Настоящее изобретение касается лечения нейродегенеративных заболеваний.
Нейродегенеративные заболевания являются заболеваниями, вызванными коллапсом нейронной сети нервной цепи, основанным на систематической дегенерации и нарушении связей нейроцитов, и среди них известны различные трудноизлечимые заболевания, такие как болезнь Альцгеймера (БА), болезнь Паркинсона, деменция с тельцами Леви, сосудистая деменция и проблемы, связанные со старением, включая (i) некогнитивную нейродегенерацию, (ii) некогнитивную нервно-мышечную дегенерацию и (iii) моторно-сенсорную нейродегенерацию, болезнь Хантингтона, заболевания двигательных нейронов, включая боковой амиотрофический склероз (БАС), синдром Дауна, кортикобазальную ганглионарную дегенерацию, множественную системную атрофию, церебральную атрофию, оливопонтоцеребеллярную атрофию, надъядерный паралич, атаксию Фридрейха, спиноцеребеллярную атаксию 2 типа, лобно-височную дегенерацию (ЛВД) и первичную прогрессирующую афазию.
Клинические симптомы нейродегенеративных заболеваний варьируются от незначительных до тяжелых, в зависимости от заболевания. К ним относятся тремор, ригидность, акинезия, гипокинезия, брадикинезия, нарушение рефлекторного изменения позы, вегетативная дистония, пропульсия, нарушение походки, депрессия, апатия, спутанность сознания, нарушение обучения, дефицит памяти, когнитивные нарушения, амиотрофия, дистрофия мышечной ткани, нарушение функции плечевого пояса, нарушение артикуляции, дисфагия, нарушение дыхания, онемение, паралич и деменция, которые представляют большие трудности для повседневной активности.
Нейродегенеративная гибель - сложный механизм, вероятно обусловленный наличием множества молекулярных групп, которые вызывают нейродегенеративные заболевания, когда возникают проблемы с их экспрессией, активностью и/или регуляцией. Это может объяснять то, почему до сих пор не было разработано эффективных методов ингибирования или обеспечения регресса нейродегенерации.
Помимо лечения для удаления причинных факторов таких заболеваний, важно также восстановить (реконструировать) нейронную сеть. Например, отмечалось, что при болезни Альцгеймера, причиной которой была признана цитотоксичность β-амилоидного пептида, как атрофия нейритов, так и сокращение синапсов запускают ухудшение нервной функции, и наоборот, даже после такого запуска нейроциты, которые не полностью разрушены или выжили без дегенерации, могут быть восстановлены, если их можно активировать с распространением нейритов для восстановления синапсов.
Таким образом, существует настоятельная потребность в новых средствах, способных защищать нейриты, но также способных увеличивать рост нейритов и стимулировать образование новых синапсов, как для предотвращения, так и для лечения нейродегенеративных заболеваний.
Настоящее изобретение удовлетворяет эту потребность.
Действительно, настоящее изобретение является результатом того неожиданного открытия авторов изобретения, что соединение Донекоприд, раскрытое в международной заявке WO 2014195593, обладает нейропротекторной активностью, поскольку оно способно, помимо снижения гиперфосфорилирования тау-белка, увеличивать рост нейритов, поддерживать частоту синапсов, а наиболее важно и неожиданно - способствовать формированию новых синапсов.
Благодаря этому общему и эффективному защитному влиянию на нейриты соединение Донекоприд является очень многообещающим нейропротекторным средством для предотвращения и/или лечения любого нейродегенеративного заболевания, включающего дегенерацию нейритов.
Этот эффект был дополнительно подтвержден большим рядом соединений общей формулы (I), приведенной ниже, на многочисленных моделях нейродегенеративных заболеваний, в частности на моделях болезни Альцгеймера, моделях болезни Паркинсона, моделях бокового амиотрофического склероза, моделях болезни Хантингтона и моделях нейродегенеративных заболеваний, вызванных ишемией или черепно-мозговой травмой.
Таким образом, настоящее изобретение относится к соединению следующей формулы (I):
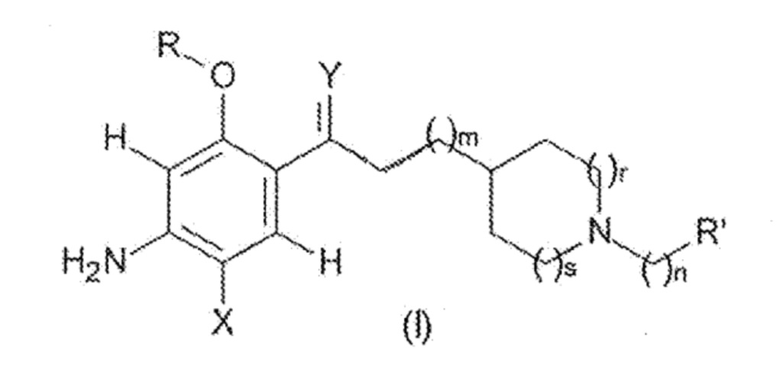
где:
X представляет собой
атом водорода, или
атом галогена (Hal), где (Hal) представляет собой фтор, хлор, бром или йод, или
Cp(Hal)2p+1 полигалогеноалкильную группу с прямой или разветвленной цепью, где p = 1, 2, 3 или 4, (Hal) имеет такое же значение, как указано выше;
Y представляет собой
атом кислорода, или
атом серы, или
радикал N-R’’, где R’’ представляет собой атом водорода, радикал -OH, радикал -OA, где A представляет собой C1-C6 алкильную группу с прямой или разветвленной цепью, в частности, где A представляет собой метильную группу, или CqH2q+1 алкильный радикал с прямой или разветвленной цепью, где q = 1, 2, 3 или 4;
m представляет собой целое число, выбранное из 1, 2 и 3;
n представляет собой целое число, выбранное из 0, 1, 2 и 3;
r и s представляют собой целые числа, значения которых: r = s = 0; или r = s = 1; или r = s = 2; или r = 0 и s = 1; или, наконец, r = 0 и s = 2;
R представляет собой
атом водорода, или
C1-C5 алкильную группу с прямой или разветвленной цепью, способную нести один или несколько атомов F;
R’ представляет собой
С1-С6 алкильный радикал с разветвленной цепью, или
C3-C10 циклоалкильную или C5-C13 бициклическую группу, способную нести одну или несколько R-групп и иметь атом кислорода, или атом азота, который может быть замещен группой R, или атом серы или радикал -SO2- или -SO-,
а также к его энантиомерам или диастереоизомерам и его рацемическим соединениям, его кислым солям, его гидратам или продуктам его сольватации;
для использования в качестве нейропротекторного агента при профилактике и/или лечении нейродегенеративного заболевания.
Подробное описание изобретения
Соединения
Соединение, используемое в контексте изобретения, представляет собой соединение следующей формулы (I):
где:
X представляет собой
атом водорода, или
атом галогена (Hal), где (Hal) представляет собой фтор, хлор, бром или йод, или
Cp(Hal)2p+1 полигалогеноалкильную группу с прямой или разветвленной цепью, где p = 1, 2, 3 или 4, (Hal) имеет такое же значение, как указано выше;
Y представляет собой
атом кислорода, или
атом серы, или
радикал N-R’’, где R’’ представляет собой атом водорода, радикал -OH, радикал -OA, где A представляет собой C1-C6 алкильную группу с прямой или разветвленной цепью, в частности, где A представляет собой метильную группу, или CqH2q+1 алкильный радикал с прямой или разветвленной цепью, где q = 1, 2, 3 или 4;
m представляет собой целое число, выбранное из 1, 2 и 3;
n представляет собой целое число, выбранное из 0, 1, 2 и 3;
r и s представляют собой целые числа, значения которых: r = s = 0; или r = s = 1; или r = s = 2; или r = 0 и s = 1; или, наконец, r = 0 и s = 2;
R представляет собой
атом водорода, или
C1-C5 алкильную группу с прямой или разветвленной цепью, способную нести один или несколько атомов F;
R’ представляет собой
С1-С6 алкильный радикал с разветвленной цепью, или
C3-C10 циклоалкильную или C5-C13 бициклическую группу, способную нести одну или несколько R-групп и иметь атом кислорода, или атом азота, который может быть замещен группой R, или атом серы или радикал -SO2- или -SO-,
а также его энантиомеры или диастереоизомеры и его рацемические соединения, его кислотные соли, его гидраты или продукты его сольватации.
В одном конкретном варианте осуществления в формуле (I) Y представляет собой
атом кислорода, или
атом серы, или
радикал N-R’’, где R’’ представляет собой атом водорода, радикал -OH, или CqH2q+1 алкильный радикал с прямой или разветвленной цепью, где q = 1, 2, 3 или 4.
В конкретном варианте осуществления в формуле (I) X представляет собой атом галогена, в частности хлор или фтор, более предпочтительно хлор, наиболее предпочтительно фтор.
В другом конкретном варианте осуществления в формуле (I) Y представляет собой атом кислорода.
В другом конкретном варианте осуществления в формуле (I) все из m, n, r и s имеют значение 1.
В другом конкретном варианте осуществления в формуле (I) R представляет собой H, CH3, CH2CH3 или CH2-CH2F. Предпочтительно R представляет собой CH3.
В другом конкретном варианте осуществления в формуле (I) R’ представляет собой радикал, выбранный из группы, состоящей из радикалов циклопропила, циклобутила, циклопентила, циклогексила, циклогептила, 4-пиперидина и –CH2NHSO2CH3. В другом конкретном варианте осуществления в формуле (I) R’ представляет собой радикал, выбранный из группы, состоящей из радикалов циклопропила, циклобутила, циклопентила, циклогексила, 4-пиперидина и –CH2NHSO2CH3. Предпочтительно в формуле (I) R’ представляет собой радикал, выбранный из группы, состоящей из радикалов циклопропила, циклобутила, циклопентила, циклогексила, циклогептила и 4-пиперидина. Предпочтительно в формуле (I) R’ представляет собой радикал, выбранный из группы, состоящей из радикалов циклопропила, циклобутила, циклопентила, циклогексила и 4-пиперидина. Более предпочтительно R’ представляет собой C4-C7 циклоалкильный радикал. Еще более предпочтительно R’ представляет собой циклопентильный радикал, циклогексильный радикал, циклогептильный радикал или 2-пиперидиновый радикал. Более предпочтительно R’ представляет собой циклопентильный радикал. Еще более предпочтительно R’ представляет собой циклогексильный радикал. Еще более предпочтительно R’ представляет собой 2-пиперидиновый радикал. Наиболее предпочтительно R’ представляет собой циклогептильный радикал.
В другом конкретном варианте осуществления в формуле (I) R представляет собой CH3, а R’ представляет собой C4-C7 циклоалкильный радикал, в частности циклогексильный радикал. В другом конкретном варианте осуществления в формуле (I) R представляет собой CH3, а R’ представляет собой C4-C7 циклоалкильный радикал, в частности циклопентильный радикал. В другом конкретном варианте осуществления в формуле (I) R представляет собой CH3, а R представляет собой C4-C7 циклоалкильный радикал, в частности радикал 2-пиперидина. В другом конкретном варианте осуществления в формуле (I) R представляет собой CH3, а R’ представляет собой C4-C7 циклоалкильный радикал, в частности циклогептильный радикал.
В особо предпочтительном варианте осуществления в формуле (I) X представляет собой хлор, Y представляет собой атом кислорода, все из m, n, r и s имеют значение 1, R представляет собой CH3, а R’ представляет собой циклогексильный радикал. Другими словами, в особо предпочтительном варианте осуществления соединение, используемое в контексте изобретения, представляет собой соединение донекоприда следующей формулы (II):
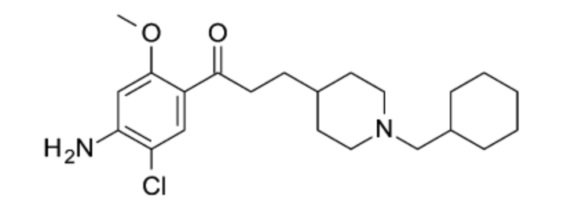 (II).
(II).
В другом особо предпочтительном варианте осуществления в формуле (I) X представляет собой фтор, Y представляет собой атом кислорода, все из m, n, r и s имеют значение 1, R представляет собой CH3, а R’ представляет собой циклогексильный радикал. Другими словами, в особо предпочтительном варианте осуществления соединение, используемое в контексте изобретения, представляет собой соединение флукоприда следующей формулы (III):
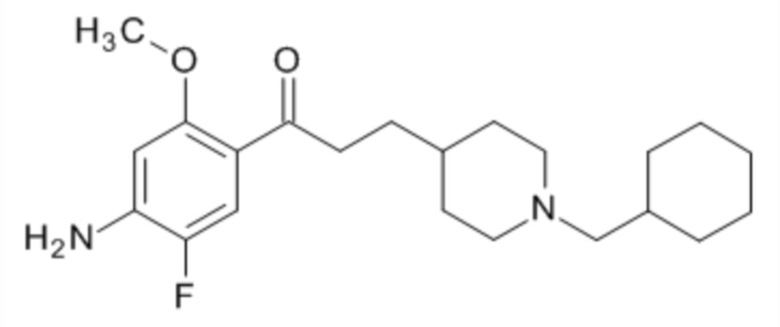 (III).
(III).
В другом особо предпочтительном варианте в формуле (I) X представляет собой хлор, Y представляет собой атом кислорода, все из m, n, r и s имеют значение 1, R представляет собой CH3, а R’ представляет собой 2-пиперидиновый радикал. Другими словами, в особо предпочтительном варианте осуществления соединение, используемое в контексте изобретения, представляет собой соединение MR31176 следующей формулы (IV):
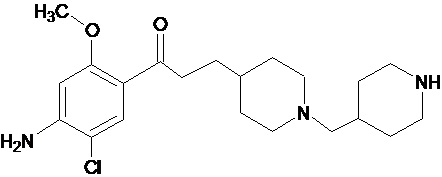 (IV).
(IV).
В другом особо предпочтительном варианте осуществления в формуле (I) X представляет собой фтор, Y представляет собой атом кислорода, все из m, n, r и s имеют значение 1, R представляет собой CH3, а R’ представляет собой циклопентильный радикал. Другими словами, в особо предпочтительном варианте осуществления соединение, используемое в контексте изобретения, представляет собой соединение MR33583 следующей формулы (V):
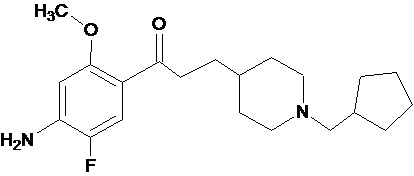 (V).
(V).
В другом особо предпочтительном варианте осуществления в формуле (I) X представляет собой хлор, Y представляет собой атом кислорода, все из m, n, r и s имеют значение 1, R представляет собой CH3, а R’ представляет собой циклопентильный радикал. Другими словами, в особо предпочтительном варианте осуществления соединение, используемое в контексте изобретения, представляет собой соединение MR31172 следующей формулы (VI):
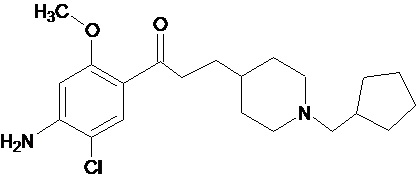 (VI).
(VI).
В другом особо предпочтительном варианте осуществления в формуле (I) X представляет собой фтор, Y представляет собой атом кислорода, все из m, n, r и s имеют значение 1, R представляет собой CH3, а R’ представляет собой циклогептильный радикал. Другими словами, в особо предпочтительном варианте осуществления соединение, используемое в контексте изобретения, представляет собой соединение MR36014 следующей формулы (VII):
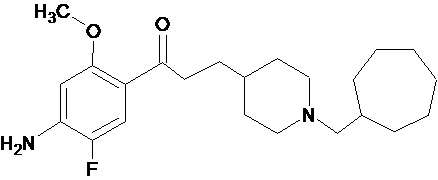 (VII).
(VII).
В другом наиболее предпочтительном варианте осуществления в формуле (I) X представляет собой хлор, Y представляет собой атом кислорода, все из m, n, r и s имеют значение 1, R представляет собой CH3, а R’ представляет собой циклогептильный радикал. Другими словами, в особо предпочтительном варианте осуществления соединение, используемое в контексте изобретения, представляет собой соединение MR31192 следующей формулы (VIII):
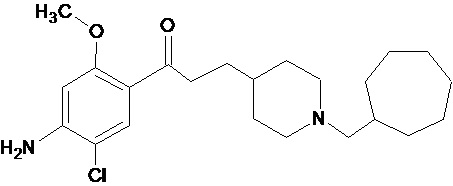 (VIII).
(VIII).
В другом конкретном варианте осуществления X представляет собой хлор, Y представляет собой радикал NR’’, где R’’ представляет собой радикал –O-A, где A представляет собой метильную группу, все из m, n, r и s имеют значение 1, R представляет собой CH3, а R’ представляет собой циклогексильный радикал. Другими словами, в конкретном варианте осуществления соединение, используемое в контексте изобретения, представляет собой соединение 2-хлор-4-[[2-[1-(циклогексилметил)-4-пиперидил]этил]-N-метоксикарбонимидоил]-5-метоксианилин.
В конкретных вариантах осуществления используются кислотные соли определенных выше соединений. В более конкретном варианте осуществления указанные кислотные соли представляют собой фумараты или дигидрохлораты.
Более конкретно в особо предпочтительном варианте осуществления соединение, используемое в контексте изобретения, представляет собой фумарат донекоприда (т.е. соединения указанной выше формулы (II)), в частности донекоприда фумарат гидрат.
В другом особо предпочтительном варианте осуществления соединение, используемое в контексте изобретения, представляет собой фумарат флуокоприда (т.е. соединения указанной выше формулы (III)).
В другом особо предпочтительном варианте осуществления соединение, используемое в контексте изобретения, представляет собой дигидрохлорат MR31176 (т.е. соединения указанной выше формулы (IV)), в частности дигидрохлорат дигидрат.
В другом особо предпочтительном варианте осуществления соединение, используемое в контексте изобретения, представляет собой фумарат MR33583 (т.е. соединения формулы (V), приведенной выше).
В другом особо предпочтительном варианте осуществления соединение, используемое в контексте изобретения, представляет собой фумарат MR31172 (т.е. соединения формулы (VI), приведенной выше).
В другом особо предпочтительном варианте осуществления соединение, используемое в контексте изобретения, представляет собой фумарат MR36014 (т.е. соединения формулы (VII), приведенной выше).
В другом особо предпочтительном варианте осуществления соединение, используемое в контексте изобретения, представляет собой фумарат MR31192 (т.е. соединения формулы (VIII), приведенной выше).
Нейродегенеративное заболевание
Под «нейродегенеративным заболеванием» в настоящем изобретении подразумевается заболевание, вызванное коллапсом нейронной сети нервной цепи, основанным на систематической дегенерации и нарушения связей нейроцитов.
Как объясняется выше, авторы настоящего изобретения продемонстрировали, что донекоприд проявляет общее и эффективное протекторное (защитное) действие в отношении нейритов. Благодаря этому общему нейропротекторному эффекту донекоприд можно использовать в качестве нейропротекторного агента для предотвращения и/или лечения любого нейродегенеративного заболевания.
Авторы изобретения дополнительно подтвердили этот эффект для большого количества соединений общей формулы (I), приведенной выше, в частности для соединений, конкретно определенных в разделе «Соединения» выше, на многочисленных моделях нейродегенеративных заболеваний, в частности на моделях болезни Альцгеймера, моделях болезни Паркинсона, моделях бокового амиотрофического склероза, моделях болезни Хантингтона и моделях нейродегенеративных заболеваний, вызванных ишемией или черепно-мозговыми травмами.
Примеры нейродегенеративных заболеваний включают болезнь Альцгеймера (БА), болезнь Паркинсона, деменцию с тельцами Леви, сосудистую деменцию и проблемы, связанные со старением, включая (i) некогнитивную нейродегенерацию, (ii) некогнитивную нейромышечную дегенерацию и (iii) моторно-сенсорную нейродегенерацию, болезнь Хантингтона, заболевания двигательных нейронов, включая боковой амиотрофический склероз (БАС), синдром Дауна, кортикобазальную ганглионарную дегенерацию, множественную системную атрофию, церебральную атрофию, оливопонтоцеребеллярную атрофию, надъядерный паралич, атаксию Фридрейха, спиноцеребеллярную атаксию 2 типа, лобно-височные дегенерации (ЛВД) и первичную прогрессирующую афазию.
В одном конкретном варианте осуществления нейродегенеративное заболевание выбрано из группы, состоящей из болезни Альцгеймера, болезни Паркинсона, болезни Хантингтона, множественной системной атрофии, заболеваний двигательных нейронов, включая боковой амиотрофический склероз, синдрома Дауна и лобно-височной дегенерации.
В одном более конкретном варианте нейродегенеративное заболевание выбрано из группы, состоящей из болезни Альцгеймера, болезни Паркинсона, болезни Хантингтона и бокового амиотрофического склероза.
В другом варианте осуществления нейродегенеративное заболевание представляет собой болезнь Альцгеймера.
В альтернативном варианте осуществления нейродегенеративное заболевание представляет собой нейродегенеративное заболевание, не являющееся болезнью Альцгеймера.
В указанном альтернативном варианте осуществления нейродегенеративное заболевание выбрано из группы, состоящей из болезни Паркинсона, болезни Хантингтона и бокового амиотрофического склероза.
Медицинские показания
Настоящее изобретение относится к соединению, определенному в разделе «Соединения» выше, предназначенному для использования в качестве нейропротекторного агента для профилактики и/или лечения нейродегенеративного заболевания, как определено в разделе «Нейродегенеративное заболевание» выше.
Настоящее изобретение также относится к применению соединения, определенного в разделе «Соединения» выше, для получения нейропротекторного агента для профилактики и/или лечения нейродегенеративного заболевания, определенного в разделе «Нейродегенеративное заболевание» выше.
Настоящее изобретение также относится к способу предотвращения и/или лечения нейродегенеративного заболевания, определенного в разделе «Нейродегенеративное заболевание» выше, включающему введение субъекту, нуждающемуся в этом, терапевтически эффективного количества соединения, определенного в разделе «Соединения» выше, в качестве нейропротекторного агента.
Настоящее изобретение также относится к способу нейропротекции у субъекта, страдающего нейродегенеративным заболеванием или подверженного риску развития нейродегенеративного заболевания, определенного в разделе «Нейродегенеративное заболевание» выше, включающему введение указанному субъекту терапевтически эффективного количества соединения, определенного в разделе «Соединения» выше.
В соответствии с изобретением термин «субъект» или «субъект, нуждающийся в этом» предназначен для обозначения человека или млекопитающего, не являющегося человеком, страдающего или вероятно страдающего нейродегенеративным заболеванием, определенным в разделе «Нейродегенеративные заболевания» выше.
В контексте изобретения термин «лечение» означает индукцию регресса, облегчение или ингибирование развития расстройства или состояния, к которому применяется такой термин, или одного или нескольких симптомов такого расстройства или состояния.
В контексте изобретения термин «предотвращение» или «профилактика» означает отсрочку или предотвращение проявления расстройства или состояния, к которому применяется такой термин, или одного или нескольких симптомов такого расстройства или состояния.
Под «нейропротекцией» в настоящей заявке подразумевается предотвращение или ингибирование дегенеративных эффектов и восстановление целостности нейронов в центральной нервной системе, в частности предотвращение или ингибирование дегенерации нейронов.
Под «нейропротекторным агентом» в настоящей заявке подразумевается агент, способный к нейропротекции, как определено выше.
Авторы изобретения, более конкретно, продемонстрировали, что соединения, используемые в контексте изобретения, способны увеличивать рост нейритов, поддерживать частоту синапсов, уменьшать гиперфосфорилирование тау-белка и/или способствовать образованию новых синапсов.
Соответственно, в конкретном варианте осуществления соединение, определенное в разделе «Соединения» выше, предназначено для увеличения роста нейритов.
Настоящее изобретение также относится к способу увеличения роста нейритов у субъекта, страдающего нейродегенеративным заболеванием или подверженного риску развития нейродегенеративного заболевания, определенного в разделе «Нейродегенеративное заболевание» выше, включающему введение указанному субъекту терапевтически эффективного количества соединения, определенного в разделе «Соединения» выше.
Под «ростом нейритов» в настоящей заявке подразумевается процесс удлинения любого отростка из тела клетки нейрона и связанный с ним генезис синапсов.
Под «усилением роста нейритов» в настоящей заявке подразумевается, что рост нейритов повышается (т.е. становится более эффективным, быстрым или длительным), предпочтительно статистически значимо повышается, когда применяют представляющее интерес соединение, по сравнению с ростом нейритов, наблюдаемым в тех же условиях, но без применения интересующего соединения. Предпочтительно рост нейритов увеличивается по меньшей мере на 120% по сравнению с ростом нейритов, наблюдаемым в тех же условиях, но без применения представляющего интерес соединения.
В другом конкретном варианте осуществления соединение, определенное в разделе «Соединения» выше, предназначено для поддержания частоты синапсов.
Другой объект по настоящему изобретению относится к способу поддержания частоты синапсов у субъекта, страдающего нейродегенеративным заболеванием или подверженного риску развития нейродегенеративого заболевания, определенного в разделе «Нейродегенеративное заболевание» выше, включающему введение указанному субъекту терапевтически эффективного количества соединения, определенного в разделе «Соединения» выше.
Под «частотой синапсов» в настоящей заявке понимается отношение количества синапсов к количеству нейронов.
Под «поддержанием частоты синапсов» в настоящей заявке подразумевается, что частота синапсов аналогична (то есть статистически значимо не отличается) при применении интересующего соединения, по сравнению с частотой синапсов, наблюдаемой в контрольных условиях, таких как те же условия, но без применения представляющего интерес соединения, или при состояниях, где нейроны не подвергаются воздействию нейродегенеративных факторов.
В другом конкретном варианте осуществления соединение, определенное в разделе «Соединения» выше, предназначено для стимуляции образования новых синапсов.
Другой объект по настоящему изобретению относится к способу стимуляции образования новых синапсов у субъекта, страдающего нейродегенеративным заболеванием или подверженного риску развития нейродегенеративного заболевания, определенного выше в разделе «Нейродегенеративное заболевание», включающему введение указанному субъекту терапевтически эффективного количества соединения, определенного в разделе «Соединения» выше.
Под «стимуляцией образования новых синапсов» в настоящей заявке подразумевается, что количество вновь образованных синапсов выше, предпочтительно статистически значимо выше, когда применяют представляющее интерес соединение, по сравнению с количеством вновь образованных синапсов, наблюдаемых в тех же условиях, но без применения интересующего соединения. Предпочтительно образование новых синапсов увеличивается по меньшей мере на 120% по сравнению с образованием новых синапсов, наблюдаемых в тех же условиях, но без применения интересующего соединения.
В другом конкретном варианте осуществления соединение, определенное в разделе «Соединения» выше, предназначено для уменьшения гиперфосфорилирования тау-белка.
Другой объект по настоящему изобретению относится к способу снижения фосфорилирования тау-белка у субъекта, страдающего нейродегенеративным заболеванием или подверженного риску развития нейродегенеративного заболевания, определенного в разделе «Нейродегенеративное заболевание» выше, включающему введение указанному субъекту терапевтически эффективного количества соединения, определенного в разделе «Соединения» выше.
Под «гиперфосфорилированием тау-белка» в настоящей заявке подразумевается аномально высокое фосфорилирование тау-белка, в частности аномальное фосфорилирование тау-белка по меньшей мере в одном из остатков Thr39, Ser46, Thr50, Ser68, Thr69, Thr71, Ser113, Thr153, Thr175, Thr181, Ser184, Ser185, Ser191, Ser198, Ser199, Ser202, Thr205, Ser208, Ser210, Thr212, Ser214, Thr217, Thr231, Ser235, Ser237, Ser238, Ser241, Ser258, Ser262, Ser285, Ser289, Ser305, Ser324, Ser352, Ser356, Tyr394, Ser396, Ser400, Thr403, Ser404, Ser409, Ser412, Ser413, Thr414, Ser416, Ser422, Ser433 и Ser435.
Под «уменьшением гиперфосфорилирования тау-белка» в настоящей заявке подразумевается, что по меньшей мере один из остатков Thr39, Ser46, Thr50, Ser68, Thr69, Thr71, Ser113, Thr153, Thr175, Thr181, Ser184, Ser185, Ser191, Ser198, Ser199, Ser202, Thr205, Ser208, Ser210, Thr212, Ser214, Thr217, Thr231, Ser235, Ser237, Ser238, Ser241, Ser258, Ser262, Ser285, Ser289, Ser305, Ser324, Ser352, Ser356, Tyr394, Ser396, Ser400, Thr403, Ser404, Ser409, Ser412, Ser413, Thr414, Ser416, Ser422, Ser433 и Ser435, в частности один из остатков Ser212 и Thr214 тау-белка, не подвергается аномальному фосфорилированию при применении представляющего интерес соединения, тогда как указанный остаток аномально фосфорилируется в тех же условиях, но без применения представляющего интерес соединения.
