Ссылка на родственные заявки
Согласно настоящей заявке испрашивается приоритет в соответствии с предварительной заявкой на выдачу патента США №62/678300, поданной 31 мая 2018 года, содержание которой настоящим включено посредством ссылки во всей своей полноте для всех целей.
Область техники, к которой относится настоящее изобретение
[0001] Настоящее раскрытие относится к слитым белкам с альбуминсвязывающим доменом (ABD) и аргиназой и к способам их получения и применения. Также предложены способы, предусматривающие истощение запасов аргинина, для лечения ожирения, метаболических нарушений и/или связанных с ними осложнений и/или сопутствующих заболеваний.
Предшествующий уровень техники настоящего изобретения
[0002] Аргиназа является гидролазой, которая катализирует катаболизм аргинина до орнитина и мочевины и присутствует у большинства живых организмов. У людей существуют две изоформы аргиназы: аргиназа I типа и II типа, которые различаются распределением в тканях и молекулярными характеристиками.
[0003] Аргиназа I встречается в основном в цитоплазме печени и представляет собой белок массой 35 кДа, который существует в виде тримера. Аргиназа II, белок с массой ~38,5 кДа, обычно локализован в митохондриях клеток и, как полагают, участвует в регуляции концентраций аргинина/орнитина в клетке.
[0004] В некоторых типах опухолей отсутствуют или вырабатываются низкие уровни аргининосукцинатсинтетазы (АСС) и/или орнитинтранскарбамоилазы (ОТС), которые необходимы для синтеза аргинина, и поэтому они являются ауксотрофными по аргинину. Депривация аргинина позволяет использовать значительную уязвимость этих опухолевых клеток и приводит к их быстрой гибели. Поэтому ферментативное истощение запасов аргинина было исследовано в качестве потенциальной стратегии избирательного разрушения опухолевых клеток.
[0005] Основным препятствием в разработке ферментативного истощения запасов аргинина в качестве средства для лечения рака является короткий период полужизни (от нескольких минут до нескольких часов) аргиназы в кровотоке из-за ее низкой молекулярной массы (<50 кДа), и поэтому она быстро выводится посредством почечного клиренса. Для поддержания эффективной терапевтической концентрации в системном кровотоке может потребоваться режим частого введения доз, что может вызвать проблемы с соблюдением пациентом режима лечения.
[0006] В связи с этой проблемой были разработаны различные стратегии по улучшению фармакокинетических свойств терапевтических белков. Было показано, что конъюгация полиэтиленгликоля (PEG) с различными терапевтическими белками, т.е. их пегилирование, увеличивает их гидродинамический радиус и, как следствие, замедляет их выведение посредством почечного клиренса. Также были исследованы стратегии слитых белков, основанные на механизмах рециркуляции неонатального рецептора Fc (FcRn), в которых было показано, что слияние кристаллизующегося фрагмента (Fc) иммуноглобулина, альбумина или альбуминсвязывающего пептида с белками увеличивает их период полужизни в кровотоке. Тем не менее, все еще существует потребность в разработке истощающих запасы аргинина белков с улучшенной фармакокинетикой.
[0007] Ожирение, метаболические нарушения и связанные с ними осложнения и сопутствующие заболевания считаются одними из наиболее серьезных глобальных проблем общественного здравоохранения в 21 веке, налагающих огромное социально-экономическое бремя. Более того, метаболические последствия ожирения являются движущими силами для других нарушений, таких как сахарный диабет 2-го типа, непереносимость глюкозы, инсулинорезистентность, гиперхолестеринемия, дислипидемия, гипертония, сердечно-сосудистые заболевания и рак. Таким образом, существует большая потребность в безопасных и эффективных способах предупреждения развития и/или лечения ожирения, метаболических нарушений и связанных с ними осложнений, а также сопутствующих заболеваний. Ожирение является результатом нарушения баланса между потреблением и расходом энергии. У некоторых людей одни лишь изменения в образе жизни могут быть неэффективными, чтобы достаточно сбросить вес или сохранить сброшенный вес для улучшения здоровья. Для снижения веса за счет снижения аппетита или уменьшения всасывания жиров можно принимать лекарственные препараты от ожирения, в том числе орлистат (ксеникал), лорказерин (белвик) и комбинацию фентермина и топирамата (ксимия). Однако такие лекарственные средства обладают различными побочными эффектами, такими как снижение всасывания жирорастворимых витаминов, головная боль, усталость, головокружение, диарея и недержание кала. Для лечения тяжелого ожирения можно применять бариатрическую хирургию, например бандажирование желудка, для достижения долгосрочной потери веса, а липосакция для удаления нежелательных очаговых жировых отложений приобрела популярность по косметическим причинам. Тем не менее, инвазивные процедуры данного типа также сопряжены с серьезными рисками. Таким образом, существует потребность в разработке усовершенствованных способов лечения ожирения, метаболических нарушений и связанных с ними осложнений, а также сопутствующих заболеваний.
Краткое раскрытие настоящего изобретения
[0008] Для удовлетворения одной или нескольких из вышеупомянутых потребностей в настоящем документе предложены слитые белки с аргиназой и ABD с улучшенным периодом полужизни in vivo и катаболизирующей аргинин активностью. Слитые белки, описываемые в настоящем документе, полезны для лечения заболеваний или патологических состояний, при которых истощение запасов аргинина оказывает терапевтический эффект, например, при лечении рака, вирусных инфекций, рассеянного склероза, ревматоидного артрита, аутоиммунных заболеваний, врожденной гипераргининемии, болезни «трансплантат против хозяина» (GvHD), воспаления, ожирения и метаболических нарушений, а также связанных с ним осложнений и сопутствующих заболеваний. Слитые белки можно легко получить и очистить из неочищенной белковой массы. Слитый белок можно применять отдельно или в комбинации по меньшей мере с одним другим средством для получения синергического эффекта при лечении или предупреждении развития заболевания.
[0009] Также предусмотрены способы лечения ожирения, метаболических нарушений и/или связанных с ними осложнений и/или сопутствующих заболеваний путем истощения уровней аргинина у субъекта, например, путем введения субъекту истощающего запасы аргинин средства.
[0010] Согласно первому аспекту, в настоящем документе предложен слитый белок, содержащий полипептид альбуминсвязывающего домена (ABD) и полипептид аргиназы.
[0011] В соответствии с первым вариантом осуществления первого аспекта, в настоящем документе предложен слитый белок по первому аспекту, причем полипептид ABD содержит полипептидную последовательность, имеющую по меньшей мере 93% гомологии последовательности с последовательностью под SEQ ID NO: 66, SEQ ID NO: 67 или SEQ ID NO: 68.
[0012] В соответствии со вторым вариантом осуществления первого аспекта, в настоящем документе предложен слитый белок по первому аспекту, причем полипептид аргиназы содержит полипептидную последовательность, имеющую по меньшей мере 95% гомологии последовательности с последовательностью под SEQ ID NO: 69, SEQ ID NO: 70, SEQ ID NO: 71 или SEQ ID NO: 72.
[0013] В соответствии с третьим вариантом осуществления первого аспекта, в настоящем документе предложен слитый белок по первому аспекту, причем полипептид ABD содержит полипептидную последовательность, имеющую по меньшей мере 93% гомологии последовательности с последовательностью под SEQ ID NO: 66, SEQ ID NO: 67 или SEQ ID NO: 68, а полипептид аргиназы содержит полипептидную последовательность, имеющую по меньшей мере 95% гомологии с последовательностью под SEQ ID NO: 69, SEQ ID NO: 70, SEQ ID NO: 71 или SEQ ID NO: 72.
[0014] В соответствии с четвертым вариантом осуществления первого аспекта, в настоящем документе предложен слитый белок по третьему варианту осуществления первого аспекта, причем полипептид ABD содержит полипептидную последовательность, имеющую по меньшей мере 93% гомологии последовательности с последовательностью под SEQ ID NO: 66, а полипептид аргиназы содержит полипептидную последовательность, имеющую по меньшей мере 95% гомологии последовательности с последовательностью под SEQ ID NO: 69.
[0015] В соответствии с пятым вариантом осуществления первого аспекта, в настоящем документе предложен слитый белок по четвертому варианту осуществления первого аспекта, дополнительно содержащий пептидный линкер, соединяющий С-конец полипептида ABD и N-конец полипептида аргиназы, причем пептидный линкер представляет собой линейный полипептид, содержащий от 1 до 20 аминокислот.
[0016] В соответствии с шестым вариантом осуществления первого аспекта, в настоящем документе предложен слитый белок по пятому варианту осуществления первого аспекта, причем пептидный линкер содержит полипептидную последовательность, имеющую по меньшей мере 90% гомологии последовательности с последовательностью под SEQ ID NO: 73 или SEQ ID NO: 74.
[0017] В соответствии с седьмым вариантом осуществления первого аспекта, в настоящем документе предложен слитый белок по шестому варианту осуществления первого аспекта, причем полипептид ABD содержит полипептидную последовательность, имеющую по меньшей мере 93% гомологии последовательности с последовательностью под SEQ ID NO: 67, а полипептид аргиназы содержит полипептидную последовательность, имеющую по меньшей мере 95% гомологии с последовательностью под SEQ ID NO: 72.
[0018] В соответствии с девятым вариантом осуществления первого аспекта, в настоящем документе предложен слитый белок по седьмому варианту осуществления первого аспекта, причем пептидный линкер представляет собой полигистидиновый линкер, содержащий от 4 до 8 аминокислот гистидин.
[0019] В соответствии с девятым вариантом осуществления первого аспекта, в настоящем документе предложен слитый белок по первому аспекту, причем слитый белок содержит полипептидную последовательность, имеющую по меньшей мере 98% гомологии последовательности с последовательностью под SEQ ID NO: 49, SEQ ID NO: 50, SEQ ID NO: 51, SEQ ID NO: 52, SEQ ID NO: 53, SEQ ID NO: 54, SEQ ID NO: 55, SEQ ID NO: 56, SEQ ID NO: 75 или SEQ ID NO: 76.
[0020] В соответствии с десятым вариантом осуществления первого аспекта, в настоящем документе предложен слитый белок по первому аспекту, причем слитый белок содержит полипептидную последовательность, имеющую по меньшей мере 98% гомологии последовательности с последовательностью под SEQ ID NO: 49, SEQ ID NO: 50, SEQ ID NO: 75 и SEQ ID NO: 76.
[0021] В соответствии с одиннадцатым вариантом осуществления первого аспекта, в настоящем документе предложен слитый белок по первому аспекту, причем слитый белок содержит полипептид, выбранный из группы, состоящей из последовательностей под SEQ ID NO: 49, SEQ ID NO: 50, SEQ ID NO: 75 и SEQ ID NO: 76.
[0022] Согласно второму аспекту, в настоящем документе предложена фармацевтическая композиция, содержащая слитый белок по первому аспекту и фармацевтически приемлемые носитель, вспомогательное вещество или их комбинацию.
[0023] Согласно третьему аспекту, в настоящем документе предложен способ лечения рака у нуждающегося в том субъекта, предусматривающий стадию введения субъекту терапевтически эффективного количества слитого белка по первому аспекту.
[0024] Согласно четвертому аспекту, в настоящем документе предложен способ лечения по меньшей мере одного патологического состояния, выбранного из группы, состоящей из ожирения, метаболического нарушения и связанного с ним осложнения, у нуждающегося в том субъекта, предусматривающий стадию введения субъекту терапевтически эффективного количества истощающего запасы аргинина средства.
[0025] В соответствии с первым вариантом осуществления четвертого аспекта, в настоящем документе предложен способ, при котором метаболическое нарушение выбрано из группы, состоящей из ожирения, непереносимости глюкозы, гипергликемии, а также сахарного диабета и связанного с ним осложнения, которое представляет собой одно или несколько патологических состояний, выбранных из группы, состоящей из диабетической нефропатии, диабетической ретинопатии, диабетической васкулопатии, диабетической невропатии, гиперхолестеринемии, дислипидемии, гипертриглицеридемии, гиперлептинемии, стеатоза, стеатогепатита, фиброза, цирроза, хронического воспаления низкой степени, гипертонии, сердечно-сосудистого заболевания и побеления бурого жира.
[0026] В соответствии со вторым вариантом осуществления четвертого аспекта, в настоящем документе предложен способ, при котором метаболическое нарушение представляет собой инсулинорезистентность.
[0027] В соответствии с третьим вариантом осуществления четвертого аспекта, в настоящем документе предложен способ, при котором лечение ожирения предусматривает по меньшей мере одно из предупреждения увеличения жировой массы и уменьшения жировой массы.
[0028] В соответствии с четвертым вариантом осуществления четвертого аспекта, в настоящем документе предложен способ, при котором метаболическое нарушение выбрано из группы, состоящей из стеатоза печени, стеатоза почек, стеатоза поджелудочной железы и стеатоза сердца.
[0029] В соответствии с пятым вариантом осуществления четвертого аспекта, в настоящем документе предложен способ, при котором концентрация аргинина в сыворотке субъекта поддерживается на уровне ниже 50 мкМ.
[0030] В соответствии с шестым вариантом осуществления четвертого аспекта, в настоящем документе предложен способ, при котором истощающее запасы аргинина средство представляет собой катаболизирующий аргинин фермент.
[0031] В соответствии с седьмым вариантом осуществления четвертого аспекта, в настоящем документе предложен способ по шестому варианту осуществления четвертого аспекта, при котором катаболизирующий аргинин фермент представляет собой белок аргиназы, белок аргининдезиминазы или белок аргининдекарбоксилазы.
[0032] В соответствии с восьмым вариантом осуществления четвертого аспекта, в настоящем документе предложен способ по седьмому варианту осуществления четвертого аспекта, при котором белок аргиназы, белок аргининдезиминазы или белок аргининдекарбоксилазы дополнительно содержит одну или несколько полиэтиленгликолевых (PEG) групп.
[0033] В соответствии с девятым вариантом осуществления четвертого аспекта, в настоящем документе предложен способ по восьмому варианту осуществления четвертого аспекта, при котором белок аргиназы содержит полипептид, имеющий последовательность под SEQ ID NO: 101, SEQ ID NO: 102, SEQ ID NO: 103 или SEQ ID NO: 104.
[0034] В соответствии с десятым вариантом осуществления четвертого аспекта, в настоящем документе предложен способ по седьмому варианту осуществления четвертого аспекта, при котором белок аргиназы, белок аргининдезиминазы или белок аргининдекарбоксилазы дополнительно содержит альбуминсвязывающий домен, или сывороточный альбумин человека, или Fc-домен IgG человека.
[0035] В соответствии с одиннадцатым вариантом осуществления четвертого аспекта, в настоящем документе предложен способ по десятому варианту осуществления четвертого аспекта, при котором катаболизирующий аргинин фермент представляет собой слитый белок, содержащий полипептид ABD и полипептид аргиназы, полипептид ABD и полипептид аргининдезиминазы или полипептид ABD и полипептид аргининдекарбоксилазы.
[0036] В соответствии с двенадцатым вариантом осуществления четвертого аспекта, в настоящем документе предложен способ по одиннадцатому варианту осуществления четвертого аспекта, при котором катаболизирующий аргинин фермент представляет собой слитый белок по первому аспекту.
[0037] В соответствии с тринадцатым вариантом осуществления четвертого аспекта, в настоящем документе предложен способ по двенадцатому варианту осуществления четвертого аспекта, при котором катаболизирующий аргинин фермент содержит полипептид, имеющий по меньшей мере 98% гомологии последовательности с последовательностью под SEQ ID NO: 49, SEQ ID NO: 50, SEQ ID NO: 51, SEQ ID NO: 52, SEQ ID NO: 53, SEQ ID NO: 54, SEQ ID NO: 55, SEQ ID NO: 56, SEQ ID NO: 75 или SEQ ID NO: 76.
[0038] В соответствии с пятым аспектом в настоящем документе предложен способ лечения по меньшей мере одного патологического состояния, выбранного из группы, состоящей из вирусных инфекций, рассеянного склероза, ревматоидного артрита, аутоиммунных заболеваний, врожденной гипераргининемии, болезни «трансплантат против хозяина» (GvHD) и воспаления, у нуждающегося в том субъекта, предусматривающий стадию введения субъекту терапевтически эффективного количества слитого белка по первому аспекту.
Краткое описание чертежей
[0039] Вышеупомянутые и другие цели и признаки настоящего раскрытия станут очевидными из последующего описания настоящего изобретения, взятого вместе с прилагаемыми чертежами.
[0040] На фиг. 1 проиллюстрирована очистка слитого белка рекомбинантной человеческой аргиназы (rhArg) альбуминсвязывающего домена (ABD) N-ABD094-rhArg (SEQ ID NO: 50) электрофорезом в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE). Дорожка 1: низкомолекулярный маркер SDS-PAGE (Bio-rad); дорожка 2: N-ABD094-rhArg (SEQ ID NO: 50) 20× разведение; дорожка 3: N-ABD094-rhArg (SEQ ID NO: 50) 10x разведение.
[0041] На фиг. 2A представлен иллюстративный ген N-ABD094-rhArg (SEQ ID NO: 38) с аннотированными местоположениями связывания праймеров.
[0042] На фиг. 2 В проиллюстрирована белковая последовательность слитого белка N-ABD094-rhArg (SEQ ID NO: 50) с теоретическим pI/Mw 6,07/42555,38.
[0043] На фиг. 3 проиллюстрирована белковая последовательность модифицированной методами инженерии аргиназы Bacillus caldovelox ВСА (S161C)-His (SEQ ID NO: 89) с односайтовой мутацией замены серина на цистеин и С-концевой гистидиновой меткой. Выделены остаток цистеина в положении 161 и 6xHis-метка.
[0044] На фиг. 4 проиллюстрированы результаты выравнивания белковой последовательности ВСА (S161C)-His (SEQ ID NO: 89), слитого белка BCA-6xHis-ABD ВНА (SEQ ID NO: 75) и слитого белка ВАН BCA-ABD-6xHis (SEQ ID NO: 76), полученные с помощью CLUSTALW. Оценки выравнивания: ВСА относительно ВНА: 99,6721; ВСА относительно ВАН: 98,0328; ВНА относительно ВАН: 97,5138.
[0045] На фиг. 5А проиллюстрирован профиль элюирования в результате очистки ВНА (SEQ ID NO: 75) из E.coli, выращенной в культуре во встряхиваемой колбе объемом 500 мл, полученный с помощью одной стадии хроматографии на загруженной никелем колонке HiTrap с хелатирующим наполнителем для HP, объемом 5 мл. mAU - миллиединица поглощения; 100% В=0,5 М имидазола.
[0046] На фиг. 5 В проиллюстрированы результаты SDS-PAGE-анализа фракций колонки, полученных в результате никель-аффинной хроматографии, для ВНА (SEQ ID NO: 75). М - маркеры молекулярной массы для SDS-PAGE, низкомолекулярные (Bio-Rad); FT - проскок; F2-F10 - фракции, элюированные из колонки.
[0047] На фиг. 5С проиллюстрирован профиль элюирования в результате очистки ВАН (SEQ ID NO: 76) из клеток E.coli, выращенной в культуре во встряхиваемой колбе объемом 500 мл, полученный с помощью одной стадии хроматографии на загруженной никелем колонке HiTrap с хелатирующим наполнителем для HP, объемом 5 мл. mAU - миллиединица поглощения; 100% В=0,5 М имидазола.
[0048] На фиг. 5D проиллюстрированы результаты SDS-PAGE-анализа фракций колонки, полученных в результате никель-аффинной хроматографии, для ВАН (SEQ ID NO: 76). M - маркеры молекулярной массы для SDS-PAGE, низкомолекулярные (Bio-Rad); FT - проскок; F2-F14 - фракции, элюированные из колонки.
[0049] На фиг. 6А проиллюстрирована хроматограмма, на которой видны результаты крупномасштабной очистки модифицированного методами инженерии ВСА (S161C)-His (SEQ ID NO: 89) с применением колонки для никель-аффинной хроматографии ХК50 с объемом слоя 196 мл. Подобно мелкомасштабной очистке, был выбран четырехсегментный градиент элюирования, а профиль элюирования отслеживали по поглощению на уровне 280 нм.
[0050] На фиг. 6 В проиллюстрированы результаты SDS-PAGE-анализа выбранных фракций после крупномасштабной очистки модифицированного методами инженерии ВСА (S161C)-His (SEQ ID NO: 89). Дорожка 1 представляет собой низкомолекулярный маркер. Дорожка 2 представляет собой общий белок из клеточного лизата. Дорожка 3 представляет собой растворимые белки, собранные после тепловой обработки. На дорожке 4 виден проскок с белком из колонки для никель-афинной хроматографии ХК50. Дорожка 5 представляет собой неспецифически связанные белки (объединенные фракции из А6-С3). На дорожках 6-10 представлены соответственно фракции Е2, Е3, F7, G3 и А'5.
[0051] На фиг. 7 проиллюстрированы результаты связывания слитого белка N-ABD-rhArg (SEQ ID NO: 49) и HSA на PAGE-геле в неденатурирующих условиях. Тридцать пикомолей HSA смешивали с 7,5, 15, 30, 60 и 120 пикомолями N-ABD-rhArg (SEQ ID NO: 49), которые представлены соответственно на дорожках 5-9. Подвижность 30 пикомолей HSA и N-ABD-rhArg (SEQ ID NO: 49) на PAGE-геле в неденатурирующих условиях показана соответственно на дорожках 2 и 3. Дорожка 1 является пустой или с холостой пробой. На дорожке 10 показано 120 пикомолей N-ABD-rhArg (SEQ ID NO: 49).
[0052] На фиг. 8 проиллюстрированы результаты связывания слитого белка ВНА (SEQ ID NO: 75) и HSA на PAGE-геле в неденатурирующих условиях. Шестьдесят пикомолей HSA смешивали с 6, 12, 60, 300 и 600 пикомолями слитого белка ВНА (SEQ ID NO: 75), которые представлены соответственно на дорожках 1-5. Подвижность 60 пикомолей HSA и слитого белка ВНА (SEQ ID NO: 75) на PAGE-геле в неденатурирующих условиях показана соответственно на дорожках 6 и 7. Бэнд, обозначенный «ВСА-ABD», представляет собой ВНА (SEQ ID NO: 75).
[0053] На фиг. 9 проиллюстрирована фармакодинамика ВСА (S161C)-His (SEQ ID NO: 89) и слитого белка ВНА (SEQ ID NO: 75) в отношении аргинина в плазме крови мышей. Каждой BALB/c мыши вводили посредством внутривенной инъекции (i.v.) или внутрибрюшинной инъекции (i.p.) 250 ед. ВСА (S161C)-His (SEQ ID NO: 89) или слитого белка ВНА (SEQ ID NO: 75). Плазму крови собирали в указанные моменты времени после инъекции. Момент времени 0 относится к образцу плазмы крови, взятому перед инъекцией. Количество аргинина в каждом образце определяли с помощью анализатора аминокислот Biochrom 30. Концентрации аргинина ниже предела детекции (3 мкМ) на этих графиках считали равными 0 мкМ. Каждая точка представляет собой среднее значение ±SD (стандартное отклонение) от трех мышей.
[0054] На фиг. 10 проиллюстрированы концентрации аргинина в плазме крови, измеренные с помощью анализатора аминокислот Biochrom 30, у BALB/c мышей после внутрибрюшинной инъекции однократной дозы (А) 125 ед., (В) 250 ед. и (С) 500 ед. N-ABD-rhArg (SEQ ID NO: 49). N-ABD-rhArg (SEQ ID NO: 49) вводили инъекцией на 0-е сутки.
[0055] На фиг. 11 проиллюстрированы результаты SDS-PAGE-анализа получения пегилированного His-rhArg (SEQ ID NO: 101). Дорожки 1 и 8: низко молекулярный маркер для SDS-PAGE (Bio-rad); дорожки 2 и 3: His-rhArg (SEQ ID NO: 101); дорожки 4 и 5: 60% элюирование; дорожки 6 и 7: 30% элюирование; дорожки 9 и 10: пегилированный His-rhArg (SEQ ID NO: 101). Молекулярная масса PEG составляет 5000. Применяемый PEG представлял собой метоксиполиэтиленгликоль-сукцинимидилпропионат (mPEG-SPA 5000). Остатки лизина на поверхности белка из His-rhArg (SEQ ID NO: 101) ковалентно присоединяют через пропионамидиловый (SPA) линкер к PEG (5000 а.е.м.), как описано в патенте США №8679810, который включен в настоящий документ посредством ссылки.
[0056] На фиг. 12 проиллюстрированы результаты получения N-ABD094-rhArg (SEQ ID NO: 50). Дорожка 1: маркер молекулярной массы для SDS-PAGE; дорожка 2: клеточный лизат; дорожка 3: после термообработки; дорожка 4: проскок; дорожка 5: 30% элюирование; дорожка 6: 60% элюирование; дорожка 7: после тангенциальной поточной фильтрации (TFF). Основной белковый бэнд на дорожке 6 или дорожке 7 представлял собой очищенный N-ABD094-rhArg (SEQ ID NO: 50).
[0057] На фиг. 13 проиллюстрированы концентрации аргинина в плазме крови у самцов C57BL/6J мышей, которых кормили стандартным рационом, измеренные с помощью трехквадрупольного масс-спектрометра Agilent 6460 для жидкостной хроматографии/с ионизацией электрораспылением с пределом детекции 0,3 мкМ L-аргинина в различные моменты времени после введения внутрибрюшинной инъекции с разовой дозой 500 ед. N-ABD094-rhArg (SEQ ID NO: 50) в солевом растворе, из которых видно, что концентрации аргинина в плазме крови могут быть снижены до <1 мкМ за 10 суток. В каждом моменте времени представлено среднее значение ±SEM от 5 мышей.
[0058] На фиг. 14 проиллюстрированы концентрации N-ABD094-rhArg (SEQ ID NO: 50) в плазме крови в различные моменты времени после инъекции самцам C57BL/6J мышей однократной дозой 500 ед., измеренные с помощью ELISA, детектирующего человеческую аргиназу (n=5). Период полужизни лекарственного средства в кровотоке составляет около 4 суток. В каждом моменте времени представлено среднее значение±SEM от 5 мышей.
[0059] На фиг. 15 проиллюстрирована значительная потеря жира у нормальных худых самцов C57BL/6 J мышей, которых кормили стандартным рационом и обрабатывали посредством N-ABD094-rhArg (SEQ ID NO: 50) [группа «Стандарт(rhArg)»]. (А) Значимое уменьшение массы висцерального и подкожного жира после обработки посредством 500 ед. N-ABD094-rhArg (SEQ ID NO: 50) с 10-суточными интервалами в течение 4 недель по сравнению с обработанным наполнителем (солевым раствором) контролем [группа «Стандарт(наполнитель)»]. *Р<0,05, независимый t-критерий. Данные выражены как ±SEM, n=5 для каждой группы. (В) На окрашенных гематоксилином и эозином (Н&Е) парафиновых срезах висцеральной белой жировой ткани (WAT) у обработанных посредством N-ABD094-rhArg (SEQ ID NO: 50) мышей видно заметное уменьшение размера адипоцитов.
[0060] На фиг. 16 проиллюстрированы уровни экспрессии нескольких важных генов, связанных с липогенезом, в различных тканях, измеренные с помощью qRT-PCR в режиме реального времени. Значимое подавление активности генов, связанных с липогенезом, в (А) висцеральной белой жировой ткани (WAT) и (В) печени после обработки посредством 500 ед. N-ABD094-rhArg (SEQ ID NO: 50) [группа «Стандарт(rhArg)»] с 10-суточными интервалами в течение 4 недель по сравнению с обработанным наполнителем (солевым раствором) контролем [группа «Стандарт(наполнитель)»]. Уровни экспрессии генов выражены относительно таковых у мышей, которых обрабатывали наполнителем [группа «Стандарт(наполнитель)»], который принимали за 1. *Р<0,05, независимый t-критерий. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0061] На фиг. 17 показано, что обработка посредством N-ABD094-rhArg (SEQ ID NO: 50) значимо улучшает чувствительность к инсулину. (А) Из результатов теста на толерантность к инсулину было видно, что у мышей, которым инъекцией вводили 500 ед. N-ABD094-rhArg (SEQ ID NO: 50) [группа «Стандарт(rhArg)»] с 10-суточными интервалами, наблюдалось значимое повышение чувствительности к понижающему уровень глюкозы в крови действию инсулина уже через 2 недели после обработки по сравнению с контрольной группой, обработанной наполнителем [группа «Стандарт(наполнитель)»]. *Р<0,05 по сравнению с группой «Стандарт(rhArg)», независимый t-критерий. Данные выражены как среднее значение±SEM, n=5 для каждой группы. (В) Из результатов анализа с применением МТТ наблюдали, что обработка первичных гепатоцитов мыши посредством 5 ед./мл N-ABD094-rhArg (SEQ ID NO: 50) в течение 48 часов не влияла на жизнеспособность клеток по сравнению с контролем без какой-либо обработки (контр.). (С) Результаты вестерн-блоттинга уровней белка фосфорилированного Akt (pAkt) (маркера передачи сигналов инсулина) в первичных гепатоцитах мыши, подвергнутых истощению запасов аргинина с помощью 5 ед./мл N-ABD094-rhArg (SEQ ID NO: 50) за 24 часа до воздействия 100 нМ инсулина в течение 20 минут. У гепатоцитов, обработанных посредством N-ABD094-rhArg (SEQ ID NO: 50), наблюдали более высокий уровень фосфорилирования Akt при стимуляции инсулином, что свидетельствовало об усилении передачи сигналов инсулина. Дорожка 1: контр.; дорожка 2: 5 ед./мл N-ABD094-rhArg (SEQ ID NO: 50); дорожка 3: 100 нМ инсулина; дорожка 4: 5 ед./мл N-ABD094-rhArg (SEQ ID NO: 50)+100 нМ инсулина.
[0062] На фиг. 18 проиллюстрировано, что в то время, как у обработанных наполнителем самцов C57BL/6J мышей, которых кормили рационом с высоким содержанием жиров (HFD), причем содержание жира достигало 60 ккал% (с 8-недельного возраста в течение 12 недель) [группа «НЕО(наполнитель)»], наблюдали заметный прирост (А) массы тела и (В) размера, одновременная обработка самцов мышей, которых кормили HFD (с возраста 8 недель в течение 12 недель) и обрабатывали 300 ед. N-ABD094-rhArg (SEQ ID NO: 50) [группа «HFD(rhArg)»] один раз в неделю, может эффективно предупреждать прирост массы. Их масса тела оставалась такой же, как и у худых мышей контрольной группы, которых обрабатывали наполнителем и кормили равноценным рационом с низким содержанием жиров (LFD), причем содержание жира достигало 10 ккал% [группа «LFD(наполнитель)»]. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0063] На фиг. 19 проиллюстрировано, что одновременная обработка самцов мышей, которых кормили HFD, посредством 300 ед. N-ABD094-rhArg (SEQ ID NO: 50) [группа «HFD(rhArg)»] один раз в неделю в течение 12 недель может эффективно предупреждать HFD-индуцированное разрастание белой жировой ткани (WAT). (А) Масса жирового тела в основных висцеральных (перигонадных, периренальных и брыжеечных) и подкожных (паховых) депо. (В) На окрашенных посредством Н&Е срезах висцерального WAT было видно, что одновременная обработка посредством N-ABD094-rhArg предупреждала HFD-индуцированную гипертрофию белых адипоцитов. (С) Уровни экспрессии нескольких важных, связанных с липогенезом генов в WAT, экспрессируемых относительно таковых у худых мышей контрольной группы [группа «LFD(наполнитель)»], уровень у которых принимали за 1. Эти гены резко подавлялись у мышей, которых кормили HFD и обрабатывали посредством N-ABD094-rhArg. *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0064] На фиг. 20 проиллюстрировано подавление HFD-индуцированной активации транскрипционных регуляторов липогенеза (Pparg и Srebplc) с сопутствующим подавлением важных липогенных ферментов (Accl и Scdl) в скелетных мышцах мышей, обработанных посредством 300 ед. N-ABD094-rhArg (SEQ ID NO: 50) [группа «HFD(rhArg)»] один раз в неделю в течение 12 недель. *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=5 для каждой группы].
[0065] На фиг. 21 проиллюстрировано, что одновременная обработка посредством 300 ед. N-ABD094-rhArg (SEQ ID NO: 50) [группа «HFD(rhArg)»] один раз в неделю может эффективно предупреждать HFD-индуцированное ожирение у самок C57BL/6J мышей. (А) Масса тела у мышей, которых кормили HFD от 8-недельного возраста в течение 12 недель, а потом обработанных посредством N-ABD094-rhArg или наполнителем. Самки мышей, которых обрабатывали наполнителем и кормили LFD, служили в качестве худых мышей контрольной группы [группа «LFD(наполнитель)»]. (В) Масса жирового тела в основных висцеральных (перигонадных, периренальных и брыжеечных) и подкожных (паховых) депо. *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0066] На фиг. 22 проиллюстрировано, что одновременная обработка посредством 300 ед. N-ABD094-rhArg (SEQ ID NO: 50) [группа «HFD(rhArg)»] один раз в неделю может эффективно предупреждать HFD-индуцированную инсулинорезистентность и непереносимость глюкозы у самок C57BL/6J мышей. (А) Тест на толерантность к инсулину (ITT), проведенный спустя 7 недель кормления HFD. (В) Тест на толерантность к глюкозе (GTT), проведенный спустя 11 недель кормления HFD. Результаты ITT и GTT выражены как площадь под кривой (AUC). *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0067] На фиг. 23 проиллюстрировано, что истощение запасов аргинина путем добавления N-ABD094-rhArg (SEQ ID NO: 50) (rhArg), бычьей аргиназы или нативной аргининдезиминазы (ADI) в культуральную среду или применение среды без аргинина ингибировало дифференцировку мышиных преадипоцитов 3T3-L1 в адипоциты. Добавление продукта метаболизма аргиназы, т.е. L-орнитина или мочевины, или продукта метаболизма ADL, т.е. L-цитруллина, в культуральную среду не оказывало ингибирующего действия на адипогенез и липогенез. (* Культуральная среда содержала 0,4 мМ L-аргинина и могла быть преобразована в 0,4 мМ L-орнитина и 0,4 мМ мочевины с помощью аргиназы или 0,4 мМ L-цитруллина с помощью ADI). Контролем служили клетки, выращенные в культуральной среде без добавления истощающих запасы аргинина средств или продуктов метаболизма аргиназы/ADI. Липиды в клетках окрашивали масляным красным О через 6 суток после воздействия специфических индуцирующих дифференцировку факторов, добавленных в среду.
[0068] На фиг. 24 проиллюстрировано, что N-ABD094-rhArg (SEQ ID NO: 50) может подавлять адипогенные и липогенные гены в преадипоцитах 3T3-L1. Мышиные преадипоциты 3T3-L1 культивировали в течение 6 суток в среде, дополненной специфическими факторами для индукции дифференцировки в адипоциты (дифференцированные). Клетки, выращенные на среде без таких факторов, оставались недифференцированными. В среду добавляли 2 ед./мл N-ABD094-rhArg (SEQ ID NO: 50) (rhArg). Контролем служили клетки, выращенные в среде без добавления N-ABD094-rhArg. Уровни экспрессии мРНК нескольких важных (А) транскрипционных регуляторов адипогенеза и (В) ферментов липогенных путей определяли с помощью qRT-PCR в режиме реального времени и выражали относительно недифференцированной (контрольной) группы, уровень у которой принимали за 1. *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=3 для каждой группы.
[0069] На фиг. 25 проиллюстрирован ответ преадипоцитов 3T3-L1 на аргинин в различных концентрациях. Мышиные преадипоциты 3T3-L1 культивировали в течение 6 суток в среде без аргинина с добавлением или без добавления L-аргинина в различных концентрациях. Клетки, культивированные в нормальной, содержащей аргинин среде с добавлением факторов, индуцирующих дифференцировку, служили положительным («+») контролем дифференцировки, в то время как клетки, культивированные в нормальной, содержащей аргинин среде без добавления факторов, индуцирующих дифференцировку, служили отрицательным («-») контролем дифференцировки. Одну группу клеток культивировали в нормальной, содержащей аргинин среде с добавлением факторов, индуцирующих дифференцировку, и обрабатывали посредством 5 ед./мл N-ABD094-rhArg (SEQ ID NO: 50) (rhArg) для ингибирования дифференцировки. (А) Клетки окрашивали на липиды масляным красным О и интенсивность окрашивания количественно оценивали путем измерения оптической плотности, которую выражали в процентах по сравнению с «+» контрольной группой. (В) Уровни мРНК нескольких важных генов, связанных с липогенезом, выражали относительно клеток, культивируемых в среде без аргинина, уровень которых принимали за 1. Данные выражены как среднее значение±SEM, n=3 для каждой группы.
[0070] На фиг. 26 проиллюстрировано, что одновременная обработка самцов мышей, которых кормили HFD, посредством 300 ед. N-ABD094-rhArg (SEQ ID NO: 50) [группа «HFD(rhArg)»] один раз в неделю может эффективно предупреждать HFD-индуцированную инсулинорезистентность. Тест на толерантность к инсулину (ITT) проводили через (А) 6 недель и (В) 11 недель после кормления HFD, а результаты выражали в виде площади под кривой (AUC). *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0071] На фиг. 27 проиллюстрировано, что одновременная обработка самцов мышей, которых кормили HFD, посредством 300 ед. N-ABD094-rhArg (SEQ ID NO: 50) [группа «HFD(rhArg)»] один раз в неделю может эффективно предупреждать HFD-индуцированную непереносимость глюкозы. Тест на толерантность к глюкозе (GTT) проводили через (А) 5 недель и (В) 10 недель после кормления HFD, а результаты выражали в виде площади под кривой (AUC). *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0072] На фиг. 28 проиллюстрировано, что одновременная обработка самцов мышей, которых кормили HFD, посредством 300 ед. N-ABD094-rhArg (SEQ ID NO: 50) [группа «HFD(rhArg)»] один раз в неделю может эффективно предупреждать HFD-индуцированное повышение натощак (А) уровня глюкозы в крови, измеренного с помощью глюкометра, и (В) концентраций инсулина в плазме крови, измеренных с помощью ELISA. (С) Показатель HOMA-IR, рассчитанный по стандартной формуле, в качестве критерия инсулинорезистентности указывал на то, что одновременная обработка посредством N-ABD094-rhArg предупреждала развитие HFD-индуцированной инсулинорезистентности. *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0073] На фиг. 29 проиллюстрировано, что одновременная обработка самцов мышей, которых кормили HFD, посредством 300 ед. N-ABD094-rhArg (SEQ ID NO: 50) [группа «HFD(rhArg)»] один раз в неделю предупреждала/уменьшала HFD-индуцированное повышение уровней в плазме натощак (А) лептина, (В) общего холестерина, (С) триглицеридов и (D) свободных жирных кислот. *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0074] На фиг. 30 проиллюстрировано, что одновременная обработка самцов мышей, которых кормили HFD, посредством 300 ед. N-ABD094-rhArg (SEQ ID NO: 50) [группа «HFD(rhArg)»] один раз в неделю в течение 12 недель предупреждала HFD-индуцированное увеличение печени. (А) Репрезентативные изображения цельной свежей печени. (В) Масса печени. *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0075] На фиг. 31 проиллюстрировано, что одновременная обработка самцов мышей, которых кормили HFD, посредством 300 ед. N-ABD094-rhArg (SEQ ID NO: 50) [группа «HFD(rhArg)»] один раз в неделю в течение 12 недель могла эффективно предупреждать HFD-индуцированный стеатоз печени. (А) По результатам окрашивания срезов печени масляным красным О было видно обширное накопление липидов у обработанных наполнителем мышей, которых кормили HFD [группа «НЕО(наполнитель)»], которое отсутствовало у мышей, обработанных посредством N-ABD094-rhArg. (В) обработка посредством N-ABD094-rhArg предупреждала повышение концентраций триглицеридов в печени мышей, которых кормили HFD. (С) N-ABD094-rhArg подавлял HFD-индуцированную активацию связанных с липогенезом генов в печени. *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0076] На фиг. 32 проиллюстрировано, что одновременная обработка самцов мышей, которых кормили HFD, посредством 300 ед. N-ABD094-rhArg (SEQ ID NO: 50) [группа «HFD(rhArg)»] один раз в неделю в течение 12 недель предупреждала HFD-индуцированное повышение сывороточных уровней аланинаминотрансферазы (ALT)), обычно измеряемой в качестве биомаркера повреждения печени. *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0077] На фиг. 33 проиллюстрировано, что одновременная обработка самцов мышей, которых кормили HFD, посредством 300 ед. N-ABD094-rhArg (SEQ ID NO: 50) [группа «HFD(rhArg)»] один раз в неделю в течение 12 недель предупреждала HFD-индуцированное увеличение массы (А) почки, (В) поджелудочной железы и (С) сердца. *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0078] На фиг. 34 проиллюстрировано, что одновременная обработка самцов мышей, которых кормили HFD, посредством 300 ед. N-ABD094-rhArg (SEQ ID NO: 50) [группа «HFD(rhArg)»] один раз в неделю в течение 12 недель может эффективно (А) подавлять HFD-индуцированное повышение уровней мРНК провоспалительных адипокинов в висцеральном WAT, выраженных относительно уровней у худых мышей контрольной группы [группы «ЬТТ^наполнитель)»], уровень у которых принимали за 1, и (В) подавлять HFD-индуцированное повышение уровней провоспалительных цитокинов в сыворотке крови. *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0079] На фиг. 35 проиллюстрировано, что одновременная обработка самцов мышей посредством 300 ед. N-ABD094-rhArg (SEQ ID NO: 50) [группа «HFD(rhArg)»] один раз в неделю в течение 12 недель может эффективно предупреждать HFD-индуцированное побеление бурой жировой ткани (ВАТ). (А) Репрезентативные изображения цельной свежей межлопаточной ВАТ. (В) Масса межлопаточной ВАТ. (С) На парафиновых срезах, окрашенных Н&Е, было видно, что клетки с большим одиночным отверстием (липидной глобулой), которые являются типичными гистологическими признаками белых адипоцитов, были многочисленны в ВАТ мышей, которых кормили HFD и обрабатывали наполнителем [группа «НРО(наполнитель)»], но редко встречались в ВАТ мышей, которых кормили HFD и обрабатывали посредством N-ABD094-rhArg. *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0080] На фиг. 36 проиллюстрировано, что одновременная обработка самцов мышей посредством 300 ед. N-ABD094-rhArg (SEQ ID NO: 50) [группа «HFD(rhArg)»] один раз в неделю в течение 12 недель значимо повышала уровни мРНК ключевых регуляторов термогенеза (Ucp1 и Pgc1a) в ВАТ и супрессировала HFD-индуцированное подавление активности гена окисления жирных кислот Acox1 в ВАТ. *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0081] На фиг. 37 проиллюстрировано, что у самцов C57BL/6J мышей с ранее существовавшим ожирением, индуцированным кормлением HFD от 5-недельного возраста в течение 12 недель, наблюдали заметное снижение массы тела после обработки посредством N-ABD094-rhArg (SEQ ID NO: 50). (А) Масса тела мышей, обработанных посредством 600 ед. N-ABD094-rhArg [группа «HFD(rhArg)»] один раз в неделю в течение 12 недель. (В) Репрезентативные изображения мышей после 12 недель обработки. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0082] На фиг. 38 проиллюстрировано, что введение 600 ед. N-ABD094-rhArg (SEQ ID NO: 50) один раз в неделю в течение 12 недель самцам мышей с ранее существовавшим HFD-индуцированным ожирением [группа «HFD(rhArg)»] может эффективно уменьшать количество белой жировой ткани. (А) Масса жирового тела в основных висцеральных (перигонадных, периренальных и брыжеечных) и подкожных (паховых) депо у страдающих ожирением мышей после обработки посредством N-ABD094-rhArg была очень похожа на массу у худых мышей контрольной группы [группы «LFD(rhArg)»]. *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=5 для каждой группы. (В) На окрашенных посредством Н&Е срезах висцерального WAT было видно, что обработка посредством N-ABD094-rhArg заметно уменьшала размер адипоцитов у страдающих ожирением мышей.
[0083] На фиг. 39 проиллюстрировано, что введение 600 ед. N-ABD094-rhArg (SEQ ID NO: 50) один раз в неделю самцам мышей с ранее существовавшим HFD-индуцированным ожирением [группа «HFD(rhArg)»] может эффективно обращать инсулинорезистентность. Тест на толерантность к инсулину (ITT) проводили до и через 4, 8 и 12 недель после обработки N-ABD094-rhArg. Результаты ITT выражены как площадь под кривой (AUC). *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0084] На фиг. 40 проиллюстрировано, что введение 600 ед. N-ABD094-rhArg (SEQ ID NO: 50) один раз в неделю самцам мышей с ранее существовавшим HFD-индуцированным ожирением [группа «HFD(rhArg)»] может эффективно обращать непереносимость глюкозы. Тест на толерантность к глюкозе (GTT) проводили до и через 3, 7 и 11 недель после обработки N-ABD094-rhArg. Результаты GTT выражены как площадь под кривой (AUC). *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0085] На фиг. 41 проиллюстрировано, что введение 600 ед. N-ABD094-rhArg (SEQ ID NO: 50) один раз в неделю самцам мышей с ранее существовавшим HFD-индуцированным ожирением [группа «HFD(rhArg)»] может эффективно обращать гипергликемию, гиперинсулинемию и инсулинорезистентность. (А) Уровень глюкозы в крови натощак, (В) уровень инсулина в плазме крови натощак и (С) показатель HOMA-IR у страдающих ожирением мышей были скорректированы до почти нормальных уровней после обработки посредством N-ABD094-rhArg. *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0086] На фиг. 42 проиллюстрировано, что введение 600 ед. N-ABD094-rhArg (SEQ ID NO: 50) один раз в неделю самцам мышей с ранее существовавшим HFD-индуцированным ожирением [группа «HFD(rhArg)»] может эффективно обращать гиперлептинемию и гиперхолестеринемию. (А) Уровень лептина в плазме крови натощак и (В) уровень общего холестерина в плазме натощак у страдающих ожирением мышей были скорректированы до почти нормальных уровней после обработки посредством N-ABD094-rhArg. *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0087] На фиг. 43 проиллюстрировано, что введение 600 ед. N-ABD094-rhArg (SEQ ID NO: 50) один раз в неделю в течение 12 недель самцам мышей с ранее существовавшим HFD-индуцированным ожирением [группа «HFD(rhArg)»] может эффективно обращать увеличение печени. (А) На репрезентативных изображениях цельной свежей печени видно, что печень у страдающих ожирением мышей вернулась к нормальному размеру после обработки N-ABD094-rhArg. (В) Масса печени у страдающих ожирением мышей резко снижалась после обработки посредством N-ABD094-rhArg и была похожа на массу у худых мышей контрольной группы [группа LFD(rhArg)]. *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0088] На фиг. 44 проиллюстрировано, что введение 600 ед. N-ABD094-rhArg (SEQ ID NO: 50) один раз в неделю в течение 12 недель самцам мышей с ранее существовавшим HFD-индуцированным ожирением [группа «HFD(rhArg)»] может эффективно обращать стеатоз печени. В печени страдающих ожирением мышей, которых обрабатывали посредством N-ABD094-rhArg, (А) в результате окрашивания срезов печени масляным красным О наблюдали резкий клиренс липидов, и (В) концентрации триглицеридов снижались до уровней, сравнимых с уровнями худых мышей контрольной группы [«ЕЕО(наполнитель)»], а также (С) снижались уровни мРНК нескольких важных генов, связанных с липогенезом. Уровни экспрессии выражены относительно худых мышей контрольной группы, уровень у которых принимали за 1. *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0089] На фиг. 45 проиллюстрировано, что введение 600 ед. N-ABD094-rhArg (SEQ ID NO: 50) один раз в неделю в течение 12 недель самцам мышей с ранее существовавшим HFD-индуцированным ожирением [группа «HFD(rhArg)»] может значительно снижать сывороточные уровни аланинтрансаминазы (ALT) и аспартаттрансаминазы (AST), которые являются традиционными биомаркерами повреждения печени. *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0090] На фиг. 46 проиллюстрировано, что введение 600 ед. N-ABD094-rhArg (SEQ ID NO: 50) один раз в неделю в течение 12 недель самцам мышей с ранее существовавшим HFD-индуцированным ожирением [группа «HFD(rhArg)»] может обращать стеатоз почек. У страдающих ожирением мышей, обработанных посредством N-ABD094-rhArg: заметно были снижены (А) масса почки; (В) концентрации триглицеридов в почках; (С) вакуолизированные структуры в почечных канальцах на срезах почек, окрашенных посредством Н&Е, и (D) эктопическое накопление липидных капель в клубочках на срезах почек, окрашенных масляным красным О. *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0091] На фиг. 47 проиллюстрировано, что введение 600 ед. N-ABD094-rhArg (SEQ ID NO: 50) один раз в неделю в течение 12 недель самцам мышей с ранее существовавшим HFD-индуцированным ожирением [группа «HFD(rhArg)»] может обращать стеатоз поджелудочной железы. У страдающих ожирением мышей, обработанных посредством N-ABD094-rhArg: заметно были снижены (А) масса поджелудочной железы; (В) чрезмерное междольковое и внутридольковое скопление белой жировой ткани, наблюдаемое на окрашенных посредством Н&Е срезах поджелудочной железы; и (С) концентрации триглицеридов в поджелудочной железе. *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0092] На фиг. 48 проиллюстрировано, что введение 600 ед. N-ABD094-rhArg (SEQ ID NO: 50) один раз в неделю в течение 12 недель самцам мышей с ранее существовавшим HFD-индуцированным ожирением [группа «HFD(rhArg)»] может обращать стеатоз сердца. У страдающих ожирением мышей, обработанных посредством N-ABD094-rhArg: заметно были снижены (А) масса сердца; (В) чрезмерное накопление липидных капель в сердце; и (С) концентрации триглицеридов в сердце. *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0093] На фиг. 49 проиллюстрировано, что введение 600 ед. N-ABD094-rhArg (SEQ ID NO: 50) один раз в неделю в течение 12 недель самцам мышей с ранее существовавшим HFD-индуцированным ожирением [группа «HFD(rhArg)»] может эффективно уменьшать концентрации триглицеридов в скелетной мышце. *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0094] На фиг. 50 проиллюстрировано, что введение 600 ед. N-ABD094-rhArg (SEQ ID NO: 50) один раз в неделю в течение 12 недель самцам мышей с ранее существовавшим HFD-индуцированным ожирением [группа «HFD(rhArg)»] может значительно (А) снижать уровни мРНК провоспалительного адипокина Мcр1 в висцеральном WAT, выраженные относительно таковых у худых мышей контрольной группы [группа «LFD(наполнитель)»], уровень у которых принимали за 1, и (В) снижать сывороточные концентрации провоспалительного цитокина Mcp1. *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0095] На фиг. 51 проиллюстрировано, что введение 600 ед. N-ABD094-rhArg (SEQ ID NO: 50) один раз в неделю в течение 12 недель самцам мышей с ранее существовавшим HFD-индуцированным ожирением [группа «HFD(rhArg)»] может уменьшать воспаление и фиброз печени. В печени страдающих ожирением мышей, которых обрабатывали посредством N-ABD094-rhArg, наблюдали значимое (А) подавление активности провоспалительных генов и (В) профибротических генов, но (С) повышение активности противовоспалительных генов. Уровни мРНК выражены относительно таковых у худых мышей контрольной группы [«LFD(наполнитель)»], уровень у которых принимали за 1. *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0096] На фиг. 52 проиллюстрировано, что введение 600 ед. N-ABD094-rhArg (SEQ ID NO: 50) один раз в неделю в течение 12 недель самцам мышей с ранее существовавшим HFD-индуцированным ожирением [«HFD(rhArg)»] может уменьшать воспаление и фиброз почки. В почках страдающих ожирением мышей, которых обрабатывали посредством N-ABD094-rhArg, наблюдали значимое (А) подавление активности провоспалительных генов и (В) профибротических генов, а также (С) уменьшение чрезмерного отложения коллагеновых волокон вокруг почечных канальцев и клубочков. Уровни мРНК выражены относительно таковых у худых мышей контрольной группы [группы «LFD(наполнитель)»], уровень у которых принимали за 1. *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0097] На фиг. 53 проиллюстрировано, что введение 600 ед. N-ABD094-rhArg (SEQ ID NO: 50) один раз в неделю в течение 12 недель самцам мышей с ранее существовавшим HFD-индуцированным ожирением [группа «HFD(rhArg)»] может уменьшать воспаление и фиброз поджелудочной железы. В поджелудочной железе страдающих ожирением мышей, которых обрабатывали посредством N-ABD094-rhArg, наблюдали значимое (А) подавление активности провоспалительного гена и (В) профибротического гена. Уровни мРНК выражены относительно таковых у худых мышей контрольной группы [группы «LFD(наполнитель)»]. *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные представляют собой среднее значение±стандартная ошибка среднего, n=5 для каждой группы.
[0098] На фиг. 54 проиллюстрировано, что введение 600 ед. N-ABD094-rhArg (SEQ ID NO: 50) один раз в неделю в течение 12 недель самцам мышей с ранее существовавшим HFD-индуцированным ожирением [группа «HFD(rhArg)»] может эффективно обращать побеление бурого жира (ВАТ). У страдающих ожирением мышей, обработанных посредством N-ABD094-rhArg, (А) масса межлопаточного ВАТ; (В) морфология и размер межлопаточного ВАТ; и (С) гистологические признаки, наблюдаемые на окрашенном посредством Н&Е срезе ВАТ, были аналогичны таковым у худых мышей контрольной группы. *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=5 для каждой группы.
[0099] На фиг. 55 проиллюстрировано, что введение 600 ед. N-ABD094-rhArg (SEQ ID NO: 50) один раз в неделю самцам мышей с ранее существовавшим HFD-индуцированным ожирением [группа «HFD(rhArg)»] может значимо снижать (А) систолическое и диастолическое артериальное давление и (В) частоту сердечных сокращений у страдающих ожирением мышей до уровня, схожего с таковым у худых мышей контрольной группы [группа «LFD(наполнитель)»] через 5 недель после обработки лекарственным средством. *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=10 для каждой группы.
[00100] На фиг. 56 проиллюстрировано, что у самцов C57BL/6J мышей, которых кормили LFD и вводили внутрибрюшинную инъекцию 500 ед. N-ABD094-rhArg (SEQ ID NO: 50) [группа «LFD(rhArg)»] один раз в неделю в течение 8 месяцев, начиная с 5-недельного возраста, вырабатывались антитела, отличные от нейтрализующих к лекарственному средству. (А) С помощью ELISA-анализа в плазме мышей, которых кормили LFD и обрабатывали посредством N-ABD094-rhArg, можно было детектировать антитела к rhArg со средним титром антител 108. (В) Плазму инкубировали с 1000 ед./мл N-ABD094-rhArg (rhArg *) в течение 1 часа с последующим измерением аргиназной активности. Снижения аргиназной активности не наблюдали, что свидетельствовало о том, что антитела, вырабатываемые у животного к данному лекарственному средству, не были нейтрализующими антителами. Данные выражены как среднее значение±SEM, n=3 для каждой группы.
[00101] На фиг. 57 проиллюстрировано, что введение 500 ед. N-ABD094-rhArg (SEQ ID NO: 50) самцам C57BL/6J мышей, которых кормили LFD [группа «LFD(rhArg)»] один раз в неделю в течение 8 месяцев, начиная с 5-недельного возраста, не вызывало побочных эффектов на печень и почки. У мышей, которых кормили LFD и обрабатывали посредством N-ABD094-rhArg, не наблюдали никаких значимых отличий от мышей, которых кормили LFD и обрабатывали наполнителем [группа «LFD(наполнитель)»], в (А) сывороточных концентрациях аланинаминотрансферазы (ALT) и аспартатаминотрансферазы (AST), которые обычно применяют в качестве биомаркеров для оценки функций или заболеваний печени; и (В) сывороточных концентрациях креатинина и альбумина/креатинина в моче, которые обычно применяют в качестве биомаркеров для оценки функций или заболеваний почек. Данные выражены как среднее значение±SEM, n=3 для каждой группы.
[00102] На фиг. 58 проиллюстрировано, что введение 500 ед. N-ABD094-rhArg (SEQ ID NO: 50) самцам C57BL/6J мышей, которых кормили LFD [группа «LFD(rhArg)»] один раз в неделю в течение 8 месяцев, начиная с 5-недельного возраста, не вызывало побочных эффектов на сосуды. У мышей, которых кормили LFD и обрабатывали посредством N- ABD094-rhArg, не наблюдали никаких значимых отличий от мышей, которых кормили LFD и обрабатывали наполнителем [группа «LFD(наполнитель)»], в (А) в сокращении гладких мышц сосудов, вызванном фенилэфрином; (В) ацетилхолин-индуцированной эндотелий-зависимой релаксации; и (С) эндотелий-независимой релаксации в ответ на нитропруссид натрия, который является донором оксида азота. Данные выражены как среднее значение±SEM, n=3 для каждой группы.
[00103] На фиг. 59 проиллюстрировано, что введение 300 ед. пегилированного His-rhArg (SEQ ID NO: 101) один раз в неделю в течение 4 недель самцам C57BL/6J мышей с ранее существовавшим HFD-индуцированным ожирением [группа «HFD(PEG-rhArg)»] может эффективно снижать массу тела мышей до массы, схожей с массой у худых мышей контрольной группы, обработанных наполнителем, которых кормили стандартным рационом [группа «Стандарт(наполнитель)»]. Данные выражены как среднее значение±SEM, n=6 для каждой группы.
[00104] На фиг. 60 проиллюстрировано, что введение 300 ед. пегилированного His-rhArg (SEQ ID NO: 101) один раз в неделю в течение 4 недель самцам мышей с ранее существовавшим HFD-индуцированным ожирением [группа «HFD(PEG-rhArg)»] может эффективно уменьшать жировую массу в основных висцеральных (перигонадных, периренальных, брыжеечных) и подкожных (паховых) депо и уменьшать массу печени, почки, поджелудочной железы и сердца. *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=4 для каждой группы.
[00105] На фиг. 61 проиллюстрировано, что введение 300 ед. пегилированного His-rhArg (SEQ ID NO: 101) раз в неделю самцам мышей с уже существующим HFD-индуцированным ожирением [группа «HFD(PEG-rhArg)»] может значимо (А) снижать уровень глюкозы в крови натощак и (В) повышать чувствительность к инсулину в тесте на толерантность к инсулину (ITT), который проводили через 2 недели после обработки лекарственным средством. Результаты ITT выражены как площадь под кривой (AUC). *Р<0,05, однофакторный ANOVA с последующим LSD Фишера. Данные выражены как среднее значение±SEM, n=6 для каждой группы.
[00106] На фиг. 62 проиллюстрировано, что введение 25-100 ед. N-ABD094-rhArg (SEQ ID NO: 50 (замещенный кобальтом)] один раз в неделю в течение 5 недель самцам C57BL/6J мышей с уже существовавшим HFD-индуцированным ожирением может эффективно снижать массу тела мышей. Данные выражены как среднее значение±SEM, n=3.
[00107] На фиг. 63 проиллюстрировано противораковое действие слитого белка ВНА (SEQ ID NO: 75) на линии клеток рака желудка MKN45. Рост MKN45 ингибировался дозозависимым образом. Жизнеспособность определяли колориметрическим анализом конечных точек с применением МТТ. Данные выражены как среднее±стандартное отклонение, n=3.
[00108] На фиг. 64 проиллюстрировано влияние, N-ABD-rhArg (SEQ ID NO:49) на объем опухоли у бестимусных мышей с ксенотрансплантатом рака молочной железы 4Т1. На кривой для «Экспериментальной группы» представлены данные для экспериментальной группы, которая получала инъекцию 500 ед. N-ABD-rhArg (SEQ ID NO: 49) один раз в неделю; на кривой для «Контрольной группы» представлены данные для контрольной группы, которая получала инъекцию PBS один раз в неделю. *Р<0,05, **Р<0,01 по сравнению с экспериментальной группой, t-критерий, n=4-6.
[00109] На фиг. 65 проиллюстрировано влияние, N-ABD-rhArg (SEQ ID NO:49) на относительный объем опухоли у бестимусных мышей с ксенотрансплантатом рака молочной железы 4Т1. На кривой для «Экспериментальной группы» представлены данные для экспериментальной группы, которая получала инъекцию 500 ед. N-ABD-rhArg (SEQ ID NO: 49) один раз в неделю. На кривой для «Контрольной группы» представлены данные для контрольной группы, которая получала инъекцию PBS один раз в неделю. * Р<0,05 по сравнению с экспериментальной группой, t-критерий, n=4-6.
[00110] На фиг. 66 проиллюстрированы значимые различия в массе опухоли между экспериментальной группой и контрольной группой. Экспериментальная группа получала инъекцию 500 ед. N-ABD-rhArg (SEQ ID NO: 49) один раз в неделю. Контрольная группа получала инъекцию PBS один раз в неделю. *Р<0,05 по сравнению с экспериментальной группой, t-критерий, n=4-6.
[00111] На фиг. 67 проиллюстрирована фотография опухолей, собранных у бестимусных мышей из контрольной группы и группы, обработанной посредством N-ABD-rhArg (SEQ ID NO: 49).
[00112] На фиг. 68 проиллюстрировано влияние, N-ABD-rhArg (SEQ ID NO:49) на относительный объем опухоли у аллотрансплантатов рака молочной железы 4Т1. На кривой для «500 ед. ABD-rhArg» представлены данные для экспериментальной группы, которая получала инъекцию 500 ед. N-ABD-rhArg (SEQ ID NO: 49) один раз в неделю. На кривой для «250 ед. ABD-rhArg» представлены данные для экспериментальной группы, которая получала инъекцию 250 ед. N-ABD-rhArg (SEQ ID NO: 49) один раз в неделю. На кривой для «500 ед. PEG-rhArg» представлены данные для экспериментальной группы, которая получала инъекцию 500 ед. пегилированного His-rhArg (SEQ ID NO: 101) один раз в неделю. На кривой для «Контрольной группы» представлены данные для контрольной группы, которая получала инъекцию PBS один раз в неделю. **Р<0,01, ***Р<0,001 по сравнению с экспериментальной группой, t-критерий, n=6-8.
[00113] На фиг. 69 проиллюстрированы значимые различия в массе опухоли между экспериментальными группами и контрольной группой. Данные для «500 ед. ABD-rhArg» представлены для экспериментальной группы, которая получала инъекцию 500 ед. N-ABD-rhArg (SEQ ID NO: 49) один раз в неделю. Данные для «250 ед. ABD-rhArg» представлены для экспериментальной группы, которая получала инъекцию 250 ед. N-ABD-rhArg (SEQ ID NO: 49) один раз в неделю. Данные для «500 ед. PEG-rhArg» представлены для группы, которая получала инъекцию 500 ед. пегилированного His-rhArg (SEQ ID NO: 101) один раз в неделю. Данные для «Контрольной группы» представлены данные для контрольной группы, которая получала инъекцию PBS один раз в неделю. *Р<0,05, **Р<0,01 по сравнению с экспериментальной группой, t-критерий, n=6-8.
[00114] На фиг. 70 проиллюстрировано среднее количество метастатических узелков из легких аллотрансплантатов 4Т1. (А) Данные для «500 ед. ABD-rhArg» представлены для экспериментальной группы, которая получала инъекцию 500 ед. N-ABD-rhArg (SEQ ID NO: 49) один раз в неделю. Данные для «250 ед. ABD-rhArg» представлены для экспериментальной группы, которая получала инъекцию 250 ед. N-ABD-rhArg (SEQ ID NO: 49) один раз в неделю. Данные для «500 ед. PEG-rhArg» представлены для группы, которая получала инъекцию 500 ед. пегилированного His-rhArg (SEQ ID NO: 101) один раз в неделю. Данные для «Контрольной группы» представлены для контрольной группы, которая получала инъекцию PBS один раз в неделю. *Р<0,05, **Р<0,01 по сравнению с экспериментальной группой, t-критерий, n=6-8. (В) График разброса количества метастатических узелков из легких аллотрансплантатов 4Т1, показанных на фиг. 70А.
Подробное раскрытие настоящего изобретения
[00115] Настоящее изобретение, в целом, относится к слитым белкам, содержащим полипептид аргиназы и полипептид ABD, и к способам их применения и получения. Слитые белки, представленные в настоящем документе, являются высокоэффективными истощающими запасы аргинина средствами и благодаря слиянию с полипептидом ABD характеризуются значительно увеличенным периодом полужизни по сравнению с известной аргиназой и известными производными аргиназы. Раскрываемые слитые белки полезны для лечения заболеваний и патологических состояний, при которых истощение запасов аргинина приводит к терапевтическому эффекту, например, при лечении рака, вирусов, рассеянного склероза, ревматоидного артрита, аутоиммунных заболеваний, врожденной гипераргининемии, болезни «трансплантат против хозяина» (GvHD), воспаления, ожирения, метаболических нарушений и связанных с ними осложнений, а также сопутствующих заболеваний.
[00116] Настоящее изобретение также, в целом, относится к способам лечения ожирения, метаболических нарушений и связанных с ними осложнений, а также сопутствующих заболеваний у нуждающегося в том субъекта путем истощения уровня аргинина у субъекта. Истощение запасов аргинина у субъекта приводит к снижению массы и жировой массы, повышению толерантности к глюкозе и чувствительности к инсулину, нормализации уровней глюкозы в крови, эндокринных и метаболических профилей и облегчению воспаления, стеатоза и фиброза. В описываемых в настоящем документе способах лечения ожирения, метаболических нарушений и связанных с ними осложнений, а также сопутствующих заболеваний можно применять любое средство, способное истощать запасы аргинина у субъекта.
Определение терминов
[00117] Подразумевают, что определения применяемых в настоящем документе терминов включают современные определения, признанные для каждого термина в области биотехнологии. В соответствующих случаях представлены примеры. Определения применимы к терминам в том виде, в котором их применяют в настоящем описании, если иное не ограничено в конкретных случаях, либо индивидуально, либо как часть более крупной группы.
[00118] В контексте настоящего документа термин «период полужизни» или
«период Уг-жизни» относится ко времени, которое необходимо для того, чтобы концентрация средства, например, слитого белка или истощающего запасы аргинина средства, которые описаны в настоящем документе, упала наполовину in vitro или in vivo, например, после инъекции млекопитающему. В некоторых случаях концентрацию аргинина в плазме после инъекции используют в настоящем документе в качестве косвенного показателя периода полужизни средства. В таких случаях термин «терапевтическая продолжительность» применяют для обозначения продолжительности времени, в течение которого заданная доза истощающего запасы аргинина средства способна поддерживать плазменную концентрацию аргинина ниже заданной пороговой концентрации, при которой наблюдают требуемый терапевтический эффект. В соответствии с определенными вариантами осуществления, пороговая концентрация аргинина в плазме крови составляет ниже 50 мкМ, ниже 40 мкМ, ниже 30 мкМ, ниже 20 мкМ, ниже 10 мкМ, ниже 5 мкМ, ниже 3 мкМ или на уровне концентрации, которая ниже предела детекции традиционных аналитических приборов. Например, истощение запасов аргинина в плазме крови до концентраций, которые ниже предела детекции анализатора аминокислот Biochrom 30 (предел детекции составляет 3 мкМ), на период 7 суток после инъекции катаболизирующего аргинин фермента, описанного в настоящем документе, указывает на терапевтическую продолжительность 7 суток и период полужизни, например, порядка 7 суток.
[00119] В контексте настоящего документа термин «присоединить» или «присоединенный» относится к соединению или объединению посредством связи или несвязывающего взаимодействия с целью удержания двух или более соединений вместе, что охватывает прямое или опосредованное присоединение, такое как, например, при связывании первого полипептида непосредственно со вторым полипептидом или другой молекулой, и варианты осуществления, в которых одно или несколько промежуточных соединений (например, линкер), таких как полипептид, расположены между первым полипептидом и вторым полипептидом или другой молекулой.
[00120] В контексте настоящего документа термин «белок» или «полипептид» означает органический полимер, состоящий из двух или более аминокислотных мономеров и/или их аналогов. Термин «полипептид» включает аминокислотные полимеры любой длины, в том числе полноразмерные белки и пептиды, а также их аналоги и фрагменты. Полипептид из трех или более аминокислот также называют олигопептидом. В контексте настоящего документа термин «аминокислота», «аминокислотный мономер» или «аминокислотный остаток» относится к любой из двадцати встречающихся в природе аминокислот, включая синтетические аминокислоты с не встречающимися в природе боковыми цепями и включая оптические D- и L-изомеры. Термин «аминокислотный аналог» относится к аминокислоте, в которой один или несколько отдельных атомов были заменены либо на другой атом, либо на изотоп, либо на другую функциональную группу, но в остальном она идентична своему природному аминокислотному аналогу
[00121] В контексте настоящего документа термин «не встречающаяся в природе аминокислота» относится к любой аминокислоте, модифицированной аминокислоте и/или аминокислотному аналогу, который не является одной из 20 обычных встречающихся в природе аминокислот, селеноцистеином или пирролизином.
[00122] В контексте настоящего документа термин «слитый белок» относится к химерному белку, содержащему белки или функциональные фрагменты белка (например, аргиназу или ее варианты), имеющее различное происхождение, которые ковалентно связаны, например, амидом, сложноэфирной группой, мочевинной группой, карбаматной группой, группой простого эфира и/или дисульфидной связью.
[00123] В контексте настоящего документа термин «вариант» относится к полинуклеотиду или нуклеиновой кислоте, отличающимся от эталонной нуклеиновой кислоты или эталонного полипептида, но сохраняющим их основные свойства. Обычно варианты, в целом, очень похожи и во многих областях идентичны эталонной нуклеиновой кислоте или эталонному полипептиду.
[00124] Вариант может, например, содержать аминокислотную последовательность исходной полипептидной последовательности по меньшей мере с одной консервативной аминокислотной заменой. В качестве альтернативы или дополнительно, вариант может содержать аминокислотную последовательность исходной полипептидной последовательности по меньшей мере с одной неконсервативной аминокислотной заменой. В этом случае предпочтительно, чтобы неконсервативная аминокислотная замена не мешала или не ингибировала биологическую активность функционального варианта. Неконсервативная аминокислотная замена может усиливать биологическую активность варианта, так чтобы биологическая активность вариантов увеличивалась по сравнению с исходным полипептидом.
[00125] Термин «функциональный фрагмент» при его применении в отношении полипептида относится к любой части или части рассматриваемого полипептида, причем эта часть или частица сохраняет биологическую активность полипептида, частью которого она является (исходного полипептида). Функциональный фрагмент может быть любым фрагментом, содержащим смежные аминокислоты полипептида, частью которого он является, при условии, что функциональный фрагмент все еще обладает по меньшей мере 40%, 50%, 60%, 70%, 80%, 90%, 95% или 99% или обладает практически такой же или даже более высокой биологической активностью исходного полипептида. Что касается исходного полипептида, функциональный фрагмент может содержать, например, приблизительно 10%, 20%, 25%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 98% или более от исходного полипептида.
[00126] Функциональный фрагмент может содержать дополнительные аминокислоты на амино- либо карбоксиконце или на обоих концах, например, аминокислоты, не встречающиеся в аминокислотной последовательности исходного полипептида.
