концентрация низкомолекулярного соединения составляет 50 мкМ или более, и агонистическую активность против CD137 в растворе, в который не добавляют низкомолекулярное соединение, каждую оценивают по количеству IL-2, IFN-y и/или IL-6, измеренным в течение 72 ч после контакта клетки, экспрессирующей CD137, и анти-CD137 антигенсвязывающей молекулы в растворе.
[19.10]
Анти-CD137 антигенсвязывающая молекула по п. [19.8], в котором агонистическую активность против CD137 в растворе, который приготовлен таким образом, что конечная концентрация низкомолекулярного соединения составляет 50 мкМ или более, и агонистическую активность против CD137 в растворе, в который не добавляют низкомолекулярное соединение, каждую оценивают по сигналу люминесценции люциферазы, который измеряют в течение 6 ч после контакта Т-клеток, экспрессирующих NF-kappaB-люцифераза репортерную конструкцию и CD137 с анти-CD137 антигенсвязывающей молекулой.
[19.11]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [19] - [19.10], в которых низкомолекулярным соединением является соединение, содержащее аденозин.
[19.12]
Анти-CD137 антигенсвязывающая молекула по любому из п.п. [19] - [19.11], в которых низкомолекулярным соединением является АТФ.
[20]
Агонистическая антигенсвязывающая молекула, содержащая измененную область Fc, причем измененная область Fc содержит по меньшей мере одно аминокислотное изменение, которое приводит к увеличению изоэлектрической точки (pi) по сравнению с изоэлектрической точкой (pi) исходной агонистической антигенсвязывающей молекулой, содержащей исходную область Fc, и где агонистическая антигенсвязывающая молекула обладает повышенной агонистической активностью по сравнению с агонистической активностью исходной агонистической антигенсвязывающей молекулой.
[20.1]
Агонистическая антигенсвязывающая молекула по п. [20], в которой по меньшей мере одно аминокислотное изменение представляет собой изменение аминокислотного остатка, который может экспонироваться на поверхности исходной области Fc.
[20.2]
Агонистическое антигенсвязывающее антитело по п.п. [20] или [20.1], в котором изменение по меньшей мере одной аминокислоты представляет собой:
(i) замену по меньшей мере одного аминокислотного остатка, имеющего отрицательный заряд в боковой цепи в исходной области Fc, на аминокислотный остаток, не имеющий заряда в боковой цепи,
(ii) замену по меньшей мере одного аминокислотного остатка, не имеющего заряда в боковой цепи в исходной области Fc, на аминокислотный остаток, имеющий положительный заряд в боковой цепи, и/или
(iii) замену по меньшей мере одного аминокислотного остатка, имеющего отрицательный заряд в боковой цепи в исходной области Fc, на аминокислотный остаток, имеющий положительный заряд в боковой цепи.
[20.3]
Агонистическая антигенсвязывающая молекула по любому из п.п. [20] - [20.2], в которой по меньшей мере одно аминокислотное изменение представляет собой комбинацию аминокислотных замен, и где аминокислотные замены расположены в положениях, конформационно близких друг к другу.
[20.4]
Агонистическая антигенсвязывающая молекула по любому из п.п. [20] - [20.3], в которой связывающая активность измененной области Fc с рецептором Fcγ существенно не снижается по сравнению со связывающей активностью исходной области Fc.
[20.5]
Агонистическая антигенсвязывающая молекула по п. [20.4], где рецептор Fcγ представляет собой FcγRIIb.
[20.6]
Антигенсвязывающая молекула-агонист по любому из п.п. [20] - [20.4], где по меньшей мере одно аминокислотное изменение представляет собой по меньшей мере одну аминокислотную замену, выбранную из группы, состоящей из Q311R, P343R и D413K, в соответствии с нумерацией EU.
[20.7]
Агонистическая антигенсвязывающая молекула по любому из п.п. [20] - [20.6], где по меньшей мере изменение одной аминокислоты представляет собой изменение аминокислоты (i) P343R/D413K, (ii) Q311R/P343R, (iii) P343R, (iv) D413K, (v) Q311R или (vi) Q311R/D413K или их комбинацию согласно нумерации EU.
[20.8]
Агонистическая антигенсвязывающая молекула по любому из п.п. [20] - [20.7], которая является анти-CD137 антигенсвязывающей молекулой.
[20.9]
Агонистическая антигенсвязывающая молекула по любому из п.п. [20] - [20.8], которая является анти-CD137 антителом.
[21]
Способ получения агонистической антигенсвязывающей молекулы, содержащей измененную область Fc, включающий:
введение в исходную Fc по меньшей мере одного изменения аминокислоты, которое приводит к увеличению изоэлектрической точки (pi) по сравнению с таковой у исходной агонистической антигенсвязывающей молекулы, содержащей исходную область Fc,
причем агонистическая активность агонистической антигенсвязывающей молекулы, содержащей измененную область Fc, повышена по сравнению с таковой у исходной агонистической антигенсвязывающей молекулы.
[21.1]
Способ по п. [21], в котором агонистическая активность агонистической антигенсвязывающей молекулы для антигена в присутствии 10 мкМ, 50 мкМ, 100 мкМ, 150 мкМ, 200 мкМ или 250 мкМ низкомолекулярного соединения в два раза или более выше, чем агонистическая для антигена в отсутствие низкомолекулярного соединения.
[21.2]
Способ по п.п. [21] или [21.1], в котором агонистическая активность агонистической антигенсвязывающей молекулы для антигена в присутствии 10 мкМ или более низкомолекулярного соединения выше в два раза или более чем агонистическая для антигена в отсутствии низкомолекулярного соединения.
[21.3]
Способ по п.п. [21] или [21.1], в котором агонистическая активность агонистической антигенсвязывающей молекулы для антигена в присутствии 50 мкМ или более низкомолекулярного соединения выше в два раза или более чем агонистическая для антигена в отсутствие низкомолекулярного соединения.
[21.4]
Способ по п.п. [21] или [21.1], в котором агонистическая активность агонистической антигенсвязывающей молекулы для антигена в присутствии 250 мкМ или более низкомолекулярного соединения выше в два раза или более, чем агонистическая для антигена в отсутствие низкомолекулярного соединения.
[21.5]
Способ по любому из п.п. [21] - [21.4], в котором агонистическую активность для антигена оценивают по количеству IL-2 и/или IFN-y, вырабатываемых клетками, экспрессирующими антиген.
[21.6]
Способ по п. [21.5], в котором антиген-экспрессирующая клетка представляет собой выделенную мононуклеарную клетку периферической крови человека (МКПК) или Т-клетку, происходящую от МКПК человека.
[21.7]
Способ по любому из п.п. [21] - [21.4], в котором агонистическую активность в отношении антигена оценивают по анализу репортерного гена.
[21.8]
Способ по любому из п.п. [21] - [21.7], дополнительно включающий:
(i) получение вектора экспрессии, который включает соответствующий промотор, оперативно связанный с геном, кодирующим агонистическую антигенсвязывающую молекулу, полученную способом по любому из п.п. [21] - [21.7],
(ii) введение вектора в клетку-хозяина и культивирование клетки-хозяина для получения агонистической антигенсвязывающей молекулы, и
(iii) выделение агонистической антигенсвязывающей молекулы из культуры клеток-хозяев.
[21.9]
Способ по любому из п.п. [21] - [21.8], в котором представлена анти-CD137 антигенсвязывающая молекула.
[21.10]
Способ по любому из п.п. [21] - [21.9], в котором представлено анти-CD137 антитело. [21.11]
Способ по любому из п.п. [21.1] - [21.10],в котором низкомолекулярное соединение является соединением, содержащим аденозин. [21.12]
Способ по любому из п.п. [21.1] - [21.11], в котором низкомолекулярное соединение является АТФ.
[22]
Способ увеличения агонистической активности агонистической антигенсвязывающей молекулы, содержащей область Fc, который включает введение в область Fc по меньшей мере одного изменения аминокислоты, которое приводит к повышению изоэлектрической точки (pi) по сравнению с агонистической активностью исходной антигенсвязывающей молекулы, содержащей исходную область Fc.
[22.1]
Способ по п. [22], в котором агонистической активностью агонистической антигенсвязывающей молекулы по отношению к антигену в присутствии 10 мкМ, 50 мкМ, 100 мкМ, 150 мкМ, 200 мкМ или 250 мкМ низкомолекулярного соединения в два или более раз выше, чем агонистическая активность по отношению к антигену в отсутствие низкомолекулярного соединения.
[22.2]
Способ по п.п. [22] или [22.1], в котором агонистической активностью агонистической антигенсвязывающей молекулы по отношению к антигену в присутствии 10 мкМ или более низкомолекулярного соединения в два или более раз выше, чем агонистическая активность по отношению к антигену в отсутствие низкомолекулярного соединения.
[22.3]
Способ по п.п. [22] или [22.1], в котором агонистической активностью агонистической антигенсвязывающей молекулы по отношению к антигену в присутствии 50 мкМ или более низкомолекулярного соединения в два или более раз выше, чем агонистическая активность по отношению к антигену в отсутствие низкомолекулярного соединения.
[22.4]
Способ по п.п. [22] или [22.1], в котором агонистической активностью агонистической антигенсвязывающей молекулы по отношению к антигену в присутствии 250 мкМ или более низкомолекулярного соединения в два или более раз выше, чем агонистическая активность по отношению к антигену в отсутствие низкомолекулярного соединения.
[22.5]
Способ по любому из п.п. [22] - [22.4], в котором агонистическую активность по отношению к антигену оценивают по количеству IL-2 и/или IFN-y, продуцируемых антиген-экспрессирующей клеткой.
[22.6]
Способ по п. [22.5], в котором клетка, экспрессирующая антиген, представляет собой выделенную мононуклеарную клетку периферической крови (МКПК) человека или Т-клетку, производную от МКПК человека.
[22.7]
Способ по любому из п.п. [22] - [22.4], в котором агонистическую активность по отношению к антигену оценивают по анализу репортерного гена.
[22.8]
Способ по любому из п.п. [22] - [22.7], который включает анти-CD137 антигенсвязывающую молекулу.
[22.9]
Способ по любому из п.п. [22] - [22.8], который включает анти-CD137 антитело.
[22.10]
Способ по любому из п.п. [22.1] - [22.9], в котором низкомолекулярное соединение является соединением, содержащим аденозин. [21.11]
Способ по любому из п.п. [22.1] - [22.10], в котором низкомолекулярным соединением является АТФ.
[23]
Способ применения по меньшей мере одного изменения аминокислоты для увеличения агонистической активности антигенсвязывающей молекулы, содержащей область Fc, причем изменение аминокислоты приводит к увеличению изоэлектрической точки (pI) по сравнению с изоэлектрической точкой исходной агонистической антигенсвязывающей молекулой, содержащая исходную область Fc.
[23.1]
Способ по п. [23], в котором агонистическая активность агонистической антигенсвязывающей молекулы по отношению к антигену в присутствии 10 мкМ, 50 мкМ, 100 мкМ, 150 мкМ, 200 мкМ или 250 мкМ низкомолекулярного соединения в два или более раз выше, чем агонистическая активность по отношению к антигену в отсутствие низкомолекулярного соединения.
[23.2]
Способ по п.п. [23] или [23.1], в котором агонистическая активность агонистической антигенсвязывающей молекулы по отношению к антигену в присутствии 10 мкМ или более низкомолекулярного соединения в два или более раз выше, чем агонистическая активность по отношению к антигену в отсутствие низкомолекулярного соединения.
[23.3]
Способ по п.п. [23] или [23.1], в котором агонистическая активность агонистической антигенсвязывающей молекулы по отношению к антигену в присутствии 50 мкМ или более низкомолекулярного соединения в два или более раз выше, чем агонистическая активность по отношению к антигену по отношению к антигену в отсутствие низкомолекулярного соединения.
[23.4]
Способ по п.п. [23] или [23.1], в котором агонистическая активность агонистической антигенсвязывающей молекулы по отношению к антигену в присутствии 250 мкМ или более низкомолекулярного соединения в два или более раз выше, чем агонистическая активность по отношению к антигену по отношению к антигену в отсутствие низкомолекулярного соединения.
[23.5]
Способ по любому из п.п. [23] - [23.4], в котором агонистическая активность по отношению к антигену оценивают по количеству IL-2 и/или IFN-γ, продуцируемых антиген-экспрессирующей клеткой.
[23.6]
Способ по п. [23.5], в котором антиген-экспрессирующая клетка представляет собой выделенную мононуклеарную клетку периферической крови (МКПК) человека или Т-клетку, происходящую от МКПК человека.
[23.7]
Способ по любому из п.п. [23] - [23.4], в котором агонистическая активность по отношению к антигену оценивают с помощью анализа репортерного гена.
[23.8]
Способ по любому из п.п. [23] - [23.7], который включает анти-CD137 антигенсвязывающую молекулу.
[23.9]
Способ по любому из п.п. [23] - [23.8], который включает анти-CD137 антитело.
[23.10]
Способ по любому из п.п. [23.1] - [23.9], в котором низкомолекулярным соединением является соединение, содержащее аденозин.
[23.11]
Способ по любому из п.п. [23.1] - [23.10], в котором низкомолекулярным соединением является АТФ.
[24]
Способ скрининга антигенсвязывающего домена или антигенсвязывающей молекулы, которая обладает антигенсвязывающей активностью, зависящей от низкомолекулярного соединения, причем указанный способ включает:
(а) контактирование антигенсвязывающего домена, или антигенсвязывающей молекулы, или библиотеки антигенсвязывающих доменов или антигенсвязывающих молекул со слитой молекулой в присутствии низкомолекулярного соединения, где в слитой молекуле две или более единицы антигена слиты с одной единицей партнера по слиянию,
(б) размещение антигенсвязывающего домена или антигенсвязывающей молекулы, связанных с антигеном, внутри молекулы, слитой на стадии (а), в отсутствие или в присутствии низкой концентрации низкомолекулярного соединения, и
(в) выделение антигенсвязывающего домена или антигенсвязывающей молекулы, диссоциированных на стадии (б).
[24.1]
Способ по [24], в котором молекула-партнер по слиянию представляет собой димерную область Fc.
[24.2]
Способ по [24.1], в котором область Fc содержит первую субъединицу Fc и вторую субъединицу Fc, причем одна единица антигена слита с каждой из первой и второй субъединиц Fc.
[24.3]
Способ по п.п. [24.1] или [24.2], в котором одна единица антигена слита с N-концом каждой из первой и второй субъединиц Fc.
[24.4]
Способ по любому из п.п. [24] - [24.3], в котором библиотека антигенсвязывающих доменов или антигенсвязывающих молекул представляет собой библиотеку фагов.
[24.5]
Способ по любому из п.п. [24] - [24.4], в котором фаги, включенные в библиотеку фагов, являются фагами, имеющими на своей поверхности два или более антигенсвязывающих домена или антигенсвязывающие молекулы.
[24.6]
Способ по любому из п.п. [24] - [24.5], в котором фаги, включенные в библиотеку фагов, являются фагами, имеющими дефект в гене pIII, производном от фага-помощника.
[25]
Способ скрининга антигенсвязывающего домена или антигенсвязывающей молекулы, которая обладает антигенсвязывающей активностью, зависящей от двух или более разных низкомолекулярных соединений, причем указанный способ включает следующие стадии:
(а) контактирование антигенсвязывающего домена, или антигенсвязывающей молекулы, или библиотеки антигенсвязывающих доменов или антигенсвязывающих молекул с антигеном в присутствии первого низкомолекулярного соединения,
(б) размещение антигенсвязывающего домена или антигенсвязывающей молекулы, связанных с антигеном со стадии (а), в отсутствие или в присутствии низкой концентрации первого низкомолекулярного соединения, и
(в) выделение антигенсвязывающего домена или антигенсвязывающей молекулы, диссоциированных на стадии (б),
(г) контактирование антигенсвязывающего домена или антигенсвязывающей молекулы, выделенной на стадии (в), с антигеном в присутствии второго низкомолекулярного соединения,
(д) размещение антигенсвязывающего домена или антигенсвязывающей молекулы, связанной с антигеном на стадии (г) в отсутствие или в присутствии низкой концентрации второго низкомолекулярного соединения, и
(е) выделение антигенсвязывающего домена или антигенсвязывающей молекулы, диссоциированных на стадии (д), причем
способ не включает между стадиями (в) и (г) амплификацию гена, кодирующего антигенсвязывающий домен или антигенсвязывающую молекулу, выделенную на стадии (в).
[25.1]
Способ по п. [25], в котором библиотека антигенсвязывающих доменов или антигенсвязывающих молекул представляет собой библиотеку фагов.
[26]
Способ скрининга антигенсвязывающего домена или антигенсвязывающей молекулы, которые обладают антигенсвязывающей активностью, зависящей от низкомолекулярного соединения, причем указанный способ включает следующие стадии:
(а) контактирование наивной библиотеки антигенсвязывающих доменов или антигенсвязывающих молекул с антигеном в присутствии низкомолекулярного соединения,
(б) размещение антигенсвязывающего домена или антигенсвязывающей молекулы, связанной с антигеном на стадии (а) в отсутствие низкомолекулярного соединения или в его присутствии, но в низкой концентрации, и
(в) выделение антигенсвязывающего домена или антигенсвязывающей молекулы, диссоциированных на стадии (б),
причем наивная библиотека представляет собой библиотеку фагов, презентирующих на своей поверхности два или более антигенсвязывающих домена или две или более антигенсвязывающие молекулы.
[27]
Способ скрининга антигенсвязывающего домена или антигенсвязывающей молекулы, которые обладают антигенсвязывающей активностью, зависящей от низкомолекулярного соединения, причем указанный способ включает следующие стадии:
(а) контактирование библиотеки антигенсвязывающих доменов или антигенсвязывающих молекул с антигеном в присутствии низкомолекулярного соединения,
(б) размещение антигенсвязывающего домена или антигенсвязывающей молекулы, связанной с антигеном на стадии (а) в отсутствие низкомолекулярного соединения или в его присутствии, но в низкой концентрации, и
(в) выделение антигенсвязывающего домена или антигенсвязывающей молекулы, диссоциированных на стадии (б),
причем библиотека представляет собой библиотеку фагов, имеющих дефект в гене pIII, производном от гена-помощника.
[28]
Способ скрининга антигенсвязывающего домена или антигенсвязывающей молекулы, которые обладают антигенсвязывающей активностью, зависящей от низкомолекулярного соединения, причем указанный способ включает следующие стадии:
(а) контактирование библиотеки антигенсвязывающих доменов или антигенсвязывающих молекул с антигеном в присутствии низкомолекулярного соединения,
(б) размещение антигенсвязывающего домена или антигенсвязывающей молекулы, связанной с антигеном на стадии (а) в отсутствие низкомолекулярного соединения или в его присутствии, но в низкой концентрации, и
(в) выделение антигенсвязывающего домена или антигенсвязывающей молекулы, диссоциированных на стадии (б),
причем библиотека представляет собой библиотеку, включающую фаги, полученные путем увеличения экспрессии антигенсвязывающего домена или антигенсвязывающей молекулы с помощью низкомолекулярной добавки, которая увеличивает уровень экспрессии с промотора, регулирующего экспрессию антигенсвязывающего домена или антигенсвязывающей молекулы.
Способ скрининга по п. [28], в котором низкомолекулярная добавка представляет собой изопропил-β-тиогалактопиранозид или арабинозу.
[28.2]
Способ по любому из п.п. [24]-[28.1], в котором низкомолекулярное соединение является соединением, содержащим аденозин.
[28.3]
Способ по любому из п.п. [24]-[28.2], в котором низкомолекулярное соединение является АТФ.
[29] Антигенсвязывающая молекула, которая обладает антигенсвязывающей активностью, зависящей от концентрации опухолевого тканеспецифичного соединения, причем антигенсвязывающая активность в присутствии 100 мкМ соединения выше в два раза или более, чем антигенсвязывающая активность в отсутствие соединения.
[29.1] Антигенсвязывающая молекула по п. [29], величина KD которой в присутствии 100 мкМ соединения равна 5×10-7 М или менее.
[29.2] Антигенсвязывающая молекула по п.п. [29] или [29.1], величина KD которой в отсутствие соединения равна 1×10-6 М или более.
[29.3] Антигенсвязывающая молекула по любому из п.п. [29]-[29.2], которая обладает нейтрализующей активностью в отношении антигена.
[29.4] Антигенсвязывающая молекула по любому из п.п. [29]-[29.3], которая обладает цитотоксической активностью в отношении клеток, экспрессирующих антиген.
[29.5] Антигенсвязывающая молекула по любому из п.п. [29]-[29.4], в которых антиген представляет собой антиген, экспрессируемый или секретируемый любой из клеток, являющихся опухолевой клеткой, иммунной клеткой или стромальной клеткой в опухолевой ткани.
[29.6] Антигенсвязывающая молекула по любому из п.п. [29]-[29.5], в которых соединение является соединением, содержащим аденозин.
[29.7] Антигенсвязывающая молекула по любому из п.п. [29]-[29.6], которая содержит область Fc.
[29.8] Антигенсвязывающая молекула по п. [29.7], в которой область Fc представляет собой мутировавшую область Fc, содержащую аминокислотное изменение, причем мутировавшая область Fc имеет повышенную связывающую активность по меньшей мере с одним рецептором Fcγ, выбранным из группы, состоящей из FcγRIa, FcγRIIa, FcγRIIb и FcγRIIIa, по сравнению с областью Fc дикого типа.
[29.9] Антигенсвязывающая молекула по любому из п.п. [29]-[29.8], в которых антигенсвязывающей молекулой является антитело или фрагмент антитела.
[30] Фармацевтический состав, включающий антигенсвязывающую молекулу по любому из п.п. [29]-[29.9] и фармацевтически приемлемый носитель.
[30.1] Фармацевтический состав по п. [30], который предназначен для лечения опухоли.
[30.2] Фармацевтический состав по п. [30.1], который обладает пониженной цитотоксической активностью в неопухолевой ткани по сравнению с фармацевтическим составом, содержащим контрольную антигенсвязывающую молекулу.
[30.3] Фармацевтический состав по п.п. [30.1] или [30.2], который обладает пониженным уровнем побочных эффектов по сравнению с фармацевтическим составом, содержащим контрольную антигенсвязывающую молекулу.
[30.4] Фармацевтический состав по п.п. [30.2] или [30.3], где контрольная антигенсвязывающая молекула представляет собой антигенсвязывающую молекулу, которая не обладает антигенсвязывающей активностью, зависящей от концентрации опухолевого тканеспецифичного соединения.
[31] Способ получения антигенсвязывающей молекулы для применения в лечении опухоли, который включает стадию выбора антигенсвязывающей молекулы, антигенсвязывающая активность которой в присутствии 100 мкМ тканеспецифичного для опухоли соединения выше в два раза или более, чем антигенсвязывающая активность при отсутствии соединения.
[32] Способ получения фармацевтического состава для применения в лечении опухоли, который включает стадию смешивания антигенсвязывающей молекулы по любому из п.п. [29]-[29.9] с фармацевтически приемлемым носителем.
[33] Антигенсвязывающая молекула, обладающая антигенсвязывающей активностью, зависящей от концентрации целевого тканеспецифичного соединения, при этом антигенсвязывающая активность в присутствии 1 мкМ такого соединения понижена в два раза или более по сравнению с антигенсвязывающей активностью в присутствии достаточного количества соединения.
[33.1] Антигенсвязывающая молекула по п. [33], причем величина KD в присутствии 1 мкМ такого соединения составляет 2×10-7 М или более.
[33.2] Антигенсвязывающая молекула по п.п. [33] или [33.1], причем величина KD в присутствии 1 мкМ такого соединения составляет 1×10-7 М или менее.
[33.3] Антигенсвязывающая молекула по любому из п.п. [33]-[33.2], в которых соединение представляет собой тканеспецифичное для опухоли соединения.
[33.4] Антигенсвязывающая молекула по п. [33.3], в котором соединение является соединением, содержащим аденозин.
[33.5] Антигенсвязывающая молекула по любому из п.п. [33]-[33.4], которая обладает повышенной удерживающей способностью в плазме и/или имеет пониженную способность к накоплению антигена в плазме по сравнению с контрольной антигенсвязывающей молекулой.
[33.6] Антигенсвязывающая молекула по п. [33.5], которая представляет собой антигенсвязывающую молекулу, не обладающую антигенсвязывающей активностью, зависящей от концентрации целевого тканеспецифичного соединения.
[33.7] Антигенсвязывающая молекула по любому из п.п. [33]-[33.6], которая представляет собой антитело или фрагмент антитела.
[34] Фармацевтический состав, содержащий антигенсвязывающую молекулу по любому из п.п. [33]-[33.7] и фармацевтически приемлемый носитель.
[35] Способ получения антигенсвязывающей молекулы, которая имеет повышенную способность удерживания в плазме и/или пониженную способность к накоплению антигена в плазме по сравнению с контрольной антигенсвязывающей молекулой, причем способ включает стадии (а) получения антигенсвязывающей молекулы, чья антигенсвязывающая активность увеличивается по мере увеличения концентрации целевого тканеспецифичного соединения, и (б) измерения способности удерживания в плазме и/или способности к накоплению антигена в плазме антигенсвязывающей молекулы, полученной по п. (а).
[35.1] Способ по п. [35], который включает стадию выбора антигенсвязывающей молекулы, антигенсвязывающая активность которой в присутствии 1 мкМ целевого тканеспецифичного соединения в два раза или более ниже, чем антигенсвязывающая активность в присутствии достаточного количества соединения.
[35.2] Способ по п.п. [35] или [35.1], в котором контрольной антигенсвязывающей молекулой является антигенсвязывающая молекула, которая не обладает антигенсвязывающей активностью, зависящей от концентрации целевого тканеспецифичного соединения.
[36] Способ получения фармацевтического состава, который включает стадию смешивания антигенсвязывающей молекулы по любому из п.п. [33]-[33.7] с фармацевтически приемлемым носителем.
[37] Способ измерения концентрации АТФ в растворе, который включает стадии (i) контакта клетки split Luc/HEK293, экспрессирующей P2Y11, с раствором, и (ii) измерения активности люциферазы в клетке.
[37.1] Способ по п. [37], который дополнительно включает стадию контакта раствора, содержащего субстрат люциферазы, с клеткой.
[37.2] Способ по п.п. [37] или [37.1], в котором раствор представляет собой межклеточную жидкость в ткани in vivo.
[37.3] Способ по п. [37.2], в котором ткань является опухолевой тканью.
[37.4] Способ по п. [37.2] или [37.3], в котором стадия (i) является стадией трансплантации клетки split Luc/HEK293, экспрессирующей P2Y11, в ткань in vivo.
Краткое описание фигур
Фиг. 1. Диаграмма, показывающая агонистическую активность различных анти-CD137 антител, исследованных с применением клеток Jurkat в присутствии или в отсутствие АТФ.
Ось X показывает концентрацию антитела (мкг/мл), ось Y показывает относительные световые единицы.
Фиг. 2. Диаграмма, показывающая агонистическую активность различных анти-CD137 антител, исследованных с применением клеток Jurkat в присутствии или в отсутствие АДФ.
Ось X показывает концентрацию антитела (мкг/мл), ось Y показывает относительные световые единицы.
Фиг. 3. Диаграмма, показывающая агонистическую активность различных анти-CD137 антител, исследованных с применением Т-клеток человека в присутствии или в отсутствие АДФ-бета-S (ADPбетаS).
Фиг. 4. Диаграмма, показывающая агонистическую активность dBBAT119-P253/dBBAT119L-LamLib (анти-CD137 антитело, переключаемое низкомолекулярным соединением) или NS1-P253 (непереключаемое анти-CD137 антитело) исследованных с применением Т-клеток человека в присутствии или в отсутствие АДФ-бета-S.
Ось X показывает концентрацию антитела (мкг/мл), ось Y показывает количество выработанного IFN-γ (нг/мл).
Фиг. 5. Диаграмма, показывающая АТФ-зависящую антигенсвязывающую активность различных анти-CD137 антител (переключаемое анти-CD137 антитело с улучшенной связывающей активностью), исследованную методом фагового иммуноферментного анализа (ELISA).
Ось Y показывает соотношение поглощения S/N (сигнал/шум) в присутствии/в отсутствие АТФ, а ось X показывает соотношение S/N в присутствии/в отсутствие антигена.
Фиг. 6. Диаграмма, показывающая связывающую активность различных вариантов анти-CD137 антитела (dBAT119H-P253/dBAT119L-LamLib) с CD137 человека в присутствии или в отсутствие АТФ.
Верхний ряд показывает связывающую активность с CD137 человека в отсутствие АТФ, а нижний ряд показывает связывающую активность с CD137 человека в присутствии АТФ.
Фиг. 7. Диаграмма, показывающая агонистическую активность dBBAT119H-P253/dBBAT119L-LamLib, dBBATk119H024-P253/dBBATk119L020-LamLib, IC17HdK-hIgGl/IC17L-k0 (контроль) или NS1-P253 (не переключаемого анти-CD137 антитела), используя Т-клетки человека в присутствии или в отсутствие АДФ-бета-S.
Фигура 7А показывает результаты тестирования в отсутствие АДФ-бета-S, фигура 7Б показывает результаты тестирования в присутствии АДФ-бета-S.
Ось X показывает концентрацию антитела (мкг/мл), ось Y показывает количество выработанного IFN-γ (нг/мл).
Фиг. 8. Диаграмма, показывающая агонистическую активность различных переключаемых анти-CD137 антител, протестированных с использованием анализа репортерного гена 4-1ВВ Jurkat в присутствии или в отсутствие АТФ.
Фигура 8А показывает результаты тестирования в отсутствие АТФ, фигура 8Б показывает результаты тестирования в присутствии АТФ.
Фиг. 9. Диаграмма, показывающая усиление агонистической активности различных анти-CD137 переключаемых антител в присутствии АТФ за счет увеличения связывающей активности константных областей тяжелой цепи с рецепторами Fcγ, протестированных с использованием мононуклеарных клеток периферической крови.
Фигура 9А показывает агонистическую активность, определенную с использованием результатов тестирования в отсутствие АТФ, фигура 9Б показывает результаты тестирования в присутствии АТФ.
Фигура 9А показывает агонистическую активность, определенную с использованием количества выработанного IL-2 в качестве показателя, фигура 9Б показывает агонистическую активность, определенную с использованием количества выработанного IFN-β в качестве показателя.
Фиг. 10. Диаграмма, показывающая усиление агонистической активности различных анти-CD137 переключаемых антител в присутствии АТФ за счет увеличения связывающей активности константных областей тяжелой цепи с рецепторами Fcγ или увеличения pI константных областей тяжелой цепи, по результатам анализа с использованием мононуклеарных клеток периферической крови человека.
Фигура 10А показывает агонистическую активность, определенную с использованием количества выработанного IL-2 в качестве показателя, фигура 10Б показывает агонистическую активность, определенную с использованием количества выработанного IFN-γ в качестве показателя.
Фиг. 11. Диаграмма, показывающая усиление агонистической активности различных переключаемых анти-CD137 антител в присутствии или в отсутствие АТФ за счет увеличения связывающей активности константных областей тяжелой цепи с рецепторами Fcγ, определенную с использованием мононуклеарных клеток периферической крови человека.
Фигура 11А показывает агонистическую активность, определенную с использованием количества выработанного IL-2 в качестве показателя, фигура 11Б показывает агонистическую активность, определенную с использованием количества выработанного IFN-γ в качестве показателя.
Фиг. 12. Диаграмма, показывающая усиление агонистической активности различных переключаемых анти-CD137 антител в присутствии или в отсутствие АТФ за счет повышения связывающей активности константных областей тяжелой цепи с рецепторами Fcγ, определенную с использованием мононуклеарных клеток периферической крови человека.
Фигура 12А показывает агонистическую активность, определенную по количеству выработанного IL-2 в качестве показателя; фигура 12Б показывает агонистическую активность, определенную по количеству выработанного IFN-γ в качестве показателя.
Фиг. 13. Диаграмма, показывающая усиление агонистической активности различных переключаемых анти-CD137 антител в присутствии или в отсутствие АТФ за счет повышения связывающей активности константных областей тяжелой цепи с рецепторами Fcγ, определенное с использованием мононуклеарных клеток периферической крови человека.
Фигура 13А показывает агонистическую активность, определенную по количеству выработанного IL-2 в качестве показателя; фигура 13Б показывает агонистическую активность, определенную по количеству выработанного IFN-γ в качестве показателя.
Фиг. 14. Диаграмма, показывающая усиление агонистической активности различных переключаемых анти-CD137 антител в присутствии или в отсутствие АТФ за счет увеличения связывающей активности константных областей тяжелой цепи с рецепторами Fcγ, по результатам исследования с использованием мононуклеарных клеток периферической крови человека.
Фигура 14А показывает агонистическую активность, определенную по количеству выработанного IL-2 в качестве показателя; фигура 14Б показывает агонистическую активность, определенную по количеству выработанного IFN-γ в качестве показателя.
Фиг. 15 Диаграмма, показывающая усиление агонистической активности различных переключаемых анти-CD137 антител в присутствии или в отсутствие АТФ за счет увеличения связывающей активности константных областей тяжелой цепи с рецепторами Fcγ, по результатам исследования с использованием мононуклеарных клеток периферической крови человека.
Фигура 15А показывает агонистическую активность, определенную по количеству выработанного IL-2 в качестве показателя; фигура 15Б показывает агонистическую активность, определенную с применением количества выработанного IFN-γ в качестве показателя.
Фиг. 16. Диаграмма, показывающая усиление агонистической активности различных переключаемых анти-CD137 антител в присутствии или в отсутствие АТФ за счет повышения величины pI константных областей тяжелой цепи, определенное с использованием мононуклеарных клеток периферической крови человека.
Фигура 16А показывает агонистическую активность, определенную по количеству выработанного IL-2 в качестве показателя; фигура 16Б показывает агонистическую активность, определенную по количеству выработанного IFN-γ в качестве показателя.
Фиг. 17. Диаграмма, показывающая усиление агонистической активности различных переключаемых анти-CD137 антител в присутствии или в отсутствие АТФ за счет повышения величины pI константных областей тяжелой цепи, определенное с использованием мононуклеарных клеток периферической крови человека.
Фигура 17А показывает агонистическую активность, определенную по количеству выработанного IL-2 в качестве показателя; фигура 17Б показывает агонистическую активность, определенную по количеству выработанного IFN-γ в качестве показателя.
Фиг. 18. Диаграмма, показывающая усиление агонистической активности различных переключаемых анти-CD137 антител в присутствии или в отсутствие АТФ за счет повышения величины pI константных областей тяжелой цепи, определенное с использованием мононуклеарных клеток периферической крови человека.
Фигура 18А показывает агонистическую активность, определенную по количеству выработанного IL-2 в качестве показателя; фигура 18Б показывает агонистическую активность, определенную по количеству выработанного IFN-γ в качестве показателя.
Фиг. 19. Диаграмма, показывающая усиление агонистической активности различных переключаемых анти-CD137 антител в присутствии или в отсутствие АТФ за счет повышения величины pI константных областей тяжелой цепи, определенное с использованием мононуклеарных клеток периферической крови человека.
Фигура 19А показывает агонистическую активность, определенную по количеству выработанного IL-2 в качестве показателя; фигура 19Б показывает агонистическую активность, определенную по количеству выработанного IFN-γ в качестве показателя.
Фиг. 20. Диаграмма, показывающая усиление агонистической активности различных переключаемых анти-CD137 антител в присутствии или в отсутствие АТФ за счет повышения величины pI константных областей тяжелой цепи, определенное с использованием мононуклеарных клеток периферической крови человека.
Фигура 20А показывает агонистическую активность, определенную по количеству выработанного IL-2 в качестве показателя; фигура 20Б показывает агонистическую активность, определенную по количеству выработанного IFN-γ в качестве показателя.
Фиг. 21. Диаграмма, показывающая усиление агонистической активности различных переключаемых анти-CD137 антител в присутствии или в отсутствие АТФ за счет повышения величины pI константных областей тяжелой цепи, определенное с использованием мононуклеарных клеток периферической крови человека.
Фигура 21А показывает агонистическую активность, определенную по количеству выработанного IL-2 в качестве показателя; фигура 21Б показывает агонистическую активность, определенную по количеству выработанного IFN-γ в качестве показателя.
Фиг. 22. Диаграмма, показывающая усиление агонистической активности различных переключаемых анти-CD137 антител в присутствии или в отсутствие АТФ за счет повышения величины р! константных областей тяжелой цепи, определенное с использованием мононуклеарных клеток периферической крови человека.
Фигура 22А показывает агонистическую активность, определенную по количеству выработанного IL-2 в качестве показателя; фигура 22Б показывает агонистическую активность, определенную по количеству выработанного IFN-γ в качестве показателя.
Фиг. 23. Диаграмма, показывающая усиление агонистической активности различных переключаемых анти-CD137 антител в присутствии или в отсутствие АТФ за счет увеличения связывающей активности константных областей тяжелой цепи с рецепторами Fcγ, по результатам исследования с использованием мононуклеарных клеток периферической крови человека.
Фигура 23А показывает агонистическую активность, определенную по количеству выработанного IL-2 в качестве показателя; фигура 23Б показывает агонистическую активность, определенную по количеству выработанного IFN-γ в качестве показателя.
Фиг. 24. Диаграмма, показывающая концентрацию в плазме различных переключаемых и не переключаемых анти-CD137 антител, тестируемых с использованием мышей с нокином CD137 человека.
Все Fc от mIgG1.
Фиг. 25. Диаграмма, показывающая концентрацию в плазме различных переключаемых и не переключаемых анти-CD137 антител, исследованных с использованием мышей с нокином CD137 человека.
Все Fc от МВ110.
Фиг. 26. Диаграмма, показывающая концентрацию в плазме различных переключаемых и не переключаемых анти-CD137 антител, проанализированных с использованием мышей с нокином CD137 человека.
Все Fc от МВ492.
Фиг. 27. Диаграмма, показывающая противоопухолевый эффект А375-mIgG1/B167-ml0r на модели мышей, трансплантированных клетками МС38.
Каждая точка показывает среднее значение объема опухоли для группы (n=5).
Фиг. 28. Диаграмма, показывающая массу органов модели мыши, трансплантированной клетками МС38 после введения антител (NO1-mIgG1 или A375-mIgG1/B167-ml0r).
Фигура 28А показывает массу лимфоузла, а фигура 28Б показывает массу селезенки.
Фиг. 29. Диаграмма, показывающая степень активации Т-клеток в лимфоузлах модели мыши, трансплантированной клетками МС38 после введения антител NO1-mIgG1 или A375-mIgG1/B167-ml0r.
Фигура 29А показывает процент PD-1-положительных Т-клеток среди CD8-положительных Т-клеток, фигура 29Б показывает процент ICOS-положительных Т-клеток среди CD8-положительных Т-клеток, фигура 29В показывает процент гранзим В-положительных Т-клеток среди CD8-положительных Т-клеток.
Фиг. 30. Диаграмма, показывающая степень активации Т-клеток в селезенке модели мыши, трансплантированной клеточной линией МС38, после введения NO1-mIgG1 или A375-mIgG1/B167-ml0r.
Фигура ЗОА показывает процент PD-1-положительных Т-клеток среди CD8-положительных Т-клеток, фигура 30Б показывает процент ICOS-положительных Т-клеток среди CD8-положительных Т-клеток, фигура 30В показывает процент гранзим В-положительных Т-клеток среди CD8-положительных Т-клеток.
Фиг. 31. Диаграмма, показывающая степень активации Т-клеток в печени модели мыши, трансплантированной клеточной линией МС38, после введения NO1-mIgG1 или A375-mIgG1/B167-ml0r.
Фигура 31А показывает процент PD-1-положительных Т-клеток среди CD8-положительных Т-клеток, фигура 31Б показывает процент гранзим В-положительных Т-клеток среди CD8-положительных Т-клеток.
Фиг. 32. Диаграмма, показывающая противоопухолевый эффект А356-МВ110/B040-ml0r в модели мыши, трансплантированный клеточной линией МС38.
Каждая точка показывает среднее значение объема опухоли для группы (n=5).
Фиг. 33. Диаграмма, показывающая массу органов в модели мыши, трансплантированной клеточной линией МС38 после введения NS2-MB110 или А356-МВ110/B040-ml0r.
Фигура 33А показывает массу лимфоузла, а фигура 33Б показывает массу селезенки.
Фиг. 34. Диаграмма, показывающая степень активации Т-клеток в печени модели мыши, трансплантированной клеточной линией МС38, после введения NS2-MB110 или А356-МВ110/В040-ml0r.
Фигура 34А показывает процент PD-1-положительных Т-клеток среди CD8-положительных Т-клеток, фигура 34Б показывает процент ICOS-положительных Т-клеток среди CD8-положительных Т-клеток.
Фиг. 35. Диаграмма, показывающая противоопухолевый эффект А372-mIgG1/B040-ml0r в модели мыши, трансплантированной клеточной линией МС38.
Каждая точка показывает среднее значение объема опухоли для группы (n=5).
Фиг. 36. Количество клеток лимфатического узла (фигура 36А) и масса селезенки (фигура Б) в модели мыши, трансплантированной клеточной линией МС38, после введения A372-mIgG1/B040-ml0r.
Фиг. 37. Диаграмма, показывающая степень активации Т-клеток в печени модели мыши, трансплантированной клеточной линией МС38, после введения A372-mIgG1/B040-ml0r (процент гранзим В-положительных Т-клеток среди CD8-положительных Т-клеток).
Фиг. 38. Диаграмма, показывающая противоопухолевый эффект А372-МВ110/B040-ml0r в модели мыши, трансплантированной клеточной линией МС38.
Каждая точка показывает среднее значение объема опухоли для группы (n=5).
Фиг. 39. Диаграмма, показывающая массу органов модели мыши, трансплантированной клетками МС38, после введения NS2-MB110 или А372-MB110/B040-ml0r.
Фигура 39А показывает массу лимфоузла, фигура 39Б показывает массу селезенки.
Фиг. 40. Диаграмма, показывающая степень активации Т-клеток в печени модели мыши, трансплантированной клеточной линией МС38, после введения NS2-MB110 или A372-MB110/B040-ml0r (процент PD-1-положительных Т-клеток среди CD8-положительных Т-клеток).
Фиг. 41. Диаграмма, показывающая противоопухолевый эффект А372-МВ110/B040-ml0r в модели мыши, трансплантированной клеточной линией МС38.
Каждая точка показывает среднее значение объема опухоли для группы (n=5).
Фиг. 42. Диаграмма, показывающая количество клеток лимфатического узла и массу органа селезенки в модели мыши, трансплантированной клеточной линией МС38, после введения NS1-MB492 или А372-МВ492/В040-ml0r.
Фигура 42А показывает количество клеток лимфоузла, фигура 42Б показывает массу селезенки.
Фиг. 43. Диаграмма, показывающая степень активации Т-клеток в печени модели мыши, трансплантированной клеточной линией МС38, после введения NS1-MB492 или A372-MB492/B040-ml0r (процент гранзим В-положительных Т-клеток среди CD8-положительных Т-клеток).
Фиг. 44. Диаграмма, показывающая противоопухолевый эффект А486-MB492/B167-ml0r или A488-MB492/B226-ml0r в модели мыши, трансплантированной клеточной линией МС38.
Каждая точка показывает среднее значение объема опухоли для группы (n=5).
Фиг. 45. Диаграмма, показывающая количество клеток в лимфоузле и массу селезенки в модели мыши, трансплантированной клеточной линией МС38, после введения NS1-MB492, А486-МВ492/В167-ml0r или A488-MB492/B226-ml0r.
Фигура 45А показывает количество клеток в лимфоузле, фигура 45Б показывает массу селезенки.
Фиг. 46. Диаграмма, показывающая уровень инфильтрации эффекторных клеток в печени модели мыши, трансплантированной клеточной линией МС38, после введения NS1-MB492, A486-MB492/B167-ml0r или А488-МВ492/В226-ml0r (процент CD3-положительных и CD8-положительных Т-клеток среди CD45-положительных Т-клеток).
Фиг. 47. Диаграмма, показывающая противоопухолевый эффект А489-MB492/B223-ml0r в модели мыши, трансплантированной клеточной линией МС38.
Каждая точка показывает среднее значение объема опухоли для группы (n=5).
Фиг. 48. Диаграмма, показывающая количество клеток лимфатического узла и количество клеток во фракции лимфоцитов селезенки в модели мыши, трансплантированной клеточной линией МС38, после введения NS1-MB492 или A489-MB492/B223-ml0r.
Фигура 48А показывает количество клеток лимфатического узла, фигура 48Б показывает количество клеток во фракции лимфоцитов селезенки.
Фиг. 49. Диаграмма, показывающая степень активации Т-клеток в печени мышиной модели, трансплантированной клеточной линией МС38, после введения NS1-MB492 или A489-MB492/B223-ml0r (процент CD8-положительных Т-клеток среди CD45-положительных Т-клеток).
Фиг. 50. Диаграмма, показывающая противоопухолевый эффект А548-mIgG1/B256-ml0r и A551-mIgG1/B256-ml0r в модели мыши, трансплантированной клеточной линией МС38.
Фигура 50А показывает противоопухолевый эффект A548-mIgG1/B256-ml0r, фигура 50Б показывает противоопухолевый эффект А551-mIgG1/B256-ml0r.
Фиг. 51. Диаграмма, показывающая массу органов в модели мыши, трансплантированной клеточной линией МС38, после введения NS1-mIgG1, A548-mIgG1/B256-ml0r или А551-mIgG1/B256-ml0r.
Фигура 51А показывает массу лимфоузла, фигура 51Б показывает массу селезенки.
Фиг. 52. Диаграмма, показывающая степень активации Т-клеток в печени мышиной модели, трансплантированной клеточной линией МС38, после введения NS1-mIgG1, A548-mIgG1/B256-ml0r или A551-mIgG1/B256-ml0r.
Фигура 52А показывает процент PD-1-положительных Т-клеток среди CD8-положительных Т-клеток, фигура 52Б показывает процент гранзим В-положительных клеток среди CD8-положительных Т-клеток.
Фиг. 53. Диаграмма, показывающая противоопухолевый эффект А551-МВ110/B379-ml0r в модели мыши, трансплантированной клеточной линией МС38.
Фиг. 54. Диаграмма, показывающая массу органов в модели мыши, трансплантированной клеточной линией МС38 после введения NS1-mIgG1 или A551-MB110/B379-ml0r.
Фигура 54А показывает массу лимфатического узла, фигура 54Б показывает массу селезенки.
Фиг. 55. Диаграмма, показывающая степень активации Т-клеток в селезенке мышиной модели, трансплантированной клеточной линией МС38, после введения NS1-mIgG1 или А551-МВ110/B379-ml0r.
Фигура 55А показывает процент PD-1-положительныхТ-клеток среди CD8-положительных Т-клеток, фигура 55Б показывает процент ICOS-положительных Т-клеток среди CD8-положительных Т-клеток, фигура 55В показывает процент гранзим В-положительных Т-клеток среди CD8-положительных Т-клеток.
Фиг. 56. Диаграмма, показывающая степень активации Т-клеток в печени мышиной модели, трансплантированной клеточной линией МС38, после введения NS1-mIgG1 или А551-МВ110/B379-ml0r.
Фигура 56А показывает процент PD-1-положительных Т-клеток среди CD8-положительных Т-клеток, фигура 56Б показывает процент ICOS-положительных Т-клеток среди CD8-положительных Т-клеток, фигура 56В показывает процент гранзим В-положительных Т-клеток среди CD8-положительных Т-клеток.
Фиг. 57. Диаграмма, показывающая агонистическую активность различных анти-CD137 антител, используя клетки Jurkat в присутствии или в отсутствие L-кинуренина.
Ось X показывает концентрацию антитела (мкг/мл), а ось Y показывает относительные световые единицы.
Фиг. 58. Диаграмма, показывающая агонистическую активность различных анти-CD137 антител, используя клетки 4-1ВВ Jurkat в присутствии или в отсутствие низкомолекулярного соединения (АТФ или ФДФ).
Ось X показывает концентрацию антитела (мкг/мл), а ось Y показывает относительные световые единицы.
Фиг. 59. Диаграмма, показывающая ответную реакцию АТФ (зависящую от концентрации АТФ люминесценцию люциферина) в клетках P2Y11 split Luc/HEK293, получаемых для определения внеклеточных уровней АТФ.
Фиг. 60. Диаграмма, показывающая in vivo ответную реакцию АТФ (зависящую от концентрации АТФ люминесценцию люциферина) в клетках P2Y11 split Luc/HEK29, которые были трансплантированы мышам подкожно.
Фиг. 61. Диаграмма, показывающая результаты люминесцентной визуализации мышей, подкожно трансплантированных клетками P2Y11 split Luc/HEK293 и предварительно определенными концентрациями АТФ, и мышей с опухолью FM3A, подкожно трансплантированных клетками P2Y11 split Luc/HEK293. Метки в вентральной части мышей указывают на выявленную люминесценцию.
Фиг. 62. Диаграмма, показывающая зависящую от концентрации АТФ связывающую активность (величину KD) анти-hIL6R антител MRAH-G4T1/MRAL-k0 (контрольное антитело) и H0002-G4T1/L1058-lam1, Н0041-G4T1/L1088-lam1, H0052-G4T1/L1083-lam1 (все являются переключаемыми антителами) против hIL6R.
Фиг. 63. Диаграмма, показывающая зависящую от концентрации АДФ связывающую активность (величину KD) анти-hIL6R антител MRAH-G4T1/MRAL-k0 (контрольное антитело) и H0002-G4T1/L1058-lam1, Н0041-G4T1/L1088-lam1, H0052-G4T1/L1083-lam1 (все являются переключаемыми антителами) против hIL6R.
Фиг. 64. Диаграмма, показывающая зависящую от концентрации АМФ связывающую активность (величину KD) анти-hIL6R антител MRAH-G4T1/MRAL-k0 (контрольное антитело) и H0002-G4T1/L1058-lam1, Н0041-G4T1/L1088-lam1, H0052-G4T1/L1083-lam1 (все являются переключаемыми антителами) против hIL6R.
Фиг. 65. Диаграмма, показывающая зависящую от концентрации АТФ активность ADCC анти-hIL6R антител MRAH-mFa55/MRAL-mk0 (контрольное антитело) и H0002-mFa55/L1058-ml0, H0041-mFa55/L1088-ml0, Н0052-mFa55/L1083-ml0 (все являются переключаемыми антителами).
Фиг. 66. Диаграмма, показывающая противоопухолевую активность in vivo анти-hIL6R антител MRAH-mFa55/MRAL-mk0 (контрольное антитело) и Н0002-mFa55/L1058-ml0, H0041-mFa55/L1088-ml0, H0052-mFa55/L1083-ml0 (все являются переключаемыми антителами). IC17Hdk-mFa55/IC17L-mk1 является отрицательным контрольным антителом.
Фиг. 67. Диаграмма, показывающая сравнение кинетики в плазме анти-hIL6R антитела MRAH-mFa55/MRAL-mk0 (контрольное антитело) в обычных мышах и в hIL6R трансгенных мышах. Вертикальная ось графика показывает концентрацию антитела в плазме.
Фиг. 68. Диаграмма, показывающая сравнение кинетики в плазме анти-hIL6R антитела H0002-mFa55/L1058-ml0 (переключаемого антитела) в обычных мышах и в hIL6R трансгенных мышах. Вертикальная ось графика показывает концентрацию антитела в плазме.
Фиг. 69. Диаграмма, показывающая сравнение кинетики в плазме анти-hIL6R антитела H0041-mFa55/L1088-ml0 (переключаемого антитела) в обычных мышах и в hIL6R трансгенных мышах. Вертикальная ось графика показывает концентрацию антитела в плазме.
Фиг. 70. Диаграмма, показывающая сравнение кинетики в плазме анти-hIL6R антитела H0052-mFa55/L1083-ml0 (переключаемого антитела) в обычных мышах и в hIL6R трансгенных мышах. Вертикальная ось графика показывает концентрацию антитела в плазме.
Фиг. 71. Диаграмма, показывающая накопление антигенов у трансгенных мышей hIL6R после введения каждого из анти-hIL6R антител: не переключаемого антитела MRAH-mFa55/MRAL-mk0 (контрольное антитело) и анти-hIL6R переключаемых антител H0002-mFa55/L1058-ml0, Н0041-mFa55/L1088-ml0, H0052-mFa55/L1083-ml0 (все являются переключаемыми антителами). Вертикальная ось графика показывает концентрацию в плазме растворимого hIL6R. IC17Hdk-mFa55/IC17L-mk1 (обозначенного на фигуре как KLH-mFa55) применяют в качестве антитела отрицательного контроля.
Фиг. 72. Диаграмма, показывающая противоопухолевую активность in vivo анти-hIL6R не переключаемые антитела MRAH-mFa55/MRAL-mk0 (контрольное антитело) и анти-hIL6R переключаемые антител H0002-mFa55/L1058-ml0 и H0041-mFa55/L1088-ml0 (оба являются переключаемыми антителами). IC17Hdk-mFa55/IC17L-mk1 является антителом отрицательного контроля.
Фиг. 73. Диаграмма, показывающая сравнение кинетики в плазме анти-hIL6R не переключаемого антитела MRAH-mFa55/MRAL-mk0 (контрольное антитело) и анти-hIL6R переключаемых антител H0002-mFa55/L1058-ml0 и H0041-mFa55/L1088-ml0 (оба являются переключаемыми антителами). Вертикальная ось графика показывает концентрацию антитела в плазме.
Фиг. 74. Диаграмма, показывающая накопление антигенов после введения каждого из анти-hIL6R антител: не переключаемого антитела MRAH-mFa55/MRAL-mk0 (контрольное антитело) и анти-hIL6R переключаемых антител H0002-mFa55/L1058-ml0 и H0041-mFa55/L1088-ml0 (оба являются переключаемыми антителами). Вертикальная ось графика показывает концентрацию растворимого hIL6R в плазме. IC17Hdk-mFa55/IC17L-mk1 (обозначенное на фигуре как KLH-mFa55) применяют в качестве отрицательного контрольного антитела.
Фиг. 75. Диаграмма, показывающая in vivo противоопухолевую активность анти-hIL6R не переключаемого антитела MRAH-mFa55/MRAL-mk0 (контрольное антитело) и анти-hIL6R переключаемых антител H0041-mFa55/L1088-ml0 и H0052-mFa55/L1083-ml0 (оба являются переключаемыми антителами). IC17Hdk-mFa55/IC17L-mk1 является антителом отрицательного контроля.
Фиг. 76. Диаграмма, показывающая сравнение в плазме кинетики анти-hIL6R не переключаемого антитела MRAH-mFa55/MRAL-mk0 (контрольное антитело) и анти-hIL6R переключаемого антитела H0052-mFa55/L1083-ml0 (переключаемое антитело). Вертикальная ось графика показывает концентрацию антитела в плазме.
Фиг. 77. Диаграмма, показывающая накопление антигенов после введения каждого из анти-hIL6R антител: не переключаемого антитела MRAH-mFa55/MRAL-mk0 (контрольное антитело) и анти-hIL6R переключаемого антитела H0052-mFa55/L1083-ml0 (переключаемое антитело). Вертикальная ось графика показывает концентрацию растворимого hIL6R в плазме. IC17Hdk-mFa55/IC17L-mk1 (обозначенного на фигуре как KLH-mFa55) используют в качестве отрицательного контрольного антитела.
Фиг. 78. Диаграмма, показывающая зависящую от концентрации АТФ активность анти-PD1 антител mPD1F2VH-mF18/mPD1F2VL-mk1 (контрольное антитело) и H5029-mFa3 l/L3021-ml0 (переключаемое антитело) по ингибированию связывания PD-1/PDL-1.
Фиг. 79. Диаграмма, показывающая зависящую от концентрации АТФ активность анти-PD1 антител mPD1F2VH-mF18/mPD1F2VL-mk1 (контрольное антитело) и H5041-mFa3 l/L3021-ml0 (переключаемое антитело) по ингибированию связывания PD-1/PDL-1.
Фиг. 80. Диаграмма, показывающая зависящую от концентрации АМФ нейтрализующую активность in vitro анти-PD1 антител mPD1F2VH-mF18/mPD1F2VL-mk1 (контрольное антитело), H5029-mFa31/L3021-ml0 и H5041-mFa31/L3021-ml0 (оба являются переключаемыми антителами).
Фиг. 81. Диаграмма, показывающая зависящую от концентрации АТФ in vitro нейтрализующую активность анти-PD1 антител mPD1F2VH-mF18/mPD1F2VL-mk1 (контрольное антитело) и H5029-mFa31/L3021-ml0 и H5041-mFa31/L3021-ml0 (оба являются переключаемыми антителами).
Фиг. 82. Диаграмма, показывающая противоопухолевую активность in vivo анти-PD1 антител mPD1F2VH-mFa55/mPD1F2VL-mk1 (контрольное антитело) и H5041-mFa55/L3023-ml0 (переключаемое антитело). IC17Hdk-mFa55/IC17L-mk1 является отрицательным контрольным антителом.
Фиг. 83. Диаграмма, показывающая активность анти-PD1 антител mPD1F2VH-mFa55/mPD1F2VL-mk1 (контрольное антитело) и Н5041-mFa55/L3023-ml0 (переключаемое антитело) в элиминировании PD-1-экспрессирующих клеток из (А) опухоли и (Б) селезенки. На фигуре «изотип» представляет отрицательное контрольное антитело (IC17Hdk-mFa55/IC17L-mk1).
Фиг. 84. Диаграмма, показывающая механизм связывания АТФ и фрагментом Fab анти-hIL6R переключаемого антитела H0041L1088. На этой фигуре показывают АТФ в виде шаростержневой модели и аминокислотные остатки, взаимодействующие с АТФ, показывают в виде стержневой модели. Пунктирные линии указывают на водородные связи между антителом и АТФ.
Фиг. 85. Диаграмма, показывающая аминокислотную последовательность внеклеточного домена hIL6R (shIL6R), картированного с эпитопом анти-hIL6R переключаемого антитела H0041L1088. На фигуре аминокислотные остатки, заштрихованные серым цветом, представляют собой остатки (остатки эпитопа) shIL6R, содержащие один или несколько атомов, не являющихся водородом и расположенных на расстоянии 4,2 ангстрема или менее от АТФ или Fab H0041L1088 в кристаллической структуре.
Фиг. 86. Диаграмма, показывающая детали связывания между shIL6R и фрагмента Fab АТР-связанного H0041L1088. На рисунке тяжелая цепь антитела показана черным цветом, легкая цепь - серым цветом, a shIL6R - белым. На фигуре АТФ показан в виде шарнирной модели, а остаток эпитопа shIL6R в пределах 4,2 ангстрем от антитела или АТФ и паратопный остаток антитела в пределах 4,2 ангстрем от остатка эпитопа показаны с помощью стержневой модели. Пунктирные линии указывают на водородные связи между антителом и shIL6R. Чтобы прояснить взаимодействие с АТФ, только F298 shIL6R показан с помощью шаровой модели.
Фиг. 87. Диаграмма, показывающая структуру, отличающуюся от структуры на фиг.86 поворотом на 180 градусов (вид сзади).
Фиг. 88. Диаграмма, показывающая агонистическую активность различных переключаемых анти-CD137 антител, протестированных с использованием анализа репортерного гена 4-1ВВ Jurkat в присутствии АТФ.
Фиг. 89. Диаграмма, показывающая сравнение в плазме показателей кинетики для каждого из анти-CD137 переключаемых антител А375-SCF041aPh/B167-Lamlib и A375-MY201aPh/B167-Lamlib. Вертикальная ось графика показывает концентрацию каждого антитела в плазме.
Фиг. 90. Диаграмма, показывающая противоопухолевый эффект каждого из A375/B167-SCF041aPh и A375/B167-MY201aPh в модели мыши, полученной путем трансплантации линии клеток LLC1/OVA/GPC3 мышам hCD137KI/mFcγR2bKO/hFcγR2bTg#90.
Каждая точка показывает среднюю величину объемов опухолей в группе (n=5).
Фиг. 91. Диаграмма, показывающая агонистическую активность различных переключаемых анти-CD3 антител, исследованных с помощью анализа репортерного гена, используя активацию Т-клеток Bioassay (NFAT) в присутствии АТФ.
Формы осуществления настоящего изобретения
I. Определения
Понятие «связывающая активность» относится к силе общей суммы нековалентных взаимодействий между одним или несколькими сайтами связывания молекулы (например, антитела) и их связывающим партнером (например, антигеном). В настоящем изобретении «связывающая активность» строго не ограничивается взаимодействием 1:1 между представителями связывающейся пары (например, антителом и антигеном). Например, если представители связывающейся пары отражают одновалентное взаимодействие 1:1, связывающую активность в частности называют внутренним связывающим сродством (аффинностью). Если член связывающейся пары способен как к одновалентному связыванию, так и к поливалентному связыванию, активность связывания является суммой каждой силы связывания. Связывающая активность молекулы X в отношении ее партнера Y обычно может быть представлена константой диссоциации (KD) или «связывающим количеством исследуемого соединения на единицу количества лиганда» (ниже может называться «связывающее количество»). Специалистам очевидно, что, как правило, более низкое значение константы диссоциации (KD) означает более высокую связывающую активность, а более высокое значение «количества связывания аналита на единицу количества лиганда» или «количества связывания» означает более высокую активность связывания. Связывающую активность можно измерить обычными методами, известными в данной области, включая описанные в настоящем изобретении. Конкретные иллюстрации и примеры вариантов осуществления настоящего изобретения для измерения связывающей активности описаны ниже.
Понятие антигенсвязывающей молекулы или антитела с «созревшей связывающей активностью» или антигенсвязывающей молекулы или антитела с «повышенной (усиленной) связывающей активностью» относится к антителу с одним или несколькими изменениями в одной или нескольких гипервариабельных областях (HVR) по сравнению к исходной антигенсвязывающей молекулой или родительским антителом, которые не несут таких изменений; такие соединения приобретают улучшенную связывающую активность антигенсвязывающей молекулы или антитела в отношении антигена.
Термины «анти-CD137 антигенсвязывающая молекула против» или «анти-CD137-антитело» и «антигенсвязывающая молекула, которая связывается с CD137» или «антитело, которое связывается с CD137» относятся к антигенсвязывающей молекуле или антителу, которые способны связываться с CD137 с достаточной связывающей активностью, в соответствии с чем антигенсвязывающая молекула или антитело может использоваться в качестве диагностического и/или терапевтического агента для нацеливания на CD137. В некоторых вариантах осуществления настоящего изобретения анти-CD137 антитело связывается с эпитопом CD137, который консервативен среди CD137 разных видов.
Понятие анти-CD137 антигенсвязывающей активности или анти-CD137 антитела, «обладающего CD137 связывающей активностью, зависящей от низкомолекулярного соединения», означает антигенсвязывающую молекулу или антитело, показывающее повышенную связывающую активность с CD137 в присутствии низкомолекулярного соединения по сравнению со связывающей активностью с CD137 в отсутствие низкомолекулярного соединения. В одном из вариантов осуществления настоящего изобретения «наличие низкомолекулярного соединения» относится к условию, при котором низкомолекулярное соединение присутствует в концентрации 10 мкМ или более, 50 мкМ или более, 100 мкМ или более, 150 мкМ или более, 200 мкМ или более или 250 мкМ или более. В одном из вариантов осуществления настоящего изобретения степень связывающей активности анти-CD137 антигенсвязывающей молекулы или антитела с неродственным, белком, не являющимся CD137, в присутствии низкомолекулярного соединения составляет примерно менее 10% связывания антигенсвязывающей молекулы или антитела с CD137 по данным измерения, например, с помощью радиоиммуноанализа (RIA, radioimmunoassay) или поверхностного плазмонного резонанса (SPR, surface plasmon resonance). В некоторых вариантах осуществления настоящего изобретения в присутствии низкомолекулярного соединения анти-CD137 антигенсвязывающая молекула или антитело имеет константу диссоциации (KD) 1 мкМ или менее, 100 нМ или менее, 10 нМ или менее, 1 нМ или менее, 0,1 нМ или менее, 0,01 нМ или менее, или 0,001 нМ или менее (например, 10-6 М или менее, 10-7 М или менее, 10-8 М или менее, 10-9 М или менее, 10-10 М или менее, например, от 10-6 М до 10-10 М, от 10-7 М до 10-9 М, например, от 10-7 М до 10-8 М).
В настоящем изобретении термин «антигенсвязывающая молекула» используют в самом широком смысле и он относится к молекуле, которая специфически связывается с антигенной детерминантой. В одном из вариантов осуществления настоящего изобретения антигенсвязывающая молекула представляет собой антитело, фрагмент антитела или производное антитела.
В контексте настоящего изобретения понятие «агонистическая антигенсвязывающая молекула» или «агонистическое антитело» в контексте настоящего изобретения представляет собой антигенсвязывающую молекулу или антитело, которые значительно индуцируют или усиливают биологическую активность антигена, с которым они связываются (например, CD137 и CD3).
Следовательно, если антигеном является, например, CD137, такая антигенсвязывающая молекула или антитело, обладающее агонистическим действием, называется «агонистической антигенсвязывающей молекулой CD137» или «агонистическим антителом CD137», соответственно.
В контексте настоящего изобретения понятие «антитело» используют в самом широком смысле и оно охватывает различные структуры антител, включая, но ими не ограничиваясь, моноклональные антитела, поликлональные антитела, мультиспецифические антитела (например, биспецифические антитела) и фрагменты антител, если они проявляют необходимую антигенсвязывающую активность.
В контексте настоящего изобретения понятие «фрагмент антитела» относится к молекуле, отличной от интактного антитела, которая включает часть интактного антитела, связывающую антиген, с которым связывается интактное антитело. Примеры фрагментов антител включают, но ими перечень не ограничивают, Fv, Fab, Fab', Fab' -SH, F(ab')2; диатела; линейные антитела; молекулы одноцепочечных антител (например, scFv); и мультиспецифические антитела, образованные из фрагментов антител.
«Антигенсвязывающая молекула, которая связывается с тем же эпитопом» или «антитело, которое связывается с тем же эпитопом», что и контрольная антигенсвязывающая молекула или контрольное антитело, относится к антителу или антигенсвязывающей молекуле, которые блокируют связывание контрольного антитела или контрольной антигенсвязывающей молекулы со своим антигеном в конкурентном анализе на 50% или более, и, наоборот, контрольное антитело блокирует связывание антитела с его антигеном в конкурентном анализе на 50% или более. Пример конкурентного анализа приводят в настоящем изобретении. В одном из вариантов осуществления настоящего изобретения в случае, если контрольная антигенсвязывающая молекула или контрольное антитело проявляет антигенсвязывающую активность в зависимости от низкомолекулярного соединения, конкурентный анализ проводят в присутствии низкомолекулярного соединения.
Понятие «химерное» антитело относится к антителу, в котором часть тяжелой и/или легкой цепи происходит из определенного источника или вида, в то время как остальная часть тяжелой и/или легкой цепи происходит из другого источника или вида.
Понятие «класс» антитела определяется типом константного домена или константной области, которой обладает его тяжелая цепь. Существует пять основных классов антител: IgA, IgD, IgE, IgG и IgM, и некоторые из них могут быть далее разделены на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелой цепи, которые соответствуют различным классам иммуноглобулинов, называются альфа, дельта, эпсилон, гамма и мю, соответственно.
Понятие «эффекторные функции» относятся к тем биологическим активностям, приписываемым области Fc антитела, которые варьируют в зависимости от изотипа антитела. Примеры эффекторных функций антитела включают: связывание C1q и комплементзависимую цитотоксичность (CDC, complement dependent cytotoxicity); связывание с рецептором Fc; антителозависимую клеточно-опосредованную цитотоксичность (ADCC, antibody-dependent cell-mediated cytotoxicity); фагоцитоз; понижающую регуляцию рецепторов на клеточной поверхности (например, рецептора В-клеток); и активацию В-клеток.
Понятие «цитотоксичность» относится к активности, которая ингибирует или препятствует клеточной функции и/или вызывает гибель или разрушение клеток. Цитотоксичность может представлять собой, например, активность антителозависимой клеточно-опосредованной цитотоксичности (ADCC), активность комплемент-зависимой цитотоксичности (CDC) и цитотоксичность, обусловленную Т-клетками; также может быть цитотоксичность, вызванная цитотоксическими агентами (например, радиоизотопами и химиотерапевтическими агентами), такими как иммуноконъюгаты.
В контексте настоящего изобретения понятие «область Fc» используют для определения С-концевой области тяжелой цепи иммуноглобулина, которая содержит, по меньшей мере, часть константной области. Понятие включает области Fc с нативной последовательностью и варианты области Fc. В одном из вариантов осуществления настоящего изобретения область Fc тяжелой цепи IgG человека располагается от Cys226 или от Pro230 до карбоксильного конца тяжелой цепи. Однако С-концевой лизин (Lys447) или глицин-лизин (остатки 446-447) области Fc могут присутствовать или их может не быть. Если в настоящем изобретении не указано иное, нумерация аминокислотных остатков в области Fc или константной области соответствует системе нумерации EU, также называемой индексом EU, как описано в книге Kabat с соавт., Sequences of Proteins of Immunological Interest, 5e изд., изд-во Национальные институты здравоохранения, Бетесда, Мэриленд, 1991.
Понятие «вариант области Fc» в настоящем изобретении включает аминокислотную последовательность, которая отличается от нативной последовательности области Fc по меньшей мере одной модификацией аминокислоты, предпочтительно одной или несколькими аминокислотными заменами. Предпочтительно вариант области Fc имеет по меньшей мере одну аминокислотную замену по сравнению с нативной последовательностью области Fc или с областью Fc исходного полипептида, например, от примерно одной до примерно десяти аминокислотных замен, и предпочтительно от примерно одной до примерно пяти аминокислотных замен в нативной последовательности области Fc или в области Fc исходного полипептида. В настоящем изобретении вариант области Fc может предпочтительно обладать по меньшей мере примерно 80% гомологии с нативной последовательностью области Fc и/или с областью Fc исходного полипептида, и наиболее предпочтительно по меньшей мере примерно 90% гомологии, более предпочтительно по меньшей мере примерно 95% гомологии.
В настоящем изобретении аминокислотные изменения или замены в области Fc или константной области могут быть представлены комбинацией аминокислот, выраженной по системе нумерации EU. Например, S424N обозначает замену в положении 424 по нумерации ЕС серина (Ser) на аспарагин (Asn). EU424N означает замену в положении 424 по нумерации EU от аминокислоты (любого типа) на аспарагин (Asn).
В настоящем изобретении понятие «антитело, содержащее область Fc» означает антитело, которое содержит область Fc. С-концевой лизин (остаток 447 по системе нумерации EU) или С-концевой глицин-лизин (остатки 446-447) области Fc можно удалить, например, во время очистки антитела или рекомбинантной инженерии нуклеиновой кислоты, кодирующей антитело. Соответственно, композиция, содержащая антитело, имеющее область Fc в соответствии с настоящим изобретением, может содержать антитело с G446-K447, с G446 и без K447, со всеми удаленными G446-K447, или смесь трех типов антител, описанных выше.
В настоящем изобретении понятия «антитело полной длины», «интактное антитело» и «целое антитело» используют взаимозаменяемо для обозначения антитела, имеющего структуру, по существу аналогичную структуре нативного антитела, или имеющего тяжелые цепи, которые содержат область Fc или вариант области Fc согласно описанию настоящего изобретения.
Понятие «антитело человека» представляет собой антитело, которое обладает аминокислотной последовательностью, соответствующей последовательности антитела, вырабатываемого организмом человека, или клеткой человека, или полученного от другого источника (не от человека), в котором используют репертуары антител человека или другие последовательности, кодирующие антитела человека. Это определение антитела человека специально исключает гуманизированное антитело, содержащее антигенсвязывающие остатки, не относящиеся к антителу человека.
Понятие «каркасный участок (FR, framework)» относится к остаткам вариабельного домена, отличным от остатков гипервариабельной области (HVR). FR вариабельного домена обычно состоит из четырех доменов FR: FR1, FR2, FR3 и FR4. Соответственно, последовательности HVR и FR обычно появляются в следующем порядке в VH (или VL): FR1-H1(L1)-FR2-H2(L2)-FR3-H3(L3)-FR4.
В настоящем изобретении понятие «акцепторная каркасная область человека» представляет собой каркас, содержащий аминокислотную последовательность каркасной области вариабельного домена легкой цепи (VL) или каркасной области вариабельного домена тяжелой цепи (VH), полученной из каркаса иммуноглобулина человека или консенсусной каркасной области человека согласно описанному ниже. Акцепторная каркасная область человека, «происходящая из» каркасной области иммуноглобулина человека или консенсусной каркасной области человека, может содержать такую же аминокислотную последовательность, или она может содержать изменения аминокислотной последовательности. В некоторых вариантах осуществления настоящего изобретения количество изменений аминокислот равно 10 или менее, 9 или менее, 8 или менее, 7 или менее, 6 или менее, 5 или менее, 4 или менее, 3 или менее или 2 или менее. В некоторых вариантах осуществления настоящего изобретения акцепторная каркасная область VL человека идентична последовательности каркасной области VL иммуноглобулина человека или последовательности консенсусной каркасной области человека.
В настоящем изобретении понятие ««консенсусная каркасная область человека» означает наиболее часто встречающиеся аминокислотные остатки при отборе последовательностей VL или VH каркасного участка иммуноглобулина человека. Как правило, выбор последовательностей VL или VH иммуноглобулина человека осуществляют из подгруппы последовательностей вариабельных доменов. Обычно подгруппа последовательностей является подгруппой, описанной в книге Kabat с соавт., Sequences of Proteins of Immunological Interest, 5e изд., изд-во NIH Publication 91-3242, 1991, тома 1-3, Бетесда, Мэриленд. В одном из вариантов осуществления настоящего изобретения для VL подгруппой является подгруппа каппа I, описанная в книге Kabat с соавт., supra. В одном из вариантов осуществления настоящего изобретения для VH подгруппой является подгруппа III, описанная в книге Kabat с соавт., supra.
Понятие «гуманизированное» антитело относится к химерному антителу, содержащему аминокислотные остатки из HVR, не относящихся к человеку, и аминокислотные остатки из FR человека. В некоторых вариантах осуществления настоящего изобретения гуманизированное антитело может включать практически все из по меньшей мере одного, обычно двух вариабельных доменов, в которых все или практически все HVR (например, CDR) соответствуют таковым из антитела, не являющегося антителом человека, и все или практически все из FR соответствуют таковым антитела человека. Гуманизированное антитело необязательно может содержать по меньшей мере часть константной области антитела, полученной из антитела человека. «Гуманизированная форма» антитела, например, не являющегося антителом человека, относится к антителу, которое было гуманизировано.
Термин «вариабельная область» или «вариабельный домен» относится к домену тяжелой или легкой цепи антитела, который участвует в связывании антитела с антигеном. Вариабельные домены тяжелой цепи и легкой цепи (VH и VL, соответственно) нативного антитела обычно имеют сходные структуры, причем каждый домен включает четыре консервативных каркасных области (FR) и три гипервариабельные области (HVR). (См., например, Kindt с соавт. Kuby Immunology, 6th ed., изд-во W.H. Freeman and Co., 2007, c. 91). Одного домена VH или VL может быть достаточно для придания антигенсвязывающей специфичности. Кроме того, антитела, которые связывают конкретный антиген, могут быть выделены с использованием домена VH или VL из антитела, которое связывает антиген, для скрининга библиотеки комплементарных доменов VL или VH, соответственно. См., например, Portolano с соавт., J. Immunol. 150, 1993, 880-887; Clarkson с соавт., Nature 352, 1991, 624-628.
Термин «гипервариабельная область» или «HVR (hypervariable region)» в контексте настоящего изобретения относится к каждой из областей вариабельного домена антитела, которые являются гипервариабельными по последовательности («области, определяющие комплементарность» или «CDR (complementarity determining regions)») и/или образуют структурно определенные петли («гипервариабельные петли») и/или содержат остатки, контактирующие с антигеном («контакты антигена»). Обычно антитела содержат шесть HVR: три в VH (H1, Н2, Н3) и три в VL (L1, L2, L3). Примеры HVR в данном документе включают:
(а) гипервариабельные петли, встречающиеся при аминокислотных остатках 26-32 (L1), 50-52 (L2), 91-96 (L3), 26-32 (H1), 53-55 (Н2) и 96-101 (Н3) (Chothia, Lesk, J. Mol. Biol. 196, 1987, 901-917);
(6) CDR, несущие аминокислотные остатки 24-34 (L1), 50-56 (L2), 89-97 (L3), 31-35b (H1), 50-65 (H2) и 95-102 (Н3) (Kabat с соавт., Sequences of Proteins of Immunological Interest, 5e изд., изд-во Национальные институты здравоохранения, Бетесда, Мэриленд, 1991);
(в) контакты антигена по аминокислотным остаткам 27 с-36 (L1), 46-55 (L2), 89-96 (L3), 30-35b (H1), 47-58 (Н2) и 93-101 (Н3) (MacCallum с соавт., J. Mol. Biol, 262, 1996, 732-745);
(г) комбинации (а), (б) и/или (в), включающие аминокислотные остатки HVR 46-56 (L2), 47-56 (L2), 48-56 (L2), 49-56 (L2), 26-35 (H1), 26-35b (H1), 49-65 (Н2), 93-102 (Н3) и 94-102 (Н3).
Если не указано иное, остатки HVR и другие остатки в вариабельном домене (например, остатки FR) пронумерованы в настоящем изобретении согласно Kabat с соавт., supra. В настоящем изобретении остатки HVR или другие остатки в вариабельном домене (например, остатки FR) и аминокислотные изменения или замены таких остатков могут быть представлены в виде комбинации по системе нумерации Kabat. Например, N99 означает аспарагин (Asn) в позиции 99 по нумерации Kabat, a N99A означает замещение в позиции 99 по нумерации Kabat аспарагина (Asn) на аланин (Ala).
В настоящем изобретении понятие «иммуноконъюгат» представляет собой антитело, конъюгированное с одной или несколькими гетерологичными молекулами (молекулой), включая, но им не ограничиваясь, цитотоксический агент.
В настоящем изобретении понятие «цитотоксический агент» относится к веществу, которое ингибирует или препятствует проявлению клеточной функции и/или вызывает гибель или разрушение клеток. Цитотоксические агенты включают, но не ограничиваются ими, радиоактивные изотопы (например, 211At, 131I, 125I, 90Y, 186Re, 188Re, 153Sm, 212Bi, 32P, 212Pb и радиоактивные изотопы Lu); химиотерапевтические агенты или лекарства (например, метотрексат, адриамицин, алкалоиды барвинка (винкристин, винбластин, этопозид), доксорубицин, мелфалан, митомицин С, хлорамбуцил, даунорубицин или другие интеркалирующие агенты); агенты, ингибирующие рост; ферменты и их фрагменты, такие как нуклеолитические ферменты; антибиотики; токсины, такие как низкомолекулярные токсины или ферментативно активные токсины бактериального, грибкового, растительного или животного происхождения, включая их фрагменты и/или варианты; и различные противоопухолевые или противораковые агенты, описанные ниже.
Понятие «выделенное» антитело - это антитело, которое было отделено от компонентов его естественного окружения. В некоторых вариантах осуществления настоящего изобретения антитело очищают до более чем 95% или 99% чистоты, что определяют, например, электрофоретическим методом (например, SDS-PAGE, изоэлектрическим фокусированием (IEF), капиллярным электрофорезом) или хроматографией (например, ионообменной или обращенно-фазовой ВЭЖХ). Для обзора методов оценки чистоты антител см., например, публикацию Flatman с соавт., J. Chromatogr. В 848, 2007, 79-87.
Понятие «выделенная» нуклеиновая кислота относится к молекуле нуклеиновой кислоты, которая была отделена от компонентов ее естественного окружения. Выделенная нуклеиновая кислота включает молекулу нуклеиновой кислоты, содержащуюся в клетках, которые обычно содержат молекулу нуклеиновой кислоты, но молекула нуклеиновой кислоты присутствует вне хромосомы или имеет хромосомную локализацию, которая отличается от ее естественной хромосомной локализации.
В настоящем изобретении понятие «вектор» относится к молекуле нуклеиновой кислоты, способной размножаться с другой нуклеиновой кислотой, с которой она связана. Понятие включает вектор как самореплицирующуюся структуру нуклеиновой кислоты, а также вектор, включенный в геном клетки-хозяина, в которую он был введен. Некоторые векторы способны управлять экспрессией нуклеиновых кислот, с которыми они функционально связаны. Такие векторы называются в настоящем изобретении «векторами экспрессии».
Понятие «кодированная нуклеиновая кислота, кодирующая анти-CD137 антигенсвязывающую молекулу» относится к одной или более молекулам нуклеиновой кислоты, которые кодируют полипептид (полипептиды), составляющие антигенсвязывающую молекулу. Понятие «выделенная нуклеиновая кислота, кодирующая анти-CD137 антитело» относится к одной или более молекулам нуклеиновой кислоты, кодирующим тяжелую и легкую цепи антитела (или их фрагменты), включая такую молекулу (молекулы) нуклеиновой кислоты в одном векторе или в отдельных векторах, и такую молекулу (молекулы) нуклеиновой кислоты, присутствующие в одном или нескольких местах локализации в клетке-хозяине.
Понятия «клетка-хозяин», «линия клеток-хозяев» и «культура клеток-хозяев» используются взаимозаменяемо и относятся к клеткам, в которые была введена экзогенная нуклеиновая кислота, включая потомство таких клеток. К клеткам-хозяевам относятся «трансформанты» и «трансформированные клетки», которые включают первичную трансформированную клетку и потомство, полученное от нее, независимо от количества пассажей. Потомство может не быть полностью идентичным родительской клетке по содержанию нуклеиновой кислоты, но может содержать мутации. В настоящем изобретении к данным понятиям также относятся мутантные потомки, которые обладают той же функцией или биологической активностью, что и проверенные или отобранные среди первоначально трансформированных клеток.
В контексте настоящего изобретения понятие «моноклональное антитело» относится к антителу, полученному из популяции по существу гомогенных антител, т.е. отдельные антитела, составляющие популяцию, идентичны и/или связываются с одним и тем же эпитопом, за исключением возможных вариантных антител, например, содержащих встречающиеся в природе мутации или возникающие во время получения препарата моноклонального антитела, такие варианты обычно присутствуют в незначительных количествах. В отличие от препаратов поликлональных антител, которые обычно включают разные антитела, направленные против разных детерминант (эпитопов), каждое моноклональное антитело в препарате моноклональных антител направлено против одной детерминанты на антигене. Таким образом, определение «моноклональное» указывает на характер антитела, как полученного по существу из гомогенной популяции антител, и не должно толковаться как требующее получения антитела каким-либо конкретным методом. Например, моноклональные антитела, которые будут использоваться в соответствии с настоящим описанием, могут быть получены различными методами, включая, но ими не ограничиваясь, гибридомный метод, методы рекомбинантной ДНК, методы фагового дисплея и методы с использованием трансгенных животных, содержащих все или часть локусов иммуноглобулина человека, а также другие методы получения моноклональных антител, примеры применения которых описаны в настоящем изобретении.
Термин «голое антитело» относится к антителу, которое не конъюгировано с гетерологичным фрагментом (например, с цитотоксическим фрагментом) или с радиоактивной меткой. Голое антитело может присутствовать в фармацевтическом составе.
Понятие «нативные антитела» относится к природным молекулам иммуноглобулинов различной структуры. Например, нативные антитела IgG представляют собой гетеротетрамерные гликопротеины массой примерно 150000 дальтон, состоящие из двух идентичных легких цепей и двух идентичных тяжелых цепей, связанных дисульфидной связью. От N- к С-концу каждая тяжелая цепь имеет вариабельную область (VH), также называемую вариабельным тяжелым доменом или вариабельным доменом тяжелой цепи, за которой следуют три константных домена (CH1, СН2 и СН3). Аналогично, от N- до С-конца каждая легкая цепь имеет вариабельную область (VL), также называемую вариабельным легким доменом или вариабельным доменом легкой цепи, за которой следует константный домен легкой цепи (CL). Легкая цепь антитела может быть отнесена к одному из двух типов, называемых каппа (κ) и лямбда (λ) на основе аминокислотной последовательности соответствующего константного домена.
Понятие «процент (%) идентичности аминокислотной последовательности» по отношению к контрольной полипептидной последовательности определяют как процент аминокислотных остатков в последовательности-кандидате, которые идентичны аминокислотным остаткам в контрольной полипептидной последовательности, после выравнивания последовательностей и введения гэпов, если необходимо, для достижения максимального процента идентичности последовательностей, и без учета каких-либо консервативных замен как части идентичности последовательностей. Выравнивание для определения процента идентичности аминокислотной последовательности может быть достигнуто различными методами, которые известны специалистам в данной области, например, с использованием общедоступного компьютерного программного обеспечения, такого как BLAST, BLAST-2, ALIGN, Megalign (DNASTAR) или GENETYX (зарегистрированная торговая марка) (фирма Genetyx Co., Ltd.). Специалисты в данной области могут определить подходящие параметры для выравнивания последовательностей, включая любые алгоритмы, необходимые для достижения максимального выравнивания по всей длине сравниваемых последовательностей.
Компьютерная программа для сравнения последовательностей ALIGN-2 была разработана фирмой Genentech, Inc., а исходный код был подан вместе с пользовательской документацией в Бюро регистрации авторских прав США, Вашингтон, округ Колумбия, 20559, где он зарегистрирован под номером регистрации авторских прав US TXU510087. Программа ALIGN-2 находится в открытом доступе от фирмы Genentech, Inc., Южный Сан-Франциско, Калифорния, или может быть скомпилирована из исходного кода. Программа ALIGN-2 может быть скомпилирована для использования в операционной системе UNIX, включая цифровую операционную систему UNIX V4.0D. Все параметры сравнения последовательностей задаются программой ALIGN-2 и не меняются. В ситуациях, когда ALIGN-2 используют для сравнения аминокислотных последовательностей, процент идентичности данной аминокислотной последовательности аминокислотной последовательности А с/против аминокислотной последовательности Б (которая альтернативно может быть сформулирована как данная аминокислота последовательность А, которая имеет или содержит определенный процент идентичности аминокислотной последовательности с данной аминокислотной последовательностью В, с ней или против нее) рассчитывают следующим образом:
100-кратная фракция X/Y
где X означает количество аминокислотных остатков, оцененных как идентичные совпадения программой выравнивания последовательностей ALIGN-2 при выравнивании А и Б этой программой, и где Y означает общее количество аминокислотных остатков в Б. Следует учитывать, что если длина аминокислотной последовательности А не равна длине аминокислотной последовательности Б, процент идентичности аминокислотной последовательности А к Б не будет равняться проценту идентичности аминокислотной последовательности Б к А. Если специально не указано иное, все значения процентов идентичности аминокислотной последовательности, используемые в настоящем изобретении, получены непосредственно по описанию в предшествующем абзаце, с использованием компьютерной программы ALIGN-2.
Понятие «фармацевтический состав» относится к препарату, который находится в такой форме, которая позволяет биологической активности содержащегося в нем активного ингредиента быть эффективной, и который не содержит дополнительных компонентов, которые являются неприемлемо токсичными для субъекта, которому состав будет введен.
Понятие «фармацевтически приемлемый носитель» относится к ингредиенту фармацевтического состава, отличному от активного ингредиента, который нетоксичен для субъекта. Фармацевтически приемлемый носитель включает, но ими не ограничивается, буфер, наполнитель, стабилизатор или консервант.
Понятие «эффективное количество» агента, например фармацевтического состава, относится к количеству, эффективному в дозировках и в течение периодов времени, необходимых для достижения желаемого терапевтического или профилактического результата.
Понятие «индивидуум» или «субъект» означает млекопитающее. Млекопитающие включают, но ими возможный перечень не ограничивают, домашних животных (например, коров, овец, кошек, собак и лошадей), приматов (например, людей и других приматов, таких как обезьяны), кроликов и грызунов (например, мышей и крысы). В некоторых вариантах осуществления настоящего изобретения индивидом или субъектом является человек.
В настоящем изобретении термин «CD137» относится к любому природному CD137 от любого из позвоночных, включая млекопитающих, таких как приматы (люди) и грызуны (например, мыши и крысы), если не указано иное. Понятие охватывает необработанный CD137«полной длины», а также любую форму CD137, которая является результатом процессинга в клетке. Термин также включает встречающиеся в природе варианты CD137, например, варианты сплайсинга или аллельные варианты. Аминокислотная последовательность примера CD137 полной длины человека показана в SEQ ID NO: 1 (контрольная последовательность NCBI: NP_001552.2), а пример аминокислотной последовательности примера внеклеточного участка CD137 человека показана в SEQ ID NO: 2. Аминокислотная последовательность типичного CD137 полной длины мыши показана в SEQ ID NO: 3 (контрольная последовательность NCBI: NP_035742.1), аминокислотная последовательность типичного внеклеточного участка CD137 мыши показана в SEQ ID NO: 4. Аминокислотная последовательность примера CD137 полной длины обезьяны показана в SEQ ID NO: 5 (контрольная последовательность NCBI: ABY47575.1), аминокислотная последовательность примера внеклеточного участка CD137 обезьяны показана в SEQ ID NO: 6.
CD137 является представителем семейства рецепторов фактора некроза опухоли (TNF). Его также обозначают как представитель 9 надсемейства рецепторов фактора некроза (TNFRSF9, tumor necrosis factor receptor superfamily member 9), 4-1BB и ILA. В дополнение к экспрессии на активированных CD4+ и CD8+ Т-клетках, CD137 экспрессируется в В-клетках, дендритных клетках, природный клетках-киллерах (NK) и NK-Т-клетках, макрофагах, моноцитах, нейтрофилах, CD4+ CD25+ регуляторных Т-клетках и эндотелиальных клетках сосудов. Экспрессия в опухолевых клетках также описана (Labiano с соавт. Oncoimmunology, 24, 2015, е1062967). Природный лиганд CD137, CD137L, представлен антигенпрезентирующими клетками, такими как В-клетки, моноциты/макрофаги и дендритные клетки (Watts с соавт., Annu. Rev. Immunol., 23, 2005, 23-68). Благодаря взаимодействию с лигандом, CD137 вызывает увеличение TCR-индуцированной пролиферации Т-клеток, выработку цитокинов, функциональное созревание, подавление апоптоза и длительное выживание CD8+ Т-клеток (Nam с соавт., Curr. Cancer Drug Targets, 5, 2005, 357-363; Watts, с соавт., Annu. Rev. Immunol., 23, 2005, 23-68).
Термины «карцинома», «рак» и «злокачественный» относятся или описывают физиологическое состояние у млекопитающих, которое обычно характеризуется нерегулируемым ростом/пролиферацией клеток.
Термин «опухоль» относится ко всем неопластическим клеткам роста и пролиферации, злокачественным или доброкачественным, и ко всем предраковым и злокачественным клеткам и тканям. Термины «карцинома», «рак», «злокачественный», «нарушение пролиферации клеток», «пролиферативное расстройство» и «опухоль» не являются взаимоисключающими в контексте настоящего изобретения.
Понятия «нарушение пролиферации клеток» и «пролиферативное расстройство» относятся к нарушениям, которые связаны с некоторой степенью аномальной пролиферации клеток. В одном из вариантов осуществления настоящего изобретения нарушение пролиферации клеток представляет собой рак. В одном из вариантов осуществления настоящего изобретения пролиферативное расстройство является раком.
В контексте настоящего изобретения понятие «лечение» (и его грамматические вариации, такие как «лечить») относится к клиническому вмешательству в попытке изменить состояние индивида, и может проводиться либо для профилактики, либо в ходе происходящей клинической патологии. Желательные эффекты лечения включают, но не ограничиваются ими, предотвращение возникновения или рецидива заболевания, облегчение симптомов, уменьшение любых прямых или косвенных патологических последствий заболевания, предотвращение метастазирования, снижение скорости прогрессирования заболевания, улучшение или паллиативное улучшение болезненного состояния и ремиссия или улучшенный прогноз. В некоторых вариантах осуществления настоящего изобретения антитела по настоящему изобретению применяют для отсрочки развития заболевания или медленного прогрессирования заболевания.
II. Композиции и методы (анти-СР137 агонистические антигенсвязывающие молекулы)
В одном из вариантов осуществления настоящее изобретение частично основано на анти-CD137 агонистических антигенсвязывающих молекулах и на их применениях. В некоторых вариантах осуществления настоящего изобретения предусматривают антитела, которые связываются с CD137. Антитела в настоящем изобретении могут проявлять активирующее действие на иммунные клетки, цитотоксичность или противоопухолевую активность и, следовательно, полезны, например, для диагностики или лечения рака.
А. Типичные анти-СР137 антигенсвязывающие молекулы или антитела
В одном из вариантов осуществления настоящего изобретения предусматривают выделенные антигенсвязывающие молекулы или антитела, которые связываются с CD137. В некоторых вариантах осуществления настоящего изобретения анти-CD137 антигенсвязывающие молекулы или антитела
- обладают CD137-связывающей активностью, зависящей от низкомолекулярного соединения;
- связываются с внеклеточной областью CD137;
- образуют тройной комплекс вместе с низкомолекулярным соединением и CD137;
- связываются с происходящим от человека CD137 и с происходящим от обезьяны CD137;
- являются агонистами по отношению к активности CD137;
- проявляют агонистическую активность по отношению к CD137 в присутствии низкомолекулярного соединения;
- обладают низкой агонистической активностью к CD137 в отсутствие низкомолекулярного соединения; и/или
- практически не проявляют агонистической активности по отношению к CD137 в отсутствие низкомолекулярного соединения.
Связывающая активность антигенсвязывающих молекул или антител В некоторых вариантах осуществления настоящего изобретения связывающая активность антигенсвязывающих молекул или антител по настоящему изобретению в присутствии низкомолекулярного соединения обладает константой диссоциации (KD) 1 мкМ или менее, 100 нМ или менее, 10 нМ или менее, 1 нМ или менее, 0,1 нМ или менее, 0,01 нМ или менее или 0,001 нМ или менее (например, 10-6 М или менее, 10-7 М или менее, 10-8 М или менее, 10-9 М или менее, 10-10 М или менее, например от 10-6 М до 10-10 М, от 10-7 М до 10-9 М, например от 10-7 М до 10-8 М).
В одном из вариантов осуществления настоящего изобретения связывающую активность антигенсвязывающих молекул или антител измеряют с помощью радиоиммуноанализа (RIA) с применением антигена с радиоактивной меткой и выражают в виде KD. В одном из вариантов осуществления настоящего изобретения осуществляют RIA с версией Fab исследуемого антитела и его антигена. Например, аффинность раствора Fab с антигеном измеряют путем уравновешивания Fab с минимальной концентрацией (125I)-меченого антигена в присутствии серии титрований немеченого антигена с последующим улавливанием связанного антигена с помощью планшета, покрытого анти-Fab антителами (см., например, Chen с соавт., J. Mol. Biol. 293, 1999, 865-881). Чтобы установить условия для анализа MICROTITER (зарегистрированная торговая марка) многолуночные планшеты (фирма Thermo Scientific) в течение ночи покрывают 5 мкг/мл захватывающего анти-Fab антитела (фирма Cappel Labs) в 50 мМ карбонате натрия (рН 9,6) и затем блокируют 2% (масса/объем) раствором бычьего сывороточного альбумина в ФСБ в течение от двух до пяти часов при комнатной температуре (примерно 23°С). В планшете без адсорбента (фирма Nunc, номер в каталоге 269620), 100 пМ или 26 пМ [125I]-антигена смешивают с серийными разведениями исследуемого фрагмента Fab (например, соответствует оценке анти-VEGF антитела, Fab-12, в публикации Presta с соавт., Cancer Res. 57, 1997, 4593-4599). Исследуемый Fab затем инкубируют в течение ночи; однако инкубирование может длиться дольше (например, примерно 65 ч), чтобы убедиться, что реакция равновесия достигнута. После этого смеси переносят в планшет с захватом для инкубации при комнатной температуре (например, в течение 1 ч). Затем раствор удаляют и планшет промывают восемь раз 0,1% полисорбатом 20 (TWEEN-20 (торговая марка)) в СФБ. После того, как планшеты высохнут, добавляют 150 мкл/лунку сцинтиллятора (MICROSCINT-20 ТМ; фирма Packard) и излучение с планшетов подсчитывают на гамма-счетчике TOPCOUNT ТМ (фирма Packard) в течение десяти мин. Концентрации каждого Fab, которые проявляют показатели менее или равные 20% максимального связывания, выбирают для использования в анализах конкурентного связывания.
В одном из вариантов осуществления настоящего изобретения для измерения связывающей активности антитела применяют методы захвата лиганда, например, используя BIACORE (зарегистрированная торговая марка) Т200 или BIACORE (зарегистрированная торговая марка) 4000 (фирма GE Healthcare, Уппсала, Швеция), которые основаны на методах анализа поверхностного плазмонного резонанса в качестве принципа измерения. Контролирующее программное обеспечение BIACORE (зарегистрированная торговая марка) используют для работы устройств. В одном из вариантов осуществления настоящего изобретения используют набор для связывания аминов (фирма GE Healthcare, Уппсала, Швеция), руководствуясь инструкциями производителя, чтобы молекула для захвата лиганда, например, антитело против метки, антитело против IgG, белка А и т.д., фиксировалась на сенсорном чипе (фирма GE Healthcare, Уппсала, Швеция), покрытом карбоксиметилдекстраном. Захватывающую лиганд молекулу разбавляют 10 мМ раствором ацетата натрия при соответствующем рН и вводят с соответствующей скоростью потока в течение соответствующего времени инъекции. Измерения связывающей активности осуществляют с использованием буфера, содержащего 0,05% полисорбат 20 (другое название Tween-20 (зарегистрированная торговая марка)), в качестве буфера для измерений при скорости потока 10-30 микролитров в минуту, при измерении температуру предпочтительно поддерживают на уровне 25°С или 37°С. Для измерения, проводимого с антигеном и/или рецептором Fc, захваченным лиганд-связывающей молекулой в качестве лиганда, вводят инъекцией антиген и/или рецептор Fc, чтобы позволить захватить их целевое количество, а затем вводят инъекцией серийное разведение антитела (исследуемого соединения), приготовленное с использованием буфера для измерения.
В одном из вариантов осуществления настоящего изобретения результаты измерений анализируют с использованием программного обеспечения BIACORE (зарегистрированная торговая марка) Evaluation Software. Расчет параметров кинетики выполняется путем подбора сенсограмм ассоциации и диссоциации одновременно с использованием модели связывания 1: 1 и скорости ассоциации (коп или ка), скорости диссоциации (koff или kd) и константы равновесной диссоциации (KD) могут быть рассчитаны. При слабой связывающей активности, в частности, для случаев, когда диссоциация является быстрой и параметры кинетики трудно вычислить, модель стационарного состояния может использоваться для расчета константы равновесной диссоциации (KD). В качестве дополнительных параметров, касающихся связывающей активности, «количество связывающего исследуемого соединения на единицу количества лиганда» можно рассчитать путем деления связывающего количества исследуемого соединения (резонансная единица: RU (resonance unit)) в конкретной концентрации на количество захваченного лиганда.
Связывающая активность, зависящая от низкомолекулярного соединения
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело обладает зависимой от низкомолекулярных соединений CD137-связывающей активностью. В одном варианте осуществления настоящего изобретения, не ограничивающего его области охвата, антигенсвязывающая молекула или анти-CD137 антитело имеет более высокую связывающую активность в отношении CD137 в присутствии низкомолекулярного соединения по сравнению со связывающей активностью в отношении CD137 в отсутствие низкомолекулярного соединения. В другом варианте осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело обладает повышенной связывающей активностью с CD137 в присутствии высокой концентрации низкомолекулярного соединения по сравнению с CD137-связывающей активностью в присутствии низкомолекулярного соединения при низкой концентрации. В одном предпочтительном варианте осуществления настоящего изобретения связывающая активность анти-CD137 антигенсвязывающей молекулы или антитела в отношении CD137 в присутствии низкомолекулярного соединения больше в 2 раза или более, в 3 раза или более, в 5 раз или более, в 10 раз или более, в 15 раз или более, в 20 раз или более, в 25 раз или более, в 30 раз или более, в 50 раз или более, в 100 раз или более, в 200 раз или более, в 300 раз или более, в 500 раз или более, кратно 1×103 или более, кратно 2×103 или более, кратно 3×103 или более, кратно 5×103 или более, кратно 1×104 или более, кратно 2×104 или более, кратно 3×104 или более, кратно 5×104 или более или кратно 1×105 или более по сравнению со связывающей активностью в отсутствие низкомолекулярного соединения. В другом предпочтительном варианте осуществления настоящего изобретения связывающая активность анти-CD137 антигенсвязывающей молекулы или антитела в отношении CD137 в присутствии низкомолекулярного соединения больше чем в 2 раза, больше чем в 3 раза, больше чем в 5 раз, больше чем в 10 раз, больше чем в 15 раз, больше чем в 20 раз, больше чем в 25 раз, больше чем в 30 раз, больше чем в 50 раз, больше чем в 100 раз, больше чем в 200 раз, больше чем в 300 раз, больше чем в 500 раз, больше чем в 1000 раз, больше чем в 2000 раз, больше чем в 3000 раз, больше чем в 5000 раз, больше чем в 10000 раз, больше чем в 20000 раз, больше чем в 30000 раз, больше чем в 50000 раз или больше чем в 100000 раз по сравнению со связывающей активностью в отсутствие низкомолекулярного соединения.
Концентрация низкомолекулярного соединения может быть любой произвольной концентрацией при условии, что обнаруживается различие в связывающей активности анти-CD137 антигенсвязывающей молекулы или антитела. В одном из вариантов осуществления настоящего изобретения концентрация низкомолекулярного соединения «в присутствии низкомолекулярного соединения» и/или «в присутствии высокой концентрации низкомолекулярного соединения» составляет, например, 100 нМ или более, 500 нМ или более, 1 мкМ или более, 3 мкМ или более, 5 мкМ или более, 10 мкМ или более, 50 мкМ или более, 100 мкМ или более, 150 мкМ или более, 200 мкМ или более, 250 мкМ или более, 300 мкМ или более, 400 мкМ или более, 500 мкМ или более или 1 мМ или более. В другом варианте концентрация может быть определена как количество, достаточное для анти-CD137 антигенсвязывающей молекулы или антитела, чтобы продемонстрировать максимальную связывающую активность. Кроме того, в одном из вариантов осуществления настоящего изобретения концентрация низкомолекулярного соединения «в присутствии низкой концентрации низкомолекулярного соединения» может составлять, например, 500 мкМ или менее, 250 мкМ или менее, 200 мкМ или менее, 150 мкМ или менее, 100 мкМ или менее, 50 мкМ или менее, 10 мкМ или менее, 1 мкМ или менее, 500 нМ или менее, 100 нМ или менее, 50 нМ или менее, или 10 нМ или 1 нМ или менее. Случай, когда концентрация низкомолекулярного соединения равна нулю, или реальная концентрация равна нулю, также может быть выбран в качестве варианта осуществления настоящего изобретения при низкой концентрации.
В настоящем изобретении термин «реальная концентрация равна нулю» означает, например, концентрацию, которая настолько мала, что ее нельзя обнаружить с помощью современных технологий, хотя низкомолекулярное соединение присутствует.
В одном из вариантов осуществления настоящего изобретения связывающая активность в отношении CD137 в присутствии
низкомолекулярного соединения в концентрации 10 мкМ, 50 мкМ, 100 мкМ, 150 мкМ, 200 мкМ или 250 мкМ больше в 2 раза или более, в 5 раз или более, в 10 раз или более, в 15 раз или более, в 16 раз или более, в 17 раз или более, в 18 раз или более, в 19 раз или более, в 20 раз или более, по сравнению со связывающей активностью в отношении CD137 в отсутствие низкомолекулярного соединения.
В одном из вариантов осуществления настоящего изобретения связывающая активность анти-CD137 антигенсвязывающей молекулы или антитела против CD137 в присутствии низкомолекулярного соединения в концентрации 10 мкМ или более больше в 2 раза или более, в 5 раз или более, в 10 раз или более, в 15 раз или более, в 16 раз или более, в 17 раз или более, в 18 раз или более, в 19 раз или более, в 20 раз или более по сравнению со связывающей активностью в отношении CD137 в отсутствие низкомолекулярного соединения. В одном из вариантов осуществления настоящего изобретения связывающая активность анти-CD137 антигенсвязывающей молекулы или антитела против CD137 в присутствии низкомолекулярного соединения в концентрации 100 мкМ или более больше в 2 раза или более, в 5 раз или более, в 10 раз или более, в 15 раз или более, в 16 раз или более, в 17 раз или более, в 18 раз или более, в 19 раз или более, в 20 раз или более по сравнению со связывающей активностью в отношении CD137 в отсутствие низкомолекулярного соединения.
В одном из вариантов осуществления настоящего изобретения связывающая активность (KD) анти-CD137 антигенсвязывающей молекулы или антитела в отношении CD137 в присутствии низкомолекулярного соединения в количестве 10 мкМ или более является константой диссоциации (KD) равной 9×10-7 М или менее, 8×10-7 М или менее, 7×10-7 М или менее, 6×10-7 М или менее, 5×10-7 М или менее, или 4×10-7 М или менее, или, предпочтительно, константой диссоциации (KD) равной 5×10-7 М или менее. В другом варианте осуществления настоящего изобретения связывающая активность (KD) анти-CD137 антигенсвязывающей молекулы или антитела в отношении CD137 в отсутствие низкомолекулярного соединения слишком большая, чтобы ее можно было подсчитать с помощью Biacore (слабая связывающая активность), или это константа диссоциации (KD) 1×10-7 М или более, 5×10-7 М или более, 7×10-7 М или более, 8×10-7 М или более, 9×10-7 М или более, 1×10-6 М или более, 2×10-6 М или более, 3×10-6 М или более, или 4×10-6 М или более, или предпочтительно, константа диссоциации (KD) 1×10-6 М или более. В другом варианте осуществления настоящего изобретения связывающая активность (KD) анти-CD137 антигенсвязывающей молекулы или антитела в отношении CD137 в присутствии низкомолекулярного соединения в количестве 100 мкМ или более составляет 9×10-7 М или менее, 8×10-7 М или менее, 7×10-7 М или менее, 6×10-7 М или менее, 5×10-7 М или менее, 4×10-7 М или менее, 3×10-7 М или менее, 2×10-7 М или менее, или 1×10-7 М или менее, или предпочтительно, константа диссоциации (KD) составляет 2×10-7 М или менее. В другом варианте осуществления настоящего изобретения связывающая активность (KD) анти-CD137 антигенсвязывающей молекулы или антитела в отношении CD137 в отсутствие низкомолекулярного соединения слишком велика для расчета с помощью Biacore (слабая связывающая активность), или представляет собой константу диссоциации (KD) 1×10-7 М или более, 5×10-7 М или более, 7×10-7 М или более, 8×10-7 М или более, 9×10-7 М или более, 1×10-6 М или более, 2×10-6 М или более, 3×10-6 М или более, или 4×10-6 М или более, или предпочтительно, константа диссоциации (KD) 1×10-6 М или более.
В одном из вариантов осуществления настоящего изобретения связывающая активность (KD) анти-CD137 антигенсвязывающей молекулы или антитела в отношении CD137 в присутствии низкомолекулярного соединения в количестве 10 мкМ или более представляет собой константу диссоциации (KD) 8×10-8 М или менее, и связывающая активность (KD) по отношению к CD137 в отсутствие низкомолекулярного соединения слишком велика, чтобы ее можно было рассчитать с помощью Biacore (слабая связывающая активность). В другом варианте осуществления настоящего изобретения связывающая активность (KD) анти-CD137 антигенсвязывающей молекулы или антитела в отношении CD137 в присутствии низкомолекулярного соединения в количестве 100 мкМ представляет собой константу диссоциации (KD) 2×10-8 М или менее, и связывающая активность по отношению к CD137 в отсутствие низкомолекулярного соединения слишком велика, чтобы ее можно было рассчитать с помощью Biacore (слабая связывающая активность).
В одном из вариантов осуществления настоящего изобретения предусматривают анти-CD137 антигенсвязывающую молекулу или антитело, у которого значение [связывающей активности (связывающей величины) по отношению к CD 137 в присутствии низкомолекулярного соединения в концентрации 10 мкМ или более]/ [связывающую активность (связывающее количество) по отношению к CD137 в отсутствие низкомолекулярного соединения)] такое же или больше, чем значение для контрольной анти-CD137 антигенсвязывающую молекулы. В другом варианте осуществления настоящего изобретения предусматривают анти-CD137 антигенсвязывающую молекулу или антитело, у которого значение [связывающая активность (количество связывания) в отношении CD137 в присутствии низкомолекулярного соединения при 100 мкМ или более]/[связывающая активность (количество связывания) в отношении CD137 в отсутствие низкомолекулярного соединения)] такое же или больше, чем значение контрольной анти-CD137 антигенсвязывающей молекулы. В любом из приведенных выше вариантов осуществления настоящего изобретения контрольная анти-CD137 антигенсвязывающая молекула может быть выбрана из анти-CD137 антител, содержащих HVR-H1, HVR-H2, HVR-H3, HVR-L1, HVR-L2 и HVR-L3, имеющих такие же аминокислотные последовательности, что и HVR-H1, HVR-H2, HVR-H3, HVR-L1, HVR-L2 и HVR-L3, содержащиеся в А375/В167, А372/В040, А356/В040, А486/В167, А487/В167, А488/В226, А489/В223, А548/В376, А551/В256, А551/В379, А555/В379, А548/В256 или А549/В167, описанных в табл. 17.
В одном из вариантов осуществления настоящего изобретения контрольная анти-CD137 антигенсвязывающая молекула представляет собой антитело, содержащее аминокислотные последовательности А375/В167, А372/В040, А356/В040, А486/В167, А487/В167, А488/В226, А489/В223, А548/В376, А551/В256, А551/В379, А555/В379, А548/В256 или А549/В167, описанные в табл. 17 в качестве комбинации вариабельной области тяжелой цепи/вариабельной области легкой цепи. В предпочтительном варианте осуществления настоящего изобретения контрольная антигенсвязывающая молекула представляет собой анти-CD137 антитело, включающее HVR-H1, HVR-Н2, HVR-H3, HVR-L1, HVR-L2 и HVR-L3, имеющие такие же аминокислотные последовательности, что и HVR-H1, HVR-H2, HVR-H3, HVR-L1, HVR-L2 и HVR-L3, содержащиеся в А375/В167. В другом варианте осуществления настоящего изобретения контрольная анти-CD137 антигенсвязывающая молекула представляет собой анти-CD137 антитело, содержащее А375/В167 в качестве комбинации вариабельной области тяжелой цепи/вариабельной области легкой цепи. В другом предпочтительном варианте осуществления настоящего изобретения контрольная антигенсвязывающая молекула представляет собой анти-CD137 антитело, содержащее HVR-H1, HVR-H2, HVR-H3, HVR-L1, HVR-L2 и HVR-L3, имеющие такие же аминокислотные последовательности, что и HVR-H1, HVR-H2, HVR-H3, HVR-L1, HVR-L2 и HVR-L3, содержащиеся в А551/В379. В другом варианте осуществления настоящего изобретения контрольная анти-CD137 антигенсвязывающая молекула представляет собой анти-CD137 антитело, содержащее А551/В379 в качестве комбинации вариабельной области тяжелой цепи/вариабельной области легкой цепи. В предпочтительном варианте осуществления настоящего изобретения контрольная антигенсвязывающая молекула содержит константные области тяжелой и легкой цепей человеческого происхождения (например, G1T3 (SEQ ID NO: 138) в качестве константной области тяжелой цепи, Х-цепи человека Lamlib (SEQ ID NO: 63) в качестве константной области легкой цепи).
В одном из вариантов осуществления настоящего изобретения предусматривают анти-CD137 антигенсвязывающую молекулу или антитело, у которого связывающая активность (связанное количество) в отношении CD137 в отсутствие низкомолекулярного соединения такое же или ниже связывающей активности контрольной анти-CD137 антигенсвязывающей молекулы, а также, связывающая активность (связанное количество) в отношении CD137 в присутствии низкомолекулярного соединения в концентрации 10 мкМ или более равно или более чем связывающая активность контрольной анти-CD137 антигенсвязывающей молекулы в отношении CD137 в тех же условиях. В другом варианте осуществления настоящего изобретения предусматривают анти-CD137 антигенсвязывающую молекулу или антитело, у которого связывающая активность в отношении CD137 в отсутствие низкомолекулярного соединения такое же или меньше связывающей активности контрольного анти-CD137 антигенсвязывающей молекулы, а также, связывающая активность (связанное количество) в отношении CD137 в присутствии низкомолекулярного соединения в концентрации 10 мкМ или более равно или более чем связывающая активность (связанное количество) контрольной анти-CD137 связывающей активности в отношении CD137 в тех же условиях. В любом из приведенных выше вариантах осуществления настоящего изобретения контрольная анти-CD137 антигенсвязывающая молекула может быть выбрана из анти-CD137 антител, содержащих HVR-H1, HVR-H2, HVR-H3, HVR-L1, HVR-L2 и HVR-L3, имеющие те же аминокислотные последовательности, что и HVR-H1, HVR-H2, HVR-H3, HVR-L1, HVR-L2 и HVR-L3, содержащиеся в А375/В167, А372/В040, А356/В040, А486/В167, А487/В167, А488/В226, А489/В223, А548/В376, А551/В256, А551/В379, А555/В379, А548/В256 или А549/В167, описанных в табл. 17.
В одном из вариантов осуществления настоящего изобретения контрольная анти-CD137 антигенсвязывающая молекула является анти-CD137 антителом, содержащим аминокислотную последовательность А375/В167, А372/В040, А356/В040, А486/В167, А487/В167, А488/В226, А489/В223, А548/В376, А551/В256, А551/В379, А555/В379, А548/В256 или А549/В167, описанных в табл. 17 в виде комбинации вариабельной области тяжелой цепи/вариабельной области легкой цепи. В другом предпочтительном варианте осуществления настоящего изобретения контрольная анти-CD137 антигенсвязывающая молекула является анти-CD137 антителом, содержащим области HVR-H1, HVR-Н2, HVR-H3, HVR-L1, HVR-L2 и HVR-L3, имеющие те же аминокислотные последовательности, что и HVR-H1, HVR-H2, HVR-H3, HVR-L1, HVR-L2 и HVR-L3, содержащиеся в А375/В167. В другом варианте осуществления настоящего изобретения контрольная анти-CD137 антигенсвязывающая молекула является анти-CD137 антителом, содержащим А375/В167 в виде комбинации вариабельной области тяжелой цепи/вариабельной области легкой цепи. В предпочтительном варианте осуществления настоящего изобретения контрольная антигенсвязывающая молекула является анти-CD137 антителом, содержащим HVR-H1, HVR-H2, HVR-H3, HVR-L1, HVR-L2 и HVR-L3, имеющие те же аминокислотные последовательности, что и HVR-H1, HVR-H2, HVR-H3, HVR-L1, HVR-L2 и HVR-L3, содержащиеся в А551/В379. В другом варианте осуществления настоящего изобретения контрольная анти-CD137 антигенсвязывающая молекула является анти-CD137 антителом, содержащим А551/В379 в виде комбинации вариабельной области тяжелой цепи/вариабельной области легкой цепи. В предпочтительном варианте осуществления настоящего изобретения контрольная антигенсвязывающая молекула содержит константные области тяжелой и легкой цепей, происходящих от человека (например, G1T3 (SEQ ID NO: 138) как константная область тяжелой цепи, % цепь Lamlib человека (SEQ ID NO: 63) как константная область легкой цепи.
В одном из вариантов осуществления настоящего изобретения предусматривают анти-CD137 антигенсвязывающую молекулу или антитело, у которого значение [связывающей активности (KD) по отношению к CD137 в присутствии низкомолекулярного соединения в концентрации 1 мкМ]/[связывающей активности (KD) по отношению к CD137 в присутствии низкомолекулярного соединения в концентрации 10 мкМ или более] такое же или больше, чем значение для контрольной анти-CD137 антигенсвязывающую молекулы. В другом варианте осуществления настоящего изобретения предусматривают анти-CD137 антигенсвязывающую молекулу или антитело, у которого значение [связывающей активности (KD) по отношению к CD137 в присутствии низкомолекулярного соединения в концентрации 1 мкМ]/[связывающей активности (KD) по отношению к CD137 в присутствии низкомолекулярного соединения в концентрации 100 мкМ или более] такое же или больше, чем значение для контрольной антигенсвязывающую молекулы. В любом из приведенных выше вариантах осуществления настоящего изобретения контрольная антигенсвязывающая молекула может быть выбрана из анти-CD137 антител, содержащих HVR-H1, HVR-H2, HVR-H3, HVR-L1, HVR-L2 и HVR-L3, имеющих такие же аминокислотные последовательности что и HVR-H1, HVR-Н2, HVR-H3, HVR-L1, HVR-L2 и HVR-L3, содержащиеся в А375/В167, А372/В040, А356/В040, А486/В167, А487/В167, А488/В226, А489/В223, А548/В376, А551/В256, А551/В379, А555/В379, А548/В256 или А549/В167, описанных в табл. 17.
В одном из вариантов осуществления настоящего изобретения контрольной антигенсвязывающей молекулой является антитело, включающее аминокислотную последовательность А375/В167, А372/В040, А356/В040, А486/В167, А487/В167, А488/В226, А489/В223, А548/В376, А551/В256, А551/В379, А555/В379, А548/В256 или А549/В167, описанные в табл. 17, в качестве комбинации вариабельной области тяжелой цепи/вариабельной области легкой цепи. В предпочтительном варианте осуществления настоящего изобретения контрольной антигенсвязывающей молекулой является антитело, включающее HVR-H1, HVR- Н2, HVR-H3, HVR-L1, HVR-L2 и HVR-L3, имеющих такие же аминокислотные последовательности что и HVR-H1, HVR-Н2, HVR-H3, HVR-L1, HVR-L2 и HVR-L3, содержащиеся в А375/В167. В другом варианте осуществления настоящего изобретения контрольной антигенсвязывающей молекулой является антитело, включающее А375/В167 в качестве комбинации вариабельной области тяжелой цепи/вариабельной области легкой цепи. В другом варианте осуществления настоящего изобретения контрольной антигенсвязывающей молекулой является антитело, включающее HVR-H1, HVR-H2, HVR-H3, HVR-L1, HVR-L2 и HVR-L3, имеющие ту же аминокислотную последовательность, что и HVR-H1, HVR-H2, HVR-H3, HVR-L1, HVR-L2 и HVR-L3, содержащиеся в А551/В379. В другом варианте осуществления настоящего изобретения контрольной антигенсвязывающей молекулой является антитело, включающее А551/В379 в качестве комбинации вариабельной области тяжелой цепи/вариабельной области легкой цепи. В предпочтительном варианте осуществления настоящего изобретения контрольная антигенсвязывающая молекула включает константные области тяжелой и легкой цепей, происходящих от человека (например, G1T3 (SEQ ID NO: 138) в качестве константной области тяжелой цепи, X цепь Lamlib человека (SEQ ID NO: 63) в качестве константной области легкой цепи.
В одном из вариантов осуществления настоящего изобретения связывающую активность анти-CD137 антитела в отношении CD137, при наличии, отсутствии, высокой концентрации и/или низкой концентрации низкомолекулярного соединения измеряют, например, методом захвата лиганда с использованием BIACORE (зарегистрированная торговая марка) Т200 поверхностного плазмонного резонанса в качестве принципа измерения.
Детали примера метода измерения связывающей активности анти-CD137 антитела в отношении CD137 описаны ниже. В одном из вариантов осуществления настоящего изобретения связывающую активности анти-CD137 антитела в отношении CD137 в отношении CD137 оценивают методом BIACORE (зарегистрированная торговая марка) Т200. В предпочтительном варианте осуществления настоящего изобретения в этом методе используют 20 мМ ACES (рН 7,4), 150 мМ NaCl, 2 мМ MgCl2 и 0,05% Tween 20 в качестве подвижного буфера и его осуществляют при 37°C. В одном из вариантов осуществления настоящего изобретения такое измерение осуществляют после захвата антитела в качестве лиганда на лиганд-захватывающей молекуле. Конкретно, необходимое количество (например, примерно 100 RU, 200 RU, 300 RU, 400 RU или 500 RU) антитела захватывается, взаимодействуя с раствором антитела, приготовленным с использованием подвижного буфера с чипом, приготовленным заранее путем иммобилизации Sure Protein А (фирма GE Healthcare) на сенсорном чипе Series S Sensor Chip СМ3 (фирма GE Healthcare).
В предпочтительном варианте осуществления настоящего изобретения примерно от 100 до 500 RU, предпочтительно примерно от 250 до 400 RU антитела захватывается. Затем связывающую активность в отношении CD137 в присутствии или в отсутствие низкомолекулярного соединения оценивают путем взаимодействия раствора CD137, приготовленного с использованием подвижного буфера, добавленного с низкомолекулярным соединением к целевой концентрации (например, 1 мкМ, 10 мкМ, 50 мкМ или 100 мкМ), или раствор CD137 приготовляют, используя подвижный буфер, который не содержит низкомолекулярного соединения. Хотя концентрация CD137 в растворе CD137 может быть определена соответствующим образом, например, когда hCD 137-HisBAP (см. пример 1-1) применяют в качестве антигена, измерение осуществляют, используя концентрации антигена 0 нМ, 15,625 нМ, 62,5 нМ, 250 нМ и 1000 нМ, соответственно. В одном из вариантов осуществления настоящего изобретения константа диссоциации (KD) анти-CD137 антитела с CD137 человека рассчитывают, используя программное обеспечение Biacore Т200 Evaluation Software 2.0. Конкретно константу скорости связывания ka (л/М/сек) и константу скорости диссоциации kd (1/сек) рассчитывают путем глобальной подгонки сенсограммы, полученной измерением с использованием модели связывания Лэнгмюра 1:1. Константу диссоциации KD (М/л) рассчитывают исходя из этих величин.
Дополнительные типичные методы анализа по измерению активности связывания анти-CD137 антитела с CD137 будут подробно описаны ниже. Связывание анти-CD137 антител с CD137 человека оценивают с помощью Biacore Т200. Связывание с CD137 человека измеряют, используя 20 мМ ACES (рН 7,4), 150 мМ NaCl, 2 мМ MgCl2, 0,05% Tween 20 в качестве подвижного буфера, измерение проводят при 37°C. Во-первых, антитело примерно от 250 до 400 RU захватывают путем взаимодействия раствора антитела, приготовленного с использованием подвижного буфера, с чипом, на котором Sure Protein А (фирма GE Healthcare) иммобилизован на сенсорном чипе Series S СМЗ (фирма GE Healthcare). Затем раствор CD137 человека, приготовленный с применением подвижного буфера с добавлением АТФ в целевой концентрации (например, 1 мкМ, 10 мкМ, 50 мкМ или 100 мкМ), или раствор CD137 человека, приготовленный с применением подвижного буфера, который не содержит АТФ, вступает во взаимодействие для оценки связывающей активности в отношении CD137 в присутствии или в отсутствие АТФ. hCD137-HisBAP получают методом, описанным в примере (1-1), и применяют в качестве CD137 человека, являющегося антигеном, измерение проводят при концентрации антигена 0 нМ, 15,625 нМ, 62,5 нМ, 250 нМ и 1000 нМ, соответственно. Чип регенерируют, используя 25 мМ NaOH и 10 мМ глицин-HCl (рН 1,5), измерение проводят путем повторяющегося захвата антител. Константа диссоциации каждого антитела в отношении CD137 человека рассчитывают, используя программное обеспечение Biacore Т200 Evaluation Software 2.0. Конкретно константу скорости связывания ka (л/М/сек) и константу скорости диссоциации kd (1/сек) рассчитывают путем глобальной подгонки сенсограммы, полученной измерением с использованием модели связывания Лэнгмюра 1:1. Константу диссоциации KD (М/л) рассчитывают, исходя из этих величин.
В одном из вариантов осуществления настоящего изобретения связывающая активность анти-CD137 антитела в отношении CD137 (предпочтительно CD137 человека) также может быть названа по-другому, а именно «количество связываемого CD137 на единицу антитела». В частности, используя сенсограммы, полученные с помощью вышеуказанного метода анализа с использованием BIACORE (зарегистрированная торговая марка) Т200, количество связывания CD137 с антителом (RU) делится на количество захваченного антитела для расчета «количества связывания CD137 на единицу количества антитела». В одном из вариантов осуществления настоящего изобретения связывающая активность анти-CD137 антитела в отношении CD137 (предпочтительно CD137 человека) также может быть измерена методом, описанным в примерах 5-3 или 6-2.
Понятия «малая молекула» и «низкомолекулярное соединение» относятся к химическому веществу природного происхождения, отличному от «биополимеров», присутствующих в живом организме, или к химическому веществу неприродного происхождения. Предпочтительно, это целевое тканеспецифическое соединение или не встречающееся в природе соединение, но ими перечень не ограничивают. В одном из вариантов осуществления настоящего изобретения «низкомолекулярное соединение» по настоящему изобретению является «соединением, специфичное в отношении раковой ткани» или «метаболитом, специфическим в отношении раковой ткани». В настоящем изобретении понятие «соединение, специфичное в отношении раковой ткани» относится к соединению, которое существует в опухолевой ткани, но не в неопухолевой ткани. В настоящем изобретении понятие «рак» применяют для описания злокачественных новообразований, которые могут быть с метастазами или без них. Термин «метаболизм» относится к химическим изменениям, которые происходят в ткани организма, включая «ассимиляцию» и «катаболизм». Ассимиляция относится к биосинтезу или накоплению молекул, а катаболизм относится к разложению молекул. «Метаболит» - это промежуточное соединение или продукт, образующийся в результате метаболизма вещества.
В настоящем изобретении понятие «целевая ткань» означает любую ткань в живом организме, к которой антигенсвязывающая молекула по настоящему изобретению должна быть доставлена. Целевая ткань может быть гистологически разной, такой как ткань различных органов или патологически отличающаяся ткань, например, нормальные ткани и больные ткани. В некоторых вариантах осуществления настоящего изобретения целевой тканью является опухолевая ткань. Напротив, «нецелевой тканью (тканями)» являются ткани в живом организме, отличные от целевой ткани.
Понятие «опухолевая ткань» означает ткань, которая включает по меньшей мере одну опухолевую клетку. Обычно опухолевая ткань производит популяцию опухолевых клеток, составляющих основное тело опухоли (паренхиму) и соединительные ткани и кровеносные сосуды, существующие между опухолевыми клетками и поддерживающей опухоль тканью (стромой). В некоторых случаях они четко различимы, но есть случаи, когда они перемешаны. В некоторых случаях, есть такие клетки, как иммунные клетки, которые инфильтруются в ткань опухоли. Напротив, понятие «не опухолевая ткань» означает ткань живого организма, отличающуюся от опухолевой ткани (тканей). Здоровые ткани/нормальные ткани представляют такие неопухолевые ткани.
Примером, который не ограничивает рамок охвата настоящего изобретения, является специфическое в отношении раковой ткани соединение или раковый тканеспецифический метаболит, применяемый в настоящем изобретении, по меньшей мере одно соединение, выбранное из соединений, подробно описанных ниже, которое можно соответствующим образом проиллюстрировать. Понятие «по меньшей мере одно соединение» включает, помимо случая, когда связывающая активность против антигена теми же антигенсвязывающими доменами, описанными ниже, зависит от одного типа ракового тканеспецифичного соединения или ракового тканеспецифичного метаболита, случай, когда связывающая активность зависит от нескольких типов, специфических в отношении раковой ткани соединений или раковых тканеспецифических метаболитов.
В настоящем изобретении термин «тканеспецифическое соединение-мишень» относится к соединению, которое специфически присутствует в ткани-мишени по сравнению с тканью, не являющейся мишенью. В некоторых вариантах осуществления настоящего изобретения тканеспецифическое соединение-мишень может быть соединением, определяемым качественной тканевой специфичностью-мишенью, такой как присутствие в ткани-мишени, но не в тканях, не являющихся мишенями, или присутствие в ткани, не являющейся мишенью, но не в ткани-мишени. В другом варианте осуществления настоящего изобретения тканеспецифическое соединение-мишень может быть соединением, определяемым количественно тканевой специфичностью-мишенью, такой как присутствие в ткани-мишени в концентрации, которая отличается (например, более высокая концентрация или более низкая концентрация) по сравнению с тканью, не являющейся мишенью. В определенных вариантах осуществления настоящего изобретения тканеспецифическое соединение-мишень присутствует в ткани-мишени в концентрации, которая больше, например, в 1,05 раз или более, в 1,1 раза или более, в 1,15 раз или более, в 1,2 раза или более, в 1,25 раз или более, в 1,3 раз или более, в 1,35 раз или более, в 1,4 раз или более, в 1,45 раз или более, в 1,5 раз или более, в 1,55 раз или более, в 1,6 раз или более, в 1,65 раз или более, в 1,7 раз или более, в 1,75 раз или более, в 1,8 раз или более, в 1,85 раз или более, в 1,9 раз или более, в 1,95 раз или более, в 2 раза или более, в 2,1 раза или более, в 2,2 раз или более, в 2,3 раза или более, в 2,4 раза или более, в 2,5 раз или более, в 3 раза или более, в 5 раз или более, в 10 раз или более, в 50 раз или более, в 100 раз или более, в 103 раз или более, в 104 раз или более, в 105 раз или более, в 106 раз или более по сравнению с тканью, не являющейся мишенью. В определенном варианте осуществления настоящего изобретения специфическое в отношении целевой ткани соединение по сравнению с нецелевой тканью, присутствует в ткани-мишени в концентрации, которая статистически значительно выше или ниже (т.е., по анализу с использованием из t-критерия Уэлча или критерия суммы рангов Вилкоксона, значение р меньше 0,05 и/или значение q меньше 0,10). В определенном варианте осуществления настоящего изобретения тканеспецифичное соединение-мишень представляет собой тканеспецифическое соединение опухоли.
В определенном варианте осуществления настоящего изобретения тканеспецифическое соединение-мишень представляет собой метаболит, специфически вырабатываемый опухолевой клеткой. Метаболит может быть продуктом, который образуется в процессе метаболизма, необходимого для жизнедеятельности (первичный метаболит), или продуктом, образующимся в результате метаболизма, необязательного для жизнедеятельности (вторичный метаболит). Примеры первичных метаболитов могут включать сахара, белки, липиды, нуклеиновые кислоты и т.п. Примеры вторичных метаболитов включают антибиотики и красители. Метаболит может быть биополимером или небольшой молекулой. В определенном варианте осуществления настоящего изобретения биополимер представляет собой молекулу с молекулярной массой около 5000 или более, которая состоит из одного или более типов повторяющихся единиц, включая, например, полисахариды, полипептиды и полинуклеотиды. В определенном варианте осуществления настоящего изобретения небольшими молекулами являются молекулы, имеющие молекулярную массу около 500 или менее, и представляют собой химические вещества, присутствующие в живом организме. В другом варианте осуществления настоящего изобретения тканеспецифическое соединение опухоли представляет собой низкомолекулярный метаболит, специфически продуцируемый в опухолевых клетках (Eva Gottfried, Katrin Peter, Marina P. Kreutz, From Molecular and Modular Tumor Therapy, 3(2), 2010, 111-132). В другом варианте осуществления настоящего изобретения тканеспецифическое соединение опухоли представляет собой представляет собой метаболит, который специфически вырабатывается клеткой, инфильтрующейся в опухолевую ткань (например, иммунной клеткой), или стромальной клеткой (например, фибробластом, связанным с раком (CAF, cancer associated fibroblast)), присутствующей в опухолевой ткани. Примерами иммунных клеток, инфильтрирующих в опухолевую ткань, являются дендритные клетки, супрессивные дендритные клетки, регуляторные Т-клетки, истощенные Т-клетки, происходящие от миеломы супрессорные клетки (MDSC, myeloma-derived suppressor cells) и т.п. В другом варианте осуществления настоящего изобретения метаболит, который продуцируется клетками, присутствующими в опухолевой ткани (например, опухолевыми клетками, иммунными клетками, стромальными клетками и т.д.), который высвобождается из клетки после их гибели в результате апоптоза или некроза или тому подобного процесса, также может быть отнесен к соединениям настоящего изобретения, специфичным для опухолевой ткани.
Чтобы идентифицировать тканеспецифическое соединение опухоли, анализ на транскриптомном уровне (см., например, Dhanasekaran с соавт., Nature, 412, 2001, 822-826; Lapointe с соавт., Proc. Natl. Acad. Sci. USA, 101, 2004, 811-816; Perou с соавт., Nature, 406, 2000, 747-752), анализ на протеомном уровне (см., например, Ahram с соавт., Mol. Carcinog., 33, 2002, 9-15; Hood с соавт., Mol. Cell. Proteomics, 4, 2005, 1741-1753) и анализ метаболизма, основанный на метаболомном профилировании (метаболомика), может быть использован надлежащим образом. То есть для идентификации метаболита в исследуемом образце используют высокоэффективную жидкостную хроматографию (ВЭЖХ), ядерный магнитный резонанс (ЯМР) (Brindle с соавт., J. Mol. Recognit., 10, 1997, 182-187)), масс-спектрометрию (GC/MS и LC/MS) (Gates, Sweeley, Clin. Chem., 24, 1978, 1663-1673) и метаболическое профилирование с применением метода ELISA, и тому подобные подходящие методы, которые можно применять отдельно и/или в комбинации.
В определенном варианте осуществления настоящего изобретения специфическое для опухолевой ткани соединение представляет собой по крайней мере одно соединение, выбранное из группы, состоящей из: нуклеозидов, имеющих структуру пуринового кольца, аминокислот и их метаболитов, липидов и их метаболитов, первичных метаболитов углеводного обмена, а также никотинамида и его метаболитов. В другом варианте осуществления настоящего изобретения специфическое для опухолевой ткани соединение представляет собой по меньшей мере одно соединение, выбранное из п.п. (1)-(6), включающих:
(1) нуклеозиды, имеющие пуриновую структуру, такие как аденозин (ADO), аденозинтрифосфат (АТФ), аденозиндифосфат (АДФ), аденозинмонофосфат (АМФ) и инозин;
(2) аминокислоты, такие как аланин, глутаминовая кислота и аспарагиновая кислота;
(3) метаболиты аминокислот, такие как кинуренин, антраниловая кислота, 3-гидроксикинуренин и кинуреновая кислота;
(4) метаболиты арахидоновой кислоты, такие как простагландин Е2;
(5) первичные метаболиты гликолитического пути или цикла Кребса, такие как молочная кислота, янтарная кислота и лимонная кислота; и,
(6) метаболиты никотинамида, такие как 1-метилникотинамид.
(1) Нуклеозиды, имеющие пуриновую структуру, такие как аденозин (ADO), аденозинтрифосфат (АТФ), аденозиндифосфат (АДФ), аденозинмонофосфат (АМФ) и инозин
Известно, что при гибели опухолевых клеток происходит утечка большого количества внутриклеточного АТФ. Следовательно, концентрация АТФ в опухолевой ткани значительно выше, чем в нормальной ткани (PLoS One. (2008) 3, е2599). АМФ метаболизируется ферментами на поверхности клетки, такими как внеклеточная 5'-нуклеотидаза (эко-5'-нуклеотидаза) (CD73) (Resta, Thompson, Immunol. Rev. 161, 1998, 95-109; Sadej с соавт., Melanoma Res. 16, 2006, 213-222). Аденозин представляет собой пуриновый нуклеозид, который постоянно присутствует во внеклеточной среде в низких концентрациях, но сообщалось о заметном повышении внеклеточной концентрации аденозина в гипоксических тканях, обнаруженных в солидных опухолях (Blay, Hoskin, Cancer Res., 57, 1997, 2602-2605). CD73 экспрессируется на поверхности опухолей и иммунных клеток (Kobie с соавт., J. Immunol. 177, 2006, 6780-6786) и повышенную активность обнаруживают при раке груди (Canbolat с соавт., Breast Cancer Res. Treat. 37, 1996, 189-193), раке желудка (Durak с соавт., Cancer Lett. 84, 1994, 199-202), раке поджелудочной железы (Flocke, Mannherz, Biochim. Biophys. Acta 1076, 1991, 273-281) и глиобластоме (Bardot с соавт., Br. J. Cancer, 70, 1994, 212-218). Было высказано предположение, что накопление аденозина в опухолевой ткани является результатом увеличения дефосфорилирования АМФ цитоплазматической 5'-нуклеотидазой (Headrick, Willis, Biochem. J., 261, 1989, 541-550). Кроме того, регуляторные Т-клетки, проникающие в опухолевую ткань, также экспрессируют АТФазу и продуцируют аденозин (Proc. Natl. Acad. Sci. USA, 103 (35), 2006, 13132-13137; Curr. Med. Chem., 18, 2011, 5217-5223). Полагают, что продуцируемый аденозин поддерживает ткани опухоли в иммуносупрессивной среде через аденозиновые рецепторы, такие как рецептор А2А (Curr. Med. Chem., 18, 2011, 5217-5223). Таким образом, АТФ, АДФ, АМФ, аденозин и т.п., которые, как считается, накапливаются в высокой концентрации в опухолевой ткани за счет метаболизма пуриновых нуклеотидов, являются примерами соединений, специфичных для опухолевой ткани, используемых в настоящем изобретении. Кроме того, поскольку аденозин разрушается до инозина под действием аденозиндезаминазы, инозин накапливается в высокой концентрации.
В определенном варианте осуществления настоящего изобретения нуклеозиды, имеющие структуру пуринового кольца, включают аденозинсодержащие соединения. В определенных вариантах осуществления настоящего изобретения аденозинсодержащие соединения включают, например, аденозин (АДО), аденозинтрифосфат (АТР), аденозиндифосфат (АДФ), аденозинмонофосфат (АМФ), циклический аденозинмонофосфат (цАМФ), дезоксиаденозин (дАДО), дезоксиаденозин трифосфат (дАТФ), дезоксиаденозин дифосфат (дАДФ), дезоксиаденозин монофосфат (дАМФ), аденозин[γ-тио]трифосфат (АТФγS) и тому подобное. В другом варианте осуществления настоящего изобретения нуклеозиды, имеющие структуру пуринового кольца, включают инозин, который является метаболитом аденозина.
Более того, в определенном варианте осуществления настоящего изобретения, нуклеозиды, имеющие структуру пуринового кольца, включают коммерчески доступные нуклеозиды, имеющие структуру пуринового кольца, такие как АДФ-бета-S (фирма Sigma Inc.) и подобные.
(2) Аминокислоты, такие как аланин, глутаминовая кислота и аспарагиновая кислота
Скорость поглощения глутамина, который действует как переносчик азота в живом организме, увеличивается в опухолевых клетках, и считается, что такое включение глутамина и связанное с этим преобразование в глутаминовую кислоту и молочную кислоту (разрушение глутамина (глутаминолиз)) являются особенностями опухолевых клеток (Mazurek, Eigenbrodt, Anticancer Res. 23, 2003, 1149-1154; Mazurek с соавт., J. cell. Physiol. 181, 1999, 136-146). У онкологических больных уровень глутамина в плазме снижается, а концентрация глутаминовой кислоты повышается (Droge с соавт., Immunobiology, 174, 1987, 473-479), и исследования метаболизма меченной по 13С глюкозы в ткани рака легких показали корреляцию между концентрациями меченной по 13С янтарной кислоты, меченного по 13С аланина, меченной по 13С глутаминовои кислоты и меченного по 13С цитрата. По этим причинам аланин, глутаминовая кислота, аспарагиновая кислота и другие подобные соединения, которые, как считается, накапливаются в высоких концентрациях в опухолевой ткани, например, из-за разрушения глутамина, являются примерами соединений, специфичных для опухолевой ткани, используемых в настоящем изобретении.
(3) Метаболиты аминокислот, такие как кинуренин, антраниловая кислота, 3-гидроксикинуренин и кинуреновая кислота
Индолеамин-2,3-диоксигеназа (IDO, Indoleamine 2,3-dioxygenase) является метаболизирующим триптофан ферментом, который высоко экспрессируется при многих раковых заболеваниях, таких как меланома, рак толстой кишки, рак почки и других подобных заболеваний (Uyttenhove с соавт., Nat. Med., 9, 2003, 1269-1274)). IDO катализирует превращение триптофана в кинуренин. В глиомах, которые не экспрессируют IDO, кинуренин вырабатывается из триптофана триптофан-2,3-диоксигеназой (TDO, tryptophan 2,3-dioxygenase) печени (Opitz с соавт., Nature, 478 (7368), 2011, 197-203). IDO также экспрессируется на дендритных клетках, проникающих в опухолевую ткань, и дендритные клетки также производят кинуренин (J. Immunol. 181, 2008, 5396-5404). Кроме того, IDO также экспрессируется в миелоидных супрессорных клетках (MDSC, myeloid-derived suppressor cell) опухолевых тканей, и MDSC также продуцирует кинуренин (Yu с соавт., J. Immunol., 190, 2013, 3783-3797). Кинуренин превращается в антраниловую кислоту с помощью кинурениназы и в 3-гидроксикинуренин с помощью кинуренин-3-гидроксилазы. Как антраниловая кислота, так и 3-гидроксикинуренин конвертируют в 3-гидроксиантраниловую кислоту, предшественник НАД. Кинуренин превращается в кинуреновую кислоту кинуренинаминотрансферазой. По этим причинам кинуренин и его метаболит, то есть антраниловая кислота, 3-гидроксикинуренин, кинурениновая кислота и т.п., являются примерами соединений, специфичных для опухолевой ткани, используемых в настоящем изобретении, в частности, метаболитов, специфичных для опухолевых клеток.
(4) Метаболиты арахидоновой кислоты, такие как простагландин Е2
Простагландин Е2 (PGE2) способствует росту раковых клеток толстой кишки и подавляет их апоптоз (Sheng с соавт., Cancer Res., 58, 1998, 362-366). Что касается синтетаз PGE2, в основном было обнаружено, что СОХ-1 конститутивно экспрессируется почти во всех тканях, тогда как СОХ-2 индуцируется некоторыми воспалительными цитокинами и онкогенами в опухолях (Warner, Mitchell, FASEB J., 18, 2004, 790-804). Сообщалось, что сверхэкспрессия СОХ-2 связана с плохим прогнозом рака груди (Denkert с соавт., Clin. Breast Cancer, 4, 2004, 428-433) и быстрым прогрессированием рака яичников (Denker с соавт., Mod. Pathol., 19, 2006, 1261-1269). Кроме того, регуляторные Т-клетки, проникающие в опухолевую ткань, также производят PGE2 (Curr. Med. Chem., 18, 2011, 5217-5223). По этим причинам метаболиты арахидоновой кислоты, такие как PGE2, являются примерами соединений, специфичных для опухолевой ткани, в частности метаболитов, специфичных для опухолевых клеток, или метаболитов, специфичных для иммунных клеток, инфильтрирующих опухолевую ткань. Помимо PGE2, выработка тромбоксана А2 (ТХА2) повышается в опухолевых тканях, например, при раке толстой кишки (J. Lab. Clin. Med., 122, 1993, 518-523).
(5) Первичные метаболиты гликолитического пути или цикла Кребса, такие как молочная кислота, янтарная кислота и лимонная кислота
Гликолитический фенотип, характеризующийся активацией гликолитических (путь Эмбдена-Мейергофа) ферментов, таких как пируваткиназа, гексокиназа и лактатдегидрогеназа (ЛДГ), традиционно известен как эффект Варбурга, характерный для солидных опухолей. Молочная кислота, которая является конечным продуктом гликолиза, а также янтарная и лимонная кислоты, образующиеся в ходе цикла Кребса, накапливаются в опухолевых тканях (Teresa с соавт., Mol. Cancer, 8, 2009, 41-59). По этим причинам молочная кислота, янтарная кислота, лимонная кислота и другие подобные соединения, которые являются первичными метаболитами, образуемыми в ходе гликолиза, являются примерами соединений, специфичных для опухолевой ткани, в частности метаболитов, специфичных для опухолевых клеток, упоминаемых в настоящем изобретении. Кроме того, известно, что из-за гибели клеток сукцинат, который присутствует в клетках в высокой концентрации, выделяется из клеток (Nature Immunology, 9, 2008, 1261-1269). Предполагают, что причиной является повышенная концентрация янтарной кислоты в опухолевых тканях, где часто происходит гибель клеток.
(б) Метаболиты никотинамида. такие как 1-метилникотинамид
Известно, что никотинамид-N-метилтрансфераза экспрессируется на высоком уровне во множестве опухолевых тканей человека. Также известно, что 1-метилникотинамид, который представляет собой стабильный метаболит никотинамида, продуцируемый этим ферментом, секретируется за пределы опухолевых клеток (Yamada с соавт., J. Nutr. Sci. Vitaminol., 56, 2010, 83-86). По этой причине 1-метилникотинамид и другие подобные соединения, которые предположительно накапливаются в опухолевой ткани в высокой концентрации в результате метаболизма никотинамида, являются примерами соединений, специфичных для опухолевой ткани, упоминаемых в настоящем изобретении.
«Антигенсвязывающая молекула» в настоящем изобретении содержит «антигенсвязывающий домен». В качестве «антигенсвязывающего домена» можно использовать домен любой структуры, если он связывается с антигеном-мишенью. В одном из вариантов осуществления настоящего изобретения антигенсвязывающие домены по настоящему изобретению включают, например, вариабельные области тяжелых цепей и/или легких цепей антител, авимеры, содержащие модуль (домен А) примерно из 35 аминокислот, содержащихся в различных белках клеточных мембран в живом организме (WO2004/044011 и WO2005/040229), аднектины, содержащие домен 10Fn3 фибронектина, который представляет собой гликопротеин, экспрессируемый на клеточной мембране (WO2002/032925), аффитела, использующие в качестве каркаса связывающий домен IgG из 58 аминокислот белка A (WO WO1995/001937), сконструированные белки с анкириновым повтором (DARPin, Designed Ankyrin Repeat protein), с использованием анкиринового повтора (AR, ankyrin repeat), который представляет собой повторяющуюся последовательность из 33 аминокислот, в качестве основания (WO2002/020565), антикалины, содержащие липокалин, такой как липокалин, связанный с желатиназой нейтрофилов (NGAL, neutrophil gelatinase-associated lipocalin) в качестве основания (WO2003/029462), вариабельные рецепторы лимфоцитов (VLR, variable lymphocyte receptor), которые являются белками, функционирующими в адаптивных иммунных системах бесчелюстных позвоночных, таких как Lampetra japonica и Eptatretus, и содержат модуль обогащенных лейцином повторов (LRR, leucine-rich-repeat) модуль (WO2008/016854) и другие подобные соединения. В определенном варианте осуществления настоящего изобретения антигенсвязывающий домен по настоящему изобретению содержит вариабельные области тяжелой и легкой цепей антитела. В другом варианте осуществления настоящего изобретения антигенсвязывающий домен по настоящему изобретению содержит, например, одноцепочечный домен scFv (single chain Fv), одноцепочечные антитела, Fv, scFv2 (single chain Fv2), Fab или F(ab')2.
HVR и вариабельная область
В одном из вариантов осуществления настоящее изобретение относится к анти-CD137 антигенсвязывающей молекуле или антителу, которые содержат по меньшей мере одну, по меньшей мере две или все три последовательности VH HVR, выбранные из: (a) HVR-H1, включающей аминокислотную последовательность SEQ ID NO: 7; (б) HVR-H2, включающей любую аминокислотную последовательность, выбранную из последовательностей SEQ ID NO: 8, 9, 10, 11, 12, 13, 14, 15 или 16; (в) HVR-H3, включающей любую аминокислотную последовательность, выбранную из последовательностей SEQ ID NO: 17, 18, 19 или 20. В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включают: (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO: 7; (б) HVR-H2, содержащую любую аминокислотную последовательность, выбранную из последовательностей SEQ ID NO: 8, 9, 10, 11, 12, 13, 14, 15 или 16; (в) HVR-H3, содержащую любую аминокислотную последовательность, выбранную из последовательностей SEQ ID NO: 17, 18, 19 или 20.
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула представляет собой антитело, содержащее аминокислотную последовательность из числа последовательностей А375/В167, А372/В040, А356/В040, А486/В167, А487/В167, А488/В226, А489/В223, А548/В376, А551/В256, А551/В379, А555/В379, А548/В256 или А549/В167, представленных в табл. 17, как комбинацию вариабельной области тяжелой цепи/вариабельной области легкой цепи. В предпочтительном варианте осуществления настоящего изобретения антигенсвязывающей молекулой является анти-CD137 антитело, включающее области HVR-H1, HVR-H2, HVR-Н3, HVR-L1, HVR-L2 и HVR-L3, имеющие такие же аминокислотные последовательности, как HVR-H1, HVR-H2, HVR-H3, HVR-L1, HVR-L2 и HVR-L3, содержащиеся в А375/В167. В другом варианте осуществления настоящего изобретения анти-CD137 антигенсвязывающей молекулой является анти-CD137 антитело, содержащее А375/В167 в виде комбинации вариабельной области тяжелой цепи/вариабельной области легкой цепи. В другом предпочтительном варианте осуществления настоящего изобретения антигенсвязывающей молекулой является анти-CD137 антитело, включающее области HVR-H1, HVR-Н2, HVR-H3, HVR-L1, HVR-L2 и HVR-L3, содержащие такие же аминокислотные последовательности, как HVR-H1, HVR-H2, HVR-H3, HVR-L1, HVR-L2 и HVR-L3, содержащиеся в А551/В379.
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включает следующие области (а) HVR-H1, включающую аминокислотную последовательность SEQ ID NO: 7; (б) HVR-H2, включающую аминокислотную последовательность SEQ ID NO: 8; (в) HVR-H3, включающей любую аминокислотную последовательность SEQ ID NO: 17.
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включает следующие области (а) HVR-H1, включающую аминокислотную последовательность SEQ ID NO: 7; (б) HVR-H2, включающую аминокислотную последовательность SEQ ID NO: 9; (в) HVR-H3, включающей любую аминокислотную последовательность SEQ ID NO: 17.
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включает следующие области (а) HVR-H1, включающую аминокислотную последовательность SEQ ID NO: 7; (б) HVR-H2, включающую аминокислотную последовательность SEQ ID NO: 10; (в) HVR-H3, включающей любую аминокислотную последовательность SEQ ID NO: 17.
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включает следующие области (а) HVR-H1, включающую аминокислотную последовательность SEQ ID NO: 7; (б) HVR-H2, включающую аминокислотную последовательность SEQ ID NO: 11; (в) HVR-H3, включающей любую аминокислотную последовательность SEQ ID NO: 18.
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включает следующие области (а) HVR-H1, включающую аминокислотную последовательность SEQ ID NO: 7; (б) HVR-H2, включающую аминокислотную последовательность SEQ ID NO: 8; (в) HVR-H3, включающей любую аминокислотную последовательность SEQ ID NO: 18.
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включает следующие области (а) HVR-H1, включающую аминокислотную последовательность SEQ ID NO: 7; (б) HVR-H2, включающую аминокислотную последовательность SEQ ID NO: 12; (в) HVR-H3, включающей любую аминокислотную последовательность SEQ ID NO: 18.
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включает следующие области (а) HVR-H1, включающую аминокислотную последовательность SEQ ID NO: 7; (б) HVR-H2, включающую аминокислотную последовательность SEQ ID NO: 13; (в) HVR-H3, включающей любую аминокислотную последовательность SEQ ID NO: 18.
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включает следующие области (а) HVR-H1, включающую аминокислотную последовательность SEQ ID NO: 7; (б) HVR-H2, включающую аминокислотную последовательность SEQ ID NO: 14; (в) HVR-H3, включающей любую аминокислотную последовательность SEQ ID NO: 19.
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включает следующие области (а) HVR-H1, включающую аминокислотную последовательность SEQ ID NO: 7; (б) HVR-H2, включающую аминокислотную последовательность SEQ ID NO: 15; (в) HVR-H3, включающей любую аминокислотную последовательность SEQ ID NO: 20.
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включает следующие области (а) HVR-H1, включающую аминокислотную последовательность SEQ ID NO: 7; (б) HVR-H2, включающую аминокислотную последовательность SEQ ID NO: 16; (в) HVR-H3, включающей любую аминокислотную последовательность SEQ ID NO: 20.
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включает следующие области (а) HVR-H1, включающую аминокислотную последовательность SEQ ID NO: 7; (б) HVR-H2, включающую аминокислотную последовательность SEQ ID NO: 14; (в) HVR-H3, включающей любую аминокислотную последовательность SEQ ID NO: 17.
В другом варианте осуществления настоящего изобретения предусматривают анти-CD137 антигенсвязывающую молекулу или антитело, включающее по меньшей мере одну, по меньшей мере две или все три последовательности VL HVR, выбранные из последовательностей: (a) HVR-L1, включающую любую аминокислотную последовательность, выбранную из SEQ ID NO: 21, 22, 23, 24 или 25; (б) HVR-L2, включающую аминокислотную последовательность SEQ ID NO: 26; (в) HVR-L3, включающую любую аминокислотную последовательность, выбранную из SEQ ID NO: 27, 28 или 29.
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включает (a) HVR-L1, включающую аминокислотную последовательность SEQ ID NO: 21; (б) HVR-L2, включающую аминокислотную последовательность SEQ ID NO: 26; (в) HVR-L3, включающую аминокислотную последовательность SEQ ID NO: 27.
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включает (a) HVR-L1, включающую аминокислотную последовательность SEQ ID NO: 22; (б) HVR-L2, включающую аминокислотную последовательность SEQ ID NO: 26; (в) HVR-L3, включающую аминокислотную последовательность SEQ ID NO: 27.
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включает (a) HVR-L1, включающую аминокислотную последовательность SEQ ID NO: 21; (б) HVR-L2, включающую аминокислотную последовательность SEQ ID NO: 26; (в) HVR-L3, включающую аминокислотную последовательность SEQ ID NO: 28.
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включает (a) HVR-L1, включающую аминокислотную последовательность SEQ ID NO: 21; (б) HVR-L2, включающую аминокислотную последовательность SEQ ID NO: 26; (в) HVR-L3, включающую аминокислотную последовательность SEQ ID NO: 29.
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включает (a) HVR-L1, включающую аминокислотную последовательность SEQ ID NO: 23; (б) HVR-L2, включающую аминокислотную последовательность SEQ ID NO: 26; (в) HVR-L3, включающую аминокислотную последовательность SEQ ID NO: 27.
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включает (a) HVR-L1, включающую аминокислотную последовательность SEQ ID NO: 24; (б) HVR-L2, включающую аминокислотную последовательность SEQ ID NO: 26; (в) HVR-L3, включающую аминокислотную последовательность SEQ ID NO: 27.
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включает (a) HVR-L1, включающую аминокислотную последовательность SEQ ID NO: 25; (б) HVR-L2, включающую аминокислотную последовательность SEQ ID NO: 26; (в) HVR-L3, включающую аминокислотную последовательность SEQ ID NO: 27.
В другом варианте осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело по настоящему изобретению включает (а) домен VH, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH HVR, выбранные из (i) HVR-H1, включающей аминокислотную последовательность SEQ ID NO: 7; (ii) HVR-H2, включающей аминокислотную последовательность, выбранную из SEQ ID NO: 8, 9, 10, 11, 12, 13, 14, 15 или 16; (iii) HVR-H3, включающей любую аминокислотную последовательность, выбранную из SEQ ID NO: 17, 18, 19 или 20; (б) домен VL, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VL HVR, выбранные из (i) HVR-L1, включающей любую аминокислотную последовательность, выбранную из SEQ ID NO: 21, 22, 23, 24 и 25; (ii) HVR-L2, включающей аминокислотную последовательность SEQ ID NO: 26; (iii) HVR-L3, включающей любую аминокислотную последовательность, выбранную из SEQ ID NO: 27, 28 или 29.
В другом варианте осуществления настоящего изобретения предусматривают анти-CD137 антигенсвязывающую молекулу или антитело, включающие (a) HVR-H1, включающую аминокислотную последовательность SEQ ID NO: 7; (б) HVR-H2, включающую аминокислотную последовательность SEQ ID NO: 8; (в) HVR-H3, включающую аминокислотную последовательность SEQ ID NO: 17; (г) HVR-L1, включающую аминокислотную последовательность SEQ ID NO: 21; (д) HVR-L2, включающую аминокислотную последовательность SEQ ID NO: 26; (е) HVR-L3, включающую аминокислотную последовательность SEQ ID NO: 27.
В другом варианте осуществления настоящего изобретения предусматривают анти-CD137 антигенсвязывающую молекулу или антитело, включающие (a) HVR-H1, включающую аминокислотную последовательность SEQ ID NO: 7; (б) HVR-H2, включающую аминокислотную последовательность SEQ ID NO: 9; (в) HVR-H3, включающую аминокислотную последовательность SEQ ID NO: 17; (г) HVR-L1, включающую аминокислотную последовательность SEQ ID NO: 22; (д) HVR-L2, включающую аминокислотную последовательность SEQ ID NO: 26; (е) HVR-L3, включающую аминокислотную последовательность SEQ ID NO: 27.
В другом варианте осуществления настоящего изобретения предусматривают анти-CD137 антигенсвязывающую молекулу или антитело, включающие (a) HVR-H1, включающую аминокислотную последовательность SEQ ID NO: 7; (б) HVR-H2, включающую аминокислотную последовательность SEQ ID NO: 10; (в) HVR-H3, включающую аминокислотную последовательность SEQ ID NO: 17; (г) HVR-L1, включающую аминокислотную последовательность SEQ ID NO: 22; (д) HVR-L2, включающую аминокислотную последовательность SEQ ID NO: 26; (е) HVR-L3, включающую аминокислотную последовательность SEQ ID NO: 27.
В другом варианте осуществления настоящего изобретения предусматривают анти-CD137 антигенсвязывающую молекулу или антитело, включающие (a) HVR-H1, включающую аминокислотную последовательность SEQ ID NO: 7; (б) HVR-H2, включающую аминокислотную последовательность SEQ ID NO: 11; (в) HVR-H3, включающую аминокислотную последовательность SEQ ID NO: 18; (г) HVR-L1, включающую аминокислотную последовательность SEQ ID NO: 21; (д) HVR-L2, включающую аминокислотную последовательность SEQ ID NO: 26; (e) HVR-L3, включающую аминокислотную последовательность SEQ ID NO: 27.
В другом варианте осуществления настоящего изобретения предусматривают анти-CD137 антигенсвязывающую молекулу или антитело, включающие (a) HVR-H1, включающую аминокислотную последовательность SEQ ID NO: 7; (б) HVR-H2, включающую аминокислотную последовательность SEQ ID NO: 8; (в) HVR-H3, включающую аминокислотную последовательность SEQ ID NO: 18; (г) HVR-L1, включающую аминокислотную последовательность SEQ ID NO: 21; (д) HVR-L2, включающую аминокислотную последовательность SEQ ID NO: 26; (е) HVR-L3, включающую аминокислотную последовательность SEQ ID NO: 27.
В другом варианте осуществления настоящего изобретения предусматривают анти-CD137 антигенсвязывающую молекулу или антитело, включающие (a) HVR-H1, включающую аминокислотную последовательность SEQ ID NO: 7; (б) HVR-H2, включающую аминокислотную последовательность SEQ ID NO: 12; (в) HVR-H3, включающую аминокислотную последовательность SEQ ID NO: 18; (г) HVR-L1, включающую аминокислотную последовательность SEQ ID NO: 21; (д) HVR-L2, включающую аминокислотную последовательность SEQ ID NO: 26; (е) HVR-L3, включающую аминокислотную последовательность SEQ ID NO: 28.
В другом варианте осуществления настоящего изобретения предусматривают анти-CD137 антигенсвязывающую молекулу или антитело, включающие (a) HVR-H1, включающую аминокислотную последовательность SEQ ID NO: 7; (б) HVR-H2, включающую аминокислотную последовательность SEQ ID NO: 13; (в) HVR-H3, включающую аминокислотную последовательность SEQ ID NO: 18; (г) HVR-L1, включающую аминокислотную последовательность SEQ ID NO: 21; (д) HVR-L2, включающую аминокислотную последовательность SEQ ID NO: 26; (е) HVR-L3, включающую аминокислотную последовательность SEQ ID NO: 29.
В другом варианте осуществления настоящего изобретения предусматривают анти-CD137 антигенсвязывающую молекулу или антитело, включающее (a) HVR-H1, включающую аминокислотную последовательность SEQ ID NO: 7; (б) HVR-H2, включающую аминокислотную последовательность SEQ ID NO: 14; (в) HVR-H3, включающую аминокислотную последовательность SEQ ID NO: 19; (г) HVR-L1, включающую аминокислотную последовательность SEQ ID NO: 23; (д) HVR-L2, включающую аминокислотную последовательность SEQ ID NO: 26; (e) HVR-L3, включающую аминокислотную последовательность SEQ ID NO: 27.
В другом варианте осуществления настоящего изобретения предусматривают анти-CD137 антигенсвязывающую молекулу или антитело, включающее (a) HVR-H1, включающую аминокислотную последовательность SEQ ID NO: 7; (б) HVR-H2, включающую аминокислотную последовательность SEQ ID NO: 14; (в) HVR-H3, включающую аминокислотную последовательность SEQ ID NO: 19; (г) HVR-L1, включающую аминокислотную последовательность SEQ ID NO: 23; (д) HVR-L2, включающую аминокислотную последовательность SEQ ID NO: 26; (е) HVR-L3, включающую аминокислотную последовательность SEQ ID NO: 27.
В другом варианте осуществления настоящего изобретения предусматривают анти-CD137 антигенсвязывающую молекулу или антитело, включающее (a) HVR-H1, включающую аминокислотную последовательность SEQ ID NO: 7; (б) HVR-H2, включающую аминокислотную последовательность SEQ ID NO: 15; (в) HVR-H3, включающую аминокислотную последовательность SEQ ID NO: 20; (г) HVR-L1, включающую аминокислотную последовательность SEQ ID NO: 25; (д) HVR-L2, включающую аминокислотную последовательность SEQ ID NO: 26; (е) HVR-L3, включающую аминокислотную последовательность SEQ ID NO: 27.
В другом варианте осуществления настоящего изобретения предусматривают анти-CD137 антигенсвязывающую молекулу или антитело, включающее (a) HVR-H1, включающую аминокислотную последовательность SEQ ID NO: 7; (б) HVR-H2, включающую аминокислотную последовательность SEQ ID NO: 16; (в) HVR-H3, включающую аминокислотную последовательность SEQ ID NO: 20; (г) HVR-L1, включающую аминокислотную последовательность SEQ ID NO: 25; (д) HVR-L2, включающую аминокислотную последовательность SEQ ID NO: 26; (е) HVR-L3, включающую аминокислотную последовательность SEQ ID NO: 27.
В другом варианте осуществления настоящего изобретения предусматривают анти-CD137 антигенсвязывающую молекулу или антитело, включающее (a) HVR-H1, включающую аминокислотную последовательность SEQ ID NO: 7; (6) HVR-H2, включающую аминокислотную последовательность SEQ ID NO: 14; (в) HVR-H3, включающую аминокислотную последовательность SEQ ID NO: 19; (г) HVR-L1, включающую аминокислотную последовательность SEQ ID NO: 24; (д) HVR-L2, включающую аминокислотную последовательность SEQ ID NO: 26; (e) HVR-L3, включающую аминокислотную последовательность SEQ ID NO: 27.
В другом варианте осуществления настоящего изобретения предусматривают анти-CD137 антигенсвязывающую молекулу или антитело, включающее (a) HVR-H1, включающую аминокислотную последовательность SEQ ID NO: 7; (б) HVR-H2, включающую аминокислотную последовательность SEQ ID NO: 14; (в) HVR-H3, включающую аминокислотную последовательность SEQ ID NO: 17; (г) HVR-L1, включающую аминокислотную последовательность SEQ ID NO: 21; (д) HVR-L2, включающую аминокислотную последовательность SEQ ID NO: 26; (е) HVR-L3, включающую аминокислотную последовательность SEQ ID NO: 27.
В определенном варианте осуществления настоящего изобретения любая одна или несколько аминокислот вышеописанного анти-CD137 антитела заменены в следующих положениях HVR:
В HVR-H2 (SEQ ID NO: 30): положения 5, 6, 7, 10, 13, 14 и/или 17
В HVR-H3 (SEQ ID NO: 31): положение 3 и/или 6
В HVR-L1 (SEQ ID NO: 32): положения 4, 5, 9 и/или 11
В HVR -L3 (SEQ ID NO: 33): положения 6, 7 и/или 8.
В определенном варианте осуществления настоящего изобретения замены, предусмотренные настоящим изобретением, являются консервативными заменами. В определенном варианте осуществления настоящего изобретения любое одно или несколько приведенных ниже замен могут быть переработаны в любой комбинации:
В HVR-H2 (SEQ ID NO: 8): К5Н или S; S6G; T7S; E10Y; D13E; S14Q; V17G или L.
В HVR-H3 (SEQ ID NO: 17): А3Р, K или I; F6E.
В HVR-L1 (SEQ ID NO: 21): R4S; Y5T; Y9F; E11N.
В HVR-L3 (SEQ ID NO: 27): E6P; H7A; Q8I.
Все возможные комбинации указанных выше замен входят в консенсусные последовательности SEQ ID NO: 30, 31, 32 и 33 для HVR-H2, HVR-H3, HVR-L1 и HVR-L3, соответственно.
В любом из указанных выше вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело являются гуманизированными. В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включают области HVR согласно любому из указанных выше вариантов осуществления настоящего изобретения, и дополнительно содержат акцепторную каркасную область человека, например, каркасную область иммуноглобулина человека или консенсусную каркасную область человека. В другом варианте осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включают области HVR как в любом из указанных выше вариантов осуществления настоящего изобретения и дополнительно содержит вариабельную область тяжелой цепи (VH) или вариабельную область легкой цепи (VL), содержащую последовательность каркасного участка (FR, framework). В одном из вариантов осуществления настоящего изобретения каркасный участок FR1 в вариабельной области тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 35, FR2 в вариабельной области тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 36, FR3 в вариабельной области тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 37 и FR4 в вариабельной области тяжелой цепи содержит аминокислотную последовательность SEQ ID No: 38. В одном из вариантов осуществления настоящего изобретения FR1 в вариабельной области легкой цепи содержит аминокислотную последовательность SEQ ID NO: 39, FR2 в вариабельной области легкой цепи содержит аминокислотную последовательность SEQ ID NO: 40, FR3 в вариабельной области легкой цепи содержит аминокислотную последовательность SEQ ID NO: 41 и FR4 в вариабельной области легкой цепи содержит аминокислотную последовательность SEQ ID NO: 42.
В другом варианте осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включают последовательность вариабельного домена тяжелой цепи (VH), которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 43, 44, 45, 46, 47, 48, 49, 50, 51, 52 или 53. В некоторых вариантах осуществления настоящего изобретения последовательность VH, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична, содержит замены (например, содержит консервативные замены), инсерции или делеции относительно контрольной последовательности, но анти-CD137 антигенсвязывающая молекула или антитело, содержащее такую последовательность, сохраняют способность связываться с CD137. В некоторых вариантах осуществления настоящего изобретения в целом от 1 до 10 аминокислот были заменены, инсертированы и/или делетированы в SEQ ID NO: 43, 44, 45, 46, 47, 48, 49, 50, 51, 52 или 53. В некоторых вариантах осуществления настоящего изобретения замены, инсерции или делеции происходят в областях вне HVR (т.е. в каркасных участках (FR)). Необязательно, анти-CD137 антитело содержит последовательность VH в SEQ ID NO: 43, 44, 45, 46, 47, 48, 49, 50, 51, 52 или 53, включая посттрансляционные модификации этой последовательности. В одном из вариантов осуществления настоящего изобретения VH содержит одну, две или три HVR, выбранных из: (а) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO: 7; (б) HVR-H2, содержащей любую одну аминокислотную последовательность, выбранную из SEQ ID NO: 8, 9, 10, 11, 12, 13, 14, 15 и 16; и (в) HVR-H3, содержащей любую одну аминокислотную последовательность, выбранную из SEQ ID NO: 17, 18, 19 или 20. Посттрансляционные модификации включают, но не ограничиваются ими, модификацию глутамина или глутамата на N-конце тяжелой цепи или легкой цепи в пироглутаминовую кислоту путем пироглутамилирования.
В другом варианте осуществления настоящего изобретения предусматривают анти-CD137 антигенсвязывающая молекулу или антитело, причем антитело включают вариабельный домен легкой цепи (VL), который по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичен аминокислотной последовательности SEQ ID NO: 54, 55, 56, 57, 58, 59 или 60. В некоторых вариантах осуществления настоящего изобретения последовательность VL, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична, содержит замены (например, содержит консервативные замены), инсерции или делеции относительно контрольной последовательности, но анти-CD137 антигенсвязывающая молекула или антитело, содержащая такую последовательность, сохраняет способность связываться с CD137. В некоторых вариантах осуществления настоящего изобретения в целом от 1 до 10 аминокислот были заменены, инсертированы и/или делетированы в SEQ ID NO: 54, 55, 56, 57, 58, 59 или 60. В некоторых вариантах осуществления настоящего изобретения замены, инсерции или делеции происходят в областях вне HVR (т.е. в каркасных участках (FR)). Необязательно, анти-CD137 антигенсвязывающая молекула или антитело содержит последовательность VL в SEQ ID NO: 54, 55, 56, 57, 58, 59 или 60, включая посттрансляционные модификации этой последовательности. В одном из вариантов осуществления настоящего изобретения VL содержит одну, две или три HVR, выбранные из: (a) HVR-L1, содержащей аминокислотную последовательность, выбранную из SEQ ID NO: 21, 22, 23, 24 и 25; (б) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO: 26; (в) HVR-L3, содержащей любую одну аминокислотную последовательность, выбранную из SEQ ID NO: 27, 28 и 29. Посттрансляционные модификации включают, но не ограничиваются ими, модификацию глутамина или глутамата на N-конце тяжелой цепи или легкой цепи в пироглутаминовую кислоту путем пироглутамилирования.
В другом варианте осуществления настоящего изобретения предусматривают анти-CD137 антигенсвязывающую молекулу или антитело, причем антигенсвязывающая молекула или антитело содержит VH, как в любом из вариантов осуществления настоящего изобретения, представленных выше, и VL, как в любом из вариантов осуществления настоящего изобретения, представленных выше.
- В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включает последовательности VH и VL в последовательностях SEQ ID NO: 43 и SEQ ID NO: 54, соответственно, включая посттрансляционные модификации этих последовательностей.
- В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включает последовательности VH и VL в последовательностях SEQ ID NO: 44 и SEQ ID NO: 55, соответственно, включая посттрансляционные модификации этих последовательностей.
- В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включает последовательности VH и VL в последовательностях SEQ ID NO: 45 и SEQ ID NO: 55, соответственно, включая посттрансляционные модификации этих последовательностей.
- В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включает последовательности VH и VL в последовательностях SEQ ID NO: 46 и SEQ ID NO: 54, соответственно, включая посттрансляционные модификации этих последовательностей.
- В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включает последовательности VH и VL в последовательностях SEQ ID NO: 47 и SEQ ID NO: 54, соответственно, включая посттрансляционные модификации этих последовательностей.
- В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включает последовательности VH и VL в последовательностях SEQ ID NO: 48 и SEQ ID NO: 56, соответственно, включая посттрансляционные модификации этих последовательностей.
- В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включает последовательности VH и VL в последовательностях SEQ ID NO: 49 и SEQ ID NO: 57, соответственно, включая посттрансляционные модификации этих последовательностей.
- В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включает последовательности VH и VL в последовательностях SEQ ID NO: 50 и SEQ ID NO: 58, соответственно, включая посттрансляционные модификации этих последовательностей.
- В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включает последовательности VH и VL в последовательностях SEQ ID NO: 51 и SEQ ID NO: 59, соответственно, включая посттрансляционные модификации этих последовательностей.
- В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включает последовательности VH и VL в последовательностях SEQ ID NO: 51 и SEQ ID NO: 60, соответственно, включая посттрансляционные модификации этих последовательностей.
- В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включает последовательности VH и VL в последовательностях SEQ ID NO: 52 и SEQ ID NO: 60, соответственно, включая посттрансляционные модификации этих последовательностей.
- В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включает последовательности VH и VL в последовательностях SEQ ID NO: 50 и SEQ ID NO: 59, соответственно, включая посттрансляционные модификации этих последовательностей.
- В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело включает последовательности VH и VL в последовательностях SEQ ID NO: 53 и SEQ ID NO: 54, соответственно, включая посттрансляционные модификации этих последовательностей.
Вышеупомянутые посттрансляционные модификации включают, но не ограничиваются ими, модификацию глутамина или глутамата на N-конце тяжелой цепи или легкой цепи в пироглутаминовую кислоту путем пироглутамилирования.
SEQ ID NO, соответствующие аминокислотным последовательностям предпочтительной вариабельной области тяжелой цепи и вариабельной области легкой цепи и их HVR1, 2 и 3 для каждой анти-CD137 антигенсвязывающей молекулы или антитела по настоящему изобретению, показаны в таблице ниже.