Под «терапевтически эффективным количеством» в настоящей заявке подразумевается достаточное количество соединения для действия в качестве нейропротекторного агента при лечении и/или предотвращении нейродегенеративного заболевания с разумным соотношением польза/риск, применимым к любому лечению. Однако следует понимать, что общее ежедневное использование соединений по настоящему изобретению будет определяться лечащим врачом в рамках здравого медицинского суждения. Конкретный терапевтически эффективный уровень дозы для любого конкретного субъекта будет зависеть от множества факторов, включая заболевание, которое лечат, и тяжесть нарушения, активность конкретных используемых соединений, возраст, массу тела, общее состояние здоровья, пол и рацион субъекта, время введения, путь введения и уровень экскреции конкретных используемых соединений, продолжительность лечения, лекарства, применяемые в комбинации или одновременно с конкретными применяемыми соединениями, и подобные факторы, хорошо известные в области медицины. Например, специалисту в данной области техники известно, что следует начинать применение соединений с уровней дозы ниже тех, которые требуются для достижения необходимого терапевтического эффекта, и постепенно увеличивать дозу до тех пор, пока необходимый эффект не будет достигнут.
Соединения, используемые в контексте изобретения, можно применять в форме фармацевтической композиции, включающей фармацевтически приемлемые вспомогательные вещества и, при необходимости, матрицы с замедленным высвобождением, такие как биоразлагаемые полимеры, для получения терапевтических композиций.
«Фармацевтически» или «фармацевтически приемлемый» относится к молекулярным объектам и композициям, которые не вызывают неблагоприятных, аллергических или других нежелательных реакций при введении млекопитающему, в частности человеку, в зависимости от ситуации. Фармацевтически приемлемый носитель или вспомогательное вещество относится к нетоксичному твердому, полутвердому или жидкому наполнителю, разбавителю, инкапсулирующему материалу или добавочному веществу любого типа.
Форма фармацевтических композиций, включая соединения, используемые в контексте изобретения, и способ введения, естественно, зависят от состояния, которое необходимо лечить, тяжести заболевания, возраста, массы тела и пола пациента и т.д.
Соединения, используемые в контексте изобретения, могут быть включены в состав для местного, перорального, парентерального, интраназального, внутривенного, внутримышечного, подкожного или внутриглазного введения и т.п. В одном конкретном варианте осуществления соединение, используемое в контексте изобретения, применяют перорально.
В альтернативном варианте фармацевтические композиции, включающие соединения, используемые в контексте изобретения, могут содержать носители, которые фармацевтически приемлемы для состава, который можно вводить путем инъекции. Это могут быть, в частности, изотонические, стерильные, солевые растворы (мононатрий- или динатрий фосфат, хлорид натрия, калия, кальция или магния и т.п., или смеси таких солей) или сухие, особенно лиофилизированные, композиции, которые при добавлении, в зависимости от необходимости, стерильной воды или физиологического раствора, позволяют получать растворы для инъекций.
Для приготовления фармацевтических композиций эффективное количество соединений, используемых в контексте изобретения, может быть растворено или диспергировано в фармацевтически приемлемом носителе или водной среде.
Фармацевтические формы, подходящие для инъекционного применения, включают стерильные водные растворы или дисперсии и стерильные порошки для немедленного приготовления стерильных инъекционных растворов или дисперсий. Во всех случаях форма должна быть стерильной и должна быть жидкой до такой степени, чтобы ее можно было легко набрать с помощью шприца. Она должна быть стабильной в условиях производства и хранения, и должна быть защищена от загрязняющего действия микроорганизмов, таких как бактерии и грибки.
Носитель может быть растворителем или дисперсионной средой, содержащей, например, воду, этанол, полиол (например, глицерин, пропиленгликоль, жидкий полиэтиленгликоль и т.п.) и их подходящие смеси. Подходящую текучесть можно поддерживать, например, путем использования покрытия, такого как лецитин, путем поддержания необходимого размера частиц в случае дисперсии и использования поверхностно-активных веществ, стабилизирующих агентов, криопротекторов или антиоксидантов. Предотвращать действие микроорганизмов можно с помощью антибактериальных и противогрибковых средств. Во многих случаях будет предпочтительно включать изотонические агенты, например сахара или хлорид натрия.
Стерильные растворы для инъекций готовят путем включения активных соединений в необходимом количестве в соответствующий растворитель с несколькими другими ингредиентами, перечисленными выше, если это требуется, с последующей стерилизацией фильтрованием. Обычно дисперсии готовят путем включения различных стерилизованных активных ингредиентов в стерильный носитель, который содержит основную дисперсионную среду и другие необходимые ингредиенты из перечисленных выше. В случае стерильных порошков для приготовления стерильных растворов для инъекций предпочтительными способами приготовления являются способы вакуумной сушки и лиофилизации, которые дают порошок активного ингредиента плюс любого дополнительного необходимого ингредиента из их раствора, предварительно стерилизованного фильтрованием.
После приготовления растворы будут вводить способом, совместимым с лекарственным препаратом, и в таком количестве, которое является терапевтически эффективным. Композиции легко вводят в различных лекарственных формах, таких как типы растворов для инъекций, описанные выше, но также можно использовать капсулы для высвобождения лекарственного средства и тому подобное.
Для парентерального введения в водном растворе, например, раствор должен быть соответствующим образом забуферен, если это необходимо, и жидкий разбавитель сначала должен быть сделан изотоническим с помощью достаточного количества физиологического раствора или глюкозы. Эти конкретные водные растворы особенно хорошо подходят для внутривенного, внутримышечного, подкожного и интраперитонеального введения. В связи с этим, стерильные водные среды, которые можно использовать, известны специалистам в данной области техники в свете настоящего изобретения. Например, одну дозу можно растворить в 1 мл изотонического раствора NaCl и либо добавить к 1000 мл жидкости для гиподермоклизиса, либо ввести в предполагаемое место инфузии (см., например, «Remington’s Pharmaceutical Sciences» 15-е издание, стр. 1035-1038 и 1570-1580). В зависимости от состояния пациента, которого лечат, обязательно будет некоторое изменение дозировки. Лицо, ответственное за введение, в любом случае определит подходящую дозу для каждого отдельного субъекта.
Соединение, используемое в контексте изобретения, обычно можно вводить перорально, например в форме таблеток, капсул, микрокапсул, порошков, гранул, сиропов, растворов или суспензий, принимаемых пероральным или сублингвальным путем.
В одном конкретном варианте осуществления соединение, используемое в контексте изобретения, подходящим образом применяют перорально в суточной дозе от 1 до 20 мг/кг массы тела, в частности в суточной дозе 2 мг/кг.
В другом конкретном варианте осуществления соединения, используемые в контексте изобретения, подходящим образом применяют перорально, два раза в неделю, в дозе от 1 до 20 мг/кг массы тела, в частности в дозе 1 мг/кг два раза в неделю.
Согласно другому конкретному варианту осуществления соединение, используемое в контексте изобретения, применяют в дозах от 10 до 1000 мг в сутки, предпочтительно в дозах от 0,2 до 2 мг 5 раз в сутки.
Настоящее изобретение будет дополнительно проиллюстрировано приведенными ниже фигурами и примерами.
Краткое описание фигур
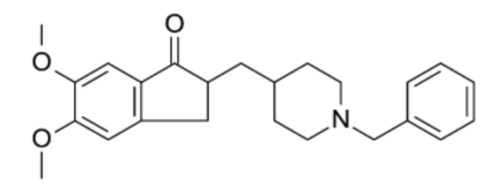
Фиг. 1: Влияние MR31193 на выживание нейронов MAP-2 (A), общую нейритную сеть (B), длину нейритов, нормализованную на количество нейронов (C), и на фосфорилирование тау-белка (AT100) (D) в первичной культуре нейронов гиппокампа, поврежденных Aβ 1-42. Результаты выражены в процентах от контроля как среднее значение ± SEM (n = 4-6 на группу). Односторонний дисперсионный анализ с последующим тестом методом наименьшей значимой разницы по Фишеру. *p <0,05 считали значимым.
Фиг. 2: Влияние MR31193 на сохранение синапсов нейронов MAP-2 в первичной культуре нейронов гиппокампа, поврежденных Aβ 1-42. Результаты выражены в процентах от контроля как среднее значение ± SEM (n = 4-6 на группу). Односторонний дисперсионный анализ с последующим тестом методом наименьшей значимой разницы по Фишеру. *p <0,05 считали значимым.
Фиг. 3: Влияние MR31176 на выживание нейронов MAP-2 (A), общую нейритную сеть (B), длину нейритов, нормализованную на количество нейронов (C), и на фосфорилирование тау-белка (AT100) (D) в первичной культуре нейронов гиппокампа, поврежденных Aβ 1-42. Результаты выражены в процентах от контроля как среднее значение ± SEM (n = 4-6 на группу). Односторонний дисперсионный анализ с последующим тестом методом наименьшей значимой разницы по Фишеру. *p <0,05 считали значимым.
Фиг. 4: Влияние MR31176 на сохранение синапсов нейронов MAP-2 в первичной культуре нейронов гиппокампа, поврежденных Aβ 1-42. Результаты выражены в процентах от контроля как среднее значение ± SEM (n = 4-6 на группу). Односторонний дисперсионный анализ с последующим тестом методом наименьшей значимой разницы по Фишеру. *p <0,05 считали значимым.
Фиг. 5: Влияние MR31192 на выживание нейронов MAP-2 (A), общую нейритную сеть (B), длину нейритов, нормализованную на количество нейронов (C), и на фосфорилирование тау-белка (AT100) (D) в первичной культуре нейронов гиппокампа, поврежденных Aβ 1-42. Результаты выражены в процентах от контроля как среднее значение ± SEM (n = 4-6 на группу). Односторонний дисперсионный анализ с последующим тестом методом наименьшей значимой разницы по Фишеру. *p <0,05 считали значимым.
Фиг. 6. Влияние MR31192 на сохранение синапсов нейронов MAP-2 в первичной культуре нейронов гиппокампа, поврежденных Aβ 1-42. Результаты выражены в процентах от контроля как среднее значение ± SEM (n = 4-6 на группу). Односторонний дисперсионный анализ с последующим тестом методом наименьшей значимой разницы по Фишеру. *p <0,05 считали значимым.
Фиг. 7: Влияние MR33583 на выживание нейронов MAP-2 (A), общую нейритную сеть (B), длину нейритов, нормализованную по количеству нейронов (C), и на фосфорилирование тау-белка (AT100) (D) в первичной культуре нейронов гиппокампа, поврежденных Aβ 1-42. Результаты выражены в процентах от контроля как среднее значение ± SEM (n = 4-6 на группу). Односторонний дисперсионный анализ с последующим тестом методом наименьшей значимой разницы по Фишеру. *p <0,05 считали значимым.
Фиг. 8: Влияние MR33583 на сохранение синапсов нейронов MAP-2 в первичной культуре нейронов гиппокампа, поврежденных Aβ 1-42. Результаты выражены в процентах от контроля как среднее значение ± SEM (n = 4-6 на группу). Односторонний дисперсионный анализ с последующим тестом методом наименьшей значимой разницы по Фишеру. *p <0,05 считали значимым.
Фиг. 9: Влияние MR31192 или донепезила на выживание нейронов MAP-2 (A), общую нейритную сеть (B), длину нейритов, нормализованную по количеству нейронов (C), и на фосфорилирование тау-белка (AT100) (D) в первичной культуре нейронов гиппокампа, поврежденных Aβ 1-42. Результаты выражены в процентах от контроля как среднее ± SEM (n = 4-6 на группу). Односторонний дисперсионный анализ с последующим тестом методом наименьшей значимой разницы по Фишеру. *p <0,05 считали значимым.
Фиг. 10: Влияние MR31192 или донепезила на сохранение синапсов нейронов MAP-2 в первичной культуре нейронов гиппокампа, поврежденных Aβ 1-42. Результаты выражены в процентах от контроля как среднее значение ± SEM (n = 4-6 на группу). Односторонний дисперсионный анализ с последующим тестом методом наименьшей значимой разницы по Фишеру. *p <0,05 считали значимым.
Фиг. 11: Влияние MR36014 или донепезила на выживание нейронов MAP-2 (A), общую нейритную сеть (B), длину нейритов, нормализованную на количество нейронов (C), и на фосфорилирование тау-белка (AT100) (D) в первичной культуре нейронов гиппокампа, поврежденных Aβ 1-42. Результаты выражены в процентах от контроля как среднее значение ± SEM (n = 4-6 на группу). Односторонний дисперсионный анализ с последующим тестом методом наименьшей значимой разницы по Фишеру. *p <0,05 считали значимым.
Фиг. 12: Влияние MR36014 или донепезила на сохранение синапсов нейронов MAP-2 в первичной культуре нейронов гиппокампа, поврежденных Aβ 1-42. Результаты выражены в процентах от контроля как среднее значение ± SEM (n = 4-6 на группу). Односторонний дисперсионный анализ с последующим тестом методом наименьшей значимой разницы по Фишеру. *p <0,05 считали значимым.
Фиг. 13: Влияние MR36014 на выживание TH-нейронов (A), на общую нейритную сеть TH-нейронов (B) и на агрегацию aSyn в TH-нейронах (C) в первичной культуре мезэнцефальных нейронов, поврежденных MPP+. Результаты выражены в процентах от контроля как среднее значение ± SEM (n = 4-6 на группу). Односторонний дисперсионный анализ с последующим тестом методом наименьшей значимой разницы по Фишеру. *p <0,05 считали значимым.
Фиг. 14: Влияние MR33583 на выживание TH-нейронов (A), на общую нейритную сеть TH-нейронов (B) и на агрегацию aSyn в TH-нейронах (C) в первичной культуре мезэнцефальных нейронов, поврежденных MPP+. Результаты выражены в процентах от контроля как среднее значение ± SEM (n = 4-6 на группу). Односторонний дисперсионный анализ с последующим тестом методом наименьшей значимой разницы по Фишеру. *p <0,05 считали значимым.
Фиг. 15: Влияние MR36014 (A-B) и MR33583 (C-D) на выживание нейронов MAP-2 (A-C)) и защиту общей нейритной сети (B-D)) в первичной культуре кортикальных нейронов, поврежденных глутаматом. Результаты выражены в процентах от контроля как среднее значение ± SEM (n = 4-6 на группу). Односторонний дисперсионный анализ с последующим тестом методом наименьшей значимой разницы по Фишеру. *p <0,05 считали значимым.
Фиг. 16: Влияние MR36014 на выживание (A) и нейритную сеть (B, C) в двигательных нейронах спинного мозга после повреждения глутаматом. Результаты выражены в процентах от контроля и показывают среднее значение ± SEM (100% = CT, без соединения). Односторонний дисперсионный анализ с последующим тестом методом наименьшей значимой разницы по Фишеру, n = 4-6. p <0,05 считали значимым.
Фиг. 17: Влияние MR33583 на выживание (A), нейритную сеть (B, C) и на цитоплазматическую транслокацию TDP-43 (D) в двигательных нейронах спинного мозга после повреждения глутаматом. Результаты выражены в процентах от контроля и показывают среднее значение ± SEM (100% = CT, без соединения). Односторонний дисперсионный анализ с последующим тестом методом наименьшей значимой разницы по Фишеру, n = 4-6. p <0,05 считали значимым.
Фиг. 18: Влияние MR33583 на выживание ГАМКергических нейронов в первичной культуре мезэнцефальных клеток после депривации факторов роста (ДФР). Результаты выражены в процентах от контроля и показывают среднее значение ± SEM (100% = CT, без соединения). Односторонний дисперсионный анализ с последующим тестом методом наименьшей значимой разницы по Фишеру, n = 4-6. p <0,05 считали значимым.
Примеры
Пример 1
В этом примере показано влияние донекоприда на первичные нейроны гиппокампа крысы, поврежденные Aβ (24-часовое воздействие).
Болезнь Альцгеймера (БА) - это нейродегенеративное заболевание, которое поражает в основном людей старше 65 лет, страдающих от различных клинических симптомов, таких как прогрессирующее снижение мышления, речи и способности к обучению. Все больше данных из различных источников указывает на олигомеры Aβ (AβO)/протофибриллы как предполагаемые токсичные частицы в патогенезе БА.
Создавая раствор AβO и варьируя концентрацию и время воздействия AβO, можно воспроизвести ранние эффекты (окислительный стресс) и долгосрочное развитие структурных изменений (гибель нейронов).
Модель, использованная в настоящем примере, с применением раствора Aβ пептида, содержащего AβO (точно измеренного с помощью автоматического вестерн-блоттинга), воспроизводила основные нейропатологические особенности БА.
Целью этого исследования была оценка эффектов одного тестируемого соединения (фумарата донекоприда в нескольких концентрациях) в отношении первичных нейронов гиппокампа крыс, поврежденных Aβ (24-часовое воздействие). Донепезил следующей формулы:
(одна концентрация) использовали в качестве эталонного тестируемого соединения.
Материалы и методы
Первичная культура нейронов гиппокампа
Нейроны гиппокампа крыс культивировали, как описано Callizot et al. (2013) J. Neurosci. Res. 91706-716. Беременных самок крыс со сроком гестации 17 дней (крысы Wistar; Janvier Labs, Франция) умерщвляли с использованием глубокой анестезии в камере CO2 с последующей цервикальной дислокацией. Затем эмбрионы извлекали из матки и немедленно помещали в ледяную среду L15 Лейбовица с 2% раствором пенициллина (10000 Ед/мл) и стрептомицина (10 мг/ мл) (ПС) и 1% бычьим сывороточным альбумином (БСА). Нейроны гиппокампа обрабатывали в течение 20 мин при 37°C раствором трипсин-ЭДТА с конечной концентрацией 0,05% трипсина и 0,02% ЭДТА. Диссоциацию останавливали добавлением модифицированной по Дульбекко среды Игла (DMEM) с 4,5 г/л глюкозы, содержащей ДНКазу I сорта II (конечная концентрация 0,5 мг/мл) и 10% эмбриональной телячьей сыворотки (ЭТС). Клетки механически диссоциировали посредством трех принудительных пропусканий через кончик 10 мл пипетки, а затем центрифугировали при 515 x g в течение 10 минут при 4°C. Надосадочную жидкость удаляли, а осадок ресуспендировали в определенной культуральной среде, состоящей из нейробазальной среды с 2% раствором добавки B27, 2 ммоль/л L-глутамина, 2% раствора ПС и 10 нг/мл нейротрофического фактора головного мозга (BDNF). Жизнеспособные клетки подсчитывали на цитометре Нойбауэра с использованием теста исключения трипанового синего. Клетки высевали с плотностью 20000 на лунку в 96-луночные планшеты, предварительно покрытые поли-L-лизином, и культивировали при 37°C в инкубаторе в среде воздух (95%) - CO2 (5%). Питательную среду меняли каждые 2 дня.
Донекоприд и воздействие Aβ1-42 человека
Нейроны гиппокампа обрабатывали растворами Aβ (см. ниже) через 17 дней культивирования. Получение Aβ1-42 осуществляли в соответствии с процедурой, описанной Callizot et al. (2013) J. Neurosci. Res. 91706-716. Вкратце, пептид Aβ1-42 растворяли в указанной выше культуральной среде при начальной концентрации 40 мкМ. Этот раствор осторожно перемешивали в течение 3 дней при 37°C в темноте и сразу же использовали после того, как он был должным образом разбавлен в культуральной среде до используемых концентраций (20 мкМ (планшет 1) или 2,5 мкМ (планшет 2), что соответствует 2 мкМ или 0,25 мкМ олигомеров (AβO), соответственно).
Донекоприда фумарат растворяли в культуральной среде и предварительно инкубировали за 1 час до нанесения Aβ. Препарат Aβ1-42 добавляли до конечной концентрации 20 или 2,5 мкМ (= 2 мкМ или 0,25 мкМ AβO, при оценке с помощью автоматического вестерн-блоттинга), при разведении в контрольной среде в присутствии донекоприда.
Организация чашек с культурой
Донекоприда фумарат (Donecopride) тестировали на одной культуре в 96-луночном планшете (6 лунок на условия). Соединение предварительно инкубировали за час до нанесения Aβ. Оценивали следующие условия:
Оценка конечной точки
Планшет 1: ВЫЖИВАНИЕ, НЕЙРИТНАЯ СЕТЬ И ОЦЕНКА ФОСФОРИЛИРОВАНИЯ ТАУ-БЕЛКА
Через 24 часа после интоксикации нейроны гиппокампа фиксировали холодным раствором этанола (95%) и уксусной кислоты (5%) в течение 5 минут при -20°C. После пермеабилизации 0,1% сапонином клетки инкубировали в течение 2 часов с:
a) куриными поликлональными антителами к белку 2, ассоциированному с микротрубочками (MAP-2), разведенными до 1/1000 в ФБР, содержащем 1% эмбриональной телячьей сыворотки и 0,1% сапонина (это антитело позволяет специфически окрашивать тела нейрональных клеток и нейритов; и позволяют изучать гибель нейронных клеток и нейритную сеть).
b) мышиным моноклональным антителом против фосфо-тау-белка (AT100) при разведении 1/400 в ФБР, содержащем 1% эмбриональной телячьей сыворотки и 0,1% сапонина.
Эти антитела были выявлены с помощью козьих антител к IgG мышей с Alexa Fluor 488 и козьих антител к куриным IgG с Alexa Fluor 568 в разведении 1/400 в ФБР, содержащем 1% ЭТС, 0,1% сапонина, в течение 1 часа при комнатной температуре.
Для каждого условия с помощью ImageXpress (Molecular Devices) при 20-кратном увеличении было сделано 30 снимков (представляющих всю площадь лунки) на лунку. Все изображения были сделаны с одинаковыми параметрами получения. Анализы проводили автоматически с применением Custom Module Editor (Molecular Devices).
Оценивали следующие конечные точки:
- Общее количество нейронов (выживание нейронов, количество МАР-2-положительных нейронов);
- Нейритная сеть (в мкм МАР-2-положительных нейронов);
- Площадь тау в нейронах (мкм², перекрывание с MAP-2-положительными нейронами).
Планшет 2: ОЦЕНКА СИНАПСОВ
Через 24 часа после интоксикации нейроны гиппокампа фиксировали холодным раствором этанола (95%) и уксусной кислоты (5%) в течение 5 минут при -20°C. После пермеабилизации 0,1% сапонином клетки инкубировали в течение 2 часов с:
а) мышиным моноклональным антителом против постсинаптического уплотнения 95 кДа (PSD95) при разведении 1/100 в ФБР, содержащем 1% эмбриональной телячьей сыворотки и 0,1% сапонина;
b) кроличьими поликлональными антителами против синаптофизина (SYN) в разведении 1/100 в ФБР, содержащем 1% эмбриональной телячьей сыворотки и 0,1% сапонина.
Эти антитела проявляли с помощью козьих антител против IgG мыши Alexa Fluor 488 и козьих антител против кроличьих IgG Alexa Fluor 568 в разведении 1/400 в ФБР, содержащем 1% ЭТС, 0,1% сапонина, в течение 1 часа при комнатной температуре.
Для каждого условия было сделано 40 снимков на лунку с помощью ImageXpress (Molecular Devices) с 40-кратным увеличением. Все изображения были сделаны с одинаковыми параметрами получения.
Оценку синапсов выполняли автоматически с помощью Custom Module Editor (Molecular Devices).
Оценивали следующие конечные точки:
- Общее количество синапсов (перекрывание между PSD95/SYN).
Статистический анализ
Все значения выражены как среднее ± SEM. Статистический анализ был выполнен с помощью однофакторного дисперсионного анализа с последующим тестом методом защищенной наименьшей значимой разницы по Фишеру, p <0,05 считали значимым.
Графики и статистический анализ для различных условий были выполнены с использованием программного обеспечения GraphPad Prism версии 7.04. *p <0,05 считали значимым.
Результаты
Влияние донекоприда на выживание и нейритную сеть МАР-2 нейронов гиппокампа и на гиперфосфорилирование тау-белка.
Выживание нейронов
Влияние донекоприда на выживание МАР-2 нейронов в первичной культуре нейронов гиппокампа показано в таблице 1 ниже.
Таблица 1. MAP-2 нейроны (% от контроля)
среднего
*: p <0,05 по сравнению с Aβ1-42, при определении с использованием однофакторного дисперсионного анализа с последующим тестом методом наименьшей значимой разницы по Фишеру.
После применения Aβ пептида имела место существенная потеря нейронов MAP-2. Донепезил, используемый здесь в качестве положительного контроля, был способен в существенной степени защищать нейроны от гибели. Две дозы донекоприда (100 нМ и 500 нМ) оказывали нейропротекторное действие в той же степени, что и донепезил.
Нейритная сеть
Влияние донекоприда на нейритную сеть МАР-2 нейронов в первичной культуре нейронов гиппокампа показано в таблице 2 ниже.
Таблица 2. Нейритная сеть MAP-2 (% от контроля)
среднего
*: p <0,05 по сравнению с Aβ1-42, определенным с использованием однофакторного дисперсионного анализа с последующим тестом методом наименьшей значимой разницы по Фишеру.
Общая нейритная сеть сильно сократилась при повреждении Aβ 1-42. Донепезил в значительной степени сохранил почти всю нейритную сеть. Четыре дозы донекоприда (50 нМ, 100 нМ, 500 нМ и 1 мкМ) также оказали защитное действие на нейритную сеть.
Нейритная сеть пропорциональна количеству нейронов в культуре. Полная нейритная сеть была нормализована на количество нейронов, чтобы определить среднюю длину нейрита на нейрон. Соответствующие результаты показаны в таблице 3 ниже.
Таблица 3. Нейритная сеть/нейроны MAP-2 (% от контроля)
среднего
*: p <0,05 по сравнению с Aβ1-42, определенным с использованием однофакторного дисперсионного анализа с последующим тестом методом наименьшей значимой разницы по Фишеру.
Эта корректировка показала, что длина нейритов на нейрон была больше в присутствии донепезила и донекоприда (1 мкМ), что позволило предположить, что эти два соединения влияли на рост нейритов.
Фосфорилирование тау-белка
Влияние донекоприда на фосфорилирование тау-белка (AT100) в нейронах MAP-2 в первичной культуре нейронов гиппокампа показано в таблице 4 ниже.
Таблица 4. Площадь тау-белка (AT100)/нейроны (% от контроля)
среднего
*: p <0,05 по сравнению с Aβ1-42, определенным с использованием однофакторного дисперсионного анализа с последующим тестом методом наименьшей значимой разницы по Фишеру.
При применении Aβ 1-42 наблюдалось гиперфосфорилирование тау-белка (на AT100). Донепезил в значительной степени уменьшал это гиперфосфорилирование. Аналогичным образом, донекоприд (50 нМ, 100 нМ, 500 нМ) в значительной степени снижал гиперфосфорилирование тау-белка дозозависимым образом.
Влияние донекоприда на синапсы в первичной культуре нейронов гиппокампа
Для оценки образования синапсов изучали распределение PSD-95, постсинаптического маркера, и синаптофизина, пресинаптического маркера. Структуру, положительную при окрашивании как на PSD-95, так и на синаптофизин, рассматривали как синапс.
Влияние донекоприда на эти синапсы показано в таблице 5 ниже.
Таблица 5. Число синапсов (% от контроля)
среднего
*: p <0,05 по сравнению с Aβ1-42, определенным с использованием однофакторного дисперсионного анализа с последующим тестом методом наименьшей значимой разницы по Фишеру.
Стресс, возникающий в результате применения Aβ 1-42, вызывал уменьшение числа синапсов (как ранее было показано Callizot et al. (2013) J. Neurosci. Res. 91: 706-716).
Донепезил уменьшал потерю синапсов. Аналогичным образом, донекоприд был способен сохранять количество синапсов дозозависимым образом (от 5 нМ до 1 мкМ). Интересно, что этот эффект наблюдался и при низкой дозе соединения (5 нМ). Этот эффект усиливался с увеличением дозы.
Из-за повреждения, вызванного пептидом Aβ 1-42, число нейронов снижалось в экспериментальных условиях, что привело к уменьшению числа синапсов. Поэтому, чтобы оценить число синапсов на нейрон, количество синапсов нормализовали на количество нейронов для групп контроля, Aβ 1-42, донекоприда (500 нМ, состоянию, показывающему наибольшее количество синапсов) и донепезила.
Эта корректировка показала, что только донекоприд в значительной степени способствовал образованию новых синапсов, в то время как донепезил не оказывал такого эффекта.