[00127] Аминокислотные замены описанных полипептидов могут быть консервативными аминокислотными заменами. Консервативные аминокислотные замены известны в настоящей области техники и включают аминокислотные замены, при которых одну аминокислоту, имеющую определенные физические и/или химические свойства, заменяют другой аминокислотой, которая имеет такие же или подобные химические или физические свойства. Например, консервативная аминокислотная замена может представлять собой кислую/отрицательно заряженную полярную аминокислоту, замененную на другую кислую/отрицательно заряженную полярную аминокислоту (например, Asp или Glu), аминокислоту с неполярной боковой цепью, замененной на другую аминокислоту с неполярной боковой цепью (например, Ala, Gly, Val, Ile, Leu, Met, Phe, Pro, Trp, Cys, Val и т.д.), основную/положительно заряженную полярную аминокислоту, замененную на другую основную/положительно заряженную полярную аминокислоту (например, Lys, His, Arg и т.д.), незаряженную аминокислоту с полярной боковой цепью, замененную на другую незаряженную аминокислоту с полярной боковой цепью (например, Asn, Gln, Ser, Thr, Tyr и т.д.), аминокислоту с бета-разветвленной боковой цепью, замененную на другую аминокислоту с бета-разветвленной боковой цепью (например, Ile, Thr и Val), аминокислоту с ароматической боковой цепью, замененную на другую аминокислоту с ароматической боковой цепью (например, His, Phe, Trp и Tyr), и т.д.
[00128] Термины «процент гомологии» и «процент идентичности последовательностей» при их применении в отношении полипептидной или полинуклеотидной последовательности в настоящем документе применяют взаимозаменяемо для обозначения результатов сравнения между полинуклеотидами и полипептидами и определяют путем сравнения двух оптимально выровненных последовательностей в интервале сравнения, причем часть полинуклеотидной или полипептидной последовательности в интервале сравнения может содержать добавления или делеции (т.е. гэпы) в сравнении с эталонной последовательностью (которая не содержит добавлений или делеций) для оптимального выравнивания этих двух последовательностей. Процент рассчитывают путем определения количества положений, в которых идентичное основание нуклеиновой кислоты или аминокислотный остаток встречается в обеих последовательностях, с получением количества совпадающих положений, потом деления количества совпадающих положений на общее количество положений в интервале сравнения и умножения результата на 100 с получением процента идентичности последовательностей. Гомологию оценивают с помощью любого из множества алгоритмов и программ сравнения последовательностей, которые известны в настоящей области техники. К таким алгоритмам и программам относятся без ограничения TBLASTN, BLASTP, FAS ТА, TFASTA и CLUSTALW [Pearson and Lipman, 1988, Proc. Natl. Acad. Sci. USA 85(8):2444-2448; Altschul et al., 1990, J. Mol. Biol. 215(3):403-410; Thompson et al., 1994, Nucleic Acids Res. 22(2):4673-4680; Higgins et al. 1996, Methods Enzymol. 266:383-402; Altschul et al., 1990, J. Mol. Biol. 215(3):403-410; Altschul et al., 1993, Nature Genetics 3:266-272]. В соответствии с определенными вариантами осуществления, гомология белковых последовательностей и последовательностей нуклеиновых кислот оценивают с помощью средства поиска основного локального выравнивания («BLAST»), которое хорошо известно в настоящей области техники (см., например, Karlin and Altschul, 1990, Proc. Natl. Acad. Sci. USA 87:2267-2268; Altschul et al., 1990, J. Mol. Biol. 215:403-410; Altschul et al., 1993, Nature Genetics 3:266-272; Altschul et al., 1997, Nuc. Acids Res. 25:3389-3402).
[00129] В контексте настоящего документа термины «лечить», «осуществление лечения», «лечение» и т.п. относятся к уменьшению или облегчению нарушения/заболевания и/или связанных с ними симптомов. Следует понимать, что лечение нарушения или патологического состояния, хотя и не исключено, не нуждается в полном устранении нарушения, патологического состояния или связанных с ними симптомов. В соответствии с определенными вариантами осуществления, лечение включает предупреждение развития нарушения или патологического состояния и/или связанных с ними симптомов. Термин «предупреждение» или «предупреждать» в контексте настоящего документа относится к любому действию, которое ингибирует или по меньшей мере задерживает развитие нарушения, патологического состояния или связанных с ними симптомов. Предупреждение может включать первичные, вторичные и третичные уровни предупреждения, причем а) первичное предупреждение позволяет избежать развития заболевания; b) мероприятия по вторичному предупреждению направлены на раннее лечение заболевания, тем самым увеличивая возможности вмешательств для предупреждения прогрессирования заболевания и появления симптомов; и с) третичное предупреждение снижает отрицательное влияние уже развившегося заболевания путем восстановления функции и уменьшения связанных с заболеванием осложнений.
[00130] В контексте настоящего документа термин «катаболизм» или «катаболический» относится к химической реакции молекулы с образованием других, например, более мелких молекул. Например, катаболизирующие аргинин ферменты относятся к любому ферменту, способному вступать в реакцию с аргинином, тем самым превращая его в другие молекулы, такие как орнитин, цитруллин и агматин.
[00131] В контексте настоящего документа термин «субъект» относится к любому животному (например, млекопитающему), включая без ограничения людей, отличных от человека приматов, собак, кошек и грызунов.
[00132] В контексте настоящего документа термин «индекс массы тела» или «BMI» означает отношение массы в кг к росту в метрах в квадрате.
[00133] В контексте настоящего документа термин «избыточная масса» относится к BMI от 25 до 30 у взрослых людей. Для людей младше 20 лет «избыточную массу» определяют как BMI между 85-м и 95-м процентилями в сравнении с людьми того же возраста.
[00134] В контексте настоящего документа термин «ожирение» относится к BMI от 30 до 40 у взрослых людей. Для людей младше 20 лет «ожирение» определяют как BMI выше 95-го процентиля в сравнении с людьми того же возраста. В контексте настоящего документа данный термин может включать как ожирение, так и патологическое ожирение.
[00135] В контексте настоящего документа термин «патологическое ожирение» относится к BMI более 40 у взрослых людей.
[00136] В контексте настоящего документа термин «стеатоз» относится к патологическому состоянию, при котором жир накапливается в тканях, таких как ткань печени, ткань почек, ткань поджелудочной железы, ткань сердечной мышцы или другие мышечные ткани.
[00137] В контексте настоящего документа термин «фиброз» относится к патологическому состоянию, при котором имеется избыточное отложение фиброзной соединительной ткани, в том числе коллагена, в органе или ткани, такой как ткань печени, ткань почек, ткань поджелудочной железы или ткань сердца.
[00138] Термин «гиперхолестеринемия» в контексте настоящего документа относится к патологическому состоянию, при котором уровни холестерина в крови повышены по сравнению с нормальным средним уровнем холестерина в крови у соответствующего эталонного субъекта того же этнического происхождения, возраста и пола. В отношении людей данный термин, в частности, относится к уровням общего холестерина в крови выше приблизительно 200, в частности выше приблизительно 240 мг/дл.
[00139] Термины «дислипидемия» и «гиперлипидемия» в контексте настоящего документа относятся к патологическому состоянию, при котором уровень липидов/жиров, в том числе холестерина, сложных эфиров холестерина, фосфолипидов, триглицеридов и/или липопротеинов, в кровотоке соответственно аномален и повышен по сравнению с нормальным средним уровнем липидов/жиров в крови у соответствующего эталонного субъекта того же этнического происхождения, возраста и пола.
[00140] Представленные в настоящем документе слитые белки содержат полипептид аргиназы. Полипептид аргиназы может быть получен из белка аргиназы, экспрессируемого любым организмом, экспрессирующим аргиназу. Примеры аргиназ включают аргиназы, которые вырабатываются бактериями, такими как бациллы, агробактерии, цианобактерии и микобактерии, и млекопитающими, такими как коровы, свиньи, овцы, козы, грызуны и люди. Если полипептид аргиназы получен из человеческой аргиназы, это может быть аргиназа 1-го типа (ARG1) или аргиназа 2-го типа (ARG2).
[00141] Полипептид аргиназы может представлять собой полноразмерный полипептид аргиназы или его функциональный фрагмент и/или вариант.
[00142] Аргиназа представляет собой марганецсодержащий фермент. Как показано в примере 32, если один или несколько ионов марганца, присутствующих в слитых белках, описанных в настоящем документе, заменены одним или несколькими двухвалентными катионными металлами, такими как Со2+ или Ni2+, каталитическая активность слитого белка может увеличиваться. Соответственно, в соответствии с определенными вариантами осуществления, слитые белки, описываемые в настоящем документе, содержат один или несколько двухвалентных металлов, отличных от марганца, таких как Со2+ или Ni2+. В соответствии с определенными вариантами осуществления, слитый белок содержит один или несколько металлов, выбранных из Со2+ и Ni2+. В соответствии с определенными вариантами осуществления, слитый белок содержит два иона Со2+ или два иона Ni2+. В соответствии с другими вариантами осуществления, слитый белок содержит два иона Mn2+.
[00143] В соответствии с определенными вариантами осуществления, полипептид аргиназы представляет собой человеческую ARG1 дикого типа. В соответствии с определенными вариантами осуществления, полипептид аргиназы содержит последовательность по меньшей мере с 95% гомологии последовательности с последовательностью под SEQ ID NO: 69. Например, полипептид аргиназы может содержать полипептидную последовательность по меньшей мере с 96%, 97%, 98%, 99%, 99,1%, 99,4% или 99,7% гомологии с последовательностью под SEQ ID NO: 69. В соответствии с определенными вариантами осуществления, последовательность полипептида аргиназы может отличаться от последовательности под SEQ ID NO: 69 по 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или 30 аминокислотным модификациям (например, вставке, замене, делеции и т.д.). В соответствии с определенными вариантами осуществления, полипептид аргиназы представляет собой полипептид с консервативными аминокислотными заменами, неконсервативными аминокислотными заменами или их комбинацией.
[00144] В соответствии с определенными вариантами осуществления, полипептид аргиназы представляет собой аргиназу Bacillus caldovelox (ВСА). В соответствии с определенными вариантами осуществления, полипептид аргиназы содержит последовательность по меньшей мере с 95% гомологии последовательности с последовательностью под SEQ ID NO: 70. Например, полипептид аргиназы может содержать полипептидную последовательность по меньшей мере с 96%, 97%, 98%, 99%, 99,3% или 99,7% гомологии с последовательностью под SEQ ID NO: 70. В соответствии с определенными вариантами осуществления, последовательность полипептида аргиназы может отличаться от последовательности под SEQ ID NO: 70 по 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или 30 аминокислотным модификациям (например, вставке, замене, делеции и т.д.). В соответствии с определенными вариантами осуществления, полипептид аргиназы представляет собой полипептид с консервативными аминокислотными заменами, неконсервативными аминокислотными заменами или их комбинацией.
[00145] В соответствии с определенными вариантами осуществления, полипептид аргиназы представляет собой ВСА, причем серии 161 заменен на цистеин, как представлено в последовательностях под SEQ ID NO: 71 и SEQ ID NO: 72. Замена серина на цистеин делает возможным сайт-направленное включение химических фрагментов, которые могут дополнительно улучшить свойства слитого белка. Например, боковая цепь цистеина 161 может вступать в реакцию с соответствующим образом активированным PEG-фрагментом, тем самым образуя пегилированную аргиназу, которая может быть включена в получаемый в результате слитый белок. Пегилирование слитого белка обладает способностью дополнительно увеличивать время удержания описываемых в настоящем документе слитых белков путем дополнительной защиты их от различных механизмов разложения, действующих внутри ткани или клетки, что, как следствие, улучшает их терапевтический потенциал. Соответственно, в соответствии с определенными вариантами осуществления, полипептид аргиназы содержит последовательность по меньшей мере с 95% гомологии последовательности с последовательностями под SEQ ID NO: 71 или SEQ ID NO: 72. Например, полипептид аргиназы может содержать полипептидную последовательность по меньшей мере с 96%, 97%, 98%, 99%, 99,3% или 99,7% гомологии с последовательностями под SEQ ID NO: 71 или SEQ ID NO: 72. В соответствии с определенными вариантами осуществления, последовательность полипептида аргиназы может отличаться от последовательности под SEQ ID NO: 71 или последовательности под SEQ ID NO: 72 по 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25 или 30 аминокислотным модификациям (например, вставке, замене, делеции и т.д.). В соответствии с определенными вариантами осуществления, полипептид аргиназы представляет собой полипептид с консервативными аминокислотными заменами, неконсервативными аминокислотными заменами или их комбинацией.
[00146] В случаях, когда описываемые в настоящем документе слитые белки являются пегилированными, PEG-группа может иметь молекулярную массу от приблизительно 5000 до приблизительно 20000 а.е.м., от приблизительно 5000 до приблизительно 15000 а.е.м., от приблизительно 5000 до приблизительно 12000 а.е.м., от приблизительно 7000 до приблизительно 12000 а.е.м. или от приблизительно 7000 до приблизительно 10000 а.е.м. В соответствии с определенными вариантами осуществления, PEG-группа имеет молекулярную массу от приблизительно 4000 до 10000 а.е.м. В соответствии с определенными вариантами осуществления, PEG-группа представляет собой PEG4000 или PEG7000.
[00147] PEG-группа может быть ковалентно непосредственно присоединена к слитому белку или присоединена через линкер. PEG-группа может быть ковалентно присоединена к слитому белку посредством реакции боковой цепи цистеина или лизина, присутствующей на белке, с реагентом для пегилирования. Альтернативно, PEG-группа может быть ковалентно присоединена к N-концевому амину белка.
[00148] В соответствии с определенными вариантами осуществления, слитый белок ковалентно присоединен к PEG через линкер на основе пропионовой кислоты. В соответствии с другими вариантами осуществления, слитый белок ковалентно присоединен к PEG через линкер на основе карбоновой кислоты с С2-С10, С2-С9, С2-С8, С2-С7, С2-С6, С2-С5 или С2-С4 прямой или разветвленной цепью. В соответствии с определенными вариантами осуществления, PEG-группа присоединена к полипептиду аргиназы.
[00149] Описываемые в настоящем документе слитые белки также содержат полипептид альбуминсвязывающего домена (ABD). В ряде исследований был продемонстрирован потенциал связывания альбумина для увеличения периода полужизни терапевтических белков. Тем не менее, разработка схемы слитых белков, включающих полипептиды ABD, может быть сложной задачей, поскольку слияние полипептида ABD с терапевтическим белком может повлиять как на эффективность терапевтического белка, так и на аффинность связывания полипептида ABD, а также растворимость слитого белка. Соответственно, выбор полипептида ABD, его сайта присоединения и конструирование любых необходимых линкеров не является незатейливым процессом и зачастую требует проб и ошибок для получения слитого продукта с требуемыми свойствами. Например, на фиг. 9, 10 и 13 показано, что продолжительность терапевтического действия (и период полужизни) N-ABD094-rhArg (SEQ ID NO: 50) неожиданно намного длиннее, чем у ВНА и ВАН. Хотя все из ВНА, ВАН и N-ABD094-rhArg характеризуются удивительно высокой продолжительностью терапевтического действия (и периодом полужизни), нельзя было спрогнозировать, что N-ABD094-rhArg будет иметь такую длительную продолжительность терапевтического действия (и период полужизни).
[00150] Обычно желательно более длительное действие лекарственного средства. Тип, длина, гибкость линкера и слияние биоактивного пептида или белка с С- или N-концом модуля для продления периода полужизни могут оказывать сильное влияние на активность слитого белка. В настоящем раскрытии различные аргиназы были слиты с молекулами ABD через подходящий линкер так, чтобы можно было сохранить как аргиназную ферментативную активность, так и альбуминсвязывающую способность ABD. Также важны хорошая стабильность и растворимость. Этого очень сложно и непросто достичь. В отличие от обычных продуктов слияния с HSA или Fc, очень мало известно о продуктах слияния с ABD. В настоящем раскрытии представлены примеры разработки схемы линкера, который можно применять для создания функциональных продуктов слияния аргиназа-ABD. Активность модифицированных методами инженерии аргиназных слитых белков (N-ABD094-rhArg, N-ABD-rhArg и т.д.) была оптимизирована в настоящем раскрытии либо посредством линкерной инженерии, либо посредством позиционирования биоактивного белка (аргиназы) относительно модуля продления периода полужизни (ABD), либо и того, и другого.
[00151] Во многих случаях для преодоления утраты активности можно применять белковую инженерию. В одном примере линкерную инженерию применяли для восстановления активности, утраченной при слиянии IFN-2b с HSA [Prot Exp Purif. 2008; 61:73-7]. Прямое слияние IFN-2b с HSA приводило к получению нестабильного белка с очень низкой биологической активностью. В формате слияния тестировали влияние различных линкеров на активность IFN-2b. Как известно, пептидные линкеры влияют на экспрессию, активность и фармакокинетику слитых белков [Adv Drug Deliv Rev. 2013; 65:1357-69]. Двумя основными типами пептидных линкеров являются: (i) гибкие линкеры (например, (G4S)n, где n=1-4); (ii) жесткие линкеры, такие как α-спиральный линкер [А(ЕАААК)nA]х(где n=2-4 и х=1 или 2) и XPn (где X представляет собой А, К или Е). Одним из преимуществ гибких линкеров является то, что гибкость может потребоваться для получения надлежащей ориентации биоактивной части молекулы по отношению к ее когнатному рецептору. Тем не менее, гибкие линкеры не оставляют большого пространства между партнером по слиянию и биоактивным белком. С другой стороны, жесткие линкеры предоставляют больше пространства, но не обладают гибкостью. В случае слитого белка IFN-2b-HSA гибкий линкер давал примерно 39% активности по сравнению с активностью нативного IFN-2b, тогда как жесткий линкер ХР и α-спиральный линкер давали соответственно 68% и 115% активности нативного IFN-2b [Prot Exp Purif. 2008; 61:73-7].
[00152] Определенные линкеры могут отрицательно влиять на свойства слитого белка. Например, гранулоцитарный колониестимулирующий фактор (G-CSF) был слит с трансферрином (Tf). Применение короткого лейцин-глутаматного (LE) линкера давало лишь примерно 10% активности нативного G-CSF. Вставка (G4S)3 или α-спирального [A(EAAAK)nA]m (n=2-4, m=1 или 2) линкера значительно увеличивала активность слитых белков по сравнению с G-CSF-LE-Tf. Слитый белок, сконструированный с линкером (A(EAAAK)4ALEA-(EAAAK)4A), давал биологическую активность, близкую к таковой у нативного G-CSF [Pharm Res. 2006; 23:2116-21]. В соответствии с другим примером, хотя С-конец Fc-фрагмента может быть напрямую связан с N-концом IFN-□-фрагмента через пептидную связь, Gillies et al. [US 7670595 B2] дополнительно соединяют Fc-фрагмент и IFN-□-фрагмент через линкерный пептид. Линкерный пептид расположен между С-концом Fc-фрагмента и N-концом зрелого IFN-□-фрагмента. В этом случае линкерный пептид предпочтительно состоит из остатков серина и глицина, например, с аминокислотной последовательностью G4SG4SG3SG. Все эти результаты демонстрируют важность тестирования линкерной технологии для достижения успеха в программах исследований и разработок слитых белков.
[00153] С помощью другого подхода была продемонстрирована важность позиционирования у продукта слияния для его активности [Curr Pharmaceut Biotechnol. 2014;15:856-63]. Натрийуретический пептид головного мозга (BNP) сливали либо с N, либо с С-концом HSA в различных форматах. Из результатов было видно, что BNP-HSA, BNP2-HSA (две копии BNP) и BNP4-HSA, все из которых слиты с N-концом HSA, не обладали активностью. Тем не менее, HSA-BNP2, слитый с С-концом HSA, обладал такой же активностью, что нативный BNP, и характеризовался длительным действием.
[00154] Такие примеры демонстрируют важность приложения значительных усилий по оптимизации слитых белков для оптимизации активности либо посредством линкерной инженерии, либо посредством позиционирования биоактивного белка или пептида по отношению к модулю для продления периода полужизни, либо и того, и другого. Важно отметить, что в настоящем изобретении был использован новый подход по слиянию ABD для совместного соединения ABD и аргиназы, так чтобы можно было сохранить аргиназную активность. Были успешно созданы стабильные и растворимые слитые молекулы аргиназа-ABD, которые могут связываться с FcRn в зависимости от рН, что обеспечивает эффективный эндосомальный рециклинг. Например, в настоящем раскрытии неожиданно было обнаружено, что в случае rhArg, слитого с ABD, конечный период полужизни белка в кровотоке резко увеличивается с нескольких минут до 4 суток у мышей. В совокупности, для различных болезненных состояний доступны различные подходы для точной настройки фармакокинетических свойств аргиназ, обеспечивающие соответствующее время пребывания в кровотоке. Другой ключевой вопрос при разработке любого терапевтического белка заключается в минимизации иммуногенности во избежании побочных эффектов. Таким образом, важны наблюдаемая низкая иммуногенность человеческой аргиназы и успешная деиммунизация ABD (например, ABD094), которая также была использована в настоящем раскрытии в качестве генного партнера по слиянию для увеличения периода полужизни in vivo.
[00155] Альбуминсвязывающий белок представляет собой трехспиральный белковый домен, встречающийся в различных поверхностных белках, экспрессируемых грамположительными бактериями. Альбуминсвязывающий белок, полученный из стрептококкового белка G, имеет 214 аминокислот и содержит три альбуминсвязывающих домена (ABD1-3), которые используются для связывания с сывороточным альбумином человека и уклонения от надзора иммунной системы хозяина. ABD3 соответствует последовательности из 46 аминокислот, для которой было продемонстрировано, что она связывается с сывороточным альбумином человека и является предметом ряда исследований, а также созревания аффинности сывороточного альбумина человека для разработки полипептидов ABD с различными свойствами, такими как аффинность связывания и селективность связывания. В результате таких исследований было создано значительное количество полипептидов ABD с широко варьирующими свойствами.
[00156] Альбуминсвязывающие белки встречаются и у других бактерий. Например, к природным альбуминсвязывающим белкам относятся определенные поверхностные белки грамположительных бактерий, такие как белки стрептококковые белки М (например, M1/Emm1, М3 Emm3, М12/Emm12, EmmL55/Emm55, Emm49/EmmL49 и белок Н), стрептококковые белки G, MAG и ZAG, а также PPL и РАВ из определенных штаммов Finegoldia magna.
[00157] В соответствии с определенными вариантами осуществления, описываемые в настоящем документе слитые белки содержат полипептид ABD, который получен из альбуминсвязывающего домена стрептококкового белка G. В соответствии с определенными вариантами осуществления, полипептид ABD представляет собой полный альбуминсвязывающий домен 3 стрептококкового белка G или его функциональный фрагмент и/или вариант.
[00158] В соответствии с определенными вариантами осуществления, слитый белок содержит полипептид ABD, содержащий полипептидную последовательность, имеющую по меньшей мере 93% гомологии последовательности с последовательностью под SEQ ID NO: 66. Например, полипептид ABD может содержать полипептидную последовательность по меньшей мере с 94%, 96% или 98% гомологии с последовательностью под SEQ ID NO: 66. В соответствии с определенными вариантами осуществления, последовательность полипептида ABD может отличаться от последовательности под SEQ ID NO: 66 по 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислотным модификациям (например, вставке, замене, делеции и т.д.). В соответствии с определенными вариантами осуществления, полипептид ABD представляет собой полипептид с консервативными аминокислотными заменами, неконсервативными аминокислотными заменами или их комбинацией.
[00159] В соответствии с определенными вариантами осуществления, слитый белок содержит полипептид ABD, содержащий полипептидную последовательность, имеющую по меньшей мере 93% гомологии последовательности с последовательностью под SEQ ID NO: 67. Например, полипептид ABD может содержать полипептидную последовательность по меньшей мере с 94%, 96% или 98% гомологии с последовательностью под SEQ ID NO: 67. В соответствии с определенными вариантами осуществления, последовательность полипептида ABD может отличаться от последовательности под SEQ ID NO: 67 по 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислотным модификациям (например, вставке, замене, делеции и т.д.). В соответствии с определенными вариантами осуществления, полипептид ABD представляет собой полипептид с консервативными аминокислотными заменами, неконсервативными аминокислотными заменами или их комбинацией.
[00160] В соответствии с определенными вариантами осуществления, слитый белок содержит полипептид ABD, содержащий полипептидную последовательность, имеющую по меньшей мере 93% гомологии последовательности с последовательностью под SEQ ID NO: 68. Например, полипептид ABD может содержать полипептидную последовательность по меньшей мере с 93%, 95% или 97% гомологии с последовательностью под SEQ ID NO: 68. В соответствии с определенными вариантами осуществления, последовательность полипептида ABD может отличаться от последовательности под SEQ ID NO: 68 по 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислотным модификациям (например, вставке, замене, делеции и т.д.). В соответствии с определенными вариантами осуществления, полипептид ABD представляет собой полипептид с консервативными аминокислотными заменами, неконсервативными аминокислотными заменами или их комбинацией.
[00161] Относительное положение полипептида ABD и полипептида аргиназы может варьировать. Например, полипептид ABD может предшествовать полипептиду аргиназы (например, полипептид аргиназы может быть присоединен напрямую или опосредовано от С-конца полипептида ABD) или полипептид аргиназы может предшествовать полипептиду ABD (например, полипептид ABD может быть присоединенным напрямую или опосредовано от С-конца полипептида аргиназы).
[00162] В соответствии с определенными вариантами осуществления, слитый белок может включать один или несколько полипептидов аргиназы и/или один или несколько полипептидов ABD. Например, слитый белок может иметь общую структуру ABD-rhArg-ABD, ABD094-rhArg-ABD094, ABD-BCA-ABD, ABD094-BCA-ABD094, rhArg-ABD-rhArg, rhArg-ABD094-rhArg, BCA-ABD-BCA или BCA-ABD094-BCA.
[00163] Полипептид ABD и полипептид аргиназы могут быть присоединены путем прямого ковалентного присоединения или опосредовано через пептидный линкер.
[00164] Пептидный линкер или линкер представляет собой полипептид, обычно с длиной в диапазоне от приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 или более аминокислот, который сконструирован для облегчения функционального соединения полипептида ABD и полипептида аргиназы в связанный слитый белок. Термин функциональная связь обозначает соединение, которое способствует надлежащему сворачиванию полипептидов в трехмерную структуру, которая позволяет связанному слитому белку проявлять некоторые или все функциональные аспекты или биологические активности белка(-ов), из которого(-ых) получены его полипептидные составляющие.
[00165] Полипептидный линкер может быть расположен между N-концом полипептида ABD и С-концом полипептида аргиназы или, альтернативно, расположен между N-концом полипептида аргиназы и С-концом полипептида ABD.
[00166] Пептидный линкер может содержать встречающиеся в природе аминокислоты, не встречающиеся в природе аминокислоты и их комбинации.
[00167] В соответствии с определенными вариантами осуществления, пептидный линкер может содержать глицин, серии, аспарагин или их комбинацию. К иллюстративным пептидным линкерам относятся линкеры, содержащие полиглицин, (GS)n и (GGS)n, где n равно 1-30. К дополнительным иллюстративным пептидным линкерам относятся гибкие линкеры (например, (G4S)n, где n=1-4) или жесткие линкеры (например, альфа-спиральный линкер [А(ЕАААК)nA]x, где n=2-4 и х=1 или 2; и XPn, где X представляет собой либо А, либо К, либо Е, а n=1-10). В соответствии с определенными вариантами осуществления, пептидный линкер представляет собой (A(EAAAK)4ALEA-(EAAAK)4A) (SEQ ID NO: 105), G4SG4SG3SG (SEQ ID NO: 106), GS(N)nGSG, где n=1-10 и GS(Q)nGSG, где n=1-10 и тому подобное. В соответствии с определенными вариантами осуществления, пептидный линкер содержит полипептидную последовательность, имеющую по меньшей мере 90% гомологии последовательности с последовательностью под SEQ ID NO: 73. Например, пептидный линкер может содержать полипептид, имеющий по меньшей мере 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% гомологии последовательности или являющийся идентичным последовательности под SEQIDNO: 73.
[00168] В соответствии с определенными вариантами осуществления, пептидный линкер может содержать глицин, серии, аспарагин или их комбинацию. В соответствии с определенными вариантами осуществления, пептидный линкер содержит полипептидную последовательность, имеющую по меньшей мере 90% гомологии последовательности с последовательностью под SEQ ID NO: 74. Например, пептидный линкер может содержать полипептид, имеющий по меньшей мере 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% гомологии последовательности или являющийся идентичным последовательности под SEQ ID NO: 74.
[00169] Для облегчения очистки слитого белка, например, с помощью аффинной хроматографии, можно применять метки для очистки. Хорошо известной меткой для очистки является гексагистидиновая (6xHis) метка, которая представляет собой последовательность из шести остатков гистидина. Таким образом, в соответствии с определенными вариантами осуществления, слитый белок дополнительно содержит полигистидин, содержащий 4-8 аминокислот гистидина, например, 6xHis-метку. Полигистидин может присутствовать на С-конце слитого белка, на N-конце слитого белка или располагаться между полипептидом ABD и полипептидом аргиназы.
[00170] Если полигистидин расположен между полипептидом ABD и полипептидом аргиназы, он может играть роль пептидного линкера или может быть включен в дополнение к пептидному линкеру. Например, слитый белок SEQ ID NO: 75 включает шестигистидиновый полипептидный линкер в положении 300-305, который служит для связывания полипептида ABD и полипептида аргиназы и может быть успешно использован для очистки слитого белка с помощью аффинной хроматографии.
[00171] После завершения очистки с помощью методик, обычно известных в настоящей области техники, полигистидиновую метку необязательно можно удалить. Например, для удаления N-концевых полигистидиновых меток можно применять экзопептидазы (например, Qiagen TAGZyme), а С-концевым полигистидиновым меткам может предшествовать подходящая аминокислотная последовательность, которая облегчает удаление полигистидиновых меток с помощью эндопептидаз. Таким образом, объемом настоящего изобретения охватываются слитые белки, за исключением N-концевой и/или С-концевой полигистидиновой метки.
[00172] В соответствии с определенными вариантами осуществления, полипептид ABD содержит полипептидную последовательность, имеющую по меньшей мере 93% гомологии последовательности с последовательностью под SEQ ID NO: 66, SEQ ID NO: 67 или SEQ ID NO: 68, а полипептид аргиназы содержит полипептидную последовательность, имеющую по меньшей мере 95% гомологии с последовательностью под SEQ ID NO: 69, SEQ ID NO: 70, SEQ ID NO: 71 или SEQ ID NO: 72. Например, полипептид ABD содержит полипептидную последовательность, имеющую по меньшей мере 94%, 95%, 96%, 97%, 98% или 99% гомологии последовательности или являющуюся идентичной последовательности под SEQ ID NO: 66, SEQ ID NO: 67 или SEQ ID NO: 68, a полипептид аргиназы содержит полипептидную последовательность, имеющую по меньшей мере 96%, 97%, 98% или 99% гомологии последовательности или являющуюся идентичной последовательности под SEQ ID NO: 69, SEQ ID NO: 70, SEQ ID NO: 71 или SEQ ID NO: 72.
[00173] В соответствии с определенными вариантами осуществления, полипептид ABD содержит полипептидную последовательность, имеющую по меньшей мере 93%, 94%, 95%, 96%, 97%, 98% или 99% гомологии последовательности или являющуюся идентичной последовательности под SEQ ID NO: 66, а полипептид аргиназы содержит полипептидную последовательность, имеющую по меньшей мере 95%, 96%, 97%, 98% или 99% гомологии последовательности или являющуюся идентичной последовательности под SEQ ID NO: 69.
[00174] В соответствии с определенными вариантами осуществления, слитый белок дополнительно содержит пептидный линкер, содержащий полипептидную последовательность, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% гомологии последовательности или являющуюся идентичной последовательности под SEQ ID NO: 73 или последовательности под SEQ ID NO: 74.
[00175] К иллюстративным слитым белкам относятся слитые белки, имеющие по меньшей мере 95%, 96%, 97%, 98% или 99% гомологии или являющиеся идентичными последовательности под SEQ ID NO: 49, SEQ ID NO: 50, SEQ ID NO: 51, SEQ ID NO: 52, SEQ ID NO: 53, SEQ ID NO: 54, SEQ ID NO: 55, SEQ ID NO: 56, SEQ ID NO: 75 и SEQ ID NO: 76.
[00176] Продолжительность терапевтического действия слитого белка на концентрацию аргинина в плазме зависит от количества введенного гибридного белка и может составлять приблизительно 1, приблизительно 2, приблизительно 3, приблизительно 4, приблизительно 5, приблизительно 6, приблизительно 7, приблизительно 8, приблизительно 9, приблизительно 10, приблизительно 11, приблизительно 12, приблизительно 13, приблизительно 14 или приблизительно 15 суток или более. В соответствии с определенными вариантами осуществления, продолжительность терапевтического действия слитого белка составляет от приблизительно 5 суток до приблизительно 20 суток, от приблизительно 5 суток до приблизительно 19 суток, от приблизительно 5 суток до приблизительно 18 суток, от приблизительно 5 суток до приблизительно 17 суток, от приблизительно 5 суток до приблизительно 16 суток, от приблизительно 5 суток до приблизительно 15 суток, от приблизительно 6 суток до приблизительно 15 суток, от приблизительно 7 суток до приблизительно 15 суток, от приблизительно 7 суток до приблизительно 14 суток, от приблизительно 7 суток до приблизительно 13 суток, от приблизительно 7 суток до приблизительно 12 суток, от приблизительно 7 суток до приблизительно 11 суток или от приблизительно 8 суток до приблизительно 11 суток.
[00177] В соответствии с определенными вариантами осуществления, период полужизни слитого белка составляет от приблизительно 1 суток до приблизительно 10 суток, от приблизительно 1 суток до приблизительно 9 суток, от приблизительно 1 суток до приблизительно 8 суток, от приблизительно 1 суток до приблизительно 7 суток, от приблизительно 1 суток до приблизительно 6 суток, от приблизительно 1 суток до приблизительно 5 суток, от приблизительно 1 суток до приблизительно 4 суток, от приблизительно 1 суток до приблизительно 3 суток или от приблизительно 1 суток до приблизительно 2 суток. В соответствии с другими вариантами осуществления, период полужизни слитого белка составляет от приблизительно 6 часов до приблизительно 30 часов.
[00178] Аргиназная активность описываемых в настоящем документе слитых белков может быть практически такой же, более низкой или более высокой, чем активность полипептида аргиназы, из которого он получен. За одну единицу аргиназной активности принимают количество слитого белка [например, ВНА (SEQ ID NO: 75), ВАН (SEQ ID NO: 76), N-ABD-rhArg (SEQ ID NO: 49) или N-ABD094-rhArg (SEQ ID NO: 50)] или аргиназы [например, ВСА (SEQ ID NO: 70)], которое катализирует выработку 1 мкмоль мочевины в минуту в стандартных условиях анализа. Специфическую активность фермента выражают в единицах активности на мг белка. В стандартных условиях анализа с применением диацетилмоноксима (DAMO) (37°С, рН 7,4) слитые белки могут обладать специфической активностью, которая приблизительно на 5%, приблизительно на 10%, приблизительно на 15%, приблизительно на 20%, приблизительно на 25%, приблизительно на 30%, приблизительно на 35% или приблизительно на 40% ниже или выше, чем у соответствующего полипептида аргиназы, который ею обладает. В соответствии с определенными вариантами осуществления, слитые белки могут обладать специфической активностью, которая ниже или выше на величину, которая варьирует от приблизительно 5% до приблизительно 40%, от приблизительно 10% до приблизительно 40%, от приблизительно 10% до приблизительно 35%, от приблизительно 10% до приблизительно 30%, от приблизительно 20% до приблизительно 30%, от приблизительно 20% до приблизительно 35%, от приблизительно 15% до приблизительно 30%, от приблизительно 15% до приблизительно 25% или от приблизительно 10% до приблизительно 20%, чем у соответствующего полипептида аргиназы, который ею обладает.
[00179] В соответствии с определенными вариантами осуществления, на аргиназную активность слитых белков практически не влияет присутствие HSA. Это является преимуществом, поскольку связывание слитых белков ABD с HSA может оказывать отрицательное влияние на активность слитого белка.
[00180] Также представлены полинуклеотидные последовательности, кодирующие описываемые в настоящем документе слитые белки, в виде выделенных полинуклеотидов, или в виде частей векторов экспрессии, или в виде частей линейных последовательностей ДНК, включая линейные последовательности ДНК, применяемые для транскрипции/трансляции in vitro, векторы, совместимые с экспрессией, секрецией и/или дисплеем конструкций у прокариот или эукариот. В настоящем документе раскрыты определенные иллюстративные полинуклеотиды, однако в объем настоящего раскрытия также входят и другие полинуклеотиды, которые, с учетом вырожденности генетического кода или предпочтений по использованию кодонов в данной системе экспрессии, кодируют описываемые в настоящем документе слитые белки.
[00181] Описываемые в настоящем документе полинуклеотиды можно получить путем химического синтеза, такого как твердофазный синтез полинуклеотидов на автоматическом синтезаторе полинуклеотидов, и собрать в полные одно- или двухцепочечные молекулы. Альтернативно, полинуклеотиды по настоящему изобретению можно получить с помощью других методик, таких как ПЦР с последующим стандартным клонированием. Способы производства или получения полинуклеотидов с заданной известной последовательностью хорошо известны в настоящей области техники.
[00182] Описываемые в настоящем документе полинуклеотиды могут содержать по меньшей мере одну некодирующую последовательность, такую как промоторную или энхансерную последовательность, интрон, сигнал полиаденилирования и тому подобное. Полинуклеотидные последовательности также могут содержать дополнительные последовательности, кодирующие дополнительные аминокислоты, которые кодируют, например, маркерную последовательность или последовательность метки, такую как полигистидиновую (6XHis) метку или метку НА, для облегчения очистки или детекции белка, сигнальную последовательность, партнер для получения слитого белка, например, кДНК, кодирующую биоактивное средство, и др.
[00183] В соответствии с другим вариантом осуществления, в настоящем документе представлен вектор, содержащий по меньшей мере один из описываемых в настоящем документе полинуклеотидов. Такие векторы могут быть плазмидными векторами, вирусными векторами, векторами для экспрессии бакуловируса, векторами на основе транспозонов или любым другим вектором, подходящим для введения каким-либо способом полинуклеотидов по настоящему изобретению в заданный организм или генетический фон. Такие векторы могут быть векторами экспрессии, содержащими элементы последовательности нуклеиновой кислоты, которые могут контролировать, регулировать, запускать или делать возможной экспрессию полипептида, кодируемого таким вектором. К таким элементам можно отнести сайты связывания энхансеров транскрипции, сайты инициации РНК-полимеразы, сайты связывания рибосом и другие сайты, которые способствуют экспрессии кодируемых полипептидов в заданной системе экспрессии. Такие системы экспрессии могут быть клеточными или бесклеточными системами, хорошо известными в настоящей области техники.
[00184] Во многих бактериальных системах экспрессии стартовый кодон обычно кодирует метионин, что, как следствие, в этих системах экспрессии приводит к получению белков, начинающихся с N-концевого метионина. Однако хорошо известно, что определенные бактериальные ферменты, такие как метионинаминопептидаза (MetAP) и др., могут катализировать гидролитическое отщепление N-концевого метионина у свежесинтезированных полипептидов. Это обычно наблюдается в случаях, когда следующей аминокислотой является, например, Gly, Ala, Ser или Thr [In vivo processing of N-terminal methionine in E.coli, FEBS Lett. 1990 Jun 18; 266(1-2): 1-3]. Соответственно, в соответствии с определенными вариантами осуществления, к описываемым в настоящем документе слитым белкам относятся варианты, у которых отсутствует N-концевой метионин белка.
[00185] Описываемые в настоящем документе слитые белки можно выделить с помощью процедур разделения, хорошо известных в настоящей области техники для захвата, иммобилизации, разделения или осаждения, и очистить до степени, необходимой для коммерческого применения.
[00186] Для терапевтического применения описываемые в настоящем документе слитые белки можно приготовить в виде фармацевтических композиций, содержащих терапевтически эффективное количество описанного в настоящем документе слитого белка в качестве активного ингредиента в фармацевтически приемлемом носителе. Термин «носитель» относится к разбавителю, адъюванту, вспомогательному веществу или наполнителю, с которым вводят активное соединение. Такие наполнители могут быть жидкостями, такими как вода и масла, включая масла нефтяного, животного, растительного или синтетического происхождения, такие как арахисовое масло, соевое масло, минеральное масло, кунжутное масло и др. Например, можно применять 0,9% солевой раствор и 0,3% глицин. Эти растворы являются стерильными и обычно не содержат твердых частиц. Простерилизовать их можно обычными, хорошо известными методиками стерилизации (например, фильтрацией). Композиции могут содержать фармацевтически приемлемые вспомогательные материалы, необходимые для приближения к физиологическим условиям, такие как регуляторы рН и буферные средства, стабилизирующие, загущающие, смазывающие и окрашивающие средства и т.д. Концентрация слитого белка в таком фармацевтическом составе может широко варьировать, например, от менее приблизительно 0,5%, обычно на уровне или по меньшей мере приблизительно от 1% вплоть до 15 или 20% по массе, и ее будут выбирать, в первую очередь, исходя из необходимой дозы, объемов жидкости, вязкости и т.д., в зависимости от конкретного выбранного способа введения. Подходящие наполнители и составы описаны, например, в публикации Remington: The Science and Practice of Pharmacy, 21st Edition, Troy, D.B. ed., Lipincott Williams and Wilkins, Philadelphia, Pa. 2006, Part 5, Pharmaceutical Manufacturing pp. 691-1092, см., в частности, страницы 958-989.
[00187] Способ введения для терапевтического применения описываемого в настоящем документе слитого белка можно реализовать любым подходящим путем, который позволяет доставить средство реципиенту, таким как парентеральное введение, например внутрикожное, внутримышечное, внутрибрюшинное, внутривенное или подкожное, легочное; трансмукозальное (пероральное, интраназальное, интравагинальное, ректальное); с помощью состава в форме таблетки, капсулы, раствора, суспензии, порошка, геля, частицы; и содержащегося в шприце, имплантируемом устройстве, осмотической помпе, картридже, микропомпе; или других признанных специалистом средств, которые хорошо известны в настоящей области техники. Введение в конкретное место можно осуществлять, например, внутрисуставным, внутрибронхиальным, внутрибрюшным, внутрикапсулярным, внутрихрящевым, внутриполостным, внутрицелиальным, внутриспиномозговым, внутрицеребровентрикулярным, внутрикишечным, внутрицервикальным, внутрижелудочным, внутрипеченочным, внутрисердечным, внутрикостным, внутритазовым, внутриперикардиальным, внутрибрюшинным, внутриплевральным, внутрипростатическим, внутрилегочным, внутриректальным, внутрипочечным, внутриретинальным, внутриспинальным, внутрисиновиальным, внутригрудным, внутриматочным, внутрисосудистым, внутрипузырным, внутриочаговым, вагинальным, ректальным, трансбуккальным, сублингвальным, интраназальным или трансдермальным введением.
[00188] Описываемые в настоящем документе слитые белки можно применять в качестве истощающих запасы аргинина средств. Такие средства полезны при лечении любого заболевания или патологического состояния, при котором истощение запасов аргинина у субъекта приводит к терапевтическому эффекту, например, при лечении рака, определенных вирусов, рассеянного склероза, ревматоидного артрита, аутоиммунных заболеваний, врожденной гипераргининемии, болезни «трансплантат против хозяина» (GvHD) и, как подробно описано в настоящем документе, ожирения, метаболических нарушений и связанных с ними осложнений, а также сопутствующих заболеваний.
[00189] Ауксотрофные по аргинину опухоли зависят от внеклеточного аргинина из-за подавления ASS или ОТС и зависят от внеклеточных источников аргинина для выживания. Было показано, что средства, истощающие запасы аргинина, являются цитотоксическими по отношению к ауксотрофным по аргинину опухолям, и их применение при лечении рака в настоящее время исследуют в клинических испытаниях.
[00190] Таким образом, описываемые в настоящем документе слитые белки можно применять для лечения рака, такого как рак поджелудочной железы, лейкоз, меланома, рак головы и шеи, колоректальный рак, рак легких, рак молочной железы, рак предстательной железы, рак шейки матки, рак печени, рак носоглотки, рак пищевода, саркома, рак желудка, мезотелиома, лимфома, рак мочевого пузыря, рак костей, рак эндометрия, рак яичников, рак почек, рак глаза, нейробластома, глиобластома, злокачественная опухоль оболочки периферических нервов (MPNST) и рак головного мозга. Рак может представлять собой ауксотрофный по аргинину рак, например рак, характеризующийся подавлением активности по меньшей мере одного из ASS и ОТС.
[00191] Описываемые в настоящем документе слитые белки также можно применять для лечения вирусных инфекций, таких как вирус иммунодефицита человека (ВИЧ), вирус гепатита В (HBV), вирус гепатита С (HCV), вирус простого герпеса (HSV), вирус папилломы человека (HPV), вирус Эпштейна - Барра (EBV), вирус герпеса, вирус ветряной оспы и грипп.
[00192] Как подробно описано в настоящем документе, представленные в настоящем документе слитые белки также можно применять при лечении по меньшей мере одного патологического состояния, выбранного из группы, состоящей из ожирения, метаболического нарушения и связанного с ним осложнения и/или сопутствующего заболевания.
[00193] Как продемонстрировано в представленных ниже экспериментах, истощение запасов аргинина может значительно уменьшать ожирение. В настоящем документе авторами настоящего изобретения был исследован терапевтический потенциал депривации аргинина с помощью слитых белков из аргиназы и ABD или пегилированной аргиназы, такой как N-ABD094-rhArg (SEQ ID NO: 50), пегилированной His-rhArg (SEQ ID NO: 101) и N-ABD094-rhArg-Co2+ [SEQ ID NO: 50 (замещенная кобальтом)] при лечении ожирения и связанных с ним метаболических нарушений. Из представленных в настоящем документе данных видно, что истощение запасов аргинина с помощью, например, N-ABD094-rhArg (SEQ ID NO: 50), может значимо уменьшать жировую массу, подавлять липогенез и повышать чувствительность к инсулину у нормальных худых мышей и страдающих ожирением мышей. Было обнаружено, что депривация аргинина, например, с помощью N-ABD094-rhArg (SEQ ID NO: 50), может предупреждать развитие и уменьшать ожирение, метаболические нарушения и связанные с ними осложнения и сопутствующие заболевания, в том числе инсулинорезистентность.
[00194] Представленные в настоящем документе эффекты истощения запасов аргинина являются удивительными и неожиданными, поскольку в многочисленных исследованиях было показано, что увеличение потребления с пищей некоторых аминокислот (АА), в том числе аргинина, оказывает значительное влияние на метаболизм глюкозы и липидов, а добавки АА можно применять для снижения массы тела. Более того, поскольку аргинин является полузаменимой аминокислотой, его истощение не приводит к пагубным последствиям для субъекта.
[00195] Использовали мышиную модель ожирения, вызванного рационом с высоким содержанием жиров (DIO). Худых мышей, начинающих питаться рационом с высоким содержанием жиров, одновременно обрабатывали посредством N-ABD094-rhArg (SEQ ID NO: 50), чтобы протестировать его эффективность в предупреждении развития DIO и связанных с ним метаболических нарушений. Мышей с ранее существовавшим DIO обрабатывали посредством N-ABD094-rhArg (SEQ ID NO: 50), чтобы проверить его эффективность в уменьшении ожирения, метаболических нарушений и связанных с ними осложнений, а также сопутствующих заболеваний.
[00196] Как продемонстрировано в представленных ниже примерах, обработка посредством N-ABD094-rhArg (SEQ ID NO: 50) приводила к снижению массы тела и жировой массы, улучшению толерантности к глюкозе и чувствительности к инсулину, нормализации эндокринных и метаболических профилей и облегчению воспаления, стеатоза и фиброза, а также к предупреждению развития или обращению побеления бурого жира.
[00197] В настоящем документе предложен способ лечения ожирения и/или метаболического нарушения у нуждающегося в том субъекта, предусматривающий стадию введения субъекту терапевтически эффективного количества истощающего запасы аргинина средства.
[00198] Истощающее запасы аргинина средство может быть любым известным из уровня техники истощающим запасы аргинина средством, которое способно снижать уровни аргинина в плазме и/или клетках у субъекта. Истощающее запасы аргинина средство может быть низкомолекулярным соединением или белком.
[00199] Белок может быть слитым белком и/или химически модифицированным белком, таким как пегилированный белок. К иллюстративным белкам относятся белки, которые способны катализировать катаболизм аргинина в другие продукты, такие как белки, обладающие аргиназной, аргининдезиминазной, аргининдекарбоксилазной или ринин-2-монооксигеназной активностью.
[00200] Аргиназа может быть любой известной из уровня техники аргиназой, такой как аргиназа, продуцируемая бактериями, грибами, рыбой, человеком, коровами, свиньей, кроликом, грызуном, приматом, овцой и козой. Например, аргиназа Bacillus caldovelox, аргиназа Thermus thermophilus, аргиназа I Capra hircus, аргиназа I Heterocephalus glaber, аргиназа I Bos taurus, аргиназа I Sus scrofa, аргиназа I Plecoglossus altivelis, аргиназа I Salmo salar, аргиназа I Oncorhynchus mykiss, аргиназа I Osmerus mordax, аргиназа I Hyriopsis cumingii, аргиназа I Rattus norvegicus, аргиназа I Mus musculus, (человеческая) аргиназа I Homo sapiens, аргиназа I Pan troglodytes, аргиназа I Oryctolagus cuniculus, аргиназа II Rattus norvegicus, аргиназа II Mus musculus, (человеческая) аргиназа II Homo sapiens, аргиназа II Bostaurus, аргиназа II Heterocephalus glaber, аргиназа II Pan troglodytes, аргиназа II Oryctolagus cuniculus, аргиназа Delftia, аргиназа Bacillus coagulans, аргиназа Hoeflea phototrophica и аргиназа Roseiflexus castenholzii. Другие примеры включают аргиназы от Bacillus methanolicus, Bacillus sp.NRRL B-14911, Planococcus donghaensis, Paenibacillus dendritiformis, Desmospora sp., Methylobacter tundripaludum, Stenotrophomonas sp., Microbacterium laevaniformans, Porphyromonas uenonis, Agrobacterium sp., Octadecabacter arcticus, Agrobacterium tumefaciens, Anoxybacillus flavithermus, Bacillus pumilus, Geobacillus thermoglucosidasius, Geobacillus thermoglucosidans, Brevibacillus laterosporus, Desulfotomaculum ruminis, Geobacillus kaustophilus, Geobacillus thermoleovorans, Geobacillus thermodenitrificans, Staphylococcus aureus, Halophilic archaeon DL31, Halopigerxanaduensis, Natrialba magadii, Plasmodium falciparum, Helicobacter pylori и тому подобных.
[00201] Аргининдезиминаза может быть любой известной из уровня техники аргининдезиминазой, например, получаемой из микроорганизмов Mycoplasma, Lactococcus, Pseudomonas, Steptococcus, Escherichia, Mycobacterium или Bacillus. К иллюстративной аргининдезиминазе относятся без ограничения аргининдезиминазы, которые продуцируются у Mycoplasma hominis, Mycoplasma arginini, Mycoplasma arthritidis, Clostridium perfringens, Bacillus licheniformis, Borrelia burgdorferi, Borrelia afzellii, Enterococcus faecalis, Lactococcus lactis, Bacillus cereus, Streptococcus pyogenes, Steptococcus pneumoniae, Lactobacillus sake, Giardia intestinalis, Mycobacterium tuberculosis, Pseudomonas plecoglossicida, Pseudomonas putida, Pseudomonas aeruginosa и тому подобных.
[00202] Аргининдекарбоксилаза может быть любой известной из уровня техники аргининдекарбоксилазой, например, продуцируемой у Escherichia coll., Salmonella typhimurium, Chlamydophila pneumoniae, Methanocaldococcus jannaschii, Paramecium bursaria Chlorella virus 1, Vibrio vulnificus YJ016, Campylobacter jejuni subsp., Trypanosoma cruzi, Sulfolobus solfataricus, Bacillus licheniformis, Bacillus cereus, Carica papaya, Nicotianatobacum, Glycine max, Lotus coniculata, Vibrio vulnificus, Vibrio cholerae, Mus musculus, Thermotoga, Rattus norvegicus, Homo sapiens, Bos taurus, Susscrofa, Thermus thermophiles, Thermus parvatiensis, Thermus aquaticus, Thermus thermophilus, Thermus islandicus, Arabidopsis thaliana, Avena sativa и тому подобных.
[00203] Аргинин-2-монооксигеназа может представлять собой любую известную из уровня техники аргинин-2-монооксигеназу, например, получаемую от Arthrobacter globiformis IFO 12137, Arthrobacter simplex IFO 12069, Brevibacterium helvolum IFO 12073, Helicobacter cinaedi CCUG 18818, Streptomyces griseus и тому подобных.
[00204] Аргининдекарбоксилаза, аргининдезиминаза, аргинин-2-монооксигеназа и аргиназа могут быть полноценным белком или его функциональным фрагментом и/или вариантом. Аргининдекарбоксилаза, аргининдезиминаза, аргинин-2-монооксигеназа и аргиназа можно модифицировать для улучшения их фармакокинетических свойств, например, путем слияния белка или его функционального фрагмента и/или варианта с человеческим сывороточным альбумином, альбуминсвязывающим доменом, Fc-участком иммуноглобулина, PEG-группой или их комбинацией.
[00205] Катаболизирующие аргинин ферменты, описанные в настоящем документе, можно модифицировать методами инженерии таким образом, чтобы они включали определенные сайты фермента, к которым может быть избирательно присоединен PEG. Выбранные сайты пегилирования предпочтительно расположены на участке, удаленном от активного центра фермента, и, как правило, доступны для взаимодействия с растворителем, что делает возможной реакцию с реагентами для пегилирования.
[00206] Например, можно получить Cys45-аргиназу I человека (HAI) и Cys161-аргиназу Bacillus caldovelox (ВСА), которые вступают в реакцию с тиол-специфичными PEG-молекулами. Конъюгация между одиночным остатком свободного цистеина модифицированной аргиназы и малеимидной группой (MAL), присоединенной к PEG-соединению, может приводить к образованию ковалентной связи между PEG-соединением и свободным цистеином модифицированной аргиназы. К последовательностям под SEQ ID NO: 102 и 104 относится мутант (C168S/C303S), сконструированный для сайт-направленного пегилирования Cys45, и поэтому необязательно они могут быть пегилированными. К последовательности под SEQ ID NO: 89 также относится мутант (S161C), сконструированный для сайт-направленного пегилирования Cys161, и поэтому необязательно она может быть пегилированнлй.
[00207] В соответствии с определенными вариантами осуществления, аргиназа может содержать последовательность под SEQ ID NO: 101, SEQ ID NO: 102, SEQ ID NO: 103 или SEQ ID NO: 104, причем SEQ ID NO: 102 и SEQ ID NO: 104 необязательно содержат полиэтиленгликолевую группу (PEG).
[00208] Для ковалентного присоединения PEG к описанным в настоящем документе катаболизирующим аргинин ферментам можно применять любой известный из уровня техники реагент для пэгилирования. К иллюстративным реагентам для пэгилирования относятся без ограничения mPEG-ALD (метоксиполиэтиленгликоль-пропиональдегид), mPEG-MAL (метоксиполиэтиленгликольмалеимид), mPEG-NHS (метоксиполиэтиленгликоль-N-гидроксисукцинимид), mPEG-SPA (метоксиполиэтиленгликоль-сукцинимидилпропионат) и mPEG-CN (метоксиполиэтиленгликоль-цианурхлорид).
[00209] PEG-группа может иметь молекулярную массу от приблизительно 5000 до приблизительно 20000 а.е.м., от приблизительно 5000 до приблизительно 15000 а.е.м., от приблизительно 5000 до приблизительно 12000 а.е.м., от приблизительно 7000 до приблизительно 12000 а.е.м. или от приблизительно 7000 до приблизительно 10000 а.е.м. В соответствии с определенными вариантами осуществления, PEG-группа имеет молекулярную массу от приблизительно 2000 до 10000 а.е.м. В соответствии с определенными вариантами осуществления, PEG-группа представляет собой PEG4000, PEG5000, PEG6000 или PEG7000.
[00210] PEG-группа может быть ковалентно присоединена к аргиназе непосредственно или через линкер. В соответствии с определенными вариантами осуществления, аргиназа ковалентно присоединена к PEG через линкер на основе пропионовой кислоты. В соответствии с другими вариантами осуществления, аргиназа ковалентно присоединена к PEG через линкер на основе карбоновой кислоты с С2-С10, С2-С9, С2-С8, С2-С7, С2-С6, С2-С5 или С2-С4 прямой или разветвленной цепью.
[00211] В соответствии с альтернативными вариантами осуществления, способ лечения по меньшей мере одного патологического состояния, выбранного из группы, состоящей из ожирения, метаболического нарушения и связанного с ним осложнения, а также/или сопутствующего заболевания, у нуждающегося в том субъекта предусматривает назначение субъекту рациона с низким содержанием аргинина или практически без аргинина.
[00212] Способ лечения по меньшей мере одного патологического состояния, выбранного из группы, состоящей из ожирения, метаболического нарушения и связанного с ним осложнения и/или сопутствующего заболевания, также может предусматривать предупреждение развития по меньшей мере одного патологического состояния, выбранного из группы, состоящей из ожирения, метаболического нарушения и связанного с ним осложнения, а также/или сопутствующего заболевания.
[00213] К метаболическому нарушению можно отнести ожирение, гиперхолестеринемию, дислипидемию, стеатоз, инсулинорезистентность, непереносимость глюкозы, гипергликемию и сахарный диабет или их комбинации, а также связанные с ними осложнения и/или сопутствующие заболевания, которые могут включать диабетическую ретинопатию, диабетическую нефропатию, диабетическую васкулопатию, диабетическую невропатию, стеатогепатит, фиброз, цирроз, воспаление, гипертонию, сердечно-сосудистое заболевание и побеление бурого жира или их комбинацию.
[00214] В соответствии с определенными вариантами осуществления, лечение метаболического нарушения представляет собой лечение и/или предупреждение развития инсулинорезистентности, например, у субъектов с сахарным диабетом I типа или особенно II типа. Как продемонстрировано в приведенных ниже примерах, истощение запасов аргинина приводит к увеличению чувствительности к инсулину у субъекта.
[00215] Способ лечения ожирения может предусматривать уменьшение жировой массы или предупреждение увеличения жировой массы у субъекта. Лечение субъекта средством, истощающим запасы аргиниа, может привести к снижению уровней экспрессии мРНК нескольких адипогенных факторов транскрипции и липогенных ферментов в висцеральной белой жировой ткани (WAT), печени и скелетных мышцах, таких как Pparg, Srebplc, Асе и Scdl, по сравнению с субъектами без лечения на рационе с высоким содержанием жиров. В соответствии с определенными вариантами осуществления, уровень экспрессии при снижении уровней экспрессии мРНК регуляторов липогенеза снижается, например, по меньшей мере приблизительно на 10%, приблизительно на 20%, приблизительно на 30%, приблизительно на 40%, приблизительно на 50%, приблизительно на 60%, приблизительно на 70%, приблизительно на 80% или приблизительно на 90% от соответствующих уровней экспрессии мРНК у субъекта без лечения на рационе с высоким содержанием жиров. В соответствии с определенными вариантами осуществления, лечение субъекта средством, истощающим запасы аргинина, приводит к снижению экспрессии мРНК одного или нескольких генов, выбранных из группы, состоящей из Pparg, Srebplc, Асе и Scdl, в WAT, печени и/или скелетных мышцах субъекта.
[00216] Ожирение может быть результатом и/или может быть связано с метаболическим нарушением (например, гипергликемией, гиперинсулинемией) и/или другими факторами, такими как переедание, недостаток физической нагрузки и т.д.
[00217] Метаболическое нарушение может также включать лечение и/или предупреждение развития у субъекта стеатоза, такого как стеатоз сердца, стеатоз печени, стеатоз почек, стеатоз поджелудочной железы и стеатоз мышц.
[00218] Метаболическое нарушение может также включать лечение и/или предупреждение развития у субъекта фиброза, такого как фиброз печени, фиброз почек, фиброз поджелудочной железы и фиброз сердца.
[00219] Метаболическое нарушение может также включать лечение гиперхолестеринемии, дислипидемии, например, путем снижения уровней общего холестерина в плазме крови, холестерина LDL, аполипопротеина В и/или триглицеридов.
[00220] Лечение субъекта средством, истощающим запасы аргинина, может снизить концентрацию лептина в плазме крови, например, по меньшей мере приблизительно на 10%, приблизительно на 20%, приблизительно на 30%, приблизительно на 40%, приблизительно на 50%, приблизительно на 60% или приблизительно на 70% от соответствующей концентрации лептина в плазме крови у субъекта без лечения на HFD. В соответствии с определенными вариантами осуществления, концентрация лептина в плазме крови можно снизить на величину, составляющую от приблизительно 10% до приблизительно 70%, от приблизительно 10% до приблизительно 60%, от приблизительно 10% до приблизительно 50%, от приблизительно 10% до приблизительно 40%, от приблизительно 10% до приблизительно 30%, от соответствующей концентрации лептина в плазме крови у субъекта без лечения на HFD.
[00221] Лечение субъекта средством, истощающим запасы аргинина, может снизить концентрацию триглицеридов в плазме крови, например, по меньшей мере приблизительно на 20%, приблизительно на 30% или приблизительно на 40% от соответствующей концентрации триглицеридов в плазме крови у субъекта без лечения на HFD. В соответствии с определенными вариантами осуществления, концентрацию триглицеридов в плазме крови можно снизить на величину, составляющую от приблизительно 5% до приблизительно 40%, от приблизительно 5% до приблизительно 30% или от приблизительно 5% до приблизительно 20% от соответствующей концентрации триглицеридов в плазме крови у субъекта без лечения на HFD.
[00222] Лечение субъекта средством, истощающим запасы аргинина, может снизить концентрацию общего холестерина в плазме крови, например, по меньшей мере приблизительно на 5%, приблизительно на 10% или приблизительно на 15% от соответствующей концентрации общего холестерина в плазме крови у субъекта без лечения на HFD. В соответствии с определенными вариантами осуществления, концентрацию общего холестерина в плазме крови можно снизить на величину, составляющую от приблизительно 5% до приблизительно 15%, от приблизительно 5% до приблизительно 10% или от приблизительно 10% до приблизительно 15% от соответствующей концентрации общего холестерина в плазме крови у субъекта без лечения на HFD.
[00223] Лечение субъекта средством, истощающим запасы аргинина, может снизить концентрацию свободных жирных кислот в плазме крови, например, по меньшей мере приблизительно на 5%, приблизительно на 10% или приблизительно на 20% от соответствующей концентрации свободных жирных кислот в плазме крови у субъекта без лечения на HFD. В соответствии с определенными вариантами осуществления, концентрацию свободных жирных кислот в плазме крови можно снизить на величину, составляющую от приблизительно 5% до приблизительно 20%, от приблизительно 5% до приблизительно 15% или от приблизительно 5% до приблизительно 10% от соответствующей концентрации свободных жирных кислот в плазме крови у субъекта без лечения на HFD.
[00224] Концентрация аргинина в плазме крови у субъекта, необходимая для наблюдения терапевтического результата, может варьировать в зависимости от ряда факторов, в том числе общего состояния субъекта и типа и тяжести заболевания и/или медицинского состояния и/или состава рациона. Выбор целевых уровней аргинина в плазме крови будет хорошо известен рядовому специалисту в настоящей области техники. В соответствии с определенными вариантами осуществления, концентрация аргинина в плазме крови ниже приблизительно 100 мкМ, приблизительно 90 мкМ, приблизительно 80 мкМ, приблизительно 70 мкМ, приблизительно 60 мкМ, приблизительно 50 мкМ, приблизительно 40 мкМ, приблизительно 30 мкМ, приблизительно 20 мкМ, приблизительно 10 мкМ или приблизительно 5 мкМ. В соответствии с определенными вариантами осуществления, концентрация аргинина в плазме крови составляет от приблизительно 0,1 мкМ до приблизительно 100 мкМ, от приблизительно 0,1 мкМ до приблизительно 90 мкМ, от приблизительно 0,1 мкМ до приблизительно 80 мкМ, от приблизительно 0,1 мкМ до приблизительно 70 мкМ, от приблизительно 0,1 мкМ до приблизительно 60 мкМ, от приблизительно 0,1 мкМ до приблизительно 50 мкМ, от приблизительно 0,1 мкМ до приблизительно 40 мкМ, от приблизительно 0,1 мкМ до приблизительно 30 мкМ, от приблизительно 0,1 мкМ до приблизительно 20 мкМ или от приблизительно 0,1 мкМ до приблизительно 10 мкМ. В соответствии с определенными вариантами осуществления, уровень аргинина ниже предела детекции у анализатора аминокислот Biochrom 30 (например, ниже приблизительно 3 мкМ) и/или ниже предела детекции у трехквадрупольного масс-спектрометра Agilent 6460 для жидкостной хроматографии/с ионизацией электрораспылением (например, менее приблизительно 0,3 мкМ).
[00225] Определение продолжительности лечения, например, продолжительности времени, в течение которого концентрации аргинина в плазме крови у субъекта сохраняются в истощенном состоянии, будет хорошо известно рядовому специалисту в настоящей области техники. В соответствии с определенными вариантами осуществления, продолжительность лечения составляет приблизительно 1, приблизительно 2, приблизительно 3, приблизительно 4, приблизительно 8, приблизительно 12, приблизительно 16, приблизительно 20, приблизительно 24, приблизительно 28, приблизительно 32, приблизительно 36, приблизительно 40, приблизительно 44, приблизительно 48, приблизительно 52, приблизительно 56 недель или дольше.
[00226] Было показано, что истощение запасов аргинина является эффективным лечением ряда патологических состояний, в том числе без ограничения вирусных инфекций (патент США №8507245, включенный в настоящий документ посредством ссылки во всей своей полноте), рассеянного склероза, ревматоидного артрита, аутоиммунных заболеваний (патент США №9789169, включенный в настоящий документ посредством ссылки во всей своей полноте), врожденной гипераргининемии (идентификатор на сайте ClinicalTrials.gov: NCT02488044), болезни «трансплантат против хозяина» (GvHD) (Hallet W., et al., Exploiting arginase to prevent GVHD, Blood 2010 116:5440-5441), воспаления (Sahin E., et al., Macrophage PTEN regulates expression and secretion of arginase I modulating innate and adaptive immune responses, J Immunol. 2014 Aug 15;193(4): 1717-27) и нейроваскулярной дегенерации сетчатки (Fouda AY. et al., Arginase 1 promotes retinal neurovascular protection from ischemia through suppression of macrophage inflammatory responses Cell Death Dis. 2018 Sep 25;9(10):1001).
[00227] Соответственно, в настоящем документе также предложен способ лечения у нуждающегося в том субъекта по меньшей мере одного патологического состояния, выбранного из группы, состоящей из вирусных инфекций, рассеянного склероза, ревматоидного артрита, аутоиммунных заболеваний, врожденной гипераргининемии, болезни «трансплантат против хозяина» (GvHD) и воспаления, предусматривающий стадию введения субъекту терапевтически эффективного количества описываемого в настоящем документе слитого белка.
[00228] Дополнительные цели, преимущества и новые признаки настоящего раскрытия станут очевидны для специалистов в настоящей области техники после изучения приведенных далее примеров, которые не подразумеваются ограничивающими.
[00229] Пример 1. Конструирование слитых генов ABD и ABD094
[00230] Ген N-ABD для N-концевого слияния конструировали путем смешивания праймеров 01-08 (SEQ ID NO: 1-8), перечисленных в таблице 1, в ПЦР-реакции с каждым праймером до концентрации 20 нМ, вместе с 1× реакционным буфером для ДНК-полимеразы iProof (Bio-rad), 0,2 мМ dNTP и 0,5 единицы ДНК-полимер азы iProof (Bio-rad) в реакционном объеме 50 мкл. Задавали следующую программу ПЦР: (1) 98°С, 10 с; (2) 50°С, 20 с; (3) 72°С, 15 с; (4) повторение (1) - (3) 35 раз. Затем 1 мкл продукта ПЦР-реакции вносили в новую смесь для ПЦР-реакции вместе с 1× реакционным буфером для ДНК-полимеразы iProof (Bio-rad), 0,2 мМ dNTP, 0,2 мкМ праймера (N-ABDNde-F): 5'-GATCTTAAGCATATGCATCATCACCATC-3' (SEQ ID NO: 9), праймера (N-ABDHind-R): 5'-ACAGCTAAAAGCTTATCAGCCTAGGATCCGCCGCTACCATTG-3' (SEQ ID NO: 93) и 0,5 единицы ДНК-полимеразы iProof в реакционном объеме 50 мкл. Задавали следующую программу ПЦР: (1) 98°С, 10 с; (2) 50°С, 20 с; (3) 72°С, 15 с; (4) повторение (1) - (3) 35 раз; (5) 72°С, 5 минут. Конструирование гена N-ABD094 было таким же, как и процедуры конструирования гена N-ABD, за исключением того, что использовали праймеры 09-16 (SEQ ID NO: 9-16), перечисленные в таблице 1, вместо праймеров 01-08 (SEQ ID NO: 1-8), перечисленных в таблице 1.
[00231] Ген C-ABD для С-концевого слияния конструировали путем смешивания праймеров 17-24, перечисленных в таблице 1, в ПЦР-реакции с каждым праймером до концентрации 20 нМ, вместе с 1× реакционным буфером для ДНК-полимеразы iProof, 0,2 мМ dNTP и 0,5 единицы ДНК-полимеразы iProof в реакционном объеме 50 мкл. Задавали следующую программу ПЦР: (1) 98°С, 10 с; (2) 50°С, 20 с; (3) 72°С, 15 с; (4) повторение (1) - (3) 35 раз. Затем 1 мкл продукта ПЦР-реакции вносили в новую смесь для ПЦР-реакции вместе с 1× реакционным буфером для ДНК-полимеразы iProof, 0,2 мМ dNTP, 0,2 мкМ праймера (C-ABDNde-F): 5'-TAGCTGATCATATGTTATGCGATGGATCCAGTAACAAC-3' (SEQ ID NO: 91), 0,2 мкМ праймера (C-ABDHind-R): 5'- GACCCTAAAAGCTTAATGGTGATGGTGATGATG-3' (SEQ ID NO: 92) и 0,5 единицы ДНК-полимеразы iProof в реакционном объеме 50 мкл. Задавали следующую программу ПЦР: (1) 98°С, 10 с; (2) 50°С, 20 с; (3) 72°С, 15 с; (4) повторение (1) - (3) 35 раз; (5) 72°С, 5 минут. Конструирование гена C-ABD094 было таким же, как и процедуры конструирования гена C-ABD, за исключением того, что использовали праймеры 25-32 (SEQ ID NO: 25-32), перечисленные в таблице 1, вместо праймеров 17-24 (SEQ ID NO: 17-24), перечисленных в таблице 1.
[00232] Любой из ПЦР-фрагмента N-ABD, N-ABD094, C-ABD или С-ABD094 расщепляли рестрикционными ферментами Nde I и Hind III и лигировали в вектор pRSET-lac, предварительно расщепленный теми же ферментами, и вносили путем трансформации в Escherichia coli Top 10 с получением соответственно плазмиды pN-ABD, pN-ABD094, pC-ABD и pC-ABD094. Плазмиду pRSET-lac получали путем модификации из pRSET-A (Invitrogen) посредством замены промотора Т7 на промотор lac из вектора pGEMT-Easy (Promega).