Если анти-CD137 антигенсвязывающая молекула или антитело по настоящему изобретению содержит глутамин в качестве N-концевой аминокислоты тяжелой цепи или легкой цепи, эта аминокислота может быть заменена глутаминовой кислотой. Если анти-CD137 антитело, представленное в настоящем изобретении, содержит глутаминовую кислоту в качестве аминокислоты на N-конце тяжелой цепи или легкой цепи, эта аминокислота может быть заменена глутамином.
В предпочтительном варианте осуществления настоящего изобретения анти-CD137 антигенсвязывающие молекулы или антитела, содержащие описанные выше HVR, вариабельные области тяжелой цепи и/или вариабельные области легкой цепи, все обладают зависимой от низкомолекулярного соединения связывающей активностью по отношению к CD137 согласно описанному выше.
Константные области
В другом варианте осуществления настоящего изобретения анти-CD137 антигенсвязывающие молекулы или антитела содержат константную область. Константная область может быть константной областью тяжелой цепи (включая область Fc), константной областью легкой цепи или обеими. В другом варианте осуществления настоящего изобретения анти-CD137 антигенсвязывающие молекулы или антитела содержат область Fc. В некоторых вариантах осуществления настоящего изобретения константная область является областью с нативной последовательностью. Примеры константных областей тяжелой цепи, полученных из нативных антител, включают, например, константную область тяжелой цепи IgG1 человека (SEQ ID NO: 61, 62), IgG2 человека, IgG3 человека, IgG4 человека и другие подобные. Примеры константных областей легкой цепи, полученных из нативных антител, включают, например, каппа-цепь человека, лямбда-цепь человека (например, SEQ ID NO: 63) и другие подобные.
В настоящем изобретении понятие «исходная константная область» или «исходная область Fc» относятся к константной области или области Fc до внедрения аминокислотных изменений, описанных в настоящем изобретении. Понятие «исходная антигенсвязывающая молекула» относится к антигенсвязывающей молекуле, которая содержит исходную константную область или исходную область Fc. В некоторых вариантах осуществления настоящего изобретения исходная область Fc является областью Fc с нативной последовательностью (или областью Fc нативного антитела). Антитела включают, например, IgA (IgA1, IgA2), IgD, IgE, IgG (IgG1, IgG2, IgG3, IgG4), IgM и др. Антитела могут происходить от человека или от обезьяны (например, макаки-крабоеда, макаки резус, мармозетки, шимпанзе или бабуина). Нативные антитела также могут иметь природные мутации. Множество аллотипов последовательностей IgG из-за генетического полиморфизма описано в публикации «Sequences of proteins of immunological interest», NIH Publication No. 91-3242, каждый из которых может применяться в настоящем изобретении. В одном из вариантов осуществления настоящего изобретения исходная область Fc является областью Fc, происходящей от константной области IgG1 человека, представленной последовательностями SEQ ID NO: 61, 62 или 182.
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающие молекулы или антитела обладают повышенной изоэлектрической точкой (pi, isoelectric point) по сравнению с анти-CD137 антигенсвязывающими молекулами или антителами, которые содержат нативную последовательность области Fc или исходной области Fc. В некоторых вариантах осуществления настоящего изобретения варианты области Fc включают по меньшей мере одно аминокислотное изменение. В других вариантах осуществления настоящего изобретения аминокислотное изменение приводит к повышению изоэлектрической точки (pI) варианта области Fc по сравнению с исходной областью Fc. Не опираясь на какую-либо теорию, предполагают, что рН биологических жидкостей (например, плазмы) находится в нейтральном диапазоне рН. В биологических жидкостях общий положительный заряд антигенсвязывающей молекулы или антитела с повышенной величиной pI увеличивается из-за увеличения pI, и в результате антигенсвязывающая молекула или антитело сильнее притягивается физико-химическим кулоновским взаимодействием к поверхности эндотелиальной клетки, которая имеет общий отрицательный заряд по сравнению с антигенсвязывающей молекулой или антителом, не имеющим повышенной величины pI. Таким образом, агонистические антигенсвязывающие молекулы (или антитела) или антигенсвязывающие агонистические антигенсвязывающие молекулы (или антитела) могут приближаться к поверхности клеток, которые экспрессируют рецептор Fc-гамма, что приводит к усилению связывания антигенсвязывающих молекул или антител с клетками, экспрессирующим Fc-гамма-рецептор. Для тех анти-CD137 агонистических антигенсвязывающих молекул или антител, которые проявляют агонистическую активность против CD137, основанную на вкладе за счет связывающей активности связывания с Fc-гамма-рецептором, анти-CD137-агонистические антигенсвязывающие молекулы или антитела, имеющие повышенное связывание с экспрессирующими Fc-гамма рецептор клетками из-за повышающих pI аминокислотных изменений могут проявлять более сильную агонистическую активность против CD137 по сравнению с анти-CD137 агонистическими антигенсвязывающими молекулами или антителами, не имеющими изменений аминокислот, которые повышают изоэлектрическую точку (pI).
В настоящем изобретении pI может быть теоретическим или экспериментально определенной величиной. Значение pI можно определить, например, с помощью изоэлектрического фокусирования, известного специалистам в данной области. Значение теоретической pI можно рассчитать, например, с помощью программного обеспечения для анализа генной и аминокислотной последовательности (Genetyx и т.д.). При расчете свойства антитела могут быть отражены в формуле расчета. Например, (i) обычно Cys, консервативный в антителе, образует дисульфидную связь и не несет электрического заряда в боковой цепи; поэтому такой Cys может быть исключен из расчета, и только Cys в свободной форме, который не образует дисульфидную связь, может быть включен в расчет. В другом варианте (ii) состояние заряда или изоэлектрическая точка антител может изменяться из-за посттрансляционных модификаций; поэтому формула расчета может быть изменена следующим образом с учетом таких посттрансляционных модификаций: (а) когда N-концом тяжелой цепи является Q (глутамин), N-концевая аминогруппа исключается из расчета, предполагая, что происходит пироглутамилирование, (б) когда С-концом тяжелой цепи является K (лизин), K (только один остаток) исключается из расчета, предполагая, что происходит усечение; и (в) боковые цепи всех С (цистеинов), присутствующих в обычно консервативных положениях, исключаются из расчета, предполагая, что все эти С образуют дисульфидные связи внутри молекулы. В одном предпочтительном варианте осуществления настоящего изобретения оба вышеописанных положения (i) и (ii) могут быть отражены в формуле расчета.
В одном из вариантов осуществления настоящего изобретения величина pI может быть увеличена, например, по меньшей мере на 0,01, 0,03, 0,05, 0,1, 0,2, 0,3, 0,4, 0,5, или более, по меньшей мере на 0,6, 0,7, 0,8, 0,9, или более, по меньшей мере на 1,0, 1,1, 1,2, 1,3, 1,4, 1,5, или более, или по меньшей мере на 1,6, 1,7, 1,8, 1,9, 2,0, 2,1, 2,2, 2,3, 2,4, 2,5, 3,0 или более, по сравнению с величиной pI до модификации.
В одном из вариантов осуществления настоящего изобретения аминокислотные изменения, относящиеся к увеличению и способы увеличения pI антигенсвязывающей молекулы или антитела, подробно описаны в настоящем изобретении в разделе «III. Композиции и способы (агонистические антигенсвязывающие молекулы, содержащие вариант области Fc с повышенной изоэлектрической точкой (pI))». Специалисты в данной области поймут, что любые аминокислотные изменения и способы увеличения pI, описанные в разделе «III. Композиции и способы (агонистические антигенсвязывающие молекулы, содержащие вариант области Fc с повышенной изоэлектрической точкой (pI))» могут применяться к анти-CD137 антигенсвязывающим молекулам или антителам.
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающие молекулы или антитела имеют вариант области Fc с увеличенной величиной pI, и вариант области Fc содержит по меньшей мере одно аминокислотное изменение по меньшей мере в одном положении, выбранном из группы, состоящей из положений 285, 311, 312, 315, 318, 333, 335, 337, 341, 342, 343, 384, 385, 388, 390, 399, 400, 401, 402, 413, 420, 422 и 431 по нумерации EU. В других вариантах осуществления настоящего изобретения варианты области Fc с повышенной величиной pI включают Arg или Lys в каждом выбранном положении.
В других вариантах осуществления настоящего изобретения анти-CD137 антигенсвязывающие молекулы или антитела имеют вариант области Fc с повышенной величиной pI, а вариант области Fc содержит по меньшей мере одно аминокислотное изменение по меньшей мере в одном положении, выбранном из группы, состоящей из положений 311, 343 и 413 по нумерации EU. В других вариантах осуществления настоящего изобретения варианты области Fc с повышенной величиной pI включают аминокислотное изменение в положении 311, 343 или 413 по нумерации EU. В другом варианте осуществления настоящего изобретения варианты области Fc с повышенной величиной pI включают Arg или Lys в каждом выбранном положении.
В другом варианте осуществления настоящего изобретения предусматривают анти-CD137 антигенсвязывающие молекулы или антитела, содержащие вариант области Fc с повышенной величиной pI, который содержит аминокислотные изменения любого из следующих (1) - (3): (1) в положениях 311 и 343; (2) в положениях 311 и 413; и (3) в положениях 343 и 413 в соответствии с нумерацией EU. В других вариантах осуществления настоящего изобретения варианты области Fc с повышенной величиной pI содержат Arg или Lys в каждом выбранном положении.
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающие молекулы или антитела по настоящему изобретению включают вариант области Fc, которая содержит аминокислотное изменение (изменения), представленные в табл. 2 ниже.

В одном из вариантов осуществления настоящего изобретения предусматривают анти-CD137 антигенсвязывающие молекулы или антитела, содержащие вариант области Fc, полученный путем внесения аминокислотных изменений в область Fc, имеющую нативную последовательность. В одном из вариантов осуществления настоящего изобретения вариантные области Fc обладают повышенной связывающей активностью по меньшей мере с одним рецептором Fc-гамма, выбранным из группы, состоящей из Fc-гамма RIa, Fc-гамма RIIa, Fc-гамма RIIb, Fc-гамма RIIIa и Fc-гамма RIIIb, по сравнению с областью Fc, имеющей нативную последовательность или исходную областью Fc. Предпочтительно, вариантные области Fc обладают повышенной связывающей активностью в отношении Fc-гамма RIIb по сравнению с областью Fc, имеющей нативную последовательность или исходную область Fc. Сообщают, что анти-CD137 антитело, содержащее вариант области Fc с повышенной связывающей активностью в отношении Fc-гамма RIIb, обладает повышенной агонистической активностью по сравнению с анти-CD137 антителом, содержащим область Fc с нативной последовательностью. В одном из вариантов осуществления настоящего изобретения в качестве аминокислотных изменений для увеличения связывающей активности по отношению к Fc-гамма RIIb, например, можно использовать аминокислотные изменения, описанные в WO2012/115241, WO2014/030728, WO2014/163101 и/или WO2017/104783. В одном предпочтительном варианте осуществления настоящего изобретения изменения для увеличения связывающей активности по отношению к Fc-gamma RIIb представляют собой аминокислотные изменения по меньшей мере в одном положении, выбранном из группы, состоящей из положений 234, 235, 236, 237, 238, 264, 268, 295, 326 и 330 по нумерации EU.
«Рецепторы Fc-гамма» (называемые в настоящем изобретении рецепторами Fc-гамма, Fc гамма R или FcgR) относятся к рецепторам, которые могут связываться с областью Fc моноклональных антител IgG1, IgG2, FgG3 и IgG4, и практически означает любого представителя группы семейства белков, кодируемых генами рецепторов Fc-гамма. У людей это семейство включает Fc-гамма RI (CD64), включая изоформы Fc-гамма RIa, Fc-гамма RIb и Fc-гамма RIc; Fc-гамма RII (CD32), включая изоформы Fc-гамма RIIa (включая аллотипы Н131 (тип Н) и R131 (тип R)), Fc-гамма RIIb (включая Fc-гамма RIIb-1 и Fc-гамма RIIb-2) и Fc-гамма RIIc; и Fc-гамма RIII (CD16), включая изоформы Fc-гамма RHIa (включая аллотипы V158 и F158), и Fc-гамма RIIIb (включая аллотипы Fc-гамма RIIIb-NA1 и Fc-гамма RIIIb-NA2) и любые Fc-гамма Rs человека, Fc-гамма изоформы или аллотипы, которые еще предстоит открыть. Установлено, что Fc-гамма RIIb1 и Fc-гамма RIIb2 являются вариантами сплайсинга Fc-гамма RIIb человека. Кроме того, сообщалось о варианте сплайсинга под названием Fc-гамма RIIb3 (J Exp Med, 170, 1989, 1369-1385). Помимо этих вариантов сплайсинга, Fc-гамма Rllb человека включает AAI46679.1, зарегистрированный в NCBI, и все варианты сплайсинга, зарегистрированные в NCBI, а именно NP 001002273.1, NP 001002274.1, NP 001002275.1, NP 001177757.1 и NP 003992.3. Кроме того, Fc-гамма RIIb человека включает все ранее описанные генетические полиморфизмы, а также Fc-гамма RIIb (Arthritis Rheum. 48, 2003, 3242-3252; Kono с соавт., Hum. Mol. Genet. 14, 2005, 2881-2892; Kyogoju с соавт., Arthritis Rheum. 46, 2002, 1242-1254) и каждый генетический полиморфизм, о котором будет известно в будущем.
В Fc-гамма RIIa существует два аллотипа, в одном из которых аминокислота в положении 131 Fc-гамма RIIa представляет собой гистидин (тип Н), а в другом аминокислота в положении 131 замещена аргинином (тип R) (Warrmerdam, J. Exp. Med. 172, 1990, 19-25).
Fc-гамма R включает Fc-гамма R человека, мыши, крысы, кролика и обезьяны, но ими не ограничивается и может происходить из любого организма. Fc-гамма R мыши включают Fc-гамма RI (CD64), Fc-гамма RII (CD32), Fc-гамма RIII (CD16) и Fc-гамма RIII-2 (CD16-2), а также любые Fc-гамма R мыши или изоформы Fc-гамма R, но ими перечень не ограничивают.
В другом варианте осуществления настоящего изобретения предусматривают анти-CD137 антигенсвязывающие молекулы или антитела, содержащие вариант области Fc с повышенной связывающей активностью в отношении Fc-гамма RIIb, которая включает аминокислотные изменения любого из следующих пунктов (1)-(8): (1) в положениях 234, 238, 264 и 330; (2) в положениях 234, 238 и 330; (3) в положениях 234, 237, 238 и 330; (4) в положениях 236, 268 и 330; (5) в положениях 235, 236, 268, 295, 326 и 330; согласно нумерации EU.
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающие молекулы или антитела по настоящему изобретению содержат вариант области Fc, содержащей изменения аминокислот, представленные ниже в табл. 3. В другом варианте осуществления настоящего изобретения анти-CD137 антигенсвязывающие молекулы или антитела по настоящему изобретению содержат вариант области Fc, который дополнительно включает, помимо аминокислотного изменения (изменений), описанных в табл. 2 (аминокислотные изменения, включающие увеличение pI области Fc), любую комбинацию аминокислотных изменений, указанных в табл. 3 ниже.

В одном из вариантов осуществления настоящего изобретения предусматривают варианты области Fc, которые имеют изменение по меньшей мере одной аминокислоты и обладают связывающей активностью по отношению к Fc-гамма RIIb, эквивалентной или более высокой, чем у контрольной области Fc. В одном из вариантов осуществления настоящего изобретения контрольная область Fc представляет собой область Fc, содержащую любую комбинацию аминокислотных изменений, указанных выше в табл. 3. В одном предпочтительном варианте осуществления настоящего изобретения контрольная область Fc представляет собой область Fc, содержащуюся в константной области тяжелой цепи ТТ14 (SEQ ID NO: 149), ТТ16 (SEQ ID NO: 150), MY201 (SEQ ID NO: 153) или MY518 (SEQ ID NO: 154). В одном предпочтительном варианте осуществления настоящего изобретения контрольная область Fc представляет собой область Fc, содержащуюся в константной области тяжелой цепи MY201 (SEQ ID NO: 153) или MY518 (SEQ ID NO: 154).
В другом варианте осуществления настоящего изобретения в настоящем изобретении предусматривают выделенные агонистические антигенсвязывающие молекулы или антитела, которые содержат вариант области Fc с повышенной связывающей активностью в отношении рецептора Fc-гамма (предпочтительно, Fc-гамма RIIb) и увеличенной pI. В одном из вариантов осуществления настоящего изобретения варианты областей Fc содержат по меньшей мере два аминокислотных изменения по сравнению с исходными областями Fc. Выше описано, что антигенсвязывающая молекула или антитело с повышенной pI более сильно притягивается физико-химическим кулоновским взаимодействием к поверхности эндотелиальной клетки, которая имеет общий отрицательный заряд, по сравнению с антигенсвязывающей молекулой или антителом, не имеющим повышенной pI. Следовательно, для тех агонистических антигенсвязывающих молекул или антител, которые проявляют агонистическую активность, основанную на вкладе связывающей активности в отношении рецептора Fc-гамма (предпочтительно Fc-гамма RIIb), агонистическая активность антигенсвязывающих молекул или антител может быть увеличена путем комбинирования аминокислотного изменения (изменений) для увеличения рецептора Fc-гамма (предпочтительно Fc-гамма RIIb) и аминокислотное изменение (изменений) для увеличения pI.
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающие молекулы или антитела содержат вариант области Fc, который включает как аминокислотные изменения для увеличения связывающей активности в отношении Fc-гамма-рецептора (например, Fc-gamma RIIb), так и аминокислотные изменения для увеличения изоэлектрической точки (pI), как описано выше. Выше описано, что антигенсвязывающая молекула или антитело с повышенной pI более сильно притягивается физико-химическим кулоновским взаимодействием к поверхности эндотелиальной клетки, которая имеет общий отрицательный заряд, по сравнению с антигенсвязывающей молекулой или антителом, не имеющим повышенной изоэлектрической точки (pI). Следовательно, для тех анти-CD137 агонистических антигенсвязывающих молекул или антител, которые проявляют агонистическую активность против CD137 на основе вклада за счет связывающей активности с рецептором Fc-гамма (предпочтительно Fc-гамма RIIb), агонистическая активность анти-CD137 антигенсвязывающих молекул против или антитела могут быть увеличены путем комбинирования аминокислотных изменений для увеличения рецептора Fc-гамма (предпочтительно Fc-гамма RIIb) и аминокислотных изменений для увеличения pI.
В одном из вариантов осуществления настоящего изобретения в настоящем изобретении предусматривают полипептиды, включающие вариант области Fc с повышенной связывающей активностью в отношении Fc-гамма RIIb и с повышенной величиной pI, который включает по меньшей мере три аминокислотных изменения, включая (а) по меньшей мере одно аминокислотное изменение по меньшей мере в одном положении, выбранном из группы, состоящей из положений 234, 235, 236, 237, 238, 264, 268, 295, 326 и 330 в соответствии с нумерацией EU, и (б) по меньшей мере два аминокислотных изменения по меньшей мере в двух положениях, выбранных из группы, состоящей из положений 311, 343 и 413, согласно нумерации EU.
В другом варианте осуществления настоящего изобретения предусматривают полипептиды, содержащие вариант области Fc с повышенной связывающей активностью по отношению к Fc-gamma RIIb и повышенной величиной pI, которые включают аминокислотные изменения по любому из следующих пунктов (1) - (26):
(1) положения 235, 236, 268, 295, 326, 330, 343 и 413;
(2) положения 214, 235, 236, 268, 295, 326, 330, 343 и 413;
(3) положения 234, 238, 250, 264, 307, 330, 343 и 413;
(4) положения 234, 238, 264, 330, 343 и 413;
(5) положения 234, 237, 238, 250, 307, 330, 343 и 413;
(6) положения 234, 237, 238, 330, 343 и 413;
(7) положения 235, 236, 268, 295, 326, 330, 311 и 343;
(8) положения 234, 238, 250, 264, 307, 330, 311 и 343;
(9) положения 234, 238, 264, 330, 311 и 343;
(10) положения 234, 237, 238, 250, 307, 330, 311 и 343;
(11) положения 234, 237, 238, 330, 311 и 343;
(12) положения 235, 236, 268, 295, 326, 330 и 343;
(13) положения 214, 235, 236, 268, 295, 326, 330 и 343;
(14) положения 235, 236, 268, 295, 326, 330 и 413;
(15) положения 214, 236, 268, 330 и 343;
(16) положения 214, 235, 236, 268, 330 и 343;
(17) положения 214, 236, 268, 330 и 413;
(18) положения 214, 236, 268, 330, 343 и 413;
(19) положения 214, 235, 236, 268, 330, 343 и 413;
(20) положения 214, 236, 268, 330 и 311;
(21) положения 214, 235, 236, 268, 330 и 311;
(22) положения 214, 236, 268, 330, 311 и 343;
(23) положения 214, 235, 236, 268, 330, 311 и 343;
(24) положения 214, 236, 268, 330, 311 и 413;
(25) положения 214, 235, 236, 268, 330, 311 и 413;
(26) положения 214, 235, 236, 268, 295, 326, 330 и 311,
в соответствии с нумерацией EU.
В одном из вариантов осуществления настоящего изобретения вариантные области Fc по настоящему изобретению включают любую комбинацию аминокислотных изменений, представленных ниже в табл. 4.
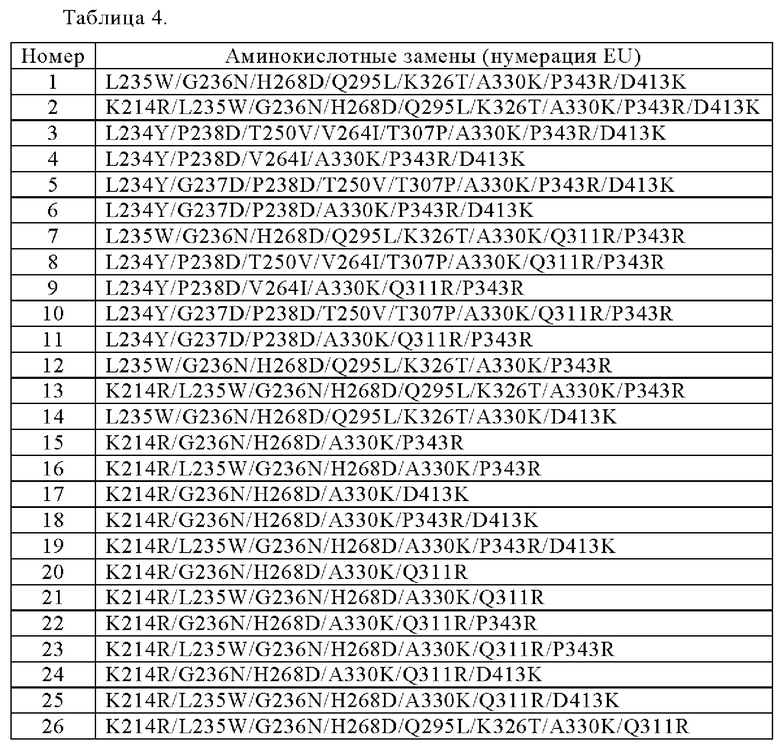
В одном из вариантов осуществления настоящего изобретения варианты области Fc, содержащие любую комбинацию аминокислотных изменений, описанных выше в табл. 4, не содержат аминокислоту в положении 447 согласно нумерации EU.
Специалистам в данной области известно, что по крайней мере одно изменение аминокислоты для увеличения связывающей активности в отношении Fc-гамма R (включая Fc-гамма RIIb) по сравнению с исходной областью Fc, как описано или предложено, например, в WO 2013/047752, WO 2013/125667, WO 2014/030728, WO 2014/163101 или WO 2017104783, и по меньшей мере одно аминокислотное изменение для увеличения pI по сравнению с исходной областью Fc, как описано или предложено, например, в WO 2017/104783, WO 2017/046994 и любой комбинации этих аминокислотных изменений могут быть использованы в дополнение к изменениям, представленным в качестве иллюстраций выше.
Кроме того, аминокислотные изменения, выполненные для других целей, могут быть объединены в варианте области Fc, описанной в настоящем изобретении. Например, аминокислотные замены, повышающие FcRn-связывающую активность (Hinton с соавт., J. Immunol. 176(1), 2006, 346-356; Dall'Acqua с соавт., J. Biol. Chem. 281(33), 2006, 23514-23524; Petkova с соавт., Intl. Immunol. 18(12), 2006, 1759-1769; Zalevsky с соавт., Nat. Biotechnol. 28(2), 2010, 157-159; WO 2006/019447; WO 2006/053301; WO 2009/086320), и могут быть добавлены аминокислотные замены для улучшения гетерогенности или стабильности антител (WO 2009/041613). В другом варианте полипептиды со свойством стимулирования клиренса антигена, которые описаны в WO 2011/122011, WO 2012/132067, WO 2013/046704 или WO 2013/180201, полипептиды со свойством специфического связывания с тканью-мишенью, которые описаны в WO 2013/180200, полипептиды со свойством многократного связывания с множеством молекул антигена, которые описаны в WO 2009/125825, WO 2012/073992 или WO 2013/047752, можно комбинировать с вариантом области Fc, описанным в настоящем изобретении. В другом варианте с целью придания связывающей активности другим антигенам, аминокислотные изменения, описанные в ЕР 752471 и ЕР 1772465, могут быть объединены в СНЗ варианта области Fc, описанного в настоящем изобретении.
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающие молекулы или антитела по настоящему изобретению содержат константную область тяжелой цепи, содержащую любую одну аминокислотную последовательность, выбранную из SEQ ID NO: 64-85. Предпочтительно анти-CD137 антигенсвязывающие молекулы или антитела по настоящему изобретению содержат константную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 75 или 82.
В одном предпочтительном варианте осуществления настоящего изобретения анти-CD137 антигенсвязывающие молекулы или антитела по настоящему изобретению содержащие описанный выше вариант области Fc, обладают описанной выше СВ137-связывающей активностью, зависящей от низкомолекулярного соединения.
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающие молекулы или антитела по настоящему изобретению содержат следующую вариабельную область и константную область: вариабельную область, содержащую описанную выше область HVR, вариабельную область тяжелой цепи и/или вариабельную область легкой цепи; и описанный выше вариант области Fc. В одном предпочтительном варианте осуществления настоящего изобретения анти-CD137 антигенсвязывающие молекулы или антитела по настоящему изобретению могут быть любым анти-CD137 антителом, выбранным из антител, описанных в табл. 52.
В другом варианте осуществления настоящего изобретения предусматривают антигенсвязывающие молекулы или антитела, связывающиеся с тем же эпитопом на CD137, что и анти-CD137 антигенсвязывающие молекулы или антитела, представленные в настоящем изобретении, в присутствии низкомолекулярного соединения (например, в присутствии 10 мкМ или более, 50 мкМ или более, 100 мкМ или более, 150 мкМ или более, 200 мкМ или более, или 250 мкМ или более низкомолекулярного соединения). Например, в одном из вариантов осуществления настоящего изобретения предусматривают те антигенсвязывающие молекулы или антитела, которые связываются с одним и тем же эпитопом с антигенсвязывающими молекулами или антителами против CD137, содержащими А375/В167, А372/В040, А356/В040, А486/В167, А487/В167, А488/В226, А489/В223, А548/В376, А551/В256, А551/В379, А555/В379, А548/В256 и/или А549/В167, описанными в табл. 17, в виде комбинации вариабельной области тяжелой цепи/вариабельной области легкой цепи. В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающие молекулы или антитела по настоящему изобретению, обладающие CD137-связывающей активностью, которая зависит от антигенсвязывающей активности, зависящей от низкомолекулярного соединения, распознают эпитоп, образованный комплексом, сформированным из антигена (например, CD137) и низкомолекулярного соединения (например, АТФ).
В другом варианте осуществления настоящего изобретения предусматривают антигенсвязывающие молекулы или антитела, которые конкурируют за связывание с CD137 с анти-CD137 антигенсвязывающими молекулами или антителами по настоящему изобретению в присутствии низкомолекулярного соединения (например, в присутствии 10 мкМ или более, 50 мкМ или более, 100 мкМ или более, 150 мкМ или более, 200 мкМ или более или 250 мкМ или более низкомолекулярного соединения). Например, в одном из вариантов осуществления настоящего изобретения они конкурируют за сайт связывания с CD137 с анти-CD137 антигенсвязывающими молекулами или антителами, содержащими А375/В167, А372/В040, А356/В040, А486/В167, А487/В167, А488/В226, А489/В223, А548/В376, А551/В256, А551/В379, А555/В379, А548/В256 и/или А549/В167, описанные в табл. 17, в виде комбинации вариабельной области тяжелой цепи/вариабельной области легкой цепи.
В другом варианте осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело по любому из указанных выше вариантов осуществления настоящего изобретения является моноклональным антителом, в том числе химерным, гуманизированным или антителом человека. В одном из вариантов осуществления настоящего изобретения анти-CD137 антитело является фрагментом антитела, например, Fv, Fab, Fab', scFv, диателом или фрагментом F(ab')2. В другом варианте осуществления настоящего изобретения антитело является антителом полной длины, например, интактным антителом IgG1 или антителом другого класса или изотипа согласно настоящему описанию.
В другом варианте осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело по любому из указанных выше вариантов осуществления настоящего изобретения может включать любую из функций, по отдельности или в комбинации, как описано в разделах 1-7 ниже:
1. Агонистическая активность анти-CD137 антигенсвязывающей молекулы или антитела
В определенном варианте осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело по настоящему изобретению обладает агонистической активностью CD137. Передача сигнала CD137 не только стимулирует секрецию IFN-γ и пролиферацию клеток NK (Buechele с соавт., 2012; Lin с соавт., 2008; Melero с соавт., 1998), но также повышает их выживаемость и активацию дендритных клеток (DC), на что указывает активация костимулирующих молекул и секреции цитокинов (Choi с соавт., 2009; Futagawa с соавт., 2002; Wilcox с соавт., 2002). Однако CD137 лучше всего охарактеризовать как костимулирующую молекулу, которая регулирует TCR-индуцированную активацию как в CD4+, так и в CD8+ подмножеств Т-клеток. В комбинации с активацией TCR анти-CD137 агонистические антитела усиливают пролиферацию Т-клеток, стимулируют секрецию лимфокинов и снижают чувствительность Т-лимфоцитов к гибели клеток, вызванной активацией (рассмотрено в публикации Snelc соавт., 2011). Из этих явлений физиологические явления, наблюдаемые после передачи сигнала CD137 на Т-клетки, опосредуются расположенными ниже по цепи сигналами, активируемыми передачей сигналов CD137, такими как TRAF2, TRAF1, в частности NF-kappaB, JNK, Erk, Akt, сурвивин, Bcl-XL и/или Bcl-2 (Ward-Kavanagh с соавт., Immunity, 44, 2016, 1005).
В одном из вариантов осуществления настоящего изобретения понятия «анти-CD137 агонистическая антигенсвязывающая молекула» или «анти-CD137 агонистическое антитело» означают антигенсвязывающую молекулу или антитело, которое, связываясь с CD137, трансдуцирует сигнал CD137 и значительно индуцирует или усиливает секрецию IFN-гамма, пролиферацию и повышение выживаемости клеток NK; активация DC указывает на повышенную регуляцию секреции цитокинов и костимулирующих молекул; индукцию TCR; пролиферацию Т-клеток; и/или секрецию лимфокинов. В другом варианте осуществления настоящего изобретения понятия «анти-CD137 агонистическая антигенсвязывающая молекула» или «анти-CD137 агонистическое антитело» означают антигенсвязывающую молекулу или антитело, которое трансдуцирует сигнал CD137 путем связывания с CD137 на Т-клетках и существенно индуцирует активацию NF-kappaB Т-клеток. Кроме того, антигенсвязывающая молекула или антитело «проявляет агонистическую активность против CD137» означает, что наблюдается любое из вышеупомянутых физиологических явлений, когда антигенсвязывающая молекула или антитело связывается с CD137. Метод измерения агонистической активности CD137 подробно описан ниже в разделе «В. Анализы».
В определенном варианте осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело по настоящему изобретению обладает зависимой от низкомолекулярных соединений агонистической активностью по отношению к CD137. В одном из вариантов осуществления настоящего изобретения, не ограничивающем рамок охвата настоящего изобретения, агонистическая активность CD137 анти-CD137 антигенсвязывающей молекулы или антитела против CD137 в присутствии низкомолекулярного соединения выше, чем агонистическая активность CD137 в отсутствие низкомолекулярного соединения. В другом варианте осуществления настоящего изобретения агонистическая активность в отношении CD137 анти-CD137 антигенсвязывающей молекулы или антитела в присутствии высокой концентрации низкомолекулярного соединения выше по сравнению с агонистической активностью CD137 в присутствии низкой концентрации низкомолекулярного соединения. В другом варианте осуществления настоящего изобретения агонистическая активность в отношении CD137 анти-CD137 антигенсвязывающей молекулы или антитела в присутствии низкомолекулярного соединения больше в 2 раза или более, в 3 раза или более, в 5 раз или более, в 10 раз или более, в 20 раз или более, в 30 раз или более, в 50 раз или более, в 100 раз или более, в 200 раз или более, в 300 раз или более, в 500 раз или более, в 1×103 раз или более, 2×103 раз или более, в 3×103 раз или более, в 5×103 раз или более, в 1×104 раз или более, в 2×104 раз или более, в 3×104 раз или более, в 5×104 раз или более, или в 1×105 раз или более по сравнению с агонистической активностью в отношении CD137 в отсутствие низкомолекулярного соединения.
Любая подходящая концентрация может быть выбрана в качестве концентрации низкомолекулярного соединения при условии, что обнаруживают различие в связывающей активности анти-CD137 антигенсвязывающей молекулы или антитела. В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело трансдуцирует сигнал CD137, связываясь с CD137 на поверхности клетки. Следовательно, специалисту в данной области очевидно, что анти-CD137 антигенсвязывающая молекула или антитело, которое обладает зависимой от низкомолекулярного соединения связывающей активностью с CD137, обладает агонистической активностью по отношению к CD137, зависящей от низкомолекулярного соединения. Однако, с другой стороны, поскольку методы измерения связывающей активности и агонистической активности различны, специалист в данной области поймет, что концентрация низкомолекулярного соединения, при которой обнаруживают различия в связывающей активности, может отличаться от концентрации низкомолекулярного соединения, для которой обнаружено различие в агонистической активности (например, для анти-CD137 антигенсвязывающей молекулы или антитела, чья CD137-связывающая активность в присутствии низкомолекулярного соединения в концентрации 10 мкМ выше в 2 раза или более по сравнению со связывающей активностью CD137 в отсутствие низкомолекулярного соединения, агонистическая активность CD137 (исследуемое значение) в присутствии низкомолекулярного соединения в концентрации 10 мкМ может быть менее чем в 2 раза по сравнению с агонистической активностью CD137 (исследуемое значение) в отсутствие низкомолекулярного соединения). Кроме того, специалистам в данной области понятно, что определение агонистической активности может варьировать в зависимости от анализа агонистической активности в отношении CD137 (см. раздел «В. Анализы»).
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело (i) проявляет агонистическую активность в отношении CD137 в присутствии низкомолекулярного соединения в количестве 10 мкМ, 50 мкМ, 100 мкМ, 150 мкМ, 200 мкМ или 250 мкМ, и (ii) по существу не проявляет агонистической активности в отношении CD137 в отсутствие низкомолекулярного соединения или имеет низкую агонистическую активность в отношении CD137 в отсутствие низкомолекулярного соединения (по сравнению с присутствием низкомолекулярного соединения).
В одном из вариантов осуществления настоящего изобретения если агонистическую активность анти-CD137 антигенсвязывающей молекулы или антитела оценивают с помощью «а) Анализа агонистической активности (МКПК)», подробно описанного в разделе «В. Анализы», анти-CD137 агонистическая антигенсвязывающая молекула или антитело (i) проявляет агонистическую активность в отношении CD137 в присутствии низкомолекулярного соединения в концентрации 250 мкМ, и (ii) имеет низкую агонистическую активность в отношении CD137 в отсутствие низкомолекулярное соединение (по сравнению с наличием низкомолекулярного соединения). В другом варианте осуществления настоящего изобретения анти-CD137 агонистическая антигенсвязывающая молекула или антитело (i) проявляет агонистическую активность по отношению к CD137 в присутствии низкомолекулярного соединения в концентрации 250 мкМ, и (ii) по существу не проявляет агонистической активности в отношении CD137 в отсутствие низкомолекулярного соединения.
В одном из вариантов осуществления настоящего изобретения если агонистическую активность анти-CD137 антигенсвязывающей молекулы или антитела оценивают с помощью пункта «б) Анализ агонистической активности (анализ репортерного гена)», подробно рассмотренного в разделе «В. Анализы», анти-CD137 антигенсвязывающая молекула или антитело (i) проявляют агонистическую активность в отношении CD137 в присутствии 10 мкМ, 50 мкМ, 100 мкМ, 150 мкМ, 200 мкМ или 250 мкМ низкомолекулярного соединения, и (ii) по существу не имеет агонистической активности в отношении CD137 или имеет более пониженную агонистическую активность в отсутствие низкомолекулярного соединения (по сравнению с таковой в присутствии низкомолекулярного соединения). Концентрации антител в анализе репортерного гена могут быть выбраны произвольно, например, конечная концентрация антитела составляет 0, 0,001, 0,01, 0,1, 1 или 10 мкг/мл. В предпочтительном варианте осуществления настоящего изобретения конечная концентрация антитела составляет 0,1 мкг/мл или 1 мкг/мл.
В одном из вариантов осуществления настоящего изобретения, когда конечная концентрация антитела составляет 0,1 мкг/мл в пункте «б) Анализ агонистической активности (анализ репортерного гена)», подробно рассмотренного в разделе «В. Анализы», (i) агонистическая активность (относительная световая единица (RLU)) анти-CD137 антигенсвязывающей молекулы или антитела против CD137 в присутствии 10 мкМ низкомолекулярного соединения является 2-кратной или более, 3-кратной или более, 5 -кратной или более, 10-кратной или более, 20-кратной или более, 30-кратной или более, 50-кратной или более, 60-кратной или более, 70-кратной или более, 80-кратной или более или 90-кратной или более по сравнению с (ii) агонистической активностью в отношении CD137 (относительная световая единица) в отсутствие низкомолекулярного соединения. В одном из вариантов осуществления настоящего изобретения если конечная концентрация антитела составляет 0,1 мкг/мл в пункте «б) Анализ агонистической активности (анализ репортерного гена)», подробно рассмотренного в разделе «В. Анализы», (i) агонистическая активность в отношении CD137 (относительная световая единица) антигенсвязывающей молекулы или антитела в отношении CD137 в присутствии 100 мкМ низкомолекулярного соединения является 2-кратной или более, 3-кратной или более, 5 -кратной или более, 10-кратной или более, 20-кратной или более, 30-кратной или более, 50-кратной или более, 60-кратной или более, 70-кратной или более, 80-кратной или более или 90- кратной или более по сравнению с (ii) агонистической активностью CD137 (относительная световая единица) в отсутствие низкомолекулярного соединения. В одном из вариантов осуществления настоящего изобретения, когда конечная концентрация антител составляет 0,1 мкг/мл в пункте «б) Анализ агонистической активности (анализ репортерного гена)», подробно рассмотренного в разделе «В. Анализы»,_(1) агонистическая активность в отношении CD137 (в относительных световых единицах (RLU)) анти-CD137 агонистической антигенсвязывающей молекулы или антитела в присутствии 250 мкМ низкомолекулярного соединения больше в 2 раза или более, в 3 раза или более, в 5 раз или более, в 10 раз или более, в 20 раз или более, в 30 раз или более, в 50 раз или более, в 60 раз или более, в 70 раз или более, в 80 раз или более, или в 90 раз или более по сравнению с (ii) агонистической активностью в отношении CD137 (в относительных световых единицах) в отсутствие низкомолекулярного соединения. В любом из указанных выше вариантов осуществления настоящего изобретения, дополнительно, 0,1 мкг/мл анти-CD137 антигенсвязывающей молекулы или антитела по существу не проявляет агонистической активности в отношении CD137 в отсутствие низкомолекулярного соединения.
В одном из вариантов осуществления настоящего изобретения когда конечная концентрация антител составляет 1 мкг/мл в пункте «б) Анализ агонистической активности (анализ репортерного гена)», подробно рассмотренного в разделе «В. Анализы», (i) агонистическая активность в отношении CD137 (в относительных световых единицах (RLU)) анти-CD137 агонистической антигенсвязывающей молекулы или антитела в присутствии 10 мкМ низкомолекулярного соединения больше в 2 раза или более, в 3 раза или более, в 5 раз или более, в 10 раз или более, в 20 раз или более, в 30 раз или более, в 50 раз или более, в 60 раз или более, в 70 раз или более, в 80 раз или более, или в 90 раз или более по сравнению с (ii) агонистической активностью в отношении CD137 (в относительных световых единицах) в отсутствие низкомолекулярного соединения. В одном из вариантов осуществления настоящего изобретения если итоговая концентрация антитела составляет 0,1 мкг/мл в пункте «б) Анализ агонистической активности (анализ репортерного гена)», подробно рассмотренного в разделе «В. Анализы», (i) агонистическая активность в отношении CD137 (в относительных световых единицах (RLU)) анти-CD137 антигенсвязывающей молекулы или антитела в присутствии 100 мкМ низкомолекулярного соединения больше в 2 раза или более, в 3 раза или более, в 5 раз или более, в 10 раз или более, в 20 раз или более, в 30 раз или более, в 50 раз или более, в 60 раз или более, в 70 раз или более, в 80 раз или более, или в 90 раз или более по сравнению с (ii) агонистической активностью в отношении CD137 (в относительных световых единицах) в отсутствие низкомолекулярного соединения. В одном из вариантов осуществления настоящего изобретения если итоговая концентрация антитела составляет 0,1 мкг/мл в пункте «б) Анализ агонистической активности (анализ репортерного гена)», подробно рассмотренного в разделе «В. Анализы», (i) агонистическая активность в отношении CD137 (в относительных световых единицах (RLU)) анти-CD137 антигенсвязывающей молекулы или антитела в присутствии 250 мкМ низкомолекулярного соединения больше в 2 раза или более, в 3 раза или более, в 5 раз или более, в 10 раз или более, в 20 раз или более, в 30 раз или более, в 50 раз или более, в 60 раз или более, в 70 раз или более, в 80 раз или более, или в 90 раз или более по сравнению с (ii) агонистической активностью в отношении CD137 (в относительных световых единицах) в отсутствие низкомолекулярного соединения. В любом из указанных выше вариантов осуществления настоящего изобретения, дополнительно, 1 мкг/мл анти-CD137 антигенсвязывающей молекулы или антитела по существу не проявляет агонистической активности в отношении CD137 в отсутствие низкомолекулярного соединения.
2. Фрагменты антител
В некоторых вариантах осуществления настоящего изобретения предусматриваемое антитело является фрагментом антитела. К фрагментам антител относятся, но ими перечень не ограничивается, фрагменты Fab, Fab', Fab'-SH, F(ab')2, Fv и scFv, а также другие фрагменты, описанные ниже. Обзор по фрагментам антител опубликован Hudson с соавт.Nat. Med. 9, 2003, 129-134. Обзор фрагментов scFv см., например, в публикации Pluckthun в «The Pharmacology of Monoclonal Antibodies», под ред. Rosenburg и Moore, (изд-во Springer-Verlag, Нью-Йорк), 113, 1994, 269-315; см. также WO 93/16185; US 5571894 и US 5587458. Для обсуждения фрагментов Fab и F(ab')2, содержащих остатки эпитопа, связывающего рецептор спасения, и имеющих увеличенный период полужизни in vivo, см. US 5869046.
Диатела и фрагменты антитела с двумя сайтами связывания антигена могут быть бивалентными или биспецифичными. См., например, ЕР 404097; WO 1993/01161; Hudson с соавт., Nat. Med. 9, 2003, 129-134; Hollinger с соавт., Proc. Natl. Acad. Sci. USA 90, 1993, 6444-6448. Триатела и тетратела также описаны в публикации Hudson с соавт., Nat. Med. 9, 2003, 129-134.
Однодоменные антитела представляют собой фрагменты антител, содержащие весь или часть вариабельного домена тяжелой цепи или весь или часть вариабельного домена легкой цепи антитела. В некоторых вариантах осуществления настоящего изобретения однодоменное антитело является однодоменным антителом человека (фирма Domantis, Inc., Уолтем, Массачусетс; см., например, US 6248516).
Фрагменты антител могут быть получены различными методами, включая, но ими не ограничиваясь, протеолитическое расщепление интактного антитела, а также выработку рекомбинантными клетками-хозяевами (например, Е. coli или фаг) по описанию настоящего изобретения.
3. Химерные и гуманизированные антитела
В некоторых вариантах осуществления настоящего изобретения предусмотренное антитело является химерным антителом. Некоторые химерные антитела описаны, например, в US 4816567; Morrison с соавт., Proc. Natl. Acad. Sci. USA, 81, 1984, 6851-6855. В одном из примеров химерное антитело содержит вариабельную область, не относящуюся к антителу человека (например, вариабельную область, производную от мыши, крысы, хомяка, кролика или приматов, но не от человека, таких как обезьяна), и константную область человека. В другом примере химерное антитело представляет собой антитело «переключенного класса», в котором класс или подкласс был изменен по сравнению с классом родительского антитела. Химерные антитела включают их антигенсвязывающие фрагменты.
В некоторых вариантах осуществления настоящего изобретения химерное антитело представляет собой гуманизированное антитело. Обычно антитело, не являющееся антителом человека, гуманизируют для снижения иммуногенности для человека, сохраняя при этом специфичность и аффинность исходного, не являющегося антителом человека, антитела. Обычно гуманизированное антитело включает один или более вариабельных доменов, в которых HVR, например, CDR (или их части) происходят из антитела, не являющегося антителом человека, a FR (или их части) происходят из последовательностей антитела человека. Гуманизированное антитело необязательно также может содержать по меньшей мере часть константной области человека. В некоторых вариантах осуществления настоящего изобретения некоторые остатки FR в гуманизированном антителе заменены соответствующими остатками антитела, не являющегося антителом человека (например, антитела, из которого получены остатки HVR), например, для восстановления или улучшения специфичности или аффинности антитела.
Гуманизированные антитела и методы их получения рассмотрены, например, Almagro и Fransson, Front. Biosci. 13, 2008, 1619-1633, а также описаны, например, в публикациях Riechmann с соавт., Nature 332, 1988, 323-329; Queen с соавт., Proc. Nat 7 Acad. Sci. USA 86, 1989, 10029-10033; US 5821337, US 7527791, US 6982321, US 7087409; Kashmiri с соавт., Methods 36, 2005, 25-34 (описание трансплантации области, определяющей специфичность (SDR)); Padlan, Mol. Immunol. 28, 1991, 489-498 (описание «перекладки»); Dall'Acqua с соавт., Methods 36, 2005, 43-60 (описание «перетасовки FR»); Osbourn с соавт., Methods 36, 2005, 61-68; Klimka с соавт., Br. J. Cancer, 83, 2000, 252-260 (описание подхода «направленного отбора» к перетасовке FR).
Области каркасного участка человека, которые можно использовать для гуманизирования, включают, но ими не ограничиваются: каркасные области, выбранные с использованием метода «наилучшего соответствия» (см., например, Sims с соавт. J. Immunol. 151, 1993, 2296); области каркасного участка, производного от консенсусной последовательности антител человека определенной подгруппы вариабельных областей легкой или тяжелой цепи (см., например, Carter с соавт. Proc. Natl. Acad. Sci. USA, 89, 1992, 4285; and Presta с соавт. J. Immunol., 151: 2623 (1993)); зрелые (соматически мутировавшие) каркасные области человека или каркасные области зародышевой линии человека (см., например, Almagro и Fransson, Front. Biosci. 13, 2008, 1619-1633); и области каркасного участка, полученные из скрининговых библиотек FR (См., например, Васа с соавт., J. Biol. Chem. 272, 1997, 10678-10684; Rosok с соавт., J. Biol. Chem. 271, 1996, 22611-22618).
4. Антитела человека
В некоторых вариантах осуществления настоящего изобретения предусмотренное в настоящем изобретении антитело является антителом человека. Антитела человека можно получить с использованием различных методик, известных в данной области. Антитела человека в общем описаны в публикациях van Dijk, van de Winkel, Curr. Opin. Pharmacol. 5, 2001, 368-374, и Lonberg, Curr. Opin. Immunol. 20, 2008, 450-459.
Антитела человека могут быть получены путем введения иммуногена трансгенному животному, которое было модифицировано для выработки интактных антител человека или интактных антител с вариабельными областями человека в ответ на антигенную пробу. Такие животные обычно содержат все или часть локусов иммуноглобулина человека, которые заменяют эндогенные локусы иммуноглобулина, или которые присутствуют вне хромосом или случайным образом интегрированы в хромосомы животного. У таких трансгенных мышей локусы эндогенного иммуноглобулина обычно инактивированы. Для обзора методов получения антител человека от трансгенных животных см. Lonberg, Nat. Biotech. 23, 2005, 1117-1125. См. также, например, патенты US 6075181 и US 6150584, описывающие технологию XENOMOUSE™; патент US 5770429, описывающий технологию HuMab (зарегистрированная торговая марка); патент US 7041870, описывающий технологию K-М MOUSE (зарегистрированная торговая марка), и публикацию патентной заявки US 2007/0061900, описывающую технологию VelociMouse (зарегистрированная торговая марка). Вариабельные области человека из интактных антител, вырабатываемые такими животными, можно дополнительно модифицировать, например, путем комбинирования с другой константной областью человека.
Антитела человека также можно получить методами на основе гибридом. Описаны клеточные линии миеломы человека и гетеромиеломы мыши-человека для выработки моноклональных антител человека. (См., например, Kozbor j. Immunol., 133, 1984, 3001); Brodeur с соавт. в кн.: Monoclonal Antibody Production Techniques and Applications, 1987, 51-63, изд-во Marcel Dekker, Inc., Нью-Йорк); Boerner с соавт., J. Immunol., 147, 1991, 86.) Антитела человека, полученные с помощью технологии гибридом В-клеток человека, также описаны Li с соавт., Proc. Natl. Acad. Sci. USA, 103, 2006, 3557-3562. Дополнительные методы описаны, например, в US 7189826 (описание выработки моноклональных антител IgM человека линиями гибридомных клеток); Ni, Xiandai Mianyixue, 26(4), 2006, 265-268 (описание гибридом клеток человека с клетками человека). Технология гибридомы человека (технология Trioma) также описана в работах Vollmers, Brandlein, Histology andHistopathology, 20(3), 2005, 927-937, и Vollmers, Brandlein, Methods and Findings in Experimental and Clinical Pharmacology, 27(3), 2005, 185-191.
Антитела человека также могут быть получены путем выделения последовательностей вариабельного домена клона Fv, выбранных из полученных от человека библиотек фагового дисплея. Такие последовательности вариабельного домена затем можно комбинировать с желаемым константным доменом человека. Методы отбора человеческих антител из библиотек антител описаны ниже.
5. Антитела, производные от библиотек
Антитела по настоящему изобретению могут быть выделены путем скрининга комбинаторных библиотек на антитела с требуемой активностью или активностями. Например, в данной области известны различные методы создания библиотек фагового дисплея и скрининга таких библиотек на антитела, обладающие требуемыми характеристиками связывания. Такие методы рассмотрены, например, в работе Hoogenboom с соавт., Methods in Molecular Biology 178, 2001, 1-37, под ред. O'Brien с соавт., изд-во Human Press, Тотова, Нью-Джерси, а также описаны, например, McCafferty с соавт., Nature 348, 552-554; Clackson с соавт., Nature 352, 1991, 624-628; Marks с соавт., J. Mol. Biol. 222, 1992, 581-597; Marks, Bradbury, Methods in Molecular Biology 248, 2003, 161-175 (под ред. Lo, изд-во Human Press, Тотова, Нью-Джерси); Sidhu с соавт., J. Mol. Biol. 338(2), 2004, 299-310; Lee с соавт., J. Mol. Biol. 340(5), 2004, 1073-1093; Fellouse, Proc. Natl. Acad. Sci. USA 101(34), 2004, 12467-12472; Lee с соавт., J. Immunol. Methods 284(1-2), 2004, 119-132.
В некоторых методах фагового дисплея репертуары генов VH и VL отдельно клонируют с помощью полимеразной цепной реакции (ПЦР) и случайным образом рекомбинируют в фаговых библиотеках, которые затем могут быть проверены на антигенсвязывающий фаг, как описано Winter с соавт., Ann. Rev. Immunol, 12, 1994, 433-455. Фаг обычно отображает фрагменты антител либо в виде одноцепочечных фрагментов Fv (scFv), либо в виде фрагментов Fab. Библиотеки из иммунизированных источников предоставляют антитела с высоким сродством к иммуногену без необходимости конструирования гибридом. В другом варианте наивный репертуар можно клонировать (например, из человека) для обеспечения единого источника антител к широкому спектру чужеродных, а также аутоантигенов без какой-либо иммунизации, как описано Griffiths с соавт., EMBO J, 12, 1993, 725-734. Наконец, наивные библиотеки также могут быть получены синтетически путем клонирования неупорядоченных сегментов V-гена из стволовых клеток и использования праймеров ПЦР, содержащих случайную последовательность, для кодирования высоко вариабельных участков CDR3 и для выполнения реаранжировки in vitro, как описано Hoogenboom and Winter, J. Мол. Biol., 227, 1992, 381-388. Патентные публикации, описывающие фаговые библиотеки человеческих антител, включают, например, US 5750373 и публикации патентных заявок 2005/0079574, 2005/0119455, 2005/0266000, 2007/0117126, 2007/0160598, 2007/0237764, 2007/0292936 и 2009/0002360.
Антитела или фрагменты антител, выделенные из библиотек антител человека, в настоящем изобретении расценивают как антитела человека или фрагменты антител человека.
Антигенсвязывающие молекулы или антитела, обладающие антигенсвязывающей активностью, зависящей от низкомолекулярного соединения по настоящему изобретению, могут быть выбраны путем проведения скрининга библиотеки антигенсвязывающих молекул. В качестве такой библиотеки можно использовать описанные выше комбинаторные библиотеки. Библиотека антигенсвязывающих молекул может быть с непредвзятым репертуаром антигенсвязывающих молекул (наивная библиотека) или может иметь необъективный репертуар антигенсвязывающих молекул. Примеры библиотеки последнего типа включают библиотеку антигенсвязывающих молекул, которым заранее придается связывающая активность по отношению к определенному соединению. В одном из вариантов осуществления настоящего изобретения библиотека антигенсвязывающих молекул представляет собой библиотеку антигенсвязывающих молекул, в которую заранее вносятся аминокислотные изменения для придания связывающей активности по отношению к определенному соединению. Примеры библиотек такого типа включают библиотеки, описанные, например, в публикации международной патентной заявки WO 2015/083764.
б. Мультиспецифические антитела
В некоторых вариантах осуществления настоящего изобретения предусматривают антитело, которое является мультиспецифическим, например, биспецифическим антителом. Мультиспецифические антитела это моноклональные антитела, которые обладают специфичностью связывания по крайней мере с двумя разными сайтами. В некоторых вариантах осуществления настоящего изобретения одна из специфичностей связывания предназначена для CD137, а другая для любого другого антигена. В некоторых вариантах осуществления настоящего изобретения биспецифические антитела могут связываться с двумя разными эпитопами CD137. Биспецифические антитела также можно использовать для локализации цитотоксических агентов в клетках, экспрессирующих CD137. Биспецифические антитела можно получить в виде полноразмерных антител или фрагментов антител.
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающие молекулы или антитела по настоящему изобретению являются биспецифическими антителами, одна составляющая которых связывается с CD137, зависящая от низкомолекулярного соединения, а другая составляющая связывается с антигеном, отличным от CD137. «Антиген», отличный от CD137, особенно не ограничен по структуре. Другими словами, антиген может быть неорганическим или органическим соединением. Типичные антигены описывают в настоящем изобретении (например, в разделе «IV. Композиции и способы (антигенсвязывающие молекулы, активность связывания которых с антигенами изменяется в зависимости от концентрации низкомолекулярного соединения)», «Б. Антиген»). В одном из вариантов осуществления настоящего изобретения антигены предпочтительно представляют собой антигены, экспрессируемые в раковых клетках, иммунных клетках, стромальных клетках или подобных клетках в раковых тканях или воспалительных тканях.
Методы получения мультиспецифических антител включают, но не ограничиваются ими, рекомбинантную совместную экспрессию двух пар тяжелая цепь-легкая цепь иммуноглобулина, имеющих разные специфичности (см. Milstein, Cuello, Nature 305, 1983, 537; WO 93/08829; Traunecker с соавт., EMBO J. 10, 1991, 3655), конструирование по типу «выступ-во-впадину» (см., например, US 5731168). Мультиспецифические антитела также могут быть получены путем создания электростатических управляющих эффектов для создания Fc-гетеродимерных молекул антитела (WO 2009/089004А1); перекрестным сшиванием двух или более антител или фрагментов (см., например, US 4676980; Brennan с соавт., Science, 229, 1985, 81); использованием лейциновых молний для выработки биспецифических антител (см., например, Kostelny с соавт., J. Immunol., 148(5), 1992, 1547-1553); использованием технологии «диател» для получения фрагментов биспецифических антител (см., например, Hollinger с соавт., Proc. Natl. Acad. Sci. USA, 90, 1993, 6444-6448); использованием димеров одноцепочечных Fv (scFv) (см., например, Gruber с соавт., J. Immunol., 152, 1994, 5368); и получением триспецифических антител, как описано, например, Tutt с соавт.J. Immunol. 147, 1991, 60.
Сконструированные антитела с тремя или более функциональными антигенсвязывающими сайтами, включая мультивалентные «антитела-осьминоги», также включены в настоящее изобретение (см., например US 2006/0025576 А1).
В настоящем изобретении также предусматривают «Fab двойного действия» или «DAF (Dual acting Fab)», содержащий антигенсвязывающий сайт, который связывается с CD137, а также с другим отличающимся антигеном (см., например, US 2008/0069820).
7. Варианты антител
В некоторых вариантах осуществления настоящего изобретения предусматривают варианты аминокислотной последовательности представленных антител. Например, может быть желательно улучшить аффинность и/или другие биологические свойства антитела. Варианты аминокислотной последовательности антитела могут быть получены путем внесения соответствующих модификаций в нуклеотидную последовательность, кодирующую антитело, или путем пептидного синтеза. Такие модификации включают, например, делеции и/или вставки и/или замены остатков в аминокислотных последовательностях антитела. Любая комбинация делеции, вставки и замены может быть использована для получения конечной конструкции при условии, что конечная конструкция обладает желаемыми характеристиками, например связыванием антигена.
а) Варианты замен, инсерций и делеции
В некоторых вариантах осуществления настоящего изобретения предусматривают варианты антител, содержащие одну или несколько аминокислотных замен. Сайты, представляющие интерес для замещающего мутагенеза, включают области HVR и FR. Консервативные замены показаны в табл. 1 под заголовком «предпочтительные замены». Более существенные изменения представлены в табл. 1 под заголовком «примеры замен» и дополнительно описаны ниже со ссылкой на классы боковых цепей аминокислот. Аминокислотные замены могут быть введены в представляющее интерес антитело, и продукты могут быть проверены на желаемую активность, например, сохранение / улучшение связывания антигена, снижение иммуногенности или улучшение ADCC или CDC.
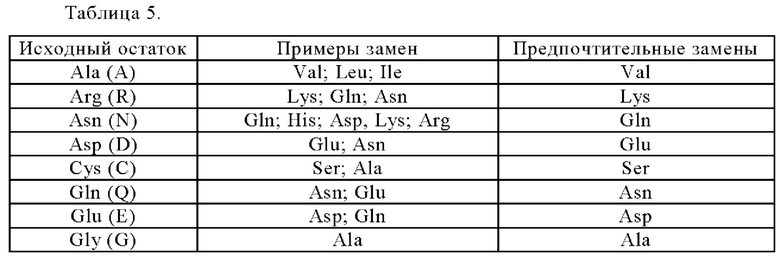
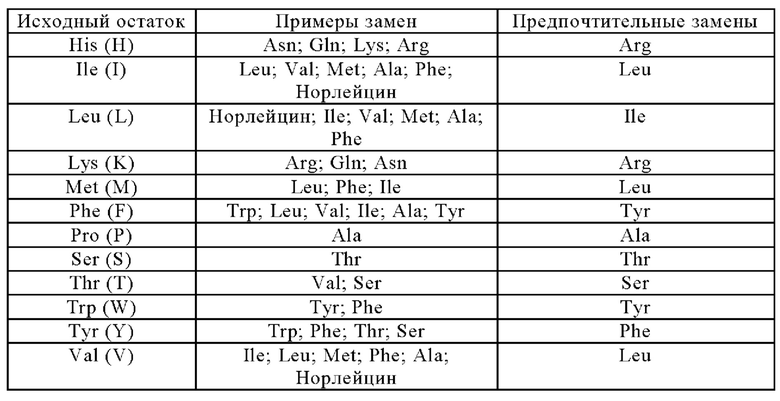
Аминокислоты могут быть сгруппированы в соответствии со свойствами боковой цепи:
(1) гидрофобные: Norleucine, Met, Ala, Val, Leu, Ile;
(2) нейтральные гидрофильные: Cys, Ser, Thr, Asn, Gln;
(3) кислые: Asp, Glu;
(4) основные: His, Lys, Arg;
(5) остатки, влияющие на ориентацию боковой цепи: Gly, Pro;
(6) ароматические: Trp, Tyr, Phe.
Неконсервативные замены приводят к замене представителя одного из этих классов на другой класс.
Один тип варианта замены включает замену одного или более остатков гипервариабельных областей родительского антитела (например, гуманизированного антитела или антитела человека). Как правило, полученный вариант (варианты), выбранные для дальнейшего исследования, могут иметь модификации (например, улучшения) определенных биологических свойств (например, повышенную аффинность, пониженную иммуногенность) по сравнению с исходным антителом и/или в значительной степени сохранят определенные биологические свойства родительского антитела. Типичный вариант замены представляет собой антитело с созревшей аффинностью, которое может быть легко получено, например, с использованием методов формирования аффинности на основе фагового дисплея, таких как описанные в настоящем изобретении. Вкратце, один или несколько остатков HVR мутируют, и варианты антитела отображаются на фаге и подвергаются скринингу на определенную биологическую активность (например, аффинность).
Изменение (например, замены) могут быть сделаны в HVR, например, для улучшения аффинности антитела. Такие изменения могут быть сделаны в «горячих точках» HVR, то есть в остатках, кодируемых кодонами, которые подвергаются мутациям с высокой частотой в процессе соматического созревания (см., например, Chowdhury, Methods Mol. Biol. 207, 2008, 179-196), и/или в остатках, которые контактируют с антигеном, при этом полученный вариант VH или VL тестируют на аффинность. Описано созревание аффинности путем конструирования и повторного отбора из вторичных библиотек, например, в публикации Hoogenboom с соавт. в кн.: Methods in Molecular Biology 178, 2001, 1-37, под ред. O'Brien с соавт., изд-во Human Press, Тотова, Нью-Джерси. В некоторых вариантах осуществления настоящего изобретения, связанных с созреванием аффинности, вводят разнообразие в вариабельные гены, выбранные для созревания, любым из множества методов (например, подверженной ошибкам ПЦР, перетасовкой цепей или олигонуклеотид-направленным мутагенезом). Затем создают вторичную библиотеку. Эту библиотеку подвергают скринингу для выявления любых вариантов антител с желаемой аффинностью. Другой метод введения разнообразия включает подходы, направленные на HVR, в которых несколько остатков HVR (например, 4-6 остатков за раз) рандомизируют. Остатки HVR, участвующие в связывании антигена, могут быть специфически идентифицированы, например, с использованием мутагенеза или моделирования аланинового сканирования. CDR-H3 и CDR-L3, в частности, часто становятся мишенью.
В некоторых вариантах осуществления настоящего изобретения замены, вставки или делеции могут происходить в одной или нескольких HVR, если такие изменения существенно не снижают способность антитела связывать антиген. Например, консервативные изменения (например, консервативные замены, как предусмотрено в данном документе), которые существенно не снижают аффинности, могут быть внесены в HVR. Такие изменения могут, например, происходить за пределами контактирующих с антигеном остатков в HVR. В некоторых вариантах осуществления настоящего изобретения из вариантов последовательностей VH и VL, представленных выше, каждая HVR либо не изменена, либо содержит не более одной, двух или трех аминокислотных замен.
Полезный метод идентификации остатков или областей антитела, которые могут быть мишенью для мутагенеза, называется «аланиновый сканирующий мутагенез», который описан Cunningham и Wells, Science, 244, 1989, 1081-1085. По этому методу остаток или группу целевых остатков (например, заряженных остатков, таких как arg, asp, his, lys и glu) идентифицируют и заменяют нейтральной или отрицательно заряженной аминокислотой (например, аланином или полиаланином) чтобы определить, есть ли воздействие на взаимодействие антитела с антигеном. Дальнейшие замены могут быть введены в аминокислотные положения, демонстрирующие функциональную чувствительность к начальным заменам. В другом варианте или дополнительно может быть проанализирована кристаллическая структура комплекса антиген-антитело для определения точек контакта между антителом и антигеном. Такие контактные остатки и соседние остатки могут быть нацелены или исключены как кандидаты на замену. Варианты могут быть проверены, чтобы определить, содержат ли они требуемые свойства.
Инсерции аминокислотной последовательности включают амино- и/или карбокси-концевые слияния длиной от одного остатка до полипептидов, содержащих сто или более остатков, а также вставки внутри последовательности одного или нескольких аминокислотных остатков. Примеры концевых вставок включают антитело с N-концевым метионильным остатком. Другие инсерционные варианты молекулы антитела включают слияние фермента (например, для ADEPT) или полипептида, который увеличивает период полужизни антитела в плазме, по N- или С-концу антитела.
б) Варианты гликозилирования
В некоторых вариантах осуществления настоящего изобретения предусмотренное антитело изменено для увеличения или уменьшения степени гликозилирования антитела. Добавление или удаление сайтов гликозилирования к антителу может быть выполнено апробированным методом изменения аминокислотной последовательности таким образом, что создают или удаляют один или несколько сайтов гликозилирования.
Если антитело включает область Fc, присоединенный к ней углевод может быть изменен. Нативные антитела, вырабатываемые клетками млекопитающих, обычно содержат разветвленный двухантенарный олигосахарид, который обычно присоединен N-связью к Asn297 домена СН2 области Fc. См., например, Wright с соавт., TIBTECH 15, 1997, 26-32. В составе могут быть разные олигосахариды, например, манноза, N-ацетилглюкозамин (GlcNAc), галактоза и сиаловая кислота, а также фукоза, присоединенная к GlcNAc в «стволе» двухантенарной структуры олигосахарида. В некоторых вариантах осуществления настоящего изобретения модификации олигосахарида в антителе по настоящему изобретению могут быть сделаны для создания вариантов антител с определенными улучшенными свойствами.
В одном из вариантов осуществления настоящего изобретения предусматривают варианты антител, имеющие углеводную структуру, в которой отсутствует фукоза, присоединенная (прямо или опосредованно) к области Fc. Например, количество фукозы в таком антителе может составлять от 1% до 80%, от 1% до 65%, от 5% до 65% или от 20% до 40%. Количество фукозы определяют путем расчета среднего количества фукозы в цепи сахара при Asn297 относительно суммы всех гликоструктур, присоединенных к Asn 297 (например, сложных, гибридных и высокоманнозных структур) по данным измерения методом масс-спектрометрии MALDI-TOF по описанию в WO 2008/077546, например при Asn297, относится к остатку аспарагина, расположенному около положения 297 в области Fc (нумерация остатков области Fc по EU); однако Asn297 может также располагаться примерно на +/- 3 аминокислоты выше или ниже по цепи от положения 297, то есть между положениями 294 и 300, из-за незначительных вариаций последовательности в антителах. Такие фукозилированные варианты могут обладать улучшенной функцией ADCC. См., например, US 2003/0157108 (Presta, L.); US 2004/0093621 (Kyowa Hakko Kogyo Co., Ltd). Примеры публикаций, относящихся к «дефукозилированным» или «дефицитным по фукозе» вариантам антител, включают: US 2003/0157108; WO 2000/61739; WO 2001/29246; US 2003/0115614; US 2002/0164328; US 2004/0093621; US 2004/0132140; US 2004/0110704; US 2004/0110282; US 2004/0109865; WO 2003/085119; WO 2003/084570; WO 2005/035586; WO 2005/035778; WO2005/053742; WO 2002/031140; Okazaki с соавт. J. Mol. Biol. 336, 2004, 1239-1249; Yamane-Ohnuki с соавт., Biotech. Bioeng. 87, 2004, 614. Примеры клеточных линий, способных вырабатывать дефукозилированные антитела, включают клетки Lec13 СНО, дефицитные по фукозилированию белка (Ripka с соавт. Arch. Biochem. Biophys. 249, 1986, 533-545; US 2003/0157108 A1; WO 2004/056312 A1, Adams с соавт., особенно в примере 11), и линии клеток с нокаутом, такие как ген альфа-1,6-фукозилтрансферазы, FUT8, клетки с нокаутом СНО (см., например, Yamane-Ohnuki с соавт. Biotech. Bioeng. 87, 2004, 614; Kanda Y. с соавт., Biotechnol. Bioeng., 94(4), 2006, 680-688; WO 2003/085107).
Варианты антител дополнительно содержат олигосахариды, разделенные пополам, например, в которых двухантенный олигосахарид, присоединенный к области Fc антитела, разделен пополам GlcNAc. Такие варианты антител могут иметь пониженное фукозилирование и/или улучшенную функцию ADCC. Описаны примеры таких вариантов антител, например, в WO 2003/011878 (Jean-Mairet с соавт.); US 6602684 (Umana с соавт.); US 2005/0123546 (Umana с соавт.). Также предусмотрены варианты антител по крайней мере с одним остатком галактозы в олигосахариде, присоединенном к области Fc. Такие варианты антител могут иметь улучшенную функцию CDC. Такие варианты антител описаны, например, в WO 1997/30087 (Patel с соавт.); WO 1998/58964 (Raju, S.); WO 1999/22764 (Raju, S.).
в) Варианты области Fc
В некоторых вариантах осуществления настоящего изобретения одна или более аминокислотных модификаций могут быть введены в область Fc антитела, предусмотренного в настоящем изобретении, тем самым создавая вариант области Fc (который также может называться «измененной областью Fc»). Вариант области Fc может содержать последовательность области Fc человека (например, области Fc IgG1, IgG2, IgG3 или IgG4 человека), включающей модификацию аминокислоты (например, замену) в одном или нескольких аминокислотных положениях.
В некоторых вариантах осуществления настоящего изобретения рассматривают вариант антитела, который обладает некоторыми, но не всеми эффекторными функциями, что делает его желательным кандидатом для применений, в которых важен период полужизни антитела in vivo, однако некоторые эффекторные функции (такие как комплемент и ADCC) являются ненужными или вредными. In vitro и/или in vivo могут быть проведены анализы цитотоксичности для подтверждения снижения/истощения активности CDC и/или ADCC. Например, анализы связывания рецептора Fc (FcR) могут быть проведены, чтобы гарантировать, что антитело утратило способность связывать Fc гамма R (следовательно, вероятно, не обладает активностью ADCC), но сохраняет связывающую активность с FcRn. Первичные клетки для опосредования ADCC, клетки NK, экспрессируют только Fc гамма RIII, тогда как моноциты экспрессируют Fc гамма RI, Fc гамма RII и Fc гамма RIII. Экспрессия FcR на гемопоэтических клетках суммирована в табл. 3 на странице 464 публикации Ravetch и Kinet, Аппи. Rev. Immunol. 9, 1991, 457-492. Примеры методов анализа активности ADCC in vitro, перечень которых не ограничивается данными примерами, приведены в US 5500362 (см., например, Hellstrom I. с соавт. Proc. Nat'l Acad. Sci. USA 83, 1986, 7059-7063; Hellstrom I с соавт., Proc. Nat'l Acad. Sci. USA 82, 1985, 1499-1502; US 5821337 (см. Bruggemann M. с соавт., J. Exp. Med. 166, 1987, 1351-1361). В другом варианте можно использовать методы нерадиоактивных анализов (см., например, анализ нерадиоактивнои цитотоксичности АСТ1™ методом жидкостной цитометрии (CellTechnology, Inc., Маунтин-Вью, Калифорния; и CytoTox 96 (зарегистрированная торговая марка) анализ нерадиоактивной цитотоксичности (фирма Promega, Мэдисон, Висконсин)). Эффекторные клетки, применимые для таких анализов, включают мононуклеарные клетки периферической крови (МКПК) и природные клетки-киллеры (NK). В другом варианте или дополнительно активность ADCC исследуемой молекулы можно оценить in vivo, например, в животной модели, такой как описанная в публикации Clynes с соавт. Proc. Nat'l Acad. Sci. USA 95, 1998, 652-656. Анализы связывания C1q также могут быть выполнены для подтверждения того, что антитело неспособно связывать C1q и, следовательно, не обладает активностью CDC. См., например, связывание C1q и С3с методом ELISA в WO 2006/029879 и WO 2005/100402. Для оценки активации комплемента может быть выполнен анализ CDC (см., например, Gazzano-Santoro с соавт., J. Immunol. Methods 202, 1996, 163; Cragg M.S. с соавт., Blood 101, 2003, 1045-1052; Cragg M.S., Glennie M.J., Blood 103, 2004, 2738-2743). Связывание FcRn и определение клиренса/периода полужизни in vivo также можно проводить с использованием методов, известных в данной области (см., например, Petkova S.B. с соавт., Int'l. Immunol. 18(12), 2006, 1759-1769).
К антителам с пониженной эффекторной функцией относят антитела с заменой одного или более из остатков 238, 265, 269, 270, 297, 327 и 329 в области Fc (US 6737056). К таким мутантам Fc относятся мутанты Fc с заменами в двух или более положениях аминокислот 265, 269, 270, 297 и 327, включая так называемый мутант Fc «DANA» с заменой остатков 265 и 297 на аланин (US 7332581).
Описаны некоторые варианты антител с повышенным или пониженным связыванием с FcR. (См., например, US 6737056; WO 2004/056312; Shields с соавт., J. Biol. Chem. 9(2), 2001, 6591-6604).
В некоторых вариантах осуществления настоящего изобретения вариант антитела включает область Fc с одной или более аминокислотными заменами, которые улучшают ADCC, например, замены в положениях 298, 333 и/или 334 области Fc (нумерация остатков по EU).
В некоторых вариантах осуществления настоящего изобретения в область Fc вносят изменения, которые приводят к изменению (т.е. к увеличению или уменьшению) связывания C1q и/или комплемент-зависимой цитотоксичности (CDC), например, как описано в US 6194551, WO 99/51642 и Idusogie с соавт. J. Immunol. 164, 2000, 4178-4184.
Антитела с увеличенной полужизнью и повышенным связыванием с неонатальным рецептором Fc (FcRn), который ответственен за передача материнских IgG плоду (Guyer с соавт., J. Immunol. 117, 1976, 587; Kim с соавт., J. Immunol. 24, 1994, 249), описаны в US 2005/0014934 A1 (Hinton с соавт.). Эти антитела содержат область Fc с одной или несколькими заменами в ней, которые увеличивают связывание области Fc с FcRn. Такие варианты Fc включают варианты с заменами в одного или нескольких остатков в области Fc: 238, 256, 265, 272, 286, 303, 305, 307, 311, 312, 317, 340, 356, 360, 362, 376, 378, 380, 382, 413, 424 или 434, например, замена остатка 434 в области Fc (US 7371826).
См. также публикации Duncan, Winter, Nature 322, 1988, 738-740; US 5648260; US 5624821; WO 94/29351, касающиеся примеров других вариантов области Fc.
В одном из вариантов осуществления настоящего изобретения связывающая активность по отношению к каждому рецептору Fc-гамма человека (Fc-gamma R) области Fc антитела (включая вариант области Fc (то же самое применимо и далее)) может быть измерена методом захвата лиганда с использованием, например, BIACORE (зарегистрированная торговая марка) Т200, в основе которых лежат методы анализа поверхностного плазмонного резонанса в качестве принципа измерения.
Подробности примера метода измерения связывающей активности области Fc антитела в отношении различных рецепторов Fc-гамма человека (Fc-гамма Rs) описаны ниже. В одном из вариантов осуществления настоящего изобретения связывающей активности области Fc по отношению к Fc-гамма R оценивают с использованием BIACORE (зарегистрированная торговая марка) Т200. В предпочтительном варианте осуществления настоящего изобретения это измерение проводят при 25°С с использованием буфера для измерений (50 мМ фосфат, 150 мМ NaCl, Р20 0,05 мас./об. %, рН 7,4. В частности, около 1000 RU антитела, содержащего вариант области Fc, сначала захватывается сенсорным чипом с помощью CaptureSelect (товарный знак) конъюгата Fab-лямбда человека-биотин (фирма ThermoFisher Scientific), иммобилизованного в качестве молекулы, захватывающей лиганд. Fc-гамма R человека разбавляют буфером для измерения до 8 нМ для Fc-гамма RIa и до 1000 нМ для других Fc-гамма R, и дают возможность связываться с захваченным антителом. Связывающую активность каждого антитела по отношению к каждому Fc-гамма R оценивают путем расчета количества связанного Fc-гамма R на единицу количества антитела (RU) с использованием программного обеспечения Biacore Т200 Evaluation Software 2.0. В одном из вариантов осуществления настоящего изобретения связывающая активность области Fc антитела в отношении различных рецепторов Fc-гамма человека (Fc-гамма R) может быть измерена методом, описанным в примере 7-4.
В одном предпочтительном варианте осуществления настоящего изобретения Fc-гамма R, используемые для описанного выше способа измерения, могут быть внеклеточным доменом Fc-гамма R, полученным методом, описанным ниже. Во-первых, синтез гена внеклеточного домена Fc-гамма R проводят методом, известным специалистам в данной области. Для этого синтеза последовательности каждого Fc-гамма R готовят на основе информации, зарегистрированной в NCBI. Более конкретно, последовательность Fc-gamma RI получают на основе последовательности NCBI (номер в базе данных NM_000566.3), последовательность Fc-gamma RIIa получают на основе последовательности NCBI (номер в базе данных NM_001136219.1), последовательности для Fc- гамма RIIb получают на основе последовательности NCBI (номер в базе данных NM_004001.3), последовательность для Fc-гамма RHIa получают на основе последовательности NCBI (номер в базе данных NM_001127593.1); к С-концу добавляют His-метку. Полиморфные сайты для Fc-гамма RIIa получают со ссылкой на J. Exp. Med., 172, 1990, 19-25, и полиморфные сайты для Fc-гамма RIIIa получают со ссылкой на J. Clin. Invest., 100, 1997, 1059-1070. Полученные фрагменты генов инсертируют в вектор экспрессии для клеток животных для получения векторов экспрессии. Такие векторы экспрессии временно вводят в клетки FreeStyle293 (фирма Invitrogen), полученные из клеток почки эмбрионального рака человека, и дают возможность экспрессироваться белку интереса. Супернатант культуры собирают и фильтруют через фильтр с диаметром пор 0,22 мкм, затем очищают в основном с помощью описанных ниже четырех этапов. На первом этапе проводят катионообменную колоночную хроматографию (SP Sepharose FF); на втором этапе колоночная аффинная хроматография к His-меткам (HisTrap HP); на третьем этапе - гель-фильтрационная колоночная хроматография (Superdex200); и, четвертый этап - асептическая фильтрация. Следует отметить, что для Fc-гамма RI в качестве первой стадии проводят анионообменную колоночную хроматографию с использованием Q-сефарозы FF. Концентрацию очищенного белка рассчитывают на основе коэффициента поглощения, рассчитанного путем измерения поглощения при 280 нм с использованием спектрофотометра и с использованием РАСЕ или подобного метода для получения значений (Protein Science, 4, 1995, 2411-2423).
В одном из вариантов осуществления настоящего изобретения связывающая активность области Fc антитела по отношению к FcRn человека может быть измерена методом захвата лиганда с использованием, например, BIACORE (зарегистрированная торговая марка) Т200, принцип измерения которого основан на методах анализа поверхностного плазмонного резонанса.
Детали примера метода измерения связывающей активности области Fc антитела с FcRn человека описаны ниже. В одном из вариантов осуществления настоящего изобретения связывающую активность области Fc антитела в отношении FcRn оценивают, используя BIACORE (зарегистрированная торговая марка) Т200. В предпочтительном варианте осуществления настоящего изобретения это измерение проводят при 25°С с использованием буфера для измерений (50 мМ фосфат, 150 мМ NaCl, Р20 - 0,05 мас./об.%; рН 6,0). В частности, около 400 RU антитела, содержащего область Fc, сначала захватывается на сенсорном чипе, на котором CaptureSelect (торговая марка) Human Fab-lambda Kinetics Biotin Conjugate (фирма ThermoFisher Scientific) иммобилизован как молекула, захватывающая лиганд, и затем FcRn человека, разбавленный буфером для измерений, позволяется связываться с ним. Связывающую активность каждого антитела по отношению к FcRn оценивают путем расчета KD (М) с использованием модели устойчивого состояния Biacore Т200 Evaluation Software 2.0. В одном предпочтительном варианте осуществления настоящего изобретения белок FcRn человека, используемый в этом измерении, получают в соответствии со способом, описанным в примере 2 WO 2010107110, используемым в качестве образца. В одном из вариантов осуществления настоящего изобретения связывающая активность области Fc антитела в отношении различных FcRn человека может быть измерена способом, описанным в примере 7-5.
г) Варианты антител, разработанных на основе цистеина
В некоторых вариантах осуществления настоящего изобретения может потребоваться разработать сконструированные на основе цистеина антитела, например, «тиоМАb», в которых один или несколько остатков антитела заменены остатками цистеина. В одном из вариантов осуществления настоящего изобретения замещенные остатки находятся в доступных сайтах антитела. За счет замены этих остатков цистеином реактивные тиоловые группы тем самым размещают в доступных сайтах антитела и их можно использоваться для конъюгирования антитела с другими фрагментами, такими как фрагменты лекарственного средства или фрагменты линкер-лекарственное средство, для создания иммуноконъюгата, как описано ниже в настоящем изобретении. В некоторых вариантах осуществления настоящего изобретения любой один или несколько из следующих остатков могут быть заменены цистеином: V205 (нумерация Kabat) легкой цепи; А118 (нумерация EU) тяжелой цепи; и S400 (EU нумерация) области Fc тяжелой цепи. Сконструированные антитела с применением цистеина могут быть получены как описано, например, в US 7521541.
д) Производные антител
В некоторых вариантах осуществления настоящего изобретения предусматривают антитело, которое может быть дополнительно модифицировано за счет включения дополнительных небелковых фрагментов, которые известны в данной области и легко доступны. Фрагменты, подходящие для дериватизации антитела, включают, но ими не ограничиваются, водорастворимые полимеры. Примеры водорастворимых полимеров включают, но ими перечень не ограничивается, полиэтиленгликоль (ПЭГ), сополимеры этиленгликоля/пропиленгликоля, карбоксиметилцеллюлозу, декстран, поливиниловый спирт, поливинилпирролидон, поли-1,3-диоксолан, поли-1,3,6-триоксан, сополимер этилена и малеинового ангидрида, полиаминокислоты (гомополимеры или статистические сополимеры) и декстран или поли (н-винилпирролидон) полиэтиленгликоль, гомополимеры полипропиленгликоля, сополимеры полипропиленоксида и этиленоксида, полиоксиэтилированные полиолы (например, глицерин), поливиниловый спирт и их смеси. Пропиональдегид полиэтиленгликоля может иметь преимущества при получении из-за его стабильности в воде. Полимер может иметь любую молекулярную массу и может быть разветвленным или неразветвленным. Количество полимеров, присоединенных к антителу, может варьировать, и если присоединено более одного полимера, они могут быть одинаковыми или разными молекулами. В общем, количество и/или тип полимеров, используемых для дериватизации, может быть определен с учетом ряда обстоятельств, например, конкретных свойств или функций подлежащего улучшению антитела, планируется ли использование производного антитела в терапии при определенные условия и т.д.
В другом варианте осуществления настоящего изобретения предусматривают конъюгаты антитела и небелкового фрагмента, которые можно избирательно нагревать под воздействием излучения. В одном из вариантов осуществления настоящего изобретения небелковый фрагмент представляет собой углеродную нанотрубку (Kam с соавт., Proc. Natl. Acad. Sci. USA 102, 2005, 11600-11605). Излучение может иметь любую длину волны и включает, помимо прочего, длины волн, которые не повреждают обычные клетки, но нагревают небелковый фрагмент до температуры, при которой гибнут клетки, проксимальные к небелковому фрагменту, не содержащему антитела.
Б. Методы рекомбинации и композиции
Антитела могут быть получены с использованием методов рекомбинации и композиции, например, описанные в US 4816567. В одном из вариантов осуществления настоящего изобретения предусматривают выделенную нуклеиновую кислоту, кодирующую анти-CD137 антигенсвязывающую молекулу или антитело, описанные в настоящем изобретении. Такая нуклеиновая кислота может кодировать аминокислотную последовательность, содержащую VL и/или аминокислотную последовательность, содержащую VH антитела (например, легкую и/или тяжелую цепи антитела). В другом варианте осуществления настоящего изобретения предусматривают один или более векторов (например, векторов экспрессии), включающих такую нуклеиновую кислоту. В другом варианте осуществления настоящего изобретения предусматривают клетку-хозяина, включающую такую нуклеиновую кислоту. В одном из подобных вариантов осуществления настоящего изобретения клетка-хозяин содержит (например, была трансформирована): (1) вектор, содержащий нуклеиновую кислоту, кодирующую аминокислотную последовательность, содержащую VL антитела, и аминокислотную последовательность, содержащую VH антитела, или (2) первый вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VL антитела, и второй вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VH антитела. В одном из вариантов осуществления настоящего изобретения клетка-хозяин является эукариотической, например клеткой яичника китайского хомячка (СНО) или лимфоидной клеткой (например, клетками Y0, NS0, Sp2/0). В одном из вариантов осуществления настоящего изобретения предусматривают способ получения анти-CD137 антигенсвязывающей молекулы или антитела, который включает культивирование клетки-хозяина, содержащей нуклеиновую кислоту, кодирующую антитело, как указано выше, в условиях, подходящих для экспрессии анти-CD137 антигенсвязывающей молекулы или антитела, и необязательно извлечение антитела из клетки-хозяина (или среды для культивирования клеток-хозяев).
Для рекомбинантного получения анти-CD137 антигенсвязывающей молекулы или антитела нуклеиновую кислоту, кодирующую антитело, например, как описано выше, выделяют и вставляют в один или несколько векторов для дальнейшего клонирования и/или экспрессии в клетке-хозяине. Такую нуклеиновую кислоту можно легко выделить и секвенировать с использованием обычных процедур (например, с использованием олигонуклеотидных зондов, которые способны специфически связываться с генами, кодирующими тяжелую и легкую цепи антитела).
Подходящие клетки-хозяева для клонирования или экспрессии векторов, кодирующих антитела, включают прокариотические или эукариотические клетки, описанные в настоящем изобретении. Например, антитела могут вырабатываться в бактериях, в частности, когда гликозилирование и эффекторная функция Fc не нужны. Для экспрессии фрагментов антител и полипептидов в бактериях см., например, US 5648237, US 5789199, US 5840523 (см. также Charlton, Methods in Molecular Biology, 2003, том 248, 245-254 (под ред. В.K.С. Lo, изд-во Humana Press, Тотова, Нью-Джерси), в которых описывают экспрессию фрагментов антител в Е. coli). После экспрессии антитело может быть выделено из биомассы бактериальных клеток в растворимую фракции и может быть дополнительно очищено.
Помимо прокариот, подходящими хозяевами для клонирования или экспрессии векторов, кодирующих антитела, являются эукариотические микроорганизмы, такие как мицелиальные грибы или дрожжи, включая штаммы грибов и дрожжей, чьи пути гликозилирования были «гуманизированы», что приводит к выработке антитела с частично или полностью гликозилированным образцом человека. См. публикации Nat. Biotech., 22, 2004, 1409-1414; Li с соавт., Nat. Biotech. 24, 2006, 210-215.
Клетки-хозяева, применимые для экспрессии гликозилированного антитела, также происходят из многоклеточных организмов (беспозвоночных и позвоночных). Примеры клеток беспозвоночных включают клетки насекомых, также применимы клетки растений. Идентифицированы многочисленные штаммы бакуловирусов, которые можно использовать в сочетании с клетками насекомых, в частности, для трансфекции клеток Spodoptera frugiperda.
В качестве клеток-хозяев также можно использовать клетки растений. См., например, US 5959177, US 6040498, US 6420548, US 7125978 и US 6417429 (описание технологии PLANTIBODIES™ по получению антител в клетках трансгенных растений).
В качестве клеток-хозяев также можно использовать клетки животных. Например, можно использовать линии клеток млекопитающих, которые адаптированы к росту в виде суспензии. К другим примерам относятся линии клеток-хозяев млекопитающих, а именно линия клеток почки обезьяны CV1, трансформированная вирусом SV40 (COS-7); линия почки эмбриона человека (293 или 293 по описанию, например, в публикации Graham с соавт., J. Gen Virol. 36, 1977, 59); клетки почки детеныша хомяка (BHK, baby hamster kidney); клетки Сертоли мыши (клетки ТМ4 по описанию, например, в публикации Mather, Biol. Reprod. 23, 1980, 243-251); клетки почки обезьяны (CV1); клетки почки африканской зеленой мартышки (VERO-76); клетки карциномы шейки матки человека (HELA); клетки почки собаки (MDCK); клетки печени буйволовой крысы (BRL 3А); клетки легкого человека (W138); клетки печени человека (Hep G2); клетки опухоли молочной железы мыши (ММТ 060562); клетки TRI согласно описанию, например, в публикации Mather с соавт., Annals N.Y. Acad. Sci. 383, 1982, 44-68; клетки MRC 5; клетки FS4. К другим применимым линиям клеток-хозяев млекопитающих относятся клетки яичника китайского хомячка (СНО, Chinese hamster ovary), включая клетки DHFR-CHO (Urlaub с соавт., Proc. Natl. Acad. Sci. USA 77, 1980, 4216); клеточные линии миеломы, такие как Y0, NS0 и Sp2/0. Для обзора некоторых линий клеток-хозяев млекопитающих, применимых для выработки антител, см., например, Yazaki и Wu, Methods in Molecular Biology, том 248, 2003, 255-268, под ред. В.K.С. Lo, изд-во Humana Press, Тотова, Нью-Джерси.
В. Методы исследования
Анти-CD137 антигенсвязывающие молекулы или антитела, предусмотренные в настоящем изобретении, могут быть идентифицированы, подвергнуты скринингу или охарактеризованы по их физическим/химическим свойствам и/или биологической активности с помощью различных методов исследования, известных в данной области.
1. Анализ связывания и другие методы
В одном из вариантов осуществления настоящего изобретения антигенсвязывающая молекула или антитело по настоящему описанию тестируют на антигенсвязывающую активность, например, известными методами, такими как ELISA, вестерн-блоттинг и т.д.
В другом варианте осуществления настоящего изобретения конкурентный анализ в присутствии низкомолекулярного соединения можно использовать для идентификации антигенсвязывающей молекулы или антитела, которые конкурируют за связывание с CD137 с анти-CD137 антигенсвязывающими молекулами или антителами, включая антитела А375/В167, А372/В040, А356/В040, А486/В167, А487/В167, А488/В226, А489/В223, А548/В376, А551/В256, А551/В379, А555/В379, А548/В256 и/или А549/В167, описанные в табл. 17, в качестве комбинации вариабельной области тяжелой цепи/вариабельной области легкой цепи в присутствии низкомолекулярного соединения (например, в присутствии 10 мкМ или более, 50 мкМ или более, 100 мкМ или более, 150 мкМ или более, 200 мкМ или более или 250 мкМ или более низкомолекулярного соединения). В некоторых вариантах осуществления настоящего изобретения такие конкурирующие антигенсвязывающие молекулы или антитела связываются с одним и тем же эпитопом (например, линейным эпитопом или конформационным эпитопом), который связывается анти-CD137 антигенсвязывающими молекулами, включая А375/В167, А372/В040, А356/В040, А486/В167, А487/В167, А488/В226, А489/В223, А548/В376, А551/В256, А551/В379, А555/В379, А548/В256 и/или А549/В167, описанные в табл. 17 в качестве комбинации вариабельной области тяжелой цепи/вариабельной области легкой цепи. Подробное описание примеров методов картирование эпитопа, с которым связывается антигенсвязывающая молекула или антитело, представлены в работе Morris "Epitope Mapping Protocols," in Methods in Molecular Biology том 66, 1966, изд-во Humana Press, Тотова, Нью-Джерси). В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающие молекулы или антитела в настоящем описании, обладающие СВ137-связывающей активностью, которая зависит от антигенсвязывающей активности, которая в свою очередь зависящей от низкомолекулярного соединения, распознает эпитоп, образованный комплексом, сформированным из антигена (например, CD137) и низкомолекулярного соединения (например, АТФ).
В типичном конкурентном анализе с использованием антитела иммобилизованный CD137 инкубируют в присутствии низкомолекулярного соединения (например, в присутствии 10 мкмоль или более, 50 мкмоль или более, 100 мкмоль или более, 150 мкМ или более, 200 мкМ или более, или 250 мкМ или большего количества низкомолекулярного соединения) в растворе, содержащем первое меченое антитело, связывающееся с CD137 (например, анти-CD137 антитела, включающие А372/В040, А356/В040, А486/В167, А487/В167, А488/В226, А489/В223, А548/В376, А551/В256, А551/В379, А555/В379, А548/В256 и/или А549/В167, описанные в табл.17, в качестве комбинации вариабельной области тяжелой цепи/вариабельной области легкой цепи), и второе немеченое антитело, которое тестируют на способность конкурировать за связывание с CD137 с первым антителом. Второе антитело может присутствовать в супернатанте гибридомы. В качестве контроля иммобилизованный CD137 инкубируют в растворе, содержащем первое меченое антитело, но не второе немеченое антитело. После инкубации в условиях, допускающих связывание первого антитела с CD137, избыток несвязавшегося антитела удаляют и измеряют количество метки, связанной с иммобилизованным CD137. Если количество метки, связанной с иммобилизованным CD137, существенно снижено в тестируемом образце по сравнению с контрольным образцом, это указывает на то, что второе антитело конкурирует с первым антителом за связывание с CD137. См. публикацию Harlow и Lane, Antibodies: A Laboratory Manual, глава 14, 1988, изд-во Cold Spring Harbor Laboratory, Нью-Йорк). Специалистам в данной области очевидно, что анализ можно проводить аналогично с исследованием антигенсвязывающих молекул, отличных от антител.
2. Анализы активности
В одном из вариантов осуществления настоящего изобретения предусматривают анализы для идентификации биологической активности анти-CD137 антигенсвязывающих молекул или антител, обладающих биологической активностью. Биологическая активность может включать, например, агонистическую активность CD137; период полужизни в плазме; противоопухолевую активность; и слабая или подавленная системная реакция в тканях, отличных от опухолей. Также предусматривают антигенсвязывающие молекулы или антитела, обладающие такой биологической активностью in vivo и/или in vitro. В некоторых вариантах осуществления настоящего изобретения антигенсвязывающая молекула (например, анти-CD137 антигенсвязывающая молекула) или антитело по настоящему изобретению тестируют на такую биологическую активность.
а) Анализ агонистической активности (МКПК)
В одном из вариантов осуществления настоящего изобретения агонистическую активность по отношению к CD137 измеряют путем контакта CD137-экспрессирующих клеток с анти-CD137 антигенсвязывающей молекулой или антителом в растворе, в который добавляют или не добавляют низкомолекулярное соединение. В одном из вариантов осуществления настоящего изобретения агонистическую активность по отношению к CD137 в растворе, в который добавлено низкомолекулярное соединение, и агонистическую активность по отношению к CD137 в растворе, в который не добавлено низкомолекулярное соединение, оценивают по количеству цитокина, вырабатываемого (например, количества вырабатываемого IL-2, IFN-γ и/или IL-6), которое измеряют в течение 18 ч, 24 ч, 36 ч, 48 ч или 72 ч после контакта в растворе CD137-экспрессирующих клеток и анти-CD137 антигенсвязывающей молекулы или антитела. В одном из вариантов осуществления настоящего изобретения раствор, в который добавляют низкомолекулярное соединение, регулируют таким образом, чтобы концентрация низкомолекулярного соединения после корректировки составляет 10 мкМ, 50 мкМ, 100 мкМ, 150 мкМ, 200 мкМ или 250 мкМ. В другом варианте осуществления настоящего изобретения используемые клетки, экспрессирующие CD 137, представляют собой выделенные мононуклеарные клетки периферической крови (МКПК) человека или Т-клетки, размноженные из выделенных МКПК человека.
В одном из вариантов осуществления настоящего изобретения используют МКПК человека, выделенные из крови здорового донора путем центрифугирования в режиме 400×g в течение 30 мин при комнатной температуре. Предпочтительно используют МКПК человека, выделенные на последующих двух этапах. На первом этапе Leucosep (Greiner Bio-One), обогащенный Ficoll-Paque PLUS (фирма GE Healthcare), центрифугируют в режиме 1000×g в течение 1 мин при комнатной температуре, затем кровь разбавляют ФСБ и центрифугируют в режиме 400×g в течение 30 мин при комнатной температуре. На втором этапе лейкоцитарную пленку из пробирки после центрифугирования собирают и промывают 60 мл ФСБ (фирма Wako).
Подробное описание примера метода измерения CD137 агонистической активности с применением МКПК человека описано ниже. Следует отметить, что даже несмотря на то, что в следующем примере для иллюстрации АТФ используют в качестве низкомолекулярного соединения, это не исключает других низкомолекулярных соединений. В одном из вариантов осуществления настоящего изобретения выделенные МКПК человека разводят до плотности 5×106/мл культуральной средой (5% сыворотки человека (фирма SIGMA), 95% AIM-V (фирма Thermo Fischer Scientific)). Затем выделенные МКПК человека контактируют с антителом против CD3ε человека и/или с антителом против CD28 человека, посредством чего экспрессия CD137 индуцируется в МКПК человека. Предпочтительно 50 мкл 0,04 мкг/мл антитела против CD3ε человека (фирма BD Co., клон SP34) и 20 мкг/мл антитела против CD28 человека (фирма BD, клон: CD28.2), разведенные культуральной средой, добавляют к выделенным МКПК человека (100 мкл с плотностью клеток 5×106/мл).
К МКПК человека, к которым добавлены антитела против CD3ε человека и/или антитела против CD28 человека, затем дополнительно добавляют (i) культуральную среду с или без АТФ; и (ii) анти-CD137 антигенсвязывающую молекулу или антитело. Предпочтительно добавляют 25 мкл питательной среды с АТФ или без него. Предпочтительно добавляют 25 мкл анти-CD137 антигенсвязывающей молекулы или антитела в концентрации 40 мкг / мл. Более предпочтительно, указанные выше (i) и (ii) добавляют примерно через 6 ч после контакта МКПК человека с антителом против CD3ε человека и/или антителом против CD28 человека. В одном из вариантов осуществления настоящего изобретения, предпочтительно количество выработанного IL-2 измеряют до выработки количества IFN-γ. В одном из вариантов осуществления настоящего изобретения количество выработки IL-2 измеряют в течение примерно 24 ч после контакта МКПК человека с антителом против CD3ε человека и/или антителом против CD28 человека. Предпочтительно количество выработки IL-2 измеряют примерно через 24 ч после контакта МКПК человека с антителом против CD3ε человека и/или антителом против CD28 человека и примерно через 18 ч после добавления анти-CD137 антигенсвязывающей молекулы или антитела.
В другом варианте осуществления настоящего изобретения количество выработанного IFN-γ измеряют примерно в течение 48 ч после контакта МКПК человека с антителом против CD3ε человека и/или с антителом против CD28 человека. Предпочтительно количество выработанного IFN-γ измеряют примерно в течение 48 ч после контакта МКПК человека с антителом против CD3ε человека и/или с антителом против CD28 человека и примерно 42 ч после добавления анти-CD137 антигенсвязывающей молекулы или антитела. В одном из вариантов осуществления настоящего изобретения количество выработанного IL-2 и/или количество выработанного IFN-γ определяют путем измерения количества выработанного IL-2 и/или количества выработанного IFN-γ в собранном культуральном супернатанте. В одном из вариантов осуществления настоящего изобретения МКПК человека, добавленные с антителом против CD3ε человека и/или антителом против CD28 человека, оставляют в инкубаторе в атмосфере 5% СО2 при 37°С пока все измерения не будут завершены.
Подробности еще одного примера метода измерения агонистической активности CD137 с использованием МКПК человека описаны ниже. Выделенный МКПК человека разводят до плотности клеток 5×106/мл культуральной средой (5% сыворотки человека (фирма SIGMA), 95% AIM-V (фирма Thermo Fischer Scientific)). Затем плотность МКПК человека доводят до 5×106/мл и по 100 мкл высевают в лунки 96-луночного планшета (плоскодонного с крышкой) (фирма Corning). Затем воздействуют на МКПК человека для индукции экспрессии CD137. Например, экспрессию CD137 в МКПК человека индуцируют добавлением 50 мкл антитела против CD3ε человека в концентрации 0,04 мкг/мл (фирма BD Co., клон SP34) и антитела против CD28 человека в концентрации 20 мкг/мл (фирма BD, клон CD28.2), разведенных культуральной средой.
Затем экспрессию CD137 индуцируют в МКПК человека, планшет встряхивают и оставляют на 6 ч при 37°С в инкубаторе в атмосфере 5% СО2. Затем по 25 мкл 2 мМ АТФ (фирма SIGMA), разведенной в среде, или только среду без АТФ, и 25 мкл каждого антитела в концентрации 40 мкг/мл вносят в каждую лунку, и планшет встряхивают и оставляют на 18 ч при 37°С в инкубаторе в атмосфере 5% CO2. Затем собирают часть культурального супернатанта, и с его помощью количество ИЛ-2, содержащегося в культуральном супернатанте, количественно определяют с помощью набора для иммуноферментного анализа Human IL-2 DuoSet ELISA (фирма R&D systems) или набора Human IL-2 ELISA Set (фирма BD Biosciences). После сбора культурального супернатанта планшет снова оставляют на 24 ч при 37°С в инкубаторе в атмосфере 5% СО2. Затем собирают часть культурального супернатанта и IFN-γ, содержащийся в культуральном супернатанте, количественно определяют с использованием набора для иммуноферментного анализа IFN-γ DuoSet ELISA (фирма R&D systems) или набора Human IFN-y ELISA Development (фирма PeproTech). Метод ELISA в основном применяют в соответствии с протоколом, прилагаемым к набору. При работе с набором Human IL-2 DuoSet ELISA (фирма R&D systems) и набором Human IFN-γ ELISA Development (фирма R&D systems) появление и прекращение окрашивания выполняют в соответствии с протоколом с использованием раствора субстрата (фирма R&D systems), содержащего Н2О2, тетраметилбензидин и 1н H2SO4 (фирма Wako). При работе с набором Human IL-2 ELISA Set (фирма BD Biosciences) прекращение окрашивания выполняют с применением 1н H2SO4 (фирма Wako).
При работе с набором IFN-γ ELISA Development (фирма PeproTech) появление окрашивания и прекращение окрашивания проводят с использованием раствора ТМВ Chromogen Solution (фирма Thermo Fischer Scientific) и 1н H2SO4 (фирма Wako). Затем измерение абсорбции выполняют с помощью En Vision (фирма PerkinElmer), и количества (пг/мл) IL-2 и IFN-γ в культуральном супернатанте соответственно рассчитывают с использованием калибровочной кривой, построенной в соответствии с протоколом. В этом анализе МКПК агонистическую активность CD137 может быть выражена как кратное изменение количества IL-2 и IFN-γ в супернатанте культуры по сравнению с отрицательным контрольным антителом (антителом, которое не связывается с CD137). В одном из вариантов осуществления настоящего изобретения агонистическую активность CD137 измеряют в соответствии с методами, описанными в примерах 5-5-1 и 5-5-2.
В одном из вариантов осуществления настоящего изобретения если агонистическую активность в отношении CD137 оценивают по количеству выработанного цитокина (например, по количеству выработанного IL-2, IFN-γ и/или IL-6) в МКПК человека, анти-CD137 антигенсвязывающую молекулу или антитело можно расценить как проявление агонистической активности в отношении CD137 в присутствии низкомолекулярного соединения, если количество выработанного цитокина в присутствии 10 мкМ, 50 мкМ, 100 мкМ, 150 мкМ, 200 мкМ или 250 мкМ низкомолекулярного соединения при добавлении анти-CD137 антигенсвязывающей молекулы или антитела больше в 1,01 раз или более, в 1,02 раз или более, в 1,03 раз или более, в 1,05 раз или более, в 1,06 раз или более, в 1,07 раз или более, в 1,08 раз или более, в 1,09 раз или более, в 1,1 раза или более, в 1,11 раз или более, в 1,12 раз или более, в 1,13 раз или более, в 1,14 раз или более или в 1,15 раз или более, в 1,5 раза или более, в 2 раза или более, или в 3 раза или более по сравнению с количеством выработанного цитокина при добавлении антитела отрицательного контроля.
В одном из вариантов осуществления настоящего изобретения агонистическую активность в отношении CD137 оценивают по количеству выработанного IL-2 в МКПК человека, когда анти-CD137 антигенсвязывающую молекулу или антитело можно оценить как проявляющую агонистическую активность в отношении CD137 в присутствии низкомолекулярного соединения, если количество вырабатываемого IL-2 в присутствии 10 мкМ, 50 мкМ, 100 мкМ, 150 мкМ, 200 мкМ или 250 мкМ низкомолекулярного соединения, если добавленные анти-CD137 антигенсвязывающая молекула или антитело больше в 1,01 раза или более, в 1,02 раза или более, в 1,03 раза или более, или в 1,05 раза или более, в 1,05 раза или более в предпочтительном варианте осуществления настоящего изобретения по сравнению с количеством выработанного IL-2 при добавлении антитела отрицательного контроля.
В одном из вариантов осуществления настоящего изобретения, когда агонистическую активность в отношении CD137 оценивают по количеству выработанного IFN-γ в анализе с МКПК человека, анти-CD137 антигенсвязывающую молекулу или антитело можно оценить как проявляющую агонистическую активность в отношении CD137 в присутствии низкомолекулярного соединения, когда количество вырабатываемого IFN-γ в присутствии 10 мкМ, 50 мкМ, 100 мкМ, 150 мкМ, 200 мкМ или 250 мкМ низкомолекулярного соединения при добавлении анти-CD137 антигенсвязывающей молекулы или антитела больше в 1,1 раза или более, в 1,11 раз или более, в 1,12 раз или более, в 1,13 раз или более, в 1,14 раз или более, в 1,15 раз или более, в 1,15 раз или более в предпочтительном варианте осуществления настоящего изобретения по сравнению с количеством выработанного IFN-γ при добавлении антитела - отрицательного контроля.
В другом варианте осуществления настоящего изобретения среди анти-CD137 антигенсвязывающих молекул или антител, для которых установлено, что они проявляют агонистическую активность в отношении CD137 в присутствии низкомолекулярного соединения в вышеупомянутом сравнении с отрицательным контролем, дополнительно определяют антитела, которые оценивают как не проявляющие агонистической активности в отношении CD137 или обладающие пониженной агонистической активностью CD137 в отсутствие низкомолекулярного соединения (по сравнению с таковой в присутствии низкомолекулярного соединения). В частности, когда указанные ниже (ii) больше, чем (i), анти-CD137 антигенсвязывающую молекулу или антитело оценивают как не проявляющую активности агониста CD137 или имеющую более пониженную активность агониста CD137 в отсутствие низкомолекулярного соединения:
(i) [выработанное количество цитокина в отсутствие низкомолекулярного соединения при добавлении анти-CD137 антигенсвязывающей молекулы или антитела]/ [выработанное количество цитокина в отсутствие низкомолекулярного соединения при добавлении антитела отрицательного контроля]
(ii) [выработанное количество цитокина в присутствии низкомолекулярного соединения при добавлении анти-CD137 антигенсвязывающей молекулы или антитела]/[выработанное количество цитокина в присутствии низкомолекулярного соединения при добавлении антитела отрицательного контроля].
В другом варианте осуществления настоящего изобретения когда агонистическая активность CD137 первой антигенсвязывающей молекулы или антитела против CD137 и второй антигенсвязывающей молекулы или антитела против CD137 сравнивают, используя количества выработанных цитокинов (например, количества IL-2, IFN-γ, и/или продуцирование IL-6) в анализе МКПК человека, первая антигенсвязывающая молекула или антитело против CD137 могут быть оценена как обладающие более высокой агонистической активностью, чем вторая антигенсвязывающая молекула или антитело против CD137, если (i) количество выработанных цитокинов при добавлении первой антигенсвязывающей молекулы или антитела против CD137 в присутствии 10 мкМ, 50 мкМ, 100 мкМ, 150 мкМ, 200 мкМ или 250 мкМ низкомолекулярного соединения больше в 1,01 раза или более, в 1,02 раза или более, в 1,03 раза или более, в 1,04 раз или более, в 1,05 раз или более, в 1,06 раз или более, в 1,07 раз или более, в 1,08 раза или более, в 1,09 раза или более, в 1,1 раз или более, в 1,11 раз или более, в 1,12 раз или более, в 1,13 раз или более, в 1,14 раз или более или в 1,15 раз или более, 1,5 раза или более, в 2 раза или более или в 3 раза или более по сравнению с (ii) количеством выработанных цитокинов при добавлении второй антигенсвязывающей молекулы или антитела против CD137 в тех же условиях.
В одном из вариантов осуществления настоящего изобретения если агонистическую активность в отношении CD137 оценивают по количеству выработанного IL-2 в анализе МКПК человека, первая антигенсвязывающая молекула или антитело против CD137 может быть оценена как имеющая более высокую агонистическую активность, чем вторая антигенсвязывающая анти-CD137 молекула или антитело, если (i) количество выработки цитокинов при добавлении первой антигенсвязывающей молекулы или антитела к CD137 в присутствии 10 мкМ, 50 мкМ, 100 мкМ, 150 мкМ, 200 мкМ или 250 мкМ низкомолекулярного соединения больше в 1, 01 раза или более, в 1,02 раза или более, в 1,03 раза или более или в 1,04 раза или более, или в 1,04 раза или более предпочтительном варианте осуществления настоящего изобретения по сравнению с (ii) количеством вырабатываемого IL-2 при добавлении второй антигенсвязывающей молекулы или антитела против CD137 в тех же условиях.
В одном из вариантов осуществления настоящего изобретения если агонистическую активность в отношении CD137 оценивают по количеству выработки IFN-γ в анализе МКПК человека, первая антигенсвязывающая молекула или антитело против CD137 может быть оценена как имеющая более высокую агонистическую активность, чем вторая антигенсвязывающая анти-CD137 молекула или антитело, если (i) количество выработанных цитокинов при добавлении первой антигенсвязывающей молекулы или антитела против CD137 в присутствии 10 мкМ, 50 мкМ, 100 мкМ, 150 мкМ, 200 мкМ или 250 мкМ низкомолекулярного соединения больше в 1,05 раза или более, в 1,06 раза или более, в 1,07 раза или более, в 1,08 раза или более, в 1,09 раза или более или в 1,1 раза или более или в 1,1 раза или более в предпочтительном варианте осуществления настоящего изобретения по сравнению с (ii) добавлением второй антигенсвязывающей молекулы или антитела против CD137 в тех же условиях. В одном из вариантов осуществления настоящего изобретения первая антигенсвязывающая молекула или антитело против CD137 представляет собой антигенсвязывающую молекулу или антитело, содержащее исходную область Fc, а вторая антигенсвязывающая молекула анти-CD137 представляет собой антигенсвязывающую молекулу или антитело, содержащее вариант области Fc.
В одном из вариантов осуществления настоящего изобретения метод, описанный в примерах 5-5-1 и 5-5-2, может использоваться для измерения агонистической активности CD137 с использованием МКПК человека, а метод, описанный в примере 2-6, может использоваться для измерения агонистической активности CD137 с использованием Т-клеток, размноженных из выделенных МКПК человека.
В одном из вариантов осуществления настоящего изобретения агонистическая активность в отношении CD137 может быть измерена путем контактирования CD8-положительных или CD4-положительных Т-клеток, выделенных из МКПК человека, с антигенсвязывающей молекулой или антителом против CD137 в растворе, в который добавлено или не добавлено низкомолекулярное соединение. В это время к раствору могут быть дополнительно добавлены клетки, экспрессирующие FcγRIIb. В другом варианте агонистическая активность в отношении CD137 может быть измерена путем контактирования В-клеток (которые могут быть выделены из МКПК человека или может использоваться известная линия В-клеток) с антигенсвязывающей молекулой или антителом против CD137. В одном из вариантов осуществления настоящего изобретения агонистическая активность в отношении CD137 может быть оценена по количеству выработки цитокинов (например, по количеству IL-2, IFN-γ и/или по количеству выработанного IL-6), которое измеряют после контакта в растворе CD8-положительных Т-клеток, CD8-положительных Т-клеток или В-клеток с антигенсвязывающей молекулой или антителом против CD137.
В одном из вариантов осуществления настоящего изобретения анализы агонистической активности с использованием мононуклеарных клеток периферической крови человека (МКПК) выделяют из крови, взятой у здоровых индивидов. Таким образом, в любом из указанных выше вариантов осуществления настоящего изобретения специалистам в данной области должно быть понятно, что результаты анализов с использованием анализов могут различаться для каждого донора образцов крови. Принимая во внимание это обстоятельство, антитела, агонистическая активность которых не проявляется в отношении части или большинства МКПК человека, выделенных от нескольких доноров, не могут считаться проявляющими агонистическую активность CD137, даже если критерии агонистической активности удовлетворяются у другой части МКПК человека. Кроме того, когда анализ в соответствии с описанным выше методом проводят на МКПК человека, выделенных от многих доноров, и критерии агонистической активности CD137 у большинства доноров соответствуют, можно сделать вывод, что проявляется агонистическая активность CD137. В другом варианте осуществления настоящего изобретения, когда анализ в соответствии с описанным выше методом проводят на МКПК человека, выделенных от многих доноров, среднее или медианное значение анализа (например, количества IL-2, IFN-γ, и/или IL-6) может использоваться для определения наличия или отсутствия агонистической активности CD137. В одном из вариантов осуществления настоящего изобретения когда анализ проводят согласно вышеуказанному методу на МКПК человека, выделенных от множества доноров, количество доноров составляет, например, 2 или более, 3 или более, 4 или более, 5 или более, 10 или более, 15 или более, или 20 или более.
б) Анализ агонистической активности (анализ репортерного гена) В одном из вариантов осуществления настоящего изобретения агонистическую активность по отношению к CD137 оценивают с помощью анализа репортерного гена в растворе, в который добавляют или не добавляют низкомолекулярное соединение. В одном из вариантов осуществления настоящего изобретения агонистическую активность по отношению к CD137 в растворе, в который добавлено низкомолекулярное соединение, и агонистическую активность по отношению к CD137 в растворе, в который не добавлено низкомолекулярное соединение, соответственно оценивают по сигналу люминесценции люциферазы, измеренному после контакта Т-клеток, экспрессирующих репортерную конструкцию NF-каппаВ-люциферазы и CD137 с антигенсвязывающей молекулой против CD137, и оставляют на определенное время в соответствующем растворе. В одном из вариантов осуществления настоящего изобретения раствор, в который добавляют низкомолекулярное соединение, регулируют таким образом, чтобы концентрация низкомолекулярного соединения после корректировки составляла 10 мкМ, 50 мкМ, 100 мкМ, 150 мкМ, 200 мкМ или 250 мкМ. Т-клетки, экспрессирующие репортерную конструкцию NF-kappaB-luciferase и CD137, предпочтительно являются клетками линии GloResponse™ NF-kappaB-Luc2/4-1BB Jurkat (фирма Promega, CS196004).
Подробности типичного метода измерения агонистической активности CD137 с использованием анализа репортерного гена описаны ниже. В приводимых ниже примерах АТФ используют в качестве примера низкомолекулярного соединения, но не исключаются и другие низкомолекулярные соединения. В одном из вариантов осуществления настоящего изобретения сначала концентрацию FcγRIIB-экспрессирующих клеток доводят до 5×104 мл средой (культуральная среда СНО (90% Ham's F12, 10% ФСБ)), и оставляют на ночь при 37°С в инкубаторе в атмосфере 5% СО2. В одном из вариантов осуществления настоящего изобретения в качестве клеток, экспрессирующих FcgRIIB, можно использовать не только клетки, усиленно экспрессирующие FcgRIIB, но также клеточные линии, которые эндогенно экспрессируют FcgRIIB, такие как линии В-клеток, В-клетки, выделенные из живого организма, и т.п. В предпочтительном варианте осуществления настоящего изобретения FcγRIIB-экспрессирующими клетками являются клетки FcγRIIB СНО-K1 (фирма Promega). Затем, после удаления среды аспирацией, концентрацию клеток линии GloResponseTM NF-kappaB-Luc2/4-1B Jurkat (далее «4-1BB Jurkat») доводят до 2×106/мл другой средой (99% RPMI, 1% ФСБ). В предпочтительном варианте осуществления настоящего изобретения на 200 мкл скорректированных средой FcγRIIB-экспрессирующих клеток добавляют 25 мкл GloResponse™ NF-kappaB-Luc2/4-1BB клеток линии Jurka, скорректированных средой. Затем по 25 мкл каждой антигенсвязывающей молекулы против CD137 разбавляют вышеупомянутой средой (99% RPMI, 1% ФСБ) до достижения желаемой концентрации (например, конечной концентрации 0, 0,001, 0,01, 0, 1, 1, 10 мкг/мл), а затем добавляют по 25 мкл раствора АТФ, разбавленного вышеупомянутой средой (99% RPMI, 1% ФСБ), чтобы получить желаемую концентрацию (например, конечная концентрация равна 0, 10, 50, 100, 150, 200, 250 мкМ). В одном из вариантов осуществления настоящего изобретения сигнал люминесценции люциферазы измеряют после пребывания в течение 2 ч или менее, 4 ч или менее, 6 ч или менее, 24 ч или менее после добавления анти-CD137 антигенсвязывающей молекулы к клеткам 4-1ВВ Jurkat. В предпочтительном варианте осуществления настоящего изобретения 4-1ВВ Jurkat выдерживают при 37°С в течение 6 ч в инкубаторе в атмосфере 5% CO2. После этого добавляют 75 мкл реагента Bio-Glo и измеряют люминесценцию с помощью планшетного ридера. В одном предпочтительном варианте осуществления настоящего изобретения реагентом Bio-Glo является Bio-Glo Luciferase Assay System (буфер и субстрат). В одном из вариантов осуществления настоящего изобретения чтобы поддерживать постоянную температуру во время реакции клетки 4-1ВВ Jurkat можно оставить при комнатной температуре в течение 5 мин, 10 мин, 15 мин или 20 мин после извлечения из инкубатора. В предпочтительном варианте осуществления настоящего изобретения 4-1ВВ Jurkat оставляют при комнатной температуре в течение 15 мин после извлечения из инкубатора. В одном из вариантов осуществления настоящего изобретения значение люминесценции клеток 4-1ВВ Jurkat, к которым добавлена антигенсвязывающая молекула против CD137, делят на значение люминесценции клеток 4-1ВВ Jurkat, к которым не была добавлена антигенсвязывающая молекула против CD137, и это значение принимают за показатель кратности индукции (относительная световая единица) и используют в качестве показателя для оценки агонистической активности CD137 каждой антигенсвязывающей молекулы.
Еще один пример способа измерения агонистической активности CD137 с использованием анализа репортерного гена описан ниже. В каждую лунку 96-луночного планшета добавляют 200 мкл клеток FcγRIIB СНО-K1 (фирма Promega), доведенных добавлением среды до концентрации 5×104/мл и оставляют на ночь при 37°С в инкубаторе в атмосфере 5% CO2. В качестве культуральной среды применяют среду СНО (90% Ham's F12, 10% ФСБ). Затем среду полностью удаляют отсасыванием, 25 мкл GloResponse™ NF-κB-Luc2/4-1ВВ клеток линии Jurkat с откорректированной концентрацией 2×106/мл в среде для анализа (99% RPMI, 1% ФСБ) добавляют в каждую лунку. Затем добавляют по 25 мкл каждого раствора антигена, разбавленного средой для анализа до конечной концентрации 0, 0,001, 0,01, 0,1, 1 или 10 мкг/мл, и, в итоге добавляют 25 мкл раствора АТФ, разбавленного средой для анализа до 0 или 250 мкМ. После того, как планшет оставляют на 6 ч при 37°С в инкубаторе с 5% СО2 в атмосфере, его оставляют при комнатной температуре на 15 мин и затем добавляют по 75 мкл реагента Bio-Glo в каждую лунку. В качестве реагента Bio-Glo, например, можно использовать систему анализа люциферазы Bio-Glo Luciferase Assay System (буфер и субстрат). После этого люминесценцию в каждой лунке измеряют с помощью планшет-ридера. Показатель, полученный делением значения люминесценции в каждой лунке на значение люминесценции в лунке, в которую не добавлено какое-либо антитело, определяют как показатель кратности индукции. В анализе репортерного гена агонистическая активность CD137 может быть оценена по кратному изменению (как относительная световая единица) люминесценции в лунках с каждым добавленным антителом, по сравнению с количеством люминесценции в лунках, в которые не было добавлено каких-либо антител.
Еще один пример способа измерения агонистической активности CD137 с использованием анализа репортерного гена описан ниже. В каждую лунку 384-луночного планшета добавляют 10 мкл клеток FcγRIIB СНО-K1 (фирма Promega), доведенных до концентрации 2×106/мл с помощью среды для анализа (99% RPMI, 1% ФСБ). Затем в каждую лунку добавляют 10 мкл раствора антитела, содержащего АДФ, или раствора антитела, содержащего АТФ, или раствора антитела, не содержащего АТФ или АДФ. После этого в каждую лунку добавляют 10 мкл линии клеток GloResponse™ NF-κB-Luc2/4-1BB Jurkat, доведенной до 2×106/мл с помощью среды для анализа (99% RPMI, 1% ФСБ). Конечная концентрация АДФ составляет 50 мкМ и конечная концентрация АТФ составляет 50 мкМ. После того, как планшет оставляют при 37°С в течение 6 ч в инкубаторе с 5% СО2 в атмосфере, его оставляют при комнатной температуре на 15 мин, и в каждую лунку добавляют 30 мкл реагента Bio-Glo. В качестве реагента Bio-Glo используют систему анализа люциферазы Bio-Glo (буфер и субстрат). После этого люминесценцию в каждой лунке измеряют с помощью планшет-ридера. Агонистическая активность CD137 может быть оценена как относительная световая единица (также оценку называют кратностью люминесценции или кратным изменением) люминесценции в лунках с каждым добавленным антителом, по сравнению с количеством люминесценции в лунках, в которые не было добавлено каких-либо антител.
Другой пример метода измерения агонистической активности в отношении CD137 с использованием анализа репортерного гена приведен ниже. В каждую лунку 384-луночного планшета добавляют 10 мкл клеток FcγRIIB СНО-K1 (фирма Promega), доведенных до концентрации 4×105/мл с помощью среды для анализа (99% RPMI, 1% ФСБ). Затем в каждую лунку добавляют 10 мкл раствора антитела, содержащего АДФ, или раствора антитела, содержащего АТФ, или раствора антитела, не содержащего АТФ или АДФ. После этого в каждую лунку добавляют 10 мкл линии клеток GloResponse™ NF-κB-Luc2/4-1ВВ Jurkat, доведенной до 2×106/мл с помощью среды для анализа (99% RPMI, 1% ФСБ). Конечная концентрация АДФ составляет 10 мкМ и конечная концентрация АТФ составляет 10 мкМ. После того, как планшет оставляют при 37°С в течение 6 ч в инкубаторе с 5% СО2 в атмосфере, его оставляют при комнатной температуре на 15 мин, и в каждую лунку добавляют 30 мкл реагента Bio-Glo. В качестве реагента Bio-Glo используют систему анализа люциферазы Bio-Glo Luciferase Assay System (буфер и субстрат). После этого люминесценцию в каждой лунке измеряют с помощью планшет-ридера. Агонистическая активность CD137 может быть оценена как относительная световая единица (также оценку называют кратностью люминесценции или кратным изменением) люминесценции в лунках с каждым добавленным антителом, по сравнению с количеством люминесценции в лунках, в которые не было добавлено каких-либо антител.
В одном из вариантов осуществления настоящего изобретения когда агонистическую активность в отношении CD137 измеряют с использованием анализа репортерного гена, антигенсвязывающая молекула или антитело против CD137 может быть оценена как проявляющая агонистическую активность в отношении CD137 в присутствии низкомолекулярного соединения, если (i) агонистическая активность в отношении CD137 (относительная световая единица) в присутствии 10 мкМ, 50 мкМ, 100 мкМ, 150 мкМ, 200 мкМ или 250 мкМ низкомолекулярного соединения больше в 1,1 раза или более, в 1,2 раза или более, в 1,3 раза или более, в 1,5 раз или более, в 2 раза или более, в 3 раза или более, в 5 раз или более, в 10 раз или более, в 20 раз или более, в 30 раз или более, в 50 раз или более, в 60 раз или более, в 70 раз или более, в 80 раз или более или в 90 раз или более по сравнению с (ii) агонистической активностью CD137 (относительная световая единица) в отсутствие низкомолекулярного соединения. В одном из вариантов осуществления настоящего изобретения итоговая концентрация антитела в (i) и (ii), указанная выше, составляет 0, 0,001, 0,01, 0,1, 1 или 10 мкг/мл, в предпочтительном варианте осуществления настоящего изобретения она составляет 0,1 мкг/мл или 1 мкг/мл.
В другом варианте осуществления настоящего изобретения если антигенсвязывающая молекула или антитело против CD137 добавляют в определенной концентрации (например, при конечной концентрации 0,001, 0,01, 0,1, 1 или 10 мкг/мл, или 0,1 мкг/мл или 1 мкг/мл), антигенсвязывающую молекулу или антитело против CD137 оценивают как по существу не проявляющую агонистической активности против CD137 в отсутствие низкомолекулярного соединения, если индукция кратности (относительная световая единица) в отсутствие низкомолекулярного соединения составляет 10 или менее, 9 или менее, 8 или менее, или 5 или менее в предпочтительном варианте осуществления настоящего изобретения, и существенно 1 в более предпочтительном варианте осуществления настоящего изобретения. В одном из вариантов осуществления настоящего изобретения «кратность индукции (относительная световая единица) в отсутствие низкомолекулярного соединения по существу равна 1» означает те случаи, когда кратная индукция (относительная световая единица) в отсутствие низкомолекулярного соединения составляет менее 2 раз, менее 1, 9 раз, менее 1,8 раз, менее 1,7 раз, менее 1,6 раз, менее 1,5 раз, менее 1,4 раз, менее 1,3 раз, менее 1,2 раз или менее 1,1 раза.
в) Концентрация в плазме
В одном из вариантов осуществления настоящего изобретения кинетику в крови антигенсвязывающей молекулы или антитела против CD137 по настоящему раскрытию измеряют и/или сравнивают с использованием мышей с нокином CD137 человека. Мышь с нокином CD137 человека получают, например, путем замены гена CD137 мыши геном CD137 человека путем введения вектора замены с геном CD137 человека в эмбриональные стволовые клетки мыши (ES-клетки). В одном из вариантов осуществления настоящего изобретения антигенсвязывающую молекулу или антитело CD137 по настоящему изобретению вводят путем однократного внутривенного введения мышам с нокином CD137 человека, и кровь собирают несколько раз с течением времени, начиная сразу после введения до примерно 5 дней, 10 дней, 15 дней, 20 дней, 25 дней или 30 дней после введения. В предпочтительном варианте осуществления настоящего изобретения антигенсвязывающую молекулу или антитело против CD137 по настоящему изобретению вводят путем однократного внутривенного введения мышам с нокином CD137 человека, кровь собирают несколько раз в течение периода времени от 5 мин до 28 дней после введения. Плазму быстро отделяют от собранной крови, и концентрацию антител в плазме измеряют электрохемилюминесценцией (ECL). В одном из вариантов осуществления настоящего изобретения концентрацию антитела в плазме можно измерить способом, описанным в примере 6-3-2.
При измерении периода полужизни в плазме с использованием мыши с нокином CD137 человека, если анти-CD137 антигенсвязывающая молекула или антитело исчезает из плазмы медленнее, чем контрольная молекула, анти-С0137 антигенсвязывающую молекулу или антитело оценивают как имеющее улучшенную кинетику в крови, чем у контрольной молекулы. Кроме того, если анти-CD137 антигенсвязывающая молекула или антитело, имеющие связывающую активность, зависящую от низкомолекулярного соединения (переключаемая молекула или переключаемое антитело), исчезает из плазмы медленнее, чем анти-CD137 антигенсвязывающая молекула или антитело, не зависящие от низкомолекулярного соединения (непереключаемая молекула или непереключаемое антитело), можно прийти к заключению, что переключаемая молекула (или переключаемое антитело) не связывается с CD137, экспрессируемым в неопухолевой ткани, по сравнению с непереключаемой молекулой (непереключаемым антителом).
г) Противоопухолевая активность
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело тестируют по способности ингибировать рост или пролиферацию клеток in vivo. В некоторых вариантах осуществления настоящего изобретения анти-CD137 антигенсвязывающая молекула или антитело тестируют по способности ингибировать рост опухоли in vivo. Системы моделей in vivo, такие как модели аллотрансплантата или модели ксенотрансплантата, могут использоваться для такого тестирования. В примере системы ксенотрансплантата опухолевые клетки человека вводят животному (не человеку) с соответствующим иммунодефицитом, например, бестимусной «голой» мыши. Антитело по настоящему изобретению вводят животному. Измеряют способность антитела ингибировать или уменьшать рост опухоли. В некоторых вариантах осуществления настоящего изобретения в указанной выше системе ксенотрансплантата опухолевые клетки человека представляют собой опухолевые клетки пациента-человека. Такие модели ксенотрансплантатов коммерчески доступны от фирмы Oncotest GmbH (Фрейбург, Германия). В некоторых вариантах осуществления настоящего изобретения опухолевые клетки человека вводят животному (но не человеку) с соответствующим иммунодефицитом путем подкожной инъекции или трансплантации в подходящее место, такое как жировая прослойка молочной железы.
В одном из вариантов осуществления настоящего изобретения противоопухолевую активность анти-CD137 антигенсвязывающих молекул или антител по настоящему изобретению может быть измерена и/или сопоставлена с использованием модели трансплантата сингенных опухолевых клеток, основанной на вышеописанных мышах с нокином CD137 человека. Подходящим образом может быть выбрана линия раковых клеток для использования в тесте, предпочтительно линия клеток рака толстой кишки мыши МС38. В одном из вариантов осуществления настоящего изобретения клеточную линия МС38 трансплантируют под кожу в брюшную область мыши, и модель считают сформированной, когда объем опухоли достигает примерно 50-300 мм3. После формирования модели мышей с трансплантированными клетками МС38 распределяют по группам и затем им вводят анти-CD137 антигенсвязывающую молекулу или антитело. В одном из вариантов осуществления настоящего изобретения измерение объема опухоли проводят с частотой от одного до двух раз в неделю, чтобы оценить противоопухолевую активность. Объем опухоли рассчитывают с использованием следующего уравнения: объем опухоли=(большая ось × малая ось × малая ось)/2. В одном из вариантов осуществления настоящего изобретения противоопухолевую активность анти-CD137 антигенсвязывающей молекулы или антитела может быть протестирована и оценена в соответствии с методом, описанным в примере 6-4.
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающие молекулы или антитела распознают как проявляющие противоопухолевую активность, когда объем опухоли в группе введения животным анти-CD137 антигенсвязывающих молекул или антител меньше, чем в группе введения животным растворителя, или увеличение объема опухоли меньше в группе введения животным анти-CD137 антигенсвязывающих молекул или антител меньше, чем в группе введения животным растворителя.
д) Системная реакция
В одном из вариантов осуществления настоящего изобретения системная реакция на анти-CD137 антигенсвязывающие молекулы или антитела по настоящему изобретению измеряют и/или сравнивают с использованием модели трансплантата сингенных опухолевых клеток на основе описанных выше мышей с нокином CD137 человека. Органы для измерения системной реакции могут быть выбраны соответствующим образом, это могут быть печень, селезенка и/или лимфатические узлы. В одном из вариантов осуществления настоящего изобретения системную реакцию оценивают путем удаления печени, селезенки и/или лимфатических узлов у мышей с нокином CD137 человека в соответствующее время после введения анти-CD137 антигенсвязывающих молекул или антител. В случае, если подлежащим удалению органом является селезенка и/или лимфатические узлы, этот орган взвешивают и/или подсчитывают клетки во фракции лимфоцитов. При подсчете предпочтительно фракцию лимфоцитов после гемолиза используют для селезенки, а фракция лимфоцитов, полученная путем гомогенизации, используется для лимфатических узлов. В случае, если подлежащим удалению органом является печень, подсчитываю клетки во фракции лимфоцитов, полученные с помощью набора Liver dissociation kit, mouse (Milteny Biotec). Кроме того, анализ Т-клеток с использованием жидкостной цитометрии (FCM, flowcytometry) можно проводить на фракции лимфоцитов из каждого органа (печени, селезенки и/или лимфатических узлов). В анализе FCM используют экспрессию гранзима В, или экспрессию PD-1, или экспрессию ICOS в CDS-альфа-положительных Т-клетках или соотношение CD8-альфа-положительных Т-клеток к CD45 положительным клеткам. В одном из вариантов осуществления настоящего изобретения системная реакция на анти-CD137 антигенсвязывающие молекулы или антитела может быть протестирована и оценена в соответствии с методом, описанным в примере 6-4.
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающие молекулы или антитела распознают как вызывающие подавленную системную реакцию и/или подавленную активацию иммунных клеток в неопухолевых тканях (например, печени, селезенке и/или лимфатических узлах) по сравнению с контрольной молекулой, когда в группе, в которой мышам вводили анти-CD137 антигенсвязывающие молекулы или антитела, отмечают более низкое значение при оценке вышеописанных различных показателей по сравнению с группой, в которой мышам вводили такое же количество контрольной молекулы. Кроме того, если группа, получающая введение анти-CD137 антигенсвязывающей молекулы или антитела, обладающего связывающей активностью, зависящей от низкомолекулярного соединения (переключаемой молекулы или переключаемого антитела), показывает пониженное значение при оценке вышеописанных различных показателей, чем что для группы, в которой мышам вводили анти-CD137 антигенсвязывающие молекулы или антитела, которое не обладает связывающей активностью, зависящей от низкомолекулярного соединения (непереключаемая молекула или непереключаемое антитело), переключаемая молекула (или переключаемое антитело) распознаются как вызывающие подавленную системную реакцию и/или подавленную активацию иммунных клеток в неопухолевых тканях (например, в печени, селезенке и/или лимфатических узлах) по сравнению с непереключаемой молекулой (или непереключаемым антителом).
Следует принять во внимание, что любой из описанных выше способов измерения может быть проведен с использованием иммуноконъюгатов по настоящему изобретению вместо или в дополнение к анти-CD137 антигенсвязывающим молекулам или антителам.
Г. Иммуноконъюгаты
В настоящем изобретении также предусматривают иммуноконъюгаты, содержащие анти-CD137 антигенсвязывающие молекулы или антитела, конъюгированные с одним или несколькими цитотоксическими агентами, такими как химиотерапевтические агенты или лекарственные препараты, агенты, ингибирующие рост, токсины (например, белковые токсины, ферментативно активные токсины бактерий, а также грибкового, растительного или животного происхождения, или их фрагменты) или радиоактивные изотопы.
В одном из вариантов осуществления настоящего изобретения иммуноконъюгат представляет собой конъюгат антитело-лекарственное средство (ADC, antibody-drug conjugate), где антитело конъюгировано с одним или несколькими лекарственными средствами, включая, но ими не ограничиваясь, майтансиноид (см. US 5208020, US 5416064, ЕР 0425235 В1); ауристатин, такой как лекарственные части монометилауристатина DE и DF (ММАЕ и MMAF) (см. US 5635483, US 5780588, US 7498298); доластатин; калихеамицин или его производное (см. US 5712374, US 5714586, US 5739116, US 5767285, US 5770701, US 5770710, US 5773001, US 5877296; Hinman с соавт., Cancer Res. 53, 1993, 3336-3342; Lode с соавт., Cancer Res. 58, 1998, 2925-2928); антрациклин, такой как дауномицин или доксорубицин (см. Kratz с соавт., Current Med. Chem. 13, 2006, 477-523; Jeffrey с соавт., Bioorganic & Med. Chem. Letters 16, 2006, 358-362; Torgov с соавт., Bioconj. Chem. 16, 2005, 717-721; Nagy с соавт., Proc. Natl. Acad. Sci. USA 97, 2000, 829-834; Dubowchik с соавт., Bioorg. & Med. Chem. Letters 12, 2002, 1529-1532; King с соавт., J. Med. Chem. 45, 2002, 4336-4343; US 6630579); метотрексат; виндезин; таксан, такой как доцетаксел, паклитаксел, ларотаксел, тезетаксел и ортатаксел; трихотецин; СС1065.
В другом варианте осуществления настоящего изобретения иммуноконъюгат включает антитело, как описано в настоявшем изобретении, конъюгированное с ферментативно активным токсином или его фрагментом, включая, помимо прочего, цепь А дифтерийного токсина, несвязывающие активные фрагменты дифтерийного токсина, цепь экзотоксина А (из Pseudomonas aeruginosa), цепь рицина А, цепь абрина А, цепь А модекцина, альфа-сарцин, белки Aleurites fordii, белки диантина, белки Phytolacca americana (PAPI, PAPII и PAP-S), ингибитор момордика харантии, курцин, кротин, ингибитор Saponaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трикотецины.
В другом варианте осуществления настоящего изобретения иммуноконъюгат включает антитело, как описано в настоявшем изобретении, конъюгированное с радиоактивным атомом для формирования радиоактивного иммуноконъюгата. Для получения радиоконъюгатов доступны различные радиоактивные изотопы. Примерами являются 211At, 131I, 125I, 90Y, 186Re, 188Re, 153Sm, 212Bi, 32P, 212Pb и радиоактивные эпитопы Lu. Когда радиоконъюгат используют для обнаружения, он может содержать радиоактивный атом для сцинтиграфических исследований, например Тс-99m или 123I, или спиновую метку для визуализации методом ядерно-магнитного резонанса (ЯМР) (также известного как магнитно-резонансная томография, МРТ), например, йод-123, йод-131, индий-111, фтор-19, углерод-13, азот-15, кислород-17, гадолиний, марганец или железо.
Конъюгаты антитела и цитотоксического агента могут быть получены с использованием различных бифункциональных белковых связывающих агентов, таких как N-сукцинимидил-3- (2-пиридилдитио) пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональные производные имидоэфиров (такие как диметиладипимидат НС1), активные сложные эфиры (такие как дисукцинимидил суберат), альдегиды (такие как глутаральдегид), бис-азидосоединения (такие как бис(п-азидобензоил)гександиамин), производные бис-диазония (такие как бис-(п-диазонийбензоил)этилендиамин), диизоцианаты (такие как толуол-2,6-диизоцианат) и бис-активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол). Например, иммунотоксин рицин можно получить, как описано в публикации Vitetta с соавт., Science 238, 1987, 1098. Меченая углеродом-14 1-изотиоцианатобензил-3-метилдиэтилентриаминпентауксусная кислота (МХ-DTPA) является типичным хелатирующим агентом для конъюгации радионуклида с антителом. См. WO 94/11026. Линкер может быть «расщепляемым линкером», способствующим высвобождению цитотоксического лекарственного средства в клетке. Например, можно использовать кислотолабильный линкер, чувствительный к пептидазе линкер, фотолабильный линкер, диметиловый линкер или дисульфидсодержащий линкер (Chari с соавт., Cancer Res. 52, 1992, 127-131; US 5208020).
Иммуноконъюгаты или ADC в настоящем изобретении явно предполагают, но не ограничиваются ими, такие конъюгаты, как полученные с помощью перекрестно-линкерных реагентов, включая, но ими не ограничиваясь, BMPS, EMCS, GMBS, HBVS, LC-SMCC, MBS, МРВН, SBAP, SIA, SIAB, SMCC, SMPB, SMPH, сульфо-EMCS, сульфо-GMBS, сульфо-KMUS, сульфо-MBS, сульфо-SIAB, сульфо-SMCC и сульфо-SMPB, а также SVSB (сукцинимидил-(4-винилсульфон)бензоат), которые являются коммерческими доступными (например, от фирмы Pierce Biotechnology, Inc., Рокфорд, Иллинойс, США).
Е. Способы и композиции для диагностики и обнаружения
В некоторых вариантах осуществления настоящего изобретения любые из предусмотренных в настоящем изобретении анти-CD137 антигенсвязывающих молекул или антител применимы для обнаружения присутствия анти-CD137 антигенсвязывающей молекулы в биологическом образце. В настоящем изобретении термин «обнаружение» означает количественное или качественное обнаружение. В некоторых вариантах осуществления настоящего изобретения биологический образец включает клетку или ткань.
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающую молекулу или антитело предусматривают для применения в способе диагностики и обнаружения. В другом варианте осуществления настоящего изобретения предусматривают метод обнаружения CD137 в биологическом образце. В некоторых вариантах осуществления настоящего изобретения способ включает приведение биологического образца в контакт с анти-CD137 антигенсвязывающей молекулой или антителом, как описано в настоящем изобретении, в условиях, допускающих связывание анти-CD137 антигенсвязывающей молекулы или антитела с CD137, и выявление того, образуется ли комплекс между анти-CD137 антигенсвязывающей молекулой или антителом с CD137. Такой метод может быть методом in vitro или in vivo. В одном из вариантов осуществления настоящего изобретения антигенсвязывающую молекулу или антитело против CD137 используют для отбора субъектов, пригодных для терапии анти-CD137 антигенсвязывающей молекулой или антителом, например, если CD137 является биомаркером для отбора пациентов.
К типичным расстройствам, которые можно диагностировать с использованием антитела по настоящему изобретению, относят рак.
В некоторых вариантах осуществления настоящего изобретения предусматривают меченые анти-CD137 антигенсвязывающие молекулы или антитела. К меткам относят, но ими перечень не ограничивается, метки или фрагменты, которые обнаруживают напрямую (такие как флуоресцентные, хромофорные, электронно-плотные, хемилюминесцентные и радиоактивные метки), а также фрагменты, такие как ферменты или лиганды, которые обнаруживают опосредованно, например, через ферментативную реакцию или молекулярное взаимодействие. К примерам меток относят, но ими перечень не ограничивается, радиоизотопы 32Р, 14С, 125I, 3Н и 131I, флуорофоры, например, такие как хелаты редкоземельных элементов или флуоресцеин и его производные, родамин и его производные, дансил, умбеллиферон, люцериферазы, например, люцифераза светлячков и бактериальная люцифераза (US 4737456), люциферин, 2,3-дигидрофталазиндионы, пероксидаза хрена (HRP), щелочная фосфатаза, бета-галактозидаза, глюкоамилаза, лизоцим, сахаридоксидазы, например, глюкозооксидаза, галактозооксидаза и глюкозо-6-фосфатдегидрогеназа, гетероциклические оксидазы, такие как уриказа и ксантиноксидаза, соединенные с ферментом, который использует перекись водорода для окисления предшественника красителя, такого как HRP, лактопероксидаза или микропероксидаза, биотин/авидин, спиновые метки, бактериофаговые метки, стабильные свободные радикалы и др.
Е. Фармацевтические составы
Фармацевтические составы анти-CD137 антигенсвязывающей молекулы или антитела по настоящему изобретению получают путем смешивания такого антитела, имеющего желаемую степень чистоты, с одним или более необязательными фармацевтически приемлемыми носителями (Remington's Pharmaceutical Sciences, 16е изд., под ред. Osol А., 1980) в виде лиофилизированных составов или водных растворов. Фармацевтически приемлемые носители обычно нетоксичны для реципиентов в используемых дозировках и концентрациях и включают, но ими не ограничиваются: буферы, такие как фосфатный, цитратный и других органических кислот; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония; хлорид бензетония; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцин; циклогексанол; 3-пентанол и м-крезол); низкомолекулярные полипептиды (менее примерно 10 остатков); белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как EDTA; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы металлов (например, комплексы Zn-белок); и/или неионные поверхностно-активные вещества, такие как полиэтиленгликоль (PEG). Примерами фармацевтически приемлемых носителей в настоящем изобретении также являются интерстициальные агенты для диспергирования лекарственных средств, такие как растворимые нейтрально-активные гликопротеины гиалуронидазы (sHASEGP), например, растворимые гликопротеины гиалуронидазы РН-20 человека, такие как rHuPH20 (HYLENEX (зарегистрированная торговая марка), Baxter International, Inc.). Некоторые примеры sHASEGP и способы их использования, включая rHuPH20, описаны в публикациях патентов US №2005/0260186 и US №2006/0104968. В одном из вариантов осуществления sHASEGP комбинируют с одной или несколькими дополнительными гликозаминогликаназами, такими как хондроитиназы.
Примеры составов лиофилизированных антител описаны в US 6267958. Водные составы антител включают те, которые описаны в US 6171586 и WO 2006/044908, причем последние составы включают гистидин-ацетатный буфер.
Активные ингредиенты могут быть заключены в микрокапсулы, приготовленные, например, методами коацервации или межфазной полимеризацией, например, микрокапсулы из гидроксиметилцеллюлозы или желатина и микрокапсулы из поли(метилметакрилата), соответственно, в коллоидных системах доставки лекарств (например, липосомы, микросферы альбумина, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсиях. Такие методы описаны в Remington's Pharmaceutical Sciences, 16е изд., под ред. Osol А., 1980.
Могут быть получены препараты с замедленным высвобождением. Подходящие примеры препаратов с замедленным высвобождением включают полупроницаемые матрицы твердых гидрофобных полимеров, содержащие антитело, причем эти матрицы имеют форму изделий определенной формы, например пленок или микрокапсул.
Составы, используемые для введения in vivo, обычно стерильны. Стерильность может быть легко достигнута, например, путем фильтрации через стерильные фильтрующие мембраны.
Ж. Способы терапии и лечебные композиции
Любая из антигеневязывающих молекул или антител против CD137, представленных в настоящем изобретении, может использоваться в терапевтических способах.
В одном из объектов настоящего изобретения предусматривают антигенсвязывающие молекулы или антитела против CD137 для использования в качестве лекарственного средства. В настоящем изобретении примеры лекарственного средства, в частности, включают лекарственные средства для индукции противоопухолевого действия, например, ингибирования неоваскуляризации в опухолях, ингибирование пролиферации опухолевых клеток, истощение опухолевых В-клеток и т.д., посредством активации клеток, возникающих в результате связывания анти-CD137 антигенсвязывающих молекул или антител с CD137, экспрессируемым на иммунных клетках, таких как Т-клетки. В других вариантах осуществления настоящего изобретения предусматривают анти-CD137 антигенсвязывающие молекулы или антитела для применения в лечении опухолей. В некоторых вариантах осуществления настоящего изобретения предусматривают некоторые анти-CD137 антигенсвязывающие молекулы или антитела для применения в методе лечения. В некоторых вариантах осуществления настоящего изобретения предусматривают некоторые анти-CD137 антигенсвязывающие молекулы или антитела для применения в методе лечения больного раком индивида, включающего введение индивиду эффективного количества анти-CD137 антигенсвязывающей молекулы или антитела. В других вариантах осуществления настоящего изобретения примеры опухоли включают солидную опухоль, в которую проникли В-клетки, дендритные клетки, естественные клетки-киллеры, макрофаги, CD8-положительные Т-клетки и/или регуляторные Т-клетки (Treg-клетки).
В других вариантах осуществления настоящего изобретения предусматривают анти-CD137 антигенсвязывающие молекулы или антитела для применения в методе активации иммунных клеток у индивида, включающем введение индивиду эффективного количества анти-CD137 антигенсвязывающей молекулы или антитела для активации иммунных клеток, таких как В-клетки, дендритные клетки, естественные клетки-киллеры, макрофаги и/или CD8-положительные Т-клетки (более конкретно, эти иммунные клетки, проникшие в опухолевую ткань).
В других вариантах осуществления настоящего изобретения предусматривают анти-CD137 антигенсвязывающие молекулы или антитела для применения в методе повреждения клеток (например, опухолевых клеток) у индивида, включающем введение индивиду эффективного количества анти-CD137 антигенсвязывающей молекулы или антитела. В некоторых вариантах осуществления настоящего изобретения примеры опухолей включают солидную опухоль, в которую проникли В-клетки, дендритные клетки, природные клетки-киллеры, макрофаги, CD8-положительные Т-клетки и/или регуляторные Т-клетки (Treg-клетки). В любом из указанных выше вариантов осуществления настоящего изобретения «индивидом» предпочтительно является человек.
В других вариантах осуществления настоящего изобретения предусматривают применение анти-CD137 антигенсвязывающих молекул или антител для получения лекарственного средства. В одном из вариантов осуществления настоящего изобретения лекарство предназначено для лечения опухоли (в зависимости от контекста может быть более подходящим называть это «раком» (то же самое применимо и в дальнейшем)). В других вариантах осуществления настоящего изобретения лекарственное средство предназначено для использования в способе лечения опухоли (или рака в зависимости от контекста), причем способ включает введение индивиду с опухолью (или раком в зависимости от контекста), эффективного количества лекарственного средства. В других вариантах осуществления настоящего изобретения лекарство предназначено для индукции противоопухолевого действия, например, ингибирования неоваскуляризации в опухолях, ингибирования пролиферации опухолевых клеток, для истощения опухолевых В-клеток и т.д., посредством активации клеток, возникающих в результате связывания анти-CD137 антигенсвязывающей молекулы или антитела с CD137, экспрессируемым на иммунных клетках, таких как Т-клетки. В других вариантах осуществления настоящего изобретения лекарство предназначено для использования в способе, например, ингибирования неоваскуляризации в опухолях, ингибирования пролиферации опухолевых клеток, истощения опухолевых В-клеток и т.д. посредством активации клеток в результате связывания анти-CD137 антигенсвязывающей молекулы или антитела с CD137, экспрессируемым на иммунных клетках, таких как Т-клетки, у индивида, причем способ включает введение для этого индивиду эффективного количества лекарственного средства. В любом из указанных выше вариантов осуществления настоящего изобретения «индивидом» предпочтительно является человек.
В других вариантах осуществления настоящего изобретения предусматривают способ лечения рака. В одном из вариантов осуществления настоящего изобретения способы включают введение индивиду с опухолью эффективного количества анти-CD137 антигенсвязывающей молекулы или антитела. В других вариантах осуществления настоящего изобретения примеры опухоли включают солидную опухоль, в которую инфильтруются В-клетки, дендритные клетки, природные клетки-киллеры, макрофаги, CD8-положительные Т-клетки и/или регуляторные Т-клетки (Treg-клетки).
В других вариантах осуществления настоящего изобретения предусматривают методы активации иммунных клеток у индивида. В одном из вариантов осуществления настоящего изобретения способ включает введение индивиду эффективного количества анти-CD137 антигенсвязывающей молекулы или антитела. В других вариантах осуществления настоящего изобретения к иммунным клеткам относят такие иммунные клетки, как В-клетки, дендритные клетки, природные клетки-киллеры, макрофаги и/или CD8-положительные Т-клетки (более конкретно, эти иммунные клетки, проникшие в опухолевую ткань).
В других вариантах осуществления настоящего изобретения предусматривают методы повреждения клеток (в частности, опухолевых клеток) у индивида. В одном из вариантов осуществления настоящего изобретения предусматривают методы, которые включают введение индивиду эффективного количества анти-CD137 антигенсвязывающей молекулы или антитела. В некоторых вариантах осуществления настоящего изобретения примеры опухоли включают солидную опухоль, в которую инфильтруются В-клетки, дендритные клетки, природные клетки-киллеры, макрофаги, CD8-положительные Т-клетки и/или регуляторные Т-клетки (Treg-клетки). В любом из указанных выше вариантов осуществления настоящего изобретения «индивидом» может быть человек.
В других вариантах осуществления настоящего изобретения фармацевтические препараты, содержащие анти-CD137 антигенсвязывающие молекулы или антитела, которые предназначены для использования в описанных выше способах лечения, применяют в лечении, и лекарственных средств могут содержать эффективное количество анти-CD137 антигенсвязывающей молекулы или антитела, которое имеет пониженный уровень иммунологической активации в неопухолевой ткани по сравнению с анти-CD137 антигенсвязывающей молекулой, которая не обладает CD137 антигенсвязывающей активностью, зависящей от низкомолекулярного соединения.
В одном из вариантов осуществления настоящего изобретения неопухолевые ткани включают лимфатические узлы, селезенку и/или печень.
В других вариантах осуществления настоящего изобретения фармацевтические препараты, содержащие анти-CD137 антигенсвязывающую молекулу или антитело для применения в описанных выше способах лечения, применения в лечении и лекарственных средствах, могут содержать эффективное количество анти-CD137 антигенсвязывающей молекулы или антитела, которое по существу не связывается с CD137, экспрессируемым в неопухолевой ткани.
В других вариантах осуществления настоящего изобретения фармацевтические препараты, содержащие анти-CD137 антигенсвязывающую молекулу или антитело для применения в описанных выше способах лечения, применения в лечении, причем лекарственные препараты могут содержать эффективное количество анти-CD137 антигенсвязывающей молекулы или антитела, которое имеет увеличенный период полужизни в крови по сравнению с анти-CD137 антигенсвязывающей молекулы или антитела, которая не обладает связывающей активностью CD137, зависящей от низкомолекулярного соединения.
В других вариантах осуществления настоящего изобретения фармацевтические препараты, содержащие анти-CD137 антигенсвязывающую молекулу или антитело для применения в описанных выше способах лечения, применения в лечении, причем лекарственные препараты могут содержать эффективное количество анти-CD137 антигенсвязывающей молекулы или антитела, которое имеет низкий уровень побочных эффектов по сравнению с анти-CD137 антигенсвязывающей молекулой или антителом, которое не обладает связывающей активностью с CD137, зависящей от низкомолекулярного соединения.
В других вариантах осуществления настоящего изобретения к побочным явлениям относят повышенный AST, повышенный ALT, лихорадку, тошноту, острый гепатит, повреждение печени, спленомегалию, энтерит, гнойное воспаление кожи, нейтропению, лимфопению, тромбопению, экспрессию трансаминаз и/или гипербилирубинемию.
В одном из вариантов осуществления настоящего изобретения анти-CD137 антигенсвязывающие молекулой по настоящему изобретению имеют низкие побочные эффекты, и, таким образом, дозировка может быть увеличена без опасения по поводу побочных эффектов, поэтому в результате они могут проявлять более высокую лекарственную эффективность (цитотоксическую активность или противоопухолевую активность).
В других вариантах осуществления настоящего изобретения предусматривают фармацевтические составы, содержащие любую из анти-CD137 антигенсвязывающих молекул или антител, предусмотренных в настоящем изобретении, например, для использования в любом из вышеуказанных терапевтических методов. В одном из вариантов осуществления настоящего изобретения фармацевтический состав содержит любую из анти-CD137 антигенсвязывающих молекул или антител, предусмотренных в настоящем изобретении, и фармацевтически приемлемый носитель.
Антигенсвязывающую молекулу или антитело по настоящему изобретению можно вводить любыми подходящим способом, включая парентеральный, внутрилегочный и интраназальный, и, если необходимо местное лечение, внутриочаговое введение. Парентеральные инфузии включают внутримышечное, внутривенное, внутриартериальное, внутрибрюшинное или подкожное введение. Дозирование может осуществляться любым подходящим путем, например, инъекциями, такими как внутривенные или подкожные инъекции, частично в зависимости от того, является ли введение кратковременным или хроническим. В настоящем изобретении предусматривают различные схемы дозирования, включая, но не ограничиваясь ими, однократное или многократное введение в различные моменты времени, болюсное введение и пульсовую инфузию.
Антитела по настоящему изобретению могут быть сформированы, дозированы и введены способом, соответствующим надлежащей медицинской практике. Факторы, которые следует учитывать в этом контексте, включают конкретное заболевание, которое лечат, конкретное млекопитающее, которое лечат, клиническое состояние отдельного пациента, причину расстройства, место доставки агента, метод введения, график введения и другие факторы, известные практикующим врачам. Антитело необязательно, но может быть составлено с одним или несколькими агентами, которые в настоящее время используются для предотвращения или лечения рассматриваемого нарушения. Эффективное количество таких других агентов зависит от количества антитела, присутствующего в составе, типа расстройства или лечения и других факторов, обсуждаемых выше. Их обычно используют в тех же дозах и вводят способами, которые описаны в настоящем изобретении, или примерно от 1 до 99% дозировок, описанных в настоящем изобретении, или в любой дозировке и любым способом, который эмпирически/клинически определен как применимый.
Для профилактики или лечения заболевания подходящая доза антитела по настоящему изобретению может зависеть от типа заболевания, подлежащего лечению, типа антитела, тяжести и течения заболевания, от того, вводят ли антитело для профилактики или в целях терапии, от предыдущей терапии, истории болезни пациента и реакции на антитело, а также на усмотрение лечащего врача. Антитело обычно вводят пациенту за один раз или в течение ряда курсов лечения. В зависимости от типа и тяжести заболевания от 1 мкг/кг до 15 мг/кг (например, 0,1 мг/кг - 10 мг/кг) антитела может быть начальной дозой антитела-кандидата для введения пациенту, например, в виде одного или нескольких отдельных введений, или путем непрерывной инфузии. Одна типичная суточная доза может составлять примерно от 1 микрограмма/кг до 100 мг/кг или более, в зависимости от факторов, упомянутых выше. При повторных введениях в течение нескольких дней или дольше, в зависимости от состояния, лечение обычно будет продолжаться до тех пор, пока не произойдет желаемое подавление симптомов заболевания. Одна примерная дозировка антитела может находиться в диапазоне от примерно 0,05 мг/кг до примерно 10 мг/кг. Таким образом, пациенту можно вводить одну или более доз, составляющих примерно 0,5 мг/кг, 2,0 мг/кг, 4,0 мг/кг или 10 мг/кг (или любую их комбинацию). Такие дозы можно вводить с перерывами, например, каждую неделю или каждые три недели (например, когда пациент получает от примерно двух до примерно двадцати, или например, примерно шести доз антитела). Может быть введена начальная более высокая ударная доза, а затем одна или несколько более низких доз. Прогресс этой терапии легко подвергать мониторингу с помощью обычных методов и анализов.
Очевидно, что любой из вышеуказанных составов или терапевтических методов может быть осуществлен с использованием иммуноконъюгата по настоящему описанию вместо или в дополнение к анти-CD137 антигенсвязывающей молекуле или антителу.
3. Получаемые изделия
В другом аспекте настоящего раскрытия предусмотрено получение изделия, содержащего материалы, полезные для лечения, профилактики и/или диагностики нарушений, описанных выше. Изделие включает контейнер и связанную с ним этикетку или вкладыш в упаковку, связанный с контейнером. Подходящие контейнеры включают, например, бутылки, флаконы, шприцы, пакеты с раствором для внутривенного введения и т.д. Контейнеры могут быть изготовлены из различных материалов, таких как стекло или пластик. Контейнер содержит состав, который сам по себе и может иметь стерильный порт доступа (например, контейнер может быть пакетом с раствором для внутривенного введения или флаконом с пробкой, которую можно проткнуть иглой для подкожных инъекций). По крайней мере, один активный ингредиент в композиции представляет собой антитело по настоящему изобретению. Этикетка или вкладыш в упаковку указывает, что композицию используют для лечения выбранного состояния. Изделие в этом варианте осуществления настоящего изобретения может дополнительно содержать вкладыш в упаковку, указывающий, что композиции можно использовать для лечения определенного состояния. В другом варианте или дополнительно изделие может дополнительно содержать второй (или третий) контейнер, содержащий фармацевтически приемлемый буфер, такой как бактериостатическая вода для инъекций (BWFI, bacteriostatic water for injection), фосфатно-солевой буфер, раствор Рингера и раствор декстрозы. Он может дополнительно включать другие материалы, желательные с коммерческой точки зрения и с точки зрения пользователя, включая другие буферы, разбавители, фильтры, иглы и шприцы.
Термин «вкладыш в упаковку» используют для обозначения инструкций, обычно включаемых в коммерческие упаковки терапевтических продуктов, которые содержат информацию о показаниях, применении, дозировке, введении, комбинированной терапии, противопоказаниях и/или предупреждениях относительно использования таких терапевтических продуктов.
Понятно, что любое из вышеуказанных промышленных изделий может включать иммуноконъюгат по настоящему изобретению вместо или в дополнение к анти-CD137 антигенсвязывающей молекуле или антителу.
III. Композиции и способы (агонистические антигенсвязывающие молекулы и цитотоксические антигенсвязывающие молекулы, содержащие вариант области Fc с повышенной изоэлектрической точкой (pI))
В одном из вариантов осуществления настоящего изобретения предусматривают агонистические антигенсвязывающие молекулы или антитела, содержащие вариант области Fc с увеличенной изоэлектрической точкой (pI), и способы их применения. В некоторых вариантах осуществления настоящего изобретения полипептиды, содержащие вариант области Fc с увеличенной изоэлектрической точкой (pI), содержат по меньшей мере одно аминокислотное изменение в исходной области Fc. В других вариантах осуществления настоящего изобретения каждое такое аминокислотное изменение вызывает повышение изоэлектрической точки (pI) вариантной области Fc по сравнению с исходной областью Fc. Не опираясь на какую-либо теорию, авторы изобретения высказывают предположение, что рН биологических жидкостей (например, плазмы) находится в нейтральном диапазоне рН. В биологических жидкостях суммарный положительный заряд антигенсвязывающей молекулы или антитела с повышенной pI увеличивается из-за увеличения pI, и в результате антигенсвязывающая молекула или антитело сильнее притягивается физико-химическим кулоновским взаимодействием к поверхности эндотелиальной клетке., которая имеет общий отрицательный заряд по сравнению с антигенсвязывающей молекулой или антителом, не имеющим повышенной pI. Таким образом, агонистические антигенсвязывающие молекулы (или антитела) или антигенсвязывающие агонистические антигенсвязывающие молекулы (или антитела) могут приближаться к поверхности клеток, которые экспрессируют рецептор Fc-гамма, что приводит к усилению связывания антигенсвязывающих молекул или антител с клетками, экспрессирующими рецептор Fc-гамма. Для тех агонистических антигенсвязывающих молекул или антител, которые проявляют агонистическую активность за счет связывающей активности с рецептором Fc-гамма, агонистические антигенсвязывающие молекулы или антитела, имеющие повышенное связывание с клетками, экспрессирующими рецептор Fc-гамма, из-за увеличивающих pI аминокислотных изменений могут проявлять более сильную агонистическую активность по сравнению с агонистическими антигенсвязывающими молекулами или антителами, не имеющими аминокислотных изменений, повышающих pI. В одном из вариантов осуществления настоящего изобретения агонистические антигенсвязывающие молекулы являются анти-CD137 антигенсвязывающими молекулами или анти-CD3 антигенсвязывающими молекулами. В другом варианте осуществления настоящего изобретения агонистические антигенсвязывающие молекулы являются анти-CD137 антителами или анти-CD3 антителами.
В другом варианте осуществления настоящего изобретения предусматривают цитотоксические антигенсвязывающие молекулы или антитела, содержащие вариант области Fc с повышенной изоэлектрической точкой (pI), и способы их использования. В биологических жидкостях суммарный положительный заряд антигенсвязывающей молекулы или антитела с повышенной pI увеличивается из-за увеличения pI, и в результате антигенсвязывающая молекула или антитело сильнее притягивается физико-химическим кулоновским взаимодействием к поверхности эндотелиальной клетки, которая имеет общий отрицательный заряд по сравнению с антигенсвязывающей молекулой или антителом, не имеющим повышенной pI. Таким образом, цитотоксические антигенсвязывающие молекулы (или антитела) или антиген-связанные цитотоксические антигенсвязывающие молекулы (или антитела) могут приближаться к поверхности клеток, которые экспрессируют рецептор Fc-гамма, что приводит к усилению связывания антигенсвязывающих молекул или антител с клеткам, экспрессирующим рецептор Fc-гамма. Для тех цитотоксических антигенсвязывающих молекул или антител, которые проявляют цитотоксическую активность за счет связывающей активности в отношении рецептора Fc-гамма, цитотоксические антигенсвязывающие молекулы или антитела, имеющие повышенное связывание из-за увеличения pI с клетками, экспрессирующими рецептор Fc-гамма, аминокислотные изменения могут проявлять более сильную агонистическую активность по сравнению с цитотоксическими антигенсвязывающими молекулами или антителами, не имеющими аминокислотных изменений, повышающих pI.
В одном из вариантов осуществления настоящего изобретения цитотоксичность, которой обладают цитотоксические антигенсвязывающие молекулы, включает, например, антителозависимую клеточную цитотоксичность (ADCC) и антителозависимый клеточный фагоцитоз (ADCP), которые вызываются эффекторными клетками.
В настоящем изобретении pI может быть определена теоретически или экспериментально. Значение pI можно определить, например, с помощью изоэлектрического фокусирования, известного специалистам в данной области. Теоретически значение pI можно рассчитать, например, с помощью программного обеспечения для анализа генной и аминокислотной последовательности (Genetyx и др.). В настоящем изобретении pI может быть либо теоретическим, либо экспериментально определенным. Значение pI можно определить, например, с помощью изоэлектрического фокусирования, известного специалистам в данной области. Значение теоретически pI можно рассчитать, например, с помощью программного обеспечения для анализа генной и аминокислотной последовательности (Genetyx и т.д.). При расчете свойства антитела могут быть отражены в формуле расчета. Например, (i) как правило, Cys, консервативный в антителе, образует дисульфидную связь и не несет электрического заряда в боковой цепи; поэтому такой Cys может быть исключен из расчета, и только Cys в свободной форме, который не образует дисульфидную связь, может быть включен в расчет. В другом варианте (ii) состояние заряда или изоэлектрическая точка антител могут быть изменены из-за посттрансляционных модификаций; поэтому формула расчета может быть изменена с учетом таких посттрансляционных модификаций: (а) когда N-концом тяжелой цепи является Q (глутамин), N-концевая аминогруппа исключается из расчета, предполагая, что происходит пироглутамилирование, (б) когда С-концом тяжелой цепи является K (лизин), K (только один остаток) исключают из расчета, предполагая, что происходит усечение; (в) боковые цепи всех С (цистеина), присутствующих в обычно консервативных положениях, исключают из расчета, предполагая, что все эти С образуют дисульфидные связи внутри молекулы. В одном предпочтительном варианте осуществления настоящего изобретения оба описанных выше пункта (i) и (ii) могут быть отражены в формуле расчета.
В одном из вариантов осуществления настоящего изобретения величина pI может быть повышена, например, по меньшей мере на 0,01, 0,03, 0,05, 0,1, 0,2, 0,3, 0,4, 0,5 или более, по меньшей мере на 0,6, 0,7, 0,8, 0,9, или более, по меньшей мере на 1,0, 1,1, 1,2, 1,3, 1,4, 1,5, или более, или по меньшей мере на 1,6, 1,7, 1,8, 1,9, 2,0, 2,1, 2,2, 2,3, 2,4, 2,5, 3,0 или более, по сравнению с тем, что было до модификации.
В некоторых вариантах осуществления настоящего изобретения аминокислота для увеличения pI может быть экспонирована на поверхности вариантной области Fc. В настоящем изобретении аминокислота, которая может экспонироваться на поверхности, обычно относится к аминокислотному остатку, расположенному на поверхности полипептида, составляющего вариант области Fc. Аминокислотный остаток, расположенный на поверхности полипептида, относится к аминокислотному остатку, боковая цепь которого может контактировать с молекулами растворителя (которые, как правило, представляют собой молекулы воды). Однако боковая цепь не обязательно должна полностью контактировать с молекулами растворителя, и когда даже часть боковой цепи находится в контакте с молекулами растворителя, аминокислоту определяют как «аминокислотный остаток, расположенный на поверхности». Аминокислотные остатки, расположенные на поверхности полипептида, также включают аминокислотные остатки, расположенные близко к поверхности, и, таким образом, могут подвергаться влиянию электрического заряда от другого аминокислотного остатка, боковая цепь которого, даже частично, находится в контакте с молекулами растворителя. Специалисты в данной области могут создать гомологичную модель полипептида, например, с использованием коммерчески доступного программного обеспечения. В другом варианте можно использовать методы, известные специалистам в данной области, такие как рентгенокристаллография. Аминокислотные остатки, которые могут быть обнаружены на поверхности, определяют, например, с использованием координат трехмерной модели и компьютерной программы, такой как программа Insightll (фирма Accelrys). Открытые участки могут быть определены с использованием алгоритмов, известных в этой области (например, Lee и Richards (J. Mol. Biol. 55, 1971, 379-400); Connolly (J. Appl. Cryst. 16, 1983, 548-558)). Участки на поверхности, подверженные воздействию, можно определить с помощью программного обеспечения по моделированию белков и получению информации о трехмерной структуре. Программное обеспечение, доступное для таких целей, включает, например, программное обеспечение SYBYL Biopolymer Module (фирма Tripos Associates). Когда алгоритм требует вводимого пользователем параметра размера, «размер» зонда, который используют в вычислении, может быть установлен примерно на 1,4 Ангстрема  или меньше по радиусу. Кроме того, Pacios описал методы определения открытых участков поверхности с использованием программного обеспечения для персональных компьютеров (Comput. Chem. 18(4), 1994, 377-386; J. Mol. Model. 1, 1995, 46-53). На основании такой информации, как описано выше, могут быть выбраны соответствующие аминокислотные остатки, расположенные на поверхности полипептида, который составляет вариант области Fc.
или меньше по радиусу. Кроме того, Pacios описал методы определения открытых участков поверхности с использованием программного обеспечения для персональных компьютеров (Comput. Chem. 18(4), 1994, 377-386; J. Mol. Model. 1, 1995, 46-53). На основании такой информации, как описано выше, могут быть выбраны соответствующие аминокислотные остатки, расположенные на поверхности полипептида, который составляет вариант области Fc.
Способы увеличения pI путем введения одной замены аминокислоты в константной области антитела особенно не ограничиваются и, например, могут быть реализованы на основе методов, описанных в WO 2014/145159. В качестве аминокислотных замен для введения в константную область предпочтительно вводить аминокислотные замены (замену) для уменьшения количества аминокислот, имеющих отрицательный заряд (например, аспарагиновой кислоты или глутаминовой кислоты), при увеличении количества аминокислот, имеющих положительный заряд (например,, аргинин или лизин), как и в случае вариабельной области.
Хотя нет особенных ограничений, в качестве сайтов для введения аминокислотных замен в константную область предпочтительны такие сайты, в которых боковые цепи аминокислот могут быть экспонированы на поверхности молекулы антитела. Предпочтительные примеры включают способы введения множества аминокислотных замен в комбинации в те сайты, которые могут экспонироваться на поверхности молекулы антитела. В другом варианте предпочтительно, чтобы множество производимых аминокислотных замен находилось в сайтах, проксимальных друг к другу в трехмерной структуре. Множественные аминокислотные замены, которые вводят, предпочтительно являются положительно заряженными аминокислотами, которые могут в некоторых случаях привести к состоянию, когда множественные положительные заряды переносятся на проксимальные участки трехмерной структуры, хотя они этим не ограничиваются. Определение «проксимальные сайты в трехмерной структуре» в настоящем изобретении строго не ограничивают, но, например, это может означать состояние, при котором замещение одной аминокислоты или замещение нескольких аминокислот вводят на расстоянии друг от друга 45 ангстрем или меньше, 40 ангстрем или меньше, 30 Ангстрем или меньше, 20 Ангстрем или меньше, лучше 15 Ангстрем или меньше, или более предпочтительно 10 Ангстрем или меньше. Установить, находятся ли представляющие интерес аминокислотные замены в сайтах, экспонированных на поверхности молекулы антитела, или множественные сайты аминокислотных замен расположены проксимально, может известными методами, такими как рентгенокристаллография.
В одном из вариантов осуществления настоящего изобретения варианты областей Fc с повышенной pI содержат по меньшей мере одно положение, выбранное из группы, включающей положения 285, 311, 312, 315, 318, 333, 335, 337, 341, 342, 343, 384, 385, 388, 390, 399, 400, 401, 402, 413, 420, 422 и 431, нумерация EU. В других вариантах осуществления настоящего изобретения варианты областей Fc с повышенной pI содержат Arg или Lys в каждом выбранном положении.
В других вариантах осуществления настоящего изобретения варианты областей Fc с повышенной pI содержат по меньшей мере одно аминокислотное изменение по меньшей мере в одном положении, выбранном из группы, включающей положения 311, 343 и 413 по нумерации EU. В других вариантах осуществления настоящего изобретения варианты областей Fc с повышенной pI содержат аминокислотное изменение в положениях 311, 343 или 413 по нумерации EU. В других вариантах осуществления настоящего изобретения варианты областей Fc с повышенной pI содержат Arg или Lys в каждой выбранной позиции.
В другом варианте осуществления настоящего изобретения предусматривают полипептиды, содержащие вариант области Fc с повышенной pI, который включает аминокислотные изменения по любому из следующих пунктов (1) - (3): (1) в позициях 311 и 343; (2) в позициях 311 и 413; (3) в позициях 343 и 413 по нумерации EU. В других вариантах осуществления настоящего изобретения варианты областей Fc с повышенной pI содержат Arg или Lys в каждой выбранной позиции.
Метод увеличения pI белка представляет, например, уменьшение количества аминокислот с отрицательно заряженной боковой цепью (например, аспарагиновой кислоты и глутаминовой кислоты) и/или увеличение количества аминокислот с положительно заряженной стороной боковой цепью (например, аргинина, лизина и гистидина) при нейтральном рН. Аминокислоты с отрицательно заряженной боковой цепью имеют отрицательный заряд, представленный как -1, в условиях рН, которые значительно выше, чем рКа их боковой цепи, что является теорией, хорошо известной специалистам в данной области. Например, теоретическое значение pKa для боковой цепи аспарагиновой кислоты составляет 3,9, а боковая цепь имеет отрицательный заряд, представленный как -1 в условиях нейтрального рН (например, в растворе с рН 7,0). И наоборот, аминокислоты с положительно заряженной боковой цепью имеют положительный заряд, представленный как +1, при условии, что рН существенно ниже, чем рКа их боковой цепи. Например, теоретическое значение рКа для боковой цепи аргинина составляет 12,5, а боковая цепь имеет положительный заряд, представленный как +1 в условиях нейтрального рН (например, в растворе с рН 7,0). Между тем известно, что аминокислоты, боковая цепь которых не имеет заряда при нейтральном рН (например, в растворе с рН 7,0), включают 15 типов природных аминокислот, то есть аланин, цистеин, фенилаланин, глицин, изолейцин, лейцин, метионин, аспарагин, пролин, глутамин, серии, треонин, валин, триптофан и тирозин. Авторы отмечают, что аминокислоты для увеличения pI могут быть неприродными аминокислотами.
Исходя из вышеизложенного, способ увеличения pI белка в условиях нейтрального рН (например, в растворе с рН 7,0) может придавать изменение заряда +1 белку интереса, например, путем замены аминокислот на незаряженные боковые цепи аспарагиновой кислоты или глутаминовой кислоты (боковая цепь которых имеет отрицательный заряд -1) в аминокислотной последовательности белка. Кроме того, изменение заряда +1 может быть придано белку, например, заменой аргинина или лизина (боковая цепь которого имеет положительный заряд +1) на аминокислоты, боковая цепь которых не имеет заряда. Более того, изменение заряда +2 может быть придано белку за один раз путем замены аргинина или лизина (чья боковая цепь имеет положительный заряд +1) на аспарагиновую кислоту или глутаминовую кислоту (чья боковая цепь имеет отрицательный заряд - 1). В другом варианте для увеличения pI белка, аминокислоты с боковой цепью, не имеющей заряда, и/или аминокислоты с положительно заряженной боковой цепью могут быть добавлены в аминокислотную последовательность белка, или аминокислоты с боковой цепью, не имеющей положительного заряда, и/или предпочтительно аминокислоты с отрицательно заряженной боковой цепью, присутствующая в аминокислотной последовательности белка, могут быть делетированы. Понятно, что, например, N-концевые и С-концевые аминокислотные остатки белка имеют заряд, производный от основной цепи (NH3+ аминогруппы на N-конце и СОО- карбонильной группы на С-конце) в дополнение к зарядам от их боковых цепей. Таким образом, pI белка также может быть увеличена путем выполнения над функциональными группами основной цепи некоторого добавления, делеции, замены или инсерции.
Замена аминокислоты для увеличения pI включает, например, замену аминокислоты, боковая цепь которой не имеет заряда, на аминокислоту с отрицательно заряженной боковой цепью, замену аминокислоты, имеющей положительно заряженную боковую цепь, на аминокислоту, боковая цепь которой не имеет заряда, и замену аминокислоты, имеющей положительно заряженную боковую цепь, на аминокислоту с отрицательно заряженной боковой цепью в аминокислотной последовательности родительской области Fc, которые выполняют отдельно или в соответствующих комбинациях.
Инсерция или добавление аминокислоты для увеличения pI включает, например, инсерцию или добавление аминокислоты, боковая цепь которой не имеет заряда, и/или инсерцию или добавление аминокислоты, имеющей положительно заряженную боковую цепь в аминокислотной последовательности родительской области Fc, которые выполняются отдельно или в соответствующих комбинациях.
Делеция аминокислоты для увеличения pI включает, например, делецию аминокислоты, боковая цепь которой не имеет заряда, и/или делецию аминокислоты, имеющей отрицательно заряженную боковую цепь, в аминокислотной последовательности родительской области Fc, которые выполняются по отдельности или в соответствующих комбинациях.
В одном из вариантов осуществления настоящего изобретения природные аминокислоты, используемые для увеличения pI, можно классифицировать следующим образом: (а) аминокислотой с отрицательно заряженной боковой цепью может быть Glu (Е) или Asp (D); (б) аминокислотой, боковая цепь которой не имеет заряда, может быть Ala (A), Asn (N), Cys (С), Gin (Q), Gly (G), His (H), Ile (I), Leu (L), Met (M), Phe (F), Pro (P), Ser (S), Thr (T), Trp (W), Tyr (Y) или Val (V); (в) аминокислотой с положительно заряженной боковой цепью может быть His (Н), Lys (K) или Arg (R). В одном из вариантов осуществления настоящего изобретения аминокислотной инсерцией или заменой после модификации является Lys (K) или Arg (R).
В другом варианте осуществления настоящего изобретения предусматривают выделенные агонистические антигенсвязывающие молекулы или антитела, содержащие последовательности варианта области Fc с повышенной связывающей активностью с рецептором Fc-гамма (предпочтительно, Fc gamma RIIb-связывающей активностью) и повышенной изоэлектрической точкой pI. В некоторых вариантах осуществления настоящего изобретения вариант области Fc по настоящему изобретению содержит по меньшей мере два аминокислотных изменения в исходной области Fc. Выше описано, что антигенсвязывающая молекула или антитело, имеющее повышенную pI, более сильно притягивается физико-химическим кулоновским взаимодействием к поверхности эндотелиальной клетки, которая имеет общий отрицательный заряд, по сравнению с антигенсвязывающей молекулой или антителом, не имеющими повышенной pI. Следовательно, для агонистических антигенсвязывающих молекул или антител, которые проявляют агонистическую активность на основе влияния связывающей активности в отношении рецептора Fc-гамма (предпочтительно Fc-gamma RIIb), можно увеличить агонистическую активность антигенсвязывающих молекул или антител путем комбинирования аминокислотных изменений для увеличения рецептора Fc-гамма (предпочтительно Fc-gamma RIIb) и аминокислотных изменений (изменения) для увеличения pI. Определение термина «рецептор Fc-гамма» приводят в разделе «II. Композиции и методы (агонистические антигенсвязывающие молекулы против CD137)» применяют сходным образом.
В одном из вариантов осуществления настоящего изобретения предусматривают полипептиды, содержащие варианты области Fc с повышенной FcγRIIb-связывающей активностью и увеличенной pI, содержащие по меньшей мере три аминокислотных изменения, включающие: (а) по меньшей мере одно аминокислотное изменение по меньшей мере в одном положении, выбранном из группы, состоящей из: 231, 232, 233, 234, 235, 236, 237, 238, 239, 264, 266, 267, 268, 271, 295, 298, 325, 326, 327, 328, 330, 331, 332, 334 и 396, EU нумерация, и (б) по меньшей мере два аминокислотных изменения по меньшей мере в двух положениях, выбранных из группы, состоящей из: 285, 311, 312, 315, 318, 333, 335, 337, 341, 342, 343, 384, 385, 388, 390, 399, 400, 401, 402, 413, 420, 422 и 431, EU нумерация.
В одном из вариантов осуществления настоящего изобретения предусматривают полипептид, содержащий вариант области Fc, который приводит к повышенной FcγRIIb-связывающей активности и увеличению pI, причем полипептид содержит по меньшей мере 3 аминокислотных изменения, включающих (а) по меньшей мере одно аминокислотное изменение по меньшей мере в одном положении, выбранном из группы, состоящей из положений 234, 235, 236, 237, 238, 264, 268, 295, 326 и 330 по нумерации EU, и (б) по меньшей мере 2 аминокислотные мутации по меньшей мере в 2 положениях, выбранных из группы, состоящей из положений 311, 343 и 413 по нумерации EU.
В другом варианте осуществления настоящего изобретения предусматривают полипептид, содержащий вариант области Fc, который приводит к повышенной FcγRIIb-связывающей активности и увеличению pI, включающий по меньшей мере одно аминокислотное изменение в любом из положений от (1) до (26) (по нумерации EU):
(1) положения 235, 236, 268, 295, 326, 330, 343 и 413;
(2) положения 214, 235, 236, 268, 295, 326, 330, 343 и 413;
(3) положения 234, 238, 250, 264, 307, 330, 343 и 413;
(4) положения 234, 238, 264, 330, 343 и 413;
(5) положения 234, 237, 238, 250, 307, 330, 343 и 413;
(6) положения 234, 237, 238, 330, 343 и 413;
(7) положения 235, 236,268, 295, 326, 330, 311 и 343;
(8) положения 234, 238, 250, 264, 307, 330, 311 и 343;
(9) положения 234, 238, 264, 330, 311 и 343;
(10) положения 234, 237, 238, 250, 307, 330, 311 и 343;
(11) положения 234, 237, 238, 330, 311 и 343;
(12) положения 235, 236, 268, 295, 326, 330 и 343;
(13) положения 214, 235, 236, 268, 295, 326, 330 и 343;
(14) положения 235, 236, 268, 295, 326, 330 и 413;
(15) положения 214, 236, 268, 330 и 343;
(16) положения 214, 235, 236, 268, 330 и 343;
(17) положения 214, 236, 268, 330 и 413;
(18) положения 214, 236, 268, 330, 343 и 413;
(19) положения 214, 235, 236, 268, 330, 343 и 413;
(20) положения 214, 236, 268, 330 и 311;
(21) положения 214, 235, 236, 268, 330 и 311;
(22) положения 214, 236, 268, 330, 311 и 343;
(23) положения 214, 235, 236, 268, 330, 311 и 343;
(24) положения 214, 236, 268, 330, 311 и 413;
(25) положения 214, 235, 236, 268, 330, 311 и 413;
(26) положения 214, 235, 236, 268, 295, 326, 330 и 311.
В одном из вариантов осуществления настоящего изобретения вариант области Fc по настоящему изобретению включает аминокислотные изменения, представленные ниже в табл. 6.
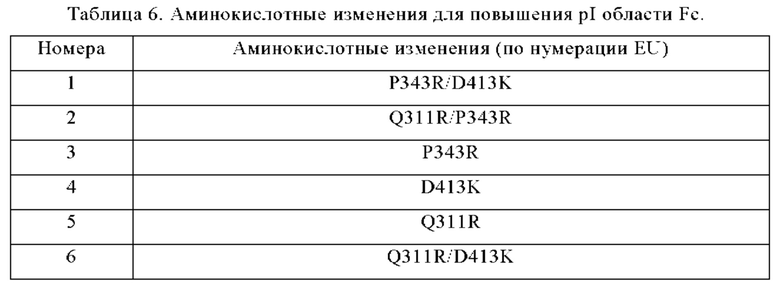
В одном из вариантов осуществления настоящего изобретения в дополнение к аминокислотным изменениям, описанным в табл. 6 (аминокислотные изменения, которые увеличивают pI Fc), вариантная область Fc по настоящему изобретению включает аминокислотные изменения, описанные в табл. 7 ниже.