Заключение
Настоящее исследование показывает, что донекоприд оказывает благотворное влияние на выживание нейронов и нейритную сеть нейронов гиппокампа, поврежденных пептидом Aβ 1-42, на модели болезни Альцгеймера in vitro.
Более того, Донекоприд был способен снижать гиперфосфорилирование тау-белка на участке AT100 и был способен в значительной степени стимулировать образование новых синапсов.
Эффект донекоприда был сильнее, чем у донепезила, что позволяет предположить, что это соединение является интересным лекарственным средством-кандидатом для использования в качестве нейропротекторного агента, в частности для лечения нейродегенеративных заболеваний.
Пример 2
Настоящий пример демонстрирует, что соединения по настоящему изобретению могут улучшать когнитивные функции мышей на модели БА in vivo.
Для этой цели эффективность двух соединений (фумарата донекоприда и фумарата флукоприда) исследовали с использованием мышей C57Bl6/J дикого типа, у которых выполняли интрацеребровентрикулярную инъекцию олигомеров Aβ1-42 (AβO).
Соединения вводили перорально (через желудочный зонд) в трех различных дозах. Когнитивные способности оценивали с помощью теста Y-лабиринта, теста распознавания новых объектов и теста водного лабиринта Морриса. В конце когнитивных испытаний всех животных умерщвляли, брали образцы крови и головного мозга для дальнейших анализов ex-vivo.
Схематическое изображение эксперимента
Вкратце, 144 самцов мышей C57Bl6/J (в возрасте 3 месяцев) получали и помещали (от 4 до 5 на клетку) в карантин в течение 7 дней или более. Испытуемые соединения вводили перорально (через желудочный зонд) один раз в день в различных концентрациях от дня -1 до дня +17.
В день 0 мыши получали однократную интрацеребровентрикулярную (ИЦВ) инъекцию контрольного растворителя или AβO.
На день +4 (то есть через четыре дня после индукции заболевания) пространственную рабочую память оценивали с помощью теста Y-лабиринта.
С дня +3 до дня +14 с помощью теста MWM (водного лабиринта Морриса) исследовали способность к обучению и долговременную память.
В дни +15 / +16 опознающую память исследовали с помощью теста распознавания новых объектов (NOR).
Животных умерщвляли на +17 день, и ткани готовили для дальнейших анализов ex-vivo.
Описание экспериментальных групп
В исследовании приняли участие 144 мыши, разделенные на 12 экспериментальных групп, по 12 мышей в каждой экспериментальной группе. Все животные получали однократную (и одностороннюю) внутривенную инъекцию растворителя или AβO в общем объеме 1 мкл.
Различные экспериментальные группы определяли следующим образом:
- ГРУППА A (Контроль с растворителем): растворитель перорально и инъекция растворителя методом ИЦВ (n = 12)
- ГРУППА B (Контроль с AβO): растворитель перорально и инъекция AβO методом ИЦВ (n = 12)
- ГРУППА C (Положительный контроль): субхроническое пероральное применение донепезила и инъекция AβO методом ИЦВ (n = 12)
- ГРУППА D (Контроль с донекопридом): перорально донекоприд (9 мг/кг) и инъекция растворителя методом ИЦВ (n = 12)
- ГРУППА E (Контроль с флукопридом): перорально флукоприд (9 мг/кг) и инъекция растворителя методом ИЦВ (n = 12)
- ГРУППА F (Лечение донекопридом): перорально донекоприд (1 мг/кг) и инъекция AβO методом ИЦВ (n = 12)
- ГРУППА G (Лечение донекопридом): перорально донекоприд (3 мг/кг) и инъекция AβO методом ИЦВ (n = 12)
- ГРУППА H (Лечение донекопридом): перорально донекоприд (9 мг/кг) и инъекция AβO методом ИЦВ (n = 12)
- ГРУППА I (Лечение флукопридом): перорально флукоприд (1 мг/кг) и инъекция AβO методом ИЦВ (n = 12)
- ГРУППА J (Лечение флукопридом): перорально флукоприд (3 мг/кг) и инъекция AβO методом ИЦВ (n = 12)
- ГРУППА K (Лечение флукопридом): перорально флукоприд (9 мг/кг) и инъекция AβO методом ИЦВ (n = 12)
- ГРУППА L (Положительный контроль): интраперитонеальная инъекция донепезила в дни испытаний и инъекция AβO методом ИЦВ (n = 12)
Поскольку в день можно было осуществить интрацеребровентрикулярные инъекции максимум 24 мышам, исследование было проведено в шести независимых циклах. В каждый цикл включал по две мыши из всех групп.
Экспериментальные процедуры
Содержание животных
Получение 144 мышей-самцов C57Bl6/J (от Janvier, Франция) (возраст 3 месяца). При получении животных помещали на карантин на срок 7 дней и более.
Мышей содержали при стандартной температуре (22±2°C) и в условиях с контролируемой освещенностью и влажностью (свет включали с 8:00 до 20:00; влажность 55 ± 10%) с неограниченным доступом к пище и воде. Мышей помещали в группы и наблюдали дважды в день силами лабораторного персонала (8:00 и 16:00). В случае 2 или более серьезных проблем общего состояния здоровья пораженную мышь умерщвляли. Критерии неприемлемого состояния здоровья были следующими: отсутствие спонтанных движений и неспособность пить или есть в течение 24-часового периода наблюдения, потеря массы тела более 12%, массивное кровотечение, спонтанное воспаление (общие наблюдения за кожей и глазами мышей) или отсутствие частей тела.
В течение всей процедуры мышей взвешивали 3 раза (т.е. до лечения и на 8-й день и 17-й день).
Хранение, приготовления состава и дозировка соединений
Для перорального применения донекоприд и флукоприд готовили путем солюбилизации в 0,9% NaCl. Дозирование соединения (общий объем 200 мкл) начинали за день до индукции заболевания (посредством инъекции AβO методом ИЦВ) и проводили в течение 19 дней подряд.
Стереотаксическая инъекция
Однократная интрацеребровентрикулярная инъекция 144 мышам растворителя или AβO (максимум 24 мыши в день, все мыши были рандомизированы): под анестезией (интраперитонеальная инъекция смеси кетамина/ксилазина в дозе 110 и 15 мг/кг соответственно) AβO (1 мкл) или растворитель (1 мкл) вводили в правый боковой желудочек. Инъекции производили с помощью 10 мкл микрошприцев Hamilton, снабженных иглой 26-го размера. Процедуру завершали подкожной инъекцией метакама (обезболивание) в дозе 5 мг/кг. Затем животных по отдельности помещали в их домашнюю клетку, и клетку помещали в обогреваемый шкаф до полного выздоровления животного. За животными тщательно наблюдали, чтобы контролировать выздоровление после анестезии.
Приготовление AβO: Aβ1-42 получали от Bachem (ссылка H1368) (номер серии: 1052301). Получение стабильных олигомеров Aβ1-42 выполняли, как описано у Garcia et al. (2010) J. Neurosci. 30: 7516-7527. Олигомерный препарат (обозначенный как серия № 2016-14) содержал смесь стабильных тримеров и тетрамеров Aβ1-42, а также мономерных форм пептида. Все используемые препараты олигомеров ранее были охарактеризованы с точки зрения состава олигомеров и нейротоксичности in vitro.
Когнитивный тест, тест Y-лабиринта
6 независимых испытаний с участием в каждом 24 мышей (т.е. по 2 мыши из каждой экспериментальной группы) всего для 144 мышей в день +4 - поведенческие тесты были выполнены через 1 час после введения дозы.
Этот тест оценивает пространственную рабочую память, которая в основном обеспечивается префронтальной корой (рабочая память) и гиппокампом (пространственный компонент). См. Yoon et al. (2008) Learn Mem. 15: 97-105 и Spellman et al. (2015) Nature 522: 309-314 для различных компонентов.
Префронтальная кора отвечает за рабочую память грызунов, приматов и человека.
Измерение спонтанного чередования поведения широко используется поведенческими фармакологами для оценки пространственной рабочей памяти, компонента кратковременной памяти. В своей простейшей форме спонтанное чередование поведения включает тенденцию мышей чередовать их обычный не закрепленный в памяти выбор конечностей при последовательных возможностях.
Этот тест успешно применялся при использовании инфузии AβO методом ИЦВ для индукции когнитивного дефицита (Garcia et al. (2010) J. Neurosci. 30: 7516-7527).
Подробный протокол: Эффективность непосредственной пространственной рабочей памяти оценивали путем регистрации спонтанного чередующегося поведения в Y-лабиринте. Лабиринт был сделан из непрозрачного оргстекла, и каждый из трех рукавов имел длину 40 см, высоту 14 см, ширину 10 см, и они были расположены под равными углами. Аппарат помещают в испытательную комнату с однородным освещением, чтобы получить 12-15 люкс во всех рукавах, а также в центральной зоне. Мышей помещают в середину одного рукава и позволяют свободно исследовать лабиринт в течение 5-минутного сеанса. Серии входов в рукава регистрируют на видео (программное обеспечение Smart v3.0, Bioseb), и вход в рукав считают завершенным, когда задние лапы мыши полностью помещаются в рукаве. Чередование определяют как последовательные входы в 3 рукава на перекрывающихся наборах триплетов. Процент чередования рассчитывают как отношение фактического общего количества чередований к возможным чередованиям, определяемое как количество входов в рукав минус 2, умноженное на 100. Двигательную активность также регистрировали и оценивали на этом этапе (например, мотивационный индекс) путем мониторинга средней скорости и общего расстояния.
Анализ данных: для статистического анализа использовали компьютерное программное обеспечение Graphpad/Prism. Был проведен непараметрический дисперсионный анализ (критерий Краскела-Уоллиса), за которым применяли непараметрические U-критерии Манна-Уитни для сравнения между группами. Статистически значимыми считали значения с р <0,05. Данные представлены как среднее значение ± стандартная ошибка среднего.
Когнитивное тестирование, анализ NOR
6 независимых испытаний с участием 24 мышей в каждом (т.е. по 2 мыши из каждой экспериментальной группы), всего 144 мыши в день +4 - поведенческие тесты были выполнены через 1 час после введения дозы.
Этот испытание оценивает опознающую память для предметов, и его человеческим эквивалентом является визуальное парное сравнение (VPC) (Wallace et al. (2015) Handb. Exp. Pharmacol. 228: 27-57). Распознавание объекта опосредуется периринальной корой у грызунов (Albasser et al. (2009) Behav. Neurosci. 123: 115-124), приматов (Zeamer et al. (2015) Dev. Cogn. Neurosci. 11: 31-41) и людей (Watson & Lee (2013) J. Neurosci. 33: 4192-4200).
Ранее было продемонстрировано, что интрацеребровентрикулярная инъекция AβO у мышей приводит к нарушениям в задаче распознавания нового объекта, и этот эффект является обратимым при использовании ингибитора ацетилхолинэстеразы донепезила, который показал защитный эффект у человека в отношении цитопротективного и нейропротекторного пептида митохондриального происхождения.
Подробный протокол: за день до когнитивного теста (то есть в день +15) мышей приучали в течение 10-минутного испытания, в течение которого их помещали в пустое открытое поле. В день когнитивного теста (то есть день +16) животных помещали в одно и то же открытое поле, и им позволяли свободно исследовать два идентичных объекта в течение пяти минут (испытание запоминания). Затем животных возвращали в домашнюю клетку на пять минут между испытаниями. Во время испытания запоминания животным позволяли исследовать два разных объекта: один знакомый и один новый объект. В течение этого времени экспериментатор, вне зависимости от лечения, регистрировал время, в течение которого мышь активно исследует каждый объект. Все испытания записывали на видео (программное обеспечение Smart v3.0, Bioseb). Затем генерировали индекс различения: Индекс различения = (время исследования нового объекта - время исследования знакомого объекта) / общее время исследования.
Анализ данных: для статистического анализа использовали компьютерное программное обеспечение Graphpad/Prism. Для сравнения между группами использовали непараметрический дисперсионный анализ (критерий Краскела-Уоллиса), после чего применяли непараметрический U-критерий Манна-Уитни. Значения p<0,05 считали статистически значимыми. Данные представлены как среднее значение ± стандартная ошибка среднего.
Когнитивное тестирование, анализ MWM
Проводили 6 независимых испытаний с участием по 24 мыши в каждом (т.е. по 2 мыши из каждой экспериментальной группы), всего 144 мыши в день +4.
Память пространственного ориентирования - это процесс, который проверяет способность запоминать пространственные ориентиры для определения местоположения. Она опосредуется гиппокампом у грызунов (Broadbent et al. (2006) Learn Mem. 13: 187-191) и у человека (Bartsch et al. (2010) Science 328: 1412-1415). Использовалась задача виртуального водного лабиринта. Этот тест также включает обучающий компонент.
Инъекция AβO методом ИЦВ у мышей вызывает дефицит памяти пространственного ориентирования в водном лабиринте Морриса (Garcia et al. (2010) J. Neurosci. 30: 7516-7527).
Подробный протокол: испытания приучения (видимая платформа) - водный лабиринт Морриса (MWM) выполняли, как описано у Garcia et al. (2010) J. Neurosci. 30: 7516-7527. Экспериментальное устройство состоит из круглого резервуара для воды (диаметр = 100 см; высота = 50 см), содержащего воду при 21°C с глубиной 25 см, сделанную непрозрачной путем добавления водной акриловой эмульсии, чтобы закрыть вид под поверхностью воды. Используют платформу (диаметр = 10 см), помещаемую в середине квадранта. Бассейн располагают в испытательной комнате, равномерно освещенной на уровне 100 люкс. Путь плавания животных фиксируют с помощью системы видеонаблюдения. Мышей помещают в экспериментальную комнату не менее чем на 30 минут до тестирования, чтобы они могли адаптироваться к условиям экспериментальной комнаты. Перед навигацией по местоположению осуществляют переход к видимой платформе для оценки зрительных и двигательных способностей всех мышей. Мышей подвергают 4 испытаниям по 60 секунд в день (в течение 2 дней подряд) с интервалом между испытаниями не менее 1 часа. Как только мыши находят платформу, их оставляют одних на платформе еще на 30 секунд. В комнате нет дополнительных ориентиров. Положение платформы и начальные точки случайным образом распределяют по всем 4 квадрантам бассейна. Мышей, которые не могут найти платформу через 60 секунд, направляют к ее местоположению и помещают на платформу на 30 секунд.
Испытания запоминания (обучающие испытания со скрытой платформой) - их выполняют в течение 5 дней подряд и используют для достижения устойчивых показателей латентности спасения (т.е. требуемого для спасения времени). Мышей помещают в экспериментальную комнату не менее чем на 30 минут до тестирования, чтобы они могли адаптироваться к условиям экспериментальной комнаты. Скрытую платформу погружают на 1 см ниже поверхности воды и помещают в середине одного квадранта. Бассейн располагают в испытательной комнате, равномерно освещенной 100 люкс и содержащей различные заметные визуальные подсказки. Пути плавания, расстояние плавания, скорость плавания и тигмотаксис регистрируют с помощью системы видеонаблюдения. Мышей подвергают 4 испытаниям по 60 секунд в день с интервалом между испытаниями не менее 1 часа. Мышам позволяют свободно плавать в течение 60 секунд, оставляют одних еще на 30 секунд на скрытой платформе, а затем возвращают в их домашнюю клетку в течение интервала между испытаниями. Начальные позиции (установленные на границе квадрантов) выбирают случайным образом для каждого животного. В каждом испытании регистрируют время, необходимое для спасения на скрытую платформу. Мышей, которые не могут найти платформу через 60 секунд, направляют на платформу и помещают на нее на 30 секунд, прежде чем их возвращают в домашнюю клетку.
Испытания на сохранение в памяти (поисковые испытания, без платформы) - их проводят через два дня после последней тренировки. Мышей снова акклиматизируют в экспериментальной комнате не менее чем за 30 минут до тестирования. Платформу удаляют, и каждому животному позволяют свободно плавать в течение 60 секунд. Во время испытания время, проведенное в целевом квадранте, время, проведенное в противоположном квадранте, и пересечения места прежнего расположения платформы измеряют и контролируют с помощью видеорегистратора.
Анализ данных: для статистического анализа используют компьютерное программное обеспечение Graphpad/Prism. Для сравнения между группами используют непараметрический дисперсионный анализ (критерий Краскела-Уоллиса), а затем непараметрический U-критерий Манна-Уитни. Значения с p <0,05 считают статистически значимыми. Данные представлены как среднее значение ± стандартная ошибка среднего.
Отбор образцов ткани
Мышей умерщвляли на +17 день.
Сбор образцов крови: под анестезией у мышей брали кровь. Приблизительно 600 мкл цельной крови забирали из полой вены с помощью шприца на 1 мл. Кровь собирали в пробирки BD Microtainer K2E (каталожный номер 365975) или микропробирки Sarstedt LiHe (каталожный номер 41.1503.005). Пробирки осторожно переворачивали, и образцы помещали на лед. Микроконтейнеры центрифугировали 5 мин при 4°C, 8600 g. Приблизительно 250 мкл плазмы переносили в пробирки Эппендорфа и замораживали при -80°C для дальнейших анализов.
Сбор образцов головного мозга: после сбора крови у мышей проводили внутрисердечную флэш-перфузию с 0,9% физиологическим раствором. Удаляли весь мозг и разделяли полушария. Контралатеральное левое полушарие мгновенно замораживали в жидком азоте и хранили при -80°C для дальнейшего анализа (т.е. оценки воздействия соединения). Ипсилатеральное правое полушарие использовали для выделения гиппокампа, префронтальной коры, и коры. Все структуры головного мозга изолировали на охлажденной подложке и мгновенно замораживали в жидком азоте.
Результаты
Влияние соединений по изобретению на пространственную рабочую память.
Чтобы определить влияние донекоприда и флукоприда на пространственную рабочую память мышей, через четыре дня после инъекции растворителя или AβO методом ИЦВ выполняли испытание с Y-лабиринтом. В течение 5 минут испытания животным позволяли исследовать устройство Y-образного лабиринта, и оценивали чередующееся поведение, как описано в экспериментальных процедурах.
Мыши из группы с контрольным растворителем (Группа A) показали нормальное поведение при исследовании Y-лабиринта с чередующимся поведением 68,0 ± 3,0%. Эти результаты согласуются с предыдущими наблюдениями аналогичных контрольных групп (см. Garcia et al. (2010) J. Neurosci. 30: 7516-7527).
Как и ожидалось, однократная инъекция AβO методом ИЦВ (Группа B) привела к значительному ухудшению (p = 0,0026) когнитивных функций в тесте Y-лабиринта по сравнению с контрольными мышами, которым был введен растворитель, лишь с 54,0 ± 2,0% чередований.
Мыши в группе положительного контроля, которым перорально вводили донепезил и осуществляли инъекцию AβO методом ИЦВ (группа C), были неотличимы от контрольных мышей (чередующееся поведение 66,1 ± 1,8%) и статистически отличались (p = 0,0011) от мышей, которым вводили AβO. Мыши в группе положительного контроля, получавшие интраперитонеально (остро) дозу донепезила и инъекцию AβO методом ИЦВ (Группа L), статистически отличались от контрольных мышей (p = 0,0278; чередующееся поведение 60,8 ± 1,9%), а также статистически отличались (p = 0,0421) от мышей, которым вводили AβO.
Мыши, у которых применяли донекоприд (9 мг/кг/сутки) и вводили растворитель методом ИЦВ (группа D), демонстрировали чередующееся поведение 59,3 ± 4,0%, не отличающееся от контрольных мышей (p = 0,2640) и от мышей, которым вводили AβO (р = 0,5371).
Мыши, у которых применяли флукоприд (9 мг/кг/сутки) и вводили растворитель методом ИЦВ (группа E), демонстрировали чередующееся поведение 63,1 ± 2,3%, не отличающееся от контрольных мышей (p = 0,1166) и отличающееся от мышей, которым вводили AβO (p = 0,0111).
Мышам вводили возрастающие дозы донекоприда (1, 3 и 9 мг/кг/сутки) и вводили AβO методом ИЦВ (группы F, G и H соответственно). В дозе 3 мг/кг/сутки донекоприд подавлял AβO-индуцированное нарушение пространственной рабочей памяти. Действительно, мыши из группы G показали чередующееся поведение 65,5 ± 4,4%, не отличающееся от контрольных мышей (p = 0,5543) и отличающееся от мышей, которым вводили AβO (p = 0,0137). В дозе 1 мг/кг/сутки донекоприд не оказывал влияния на AβO-индуцированное нарушение пространственной рабочей памяти, поскольку эти мыши демонстрировали чередующееся поведение на 56,6 ± 3,7%, отличающееся от контрольных мышей (p = 0,0488) и не отличающееся от мышей, которым вводили AβO (p = 0,6859). Из-за более высокой гетерогенности мыши из Группы H (получавшие донекоприд в дозе 9 мг/кг/сутки) демонстрировали промежуточный фенотип с чередующимся поведением 64,3 ± 5,1%, не отличающимся от поведения контрольных мышей (p = 0,5994) и также не отличающимся от поведения мышей, которым вводили AβO (p = 0,1316).
Мышам вводили возрастающие дозы флукоприда (1, 3 и 9 мг/кг/сутки) и вводили AβO методом ИЦВ (группы I, J и K, соответственно). В дозах 1 и 3 мг/кг/сутки флукоприд ингибировал AβO-индуцированное нарушение пространственной рабочей памяти. Действительно, мыши из групп I и J демонстрировали чередующееся поведение 67,1 ± 2,7% и 65,3 ± 4,1% соответственно, не отличаясь от контрольных мышей (p = 0,9754 и p = 0,6936 для групп I и J, соответственно) и отличаясь от мышей, которым вводили AβO (p = 0,0029 и p = 0,0454, для групп I и J, соответственно). Напротив, в дозе 9 мг/кг/сутки флукоприд не обеспечивал защиту мышей от индуцированного AβO нарушения пространственной рабочей памяти. Действительно, мыши из группы K демонстрировали чередующееся поведение 59,1 ± 3,0%, отличающееся от контрольных мышей (p = 0,0290), но не отличающееся от мышей, которым вводили AβO (р = 0,2346).
Влияние соединений по изобретению на способность к обучению и долговременную память.
Чтобы определить влияние соединений по настоящему изобретению на способность к обучению и долговременную память, тест MWM выполняли с дня +3 до дня +14 после инъекции растворителя или AβO методом ИЦВ.
Испытание адаптации
На первом этапе (испытания приучения, проведенные в дни +3 и +4 после инфузии растворителя или AβO методом ИЦВ) мышей обучали выходить из воды на видимую платформу при отсутствии других визуальных подсказок. Во время этих испытаний регистрировали латентность спасения, расстояние плавания и скорость плавания. Не было существенной разницы между группами по латентности спасения. Не выявлено никакой разницы в скорости или расстоянии плавания.
В целом, эти данные позволяют предположить, что лечение не повлияло ни на физические возможности, ни на мотивацию мышей к спасению из воды.
Испытания обучения
Способность к обучению мышей контролировали в течение пяти дней подряд (от дня +7 до дня +11 после инфузии растворителя или AβO методом ИЦВ). Животные выполняли 4 испытания в день и были обучены определять местонахождение скрытой платформы с помощью визуальных подсказок. Во время этих испытаний обучения регистрировали латентность спасения, расстояние плавания и скорость плавания.
Статистически значимой разницы в средней латентности спасения между группами в начале испытаний не было выявлено. Все группы тогда проявили некоторую степень обучения.
Мыши из группы контрольного растворителя (Группа A) эффективно выучили локализацию платформы, и после пяти дней обучения (день 5) у них обнаруживалась средняя латентность спасения 11,7 ± 1,5 секунд (p = 0,00031 по сравнению с днем 1). Напротив, мыши, которым вводили AβO (Группа B), продемонстрировали нарушенное поведение, в результате которого средняя латентность спасения составила 17,3 ± 2,0 секунд после пяти дней обучения (p = 0,4537, не отличается от дня 1, и p = 0,00293, значительно отличается от контрольных мышей).
Мыши, которым субхронически вводили донепезил (перорально) и вводили AβO методом ИЦВ (Группа C), эффективно выучивали локализацию платформы. После пяти дней обучения (5-й день) средняя латентность спасения составила 14,7 ± 1,9 секунд (p = 0,09593 по сравнению с 1-м днем). На 5 день их показатели не отличались от контрольных мышей (p = 0,2188) и не отличались от мышей, которым вводили AβO (p = 0,3477).
Мыши, получившие острую дозу (в день тестирования) донепезила (итраперитонеально) и инъекцию AβO методом ИЦВ (Группа L), эффективно выучивали локализацию платформы. После пяти дней тренировок (5-й день) средняя латентность спасения составила 13,4 ± 1,5 секунд (p = 0,01573 по сравнению с 1-м днем). На 5 день их показатели не отличались от контрольных мышей (p = 0,4187) и не отличались от мышей, которым вводили AβO (p = 0,1272).
Мыши, которым вводили дозу донекоприда (9 мг/кг/сутки) или флукоприда (9 мг/кг/сутки день), и вводили растворитель методом ИЦВ (Группы D и E, соответственно), демонстрировали нормальные когнитивные способности. После пяти дней обучения (день 5) у мышей, которым вводили дозу донекоприда, наблюдалась средняя латентность спасения 13,0 ± 1,7 секунд (p = 0,01778 по сравнению с днем 1). На 5 день их показатели не отличались от контрольных мышей (p = 0,5476) и не отличались от мышей, которым вводили AβO (p = 0,10731). После пяти дней обучения (день 5) у мышей, которым вводили флукоприд, средняя латентность спасения составила 16,3 ± 2,0 секунд (p = 0,04439 по сравнению с днем 1). На 5 день их показатели (по неизвестным причинам) отличались от контрольных мышей (p = 0,06709) и не отличались от мышей, которым вводили AβO (p = 0,7235). Следует отметить, что на 4-й день группа флукоприда продемонстрировала среднюю латентность спасения 14,2 ± 1,9 секунд, не отличаясь от контрольных мышей (p = 0,7933) и не отличаясь от мышей, которым вводили AβO (p = 0,1225).
Мышам вводили возрастающие дозы донекоприда, и вводили AβO методом ИЦВ (Группы F, G и H). Присутствие донекоприда (в дозе 3 мг/кг/сутки) приводило к улучшению способности к обучению мышей по сравнению с мышами, которым вводили AβO. Действительно, у мышей из Группы G средняя латентность спасения составила 12,7 ± 1,5 секунд (p = 0,02933 по сравнению с днем 1) после пяти дней обучения. Их характеристики не отличались от контрольных мышей (p = 0,6284), но отличались от мышей, которым вводили AβO (p = 0,07381).
Мышам вводили возрастающие дозы флукоприда, и вводили AβO методом ИЦВ (Группы I, J и K). Присутствие флукоприда (в дозе 1 и 3 мг/кг/сутки) привело к улучшению обучаемости мышей по сравнению с мышами, которым вводили AβO, тогда как соединение в дозе 9 мг/кг/сутки не оказывало влияния. У мышей, которым вводили флукоприд в дозе 1 мг/кг/сутки, средняя латентность спасения составила 13,1 ± 1,1 секунд (p = 0,03759 по сравнению с днем 1) после пяти дней обучения. Их характеристики не отличались от контрольных мышей (p = 0,4367), но отличались от мышей, которым вводили AβO (p = 0,07422). Аналогичным образом, мыши, которым вводили флукоприд в дозе 3 мг/кг/сутки, показали среднюю латентность спасения 12,5 ± 1,1 секунд (p = 0,00146 по сравнению с днем 1) после пяти дней обучения. Их характеристики не отличались от контрольных мышей (p = 0,6477), но отличались от мышей, которым вводили AβO (p = 0,04328).
Поисковые испытания
Эффективность долговременной памяти определяли через четыре дня после последней тренировки и через 14 дней после инъекции растворителя или AβO методом ИЦВ. Платформу удаляли, и каждому животному давали свободно плавать в течение 60 секунд. Во время поискового испытания определяли время, проведенное в различных квадрантах бассейна, количество пересечений прежнего местоположения платформы и время первого такого пересечения (то есть латентность до цели). Во время поисковых испытаний не наблюдалось разницы в скорости плавания между группами, что свидетельствует о нормальных физических возможностях и мотивации.
Количество пересечений - Мыши из группы контроля с растворителем (Группа А) продемонстрировали нормальное поведение со средним числом пересечений 5,3 ± 0,6 (фиг. 3). Эти результаты согласуются с предыдущими наблюдениями аналогичных контрольных групп. Как и ожидалось, однократная инъекция AβO методом ИЦВ (Группа B) привела к значительному ухудшению когнитивных функций по сравнению с мышами, которым вводили контрольный растворитель, со снижением среднего числа пересечений до 2,2 ± 0,4 (p = 0,0010 по сравнению с контрольными мышами).
