Область техники, к которой относится настоящее изобретение
Настоящее изобретение относится к способам модуляции или стабилизации кишечной микробиоты путем перорального введения одного или нескольких дефензинов. Способы могут быть использованы для лечения или предупреждения воспаления кишечника, злокачественной опухоли толстой и прямой кишки, метаболического синдрома, ожирения, преддиабета, сахарного диабета и сердечно-сосудистого заболевания, а также для стимуляции рост мышц при производстве мяса.
Предшествующий уровень техники настоящего изобретения
Кишечная микробиота
Возрастающая распространенность общих нарушений, таких как ожирение и связанные с ожирением заболевания, тесно связана с нашим вестернизированным образом жизни и рационом. Наиболее распространенными заболеваниями, связанными с ожирением, являются инсулиновая резистентность, манифестный сахарный диабет 2 типа (T2D) и некоторые злокачественные опухоли (Faulds & Dahlman-Wright, 2012). Хотя этиология этих заболеваний сложна, многие из них характеризуются общим состоянием слабо выраженного воспаления, которое может возникать из-за разрегулированной кишечной микробиоты (Everard & Cani, 2013; Belkaid & Hand, 2014). Несмотря на то, что проблемы, связанные с образом жизни современного человека и производством мяса животных, могут показаться далекими друг от друга, предполагается, что нарушение здорового состояния кишечника является объединяющим фактором. Разрегулированное здоровое состояние кишечника действительно связано с множеством различных заболеваний, таких как ожирение (Ridaura et al., 2013), сахарный диабет 2 типа (Qin et al., 2012), ревматоидный артрит (Zhang et al., 2015) и злокачественная опухоль толстой и прямой кишки (Feng et al., 2015). Недавно сообщалось о связи между кишечной микробиотой и, в частности, присутствием определенных липополисахаридов из Bacteroides, и более высокой частотой возникновения сахарного диабета 1 типа в Финляндии по сравнению с соседними регионами (Leviten, 2016).
Ожирение и сопутствующее ему слабо выраженное воспаление образуют мощный фактор разрегулированного метаболического гомеостаза. Turnbaugh et al. (2006) обнаружили, что ассоциированная с ожирением микробиота имела повышенную способность накопления энергии, а через 2 недели после трансплантации микробиоты от тучных мышей безмикробные мыши показали значительно большее увеличение массы жира, чем при подобной трансплантации от тощих мышей. Кроме того, и что особенно важно, Turnbaugh et al. (2008) открыли, что изменение микробного состава кишечника полностью обращалось после возврата к исходному рациону у мышей, которых какое-то время кормили «западным» рационом с высоким содержанием жира/сахара. Эти данные Vrieze et al. (2012) подтвердили на человеке, продемонстрировав, что перенос кишечной микробиоты от худых доноров-людей повышал чувствительность к инсулину у индивидуумов с метаболическим синдромом.
Манипуляцию с кишечной микробиотой для повышения массы и интенсивности прироста массы использовали на протяжении многих лет в животноводстве посредством применения низких доз антибиотиков и пробиотиков, таких как Lactobacillus ingluviei. Манипуляцию с кишечной микробиотой для прироста массы продемонстрировали на цыплятах (Khan et al., 2007), утках (Angelakis & Raoult, 2010) и мышах (Angelakis et al., 2012). У людей обнаружили, что младенцы, получавшие антибиотики, были крупнее, чем их контроли (Trasande et al., 2012), поскольку воздействие в раннем возрасте пероральных антибиотиков ассоциируется с избыточной массой у детей (Ajslev et al., 2014). А у беременных женщин физиологическое увеличение ожирения и потенциальное развитие гестационного диабета в третьем триместре, по-видимому, связано с глубоким изменением кишечной микробиоты (Koren et al., 2012).
Слизистая оболочка кишечника, несомненно, является самой большой поверхностью в организме (приблизительно 200 м2), подверженной воздействию внешней среды. Следовательно, поверхность кишечника находится в тесном контакте с посторонним материалом, метаболитами, полученными из нашего рациона, и, по оценкам, с 1014 бактериями - кишечной микробиотой, которые обитают в нашем кишечнике. Таким образом, кишечный барьер находится под постоянным и интенсивным иммунологическим надзором, что требует динамического взаимовлияния между иммунной системой, компонентами рациона и кишечной микробиотой. Воздействия на рацион оказывают огромное влияние на иммунную регуляцию (Mowat & Agace, 2014) и состав кишечной микробиоты (Walter, 2015), которые независимо и синергетически влияют на метаболический гомеостаз. В связи с этим в двух последних статьях подчеркивается (неблагоприятный) эффект пищевых добавок в отношении изменения посредством модуляции микробиоты метаболического гомеостаза. В недавней статье (Chassaing et al., 2015) показали, как пищевые эмульгаторы нарушают толерантность к глюкозе, повышая тем самым прирост массы, а также склонность к колиту, за счет индуцирования разрегулированной кишечной микробиоты. Наблюдения не могли воспроизвести на безмикробных (GF) мышах, что указывает на ключевую роль кишечной микробиоты. Подобным образом, Suez et al. (2014) недавно показали, как некалорийные искусственные подсластители вызывают метаболическую дисфункцию посредством изменений кишечной микробиоты. Авторы подтвердили свои результаты с помощью фекального переноса GF мышам, после чего у GF мышей быстро развивалось нарушение толерантности к глюкозе. Эти наблюдения повторяют новаторское исследование на GF мышах (Bäckhed et al., 2007), в котором выясняется роль кишечных микробов в поддержании метаболического здоровья. Это исследование показало, что при отсутствии комменсальных микробов, что тем самым вызывает дисбаланс иммунного гомеостаза слизистой оболочки, снижаются размер жировых тканей и функция в ответ на рацион с высоким содержанием жира. Несмотря на отсутствие прироста массы, что обычно выглядит как здоровый фенотип, эктопическое накопление липидов (стеатоз печени и повышенное содержание триглицеридов в сыворотке крови) приводило к тяжелым метаболическим нарушениям. Было показано, что у человека многообразие генов микробиоты ассоциируется со здоровым фенотипом, тогда как скудность генов (низкие количества генов) коррелирует с повышенным риском метаболических нарушений (Le Chatelier et al., 2013).
Дефензины
Дефензины представляют собой одно из доминантных врожденных средств защиты хозяина, которые служат поддержанию здорового микробиома и блокированию потенциальных патогенов (Wehkamp et al., 2002, и Salzman et al., 2007). Дефензины представляют собой пептиды, обладающие противомикробной активностью в отношении грамположительных и грамотрицательных бактерий, грибков и архебактерий, а также противовоспалительной активностью, повышающей противовоспалительные цитокины и уменьшающей воспалительные цитокины.
Дефензины человека представляют собой небольшие катионные пептиды, которые можно разделить на α- и β-дефензины на основании топологии трех их внутримолекулярных цистеиновых дисульфидных связей. α-дефензины человека можно далее разделить на те, которые были впервые выделены из гранулы нейтрофилов (HNP1-4), и кишечные α-дефензины, которые экспрессируются клетками Панета в криптах тонкого кишечника (HD5 и HD6 или DEFA5 и DEFA6). β-дефензины (DEFBn) в основном продуцируются эпителиальными клетками в различных тканях и органах, в том числе в коже, глазу, среднем ухе, рту, трахеи, легком, желудочно-кишечном тракте, печени, мочеполовой системе, почках, влагалище, поджелудочной железе и молочных железах. Наилучшим образом охарактеризованными представителями семейства β-дефензинов человека являются hBD1-4. Некоторые из дефензинов человека продуцируются конститутивно, тогда как другие индуцируются провоспалительными цитокинами или продуктами экзогенных микробов. Некоторые из дефензинов человека экспрессируются даже в амниотической жидкости при уровнях, повышающихся с гестационным сроком, защищая плод в матке. Грудное молоко и особенно первое молоко, молозиво, содержит как α-, так и β-дефензины, но только некоторые из них находятся в значительных концентрациях в материнском молоке (Armogida et al., 2004).
Liu et al. (2008) обнаружили, что HNP-1 и HNP-2, оба продуцируемые лейкоцитами и принадлежащие подгруппе α-дефензинов в крови, были способны ингибировать гликогенолиз и глюконеогенез в выделенных гепатоцитах посредством внутримолекулярного механизма, совершенно отличного от классического инсулинового пути передачи сигнала.
Краткое раскрытие настоящего изобретения
Настоящее раскрытие демонстрирует, что перорально вводимые кишечные α- и β-дефензины млекопитающих обладают способностью поддерживать нормальный состав микробиоты в кишечнике мыши, получающей рацион с высоким содержанием жира. Данные в примерах демонстрируют, что пероральное введение α- и/или β-дефензинов млекопитающих приводит к стабилизации или нормализации дисбиотической микробиоты. Дефензины, поэтому, применимы в лечении или предупреждении злокачественной опухоли толстой и прямой кишки, эндокринного, связанного с питанием, метаболического или сердечно-сосудистого заболевания или в качестве стимуляторов роста мышц в производстве мяса.
Как показано в примерах 1 и 3, пероральная дозировка человеческого альфа-дефензина 5 (HD5) или человеческого бета-дефензина 2 (hBD2) предупреждает или уменьшает прирост массы у мышей, содержащихся на рационе с высоким содержанием жира. Животная модель представляет собой модель метаболического синдрома и раннего сахарного диабета 2 типа. При отсутствии лечения у мышей, выкармливаемых рационом с высоким содержанием жира, развивается ожирение за счет накопления жира, особенно абдоминального жира и жира печени. У мышей, кроме того, развиваются признаки сахарного диабета, такие как инсулиновая резистентность и ухудшенная толерантность к глюкозе.
Без HD5 или hBD2 в рационе животные значительно сильнее прибавляют в массе на рационе с высоким содержанием жира, и в конечном итоге у них развиваются признаки сахарного диабета и ожирения. Уменьшение прироста массы происходит по мере снижения накопления массы жира. Животные, получающие дозировку HD5 или hBD2, демонстрируют повышенную толерантность к глюкозе и пониженную инсулиновую резистентность по сравнению с необработанными животными на рационе с высоким содержанием жира.
Подобным образом, примеры демонстрируют, что пероральное введение HD5 животным с дисбиотической микрофлорой может по меньшей мере частично нормализовать микрофлору. Поэтому, HD5 и другие дефензины могут быть использованы для нормализации дисбиотической микрофлоры или для нормализации микрофлоры. Рацион с высоким содержанием жира и рацион с высоким содержанием сахара, как известно, индуцируют дисбиотическую микрофлору. Таким образом, дефензины могут быть использованы для лечения такой дисбиотической микрофлоры. Дефензины также могут быть использованы профилактически для субъектов, подвергающихся методам лечения, которые, как предполагается, негативно влияют на микрофлору, например, лечение антибиотиками, иммуносупрессивное лечение, химиотерапия, иммунотерапия или радиационная терапия.
Один аспект относится к способу лечения воспаления кишечника у отличных от человека животных, при этом способ предусматривает введение эффективного количества дефензина, α- и/или β-дефензина млекопитающего или птицы субъекту при необходимости этого.
Один аспект относится к способу лечения воспаления кишечника у людей, при этом способ предусматривает введение эффективного количества дефензина, α- и/или β-дефензина млекопитающего или птицы субъекту при необходимости этого.
Один аспект относится к способу лечения воспаления кишечника, причем воспаление локализуется во рту, пищеводе, желудке, двенадцатиперстной кишке, тонкой кишке, подвздошной кишке, слепой кишке, прямой кишке и/или анальном канале животного, при этом способ предусматривает введение эффективного количества дефензина, α- и/или β-дефензина млекопитающего или птицы субъекту при необходимости этого.
Один аспект относится к способу поддержания нормального состава микробиоты в кишечнике, при этом способ предусматривает введение эффективного количества дефензина, α- и/или β-дефензина млекопитающего или птицы и/или глюкагоноподобного пептида 1 (GLP-1)/аналога GLP-1 субъекту при необходимости этого.
Один аспект относится к способу лечения дисбиотической микробиоты в кишечнике, при этом способ предусматривает введение эффективного количества дефензина, α- и/или β-дефензина млекопитающего или птицы и/или глюкагоноподобного пептида 1 (GLP-1)/аналога GLP-1 субъекту при необходимости этого.
Один аспект относится к способу повышения многообразия генов кишечной микробиоты, при этом способ предусматривает введение эффективного количества дефензина, α- и/или β-дефензина млекопитающего или птицы и/или глюкагоноподобного пептида 1 (GLP-1)/аналога GLP-1.
Один аспект относится к способу повышения числа филумов кишечной микробиоты, при этом указанный способ предусматривает введение эффективного количества дефензина, α- и/или β-дефензина млекопитающего или птицы и/или глюкагоноподобного пептида 1 (GLP-1)/аналога GLP-1.
Один аспект относится к способу повышения продуцирования короткоцепочечных жирных кислот кишечной микробиотой, при этом указанный способ предусматривает введение эффективного количества дефензина, α- и/или β-дефензина млекопитающего или птицы и/или глюкагоноподобного пептида 1 (GLP-1)/аналога GLP-1.
Один аспект относится к способу повышения продуцирования бутирата или повышения продуцирования ацетата кишечной микробиоты, при этом указанный способ предусматривает введение эффективного количества дефензина, α- и/или β-дефензина млекопитающего или птицы и/или глюкагоноподобного пептида 1 (GLP-1)/аналога GLP-1.
Один аспект относится к способу повышения числа бактерий, принадлежащих роду, выбранному из группы, состоящей из Bacterioidetes, Faecalibacterium, Roseburia, Blautia, Ruminococcus, Bifidobacterium, Methanobrevibacter, Lactobacillus, Coprococcus, Clostridium, Allobaculum, Alloprevotella, Akkermansia, Eubacterium, в кишечнике, при этом указанный способ предусматривает введение эффективного количества α- и/или β-дефензина млекопитающего или птицы и/или глюкагоноподобного пептида 1 (GLP-1)/аналога GLP-1. Предпочтительно, род бактерии включает в себя один или несколько из Allobaculum, Alloprevotella, Akkermansia и Lactobacillus.
Один аспект относится к способу снижения числа бактерий, выбранных из группы, состоящей из Bacteroidetes fragilis, Sutturella wadsworthia, Veillonella parvula, Escherichi coli, Haemophilus parainfluenzae, Fusobacterium nucleatum, Eikenella corodens, Gemella moribillum, в кишечнике, при этом указанный способ предусматривает введение эффективного количества α- и/или β-дефензина млекопитающего или птицы и/или глюкагоноподобного пептида 1 (GLP-1)/аналога GLP-1.
Один аспект относится к способу лечения злокачественной опухоли толстой и прямой кишки, эндокринного, связанного с питанием, метаболического или сердечно-сосудистого заболевания, при этом указанный способ предусматривает введение эффективного количества дефензина, α- и/или β-дефензина млекопитающего или птицы субъекту при необходимости этого. Поскольку дефензины могут быть использованы для лечения ожирения и различных симптомов сахарного диабета 2 типа, а также могут быть использованы для нормализации микрофлоры, они также могут снижать риск возникновения одного или нескольких из упомянутых нарушений.
Один аспект относится к способу стимуляции роста мышц при производстве мяса животных, при этом указанный способ предусматривает введение эффективного количества дефензина, α- и/или β-дефензина млекопитающего или птицы субъекту при необходимости этого. Этот аспект подтверждается демонстрацией того, что введение HD5 или hBD2 тучным мышам ведет к снижению процента жира, т.e. дефензины способствуют преобладанию роста мышц над наращиванием жира.
Один аспект относится к композиции, содержащей по меньшей мере один α-дефензин млекопитающего или птицы и по меньшей мере один β-дефензин млекопитающего или птицы.
Один аспект относится к композиции, содержащей по меньшей мере один α- или β-дефензин млекопитающего или птицы в комбинации или с инсулином/аналогами инсулина, и/или с глюкагоноподобным пептидом-1 (GLP-1)/аналогами GLP-1, и/или с глюкагоноподобным пептидом-2 (GLP-2)/аналогами GLP-2, и/или с ингибитором дипептидилпептидазы IV (DPP-IV), и/или с метформином, и/или с ингибитором натрий-зависимого переносчика глюкозы-2 (SGLT-2), и/или с антагонистом рецептора глюкагона, и/или с антагонистом представителя 1 семейства V катионного канала транзиторного рецепторного потенциала (TRPV1), или с их комбинацией. В одном аспекте дефензином является HD5 или hBD-2.
Один аспект относится к композиции, содержащей по меньшей мере один α- или β-дефензин млекопитающего или птицы, для применения в комбинации с химиотерапией, иммунотерапией, радиотерапией или их комбинацией. В одном аспекте дефензином является HD5 или hBD-2, предпочтительно, если дефензин вводят перорально.
В уровне техники известно, что дефензины, в том числе человеческий бета-дефензин 2, являются сильными противовоспалительными средствами (WO 2010/007165). Авторы настоящего изобретения продемонстрировали эффекты перорально вводимого человеческого бета-дефензина 2 против ожирения и против сахарного диабета. Другой кишечный гормон GLP-1 и аналоги GLP-1, такие как лираглутид, также могут быть использованы для лечения ожирения и сахарного диабета. Авторы настоящего изобретения демонстрируют в примере 4, что парентерально вводимый лираглутид не влияет на различные воспалительные и противовоспалительные цитокины. Поэтому, GLP-1 и аналоги GLP-1 обладают другим способом действия в отличие от дефензинов. Поэтому, авторы настоящего изобретения рассматривают введение комбинации по меньшей мере одного дефензина по меньшей мере с одним GLP-1 или аналогом GLP-1 для лечения показаний, описанных в настоящем документе.
Краткое описание графических материалов
Фиг. 1A. Схематический план экспериментальной установки для исследования эффектов альфа- и/или бета-дефензинов млекопитающих на метаболизм мышей. На неделе -1 мышей C57/BL/6J делили на группы и клетки так, что на клетку приходилось 3 мыши и клетки на группу. От недели -1 до 0 мышей клинически обследовали с помощью магнитно-резонансного сканирования для оценки распределения жира. На неделях 0, 1 и 4 анализировали микробиом фекалий. На неделе 4 помимо анализа микробиоты мышей сканировали и измеряли содержание в крови глюкозы и инсулина. На неделе 6 оценивали потребление энергии путем анализа содержания азота и липида в фекалиях. На неделе 7 проводили тест на толерантность к инсулину (ITT). На неделе 8 проводили пероральный тест на толерантность к глюкозе (OGTT) и стимулированную глюкозой секрецию инсулина (GSIS). На неделе 9 (завершение) выполняли несколько анализов, в частности, мышей взвешивали и сканировали, а также оценивали состав плазмы и состав микробиоты толстой кишки, слепой кишки и тонкой кишки. Кроме того, выполняли гистологический анализ и анализ белка/РНК на мышечной ткани (квадрицепсе), iWAT, eWAT, iBAT, печени, толстой кишке, тонкой кишке, подвздошной кишке и двенадцатиперстной кишке.
Фиг. 1B. Схематический план экспериментальной установки для исследования эффектов альфа- и/или бета-дефензинов млекопитающих на метаболизм мышей. На неделе -1 получали мышей C57/BL/6J. На неделе 0 собирали фекалии. На протяжении вводного периода от недели 0 до недели 12 мышей кормили рационом с высоким содержанием жира. На неделе 12 мышей клинически обследовали с помощью магнитно-резонансного сканирования для оценки распределения жира, собирали фекалии и выполняли пероральный тест на толерантность к глюкозе (OGTT) и стимулированную глюкозой секрецию инсулина (GSIS). На неделе 13-0 мышей делили на группы и клетки по 4 мыши на клетку и 3 клетки на группу. На неделях 0, 12 и 13-10 анализировали микробиом фекалий. На неделях 13-2, 13-4, 13-6, 13-8 и 13-10 мышей сканировали и измеряли содержание в крови глюкозы и инсулина. На неделе 13-9 проводили тест на толерантность к инсулину (ITT). На неделе 13-10 (завершение) выполняли несколько анализов, в частности, мышей взвешивали и сканировали, а также оценивали состав плазмы и состав микробиоты толстой кишки, слепой кишки и тонкой кишки. Кроме того, измеряли iWAT, eWAT и массу печени.
Фиг. 2. Множественное выравнивание последовательностей Clustal W (2,1) человеческого бета-дефензина 1-4.
Фиг. 3. Множественное выравнивание последовательностей Clustal W (2,1) человеческого альфа-дефензина 5 и 6.
Фиг. 4. Множественное выравнивание последовательностей Clustal W (2,1) человеческого нейтрофильного пептида 1-3.
Фиг. 5. Множественное выравнивание последовательностей Clustal W (2,1) бета-дефензина 2 человека, макака-резус, шимпанзе и орангутана.
В выравниваниях Clustal W
* указывает положения, которые занимает отдельный полностью консервативный остаток;
: указывает на то, что одна из следующих «сильных» групп является полностью консервативной:
S,T,A; N,E,Q,K; N,H,Q,K; N,D,E,Q; Q,H,R,K; M,I,L,V; M,I,L,F; H,Y; F,Y,W.
. указывает на то, что одна из следующих «более слабых» групп является полностью консервативной:
C,S,A; A,T,V; S,A,G; S,T,N,K; S,T,P,A; S,G,N,D; S,N,D,E,Q,K; N,D,E,Q,H,K; N,E,Q,H,R,K; V,L,I,M; H,F,Y.
Фиг. 6. Изменение массы (A), наращивание массы (B) и суммарное потребление корма (C) на протяжении 7 недель у мышей, получавших рацион с низким содержанием жира (LFD), рацион с высоким содержанием жира (HFD) или HFD и дефензин hBD2 (HFD + P2).
A. Изменение массы. N = 12. Обе группы на HFD существенно отличаются от эталонной группы на LFD. Звездочки означают различия между группами на HFD и HFD + P2. Обе группы на HFD существенно отличаются от группы на LFD с недели w (p < 0,001). 2-факторный ANOVA, апостериорный критерий Тьюки.
B. Наращивание массы. N = 12. Обе группы на HFD существенно отличаются от эталонной группы на LFD. Звездочки означают различия между группами на HFD и HFD + P2. Обе группы на HFD существенно отличаются от группы на LFD с недели w (p < 0,001). 2-факторный ANOVA, апостериорный критерий Тьюки.
C. Суммарное потребление корма. Эталонная группа на LFD имеет одинаковое количество как сахарозы, так и белка на грамм корма по сравнению с группами на HFD.
Фиг. 7. Наращивание массы мышц/жира на протяжении 7 недель у мышей, получавших рацион с низким содержанием жира (LFD), рацион с высоким содержанием жира (HFD) или HFD и дефензин hBD2 (HFD + P2). (A) Наращивание массы мышц на неделе 1 и на неделе 7. (B) Наращивание массы жира на неделе 1 и на неделе 7. A: Масса мышц на неделе -1 и неделе 7. N = 12. Однофакторный ANOVA, апостериорный критерий Тьюки. B: Масса жира на неделе -1 и неделе 7. N = 12. Однофакторный ANOVA, апостериорный критерий Тьюки.
Фиг. 8. Гомеостаз глюкозы у мышей, получавших на протяжении 7 недель рацион с низким содержанием жира (LFD), рацион с высоким содержанием жира (HFD) или HFD и дефензин hBD2 (HFD + P2). (A) Тест на толерантность к инсулину (ITT). (B) Пероральный тест на толерантность к глюкозе. (C) Стимулированная глюкозой секреция инсулина (GSIS). (D) Тест на инсулин при 5-часовом воздержании от пищи.
A. Тест на толерантность к инсулину через 7 недель после начала получения рациона. N = 6 на группу. Группы на HFD сравниваются с эталонной группой на LFD. Отображаются только статистически значимые изменения. Звездочки над верхней кривой указывают различия между группами на HFD и LFD.
B. Тест на толерантность к глюкозе через 7 недель после начала получения рациона. N = 11-12 на группу. Группы на HFD сравниваются с эталонной группой на LFD. Отображаются только статистически значимые изменения. Звездочки над верхней кривой указывают различия между группами на HFD и LFD, а звездочки ниже средней кривой указывают различия между группами на HFD +P2 и LFD.
C. Стимулированная глюкозой секреция инсулина (в ходе GTT) через 7 недель после начала получения рациона. N = 11-12 на группу. Группы на HFD сравниваются с эталонной группой на LFD. Отображаются только статистически значимые изменения. Звездочки над верхней кривой указывают различия между группами на HFD и LFD. Показатели групп на HFD + P2 и LFD не являются статистически значимыми ни в какой момент времени.
D. Инсулин при 5-часовом воздержании от пищи через 7 недель после начала получения рациона. N = 11-12 на группу.
A-C. 2-Факторный ANOVA, апостериорный критерий Даннетта.
D. 1-Факторный ANOVA, апостериорный критерий Тьюки.
Фиг. 9A, 9B и 9C. Наращивание массы (a), эффективность корма (b) и потребление энергии (c) за 10 недель у мышей, получавших рацион с низким содержанием жира (низкое содержание жира), рацион с высоким содержанием жира (высокое содержание жира) или рацион с высоким содержанием жира и с превентивной обработкой дефензином hBD2 (высокое содержание жира + hBD-2).
Значение: низкое содержание жира по сравнению с высоким содержанием жира = A; низкое содержание жира по сравнению с высоким содержанием жира + hBD-2 = B; высокое содержание жира по сравнению с высоким содержанием жира + hBD-2 = C.
9A. Наращивание массы. Двухфакторный ANOVA с поправкой Тьюки (собранные сопоставляемые значения).
9B. Эффективность корма (грамм набранной массы с учетом среднего потребления пищи в клетке). Однофакторный ANOVA с поправкой Тьюки NB(!), n = 4, вследствие совместного содержания в клетке.
9C. Потребление энергии. Двухфакторный ANOVA с поправкой Тьюки (собранные сопоставляемые значения).
Фиг. 10A, 10B и 10C. Жир в виде процента от общей массы тела (a), масса печени в граммах (b) и масса эпидидимального жира (eWAT) в граммах (c) за 10 недель у мышей, получавших рацион с низким содержанием жира (низкое содержание жира), рацион с высоким содержанием жира (высокое содержание жира) или рацион с высоким содержанием жира и с превентивной обработкой дефензином hBD2 (hBD-2).
Значение: низкое содержание жира по сравнению с высоким содержанием жира = A; низкое содержание жира по сравнению с высоким содержанием жира + hBD-2 = B; высокое содержание жира по сравнению с высоким содержанием жира + hBD-2 = C.
10A. Процентное отношение жира от общей массы тела на разных неделях. Двухфакторный ANOVA с поправкой Тьюки (собранные сопоставляемые значения).
10B. Масса эпидидимальной жировой ткани (висцеральной AT) при завершении. Однофакторный ANOVA с поправкой Тьюки.
10C. Масса при завершении. Однофакторный ANOVA с поправкой Тьюки.
Фиг. 11A и 11B. Гомеостаз глюкозы у мышей, получавших в течение 10 недель рацион с низким содержанием жира (низкое содержание жира), рацион с высоким содержанием жира (высокое содержание жира) или рацион с высоким содержанием жира и с превентивной обработкой дефензином hBD2 (высокое содержание жира + hBD-2). (a) Пероральный тест на толерантность к глюкозе. (b) Стимулированная глюкозой секреция инсулина (GSIS).
Значение: низкое содержание жира по сравнению с высоким содержанием жира = A; низкое содержание жира по сравнению с высоким содержанием жира + hBD-2 = B; высокое содержание жира по сравнению с высоким содержанием жира + hBD-2 = C.
11A. Пероральный тест на толерантность к глюкозе на неделе 7. Двухфакторный ANOVA с поправкой Тьюки (собранные сопоставляемые значения).
11B. Стимулированная глюкозой секреция инсулина на неделе 7, рассматриваемая в ходе oGTT. Двухфакторный ANOVA с поправкой Тьюки (собранные сопоставляемые значения).
Фиг. 12A и 12B Гомеостаз глюкозы у мышей, получавших в течение 10 недель рацион с низким содержанием жира (низкое содержание жира), рацион с высоким содержанием жира (высокое содержание жира) или рационе с высоким содержанием жира и с превентивной обработкой дефензином hBD2 (высокое содержание жира + hBD-2). (a) Тест на толерантность к инсулину (ITT). (b) HOMA-IR.
Значение: низкое содержание жира по сравнению с высоким содержанием жира = A; низкое содержание жира по сравнению с высоким содержанием жира + hBD-2 = B; высокое содержание жира по сравнению с высоким содержанием жира + hBD-2 = C.
12a. Тест на толерантность к инсулину на неделе 8. Двухфакторный ANOVA с поправкой Тьюки (собранные сопоставляемые значения).
12b. Оценка модели гомеостаза (HOMA) на неделе 9. Однофакторный ANOVA с поправкой Тьюки.
Фиг. 13A и 13B. Наращивание массы (a) и изменение массы (b) за 10 недель у мышей, получавших рацион с низким содержанием жира (низкое содержание жира), рацион с высоким содержанием жира (высокое содержание жира) или рационе высоким содержанием жира и с применением обработки дефензином hBD2 (высокое содержание жира + hBD-2).
Значение: низкое содержание жира по сравнению с высоким содержанием жира = A; низкое содержание жира по сравнению с высоким содержанием жира + hBD-2 = B; высокое содержание жира по сравнению с высоким содержанием жира + hBD-2 = C.
13A. Наращивание массы. Двухфакторный ANOVA с поправкой Тьюки (собранные сопоставляемые значения).
13B. Изменение массы от недели 13 до конца вводного периода и следующие 10 недель на экспериментальных рационах. Двухфакторный ANOVA с поправкой Тьюки (собранные сопоставляемые значения).
Фиг. 14A и 14B. Жир в виде процента от общей массы тела (a) и изменение % жира от недели 0-4 в граммах (b) за 10 недель у мышей, получавших рацион с низким содержанием жира (низкое содержание жира), рацион с высоким содержанием жира (высокое содержание жира) или рацион с высоким содержанием жира и с применением обработки дефензином hBD2 (hBD-2).
Значение: низкое содержание жира по сравнению с высоким содержанием жира = A; низкое содержание жира по сравнению с высоким содержанием жира + hBD-2 = B; высокое содержание жира по сравнению с высоким содержанием жира + hBD-2 = C.
14A. Процентное отношение жира от общей массы тела на разных неделях. Двухфакторный ANOVA с поправкой Тьюки (собранные сопоставляемые значения).
14B. Изменение процентного отношения жира от конца водного периода и на протяжении 4 недель на экспериментальных рационах. Однофакторный ANOVA с поправкой Тьюки.
Фиг. 15A и 15B. Масса печени в граммах (a) и масса эпидидимального жира (eWAT) в граммах (b) за 10 недель у мышей, получавших рацион с низким содержанием жира (низкое содержание жира), рацион с высоким содержанием жира (высокое содержание жира) или рацион с высоким содержанием жира и с применением обработки дефензином hBD2 (hBD-2).
Значение: низкое содержание жира по сравнению с высоким содержанием жира = A; низкое содержание жира по сравнению с высоким содержанием жира + hBD-2 = B; высокое содержание жира по сравнению с высоким содержанием жира + hBD-2 = C.
15A. Масса печени при завершении. Однофакторный ANOVA с поправкой Тьюки.
15B. Масса эпидидимальной жировой ткани (висцерального жира) при завершении. Однофакторный ANOVA с поправкой Тьюки.
Фиг. 16A, 16B и 16C. Гомеостаз глюкозы у мышей, получавших в течение 10 недель рацион с низким содержанием жира (низкое содержание жира), рацион с высоким содержанием жира (высокое содержание жира) или рацион с высоким содержанием жира и с применением обработки дефензином hBD2 (высокое содержание жира + hBD-2). (a) Пероральный тест на толерантность к глюкозе из клетки 1. (b) Пероральный тест на толерантность к глюкозе мыши D1. (c) Тест на толерантность к инсулину (ITT).
Значение: низкое содержание жира по сравнению с высоким содержанием жира = A; низкое содержание жира по сравнению с высоким содержанием жира + hBD-2 = B; высокое содержание жира по сравнению с высоким содержанием жира + hBD-2 = C.
16A. Пероральные тесты на толерантность к глюкозе, повторяемые один раз в две недели от конца вводного периода (неделя 13-0), демонстрирующие первую клетку группы на высоком содержании жира + hBD-2.
16B. Пероральные тесты на толерантность к глюкозе, повторяемые один раз в две недели от конца вводного периода (неделя 13-0), демонстрирующие ТОЛЬКО мышь D1 из группы на высоком содержании жира + hBD-2.
16C. Тест на толерантность к инсулину на неделе 9. Двухфакторный ANOVA с поправкой Тьюки (собранные сопоставляемые значения).
Фиг. 17A, 17B и 17C. Наращивание массы (a), эффективность корма (b) и потребление энергии (c) за 10 недель у мышей, получавших рацион с низким содержанием жира (низкое содержание жира), рацион с высоким содержанием жира (высокое содержание жира) или рацион с высоким содержанием жира и с превентивной обработкой дефензином HD5 (высокое содержание жира + HD5).
Значение: низкое содержание жира по сравнению с высоким содержанием жира = A; низкое содержание жира по сравнению с высоким содержанием жира + НD-5 = B; высокое содержание жира по сравнению с высоким содержанием жира + НD-5 = C.
17A. Наращивание массы. Двухфакторный ANOVA с поправкой Тьюки (собранные сопоставляемые значения).
17B. Эффективность корма (грамм набранной массы с учетом среднего потребления пищи в клетке). Однофакторный ANOVA, поправка Тьюки NB(!), N = 4, вследствие совместного содержания в клетке.
17C. Потребление энергии. Двухфакторный ANOVA с поправкой Тьюки (собранные сопоставляемые значения).