В одном из вариантов осуществления настоящего изобретения вариант области Fc по настоящему изобретению включает аминокислотные изменения, описанные в табл. 8 ниже. В предпочтительном варианте осуществления настоящего изобретения вариант области Fc по настоящему изобретению включает аминокислотную делецию в положении 447 по нумерации EU помимо аминокислотных изменений, описанных ниже в табл. 8. В другом предпочтительном варианте осуществления настоящего изобретения вариант области Fc по настоящему изобретению включает аминокислотные делеции в положениях 446 и 447 по нумерации EU помимо аминокислотных изменений, описанных ниже в табл. 8.

Помимо изменений, приведенных в качестве примеров выше, специалисты в данной области поймут, что по крайней мере одно аминокислотное изменение, которое увеличивает активность связывания с FcγR, включая FcγRIIb, по сравнению с исходной областью Fc, например, как описано или предложено в WO 2013/047752, WO 2013/125667, WO 2014/030728, WO 2014/163101 или WO 2017104783, можно использовать по меньшей мере одно изменение аминокислоты, которое увеличивает pI по сравнению с исходной областью Fc, например, как описано или предложено в WO 2017/104783 или WO 2017/046994, и комбинации этих аминокислотных изменений.
В одном из вариантов осуществления настоящего изобретения агонистическая антигенсвязывающая молекула или антитело содержит как вариантную область Fc, так и антигенсвязывающий домен. В другом варианте осуществления настоящего изобретения антиген является мембранным антигеном. В другом варианте осуществления настоящего изобретения антиген представляет собой рецептор, экспрессируемый на поверхности клетки. В некоторых вариантах осуществления настоящего изобретения родительская область Fc происходит от IgG1 человека. В другом варианте осуществления настоящего изобретения полипептид представляет собой антитело. В другом варианте осуществления настоящего изобретения полипептид представляет собой слитый белок Fc.
В одном из вариантов осуществления настоящего изобретения предусматривают способ получения агонистической антигенсвязывающей молекулы, которая включает подробно описанный выше вариант области Fc (вариант области Fc с увеличенной изоэлектрической точкой (pI)).
В одном из вариантов осуществления настоящего изобретения способ по настоящему изобретению имеет следующие особенности: введение в исходную область Fc по меньшей мере одного аминокислотного изменения (как описано выше), которое приводит к повышению изоэлектрической точки (pI) по сравнению с изоэлектрической точкой (pI) родительской агонистической антигенсвязывающей молекулы, содержащей исходную область Fc; идентификация и выделение агонистической антигенсвязывающей молекулы, в которой агонистическая активность агонистической антигенсвязывающей молекулы, включающей вариант области Fc, полученный в результате вышеуказанной интродукции, повышена по сравнению с таковой у исходной агонистической антигенсвязывающей молекулы.
В одном из вариантов осуществления настоящего изобретения способ по настоящему изобретению имеет особенности получения вектора экспрессии, содержащего подходящий промотор, функционально связанный с геном, кодирующим вышеупомянутую агонистическую антигенсвязывающую молекулу, идентифицированную и выделенную, как указано выше, предусматривает введение вектора в клетку-хозяин, культивирование клеток-хозяев для выработки агонистической антигенсвязывающей молекулы и выделения вышеупомянутой агонистической антигенсвязывающей молекулы из культуры клеток-хозяев.
В одном из вариантов осуществления настоящего изобретения способ по настоящему изобретению позволяет получать агонистическую антигенсвязывающую молекулу, агонистическая активность которой в отношении антигена в присутствии 10 мкМ, 50 мкМ, 100 мкМ, 150 мкМ, 200 мкМ или 250 мкМ низкомолекулярного соединения выше в 2 раза или более, в 3 раза или более, в 5 раз или более, в 10 раз или более, в 20 раз или более, в 30 раз или более, в 50 раз или более, в 60 раз или более, в 70 раз или более, в 80 раз или более или в 90 раз или более по сравнению с агонистической активностью по отношению к антигену в отсутствие низкомолекулярного соединения.
В одном из вариантов осуществления настоящего изобретения способом по настоящему изобретению можно получить агонистическую антигенсвязывающую молекулу, агонистическая активность которой по отношению к антигену в присутствии 10 мкМ или более низкомолекулярного соединения выше в 2 раза или более, в 3 раза или более, в 5 раз или более, в 10 раз или более, в 20 раз или более, в 30 раз или более, в 50 раз или более, в 60 раз или более, в 70 раз или более, в 80 раз или более или в 90 раз или более по сравнению с агонистической активностью по отношению к антигену в отсутствие низкомолекулярного соединения.
В одном из вариантов осуществления настоящего изобретения способом по настоящему изобретению можно получить агонистическую антигенсвязывающую молекулу, агонистическая активность которой по отношению к антигену в присутствии 50 мкМ или более низкомолекулярного соединения выше в 2 раза или более, в 3 раза или более, в 5 раз или более, в 10 раз или более, в 20 раз или более, в 30 раз или более, в 50 раз или более, в 60 раз или более, в 70 раз или более, в 80 раз или более или в 90 раз или более по сравнению с агонистической активностью по отношению к антигену в отсутствие низкомолекулярного соединения.
В одном из вариантов осуществления настоящего изобретения способом по настоящему изобретению можно получить агонистическую антигенсвязывающую молекулу, агонистическая активность которой по отношению к антигену в присутствии 250 мкМ или более низкомолекулярного соединения выше в 2 раза или более, в 3 раза или более, в 5 раз или более, в 10 раз или более, в 20 раз или более, в 30 раз или более, в 50 раз или более, в 60 раз или более, в 70 раз или более, в 80 раз или более или в 90 раз или более по сравнению с агонистической активностью по отношению к антигену в отсутствие низкомолекулярного соединения.
В одном из вариантов осуществления настоящего изобретения в таком способе агонистическая активность агонистической антигенсвязывающей молекулы по отношению к антигену может быть оценена по количеству IL-2 и/или IFN-γ, вырабатываемых антиген-экспрессирующими клетками.
В одном из вариантов осуществления настоящего изобретения в таком способе агонистическая активность агонистической антигенсвязывающей молекулы по отношению к антигену может быть оценена по количеству IL-2 и/или IFN-γ, вырабатываемых выделенными мононуклеарными клетками периферической крови человека (МКПК) или Т-клетками, производными от мононуклеарных клеток периферической крови человека (МКПК).
В одном из вариантов осуществления настоящего изобретения в таком способе агонистическая активность агонистической антигенсвязывающей молекулы по отношению к антигену может быть оценена путем анализа репортерного гена.
IV. Композиции и способы (антигенсвязывающие молекулы, связывающая активность которых с антигенами изменяется в зависимости от концентрации низкомолекулярного соединения)
А. Антигенсвязывающие молекулы, связывающая активность которых с антигенами изменяется в зависимости от концентрации низкомолекулярного соединения
В одном из вариантов осуществления настоящего изобретения предусматривают антигенсвязывающие молекулам, антигенсвязывающая активность которых изменяется в зависимости от концентрации низкомолекулярного соединения. Молекулы также могут называться зависимыми от низкомолекулярного соединения антигенсвязывающими молекулами (обладающими антигенсвязывающей активностью, зависящей от концентрации низкомолекулярного соединения). В некоторых вариантах осуществления настоящего изобретения антигенсвязывающие молекулы представляют собой антитела. В некоторых вариантах осуществления настоящего изобретения антигенсвязывающие молекулы по настоящему изобретению обладают антигенсвязывающей активностью, которая увеличивается по мере того, как концентрация низкомолекулярного соединения становится выше. В некоторых вариантах осуществления настоящего изобретения антигенсвязывающие молекулы по настоящему изобретению обладают антигенсвязывающей активностью, которая снижается по мере того, как концентрация низкомолекулярного соединения становится выше. В одном из вариантов осуществления настоящего изобретения когда связывающую активность антигенсвязывающей молекулы в присутствии низкомолекулярного соединения сравнивают со связывающей активностью антигенсвязывающей молекулы в отсутствие низкомолекулярного соединения, одно из двух значений выше, чем другое значение. В другом варианте осуществления настоящего изобретения когда связывающая активность антигенсвязывающей молекулы в присутствии высокой концентрации низкомолекулярного соединения сравнивают со связывающей активностью антигенсвязывающей молекулы в присутствии низкой концентрации низкомолекулярного соединения, одно из двух значений выше чем другое значение. В некоторых вариантах осуществления настоящего изобретения низкомолекулярное соединение в настоящем изобретении представляет собой целевое тканеспецифичное соединение. В других вариантах осуществления настоящего изобретения низкомолекулярное соединение в настоящем изобретении представляет собой соединение, специфичное для опухолевой ткани.
В другом варианте осуществления настоящего изобретения предусматривают антигенсвязывающие молекулы, обладающие высокой способностью удерживания в плазме. В другом варианте осуществления настоящего изобретения предусматривают антигенсвязывающие молекулы, обладающие антигенсвязывающей активностью, которая увеличивается по мере увеличения концентрации низкомолекулярного соединения. В некоторых вариантах осуществления настоящего изобретения низкомолекулярное соединение представляет собой тканеспецифичное соединение-мишень. В других вариантах осуществления настоящего изобретения антигенсвязывающие молекулы обладают повышенной антигенсвязывающей активностью в ткани-мишени по сравнению с антигенсвязывающей активностью в ткани, не являющейся мишенью. В одном из вариантов осуществления настоящего изобретения антигенсвязывающие молекулы представляют собой антитела. Не ограничиваясь какой-либо конкретной теорией, описанное выше изменение кинетики плазмы можно объяснить следующим образом. Когда антигенсвязывающая активность антигенсвязывающей молекулы высока в зависимости от концентрации тканеспецифичного соединения-мишени, антигенсвязывающая активность этой антигенсвязывающей молекулы в тканях, отличных от ткани-мишени, будет ниже. В результате антиген-зависимая элиминация (клиренс) этой антигенсвязывающей молекулы в тканях, отличных от ткани-мишени, также станет ниже. Более низкая антиген-зависимая элиминация (клиренс) в большинстве тканей живого организма (то есть в тканях, отличных от ткани-мишени) в целом приводит к повышенной способности удерживать антигенсвязывающую молекулу в плазме. Определение того, обладает ли антигенсвязывающая молекула по настоящему изобретению высокой способностью удерживания в плазме, может быть выполнено путем сравнения с контрольной антигенсвязывающей молекулой. В некоторых вариантах осуществления настоящего изобретения антигенсвязывающие молекулы, которые обладают повышенной антигенсвязывающей активностью, когда концентрация целевого тканеспецифичного соединения становится выше, имеют более высокую способность удерживания в плазме по сравнению с контрольной антигенсвязывающей молекулой. В одном из вариантов осуществления настоящего изобретения контрольная антигенсвязывающая молекула не обладает антигенсвязывающей активностью, зависящей от концентрации низкомолекулярного соединения. В некоторых вариантах осуществления настоящего изобретения антигенсвязывающая молекула, которая не обладает антигенсвязывающей активностью, зависящей от концентрации низкомолекулярного соединения, означает антигенсвязывающую молекулу, для которой разница в антигенсвязывающей активности в присутствии низкомолекулярного соединения и в отсутствие низкомолекулярного соединения отличается в 2 раза или меньше, в 1,8 раза или меньше, в 1,5 раза или меньше, в 1,3 раза или меньше, в 1,2 раза или меньше, или в 1,1 раза или меньше. Для сравнения предпочтительно, чтобы антиген связывающая активность антигенсвязывающих молекул по настоящему изобретению и контрольных антигенсвязывающих молекул в присутствии достаточного количества низкомолекулярного соединения была по существу одинаковой.
Авторы настоящего изобретения полагают, что величина антиген-зависимой элиминации антигенсвязывающей молекулы, обнаруживаемой in vivo, изменяется в соответствии с количественным балансом антигенов и антигенсвязывающих молекул, присутствующих в плазме. В целом полагают, что чем больше антигенов присутствует в плазме / чем меньше антигенсвязывающих молекул присутствует в плазме, тем более выявляемой становится антиген-зависимая элиминация антигенсвязывающей молекулы. Напротив, считают, что чем меньше антигенов присутствует в плазме / чем больше антигенсвязывающих молекул присутствует в плазме, тем менее выявляемой становится антиген-зависимая элиминация антигенсвязывающей молекулы. Антигенсвязывающие молекулы по настоящему изобретению не обязательно должны проявлять высокую способность удерживания в плазме при любых условиях, но достаточно, чтобы антигенсвязывающие молекулы проявляли высокую способность удерживания в плазме при подходящих условиях, когда обнаруживают достаточную антиген-зависимую элиминацию. Если количество антигена в плазме слишком мало, способность удерживания в плазме может быть оценена после увеличения количества антигена любыми искусственными методами.
В другом варианте осуществления настоящее изобретение предусматривает антигенсвязывающие молекулы, имеющие низкую способность накапливать антиген в плазме. В другом варианте осуществления настоящего изобретения антигенсвязывающие молекулы обладают антигенсвязывающей активностью, которая увеличивается по мере того, как концентрация низкомолекулярного соединения становится выше. В некоторых вариантах осуществления настоящего изобретения низкомолекулярное соединение представляет собой целевое тканеспецифичное соединение. В других вариантах осуществления настоящего изобретения антигенсвязывающие молекулы обладают повышенной антигенсвязывающей активностью в ткани-мишени по сравнению с антигенсвязывающей активностью в ткани, не являющейся мишенью. В одном из вариантов осуществления настоящего изобретения антигенсвязывающие молекулы представляют собой антитела. Не ограничиваясь какой-либо конкретной теорией, описанное выше изменение кинетики плазмы можно понять следующим образом. Когда антигенсвязывающая активность антигенсвязывающей молекулы высока в зависимости от концентрации тканеспецифичного соединения-мишени, антигенсвязывающая активность этой антигенсвязывающей молекулы в тканях, отличных от ткани-мишени, будет ниже. В результате эта антигенсвязывающая молекула будет иметь пониженную способность образовывать комплекс антиген-антитело в тканях, отличных от ткани-мишени. В целом известно, что когда антиген связывается антигенсвязывающей молекулой, такой как антитело, клиренс антигена становится ниже, а концентрация антигена в плазме увеличивается (антиген накапливается). Пониженная способность образовывать комплекс антиген-антитело в большинстве тканей живого организма (то есть в тканях, отличных от ткани-мишени) в целом приводит к пониженному накоплению антигена (другими словами, к пониженной способности антигенсвязывающей молекулы к накоплению антигена). Определение того, обладает ли антигенсвязывающая молекула в настоящем изобретении пониженной способностью к накоплению антигена в плазме, может быть выполнено путем сравнения с контрольной антигенсвязывающей молекулой. В некоторых вариантах осуществления настоящего изобретения антигенсвязывающие молекулы, которые обладают повышенной антигенсвязывающей активностью, когда концентрация целевого тканеспецифичного соединения становится выше, имеют пониженную способность к накоплению антигена в плазме по сравнению с контрольной антигенсвязывающей молекулой. В одном из вариантов осуществления настоящего изобретения контрольная антигенсвязывающая молекула не обладает антигенсвязывающей активностью, зависящей от концентрации низкомолекулярного соединения. В некоторых вариантах осуществления настоящего изобретения антигенсвязывающая молекула, которая не обладает антигенсвязывающей активностью, зависящей от концентрации низкомолекулярного соединения, означает антигенсвязывающую молекулу, для которой разница в антигенсвязывающей активности в присутствии низкомолекулярного соединения и в отсутствие низкомолекулярного соединения отличается, например, в 2 раза или меньше, в 1,8 раз или меньше, в 1,5 раз или меньше, в 1,3 раза или меньше, в 1,2 раза или меньше, или в 1,1 раза или меньше. При сравнении предпочтительно, чтобы антигенсвязывающая активность антигенсвязывающих молекул по настоящему изобретению и контрольных антигенсвязывающих молекул в присутствии достаточного количества соединения были по существу одинаковыми.
В настоящем изобретении авторы отмечают, что количество комплекса антиген-антитело, образованного in vivo, зависит от количества антигена и антигенсвязывающей молекулы, присутствующих в плазме. Обычно считают, что чем больше антигенов / антител присутствует в плазме, тем больше образуется комплексов антиген-антитело. И наоборот, считают, что чем меньше антигенов / антител присутствует в плазме, тем меньше образуется комплексов антиген-антитело. Антигенсвязывающие молекулы по настоящему изобретению не обязательно должны демонстрировать низкую способность к накоплению антигена в плазме при любых условиях, но достаточно, чтобы антигенсвязывающие молекулы проявляли низкую способность к накоплению антигена в плазме при подходящих условиях, когда формируется достаточное количество комплекса антиген-антитело. Если количество антигена в плазме слишком мало, способность к накоплению антигена в плазме можно оценить после увеличения количества антигена любыми искусственными методами.
В некоторых вариантах осуществления настоящего изобретения разница в антигенсвязывающей активности, зависящая от концентрации низкомолекулярного соединения антигенсвязывающих молекул по настоящему изобретению, больше например, в два раза или более, в три раза или более, в пять раз или более, в десять раз или более, в 20 раз или более, в 30 раз или более, в 50 раз или более, в 100 раз или более, в 200 раз или более, в 300 раз или более, в 500 раз или более, в 1×103 раз или более, в 2×103 раз или более, в 3×103 раз или более, в 5×103 раз или более, в 1×104 раз или более, в 2×104 раз или более, в 3×104 раз или более, в 5×104 раз или более или в 1×105 раз или более.
В некоторых вариантах осуществления настоящего изобретения связывающая активность антигенсвязывающих молекул может быть выражена значением KD (константа диссоциации). В других вариантах осуществления настоящего изобретения когда сравнивают значение KD для антигенсвязывающей молекулы в присутствии низкомолекулярного соединения и значение KD для антигенсвязывающей молекулы в отсутствие низкомолекулярного соединения, одно значение меньше, чем другое значение. В другом варианте осуществления настоящего изобретения если значение KD антигенсвязывающей молекулы в присутствии высокой концентрации низкомолекулярного соединения сравнивают со значением KD антигенсвязывающей молекулы в присутствии низкой концентрации низкомолекулярного соединения, одно значение меньше чем другое значение. В других вариантах осуществления настоящего изобретения разница в значениях KD антигенсвязывающих молекул больше, например, в два раза или более, в три раза или более, в пять раз или более, в 10 раз или более, в 20 раз или более, в 30 раз или более, в 50 или более, в 100 раз или более, в раз или более, в 300 раз или более, в 500 раз или более, в 1×103 раз или более, в 2×103 раз или более, в 3×103 раз или более, в 5×103 раз или более, в 1×104 раз или более, в 2×104 раз или более, в 3×104 раз или более, в 5×104 раз или более или в 1×105 раз или более. Величина KD может быть меньше, например, 9×10-7 М или менее, 8×10-7 М или менее, 7×10-7 М или менее, 6×10-7 М или менее, 5×10-7 М или менее, 4×10-7 М или менее, 3×10-7 М или менее, 2×10-7 М или менее, 1×10-7 М или менее, 9×10-8 М или менее, 8×10-8 М или менее, 7×10-8 М или менее, 6×10-8 М или менее, 5×10-8 М или менее, 4×10-8 М или менее, 3×10-8 М или менее, 2×10-8 М или менее, 1×10-8 М или менее, 9×10-9 М или менее, 8×10-9 М или менее, 7×10-9 М или менее, 6×10-9 М или менее, 5×10-9 М или менее, 4×10-9 М или менее, 3×10-9 М или менее, 2×10-9 М или менее, 1×10-9 М или менее, 9×10-10 М или менее, 8×10-10 М или менее, 7×10-10 М или менее, 6×10-10 М или менее, 5×10-10 М или менее, 4×10-10 М или менее, 3×10-10 М или менее, 2×10-10 М или менее, 1×10-10 М или менее. Величина KD может быть больше, например, 1×10-8 М или более, 2×10-8 М или более, 3×10-8 М или более, 4×10-8 М или более, 5×10-8 М или более, 6×10-8 М или более, 7×10-8 М или более, 8×10-8 М или более, 9×10-8 М или более, 1×10-7 М или более, 2×10-7 М или более, 3×10-7 М или более, 4×10-7 М или более, 5×10-7 М или более, 6×10-7 М или более, 7×10-7 М или более, 8×10-7 М или более, 9×10-7 М или более, 1×10-6 М или более, 2×10-6 М или более, 3×10-6 М или более, 4×10-6 М или более, 5×10-6 М или более, 6×10-6 М или более, 7×10-6 М или более, 8×10-6 М или более, 9×10-6 М или более.
В другом варианте осуществления настоящего изобретения связывающая активность антигенсвязывающих молекул может быть выражена значением kd (константа скорости диссоциации) вместо значения KD.
В другом варианте осуществления настоящего изобретения связывающая активность антигенсвязывающих молекул может быть выражена количеством антигена, который связался с антигенсвязывающей молекулой. Например, методами поверхностного плазмонного резонанса связываемое количество антигенсвязывающей молекулы, иммобилизованной на сенсорном чипе, и связываемое количество антигена, которое связывается с ним, измеряют как резонансную единицу (RU). Антигенсвязывающая активность может быть выражена с использованием связываемого количества антигена, полученного из него, в качестве индикатора, или, альтернативно, может быть выражена с использованием значения, полученного путем деления количества связанного антигена на количество связанной антигенсвязывающей молекулы (т.е., количество связанного антигена на единицу количества антигенсвязывающей молекулы) в качестве индикатора. Конкретные методы измерения и расчета количества связывания описаны ниже в примерах. В некоторых вариантах осуществления настоящего изобретения если количество связанного антигена в присутствии низкомолекулярного соединения сравнивается с количеством связанного антигена в отсутствие низкомолекулярного соединения, одно из значений больше, чем другое. В другом варианте осуществления настоящего изобретения если количество связанного антигена в присутствии высокой концентрации низкомолекулярного соединения сравнивают с количеством связанного антигена в присутствии низкой концентрации низкомолекулярного соединения, либо одно значение больше, чем другое значение. В других вариантах осуществления настоящего изобретения разница в связываемое количестве антигена больше, например, в два раза или более, в три раза или более, в пять раз или более, в 10 раз или более, в 20 раз или более, в 30 раз или более, в 50 раз или более, в 100 раз или более, в 200 раз или более, в 300 раз или более, в 500 раз или более, в 1×103 раз или более, в 2×10×103 раз или более, в ×103 раз или более, в 5×103 раз или более, в 1×104 раз или более, в 2×104 раз или более, в 3×104 раз или более, в 5×104 раз или более или в 1×105 раз или более. Связываемое количество антигена большего значения может быть, например, 0,01 или более, 0,02 или более, 0,03 или более, 0,04 или более, 0,05 или более, 0,06 или более, 0,07 или более, 0,08 или более, 0,09 или более, 0,1 или более, 0,2 или более, 0,3 или более, 0,4 или более, 0,5 или более, 0,6 или более, 0,7 или более, 0,8 или более, 0,9 или более, 1 или более. Связываемое количество антигена меньшего значения может составлять, например, 0,5 или менее, 0,4 или менее, 0,3 или менее, 0,2 или менее, 0,1 или менее, 0,09 или менее, 0,08 или менее, 0,07 или менее, 0,06 или менее, 0,05 или менее, 0,04 или менее, 0,03 или менее, 0,02 или менее, 0,01 или менее, 0,009 или менее, 0,008 или менее, 0,007 или менее, 0,006 или менее, 0,005 или менее, 0,004 или менее, 0,003 или менее, 0,002 или менее, 0,001 или менее.
В некоторых вариантах осуществления настоящего изобретения значения KD, значения kd или значения связанных количеств, представленные в настоящем изобретении, измеряют или вычисляют путем проведения анализа поверхностного плазмонного резонанса при 25°С или 37°С (например, см. примеры в настоящем изобретении).
Концентрация низкомолекулярного соединения может быть выбрана произвольно, при условии, что может быть обнаружено различие в связывающей активности в отношении антигенсвязывающей молекулы. В некоторых вариантах осуществления настоящего изобретения высокая концентрация включает, например, 1 нМ или более высокую концентрацию, 3 нМ или более высокую концентрацию, 10 нМ или более высокую концентрацию, 30 нМ или более высокую концентрацию, 100 нМ или более высокую концентрацию, 300 нМ или более высокую концентрацию, 1 мкМ или более высокую концентрацию, 3 мкМ или более высокую концентрацию, 10 мкМ или более высокую концентрацию, 30 мкМ или более высокую концентрацию, 100 мкМ или более высокую концентрацию, 300 мкМ или более высокую концентрацию, 1 мМ или более высокую концентрацию, 3 мМ или более высокую концентрацию, 10 мМ или более высокую концентрацию, 30 мМ или более высокую концентрацию, 100 мМ или более высокую концентрацию, 300 мМ или более высокую концентрацию, 1 М или более высокую концентрацию. В другом варианте настоящего изобретения высокая концентрация может быть такой концентрацией, которая обеспечивает достаточное количество, чтобы допустить каждой антигенсвязывающей молекуле проявить максимальную связывающую активность. В одном из вариантов осуществления настоящего изобретения 1 микромоль, 10 мкМ, 100 мкМ, 1 мМ или количество, достаточное для того, чтобы каждая антигенсвязывающая молекула проявляла максимальную связывающую активность, в настоящем изобретении могут быть выбраны в качестве высокой концентрации. В некоторых вариантах осуществления настоящего изобретения низкая концентрация включает, например, 1 мМ или более низкую концентрацию, 300 мкМ или более низкую концентрацию, 100 мкМ или более низкую концентрацию, 30 мкМ или более низкую концентрацию, 10 мкМ или более низкую концентрацию, 3 мкМ или более низкую концентрацию, 1 мкМ или более низкую концентрацию, 300 нМ или более низкую концентрацию, 100 нМ или более низкую концентрацию, 30 нМ или более низкую концентрацию, 10 нМ или более низкую концентрацию, 3 нМ или более низкую концентрацию, 1 нМ или более низкую концентрацию, 300 пМ или более низкую концентрацию, 100 пМ или более низкую концентрацию, 30 пМ или более низкую концентрацию, 10 пМ или более низкую концентрацию, 3 пМ или более низкую концентрацию, 1 пМ или более низкую концентрацию и т.д. В качестве альтернативы, низкая концентрация в настоящем изобретении может быть такой концентрацией, при которой каждая антигенсвязывающая молекула проявляет минимальную связывающую активность. Случай практически «нулевой» концентрации (т.е. отсутствие низкомолекулярного соединения) может быть выбран в качестве одного из вариантов низкой концентрации. В одном из вариантов осуществления настоящего изобретения концентрация 1 мМ, 100 мкМ, 10 мкМ, 1 мкМ является такой концентрацией, при которой каждая антигенсвязывающая молекула по настоящему изобретению проявляет минимальную связывающую активность, или отсутствие низкомолекулярного соединения может быть выбрано в настоящем изобретении в качестве низкой концентрации. В другом варианте осуществления настоящего изобретения может быть выбрано соотношение высокой концентрации к низкой концентрации, например, составляющее трехкратную величину или больше, 10-кратную величину или больше, 30-кратную величину или больше, 100-кратную величину или больше, 300-кратную величину или больше, 1×103-кратную величину или больше, 1×104-кратную величину или больше, 3×104-кратную величину или больше, 1×105-кратную величину или больше, 3×105-кратную величину или больше, 1×106-кратную величину или больше, 3×106-кратную величину или больше, 1×107-кратную величину или больше, 3×107-кратную величину или больше, 1×108 -кратную величину или больше, 3×108-кратную величину или больше, 1×109-кратную величину или больше, 3×109-кратную величину или больше, 1×1010-кратную величину или больше, 3×1010-кратную величину или больше, 1×1011кратную величину или больше, 3×1011-кратную величину или больше, 1×1012-кратную величину или больше.
В другом варианте осуществления настоящего изобретения антигенсвязывающие молекулы по настоящему изобретению проявляют цитотоксичность в отношении клеток, экспрессирующих свои антигены. Когда клетка-мишень экспрессирует антиген на своей поверхности и антигенсвязывающая молекула по настоящему изобретению связывается с ним, клетка может быть повреждена. Повреждение клеток может быть таким повреждением, вызванным эффекторными клетками, например, антителозависимой клеточной цитотоксичностью (ADCC, antibody-dependent cellular cytotoxicity) и антителозависимым клеточным фагоцитозом (ADCP, antibody-dependent cellular phagocytosis), и могут быть такие повреждения, вызванные комплементом, как, например, комплемент-зависимая цитотоксичность (CDC, complement-dependent cytotoxicity). В другом варианте повреждение может быть таким, как повреждения, вызванные цитотоксическим агентом (например, радиоизотопами или химиотерапевтическими агентами) в качестве иммуноконъюгатов. Цитотоксичность в настоящем изобретении может включать действия по индукции гибели клеток, действия по подавлению пролиферации клеток, действия по повреждению функций клеток и т.д. Цитотоксичность по настоящему изобретению может включать действия по индукции гибели клеток, действия по подавлению пролиферации клеток, действия по повреждению функций клеток и т.д. В присутствии достаточного количества антигенсвязывающей молекулы по настоящему изобретению повреждение может быть вызвано, например, 10% или более, 15% или более, 20% или более, 25% или более, 30% или более, 35% или более, 40% или более, 45% или более, 50% или более, 55% или более, 60% или более, 65% или более, 70% или более, 75% или более, 80% или более, 85% или более, 90%, или более, или 95%, или более клеток, экспрессирующих антиген. Измерение такой цитотоксической активности может быть выполнено как сравнение с измерением в отсутствие антигенсвязывающей молекулы по настоящему изобретению или в присутствии антигенсвязывающей молекулы отрицательного контроля. В настоящем изобретении представлены примеры анализов цитотоксичности.
В другом варианте осуществления настоящего изобретения антигенсвязывающие молекулы по настоящему изобретению проявляют нейтрализующую активность по отношению к своим антигенам. Нейтрализующая активность означает активность по нейтрализации (или ингибированию, блокированию) биологической активности, связанной с антигенами. В одном из вариантов осуществления настоящего изобретения биологическая активность обеспечивается связыванием между лигандом и рецептором. В некоторых вариантах осуществления настоящего изобретения антигенсвязывающие молекулы по настоящему изобретению ингибируют такое связывание между лигандом и рецептором. В присутствии достаточного количества антигенсвязывающей молекулы по настоящему изобретению биологическая активность может подавляться, например, на 10% или более, 15% или более, 20% или более, 25% или более, 30% или более, 35%. или более, 40% или более, 45% или более, 50% или более, 55% или более, 60% или более, 65% или более, 70% или более, 75% или более, 80% или более, 85% или более, 90% или более, или 95% или более. Измерение такой ингибирующей активности может быть выполнено как сравнение с измерением в отсутствие антигенсвязывающей молекулы по настоящему изобретению или в присутствии антигенсвязывающей молекулы отрицательного контроля. В настоящем изобретении приводят конкретные методы измерения нейтрализующей активности.
В другом варианте осуществления настоящего изобретения антигенсвязывающие молекулы по настоящему изобретению включают область Fc. В других вариантах осуществления настоящего изобретения антигенсвязывающие молекулы по настоящему изобретению включают константную область. Постоянная область может быть константной областью тяжелой цепи (включающей область Fc), константной областью легкой цепи или и тем, и другим. В некоторых вариантах осуществления настоящего изобретения область Fc является областью с нативной последовательностью. Примеры константной области тяжелой цепи, полученной из антител с нативной последовательностью, включают, например, константную область тяжелой цепи IgG1 человека (SEQ ID NO: 219), IgG2 человека (SEQ ID NO: 220), IgG3 человека (SEQ ID NO: 221), IgG4 человека (SEQ ID NO: 222) и т.п. Другие примеры константной области тяжелой цепи включают константные области тяжелой цепи, такие как SEQ ID NO: 215, SEQ ID NO: 217. Примеры константной области легкой цепи, полученной из антител с нативной последовательностью, включают, например, константные области легкой цепи каппа-цепи человека. (SEQ ID NO: 186, SEQ ID NO: 199, SEQ ID NO: 218), лямбда-цепь человека (SEQ ID NO: 189, SEQ ID NO: 216) и т.п.
В другом варианте осуществления настоящего изобретения область Fc может быть вариантом области Fc, полученным путем введения аминокислотного изменения (изменений) в область Fc нативной последовательностью. В некоторых вариантах осуществления настоящего изобретения вариант области Fc обладает повышенной связывающей активностью по меньшей мере с одним гамма-рецептором Fc, выбранным из группы, состоящей из Fc гамма RIa, Fc гамма RIIa, Fc гамма RIIb и Fc гамма RIIIa, по сравнению с областью Fc с нативной последовательностью.
В других вариантах осуществления настоящего изобретения антигенсвязывающие молекулы по настоящему изобретению являются антителами. В некоторых вариантах осуществления настоящего изобретения, антитела представляют собой моноклональные антитела, включая химерные антитела, гуманизированные антитела или антитела человека. В одном из вариантов осуществления настоящего изобретения, антитела представляют собой фрагменты антител, например, Fv, Fab, Fab', scFv, диатела или фрагменты F(ab')2 и т.п. В другом варианте осуществления настоящего изобретения антитела представляют собой полноразмерные антитела, например, интактные антитела IgG1 , интактные антитела IgG4 или другой класс или изотип антител, как описано в настоящем изобретении.
В другом варианте осуществления настоящего изобретения антигенсвязывающие молекулы по настоящему изобретению образуют тройной комплекс вместе с низкомолекулярным соединением и антигеном. В некоторых вариантах осуществления настоящего изобретения антигенсвязывающими молекулами являются антитела. В одном из вариантов осуществления настоящего изобретения антигенсвязывающие молекулы по настоящему изобретению связываются с низкомолекулярным соединением через CDR1, CDR2, CDR3 тяжелой цепи. В одном из вариантов осуществления настоящего изобретения антигенсвязывающие молекулы по настоящему изобретению имеют связывающий мотив по отношению к низкомолекулярному соединению. Связывающий мотив к низкомолекулярному соединению получают, например, по меньшей мере, из одной аминокислоты, находящейся в положении 33, в положении 52, в положении 52а, в положении 53, в положении 55, в положении 56, в положении 58, в положении 95, в положении 96, в положении 98, в положении 100а, в положении 100b, в положении 100 с по нумерации Kabat. В других вариантах осуществления настоящего изобретения антигенсвязывающие молекулы по настоящему изобретению связываются с низкомолекулярным соединением через, по меньшей мере, одну аминокислоту через группу, находящуюся в положении 33, в положении 52, в положении 52а, в положении 53, в положении 55, в положении 56, в положении 58, в положении 95, в положении 96, в положении 98, в положении 100а, в положении 100b, в положении 100 с по нумерации Kabat Антиген может дополнительно связываться с комплексом, образованным в результате связывания антигенсвязывающей молекулы по настоящему изобретению и низкомолекулярного соединения. Кроме того, низкомолекулярное соединение может присутствовать на границе взаимодействия между антигенсвязывающей молекулой и антигеном и может связываться с ними обоими. Образование тройного комплекса антигенсвязывающей молекулой по настоящему изобретению вместе с низкомолекулярным соединением и антигеном может быть подтверждено, например, кристаллографией, как описано ниже, или такими методами. В некоторых вариантах осуществления настоящего изобретения низкомолекулярное соединение является соединением, содержащим аденозин.
В одном из вариантов осуществления настоящего изобретения низкомолекулярное соединение по настоящему изобретению представляет собой «тканеспецифичное соединение-мишень». В других вариантах осуществления настоящего изобретения «тканеспецифичное соединение-мишень» представляет собой тканеспецифичное соединение опухоли. Типичные тканеспецифичные соединения-мишени и опухолевые тканеспецифические соединения описывают в настоящем изобретении (например, «II. Композиции и методы (агонистические антигенсвязывающие молекулы против CD137)», «А. Типичные антигенсвязывающие молекулы или антитела против CD137», «[Связывающая активность, зависящая от низкомолекулярных соединений]»).
Б. Антиген
В настоящем изобретении «антигены» существенным образом не ограничиваются по своей структуре, при условии, что они содержат эпитопы, с которыми связываются антигенсвязывающие молекулы по настоящему изобретению. Антигены могут быть неорганическими или органическими веществами. В некоторых вариантах осуществления настоящего изобретения к примерам антигенов относятся: 17-IA, 4-1BB, 4Dc, 6-кето-PGF1a, 8-изо-PGF2a, 8-оксо-dG, рецептор аденозина A1, А33, АСЕ, АСЕ-2, активин, активин А, активин АВ, активин В, активин С, активин RIA, активин RIA ALK-2, активин RIB ALK-4, активин RIIA, активин RIIB, ADAM, ADAM 10, ADAM 12, ADAM 15, ADAM 17/TACE, ADAM8, ADAM9, ADAMTS, ADAMTS4, ADAMTS5, аддрессин, aFGF, ALCAM, ALK, ALK-1, ALK-7, альфа-1-антитрипсин, антагонист альфа-V/бета-1, ANG, Ang, APAF-1, АРЕ, APJ, АРР, APRIL, AR, ARC, ART, артемин, анти-Id, ASPARTIC, предсердный натрийуретический пептид, av/v3 интегрин, Axl, b2M, В7-1, В7-2, В7-Н, фактор стимуляции В-лимфоцитов (BlyS), ВАСЕ, ВАСЕ-1, Bad, BAFF, BAFF-R, Bag-1, BAK, Вах, ВСА-1, ВСАМ, Bel, ВСМА, BDNF, b-ECGF, bFGF, BID, Bik, BIM, BLC, BL-CAM, BLK, BMP, BMP-2 BMP-2a, BMP-3 остеогенин, BMP-4 BMP-2b, BMP-5, BMP-6 Vgr-1, BMP-7 (OP-1), BMP-8 (BMP-8a, OP-2), BMPR, BMPR-IA (ALK-3), BMPR-IB (ALK-6), BRK-2, RPK-1, BMPR-II (BRK-3), BMP, b-NGF, BOK, бомбезин, костный нейротрофический фактор, BPDE, BPDE-ДНК, ВТС, фактор комплемента 3 (С3), С3а, С4, С5, С5а, С10, СА125, CAD-8, кальцитонин, цАМФ, карциноэмбриональный антиген (СЕА), рак связанный антиген, катепсин А, катепсин В, катепсин С / DPPI, катепсин D, катепсин Е, катепсин Н, катепсин L, катепсин О, катепсин S, катепсин V, катепсин X / Z / Р, CBL, CCI, CCK2, CCL, CCL1, CCL11, CCL12, CCL13, CCL14, CCL15, CCL16, CCL17, CCL18, CCL19, CCL2, CCL20, CCL21, CCL22, CCL23, CCL24, CCL25, CCL26, CCL27, CCL28, CCL3, CCL4, CCL5, CCL6, CCL7, CCL8, CCL9/10, CCR, CCR1, CCR10, CCR10, CCR2, CCR3, CCR4, CCR5, CCR6, CCR7, CCR8, CCR9, CD1, CD2, CD3, CD3E, CD4, CD5, CD6, CD7, CD8, CD10, CD11a, CD11b, CD11c, CD13, CD14, CD15, CD16, CD18, CD19, CD20, CD21, CD22, CD23, CD25, CD27L, CD28, CD29, CD30, CD30L, CD32, CD33 (белок р67), CD34, CD38, CD40, CD40L, CD44, CD45, CD46, CD49a, CD52, CD54, CD55, CD56, CD61, CD64, CD66e, CD74, CD80 (В7-1), CD89, CD95, CD123, CD137, CD138, CD140a, CD146, CD147, CD148, CD152, CD164, CEACAM5, CFTR, cGMP, CINC, ботулиновый токсин, токсин Clostridium perfringens, CKb8-l, CLC, CMV, CMV UL, CNTF, CNTN-1, COX, C-Ret, CRG-2, CT-1, CTACK, CTGF, CTLA-4, PD1, PDL1, LAG3, TIM3, галектин-9, CX3CL1, CX3CR1, CXCL, CXCL1, CXCL2, CXCL3, CXCL4, CXCL5, CXCL6, CXCL7, CXCL8, CXCL9, CXCL10, CXCL11, CXCL12, CXCL13, CXCL14, CXCL15, CXCL16, CXCR, CXCR1, CXCR2, CXCR3, CXCR4, CXCR5, CXCR6, цитокератин, ассоциированный с опухолью антиген, DAN, DCC, DcR3, DC-SIGN, регуляторный фактор комплемента (фактор ускорения распада), des (1-3)-IGF-I (IGF-1 мозга), Dim, дигоксин, DNAM-1, Dnase, Dpp, DPPIV/CD26, Dtk, ECAD, EDA, EDA-A1, EDA-A2, EDAR, EGF, EGFR (ErbB-1), EMA, EMMPRIN, ENA рецептор эндотелина, энкефалиназа, eNOS, Eot, эотаксин 1, EpCAM, эфрин B2 / EphB4, EPO, ERCC, Е-селектин, ET-1, фактор Па, фактор VII, фактор VIIIc, фактор IX, белок активации фибробластов (FAP), Fas, FcR1, FEN-1, ферритин, FGF, FGF-19, FGF-2, FGF-3, FGF-8, FGFR, FGFR-3, фибрин, FL, FLIP, Flt-3, Flt-4, фолликул стимулирующий гормон, фракталкин, FZD1, FZD2, FZD3, FZD4, FZD5, FZD6, FZD7, FZD8, FZD9, FZD10, G250, Gas6, GCP-2, GCSF, GD2, GD3, GDF, GDF-1, GDF-3 (Vgr-2), GDF-5 (BMP-14, CDMP-1), GDF-6 (BMP-13, CDMP-2), GDF-7 (BMP-12, CDMP-3), GDF-8 (миостатин), GDF-9, GDF-15 (MIC-1), GDNF, GDNF, GFAP, GFRa-1, GFR-альфа1, GFR-альфа2, GFR-альфа3, GITR, глюкагон, Glut4, гликопротеин IIb/IIIa (GPIIb/IIIa), GM-CSF, gp130, gp72, GRO, гормон высвобождения гормона роста, гаптен (NP-cap или NIP-cap), HB-EGF, НСС, гликопротеин оболочки gB HCMV, гликопротеин оболочки gH HCMV, UL HCMV, гематопоэтический фактор роста (HGF), Hep В gpl20, гепараназа, Her2, Her2 / neu (ErbB-2), Her3 (ErbB-3), Her4 (ErbB-4), гликопротеин gB вируса простого герпеса (HSV), гликопротеин gD HSV, HGF А, высокомолекулярный антиген, связанный с меланомой (HMW-MAA), HIV gpl20, HIV ШВ gp 120 V3 петля, HLA, HLA-DR, HM1.24, HMFG РЕМ, HRG, Hrk, сердечный миозин человека, цитомегаловирус человека (HCMV), гормон роста человека (HGH), HVEM, 1-309, IAP, ICAM, ICAM-1, ICAM-3, ICE, ICOS, IFNg, Ig, IgA receptor, IgE, IGF, IGF-связывающий белок, IGF-1R, IGFBP, IGF-I, IGF-II, IL, IL-1, IL-1R, IL-2, IL-2R, IL-4, IL-4R, IL-5, IL-5R, IL-6, IL-6R, IL-8, IL-9, IL-10, IL-12, IL-13, IL-15, IL-18, IL-18R, IL-21, IL-23, IL-27, интерферон (INF) - альфа, INF-бета, INF-гамма, ингибин, iNOS, цепь А инсулина, цепь В инсулина, инсулиноподобный фактор роста 1, интегрин альфа 2, интегрин альфа 3, интегрин альфа 4, интегрин альфа4/бета1, интегрин альфа4/бета7, интегрин альфа5 (альфа V), интегрин альфа5/бета1, интегрин альфа5/бетаЗ, интегрин альфаб, интегрин бета1, интегрин бета2, интерферон гамма, IP-10, I-TAC, JE, калликреин 2, калликреин 5, калликреин 6, калликреин 11, калликреин 12, калликреин 14, калликреин 15, калликреин L1, калликреин L2, калликреин L3, калликреин, калликреин L3, калликреин фактор роста кератиноцитов (KGF), ламинин 5, LAMP, LAP, LAP (TGF-1), латентный TGF-1, латентный TGF-1 bpl, LBP, LDGF, LECT2, левша, антиген Lewis-Y, антиген Lewis-Y, LFA-1, LFA-3, Lfo, LIF, LIGHT, липопротеин, LIX, LKN, Lptn, L-селектин, LT-a, LT-b, LTB4, LTBP-1, поверхность легких, лютеинизирующий гормон, рецептор лимфотоксина бета, Мас-1, MAdCAM, MAG, МАР2, MARC, МСАМ, МСАМ, МСК-2, МСР, M-CSF, MDC, Mer, METALLOPROTEASES, MGDF receptor, MGMT, МНС (HLA-DR), MIF, MIG, MIP, MIP-1-альфа, МК, ММАС1, ММР, ММР-1, ММР-10, ММР-11, ММР-12, ММР-13, ММР-14, ММР-15, ММР-2, ММР-24, ММР-3, ММР-7, ММР-8, ММР-9, MPIF, Мро, MSK, MSP, муцин (Muc1), MUC18, вещество, ингибирующее мюллериан, Mug, MuSK, NAIP, NAP, NCAD, N-C адгерин, NCA 90, NCAM, NCAM, неприлизин, нейротрофин-3, -4 или -6, нейротрофин, фактор роста нервов (NGF), NGFR, NGF-бета, nNOS, NO, NOS, Npn, NRG-3, NT, NTN, OB, OGG1, OPG, OPN, OSM, OX40L, OX40R, p150, p95, PADPr, паратироидный гормон, PARC, PARP, PBR, PBSF, PCAD, Р-кадгерин, PCNA, PDGF, PDGF, PDK-1, PECAM, РЕМ, PF4, PGE, PGF, PGI2, PGJ2, PIN, PLA2, щелочная фосфатаза плаценты (PLAP), P1GF, PLP, PP14, проинсулин, прорелаксин, белок С, PS, PSA, PSCA, простатоспецифический мембранный антиген (PSMA), PTEN, PTHrp, Ptk, PTN, R51, RANK, RANKL, RANTES, RANTES, цепь релаксина А, цепь релаксина В, ренин, респираторный синцитиальный вирус (RSV) F, RSV Fgp, Ret, ревматоидный фактор, RLIP76, RPA2, RSK, S100, SCF/KL, SDF-1, SERINE, сывороточный альбумин, sFRP-3, Shh, SIGIRR, SK-1, SLAM, SLPI, SMAC, SMDF, SMOH, SOD, SPARC, Stat, STEAP, STEAP-II, TACE, TACI, TAG-72 (опухоль-ассоциированный гликопротеин-72), TARC, TCA-3, рецептор Т-клеток (например, рецептор Т-клеток альфа/бета), TdT, TECK, ТЕМ1, ТЕМ5, ТЕМ7, ТЕМ8, TERT, PLAP-подобная щелочная фосфатаза семенников, TfR, TGF, TGF-альфа, TGF-бета, TGF-бета Pan специфический, TGF-бeтaRI (ALK-5), TGF-бетаRII, TGF-бетаRIIb, TGF-бетаRIII, TGF-бета1, TGF-бета2, TGF-бета3, TGF-бета4, TGF-бета5, тромбин, тимус Ck-1, тиреотропный гормон, Tie, TIMP, TIQ, тканевой фактор, TMEFF2, Tmpo, TMPRSS2, TNF, TNF-альфа, TNF-альфабета, TNF-бета2, TNFc, TNF-RI, TNF-RII, TNFRSF10A (TRAIL R1 Apo-2, DR4), TNFRSF10B (TRAIL R2 DR5, KILLER, TRICK-2A, TRICK-B), TNFRSF10C (TRAIL R3 DcR1, LIT, TRID), TNFRSFIOD (TRAIL R4 DcR2, TRUNDD), TNFRSF11A (RANK ODF R, TRANCE R), TNFRSF1 IB (OPG OCIF, TR1), TNFRSF12 (TWEAK R FN14), TNFRSF13B (TACI), TNFRSF13C (BAFF R), TNFRSF14 (HVEM ATAR, HveA, LIGHT R, TR2), TNFRSF16 (NGFR p75NTR), TNFRSF17 (BCMA), TNFRSF18 (GITR AITR), TNFRSF19 (TROY TAJ, TRADE), TNFRSF19L (RELT), TNFRSF1A (TNF RI CD120a, p55-60), TNFRSF1B (TNF RII CD120b, p75-80), TNFRSF26 (TNFRH3), TNFRSF3 (LTbR TNF RIII, TNFC R), TNFRSF4 (OX40 ACT35, TXGP1 R), TNFRSF5 (CD40 p50), TNFRSF6 (Fas Apo-1, APT1, CD95), TNFRSF6B (DcR3 M68, TR6), TNFRSF7 (CD27), TNFRSF8 (CD30), TNFRSF9 (4-IBB CD 137, ILA), TNFRSF21 (DR6), TNFRSF22 (DcTRAIL R2 TNFRH2), TNFRST23 (DcTRAILR1 TNFRH1), TNFRSF25 (DR3 Apo-3, LARD, TR-3, TRAMP, WSL-1), TNFSF10 (TRAIL Apo-2 лиганд, TL2), TNFSF11 (TRANCE/RANK лиганд ODF, OPG лиганд), TNFSF12 (TWEAK Apo-3 лиганд, DR3 лиганд), TNFSF13 (APRIL TALL2), TNFSF13B (BAFF BLYS, TALL1, THANK, TNFSF20), TNFSF14 (LIGHT HVEM лиганд, LTg), TNFSF15 (TL1A/VEGI), TNFSF18 (GITR лиганд AITR лиганд, TL6), TNFSF1A (TNF-a конектин, DIF, TNFSF2), TNFSF1B (TNF-b LTa, TNFSF1), TNFSF3 (LTb TNFC, p33), TNFSF4 (OX40 лиганд gp34, TXGP1), TNFSF5 (CD40 лиганд CD154, gp39, HIGM1, IMD3, TRAP), TNFSF6 (Fas лиганд Apo-1 лиганд, APT1 лиганд), TNFSF7 (CD27 лиганд CD70), TNFSF8 (CD30 лиганд CD153), TNFSF9 (4-1BB лиганд CD137 лиганд), ТР-1, t-PA, Тро, TRAIL, TRAIL R, TRAIL-R1, TRAIL-R2, TRANCE, рецептор трансферрина, TRF, Trk, TROP-2, TLR1 (Толл-подобный рецептор 1), TLR2, TLR3, TLR4, TLR5, TLR6, TLR7, TLR8, TLR9, TLR10, TSG, TSLP, опухоль-ассоциированный антиген CA125, опухольассоциированный антигенэкспрессирующий Lewis-Y ассоциированные углеводы, TWEAK, ТХВ2, Ung, uPAR, uPAR-1, урокиназа, VCAM, VCAM-1, VECAD, VE- кадгерин, VE- кадгерин -2, VEFGR-1 (fit-1), VEGF, VEGFR, VEGFR-3 (flt-4), VEGI, VIM, вирусный антиген, VLA, VLA-1, VLA-4, VNR интегрин, фактор фон Виллебранда, WIF-1, WNT1, WNT2, WNT2B/13, WNT3, WNT3A, WNT4, WNT5A, WNT5B, WNT6, WNT7A, WNT7B, WNT8A, WNT8B, WNT9A, WNT9A, WNT9B, WNT10A, WNT10B, WNT11, WNT16, XCL1, XCL2, XCR1, XCR1, XEDAR, XIAP, XPD, HMGB1, IgA, А бета, CD81, CD97, CD98, DDR1, DKK1, EREG, Hsp90, IL-17/IL-17R, IL-20/IL-20R, окисленный LDL, PCSK9, прекалликреин, RON, TMEM16F, SOD1, хромогранин А, хромогранин В, тау, VAP1, высокомолекулярный кининоген, IL-31, IL-31R, Nav1.1, Nav1.2, Nav1.3, Nav1.4, Nav1.5, Nav1.6, Nav1.7, Nav1.8, Nav1.9, EPCR, C1, C1q, C1r, C1s, C2, C2a, C2b, С3, С3а, C3b, C4, C4a, C4b, C5, C5a, C5b, C6, C7, C8, C9, фактор В, фактор D, фактор H, пропердин, склеростин, фибриноген, фибрин, протромбин, тромбин, тканевой фактор, фактор V, фактор Va, фактор VII, фактор VIIa, фактор VIII, фактор VIIIa, фактор IX, фактор IXa, фактор X, фактор Ха, фактор XI, фактор XIa, фактор XII, фактор ХПа, фактор XIII, фактор ХШа, TFPI, антитромбин III, EPCR, тромбомодулин, TAPI, tPA, плазминоген, плазмин, PAI-1, PAI-2, GPC3, синдекан-1, синдекан-2, синдекан-3, синдекан-4, LPA и S1P. В некоторых вариантах осуществления настоящего изобретения примеры антигенов включают рецепторы гормонов и факторов роста. В некоторых вариантах осуществления настоящего изобретения антигенами являются те антигены, которые экспрессируются или секретируются клетками, присутствующими в опухолевых тканях (например, опухолевыми клетками, иммунными клетками, стромальными клетками или тому подобными).
В. Активности, композиции и методы
В одном из вариантов осуществления настоящего изобретения цитотоксическая активность по настоящему изобретению включает, например, активность антителозависимой клеточно-опосредованной цитотоксичности (ADCC), антителозависимый клеточный фагоцитоз (ADCP), комплемент-зависимую цитотоксичность (CDC) и цитотоксическую активность Т-клеток. Активность CDC относится к цитотоксической активности системы комплемента. С другой стороны, активность ADCC относится к активности повреждения клеток-мишеней через эффекторные клетки, что происходит, когда антигенсвязывающие молекулы связываются с антигенами, присутствующими на клеточной поверхности клеток-мишеней, а эффекторные клетки дополнительно связываются с антигенсвязывающими молекулами. Обладает ли представляющая интерес антигенсвязывающая молекула активностью ADCC или имеет ли она активность CDC, можно определить с помощью известных методов (например, Current Protocols in Immunology, глава 7, 1993. Immunologic studies in humans, под ред. Coligan с соавт.).
В одном из вариантов осуществления настоящего изобретения «нейтрализующая активность» в настоящем изобретении относится к активности антигенсвязывающей молекулы по ингибированию биологической активности посредством ее связывания с молекулой, участвующей в этой биологической активности. В некоторых вариантах осуществления настоящего изобретения биологическая активность обеспечивается связыванием лиганда с рецептором. В некоторых вариантах осуществления настоящего изобретения антигенсвязывающая молекула связывается с лигандом или рецептором, тем самым ингибируя связывание между лигандом и рецептором. Когда антигенсвязывающая молекула представляет собой антитело, антигенсвязывающая молекула, обладающая такой нейтрализующей активностью, называется нейтрализующим антителом. Нейтрализующая активность тестируемого вещества может быть измерена путем сравнения биологической активности в присутствии лиганда при условиях, когда тестируемое вещество присутствует или отсутствует.
В одном варианте осуществления настоящее изобретение обеспечивает способы получения антигенсвязывающей молекулы, антигенсвязывающая активность которой изменяется в зависимости от концентрации соединения. Когда соединение представляет собой тканеспецифичное соединение-мишень, настоящее изобретение обеспечивает способы получения антигенсвязывающей молекулы, которая специфически проявляет свое действие в ткани-мишени. В некоторых вариантах осуществления настоящего изобретения целевая ткань представляет собой опухолевую ткань. Когда соединение представляет собой соединение, специфичное для опухолевой ткани, настоящее изобретение предусматривает способы получения антигенсвязывающей молекулы для использования при лечении опухолей. В некоторых вариантах осуществления настоящего изобретения эти способы получения включают стадию выбора антигенсвязывающей молекулы, антигенсвязывающая активность которой в присутствии соединения отличается от ее антигенсвязывающей активности в отсутствие соединения. В некоторых вариантах осуществления настоящего изобретения эти способы получения включают стадию выбора антигенсвязывающей молекулы, антигенсвязывающая активность которой в присутствии высокой концентрации соединения отличается от ее антигенсвязывающей активности в присутствии низкой концентрации В некоторых вариантах осуществления настоящего изобретения эти методы получения включают стадию выбора антигенсвязывающей молекулы, антигенсвязывающая активность которой в присутствии высокой концентрации соединения отличается от ее антигенсвязывающей активности в присутствии низкой концентрации соединения. В некоторых вариантах осуществления настоящего изобретения эти методы получения включают стадии (а) получения антигенсвязывающей активности антигенсвязывающей молекулы в присутствии соединения, (б) получения антигенсвязывающей активности антигенсвязывающей молекулы в отсутствие соединение, и (в) выбор антигенсвязывающей молекулы, антигенсвязывающая активность которой в присутствии соединения отличается от ее антигенсвязывающей активности в отсутствие соединения. В некоторых вариантах осуществления настоящего изобретения эти методы получения включают стадии (а) получения антигенсвязывающей активности антигенсвязывающей молекулы в присутствии высокой концентрации соединения, (б) получения антигенсвязывающей активности антигенсвязывающей молекулы при наличии низкой концентрации соединения, и (в) выбор антигенсвязывающей молекулы, антигенсвязывающая активность которой в присутствии высокой концентрации соединения отличается от ее антигенсвязывающей активности в присутствии низкой концентрации соединения.
В другом варианте осуществления настоящего изобретения предусматривают методы получения антигенсвязывающей молекулы, антигенсвязывающая активность которой увеличивается по мере того, как концентрация соединения становится выше. В другом варианте настоящее изобретение предусматривает способы получения антигенсвязывающей молекулы, антигенсвязывающая активность которой снижается по мере того, как концентрация соединения становится выше. Способы получения включают стадию выбора антигенсвязывающей молекулы, антигенсвязывающая активность которой в присутствии соединения выше, чем ее антигенсвязывающая активность в отсутствие соединения. В другом варианте осуществления настоящего изобретения способы получения включают стадию выбора антигенсвязывающей молекулы, антигенсвязывающая активность которой в отсутствие соединения ниже, чем ее антигенсвязывающая активность в присутствии соединения. В другом варианте осуществления настоящего изобретения способы получения включают стадию выбора антигенсвязывающей молекулы, антигенсвязывающая активность которой в отсутствие соединения выше, чем ее антигенсвязывающая активность в присутствии соединения. В другом варианте осуществления настоящего изобретения способы получения включают стадию выбора антигенсвязывающей молекулы, антигенсвязывающая активность которой в присутствии соединения ниже, чем ее антигенсвязывающая активность в отсутствие соединения. В еще одном варианте осуществления настоящего изобретения способы получения включают стадию выбора антигенсвязывающей молекулы, антигенсвязывающая активность которой в присутствии высокой концентрации соединения выше, чем ее антигенсвязывающая активность в присутствии низкой концентрации соединения. В другом варианте осуществления настоящего изобретения способы получения включают стадию выбора антигенсвязывающей молекулы, антигенсвязывающая активность которой в присутствии низкой концентрации соединения ниже, чем ее антигенсвязывающая активность в присутствии высокой концентрации соединения. В еще одном варианте осуществления настоящего изобретения способы получения включают стадию выбора антигенсвязывающей молекулы, антигенсвязывающая активность которой в присутствии низкой концентрации соединения выше, чем ее антигенсвязывающая активность в присутствии высокой концентрации соединения. В другом варианте осуществления настоящего изобретения способы получения включают стадию выбора антигенсвязывающей молекулы, антигенсвязывающая активность которой в присутствии высокой концентрации соединения ниже, чем ее антигенсвязывающая активность в присутствии низкой концентрации соединения.
В одном из вариантов осуществления настоящее изобретение предусматривает способы получения антигенсвязывающей молекулы, обладающей высокой способностью удерживания в плазме. В некоторых вариантах осуществления настоящего изобретения способы получения включают стадию получения антигенсвязывающей молекулы, антигенсвязывающая активность которой увеличивается по мере того, как концентрация соединения становится выше. В других вариантах осуществления настоящего изобретения способы получения включают стадии (а) получения антигенсвязывающей молекулы, антигенсвязывающая активность которой увеличивается по мере того, как концентрация соединения становится выше; и (б) измерения удерживающей способности в плазме антигенсвязывающей молекулы, полученной на этапе (а). В некоторых вариантах осуществления настоящего изобретения соединение представляет собой целевое тканеспецифичное соединение. В других вариантах осуществления настоящего изобретения соединение представляет собой соединение, специфичное для опухолевой ткани. Определение того, обладает ли антигенсвязывающая молекула в настоящем изобретении высокой способностью удерживания в плазме, может быть выполнено путем сравнения с контрольной антигенсвязывающей молекулой. В некоторых вариантах осуществления настоящего изобретения те антигенсвязывающие молекулы, антигенсвязывающая активность которых увеличивается по мере того, как концентрация целевого тканеспецифичного соединения становится выше, имеют более высокую способность удерживания в плазме по сравнению с контрольной антигенсвязывающей молекулой. В одном из вариантов осуществления настоящего изобретения контрольная антигенсвязывающая молекула не обладает антигенсвязывающей активностью, зависящей от концентрации соединения. В некоторых вариантах осуществления настоящего изобретения антигенсвязывающая молекула, которая не обладает антигенсвязывающей активностью, зависящей от концентрации соединения, означает антигенсвязывающую молекулу, для которой разница в антигенсвязывающей активности в присутствии соединения и в отсутствие соединения меньше, например, в 2 раза или меньше, в 1,8 раз или меньше, в 1,5 раз или меньше, в 1,3 раз или меньше, в 1,2 раза или меньше или в 1,1 раза или меньше. С точки зрения сравнения, предпочтительно, чтобы антигенсвязывающая активность антигенсвязывающих молекул по настоящему изобретению и контрольных антигенсвязывающих молекул в присутствии достаточного количества соединения была по существу одинаковой.
В другом варианте осуществления настоящее изобретение обеспечивает способы получения антигенсвязывающей молекулы, имеющей низкую способность к накоплению антигена в плазме. В некоторых вариантах осуществления настоящего изобретения способы получения включают стадию получения антигенсвязывающей молекулы, антигенсвязывающая активность которой увеличивается по мере того, как концентрация соединения становится выше. В других вариантах осуществления настоящего изобретения способы получения включают стадии (а) получения антигенсвязывающей молекулы, антигенсвязывающая активность которой увеличивается по мере того, как концентрация соединения становится выше; и (б) измерение способности к накоплению антигена в плазме для антигенсвязывающей молекулы, получаемой в пункте (а). В некоторых вариантах осуществления настоящего изобретения соединение представляет собой целевое тканеспецифичное соединение. В других вариантах осуществления настоящего изобретения соединение представляет собой соединение, специфичное для опухолевой ткани. Определение того, обладает ли антигенсвязывающая молекула в настоящем изобретении низкой способностью к накоплению антигена в плазме, может быть выполнено путем сравнения с контрольной антигенсвязывающей молекулой. В некоторых вариантах осуществления настоящего изобретения те антигенсвязывающие молекулы, антигенсвязывающая активность которых увеличивается по мере повышения концентрации целевого тканеспецифичного соединения, имеют более низкую способность к накоплению антигена в плазме по сравнению с контрольной антигенсвязывающей молекулой. В одном из вариантов осуществления настоящего изобретения контрольная антигенсвязывающая молекула не обладает антигенсвязывающей активностью, зависящей от концентрации соединения. В некоторых вариантах осуществления настоящего изобретения антигенсвязывающая молекула, которая не обладает антигенсвязывающей активностью, зависящей от концентрации соединения, означает антигенсвязывающую молекулу, для которой разница в антигенсвязывающей активности в присутствии соединения и в отсутствие соединения меньше, например, в 2 раза или меньше, в 1,8 раз или меньше, в 1,5 раз или меньше, в 1,3 раза или меньше, в 1,2 раза или меньше или в 1,1 раза или меньше. С точки зрения сравнения, предпочтительно, чтобы антигенсвязывающая активность антигенсвязывающих молекул по настоящему изобретению и контрольных антигенсвязывающих молекул в присутствии достаточного количества соединения была по существу одинаковой.
В некоторых вариантах осуществления настоящего изобретения для антигенсвязывающих молекул, получаемых способами по настоящему изобретению, разница в антигенсвязывающей активности, зависящей от концентрации соединения, больше, например, в два раза или более, в три раза или более, в пять раз или более, в 10 раз или более, в 20 раз или более, в 30 раз или более, в 50 раз или более, в 100 раз или более, в 200 раз или более, в 300 раз или более, в 500 раз или более, в 1000 раз или более, в 2000 раз или более, в 3000 раз или более, в 5000 раз или более, в 10000 раз или более, 20000 раз или более, 30000 раз или более, 50000 раз или более или в 100000 раз или более.
В некоторых вариантах осуществления настоящего изобретения антигенсвязывающая активность может быть выражена с помощью величины константы диссоциации (KD) или величины константы скорости диссоциации (kd). В другом варианте она может быть выражена с использованием количества антигена, связавшегося с антигенсвязывающей молекулой, как описано выше. В другом варианте осуществления настоящего изобретения цитотоксическая активность или нейтрализующая активность антигенсвязывающей молекулы может быть использована в качестве альтернативного индикатора антигенсвязывающей активности.
В некоторых вариантах осуществления настоящего изобретения повышенная антигенсвязывающая активность составляет, например, величину KD 9×10-7 М или менее, 8×10-7 М или менее, 7×10-7 М или менее, 6×10-7 М или менее, 5×10-7 М или менее, 4×10-7 М или менее, 3×10-7 М или менее, 2×10-7 М или менее, 1×10-7 М или менее, 9×10-8 М или менее, 8×10-8 М или менее, 7×10-8 М или менее, 6×10-8 М или менее, 5×10-8 М или менее, 4×10-8 М или менее, 3×10-8 М или менее, 2×10-8 М или менее, 1×10-8 М или менее, 9×10-9 М или менее, 8×10-9 М или менее, 7×10-9 М или менее, 6×10-9 М или менее, 5×10-9 М или менее, 4×10-9 М или менее, 3×10-9 М или менее, 2×10-9 М или менее, 1×10-9 М или менее, или, например, количество связанного антигена составляет 0,01 или более, 0,02 или более, 0,03 или более, 0,04 или более, 0,05 или более, 0,06 или более, 0,07 или более, 0,08 или более, 0,09 или более, 0,1 или более, 0,2 или более, 0,3 или более, 0,4 или более, 0,5 или более, 0,6 или более, 0,7 или более, 0,8 или более, 0,9 или более, 1 или более.
В некоторых вариантах осуществления настоящего изобретения пониженная антигенсвязывающая активность составляет, например, величину KD 1×10-8 М или более, 2×10-8 М или более, 3×10-8 М или более, 4×10-8 М или более, 5×10-8 М или более, 6×10-8 М или более, 7×10-8 М или более, 8×10-8 М или более, 9×10-9 М или более, 1×10-7 М или более, 2×10-7 М или более, 3×10-7 М или более, 4×10-7 М или более, 5×10-7 М или более, 6×10-7 М или более, 7×10-7 М или более, 8×10-7 М или более, 9×10-7 М или более, 1 ×10-6 M или более, 2×10-6 М или более, 3×10-6 М или более, 4×10-6 М или более, 5×10-6 М или более, 6×10-6 М или более, 7×10-6М или более, 8×10-6 М или более, 9×10-6 М или более, или, например, количество связанного антигена составляет 0,5 или менее, 0,4 или менее, 0,3 или менее, 0,2 или менее, 0,1 или менее, 0,09 или менее, 0,08 или менее, 0,07 или менее, 0,06 или менее, 0,05 или менее, 0,04 или менее, 0,03 или менее, 0,02 или менее, 0,01 или менее, 0,009 или менее, 0,008 или менее, 0,007 или менее, 0,006 или менее, 0,005 или менее, 0,004 или менее, 0,003 или менее, 0,002 или менее, 0,001 или менее.
Концентрация соединения может быть выбрана в широком диапазоне, при условии, что различие в связывающей активности для антигенсвязывающей молекулы может быть обнаружено. Примеры концентрации (высокой и низкой) представлены в настоящем изобретении. В некоторых вариантах осуществления настоящего изобретения к примерам высокой концентрации относятся, например, концентрация 1 мкМ или выше, концентрация 3 мкМ или выше, концентрация 10 мкМ или выше, концентрация 30 мкМ или выше, концентрация 100 мкМ или выше, концентрация 300 мкМ или выше, концентрация 1 мМ или выше. В другом варианте по настоящему изобретению высокая концентрация может быть такой концентрацией, которая обеспечивает достаточное количество, позволяющее каждой антигенсвязывающей молекуле проявить максимальную связывающую активность. В некоторых вариантах осуществления настоящего изобретения примерами низких концентраций являются, например, концентрация 1 мМ или ниже, концентрация 300 мкМ или ниже, концентрация 100 мкМ или ниже, концентрация 30 мкМ или ниже, концентрация 10 мкМ или ниже, концентрация 3 мкМ или ниже, концентрация 1 мкМ или ниже. В другом варианте осуществления настоящего изобретения низкая концентрация может быть такой концентрацией, при которой каждая антигенсвязывающая молекула проявляет минимальную связывающую активность. Случай по существу «нулевой» концентрации (т.е. отсутствие соединения) может быть одним из вариантов низкой концентрации.
В другом варианте осуществления настоящего изобретения эти способы получения включают стадию выбора антигенсвязывающей молекулы, антигенсвязывающая активность которой в присутствии соединения отличается от ее антигенсвязывающей активности в отсутствие соединения, из библиотеки антигенсвязывающих молекул. Библиотека антигенсвязывающих молекул с может быть со свободным репертуаром антигенсвязывающих молекул (подбор молекул не подвергавшийся воздействию) или может быть с репертуаром определенным образом отобранных антигенсвязывающих молекул. К примерам библиотек такого типа относится библиотека антигенсвязывающих молекул, которым заранее придается связывающая активность с определенным соединением. В некоторых вариантах осуществления настоящего изобретения библиотека антигенсвязывающих молекул представляет собой библиотеку антигенсвязывающих молекул, в которые вводят аминокислотные изменения для придания связывающей активности определенному соединению заранее. Примеры библиотек такого типа включают библиотеки, описанные, например, в WO 2015/083764.
В некоторых вариантах осуществления настоящего изобретения способы получения дополнительно включают стадии (г), на которой получают нуклеиновую кислоту, которая кодирует антигенсвязывающую молекулу, выбранную на стадии (в), стадию (е) по введению нуклеиновой кислоты со стадии (г) в клетку-хозяина, и (е) культивирование клетка-хозяин по пункту (д), так что антигенсвязывающая молекула экспрессируется. Нуклеиновая кислота по п. (г) может быть одной или несколькими нуклеиновыми кислотами и может быть включена в один или несколько векторов (например, в вектор экспрессии). В некоторых вариантах осуществления настоящего изобретения способы получения дополнительно включают стадию по п. (ж), на которой выделяют антигенсвязывающую молекулу из культуры клеток по п. (е).
Антигенсвязывающие молекулы, полученные способами получения по настоящему изобретению, также включены в настоящее изобретение.
В другом варианте осуществления настоящего изобретения предусматривают фармацевтические препараты, включающие антигенсвязывающую молекулу, предусмотренную в настоящем изобретении. В одном из вариантов осуществления настоящего изобретения вышеупомянутые фармацевтические препараты дополнительно содержат фармацевтически приемлемый носитель. В других вариантах осуществления настоящего изобретения предусматривают фармацевтические препараты для применения в лечении опухолей.
В другом варианте осуществления настоящего изобретения предусматривают способы получения фармацевтического препарата, включающие смешивание антигенсвязывающей молекулы по настоящему изобретению с фармацевтически приемлемым носителем. В других вариантах осуществления настоящего изобретения предусматривают способы получения фармацевтического препарата для лечения опухолей.
В одном из вариантов осуществления настоящего изобретения предусматривают антигенсвязывающую молекулу, которая при введении в живой организм проявляет более сильную антигенсвязывающую активность в опухолевой ткани, чем в неопухолевой ткани. Такую разницу в ответе необязательно наблюдать для каждой дозы антигенсвязывающей молекулы, и достаточно, чтобы разница наблюдалась для определенного диапазона доз. В другом варианте осуществления настоящего изобретения, клетки-мишени (клетки, экспрессирующие антиген) более сильно повреждаются в опухолевой ткани, чем в неопухолевой ткани. В другом варианте осуществления настоящего изобретения клетки-мишени повреждаются при более низкой дозе в опухолевой ткани, чем в неопухолевой ткани. В другом варианте осуществления настоящего изобретения терапевтический эффект наблюдают при более низкой дозе, чем доза, при которой наблюдают побочный эффект. В настоящем изобретении терапевтический эффект представляет собой развитие противоопухолевого эффекта (например, регресс опухоли, индукция гибели опухолевых клеток или ингибирование пролиферации опухолевых клеток и т.д.), причем побочным эффектом является возникновение нежелательного явления в нормальных тканях (например, повреждение нормальных тканей).
В одном из вариантов осуществления настоящего изобретения степень эффективности лекарственного средства, обеспечиваемого антигенсвязывающей молекулой, представленной в настоящем изобретении, различна в зависимости от того, обладает ли антигенсвязывающая молекула антигенсвязывающей активностью, зависящей от концентрации соединения (т.е. изменяющейся в зависимости от концентрации соединения). В некоторых вариантах осуществления настоящего изобретения антигенсвязывающие молекулы по настоящему изобретению представляют собой антигенсвязывающую молекулу, антигенсвязывающая активность которой увеличивается по мере того, как концентрация соединения становится выше. В некоторых вариантах осуществления настоящего изобретения контрольная антигенсвязывающая молекула представляет собой антигенсвязывающую молекулу, которая не обладает антигенсвязывающей активностью, зависящей от концентрации соединения. В некоторых вариантах осуществления настоящего изобретения соединение представляет собой соединение, специфичное для опухолевой ткани. В некоторых вариантах осуществления настоящего изобретения антигенсвязывающая молекула, которая не обладает антигенсвязывающей активностью, зависящей от концентрации соединения, означает антигенсвязывающую молекулу, для которой разница в антигенсвязывающей активности в присутствии соединения и в отсутствие соединения, например, отличается в 2 раза или меньше, в 1,8 раза или меньше, в 1,5 раза или меньше, в 1,3 раза или меньше, в 1,2 раза или меньше, или в 1,1 раза или меньше. Предпочтительно, чтобы антигенсвязывающая активность антигенсвязывающих молекул по настоящему изобретению и контрольных антигенсвязывающих молекул в присутствии достаточного количества соединения была по существу одинаковой.
В одном из вариантов осуществления настоящего изобретения для антигенсвязывающей молекулы по настоящему изобретению и контрольной антигенсвязывающей молекулы вызываемые ими эффекты лекарственного средства различны. В еще одном из вариантов осуществления настоящего изобретения эффекты в качестве лекарственного средства различны в тканях, где концентрация соединения низкая. Примеры ткани с низкой концентрацией соединения включают неопухолевые ткани, такие как нормальные ткани. Антигенсвязывающая молекула может быть представлена в виде фармацевтического препарата, содержащего антигенсвязывающую молекулу. В некоторых вариантах осуществления настоящего изобретения в ткани с низкой концентрацией соединения антигенсвязывающая молекула настоящего изобретения имеет пониженную активность по повреждению клетки-мишени (клетки, экспрессирующей антиген) по сравнению с контрольной антигенсвязывающей молекулой. В некоторых вариантах осуществления настоящего изобретения в ткани с низкой концентрацией соединения антигенсвязывающая молекула по настоящему изобретению требует более высокой дозы для повреждения клетки по сравнению с контрольной антигенсвязывающей молекулой. В некоторых вариантах осуществления настоящего изобретения в ткани с низкой концентрацией соединения антигенсвязывающая молекула настоящего изобретения демонстрирует пониженный уровень побочного эффекта по сравнению с контрольной антигенсвязывающей молекулой. В некоторых вариантах осуществления настоящего изобретения в ткани с низкой концентрацией соединения доза, при которой наблюдают побочный эффект, выше для антигенсвязывающей молекулы по настоящему изобретению по сравнению с контрольной антигенсвязывающей молекулой. Эти различия в ответе необязательно наблюдают для каждой ткани (например, для всех тканей, демонстрирующих низкую концентрацию соединения), и достаточно, чтобы различия наблюдались для некоторых тканей. В некоторых вариантах осуществления настоящего изобретения побочный эффект является нежелательным явлением в нормальных тканях (например, повреждение нормальных тканей).
В одном из вариантов осуществления настоящего изобретения для антигенсвязывающей молекулы по настоящему изобретению и контрольной антигенсвязывающей молекулы эффекты, вызываемые лекарственным средством, по существу одинаковы. В некоторых вариантах осуществления настоящего изобретения эффекты в качестве лекарственного средства практически такие же в ткани, где концентрация соединения высока. Примеры ткани с высокой концентрацией соединения включают опухолевую ткань. Антигенсвязывающая молекула может быть представлена в виде фармацевтического препарата, содержащего антигенсвязывающую молекулу. В некоторых вариантах осуществления настоящего изобретения в ткани с высокой концентрацией соединения активность повреждения клетки-мишени (клетки, экспрессирующей антиген) по существу одинакова для антигенсвязывающей молекулы по настоящему изобретению и контрольной антигенсвязывающей молекулы. В некоторых вариантах осуществления настоящего изобретения в ткани с высокой концентрацией соединения доза, необходимая для повреждения клетки, по существу одинакова для антигенсвязывающей молекулы по настоящему изобретению и контрольной антигенсвязывающей молекулы. В некоторых вариантах осуществления настоящего изобретения в ткани с высокой концентрацией соединения уровень проявления терапевтического эффекта по существу одинаков для антигенсвязывающей молекулы по настоящему изобретению и контрольной антигенсвязывающей молекулы. В некоторых вариантах осуществления настоящего изобретения в ткани с высокой концентрацией соединения доза, при которой наблюдают терапевтический эффект, по существу одинакова для антигенсвязывающей молекулы по настоящему изобретению и контрольной антигенсвязывающей молекулы. В некоторых вариантах осуществления настоящего изобретения терапевтический эффект представляет противоопухолевый эффект (например, регресс опухоли, индукцию гибели опухолевых клеток или ингибирование пролиферации опухолевых клеток и т.д.).
В другом варианте осуществления настоящего изобретения предусматривают антигенсвязывающую молекулу для использования в качестве лекарственного средства. В еще одном варианте осуществления настоящего изобретения предусматривают антигенсвязывающую молекулу для использования при лечении опухоли. В другом варианте осуществления настоящего изобретения предусматривают использование антигенсвязывающей молекулы при получении лекарственного средства. В еще одном варианте осуществления настоящего изобретения предусматривают использование антигенсвязывающей молекулы для лечения опухоли. В дополнительном варианте осуществления настоящего изобретения предусматривают способ лечения опухоли, включающий введение эффективного количества антигенсвязывающей молекулы индивиду с опухолью. «Индивидом» по любому из указанных выше вариантов осуществления настоящего изобретения предпочтительно является человек.
<Методы измерения концентрации АТФ>
В другом варианте осуществления настоящего изобретения предусматривают методы измерения концентрации АТФ в растворе. В некоторых вариантах осуществления настоящего изобретения вышеупомянутые методы включают стадии (а) контактирования расщепленной клетки Luc / HEK293, экспрессирующей рецептор P2Y, с раствором и (б) измерение активности люциферазы в клетке. Рецептор P2Y представляет собой рецептор на клеточной поверхности и семимембранный, связанный с G-белком (GPCR) рецептор, эндогенным лигандом которого являются внеклеточные пуриновые нуклеотиды (АТФ, АДФ), пиримидиновые нуклеотиды (УТФ, УДФ), сахара нуклеотидов и т.д. В других вариантах осуществления настоящего изобретения рецептором P2Y является P2Y1 (номер в каталоге U42029), P2Y2 (номер в каталоге U07225), P2Y4 (номер в каталоге Х91852), P2Y6 (номер в каталоге Х97058), P2Y11 (номер в каталоге AF030335), P2Y12 (номер в каталоге AJ320495), P2Y13 (номер в каталоге АР295368)или P2Y14 (номер в каталоге D13626). В некоторых вариантах осуществления настоящего изобретения рецептором P2Y является P2Y11 (номер в каталоге AF030335). Расщепленные клетки Luc / HEK293 представляют собой рекомбинантные клетки, полученные с использованием технологии расщепленной люциферазы, принадлежащей ProbeX (Misawa с соавт., Anal. Chem., 82, 2010, 2552-2560), и экспрессирует следующие два белка: (i) слитый белок, в котором С-концевой фрагмент из двух фрагментов молекулы люциферазы, разделенной на два, присоединен к С-концу рецептора P2Y, и (ii) слитый белок, в котором N-концевой фрагмент из двух фрагментов молекулы люциферазы, разделенной на два, присоединены к N-концу бета-аррестина (номер в каталоге NM 004313).
Когда АТФ добавляют к расщепленным клеткам Luc / HEK293, экспрессирующим рецептор P2Y, взаимодействие между рецептором P2Y и бета-аррестином происходит в непосредственной близости от клеточной мембраны, и в результате фрагменты расщепленной люциферазы связываются, чтобы восстановить активную молекулу люциферазы внутри клетки. Активность люциферазы можно измерить с использованием субстрата люциферазы, такого как, например, люциферина. В некоторых вариантах осуществления настоящего изобретения вышеописанные способы дополнительно включают стадию контактирования клетки с раствором, содержащим субстрат люциферазы. Раствор, содержащий субстрат люциферазы, может контактировать с клеткой до стадии (а) или после стадии (а).
В некоторых вариантах осуществления настоящего изобретения раствор, для которого измеряют концентрацию АТФ, может быть раствором in vitro или жидкостью организма в ткани in vivo. В других вариантах осуществления настоящего изобретения жидкость организма может быть выбрана из группы, состоящей из крови, лимфатической жидкости, тканевой жидкости (межтканевой жидкости, межклеточной жидкости, интерстициальной жидкости), жидкости полости тела (жидкости серозной полости, плевральной жидкости, асцитной жидкости, перикардиальной жидкости), спинномозговой жидкости, суставной жидкости (синовиальной жидкости) и внутриглазной жидкости. В некоторых вариантах осуществления настоящего изобретения жидкость тела является межклеточной жидкостью. В другом варианте осуществления настоящего изобретения ткань может быть здоровой тканью / нормальной тканью и может быть болезненной тканью (например, опухолевой тканью). В одном из вариантов осуществления настоящего изобретения когда описанными выше способами измеряют концентрацию АТФ в жидкости тела в ткани in vivo, этап (i) может быть этапом трансплантации расщепленной клетки Luc / HEK293, экспрессирующей рецептор P2Y, в ткань in vivo.
В некоторых вариантах осуществления настоящего изобретения люциферазную активность можно измерить, фиксируя свет, излучаемый, когда люцифераза реагирует со своим субстратом, таким как люциферин. В случае, если раствор представляет собой раствор in vitro, такое свечение может быть измерено с использованием устройства для обнаружения люминесценции, такого как планшет-ридер. В случае, если раствор представляет собой жидкость организма в ткани in vivo, люминесценцию можно измерить с использованием устройства формирования изображения люминесценции in vivo и т.п. Количественная оценка на основе измерений может быть проведена методами, известными в данной области, например, путем измерения растворов, содержащих известную концентрацию АТФ, и построения калибровочной кривой, показывающей корреляцию между концентрацией АТФ и интенсивностью люминесценции.
V. Способы скрининга
(1) Способ скрининга антигенсвязывающего домена или антигенсвязывающей молекулы, которая обладает антигенсвязывающей активностью, зависящей от молекулярного соединения, с использованием поливалентных антигенов.
В одном варианте осуществления настоящего изобретения предусматривают способ высокоэффективного и высокоточного скрининга, идентификации и получения антигенсвязывающего домена или антигенсвязывающей молекулы, которая обладает антигенсвязывающей активностью, зависящей от молекулярного соединения, с использованием слитой молекулы, полученной путем слияния двух или более молекул (двух или более единиц) антигена с одной молекулой (одной единицей) молекулы-партнера слияния.
В одном из вариантов осуществления настоящего изобретения в способе по настоящему изобретению, способ скрининга антигенсвязывающего домена или антигенсвязывающей молекулы, которая обладает антигенсвязывающей активностью, зависящей от молекулярного соединения, включает:
(а) контактирование, в присутствии низкомолекулярного соединения, антигенсвязывающего домена или антигенсвязывающей молекулы или библиотеки антигенсвязывающих доменов или антигенсвязывающих молекул со слитой молекулой, в которой две или более единиц антигена слиты с одной единицей партнера по слиянию,
(б) размещение антигенсвязывающего домена или антигенсвязывающей молекулы, связанной с антигеном в составе слитой молекулы на стадии (а), в отсутствие или в присутствии низкой концентрации соединения, и
(в) выделение антигенсвязывающего домена или антигенсвязывающей молекулы, диссоциированной на стадии (б).
В одном из вариантов осуществления настоящего изобретения в способе по настоящему изобретению молекула-партнер слияния, с которой слиты две или более молекул (две или более единиц) антигена, представляет собой димерную область Fc.
В одном из вариантов осуществления настоящего изобретения в способе по настоящему изобретению область Fc включает первую субъединицу Fc и вторую субъединицу Fc, и один антиген слит как с первой, так и со второй субъединицей Fc.
В одном из вариантов осуществления настоящего изобретения в способе по настоящему изобретению один из антигенов слит с N-концом как первой, так и второй субъединиц Fc.
В одном из вариантов осуществления настоящего изобретения в способе по настоящему изобретению библиотека антигенсвязывающих доменов или антигенсвязывающих молекул представляет собой фаговую библиотеку.
В одном из вариантов осуществления настоящего изобретения в способе по настоящему изобретению фаги, включенные в фаговую библиотеку, являются фагами, имеющими на своей поверхности два или более антигенсвязывающих домена или антигенсвязывающие молекулы.
В одном из вариантов осуществления настоящего изобретения в способе по настоящему изобретению фаги, включенные в фаговую библиотеку, являются фагами, имеющими дефект в гене pIII, производном от фага-помощника.
В одном из вариантов осуществления настоящего изобретения в способе по настоящему изобретению антиген может быть любым антигеном, как описано выше, и примеры могут включать различные типы «антигенов», описанные в разделе «IV. Композиции и способы (антигенсвязывающие молекулы, связывающая активность которых с антигенами изменяется в зависимости от концентрации низкомолекулярного соединения)», «Б. Антиген». В одном из вариантов осуществления настоящего изобретения в качестве антигена предпочтительно могут быть предоставлены антигены мембранного типа (например, костимулирующие молекулы, такие как CD137, CTLA4, CD40, ОХ40, RANK, GITR и ICOS).
В одном из вариантов осуществления настоящего изобретения в способе по настоящему изобретению слияние области Fc (субъединицы Fc) и антигена может быть получено обычным методом с использованием технологии генетической рекомбинации, как описано выше.
В одном из вариантов осуществления настоящего изобретения в способе по настоящему изобретению предусматривают в качестве низкомолекулярного соединения различные соединения, как описано выше (например, соединения, специфичные для опухолевой ткани, такие как нуклеозиды, имеющие структуру пуринового кольца, такие как аденозин (ADO), аденозинтрифосфат (АТР), аденозиндифосфат (ADP), аденозинмонофосфат (AMP), инозин, а также коммерчески доступный АДФ-бета-S (фирма Sigma); аминокислоты, такие как аланин, глутаминовая кислота и аспарагиновая кислота; метаболиты аминокислот, такие как кинуренин, антраниловая кислота, 3-гидроксикинуренин и кинуреновая кислота; метаболиты арахидоновой кислоты, такие как простагландин Е2; первичные метаболиты гликолитического пути или цикла Кребса, такие как молочная кислота, янтарная кислота и лимонная кислота; и метаболиты никотинамида, такие как 1-метилникотинамид), могут применяться.
В одном из вариантов осуществления настоящего изобретения в способе по настоящему изобретению предусматривают контакт антигена (например, слитой молекулы из области Fc и антигена) с фаговой библиотекой антигенсвязывающих доменов или антигенсвязывающих молекул, выделение антигенсвязывающего домена или антигена -связывающей молекулы, связанной с антигеном, анализ выделенного антигенсвязывающего домена или антигенсвязывающей молекулы в присутствии или в отсутствие низкомолекулярного соединения можно проводить в соответствии с описанным выше способом и способом, описанным ниже в примерах, ссылаясь на известные методы в области техники, к которой относится настоящее изобретение.
В одном из вариантов осуществления настоящего изобретения способ по настоящему изобретению может быть способом, описанным в примере 2.
(2) Способ скрининга антигенсвязывающего домена или антигенсвязывающей молекулы, которая обладает антигенсвязывающей активностью, зависящей от двух или более различных низкомолекулярных соединений.
В одном варианте осуществления настоящего описания предусматривают способ скрининга антигенсвязывающего домена или антигенсвязывающей молекулы, которая обладает антигенсвязывающей активностью, зависящей от двух или более различных низкомолекулярных соединений.
В одном из вариантов осуществления настоящего изобретения способ по настоящему изобретению включает:
(а) контактирование антигенсвязывающего домена, или антигенсвязывающей молекулы, или библиотеки антигенсвязывающих доменов, или антигенсвязывающих молекул с антигеном в присутствии первого низкомолекулярного соединения,
(б) размещение антигенсвязывающего домена или антигенсвязывающей молекулы, связанной с антигеном на стадии (а), в отсутствие или в присутствии низкой концентрации первого соединения,
(в) выделение антигенсвязывающего домена или антигенсвязывающей молекулы, диссоциированных на стадии (б),
(г) контактирование антигенсвязывающего домена или антигенсвязывающей молекулы, выделенных на стадии (в), с антигеном в присутствии низкомолекулярного соединения,
(д) размещение антигенсвязывающего домена или антигенсвязывающей молекулы, связанной с антигеном на стадии (в), в отсутствие или в присутствии низкой концентрации второго соединения, и
(е) выделение антигенсвязывающего домена или антигенсвязывающей молекулы, диссоциированной на стадии (д),
причем способ не включает между этапами (в) и (г) амплификацию гена, кодирующего антигенсвязывающий домен или антигенсвязывающую молекулу, выделенную на этапе (в).
В одном из вариантов осуществления настоящего изобретения в способе по настоящему изобретению библиотека антигенсвязывающих доменов или антигенсвязывающих молекул представляет собой фаговую библиотеку.
В одном из вариантов осуществления настоящего изобретения в способе по настоящему изобретению антиген может быть любым антигеном, как описано выше, предпочтительно антигеном мембранного типа (например, костимулирующими молекулами, такими как CD137, CTLA4, CD40, ОХ40, RANK, GITR и ICOS).
В одном из вариантов осуществления настоящего изобретения в способе по настоящему изобретению в качестве низкомолекулярных соединений могут применяться различные описанные выше соединения, например, соединения, специфичные для опухолевой ткани, такие как нуклеозиды, имеющие структуру пуринового кольца, такие как аденозин (ADO), аденозинтрифосфат (АТР), аденозин дифосфат (АДФ), аденозинмонофосфат (АМФ) и инозин; аминокислоты, такие как аланин, глутаминовая кислота и аспарагиновая кислота; метаболиты аминокислот, такие как кинуренин, антраниловая кислота, 3-гидроксикинуренин и кинурениновая кислота; метаболиты арахидоновой кислоты, такие как простагландин Е2; первичные метаболиты гликолитического пути или цикла Кребса, такие как молочная кислота, янтарная кислота и лимонная кислота; и метаболиты никотинамида, такие как 1-метилникотинамид.
В одном из вариантов осуществления настоящего изобретения в способе по настоящему изобретению первое низкомолекулярное соединение и второе низкомолекулярное соединение могут быть любыми двумя типами различных соединений, выбранными из вышеупомянутых различных низкомолекулярных соединений.
В одном из вариантов осуществления настоящего изобретения в способе по настоящему изобретению контакт антигена с фаговой библиотекой антигенсвязывающих доменов или антигенсвязывающих молекул, выделение антигенсвязывающего домена или антигенсвязывающей молекулы, связанной с антигеном, анализ выделенного антигенсвязывающего домена связывающий домен или антигенсвязывающая молекула в присутствии или в отсутствие первого низкомолекулярного соединения, и анализ в присутствии или отсутствие второго низкомолекулярного соединения можно проводить в соответствии с описанным выше способом и способом, описанным ниже в примерах, ссылаясь на известные методы в области, к которому относится настоящее изобретение.
В одном из вариантов осуществления настоящего изобретения в качестве способа настоящего изобретения можно указать способ, описанный в примере 9.
(3) Способ скрининга антигенсвязывающего домена или антигенсвязывающей молекулы, которая обладает антигенсвязывающей активностью, зависящей от низкомолекулярного соединения, с использованием наивной библиотеки.
В одном объекте настоящего изобретения предусматривают способ скрининга антигенсвязывающего домена или антигенсвязывающей молекулы, которые обладают антигенсвязывающей активностью, зависящей от низкомолекулярного соединения, с использованием наивной библиотеки, содержащей множество фагов, презентирующих антигенсвязывающие домены или антигенсвязывающие молекулы (например, домены Fab) различных последовательностей антител человека, полученных из лимфоцитов человека (например, МКПК человека), которые не получили антигенной стимуляции.
В одном из вариантов осуществления настоящего изобретения способ по настоящему изобретению включает:
(а) контактирование наивной библиотеки антигенсвязывающих доменов или антигенсвязывающих молекул с антигеном в присутствии низкомолекулярного соединения,
(б) размещение антигенсвязывающего домена или антигенсвязывающей молекулы, связанной с антигеном на стадии (а), в отсутствие или в присутствии низкой концентрации соединения, и
(в) выделение антигенсвязывающего домена или антигенсвязывающей молекулы, диссоциированной на стадии (б).
В одном из вариантов осуществления настоящего изобретения в способе настоящего изобретения наивная библиотека представляет собой фаговую библиотеку, содержащую фаги, презентирующие на своей поверхности два или более антигенсвязывающих домена или антигенсвязывающих молекул.
В одном из вариантов осуществления настоящего изобретения в способе настоящего изобретения наивная библиотека представляет собой библиотеку, содержащую фаги, имеющие дефект в гене pIII, производном от фага-помощника.
В одном из вариантов осуществления настоящего изобретения в способе настоящего изобретения наивная библиотека представляет собой библиотеку, содержащую фаги, полученные путем увеличения экспрессии антигенсвязывающего домена или антигенсвязывающей молекулы с помощью небольшой молекулярной добавки, которая увеличивает уровень экспрессии с промотора, регулирующего экспрессию антигенсвязывающего домена или антигенсвязывающей молекулы.
В одном из вариантов осуществления настоящего изобретения в способе настоящего изобретения низкомолекулярная добавка представляет собой изопропил-β-тиогалактопиранозид (IPTG, isopropyl-β-thiogalactopyranoside) или арабинозу.
В одном из вариантов осуществления настоящего изобретения в способе настоящего изобретения антиген может быть любым антигеном, как описано выше, предпочтительно антигеном мембранного типа (например, костимулирующей молекулой, такой как CD 137, CTLA4, CD40, ОХ40, RANK, GITR или ICOS).
В одном из вариантов осуществления настоящего изобретения в способе по настоящему изобретению в качестве низкомолекулярного соединения могут применяться различные соединения согласно описанному выше (например, соединения, специфичные для опухолевой ткани, такие как нуклеозиды, имеющие структуру пуринового кольца, такие как аденозин (ADO), аденозинтрифосфат (АТФ), аденозиндифосфат (АДФ), аденозинмонофосфат (АМФ) и инозин; аминокислоты, такие как аланин, глутаминовая кислота и аспарагиновая кислота; метаболиты аминокислот, такие как кинуренин, антраниловая кислота, 3-гидроксикинуренин и кинуреновая кислота, метаболиты арахидоновой кислоты, такие как простагландин Е2; первичные метаболиты гликолитического пути или цикла Кребса, такие как молочная кислота, янтарная кислота и лимонная кислота, и метаболиты никотинамида, такие как 1 -метилникотинамид).
В одном из вариантов осуществления настоящего изобретения в способе по настоящему изобретению контакт антигена с фаговой библиотекой антигенсвязывающих доменов или антигенсвязывающих молекул, выделение антигенсвязывающего домена или антигенсвязывающей молекулы, связанной с антигеном, анализ выделенного антигенсвязывающего домена или антигенсвязывающей молекулы в присутствии или в отсутствие низкомолекулярного соединения может быть осуществлен в соответствии с описанным выше способом и способом, описанным ниже в примерах, со ссылкой на хорошо известные методы в области техники, к которой относится настоящее изобретение.
В одном из вариантов осуществления настоящего изобретения в качестве способа настоящего изобретения может быть представлен способ, описанный в примере 10.
ПРИМЕРЫ
Ниже приведены примеры способов и композиций по настоящему изобретению. Учитывая приведенное выше описание изобретения, очевидно, что могут быть реализованы различные другие варианты его осуществления.
Пример 1. Получение антигена
(1-1) Получение внеклеточной области CD137 человека
Внеклеточный домен CD137 человека (также называемый hCD137) получают способами, известными специалистам в данной области. В частности, связаны расположенный ниже по цепи генный фрагмент, кодирующий внеклеточную область CD 137 человека, генный фрагмент, кодирующий гистидиновую метку, и фрагмент гена, кодирующий конкретную последовательность, к которой добавлен биотин (последовательность AviTag, SEQ ID NO: 86). Фрагмент гена, кодирующий белок, в котором связаны внеклеточная область CD137 человека, гистидиновая метка и Avitag (CD137 человека или hCD137-HisBAP, SEQ ID NO: 87), включают в вектор экспрессии для животных клеток. Сконструированный плазмидный вектор трансфицируют в клетки FreeStyle293 (фирма Invitrogen), используя 293-фектин (фирма Invitrogen). Одновременно трансфицируют плазмидный вектор, содержащий ген, экспрессирующий EBNA1 (SEQ ID NO: 88). Клетки, трансфицированные генами в соответствии с вышеупомянутой процедурой, культивируют при 37°С, в атмосфере 8% СО2; и представляющий интерес белок (внеклеточная область CD137 человека) секретируется в супернатант культуры. Эту среду для культивирования клеток фильтруют через фильтр с порами 0,22 мкм для получения супернатанта культуры. Супернатант культуры вносят в колонку HisTrap-HP (фирма GE Healthcare) и внеклеточную область CD137 человека связывают с колонкой.
Внеклеточную область CD137 человека элюируют раствором 20 мМ фосфата натрия, 500 мМ хлорида натрия и 500 мМ имидазола, рН 7,5. Затем агрегаты удаляют гель-фильтрационной хроматографией с использованием колонки Superdex 200 26/600 (фирма GE Healthcare) для получения очищенной внеклеточной области CD137 человека. Концентрацию CD137 человека рассчитывают по методу Расе с соавт.на основе аминокислотной последовательности, исключая сигнальную последовательность, удаленную из SEQ ID NO: 87 (Расе C.N., с соавт. Protein Science, 4, 1995, 2411-2423).
(1-2) Получение внеклеточного домена CD 137 человека (hCD137 (расщепленный FXa))
Внеклеточный домен CD137 человека (также называемый hCD137 (расщепленный протеазой FXa)) получают способом, известным специалистам в данной области. В частности, фрагмент гена, кодирующий последовательность расщепления фактора Ха, и фрагмент гена, кодирующий константную область антитела, соединяют ниже по цепи от фрагмента гена, кодирующего внеклеточную область CD 137 человека. Фрагмент гена, кодирующий белок, в котором связаны внеклеточная область CD137 человека, расщепляемая протеазой FactorXa последовательность и константная область антитела (hCD137-F-Fc, SEQ ID NO: 89), включен в вектор экспрессии животного. Сконструированный плазмидный вектор трансфицируют в клетки FreeStyle293 (фирма Invitrogen) с использованием 293-фектина (фирма Invitrogen). Одновременно трансфицируют плазмидный вектор, содержащий ген, кодирующий EBNA1 (SEQ ID NO: 88). Клетки, трансфицированные генами в соответствии с вышеупомянутыми процедурами, культивируют при 37°С в атмосфере 8% CO2, при этом hCD137-F-Fc секретируется в супернатант культуры. Среду для культивирования клеток фильтруют через фильтр с порами 0,22 мкм и собирают культуральный супернатант.
Супернатанты культур клеток наносят на колонку, заполненную белком А (продукт MabSelect SuRe, фирма GE Healthcare). hCD137-F-Fc связывается с колонкой, после чего его элюируют 50 мМ раствором уксусной кислоты. После нейтрализации элюата с помощью 1 М трис-HCl, рН 8,0, растворитель hCD137-F-Fc заменяют раствором 20 мМ Трис, 100 мМ хлорид натрия, 2 мМ хлорид кальция, рН 8,0. Затем добавляют протеазу Factor Ха (фирма NEW ENGLAND BioLabs, номер в каталоге P8010L) и расщепляют hCD137-F-Fc. Затем добавляют ингибитор протеазы (продукт DNS-GGACK, фирма Calbiochem, номер в каталоге 251700), чтобы остановить реакцию. К раствору, в котором действует протеаза, добавляют 5М хлорида натрия и весь объем приготовленного раствора вносят в колонку HiTrap Benzamidine (фирма GE Healthcare, номер в каталоге 17-5143-01). Пропущенный через колонку раствор затем наносят на колонку с белком А (продукт MabSelect SuRe, фирма GE Healthcare) и собирают прошедшие сквозь колонку фракции. Из пропущенных фракций удаляют агрегаты с помощью гель-фильтрационной хроматографии, используя Superdex 200 Increase, 10/30 (фирма GE Healthcare, номер в каталоге 28990944), получая очищенную внеклеточную область CD137 человека.
(1-3) Получение биотинилированного слитого с Fc CD137 человека
Биотинилированный слитый с Fc CD137 человека (также называемый «биотинилированный hCD137-Fc», или «био-hCD137-Fc», или hCD137-Fc-Bio) получают способами, известными специалистам в данной области. В частности, фрагмент гена, кодирующий константную область антитела, и фрагмент гена, кодирующий специфическую последовательность, к которой добавлен биотин (последовательность AviTag, SEQ ID NO: 86), присоединяют ниже по цепи от фрагмента гена, кодирующего внеклеточную область CD137 человека. Фрагмент гена, кодирующий белок (Fc, слитый с CD137 человека, SEQ ID NO: 90), в котором связаны внеклеточная область CD137 человека, константная область антитела и Avitag, включен в вектор экспрессии животного. Сконструированный плазмидный вектор трансфицируют в клетки FreeStyle293 (фирма Invitrogen) с использованием 293-фектина (фирма Invitrogen). Одновременно трансфицируют ген, экспрессирующий EBNA1 (SEQ ID NO: 88), и ген, экспрессирующий биотинлигазу (BirA, SEQ ID NO: 91), добавляют биотин с целью биотинилирования Fc, слитого с CD 137 человека. Клетки, трансфицированные генами в соответствии с вышеупомянутыми процедурами, культивируют при 37°С в атмосфере 8% CO2; белок интереса (биотинилированный слитый с Fc CD137 человека) секретируется в супернатант культуры. Среду для культивирования клеток фильтруют через фильтр с порами 0,22 мкм и собирают культуральный супернатант.
Супернатант культуры вносят в колонку, заполненную белком A (MabSelect SuRe, GE Healthcare), и биотинилированный слитый с Fc CD137 человека связывается с колонкой. Биотинилированный слитый с Fc CD 137 человека элюируют 50 мМ раствором уксусной кислоты. Затем агрегаты удаляют гель-фильтрационной хроматографией с использованием Superdex 200, 26/600 (фирма GE Healthcare) и получают очищенный биотинилированный слитый с Fc CD137 человека.
(1-4) Получение слитого с Fc CD137 человека
Слитый с Fc CD137 человека (также называемый hCD137-Fc) получают способами, известными специалистам в данной области. В частности, фрагмент гена, кодирующий константную область антитела, и фрагмент гена, кодирующий специфическую последовательность, к которой добавлен биотин (последовательность AviTag, SEQ ID NO: 86), присоединяют ниже по цепи от фрагмента гена, кодирующего внеклеточную область CD137 человека. Фрагменты гена, кодирующие белок (слитый с Fc человека CD137, SEQ ID NO: 90), в котором внеклеточная область CD137 человека, константная область антитела и Avitag включены в вектор экспрессии животного. Сконструированный плазмидный вектор трансфицируют в клетки FreeStyle293 (фирма Invitrogen) с использованием 293-фектина (фирма Invitrogen). Одновременно трансфицируют ген, экспрессирующий EBNA1 (SEQ ID NO: 88). Клетки, трансфицированные генами в соответствии с вышеупомянутыми процедурами, культивируют при 37°С в атмосфере 8% СО2; белок интереса (слитый с Fc CD137 человека) секретируется в супернатант культуры. Среду для культивирования клеток фильтруют через фильтр с порами 0,22 мкм и собирают культуральный супернатант.
Супернатант культуры вносят в колонку, заполненную белком A (MabSelect SuRe, GE Healthcare), и слитый с Fc CD137 человека связывается с колонкой. Слитый с Fc CD137 человека элюируют 50 мМ раствором уксусной кислоты. Затем агрегаты удаляют гель-фильтрационной хроматографией с использованием Superdex 200, 26/600 (фирма GE Healthcare) и получают очищенный слитый с Fc CD137 человека.
(1-5) Получение биотинилированного слитого с Fc CD137 обезьяны
Биотинилированный слитый с Fc CD 137 обезьяны (также называемый cyCD137-Fc-BAP) получают способами, известными специалистам в данной области. В частности, фрагмент гена, кодирующий константную область антитела, и фрагмент гена, кодирующий специфическую последовательность, к которой добавлен биотин (последовательность AviTag, SEQ ID NO: 86), присоединяют ниже по цепи от фрагмента гена, кодирующего внеклеточную область CD 137 обезьяны. Фрагмент гена, кодирующий белок, в котором внеклеточная область CD 137 обезьяны, константная область антитела и Avitag связаны (слитый с Fc обезьяны CD137, SEQ ID NO: 92), включена в вектор экспрессии животного.
Сконструированный плазмидный вектор трансфицируют в клетки FreeStyle293 (фирма Invitrogen) с использованием 293-фектина (фирма Invitrogen). Одновременно трансфицируют ген, экспрессирующий EBNA1 (SEQ ID NO: 88), и ген, экспрессирующий биотинлигазу (BirA, SEQ ID NO: 91); биотин добавляют с целью нанесения метки в виде биотина для слитого с Fc CD137 обезьяны. Клетки, трансфицированные генами в соответствии с вышеупомянутыми процедурами, культивируют при 37°С в атмосфере 8% CO2; белок интереса (биотинилированный слитый с Fc CD137 обезьяны) секретируется в супернатант культуры. Среду для культивирования клеток фильтруют через фильтр с порами 0,22 мкм и собирают культуральный супернатант.
Супернатант культуры вносят в колонку, заполненную белком А (MabSelect SuRe, GE Healthcare), и биотинилированный слитый с Fc CD137 обезьяны связывается с колонкой. Биотинилированный слитый с Fc CD 137 обезьяны элюируют 50 мМ раствором уксусной кислоты. Затем агрегаты удаляют гель-фильтрационной хроматографией с использованием Superdex 200, 26/600 (фирма GE Healthcare) и получают очищенный биотинилированный слитый с Fc CD 137 обезьяны.
Пример 2. Получение АТФ-зависимых анти-СР137 антител
(2-1) Получение антител с антигенсвязывающей активностью, зависящей от низкомолекулярных соединений (антитела, переключаемые низкомолекулярными соединениями), из рационально разработанной библиотеки, использующей АТФ (1)
(2-1-1) Пэннинг
Антитела, которые проявляют связывающую активность по отношению к антигенам в присутствии аденозинтрифосфата (аденозин-5'-трифосфат; АТФ), были получены из рационально разработанной библиотеки фагового дисплея антител, созданной в предыдущем патенте WO 2015/083764. Следует отметить, что антитела с антигенсвязывающей активностью (антиген, например, CD 137), зависящей от низкомолекулярных соединений, могут называться «переключаемыми антителами» или «антителами, переключаемыми низкомолекулярными соединениями», и антитела с антигенсвязывающей активностью (антиген CD137), зависящей от АТФ, могут называться «переключаемыми антителами» или «АТФ-переключаемыми антителами». Для получения антител собирают фаги, презентирующие антитела, которые проявляют связывающую активность в присутствии АТФ по отношению к антигену, захваченному на гранулах. Затем фаги собирают из элюатов, элюированных с гранул в отсутствие АТФ.
Фаги получают обычным методом из бактерии Е. coli, несущей фагмиду, сконструированную методом фагового дисплея. В частности, Е. coli, несущую сконструированный фагмидный вектор, инфицируют фагом M13KO7ΔpIII (обозначенным как «гиперфаг») (фирма PROGEN Biotechnik), и фаги собирают из супернатантов, культивируемых в течение ночи при 30°С. Суспензию фаговой библиотеки получают путем разбавления трис-буферным физиологическим раствором (TBS, Tris-Buffered Saline) популяции фагов, которых осаждают путем добавления 2,5 М NaCl/10% PEG к культуре Е. coli, в которой вырабатывается фаг. Затем к суспензии фаговой библиотеки добавляют БСА до конечной концентрации 4%. Пэннинг проводят с использованием антигена, иммобилизованного на магнитных гранулах. В качестве магнитных гранул используют гранулы Sera-Mag NeutrAvidin (фирма Thermo Fisher Scientific) или Dynabeads M-280 StreptAvidin (фирма Life Technologies). В качестве антигена применяют биотинилированный аденозинтрифосфат (аденозин-5'-трифосфат; АТФ) от фирмы Jena Bioscience, или hCD137-Fc-Bio (SEQ ID NO: 90) или био-hCD137 (SEQ ID NO: 89), которые биотинилированы из hCD137 (расщепление FXa), с использованием предварительно измеренного NHS-PEO4-биотина без взвешивания (фирма PIERCE), полученного в примерах 1-2 и 1-3.
Пэннинг проводят для эффективного получения небольшой молекулы, которая может играть роль переключателя в зависимых от раковых тканях переключаемых антителах. В частности, пэннинг для обогащения антителами, которые связываются с антигеном в присутствии АТФ, который представляет собой небольшую молекулу, и не связываются с антигеном в отсутствие АТФ, выполняют со ссылкой на методы, показанные в предыдущем патенте WO 2015/083764. В первом раунде для всех биотинилированных hCD137 (bio-hCD137), hCD137-Fc-Bio и биотинилированного АТФ (био-АТФ) пэннинг выполняют с использованием метода, в котором твердофазная иммобилизация биотинилированных антигенов на магнитных гранулах (называемая «твердофазным методом на основе гранул») предшествует добавлению приготовленной суспензии фаговой библиотеки. Для Био-АТФ пэннинг для обогащения антителами, которые могут связываться с антигеном (Био-АТФ) в отсутствие АТФ в качестве низкомолекулярного соединения, выполняют со ссылкой на методы, описанные в предыдущем патенте WO 2015/083764, отмеченном выше. Для hCD137-Fc-Bio добавляют 4 нМ небиотинилированной области Fc IgG1 человека для удаления антител, которые связываются с областью Fc. Собранный фаг добавляют к штамму Е. coli ER2738 для заражения фагом Е. coli, затем собранные клетки Е. coli инфицируют гиперфагом и фаг собирают из супернатанта после культивирования в течение ночи при 30°С.
Начиная со 2 раунда, пэннинг выполняют только на биотинилированном hCD137 (bio-hCD137) и hCD137-Fc-Bio для обогащения антител, которые связываются с антигеном в присутствии АТФ, и не связываются с антигеном в отсутствие АТФ, используя твердофазный метод с гранулами, ссылаясь на метод, показанный в предыдущем патенте WO 2015/083764. Для hCD137-Fc-Bio в обоих случаях добавляют 4 нМ небиотинилированной области Fc IgG1 человека для удаления антител, которые связываются с областью Fc. Подобный пэннинг повторяют до 5 раунда для обогащения представляющих интерес последовательностей антител.
(2-1-2) Оценка связывающей активности в присутствии и в отсутствие АТФ с помощью метода фагового иммуноферментного анализа ELIS А
Из отдельных колоний Е. coli, полученных описанными выше методами, супернатант фагосодержащей культуры собирают обычным методом {Methods Mol. Biol, 178, 2002, 133-145). NucleoFast 96 (фирма MACHEREY-NAGEL) используют для ультрафильтрации собранного супернатанта культуры. Элюат удаляют центрифугированием (4500 g, 45 мин) планшета NucleoFast 96, в который вносили по 100 мкл супернатанта культуры в каждую лунку. NucleoFast 96 снова промывают центрифугированием (4500 g в течение 30 мин), для чего добавляют 100 мкл H2O в каждую лунку. В итоге добавляют 100 мкл TBS, и суспензию фага, содержащуюся в супернатантах в лунках планшета NucleoFast 96, которые оставляли при комнатной температуре на 5 мин, извлекают.
Очищенные фаги с добавлением TBS или АТФ/TBS подвергают иммуноферментномуанализу (ELISA) по следующей процедуре. 384-луночные микропланшеты, покрытые стрептавидином (фирма Greiner), покрывают в течение ночи 10 мкл буфера TBS, содержащего биотинилированные антигены (bio-hCD137, hCD137-Fc-Bio и bio-Fc), полученные в примере 1. После удаления биотинилированных антигенов, которые не связались с планшетом, путем промывания каждой лунки планшета трис-буферным солевым раствором с Tween-20 (TBST), лунки блокируют 80 мкл 2% обезжиренного молока-TBS в течение 1 ч или более. 2% обезжиренное молоко-TBS удаляют промывками TBST, после чего допускают, чтобы презентирующие антитела фаги связывались с биотинилированным антигеном, присутствующим в каждой лунке в отсутствие и в присутствии АТФ, оставляя планшеты с подготовленными очищенными фагами, добавленными в каждую лунку, на 1 ч при комнатной температуре. В каждую лунку, промытую TBST или АТФ/TBST, добавляют HRP-конъюгированные анти-М13 антитела (фирма GE Healthcare, номер в каталоге 27-9421-01), разведенные TBS или АТФ/TBS, и планшет инкубируют в течение одного часа. После промывки TBST или АТФ/TBST проявление цвета раствора в каждой лунке, в которую был добавлен единый раствор ТМВ (фирма ZYMED), останавливают добавлением серной кислоты, а затем проявление цвета измеряют по поглощению при 450 нм.
В результате идентифицировано несколько антител с измененной связывающей активностью с bio-hCD137 или hCD137-Fc-Bio в присутствии и в отсутствие АТФ.
Результаты фагового ELISA с использованием клонов после 4 и 5 раундов пэннинга показаны в табл.9.
В настоящем изобретении клоны с абсорбцией 0,2 или более в присутствии АТФ и отношением S/N (сигнал/шум) абсорбции выше 2 в
присутствии/отсутствие антигена определяют как положительные клоны. Кроме того, среди положительных клонов клоны с коэффициентом абсорбцией S/N выше 2 раза в присутствии/отсутствие АТФ считают клонами с АТФ-зависимой антигенсвязывающей активностью (переключаемые клоны).
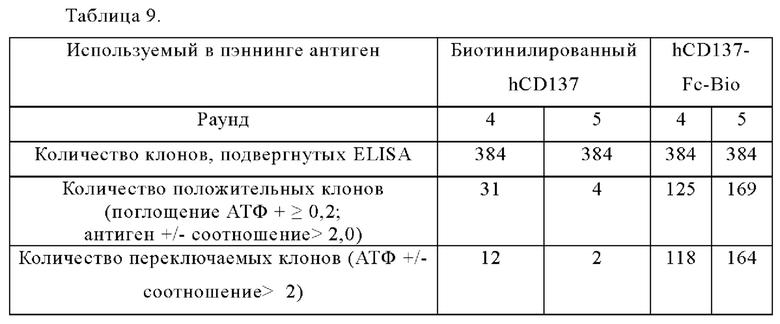
(2-1-3) Анализ последовательности переключаемых антител, антигенсвязывающая активность которых изменяется в зависимости от присутствия/отсутствия АТФ
Нуклеотидные последовательности генов, амплифицированных с использованием специфических праймеров pBAD-F, Glseq-R, из клонов с АТФ-зависимой антигенсвязывающей активностью (переключаемые клоны) анализируют на основе результатов фагового ELISA. В результате анализа получают нуклеотидные последовательности клонов, которые, как установлено, связываются с CD137 человека в присутствии АТФ, но не с CD137 человека в отсутствие АТФ.
(2-2) Получение антител, которые связываются с антигенами в присутствии небольшой молекулы, из рационально разработанной библиотеки, использующей АТФ (2)
(2-2-1) Пэннинг
Антитела, которые проявляют активность связывания с антигенами в присутствии АТФ, были получены из рационально разработанной библиотеки фагового дисплея антител, сконструированной в предыдущем патенте WO 2015/083764. Для получения антител собирают фаги, презентирующие антитела, которые проявляют связывающую активность в присутствии АТФ по отношению к антигенам, с последующим выделением фагов из элюата, элюированного с гранул в отсутствие АТФ.
Фаги получают обычным методом из бактерии Е. coli, несущей фагмиду, сконструированную методом фагового дисплея. В частности, Е. coli, несущую сконструированный фагмидный вектор, инфицируют фагом M13KO7TC (WO 2015046554A1) или M13KO7ΔpIII (обозначенным как «гиперфаг») (фирма PROGEN Biotechnik), и фаги выделяют из супернатантов, культивируемых в течение ночи при 30°С.Суспензию фаговой библиотеки получают путем разбавления трис-буферным физиологическим раствором (TBS, Tris-Buffered Saline) популяции фагов, которых осаждают путем добавления 2,5 М NaCl/10% PEG к культуре Е. coli, в которой вырабатывается фаг. Затем к суспензии фаговой библиотеки добавляют БСА до конечной концентрации 4%. Пэннинг проводят с использованием антигенов, иммобилизованных на магнитных гранулах. В качестве магнитных гранул используют гранулы с покрытием NeutrAvidin (Sera-Mag SpeedBeads NeutrAvidin-покрытие), гранулы NeutrAvidin (фирма TAMAGAWA SEIKI) или Dynabeads MyOne StreptAvidin T1 (фирма Thermo Fisher Scientific). В качестве антигена применяют hCD137-Fc-Bio или bio-hCD137, полученные в примере 1.
Пэннинг проводят для эффективного получения антител, переключаемых небольшой молекулой, которая может играть роль переключателя в раковых тканях. В частности, пэннинг для обогащения антителами, которые связываются с антигеном в присутствии небольшой молекулы аденозинтрифосфата (аденозин-5'-трифосфат; АТФ) и не связываются с антигеном в отсутствие АТФ, выполняют со ссылкой на методы, описанные в предыдущем патенте WO 2015/083764. Для био-hCD137 применяют метод, в котором добавляют приготовленную суспензию библиотеки фагов после заранее произведенной твердофазной иммобилизации биотинилированных антигенов на магнитных гранулах (так называемый «твердофазный метод гранул»), и метод, в котором добавляют магнитные гранулы после смешивания приготовленной суспензии фаговой библиотеки с заранее биотинилированными антигенами (так называемый «жидкофазный метод»). Для hCD137-Fc-Bio добавляют 4 нМ небиотинилированной области Fc IgG1 человека для удаления антител, которые связываются с областью Fc, и пэннинг осуществляют только жидкофазным методом. Собранный фаг добавляют к штамму Е. coli ER2738 для заражения фагом Е. coli, затем собранные клетки Е. coli инфицируют M13KO7TC (WO 2015046554A1) или M13KO7ΔpIII (гиперфагом) (фирма PROGEN Biotechnik) и после культивирования в течение ночи при 30°С фаг собирают из супернатанта. Подобный пэннинг повторяют до 5 раунда.
Пример (2-2-2). Оценка связывающей активности в присутствии и в отсутствие АТФ с помощью фагового ELISA
Из отдельных колоний Е. coli, полученных после каждого раунда описанными выше методами, супернатанты фагосодержащих культур собирают с использованием обычного метода (Methods Mol. Biol., 178, 2002, 133-145). NucleoFast 96 (фирма MACHEREY-NAGEL) применяют для ультрафильтрации собранных культуральных супернатантов. Элюат удаляют центрифугированием (4500 g, 45 мин) планшетов NucleoFast 96, в которые вносили по 100 мкл супернатанта культуры в каждую лунку. NucleoFast 96 снова промывают центрифугированием (4500 g в течение 30 мин), для чего добавляют 100 мкл H2O в каждую лунку. В итоге добавляют 100 мкл TBS, и суспензию фага, содержащуюся в супернатантах в лунках планшета NucleoFast 96, которые оставляли при комнатной температуре на 5 мин, извлекают.
Очищенные фаги с добавлением TBS или АТФ/TBS подвергают иммуноферментному анализу (ELISA) по следующей процедуре. Планшеты для микротитрования StreptaWell 96 (фирма Roche) иммобилизуют в течение ночи 100 мкл TBS, содержащего биотинилированные антигены (hCD137-Fc-Bio или bio-hCD137), полученные в Примере 1. После удаления биотинилированных антигенов, которые не связались с планшетом, путем промывания каждой лунки планшета трис-буферным солевым раствором (TBST), лунки блокируют 250 мкл 2% обезжиренного молока-TBS в течение 1 ч или более. 2% обезжиренное молоко-TBS удаляют, после чего фаги, презентирующие антитела, связываются с биотинилированным антигеном, присутствующим в каждой лунке в отсутствие и в присутствии АТФ, оставляя планшеты с подготовленными очищенными фагами, добавленными в каждую лунку при 37°С на 1 ч. В каждую лунку, промытую TBST или АТФ/TBST, добавляют HRP-конъюгированные анти-М13 антитела (фирма GE Healthcare, номер в каталоге 27-9421-01), разведенные TBS или АТФ/TBS, и планшет инкубируют в течение одного часа. После промывки TBST или АТФ/TBST проявление цвета раствора в каждой лунке, в которую был добавлен единый раствор ТМВ (фирма ZYMED), останавливают добавлением серной кислоты, а затем проявление цвета измеряют по поглощению при 450 нм.
В результате идентифицировано несколько антител с измененной связывающей активностью с bio-hCD137 или hCD137-Fc-Bio в присутствии и в отсутствие АТФ.
В качестве примера, результаты фагового ELISA с использованием клонов после 4 и 5 раундов пэннинга показаны в табл.10.
В настоящем изобретении клоны с абсорбцией 0,2 или более в присутствии АТФ и отношением S/N (сигнал/шум) абсорбции выше 2 в
присутствии/отсутствие антигена определяют как положительные клоны. Кроме того, среди положительных клонов клоны с коэффициентом абсорбцией S/N выше 2 раза в присутствии/отсутствие АТФ считают клонами с АТФ-зависимой антигенсвязывающей активностью (переключаемые клоны).
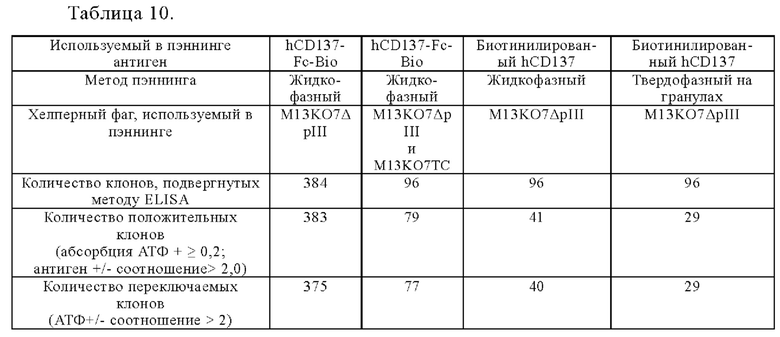
(2-2-3) Анализ последовательности переключаемых антител, антигенсвязывающая активность которых изменяется присутствием/отсутствием АТФ
Нуклеотидные последовательности генов, амплифицированных из клонов с АТФ-зависимой антигенсвязывающей активностью (переключаемые клоны) на основе результатов ELISA фага, анализируют с использованием специфических праймеров pBAD-F, Glseq-R. В результате анализа получены нуклеотидные последовательности клонов, которые, как установлено, связываются с CD 137 человека в присутствии АТФ, но не с CD137 человека в отсутствие АТФ.
(2-3) Отбор переключаемых антител
Семнадцать образцов отобраны из клонов, которые, как было установлено, обладают АТФ-зависимой антигенсвязывающей активностью, полученной в результате анализов в примерах 2-1-3 и 2-2-3; названия клонов повторно присвоены, как описано в таблице 11.

(2-4) Экспрессия и очистка переключаемых антител, антигенсвязывающая активность которых изменяется присутствием/отсутствием АТФ
Гены, кодирующие вариабельные области антител, описанных в табл.11, которые получают из рационально разработанной фаговой библиотеки, инсертируют в плазмиды IgG1 человека/Lambda для экспрессии у животных. Антитела экспрессируют с использованием описанных ниже методов. Приготовленные плазмиды вводят методом липофекции в производную от клеток линию клеток FreeStyle 293-F, полученную от почки плода человека (фирма Invitrogen), которую суспендируют в среде для экспрессии FreeStyle 293 (фирма Invitrogen) при плотности клеток 1,33×106 клеток/мл, полученную суспензию высевают по 3 мл в каждую лунку 6-луночных планшетов. Антитела очищают с использованием rProtein A Sepharose™ Fast Flow (фирма Amersham Biosciences) из супернатантов культур, культивированных в течение четырех дней в инкубаторе с СО2 (37°С, 8% СО2, 90 об/мин) методом, известным специалистам в данной области. Спектрофотометры используют для измерения абсорбции очищенных растворов антител при 280 нм. По полученным результатам измерений концентрации очищенных антител были рассчитывают с использованием коэффициентов экстинкции, рассчитанных методом РАСЕ (Protein Science, 4, 1995, 2411-2423).
(2-5) Оценка анти-CD137 антител, идентифицированных методом фагового дисплея
(2-5-1) Экспрессия и очистка переключаемых антител, которые связываются с антигенами, в зависимости от наличия или отсутствия АТФ и его метаболитов
Гены, кодирующие вариабельные области антител, полученных из рационально сконструированной фаговой библиотеки человека, инсертируют в плазмиды экспрессии животных, имеющие константную область тяжелой цепи, слитую с геном, кодирующим Gly и Lys ниже по цепи модифицированного IgG1 человека (Р253) (SEQ ID NO: 93), и константную область легкой цепи, лямбда-цепи Lamlib (SEQ ID NO: 63). Обозначения и порядковые номера клонов перечислены в табл. 12.
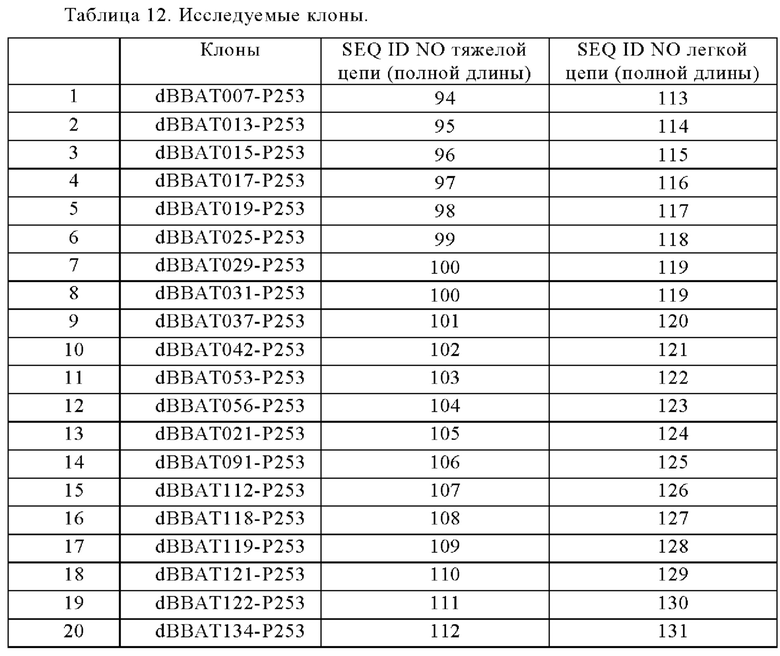
После экспрессирования антитела очищают методами, известными специалистам в данной области. Спектрофотометры применяют для измерения оптической плотности очищенных растворов антител при 280 нм. По установленным значениям рассчитывают концентрации очищенных антител, используя коэффициенты экстинкции, рассчитанные методом РАСЕ (Protein Science, 4, 1995, 2411-2423).
(2-5-2) Оценка методом поверхностного плазмонного резонанса влияния АТФ, АДФ и АМФ на связывание CD137 человека
Biacore Т200 (фирма GE Healthcare) используют для анализа взаимодействия антиген-антитело между анти-СD137антителами и hCD137 (расщепление FXa). Анти-CD137 антитела захватывают на сенсорном чипе СМ3 (фирма GE Healthcare), на котором был иммобилизован белок G (фирма CALBIOCHEM) в соответствующих количествах с помощью метода связывания с амином, и hCD137 (расщепление FXa), полученный в примере 1-2, может вступать в реакцию взаимодействовать. В качестве подвижного буфера используют 20 мМ ACES, 150 мМ NaCl, 2 мМ MgCl2, 0,05% Tween20 (рН 7,4) и 10 мМ глицин-HCl (рН 1,5) в качестве раствора для регенерации.
После улавливания антител к CD 137, суспендированных в TBS, в каждую проточную кювету вводят 500 нМ hCD137 (расщепление FXa) при скорости потока 10 мкл/мин в течение трех минут. Этот трехминутный период служит фазой связывания для hCD137 (расщепление FXa), а после окончания фазы связывания двухминутный период переключения на рабочий буфер служит фазой диссоциации для hCD137 (расщепление FXa). После завершения фазы диссоциации регенерирующий раствор вводят со скоростью 30 мкл/мин в течение 30 сек. Выше представлен цикл измерения связывающей активности анти-CD137 антител. Связывающее количество hCD137 (расщепленный FXa), которое взаимодействуете анти-С0137-антителом в фазе связывания, регулируют количеством захваченного антитела. Программное обеспечение Biacore Т200 Evaluation Software Version 2,0 используют для отображения количества связывания (RU) на 1RU захватываемого лиганда, и получают значения количества захваченного антитела и количества связываемого антигена. Количество связанного антигена показано в табл. 13. Поскольку количество связанного антигена отражает связывающую активность, можно сказать, что зависимость от низкомолекулярного соединения распознается, когда значение в присутствии низкомолекулярного соединения (АТФ, АДФ или АМФ) выше, чем значение без низкомолекулярного соединения. В частности, чем больше разница, тем выше зависимость от низкомолекулярного соединения.
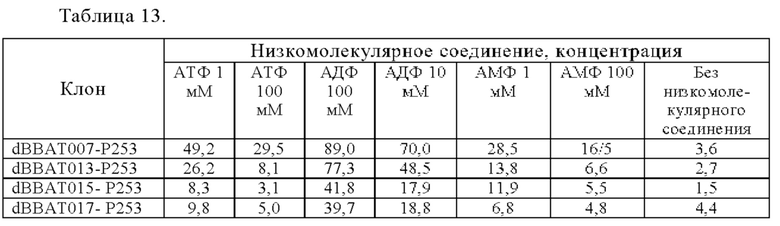
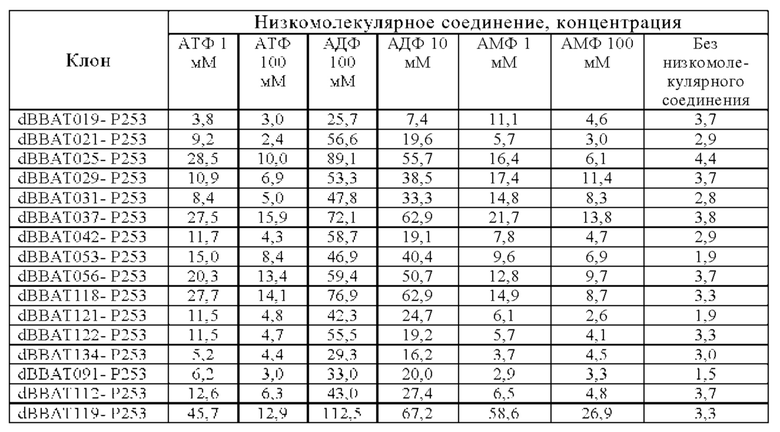
(2-5-3) Оценка связывающей активности с антигеном CD137 обезьяны
Полученные антитела оценивают по связыванию с CD137 обезьяны методом ELISA. Полученный в примере 1 cyCD137-Fc-BAP иммобилизуют на планшетах для микротитрования Streptawell. После удаления несвязавшегося антигена из планшетов промыванием каждой лунки планшетов промывочным буфером лунки блокируют 150 мкл блокирующего буфера (TBS с 2% БСА) в течение 1 ч или более. Блокирующий буфер удаляют из каждой лунки и в каждую лунку добавляют 100 мкл очищенных антител, разведенных в TBS с конечной концентрацией 1 мМ АДФ или TBS с конечной концентрацией 1 мМ АДФ. Планшет, в который вносили антитело, встряхивают при 600 об/мин в течение 1 ч. АР-конъюгированные антитела против лямбда человека (фирма BETHYL), разведенные в TBS с конечной концентрацией 1 мМ АДФ, добавляют в каждую лунку после промывания промывочным буфером (TBS с 0,1% Tween20), содержащим конечную концентрацию 1 мМ АДФ. После инкубации в течение одного часа и промывки промывочным буфером с конечной концентрацией 1 мМ АТФ, добавляют фосфатный субстрат BluePhos (KPL). Развитие цвета измеряют по абсорбции при 600 нм. Скорость увеличения абсорбции при концентрации антитела 0 мкг/мл показана в табл.14. Можно сказать, что образцы с зависимым от концентрации увеличением коэффициента абсорбции связаны с CD 137 обезьяны.
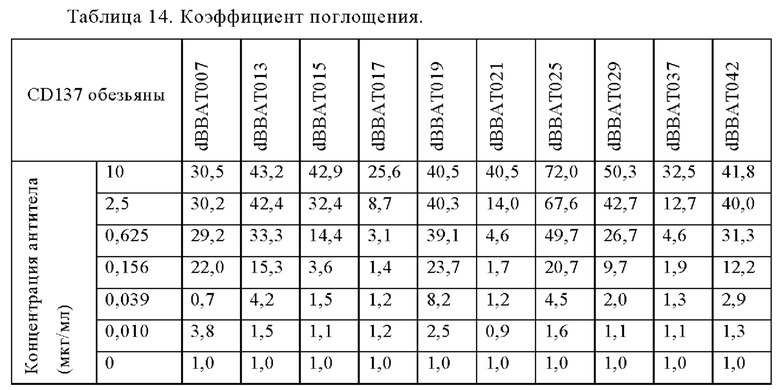

(2-5-4) Оценка агонистической активности CD137 с использованием клеток Jurkat
Клеточную линию GloResponse™ NF-kappa B-Luc2/4-lBB Jurkat (фирма Promega) используют для измерения активности антител in vitro. В каждую лунку 384-луночных планшетов добавляют 10 мкл клеток FcγRIIB СНО-К1 (фирма Promega), приготовленных в концентрации 2 × 106/мл в среде для анализа (99% RPMI, 1% ФСБ). Затем в каждую лунку добавляют 10 мкл раствора антитела, содержащего АДФ, или раствора антитела, содержащего АТФ, или раствора антитела, не содержащего АТФ или АДФ. Затем в каждую лунку добавляют 10 мкл клеточной линии GloResponseTM NF-κB-Luc2/4-1BB Jurkat, приготовленной в концентрации 2 × 10б/мл в среде для анализа (99% RPMI, 1% ФСБ). Конечная концентрация АДФ составляет 50 мкМ; конечная концентрация АТФ составляет 50 мкМ. Планшеты оставляют на шесть часов при 37°С в инкубаторе в атмосфере 5% CO2, затем 15 мин при комнатной температуре и добавляют 30 мкл реагента Bio-Glo в каждую лунку. Систему анализа люциферазы Bio-Glo (буфер и субстрат) применяют для реагента Bio-Glo. Впоследствии люминесценцию каждой лунки измеряют с помощью планшет-ридера. Значение люминесценции каждой лунки, деленное на величину люминесценции лунки без добавленных антител, представляет кратность люминесценции и служит индикатором для оценки активности каждого антитела.
Полученные результаты показаны на фиг. 1 и 2. Фиг. 1 и 2 подтверждают, что все антитела, кроме NS1-P253, которое не является переключаемым антителом, проявляют агонистическую активность CD137 человека АТФ- и АДФ-зависимым образом.
(2-6) Оценка агонистической активности CD137 in vitro переключаемых антител, полученных методом пэннинга. с использованием Т-клеток человека
(2-6-1) Размножение культур Т-клеток человека
Используют мононуклеарные клетки периферической крови человека, выделенные из образцов крови здоровых добровольцев. 50 мл крови смешивают с 0,5 мл гепарина и дополнительно разбавляют 50 мл ФСБ. Мононуклеарные клетки периферической крови человека выделяют в два этапа, описанные ниже. На первом этапе в пробирки Leucosep (фирма greiner bio-one) вносят Ficoll-Paque PLUS (фирма GE Healthcare) и центрифугируют 1000×g в течение одной минуты при комнатной температуре с последующим добавлением крови, разбавленной ФСБ, и центрифугируют при комнатной температуре в режиме 400×g в течение 30 мин. На втором этапе лейкоцитарный слой собирают из пробирок после центрифугирования и затем промывают 60 мл ФСБ (фирма Wako). Затем используют набор для активации/размножения Т-клеток человека (фирма MACS Miltenyi biotec) для размножения культур Т-клеток.
(2-6-2) Оценка агонистической активности CD137 in vitro с использованием Т-клеток человека
2×104 Т-клеток, происходящих из мононуклеарных клеток периферической крови, и 2×104 клеток REC-1 суспендируют в 100 мкл среды RPMI1640, содержащей 30 ЕД/мл IL-2 человека (фирма SIGMA), 10 нг/мл РМА (фирма SIGMA), 0,5 мкг/мл иономицина, 500 мкМ АДФ-бета-S (фирма Sigma) и 10% ФСТ (фирма Sigma), высевают в 96-луночные многолуночные планшеты с плоским дном (фирма Corning) с контрольным антителом IC17HdK-hIgG1/IC17L-k0, NS1-P253, которое является антителом с АТФ-независимой связывающей активностью в отношении CD137-человека (называемым во всех приводимых в настоящем описании примерах «непереключаемым антителом» или «непереключаемым анти-CD137 антителом»), или с любым из клонов, описанных в табл.12. NS1-P253, IC17HdK-hIgG1/IC17L-k0 и клоны, описанные в табл.12, оценивают в концентрации 10 мкг/мл. Также оценивают выработку IFN-γ в АДФ-бета-S-минус среде. Планшеты встряхивают и затем оставляют при 37°С в течение 72 ч в инкубаторе в атмосфере 5% CO2. Затем культуральные супернатанты собирают и определяют количество IFN-γ, содержащегося в культуральных супернатантах, используя набор Human IFN-γ ELISA Development Kit (фирма PeproTech). Метод ELISA осуществляют в соответствии с инструкциями производителя (фирма PeproTech). Измерения абсорбции проводят на приборе EnVision (фирма PerkinElmer).
Результаты показаны на фиг. 3.
Было подтверждено, что dBBAT007-P253, dBBAT013-P253, dBBAT015-Р253, dBBAT019-P253, dBBAT021-P253, dBBAT025-P253, dBBAT031-P253, dBBAT042-P253, dBBAT056-P253, dBBAT118-P253, dBBAT121-P253, dBBAT122-P253 и dBBAT119-P253 проявляют агонистическую активность в отношении CD137 человека в АДФ-бета-S-зависимым образом. АДФ-бета-S является аналогом АДФ, который менее подвержен гидролизу, чем АДФ. Это указывает на возможность того, что эти анти-CD137 человека переключаемые антитела проявляют агонистическую активность в отношении CD 137 человека, зависящую от небольшой молекулы, такой как АТФ, АДФ или АМФ.
(2-6-3) Оценка агонистической активности CD137 in vitro с использованием Т-клеток человека (2)
2×104 мононуклеарные клетки периферической крови человека, полученные из Т-клеток, и 2×104 клеток REC-1 суспендируют в 100 мкл среды RPMI1640, содержащей 30 ЕД/мл IL-2 человека (фирма SIGMA), 10 нг/мл РМА (фирма SIGMA), 0,5 мкг/мл иономицина, 500 мкМ АДФ-бета-S (фирма Sigma) и 10% ФСТ (фирма Sigma), и высевают в 96-луночные плоскодонные планшеты (фирма Corning), либо с NS1-P253 (непереключаемое антитело), либо с dBBAT119-Р253. NS1-P253 и dBBAT119-Р253 оценивают в концентрации 10, 2, 0,4, 0,08, 0,016, 0,0032 и 0,00064 мкг/мл. Планшеты встряхивают и оставляют при 37°С на 72 ч в инкубаторе в атмосфере 5% CO2. Затем собирают культуральные супернатанты и определяют количество IFN-γ, содержащегося в культуральных супернатантах, с использованием набора для метода ELISA для IFN-γ человека (фирма PeproTech). Измерения абсорбции проводят с помощью En Vision (фирма PerkinElmer).
Результаты представлены на фиг. 4.
Было подтверждено, что dBBAT119-Р253 проявляет агонистическую активность в отношении CD137 человека в присутствии АДФ-бета-S. Это указывает на возможность того, что dBBAT119-Р253 проявляет агонистическую активность в отношении CD137 человека в зависимости от небольшой молекулы, такой как АТФ, АДФ или АМФ.
Пример 3. Повышение связывающей активности антител, связывающих антигены в присутствии малых молекул, с использованием рационально разработанных библиотек легких и тяжелых цепей
(3-1) Создание библиотеки для повышения связывающей активности с использованием рационально разработанной библиотеки легких цепей
Для библиотеки антител, содержащей большое количество антител, собранных в примере 2-2-1, с АТФ-зависимой антигенсвязывающей активностью, усиление связывающей активности осуществляют путем повторной библиотекаризации легких цепей антитела.
Области легкой цепи и тяжелой цепи рационально сконструированной библиотеки фагового дисплея антитела, сконструированной в предыдущем патенте WO 2015/083764, используют для конструирования библиотеки легкой цепи антитела и библиотеки тяжелой цепи антитела для усиления связывающей активности. Их вводят в указанную выше библиотеку легких цепей или в области легкой цепи или тяжелой цепи библиотеки фагмидных векторов, собранные в примере 2-2-1, и вводят в штамм Е. coli ER2738 путем электропорации.
(3-2) Повышение связывающей активности антител с АТФ-зависимой антигенсвязывающей активностью с использованием рационально разработанной библиотеки
Фаги получают обычным способом из Е. coli, несущий сконструированную фагмиду фагового дисплея. В частности, штамм Е. coli, несущую сконструированный фагмидный вектор, инфицируют M13KO7TC (WO 2015/046554) или M13KO7ΔpIII (гиперфаг) (фирма PROGEN Biotechnik), и фаги собирают из супернатантов после культивирования в течение ночи при 30°С. Суспензию фаговой библиотеки получают разбавлением TBS популяции фагов, осажденных путем добавления 2,5 М NaCl/10% PEG к культуре Е. coli, в которой осуществляли выработку фага. Затем к раствору фаговой библиотеки добавляют БСА до конечной концентрации 4%. Пэннинг проводят, используя антиген, иммобилизованный на магнитных гранулах. Покрытые NeutrAvidin гранулы (Sera-Mag SpeedBeads NeutrAvidin-coated), гранулы NeutrAvidin (фирма TAMAGAWA SEIKI) или гранулы Dynabeads MyOne StreptAvidin T1 (фирма Thermo Fisher Scientific) применяют в качестве магнитных гранул. В качестве антигена применяют биотинилированный hCD137-Fc.
Пэннинг проводят для эффективного получения низкомолекулярных переключаемых антител, которые зависят от малой молекулы, которая может играть роль переключателя в раковых тканях. В частности, пэннинг для обогащения антител, которые связываются с антигенами в присутствии аденозинтрифосфата (аденозин-5'-трифосфата; АТФ) небольшую молекулы, и не связывается с антигенами в отсутствие АТФ, выполняют со ссылкой на методы, изложенные в предыдущем патенте WO 2015/083764.
(3-3) Оценка связывающей активности в присутствии и в отсутствие АТФ с помощью фагового ELISA
Из отдельных колоний Е. coli, полученных после каждого раунда описанными выше методами, супернатанты фагосодержащих культур собирают с использованием обычного метода {Methods Mol. Biol., 178, 2002, 133-145). Связывающую активность с CD 137 человека в присутствии и в отсутствие АТФ затем подтверждают методами фагового ELISA, описанными в примере 2-2-2.
Результаты представлены на фиг. 5.
Был получен ряд клонов, которые были признаны клонами, обладающими АТФ-зависимой антигенсвязывающей активностью, с отношением поглощения S/N более 2 в присутствии/в отсутствие антигена и отношением S/N поглощения более 2 в присутствии/отсутствии АТФ (переключенные клоны).
Пример 4. Выработка модифицированных анти-СР137 антител и оценка их активности
(4-1) Повышение связывающей активности за счет изменения dBBAT119H-P253/dBBAT119L-LamLib
Варианты вариабельной области тяжелой цепи, dBAT119H, и вариабельной области легкой цепи, dBAT119L, анти-CD137 антитела (название клона: dBBAT119H-P253/dBAT119L-LamLib), полученные в примере 2-4, получают методами, известными специалистам в данной области, например ПЦР. Для вариабельной области тяжелой цепи создают варианты, в которых D10 (относится к аспарагиновой кислоте (Asp) в положении 10 (нумерация по Kabat)) и G17 (относится к глицину (Gly) в положении 17 (нумерация по Kabat)) dBBAT119H заменены глицином (Gly) и серином (Ser), соответственно, с получением dBBAT119H010; и N99 (в отношении аспарагина (Asn) в позиции 99 (нумерация по Kabat)), М100а (в отношении метионина (Met) в позиции 100а (нумерация по Kabat)) и N100b (в отношении аспарагина (Asn) в позиции 100b (нумерация по Kabat))) dBBAT119H010 заменены другими аминокислотами. Для вариабельной области легкой цепи были созданы варианты, в которых F87 (относящийся к фенилаланину (Phe) в положении 87 (нумерация по Kabat)) dBBAT119L заменен тирозином (Tyr) с образованием dBBAT119L010; и D27b (относится к аспартату (Asp) в позиции 27b (нумерация по Kabat)), N31 (относится к аспарагину (Asn) в позиции 31 (нумерация по Kabat)) и D94 (относится к аспартату (Asp) в позиции 94 (нумерация по Kabat))) в dBBATl 19L010 заменены другими аминокислотами.
Для вариантов вариабельной области тяжелой цепи связывающую активность с CD 137 человека измеряют с помощью анализатора поверхностного плазмонного резонанса BiacoreT200 (фирма GE Healthcare). Антитела захвачены путем взаимодействия очищенных вариантов с иммобилизованным белком G (фирма CALBIOCHEM) на сенсорном чипе СМ3 серии S (фирма GE Healthcare). Затем раствор CD137 человека (расщепленного FXa) с добавлением АТФ или раствор CD137 человека (расщепленного FXa) без АТФ взаимодействует в присутствии АТФ и в отсутствие АТФ для оценки связывающей активности вариантов по отношению к CD137 человека (расщепленного FXa). Измерения проводят при 25°С с использованием подвижного буфера, включающего 20 мМ ACES, 150 мМ NaCl, 0,05% Tween20 и 2 мМ MgCl2, рН 7,4. Результаты измерений показывают, что вариант L100a (вариант, в котором метионин (Met) в положении 100а (нумерация по Kabat) заменен лейцином (Leu)) имеет повышенную связывающую активность с CD 137 человека только в присутствии АТФ (фиг. 6). Вариабельная область тяжелой цепи варианта L100a представляет собой dBATk119H024 (SEQ ID NO: 132). Связывающее количество CD137 человека выверяют по захваченному количеству каждого варианта (1000 RU).
Для вариантов вариабельной области легкой цепи связывающую активность с CD137 человека (расщепление FXa) определяют с помощью BiacoreT200 в условиях, аналогичных описанным выше. Результаты измерений показывают, что вариант Е94 (вариант, в котором аспартат (Asp) в положении 94 (нумерация по Kabat) заменен глутаматом (Glu)) имеет повышенную связывающая активность только с CD137 человека в присутствии АТФ (фиг. 6). Вариабельной областью легкой цепи варианта Е94 является dBBATk119L020 (SEQ ID NO: 133).
Такие варианты, сочетающие указанные варианты тяжелой цепи и легкой цепи, сокращенно обозначают аббревиатурами dBBATk119H024-P253/dBATk119L020-LamLib (вариабельная область тяжелой цепи SEQ ID NO: 132; вариабельная область легкой цепи SEQ ID NO: 133; константная область тяжелой цепи SEQ ID NO: 93 и константная область легкой цепи SEQ ID NO: 63).
Антитела в настоящем изобретении обозначают в соответствии со следующим правилом: (вариабельная область тяжелой цепи) - (константная область тяжелой цепи)/(вариабельная область легкой цепи) - (константная область легкой цепи).
Например, название антитела dBBAT119H-P253/dBAT119L-LamLib означает, что оно состоит из вариабельной области тяжелой цепи антитела - dBAT119H, константной области тяжелой цепи антитела - Р253, вариабельной области легкой цепи антитела - dBBAT119L, и константной области легкой цепи - LamLib.
(4-2) Оценка агонистической активности CD137 модифицированных анти-CD137 антител человека in vitro, используя Т-клетки человека
(4-2-1) Размножение культур Т-клеток человека
Т-клетки человека размножают и культивируют согласно описанному в примерах 2-6-1.
(4-2-2) Оценка агонистической активности CD 137 in vitro, используя Т-клетки человека
Методами, описанными в примере 2-6-2, оценивают выработку IFN-γ в присутствии NS1-P253 (непереключаемого антитела), dBBAT119H-P253/dBAT119L-LamLib, dBBATk119H024-P253/dBATk119L020-LamLib или контрольного антитела, IC17HdK-hIgG1/IC17L-k0 при 10, 2, 0,4, 0,08 и 0,016 мкг/мл, соответственно. Также оценивают выработку IFN-γ в среде АДФ-бета-S-минус.
Оба результата показаны на фиг. 7.
Модифицированное антитело dBBATk119H024-P253/dBBATk119L020-LamLib проявляет усиленную АДФ-бета-S-зависимую агонистическую активность по сравнению с dBBAT119H-P253/dBBAT119L-LamLib. Это указывает на возможность того, что dBBATk119H024-P253/dBATk119L020-LamLib проявляет более сильную АТФ-, АДФ- и АМФ-зависимую агонистическую активность в отношении CD137 человека по сравнению с dBBAT119H-P253/dBBAT119L-LamLib.
Пример 5. Дополнительные модификации анти-СР137 антител
(5-1) Поиск модификаций, которые повышают связывающую активность, путем внесения комплексных модификаций
Для получения высококачественных анти-CD137 антител аминокислотные модификации четко интродуцируют в вариабельную область тяжелой цепи, dBBATk119H024, и ввариабельную область легкой цепи, dBBATk119L020, анти-CD137 антител, полученных в примерах 4-1. С помощью методов, известных специалистам в данной области, таких как ПЦР, созданы варианты, в которых каждый из всех аминокислотных остатков, составляющих CDR dBBATk119H024 и dBBATk119L020, заменены всеми 18 аминокислотами, кроме цистеина. Измерения приблизительно 1200 вариантов, вырабатываемых для связывания CD137 человека, выполняют с использованием Biacore4000. Антитела захвачены взаимодействующими культуральными супернатантами вариантов с иммобилизованным белком G (фирма CALBIOCHEM) на сенсорном чипе СМ3 серии S (фирма GE Healthcare). Затем раствор CD137 человека с добавлением малых молекул (АТФ) или раствор CD 137 человека без добавления малых молекул подвергают взаимодействию в присутствии малой молекулы или в отсутствие малой молекулы для оценки связывающей активности антител по отношению к CD 137 человека. Измерения проводят при 25°С с использованием подвижного буфера с добавлением CaCl2, содержащего 20 мМ ACES, 150 мМ NaCl, 0,02% Tween20 и 2 мМ MgCl2, рН 7,4.
(5-2) Повышение связывания АТФ
Ген тяжелой цепи антитела А002-Р253 (SEQ ID NO: 134) создан путем объединения модификаций, выявленных в примере 5-1, которые увеличивают связывающую активность с CD137 человека в присутствии небольшой молекулы, в гене dBBATk119H024-Р253, имеющем dBBATk119H024 (SEQ ID NO: 132) в качестве вариабельной области тяжелой цепи, и Р253 (SEQ ID NO: 93) в качестве константной области тяжелой цепи, который получают путем введения модификации S267E/L328F и удалением С-концевого Gly и Lys из IgG1 человека. Ген легкой цепи антитела B040-Lamlib (SEQ ID NO: 135) также создают путем объединения модификаций, обнаруженных в примере 5-1, которые увеличивают связывающую активность с CD137 человека в присутствии небольшой молекулы, в легкой цепи антитела dBBATk119L020-Lamlib, имеющий dBBATk119L020 (SEQ ID NO: 133) в качестве вариабельной области легкой цепь X человека Lamlib (SEQ ID NO: 63) в качестве константной области легкой цепи. Эти гены объединяют для экспрессии и очистки антител способами, известными специалистам в данной области, для получения анти-CD137 антитела интереса, A002-P253/B040-Lamlib. Вариабельная область тяжелой цепи A002-P253/B040-Lamlib - это А002 (SEQ ID NO: 136), вариабельная область легкой цепи - В040 (SEQ ID NO: 137), константная область тяжелой цепи - Р253 (SEQ ID NO: 93), а константная область легкой цепи представляет собой цепь λ Lamlib человека (SEQ ID NO: 63).
Для вариабельной области тяжелой цепи A002-P253/B040-Lamlib, полученной в этом разделе, создают различные варианты, в которых 53-я, 54-я или 55-я аминокислоты по нумерации Kabat заменены другими аминокислотами с целью повышения уровня АТФ-связывающей активности. Табл. 15 показывает модификации аминокислот (нумерация по Kabat) из А002 в вариабельных областях разработанных антител.
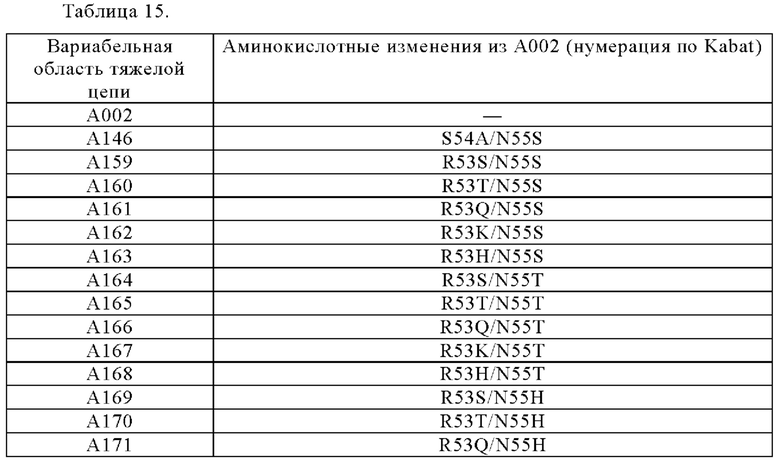
Связывающую активность вырабатываемых вариантов по отношению к АТФ и CD137 человека оценивают с помощью Biacore Т200.
Измерения АТФ-связывающей активности проводят при 37°С с использованием подвижного буфера, содержащего 20 мМ ACES (рН 7,4), 150 мМ NaCl, 2 мМ MgCl2 и 0,05% Tween20. Сначала антитела захватываются путем взаимодействия растворов антител в подвижном буфере с иммобилизованным Sure Protein А (фирма GE Healthcare) на сенсорном чипом СМ3 Series S (фирма GE Healthcare). Затем связывающую активность антител оценивают по взаимодействию с раствором АТФ, приготовленным в подвижном буфере. Чип регенерируют, используя 25 мМ NaOH и 10 мМ глицин-HCl (рН 1,5), измерения проводят путем многократного захвата антител. Количество связывания АТФ для каждого антитела рассчитывают путем корректировки количества связывания АТФ при инъекции в концентрации 100 нМ на количество антитела, захваченного на поверхности чипа, как количество АТФ на единицу количества антитела.
Измерение связывающей активности по отношению к CD137 человека проводят при 37°С с использованием 20 мМ ACES (рН 7,4), 150 мМ NaCl, 2 мМ MgCl2 и 0,05% Tween20 в качестве подвижного буфера. Сначала антитела улавливают путем взаимодействия растворов антител, приготовленных в подвижном буфере, с иммобилизованным Sure Protein А (фирма GE Healthcare) на сенсорным чипе СМ3 Series S (фирма GE Healthcare). Затем оценивают активность связывания с CD 137 человека путем взаимодействия с раствором CD137 человека, дополненным 100 мкМ АТФ в качестве небольшой молекулы. Для антигена CD137 человека используют hCD137-HisBAP, полученный в примере (1-1), и измерения проводят при концентрации антигена 0, 15 625, 62,5, 250 или 1000 нМ. Чип регенерируют, используя 25 мМ NaOH и 10 мМ глицин-HCl (рН 1,5), измерения проводят путем многократного захвата антител. Константу диссоциации (KD) соответствующих антител для CD137 человека рассчитывают с использованием программного обеспечения Biacore Т200 Evaluation Software 2.0. В частности, константу скорости ассоциации ka (л/моль/сек) и константу скорости диссоциации kd (1/с) рассчитывают путем глобальной подгонки сенсограмм, полученных путем измерения с помощью модели связывания Ленгмюра 1:1, и константу диссоциации KD (моль/л) рассчитывают по этим значениям.
В табл.16 представлены результаты таких измерений.
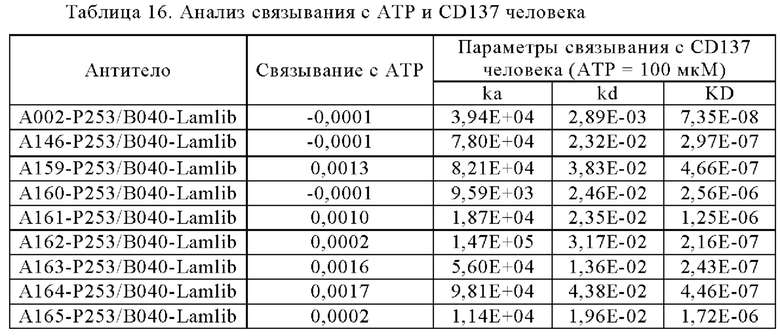
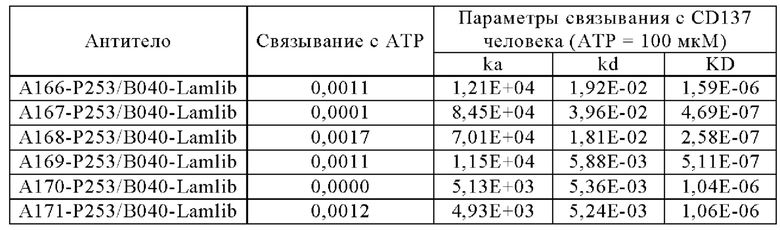
Как показано в таблице 16, варианты, в которых 53-я, 54-я или 55-я аминокислоты (нумерация по Kabat) вариабельной области тяжелой цепи заменены другими аминокислотами, имеют повышенную АТФ-связывающую активность по сравнению с антителом A002-P253/B040-Lamlib до модификации, за исключением A146-P253/B040-Lamlib и A160-P253/B040-Lamlib. Сравнение константы скорости ассоциации ka (л/моль/сек) также показывает, что константа скорости ассоциации для CD 137 человека увеличена для антител А146-P253/B040-Lamlib, A159-P253/B040-Lamlib, A162-P253/B040-Lamlib., А163-P253/B040-Lamlib, A164-P253/B040-Lamlib, A167-P253/B040-Lamlib и A168-P253/B040-Lamlib среди антител, оценка которым дана выше.
(5-3) Повышение связывающей активности путем внесения комплексных модификаций
Для создания лучших анти-CD137 антител, модификации, выявленные в примере 5-1, которые увеличивают активность связывания CD 137 человека в присутствии малой молекулы и уменьшают связывание с CD 137 человека в условиях, когда эта малая молекула отсутствует, и модификации, выявленные в примерах 5-2, которые увеличивают АТФ-связывающую активность и увеличивают константу скорости ассоциации для CD137 человека, были объединены для получения антител против CD 137 человека, которые проявляют лучший профиль. Были разработаны гены тяжелой цепи антитела, которые объединяют модификации, обнаруженные в примерах 5-1 и 5-2 в гене тяжелой цепи антитела A002-G1T3, имеющем последовательность А002 (SEQ ID NO: 136) в качестве вариабельной области тяжелой цепи, и последовательность G1T3 (SEQ ID NO: 138) в качестве константной области тяжелой цепи, которая была получена путем введения модификации K214R/Q419E и удаления С-концевых Gly и Lys из IgG1 человека. Гены легкой цепи антитела получены путем объединения модификаций, обнаруженных в примере 5-1, в легкой цепи антитела B040-Lamlib, имеющей последовательность В040 (SEQ ID NO: 137) в качестве вариабельной области легкой цепи и Lamlib λ-цепи человека в качестве константной области легкой цепи.
В качестве объекта для сравнения используют ген тяжелой цепи антитела 20Н4.9-Р253, имеющий вариабельную область тяжелой цепи 20Н4.9 (SEQ ID NO: 139) существующего анти-С0137 антитела, описанного в US8137667 и Р253 (SEQ ID NO: 93) как константная область тяжелой цепи; и ген легкой цепи антитела, который объединяет вариабельную область легкой цепи 20H4.9LC (SEQ ID NO: 140) с цепью k человека κ0 (SEQ ID NO: 141) в качестве константной области легкой цепи. В качестве другого объекта сравнения, ген тяжелой цепи антитела MOR-7480.1Н-Р253, имеющий вариабельную область тяжелой цепи MOR-7480.1H (SEQ ID NO: 142), который составляет существующее анти-CD137MOR-7480.1 антитело, описанное в US8337850, и Р253 (SEQ ID NO: 93) в качестве константной области тяжелой цепи; и легкая цепь антитела MOR-7480.1L-lam, объединяющая вариабельную область легкой цепи MOR-7480.1L (SEQ ID NO: 143) с lam λ-цепи человека (SEQ ID NO: 63) в качестве константной области легкой цепи (примечание: и Lamlib λ-цепи человека, и lam человека имеют одинаковую аминокислотную последовательность (SEQ ID NO: 63)).
Эти гены объединяют для экспрессии и очистки антител с использованием методов, известных специалистам в данной области, для получения анти-С0137 антител интереса. Табл. 17 представляет собой список номеров последовательностей вариабельной области тяжелой цепи, вариабельной области легкой цепи, константной области тяжелой цепи, константной области легкой цепи и гипервариабельной области (гипервариабельная область; также называемая HVR или CDR) разработанных антител.
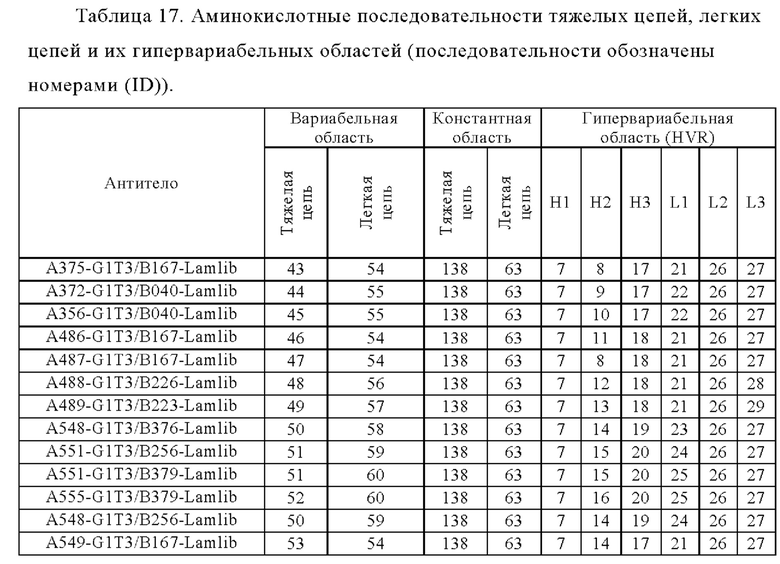
Связывание разработанных вариантов с АТФ и CD137 человека оценивают с помощью Biacore Т200. Измерения связывания CD137 человека проводят при 37°С с использованием 20 мМ ACES (рН 7,4), 150 мМ NaCl, 2 мМ MgCl2 и 0,05% Tween20 в качестве подвижного буфера. Сначала 250-400 RU антител захватывают путем взаимодействия растворов антител, приготовленных в подвижном буфере, с иммобилизованным Sure Protein А (фирма GE Healthcare) на сенсорным чипе СМ3 Series S (фирма GE Healthcare). Связывающую активность по отношению к CD137 человека в присутствии АТФ или в отсутствие АТФ затем оценивают путем взаимодействия с раствором CD137 человека, приготовленным в подвижном буфере, дополненном требуемой концентрацией АТФ, или с раствором CD137 человека, приготовленным в подвижном буфере без АТФ. Для антигена CD137 человека используют hCD137-HisBAP, полученный в примере (1-1), и измерения значения KD проводят при концентрации антигена 0, 15 625, 62,5, 250 и 1000 нМ. Для оценки связанного количества проводят измерения при концентрации антигена 0 и 1000 нМ. Чип регенерируют с помощью 25 мМ NaOH и 10 мМ глицин-HCl (рН 1,5), и измерения проводят путем многократного захватываемых антител. Константу диссоциации соответствующих антител для CD 137 человека рассчитывают с использованием программного обеспечения Biacore Т200 Evaluation Software 2.0. В частности, константу скорости ассоциации ка (л/моль/с) и константу скорости диссоциации kd (1/с) рассчитывают путем глобальной подгонки сенсограмм, полученных путем измерения с помощью модели связывания Ленгмюра 1:1, и константу диссоциации KD (моль/л) рассчитывают по этим значениям.
В табл.18 представлены результаты таких измерений.
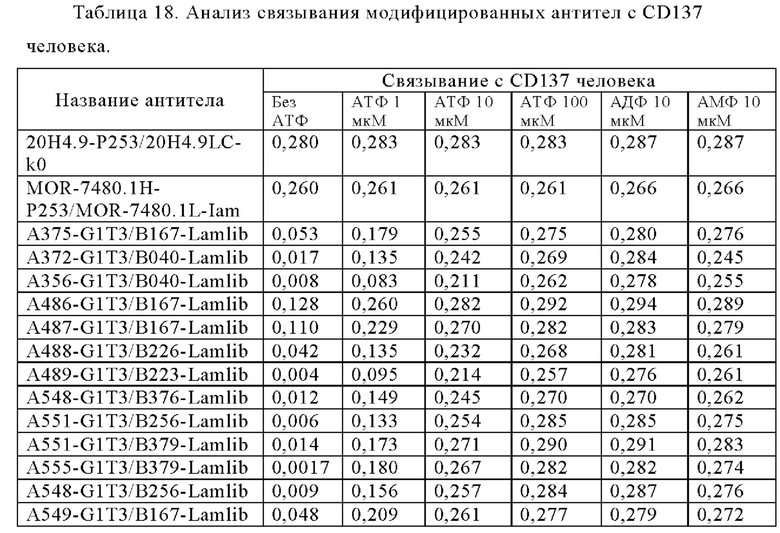
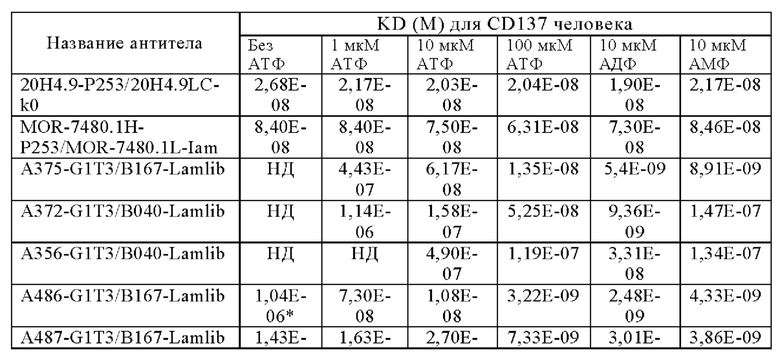
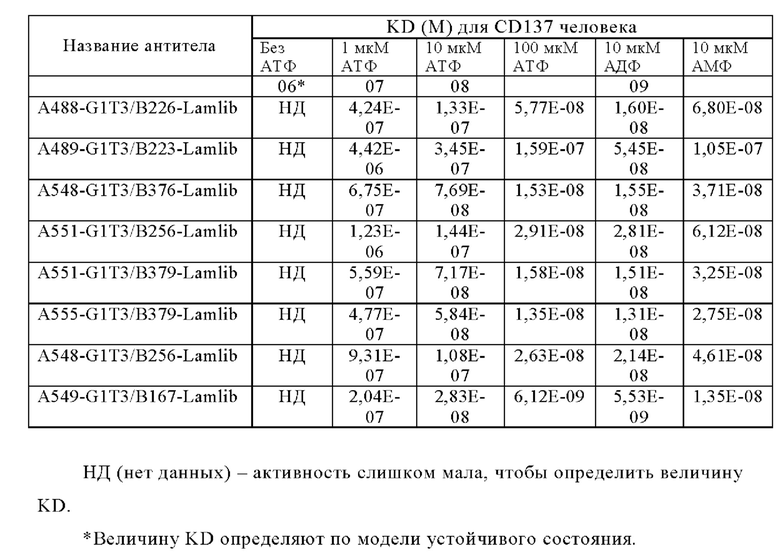
Значения «связывание с CD137 человека» в табл.18 выше указывают количество связывания CD 137 человека на единицу количества антитела, когда CD137 человека допускает взаимодействовать при 1000 нМ при каждой концентрации АТФ, указанной выше, и «KD (М) для CD 137 человека «указывает константу диссоциации CD 137 человека при каждой концентрации АТФ. Значения KD, отмеченные в таблице знаком «*», рассчитаны с использованием модели устойчивого состояния. Количество связывания всех разработанных вариантов в присутствии 10 мкМ АТФ больше, чем при концентрации АТФ 1 мкМ, и даже больше в присутствии 100 мкМ, что позволяет предположить, что они связываются с CD137 человека в зависимости от концентрации АТФ. С другой стороны, объекты сравнения, 20H4.9-P253/20H4.9LC-k0 и MOR-7480.1 Н-P253/MOR-7480.1L-lam, не проявляют связывания с CD137 человека зависимым от концентрации АТФ образом.
(5-4) Оценка in vitro АТФ-зависимой агонистической активности в отношении CD137 модифицированных антител против CD137 человека с использованием анализа репортерного гена 4-1 ВВ Jurkat
Клеточную линию GloResponse™ NF-κ B-Luc2/4-lBB Jurkat (фирма Promega, CS196004) используют для измерения in vitro активности разработанных вариантов. В каждую лунку 96-луночных планшетов добавляют по 200 мкл суспензии клеток FcγRIIB СНО-К1 (фирма Promega), приготовленных в концентрации 5 × 104/мл в среде, и оставляют на ночь при 37°С в инкубаторе в атмосфере 5% CO2. В качестве питательной среды используют культуральную среду СНО (90% Ham's F12, 10% ФСБ). Затем, после того, как всю среду удаляют, по 25 мкл суспензии клеточной линии GloResponse NF-κB-Luc2/4-1BB Jurkat с концентрацией 2 × 106/мл в среде для исследования (99% RPMI, 1% ФСБ) вносят в каждую лунку. Затем добавляют 25 мкл каждого раствора антител, разбавленного средой для исследования таким образом, чтобы конечная концентрация составляла 0, 0,001, 0,01, 0,1, 1 и 10 мкг/мл; и, наконец, добавляли 25 мкл раствора АТФ, разбавленного средой для исследования, таким образом, чтобы конечная концентрация стала 0 и 250 мкМ. Планшеты оставляют на 6 ч при 37°С в инкубаторе в атмосфере 5% CO2 и на 15 мин при комнатной температуре; по 75 мкл реагента Bio-Glo вносят в каждую лунку. Для реагента Bio-Glo используют систему анализа люциферазы Bio-Glo (буфер и субстрат). Впоследствии люминесценцию каждой лунки измеряют с помощью планшет-ридера. Величина люминесценции каждой лунки, деленная на величину люминесценции лунки без добавления антител, является относительной световой единицей (кратная индукция) и служит индикатором для оценки агонистической активности в отношении CD 137 каждого антитела.
Результаты показаны на фиг. 8.
(5-5) Оценка in vitro АТФ-зависимой агонистической активности в отношении CD137 модифицированных антител против CD137 человека с использованием мононуклеарных клеток периферической крови человека
(5-5-1) Выделение мононуклеарных клеток периферической крови человека
Мононуклеарные клетки периферической крови человека (МКПК) выделяют из образцов крови здоровых добровольцев в компании Заявителя по технологии, описанной в примере 2-6-1. Впоследствии клетки разводят в среде (5% сыворотки человека (фирма SIGMA), 95% AIM-V (фирма Thermo Fischer Scientific) до плотности клеток 5×106/мл.
(5-5-2) Оценка агонистической активности в отношении CD 137 с использованием мононуклеарных клеток периферической крови человека
Мононуклеарные клетки периферической крови человека доводят до плотности 5 × 106/мл и высевают в 96-луночные планшеты с плоским дном (фирма Corning) по 100 мкл в каждую лунку. Затем добавляют по 50 мкл раствора с концентрацией 0,04 мкг/мл антитела против CD3ε человека (фирма BD, клон SP34) и по 20 мкг/мл антитела против CD28 человека (фирма BD, клон: CD28.2), разведенных в среде. Планшеты встряхивают и оставляют в инкубаторе в атмосфере 5% CO2 при 37°С на 6 ч. Затем в каждую лунку добавляют по 25 мкл среды с 2 мМ АТФ (фирма SIGMA) или среды без АТФ и по 25 мкл каждого антитела в концентрации 40 мкг/мл, и планшеты встряхивают перед тем, как оставить в инкубаторе в атмосфере с5% CO2 при 37°С на 18 ч. Затем часть культурального супернатанта собирают, и культуральный супернатант используют для подсчета содержания IL-2 в культуральном супернатанте с Human IL-2 DuoSet ELISA kit (фирма R&D systems) или Human IL-2 ELISA Set (фирма BD Biosciences). После сбора культурального супернатанта планшеты снова оставляют в инкубаторе в атмосфере с5% CO2 при 37°С на 24 ч. Затем собирают часть культурального супернатанта, и количество IFN-γ, содержащегося в культуральном супернатанте, определяют с использованием набора Human IFN-γ DuoSet ELISA kit (фирма R&D systems) или Human IFN-γ ELISA Development Kit (фирма PeproTech). Анализ ELISA осуществляют в основном по протоколу поставщика набора. Для набора Human IL-2 DuoSet ELISA (фирма R&D systems) и набора Human IFN-γ DuoSet ELISA (фирма R&D systems) появление окраски и прекращение проявления окраски выполняют в соответствии с протоколами с использованием раствора субстрата (фирма R&D systems), содержащего H2O2 и тетраметилбензидин, и 1н H2SO4 (фирма Wako). Для набора Human IL-2 ELISA Set (фирма BD Biosciences) развитие окраски прекращают с использованием 1 н H2SO4 (фирма Wako). Для набора IFN-γ ELISA Development Kit (фирма PeproTech) используют раствор хромогена ТМВ (фирма Thermo Fischer Scientific) и 1 н H2SO4 (фирма Wako) для проявления окраски и ее прекращения. Затем проводят измерения поглощения с помощью ридера EnVision (фирма PerkinElmer).
Результаты подробно описаны в примере 5-5-3 и в дальнейшем описании.
(5-5-3) Оценка усиления агонистической активности константной области тяжелой цепи путем увеличения связывающей активности с рецептором Fcγ
In vitro с применением моноцитов периферической крови человека согласно описанию в примерах 5-5-1 и 5-5-2 оценивают эффект применения Р587, MY518, ТТ14 и ТТ16, которые по-разному увеличивают FcγRIIb-связывающую активность константной области тяжелой цепи, по агонистической активности в отношении CD137.
Оцениваемые антитела представлены в табл. 19.
Получение антител, перечисленных в табл. 19, описано в примере 7-1. По сравнению с количествами IL-2 и IFN-γ в супернатанте культуры, когда антитело отрицательного контроля (IC17HdK-MY518/IC17L-k0, IC17HdK-G4d/IC17L-k0) добавляют в присутствии 250 мкМ АТФ, если добавление антитела увеличивает количество IL-2 в супернатанте культуры в 1,05 раза или более и количество IFN-γ в 1,15 раза или более, установлено, что антитело демонстрирует агонистическую активность в отношении CD137. Измерение агонистической активности с использованием мононуклеарных клеток периферической крови человека (МКПК) может отличаться в зависимости от доноров крови. В связи с этим было определено, что антитела, которые не проявляют агонистической активности в отношении CD137 для некоторой части или более чем для половины МКПК человека, выделенных от нескольких доноров, можно расценивать как не проявляющие агонистической активности в отношении CD137, даже если они соответствуют стандартам агонистической активности для другой части МКПК человека.
Антитела, которые рассматривают в качестве проявляющих агонистическую активность в отношении CD137 в присутствии 250 мкМ АТФ, показаны в табл. 19. Это указывает на то, что агонистическая активность в отношении CD137 была усилена путем комбинирования ТТ16 с Р587, который увеличивает связывающую активность с FcγRIIb константной области тяжелой цепи (фиг. 9 и 10). Предполагают, что агонистическая активность усилена за счет увеличения связывающей активности в отношении клеток, экспрессирующих FcγRIIb, которые представляют собой перекрестно-связывающие каркасы, необходимые для того, чтобы антитело проявляло агонистическую активность в отношении клеток, экспрессирующих CD137 (см. Protein Eng Des Sel, 26(10), 2013, 589-598, doi: 10.1093/protein).
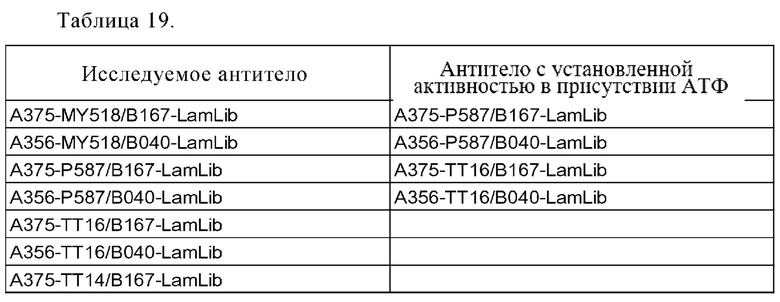
(5-5-4) Оценка АТФ-зависимой агонистической активности в отношении CD137 вариабельных областей
АТФ-зависимая агонистическая активность в отношении CD137 каждой вариабельной области путем комбинирования константных областей тяжелой цепи Р587 и Р253 с различными вариабельными областями оценена in vitro с использованием моноцитов периферической крови человека, как описано в примерах 5-5-1 и 5-5-2.
Оцениваемые антитела представлены в Таблице 20. Выработка антител, перечисленных в таблице 20, описана в примере 7-1. По сравнению с количествами IL-2 и IFN-γ в супернатанте культуры, когда отрицательное контрольное антитело (IC17HdK-P253/IC17L-k0, IC17HdK-P587/IC17L-k0) добавляют в присутствии 250 мкМ АТФ, и когда добавление антитела увеличивает количество IL-2 в культуральном супернатанте в 1,05 раза или более и количество IFN-γ в 1,15 раза или более, делают вывод, что антитело проявляет агонистическую активность в отношении CD137. Измерение агонистической активности с использованием мононуклеарных клеток периферической крови человека (МКПК) может отличаться от донора к донору в зависимости от образца крови. В связи с этим определено, что антитела, которые не проявляют агонистической активности в отношении CD137 для некоторой или более половины МКПК человека, выделенного от нескольких доноров, можно расценивать как не проявляющие агонистической активности в отношении CD137, даже если они соответствуют стандартам агонистической деятельности по другой части МКПК человека. Антитела, которые согласно оценке проявляют агонистическую активность в отношении CD137 в присутствии 250 мкМ АТФ, представлены в табл. 20 (фиг. 11, фиг. 12, фиг. 13, фиг. 14 и фиг. 15).
Кроме того, среди антител, которые согласно оценке проявляют агонистическую активность в отношении CD137 в присутствии 250 мкМ АТФ, антитела, кратность которых изменяется относительно группы, в которую добавлено отрицательное контрольное антитело (IC17HdK-P253/IC17L-k0, IC17HdK-P587/IC17L-k0) по отношению к количеству IL-2 и IFN-γ в супернатанте культуры в условиях отсутствия АТФ, меньше, чем кратное изменение по сравнению с отрицательным контролем в условиях в присутствии 250 мкМ АТФ, обладают пониженной агонистической активностью в отношении CD137 при отсутствии АТФ. Антитела, которые проявляют агонистическую активность в отношении CD137 в присутствии 250 мкМ АТФ и которые оценивают как имеющие агонистическую активность в отношении CD137 в отсутствие АТФ, представлены в табл. 20 (фиг. 11, фиг. 12, фиг. 13, фиг. 14 и фиг. 15). Было установлено, что эти антитела проявляют АТФ-зависимую агонистическую активность в отношении CD137.
На основании вышеизложенного подтверждено, что для вариабельных областей комбинации вариабельная область тяжелой цепи/вариабельная область легкой цепи (А375/В167), (А356/В040), (А372/В040), (А486/В167), (А488/В226), (А489/В223), (А551/В256), (А548/В256) и (А551/В379) проявляют агонистическую активность в отношении CD137 АТФ-зависимым образом.
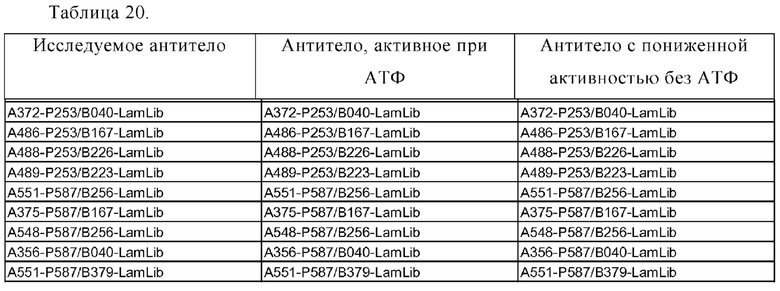
(5-5-5) Оценка усиливающего воздействия на агонистическую активность в отношении CD 137 комбинированных изменений, которые увеличивают pI константной области тяжелой цепи с константной областью тяжелой цепи с повышенной связывающей активностью с рецептором Fcγ
Влияние на агонистическую активность в отношении CD137 интродуцированных различных модификаций аминокислот, которые увеличивают pI константной области тяжелой цепи в константную область тяжелой цепи с различной повышенной FcγRIIb-связывающей активностью, оценивают путем оценки in vitro с использованием моноцитов периферической крови человека согласно описанному в примерах 5-5-1 и 5-5-2. Аминокислотные изменения, которые увеличивают pI, вводят в константные области тяжелой цепи ТТ16, MY518 и ТТ14 и, таким образом, в константные области тяжелой цепи ТТ16+P343R/D413K (SCF028), TT16+Q311 R/P343R (SCF033), MY518+P343R/D413K (SCF025), MY518+Q311R/P343R (SCF030), TT14+P343R/D413K(SCF027) и TT14+Q311R/P343R (SCF032). Названия оцениваемых антител, в которые внесены аминокислотные изменения, увеличивающие pI, и названия соответствующих антител до введения аминокислотных изменений, которые увеличивают pI, показаны в табл. 21. Выработка антител, перечисленных в табл. 21 описана в примере 7-2.
Что касается оценки усиления агонистической активности путем интродукции модификаций аминокислот, повышающих pI, когда добавление антитела увеличивает количество IL-2 в 1,04 раза или более и количество IFN-γ в 1,1 раза или более в супернатанте культуры, относительно групп антител, содержащих константную область тяжелой цепи, которая не содержит этих аминокислотных модификаций, A375-TT16/B167-LamLib, A375-MY518/B167-LamLib и A375-TT14/B167-LamLib, установлено, что агонистическая активность в отношении CD137 усиливается. По сравнению с антителами, содержащими константную область тяжелой цепи без повышающих pI аминокислотных модификаций, антитела, содержащие константную область тяжелой цепи, в которой модификации аминокислот, как было показано, обладают повышенной агонистической активностью в отношении CD137 в присутствии 250 мкМ АТФ, показаны в табл. 22 (фиг. 10).
Это указывает на то, что независимо от степени увеличения FcγRIIb-связывающей активности константной области тяжелой цепи, введение модификаций, которые увеличивают pI константной области тяжелой цепи, усиливает агонистическую активность в отношении CD137 антитела против CD137 человека. Было высказано предположение, что: сочетание модификаций, которые увеличивают pI константной области тяжелой цепи, с модификациями, которые увеличивают связывание FcγRIIb константной области тяжелой цепи с константной областью тяжелой цепи, усиливает взаимодействие с отрицательно заряженной FcγRIIb-экспрессирующей поверхностью клетки; иммунные комплексы антител или связанных с антигеном антител ближе подходят к поверхности клеток, экспрессирующих FcγRIIb; это дополнительно увеличивает связывание с клетками, экспрессирующими FcγRIIb, которые представляют собой перекрестно-сшивающие каркасы, необходимые для того, чтобы антитела проявляли агонистическую активность в отношении клеток, экспрессирующих CD137; и, таким образом, агонистическая активность была дополнительно усилена.
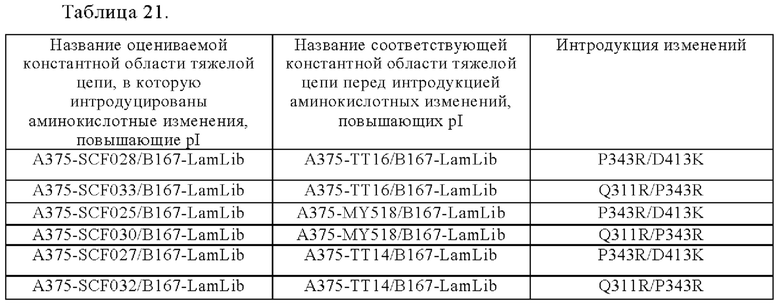