[00233] Продукты слияния N-ABD или N-ABD094 с геном аргиназы I человека конструировали путем смешивания 0,2 мкМ праймера HARGBam-F (5'-TTAGCTGGGGATCCGCCAAGTCCAGAACCATAGG-3') (SEQ ID NO: 93), 0,2 мкМ праймера HARGHind-R (5 '-GAGATCAAAGCTTACTTAGGTGGGTTAAGGTAGTC-3') (SEQ ID NO: 94) вместе с 1× реакционным буфером для ДНК-полимеразы iProof, 0,2 мМ dNTP, 100 нг кДНК аргиназы I человека и 0,5 единицы ДНК-полимеразы iProof в реакционном объеме 50 мкл. Задавали следующую программу ПЦР: (1) 98°С, 10 с; (2) 50°С, 20 с; (3) 72°С, 30 с; (4) повторение (1) - (3) 35 раз; (5) 72°С, 5 минут. ПЦР-фрагмент гена аргиназы I человека расщепляли рестрикционными ферментами BamH I и Hind III и лигировали в вектор pN-ABD или pN-ABD094, предварительно расщепленный теми же ферментами, и вносили путем трансформации в Escherichia coli ER2566 (NEB) с получением соответственно плазмиды pN-ABD -rhArg и pN-ABD094-rhArg.
[00234] Продукты слияния C-ABD или C-ABD094 с геном аргиназы I человека конструировали путем смешивания 0,2 мкМ праймера HARGNde-F (5'-TAGGCTGCATATGAGCGCCAAGTCCAGAACCATAG-3') (SEQ ID NO: 95), 0,2 мкМ праймера HARGBam-R (5'-ATCAGCTAGGATCCCTTAGGTGGGTTAAGGTAGTCAATAGG-3') (SEQ ID NO: 96) вместе с 1× реакционным буфером для ДНК-полимеразы iProof (Bio-rad), 0,2 мМ dNTP, 100 нг кДНК аргиназы I человека и 0,5 единицы ДНК-полимеразы iProof (Bio-rad) в реакционном объеме 50 мкл. Задавали следующую программу ПЦР: (1) 98°С, 10 с; (2) 50°С, 20 с; (3) 72°С, 30 с; (4) повторение (1) - (3) 35 раз; (5) 72°С, 5 минут. ПЦР-фрагмент гена аргиназы I человека расщепляли рестрикционными ферментами Nde I и BamH I и лигировали в вектор pC-ABD или pC-ABD094, предварительно расщепленный теми же ферментами, и вносили путем трансформации в Escherichia coli ER2566 (NEB) с получением соответственно плазмиды pC-ABD-rhArg и pC-ABD094-rhArg.
[00235] Продукты слияния N-ABD или N-ABD094 с геном аргиназы Bacillus caldovelox (ВСА) конструировали путем смешивания 0,2 мкМ праймера BCABam-F (5'-ATGC Т AGTGG АТСС ATG A AGCC А ATTTC A ATT ATCG -3') (SEQ ID NO: 97), 0,2 мкМ праймера BCAHind-R (5'-TCAGCCTAAAGCTTACATGAGTTTTTCACCAAACAACG-3') (SEQ ID NO:) вместе с 1× реакционным буфером для ДНК-полимеразы iProof, 0,2 мМ dNTP, 100 нг геномной ДНК Bacillus caldovelox и 0,5 единицы ДНК-полимеразы iProof в реакционном объеме 50 мкл. Задавали следующую программу ПЦР: (1) 98°С, 10 с; (2) 50°С, 20 с; (3) 72°С, 30 с; (4) повторение (1) - (3) 35 раз; (5) 72°С, 5 минут. ПЦР-фрагмент гена аргиназы I Bacillus caldovelox расщепляли рестрикционными ферментами BamH I и Hind III и лигировали в вектор pN-ABD или pN-ABD094, предварительно расщепленный теми же ферментами, и вносили путем трансформации в Escherichia coli ER2566 (NEB) с получением соответственно плазмиды pN-ABD-BCA и pN-ABD094-BCA.
[00236] Продукты слияния C-ABD или C-ABD094 с геном аргиназы Bacillus caldovelox (ВСА) конструировали путем смешивания 0,2 мкМ праймера BCANde-F (5'-ATGCTAGCCATATGAAGCCAATTTCAATTATCGGG -3') (SEQ ID NO: 99), 0,2 мкМ праймера BCABam-R (5'-TCAGCTAAGGATCCCATGAGTTTTTCACCAAACAACG-3') (SEQ ID NO: 100) вместе с 1× реакционным буфером для ДНК-полимеразы iProof, 0,2 мМ dNTP, 100 нг геномной ДНК Bacillus caldovelox и 0,5 единицы ДНК-полимеразы iProof в реакционном объеме 50 мкл. Задавали следующую программу ПЦР: (1) 98°С, 10 с; (2) 50°С, 20 с; (3) 72°С, 30 с; (4) повторение (1) - (3) 35 раз; (5) 72°С, 5 минут. ПЦР-фрагмент гена аргиназы I Bacillus caldovelox расщепляли рестрикционными ферментами Nde I и BamH I и лигировали в вектор ppC-ABD или pC-ABD094, предварительно расщепленный теми же ферментами, и вносили путем трансформации в Escherichia coli ER2566 (NEB) с получением соответственно плазмиды pC-ABD-BCA и pC-ABD094-BCA.



[00238] Пример 2. Экспрессия и очистка слитого белка ABD-rhArg или ABD094-rhArg путем ферментации
[00239] Для получения слитого белка N-ABD-rhArg (SEQ ID NO: 49) Escherichia coli ER2566, трансформированную посредством pN-ABD-rhArg, выращивали в 50 мл посевной среды (1,5 г дрожжевого экстракта, 0,25 г NaCl и 5 мг ампициллина) при 30°С в орбитальном шейкере на скорости 250 об/мин в течение 16 часов. Затем посевную культуру переносили в 1 л ферментационной среды (10 г дрожжевого экстракта, 16 г триптона, 6,7 г Na2HPO4⋅7H2O, 3,41 г KH2PO4, 2,41 г NH4Cl, 0,67 г (NH4)2SO4, 10 г глицерина, 1 г глюкозы, MgSO4.7H2O, 1 мМ CaCl2 и 0,1 г ампициллина) и выращивали при 28°С с подачей воздуха 1 л/мин. и с минимальной скоростью 500 об/мин, рН поддерживали на уровне 7,4. Скорость перемешивания увеличивалась автоматически для поддержания минимального содержания растворенного кислорода pO2 на уровне по меньшей мере 20%. При достижении оптической плотности (OD) 15 при длине волны 600 нм у культуры подавали 500 мл среды (10 г дрожжевого экстракта, 16 г триптона, 2,41 г NH4Cl, 0,67 г (NH4)2SO4, 20 г глицерина) со скоростью 31,25 мл/час в течение непрерывных 16 часов. Клетки собирали центрифугированием, а белок очищали на колонке с Ni-IDA-сефарозой. Обычно очищенного белка получали более 1 г/л культуры. Затем из очищенного белка удаляли эндотоксин по способу с применением Triton Х-114 [Aida Y. and Pabst M.J. (1990) Journal of Immunological Methods. 132: 191-195, которая включена в настоящий документ посредством ссылки]. Процедуры получения слитых белков N-ABD094-rhArg (SEQ ID NO: 50), С-ABD-rhArg (SEQ ID NO: 51), C-ABD094-rhArg (SEQ ID NO: 52), N-ABD-BCA (SEQ ID NO: 53), N-ABD094-BCA (SEQ ID NO: 54), C-ABD-BCA (SEQ ID NO: 55) и C-ABD094-BCA (SEQ ID NO: 56) были такими же, как и процедуры в случае получения N-ABD-rhArg (SEQ ID NO: 49) за исключением того, что клетки соответственно трансформировали соответствующей плазмидой. На фиг. 1 и в таблице 2 показан пример очистки белка вышеупомянутой рекомбинантной аргиназы человека с альбуминсвязывающим доменом (rhArg), т.е. N-ABD094-rhArg (SEQ ID NO: 50).
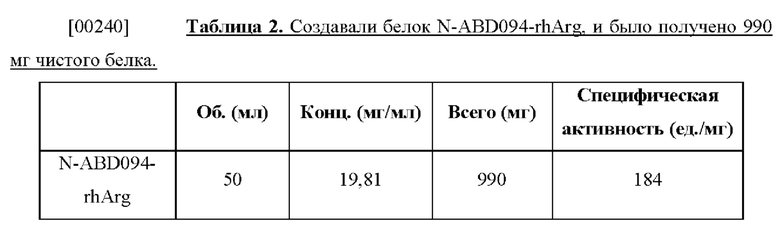
На фиг. 12 показаны результаты создания. Было получено 709 мг N-ABD094-rhArg (SEQ ID NO: 50) со специфической активностью 205 ед./мг и концентрацией белка 10,5 мг/мл.
[00241] Пример 3. Конструирование и клонирование гена N-ABD094-rhArg (SEP ID NO: 38)
[00242] Ген N-ABD094-rhArg (SEQ ID NO: 38) синтезировали путем ПЦР-элонгации с перекрывающимися праймерами на основе гена N-ABD-rhArg (SEQ ID NO: 37). Проводили две ПЦР-реакции. Прежде всего, использовали плазмиду pRSET/lac с геном N-ABD094-rhArg (SEQ ID NO: 38). Выделяли участок rhArg и амплифицировали его в первой ПЦР-реакции с применением ABD094-0F (SEQ ID NO: 57) и HuArgHinBam-R (SEQ ID NO: 64). ПЦР-продукт выделяли и очищали с помощью 0,7% агарозного геля (Biorad) при напряжении 100 В в течение 30 мин. и набора для очистки бэндов от геля (GE Healthcare Life Science). Затем к ПЦР-продукту добавляли ABD094 посредством второй ПЦР-реакции. Сначала получали смесь праймеров, разведенных в 20 раз, включавшую ABD094-0F, ABD094-1F, ABD094-2F, ABD094-3F, ABD094-4F и ABD094-5F (SEQ ID NO: 57-62), со стерилизованной водой milli-Q. Праймеры в смеси и abdNde-F перекрывали друг друга. Проводили вторую ПЦР-реакцию с применением abdNde-F (SEQ ID NO: 63), HuArgHinBam-R (SEQ ID NO: 64) и 20× разведенной смеси праймеров. ПЦР-продукт выделяли и очищали теми же способы, что были описаны выше. Последовательности праймеров показаны в таблице 3. Местоположения связывания праймеров проиллюстрированы на фиг. 2А. Полученная последовательность ДНК слитого гена N-ABD094-rhArg (SEQ ID NO: 38) и его белковая последовательность показаны на фиг. 2 В.
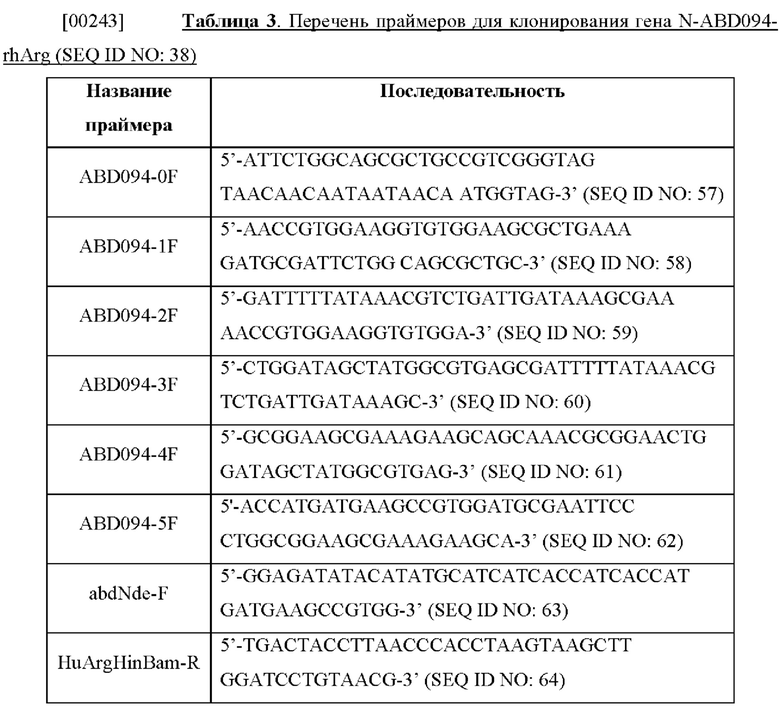
[00244] ПЦР-продукт и вектор pRSET/lac дважды расщепляли посредством BamHI-HF и Ndel (New England BioLabs) при температуре 37°С в течение 2 часов. Как вставку, так и вектор выделяли и очищали с помощью 0,7% агарозного геля (Biorad) при напряжении 100 В в течение 30 мин. и с применением набора для очистки бэндов от геля (GE Healthcare Life Science). Затем вставку и вектор лигировали лигазой Т4 (New England BioLabs) при комнатной температуре в течение 20 мин. В компетентные клетки трансформацией вводили рекомбинантную ДНК и наносили на среду LB с 50 мкг/мл канамицина. Среду LB инкубировали при 37°С в течение ночи. Отбирали 8 колоний и проверяли на наличие N-ABD094-rhArg с помощью ПЦР с abdNde-F и HuArgHinBam-R. Колонии с положительным результатом в ПЦР дополнительно растили в 5 мл 2× TY с 50 мкг/мл канамицина при 30°С в течение ночи. Бактериальный бульон центрифугировали в микроцентрифужной пробирке объемом 1,7 мл на ускорении 16100 × g в течение 1 мин. Осадки бактерий ресуспендировали в буфере для солюбилизации (50 мМ Tris-HCl, 0,1 М NaCl, 1 мМ MnCl2, рН 7,4) и разбивали путем цикличного повторения замораживания и оттаивания в жидком азоте. Лизат клеток центрифугировали с получением раствора белка. Для проверки присутствия аргиназы проводили пробу на аргиназную активность с применением DAMO.
[00245] Пример 4. Конструирование вектора экспрессии ВСА
[00246] Ген, кодирующий ВСА (номер доступа в Pubmed: U48226), заказывали у GenScript, где его синтезировали de novo и вставляли в вектор для клонирования pUC57. Одиночный остаток цистеина вводили посредством сайт-направленного мутагенеза, при этом для облегчения процесса очистки белка к С-концу добавляли шестигистидиновую метку. Модифицированный методами инженерии ген ВСА субклонировали в вектор рЕТ-3а и вводили путем трансформации в компетентные клетки BL21 (DE3) для экспрессии ВСА. Кристаллическую структуру ВСА (PDB: 2CEV) можно получить из банка данных по белкам. Белковая последовательность модифицированной методами инженерии ВСА (S161C)-His (SEQ ID NO: 89) показана на фиг. 3.
[00247] Пример 5. Конструирование вектора экспрессии BCA-ABD
[00248] Пару праймеров, кодирующих альбуминсвязывающий домен (ABD), конструировали и амплифицировали с помощью ПЦР с перекрывающимися праймерами. Вкратце: 200 нг праймеров использовали в качестве матрицы и амплифицировали с применением 1 единицы высокоточной ДНК-полимеразы iProof (Bio-Rad). Праймеры инкубировали при 98°С в течение 30 с в ходе 15 циклов амплификации, включающих тепловую денатурацию при 98°С в течение 10 с, гибридизацию при 60°С в течение 30 с и элонгацию при 72°С в течение 15 с.
[00249] 5'смысловая цепь-ABD
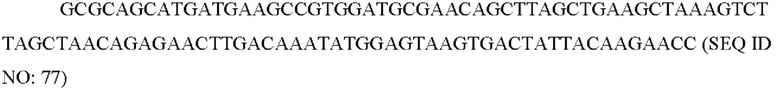
[00250] 5'-антисмысловая цепь-ABD

[00251] Кодирующий ABD ген, сконструированный с помощью этой пары праймеров, амплифицировали с применением 2 различных наборов праймеров с целью конструирования BCA-6xHis-ABD (также называемой ВНА) (SEQ ID NO: 75) и ВСА-ABD-6xHis (также называемой ВАН) (SEQ ID NO: 76). Для конструирования ВНА сначала ABD амплифицировали с помощью представленной далее пары праймеров, в случае которой был разработан прямой праймер для ПЦР с перекрывающимися праймерами с ВСА через шестигистидиновую метку, при этом были включены сайт рестрикции BamH I и стоп-кодон для лигирования в вектор рЕТ-За с помощью обратного праймера:
[00252] Прямой праймер для клонирования ABD:
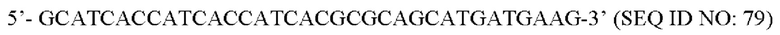
[00253] Обратный праймер для клонирования ABD:
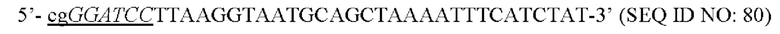
[00254] ПЦР-продукт, полученный в результате амплификации ABD,
подвергали электрофорезу в 1,5% агарозном геле и очищали ПЦР-продукты. Тогда как модифицированную методами инженерии ВСА с дополнительной мутацией (V20P) клонировали с применением следующей пары праймеров, для данного случая был удален стоп-кодон:
[00255] Прямой праймер для клонирования ВСА:

[00256] Обратный праймер для клонирования ВСА:
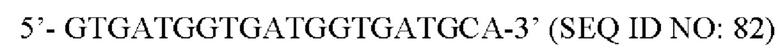
[00257] ВСА, амплифицированную с помощью описанных выше праймеров, подвергали электрофорезу в 1,5% агарозном геле и очищали ПЦР-продукты. Вышеупомянутые ПЦР-продукты ВСА и ABD связывали с помощью ПЦР с перекрывающимися праймерами. Аналогично, 200 нг ВСА и ABD использовали в качестве матрицы и амплифицировали с применением 1 единицы высокоточной ДНК-полимеразы iProof. ВСА и ABD инкубировали при 98°С в течение 30 секунд в ходе 15 циклов амплификации, включающих тепловую денатурацию при 98°С в течение 10 секунд, гибридизацию при 60°С в течение 30 секунд и элонгацию при 72°С в течение 30 секунд. ПЦР-продукт с перекрывающимися праймерами использовали в качестве матрицы для дальнейшей амплификации с применением следующей пары праймеров: [00258] Прямой праймер для клонирования ВНА:

[00259] Обратный праймер для клонирования ВНА:
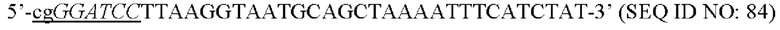
[00260] ПЦР-продукт подвергали электрофорезу в 1% агарозном геле и вырезали из геля бэнд, соответствующий ВНА, для очистки.
[00261] Для конструирования ВАН сначала амплифицировали ABD с применением представленной далее пары праймеров, в случае которой был разработан прямой праймер для ПЦР с перекрывающимися праймерами непосредственно с ВСА, при этом сайт рестрикции BamHI, шестигистидиновая метка и стоп-кодон были включены с помощью обратного праймера:
[00262] Прямой праймер для клонирования ABD:
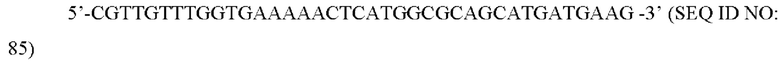
[00263] Обратный праймер для клонирования ABD:
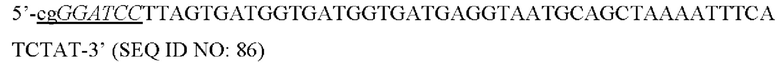
[00264] ПЦР-продукт, полученный в результате амплификации ABD, подвергали электрофорезу в 1,5% агарозном геле и очищали ПЦР-продукты. Модифицированную методами инженерии ВСА с дополнительной мутацией (V20P) клонировали с применением следующей пары праймеров, для данного случая были удалены стоп-кодон и шестигистидиновая метка:
[00265] Прямой праймер для клонирования ВСА:

[00266] Обратный праймер для клонирования ВСА:
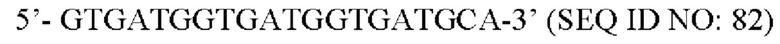
[00267] ВСА, амплифицированную с помощью описанного выше праймера, подвергали электрофорезу в 1,5% агарозном геле и очищали ПЦР-продукты. Вышеупомянутый ПЦР-продукт ВСА и ABD связывали с помощью ПЦР с перекрывающимися праймерами. Аналогично, 200 нг ВСА и ABD использовали в качестве матрицы и амплифицировали с применением 1 единицы высокоточной ДНК-полимеразы iProof. ВСА и ABD инкубировали при 98°С в течение 30 секунд в ходе 15 циклов амплификации, включающих тепловую денатурацию при 98°С в течение 10 секунд, гибридизацию при 60°С в течение 30 секунд и элонгацию при 72°С в течение 30 секунд. ПЦР-продукт с перекрывающимися праймерами использовали в качестве матрицы для дальнейшей амплификации с применением следующей пары праймеров:
[00268] Прямой праймер для клонирования ВАН:

[00269] Обратный праймер для клонирования ВАН:
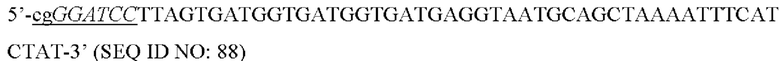
[00270] ПЦР-продукт подвергали электрофорезу в 1% агарозном геле и вырезали из геля бэнд, соответствующий ВНА, для очистки. ПЦР-продукт, кодирующий ВНА и ВАН, и вектор рЕТ-За дважды расщепляли с помощью Ndel и BamHI. ВНА и ВАН лигировали в вектор рЕТ-3а и вводили путем трансформации в компетентные клетки Е. coli ТОР10. Нуклеотидные последовательности ВНА и ВАН подтверждали с помощью секвенирования ДНК. Вектор рЕТ-3а, содержащий вставку ВНА или ВАН, вводили путем трансформации в вектор BL21(DE3) для экспрессии белка.
[00271] Пример 6. Белковые конструкции ВНА и ВАН
[00272] Белковая последовательность ВСА, используемая в данном исследовании, была такой же, как и у Bewley et al. (Structure 7, 435-448, 1999) (PDB ID: 2CEV, которая включена в настоящий документ посредством ссылки), за исключением того, что она содержала шестигистидиновую метку на С-конце и мутацию S161C. Разрабатывали конструкции как BCA-6xHis-ABD (ВНА), так и BCA-ABD-6xHis (ВАН), а экспрессировать ВНА и ВАН можно было с помощью биомасс клеток Е. coli. Как у ВНА, так и у ВАН ВСА-часть представляла собой мутантную версию ВСА с остатком валина в положении 20, который был мутирован в пролин (V20P), тогда как ABD-часть представляла собой версию ABD дикого типа с линкерной последовательностью (AQHDEAVDANS) (Jonsson et al., 2008, Protein Eng. Des. Sel. 21, 515-527). ABD-часть либо была расположена на самом С-конце слитого белка (в случае ВНА), либо были слита между ВСА и 6-гестидиновой меткой (в случае ВАН). Результаты выравнивания белковых последовательностей ВСА, ВНА и ВАН, использованных в настоящем исследовании, представлены на фиг. 4.
[00273] Аминокислотная последовательность ВСА (дикого типа) (PDB ID: 2CEV):

[00274] Аминокислотная последовательность ВСА (S161C)-His:

[00275] Аминокислотная последовательность ВНА:

[00276] Аминокислотная последовательность ВАН:

[00277] Пример 7. Получение ВНА и ВАН
[00278] Использовали биомассы клеток Е. coli, которые экспрессировали ВНА и ВАН. Для получения инокуляционной культуры небольшое количество биомассы E.coli BL21 (DE3) в глицерине инокулировали в 10 мл среды LB с добавлением 100 мкг/мл ампициллина. Инокуляционную культуру выращивали при 37°С со встряхиванием на скорости 700 об/мин в течение ночи (~16 ч). Затем полученную за ночь инокуляционную культуру в объеме 2,5 мл использовали для инокуляции 250 мл среды LB с 100 мкг/мл ампициллина. Инокулированные клетки выращивали при 37°С со встряхиванием на скорости 280 об/мин в течение 2-4 часов до оптической плотности 0,6-0,8 при длине волны 600 нм (OD600). Затем для индукции сверхэкспрессии целевого белка в культур ал ьную среду добавляли IPTG (0,2 мМ). Клетки выращивали дополнительные 4 часа и собирали центрифугированием на скорости 4500 об/мин.
[00279] Очистка белков методом никель-аффинной хроматографии
[00280] Осадки клеток Е. coli ресуспендировали в буфере, содержащем 50 мМ Tris-HCl, 100 мМ NaCl (рН 7,4), и разрушали посредством обработки ультразвуком. Затем удаляли клеточный дебрис центрифугированием на скорости 10000 об/мин в течение 40 мин. Надосадочную жидкость наносили на 5 мл хелатирующую колонку HiTrap (GE Healthcare), которая была загружена Ni2+ и уравновешена буфером А (20 мМ фосфат натрия, 0,5 М NaCl, рН 7,4) с применением очистителя АКТА. Белки элюировали с помощью 0-0,5 М градиента имидазола, что достигали с помощью комбинации буфера А с буфером В (0,5 М имидазола в буфере А) в различных соотношениях. Фракции анализировали с помощью электрофореза в полиакриламидном геле с добавлением додецилсульфата натрия (SDS-PAGE) с помощью 12% полиакриламидных гелей и объединяли фракции, содержащие целевой белок. Объединенные фракции концентрировали и заменяли буфер из концентрированных объединенных фракций на 20 мМ фосфат натрия (рН 7,4) с помощью центробежного фильтрующего устройства (Amicon) или устройства для тангенциальной поточной фильтрации (TFF) (Millipore). Пределом фильтрации у мембраны для ВНА и ВАН была молекулярная масса 30000. Затем ретентат разводили до требуемой концентрации, и подвергали стерилизующей фильтрации через 0,2-мкм шприц-фильтр, и хранили при 4°С.
[00281] Согласно хромато грамме результатов очистки ВНА, во время процесса элюирования с использованием возрастающих концентраций имидазола наблюдали три пика (фиг. 5А). Первые два пика были слиты вместе, за ними шел третий пик, который возникал при достижении концентрации имидазола ~0,13 М (фиг. 5А). При анализе с помощью SDS-PAGE было обнаружено, что третий пик в основном состоял из целевого белка массой около 40 кДа (фиг. 5 В, дорожки F4-F8). Этим целевым белком, вероятно, был ВНА, поскольку рассчитанная молекулярная масса мономера ВНА на основе его аминокислотной последовательности составляла 39,44 кДа. Поэтому был сделан вывод, что ВНА успешно экспрессировался и очищался. Фракции, содержащие относительно чистый ВНА, объединяли для дальнейших процессов.
[00282] Согласно хроматограмме результатов очистки ВАН, во время процесса элюирования с использованием возрастающих концентраций имидазола наблюдали три пика (фиг. 5А). 5С). Второй и третий пики были слиты вместе и возникали при достижении концентрации имидазола ~0,18 М (фиг. 5С). При анализе с помощью SDS-PAGE было обнаружено, что данный слитый пик в основном состоял из целевого белка массой около 40 кДа (фиг. 5D, дорожки F6-F14), который, скорее всего, представлял собой ВАН. По сравнению с ВНА, который элюировался при ~0,13 М имидазола, ВАН, по-видимому, более прочно связывался с никель-аффинной колонкой. Вероятно, это связано с тем, что ABD-домен, вставленный между ВСА и шестигистидиновой меткой, помогает сделать шестигистидиновую метку более доступной, а значит ВАН может эффективнее взаимодействовать с ионами никеля в колонке, чем ВНА. Фракции, содержащие относительно чистый ВАН, объединяли вместе и заменяли у них буфер с использованием устройства для тангенциальной поточной фильтрации (TFF) (Millipore) вместо центробежного фильтрующего устройства.
[00283] За одну единицу ферментативной активности принимали количество фермента (ВСА, ВНА или ВАН), которое катализирует получение 1 мкмоля мочевины за минуту при стандартных условиях анализа. Специфическую активность фермента выражали в единицах активности на мг белка. В стандартных условиях анализа с применением DAMO (37°С, рН 7,4) свежеочищенный ВНА (SEQ ID NO: 75) имел значение специфической активности 220,6±15,8 ед./мг, что близко к значению ВСА (S161C)-His (SEQ ID NO: 89) (243,30 ед./мг).
[00284] Для слитых белков, содержащих полипептид ВСА и полипептид ABD, в случае ВНА был получен чрезвычайно высокий выход белка, составляющий приблизительно 290 мг белка на литр культуры во встряхиваемой колбе (SEQ ID NO: 75). Выходы слитых белков ABD сильно зависели от используемых систем экспрессии. Сообщалось, что выход ABD, слитого с sCR1, варьировал в диапазоне от 0,4 до 1,1 мкг за 24 ч на мл культуры клеток яичника китайского хомячка (Makrides et al., 1996, J. Pharmacol. Exp.Ther. 277, 534-542). Из каждого л культуры клеток НЕК293 можно получить около 2-15 мг слитых белков одноцепочечного диатела с ABD (Норр et al., 2010, Protein Eng. Des. Sel. 23, 827-834; Stork et al., 2007, Protein Eng. Des. Sel. 20, 569-576). Выход слитого белка ABD-TolA, экспрессируемого в хозяине Е. coli BL21, составлял 20 мг/л питательной среды LB (Ahmad et al., 2012, Proteins 80, 774-789). Для сравнения, выход ВНА был намного выше. Из наших результатов видно, что получение ВНА (SEQ ID NO: 75) является простым и экономичным.
[00285] Пример 8. Экспрессия и очистка ВСА (S161CVHis (SEP ID NO: 89)
[00286] В 5 мл среды LB, содержащей 100 мкг/мл ампицилина, получали ночную инокуляционную культуру Е. coli штамма BL21 (DE3)pLysS, трансформированную плазмидой pET-3a-BCA(S161C)-His, и оставляли ее расти во встряхиваемой колбе при температуре 37°С в течение 16 ч со скоростью встряхивания 280 об/мин. На 250 мл среды LB, содержащей 100 мкг/мл ампицилина, добавляли 2,5 мл ночной культуры и культивировали при температуре 37°С со встряхиванием на скорости 280 об/мин до достижения оптической плотности 0,8-0,9 (OD600) на длине волны 600 нм. Добавляли изопропил-β-D-тиогалактопиранозид (IPTG) до конечной концентрации 0,2 мМ. Спустя 4 ч индукции клетки собирали путем центрифугирования на ускорении 12000 × g в течение 20 минут при температуре 4°С. Клеточный осадок можно было либо разрушить ультразвуком и подвергнуть очистке, либо сохранить при температуре -20°С до необходимости. Клеточный осадок суспендировали в 20 мл солюбилизирующего буфера (50 мМ Tris-HCl, 100 мМ NaCl, рН 7,4), содержащего 10 мМ MnCl. Сначала клетки разрушали путем обработки ультразвуком на льду в течение 10 минут с импульсом каждые 30 секунд с 40% амплитудой. Клеточный дебрис удаляли путем центрифугирования на ускорении 12000 × g в течение 20 минут при температуре 4°С. Надосадочную жидкость инкубировали при температуре 70°С в течение 15 минут, а осадки, образовавшиеся во время стадии термообработки, удаляли путем центрифугирования. Растворимую фракцию фильтровали через 0,22-мкм фильтр перед загрузкой в 5-мл колонку HisTrap Chelating HP. Фракцию проскока собирали во время загрузки образца, а несвязанные белки вымывали 5 объемами колонки (CV) стартового буфера или до тех пор, пока уровень не снижался ниже исходного. Целевые белки, связавшиеся с колонкой, элюировали путем конкурентного градиентного элюирования с применением имидазола (0,5 М имидазола в стартовом буфере). Целевые фракции подвергали идентификации и объединяли друг с другом, у которых заменяли буфер на 20 мМ фосфат натрия при рН 7,4 с использованием центробежного фильтра Amicon Ultra-15 с молекулярной массой отсечки 10 кДа в соответствии с инструкциями производителя. Образец фильтровали в стерильных условиях через 0,22-мкм фильтр и хранили при 4°С до необходимости.
[00287] Пример 9. Очистка белка ВСА (S161C)-His (SEQ ID NO: 89), полученного в результате периодической ферментации с подпиткой
[00288] Ферментация клеток Е. coli, экспрессирующих ВСА (S161C)-His, в 5-литровом биореакторе давала 248,86 г массы влажных клеток. Клеточный осадок ресуспендировали в солюбилизирующем буфере, содержащем ДНКазу и РНКазу, которые разрушали огромное количество ДНК и РНК, высвобожденных при гомогенизации, тем самым снижая вязкость клеточного лизата. Клеточный лизат после трехкратного пропускания через гомогенизатор инкубировали при температуре 37°С в течение 15 минут для дополнительного снижения вязкости с последующим центрифугированием для получения растворимой фракции. На первой стадии очистки удаляли нецелевые примеси путем термообработки при 70°С в течение 15 минут, в результате чего осаждались термолабильные белки, и посредством центрифугирования выделяли термостойкий белок ВСА (S161C)-His (SEQ ID NO: 89).
[00289] Растворимую фракцию, содержащую целевой белок, дополнительно очищали на Ni-аффинной колонке с объемом слоя 196 мл, заполненным в колонку ХК50. Вкратце: для элюирования ВСА (S161C)-His (SEQ ID NO: 89) из Ni-аффинной колонки использовали 4-сегментную программу элюирования. Первый градиент элюирования от 0 до 30% элюирующего буфера был предназначен для удаления белка, который неспецифично связался с колонкой. Во втором сегменте поддерживали уровень 30%, что позволяло полностью элюировать неспецифически связанные белки. Целевой белок элюировали в третьем сегменте, увеличивая концентрацию элюирующего буфера с 30% до 70%. В четвертом сегменте, который заключался в увеличении с 70 до 100% элюирующего буфера, элюировали весь оставшийся белок, который все еще был связан на колонке. Фракции, которые давали поглощение при длине УФ-волны 280 нм, анализировали с помощью SDS-PAGE, тогда как 24 фракции Е1 - А'5 (по 50 мл каждая), содержащие целевой белок, объединяли друг с другом (фиг. 6А и фиг. 6 В). Объединенные фракции, содержащие высокую концентрацию имидазола, подвергали обессоливанию с помощью системы для тангенциальной поточной фильтрации (TFF), исходя из нашего понимании гексамерной структуры ВСА (дикого типа) (SEQ ID NO: 70), для TFF применяли мембраны с NMWC (номинальной молекулярной массой отсечки) 30 кДа. Очищенный ВСА (S161C)-His (SEQ ID NO: 89) хранили в PBS-буфере, фильтровали через 0,2-мкм фильтр в стерильных условиях и хранили при 4°С до дальнейшего применения. Путем 2-стадийной очистки в исследовательском масштабе получали приблизительно 1096 мг белка ВСА (S161C)-His (SEQ ID NO: 89) и определяли, что специфическая активность составляла 264 ед./мг.
[00290] Пример 10. Определение аргиназной ферментативной активности
[00291] Активность ВСА (S161C)-His (SEQ ID NO: 89) анализировали прямым колориметрическим способом с применением диацетилмоноксима (DAMO), который позволяет определять количество такого продукта, как мочевина, в ферментативной реакции. Вкратце: серийные разведения 0,3 мл растворов ВСА вместе с 0,15 мл воды и 720 мМ аргининового субстрата при рН 7,4 предварительно инкубировали отдельно при 37°С в течение 10 мин. Ферментативную реакцию проводили путем добавления 0,3 мл аргининового субстрата к предварительно инкубированным растворам ферментов. Реакционную смесь инкубировали при 37°С ровно 5 минут и добавляли гасящий реагент [50% трихлоруксусную кислоту (ТСА)] для остановки реакции. Холостые пробы готовили аналогичным образом, но такой же объем растворов ферментов добавляли после раствора ТСА.
[00292] Количество мочевины, образовавшейся в результате указанной выше реакции, определяли с помощью прямого колориметрического способа с применением DAMO, описанного Всемирной организацией здравоохранения (WHO). Данный протокол был принят WHO в качестве стандартных операционных процедур (SOP) для измерения концентрации мочевины в крови. Готовили несколько реагентов, относящихся к SOP, смешанный кислотный реагент готовили путем медленного добавления 100 мл концентрированной серной кислоты к 400 мл дистиллированной воды с последующим добавлением 0,3 мл стокового кислотного реагента (0,5 г гексагидрата хлорида железа в 15 мл дистиллированной воды с доведением до 25 мл концентрированной фосфорной кислотой). Смешанный цветной реагент получали путем смешивания 35 мл исходного цветного реагента А (1 г DAMO в 50 мл дистиллированной воды) и 35 мл стокового цветного реагента В (0,25 г тиосемикарбазида в 50 мл дистиллированной воды) и доводили до 500 мл дистиллированной водой. Рабочий стандарт мочевины получали путем добавления 5 мл стокового стандарта мочевины (0,5 г мочевины в 50 мл бензойной кислоты) к 45 мл бензойной кислоты (1 г/дл). Анализ с применением DAMO проводили путем добавления 100 мкл 20-кратно разведенной реакционной смеси в стеклянную пробирку для кипячения с последующим добавлением 3 мл реагента для проявления окраски, который готовили свежим путем смешивания дистиллированной воды, смешанного цветного реагента и смешанного кислотного реагента в соотношении 1:1:1. Пробирки инкубировали при 100°С в течение 15 минут и давали им остыть до комнатной температуры в течение 5 минут. Оптическую плотность измеряли на 540 нм. За одну единицу аргиназы принимали количество фермента, который превращает микромоль аргинина в орнитин и мочевину за минуту при 37°С, рН 7,4. Ферментативную активность рассчитывали по приведенным далее уравнениям, из значения оптической плотности образца вычитали значение его холостой пробы:
ΔА540нм образца=А540нм образца - А540нм холостого образца
Количество полученной мочевины рассчитывали из стандартной кривой:
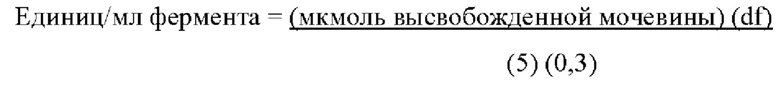
df=кратность разведения фермента
0,3=объем (в миллилитрах) использованного фермента
5 = время (в минутах) анализа, затрачиваемое для определения одной единицы
[00293] Пример 11. Определение концентрации белка
[00294] Концентрацию белка определяли с помощью анализа методом Брэдфорда. Концентрат красителя для анализа белка (Bio-rad) перед применением разводили в 5 раз. 20 мкл образца смешивали с 1 мл разведенного красителя и инкубировали при комнатной температуре в течение 10 минут. Для определения концентрации белка смесь измеряли на 595 нм с помощью набора стандартов на основе бычьего сывороточного альбумина Quick Start (Bio-rad) в качестве стандарта в диапазоне концентраций 0-1 мг/мл.
[00295] Пример 12. Ферментативная активность слитого с ВНА белка (SEQ ID NO: 75)
[00296] Анализировали катаболизирующую аргинин активность у ВНА (SEQ ID NO: 75) с целью изучения, будет ли затронута ферментативная активность в присутствии HSA. Из результатов было видно, что на активность ВНА (SEQ ID NO: 75) не влияли различные количества HSA, добавляемые в ходе анализа активности. Мольное соотношение ВНА: HSA, протестированное в исследованиях по связыванию, составляло 1:10, 1:5, 1:1, 5:1 и 10:1. Во всех данных условиях измеренные специфические активности отдельно у ВНА (SEQ ID NO: 75) и у комплексов BHA-HSA были очень схожими и составляли приблизительно 200 ед./мг.
[00297] Пример 13. Связывание N-ABD-rhArg (SEQ ID NO: 49) или ВНА (SEQ ID NO: 75) с HSA, выявленное с помощью анализа PAGE в неденатурирующих условиях
[00298] При слиянии ABD с rhArg или ВСА важно, чтобы слитый белок сохранял как альбуминсвязывающую способность, так и ферментативную активность. Для демонстрации связывающей способности ABD в слитых белках инкубировали фиксированное количество сывороточного альбумина человека (HSA) с N-ABD-rhArg (SEQ ID NO: 49) или ВНА (SEQ ID NO: 75) в различных молярных отношениях. Реакционные смеси анализировали в их нативном состоянии на PAGE-геле в неденатурирующих условиях, где белки разделялись по размеру и конформации.
[00299] На фиг. 7 подвижность каждого из отдельно HSA и отдельно N-ABD-rhArg (SEQ ID NO: 49) показана соответственно на дорожках 2 и 3 PAGE в неденатурирующих условиях (фиг. 7). Фиксированное количество HSA (30 пмоль) титровали против N-ABD-rhArg (SEQ ID NO: 49) в различных молярных соотношениях, а результаты проиллюстрированы на дорожках 5-9. При увеличении количества N-ABD-rhArg (SEQ ID NO: 49) в реакции связывания количество свободного HSA пропорционально уменьшалось, что указывало на специфическое связывание N-ABD-rhArg (SEQ ID NO: 49) с HSA. На фиг. 8 подвижность каждого из отдельно HSA и отдельно ВНА (SEQ ID NO: 75) показана соответственно на дорожках 6 и 7 PAGE в неденатурирующих условиях (фиг. 8). Фиксированное количество HSA (60 пмоль) титровали против ВНА (SEQ ID NO: 75) в различных молярных соотношениях, а результаты проиллюстрированы на дорожках 1-5. При увеличении количества ВНА (SEQ ID NO: 75) в реакции связывания количество свободного HSA пропорционально уменьшалось, что указывало на специфическое связывание ВНА (SEQ ID NO: 75) с HSA.
[00300] Пример 14. In vivo исследование ВНА (SEP ID NO: 75)
[00301] Для in vivo исследования ВНА (SEQ ID NO: 75) использовали шесть BALB/c мышей в возрасте 8 недель. Перед инъекцией ВНА (SEQ ID NO: 75) у каждой мыши брали образец крови и использовали его в качестве контроля (0 ч). Мышей случайным образом разделяли на две группы для соответственно внутрибрюшинной (i.p.) и внутривенной (i.v.) инъекции. Каждой мыши вводили ВНА (SEQ ID NO: 75) (250 ед.), а через 6, 24, 72 и 120 ч после инъекции брали образцы крови. Плазму каждого образца крови в количестве 16 мкл анализировали с помощью анализатора аминокислот (Biochrom) на содержание аргинина и орнитина.
[00302] Истощающее запасы аргинина in vivo влияние ВНА (ВНА SEQ ID NO: 75) изучали на BALB/c мышах и сравнивали с аналогичным влиянием ВСА (S161C)-His (SEQ ID NO: 89), результаты чего показаны на фиг. 9. Согласно результатам, уровень аргинина в плазме крови быстро возвращался к норме в течение 24 часов после инъекции ВСА (S161C)-His (SEQ ID NO: 89) (фиг. 9). В отличие от этого, в плазме крови сохранялся недетектируемый уровень аргинина в течение по меньшей мере 24 часов после инъекции ВНА (SEQ ID NO: 75) (фиг. 9). Данный результат свидетельствовал о том, что слияние ABD характеризовалось пролонгированным периодом полужизни ВСА (S161C)-His в кровотоке (SEQ ID NO: 89) in vivo. Из результатов было видно, что слияние с ABD является осуществимой и многообещающей стратегией улучшения фармакодинамических свойств истощающих запасы аргинина ферментов. Судя по всему, путь введения не оказывал влияния на истощающее запасы аргинина действие ВНА (SEQ ID NO: 75) in vivo (фиг. 9). Кроме того, у всех мышей наблюдали хорошую переносимость при введении ВНА (SEQ ID NO: 75), поскольку не наблюдали никаких явных побочных эффектов. В своей совокупности, для описанных в настоящем документе слитых белков, таких как ВНА (SEQ ID NO: 75), была доказана осуществимость стратегии слияния с ABD по снижению уровня аргинина в плазме крови до недетектируемого уровня на гораздо более длительный период (по меньшей мере 24 часа), чем при применении нативного белка ВСА (S161C)-His (SEQ ID NO: 89) (8 часов) без слияния с ABD.
[00303] Пример 15. In vivo исследование фармакодинамики N-ABD-rhArg (SEP ID NO: 49)
[00304] Противораковые действия аргиназы уже давно исследованы (Greenberg and Sassenrath, 1953, Cancer Res. 13, 709-715). Было обнаружено, что аргиназа обладает очень хорошим противораковым действием in vitro. Тем не менее, in vivo практически не наблюдали никакого действия из-за ее короткого периода полужизни в плазме. Уровень аргинина в плазме падал до наиболее низкого уровня через 3 часа после инъекции нативной аргиназы мышам С3Н. У того же модельного животного уровень аргинина возрастал до нормального исходного уровня через 5 часов после инъекции (Greenberg and Sassenrath, 1953, Cancer Res. 13, 709-715). Поэтому аргиназу считали неэффективным противораковым средством из-за короткого периода полужизни в плазме. Это позволило предположить, что период полужизни в плазме может влиять на эффективность ферментного лекарственного средства in vivo. У N-ABD-rhArg (SEQ ID NO: 49) и N-ABD094-rhArg (SEQ ID NO: 50) наблюдали схожую специфическую активность. Авторами настоящего изобретения было обнаружено, что эффективным способом является продление периода полужизни rhArg (SEQ ID NO: 103) в плазме крови за счет слияния с ABD. Для снижения стоимости получения rhArg (SEQ ID NO: 103) с увеличенным периодом полужизни в плазме были разработаны N-ABD-rhArg (SEQ ID NO: 49) и N-ABD094-rhArg (SEQ ID NO: 50).
[00305] В данном исследовании использовали пятнадцать самок BALB/c мышей (возрастом 10 недель). Их держали группами по пять животных в клетке. Мышам вводили разовую дозу N-ABD-rhArg (SEQ ID NO: 49) (500 ед., 250 ед. и 125 ед. соответственно) посредством внутрибрюшинной инъекции. Образцы крови собирали из подкожной вены с применением гепаринизированных капиллярных пробирок и центрифугировали с ускорением 800 × g для получения плазмы крови. С помощью анализатора аминокислот Biochrom 30 у шестнадцати микролитров образца плазмы крови измеряли концентрацию L-аргинина. Результаты представлены на фиг. 10. Из результатов измерения концентрации аргинина в плазме крови было видно, что у BALB/c мышей 500 ед. N-ABD-rhArg (SEQ ID NO: 49) могут истощать запасы аргинина в плазме крови до уровней, которые ниже предела детекции анализатора аминокислот Biochrom 30 (3 мкМ), на срок по меньшей мере 9 суток, 250 ед. N-ABD-rhArg (SEQ ID NO: 49) могут истощать запасы аргинина в сыворотке крови до уровней, которые ниже предела детекции анализатора аминокислот Biochrom 30, на срок по меньшей мере 7 суток, а 125 ед. N-ABD-rhArg (SEQ ID NO: 49) могут истощать запасы аргинина в сыворотке крови до уровней, которые ниже предела детекции анализатора аминокислот Biochrom 30, на срок по меньшей мере 3 суток. Эти результаты ясно свидетельствовали, что период полужизни rhArg в плазме крови можно продлить за счет слияния с ABD. По сравнению с ВНА (SEQ ID NO: 75) (фиг. 9) N-ABD-rhArg (SEQ ID NO: 49) может истощать запасы аргинина до очень низкого уровня у мышей на исключительно длительный период, что подчеркивает важность необходимости рационального конструирования слитого белка.
[00306] Пример 16. In vivo исследование фармакодинамики и Фармакокинетики у N-ABD094-rhArg (SEO ID NO: 50)
[00307] В данном исследовании использовали пятьдесят самцов C57BL/6J мышей (возрастом 10-12 недель). Мыши вводили разовую дозу 500 ед. N-ABD094-rhArg (SEQ ID NO: 50) посредством внутрибрюшинной инъекции. Образцы крови собирали из подкожной вены с применением гепаринизированных капиллярных пробирок и центрифугировали с ускорением 800 × g для получения плазмы крови. В пяти микролитрах образца плазмы крови производили измерение концентрации L-аргинина способом жидкостной хроматографии-тандемной масс-спектрометрии с применением трехквадрупольного масс-спектрометра Agilent 6460 для жидкостной хроматографии/с ионизацией электрораспылением с пределом детекции 0,3 мкМ L-аргинина. В пяти микролитрах образца плазмы крови производили измерение концентраций N-ABD094-rhArg (SEQ ID NO: 50) с применением набора для ELISA (твердофазного иммуноферментного анализа) человеческой аргиназы I (Abcam), который позволяет специфически определять человеческую аргиназу и не реагирует с мышиной аргиназой.
[00308] После инъекции сыворотку собирали в моменты времени 2-й час, 4-й час, 8-й час, 16-й час, 24-й час (1-е сутки) и со 2-х по 12-е сутки с 24-часовыми интервалами, а также с 12-х по 30-е сутки с 72-часовыми интервалами. Для достижения баланса между количеством используемых животных и для минимизации влияния на результаты исследований фармакокинетики и фармакодинамики, вызванного удалением N-ABD094-rhArg (SEQ ID NO: 50) из мыши в результате забора крови, мышей разделяли на 10 групп, n=5 в каждой группе, и забор крови производили в указанные моменты времени, которые представлены в скобках: группа 1 (2-й час), группа 2 (4-й час), группа 3 (8-й час), группа 4 (16-й час), группа 5 (24-й час), группа 6 (2-е сутки, 7-е сутки, 12-е сутки), группа 7 (3-е сутки, 8-е сутки, 18-е сутки), группа 8 (4-е сутки, 9-е сутки, 21-е сутки), группа 9 (5-е сутки, 10-е сутки, 15-е сутки, 24-е сутки), группа 10 (0-е сутки, 6-е сутки, 11-е сутки, 27-е сутки, 30-е сутки). Результаты изучения фармакодинамики и фармакокинетики соответственно показаны на фиг. 13 и фиг. 14.
[00309] По результатам измерения концентраций аргинина в плазме крови было видно, что 500 ед. N-ABD094-rhArg (SEQ ID NO: 50), вводимые внутрибрюшинно, могут снижать концентрации аргинина в плазме крови до 1 мкМ в течение 2 часов после инъекции (фиг. 13). Концентрация аргинина в плазме крови сохранялась на уровне ≤1 мкМ в течение 10 суток до постепенного повышения в последующие сутки (таблица 4). Концентрации аргинина в плазме возвращалась к половине от исходного уровня в период с 18-е сутки по 21-е сутки. Эти результаты позволяли предположить, что N-ABD094-rhArg (SEQ ID NO: 50) может поддерживать уровень аргинина в плазме крови на очень низком уровне (≤1 мкМ) у мышей в течение исключительно длительного периода.

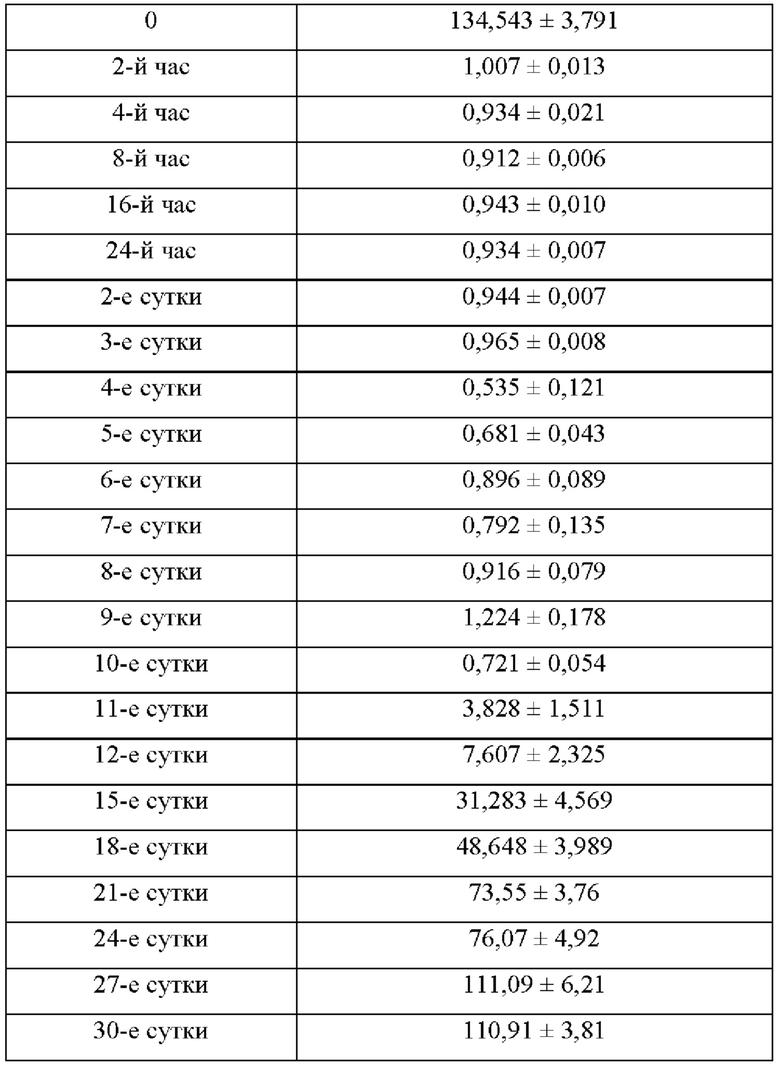
[00311] Фармакокинетические параметры рассчитывали с помощью двухкомпонентной модели.
Константу скорости элиминации ke рассчитывали с помощью следующего уравнения:
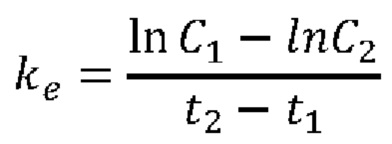
Период полужизни в плазме крови t1/2 рассчитывали с помощью следующего уравнения:
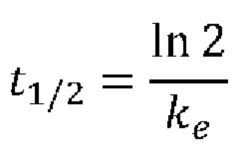
Площадь под кривой (AUC) рассчитывали по правилу трапеций с помощью следующего уравнения:
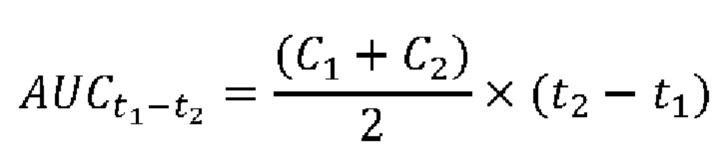
Для сравнения авторами настоящего изобретения были использованы данные о периоде полужизни нативной rhArg, которые доступны в литературе. Слитый белок N-ABD094-rhArg (SEQ ID NO: 50) характеризовался периодом полужизни 3,9±0,3 суток (фиг. 14), что означало, что молекулы выводились намного медленнее, чем в случае неслитого белка rhArg, который характеризовался гораздо более коротким периодом полужизни. Для конкретного сравнения, период полужизни человеческой аргиназы в кровотоке составляет всего несколько минут (Georgiou et al., US 8440184 B2), что указывает на то, что конъюгация значительно увеличивала время ее пребывания в кровотоке. Эти результаты ясно свидетельствовали, что период полужизни rhArg в плазме крови можно значительно продлить за счет слияния с ABD. Период полужизни нативного ADI (несколько часов) в кровотоке был продлен за счет пегилирования (Ensor et al., Cancer Res 2002, 62: 5443-5450). Период полужизни пегилированного ADI в кровотоке составлял 2-3 суток, и только пегилированный ADI (не нативный ADI) был эффективным в ингибировании роста опухолей in vivo. При сравнении нативного rhArg и пегилированного rhArg, вводимого крысам с помощью внутрибрюшинной инъекции, 1500 ед. пегилированного rhArg на крысу (6 ед./г крысы) могли истощить запасы аргинина в кровотоке до уровней, которые ниже предела детекции у анализатора аминокислот (3 мкМ), на период 6 суток, тогда как высокая доза нативного rhArg (7500 ед./крыса) уменьшала уровень аргинина в кровотоке лишь до ~20 мкМ в течение 3 часов после инъекции, при этом аргинин возвращался к исходному уровню примерно через 2 суток после инъекции (Tsui et al., Cancer Cell Int. 2009 Apr 17;9:9). Нативная аргиназа выводится из кровотока в течение нескольких минут (Savoca et al., Cancer Biochem. Biophy's. 7:261-268, 1984). Для клинического применения важно, чтобы аргиназа была модифицирована методами инженерии так, чтобы она была способна оставаться в кровотоке в течение многих суток или недель. В отсутствие каких-либо модификаций человеческая аргиназа имеет период полужизни всего несколько минут в кровотоке, поскольку ее размер недостаточно велик, чтобы избежать фильтрации почками (Georgiou et al., US 8440184 B2). Таким образом, в соответствии с настоящим изобретением, были разработаны совершенно новые и значительно улучшенные формы аргиназы со значительно улучшенной устойчивостью в кровотоке для терапевтического применения.
Пример 17. Обработка посредством N-ABD094-rhArg (SEQ ID NO: 50) снижала жировую массу, подавляла липогенез и улучшала чувствительность к инсулину у нормальных худых мышей
[00312] Из наших данных, полученных на мышиной модели, видно, что однократная внутрибрюшинная инъекция 500 ед. N-ABD094-rhArg (SEQ ID NO: 50) 8-недельным нормальным худым самцам C57BL/6J мышей могла сохранять концентрацию аргинина в плазме крови на уровне ≤1 мкМ в течение по меньшей мере 10 суток, при этом характеризуясь длительным периодом полужизни N-ABD094-rhArg (SEQ ID NO: 50) в кровотоке (фиг. 13). Поэтому мы обрабатывали нормальных худых самцов C57BL/6J мышей, получавших стандартный рацион питания, путем внутрибрюшинной инъекции 500 ед. N-ABD094-rhArg (SEQ ID NO: 50) с 10-суточными интервалами в течение 4 недель. Спустя 4 недели после обработки имело место снижение массы висцеральной (перигонадного, периренального и брыжеечного жирового тела) и подкожной (пахового жирового тела) жировой массы (фиг. 15 А) по сравнению с контрольными мышами, которым вводили наполнитель (солевой раствор). По результатам гистологического исследования висцеральной белой жировой ткани (WAT) было видно заметное уменьшение размера адипоцитов (фиг. 15 В). Дополнительно был проанализирован липогенез и было обнаружено, что у мышей, которых обрабатывали посредством N-ABD094-rhArg (SEQ ID NO: 50), уровни мРНК нескольких ключевых липогенных ферментов, в том числе ацетил-СоА-карбоксилазы 1 (Acc1), синтазы жирных кислот (Fasn) и стеароил-СоА-десатуразы 1 (Scd1), резко подавлялись в висцеральном WAT (фиг. 16А) и печени (фиг. 16 В).
[00313] Ожирение тесно связано с инсулинорезистентностью. Поэтому было проведено как in vivo, так и in vitro исследование с целью определения, будет ли обработка посредством N-ABD094-rhArg (SEQ ID NO: 50) влиять на чувствительность к инсулину. Из результатов теста на толерантность к инсулину (ITT) было видно, что у мышей, обработанных посредством N-ABD094-rhArg (SEQ ID NO: 50), наблюдали значимое повышение чувствительности к инсулину через 2 недели после обработки лекарственным средством (фиг. 17 А). Затем отдельно исследовали чувствительность печени к инсулину при обработке первичных гепатоцитов, полученных от самцов C57BL/6J мышей, средой, содержащей 5 ед./мл N-ABD094-rhArg (SEQ ID NO: 50), которая при этой концентрации не влияла на жизнеспособность клеток (фиг. 17 В). Через 24 часа после культивирования в среде с N-ABD094-rhArg (SEQ ID NO: 50) или контрольной среде (без добавления N-ABD094-rhArg) первичные гепатоциты стимулировали посредством 100 нМ инсулина в течение 20 минут. Затем клетки собирали для вестерн-блоттинга, который проводили с целью детекции фосфорилированной протеинкиназы В (pAkt), которую обычно используют в качестве маркера передачи сигналов с участием инсулина. Из результатов видно, что имело место заметное увеличение фосфорилирования Akt в группе, обработанной посредством N-ABD094-rhArg (SEQ ID NO: 50), по сравнению с контрольной группой, что свидетельствовало об усилении передачи сигналов в клетках печени с участием инсулина (фиг. 17 С).
[00314] Из результатов, полученных на нормальных худых мышах, было видно, что N-ABD094-rhArg (SEQ ID NO: 50) имел длительный период полужизни в кровотоке и мог уменьшать жировую массу, подавлять липогенез и улучшать чувствительность к инсулину как in vivo, так и in vitro. Данные результаты подтверждали, что слитый белок ABD-rhArg можно применять для предупреждения развития и лечения ожирения и связанных с ним метаболических нарушений, таких как инсулинорезистентность и стеатоз печени (жировая дистрофия печени).
[00315] Пример 18. Обработка посредством N-ABD094-rhArg (SEQ ID NO: 50) эффективно предупреждала развития HFD-индуцированного ожирения
[00316] Обработка посредством рекомбинантной человеческой аргиназы [N-ABD094-rhArg (SEQ ID NO: 50)] могла значимо снижать жировую массу, подавлять липогенез и улучшать чувствительность к инсулину у нормальных худых мышей.
[00317] Тестировали влияние N-ABD094-rhArg (SEQ ID NO: 50) на ожирение и различные метаболические параметры у страдающих ожирением животных. Мышиная модель ожирения, индуцированного рационом с высоким содержанием жиров (DIO) [Wang and Liao, 2012, Methods Mol. Биол. 821: 421-433], имеет много общих черт с человеческим ожирением и широко используется для тестирования потенциальных средств против ожирения [Vickers et al., 2011, Br. J. Pharmacol. 164 (4): 1248-1262]. В частности, в настоящем исследовании применяли C57BL/6J, которая является наиболее часто используемой линией мышей для моделирования полигенного ожирения. Хорошо известно, что C57BL/6J мыши, питающиеся рационом с высоким содержанием жиров, будут значительно прибавлять в массе, станут страдать ожирением и у них разовьются гиперинсулинемия, гипергликемия, нарушение толерантности к глюкозе и инсулинорезистентность [Gallou-Kabani et al., 2007, Obesity 15(8): 1996-2005].
[00318] Для определения эффективности N-ABD094-rhArg (SEQ ID NO: 50) при предупреждении развития индуцированного рационом ожирения и связанных с ним метаболических нарушений самцов C57BL/6J мышей кормили рационом с высоким содержанием жиров, причем содержание жира достигало 60 ккал% (HFD; Research Diets, D12492), от 8-недельного возраста в течение 12 недель, и одновременно они получали внутрибрюшинную инъекцию 200-300 ед. N-ABD094-rhArg (SEQ ID NO: 50) или солевого раствора в качестве контрольного наполнителя с 7-суточными интервалами. C57BL/6J мыши, которых кормили равноценным рационом с низким содержанием жира, причем содержание жира достигало 10 ккал% (LFD; Research Diets, D12450J), и которым вводили солевой раствор, служили при различных измерениях в качестве контрольной группы худых мышей.
[00319] Опыт проводили раздельно на двух когортах, по n=5 в каждой группе. Мышей размещали группой по 5 животных. В двух независимых когортах были получены схожие результаты. Данные по одной когорте мышей суммированы в примерах 18-22.
[00320] По сравнению с контрольной группой худых мышей, которых кормили LFD и обрабатывали наполнителем [называемой группой «LFD(наполнитель)»], самцы мышей, которых обрабатывали наполнителем и кормили HFD [называемой группой «НFD(наполнитель)»], характеризовались заметным увеличением массы тела (фиг. 18А) и размера (фиг. 18 В) со значимым увеличением массы белой жировой ткани (WAT) в основных висцеральных (перигонадных, периренальных и брыжеечных) и подкожных (паховых) депо (фиг. 19А). Тем не менее, у мышей, которых кормили HFD, но одновременно обрабатывали посредством N-ABD094-rhArg (SEQ ID NO: 50) [называемых группой «HFD(rhArg)»], можно было эффективно предупредить увеличение массы и увеличение WAT, вызванное потреблением HFD. По результатам гистологического исследования WAT было видно, что обработка посредством N-ABD094-rhArg (SEQ ID NO: 50) заметно подавляла гипертрофию адипоцитов (фиг. 19 В). Из результатов анализа уровней экспрессии мРНК с помощью qRT-PCR в режиме реального времени в gWAT дополнительно было видно резкое снижение уровня нескольких важных адипогенных факторов транскрипции, в том числе гамма-рецептора, активируемого пролифератором пероксисом (Pparg), и белка 1 с, связывающего стеролрегулирующие элементы (Srebplc), а также липогенных ферментов (Асc1 и Scd1) (фиг. 19С). Данные гены также значимо снижали уровень активности в скелетных мышцах мышей, которых кормили HFD и обрабатывали посредством N-ABD094-rhArg (SEQ ID NO: 50) (фиг. 20), что позволяло предположить, что антилипогенезное влияние N-ABD094-rhArg (SEQ ID NO: 50) не ограничивалось WAT.
[00321] После подтверждения влияния N-ABD094-rhArg (SEQ ID NO: 50) против ожирения на самцах мышей изучали влияние N-ABD094-rhArg (SEQ ID NO: 50) на самках мышей (n=5 в каждой группе) с целью определения, есть ли какие-либо половые различия. Из результатов было видно, что одновременная обработка посредством N-ABD094-rhArg (SEQ ID NO: 50) была аналогично эффективной при защите самок мышей, которых кормили HFD с 8-недельного возраста в течение 12 недель, от увеличения массы (фиг. 21 А) и чрезмерного накопления WAT в основном висцеральном (перигонадном, периренальном и брыжеечном) и подкожном (паховом) депо (фиг. 21 В) и могла улучшать чувствительность к инсулину (фиг. 22А) и толерантность к глюкозе (фиг. 22 В).
[00322] Для тестирования влияния N-ABD094-rhArg (SEQ ID NO: 50) на жировые клетки использовали преадипоциты 3T3-L1 мышей, которые обычно используют в качестве модельных клеток для изучения адипогенеза [Green and Meuth, 1974, Cell 3:127-133]. Было обнаружено, что N-ABD094-rhArg (SEQ ID NO: 50) мог эффективно ингибировать адипогенез и липогенез в клетках 3T3-L1 (фиг. 23) и значимо подавлять уровни мРНК некоторых адипогенных факторов транскрипции (фиг. 24А) и липогенных ферментов (фиг. 24 В). Подобное ингибирующее влияние наблюдали при культивации преадипоцитов 3T3-L1 в бычьей аргиназе или другом истощающем запасы аргинина ферменте, т.е. аргининдезиминазе (ADI), или не содержащей аргинина среде (фиг. 23). Аргиназа превращает L-аргинин в L-орнитин и мочевину, тогда как ADI превращает L-аргинин в L-цитруллин. Тем не менее, L-орнитин, мочевина или L-цитруллин не могли ингибировать адипогенез и липогенез в клетках 3T3-L1 (фиг. 23).
[00323] Более того, в результате дополнения среды без аргинина L-аргинином в различных концентрациях наблюдали дозозависимое влияние аргинина на адипогенез и липогенез (фиг. 25А) с пороговой концентрацией аргинина 10-40 мкМ по активации важных адипогенных факторов транскрипции, в том числе Cebpa и Pparg и липогенного фермента Scd1 (фиг. 25 В).
[00324] В совокупности из этих данных видно, что антиадипогенезное и липогенезное влияние N-ABD094-rhArg (SEQ ID NO: 50) опосредовалось депривацией аргинина.
[00325] Совместно эти данные подтверждали, что истощение запасов аргинина слитым белком ABD-rhArg, или другими формами аргиназы (например, rhArg-PEG, BCA-PEG или BCA-ABD), или истощающими запасы аргинина ферментами (например, ADI-PEG, ADI-ABD, ADC-PEG или ADC-ABD) можно использовать в качестве превентивной терапии против ожирения.
[00326] Пример 19. Обработка посредством N-ABD094-rhArg (SEQ ID NO:50) предупреждала HFD-индуцпрованную инсулинорезистентность, нарушение толерантности к глюкозе и метаболические расстройства
[00327] Ожирение обычно связано с инсулинорезистентностью, нарушением толерантности к глюкозе и метаболическими расстройствами. Действительно, по сравнению с худыми мышами контрольной группы, которых обрабатывали наполнителем и кормили посредством LFD, мыши, которых обрабатывали наполнителем и кормили HFD, характеризовались значимым снижением чувствительности к глюкозоснижающему влиянию вводимого инсулина в тесте на толерантность к инсулину (ITT), проведенном через 6 недель после периода кормления HFD (фиг. 26 А), которое еще больше ухудшилось через 11 недель (фиг. 26 В). Одновременно со сниженной чувствительностью к инсулину эти мыши были значимо менее эффективны в возвращении глюкозы в крови к исходным уровням после инъекции D-глюкозы в тесте на толерантность к глюкозе (GTT) спустя 5 недель кормления HFD (фиг. 27 А), что означало развитие нарушения толерантности к глюкозе, которое еще больше ухудшалось через 10 недель употребления HFD (фиг. 27 В). Действительно, уровни глюкозы в крови натощак (в течение 5 часов) (фиг. 28 А) и уровни инсулина в плазме крови натощак (фиг. 28 В) были значимо повышены, а показатель HOMA-IR (фиг. 28 С), т.е. показатель инсулинорезистентности, был выше у мышей, которых кормили HFD и обрабатывали наполнителем, по сравнению с худыми мышами контрольной группы. Уровни лептина в плазме крови натощак (фиг. 29 А), общего холестерина (фиг. 29 В), триглицеридов (фиг. 29 С) и свободных жирных кислот (фиг. 29 D) были значимо увеличены при измерении через 12 недель после кормления HFD. Примечательно, что одновременная обработка посредством N-ABD094-rhArg (SEQ ID NO: 50) могла полностью предупредить развитие инсулинорезистентности, нарушение толерантности к глюкозе, гипергликемию, гиперинсулинемию, гиперлептинемию, гиперхолестеринемию, гипертриглицеридемию и гиперацидемию свободных жирных кислот, индуцированную посредством HFD.
[00328] Совместно из этих результатов видно, что N-ABD094-rhArg (SEQ ID NO: 50) может эффективно предупреждать развитие инсулинорезистентности и непереносимости глюкозы, которые являются предшественниками развития сахарного диабета 2-го типа. Он также может предупреждать развитие резистентности к лептину и других метаболических расстройств, в том числе связанной с ожирением дислипидемии.
[00329] Совместно эти данные подтверждали, что истощение запасов аргинина слитым белком ABD-rhArg, или другими формами аргиназы (например, rhArg-PEG, BCA-PEG или BCA-ABD), или истощающими запасы аргинина ферментами (например, ADI-PEG, ADI-ABD, ADC-PEG или ADC-ABD) можно использовать в качестве превентивной терапии против диабета, против дислипидемии, против атеросклероза и против метаболических расстройств.