Мыши, которым субхронически вводили донепезил (перорально) и вводили AβO методом ИЦВ (группа C), демонстрировали улучшенное поведение со средним числом пересечений 3,7 ± 0,4. Эти мыши не отличались от мышей с контрольным растворителем (p = 0,0502) и значимо отличались от мышей, которым вводили AβO (p = 0,0214). Аналогичным образом, мыши, получавшие острую дозу (в день испытания) донепезила (интраперитонеально) и инъекцию AβO методом ИЦВ (группа L), продемонстрировали улучшенное поведение со средним числом пересечений 3,8 ± 0,4. Эти мыши не отличались от мышей с контрольным растворителем (p = 0,0857) и значимо отличались от мышей, которым вводили AβO (p = 0,0159).
Мыши, получавшие дозу донекоприда (9 мг/кг/сутки) или флукоприда (9 мг/кг/сутки) при введении растворителя (группы D и E, соответственно), продемонстрировали нормальные когнитивные способности со средним числом пересечений 4,6 ± 0,5 и 4,6 ± 0,7. Эти мыши не отличались от контрольных мышей с применением растворителя (p = 0,4003 и p = 0,3945 для донекоприда и флукоприда, соответственно) и значимо отличались от мышей, которым вводили AβO (p = 0,0026 и p = 0,0077 для донекоприда и флукоприда, соответственно).
Мышам вводили возрастающие дозы донекоприда и вводили AβO методом ИЦВ (Группы F, G и H). Присутствие донекоприда (во всех дозах) приводило к ингибированию AβO-индуцированного нарушения долговременной памяти, оцениваемого по количеству пересечений. Максимальный эффект достигался при дозе 3 мг/кг/сутки (Группа G). Действительно, эти мыши демонстрировали нормальное поведение со средним числом пересечений 5,1 ± 0,6, не отличаясь от контрольных мышей с растворителем (p = 0,6575) и значимо отличаясь от мышей, которым вводили AβO (p = 0,0018).
Мышам вводили увеличивающиеся дозы флукоприда и вводили AβO методом ИЦВ (группы I, J и K). Присутствие флукоприда (во всех дозах) приводило к ингибированию AβO-индуцированного нарушения долговременной памяти, оцениваемого по количеству пересечений. Максимальный эффект достигался при дозе 3 мг/кг/сутки (Группа J). Действительно, эти мыши демонстрировали нормальное поведение со средним числом пересечений 4,1 ± 0,6, не отличаясь от контрольных мышей с растворителем (p = 0,1668) и значимо отличаясь от мышей, которым вводили AβO (p = 0,0242).
Латентность до цели. Мыши из контрольной группы с растворителем (Группа A) демонстрировали нормальное поведение со средней латентностью до цели (т.е. временем достижения цели) 12,0 ± 1,9 секунд. Эти результаты согласуются с предыдущими наблюдениями аналогичных контрольных групп. Как и ожидалось, однократная инъекция AβO методом ИЦВ (Группа B) привела к значительному ухудшению когнитивных функций по сравнению с контрольными мышами, которым вводили растворитель, с увеличенной средней латентностью до цели 24,4 ± 4,7 с (p = 0,0074 по сравнению с контрольными мышами).
Мыши, которым субхронически вводили донепезил (перорально) и вводили AβO методом ИЦВ (группа C), демонстрировали улучшенное поведение со средней латентностью до цели 14,6 ± 2,7 секунд. Эти мыши не отличались от контрольных мышей с растворителем (p = 0,6860) и значимо отличались от мышей, которым вводили AβO (p = 0,0272). Аналогичным образом, мыши, получившие острую дозу (в день тестирования) донепезила (интраперитонеально) и интрацеребровентрикулярную инъекцию AβO (группа L), показали улучшенное поведение со средней латентностью до цели 13,3 ± 2,3 секунд. Эти мыши не отличались от контрольных мышей с растворителем (p = 0,9264) и значимо отличались от мышей, которым вводили AβO (p = 0,0028).
Было обнаружено, что мыши, которым вводили дозу донекоприда (9 мг/кг/сутки) или флукоприда (9 мг/кг/день) с использованием растворителя (Группы D и E, соответственно), демонстрировали нормальные когнитивные способности со средней латентностью до цели 7,2 ± 1,5 секунд и 12,0 ± 2,1 секунд. Эти мыши не отличались от контрольных мышей с растворителем (p = 0,0524 и p = 0,7582 для донекоприда и флукоприда, соответственно) и значимо отличались от мышей, которым вводили AβO (p = 0,0017 и p = 0,0082 для донекоприда и флукоприда, соответственно).
Мышам вводили возрастающие дозы донекоприда и вводили AβO методом ИЦВ (группы F, G и H). Присутствие донекоприда (во всех дозах) приводило к ингибированию AβO-индуцированного нарушения долговременной памяти, оцениваемого с использованием показателя латентности до цели. Максимальный эффект достигался при дозе 1 мг/кг/сутки (Группа F). Действительно, эти мыши демонстрировали нормальное поведение со средней латентностью до цели 9,3 ± 1,4 секунд, не отличаясь от контрольных мышей с растворителем (p = 0,2548) и значимо отличаясь от мышей, которым вводили AβO (p = 0,0005).
Мышам вводили увеличивающиеся дозы флукоприда и вводили AβO методом ИЦВ (Группы I, J и K). Присутствие флукоприда (только в дозе 9 мг/кг) приводило к ингибированию AβO-индуцированного нарушения долговременной памяти, оцениваемого с использованием показателя латентности до цели. Действительно, мыши из Группы К показали обычное поведение со средней латентностью до цели 10,7 ± 1,9 секунд, не отличаясь от контрольных мышей с растворителем (p = 0,6891) и значимо отличаясь от мышей, которым вводили AβO (p = 0,0031).
Время в квадранте. Мыши из контрольной группы с растворителем (Группа A) провели значительно больше времени в целевом квадранте (p <0,0001 по сравнению с временем, проведенным в противоположном квадранте). Эти результаты согласуются с предыдущими наблюдениями аналогичных контрольных групп. Как и ожидалось, однократная инъекция AβO методом ИЦВ (Группа B) привела к значительному ухудшению когнитивных функций по сравнению с контрольными мышами с растворителем, при этом в противоположном квадранте было проведено больше времени, чем в целевом квадранте (p = 0,0779, при сравнении времени, проведенного в целевом и противоположном квадранте). Субхроническое или острое введение донепезила (группы C и L) не помогло улучшить AβO-индуцированное нарушение долговременной памяти при оценке времени, проведенного в целевом и противоположном квадрантах. Аналогичным образом, донекоприд и флукоприд не улучшили разницу для квадрантов при поисковом испытании.
Влияние соединений по изобретению на опознающую память
Чтобы определить влияние тестовых соединений на опознающую память, выполняли тест NOR через 16 дней после инъекции растворителя или AβO методом ИЦВ. Поведенческие испытания выполняли, как описано в экспериментальных процедурах.
Мыши из контрольной группы с растворителем (Группа А) демонстрировали нормальное поведение со средним индексом различения 0,35 ± 0,04. Эти результаты согласуются с предыдущими наблюдениями аналогичных контрольных групп. Как и ожидалось, однократная инъекция AβO методом ИЦВ (Группа B) привела к значительному ухудшению (p <0,0001) когнитивных функций по сравнению с контрольными мышами с растворителем; со средним индексом различения -0,01 ± 0,03; мыши, которым вводили AβO, не могли различать новые и знакомые объекты.
Мыши, которым субхронически вводили донепезил (перорально) и AβO (ИЦВ) (группа C), проявляли нормальное поведение со средним индексом различения 0,29 ± 0,09. Эти мыши не отличались от контрольных мышей с растворителем (p = 0,7439) и значимо отличались от мышей, которым вводили AβO (p = 0,0192).
Мыши, получившие острую дозу (в день испытания) донепезила (интраперитонеально) и инъекцию AβO методом ИЦВ (Группа L), показали нормальное поведение со средним индексом различения 0,30 ± 0,07. Эти мыши не отличались от контрольных мышей с растворителем (p = 0,9158) и значимо отличались от мышей, которым вводили AβO (p = 0,0030).
Было обнаружено, что мыши, которым вводили дозу донекоприда (9 мг/кг/сутки) или флукоприда (9 мг/кг/сутки) в присутствии растворителя (Группы D и E, соответственно), демонстрировали нормальные когнитивные способности со средним индексом различения 0,29 ± 0,04 и 0,36 ± 0,06. Эти мыши не отличались от контрольных мышей с растворителем (p = 0,3071 и p = 0,8205 для донекоприда и флукоприда, соответственно) и значимо отличались от мышей, которым вводили AβO (p = 0,0007 и p = 0,0003 для донекоприда и флукоприда, соответственно).
Мышам вводили возрастающие дозы донекоприда и вводили AβO методом ИЦВ (группы F, G и H). Присутствие донекоприда (во всех дозах) приводило к дозозависимому ингибированию AβO-индуцированного нарушения опознающей памяти. Максимальный эффект достигался при дозе 9 мг/кг/сутки (Группа H). Действительно, эти мыши демонстрировали нормальное поведение со средним индексом различения 0,43 ± 0,06, не отличаясь от контрольных мышей с растворителем (p = 0,3440) и значительно отличаясь от мышей, которым вводили AβO (p = 0,0003).
Мышам вводили возрастающие дозы флукоприда и вводили AβO методом ИЦВ (Группы I, J и K). Присутствие флукоприда (во всех дозах) приводило к дозозависимому ингибированию (колоколообразная кривая) AβO-индуцированного нарушения опознающей памяти. Максимальный эффект достигался при дозе 3 мг/кг/сутки (Группа J). Действительно, эти мыши демонстрировали нормальное поведение со средним индексом различения 0,46 ± 0,05, не отличавшимся от контрольных мышей с растворителем (p = 0,1039) и значительно отличавшимся от мышей, которым вводили AβO (p = 0,0003).
Заключение
В целом, эти данные убедительно свидетельствуют о том, что и Донекоприд, и Флукоприд улучшают когнитивные способности мышей, которым вводили AβO.
Пример 3
В этом примере показаны антиамнестические, антиамилоидные и противовоспалительные эффекты донекоприда и флукоприда при хроническом введении на модели болезни Альцгеймера у мышей 5XFAD.
Обоснование исследования
Целью исследования была оценка потенциального антиамнестического, антиамилоидного и противовоспалительного эффекта донекоприда и флукоприда при хроническом введении на мышиной модели болезни Альцгеймера.
Используемая мышиная модель представляла собой линию 5XFAD (обозначаемый в лаборатории Джексона как «B6SJL-Tg(APPSwFlLon,PSEN1*M146L*L286V)6799Vas/Mmjax», инвентарный номер MMRRC: 34840-JAX или «B6.Cg-Tg(APPSwFlLon,PSEN1*M146L*L286V)6799Vas/Mmjax», инвентарный номер MMRRC: 34848-JAX).
Эти трансгенные мыши 5XFAD сверхэкспрессируют мутантный человеческий APP (695) со шведской (K670N, M671L), флоридской (I716V) и лондонской (V717I) мутациями семейной болезни Альцгеймера (FAD) вместе с человеческим PS1, несущим две мутации FAD, M146L и L286V. Оба трансгена регулируются мышиным промотором Thy1, обеспечивая сверхэкспрессию в головном мозге. Мыши 5XFAD воспроизводят основные черты амилоидной патологии при болезни Альцгеймера и представляют собой полезную модель нейродегенерации, индуцированной внутринерональным Aβ42, и образования амилоидных бляшек.
Методы и дизайн эксперимента
Мыши
Эксперименты на животных проводили в соответствии с Директивой Совета Европейских Сообществ от 24 ноября 1986 (86/609/EEC). Были предприняты все усилия, чтобы свести к минимуму страдания животных и уменьшить количество используемых мышей. Получение мышей 5XFAD описано у Oakley et al. (2006) J. Neurosci. 26: 10129-10140. Эти трансгенные мыши сверхэкспрессируют как человеческий APP (695), несущий шведскую (K670N, M671L), флоридскую (I716V) и лондонскую (V717I) мутации семейной болезни Альцгеймера (FAD), так и человеческий пресенилин 1 (PS1), несущий две мутации FAD M146L и L286V. Экспрессия обоих трансгенов регулируется нейронально-специфическими элементами мышиного промотора Thy1. Линию 5XFAD (генетический фон B6/SJL) поддерживали путем скрещивания гемизиготных трансгенных мышей с производителями B6/SJL F1. Линию 5XFAD (генетический фон C57BL/6J) поддерживали путем скрещивания гемизиготных трансгенных мышей с производителями C57BL/6J F1. Все животные были генотипированы с помощью ПЦР с использованием геномной ДНК хвоста. Трансгенных мышей и мышей дикого типа разводили в собственном виварии, они имели доступ к пище и воде без ограничений и содержались при 12-часовом цикле свет-темнота при 22–24°C. В этом исследовании использовали самок мышей 5XFAD. Для каждого эксперимента возраст мышей указан в месяцах (с двухнедельным диапазоном).
Лекарства
Донекоприд (донекоприда фумарат, MR 31155) и флукоприд (флукоприда фумарат, MR 31193) были синтезированы в CERMN, Кан, Франция.
RS 67333 (1-(4-амино-5-хлор-2-метоксифенил)-3-(1-бутил-4-пиперидинил)-1-пропанон) был использован в качестве эталонного агониста рецептора 5-HT4 и был получен у Tocris Bioscience (R&D Systems Europe, Лилль, Франция).
Соединения ресуспендировали в ДМСО (37,5 мкг/мкл, хранили при -20°C) и перед введением готовили свежий раствор 1:250 в 0,9% NaCl. Раствор носителя (0,9% NaCl, 0,4% ДМСО) использовали в качестве контроля.
Лечение животных
Мыши 5XFAD получали хроническое лечение лекарствами или растворителем в соответствии с 2 различными протоколами. В каждом эксперименте лекарства или растворитель вводили интраперитонеально два раза в неделю (1 мг/кг). В «Протоколе 1» мышей (n = 6-7 мышей в группе) лечили соединениями в течение двух месяцев в возрасте с 2 до 4 месяцев. В «Протоколе 2» мышей (n = 7) лечили соединениями в течение трех месяцев в возрасте с 1 до 4 месяцев. В конце каждого протокола мышей анестезировали смесью 100 мг/кг кетамина и 10 мг/кг ксилазина в физиологическом растворе и транскардиально перфузировали ФБР. Мозг быстро изолировали на льду, удаляли обонятельные луковицы и мозжечок, и разделяли два полушария. Одно полушарие замораживали на сухом льду и хранили при -80°C для биохимического анализа, а другое полушарие фиксировали в 4% ПФА для иммуногистохимии (IHC).
Сбор цереброспинальной жидкости (ЦСЖ)
Мышей анестезировали и помещали на стереотаксический инструмент. Кожу шеи разрезали, подкожную клетчатку и мышцы отделяли с помощью микро-ретракторов (Fine Science Tools, Гейдельберг, Германия). Затем мышей укладывали так, чтобы голова составляла угол около 135° с телом (Liu и Duff (2008) J. Vis. Exp. 10: 960). Для прокалывания твердой мозговой оболочки большой цистерны использовали капиллярную трубку (боросиликатное стекло, B100-75-10, Sutter Instruments, Новато, Калифорния, США). ЦСЖ собирали за счет капиллярных сил и переносили в микропробирки на 0,5 мл, немедленно замораживали на сухом льду и хранили при -80°C до использования. После размораживания образцы нагревали при 60°C в течение 5 минут, как описано в международной заявке WO 2008/009868, и анализировали без дальнейших циклов замораживания-размораживания.
Приготовление экстракта головного мозга
Полушария головного мозга, лобную кору и гиппокамп мышей 5XFAD и контрольных мышей размораживали, взвешивали и гомогенизировали в 4 объемах трис-физиологического раствора (50 мМ трис-HCl pH = 7,4, 150 мМ NaCl) с коктейлем ингибиторов протеазы (Roche Applied Science, Мелан, Франция). Полученные гомогенаты центрифугировали при 54000 x g в течение 20 минут, надосадочную жидкость («растворимую фракцию») собирали и разделяли на аликвоты для хранения при -80°C. Осадок ресуспендировали кратковременной обработкой ультразвуком в 10 объемах 6 М гуанидин-HCl в 50 мМ трис-HCl, pH = 7,6 и снова центрифугировали при 26 500 x g в течение 20 минут. Надосадочную жидкость («нерастворимую фракцию») разделяли на аликвоты и хранили при -80°C (Morishima-Kawashima et al. (2000) Am. J. Pathol. 157: 2093-2099).
Количественное определение Aβ40 и Aβ42
Наборы ИФА от IBL International (Гамбург, Германия) для дозировки Aβ40 (набор для анализа на β-амилоид (1-40) человека, #27713) или Aβ42 (набор для анализа на β-амилоид (1-42) человека, #27719) использовали в соответствии с инструкциями производителя. Реакции регистрировали при 620 нм и 450 нм с использованием счетчика люминесценции Infinite 2000. Полученные значения были нормализованы к концентрации белка в каждом образце, измеренной бицинхониновым методом (Sigma-Aldrich).
Иммуногистохимия
Срезы толщиной 30 микрометров готовили с помощью вибратома (Microm HM 650V, Thermo Scientific, Сен-Эрблен, Франция) и хранили в криопротекторной среде при -20°C. Для маркировки амилоидных бляшек свободно плавающие срезы тканей лобной коры, гиппокампа и энторинальной коры (координаты от брегмы: лобная кора = 1,98 мм, гиппокамп = -1,94 мм, энторинальная кора = -3,08 мм) тщательно промывали в ФБР, а затем инкубировали в блокирующем растворе (фБР; 3% БСА; 0,1% Triton X-100) в течение 1 часа. Срезы окрашивали красителем Хехста (1:1000, Life Technologies, Saint Aubin, Франция) в течение 15 минут для обнаружения ядер клеток, а затем свежеприготовленным раствором тиофлавина T (# T3516-5G, Sigma-Aldrich) (конечная концентрация: 0,01 мг/мл в блокирующем буфере) в течение 15 минут. После промывания в 70% этаноле в течение 5 минут образцы помещали на полилизиновые предметные стекла с покровными стеклами. Для окрашивания GFAP (глиальный фибриллярный кислый белок) свободно плавающие срезы мозга блокировали, как и раньше, и инкубировали с поликлональными кроличьими анти-GFAP (1:1000, Z0334, Dako, Лез-Юлис, Франция) при 4°C в течение ночи. После окрашивания тиофлавином Т и промывания 70% этанолом на 2 часа добавляли вторичные козьи антитела к IgG кролика с Alexafluor 594 (1: 1000, A11012, Life Technologies). Промывание ФБР и установку выполняли, как описано ранее.
Получение и анализ изображений
Изображения получали с помощью микроскопа AxioImager Z1 (Carl Zeiss S.A.S., Марли-ле-Руа, Франция). Анализ окрашивания тиофлавином Т проводили вслепую, и данные представлены как среднее количество частиц на мм2 в двух-трех срезах ткани из одной и той же области мозга/животного (программное обеспечение Image J). Экспрессия GFAP была количественно определена с использованием программного обеспечения Image J, и результаты выражены в долях площади.
Тест распознавания нового объекта
Когнитивные способности мышей проверяли с помощью теста на распознавание новых объектов (NOR) (Bevins и Besheer (2006) Nat. Protoc. 1: 1306-1311). Во время медикаментозного лечения до начала теста животных интенсивно обрабатывали. Каждый день мышам позволяли знакомиться с испытательной комнатой в течение как минимум 1 часа перед испытанием. Испытание проводили в боксе из оргстекла (ширина 35 см, длина 20 см, высота 20 см), помещенном в тускло освещенную комнату. В день 1 и 2 каждую мышь приучали к пустому боксу в течение 10 минут в сутки. На 3-й день два объекта (сконструированные из пластиковых игрушек) были помещены в клетку на расстоянии 5 см от противоположных стен. Во время тренировки каждое животное помещали между двумя объектами лицом к стене, а затем им позволяли исследовать объекты в течение 5 минут. Затем мышей возвращали в их домашнюю клетку и через 24 часа проходили 5-минутную экспериментальную сессию, в которой один из двух (знакомых) объектов был заменен новым (незнакомым). Весь эксперимент регистрировали на видео, и исследование объекта [время контактирования носа мыши с объектом или время обнюхивания объекта на расстоянии ≤ 1 см] измеряли вслепую. Учитывали два параметра: (1) время исследования (в секундах), затраченное животным при взаимодействии с двумя знакомыми объектами во время сеанса обучения; и (2) время исследования, потраченное животным при взаимодействии с новым объектом, по отношению к общему времени исследования ([новый/(знакомый + новый)] × 100) во время испытания. Также был рассчитан индекс различения ([новый - знакомый]/[знакомый + новый]).
Статистический анализ
Все значения выражены как среднее значение ± SEM. Значимые эффекты лечения определяли с помощью анализа ANOVA с последующим апостериорным тестом Тьюки в случае нескольких групп сравнения. Во всех остальных случаях использовали непарный t-критерий Стьюдента. Для всех статистических тестов достоверными считали значения с р<0,05. Анализ выполняли с помощью GraphPad Prism 7 (GraphPad Software, Ла-Хойя, Калифорния).
Результаты
Влияние донекоприда и флукоприда на амилоидную нагрузку в мозге 5XFAD
В этом исследовании использовали 27 самок мышей 5XFAD (генетический фон B6/SJL). Мышам вводили растворитель или соединения (1 мг/кг) интраперитонеально дважды в неделю в течение двух месяцев в возрасте с 2 до 4 месяцев.
Aβ40 и Aβ42 определяли количественно в экстракте головного мозга в растворимой и нерастворимой фракциях. Результаты выражены в % от растворителя.
Полученные результаты представлены в Таблице 6 ниже:
Таблица 6. Уровни Aβ40 и Aβ42 (% от контроля)
*: p <0,05 по сравнению с растворителем, определено односторонним дисперсионным анализом с последующим тестом Тьюки.
Значения групп растворителя, представленные в таблице 6, были следующими:
растворимая фракция:
- Aβ40 = 9,48 ± 1,08 мкг/мг белка,
- Aβ42 = 0,24 ± 0,03 мкг/мг белка;
нерастворимая фракция:
- Aβ40 = 0,68 ± 0,10 мкг/мг белка,
- Aβ42 = 10,84 ± 0,73 мкг/мг белка.
Антиамилоидный эффект. В растворимой фракции мозга мышей 5XFAD хроническое введение донекоприда и флукоприда (1 мг/кг два раза в неделю) позволило значительно снизить уровни Aβ42 (*p <0,05 по сравнению с растворителем для донекоприда и ****p <0,0001 по сравнению с растворителем для флукоприда, при определении односторонним дисперсионным анализом с последующим тестом Тьюки) без каких-либо изменений уровней Aβ40. Аналогичным образом, в нерастворимой фракции донекоприд снижал нагрузку Aβ42 в мозге мышей (*p <0,05 по сравнению с растворителем, при определении с помощью однофакторного дисперсионного анализа с последующим тестом Тьюки). Оба препарата не повлияли на уровни нерастворимого Aβ40.
Влияние донекоприда и флукоприда на память, амилоидную нагрузку в цереброспинальной жидкости, амилоидные бляшки и астроглиоз у мышей 5XFAD.
В этом исследовании использовали 28 самок мышей 5XFAD (генетический фон C57BL/6J). Мышам вводили растворитель или соединения (1 мг/кг) интраперитонеально дважды в неделю в течение трех месяцев в возрасте с 1 до 4 месяцев.
В конце лечения у мышей оценивали долговременную память в испытании распознавания нового объекта (NOR). Межсессионный интервал составлял 24 часа. Поскольку ни одна мышь не смогла исследовать любой из двух объектов или исследовать оба объекта менее чем за 10 секунд во время сеанса обучения, все мыши были включены в анализ данных.
Антиамнестический эффект. Самкам 5XFAD и однопометникам дикого типа в возрасте одного месяца вводили RS 67333, растворитель, донекоприд или флукоприд (1 мг/ кг, два раза в неделю в течение 3 месяцев), а затем оценивали опознающую память с помощью теста распознавания нового объекта (NOR). Сеансы приучения начинали через 3 дня после окончания лечения, чтобы избежать влияния острых эффектов. Интервал между тренировкой и испытанием составлял 24 часа. У мышей, которым вводили растворитель, наблюдались нарушения памяти, и они не могли отличить новый объект от знакомого, о чем свидетельствуют сходное время исследования и незначительный индекс различения. Лечение RS 67333 устраняло когнитивные нарушения, на что указывает более высокий процент времени, посвященный исследованию нового объекта по сравнению с мышами, получавшими растворитель (** p <0,01 по сравнению с знакомым объектом, при определении двухфакторным дисперсионным анализом ANOVA с последующим тестом Сидака). Аналогичным образом, хроническое применение донекоприда и флукоприда позволяло предотвратить изменение памяти (**p<0,01 по сравнению со знакомым объектом для донекоприда, *p<0,05 по сравнению со знакомым объектом для флукоприда, при определении с помощью двухфакторного дисперсионного анализа ANOVA с последующим тестом Сидака). Индекс различения в 3 группах, получавших соединение, отличался от нуля, т.е. случайного уровня (определяемого непарным t-критерием Стьюдента). Общее время исследования во время сеанса обучения существенно не различалось в четырех группах, что указывает на то, что улучшение опознающей памяти у мышей 5XFAD, получавших донекоприд или флукоприд, не является следствием увеличения исследовательской активности.
Амилоидная нагрузка в цереброспинальной жидкости. Концентрация Aβ42 в цереброспинальной жидкости (ЦСЖ) мышей 5XFAD, получавших RS 67333, донекоприд или флукоприд (1 мг/кг, два раза в неделю в течение 3 месяцев), существенно не отличалась по сравнению с контролем, что указывает на то, что хроническая стимуляция рецептора 5-HT4 не влияла на уровни амилоида в цереброспинальной жидкости (при определении односторонним дисперсионным анализом ANOVA с последующим тестом Тьюки).
Антиамилоидный эффект. Фронтальная кора, гиппокамп и энторинальная кора были проанализированы как репрезентативные области мозга мышей 5XFAD (фронтальный, средний и каудальный срезы) и областей мозга человека, которые на ранней стадии поражены болезнью Альцгеймера и сильно обогащены амилоидными отложениями.
Лечение донекопридом (1 мг/кг, два раза в неделю в течение 3 месяцев) значительно уменьшило количество амилоидных бляшек по сравнению с растворителем во фронтальной коре головного мозга (уменьшение на 16 ± 3%, **p <0,01 по сравнению с растворителем) и в энторинальной коре головного мозга (снижение на 24 ± 4%, **p <0,01 по сравнению с растворителем). После лечения флукопридом (1 мг/кг, дважды в неделю в течение 3 месяцев) в энторинальной коре также наблюдалось значительное снижение количества бляшек с уменьшением на 28 ± 6% по сравнению с контролем (**p <0,01 по сравнению с растворителем). После лечения RS 67333 (1 мг/кг дважды в неделю в течение 3 месяцев) значительного эффекта не наблюдалось ни в лобной, ни в энторинальной коре. Число амилоидных бляшек в гиппокампе мышей 5XFAD, получавших RS 67333, донекоприд или флукоприд (1 мг/кг, два раза в неделю в течение 3 месяцев), существенно не отличалось от контроля. Статистические данные определяли с помощью однофакторного дисперсионного анализа с последующим тестом Тьюки.
Противовоспалительный эффект. Иммуногистохимическая оценка астроглии (с антителом против GFAP, глиального фибриллярного кислого белка) показала, что длительное лечение донекопридом и RS 67333 (1 мг/кг, два раза в неделю в течение 3 месяцев) значительно уменьшало астроглиоз (54 ± 10% и 62 ± 6% снижение окрашивания GFAP в срезах энторинальной коры головного мозга получавших лечение животных по сравнению с контрольными, получавшими растворитель, соответственно; *p <0,01 по сравнению с растворителем). В энторинальной коре лечение флукопридом (1 мг/кг, дважды в неделю в течение 3 месяцев) вызвало снижение окрашивания GFAP на 51 ± 5%, что было близко к значимости (p = 0,06). Однако окрашивание GFAP в лобной коре или гиппокампе мышей 5XFAD, получавших RS 67333, донекоприд или флукоприд (1 мг/кг, два раза в неделю в течение 3 месяцев), существенно не отличалось по сравнению с контролем. Статистические данные определяли с помощью однофакторного дисперсионного анализа с последующим тестом Тьюки.
Заключение
Исследование демонстрирует, что донекоприд и флукоприд проявляют антиамилоидные эффекты (снижение уровней Aβ42 и амилоидных бляшек) и противовоспалительные эффекты (уменьшение астроглиоза) при хроническом введении мышам 5XFAD, на трансгенной мышиной модели болезни Альцгеймера. Эти эффекты наиболее заметны в энторинальной и лобной коре. Более того, хроническое введение донекоприда и флукоприда может оказывать антиамнестический эффект и предотвращать появление дефицита памяти у мышей 5XFAD.