(5-5-6) Оценка воздействия на агонистическую активность путем внесения модификаций, которые увеличивают pI константной области тяжелой цепи
Влияние на агонистическую активность в отношении CD137 введения различных модификаций аминокислот, которые увеличивают pI константных областей тяжелой цепи MY201aPh и MY518, оценивают in vitro с использованием моноцитов периферической крови человека согласно описанию в примерах 5-5-1 и 5-5-2. В константные области тяжелой цепи MY518, MY518a и MY201aPh введены аминокислотными модификациями, которые увеличивают pI; таким образом были созданы MY518+P343R/D413K (SCF025), MY518+Q311R/P343R(SCF030), MY518+P343R(SCF039), MY518+D413K (SCF040), MY518a+Q311R (SCF060a), MY201aPh+P343R(SCF041aPh), MY201aPh+P343R/D413K (SCF043aPh), MY201aPh+Q311R (SCF056aPh), MY201aPh+Q311R/P343Rb(SCF057aPh) и MY201aPh+Q311R/D413K (SCF059aPh). Названия рассматриваемых антител, в которые внесены аминокислотные изменения, которые увеличивают pI, и названия антител до внесения аминокислотных изменений, которые увеличивают соответствующую величину pI, показаны в таблице 23. Выработка антител, перечисленных в табл. 23, описана в примере 7-2.
Что касается оценки усиления агонистической активности путем введения модификаций аминокислот, повышающих pI, когда добавление антитела увеличивает количество IL-2 в 1,04 раза или более и количество IFN-γ в 1,1 раза или более в супернатанте культуры, относительно группы, в которую добавлены антитела, содержащие константную область тяжелой цепи, которая не содержит таких аминокислотных модификаций, A375-MY518a/B167-LamLib и А375-MY201aPh/B167-LamLib, было подтверждено, что агонистическая активность в отношении CD137 улучшена. По сравнению с антителами, содержащими константную область тяжелой цепи без повышающих pI аминокислотных модификаций, антитела, содержащие константную область тяжелой цепи, в которой аминокислотные изменения, как показано, обладают повышенной агонистической активностью в отношении CD137 в присутствии 250 мкМ АТФ, показаны в табл. 24 (фиг. 16 и 17).
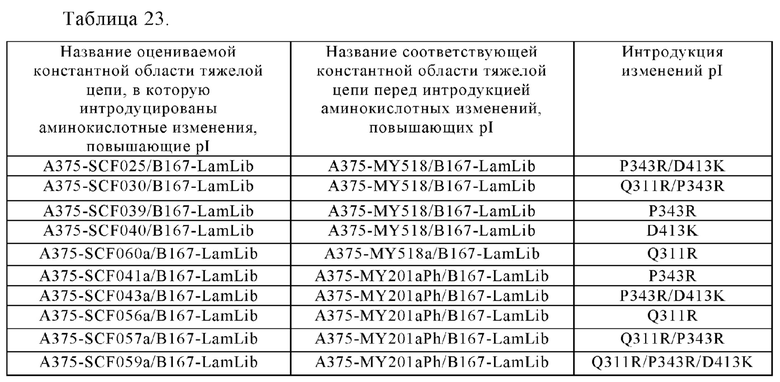
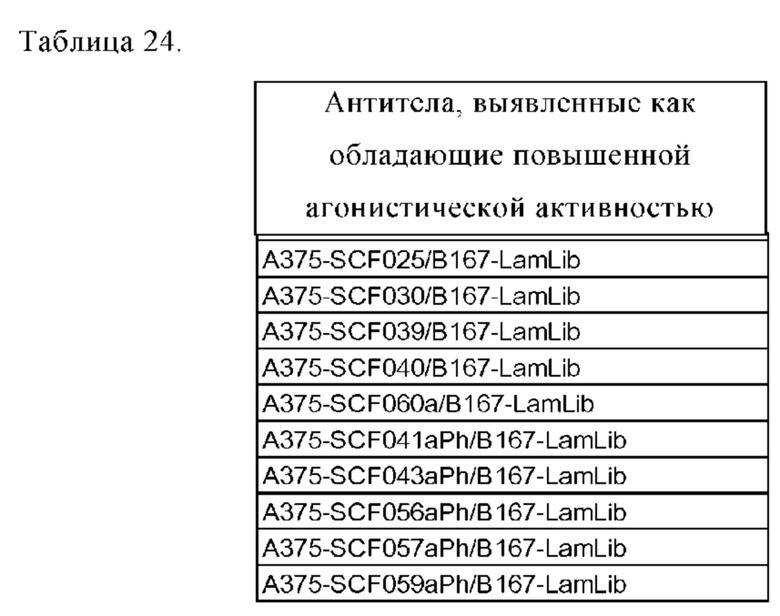
(5-5-7) Оценка АТФ-зависимой агонистической активности в отношении CD137 антител против CD137 человека, которые объединяют разработанные вариабельные области и константные области
АТФ-зависимая агонистическая активность в отношении CD 137 антител против CD137 человека, полученных комбинированием описанных выше вариабельных областей с константными областями тяжелой цепи, повышающими pI, оценивают in vitro с использованием моноцитов периферической крови человека согласно описанию в примерах 5-5-1 и 5-5-2. Исследованные антитела представлены в табл. 25.
По сравнению с количествами IL-2 и IFN-γ в супернатанте культуры при добавлении отрицательного контрольного антитела (IC17HdK-MY518a/IC17L-k0, IC17HdK-MY201aPh/IC17L-k0, IC17HdK-MY201/IC17L-k0, IC17HdK-G4d/IC17L-k0, IC17HdK-MY518/IC17L-k0, или IC17HdK-TT16/IC17L-k0) в присутствии 250 мкМ АТФ, когда добавление антитела увеличивает количество IL-2 в супернатанте культуры в 1,05 раза или более и количество IFN-γ в 1,15 раза или более, согласно оценке антитело демонстрирует агонистическую активность в отношении CD137. Измерение агонистической активности с использованием мононуклеарных клеток периферической крови человека (МКПК) может отличаться от донора к донору по образцу крови. В связи с этим определено, что антитела, которые не проявляют агонистической активности в отношении CD137 некоторой или более половины МКПК человека, выделенного от нескольких доноров, можно расценивать как не проявляющие активности агониста CD137, даже если они соответствовали стандартам для агонистов для другой части МКПК человека. Антитела, которые, как установлено, проявляют агонистическую активность в отношении CD137 в присутствии 250 мкМ АТФ, показаны в таблице 25 (фиг. 16, фиг. 17, фиг. 18, фиг. 19, фиг. 20, фиг. 21, фиг. 22 и фиг. 23).
Кроме того, среди антител, которые, как установлено, проявляют агонистическую активность в отношении CD137 в присутствии 250 мкМ АТФ, антитела, кратность изменения которых по сравнению с группой, в которую добавлено отрицательное контрольное антитело (IC17HdK-MY518a/IC17L-k0, IC17HdK-MY201aPh/IC17L-k0, IC17HdK-MY201/IC17L-k0, IC17HdK-G4d/IC17L-kO, IC17HdK-MY518/IC17L-k0, или IC17HdK-TT16/IC17L-k0), относительно количеств IL-2 и IFN-γ в супернатанте культуры при отсутствии АТФ меньше, чем кратное изменение относительно отрицательного контроля при наличии 250 мкМ АТФ, как установлено, проявляют пониженную агонистическую активность в отношении CD137 при отсутствии АТФ. Антитела, которые, как установлено, обладают пониженной агонистической активностью в отношении CD137 в отсутствие АТФ, показаны в табл.25 (фиг. 16-23). Показано, что эти антитела проявляют АТФ-зависимую агонистическую активность в отношении CD137.
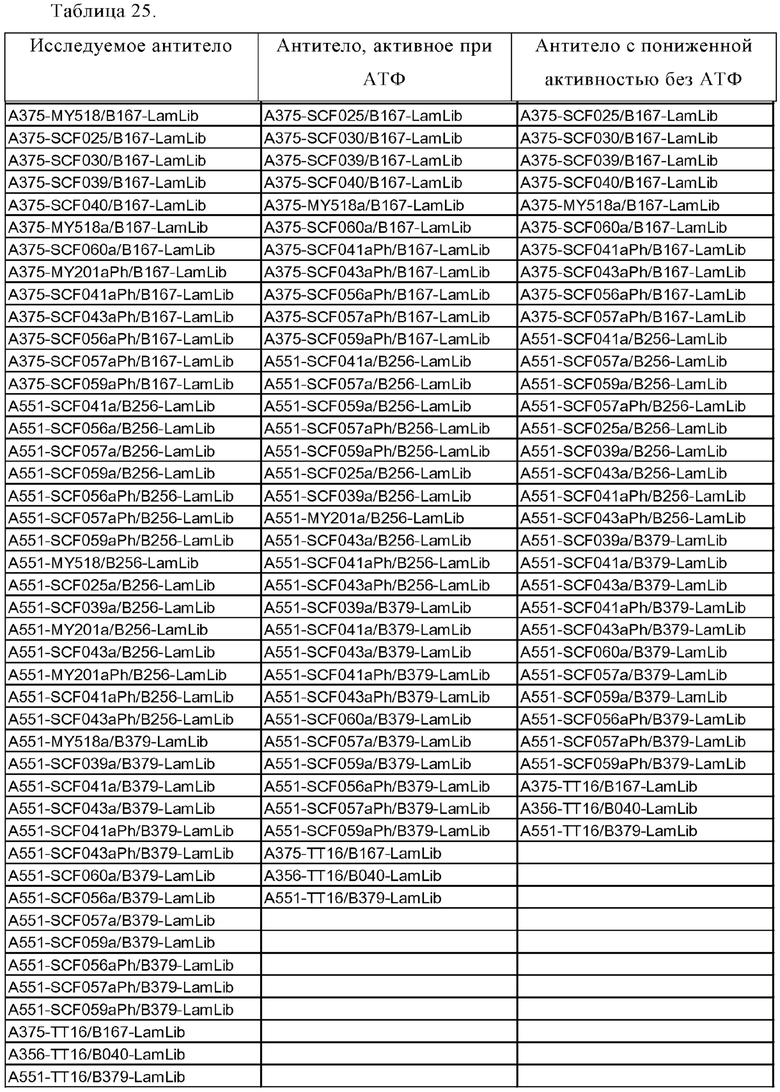
Пример 6. Мыши с нокином CD137 человека в качестве теста для введения переключаемых антител против CD137 человека
(6-1) Разработка антител для исследования введения мышам с нокином CD137 человека
Переключаемые и непереключаемые антитела против CD137 человека, содержащие константную область мыши, разработаны для исследования введения мышам с нокином CD137 человека. В частности получают непереключаемые антитела против CD137 человека (20H4.9-mIgG1/20H4.9LC-mk0 аббревиатура: NS1-mIgG1, 20Н4.9-МВ110/20H4.9LC-mk0 аббревиатура: NS1-MB110, 20H4.9-MB492/20H4.9LC-mk0 аббревиатура: NS1-MB492, MOR-7480.1 H-MB110/MOR-7480.1 L-ml0r аббревиатура: NS2-MB110, MOR-7480.1 H-MB492/MOR-7480.1 L-ml0r аббревиатура: NS2-MB492) и переключаемые антитела против CD137 человека (A375-mIgG1/B167-ml0r, A372-mIgG1/B040-ml0r, A372-MB110/B040-ml0r, A372-MB492/B040-ml0r, A356-MB110/B040-ml0r, A486-MB492/B167-ml0r, A488-MB492/B226-ml0r, A489-MB492/B223-ml0r, A551-mIgG1/B256-ml0r, A551-MB110/B379-ml0r и A548-mIgGl/B256-ml0r).
Для тяжелых цепей антител NS1-mIgG1, NS 1-MB110 и NS1-MB492 гены тяжелых цепей антитела разрабатывают комбинированием вариабельной области тяжелой цепи 20Н4.9 (SEQ ID NO: 139) с любой из последовательностей:
(i) mIgG1, константной области тяжелой цепи IgG1 мыши (SEQ ID NO: 144),
(ii) МВ110, описанной в WO 2014030750 (SEQ ID NO: 145), и
(iii) MB492, описанной в WO 2014030750 (SEQ ID NO: 146),
как константной областью тяжелой цепи. То есть были разработаны гены 20H4.9-mIgG1, 20Н4.9-МВ110 и 20Н4.9-МВ492.
Для легких цепей антител NS1-mlgGl, NS1-MB110 и NS1-MB492 разрабатывают ген легкой цепи антитела 20H4.9LC-mk0 путем комбинирования вариабельной области легкой цепи 20H4.9LC (SEQ ID NO: 140) с цепью к мыши mk0 (SEQ ID NO: 147) в качестве константной области легкой цепи. Комбинируя эти гены тяжелой цепи и легкой цепи, каждое антитело получают экспрессией и очищают методами, известными специалистам в данной области.
Для тяжелых цепей антител NS2-MB110 и NS2-MB492 гены тяжелых цепей антитела были созданы путем объединения вариабельной области тяжелой цепи MOR-7480.1 Н (SEQ ID NO: 142) с одной из последовательностей:
(i) MB110 (SEQ ID NO: 145), или
(ii) MB492 (SEQ ID NO: 146),
в качестве константной области тяжелой цепи. То есть созданы гены MOR-7480.1Н-МВ110 и MOR-7480.1H-MB492.
Для легких цепей антител NS2-MB110 и NS2-MB492 ген легкой цепи антитела MOR-7480. 1L-ml0r получают путем объединения вариабельной области легкой цепи MOR-7480.1L (SEQ ID NO: 143) с цепью λ мыши ml0r (SEQ ID NO: 148) в качестве константной области легкой цепи. Комбинируя указанные гены тяжелой цепи и легкой цепи, каждое антитело экспрессируют и очищают методами, известными специалистам в данной области.
Для анти-CD137 переключаемых антител получают гены тяжелой цепи антитела и легкой цепи антитела, указанные в табл. 26 и 27; каждое антитело экспрессируют и очищают путем объединения этих генов методами, известными специалистам в данной области.
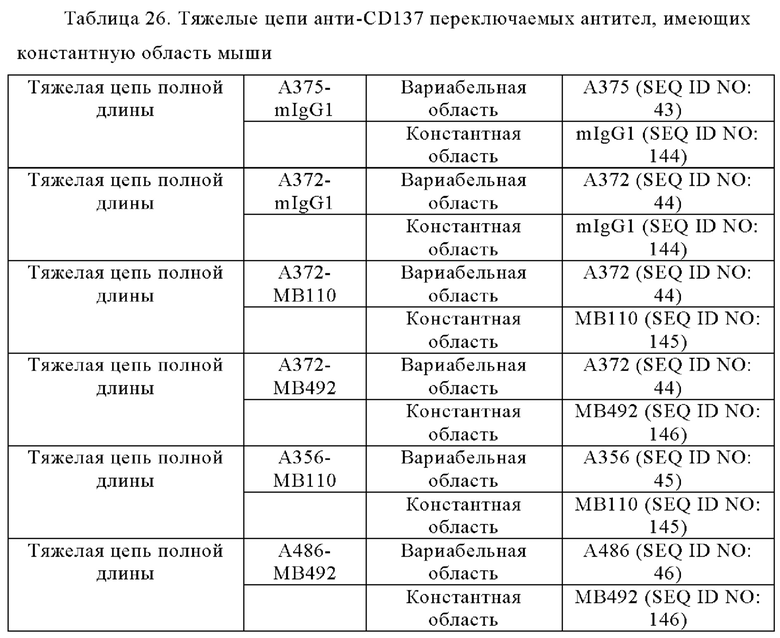
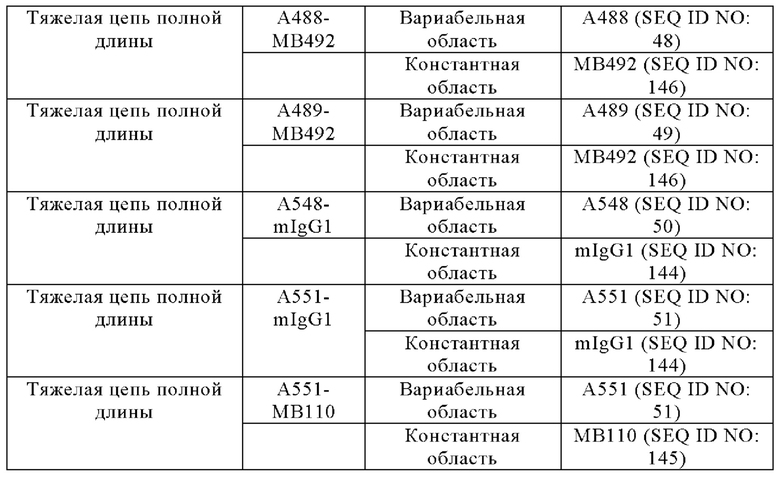
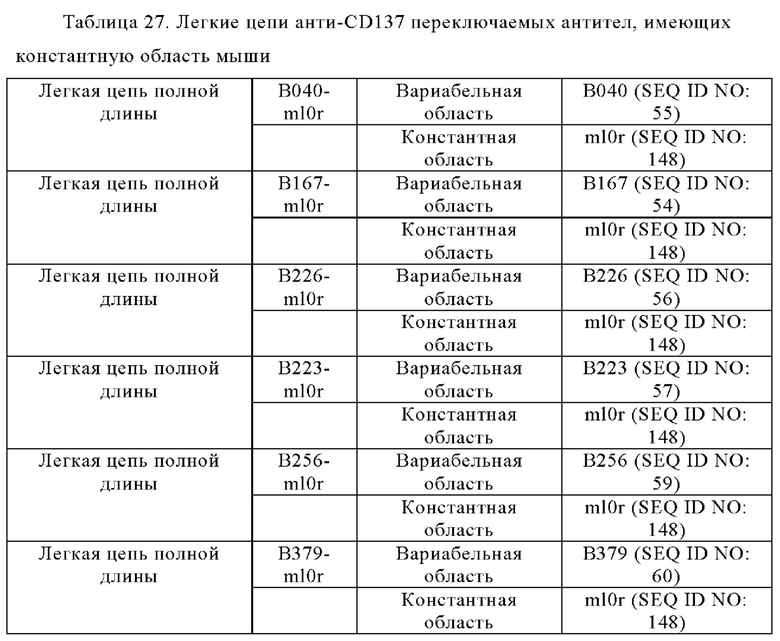
(6-2) Оценка связывающей активности антител против CD137 человека, приготовленных для исследования введения мышам с нокином CD137 человека
Оценивают связывающую активность непереключаемого антитела против CD137 человека и переключаемого антитела против CD 137, полученного в примере 6-1, по отношению к CD 137 человека. Измерения связывания CD137 человека проводят при 37°С с использованием 20 мМ ACES (рН 7,4), 150 мМ NaCl, 2 мМ MgCl2 и 0,05% Tween20 в качестве подвижного буфера. Сначала антитела улавливают с помощью взаимодействующих растворов антител, приготовленных в подвижном буфере с иммобилизованным антителом кролика против IgG мыши (фирма Thermo Scientific) на сенсорном чипе Series S Sensor Chip СМ5 (фирма GE Healthcare). Затем оценивают связывающую активность в отношении CD137 человека в присутствии АТФ и в отсутствие АТФ путем взаимодействия с раствором CD137 человека в подвижном буфере без АТФ. Для антигена CD137 человека используют hCD137 (расщепленный FXa), полученный в примере (1-2), и измерения проводят при концентрации антигена 0, 15 625, 62,5, 250 и 1000 нМ. Чип регенерируют, используя 25 мМ NaOH и 10 мМ глицин-HCl (рН 1,5), измерения проводят путем многократного захвата антител.
Константу диссоциации (KD) соответствующих антител в отношении CD137 человека рассчитывают с использованием программного обеспечения Biacore Т200 Evaluation Software 2.0. В частности, константу скорости ассоциации ka (л/моль/с) и константу скорости диссоциации kd (1/с) рассчитывают путем глобальной подгонки сенсограмм, полученных путем измерения с помощью модели связывания Ленгмюра 1:1, и константы диссоциации KD (моль/л) рассчитывают по этим значениям.
В таблице 28 представлены результаты таких измерений.
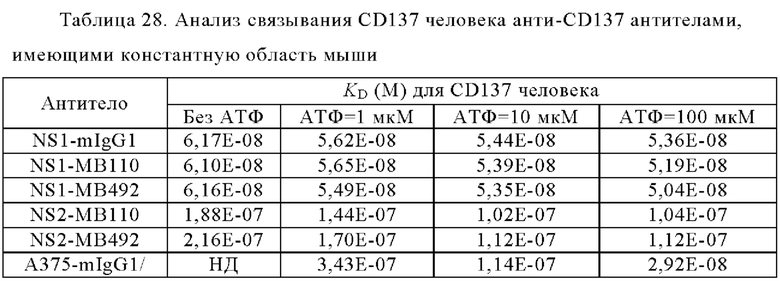
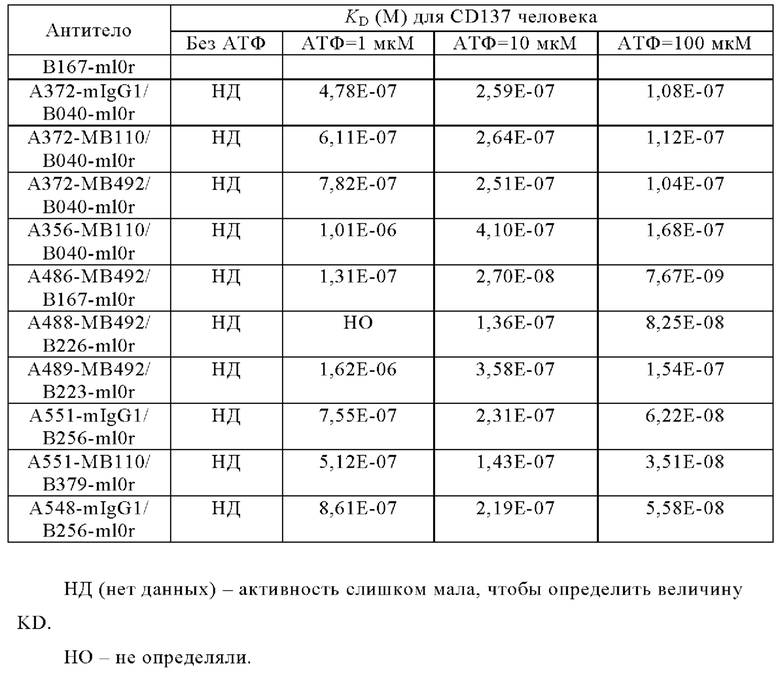
Было подтверждено, что как непереключаемое антитело против CD137 человека, так и полученное выше переключаемое антитело против CD137 человека, которые содержат константную область мыши, связываются с CD137 человека. Также было показано, что все переключаемые антитела против CD137 человека связываются с CD137 человека зависимым от концентрации АТФ образом. С другой стороны, для непереключаемых антител не наблюдают такого зависимого от концентрации АТФ связывания с CD137 человека, причем связывание было почти равным при любой концентрации АТФ.
(6-3) Оценка кинетики модифицированных антител в плазме мышей с нокином CD137 человека
(6-3-1) Получение мышей с нокином CD137 человека
Путем интродукции вектора для замены гена CD137 человека в эмбриональные стволовые клетки мыши (ES-клетки) получают мышь с нокином CD137 человека, у которой ген CD137 мыши заменен геном CD 137 человека. Такую мышь обозначают «hCD137 KI».
(6-3-2) Измерение концентрации антитела против CD137 человека в плазме модели мыши hCD137KI
После создания мыши hCD137KI в примере 6-3-1, соответствующие антитела вводят внутривенно в виде однократной дозы мыши hCD137KI, как показано в табл. 29. В табл. 29 антитела NS1-mIgG1, NS1-MB110 HNS1-MB492 все являются непереключаемыми антителами, а другие - переключаемыми антителами. Кровь собирают несколько раз, начиная с 5 мин до 28 дней после введения. Полученную кровь центрифугируют для отделения плазмы. Плазму хранят в морозильной камере при температуре ниже -20°С до проведения измерения.
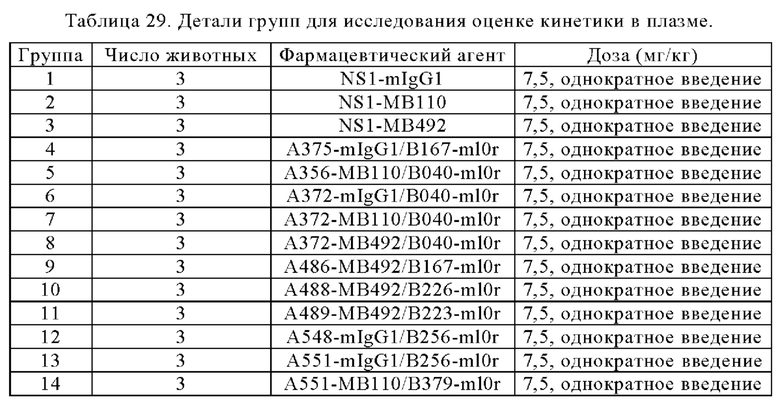
Концентрацию каждого переключаемого антитела в плазме измеряют методом электрохемилюминесценции (ECL, electrochemiluminescence). В частности, hCD137 (фирма Sino Biological Inc.) разводят в ФСБ (-) и вносят в MULTI-ARRAY 96-луночный планшет (фирма Meso Scale Diagnostics, LLC). В планшет добавляют hCD137 и встряхивают при комнатной температуре в течение 1 ч, hCD137 иммобилизуют в планшете. Затем для блокирования добавляют раствор ФСБ, содержащий 1% БСА и 0,05% Tween 20, и встряхивают при комнатной температуре в течение 1 ч. Калибровочные кривые для индивидуальных переключаемых антител построены при концентрациях в плазме 64, 32, 16, 8, 4, 2 и 1 нг/мл. После добавления раствора ФСБ, содержащего 3 мМ АДФ, 1% БСА и 0,05% Tween 20, в планшет с иммобилизованным hCD137, двукратный объем образцов плазмы разбавляют в растворе ФСБ, содержащем 1% БСА и 0,05% Tween20, а также образцы калибровочной кривой. После встряхивания планшета при комнатной температуре в течение 1 ч добавляют биотинилированное антитело против IgG мыши (фирма Jackson ImmunoResearch Laboratories, Inc.). Кроме того, после встряхивания планшета при комнатной температуре в течение 1 ч добавляют меченый стрептавидином SULFO-TAG (фирма Meso Scale Diagnostics, LLCs). Кроме того, после встряхивания планшета при комнатной температуре в течение 1 ч добавляют буфер Read buffer Т (фирма Meso Scale Diagnostics, LLCs), разведенный в два раза раствором ФСБ, содержащим 2 мМ АДФ, 1% БСА и 0,05% Tween 20. Измерение концентраций антител в плазме мышей hCD137KI проводят путем обнаружения с использованием SULFO-TAG на SECTOR Imager (фирма Meso Scale Diagnostics, LLC). Расчет индивидуальных концентраций антител в плазме мыши производят с помощью программы SOFTmax PRO (фирма Molecular Devices).
Концентрацию непереключаемых антител в плазме измеряют методом электрохемилюминесценции (ECL). В частности, hCD137 (фирма Sino Biological Inc.) разводят в ФСБ (-) и добавляют в 96-луночный планшет MULTI-ARRAY (фирма Meso Scale Diagnostics, LLC). В планшет добавляют hCD137 и встряхивают при комнатной температуре в течение одного часа для иммобилизации hCD137 в планшете. Затем для блокирования добавляют раствор ФСБ, содержащий 1% БСА и 0,05% Tween 20, встряхивают при комнатной температуре в течение одного часа. Калибровочные кривые для индивидуальных непереключаемых антител построены при концентрациях в плазме 32, 16, 8, 4, 2, 1 и 0,5 нг/мл. Образцы плазмы, разведенные в растворе ФСБ, содержащем 1% БСА, 0,05% Tween 20, а также образцы калибровочной кривой, добавляют в планшет с иммобилизованным hCD137. После встряхивания планшета при комнатной температуре в течение одного часа добавляют биотинилированное антитело против IgG мыши (фирма Jackson ImmunoResearch Laboratories, Inc.). Кроме того, после встряхивания планшета при комнатной температуре в течение одного часа добавляют SULFO-TAG Labeled Streptavidin (фирма Meso Scale Diagnostics, LLC). Добавляют буфер для считывания Т (фирма Meso Scale Diagnostics, LLCs), разведенный в два раза. Измерение концентраций антител в плазме мышей hCD137KI выполняют аналогично описанному выше для переключаемых антител.
Результаты показаны на фиг. 24, 25 и 26.
На фиг. 24 показана динамика средних концентраций в плазме мыши hCD137 KI антител, имеющих mIgG1 в качестве Fc (NS1-mIgG1 в качестве непереключаемого антитела и A375-mIgG1/B167-ml0r, A372-mIgG1/B040-ml0r, A548-mIgGl/B256-ml0r или A551-mIgG1/B256-ml0r в качестве переключаемого антитела) среди антител, показанных в таблице 29. На фиг. 25 показана динамика средних концентраций в плазме мыши hCD137 KI антител, имеющих МВ110 в качестве Fc (NS1-MB110 в качестве не переключаемого антитела и A356-MB110/B040-ml0r, А372-МВ110/B040-ml0r, и А551-МВ110/B379-ml0r в качестве переключаемых антител). На фиг. 26 показана динамика средних концентраций в плазме мыши CD 137 KI антител, имеющих МВ492 в качестве Fc (NS1-MB492 в качестве непереключаемого антитела и A372-MB492/B040-ml0r, A486-MB492/B167-ml0r, A488-MB492/B226-ml0r или A489-MB492/B223-ml0r в качестве переключаемого антитела). На фиг. 26, что касается А488-МВ492/В226-ml0r, резкое снижение во времени концентрации в плазме, предположительно вызванное ADA, подтверждено у одного субъекта, а среднее значение было взято по данным двух субъектов.
У мыши hCD137 KI с любым Fc переключающее антитело демонстрирует более медленное выведение из плазмы, чем непереключаемое антитело. Предполагают, что это связано с тем, что по сравнению с переключаемым антителом непереключаемое антитело связывается с CD137, экспрессируемым в организме, и быстрее элиминирует в плазме. Кроме того, высказывают предположение, что концентрация внеклеточного АТФ в нормальной ткани до такой степени низкая, что переключаемые антитела, используемые в этом исследовании, не связывались с CD137 человека. Эти результаты позволяют предположить, что связывание с антигеном, экспрессируемым системно, можно уменьшить с помощью переключаемых антител, и, таким образом, можно получить антитело с улучшенной кинетикой крови.
(6-4) Движение маркеров эффективности лекарственного средства и системного ответа антител к CD137 на модели трансплантации сингенных опухолевых клеток с использованием мышей hCD137KI
(6-4-1) Получение клеточных линий и мышиная модель трансплантации сингенной опухоли, а также метод оценки противоопухолевого эффекта и системного ответа
Клетки линии клеток колоректального рака МС38 мыши, лицензированной Национальным институтом рака, и мышей hCD137 KI, описанных в примере 6-3-1, используют для различных исследований. Модель формируется, когда объем опухоли достигает приблизительно 50-300 мм3, путем подкожной имплантации линии клеток МС38 в брюшную полость мышей. После создания модели мышей с трансплантированной клеточной линией МС38 группируют, а затем вводят различные анти-CD137 антитела.
Для оценки противоопухолевого эффекта измерение объема опухоли проводят 1-2 раза в неделю. Объем опухоли рассчитывают по следующей формуле:
Объем опухоли (мм3) = длина (мм) × ширина (мм) × ширина (мм)/2
Путем удаления печени и селезенки или лимфатических узлов в соответствующее время после введения антитела системные ответы оценивают по массе органа, по количеству клеток во фракциях лимфоцитов или по анализу Т-клеток с использованием жидкостной цитометрии (FCM). При оценке этих показателей установлено, что переключаемое антитело субъекта подавляет системные ответы и/или активацию Т-клеток в неопухолевых тканях (например, лимфатических узлах, селезенке и печени), когда показатель системного ответа был низким по сравнению с таким же количество непереключаемого антитела (контрольное антитело против CD137 человека без АТФ-зависимой связывающей активности в отношении CD137 человека).
Антитела, полученные в примере 6-1 (вырабатывание антител для теста по введению мышам с нокином CD137 человека), используют в различных тестах.
(6-4-2) Получение различных органов от мыши с пересаженной клеточной линией МС38 и подготовка фракций лимфоцитов
Проводят измерение массы собранных селезенок и лимфатических узлов, а также подсчет клеток фракций лимфоцитов. Фракции лимфоцитов, полученные с использованием набора для диссоциации печени, мыши (фирма Milteny Biotec), также используют для оценки активации печени. Фракции лимфоцитов после лизиса эритроцитов используют для оценки активации в селезенке, а фракции лимфоцитов, полученные путем измельчения, используют для оценки активации в лимфатических узлах.
(6-4-3) Анализ FCM с использованием фракций лимфоцитов из различных органов
Для оценки активации маркеров методом FCM применяют экспрессию гранзима В или экспрессию ICOS на CD8α-положительных Т-клетках или процент CD8α-положительных Т-клеток среди CD45-положительных клеток. По этой причине в анализе FCM используют флуоресцентно меченые антитела против гранзима В, PD-1, ICOS, CD8α и CD45. Измерения проводят с помощью BD LSRFortessa™ Х-20 (фирма BD Biosciences).
(6-4-4) Исследования лекарственной эффективности (противоопухолевого эффекта) антитела A375-mIgG1/B167-ml0r
После формирования у мышей опухоли в результате трансплантации раковых клеток МС38 в примерах 6-4-1, вводят растворитель и А375-mIgG1/B167-ml0r в дозах, показанных в табл. 30. Введение антител проводят дважды через хвостовую вену в день формирования групп и через четыре дня после этого. В качестве растворителя применяют ФСБ, содержащий 0,05% Tween-20.
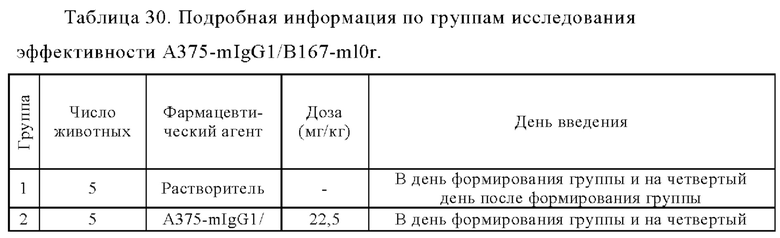
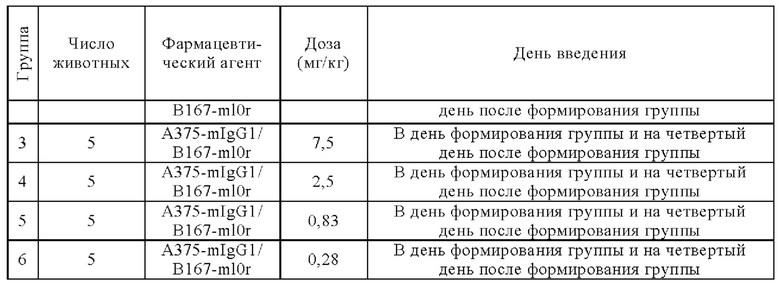
Противоопухолевый эффект в каждой группе оценивают по объему опухоли, рассчитанной согласно описанию в примере 6-4-1. В результате подтверждают доза-зависимый противоопухолевый эффект антитела А375-mIgG1/B167-ml0r (фиг. 27).
(6-4-5) Введение антител и отбор образцов, а также анализ FCM для оценки системных эффектов A375-mIgG1/B167-ml0r
После создания мышиной модели после трансплантации МС38 в примере 6-4-1, вводят растворитель, NS1-mIgG1 (непереключаемое антитело) и А375-mIgG1/B167-ml0r, как показано в табл. 31. Введение антител вводят дважды в хвостовую вену в день формирования групп и через три дня после формирования групп. В качестве растворителя применяют ФСБ, содержащий 0,05% Tween-20.
Через пять дней после первоначального введения собирают печень, лимфатические узлы и селезенку, как показано в примере 6-4-2 (сбор различных органов от мыши с трансплантированной линией клеток МС38 и получение фракций лимфоцитов). Кроме того, для селезенки и лимфатических узлов органы трех мышей объединяют и проводят анализ FCM, как показано в примере 6-4-3 (анализ FCM с использованием фракций лимфоцитов из различных органов).
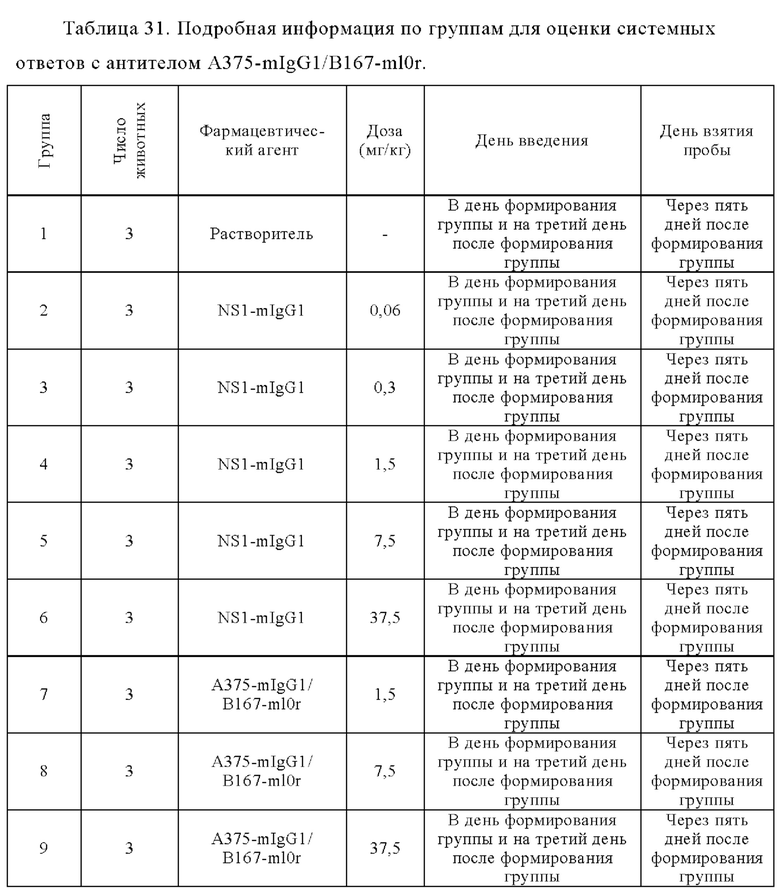
Оценивают системные эффекты от введения антител. В результате оценки веса органов лимфатических узлов и селезенок установлено увеличение веса этих органов при введении NS1-mIgG1 в дозе 0,3 мг/кг и более выраженная гипертрофия органов при 1,5 мг/кг и 7,5 мг/кг. С другой стороны, увеличения органа не наблюдают при введении A375-mIgG1/B167-ml0r ни при одной из доз 1,5 мг/кг, 7,5 мг/кг или 37,5 мг/кг (фиг. 28).
Кроме того, в результате оценки активации Т-клеток с помощью оценки FCM наблюдают повышение экспрессии всех маркеров PD-1, ICOS и гранзима В при 0,3 мг/кг после введения NS1-mIgG1. С другой стороны, при введении А375-mIgGl/B167-ml0r наблюдают заметное ингибирование экспрессии всех маркеров PD-1, ICOS и гранзима В в дозах 1,5 мг/кг, 7,5 мг/кг и 37,5 мг/кг (фиг. 29 и фиг. 30). Поскольку результаты по A375-mIgG1/B167-ml0r показали, что вес органа коррелирует с маркерами активации Т-клеток в лимфатических узлах и селезенке, вес органа используют в основном в качестве индикатора активации иммунных клеток в лимфатических узлах и селезенке при последующей оценке антител.
В результате оценки активации Т-клеток в печени наблюдают увеличение экспрессии PD-1 и гранзима В при введении NS1-mIgG1 в дозе 0,3 мг/кг. С другой стороны, при введении A375-mIgG1/B167-ml0r, экспрессии подавлялись при всех дозах 1,5 мг/кг, 7,5 мг/кг и 37,5 мг/кг (фиг. 31).
Таким образом, переключающее антитело ингибирует активацию Т-клеток в лимфатических узлах, селезенке и печени по сравнению с таким же количеством непереключаемого антитела.
(6-4-6) Исследование лекарственной эффективности (противоопухолевого эффекта) антитела А356-МВ110/B040-ml0r
После формирования мышиной модели после трансплантации МС38 в примере 6-4-1 растворитель и А356-МВ110/B040-ml0r вводят в дозах, показанных в табл. 32. Введение антитела проводят дважды в хвостовую вену в день формирования группы и через три дня после формирования группы. В качестве растворителя применяют ФСБ, содержащий 0,05% Tween-20.
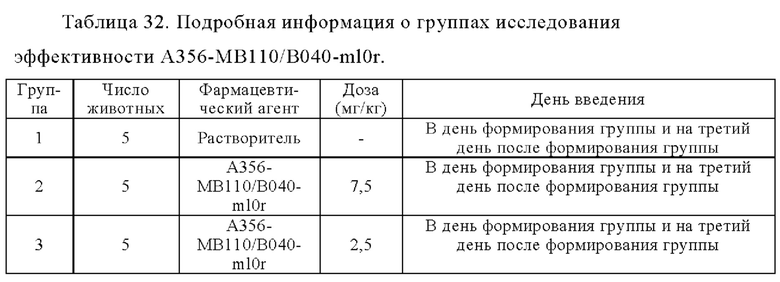
Противоопухолевый эффект в каждой группе оценивают по объему опухоли по расчетам, описанным в примере 6-4-1. Результаты подтверждают дозозависимый противоопухолевый эффект А356-МВ110/B040-ml0r (фиг. 32).
(6-4-7) Введение антител и отбор образцов, а также FCM-анализ для оценки системных эффектов А356-МВ110/B040-ml0r
После создания модели мышей с трансплантацией МС38 в примере 6-4-1 вводят растворитель, NS2-MB110 (непереключаемое антитело) и А356-МВ110/B040-ml0r, как показано в табл. 33. Введение антител проводят дважды в хвостовую вену в день формирования групп и через три дня после г формирования групп. В качестве растворителя применяют ФСБ, содержащий 0,05% Tween-20.
Через семь дней после первого введения печень, лимфатические узлы и селезенку собирали, как показано в примере 6-4-2 (сбор различных органов у мышей, трансплантированных клеточной линией МС38, и подготовка фракций лимфоцитов). Для селезенки и лимфатических узлов проводят только измерение веса органов.
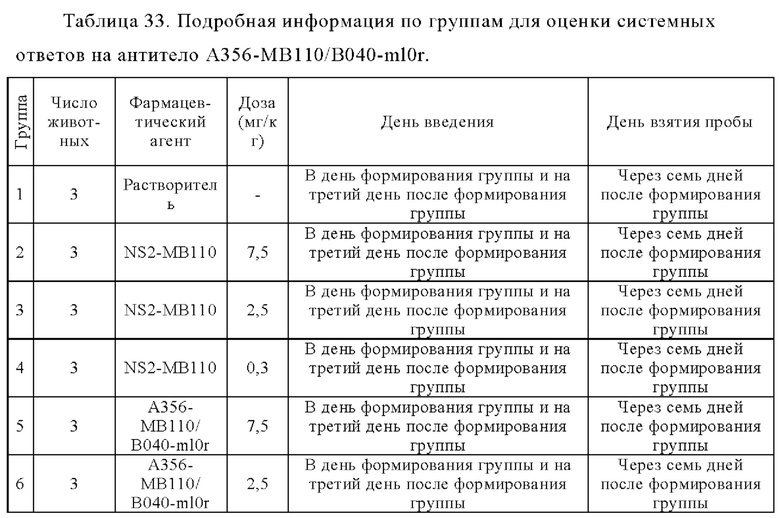
Оценивают системные эффекты от введенных антител. Результат оценки массы органов лимфатических узлов и селезенки при введении NS2-MB110 и А356-МВ110/B040-ml0r показывает, что гипертрофия наблюдается также при введении NS2-MB110 в дозе 2,5 мг/кг. С другой стороны, на момент введения А356-МВ110/В040-мл0r лимфаденопатия подавляется даже при 7,5 мг/кг, а спленомегалия подавляется при 2,5 мг/кг (фиг. 33).
Кроме того, результат оценки активации Т-клеток в печени показывает, что увеличение экспрессии PD-1 в CD8α-положительных клетках также наблюдают при введении NS2-MB110 в дозе 0,3 мг/кг. Между тем, когда вводят А356-МВ110/B040-ml0r, экспрессия PD-1 подавляется при дозе 7,5 мг/кг и существенно подавляется при 2,5 мг/кг. Небольшое увеличение экспрессии ICOS в CD8α-положительных клетках также наблюдают при введении антитела NS2-МВ110 в дозе 0,3 мг/кг. С другой стороны, наблюдают, что экспрессия растворителя подавляется при 2,5 мг/кг при введении А356-МВ110/B040-ml0r (фиг. 34).
Таким образом, переключаемое антитело ингибирует активацию Т-клеток в лимфатических узлах, селезенке и печени по сравнению с непереключаемым антителом.
(6-4-8) Исследование лекарственной эффективности (противоопухолевого эффекта) антитела A372-mIgG1/B040-ml0r
После создания модели мышей с трансплантацией МС38 в примере 6-4-1 вводят растворитель и A372-mIgG1/B040-ml0r в дозах, показанных в табл. 34. Введение антитела проводят четыре раза в хвостовую вену в день формирования группы и через 4 дня, 7 дней и 10 дней после формирования группы. В качестве растворителя применяют ФСБ, содержащий 0,05% Tween-20.

Противоопухолевый эффект в каждой группе оценивают по объему опухоли по расчетам, описанным в примере 6-4-1. Результаты подтверждают противоопухолевый эффект от введения антитела A372-mIgG1/B040-ml0r (фиг. 35).
(6-4-9) Введение антител и отбор образцов, а также анализ FCM для оценки системных эффектов A372-mIgG1/B040-ml0r
После создания модели мышей с трансплантацией МС38 в примере 6-4-1 вводят растворитель и A372-mIgG1/B040-ml0r, как показано в табл. 35. Введение антитела проводят три раза в хвостовую вену в день формирования группы и через 3 дня и 6 дней после формирования группы. В качестве растворителя применяют ФСБ, содержащий 0,05% Tween-20.
Через семь дней после первого введения печень, лимфатические узлы и селезенку собирают, как показано в примере 6-4-2 (сбор различных органов у мышей, трансплантированных клеточной линией МС38, и подготовка фракций лимфоцитов). Для селезенки проводят только измерение массы органов; для лимфатических узлов проводят только подсчет клеток фракций лимфоцитов, полученных из лимфоцитов, объединенных от трех мышей.

Оценивают системные эффекты от введения антител. В результате измерения количества клеток в лимфатических узлах и массы селезенок было подтверждено, что нет увеличения количества клеток в лимфатических узлах и увеличения массы селезенки при введении A372-mIgG1/B040-ml0r, и что масса этого органа эквивалентна массе в группе введения растворителя (фиг. 36).
В результате оценки активации Т-клеток в печени показано, что уровни экспрессии гранзима В подобны аналогичным уровням в группе введения растворителя когда вводили A372-mIgG1/B040-ml0r (фиг. 37).
Таким образом, подтверждено, что переключаемое антитело ингибирует активацию Т-клеток в лимфатических узлах, селезенке и печени.
(6-4-10) Исследование лекарственной эффективности (противоопухолевого эффекта) антитела А372-МВ110/B040-ml0r
После создания модели мышей с трансплантацией рака МС38 в примере 6-4-1 растворитель и А372-МВ110/B040-ml0r вводят в дозах, представленных в табл. 36. Введение антител осуществляют дважды в хвостовую вену в день формирования группы и через четыре дня после этого. В качестве растворителя используют ФСБ, содержащий 0,05% Tween-20.
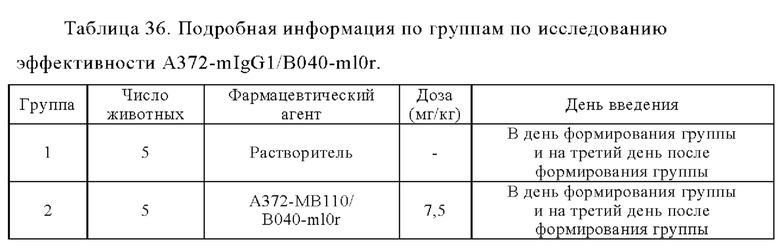
Противоопухолевый эффект в каждой группе оценивают по объему опухоли по расчетам, описанным в примере 6-4-1. Результаты подтверждают противоопухолевый эффект A372-mIgG1/B040-ml0r (фиг. 38).
(6-4-11) Введение антител и отбор образцов, а также анализ FCM для оценки системных эффектов А372-МВ110/B040-ml0r
После создания модели мышей с трансплантацией рака МС38 в примере 6-4-1 растворитель, NS2-MB110 (непереключаемое антитело) и А372-МВ110/В040-ml0r вводят в дозах, представленных в табл. 37. Введение антител осуществляют дважды в хвостовую вену, а именно в день формирования группы и через три дня после этого. В качестве растворителя используют ФСБ, содержащий 0,05% Tween-20.
Через семь дней после первоначального введения собирают печень, лимфатические узлы и селезенку, как показано в примере 6-4-2 (сбор различных органов у мышей, трансплантированных клеточной линией МС38, и получение фракций лимфоцитов). Для селезенки и лимфатических узлов проводят только измерение массы органов.
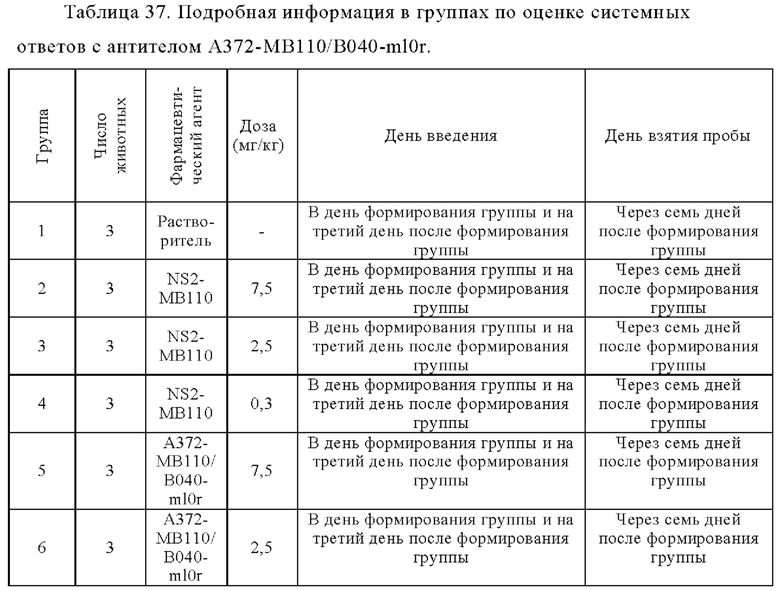
Оценивают системные эффекты от введения антител. В результате измерения массы лимфатических узлов и селезенок подтверждено, что масса этих органов увеличивается при введении NS2-MB110 в дозах 2,5 мг/кг и 7,5 мг/кг. С другой стороны наблюдают ингибирование органомегалии при обеих дозах 2,5 мг/кг и 7,5 мг/кг при введении А372-МВ110/B040-ml0r (фиг. 39).
В результате оценки активации Т-клеток в печени наблюдают увеличение экспрессии PD-1 при введении NS2-MB110. Это также наблюдают при введении A372-MB110/B040-ml0r (фиг. 40).
Таким образом, переключаемое антитело ингибирует активацию Т-клеток в лимфатических узлах и селезенке в отличие от непереключаемого антитела.
(6-4-12) Исследование лекарственной эффективности (противоопухолевого эффекта) антитела A372-MB492/B040-ml0r
После создания модели мышей с трансплантацией рака МС38 в примере 6-4-1 растворитель и А372-МВ110/B040-ml0r вводят в дозах, представленных в табл. 38. Введение антител осуществляют дважды в хвостовую вену в день формирования группы и через три дня после этого. В качестве растворителя используют ФСБ, содержащий 0,05% Tween-20.
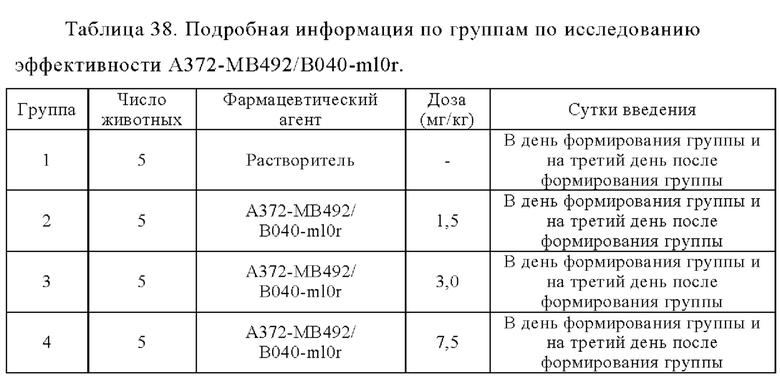
Противоопухолевый эффект в каждой группе оценивают по объему опухоли по расчетам, описанным в примере 6-4-1. В результате наблюдают противоопухолевый эффект антитела A372-MB492/B040-ml0r для всех доз (фиг. 41).
(6-4-13) Введение антител и отбор образцов, а также анализ FCM для оценки системных эффектов антитела A372-MB492/B040-ml0r
После получения модели мышей с трансплантацией рака МС38 в примере 6-4-1 растворитель, NS1-MB492 (непереключаемое антитело) и А372-MB492/B040-ml0r вводят в дозах, представленных в табл. 39. Введение антител осуществляют дважды в хвостовую вену, а именно в день формирования группы и через три дня после этого. В качестве растворителя используют ФСБ, содержащий 0,05% Tween-20.
Через восемь дней после первоначального введения собирают печень, лимфатические узлы и селезенку, как показано в примере 6-4-2 (сбор различных органов у мышей, трансплантированных клеточной линией МС38, и получение фракций лимфоцитов). Измеряют массу отобранных селезенок, а также подсчитывают лимфоциты во фракциях из лимфоузлов.
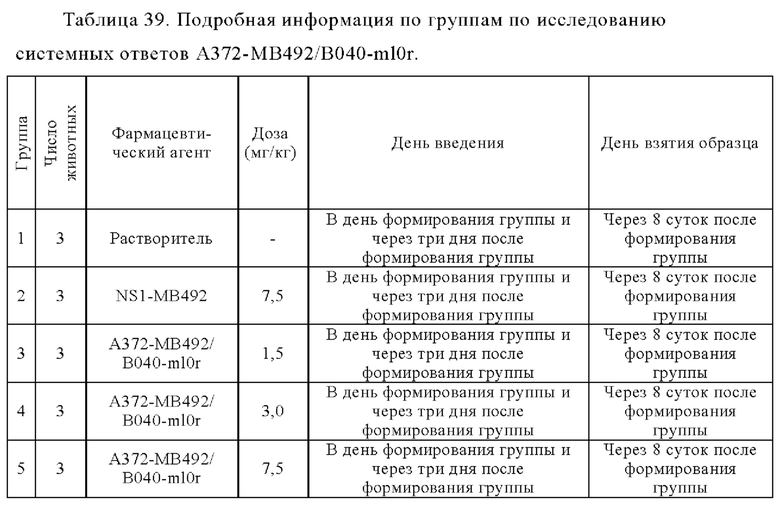
Оценивают системные эффекты от введенных антител. Наблюдают увеличение массы селезенок при введении NS1-MB492 и увеличение клеток во фракциях лимфоцитов при введении NS1-MB492. С другой стороны, при введении A372-MB492/B040-ml0r в дозах 1,5 мг/кг, 3,0 мг/кг и 7,5 мг/кг наблюдают ингибирование органомегалии по сравнению с введением NS1-МВ492 (фиг. 42).
В результате оценки активации Т-клеток в печени увеличение экспрессии гранзима В наблюдают при введении NS1-MB492 и A372-MB492/B040-ml0r по сравнению с введением растворителя (фиг. 43).
Таким образом, переключаемое антитело ингибирует активацию Т-клеток в лимфатических узлах и селезенке по сравнению с непереключаемым антителом.
(6-4-14) Исследование лекарственной эффективности (противоопухолевого эффекта) антител A486-MB492/B167-ml0r и А488-МВ492/В226-ml0r
После создания модели мышей с трансплантацией рака МС38 в примере 6-4-1 растворитель и антитела A486-MB492/B167-ml0r и A488-MB492/B226-ml0r вводят в дозах, представленных в табл. 40. Формирование групп проводят через 10 дней после трансплантации клеток, а введение антител осуществляют в хвостовую вену через 11, 15, 18 и 22 дня после трансплантации клеток, в общей сложности четыре раза.
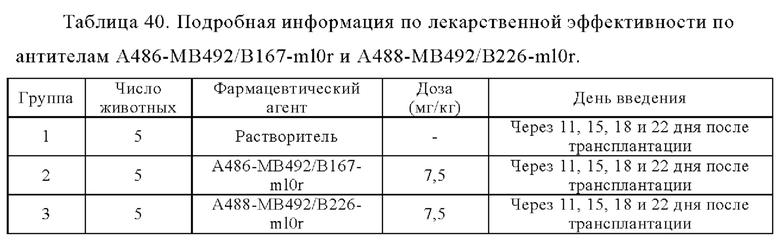
Противоопухолевый эффект в каждой группе оценивают по объему опухоли по расчетам, описанным в примере 6-4-1. Результаты подтверждают противоопухолевый эффект A486-MB492/B167-ml0r и A488-MB492/B226-ml0r (фиг. 44).
(6-4-15) Введение антител и отбор образцов, а также анализ FCM для оценки системных эффектов антител А48б-МВ492/В167-ml0r и А488-МВ492/В226-ml0r
После создания модели мышей с трансплантацией рака МС38 в примере 6-4-1 растворитель, NS1-MB492 (непереключаемое антитело), А486-МВ492/В167-ml0r и A488-MB492/B226-ml0r вводят в дозах, представленных в табл. 41. Формирование групп проводят через 10 дней после трансплантации клеток, а введение антител осуществляют в хвостовую вену через 11, 15, 18 и 22 дня после трансплантации клеток, в общей сложности четыре раза. В качестве растворителя применяют ФСБ, содержащий 0,05% Tween-20.
Печень, лимфатические узлы и селезенку собирают через 13 дней после первоначального введения, как показано в примере 6-4-2 (сбор различных органов у мышей, трансплантированных клеточной линией МС38, и подготовка фракций лимфоцитов). Проводят измерение массы селезенок, а также подсчитывают лимфоциты для фракций из лимфоузлов.
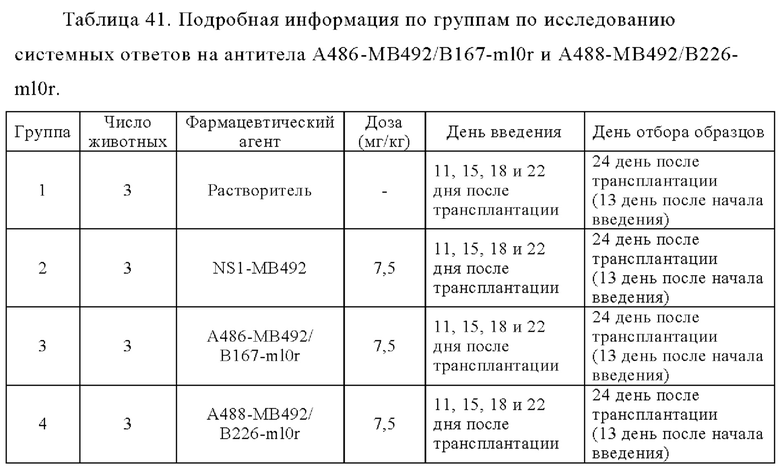
В результате измерения количества клеток во фракциях лимфоцитов из лимфатических узлов и измерения массы селезенки при введении NS1-MB492 наблюдают увеличение количества лимфоцитов и массы селезенки. С другой стороны, замечено, что увеличение количества лимфоцитов и массы селезенки подавляются при введении A486-MB492/B167-ml0r и A488-MB492/B226-ml0r по сравнению с таковыми при введении NS1-MB492 (фиг. 45).
В результате оценки активации Т-клеток в печени увеличение процента CD8α-положительных клеток наблюдают при введении NS1-MB492, А486-MB492/B167-ml0r и A488-MB492/B226-ml0r по сравнению с введением растворителя (фиг. 46).
На основании вышеизложенного подтверждено, что по сравнению с непереключаемым антителом переключаемое антитело ингибирует активацию Т-клеток в лимфатических узлах и селезенке.
(6-4-16) Исследование лекарственной эффективности (противоопухолевого эффекта) антитела A489-MB492/B223-ml0r
После создания модели мышей с трансплантацией рака МС38 в примере 6-4-1 растворитель и антитело A489-MB492/B223-ml0r вводят в дозах, представленных в табл. 42. Введение антитела осуществляют четыре раза через хвостовую вену на следующий день, через четыре, семь и десять дней после формирования группы. В качестве растворителя применяют ФСБ, содержащий 0,05% Tween-20.
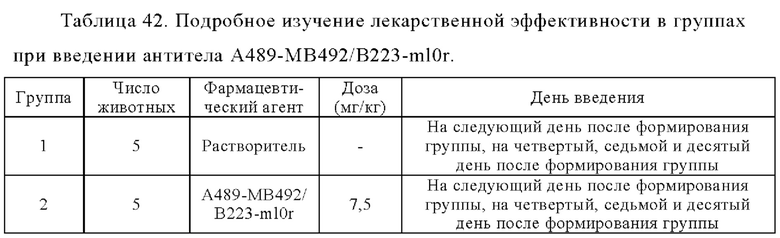
Противоопухолевый эффект в каждой группе оценивают по объему опухоли по расчетам, описанным в примере 6-4-1. Результаты подтверждают противоопухолевый эффект от введения дозы 7,5 мг/кг (фиг. 47).
(6-4-17) Введение антител и отбор образцов, а также анализ FCM для оценки системных эффектов антитела A489-MB492/B223-ml0r
После создания модели мышей с трансплантацией рака МС38 в примере 6-4-1 растворитель, NS1-MB492 (непереключаемое антитело) и А489-МВ492/В223-ml0r вводят в дозах, представленных в табл. 43. Введение антител осуществляют через хвостовую вену в день формирования группы, а также через 3 дня и 6 дней после формирования группы, в общей сложности три раза. В качестве растворителя применяют ФСБ, содержащий 0,05% Tween-20.
Через семь дней после первоначального введения печень, лимфатические узлы и селезенку собирают, как показано в примере 6-4-2 (сбор различных органов у мышей, трансплантированных клеточной линией МС38, и подготовка фракций лимфоцитов). В селезенках и лимфоузлах проводят только измерение количества клеток только во фракциях лимфоцитов.
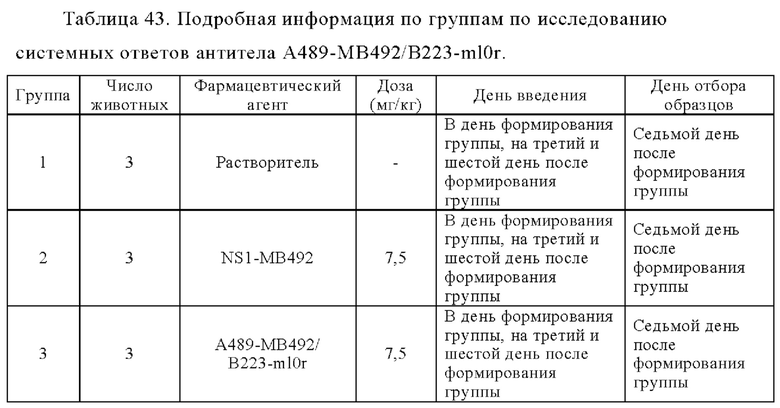
В результате измерения количества клеток во фракциях лимфоцитов из лимфатических узлов и селезенки наблюдают увеличение количества клеток при введении NS1-MB492. С другой стороны, подавление увеличения количества лимфоцитов наблюдают при введении A489-MB492/B223-ml0r по сравнению с таковым при введении NS1-MB492 (фиг. 48).
В результате оценки активации Т-клеток в печени наблюдают увеличение процента CD8α-положительных клеток при введении NS1-MB492 и А489-MB492/B223-ml0r по сравнению с растворителем (фиг. 49).
На основании вышеизложенного подтверждено, что по сравнению с непереключаемым антителом переключаемое антитело ингибирует активацию Т-клеток в лимфатических узлах и селезенке.
(6-4-18) Исследование лекарственной эффективности (противоопухолевого эффекта) антител A548-mIgG1/B256-ml0r и A551-mIgG1/B256-ml0r
После создания модели мышей с трансплантацией рака МС38 в примере 6-4-1 растворитель и антитела A548-mIgG1/B256-ml0r и А551-mIgG1/B256-ml0r вводят в дозах, представленных в табл. 44. Введение антитела осуществляют через хвостовую вену два раза: в день формирования группы и через три дня. В качестве растворителя применяют ФСБ, содержащий 0,05% Tween-20.
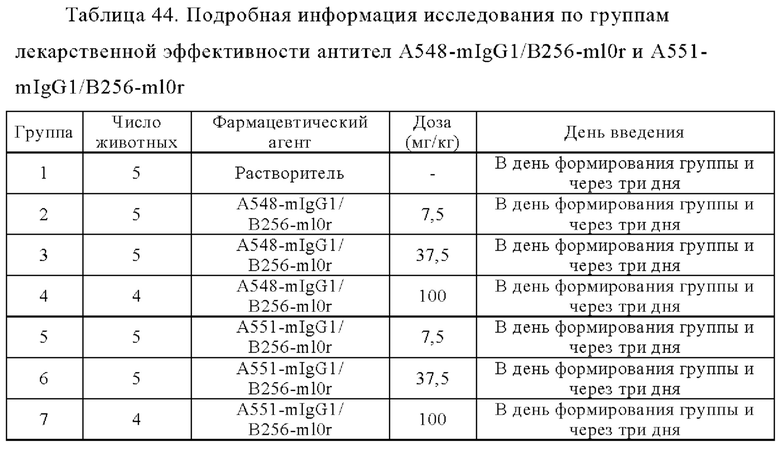
Противоопухолевый эффект в каждой группе оценивают по объему опухоли по расчетам, описанным в примере 6-4-1. Результаты подтверждают противоопухолевый эффект от введения A548-mIgG1/B256-ml0r в дозах 37,5 мг/кг и 100 мг/кг. A551-mIgG1/B256-ml0r также проявляет существенный противоопухолевый эффект в дозе 100 мг/кг (фиг. 50).
(6-4-19) Введение антител и отбор образцов, а также анализ FCM для оценки системных эффектов антител A548-mIgG1/B256 и A551-mIgG1/B256
После создания модели мышей с трансплантацией рака МС38 в примере 6-4-1 растворитель, NS1-mIgG1 (непереключаемое антитело), A548-mIgGl/B256-ml0r и A551-mIgG1/B256-ml0r вводят в дозах, представленных в табл.45. Введение антител осуществляют через хвостовую вену в день формирования группы, а также через 3 дня после формирования группы, в общей сложности два раза. В качестве растворителя применяют ФСБ, содержащий 0,05% Tween-20.
Через семь дней после первоначального введения печень, лимфатические узлы и селезенку собирают, как показано в примере 6-4-2 (сбор различных органов у мышей, трансплантированных клеточной линией МС38, и подготовка фракций лимфоцитов). У образцов селезенок и лимфоузлов определяют только массу органов.
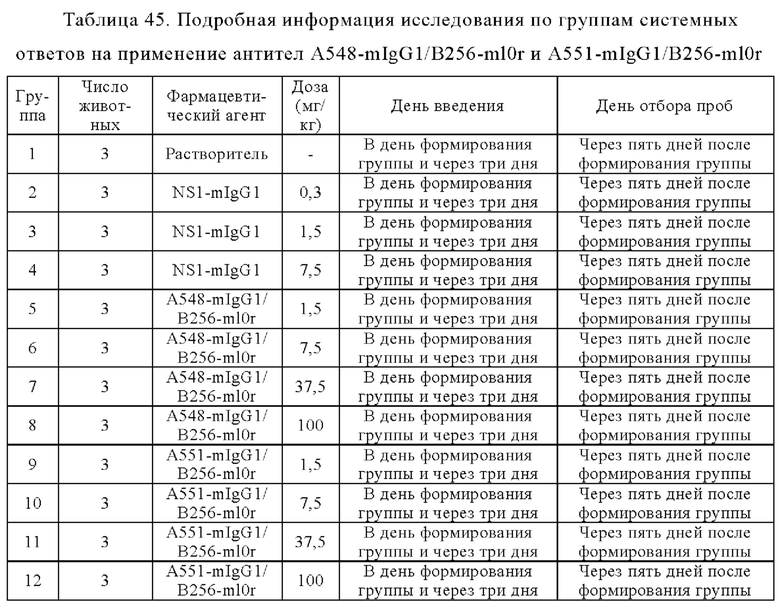
В результате оценки массы органов лимфатических узлов и селезенки наблюдают увеличение массы органа при введении NS1-mIgG1 в дозе 0,3 мг/кг, 1,5 мг/кг и 7,5 мг/кг. С другой стороны не наблюдают гипертрофии органов при введении A548-mIgGl/B256-ml0r или А551-mIgG1/B256-ml0r в любой дозе из доз 1,5 мг/кг, 7,5 мг/кг, 37,5 мг/кг и 100 мг/кг (фиг. 51).
В результате оценки активации Т-клеток в печени наблюдают увеличение экспрессии как гранзима В, так и PD-1 при введении NS1-mIgG1 в дозе 0,3 мг/кг. С другой стороны, экспрессия подавляется при введении A548-mIgG1/B256-ml0r и А551-mIgG1/B256-ml0r дозе из доз 1,5 мг/кг, 7,5 мг/кг, 37,5 мг/кг и 100 мг/кг (фиг. 52).
Таким образом, по сравнению с не переключаемым антителом переключаемое антитело ингибирует активацию Т-клеток в лимфатических узлах, селезенке и печени.
(6-4-20) Исследование лекарственной эффективности (противоопухолевого эффекта) антитела А551-МВ110/B379-ml0r
После создания модели мышей с трансплантацией МС38в примере 6-4-1, растворитель и антитело А551-МВ110/B379-ml0r вводят в дозах, указанных в табл. 46. Антитела вводят дважды в хвостовую вену и через три дня после формирования группы. В качестве растворителя используют ФСБ с 0,05% Tween-20.
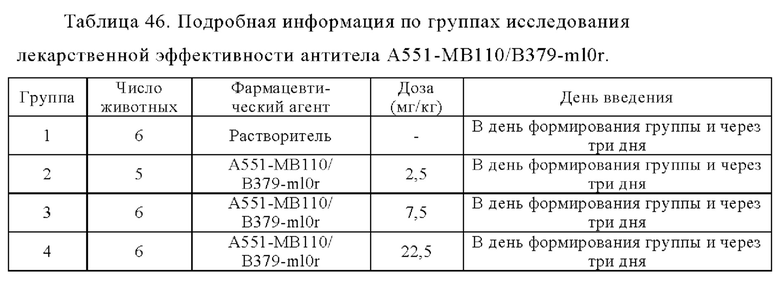
Противоопухолевый эффект в каждой группе оценивают по объему опухоли, рассчитанной ка описано в примере 6-4-1. В результате противоопухолевый эффект подтвержден для каждой дозировки (фиг. 53).
(6-4-21) Введение антител и отбор образцов, а также анализ FCM для оценки системных эффектов
После создания модели мышей с трансплантацией МС38 в примере 6-4-1 вводят растворитель, NS1-mIgG1 (непереключаемое антитело) и А551-МВ110/B379-ml0r вводят как показано в табл. 47. Введение антитела проводят два раза в хвостовую вену в день формирования группы и через 3 дня после формирования группы. В качестве растворителя применяют ФСБ, содержащий 0,05% Tween-20.
Печень, лимфатические узлы и селезенки собирают через 5 дней после первоначального введения, как показано в примере 6-4-2 (сбор различных органов у мышей с трансплантированной клеточной линией МС38 и подготовка фракций лимфоцитов) и анализ FCM выполняют, как показано в Примере 6-4-3 (анализ FCM с использованием фракций лимфоцитов различных органов).
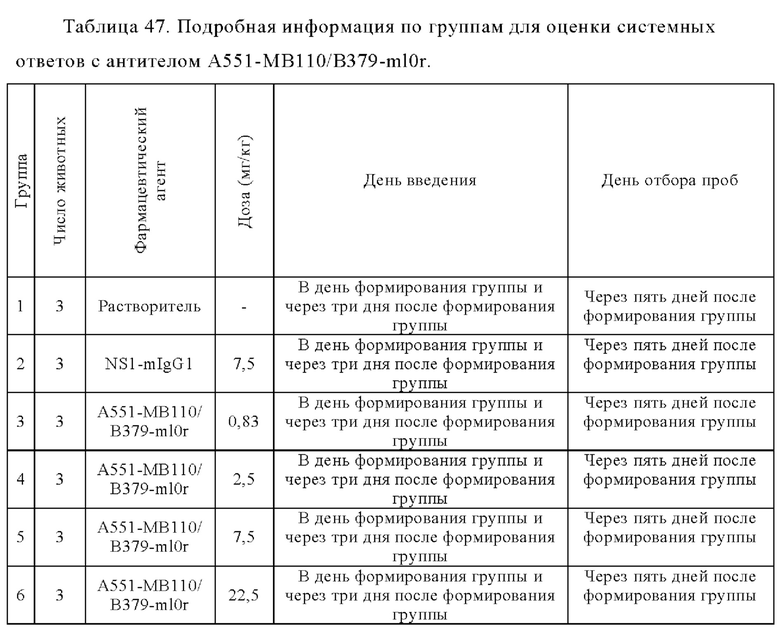
В результате оценки массы органов лимфатических узлов и селезенки наблюдают увеличение массы органов при введении NS1-mlgG1 в дозе 7,5 мг/кг. С другой стороны, увеличения органов не наблюдают при введении А551-МВ110/B379-ml0r в любой дозе, а именно 0,83 мг/кг, 2,5 мг/кг, 7,5 мг/кг и 22,5 мг/кг (фиг. 54). Кроме того, при оценке активации Т-клеток в селезенке методом FCM, увеличение маркеров активации (PD-1, ICOS, гранзима В), наблюдаемое при введении NSl-mIgG1, не подтверждалось при введении А551-МВ110/В379-ml0r в любой дозе (фиг. 55).
При оценке активации Т-клеток в печени увеличение экспрессии как PD-1, так и ICOS наблюдают при введении NS1-mIgG1 в дозе 7,5 мг/кг. С другой стороны, подавление экспрессии на уровне растворителя, когда А551-МВ110/B379-ml0r вводят в дозах 0,83 мг/кг и 2,5 мг/кг, и даже большее подавление экспрессии, чем от NS1-mIgG1, при дозе 7,5 мг/кг (фиг. 56).
Таким образом, по сравнению с непереключаемым антителом переключаемое антитело ингибирует активацию Т-клеток в лимфатических узлах, селезенке и печени.
Пример 7. Получение модифицированного Fc, способного усиливать агонистическую активность
(7-1) Получение вариантов с повышенной FcγR-связывающей активностью.
Константная область тяжелой цепи ТТ14 (SEQ ID NO: 149), которая увеличивает FcγRIIb-связывающую активность, как описано в WO 2017104783 (константная область тяжелой цепи, содержащая T250V/T307P в качестве аминокислотных модификаций, и L234Y/P238D/V264I/A330K в качестве аминокислотных модификаций для увеличения FcγRIIb-связывающей активности в области Fc, содержащей SEQ ID NO: 182 (оба модифицированных положения приведены в соответствии с нумерацией EU)), область ТТ16 (SEQ ID NO: 150), содержащую мутацию аминокислоты G237D, введенную в константную область тяжелой цепи ТТ11, которая увеличивает FcγRIIb-связывающую активность, как описано в WO 2017104783 (константная область тяжелой цепи, содержащую аминокислотные модификации T250V/T307P и L234Y/P238D/A330K в качестве модификаций для увеличения FcγRIIb-связывающей активности в области Fc, содержащей SEQ ID NO: 182 (оба модифицированных положения приведены в соответствии с нумерацией EU)), константную область тяжелой цепи Р587 (SEQ ID NO: 151), которая увеличивает FcγRIIb-связывающую активность, как описано в WO 2014163101; и последовательность Р253 (SEQ ID NO: 93), являющуюся предшествующим вариантом константной области тяжелой цепи с повышенной связываемостью с FcγRIIb, объединенную с вариабельной областью тяжелой цепи переключаемого антитела (А375; А372; А356; А486; А488; А489; А548; и А551) или с вариабельной областью тяжелой цепи антитела - отрицательного контрольного (IC17HdK (SEQ ID NO: 152)). В результате были созданы гены со следующими комбинациями каждой константной области и вариабельной области тяжелой цепи.
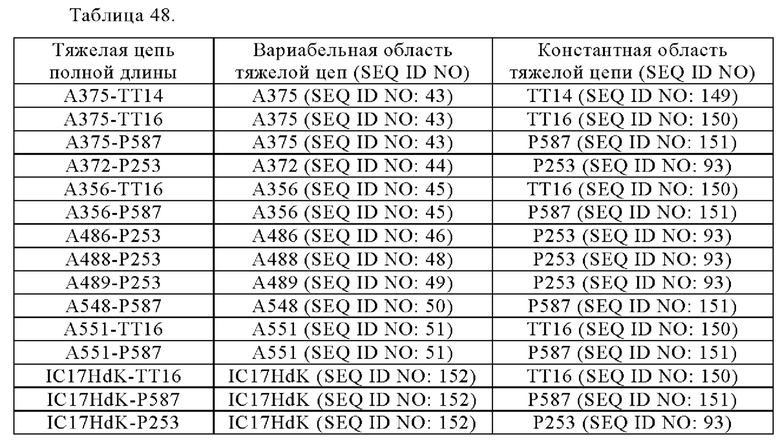
Вариабельную область тяжелой цепи переключаемого антитела (А375, A356 и А551) или вариабельную область тяжелой цепи антитела - отрицательного контроля (IC17HdK (SEQ ID NO: 152)) объединяют с константной областью тяжелой цепи MY201 (SEQ ID НЕТ: 153) (константной областью тяжелой цепи, содержащей G236N/H268D/A330K в качестве аминокислотных модификаций, которые повышают связывающую активность с FcγRIIb в области Fc, включающей аминокислотную последовательность SEQ ID NO: 182 (все модифицированные положения указаны по нумерации EU)) или MY518 (SEQ ID NO: 154) (константной областью тяжелой цепи, содержащей L235W/G236N/H268D/Q295L/K326T/A330K в качестве аминокислотных модификаций, которые повышают связывающую активность с FcγRIIb в области Fc, содержащей аминокислотную последовательность SEQ ID NO: 182 (все модифицированные положения указаны по нумерации EU), которые описаны в WO 2017104783 и имеют повышенную связывающую активность с FcγR, константную область тяжелой цепи MY201a (SEQ ID NO: 155) или MY518a (SEQ ID NO: 156), в которой модификация K214R интродуцирована в MY201 или MY518, или константную область тяжелой цепи MY201aPh (SEQ ID NO: 157), в которой модификация L235W интродуцирована в MY201a. В результате созданы гены с комбинациями каждой константной области и вариабельной области тяжелой цепи, представленные ниже в табл.49.
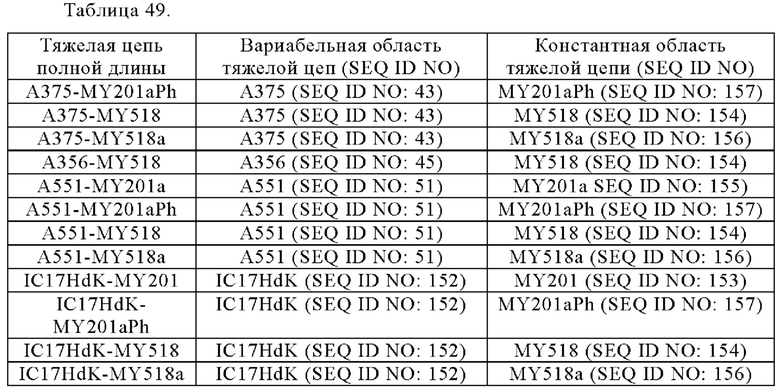
В табл.50 показано, что представляющие интерес антитела экспрессированы и очищены методами, известными специалистам в данной области техники, путем комбинирования указанных выше генов тяжелой цепи антитела с геном легкой цепи переключаемого антитела (B040-Lamlib, В167-Lamlib, B226-Lamlib, B223-Lamlib, B256-Lamlib и B379-Lamlib, полученные в примерах 5-2 и 5-3) или геном легкой цепи антитела - отрицательного контроля (IC17L-k0, полученного путем объединения легкой цепи вариабельная область IC17L (SEQ ID NO: 158) с цепью к человека k0 (SEQ ID NO: 141) в качестве константной области легкой цепи).
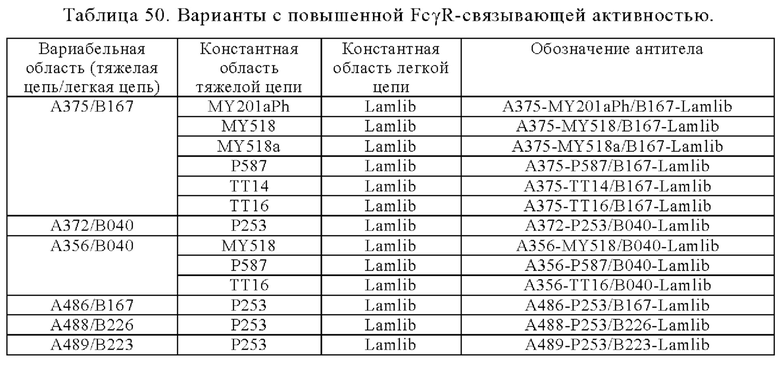
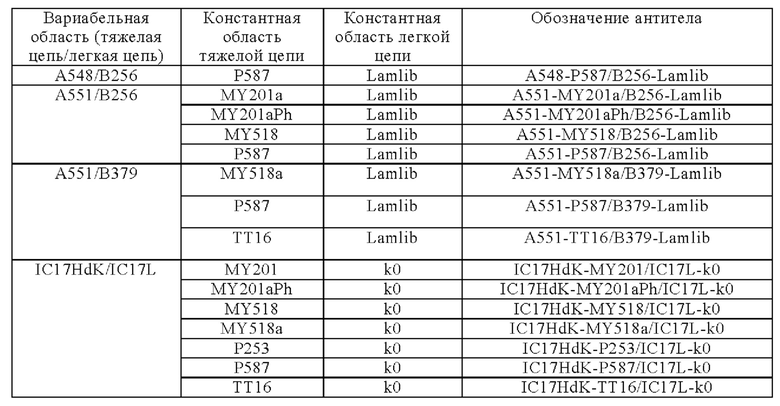
(7-2) Получение вариантов с увеличенной pI путем аминокислотной модификации константной области
Константную область тяжелой цепи получают путем объединения константной области тяжелой цепи с повышенной FcγR-связывающей активностью, полученной в примере 7-1, с Q311R, P343R и D413K, которые представляют собой аминокислотные мутации, увеличивающие pI без значительного изменения FcγR-связывающей активности, как описано в WO 2017046994.
В частности, получают гены константной области тяжелой цепи с аминокислотными мутациями, введенными в каждый из генов константной области тяжелой цепи антитела, как показано ниже.
- Введение P343R/D413K в MY518 (SEQ ID NO: 154): SCF025 (SEQ ID NO:64)
- Введение P343R/D413K в MY518a (SEQ ID NO: 156): SCF025a (SEQ ID NO: 65)
- Введение P343R/D413K в TT14 (SEQ ID NO: 149): SCF027 (SEQ ID NO: 66)
- Введение P343R/D413K в TT16 (SEQ ID NO: 150): SCF028 (SEQ ID NO: 67)
- Введение Q311R/P343R в MY518 (SEQ ID NO: 154): SCF030 (SEQ ID NO:68)
- Введение Q311R/P343R в ТТ14 (SEQ ID NO: 149): SCF032 (SEQ ID NO: 69)
- Введение Q311R/P343R в TT16 (SEQ ID NO: 150): SCF033 (SEQ ID NO: 70)
- Введение P343R в MY518 (SEQ ID NO: 154): SCF039 (SEQ ID NO: 71)
- Введение P343R в MY518a (SEQ ID NO: 156): SCF039a (SEQ ID NO: 72)
- Введение D413K в MY518 (SEQ ID NO: 154): SCF040 (SEQ ID NO: 73)
- Введение P343R в MY201a (SEQ ID NO: 155): SCF041a (SEQ ID NO: 74)
- Введение P343R в MY201aPh (SEQ ID NO: 157): SCF041aPh (SEQ ID NO: 75)
- Введение D413K в MY201a (SEQ ID NO: 155): SCF042a (SEQ ID NO: 76)
- Введение P343R/D413K в MY201a (SEQ ID NO: 155): SCF043a (SEQ ID NO: 77)
- Введение P343R/D413K в MY201aPh (SEQ ID NO: 157): SCF043aPh (SEQ ID NO: 78)
- Введение Q311R в MY201a (SEQ ID NO: 155): SCF056a (SEQ ID NO: 79)
- Введение Q311R в MY201aPh (SEQ ID NO: 157): SCF056aPh (SEQ ID NO: 80)
- Введение Q311R/P343R в MY201a (SEQ ID NO: 155): SCF057a (SEQ ID NO: 81)
- Введение Q311R/P343R в MY201aPh (SEQ ID NO: 157): SCF057aPh (SEQ ID NO: 82)
- Введение Q311R/D413K в MY201a (SEQ ID NO: 155): SCF059a (SEQ ID NO: 83)
- Введение Q311R/D413K в MY201aPh (SEQ ID NO: 157): SCF059aPh (SEQ ID NO: 84)
- Введение Q311R в MY518a (SEQ ID NO: 156): SCF060a (SEQ ID NO: 85)
Гены тяжелой цепи антитела получают, как показано в табл.51 ниже, путем комбинирования гена константной области тяжелой цепи, полученного в настоящем изобретении, с геном вариабельной области тяжелой цепи А375 (SEQ ID NO: 43) или А551 (SEQ ID NO: 51).
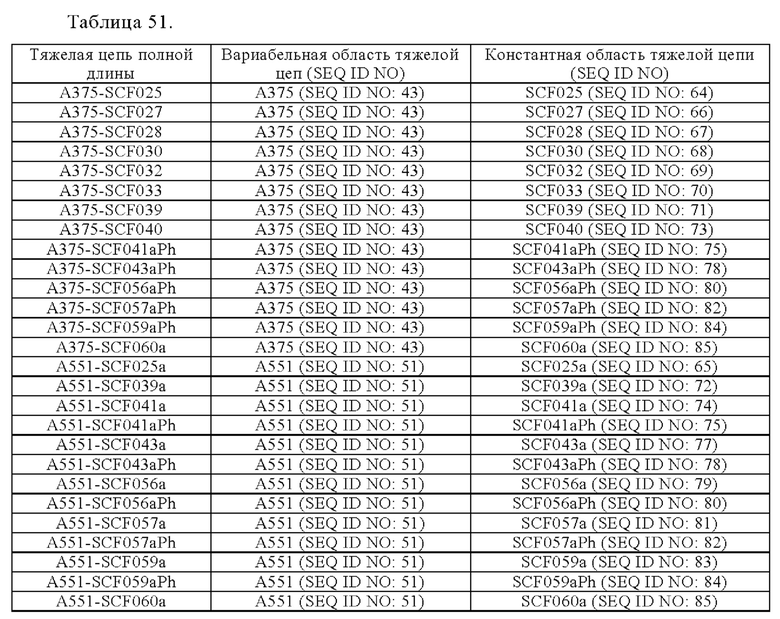
Кроме того, путем объединения этих генов тяжелой цепи антитела с генами B167-Lamlib, B256-Lamlib и B379-Lamlib, полученными в примере 5-3 в качестве генов легкой цепи антитела, требуемое антитело было экспрессировано и очищено, как показано в табл.52 методами, известными специалистам в данной области.
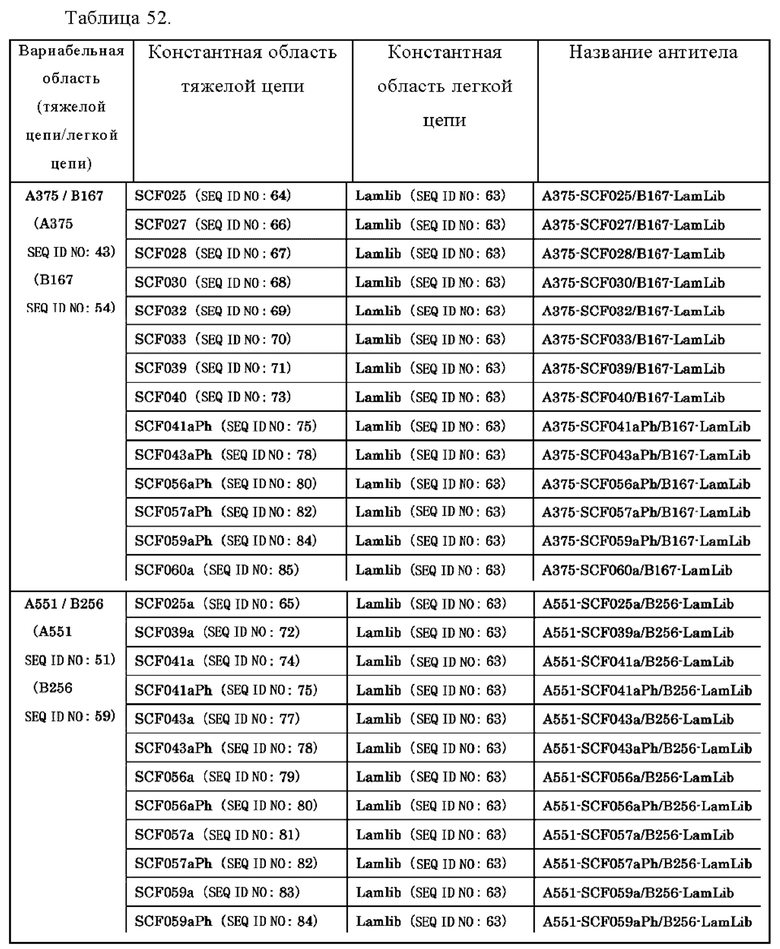
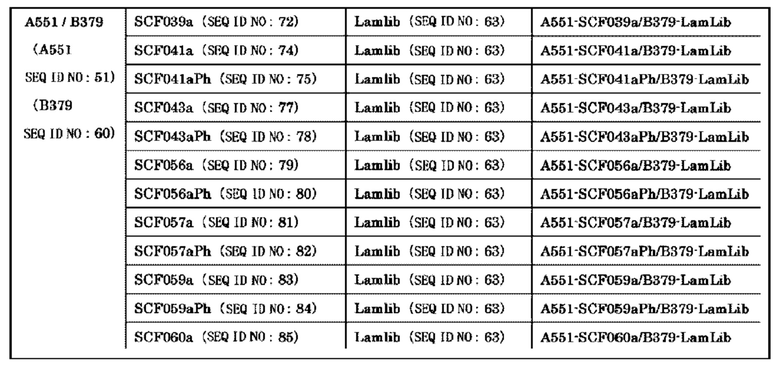
(7-3) Выработка контрольных антител для сравнения агонистической активности
Получают антитела (IC17HdK-MY201/IC17L-k0-IC17HdK-MY201aPh/IC17L-k0; 1С 17HdK-MY518/IC17L-k0; IC17HdK-MY5 18a/IC17L-k0; IC17HdK-P253/IC17L-k0; IC17HdK-P587/IC17L-k0; IC17HdK-TTl6/IC17L-k0) с вариабельной областью тяжелой цепи IC17HdK (SEQ ID NO: 152), получение которых описано в примере 7-1 в качестве контрольного антитела для сравнения агонистической активности. Кроме того, представляющее интерес антитело (IC17HdK-G4d/IC17L-k0) экспрессируют и очищают способами, известными в данной области, путем комбинирования гена легкой цепи антитела IC17L-k0 (SEQ ID NO: 158, вариабельная область; SEQ ID NO: 141, константная область) с геном тяжелой цепи антитела IC17HdK-G4d, который получают путем комбинирования гена вариабельной области тяжелой цепи IC17HdK (SEQ ID NO: 152) с геном G4d (SEQ ID NO: 159), из которого удалены Gly и Lys на C-конце константной области тяжелой цепи IgG4 человека. Эти контрольные антитела использовали в Примере 5.
(7-4) Оценка связывания рецептора Fcγ человека вариантов с повышенным показателем pI путем аминокислотной модификации константной области
Программное обеспечение Biacore Т200 (фирма GE Healthcare) используют для оценки связывающей активности с соответствующим рецептором Fcγ человека (далее именуемым FcγR). Измерение выполняют при 25°С с использованием буфера, включающего 50 мМ фосфата, 150 мМ NaCl, 0,05 мае./об.% -Р20, рН 7,4. Около 1000 RU антитела захвачено сенсорным чипом, иммобилизованным с Capture Select™ Human Fab-lambda Kinetics Biotin Conjugate (фирма Thermo Fisher Scientific) в качестве молекулы для захвата лиганда. FcγR человека разбавляют до 8 нМ для FcγRIa и 1000 нМ для другого FcγR в буфере для измерения, после чего он взаимодействует с захваченным антителом. Связывающую активность каждого антитела с FcγR оценивают путем расчета связывающего количества FcγR (RU) на единицу количества антитела с использованием Biacore Т200 Evaluation Software 2,0.
Внеклеточный домен FcγR получают описанными ниже методами. Во-первых, генный синтез внеклеточного домена FcγR осуществляют методом, известным специалистам в данной области. При этом последовательности FcγR получают на основе данных, зарегистрированных NCBI. В частности, последовательность FcγRI получают на основе последовательности из базы данных NCBI NM_000566.3, последовательность FcγRIIa на основе последовательности из базы данных NCBI NM_001136219.1, последовательности FcγRIIb на основе последовательности из базы данных NCBI NM_004001.3 и последовательности FcγRIIIa на основе последовательности из базы данных NCBI NM_001127593.1; и его тег был к их С-концам добавлена метка His. Полиморфный сайт FcγRIIa получают со ссылкой на публикацию J. Exp.Med., 172, 1990, 19-25; и полиморфный сайт FcγRIIIa получают со ссылкой на публикацию J. Clin. Invest., 100, 1997, 1059-1070. Полученные в результате фрагменты генов инсертируют в вектор экспрессии клеток животных для получения векторов экспрессии. Полученные векторы экспрессии временно вводят в клетки FreeStyle293, полученные из клеток эмбриональной почки человека (фирма Invitrogen), после чего экспрессируются представляющие интерес белки. Культуральные супернатанты собирают, пропускают через фильтр с порами 0,22 мкм и очищали в основном за четыре этапа, описанные ниже. Первый этап выполняют с помощью катионообменной колоночной хроматографии (SP Sepharose FF), второй этап - с помощью аффинной колоночной хроматографии (HisTrap HP) для His-метки, третий этап - с помощью гель-фильтрационной колоночной хроматографии (Superdex200), а четвертый этап - с помощью стерилизующей фильтрации. Однако для FcγRI на первом этапе проводят анионообменную колоночную хроматографию с Q Sepharose FF. Концентрации очищенных белков рассчитывают путем измерения адсорбции при 280 нм с помощью спектрофотометра, а коэффициенты адсорбции рассчитывают из полученных значений методами РАСЕ с соавт. (Protein Science, 4, 1995, 2411-2423).
Табл. 53 показывает связывающееся количество на единицу количества антитела, а также количество относительно связывающегося количества А375-G1T3/B167-Lamlib. Все антитела в табл.53 показывают большее связывающее количество с FcγRIIb человека на единицу количества антитела, чем у А375-G1T3/B167-Lamlib, и относительное значение составляет 2,85-14,26, тогда как связывающееся количество А375-С1Т3/Значение B167-Lamlib с FcγRIIb человека установлено равным 1,0. Количество FcγRIIb человека, связывающееся с единицей количества антитела, и относительные связывающиеся количества были аналогичными в присутствии или в отсутствие аминокислотных модификаций для увеличения pI константной области. Кроме того, связывающееся количество антител, содержащих модификацию L235W, с hFcgRIa снижено по сравнению с таковым для A375-TT14/B167-Lamlib.
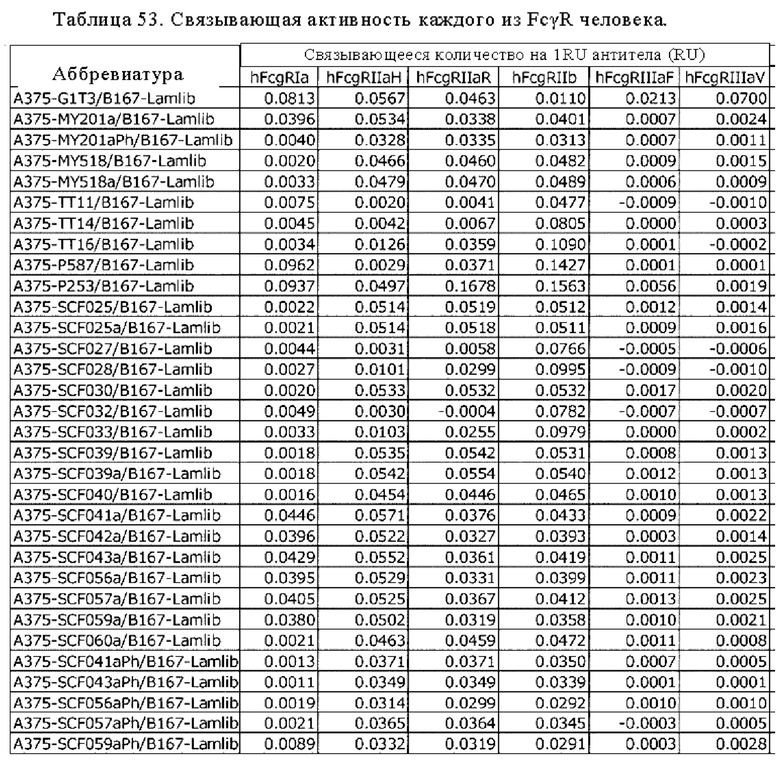
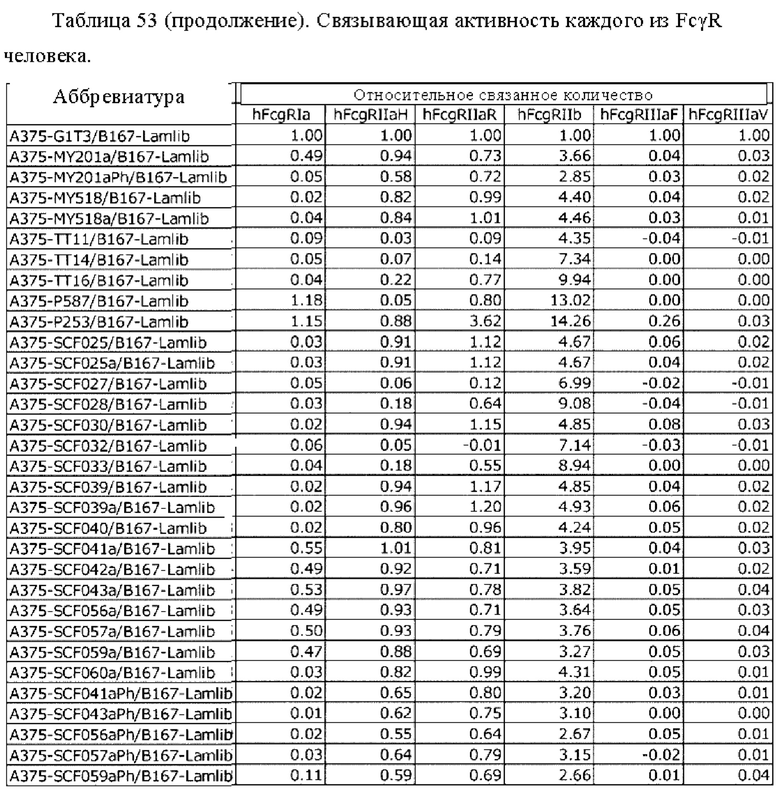
(7-5) Оценка связывания FcRn человека у вариантов с увеличенной pI путем аминокислотной модификации константной области
Программное обеспечение Biacore Т200 (фирма GE Healthcare) используют для оценки связывающей активности с FcRn человека. Измерения выполняют при 25°С с использованием буфера, включающего 50 мМ фосфата, 150 мМ NaCl, 0,05 мае./Об.% -Р20, рН 6,0. Белки FcRn человека, используемые для этого определения, получены в соответствии с процедурой, описанной в WO 2010107110. Около 400 RU антитела захвачено с использованием иммобилизованного CaptureSelect™ Human Fab-lambda Kinetics Biotin Conjugate сенсорного чипа (фирма Thermo Fisher Scientific) в качестве молекулы для захвата лиганда с последующим связыванием FcRn человека, разведенного в буфере для измерений. Активность соответствующих антител по связыванию FcRn оценивают путем расчета KD (моль/л) с помощью стационарной модели с использованием Biacore Т200 Evaluation Software 2,0. Значения KD были близкими в присутствии или в отсутствие аминокислотных модификаций для увеличения pI константной области. В табл. 54 представлена величина KD (моль/л) соответствующих антител к FcRn человека.
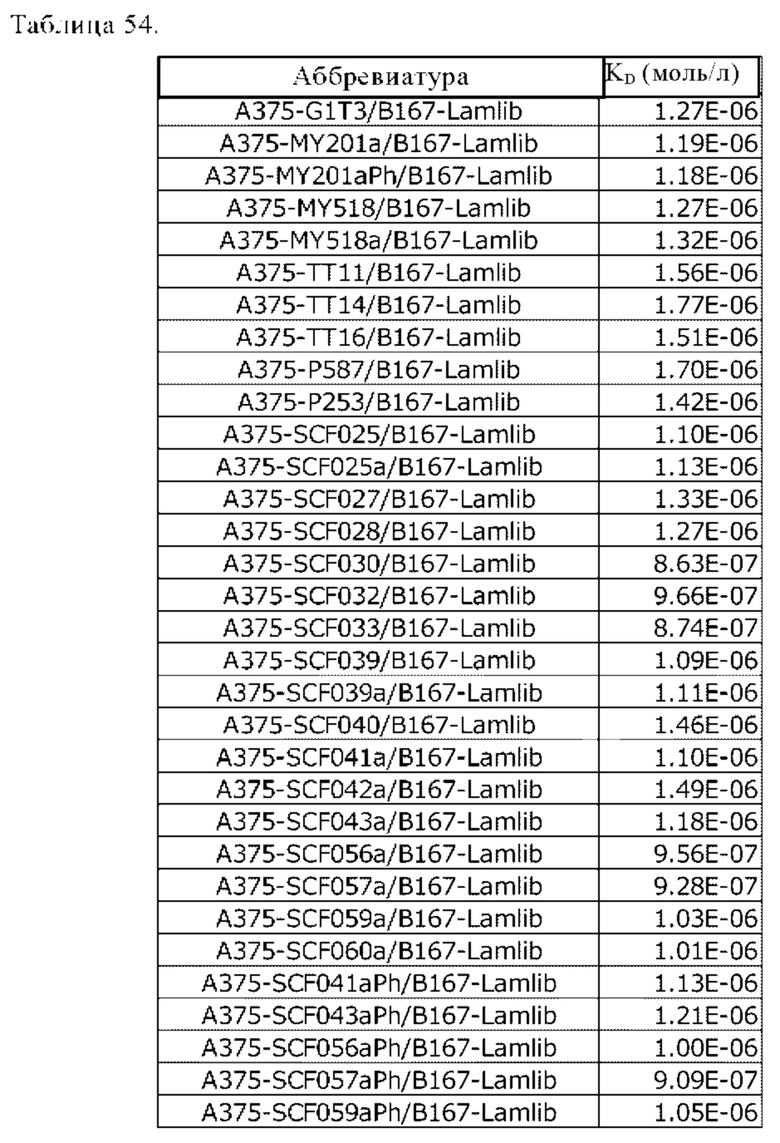
Пример 8. Получение кинуренин (Kyn)-зависимых антител против CD137
(8-1) Получение антител с кинуренин-зависимой связывающей активности в отношении CD137 человека из рационально разработанной библиотеки антител.
Антитела, проявляющие связывающую активность с CD 137 человека в присутствии кинуренина и не проявляющие связывающей активности в отсутствие кинуренина, получены из рационально разработанной библиотеки фагового дисплея антител (библиотеки кинуренина) для получения кинуренин-переключаемых антител, используя пэннинг против кинуренина, которые сконструированы, как описано в WO 2015/083764. Для получения биотинилированные антигены, связанные фагами в присутствии кинуренина, улавливаются гранулами, и извлекают фаги, презентирующие антитела, которые проявляют связывающую активность по отношению к антигенам в присутствии кинуренина. Затем фаги собирают из элюатов, полученных с гранул.
Фаги получают обычным образом из клеток Е. coli, несущих сконструированную методом фагового дисплея фагмиду. В частности, клетки E.coli, несущие сконструированный фагмидный вектор, инфицируют фагом M13KO7ΔpIII (обозначенным как «гиперфаг») (фирма PROGEN Biotechnik) и после культивирования в течение ночи при 30°С фаги выделяют из супернатантов. Раствор фаговой библиотеки получают разбавлением популяции фагов, осажденных добавлением 2,5 М NaCl/10% PEG к культуре E.coli, в которой выработку фагов осуществляют с помощью трис-буферного солевого раствора (TBS, Tris Buffered Saline). Затем к раствору фаговой библиотекой добавляют БСА до конечной концентрации 4%. Пэннинг проводят с использованием антигена, иммобилизованного на магнитных гранулах. В качестве магнитных шариков использовали Sera-Mag SpeedBeads с покрытием NeutrAvidin (фирма Thermo Fisher Scientific), шарики FG NeutrAvidin (фирма Tamakawa Seminator) или Dynabeads MyOne Streptavidin T1 (фирма Life Technologies). Гранулы с покрытием Sera-Mag SpeedBeads NeutrAvidin-coated (фирма Thermo Fisher Scientific), FG гранулы NeutrAvidin (фирма Tamakawa Seminator) или гранулы Dynabeads MyOne Streptavidin T1 (фирма Life Technologies) применяют в качестве магнитных гранул. В качестве антигена используют биотинилированный hCD137-Fc (hCD137-Fc-Bio), полученный, как описано в примере 1-3 (получение биотинилированного Fc-слитого CD137 человека).
Пэннинг проводят для эффективного получения низкомолекулярных переключаемых антител, которые зависят от небольшой молекулы, которая может играть переключающую роль в раковых тканях. В частности, пэннинг для обогащения антителами, которые связываются с антигенами в присутствии кинуренина, который является небольшой молекулой, и которые не связывают антигены в отсутствие кинуренина, выполняют со ссылкой на методы, описанные в предыдущем патенте WO 2015/083764. В 1 раунде пэннинг выполняют на hCD137-Fc-Bio или биотинилированных кинуренинах (bio-Kyn) (соединение 028 (представлено [формулой 28]), 029 (представлено [формулой 29]) и 036 (представлено [формулой 36]), как описано в WO 2015/083764). Пэннинг для hCD137-Fc-Bio выполняют путем добавления раствора фага к антигену с последующим выделением фага, связанного с антигеном, с помощью магнитных гранул; и пэннинг для bio-Kyn выполняют таким образом, что добавляют приготовленный раствор фаговой библиотеки после первой иммобилизации биотинилированного антигена на магнитных гранулах. Для Bio-Kyn пэннинг для обогащения антител, способных связываться с антигеном в отсутствие кинуренина в виде небольшой молекулы, выполняют со ссылкой на методы, описанные в вышеупомянутом патенте WO 2015/083764. Для hCD137-Fc-Bio добавляют 4 нмоль небиотинилированной области Fc IgG1 человека для удаления антител, которые связываются с областью Fc. Восстановленные фаги добавляют к штамму Е. coli ER2738 для заражения фагами Е. coli, затем выделенные клетки Е. coli инфицируют гиперфагами, и фаги выделяют из супернатантов после культивирования в течение ночи при 30°С.
После второго раунда пэннинг для обогащения антител, которые связываются с антигенами в присутствии кинуренина и не связываются с антигенами в отсутствие кинуренина, выполняют на hCD137-Fc-Bio со ссылкой на методы, показанные в предыдущем патенте WO 2015/083764. В обоих случаях добавляют 4 нМ небиотинилированной области Fc IgG1 человека для удаления антител, которые связываются с областью Fc для hCD137-Fc-Bio. Подобный пэннинг повторяют до пятого раунда для обогащения представляющих интерес последовательностей антител.
(8-2) Получение антител, связывающих hCD137 в присутствии кинуренина, из библиотеки антител.
(8-2-1) Получение hCD137p12-FX-Fc-Bio
Получение hCD137p12-FX-Fc-Bio осуществляют методом, известным специалистам в данной области. В частности, фрагмент гена, кодирующий расщепляемую фактором Ха последовательность, и фрагмент гена, кодирующий константную область антитела, присоединены ниже по цепи от фрагмента гена, кодирующего область внеклеточного домена CD137 человека. Фрагмент гена, кодирующий белок (hCD137p12-FX-Fc-Bio, SEQ ID NO: 160), в котором связаны внеклеточная часть CD137 человека, расщепляемая фактором Ха последовательность и константная область антитела, включают в вектор экспрессии животного. Сконструированный плазмидный вектор вводят в клетки FreeStyle293 (фирма Invitrogen) с использованием 293-фектина (фирма Invitrogen). Одновременно вводят плазмидный вектор, содержащий гены, экспрессирующие BirA и EBNA1 (SEQ ID NO: 88). В культуру добавляют биотин. Клетки, трансдуцированные генами в соответствии с вышеупомянутыми процедурами, культивируют при 37°С, 8% СО2, и hCD137p12-FX-Fc-Bio секретируется в супернатант культуры. Этот культуральный супернатант собирают. Белок А используют для очистки hCD137p12-FX-Fc-Bio из культурального супернатанта.
При использовании рационально разработанной библиотеки считают, что путем пэннинга антител, проявляющих антигенсвязывающую активность только в присутствии небольшой молекулы, можно получить переключаемые низкомолекулярными соединениями антитела, проявляющие антигенсвязывающую активность в присутствии малых молекул. То есть из библиотеки кинуренина, созданной способом, описанным в международной публикации WO 2015/083764, были извлечены фаги, презентирующие антитела, которые проявляют связывающую активность в присутствии кинуренина по отношению к hCD137, захваченному на гранулах. В настоящем методе получения биотинилированный hCD137-Fc (hCD137-Fc-Bio), полученный, как описано в примере 1-3 (получение меченного биотином Fc-слитого CD137 человека), или hCD137p12-FX-Fc-Bio используют в качестве антигенов.
Бактерию Е. coli, несущую фагмидный вектор для фагового дисплея, сконструированный, как описано в международной публикации WO 2015/083764, инфицируют M13KO7ΔpIII (гиперфагом) (фирма PROGEN Biotechnik), и фаги выделяют из супернатантов после культивирования в течение ночи при 30°С. Раствор библиотеки фагов с поливалентным дисплеем антител получают разбавлением TBS популяции фагов, осажденных путем добавления 2,5 М NaCl/10% PEG к культуре Е. coli, в которой размножали фаг. Затем к раствору фаговой библиотеки добавляют БСА до конечной концентрации 4%. Проводят пэннинг с иммобилизованным на магнитных гранулах антигеном. Гранулы FG NeutrAvidin (фирма Tamakawa Seminator) или Dynabeads MyOne Streptavidin T1 (фирма Life Technologies) используют в качестве магнитных гранул.
В первом раунде пэннинга проводят обогащение фагов, способных связываться с антигеном, в присутствии кинуренина. В частности, фаговую библиотеку контактируют с антигеном и кинуренином в течение одного часа при комнатной температуре путем добавления 0,8 мл приготовленного раствора фаговой библиотеки, при конечной концентрации кинуренина 500 мкМ и 4 нмоль небиотинилированной области Fc IgG1 человека (hFC) с 125 пМ меченного биотином антигена. Добавляют блокированные БСА магнитные гранулы (гранулы FG NeutrAvidin) и позволяют антиген-фаговым комплексам связываться с магнитными гранулами в течение 15 мин при комнатной температуре. Гранулы дважды промывают 0,5 мл кинуренина/TBST и один раз кинуренином/TBS. Затем к смеси добавляют 0,5 мл раствора трипсина с концентрацией 1 мг/мл. После суспендирования смеси при комнатной температуре в течение 15 мин раствор фага выделяют из гранул, которые сразу же отделяют с помощью магнитной платформы. Восстановленные фаги добавляют к 20 мл штамма Е. coli ER2738 в логарифмической фазе роста (OD600 0,4-0,7). Е. coli инфицируют фагами путем культивирования Е. coli при 37°С в течение одного часа при осторожном перемешивании. Инфицированные клетки Е. coli высевают в чашки размером 225 мм × 225 мм. Затем готовят библиотеки фагов, презентирующих моновалентные антитела, путем инфицирования культур диссеминированных Е. coli фагом M13KO7TC (WO 2015046554A1) (фагом-помощником) (фирма Takara) и собирают фаги из супернатантов, культивируемых в течение ночи при 30°С.
Для второго раунда пэннинга фаговую библиотеку контактируют с антигеном и кинуренином в течение одного часа при комнатной температуре путем добавления 0,8 мл приготовленного раствора фаговой библиотеки до конечной концентрации 500 мкМ кинуренина и 4 нМ немеченной биотином области Fc IgG1 человека вместе с 40 пМ меченного биотином антигена. Добавляют блокированные БСА магнитные гранулы (Dynabeads MyOne Streptavidin T1) и допускают антиген-фаговым комплексам связываться с магнитными гранулами в течение 15 мин при комнатной температуре. Гранулы дважды промывают 0,5 мл кинуренина/TBST и один раз кинуренином/TBS. Затем к смеси добавляют 0,5 мл раствора трипсина с концентрацией 1 мг/мл. После суспендирования смеси при комнатной температуре в течение 15 мин раствор фага выделяют из гранул, которые сразу же отделяют с помощью магнитной платформы. Затем готовят раствор библиотеки фагов для моновалентного дисплея антител таким же образом, как и в первом раунде, описанном выше.
Для раунда 3 пэннинга фаговую библиотеку контактируют с антигеном и кинуренином в течение одного часа при комнатной температуре путем добавления 0,8 мл приготовленного раствора фаговой библиотеки, при конечной концентрации кинуренина 500 мкМ и 4 нМ немеченной биотином области Fc IgG1 человека наряду с 10 пМ меченного биотином антигена. Добавляют блокированные БСА магнитные гранулы (гранулы FG NeutrAvidin) и допускают антиген-фаговым комплексам связываться с магнитными гранулами в течение 15 мин при комнатной температуре. Гранулы трижды промывают 0,5 мл кинуренина/TBST и дважды - кинуренина/TBS. Затем к смеси добавляют 0,5 мл раствора трипсина с концентрацией 1 мг/мл. После суспендирования смеси при комнатной температуре в течение 15 мин раствор фага выделяли из гранул, которые сразу же отделяют с помощью магнитной платформы. Затем готовят раствор библиотеки фагового дисплея моновалентных антител таким же образом, как и в первом раунде, описанном выше. В течение 4-го раунда пэннинг осуществляют в условиях, сходных с условиями 3-го раунда пэннинга. Однако в качестве магнитных гранул применяют Dynabeads MyOne Streptavidin T1.
(8-3) Получение антител, которые связываются с CD137 человека в присутствии кинуренина. из библиотек антител с использованием SS-биотинилированных антигенов и методов отрицательной селекции.
(8-3-1) Получение hCD137p12-FX-Fc
hCD137p12-FX-Fc получают способом, известным специалистам в данной области. В частности, фрагмент гена, кодирующий последовательность расщепления фактором Ха, и фрагмент гена, кодирующий константную область антитела, присоединены ниже по цепи от фрагмента гена, кодирующего область внеклеточного домена CD137 человека. Фрагмент гена, кодирующий белок, в котором связаны внеклеточная часть CD137 человека, последовательность расщепления FactorXa и константная область антитела (hCD137p12-FX-Fc, SEQ ID NO: 160), включен в вектор экспрессии животного. Сконструированный плазмидный вектор вводят в клетки FreeStyle293 (фирма Invitrogen) с использованием 293-фектина (фирма Invitrogen). Одновременно вводят плазмидный вектор, содержащий гены, экспрессирующие EBNA1 (SEQ ID NO: 88). Клетки, трансдуцированные генами в соответствии с вышеупомянутыми процедурами, культивируют при 37°С, 8% СО2, и hCD137p12-FX-Fc-Bio секретируется в супернатант культуры. Белок А используют для очистки hCD137p12-FX-Fc-Bio из этого супернатанта клеточной культуры.
(8-3-2) Получение антител, связывающих CD137 человека в присутствии кинуренина. из библиотеки антител с использованием SS-биотинилированных антигенов и метода отрицательной селекции.
При получении низкомолекулярных переключаемых антител, которые проявляют антигенсвязывающую активность в присутствии небольшой молекулы, считалось, что после фагового дисплея антител, которые связываются с антигеном с помощью магнитных гранул, захватываются с использованием антигена, биотинилированного (SS-биотинилированного) через линкер с дисульфидной связью (SS-связь), только антигенсвязывающее антитело по методу фагового дисплея может быть элюировано с гранул с использованием дитиотреитола (DTT) или подобного соединения для расщепления SS-связей в восстанавливающих условиях. Библиотеку кинуренина, сконструированную, как описано в международной публикации WO 2015/083764, подвергают скринингу на антитела, которые проявляют связывающую активность с антигенами в присутствии кинуренина. Для скрининга библиотеку фагового дисплея антител сначала приводят в контакт с меченными биотином антиген-магнитными гранулами в отсутствие кинуренина, и фаги, демонстрирующие антитела, которые обладают антигенсвязывающей активностью даже в отсутствие кинуренина, удаляют. Последующий скрининг на антитела с антигенсвязывающей активностью проводят только в присутствии кинуренина, аналогичным образом проводя пэннинг в присутствии кинуренина. В этом методе меченный биотином антиген (SS-биотинилированный антиген) используют в качестве антигена в соответствии с прилагаемыми протоколами с использованием hCD137-Fc, полученного согласно описанию в примере 1-4, или hCD137p12-FX-Fc, полученного в (8-3-1), и набор Sulfo-NHS-SS-Biotinylation EZ-Link (фирма Thermo Fisher Scientific). Следует отметить, что hCD137-Fc-Bio, полученный по описанию примера 1-3, или hCD137p12-FX-Fc-Bio, полученный в (8-2-1), используют для отрицательной селекции.
Клетки Е. coli, несущие фагмидный вектор фагового дисплея, сконструированный согласно описанию WO 2015/083764, инфицированы M13KO7ΔpIII (гиперфагом) (фирма PROGEN Biotechnik), и после культивирования в течение ночи при 30°С фаги выделяют из супернатантов. Библиотеки фагового дисплея поливалентных антител получают разбавлением TBS популяции фагов, осажденных путем добавления 2,5 М NaCl/10% PEG к культуре Е. coli, в которой производили наработку фага. Затем к раствору библиотеки фагов добавляют БСА до конечной концентрации 4%. Осуществляют пэннинг с иммобилизованным на магнитных гранулах антигеном. В качестве магнитных гранул используют Sera-Mag SpeedBeads NeutrAvidin-coated (фирма Thermo Fisher Scientific), гранулы FG NeutrAvidin (фирма Tamakawa Seminator) или гранулы Dynabeads MyOne Streptavidin Tl (фирма Life Technologies).
В первом раунде пэннинга проводят обогащение фагов, способных связываться с антигеном в присутствии кинуренина, применяя метод отрицательной селекции. В частности, 800 пмоль hCD137-Fc-Bio или hCD137p12-FX-Fc-Bio добавляют к гранулам Sera-Mag SpeedBeads, покрытым нейтравидином, блокированным обезжиренным молоком, которое предварительно инкубировали со стрептавидином (фирма Roche) и допускают связывание в течение 15 мин при комнатной температуре. Гранулы трижды промывают TBST и добавляют 0,8 мл раствора библиотеки фагов, блокированных БСА чтобы была возможность связаться в течение одного часа при комнатной температуре. Разделяя гранулы на магнитной платформе, собирают антигены и фаги, которые не связались с гранулами.
Затем 225 пМ SS-биотинилированного hCD137-Fc или 450 пМ SS-биотинилированного hCD137p12-FX-Fc-Bio добавляют к гранулам FG NeutrAvidin, блокированным обезжиренным молоком, которое инкубировали со стрептавидином (фирма Roche), и допускают связывание в течение 15 мин при комнатной температуре. Гранулы трижды промывают TBST, и вместе с выделенными фагами добавляют до конечной концентрации 0,2 мМ кинуренина и 4 нМ hFc; фаговую библиотеку приводят в контакт с антигеном, а также с кинуренином, в течение одного часа при комнатной температуре. Гранулы дважды промывают 0,5 мл кинуренина/TBST (0,2 мМ кинуренин, 0,1% Tween20, TBS, рН 7,4) и один раз кинуренином/TBS (0,2 мМ кинуренин, TBS, рН 7,4). Затем добавляют 500 мкл раствора TBS, содержащего в конечной концентрации 25 мМ DTT, и после пяти минут перемешивания при комнатной температуре фаги выделяют из отделенных гранул с использованием магнитной платформы. Затем к смесям добавляют трипсин до конечной концентрации 1 мг/мл. Смесь перемешивают при комнатной температуре в течение 15 мин перед извлечением фагов из отделенных шариков с помощью магнитной платформы. Восстановленные фаги добавляют к 20 мл штамма Е. coli ER2738 в логарифмической фазе роста (OD600 0,4-0,7). Е. coli инфицируют фагами путем культивирования Е. coli при 37°С в течение одного часа при осторожном перемешивании. Инфицированные клетки Е. coli высевают в чашки размером 225 мм × 225 мм. Затем получают библиотеку фагов с дисплеем поливалентных антител путем инфицирования культур диссеминированных Е. coli гиперфагами и извлечения фагов из супернатантов после культивирования в течение ночи при 30°С.
Для второго раунда пэннинга 400 пМ hCD137-Fc-Bio или hCD137p12-FX-Fc-Bio добавляют к гранулам к Sera-Mag SpeedBeads NeutrAvidin, покрытым нейтральным авидином, блокированным обезжиренным молоком, предварительно инкубированным со стрептавидином (фирма Roche), и дают возможность связываться в течение 15 мин при комнатной температуре. К гранулам, трижды промытым TBST, добавляют 0,4 мл раствора библиотеки фагов, блокированных БСА, и дают возможность связываться в течение одного часа при комнатной температуре. Путем разделения гранул с помощью магнитной платформы, извлекают антиген и фаги, которые не связались с гранулами. Извлеченные фаги снова инкубируют с гранулами Sera-Mag SpeedBeads NeutrAvidin, покрытыми нейтральным авидином, которые инкубировали с 400 пМ hCD137-Fc-Bio или hCD137p12-FX-Fc-Bio по аналогичной процедуре, а затем антигены и фаги, которые не связывались с гранулами, собирают на магнитной платформе.
56,3 пМ SS-биотинилированного hCD137-Fc или 113 пМ SS-биотинилированного hCD137p12-FX-Fc добавляют к гранулам FG NeutrAvidin, блокированным обезжиренным молоком, которое предварительно инкубируют со стрептавидином (фирма Roche), и дают возможность связать в течение 15 мин при комнатной температуре. Гранулы трижды промывают TBST, и вместе с выделенными фагами добавляют до конечной концентрации 0,2 мМ кинуренина и 4 нМ небиотинилированной области Fc IgG1 человека; фаговую библиотеку приводят в контакт с антигеном, а также кинуренином, в течение одного часа при комнатной температуре. Гранулы трижды промывают 0,5 мл кинуренина/TBST (0,2 мМ кинуренин, 0,1% Tween20, TBS, рН 7,4) и дважды кинуренином/TBS (0,2 мМ кинуренин, TBS, рН 7,4). Затем добавляют 500 мкл раствора TBS, содержащего в конечной концентрации 25 мМ DTT, и после 5 мин перемешивания при комнатной температуре фаги выделяют из отделенных гранул с использованием магнитной платформы. Затем готовят раствор библиотеки фагов с многовалентным дисплеем антител таким же образом, как в первом раунде, описанном выше.
Для третьего раунда пэннинга 200 пМ hCD137-Fc-Bio или hCD137p12-FX-Fc-Bio добавляют к покрытым нейтральным авидином гранулам Sera-Mag SpeedBeads NeutrAvidin, блокированным обезжиренным молоком Sera-Mag SpeedBeads, предварительно инкубированным со стрептавидином (фирма Roche), и дают возможность связать при комнатной температуре в течение 15 мин. Гранулы трижды промывают TBST и добавляют 0,2 мл раствора библиотеки фагов, блокированных БСА, и дают возможность связываться в течение одного часа при комнатной температуре. Разделив гранулы с помощью магнитной платформы извлекают антигены и фаги, которые не связались с гранулами. Извлеченные фаги снова инкубируют с Sera-Mag SpeedBeads NeutrAvidin, которые инкубировали с 200 пМ hCD137-Fc-Bio или hCD137p12-FX-Fc-Bio по аналогичной процедуре, затем антигены и фаги, которые не связывались с гранулами, восстанавливают с помощью магнитной платформы.
12,5 пМ SS-биотинилированного hCD137-Fc или 25 пмоль SS-биотинилированного hCD137p12-FX-Fc добавляют к гранулам FG NeutrAvidin NeutrAvidin, блокированным обезжиренным молоком, который инкубировали со стрептавидином (фирма Roche), и дают возможность связываться при комнатной температуре в течение 15 мин. Гранулы трижды промывают TBST, и вместе с выделенными фагами добавляют до конечной концентрации 0,2 мМ кинуренина и 4 нМ небиотинилированной области Fc IgG1 человека; фаговую библиотеку приводят в контакт с антигеном, а также кинуренином, в течение одного часа при комнатной температуре.
Гранулы пять раз промывают 0,5 мл кинуренина/TBST (0,2 мМ кинуренин, 0,1% Tween20, TBS, рН 7,4) и пять раз кинуренином/TBS (0,2 мМ кинуренин, TBS, рН 7,4). Затем добавляют 500 мкл раствора TBS, содержащего в конечной концентрации 25 мМ DTT, и после 5 мин перемешивания при комнатной температуре фаги выделяют из отделенных гранул с использованием магнитной платформы. Затем готовят раствор библиотеки фагов с многовалентным дисплеем антител таким же образом, как в первом раунде, описанном выше. В течение четвертого раунда пэннинг осуществляют в условиях, аналогичных условиям третьего раунда пэннинга. Однако применяют в качестве магнитных гранул гранулы Dynabeads MyOne Streptavidin Т1 вместо гранул FG beads NeutrAvidin. For пятого раунда пэннинг осуществляют в условиях, близких к условиям третьего раунда.
(8-4) Оценка кинуренин-зависимых анти-СР137 антител, идентифицированных с помощью фагового дисплея
(8-4-1) Экспрессия и очистка переключаемых антител, которые изменяют свое связывание с антигенами за счет присутствия/отсутствия кинуренина
Гены, кодирующие вариабельные области антител, полученных из кинурениновых библиотек, описанных в настоящем изобретении, инсертируют в экспрессирующие плазмиды животных, имеющие IgG1 человека или модифицированный IgG1 человека (Р253) и цепи каппа. Обозначения клонов и порядковые номера перечислены в табл. 55.

Антитела экспрессируют и очищают с использованием методов, известных специалистам в данной области. Спектрофотометры использовали для измерения абсорбции раствора очищенных антител при 280 нм. Из полученных измеренных значений рассчитывают концентрации очищенных антител с использованием коэффициентов экстинкции, рассчитанных методом РАСЕ (Protein Science, 4, 1995, 2411-2423).
(8-4-2) Оценка влияния кинуренина на связывание CD137 человека с помощью метода поверхностного плазмонного резонанса
Программное обеспечение Biacore Т200 (фирма GE Healthcare) используют для оценки взаимодействия по связывающей активности анти-CD137 антитела и hCD137-His-BAP (SEQ ID NO: 87). Чип сенсора СМ3 (фирма GE Healthcare), иммобилизованный с соответствующими количествами белка А путем иммобилизации по аминогруппе, захватывается анти-С0137 антителом для взаимодействия с hCD137-His-BAP, полученным в примере 1-1. TBS используют в качестве подвижного буфера, а 10 мМ глицин-HCl (рН 1,5) и 25 мМ NaOH используют в качестве раствора для регенерации.
После захвата анти-CD137 антитела, суспендированного в TBS, 500 нМ hCD137-His-BAP вводят в каждую проточную ячейку при скорости потока 10 мкл/мин в течение 3 мин. Этот 3-минутный период является фазой связывания для hCD137-His-BAP, а после завершения фазы связывания наступает 5-минутный период переключения на подвижный буфер для фазы диссоциации для hCD137-His-BAP. После завершения фазы диссоциации регенерирующий раствор впрыскивают со скоростью 30 мкл/мин в течение 10 сек. Приведенное выше описание является циклом измерения связывающей активности анти-CD137 антител. Связывающееся количество hCD137-His-BAP, которое взаимодействовало с анти-CD137 антителом в фазе связывания, анализируют с помощью Biacore Т200 Evaluation Software Version 2.0, как показано в табл.56.
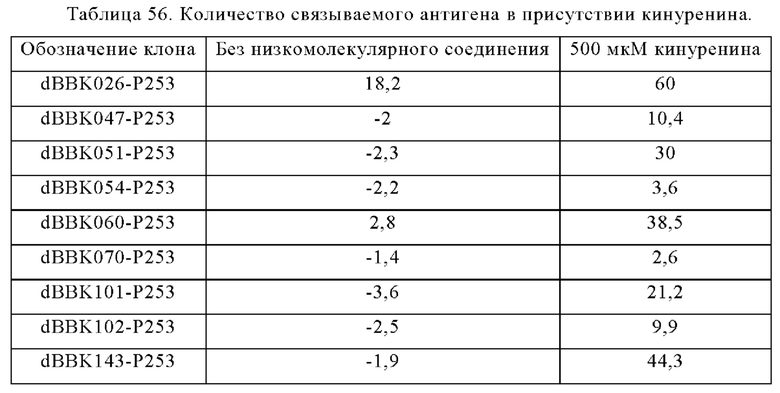
(8-4-3) Оценка связывающей активности полученных антител по отношению к частям последовательностей CD137.
(8-4-3-1) Получение антигенов с частями последовательностей CD137.
Антигены с частями последовательностей CD 137 получают методами, известными специалистам в данной области. В частности, фрагмент гена, кодирующий константную область антитела, был связан ниже по цепи после фрагмента гена, кодирующего часть внеклеточной области CD137 человека. Фрагмент гена, кодирующий белок (табл.57), в котором часть внеклеточного домена CD 137 человека и константная область антитела связаны, включают в экспрессирующий вектор животного. Сконструированный плазмидный вектор вводят в клетки Expi293 (фирма Invitrogen) с использованием ExpiFectamine293 (фирма Invitrogen), и представляющий интерес белок секретируется в супернатант культуры. Белок А используют для очистки представляющего интерес белка из супернатанта культуры.