Фиг. 18A, 18B и 18C. Жир в виде процента от общей массы тела (a), масса печени в граммах (b) и масса эпидидимального жира (eWAT) в граммах (c) за 10 недель у мышей, получавших рацион с низким содержанием жира (низкое содержание жира), рацион с высоким содержанием жира (высокое содержание жира) или рацион с высоким содержанием жира и с превентивной обработкой дефензином HD5 (HD5).
Значение: низкое содержание жира по сравнению с высоким содержанием жира = A; низкое содержание жира по сравнению с высоким содержанием жира + НD-5 = B; высокое содержание жира по сравнению с высоким содержанием жира + НD-5 = C.
18A. Процентное отношение жира от общей массы тела на разных неделях. Двухфакторный ANOVA с поправкой Тьюки (собранные сопоставляемые значения).
18B. Масса печени при завершении. Однофакторный ANOVA с поправкой Тьюки.
18C. Масса эпидидимальной жировой ткани (висцеральной AT) при завершении. Однофакторный ANOVA с поправкой Тьюки.
Фиг. 19A и 19B. Гомеостаз глюкозы у мышей, получавших в течение 10 недель рацион с низким содержанием жира (низкое содержание жира), рацион с высоким содержанием жира (высокое содержание жира) или рацион с высоким содержанием жира и с превентивной обработкой дефензином HD5 (высокое содержание жира + HD5). (a) Пероральный тест на толерантность к глюкозе. (b) Стимулированная глюкозой секреция инсулина (GSIS).
Значение: низкое содержание жира по сравнению с высоким содержанием жира = A; низкое содержание жира по сравнению с высоким содержанием жира + НD-5 = B; высокое содержание жира по сравнению с высоким содержанием жира + НD-5 = C.
19A. Пероральный тест на толерантность к глюкозе на неделе 7. Двухфакторный ANOVA с поправкой Тьюки (собранные сопоставляемые значения).
19B. Стимулированная глюкозой секреция инсулина на неделе 7 в ходе oGTT. Двухфакторный ANOVA с поправкой Тьюки (собранные сопоставляемые значения).
Фиг. 20A и 20B. Гомеостаз глюкозы у мышей, получавших в течение 10 недель рацион с низким содержанием жира (низкое содержание жира), рацион с высоким содержанием жира (высокое содержание жира) или рацион с высоким содержанием жира и с превентивной обработкой дефензином HD5 (высокое содержание жира + HD5). (a) Тест на толерантность к инсулину (ITT). (b) HOMA-IR.
Значение: низкое содержание жира по сравнению с высоким содержанием жира = A; низкое содержание жира по сравнению с высоким содержанием жира + НD-5 = B; высокое содержание жира по сравнению с высоким содержанием жира + НD-5 = C.
20A. Тест на толерантность к инсулину на неделе 8. Двухфакторный ANOVA с поправкой Тьюки (собранные сопоставляемые значения).
20B. Оценка модели гомеостаза (HOMA) на неделе 9. Однофакторный ANOVA с поправкой Тьюки.
Фиг. 21A и 21B. На данных фиг. показаны изменения от недели 1 (Wk1) до недели 10 (Wk10) для четырех обработок в примере 1: рацион с высоким содержанием жира с HD5 – HF HD5; рацион с высоким содержанием жира с hBD2 – HD hBD2; рацион с высоким содержанием жира – без обработки: HF; рацион с низким содержанием жира: LF.
Взвешенный анализ UniFrac микробиоты на неделе 1 (21A) и неделе 10 (21B), т.e. относительное содержание видов бактерий. Микробиота от мышей, получавших HFD плюс HD5, постепенно приближалась к бактериальной флоре мышей, получавших LFD, т.e. наблюдалась нормализация микробиоты. Сокращения: Wk1 – неделя 1; W10 – неделя 10.
Фиг. 21C. Иллюстрация видов микробиоты, способствующей изменению. Изменения микробиоты в основном были обусловлены повышенным содержанием Allobaculum и Lactobacillus и понижением содержания Clostridium. Allobaculum является продуцирующим короткоцепочечную жирную кислоту видом. Lactobacillus является бактерией с противовоспалительными свойствами.
Фиг. 22A и 22B. Наращивание массы (a) и изменение массы (b) за 10 недель у мышей, получавших рацион с низким содержанием жира (низкое содержание жира), рацион с высоким содержанием жира (высокое содержание жира) или рацион с высоким содержанием жира и с применением обработки дефензином HD5 (высокое содержание жира + HD5).
Значение: низкое содержание жира по сравнению с высоким содержанием жира = A; низкое содержание жира по сравнению с высоким содержанием жира + НD-5 = B; высокое содержание жира по сравнению с высоким содержанием жира + НD-5 = C.
22A. Наращивание массы. Двухфакторный ANOVA с поправкой Тьюки (собранные сопоставляемые значения).
22B. Изменение массы от недели 13 до конца вводного периода и следующие 10 недель на экспериментальных рационах. Двухфакторный ANOVA с поправкой Тьюки (собранные сопоставляемые значения).
Фиг. 23A и 23B. Жир в виде процента от общей массы тела (a) и изменение % жира от недели 0-4 в граммах (b) за 10 недель у мышей, получавших рацион с низким содержанием жира (низкое содержание жира), рацион с высоким содержанием жира (высокое содержание жира) или рацион с высоким содержанием жира и с применением обработки дефензином HD5 (HD-5).
Значение: низкое содержание жира по сравнению с высоким содержанием жира = A; низкое содержание жира по сравнению с высоким содержанием жира + НD-5 = B; высокое содержание жира по сравнению с высоким содержанием жира + НD-5 = C.
23A. Процентное отношение жира от общей массы тела на разных неделях. Двухфакторный ANOVA с поправкой Тьюки (собранные сопоставляемые значения).
23B. Изменение процентного отношения жира от конца водного периода и до недели 4 на экспериментальных рационах. Однофакторный ANOVA с поправкой Тьюки.
Фиг. 24A и 24B. Масса печени в граммах (a) и масса эпидидимального жира (eWAT) в граммах (b) за 10 недель у мышей, получавших рацион с низким содержанием жира (низкое содержание жира), рацион с высоким содержанием жира (высокое содержание жира) или рацион с высоким содержанием жира и с применением обработки дефензином HD5 (HD-5).
Значение: низкое содержание жира по сравнению с высоким содержанием жира = A; низкое содержание жира по сравнению с высоким содержанием жира + НD-5 = B; высокое содержание жира по сравнению с высоким содержанием жира + НD-5 = C.
24A. Масса печени при завершении. Однофакторный ANOVA с поправкой Тьюки.
24B. Масса эпидидимальной жировой ткани (висцеральной AT) при завершении. Однофакторный ANOVA с поправкой Тьюки.
Фиг. 25A и 25B. Гомеостаз глюкозы у мышей, получавших в течение 10 недель рацион с низким содержанием жира (низкое содержание жира), рацион с высоким содержанием жира (высокое содержание жира) или рацион с высоким содержанием жира и с применением обработки дефензином HD5 (высокое содержание жира + HD5). (a) Пероральный тест на толерантность к глюкозе из клетки 2 (b) Тест на толерантность к инсулину (ITT).
Значение: низкое содержание жира по сравнению с высоким содержанием жира = A; низкое содержание жира по сравнению с высоким содержанием жира + НD-5 = B; высокое содержание жира по сравнению с высоким содержанием жира + НD-5 = C.
25A. Пероральные тесты на толерантность к глюкозе, повторяемые один раз в две недели от конца вводного периода (неделя 13-0), демонстрирующие вторую клетку группы на высоком содержании жира + HD-5.
25B. Тест на толерантность к инсулину на неделе 9. Двухфакторный ANOVA с поправкой Тьюки (собранные сопоставляемые значения).
Фиг. 26. Схематический план экспериментальной установки для исследования эффектов аналога GLP-1 (лираглутида) в отношении воспаления кишечника и микробиоты мыши. На неделе -40 получали мышей C57/BI/6J DIO. Мышей кормили рационом с высоким содержанием жира 60%, SSNIFF (рационом № D12492) или Рurina Сhow в течение 38 недель до достижения средней массы тела 55 грамм. С недели -2 мышей размещали по отдельности. Образцы фекалий собирали в дни -1 и 27 для анализа 16S РНК. Образцы из подвздошной кишки собирали в 2 см от слепой кишки в день 28.
Фиг. 27A. Результаты микробиомного анализа из примера 4. Мышей обрабатывали в течение четырех недель лираглутидом или средой-носителем (средой-носителем DIO).
Невзвешенный анализ UniFrac микробиоты в день -1 и день 28 показывает изменения в микробиоме двух групп с обработкой от начала до конца эксперимента. Микробиота от мышей, получавших HFD плюс лираглутид, постепенно приближалась к бактериальной флоре мышей, получавших LFD, тогда как микробиота обработанных средой-носителем мышей не изменялась на протяжении исследования.
Фиг. 27B. Изменения микробиоты с видами добавленной микробиоты. Изменения в основном были обусловлены повышенным содержанием Akkermansia и Alloprevotella. Akkermansia включает в себя продуцирующие короткоцепочечную жирную кислоту виды.
Фиг. 28. Фармакокинетические данные после перорального введения 4 мг/кг hBD-2 самкам мышей NMRI. По оси Y показан hBD2 в мкг/г ткани. Результаты приводятся как среднее по группе +/- SEM.
Фиг. 29. Фармакокинетические данные для hBD-2 после подкожного (SC) и внутривенного (IV) введения 1 мг/кг, соответственно. По оси Y показан hBD2 в мкг/мл. Разные кривые представляют разные эксперименты и способы выявления (HPLC и ELISA).
Фиг. 30. Фармакокинетические данные для «N-концевого слияния hBD-2-альбумин» после подкожного и внутривенного введения 16,5 мг/кг, соответственно. По оси Y показана концентрация белка слияния в мкг/мл. Результаты представляют собой среднее от 4 мышей/время отбора образцов +/- SD.
Фиг. 31. Фармакокинетические данные для «С-концевого слияния hBD-2-альбумин» после подкожного и внутривенного введения 16,5 мг/кг, соответственно. По оси Y показана концентрация белка слияния в мкг/мл. Результаты представляют собой среднее от 4 мышей/время отбора образцов +/- SD.
Подробное раскрытие настоящего изобретения
Определения
Дефензин. Используемый в настоящем документе термин «дефензин» относится к полипептидам, известным специалисту в данной области как принадлежащие дефензиновому классу противомикробных пептидов. Дефензины принадлежат классу альфа-дефензинов или классу бета-дефензинов. Примеры дефензинов включают в себя человеческий кишечный альфа-дефензин 5 (HD5; SEQ ID NO. 8); человеческий альфа-дефензин 6 (HD6; SEQ ID NO. 9); человеческий нейтрофильный пептид 1 (HNP-1); человеческий нейтрофильный пептид 2 (HNP-2); человеческий нейтрофильный пептид 3 (HNP-3), все из которых принадлежат классу альфа-дефензинов; а также человеческий бета-дефензин 1 (hBD1; SEQ ID NO. 4); человеческий бета-дефензин 2 (hBD2; SEQ ID NO. 5); человеческий бета-дефензин 3 (hBD3; SEQ ID NO. 6); человеческий бета-дефензин 4 (hBD4; SEQ ID NO. 7), бета-дефензин 2 шимпанзе (SEQ ID NO: 10), бета-дефензин 2 макака (SEQ ID NO: 11), бета-дефензин 2 орангутана (SEQ ID NO: 3), бета-дефензин 3 мыши (SEQ ID NO: 12), бета-дефензин 2 лошади (SEQ ID NO: 13), бета-дефензин 1 свиньи (SEQ ID NO: 14), бета-дефензин 2 козла (SEQ ID NO: 15), бета-дефензин 2 быка (SEQ ID NO: 1), бета-дефензин 2 курицы (SEQ ID NO: 2), LL37 человека (SEQ ID NO: 16), полученный из человеческого кателицидина, и усеченный hBD2 (SEQ ID NO: 17), принадлежащий классу бета-дефензинов. Дефензины могут быть гликозилированы, и дефензины могут быть протеолитически расщеплены на более мелкие биоактивные фрагменты. Гликозилированные дефензины и фрагменты дефензина также включены в объем настоящего раскрытия.
Дефензины экспрессируются как предшественники и процессируются расщеплением сигнального пептида, а в некоторых случаях пропептидов, а также перед секрецией во внеклеточное пространство. Указанные выше последовательности представляют прогнозированные зрелые биоактивные дефензины. Специалисту в данной области будет понятно, что процессирование может отличаться от клетки к клетке, и что полученный в результате секретированный зрелый пептид может отличаться одним или обеими C- или N-концевыми аминокислотами от прогнозированных последовательностей и все же сохранять свою биоактивность.
Кишечник. Кишечник представляет собой трубку, используемую животными для перемещения пищи в пищеварительные органы, и включает в себя собственно пищеварительные органы. Используемый в настоящем документе термин «кишечник человека» относится к пищеварительной системе, состоящей из рта, пищевода, желудка, двенадцатиперстной кишки, тонкой кишки, подвздошной кишки, слепой кишки, толстой кишки, прямой кишки и анального канала. Некоторые варианты осуществления относятся к частям человеческого кишечника и, в частности, ко рту, пищеводу, желудку, двенадцатиперстной кишке, тонкой кишке, подвздошной кишке, слепой кишке, толстой кишке, прямой кишке и анальному каналу. Другие варианты осуществления относятся ко всем эти частям за исключением толстой кишки. Кишечник жвачных животных, описываемый в настоящем документе, представляет собой кишечник, состоящий из рта, пищевода, желудка, двенадцатиперстной кишки, тонкой кишки, подвздошной кишки, слепой кишки, толстой кишки, прямой кишки и анального канала, но характеризуется тем фактом, что желудок делится на четыре отдела - рубец, сетку, книжку и сычуг. Кишечник птиц, описываемый в настоящем документе, представляет собой кишечник, состоящий из пищевода, желудка, двенадцатиперстной кишки, тонкой кишки, подвздошной кишки, слепой кишки, толстой кишки, прямой кишки и анального канала, но характеризуется тем фактом, что желудок делится на железистый желудок или истинный желудок и мускульный желудок. В некоторых случаях имеется мускульный мешок вдоль пищевода, называемый зобом.
Глюкагоноподобный пептид-1. GLP-1 представляет собой нейропептид и инкретин, полученный из продукта транскрипции гена проглюкагона. Основным источником GLP-1 в периферии является L-клетка кишечника, которая секретирует GLP-1 как гормон кишечника. Биологически активными формами GLP-1 являются GLP-1-(7-37) и GLP-1-(7-36)NH2. Эти пептиды являются результатом селективного расщепления молекулы проглюкагона.
Секреция GLP-1 L-клетками кишечника зависит от присутствия питательных веществ в просвете тонкого кишечника. Стимуляторы секреции (средства, которые вызывают или стимулируют секрецию) этого гормона включают в себя основные питательные вещества, такие как углеводы, белки и липиды. GLP-1 характеризуется периодом полувыведения из кровотока менее 2 минут из-за быстрого разложения ферментом дипептидилпептидазой-4.
GLP-1 является мощным антигипергликемическим гормоном, индуцирующим высвобождение из β-клеток поджелудочной железы гормона инсулина в ответ на повышение содержания глюкозы с подавлением тем самым секреции глюкагона. Такое зависимое от глюкозы действие является особенно привлекательным, поскольку нерегулируемое высвобождение инсулина, если концентрация глюкозы в плазме находится в нормальном диапазоне натощак, или при несвоевременных инъекциях инсулина, может вызывать опасное падение содержания глюкозы в крови - гипогликемию. Это не происходит в результате действия GLP-1, потому что GLP-1 более не стимулирует высвобождение из β-клеток большего количества инсулина, если содержание глюкозы в крови находится в диапазоне натощак. Кроме того, GLP-1 ингибирует секрецию и моторику желудка. Это задерживает и затягивает всасывание углеводов и способствует эффекту насыщения.
Идентичность. Сходство между двумя аминокислотными последовательностями или между двумя нуклеотидными последовательностями описывается параметром «идентичность».
Степень идентичности между двумя аминокислотными последовательностями определяют с использованием алгоритма Нидлмана-Вунша (Needleman and Wunsch, 1970, J. Mol. Biol. 48: 443-453), внедренного в программу Needle пакета EMBOSS (Rice et al., 2000, http://emboss.org), предпочтительно версии 3,0,0 или более поздней. Используемыми опционными параметрами являются штраф за открытие гэпа 10, штраф за продление гэпа 0,5 и матрица замен EBLOSUM62 (версия EMBOSS от BLOSUM62). Выходные данные, обозначенные в Needle как «наиболее длинная идентичность» (полученные с использованием опции –nobrief), используют как процент идентичности и рассчитывают следующим образом:
(идентичные остатки × 100)/(длина выравнивания – общее количество гэпов в выравнивании).
Стимуляция роста мышц. Используемый в настоящем документе термин «стимуляция роста мышц» или «усиление роста мышц» относится к откармливанию домашнего скота или домашних животных, например, коров, свиней, овец, коз, лошадей, уток, гусей, голубей, индюков, фазанов и цыплят, при производстве мяса, если целью является быстрое повышение массы тела за счет мышц.
Домашний скот. Крупный рогатый скот, лошади, птицы и подобные животные, содержащиеся для домашнего применения.
Нормальная микробиота. Используемый в настоящем документе термин «нормальная микробиота» означает микробиоту, которая не является дисбиотической. Нормальная микробиота характеризуется наличием большого разнообразия генов и филумов. Нормальная микробиота характеризуется наличием бактерий, принадлежащих родам Bacterioidetes, Faecalibacterium, Roseburia, Blautia, Ruminococcus, Coprococcus, Bifidobacterium, Methanobrevibacter, Lactobacillus, Coprococcus, Clostridium, Akkermansia, Eubacterium.
Лечение. Используемые в настоящем документе термины «лечение» и «процесс лечения» относятся к наблюдению больного и уходу за ним с целью борьбы с состоянием, заболеванием или нарушением. Данные термины включают в себя полный спектр методов лечения данного состояния, которым страдает больной, например, введение активного соединения с целью ослабления или облегчения симптомов или осложнений; замедления прогрессирования состояния, заболевания или нарушения; излечения или устранения состояния, заболевания или нарушения; и/или предупреждения состояния, заболевания или нарушения, при этом термин «предупреждения» или «профилактика» относится к наблюдению больного и уходу за ним с целью торможения, снижения или замедления развития состояния, заболевания или нарушения и включает в себя введение активных соединений для предупреждения или уменьшения риска возникновения симптомов или осложнений. Больным, подлежащим лечению, предпочтительно является млекопитающее, в частности, человек. Больные, подлежащие лечению, могут быть разного возраста.
Субъект, больной. Субъект представляет собой индивидуума одного из видов млекопитающих или птиц, раскрываемых в настоящем документе. Больной представляет собой субъекта, у которого диагностировали конкретное нарушение.
Альфа-дефензины млекопитающего и птицы и бета-дефензины млекопитающего и птицы
Настоящее раскрытие относится к применениям дефензинов, альфа- и/или бета-дефензинов млекопитающего и птицы, таких как бета-дефензины быка, свиньи, овцы, мыши, обезьяны, лошади и птицы, например, цыпленка, индюка, утки, гуся, фазана, голубя, мыши, обезьяны или человека, более предпочтительно Hominidae, более предпочтительно человеческих альфа- и/или бета-дефензинов, в лечении показаний, раскрываемых в настоящем документе, в том числе без ограничения воспаления кишечника, или злокачественной опухоли толстой и прямой кишки, или эндокринного, связанного с питанием, метаболического или сердечно-сосудистого заболевания.
Фрагмент LL37 кателицидина также предусматривается для применений в соответствии с настоящим изобретением. LL37 имеет последовательность из SEQ ID NO: 16.
Согласно конкретным предпочтительным вариантам осуществления дефензинами являются альфа- или бета-дефензины.
Согласно варианту осуществления LL37 - альфа- и/или бета-дефензины млекопитающих – имеют степень идентичности по меньшей мере 80%, предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95% по отношению к любой из аминокислотных последовательностей из SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 14, SEQ ID NO: 15, SEQ ID NO: 16 и/или SEQ ID NO 17. Согласно другому варианту осуществления дефензин отличается от одной из SEQ ID NO: 1-17 менее чем 10, например, менее чем 8, например, менее чем 5, например, менее чем 4, например, менее чем 3, например, менее чем 2 аминокислотами.
Согласно предпочтительному варианту осуществления человеческие альфа-дефензины включают в себя альфа-дефензин 5 (SEQ ID NO: 8) и/или альфа-дефензин 6 (SEQ ID NO: 9). Согласно предпочтительному варианту осуществления бета-дефензины млекопитающего включают в себя человеческий бета-дефензин 1 (SEQ ID NO: 4), человеческий бета-дефензин 2 (SEQ ID NO: 5), человеческий бета-дефензин 3 (SEQ ID NO: 6), человеческий бета-дефензин 4 (SEQ ID NO: 7) и/или усеченный человеческий бета-дефензин 2 (SEQ ID NO 17).
Согласно предпочтительному варианту осуществления человеческий альфа-дефензин имеет степень идентичности по меньшей мере 80%, предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95% по отношению к аминокислотной последовательности из SEQ ID NO: 8. Согласно предпочтительному варианту осуществления человеческие альфа-дефензины включают в себя альфа-дефензин 5 (SEQ ID NO: 8). Согласно предпочтительному варианту осуществления человеческий бета-дефензин имеет степень идентичности по меньшей мере 80%, предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95% по отношению к аминокислотной последовательности из SEQ ID NO: 5. Согласно предпочтительному варианту осуществления человеческий бета-дефензин включает в себя человеческий бета-дефензин 2 (SEQ ID NO: 5). Другим предпочтительным человеческим бета-дефензином является усеченный человеческий бета-дефензин 2 (SEQ ID NO: 17). Усеченный hBD2 (SEQ ID NO: 17) обладает противовоспалительными эффектами в отличие от hBD2 (SEQ ID NO: 5) (WO 2013/026794).
Что касается отличных от человека видов, субъектов предпочтительно лечат дефензином, полученным от тех же или родственных видов, или дефензином, общим по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90% и наиболее предпочтительно по меньшей мере на 95% по отношению к аминокислотной последовательности дефензина от тех же видов (например, дефензин имеет аминокислотную последовательность, выбранную из SEQ ID NO: 1-3 и 10-15). Например, можно предположить, что птицу можно лечить ортологичным дефензином от тех же или других видов птиц.
Согласно еще одному варианту осуществления альфа-дефензины млекопитающего включают в себя человеческие альфа-дефензины и/или мышиные альфа-дефензины, а также их функционально эквивалентные варианты. Предпочтительно, альфа-дефензином млекопитающего является человеческий альфа-дефензин, который может включать в себя человеческий альфа-дефензин 5, человеческий альфа-дефензин 6, а также их функционально эквивалентные варианты. Более предпочтительно альфа-дефензины млекопитающего включают в себя человеческий альфа-дефензин 5 и его функционально эквивалентные варианты или ортологи.
Согласно следующему варианту осуществления бета-дефензины млекопитающего включают в себя человеческие бета-дефензины и/или мышиные бета-дефензины, а также их функционально эквивалентные варианты. Предпочтительно, бета-дефензины млекопитающего или птицы включают в себя человеческий бета-дефензин 1, человеческий бета-дефензин 2, человеческий бета-дефензин 3, человеческий бета-дефензин 4, бета-дефензин 2 шимпанзе, бета-дефензин 2 макака и бета-дефензин 3 мыши, бета-дефензин 2 орангутана, бета-дефензин 2 лошади, бета-дефензин 1 свиньи, бета-дефензин 2 козла, бета-дефензин 2 быка, бета-дефензин 2 цыпленка, а также их функционально эквивалентные варианты. Более предпочтительно бета-дефензины млекопитающего включают в себя человеческий бета-дефензин 1, человеческий бета-дефензин 2, человеческий бета-дефензин 3, человеческий бета-дефензин 4, а также их функционально эквивалентные варианты. Еще более предпочтительно бета-дефензины млекопитающего включают в себя человеческий бета-дефензин 2 и его функционально эквивалентные варианты или ортологи.
Согласно одному варианту осуществления способы предусматривают введение эффективного количества по меньшей мере одного α-дефензина млекопитающего или птицы субъекту при необходимости такого лечения. Согласно другим вариантам осуществления представленные способы предусматривают введение эффективного количества по меньшей мере одного β-дефензина млекопитающего или птицы субъекту при необходимости такого лечения. Согласно следующему варианту осуществления представленные способы предусматривают введение эффективного количества по меньшей мере одного α-дефензина млекопитающего или птицы и по меньшей мере одного β-дефензина млекопитающего или птицы субъекту при необходимости такого лечения. Предпочтительный вариант осуществления относится к введению альфа-дефензина млекопитающего HD5 и/или бета-дефензина млекопитающего hBD-2
«Функционально эквивалентным вариантом» альфа- или бета-дефензина млекопитающего (например, человека) или птицы является модифицированный альфа- или бета-дефензин млекопитающего (например, человека) или птицы, оказывающий аппроксимативно такой же эффект в отношении микробиоты в кишечнике, как и исходные альфа- и/или бета-дефензины млекопитающего (например, человека) или птицы. Функционально эквивалентный вариант дефензина млекопитающего (например, человека) или птицы может содержать 1-5 аминокислотных модификаций, предпочтительно 1-4 аминокислотных модификации, более предпочтительно 1-3 аминокислотных модификации, наиболее предпочтительно 1-2 аминокислотных модификации и, в частности, одну аминокислотную модификацию, по сравнению с аминокислотной последовательностью дефензина млекопитающего (например, человека) или птицы. Предпочтительно бета-дефензины млекопитающего сравнивать с человеческим бета-дефензином 2, имеющим SEQ ID NO 5.
Термин «модификация» в настоящем документе означает любую химическую модификацию дефензина млекопитающего (например, человека) или птицы. Модификацией(ями) может быть замена(ы), делеция(и) и/или вставка(и) аминокислоты(аминокислот), а также замещение(ия) аминокислотной боковой цепи(ей); или применение ненатуральных аминокислот с подобными характеристиками в аминокислотной последовательности. В частности, модификацией(ями) могут быть амидирования, например, амидирование C-конца.
Предпочтительно аминокислотные модификации являются не существенными, то есть консервативными аминокислотными заменами или вставками, которые существенно не влияют на укладку и/или активность полипептида; отдельными делециями; небольшими амино- или карбокси-концевыми удлинениями или небольшим удлинением, которое облегчает очистку путем изменения суммарного заряда или другой функции, например, полигистидиновая метка, антигенный эпитоп или связывающий домен. Согласно одному варианту осуществления небольшое удлинение, такое как полигистидиновая метка, антигенный эпитоп или связывающий домен, присоединяют к альфа- или бета-дефензину млекопитающего (например, человека) или птицы через небольшой линкерный пептид до приблизительно 20-25 остатков, и указанный линкер может содержать сайт расщепления ферментом рестрикции. Могут быть использованы выравнивания Clustal W, как на фиг. 2-5, для прогнозирования того, какие аминокислотные остатки могут быть замещены без существенного влияния на биологическую активность белка. Последовательности выравнивали с использованием Clustal W 2,1 (http://www.geno,me.jp/tools/clustalw/) и следующих установок: штраф за открытие гэпа: 10, штраф за продление гэпа: 0,05, вес транзиции: NO, гидрофильные остатки для белков: GPSNDQE, гидрофильные гэпы: YES, матрица сравнения: BLOSUM (для белка). Замены в следующей группе (Clustal W, группа «сильной» консервативности) рассматривают как консервативные замены:
- S,T,A; N,E,Q,K; N,H,Q,K; N,D,E,Q; Q,H,R,K; M,I,L,V; M,I,L,F; H,Y; F,Y,W.
Замены в следующей группе (Clustal W, группа «слабой» консервативности) рассматривают как полуконсервативные замены:
- C,S,A; A,T,V; S,A,G; S,T,N,K; S,T,P,A; S,G,N,D; S,N,D,E,Q,K; N,D,E,Q,H,K; N,E,Q,H,R,K; V,L,I,M; H,F,Y.
Примеры консервативных замен являются замены, выполненные в группе оснóвных аминокислот (аргинин, лизин и гистидин), кислых аминокислот (глутаминовая кислота и аспарагиновая кислота), полярных аминокислот (глутамин и аспарагин), гидрофобных аминокислот (лейцин, изолейцин и валин), ароматических аминокислот (фенилаланин, триптофан и тирозин) и малых аминокислот (глицин, аланин, серин, треонин и метионин). Аминокислотные замены, которые обычно не изменяют специфическую активность, известны в уровне техники и описаны, например, у Neurath and Hill (1979). Наиболее часто встречающимися заменами являются Ala/Ser, Val/Ile, Asp/Glu, Thr/Ser, Ala/Gly, Ala/Thr, Ser/Asn, Ala/Val, Ser/Gly, Tyr/Phe, Ala/Pro, Lys/Arg, Asp/Asn, Leu/Ile, Leu/Val, Ala/Glu и Asp/Gly.
В дополнение к 20 стандартным аминокислотам нестандартными аминокислотами (такими как 4-гидроксипролин, 6-N-метиллизин, 2-аминоизомасляная кислота, изовалин и альфа-метилсерин) можно заменить аминокислотные остатки в порлипептиде дикого типа. Аминокислотные остатки можно заменять ограниченным числом неконсервативных аминокислот, аминокислот, которые не кодируются генетическим кодом, и ненатуральных аминокислот. «Ненатуральные аминокислоты» были модифицированы после синтеза белков и/или имеют химическую структуру в своей боковой цепи(ях), отличную от таковой у стандартных аминокислот. Ненатуральные аминокислоты могут быть химически синтезированы, предпочтительно являются коммерчески доступными и включают в себя пипеколиновую кислоту, тиазолидиновую карбоновую кислоту, дегидропролин, 3- и 4-метилпролин и 3,3-диметилпролин.
Незаменимые аминокислоты в альфа- и/или бета-дефензине млекопитающего или птицы могут быть идентифицированы согласно процедурам, известным в уровне техники, таким как сайт-направленный мутагенез или аланин-сканирующий мутагенез (Cunningham and Wells, 1989, Science 244: 1081-1085). В последней методике отдельные мутации аланина вводят на каждый остаток в молекуле и полученные в результате мутантные молекулы тестируют на предмет биологической активности (т.e. активности в отношении воспалительного заболевания кишечника и/или подавления активности TNF-альфа) для идентификации аминокислотных остатков, которые важны для активности молекулы. См. также, Hilton et al., 1996, J. Biol. Chem. 271: 4699-4708. Идентичности незаменимых аминокислот также могут быть определены с помощью анализа идентичностей с полипептидами, которые являются родственными по отношению к альфа- и/или бета-дефензинам млекопитающего или птицы (см. выравнивание Clustal W на фиг. 2-5).
Замены одной или нескольких аминокислот могут быть выполнены и протестированы с использованием известных способов мутагенеза, рекомбинации и/или шаффлинга с последующей процедурой соответствующего скрининга, такой как раскрываемые в Reidhaar-Olson and Sauer, 1988, Science 241: 53-57; Bowie and Sauer, 1989, Proc. Natl. Acad. Sci. USA 86: 2152-2156; WO 95/17413 или WO 95/22625. Другие способы, которые могут быть использованы, включают в себя ПЦР с внесением ошибок, фаговый дисплей (например, Lowman et al., 1991, Biochem. 30:10832-10837; патент США №5223409; WO 92/06204) и направленный на участок мутагенез (Derbyshire et al., 1986, Gene 46:145; Ner et al., 1988, DNA 7:127).
Если результат данной замены не может быть прогнозирован с определенностью, можно быстро проанализировать производные согласно способам, описанным в настоящем документе выше, для определения присутствия или отсутствия биологической активности.
Дефензины, раскрываемые в настоящем документе, могут быть подвергнуты гликозилированию. Кроме того, в уровне техники известно, что встречающиеся в природе дефензины могут быть подвергнуты протеолитическому процессированию и могут быть расщеплены на более мелкие биоактивные фрагменты. Гликозилированные дефензины и биоактивные фрагменты дефензинов включены в настоящее раскрытие.
Кроме того, в объем настоящего раскрытия включены индукторы дефензинов. В уровне техники известно, что, например, витамин D и E. coli Nissle могут индуцировать секрецию дефензинов и, таким образом, могут быть использованы для лечения показаний, описанных в настоящем документе.
Комбинации альфа- и бета-дефензинов
В одном аспекте представлена композиция, содержащая по меньшей мере один α-дефензин млекопитающего или птицы и по меньшей мере один β-дефензин млекопитающего или птицы. Как показано в примерах 1 и 3, альфа- и бета-дефензины могут быть введены перорально. Типичный альфа-дефензин HD5, в частности, снижает эктопическое накопление липидов, и типичный бета-дефензин hBD2, в частности, улучшает глюкорегуляторный путь. Поэтому, комбинация альфа- и бета-дефензинов может обеспечивать особенно эффективное лечение ожирения и эндокринных показаний, описанных в настоящем документе.
α-дефензин млекопитающего может быть выбран из группы, состоящей из HD5 и HD6, а по меньшей мере один β-дефензин млекопитающего может быть выбран из hBD-1, hBD-2, hBD-3 и hBD-4.
Предпочтительно композиция содержит HD5 и hBD-2.
Композиция дополнительно может содержать фармацевтически приемлемое вспомогательное средство, является стерильной и может быть составлена в виде стерильного и изотонического раствора.