[00330] Пример 20. Обработка посредством N-ABD094-rhArg (SEQ ID NO:50) предупреждала развитие HFD-индуцированного стеатоза
[00331] Стеатоз относится к аномальному удержанию липидов в клетке. Неалкогольный стеатоз печени (неалкогольная жировая дистрофия печени) является частым сопутствующим заболеванием, связанным с ожирением. Хотя на ранней стадии он может протекать бессимптомно, при воспалении печени он перерастает в стеатогепатит, который может прогрессировать до фиброза и далее развиться в цирроз на более поздних стадиях, что значительно увеличивает риск возникновения гепатоклеточной карциномы. Действительно, было обнаружено, что у мышей, которых обрабатывали наполнителем и кормили HFD, печень была заметно увеличена и имела бледный цвет (фиг. 30 А), при этом ее масса почти вдвое превышала ее массу у худых мышей контрольной группы (фиг. 30 В). По результатам окрашивания срезов печени масляным красным О было обнаружено обширное скопление крупных липидных капель (фиг. 31 А) с соответствующим десятикратным увеличением концентрации триглицеридов (фиг. 31 В). Одновременно с этими полученными результатами были значимо повышены уровни аланинаминотрансферазы (ALT) в сыворотке крови (фиг. 32), которые обычно измеряют в качестве биомаркеров повреждения печени. Примечательно, что одновременная обработка посредством N-ABD094-rhArg (SEQ ID NO: 50) могла эффективно предупреждать чрезмерное накопление липидов в печени мышей, которых кормили HFD, так что масса и содержание триглицеридов в печени, а также уровни ALT в сыворотке крови были схожи с таковыми у худых мышей контрольной группы. Наряду с этими результатами было обнаружено, что, хотя HFD индуцировал значимое повышение уровня нескольких важных адипогенной факторов транскрипции и лиогенных ферментов в печени, обработка посредством N-ABD094-rhArg (SEQ ID NO: 50) могла значимо подавлять активацию этих генов (фиг. 31 С). Совместно из этих результатов видно, что N-ABD094-rhArg (SEQ ID NO: 50) очень эффективен в предупреждении развития стеатоза печени.
[00332] Помимо печени ожирение также связано с чрезмерным накоплением липидов в почках (стеатоз почек), поджелудочной железе (стеатоз поджелудочной железы) и сердце (стеатоз сердца), обуславливая липотоксичность и дисфункцию этих органов. Действительно, было обнаружено, что у мышей, которых обрабатывали наполнителем и кормили HFD, наблюдали значимое увеличение массы почки (фиг. 33 А), поджелудочной железы (фиг. 33 В) и сердца (фиг. 33 С). Одновременное обработка посредством N-ABD094-rhArg (SEQ ID NO: 50) могла эффективно предупреждать увеличение массы этих органов, что позволяло предположить, что N-ABD094-rhArg (SEQ ID NO: 50) может предупреждать чрезмерное накопление липидов и липотоксичность в этих органах.
[00333] Совместно эти данные подтверждали, что слитый белок ABD-rhArg может эффективно предупреждать развитие стеатоза, а истощение запасов аргинина слитым белком N-ABD094-rhArg (SEQ ID NO: 50), или другими формами аргиназы (например, rhArg-PEG, BCA-PEG или BCA-ABD), или истощающими запасы аргинина ферментами (например, ADI-PEG, ADI-ABD, ADC-PEG или ADC-ABD) можно использовать в качестве превентивной терапии против стеатоза.
[00334] Пример 21. Обработка посредством N-ABD094-rhArg (SEP ID NO:50) предупреждала развитие HFD-индуцированного хронического воспаления
[00335] Чрезмерное увеличение количества WAT будет увеличиватьпродуцирование ряда адипокинов, которые запускают развитие слабого хронического воспаления, которое способствует развитию инсулинорезистентности. Хемоаттрактантный белок-1 моноцитов (Mcp1) является одним из ключевых провоспалительных адипокинов, секретируемых WAT, который регулирует инфильтрацию макрофагов в WAT, инсулинорезистентность и стеатоз печени при ожирении. Действительно, авторами настоящего изобретения было обнаружено, что уровень мРНК Mcp1 значимо повышался в перигонадном WAT у мышей, которых обрабатывали наполнителем и кормили HFD (фиг. 34 А). Одновременно с этим сывороточные уровни Mcp1 в кровотоке, измеренные с помощью твердофазного иммуноферментного анализа (ELIS А), были значимо повышены по сравнению с таковыми у худых мышей контрольной группы (фиг. 34 В). Другие провоспалительные адипокины, к которым относятся интерлейкин 1 бета (IIIb) и фактор некроза опухоли альфа (Tnfa), также характеризовались повышенной экспрессией мРНК в перигонадном WAT (фиг. 34 А). Следует отметить, что одновременная обработка посредством N-ABD094-rhArg (SEQ ID NO: 50) может подавлять активацию этих адипокинов в перигонадном WAT и подавлять повышение сывороточных уровней Mcp1.
[00336] Из этих результатов видно, что N-ABD094-rhArg (SEQ ID NO: 50) может предупреждать развитие слабого хронического воспаления, связанного с ожирением, и поддерживать потенциал истощения запасов аргинина слитым белком ABD-rhArg, или другими формами аргиназы (например, rhArg-PEG, BCA-PEG или BCA-ABD), или истощающем запасы аргинина ферментом (например, ADI-PEG, ADI-ABD, ADC-PEG или ADC-ABD) в качестве превентивной терапии против воспаления.
[00337] Пример 22. Обработка посредством N-ABD094-rhArg (SEQ ID NO: 50) предупреждала развитие HFD-индуцированного побеления и дерегуляции окисления жирных кислот бурого жира и усиливала недрожательный термогенез
[00338] Бурый жир (ВАТ) служит для растрачивания окислительного топлива в тепло (недрожательный термогенез). Из результатов недавних исследований видно, что он обладает большой способностью к расходу энергии для борьбы с ожирением. Напротив, побеление ВАТ связано с ожирением и нарушает функционирование ВАТ. Действительно, авторами настоящего изобретения было обнаружено, что у мышей, которых обрабатывали наполнителем и кормили HFD, было заметно увеличено количество межлопаточного ВАТ (фиг. 35 А), при этом его масса почти вдвое превышала его массу у худых мышей контрольной группы (фиг. 35 В). По результатам гистологического исследования было выявлено обширное скопление крупных однодольковых клеток, встречающихся между многодольковыми бурыми адипоцитами (фиг. 35 С), что свидетельствовало о превращении коричневых адипоцитов в бело-подобные клетки. Тем не менее, у мышей, которых кормили HFD и одновременно обрабатывали N-ABD094-rhArg (SEQ ID NO: 50), масса, размер и гистологическая картина ВАТ были такими же, как и у худых мышей контрольной группы. Авторами настоящего изобретения дополнительно было обнаружено, что обработка посредством N-ABD094-rhArg (SEQ ID NO: 50) значительно увеличивала активность ВАТ-специфического гена разобщающего белка 1 (Ucp1), который имеет решающее значение для термогенеза, а также увеличивает активность коактиватора гамма-рецептора 1-альфа, активируемого пролифератором пероксисом (Pgcla), ключевого регулятора транскрипции Ucp1 в бурых адипоцитах, при этом подавляя HFD-индуцированное снижение активности гена ацил-СоА-оксидазы 1 (Acoxl), главного гена, задействованного в окислении жирных кислот (фиг. 36). Было обнаружено, что обработка посредством N-ABD094-rhArg (SEQ ID NO: 50) может блокировать снижение активности Acoxl, что указывает на потенциал N-ABD094-rhArg (SEQ ID NO: 50) для нормализации окисления жирных кислот.
[00339] Совместно из этих результатов видно, что обработка посредством N-ABD094-rhArg (SEQ ID NO: 50) может эффективно предупреждать развитие побеления бурого жира и дерегуляции окисления жирных кислот в ВАТ, связанном с ожирением, и усиливает недрожательный термогенез для борьбы с эффектом ожирения в результате HFD.
[00340] Эти результаты подтверждали потенциал истощения запасов аргинина слитым белком ABD-rhArg, или другими формами аргиназы (например, ABD094-rhArg, BCA-PEG или BCA-ABD), или истощающим запасы аргинина ферментом (например, ADI-PEG, ADI-ABD, ADC-PEG или ADC-ABD) в качестве превентивной терапии против ожирения.
[00341] Пример 23. Обработка посредством N-ABD094-rhArg fSEO ID NO:50) эффективно уменьшала HFD-индуцированное ожирение
[00342] После подтверждения того, что одновременная обработка посредством N-ABD094-rhArg (SEQ ID NO: 50) может эффективно предупреждать развитие HFD-индуцированного ожирения и связанных с ним осложнений, а также сопутствующих заболеваний, в том числе инсулинорезистентности, непереносимости глюкозы, гипергликемии, дислипидемии, стеатоза печени, хронического воспаления и побеления ВАТ, авторами настоящего изобретения была изучена эффективность N-ABD094-rhArg (SEQ ID NO: 50) в снижении ожирения и обращении этих нарушений у мышей с уже существовавшим, индуцированным рационом ожирением (DIO). Самцов C57BL/6J мышей кормили HFD с 5-недельного возраста в течение 12 недель для индукции у них развития ожирения и связанных с ним осложнений, а также сопутствующих заболеваний. Затем их стратифицировали в соответствии с массой тела на 2 группы и обрабатывали либо 600-700 ед. N-ABD094-rhArg (SEQ ID NO: 50), либо солевым раствором в качестве контрольного наполнителя с 7-суточными интервалами в течение 12 недель, при этом их продолжали кормить HFD. Одну группу мышей кормили равноценным рационом с низким содержанием жиров (LFD) и на протяжении всего исследования вводили солевой раствор, что служило в качестве контрольной группы худых мышей при различных измерениях. Опыт был проведен раздельно на двух когортах, по n=5 в каждой группе. Мышей размещали группами по 5 животных. В двух независимых когортах были получены схожие результаты. Данные по одной когорте мышей представлены в примерах 23-28.
[00343] Двенадцать недель кормления HFD могли увеличивать массу тела самцов мышей до более 50 г, что было заметно больше, чем у худых самцов мышей контрольной группы, которых кормили LFD (фиг. 37 А и 37 В). После обработки посредством N-ABD094-rhArg (SEQ ID NO: 50) масса тела у мышей, страдающих HFD-индуцированным ожирением, постепенно снижалась. К 5 неделям масса их тела снижалась более чем на 30% и была схожей с массой у мышей из контрольной группы. Масса тела сохранялась на том же уровне до завершения 12 недель обработки. Масса жирового тела в основных висцеральных депо (перигонадном, периренальном и мезентериальном WAT) и подкожных депо (паховом WAT) у мышей с DIO, обрабатываемых посредством N-ABD094-rhArg (SEQ ID NO: 50), значимо снижалась и была очень похожа на таковую у худых мышей контрольной группы (фиг. 38 А). По результатам гистологического исследования WAT было выявлено заметное уменьшение количества адипоцитов у мышей с DIO, обработанных посредством N-ABD094-rhArg (SEQ ID NO: 50) (фиг. 38 В).
[00344] Вместе с результатами, представленными в примере 18, эти данные подтверждали, что слитый белок ABD-rhArg, или другие формы аргиназы (например, rhArg-PEG, BCA-PEG или BCA-ABD), или истощающий запасы аргинина фермент (например, ADI-PEG, ADI-ABD, ADC-PEG или ADC-ABD) можно применять для терапевтического лечения ожирения.
[00345] Пример 24. Обработка посредством N-ABD094-rhArg (SEQ ID NO:50) обращала HIT) им tvunp она иную инсулинорезистентность и нарушение толерантности к глюкозе, а также исправляла и метаболическое расстройство
[00346] До обработки у мышей, которых кормили HFD в течение 12 недель, развивалась периферическая инсулинорезистентность (фиг. 39) и нарушенная толерантность к глюкозе (фиг. 40), которые являются характеристиками преддиабета, по сравнению с худыми мышами контрольной группы. Тем не менее, к 4-м неделям после обработки посредством N-ABD094-rhArg (SEQ ID NO: 50) мыши с DIO характеризовались заметным повышением чувствительности к инсулину, которая стала сравнимой с таковой у худых мышей контрольной группы. Нормализация чувствительности к инсулину у мышей, получавших N-ABD094-rhArg (SEQ ID NO: 50) при непрерывном кормлении HFD, могла сохраняться до конца 12 недель обработки лекарственным средством. Более того, мыши, которых обрабатывали посредством N-ABD094-rhArg (SEQ ID NO: 50), к 3 неделям обработки характеризовались значимым улучшением толерантности к глюкозе и к 7 неделям обработки полностью восстанавливались до уровня, схожего с таковым у мышей из контрольной группы. Наряду с этими улучшениями уровни глюкозы в крови натощак (фиг. 41 А) и инсулина в плазме крови (фиг. 41 В) у мышей с DIO, которых обрабатывали посредством N-ABD094-rhArg (SEQ ID NO: 50), к 4-й неделе обработки rhArg восстанавливались до уровней, схожих с уровнями у худых мышей контрольной группы, и сохранялись в пределах нормального диапазона до конца периода обработки. Это приводило к резкому снижению показателя HOMA-IR с 35,1 у мышей до обработки и до 1,2 у мышей спустя 4 недели обработки (фиг. 41 С), что приводило к обращению инсулинорезистентности. Уровни лептина (фиг. 42 А) и общего холестерина (фиг. 42 В) в плазме крови натощак также заметно снижались до нормальных уровней.
[00347] В совокупности из наших результатов видна высокая эффективность N-ABD094-rhArg (SEQ ID NO: 50) при обращении инсулинорезистентности, нарушения толерантности к глюкозе, гипергликемии, гиперинсулинемии, гиперлептинемии и гиперхолестеринемии в пределах 4 недель обработки.
[00348] В совокупности эти результаты подтверждали, что истощение запасов аргинина при помощи слитого белка ABD-rhArg, или других форм аргиназы (например, rhArg-PEG, BCA-PEG или BCA-ABD), или истощающего запасы аргинина фермента (например, ADI-PEG, ADI-ABD, ADC-PEG или ADC-ABD) можно применять в качестве средства, повышающего чувствительность к инсулину, и для терапевтического лечения преддиабета и сахарного диабета 2-го типа, характеризующихся инсулинорезистентностью, непереносимостью глюкозы и гипергликемией, а также других нарушений, связанных с инсулинорезистентностью. Его также можно применять для терапевтического лечения гиперлептинемии и гиперхолестеринемии.
[00349] Пример 25. Обработка посредством N-ABD094-rhArg (SEQ ID NO: 50) обращала развитие HFD-индуцированного стеатоза
[00350] Двенадцать недель кормления посредством HFD индуцировали стеатоз печени у самцов мышей (фиг. 30 и 31). После продления кормления посредством HFD на 12 недель печень еще больше увеличивалась в размерах и становилась бледной (фиг. 43 А). Масса печени была более чем в два раза больше, чем у худых мышей контрольной группы (фиг. 43 В). Массивное накопление липидов распространялось на всю печень (фиг. 44 А) с соответствующим увеличением содержания триглицеридов (фиг. 44 В). Уровни экспрессии важных адипогенных факторов транскрипции Pparg и Srebplc были подавлены до уровней, схожих с таковыми у худых мышей контрольной группы, при этом уровни экспрессии липогенных ферментов Асc1 были снижены еще больше, а уровни экспрессии Scdl были коренным образом подавлены (фиг. 44 С). При сравнении с мышами, которых кормили HFD в течение 12 недель (пример 20, фиг. 32), наблюдали дополнительное повышение уровней сывороточной аланинаминотрансферазы (ALT) у мышей, которых кормили HFD в течение 24 недель, а также значимо увеличивались уровни сывороточной аспартатаминотрансферазы (AST) (фиг. 45), что свидетельствовало о дальнейшем прогрессировании поражения печени. Однако у мышей с DIO, которых обрабатывали посредством N-ABD094-rhArg (SEQ ID NO: 50), наблюдали заметное уменьшение размеров печени до такой степени, что масса была схожей с таковой у худых мышей контрольной группы (фиг. 43 В). Более того, печень восстанавливалась до красноватого цвета (фиг. 43 А), что хорошо коррелировало с резким клиренсом липидных капель (фиг. 44 А), при этом концентрации триглицеридов снижались до уровня, схожего с таковым у худых мышей контрольной группы (фиг. 44 В). Следует отметить, что концентрации ALT и AST в плазме крови были сравнимы с таковыми у худых мышей контрольной группы, что позволяло предположить, что обработка посредством N-ABD094-rhArg (SEQ ID NO: 50) может позволить предупредить дальнейшее прогрессирование и исправить существующее повреждение печени. В совокупности из наших результатов видно выраженное влияние N-ABD094-rhArg (SEQ ID NO: 50) на обращение стеатоза печени.
[00351] Помимо печени ожирение также связано с чрезмерным накоплением липидов в почках (стеатоз почек), поджелудочной железе (стеатоз поджелудочной железы) и сердце (стеатоз сердца), обуславливая липотоксичность и дисфункцию этих жизненно важных органов. Действительно, у мышей с DIO, получавших наполнитель, наблюдали значимое увеличение массы почки (фиг. 46 А), поджелудочной железы (фиг. 47 А) и сердца (фиг. 48 А). По результатам гистологического исследования было выявлено скопление вакуолей, которые напоминают липидные капли в почечных канальцах (фиг. 46 С). По результатам окрашивания срезов почек масляным красным О было видно, что в клубочках имело место эктопическое накопление липидов (фиг. 46 D). В поджелудочной железе имело место обширное внутридольковое и междольковое накопление пятен WAT (фиг. 47 В), а среди волокон сердечной мышцы были обнаружены островки белых адипоцитов (фиг. 48 В). Концентрации триглицеридов значимо возрастали в почке (фиг. 46 В), поджелудочной железе (фиг. 47 С), сердце (фиг. 48 С). Более того, значимое повышение концентраций триглицеридов также было обнаружено в скелетной мышце (фиг. 49). Эти данные указывали на развитие стеатоза в этих органах мышей с DIO, которых обрабатывали наполнителем. Однако обработка посредством N-ABD094-rhArg (SEQ ID NO: 50) могла значимо снижать массу почки (фиг. 46 А), поджелудочной железы (фиг. 47 А) и сердца (фиг. 48 А) мышей с DIO до уровня, схожего с таковым у худых мышей контрольной группы. Результаты гистологического исследования подтверждали заметное уменьшение количества вакуолей в почечных канальцах (фиг. 46 С) и липидов в клубочках почек, выявленное при окрашивании масляным красным О (фиг. 46 D). Имело место заметное снижение количества WAT в поджелудочной железе (фиг. 47 В) и в сердце (фиг. 48 В). Концентрации триглицеридов в почке (фиг. 46 В), поджелудочной железе (фиг. 47 С), сердце (фиг. 48 С) и скелетной мышце (фиг. 49) у мышей с DIO, которых обрабатывали посредством N-ABD094-rhArg (SEQ ID NO: 50), восстанавливались до уровня, схожего с таковым у худых мышей контрольной группы. Из этих результатов было видно, что N-ABD-rhArg (SEQ ID NO: 50) может эффективно обращать развитие стеатоза в почке, поджелудочной железе и сердце, а также, возможно, и в других органах.
[00352] Совместно эти данные подтверждали, что слитый белок ABD-rhArg, или другие формы аргиназы (например, rhArg-PEG, BCA-PEG или BCA-ABD), или истощающий запасы аргинина фермент (например, ADI-PEG, ADI-ABD, ADC-PEG или ADC-ABD) можно применять для терапевтического лечения стеатоза.
[00353] Пример 26. Обработка посредством N-ABD094-rhArg fSEO ID NO:50) уменьшала HFD-индуцированное хроническое воспаление и фиброз
[00354] Как проиллюстрировано в примере 21, кормление HFD индуцировало развитие хронического воспаления. У мышей с DIO, которых обрабатывали наполнителем, одновременно с повышенной экспрессией провоспалительного адипокина Mcp1 в перигонадном WAT (фиг. 50 А) были значимо повышены сывороточные концентрации Mcp1 (фиг. 50 В). С другой стороны, у мышей с DIO, которых обрабатывали посредством N-ABD094-rhArg (SEQ ID NO: 50), значения экспрессии МСР1 в перигонадном WAT были значимо снижены, а значения сывороточной концентрации МСР1 скорректированы до уровня, схожего с таковым у худых мышей контрольной группы (фиг. 50 В), что позволяло предположить, что обработка посредством N-ABD094-rhArg может облегчать HFD-индуцированное хроническое воспаление в WAT.
[00355] В печени мышей с DIO, которых обрабатывали наполнителем, наблюдали значимую активацию нескольких провоспалительных генов, в том числе генов интерлейкина 1 альфа и 1 бета (IIIa и IIIb) (фиг. 51 А). Хроническое воспаление связано с фиброзом. И действительно, наблюдали значимую активацию важного профибротического гена трансформирующего ростового фактора бета 1 (Tgfb) (фиг. 51 В). Обработка посредством N-ABD094-rhArg (SEQ ID NO: 50) могла значимо подавлять активность этих провоспалительных генов и заметно активировать несколько противовоспалительных генов, в том числе гены интерлейкина 10 (1110), интерлейкина 22 (1122) и интерферона гамма (Infg) (фиг. 51 С). Экспрессия профибротического гена Tgfb восстанавливалась до уровня, схожего с таковым у худых мышей контрольной группы. Как и в случае с печенью, в почке мышей с DIO, которых обрабатывали наполнителем, наблюдали значимую активацию нескольких провоспалительных генов, в том числе Mcp1, гена фактора некроза опухоли альфа (Tnfa) и IIIb (фиг. 52 А). Профибротический ген цепи альфа-1 коллагена 1-го типа (Collal) также был значимо активирован, что согласовывалось с фактами избыточного отложения коллагеновых волокон, выявленных в почке при окрашивании сириусом красным (фиг. 52 С). Следует отметить, что обработка мышей с DIO посредством N-ABD094-rhArg (SEQ ID NO: 50) могла подавлять экспрессию этих провоспалительных генов до уровней, схожих с таковыми у худых мышей контрольной группы. Был значительно подавлен профибротический ген Collal, и заметно уменьшалось количество коллагеновых волокон. Аналогичным образом, в поджелудочной железе обработка посредством N-ABD094-rhArg (SEQ ID NO: 50) могла значимо подавлять такой провоспалительный ген, как Mcp1 (фиг. 52 А), и важный профибротический ген Tgfb (фиг. 53 В).
[00356] В своей совокупности эти данные свидетельствовали, что обработка посредством N-ABD094-rhArg могла эффективно облегчать хроническое воспаление и фиброз печени, почки, поджелудочной железы и, возможно, других органов.
[00357] Жировая дистрофия печени (стеатоз печени) на ранней стадии может протекать бессимптомно. При воспалении печени он перерастает в стеатогепатит, который может прогрессировать до фиброза и далее развиться в цирроз на более поздних стадиях, что значительно увеличивает риск возникновения гепатоклеточной карциномы. Наши результаты подтверждали, что обработка посредством N-ABD094-rhArg может останавливать прогрессирование заболевания печени и обращать ее повреждение.
[00358] Все вместе эти результаты подтверждали, что истощение запасов аргинина слитым белком ABD-rhArg, или другими формами аргиназы (например, rhArg-PEG, BCA-PEG, BCA-ABD), или истощающим запасы аргинина ферментом (например, ADI-PEG, ABD-ADI, ADC-PEG или ADC-ABD) можно применять для терапевтического лечения хронического воспаления или фиброза.
[00359] Пример 27. Обработка посредством N-ABD094-rhArg (SEQ ID NO:50) обращала побеление бурого жира
[00360] После продолжительного кормления на HFD наблюдали дополнительное увеличение размера (фиг. 54 А) и массы межлопаточного ВАТ (фиг. 54 В) у мышей с DIO, которых обрабатывали наполнителем, по сравнению с худыми мышами контрольной группы, что коррелировало с присутствием большого количества увеличенных белесых однодольковых клеток (фиг. 54 С). Напротив, обработка мышей с DIO посредством N-ABD094-rhArg (SEQ ID NO: 50) могла резко восстановить межлопаточный ВАТ до размера и массы, схожих с таковыми у худых мышей контрольной группы, с почти полной ремиссией белесых однодольковых клеток. Совместно эти результаты свидетельствовали о значительной способности N-ABD094-rhArg (SEQ ID NO: 50) обращать побеление бурого жира.
[00361] Совместно эти результаты подтверждали, что слитый белок ABD-rhArg, или другие формы аргиназы (например, rhArg-PEG, BCA-PEG или BCA-ABD), или истощающий запасы аргинина фермент (например, ADI-PEG, ADI-ABD, ADC-PEG или ADC-ABD) можно применять для терапевтического лечения ожирения путем обращения побеления бурого жира.
[00362] Пример 28. Обработка посредством N-ABD094-rhArg (SEQ ID NO:50) снижала кровяное давление
[00363] Ожирение связано с сердечно-сосудистым заболеванием и гипертонией. У мышей с DIO, которых обрабатывали посредством 600 ед. N-ABD094-rhArg (SEQ ID NO: 50) с 7-суточными интервалами в течение 5 недель, измеряли кровяное давление способом хвостовой манжеты с применением неинвазивной системы измерения кровяного давления CODA (Kents Scientific Corporation). Было обнаружено, что систолическое и диастолическое кровяное давление (фиг. 55 А), а также частота сердечных сокращений (фиг. 55 В) у мышей с DIO, которых обрабатывали посредством N-ABD094-rhArg (SEQ ID NO: 50), были значимо ниже, чем у мышей с DIO, которых обрабатывали наполнителем, и были очень сопоставимы с таковыми у худых мышей контрольной группы. В примере 25 продемонстрировано, что N-ABD094-rhArg (SEQ ID NO: 50) может эффективно обращать стеатоз почек и сердца, что может оказывать положительное влияние на снижение кровяного давления.
[00364] Совместно эти данные подтверждали, что слитый белок ABD-rhArg, или другие формы аргиназы (например, rhArg-PEG, BCA-PEG или BCA-ABD), или истощающий запасы аргинина фермент (например, ADI-PEG, ADI-ABD, ADC-PEG или ADC-ABD) можно применять для терапевтического лечения гипертонии.
[00365] Пример 29. N-ABD0094-rhArg (SEP ID NO: 50) не индуцировал выработку нейтрализующих антител и оставался эффективным при продолжительном применении
[00366] Образование нейтрализующих антител может влиять на фармакодинамику, фармакокинетику, стабильность и эффективность лекарственного средства. Для определения иммуногенного ответа 8-недельным самцам C57BL/6J мышей, которых кормили LFD, делали внутрибрюшинную инъекцию 500 ед. N-ABD094-rhArg (SEQ ID NO: 50) или солевого раствора в качестве контрольного наполнителя с 7-суточными интервалами в течение 8 месяцев. Было обнаружено, что несмотря на наличие антител к лекарственному средству, т.е. к N-ABD094-rhArg (SEQ ID NO: 50) (фиг. 56A), данные антитела не оказывали нейтрализующего действия на аргиназную активность (фиг. 56 В), что согласовывалось выводами о том, что концентрации аргинина в плазме крови можно поддерживать на уровнях, которые ниже предела детекции анализатора аминокислот Biochrom 30 (3 мкМ), на протяжении всего периода исследования.
[00367] Для оценки безопасности длительного применения образцы сыворотки крови и мочи мышей, которых кормили LFD и обрабатывали посредством N-ABD094-rhArg (SEQ ID NO: 50), собирали в течение 8 месяцев для анализа на ALT и AST, которые являются традиционно используемыми биомаркерами повреждения печени, а также на сывороточный креатинин и соотношение альбумин/креатинин в моче для оценки функции почек. Было обнаружено, что мыши, которых кормили LFD и обрабатывали лекарственным средством, не характеризовались по всем этим параметрам какими-либо значимыми отличиями от мышей, которых кормили LFD и обрабатывали наполнителем (фиг. 57 А и 57 В). Более того, при изучении функциональных ответов и сосудистой реактивности изолированных артерий с помощью проволочной миографии между мышами, которых кормили LFD и обрабатывали посредством N-ABD094-rhArg (SEQ ID NO: 50), и мышами, которых кормили LFD и обрабатывали наполнителем, не было обнаружено значимых различий в сокращении гладких мышц сосудов, вызванном фенилэфрином (фиг. 58 А), ацетилхолин-индуцированной эндотелий-зависимой релаксации (фиг. 58 В) и эндотелий-независимой релаксации в ответ на нитропруссид натрия, который является донором оксида азота (фиг. 58 С).
[00368] В отдельном исследовании, в котором самцов C57BL/6J мышей с индуцированным рационом ожирением обрабатывали посредством 600 ед. N-ABD094-rhArg (SEQ ID NO: 50) в течение 20 недель, при этом продолжали кормление HFD, к 5 неделе масса тела снижалась до уровня, схожего с таковым у худых мышей контрольной группы, и продолжала поддерживаться на том же уровне без повторного увеличения. В результате регулярного отслеживания концентраций аргинина в плазме наблюдали, что уровень аргинина оставался ниже предела детекции анализатора аминокислот Biochrom 30 (3 мкМ). Эти результаты подтверждали, что N-ABD094-rhArg (SEQ ID NO: 50) оставался эффективным в течение всего периода обработки. Совместно эти данные подтверждали, что N-ABD094-rhArg (SEQ ID NO: 50) безопасен и эффективен для длительного применения.
[00369] Пример 30. Протокол пегилирования рекомбинантной человеческой аргиназы (rhArg)
[00370] Рекомбинантную человеческую аргиназу (His-rhArg SEQ ID NO: 101) получали путем очистки с применением аффинной хроматографии. Перед пегилированием очищенный His-rhArg (SEQ ID NO: 101) подвергали диафильтрации против 20 мМ фосфата натрия (рН 8,0), а концентрацию белка доводили до 2 мг/мл. Реагенты для пегилирования готовили в 20 мМ фосфате натрия (рН 8,0). Использовали молярное соотношение PEG-реагента к rhArg 20:1. PEG-реагент добавляли к His-rhArg (SEQ ID NO: 101), а затем инкубировали в течение 4 часов при комнатной температуре с перемешиванием. Затем реакционную смесь подвергали диафильтрации против 20 мМ фосфата натрия (рН 7,4) с помощью тангенциальной поточной фильтрации (TFF). Избыток PEG в проскоке удаляли с помощью аффинной хроматографии и выделяли пегилированный His-rhArg (SEQ ID NO: 101), элюируя белок 0,5 М имидазолом. Весь недериватизированный His-rhArg (SEQ ID NO: 101) отделяли с помощью еще одного раунда аффинной хроматографии. Затем реакционную смесь анализировали методом SDS-PAGE с применением 12% полиакриламидного геля. Гель красили кумасси синим для определения молекулярной массы или 0,1 М раствором йода для детекции свободного PEG.
[00371] Например, в препарате получали 150 мг пегилированного His-rhArg (SEQ ID NO: 101) со специфической активностью 245 ед./мг и концентрацией 7,5 мг/мл. На фиг. 11 и в таблице 5 представлен пример получения PEG-rhArg и результаты его анализа.
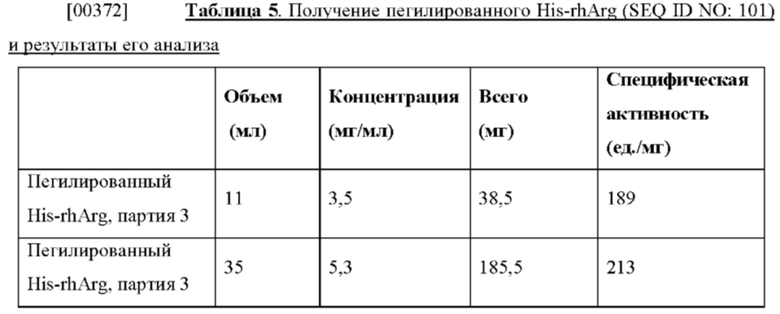
[00373] Пример 31. Обработка пегилированным His-rhArg [SEQ ID NO:1011 уменьшала ожирение и улучшала чувствительность к инсулину
[00374] Получение пегилированного His-rhArg (SEQ ID NO: 101) описано в примере 30. Самцам C57BL/6J мышей с уже существовавшим ожирением, индуцированным рационом (DIO), вводили внутрибрюшинной инъекцией 300 ед. пегилированного His-rhArg [SEQ ID NO: 101] или солевого раствора в качестве контрольного наполнителя с 7-суточными интервалами в течение 5 недель при непрерывном кормлении HFD. Масса тела мышей, которых обрабатывали пегилированным His-rhArg (SEQ ID NO: 101), постепенно снижалась до 30 г, что было похоже на массу тела у худых мышей контрольной группы, которых обрабатывали наполнителем и кормили стандартным рационом (фиг. 59). Масса жирового тела в основных висцеральных депо (перигонадном, периренальном и мезентериальном WAT) и подкожных депо (паховом WAT) значимо снижалась и в конце исследования была схожа с таковой у худых мышей контрольной группы (фиг. 60). Масса печени, почки, поджелудочной железы и сердца была значимо ниже, чем у мышей с DIO, которых обрабатывали наполнителем, и была сравнима с таковой у худых мышей контрольной группы (фиг. 60). В результате регулярного отслеживания концентраций аргинина в плазме подтверждали, что уровень аргинина оставался ниже предела детекции анализатора аминокислот Biochrom 30 (3 мкМ) на протяжении всего исследования.
[00375] Через две недели после обработки пегилированным His-rhArg [SEQ ID NO: 101] мыши с DIO уже имели более низкие уровни глюкозы в крови натощак (фиг. 61 А) и значимое увеличение чувствительности к инсулину по сравнению с мышами с DIO, которых обрабатывали наполнителем (фиг. 61 В).
[00376] В своей совокупности эти данные подтверждали, что пегилированный His-rhArg [SEQ ID NO: 101] обладает противожировым и инсулин-сенсибилизирующим действием, схожим с таковым у N-ABD094-rhArg [SEQ ID NO: 50]. Депривация аргинина с помощью пегилированного His-rhArg (SEQ ID NO: 101) оказывала терапевтическое действие при лечении ожирения и заболеваний, связанных с инсулинорезистентностью.
[00377] Пример 32. Замещение иона металлов
[00378] Аргиназа дикого типа обычно включает ион металла Mn2+ (D'Antonio & Christianson, 2011, Biochemistry, 50:8018-27). Было обнаружено, что замещение иона металла представляет собой способ получения гораздо более сильнодействующей формы ABD094-rhArg (с низким значением Km и высоким значением kcat/Km). Очищенный N-ABD094-rhArg (SEQ ID NO: 50) в 20 мМ Tris-HCl буфере (рН 7,4) смешивали с 20 мМ различных двухвалентных ионов металлов (Mn2+, Ni2+ или Со2+) и подвергали инкубации при 52-55°С в течение 15-60 минут, а затем заменяли буфер на PBS-буфер (рН 7,4).
[00379] У образцов изучали ферментативную активность и проводили над ними ферментно-кинетические исследования для определения специфической активности и каталитической эффективности (kcat/Km). Образцы использовали для атомно-эмиссионной спектроскопии с индуктивно связанной плазмой (ICP-OES, спектрометр для атомно-эмиссионной спектроскопии с ICP марки Agilent 700 Series) для количественного анализа содержания металлов.
[00380] Каждый образец белка (100-200 мкл) добавляли к 10 мл 1% (масс/об) HNO3, а затем выбирали различные элементы и значения длины волны для ICP-OES-анализа. Соотношение металла к белку определяли делением концентрации металла на концентрацию белка. Для оценки концентрации белка использовали анализ белка Pierce™ ВСА в соответствии с инструкциями производителя (Thermo Fisher Scientific).
[00381] Добавление иона металла Со2+ или Ni2+ к N-ABD094-rhArg (SEQ ID NO: 50) приводило к надлежащему удалению Mn2+ и включению Со2+ или Ni2+.
[00382] Было обнаружено, что замещенный кобальтом фермент (N-ABD094-rhArg-Co2+) обладал гораздо меньшей Km (0,27-0,35 мМ), более высокой kcat (250-316 с-1) и более высокой каталитической эффективностью kcat/Km (905-917 с-1 мМ-1). В случае исходного фермента (N-ABD094-rhArg-Mn2+) он имел Km 1,37-1,89 мМ, kcat 82-98 с-1 и kcat/Km 44-72 с-1 мМ-1. С помощью способа замещения металла успешно заменяли Mn2+ на Со2+, что приводило к получению соотношения насыщенного Со2+ к белку примерно 2-4:1. Эти данные подтверждали, что путем замещения ионами других двухвалентных металлов (например, Ni2+ или Со2+) можно получать более эффективную форму N-ABD094-rhArg (SEQ ID NO: 50) или других слитых белков, описанных в настоящем документе.
[00383] Пример 33. Обработка посредством N-ABD094-rhArg-Co2+ [SEP ID NO: 50 (с кобальтовым заместителем)] уменьшала ожирение
[00384] Получение N-ABD094-rhArg-Co2+ [SEQ ID NO: 50 (с кобальтовым заместителем)] описано в примере 32. Самцам C57BL/6J мышей с уже существовавшим ожирением, индуцированным рационом (DIO), вводили внутрибрюшинной инъекцией 25-100 ед. N-ABD094-rhArg-Co2+ [SEQ ID NO: 50 (с кобальтовым заместителем)] в течение э недель при непрерывном кормлении HFD. Масса тела у мышей постепенно снижалась с 55 г до 30 г за 5 недель обработки (фиг. 62), что было схожим с результатом воздействия 600-700 ед. N-ABD094-rhArg (фиг. 37). Данный результат подтверждал, что замещение Мп2+ на Со2+ заметно увеличивало эффективность слитого белка ABD-rhArg в отношении снижения массы тела.
[00385] Пример 34. Культура клеток млекопитающих и анализ их пролиферации
[00386] Линии раковых клеток (например, MKN45, AGS, BGC823, НСТ-15 и НСТ-116) поддерживали в среде RPMI с 10% FBS и 100 ед./мл смеси пенициллина/стрептомицина по стандартному протоколу.
[00387] Для анализа пролиферации клеток раковые клетки (5×103) в объеме 100 мкл ростовой среды высевали в каждую лунку 96-луночного планшета и инкубировали в течение 24 часов. Культуральную среду дополняли средой без аргинина (AFM), дополненной аргинином в различных концентрациях, свежей средой, содержащей в различной концентрации различные ферменты, истощающие запасы аргинина, а также с применением этих ферментов в сочетании с цитруллином. Планшет инкубировали в течение дополнительных 1-3 суток при 37°С в термостате с влажной атмосферой, состоящей из 95% воздуха и 5% СО2.
[00388] Количественные анализы пролиферации клеток проводили с помощью МТТ-проб (3-[4,5-диметилтиазол-2-ил]-2,5-дифенилтетразолия бромида), как подробно описано в других документах. За IC50 принимали количество истощающего запасы аргинина фермента, необходимое для уничтожения 50% клеток в культуре.
[00389] Пример 35. Ингибирование пролиферации различных линий раковых клеток человека различными истощающими запасы аргинина ферментами (API, ВСА и АРС)
[00390] Цитотоксичность аргининдезиминазы (ADI), бактериальной аргиназы (ВСА) и аргининдекарбоксилазы (ADC) определяли в сравнительном исследовании на пяти линиях опухолевых клеток: А375, HeLa, ВхРС-3, PANC-1 и НСТ116.
[00391] Поскольку реакции, катализируемые как ADI, так и ВСА, являются стадиями цикла мочевины, противоопухолевое действие как ADI, так и ВСА связано с уровнем клеточной ASS. В отличие от этого, на противоопухолевое действие ADC может относительно меньше влиять присутствие ASS, поскольку ни один из каталитических продуктов ADC не является промежуточным звеном цикла мочевины. Нашими результатами было продемонстрировано, что все из ADI, ВСА и ADC оказывали ингибирующее действие на исследуемые клеточные линии. ADI была эффективной против А375, НСТ116, ВхРС-3, PANC-1, но не против клеток HeLa, в то время как ВСА и ADC были эффективны против всех этих пяти протестированных линий раковых клеток (таблица 6). ADC является наименее активным истощающим запасы аргинина ферментом, но обладает преимуществом перед ADI и ВСА с точки зрения эффективности, особенно в отношении ASS-положительных раковых клеток (таблица 6).
[00392] Чувствительность опухолевых клеток к ADI свидетельствовала о четкой взаимосвязи с клеточным уровнем ASS (таблица 6). Взятые совместно, ADC и ВСА, в отличие от ADI, могут ингибировать пролиферацию линий раковых клеток человека независимым от ASS образом.
[00393] Таблица 6. IC50 и максимальная цитотоксичность ADI, ВСА и ADC на пяти линиях раковых клеток человека. Перед анализом с применением МТТ клетки обрабатывали в течение 72 часов. Данные представлены в средних значениях по результатам трех экспериментов, проведенных в трех повторностях. Значение IC50 определяли как количество фермента, необходимое для достижения 50% ингибирования жизнеспособности клеток. Максимальную цитотоксичность выражали как максимальный процент нежизнеспособных клеток, достигаемый с каждым ферментом. Устойчивые к ADI клетки определяли как клетки, более 50% которых выживали при максимальной протестированной дозе ADI.
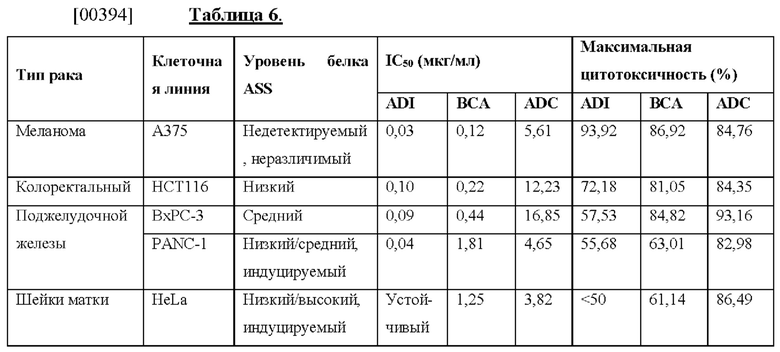
[00395] Пример 36. Цитотоксичность ВНА (SEQ ID NO: 75) в отношении клеток рака желудка
[00396] In vitro эффективность альбуминсвязывающей аргиназы ВНА (SEQ ID NO: 75) тестировали путем инкубации линии клеток рака желудка MKN45 с ВНА (SEQ ID NO: 75) в различных концентрациях в течение 72 часов и определения жизнеспособности MKN45 с помощью анализа с применением МТТ. Как видно на фиг. 63, рост раковых клеток MKN45 ингибировался посредством ВНА (SEQ ID NO: 75) дозозависимым образом. Это дополнительно подтверждало цитотоксичность истощающего запасы аргинина слитого белка в культуральных условиях, в которых среда была дополнена фетальной бычьей сывороткой (FBS) и, в том числе, альбумином, а также другими различными белками.
[00397] Пример 37. Оценка апоптоза с помощью двойного окрашивания аннексином V-FITC и PI
[00398] Для оценки индукции апоптоза аргиназой ВСА (SEQ ID NO: 71) 3×105 клеток высеивали в 6-луночный планшет за одни сутки до различных условий обработки. Удаляли среду и повторно заполняли полной средой с ВСА (SEQ ID NO: 71) в различных концентрациях. Как плавающие, так и прикрепленные клетки собирали после воздействия в течение 24, 48 и 72 часов посредством ВСА. Вкратце: плавающие клетки в культуральной среде собирали центрифугированием в количестве 100 мкг в течение 3 минут при комнатной температуре, при этом прикрепленные клетки промывали PBS и поднимали на поверхность путем инкубирования с 0,25% раствором трипсина в течение 3 минут при 37°С. После добавления полной среды клетки собирали центрифугированием. Клетки промывали PBS и ресуспендировали в 0,1 мл связывающего буфера (0,01 М HEPES, рН 7,4; 0,14 М NaCl; 2,5 мМ CaCl2), содержащего йодид пропидия (PI) и аннексии V-FITC. Клетки окрашивали в темноте при комнатной температуре в течение 15 минут и перед анализом методом проточной цитометрии с применением FACSAria (BD biosciences) в каждую пробирку добавляли 0,4 мл связывающего буфера. Питометр настраивали на получение данных от 10000 клеток для каждого образца, а данные анализировали с помощью программного обеспечения FACSAria.
[00399] Согласно результатам проточной цитометрии, при обработке посредством ВСА (SEQ ID NO: 71) (50 мкг/мл ВСА) апоптоз индуцировался в клетках рака толстой кишки человека НСТ116 в зависимости от времени. Через 24, 48 и 72 часа после обработки ВСА апоптозу подвергались соответственно ~16%, 21% и 25% раковых клеток НСТ116.
[00400] Пример 38. Анализ жизнеспособности клеток и воздействие комбинированных лекарственных средств
[00401] В данных исследованиях использовали линии клеток рака молочной железы (например, человеческие клетки MCF-7 и MDA-MB-231). Раковые клетки (5×103 клеток) высевали в 96-луночные планшеты (ТРР) со 100 мкл полной среды. Планшеты инкубировали в термостате в течение ночи и позволяли клеткам приклеиться к стенкам планшета. Затем в 96-луночный планшет добавляли аргиназу His-rhArg (SEQ ID NO: 101) (в количестве 2 ед./мл до 0,0032 ед./мл) и ингибитор аутофагии хлорохин (CQ, в количестве от 100 мкМ до 0,16 мкМ). В случае группы, получавшей комбинацию лекарственных средств, His-rhArg (SEQ ID NO: 101) (в количестве от 2 ед./мл до 0,0032 ед./мл), в лунки добавляли с 20 мкМ CQ (способ непостоянного соотношения). Каждую дозу повторяли в трех повторностях в 3 лунках для получения среднего значения изменения жизнеспособности клеток. Спустя 68 ч инкубации добавляли 10 мкл 5 мг/мл раствора МТТ [3-(4,5-диметилтиазол-2-ил)-2,5-дифенилтетразолийбромида]. Планшеты снова помещали в термостат и инкубировали дополнительные 4 часа. Для растворения нерастворимого соединения формазана, образованного под действием МТТ, добавляли 100 мкл 10% SDS в 0,01 М HCl. Планшеты инкубировали при 37°С в течение ночи. Изменение цвета измеряли спектрофотометром на длине волны 570 нм. Результаты контрольного эксперимента (без какого-либо лекарственного средства) использовали в качестве основы для расчета процента жизнеспособности клеток, который использовали для построения кривой зависимости дозы от ответа с помощью программного обеспечения Prism 6.0 (GraphPad) для определения значений IC50. Эффекты взаимодействия или аддитивности между His-rhArg (SEQ ID NO: 101) и CQ можно рассчитать с помощью показателя аддитивности (CI). Для расчета значений CI применяли программное обеспечение CalcuSyn (Biosoft).
[00402] Раковые клетки MCF-7 были чувствительны к обусловленному аргиназой аргининовому голоданию в течение 72 часов (таблица 7). Для CQ также наблюдали его цитотоксическое действие на MCF-7 (таблица 8). У группы комбинации лекарственных средств наблюдали синергические эффекты (CI<1) между His-rhArg (SEQ ID NO: 101) и CQ (таблица 9). В таблице 10 показаны IC50 для His-rhArg (SEQ ID NO: 101), CQ и rhArg плюс CQ против раковых клеток MCF-7.
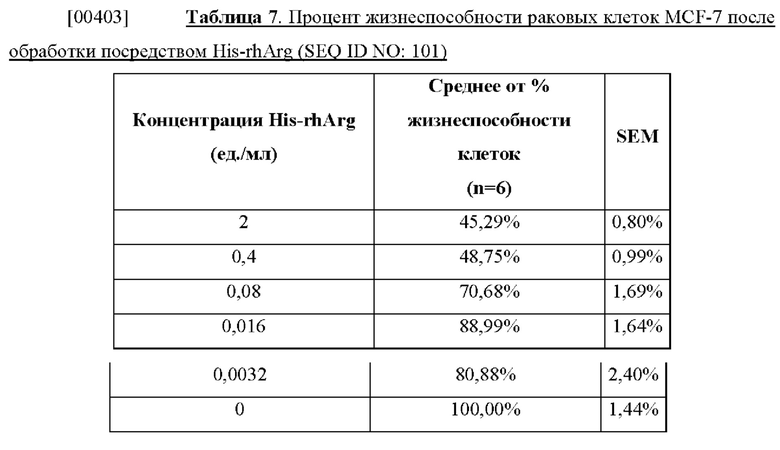
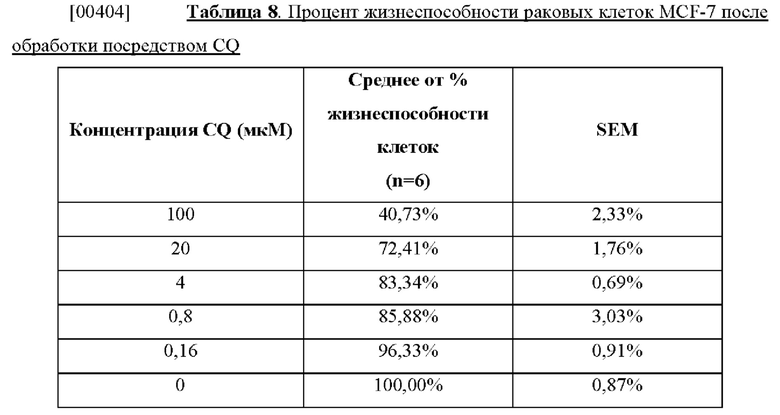
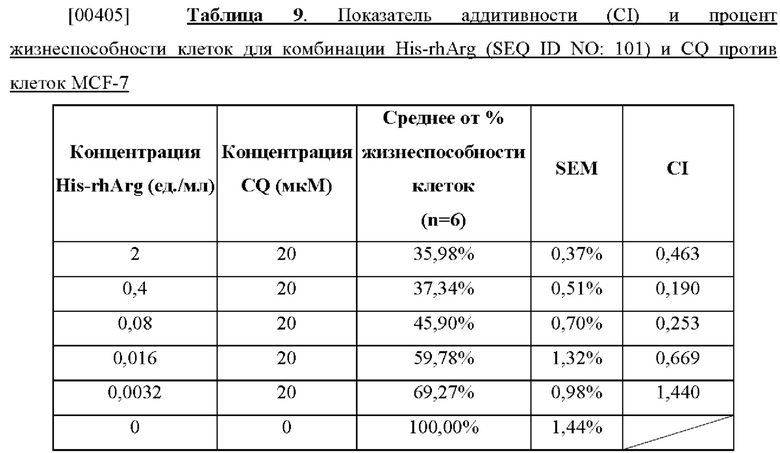
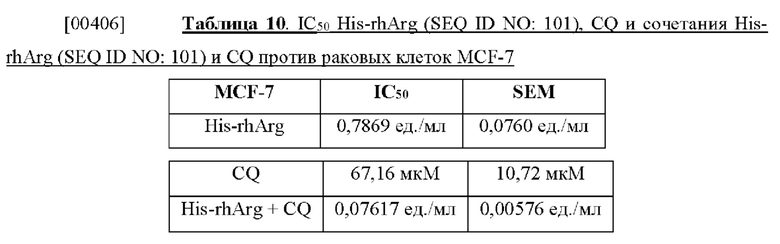
[00407] Из экспериментальных данных было видно, что клетки рака
молочной железы MDA-MB-231 были чувствительны к аргининовому голоданию, обусловленному посредством His-rhArg (SEQ ID NO: 101) и CQ (таблица 11 и таблица 12). Комбинация His-rhArg (SEQ ID NO: 101) и CQ также обладала синергической цитотоксичностью в отношении клеток MDA-MB-231. Значение CI было меньше 1 (таблица 13). В таблице 14 показаны IC50 His-rhArg (SEQ ID NO: 101), CQ и His-rhArg (SEQ ID NO: 101) плюс CQ против клеток рака молочной железы MDA-MB-231.
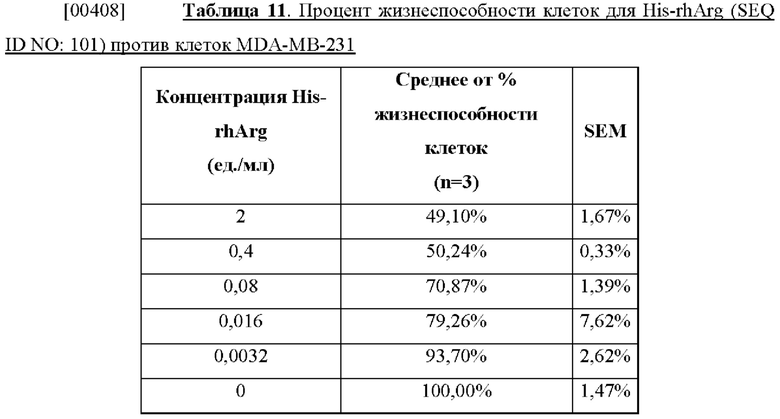
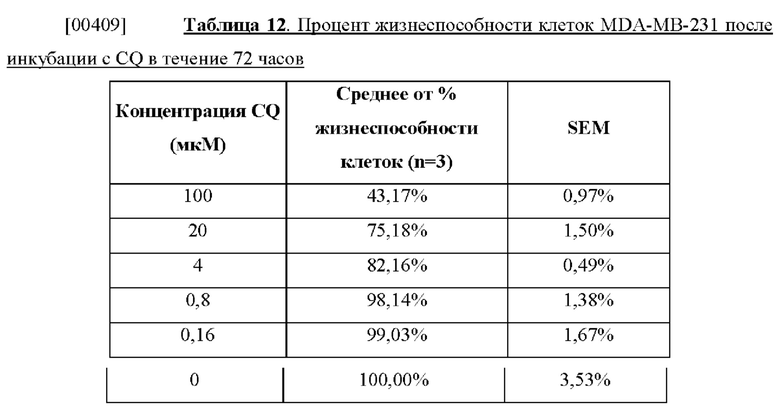
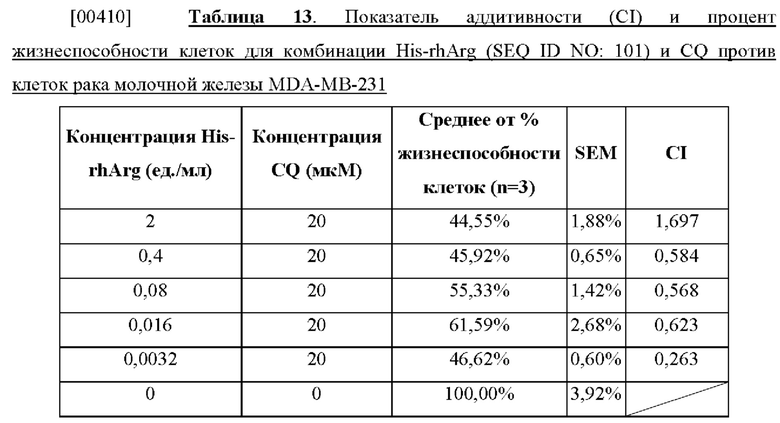
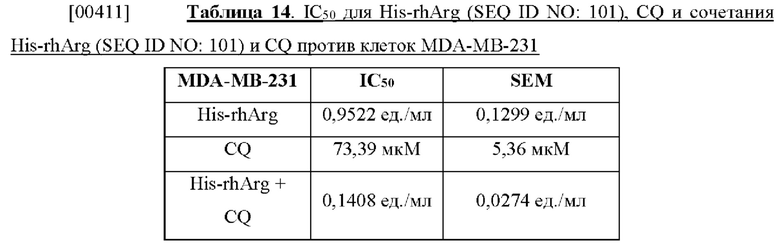
[00412] Авторами настоящего изобретения также была протестирована лекарственная комбинация His-rhArg (SEQ ID NO: 101) (0,016, 0,08 или 0,4 ед./мл) и CQ (20 мкМ) на линии клеток рака печени человека «Red Flu HepG2». В данном случае также наблюдали синергические эффекты (значения CI менее 1).
[00413] Пример 39. Анализ миграции и анализ инвазии Transwell
[00414] В данных исследованиях использовали линии клеток рака молочной железы (человеческие клетки MDA-MB-231 и мышиные клетки 4Т1). Как анализ миграции, так и анализ инвазии проводили в 24-луночной системе Transwell от Costar (Corning) (поликарбонатные мембранные вставки с размером пор 8,0 мкм). Перед анализом клетки культивировали в соответствующей бессывороточной среде в течение по меньшей мере 24 часов. За сутки до посева клеток в верхние камеры для анализа инвазии в верхние камеры вставок добавляли 100 мкл разведенного матригеля. В верхнюю камеру, используемую для анализа миграции, добавляли бессывороточную среду (100 мкл). 24-луночные планшеты инкубировали в термостате в течение ночи. На следующий день клетки обрабатывали трипсином и подсчитывали. В данных исследованиях в верхние камеры засевали 3×104 клеток 4Т1 или 5×104 клеток MDA-MB-231 с 200 мкл соответствующей бессывороточной среды. В нижние камеры добавляли соответствующую полную среду (750 мкл). В случае группы обработки лекарственным средством в обе камеры добавляли 1 ед./мл His-rhArg (SEQ ID NO: 101). 24-луночные планшеты инкубировали при 37°С во влажной атмосфере с 5% СО2 в течение 20 часов (клетки 4Т1) или 24 часов (клетки MDA-MB-231).
[00415] После инкубации среду из верхних камер удаляли. Вставки Transwell дважды промывали 1× PBS. Как в верхние, так и в нижние камеры на 10 минут добавляли холодный 100% метанол для фиксации клеток. Вставки дважды промывали 1× PBS. После этого для окрашивания клеток в обе камеры на 15 минут добавляли 1% кристаллический фиолетовый в 1× PBS. Вставки дважды промывали 1× PBS. Клетки на поверхности верхней камеры удаляли с помощью турунды. Вставки сушили на воздухе в течение ночи. Делали фотографии с помощью конфокального микроскопа. Из результатов видно, что количество раковых клеток, которые могут мигрировать и инвазировать через вставки Transwell, уменьшалось, когда клетки были обработаны 1 ед./мл His-rhArg (SEQ ID NO: 101), что указывало на то, что истощение запасов аргинина аргиназой может ингибировать миграцию и инвазию раковых клеток.
[00416] Пример 40. N-ABD-rhArg (SEQ ID NO: 49) ингибировал рост и метастазирование клеток агрессивного рака молочной in vivo
[00417] Для исследований модели ксенотрансплантата клеток агрессивного рака молочной железы 4Т1 использовали 10 самок бестимусных мышей (возрастом 11-12 недель). Их держали группами по пять животных в клетке, корм и воду давали без ограничений. Раковые клетки 4Т1 собирали трипсинизацией, дважды промывали 1× PBS и ресуспендировали в 1× PBS для инокуляции. Раковые клетки (1×106 клеток 4Т1) вводили подкожно в левый бок мышей. Мышей случайным образом разделяли на две группы при достижении средней длины опухоли 6 мм спустя 3 суток после инокуляции. Один раз в неделю им вводили либо 500 ед. (3,2 мг, 230 мкл) N-ABD-rhArg (SEQ ID NO: 49), либо 1× PBS. Образцы крови брали из подкожной вены. Регулярно отслеживали размер опухоли и массу тела мышей. Объем опухоли оценивали с помощью следующего уравнения:
[00418] 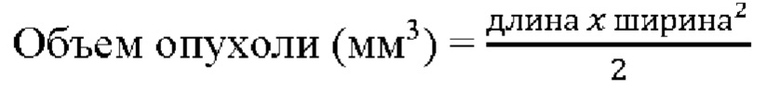
[00419] Мышей умерщвляли спустя 16 суток после инокуляции раковых клеток (т.е. спустя 13 суток после начала эксперимента) из-за некроза опухоли в контрольной группе, получавшей инъекцию PBS. После умерщвления у мышей собирали опухоли и измеряли массу опухоли. Также собирали легкие и промывали их посредством 1× PBS. Легкие фиксировали в растворе Боуина в течение ночи и после фиксации промывали 1× PBS. Подсчитывали метастатические узелки в легких.
[00420] Важно отметить, что 500 ед. альбуминсвязывающей аргиназы N-ABD-rhArg (SEQ ID NO: 49) истощали запасы аргинина до уровня, который был ниже предела детекции у анализатора аминокислот Biochrom 30 (3 мкМ), в течение по меньшей мере 10 суток у BALB/c мышей (фиг. 10). Поэтому для исследования эффективности бестимусным мышам с ксенотрансплантатом рака молочной железы 4Т1 один раз в неделю вводили 500 ед. N-ABD-rhArg (SEQ ID NO: 49) посредством внутрибрюшинной инъекции. В таблице 15 показано, что 500 ед. N-ABD-rhArg (SEQ ID NO: 49) в ходе эксперимента истощали запасы аргинина до недетектируемого уровня.
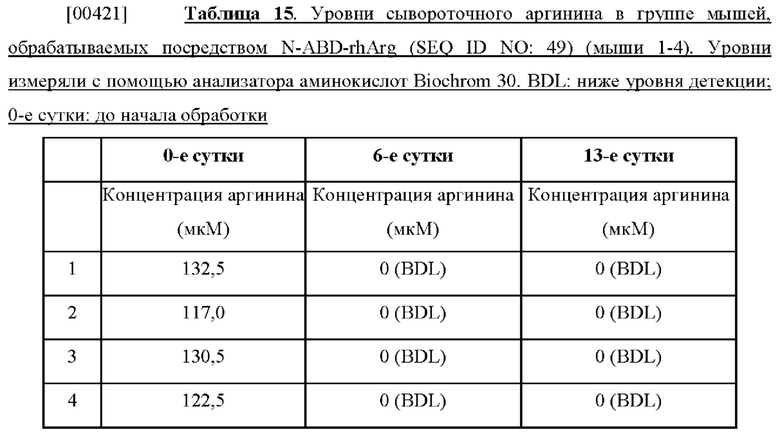
[00422] На фиг. 64-67 видно, что 500 ед. N-ABD-rhArg (SEQ ID NO: 49) были способны ингибировать рост ксенотрансплантата рака молочной железы 4Т1 у бестимусных мышей. Через 12 суток и 13 суток после начала эксперимента наблюдали значимую разницу в расчетном объеме опухоли между контрольной группой и экспериментальной группой (фиг. 64 и 65). Также между двумя группами наблюдали заметные различия в размере опухоли (фиг. 66 и 67).
[00423] Данные результаты свидетельствовали, что истощение запасов аргинина при применении 500 ед. N-ABD-rhArg (SEQ ID NO: 49), вводимого инъекцией один раз в неделю, может у бестимусных мышей значимо ингибировать рост ксенотрансплантата агрессивного рака молочной железы 4Т1. Из результатов in vivo исследования было видно, что N-ABD-rhArg (SEQ ID NO: 49) может безопасно и эффективно ингибировать рост рака у бестимусных мышей.
[00424] Более того, авторами настоящего изобретения было обнаружено, что аргининовое голодание может ингибировать метастазирование клеток рака молочной железы 4Т1, что согласовывалось с нашими результатами, полученными в условиях in vitro. После умерщвления мышей собирали легкие мышей из группы обработки и тщательно их исследовали на наличие признаков метастазирования. В 3 разных легких контрольной группы были обнаружены три метастатических узелка в легких; тогда как в группе, обработанной посредством N-ABD-rhArg (SEQ ID NO: 49), метастатических узелков обнаружено не было (таблица 16). Из результатов убедительно видно, что аргининовое голодание, обусловленное N-ABD-rhArg (SEQ ID NO: 49), может ингибировать метастазирование опухоли. Это свидетельствовало о том, что аргинин может играть важную роль в миграции клеток и метастазировании опухолей. Истощение запасов аргинина, обусловленное N-ABD-rhArg (SEQ ID NO: 49), может ингибировать некоторые пути, которые важны для метастазирования раковых клеток (Knott et al., 2018, Nature, 554:378-381).
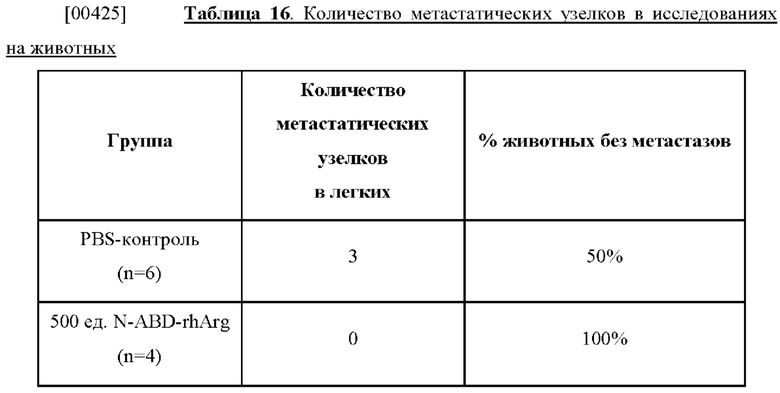
[00426] Пример 41. Цитотоксичность на линиях клеток рака толстой кишки человека при применении слитого фермента N-ABD094-rhArg (SEQ ID NO: 50)
[00427] Эффективность слитого белка ABD и rhArg in vitro тестировали на линиях клеток рака толстой кишки человека. Линии клеток рака толстой кишки человека (НСТ116, LOVO и НТ29) высевали в 96-луночный планшет плотностью 5×103 клеток/лунка. Спустя 1 сутки после инкубации культуральную среду заменяли средой, содержащей N-ABD094-rhArg (SEQ ID NO: 50) в различных концентрациях (2, 0,4, 0,08, 0,016 и 0,0032 ед./мл). На 3-й сутки [спустя 3 суток после обработки посредством N-ABD094-rhArg (SEQ ID NO: 50)] проводили анализ с применением МТТ для определения значений IC50. Культуральную среду заменяли раствором МТТ (1 мг/мл) (Invitrogen) и инкубировали при 37°С в течение 4 часов. После инкубации раствор МТТ заменяли диметилсульфоксидом (DMSO) и измеряли оптическую плотность на 570 нм относительно 650 нм. Жизнеспособность клеток определяли путем деления оптической плотности у клеток, обработанных посредством N-ABD094-rhArg, на среднюю оптическую плотность у клеток без обработки. Значения IC50 для N-ABD094-rhArg (SEQ ID NO: 50) на линиях раковых клеток (при 95% доверительном интервале) анализировали с помощью программного обеспечения Prism 6.0. Для каждой клеточной линии проводили три независимых серии экспериментов (n=3). Результаты представлены в таблице 17.
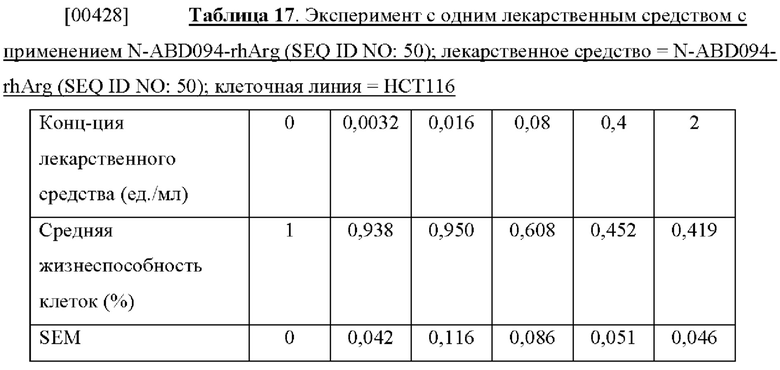
[00429] Клеточная линия=LOVO
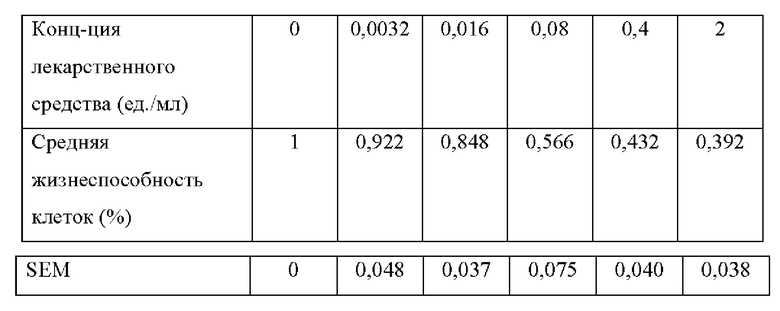
[00430] Клеточная линия=HT29
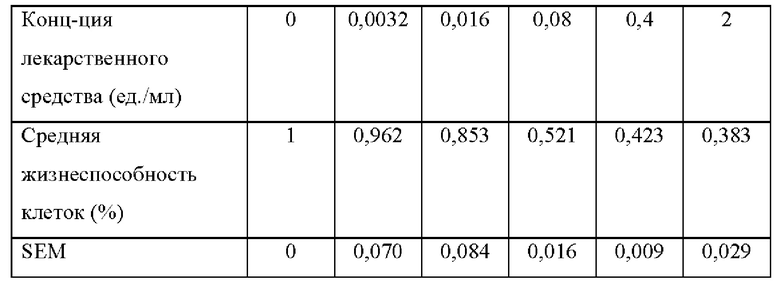
[00431] Все протестированные линии клеток рака толстой кишки были очень чувствительны к слитому ферменту N-ABD094-rhArg (SEQ ID NO: 50) in vitro со значениями IC50 от 0,22 до 0,58 ед./мл. НСТ116 была наиболее чувствительной опухолью с IC50 0,22 ед./мл. Значения IC50 в случае LOVO и НТ29 составляли соответственно 0,38 ед./мл и 0,58 ед./мл.
[00432] Пример 42. Комбинация N-ABD094-rhArg (SEQ ID NO: 50) и хлорохина оказывала синергическое цитотоксическое действие на линии клеток рака толстой кишки человека
[00433] Значения IC50 у ингибитора аутофагии хлорохин (CQ) определяли следующим образом: линии клеток рака толстой кишки НСТ116, LOVO и НТ29 высевали отдельно и спустя 1 сутки инкубации культуральную среду заменяли средой, содержащей CQ в различных концентрациях (100, 20, 4, 0,8, 0,16 мкМ). На 3 сутки проводили анализ с применением МТТ. Для каждой клеточной линии проводили три независимых серии экспериментов (n=3). Результаты представлены в таблице 18.