(8-4-3-2) Подтверждение связывания
Связываются ли полученные антитела с антигенами, имеющими частичную последовательность CD137, оценивают методом ELISA. Сначала различные антигены, полученные в примере 1 или примере 8-4-3-1, иммобилизуют на планшетах для микротитрования Maxisorp 384. После удаления из планшетов несвязавшихся антигенов промыванием каждой лунки промывочным буфером лунки блокируют 50 мкл блокирующего буфера (TBS, содержащего BlockAce) в течение одного часа или более. В каждую из лунок, из которых удален блокирующий буфер, добавляют 10 мкл очищенных антител, приготовленных в концентрации 10 мкг/мл в TBS с конечной концентрацией 500 мкМ L-кинуренина или TBS с конечной концентрацией 500 мкМ кинуренина, и планшет оставляют в покое на один час. АР-конъюгированные антитела против каппа человека (фирма BIOSOURCE), разведенные в TBS с конечной концентрацией 500 мкМ кинуренина, добавляют в каждую лунку после промывания промывочным буфером, содержащим конечную концентрацию кинуренина 500 мкМ (TBS с 0,1% Tween20). После 1 ч инкубации добавляют фосфатный субстрат BluePhos (KPL) после промывания промывочным буфером, содержащим 500 мкМ кинуренина. Развитие цвета раствора в каждой лунке измеряют по абсорбции при 620 нм после того, как реакцию проявления цвета раствора в каждой лунке останавливают добавлением стоп-раствора BluePhos. Показан коэффициент абсорбции путем установки значений в лунках с концентрацией антител от 0 до 1 мкг/мл. Судя по связыванию с коэффициентом абсорбции больше или равным 2, предполагают, что оцениваемые клоны включают антитела, которые проявляют отличающиеся связывающие свойства.
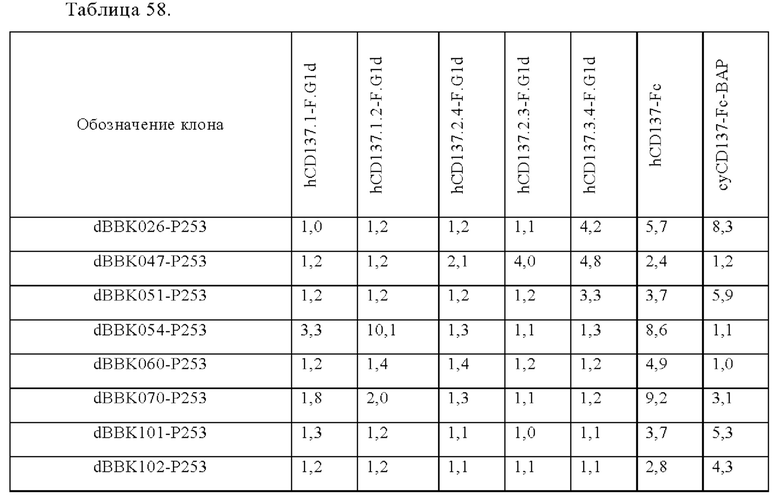
(8-5) In vitro оценка активности полученных антител
Клеточную линию GloResponseTM NF-κB-Luc2/4-1BB Jurkat (фирма Promega) используют для измерения активности антител in vitro. В каждую лунку 384-луночных планшетов добавляют 10 мкл клеток FcγRIIB СНО-К1, приготовленных в концентрации 2 × 106/мл со средой для анализа (99% RPMI, 1% ФСБ). Растворы антител с кинуренином или без кинуренина добавляют в каждую лунку по 10 мкл. Затем в каждую лунку добавляли 10 мкл клеточной линии GloResponseTM NF-κB-Luc2/4-1BB Jurkat, приготовленной в концентрации 2 × 106/мл в среде для анализа (99% RPMI, 1% ФСБ). Конечная концентрация кинуренина составляет 500 мкМ. Планшеты оставляют на шесть часов при 37°С в инкубаторе в атмосфере 5% СО2, затем выдерживают 15 мин при комнатной температуре и добавляют 30 мкл реагента Bio-Glo в каждую лунку. Для реагента Bio-Glo используют систему анализа люциферазы Bio-Glo (буфер и субстрат). Впоследствии люминесценцию каждой лунки измеряют с помощью планшет-ридера. Значение люминесценции каждой лунки, деленное на величину люминесценции лунки без добавленного антитела, представляет кратность люминесценции и служит индикатором для оценки активности каждого антитела.
Полученные результаты показаны на фиг. 57.
Кратность люминесценции была выше, когда концентрация L-кинуренина составляла 500 мкМ, чем когда она составляла 0 мкМ, что указывает на то, что активность агониста изменялась в зависимости от концентрации или присутствия кинуренина.
Пример 9. Получение АТФ- и АМФ-зависимых CD137-связывающих антител путем двойного раунда отбора
(9-1) Получение антител, связывающихся с антигенами в присутствии АТФ и АМФ. соответственно, из рационально разработанной библиотеки методом двойного раунда отбора
Получены антитела, которые проявляют связывающую активность с антигеном в присутствии определенной небольшой молекулы, но никогда не сообщалось о получении антител, которые проявляют связывающую активность с антигеном в присутствии множества различных малых молекул. Один из путей решения этой проблему является рассмотрение возможности чередования пэннинга, который меняет небольшую молекулу, используемую для каждого раунда пэннинга, например, АТФ и АМФ. Однако в этом случае существует проблема, заключающаяся в том, что давление отбора применяют в отношении небольшой молекуле в каждом раунде, и, таким образом, давление отбора не может применяться одинаково для обеих небольших молекул одновременно. Для решения этой проблемы был реализован метод двойного раунда отбора. Метод двойного раунда отбора является процедурой, при которой два цикла пэннинга выполняют на одном антигене за один раунд пэннинга (J Mol Biol., 226(3), 1992, 889-896). В настоящем изобретении, например, в случае метода фагового дисплея, один раунд относится к этапу выделения фагов из Е. coli для повторного инфицирования Е. coli фагами после пэннинга; а в случае метода рибосомального дисплея один раунд относится к этапу от транскрипции in vitro до амплификации гена с методом ПЦР. Хотя сообщалось о двойном цикле отбора для одного типа антигена, нет сообщений о применении метода для получения антител, зависимых от малых молекул, и, соответственно, нет сообщений о применении метода для получения антител, которые проявляют зависимость от нескольких отдельных разных малых молекул. Модифицируя метод двойного раунда отбора и применяя селективное давление как на АТФ, так и на АМФ в одном цикле, авторы настоящего изобретения полагают, что можно получить антитела, которые проявляют антигенсвязывающую активность АТФ- и АМФ-зависимым образом более эффективно.
Антитела, проявляющие антигенсвязывающую активность в присутствии АТФ и AMP, получают из рационально разработанной библиотеки фагового дисплея антител, созданной в предыдущем патенте WO 2015 / 083764. Условия пэннинга показаны в табл.59. Условия 1 и 2 применены к условию обычного пэннинга и вышеуказанному переменному условию пэннинга; в то время как условие 3 применено к вышеупомянутому методу двойного раунда отбора. В настоящем изобретении двойной раунд отбора проведен в раунде 2 и раунде 3. Биотинилированный hCD137-Fc используют в качестве антигена.
В табл. 59 АТФ обозначает пэннинг с использованием АТФ, АМФ обозначает пэннинг с использованием АМФ, обозначение «Двойной» означает пэннинг с использованием двойного раунда отбора.
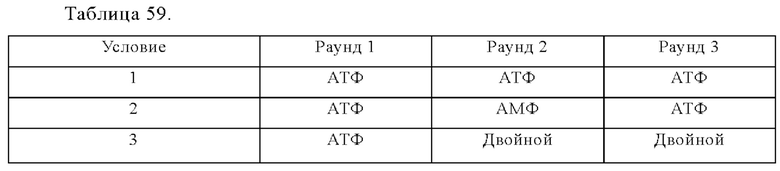
Фаги получают обычным образом из клеток Е. coli, несущих сконструированную фагмиду фагового дисплея. В частности, штамм Е. coli, несущий сконструированный фагмидный вектор, инфицируют фагами M13KO7TC (WO 2015046554A1) или M13KO7ΔpIII (гиперфагом) (фирма PROGEN Biotechnik), и фаги выделяют из супернатантов после культивирования в течение ночи при 30°С. Библиотеки фаговых дисплеев антител получают разбавлением TBS, популяции фагов, осажденных добавлением 2,5 М NaCl / 10% PEG к культуре Е. coli, в которой производили получение фагов.
Затем к раствору фаговой библиотеки добавляют БСФ до конечной концентрации 4%. Пэннинг проводят с использованием антигена, иммобилизованного на магнитных гранулах. В качестве магнитных гранул используют гранулы NeutrAvidin (фирма TAMAGAWA SEIKI) или Dynabeads MyOne StreptAvidin T1 (фирма Thermo Fisher Scientific).
В приготовленный раствор фаговой библиотеки добавляют биотинилированный hCD137-Fc и АТФ в конечной концентрации 1 мМ и проводят реакцию при комнатной температуре в течение 60 мин. Блокированные магнитные гранулы добавляют к реакционному раствору фага и антигена, и они реагируют при комнатной температуре в течение 15 мин. Гранулы дважды или трижды промывают буфером TBS / 0,1% Tween20, содержащим 1 мМ АТФ, и один или два раза TBS, содержащим 1 мМ АТФ. Гранулы, к которым впоследствии добавляют TBS, суспендируют при комнатной температуре, и раствор фага выделяют из гранул, разделенных с использованием магнитной платформы. После двух повторений этой процедуры элюированный раствор фага перемешивают. К полученному раствору фага добавляют трипсин в конечной концентрацию 1 мг / мл. Восстановленные фаги добавляют к 20 мл штамма Е. coli ER2738 в логарифмической фазе роста (OD600 0,4-0,7). Культуру Е. coli инфицируют фагами путем культивирования Е. coli в течение одного часа при 37°С при перемешивании. Е. coli высевают в чашки размером 225 мм × 225 мм. Е. coli инфицируют M13KO7TC (WO 2015046554A1) или M13KO7ΔpIII (называемым гиперфагом) (фирма PROGEN Biotechnik) и культивируют в течение ночи при 30°С; фаги выделяют из супернатантов. При условии 1 эту процедуру повторяют три раза. При условии 2, после того, как вышеупомянутая процедура была выполнена один раз, АТФ заменяют на АМФ, и вышеупомянутую процедуру была выполняют снова (раунд 2), а затем вышеупомянутую процедура выполняют заново с АТф (раунд 3).
С другой стороны, в соответствии с условием 3, после того, как описанную выше процедуру выполняют один раз, двойной раунд отбора выполняют в раундах 2 и 3. В частности, в приготовленный раствор фаговой библиотеки сначала добавляют биотинилированный hCD137-Fc и AMP в конечной концентрации 1 мМ, и они реагируют при комнатной температуре в течение 60 мин. Блокированные магнитные гранулы добавляют к реакционному раствору фага и антигена, и они реагируют при комнатной температуре в течение 15 мин. Гранулы дважды или трижды промывают буфером TBS / 0,1% Tween20, содержащим 1 мМ АМФ, и один или два раза буфер TBS, содержащий 1 мМ АМФ. Гранулы, к которым впоследствии добавляют TBS, суспендируют при комнатной температуре, и раствор фага выделяют из гранул, разделенных с использованием магнитной платформы. После двух повторений этой процедуры элюированный раствор фага перемешивают. Кроме того, в раствор восстановленной фаговой библиотеки затем добавляют биотинилированный hCD137-Fc и АТФ в конечной концентрации 1 мМ, и они реагируют при комнатной температуре в течение 60 мин. Блокированные магнитные гранулы добавляют к реакционному раствору фага и антигена, и они реагируют при комнатной температуре в течение 15 мин. Гранулы дважды или трижды промывают буфером TBS / 0,1% Tween20, содержащим 1 мМ АТФ, и один или два раза TBS, содержащим 1 мМ АТФ. Гранулы, к которым впоследствии добавляют TBS, суспендируют при комнатной температуре, и раствор фага выделяют из гранул, разделенных с использованием магнитной платформы. После двух повторений этой процедуры элюированный раствор фага перемешивают. К полученному раствору фага добавляют трипсин в конечной концентрации 1 мг / мл. Восстановленные фаги добавляют к 20 мл суспензии штаммаЕ. coli ER2738 в логарифмической фазе роста (OD600 0,4-0,7). Е. coli инфицируют фагами путем культивирования Е. coli в течение одного часа при 37°С при перемешивании. Е. coli высевают в чашки размером 225 мм × 225 мм. Е. coli инфицируют фагами M13KO7TC (WO 2015046554A1) или М13КO7ΔрIII (называемым гиперфагом) (фирма PROGEN Biotechnik) и культивируют в течение ночи при 30°С; фаги выделяют из супернатантов.
(9-2) Оценка связывающей активности в присутствии и в отсутствие АТФ или АМФ методом фагового ELISA
Из отдельных колоний Е. coli, полученных описанными выше методами, отбирают супернатанты фагосодержащих культур с использованием обычного метода (Methods Mol. Biol, 178, 2002, 133-145). NucleoFast 96 (фирма MACHEREY-NAGEL) используют для ультрафильтрации собранного культурального супернатанта. Проток удаляют центрифугированием (4500 g, 45 мин) NucleoFast 96, в который вносят по 100 мкл супернатанта культуры в каждую лунку. NucleoFast 96, в котором по 100 мкл Н2О добавляют в каждую лунку, снова промывают центрифугированием (4500 g, 30 мин). В итоге добавляют от по 100 мкл TBS и собирают раствор фага, содержащийся в супернатантах 96-луночных планшетов NucleoFast, которые оставляли стоять при комнатной температуре в течение 5 мин.
TBS, 1 мМ АМФ / TBS или 1 мМ АТФ / TBS добавляют к очищенным фагам, которые затем подвергают методу ELISA с помощью процедуры, описанной ниже. 384-луночные микропланшеты, покрытые стрептавидином (greiner, 781990), покрывают в течение ночи 100 мкл TBS, содержащего биотинилированный hCD137-Fc, полученный в примере 1. После удаления несвязавшихся антигенов из планшетов промыванием каждой лунки TBST лунки блокируют 250 мкл 0,4% Block Асе, 1% БСА, 0,02% Tween, 0,05% ProClin 300-TBS в течение одного часа или более. Затем допускают, чтобы фаги, презентирующие антитела, связывались с антигеном, присутствующим в каждой лунке, в отсутствие и в присутствии АТФ или АМФ, выдерживая планшеты с подготовленными очищенными фагами, добавленными в каждую лунку при 37°С в течение одного часа. Каждую лунку промывают TBST, 1 мМ ATP / TBST или 1 мМ AMP / TBST и дополняют HRP-конъюгированным антителом против М13 (фирма Amersham Pharmacia Biotech), разведенным в TBS, 1 мМ АМФ / TBS или 1 мМ АТФ / TBS и инкубируют в течение одного часа. После промывания TBST, 1 мМ АТФ / TBST или 1 мМ АМФ / TBST, окрашивание раствора в каждой лунке, в которую был добавлен единственный раствор ТМВ (фирма ZYMED), останавливают добавлением серной кислоты, а затем проявление окрашивания измеряют по абсорбции при 450 нм.
Результаты фагового ELISA представлены в табл.60.
В настоящем изобретении клоны с отношением поглощения S/N (сигнал / шум) больше 2 в присутствии АТФ (отношение поглощения в присутствии антигена к поглощению в отсутствие антигена), были оценены как положительные по связыванию, а клоны с коэффициентом поглощения 3 или выше в присутствии / отсутствии АТФ или АМФ были расценены как клоны с антигенсвязывающей активностью, зависящей от АТФ или АМФ (переключаемые клоны). В результате для hCD137-Fc большинство клонов, полученных по условию 1, показывают связывание в присутствии АТФ, но не показывают связывания в присутствии АМФ. Среди клонов, которые проявляют связывание в присутствии АТФ и в отсутствие связывания в отсутствие АТФ, полученных по условию 2, есть несколько антител, которые показывают связывание даже в присутствии АМФ, но процентное соотношение равно 50% или менее. С другой стороны, по условию 3, согласно которому проведен двойной отбор, 65% или более клонов, показывающих связывание в присутствии АТФ и отсутствие связывания в отсутствие АТФ, способны проявлять связывание даже в присутствии АМФ.
Для полученных клонов амплифицированные гены тяжелой цепи секвенируют с использованием специфических праймеров pBAD-F и Glseq-R.
В результате при условии 3, при котором выполняют двойной раунд отбора, полученные последовательности тяжелой цепи антитела, показывающие связывание CD137 человека, зависящее как от АТФ, так и от АМФ, которое в 3 раза больше, чем при условии 1, и которое выполняли только с АТФ, и примерно в 1,5 раза раз больше, чем при условии 2, при котором поочередно используют АТФ и АМФ. Это указывает на то обстоятельство, что двойной круговой отбор является более ценным подходом к получению антител, которые проявляют антигенсвязывающую активность, зависящую от множества отдельных небольших молекул.
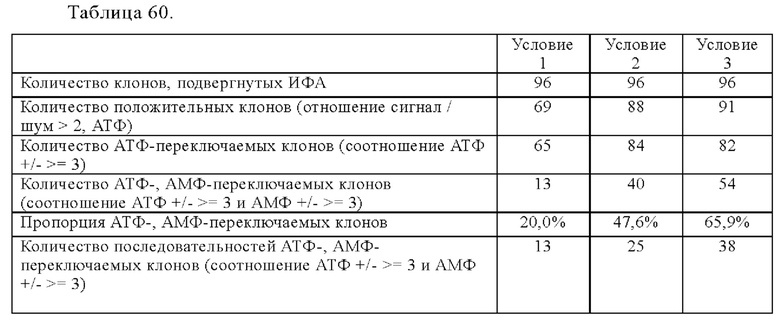
Пример 10. Получение АТФ-зависимых антител против CTLA4 мыши из наивной библиотеки
(10-1) Получение антител, которые связываются с CTLA4 мыши в присутствии малых молекул, из библиотеки наивных антител человека с помощью пэннинга на гранулах
Библиотека фагового дисплея антител человека, состоящая из множества фагов, презентирующих домены Fab различных последовательностей антител человека, была сконструирована в соответствии с методами, известными специалистам в данной области, с использованием поли(А) РНК, полученной из МКПК человека, коммерчески доступной поли(А) человека РНК, или такой как матрица.
Из созданной библиотеки фагового дисплея наивных антител человека проводят скрининг антител, проявляющих связывающую активность с внеклеточным доменом CTLA4 мыши (mCTLA4) в присутствии малых молекул. То есть собирают фаги, презентирующие антитела, которые проявляют связывающую активность с mCTLA4, захваченными гранулами, в присутствии малых молекул. Фаги выделяют из фаговых элюатов, элюированных с гранул, в отсутствие малых молекул. Биотинилированный mCTLA4 (mCTLA4-His-Biotin) в этом методе используют в качестве антигена. Получают mCTLA4-His-Biotin путем биотинилирования (фирма PIERCE, номер в каталоге 21329) mCTLA4-His (фирма Sino Biologies Inc. 50503-М08Н, номер в каталоге NP_033973.2) методом соединения через амин, которое осуществляют слиянием His-метки с внеклеточным доменом mCTLA4.
Фаги, полученные из клеток Е. coli, несущих сконструированные фагмиды фагового дисплея, очищают известными методами. Затем раствор фаговой библиотеки получают диализом против TBS. Проводят пэннинг с иммобилизованным на магнитных гранулах антигеном. В качестве магнитных гранул используют гранулы, покрытые нейтравидином (Sera-Mag SpeedBeads NeutrAvidin) или гранулы, покрытые стрептавидином (Dynabeads М-280 Streptavidin).
Для эффективного получения переключаемых низкомолекулярными соединениями антител, которые зависят от малой молекулы, способной играть переключающую роль в раковых тканях, пэннинг для обогащения антител, которые связываются с антигенами в присутствии аденозинтрифосфата (аденозин-5'-трифосфат; АТФ) и метаболитами АТФ, которые не связывают антигены в отсутствие АТФ, выполняют со ссылкой на методы, описанные в предыдущем патенте WO 2013/180200.
В частности, в приготовленный раствор фаговой библиотеки добавляют биотинилированный mCTLA4 и АТФ, а также метаболиты АТФ, в конечной концентрации 1 мМ для осуществления реакции при комнатной температуре в течение 60 мин. Блокированные магнитные гранулы добавляют к реакционному раствору фага и антигена, и они реагируют при комнатной температуре в течение 15 мин. Гранулы дважды или трижды промывают буфером TBS / 0,1% Tween20, содержащим 1 мМ АТФ, и один или два раза TBS, содержащим 1 мМ АТФ. Гранулы, к которым впоследствии добавляют TBS, суспендируют при комнатной температуре, и раствор фага выделяют из гранул, разделенных с использованием магнитной платформы. Затем к гранулам добавляют трипсин в конечной концентрации 1 мг / мл в 1-м раунде и TBS во 2-м раунде и далее, и их суспендируют при комнатной температуре; растворы фагов извлекают из гранул, разделенных с помощью магнитной платформы. Если используют TBS, эту процедуру повторяют дважды, после чего перемешивают элюированный раствор фага и добавляют к полученному раствору фага трипсин в конечной концентрации 1 мг / мл. Извлеченные фаги добавляют к 10 мл штамма Е. coli ER2738 в логарифмической фазе роста (OD600 0,4-0,7). Е. coli инфицируют фагами путем культивирования Е. coli в течение одного часа при 37°С при перемешивании. Е. coli высевают в чашки размером 225 мм × 225 мм. Е. coli инфицируют М13К07ТС (WO 2015046554A1) или M13KO7ΔpIII (называемым гиперфагом) (фирма PROGEN Biotechnik) и культивируют в течение ночи при 30°С; фаги выделяют из супернатантов. Во время культивирования для индукции экспрессии генов Fab с промотора lac используют условие добавления 100 мкМ IPTG и условие отсутствия добавления IPTG. Эту процедуру повторяют четыре раза.
(10-2) Оценка связывающей активности в присутствии и в отсутствие малых молекул методом фагового иммуноферментного анализа (ELISA)
Из отдельных колоний Е. coli, полученных в примере 10-1, отбирают супернатанты фагосодержащих культур с использованием обычного метода {Methods Mol. Biol, 178, 2002, 133-145). NucleoFast 96 (фирма MACHEREY-NAGEL) используют для ультрафильтрации собранных культуральных супернатантов. По 100 мкл каждого культурального супернатанта вносят в каждую лунку планшета NucleoFast 96 и центрифугируют в режиме 4500 g в течение 45 мин для удаления протока. По 100 мкл H2O добавляют в каждую лунку, снова промывают центрифугированием (4500 g, 30 мин). В итоге добавляют по 100 мкл TBS и оставляют при комнатной температуре в течение 5 мин, затем выделяют раствор фага из супернатантов.
TBS добавляют к очищенным фагам, которые затем подвергают анализу методом ELISA с помощью процедуры, описанной ниже. Планшеты для микротитрования StreptaWell 96 (фирма Roche) покрывают в течение ночи 100 мкл TBS, содержащего mCTLA4-His-Biotin. После удаления несвязанного mCTLA4-His- Biotin из планшетов промыванием каждой лунки TBST, лунки блокируют 250 мкл 2% обезжиренного молока-TBS в течение более чем одного часа. Затем допускают связывание антитело-презентирующих фагов с mCTLA4-His-Biotin, присутствующим в каждой лунке в отсутствие и в присутствии АТФ, путем удаления 2% обезжиренного молока-TBS с последующим хранением планшетов с подготовленными очищенными фагами, добавленными в каждую лунку, при комнатной температуре в течение одного часа. Каждую лунку промывают TBST или ATP / TBST и дополняют HRP-конъюгированным антителом против М13 (фирма Amersham Pharmacia Biotech), разведенным в TBS или ATP / TBS, и инкубируют в течение одного часа. После промывания TBST или ATP / TBST развитие окраски раствора в каждой лунке, в которую был добавлен отдельный раствор ТМВ (фирма ZYMED), остановлено добавлением серной кислоты, а затем проявление цвета измеряют по абсорбции при 450 нм. В результате подтверждают, что несколько антител связываются с mCTLA4 только в присутствии АТФ.
Результаты анализа методом фагового ELISA представлены в табл. 61.
В настоящем изобретении клоны с абсорбцией более 0,2 в присутствии АТФ признают положительными, а клоны с абсорбцией более 2 в присутствии /в отсутствие АТФ оценивают как клоны с АТФ-зависимой антигенсвязывающей активностью (переключаемые клоны).
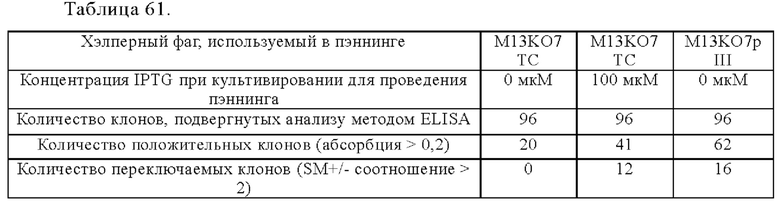
Вышеупомянутое изобретение подробно описано с примерами и иллюстрациями в целях содействия ясному пониманию, и приведенное описание и иллюстрации не следует истолковывать как ограничивающие объем настоящего изобретения. Сущность всех патентов и научной литературы, цитируемых в настоящем изобретении, включена в него в виде ссылок.
Пример 11. Оценка in vitro АТФ- и АДФ-зависимой CD137 агонистической активности модифицированных антител против CD 137 человека с использованием анализа репортерного гена 4-1 ВВ Jurkat
GloResponse™ NF-κВ Luc2/4-1BB Jurkat линию клеток (фирма Promega, CS196004) используют для применения in vitro активности вариантов, полученных в примере 7-2 (табл. 52). В каждую лунку 384-луночных планшетов добавляют по 10 мкл каждой из клеток FcγRIIB СНО-К1 (фирма Promega), приготовленных в концентрации 4 × 105 / мл в среде для анализа (99% RPMI, 1% ФСБ). Затем в каждую лунку добавляют 10 мкл раствора антитела, содержащего АДФ, или раствора антитела, содержащего АТФ, или раствора антитела без АТФ или без АДФ. Затем в каждую лунку добавляют 10 мкл клеточной линии GloResponseTM NF-κB-Luc2 / 4-1 ВВ Jurkat, приготовленной в концентрации 2 × 106 / мл в среде для анализа (99% RPMI, 1% ФСБ). Конечная концентрация АДФ составляет 10 мкМ, конечная концентрация АТФ составляет 10 мкМ. Планшеты оставляют на 6 ч при 37°С в инкубаторе с 5% СО2 и 15 мин при комнатной температуре; 30 мкл реагента Bio-Glo добавляют в каждую лунку. Для реагента Bio-Glo используют систему анализа люциферазы Bio-Glo (буфер и субстрат). Затем количество люминесценции в каждой лунке измеряют с помощью планшет-ридера. Величину люминесценции каждой лунки, деленную на величину люминесценции лунки без добавления антитела, определяют как «относительную световую единицу» (кратная индукция), которая служит индикатором оценки агонистической активности в отношении CD 137 каждого антитела.
Результаты представлены на фиг. 58. На фиг. 58 приводят подтверждение, что A375-P587/B167-LamLib, А551-P587/B256-LamLib, А551-P587/B379-LamLib, A375-SCF041aPh/B167-LamLib, A551-SCF041aPh/B256-LamLib, A551-SCF041aPh/B379-LamLib, A375-SCF043aPh/B167-LamLib, A551-SCF043aPh/B256-LamLib, A551-SCF043aPh/B379-LamLib, A375-SCF057aPh/B167-LamLib, A551-SCF057aPh/B256-LamLib и A551-SCF057aPh/B379-LamLib проявляют агонистическую активность в отношении CD137 человека АТФ- и АДФ-зависимым образом.
Пример 12. Измерение уровня внеклеточного АТФ в участках опухоли
Антитело, переключаемое АТФ, связывается с молекулой-мишенью в присутствии АТФ и проявляет лекарственную эффективность. Оценку внеклеточного уровня АТФ в участках опухолевой ткани проводят с использованием клеток P2Y11 split Luc/HEK293. Клетки получают с применением ProbeX с использованием технологии компании сплит-люциферазы. Клетки экспрессируют С-концевой белок люциферазы, связанный с пуринергическим рецептором P2Y11 (номер доступа AF030335) с АТФ в качестве лиганда, и N-концевой белок люциферазы, связанный с изоформой β-аррестина-2 (номер доступа NM_004313). Активную люциферазу вырабатывают путем объединения двух фрагментов люциферазы, которые отдельно присутствуют в клетках, путем стимуляции АТФ, и может быть получена люминесценция с использованием люциферина в качестве субстрата.
(12-1) Чувствительность к АТФ расщепленных P2Y11 клеток Luc / НЕК293
Клетки P2Y11 split Luc/HEK293 культивируют и поддерживают в среде Игла, модифицированной Дульбекко (Dulbecco's Modified Eagle's Medium с высоким содержанием глюкозы: D5796 / Sigma-Aldrich), содержащей 10% фетальной сыворотки теленка, 0,8 мг / мл G418 (Geneticin™) и 0,05 мг / мл флеомицина Dl (Zeocin™) для пассажей. Клетки P2Y11 split Luc/HEK293 помещают в 96-луночный микропланшет (jiCLEAR™, WHITE, CELLSTAR/фирма Greiner Bio-One) в количестве 30000 клеток/лунку, культивируют в течение ночи и затем раствор культуры забирают из каждой лунки и заменяют 0,18 мл раствора культуры, содержащего 0,5 мг/мл люциферина (VivoGlo™ Luciferin, In Vivo Grade /фирма Promega). Для достижения конечных концентраций 0, 0,125, 0,25, 0,5, 1,0 и 2,0 мМ добавляют 0,02 мл раствора аденозина, АМФ, АДФ или АТФ и измеряют значения люминесценции на планшет-ридере En Vision (фирма Perkin Elmer) после 30 мин инкубации в инкубаторе с CO2 (37°С, в атмосфере 5% CO2).
Результаты показаны на фиг. 59. Клетки P2Y11 split Luc/HEK293 увеличивают свою люминесценцию АТФ-специфическим образом в зависимости от концентрации лиганда.
(12-2) Оценка концентрации внеклеточного АТФ в участках опухолевой ткани
Клетки P2Y11 split Luc/HEK293 отделяют от культуральных колб с помощью буфера для диссоциации клеток, не содержащего ферментов, и ФСБ (фирма Thermo Fisher Scientific), промывают HBSS и затем получают суспензию клеток в HBSS, которую используют для измерения уровня внеклеточного АТФ in vivo.
(12-2-1) Получение калибровочной кривой АТФ in vivo
0,2 мл суспензии клеток, содержащей фиксированную концентрацию АТФ, 1 мг / мл люциферина (VivoGlo, фирма Promega) и 1,0×107 клеток / мл (клеток P2Y11 split Luc/HEK293), трансплантируют подкожно в вентральную область мышей С3Н / HeN, и через 20 минут измеряют люминесцентную визуализацию под анестезией изофлураном с помощью устройства визуализации in vivo IVIS Spectrum СТ. Полученные изображения проанализированы с помощью программного обеспечения Living Image Software, и выстраивают калибровочную кривую АТФ в условиях измерения in vivo на основе рассчитанных значений люминесценции и концентраций добавленного АТФ.
На фиг. 60 показаны результаты. Клетки P2Y1 lsplit Luc/HEK293 обладают люминесценцией, зависящей от концентрации АТФ, даже в условиях in vivo при подкожной трансплантации мышам С3Н / HeN; эти результаты можно преобразовать в калибровочную кривую для оценки уровней АТФ in vivo.
(12-2-2) Оценка уровня внеклеточного АТФ в участках опухолевой ткани
0,2 мл суспензии клеток, содержащей 1 мг / мл люциферина (VivoGlo, фирма Promega) и 1,0 × 107 клеток / мл (клеток P2Y11 split Luc/HEK293), трансплантируют подкожно в участок опухоли мышей с опухолью FM3A (объем опухоли 200-300 мм) и измеряют люминесцентную визуализацию через 20 мин под анестезией изофлураном с помощью устройства визуализации in vivo IVIS Spectrum СТ. Полученные изображения анализируют с помощью программного обеспечения Living Image Software, уровень АТФ рассчитывают по величине люминесценции участка опухоли с использованием калибровочной кривой АТФ, полученной in vivo, который рассматривают как уровень внеклеточного АТФ в участке опухоли.
Результаты измерения люминесцентной визуализации показаны на фиг. 61, а уровни внеклеточного АТФ на участке опухоли, рассчитанные по калибровочной кривой АТФ, показаны в табл.62. Уровень люминесценции на участке ткани опухоли (мышь с опухолью FM3A) выше, чем на фоне (нормальный участок), и уровень внеклеточного АТФ на участке опухолевой ткани в модели трансплантации FM3A рассчитывают как среднее значение 1,31 мМ на подготовленной in vivo калибровочной кривой АТФ.
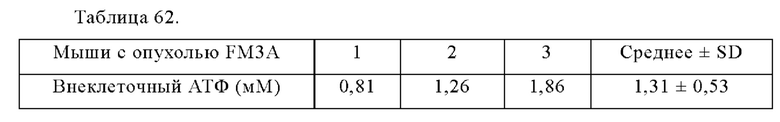
Пример 13. АТФ-зависимая противоопухолевая активность анти-ЬЛЬбРу антител, обладающих АТФ-зависимыми связывающими свойствами
(13-1) Получение анти-hIL6R антител, имеющих АТФ-зависимые связывающие свойства
Ссылаясь на методы, описанные в WO 2015/083764 и WO 2013/180200, полученные антитела оптимизируют для получения антител против рецептора интерлейкина 6 человека (hIL6R), обладающих АТФ-зависимыми связывающими свойствами. Тяжелые цепи и легкие цепи объединяют, как показано в табл. 63, для экспрессии и очистки антител против hIL6R методом, известным специалистам в данной области.
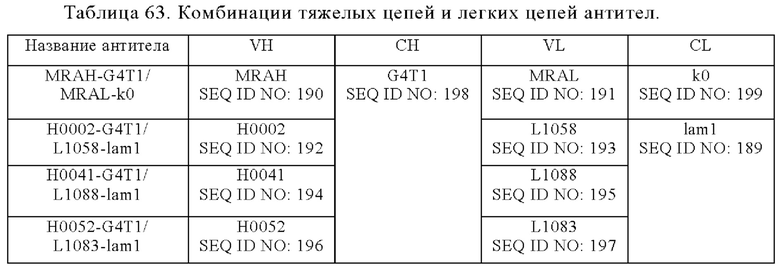
hIL6R (SEQ ID NO: 200) синтезируют генетически в качестве антигена и инсертируют в плазмиду для экспрессии в животных, затем его вводят в клетки СНО, и, таким образом, в результате клонирования получают стабильную линию экспрессии (hIL6R-CHO), которая конститутивно экспрессирует антиген. Антигенный белок экспрессируют и очищают с использованием описанных ниже методов. Линию клеток hIL6R-CHO суспендируют при подходящей плотности клеток, высевают и культивируют в колбах, а антиген очищали из супернатанта культуры способом, известным специалистам в данной области. Спектрофотометры используют для измерения абсорбции очищенного раствора антигена при 280 нм. Концентрации очищенного антигена рассчитывают по полученным значениям измерений с использованием коэффициентов абсорбции, рассчитанных методом РАСЕ (Protein Science 4, 1995, 2411-2423).
(13-2) Расчет зависимых от концентрации АТФ значений KD антител против hIL6R
Анализ аффинности для hIL6R в присутствии каждой из концентраций АТФ 0, 1, 10 и 100 мкМ проводят с помощью Biacore Т200 с использованием антител, описанных в табл.63. Измерения проводят при скорости потока 10 мкл / мин, время взаимодействия 1 мин, время диссоциации 1 мин и кинетика одного цикла при 37°С. Используют буфер 20 мМ ACES, 150 мМ NaCl, 2 мМ MgCl2 и 0,05% Tween20, рН 7,4, содержащие различные концентрации (0, 1, 10 и 100 мкМ) АТФ. Дополнительное программное обеспечение Biacore Evaluation Software от Biacore Т200 используют для анализа, а модель связывания Ленгмюра 1:1 используют для модели соразмерности. Результаты анализа представлены в табл.64 и на фиг. 62.
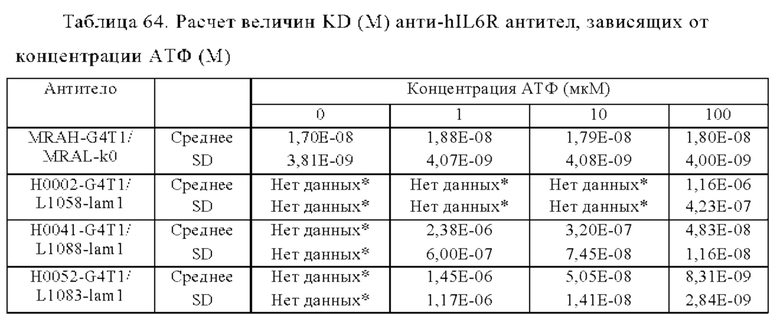

Результаты показывают, что KD для hIL6R MRAH-G4T1 / MRAL-k0 не зависит от концентрации АТФ в анализе, но KD для hIL6R H0002-G4T1 / L1058-lam1, H0041-G4T1 / L1088-lam1 и Н0052 -G4T1 / L1083-lam1 становится меньше при более высокой концентрацией АТФ в анализе. То есть, чем выше концентрация АТФ в анализе, тем сильнее становится связывающая активность. Результаты подтверждают, что H0002-G4T1 / L1058-lam1, H0041-G4T1 / L1088-lam 1 и H0052-G4T1 / L1083-lam1 обладают АТФ-зависимой hIL6R-связывающей активностью.
В дальнейшем антитело, антигенсвязывающая активность которого не зависит от концентрации АТФ, такое как MRAH-G4T1 / MRAL-k0, может называться непереключаемым антителом, а антитело, антигенсвязывающая активность которого зависит от концентрации АТФ, такое как H0002-G4T1 / L1058-lam1, H0041-G4T1 / L1088-lam1 и H0052-G4T1 / L1083-lam1, может быть обозначена как переключаемое антитело.
(13-3) Расчет зависимых от концентрации АДФ и АМФ значений KD антител против hIL6R
Затем антитела, описанные в табл. 63, используют для анализа аффинности hIL6R в присутствии 10 и 100 мкМ АДФ или АМФ вместо АТФ с использованием Biacore Т200 в соответствии с условиями, описанными выше. Результаты анализа измерения с использованием АДФ показаны в табл.65 и на фиг. 63, а результаты анализа измерения с использованием АМФ показаны в табл. 66 и на фиг. 64. Антитела с ответом 10 RU или менее во время измерения рассматривают как не анализируемые и обозначают в таблице как «нет данных».
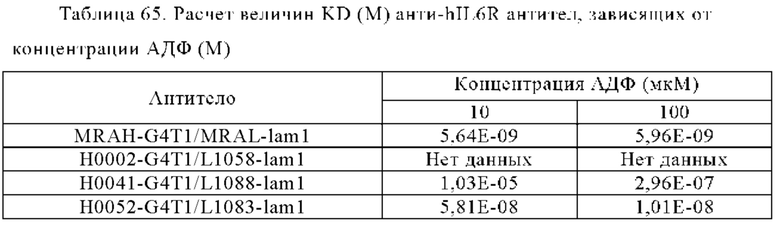
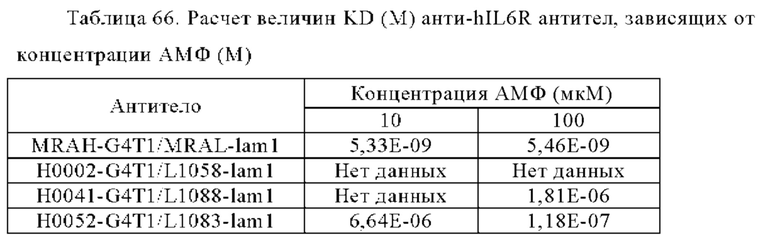
Результаты показывают, что в анализе на KD для hIL6R MRAH-G4T1 / MRAL-k0 не влияют концентрации АДФ и АМФ, но, как и в случае АТФ, более высокие концентрации АДФ или АМФ в анализе сопровождаются меньшими значениями KD для hIL6R H0002-G4T1 / L1058-lam1, H0041-G4T1 / L1088-lam1 и H0052-G4T1 / L1083-lam1, т.е. они обладают более сильной связывающей активностью. Результаты подтверждают, что H0002-G4T1 / L1058-lam1, Н0041-G4T1 / L1088-lam1 и H0052-G4T1 / L1083-lam1 обладают АДФ- и АМФ-зависимой связывающей активностью по отношению к hIL6R.
(13-4) Оценка активности ADCC антител против hIL6R. зависящей от концентрации АТФ
Антитела получают методом, известным специалистам в данной области, как показано в табл. 67, путем изменения константных областей тяжелой цепи указанных выше антител на mFa55 (SEQ ID NO: 184), в которых активность ADCC усиливают путем добавления модификаций mIgG2a, усиливающих связывание FcγRIV, и путем изменения константной области легкой цепи с цепи человека на цепь мыши. Кроме того, KLH-mFa55 (IC17Hdk-mFa55 / IC17L-mk1) получают в качестве отрицательного контроля.
Эти антитела используют для оценки ADCC против Hepal-6 (hIL6R-Hepal-6), линии гепатомы мыши, которая экспрессирует полноразмерный hIL6R (SEQ ID NO: 213), полученный методами, известными специалистам в данной области, описанными ниже.
Для измерений, приведенных ниже, используют mFcγRIV ADCC Reporter Bioassay, Core Kit (фирма Promega). В каждую лунку 96-луночного планшета добавляют 100 мкл hIL6R-Hepal-6, приготовленного в среде с концентрацией клеток 2хЮ5/мл, и оставляют планшет при 37°С в течение 21 ч. В качестве питательной среды используют 90% DMEM, 10% ФСБ, 600 мкг генетицина и 500 мкг зеоцина. После центрифугирования планшета в течение пяти мин в режиме 200 × g, 4°С, супернатант из каждой лунки удаляют. Раствор антител, описанных в табл. 67, разбавленный в буфере для анализа до конечной концентрации 0,2 мкг/мл, раствор АТФ, разбавленный в буфере для анализа до конечной концентрации 0, 1, 10 и 100 мкМ, и 25 мкл раствора, приготовленного путем добавления 0,69 мл набора вспомогательных эффекторных клеток к 3,82 мл буфера для анализа, смешивают до общего объема 75 мкл, смеси добавляют в каждую лунку и оставляют выстаиваться при 37°С в течение шести часов. В качестве буфера для анализа используют 1,5 мл вспомогательной сыворотки с низким уровнем IgG из набора, которые добавляют к 36 мл среды RPMI 1640. Затем планшет оставляют при комнатной температуре на 15 мин, и в каждую лунку добавляют 75 мкл реагента Bio-Glo. Для реагента Bio-Glo используют систему анализа люциферазы Bio-glo (буфер и субстрат). Затем измеряют люминесценцию в каждой лунке с помощью планшет-ридера. Значение люминесценции в каждой лунке, деленное на величину люминесценции в лунке без добавления антител, является показателем кратности индукции, и она служила индикатором для оценки ADCC каждого антитела.
Полученные результаты показаны на фиг. 65. На этой фигуре кратность индукции обозначена как относительная световая единица (RLU, relative light unit).
Результаты показывают, что на ADCC непереключаемого антитела MRAH-mFa55 / MRAL-mk0 не влияет на экспрессирующие hIL6R клеток в зависимости от концентрации АТФ, a ADCC переключаемых антител H0002-mFa55/L1058-ml0, H0041-mFa55/L1088-ml0 и H0052-mFa55/L1083-ml0 становится сильнее при более высокой концентрацией АТФ в анализе. Результаты подтверждают, что H0002-mFa55 / L1058-ml0, H0041-mFa55 / L1088-ml0 и H0052-mFa55 / L1083-ml0 обладают АТФ-зависимой цитотоксической активностью в отношении hIL6R.
(13-5) Оценка противоопухолевой активности антител против hIL6R in vivo
После подтверждения ADCC каждого антитела проводят тест для оценки противоопухолевой активности этих антител in vivo на несущей опухоль модели мышей C57BL /6 с hIL6R-Hepa1-6 согласно приведенным ниже методам.
Анти-asialo GM1 антитело (фирма Wako Pure Chemical Industries) растворяют в 1 мл дистиллированной воды (фирма Otsuka Pharmaceutical Factory), и затем разбавляют 4 мл ФСБ. Полученное таким образом анти-asialo GM1 антитело вводят внутрибрюшинно 100 мышам C57BL / 6 в дозе 0,1 мл на мышь. Через день готовят смешанный раствор, к которому добавлен равный объем Matrigel Matrix (фирма Corning), вносят в клеточную суспензию hIL6R-Нера1-6 ститром 1×108/ мл в сбалансированном солевом растворе Хэнкса (фирма Sigma). Смешанный раствор вводят подкожно 100 мышам C57BL / 6 в дозе 0,2 мл на мышь и имплантируют клетки. Диаметр опухоли мышей измеряют штангенциркулем, а объем опухоли определяют как (большая ось) × (малая ось) × (малая ось) × (1/2). Измерение диаметра опухоли проводит индивидуум, не знающий о составе раствора лекарственного средства, вводимого каждой мыши. Диаметр опухоли и вес тела 100 из указанных выше мышей C57BL / 6 измеряют на 11-й день после трансплантации клеток. На основании полученных значений рандомизируют мышей, которых разделяют в общей сложности на пять групп: четыре группы по 10 животных в каждой и одна группа из 8 животных; в каждой группе животным вводят через хвостовую вену раствор антител, показанных в табл.67. В группе из 8 животных животные получают H0002-mFa55 / L1058-ml0. Доза (мл) введенного раствора антитела составляет (0,01 (мл/г)) × (масса тела (г) каждой мыши). Диаметр опухоли и массу тела мышей C57BL / 6, сгруппированных выше, измеряют таким же образом на 14, 18, 21 и 25 день после трансплантации. На 18-й день после трансплантации растворы антител, показанных в табл. 67, вводят животным в соответствующей группе через хвостовую вену в дозировке, определенной с помощью той же формулы расчета, которая описана выше, на основе данных измерения массы тела. Был рассчитан средний объем опухоли в каждой группе лечения антителом в каждый день измерения, и он является показателем для оценки противоопухолевой активности in vivo каждого антитела.
Полученные результаты показаны на фиг. 66.
Среди переключаемых антител, для которых подтверждена АТФ-зависимая ADCC, подтверждено, что H0041-mFa55 / L1088-ml0 и H0052-mFa55 / L1083-ml0 проявляют противоопухолевую активность в отношении клеток, экспрессирующих hIL6R, в той же степени, что и MRAH-mFa55 / MRAL- mk0, которое не является непереключаемым антителом согласно приведенным выше результатов. Результаты показывают, что внеклеточный АТФ в непосредственной близости от опухолей присутствовал на уровне концентрации, при котором H0041-mFa55 / L1088-ml0 и H0052-mFa55 / L1083-ml0 функционируют схожим образом с MRAH-mFa55 / MRAL-mk0.
Пример 14. Оценка системных эффектов анти-hIL6R антител, обладающих АТФ-зависимыми связывающими свойствами
Чтобы оценить в нормальных тканях эффекты антител против hIL6R, обладающих АТФ-зависимыми связывающими свойствами, нормальных мышей (C57BL / 6) и трансгенных мышей с системной сверхэкспрессией hIL6R (hIL6R-tgm) (Proc Natl Acad Sci USA, 92 (11), 1995, 4862-4866) используют для оценки кинетики антител в плазме. Для этой оценки антитела, описанные в табл. 67, вводят нормальным мышам (C57BL / 6) и трансгенным мышам hIL6R-tgm, и сравнивают их кинетику в плазме. Сообщалось, что в hIL6R-tgm обычные антитела, которые связываются с hIL6R, исчезают через hIL6R мембранного типа в результате связывания с системно экспрессируемым hIL6R мембранного типа hIL6R (Nature Biotechnology, 28, 2010, 1203-1207).
2 мг/мл MRAH-mFa55 / MRAL-mk0, H0002-mFa55 / L1058-ml0, H0041-mFa55 / L1088-ml0 или H0052-mFa55 / L1083-ml0 вводят внутривенно нормальным мышам и hIL6R-tgm за один раз в дозе 10 мл / кг. У нормальных мышей кровь собирают через 5 мин, 7, 24, 48, 72 и 168 ч после введения во всех группах: группе MRAH-mFa55/MRAL-mk0, группе H0002-mFa55/L1058-ml0, группе H0041-mFa55 / L1088-ml0 и группе H0052-mFa55 / L1083-ml0. У мышей hIL6R-tgm кровь собирают через 5 мин, 7, 24, 48, 52, 55, 72 и 168 ч после введения в группе MRAH-mFa55 / MRAL-mk0 и через 5 мин, 7, 24, 48, 72. и через 168 ч после введения в группе H0002-mFa55 / L1058-ml0, группе Н0041 mFa55 / L1088-ml0 и группе H0052-mFa55 / L1083-ml0. Полученную кровь быстро центрифугировали при 4°С в режиме 12000 об / мин, 10 мин; плазму отделяют. Плазму хранят в морозильной камере, установленной на -20°С или ниже, до проведения измерения.
Концентрацию каждого антитела в плазме измеряют методом электрохемилюминесценции (ECL, electrochemiluminescence). Раствор hIL6R добавляют в планшет MULTI-ARRAY PR (фирма Meso Scale Diagnostics, LLCs), оставляют выстаиваться при 37°С в течение одного часа, и hIL6R иммобилизуют на планшете. Для калибровочной кривой получают MRAH-mFa55 / MRAL-mk0, H0041-mFa55 / L1088-ml0 и H0052-mFa55 / L1083-ml0 при концентрациях в плазме 8, 4, 2, 1, 0,5, 0,25 и 0,125 мкг/мл; и H0002-mFa55 / L1058-ml0 получают при концентрациях в плазме 64, 32, 16, 8, 4, 2 и 1 мкг/мл. После добавления 1 М раствора трис-HCl, рН 8,0, содержащего 0,05% Tween20 и 1% БСА, на планшет с иммобилизованным hIL6R, образцы плазмы, разбавленные в 250 раз в 0,3 М растворе уксусной кислоты, а также образцы калибровочной кривой быстро добавляют, а затем добавляют раствор АТФ и перемешивают. После того, как планшет оставляют выстаиваться в течение одного часа при комнатной температуре, добавляют меченное биотином антитело против IgG мыши (фирма Southern Biotechnology Associates, Inc) и оставляют при комнатной температуре на один час. Затем добавляют стрептавидин SULFO-TAG Labeled (фирма Meso Scale Diagnostics, LLC) и оставляют при комнатной температуре на один час, после чего добавляют буфер считывания Т (фирма Meso Scale Diagnostics, LLC), и реакцию измеряют с помощью считывателя электрохимической люминесценции SECTOR Imager 6000 (фирма Meso Scale Diagnostics, LLC). Программу SOFTmax PRO (фирма Molecular Devices) используют для расчета концентрации каждого антитела в плазме мыши.
Концентрацию растворимого антигена в плазме измеряют методом ECL. Разбавленные в 50 раз образцы плазмы мышей смешивают с SULFO-TAG (фирма Meso Scale Diagnostics, LLC)-меченным антителом против IL6R человека (фирма R&D Systems), биотин-меченным антителом против IL6R человека (фирма R&D Systems) и избыточным количеством антитела против IL6R человека тоцилизумаба (продукт получают сами авторы настоящего изобретения) и смесь инкубируют в течение ночи при 37°С. После добавления смеси в стандартный 96-луночный блокированный планшет, покрытый стрептавидином (фирма Meso Scale Diagnostics, LLC), с последующим добавлением буфера для считывания результатов, реакцию измеряют на SECTOR Imager 6000 (фирма Meso Scale Diagnostics, LLC). Программное обеспечение SOFTmax PRO (фирма Molecular Devices) используют для расчета концентраций растворимого антигена в плазме мышей.
Результаты сравнения уровней MRAH-mFa55 / MRAL-mk0, H0002-mFa55 / L1058-ml0, H0041-mFa55 / L1088-ml0 и H0052-mFa55 / L1083-ml0 в крови у нормальных мышей и hIL6R-tgm представлены на фиг. 67, фиг. 68, фиг. 69 и фиг. 70, соответственно.
Непереключаемое антитело MRAH-mFa55 / MRAL-mk0 быстрее выводится у мышей hIL6R-tgm, которые системно экспрессируют hIL6R, чем у нормальных мышей (фиг. 67). Этот результат, по-видимому, вызван тем обстоятельством, что непереключаемое антитело связывается с hIL6R мембранного типа, экспрессируемым в нормальных тканях всего организма, и исчезает через hIL6R.
С другой стороны, для переключаемых антител H0002-mFa55 / L1058-ml0, H0041-mFa55 / L1088-ml0 и H0052-mFa55 / L1083-ml0 в трансгенных мышах hIL6R-tgm не наблюдают резкого снижения концентраций антител по сравнению с нормальными мышами. Результаты означают, что, в отличие от результата с непереключаемым антителом, переключаемые антитела не связываются с системно экспрессируемым hIL6R мембранного типа и не вызывают опосредованную hIL6R элиминацию.
По сравнению с переключаемыми антителами H0002-mFa55 / L1058-ml0, H0041-mFa55 / L1088-ml0 и H0052-mFa55 / L1083-ml0, только антитело H0052-mFa55 / L1083-ml0, демонстрирующее более сильное связывание при низких концентрациях АТФ, приводит к ускоренной потере антитела в hIL6R-tgm по сравнению с нормальными мышами.
Эти результаты позволяют предположить, что внеклеточные концентрации АТФ в нормальных тканях являются недостаточными для переключения антител на связывание с hIL6R.
Затем концентрации в плазме растворимого hIL6R у мышей hIL6R-tgm сравнивают после введения соответствующих антител. Результаты показаны на фиг. 71.
По сравнению с концентрациями растворимого hIL6R у мышей, получающих изотипический контроль IC17HdK-mFa55 / IC17L-mk1, концентрации растворимого hIL6R у мышей, получающих непереключаемое антитело, MRAH-mFa55 / MRAL-mk0, по данным наблюдений накапливается. Считают, что это вызвано связыванием непереключаемого антитела с растворимой формой hIL6R, что препятствует нормальным метаболическим путям.
С другой стороны, у мышей, получавших переключаемые антитела Н0002-mFa55 / L1058-ml0, H0041-mFa55 / L1088-ml0 и H0052-mFa55 / L1083-ml0, не наблюдают значительного накопления антигена, подобного накоплению в случае введения непереключаемого антитела. При сравнении переключаемых антител H0002-mFa55 / L1058-ml0, H0041-mFa55 / L1088-ml0 и H0052-mFa55 / L1083-ml0 наблюдают тенденцию по накоплению антигена для антител, демонстрирующих более сильное связывание при низких концентрациях АТФ.
Эти результаты позволяют предположить, что внеклеточные концентрации АТФ в нормальных тканях и крови являются низкими для переключения антител на связывание с мембраной и растворимыми формами hIL6R.
Пример 15: Одновременная оценка противоопухолевых эффектов и системных эффектов антител против hIL6R, обладающих АТФ-зависимыми связывающими свойствами
Противоопухолевые эффекты и системные эффекты одновременно оценивают для выявления системных эффектов aHTH-hIL6R антител, обладающих АТФ-зависимыми связывающими свойствами, в присутствии опухоли. Для этой оценки используют модель hIL6R-tgm с трансплантированным hIL6R-Hepal-6 и вводят антитела, описанные в табл. 67.
Анти-asialo GM1 антитело (фирма Wako Pure Chemical Industries) растворяют в 1 мл дистиллированной воды (фирма Otsuka Pharmaceutical Factory), а затем разбавляют 4 мл ФСБ. Полученное таким образом анти-asialo GM1 антитело вводят внутрибрюшинно 83 мышам hIL6R-tgm в дозе 0,1 мл на мышь. Через день получают смешанный раствор, в который добавляют такой же объем Matrigel Matrix (фирма Corning) к клеточной суспензии hIL6R-Hepal-6, суспендированной до титра 1 × 108 / мл в сбалансированном солевом растворе Хэнкса (фирма Sigma). Смешанный раствор вводят подкожно 83 трансгенным мышам hIL6R-tgm в дозе 0,2 мл на мышь, в результате чего клетки имплантируются. Диаметр опухоли мышей измеряют штангенциркулем, а объем опухоли определяют как (большая ось) × (малая ось) × (малая ось) × (1/2). Измерение диаметра опухоли проводит индивидуум, не знающий о составе раствора лекарственного средства, вводимого каждой мыши. Диаметр опухоли и вес тела 83 указанных выше мышей hIL6R-tgm измеряют на 10-й день после трансплантации клеток. На основании полученных значений рандомизируют мышей, которых разделяют в общей сложности на четыре группы по 10 животных, в каждой группе животным вводят через хвостовую вену соответствующий раствор антитела из числа антител, показанных в табл. 67, за исключением H0052-mFa55/L1083-ml0. Доза (мл) введенного раствора антитела составляет (0,01 (мл/г)) × (масса тела (г) каждой мыши). Впоследствии диаметры опухоли и массу тела hIL6R-tgm, сгруппированных указанным выше образом, измеряют таким же образом на 13, 17 и 20 день после трансплантации. На 17-й день после трансплантации растворы четырех антител, показанных в табл. 67, за исключением H0052-mFa55/L1083-ml0, вводят животным в соответствующей группе через хвостовую вену в дозировке, определенной с помощью той же формулы расчета, которая описана выше, на основе данных по измерению массы тела. Рассчитывали средний объем опухоли в каждой группе лечения антителом и в каждый день измерения, которые является показателем для оценки противоопухолевой активности in vivo для каждого антитела.
В вышеупомянутых экспериментах берут на анализ кровь у мышей hIL6R-tgm, разделенных на указанные выше группы, под анестезией изофлураном через один час, 1, 2, 3 и 7 дней после первоначального введения растворов антител. Образцы крови переносят в серии из восьми пробирок с индивидуальным номером и оставляют на льду. После центрифугирования серий из восьми пробирок в режиме 1900 × g в течение 10 мин при 4°С супернатант после центрифугирования получают в виде компонента плазмы и переносят в другую серию из восьми пробирок с индивидуальным номером, которые хранят при -30°С.
Концентрацию каждого антитела в плазме измеряют методом электрохемилюминесценции (ECL). Раствор hIL6R добавляют в планшет MULTI-ARRAY PR (фирма Meso Scale Diagnostics, LLC) и оставляют при 37°С в течение одного часа, hIL6R иммобилизуется на планшете. Для калибровочной кривой готовят растворы MRAH-mFa55 / MRAL-mk0 и H0041-mFa55 / L1088-ml0 в плазме с концентрациями 8, 4, 2, 1, 0,5, 0,25 и 0,125 мкг/мл; H0002-mFa55 / L1058-ml0 получают с концентрациях в плазме 64, 32, 16, 8, 4, 2 и 1 мкг/мл. После добавления 1 М раствора трис-HCl, рН 8,0, содержащего 0,05% Tween20 и 1% БСА, в планшет с иммобилизованным hIL6R, образцы плазмы разбавляют в 250 раз в 0,3 М растворе уксусной кислоты, а также быстро добавляют образцы для калибровочной кривой и раствор АТФ и перемешивают. После того, как планшет оставляли в течение одного часа при комнатной температуре, добавляют меченое биотином антитело против IgG мыши (фирма Southern Biotechnology Associates, Inc) и оставляли стоять при комнатной температуре в течение одного часа. Впоследствии добавляют меченый стрептавидином SULFO-TAG (фирма Meso Scale Diagnostics, LLC), и планшет оставляют при комнатной температуре в течение одного часа с последующим добавлением буфера для считывания результатов Т (фирма Meso Scale Diagnostics, LLC) и реакцию измеряют с помощью SECTOR Imager 6000 (фирма Meso Scale Diagnostics, LLC). SOFTmax PRO (фирма Molecular Devices) используют для расчета уровней антител в плазме мышей.
Аналогичным образом анти-asialo GM1 антитело (фирма Wako Pure Chemical Industries) растворяют в 1 мл дистиллированной воды (фирма Otsuka Pharmaceutical Factory), а затем разбавляют 4 мл ФСБ. Полученное таким образом анти-asialo GM1 антитело вводят внутрибрюшинно в 91 мыши hIL6R-tgm в дозе 0,1 мл на мышь. Через день получают смешанный раствор, в котором такой же объем Matrigel Matrix (фирма Corning) добавлен к клеточной суспензии hIL6R-Hepal-6, суспендированной до 1 × 108 клеток/мл в сбалансированном солевом растворе Хэнкса (фирма Sigma). Смешанный раствор вводят подкожно в 91 мышь hIL6R-tgm в дозе 0,2 мл на мышь и клетки имплантируют. Диаметр опухоли мышей измеряли штангенциркулем, а объем опухоли определяли как (большая ось) × (малая ось) × (малая ось) × (1/2). Измерение диаметра опухоли проводит индивид, не знающий о составе раствора лекарственного средства, вводимого каждой мыши. Диаметр опухоли и вес тела 91 указанной выше мыши hIL6R-tgm измеряют на 10-й день после трансплантации клеток. На основании полученных значений рандомизируют мышей, которых разделяют в общей сложности на четыре группы по 6 животных, в каждой группе животным вводят через хвостовую вену соответствующий раствор антитела из числа антител, показанных в табл. 67, за исключением H0052-mFa55/L1083-ml0. Доза (мл) введенного раствора антитела составляет (0,01 (мл/г)) × (масса тела (г) каждой мыши). Впоследствии диаметры опухоли и массу тела мышей hIL6R-tgm, сгруппированных указанным выше образом, измеряют таким же образом на 13, 17 и 20 день после трансплантации. На 17-й день после трансплантации четыре раствора антител из растворов антител, представленных в табл.67, за исключением H0002-mFa55 / L1058-ml0, вводят мышам соответствующих групп через хвостовую вену в дозировке, определенной по формуле расчета, описанной выше, на основе измеренных данных о массе тела. Рассчитан средний объем опухоли в каждой группе лечения антителом в каждый день измерения, и на основе полученных данных оценивают противоопухолевую активность in vivo каждого антитела.
В вышеупомянутых экспериментах кровь собирают путем взятия пробы крови из орбитального синуса мышей hIL6R-tgm, разделенных на указанные выше группы, под анестезией изофлураном через один час, 3 и 7 дней после первоначального введения растворов антител. Собранную кровь переносят в серию из восьми пробирок с индивидуальным номером и оставляют на льду. После центрифугирования серий из восьми пробирок с индивидуальным номером в режиме 1900×g в течение 10 мин при 4°С получают супернатант в качестве компонента плазмы и переносят в другие серии из восьми пробирок с индивидуальным номером и хранят при -30°С.
Концентрацию каждого антитела в плазме измеряют методом электрохемилюминесценции (ECL). Раствор hIL6R добавляют в планшет MULTI-ARRAY PR (фирма Meso Scale Diagnostics, LLC) и выстаивают при 37°С в течение одного часа, в результате чего hIL6R иммобилизуется на планшете. Для калибровочной кривой готовят MRAH-mFa55/MRAL-mk0, Н0041-mFa55/L1088-ml0 и H0052-mFa55/L1083-ml0 в плазме с концентрациями 8, 4, 2, 1, 0,5, 0,25 и 0,125 мкг/мл. После добавления 1 М раствора трис-HCl, рН 8,0, содержащего 0,05% Tween20 и 1% БСА, в планшет с иммобилизованным hIL6R, образцы плазмы разбавляют в 250 раз в 0,3 М растворе уксусной кислоты, а также быстро добавляют образцы для калибровочной кривой и затем раствор АТФ и перемешивают. После того, как планшет оставляли в течение одного часа при комнатной температуре, добавляют меченое биотином антитело против IgG мыши (фирма Southern Biotechnology Associates, Inc) и оставляют стоять при комнатной температуре в течение одного часа. Впоследствии добавляют меченый стрептавидином SULFO-TAG (фирма Meso Scale Diagnostics, LLC), и планшет оставляют при комнатной температуре в течение одного часа с последующим добавлением буфера для считывания результатов Т (фирма Meso Scale Diagnostics, LLC) и реакцию измеряют с помощью SECTOR Imager 6000 (фирма Meso Scale Diagnostics, LLC). SOFTmax PRO (фирма Molecular Devices) используют для расчета уровней антител в плазме мышей.
Концентрацию растворимого антигена в плазме измеряют методом электрохемилюминесценции (ECL). Образцы плазмы мыши, разведенные в 50 раз, смешивают с SULFO-TAG (фирма Meso Scale Diagnostics, LLC)-меченым антителом против IL6R человека (фирма R&D Systems), биотин-меченым антителом против IL6R человека (фирма R&D Systems) и избытком антитела против IL6R человека толизумамом (полученным авторами настоящего изобретения), смесь инкубируют в течение ночи при 37°С. После добавления смеси в стандартный 96-луночный блокированный планшет, покрытый стрептавидином (фирма Meso Scale Diagnostics, LLC), с последующим добавлением буфера для считывания результатов, реакцию измеряют на SECTOR Imager 6000 (фирма Meso Scale Diagnostics, LLC). Программное обеспечение SOFTmax PRO (фирма Molecular Devices) используют для расчета концентраций растворимого антигена в плазме мышей.
Сначала сравнивают противоопухолевые эффекты, концентрации антител в плазме и концентрации антигена в плазме IC17HdK-mFa55 / IC17L-mk1, MRAH-mFa55 / MRAL-mk0, H0002-mFa55 / L1058-ml0 и H0041-mFa55 / L1088-ml0. Результаты показаны на рисунках 72, 73 и 74, соответственно.
В настоящем изобретении по сравнению с группой, получавшей изотипический контроль IC17HdK-mFa55 / IC17L-mkl, группы, получавшие MRAH-mFa55 / MRAL-mk0 и H0002-mFa55 / L1058-ml0, не показывают более высоких показателей ингибирования роста опухоли (TGI). С другой стороны, больший показатель TGI наблюдают в группе, получавшей переключаемое антитело H0041-mFa55 / L1088-ml0 (фиг.72). Слабее эффективность препарата наблюдают в группе мышей, получавших MRAH-mFa55 / MRAL-mk0, по сравнению с таковой H0041-mFa55 / L1088-ml0, что позволяет предположить, что MRAH-mFa55 / MRAL-mk0 выводится быстрее, чем H0041-mFa55 / L1088-ml0, и он не может поддерживать концентрацию, достаточную для демонстрации эффективности лекарственного средства (фиг. 73).
Также накопления растворимого hIL6R не наблюдают в группе, обработанной H0002-mFa55 / L1058-ml0. В группе, обработанной H0041-mFa55 / L1083-ml0, накопление растворимого IL6R происходит в меньшем количестве, чем в группе, обработанной MRAH-mFa55 / MRAL-mk0 (фиг. 74).
Затем сравнивают противоопухолевые эффекты, концентрации антител в плазме и концентрации антигена в плазме IC17HdK-mFa55 / IC17L-mk1, MRAH-mFa55 / MRAL-mk0, H0041-mFa55 / L1088-ml0 и H0052-mFa55 / L1083-ml0. Результаты показаны на фиг. 75, 76 и 77 соответственно.
В настоящем исследовании по сравнению с группой, получавшей изотипический контроль IC17HdK-mFa55 / IC17L-mkl, группы, получавшие MRAH-mFa55 / MRAL-mk0, H0041-mFa55 / L1088-ml0 и H0052-mFa55 / L1083-ml0, показывают более высокие показатели ингибирования роста опухоли (TGI) (фиг. 75). С другой стороны, при сравнении их концентраций в плазме MRAH-mFa55 / MRAL-mk0 демонстрирует более быстрое выведение, чем Н0052-mFa55 / L1083-ml0, и пониженную концентрацию в крови (фиг. 76).
Кроме того, по сравнению с MRAH-mFa55 / MRAL-mk0, H0052-mFa55 / L1083-ml0 только накапливает меньше растворимого IL6R (фиг. 77).
Эти результаты позволяют предположить, что у мышей с опухолью переключаемые антитела обладают аналогичным или более сильным противоопухолевым действием, чем непереключаемые антитела; однако, в отличие от непереключаемых антител, переключаемые антитела не связываются с hIL6R в нормальных тканях. Результаты показывают, что даже если рак присутствовал, внеклеточные концентрации АТФ в нормальных тканях и крови недостаточно повышены, чтобы стимулировать связывание переключаемых антител.
Пример 16. АТФ-зависимая нейтрализующая активность анти-PD1 антител, обладающих АТФ-зависимыми связывающими свойствами
Тяжелые цепи и легкие цепи объединяют, как показано в табл. 68, для экспрессии и очистки анти-PD-1 антител методами, известными специалистам в данной области.

Нейтрализующую активность антител, описанных в табл. 68, для взаимодействия PDL1-PD1 оценивают в соответствии с методами, приведенными ниже.
Первоначально антигены mPD1-G1CH2 (SEQ ID NO: 209) и mPDL1-G1dCH2His (SEQ ID NO: 210) были генетически синтезированы и инсертированы в плазмиды для экспрессии в животных. Антигенные белки экспрессируют и очищают описанными ниже методами. Полученные плазмиды вводят методом липофекции в линию FreeStyle 293-F (фирма Invitrogen), полученную из фетальных клеток почки человека (фирма Invitrogen), суспендированную в среде для экспрессии FreeStyle 293 (фирма Invitrogen) при соответствующей плотности, и высевают в колбы. Антигены очищают из культуральных супернатантов клеток, культивируемых в CO2-инкубаторе (37°С, 8% CO2, 125 об/мин) в течение четырех дней методом, известным специалистам в данной области. Спектрофотометры используют для измерения абсорбции очищенных растворов антигенов при 280 нм. Концентрации очищенных антигенов рассчитывают с использованием коэффициентов абсорбции, рассчитанных методом РАСЕ на основе полученных измеренных значений (Protein Science 4, 1995, 2411-2423).
mPD1-G1CH2 (SEQ ID NO: 209), который представляет собой Fc-слитый PD1 мыши, разбавляют до 5 мкг/мл (55 нМ) раствором 0,1 М NaHCO3, 0,05% NaN3, и 100 мкл разбавленного раствора добавляют в 96-луночный планшет, оставляют на ночь при 4°С и иммобилизуют на поверхности планшета. После трехкратной промывки лунок ФСБ и 0,1% Tween20 в каждую лунку добавляли 250 мкл раствора БСА, разбавленного до 2% ФСБ, для блокирования поверхностей планшета. Затем трижды промывают.mPDLl-G1dCH2His (SEQ ID NO: 210), который представляет собой PDL1 мыши, слитый с Fc антитела и His-меткой, разводят в ФСБ до конечной концентрации 55 нМ; раствор антител, описанных в табл. 68, разбавленный до конечных концентраций 6,25, 1,56, 0,390, 0,0977, 0,0061 и 0 мкг/мл; и раствор АТФ, разбавленный до конечных концентраций 0, 1, 10 и 100 мкМ, смешивают до общего количества 100 мкл. Затем смесь добавляют в каждую лунку и оставляют при 37°С в течение одного часа. Затем каждую лунку трижды промывают 0,1% Tween20 и ФСБ, которые приготовлены с такой же концентрацией АТФ, что и концентрация АТФ в растворах, добавленных в каждую лунку. Анти-His-tag mAb-HRP-Direct (фирма MBL Life Science) разводят в 10000 раз в блокирующем буфере, чтобы он содержал ту же концентрацию АТФ, что и концентрация АТФ в растворах, добавленных в каждую лунку, из которых 100 мкл добавляют в каждую лунку, и оставляют при 37°С в течение одного часа. Затем каждую лунку трижды промывают 0,1% Tween20 и ФСБ, которые приготовлены с такой же концентрацией АТФ, что и концентрация АТФ в растворах, добавленных в каждую лунку. Затем в каждую лунку добавляли 100 мкл раствора ТМВ, и планшет оставляют при 37°С в течение одного часа, и в каждую лунку добавляют 50 мкл 1 М H2SO4 для остановки реакции. Затем определяют абсорбцию в каждой лунки при 450 нм на микропланшетном ридере оптической плотности (фирма Wako Sunrise).
Величину абсорбции в лунке, свободной от антител, при тех же условиях концентрации АТФ устанавливают как 100% связывания PD1-PDL1, и оценивают степень, до которой скорость связывания снижается при добавлении антитела. Результаты показаны на фиг. 78 и 79.
Результаты показывают, что на нейтрализующую активность mPD1F2VH-mF18 / mPD1F2VL-mkl при взаимодействии PD1 / PDL-1 не влияют концентрации АТФ, тогда как нейтрализующая активность H5029-mFa31 / L3021-ml0 и H5041-mFa31 / L3021-ml0 становится сильнее при более высокой концентрации АТФ в анализах. Результаты подтвердили, что H5029-mFa31 / L3021-ml0 и H5041-mFa31 / L3021-ml0 обладают АТФ-зависимой связывающей активностью и нейтрализующей активностью.
Систему анализа люциферазы (фирма Promega) используют для измерения нейтрализующей активности in vitro. Для эффекторной клетки используют вспомогательные клетки hPDL1-CHO для биотеста блокады PD-1 / PD-L1, Core Kit (фирма Promega). В качестве клетки-мишени используют клетки Jurkat-NFAT-Luc2-mPD1, полученные авторами настоящего изобретения. Сначала ген гибридного белка внеклеточного mPD1 и внутриклеточного hPD1 (SEQ ID NO: 214) вводят в клетки Jurkat-NFAT-Luc2 (фирма Promega) методом, известным специалистам в данной области, для получения клеток Jurkat-NFAT-Luc2-mPD1. Взаимодействие hPDL1 на hPDL1-CHO со слитым белком внеклеточного mPD1 и внутриклеточного hPD1 на Jurkat-NFAT-Luc2-mPD1 используют в качестве взаимодействия PD-1 / PDL-1 для оценки нейтрализующей активности in vitro.
В каждую лунку 96-луночного планшета добавляют 100 мкл из вспомогательного набора hPDL1-CHO, который приготовлен до концентрации клеток 4 × 105/ мл в среде, и планшет оставляют на ночь при 37°С. Для культуральной среды, 90% Ham's F-12, 10% ФСБ, применяют 250 мкг генетицина и 200 мкг гигромицина. Супернатанты удаляют из каждой лунки; 20 мкл раствора антител, описанных в табл. 68, разбавляют буфером для анализа до конечной концентрации 7,5 мкг/мл, смешивают с 20 мкл раствора АМФ, разбавленного буфером для анализа до конечных концентраций 0, 1, 10 и 100 мкМ, и 40 мкл раствора клеток NFAT-Luc2-hPD1-mPD1-Jurkat, приготовленного с буфером для анализа, для достижения количества клеток 5хЮ4/лунку. Смесь добавляют в каждую лунку и оставляют при 37°С в течение четырех часов. В качестве буфера для анализа используют 98% RPMI 1640 и 2% ФСТ. Затем планшеты оставляют при комнатной температуре в течение 10 мин и в каждую лунку добавляют по 80 мкл реагента Bio-Glo. Для реагента Bio-Glo используют систему анализа люциферазы Bio-glo (буфер и субстрат). Затем измеряют люминесценцию в каждой лунке с помощью планшет-ридера.
Величину люминесценции в каждой лунке при каждой концентрации АМФ, деленное на величину люминесценции лунки, свободной от антител, представляет собой кратность индукции, и она служит показателем активности каждого антитела по нейтрализации взаимодействия PD- 1 / PDL-1. Результаты показаны на фиг. 80. На фигуре кратность индукции обозначена как относительная световая единица (RLU).
АТФ тестируют так же, как АМФ, и 100 мл hPDL1-CHO, приготовленного в концентрации 4 × 105 / мл в среде, добавляют в каждую лунку 96-луночного планшета, и планшет оставляют при 37°С на ночь. В качестве питательной среды используют 90% Ham's F-12, 10% ФСБ, 250 мкг генетицина и 200 мкг гигромицина. Супернатанты удаляют из каждой лунки аспирацией; 20 мкл раствора антител, описанных в табл. 68, разбавляют буфером для анализа до конечной концентрации 10 мкг/мл, смешивают с 20 мкл раствора АТФ, разбавленного буфером для анализа до конечных концентраций 0, 12,5 и 125 мкМ, и 40 мкл суспензии клеток NFAT-Luc2-hPD1-mPD1-Jurkat, приготовленной с буфером для анализа для достижения количества клеток 5×104/лунку. Смесь добавляют в каждую лунку и оставляют при 37°С на четыре часа. В качестве буфера для анализа используют 98% RPMI 1640 и 2% ФСТ. Затем планшеты оставляют при комнатной температуре в течение 10 мин и в каждую лунку добавляют по 80 мкл реагента Bio-Glo. Для реагента Bio-Glo используют систему анализа люциферазы Bio-glo (буфер и субстрат). Затем измеряют люминесценцию в каждой лунке с помощью планшет-ридера.
Значение люминесценции в каждой лунке при каждой концентрации АМФ, деленное на величину люминесценции в лунке, свободной от антител, представляет собой кратность индукции, и она служит в качестве показателя для оценки активности каждого антитела по нейтрализации взаимодействия PD-1 / PDL-1. Результаты показаны на фиг. 81. На рисунке кратность индукции обозначена как относительная световая единица (RLU).
Пример 17. АТФ-зависимая ADCC-опосредованная противоопухолевая активность анти-PDl антител, обладающих АТФ-зависимыми связывающими свойствами
Тяжелые цепи и легкие цепи объединяют, как показано в табл.69, для экспрессии и очистки анти-PD-1 антител методами, известными специалистам в данной области.
Константные области тяжелой цепи этих антител модифицированы до mFa55, который усиливает активность ADCC путем добавления модификаций, усиливающих связывание FcγRIV с mIgG2a. Следовательно, считают возможным проявить активность ADCC в отношении клеток, экспрессирующих PD-1, и удалить клетки, экспрессирующие PD-1. Антитела используют для оценки противоопухолевой активности in vivo и для оценки элиминации клеток, экспрессирующих PD-1, в соответствии с приведенными ниже методами.
Линию колоректального рака мыши Colon38 (NCI) имплантируют в количестве 5 × 106 с Matrigel (фирма Corning) подкожно в правую часть живота мышей C57BL / 6J (фирма Japan Charles River) для образования солидных опухолей. На четырнадцатый день после имплантации IC17HdK-mFa55 / IC17L-mkl вводят внутривенно (в / в) в дозе 15 мг/кг, mPD1F2VH-mFa55 / mPD1F2VL-mkl в дозе 1,5, 5 и 15 мг/кг и H5041-mFa55/L3023-ml0 в дозе 25,5 и 100 мг/кг, соответственно (п=7 на группу). Кроме того, на 17-й день после трансплантации в группе F£5041-mFa55 / L3023-ml0 мышам вводят те же антитела внутривенно (в / в) в дозах 25,5 и 100 мг/кг, соответственно. В результате противоопухолевый эффект наблюдают во всех группах введения антител, за исключением группы отрицательного контроля (фиг. 82).
Элиминацию PD1-экспрессирующих клеток в опухолевой ткани после введения соответствующих антител оценивают с помощью жидкостной цитометрии. Солидным опухолям Со1оп38 дают возможность сформироваться, как описано выше, и на 14-й день после имплантации IC17HdK-mFa55 / IC17L-mk1 (изотипический контроль) вводят внутривенно (в/в) в дозе 15 мг/кг, mPD1F2VH-mFa55 / mPD1F2VL-mkl в дозе 1,5, 5 и 15 мг/кг, и H5041-mFa55 / L3023-ml0 в дозе 25,5 и 100 мг/кг, соответственно (n=3 на группу). Ткани селезенки и опухоли каждой мыши собирают через три дня после введения антитела. Селезенки измельчают пинцетом, добавляют 10% ФСБ-дополненную среду RPMI 1640 и центрифугируют при 400 × g в течение 5 мин, после чего супернатант удаляют. Добавляют 2 мл лизирующего буфера АСК (фирма Thermo Fisher Scientific) и оставляют при комнатной температуре на 2,5 мин, гемолизированный образец используют в качестве клеток селезенки для анализа методом жидкостной цитометрии. Ткань опухоли измельчают ножницами с последующим добавлением ферментов в соответствии с протоколом набора для диссоциации опухолей Tumor Dissociation Kit (фирма Miltenyi), а затем измельчают с помощью мягкого диссоциатора MACS (фирма Miltenyi Biotec). С добавлением 10% ФСБ-дополненной среды RPMI 1640 ее центрифугируют при 400 × g в течение 5 мин, супернатант удаляют и используют для анализа методом жидкостной цитометрии в качестве опухолевых клеток. Клетки селезенки и опухолевые клетки переносят в планшет с V-образным дном (фирма Corning), центрифугируют при 400 × g в течение 5 мин и супернатант удаляют. Клетки ресуспендируют в 100 мкл реагента, блокирующего FcR (фирма Miltenyi Biotec), разведенного в 10 раз в ФСБ (буфер FACS), содержащем 1% ФСТ и 2 мМ EDTA (фирма Sigma). После инкубирования клеток при комнатной температуре в течение 10 мин, 0,4 мкл BUV737 Anti-Mouse CD3e (фирма BD), 0,1 мкл Zombie Aqua (фирма Biolegend), 0,4 мкл PerCP / Су5.5 Anti-Mouse CD8a (фирма Biolegend), 0,4 мкл РЕ / Су7 Anti-Mouse CD4 (фирма Biolegend), 0, 2 мкл APC-R700 против CD45 мыши (фирма BD), 0,4 мкл FITC против CD278 человека / мыши / крысы (фирма Biolegend), 0,4 мкл АРС / FireTM 750 против CD25 мыши (фирма Biolegend) и 0,4 мкл АРС антимышиного CD279 (фирма Biolegend), который, как уже было подтверждено, не конкурирует с каждым из введенных антител PD1, добавляют в каждую лунку, и буфер FACS добавляют по 20 мкл /лунку. Через 30 мин инкубирования при 4°С добавляют 100 мкл буфера FACS, центрифугируют в режиме 400×g в течение 5 мин и удаляют супернатант. В соответствии с протоколом из набора буферов для окрашивания Foxp3 / транскрипционного фактора (фирма eBioscience), концентрат фиксации / проницаемости и разбавитель фиксации / проницаемости смешивают и по 200 мкл смеси вносят в каждое лунку. После инкубации при 4°С в течение 30 мин и центрифугирования при 400 × g в течение 5 мин супернатант удаляют.Добавляют буфер для пермеабилизации в количестве 200 мкл и после центрифугирования в режиме 400xg в течение 5 мин супернатант удаляют. Эту процедуру промывки повторяют еще раз. eFluor450 Anti-Mouse / Rat Foxp3 (фирма eBioscience) в количестве 0,4 мкл и РЕ Anti-Mouse CD152 (фирма BD) в концентрации 0,4 мкл добавляют в каждую лунку, буфер FACS добавляют в количестве 20 мкл / лунку. После инкубации при 4°С в течение 30 мин добавляют 100 мкл буфера FACS и после центрифугирования при 400 × g в течение 5 мин супернатант удаляют. Буфер для пермеабилизации добавляют в каждую лунку на 200 мкл, и после центрифугирования при 400 × g в течение 5 мин супернатант удаляют. Образцы ресуспендируют в 200 мкл буфера FACS и измеряют в жидкостном цитометре FACS Fortessa (фирма BD). Для анализа экспрессии используют программное обеспечение FlowJo. С04-положительные клетки, проходящие в популяции клеток, подлежат анализу, для анализа экспрессии Foxp3 и PD1. Уровень экспрессии PD1 в популяции клеток CD4+ Foxp3- рассчитывают по интенсивности флуоресценции. В результате уровень экспрессии PD1 как в клетках селезенки, так и в опухолевых клетках проявляет тенденцию к снижению в группах, в которых мышам вводили 5 и 15 мг/кг mPD1F2VH-mFa55 / mPD1F2VL-mk1, но не в группах, в которых вводили 25,5 и 100 мг/кг H5041-mFa55 / L3023-ml0, где уровень экспрессии PD1 имел тенденцию к снижению только в опухолевых клетках (фиг. 83А и Б). Подтверждают, что все переключаемые антитела, которые связываются с PD1 АТФ-зависимым образом, проявляют лекарственную эффективность против опухоли, в то время как системный ответ (снижение уровня экспрессии PD1) не происходит, и было подтверждено, что переключаемые антитела способны проявлять специфическую активность в участке опухоли.
Пример 18. Анализ кристаллической структуры hIL6R-связывающих антител, обладающих АТФ-зависимыми связывающими свойствами
Проводят рентгеноструктурный анализ комплекса АТФ, внеклеточного домена hIL6R (shIL6R) и фрагмента Fab антитела H0041L1088, связывающегося с рецептором человека IL6 (hIL6R) (переключатель АТФ), полученного в примере 13.
(18-1) Получение антител H0041L1088 полной длины
Получение и очистка антител полной длины H0041L1088 (VH: SEQ ID NO: 194; СН: SEQ ID NO: 217, VL: SEQ ID NO: 195, CL: SEQ ID NO: 189), имеющих формат IgG1 человека, выполняют методами, известные специалистам в данной области.
(18-2) Получение фрагментов Fab H0041L1088
Образец антитела полной длины H0041L1088 фрагментируют на Fab и Fc с использованием папаина (фирма SIGMA-ALDRICH, 10108014001) в условиях 35°С, примерно в течение 18 ч, а затем подвергают очистке на колонке на HiTrap SP HP 1 мл (фирма GE Healthcare) и HiTrap MabSelect SuRe 1 мл (фирма GE Healthcare), а также очистке SEC с помощью HiLoad 16/600 Superdex 200 пг (фирма GE Healthcare) для получения образцов Fab.
(18-3) Получение shIL6R
Основываясь на аминокислотной последовательности UniProtKB: Р08887 (IL6RA HUMAN) (SEQ ID NO: 213), его домены 2 и 3 (аминокислоты 111-320) были получены генетически и включены в векторы экспрессии. Для синтеза гена сигнальная последовательность для экспрессии секреции добавлена на N-конце, метки FLAG tag + His×8 tag добавлены на С-конце для очистки, и была введена модификация C193S для устранения несовершенства пар Cys-Cys. который является результатом удаления домена 1. Экспрессию белка с помощью системы экспрессии Expi293TM (фирма Thermo Fisher Scientific) выполняют в присутствии кифуненсина (фирма Santa Cruz Biotechnology) с использованием полученных плазмид экспрессии и супернатанта культуры экспрессии, содержащего исследуемый белок. Из этого образца shIL6R получают путем аффинной очистки с помощью колонки для очистки cOmplete™ His-Tag Purification объемом 5 мл (фирма SIGMA-ALDRICH), а также очистки SEC с помощью HiLoad 16/600 Superdex 200 пг (фирма GE Healthcare).
(18-4) Получение тройного комплекса H0041L1088 Fab-shIL6R-ATO
В образец shIL6R добавляют эндогликозидазу F1 (EndoF1) для расщепления цепи сахара N-типа и энтерокиназу (фирма SIGMA-ALDRICH, 11334115001) для отщепления метки Hisx8 tag, оставляют стоять при комнатной температуре в течение одного дня и затем проводят сквозную обработку в колонке HisTrap EXCEL объемом 1 мл (фирма GE Healthcare) и SEC очистку Superdex 200 Increase 10/300 GL (фирма GE Healthcare). После добавления образца Fab H0041L1088 полученную очищенную фракцию обогащают ультрафильтрацией и очисткой SEC с помощью Superdex 200 Increase 10/300 GL (фирма GE Healthcare) с использованием 20 мМ HEPES, рН 7,3, 100 мМ NaCl, 0,5 мМ АТФ в качестве буфера для подготовки сложных (комплексных) образцов. Комплексные образцы для кристаллизации получают путем обогащения ультрафильтрацией полученных очищенных фракций.
(18-5) Получение кристаллов тройного комплекса H0041L1088 Fab-shIL6R-АТФ
Кристаллизацию методом диффузии паров в сидячей капле проводят при температуре 21°С с использованием комплексных образцов для кристаллизации в резервуарных условиях в растворе 60 мМ Трис, рН 7,5, 12,0% мас./об. полиэтиленгликоля 1500 (полиэтиленгликоль 1500), 60 мМ сульфата аммония; были получены кристаллы, пригодные для рентгеноструктурного анализа.
(18-6) Измерение данных дифракции рентгеновских лучей от кристаллов тройного комплекса Fab-shIL6R-ATP H0041L1088 и определение кристаллических структур
Полученные кристаллы погружают в раствор 68 мМ Трис, рН 7,5, 13,6% мас./об. полиэтиленгликоля 1500 (полиэтиленгликоль 1500), 68 мМ сульфата аммония, 14,6% этиленгликоля и 0,375 мМ АТФ, и затем замораживали в жидком азоте. Данные рентгеновской дифрактометрии измеряют в установке радиационного света Научно-исследовательской организации по ускорителям высоких энергий (High Energy Accelerator Research Organization), Photon Factory BL-17A. Во время измерения кристаллы постоянно помещали в поток азота с температурой -178°С для поддержания замороженного состояния. Полученные дифрагированные изображения обрабатывают с помощью autoPROC ((Acta Cryst. D67, 2011, 293-302) и получают данные по интенсивности дифракции с разрешением до 2,76 Ангстрем.
Используя полученные данные интенсивности дифракции рентгеновских лучей, исходную структуру определяют методом молекулярного замещения с использованием Phaser (J. Appl. Cryst., 40, 2007, 658-674), используя кристаллические структуры известных Fab и shIL6R PDB. ID=1N26 как поисковые модели. Впоследствии построение и уточнение модели с помощью coot (Acta Cryst. D66, 2010, 486-501) и refmacS (Acta Cryst. D67, 2011, 355-367), a также phenix.refine (Acta Cryst. D68, 2012, 352- 367) повторяют, что приводит к окончательному уточнению координат. Кристаллографическая статистика представлена в табл.70. Следует отметить, что среди координат кристаллической структуры, номера остатков аминокислот в Fab даны по схеме нумерации Kabat, а номера остатков аминокислот в антигене, shIL6R, даны, чтобы соответствовать номерам остатков аминокислот в UniProtKB: Р08887 (I6TRA_HUMAN). Хотя есть два комплекса в асимметричных единицах этого кристалла, в домене 2 shIL6R наблюдают специализированную димеризацию, называемую перестановкой доменов. Такие димеры не обнаруживаются в кристаллических структурах shIL6R PDB ID=1N26, что делает их уникальными для используемых в настоящем изобретении конструкций shIL6R.
(18-71 Взаимодействие между H0041L1088 и АТФ
На фиг. 84, АТФ распознается в первую очередь тяжелой цепью антитела. В частности, часть аденинового кольца АТФ распознается каждой боковой цепью CDR1 тяжелой цепи антитела: Т33, CDR3: Y95, L98, N100B и W100C, а также каждой основной цепью G96, L100A и W100C. В частности, водородные связи образуются между карбонильным кислородом основной цепи G96 и L100A и NH2 в положении 6 NH2 АТФ, а также между NH группой амида основной цепи W100C и N положением 1 АТФ; и взаимодействия, такие как СН-пи (π) и π-π образуется между боковыми цепями Y95, L98 и W100C и остатком аденинового кольца. Антитело надежно распознает фрагмент аденинового кольца АТФ. Фрагмент рибозы распознается ван-дер-ваальсовыми взаимодействиями с соответствующими боковыми цепями CDR1 тяжелой цепи: Т33 и CDR2: Н56 и Y58. Фрагмент трифосфорной группы распознается боковыми цепями CDR2 тяжелой цепи: S52, S52A, Y55 и Н56, а также основной цепью S52A и Q53. В частности, боковая цепь S52 и боковая цепь S52A, а также NH-группа основной цепи и NH-группа основной цепи Q53 образуют плотную сеть водородных связей с фрагментом трифосфорной группы, которые играют решающую роль в распознавании фрагментов трифосфорной группы.
(18-8) Взаимодействие между H0041L1088 и hIL6R
На основании настоящей кристаллической структуры аминокислотные остатки shIL6R, содержащие один или несколько неводородных атомов, расположенных на расстояниях в пределах 4,2 Ангстрем от Fab или АТФ H0041L1088, выбраны в качестве остатков эпитопа, и они показаны на аминокислотной последовательности shIL6R. (фиг. 85). Детали взаимодействий этих остатков эпитопа с антителом показаны на фиг. 86 и 87. Эти остатки эпитопа образуют ван-дер-ваальсовы взаимодействия, водородные связи, электростатические взаимодействия и т.д. с остатками CDR1 легкой цепи: D27B, G28, D29, А31 и Y32, и CDR3: R91, S92, Р93, G94 и Р95, и CDR2 тяжелой цепи: Н56 и Y58 и CDR3: L98, Y99, N100B и W100C антитела; и они прочно связаны с антителом.
(18-9) АТФ-зависимый антигенсвязывающий механизм
Кроме того, при связывании Fab-shIL6R H0041L1088, как показано на фиг. 86 и 87, образуется большой межмолекулярный контакт между F298 shIL6R и АТФ, который связывается с Fab H0041L1088. Поскольку это взаимодействие теряется в АТФ-несвязанном состоянии, предполагают, что это прямое взаимодействие между антигеном и АТФ, который связывается с Fab H0041L1088, является основным фактором АТФ-зависимого связывания. Полагают, что структурно АТФ способствует структурной стабилизации CDR3 тяжелой цепи за счет водородных связей и ван-дер-ваальсовых взаимодействий с остатками CDR3 тяжелой цепи. Полагают, что стабилизация структуры CDR3 тяжелой цепи в ее антиген-связанной форме путем связывания АТФ приводит к значительному усилению прямого взаимодействия CDR3: L98 или Y99 тяжелой цепи с shIL6R, тем самым внося вклад в АТФ-зависимое связывание.
Пример 19. Повышение агонистической активности модифицированными анти-СР137 антителами
(19-1) Получение антител для оценки
Что касается случая, когда различные модификации аминокислот для увеличения pi были введены в константную область тяжелой цепи с повышенной активностью связывания с FcγRIIb, оценивают влияние таких модификаций аминокислот на активность агониста CD137 и на кинетику плазмы и эффективность лекарственного средства в hCD137KI / mFcγR2bKO/hFcγR2bTg # 90. Во-первых, как описано в примере 7-1, примере 7-2 и примере 7-3, получают A375-MY201aPh / B167-Lamlib, A375-SCF041aPh / B167-Lamlib, A375-SCF057aPh / B167-Lamlib и IC17HdK-MY201aPh / IC17L-k0.
(19-2) Оценка in vitro АТФ-зависимой агонистической активности CD137 модифицированных антител против CD137 человека с использованием анализа репортерного гена 4-1 ВВ Jurkat
Что касается случая, когда различные модификации аминокислот для увеличения pi введены в константную область тяжелой цепи с повышенной активностью связывания с FcγRIIb, чтобы оценить влияние таких модификаций аминокислот на активность агониста CD137 оценивают активность агониста CD137 A375-MY201aPh / B167-Lamlib, A375-SCF041aPh / B167-Lamlib, A375-SCF057aPh / B167-Lamlib и IC17HdK-MY201aPh / IC17L-k0.
Клеточную линию GloResponseTM NF-kB-Luc2 / 4-IBB Jurkat (фирма Promega, CS196004) используют для измерения активности вырабатываемых антител in vitro. В каждую лунку 96-луночного планшета добавляют 200 мкл клеток FcγRIIB СНО-К1 (фирма Promega), приготовленных в концентрации 5 × 104 / мл с культуральной средой, и планшет оставляют на ночь в инкубаторе в атмосфере 5% CO2 при 37°С. В качестве питательной среды используют культуральную среду СНО (90% Ham's F12, 10% ФСБ). Затем всю культуральную среду удаляют отсасыванием, после чего 25 мкл клеточной линии GloResponseTM NF-kB-Luc2 / 4-1 ВВ Jurkat, приготовленной с концентрацией 2 × 106 клеток/мл со средой для анализа (99% RPMI, 1% ФСБ) добавляют в каждую лунку. Затем добавляют 25 мкл каждого из растворов антител, разбавленных аналитической средой до конечной концентрации 0, 0,001, 0,01, 0,1, 1 и 10 мкг/мл. Потом добавляют 25 мкл раствора АТФ, разбавленного аналитической средой до конечной концентрации 250 мкМ. Планшет оставляют на 6 ч в инкубаторе в атмосфере 5% CO2 при 37°С, а затем оставляют на 15 мин при комнатной температуре, и в каждую лунку добавляют 75 мкл реагента Bio-Glo. Для реагента Bio-Glo используют систему анализа люциферазы Bio-Glo (буфер и субстрат). После этого с помощью планшет-ридера определяют относительную световую единицу каждой лунки. Величину люминесценции в каждой лунке, деленную на величину люминесценции в лунке без добавления антител, определяют как «относительную световую единицу», и она служит индикатором для оценки активности агониста CD137 каждого антитела.
Результаты показаны на фиг. 88. Результаты показывают, что агонистическая активность CD 137 увеличивается путем введения различных модификаций аминокислот для увеличения pI
(19-3) Фармакокинетическое исследование на мышах модифицированных антител против CD137 человека
(Т9-3-П Получение мыши hCD137KI/mFcγR2bKO/hFcγR2bTg # 90
Сначала получают мышь с нокином CD137 человека, в которой ген CD137 мыши заменен геном CD137 человека, путем введения вектора замены гена CD137 человека в эмбриональные стволовые клетки мыши (ES-клетки) вместе с нуклеазой цинкового пальца (ZFN), который нацелен на CD137 мыши. Затем мРНК ZFN, нацеленную на ген Fcgr2b мыши, путем микроинъекции вводят оплодотворенным яйцеклеткам мышей, и мышей с нокаутом Fcgr2b получают путем отбора мышей, которым была введена мутация в целевой сайт. роме того, вектор ВАС, в котором клонирован ген FCGR2B человека, путем микроинъекции вводят оплодотворенным яйцеклеткам мышей, и путем отбора из мышей, полученных из этих клеток, тех, у которых была введена область генома гена FCGR2B человека, получают трансгенную мышь с FCGR2B человека (Iwayanagi. с соавт., J Immunol, 195, 2015, 3198-3205).
Путем скрещивания мышей трех вышеуказанных линий получают «трансгенную мышь с нокином Fcgr2b CD137 человека и нокаутом FCGR2B человека». Эту мышь обозначают как мышь hCD137KI / mFcγR2bKO / hFcγR2bTg # 90.
(19-3-2) Измерение концентрации антитела против CD137 человека в плазме модельных мышей hCD137KI/mFcγR2bKO/hFcγR2bTg#90
Соответствующие антитела против CD137 человека вводят внутривенно в виде однократной дозы мышам hCD137KI/mFcγR2bKO/hFcγR2bTg#90, как показано в табл. 1. Кровь собирают несколько раз в течение времени от 5 мин до 28 дней после введения. Полученную кровь центрифугируют для отделения плазмы. Плазму хранят в морозильной камере при температуре ниже -20°С до проведения измерения.

Концентрацию в плазме каждого антитела против CD137 человека измеряют методом электрохемилюминесценции (ECL, electrochemiluminescence). hCD137 (фирма Sino Biological Inc.) разбавляют ФСБ (-) и добавляют в 96-луночный планшет MULTI-ARRAY (фирма Meso Scale Diagnostics, LLC). Планшет с добавлением hCD137 встряхивают в течение 1 ч при комнатной температуре, и hCD137 иммобилизуют на планшете. Затем для блокирования добавляют раствор ФСБ, содержащий 1 мМ АДФ, 1% БСА и 0,05% Tween-20, и планшет встряхивают в течение 1 ч при комнатной температуре. Калибровочные кривые для соответствующих антител против CD137 человека были построены при концентрациях в плазме 640, 320, 160, 80, 40, 20 и 10 нг/мл. Образцы плазмы, разбавленные раствором ФСБ, содержащим 1 мМ АДФ, 1% БСА и 0,05% Tween-20, и образцы для калибровочной кривой добавляют в планшет с иммобилизованным hCD137. Затем планшет встряхивают в течение 1 ч при комнатной температуре, а затем в качестве вторичного антитела добавляют антитело, описанное в WO 2019112027, которое специфически распознает константные области антител против CD137 человека. Планшет дополнительно встряхивают в течение 1 ч при комнатной температуре и затем добавляют в него меченые SULFO-TAG козьи анти-кроличьи антитела (фирма Meso Scale Diagnostics, LLC). Планшет дополнительно встряхивают в течение 1 ч при комнатной температуре и затем добавляют к нему меченые SULFO-TAG козьи анти-кроличьи антитела (фирма Meso Scale Diagnostics, LLC), разведенные двукратно и добавляют 1 мМ АДФ. Концентрацию каждого антитела в плазме мыши измеряют путем обнаружения SULFO-TAG с помощью SECTOR Imager (фирма Meso Scale Diagnostics, LLC). Концентрацию каждого антитела в плазме мыши рассчитывали с помощью SOFTmax PRO (фирма Molecular Devices).
Результаты показаны на фиг. 89. A375-SCF041aPh/B167-Lamlib демонстрирует более быструю элиминацию чем A375-MY201aPh/B167-Lamlib. Считается, что это связано с тем, что константная область тяжелой цепи А375-SCF041aPh/B167-Lamlib введена с аминокислотными модификациями, которые увеличивают pi.
(19-4) Оценка лекарственной эффективности модифицированных анти-CD137 человека антител у мышей
(19-4-1) Получение клеточных линий и мышиной модели, трансплантированной сингенной опухолевой линией
Используют клон LLC1/OVA/GPC3 линии клеток С5 (LLC1/OVA/GPC3), полученных путем интродукции плазмид экспрессии куриного овальбумина (OVA) и человеческого глипикана-3 (GPC3) в клетки рака легких мыши, полученные из клеток клеточной линии LLC1 [LL/2 (обозначение: LLC1), дистрибьютор: фирма АТСС, номер в каталоге: CRL-1642]. Использовали мышь hCD137KI/mFcγR2bKO/hFcγR2bTg#90 (возраст 11 недель, самка), описанную выше в (19-3-1). Клеточную линию LLC1/OVA/GPC3 поддерживают и пассируют в среде RPMI1640 (фирма Sigma-Aldrich, Co. LLC), содержащей 9,8% фетальной бычьей сыворотки (фирма Sigma-Aldrich, Co. LLC), 0,44 мг/мл G418 (фирма Nacalai Tesque, Inc.) и 0,88 мг/мл зеоцина (фирма Thermo Fisher Scientific, Inc.). Клеточную линию LLC1/OVA/GPC3 трансплантируют подкожно в брюшную полость мыши, и считают, что модель сформировалась, когда объем опухоли достигал примерно 250-500 мм3 После формирования модели мышей с трансплантированной клеточной линией LLC1/OVA/GPC3 группируют и затем вводят носитель и соответствующие антит-CD137 человека антитела.
(19-4-2) Приготовление и введение фармацевтических агентов для введения и измерения опухолей
A375-MY201aPh/B167-Lamlib или A375-SCF041aPh /B167-Lamlib, приготовленные в дозировках, указанных в таблице 72, с ФСБ, содержащим 0,05% Tween-20, вводят в модель с трансплантированной клеточной линией LLC1/OVA/GPC3 на 11-е и 14-е сутки после трансплантации опухоли. ФСБ, содержащий 0,05% Tween-20, вводят в группе контроля. Приготовленную жидкость для введения вводят в дозе 10 мл/кг в хвостовую вену.
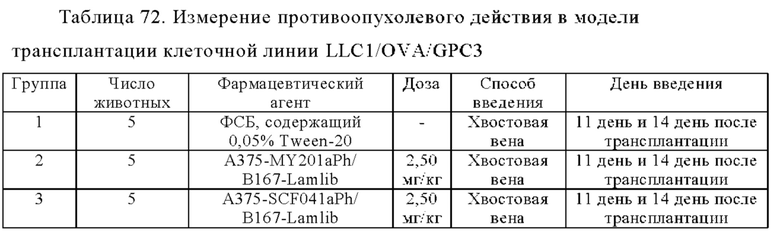
Для оценки противоопухолевого эффекта объем опухоли измеряют 1-2 раза в неделю. Объем опухоли рассчитывают по следующему уравнению. Объем опухоли (мм3) = длина (мм) × ширина (мм) × ширина (мм)/2
Таким образом, результаты примера (19-3-2) показывают, что изменение концентрации в крови у мышей, которым вводили A375-SCF041aPh/B167-Lamlib, меньше по сравнению с таковым для A375-MY201aPh/B167-Lamlib; однако A375-SCF041aPh/B167-Lamlib проявляет более сильный противоопухолевый эффект, чем A375-MY201aPh/B167-Lamlib (фиг. 90).
Основываясь на приведенных выше результатах на данной мышиной модели, противоопухолевый эффект анти-CD137 человека антитела увеличивают путем введения модификаций аминокислот, которые увеличивают pI в константной области тяжелой цепи.
Пример 20. Повышение агонистической активности модифицированными антителами против CD3 человека
Исследуют повышение агонистической активности анти-C3 антитела путем комбинирования аминокислотных модификаций для увеличения pi и константной области тяжелой цепи с повышенной связывающей активностью в отношении FcγRIIb.
Используют SCF057aPh, который получают путем введения аминокислотной модификации Q311R/P343R для увеличения pI до MY201aPh и MY201aPh, которые представляют собой константные области тяжелой цепи с повышенной связывающей активностью в отношении FcγRIIb, полученного в примере 7-1. Кроме того, в качестве константной области нативного IgG1 человека используют G1T6 (SEQ ID NO: 223). Соответствующие анти-CD3 человека антитела получают с использованием TR01H113 (SEQ ID NO: 224) в качестве вариабельной области тяжелой цепи; MY201aPh, SCF057aPh или G1T6 в качестве константной области тяжелой цепи; и L0011-k0 (SEQ ID NO: 225) в качестве легкой цепи анти-CD3 человека антитела. IC17HdK-MY201aPh/IC17L-k0 используют в качестве отрицательного контроля.
Bioassay (NFAT) (фирма Promega, CS176401) используют для активации Т-клеток для измерения in vitro активности продуцируемых антител. В каждую лунку 96-луночного планшета добавляют по 200 мкл клеток FcγRIIB СНО-K1 (фирма Promega), приготовленных в концентрации 5 × 104/мл в культуральной среде, и планшет оставляют на ночь в инкубаторе в атмосфере 5% CO2 при 37°С. В качестве питательной среды используют культуральную среду СНО (90% Ham's F12, 10% ФСБ). Затем всю культуральную среду удаляют отсасыванием, затем в каждую лунку добавляют по 25 мкл суспензии клеток NFAT-RE-Luc2, приготовленных в концентрацией 2 × 106/мл со средой для анализа (99% RPMI, 1% ФСБ). Затем добавляют по 25 мкл каждого раствора антител, разбавленного аналитической средой, до конечной концентрации 0, 0,0001, 0,001, 0,01, 0,1, 1 и 10 мкг/мл. Планшет оставили на 6 ч в инкубаторе в атмосфере 5% CO2 при 37°С, затем оставляли на 15 мин при комнатной температуре, потом в каждую лунку добавляли 75 мкл реагента Bio-Glo. Для реагента Bio-Glo используют систему анализа люциферазы Bio-Glo (буфер и субстрат). После этого с помощью планшет-ридера определяют относительную световую единицу каждой лунки. Величину люминесценции каждой лунки, деленную на величину люминесценции лунки без добавления антител, определяют как «относительную световую единицу», и она служит индикатором для оценки активности агониста CD3 каждого антитела.
Результаты показаны на фиг. 91. По сравнению с TR01H113-G1T6/L0011-k0, имеющим константную область нативного IgG1 человека, TR01H113-MY201aPh/L0011-k0, который имеет повышенное связывание с FcγRIIb, показал более высокую активность агониста CD3. TR01H113-SCF057aPh/L0011-k0 с произведенными аминокислотными модификациями, которые увеличивают pi, показал более высокую агонистическую активность CD3 по сравнению с TR01H113-MY201aPh/L0011-k0 до введения аминокислотных модификаций.
Промышленное применение
Описанные анти-CD137 антигенсвязывающие молекулы и способы их использования применимы для разработки, получения, поставки и применения фармацевтических препаратов, которые обладают эффектом активации иммунных клеток, цитотоксическим действием или противоопухолевым действием, но оказывают слабое воздействие на неопухолевые ткани, такие как нормальные ткани, и вызывают незначительное число побочных эффектов.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ЧУГАЙ СЕЙЯКУ КАБУСИКИ КАЙСЯ
<120> АНТИ-CD137 АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ И ИХ ПРИМЕНЕНИЕ
<130> C1-A1821P
<150> JP 2018-152126
<151> 2018-08-10
<160> 225
<170> PatentIn version 3.5
<210> 1
<211> 255
<212> PRT
<213> Homo sapiens
<400> 1
Met Gly Asn Ser Cys Tyr Asn Ile Val Ala Thr Leu Leu Leu Val Leu
1 5 10 15
Asn Phe Glu Arg Thr Arg Ser Leu Gln Asp Pro Cys Ser Asn Cys Pro
20 25 30
Ala Gly Thr Phe Cys Asp Asn Asn Arg Asn Gln Ile Cys Ser Pro Cys
35 40 45
Pro Pro Asn Ser Phe Ser Ser Ala Gly Gly Gln Arg Thr Cys Asp Ile
50 55 60
Cys Arg Gln Cys Lys Gly Val Phe Arg Thr Arg Lys Glu Cys Ser Ser
65 70 75 80
Thr Ser Asn Ala Glu Cys Asp Cys Thr Pro Gly Phe His Cys Leu Gly
85 90 95
Ala Gly Cys Ser Met Cys Glu Gln Asp Cys Lys Gln Gly Gln Glu Leu
100 105 110
Thr Lys Lys Gly Cys Lys Asp Cys Cys Phe Gly Thr Phe Asn Asp Gln
115 120 125
Lys Arg Gly Ile Cys Arg Pro Trp Thr Asn Cys Ser Leu Asp Gly Lys
130 135 140
Ser Val Leu Val Asn Gly Thr Lys Glu Arg Asp Val Val Cys Gly Pro
145 150 155 160
Ser Pro Ala Asp Leu Ser Pro Gly Ala Ser Ser Val Thr Pro Pro Ala
165 170 175
Pro Ala Arg Glu Pro Gly His Ser Pro Gln Ile Ile Ser Phe Phe Leu
180 185 190
Ala Leu Thr Ser Thr Ala Leu Leu Phe Leu Leu Phe Phe Leu Thr Leu
195 200 205
Arg Phe Ser Val Val Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe
210 215 220
Lys Gln Pro Phe Met Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly
225 230 235 240
Cys Ser Cys Arg Phe Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu
245 250 255
<210> 2
<211> 164
<212> PRT
<213> Homo sapiens
<400> 2
Ser Leu Gln Asp Pro Cys Ser Asn Cys Pro Ala Gly Thr Phe Cys Asp
1 5 10 15
Asn Asn Arg Asn Gln Ile Cys Ser Pro Cys Pro Pro Asn Ser Phe Ser
20 25 30
Ser Ala Gly Gly Gln Arg Thr Cys Asp Ile Cys Arg Gln Cys Lys Gly
35 40 45
Val Phe Arg Thr Arg Lys Glu Cys Ser Ser Thr Ser Asn Ala Glu Cys
50 55 60
Asp Cys Thr Pro Gly Phe His Cys Leu Gly Ala Gly Cys Ser Met Cys
65 70 75 80
Glu Gln Asp Cys Lys Gln Gly Gln Glu Leu Thr Lys Lys Gly Cys Lys
85 90 95
Asp Cys Cys Phe Gly Thr Phe Asn Asp Gln Lys Arg Gly Ile Cys Arg
100 105 110
Pro Trp Thr Asn Cys Ser Leu Asp Gly Lys Ser Val Leu Val Asn Gly
115 120 125
Thr Lys Glu Arg Asp Val Val Cys Gly Pro Ser Pro Ala Asp Leu Ser
130 135 140
Pro Gly Ala Ser Ser Val Thr Pro Pro Ala Pro Ala Arg Glu Pro Gly
145 150 155 160
His Ser Pro Gln
<210> 3
<211> 256
<212> PRT
<213> Mus musculus
<400> 3
Met Gly Asn Asn Cys Tyr Asn Val Val Val Ile Val Leu Leu Leu Val
1 5 10 15
Gly Cys Glu Lys Val Gly Ala Val Gln Asn Ser Cys Asp Asn Cys Gln
20 25 30
Pro Gly Thr Phe Cys Arg Lys Tyr Asn Pro Val Cys Lys Ser Cys Pro
35 40 45
Pro Ser Thr Phe Ser Ser Ile Gly Gly Gln Pro Asn Cys Asn Ile Cys
50 55 60
Arg Val Cys Ala Gly Tyr Phe Arg Phe Lys Lys Phe Cys Ser Ser Thr
65 70 75 80
His Asn Ala Glu Cys Glu Cys Ile Glu Gly Phe His Cys Leu Gly Pro
85 90 95
Gln Cys Thr Arg Cys Glu Lys Asp Cys Arg Pro Gly Gln Glu Leu Thr
100 105 110
Lys Gln Gly Cys Lys Thr Cys Ser Leu Gly Thr Phe Asn Asp Gln Asn
115 120 125
Gly Thr Gly Val Cys Arg Pro Trp Thr Asn Cys Ser Leu Asp Gly Arg
130 135 140
Ser Val Leu Lys Thr Gly Thr Thr Glu Lys Asp Val Val Cys Gly Pro
145 150 155 160
Pro Val Val Ser Phe Ser Pro Ser Thr Thr Ile Ser Val Thr Pro Glu
165 170 175
Gly Gly Pro Gly Gly His Ser Leu Gln Val Leu Thr Leu Phe Leu Ala
180 185 190
Leu Thr Ser Ala Leu Leu Leu Ala Leu Ile Phe Ile Thr Leu Leu Phe
195 200 205
Ser Val Leu Lys Trp Ile Arg Lys Lys Phe Pro His Ile Phe Lys Gln
210 215 220
Pro Phe Lys Lys Thr Thr Gly Ala Ala Gln Glu Glu Asp Ala Cys Ser
225 230 235 240
Cys Arg Cys Pro Gln Glu Glu Glu Gly Gly Gly Gly Gly Tyr Glu Leu
245 250 255
<210> 4
<211> 165
<212> PRT
<213> Mus musculus
<400> 4
Ala Val Gln Asn Ser Cys Asp Asn Cys Gln Pro Gly Thr Phe Cys Arg
1 5 10 15
Lys Tyr Asn Pro Val Cys Lys Ser Cys Pro Pro Ser Thr Phe Ser Ser
20 25 30
Ile Gly Gly Gln Pro Asn Cys Asn Ile Cys Arg Val Cys Ala Gly Tyr
35 40 45
Phe Arg Phe Lys Lys Phe Cys Ser Ser Thr His Asn Ala Glu Cys Glu
50 55 60
Cys Ile Glu Gly Phe His Cys Leu Gly Pro Gln Cys Thr Arg Cys Glu
65 70 75 80
Lys Asp Cys Arg Pro Gly Gln Glu Leu Thr Lys Gln Gly Cys Lys Thr
85 90 95
Cys Ser Leu Gly Thr Phe Asn Asp Gln Asn Gly Thr Gly Val Cys Arg
100 105 110
Pro Trp Thr Asn Cys Ser Leu Asp Gly Arg Ser Val Leu Lys Thr Gly
115 120 125
Thr Thr Glu Lys Asp Val Val Cys Gly Pro Pro Val Val Ser Phe Ser
130 135 140
Pro Ser Thr Thr Ile Ser Val Thr Pro Glu Gly Gly Pro Gly Gly His
145 150 155 160
Ser Leu Gln Val Leu
165
<210> 5
<211> 254
<212> PRT
<213> Macaca fascicularis
<400> 5
Met Gly Asn Ser Cys Tyr Asn Ile Val Ala Thr Leu Leu Leu Val Leu
1 5 10 15
Asn Phe Glu Arg Thr Arg Ser Leu Gln Asp Leu Cys Ser Asn Cys Pro
20 25 30
Ala Gly Thr Phe Cys Asp Asn Asn Arg Ser Gln Ile Cys Ser Pro Cys
35 40 45
Pro Pro Asn Ser Phe Ser Ser Ala Gly Gly Gln Arg Thr Cys Asp Ile
50 55 60
Cys Arg Gln Cys Lys Gly Val Phe Lys Thr Arg Lys Glu Cys Ser Ser
65 70 75 80
Thr Ser Asn Ala Glu Cys Asp Cys Ile Ser Gly Tyr His Cys Leu Gly
85 90 95
Ala Glu Cys Ser Met Cys Glu Gln Asp Cys Lys Gln Gly Gln Glu Leu
100 105 110
Thr Lys Lys Gly Cys Lys Asp Cys Cys Phe Gly Thr Phe Asn Asp Gln
115 120 125
Lys Arg Gly Ile Cys Arg Pro Trp Thr Asn Cys Ser Leu Asp Gly Lys
130 135 140
Ser Val Leu Val Asn Gly Thr Lys Glu Arg Asp Val Val Cys Gly Pro
145 150 155 160
Ser Pro Ala Asp Leu Ser Pro Gly Ala Ser Ser Ala Thr Pro Pro Ala
165 170 175
Pro Ala Arg Glu Pro Gly His Ser Pro Gln Ile Ile Phe Phe Leu Ala
180 185 190
Leu Thr Ser Thr Val Val Leu Phe Leu Leu Phe Phe Leu Val Leu Arg
195 200 205
Phe Ser Val Val Lys Arg Ser Arg Lys Lys Leu Leu Tyr Ile Phe Lys
210 215 220
Gln Pro Phe Met Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys
225 230 235 240
Ser Cys Arg Phe Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu
245 250
<210> 6
<211> 165
<212> PRT
<213> Macaca fascicularis
<400> 6
Ser Leu Gln Asp Leu Cys Ser Asn Cys Pro Ala Gly Thr Phe Cys Asp
1 5 10 15
Asn Asn Arg Ser Gln Ile Cys Ser Pro Cys Pro Pro Asn Ser Phe Ser
20 25 30
Ser Ala Gly Gly Gln Arg Thr Cys Asp Ile Cys Arg Gln Cys Lys Gly
35 40 45
Val Phe Lys Thr Arg Lys Glu Cys Ser Ser Thr Ser Asn Ala Glu Cys
50 55 60
Asp Cys Ile Ser Gly Tyr His Cys Leu Gly Ala Glu Cys Ser Met Cys
65 70 75 80
Glu Gln Asp Cys Lys Gln Gly Gln Glu Leu Thr Lys Lys Gly Cys Lys
85 90 95
Asp Cys Cys Phe Gly Thr Phe Asn Asp Gln Lys Arg Gly Ile Cys Arg
100 105 110
Pro Trp Thr Asn Cys Ser Leu Asp Gly Lys Ser Val Leu Val Asn Gly
115 120 125
Thr Lys Glu Arg Asp Val Val Cys Gly Pro Ser Pro Ala Asp Leu Ser
130 135 140
Pro Gly Ala Ser Ser Ala Thr Pro Pro Ala Pro Ala Arg Glu Pro Gly
145 150 155 160
His Ser Pro Gln Ile
165
<210> 7
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 7
Thr Phe Thr Met Asn
1 5
<210> 8
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 8
Ser Ile Ser Ser Lys Ser Thr Tyr Ile Glu Tyr Ala Asp Ser Phe Lys
1 5 10 15
Val
<210> 9
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 9
Ser Ile Ser Ser His Ser Ser Tyr Ile Glu Tyr Ala Asp Ser Phe Lys
1 5 10 15