Соотношением альфа-дефензина к бета-дефензину может быть любое соотношение. Согласно некоторым вариантам осуществления композиция содержит, по сути, равные количества по меньшей мере одного α-дефензина млекопитающего или птицы и по меньшей мере одного β-дефензина млекопитающего или птицы, выраженные в молях, или в массе, или в мг/мл.
Длительно действующие дефензины
Период полувыведения α- или β-дефензина может быть продлен с помощью слияния или конъюгации α- или β-дефензина с другой молекулой, т.e. конструирования длительно действующего биологически активного α- или β-дефензина, связанного с фармацевтически приемлемой молекулой, что обеспечивает in vivo период полувыведения α- или β-дефензина из плазмы, который существенно продлен по сравнению с in vivo периодом полувыведения из плазмы α- или β-дефензина, введенного тем же способом, что и α- или β-дефензин.
Длительно действующий биологически активный α- или β-дефензин включает в себя α-дефензин млекопитающего или его аналог или β-дефензин млекопитающего или его аналог, связанный с фармацевтически приемлемой молекулой, выбранной из молекулы, связывающейся с неонатальным Fc-рецептором млекопитающего, трансферрином или CH3(CH2)nCO-, в котором n равняется 8-22, или с полимером.
α- или β-дефензиновый агонист также может иметь отличное от млекопитающего происхождение и может быть выбран из малых органических молекул, пептидов, полипептидов и белков.
α- или β-дефензиновый агонист может быть связан с фармацевтически приемлемой молекулой различными путями, описанными в имеющейся в уровне техники литературе, такими как без ограничения химическое сочетание посредством бифункционального линкера, генной технологией с помощью сочетания N-конца или C-конца дефензина, такого как α-дефензин или β-дефензин, с фармацевтически приемлемой молекулой, такой как альбумин или аналог альбумина. В частности, N-конец альбумина или аналога альбумина, например, человеческого альбумина, может быть введен в реакцию сочетания с C-концом α-дефензина или β-дефензина или N-концом α- или β-дефензина; или C-конец альбумина, например, человеческого альбумина, может быть введен в реакцию сочетания с C-концом α-дефензина или β-дефензина или N-концом α- или β-дефензина. Может быть вставлена линкерная последовательность между альбумином и цепью α- или β-дефензина.
α- или β-дефензиновый агонист может быть связан с фармацевтически приемлемой молекулой через стабильный линкер или более лабильный линкер. В уровне техники известно несколько линкеров, в том числе бифункциональные молекулы PEG (см., например, Paige et.al. Pharmaceutical Research, vol. 12, no. 12, 1995), гидролизируемые линкеры (Shechter et al. Bioconjugate Chem. 2005,16: 913- 920, International Journal of Peptide Research and Therapeutics, Vol. 13, Nos. 1-2, June 2007, и W02009095479), PDPH и EMCH, см., например, WО 2010092135. В особом случае, если химическая конъюгация (связывание двух или более молекул) α- или β-дефензинового агониста с фармацевтически приемлемой молекулой сильно снижает функциональную активность α- или β-дефензина, может быть предпочтительным применение более лабильного линкера, который может высвобождать функциональный α- или β-дефензиновый агонист.
Удлинение периода полувыведения также может быть выполнено с помощью ацилирования пептидного остова со спейсером, например, γ-L-глутамиловым спейсером, и цепи C-18 жирной двухосновной кислоты с лизином. Боковая цепь жирной двухосновной кислоты и спейсер опосредуют прочное, но обратимое, связывание с альбумином, замедляющее высвобождение из участка инъекции и снижение почечного выведения.
Способы и применения
Как показано в примере 4, введение лираглутида - аналога GLP-1 – ведет к изменениям в микрофлоре у тучных мышей, получающих рационы высоким содержанием жира. Изменения касаются оздоровления или нормализации микрофлоры inter alia при повышении числа видов бактерий, которые способствуют продуцированию короткоцепочечных жирных кислот. Поэтому, авторы настоящего изобретения рассматривают лечение дисбиотической микрофлоры и другие применения, описанные в настоящем документе, с помощью введения GLP-1 или аналога GLP-1.
Предпочтительно GLP-1 или аналоги GLP-1 вводят парентерально посредством либо подкожного, либо внутримышечного введения. Аналог GLP-1 может быть выбран из эксенатида, лираглутида, ликсисенатида, албиглутида и дулаглутида.
Выяснили, что человеческий альфа-дефензин 5 и человеческий бета-дефензин 2 способны поддерживать или стабилизировать нормальную микробиоту в кишечнике и даже лечить или нормализовать дисбиотическую микробиоту в кишечнике, демонстрируя, таким образом, мощную активность в качестве лекарственного препарата для лечения злокачественной опухоли толстой и прямой кишки, воспаления кишечника, эндокринного, связанного с питанием, метаболического или сердечно-сосудистого заболеваний или в качестве стимуляторов роста мышц. Поэтому, один аспект относится к способам лечения воспаления кишечника в целом или лечения злокачественной опухоли толстой и прямой кишки, эндокринного, связанного с питанием, метаболического или сердечно-сосудистого заболеваний путем введения эффективного количества α-дефензина и/или β-дефензина млекопитающего субъекту при необходимости такого лечения. Примерами таких заболеваний являются сахарный диабет 1 типа, сахарный диабет 2 типа, метаболический синдром, системное неспецифическое воспаление, ожирение, инсулиновая резистентность, нарушение толерантности к глюкозе и сердечно-сосудистое заболевание.
В частности, продемонстрировали, что HD5 и hBD2 могут быть использованы для лечения инсулиновой резистентности путем улучшения чувствительности к инсулину и толерантности к глюкозе, а также для лечения или предупреждения ожирения. HD5, в частности, может снижать эктопическое накопление липидов, тогда как hBD2, в частности, может улучшать глюкорегуляторную эффективность.
Предупреждение ожирения, или индуцирование потери массы, или предупреждение прироста массы предпочтительно предусматривают снижение или предупреждение накопления висцерального жира, снижение или предупреждение повышения процентного отношения жира, или снижение, или предупреждение увеличения окружности талии.
Представленными способами можно лечить или предупреждать воспаление кишечника за счет изменения бактериальных фенотипов посредством изменения на уровне транскрипции, а также структуры и состава кишечной бактериальной флоры у субъекта, пораженного одним из указанных состояний, описанных в настоящем документе.
Представленными способами можно лечить злокачественную опухоль толстой и прямой кишки, эндокринное, связанное с питанием, метаболическое или сердечно-сосудистое заболевания путем изменения структуры и состава кишечной микробиоты и, таким образом, метаболома у субъекта, пораженного одним из указанных состояний, описанных в настоящем документе.
Один аспект относится к способам лечения воспаления кишечника у человека, при этом воспаление локализуется во рту, пищеводе, желудке, двенадцатиперстной кишке, тонкой кишке, подвздошной кишке, слепой кишке, толстой кишке, прямой кишке и/или анальном канале животного, путем введения эффективного количества α-дефензина и/или β-дефензина млекопитающего субъекту при необходимости такого лечения. Предпочтительно дефензином является человеческий альфа-дефензин. Согласно другим предпочтительным вариантам осуществления дефензином является человеческий бета-дефензин, предпочтительно hBD2, и воспаление снижают во рту, пищеводе, желудке, двенадцатиперстной кишке, тонкой кишке, подвздошной кишке, слепой кишке, прямой кишке и/или анальном канале.
Один аспект относится к способам лечения воспаления кишечника путем введения эффективного количества α-дефензина и β-дефензина млекопитающего или птицы субъекту при необходимости такого лечения.
Один аспект относится к способам стабилизации или поддержания нормальной микробиоты в кишечнике. Другой аспект относится к способам лечения или нормализации дисбиотической микробиоты в кишечнике путем введения эффективного количества α-дефензина и/или β-дефензина млекопитающего или птицы, и/или GLP-1/аналога GLP-1 субъекту при необходимости такого лечения.
Следующий аспект относится к способам повышения многообразия генов кишечной микробиоты путем введения эффективного количества α-дефензина и/или β-дефензина млекопитающего или птицы, и/или GLP-1/аналога GLP-1 субъекту при необходимости такого лечения.
Один аспект относится к способам повышения числа филумов кишечной микробиоты путем введения эффективного количества α-дефензина и/или β-дефензина млекопитающего или птицы, и/или GLP-1/аналога GLP-1 субъекту при необходимости такого лечения.
Один аспект относится к способам повышения продуцирования бутирата и/или повышения продуцирования ацетата из кишечной микробиоты путем введения эффективного количества α-дефензина и/или β-дефензина млекопитающего или птицы, и/или GLP-1/аналога GLP-1 субъекту при необходимости такого лечения.
Один аспект относится к способам повышения продуцирования короткоцепочечных жирных кислот из кишечной микробиоты путем введения эффективного количества α-дефензина и/или β-дефензина млекопитающего или птицы, и/или GLP-1/аналога GLP-1 субъекту при необходимости такого лечения.
Некоторые аспекты относятся к способам повышения числа бактерий, принадлежащих роду, выбранному из группы, состоящей из Bacterioidetes, Faecalibacterium, Roseburia, Blautia, Ruminococcus, Coprococcus, Bifidobacterium, Methanobrevibacter, Lactobacillus, Clostridium, Allobaculum, Alloprevotella, Akkermansia, Eubacterium, в кишечной микробиоте путем введения эффективного количества α-дефензина и/или β-дефензина млекопитающего или птицы, и/или GLP-1/аналога GLP-1 субъекту при необходимости такого лечения. Предпочтительными бактериями являются Allobaculum, Alloprevotella, Akkermansia или Lactobacillus.
Согласно предпочтительному варианту осуществления представлены способы повышения числа бактерий, выбранных из группы, состоящей из Bacteroides vulgatus, Bacteroides caccae, Faecalibacterium prausnitzii, Roseburia intestinalis, Blautia hansenii, Ruminococcus gnavus, Coprococcus comes, Clostridium nexile, Clostridium bolteae, Bifidobacterium longum, Bifidobacterium bifidum, Bifidobacterium adolescentis, Bifidobacterium dentum, Lactobacillus gasseri, Lactobacillus plantarum, Akkermansia muciniphila, Eubacterium rectale. Представленные способы повышают число бактерий, которые являются типичными для микробиоты здорового кишечника.
Один аспект относится к способам снижения числа бактерий, принадлежащих роду, выбранному из группы, состоящей из Bacteroidetes fragilis, Sutturella wadsworthia, Veillonella parvula, Escherichi coli, Haemophilus parainfluenzae, Fusobacterium nucleatum, Eikenella corodens, Gemella moribillum, в кишечной микробиоте путем введения эффективного количества α-дефензина и/или β-дефензина млекопитающего или птицы, и/или GLP-1/аналога GLP-1 субъекту при необходимости такого лечения. Представленные способы снижают число бактерий, которые являются типичными для дисбиотической микробиоты в кишечнике субъекта, нуждающегося в лечении.
Поэтому, описанные воспаление кишечника, злокачественная опухоль толстой и прямой кишки, эндокринное, связанное с питанием, метаболическое или сердечно-сосудистое заболевания, которые можно лечить с использованием способов, раскрываемых согласно предпочтительным вариантам осуществления, характеризуются дисбиотической микробиотой в кишечнике субъектов, нуждающихся в лечении. Согласно некоторым вариантам осуществления дисбиотическая микробиота в кишечнике субъекта, нуждающегося в лечении, обеспечиваемом раскрываемыми способами, имеет низкое многообразие генов. Согласно другим вариантам осуществления дисбиотическая микробиота в кишечнике субъекта, нуждающегося в лечении, обеспечиваемом раскрываемыми способами, имеет низкое число филумов. Согласно другим вариантам осуществления дисбиотическая микробиота в кишечнике субъекта, нуждающегося в лечении, обеспечиваемом раскрываемыми способами, характеризуется повышенным продуцированием ацетата из микробиоты. С помощью раскрываемых способов повышенное продуцирование ацетата может быть снижено в пользу продуцирования бутирата.
Согласно предпочтительным вариантам осуществления дисбиотическая микробиота в кишечнике субъекта, нуждающегося в лечении, обеспечиваемом раскрываемыми способами, имеет низкое число бактерий, принадлежащих роду, выбранному из группы, состоящей из Bacterioidetes, Faecalibacterium, Roseburia, Blautia, Ruminococcus, Coprococcus, Bifidobacterium, Methanobrevibacter, Lactobacillus, Clostridium, Allobaculum, Alloprevotella Akkermansia и Eubacterium. Согласно более предпочтительным вариантам осуществления дисбиотическая микробиота в кишечнике субъекта, нуждающегося в лечении, обеспечиваемом раскрываемыми способами, имеет низкое число бактерий, выбранный из группы, состоящей из Bacteroidetes vulgatus, Bacteroides caccae, Faecalibacterium prausnitzii, Roseburia intestinalis, Blautia hansenii, Ruminococcus gnavus, Coprococcus comes, Clostridium nexile, Clostridium bolteae, Bifidobacterium longum, Bifidobacterium bifidum, Bifidobacterium adolescentis, Bifidobacterium dentum, Lactobacillus gasseri, Lactobacillus plantarum, Akkermansia muciniphila, Eubacterium rectale. Согласно следующим вариантам осуществления дисбиотическая микробиота в кишечнике субъекта, нуждающегося в лечении, имеет высокое число бактерий, выбранных из группы, состоящей из Bacteroides fragilis, Sutturella wadsworthia, Veillonella parvula, Escherichi coli, Haemophilus parainfluenzae, Fusobacterium nucleatum, Eikenella corodens, Gemella moribillum.
Способами, раскрываемыми согласно предпочтительным вариантам осуществления, можно лечить дисбиотическую микробиоту и метаболом путем введения по меньшей мере альфа-дефензина млекопитающего или птицы, и/или по меньшей мере бета-дефензина млекопитающего или птицы, и/или по меньшей мере GLP-1/аналога GLP-1.
Раскрываемые способы могут быть использованы для лечения, предупреждения или нормализации дисбиотической микробиоты и/или метаболома в кишечнике субъекта, который получал лечение и/или получает лечение антибиотиком, или химиотерапию, или иммунотерапию, или иммуносупрессивную терапию, или другой метод лечения, отрицательно влияющие на кишечную микробиоту.
Раскрываемые способы также могут быть использованы для лечения, предупреждения или нормализации дисбиотической микробиоты во рту субъекта, такого как субъект, страдающий периодонтитом, в том числе гингивитом. Периодонтит может быть вызван курением и стрессом, а также может быть индуцирован лекарственным средством, например, химиотерапией, иммунотерапией и иммуносупрессивной терапией.
Субъект, нуждающийся в лечении, обеспечиваемом раскрываемыми способами, страдает от воспаления кишечника, или злокачественной опухоли толстой и прямой кишки, или эндокринного, связанного с питанием, метаболического, или сердечно-сосудистого заболевания. Согласно одному варианту осуществления субъект, нуждающийся в лечении, имеет BMI 25 или больше, например, 30 или больше, например, 35 или больше, например, 40 или больше. Согласно другому варианту осуществления субъект, нуждающийся в лечении, имеет соотношение окружностей талии и бедра по меньшей мере 0,80, например, 0,80-0,84, например, по меньшей мере 0,85 (у женского пола) или по меньшей мере 0,90, например, 0,9-0,99, например, приблизительно 1,00 (у мужского пола). Согласно следующему варианту осуществления субъект, нуждающийся в лечении, имеет содержание глюкозы в крови натощак по меньшей мере 6,1 ммоль/л, например, по меньшей мере 7,0 ммоль/л. Согласно следующему варианту осуществления субъект, нуждающийся в лечении, имеет содержание гликированного гемоглобина (HbA1C) по меньшей мере 42 ммоль/моль Hb, например, от 42 до 46 ммоль/моль Hb, например, по меньшей мере 48 ммоль/моль Hb.
У субъекта, нуждающегося в лечении, обеспечиваемом раскрываемыми способами, может иметься один или несколько из следующих симптомов:
повышенное давление крови: ≥ 140/90 мм ртутного столба;
дислипидемия: триглицериды (TG) ≥ 1,695 ммоль/л и холестерин липопротеинов высокой плотности (HDL-C) ≤ 0,9 ммоль/л (у мужского пола), ≤ 1,0 ммоль/л (у женского пола);
центральный тип ожирения: соотношение окружностей талии и бедра > 0,90 (у мужского пола); > 0,85 (у женского пола) или индекс массы тела > 30 кг/м2; и
микроальбуминурия: показатель выведения альбумина с мочой ≥ 20 мкг/минута или соотношение альбумин/креатинин ≥ 30 мг/г.
Согласно одному варианту осуществления введение по меньшей мере одного α-дефензина млекопитающего или птицы и/или по меньшей мере одного β-дефензина млекопитающего или птицы согласно раскрываемым способам, как правило, является пероральным.
Альфа- и бета-дефензины млекопитающего или птицы могут быть использованы терапевтически в композициях, составленных для введения любым удобным способом. Согласно одному варианту осуществления альфа- и/или бета-дефензины млекопитающего или птицы вводят перорального. Известно, что человеческий бета-дефензин 2 может быть введен перорально для лечения воспалительного заболевания кишечника в толстой кишке. Авторы настоящего изобретения неожиданно продемонстрировали, что подобно человеческому альфа-дефензину HD5 может быть введен перорально, и что он сохраняет свою биоактивность в кишечнике несмотря на прохождение через кислотную среду желудка.
Пероральное введение является нормальным для парентеральной доставки лекарственного средства, при этом средство доставляется через слизистую оболочку кишечника. Однако как показали авторы настоящего изобретения, hBD2 не абсорбируется из кишечника ни в какой выявляемой степени. Предполагается, что другие дефензины также не абсорбируются из кишечника.
Согласно одному варианту осуществления альфа- и/или бета-дефензины млекопитающего или птицы вводят подкожно. В частности, рассматривают, что hBD2 и HD5 могут быть введены подкожно.
Согласно некоторым вариантам осуществления композиции согласно предпочтительным вариантам осуществления могут быть составлены как лиофилизат с использованием подходящих вспомогательных средств, которые обеспечивают стабильность в виде лиофилизата и после регидратации.
Фармацевтические или животное кормовые композиции, содержащие альфа-дефензин млекопитающего и/или бета-дефензин млекопитающего, такой как человеческий альфа-дефензин и/или человеческий бета-дефензин, могут быть изготовлены согласно традиционным способам, например, с помощью процессов смешивания, гранулирования, покрытия, растворения или лиофилизации. Согласно предпочтительному варианту осуществления фармацевтические композиции, содержащие альфа-дефензин млекопитающего и/или бета-дефензин млекопитающего, составляют в виде стерильного и изотонического раствора.
Представленные фармацевтические композиции содержат согласно одному варианту осуществления по меньшей мере один альфа-дефензин млекопитающего. Примерами альфа-дефензинов млекопитающего являются HD5 и HD6. Согласно предпочтительному варианту осуществления композиции содержат альфа-дефензин млекопитающего HD5. Фармацевтические композиции содержат согласно другому варианту осуществления по меньшей мере один бета-дефензин млекопитающего. Примерами бета-дефензинов млекопитающего являются hBD1, hBD2, hBD3 и hBD4. Согласно предпочтительному варианту осуществления композиции содержат бета-дефензин млекопитающего hBD2. Фармацевтические композиции содержат согласно следующему варианту осуществления по меньшей мере один альфа-дефензин млекопитающего и по меньшей мере один бета-дефензин млекопитающего. Примерами альфа-дефензинов млекопитающего являются HD5 и HD6. Примерами бета-дефензинов млекопитающего являются hBD1, hBD2, hBD3 и hBD4. Согласно предпочтительному варианту осуществления композиции содержат альфа-дефензин млекопитающего HD5 и бета-дефензин млекопитающего hBD2. Согласно другим вариантам осуществления композиции или кормовые композиции содержат один или несколько отличных от человеческого дефензинов, выбранных из дефензинов, имеющих аминокислотную последовательность, выбранную из SEQ ID NO: 1-3 и 10-17, а также варианты и фрагменты последовательности, определенные в настоящем документе.
Фармацевтические композиции согласно предпочтительным вариантам осуществления содержат альфа-дефензин млекопитающего и/или бета-дефензин млекопитающего, например, человеческий альфа-дефензин и человеческий бета-дефензин, и фармацевтически приемлемый носитель и/или разбавитель.
Фармацевтически приемлемые и приемлемые для корма животных носители и/или разбавители известны специалистам в данной области. Для композиций, составленных в виде жидких растворов, приемлемые носители и/или разбавители включают в себя солевой раствор и стерильную воду и необязательно могут включать в себя антиоксиданты, буферы, бактериостатики и другие распространенные добавки.
Раскрываемое соединение может быть составлено в виде разнообразных составов для перорального введения. Препараты в твердой форме могут включать в себя порошки, таблетки, капли, капсулы, саше, пастилки и диспергируемые гранулы. Другие формы, подходящие для перорального введения, могут включать в себя препараты в жидкой форме, в том числе эмульсии, сиропы, эликсиры, водные растворы, водные суспензии, зубную пасту, гель для чистки зубов, жевательную резинку, или препараты в твердой форме, которые предназначены для превращения непосредственно перед применением в препараты в жилкой форме, такие как растворы, суспензии и эмульсии.
Раскрываемое соединение может быть составлено в виде ряда составов для подкожного введения.
Состав может содержать (в дополнение к альфа-дефензину млекопитающего и/или бета-дефензину млекопитающего и другим необязательным активным ингредиентам) носители, наполнители, разрыхлители, средства для улучшения сыпучести, сахара и подсластители, ароматизаторы, консерванты, стабилизаторы, увлажнители, эмульгаторы, солюбилизаторы, соли для регулирования осмотического давления, буферы, разбавители, диспергаторы и поверхностно-активные средства, связующие, смазки и/или другие фармацевтические вспомогательные средства, известные в уровне техники.
Специалист в данной области, кроме того, сможет составить альфа-дефензин млекопитающего или птицы и бета-дефензины млекопитающего или птицы соответствующим образом и в соответствии с принятой технологией, например, как описано в Remington's Pharmaceutical Sciences, Gennaro (1990).
Альфа-дефензин млекопитающего и бета-дефензин млекопитающего, такие как человеческий альфа-дефензин и человеческий бета-дефензин, могут быть использованы отдельно или в комбинированных терапевтических средствах с одним, двумя или более другими фармацевтическими соединениями или лекарственными веществами, например, с инсулином/аналогами инсулина, и/или с глюкагоноподобным пептидом-1 (GLP-1)/аналогами GLP-1, и/или с глюкагоноподобным пептидом-2 (GLP-2)/аналогами GLP-2, и/или с ингибитором дипептидилпептидазы IV (DPP-IV), и/или с метформином, и/или с ингибитором натрий-зависимого переносчика глюкозы-2 (SGLT-2), и/или с антагонистом рецептора глюкагона, и/или с антагонистом представителя 1 семейства V катионного канала транзиторного рецепторного потенциала (TRPV1), и/или с одним или несколькими фармацевтически приемлемыми вспомогательными средствами. Предпочтительно инсулин, или аналоги инсулина, или GLP-1, или аналоги GLP-1 вводят парентерально посредством либо подкожного, либо внутримышечного введения. Аналог GLP-1 может быть выбран из эксенатида, лираглутида, ликсисенатида, албиглутида и дулаглутида, а аналог инсулина может быть выбран из лизпро, аспарта, глулизина, инсулина детемира, инсулина деглудека и инсулина гларгина.
Альфа-дефензин млекопитающего и бета-дефензин млекопитающего, такие как человеческий альфа-дефензин и человеческий бета-дефензин, также могут быть использованы в комбинированных терапевтических средствах либо с химиотерапией, иммунотерапией, радиотерапией, либо с их комбинацией.
Способы и применения для домашнего скота
Способы, раскрываемые в настоящем документе, могут быть использованы для домашнего скота с целью улучшения скорости их роста и эффективности корма. Способы, например, могут быть использованы как альтернатива антибиотикам.
Один аспект относится к способам лечения воспаления кишечника у отличных от человека животных путем введения эффективного количества α-дефензина и/или β-дефензина млекопитающего или птицы субъекту при необходимости такого лечения.
Другой аспект относится к способу стимуляции роста мышц при производстве мяса животных, при этом указанный способ предусматривает введение эффективного количества α- и/или β-дефензина млекопитающего или птицы субъекту при необходимости этого. Дефензины могут быть использованы как альтернатива гормонам, стероидам и антибиотикам. Как показано в примерах 1 и 3, введение HD5 и hBD2 мышам, получавшим рацион с высоким содержанием жира, стимулирует рост мышц, поскольку оно предупреждает накопление массы жира.
In vitro синтез
Альфа-дефензины млекопитающего и птицы и бета-дефензины млекопитающего и птицы могут быть получены с помощью in vitro синтеза с использованием традиционных способов, известных в уровне техники. Доступны различные коммерческие устройства для синтеза, например, автоматизированные синтезаторы от Applied Biosystems Inc., Beckman и т.д. С использованием синтезаторов встречающиеся в природе аминокислоты могут быть замещены не встречающимися в природе аминокислотами, в частности, D-изомерами (или D-формами), например, D-аланином и D-изолейцином, диастереоизомерами, боковыми цепями с разной длиной или функциональными группами и т.п. Конкретные последовательность и способы получения будут определяться удобством, экономией, требуемой чистотой и т.п.
Химическое связывание может быть обеспечено для различных пептидов или белков, содержащих удобные для связи функциональные группы, такие как аминогруппы для образования амида или замещенного амина, например, восстановительное аминирование, тиольные группы для образования тиоэфира или дисульфида, карбоксильные группы для образования амида и т.п.
При необходимости различные группы могут быть введены в пептид в ходе синтеза или в ходе экспрессии, что обеспечивает связывание с другими молекулами или с поверхностью. Таким образом, цистеины могут быть использованы для получения тиоэфиров, гистидины для связывания с комплексом ионов металла, карбоксильные группы для образования амидов или сложных эфиров, аминогруппы для образования амидов и т.п.
Альфа-дефензины млекопитающего и птицы и бета-дефензины млекопитающего и птицы или их функциональные эквиваленты также могут быть выделены и очищены в соответствии с традиционными способы рекомбинантного синтеза. Рекомбинантный синтез может быть выполнен с использованием соответствующих векторов экспрессии и эукариотической системы экспрессии. Может быть получен раствор хозяина экспрессии, среды и дефензинов, очищенных с использованием HPLC, эксклюзионной хроматографии, электрофореза в геле, аффинной хроматографии или других методик очистки. Способы рекомбинантной экспрессии человеческого бета-дефензина 2 в E. coli раскрываются в WO 2010/007166 (Novozymes).
Дозировки
Альфа-дефензин млекопитающего и бета-дефензин млекопитающего, такие как человеческий альфа-дефензин и человеческий бета-дефензин, предпочтительно используют в фармацевтических композициях в количестве, которое является эффективным для лечения воспаления кишечника, злокачественной опухоли толстой и прямой кишки, эндокринного, связанного с питанием, метаболического или сердечно-сосудистого заболевания, предпочтительно с приемлемой токсичностью, для больного. Альфа-дефензин млекопитающего или птицы и бета-дефензин млекопитающего или птицы предпочтительно применяют в корме животных для стимуляции роста мышц. Альфа-дефензин млекопитающего или птицы и бета-дефензин млекопитающего или птицы, такие как человеческий альфа-дефензин и человеческий бета-дефензин, также предпочтительно используют в фармацевтических композициях или в корме животных в количестве, которое является эффективным для поддержания нормального состава микробиоты в кишечнике или для лечения или нормализации дисбиотической микробиоты в кишечнике, предпочтительно с приемлемой токсичностью, для больного или животного при необходимости лечения.
Для таких методов лечения соответствующая дозировка, конечно, будет варьировать в зависимости, например, от химической природы и фармакокинетических данных используемого соединения, отдельного хозяина, пути введения, а также природы и тяжести подлежащих лечению состояний. Используемый в настоящем документе термин «мг-эквиваленты HD5» относится к эквимолярной концентрации человеческих альфа- и бета-дефензинов по сравнению с концентрацией HD5. Используемый в настоящем документе термин «мг-эквиваленты hBD-2» относится к эквимолярной концентрации человеческих альфа- и бета-дефензинов в сравнении с концентрацией hBD-2. Поскольку молекулярные массы раскрываемых дефензинов являются сопоставимыми, термины «мг-эквиваленты HD5» и «мг-эквиваленты hBD2» могут просто означать мг используемого дефензина.
Однако, в целом, для удовлетворительных результатов у млекопитающих или птицы, например, у людей, указанная суточная дозировка человеческого альфа-дефензина составляет предпочтительно от приблизительно 0,1 мг-эквивалента HD5/кг массы тела до приблизительно 10 мг-эквивалентов HD5/кг массы тела, более предпочтительно от приблизительно 0,5 мг-эквивалента HD5/кг массы тела до приблизительно 10 мг-эквивалентов HD5/кг массы тела; например, от 1 мг-эквивалента HD5/кг массы тела до 10 мг-эквивалентов HD5/кг массы тела, более предпочтительно от приблизительно 1,2 мг-эквивалента HD5/кг массы тела до приблизительно 10 мг-эквивалентов HD5/кг массы тела, предпочтительно от приблизительно 1,2 мг-эквивалента HD5/кг массы тела до приблизительно 5 мг-эквивалентов HD5/кг массы тела, еще более предпочтительно 1,2 мг-эквивалента HD5/кг массы тела, например, вводят поделенными дозами до одного, двух или трех раз в сутки.
Согласно одному варианту осуществления указанная суточная дозировка человеческого бета-дефензина составляет предпочтительно от приблизительно 0,1 мг-эквивалента hBD-2/кг массы тела до приблизительно 10 мг-эквивалентов hBD-2/кг массы тела, более предпочтительно от приблизительно 0,5 мг-эквивалента hBD-2/кг массы тела до приблизительно 10 мг-эквивалентов hBD-2/кг массы тела; например, от 1 мг-эквивалента hBD-2/кг массы тела до 10 мг-эквивалентов hBD-2/кг массы тела, более предпочтительно от приблизительно 1,2 мг-эквивалента hBD-2/кг массы тела до приблизительно 10 мг-эквивалентов hBD-2/кг массы тела, предпочтительно от приблизительно 1,2 мг-эквивалента hBD-2/кг массы тела до приблизительно 5 мг-эквивалентов hBD-2/кг массы тела, еще более предпочтительно 1,2 мг-эквивалента hBD-2/кг массы тела, например, вводят поделенными дозами до одного, двух или трех раз в сутки.
Согласно одному варианту осуществления указанная суточная дозировка человеческого альфа-дефензина вместе с человеческим бета-дефензином составляет предпочтительно от приблизительно 0,1 мг-эквивалента hBD-2/кг массы тела до приблизительно 10 мг-эквивалентов hBD-2/кг массы тела, более предпочтительно от приблизительно 0,5 мг-эквивалента hBD-2/кг массы тела до приблизительно 10 мг-эквивалентов hBD-2/кг массы тела; например, от 1 мг-эквивалента hBD-2/кг массы тела до 10 мг-эквивалентов hBD-2/кг массы тела, более предпочтительно от приблизительно 1,2 мг-эквивалента hBD-2/кг массы тела до приблизительно 10 мг-эквивалентов hBD-2/кг массы тела, предпочтительно от приблизительно 1,2 мг-эквивалента hBD-2/кг массы тела до приблизительно 5 мг-эквивалентов hBD-2/кг массы тела, еще более предпочтительно 1,2 мг-эквивалента hBD-2/кг массы тела, например, вводят поделенными дозами до одного, двух или трех раз в сутки.
В целом, для млекопитающих, например, людей, указанная суточная дозировка человеческого альфа-дефензина составляет предпочтительно от приблизительно 0,1 мг HD5/кг массы тела до приблизительно 10 мг HD5/кг массы тела, более предпочтительно от приблизительно 0,5 мг HD5/кг массы тела до приблизительно 10 мг HD5/кг массы тела; например, от 1 мг HD5/кг массы тела до 10 мг HD5/кг массы тела, более предпочтительно от приблизительно 1,2 мг HD5/кг массы тела до приблизительно 10 мг HD5/кг массы тела, предпочтительно от приблизительно 1,2 мг HD5/кг массы тела до приблизительно 5 мг HD5/кг массы тела, еще более предпочтительно 1,2 мг HD5/кг массы тела, например, вводят поделенными дозами до одного, двух или трех раз в сутки. Подобные дозировки могут быть использованы для других альфа-дефензинов.
Согласно одному варианту осуществления указанная суточная дозировка человеческого бета-дефензина составляет предпочтительно от приблизительно 0,1 мг hBD-2/кг массы тела до приблизительно 10 мг hBD-2/кг массы тела, более предпочтительно от приблизительно 0,5 мг hBD-2/кг массы тела до приблизительно 10 мг hBD-2/кг массы тела; например, от 1 мг hBD-2/кг массы тела до 10 мг hBD-2/кг массы тела, более предпочтительно от приблизительно 1,2 мг hBD-2/кг массы тела до приблизительно 10 мг hBD-2/кг массы тела, предпочтительно от приблизительно 1,2 мг hBD-2/кг массы тела до приблизительно 5 мг hBD-2/кг массы тела, еще более предпочтительно 1,2 мг hBD-2/кг массы тела, например, вводят поделенными дозами до одного, двух или трех раз в сутки. Подобные дозировки могут быть использованы для других бета-дефензинов.
Указанная суточная дозировка человеческого альфа-дефензина вместе с человеческим бета-дефензином составляет предпочтительно от приблизительно 0,1 мг дефензина/кг массы тела до приблизительно 10 мг дефензина/кг массы тела, более предпочтительно от приблизительно 0,5 мг дефензина/кг массы тела до приблизительно 10 мг дефензина/кг массы тела; например, от 1 мг дефензина/кг массы тела до 10 мг дефензина/кг массы тела, более предпочтительно от приблизительно 1,2 мг дефензина/кг массы тела до приблизительно 10 мг дефензина/кг массы тела, предпочтительно от приблизительно 1,2 мг дефензина/кг массы тела до приблизительно 5 мг дефензина/кг массы тела, еще более предпочтительно 1,2 мг дефензина/кг массы тела, например, вводят поделенными дозами до одного, двух или трех раз в сутки.