Все эти эффекты сравнимы, а иногда и более выражены, чем эффекты RS 67333 (агониста рецептора 5-HT4, выбранного в качестве препарата сравнения), что указывает на то, что донекоприд и флукоприд являются многообещающими препаратами против болезни Альцгеймера.
Пример 4
В этом примере раскрывается нейропротекторный эффект множества соединений общей формулы (I), определенной выше, в отношении нейронов гиппокампа, интоксицированных олигомерами амилоидных пептидов.
Материалы и методы
Первичная культура нейронов гиппокампа
Нейроны гиппокампа крыс культивировали, как описано Callizot et al. (2013) J. Neurosci. Res. 91: 706-716. Вкратце, беременных самок крыс со сроком беременности 17 дней (Rats Wistar; Janvier Labs France) умерщвляли с использованием глубокой анестезии в камере с CO2 и цервикальной дислокации. Затем эмбрионы извлекали из матки и немедленно помещали в ледяную среду L15 Лейбовица с 2% раствором пенициллина (10000 Ед/мл) и стрептомицина (10 мг/мл) (ПС) и 1% бычьим сывороточным альбумином (БСА). Гиппокамп обрабатывали в течение 20 мин при 37°C раствором трипсин-ЭДТА с конечной концентрацией 0,05% трипсина и 0,02% ЭДТА. Диссоциацию останавливали добавлением модифицированной по Дульбекко среды Игла (DMEM) с 4,5 г/л глюкозы, содержащей ДНКазу I сорта II (конечная концентрация 0,5 мг/мл) и 10% эмбриональной телячьей сыворотки (ЭТС). Клетки механически диссоциировали тремя принудительными пропусканиями через кончик 10 мл пипетки.
Затем клетки центрифугировали при 515 x g в течение 10 минут при 4°C. Надосадочную жидкость удаляли, а осадок ресуспендировали в определенной культуральной среде, состоящей из нейробазальной среды с 2% раствором добавки B27, 2 ммоль/л L-глутамина, 2% раствором ПС и 10 нг/мл нейротрофического фактора головного мозга (BDNF). Жизнеспособные клетки подсчитывали на цитометре Нойбауэра с использованием теста исключения трипанового синего. Клетки высевали с плотностью 20000 на лунку в 96-луночные планшеты, предварительно покрытые поли-L-лизином, и культивировали при 37°C в инкубаторе в среде воздух (95%) - CO2 (5%).
Питательную среду меняли каждые 2 дня. Для 96-луночных планшетов использовали только 60 лунок. Лунки первой и последней линий и рядов не использовали (во избежание краевого эффекта) и заполняли стерильной водой.
Тестируемые соединения и воздействие Aβ1-42 человека
Тестируемые соединения: Были протестированы следующие соединения:
- MR31193
- MR31176
- MR31192, и
- MR33583
Предварительная инкубация: на 17 день культивирования соединения растворяли в культуральной среде и предварительно инкубировали с первичными нейронами гиппокампа в течение 1 часа перед воздействием Aβ1-42.
Повреждение: нейроны гиппокампа подвергали воздействию растворов Aβ (см. ниже) через 17 дней культивирования. Получение Aβ1-42 было выполнено в соответствии с процедурой, описанной Callizot et al. (2013) J. Neurosci. Res. 91: 706-716. Партия Aβ1-42, использованная в этом исследовании, содержит 10% олигомеров (количественная оценка по WB, проведенная в Combes et al. (2015) J. Neurosci. Res. 93: 633-643). Вкратце, пептид Aβ1-42 растворяли в указанной выше культуральной среде при начальной концентрации 40 мкмоль/л. Этот раствор осторожно перемешивали в течение 3 дней при 37°C в темноте и сразу же использовали после того, как он был должным образом разбавлен в культуральной среде до используемых концентраций (20 мкмоль/л или 2,5 мкмоль/л, что соответствует 2 мкмоль/л или 0,25 мкмоль/л олигомеров (AβO), соответственно). Препарат Aβ1-42 добавляли до конечной концентрации 20 или 2,5 мкмоль/л (2 мкмоль/л или 0,25 мкмоль/л AβO, соответственно, при определении автоматическим WB), при разведении в контрольной среде в присутствии соединений.
Конечная оценка
Через 24 часа после интоксикации надосадочную жидкость отбирали и немедленно замораживали для будущего анализа. Её хранили при -20°C до 6 месяцев. Затем нейроны гиппокампа фиксировали холодным раствором этанола (95%) и уксусной кислоты (5%) в течение 5 мин при -20°C. Их снова дважды промывали ФБР, затем повышали проницаемость и блокировали неспецифические участки с использованием раствора ФБР, содержащего 0,1% сапонина и 1% ЭТС, в течение 15 мин при комнатной температуре.
Иммуноокрашивание: MAP-2/AT100 - После фиксации и пермеабилизации клетки инкубировали в течение 2 часов с:
- куриным поликлональным антителом к белку 2, ассоциированному с микротрубочками (MAP-2), при разведении 1/1000 в ФБР, содержащем 1% эмбриональной телячьей сыворотки и 0,1% сапонина (это антитело специфически окрашивает тела клеток и нейриты, что позволяет изучать гибель нейрональных клеток и нейритную сеть); и
- мышиным моноклональным антителом против фосфо-тау-белка (AT100) при разведении 1/100 в ФБР, содержащем 1% эмбриональной телячьей сыворотки и 0,1% сапонина.
Эти антитела выявляли с помощью козьих антител к IgG мыши с Alexa Fluor 488 и козьих антител к куриным IgG с Alexa Fluor 568 в разведении 1/400 в ФБР, содержащем 1% ЭТС, 0,1% сапонина, в течение 1 часа при комнатной температуре.
Для каждого условия с помощью ImageXpress® (Molecular Devices) с 20-кратным увеличением было сделано 30 снимков (представляющих всю площадь лунки) на лунку. Все изображения были созданы с использованием одних и тех же параметров сбора данных. Анализ изображений проводили напрямую и автоматически с помощью Custom Module Editor® (Molecular Devices). Были исследованы следующие показания:
- Общее количество нейронов (выживание нейронов, количество МАР-2-положительных нейронов)
- Нейритная сеть (в мкм MAP-2-положительных нейронов)
- Площадь AT100 (мкм², перекрывание с МАР-2-положительными нейронами).
Иммуноокрашивание: PSD95 и SYN - после фиксации и пермеабилизации клетки инкубировали в течение 2 часов с:
- мышиным моноклональным антителом против постсинаптического уплотнения 95 кДа (PSD95) при разведении 1/200 в ФБР, содержащем 1% эмбриональной телячьей сыворотки и 0,1% сапонина;
- кроличьими поликлональными антителами против синаптофизина (SYN) в разведении 1/100 в ФБР, содержащем 1% эмбриональной телячьей сыворотки и 0,1% сапонина.
Эти антитела выявляли с помощью козьих антител против IgG мыши с Alexa Fluor 488 и козьих антител против кроличьих IgG с Alexa Fluor 568 в разведении 1/400 в ФБР, содержащем 1% ЭТС, 0,1% сапонина, в течение 1 часа при комнатной температуре.
Для каждого условия было сделано 40 снимков на лунку с помощью ImageXpress (Molecular Devices) с 40-кратным увеличением. Все изображения были сделаны в одинаковых условиях. Оценку синапсов выполняли автоматически с помощью Custom module editor® (Molecular Devices). Были оценены следующие конечные точки:
- Количество синапсов (перекрывание между PSD95/SYN)
Статистический анализ
Все значения выражены как среднее значение ± SEM (стандартная ошибка среднего). Статистический анализ был выполнен с помощью однофакторного дисперсионного анализа с последующим тестом Даннета или наименьшей значимой разницы по Фишеру. p <0,05 считали значимым.
Результаты
Были получены следующие результаты.
- MR31193 продемонстрировал сильный нейропротекторный эффект (выживание, нейритная и синаптическая сеть). Как видно на фиг. 1 и 2, этот эффект соответствует колоколообразной кривой с максимальным эффектом, наблюдаемым при концентрации 100 нМ для выживания и нейритной сети (фиг. 1A-C). Максимальное влияние на сеть синапсов было получено при более низких концентрациях (от 5 до 100 нМ, фиг. 2). Также наблюдался значительный эффект в отношении гиперфосфорилирования тау-белка с максимальным эффектом при 50 нМ (см. фиг. 1D).
- Как видно на фиг. 3 и 4, MR31176 демонстрирует нейропротекторный эффект при высоких концентрациях (выживание, нейритная и синаптическая сеть). Значительное влияние на гиперфосфорилирование тау-белка также наблюдалось при высоких концентрациях (см. фиг. 3D).
- Как видно на фиг. 5 и 6, MR31192 демонстрирует очень сильный нейропротекторный эффект при очень низких дозах (например, 1-5 нМ для выживания, см. фиг. 5A). Очень сильный эффект также наблюдался в отношении нейритной сети при каждой испытанной дозе (см. фиг. 5B). Это соединение даже показало нейротрофический эффект (оно вызывало рост нейритов), как это видно на фиг. 5C. Количество синапсов также увеличивалось при низких дозах (1-5 нМ, см. фиг. 6). Также наблюдался важный эффект в отношении гиперфосфорилирования тау-белка (см. фиг. 5D).
- Нейропротекторный эффект, соответствующий колоколообразной кривой, также наблюдался у MR33583 (см. фиг. 7 и 8). Активные концентрации были выше 50-500 нМ. Сильный эффект на нейритную сеть наблюдался почти при каждой испытанной дозе (см. фиг. 7B). Колоколообразный эффект также наблюдался в отношении гиперфосфорилирования тау-белка с максимальным эффектом при 50-100 нМ (см. фиг. 7D).
Эти результаты подтверждают, что все испытанные соединения общей формулы (I) являются нейропротекторными агентами, причем соединения MR31193, MR31192 и MR33583 особенно эффективны.
Пример 5
В этом примере показаны нейропротекторные эффекты соединений MR31192 и MR36014.
Материалы и методы
Первичная культура нейронов гиппокампа
Нейроны гиппокампа крыс культивировали, как описано Callizot et al. (2013) J. Neurosci. Res. 91: 706-716. Вкратце, беременных самок крыс со сроком беременности 17 дней (Rats Wistar; Janvier Labs France) умерщвляли с использованием глубокой анестезии в камере с CO2 и цервикальной дислокации. Затем эмбрионы извлекали из матки и немедленно помещали в ледяную среду L15 Лейбовица с 2% раствором пенициллина (10000 Ед/мл) и стрептомицина (10 мг/мл) (ПС) и 1% бычьим сывороточным альбумином (БСА). Гиппокамп обрабатывали в течение 20 мин при 37°C раствором трипсин-ЭДТА с конечной концентрацией 0,05% трипсина и 0,02% ЭДТА. Диссоциацию останавливали добавлением модифицированной по Дульбекко среды Игла (DMEM) с 4,5 г/л глюкозы, содержащей ДНКазу I сорта II (конечная концентрация 0,5 мг/мл) и 10% эмбриональной телячьей сыворотки (ЭТС). Клетки механически диссоциировали тремя принудительными пропусканиями через кончик пипетки на 10 мл.
Затем клетки центрифугировали при 515 x g в течение 10 минут при 4°C. Надосадочную жидкость удаляли, а осадок ресуспендировали в определенной культуральной среде, состоящей из нейробазальной среды с 2% раствором добавки B27, 2 ммоль/л L-глутамина, 2% раствором ПС и 10 нг/мл нейротрофического фактора головного мозга (BDNF). Жизнеспособные клетки подсчитывали на цитометре Нойбауэра с использованием теста исключения трипанового синего. Клетки высевали с плотностью 20000 на лунку в 96-луночные планшеты, предварительно покрытые поли-L-лизином, и культивировали при 37°C в инкубаторе в среде воздух (95%) - CO2 (5%).
Питательную среду меняли каждые 2 дня. Для 96-луночных планшетов использовали только 60 лунок. Лунки первой и последней линий и рядов не использовали (во избежание краевого эффекта), и их заполняли стерильной водой.
Тестируемые соединения и воздействие Aβ1-42 человека
Тестируемые соединения: Были протестированы следующие соединения:
- MR31192,
- MR36014, и
- Донепезил.
Предварительная инкубация: на 17 день культивирования соединения растворяли в питательной среде и предварительно инкубировали с первичными нейронами гиппокампа в течение 1 часа перед воздействием Aβ1-42.
Повреждение: Нейроны гиппокампа подвергали воздействию растворов Aβ (см. ниже) через 17 дней культивирования. Aβ1-42 получали в соответствии с процедурой, описанной Callizot et al. (2013) J. Neurosci. Res. 91: 706-716. Партия Aβ1-42, использованная в этом исследовании, содержит 10% олигомеров (количественная оценка WB, проведенная в Combes et al. (2015) J. Neurosci. Res. 93: 633-643). Вкратце, пептид Aβ1-42 растворяли в указанной выше культуральной среде при начальной концентрации 40 мкмоль/л. Этот раствор осторожно перемешивали в течение 3 дней при 37°C в темноте и сразу же использовали после того, как он был должным образом разбавлен в культуральной среде до используемых концентраций (20 мкмоль/л или 2,5 мкмоль/л, что соответствует 2 мкмоль/л или 0,25 мкмоль/л олигомеров (AβO), соответственно). Препарат Aβ1-42 добавляли до конечной концентрации 20 или 2,5 мкмоль/л, (соответственно 2 мкмоль/л или 0,25 мкмоль/л AβO, при определении автоматическим WB), при разведении в контрольной среде в присутствии соединений.
Конечная оценка
Через 24 часа после интоксикации надосадочную жидкость отбирали и немедленно замораживали для будущего анализа. Её хранили при -20°C до 6 месяцев. Затем нейроны гиппокампа фиксировали холодным раствором этанола (95%) и уксусной кислоты (5%) в течение 5 мин при -20°C. Их снова дважды промывали ФБР, затем повышали проницаемость и блокировали неспецифическое связывание с использованием раствора ФБР, содержащего 0,1% сапонина и 1% ЭТС, в течение 15 мин при комнатной температуре.
Иммуноокрашивание: MAP-2 / AT100 - После фиксации и пермеабилизации клетки инкубировали в течение 2 часов с:
- куриным поликлональным антителом к белку 2, ассоциированному с микротрубочками (MAP-2), при разведении 1/1000 в ФБР, содержащем 1% эмбриональной телячьей сыворотки и 0,1% сапонина (это антитело специфически окрашивает тела клеток и нейриты, что позволяет изучать гибель нейрональных клеток и нейритную сеть); и
- мышиным моноклональным антителом против фосфо-тау-белка (AT100) при разведении 1/100 в ФБР, содержащем 1% эмбриональной телячьей сыворотки и 0,1% сапонина.
Эти антитела были выявлены с помощью козьих антител к IgG мыши с Alexa Fluor 488 и козьих антител к куриным IgG с Alexa Fluor 568 в разведении 1/400 в ФБР, содержащем 1% ЭТС, 0,1% сапонина, в течение 1 часа при комнатной температуре.
Для каждого условия с помощью ImageXpress® (Molecular Devices) с 20-кратным увеличением было сделано 30 снимков (представляющих всю площадь лунки) на лунку. Все изображения были созданы с использованием одних и тех же параметров сбора данных. Анализ изображений проводили напрямую и автоматически с помощью Custom Module Editor® (Molecular Devices). Были исследованы следующие показания:
- Общее количество нейронов (выживание нейронов, количество МАР-2-положительных нейронов);
- Нейритная сеть (в мкм MAP-2-положительных нейронов);
- Площадь AT100 (мкм², перекрывание с МАР-2-положительными нейронами).
Иммуноокрашивание: PSD95 и SYN - после фиксации и пермеабилизации клетки инкубировали в течение 2 часов с:
- мышиным моноклональным антителом против постсинаптического уплотнения 95 кДа (PSD95) при разведении 1/200 в ФБР, содержащем 1% эмбриональной телячьей сыворотки и 0,1% сапонина;
- кроличьими поликлональными антителами против синаптофизина (SYN) в разведении 1/100 в ФБР, содержащем 1% эмбриональной телячьей сыворотки и 0,1% сапонина.
Эти антитела были выявлены с помощью козьих антител против IgG мыши с Alexa Fluor 488 и козьих антител против кроличьих IgG с Alexa Fluor 568 в разведении 1/400 в ФБР, содержащем 1% ЭТС, 0,1% сапонина, в течение 1 часа при комнатной температуре.
Для каждого условия было сделано 40 снимков на лунку с помощью ImageXpress (Molecular Devices) с 40-кратным увеличением. Все изображения были сделаны в одинаковых условиях. Оценку синапсов выполняли автоматически с помощью Custom module editor® (Molecular Devices). Оценивали следующие конечные точки:
- Количество синапсов (перекрывание между PSD95/SYN)
Статистический анализ
Все значения выражены как среднее значение ± SEM (стандартная ошибка среднего). Статистический анализ был выполнен с помощью однофакторного дисперсионного анализа с последующим тестом Даннета или наименьшей значимой разницы по Фишеру. p <0,05 считали значимым.
Результаты
Как видно на фиг. 9 и 10, соединение MR31192 является активным до концентрации 500 пМ (выживание и нейритная сеть, см. фиг. 9A, B, C), а также наблюдалось значительное снижение гиперфосфорилирования тау-белка (см. фиг. 9D) при 100 пМ. Нейропротекторное влияние на синапсы далее наблюдалось при 500 пМ (см. фиг. 10). Соответственно, соединение MR31192 эффективно при очень низких дозах, в частности, при дозах ниже 100 нМ (от 100 до 500 пМ). В этих очень низких дозах MR31192 проявляет нейропротекторный эффект, аналогичный тому, который наблюдается у донепезила, но при 1 мкМ.
Как видно на фиг. 11 и 12, соединение MR36014 (фтористый аналог MR31192) демонстрирует нейропротекторный профиль, аналогичный тому, который получен с MR31192 при дозах в области нМ. Нейропротекторный эффект также наблюдался при дозах в области пМ. Этот эффект особенно важен для выживания и нейритной сети (см. фиг. 11A, B, C) и для гиперфосфорилирования тау-белка (см. фиг. 11D). Влияние на синапсы является значимым до концентрации 50 пМ (см. фиг. 12). В этих очень низких дозах MR36014 проявляет нейропротекторный эффект, почти эквивалентный тому, который наблюдается у донепезила, но при 1 мкМ.
Соответственно, оба соединения активны в очень низких дозах (как в дозах в области нМ, так и в дозах в области пМ), что особенно интересно для предотвращения потенциальной токсичности и/или побочных эффектов.
Пример 6
В этом примере показаны нейропротекторные эффекты MR36014 и MR33583 в отношении дофаминергических TH-положительных нейронов, поврежденных MPP+.
Болезнь Паркинсона (БП) - распространенное нейродегенеративное двигательное расстройство, которым страдает около 1% населения старше 70 лет. Это второе по распространенности нейродегенеративное заболевание после болезни Альцгеймера. У пациентов, страдающих БП, в 70% случаев в качестве первого симптома проявляются симптомы двигательной нестабильности с тремором покоя. Другие клинические симптомы включают ригидность, брадикинезию и постуральную нестабильность, и часто включают когнитивные нарушения, депрессию и расстройства сна.
Хотя патологические механизмы, приводящие к дегенерации черной субстанции (SN), могут быть множественными, несколько путей считаются центральными: агрегация белков, связанная с протеасомными нарушениями, дисфункция митохондрий (также вызванная специфическими DA-токсинами) и нарушение высвобождения дофамина (DA).
Как подтверждают Dauer и Przedborski (2003) Neuron 39: 889-909, токсины, специфичные для дофаминергических нейронов (DA-токсины), в частности 1-метил-4-фенил-1,2,3,6-тетрагидропиридин (MPTP, пролекарство нейротоксина MPP+), 6-гидроксидофамин (6-OHDA) или ротенон, вызывают паркинсонизм у людей. Эти токсины в основном используются учеными для экспериментального моделирования патологии у животных. Любые вещества, снижающие нейротоксичность DA-токсинов, могут быть полезны в качестве нового терапевтического агента для лечения или профилактики БП.
Целью настоящего исследования было изучение нейропротекторного влияния MR36014 и MR33583 (в 4 концентрациях) на дофаминергические TH-положительные нейроны, поврежденные MPP+. Тестируемые соединения предварительно инкубировали в течение 1 часа перед применением MPP+ (4 мкМ в течение 48 часов). Исследовали нейропротекторный эффект (выживание и нейритная сеть) тестируемых соединений. Кроме того, было изучено влияние на накопление альфа-синуклеина в цитоплазме.
Материалы и методы
Первичная культура мезэнцефальных нейронов
Все эксперименты проводили в соответствии с Руководством по уходу и использованию лабораторных животных Национального института здравоохранения и действующими правилами Европейского Союза (Директива 2010/63/EU). Номер договора: A1301310.
Дофаминергические нейроны крыс культивировали, как описано Callizot et al. (2019) PLoS One 14: e0215277 и Visanji et al. (2008) FASEB J. 22: 2488-2497. Вкратце, беременных крыс-самок (Wistar) с 15-дневным сроком беременности умерщвляли с использованием глубокой анестезии в камере с CO2 и цервикальной дислокации. Средний мозг, полученный от 15-дневных эмбрионов крыс (Janvier, Франция), препарировали под микроскопом. Эмбриональный средний мозг удаляли и помещали в ледяную среду Лейбовица (L15), содержащую 2% пенициллин-стрептомицин (ПС) и 1% бычий сывороточный альбумин (БСА). Вентральная часть мезэнцефалического изгиба, область развивающегося мозга, богатая дофаминергическими нейронами, была использована для клеточных препаратов.
Средний мозг диссоциировали трипсинизацией в течение 20 минут при 37°C (раствор с конечной концентрацией 0,05% трипсина и 0,02% ЭДТА). Реакцию останавливали добавлением модифицированной по Дульбекко среды Игла (DMEM), содержащей ДНКазу I сорта II (0,5 мг/мл) и 10% эмбриональную телячью сыворотку (ЭТС). Затем клетки механически диссоциировали путем 3 пропусканий через пипетку на 10 мл. Затем клетки центрифугировали при 180 x g в течение 10 мин при 4°C на слое БСА (3,5%) в среде L15. Надосадочную жидкость удаляли, а осадок клеток ресуспендировали в определенной культуральной среде, состоящей из нейробазальной среды с добавлением B27 (2%), L-глутамина (2 мМ), 2% раствора ПС, 10 нг/мл нейротрофического фактора головного мозга (BDNF) и 1 нг/мл глиального нейротрофического фактора (GDNF). Жизнеспособные клетки подсчитывали на цитометре Нейбауэра с использованием теста исключения трипанового синего. Клетки высевали при плотности 40 000 клеток/лунку в 96-луночные планшеты (предварительно покрытые поли-L-лизином) и выдерживали в увлажненном инкубаторе при 37°C в атмосфере 5% CO2/95% воздуха. Половину среды меняли каждые 2 дня на свежую среду.
Вкратце, на 6 день культивирования среду удаляли и добавляли свежую среду с тестируемыми соединениями или без них, и с MPP+ или без него на 48 часов, при разведении в контрольной среде. Было оценено 6 лунок на одно условие.
Испытуемые соединения и воздействие MPP+
Предварительная инкубация: на 6 день культивирования соединения разбавляли в культуральной среде и предварительно инкубировали с первичными дофаминергическими нейронами в течение 1 часа перед воздействием MPP+.
Повреждение: через 1 час после инкубации испытуемых соединений добавляли MPP+ до конечной концентрации 4 мкМ, при разбавлении в культуральной среде в присутствии соединений в течение 48 часов.
Организация планшетов с культурой
Испытуемые соединения тестировали в 96-луночном планшете (n = 6 лунок для культивирования на условие). MR36014 и MR33583 предварительно инкубировали в течение 1 часа перед интоксикацией MPP +. Оценивали следующие условия:
Конечная оценка
После 48 часов интоксикации надосадочную жидкость клеточных культур отделяли и немедленно замораживали для дальнейшего анализа. Клетки фиксировали раствором 4% параформальдегида в ФБР, pH = 7,3, в течение 20 минут при комнатной температуре. Клетки дважды промывали ФБР. Проницаемость клеточной мембраны и блокирование неспецифического связывания проводили с использованием раствора ФБР, содержащего 0,1% сапонина и 1% ЭТС, в течение 15 минут при комнатной температуре.
Иммуноокрашивание: TH и α-syn. Культуры инкубировали с:
- моноклональными антителами против тирозингидроксилазы (TH), полученными у мышей, при разведении 1/10 000 в ФБР, содержащем 1% ЭТС, 0,1% сапонина, в течение 2 часов при комнатной температуре.
- поликлональными антителами против альфа-синуклеина (α-syn), полученными у кроликов при разведении 1/200 в ФБР, содержащем 1% ЭТС, 0,1% сапонина, в течение 2 часов при комнатной температуре.
Эти антитела были выявлены с помощью козьих антител против IgG мыши с Alexa Fluor 488 в разведении 1/800 и с помощью козьих антител против кроличьих IgG с Alexa Fluor 568 в разведении 1/400 в ФБР, содержащем 1% ЭТС, 0,1% сапонина, в течение 1 часа при комнатной температуре.
Автоматический компьютерный анализ - для каждого условия автоматически делали 20 снимков (представляющих всю область лунки) с помощью ImageXpress® (Molecular Devices) при 10-кратном увеличении (20 снимков) с использованием тех же параметров сбора данных. Анализ изображений проводили напрямую и автоматически с помощью Custom Module Editor® (Molecular Devices). Оценивали следующие показания:
- Анализ общего количества TH-нейронов (TH-положительные нейроны),
- общая нейритная сеть TH-положительных нейронов (в мкм)
- агрегация α-syn (площадь перекрывания между окрашиванием TH и α-syn, в мкм²).
Статистический анализ
Все значения выражены как среднее значение ± SEM (стандартная ошибка среднего). Статистический анализ был выполнен с помощью однофакторного дисперсионного анализа с последующим тестом Даннета или наименьшей значимой разницы по Фишеру. p <0,05 считали значимым.
Результаты
Влияние MR36014 на выживание, нейритную сеть и агрегацию α-syn TH-нейронов после повреждения MPP+.
Применение MPP+ вызвало явную нейротоксичность для дофаминергических нейронов, о чем свидетельствует снижение выживания (фиг. 13A), потеря нейритной сети (фиг. 13B) и патологическое накопление α-syn (фиг. 13C), как показано ранее (Callizot et al. др. (2019) PLoS One 14: e0215277).
Применение MR36014 оказало сильное общее нейропротекторное влияние на TH-нейроны, поскольку это соединение улучшило выживание TH-нейронов (с 1 нМ до 10 нМ), защитило их нейритную сеть (все исследованные дозы) и уменьшило накопление α- syn (все исследованные дозы).
Влияние MR33583 на выживание, нейритную сеть и агрегацию α-syn TH-нейронов после повреждения MPP+.
После повреждения MPP+ MR33583 улучшил выживание нейронов (100 нМ, см. фиг. 14A) и частично защитил нейритную сеть (от 10 нМ до 100 нМ, см. фиг. 14B). Положительный эффект MR33583 также наблюдался в отношении агрегации α-syn (50 нМ и 100 нМ, см. фиг. 14C).
Обсуждение
Целью исследования было изучение нейропротекторных эффектов MR36014 и MR33583 на модели болезни Паркинсона in vitro.
Это исследование показывает, что:
- оба соединения противодействуют токсичности MPP+ для дофаминергических нейронов;
- MR36014 более эффективен, чем MR33583, так как это соединение показало более высокую эффективность в повышении выживания TH-нейронов;
- MR36014 был активен при более низких дозах, чем MR33583.
В целом, это исследование показало, что MR36014 и MR33583 являются интересными кандидатами в качестве лекарств от болезни Паркинсона, и что MR36014 более активен, чем MR33583.
Пример 7
В этом примере раскрыта оценка нейропротекторных эффектов соединений MR36014 и MR33583 в отношении первичных корковых нейронов крыс после глутаматного повреждения.
Глутаматергическая система и, в частности, рецепторы NMDA (глутаматергические рецепторы) играют важную роль в процессах обучения и памяти. Синаптическая пластичность может регулироваться передачей сигналов рецептора NMDA. Однако чрезмерная активация рецепторов NMDA и глутаматергическая эксцитотоксичность являются обычным патологическим признаком нейродегенеративных заболеваний (Lewerenz и Maher (2015) Front Neurosci. 9: 469).