Val
<210> 10
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 10
Ser Ile Ser Ser Ser Ser Ser Tyr Ile Glu Tyr Ala Asp Ser Phe Lys
1 5 10 15
Gly
<210> 11
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 11
Ser Ile Ser Ser Lys Ser Ser Tyr Ile Glu Tyr Ala Asp Ser Phe Lys
1 5 10 15
Val
<210> 12
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 12
Ser Ile Ser Ser Ser Ser Ser Tyr Ile Glu Tyr Ala Asp Ser Phe Lys
1 5 10 15
Leu
<210> 13
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 13
Ser Ile Ser Ser Lys Ser Ser Tyr Ile Tyr Tyr Ala Asp Ser Phe Lys
1 5 10 15
Gly
<210> 14
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 14
Ser Ile Ser Ser Lys Gly Ser Tyr Ile Glu Tyr Ala Glu Gln Phe Lys
1 5 10 15
Val
<210> 15
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 15
Ser Ile Ser Ser Lys Gly Ser Tyr Ile Glu Tyr Ala Glu Ser Phe Lys
1 5 10 15
Val
<210> 16
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 16
Ser Ile Ser Ser Lys Gly Ser Tyr Ile Glu Tyr Ala Asp Ser Phe Lys
1 5 10 15
Val
<210> 17
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 17
Tyr Gly Ala Lys Asn Phe Leu Asn Trp Val Phe Asp Tyr
1 5 10
<210> 18
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 18
Tyr Gly Pro Lys Asn Glu Leu Asn Trp Val Phe Asp Tyr
1 5 10
<210> 19
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 19
Tyr Gly Lys Lys Asn Glu Leu Asn Trp Val Phe Asp Tyr
1 5 10
<210> 20
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 20
Tyr Gly Ile Lys Asn Glu Leu Asn Trp Val Phe Asp Tyr
1 5 10
<210> 21
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 21
Thr Gly Thr Arg Tyr Asp Val Gly Tyr Tyr Glu Tyr Val Ser
1 5 10
<210> 22
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 22
Thr Gly Thr Ser Thr Asp Val Gly Phe Tyr Asn Tyr Val Ser
1 5 10
<210> 23
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 23
Thr Gly Thr Arg Thr Asp Val Gly Phe Tyr Glu Tyr Val Ser
1 5 10
<210> 24
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 24
Thr Gly Thr Ser Thr Asp Val Gly Phe Tyr Glu Tyr Val Ser
1 5 10
<210> 25
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 25
Thr Gly Thr Ser Thr Asp Val Gly Tyr Tyr Glu Tyr Val Ser
1 5 10
<210> 26
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 26
Glu Thr Ser Lys Arg Leu Ser
1 5
<210> 27
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 27
Ser Ser Tyr Arg Tyr Glu His Gln Val Ser
1 5 10
<210> 28
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 28
Ser Ser Tyr Arg Tyr Pro His Ile Val Ser
1 5 10
<210> 29
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 29
Ser Ser Tyr Arg Tyr Glu Ala Gln Val Ser
1 5 10
<210> 30
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<220>
<221> misc_feature
<222> (5)..(5)
<223> Xaa can be Lys, His or Ser
<220>
<221> misc_feature
<222> (6)..(6)
<223> Xaa can be Ser or Gly
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa can be Thr or Ser
<220>
<221> misc_feature
<222> (10)..(10)
<223> Xaa can Glu or Tyr
<220>
<221> misc_feature
<222> (13)..(13)
<223> Xaa can be Asp or Glu
<220>
<221> misc_feature
<222> (14)..(14)
<223> Xaa can be Ser or Gln
<220>
<221> misc_feature
<222> (17)..(17)
<223> Xaa can be Val, Gly or Leu
<400> 30
Ser Ile Ser Ser Xaa Xaa Xaa Tyr Ile Xaa Tyr Ala Xaa Xaa Phe Lys
1 5 10 15
Xaa
<210> 31
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<220>
<221> misc_feature
<222> (3)..(3)
<223> Xaa can be Ala, Pro, Lys or Ile
<220>
<221> misc_feature
<222> (6)..(6)
<223> Xaa can be Phe or Glu
<400> 31
Tyr Gly Xaa Lys Asn Xaa Leu Asn Trp Val Phe Asp Tyr
1 5 10
<210> 32
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<220>
<221> misc_feature
<222> (4)..(4)
<223> Xaa can be Arg or Ser
<220>
<221> misc_feature
<222> (5)..(5)
<223> Xaa can be Tyr or Thr
<220>
<221> misc_feature
<222> (9)..(9)
<223> Xaa can be Tyr or Phe
<220>
<221> misc_feature
<222> (11)..(11)
<223> Xaa can be Glu or Asn
<400> 32
Thr Gly Thr Xaa Xaa Asp Val Gly Xaa Tyr Xaa Tyr Val Ser
1 5 10
<210> 33
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<220>
<221> misc_feature
<222> (6)..(6)
<223> Xaa can be Glu or Pro
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa can be His or Ala
<220>
<221> misc_feature
<222> (8)..(8)
<223> Xaa can be Gln or Ile
<400> 33
Ser Ser Tyr Arg Tyr Xaa Xaa Xaa Val Ser
1 5 10
<210> 34
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 34
Ser Ile Ser Ser Arg Ser Asn Tyr Lys Glu Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 35
<211> 30
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 35
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser
20 25 30
<210> 36
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 36
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser
1 5 10
<210> 37
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 37
Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln
1 5 10 15
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg
20 25 30
<210> 38
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 38
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
1 5 10
<210> 39
<211> 22
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 39
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys
20
<210> 40
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 40
Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu Met Ile Tyr
1 5 10 15
<210> 41
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 41
Gly Val Pro Asp Arg Phe Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser
1 5 10 15
Leu Thr Val Ser Gly Leu Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys
20 25 30
<210> 42
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 42
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
1 5 10
<210> 43
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 43
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Phe
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Lys Ser Thr Tyr Ile Glu Tyr Ala Asp Ser Phe
50 55 60
Lys Val Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Ala Lys Asn Phe Leu Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 44
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 44
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Phe
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser His Ser Ser Tyr Ile Glu Tyr Ala Asp Ser Phe
50 55 60
Lys Val Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Ala Lys Asn Phe Leu Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 45
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 45
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Phe
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Ser Ser Ser Tyr Ile Glu Tyr Ala Asp Ser Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Ala Lys Asn Phe Leu Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 46
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 46
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Phe
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Lys Ser Ser Tyr Ile Glu Tyr Ala Asp Ser Phe
50 55 60
Lys Val Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Pro Lys Asn Glu Leu Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 47
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 47
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Phe
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Lys Ser Thr Tyr Ile Glu Tyr Ala Asp Ser Phe
50 55 60
Lys Val Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Pro Lys Asn Glu Leu Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 48
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 48
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Phe
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Ser Ser Ser Tyr Ile Glu Tyr Ala Asp Ser Phe
50 55 60
Lys Leu Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Pro Lys Asn Glu Leu Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 49
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 49
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Phe
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Lys Ser Ser Tyr Ile Tyr Tyr Ala Asp Ser Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Pro Lys Asn Glu Leu Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 50
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 50
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Phe
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Lys Gly Ser Tyr Ile Glu Tyr Ala Glu Gln Phe
50 55 60
Lys Val Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Lys Lys Asn Glu Leu Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 51
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 51
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Phe
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Lys Gly Ser Tyr Ile Glu Tyr Ala Glu Ser Phe
50 55 60
Lys Val Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Ile Lys Asn Glu Leu Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 52
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 52
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Phe
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Lys Gly Ser Tyr Ile Glu Tyr Ala Asp Ser Phe
50 55 60
Lys Val Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Ile Lys Asn Glu Leu Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 53
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 53
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Phe
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Lys Gly Ser Tyr Ile Glu Tyr Ala Glu Gln Phe
50 55 60
Lys Val Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Ala Lys Asn Phe Leu Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 54
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 54
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Arg Tyr Asp Val Gly Tyr Tyr
20 25 30
Glu Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Thr Ser Lys Arg Leu Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Arg Tyr Glu
85 90 95
His Gln Val Ser Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 55
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 55
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Phe Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Thr Ser Lys Arg Leu Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Arg Tyr Glu
85 90 95
His Gln Val Ser Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 56
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 56
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Arg Tyr Asp Val Gly Tyr Tyr
20 25 30
Glu Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Thr Ser Lys Arg Leu Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Arg Tyr Pro
85 90 95
His Ile Val Ser Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 57
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 57
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Arg Tyr Asp Val Gly Tyr Tyr
20 25 30
Glu Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Thr Ser Lys Arg Leu Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Arg Tyr Glu
85 90 95
Ala Gln Val Ser Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 58
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 58
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Arg Thr Asp Val Gly Phe Tyr
20 25 30
Glu Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Thr Ser Lys Arg Leu Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Arg Tyr Glu
85 90 95
His Gln Val Ser Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 59
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 59
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Phe Tyr
20 25 30
Glu Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Thr Ser Lys Arg Leu Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Arg Tyr Glu
85 90 95
His Gln Val Ser Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 60
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 60
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Tyr Tyr
20 25 30
Glu Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Thr Ser Lys Arg Leu Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Arg Tyr Glu
85 90 95
His Gln Val Ser Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 61
<211> 330
<212> PRT
<213> Homo sapiens
<400> 61
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 62
<211> 330
<212> PRT
<213> Homo sapiens
<400> 62
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 63
<211> 106
<212> PRT
<213> Homo sapiens
<400> 63
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
35 40 45
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 64
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 64
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Trp Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Leu Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Thr Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 65
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 65
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Trp Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Leu Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Thr Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 66
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 66
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 67
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 67
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Asp Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 68
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 68
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Trp Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Leu Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Thr Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 69
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 69
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 70
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 70
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Asp Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 71
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 71
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Trp Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Leu Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Thr Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 72
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 72
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Trp Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Leu Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Thr Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 73
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 73
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Trp Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Leu Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Thr Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 74
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 74
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 75
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 75
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Trp Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 76
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 76
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 77
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 77
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 78
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 78
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Trp Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 79
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 79
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 80
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 80
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Trp Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 81
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 81
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 82
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 82
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Trp Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 83
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 83
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 84
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 84
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Trp Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 85
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 85
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Trp Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Leu Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Thr Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 86
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 86
Gly Leu Asn Asp Ile Phe Glu Ala Gln Lys Ile Glu Trp His Glu
1 5 10 15
<210> 87
<211> 212
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 87
Met Gly Asn Ser Cys Tyr Asn Ile Val Ala Thr Leu Leu Leu Val Leu
1 5 10 15
Asn Phe Glu Arg Thr Arg Ser Leu Gln Asp Pro Cys Ser Asn Cys Pro
20 25 30
Ala Gly Thr Phe Cys Asp Asn Asn Arg Asn Gln Ile Cys Ser Pro Cys
35 40 45
Pro Pro Asn Ser Phe Ser Ser Ala Gly Gly Gln Arg Thr Cys Asp Ile
50 55 60
Cys Arg Gln Cys Lys Gly Val Phe Arg Thr Arg Lys Glu Cys Ser Ser
65 70 75 80
Thr Ser Asn Ala Glu Cys Asp Cys Thr Pro Gly Phe His Cys Leu Gly
85 90 95
Ala Gly Cys Ser Met Cys Glu Gln Asp Cys Lys Gln Gly Gln Glu Leu
100 105 110
Thr Lys Lys Gly Cys Lys Asp Cys Cys Phe Gly Thr Phe Asn Asp Gln
115 120 125
Lys Arg Gly Ile Cys Arg Pro Trp Thr Asn Cys Ser Leu Asp Gly Lys
130 135 140
Ser Val Leu Val Asn Gly Thr Lys Glu Arg Asp Val Val Cys Gly Pro
145 150 155 160
Ser Pro Ala Asp Leu Ser Pro Gly Ala Ser Ser Val Thr Pro Pro Ala
165 170 175
Pro Ala Arg Glu Pro Gly His Ser Pro Gln His His His His His His
180 185 190
Gly Gly Gly Gly Ser Gly Leu Asn Asp Ile Phe Glu Ala Gln Lys Ile
195 200 205
Glu Trp His Glu
210
<210> 88
<211> 641
<212> PRT
<213> Human gammaherpesvirus 4
<400> 88
Met Ser Asp Glu Gly Pro Gly Thr Gly Pro Gly Asn Gly Leu Gly Glu
1 5 10 15
Lys Gly Asp Thr Ser Gly Pro Glu Gly Ser Gly Gly Ser Gly Pro Gln
20 25 30
Arg Arg Gly Gly Asp Asn His Gly Arg Gly Arg Gly Arg Gly Arg Gly
35 40 45
Arg Gly Gly Gly Arg Pro Gly Ala Pro Gly Gly Ser Gly Ser Gly Pro
50 55 60
Arg His Arg Asp Gly Val Arg Arg Pro Gln Lys Arg Pro Ser Cys Ile
65 70 75 80
Gly Cys Lys Gly Thr His Gly Gly Thr Gly Ala Gly Ala Gly Ala Gly
85 90 95
Gly Ala Gly Ala Gly Gly Ala Gly Ala Gly Gly Gly Ala Gly Ala Gly
100 105 110
Gly Gly Ala Gly Gly Ala Gly Gly Ala Gly Gly Ala Gly Ala Gly Gly
115 120 125
Gly Ala Gly Ala Gly Gly Gly Ala Gly Gly Ala Gly Gly Ala Gly Ala
130 135 140
Gly Gly Gly Ala Gly Ala Gly Gly Gly Ala Gly Gly Ala Gly Ala Gly
145 150 155 160
Gly Gly Ala Gly Gly Ala Gly Gly Ala Gly Ala Gly Gly Gly Ala Gly
165 170 175
Ala Gly Gly Gly Ala Gly Gly Ala Gly Ala Gly Gly Gly Ala Gly Gly
180 185 190
Ala Gly Gly Ala Gly Ala Gly Gly Gly Ala Gly Ala Gly Gly Ala Gly
195 200 205
Gly Ala Gly Gly Ala Gly Ala Gly Gly Ala Gly Ala Gly Gly Gly Ala
210 215 220
Gly Gly Ala Gly Gly Ala Gly Ala Gly Gly Ala Gly Ala Gly Gly Ala
225 230 235 240
Gly Ala Gly Gly Ala Gly Ala Gly Gly Ala Gly Gly Ala Gly Ala Gly
245 250 255
Gly Ala Gly Gly Ala Gly Ala Gly Gly Ala Gly Gly Ala Gly Ala Gly
260 265 270
Gly Gly Ala Gly Gly Ala Gly Ala Gly Gly Gly Ala Gly Gly Ala Gly
275 280 285
Ala Gly Gly Ala Gly Gly Ala Gly Ala Gly Gly Ala Gly Gly Ala Gly
290 295 300
Ala Gly Gly Ala Gly Gly Ala Gly Ala Gly Gly Gly Ala Gly Ala Gly
305 310 315 320
Gly Ala Gly Ala Gly Gly Gly Gly Arg Gly Arg Gly Gly Ser Gly Gly
325 330 335
Arg Gly Arg Gly Gly Ser Gly Gly Arg Gly Arg Gly Gly Ser Gly Gly
340 345 350
Arg Arg Gly Arg Gly Arg Glu Arg Ala Arg Gly Gly Ser Arg Glu Arg
355 360 365
Ala Arg Gly Arg Gly Arg Gly Arg Gly Glu Lys Arg Pro Arg Ser Pro
370 375 380
Ser Ser Gln Ser Ser Ser Ser Gly Ser Pro Pro Arg Arg Pro Pro Pro
385 390 395 400
Gly Arg Arg Pro Phe Phe His Pro Val Gly Glu Ala Asp Tyr Phe Glu
405 410 415
Tyr His Gln Glu Gly Gly Pro Asp Gly Glu Pro Asp Val Pro Pro Gly
420 425 430
Ala Ile Glu Gln Gly Pro Ala Asp Asp Pro Gly Glu Gly Pro Ser Thr
435 440 445
Gly Pro Arg Gly Gln Gly Asp Gly Gly Arg Arg Lys Lys Gly Gly Trp
450 455 460
Phe Gly Lys His Arg Gly Gln Gly Gly Ser Asn Pro Lys Phe Glu Asn
465 470 475 480
Ile Ala Glu Gly Leu Arg Ala Leu Leu Ala Arg Ser His Val Glu Arg
485 490 495
Thr Thr Asp Glu Gly Thr Trp Val Ala Gly Val Phe Val Tyr Gly Gly
500 505 510
Ser Lys Thr Ser Leu Tyr Asn Leu Arg Arg Gly Thr Ala Leu Ala Ile
515 520 525
Pro Gln Cys Arg Leu Thr Pro Leu Ser Arg Leu Pro Phe Gly Met Ala
530 535 540
Pro Gly Pro Gly Pro Gln Pro Gly Pro Leu Arg Glu Ser Ile Val Cys
545 550 555 560
Tyr Phe Met Val Phe Leu Gln Thr His Ile Phe Ala Glu Val Leu Lys
565 570 575
Asp Ala Ile Lys Asp Leu Val Met Thr Lys Pro Ala Pro Thr Cys Asn
580 585 590
Ile Arg Val Thr Val Cys Ser Phe Asp Asp Gly Val Asp Leu Pro Pro
595 600 605
Trp Phe Pro Pro Met Val Glu Gly Ala Ala Ala Glu Gly Asp Asp Gly
610 615 620
Asp Asp Gly Asp Glu Gly Gly Asp Gly Asp Glu Gly Glu Glu Gly Gln
625 630 635 640
Glu
<210> 89
<211> 399
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 89
Leu Gln Asp Pro Cys Ser Asn Cys Pro Ala Gly Thr Phe Cys Asp Asn
1 5 10 15
Asn Arg Asn Gln Ile Cys Ser Pro Cys Pro Pro Asn Ser Phe Ser Ser
20 25 30
Ala Gly Gly Gln Arg Thr Cys Asp Ile Cys Arg Gln Cys Lys Gly Val
35 40 45
Phe Arg Thr Arg Lys Glu Cys Ser Ser Thr Ser Asn Ala Glu Cys Asp
50 55 60
Cys Thr Pro Gly Phe His Cys Leu Gly Ala Gly Cys Ser Met Cys Glu
65 70 75 80
Gln Asp Cys Lys Gln Gly Gln Glu Leu Thr Lys Lys Gly Cys Lys Asp
85 90 95
Cys Cys Phe Gly Thr Phe Asn Asp Gln Lys Arg Gly Ile Cys Arg Pro
100 105 110
Trp Thr Asn Cys Ser Leu Asp Gly Lys Ser Val Leu Val Asn Gly Thr
115 120 125
Lys Glu Arg Asp Val Val Cys Gly Pro Ser Pro Ala Asp Leu Ser Pro
130 135 140
Gly Ala Ser Ser Val Thr Pro Pro Ala Pro Ala Arg Glu Pro Gly His
145 150 155 160
Ser Pro Gln Asp Ile Glu Gly Arg Met Asp Pro Lys Ser Cys Asp Lys
165 170 175
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
180 185 190
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
195 200 205
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
210 215 220
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
225 230 235 240
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
245 250 255
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
260 265 270
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
275 280 285
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
290 295 300
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
305 310 315 320
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
325 330 335
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
340 345 350
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
355 360 365
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
370 375 380
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
385 390 395
<210> 90
<211> 442
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 90
Met Gly Asn Ser Cys Tyr Asn Ile Val Ala Thr Leu Leu Leu Val Leu
1 5 10 15
Asn Phe Glu Arg Thr Arg Ser Leu Gln Asp Pro Cys Ser Asn Cys Pro
20 25 30
Ala Gly Thr Phe Cys Asp Asn Asn Arg Asn Gln Ile Cys Ser Pro Cys
35 40 45
Pro Pro Asn Ser Phe Ser Ser Ala Gly Gly Gln Arg Thr Cys Asp Ile
50 55 60
Cys Arg Gln Cys Lys Gly Val Phe Arg Thr Arg Lys Glu Cys Ser Ser
65 70 75 80
Thr Ser Asn Ala Glu Cys Asp Cys Thr Pro Gly Phe His Cys Leu Gly
85 90 95
Ala Gly Cys Ser Met Cys Glu Gln Asp Cys Lys Gln Gly Gln Glu Leu
100 105 110
Thr Lys Lys Gly Cys Lys Asp Cys Cys Phe Gly Thr Phe Asn Asp Gln
115 120 125
Lys Arg Gly Ile Cys Arg Pro Trp Thr Asn Cys Ser Leu Asp Gly Lys
130 135 140
Ser Val Leu Val Asn Gly Thr Lys Glu Arg Asp Val Val Cys Gly Pro
145 150 155 160
Ser Pro Ala Asp Leu Ser Pro Gly Ala Ser Ser Val Thr Pro Pro Ala
165 170 175
Pro Ala Arg Glu Pro Gly His Ser Pro Gln Asp Ile Glu Gly Arg Met
180 185 190
Asp Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
195 200 205
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
210 215 220
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
225 230 235 240
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
245 250 255
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
260 265 270
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
275 280 285
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
290 295 300
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
305 310 315 320
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
325 330 335
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
340 345 350
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
355 360 365
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
370 375 380
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
385 390 395 400
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
405 410 415
Ser Leu Ser Leu Ser Pro Gly Gly Gly Gly Ser Gly Leu Asn Asp Ile
420 425 430
Phe Glu Ala Gln Lys Ile Glu Trp His Glu
435 440
<210> 91
<211> 347
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 91
Met Val Leu Ala Ser Ser Thr Thr Ser Ile His Thr Met Leu Leu Leu
1 5 10 15
Leu Leu Met Leu Ala Gln Pro Ala Met Ala Met Lys Asp Asn Thr Val
20 25 30
Pro Leu Lys Leu Ile Ala Leu Leu Ala Asn Gly Glu Phe His Ser Gly
35 40 45
Glu Gln Leu Gly Glu Thr Leu Gly Met Ser Arg Ala Ala Ile Asn Lys
50 55 60
His Ile Gln Thr Leu Arg Asp Trp Gly Val Asp Val Phe Thr Val Pro
65 70 75 80
Gly Lys Gly Tyr Ser Leu Pro Glu Pro Ile Gln Leu Leu Asn Ala Lys
85 90 95
Gln Ile Leu Gly Gln Leu Asp Gly Gly Ser Val Ala Val Leu Pro Val
100 105 110
Ile Asp Ser Thr Asn Gln Tyr Leu Leu Asp Arg Ile Gly Glu Leu Lys
115 120 125
Ser Gly Asp Ala Cys Ile Ala Glu Tyr Gln Gln Ala Gly Arg Gly Arg
130 135 140
Arg Gly Arg Lys Trp Phe Ser Pro Phe Gly Ala Asn Leu Tyr Leu Ser
145 150 155 160
Met Phe Trp Arg Leu Glu Gln Gly Pro Ala Ala Ala Ile Gly Leu Ser
165 170 175
Leu Val Ile Gly Ile Val Met Ala Glu Val Leu Arg Lys Leu Gly Ala
180 185 190
Asp Lys Val Arg Val Lys Trp Pro Asn Asp Leu Tyr Leu Gln Asp Arg
195 200 205
Lys Leu Ala Gly Ile Leu Val Glu Leu Thr Gly Lys Thr Gly Asp Ala
210 215 220
Ala Gln Ile Val Ile Gly Ala Gly Ile Asn Met Ala Met Arg Arg Val
225 230 235 240
Glu Glu Ser Val Val Asn Gln Gly Trp Ile Thr Leu Gln Glu Ala Gly
245 250 255
Ile Asn Leu Asp Arg Asn Thr Leu Ala Ala Met Leu Ile Arg Glu Leu
260 265 270
Arg Ala Ala Leu Glu Leu Phe Glu Gln Glu Gly Leu Ala Pro Tyr Leu
275 280 285
Ser Arg Trp Glu Lys Leu Asp Asn Phe Ile Asn Arg Pro Val Lys Leu
290 295 300
Ile Ile Gly Asp Lys Glu Ile Phe Gly Ile Ser Arg Gly Ile Asp Lys
305 310 315 320
Gln Gly Ala Leu Leu Leu Glu Gln Asp Gly Ile Ile Lys Pro Trp Met
325 330 335
Gly Gly Glu Ile Ser Leu Arg Ser Ala Glu Lys
340 345
<210> 92
<211> 442
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 92
Met Gly Asn Ser Cys Tyr Asn Ile Val Ala Thr Leu Leu Leu Val Leu
1 5 10 15
Asn Phe Glu Arg Thr Arg Ser Leu Gln Asp Leu Cys Ser Asn Cys Pro
20 25 30
Ala Gly Thr Phe Cys Asp Asn Asn Arg Ser Gln Ile Cys Ser Pro Cys
35 40 45
Pro Pro Asn Ser Phe Ser Ser Ala Gly Gly Gln Arg Thr Cys Asp Ile
50 55 60
Cys Arg Gln Cys Lys Gly Val Phe Lys Thr Arg Lys Glu Cys Ser Ser
65 70 75 80
Thr Ser Asn Ala Glu Cys Asp Cys Ile Ser Gly Tyr His Cys Leu Gly
85 90 95
Ala Glu Cys Ser Met Cys Glu Gln Asp Cys Lys Gln Gly Gln Glu Leu
100 105 110
Thr Lys Lys Gly Cys Lys Asp Cys Cys Phe Gly Thr Phe Asn Asp Gln
115 120 125
Lys Arg Gly Ile Cys Arg Pro Trp Thr Asn Cys Ser Leu Asp Gly Lys
130 135 140
Ser Val Leu Val Asn Gly Thr Lys Glu Arg Asp Val Val Cys Gly Pro
145 150 155 160
Ser Pro Ala Asp Leu Ser Pro Gly Ala Ser Ser Ala Thr Pro Pro Ala
165 170 175
Pro Ala Arg Glu Pro Gly His Ser Pro Gln Asp Ile Glu Gly Arg Met
180 185 190
Asp Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
195 200 205
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
210 215 220
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
225 230 235 240
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
245 250 255
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
260 265 270
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
275 280 285
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
290 295 300
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
305 310 315 320
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
325 330 335
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
340 345 350
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
355 360 365
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
370 375 380
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
385 390 395 400
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
405 410 415
Ser Leu Ser Leu Ser Pro Gly Gly Gly Gly Ser Gly Leu Asn Asp Ile
420 425 430
Phe Glu Ala Gln Lys Ile Glu Trp His Glu
435 440
<210> 93
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 93
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Glu His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Phe Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 94
<211> 452
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 94
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Gly Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Asn Tyr Ala His Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Gly Leu Lys Asn Phe Met Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Glu
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Phe Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 95
<211> 452
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 95
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser His
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Asn Tyr Ile Asp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Gly Met Lys Asn Lys Met Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Glu
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Phe Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 96
<211> 452
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 96
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Thr Tyr Ala Thr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Leu Lys Asn Ser Leu Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Glu
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Phe Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 97
<211> 452
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 97
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Gly Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Asn Tyr Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Thr Lys Asn Ser Arg Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Glu
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Phe Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 98
<211> 452
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 98
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Arg Tyr Ile Ser Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Lys Lys Asn Tyr His Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Glu
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Phe Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 99
<211> 452
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 99
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser His
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Asn Tyr Lys Asp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Gly Ile Lys Asn His Met Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Glu
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Phe Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 100
<211> 452
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 100
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Thr Tyr Ala Ser Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Lys Lys Asn Met Met Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Glu
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Phe Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 101
<211> 452
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 101
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Gly Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Asn Tyr Ile Asp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Gly Leu Lys Asn Ser Leu Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Glu
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Phe Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 102
<211> 452
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 102
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser His
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Ser Ser Ile Ser Ser Arg Ser Asn Tyr Lys Ser Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly His Lys Asn Met His Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Glu
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Phe Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 103
<211> 452
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 103
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Phe
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Asn Tyr Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Gly Leu Lys Asn Glu Met Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Glu
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Phe Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 104
<211> 452
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 104
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Asn Tyr Ile Asp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Ile Lys Asn Glu Met Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Glu
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Phe Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 105
<211> 452
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 105
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Asn Tyr Arg Asp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Gly Lys Asn Ile Leu Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Glu
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Phe Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 106
<211> 452
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 106
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Asn Tyr Ile Asp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Gly Lys Lys Asn Asp Met Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Glu
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Phe Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 107
<211> 452
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 107
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Asn Tyr Lys Phe Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Gly Val Lys Asn Asn Met Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Glu
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Phe Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 108
<211> 452
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 108
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Gly Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Asn Tyr Ala Asp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Ala Lys Asn Val Met Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Glu
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Phe Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 109
<211> 452
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 109
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Phe
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Asn Tyr Lys Ser Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Ala Lys Asn Phe Met Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Glu
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Phe Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 110
<211> 452
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 110
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Gly Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Asn Tyr Lys His Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Gly Leu Lys Asn Glu Arg Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Glu
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Phe Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 111
<211> 452
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 111
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Asn Tyr Lys Phe Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Gly Gly Lys Asn Asn Met Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Glu
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Phe Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 112
<211> 452
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 112
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Asn Tyr Ala Asp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Ala Lys Asn Glu Met Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Glu
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Phe Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 113
<211> 216
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 113
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Phe Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Gly Thr Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Ser Tyr Ala Tyr Asp
85 90 95
Ala Gln Leu Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 114
<211> 216
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 114
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Asn Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Phe Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Thr Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Thr Tyr Arg Tyr Asn
85 90 95
Asn Leu Leu Ala Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 115
<211> 216
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 115
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Phe Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Val Ser Lys Lys Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Thr Tyr Ser Tyr Asp
85 90 95
Glu Ser Val Ala Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 116
<211> 216
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 116
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Phe Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Gly Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Ser Tyr Ala Tyr Thr
85 90 95
Lys Asn Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 117
<211> 216
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 117
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Phe Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Thr Ser Lys Lys Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Val Tyr Arg Tyr Asn
85 90 95
Asp Gln Val Ala Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 118
<211> 216
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 118
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Phe Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Val Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Thr Tyr Ala Tyr Ser
85 90 95
Thr Ile Val Met Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 119
<211> 216
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 119
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Phe Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Val Ser Lys Lys Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Thr Tyr Arg Tyr Arg
85 90 95
Asp Tyr Val Gly Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 120
<211> 216
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 120
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Phe Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Thr Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Ser Tyr Ser Gly Ala
85 90 95
Ser Ile Leu Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 121
<211> 216
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 121
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Phe Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Val Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Thr Tyr Arg Tyr Asp
85 90 95
Gln Asn Val Ala Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 122
<211> 216
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 122
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Phe Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Gly Ser Lys Lys Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Val Tyr Ser Tyr Ile
85 90 95
Tyr Tyr Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 123
<211> 216
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 123
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Phe Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Thr Ser Lys Lys Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Val Tyr Arg Tyr Ser
85 90 95
Leu Gly Val Gly Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 124
<211> 216
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 124
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Phe Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Val Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Ser Tyr Ser Ser Asp
85 90 95
Thr His Val Gly Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 125
<211> 216
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 125
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Phe Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Gly Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Ser Tyr Glu Tyr Thr
85 90 95
Glu Ser Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 126
<211> 216
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 126
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Phe Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Val Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Ser Tyr Glu Tyr Ile
85 90 95
Glu Ser Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 127
<211> 216
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 127
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Phe Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Gly Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Ser Tyr Ala Tyr Thr
85 90 95
Lys Asn Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 128
<211> 216
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 128
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Phe Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Tyr Thr Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Ser Tyr Arg Tyr Asp
85 90 95
His Gln Val Ser Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 129
<211> 216
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 129
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Phe Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Val Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Thr Tyr Thr Tyr Asn
85 90 95
Ala Leu Leu Ala Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 130
<211> 216
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 130
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Phe Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Val Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Ser Tyr Glu Tyr Ile
85 90 95
Glu Ser Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 131
<211> 216
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 131
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Phe Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Val Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Ser Tyr Thr Tyr Asp
85 90 95
Gln Gln Leu Met Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 132
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 132
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Phe
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Asn Tyr Lys Ser Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Ala Lys Asn Phe Leu Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 133
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 133
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Phe Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Tyr Thr Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Arg Tyr Glu
85 90 95
His Gln Val Ser Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 134
<211> 450
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 134
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Phe
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Asn Tyr Lys Glu Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Ala Lys Asn Phe Leu Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Glu
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Phe Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro
450
<210> 135
<211> 216
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 135
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Phe Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Thr Ser Lys Arg Leu Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Arg Tyr Glu
85 90 95
His Gln Val Ser Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 136
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 136
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Phe
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Asn Tyr Lys Glu Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Ala Lys Asn Phe Leu Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 137
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 137
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Phe Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Thr Ser Lys Arg Leu Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Arg Tyr Glu
85 90 95
His Gln Val Ser Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 138
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 138
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 139
<211> 121
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 139
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn His Gly Gly Tyr Val Thr Tyr Asn Pro Ser Leu Glu
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Tyr Gly Pro Gly Asn Tyr Asp Trp Tyr Phe Asp Leu Trp Gly
100 105 110
Arg Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 140
<211> 109
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 140
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Pro
85 90 95
Ala Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 141
<211> 107
<212> PRT
<213> Homo sapiens
<400> 141
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 142
<211> 116
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 142
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Arg Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Ser Thr Tyr
20 25 30
Trp Ile Ser Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Lys Ile Tyr Pro Gly Asp Ser Tyr Thr Asn Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Gly Tyr Gly Ile Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 143
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 143
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly Asp Asn Ile Gly Asp Gln Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Lys Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Tyr Thr Gly Phe Gly Ser Leu
85 90 95
Ala Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 144
<211> 324
<212> PRT
<213> Mus musculus
<400> 144
Ala Lys Thr Thr Pro Pro Ser Val Tyr Pro Leu Ala Pro Gly Ser Ala
1 5 10 15
Ala Gln Thr Asn Ser Met Val Thr Leu Gly Cys Leu Val Lys Gly Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ser Leu Ser Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Asp Leu Tyr Thr Leu
50 55 60
Ser Ser Ser Val Thr Val Pro Ser Ser Thr Trp Pro Ser Glu Thr Val
65 70 75 80
Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys Lys
85 90 95
Ile Val Pro Arg Asp Cys Gly Cys Lys Pro Cys Ile Cys Thr Val Pro
100 105 110
Glu Val Ser Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Val Leu
115 120 125
Thr Ile Thr Leu Thr Pro Lys Val Thr Cys Val Val Val Asp Ile Ser
130 135 140
Lys Asp Asp Pro Glu Val Gln Phe Ser Trp Phe Val Asp Asp Val Glu
145 150 155 160
Val His Thr Ala Gln Thr Gln Pro Arg Glu Glu Gln Phe Asn Ser Thr
165 170 175
Phe Arg Ser Val Ser Glu Leu Pro Ile Met His Gln Asp Trp Leu Asn
180 185 190
Gly Lys Glu Phe Lys Cys Arg Val Asn Ser Ala Ala Phe Pro Ala Pro
195 200 205
Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Arg Pro Lys Ala Pro Gln
210 215 220
Val Tyr Thr Ile Pro Pro Pro Lys Glu Gln Met Ala Lys Asp Lys Val
225 230 235 240
Ser Leu Thr Cys Met Ile Thr Asp Phe Phe Pro Glu Asp Ile Thr Val
245 250 255
Glu Trp Gln Trp Asn Gly Gln Pro Ala Glu Asn Tyr Lys Asn Thr Gln
260 265 270
Pro Ile Met Asp Thr Asp Gly Ser Tyr Phe Val Tyr Ser Lys Leu Asn
275 280 285
Val Gln Lys Ser Asn Trp Glu Ala Gly Asn Thr Phe Thr Cys Ser Val
290 295 300
Leu His Glu Gly Leu His Asn His His Thr Glu Lys Ser Leu Ser His
305 310 315 320
Ser Pro Gly Lys
<210> 145
<211> 324
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 145
Ala Lys Thr Thr Pro Pro Ser Val Tyr Pro Leu Ala Pro Gly Ser Ala
1 5 10 15
Ala Gln Thr Asn Ser Met Val Thr Leu Gly Cys Leu Val Lys Gly Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ser Leu Ser Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Asp Leu Tyr Thr Leu
50 55 60
Ser Ser Ser Val Thr Val Pro Ser Ser Thr Trp Pro Ser Glu Thr Val
65 70 75 80
Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys Lys
85 90 95
Ile Val Pro Arg Asp Cys Gly Cys Lys Pro Cys Ile Cys Thr Val Pro
100 105 110
Glu Val Ser Asp Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Val Leu
115 120 125
Thr Ile Thr Leu Thr Pro Lys Val Thr Cys Val Val Val Asp Ile Ser
130 135 140
Lys Asp Asp Pro Glu Val Gln Phe Ser Trp Phe Val Asp Asp Val Glu
145 150 155 160
Val His Thr Ala Gln Thr Gln Pro Arg Glu Glu Gln Phe Asn Ser Thr
165 170 175
Phe Arg Ser Val Ser Glu Leu Pro Ile Met His Gln Asp Trp Leu Asn
180 185 190
Gly Lys Glu Phe Lys Cys Arg Val Asn Ser Ala Ala Phe Pro Ala Pro
195 200 205
Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Arg Pro Lys Ala Pro Gln
210 215 220
Val Tyr Thr Ile Pro Pro Pro Lys Glu Gln Met Ala Lys Asp Lys Val
225 230 235 240
Ser Leu Thr Cys Met Ile Thr Asp Phe Phe Pro Glu Asp Ile Thr Val
245 250 255
Glu Trp Gln Trp Asn Gly Gln Pro Ala Glu Asn Tyr Lys Asn Thr Gln
260 265 270
Pro Ile Met Asp Thr Asp Gly Ser Tyr Phe Val Tyr Ser Lys Leu Asn
275 280 285
Val Gln Lys Ser Asn Trp Glu Ala Gly Asn Thr Phe Thr Cys Ser Val
290 295 300
Leu His Glu Gly Leu His Asn His His Thr Glu Lys Ser Leu Ser His
305 310 315 320
Ser Pro Gly Lys
<210> 146
<211> 324
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 146
Ala Lys Thr Thr Pro Pro Ser Val Tyr Pro Leu Ala Pro Gly Ser Ala
1 5 10 15
Ala Gln Thr Asn Ser Met Val Thr Leu Gly Cys Leu Val Lys Gly Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ser Leu Ser Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Asp Leu Tyr Thr Leu
50 55 60
Ser Ser Ser Val Thr Val Pro Ser Ser Thr Trp Pro Ser Glu Thr Val
65 70 75 80
Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys Lys
85 90 95
Ile Val Pro Arg Asp Cys Gly Cys Lys Pro Cys Ile Cys Glu Pro Asn
100 105 110
Glu Val Glu Asp Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Val Leu
115 120 125
Thr Ile Thr Leu Thr Pro Lys Val Thr Cys Val Val Val Asp Ile Ser
130 135 140
Lys Asp Asp Pro Glu Val Gln Phe Ser Trp Phe Val Asp Asp Val Glu
145 150 155 160
Val His Thr Ala Gln Thr Gln Pro Arg Glu Glu Gln Phe Asn Ser Thr
165 170 175
Phe Arg Ser Val Ser Glu Leu Pro Ile Met His Gln Asp Trp Leu Asn
180 185 190
Gly Lys Glu Phe Lys Cys Arg Val Asp Ser Ala Ala Phe Pro Ala Pro
195 200 205
Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Arg Pro Lys Ala Pro Gln
210 215 220
Val Tyr Thr Ile Pro Pro Pro Lys Glu Gln Met Ala Lys Asp Lys Val
225 230 235 240
Ser Leu Thr Cys Met Ile Thr Asp Phe Phe Pro Glu Asp Ile Thr Val
245 250 255
Glu Trp Gln Trp Asn Gly Gln Pro Ala Glu Asn Tyr Lys Asn Thr Gln
260 265 270
Pro Ile Met Asp Thr Asp Gly Ser Tyr Phe Val Tyr Ser Lys Leu Asn
275 280 285
Val Gln Lys Ser Asn Trp Glu Ala Gly Asn Thr Phe Thr Cys Ser Val
290 295 300
Leu His Glu Gly Leu His Asn His His Thr Glu Lys Ser Leu Ser His
305 310 315 320
Ser Pro Gly Lys
<210> 147
<211> 107
<212> PRT
<213> Mus musculus
<400> 147
Arg Ala Asp Ala Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu
1 5 10 15
Gln Leu Thr Ser Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe
20 25 30
Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg
35 40 45
Gln Asn Gly Val Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Met Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu
65 70 75 80
Arg His Asn Ser Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser
85 90 95
Pro Ile Val Lys Ser Phe Asn Arg Asn Glu Cys
100 105
<210> 148
<211> 106
<212> PRT
<213> Mus musculus
<400> 148
Gly Gln Pro Lys Ser Ser Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Glu Thr Asn Lys Ala Thr Leu Val Cys Thr Ile Thr Asp
20 25 30
Phe Tyr Pro Gly Val Val Thr Val Asp Trp Lys Val Asp Gly Thr Pro
35 40 45
Val Thr Gln Gly Met Glu Thr Thr Gln Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Met Ala Ser Ser Tyr Leu Thr Leu Thr Ala Arg Ala Trp Glu
65 70 75 80
Arg His Ser Ser Tyr Ser Cys Gln Val Thr His Glu Gly His Thr Val
85 90 95
Glu Lys Ser Leu Ser Arg Ala Asp Cys Ser
100 105
<210> 149
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 149
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 150
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 150
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Asp Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 151
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 151
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Asp Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala Glu Glu Asp Gly Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 152
<211> 116
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 152
Gln Val Gln Leu Gln Gln Ser Gly Pro Gln Leu Val Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Ser Tyr
20 25 30
Trp Met His Trp Val Asn Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Met Ile Asp Pro Ser Tyr Ser Glu Thr Arg Leu Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Pro Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Leu Tyr Gly Asn Tyr Phe Asp Tyr Trp Gly Gln Gly Thr Thr Leu
100 105 110
Thr Val Ser Ser
115
<210> 153
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 153
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 154
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 154
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Trp Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Leu Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Thr Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 155
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 155
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 156
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 156
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Trp Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Leu Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Thr Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 157
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 157
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Trp Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 158
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 158
Asp Ile Gln Met Thr Gln Ser Ser Ser Ser Phe Ser Val Ser Leu Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Glu Asp Ile Tyr Asn Arg
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Asn Ala Pro Arg Leu Leu Ile
35 40 45
Ser Gly Ala Thr Ser Leu Glu Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Lys Asp Tyr Thr Leu Ser Ile Thr Ser Leu Gln Thr
65 70 75 80
Glu Asp Val Ala Thr Tyr Tyr Cys Gln Gln Tyr Trp Ser Thr Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Val Lys
100 105
<210> 159
<211> 325
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 159
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu
325
<210> 160
<211> 319
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 160
Leu Gln Asp Pro Cys Ser Asn Cys Pro Ala Gly Thr Phe Cys Asp Asn
1 5 10 15
Asn Arg Asn Gln Ile Cys Ser Pro Cys Pro Pro Asn Ser Phe Ser Ser
20 25 30
Ala Gly Gly Gln Arg Thr Cys Asp Ile Cys Arg Gln Cys Lys Gly Val
35 40 45
Phe Arg Thr Arg Lys Glu Cys Ser Ser Thr Ser Asn Ala Glu Cys Asp
50 55 60
Ile Glu Gly Arg Met Asp Pro Lys Ser Cys Asp Lys Thr His Thr Cys
65 70 75 80
Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu
85 90 95
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
100 105 110
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
115 120 125
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
130 135 140
Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
145 150 155 160
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
165 170 175
Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
180 185 190
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
195 200 205
Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
210 215 220
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
225 230 235 240
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
245 250 255
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
260 265 270
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
275 280 285
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Gly Gly Gly Ser
290 295 300
Gly Leu Asn Asp Ile Phe Glu Ala Gln Lys Ile Glu Trp His Glu
305 310 315
<210> 161
<211> 451
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 161
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Glu Phe Ser Asn Tyr
20 25 30
Phe Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Met Val His Pro Ile Glu Gly Trp Ala Val Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Arg Pro Val Phe Cys Cys Pro Arg Gly Ala Phe Asp Ile
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Glu His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Phe Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro
450
<210> 162
<211> 451
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 162
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Gly Phe Ser Ser Tyr
20 25 30
Gly Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ala Leu Phe Pro Ile Ser Ser Gln Ala Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Arg Pro Val Asp Tyr Val Pro His Gly Ala Phe Asp Ile
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Glu His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Phe Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro
450
<210> 163
<211> 451
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 163
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Trp Phe Ser Tyr Tyr
20 25 30
Met Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Val Ile Val Pro Ile Asn Gly Ala Ala Gln Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Arg Pro Val Trp Arg Thr Pro Arg Gly Ala Phe Asp Ile
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Glu His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Phe Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro
450
<210> 164
<211> 451
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 164
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Asp Phe Ser Asn Tyr
20 25 30
Val Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Phe Ile Asn Pro Ile Tyr Gly Ala Ala Gly Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Glu Pro Val Ala Ser Leu Pro His Gly Ala Phe Asp Ile
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Glu His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Phe Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro
450
<210> 165
<211> 451
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 165
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Trp Phe Ser His Tyr
20 25 30
Ser Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Val Val Trp Pro Ile Arg Ser Phe Ala Thr Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Asn Pro Val Thr Ser Trp Pro His Gly Ala Phe Asp Ile
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Glu His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Phe Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro
450
<210> 166
<211> 451
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 166
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Ala Phe Ser Tyr Tyr
20 25 30
Leu Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Phe Val Asp Pro Ile Arg Gly Gly Ala Phe Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Arg Pro Val Asn Ala Ala Pro Arg Gly Ala Phe Asp Ile
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Glu His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Phe Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro
450
<210> 167
<211> 451
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 167
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Ala Phe Ser Tyr Tyr
20 25 30
Leu Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Phe Val Asp Pro Ile Arg Gly Gly Ala Phe Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Arg Pro Val Asn Ala Ala Pro Arg Gly Ala Phe Asp Ile
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Glu His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Phe Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro
450
<210> 168
<211> 451
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 168
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Trp Phe Ser Arg Tyr
20 25 30
Ile Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Thr Ile Asn Pro Ile Tyr Gly Val Ala Leu Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Ala Pro Val Asn Asp Met Pro Arg Gly Ala Phe Asp Ile
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Glu His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Phe Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro
450
<210> 169
<211> 219
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 169
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val Ser Pro
20 25 30
Gly Asp Asn Thr Phe Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Glu Ile Glu Arg Arg Phe Tyr Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Ala
85 90 95
Ser Thr Phe Pro Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 170
<211> 219
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 170
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val Ser Pro
20 25 30
Gly Asp Asn Thr Phe Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Glu Ile Glu Arg Arg Phe Tyr Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Ala
85 90 95
Ser Thr Phe Pro Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 171
<211> 219
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 171
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val Asp Ser
20 25 30
Asn Met Asn Thr Phe Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Glu Ile Gln His Arg Phe Tyr Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Ala
85 90 95
Met Gly Phe Pro Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 172
<211> 219
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 172
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val Asp Pro
20 25 30
Trp Gly Asn Thr Phe Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Glu Ile Ser Asn Arg Phe Tyr Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Ala
85 90 95
Lys Asp Phe Pro Arg Thr Phe Gly Pro Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 173
<211> 219
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 173
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val Arg Pro
20 25 30
Phe Gly Asn Thr Phe Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Glu Ile Ser Thr Arg Phe Tyr Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Ala
85 90 95
His Gln Phe Pro Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 174
<211> 219
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 174
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val Gly Asp
20 25 30
Arg Gly Asn Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Glu Ile Gly Ser Ile Phe Tyr Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Ala
85 90 95
Ala Glu Phe Pro Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 175
<211> 219
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 175
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val Trp Asn
20 25 30
Asp Arg Asn Thr Phe Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Glu Ile Arg Lys Arg Phe Tyr Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Ala
85 90 95
Gln Asp Phe Pro Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 176
<211> 219
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 176
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val Asp Gly
20 25 30
Phe Gly Asn Thr Phe Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Glu Ile Arg Lys Arg Phe Tyr Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Ala
85 90 95
Phe Ala Phe Pro Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 177
<211> 258
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 177
Leu Gln Asp Pro Cys Ser Asn Cys Pro Ala Gly Thr Phe Cys Asp Asn
1 5 10 15
Asn Arg Asn Gln Ile Cys Asp Ile Glu Gly Arg Met Asp Pro Lys Ser
20 25 30
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
35 40 45
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
50 55 60
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
65 70 75 80
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
85 90 95
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
100 105 110
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
115 120 125
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
130 135 140
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
145 150 155 160
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
165 170 175
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
180 185 190
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
195 200 205
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
210 215 220
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
225 230 235 240
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
245 250 255
Ser Pro
<210> 178
<211> 299
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 178
Leu Gln Asp Pro Cys Ser Asn Cys Pro Ala Gly Thr Phe Cys Asp Asn
1 5 10 15
Asn Arg Asn Gln Ile Cys Ser Pro Cys Pro Pro Asn Ser Phe Ser Ser
20 25 30
Ala Gly Gly Gln Arg Thr Cys Asp Ile Cys Arg Gln Cys Lys Gly Val
35 40 45
Phe Arg Thr Arg Lys Glu Cys Ser Ser Thr Ser Asn Ala Glu Cys Asp
50 55 60
Ile Glu Gly Arg Met Asp Pro Lys Ser Cys Asp Lys Thr His Thr Cys
65 70 75 80
Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu
85 90 95
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
100 105 110
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
115 120 125
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
130 135 140
Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
145 150 155 160
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
165 170 175
Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
180 185 190
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
195 200 205
Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
210 215 220
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
225 230 235 240
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
245 250 255
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
260 265 270
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
275 280 285
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
290 295
<210> 179
<211> 377
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 179
Ser Pro Cys Pro Pro Asn Ser Phe Ser Ser Ala Gly Gly Gln Arg Thr
1 5 10 15
Cys Asp Ile Cys Arg Gln Cys Lys Gly Val Phe Arg Thr Arg Lys Glu
20 25 30
Cys Ser Ser Thr Ser Asn Ala Glu Cys Asp Cys Thr Pro Gly Phe His
35 40 45
Cys Leu Gly Ala Gly Cys Ser Met Cys Glu Gln Asp Cys Lys Gln Gly
50 55 60
Gln Glu Leu Thr Lys Lys Gly Cys Lys Asp Cys Cys Phe Gly Thr Phe
65 70 75 80
Asn Asp Gln Lys Arg Gly Ile Cys Arg Pro Trp Thr Asn Cys Ser Leu
85 90 95
Asp Gly Lys Ser Val Leu Val Asn Gly Thr Lys Glu Arg Asp Val Val
100 105 110
Cys Gly Pro Ser Pro Ala Asp Leu Ser Pro Gly Ala Ser Ser Val Thr
115 120 125
Pro Pro Ala Pro Ala Arg Glu Pro Gly His Ser Pro Gln Asp Ile Glu
130 135 140
Gly Arg Met Asp Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro
145 150 155 160
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
165 170 175
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
180 185 190
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
195 200 205
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
210 215 220
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
225 230 235 240
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
245 250 255
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
260 265 270
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
275 280 285
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
290 295 300
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
305 310 315 320
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
325 330 335
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
340 345 350
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
355 360 365
Thr Gln Lys Ser Leu Ser Leu Ser Pro
370 375
<210> 180
<211> 308
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 180
Ser Pro Cys Pro Pro Asn Ser Phe Ser Ser Ala Gly Gly Gln Arg Thr
1 5 10 15
Cys Asp Ile Cys Arg Gln Cys Lys Gly Val Phe Arg Thr Arg Lys Glu
20 25 30
Cys Ser Ser Thr Ser Asn Ala Glu Cys Asp Cys Thr Pro Gly Phe His
35 40 45
Cys Leu Gly Ala Gly Cys Ser Met Cys Glu Gln Asp Cys Lys Gln Gly
50 55 60
Gln Glu Leu Thr Lys Lys Gly Cys Asp Ile Glu Gly Arg Met Asp Pro
65 70 75 80
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
85 90 95
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
100 105 110
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
115 120 125
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
130 135 140
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
145 150 155 160
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
165 170 175
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
180 185 190
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
195 200 205
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
210 215 220
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
225 230 235 240
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
245 250 255
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
260 265 270
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
275 280 285
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
290 295 300
Ser Leu Ser Pro
305
<210> 181
<211> 336
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 181
Asp Cys Thr Pro Gly Phe His Cys Leu Gly Ala Gly Cys Ser Met Cys
1 5 10 15
Glu Gln Asp Cys Lys Gln Gly Gln Glu Leu Thr Lys Lys Gly Cys Lys
20 25 30
Asp Cys Cys Phe Gly Thr Phe Asn Asp Gln Lys Arg Gly Ile Cys Arg
35 40 45
Pro Trp Thr Asn Cys Ser Leu Asp Gly Lys Ser Val Leu Val Asn Gly
50 55 60
Thr Lys Glu Arg Asp Val Val Cys Gly Pro Ser Pro Ala Asp Leu Ser
65 70 75 80
Pro Gly Ala Ser Ser Val Thr Pro Pro Ala Pro Ala Arg Glu Pro Gly
85 90 95
His Ser Pro Gln Asp Ile Glu Gly Arg Met Asp Pro Lys Ser Cys Asp
100 105 110
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
115 120 125
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
130 135 140
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
145 150 155 160
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
165 170 175
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
180 185 190
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
195 200 205
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
210 215 220
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
225 230 235 240
Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu
245 250 255
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
260 265 270
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
275 280 285
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
290 295 300
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
305 310 315 320
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
325 330 335
<210> 182
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 182
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 183
<211> 189
<212> PRT
<213> Homo sapiens
<400> 183
Leu Gln Asp Pro Cys Ser Asn Cys Pro Ala Gly Thr Phe Cys Asp Asn
1 5 10 15
Asn Arg Asn Gln Ile Cys Ser Pro Cys Pro Pro Asn Ser Phe Ser Ser
20 25 30
Ala Gly Gly Gln Arg Thr Cys Asp Ile Cys Arg Gln Cys Lys Gly Val
35 40 45
Phe Arg Thr Arg Lys Glu Cys Ser Ser Thr Ser Asn Ala Glu Cys Asp
50 55 60
Cys Thr Pro Gly Phe His Cys Leu Gly Ala Gly Cys Ser Met Cys Glu
65 70 75 80
Gln Asp Cys Lys Gln Gly Gln Glu Leu Thr Lys Lys Gly Cys Lys Asp
85 90 95
Cys Cys Phe Gly Thr Phe Asn Asp Gln Lys Arg Gly Ile Cys Arg Pro
100 105 110
Trp Thr Asn Cys Ser Leu Asp Gly Lys Ser Val Leu Val Asn Gly Thr
115 120 125
Lys Glu Arg Asp Val Val Cys Gly Pro Ser Pro Ala Asp Leu Ser Pro
130 135 140
Gly Ala Ser Ser Val Thr Pro Pro Ala Pro Ala Arg Glu Pro Gly His
145 150 155 160
Ser Pro Gln His His His His His His Gly Gly Gly Gly Ser Gly Leu
165 170 175
Asn Asp Ile Phe Glu Ala Gln Lys Ile Glu Trp His Glu
180 185
<210> 184
<211> 330
<212> PRT
<213> Mus musculus
<400> 184
Ala Lys Thr Thr Ala Pro Ser Val Tyr Pro Leu Ala Pro Val Cys Gly
1 5 10 15
Asp Thr Thr Gly Ser Ser Val Thr Leu Gly Cys Leu Val Lys Gly Tyr
20 25 30
Phe Pro Glu Pro Val Thr Leu Thr Trp Asn Ser Gly Ser Leu Ser Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Asp Leu Tyr Thr Leu
50 55 60
Ser Ser Ser Val Thr Val Thr Ser Ser Thr Trp Pro Ser Gln Ser Ile
65 70 75 80
Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys Lys
85 90 95
Ile Glu Pro Arg Gly Pro Thr Ile Lys Pro Cys Pro Pro Cys Lys Cys
100 105 110
Pro Ala Pro Asn Leu Leu Gly Gly Pro Asp Val Phe Ile Phe Pro Pro
115 120 125
Lys Ile Lys Asp Val Leu Met Ile Ser Leu Ser Pro Ile Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Glu Asp Asp Pro Asp Val Gln Ile Ser Trp
145 150 155 160
Phe Val Asn Asn Val Glu Val His Thr Ala Gln Thr Gln Thr His Arg
165 170 175
Glu Asp Tyr Asn Ser Thr Leu Arg Val Val Ser Ala Leu Pro Ile Gln
180 185 190
His Gln Asp Trp Met Ser Gly Lys Glu Phe Lys Cys Lys Val Asn Asn
195 200 205
Lys Asp Leu Pro Ala Pro Ile Glu Arg Thr Ile Ser Lys Pro Lys Gly
210 215 220
Ser Val Arg Ala Pro Gln Val Tyr Val Leu Pro Pro Pro Glu Glu Glu
225 230 235 240
Met Thr Lys Lys Gln Val Thr Leu Thr Cys Met Val Thr Asp Phe Met
245 250 255
Pro Glu Asp Ile Tyr Val Glu Trp Thr Asn Asn Gly Lys Thr Glu Leu
260 265 270
Asn Tyr Lys Asn Thr Glu Pro Val Leu Asp Ser Asp Gly Ser Tyr Phe
275 280 285
Met Tyr Ser Lys Leu Arg Val Glu Lys Lys Asn Trp Val Glu Arg Asn
290 295 300
Ser Tyr Ser Cys Ser Val Val His Glu Gly Leu His Asn His His Thr
305 310 315 320
Thr Lys Ser Phe Ser Arg Thr Pro Gly Lys
325 330
<210> 185
<211> 107
<212> PRT
<213> Mus musculus
<400> 185
Arg Ala Asp Ala Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu
1 5 10 15
Gln Leu Thr Ser Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe
20 25 30
Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg
35 40 45
Gln Asn Gly Val Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Met Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu
65 70 75 80
Arg His Asn Ser Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser
85 90 95
Pro Ile Val Lys Ser Phe Asn Arg Asn Glu Cys
100 105
<210> 186
<211> 107
<212> PRT
<213> Homo sapiens
<400> 186
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 187
<211> 116
<212> PRT
<213> Artificial Sequence
<220>
<223> IC17Hdk
<400> 187
Gln Val Gln Leu Gln Gln Ser Gly Pro Gln Leu Val Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Ser Tyr
20 25 30
Trp Met His Trp Val Asn Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Met Ile Asp Pro Ser Tyr Ser Glu Thr Arg Leu Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Pro Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Leu Tyr Gly Asn Tyr Phe Asp Tyr Trp Gly Gln Gly Thr Thr Leu
100 105 110
Thr Val Ser Ser
115
<210> 188
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> IC17VL
<400> 188
Asp Ile Gln Met Thr Gln Ser Ser Ser Ser Phe Ser Val Ser Leu Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Glu Asp Ile Tyr Asn Arg
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Asn Ala Pro Arg Leu Leu Ile
35 40 45
Ser Gly Ala Thr Ser Leu Glu Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Lys Asp Tyr Thr Leu Ser Ile Thr Ser Leu Gln Thr
65 70 75 80
Glu Asp Val Ala Thr Tyr Tyr Cys Gln Gln Tyr Trp Ser Thr Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Val Lys
100 105
<210> 189
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> lam1
<400> 189
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
35 40 45
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 190
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> MRAH
<400> 190
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Arg Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Tyr Ser Ile Thr Ser Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Arg Gly Leu Glu Trp
35 40 45
Ile Gly Tyr Ile Ser Tyr Ser Gly Ile Thr Thr Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Val Thr Met Leu Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Arg Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Ser Leu Val Thr Val Ser Ser
115
<210> 191
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> MRAL
<400> 191
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 192
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> H0002
<400> 192
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Tyr His Ala Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Lys Leu Tyr Ser Leu Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 193
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> L1058
<400> 193
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Asp Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Val Val Lys Lys Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Leu Arg Ser Ala Gly
85 90 95
Pro Tyr Pro Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 194
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> H0041
<400> 194
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Gln Ser Tyr His Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Lys Leu Tyr Ser Leu Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 195
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> L1088
<400> 195
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Asp Tyr
20 25 30
Ala Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Tyr Val Ser Lys Lys Pro Asp Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Leu Arg Ser Pro Gly
85 90 95
Pro Tyr Pro Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 196
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> H0052
<400> 196
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Tyr His Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Asp Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Lys Leu Tyr Ser Leu Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 197
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> L1083
<400> 197
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Asp Tyr
20 25 30
Ala Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Tyr Val Asp Lys Lys Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Leu Arg Ser Pro Gly
85 90 95
Pro Tyr Pro Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 198
<211> 325
<212> PRT
<213> Artificial Sequence
<220>
<223> G4T1
<400> 198
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Arg Arg Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Pro
325
<210> 199
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> k0
<400> 199
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 200
<211> 338
<212> PRT
<213> Homo sapiens
<400> 200
Leu Ala Pro Arg Arg Cys Pro Ala Gln Glu Val Ala Arg Gly Val Leu
1 5 10 15
Thr Ser Leu Pro Gly Asp Ser Val Thr Leu Thr Cys Pro Gly Val Glu
20 25 30
Pro Glu Asp Asn Ala Thr Val His Trp Val Leu Arg Lys Pro Ala Ala
35 40 45
Gly Ser His Pro Ser Arg Trp Ala Gly Met Gly Arg Arg Leu Leu Leu
50 55 60
Arg Ser Val Gln Leu His Asp Ser Gly Asn Tyr Ser Cys Tyr Arg Ala
65 70 75 80
Gly Arg Pro Ala Gly Thr Val His Leu Leu Val Asp Val Pro Pro Glu
85 90 95
Glu Pro Gln Leu Ser Cys Phe Arg Lys Ser Pro Leu Ser Asn Val Val
100 105 110
Cys Glu Trp Gly Pro Arg Ser Thr Pro Ser Leu Thr Thr Lys Ala Val
115 120 125
Leu Leu Val Arg Lys Phe Gln Asn Ser Pro Ala Glu Asp Phe Gln Glu
130 135 140
Pro Cys Gln Tyr Ser Gln Glu Ser Gln Lys Phe Ser Cys Gln Leu Ala
145 150 155 160
Val Pro Glu Gly Asp Ser Ser Phe Tyr Ile Val Ser Met Cys Val Ala
165 170 175
Ser Ser Val Gly Ser Lys Phe Ser Lys Thr Gln Thr Phe Gln Gly Cys
180 185 190
Gly Ile Leu Gln Pro Asp Pro Pro Ala Asn Ile Thr Val Thr Ala Val
195 200 205
Ala Arg Asn Pro Arg Trp Leu Ser Val Thr Trp Gln Asp Pro His Ser
210 215 220
Trp Asn Ser Ser Phe Tyr Arg Leu Arg Phe Glu Leu Arg Tyr Arg Ala
225 230 235 240
Glu Arg Ser Lys Thr Phe Thr Thr Trp Met Val Lys Asp Leu Gln His
245 250 255
His Cys Val Ile His Asp Ala Trp Ser Gly Leu Arg His Val Val Gln
260 265 270
Leu Arg Ala Gln Glu Glu Phe Gly Gln Gly Glu Trp Ser Glu Trp Ser
275 280 285
Pro Glu Ala Met Gly Thr Pro Trp Thr Glu Ser Arg Ser Pro Pro Ala
290 295 300
Glu Asn Glu Val Ser Thr Pro Met Gln Ala Leu Thr Thr Asn Lys Asp
305 310 315 320
Asp Asp Asn Ile Leu Phe Arg Asp Ser Ala Asn Ala Thr Ser Leu Pro
325 330 335
Val Gln
<210> 201
<211> 118
<212> PRT
<213> Artificial Sequence
<220>
<223> mPD1F2VH
<400> 201
Glu Val Gln Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Glu Asn Trp Gly Ser Tyr Phe Asp Leu Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser
115
<210> 202
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> mPD1F2VL
<400> 202
Asp Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Arg Ala Pro Lys Val Leu Ile
35 40 45
Tyr Lys Ala Ser Thr Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Ser Thr Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 203
<211> 324
<212> PRT
<213> Mus musculus
<400> 203
Ala Lys Thr Thr Pro Pro Ser Val Tyr Pro Leu Ala Pro Gly Ser Ala
1 5 10 15
Ala Gln Thr Asn Ser Met Val Thr Leu Gly Cys Leu Val Lys Gly Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ser Leu Ser Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Asp Leu Tyr Thr Leu
50 55 60
Ser Ser Ser Val Thr Val Pro Ser Ser Thr Trp Pro Ser Glu Thr Val
65 70 75 80
Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys Lys
85 90 95
Ile Val Pro Arg Asp Cys Gly Cys Lys Pro Cys Ile Cys Thr Val Lys
100 105 110
Glu Val Ser Lys Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Val Leu
115 120 125
Thr Ile Thr Leu Thr Pro Lys Val Thr Cys Val Val Val Asp Ile Ser
130 135 140
Lys Asp Asp Pro Glu Val Gln Phe Ser Trp Phe Val Asp Asp Val Glu
145 150 155 160
Val His Thr Ala Gln Thr Gln Pro Arg Glu Glu Gln Phe Asn Ser Thr
165 170 175
Phe Arg Ser Val Ser Glu Leu Pro Ile Met His Gln Asp Trp Leu Asn
180 185 190
Gly Lys Glu Phe Lys Cys Arg Val Asn Ser Ala Ala Phe Pro Ala Pro
195 200 205
Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Arg Pro Lys Ala Pro Gln
210 215 220
Val Tyr Thr Ile Pro Pro Pro Lys Glu Gln Met Ala Lys Asp Lys Val
225 230 235 240
Ser Leu Thr Cys Met Ile Thr Asp Phe Phe Pro Glu Asp Ile Thr Val
245 250 255
Glu Trp Gln Trp Asn Gly Gln Pro Ala Glu Asn Tyr Lys Asn Thr Gln
260 265 270
Pro Ile Met Asp Thr Asp Gly Ser Tyr Phe Val Tyr Ser Lys Leu Asn
275 280 285
Val Gln Lys Ser Asn Trp Glu Ala Gly Asn Thr Phe Thr Cys Ser Val
290 295 300
Leu His Glu Gly Leu His Asn His His Thr Glu Lys Ser Leu Ser His
305 310 315 320
Ser Pro Gly Lys
<210> 204
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> H5029
<400> 204
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Thr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Ser Ser Arg His Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Arg Pro Phe Glu Leu Leu Trp Val Leu Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 205
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> L3021
<400> 205
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Pro Ser Gly Ala Trp
20 25 30
Asp Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Gln Val Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Thr Thr Ser Ser Ala
85 90 95
Trp Asp Pro Asp Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 206
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> H5041
<400> 206
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Thr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Ser Ser Lys His Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Arg Pro Trp Glu Leu Leu Trp Val Leu Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 207
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> L3023
<400> 207
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Pro Ser Gly Ala Tyr
20 25 30
Asp Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Gln Val Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Thr Thr Asn Ser Ala
85 90 95
Trp Asp Pro Asp Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 208
<211> 330
<212> PRT
<213> Artificial Sequence
<220>
<223> mFa31
<400> 208
Ala Lys Thr Thr Ala Pro Ser Val Tyr Pro Leu Ala Pro Val Cys Gly
1 5 10 15
Asp Thr Thr Gly Ser Ser Val Thr Leu Gly Cys Leu Val Lys Gly Tyr
20 25 30
Phe Pro Glu Pro Val Thr Leu Thr Trp Asn Ser Gly Ser Leu Ser Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Asp Leu Tyr Thr Leu
50 55 60
Ser Ser Ser Val Thr Val Thr Ser Ser Thr Trp Pro Ser Gln Ser Ile
65 70 75 80
Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys Lys
85 90 95
Ile Glu Pro Arg Gly Pro Thr Ile Lys Pro Cys Pro Pro Cys Lys Cys
100 105 110
Pro Ala Pro Asn Leu Arg Arg Gly Pro Lys Val Phe Ile Phe Pro Pro
115 120 125
Lys Ile Lys Asp Val Leu Met Ile Ser Leu Ser Pro Ile Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Glu Asp Asp Pro Asp Val Gln Ile Ser Trp
145 150 155 160
Phe Val Asn Asn Val Glu Val His Thr Ala Gln Thr Gln Thr His Arg
165 170 175
Glu Asp Tyr Asn Ser Thr Leu Arg Val Val Ser Ala Leu Pro Ile Gln
180 185 190
His Gln Asp Trp Met Ser Gly Lys Glu Phe Lys Cys Lys Val Asn Asn
195 200 205
Lys Asp Leu Pro Ala Pro Ile Glu Arg Thr Ile Ser Lys Pro Lys Gly
210 215 220
Ser Val Arg Ala Pro Gln Val Tyr Val Leu Pro Pro Pro Glu Glu Glu
225 230 235 240
Met Thr Lys Lys Gln Val Thr Leu Thr Cys Met Val Thr Asp Phe Met
245 250 255
Pro Glu Asp Ile Tyr Val Glu Trp Thr Asn Asn Gly Lys Thr Glu Leu
260 265 270
Asn Tyr Lys Asn Thr Glu Pro Val Leu Asp Ser Asp Gly Ser Tyr Phe
275 280 285
Met Tyr Ser Lys Leu Arg Val Glu Lys Lys Asn Trp Val Glu Arg Asn
290 295 300
Ser Tyr Ser Cys Ser Val Val His Glu Gly Leu His Asn His His Thr
305 310 315 320
Thr Lys Ser Phe Ser Arg Thr Pro Gly Lys
325 330
<210> 209
<211> 402
<212> PRT
<213> Artificial Sequence
<220>
<223> mPD1-G1CH2
<400> 209
Trp Val Arg Gln Val Pro Trp Ser Phe Thr Trp Ala Val Leu Gln Leu
1 5 10 15
Ser Trp Gln Ser Gly Trp Leu Leu Glu Val Pro Asn Gly Pro Trp Arg
20 25 30
Ser Leu Thr Phe Tyr Pro Ala Trp Leu Thr Val Ser Glu Gly Ala Asn
35 40 45
Ala Thr Phe Thr Cys Ser Leu Ser Asn Trp Ser Glu Asp Leu Met Leu
50 55 60
Asn Trp Asn Arg Leu Ser Pro Ser Asn Gln Thr Glu Lys Gln Ala Ala
65 70 75 80
Phe Ser Asn Gly Leu Ser Gln Pro Val Gln Asp Ala Arg Phe Gln Ile
85 90 95
Ile Gln Leu Pro Asn Arg His Asp Phe His Met Asn Ile Leu Asp Thr
100 105 110
Arg Arg Asn Asp Ser Gly Ile Tyr Leu Cys Gly Ala Ile Ser Leu His
115 120 125
Pro Lys Ala Lys Ile Glu Glu Ser Pro Gly Ala Glu Leu Val Val Thr
130 135 140
Glu Arg Ile Leu Glu Thr Ser Thr Arg Tyr Pro Ser Pro Ser Pro Lys
145 150 155 160
Pro Glu Gly Arg Phe Gln Asp Ile Glu Gly Arg Met Asp Pro Lys Ser
165 170 175
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
180 185 190
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
195 200 205
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
210 215 220
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
225 230 235 240
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
245 250 255
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
260 265 270
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
275 280 285
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
290 295 300
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
305 310 315 320
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
325 330 335
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
340 345 350
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
355 360 365
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
370 375 380
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
385 390 395 400
Ser Pro
<210> 210
<211> 463
<212> PRT
<213> Artificial Sequence
<220>
<223> mPDL1-G1dCH2His
<400> 210
Phe Thr Ile Thr Ala Pro Lys Asp Leu Tyr Val Val Glu Tyr Gly Ser
1 5 10 15
Asn Val Thr Met Glu Cys Arg Phe Pro Val Glu Arg Glu Leu Asp Leu
20 25 30
Leu Ala Leu Val Val Tyr Trp Glu Lys Glu Asp Glu Gln Val Ile Gln
35 40 45
Phe Val Ala Gly Glu Glu Asp Leu Lys Pro Gln His Ser Asn Phe Arg
50 55 60
Gly Arg Ala Ser Leu Pro Lys Asp Gln Leu Leu Lys Gly Asn Ala Ala
65 70 75 80
Leu Gln Ile Thr Asp Val Lys Leu Gln Asp Ala Gly Val Tyr Cys Cys
85 90 95
Ile Ile Ser Tyr Gly Gly Ala Asp Tyr Lys Arg Ile Thr Leu Lys Val
100 105 110
Asn Ala Pro Tyr Arg Lys Ile Asn Gln Arg Ile Ser Val Asp Pro Ala
115 120 125
Thr Ser Glu His Glu Leu Ile Cys Gln Ala Glu Gly Tyr Pro Glu Ala
130 135 140
Glu Val Ile Trp Thr Asn Ser Asp His Gln Pro Val Ser Gly Lys Arg
145 150 155 160
Ser Val Thr Thr Ser Arg Thr Glu Gly Met Leu Leu Asn Val Thr Ser
165 170 175
Ser Leu Arg Val Asn Ala Thr Ala Asn Asp Val Phe Tyr Cys Thr Phe
180 185 190
Trp Arg Ser Gln Pro Gly Gln Asn His Thr Ala Glu Leu Ile Ile Pro
195 200 205
Glu Leu Pro Ala Thr His Pro Pro Gln Asn Arg Thr His Ile Glu Gly
210 215 220
Arg Met Asp Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro
225 230 235 240
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
245 250 255
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
260 265 270
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
275 280 285
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
290 295 300
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
305 310 315 320
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
325 330 335
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
340 345 350
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
355 360 365
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
370 375 380
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
385 390 395 400
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
405 410 415
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
420 425 430
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
435 440 445
Thr Gln Lys Ser Leu Ser Leu Ser Pro His His His His His His
450 455 460
<210> 211
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> mk0
<400> 211
Arg Ala Asp Ala Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu
1 5 10 15
Gln Leu Thr Ser Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe
20 25 30
Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg
35 40 45
Gln Asn Gly Val Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Met Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu
65 70 75 80
Arg His Asn Ser Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser
85 90 95
Pro Ile Val Lys Ser Phe Asn Arg Asn Glu Cys
100 105
<210> 212
<211> 105
<212> PRT
<213> Artificial Sequence
<220>
<223> ml0
<400> 212
Gln Pro Lys Ser Ser Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu
1 5 10 15
Glu Leu Glu Thr Asn Lys Ala Thr Leu Val Cys Thr Ile Thr Asp Phe
20 25 30
Tyr Pro Gly Val Val Thr Val Asp Trp Lys Val Asp Gly Thr Pro Val
35 40 45
Thr Gln Gly Met Glu Thr Thr Gln Pro Ser Lys Gln Ser Asn Asn Lys
50 55 60
Tyr Met Ala Ser Ser Tyr Leu Thr Leu Thr Ala Arg Ala Trp Glu Arg
65 70 75 80
His Ser Ser Tyr Ser Cys Gln Val Thr His Glu Gly His Thr Val Glu
85 90 95
Lys Ser Leu Ser Arg Ala Asp Cys Ser
100 105
<210> 213
<211> 468
<212> PRT
<213> Artificial Sequence
<220>
<223> hIL6R
<400> 213
Met Leu Ala Val Gly Cys Ala Leu Leu Ala Ala Leu Leu Ala Ala Pro
1 5 10 15
Gly Ala Ala Leu Ala Pro Arg Arg Cys Pro Ala Gln Glu Val Ala Arg
20 25 30
Gly Val Leu Thr Ser Leu Pro Gly Asp Ser Val Thr Leu Thr Cys Pro
35 40 45
Gly Val Glu Pro Glu Asp Asn Ala Thr Val His Trp Val Leu Arg Lys
50 55 60
Pro Ala Ala Gly Ser His Pro Ser Arg Trp Ala Gly Met Gly Arg Arg
65 70 75 80
Leu Leu Leu Arg Ser Val Gln Leu His Asp Ser Gly Asn Tyr Ser Cys
85 90 95
Tyr Arg Ala Gly Arg Pro Ala Gly Thr Val His Leu Leu Val Asp Val
100 105 110
Pro Pro Glu Glu Pro Gln Leu Ser Cys Phe Arg Lys Ser Pro Leu Ser
115 120 125
Asn Val Val Cys Glu Trp Gly Pro Arg Ser Thr Pro Ser Leu Thr Thr
130 135 140
Lys Ala Val Leu Leu Val Arg Lys Phe Gln Asn Ser Pro Ala Glu Asp
145 150 155 160
Phe Gln Glu Pro Cys Gln Tyr Ser Gln Glu Ser Gln Lys Phe Ser Cys
165 170 175
Gln Leu Ala Val Pro Glu Gly Asp Ser Ser Phe Tyr Ile Val Ser Met
180 185 190
Cys Val Ala Ser Ser Val Gly Ser Lys Phe Ser Lys Thr Gln Thr Phe
195 200 205
Gln Gly Cys Gly Ile Leu Gln Pro Asp Pro Pro Ala Asn Ile Thr Val
210 215 220
Thr Ala Val Ala Arg Asn Pro Arg Trp Leu Ser Val Thr Trp Gln Asp
225 230 235 240
Pro His Ser Trp Asn Ser Ser Phe Tyr Arg Leu Arg Phe Glu Leu Arg
245 250 255
Tyr Arg Ala Glu Arg Ser Lys Thr Phe Thr Thr Trp Met Val Lys Asp
260 265 270
Leu Gln His His Cys Val Ile His Asp Ala Trp Ser Gly Leu Arg His
275 280 285
Val Val Gln Leu Arg Ala Gln Glu Glu Phe Gly Gln Gly Glu Trp Ser
290 295 300
Glu Trp Ser Pro Glu Ala Met Gly Thr Pro Trp Thr Glu Ser Arg Ser
305 310 315 320
Pro Pro Ala Glu Asn Glu Val Ser Thr Pro Met Gln Ala Leu Thr Thr
325 330 335
Asn Lys Asp Asp Asp Asn Ile Leu Phe Arg Asp Ser Ala Asn Ala Thr
340 345 350
Ser Leu Pro Val Gln Asp Ser Ser Ser Val Pro Leu Pro Thr Phe Leu
355 360 365
Val Ala Gly Gly Ser Leu Ala Phe Gly Thr Leu Leu Cys Ile Ala Ile
370 375 380
Val Leu Arg Phe Lys Lys Thr Trp Lys Leu Arg Ala Leu Lys Glu Gly
385 390 395 400
Lys Thr Ser Met His Pro Pro Tyr Ser Leu Gly Gln Leu Val Pro Glu
405 410 415
Arg Pro Arg Pro Thr Pro Val Leu Val Pro Leu Ile Ser Pro Pro Val
420 425 430
Ser Pro Ser Ser Leu Gly Ser Asp Asn Thr Ser Ser His Asn Arg Pro
435 440 445
Asp Ala Arg Asp Pro Arg Ser Pro Tyr Asp Ile Ser Asn Thr Asp Tyr
450 455 460
Phe Phe Pro Arg
465
<210> 214
<211> 288
<212> PRT
<213> Mus musculus
<400> 214
Met Trp Val Arg Gln Val Pro Trp Ser Phe Thr Trp Ala Val Leu Gln
1 5 10 15
Leu Ser Trp Gln Ser Gly Trp Leu Leu Glu Val Pro Asn Gly Pro Trp
20 25 30
Arg Ser Leu Thr Phe Tyr Pro Ala Trp Leu Thr Val Ser Glu Gly Ala
35 40 45
Asn Ala Thr Phe Thr Cys Ser Leu Ser Asn Trp Ser Glu Asp Leu Met
50 55 60
Leu Asn Trp Asn Arg Leu Ser Pro Ser Asn Gln Thr Glu Lys Gln Ala
65 70 75 80
Ala Phe Cys Asn Gly Leu Ser Gln Pro Val Gln Asp Ala Arg Phe Gln
85 90 95
Ile Ile Gln Leu Pro Asn Arg His Asp Phe His Met Asn Ile Leu Asp
100 105 110
Thr Arg Arg Asn Asp Ser Gly Ile Tyr Leu Cys Gly Ala Ile Ser Leu
115 120 125
His Pro Lys Ala Lys Ile Glu Glu Ser Pro Gly Ala Glu Leu Val Val
130 135 140
Thr Glu Arg Ile Leu Glu Thr Ser Thr Arg Tyr Pro Ser Pro Ser Pro
145 150 155 160
Lys Pro Glu Gly Arg Phe Gln Gly Leu Val Val Gly Val Val Gly Gly
165 170 175
Leu Leu Gly Ser Leu Val Leu Leu Val Trp Val Leu Ala Val Ile Cys
180 185 190
Ser Arg Ala Ala Arg Gly Thr Ile Gly Ala Arg Arg Thr Gly Gln Pro
195 200 205
Leu Lys Glu Asp Pro Ser Ala Val Pro Val Phe Ser Val Asp Tyr Gly
210 215 220
Glu Leu Asp Phe Gln Trp Arg Glu Lys Thr Pro Glu Pro Pro Val Pro
225 230 235 240
Cys Val Pro Glu Gln Thr Glu Tyr Ala Thr Ile Val Phe Pro Ser Gly
245 250 255
Met Gly Thr Ser Ser Pro Ala Arg Arg Gly Ser Ala Asp Gly Pro Arg
260 265 270
Ser Ala Gln Pro Leu Arg Pro Glu Asp Gly His Cys Ser Trp Pro Leu
275 280 285
<210> 215
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> G1m
<400> 215
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 216
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> lam1
<400> 216
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
35 40 45
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 217
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> G1d
<400> 217
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 218
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> k0
<400> 218
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 219
<211> 330
<212> PRT
<213> Homo sapiens
<400> 219
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 220
<211> 326
<212> PRT
<213> Homo sapiens
<400> 220
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro Gly Lys
325
<210> 221
<211> 377
<212> PRT
<213> Homo sapiens
<400> 221
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Leu Lys Thr Pro Leu Gly Asp Thr Thr His Thr Cys Pro
100 105 110
Arg Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg
115 120 125
Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys
130 135 140
Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys Pro
145 150 155 160
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
165 170 175
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
180 185 190
Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Lys Trp Tyr
195 200 205
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
210 215 220
Gln Tyr Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Leu His
225 230 235 240
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
245 250 255
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln
260 265 270
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
275 280 285
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
290 295 300
Ser Asp Ile Ala Val Glu Trp Glu Ser Ser Gly Gln Pro Glu Asn Asn
305 310 315 320
Tyr Asn Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu
325 330 335
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Ile
340 345 350
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn Arg Phe Thr Gln
355 360 365
Lys Ser Leu Ser Leu Ser Pro Gly Lys
370 375
<210> 222
<211> 327
<212> PRT
<213> Homo sapiens
<400> 222
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 223
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> G1T6
<400> 223
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 224
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> TR01H113
<400> 224
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Ala
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Lys Asp Lys Ser Gln Asn Tyr Ala Thr Tyr Val Ala Glu
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Ala Asp Ser Lys Asn Ser
65 70 75 80
Ile Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Arg Tyr Val His Tyr Ala Ala Gly Tyr Gly Val Asp Ile Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 225
<211> 219
<212> PRT
<213> Artificial Sequence
<220>
<223> L0011-k0
<400> 225
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Pro Leu Val His Ser
20 25 30
Asn Arg Asn Thr Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala
35 40 45
Pro Arg Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Val Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| БИСПЕЦИФИЧНЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2800779C2 |
| МУЛЬТИСПЕЦИФИЧЕСКАЯ АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА, ОБЛАДАЮЩАЯ ЗАМЕЩАЮЩЕЙ ФУНКЦИОНАЛЬНОЙ АКТИВНОСТЬЮ КОФАКТОРА КОАГУЛИРУЮЩЕГО ФАКТОРА КРОВИ VIII, И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ УКАЗАННУЮ МОЛЕКУЛУ В КАЧЕСТВЕ АКТИВНОГО ИНГРЕДИЕНТА | 2018 |
|
RU2812909C2 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ НА ОСНОВЕ АКТИВИРУЮЩИХ Т-КЛЕТКИ БИСПЕЦИФИЧЕСКИХ АНТИГЕНСВЯЗЫВАЮЩИХ МОЛЕКУЛ ПРОТИВ CD3 И ФОЛАТНОГО РЕЦЕПТОРА 1 (FOLR1) И АНТАГОНИСТОВ, СВЯЗЫВАЮЩИХСЯ С ОСЬЮ PD-1 | 2015 |
|
RU2753902C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, СПЕЦИФИЧЕСКИ СВЯЗЫВАЮЩИЕСЯ С PD1 И LAG3 | 2018 |
|
RU2778805C2 |
| ИММУНОАКТИВИРУЮЩАЯ АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА | 2015 |
|
RU2722788C2 |
| АНТИТЕЛА ПРОТИВ СИГНАЛ-РЕГУЛЯТОРНОГО БЕЛКА АЛЬФА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2771964C2 |
| НОВЫЕ СЛИТЫЕ БЕЛКИ, СПЕЦИФИЧЕСКИЕ В ОТНОШЕНИИ CD137 И GPC3 | 2020 |
|
RU2814653C2 |
| АНТИТЕЛА К МИОСТАТИНУ, ПОЛИПЕПТИДЫ, СОДЕРЖАЩИЕ ВАРИАНТЫ FC-ОБЛАСТЕЙ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2789884C2 |
| АНТИТЕЛА К МИОСТАТИНУ, ПОЛИПЕПТИДЫ, СОДЕРЖАЩИЕ ВАРИАНТЫ FC-ОБЛАСТЕЙ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2799522C2 |
| АНТИТЕЛА К МИОСТАТИНУ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2778945C2 |