Если два разных дефензина вводят в одной дозировке, дозировка может содержать равные или приблизительно равные количества двух дефензинов, определенные в пересчете на массу или в пересчете на молярность. Соотношение также может отличаться так, что соотношение альфа-дефензина к бета-дефензину варьирует от 10:1 до 1:10, например, от 5:1 до 1:5, например, от 2:1 до 1:2, при определении пересчете на массу или молярность.
Суточная дозировка может соответствовать 0,6 мг HD5/кг массы тела плюс 0,6 мг hBD-2/кг массы тела.
Соединения согласно предпочтительным вариантам осуществления могут быть введены млекопитающим или птице, например, людям, поросятам или телятам, путями введения, подобными традиционно используемым, при дозировках, подобных традиционно используемым.
Согласно определенным вариантам осуществления фармацевтические композиции или композиции корма животных согласно предпочтительным вариантам осуществления могут включать в себя альфа-дефензин млекопитающего или птицы и/или бета-дефензин млекопитающего или птицы, такие как человеческий альфа-дефензин и/или человеческий бета-дефензин, в количестве от приблизительно 0,5 мг или меньше до приблизительно 1500 мг или больше на единичную дозированную форму, предпочтительно от приблизительно 0,1, 0 3, 0,5, 0,6, 0.7, 0,8 или 0,9 мг до приблизительно 150, 200, 250, 300, 350, 400, 450, 500, 600, 700, 800, 900 или 1000 мг и более предпочтительно от приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20 или 25 мг до приблизительно 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95 или 100 мг. Согласно определенным вариантам осуществления, однако, могут быть предпочтительными более низкие или более высокие дозировки, чем упомянутые выше. Соответствующие концентрации и дозировки могут быть легко определены специалистом в данной области. Согласно определенным вариантам осуществления фармацевтические композиции согласно предпочтительным вариантам осуществления включают в себя альфа-дефензин млекопитающего, такой как человеческий альфа-дефензин. Согласно другим вариантам осуществления фармацевтические композиции согласно предпочтительным вариантам осуществления включают в себя бета-дефензин млекопитающего, такой как человеческий бета-дефензин. Согласно следующим вариантам осуществления фармацевтические композиции согласно предпочтительным вариантам осуществления включают в себя альфа-дефензин млекопитающего и бета-дефензин млекопитающего, такие как человеческий альфа-дефензин и человеческий бета-дефензин, при этом альфа- и бета-дефензины присутствуют в равных количествах в пересчете на молярность или на мг/мл.
Согласно одному варианту осуществления альфа- и/или бета-дефензин млекопитающего или птицы вводят по меньшей мере один раз в сутки, например, по меньшей мере два раза в сутки, например, по меньшей мере 3 раза в сутки.
Далее настоящее раскрытие описывается с помощью следующих примеров, которые не следует считать ограничивающими объем настоящего изобретения.
Примеры
Пример 1
Модуляция микробиоты кишечника и предупреждение воспаления кишечника и метаболического синдрома путем профилактического лечения дефензинами
Материалы и способы
Мыши. Мышей размещали по трое, по 4 клетки на группу. Потребление корма регистрировали ежесуточно непосредственно перед выключением света (в 6 часов вечера). Отдельных мышей подвергали экспериментальным процедурам в измененном порядке как в группе, так и в клетке. Мышей держали при комнатной температуре при 12-часовом цикле свет/темнота при стандартных условиях SPF.
Рационы. Для введения дозы среднюю массу оценивали как 25 грамм на мышь. Мыши съедали приблизительно 3 грамма корма на мышь в сутки.
Режим лечения (фиг. 1A). Мышей кормили либо рационом с высоким содержанием жира (HFD), либо контрольным рационом с низким содержанием жира (LF). HFD предусматривал 4 подгруппы: 1 на hBD2, 1 на HD5, 1 на hBD2/HD5 и 1 на стандартном HFD без дополнения дефензинами. Концентрация дефензина составляла 1,2 мг hBD2 на кг массы мыши в сутки. HD5 давали в эквимолярной концентрации по отношению к hBD2. Комбинационная группа получала 50% hBD2 + 50% HD5, следовательно, суммарное количество дефензинов эквивалентно таковому в остальных тестируемых группах.
Тесты. Тест на толерантность к инсулину (ITT), тест на стимулированную глюкозой секрецию инсулина (GSIS), пероральный тест на толерантность к глюкозе (OGTT) и тест на инсулин при пятичасовом воздержании от пищи выполняли за двое суток с 50% мышей на группу в сутки, следовательно, избегали суточного колебания в виде конфаундинг-фактора.
Для исследования микробиоты кишечника осуществляли анализы микроорганизмов. Лонгитудинальную характеристику 16S проводили на 4 парных образцах от 60 мышей, в сумме на 240 образцах. У каждой мыши брали образец перед изменением рациона, через 1 неделю после изменения рациона, через 4 недели после изменения рациона и при завершении, что таким образом обеспечивало тщательную характеристику фекальной микробиоты в результате обработки дефензином. Кроме того, содержимое тонкого кишечника анализировали при завершении, что обеспечивало таким образом ценную информацию о возможных изменениях на ключевом участке поглощения питательных веществ.
Полный метаболомический профиль содержимого слепой кишки может быть проведен для обеспечения переноса изменений микроорганизмов на метаболизм всего организма. Также может быть выполнен подробный гистологический и иммуногистохимический анализ двенадцатиперстной кишки, тонкой кишки, подвздошной кишки и толстой кишки.
Результаты обработки hBD2+HD5 исключили из анализа.
Результаты
Изменение массы. В то время как потребление пищи было одинаковым во всех трех группах на экспериментальном рационе (фиг. 6C), обе группы на рационе с высоким содержанием жира (HFD) набирали значительно больше массы, чем эталонная группа на рационе с низким содержанием жира (LFD) за 10-недельный период исследования (***p < 0,0001, 2-факторный ANOVA, апостериорный критерий Тьюки). Группа на HFD плюс hBD-2, однако, набирала существенно меньше массы, чем эталонная группа на HFD (*p = 0,0028) (фиг. 6 A и B).
Наращивание массы мышц/жира. Масса мышц/жира была равномерно распределена между тремя экспериментальными группами в начале исследования (фиг. 7 A и B). В конце исследования обе группы на HFD набрали одинаковую массу мышц (фиг. 7B), которая была существенно больше, чем у группы на LFD (*p < 0,0001, однофакторный ANOVA, апостериорный критерий Тьюки), вероятно, из-за увеличения масса тела. В конце исследования группа на HFD плюс hBD-2 имела тенденцию к увеличению массы жира по сравнению с группой на LFD (фиг. 7B). Однако это не было статистически значимым (*p = 0,25). Группа на HFD набирала почти в четыре раза бóльшую массу жира по сравнению с группой на LFD и в 2 раза бóльшую массу жира по сравнению с группой на HFD плюс hBD-2 (*p < 0,0001 и *p = 0,005, соответственно) (фиг. 7B).
Тест на толерантность к инсулину. Как группа на LFD, так и группа на HFD плюс hBD-2 были существенно более чувствительными к инсулину, чем группа на HFD (p < 0,05) (фиг. 8a).
Тест на толерантность к глюкозе. Группа на HFD имела нарушенную толерантность к глюкозе с пролонгированным выведением глюкозы от пикового значения за 15 минут до полувыведения через 120 минут. Группа на LFD характеризовалась быстрым выведением глюкозы от пикового значения за 15 минут. Группа на HFD плюс hBD-2 характеризовалась слегка пролонгированным выведением глюкозы, но достигала значительно более низкого содержания глюкозы, чем группа на HFD (p < 0,05) (фиг. 8B).
Стимулированная глюкозой секреция инсулина. Группа на HFD характеризовалась ухудшенным гомеостазом глюкозы со значительно более высокой и устойчивой концентрацией инсулина после введения глюкозы (p < 0,05). У группы на LFD почти не повышалась концентрация инсулина после стимуляции глюкозой. Группа на HFD плюс hBD-2 имела более высокую концентрацию инсулина, но не значительно отличающуюся от таковой у группы на LFD (фиг. 8C).
Инсулин при пятичасовом воздержании от пищи. Группа на HFD характеризовалась тяжелым диабетом со значительно более высоким содержанием инсулина натощак, чем на LFD (*p = 0,0004), и с пограничным значительно более высоким содержанием инсулина натощак, чем группа на HFD плюс hBD-2 (*p = 0,057). Не было существенной разницы между группами на LFD и на HFD плюс hBD-2 (*p = 0,17) (фиг. 8D).
*Апостериорный критерий Тьюки, другой апостериорный критерий Даннетта.
После завершения исследования в примере 1 результаты снова были анализированы и представлены ниже и в приложенных графических материалах.
Изменение массы. В то время как потребление пищи было одинаковым во всех трех группах на экспериментальном рационе, обе группы на рационе с высоким содержанием жира (HFD) набирали значительно больше массы, чем эталонная группа на рационе с низким содержанием жира (LFD), за 10-недельный период исследования (*p < 0,0001, 2-факторный ANOVA, апостериорный критерий Тьюки). Группа на HFD плюс hBD-2, однако, набирала существенно меньше массы, чем эталонная группа на HFD (*p = 0,0028) (фиг. 9a). На фиг. 9B показана эффективность корма, а на фиг. 9C - потребление энергии.
Наращивание массы мышц/жира. Масса мышц/жира была равномерно распределена между тремя экспериментальными группами в начале исследования. В конце исследования обе группы на HFD набрали одинаковую массу мышц, которая была существенно больше, чем у группы на LFD (*p < 0,0001, однофакторный ANOVA, апостериорный критерий Тьюки), вероятно, из-за увеличения масса тела. В конце исследования группа на HFD плюс hBD-2 имела тенденцию к увеличению массы жира по сравнению с группой на LFD. Однако это не было статистически значимым (*p = 0,25). Группа на HFD набирала почти в четыре раза бóльшую массу жира в процентах от общей массы тела по сравнению с группой на LFD и в два раза бóльшую массу жира в процентах от общей массы тела по сравнению с группой на HFD плюс hBD-2 (*p < 0,0001 и *p = 0,005, соответственно) (фиг. 10a). Не было статистического различия между тремя группами в отношении массы печени (фиг. 10b), тогда как группа на HFD плюс hBD-2 имела существенно меньше висцерального жира (eWAT), чем группа на HFD (фиг. 10c).
Тест на толерантность к глюкозе. Группа на HFD имела нарушенную толерантность к глюкозе с пролонгированным выведением глюкозы от пикового значения за 15 минут до полувыведения через 120 минут. Группа на LFD характеризовалась быстрым выведением глюкозы от пикового значения за 15 минут. Группа на HFD плюс hBD-2 характеризовалась слегка пролонгированным выведением глюкозы, но достигала значительно более низкого содержания глюкозы, чем группа на HFD (p < 0,05) (фиг. 11a).
Стимулированная глюкозой секреция инсулина. Группа на HFD характеризовалась ухудшенным гомеостазом глюкозы со значительно более высокой и устойчивой концентрацией инсулина после введения глюкозы (p < 0,05). У группы на LFD почти не повышалась концентрация инсулина после стимуляции глюкозой. Группа на HFD плюс hBD-2 имела более высокую концентрацию инсулина, но не значительно отличающуюся от таковой у группы на LFD (фиг. 11b).
Тест на толерантность к инсулину. Как группа на LFD, так и группа на HFD плюс hBD-2 были существенно более чувствительными к инсулину, чем группа на HFD (p < 0,05) (фиг. 12A). Не было статистически значимого различия между эталонной группой на LFD и группой на HFD плюс hBD-2.
Оценка гомеостатической модели (HOMA-IR). Взаимосвязь между инсулиновой резистентностью и функцией бета-клеток при оценивании с помощью индекса HOMA-IR была статистически достоверно ниже для группы на HFD плюс hBD-2 по сравнению с группой на HFD (фиг. 12b).
*Апостериорный критерий Тьюки, другой апостериорный критерий Даннетта.
Заключение по hBD-2 в качестве предупреждения развития сахарного диабета и ожирения у мышей, получавших рацион с высоким содержанием жира
- Постоянный прирост массы (с низким внутригрупповым SD).
- 50% мышей, получавших HFD-hBD2, имели процент жира в организме, сходный с таковым у эталонных мышей на LFD, несмотря на то, что они получали 60% HFD. У нескольких мышей был еще более низкий % жира, чем самый низкий у эталонных мышей на LFD.
- У наилучшим образом защищенных мышей, получавших hBD2, была такая же или меньшая масса висцерального жира, чем у эталонных мышей на LFD, что очень необычно для 60% HFD(!).
- Улучшенная чувствительность к инсулину(!) у мышей, получавших hBD2, существенно не отличалась от таковой эталонных мышей на LFD; и тест на толерантность к инсулину, и HOMA-IR указывают на улучшенную инсулиновую сигнализацию.
- Толерантность к глюкозе заметно улучшилась по сравнению с контрольными мышами на HFD. Важно отметить, что и толерантность к глюкозе, и стимулированная глюкозой реакция инсулина при глюкозной нагрузке была улучшена. Это означает, что мыши, получавшие hBD2, нуждались в меньшем количестве инсулина для обработки болюса глюкозы, чем контрольные мыши на HFD.
Заключение по HD5 в качестве предупреждения развития сахарного диабета и ожирения у мышей, получавших рацион с высоким содержанием жира
- Мыши, получавшие HD5, набирали меньшую массу, чем контрольные мыши, получавшие HFD (фиг. 17A), хотя эффект не был статистически значимым. Также наблюдали тенденцию к более низким эффективности корма (фиг. 17B) и потреблению энергии (фиг. 17C).
- Мыши, получавшие HD5, характеризовались погранично меньшим % жира (фиг. 18A) и пограничным меньшим содержанием висцерального жира, чем контрольные мыши, получавшие HFD (фиг. 18C). Не наблюдали статистически значимое различие в отношении массы печени (фиг. 18B).
- Не наблюдали значительного улучшения толерантности к глюкозе (фиг. 19A), стимулированной глюкозой секреции инсулина (фиг. 19B), толерантности к инсулину (фиг. 20A) и HOMA-IR (фиг. 20B), хотя мыши, получавшие HD5, демонстрировали более хорошие результаты, чем контрольные мыши, получавшие HFD. Различие результатов, полученных с hBD2 и HD5, подтверждает различия в механизмах действия между hBD2 и HD5.
Что касается модуляции микробиоты, результаты на фиг. 21A недели 1 исследования демонстрируют, что изменение в микрофлоре произошло уже на первой неделе исследования. Три группы на HFD имеют сравнимую микрофлору, отличную от микрофлоры группы на LFD.
При завершении исследования через 10 недель микрофлора группы на HFD-HD5 изменилась и теперь стала промежуточной между таковой у группы на LFD и необработанной группы на HFD (фиг. 21B). Вывод заключается в том, что пероральное введение HD5 приводит к частичной нормализации микрофлоры так, что она больше похожа на микрофлору у группы на LFD, чем на таковую у необработанной группы.
В группах на HFD при завершении мыши, обработанные альфа-дефензином (HD5), демонстрировали нормализацию микробиоты с повышенным содержанием Allobaculum и Lactobacillus и понижением содержания Clostridium в кишечной микробиоте по сравнению с мышами, которые не были обработаны (фиг. 21C). Allobaculum представляет продуцирующие короткоцепочечную жирную кислоту виды. Lactobacillus является бактерией с противовоспалительными свойствами.
Результаты могут быть истолкованы как означающие, что в группах на HFD при завершении мыши, обработанные альфа-, бета и альфа-, а также бета-дефензинами, демонстрируют более высокое многообразие генов и более высокое число бактерий в кишечной микробиоте, чем мыши, которых не обрабатывали.
В контрольной группе на LFD при завершении мыши демонстрируют здоровую неизмененную микробиоту в кишечнике.
Пример 2
Модуляция микробиоты кишечника с помощью дефензинов
Беспозвоночные. Для доказательства концепции in vivo можно использовать модель беспозвоночных восковых огневок Galleria mellonella (G. mellonella). Фекалии можно анализировать после принудительного введения α- и/или β-дефензинов (Giannouli et al. 2014) (Favre-Godal et al. 2014).
Пример 3
Модуляция параметров микробиоты кишечника, воспаления кишечника и метаболического синдрома с помощью применения обработки дефензинами у тучных мышей
Мыши и рационы. В данном эксперименте устанавливали эффект hBD-2 и HD5 в лечении метаболического синдрома (MetS) у индуцированных рационом тучных мышей. Применению обработки предшествовал вводный период в 13 недель, при котором мышей кормили HFD в высокой степени (60% энергии из жира). В заключительные анализы включали исключительно мышей, отвечающих критерию прироста массы минимум 12 грамм (приблизительно 50% начальной массы тела) за время вводного периода. Мышей, которые не отвечали данному критерию, оставляли в соответствующих им клетках как иерархических «хранителей». Их подвергали всем экспериментальным тестам, но исключали из анализов.
Режим лечения (фиг. 1B). Перед применением обработки всех мышей сканировали с помощью MR и выполняли oGTT. Клетки с мышами распределяли по экспериментальным группам на основании массы жира у них. Все последующие измерения связывали с данными от той же мыши перед применением обработки.
Эталонную группу на LFD (рационе с низким содержанием жира) анализировали в параллельном режиме. В качестве контролей для применения обработки предусматривали 2 дополнительных группы: 1 на HFD в высокой степени и 1 на LFD. Экспериментальных мышей оставляли на HFD в высокой степени на протяжении применения обработки. Мышей держали на экспериментальном рационе на протяжении 10 недель. На протяжении эксперимента их размещали по 4 мыши на клетку, 3 клетки на группу. Все тесты выполняли за 3 суток, по 1 клетке на группу в сутки.
Тесты. Тест на толерантность к инсулину (ITT), тест на стимулированную глюкозой секрецию инсулина (GSIS), пероральный тест на толерантность к глюкозе (oGTT) и тест на инсулин при пятичасовом воздержании от пищи выполняли за трое суток с 1/3 мышей на группу в сутки, следовательно, избегали суточного колебания в виде конфаундинг-фактора.
Для исследования микробиоты кишечника можно было осуществлять анализы микроорганизмов. Лонгитудинальную характеристику 16S можно было проводить на 7 парных образцах от 60 мышей, в сумме на 240 образцах. У каждой мыши можно было брать образец перед изменением рациона, через 2 недели, 4 недели, 6 недель, 8 недель после изменения рациона и при завершении, что таким образом обеспечивало тщательную характеристику фекальной микробиоты в результате обработки дефензином.
Кроме того, содержимое тонкого кишечника можно было анализировать при завершении (с помощью 16S или глубокого секвенирования), что обеспечивало таким образом ценную информацию о возможных изменениях на ключевом участке поглощения питательных веществ.
В конце может быть проведен полный метаболомический профиль содержимого слепой кишки для обеспечения переноса изменений микроорганизмов на метаболизм всего организма. Также может быть выполнен подробный гистологический и иммуногистохимический анализ двенадцатиперстной кишки, тонкой кишки, подвздошной кишки и толстой кишки.
hBD-2 как лечение метаболического синдрома у тучных мышей, получающих рацион с высоким содержанием жира
Результаты
Изменение массы. Группы, получавшие стандартный рацион с высоким содержанием жира (HFD), равномерно потребляли пищу на протяжении всего периода исследования, характеризовались одинаковым наращиванием массы с равной массой жира и мышц первые 13 недель и, таким образом, имели одинаковую исходную точку перед применением рациона. Прирост массы значительно превышал таковой у группы, получавшей рацион с низким содержанием жира (LFD) (*p < 0,05, 2-факторный ANOVA) (фиг. 13A). После применения рациона группы на HFD продолжали наращивать массу, однако группа на HFD плюс hBD-2 характеризовалась тенденцией меньшего прироста массы первый 4 недели после применения рациона, хотя и не значительного (*p = 0,07, 2-факторный ANOVA). С недели 4 до конца периода исследования группа на HFD плюс hBD-2 набирала массу, подобную таковой у группы на стандартном HFD (*p = 0,82, 2-факторный ANOVA) (фиг. 13B).
Процентное отношение жира. Процентное отношение жира от общей массы тела было подобным у трех экспериментальных групп в начале периода исследования. На момент применения рациона процентное отношение жира у двух получавших HFD групп было одинаковым и значительно превышало таковое у получавшей LFD группы, что было постоянным на протяжении 10 недель после применения рациона (*p < 0,05, 2-факторный ANOVA) (фиг. 14A). На неделе 4 после применения рациона ~75% группы на HFD плюс hBD-2 имели процентное отношение жира, подобное таковому перед применением, что резко контрастировало с группой на стандартном HFD, при этом все мыши имели повышенное процентное отношение жира (фиг. 14B). Изменение в процентном отношении жира было значительно меньшим у группы на HFD плюс hBD-2 по сравнению с группой на стандартном HFD на данный момент времени (*p = 0,003, 2-факторный ANOVA). Масса печени при завершении была значительно больше у получавших HFD групп по сравнению с группой на LFD (*p < 0,05, однофакторный ANOVA) (фиг. 15A). Количество висцерального жира (eWAT) при завершении также было выше у групп на HFD по сравнению с группой на LFD (*p < 0,05, однофакторный ANOVA). Не наблюдали значительной разницы в висцеральном жире (eWAT) между получавшими HFD группами (фиг. 15B).
Тест на толерантность к глюкозе. Толерантность к глюкозе быстро улучшалась от момента применения рациона у группы на HFD плюс hBD-2, которая демонстрировала меньшее пиковое значение содержания глюкозы в крови, а также более быстрое выведение глюкозы уже через 2 недели (фиг. 16A). При исследовании у большинства мышей с нарушенной толерантностью к глюкозе наблюдали кардинальное улучшение в первые две недели после перевода со стандартного HFD на HFD плюс hBD-2 (фиг. 16B).
Тест на толерантность к инсулину. Группа на LFD была значительно более чувствительной к инсулину, чем обе группы на HFD (*p < 0,05, 2-факторный ANOVA). Группа на HFD плюс hBD-2 при этом была более чувствительной к инсулину по сравнению с контрольной группой на HFD, что подразумевало улучшение толерантности к инсулину после применения рациона (*p < 0,05, 2-факторный ANOVA) (фиг. 16C).
Заключение по hBD-2 в качестве применения обработки против развития сахарного диабета и ожирения у мышей, получавших рацион с высоким содержанием жира
- В целом, получавшие hBD-2 мыши набирали меньше массы первые 4 недели применения рациона, чем контрольные мыши на HFD (фиг. 143).
- У 7/8 тучных и с нарушенной толерантностью к глюкозе мышей значительно улучшалась их толерантность к глюкозе после всего лишь 2 недель применения обработки (фиг. 14a). Одна мышь была мышью с наиболее нарушенной толерантностью к глюкозе в начале исследования с массой жира приблизительно 20 грамм из массы тела 50 грамм. Несмотря на такой крайне нездоровый фенотип, мышь полностью избавилась от нарушения толерантности к глюкозе за 2 недели применения обработки (фиг. 16B).
- В отношении всего организма, получавшие hBD-2 мыши были менее устойчивы к инсулину, чем контрольные мыши на HFD (фиг. 16C). Это является ключевым моментом, поскольку тяжелую системную инсулиновую резистентность чрезвычайно сложно обратить, и она представляет основное ограничение в лечении заболеваний человека (например, сахарного диабета, CVD, некоторых злокачественных опухолей среди прочих).
HD-5 в качестве лечения метаболического синдрома у тучных мышей, получающих рацион с высоким содержанием жира
Изменение массы. Все получавшие HFD группы характеризовались одинаковым потреблением пищи не протяжении периода исследования и равным приростом массы на протяжении вводного периода в 13 недель (фиг. 22A). После применения рациона группа, получавшая HFD плюс HD-5, набирала существенно меньше массы, чем контроль на HFD (*p < 0,05, 2-факторный ANOVA) (фиг. 22B). Кроме того, наблюдали тенденцию к снижению процентного отношения жира у группы на HFD плюс HD-5 (фиг. 23A) и измеряли значительно более низкое процентное отношение жира у группы на HFD плюс HD-5 через 4 недели после изменения рациона по сравнению с контролем на HFD (*p = 0,009, 2-факторный ANOVA) (фиг. 23B). Наблюдали тенденцию к снижению массы печени при завершении у получавшей HFD плюс HD-5 группы по сравнению с контролем на HFD. В частности, у ~50% мышей, получавших стандартный HFD, был более высокий результат, чем у мыши, получавшей HFD плюс HD-5, с самым высоким результатом (фиг. 24A). Масса висцерального жира была больше у получавших HFD групп, чем у получавшей LFD группы (*p < 0,05, однофакторный ANOVA) (фиг. 24B).
Тест на толерантность к глюкозе. Толерантность к глюкозе для получавших HFD + HD5 животных в типичной клетке - клетке 2 - улучшалась со временем от начала применения обработки (неделя 13-0) до недели 13,8 (фиг. 25A).
Тест на толерантность к инсулину. Группа на LFD была значительно более чувствительной к инсулину, чем получавшие HFD группы (*p < 0,05, 2-факторный ANOVA). Группы на HFD плюс HD-5 были более чувствительными к инсулину, чем контроль на HFD, что подразумевало улучшение толерантности к инсулину после применения рациона (*p < 0,05, 2-факторный ANOVA) (фиг. 25B).
Заключение по HD5 в качестве применения обработки против развития сахарного диабета и ожирения у мышей, получавших рацион с высоким содержанием жира
- Мыши, получавшие HD5, характеризовались значительно меньшим изменением массы по сравнению с контрольными мышами, получавшими HFD (фиг. 22B).
- Наблюдается общая тенденция к снижению массы жира у тучных мышей, получавших HFD-HD5 (фиг. 23A и B).
- Наблюдается тенденция к снижению массы печени у мышей, получавших HD5, по сравнению с контрольными мышами, получавшими HFD (фиг. 24a). Поскольку висцеральное и подкожное депо отличались не значительно (фиг. 24b), это наблюдение подтверждает тот факт, что умеренное снижение % жира у мышей на HD5 ограничивается печеночным липолизом/окислением липидов.
- Толерантность к глюкозе у мышей, получавших HD5, со временем улучшалась (фиг. 25A).
- Мыши, получавшие HD5, были менее устойчивы к инсулину, чем контрольные мыши, получавшие HFD (фиг. 25b).
Наблюдаемые изменения согласуются с более высоким многообразием генов и более высоким числом филумов в кишечной микробиоте у групп на HFD, обрабатываемых дефензинами, чем у мышей, которых не обрабатывали.
Пример 4
Модуляция микробиоты кишечника и лечение воспаления кишечника путем применения лечения аналогом глюкагоноподобного пептида-1
Материалы и способы
Мыши. Самцов мышей C57Bl/6J DIO возрастом 4 недели кормили рационом с высоким содержанием жира (HFD, 60% жира, SSNIFF (рацион № D12492)) или Рurina Сhow на протяжении 36 недель. Получавшая HFD группа достигала средней массы приблизительно 55 грамм к началу применения обработки. Мышей размещали группами по 10 на клетку до недели -2. С недели -2 мышей размещали по отдельности на протяжении исследования. Потребление корма регистрировали ежесуточно непосредственно перед выключением света в 3 часа дня. Отдельных мышей подвергали экспериментальным процедурам в измененном порядке как в группе, так и в клетке. Мышей держали при комнатной температуре при 12-часовом цикле свет/темнота при стандартных условиях SPF.
Режим обработки (фиг. 26). Мышей кормили либо рационом с высоким содержанием жира (HFD), либо контрольным рационом с низким содержанием жира (LF). HFD предусматривал 2 подгруппы: 1 на GLP-1 и 1 на стандартном HFD без дополнения GLP-1. GLP-1 растворяли в PBS и добавляли 0,1% BSA. GLP-1 вводили подкожно при 0,2 мг/кг BID.
Анализ. Анализ 16S РНК микробиома выполняли в дни -1 и 27 исследования. Образец из подвздошной кишки собирали в 2 см от слепой кишки и быстро замораживали в жидком азоте для анализа концентрации цитокинов IFNγ, TNF-α, IL-1β, IL12p70, IL-2, IL-4, IL-5, IL-6, IL-8 и IL-10.
Результаты
Невзвешенный анализ UniFrac, т.e. относительное содержание видов бактерий. В начале исследования (день 1) четыре группы имели сопоставимую микрофлору, как и ожидалось. Однако после 4-недельной обработки микробиота от мышей, получавших рацион с высоким содержанием жира и применение обработки с GLP-1 (лираглутидом), значительно отличалась от микрофлоры контрольной (обработанной средой-носителем) группы (фиг. 27A).
Наблюдаемые изменения в основном были обусловлены повышенным содержанием Akkermansia и Alloprevotella. Akkermansia включает в себя продуцирующие короткоцепочечную жирную кислоту виды (фиг. 27B). Повышение количества этих бактерий считается показателем более здоровой или нормализованной микрофлоры. Известно, что короткоцепочечные жирные кислоты оказывают положительное влияние на индукцию эндогенного GLP-1.
Не наблюдали, что неожиданно, противовоспалительный эффект GLP-1 (лираглутида), поскольку не было статистического различия в концентрации цитокинов в подвздошной кишке ни для какого из цитокинов между тремя группами исследования.
Пример 5
Фармакокинетическое исследование по определению пероральной биодоступности и установлению фармакокинетического профиля hBD-2 после однократного перорального введения мышам NMRI через желудочный зонд 4 мг/кг
Материалы и способы
Режим лечения. 21 самке мышей NMRI вводили дозу 5 мл/кг пероральным введением через желудочный зонд с использованием трубки для введения через желудочный зонд и 1-мл шприца в соответствии с массой тела индивидуума, полученной в день введения дозы. В произвольные моменты времени старались отбирать образцы мочи с помощью осторожного массажа паховой области живота. Первый образец крови отбирали с использованием метода подчелюстного отбора образца. Второй образец крови отбирали у анестезированных изофлураном мышей. Кишечные образцы отбирали после эвтаназии. Живот каждой мыши вскрывали и отбирали образцы с трех отделов кишечника (тонкой кишки, двенадцатиперстной кишки и толстой кишки).
Результаты
hBD-2, по-видимому, не абсорбируется из здорового кишечника, так как он не может быть обнаружен с помощью HPLC ни в каком из образцов сыворотки или мочи, поскольку все значения ниже уровня выявления < 10 пг/мл. Это указывает на то, что hBD-2 не является системно доступным после перорального введения дозы 4 мг/кг у мышей.
Перорально вводимый hBD2 остается выявляемым в содержимом толстой кишки до 360 минут после введения (фиг. 28).
Пример 6
Исследование и сравнение фармакокинетических профилей hBD-2, слитого с C-концом (молекулярная масса 71,336 Да) или N-концом (молекулярная масса 71,666 Да) человеческого сывороточного альбумина, после подкожного или внутривенного введения молярного эквивалента 1 мг/кг hBD-2 (молекулярная масса 66437 Да) самкам мышей NMRI
Материал и способы
Режим лечения. Животным вводили дозу 10 мл/кг исходной концентрации 1,65 мг/мл в соответствии с индивидуальной массой тела (300 мкл для 30-граммовой мыши). Первый образец крови отбирали с использованием метода подчелюстного отбора образца, а второй после анестезии изофлураном и эвтаназии.
Результаты
hBD-2 демонстрировал период полувыведения 1 час, а два слитых белка - период полувыведения 12 часов. AUC изменялась критически. Почечное выведение также изменялось от 10 мл/минута для hBD-2 (фиг. 29) до 0,5-2,2 мл/мин для двух слитых молекул (фиг. 30, 31).
Пример демонстрирует, что период полувыведения hBD2 может быть заметно продлен с помощью C- или N-концевой конъюгации с альбумином.
Пример 7
Определение и оценивание противовоспалительного эффекта N-концевого слияния hBD-2-альбумин в отношении модели острого индуцированного декстрана сульфатом натрия (DSS) в течение 10 суток колита у мышей
Материал и способы
Режим лечения. N-концевое слияния hBD-2-альбумин вводили внутривенно через хвостовую вену или подкожно с применением стерильной иглы 25G при объеме вводимой дозы 10 мл/кг массы тела. Животные получали 1 дозу ежесуточно на протяжении 10 рабочих дней. Активный контроль дексаметазон (DEX) вводили подкожно дозой 1 мг/кг при объеме вводимой дозы 10 мл/кг массы тела OD.
Результаты
Обработка N-концевым слиянием hBD-2-альбумин приводила к значительному ингибированию индекса активности заболевания (DAI) при введении ежесуточно дозой 1,65 мг/кг внутривенным путем (p < 0,05). Кроме того, в день 10 значительное ингибирование значения DAI также наблюдали при введении N-концевого слияния hBD-2-альбумин ежесуточно дозой 1,65 мг/кг и дозой 125 мг/кг подкожно, соответственно (p < 0,05).
Введение декстрана сульфата натрия приводило к значительному воспалению и повреждению ткани толстой кишки, о чем свидетельствует гистологическое обследование. Обработка N-концевым слиянием hBD-2-альбумин не приводит к какому-либо статистически значимому снижению данного гистологического поражения, и, подобным образом, активный контроль DEX не смог существенно уменьшить гистологическое повреждение.