Эксайтотоксичность глутамата связана с острым неврологическим повреждением в результате ишемии и черепно-мозговой травмы, а также с хронической нейродегенерацией при болезни Альцгеймера (БА) и болезни Хантингтона (БХ). Эксайтотоксическая гибель нейронов из-за чрезмерной активации NMDAR способствует чрезмерному притоку кальция (Ca2+) в клетку. Это вызывает ряд реакций, приводящих к гибели клеток, включая повышенный окислительный стресс, неадекватную активацию протеаз, таких как кальпаин, нарушение регуляции Са2+-зависимых путей, повреждение митохондрий и апоптотический каскад.
С этой точки зрения, раннее фармакологическое лечение веществами, снижающими эксайтотоксичность глутамата, может представлять собой очень хороший вариант для улучшения выживания нейронов (и улучшения когнитивной функции), и может представлять интересную терапевтическую стратегию для пациентов с диагнозом болезни Альцгеймера (Wang и Reddy (2017) J. Alzheimers Dis. 57: 1041-1048).
В настоящем исследовании оценивали нейропротекторный эффект MR36014 и MR33583 (в 4 концентрациях) в отношении первичных корковых нейронов крыс после глутаматного повреждения. Перед применением глутамата испытуемые соединения предварительно инкубировали в течение 1 часа (20 мкМ в течение 20 минут). Исследовали нейропротекторный эффект (выживание и нейритная сеть) испытуемых соединений.
Материалы и методы
Первичная культура кортикальных нейронов
Все эксперименты проводили в соответствии с Руководством по уходу и использованию лабораторных животных Национального института здравоохранения и действующими правилами Европейского Союза (Директива 2010/63/EU). Номер договора: A1301310.
Кортикальные нейроны крыс культивировали, как описано Callizot et al. (2013) J. Neurosci. Res. 91: 706-716. Беременных крыс-самок (Wistar) с 15-дневным сроком беременности умерщвляли с использованием глубокой анестезии в камере с CO2 и цервикальной дислокации. Вкратце, эмбрионы собирали и немедленно помещали в ледяную среду L15 Лейбовица с 2% раствором пенициллина (10000 Ед/мл) и стрептомицина (10 мг/мл) (ПС) и 1% бычьим сывороточным альбумином (БСА). Кортекс обрабатывали в течение 20 мин при 37°C раствором трипсин-ЭДТА с конечной концентрацией 0,05% трипсина и 0,02% ЭДТА. Диссоциацию останавливали добавлением модифицированной по Дульбекко среды Игла (DMEM) с 4,5 г/л глюкозы, содержащей ДНКазу I сорта II (конечная концентрация 0,5 мг/мл) и 10% эмбриональной телячьей сыворотки (ЭТС). Клетки механически диссоциировали тремя принудительными пропусканиями через кончик пипетки на 10 мл. Затем клетки центрифугировали при 515 x g в течение 10 минут при 4°C. Надосадочную жидкость удаляли, а осадок ресуспендировали в определенной культуральной среде, состоящей из нейробазальной среды с 2% раствором добавки B27, 2 ммоль/л L-глутамина, 2% раствором ПС и 10 нг/мл нейротрофического фактора головного мозга (BDNF). Жизнеспособные клетки подсчитывали на цитометре Нойбауэра с использованием теста исключения трипанового синего.
Клетки высевали с плотностью 25000 на лунку в 96-луночные планшеты, предварительно покрытые поли-L-лизином, и культивировали при 37°C в инкубаторе воздух (95%) - CO2 (5%).
Чтобы избежать краевого эффекта, в исследовании не использовали первую и последнюю линии, а также первый и последний ряды планшета. Пустые лунки заполняли водой.
Среду меняли каждые 2 дня. Кортикальные нейроны повреждали растворами Aβ (см. ниже) после 11 дней культивирования.
Тестируемые соединения и воздействие глутамата
Предварительная инкубация: на 13 день культивирования соединения растворяли в культуральной среде и предварительно инкубировали с первичными кортикальными нейронами в течение 1 часа перед воздействием глутамата.
Повреждение: через 1 час после инкубации тестируемого соединения добавляли глутамат до конечной концентрации 20 мкМ, при разведении в контрольной среде, все еще в присутствии тестируемых соединений, в течение 20 минут. Через 20 минут глутамат вымывали и добавляли свежую культуральную среду с тестируемыми соединениями на дополнительные 48 часов.
Организация планшетов с культурой
Испытуемые соединения тестировали в 96-луночном планшете (n = 6 лунок для культивирования на условие). Оценивали следующие условия:
Конечная оценка
Иммуноокрашивание: MAP-2 (выживание и нейритная сеть) - через 48 часов после интоксикации надосадочную жидкость от клеточной культуры удаляли и немедленно замораживали для будущего анализа. Клетки фиксировали холодным раствором этанола (95%) и уксусной кислоты (5%) в течение 5 мин при -20°C. Клетки дважды промывали ФБР. Клетки пермеабилизировали, а неспецифическое связывание блокировали раствором ФБР, содержащим 0,1% сапонина и 1% ЭТС, в течение 15 минут при комнатной температуре.
Культуры инкубировали в течение 2 часов с мышиным моноклональным антителом против белка, ассоциированного с микротрубочками 2 (MAP-2), в разведении 1/400, в ФБР, содержащем 1% эмбриональной телячьей сыворотки и 0,1% сапонина. Это антитело выявляли с помощью козьего антитела к IgG мыши с Alexa Fluor 488 (разведение 1/400) в ФБР, содержащем 1% ЭТС, 0,1% сапонина, в течение 1 часа при комнатной температуре. MAP-2 представляет собой нейрональный маркер, присутствующий в телах клеток и нейритах нейронов, который позволяет изучать выживание нейронов и длину нейритной сети.
Автоматический компьютерный анализ - для каждого условия с помощью ImageXpress® (Molecular Devices) с 20-кратным увеличением будет выполнено 30 снимков (репрезентативных для всей площади лунки) на лунку. Все изображения будут созданы с использованием одних и тех же параметров получения. Анализ изображений будет выполняться автоматически и напрямую с помощью Custom Module Editor® (Molecular Devices).
Будут исследованы следующие показания:
- Общее выживание нейронов (MAP-2-положительные, число) (данные будут выражены в виде среднего числа нейронов из 30 изображений на лунку).
- Общая сеть нейритов (MAP-2 в мкм) (данные будут выражены в виде среднего значения нейронов из 30 изображений на лунку).
Статистический анализ
Все значения выражены как среднее значение ± SEM (стандартная ошибка среднего). Статистический анализ был выполнен с помощью однофакторного дисперсионного анализа с последующим тестом Даннета или наименьшей значимой разницы по Фишеру. p <0,05 считали значимым.
Результаты
Влияние MR36014 и MR33583 на кортикальные нейроны, поврежденные глутаматом
Глутаматергическое повреждение вызвало потерю кортикальных нейронов (фиг. 15A и 15C; 32% гибель нейронов) и сокращение общей нейритной сети (фиг. 15B и 15D; 38% потеря нейритов).
Выживание нейронов. MR36014, добавленный за 1 час до глутамата и оставленный в течение 48 часов после применения глутамата, значительно защищал корковые нейроны от гибели при концентрации 1 нМ (см. фиг. 15A). В тех же экспериментальных условиях MR33583 значительно защищал кортикальные нейроны от гибели при 50 нМ и 100 нМ (см. фиг. 15C).
Нейритная сеть. Все протестированные концентрации MR36014 (от 500 пМ до 10 нМ) в значительной степени защищали сеть нейритов с максимальным эффектом при 1 нМ (см. фиг. 15B). MR33583 в значительной степени защищал нейритную сеть от 1 до 100 нМ. Эффект соответствовал колоколообразной кривой с максимальным эффектом при 50 нМ (см. фиг. 15D).
Обсуждение
Целью исследования было изучение нейропротекторных эффектов MR36014 и MR33583 на модели нейронального стресса in vitro, вызванного глутаматергической эксайтотоксичностью. Глутаматергический стресс является патологическим признаком, присутствующим при различных неврологических заболеваниях, включая болезнь Альцгеймера и церебральную ишемию.
По результатам исследования можно сделать вывод, что:
- оба соединения противодействуют токсичности глутамата для нейронов коры головного мозга;
- MR36014 и MR33583 показали аналогичную эффективность, хотя MR36014 был активен при более низкой дозе (в отношении выживаемости нейронов).
В целом, это исследование показывает, что MR36014 и MR33583 являются интересными кандидатами в качестве лекарств от неврологических заболеваний, связанных с глутаматергическим стрессом.
Пример 8
В этом примере раскрывается оценка нейропротекторных эффектов MR36014 и MR33583 в отношении первичных кортикальных нейронов крыс после глутаматного повреждения.
Боковой амиотрофический склероз (БАС) - это фатальное заболевание, характеризующееся началом с небольшой очаговой слабостью, обычно в конечностях, но иногда и в бульбарных мышцах, которая прогрессирует до паралича почти всех скелетных мышц. Существует значительное клинико-патологическое и генетическое совпадение между БАС и лобно-височной долевой деменцией (ЛВП). При БАС смерть от паралича дыхания обычно наступает в течение пяти лет. Клеточная патология является очаговой с самого начала и распространяется по схеме, предполагающей последовательное вовлечение смежных популяций нейронов. Гибель моторных нейронов происходит в сочетании с отложением агрегированных белков в моторных нейронах и олигодендроцитах и нейровоспалением.
Сложная патофизиология БАС представляет собой множество потенциальных терапевтических мишеней. Несмотря на то, что был исследован широкий спектр агентов, только Рилузол (Рилутек®), антиглутаматергический препарат, продемонстрировал устойчивую эффективность, и является единственным одобренным препаратом для лечения этого заболевания. Однако преимущества рилузола скромны - он продлевает выживание пациентов с БАС на несколько месяцев (~7%) с минимальным влиянием на функциональные показатели.
Хотя точные молекулярные пути, вызывающие гибель моторных нейронов при БАС, остаются неизвестными, некоторые возможные механизмы включают: (а) глутамат-опосредованную эксайтотоксичность; (b) снижение нейротрофических факторов (BDNF, GDNF…); (с) митохондриальные изменения и окислительные повреждения; (d) аномалии белков цитоскелета, приводящие к атрофии и гибели нейронов.
Было показано, что TDP-43 (транзактивный ДНК-связывающий белок 43 кДа) накапливается в цитоплазме моторных нейронов в большинстве случаев БАС. TDP43 представляет собой ядерный РНК-связывающий белок, участвующий в нескольких аспектах процессинга РНК, который активно перемещается между ядром и цитоплазмой.
Целью настоящего исследования была оценка влияния MR36014 и MR33583 (в 4 концентрациях) на первичный спинномозговой моторный нейрон (MN) после повреждения глутаматом.
Материалы и методы
Первичная культура спинальных моторных нейронов
Все эксперименты проводились в соответствии с Руководством по уходу и использованию лабораторных животных Национального института здравоохранения и действующими правилами Европейского Союза (Директива 2010/63/EU). Номер договора: A1301310.
Моторные нейроны спинного мозга (MN) крыс культивировали, как описано Martinou et al. (1992) Neuron. 8:737-744 и Wang et al. (2013) Hum. Mol. Genet. 22:4706-4719. Вкратце, беременных самок крыс со сроком беременности 14 дней (Rats Wistar; Janvier Labs, France) умерщвляли с использованием глубокой анестезии в камере с CO2 и цервикальной дислокации. Затем эмбрионы извлекали из матки и немедленно помещали в ледяную среду L15 Лейбовица с 2% раствором пенициллина (10000 Ед/мл) и стрептомицина (10 мг/мл) (ПС) и 1% бычьим сывороточным альбумином (БСА).
Спинной мозг обрабатывали в течение 20 мин при 37°C раствором трипсин-ЭДТА с конечной концентрацией 0,05% трипсина и 0,02% ЭДТА. Диссоциацию останавливали добавлением модифицированной по Дульбекко среды Игла (DMEM) с 4,5 г/л глюкозы, содержащей ДНКазу I сорта II (конечная концентрация 0,5 мг/мл) и 10% эмбриональной телячьей сыворотки (ЭТС). Клетки механически диссоциировали тремя принудительными пропусканиями через кончик пипетки на 10 мл. Затем клетки центрифугировали при 515 x g в течение 10 минут при 4°C. Надосадочную жидкость удаляли, а осадок ресуспендировали в определенной культуральной среде, состоящей из нейробазальной среды с 2% раствором добавки B27, 2 ммоль/л L-глутамина, 2% раствором ПС и 10 нг/мл нейротрофического фактора головного мозга (BDNF). Жизнеспособные клетки подсчитывали на цитометре Нойбауэра с использованием теста исключения трипанового синего. Клетки высевали с плотностью 20 000 на лунку в 96-луночные планшеты, предварительно покрытые поли-L-лизином, и культивировали при 37°C в инкубаторе в среде воздух (95%) - CO2 (5%).
Среду меняли каждые 2 дня. Лунки первых линий и рядов не использовали для посева (во избежание краевого эффекта) и заполняли стерильной водой. Моторные нейроны повреждали глутаматом после 13 дней культивирования.
Тестируемые соединения и воздействие глутамата
Предварительная инкубация: на 13 день культивирования соединения растворяли в культуральной среде и предварительно инкубировали с первичными кортикальными нейронами в течение 1 часа перед воздействием глутамата.
Глутаматное повреждение: на 13 день культивирования и после 1 часа предварительной инкубации добавляли глутамат до конечной концентрации 5 мкМ, разведенной в контрольной среде, все еще в присутствии соединений, в течение 20 минут. Через 20 минут глутамат вымывали и добавляли свежую культуральную среду с соединениями еще на 24 часа.
Организация планшетов с культурой
Испытуемые соединения тестировали в 96-луночном планшете (n = 6 лунок для культивирования на условие).
Оценивали следующие условия:
Конечная оценка
Через 24 часа после интоксикации надосадочную жидкость отделяли и немедленно замораживали для дальнейшего анализа. Клетки фиксировали холодным раствором этанола (95%) и уксусной кислоты (5%) в течение 5 мин при -20°C. После пермеабилизации 0,1% сапонином клетки инкубировали в течение 2 часов с:
- мышиным моноклональным антителом против ассоциированного с микротрубочками белка 2 (MAP-2) при разведении 1/400 в ФБР, содержащем 1% эмбриональной телячьей сыворотки и 0,1% сапонина. Это антитело выявляли с помощью козьего антитела против IgG мыши с Alexa Fluor 488 в разведении 1/400 в ФБР, содержащем 1% ЭТС, 0,1% сапонина, в течение 1 часа при комнатной температуре.
- кроличьими поликлональными антителами против ядерного транзактивного ДНК-связывающего белка 43 (TDP-43) при разведении 1/100 в ФБР, содержащем 1% эмбриональной телячьей сыворотки и 0,1% сапонина. Антитело против TDP-43 было выявлено с помощью козьего антитела к IgG кролика с Alexa Fluor 568 в разведении 1/400 в ФБР, содержащем 1% ЭТС, 0,1% сапонина, в течение 1 часа при комнатной температуре.
Для каждого условия с помощью ImageXpress® (Molecular Devices) с 20-кратным увеличением с использованием тех же параметров сбора данных было автоматически сделано 30 снимков (представляющих всю площадь лунки) на лунку. Анализ изображений проводили напрямую и автоматически с помощью Custom Module Editor® (Molecular Devices).
Конечный анализ
Следующие конечные точки оценивали автоматически:
- Анализ выживания нейронов (окрашивание MAP-2, количество нейронов);
- Анализ нейритной сети (окрашивание MAP-2, общая длина нейритов в мкм);
- Анализ нейритной сети / нейронов (нейронная сеть/ количество нейронов MAP-2);
- Анализ цитоплазматического TDP-43 в MAP-2-положительных нейронах (перекрывание между MAP-2 и цитоплазматическим TDP-43 в мкм²).
Статистический анализ
Все значения выражены как среднее значение ± SEM (стандартная ошибка среднего). Статистический анализ был выполнен с помощью однофакторного дисперсионного анализа с последующим применением метода наименьшей значимой разницы по Фишеру. p <0,05 считали значимым.
Результаты
Эффекты MR36014 в первичной культуре спинальных моторных нейронов, поврежденных глутаматом
Выживание нейронов. Потеря больших MAP-2 моторных нейронов спинного мозга после применения глутамата (42% гибели нейронов) (фиг. 16A). При 500 пМ и 1 нМ MR36014 значительно улучшал выживание спинальных моторных нейронов.
Нейритная сеть. Нейритная сеть была сильно уменьшена после повреждения глутаматом (46% потери нейритов, фиг. 16B). MR36014 значительно защищал нейритную сеть при 500 пМ и 1 нМ. Длина нейритной сети пропорциональна количеству нейронов. Отношение «длина нейритной сети»/ «количество нейронов» было рассчитано для оценки средней длины нейрита на нейрон. MR36014 не изменял это соотношение (фиг. 16C).
Эффекты MR33583 в первичной культуре спинальных моторных нейронов, поврежденных глутаматом
Выживание нейронов. На фиг. 17 показано, что после применения глутамата отмечалась потеря больших MAP-2 моторных нейронов позвоночника (фиг. 17A). MR33583 (добавленный за 1 час до глутамата и оставленный на 24 часа после повреждения) проявлял дозозависимый нейропротекторный эффект (от 10 до 100 нМ).
Нейритная сеть. Как показано на фиг. 17, нейритная сеть сильно сократилась после повреждения глутаматом (фиг. 17B). MR33583 значительно защищал нейритную сеть от 10 до 100 нМ дозозависимым образом. MR36014 не изменял это соотношение (фиг. 17C).
TDP43. При наивысшей дозе MR33583 (100 нМ) снижал сигнал TDP43 в цитоплазме спинальных моторных нейронов (фиг. 17D).
Обсуждение
Целью этого исследования была оценка нейропротекторных эффектов двух соединений, MR36014 и MR33583, на модели бокового амиотрофического склероза in vitro, основанной на первичной культуре моторных нейронов спинного мозга, поврежденных глутаматом.
Результаты этого исследования показывают, что:
- MR36014 обеспечивает явный нейропротекторный эффект при 500 пМ и 1 нМ, поскольку он улучшает выживание нейронов и защищает нейритную сеть.
- MR33583 продемонстрировал явный нейропротекторный эффект в отношении всех параметров (выживание нейронов, сохранение нейритной сети и накопление цитоплазматического TDP43).
В целом, эти результаты показывают, что MR36014 и MR33583 были способны опосредовать нейропротекцию моторных нейронов спинного мозга, и они являются потенциальными кандидатами в качестве лекарств от бокового амиотрофического склероза.
Пример 9
В этом примере показана оценка нейропротекторного действия MR33583 на первичные ГАМКергические нейроны крысы в условиях депривации фактора роста.
БХ (болезнь Хантингтона) - аутосомно-доминантное наследственное нейродегенеративное заболевание, поражающее примерно 4–10 из 10 000 у представителей европеоидной расы. Головной мозг при БХ характеризуется потерей MSN (средних шипиковых нейронов) полосатого тела, увеличением желудочков и соответствующей атрофией вышележащей коры. ГАМКергические (γ-аминомасляная кислота) клетки являются наиболее многочисленным типом нейрональных клеток полосатого тела (90–95% у крыс и более 85% у людей), наряду с несколькими небольшими популяциями интернейронов (Wictorin (1992) Prog. Neurobiol. 38: 611-639). MSN характеризуются экспрессией фермента GAD64. Они принимают основную часть входных сигналов в полосатое тело, получая афференты от коры, таламуса и среднего мозга, а также являются основным выходным нейроном полосатого тела, проецируясь в различные области ЦНС (центральной нервной системы), включая бледный шар и черную субстанцию. Как таковые, они образуют сложные нейронные петли в базальных ганглиях, которые важны для контроля действий, познания и эмоций (Nakano et al. (2000) J. Neurol. 247 (Suppl. 5): v1-v15).
Важность факторов роста и их уменьшение при нейродегенеративных заболеваниях (таких как БА, БП или БХ) часто упоминается как одна из множества причин этих патологий. Генетическая, а также фармакологическая депривация BDNF или GDNF ухудшает обучение и память и приводит к гибели нейронов (Yamada et al. (2003) J. Pharmacol. Sci. 91: 267-270). Модель депривации фактора роста была охарактеризована в нескольких типах нейрональных клеток, таких как первичная культура нейронов грызунов (Atabay et al. (1996) J. Neurosci. Res. 43: 465-475), человеческих SK-N-SH и SH-SY5Y клеток нейробластомы (Ba et al. (2003) J. Neurosci. Methods 123: 11-22; Russo et al. (2004) Brain Res. 1009: 40-53).
В этих условиях с депривацией фактора роста серьезная потеря жизнеспособности происходит из-за апоптотической гибели клеток, обнаруживаемой по значительному усилению параметров, связанных с апоптозом, включая расщепленную каспазу-3, PARP и H2A.X. Кроме того, было показано, что экспрессия гена и белка антиапоптотического Bcl-2 снижается, тогда как экспрессия проапоптотического Bax повышается. Кроме того, было показано, что депривация нейротрофического фактора головного мозга (BDNF) запускает пути клеточной гибели за счет активации каспаз благодаря усилению передачи сигналов p75NTR (Yu et al. (2008) J. Neurosci. 28: 7467-7475).
Интересно, что мыши Bdnf-/- также обнаруживают структурно-специфические нейроанатомические изменения и поведенческие аномалии. Они демонстрируют пониженную сложность дендритов и плотность шипов в коре и гиппокампе и в еще большей степени в полосатом теле, где 90% пораженных клеток являются ГАМКергическими MSN (Rauskolb et al. (2010) J. Neurosci.30: 1739-1749).
Целью этого исследования было изучение нейропротекторного эффекта MR33583 (в 4 концентрациях) в отношении выживания полосатого тела после депривации фактора роста (депривации BDNF и GDNF), на модели болезни Хантингтона in vitro.
Материалы и методы
Первичная культура ГАМКергических нейронов
Все эксперименты были проведены в соответствии с Руководством по уходу и использованию лабораторных животных Национального института здравоохранения и действующими правилами Европейского Союза (Директива 2010/63/EU). Номер договора: A1301310.
MSN полосатого тела крысы культивировали, как описано Ivkovic et al. (1999) J. Neurosci. 19: 5409-5419 и Schinelli et al. (1988) J. Neurochem. 50: 1900–1907. Вкратце, беременных самок крыс с 15-дневной беременностью умерщвляли с использованием глубокой анестезии в камере с CO2 и цервикальной дислокации. Эмбрионы собирали и немедленно помещали в ледяную среду L15 Лейбовица с 2% раствором пенициллина (10000 Ед/мл) и стрептомицина (10 мг/мл) (ПС) и 1% бычьим сывороточным альбумином (БСА). Для приготовления клеток использовали только ограниченную вентральная часть мезэнцефалического изгиба, поскольку это область развивающегося мозга, богатая клетками MSN. Эти участки обрабатывали в течение 20 мин при 37°C раствором трипсин-ЭДТА с конечной концентрацией 0,05% трипсина и 0,02% ЭДТА. Диссоциацию останавливали добавлением модифицированной по Дульбекко среды Игла (DMEM) с 4,5 г/л глюкозы, содержащей ДНКазу I сорта II (конечная концентрация 0,5 мг/мл) и 10% эмбриональной телячьей сыворотки (ЭТС). Клетки механически диссоциировали тремя принудительными пропусканиями через кончик пипетки на 10 мл. Затем клетки центрифугировали при 180 x g в течение 10 минут при комнатной температуре на слое БСА (3,5%) в среде L15. Надосадочную жидкость удаляли, а осадок клеток ресуспендировали в определенной культуральной среде, состоящей из нейробазальной среды с добавлением B27 (2%), L-глутамина (2 мМ), 1% раствора ПС, BDNF (10 нг/мл) и GDNF (1 нг/мл). Жизнеспособные клетки подсчитывали на цитометре Нейбауэра с использованием теста исключения трипанового синего. Клетки высевали с плотностью 40000 клеток/лунку в 96-луночные планшеты (лунки были предварительно покрыты поли-L-лизином) и культивировали при 37°C в инкубаторе воздух (95%)/CO2 (5%).
Чтобы избежать краевого эффекта, в исследовании не использовали первую и последнюю линии, а также первый и последний ряды планшета. Пустые лунки заполняли водой. Среду меняли каждые 2 дня.
Тестируемые соединения и депривация факторов роста
Предварительная инкубация: на 13 день культивирования тестируемое соединение разводили в культуральной среде и предварительно инкубировали с ГАМКергическими нейронами в течение 1 часа перед депривацией факторов роста (BDNF и GDNF).
Депривация факторов роста: через 1 час после инкубации с тестируемым соединением половину среды заменяли средой, лишенной факторов роста. Депривацию применяли на 96 часов.
Организация планшетов с культурой
Испытуемые соединения тестировали в 96-луночном планшете (n = 6 лунок для культивирования на условие). Оценивали следующие условия:
Конечная оценка
Через 96 часов после депривации факторов роста надосадочную жидкость клеточной культуры отделяли и немедленно замораживали для будущего анализа. Клетки фиксировали холодным раствором этанола (95%) и уксусной кислоты (5%) в течение 5 мин при -20°C. Клетки дважды промывали ФБР и затем повышали проницаемость. Неспецифическое связывание блокировали раствором ФБР, содержащим 0,1% сапонина и 1% ЭТС, в течение 15 мин при комнатной температуре.
Иммуноокрашивание: GAD67 (выживание). Культуры инкубировали в течение 2 часов с мышиным моноклональным антителом против GAD67 при разведении 1/200 в ФБР, содержащем 1% эмбриональной телячьей сыворотки и 0,1% сапонина (это антитело специфически окрашивает ГАМКергические нейроны). Это антитело выявляли с помощью козьего антитела против IgG мыши с Alexa Fluor 488 (разведение 1/400) в ФБР, содержащем 1% ЭТС, 0,1% сапонина, в течение 1 часа при комнатной температуре.
Автоматический компьютерный анализ - для каждого условия с помощью ImageXpress® (Molecular Devices) с 20-кратным увеличением было сделано 30 снимков (представляющих всю площадь лунки) на лунку. Все изображения были созданы с использованием одних и тех же параметров сбора данных. Анализ изображений проводили напрямую и автоматически с помощью Custom Module Editor® (Molecular Devices).
Следующие данные были исследованы: Анализ выживания MSN (окрашивание GAD67, количество ГАМКергических нейронов).
Статистический анализ
Все значения выражены как среднее значение ± SEM (стандартная ошибка среднего). Статистический анализ был выполнен с помощью однофакторного дисперсионного анализа с последующим применением метода наименьшей значимой разницы по Фишеру. p <0,05 считали значимым.
Результаты
Влияние MR33583 на первичную культуру ГАМКергических нейронов, подвергнутых стрессу путем депривации факторов роста
Выживание нейронов. Потеря больших GAD-67-положительных ГАМКергических нейронов последовала за депривацией факторов роста (фиг. 18).
MR33583 (50 нМ) оказывал значительный нейропротекторный эффект, что подтверждалось улучшением выживания ГАМКергических нейронов.