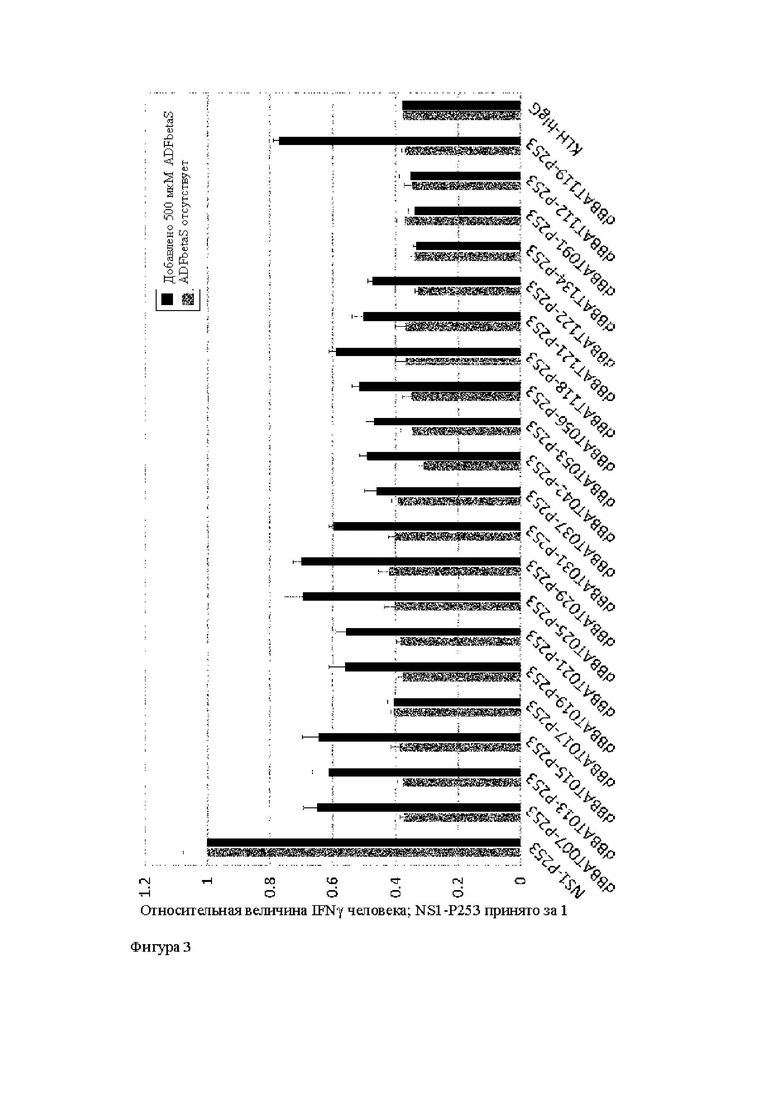
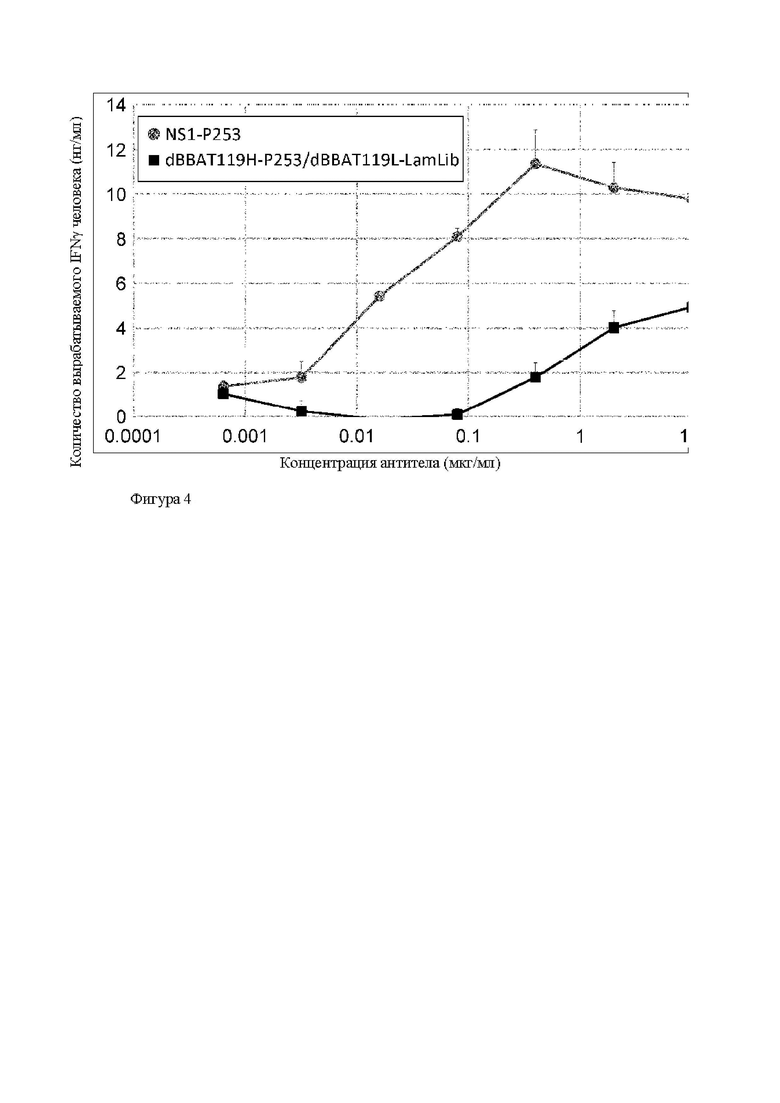
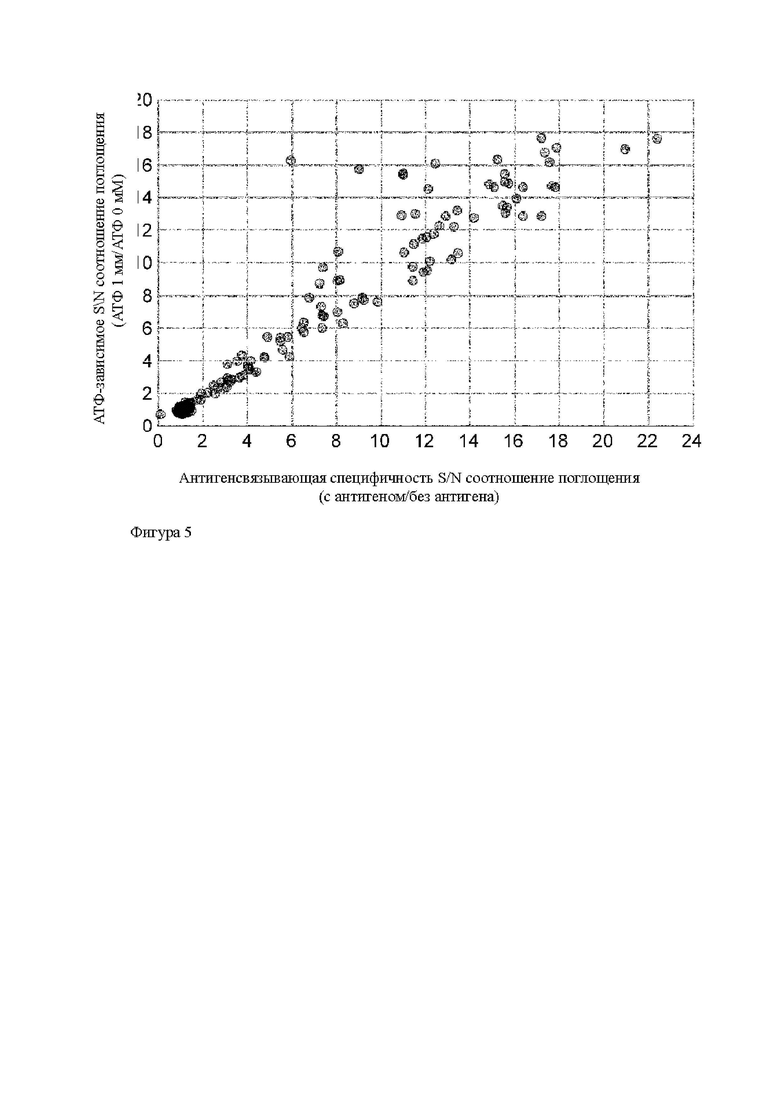
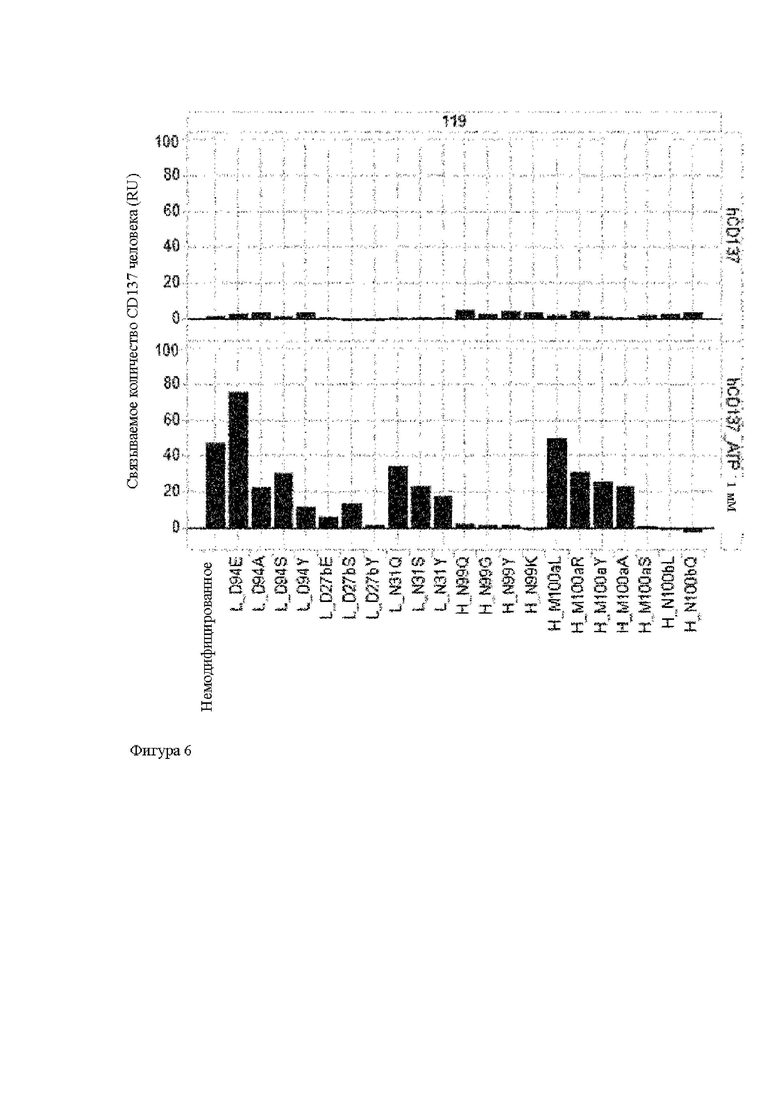
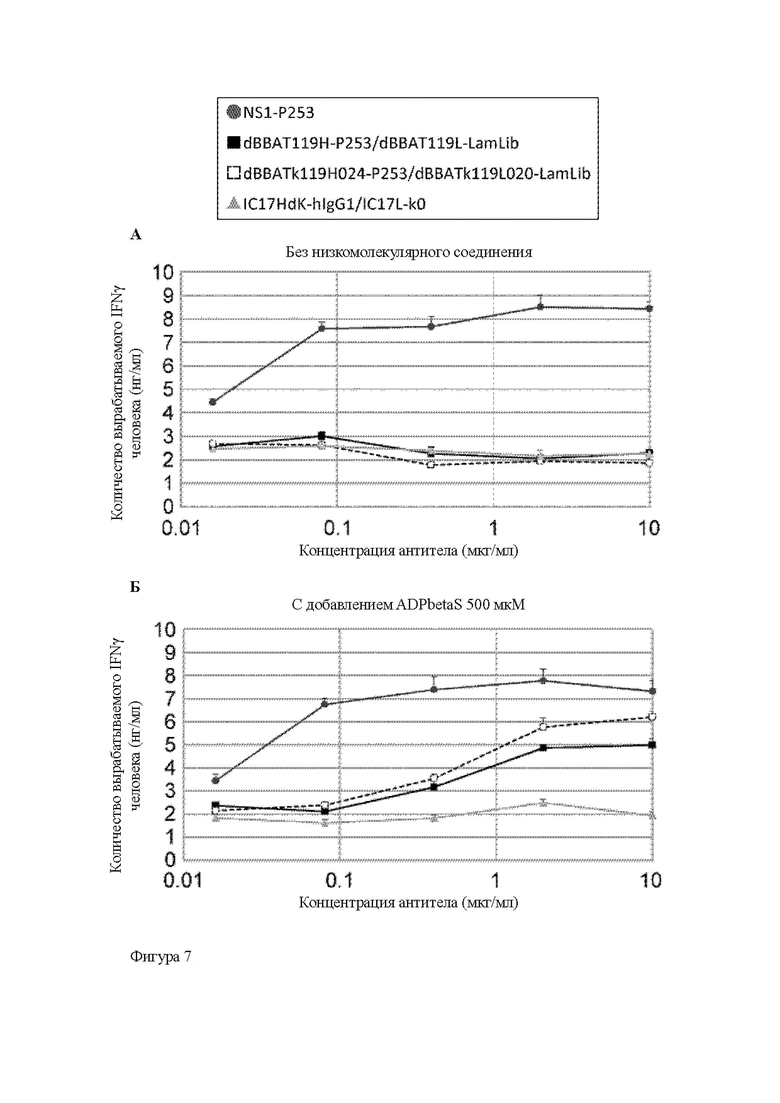
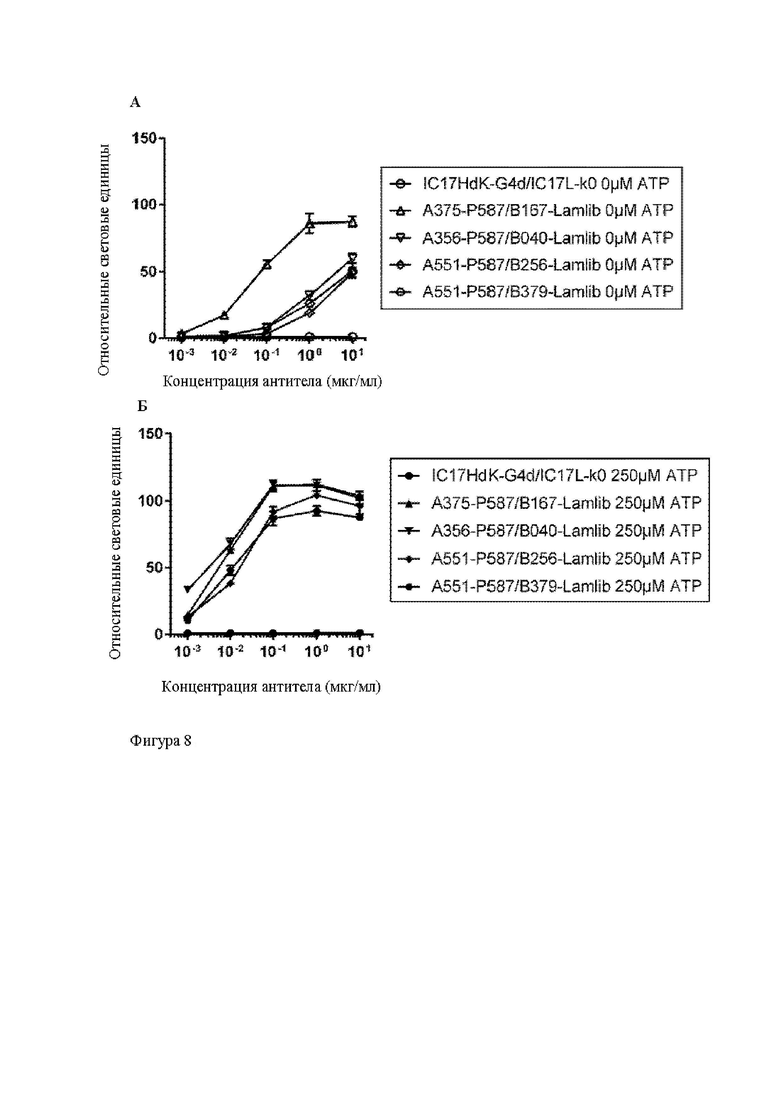
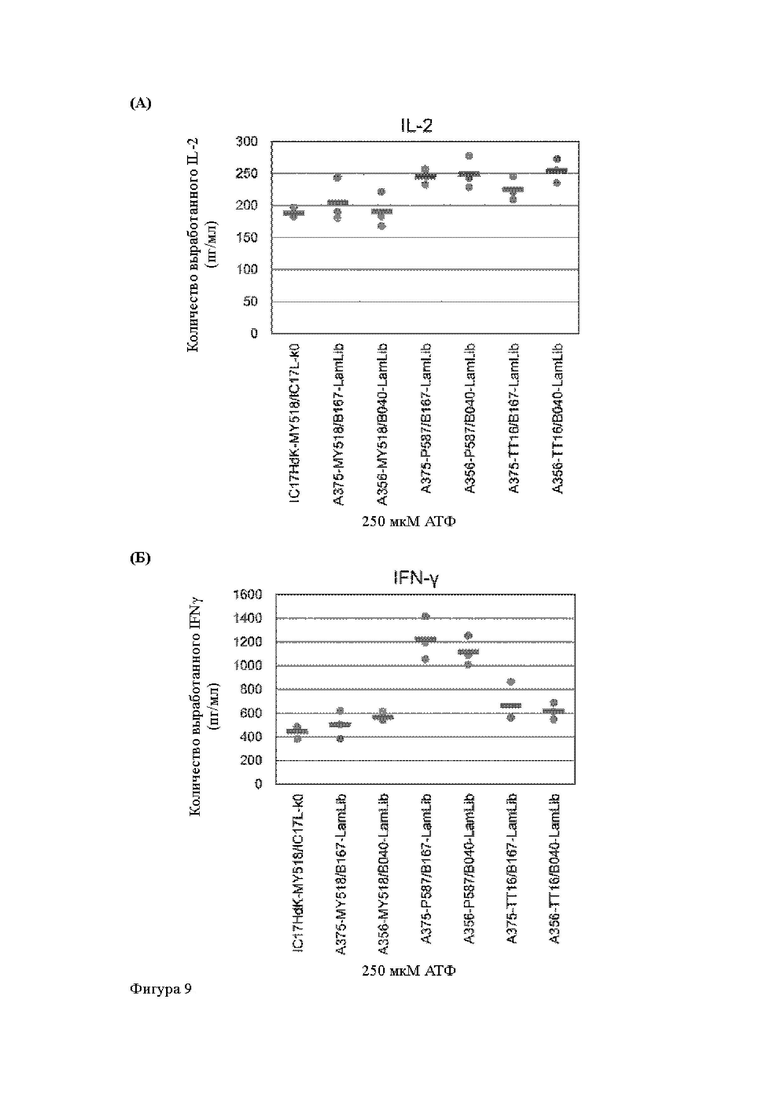
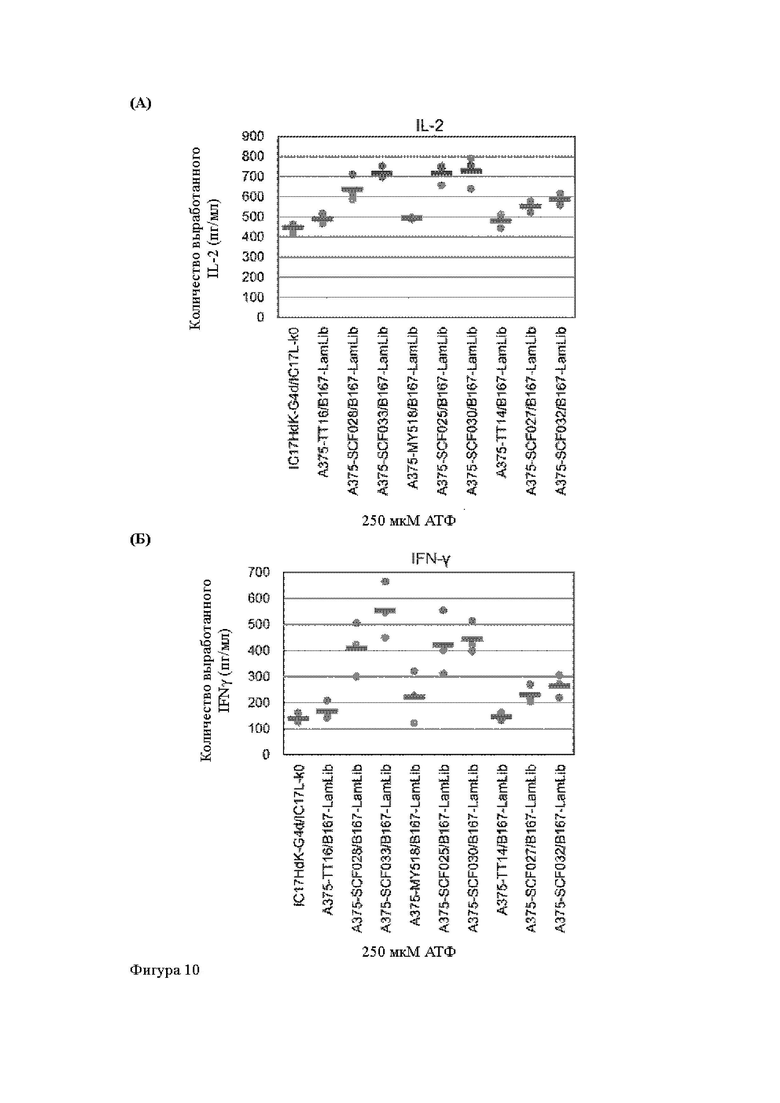
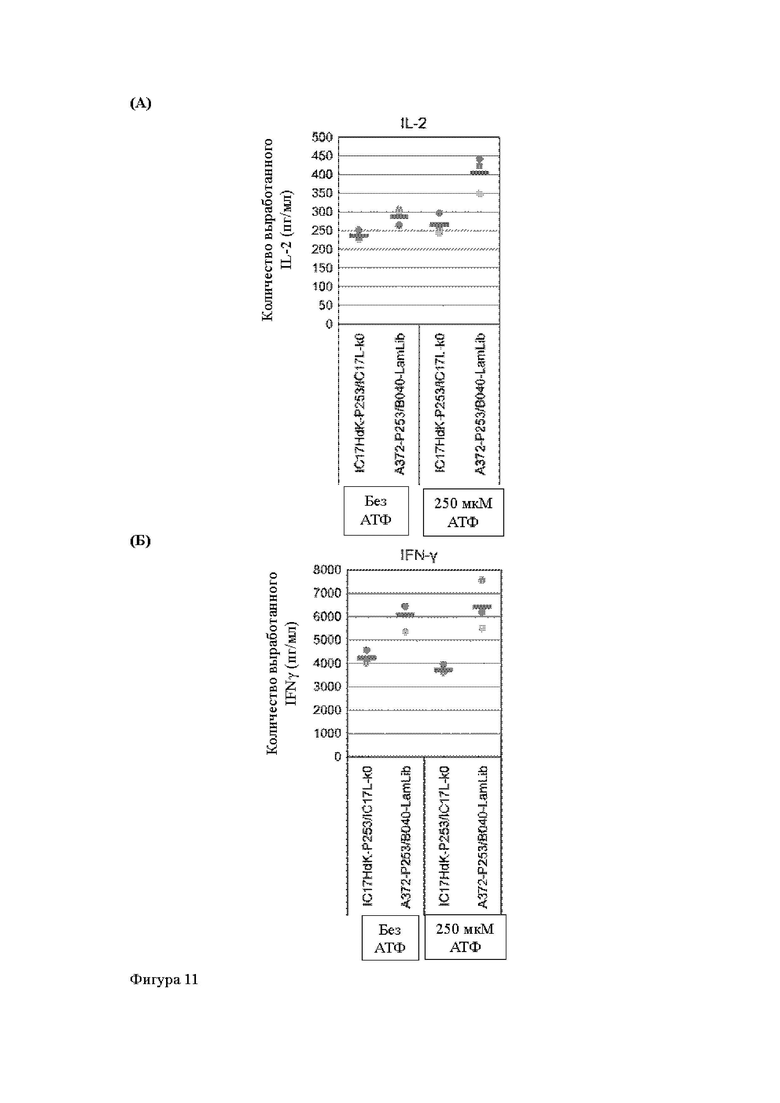
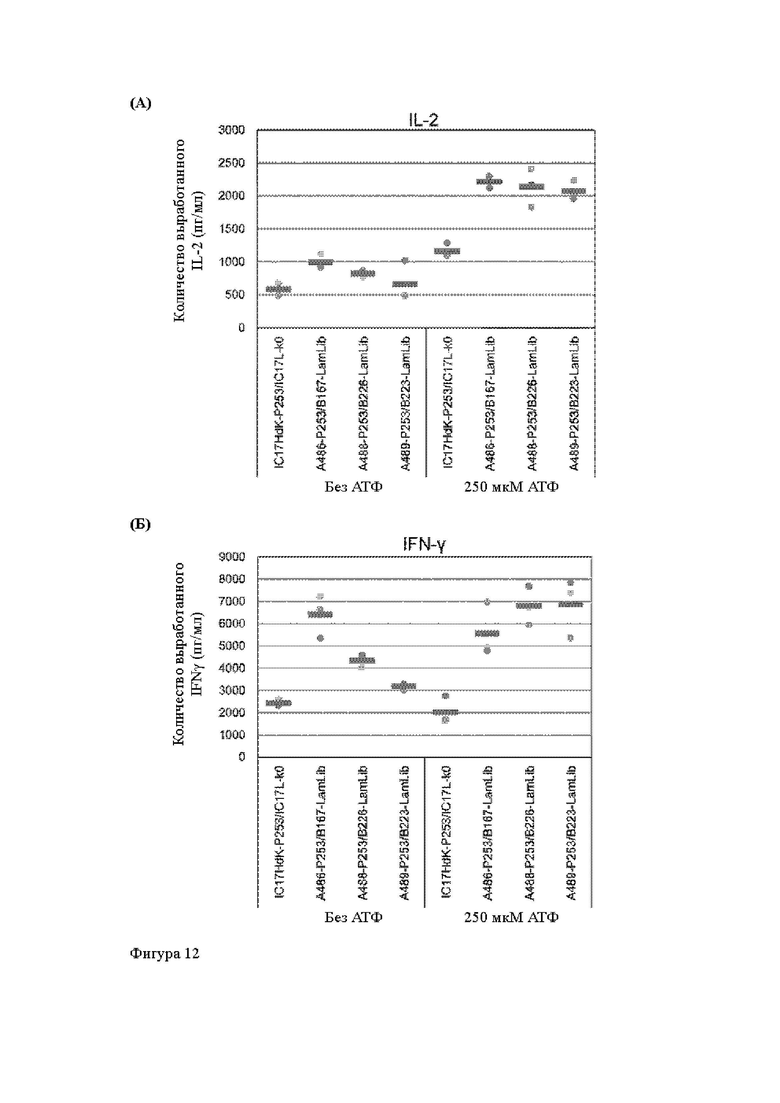
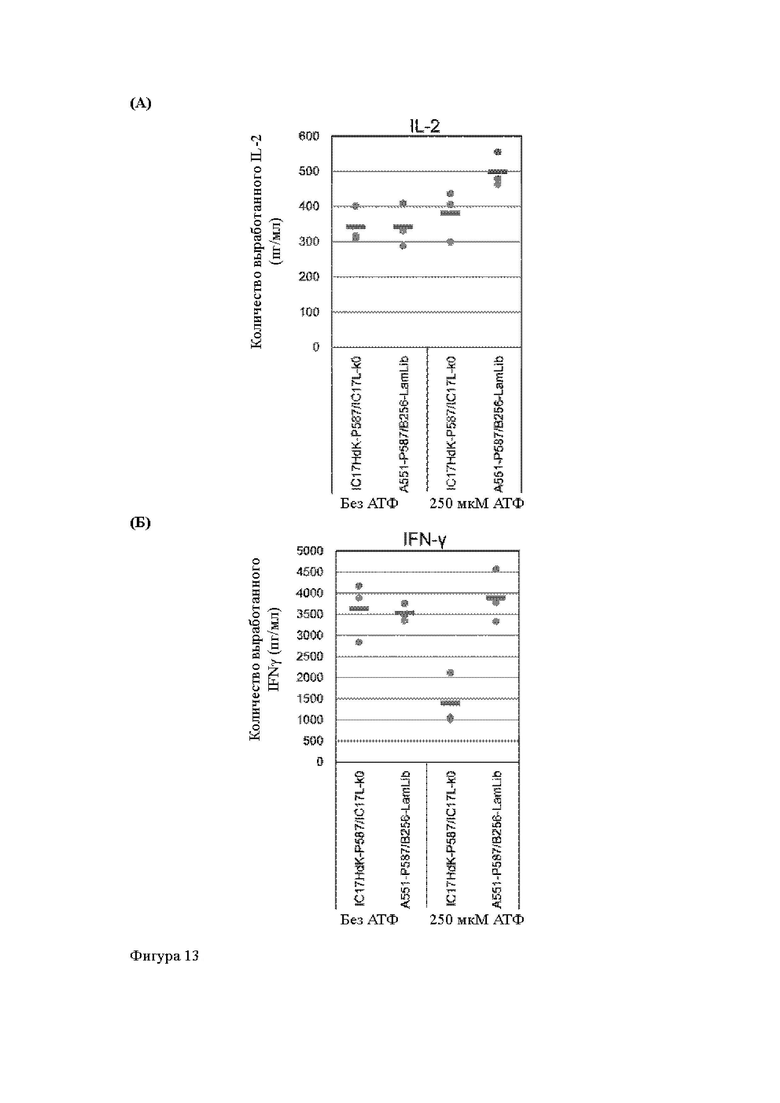
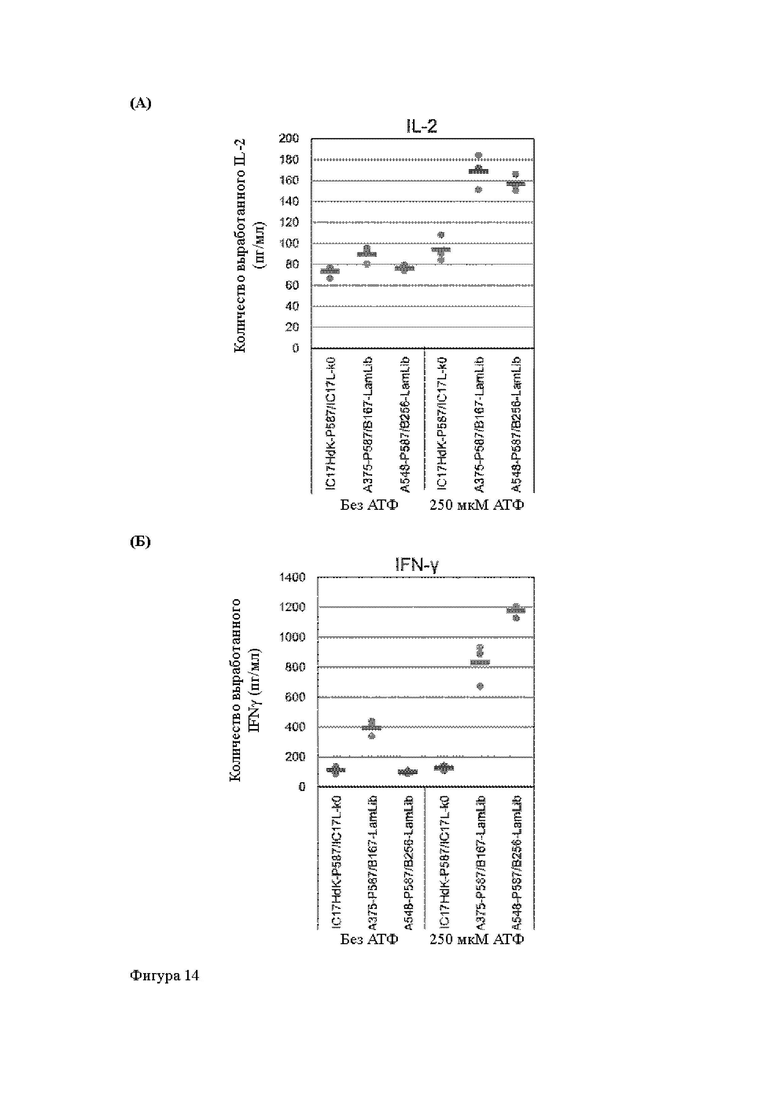
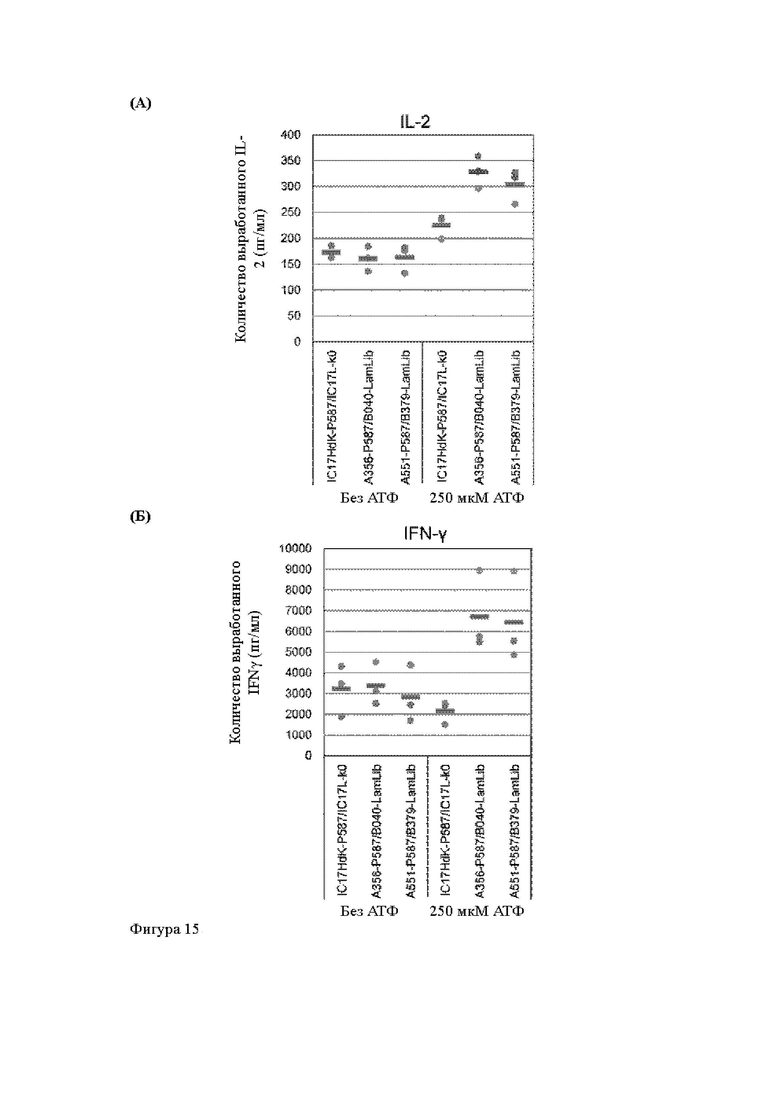

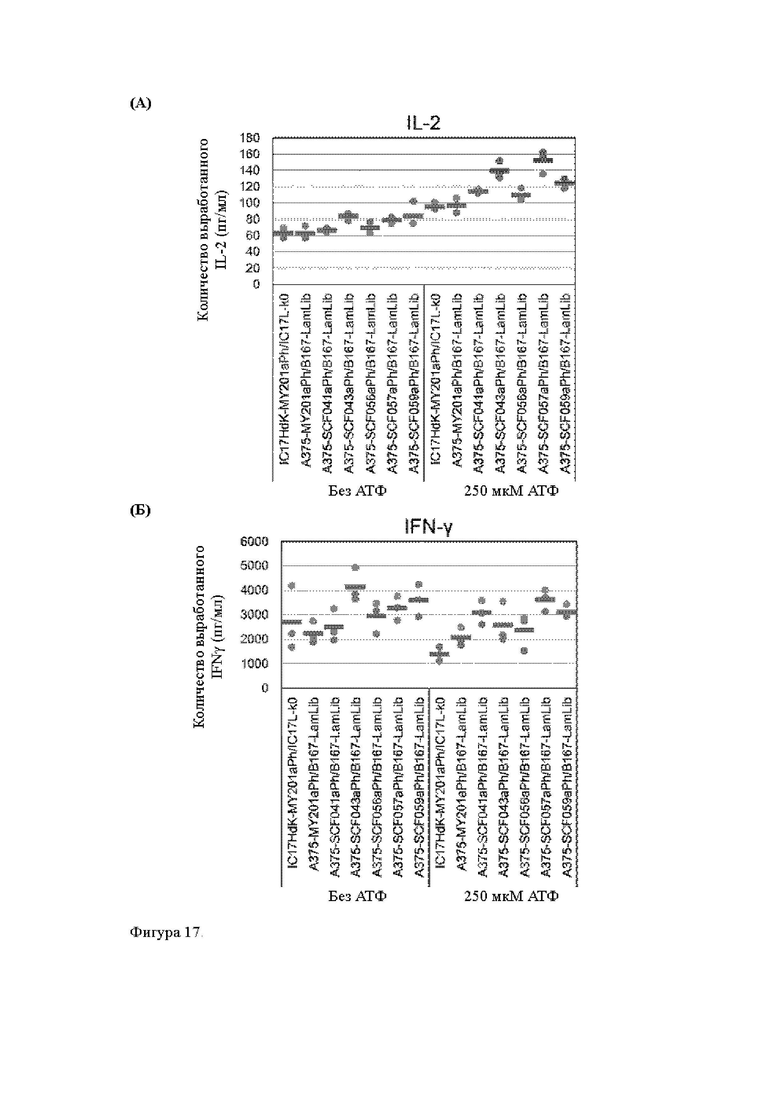
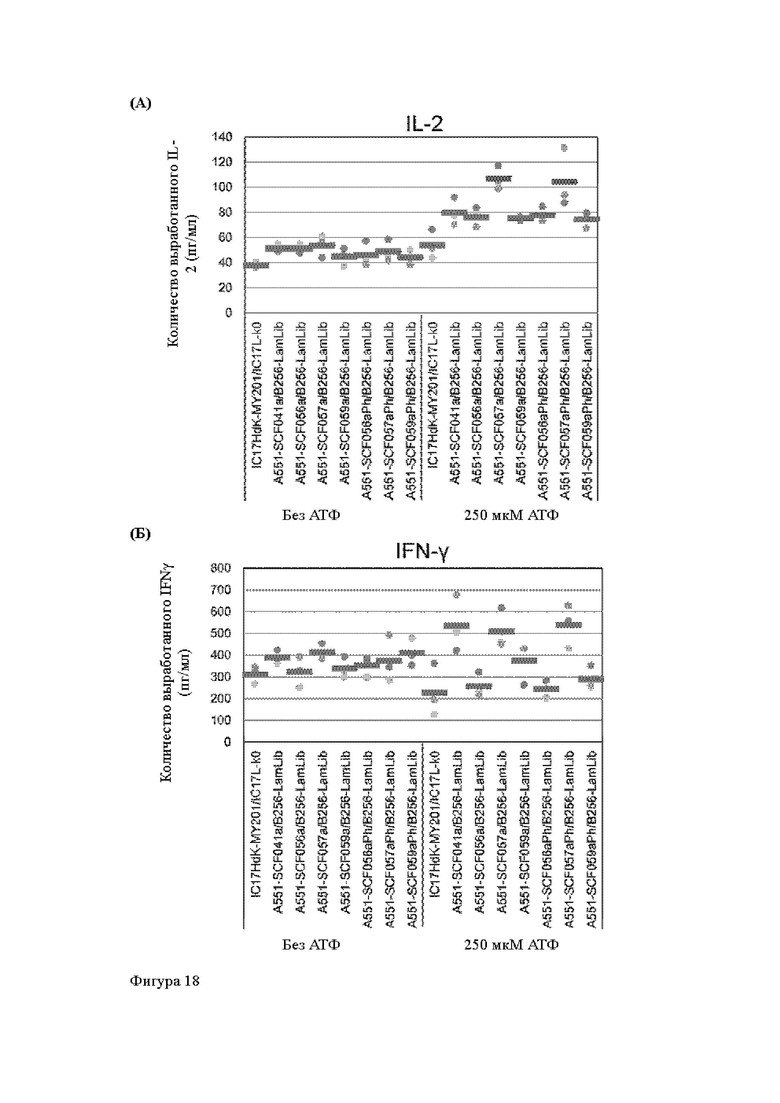
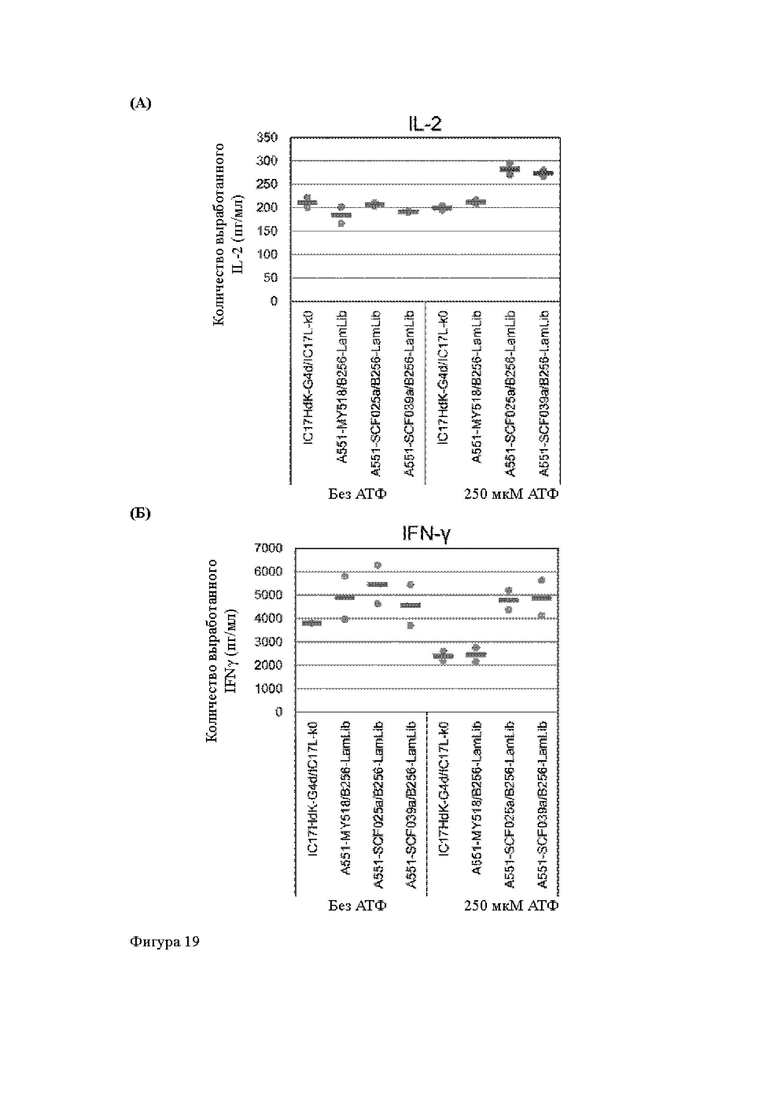
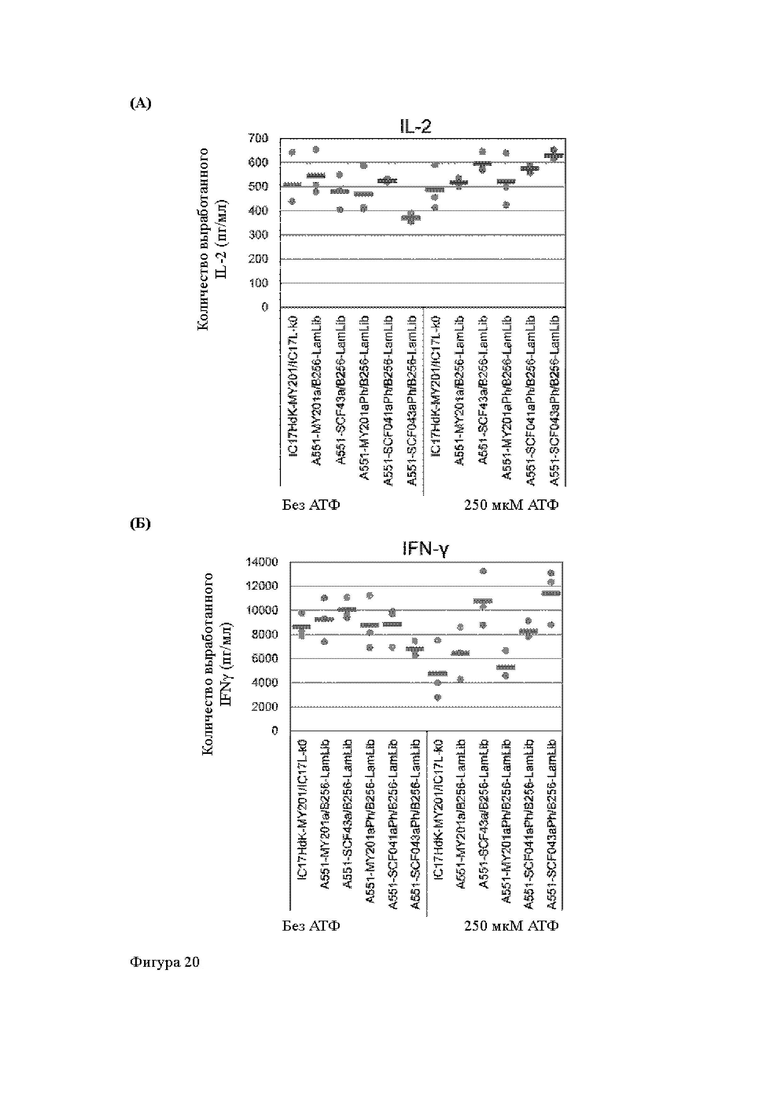
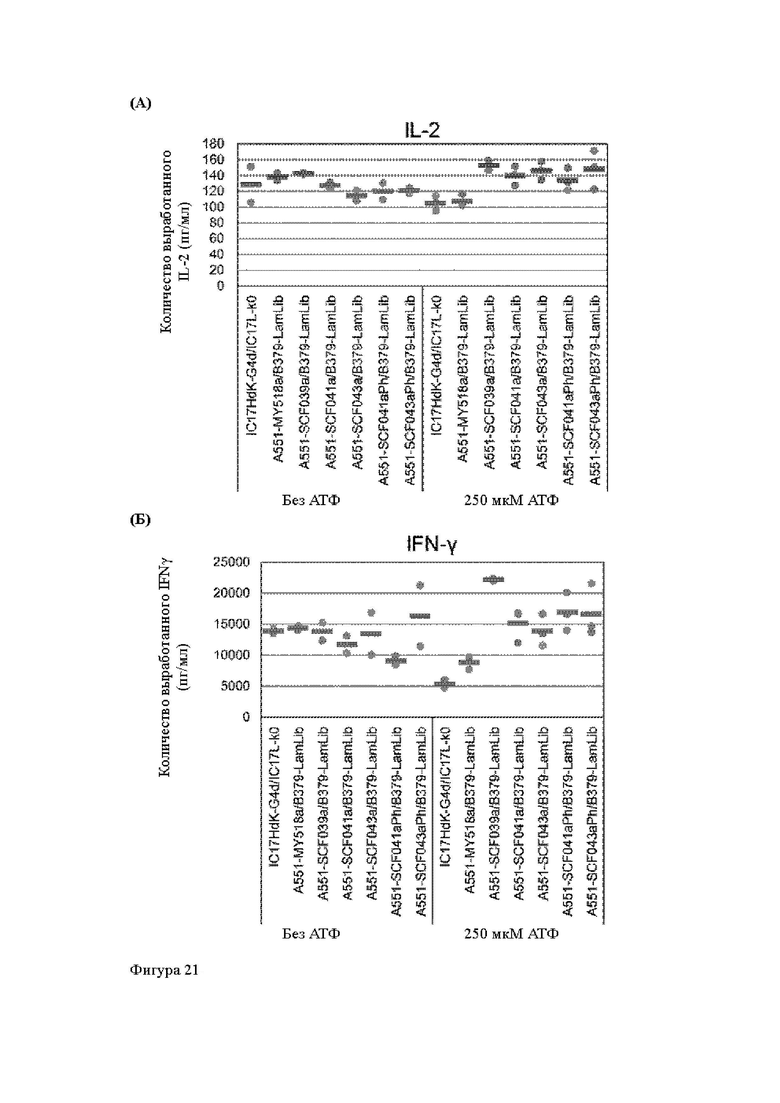
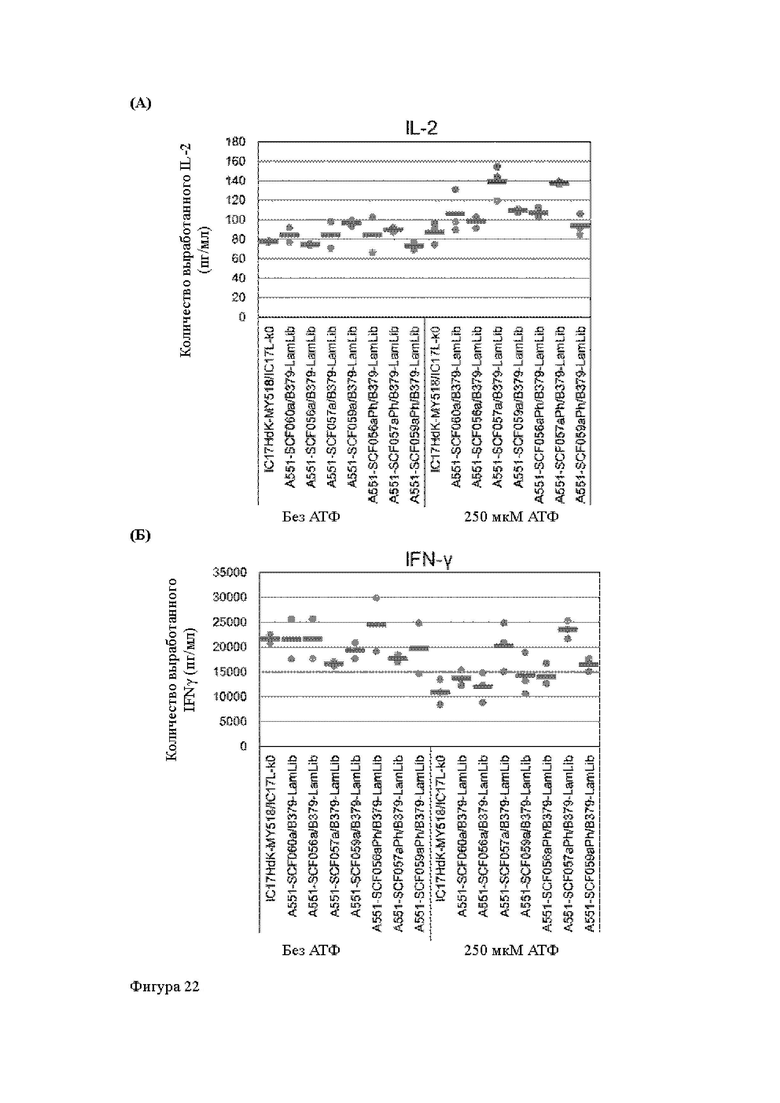
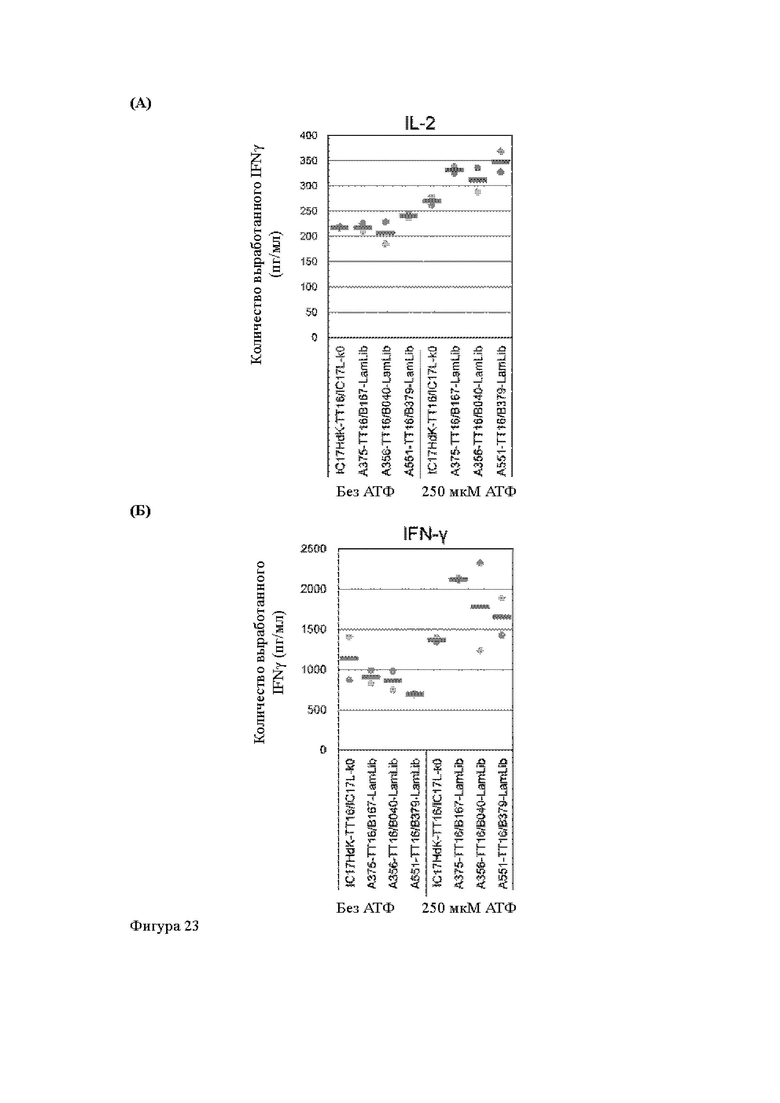
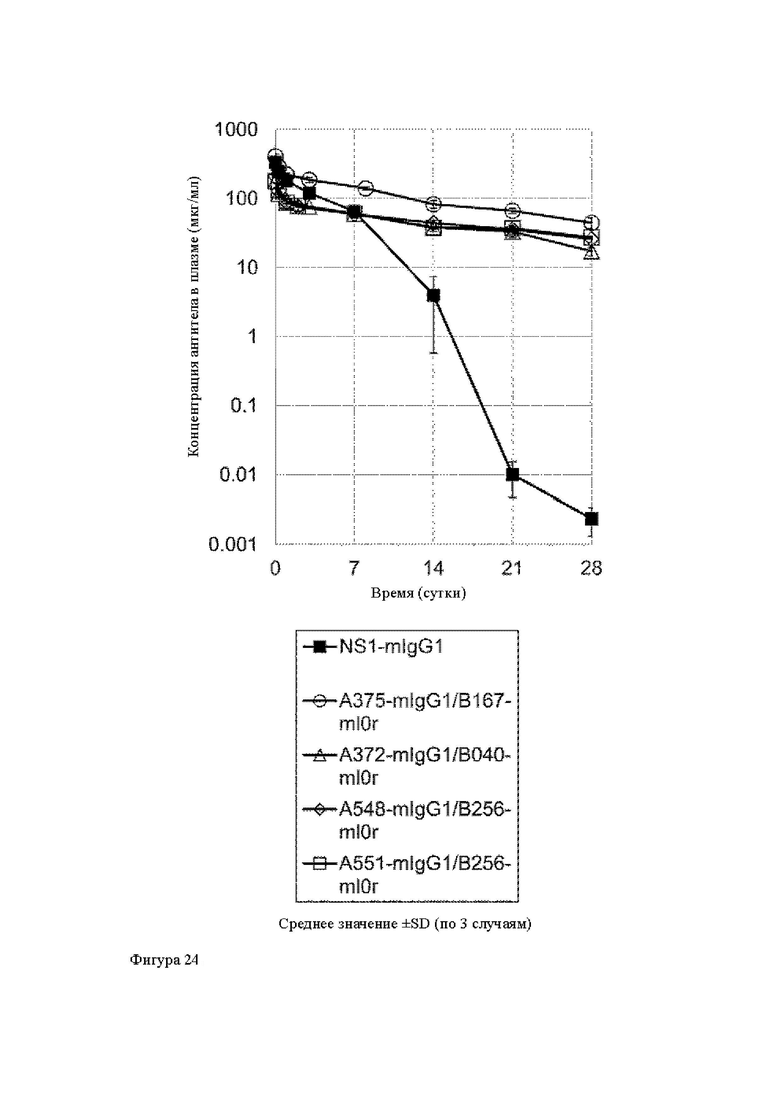
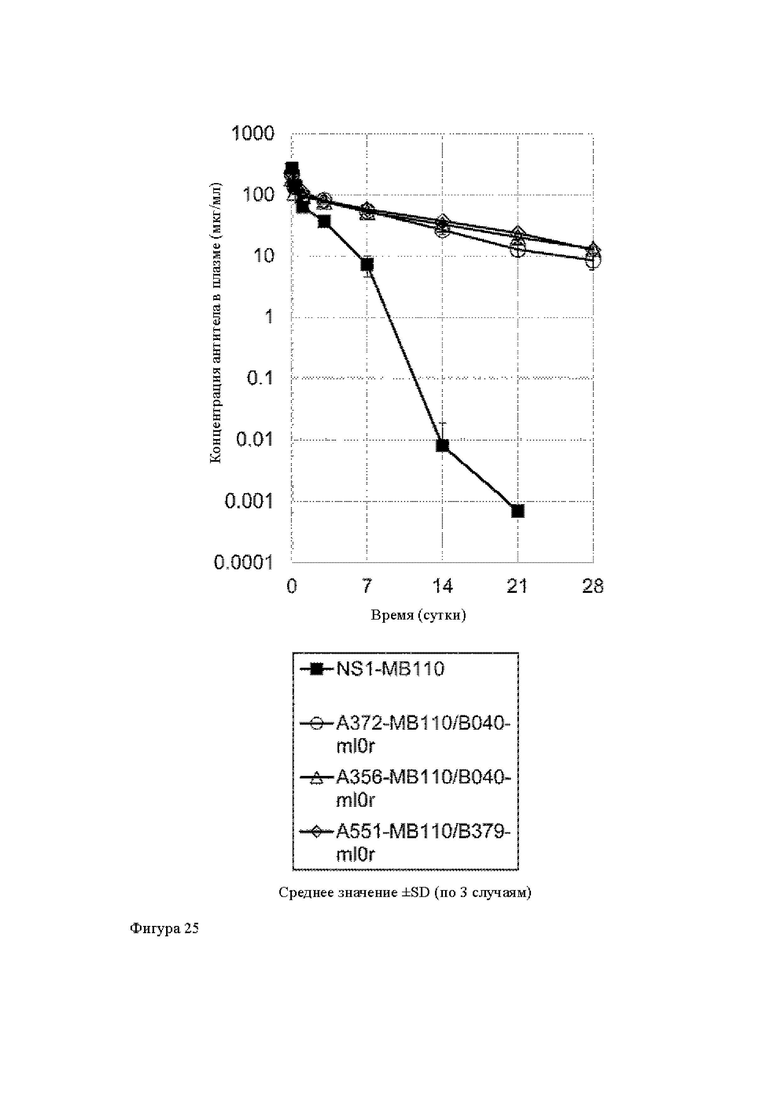

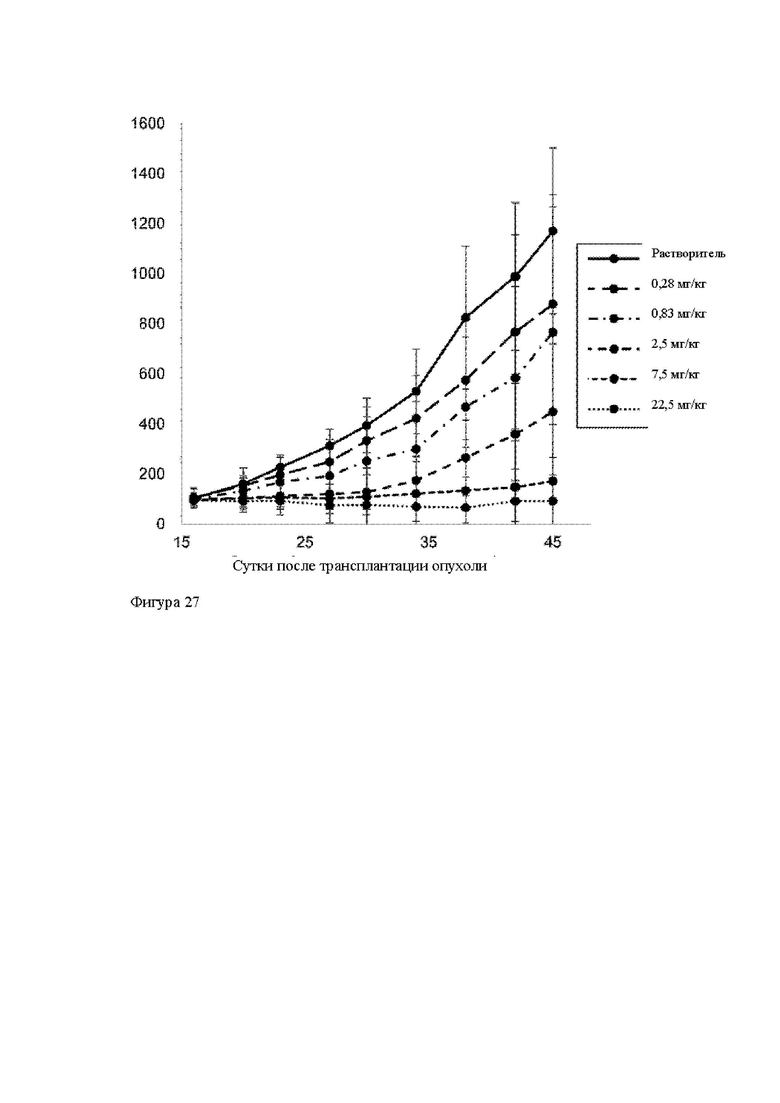
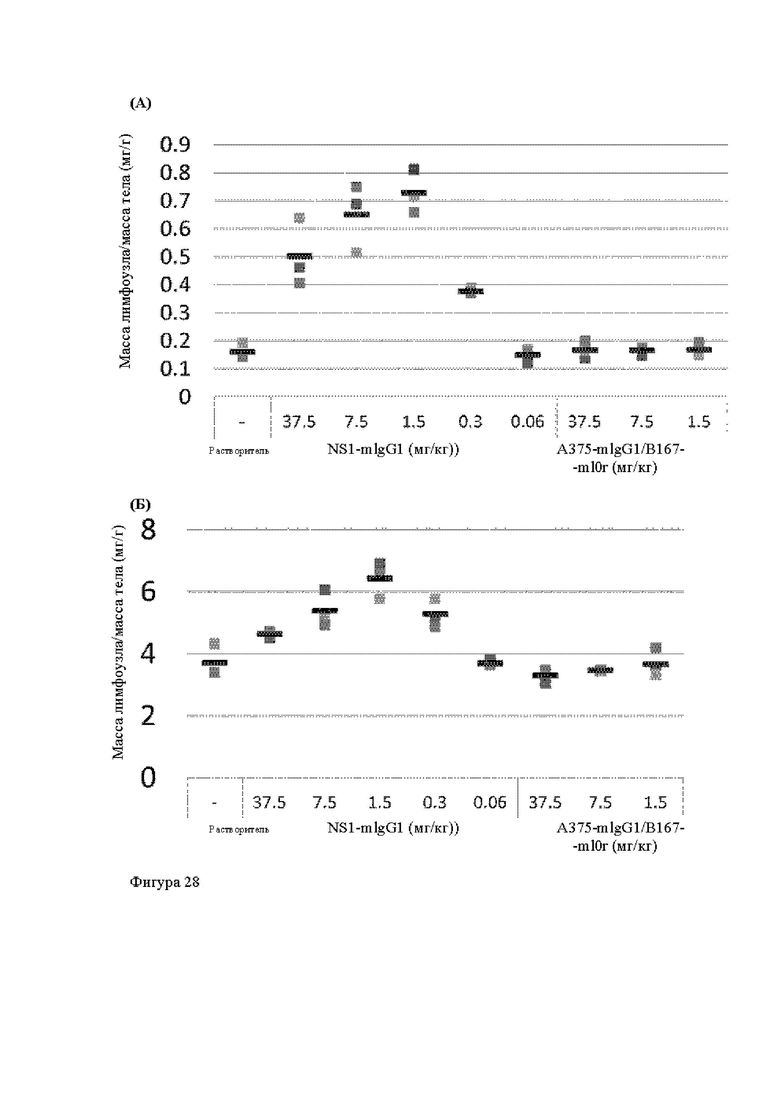
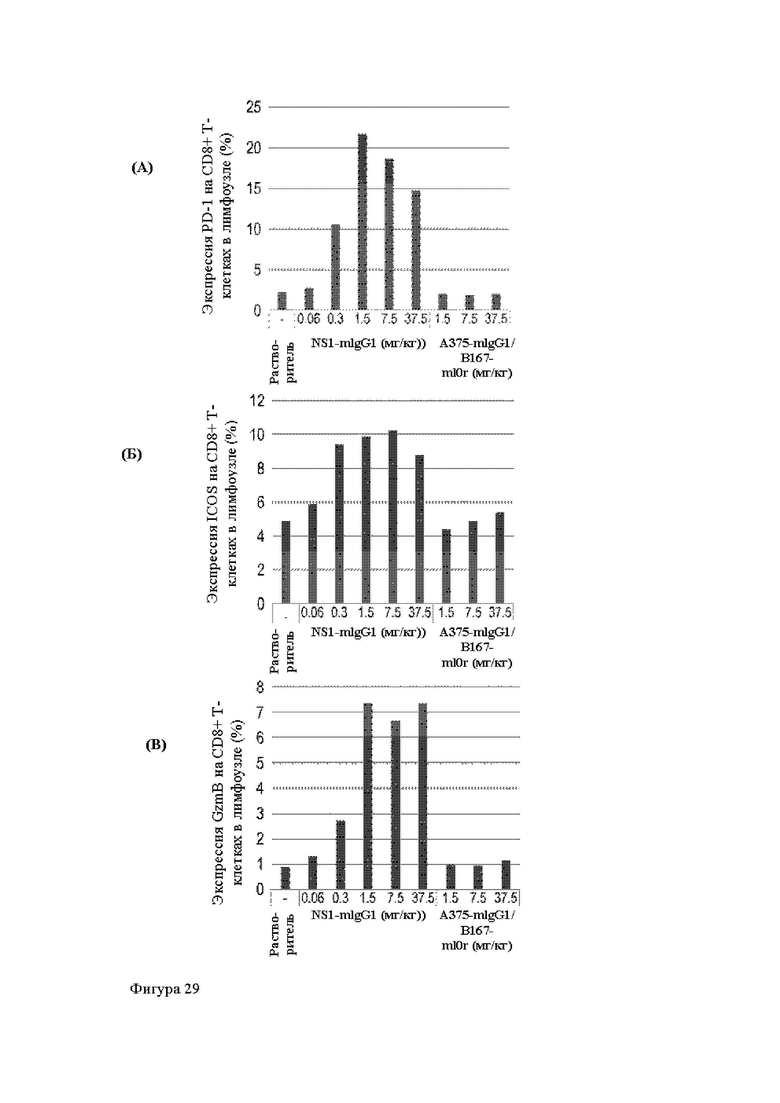
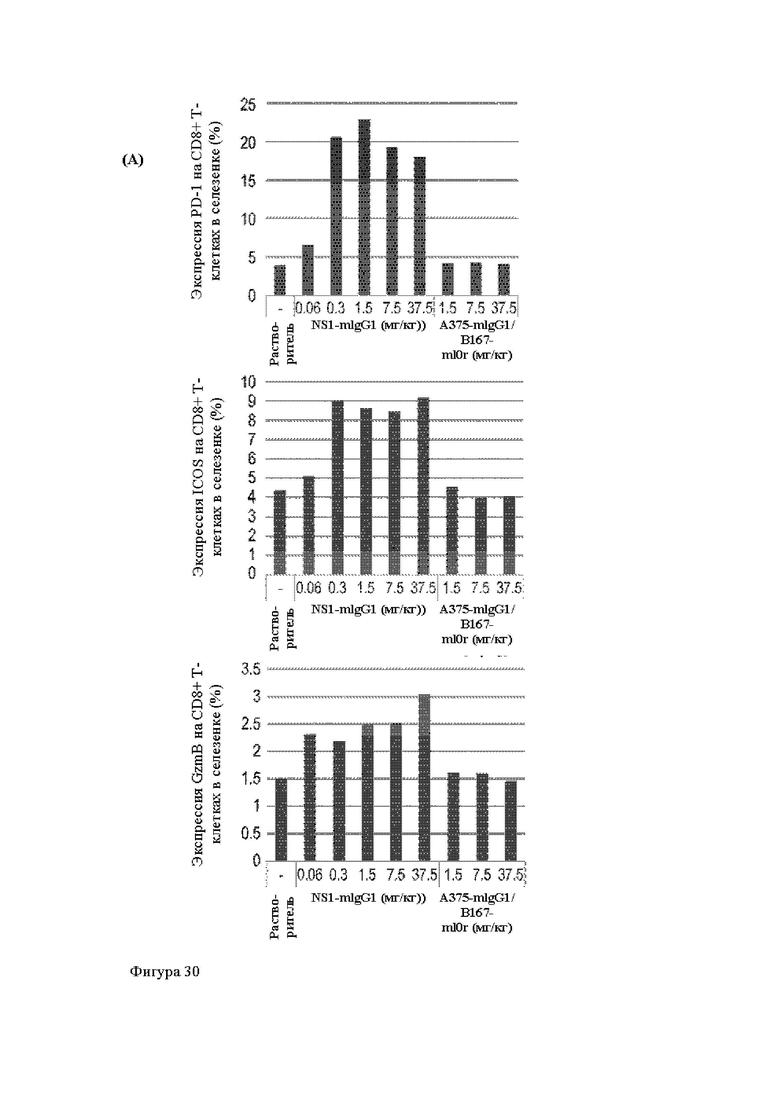
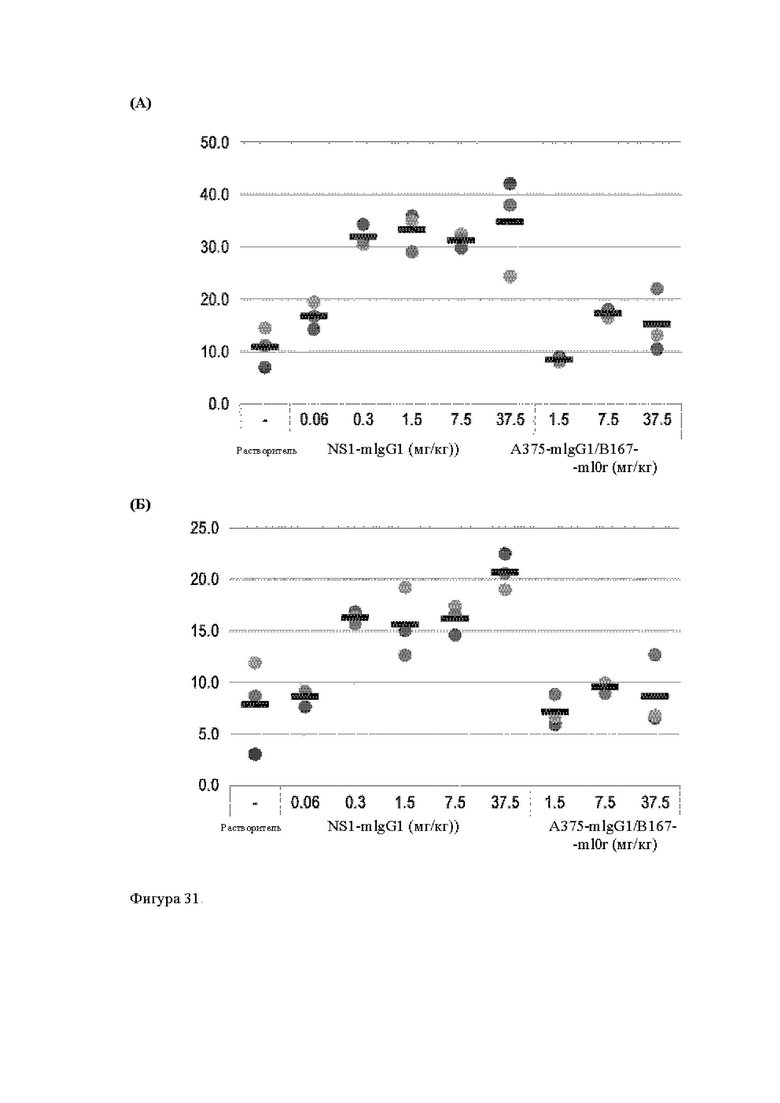
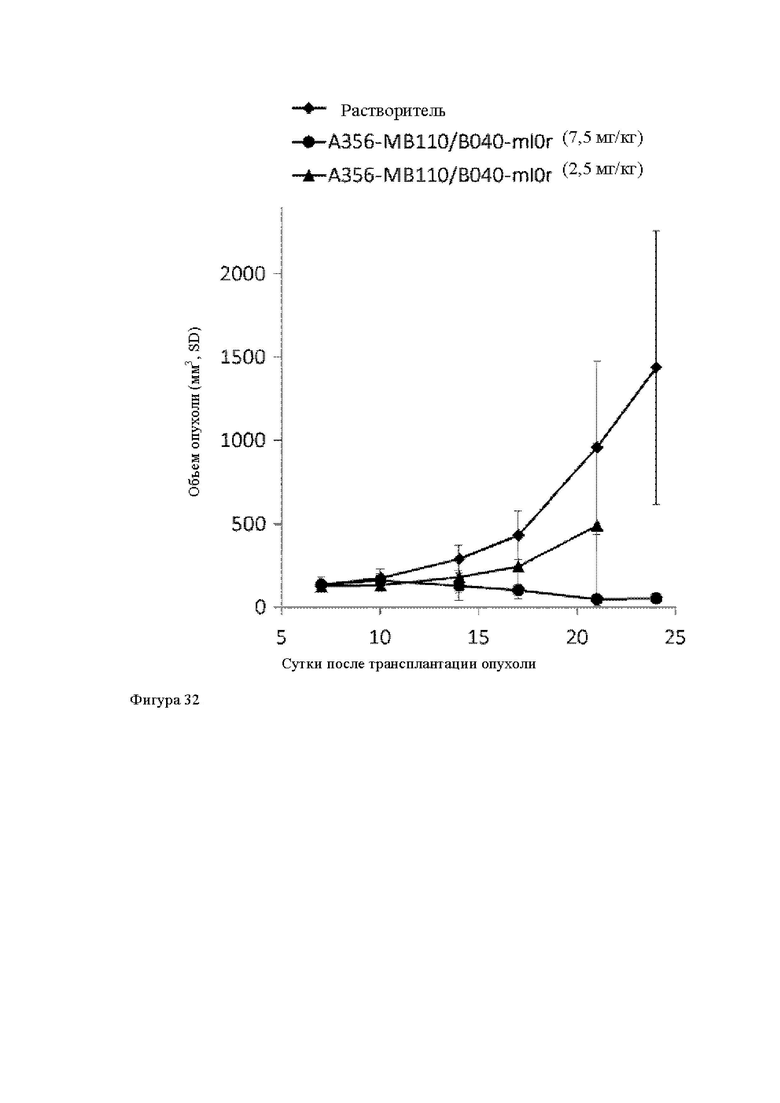
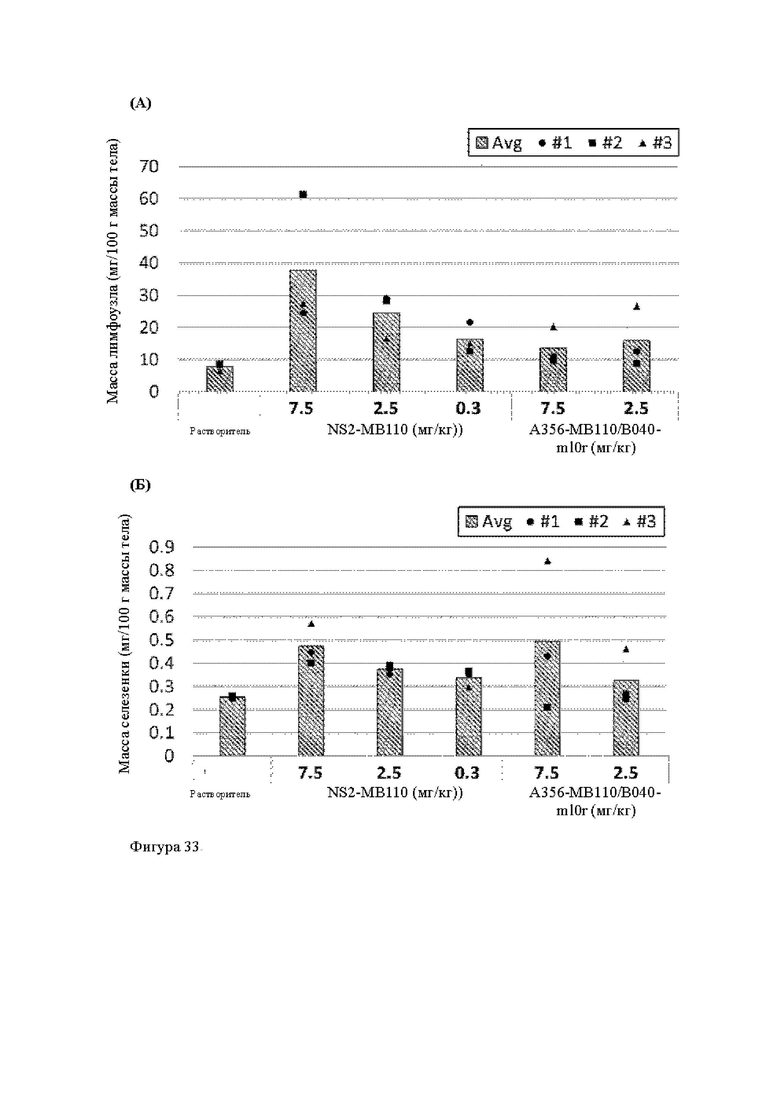
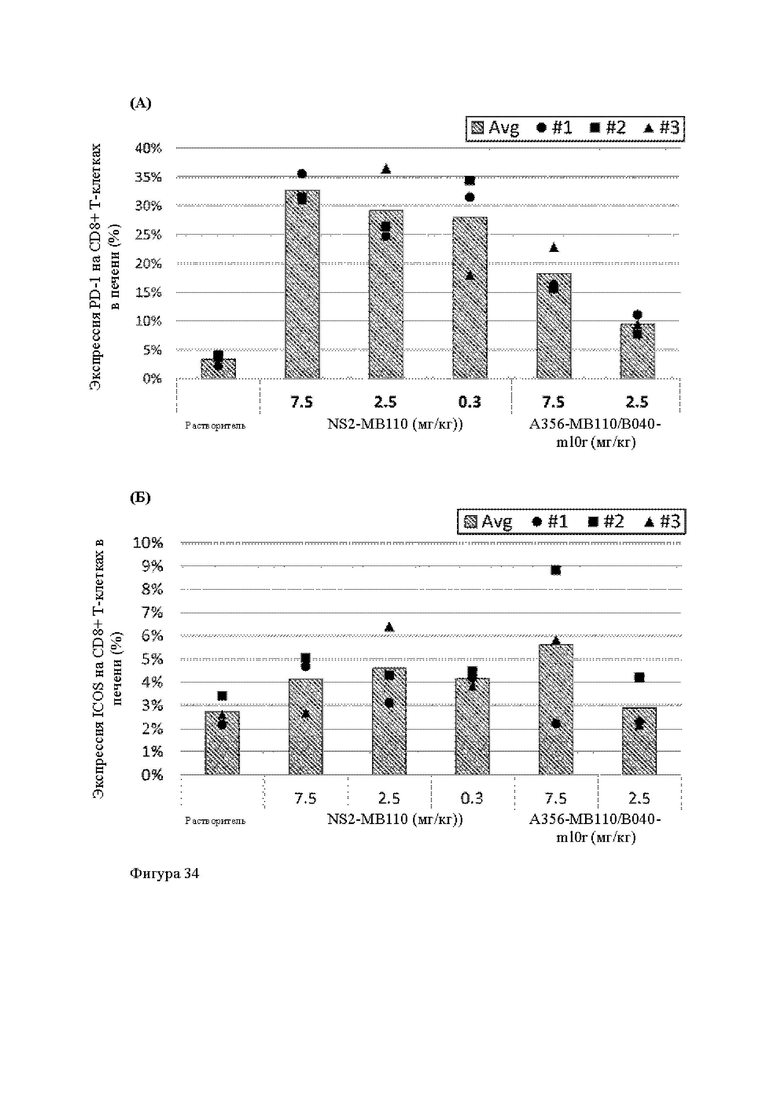
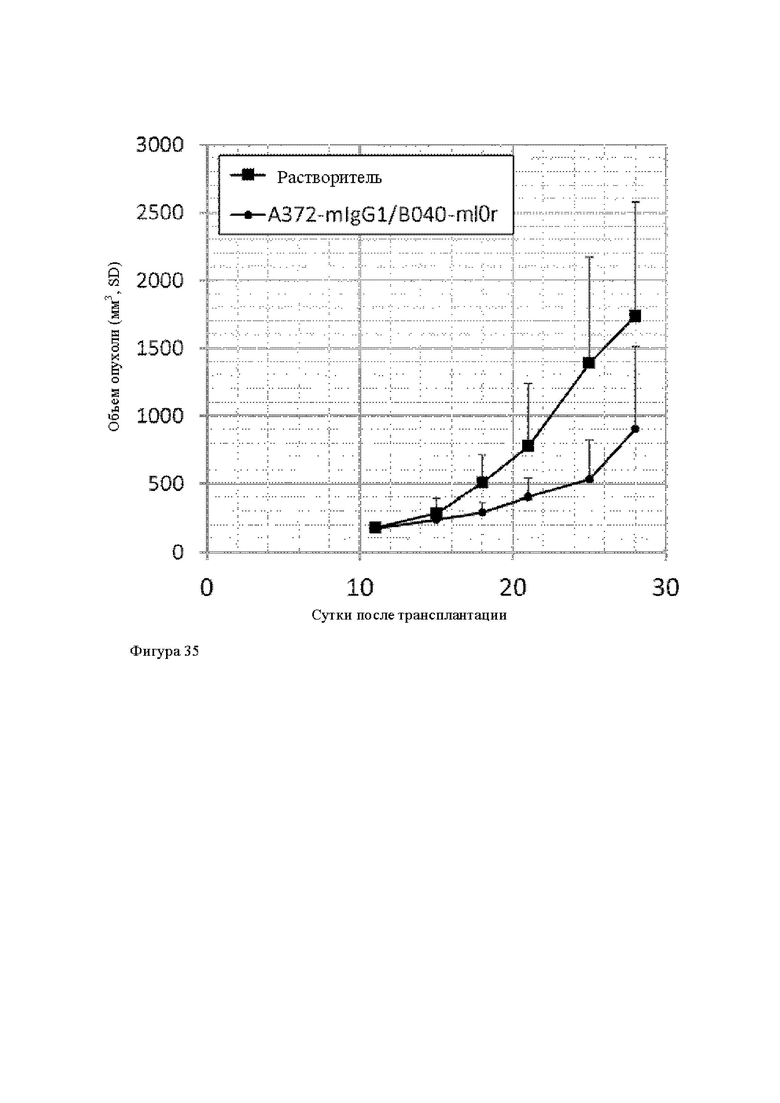
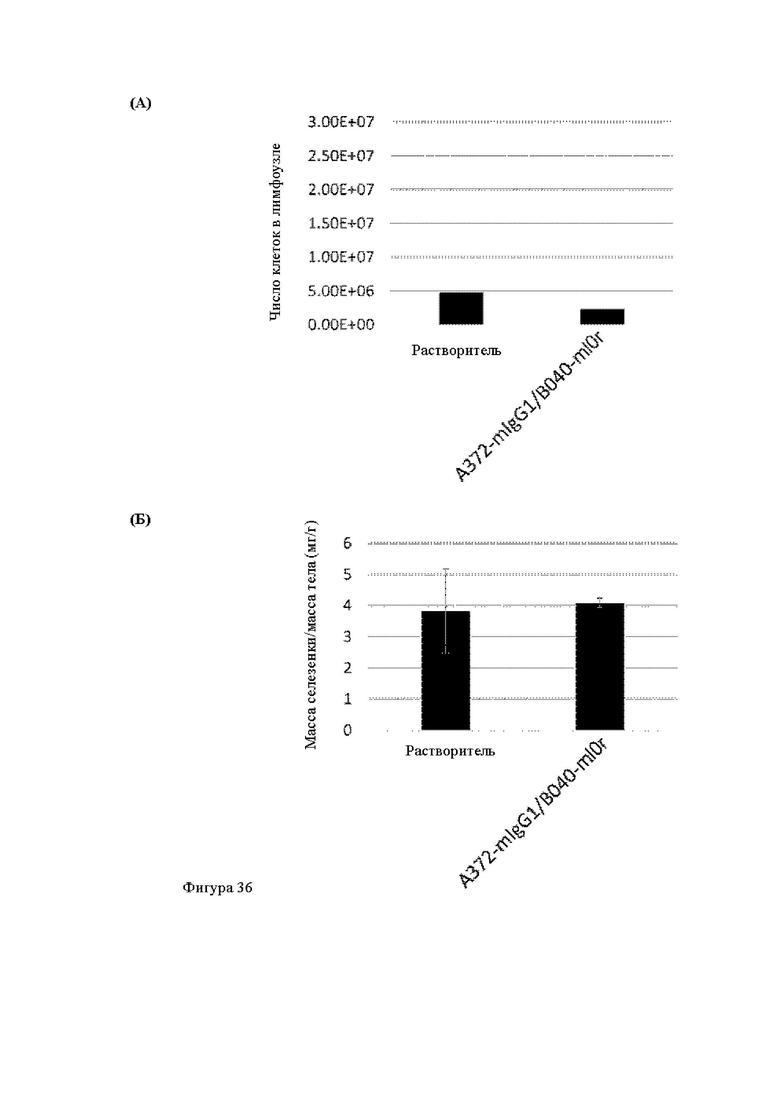
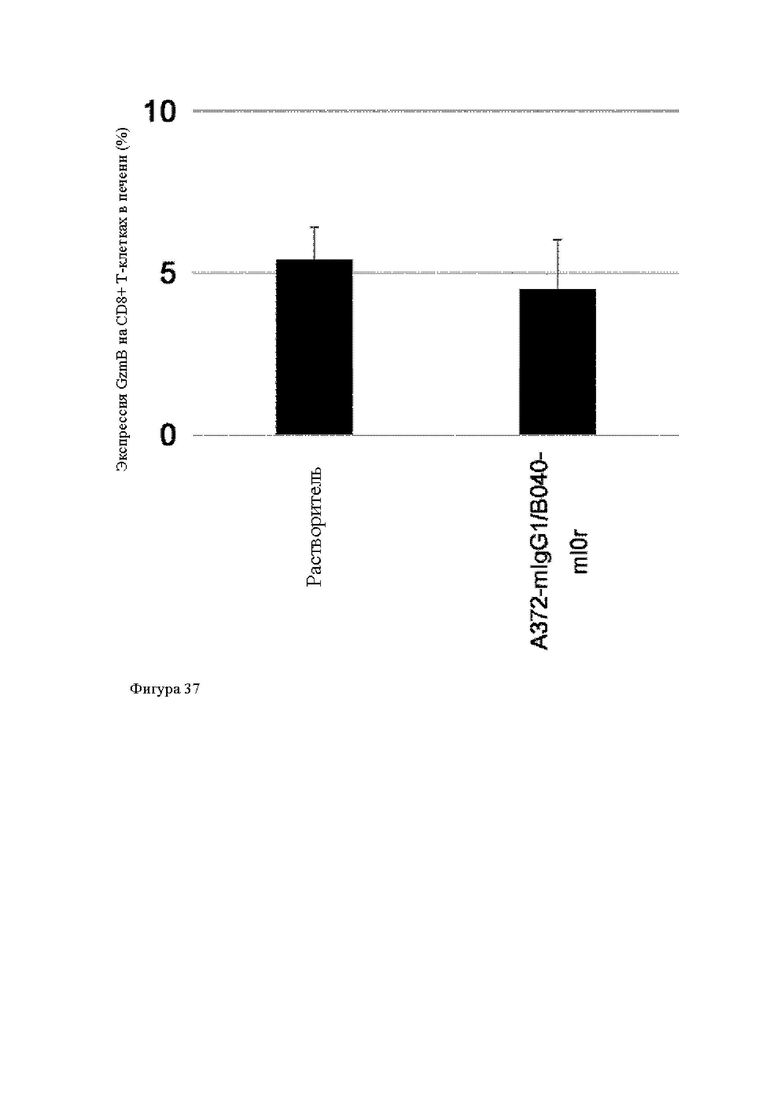