Кроме того, результаты показывают, что значительное увеличение массы тела в день 7 у животных, обработанных N-концевым слиянием hBD-2-альбумин, несмотря на временное снижение массы тела в дни 2 и 3. Напротив, животные, обработанные DEX, демонстрировали весьма значительное снижение массы тела со дня 5 и далее (p < 0,01). Это указывает на то, что N-концевое слияние hBD-2-альбумин поддерживает нормальную микробиоту и, таким образом, обладает эффектом сохранения массы.
Пример демонстрирует, что N-концевое слияние hBD-2-альбумин является биологически активным в животной модели воспалительного состояния.
Пример 8
Определение и оценивание противовоспалительного эффекта С-концевого слияния hBD-2-альбумин в отношении модели острого индуцированного декстрана сульфатом натрия (DSS) в течение 10 суток колита у мышей
Материал и способы
Режим лечения. С-концевое слияния hBD-2-альбумин вводили внутривенно через хвостовую вену или подкожно с применением стерильной иглы 25G при объеме вводимой дозы 10 мл/кг массы тела. Животные получали 1 дозу ежесуточно на протяжении 10 рабочих дней. Активный контроль преднизолон (Pred) давали перорально путем введения через желудочный зонд дозой 1 мг/кг при объеме вводимой дозы 10 мл/кг массы тела OD.
Результаты
Обработка С-концевым слиянием hBD-2-альбумин приводила к значительному ингибированию DAI при введении ежесуточно дозой 1,6 мг/кг внутривенным путем (p < 0,05). Кроме того, С-концевое слияние hBD-2-альбумин приводило к значительному ингибированию DAI при введении в альтернативные дни 0, 2, 4, 6, 8 и 10 дозой 1,6 мг/кг внутривенным путем (p < 0,05). Ежесуточная обработка преднизолоном приводила к значительному ингибированию DAI в день 9 (p < 0,05).
Введение декстрана сульфата натрия приводило к значительному воспалению и повреждению ткани толстой кишки, о чем свидетельствует гистологическое обследование. Обработка С-концевым слиянием hBD-2-альбумин дозой 1,6 мг/кг приводила к статистически значимому снижению такого гистологического поражения (p < 0,05). Подобным образом, ежесуточная обработка С-концевым слиянием hBD-2-альбумин дозой 1,6 мг/кг и 16,5 мг/кг в дни 0, 2, 4, 6, 8 и 10 приводила к значительному снижению гистологического поражения толстой кишки (p < 0,01). Обработка активным контролем преднизолоном не могла существенно уменьшить гистологическое повреждение в проксимальной части толстой кишки, но снижала повреждение в дистальной части толстой кишки (p < 0,01).
Кроме того, результаты показали значительное повышение массы тела у животных, обработанных С-концевым слиянием hBD-2-альбумин (p < 0,05). Это указывает на то, что С-концевое слияние hBD-2-альбумин поддерживает нормальную микробиоту и, таким образом, обладает эффектом сохранения массы.
Пример демонстрирует, что С-концевое слияние hBD-2-альбумин является биологически активным в отношении животной модели воспалительного состояния.
Пример 9
Последовательности
Ссылки
Ajslev TA, et al., 2014. Trends in parent-child correlations of childhood body mass index during the development of the obesity epidemic. PLoS One 9(10).
Angelakis E and Raoult D, 2010. The increase of Lactobacillus species in the gut flora of newborn broiler chicks and ducks is associated with weight gain. PLoS One 5(5).
Angelakis E., et al. 2012. An evaluation of the effects of Lactobacillus ingluviei on body weight, the intestinal microbiome and metabolism in mice. Microb Pathog 52(1):61-8.
Armogida SA, et al., 2004. Identification and quantification of innate immune system mediators in human breast milk. Allergy Asthma Proc. 25(5):297-304.
Bäckhed F, et al., 2007. Mechanisms underlying the resistance to diet-induced obesity in germ-free mice. Proc Natl Acad Sci USA 104(3):979-84.
Belkaid W and Hand TW, 2014. Role of the microbiota in immunity and inflammation. Cell 157(1):121-41.
Bowie JU and Sauer RT, 1989. Identifying determinants of folding and activity for a protein of unknown structure. Proc. Natl. Acad. Sci. USA 86: 2152-2156.
Chassaing B, et al., 2015. Dietary emulsifiers impact the mouse gut microbiota promoting colitis and metabolic syndrome. Nature 519(7541):92-6.
Cunningham BC and Wells JA, 1989. High-resolution epitope mapping of hGH-receptor interactions by alanine-scanning mutagenesis. Science 244: 1081-1085.
Derbyshire KM, Salvo JJ and Grindley ND, 1986. A simple and efficient procedure for saturation mutagenesis using mixed oligodeoxynucleotides. Gene 46:145-152.
Everard A and Cani PD, 2013. Diabetes, obesity and gut microbiota. Best Pract Res Clin Gastroenterol 27(1):73-83.
Faulds MH and Dahlman-Wright K, 2012. Metabolic diseases and cancer risk. Curr Opin Oncol. 24(1):58-61.
Favre-Godal Q, et al., 2014. Comprehensive approach for the detection of antifungal compounds using a susceptible strain of Candida albicans and confirmation of in vivo activity with the Galleria mellonella model. Phytochemistry. 105: 68-78.
Feng Q, et al., 2015. Gut microbiome development along the colorectal adenoma-carcinoma sequence. Nat Commun 6:6528.
Giannouli M, et al. Use of larvae of the wax moth Galleria mellonella as an in vivo model to study the virulence of Helicobacter pylori. 2014. BMC Microbiol14: 228.
Hilton DJ, et al., 1996. Saturation mutagenesis of the WSXWS motif of the erythropoietin receptor. J. Biol. Chem. 271: 4699-4708.
Khan M, et al., 2007. Growth-promoting effects of single-dose intragastrically administered probiotics in chickens. Br Poult Sci 48(6):732-5.
Koren O, et al., 2012. Host remodeling of the gut microbiome and metabolic changes during pregnancy. Cell 150(3):470-80.
Le Chatelier E et al. 2013. Richness of human gut microbiome correlates with metabolic markers. Nature 500(7464):541-6.
Leviten M, 2016. The Finnish connection. Biocentury Innovations, June 16.
Liu hY et al., 2008. Suppression of hepatic glucose production by human neutrophil α-defensin through a signaling pathway distinct from insulin. The Journal of Biological Chemistry 283(18):12056-12063.
Lowman HB, Bass SH, Simpson N and Wells JA, 1991. Selecting high-affinity binding proteins by monovalent phage display. Biochem 30:10832-10837.
Mowat AM and Agace WW, 2014. Regional specialization within the intestinal immune system. Nat Rev Immunol. 14(10):667-85.
Needleman SB and Wunsch CD, 1970. A general method applicable to the search for similarities in the amino acid sequence of two proteins. J. Mol. Biol. 48: 443-453.
Ner SS, Goodin DB and Smith M, 1988. A simple and efficient procedure for generating random point mutations and for codon replacements using mixed oligodeoxynucleotides. DNA 7:127-134.
Neurath H and Hill RL, 1979. The Proteins. Academic Press, New York.
Qin J, et al. 2012. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 490(7418):55-60.
Reidhaar-Olson JF and Sauer RT, 1988. Combinatorial cassette mutagenesis as a probe of the informational content of protein sequences. Science 241:53-57;
Gennaro AR, 1990. Remington 's Pharmaceutical Sciences. Ed. Mack Publishing Co., Easton, PA.