Обсуждение
В целом, эти результаты показывают, что MR33583 способен вызывать нейропротекторные эффекты (50 нМ) в ГАМКергических нейронах и что он может быть потенциальным лекарством от болезни Хантингтона.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ И ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ, СВЯЗАННЫХ С ТУБУЛИН КАРБОКСИПЕПТИДАЗАМИ (TCP) | 2018 |
|
RU2797200C1 |
| СПОСОБЫ И ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ, СВЯЗАННЫХ С ТУБУЛИН КАРБОКСИПЕПТИДАЗАМИ (TCP) | 2018 |
|
RU2812207C2 |
| ПРИМЕНЕНИЕ ЦИКЛИЧЕСКОГО ТРИПЕПТИДА ДЛЯ УЛУЧШЕНИЯ КЛЕТОЧНОГО ЭНЕРГЕТИЧЕСКОГО МЕТАБОЛИЗМА | 2016 |
|
RU2737116C2 |
| СПОСОБ ПОЛУЧЕНИЯ ОБЪЕДИНЕННОГО ЛИЗАТА ТРОМБОЦИТОВ ЧЕЛОВЕКА, ОБЪЕДИНЕННЫЙ ЛИЗАТ ТРОМБОЦИТОВ ЧЕЛОВЕКА И ЕГО ПРИМЕНЕНИЕ ДЛЯ ЛЕЧЕНИЯ НЕВРОЛОГИЧЕСКИХ НАРУШЕНИЙ | 2018 |
|
RU2802440C2 |
| 3β-(БЕНЗИЛОКСИ)-17α-МЕТИЛ-ПРЕГН-5-ЕН-20-ОН ДЛЯ ПРИМЕНЕНИЯ ПРИ ЛЕЧЕНИИ КОГНИТИВНЫХ РАССТРОЙСТВ | 2019 |
|
RU2796005C2 |
| 3-β(4-МЕТОКСИБЕНЗИЛОКСИ)ПРЕГН-5-ЕН-20-ОН ДЛЯ ПРИМЕНЕНИЯ ПРИ ЛЕЧЕНИИ НАРУШЕНИЙ, СВЯЗАННЫХ С УПОТРЕБЛЕНИЕМ КАННАБИНОИДОВ | 2019 |
|
RU2788004C2 |
| ХИМИЧЕСКИЕ СОЕДИНЕНИЯ, ВОЗДЕЙСТВУЮЩИЕ НА ГЛАЗА, И ИХ ПРИМЕНЕНИЕ ПРИ ЛЕЧЕНИИ ГЛАЗНЫХ ЗАБОЛЕВАНИЙ | 2020 |
|
RU2830634C1 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ПРОТИВ КЛАУДИНА 1 ДЛЯ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ ГЕПАТОЦЕЛЛЮЛЯРНОЙ КАРЦИНОМЫ | 2016 |
|
RU2770021C2 |
| АНТИТЕЛА ПРОТИВ BTN3A И ИХ ПРИМЕНЕНИЕ ПРИ ЛЕЧЕНИИ РАКА ИЛИ ИНФЕКЦИОННЫХ ЗАБОЛЕВАНИЙ | 2019 |
|
RU2800726C2 |
| НЕЙРОПРОТЕКТОРНОЕ ФАРМАКОЛОГИЧЕСКОЕ СРЕДСТВО | 2014 |
|
RU2561045C1 |
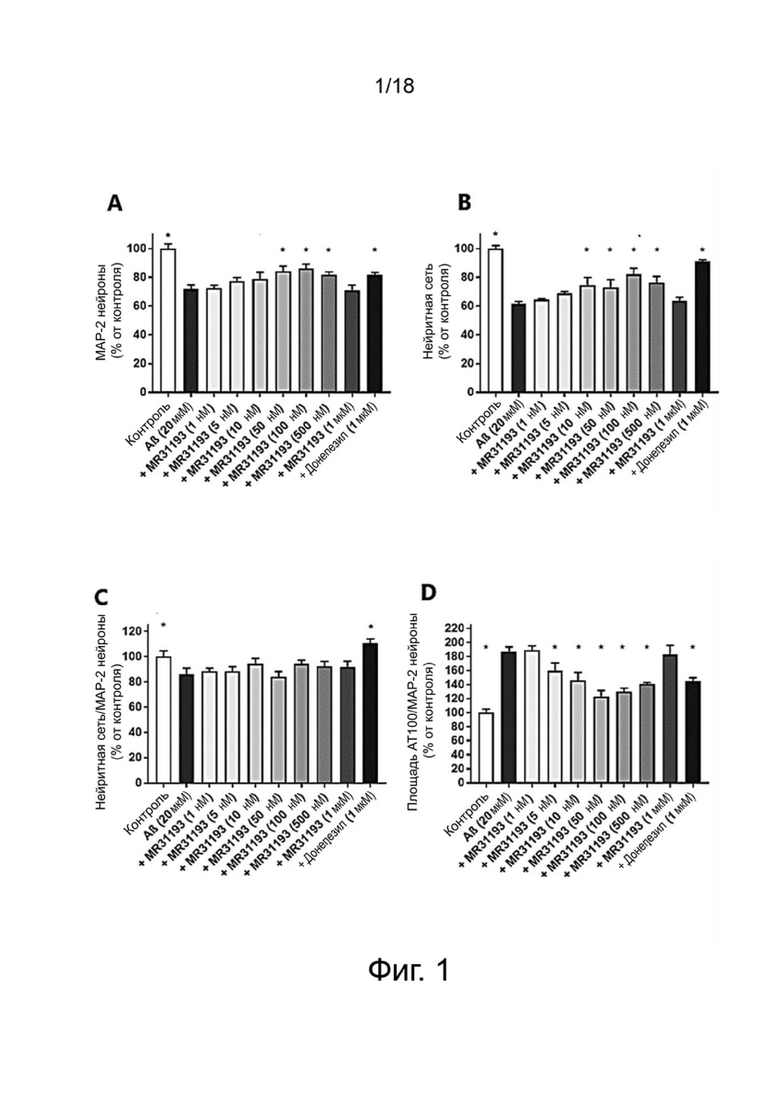
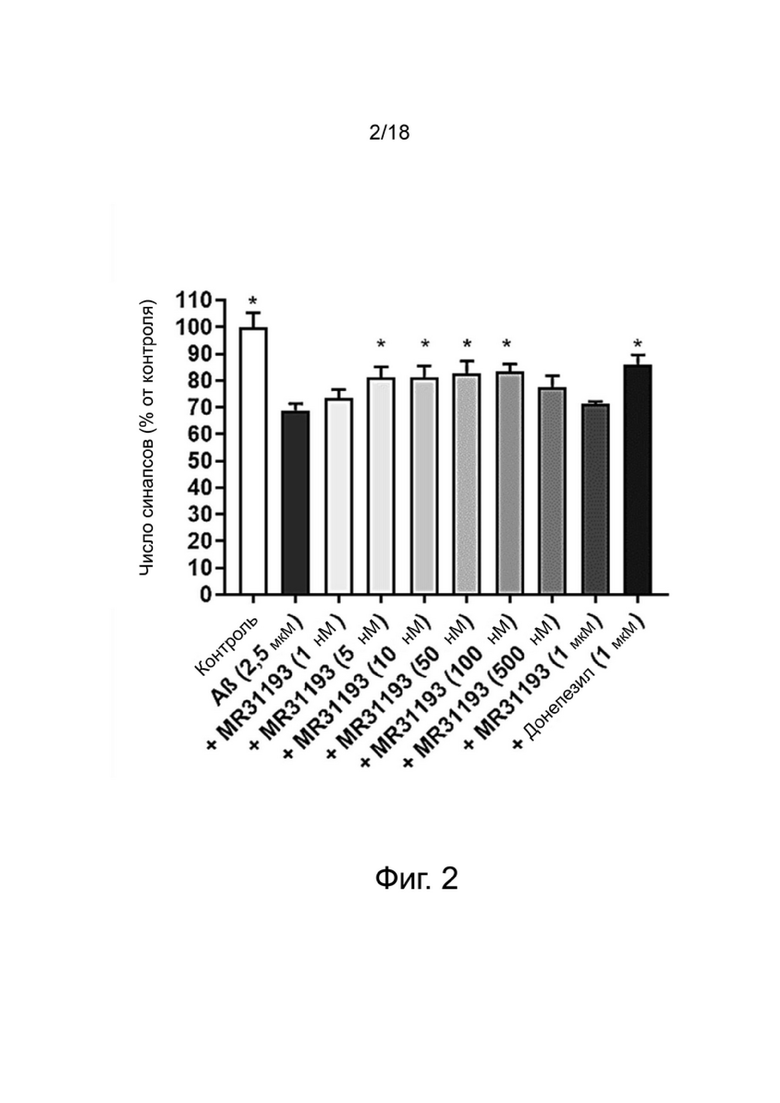
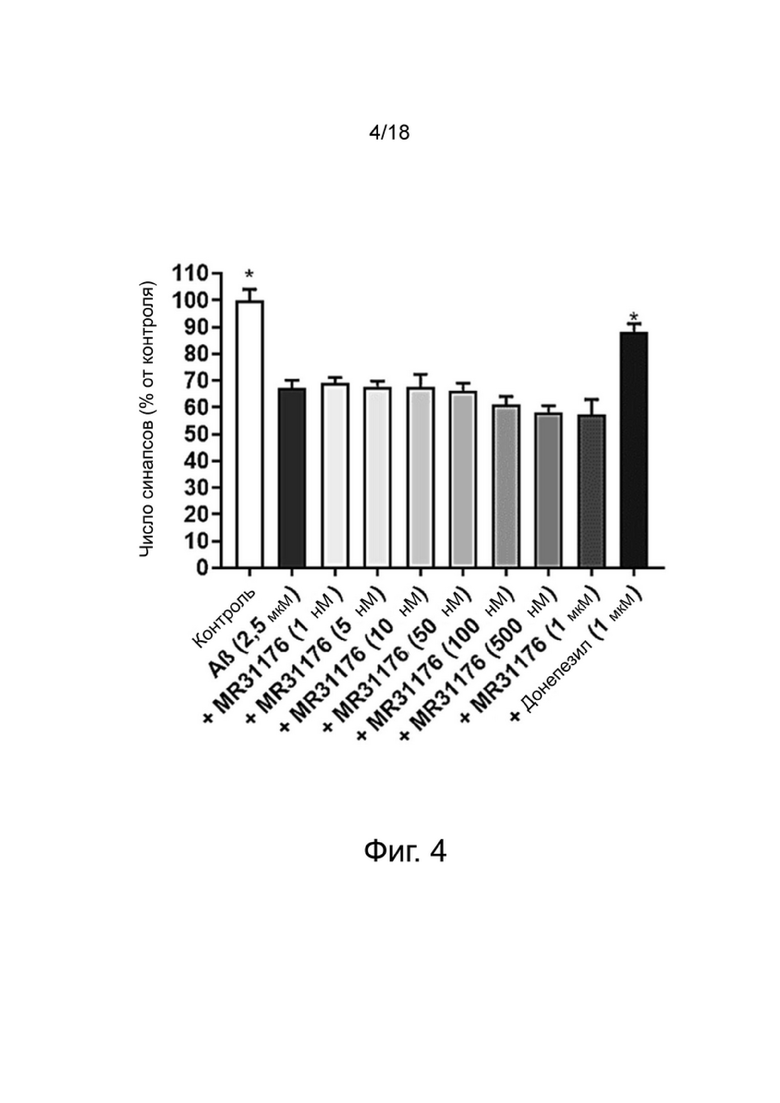
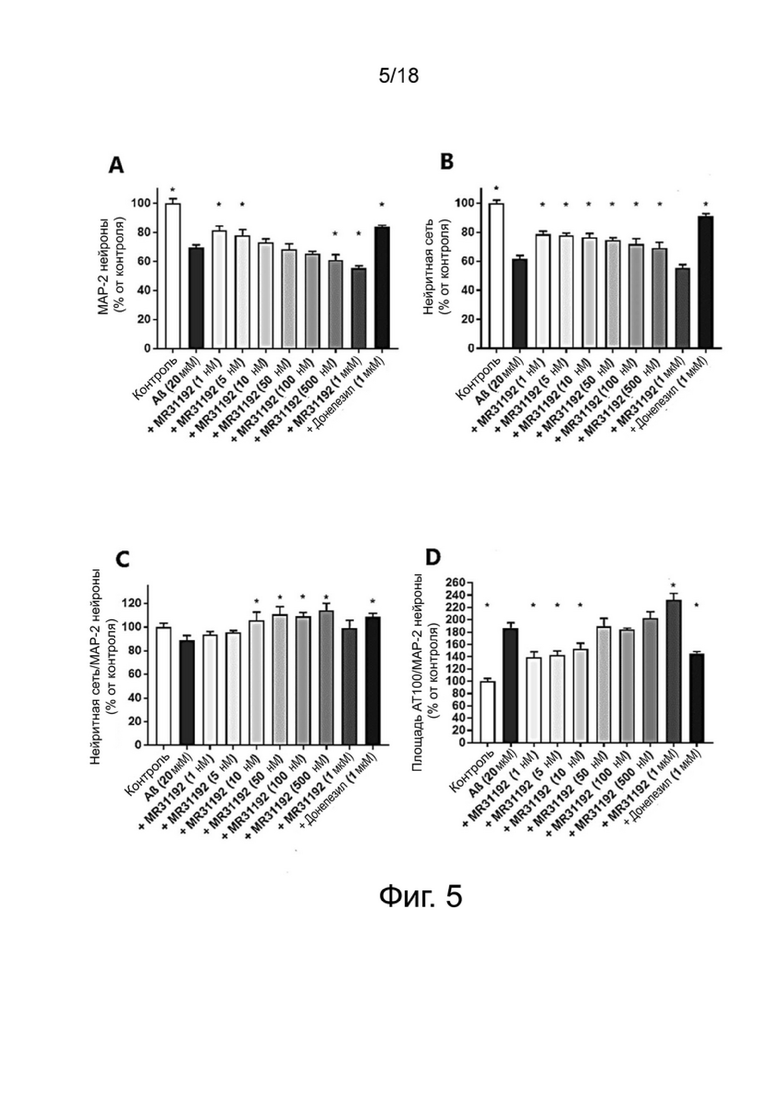
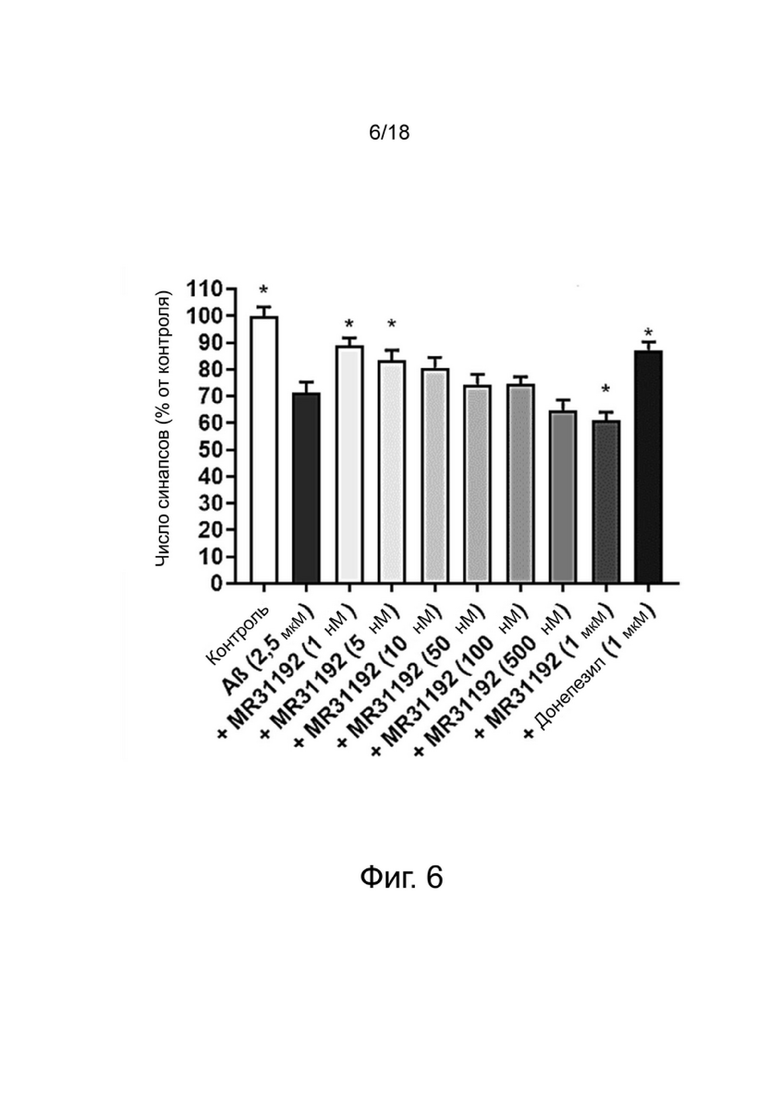
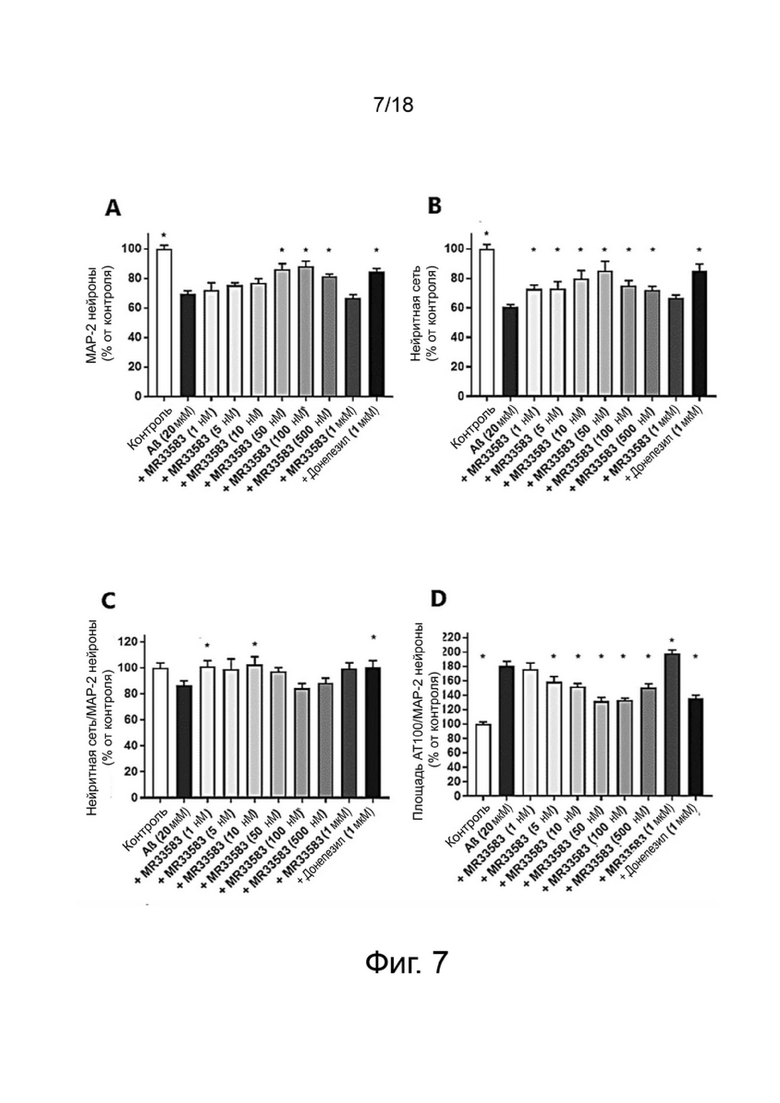
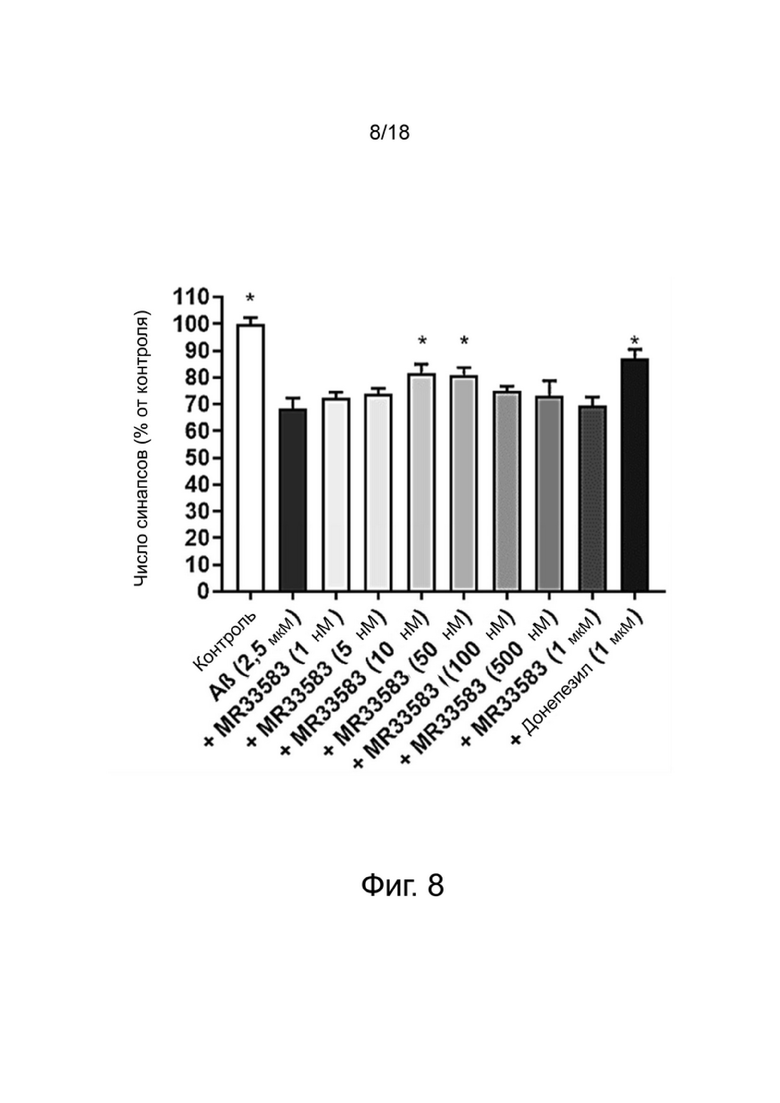
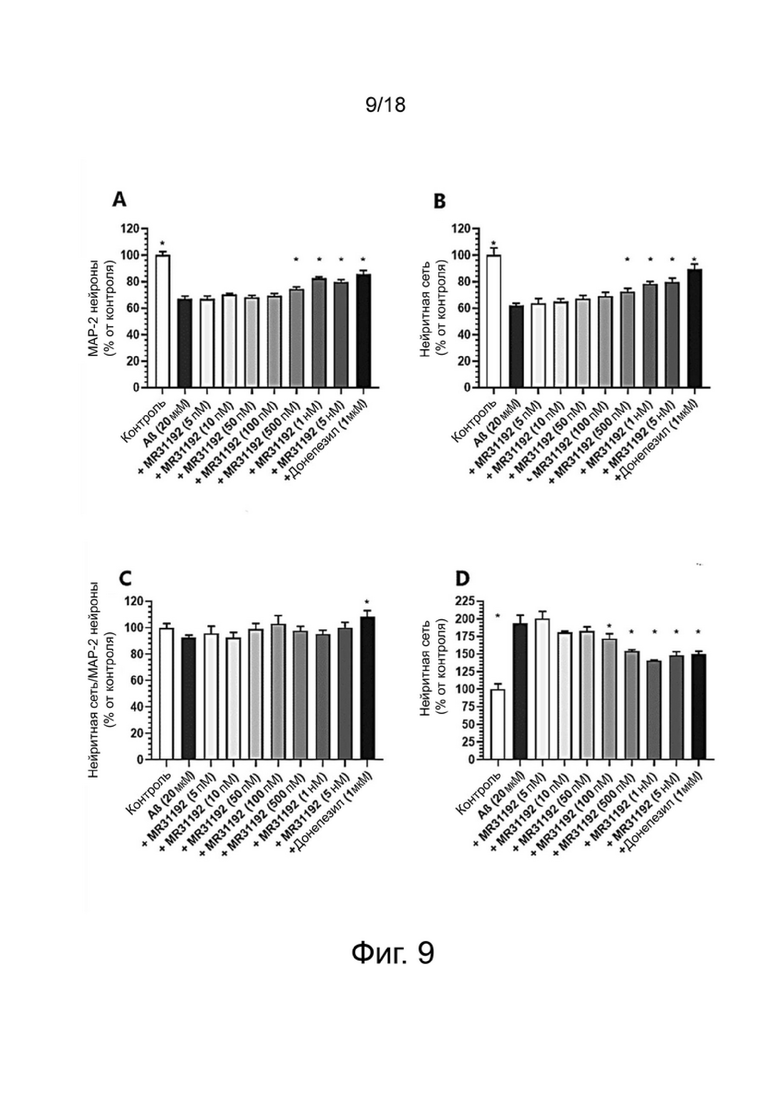
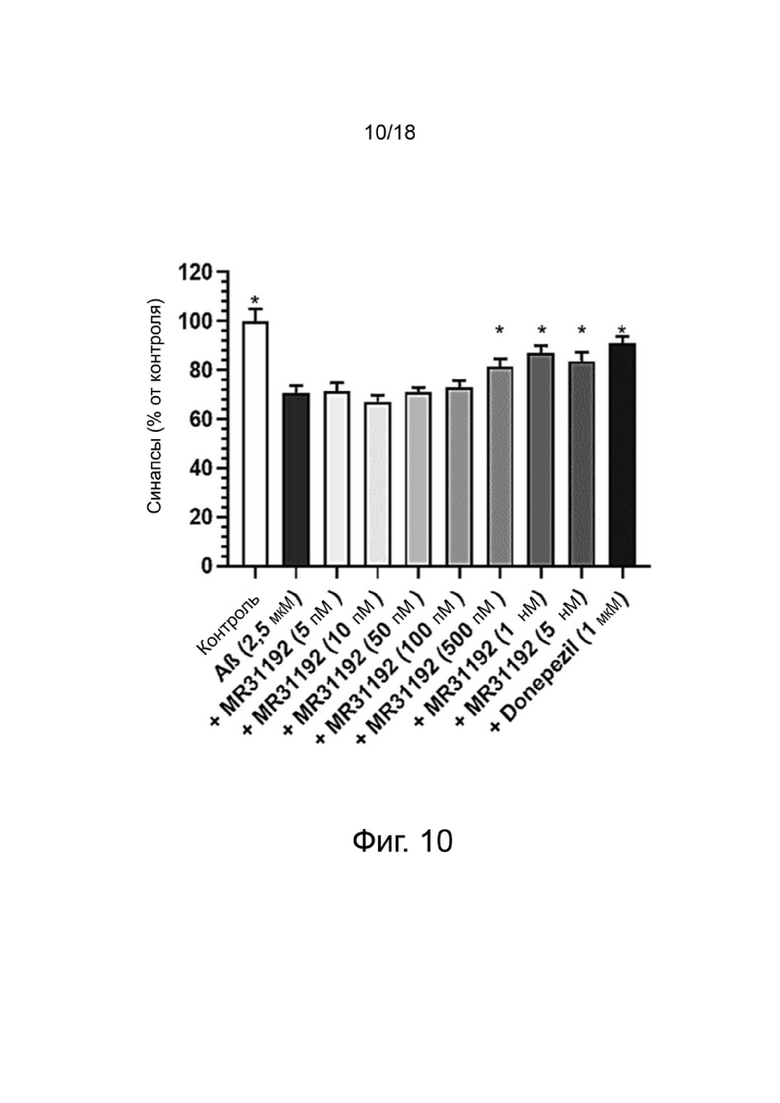
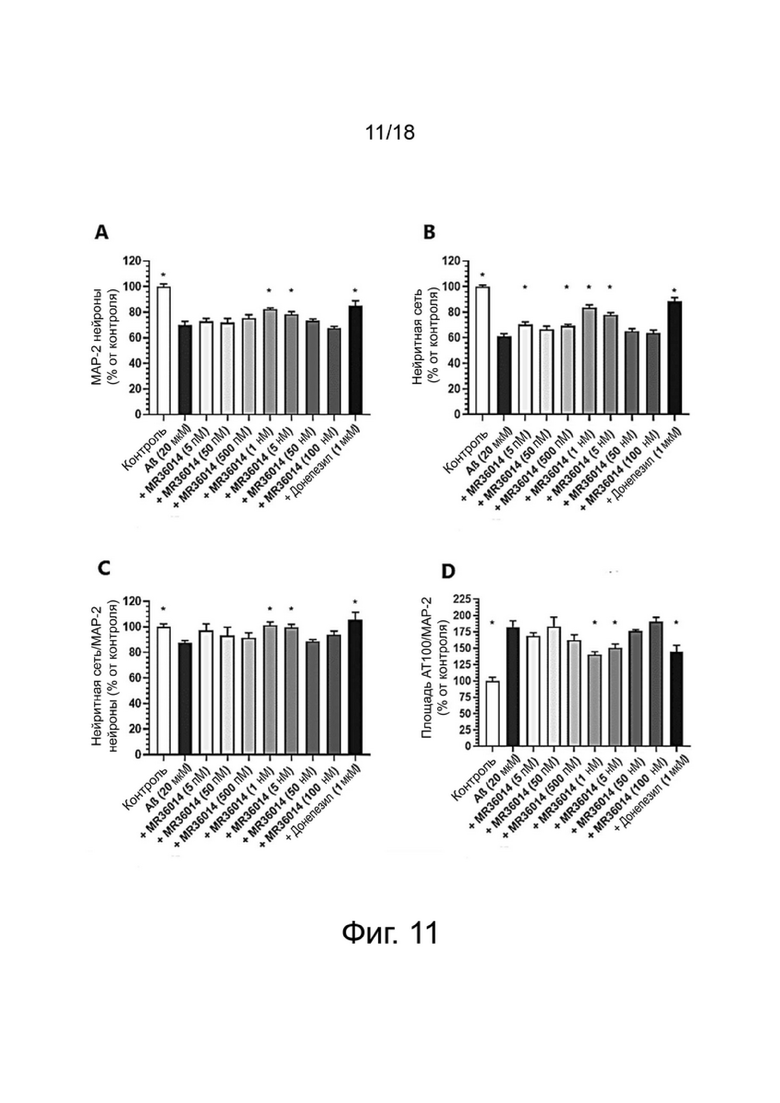
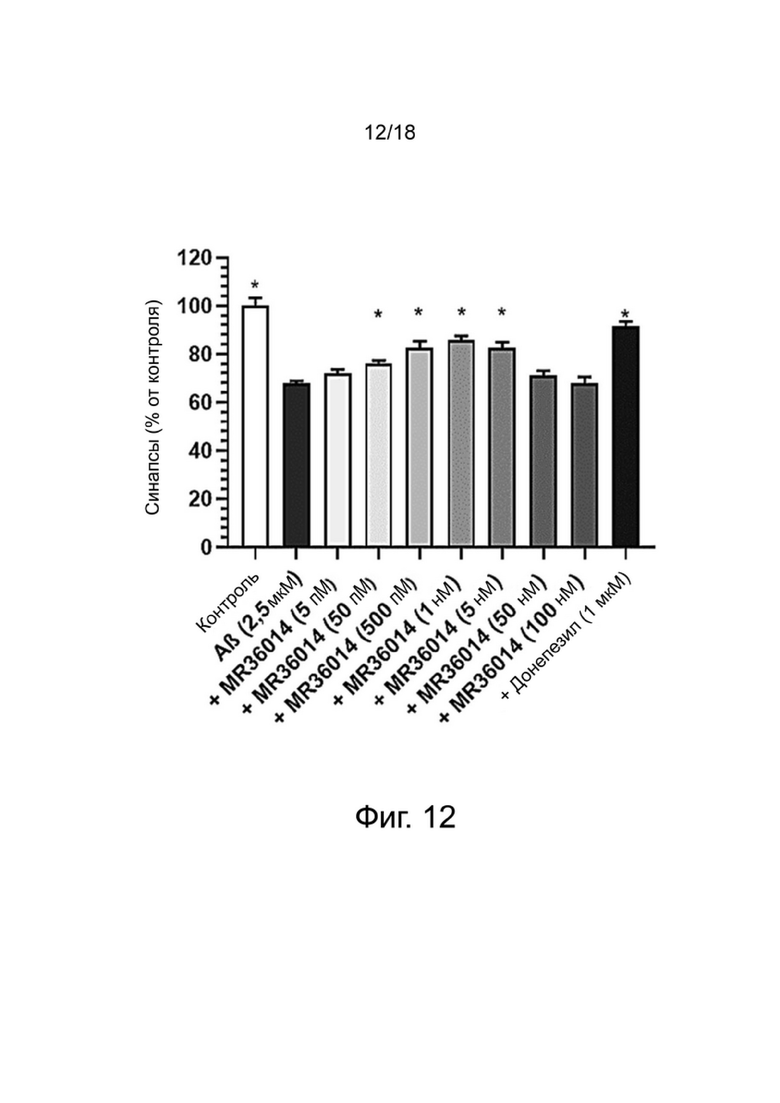
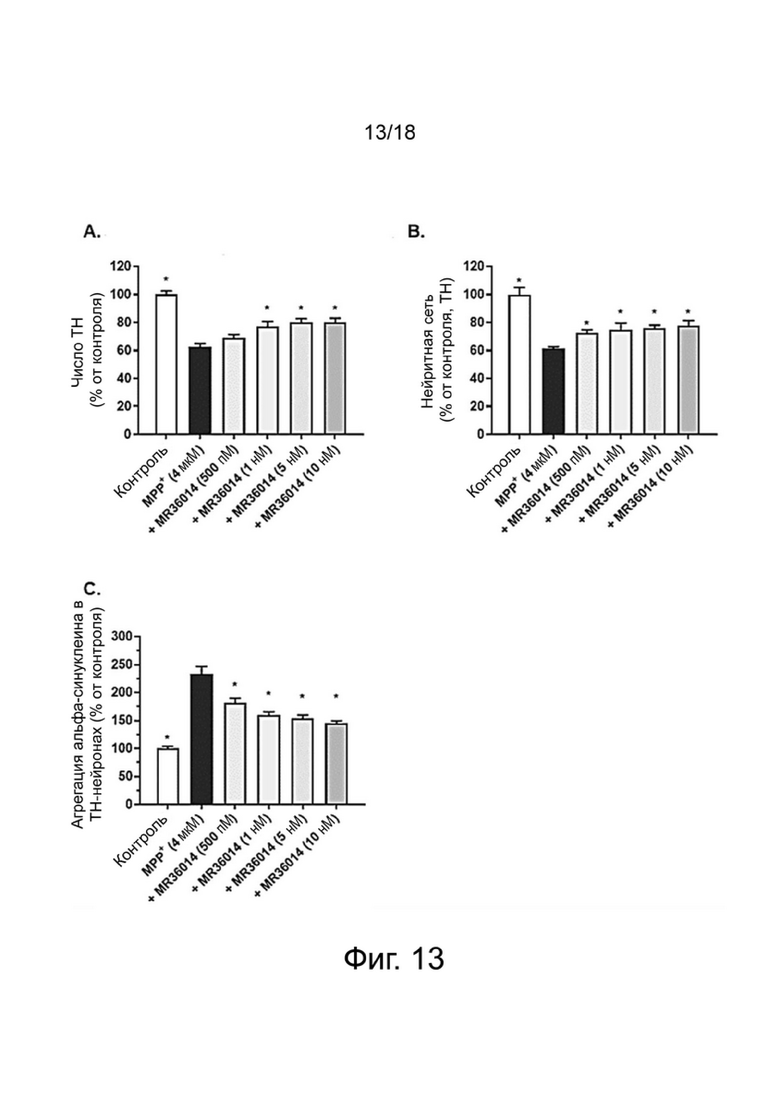
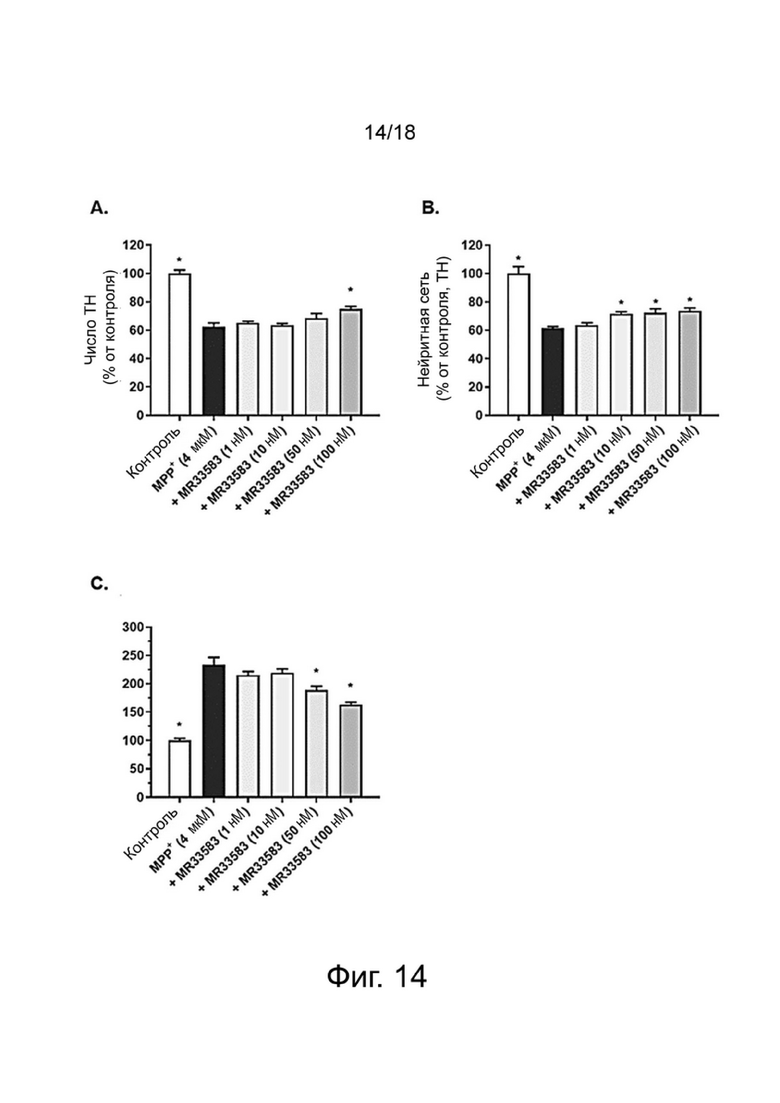
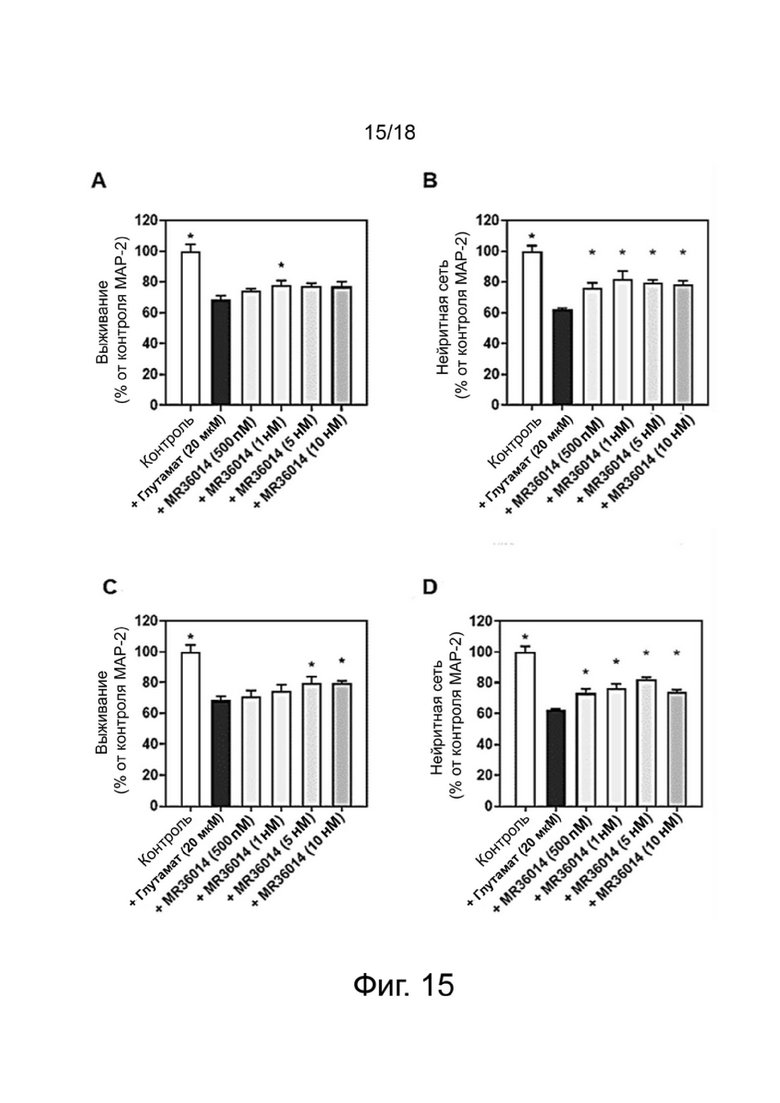
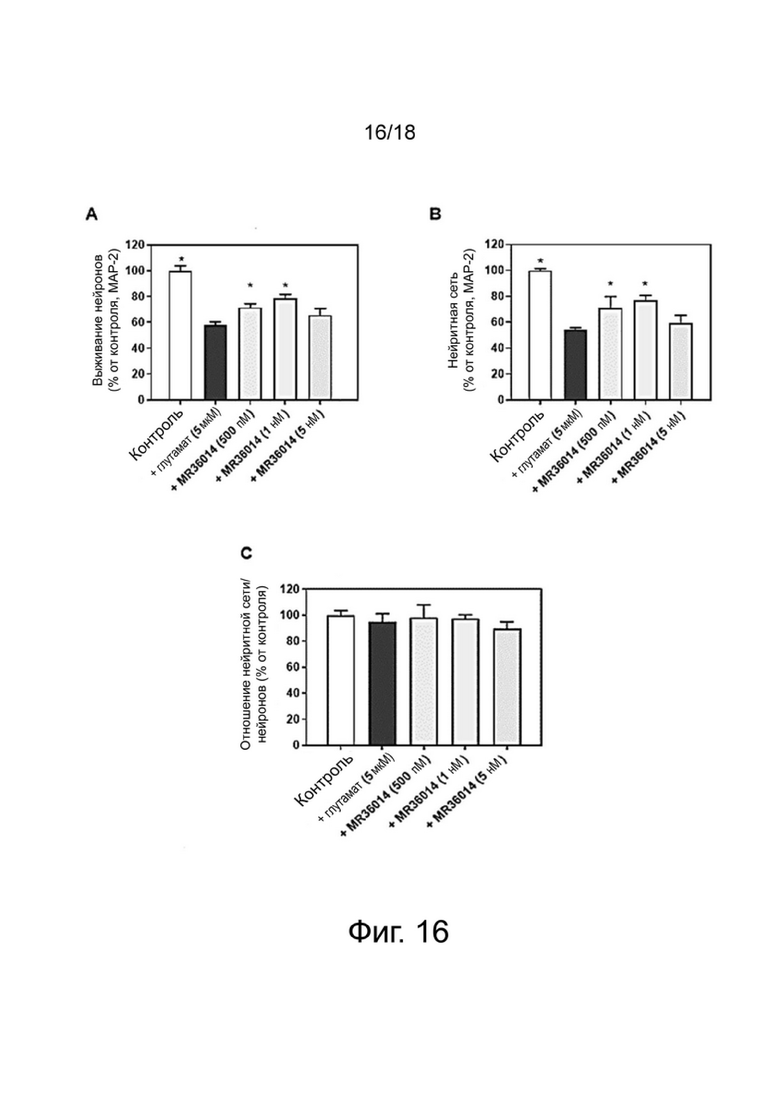
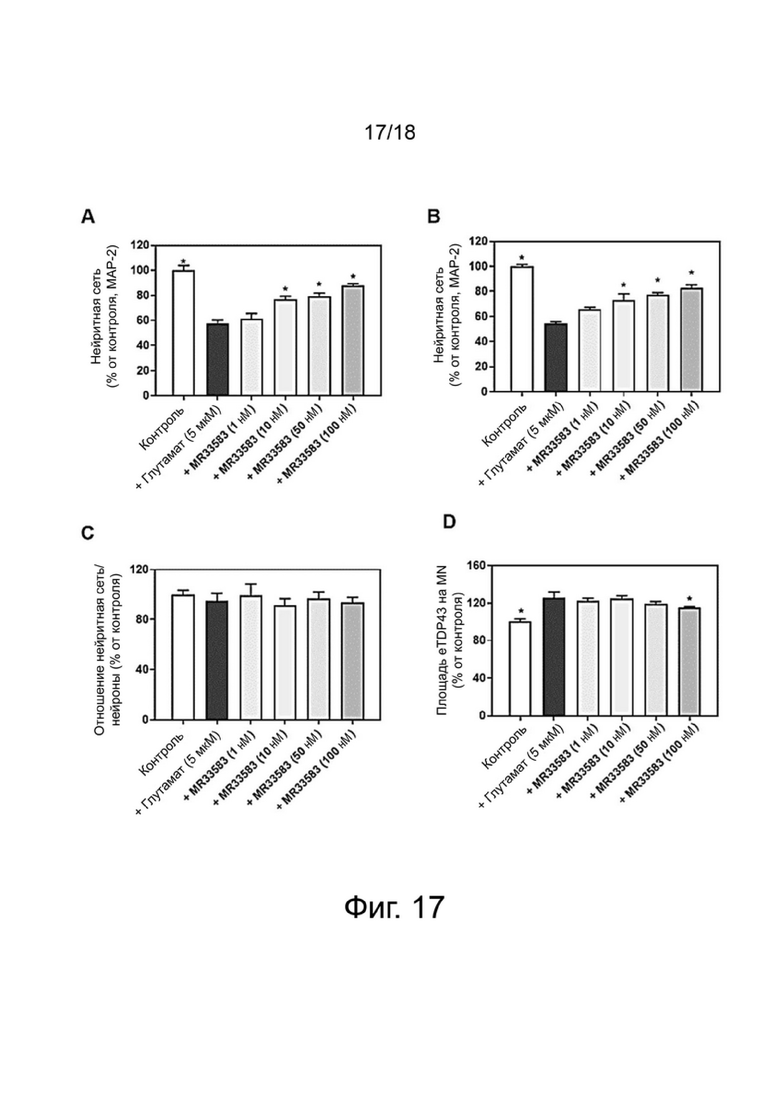
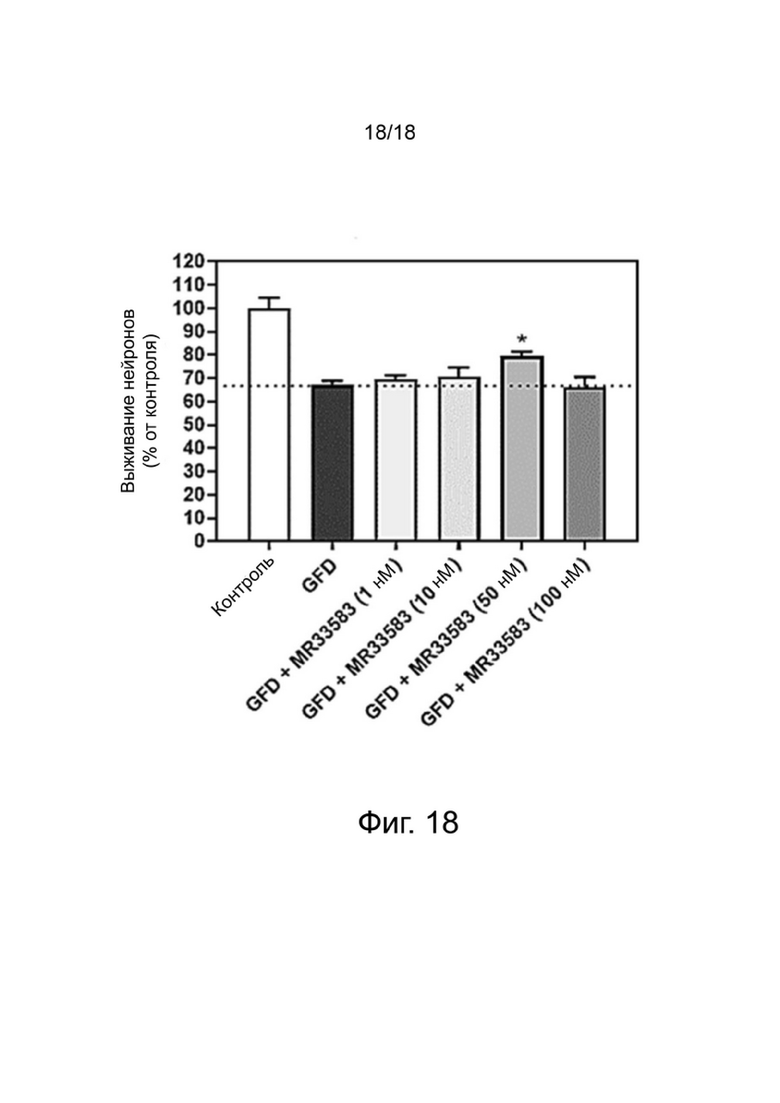
Изобретение относится к применению соединения формулы (I), а также его кислотных солей в качестве нейропротекторного агента при профилактике нейродегенеративного заболевания, выбранного из болезни Альцгеймера, болезни Паркинсона, болезни Хантингтона, бокового амиотрофического склероза и нейродегенеративного заболевания, вызванного ишемией или черепно-мозговой травмой. В общей формуле (I) X представляет собой атом галогена (Hal), где (Hal) представляет собой фтор, хлор, бром или йод; Y представляет собой атом кислорода; m, n, r и s имеют значение 1; R представляет собой С1-С5 алкильную группу с прямой или разветвленной цепью; R' представляет собой С3-С10 циклоалкил. 9 з.п. ф-лы, 18 ил., 6 табл., 9 пр.