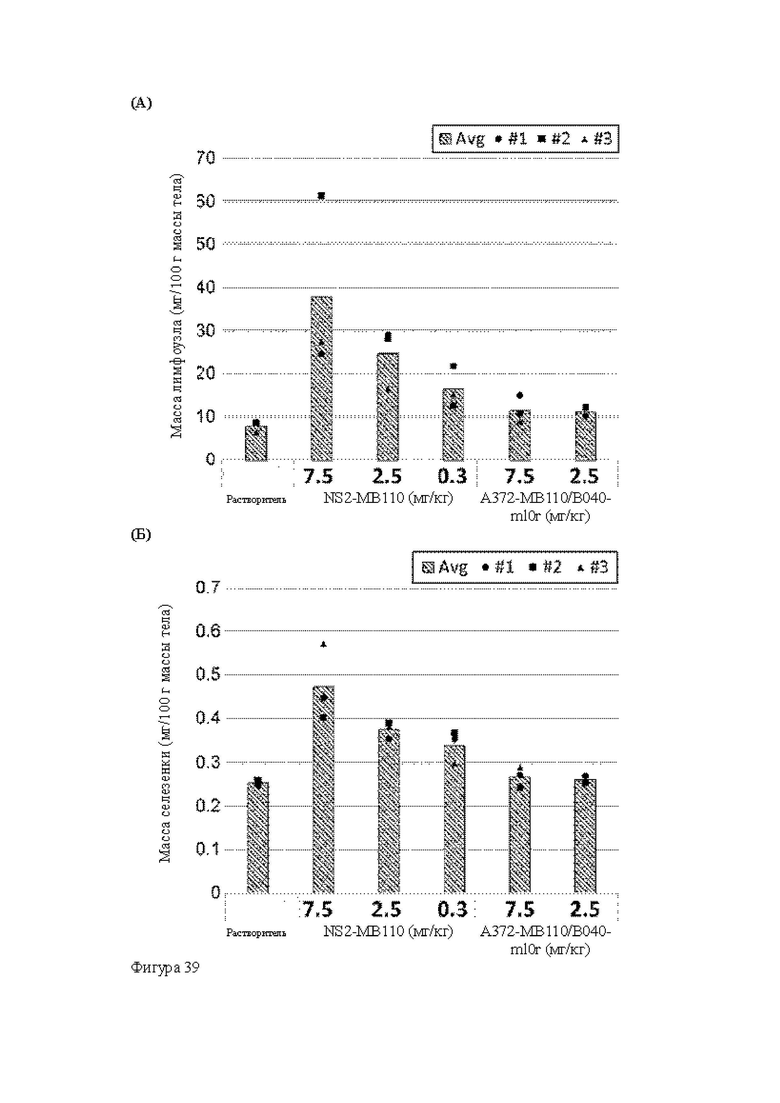
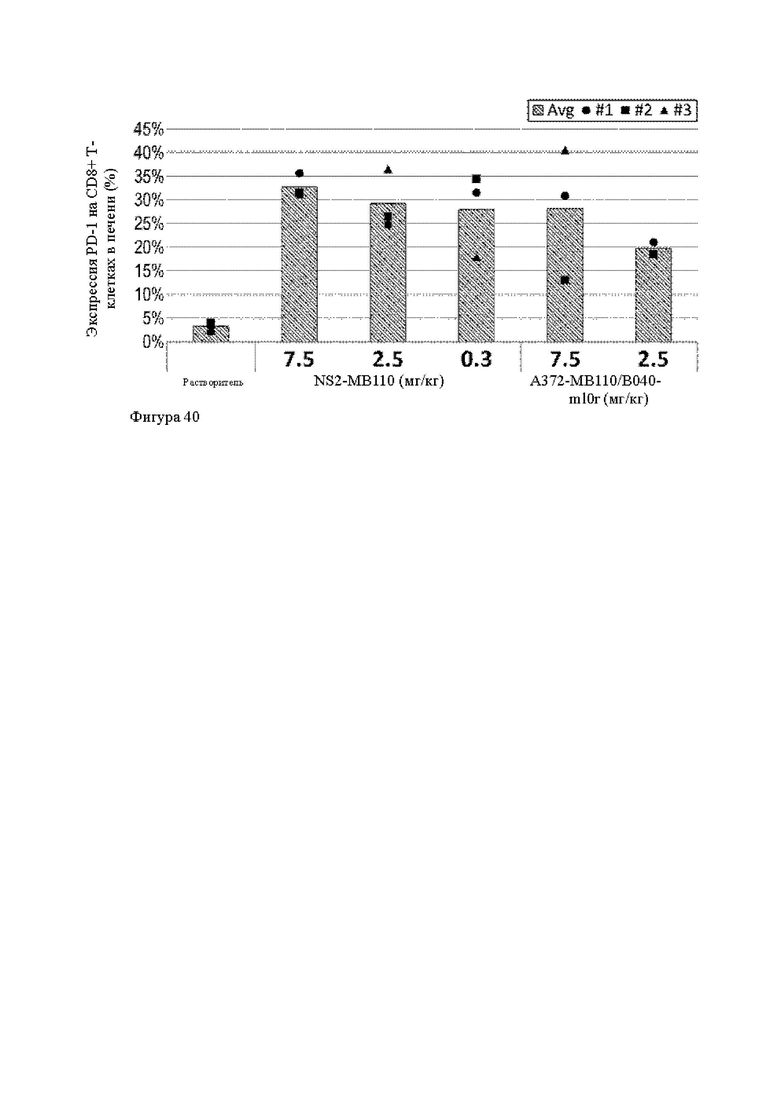
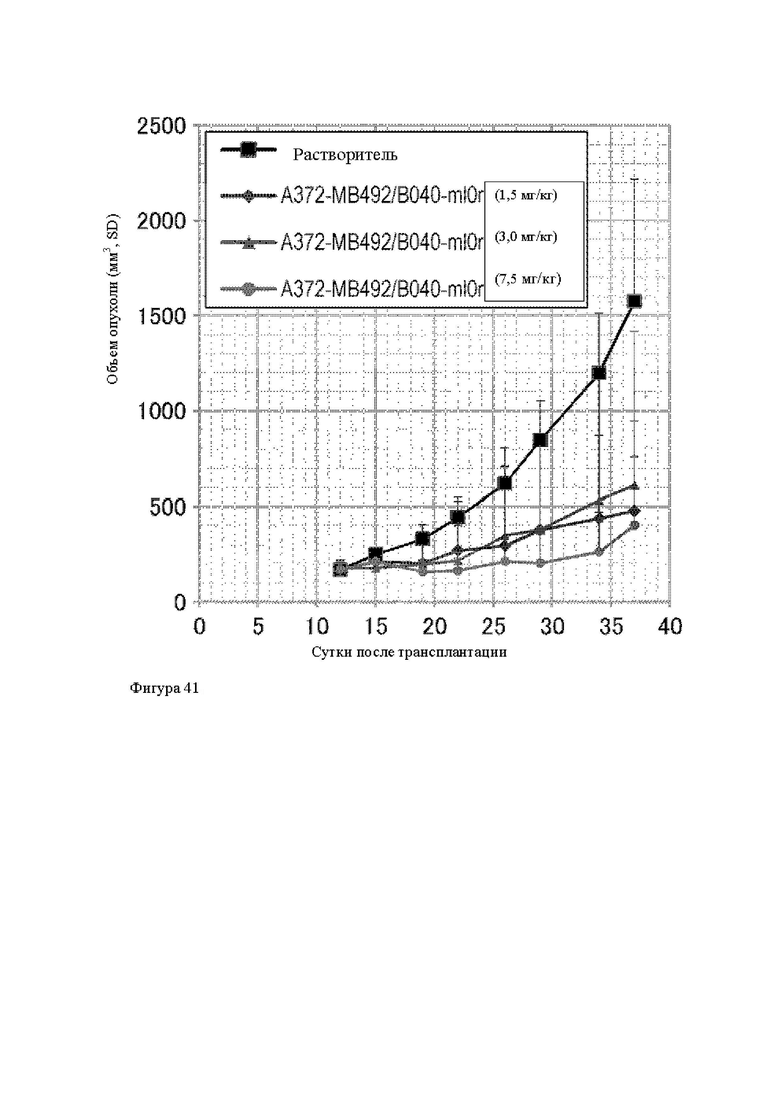
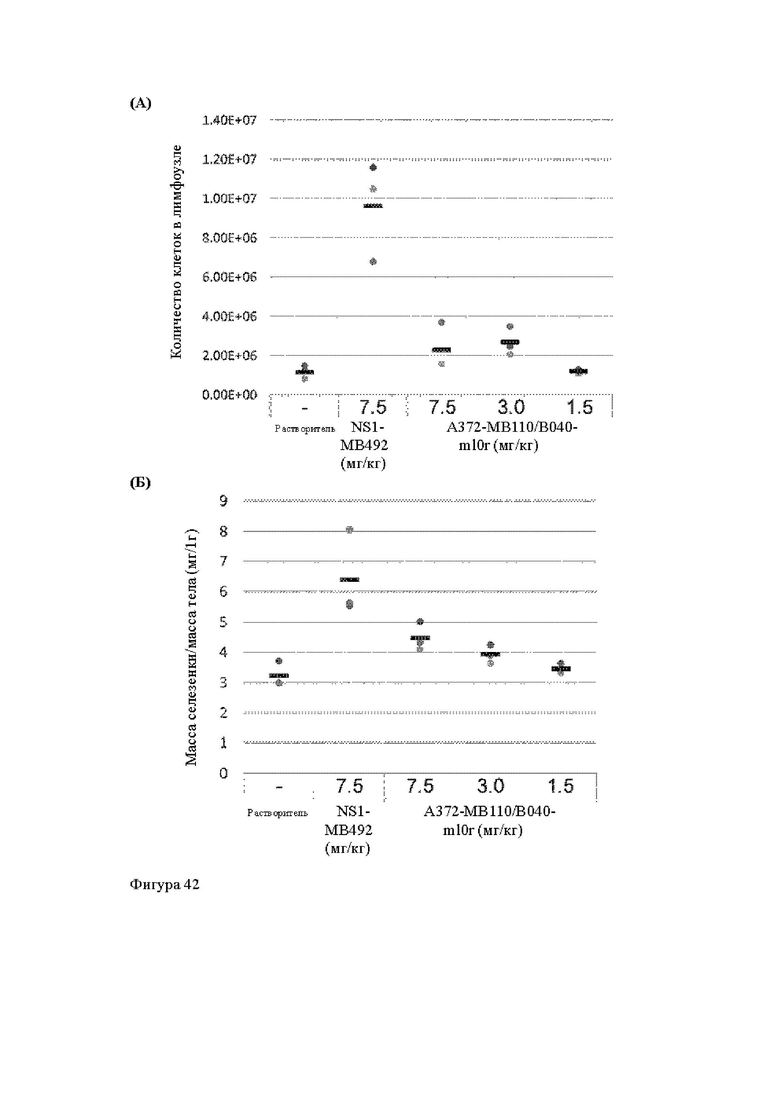
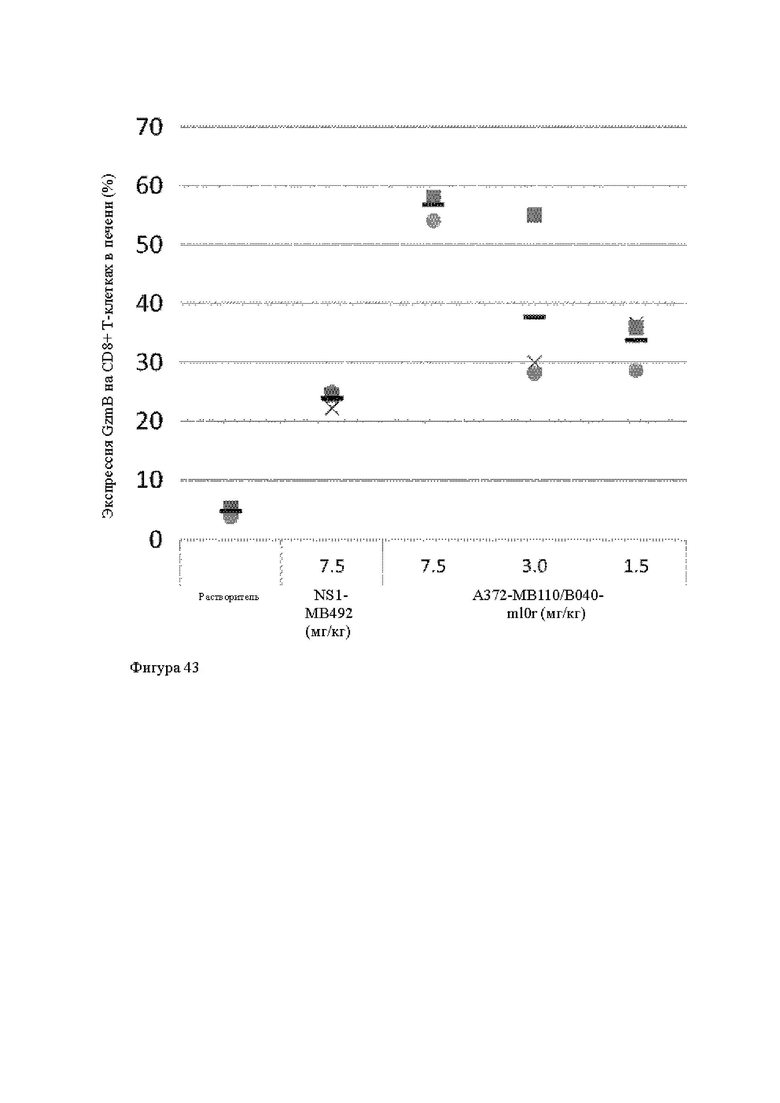
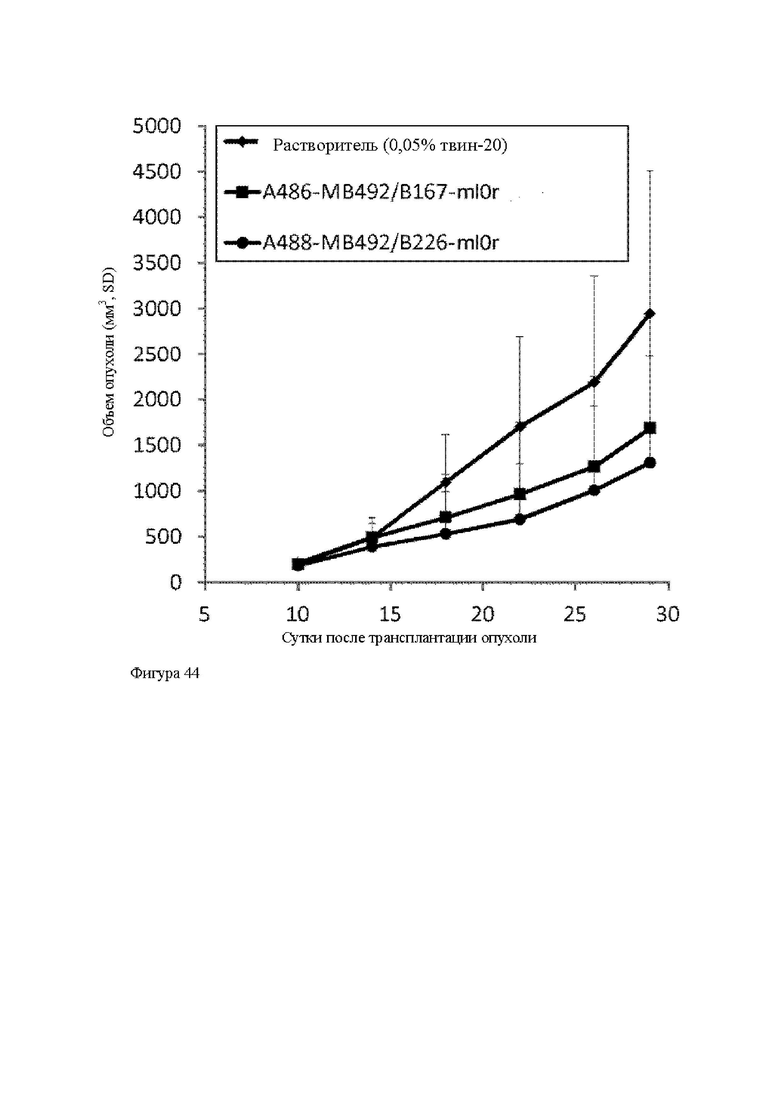
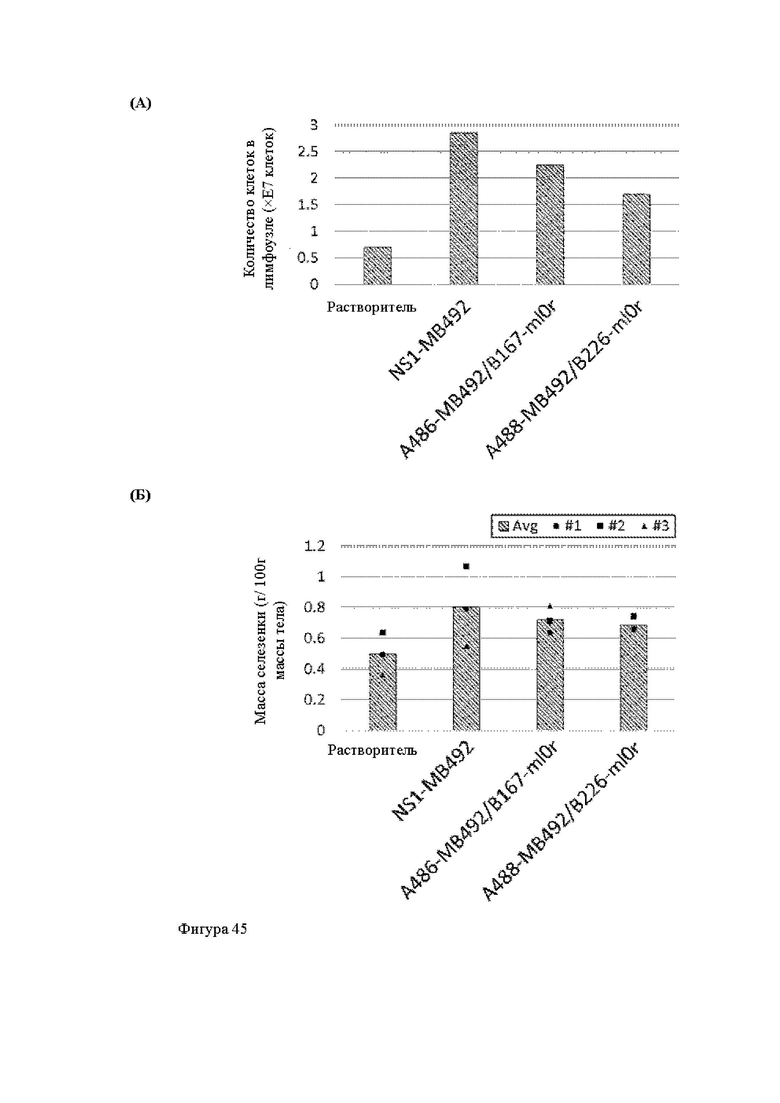
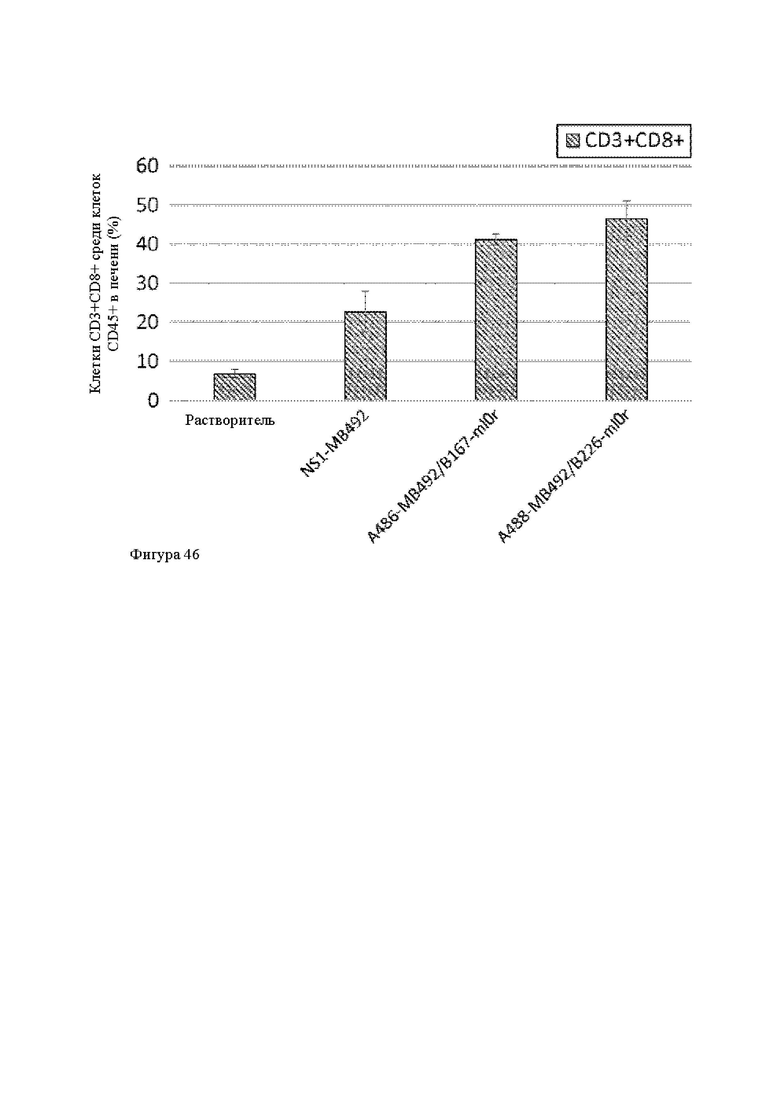
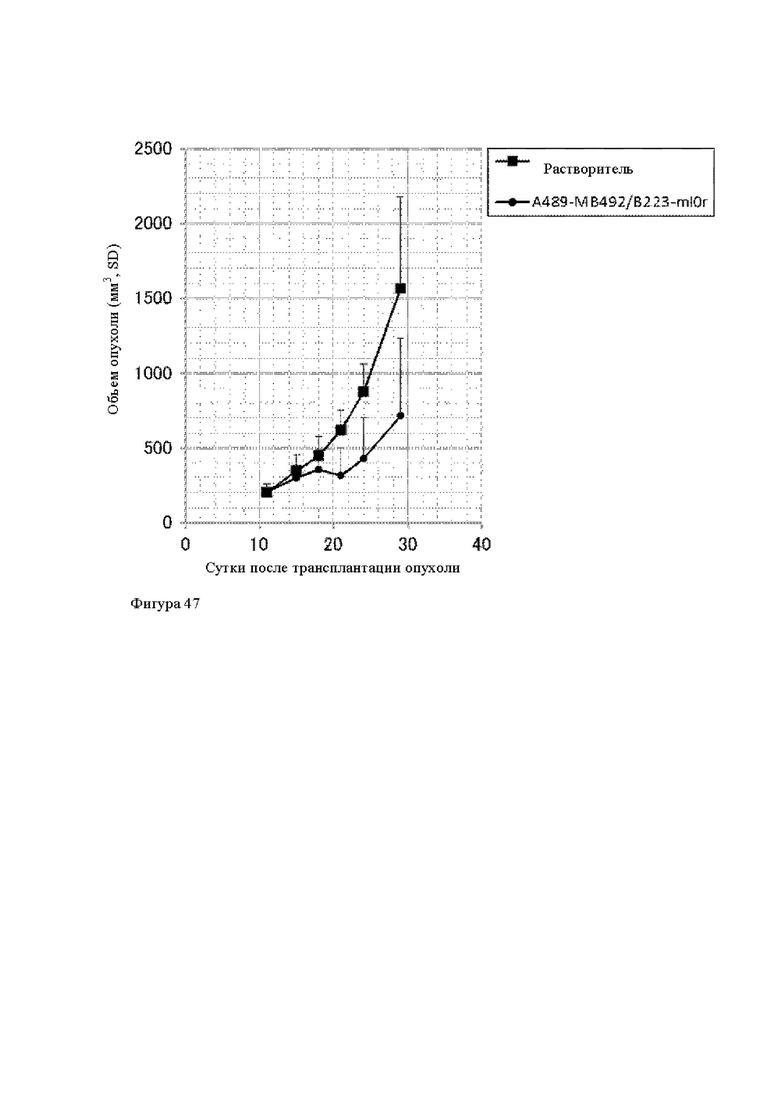
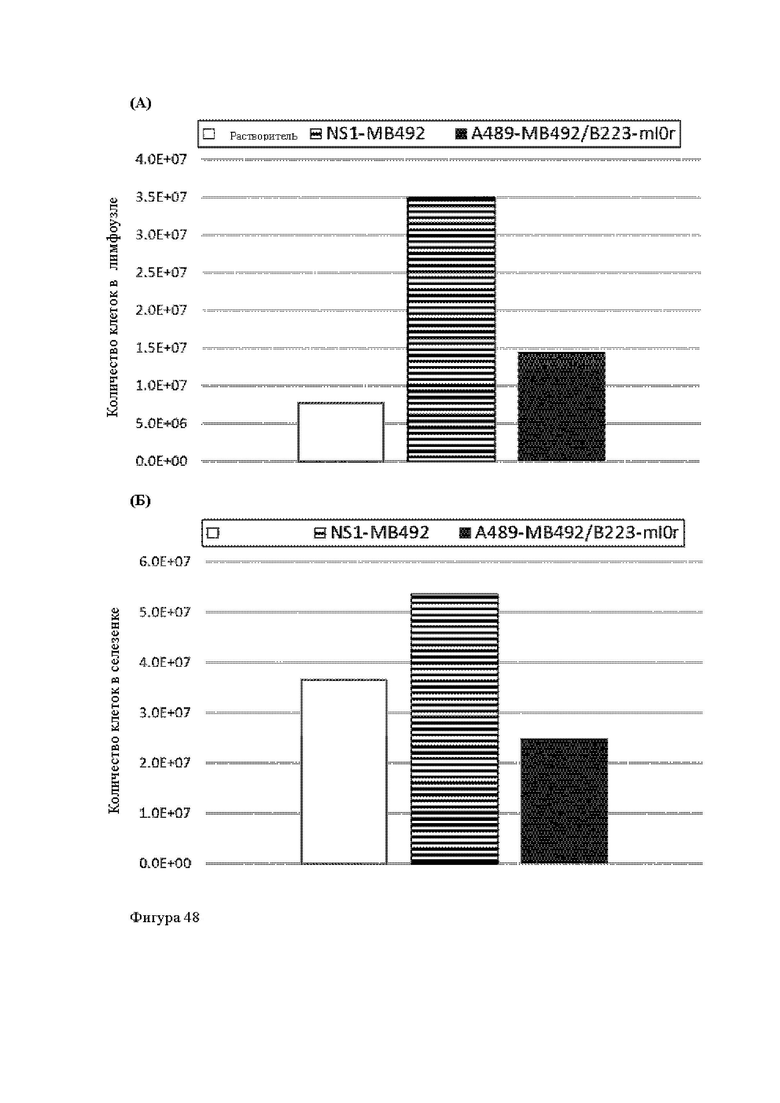
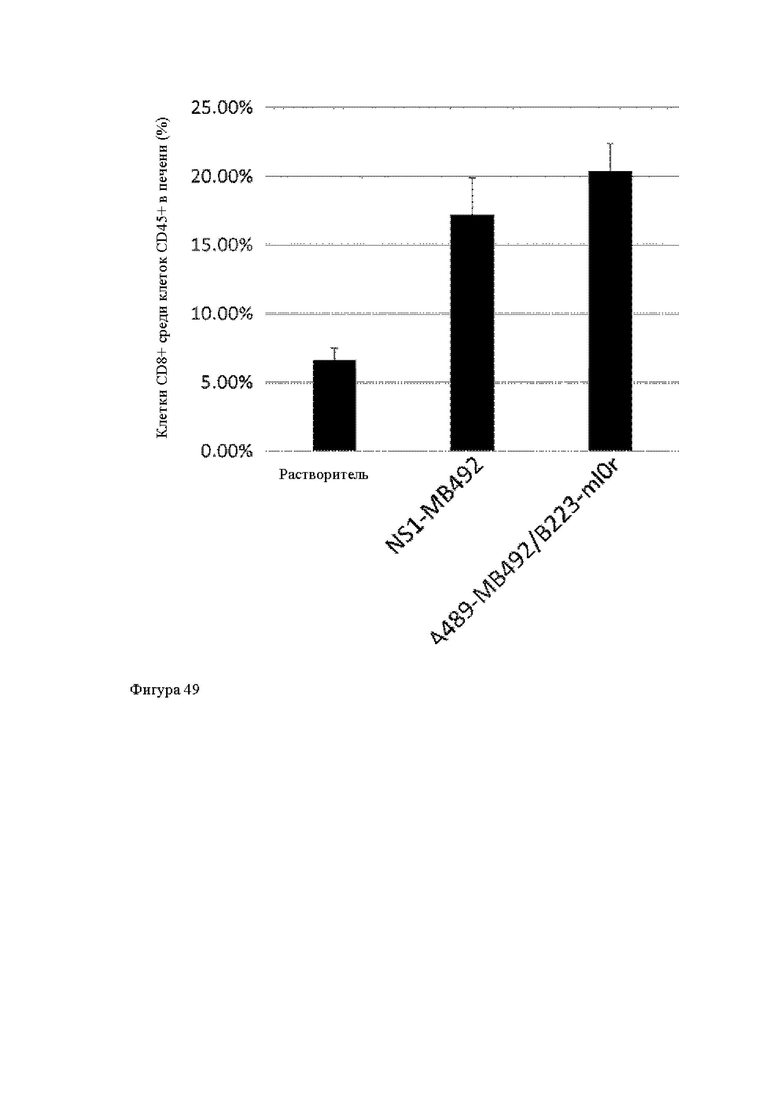
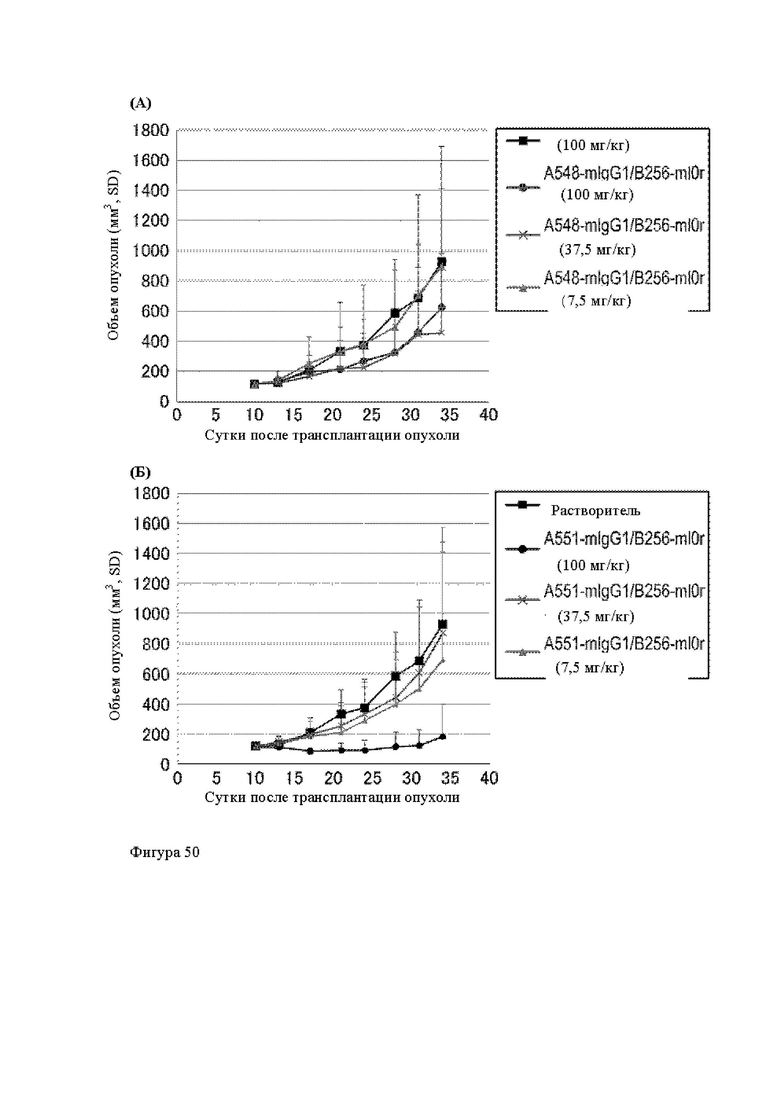
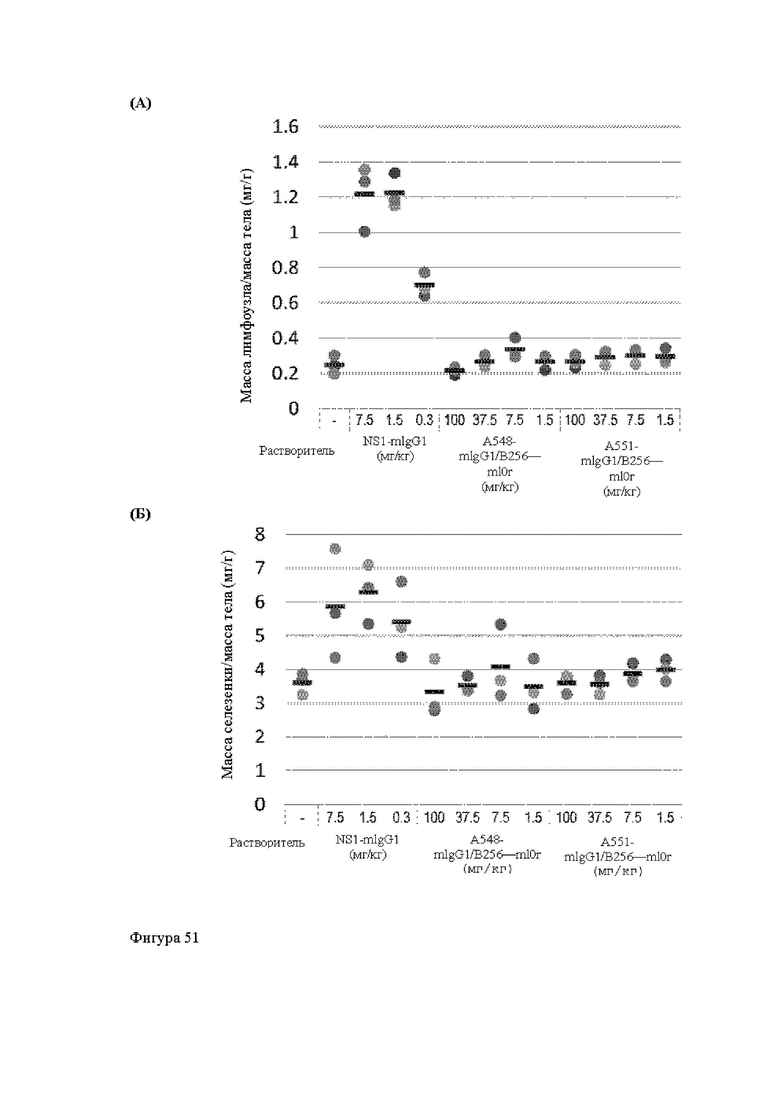
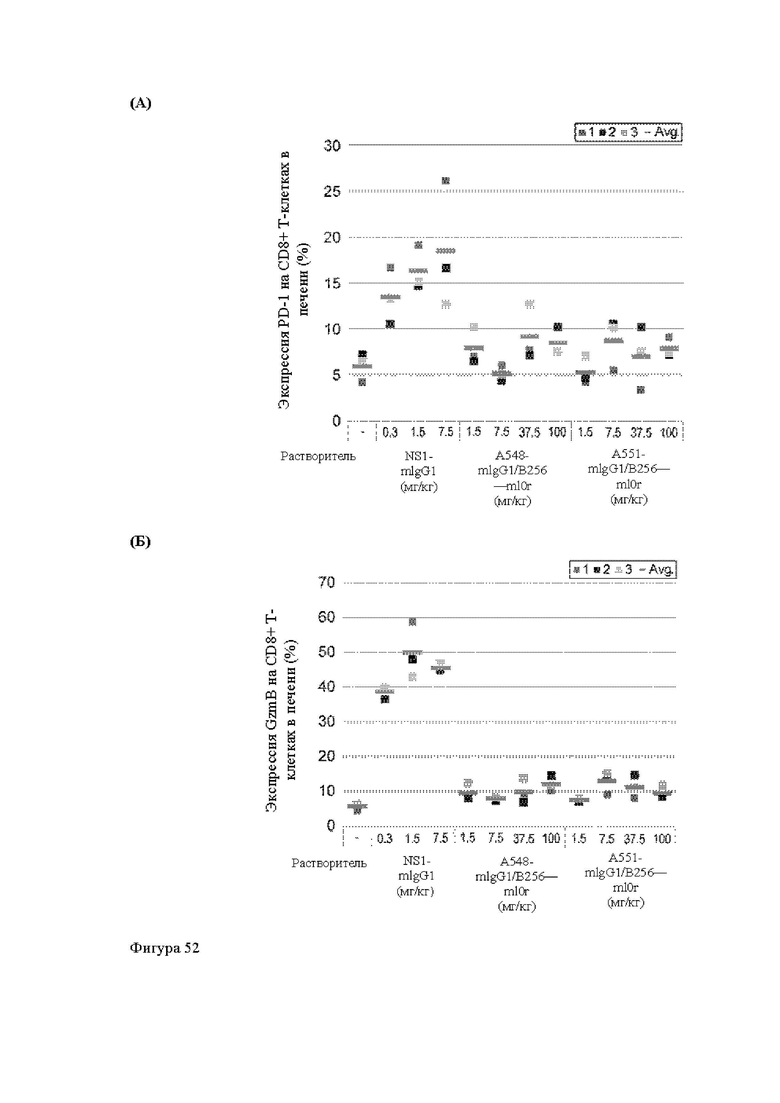
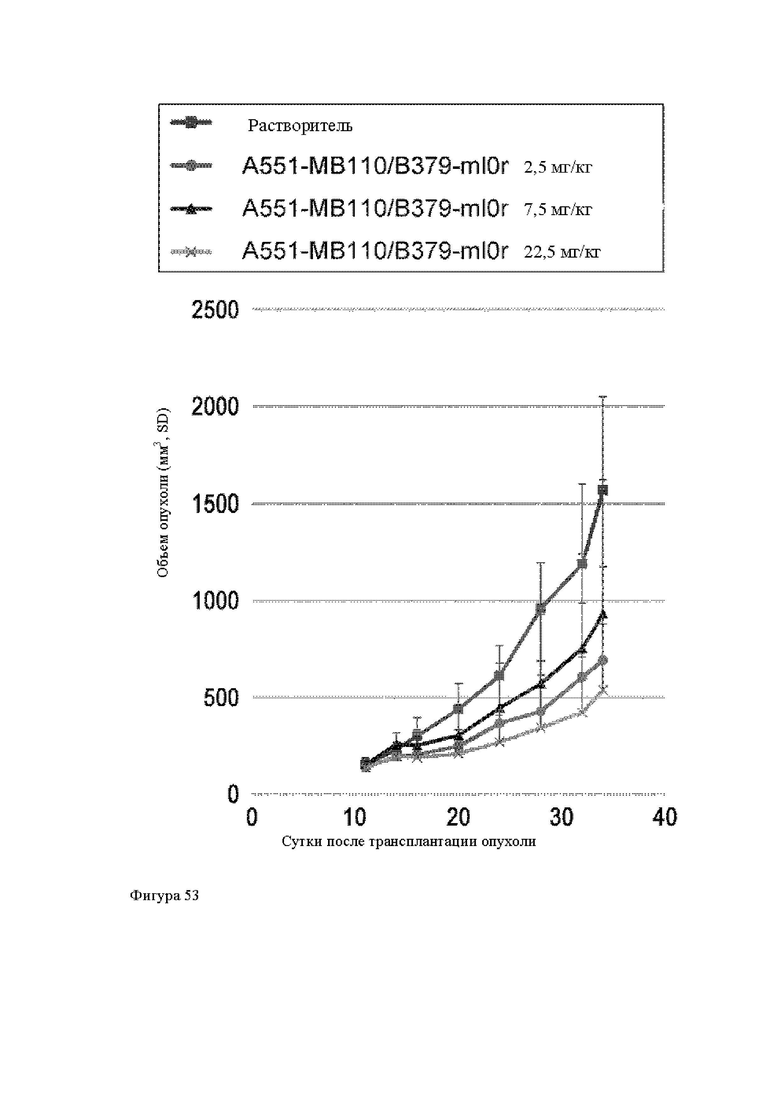
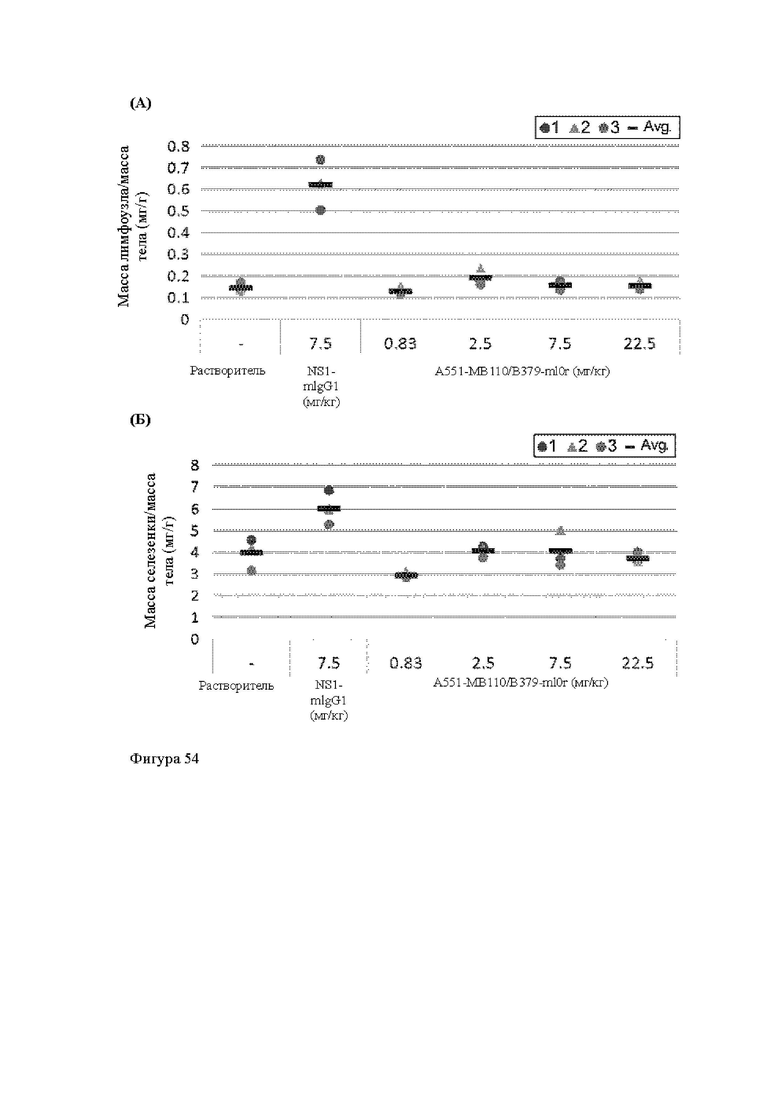
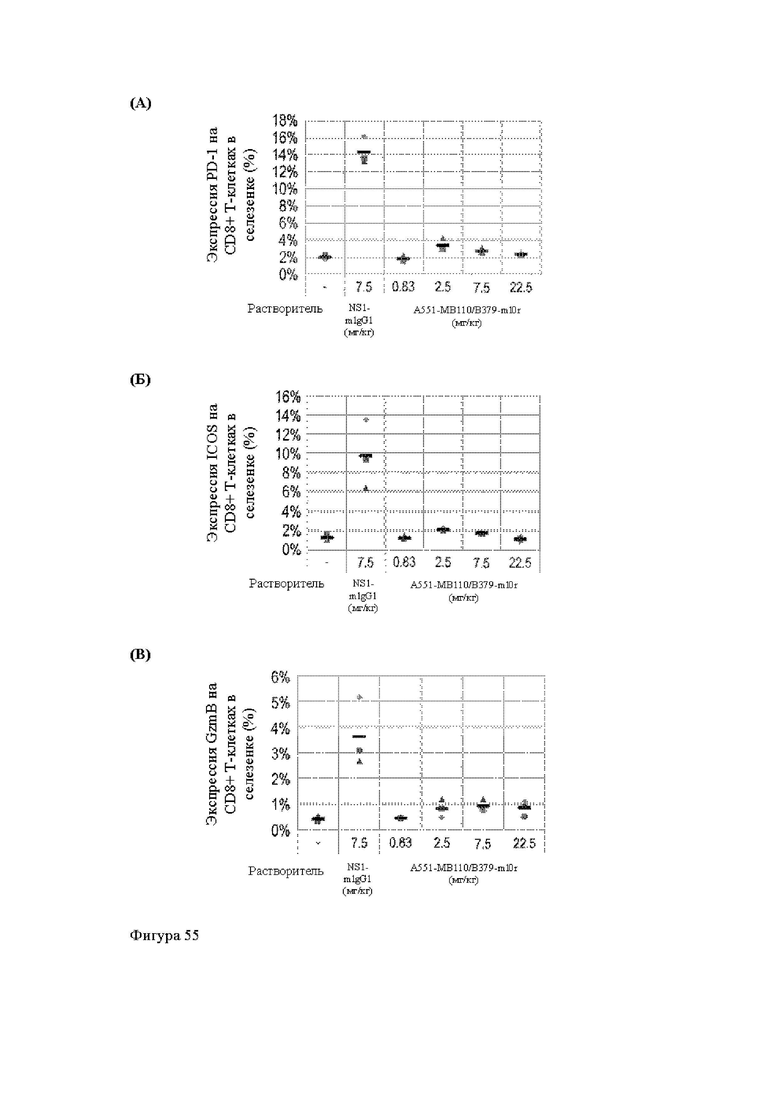
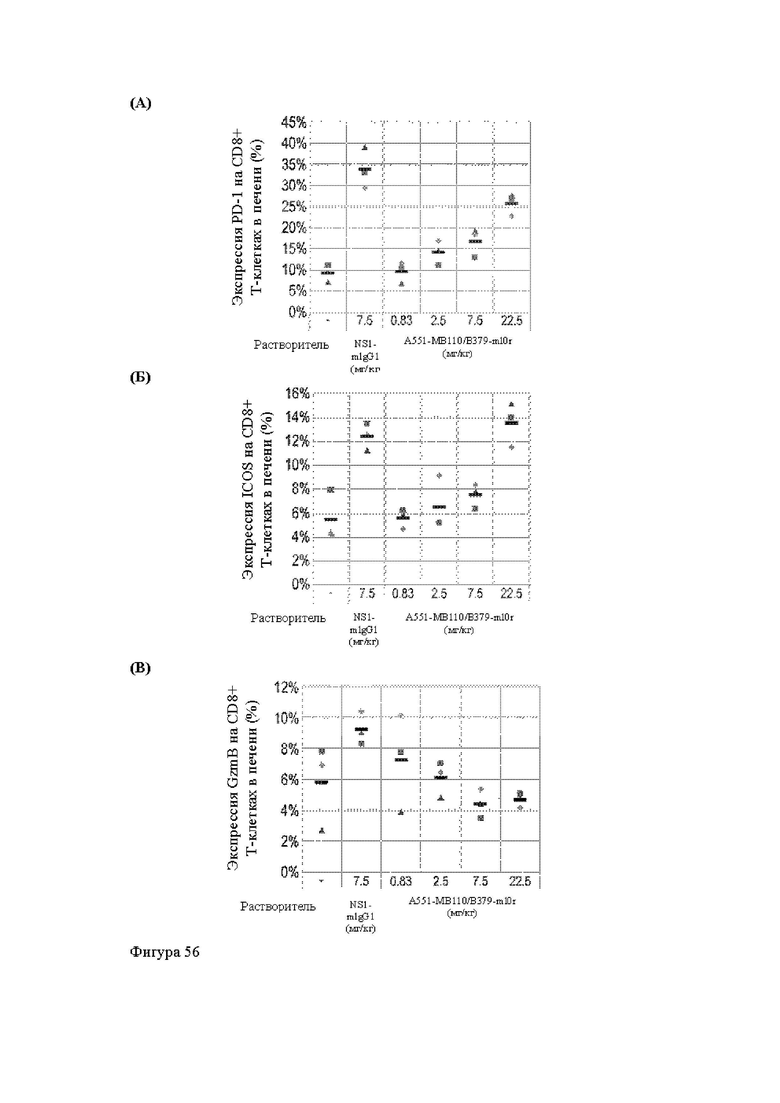
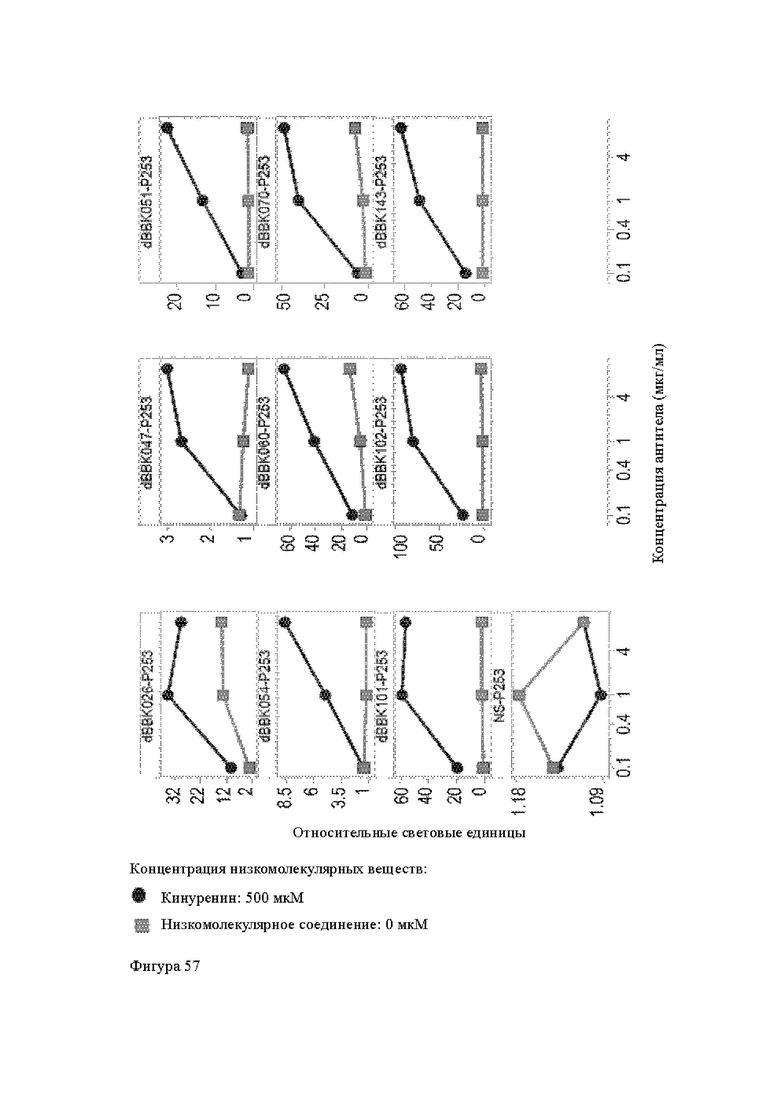
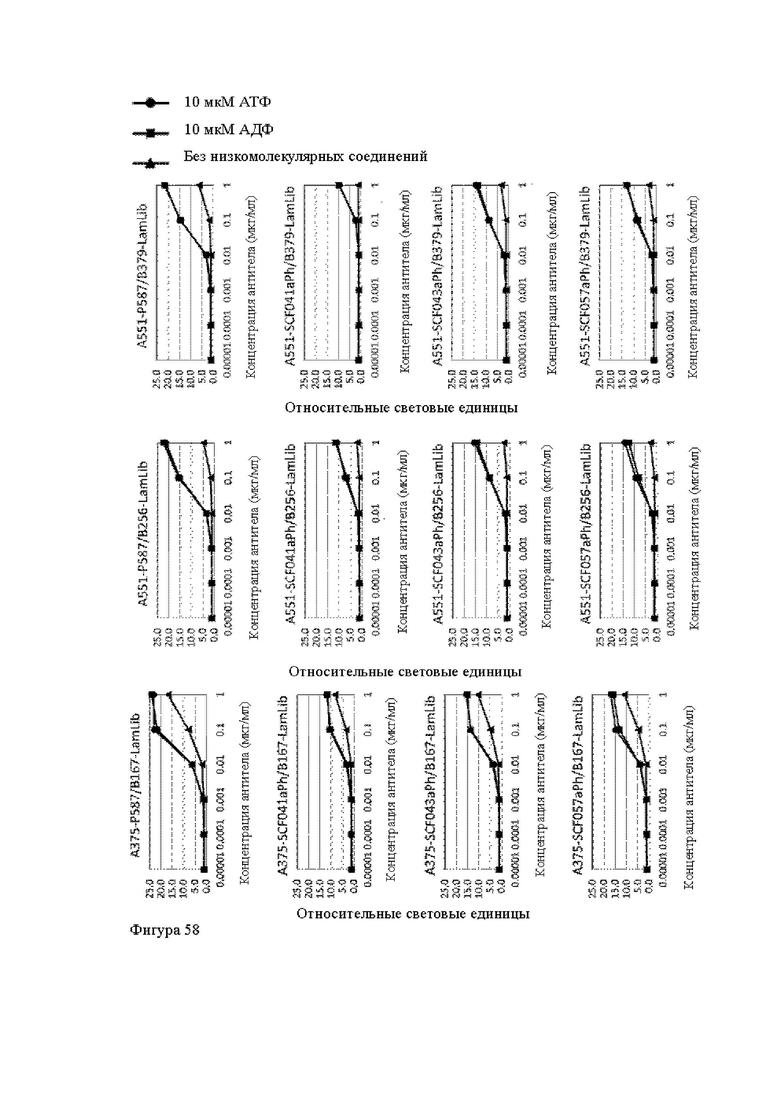
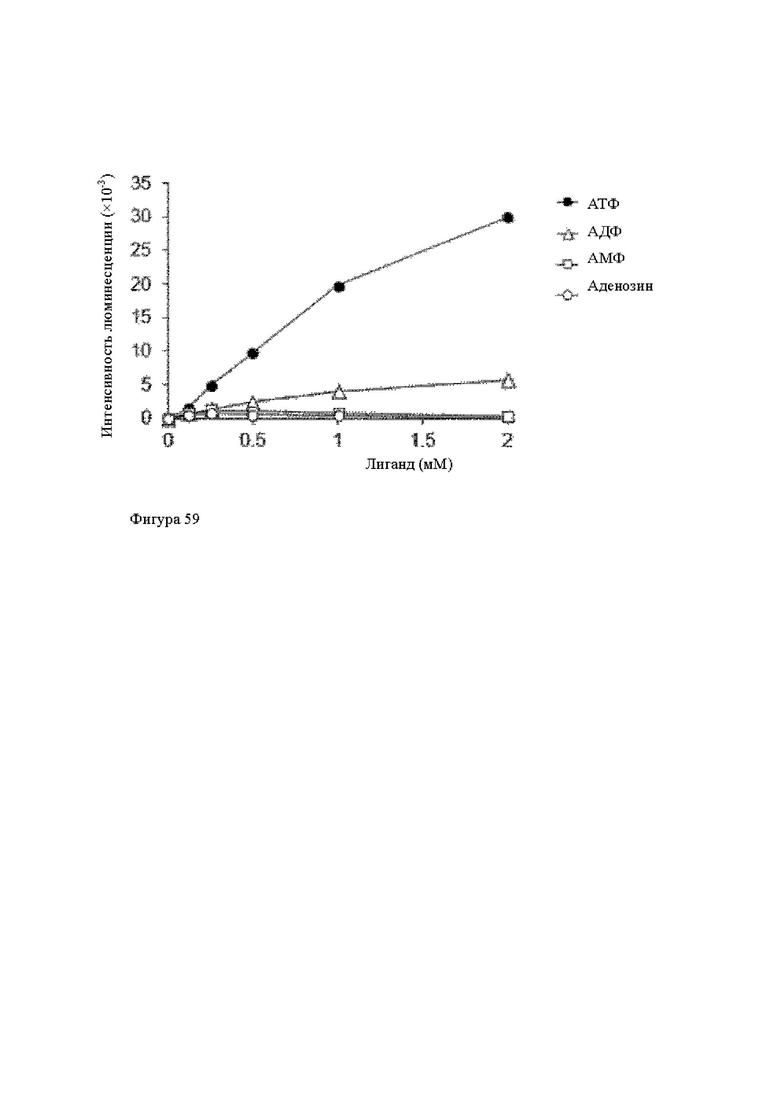
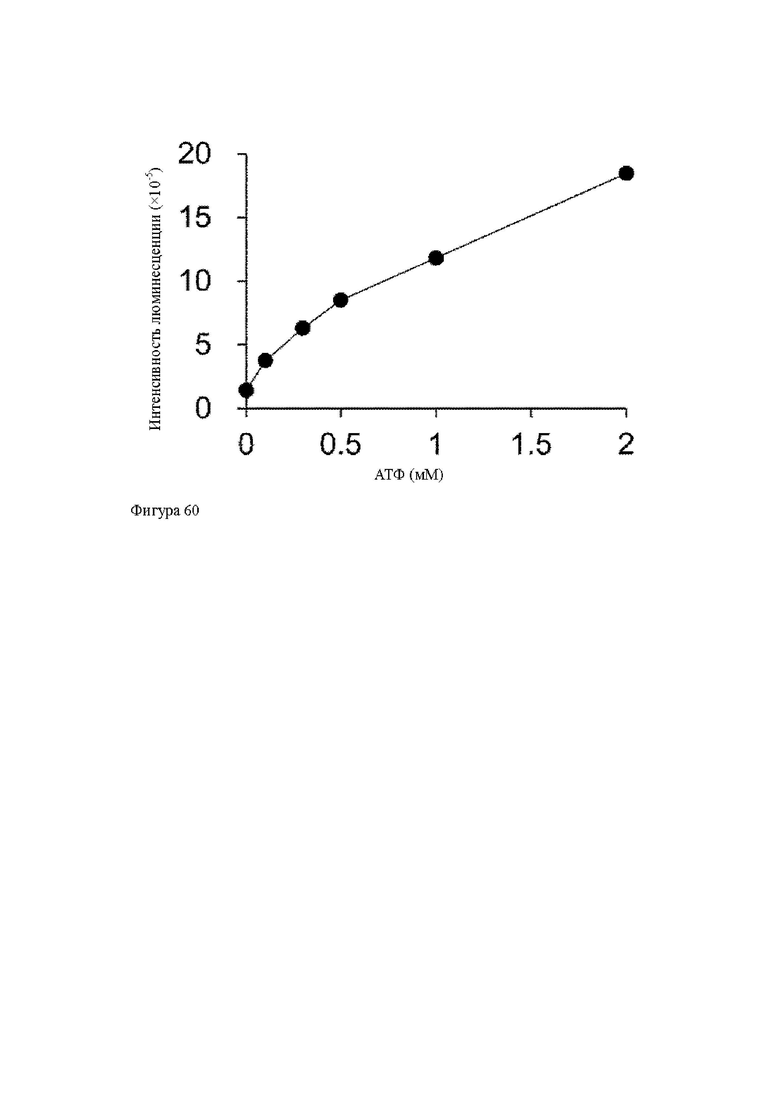


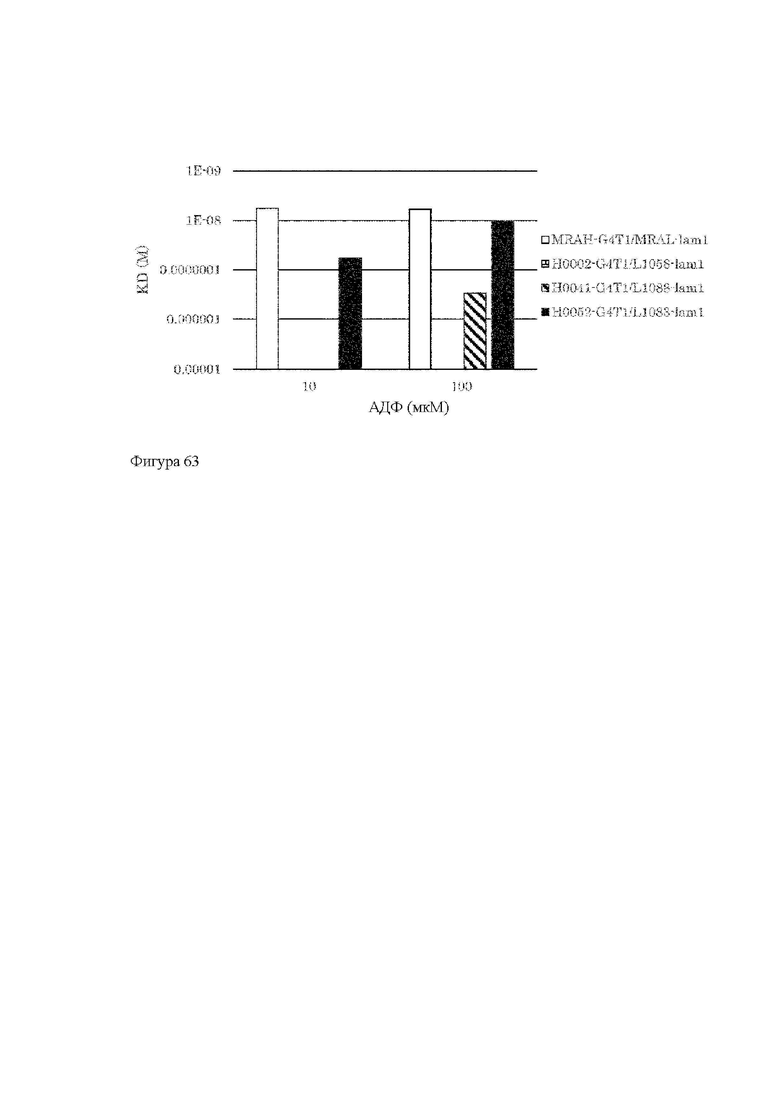
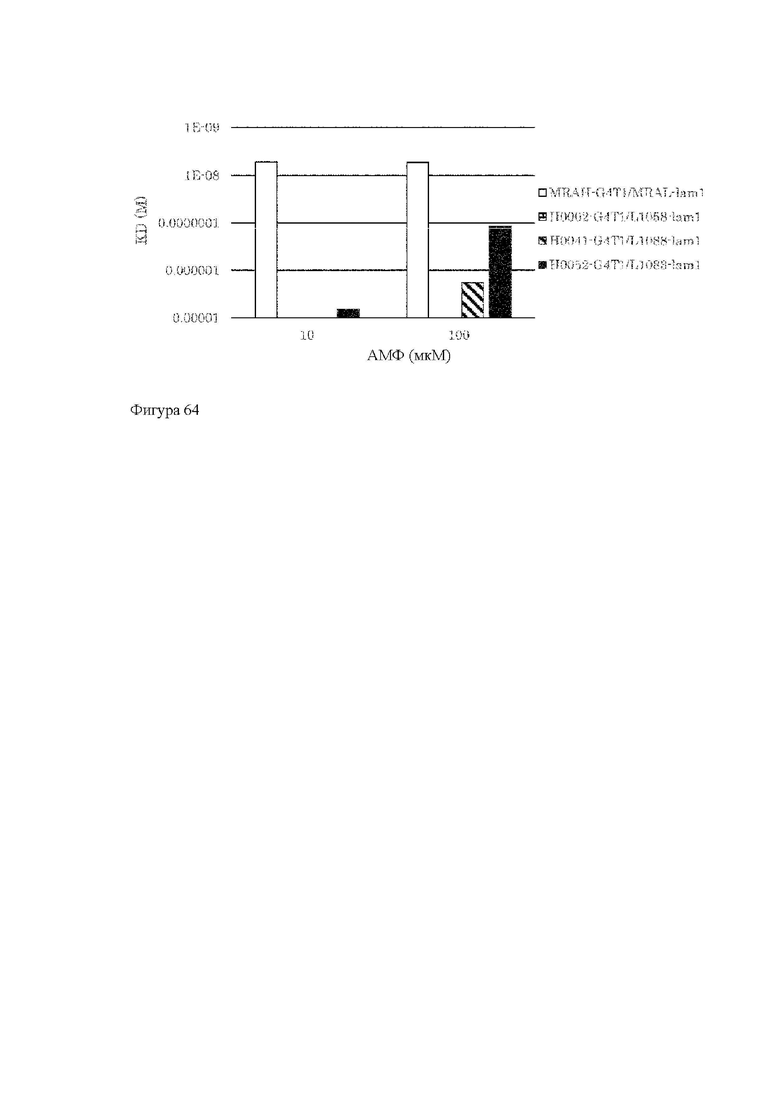
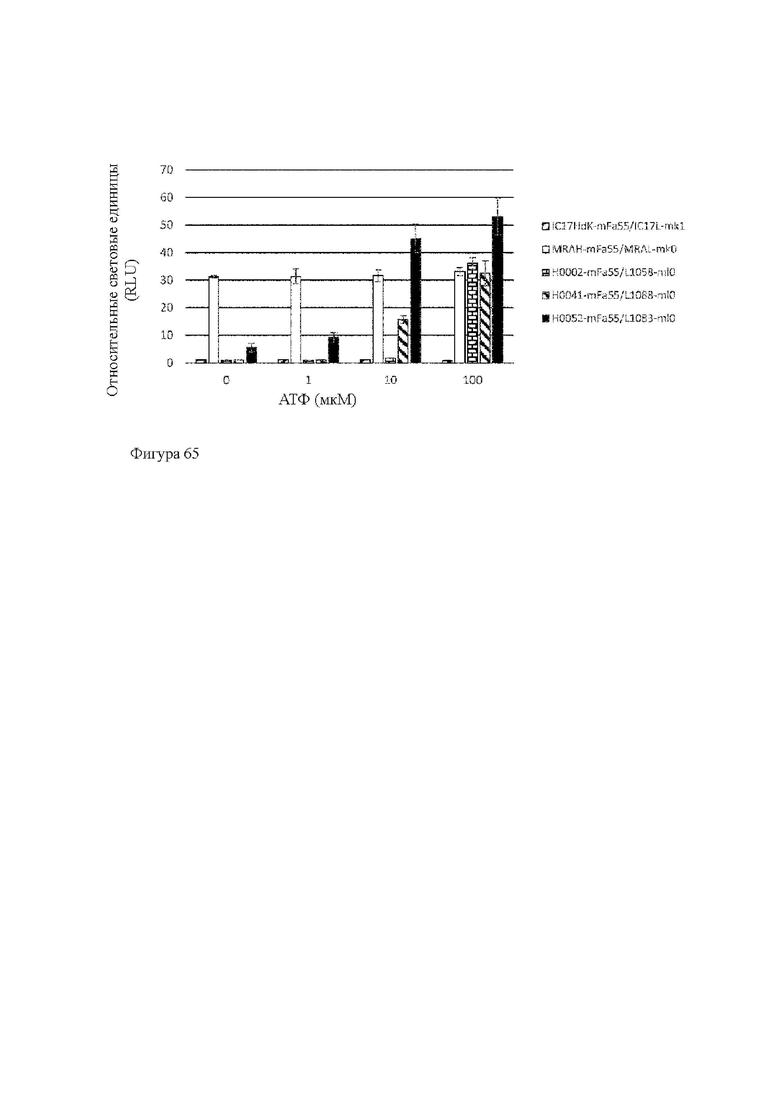
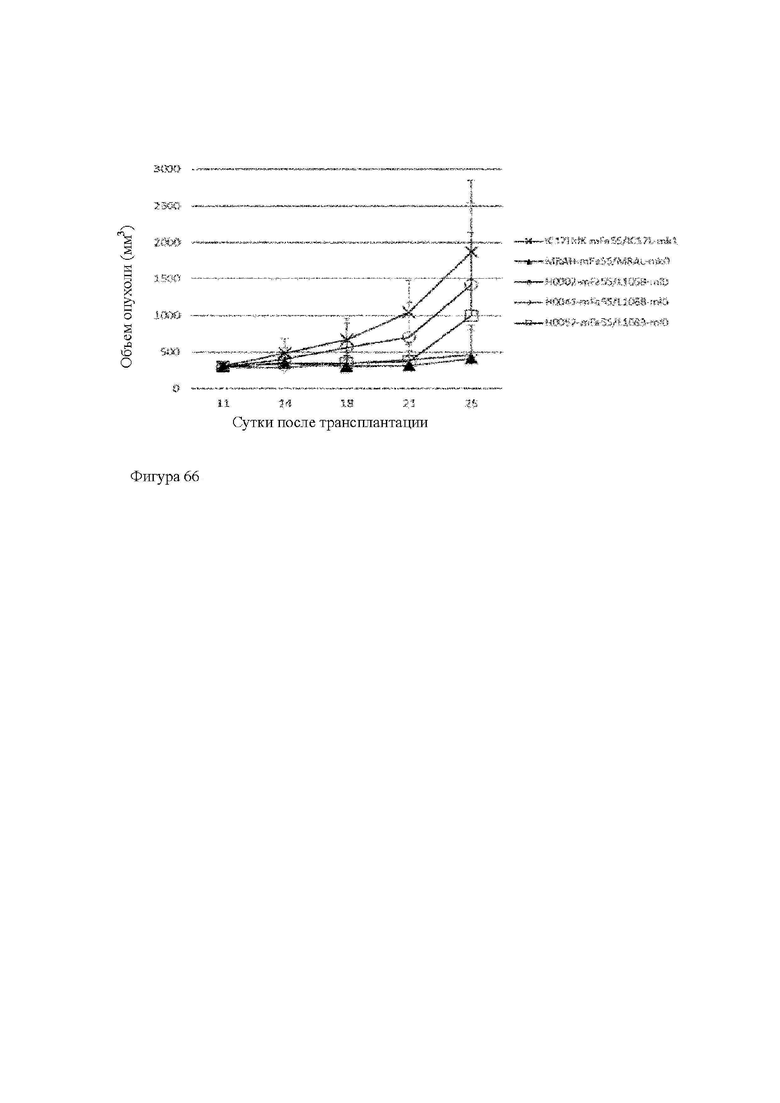
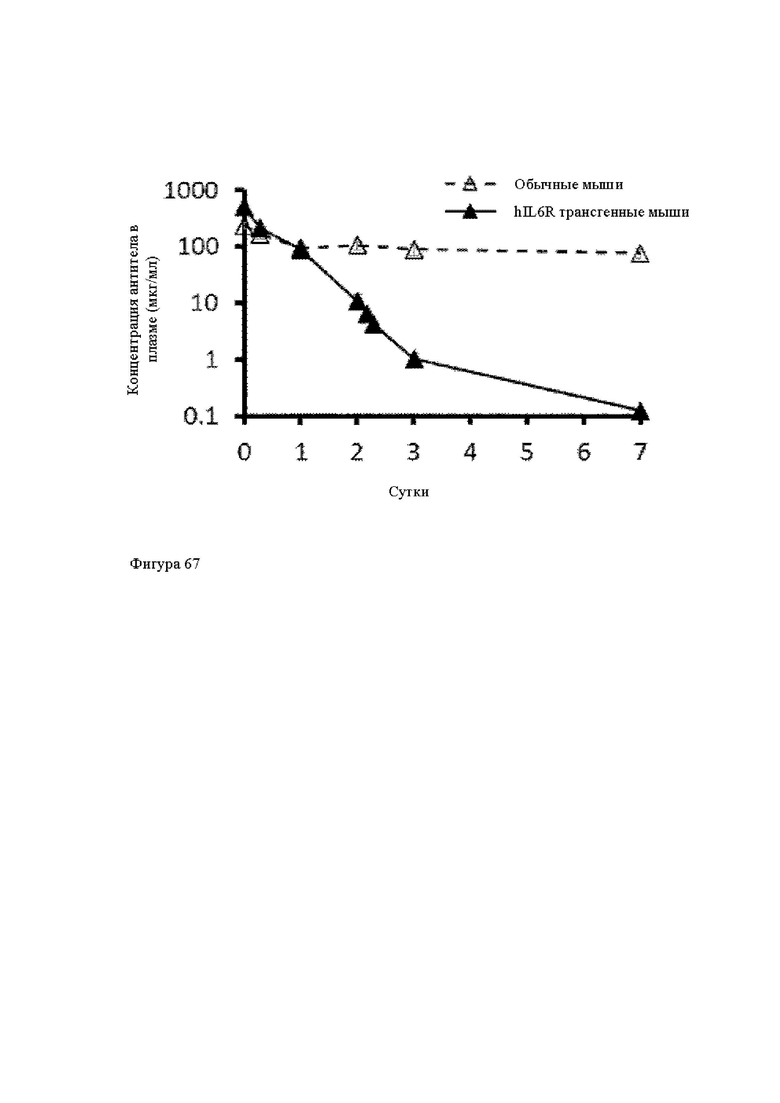
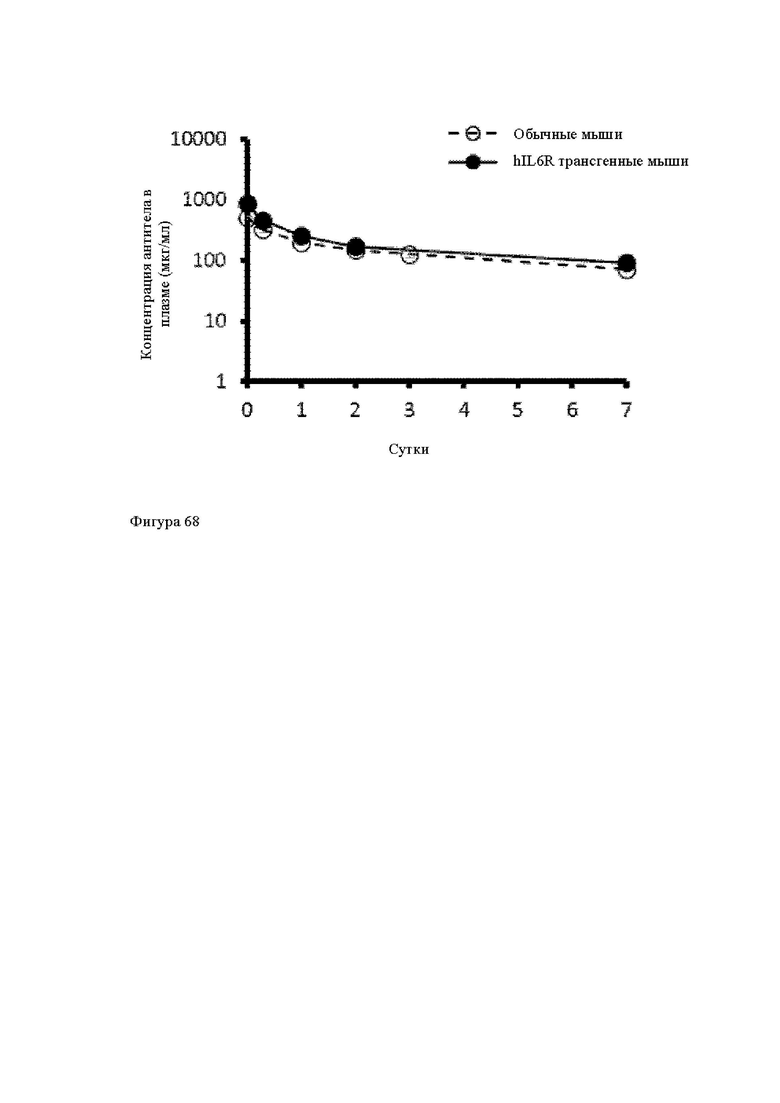

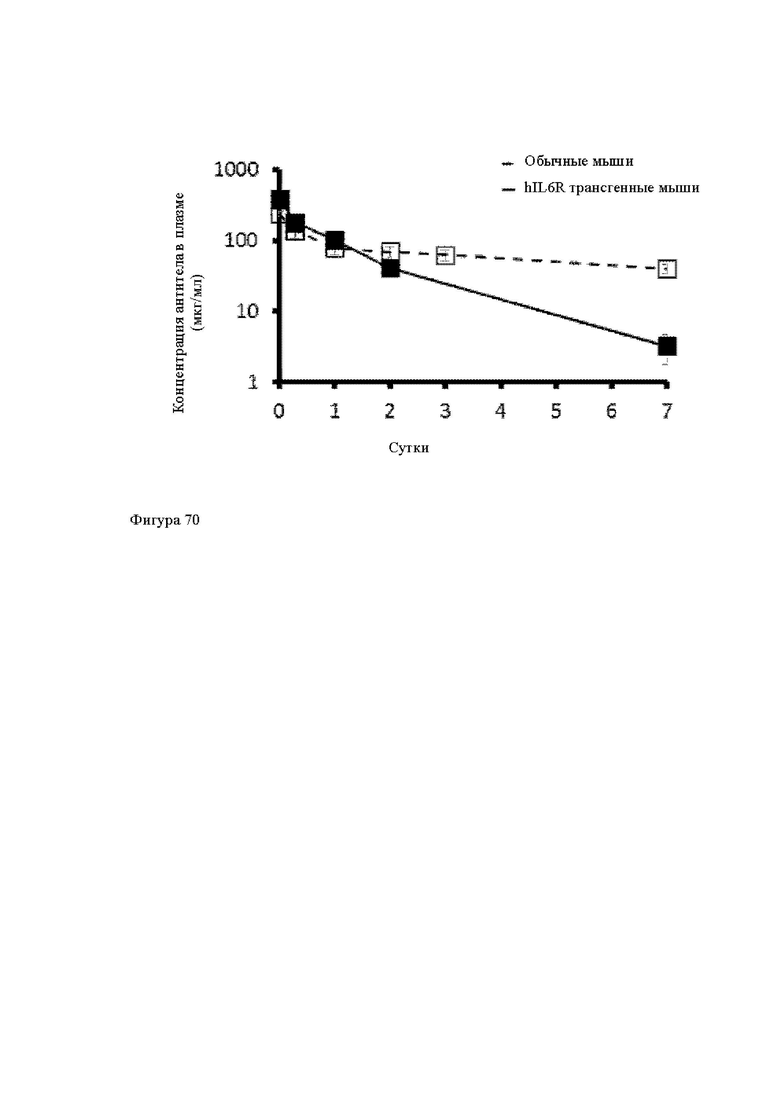
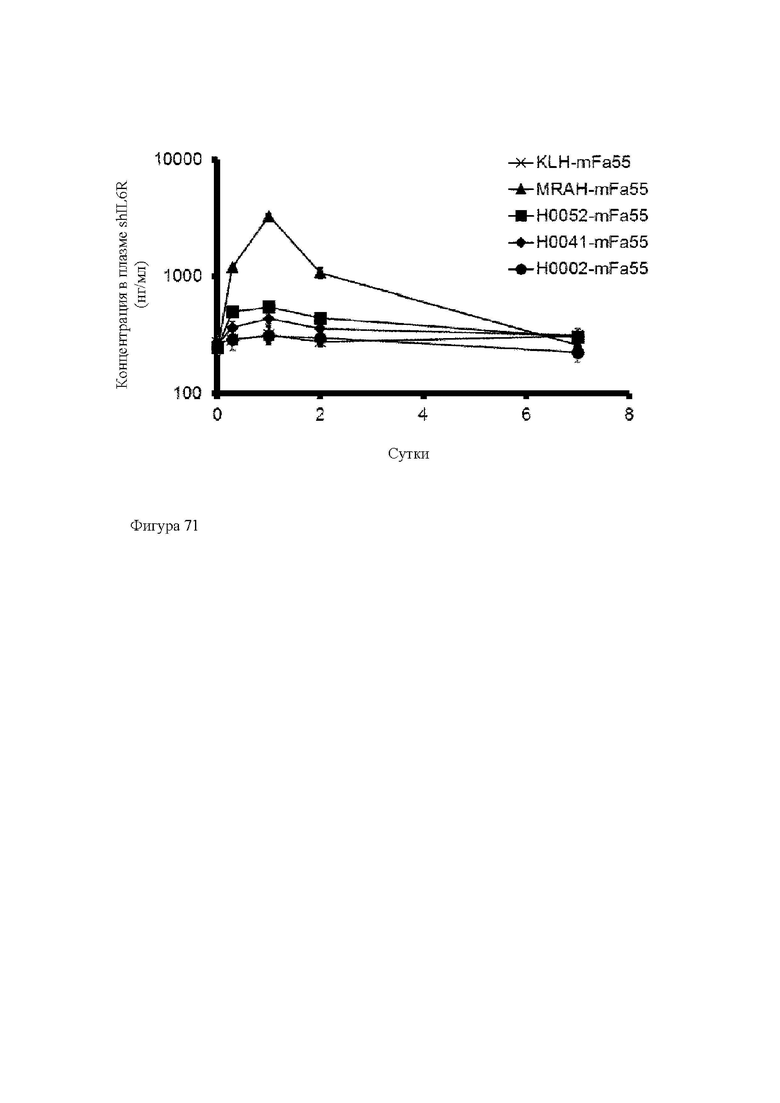
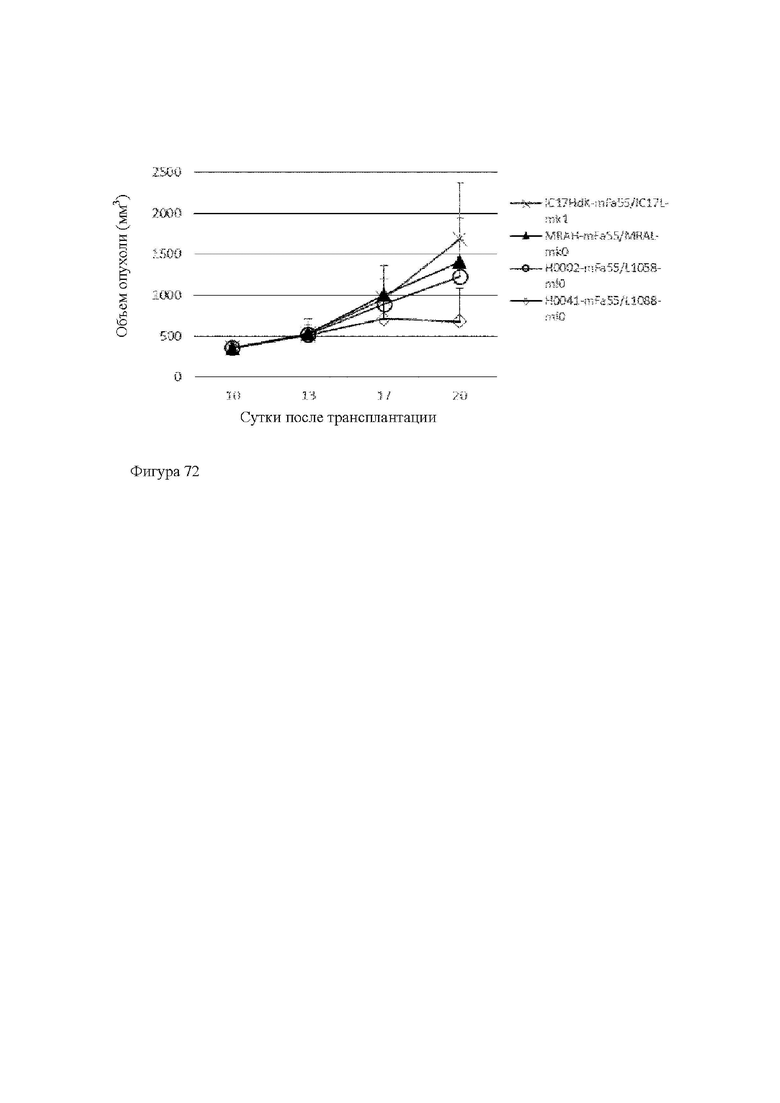
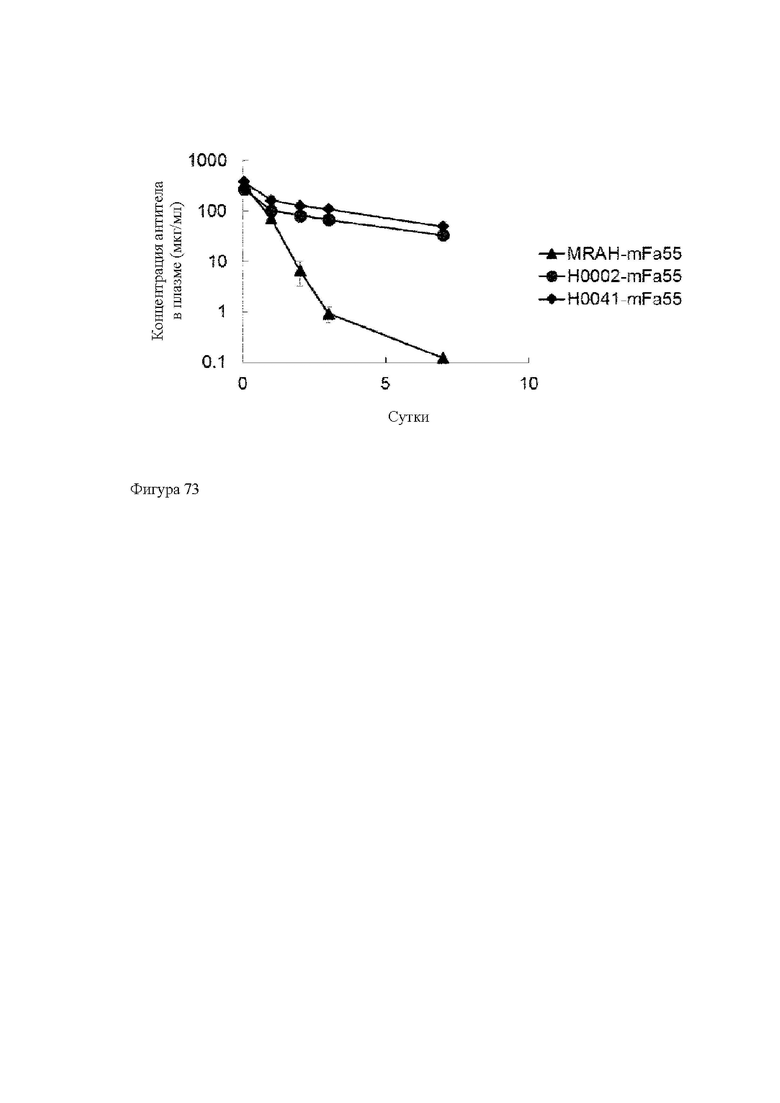
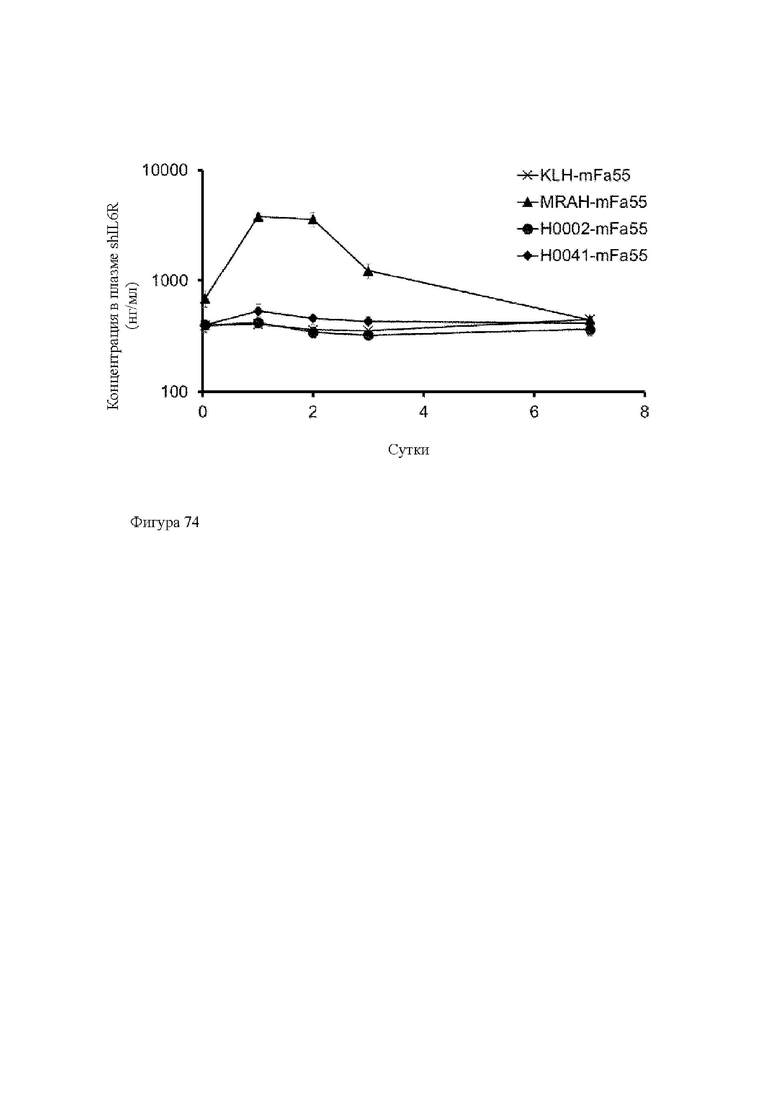
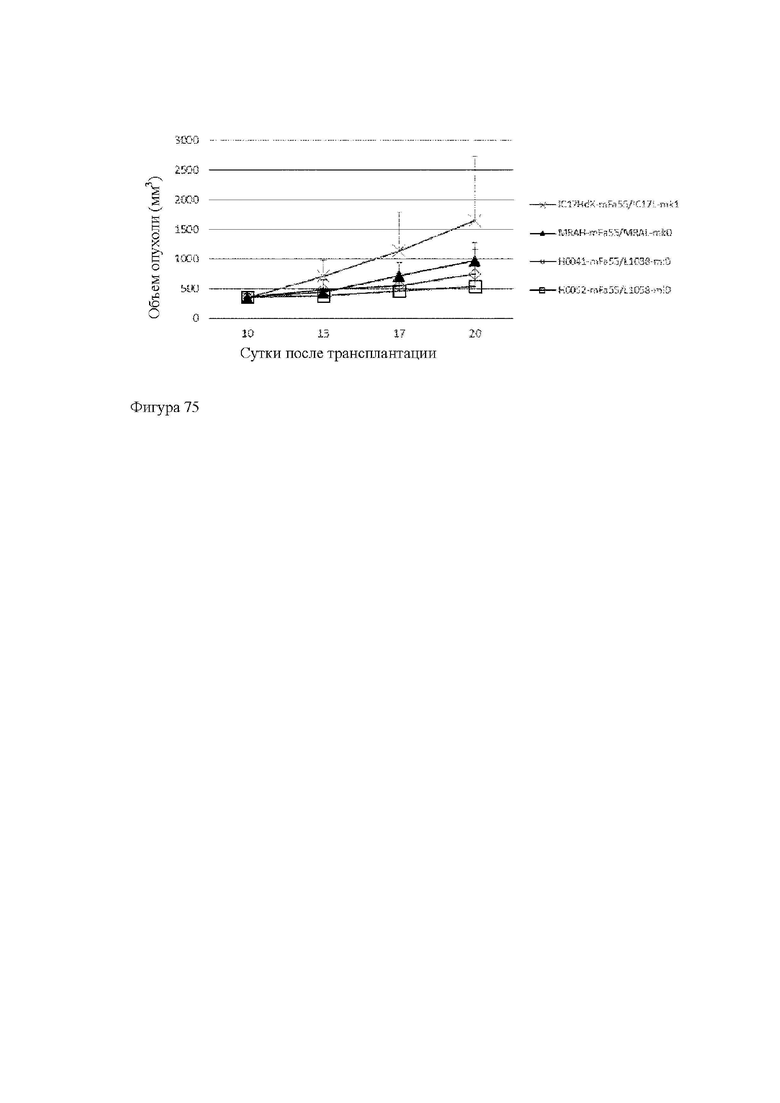
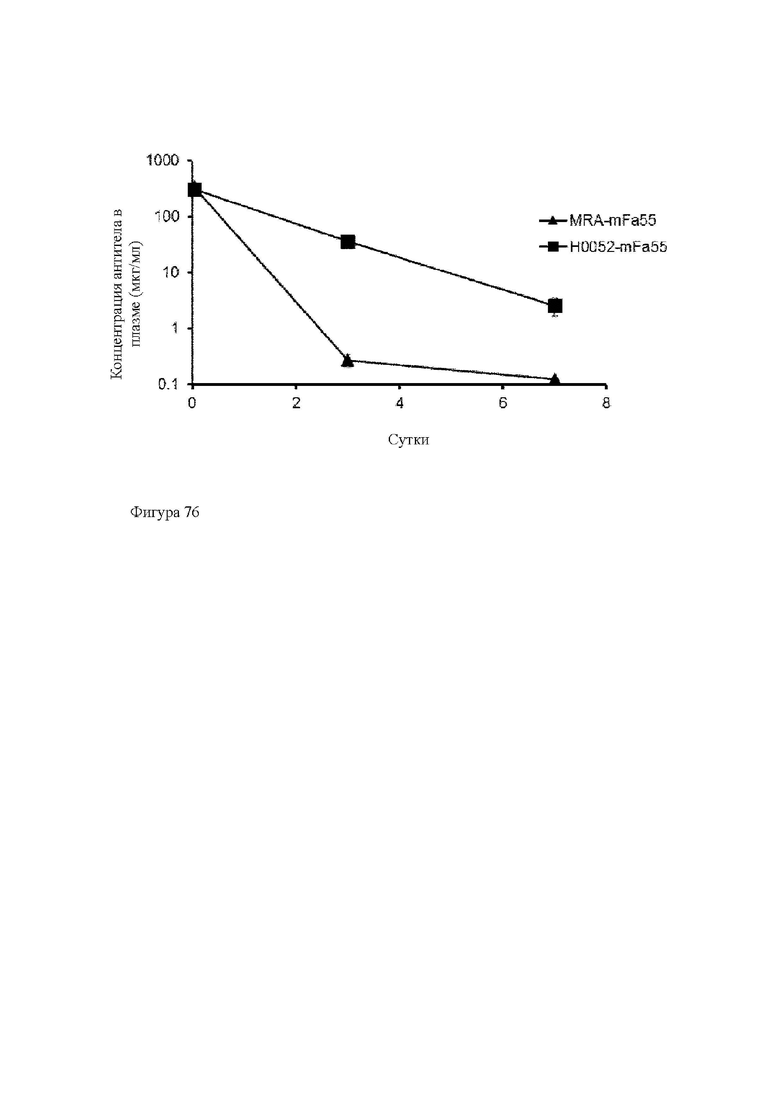
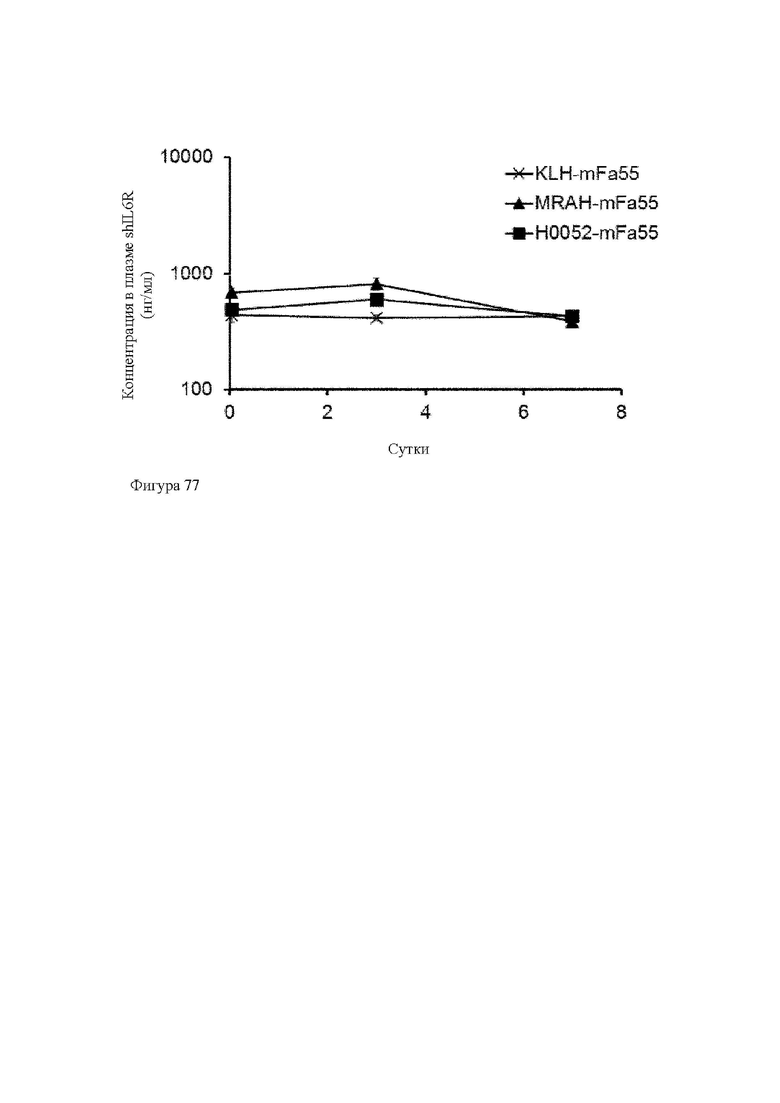
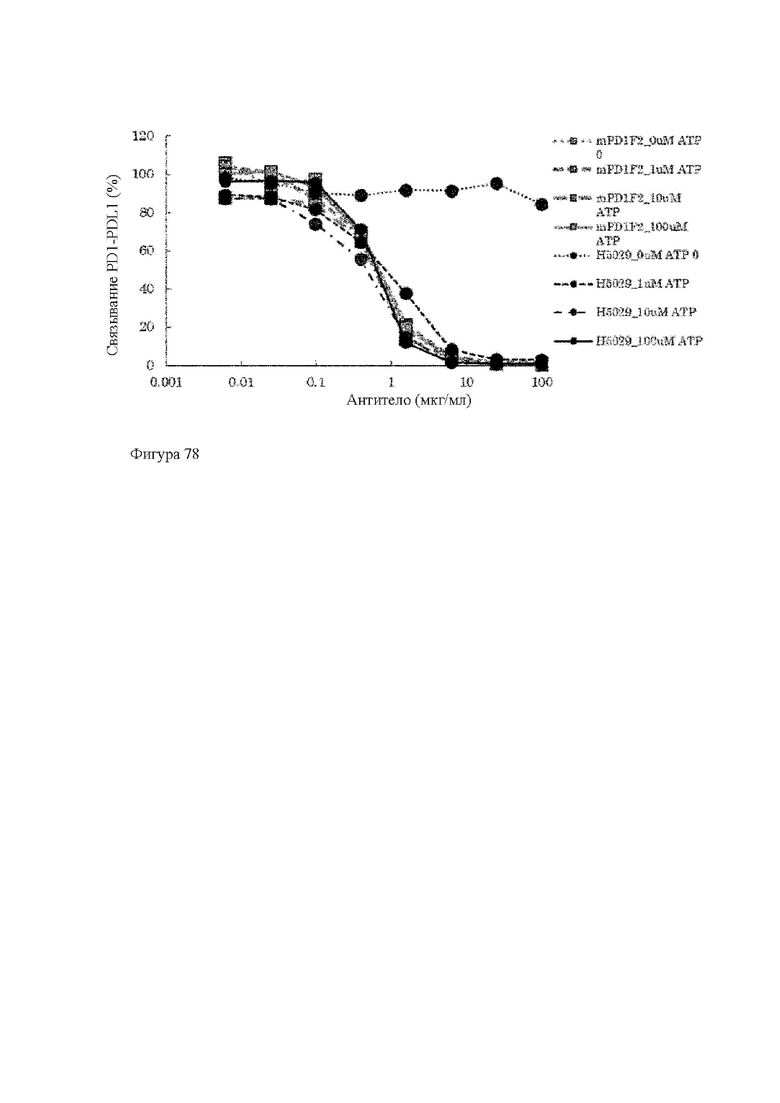
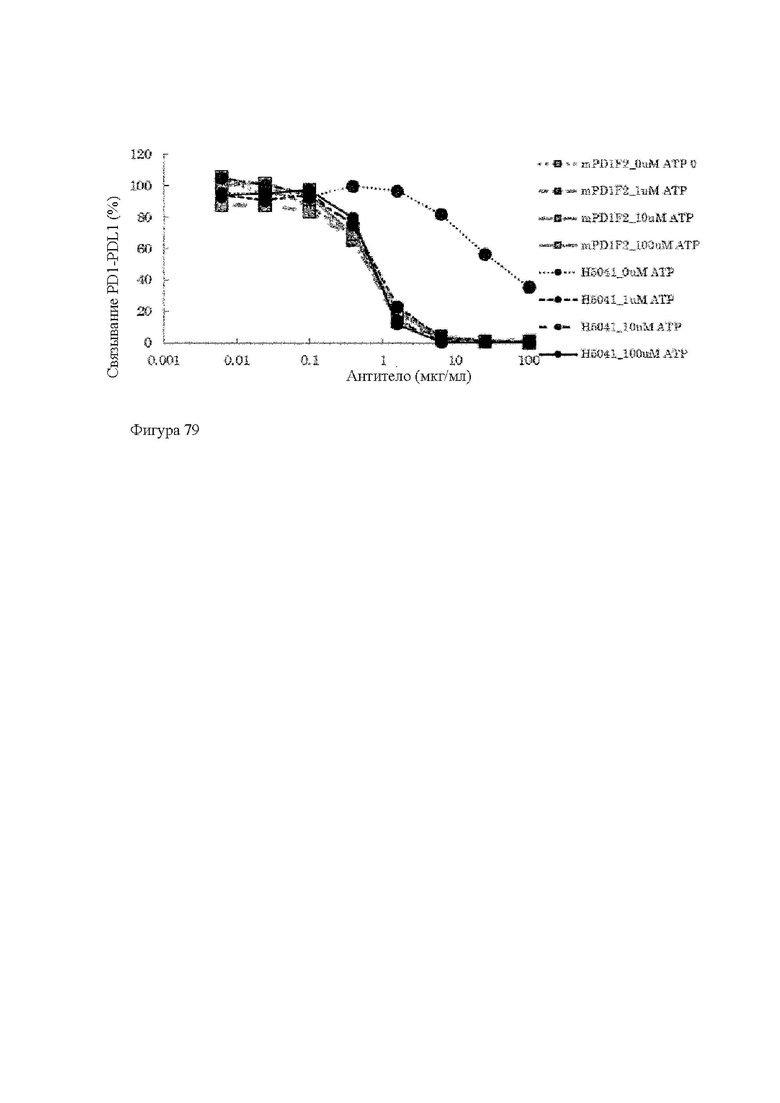
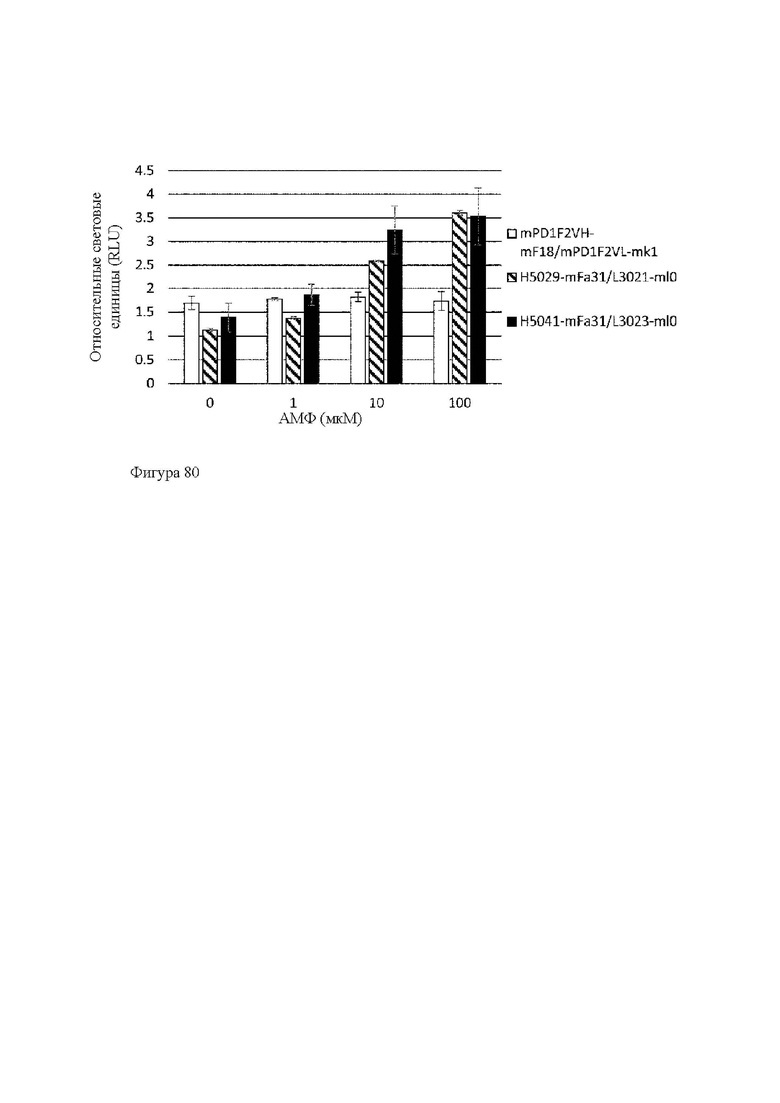
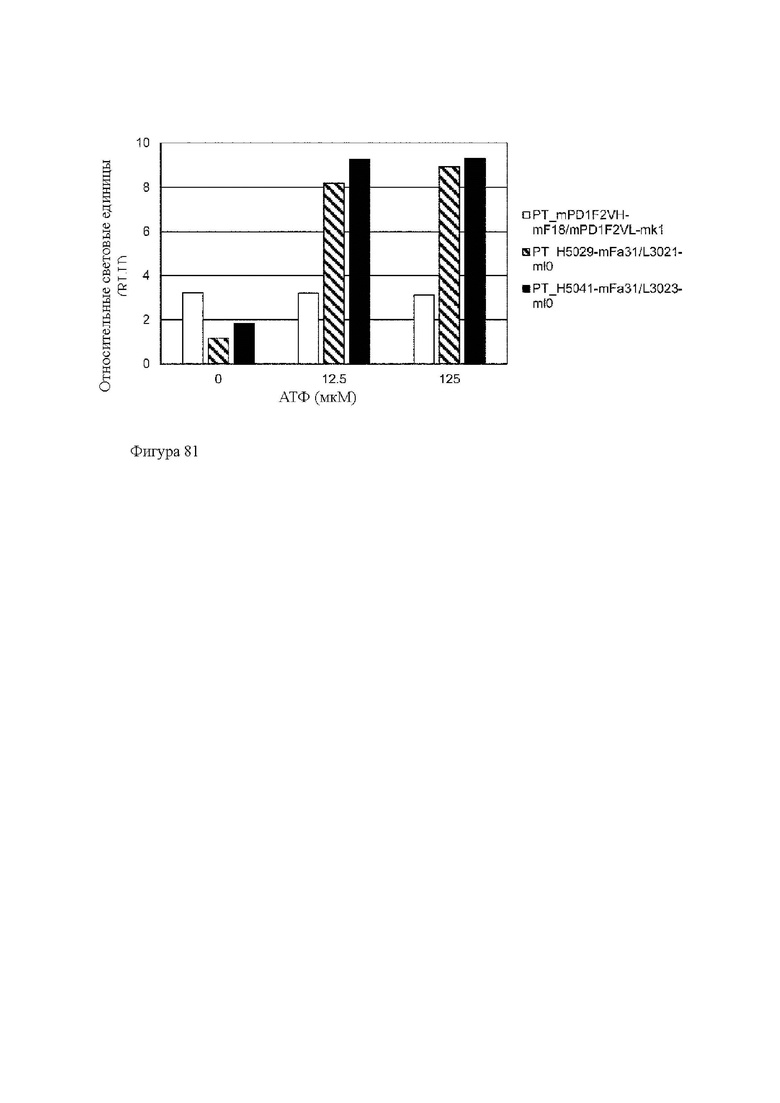
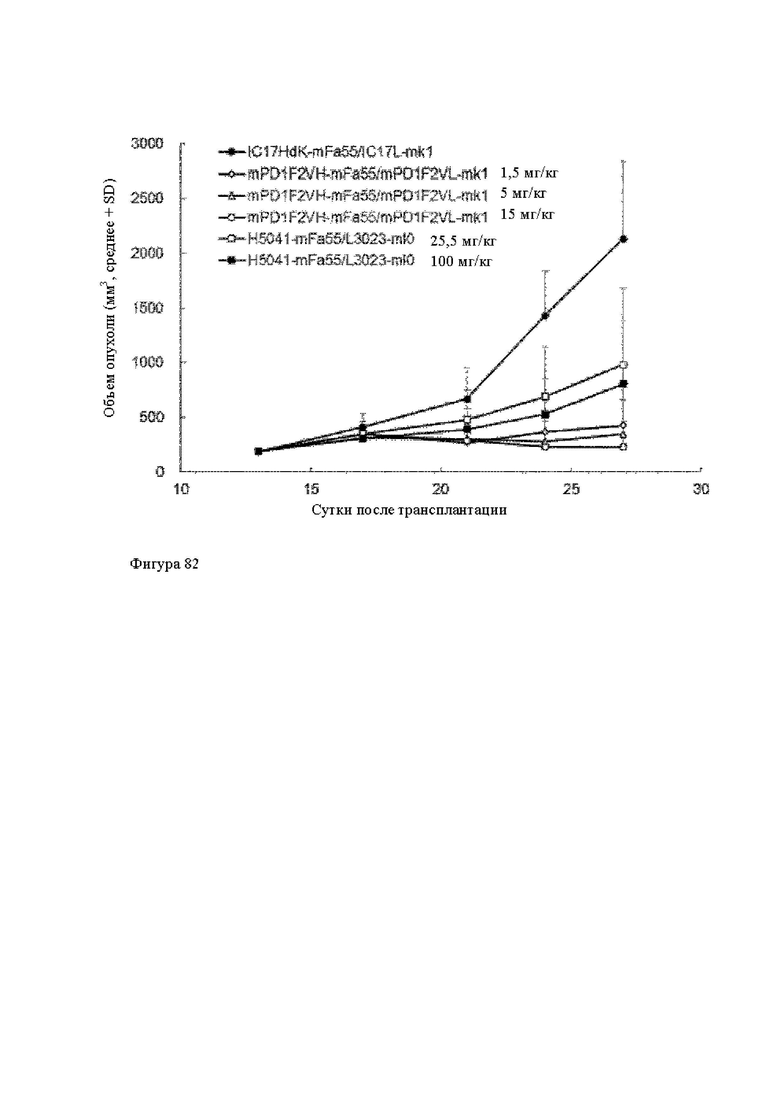
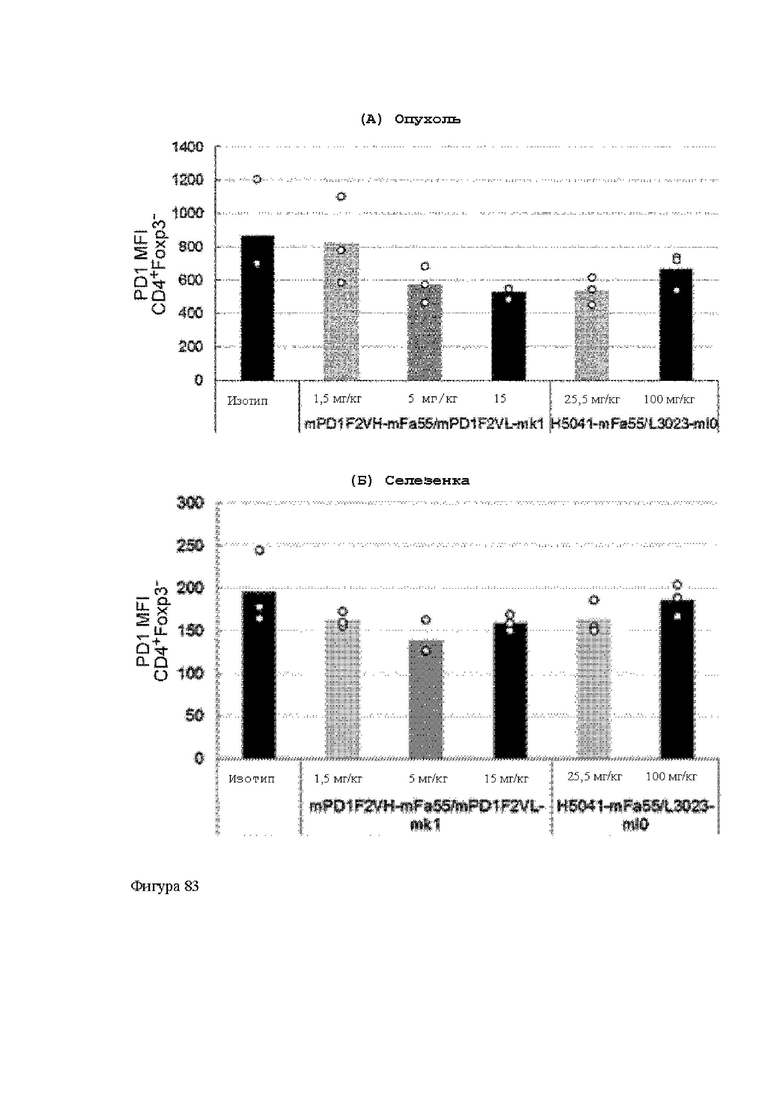
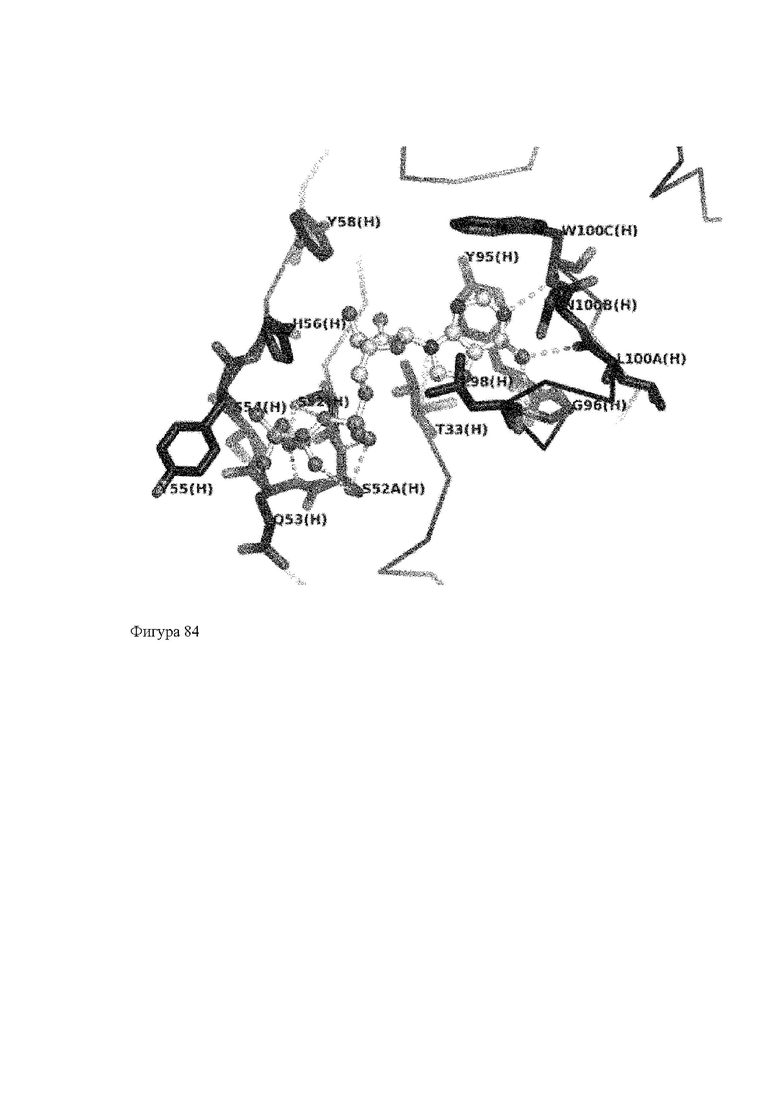

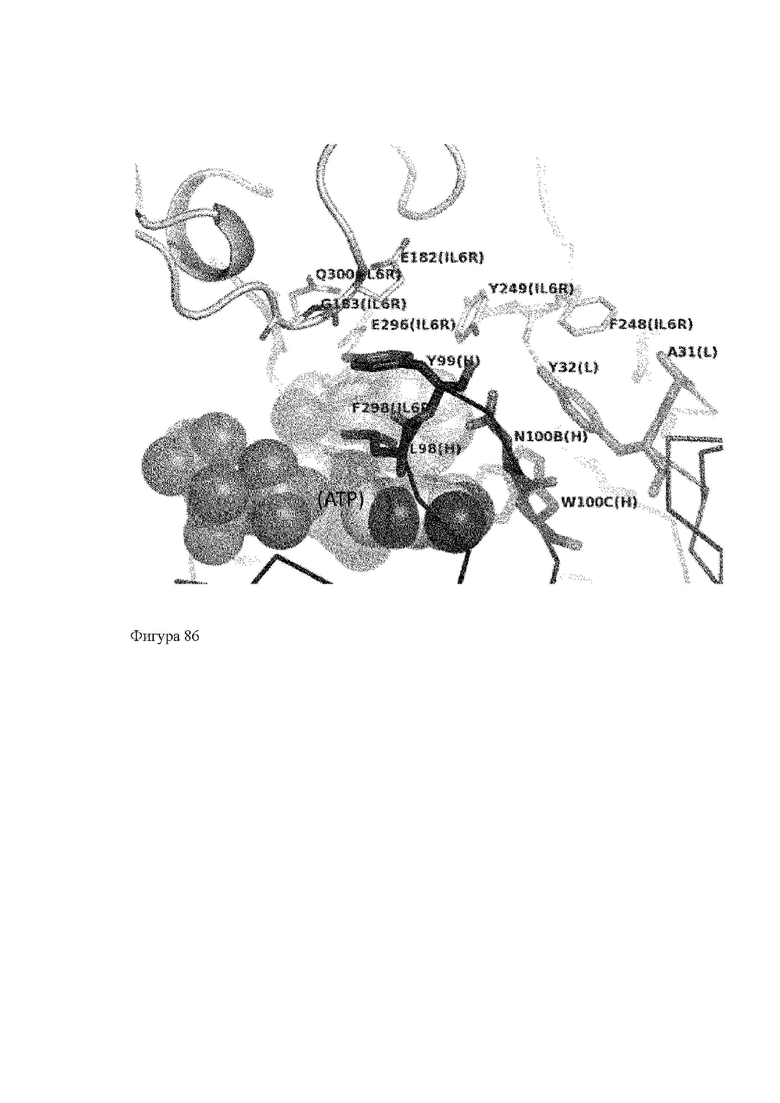
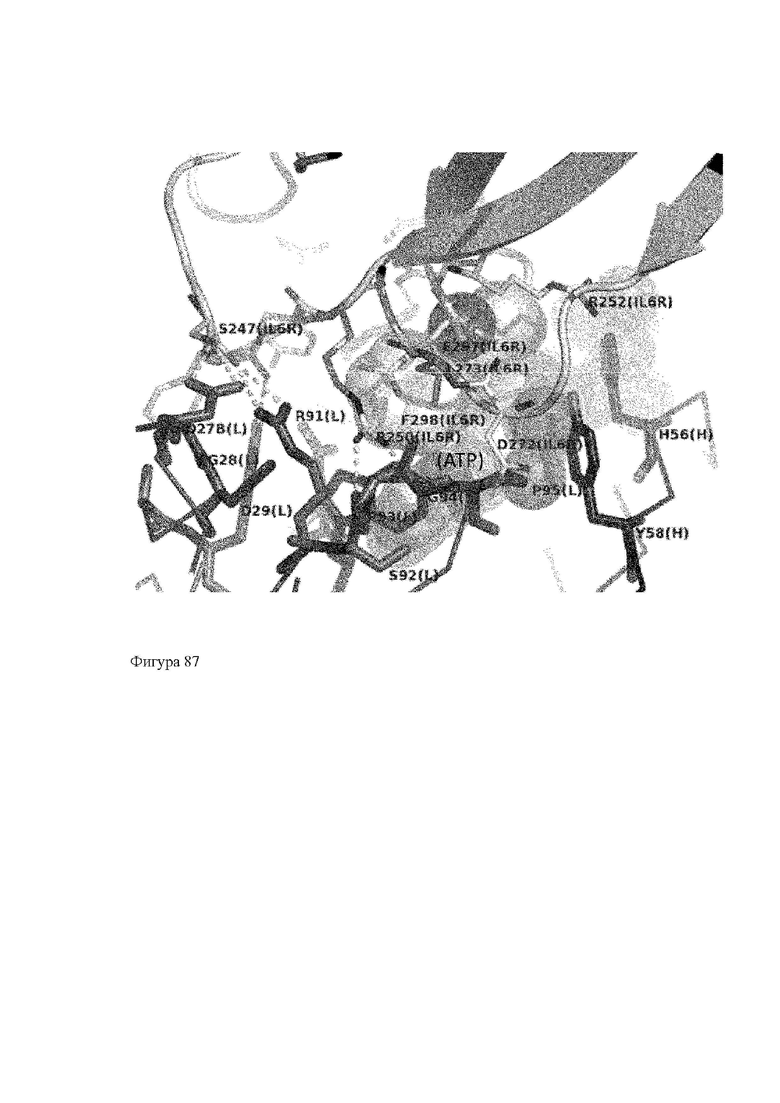
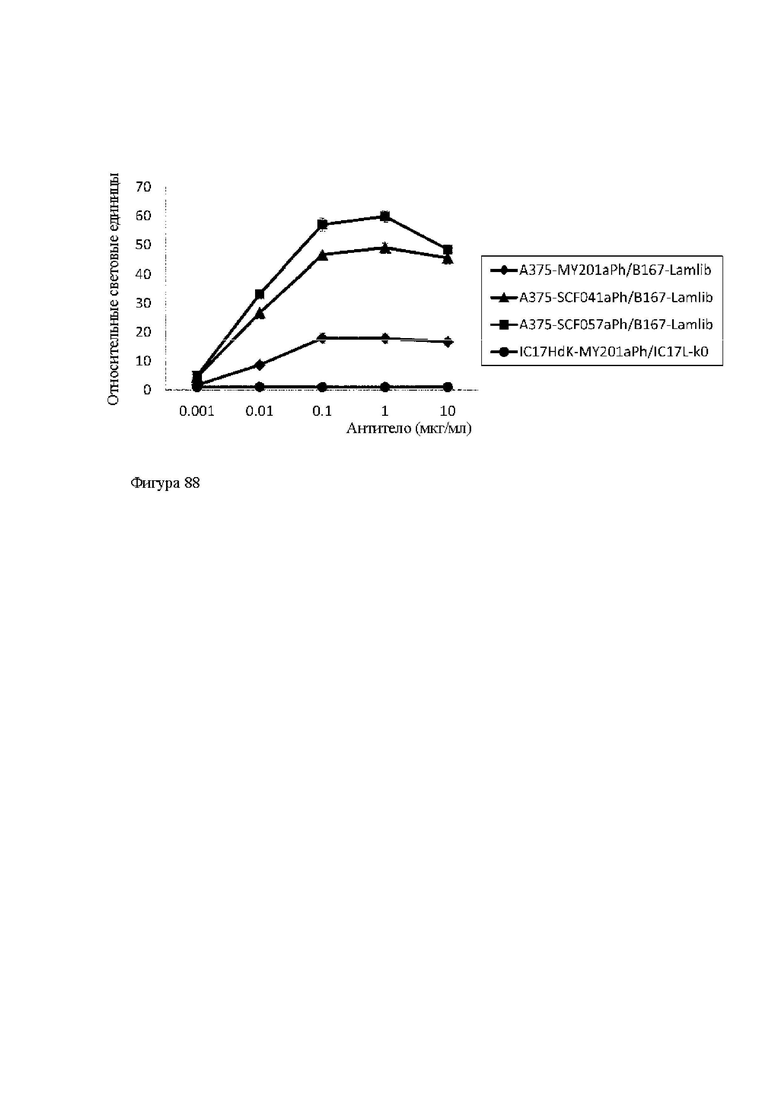
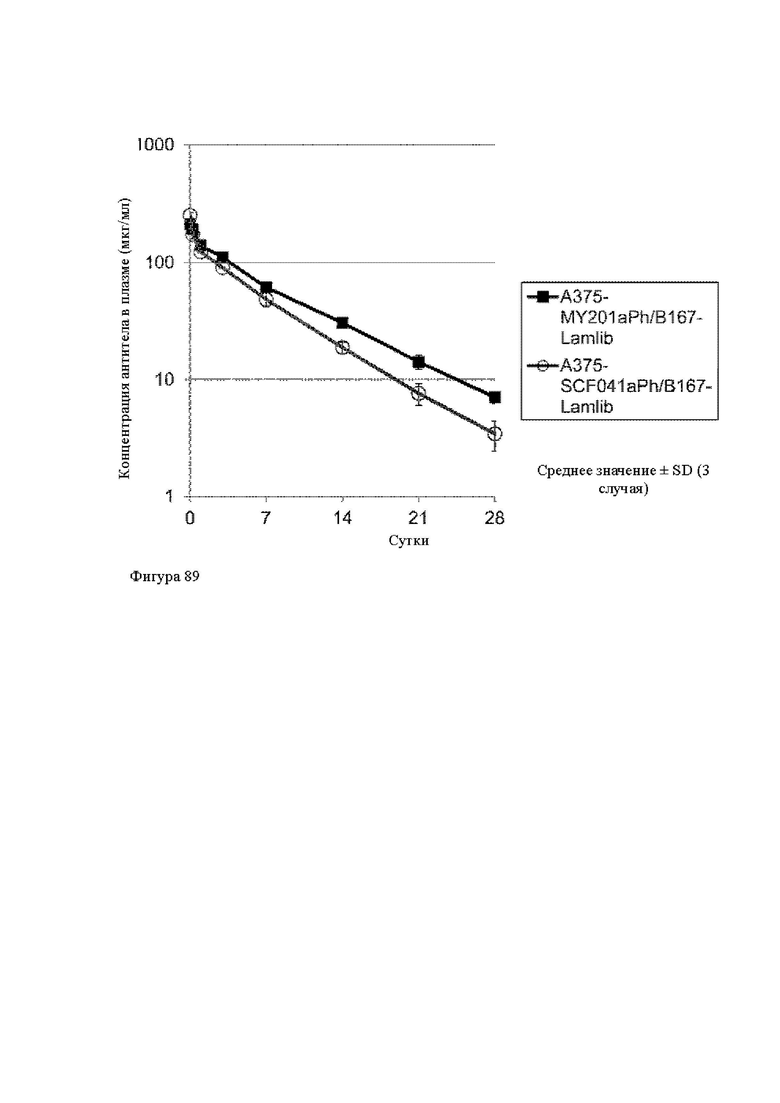
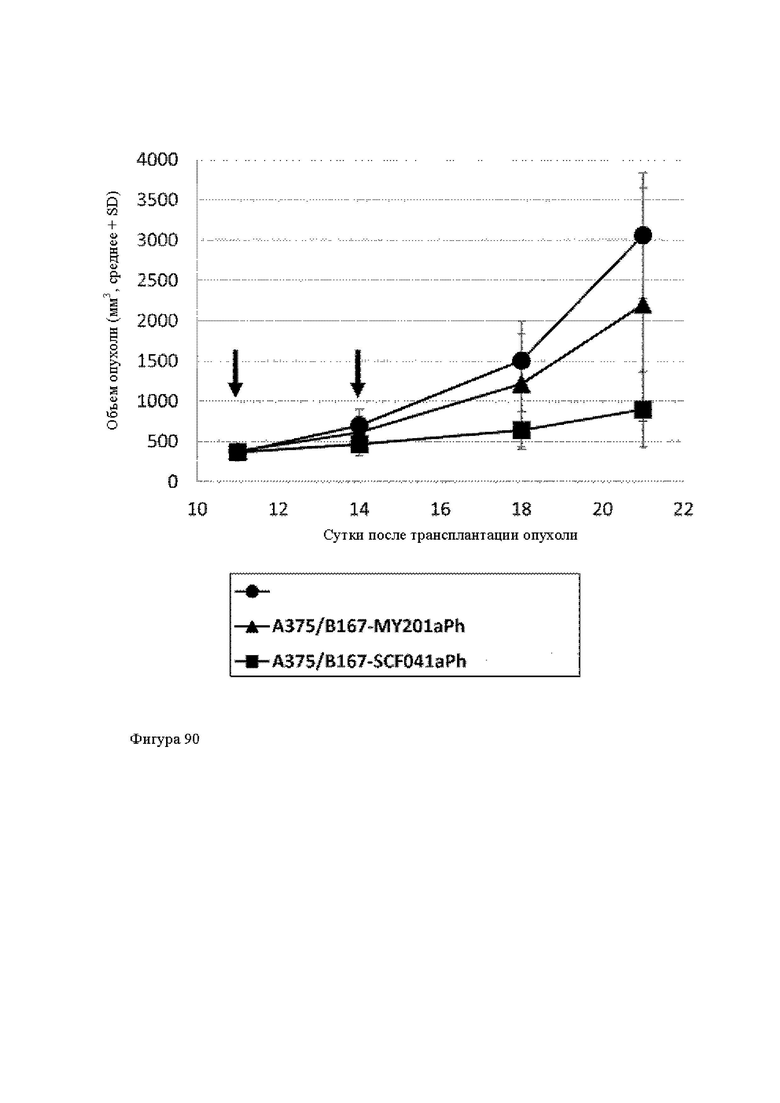
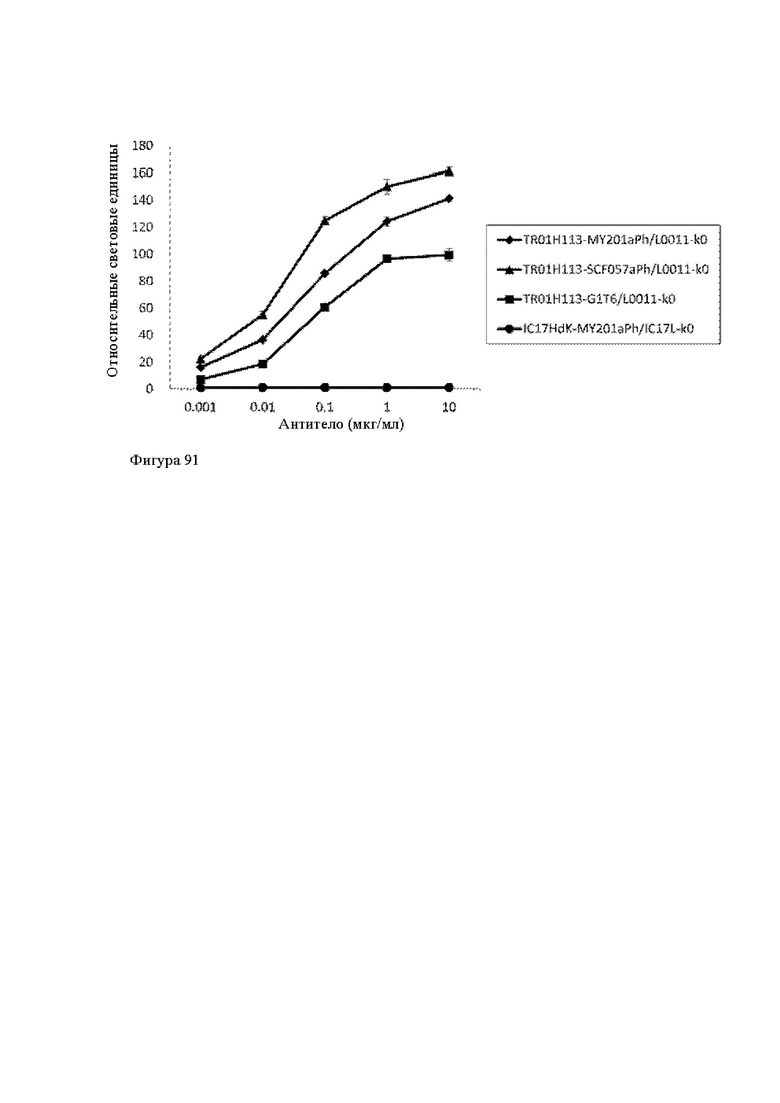
Группа изобретений относится к биотехнологии. Представлены анти-CD137 антигенсвязывающие молекулы и способ их получения. Также раскрыты выделенная нуклеиновая кислота, кодирующая анти-CD137 антигенсвязывающую молекулу, вектор экспрессии, в который интродуцирована нуклеиновая кислота и клетка-хозяин для получения анти-CD137 антигенсвязывающей молекулы. Изобретение используется для получения анти-CD137 антигенсвязывающих молекул, которые обладают эффектом активации иммуноцитов, цитотоксической активностью или противоопухолевой активностью, и в то же время обладают пониженным действием на неопухолевые ткани, такие как нормальные ткани, и вызывают меньше побочных эффектов. 6 н. и 2 з.п. ф-лы, 72 табл., 91 ил., 20 пр.
1. Анти-CD137 антигенсвязывающая молекула, включающая любую комбинацию областей HVR-H1, HVR-H2, HVR-H3, HVR-L1, HVR-L2 и HVR-L3, выбранную из приведенных ниже подпунктов от (а) до (н):
(а) HVR-H1, содержащая аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащая аминокислотную последовательность SEQ ID NO: 8, HVR-H3, содержащая аминокислотную последовательность SEQ ID NO: 17, HVR-L1, содержащая аминокислотную последовательность SEQ ID NO: 21, HVR-L2, содержащая аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащая аминокислотную последовательность SEQ ID NO: 27;
(б) HVR-H1, содержащая аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащая аминокислотную последовательность SEQ ID NO: 9, HVR-H3, содержащая аминокислотную последовательность SEQ ID NO: 17, HVR-L1, содержащая аминокислотную последовательность SEQ ID NO: 22, HVR-L2, содержащая аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащая аминокислотную последовательность SEQ ID NO: 27;
(в) HVR-H1, содержащая аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащая аминокислотную последовательность SEQ ID NO: 10, HVR-H3, содержащая аминокислотную последовательность SEQ ID NO: 17, HVR-L1, содержащая аминокислотную последовательность SEQ ID NO: 22, HVR-L2, содержащая аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащая аминокислотную последовательность SEQ ID NO: 27;
(г) HVR-H1, содержащая аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащая аминокислотную последовательность SEQ ID NO: 11, HVR-H3, содержащая аминокислотную последовательность SEQ ID NO: 18, HVR-L1, содержащая аминокислотную последовательность SEQ ID NO: 21, HVR-L2, содержащая аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащая аминокислотную последовательность SEQ ID NO: 27;
(д) HVR-H1, содержащая аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащая аминокислотную последовательность SEQ ID NO: 8, HVR-H3, содержащая аминокислотную последовательность SEQ ID NO: 18, HVR-L1, содержащая аминокислотную последовательность SEQ ID NO: 21, HVR-L2, содержащая аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащая аминокислотную последовательность SEQ ID NO: 27;
(е) HVR-H1, содержащая аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащая аминокислотную последовательность SEQ ID NO: 12, HVR-H3, содержащая аминокислотную последовательность SEQ ID NO: 18, HVR-L1, содержащая аминокислотную последовательность SEQ ID NO: 21, HVR-L2, содержащая аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащая аминокислотную последовательность SEQ ID NO: 28;
(ж) HVR-H1, содержащая аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащая аминокислотную последовательность SEQ ID NO: 13, HVR-H3, содержащая аминокислотную последовательность SEQ ID NO: 18, HVR-L1, содержащая аминокислотную последовательность SEQ ID NO: 21, HVR-L2, содержащая аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащая аминокислотную последовательность SEQ ID NO: 29;
(з) HVR-H1, содержащая аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащая аминокислотную последовательность SEQ ID NO: 14, HVR-H3, содержащая аминокислотную последовательность SEQ ID NO: 19, HVR-L1, содержащая аминокислотную последовательность SEQ ID NO: 23, HVR-L2, содержащая аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащая аминокислотную последовательность SEQ ID NO: 27;
(и) HVR-H1, содержащая аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащая аминокислотную последовательность SEQ ID NO: 15, HVR-H3, содержащая аминокислотную последовательность SEQ ID NO: 20, HVR-L1, содержащая аминокислотную последовательность SEQ ID NO: 24, HVR-L2, содержащая аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащая аминокислотную последовательность SEQ ID NO: 27;
(к) HVR-H1, содержащая аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащая аминокислотную последовательность SEQ ID NO: 15, HVR-H3, содержащая аминокислотную последовательность SEQ ID NO: 20, HVR-L1, содержащая аминокислотную последовательность SEQ ID NO: 25, HVR-L2, содержащая аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащая аминокислотную последовательность SEQ ID NO: 27;
(л) HVR-H1, содержащая аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащая аминокислотную последовательность SEQ ID NO: 16, HVR-H3, содержащая аминокислотную последовательность SEQ ID NO: 20, HVR-L1, содержащая аминокислотную последовательность SEQ ID NO: 25, HVR-L2, содержащая аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащая аминокислотную последовательность SEQ ID NO: 27;
(м) HVR-H1, содержащая аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащая аминокислотную последовательность SEQ ID NO: 14, HVR-H3, содержащая аминокислотную последовательность SEQ ID NO: 19, HVR-L1, содержащая аминокислотную последовательность SEQ ID NO: 24, HVR-L2, содержащая аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащая аминокислотную последовательность SEQ ID NO: 27; и
(н) HVR-H1, содержащая аминокислотную последовательность SEQ ID NO: 7, HVR-H2, содержащая аминокислотную последовательность SEQ ID NO: 14, HVR-H3, содержащая аминокислотную последовательность SEQ ID NO: 17, HVR-L1, содержащая аминокислотную последовательность SEQ ID NO: 21, HVR-L2, содержащая аминокислотную последовательность SEQ ID NO: 26, и HVR-L3, содержащая аминокислотную последовательность SEQ ID NO: 27.
2. Анти-CD137 антигенсвязывающая молекула, включающая:
(а) VH область, имеющую любую одну из аминокислотных последовательностей SEQ ID NO: 43-53; или
(б) VL область, имеющую любую одну из аминокислотных последовательностей SEQ ID NO: 54-60.
3. Анти-CD137 антигенсвязывающая молекула по п. 1 или 2, включающая измененную область Fc, причем измененная область Fc содержит любую комбинацию измененных аминокислот, выбранную из следующих:
L235W/G236N/H268D/Q295L/K326T/A330K/P343R/D413K;
K214R/L235W/G236N/H268D/Q295L/K326T/A330K/P343R/D413K;
L234Y/P238D/T250V/V264I/T307P/A330K/P343R/D413K;
L234Y/P238D/V264I/A330K/P343R/D413K;
L234Y/G237D/P238D/T250V/T307P/A330K/P343R/D413K;
L234Y/G237D/P238D/A330K/P343R/D413K;
L235W/G236N/H268D/Q295L/K326T/A330K/Q311R/P343R;
L234Y/P238D/T250V/V264I/T307P/A330K/Q311R/P343R;
L234Y/P238D/V264I/A330K/Q311R/P343R;
L234Y/G237D/P238D/T250V/T307P/A330K/Q311R/P343R;
L234Y/G237D/P238D/A330K/Q311R/P343R;
L235W/G236N/H268D/Q295L/K326T/A330K/P343R;
K214R/L235W/G236N/H268D/Q295L/K326T/A330K/P343R;
L235W/G236N/H268D/Q295L/K326T/A330K/D413K;
K214R/G236N/H268D/A330K/P343R;
K214R/L235W/G236N/H268D/A330K/P343R;
K214R/G236N/H268D/A330K/D413K;
K214R/G236N/H268D/A330K/P343R/D413K;
K214R/L235W/G236N/H268D/A330K/P343R/D413K;
K214R/G236N/H268D/A330K/Q311R;
K214R/L235W/G236N/H268D/A330K/Q311R;
K214R/G236N/H268D/A330K/Q311R/P343R;
K214R/L235W/G236N/H268D/A330K/Q311R/P343R;
K214R/G236N/H268D/A330K/Q311R/D413K;
K214R/L235W/G236N/H268D/A330K/Q311R/D413K;
K214R/L235W/G236N/H268D/Q295L/K326T/A330K/Q311R.
4. Анти-CD137 антигенсвязывающая молекула по любому из пп. 1-3, включающая константную область тяжелой цепи, содержащую любую из аминокислотных последовательностей из SEQ ID NO: 64-85.
5. Выделенная нуклеиновая кислота, кодирующая анти-CD137 антигенсвязывающую молекулу по любому из пп. 1-4.
6. Вектор экспрессии, в который интродуцирована нуклеиновая кислота по п. 5.
7. Клетка-хозяин для получения анти-CD137 антигенсвязывающей молекулы по любому из пп. 1-4, содержащая нуклеиновую кислоту по п. 5 или вектор экспрессии по п. 6.
8. Способ получения анти-CD137 антигенсвязывающей молекулы, который включает культивирование клетки-хозяина по п. 7 таким образом, чтобы вырабатывалась анти-CD137 антигенсвязывающая молекула.
| WO 2013180200 A1, 05.12.2013 | |||
| WO 2015083764 A1, 11.06.2015 | |||
| WO 2016194992 A1, 08.12.2016 | |||
| WO 2015190538 A1, 17.12.2011 | |||
| RU 2016129045 A, 23.01.2018. |