Rice P, Longden I and Bleasby A, 2000. EMBOSS: the European Molecular Biology Open Software Suite. Trends in Genetics 16: 276-277.
Ridaura VK, et al., 2013. Gut microbiota from twins discordant for obesity modulate metabolism in mice. Science 341(6150):1241214.
Salzman NH, Underwood MA and Bevins CL, 2007. Paneth cells, defensins, and the commensal microbiota: a hypothesis on intimate interplay at the intestinal mucosa. Semin Immunol 19(2):70-83.
Suez J, et al., 2014. Artificial sweeteners induce glucose intolerance by altering the gut microbiota. Nature 514(7521):181-6.
Trasande L, Blustein J, Liu M, Corwin E, Cox LM and Blaser MJ, 2012. Infant antibiotic exposures and early-life body mass. Int J Obes (Lond) 37(1):16-23.
Turnbaugh PJ, Ley RE, Mahowald MA, Magrini V, Mardis ER and Gordon JI, 2006. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature. 2006 Dec 21;444(7122):1027-31.
Turnbaugh PJ, Bäckhed F, Fulton L and Gordon JI, 2008. Diet-induced obesity is linked to marked but reversible alterations in the mouse distal gut microbiome. Cell Host Microbe 3(4):213-23.
Vrieze A, et al. 2012. Transfer of intestinal microbiota from lean donors increases insulin sensitivity in individuals with metabolic syndrome. Gastroent 143(4):913-6.
Walter, 2015. Murine gut microbiota-diet trumps genes. Cell Host Microbe 17(1):3-5.
Wehkamp J, et al., 2002. Innate immunity and colonic inflammation: enhanced expression of epithelial alpha-defensins. Dig Dis Sci. 47(6):1349-55.
Zhang X, et al., 2015. The oral and gut microbiomes are perturbed in rheumatoid arthritis and partly normalized after treatment. Nat Med 21(8):895-905.
Следующие патенты и заявки на выдачу патентов
WO 2010/007166
WO 92/06204
WO 95/17413
WO 95/22625
US 5223409.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ДЕФЕНСИН ТЕРАПЬЮТИКС АПС
<120> СПОСОБЫ МОДУЛЯЦИИ КИШЕЧНОЙ МИКРОБИОТЫ
<130> P4094PC00
<150> DK PA 2016 70041
<151> 2016-01-26
<150> DK PA 2016 70483
<151> 2016-07-01
<160> 17
<170> PatentIn version 3.5
<210> 1
<211> 40
<212> PRT
<213> Bos taurus
<400> 1
Gly Val Gly Asn Pro Val Ser Cys Val Arg Asn Lys Gly Ile Cys Val
1 5 10 15
Pro Ile Arg Cys Pro Gly Ser Met Lys Gln Ile Gly Thr Cys Val Gly
20 25 30
Arg Ala Val Lys Cys Cys Arg Lys
35 40
<210> 2
<211> 34
<212> PRT
<213> Gallus gallus
<400> 2
Leu Phe Cys Lys Gly Gly Ser Cys His Phe Gly Gly Cys Pro Ser His
1 5 10 15
Leu Ile Lys Val Gly Ser Cys Phe Arg Ser Cys Cys Lys Trp Pro Trp
20 25 30
Asn Ala
<210> 3
<211> 44
<212> PRT
<213> Pongo pygmaeus
<400> 3
Val Phe Gly Asp Ile Ser Asn Pro Val Thr Cys Leu Arg Ser Gly Ala
1 5 10 15
Ile Cys His Pro Gly Phe Cys Pro Arg Arg Tyr Lys His Ile Gly Thr
20 25 30
Cys Gly Leu Ser Val Ile Lys Cys Cys Lys Lys Pro
35 40
<210> 4
<211> 36
<212> PRT
<213> Homo sapiens
<400> 4
Asp His Tyr Asn Cys Val Ser Ser Gly Gly Gln Cys Leu Tyr Ser Ala
1 5 10 15
Cys Pro Ile Phe Thr Lys Ile Gln Gly Thr Cys Tyr Arg Gly Lys Ala
20 25 30
Lys Cys Cys Lys
35
<210> 5
<211> 41
<212> PRT
<213> Homo sapiens
<400> 5
Gly Ile Gly Asp Pro Val Thr Cys Leu Lys Ser Gly Ala Ile Cys His
1 5 10 15
Pro Val Phe Cys Pro Arg Arg Tyr Lys Gln Ile Gly Thr Cys Gly Leu
20 25 30
Pro Gly Thr Lys Cys Cys Lys Lys Pro
35 40
<210> 6
<211> 45
<212> PRT
<213> Homo sapiens
<400> 6
Gly Ile Ile Asn Thr Leu Gln Lys Tyr Tyr Cys Arg Val Arg Gly Gly
1 5 10 15
Arg Cys Ala Val Leu Ser Cys Leu Pro Lys Glu Glu Gln Ile Gly Lys
20 25 30
Cys Ser Thr Arg Gly Arg Lys Cys Cys Arg Arg Lys Lys
35 40 45
<210> 7
<211> 37
<212> PRT
<213> Homo sapiens
<400> 7
Glu Leu Asp Arg Ile Cys Gly Tyr Gly Thr Ala Arg Cys Arg Lys Lys
1 5 10 15
Cys Arg Ser Gln Glu Tyr Arg Ile Gly Arg Cys Pro Asn Thr Tyr Ala
20 25 30
Cys Cys Leu Arg Lys
35
<210> 8
<211> 32
<212> PRT
<213> Homo sapiens
<400> 8
Ala Thr Cys Tyr Cys Arg Thr Gly Arg Cys Ala Thr Arg Glu Ser Leu
1 5 10 15
Ser Gly Val Cys Glu Ile Ser Gly Arg Leu Tyr Arg Leu Cys Cys Arg
20 25 30
<210> 9
<211> 32
<212> PRT
<213> Homo sapiens
<400> 9
Ala Phe Thr Cys His Cys Arg Arg Ser Cys Tyr Ser Thr Glu Tyr Ser
1 5 10 15
Tyr Gly Thr Cys Thr Val Met Gly Ile Asn His Arg Phe Cys Cys Leu
20 25 30
<210> 10
<211> 41
<212> PRT
<213> Pan troglodytes
<400> 10
Gly Ile Ser Asp Pro Val Thr Cys Leu Lys Ser Gly Ala Ile Cys His
1 5 10 15
Pro Val Phe Cys Pro Arg Arg Tyr Lys Gln Ile Gly Thr Cys Gly Leu
20 25 30
Pro Gly Thr Lys Cys Cys Lys Lys Pro
35 40
<210> 11
<211> 41
<212> PRT
<213> Macaca mulatta
<400> 11
Gly Ile Gly Asp Pro Val Thr Cys Leu Lys Asn Gly Ala Ile Cys His
1 5 10 15
Pro Val Phe Cys Pro Arg Arg Tyr Lys Gln Ile Gly Thr Cys Gly Leu
20 25 30
Pro Gly Thr Lys Cys Cys Lys Lys Pro
35 40
<210> 12
<211> 40
<212> PRT
<213> Mus musculus
<400> 12
Lys Ile Asn Asn Pro Val Ser Cys Leu Arg Lys Gly Gly Arg Cys Trp
1 5 10 15
Asn Arg Cys Ile Gly Asn Thr Arg Gln Ile Gly Ser Cys Gly Val Pro
20 25 30
Phe Leu Lys Cys Cys Lys Arg Lys
35 40
<210> 13
<211> 40
<212> PRT
<213> Equus caballus
<400> 13
Gly Ile Gly Asn Pro Ile Ser Cys Ala Arg Asn Arg Gly Val Cys Ile
1 5 10 15
Pro Ile Gly Cys Leu Pro Gly Met Lys Gln Ile Gly Thr Cys Gly Leu
20 25 30
Pro Gly Thr Lys Cys Cys Arg Lys
35 40
<210> 14
<211> 40
<212> PRT
<213> Sus scrofa
<400> 14
Asn Ile Gly Asn Ser Val Ser Cys Leu Arg Asn Lys Gly Val Cys Met
1 5 10 15
Pro Gly Lys Cys Ala Pro Lys Met Lys Gln Ile Gly Thr Cys Gly Met
20 25 30
Pro Gln Val Lys Cys Cys Lys Arg
35 40
<210> 15
<211> 40
<212> PRT
<213> Capra aegagrus
<400> 15
Gly Ile Ile Asn His Arg Ser Cys Tyr Arg Asn Lys Gly Val Cys Ala
1 5 10 15
Pro Ala Arg Cys Pro Arg Asn Met Arg Gln Ile Gly Thr Cys His Gly
20 25 30
Pro Pro Val Lys Cys Cys Arg Lys
35 40
<210> 16
<211> 37
<212> PRT
<213> Homo sapiens
<400> 16
Leu Leu Gly Asp Phe Phe Arg Lys Ser Lys Glu Lys Ile Gly Lys Glu
1 5 10 15
Phe Lys Arg Ile Val Gln Arg Ile Lys Asp Phe Leu Arg Asn Leu Val
20 25 30
Pro Arg Thr Glu Ser
35
<210> 17
<211> 37
<212> PRT
<213> Homo sapiens
<400> 17
Pro Val Thr Cys Leu Lys Ser Gly Ala Ile Cys His Pro Val Phe Cys
1 5 10 15
Pro Arg Arg Tyr Lys Gln Ile Gly Thr Cys Gly Leu Pro Gly Thr Lys
20 25 30
Cys Cys Lys Lys Pro
35
<---


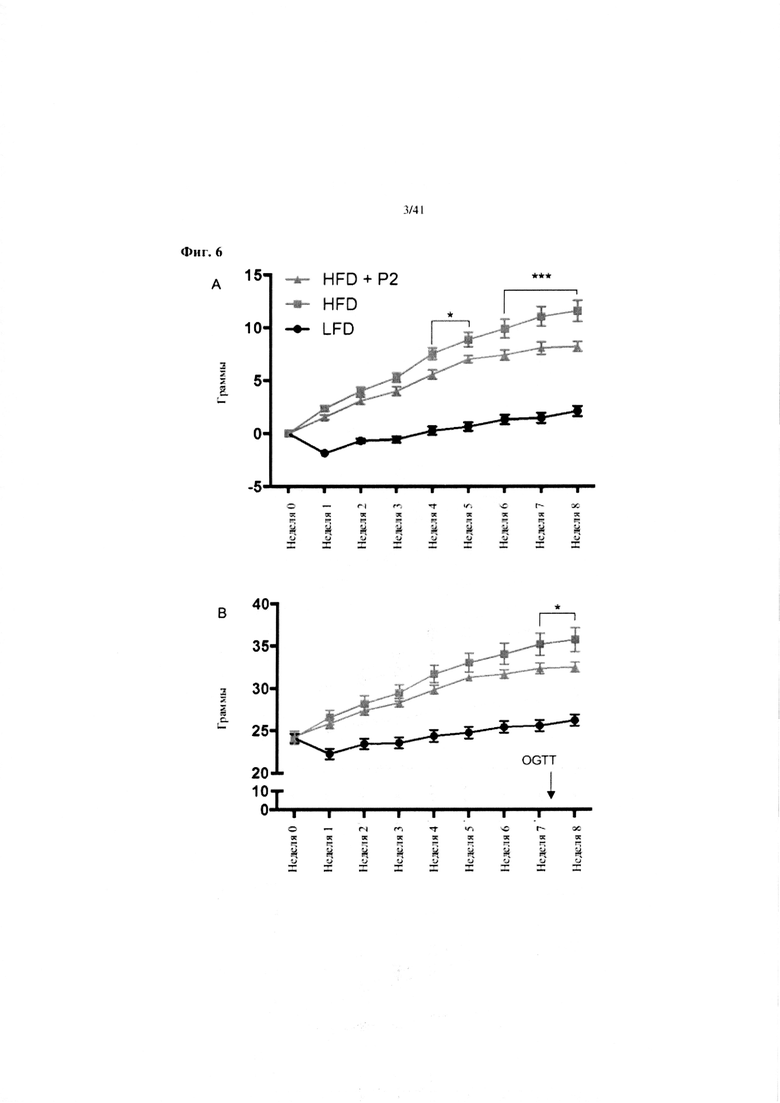
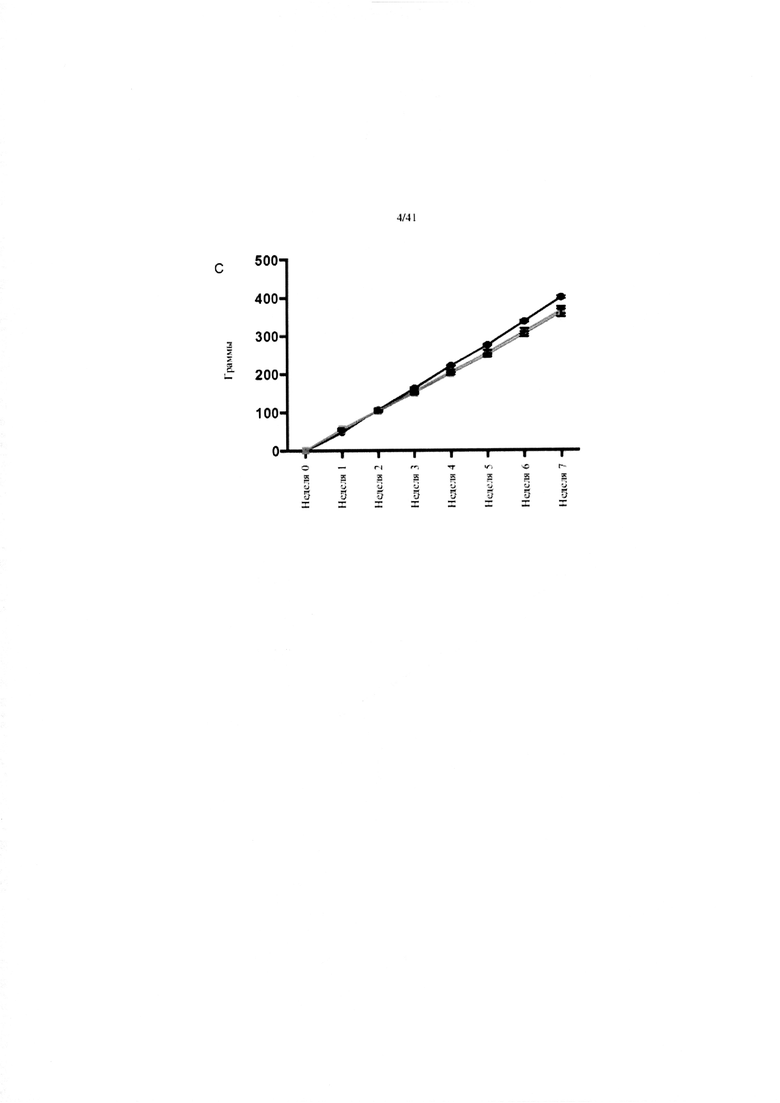
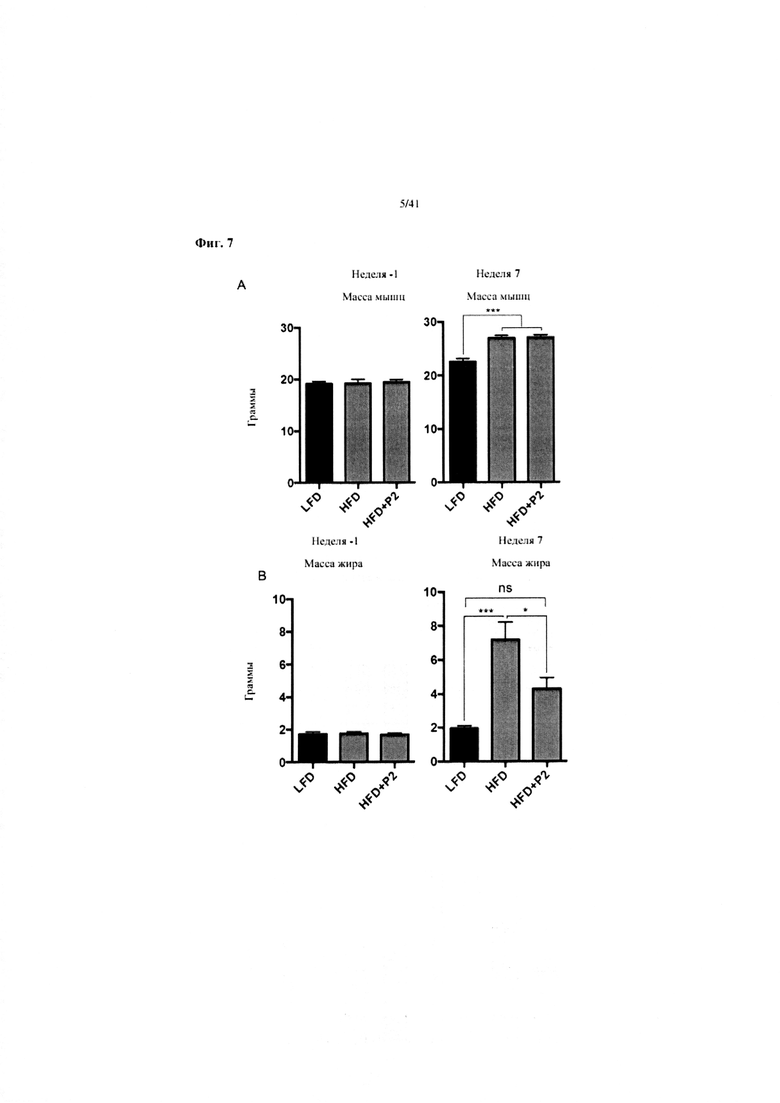
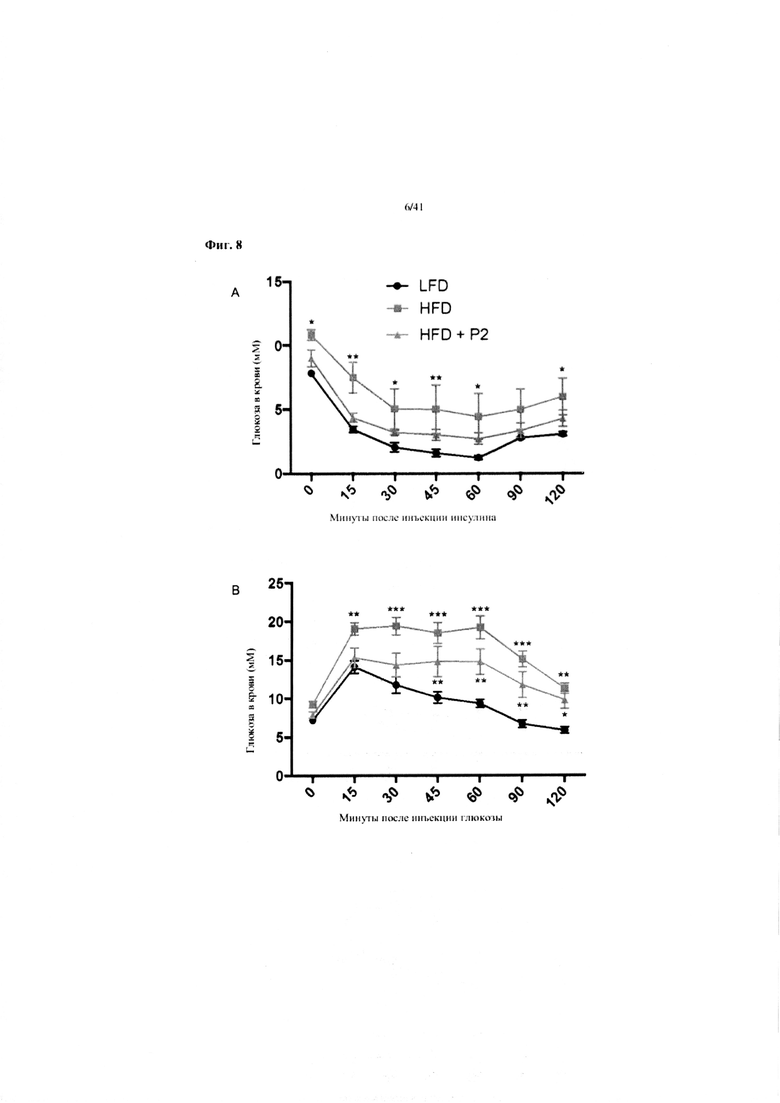
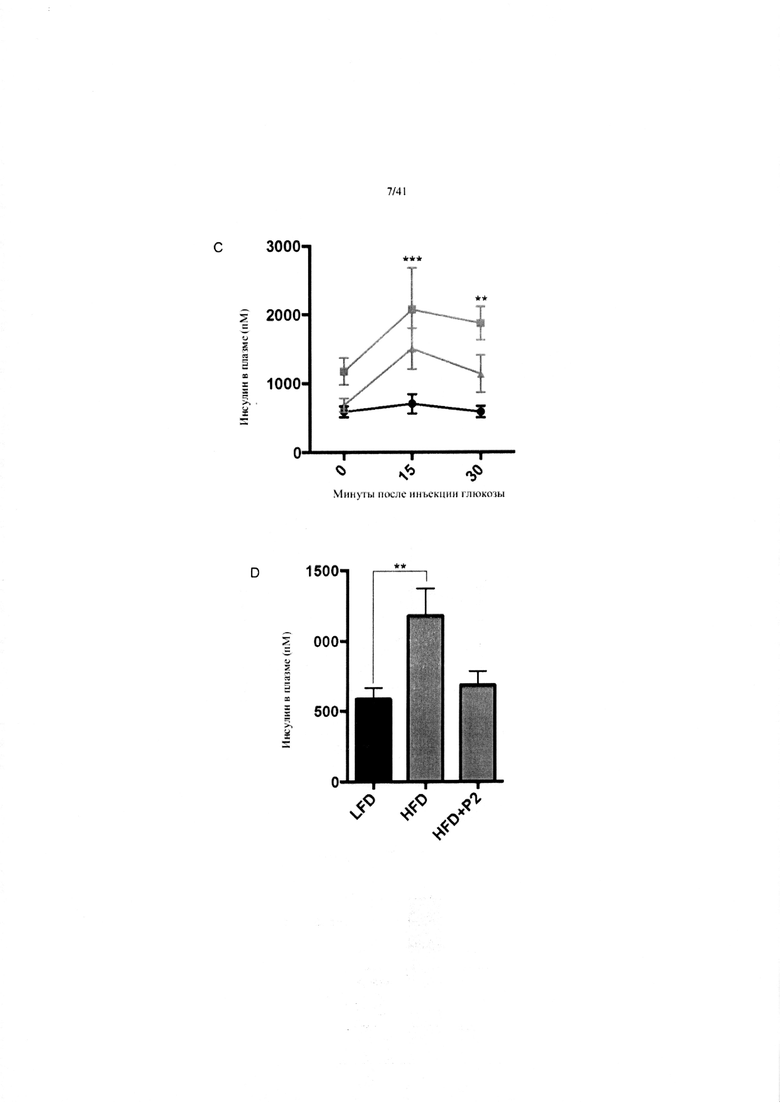
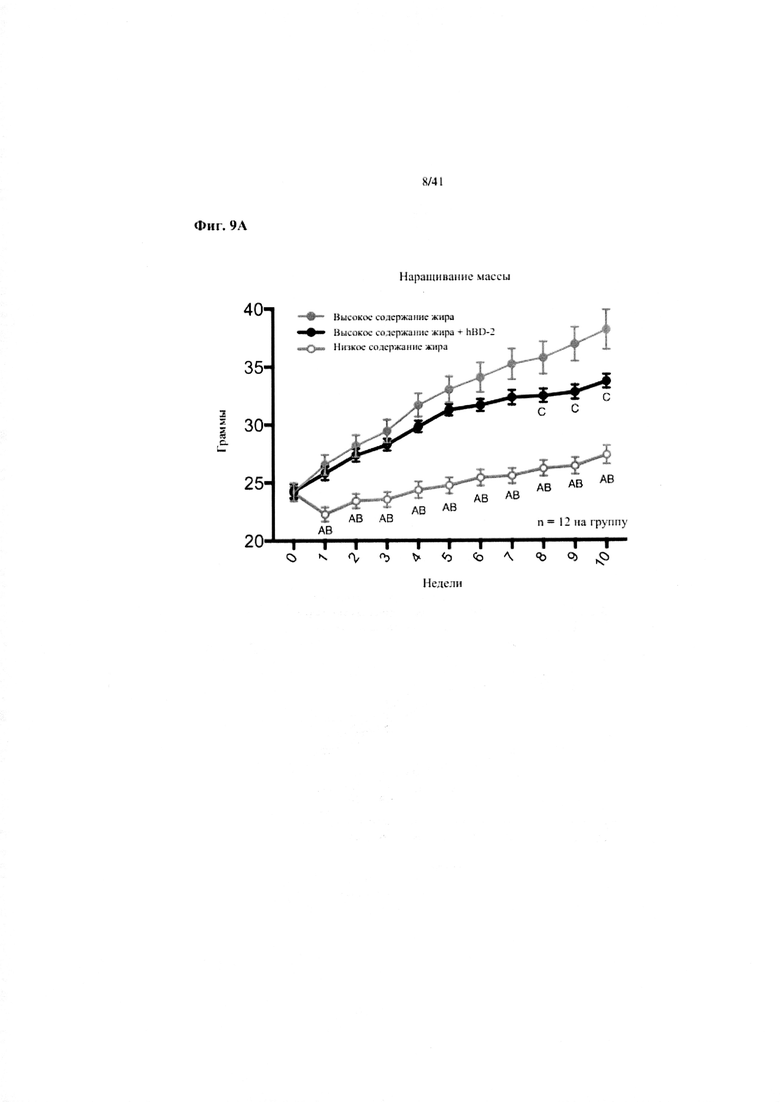
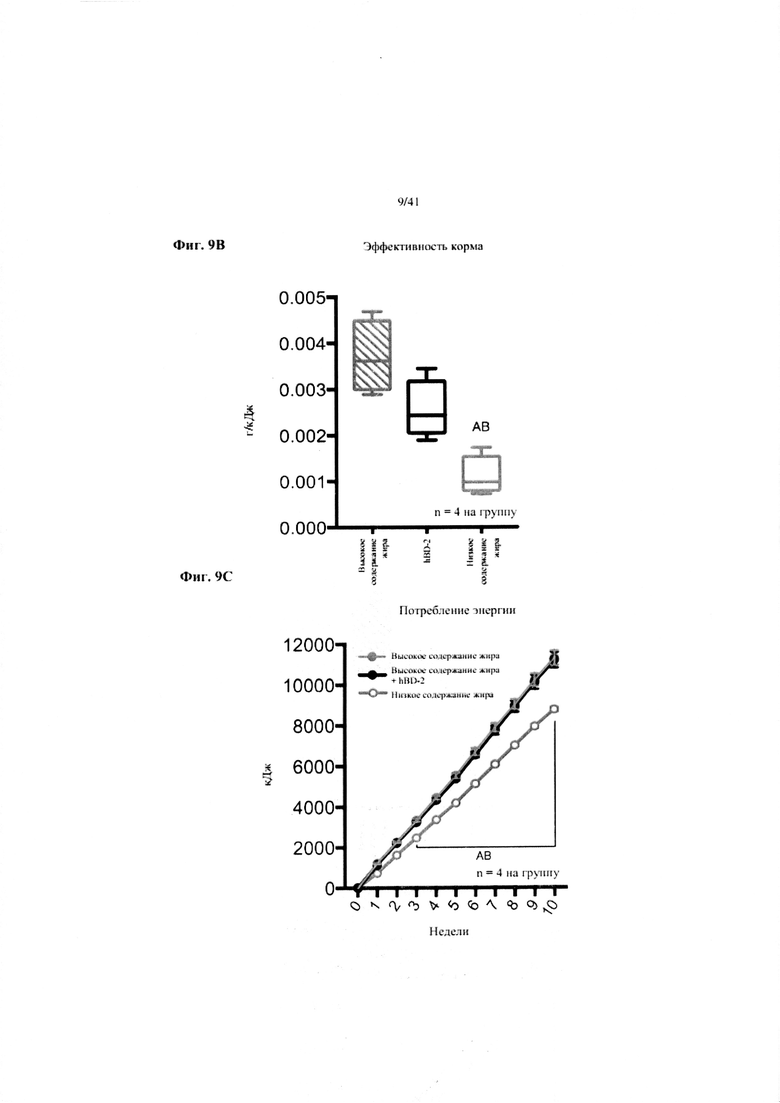
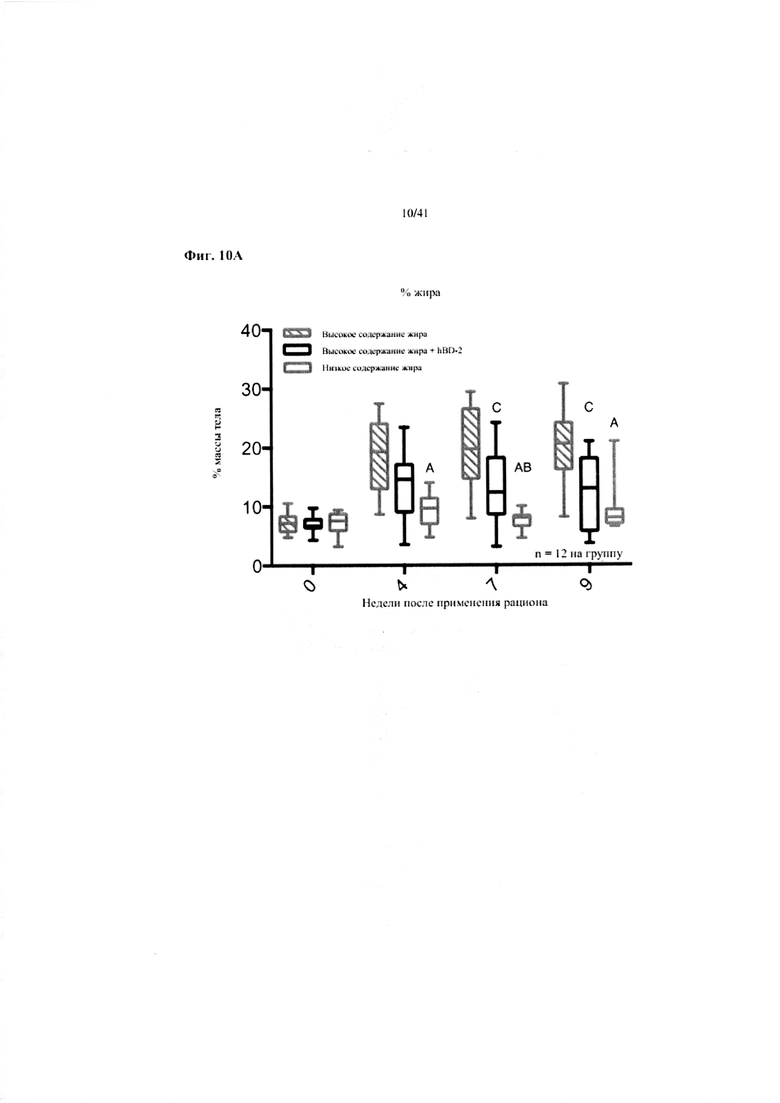
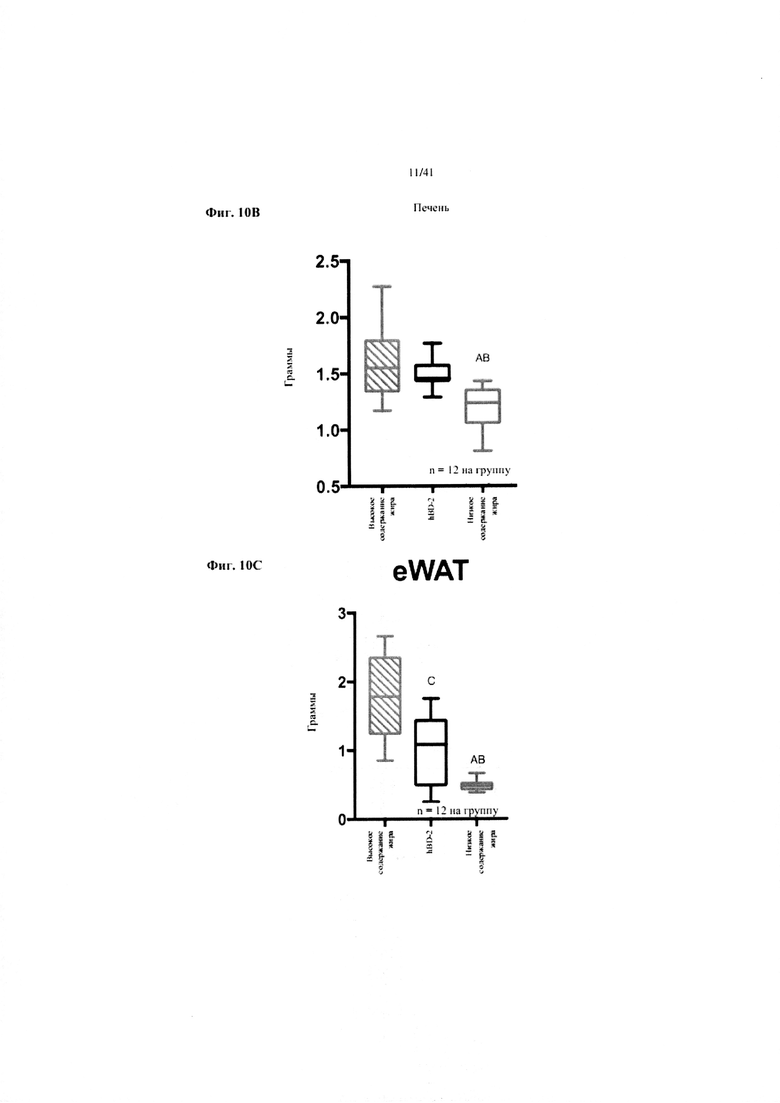
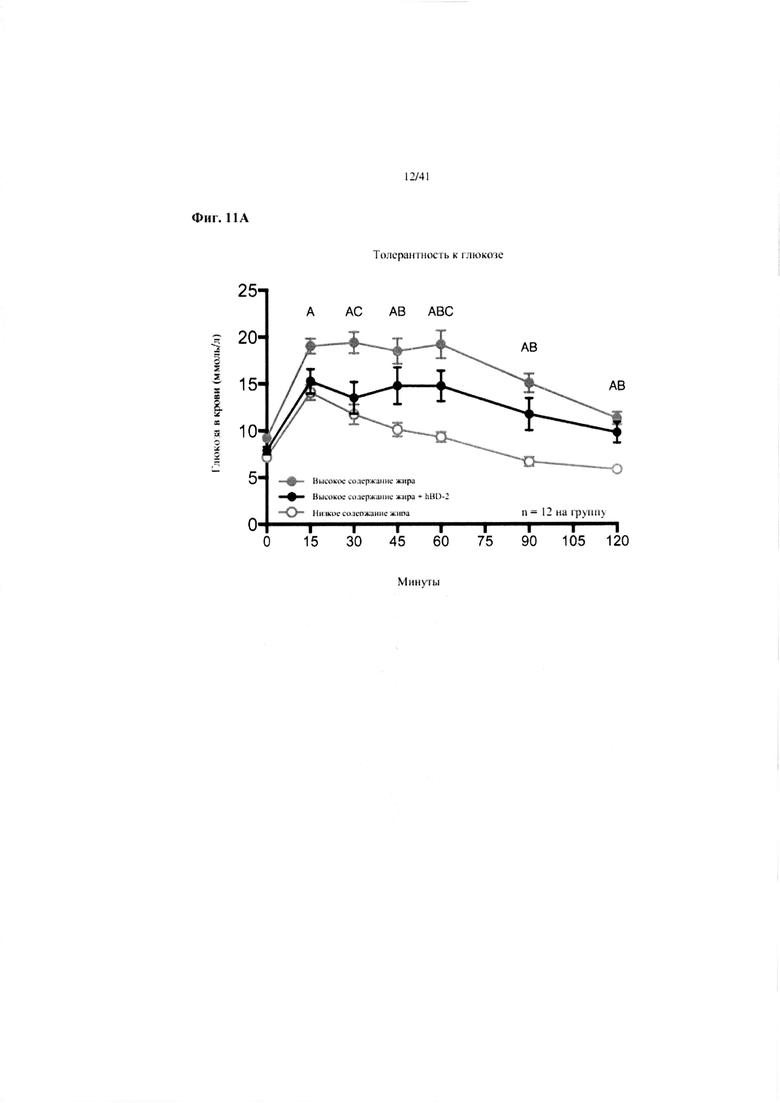
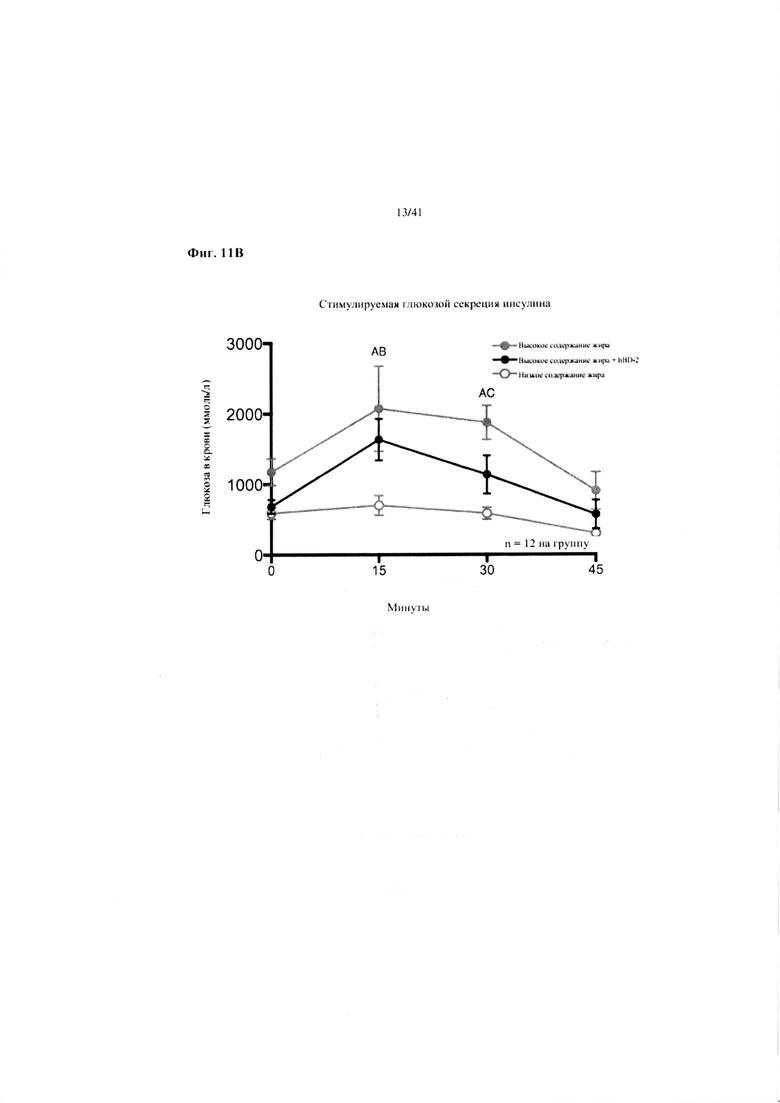
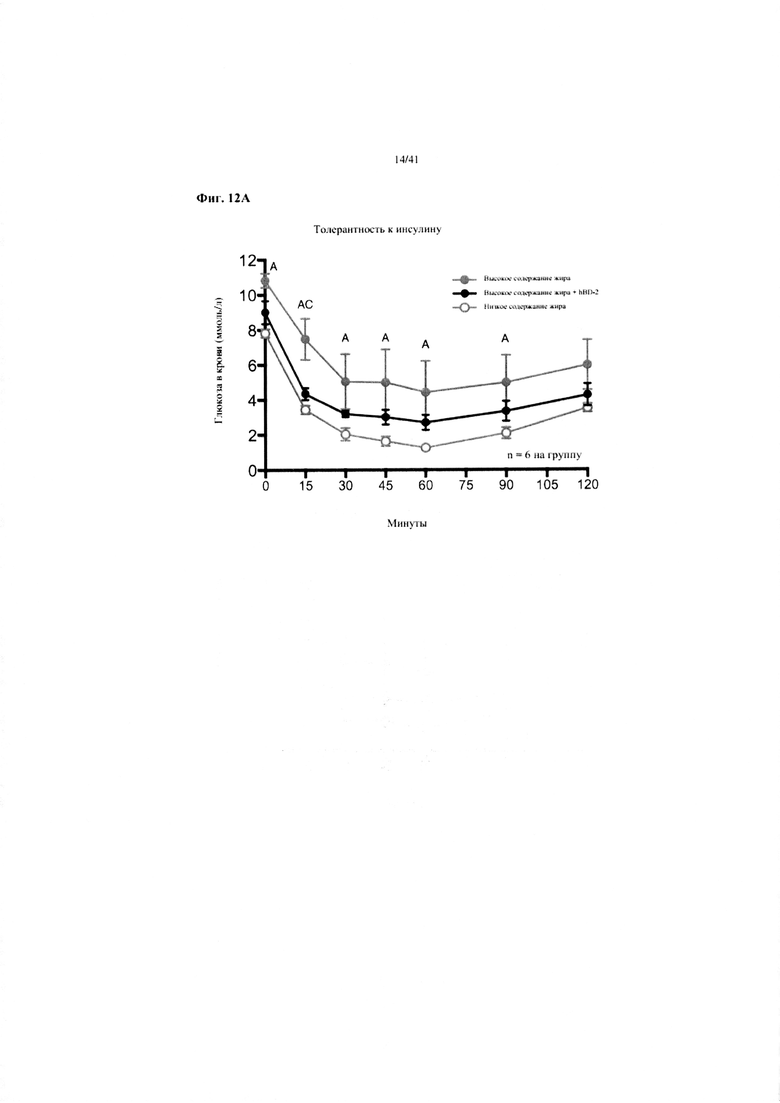

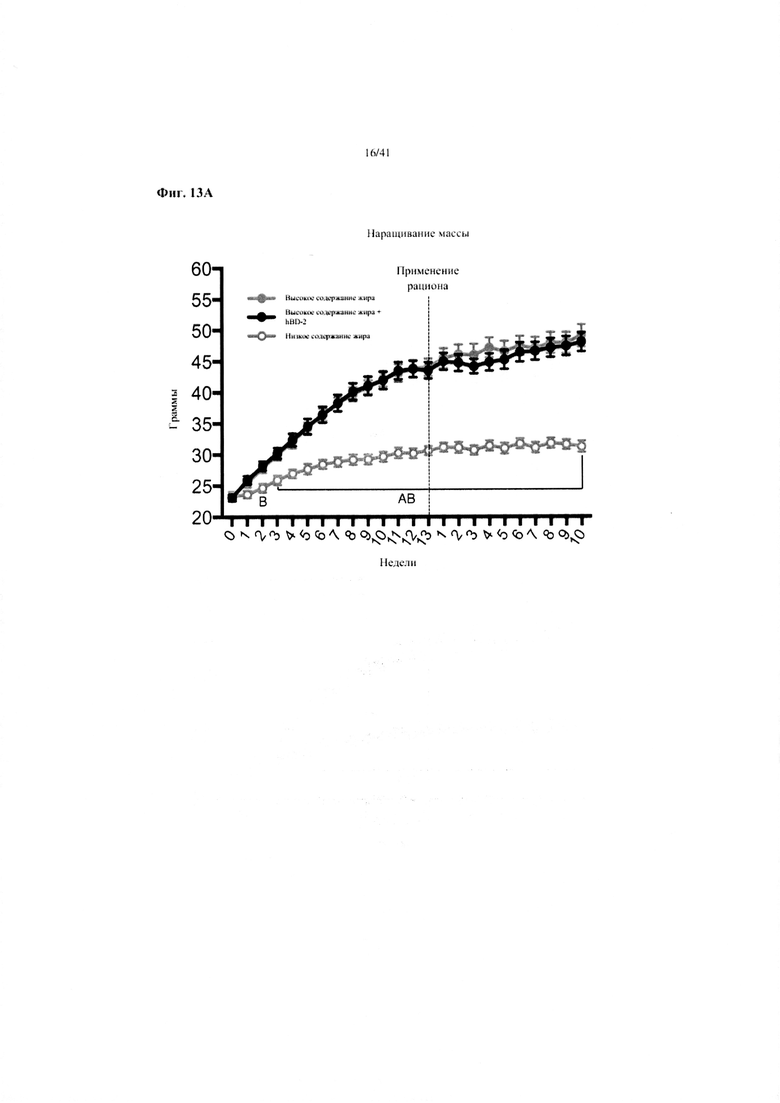
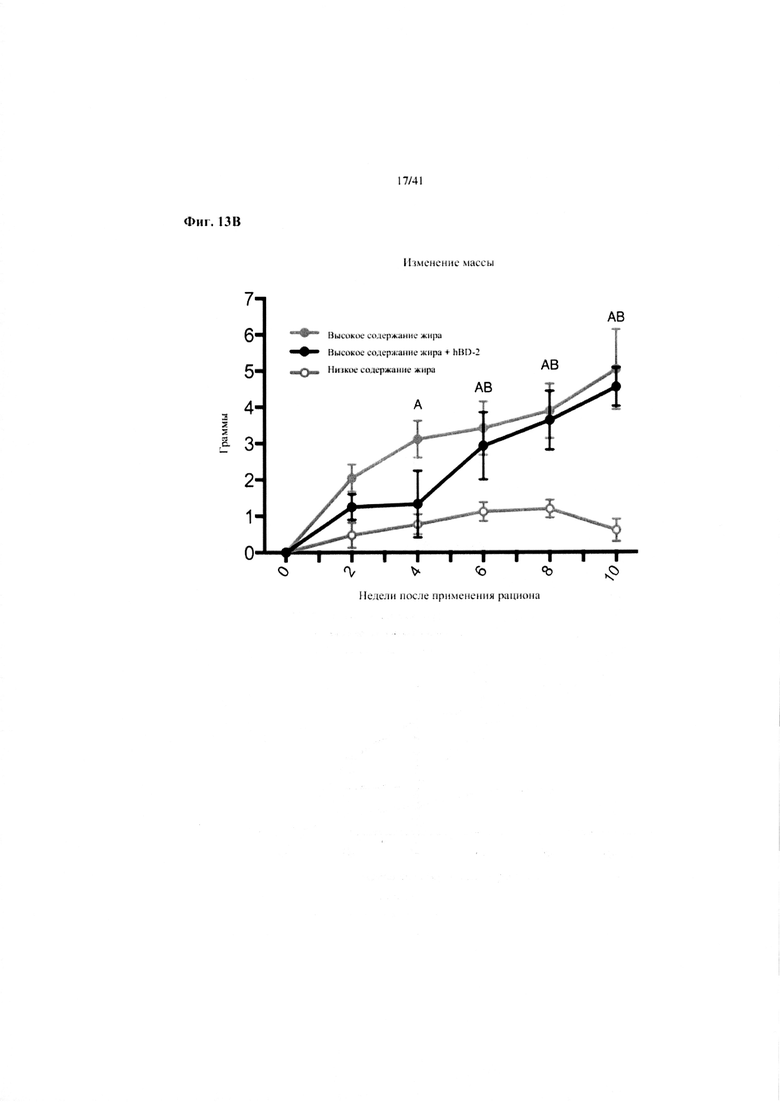
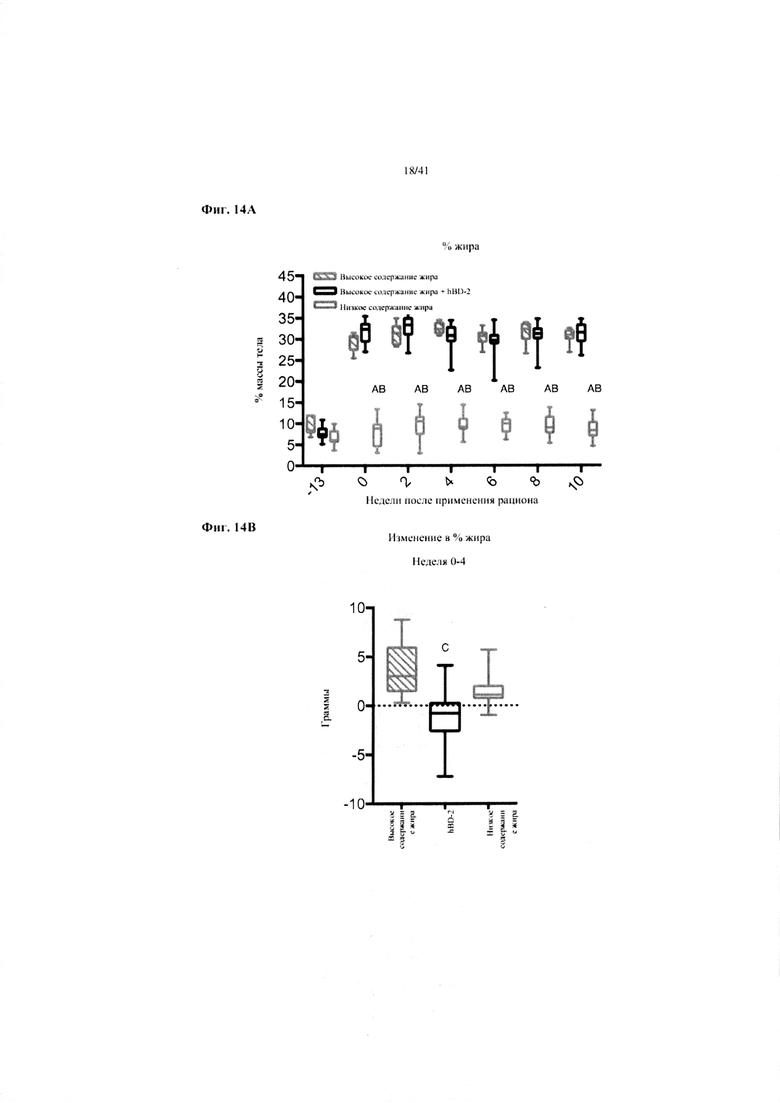
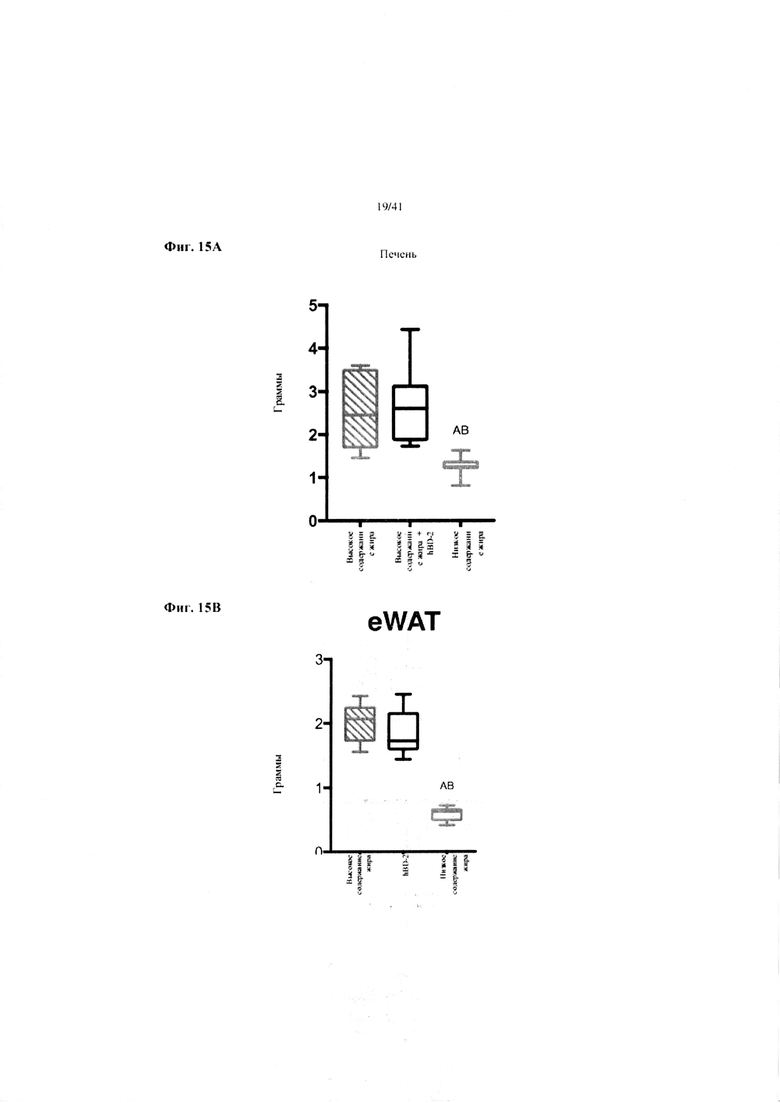
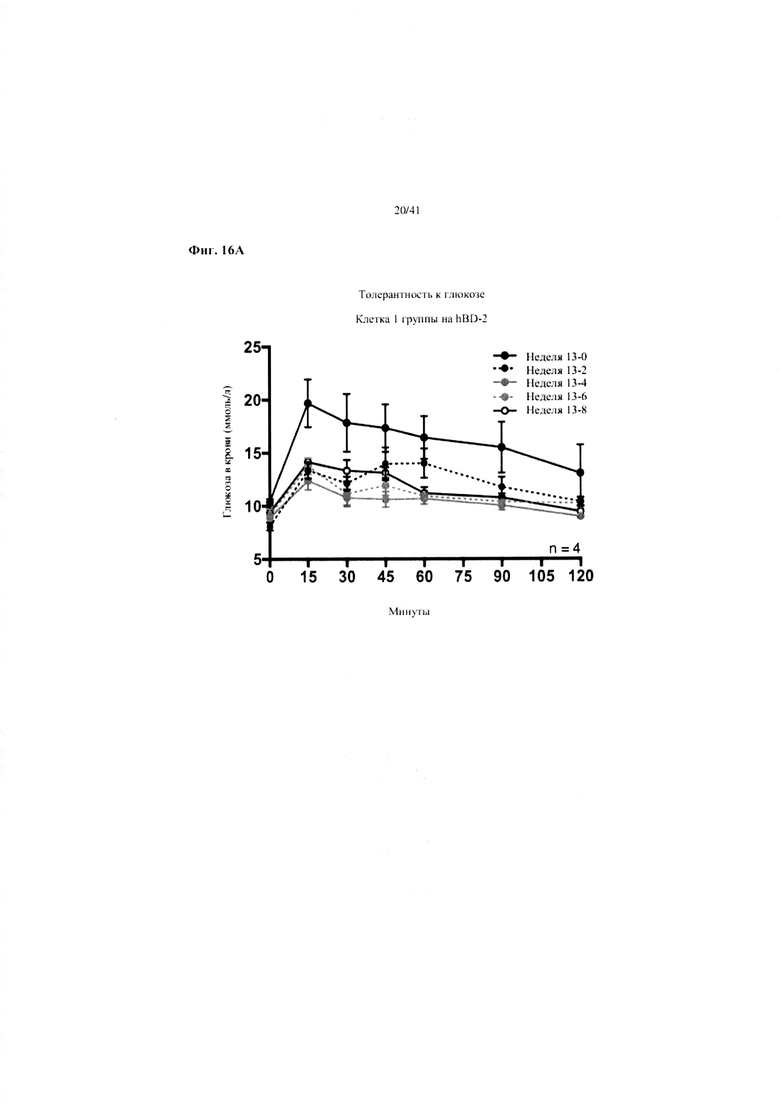
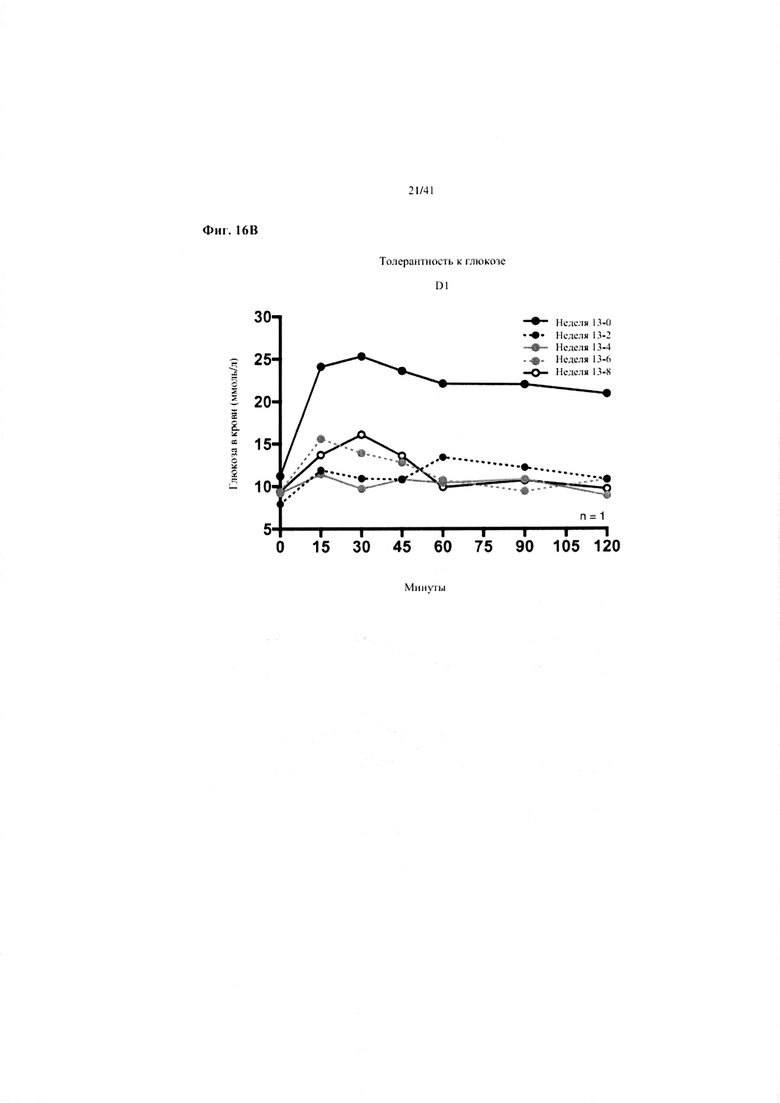
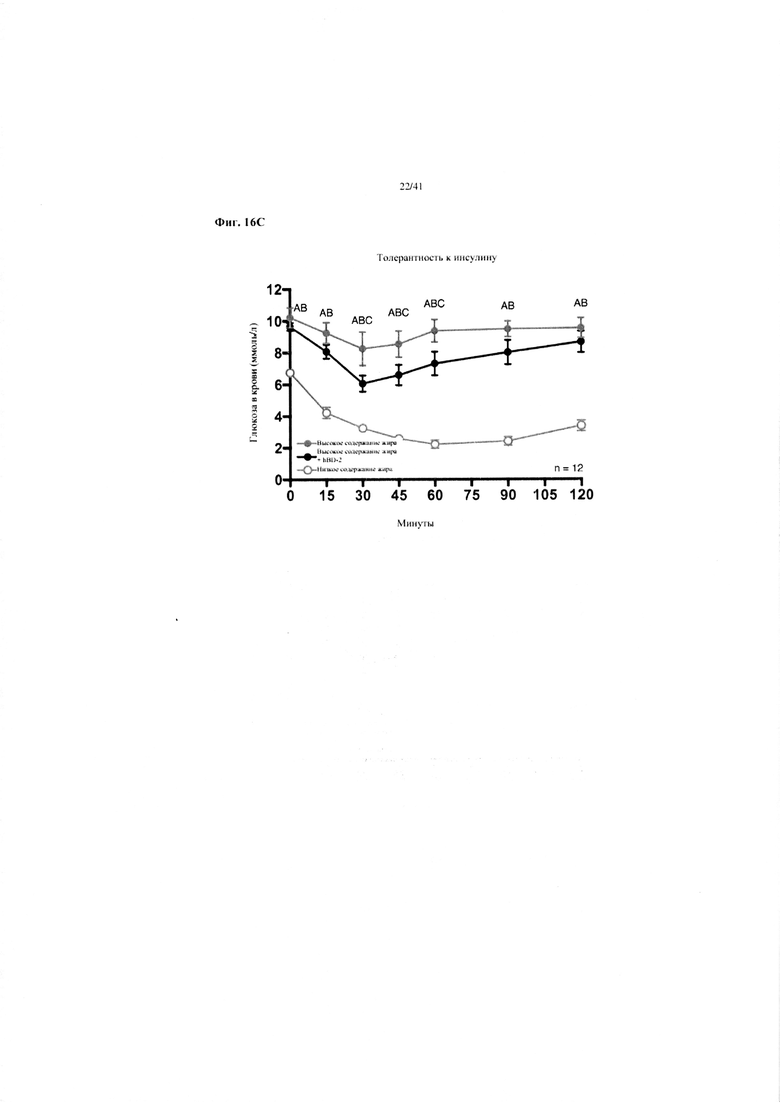
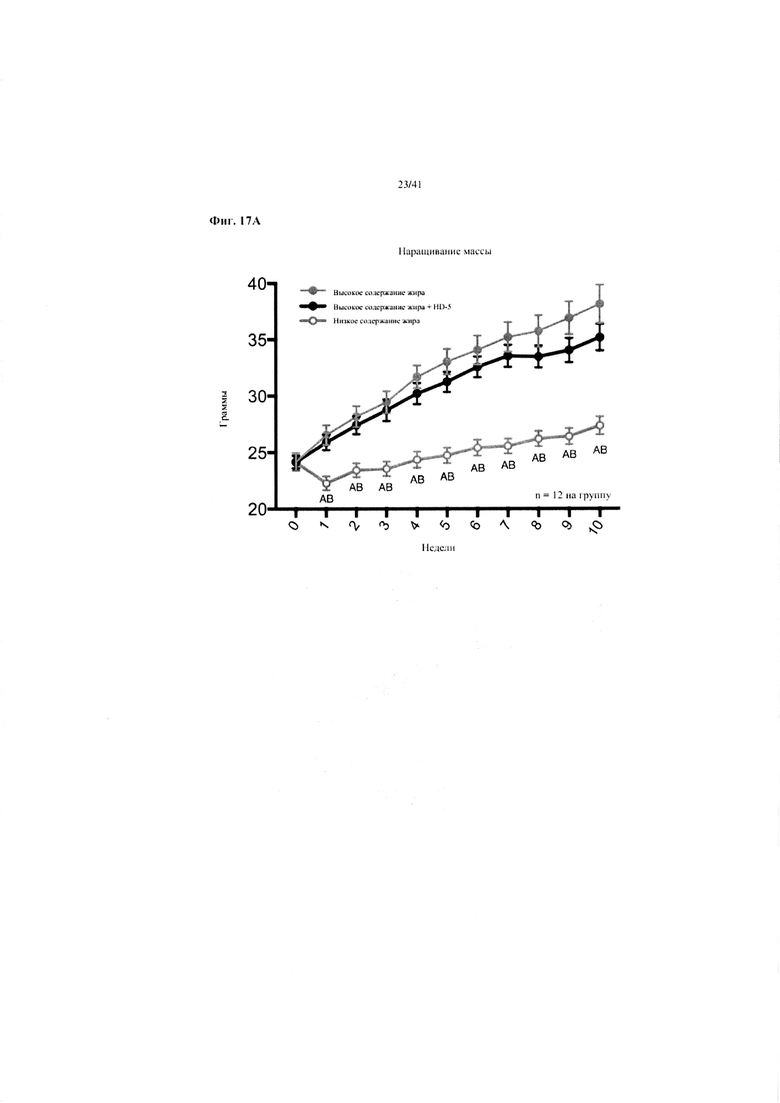
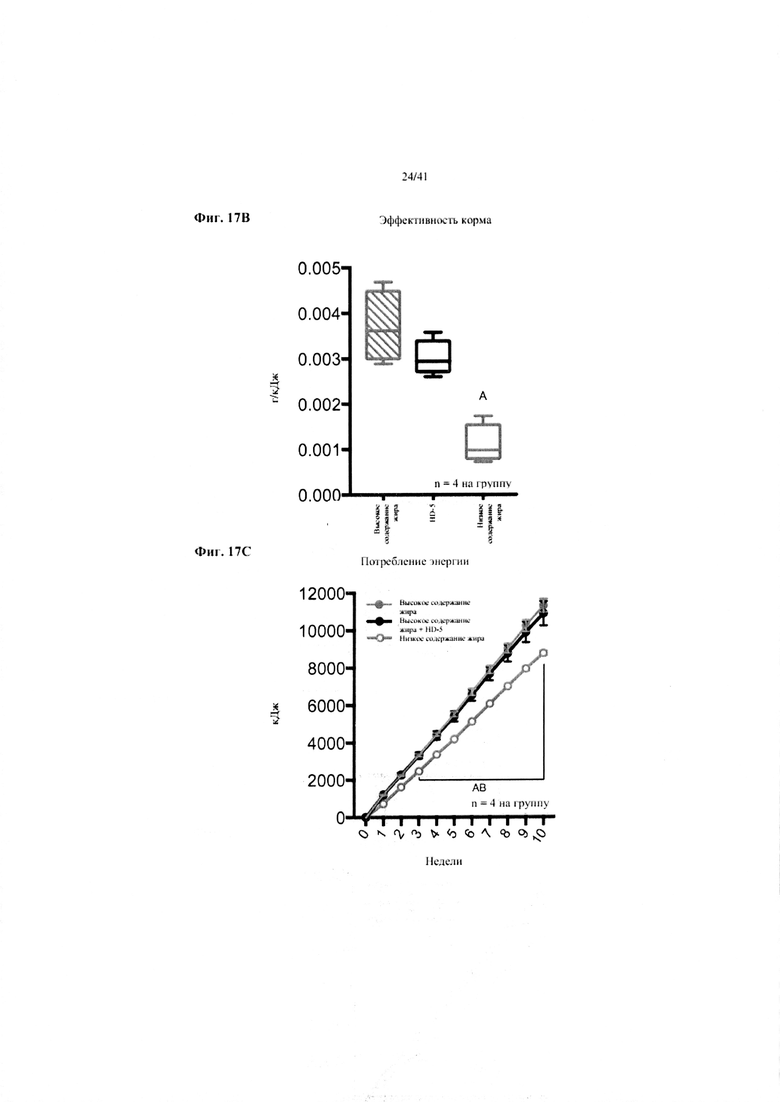
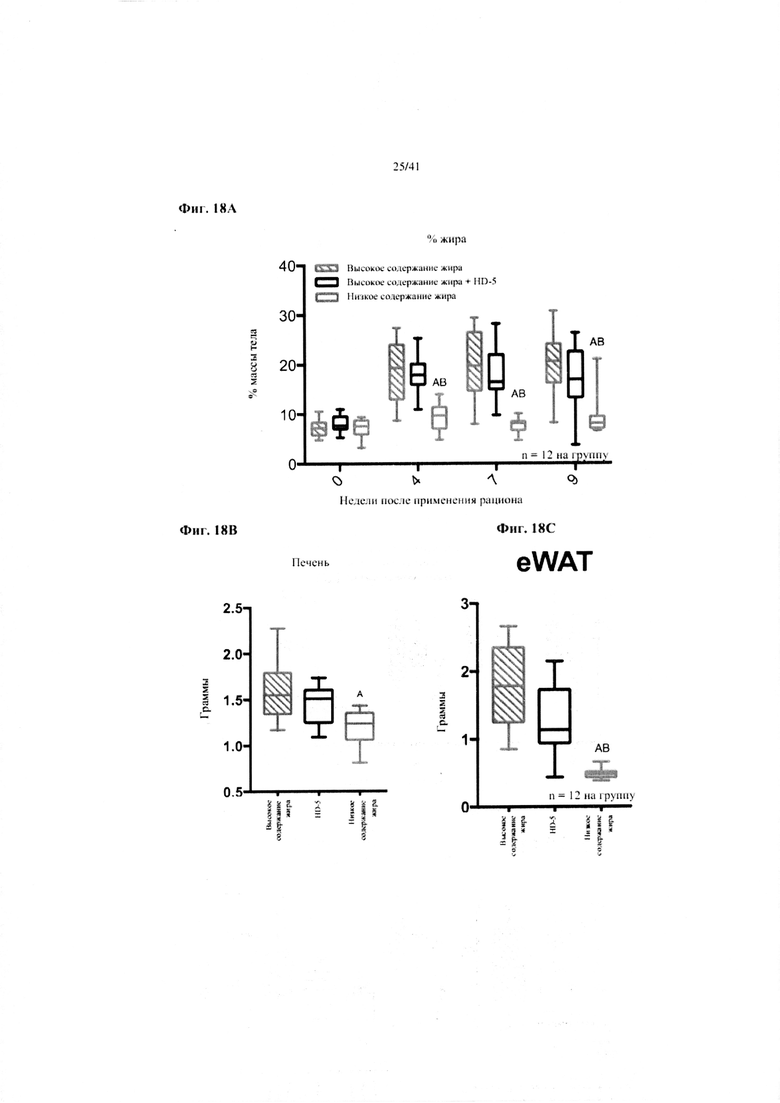
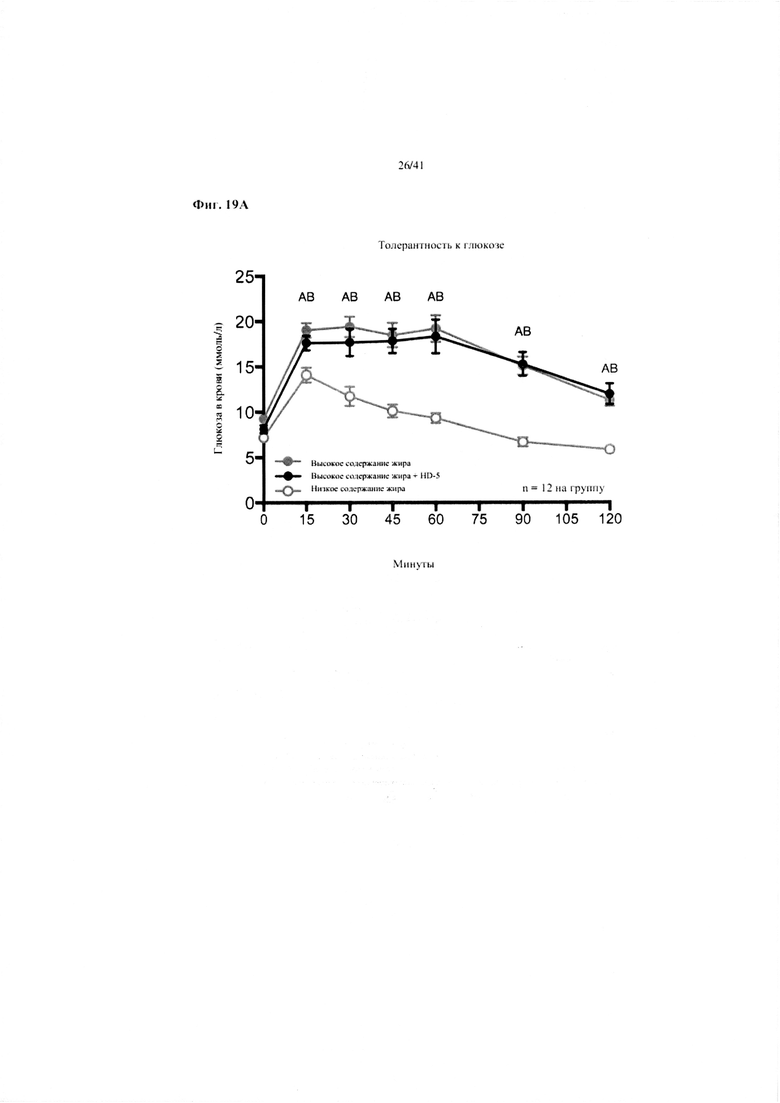
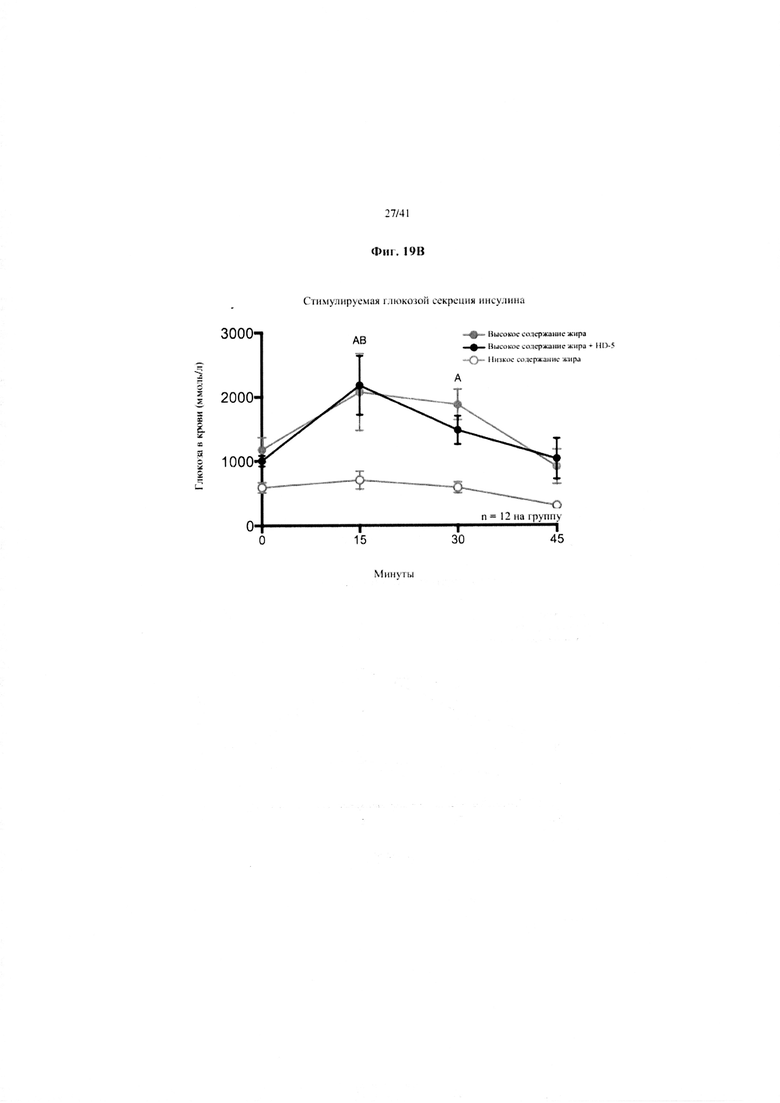
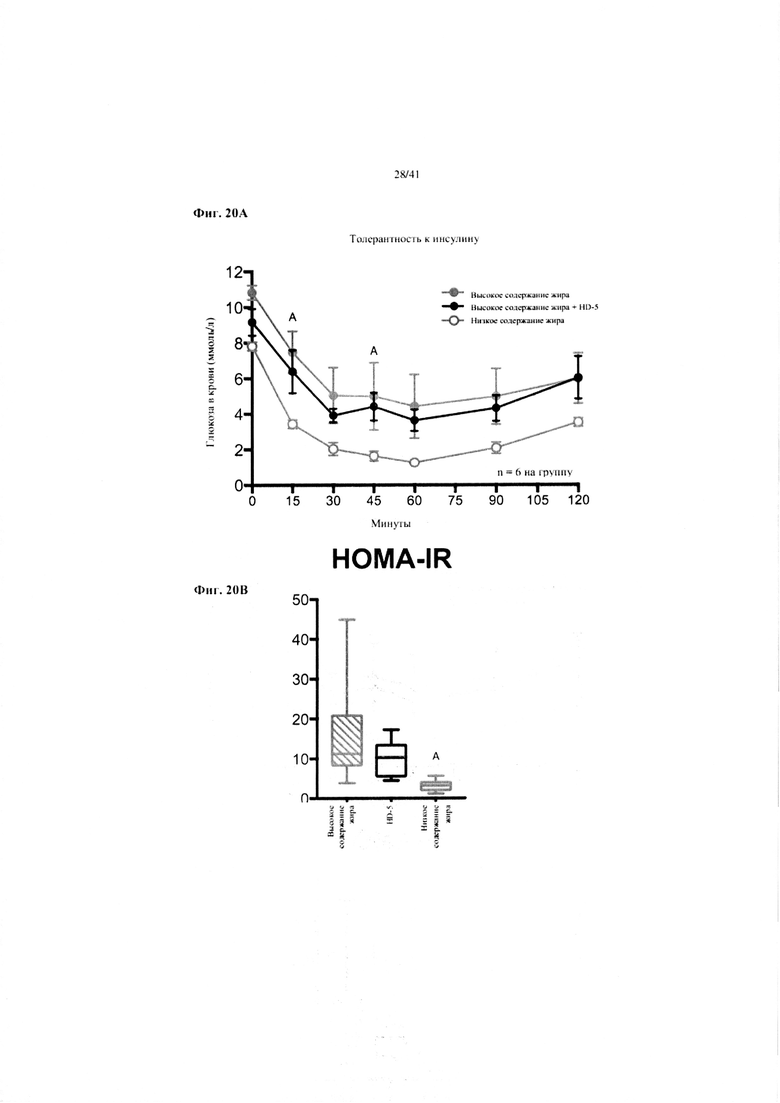
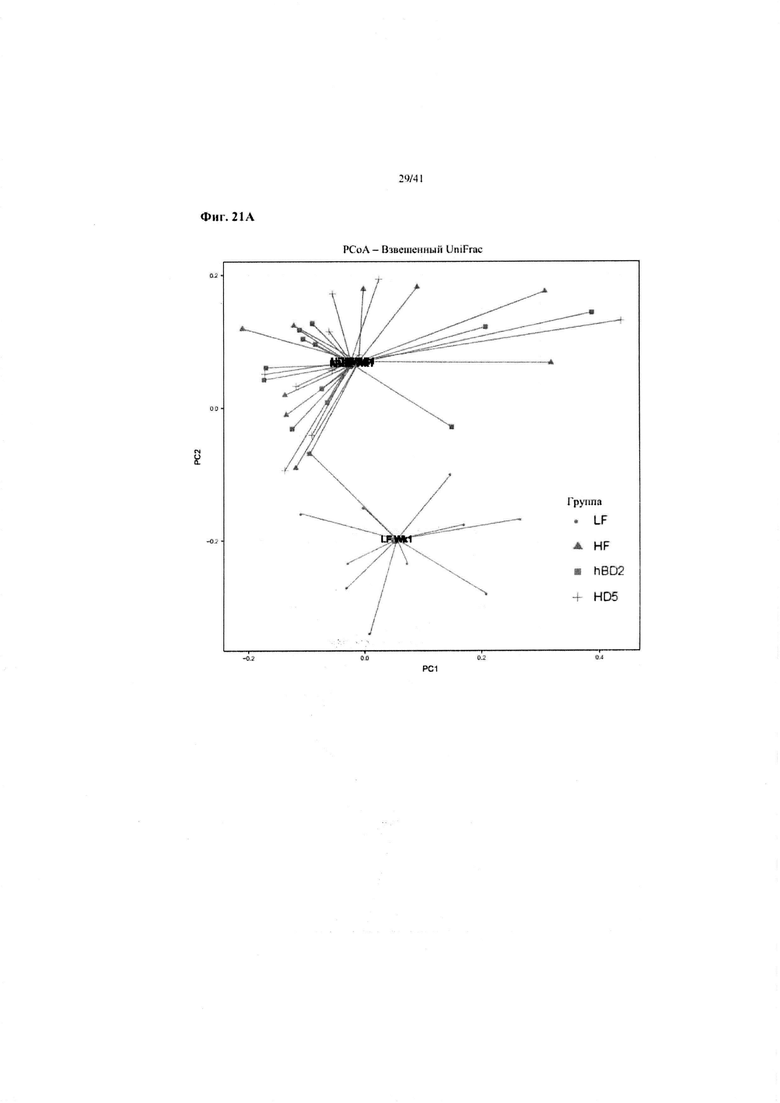