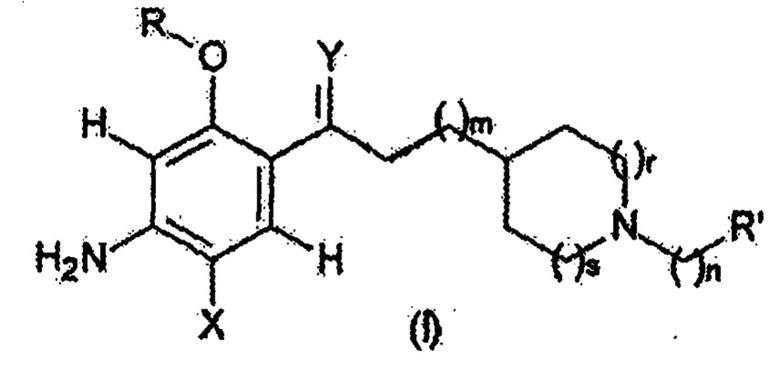
1. Применение соединения следующей формулы (I):
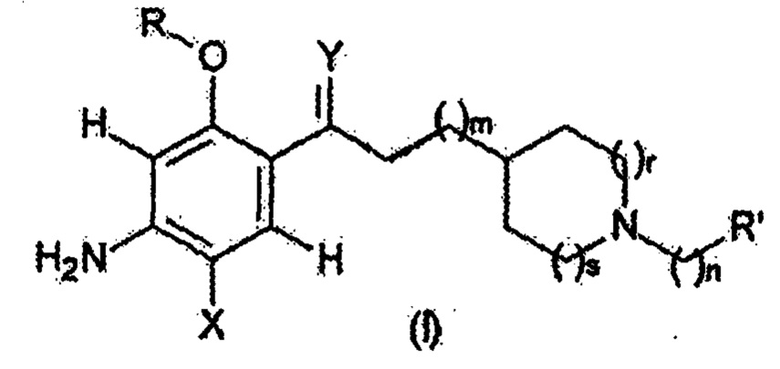
где:
X представляет собой атом галогена (Hal), где (Hal) представляет собой фтор, хлор, бром или йод;
Y представляет собой атом кислорода;
m, n, r и s имеют значение 1;
R представляет собой С1-С5 алкильную группу с прямой или разветвленной цепью;
R' представляет собой С3-С10 циклоалкил, а также его кислотных солей в качестве нейропротекторного агента при профилактике нейродегенеративного заболевания,
где указанные кислотные соли представляют собой фумараты, и
где нейродегенеративное заболевание выбрано из группы, состоящей из болезни Альцгеймера, болезни Паркинсона, болезни Хантингтона, бокового амиотрофического склероза и нейродегенеративного заболевания, вызванного ишемией или черепно-мозговой травмой.
2. Применение по п. 1, где R в формуле (I) представляет собой СН3 или СН2СН3.
3. Применение по п. 1 или 2, где R' в формуле (I) представляет собой радикал, выбранный из группы, состоящей из радикалов циклопропила, циклобутила, циклопентила, циклогексила, циклогептила и 4-пиперидина.
4. Применение по пп. 1-3, где R в формуле (I) представляет собой метильный радикал, a R' - циклоалкильный радикал С4-С7.
5. Применение по любому из пп. 1-4, где соединение представляет собой соединение донекоприд следующей формулы (II):
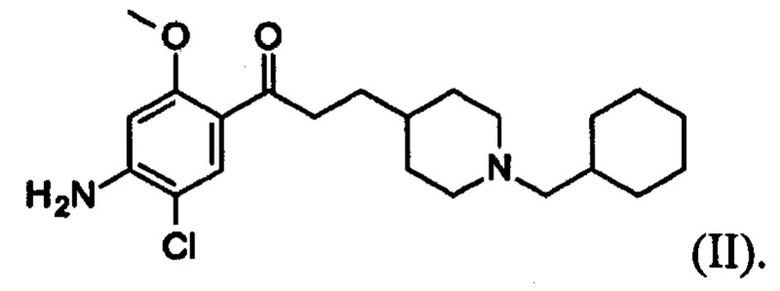
6. Применение по любому из пп. 1-4, где соединение представляет собой соединение флукоприд следующей формулы (III):
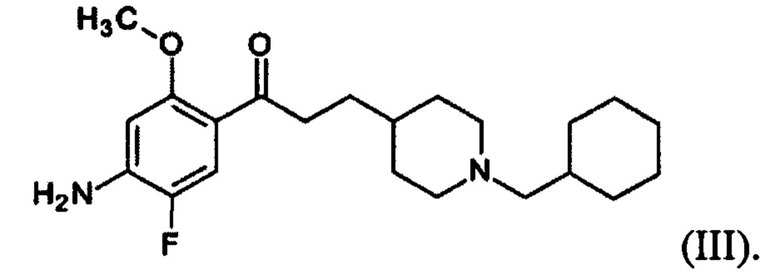
7. Применение по любому из пп. 1-4, где соединение представляет собой соединение MR31192 следующей формулы (VIII):
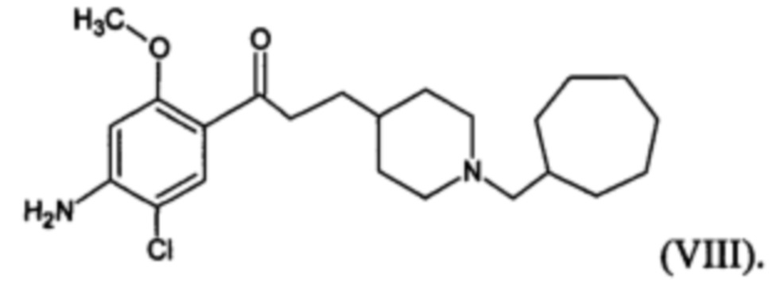
8. Применение по любому из пп. 1-4, где соединение представляет собой соединение MR33583 следующей формулы (V):
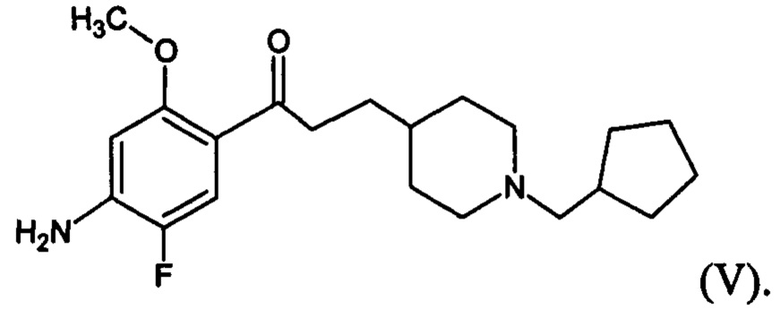
9. Применение по любому из пп. 1-4, где соединение представляет собой соединение MR36014
следующей формулы (VII):
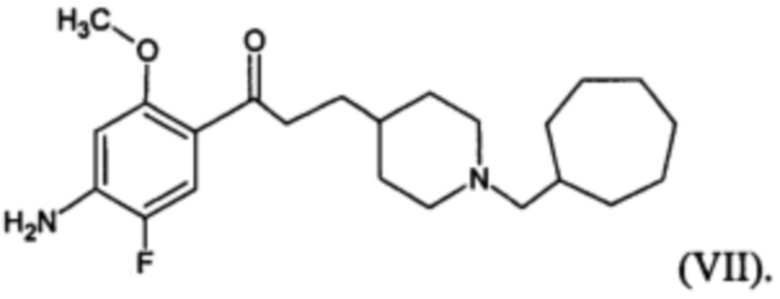
10. Применение по любому из пп. 1-4, где соединение представляет собой соединение MR31176 следующей формулы (IV):
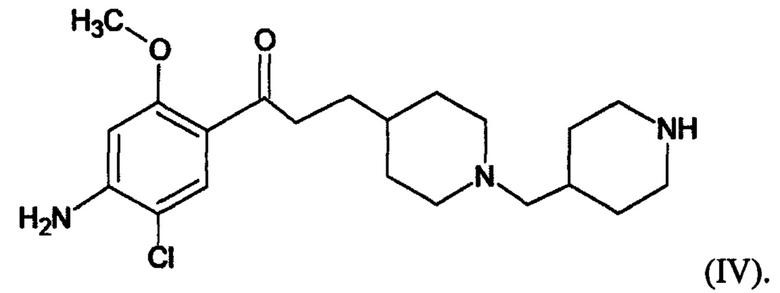
| WO 2014195593 A3, 11.12.2014 | |||
| Samir Yahiaoui "Design, synthesis, and pharmacological evaluation of multitarget-directed ligands with both serotonergic subtype 4 receptor (5-HT4R) partial agonist and 5-HT6R antagonist activities, as potential treatment of Alzheimer’s disease", European Journal of Medicinal Chemistry, Vol | |||
| Ребристый каток | 1922 |
|
SU121A1 |
| ПРИБОР ДЛЯ ОПРЕДЕЛЕНИЯ ОСАДКИ ВАЛОВ ПАРОВЫХ ТУРБИН | 1917 |
|
SU283A1 |