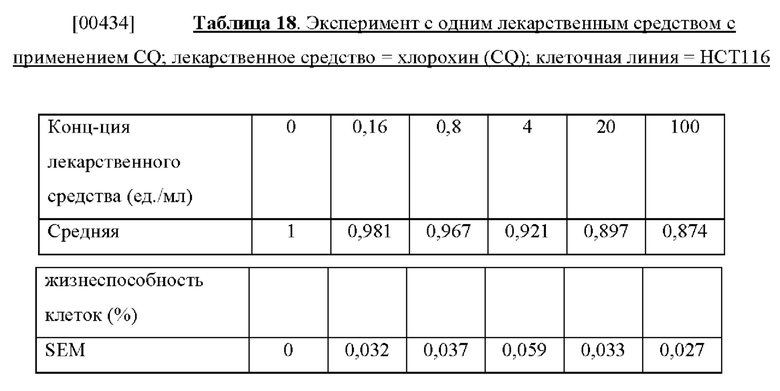
[00435] Клеточная линия=LOVO
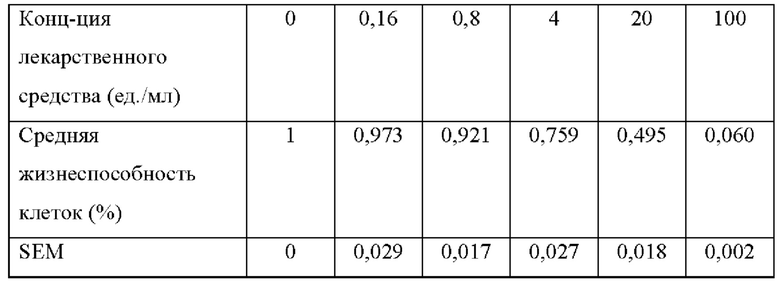
[00436] Клеточная линия=НТ29
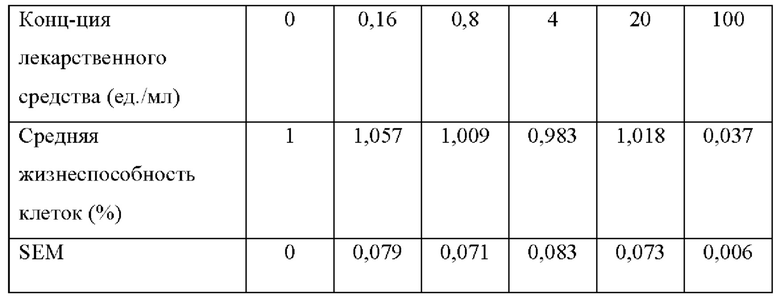
[00437] Значения IC50 CQ в случае НСТ116, LOVO и НТ29 составляли
соответственно >100 мкМ, 15 мкМ и 71 мкМ. Для анализа комбинации лекарственных средств проводили анализ способом непостоянного соотношения (Hastak et al., 2010, Cancer Res. 70(20):7970-7980). Высеивали клетки рака толстой кишки и спустя 1 сутки инкубации культуральную среду заменяли средой, содержащей N-ABD094-rhArg (SEQ ID NO: 50) в различных концентрациях (2, 0,4, 0,08, 0,016, 0,0032 ед./мл) в сочетании с фиксированным количеством CQ согласно его IC50. На 3 сутки проводили анализ с применением МТТ. Показатель аддитивности (CI) комбинированной обработки посредством N-ABD094-rhArg (SEQ ID NO: 50) и CQ анализировали с помощью программного обеспечения CalcuSyn версии 2.1. Для каждой клеточной линии проводили три независимых серии экспериментов (n=3). Результаты представлены в таблице 19.
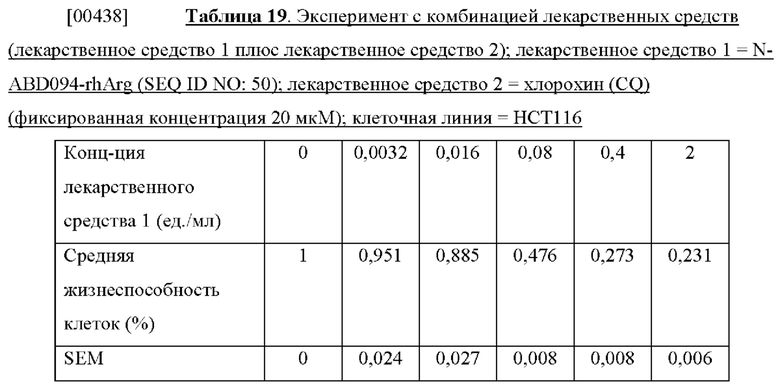
[00439] Клеточная линия=LOVO
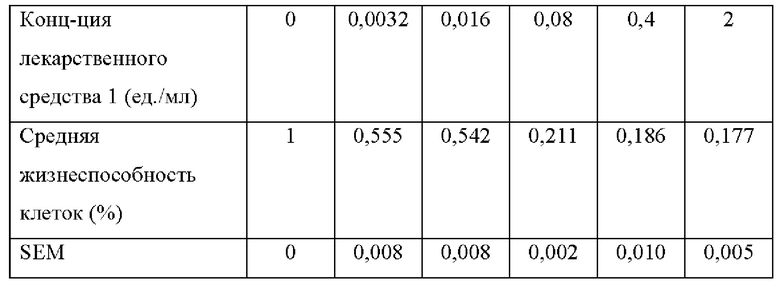
[00440] Клеточная линия=НТ29
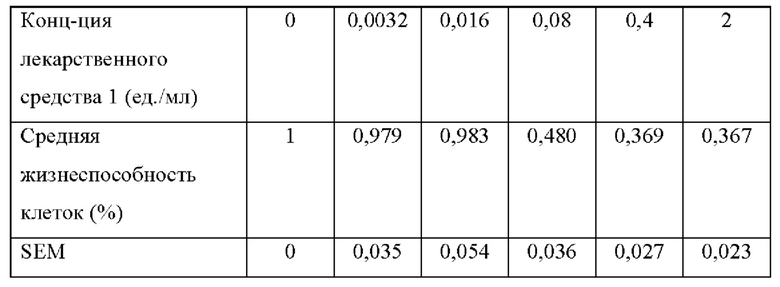
[00441] Из результатов было видно, что при объединении 0,08 или 0,4 ед./мл N-ABD094-rhArg (SEQ ID NO: 50) в комбинацию с 20 мкМ CQ у всех 3 линий клеток рака толстой кишки значения CI были <1,0. Это позволяло предположить, что N-ABD094-rhArg (SEQ ID NO: 50) активен сам по себе и синергичен в комбинации с CQ. Подводя итог вышесказанному, ингибитор аутофагии хлорохин (CQ) усиливает индуцирующее гибель клеток действие слитого белка N-ABD094-rhArg (SEQ ID NO: 50) на клетках рака толстой кишки человека.
[00442] Пример 43. Как N-ABD-rhArg (SEQ ID NO: 49), так и пегилированный His-rhArg (SEQ ID NO: 101) ингибировали рост и метастазирование клеток агрессивного рака молочной in vivo
[00443] В другом исследовании аллотрансплантата рака молочной железы 4Т1 использовали самок бестимусных мышей (возрастом 5-6 недель). Их держали группами по пять животных в клетке, корм и воду давали без ограничений. Клетки рака молочной железы 4Т1 (1×105) подкожно вводили в правый бок мышей. Мышей случайным образом разделяли на четыре группы при достижении средней длины опухоли 8 мм спустя 11 суток после инокуляции. Была одна контрольная группа (n=8) и три экспериментальные группы, в том числе группа с 500 ед. пегилированного His-rhArg (SEQ ID NO: 101) (n=8), группа с 500 ед. N-ABD-rhArg (SEQ ID NO: 49) (n=6) и группа с 250 ед. N-ABD-rhArg (SEQ ID NO: 49) (n=8). Мышам в контрольной группе ежесуточно вводили 1× PBS. В случае группы с 500 ед. ABD-rhArg и группы с 250 ед. ABD-rhArg мышам вводили соответственно 500 ед. и 250 ед. ABD-rhArg один раз в неделю; одной группе вводили 500 ед. PEG-rhArg один раз в неделю. Мышей умерщвляли спустя 24 суток после инокуляции (т.е. спустя 14 суток после начала эксперимента) из-за некроза опухоли в контрольной группе с PBS. После умерщвления мышей препарировали. У мышей собирали опухоли и измеряли их массу. Также собирали легкие и промывали их посредством 1× PBS. Легкие фиксировали в растворе Боуина в течение ночи и после фиксации промывали 1× PBS. Подсчитывали метастатические узелки в легких.
[00444] Что касается результатов, на фиг. 68 видно, что как 500 ед. ABD-rhArg, так и 500 ед. PEG-rhArg значимо ингибировали рост агрессивной солидной опухоли 4Т1. Однако более низкая доза 250 ед. ABD-rhArg не ингибировала рост опухоли. Результат также совпадал с изменением массы опухоли в различных группах (фиг. 69). Следовательно, ABD-rhArg ингибировал рост опухоли дозозависимым образом. Важно отметить, что из данных ясно видно, что аргининовое голодание (либо обусловленное посредством ABD-rhArg, либо обусловленное посредством PEG-rhArg) ингибировало или уменьшало метастазирование опухоли в легкие (фиг. 70 А и фиг. 70 В). Узелки метастазов из легких как в группе 500 ед. PEG-rhArg, так и в группе 500 ед. ABD-rhArg были весьма значимо меньше по сравнению с контрольной группой. Из гистологических срезов легких с окрашиванием гематоксилином и эозином (Н&Е) было видно, что узелки образованы рядом плотных клеток рака молочной железы. Результаты позволяли предположить, что данные узелки были метастатическими очагами. Исходя из вышеописанного, истощение запасов аргинина посредством аргиназы длительного действия ингибировало метастазирование клеток агрессивной формы рака у мышиной модели. Поскольку метастазы рака являются основной причиной смерти пациентов (Knott et al., 2018, Nature, 554: 378-381), данный результат может открыть новые возможности для более эффективной противораковой терапии.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Зе Хонг-Конг Политекник Юнивесити
Зе Чайниз Юнивесити оф Хон-Конг
<120> Композиция и применение истощающих запасы аргинина средств при
раке, ожирении, метаболических нарушениях и связанных с ними
осложнениях и сопутствующих заболеваниях
<130> P10866PCT00
<150> US 62/678,300
<151> 2018-05-31
<160> 106
<170> PatentIn version 3.5
<210> 1
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 1 ABD, синтезирован в лаборатории
<400> 1
ggagatggac atatgcatca tcaccatcac catgatgaag ccgtggatgc 50
<210> 2
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 2 ABD, синтезирован в лаборатории
<400> 2
gctaagactt tagcttcagc taaggaattc gcatccacgg cttcatcatg 50
<210> 3
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 3 ABD, синтезирован в лаборатории
<400> 3
ccttagctga agctaaagtc ttagctaaca gagaacttga caaatatgga 50
<210> 4
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 4 ABD, синтезирован в лаборатории
<400> 4
taggttcttg taatagtcac ttactccata tttgtcaagt tctctgttag 50
<210> 5
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 5 ABD, синтезирован в лаборатории
<400> 5
ggagtaagtg actattacaa gaacctaatc aacaatgcca aaactgttga 50
<210> 6
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 6 ABD, синтезирован в лаборатории
<400> 6
aaatttcatc tatcagtgct tttacacctt caacagtttt ggcattgttg 50
<210> 7
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 7 ABD, синтезирован в лаборатории
<400> 7
gtaaaagcac tgatagatga aattttagct gcattacctt cgggtagtaa 50
<210> 8
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 8 ABD, синтезирован в лаборатории
<400> 8
tccgccgcta ccattgttat tattgttgtt actacccgaa ggtaatgcag 50
<210> 9
<211> 47
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 9 ABD, синтезирован в лаборатории
<400> 9
gatcttaagc atatgcatca tcaccatcac catgatgaag cggtgga 47
<210> 10
<211> 47
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 10 ABD, синтезирован в лаборатории
<400> 10
gcttctttcg cttccgccag gctgttcgca tccaccgctt catcatg 47
<210> 11
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 11 ABD, синтезирован в лаборатории
<400> 11
gcggaagcga aagaagcggc gaacgcggaa ctggatagct atggcgtgag 50
<210> 12
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 12 ABD, синтезирован в лаборатории
<400> 12
ttatcaatca ggcgtttata aaaatcgctc acgccatagc tatccagttc 50
<210> 13
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 13 ABD, синтезирован в лаборатории
<400> 13
gatttttata aacgcctgat tgataaagcg aaaaccgtgg aaggcgtgga 50
<210> 14
<211> 51
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 14 ABD, синтезирован в лаборатории
<400> 14
cggcagcgcc gccagaatcg catctttcag cgcttccacg ccttccacgg t 51
<210> 15
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 15 ABD, синтезирован в лаборатории
<400> 15
ttctggcggc gctgccgtcg ggtagtaaca acaataataa caatggtagc 50
<210> 16
<211> 39
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 16 ABD, синтезирован в лаборатории
<400> 16
ggatccgccg ctaccattgt tattattgtt gttactacc 39
<210> 17
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 17 ABD, синтезирован в лаборатории
<400> 17
catatgttag ctgatggatc cagtaacaac aataataaca atggtagcgg 50
<210> 18
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 18 ABD, синтезирован в лаборатории
<400> 18
aattcgcatc cacggcttca tcaccgccgc taccattgtt attattgttg 50
<210> 19
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 19 ABD, синтезирован в лаборатории
<400> 19
gaagccgtgg atgcgaattc cttagctgaa gctaaagtct tagctaacag 50
<210> 20
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 20 ABD, синтезирован в лаборатории
<400> 20
cttactccat atttgtcaag ttctctgtta gctaagactt tagcttcagc 50
<210> 21
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 21 ABD, синтезирован в лаборатории
<400> 21
gagaacttga caaatatgga gtaagtgact attacaagaa cctaatcaac 50
<210> 22
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 22 ABD, синтезирован в лаборатории
<400> 22
tacaccttca acagttttgg cattgttgat taggttcttg taatagtcac 50
<210> 23
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 23 ABD, синтезирован в лаборатории
<400> 23
gccaaaactg ttgaaggtgt aaaagcactg atagatgaaa ttttagctgc 50
<210> 24
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 24 ABD, синтезирован в лаборатории
<400> 24
atggtgatgg tgatgatgag gtaatgcagc taaaatttca tctatcagtg 50
<210> 25
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 25 ABD, синтезирован в лаборатории
<400> 25
catatgttat gcgatggatc cagtaacaac aataataaca atggtagcgg 50
<210> 26
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 26 ABD, синтезирован в лаборатории
<400> 26
tgttcgcatc caccgcttca tcaccgccgc taccattgtt attattgttg 50
<210> 27
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 27 ABD, синтезирован в лаборатории
<400> 27
gaagcggtgg atgcgaacag cctggcggaa gcgaaagaag cggcgaacgc 50
<210> 28
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 28 ABD, синтезирован в лаборатории
<400> 28
aaaatcgctc acgccatagc tatccagttc cgcgttcgcc gcttctttcg 50
<210> 29
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 29 ABD, синтезирован в лаборатории
<400> 29
ctatggcgtg agcgattttt ataaacgcct gattgataaa gcgaaaaccg 50
<210> 30
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 30 ABD, синтезирован в лаборатории
<400> 30
tcgcatcttt cagcgcttcc acgccttcca cggttttcgc tttatcaatc 50
<210> 31
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 31 ABD, синтезирован в лаборатории
<400> 31
gaagcgctga aagatgcgat tctggcggcg ctgccgcatc atcaccatca 50
<210> 32
<211> 35
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 32 ABD, синтезирован в лаборатории
<400> 32
ctaaaagctt aatggtgatg gtgatgatgc ggcag 35
<210> 33
<211> 222
<212> DNA
<213> Искусственная последовательность
<220>
<223> His-ABD-линкер, синтезирован в лаборатории
<400> 33
atgcatcatc accatcacca tgatgaagcc gtggatgcga attccttagc tgaagctaaa 60
gtcttagcta acagagaact tgacaaatat ggagtaagtg actattacaa gaacctaatc 120
aacaatgcca aaactgttga aggtgtaaaa gcactgatag atgaaatttt agctgcatta 180
ccttcgggta gtaacaacaa taataacaat ggtagcggcg ga 222
<210> 34
<211> 222
<212> DNA
<213> Искусственная последовательность
<220>
<223> His-ABD094-линкер, синтезирован в лаборатории
<400> 34
atgcatcatc accatcacca tgatgaagcg gtggatgcga acagcctggc ggaagcgaaa 60
gaagcggcga acgcggaact ggatagctat ggcgtgagcg atttttataa acgcctgatt 120
gataaagcga aaaccgtgga aggcgtggaa gcgctgaaag atgcgattct ggcggcgctg 180
ccgtcgggta gtaacaacaa taataacaat ggtagcggcg ga 222
<210> 35
<211> 222
<212> DNA
<213> Искусственная последовательность
<220>
<223> Линкер-ABD-His, синтезирован в лаборатории
<400> 35
ggatccagta acaacaataa taacaatggt agcggcggtg atgaagccgt ggatgcgaat 60
tccttagctg aagctaaagt cttagctaac agagaacttg acaaatatgg agtaagtgac 120
tattacaaga acctaatcaa caatgccaaa actgttgaag gtgtaaaagc actgatagat 180
gaaattttag ctgcattacc tcatcatcac catcaccatt aa 222
<210> 36
<211> 222
<212> DNA
<213> Искусственная последовательность
<220>
<223> Линкер-ABD094-His, синтезирован в лаборатории
<400> 36
ggatccagta acaacaataa taacaatggt agcggcggtg atgaagcggt ggatgcgaac 60
agcctggcgg aagcgaaaga agcggcgaac gcggaactgg atagctatgg cgtgagcgat 120
ttttataaac gcctgattga taaagcgaaa accgtggaag gcgtggaagc gctgaaagat 180
gcgattctgg cggcgctgcc gcatcatcac catcaccatt aa 222
<210> 37
<211> 1191
<212> DNA
<213> Искусственная последовательность
<220>
<223> Ген N-ABD-rhArg, синтезирован в лаборатории
<400> 37
atgcatcatc accatcacca tgatgaagcc gtggatgcga attccttagc tgaagctaaa 60
gtcttagcta acagagaact tgacaaatat ggagtaagtg actattacaa gaacctaatc 120
aacaatgcca aaactgttga aggtgtaaaa gcactgatag atgaaatttt agctgcatta 180
ccttcgggta gtaacaacaa taataacaat ggtagcggcg gaatgtccgc caagtccaga 240
accataggga ttattggagc tcctttctca aagggacagc cacgaggagg ggtggaagaa 300
ggccctacag tattgagaaa ggctggtctg cttgagaaac ttaaagaaca agagtgtgat 360
gtgaaggatt atggggacct gccctttgct gacatcccta atgacagtcc ctttcaaatt 420
gtgaagaatc caaggtctgt gggaaaagca agcgagcagc tggctggcaa ggtggcagaa 480
gtcaagaaga acggaagaat cagcctggtg ctgggcggag accacagttt ggcaattgga 540
agcatctctg gccatgccag ggtccaccct gatcttggag tcatctgggt ggatgctcac 600
actgatatca acactccact gacaaccaca agtggaaact tgcatggaca acctgtatct 660
ttcctcctga aggaactaaa aggaaagatt cccgatgtgc caggattctc ctgggtgact 720
ccctgtatat ctgccaagga tattgtgtat attggcttga gagacgtgga ccctggggaa 780
cactacattt tgaaaactct aggcattaaa tacttttcaa tgactgaagt ggacagacta 840
ggaattggca aggtgatgga agaaacactc agctatctac taggaagaaa gaaaaggcca 900
attcatctaa gttttgatgt tgacggactg gacccatctt tcacaccagc tactggcaca 960
ccagtcgtgg gaggtctgac atacagagaa ggtctctaca tcacagaaga aatctacaaa 1020
acagggctac tctcaggatt agatataatg gaagtgaacc catccctggg gaagacacca 1080
gaagaagtaa ctcgaacagt gaacacagca gttgcaataa ccttggcttg tttcggactt 1140
gctcgggagg gtaatcacaa gcctattgac taccttaacc cacctaagta a 1191
<210> 38
<211> 1191
<212> DNA
<213> Искусственная последовательность
<220>
<223> Ген N-ABD094-rhArg, синтезирован в лаборатории
<400> 38
atgcatcatc accatcacca tgatgaagcg gtggatgcga acagcctggc ggaagcgaaa 60
gaagcggcga acgcggaact ggatagctat ggcgtgagcg atttttataa acgcctgatt 120
gataaagcga aaaccgtgga aggcgtggaa gcgctgaaag atgcgattct ggcggcgctg 180
ccgtcgggta gtaacaacaa taataacaat ggtagcggcg gaatgtccgc caagtccaga 240
accataggga ttattggagc tcctttctca aagggacagc cacgaggagg ggtggaagaa 300
ggccctacag tattgagaaa ggctggtctg cttgagaaac ttaaagaaca agagtgtgat 360
gtgaaggatt atggggacct gccctttgct gacatcccta atgacagtcc ctttcaaatt 420
gtgaagaatc caaggtctgt gggaaaagca agcgagcagc tggctggcaa ggtggcagaa 480
gtcaagaaga acggaagaat cagcctggtg ctgggcggag accacagttt ggcaattgga 540
agcatctctg gccatgccag ggtccaccct gatcttggag tcatctgggt ggatgctcac 600
actgatatca acactccact gacaaccaca agtggaaact tgcatggaca acctgtatct 660
ttcctcctga aggaactaaa aggaaagatt cccgatgtgc caggattctc ctgggtgact 720
ccctgtatat ctgccaagga tattgtgtat attggcttga gagacgtgga ccctggggaa 780
cactacattt tgaaaactct aggcattaaa tacttttcaa tgactgaagt ggacagacta 840
ggaattggca aggtgatgga agaaacactc agctatctac taggaagaaa gaaaaggcca 900
attcatctaa gttttgatgt tgacggactg gacccatctt tcacaccagc tactggcaca 960
ccagtcgtgg gaggtctgac atacagagaa ggtctctaca tcacagaaga aatctacaaa 1020
acagggctac tctcaggatt agatataatg gaagtgaacc catccctggg gaagacacca 1080
gaagaagtaa ctcgaacagt gaacacagca gttgcaataa ccttggcttg tttcggactt 1140
gctcgggagg gtaatcacaa gcctattgac taccttaacc cacctaagta a 1191
<210> 39
<211> 1188
<212> DNA
<213> Искусственная последовательность
<220>
<223> Ген C-ABD-rhArg, синтезирован в лаборатории
<400> 39
atgagcgcca agtccagaac catagggatt attggagctc ctttctcaaa gggacagcca 60
cgaggagggg tggaagaagg ccctacagta ttgagaaagg ctggtctgct tgagaaactt 120
aaagaacaag agtgtgatgt gaaggattat ggggacctgc cctttgctga catccctaat 180
gacagtccct ttcaaattgt gaagaatcca aggtctgtgg gaaaagcaag cgagcagctg 240
gctggcaagg tggcagaagt caagaagaac ggaagaatca gcctggtgct gggcggagac 300
cacagtttgg caattggaag catctctggc catgccaggg tccaccctga tcttggagtc 360
atctgggtgg atgctcacac tgatatcaac actccactga caaccacaag tggaaacttg 420
catggacaac ctgtatcttt cctcctgaag gaactaaaag gaaagattcc cgatgtgcca 480
ggattctcct gggtgactcc ctgtatatct gccaaggata ttgtgtatat tggcttgaga 540
gacgtggacc ctggggaaca ctacattttg aaaactctag gcattaaata cttttcaatg 600
actgaagtgg acagactagg aattggcaag gtgatggaag aaacactcag ctatctacta 660
ggaagaaaga aaaggccaat tcatctaagt tttgatgttg acggactgga cccatctttc 720
acaccagcta ctggcacacc agtcgtggga ggtctgacat acagagaagg tctctacatc 780
acagaagaaa tctacaaaac agggctactc tcaggattag atataatgga agtgaaccca 840
tccctgggga agacaccaga agaagtaact cgaacagtga acacagcagt tgcaataacc 900
ttggcttgtt tcggacttgc tcgggagggt aatcacaagc ctattgacta ccttaaccca 960
cctaagggat ccagtaacaa caataataac aatggtagcg gcggtgatga agccgtggat 1020
gcgaattcct tagctgaagc taaagtctta gctaacagag aacttgacaa atatggagta 1080
agtgactatt acaagaacct aatcaacaat gccaaaactg ttgaaggtgt aaaagcactg 1140
atagatgaaa ttttagctgc attacctcat catcaccatc accattaa 1188
<210> 40
<211> 1188
<212> DNA
<213> Искусственная последовательность
<220>
<223> Ген C-ABD094-rhArg, синтезирован в лаборатории
<400> 40
atgagcgcca agtccagaac catagggatt attggagctc ctttctcaaa gggacagcca 60
cgaggagggg tggaagaagg ccctacagta ttgagaaagg ctggtctgct tgagaaactt 120
aaagaacaag agtgtgatgt gaaggattat ggggacctgc cctttgctga catccctaat 180
gacagtccct ttcaaattgt gaagaatcca aggtctgtgg gaaaagcaag cgagcagctg 240
gctggcaagg tggcagaagt caagaagaac ggaagaatca gcctggtgct gggcggagac 300
cacagtttgg caattggaag catctctggc catgccaggg tccaccctga tcttggagtc 360
atctgggtgg atgctcacac tgatatcaac actccactga caaccacaag tggaaacttg 420
catggacaac ctgtatcttt cctcctgaag gaactaaaag gaaagattcc cgatgtgcca 480
ggattctcct gggtgactcc ctgtatatct gccaaggata ttgtgtatat tggcttgaga 540
gacgtggacc ctggggaaca ctacattttg aaaactctag gcattaaata cttttcaatg 600
actgaagtgg acagactagg aattggcaag gtgatggaag aaacactcag ctatctacta 660
ggaagaaaga aaaggccaat tcatctaagt tttgatgttg acggactgga cccatctttc 720
acaccagcta ctggcacacc agtcgtggga ggtctgacat acagagaagg tctctacatc 780
acagaagaaa tctacaaaac agggctactc tcaggattag atataatgga agtgaaccca 840
tccctgggga agacaccaga agaagtaact cgaacagtga acacagcagt tgcaataacc 900
ttggcttgtt tcggacttgc tcgggagggt aatcacaagc ctattgacta ccttaaccca 960
cctaagggat ccagtaacaa caataataac aatggtagcg gcggtgatga agcggtggat 1020
gcgaacagcc tggcggaagc gaaagaagcg gcgaacgcgg aactggatag ctatggcgtg 1080
agcgattttt ataaacgcct gattgataaa gcgaaaaccg tggaaggcgt ggaagcgctg 1140
aaagatgcga ttctggcggc gctgccgcat catcaccatc accattaa 1188
<210> 41
<211> 1125
<212> DNA
<213> Искусственная последовательность
<220>
<223> Ген N-ABD-BCA, синтезирован в лаборатории
<400> 41
atgcatcatc accatcacca tgatgaagcc gtggatgcga attccttagc tgaagctaaa 60
gtcttagcta acagagaact tgacaaatat ggagtaagtg actattacaa gaacctaatc 120
aacaatgcca aaactgttga aggtgtaaaa gcactgatag atgaaatttt agctgcatta 180
ccttcgggta gtaacaacaa taataacaat ggtagcggcg gatccatgaa gccaatttca 240
attatcgggg ttccgatgga tttagggcag acacgccgcg gcgttgatat ggggccgagc 300
gcaatgcgtt atgcaggcgt catcgaacgt ctggaacgtc ttcattacga tattgaagat 360
ttgggagata ttccgattgg aaaagcagag cggttgcacg agcaaggaga ttcacggttg 420
cgcaatttga aagcggttgc ggaagcgaac gagaaacttg cggcggcggt tgaccaagtc 480
gttcagcggg ggcgatttcc gcttgtgttg ggcggcgacc atagcatcgc cattggcacg 540
ctcgccgggg tggcgaaaca ttatgagcgg cttggagtga tctggtatga cgcgcatggc 600
gacgtcaaca ccgcggaaac gtcgccgtct ggaaacattc atggcatgcc gctggcggcg 660
agcctcgggt ttggccatcc ggcgctgacg caaatcggcg gatacagccc caaaatcaag 720
ccggaacatg tcgtgttgat cggcgtccgt tcccttgatg aaggggagaa gaagtttatt 780
cgcgaaaaag gaatcaaaat ttacacgatg catgaggttg atcggctcgg aatgacaagg 840
gtgatggaag aaacgatcgc ctatttaaaa gaacgaacgg atggcgttca tttgtcgctt 900
gacttggatg gccttgaccc aagcgacgca ccgggagtcg gaacgcctgt cattggagga 960
ttgacatacc gcgaaagcca tttggcgatg gagatgctgg ccgaggcaca aatcatcact 1020
tcagcggaat ttgtcgaagt gaacccgatc ttggatgagc ggaacaaaac agcatcagtg 1080
gctgtagcgc tgatggggtc gttgtttggt gaaaaactca tgtaa 1125
<210> 42
<211> 1125
<212> DNA
<213> Искусственная последовательность
<220>
<223> Ген N-ABD094-BCA, синтезирован в лаборатории
<400> 42
atgcatcatc accatcacca tgatgaagcg gtggatgcga acagcctggc ggaagcgaaa 60
gaagcggcga acgcggaact ggatagctat ggcgtgagcg atttttataa acgcctgatt 120
gataaagcga aaaccgtgga aggcgtggaa gcgctgaaag atgcgattct ggcggcgctg 180
ccgtcgggta gtaacaacaa taataacaat ggtagcggcg gatccatgaa gccaatttca 240
attatcgggg ttccgatgga tttagggcag acacgccgcg gcgttgatat ggggccgagc 300
gcaatgcgtt atgcaggcgt catcgaacgt ctggaacgtc ttcattacga tattgaagat 360
ttgggagata ttccgattgg aaaagcagag cggttgcacg agcaaggaga ttcacggttg 420
cgcaatttga aagcggttgc ggaagcgaac gagaaacttg cggcggcggt tgaccaagtc 480
gttcagcggg ggcgatttcc gcttgtgttg ggcggcgacc atagcatcgc cattggcacg 540
ctcgccgggg tggcgaaaca ttatgagcgg cttggagtga tctggtatga cgcgcatggc 600
gacgtcaaca ccgcggaaac gtcgccgtct ggaaacattc atggcatgcc gctggcggcg 660
agcctcgggt ttggccatcc ggcgctgacg caaatcggcg gatacagccc caaaatcaag 720
ccggaacatg tcgtgttgat cggcgtccgt tcccttgatg aaggggagaa gaagtttatt 780
cgcgaaaaag gaatcaaaat ttacacgatg catgaggttg atcggctcgg aatgacaagg 840
gtgatggaag aaacgatcgc ctatttaaaa gaacgaacgg atggcgttca tttgtcgctt 900
gacttggatg gccttgaccc aagcgacgca ccgggagtcg gaacgcctgt cattggagga 960
ttgacatacc gcgaaagcca tttggcgatg gagatgctgg ccgaggcaca aatcatcact 1020
tcagcggaat ttgtcgaagt gaacccgatc ttggatgagc ggaacaaaac agcatcagtg 1080
gctgtagcgc tgatggggtc gttgtttggt gaaaaactca tgtaa 1125
<210> 43
<211> 1119
<212> DNA
<213> Искусственная последовательность
<220>
<223> Ген C-ABD-BCA, синтезирован в лаборатории
<400> 43
atgaagccaa tttcaattat cggggttccg atggatttag ggcagacacg ccgcggcgtt 60
gatatggggc cgagcgcaat gcgttatgca ggcgtcatcg aacgtctgga acgtcttcat 120
tacgatattg aagatttggg agatattccg attggaaaag cagagcggtt gcacgagcaa 180
ggagattcac ggttgcgcaa tttgaaagcg gttgcggaag cgaacgagaa acttgcggcg 240
gcggttgacc aagtcgttca gcgggggcga tttccgcttg tgttgggcgg cgaccatagc 300
atcgccattg gcacgctcgc cggggtggcg aaacattatg agcggcttgg agtgatctgg 360
tatgacgcgc atggcgacgt caacaccgcg gaaacgtcgc cgtctggaaa cattcatggc 420
atgccgctgg cggcgagcct cgggtttggc catccggcgc tgacgcaaat cggcggatac 480
agccccaaaa tcaagccgga acatgtcgtg ttgatcggcg tccgttccct tgatgaaggg 540
gagaagaagt ttattcgcga aaaaggaatc aaaatttaca cgatgcatga ggttgatcgg 600
ctcggaatga caagggtgat ggaagaaacg atcgcctatt taaaagaacg aacggatggc 660
gttcatttgt cgcttgactt ggatggcctt gacccaagcg acgcaccggg agtcggaacg 720
cctgtcattg gaggattgac ataccgcgaa agccatttgg cgatggagat gctggccgag 780
gcacaaatca tcacttcagc ggaatttgtc gaagtgaacc cgatcttgga tgagcggaac 840
aaaacagcat cagtggctgt agcgctgatg gggtcgttgt ttggtgaaaa actcatggga 900
tccagtaaca acaataataa caatggtagc ggcggtgatg aagccgtgga tgcgaattcc 960
ttagctgaag ctaaagtctt agctaacaga gaacttgaca aatatggagt aagtgactat 1020
tacaagaacc taatcaacaa tgccaaaact gttgaaggtg taaaagcact gatagatgaa 1080
attttagctg cattacctca tcatcaccat caccattaa 1119
<210> 44
<211> 1119
<212> DNA
<213> Искусственная последовательность
<220>
<223> Ген C-ABD094-BCA, синтезирован в лаборатории
<400> 44
atgaagccaa tttcaattat cggggttccg atggatttag ggcagacacg ccgcggcgtt 60
gatatggggc cgagcgcaat gcgttatgca ggcgtcatcg aacgtctgga acgtcttcat 120
tacgatattg aagatttggg agatattccg attggaaaag cagagcggtt gcacgagcaa 180
ggagattcac ggttgcgcaa tttgaaagcg gttgcggaag cgaacgagaa acttgcggcg 240
gcggttgacc aagtcgttca gcgggggcga tttccgcttg tgttgggcgg cgaccatagc 300
atcgccattg gcacgctcgc cggggtggcg aaacattatg agcggcttgg agtgatctgg 360
tatgacgcgc atggcgacgt caacaccgcg gaaacgtcgc cgtctggaaa cattcatggc 420
atgccgctgg cggcgagcct cgggtttggc catccggcgc tgacgcaaat cggcggatac 480
agccccaaaa tcaagccgga acatgtcgtg ttgatcggcg tccgttccct tgatgaaggg 540
gagaagaagt ttattcgcga aaaaggaatc aaaatttaca cgatgcatga ggttgatcgg 600
ctcggaatga caagggtgat ggaagaaacg atcgcctatt taaaagaacg aacggatggc 660
gttcatttgt cgcttgactt ggatggcctt gacccaagcg acgcaccggg agtcggaacg 720
cctgtcattg gaggattgac ataccgcgaa agccatttgg cgatggagat gctggccgag 780
gcacaaatca tcacttcagc ggaatttgtc gaagtgaacc cgatcttgga tgagcggaac 840
aaaacagcat cagtggctgt agcgctgatg gggtcgttgt ttggtgaaaa actcatggga 900
tccagtaaca acaataataa caatggtagc ggcggtgatg aagcggtgga tgcgaacagc 960
ctggcggaag cgaaagaagc ggcgaacgcg gaactggata gctatggcgt gagcgatttt 1020
tataaacgcc tgattgataa agcgaaaacc gtggaaggcg tggaagcgct gaaagatgcg 1080
attctggcgg cgctgccgca tcatcaccat caccattaa 1119
<210> 45
<211> 74
<212> PRT
<213> Искусственная последовательность
<220>
<223> His-ABD-линкер, синтезирован в лаборатории
<400> 45
Met His His His His His His Asp Glu Ala Val Asp Ala Asn Ser Leu
1 5 10 15
Ala Glu Ala Lys Val Leu Ala Asn Arg Glu Leu Asp Lys Tyr Gly Val
20 25 30
Ser Asp Tyr Tyr Lys Asn Leu Ile Asn Asn Ala Lys Thr Val Glu Gly
35 40 45
Val Lys Ala Leu Ile Asp Glu Ile Leu Ala Ala Leu Pro Ser Gly Ser
50 55 60
Asn Asn Asn Asn Asn Asn Gly Ser Gly Gly
65 70
<210> 46
<211> 74
<212> PRT
<213> Искусственная последовательность
<220>
<223> His-ABD094-линкер, синтезирован в лаборатории
<400> 46
Met His His His His His His Asp Glu Ala Val Asp Ala Asn Ser Leu
1 5 10 15
Ala Glu Ala Lys Glu Ala Ala Asn Ala Glu Leu Asp Ser Tyr Gly Val
20 25 30
Ser Asp Phe Tyr Lys Arg Leu Ile Asp Lys Ala Lys Thr Val Glu Gly
35 40 45
Val Glu Ala Leu Lys Asp Ala Ile Leu Ala Ala Leu Pro Ser Gly Ser
50 55 60
Asn Asn Asn Asn Asn Asn Gly Ser Gly Gly
65 70
<210> 47
<211> 73
<212> PRT
<213> Искусственная последовательность
<220>
<223> Линкер-ABD-His, синтезирован в лаборатории
<400> 47
Gly Ser Ser Asn Asn Asn Asn Asn Asn Gly Ser Gly Gly Asp Glu Ala
1 5 10 15
Val Asp Ala Asn Ser Leu Ala Glu Ala Lys Val Leu Ala Asn Arg Glu
20 25 30
Leu Asp Lys Tyr Gly Val Ser Asp Tyr Tyr Lys Asn Leu Ile Asn Asn
35 40 45
Ala Lys Thr Val Glu Gly Val Lys Ala Leu Ile Asp Glu Ile Leu Ala
50 55 60
Ala Leu Pro His His His His His His
65 70
<210> 48
<211> 73
<212> PRT
<213> Искусственная последовательность
<220>
<223> Линкер-ABD094-His, синтезирован в лаборатории
<400> 48
Gly Ser Ser Asn Asn Asn Asn Asn Asn Gly Ser Gly Gly Asp Glu Ala
1 5 10 15
Val Asp Ala Asn Ser Leu Ala Glu Ala Lys Glu Ala Ala Asn Ala Glu
20 25 30
Leu Asp Ser Tyr Gly Val Ser Asp Phe Tyr Lys Arg Leu Ile Asp Lys
35 40 45
Ala Lys Thr Val Glu Gly Val Glu Ala Leu Lys Asp Ala Ile Leu Ala
50 55 60
Ala Leu Pro His His His His His His
65 70
<210> 49
<211> 396
<212> PRT
<213> Искусственная последовательность
<220>
<223> N-ABD-rhArg, синтезирован в лаборатории
<400> 49
Met His His His His His His Asp Glu Ala Val Asp Ala Asn Ser Leu
1 5 10 15
Ala Glu Ala Lys Val Leu Ala Asn Arg Glu Leu Asp Lys Tyr Gly Val
20 25 30
Ser Asp Tyr Tyr Lys Asn Leu Ile Asn Asn Ala Lys Thr Val Glu Gly
35 40 45
Val Lys Ala Leu Ile Asp Glu Ile Leu Ala Ala Leu Pro Ser Gly Ser
50 55 60
Asn Asn Asn Asn Asn Asn Gly Ser Gly Gly Met Ser Ala Lys Ser Arg
65 70 75 80
Thr Ile Gly Ile Ile Gly Ala Pro Phe Ser Lys Gly Gln Pro Arg Gly
85 90 95
Gly Val Glu Glu Gly Pro Thr Val Leu Arg Lys Ala Gly Leu Leu Glu
100 105 110
Lys Leu Lys Glu Gln Glu Cys Asp Val Lys Asp Tyr Gly Asp Leu Pro
115 120 125
Phe Ala Asp Ile Pro Asn Asp Ser Pro Phe Gln Ile Val Lys Asn Pro
130 135 140
Arg Ser Val Gly Lys Ala Ser Glu Gln Leu Ala Gly Lys Val Ala Glu
145 150 155 160
Val Lys Lys Asn Gly Arg Ile Ser Leu Val Leu Gly Gly Asp His Ser
165 170 175
Leu Ala Ile Gly Ser Ile Ser Gly His Ala Arg Val His Pro Asp Leu
180 185 190
Gly Val Ile Trp Val Asp Ala His Thr Asp Ile Asn Thr Pro Leu Thr
195 200 205
Thr Thr Ser Gly Asn Leu His Gly Gln Pro Val Ser Phe Leu Leu Lys
210 215 220
Glu Leu Lys Gly Lys Ile Pro Asp Val Pro Gly Phe Ser Trp Val Thr
225 230 235 240
Pro Cys Ile Ser Ala Lys Asp Ile Val Tyr Ile Gly Leu Arg Asp Val
245 250 255
Asp Pro Gly Glu His Tyr Ile Leu Lys Thr Leu Gly Ile Lys Tyr Phe
260 265 270
Ser Met Thr Glu Val Asp Arg Leu Gly Ile Gly Lys Val Met Glu Glu
275 280 285
Thr Leu Ser Tyr Leu Leu Gly Arg Lys Lys Arg Pro Ile His Leu Ser
290 295 300
Phe Asp Val Asp Gly Leu Asp Pro Ser Phe Thr Pro Ala Thr Gly Thr
305 310 315 320
Pro Val Val Gly Gly Leu Thr Tyr Arg Glu Gly Leu Tyr Ile Thr Glu
325 330 335
Glu Ile Tyr Lys Thr Gly Leu Leu Ser Gly Leu Asp Ile Met Glu Val
340 345 350
Asn Pro Ser Leu Gly Lys Thr Pro Glu Glu Val Thr Arg Thr Val Asn
355 360 365
Thr Ala Val Ala Ile Thr Leu Ala Cys Phe Gly Leu Ala Arg Glu Gly
370 375 380
Asn His Lys Pro Ile Asp Tyr Leu Asn Pro Pro Lys
385 390 395
<210> 50
<211> 396
<212> PRT
<213> Искусственная последовательность
<220>
<223> N-ABD094-rhArg, синтезирован в лаборатории
<400> 50
Met His His His His His His Asp Glu Ala Val Asp Ala Asn Ser Leu
1 5 10 15
Ala Glu Ala Lys Glu Ala Ala Asn Ala Glu Leu Asp Ser Tyr Gly Val
20 25 30
Ser Asp Phe Tyr Lys Arg Leu Ile Asp Lys Ala Lys Thr Val Glu Gly
35 40 45
Val Glu Ala Leu Lys Asp Ala Ile Leu Ala Ala Leu Pro Ser Gly Ser
50 55 60
Asn Asn Asn Asn Asn Asn Gly Ser Gly Gly Met Ser Ala Lys Ser Arg
65 70 75 80
Thr Ile Gly Ile Ile Gly Ala Pro Phe Ser Lys Gly Gln Pro Arg Gly
85 90 95
Gly Val Glu Glu Gly Pro Thr Val Leu Arg Lys Ala Gly Leu Leu Glu
100 105 110
Lys Leu Lys Glu Gln Glu Cys Asp Val Lys Asp Tyr Gly Asp Leu Pro
115 120 125
Phe Ala Asp Ile Pro Asn Asp Ser Pro Phe Gln Ile Val Lys Asn Pro
130 135 140
Arg Ser Val Gly Lys Ala Ser Glu Gln Leu Ala Gly Lys Val Ala Glu
145 150 155 160
Val Lys Lys Asn Gly Arg Ile Ser Leu Val Leu Gly Gly Asp His Ser
165 170 175
Leu Ala Ile Gly Ser Ile Ser Gly His Ala Arg Val His Pro Asp Leu
180 185 190
Gly Val Ile Trp Val Asp Ala His Thr Asp Ile Asn Thr Pro Leu Thr
195 200 205
Thr Thr Ser Gly Asn Leu His Gly Gln Pro Val Ser Phe Leu Leu Lys
210 215 220
Glu Leu Lys Gly Lys Ile Pro Asp Val Pro Gly Phe Ser Trp Val Thr
225 230 235 240
Pro Cys Ile Ser Ala Lys Asp Ile Val Tyr Ile Gly Leu Arg Asp Val
245 250 255
Asp Pro Gly Glu His Tyr Ile Leu Lys Thr Leu Gly Ile Lys Tyr Phe
260 265 270
Ser Met Thr Glu Val Asp Arg Leu Gly Ile Gly Lys Val Met Glu Glu
275 280 285
Thr Leu Ser Tyr Leu Leu Gly Arg Lys Lys Arg Pro Ile His Leu Ser
290 295 300
Phe Asp Val Asp Gly Leu Asp Pro Ser Phe Thr Pro Ala Thr Gly Thr
305 310 315 320
Pro Val Val Gly Gly Leu Thr Tyr Arg Glu Gly Leu Tyr Ile Thr Glu
325 330 335
Glu Ile Tyr Lys Thr Gly Leu Leu Ser Gly Leu Asp Ile Met Glu Val
340 345 350
Asn Pro Ser Leu Gly Lys Thr Pro Glu Glu Val Thr Arg Thr Val Asn
355 360 365
Thr Ala Val Ala Ile Thr Leu Ala Cys Phe Gly Leu Ala Arg Glu Gly
370 375 380
Asn His Lys Pro Ile Asp Tyr Leu Asn Pro Pro Lys
385 390 395
<210> 51
<211> 395
<212> PRT
<213> Искусственная последовательность
<220>
<223> C-ABD-rhArg, синтезирован в лаборатории
<400> 51
Met Ser Ala Lys Ser Arg Thr Ile Gly Ile Ile Gly Ala Pro Phe Ser
1 5 10 15
Lys Gly Gln Pro Arg Gly Gly Val Glu Glu Gly Pro Thr Val Leu Arg
20 25 30
Lys Ala Gly Leu Leu Glu Lys Leu Lys Glu Gln Glu Cys Asp Val Lys
35 40 45
Asp Tyr Gly Asp Leu Pro Phe Ala Asp Ile Pro Asn Asp Ser Pro Phe
50 55 60
Gln Ile Val Lys Asn Pro Arg Ser Val Gly Lys Ala Ser Glu Gln Leu
65 70 75 80
Ala Gly Lys Val Ala Glu Val Lys Lys Asn Gly Arg Ile Ser Leu Val
85 90 95
Leu Gly Gly Asp His Ser Leu Ala Ile Gly Ser Ile Ser Gly His Ala
100 105 110
Arg Val His Pro Asp Leu Gly Val Ile Trp Val Asp Ala His Thr Asp
115 120 125
Ile Asn Thr Pro Leu Thr Thr Thr Ser Gly Asn Leu His Gly Gln Pro
130 135 140
Val Ser Phe Leu Leu Lys Glu Leu Lys Gly Lys Ile Pro Asp Val Pro
145 150 155 160
Gly Phe Ser Trp Val Thr Pro Cys Ile Ser Ala Lys Asp Ile Val Tyr
165 170 175
Ile Gly Leu Arg Asp Val Asp Pro Gly Glu His Tyr Ile Leu Lys Thr
180 185 190
Leu Gly Ile Lys Tyr Phe Ser Met Thr Glu Val Asp Arg Leu Gly Ile
195 200 205
Gly Lys Val Met Glu Glu Thr Leu Ser Tyr Leu Leu Gly Arg Lys Lys
210 215 220
Arg Pro Ile His Leu Ser Phe Asp Val Asp Gly Leu Asp Pro Ser Phe
225 230 235 240
Thr Pro Ala Thr Gly Thr Pro Val Val Gly Gly Leu Thr Tyr Arg Glu
245 250 255
Gly Leu Tyr Ile Thr Glu Glu Ile Tyr Lys Thr Gly Leu Leu Ser Gly
260 265 270
Leu Asp Ile Met Glu Val Asn Pro Ser Leu Gly Lys Thr Pro Glu Glu
275 280 285
Val Thr Arg Thr Val Asn Thr Ala Val Ala Ile Thr Leu Ala Cys Phe
290 295 300
Gly Leu Ala Arg Glu Gly Asn His Lys Pro Ile Asp Tyr Leu Asn Pro
305 310 315 320
Pro Lys Gly Ser Ser Asn Asn Asn Asn Asn Asn Gly Ser Gly Gly Asp
325 330 335
Glu Ala Val Asp Ala Asn Ser Leu Ala Glu Ala Lys Val Leu Ala Asn
340 345 350
Arg Glu Leu Asp Lys Tyr Gly Val Ser Asp Tyr Tyr Lys Asn Leu Ile
355 360 365
Asn Asn Ala Lys Thr Val Glu Gly Val Lys Ala Leu Ile Asp Glu Ile
370 375 380
Leu Ala Ala Leu Pro His His His His His His
385 390 395
<210> 52
<211> 395
<212> PRT
<213> Искусственная последовательность
<220>
<223> C-ABD094-rhArg, синтезирован в лаборатории
<400> 52
Met Ser Ala Lys Ser Arg Thr Ile Gly Ile Ile Gly Ala Pro Phe Ser
1 5 10 15
Lys Gly Gln Pro Arg Gly Gly Val Glu Glu Gly Pro Thr Val Leu Arg
20 25 30
Lys Ala Gly Leu Leu Glu Lys Leu Lys Glu Gln Glu Cys Asp Val Lys
35 40 45
Asp Tyr Gly Asp Leu Pro Phe Ala Asp Ile Pro Asn Asp Ser Pro Phe
50 55 60
Gln Ile Val Lys Asn Pro Arg Ser Val Gly Lys Ala Ser Glu Gln Leu
65 70 75 80
Ala Gly Lys Val Ala Glu Val Lys Lys Asn Gly Arg Ile Ser Leu Val
85 90 95
Leu Gly Gly Asp His Ser Leu Ala Ile Gly Ser Ile Ser Gly His Ala
100 105 110
Arg Val His Pro Asp Leu Gly Val Ile Trp Val Asp Ala His Thr Asp
115 120 125
Ile Asn Thr Pro Leu Thr Thr Thr Ser Gly Asn Leu His Gly Gln Pro
130 135 140
Val Ser Phe Leu Leu Lys Glu Leu Lys Gly Lys Ile Pro Asp Val Pro
145 150 155 160
Gly Phe Ser Trp Val Thr Pro Cys Ile Ser Ala Lys Asp Ile Val Tyr
165 170 175
Ile Gly Leu Arg Asp Val Asp Pro Gly Glu His Tyr Ile Leu Lys Thr
180 185 190
Leu Gly Ile Lys Tyr Phe Ser Met Thr Glu Val Asp Arg Leu Gly Ile
195 200 205
Gly Lys Val Met Glu Glu Thr Leu Ser Tyr Leu Leu Gly Arg Lys Lys
210 215 220
Arg Pro Ile His Leu Ser Phe Asp Val Asp Gly Leu Asp Pro Ser Phe
225 230 235 240
Thr Pro Ala Thr Gly Thr Pro Val Val Gly Gly Leu Thr Tyr Arg Glu
245 250 255
Gly Leu Tyr Ile Thr Glu Glu Ile Tyr Lys Thr Gly Leu Leu Ser Gly
260 265 270
Leu Asp Ile Met Glu Val Asn Pro Ser Leu Gly Lys Thr Pro Glu Glu
275 280 285
Val Thr Arg Thr Val Asn Thr Ala Val Ala Ile Thr Leu Ala Cys Phe
290 295 300
Gly Leu Ala Arg Glu Gly Asn His Lys Pro Ile Asp Tyr Leu Asn Pro
305 310 315 320
Pro Lys Gly Ser Ser Asn Asn Asn Asn Asn Asn Gly Ser Gly Gly Asp
325 330 335
Glu Ala Val Asp Ala Asn Ser Leu Ala Glu Ala Lys Glu Ala Ala Asn
340 345 350
Ala Glu Leu Asp Ser Tyr Gly Val Ser Asp Phe Tyr Lys Arg Leu Ile
355 360 365
Asp Lys Ala Lys Thr Val Glu Gly Val Glu Ala Leu Lys Asp Ala Ile
370 375 380
Leu Ala Ala Leu Pro His His His His His His
385 390 395
<210> 53
<211> 374
<212> PRT
<213> Искусственная последовательность
<220>
<223> N-ABD-BCA, синтезирован в лаборатории
<400> 53
Met His His His His His His Asp Glu Ala Val Asp Ala Asn Ser Leu
1 5 10 15
Ala Glu Ala Lys Val Leu Ala Asn Arg Glu Leu Asp Lys Tyr Gly Val
20 25 30
Ser Asp Tyr Tyr Lys Asn Leu Ile Asn Asn Ala Lys Thr Val Glu Gly
35 40 45
Val Lys Ala Leu Ile Asp Glu Ile Leu Ala Ala Leu Pro Ser Gly Ser
50 55 60
Asn Asn Asn Asn Asn Asn Gly Ser Gly Gly Ser Met Lys Pro Ile Ser
65 70 75 80
Ile Ile Gly Val Pro Met Asp Leu Gly Gln Thr Arg Arg Gly Val Asp
85 90 95
Met Gly Pro Ser Ala Met Arg Tyr Ala Gly Val Ile Glu Arg Leu Glu
100 105 110
Arg Leu His Tyr Asp Ile Glu Asp Leu Gly Asp Ile Pro Ile Gly Lys
115 120 125
Ala Glu Arg Leu His Glu Gln Gly Asp Ser Arg Leu Arg Asn Leu Lys
130 135 140
Ala Val Ala Glu Ala Asn Glu Lys Leu Ala Ala Ala Val Asp Gln Val
145 150 155 160
Val Gln Arg Gly Arg Phe Pro Leu Val Leu Gly Gly Asp His Ser Ile
165 170 175
Ala Ile Gly Thr Leu Ala Gly Val Ala Lys His Tyr Glu Arg Leu Gly
180 185 190
Val Ile Trp Tyr Asp Ala His Gly Asp Val Asn Thr Ala Glu Thr Ser
195 200 205
Pro Ser Gly Asn Ile His Gly Met Pro Leu Ala Ala Ser Leu Gly Phe
210 215 220
Gly His Pro Ala Leu Thr Gln Ile Gly Gly Tyr Ser Pro Lys Ile Lys
225 230 235 240
Pro Glu His Val Val Leu Ile Gly Val Arg Ser Leu Asp Glu Gly Glu
245 250 255
Lys Lys Phe Ile Arg Glu Lys Gly Ile Lys Ile Tyr Thr Met His Glu
260 265 270
Val Asp Arg Leu Gly Met Thr Arg Val Met Glu Glu Thr Ile Ala Tyr
275 280 285
Leu Lys Glu Arg Thr Asp Gly Val His Leu Ser Leu Asp Leu Asp Gly
290 295 300
Leu Asp Pro Ser Asp Ala Pro Gly Val Gly Thr Pro Val Ile Gly Gly
305 310 315 320
Leu Thr Tyr Arg Glu Ser His Leu Ala Met Glu Met Leu Ala Glu Ala
325 330 335
Gln Ile Ile Thr Ser Ala Glu Phe Val Glu Val Asn Pro Ile Leu Asp
340 345 350
Glu Arg Asn Lys Thr Ala Ser Val Ala Val Ala Leu Met Gly Ser Leu
355 360 365
Phe Gly Glu Lys Leu Met
370
<210> 54
<211> 374
<212> PRT
<213> Искусственная последовательность
<220>
<223> N-ABD094-BCA, синтезирован в лаборатории
<400> 54
Met His His His His His His Asp Glu Ala Val Asp Ala Asn Ser Leu
1 5 10 15
Ala Glu Ala Lys Glu Ala Ala Asn Ala Glu Leu Asp Ser Tyr Gly Val
20 25 30
Ser Asp Phe Tyr Lys Arg Leu Ile Asp Lys Ala Lys Thr Val Glu Gly
35 40 45
Val Glu Ala Leu Lys Asp Ala Ile Leu Ala Ala Leu Pro Ser Gly Ser
50 55 60
Asn Asn Asn Asn Asn Asn Gly Ser Gly Gly Ser Met Lys Pro Ile Ser
65 70 75 80
Ile Ile Gly Val Pro Met Asp Leu Gly Gln Thr Arg Arg Gly Val Asp
85 90 95
Met Gly Pro Ser Ala Met Arg Tyr Ala Gly Val Ile Glu Arg Leu Glu
100 105 110
Arg Leu His Tyr Asp Ile Glu Asp Leu Gly Asp Ile Pro Ile Gly Lys
115 120 125
Ala Glu Arg Leu His Glu Gln Gly Asp Ser Arg Leu Arg Asn Leu Lys
130 135 140
Ala Val Ala Glu Ala Asn Glu Lys Leu Ala Ala Ala Val Asp Gln Val
145 150 155 160
Val Gln Arg Gly Arg Phe Pro Leu Val Leu Gly Gly Asp His Ser Ile
165 170 175
Ala Ile Gly Thr Leu Ala Gly Val Ala Lys His Tyr Glu Arg Leu Gly
180 185 190
Val Ile Trp Tyr Asp Ala His Gly Asp Val Asn Thr Ala Glu Thr Ser
195 200 205
Pro Ser Gly Asn Ile His Gly Met Pro Leu Ala Ala Ser Leu Gly Phe
210 215 220
Gly His Pro Ala Leu Thr Gln Ile Gly Gly Tyr Ser Pro Lys Ile Lys
225 230 235 240
Pro Glu His Val Val Leu Ile Gly Val Arg Ser Leu Asp Glu Gly Glu
245 250 255
Lys Lys Phe Ile Arg Glu Lys Gly Ile Lys Ile Tyr Thr Met His Glu
260 265 270
Val Asp Arg Leu Gly Met Thr Arg Val Met Glu Glu Thr Ile Ala Tyr
275 280 285
Leu Lys Glu Arg Thr Asp Gly Val His Leu Ser Leu Asp Leu Asp Gly
290 295 300
Leu Asp Pro Ser Asp Ala Pro Gly Val Gly Thr Pro Val Ile Gly Gly
305 310 315 320
Leu Thr Tyr Arg Glu Ser His Leu Ala Met Glu Met Leu Ala Glu Ala
325 330 335
Gln Ile Ile Thr Ser Ala Glu Phe Val Glu Val Asn Pro Ile Leu Asp
340 345 350
Glu Arg Asn Lys Thr Ala Ser Val Ala Val Ala Leu Met Gly Ser Leu
355 360 365
Phe Gly Glu Lys Leu Met
370
<210> 55
<211> 372
<212> PRT
<213> Искусственная последовательность
<220>
<223> C-ABD-BCA, синтезирован в лаборатории
<400> 55
Met Lys Pro Ile Ser Ile Ile Gly Val Pro Met Asp Leu Gly Gln Thr
1 5 10 15
Arg Arg Gly Val Asp Met Gly Pro Ser Ala Met Arg Tyr Ala Gly Val
20 25 30
Ile Glu Arg Leu Glu Arg Leu His Tyr Asp Ile Glu Asp Leu Gly Asp
35 40 45
Ile Pro Ile Gly Lys Ala Glu Arg Leu His Glu Gln Gly Asp Ser Arg
50 55 60
Leu Arg Asn Leu Lys Ala Val Ala Glu Ala Asn Glu Lys Leu Ala Ala
65 70 75 80
Ala Val Asp Gln Val Val Gln Arg Gly Arg Phe Pro Leu Val Leu Gly
85 90 95
Gly Asp His Ser Ile Ala Ile Gly Thr Leu Ala Gly Val Ala Lys His
100 105 110
Tyr Glu Arg Leu Gly Val Ile Trp Tyr Asp Ala His Gly Asp Val Asn
115 120 125
Thr Ala Glu Thr Ser Pro Ser Gly Asn Ile His Gly Met Pro Leu Ala
130 135 140
Ala Ser Leu Gly Phe Gly His Pro Ala Leu Thr Gln Ile Gly Gly Tyr
145 150 155 160
Ser Pro Lys Ile Lys Pro Glu His Val Val Leu Ile Gly Val Arg Ser
165 170 175
Leu Asp Glu Gly Glu Lys Lys Phe Ile Arg Glu Lys Gly Ile Lys Ile
180 185 190
Tyr Thr Met His Glu Val Asp Arg Leu Gly Met Thr Arg Val Met Glu
195 200 205
Glu Thr Ile Ala Tyr Leu Lys Glu Arg Thr Asp Gly Val His Leu Ser
210 215 220
Leu Asp Leu Asp Gly Leu Asp Pro Ser Asp Ala Pro Gly Val Gly Thr
225 230 235 240
Pro Val Ile Gly Gly Leu Thr Tyr Arg Glu Ser His Leu Ala Met Glu
245 250 255
Met Leu Ala Glu Ala Gln Ile Ile Thr Ser Ala Glu Phe Val Glu Val
260 265 270
Asn Pro Ile Leu Asp Glu Arg Asn Lys Thr Ala Ser Val Ala Val Ala
275 280 285
Leu Met Gly Ser Leu Phe Gly Glu Lys Leu Met Gly Ser Ser Asn Asn
290 295 300
Asn Asn Asn Asn Gly Ser Gly Gly Asp Glu Ala Val Asp Ala Asn Ser
305 310 315 320
Leu Ala Glu Ala Lys Val Leu Ala Asn Arg Glu Leu Asp Lys Tyr Gly
325 330 335
Val Ser Asp Tyr Tyr Lys Asn Leu Ile Asn Asn Ala Lys Thr Val Glu
340 345 350
Gly Val Lys Ala Leu Ile Asp Glu Ile Leu Ala Ala Leu Pro His His
355 360 365
His His His His
370
<210> 56
<211> 372
<212> PRT
<213> Искусственная последовательность
<220>
<223> C-ABD094-BCA, синтезирован в лаборатории
<400> 56
Met Lys Pro Ile Ser Ile Ile Gly Val Pro Met Asp Leu Gly Gln Thr
1 5 10 15
Arg Arg Gly Val Asp Met Gly Pro Ser Ala Met Arg Tyr Ala Gly Val
20 25 30
Ile Glu Arg Leu Glu Arg Leu His Tyr Asp Ile Glu Asp Leu Gly Asp
35 40 45
Ile Pro Ile Gly Lys Ala Glu Arg Leu His Glu Gln Gly Asp Ser Arg
50 55 60
Leu Arg Asn Leu Lys Ala Val Ala Glu Ala Asn Glu Lys Leu Ala Ala
65 70 75 80
Ala Val Asp Gln Val Val Gln Arg Gly Arg Phe Pro Leu Val Leu Gly
85 90 95
Gly Asp His Ser Ile Ala Ile Gly Thr Leu Ala Gly Val Ala Lys His
100 105 110
Tyr Glu Arg Leu Gly Val Ile Trp Tyr Asp Ala His Gly Asp Val Asn
115 120 125
Thr Ala Glu Thr Ser Pro Ser Gly Asn Ile His Gly Met Pro Leu Ala
130 135 140
Ala Ser Leu Gly Phe Gly His Pro Ala Leu Thr Gln Ile Gly Gly Tyr
145 150 155 160
Ser Pro Lys Ile Lys Pro Glu His Val Val Leu Ile Gly Val Arg Ser
165 170 175
Leu Asp Glu Gly Glu Lys Lys Phe Ile Arg Glu Lys Gly Ile Lys Ile
180 185 190
Tyr Thr Met His Glu Val Asp Arg Leu Gly Met Thr Arg Val Met Glu
195 200 205
Glu Thr Ile Ala Tyr Leu Lys Glu Arg Thr Asp Gly Val His Leu Ser
210 215 220
Leu Asp Leu Asp Gly Leu Asp Pro Ser Asp Ala Pro Gly Val Gly Thr
225 230 235 240
Pro Val Ile Gly Gly Leu Thr Tyr Arg Glu Ser His Leu Ala Met Glu
245 250 255
Met Leu Ala Glu Ala Gln Ile Ile Thr Ser Ala Glu Phe Val Glu Val
260 265 270
Asn Pro Ile Leu Asp Glu Arg Asn Lys Thr Ala Ser Val Ala Val Ala
275 280 285
Leu Met Gly Ser Leu Phe Gly Glu Lys Leu Met Gly Ser Ser Asn Asn
290 295 300
Asn Asn Asn Asn Gly Ser Gly Gly Asp Glu Ala Val Asp Ala Asn Ser
305 310 315 320
Leu Ala Glu Ala Lys Glu Ala Ala Asn Ala Glu Leu Asp Ser Tyr Gly
325 330 335
Val Ser Asp Phe Tyr Lys Arg Leu Ile Asp Lys Ala Lys Thr Val Glu
340 345 350
Gly Val Glu Ala Leu Lys Asp Ala Ile Leu Ala Ala Leu Pro His His
355 360 365
His His His His
370
<210> 57
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер ABD094-0F, синтезирован в лаборатории
<400> 57
attctggcag cgctgccgtc gggtagtaac aacaataata acaatggtag 50
<210> 58
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер ABD094-1F, синтезирован в лаборатории
<400> 58
aaccgtggaa ggtgtggaag cgctgaaaga tgcgattctg gcagcgctgc 50
<210> 59
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер ABD094-2F, синтезирован в лаборатории
<400> 59
gatttttata aacgtctgat tgataaagcg aaaaccgtgg aaggtgtgga 50
<210> 60
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер ABD094-3F, синтезирован в лаборатории
<400> 60
ctggatagct atggcgtgag cgatttttat aaacgtctga ttgataaagc 50
<210> 61
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер ABD094-4F, синтезирован в лаборатории
<400> 61
gcggaagcga aagaagcagc aaacgcggaa ctggatagct atggcgtgag 50
<210> 62
<211> 50
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер ABD094-5F, синтезирован в лаборатории
<400> 62
accatgatga agccgtggat gcgaattccc tggcggaagc gaaagaagca 50
<210> 63
<211> 46
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер abdNde-F, синтезирован в лаборатории
<400> 63
ggagatatac atatgcatca tcaccatcac catgatgaag ccgtgg 46
<210> 64
<211> 42
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер HuArgHinBam-R, синтезирован в лаборатории
<400> 64
tgactacctt aacccaccta agtaagcttg gatcctgtaa cg 42
<210> 65
<211> 1220
<212> DNA
<213> Искусственная последовательность
<220>
<223> Ген N-ABD094-rhArg, синтезирован в лаборатории
<400> 65
ggagatatac atatgcatca tcaccatcac catgatgaag ccgtggatgc gaattccctg 60
gcggaagcga aagaagcagc aaacgcggaa ctggatagct atggcgtgag cgatttttat 120
aaacgtctga ttgataaagc gaaaaccgtg gaaggtgtgg aagcgctgaa agatgcgatt 180
ctggcagcgc tgccgtcggg tagtaacaac aataataaca atggtagcgg cggtatgagc 240
gccaagtcca gaaccatagg gattattgga gctcctttct caaagggaca gccacgagga 300
ggggtggaag aaggccctac agtattgaga aaggctggtc tgcttgagaa acttaaagaa 360
caagagtgtg atgtgaagga ttatggggac ctgccctttg ctgacatccc taatgacagt 420
ccctttcaaa ttgtgaagaa tccaaggtct gtgggaaaag caagcgagca gctggctggc 480
aaggtggcag aagtcaagaa gaacggaaga atcagcctgg tgctgggcgg agaccacagt 540
ttggcaattg gaagcatctc tggccatgcc agggtccacc ctgatcttgg agtcatctgg 600
gtggatgctc acactgatat caacactcca ctgacaacca caagtggaaa cttgcatgga 660
caacctgtat ctttcctcct gaaggaacta aaaggaaaga ttcccgatgt gccaggattc 720
tcctgggtga ctccctgtat atctgccaag gatattgtgt atattggctt gagagacgtg 780
gaccctgggg aacactacat tttgaaaact ctaggcatta aatacttttc aatgactgaa 840
gtggacagac taggaattgg caaggtgatg gaagaaacac tcagctatct actaggaaga 900
aagaaaaggc caattcatct aagttttgat gttgacggac tggacccatc tttcacacca 960
gctactggca caccagtcgt gggaggtctg acatacagag aaggtctcta catcacagaa 1020
gaaatctaca aaacagggct actctcagga ttagatataa tggaagtgaa cccatccctg 1080
gggaagacac cagaagaagt aactcgaaca gtgaacacag cagttgcaat aaccttggct 1140
tgtttcggac ttgctcggga gggtaatcac aagcctattg actaccttaa cccacctaag 1200
taagcttgga tcctgtaacg 1220
<210> 66
<211> 54
<212> PRT
<213> Искусственная последовательность
<220>
<223> ABD на основе Streptococcus sp. G148, синтезирован в лаборатории
<400> 66
Asp Glu Ala Val Asp Ala Asn Ser Leu Ala Glu Ala Lys Val Leu Ala
1 5 10 15
Asn Arg Glu Leu Asp Lys Tyr Gly Val Ser Asp Tyr Tyr Lys Asn Leu
20 25 30
Ile Asn Asn Ala Lys Thr Val Glu Gly Val Lys Ala Leu Ile Asp Glu
35 40 45
Ile Leu Ala Ala Leu Pro
50
<210> 67
<211> 57
<212> PRT
<213> Искусственная последовательность
<220>
<223> ABD на основе Streptococcus sp. G148, синтезирован в лаборатории
<400> 67
Ala Gln His Asp Glu Ala Val Asp Ala Asn Ser Leu Ala Glu Ala Lys
1 5 10 15
Val Leu Ala Asn Arg Glu Leu Asp Lys Tyr Gly Val Ser Asp Tyr Tyr
20 25 30
Lys Asn Leu Ile Asn Asn Ala Lys Thr Val Glu Gly Val Lys Ala Leu
35 40 45
Ile Asp Glu Ile Leu Ala Ala Leu Pro
50 55
<210> 68
<211> 46
<212> PRT
<213> Искусственная последовательность
<220>
<223> ABD094 без линкера, синтезирован в лаборатории
<400> 68
Leu Ala Glu Ala Lys Glu Ala Ala Asn Ala Glu Leu Asp Ser Tyr Gly
1 5 10 15
Val Ser Asp Phe Tyr Lys Arg Leu Ile Asp Lys Ala Lys Thr Val Glu
20 25 30
Gly Val Glu Ala Leu Lys Asp Ala Ile Leu Ala Ala Leu Pro
35 40 45
<210> 69
<211> 324
<212> PRT
<213> Homo sapiens
<220>
<221> Xaa
<222> (1)..(1)
<223> Xaa в положении один либо является метионином, либо
отсутствует.