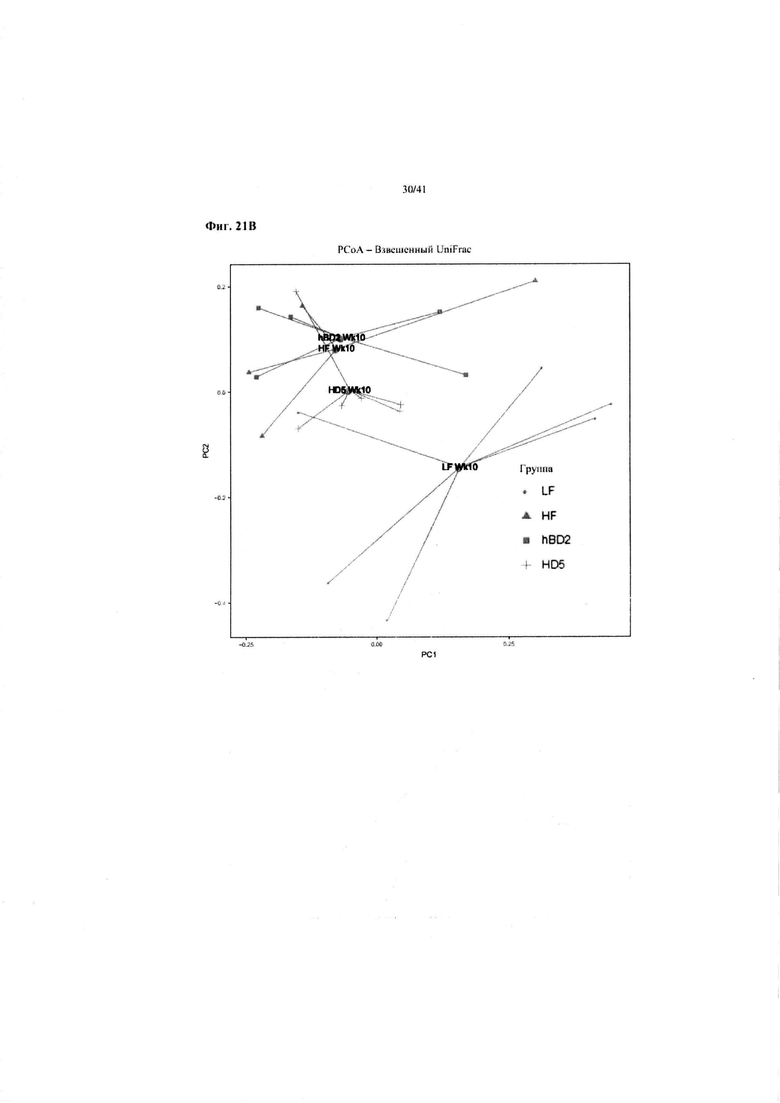
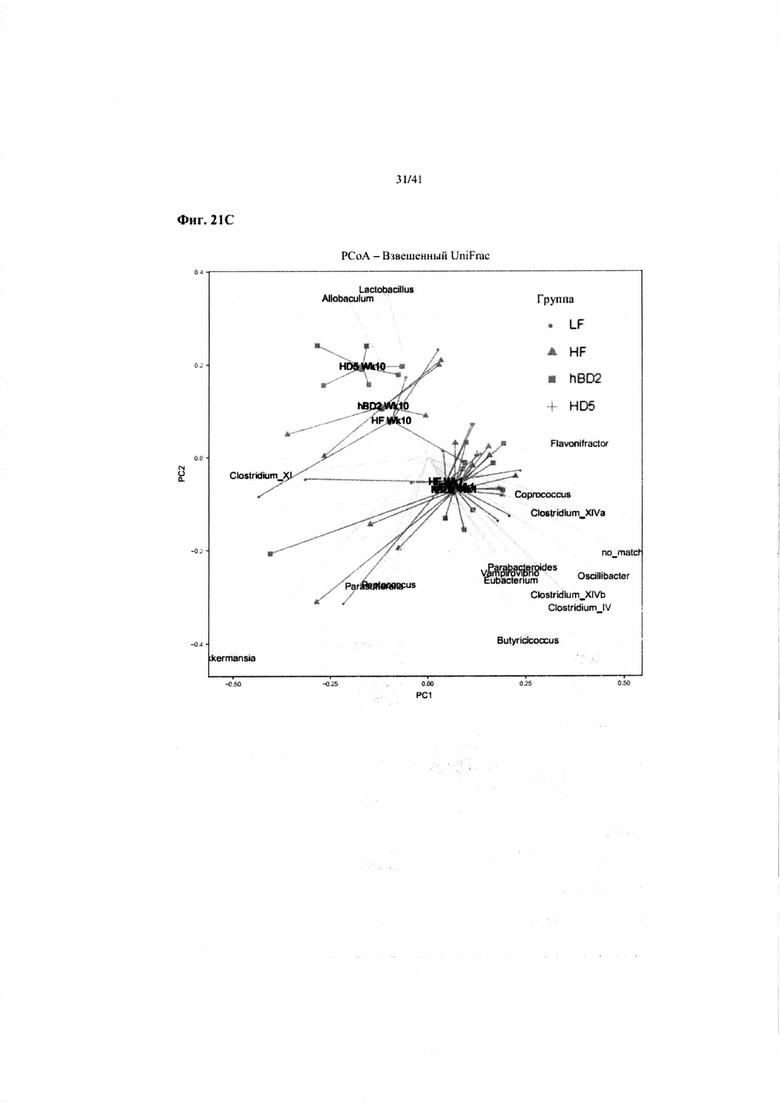
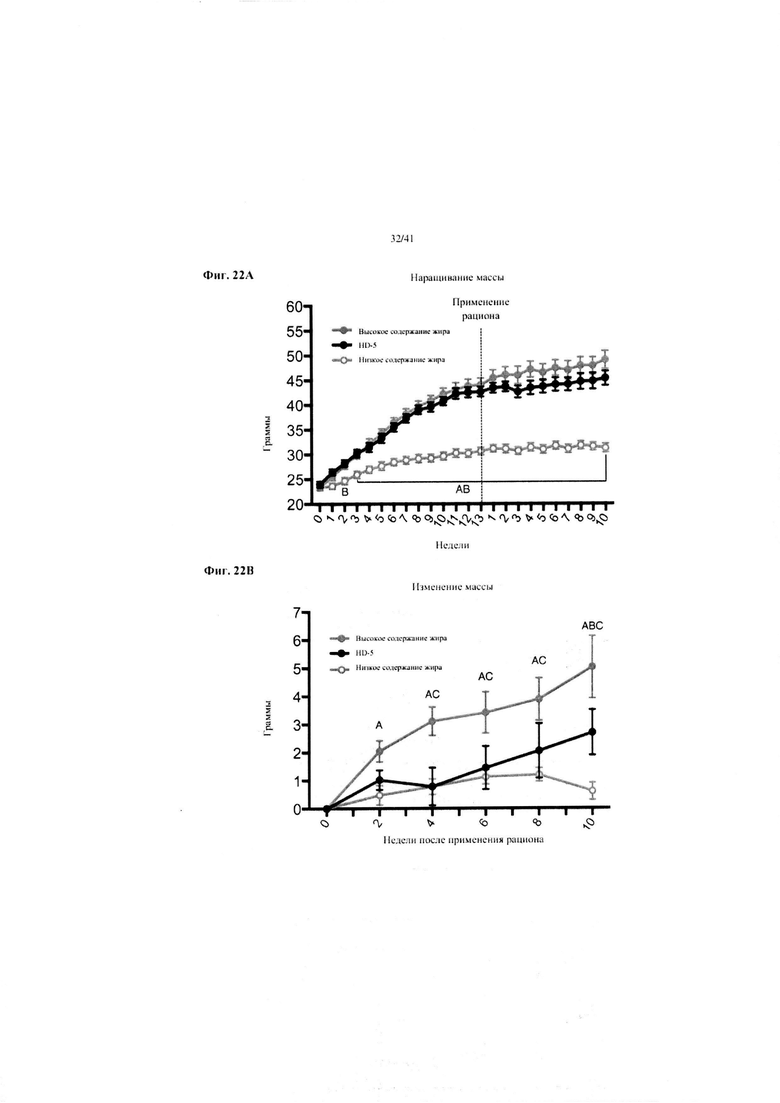
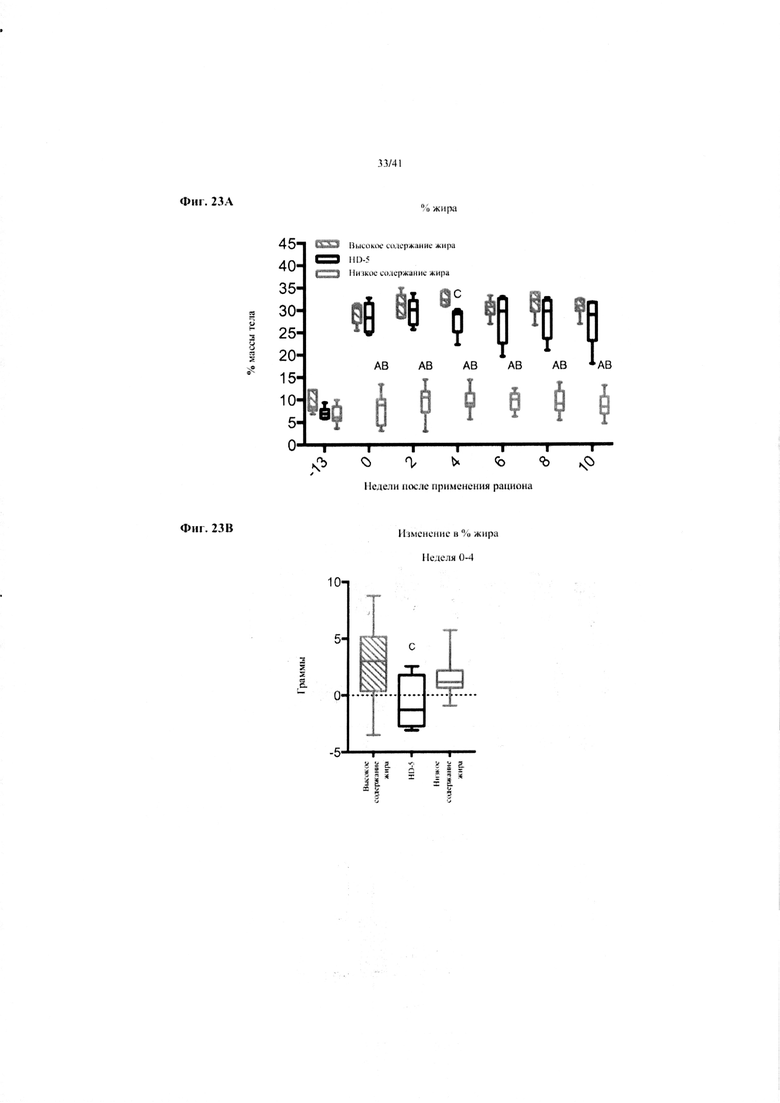
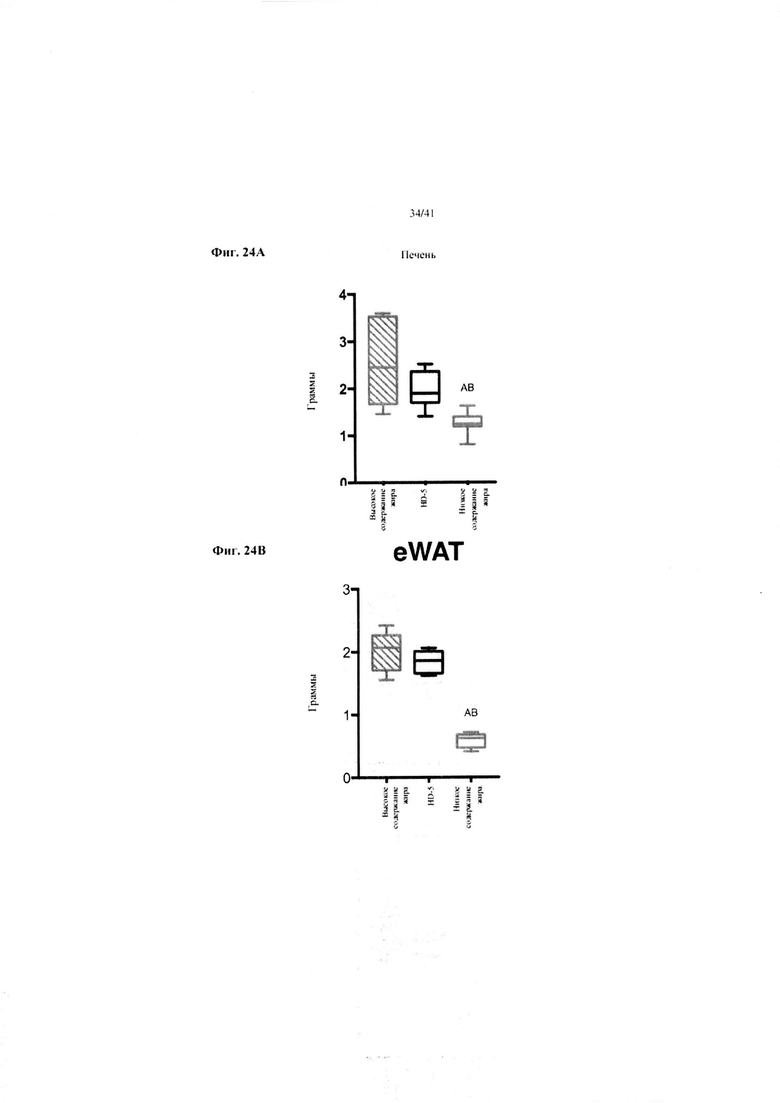
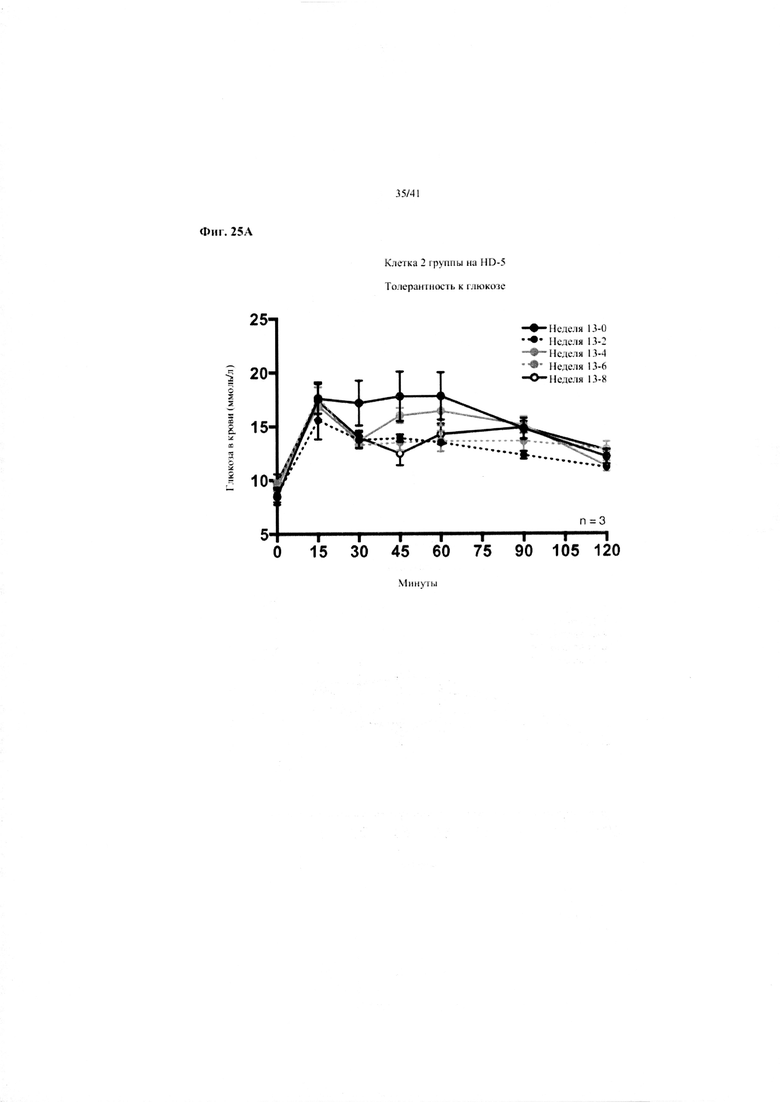
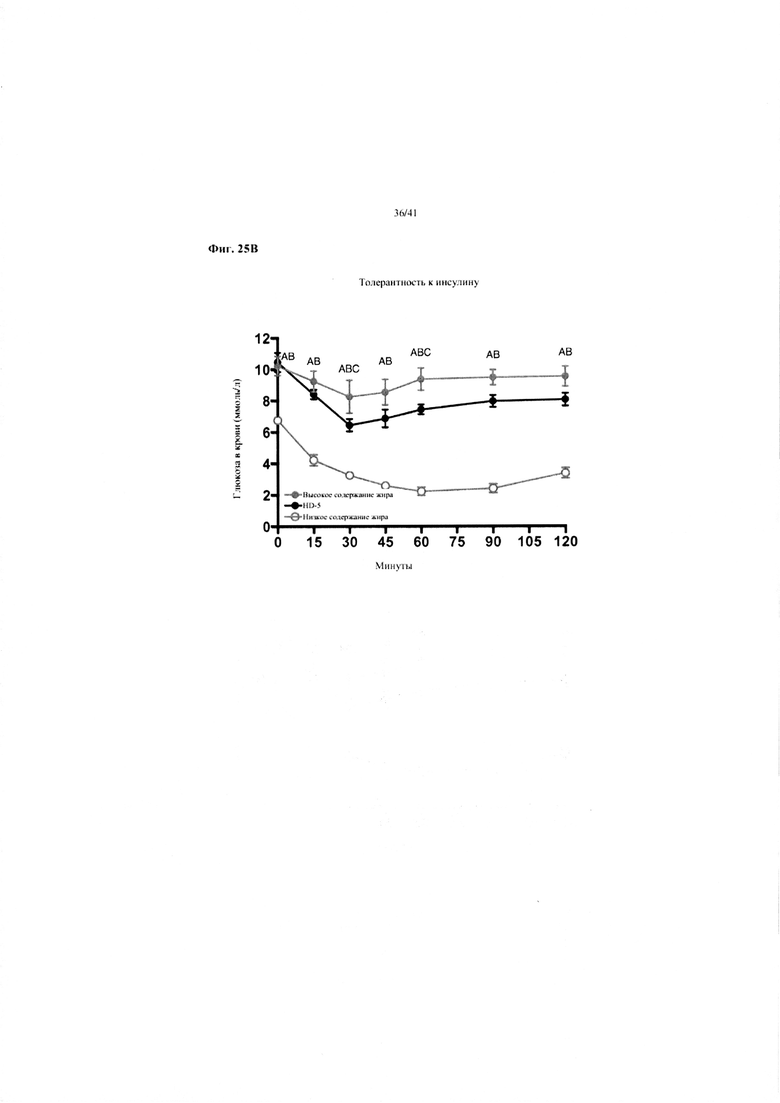
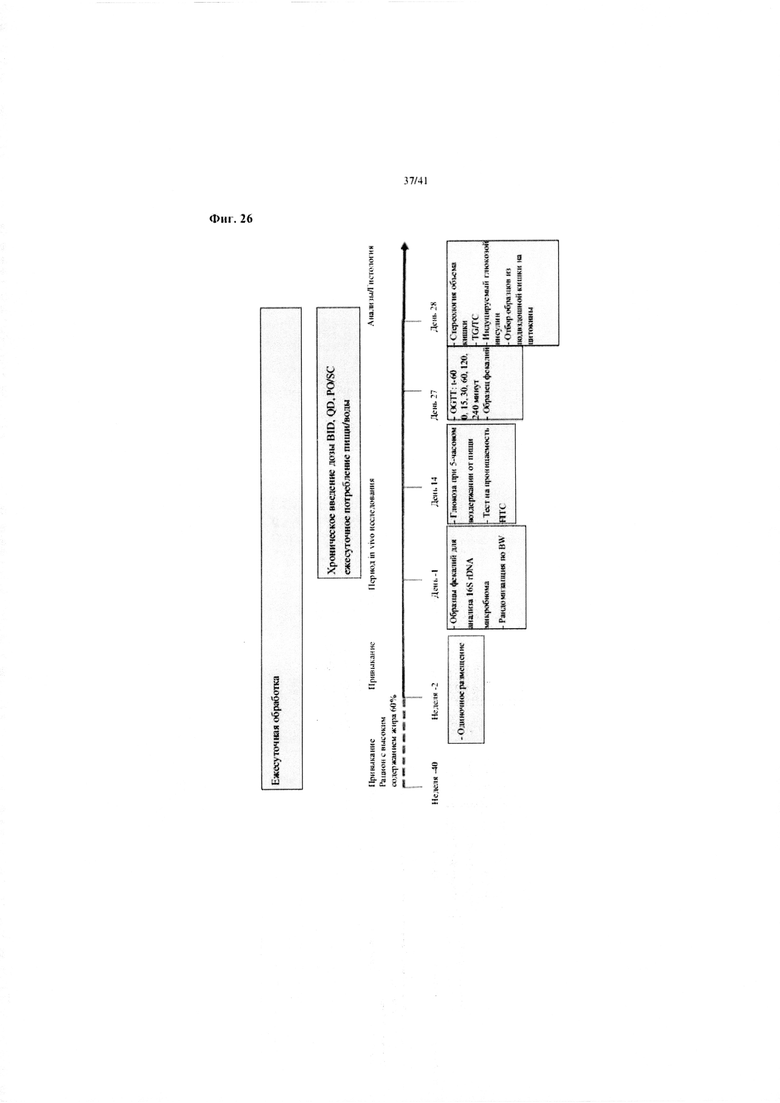
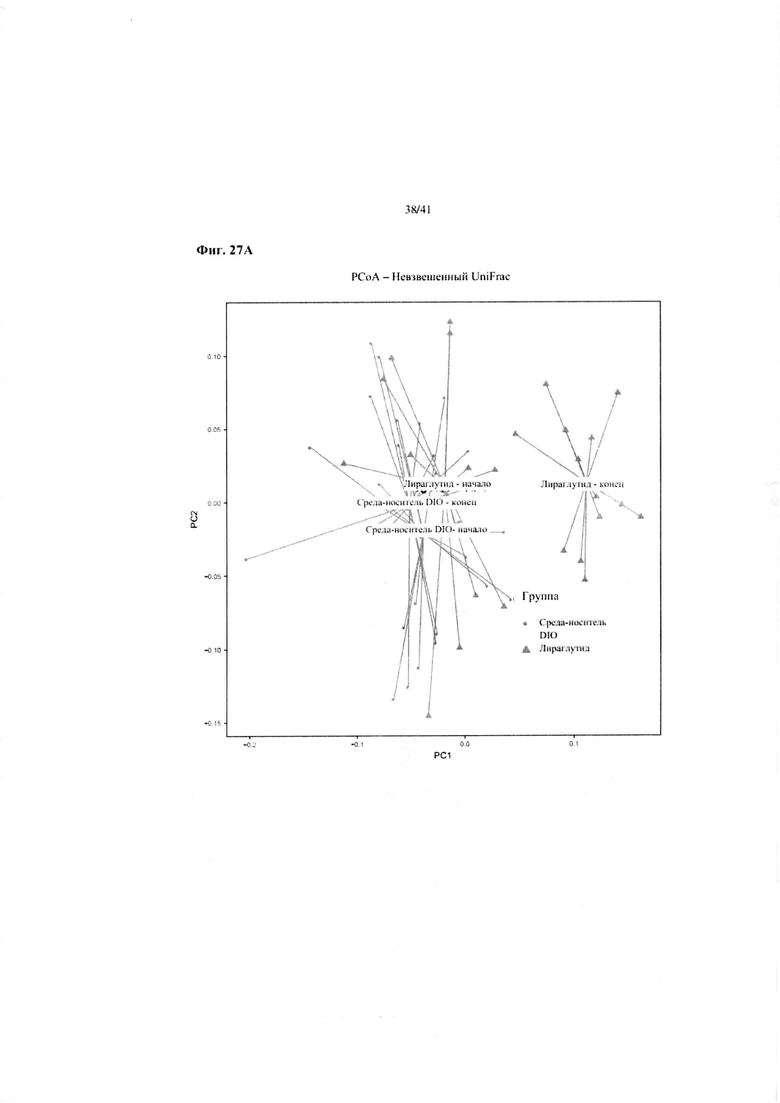
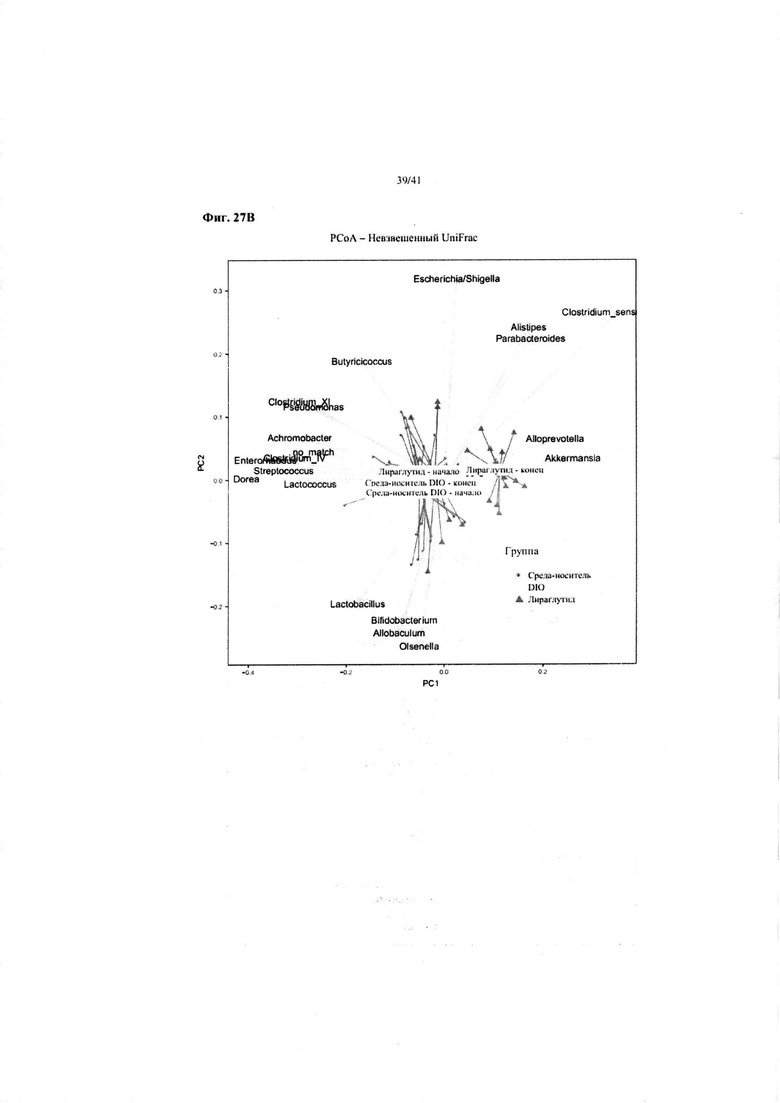
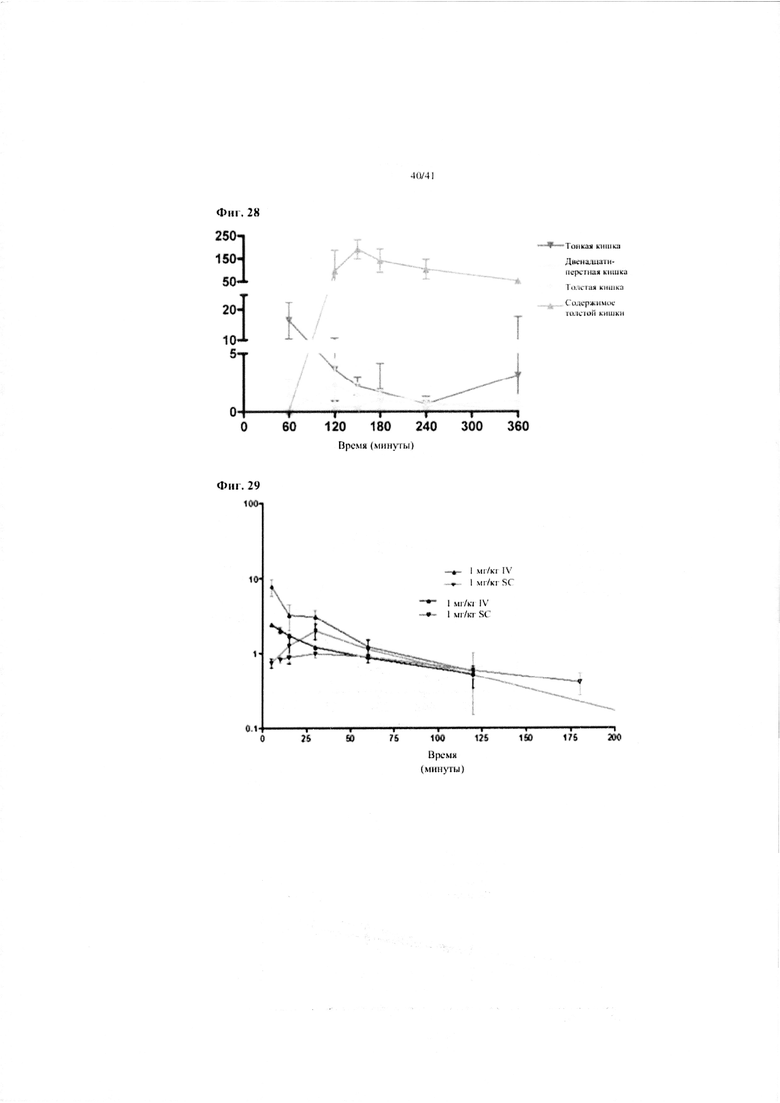
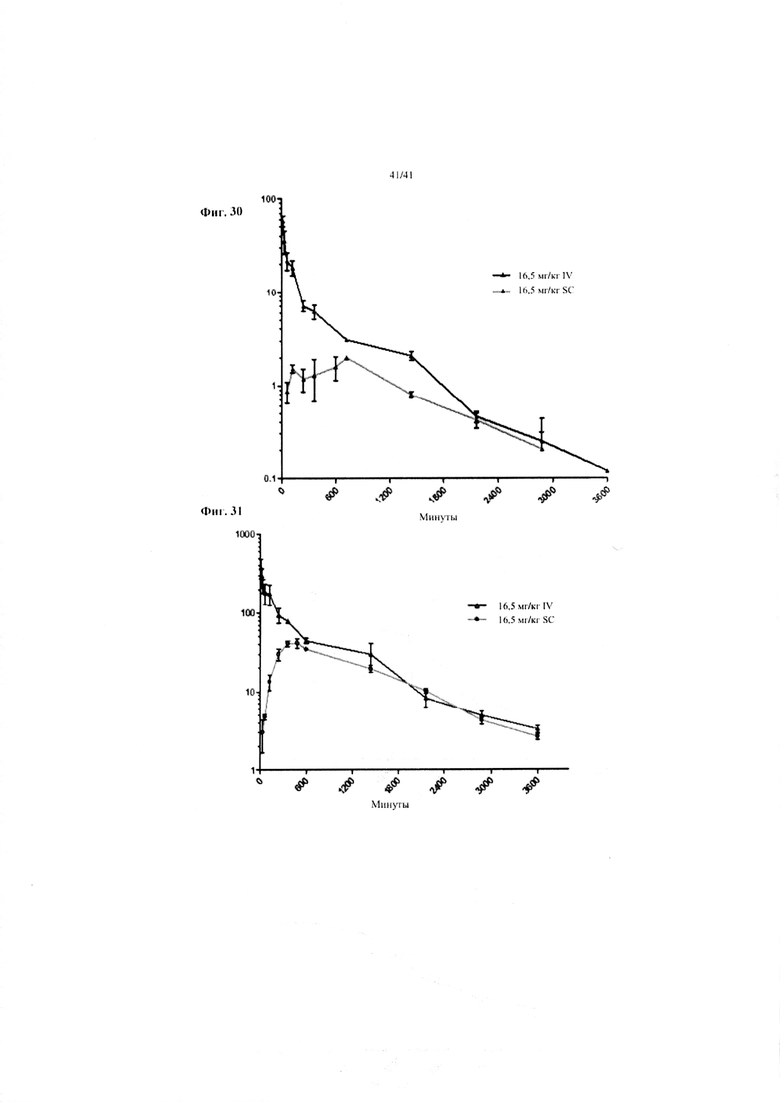
Настоящее изобретение относится к области биотехнологии, а именно к применениям α-дефензина млекопитающего, представляющего собой HD5 или HD6, и/или β-дефензина млекопитающего, представляющего собой hBD1, hBD2, усеченный hBD2, hBD3 или hBD4, или его вариант, отличающийся менее чем 5 консервативными аминокислотными заменами. α-дефензин и β-дефензин применяются для лечения сахарного диабета 2 типа, метаболического синдрома, системного неспецифического воспаления, ожирения, инсулиновой резистентности, нарушения толерантности к глюкозе, дисбиотической микробиоты в кишечнике, а также для стимуляции роста мышц у домашнего скота, повышения многообразия генов кишечной микробиоты/метболомы, повышения числа филумов кишечной микробиоты/метболомы, повышения продуцирования короткоцепочечных жирных кислот кишечной микробиоты/метболомы, повышения числа бактерий в кишечнике, принадлежащих роду, Bacterioidetes, Faecalibacterium, Roseburia, Blautia, Ruminococcus, Coprococcus, Bifidobacterium, Methanobrevibacter, Lactobacillus, Akkermansia, Alloprevotella, Allobaculum и Eubacterium и снижения числа бактерий в кишечнике, принадлежащих роду, Bacteroidetes fragilis, Sutturella wadsworthia, Veillonella parvula, Escherichi coli, Haemophilus parainfluenzae, Fusobacterium nucleatum, Eikenella corodens и Gemella moribillum. Изобретение позволяет расширить арсенал технических средств. 13 н. и 30 з.п. ф-лы, 1 табл., 9 пр., 31 ил.
1. Применение α-дефензина и/или β-дефензина млекопитающего для лечения сахарного диабета 2 типа посредством перорального или подкожного введения, причем
a) α-дефензин млекопитающего представляет собой HD5 или HD6, и/или
b) β-дефензин млекопитающего представляет собой hBD1, hBD2, усеченный hBD2, hBD3 или hBD4, или его вариант, отличающийся менее чем 5 консервативными аминокислотными заменами, нуждающемуся в этом субъекту.
2. Применение α-дефензина и/или β-дефензина млекопитающего для лечения метаболического синдрома посредством перорального или подкожного введения, причем
a) α-дефензин млекопитающего представляет собой HD5 или HD6, и/или
b) β-дефензин млекопитающего представляет собой hBD1, hBD2, усеченный hBD2, hBD3 или hBD4, или его вариант, отличающийся менее чем 5 консервативными аминокислотными заменами,
нуждающемуся в этом субъекту.
3. Применение α-дефензина и/или β-дефензина млекопитающего для лечения системного неспецифического воспаления посредством перорального или подкожного введения, причем
a) α-дефензин млекопитающего представляет собой HD5 или HD6, и/или
b) β-дефензин млекопитающего представляет собой hBD1, hBD2, усеченный hBD2, hBD3 или hBD4, или его вариант, отличающийся менее чем 5 консервативными аминокислотными заменами,
нуждающемуся в этом субъекту.
4. Применение α-дефензина и/или β-дефензина млекопитающего для лечения ожирения посредством перорального или подкожного введения, причем
a) α-дефензин млекопитающего представляет собой HD5 или HD6, и/или
b) β-дефензин млекопитающего представляет собой hBD1, hBD2, усеченный hBD2, hBD3 или hBD4, или его вариант, отличающийся менее чем 5 консервативными аминокислотными заменами,
нуждающемуся в этом субъекту.
5. Применение α-дефензина и/или β-дефензина млекопитающего для лечения инсулиновой резистентности посредством перорального или подкожного введения, причем
a) α-дефензин млекопитающего представляет собой HD5 или HD6, и/или
b) β-дефензин млекопитающего представляет собой hBD1, hBD2, усеченный hBD2, hBD3 или hBD4, или его вариант, отличающийся менее чем 5 консервативными аминокислотными заменами,
нуждающемуся в этом субъекту.
6. Применение α-дефензина и/или β-дефензина млекопитающего для лечения нарушения толерантности к глюкозе посредством перорального или подкожного введения, причем
a) α-дефензин млекопитающего представляет собой HD5 или HD6, и/или
b) β-дефензин млекопитающего представляет собой hBD1, hBD2, усеченный hBD2, hBD3 или hBD4, или его вариант, отличающийся менее чем 5 консервативными аминокислотными заменами,
нуждающемуся в этом субъекту.
7. Применение по п. 4, при котором лечение ожирения предусматривает снижение процентного отношения жира или предупреждение повышения процентного отношения жира у субъекта.
8. Применение по п. 4, при котором лечение ожирения предусматривает предупреждение или снижение количества висцерального жира и/или жира в печени и/или уменьшение окружности талии.
9. Применение по любому из предыдущих пунктов, при котором лечение предусматривает снижение массы или предупреждение прироста массы.
10. Применение по любому из предыдущих пунктов, при котором лечение предусматривает снижение содержания глюкозы в крови натощак.
11. Применение α-дефензина и/или β-дефензина млекопитающего или птицы для стимуляции роста мышц у домашнего скота, посредством введения эффективного количества указанных α- и/или β-дефензина млекопитающего или птицы нуждающемуся в этом субъекту.
12. Применение по п. 7, при котором домашний скот выбран из коров, свиней, овец, коз, лошадей, уток, гусей, голубей, индюков, фазанов и цыплят.
13. Применение по любому из предыдущих пунктов, где указанные α-дефензин и/или β-дефензин млекопитающего или плицы гликозилированы.
14. Применение по любому из предыдущих пунктов, где указанные α-дефензин и/или β-дефензин млекопитающего представляют собой HD5 и/или hBD-2.
15. Применение по любому из предыдущих пунктов, при котором указанные α-дефензин и/или β-дефензин млекопитающего дополнительно содержит по меньшей мере один дополнительный фрагмент, выбранный из группы, состоящей из проникающего в клетку пептида (СРР), фрагмента связывания с альбумином (АВМ), выявляемого фрагмента (Z) и увеличивающего период полувыведения пептида.
16. Применение по п. 15, при котором дополнительным фрагментом является увеличивающий период полувыведения пептид.
17. Применение по п. 15, при котором по меньшей мере один дополнительный фрагмент выбран из группы, состоящей из молекулы, способной связываться с неонатальным Fc-рецептором (FcRn), трансферрином, альбумином (HSA), XTEN или PEG, аминокислотным гомополимером (НАР), полимером пролин-аланин-серин (PAS) или подобным эластину пептидом (ELP), гиалуроновой кислотой, отрицательно заряженным сиалированным пептидом, таким как карбокси-концевой пептид (СТР) β-цепи хорионического гонадотропина (CG), человеческим IgG и СН3(СН2)nCO-, в котором n равно 8-22.
18. Применение по любому из предыдущих пунктов, при котором субъект имеет BMI 25 или больше, например, 30 или больше, например, 35 или больше, например, 40 или больше.
19. Применение по любому из предыдущих пунктов, при котором субъект имеет соотношение окружностей талии и бедра по меньшей мере 0,80, например, 0,80-0,84, например, по меньшей мере 0,85 (у женского пола) или по меньшей мере 0,90, например, 0,9-0,99, например, приблизительно 1,00 (у мужского пола).
20. Применение по любому из предыдущих пунктов, при котором субъект имеет содержание глюкозы в крови натощак по меньшей мере 6,1 ммоль/л, например, по меньшей мере 7,0 ммоль/л.
21. Применение по любому из предыдущих пунктов, при котором субъект имеет содержание гликированного гемоглобина по меньшей мере 42 ммоль/моль Hb, например, от 42 до 46 ммоль/моль Hb, например, по меньшей мере 48 ммоль/моль Hb.
22. Применение по любому из предыдущих пунктов, при котором субъект имеет один или несколько из следующих симптомов:
повышенное давление крови: ≥140/90 мм ртутного столба;
дислипидемию: триглицериды (TG) ≥1,695 ммоль/л и холестерин липопротеинов высокой плотности (HDL-C) ≤0,9 ммоль/л (у мужского пола), ≤1,0 ммоль/л (у женского пола);
центральный тип ожирения: соотношение окружностей талии и бедра >0,90 (у мужского пола); >0,85 (у женского пола) или индекс массы тела >30 кг/м2; и
микроальбуминурию: показатель выведения альбумина с мочой ≥20 мкг/мин или соотношение альбумин/креатинин ≥30 мг/г.
23. Применение по любому из предыдущих пунктов, при котором дефензин вводят нуждающемуся в этом субъекту в суточной дозировке от 0,1 мг hBD-2/кг до 10 мг hBD-2/кг.
24. Применение по любому из предыдущих пунктов, при котором дефензин вводят нуждающемуся в этом субъекту в суточной дозировке от 0,1 мг HD5/кг до 10 мг HD5/кг.
25. Применение по любому из предыдущих пунктов, при котором указанный дефензин вводят в комбинации с инсулином/аналогами инсулина, и/или с глюкагоноподобным пептидом-1 (GLP-1)/аналогами GLP-1, и/или с глюкагоноподобным пептидом-2 (GLP-2)/аналогами GLP-2, и/или с ингибитором дипептидилпептидазы IV (DPP-IV), и/или с метформином, и/или с ингибитором натрий-зависимого переносчика глюкозы-2 (SGLT-2), и/или с антагонистом рецептора глюкагона, и/или с антагонистом представителя 1 семейства V катионного канала транзиторного рецепторного потенциала (TRPV1) или с их комбинацией.
26. Применение по п. 25, при котором аналог GLP-1 выбирают из эксенатида, лираглутида, ликсисенатида, албиглутида и дулаглутида.
27. Применение по п. 25, при котором аналог инсулина выбирают из лизпро, аспарта, глулизина, инсулина детемира, инсулина деглудека и инсулина гларгина.
28. Применение по любому из предыдущих пунктов, при котором дефензин вводят нуждающемуся в этом субъекту один раз в сутки, два раза в сутки или три раза в сутки.
29. Применение по любому из пп. 25-27, при котором введение инсулина/аналога инсулина или GLP-1/аналога GLP-1 является подкожным или внутримышечным.
30. Применение по любому из предыдущих пунктов, при котором дефензин, вводится нуждающемуся в этом субъекту, посредством смешивания с кормом животного.
31. Применение α-дефензина и/или β-дефензина млекопитающего для лечения или нормализации дисбиотической микробиоты в кишечнике,
причем
a) α-дефензин млекопитающего представляет собой HD5 или HD6, и/или
b) β-дефензин млекопитающего представляет собой hBD1, hBD2, усеченный hBD2, hBD3 или hBD4, или его вариант, отличающийся менее чем 5 консервативными аминокислотными заменами.
32. Применение α-дефензина и/или β-дефензина млекопитающего для повышения многообразия генов кишечной микробиоты.
33. Применение α-дефензина и/или β-дефензина млекопитающего для повышения числа филумов кишечной микробиоты.
34. Применение α-дефензина и/или β-дефензина млекопитающего для повышения продуцирования короткоцепочечных жирных кислот кишечной микробиоты/метболомы.
35. Применение α-дефензина и/или β-дефензина млекопитающего для повышения числа бактерий, принадлежащих роду, выбранному из группы, состоящей из Bacterioidetes, Faecalibacterium, Roseburia, Blautia, Ruminococcus, Coprococcus, Bifidobacterium, Methanobrevibacter, Lactobacillus, Akkermansia, Alloprevotella, Allobaculum и Eubacterium, в кишечнике, причем
a) α-дефензин млекопитающего представляет собой HD5 или HD6, и/или
b) β-дефензин млекопитающего представляет собой hBD1, hBD2, усеченный hBD2, hBD3 или hBD4, или его вариант, отличающийся менее чем 5 консервативными аминокислотными заменами.
36. Применение по п. 35, при котором род выбран из Lactobacillus, Akkermansia, Alloprevotella и Allobaculum.
37. Применение α-дефензина и/или β-дефензина млекопитающего для снижения числа бактерий, выбранных из группы, состоящей из Bacteroidetes fragilis, Sutturella wadsworthia, Veillonella parvula, Escherichi coli, Haemophilus parainfluenzae, Fusobacterium nucleatum, Eikenella corodens и Gemella moribillum, в кишечнике, причем
a) α-дефензин млекопитающего представляет собой HD5 или HD6, и/или
b) β-дефензин млекопитающего представляет собой hBD1, hBD2, усеченный hBD2, hBD3 или hBD4, или его вариант, отличающийся менее чем 5 консервативными аминокислотными заменами.
38. Применение по любому из пп. 31-37, при котором дефензин вводят перорально, и при котором предпочтительно дефензином является человеческий HD5 или hBD2.
39. Применение по любому из пп. 31-38, при котором субъект имеет BMI 25 или больше, например, 30 или больше, например, 35 или больше, например, 40 или больше.
40. Применение по любому из пп. 31-39, при котором субъект имеет соотношение окружностей талии и бедра по меньшей мере 0,80, например, 0,80-0,84, например, по меньшей мере 0,85 (у женского пола) или по меньшей мере 0,90, например, 0,9-0,99, например, приблизительно 1,00 (у мужского пола).
41. Применение по любому из пп. 31-40, при котором субъект имеет содержание глюкозы в крови натощак по меньшей мере 6,1 ммоль/л, например, по меньшей мере 7,0 ммоль/л.
42. Применение по любому из пп. 31-41, при котором субъект имеет содержание гликированного гемоглобина по меньшей мере 42 ммоль/моль Hb, например, от 42 до 46 ммоль/моль Hb, например, по меньшей мере 48 ммоль/моль Hb.
43. Применение по любому из пп. 31-42, при котором субъект имеет один или несколько из следующих симптомов:
повышенное давление крови: ≥140/90 мм ртутного столба;
дислипидемию: триглицериды (TG) ≥1,695 ммоль/л и холестерин липопротеинов высокой плотности (HDL-C) ≤0,9 ммоль/л (у мужского пола), ≤1,0 ммоль/л (у женского пола);
центральный тип ожирения: соотношение окружностей талии и бедра >0,90 (у мужского пола); >0,85 (у женского пола) или индекс массы тела >30 кг/м2; и
микроальбуминурию: показатель выведения альбумина с мочой ≥20 мкг/мин или соотношение альбумин/креатинин ≥30 мг/г.
| WO 2008115390 А2, 25.09.2008 | |||
| CN 104971343A, 14.10.2015 | |||
| CN 104971343A, 14.10.2015 | |||
| LEHRER R.I., et al | |||
| "Interaction of human defensins with Escherichia coli | |||
| Mechanism of bactericidal activity", J Clin Invest | |||
| Механизм для сообщения поршню рабочего цилиндра возвратно-поступательного движения | 1918 |
|
SU1989A1 |
| WO 2008115390 А2, 25.09.2008 | |||
| КУЗНЕЦОВА Э.Э, и др | |||
| "Микробиота кишечника | |||
| Роль в развитии | |||