<400> 69
Xaa Ala Ala Ser Ala Lys Ser Arg Thr Ile Gly Ile Ile Gly Ala Pro
1 5 10 15
Phe Ser Lys Gly Gln Pro Arg Gly Gly Val Glu Glu Gly Pro Thr Val
20 25 30
Leu Arg Lys Ala Gly Leu Leu Glu Lys Leu Lys Glu Gln Glu Cys Asp
35 40 45
Val Lys Asp Tyr Gly Asp Leu Pro Phe Ala Asp Ile Pro Asn Asp Ser
50 55 60
Pro Phe Gln Ile Val Lys Asn Pro Arg Ser Val Gly Lys Ala Ser Glu
65 70 75 80
Gln Leu Ala Gly Lys Val Ala Glu Val Lys Lys Asn Gly Arg Ile Ser
85 90 95
Leu Val Leu Gly Gly Asp His Ser Leu Ala Ile Gly Ser Ile Ser Gly
100 105 110
His Ala Arg Val His Pro Asp Leu Gly Val Ile Trp Val Asp Ala His
115 120 125
Thr Asp Ile Asn Thr Pro Leu Thr Thr Thr Ser Gly Asn Leu His Gly
130 135 140
Gln Pro Val Ser Phe Leu Leu Lys Glu Leu Lys Gly Lys Ile Pro Asp
145 150 155 160
Val Pro Gly Phe Ser Trp Val Thr Pro Cys Ile Ser Ala Lys Asp Ile
165 170 175
Val Tyr Ile Gly Leu Arg Asp Val Asp Pro Gly Glu His Tyr Ile Leu
180 185 190
Lys Thr Leu Gly Ile Lys Tyr Phe Ser Met Thr Glu Val Asp Arg Leu
195 200 205
Gly Ile Gly Lys Val Met Glu Glu Thr Leu Ser Tyr Leu Leu Gly Arg
210 215 220
Lys Lys Arg Pro Ile His Leu Ser Phe Asp Val Asp Gly Leu Asp Pro
225 230 235 240
Ser Phe Thr Pro Ala Thr Gly Thr Pro Val Val Gly Gly Leu Thr Tyr
245 250 255
Arg Glu Gly Leu Tyr Ile Thr Glu Glu Ile Tyr Lys Thr Gly Leu Leu
260 265 270
Ser Gly Leu Asp Ile Met Glu Val Asn Pro Ser Leu Gly Lys Thr Pro
275 280 285
Glu Glu Val Thr Arg Thr Val Asn Thr Ala Val Ala Ile Thr Leu Ala
290 295 300
Cys Phe Gly Leu Ala Arg Glu Gly Asn His Lys Pro Ile Asp Tyr Leu
305 310 315 320
Asn Pro Pro Lys
<210> 70
<211> 299
<212> PRT
<213> Bacillus caldovelox
<400> 70
Met Lys Pro Ile Ser Ile Ile Gly Val Pro Met Asp Leu Gly Gln Thr
1 5 10 15
Arg Arg Gly Val Asp Met Gly Pro Ser Ala Met Arg Tyr Ala Gly Val
20 25 30
Ile Glu Arg Leu Glu Arg Leu His Tyr Asp Ile Glu Asp Leu Gly Asp
35 40 45
Ile Pro Ile Gly Lys Ala Glu Arg Leu His Glu Gln Gly Asp Ser Arg
50 55 60
Leu Arg Asn Leu Lys Ala Val Ala Glu Ala Asn Glu Lys Leu Ala Ala
65 70 75 80
Ala Val Asp Gln Val Val Gln Arg Gly Arg Phe Pro Leu Val Leu Gly
85 90 95
Gly Asp His Ser Ile Ala Ile Gly Thr Leu Ala Gly Val Ala Lys His
100 105 110
Tyr Glu Arg Leu Gly Val Ile Trp Tyr Asp Ala His Gly Asp Val Asn
115 120 125
Thr Ala Glu Thr Ser Pro Ser Gly Asn Ile His Gly Met Pro Leu Ala
130 135 140
Ala Ser Leu Gly Phe Gly His Pro Ala Leu Thr Gln Ile Gly Gly Tyr
145 150 155 160
Ser Pro Lys Ile Lys Pro Glu His Val Val Leu Ile Gly Val Arg Ser
165 170 175
Leu Asp Glu Gly Glu Lys Lys Phe Ile Arg Glu Lys Gly Ile Lys Ile
180 185 190
Tyr Thr Met His Glu Val Asp Arg Leu Gly Met Thr Arg Val Met Glu
195 200 205
Glu Thr Ile Ala Tyr Leu Lys Glu Arg Thr Asp Gly Val His Leu Ser
210 215 220
Leu Asp Leu Asp Gly Leu Asp Pro Ser Asp Ala Pro Gly Val Gly Thr
225 230 235 240
Pro Val Ile Gly Gly Leu Thr Tyr Arg Glu Ser His Leu Ala Met Glu
245 250 255
Met Leu Ala Glu Ala Gln Ile Ile Thr Ser Ala Glu Phe Val Glu Val
260 265 270
Asn Pro Ile Leu Asp Glu Arg Asn Lys Thr Ala Ser Val Ala Val Ala
275 280 285
Leu Met Gly Ser Leu Phe Gly Glu Lys Leu Met
290 295
<210> 71
<211> 299
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аргиназа BCA (S161C), синтезирована в лаборатории
<400> 71
Met Lys Pro Ile Ser Ile Ile Gly Val Pro Met Asp Leu Gly Gln Thr
1 5 10 15
Arg Arg Gly Val Asp Met Gly Pro Ser Ala Met Arg Tyr Ala Gly Val
20 25 30
Ile Glu Arg Leu Glu Arg Leu His Tyr Asp Ile Glu Asp Leu Gly Asp
35 40 45
Ile Pro Ile Gly Lys Ala Glu Arg Leu His Glu Gln Gly Asp Ser Arg
50 55 60
Leu Arg Asn Leu Lys Ala Val Ala Glu Ala Asn Glu Lys Leu Ala Ala
65 70 75 80
Ala Val Asp Gln Val Val Gln Arg Gly Arg Phe Pro Leu Val Leu Gly
85 90 95
Gly Asp His Ser Ile Ala Ile Gly Thr Leu Ala Gly Val Ala Lys His
100 105 110
Tyr Glu Arg Leu Gly Val Ile Trp Tyr Asp Ala His Gly Asp Val Asn
115 120 125
Thr Ala Glu Thr Ser Pro Ser Gly Asn Ile His Gly Met Pro Leu Ala
130 135 140
Ala Ser Leu Gly Phe Gly His Pro Ala Leu Thr Gln Ile Gly Gly Tyr
145 150 155 160
Cys Pro Lys Ile Lys Pro Glu His Val Val Leu Ile Gly Val Arg Ser
165 170 175
Leu Asp Glu Gly Glu Lys Lys Phe Ile Arg Glu Lys Gly Ile Lys Ile
180 185 190
Tyr Thr Met His Glu Val Asp Arg Leu Gly Met Thr Arg Val Met Glu
195 200 205
Glu Thr Ile Ala Tyr Leu Lys Glu Arg Thr Asp Gly Val His Leu Ser
210 215 220
Leu Asp Leu Asp Gly Leu Asp Pro Ser Asp Ala Pro Gly Val Gly Thr
225 230 235 240
Pro Val Ile Gly Gly Leu Thr Tyr Arg Glu Ser His Leu Ala Met Glu
245 250 255
Met Leu Ala Glu Ala Gln Ile Ile Thr Ser Ala Glu Phe Val Glu Val
260 265 270
Asn Pro Ile Leu Asp Glu Arg Asn Lys Thr Ala Ser Val Ala Val Ala
275 280 285
Leu Met Gly Ser Leu Phe Gly Glu Lys Leu Met
290 295
<210> 72
<211> 299
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аргиназа BCA (V20P, S161C), синтезирована в лаборатории
<400> 72
Met Lys Pro Ile Ser Ile Ile Gly Val Pro Met Asp Leu Gly Gln Thr
1 5 10 15
Arg Arg Gly Pro Asp Met Gly Pro Ser Ala Met Arg Tyr Ala Gly Val
20 25 30
Ile Glu Arg Leu Glu Arg Leu His Tyr Asp Ile Glu Asp Leu Gly Asp
35 40 45
Ile Pro Ile Gly Lys Ala Glu Arg Leu His Glu Gln Gly Asp Ser Arg
50 55 60
Leu Arg Asn Leu Lys Ala Val Ala Glu Ala Asn Glu Lys Leu Ala Ala
65 70 75 80
Ala Val Asp Gln Val Val Gln Arg Gly Arg Phe Pro Leu Val Leu Gly
85 90 95
Gly Asp His Ser Ile Ala Ile Gly Thr Leu Ala Gly Val Ala Lys His
100 105 110
Tyr Glu Arg Leu Gly Val Ile Trp Tyr Asp Ala His Gly Asp Val Asn
115 120 125
Thr Ala Glu Thr Ser Pro Ser Gly Asn Ile His Gly Met Pro Leu Ala
130 135 140
Ala Ser Leu Gly Phe Gly His Pro Ala Leu Thr Gln Ile Gly Gly Tyr
145 150 155 160
Cys Pro Lys Ile Lys Pro Glu His Val Val Leu Ile Gly Val Arg Ser
165 170 175
Leu Asp Glu Gly Glu Lys Lys Phe Ile Arg Glu Lys Gly Ile Lys Ile
180 185 190
Tyr Thr Met His Glu Val Asp Arg Leu Gly Met Thr Arg Val Met Glu
195 200 205
Glu Thr Ile Ala Tyr Leu Lys Glu Arg Thr Asp Gly Val His Leu Ser
210 215 220
Leu Asp Leu Asp Gly Leu Asp Pro Ser Asp Ala Pro Gly Val Gly Thr
225 230 235 240
Pro Val Ile Gly Gly Leu Thr Tyr Arg Glu Ser His Leu Ala Met Glu
245 250 255
Met Leu Ala Glu Ala Gln Ile Ile Thr Ser Ala Glu Phe Val Glu Val
260 265 270
Asn Pro Ile Leu Asp Glu Arg Asn Lys Thr Ala Ser Val Ala Val Ala
275 280 285
Leu Met Gly Ser Leu Phe Gly Glu Lys Leu Met
290 295
<210> 73
<211> 12
<212> PRT
<213> Искусственная последовательность
<220>
<223> Линкерная последовательность 1, синтезирована в лаборатории
<400> 73
Gly Ser Asn Asn Asn Asn Asn Asn Gly Ser Gly Gly
1 5 10
<210> 74
<211> 13
<212> PRT
<213> Искусственная последовательность
<220>
<223> Линкерная последовательность 2, синтезирована в лаборатории
<400> 74
Ser Gly Ser Asn Asn Asn Asn Asn Asn Gly Ser Gly Gly
1 5 10
<210> 75
<211> 362
<212> PRT
<213> Искусственная последовательность
<220>
<223> Слитый белок BHA, синтезирован в лаборатории
<400> 75
Met Lys Pro Ile Ser Ile Ile Gly Val Pro Met Asp Leu Gly Gln Thr
1 5 10 15
Arg Arg Gly Pro Asp Met Gly Pro Ser Ala Met Arg Tyr Ala Gly Val
20 25 30
Ile Glu Arg Leu Glu Arg Leu His Tyr Asp Ile Glu Asp Leu Gly Asp
35 40 45
Ile Pro Ile Gly Lys Ala Glu Arg Leu His Glu Gln Gly Asp Ser Arg
50 55 60
Leu Arg Asn Leu Lys Ala Val Ala Glu Ala Asn Glu Lys Leu Ala Ala
65 70 75 80
Ala Val Asp Gln Val Val Gln Arg Gly Arg Phe Pro Leu Val Leu Gly
85 90 95
Gly Asp His Ser Ile Ala Ile Gly Thr Leu Ala Gly Val Ala Lys His
100 105 110
Tyr Glu Arg Leu Gly Val Ile Trp Tyr Asp Ala His Gly Asp Val Asn
115 120 125
Thr Ala Glu Thr Ser Pro Ser Gly Asn Ile His Gly Met Pro Leu Ala
130 135 140
Ala Ser Leu Gly Phe Gly His Pro Ala Leu Thr Gln Ile Gly Gly Tyr
145 150 155 160
Cys Pro Lys Ile Lys Pro Glu His Val Val Leu Ile Gly Val Arg Ser
165 170 175
Leu Asp Glu Gly Glu Lys Lys Phe Ile Arg Glu Lys Gly Ile Lys Ile
180 185 190
Tyr Thr Met His Glu Val Asp Arg Leu Gly Met Thr Arg Val Met Glu
195 200 205
Glu Thr Ile Ala Tyr Leu Lys Glu Arg Thr Asp Gly Val His Leu Ser
210 215 220
Leu Asp Leu Asp Gly Leu Asp Pro Ser Asp Ala Pro Gly Val Gly Thr
225 230 235 240
Pro Val Ile Gly Gly Leu Thr Tyr Arg Glu Ser His Leu Ala Met Glu
245 250 255
Met Leu Ala Glu Ala Gln Ile Ile Thr Ser Ala Glu Phe Val Glu Val
260 265 270
Asn Pro Ile Leu Asp Glu Arg Asn Lys Thr Ala Ser Val Ala Val Ala
275 280 285
Leu Met Gly Ser Leu Phe Gly Glu Lys Leu Met His His His His His
290 295 300
His Ala Gln His Asp Glu Ala Val Asp Ala Asn Ser Leu Ala Glu Ala
305 310 315 320
Lys Val Leu Ala Asn Arg Glu Leu Asp Lys Tyr Gly Val Ser Asp Tyr
325 330 335
Tyr Lys Asn Leu Ile Asn Asn Ala Lys Thr Val Glu Gly Val Lys Ala
340 345 350
Leu Ile Asp Glu Ile Leu Ala Ala Leu Pro
355 360
<210> 76
<211> 362
<212> PRT
<213> Искусственная последовательность
<220>
<223> Слитый белок BAH, синтезирован в лаборатории
<400> 76
Met Lys Pro Ile Ser Ile Ile Gly Val Pro Met Asp Leu Gly Gln Thr
1 5 10 15
Arg Arg Gly Pro Asp Met Gly Pro Ser Ala Met Arg Tyr Ala Gly Val
20 25 30
Ile Glu Arg Leu Glu Arg Leu His Tyr Asp Ile Glu Asp Leu Gly Asp
35 40 45
Ile Pro Ile Gly Lys Ala Glu Arg Leu His Glu Gln Gly Asp Ser Arg
50 55 60
Leu Arg Asn Leu Lys Ala Val Ala Glu Ala Asn Glu Lys Leu Ala Ala
65 70 75 80
Ala Val Asp Gln Val Val Gln Arg Gly Arg Phe Pro Leu Val Leu Gly
85 90 95
Gly Asp His Ser Ile Ala Ile Gly Thr Leu Ala Gly Val Ala Lys His
100 105 110
Tyr Glu Arg Leu Gly Val Ile Trp Tyr Asp Ala His Gly Asp Val Asn
115 120 125
Thr Ala Glu Thr Ser Pro Ser Gly Asn Ile His Gly Met Pro Leu Ala
130 135 140
Ala Ser Leu Gly Phe Gly His Pro Ala Leu Thr Gln Ile Gly Gly Tyr
145 150 155 160
Cys Pro Lys Ile Lys Pro Glu His Val Val Leu Ile Gly Val Arg Ser
165 170 175
Leu Asp Glu Gly Glu Lys Lys Phe Ile Arg Glu Lys Gly Ile Lys Ile
180 185 190
Tyr Thr Met His Glu Val Asp Arg Leu Gly Met Thr Arg Val Met Glu
195 200 205
Glu Thr Ile Ala Tyr Leu Lys Glu Arg Thr Asp Gly Val His Leu Ser
210 215 220
Leu Asp Leu Asp Gly Leu Asp Pro Ser Asp Ala Pro Gly Val Gly Thr
225 230 235 240
Pro Val Ile Gly Gly Leu Thr Tyr Arg Glu Ser His Leu Ala Met Glu
245 250 255
Met Leu Ala Glu Ala Gln Ile Ile Thr Ser Ala Glu Phe Val Glu Val
260 265 270
Asn Pro Ile Leu Asp Glu Arg Asn Lys Thr Ala Ser Val Ala Val Ala
275 280 285
Leu Met Gly Ser Leu Phe Gly Glu Lys Leu Met Ala Gln His Asp Glu
290 295 300
Ala Val Asp Ala Asn Ser Leu Ala Glu Ala Lys Val Leu Ala Asn Arg
305 310 315 320
Glu Leu Asp Lys Tyr Gly Val Ser Asp Tyr Tyr Lys Asn Leu Ile Asn
325 330 335
Asn Ala Lys Thr Val Glu Gly Val Lys Ala Leu Ile Asp Glu Ile Leu
340 345 350
Ala Ala Leu Pro His His His His His His
355 360
<210> 77
<211> 103
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 5'-смысловая цепь-ABD, синтезирован в лаборатории
<400> 77
gcgcagcatg atgaagccgt ggatgcgaac agcttagctg aagctaaagt cttagctaac 60
agagaacttg acaaatatgg agtaagtgac tattacaaga acc 103
<210> 78
<211> 98
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер 5'-антисмысловая цепь-ABD, синтезирован в лаборатории
<400> 78
ttaaggtaat gcagctaaaa tttcatctat cagtgctttt acaccttcaa cagttttggc 60
attgttgatt aggttcttgt aatagtcact tactccat 98
<210> 79
<211> 35
<212> DNA
<213> Искусственная последовательность
<220>
<223> Прямой праймер для клонирования ABD, синтезирован в лаборатории
<400> 79
gcatcaccat caccatcacg cgcagcatga tgaag 35
<210> 80
<211> 38
<212> DNA
<213> Искусственная последовательность
<220>
<223> Обратный праймер для клонирования ABD, синтезирован в лаборатории
<400> 80
cgggatcctt aaggtaatgc agctaaaatt tcatctat 38
<210> 81
<211> 33
<212> DNA
<213> Искусственная последовательность
<220>
<223> Прямой праймер для клонирования BCA, синтезирован в лаборатории
<400> 81
ggaattccca tatgaagcca atttcaatta tcg 33
<210> 82
<211> 20
<212> DNA
<213> Искусственная последовательность
<220>
<223> Обратный праймер для клонирования BCA, синтезирован в лаборатории
<400> 82
gtgatggtga tggtgatgca 20
<210> 83
<211> 33
<212> DNA
<213> Искусственная последовательность
<220>
<223> Прямой праймер для клонирования BHA, синтезирован в лаборатории
<400> 83
ggaattccca tatgaagcca atttcaatta tcg 33
<210> 84
<211> 38
<212> DNA
<213> Искусственная последовательность
<220>
<223> Обратный праймер для клонирования BHA, синтезирован в лаборатории
<400> 84
cgggatcctt aaggtaatgc agctaaaatt tcatctat 38
<210> 85
<211> 39
<212> DNA
<213> Искусственная последовательность
<220>
<223> Прямой праймер для клонирования ABD, синтезирован в лаборатории
<400> 85
cgttgtttgg tgaaaaactc atggcgcagc atgatgaag 39
<210> 86
<211> 56
<212> DNA
<213> Искусственная последовательность
<220>
<223> Обратный праймер для клонирования ABD, синтезирован в лаборатории
<400> 86
cgggatcctt agtgatggtg atggtgatga ggtaatgcag ctaaaatttc atctat 56
<210> 87
<211> 33
<212> DNA
<213> Искусственная последовательность
<220>
<223> Прямой праймер для клонирования BAH, синтезирован в лаборатории
<400> 87
ggaattccca tatgaagcca atttcaatta tcg 33
<210> 88
<211> 56
<212> DNA
<213> Искусственная последовательность
<220>
<223> Обратный праймер для клонирования BAH, синтезирован в лаборатории
<400> 88
cgggatcctt agtgatggtg atggtgatga ggtaatgcag ctaaaatttc atctat 56
<210> 89
<211> 305
<212> PRT
<213> Искусственная последовательность
<220>
<223> BCA (S161C) - His, синтезирован в лаборатории
<400> 89
Met Lys Pro Ile Ser Ile Ile Gly Val Pro Met Asp Leu Gly Gln Thr
1 5 10 15
Arg Arg Gly Val Asp Met Gly Pro Ser Ala Met Arg Tyr Ala Gly Val
20 25 30
Ile Glu Arg Leu Glu Arg Leu His Tyr Asp Ile Glu Asp Leu Gly Asp
35 40 45
Ile Pro Ile Gly Lys Ala Glu Arg Leu His Glu Gln Gly Asp Ser Arg
50 55 60
Leu Arg Asn Leu Lys Ala Val Ala Glu Ala Asn Glu Lys Leu Ala Ala
65 70 75 80
Ala Val Asp Gln Val Val Gln Arg Gly Arg Phe Pro Leu Val Leu Gly
85 90 95
Gly Asp His Ser Ile Ala Ile Gly Thr Leu Ala Gly Val Ala Lys His
100 105 110
Tyr Glu Arg Leu Gly Val Ile Trp Tyr Asp Ala His Gly Asp Val Asn
115 120 125
Thr Ala Glu Thr Ser Pro Ser Gly Asn Ile His Gly Met Pro Leu Ala
130 135 140
Ala Ser Leu Gly Phe Gly His Pro Ala Leu Thr Gln Ile Gly Gly Tyr
145 150 155 160
Cys Pro Lys Ile Lys Pro Glu His Val Val Leu Ile Gly Val Arg Ser
165 170 175
Leu Asp Glu Gly Glu Lys Lys Phe Ile Arg Glu Lys Gly Ile Lys Ile
180 185 190
Tyr Thr Met His Glu Val Asp Arg Leu Gly Met Thr Arg Val Met Glu
195 200 205
Glu Thr Ile Ala Tyr Leu Lys Glu Arg Thr Asp Gly Val His Leu Ser
210 215 220
Leu Asp Leu Asp Gly Leu Asp Pro Ser Asp Ala Pro Gly Val Gly Thr
225 230 235 240
Pro Val Ile Gly Gly Leu Thr Tyr Arg Glu Ser His Leu Ala Met Glu
245 250 255
Met Leu Ala Glu Ala Gln Ile Ile Thr Ser Ala Glu Phe Val Glu Val
260 265 270
Asn Pro Ile Leu Asp Glu Arg Asn Lys Thr Ala Ser Val Ala Val Ala
275 280 285
Leu Met Gly Ser Leu Phe Gly Glu Lys Leu Met His His His His His
290 295 300
His
305
<210> 90
<211> 42
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер N-ABDHind-R, синтезирован в лаборатории
<400> 90
acagctaaaa gcttatcagc ctaggatccg ccgctaccat tg 42
<210> 91
<211> 38
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер C-ABDNde-F, синтезирован в лаборатории
<400> 91
tagctgatca tatgttatgc gatggatcca gtaacaac 38
<210> 92
<211> 33
<212> DNA
<213> Искусственная последовательность
<220>
<223> C-ABDHind-R, синтезирован в лаборатории
<400> 92
gaccctaaaa gcttaatggt gatggtgatg atg 33
<210> 93
<211> 34
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер HARGBam-F, синтезирован в лаборатории
<400> 93
ttagctgggg atccgccaag tccagaacca tagg 34
<210> 94
<211> 35
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер HARGHind-R, синтезирован в лаборатории
<400> 94
gagatcaaag cttacttagg tgggttaagg tagtc 35
<210> 95
<211> 35
<212> DNA
<213> Искусственная последовательность
<220>
<223> HARGNde-F, синтезирован в лаборатории
<400> 95
taggctgcat atgagcgcca agtccagaac catag 35
<210> 96
<211> 41
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер HARGBam-R, синтезирован в лаборатории
<400> 96
atcagctagg atcccttagg tgggttaagg tagtcaatag g 41
<210> 97
<211> 36
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер BCABam-F, синтезирован в лаборатории
<400> 97
atgctagtgg atccatgaag ccaatttcaa ttatcg 36
<210> 98
<211> 38
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер BCAHind-R, синтезирован в лаборатории
<400> 98
tcagcctaaa gcttacatga gtttttcacc aaacaacg 38
<210> 99
<211> 35
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер BCANde-F, синтезирован в лаборатории
<400> 99
atgctagcca tatgaagcca atttcaatta tcggg 35
<210> 100
<211> 37
<212> DNA
<213> Искусственная последовательность
<220>
<223> Праймер BCABam-R, синтезирован в лаборатории
<400> 100
tcagctaagg atcccatgag tttttcacca aacaacg 37
<210> 101
<211> 329
<212> PRT
<213> Искусственная последовательность
<220>
<223> His-rhArg, синтезирован в лаборатории
<400> 101
Met His His His His His His Met Ser Ala Lys Ser Arg Thr Ile Gly
1 5 10 15
Ile Ile Gly Ala Pro Phe Ser Lys Gly Gln Pro Arg Gly Gly Val Glu
20 25 30
Glu Gly Pro Thr Val Leu Arg Lys Ala Gly Leu Leu Glu Lys Leu Lys
35 40 45
Glu Gln Glu Cys Asp Val Lys Asp Tyr Gly Asp Leu Pro Phe Ala Asp
50 55 60
Ile Pro Asn Asp Ser Pro Phe Gln Ile Val Lys Asn Pro Arg Ser Val
65 70 75 80
Gly Lys Ala Ser Glu Gln Leu Ala Gly Lys Val Ala Glu Val Lys Lys
85 90 95
Asn Gly Arg Ile Ser Leu Val Leu Gly Gly Asp His Ser Leu Ala Ile
100 105 110
Gly Ser Ile Ser Gly His Ala Arg Val His Pro Asp Leu Gly Val Ile
115 120 125
Trp Val Asp Ala His Thr Asp Ile Asn Thr Pro Leu Thr Thr Thr Ser
130 135 140
Gly Asn Leu His Gly Gln Pro Val Ser Phe Leu Leu Lys Glu Leu Lys
145 150 155 160
Gly Lys Ile Pro Asp Val Pro Gly Phe Ser Trp Val Thr Pro Cys Ile
165 170 175
Ser Ala Lys Asp Ile Val Tyr Ile Gly Leu Arg Asp Val Asp Pro Gly
180 185 190
Glu His Tyr Ile Leu Lys Thr Leu Gly Ile Lys Tyr Phe Ser Met Thr
195 200 205
Glu Val Asp Arg Leu Gly Ile Gly Lys Val Met Glu Glu Thr Leu Ser
210 215 220
Tyr Leu Leu Gly Arg Lys Lys Arg Pro Ile His Leu Ser Phe Asp Val
225 230 235 240
Asp Gly Leu Asp Pro Ser Phe Thr Pro Ala Thr Gly Thr Pro Val Val
245 250 255
Gly Gly Leu Thr Tyr Arg Glu Gly Leu Tyr Ile Thr Glu Glu Ile Tyr
260 265 270
Lys Thr Gly Leu Leu Ser Gly Leu Asp Ile Met Glu Val Asn Pro Ser
275 280 285
Leu Gly Lys Thr Pro Glu Glu Val Thr Arg Thr Val Asn Thr Ala Val
290 295 300
Ala Ile Thr Leu Ala Cys Phe Gly Leu Ala Arg Glu Gly Asn His Lys
305 310 315 320
Pro Ile Asp Tyr Leu Asn Pro Pro Lys
325
<210> 102
<211> 329
<212> PRT
<213> Искусственная последовательность
<220>
<223> His-rhArg-моно-Cys, синтезирован в лаборатории
<400> 102
Met His His His His His His Met Ser Ala Lys Ser Arg Thr Ile Gly
1 5 10 15
Ile Ile Gly Ala Pro Phe Ser Lys Gly Gln Pro Arg Gly Gly Val Glu
20 25 30
Glu Gly Pro Thr Val Leu Arg Lys Ala Gly Leu Leu Glu Lys Leu Lys
35 40 45
Glu Gln Glu Cys Asp Val Lys Asp Tyr Gly Asp Leu Pro Phe Ala Asp
50 55 60
Ile Pro Asn Asp Ser Pro Phe Gln Ile Val Lys Asn Pro Arg Ser Val
65 70 75 80
Gly Lys Ala Ser Glu Gln Leu Ala Gly Lys Val Ala Glu Val Lys Lys
85 90 95
Asn Gly Arg Ile Ser Leu Val Leu Gly Gly Asp His Ser Leu Ala Ile
100 105 110
Gly Ser Ile Ser Gly His Ala Arg Val His Pro Asp Leu Gly Val Ile
115 120 125
Trp Val Asp Ala His Thr Asp Ile Asn Thr Pro Leu Thr Thr Thr Ser
130 135 140
Gly Asn Leu His Gly Gln Pro Val Ser Phe Leu Leu Lys Glu Leu Lys
145 150 155 160
Gly Lys Ile Pro Asp Val Pro Gly Phe Ser Trp Val Thr Pro Ser Ile
165 170 175
Ser Ala Lys Asp Ile Val Tyr Ile Gly Leu Arg Asp Val Asp Pro Gly
180 185 190
Glu His Tyr Ile Leu Lys Thr Leu Gly Ile Lys Tyr Phe Ser Met Thr
195 200 205
Glu Val Asp Arg Leu Gly Ile Gly Lys Val Met Glu Glu Thr Leu Ser
210 215 220
Tyr Leu Leu Gly Arg Lys Lys Arg Pro Ile His Leu Ser Phe Asp Val
225 230 235 240
Asp Gly Leu Asp Pro Ser Phe Thr Pro Ala Thr Gly Thr Pro Val Val
245 250 255
Gly Gly Leu Thr Tyr Arg Glu Gly Leu Tyr Ile Thr Glu Glu Ile Tyr
260 265 270
Lys Thr Gly Leu Leu Ser Gly Leu Asp Ile Met Glu Val Asn Pro Ser
275 280 285
Leu Gly Lys Thr Pro Glu Glu Val Thr Arg Thr Val Asn Thr Ala Val
290 295 300
Ala Ile Thr Leu Ala Ser Phe Gly Leu Ala Arg Glu Gly Asn His Lys
305 310 315 320
Pro Ile Asp Tyr Leu Asn Pro Pro Lys
325
<210> 103
<211> 322
<212> PRT
<213> Homo sapiens
<400> 103
Met Ser Ala Lys Ser Arg Thr Ile Gly Ile Ile Gly Ala Pro Phe Ser
1 5 10 15
Lys Gly Gln Pro Arg Gly Gly Val Glu Glu Gly Pro Thr Val Leu Arg
20 25 30
Lys Ala Gly Leu Leu Glu Lys Leu Lys Glu Gln Glu Cys Asp Val Lys
35 40 45
Asp Tyr Gly Asp Leu Pro Phe Ala Asp Ile Pro Asn Asp Ser Pro Phe
50 55 60
Gln Ile Val Lys Asn Pro Arg Ser Val Gly Lys Ala Ser Glu Gln Leu
65 70 75 80
Ala Gly Lys Val Ala Glu Val Lys Lys Asn Gly Arg Ile Ser Leu Val
85 90 95
Leu Gly Gly Asp His Ser Leu Ala Ile Gly Ser Ile Ser Gly His Ala
100 105 110
Arg Val His Pro Asp Leu Gly Val Ile Trp Val Asp Ala His Thr Asp
115 120 125
Ile Asn Thr Pro Leu Thr Thr Thr Ser Gly Asn Leu His Gly Gln Pro
130 135 140
Val Ser Phe Leu Leu Lys Glu Leu Lys Gly Lys Ile Pro Asp Val Pro
145 150 155 160
Gly Phe Ser Trp Val Thr Pro Cys Ile Ser Ala Lys Asp Ile Val Tyr
165 170 175
Ile Gly Leu Arg Asp Val Asp Pro Gly Glu His Tyr Ile Leu Lys Thr
180 185 190
Leu Gly Ile Lys Tyr Phe Ser Met Thr Glu Val Asp Arg Leu Gly Ile
195 200 205
Gly Lys Val Met Glu Glu Thr Leu Ser Tyr Leu Leu Gly Arg Lys Lys
210 215 220
Arg Pro Ile His Leu Ser Phe Asp Val Asp Gly Leu Asp Pro Ser Phe
225 230 235 240
Thr Pro Ala Thr Gly Thr Pro Val Val Gly Gly Leu Thr Tyr Arg Glu
245 250 255
Gly Leu Tyr Ile Thr Glu Glu Ile Tyr Lys Thr Gly Leu Leu Ser Gly
260 265 270
Leu Asp Ile Met Glu Val Asn Pro Ser Leu Gly Lys Thr Pro Glu Glu
275 280 285
Val Thr Arg Thr Val Asn Thr Ala Val Ala Ile Thr Leu Ala Cys Phe
290 295 300
Gly Leu Ala Arg Glu Gly Asn His Lys Pro Ile Asp Tyr Leu Asn Pro
305 310 315 320
Pro Lys
<210> 104
<211> 322
<212> PRT
<213> Искусственная последовательность
<220>
<223> rhArg-моно-Cys, синтезирован в лаборатории
<400> 104
Met Ser Ala Lys Ser Arg Thr Ile Gly Ile Ile Gly Ala Pro Phe Ser
1 5 10 15
Lys Gly Gln Pro Arg Gly Gly Val Glu Glu Gly Pro Thr Val Leu Arg
20 25 30
Lys Ala Gly Leu Leu Glu Lys Leu Lys Glu Gln Glu Cys Asp Val Lys
35 40 45
Asp Tyr Gly Asp Leu Pro Phe Ala Asp Ile Pro Asn Asp Ser Pro Phe
50 55 60
Gln Ile Val Lys Asn Pro Arg Ser Val Gly Lys Ala Ser Glu Gln Leu
65 70 75 80
Ala Gly Lys Val Ala Glu Val Lys Lys Asn Gly Arg Ile Ser Leu Val
85 90 95
Leu Gly Gly Asp His Ser Leu Ala Ile Gly Ser Ile Ser Gly His Ala
100 105 110
Arg Val His Pro Asp Leu Gly Val Ile Trp Val Asp Ala His Thr Asp
115 120 125
Ile Asn Thr Pro Leu Thr Thr Thr Ser Gly Asn Leu His Gly Gln Pro
130 135 140
Val Ser Phe Leu Leu Lys Glu Leu Lys Gly Lys Ile Pro Asp Val Pro
145 150 155 160
Gly Phe Ser Trp Val Thr Pro Ser Ile Ser Ala Lys Asp Ile Val Tyr
165 170 175
Ile Gly Leu Arg Asp Val Asp Pro Gly Glu His Tyr Ile Leu Lys Thr
180 185 190
Leu Gly Ile Lys Tyr Phe Ser Met Thr Glu Val Asp Arg Leu Gly Ile
195 200 205
Gly Lys Val Met Glu Glu Thr Leu Ser Tyr Leu Leu Gly Arg Lys Lys
210 215 220
Arg Pro Ile His Leu Ser Phe Asp Val Asp Gly Leu Asp Pro Ser Phe
225 230 235 240
Thr Pro Ala Thr Gly Thr Pro Val Val Gly Gly Leu Thr Tyr Arg Glu
245 250 255
Gly Leu Tyr Ile Thr Glu Glu Ile Tyr Lys Thr Gly Leu Leu Ser Gly
260 265 270
Leu Asp Ile Met Glu Val Asn Pro Ser Leu Gly Lys Thr Pro Glu Glu
275 280 285
Val Thr Arg Thr Val Asn Thr Ala Val Ala Ile Thr Leu Ala Ser Phe
290 295 300
Gly Leu Ala Arg Glu Gly Asn His Lys Pro Ile Asp Tyr Leu Asn Pro
305 310 315 320
Pro Lys
<210> 105
<211> 46
<212> PRT
<213> Искусственная последовательность
<220>
<223> Пептидный линкер A(EAAAK)4ALEA-(EAAAK)4A, синтезирован в
лаборатории
<400> 105
Ala Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys
1 5 10 15
Glu Ala Ala Ala Lys Ala Leu Glu Ala Glu Ala Ala Ala Lys Glu Ala
20 25 30
Ala Ala Lys Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys Ala
35 40 45
<210> 106
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Пептидный линкер G4SG4SG3SG, синтезирован в лаборатории
<400> 106
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Ser Gly
1 5 10 15
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ВАКЦИНА И СПОСОБЫ ОБНАРУЖЕНИЯ И ПРОФИЛАКТИКИ ФИЛЯРИОЗА | 2021 |
|
RU2832185C1 |
| ТЕРАПЕВТИЧЕСКИЕ СОЕДИНЕНИЯ И СПОСОБЫ | 2016 |
|
RU2770001C2 |
| ХИМЕРНЫЕ РЕЦЕПТОРЫ АНТИГЕНА ПРОТИВ МЕЗОТЕЛИНА ЧЕЛОВЕКА И ИХ ПРИМЕНЕНИЕ | 2014 |
|
RU2714902C2 |
| ТЕХНОЛОГИИ МУЛЬТИМЕРИЗАЦИИ | 2014 |
|
RU2714733C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ ДЛЯ ЛЕЧЕНИЯ РАКА | 2017 |
|
RU2826270C2 |
| АНТИ-αvβ8 АНТИТЕЛА И КОМПОЗИЦИИ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2812478C2 |
| КОМПОЗИЦИИ И СПОСОБЫ РЕДАКТИРОВАНИЯ ГЕНОВ | 2019 |
|
RU2804665C2 |
| БЕЛКИ С ДВОЙНОЙ ФУНКЦИЕЙ И СОДЕРЖАЩАЯ ИХ ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ | 2016 |
|
RU2741345C2 |
| АНТИТЕЛА ПРОТИВ CXCR5 И ИХ КОМПОЗИЦИИ И ПРИМЕНЕНИЕ | 2018 |
|
RU2798422C2 |
| НЕ ВСТРЕЧАЮЩИЙСЯ В ПРИРОДЕ ВИРУС РЕПРОДУКТИВНО-РЕСПИРАТОРНОГО СИНДРОМА СВИНЕЙ (BPPCC) И СПОСОБЫ ПРИМЕНЕНИЯ | 2015 |
|
RU2687150C2 |
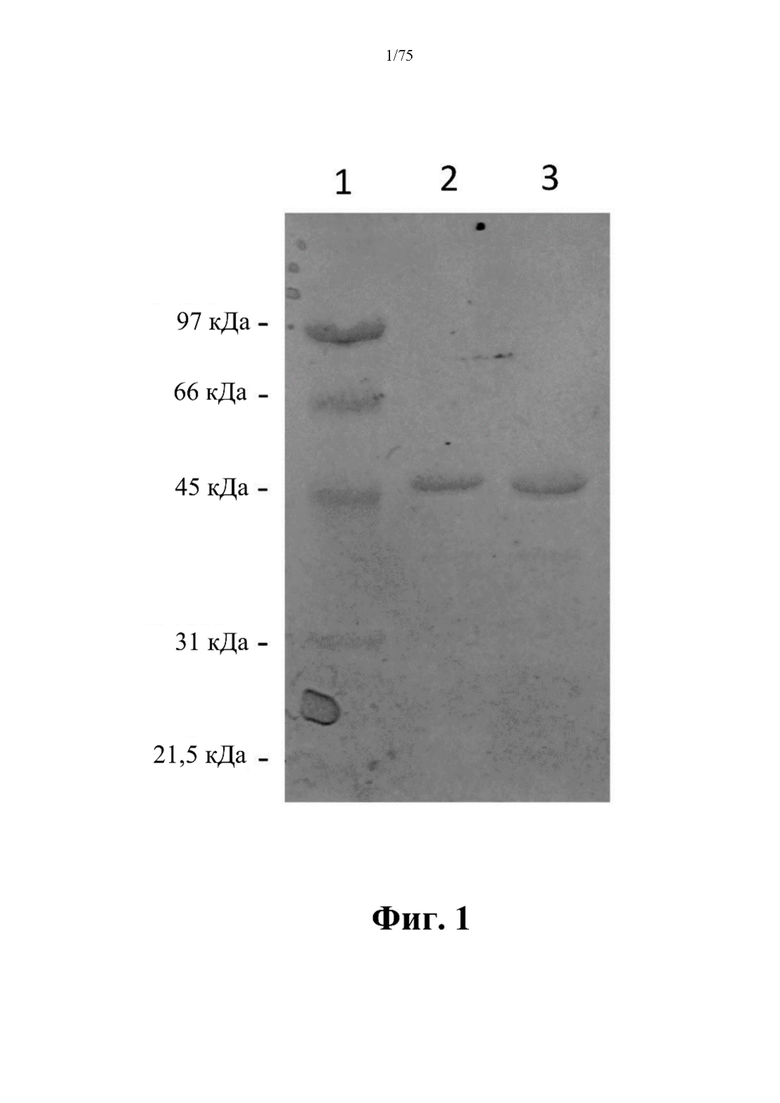
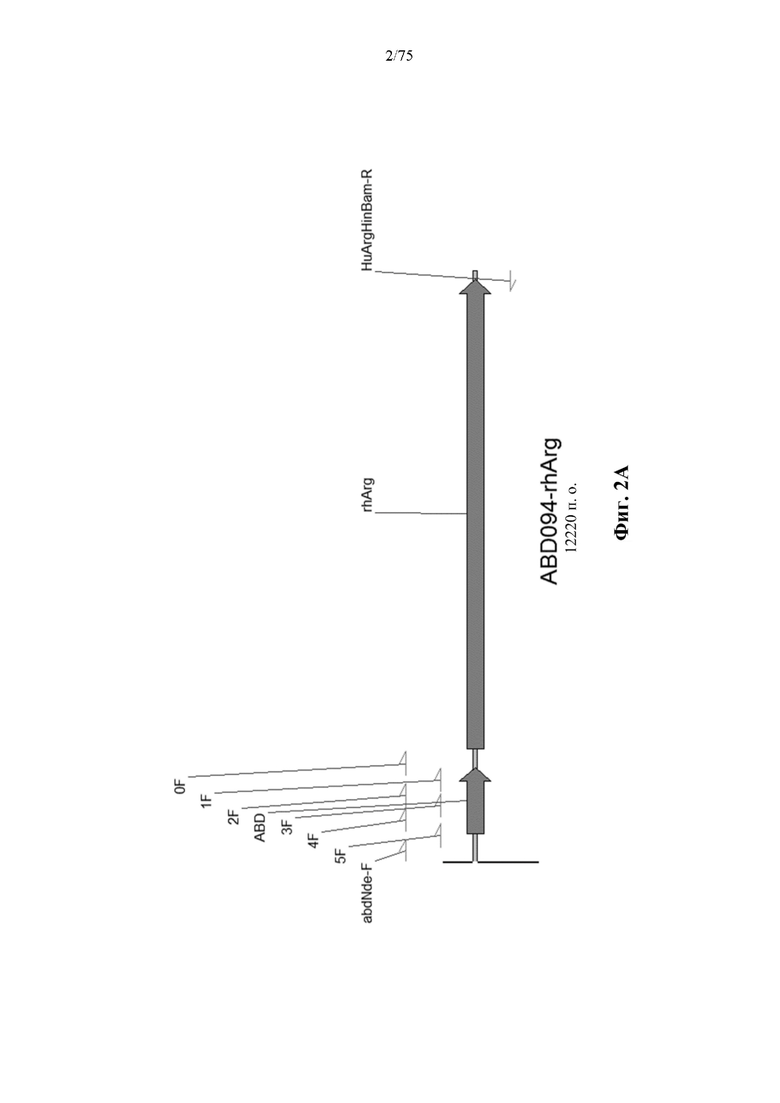



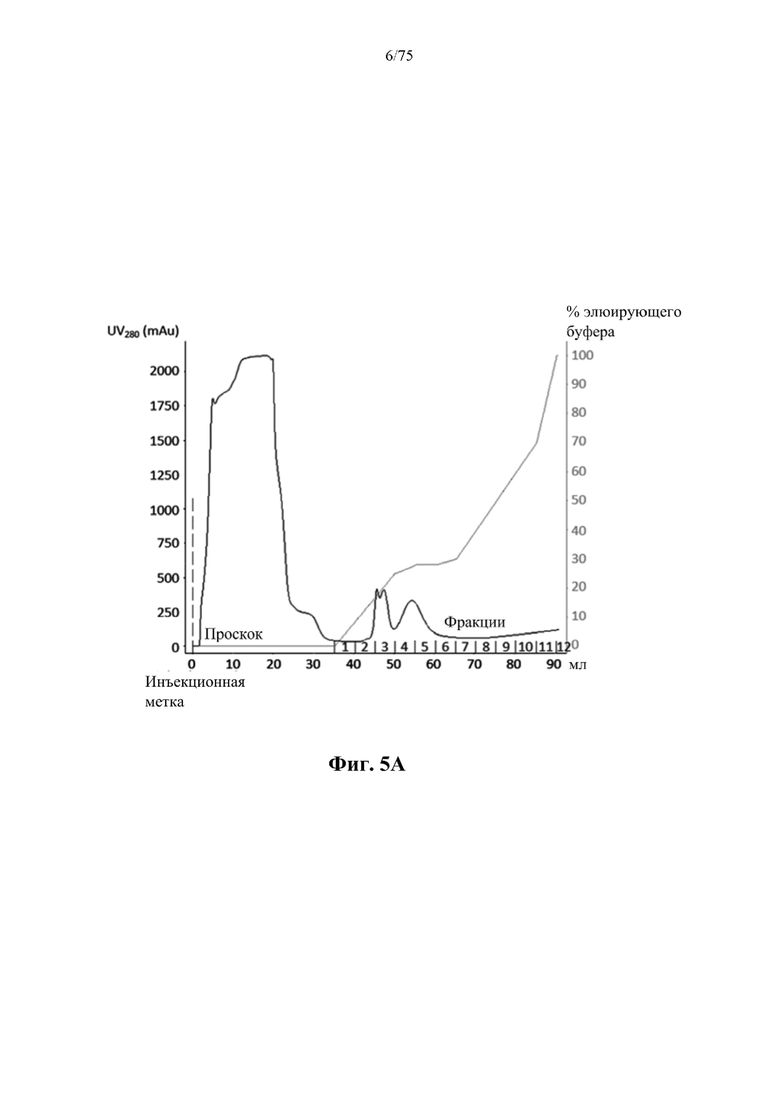
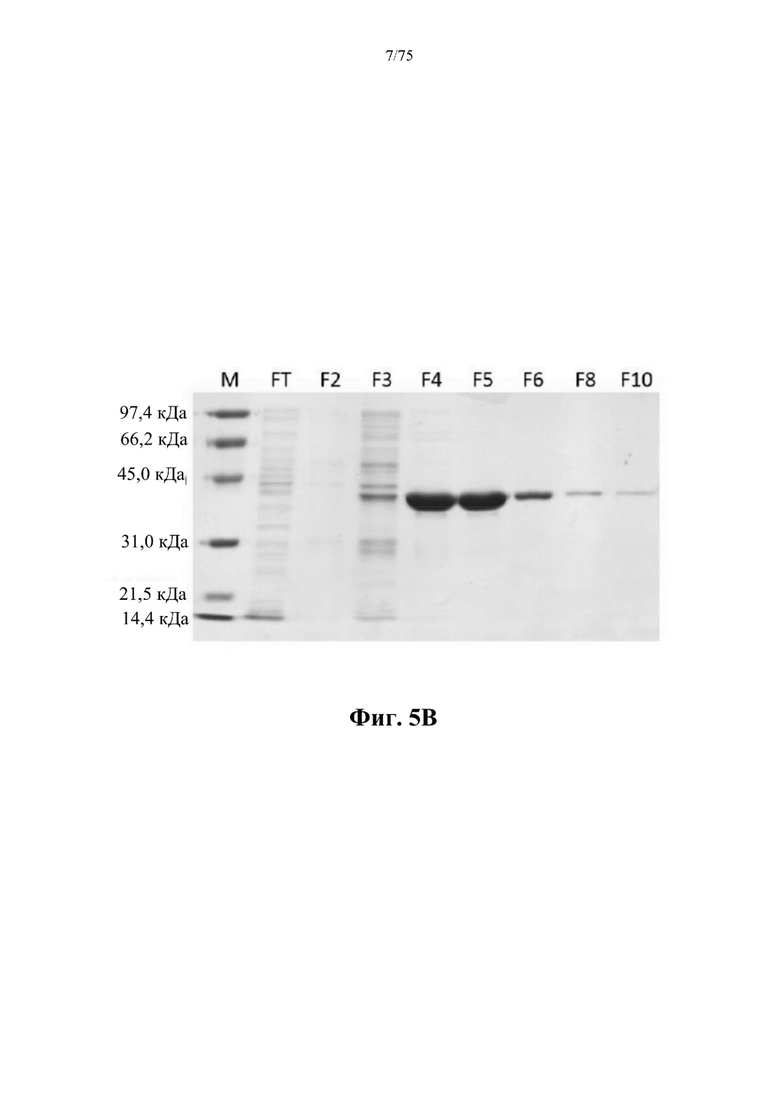
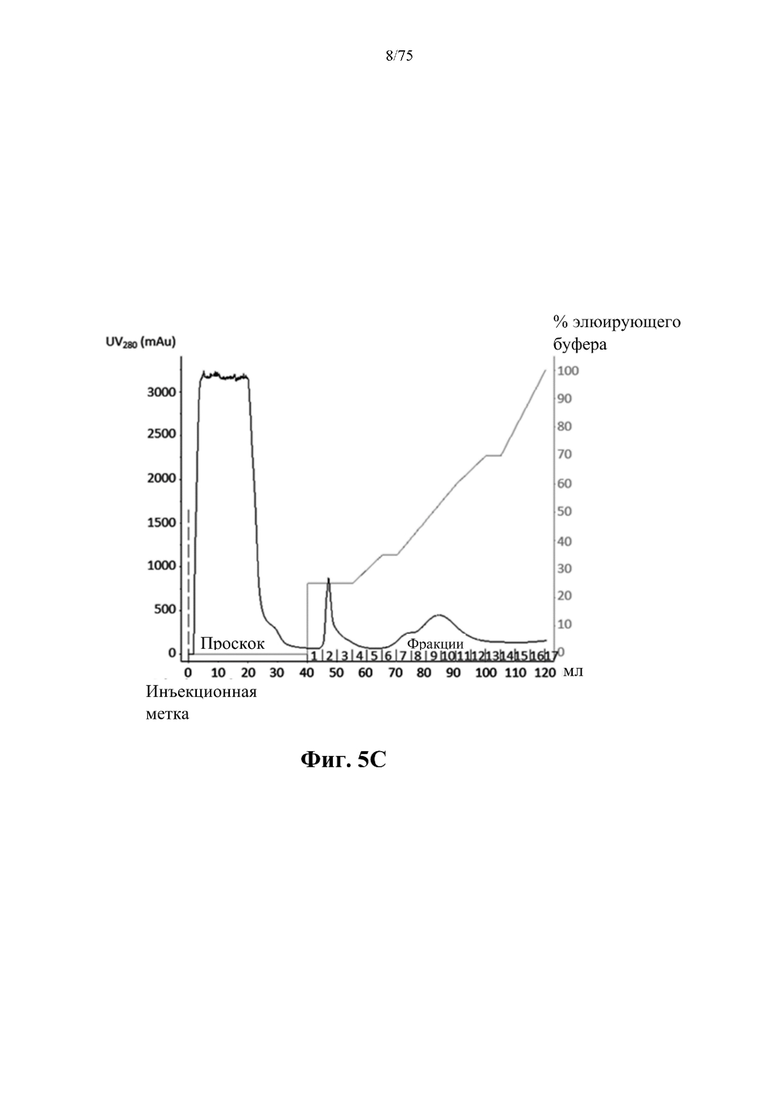
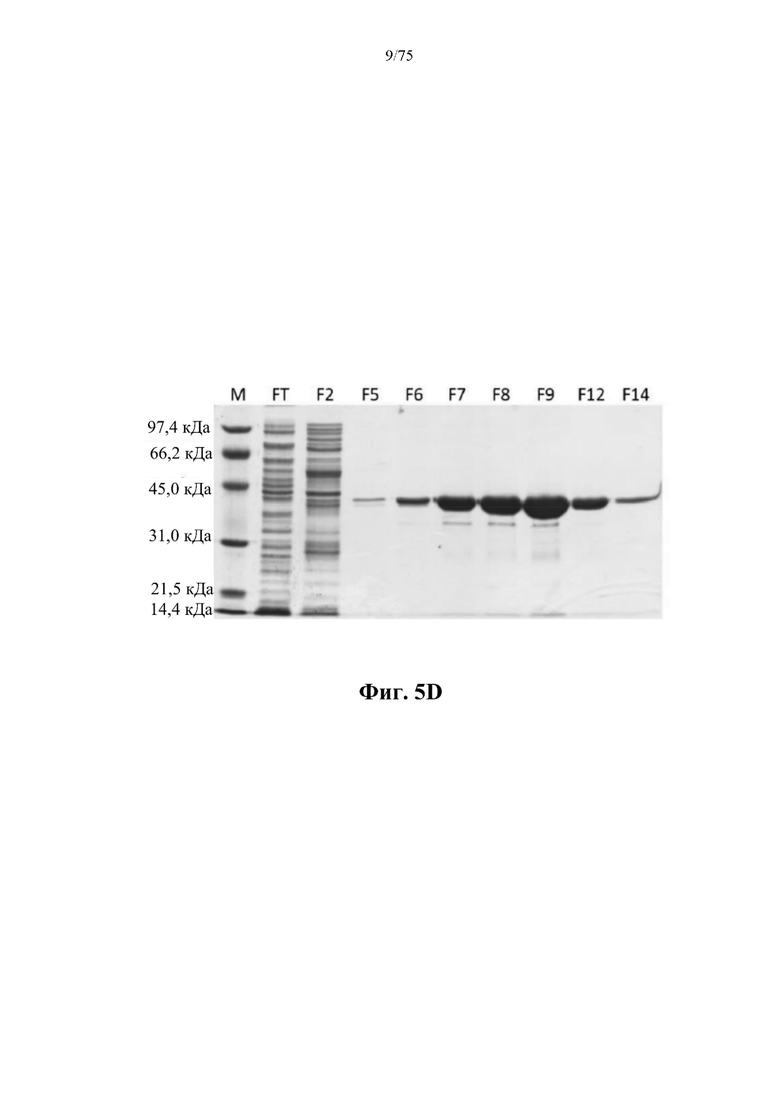
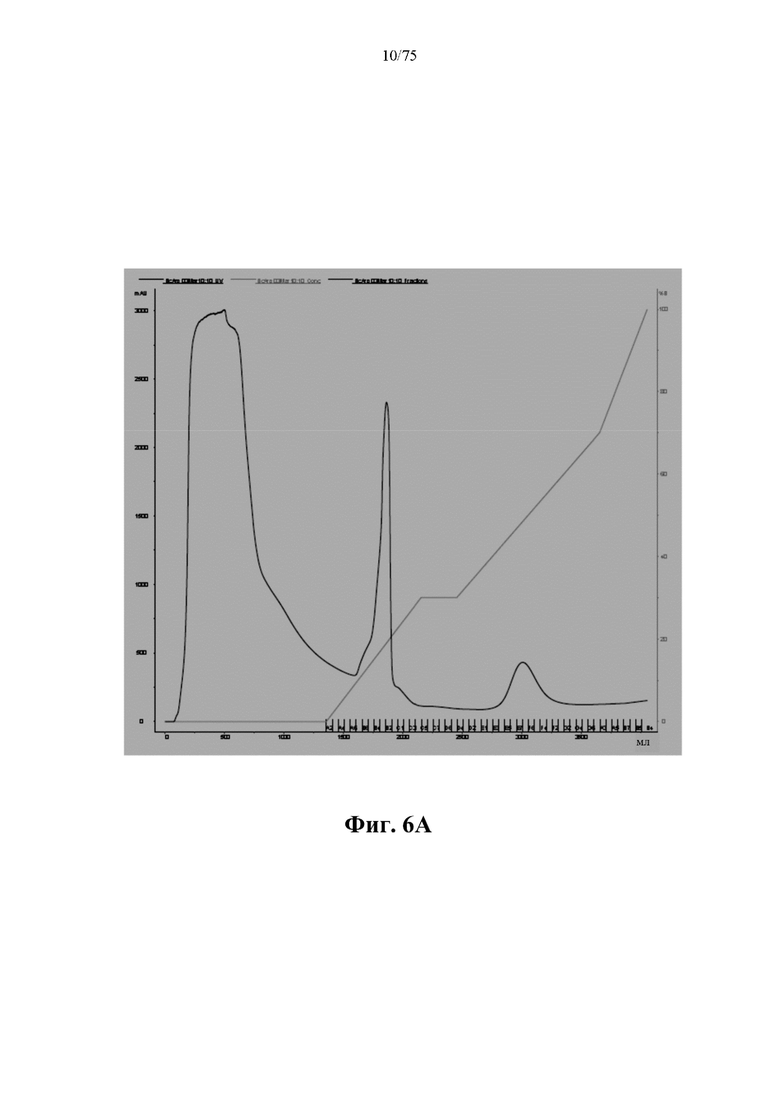
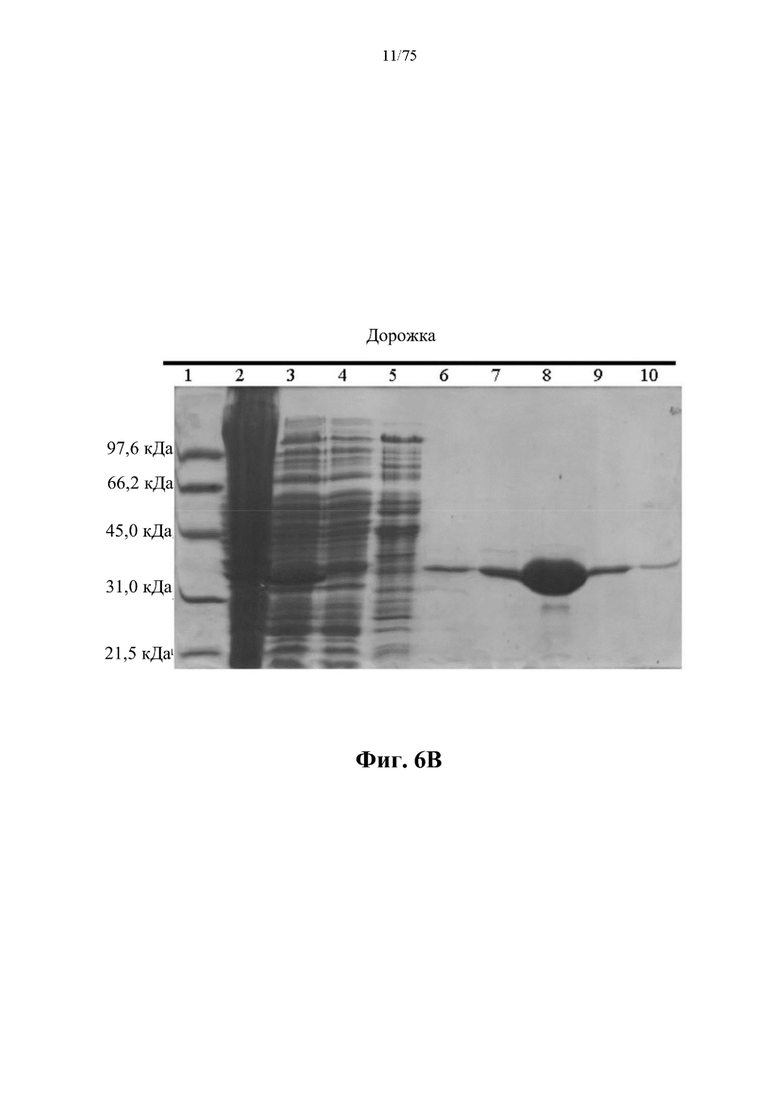
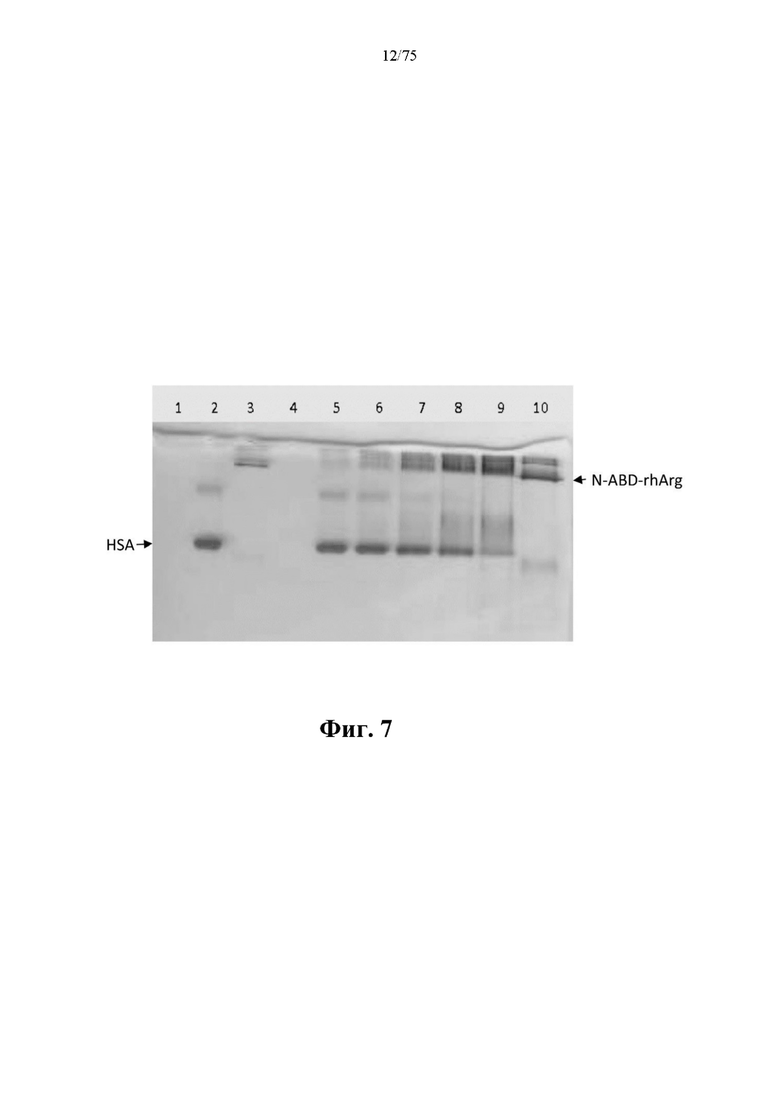
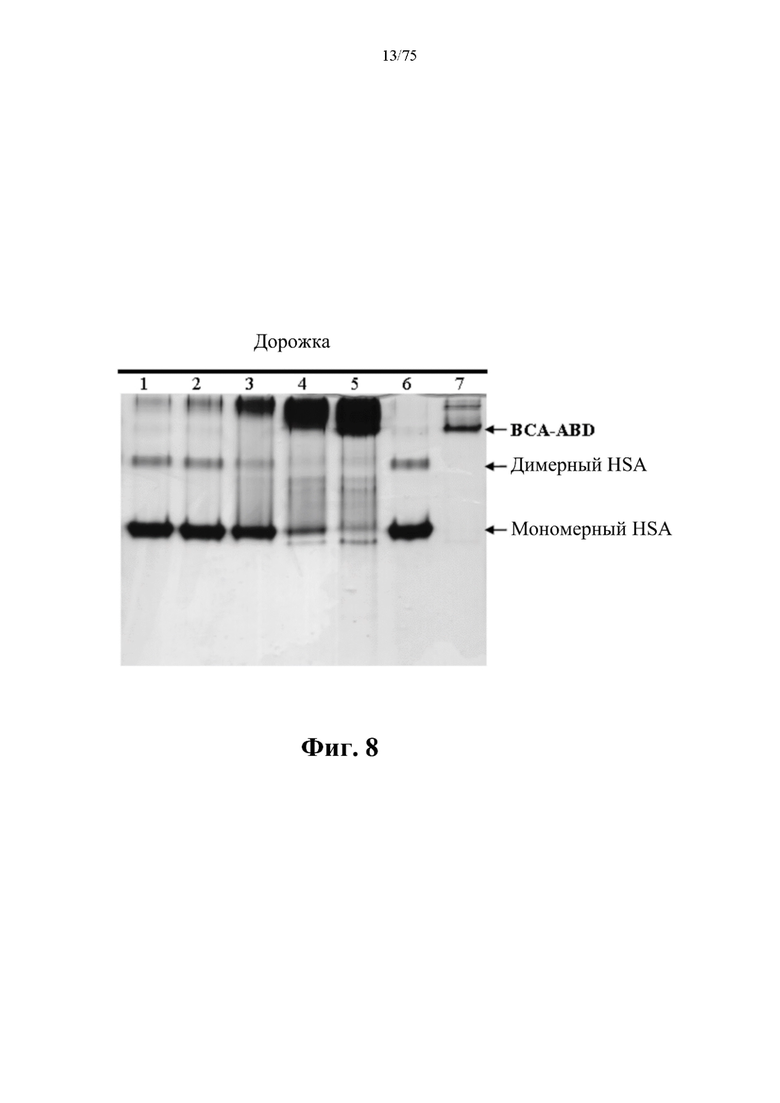
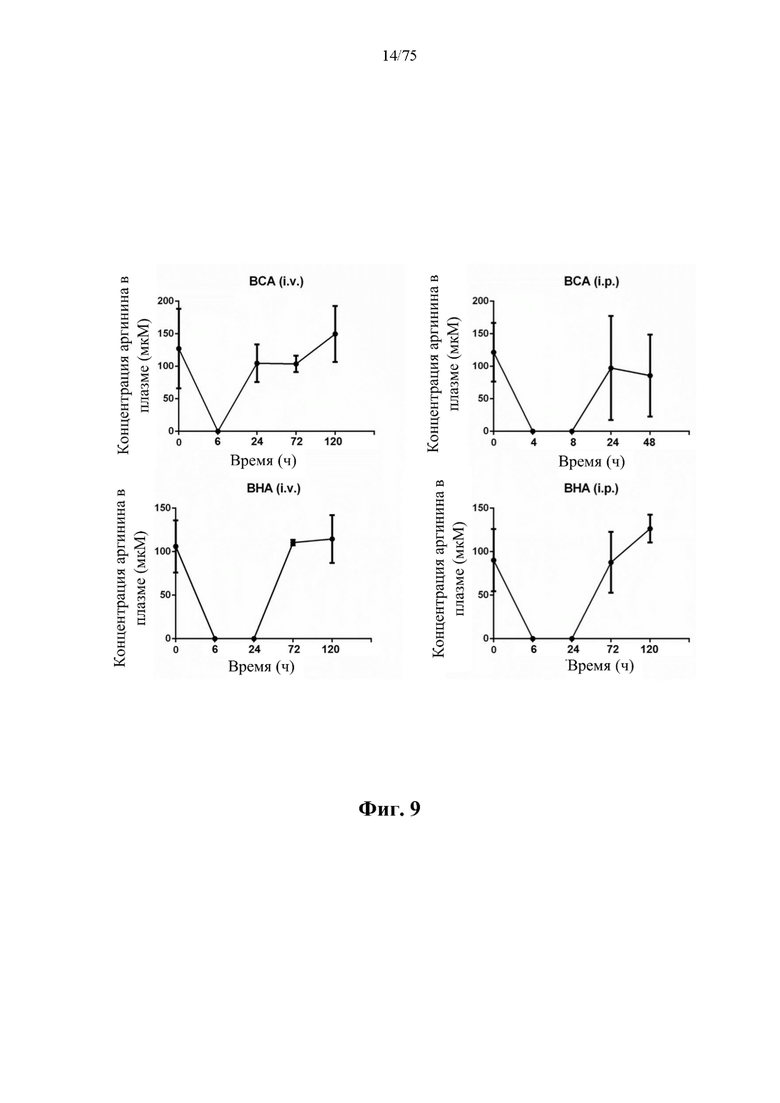
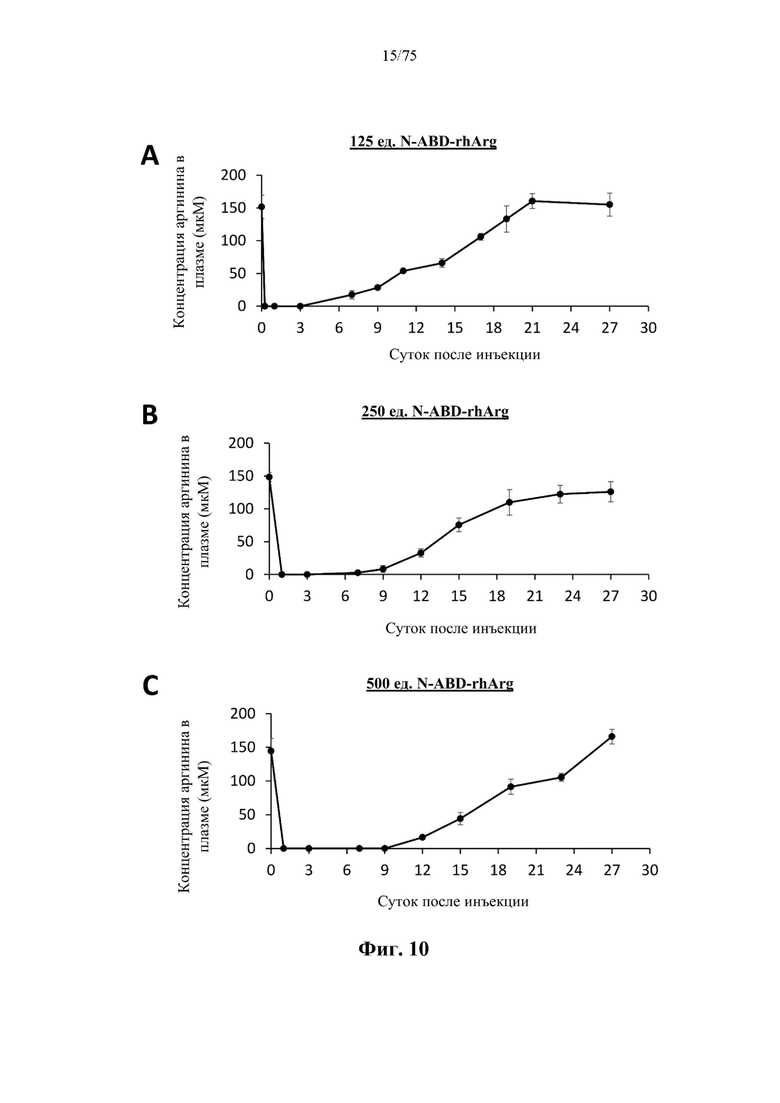
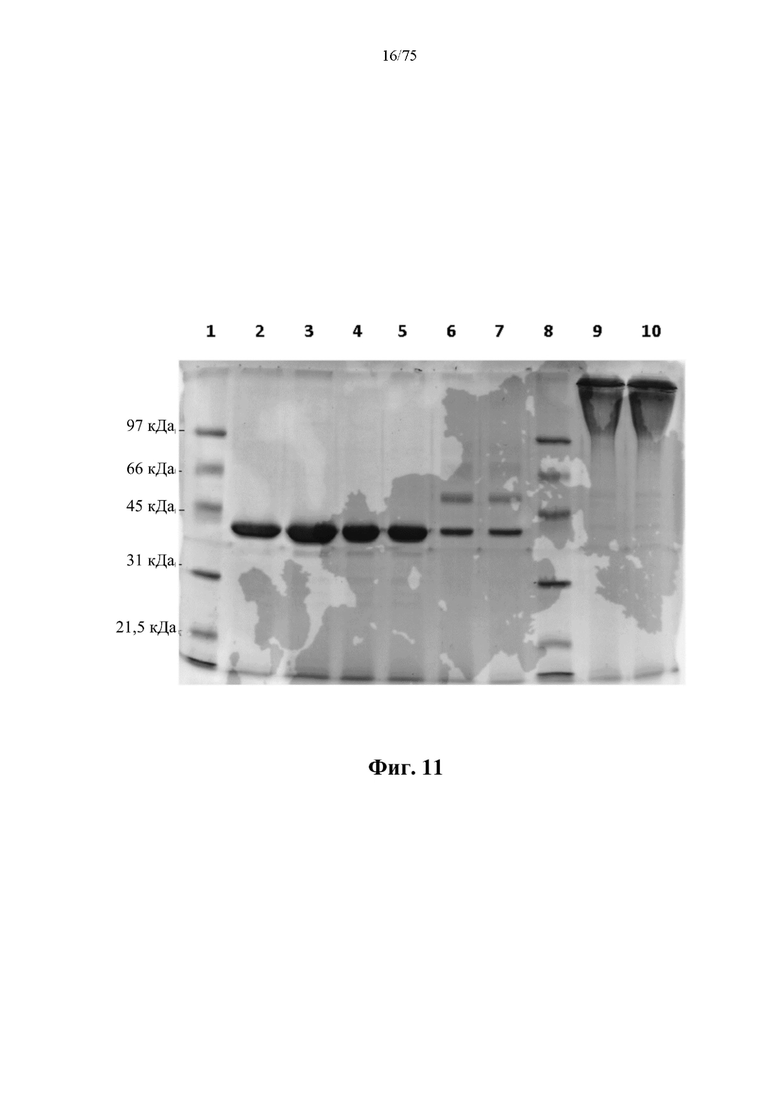
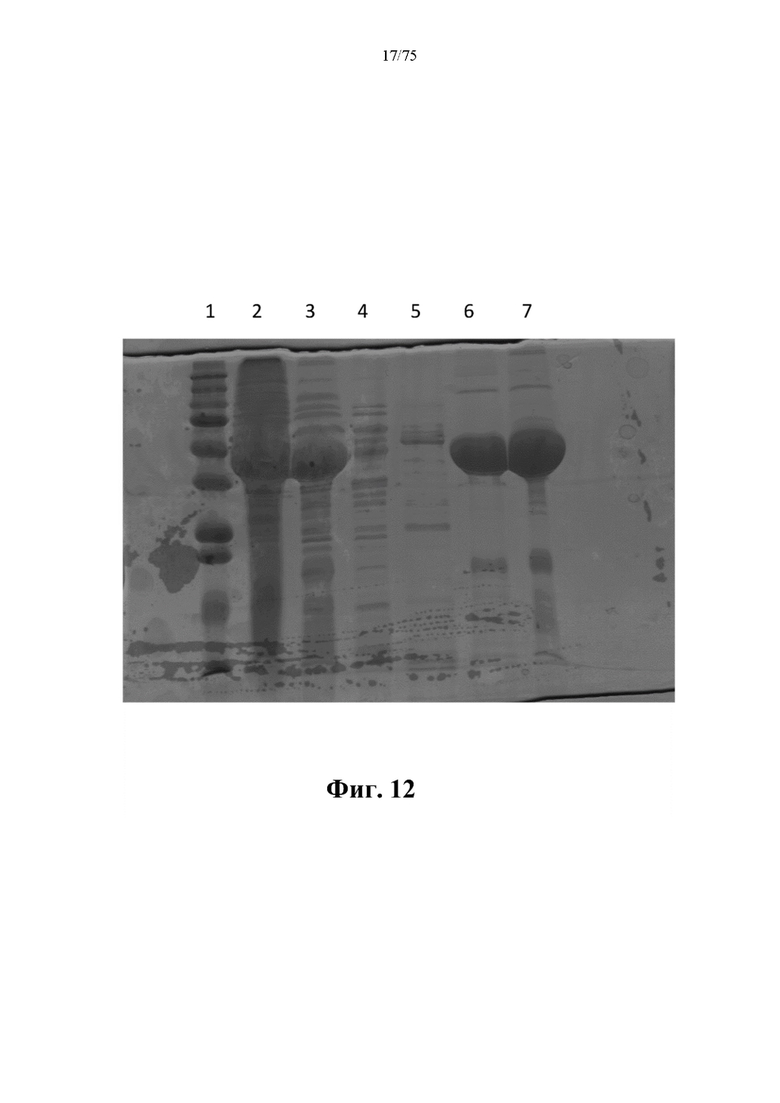
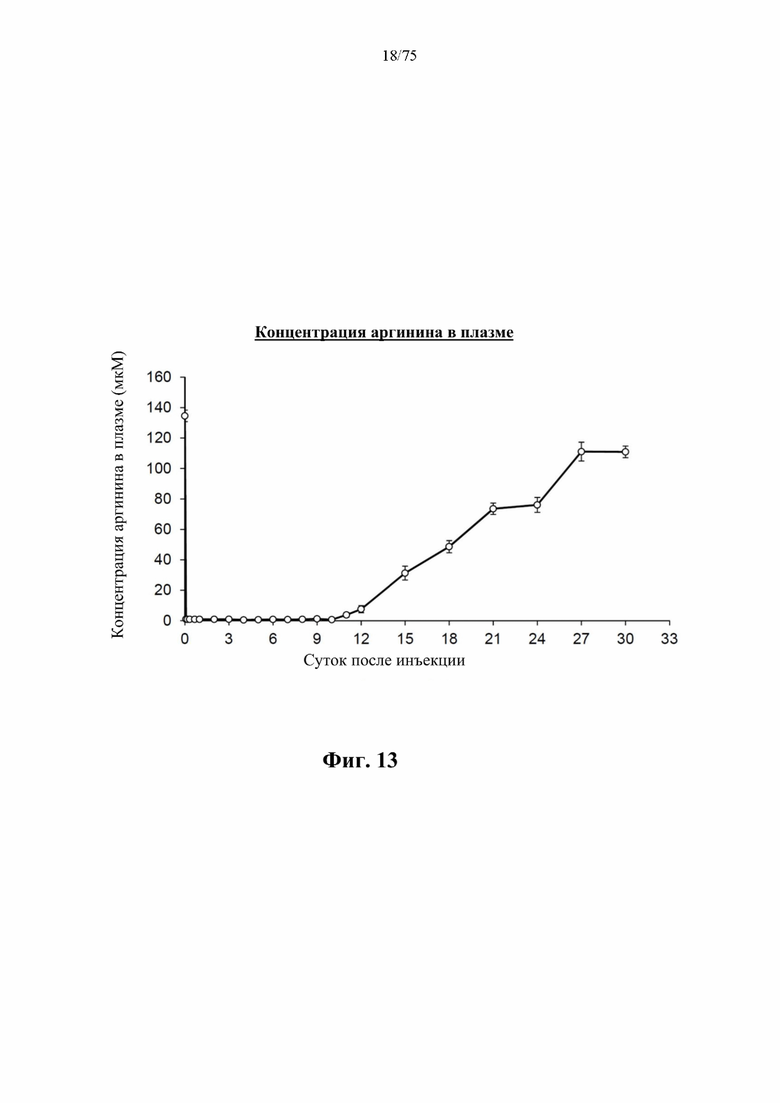
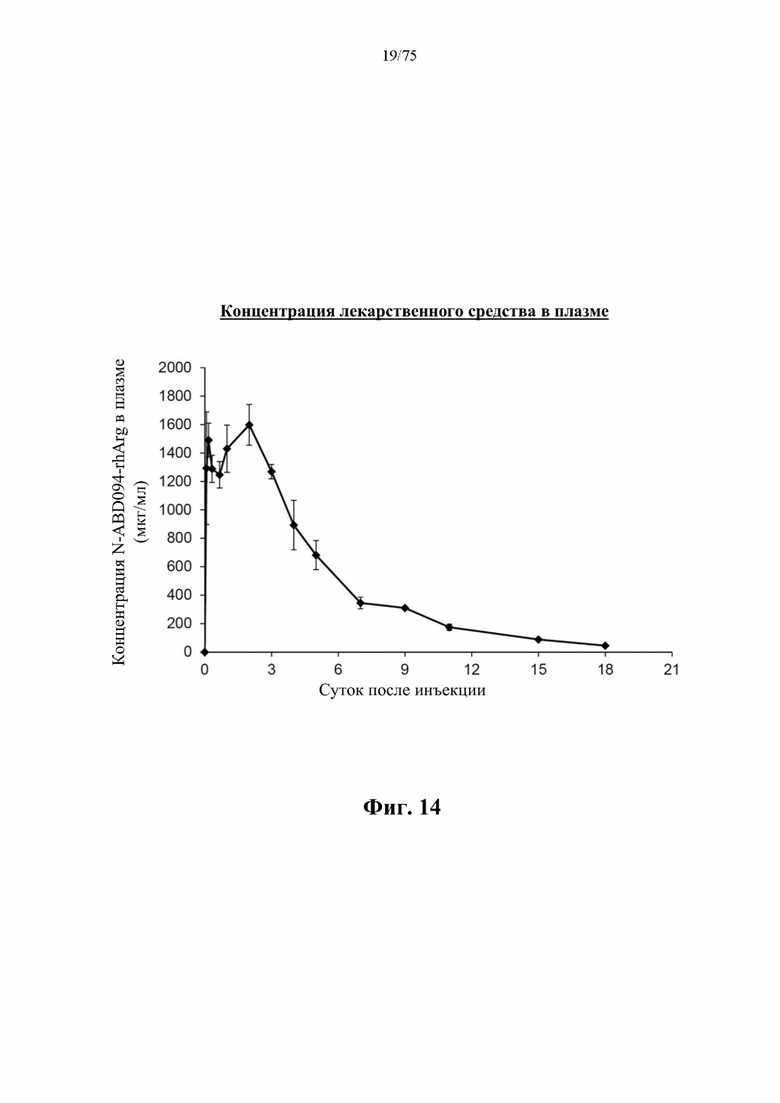
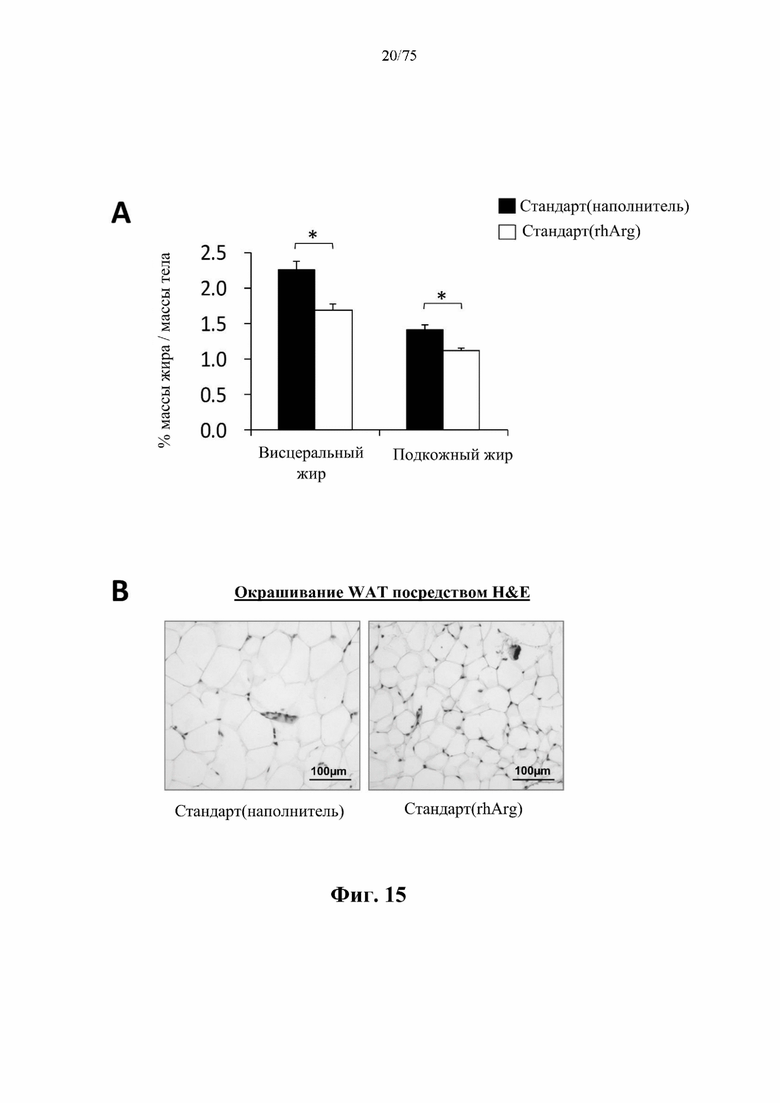
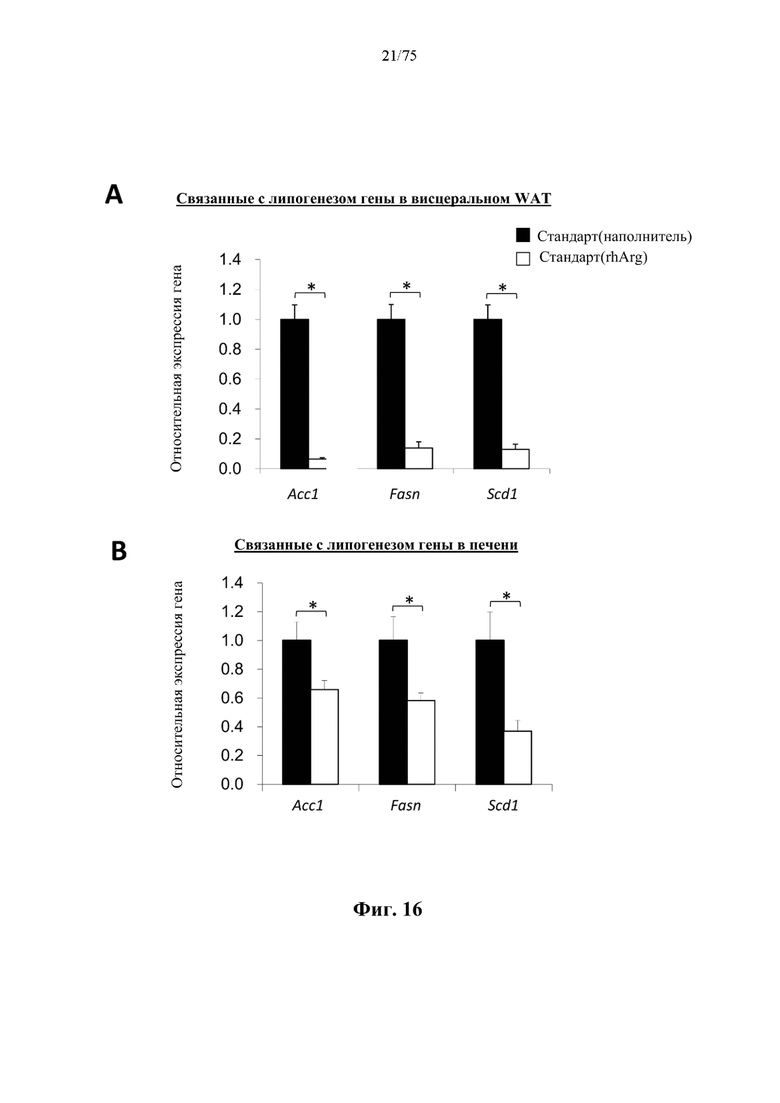
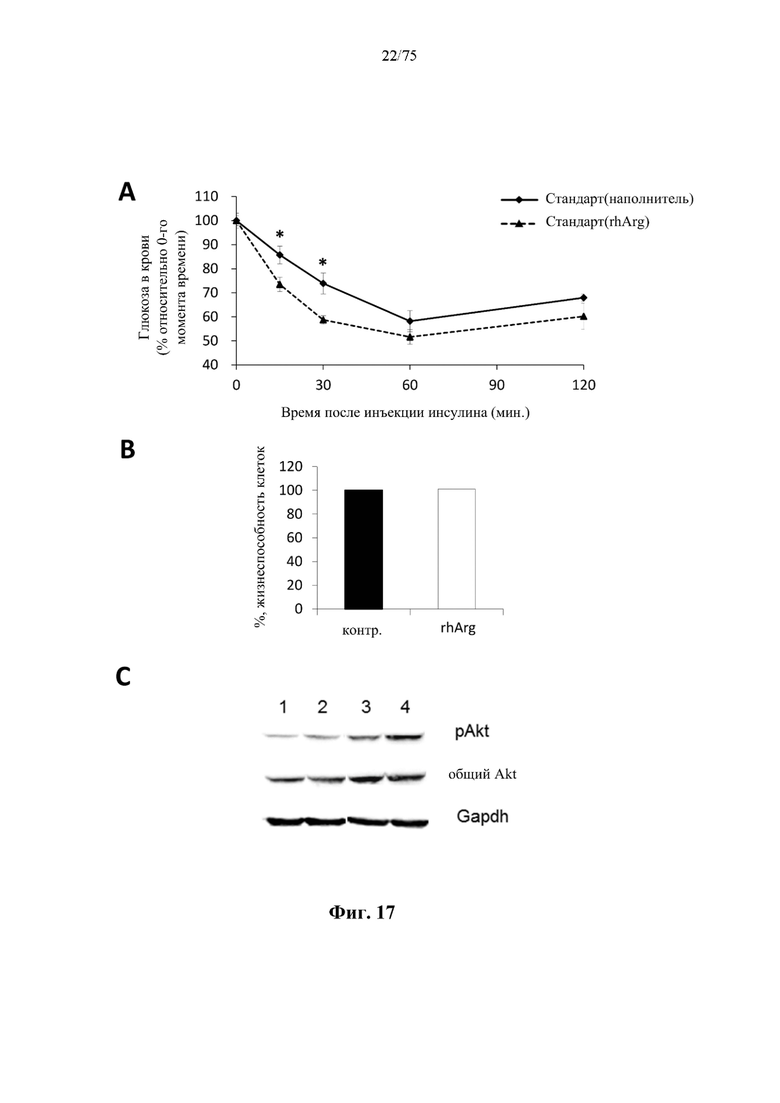
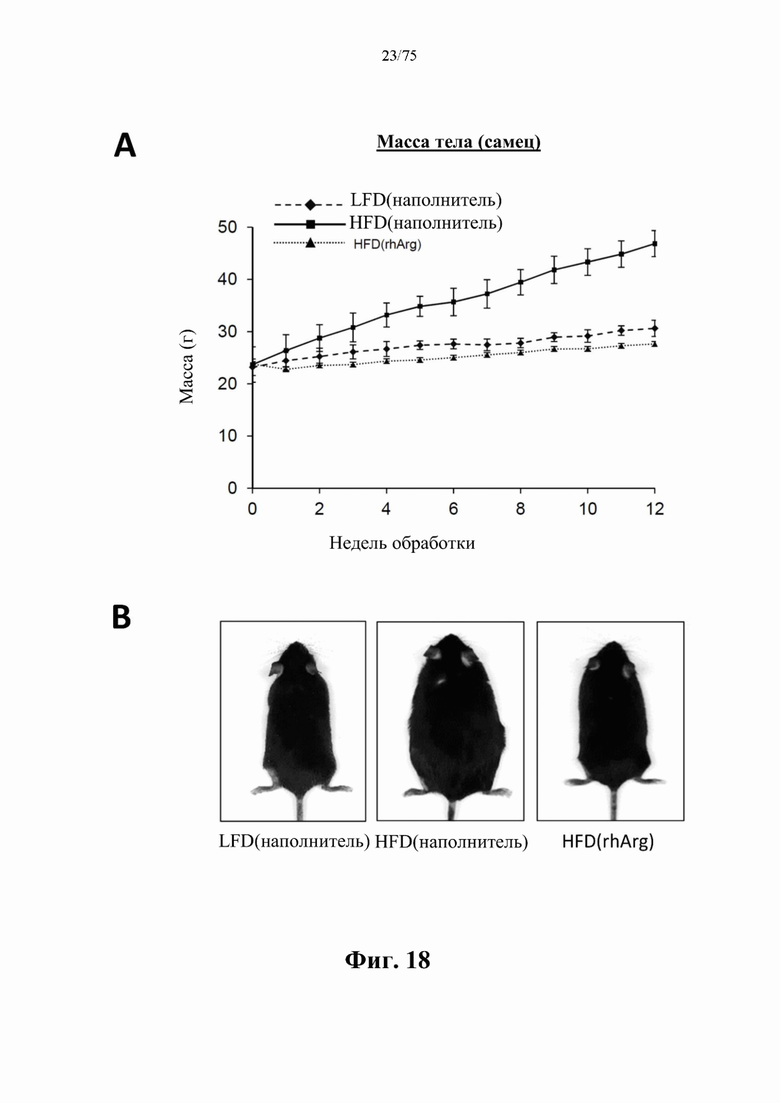
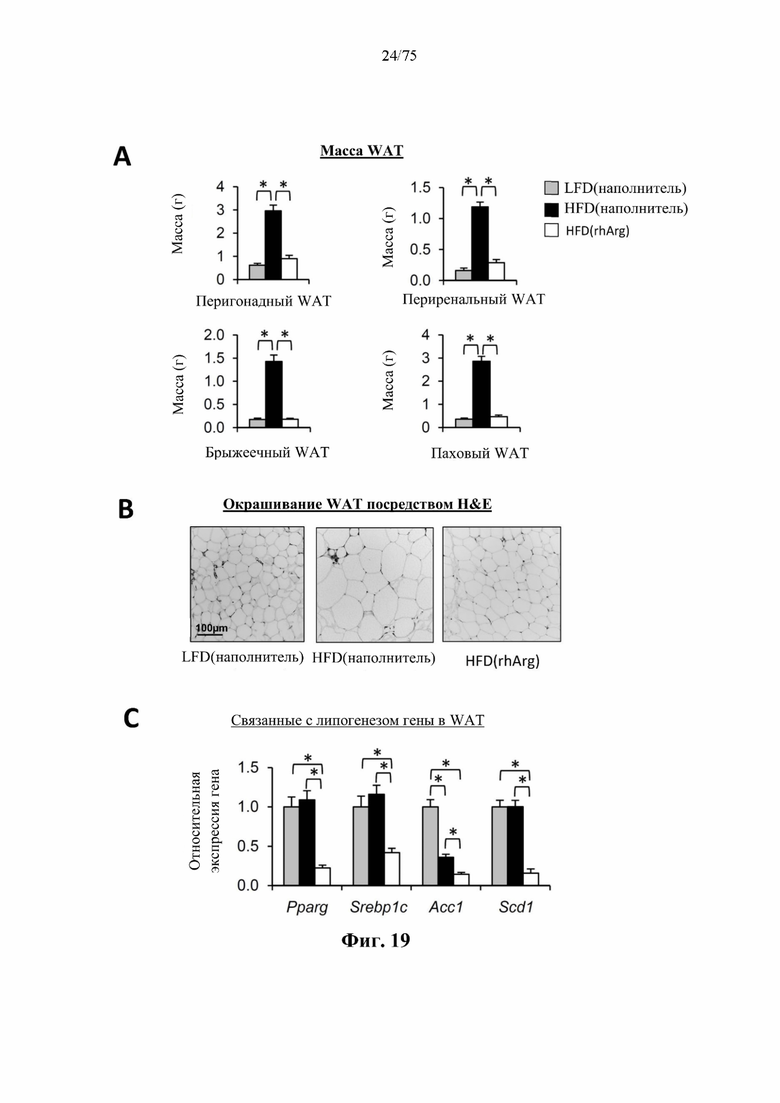
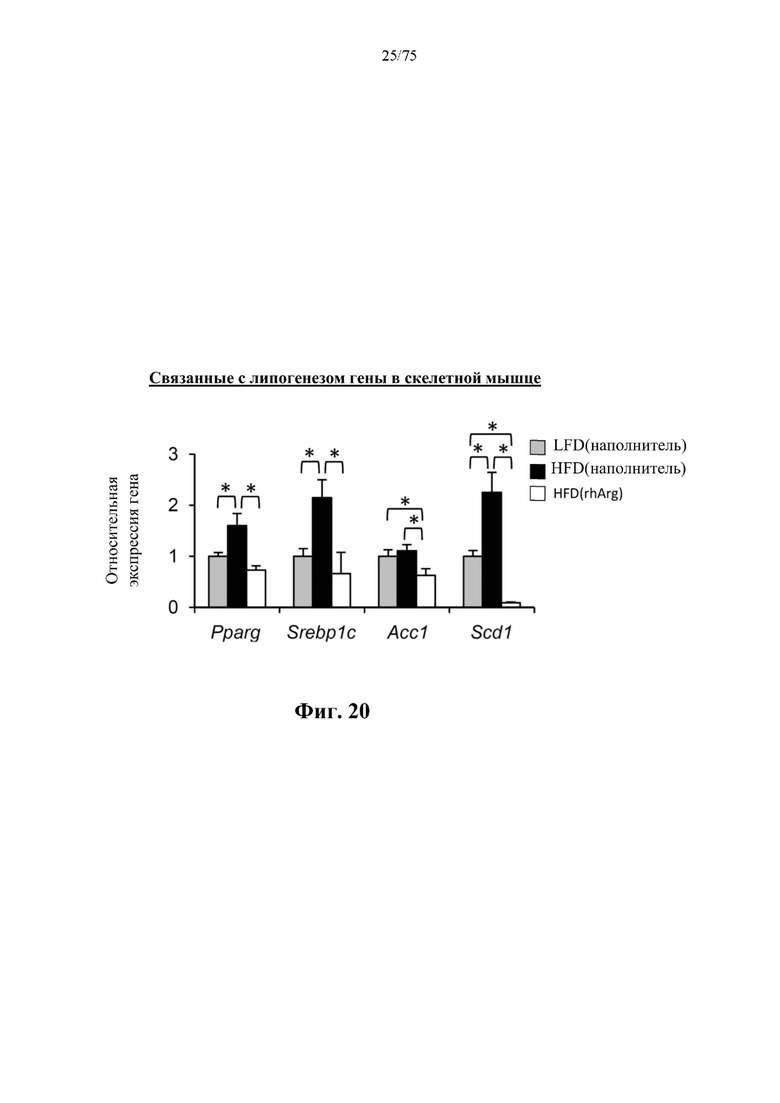
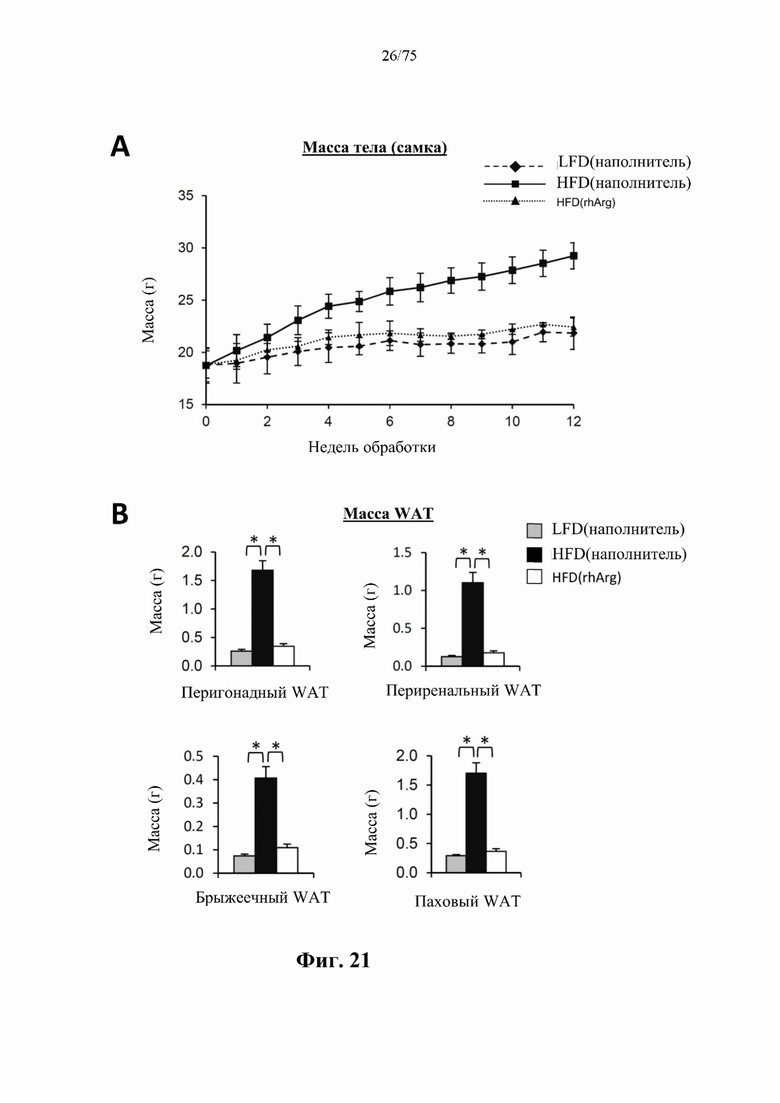
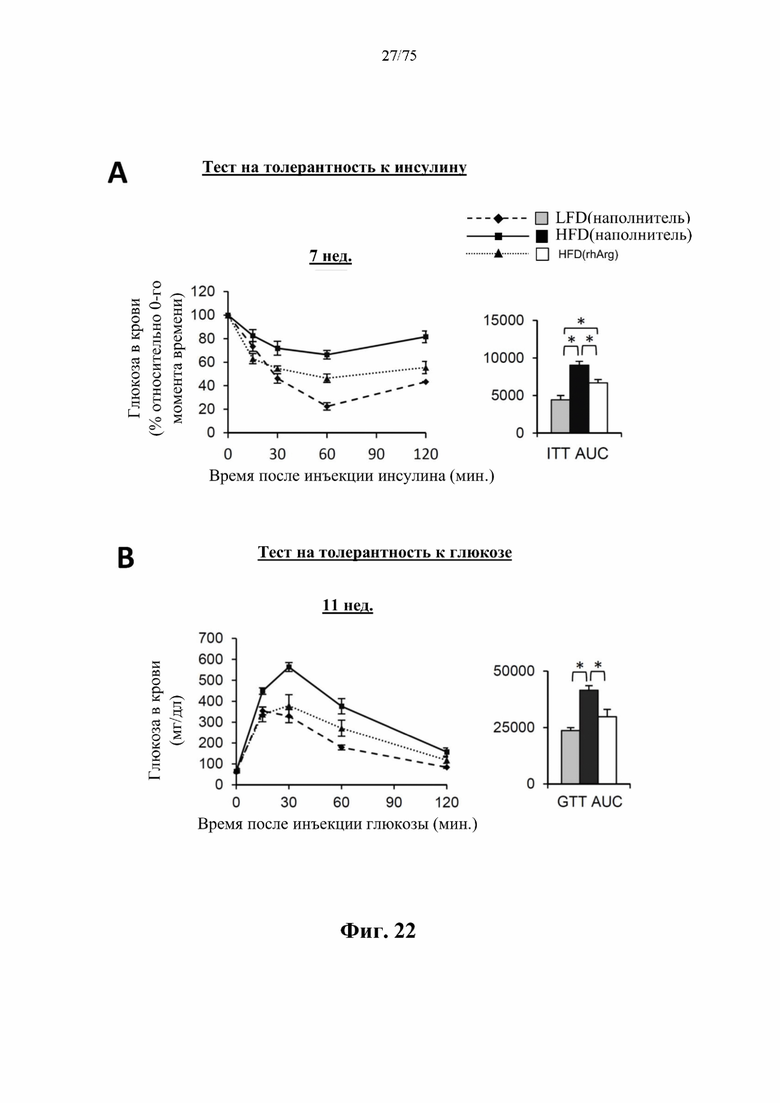
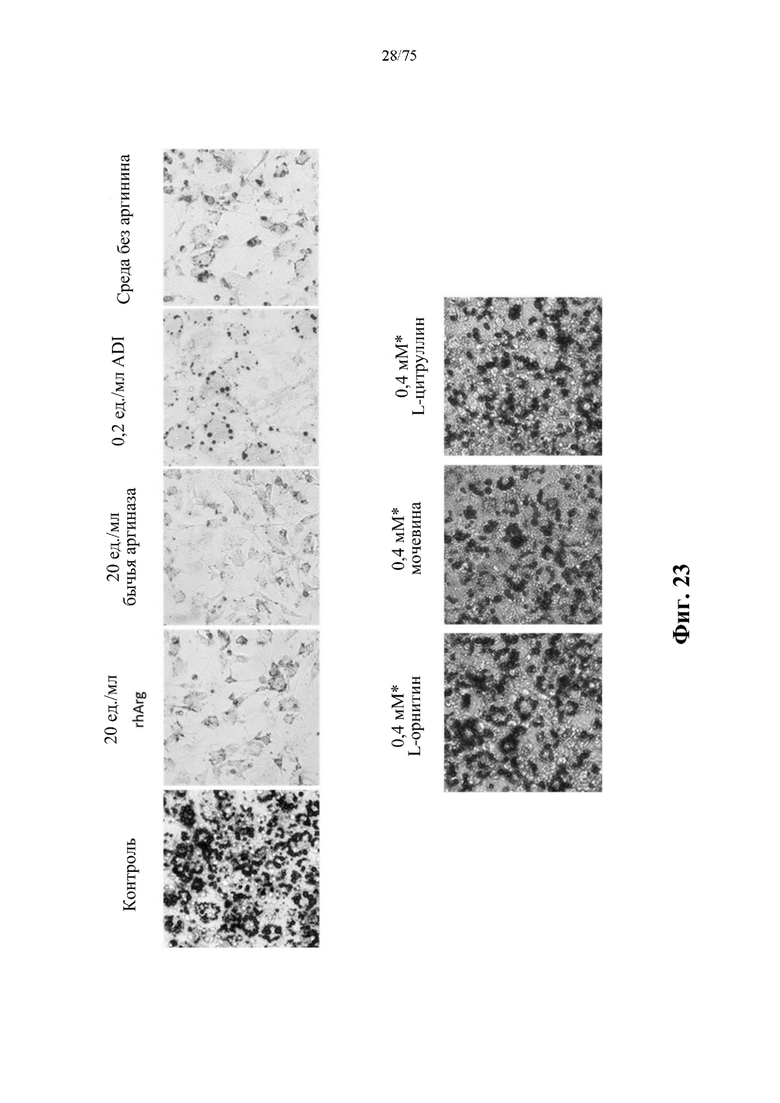
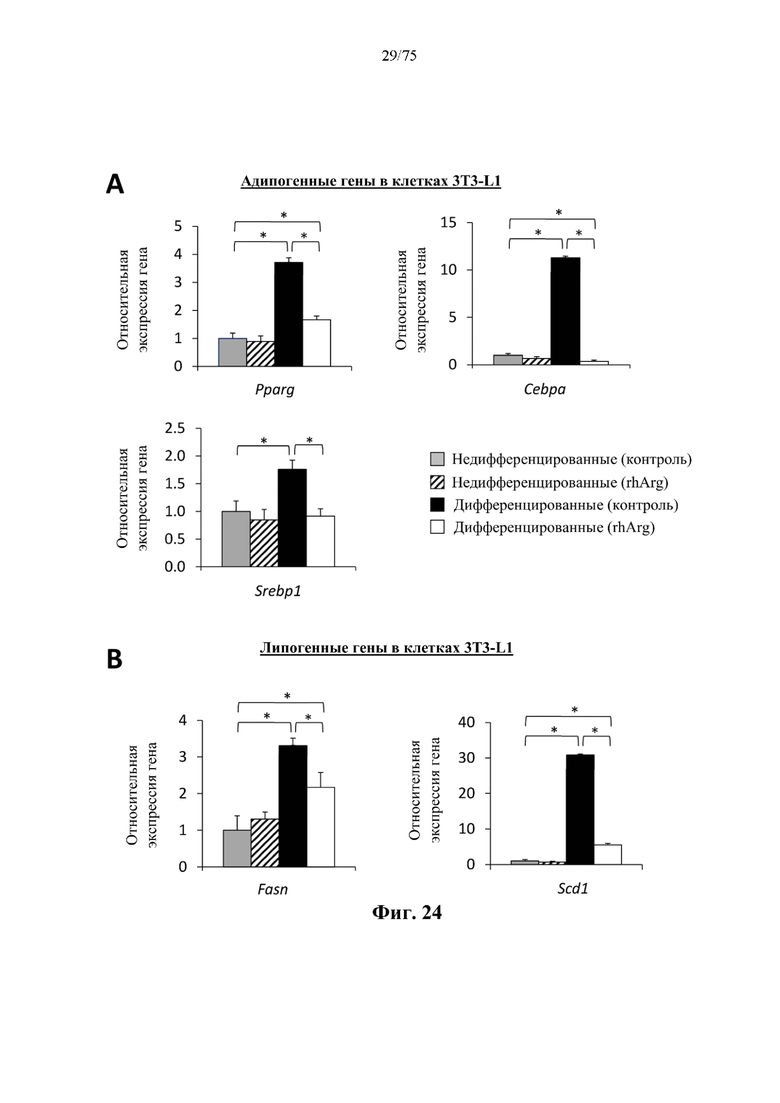
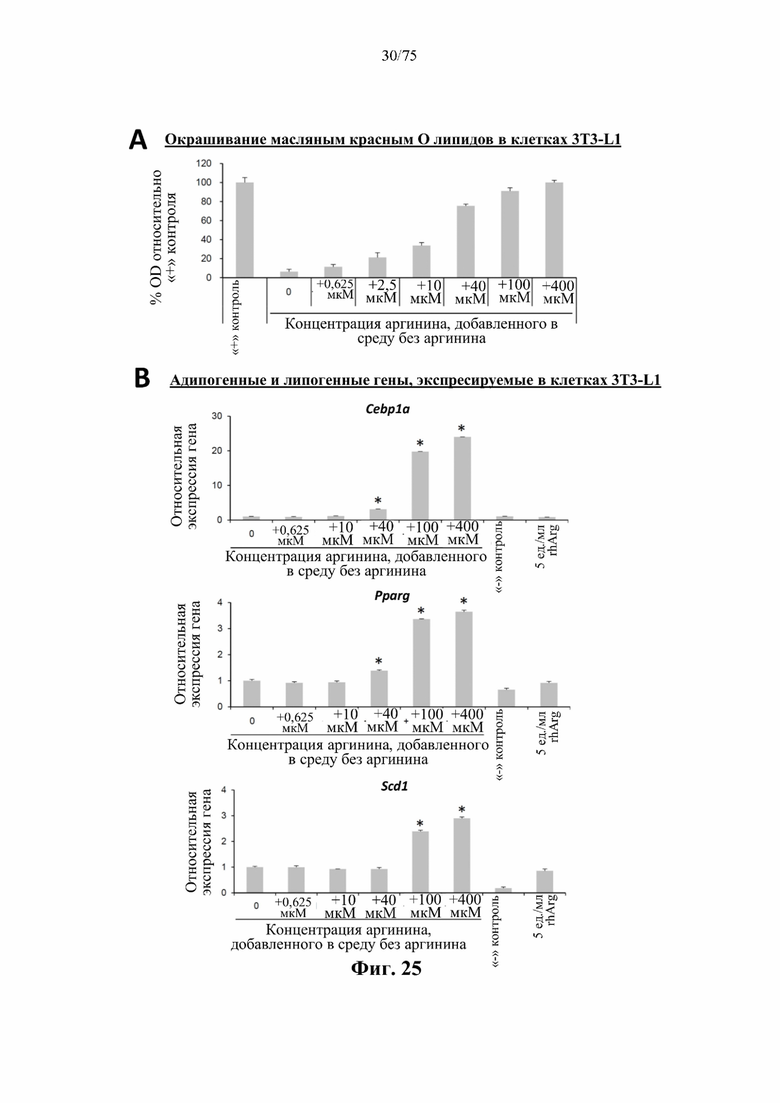
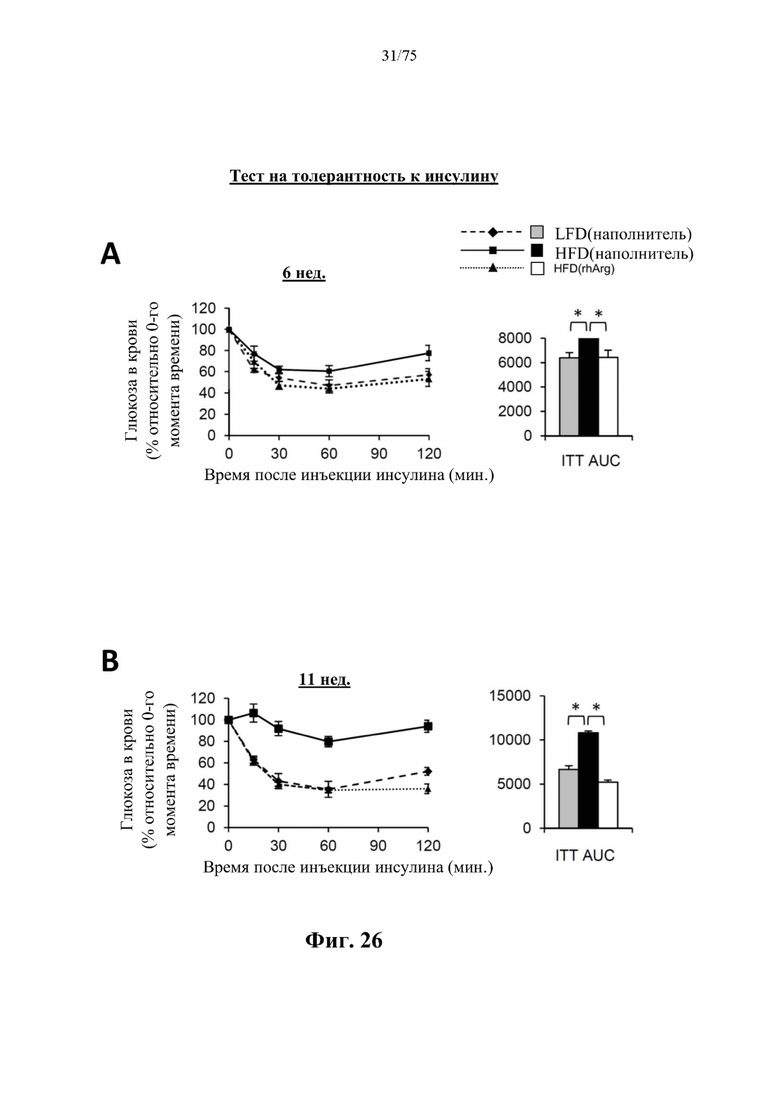
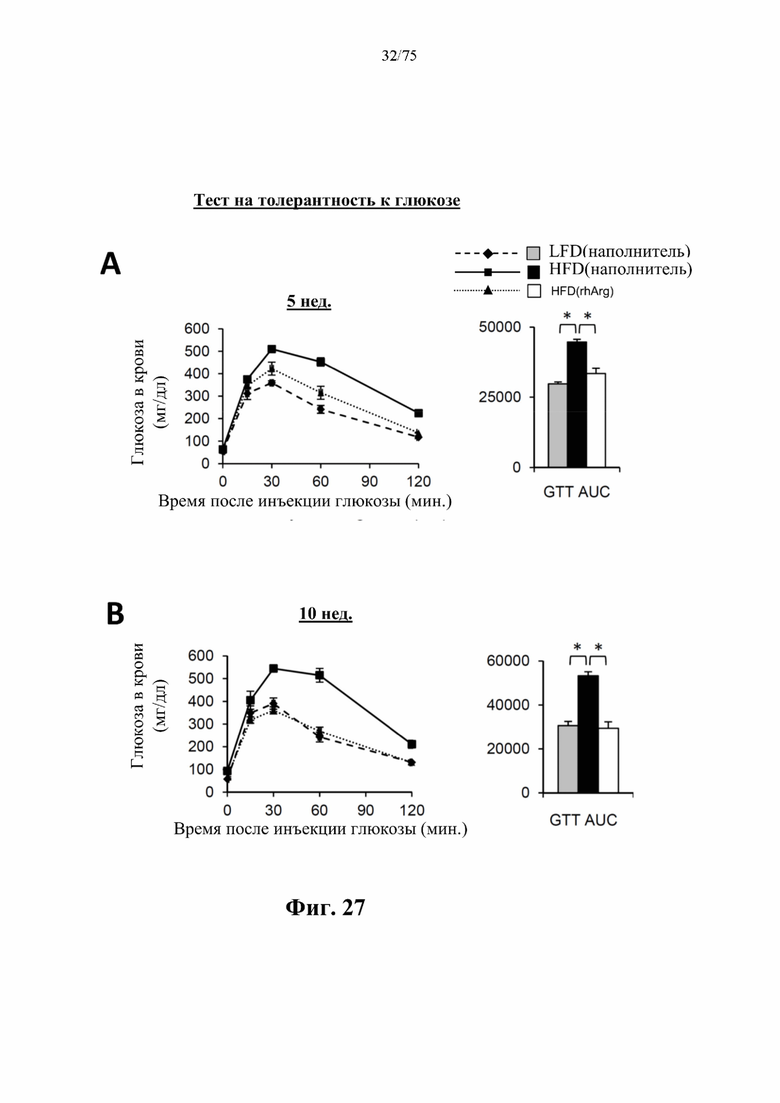
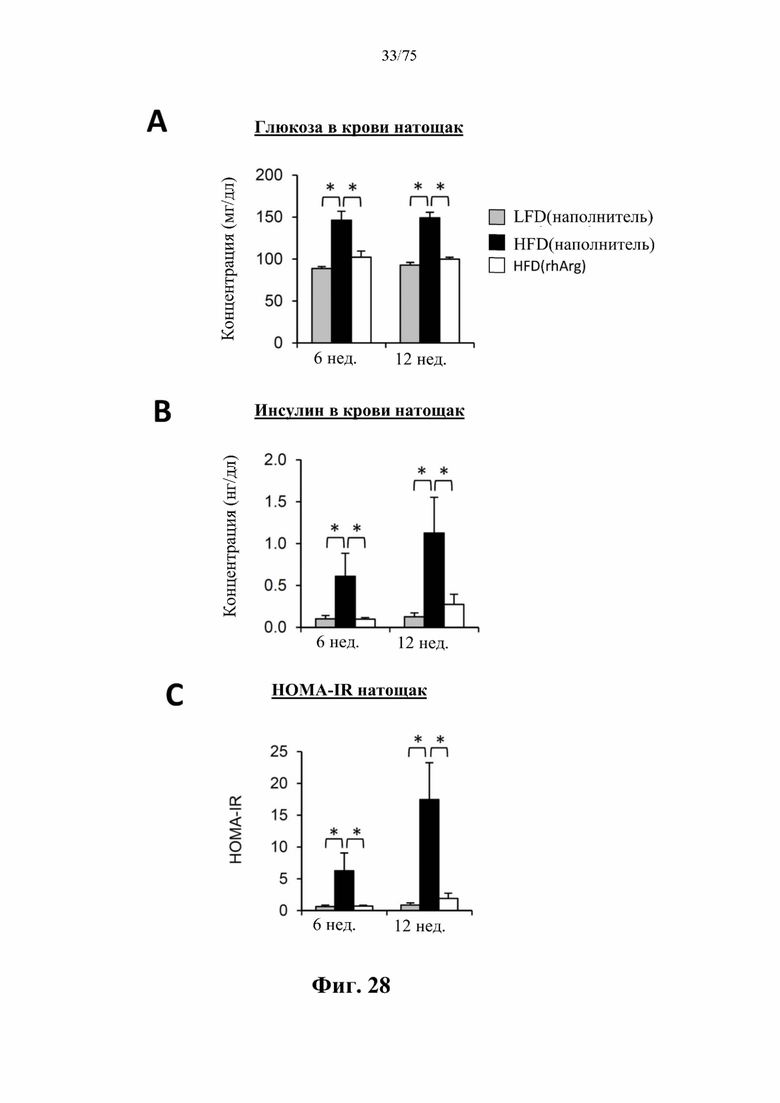
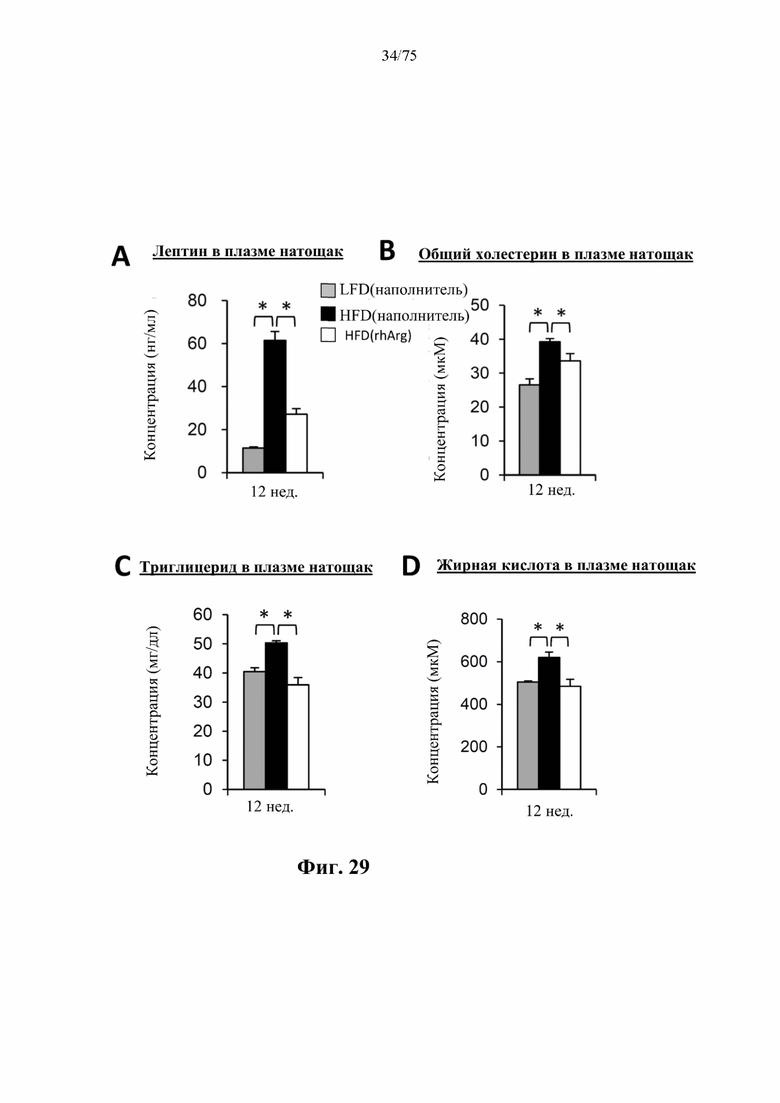
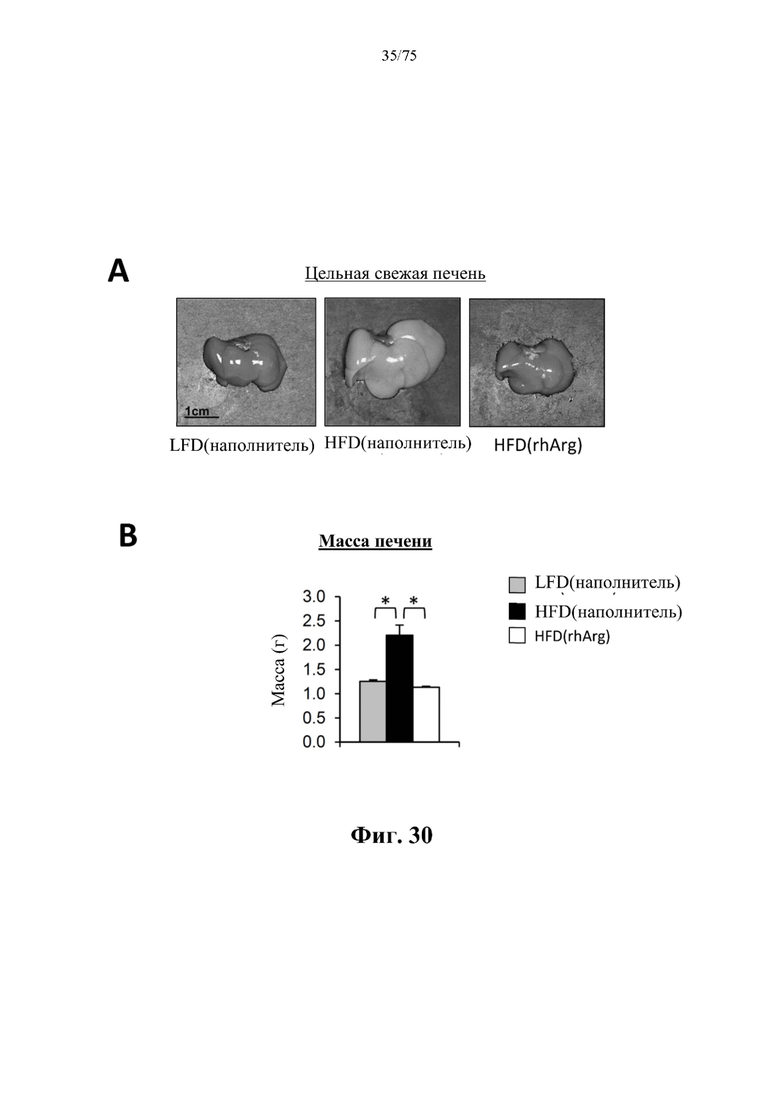
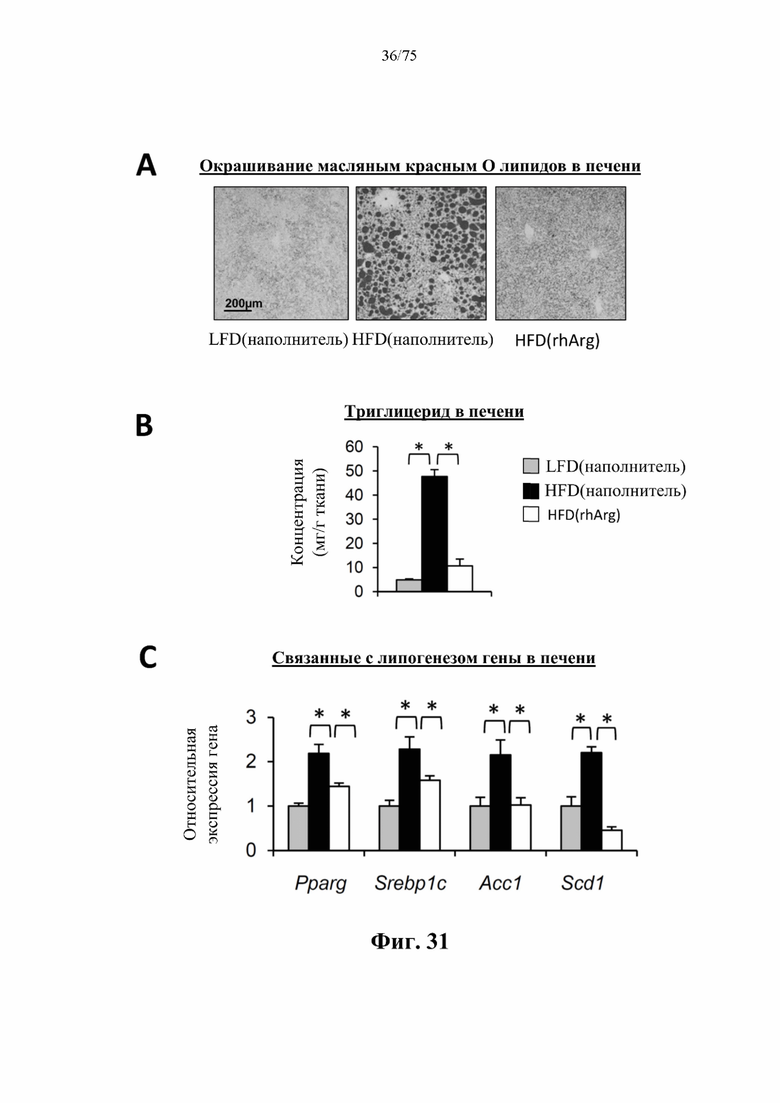
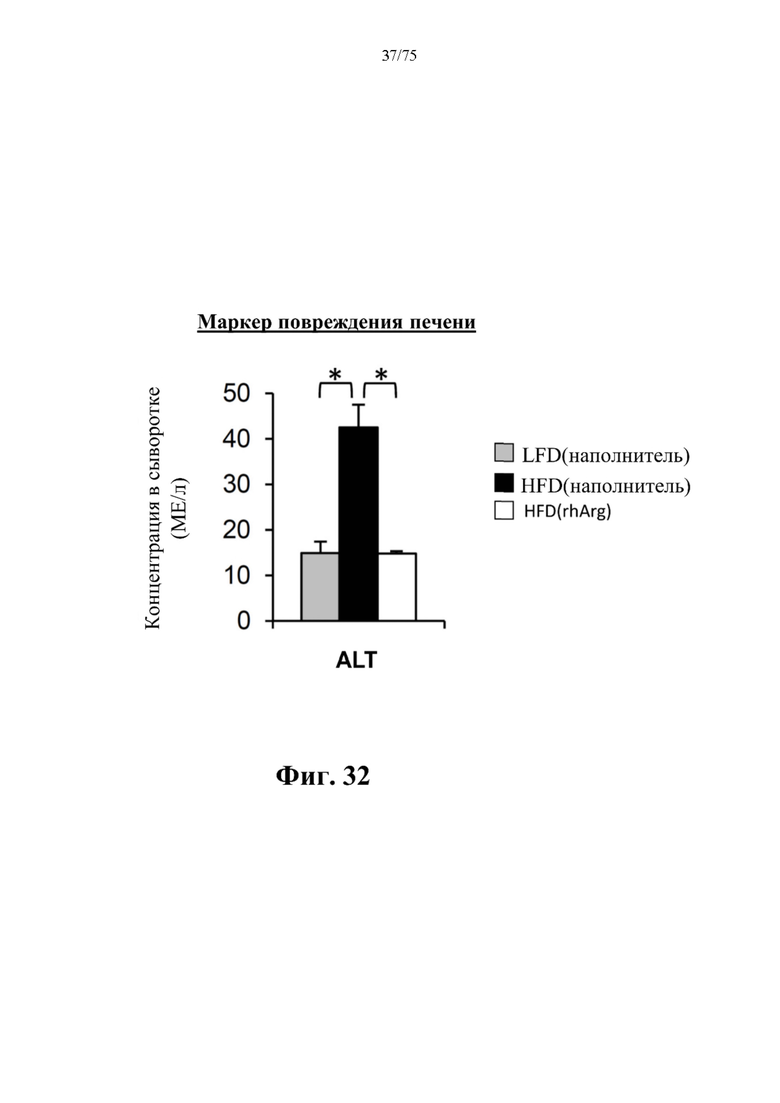
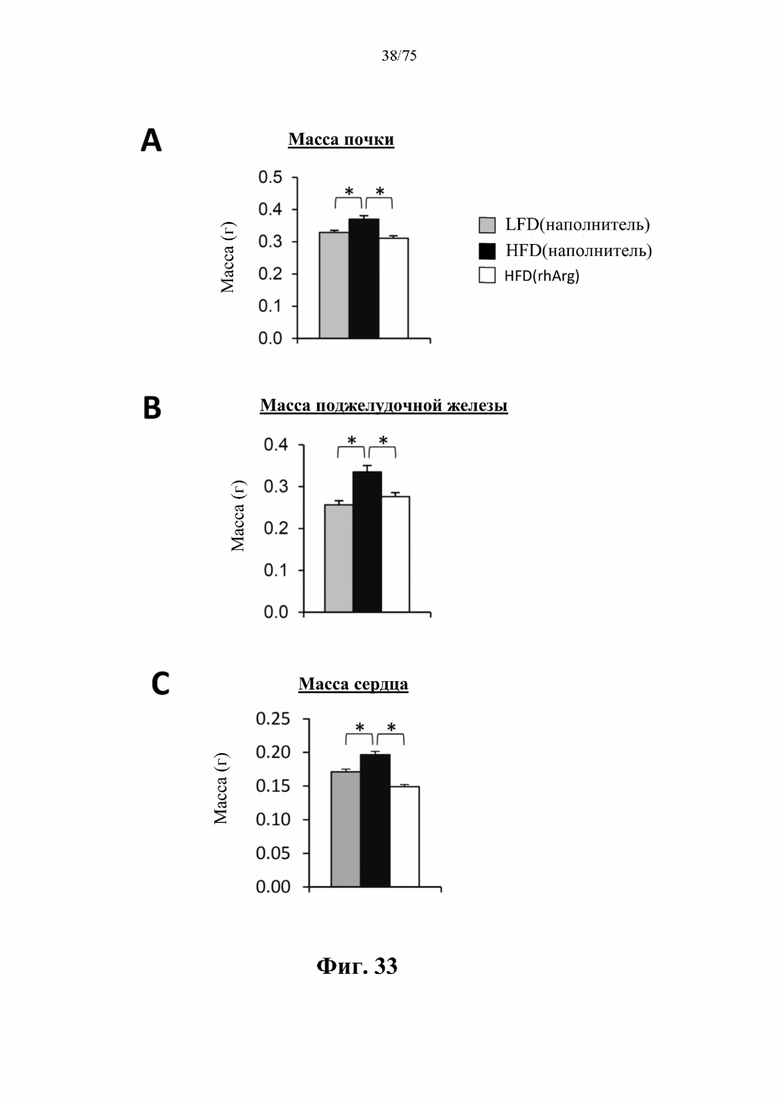
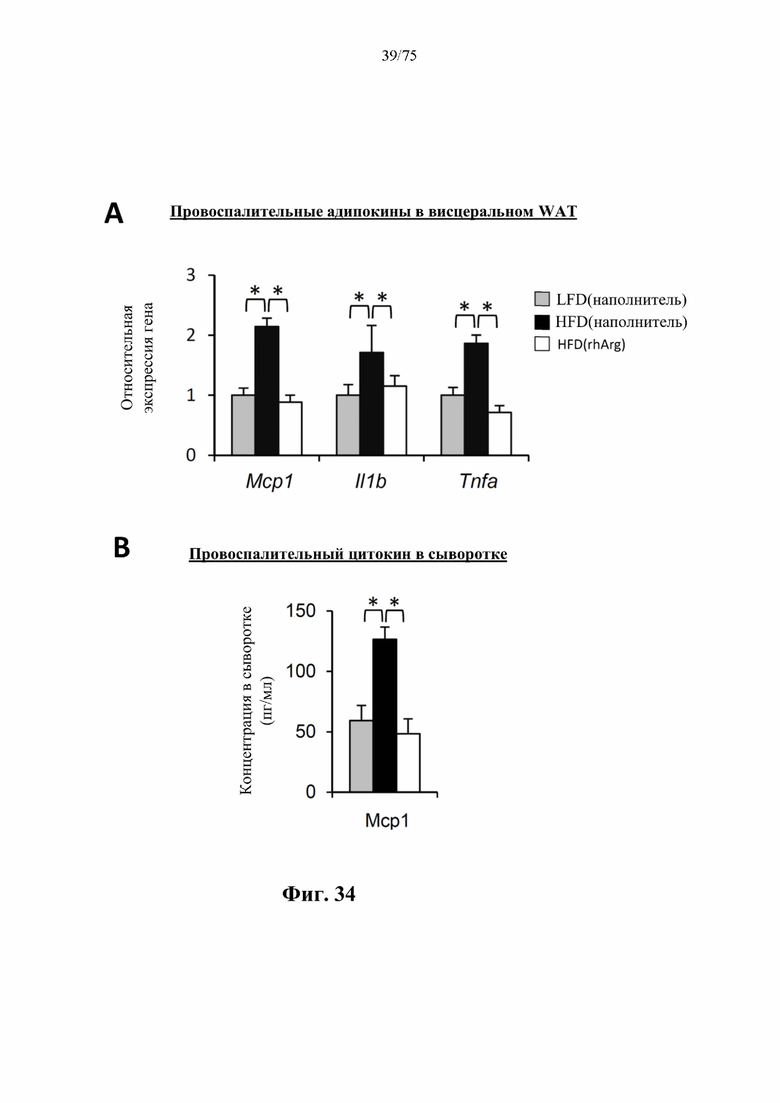
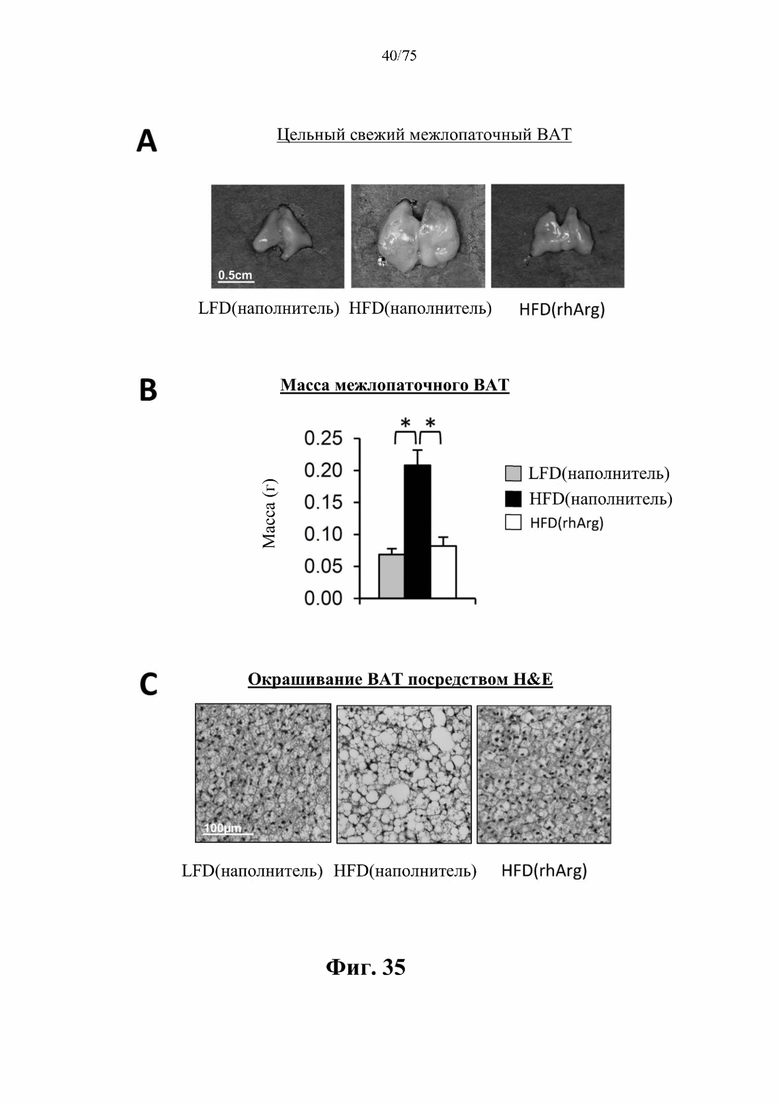
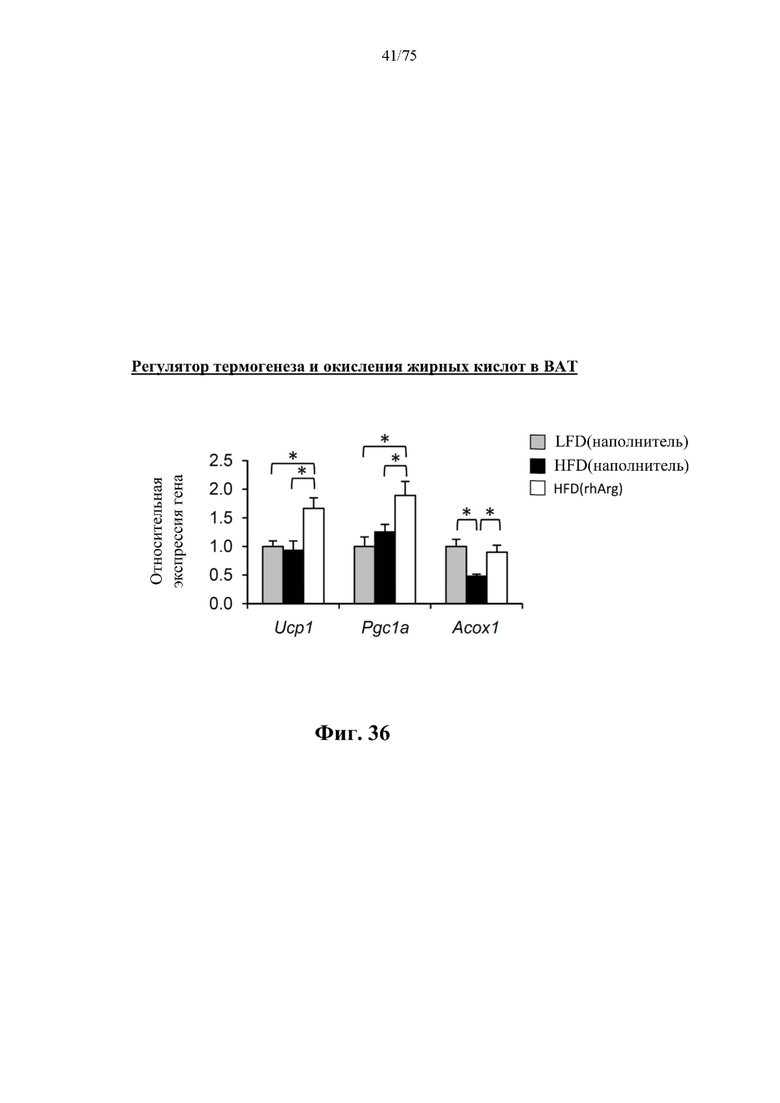
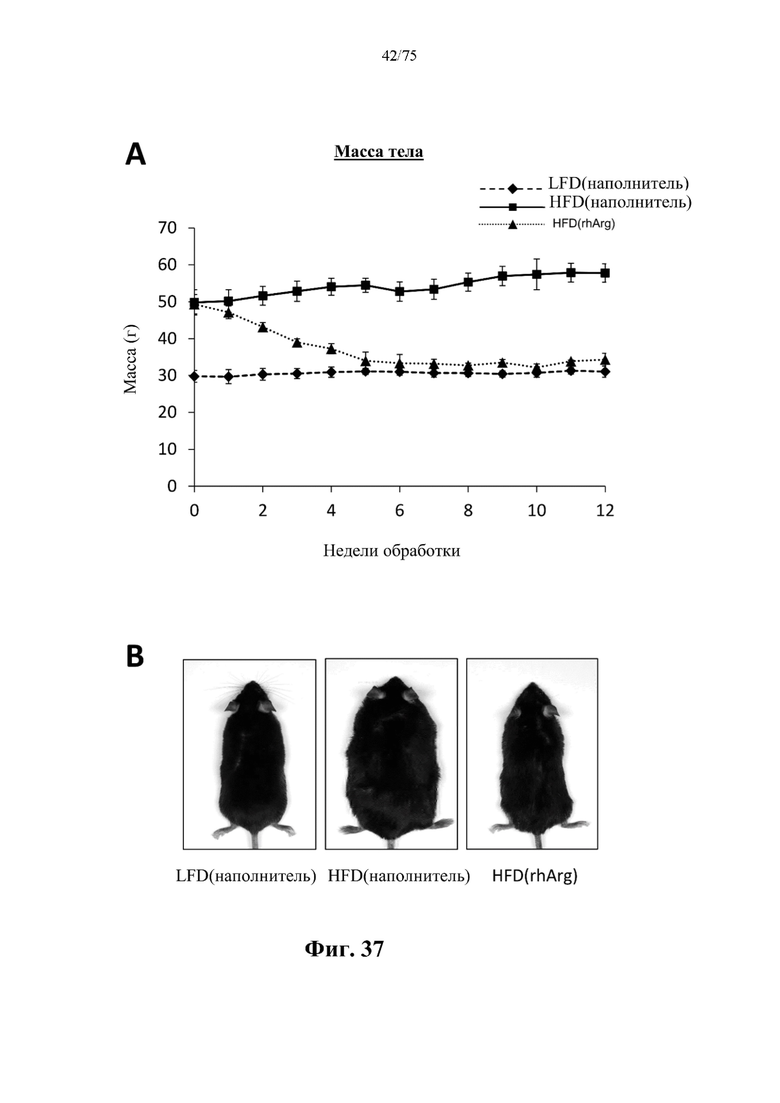
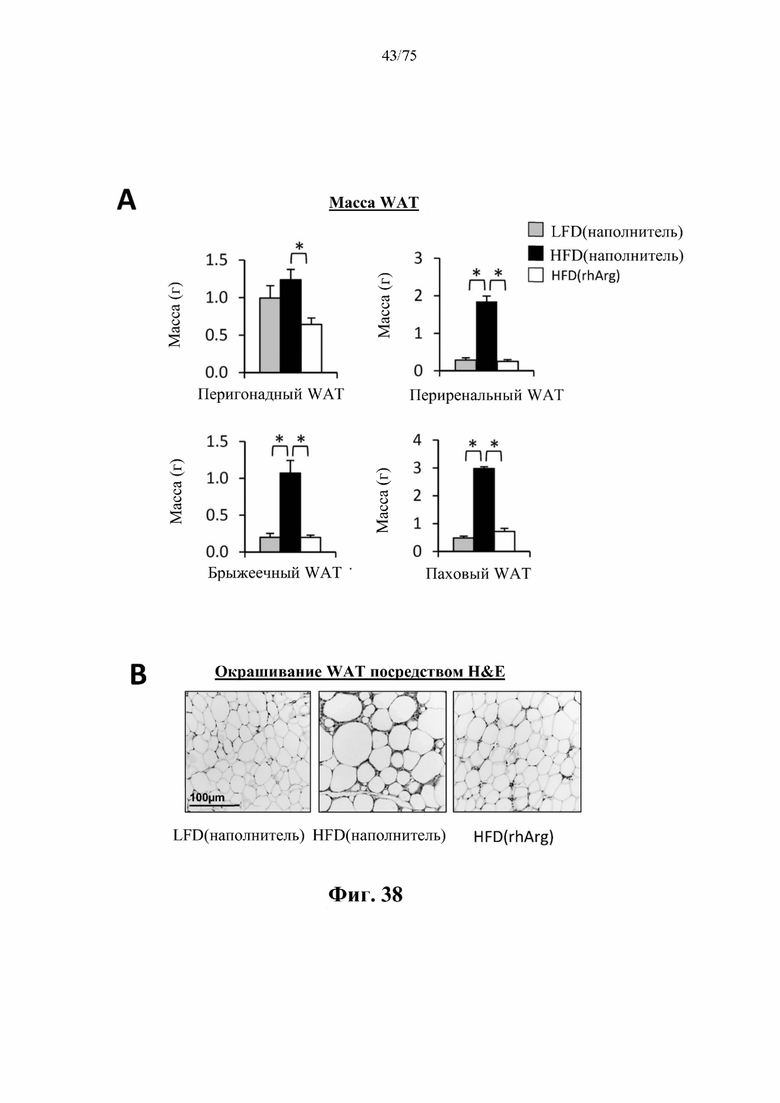
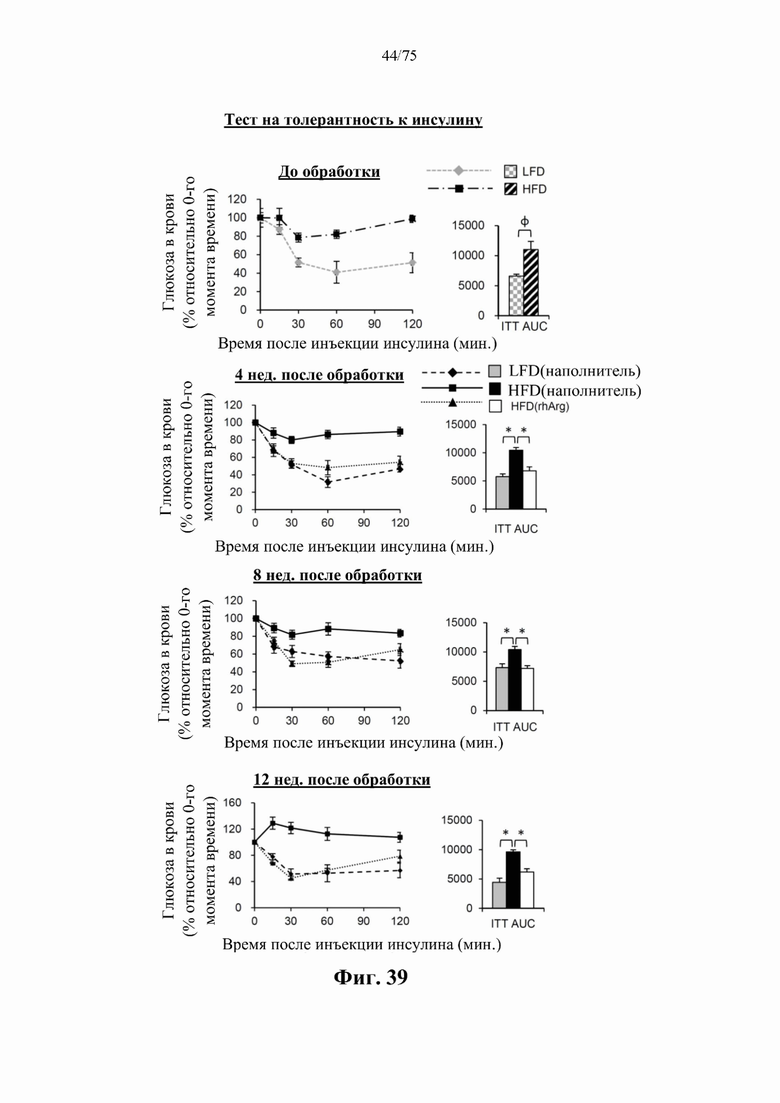
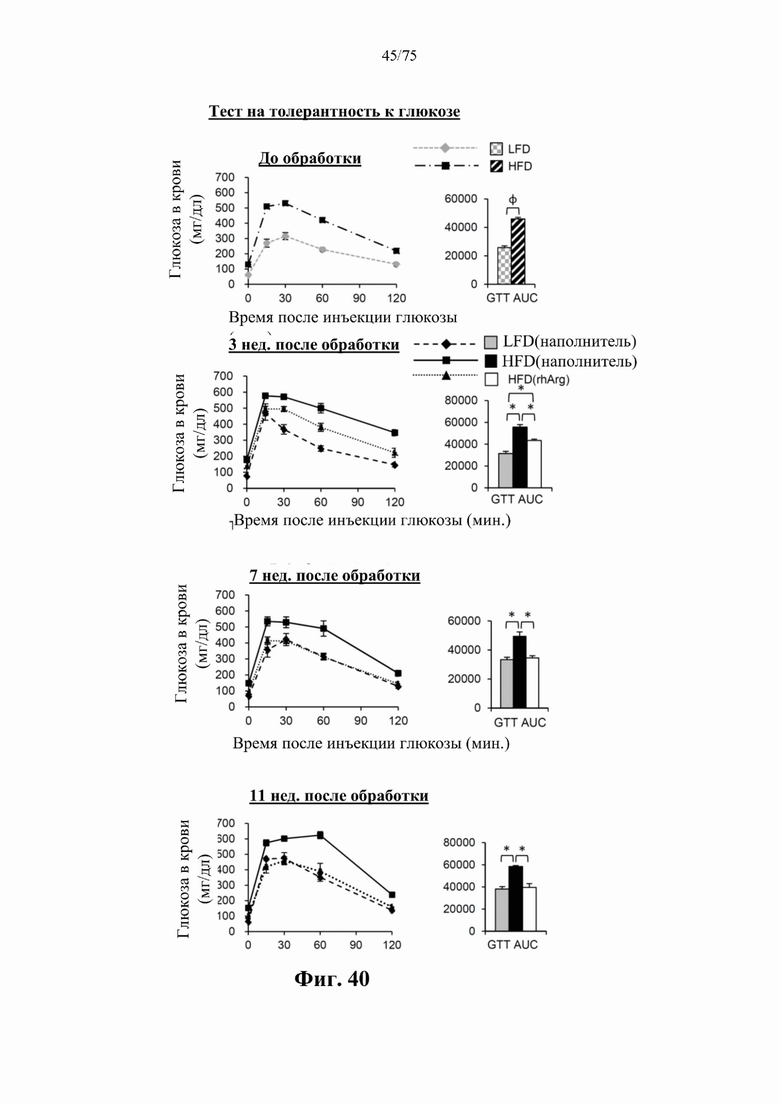
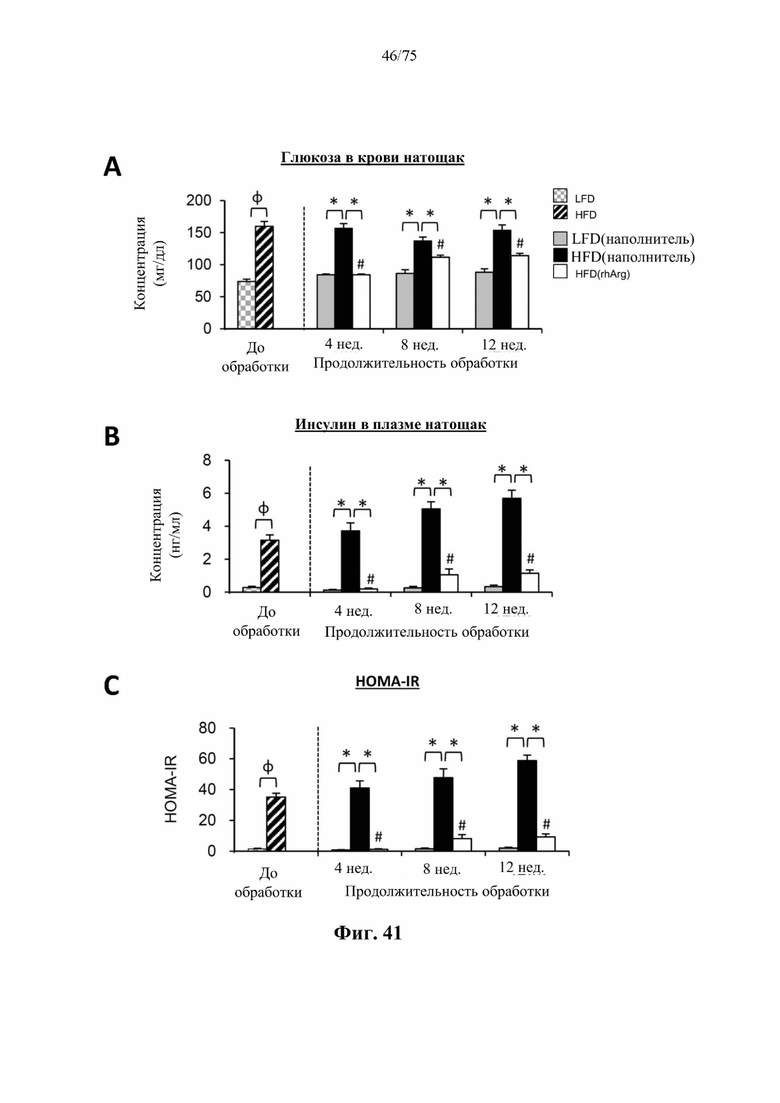
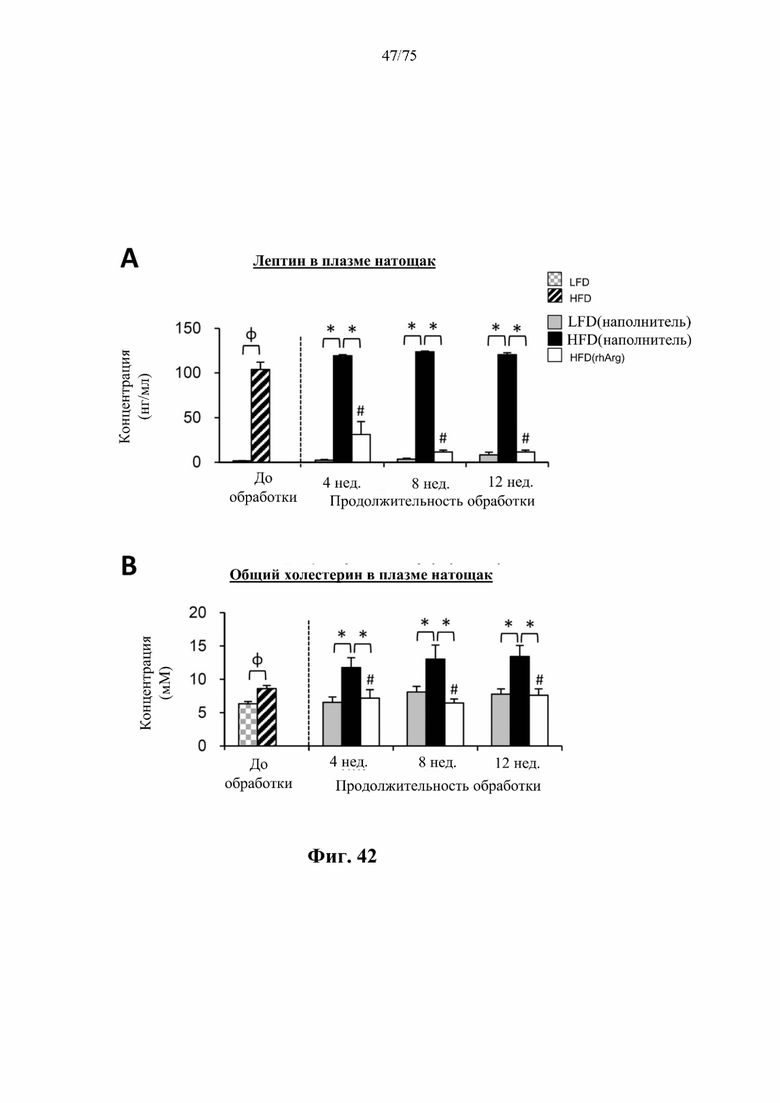
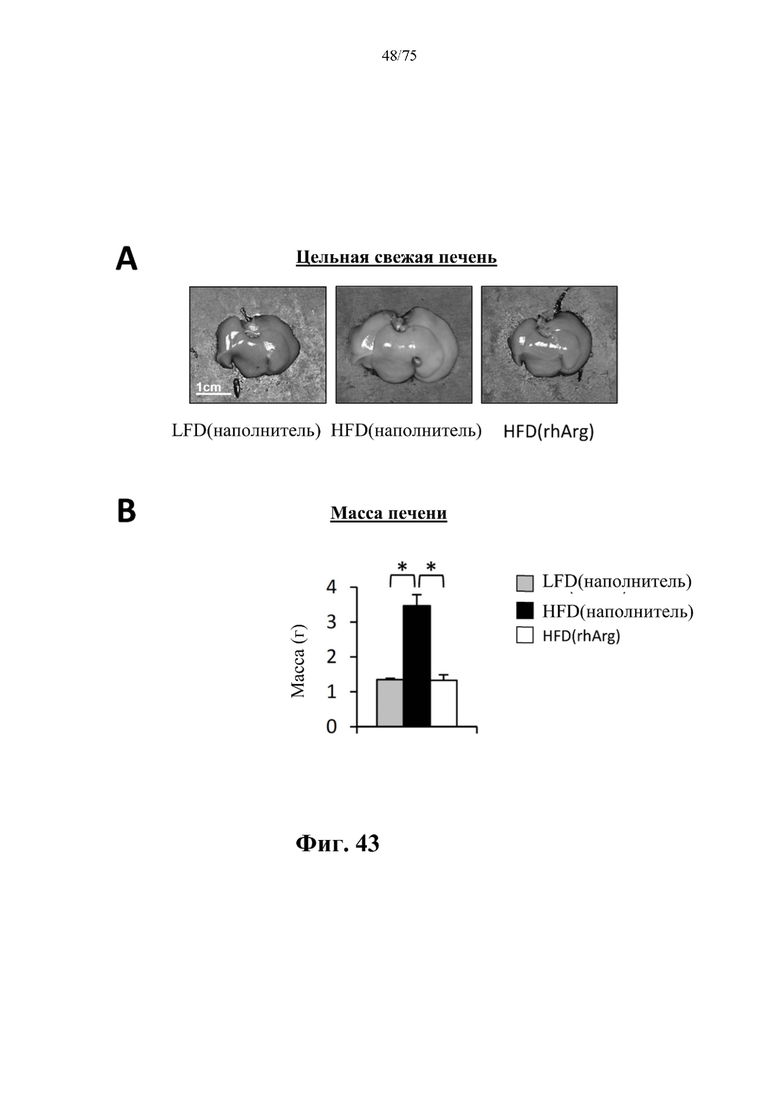
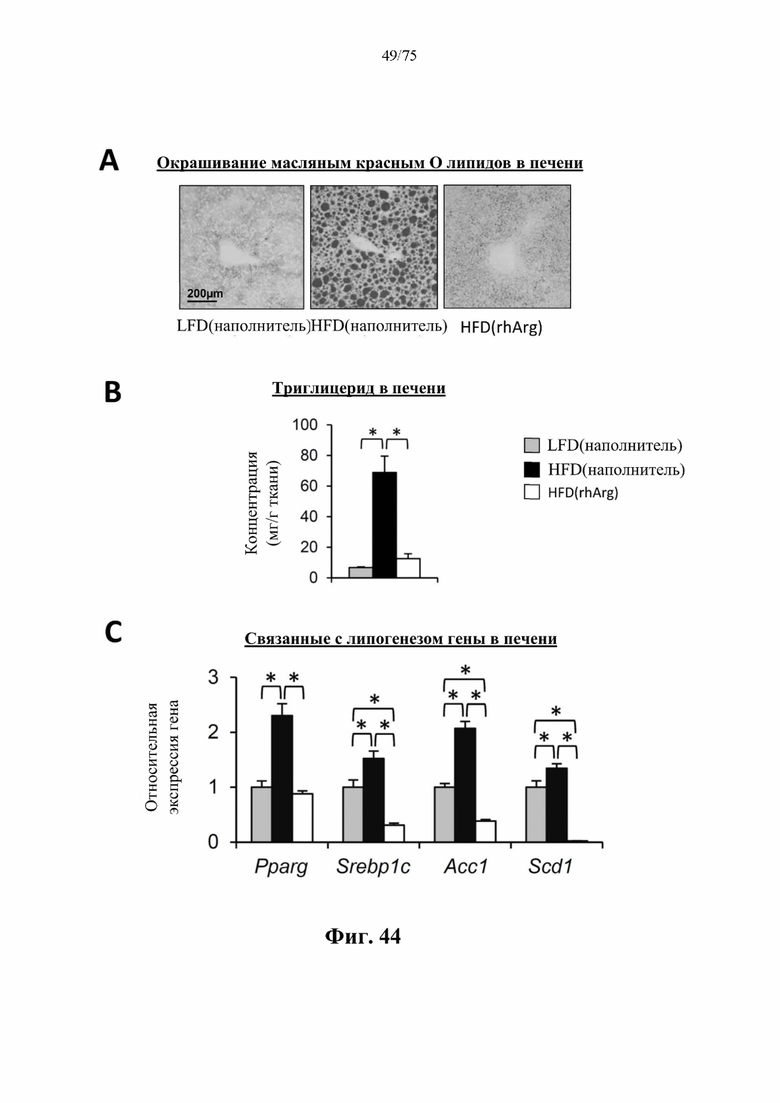
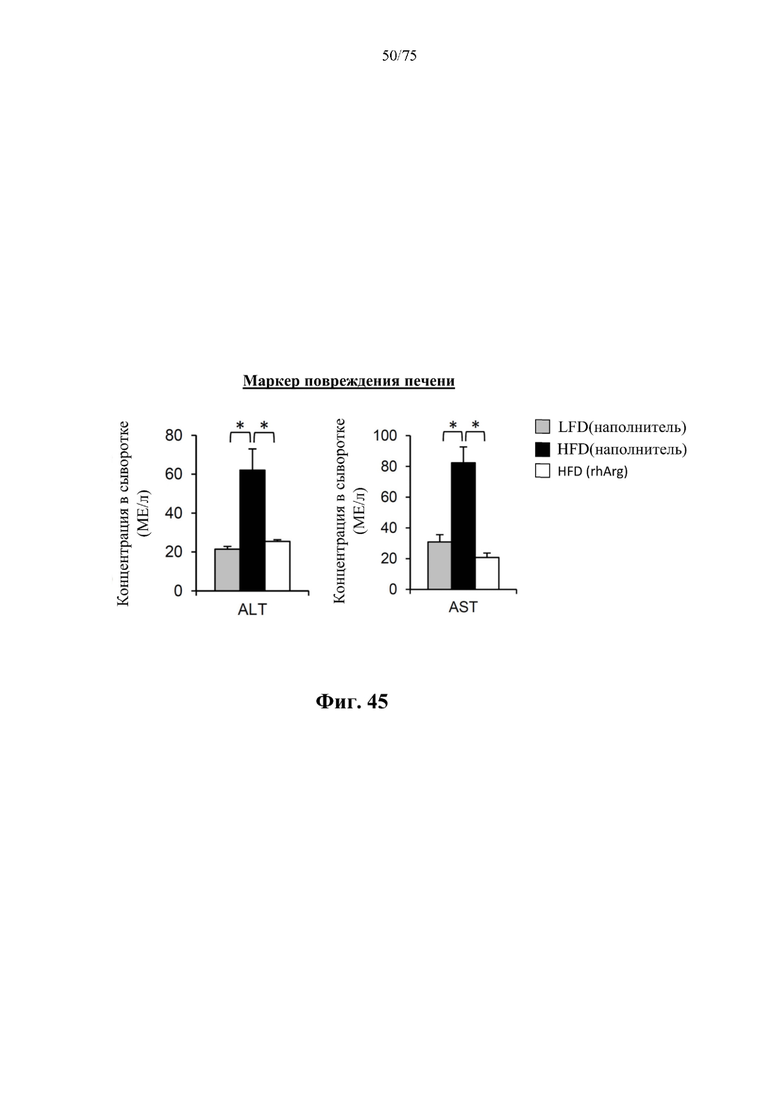
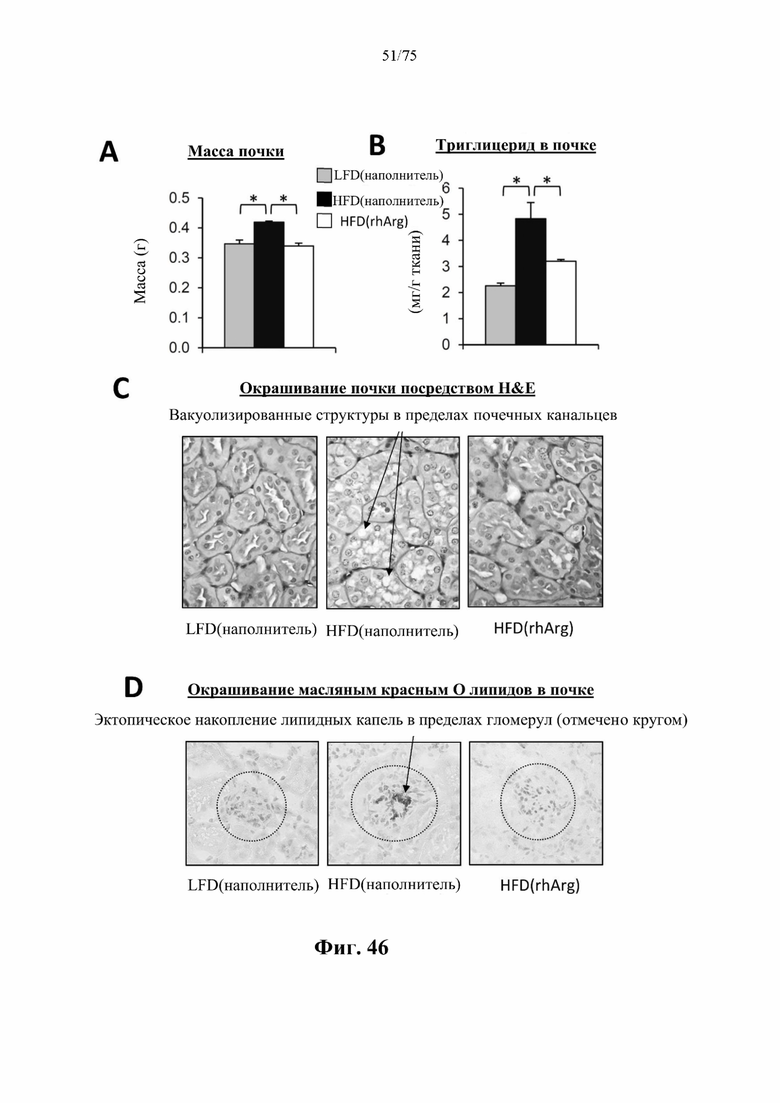
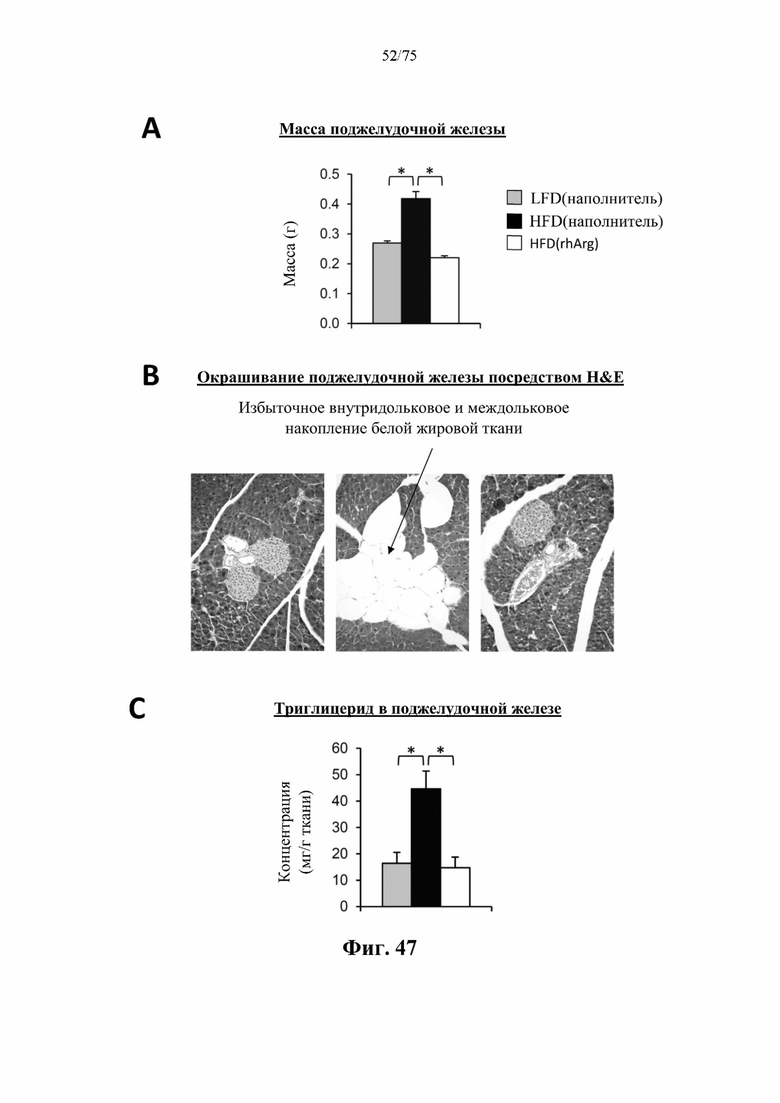
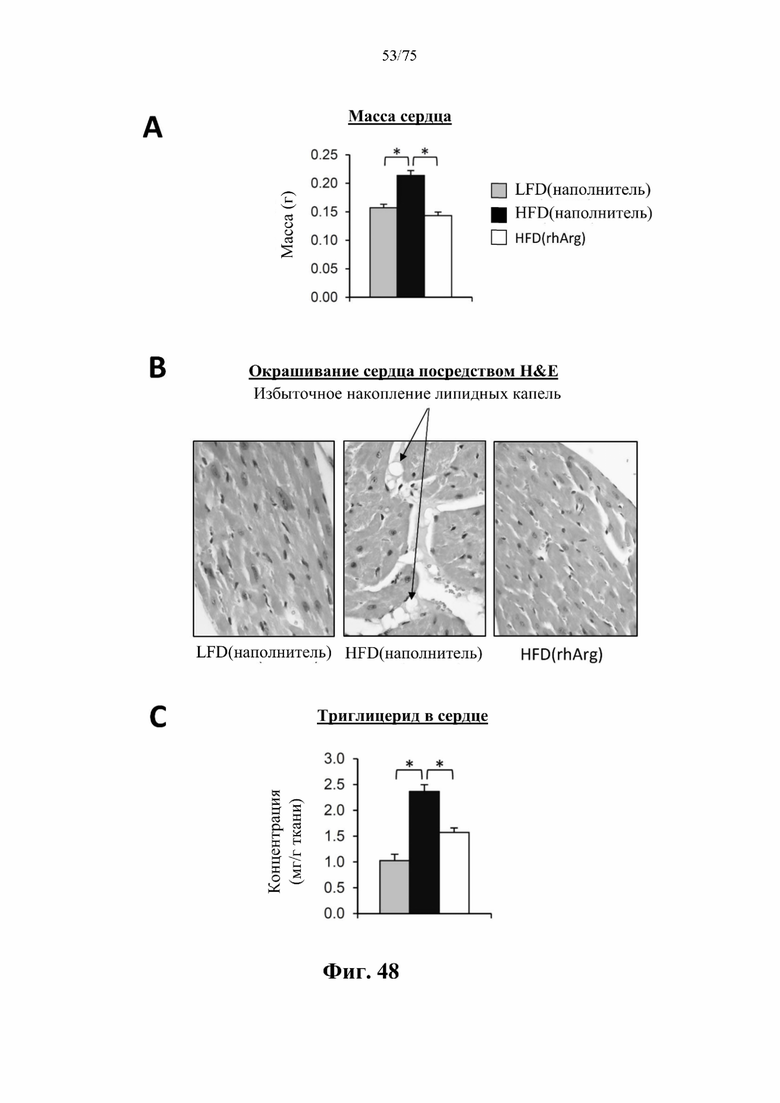
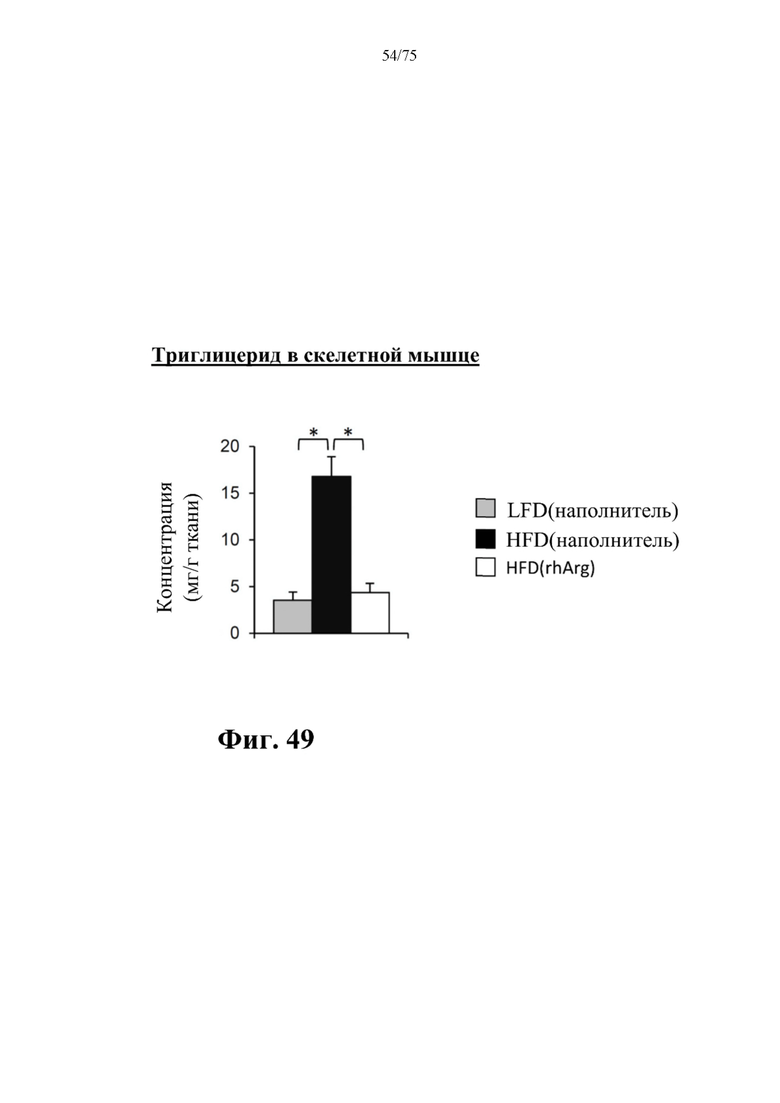
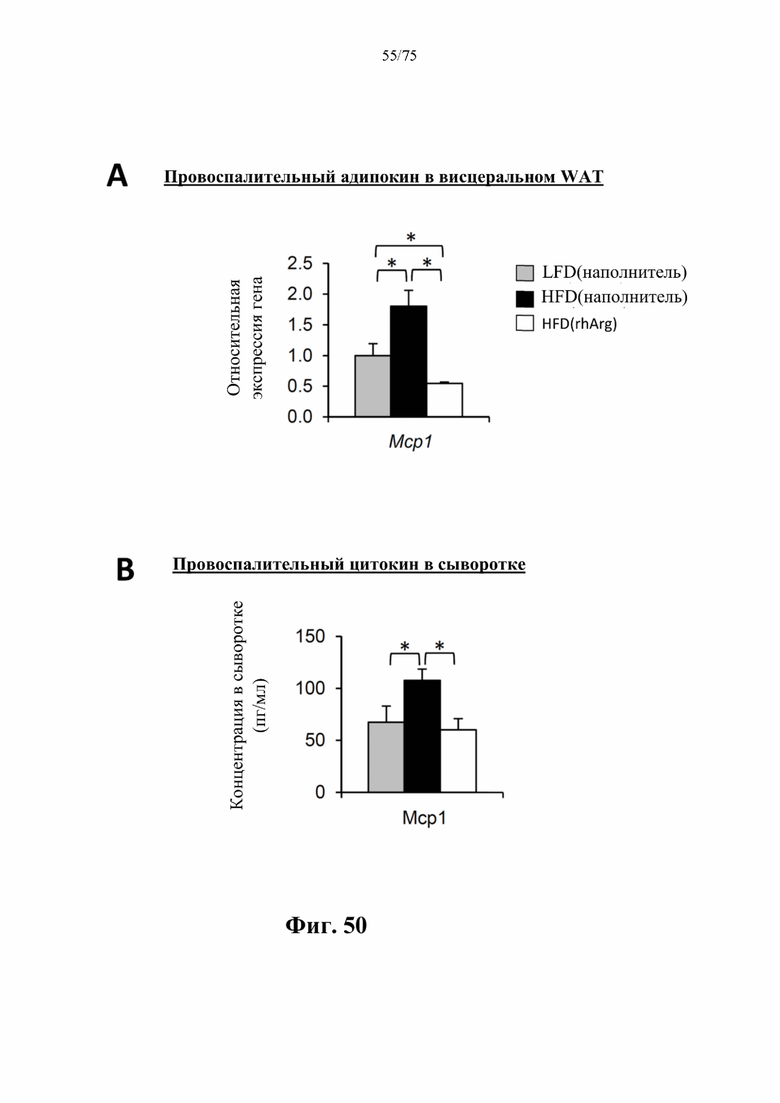
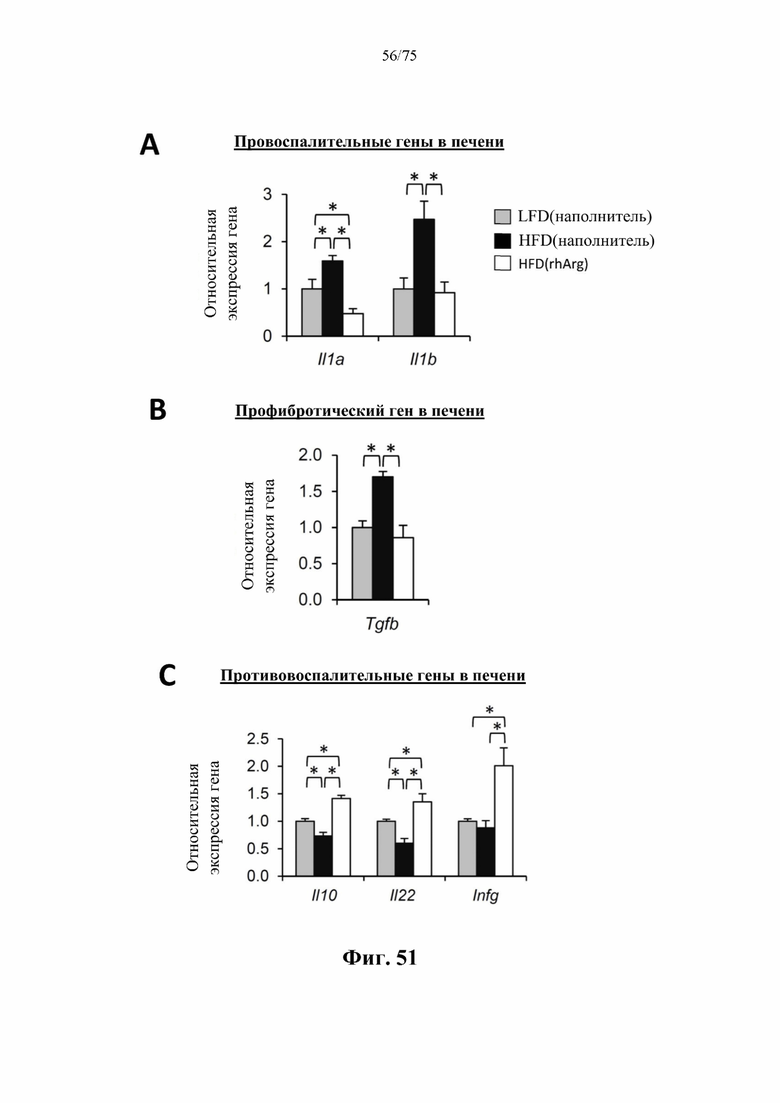
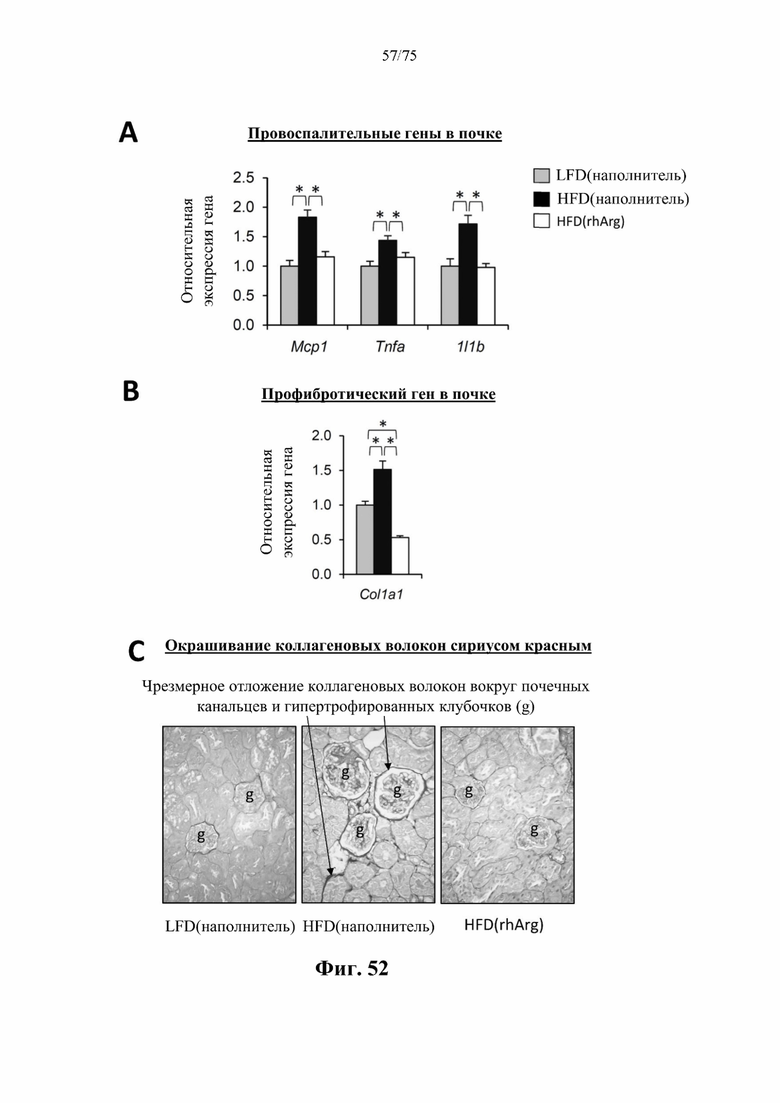
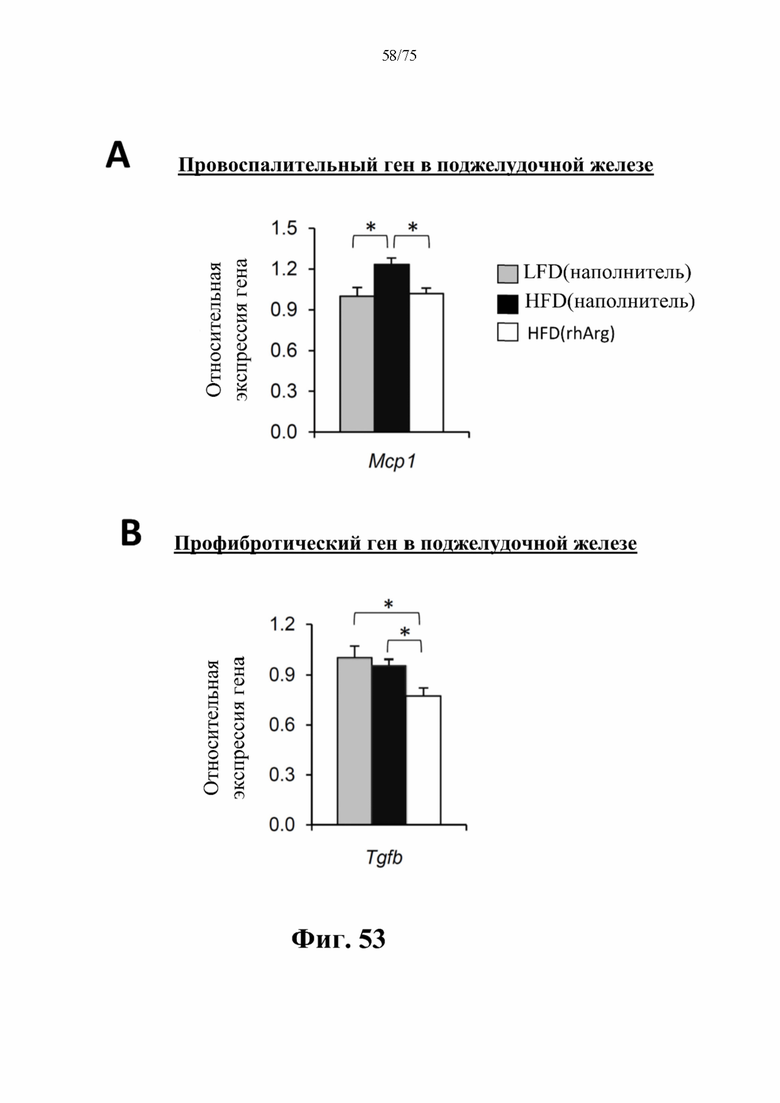
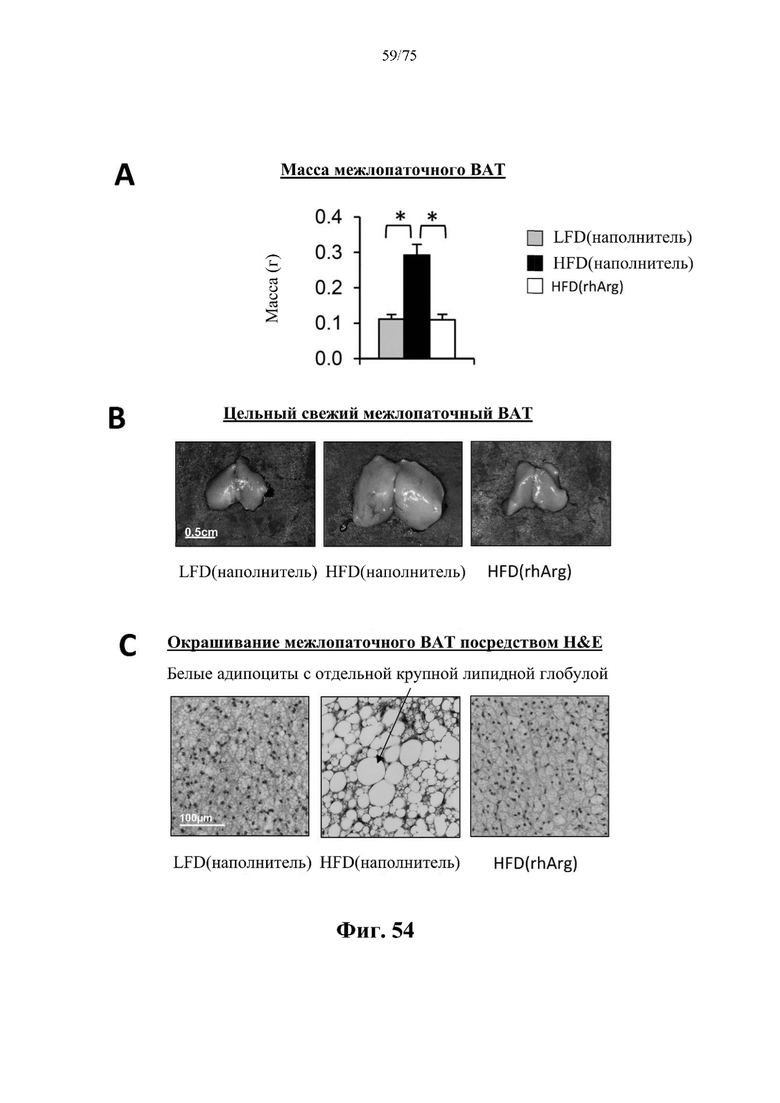
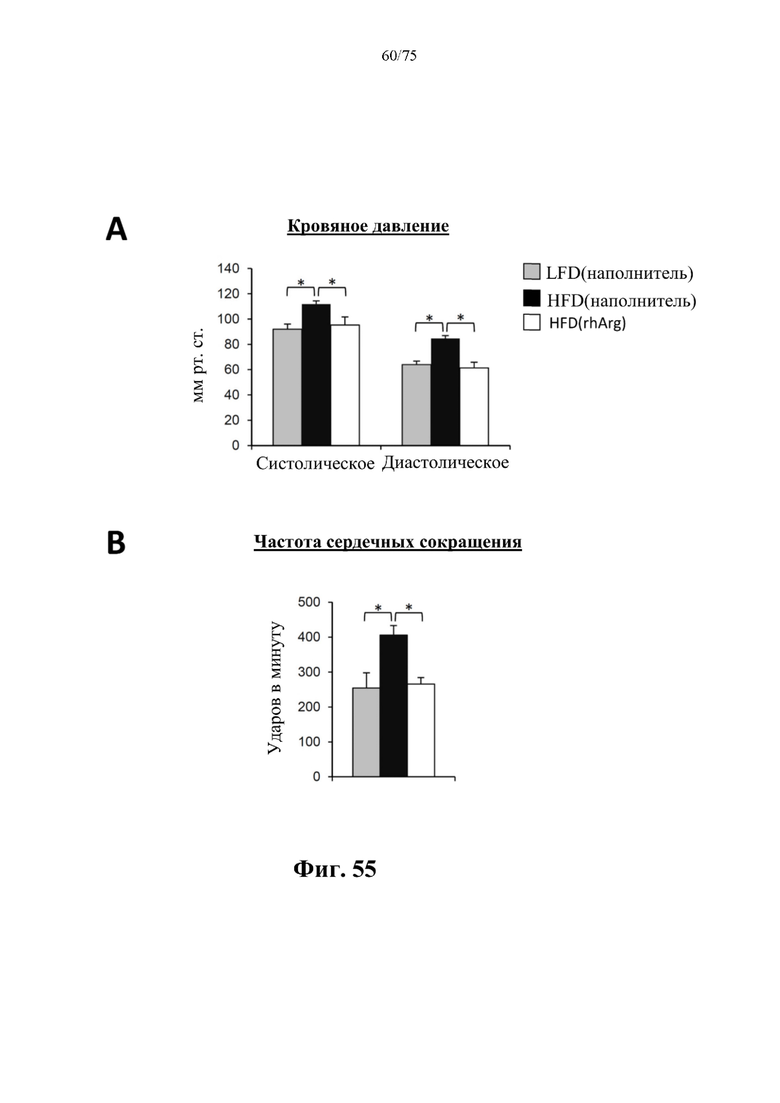
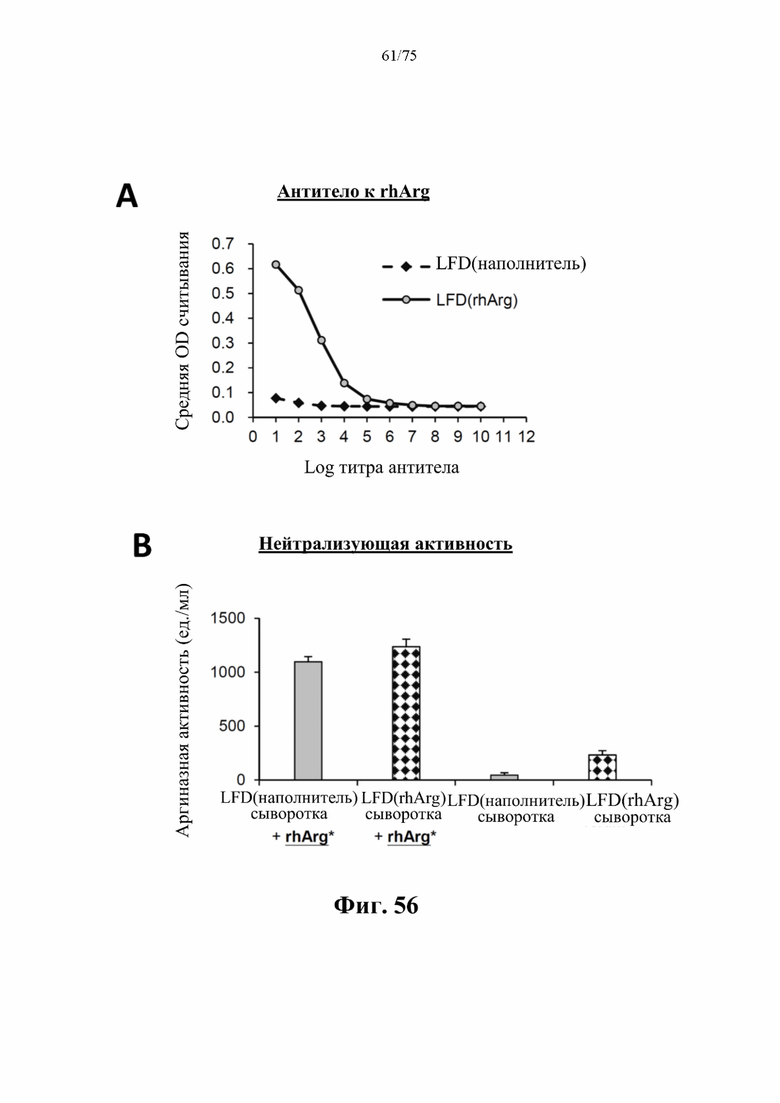
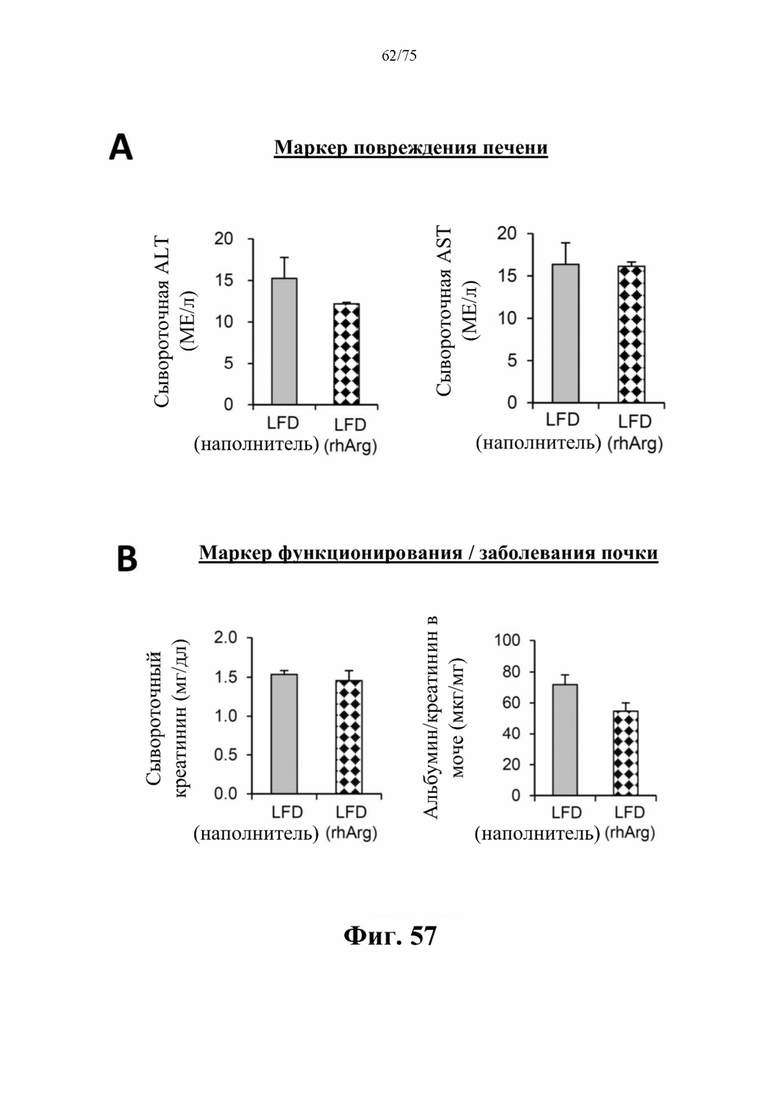
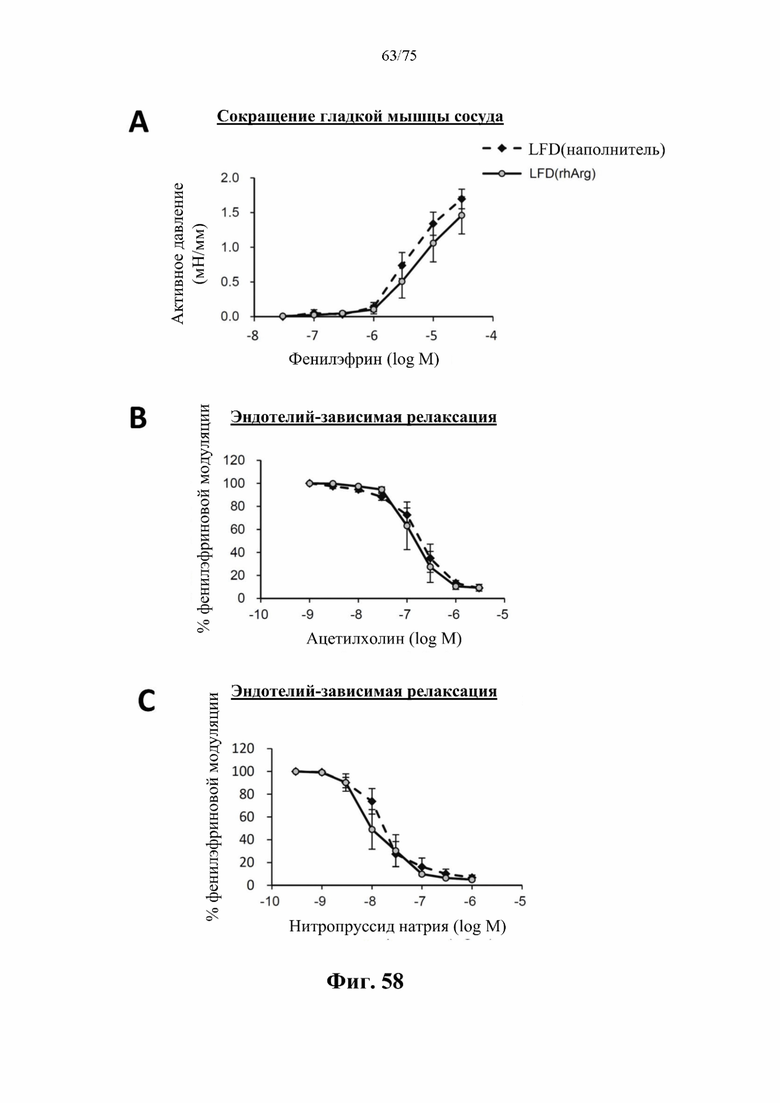
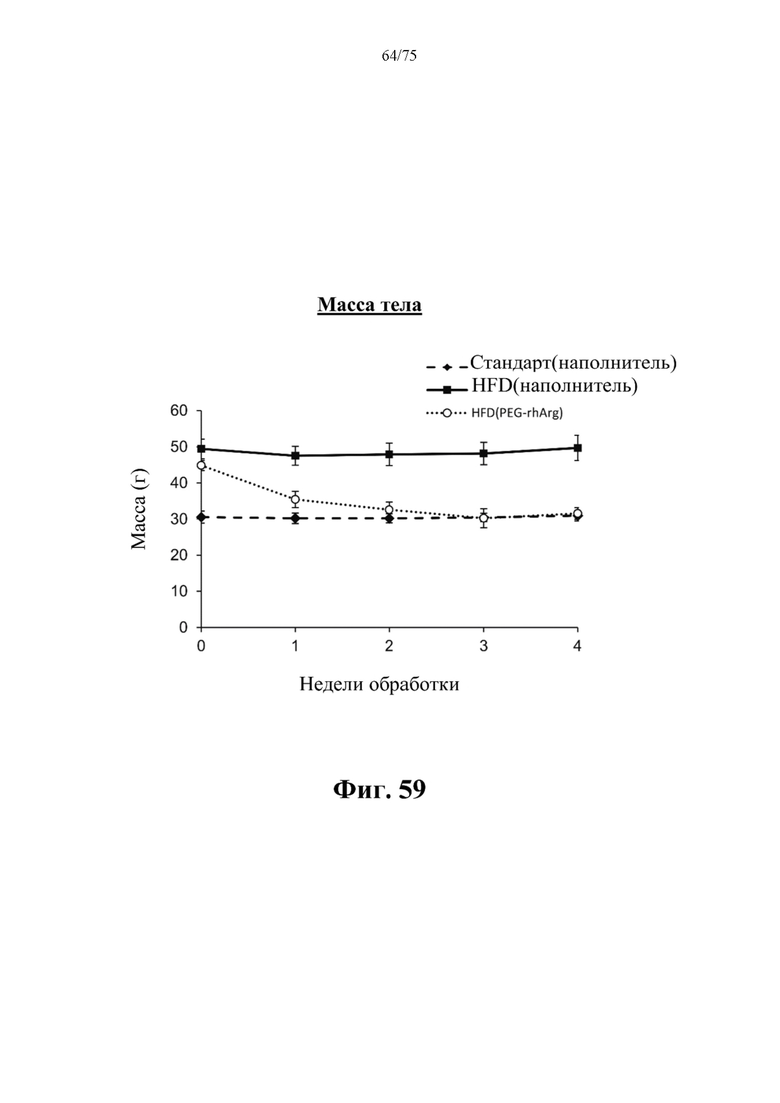
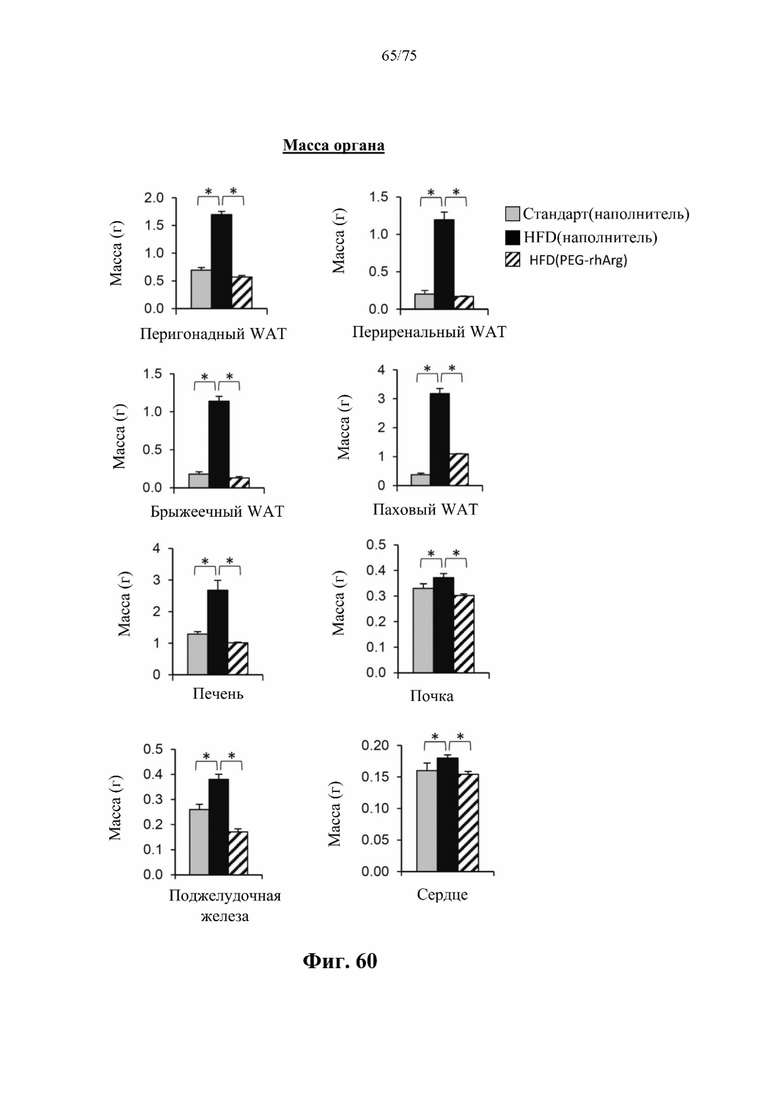
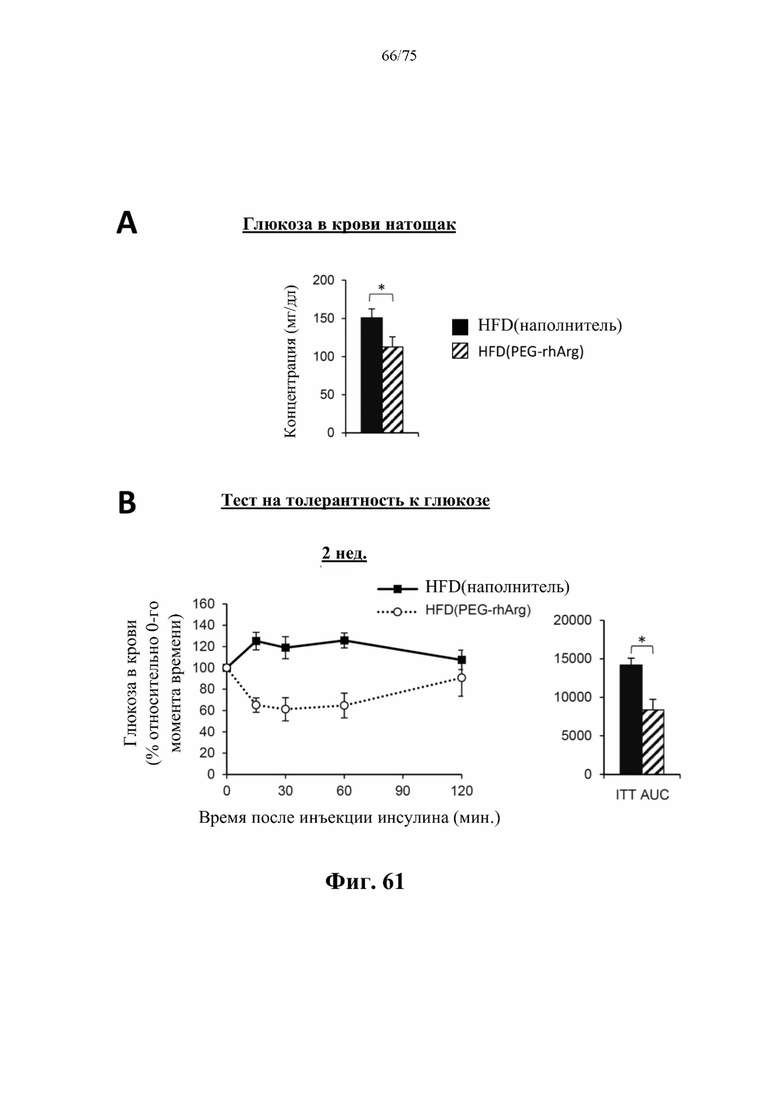
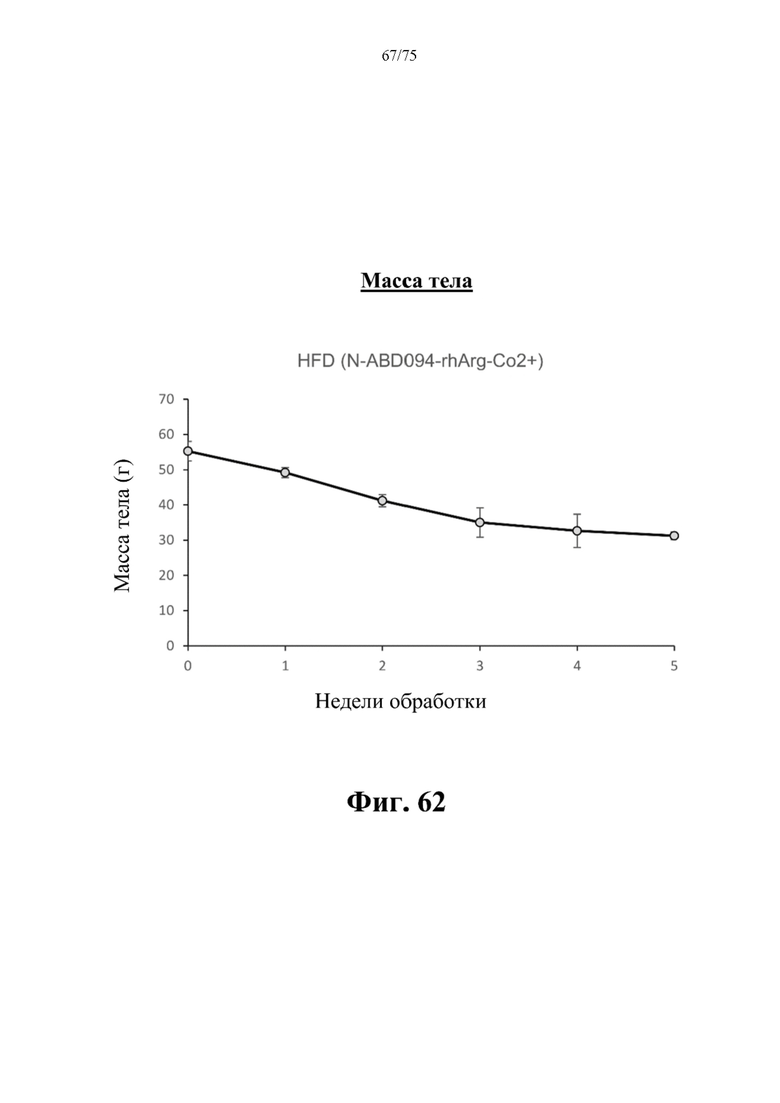
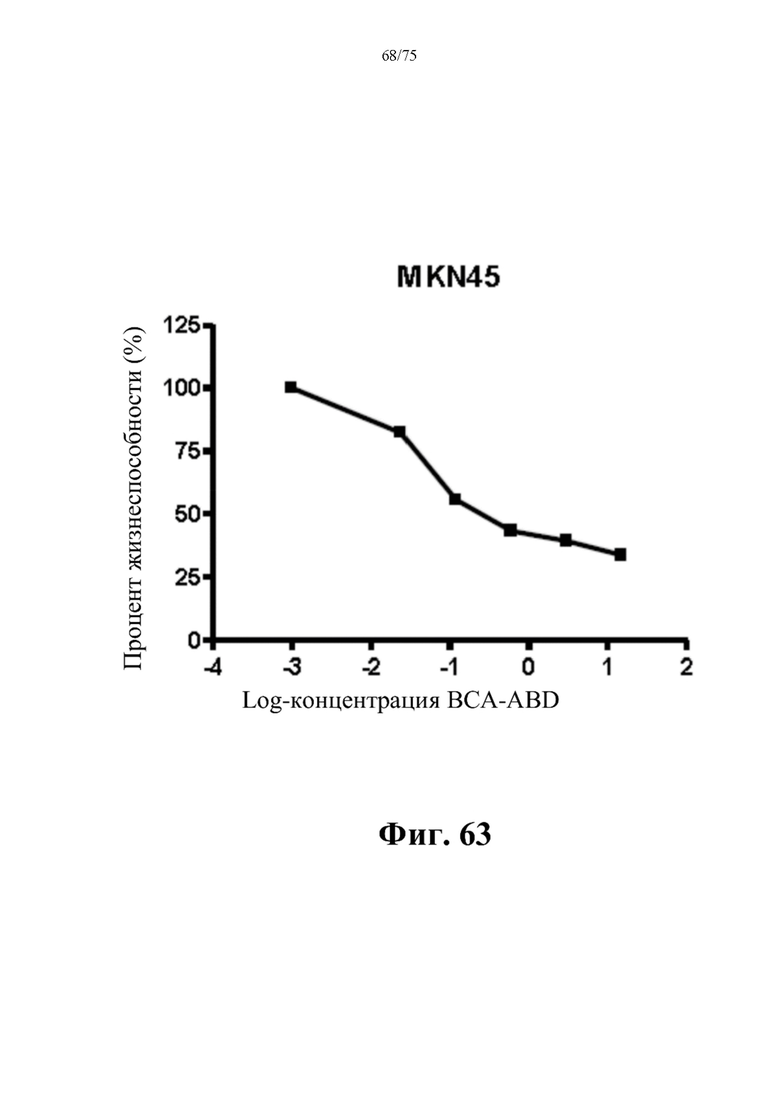
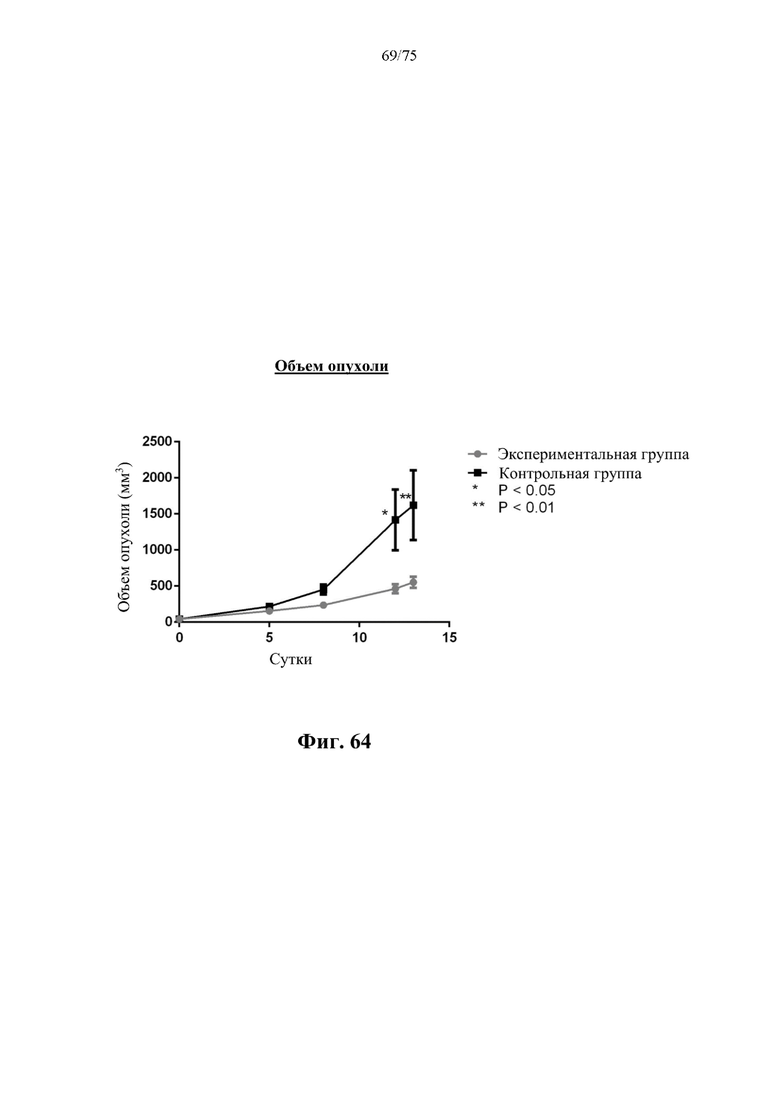
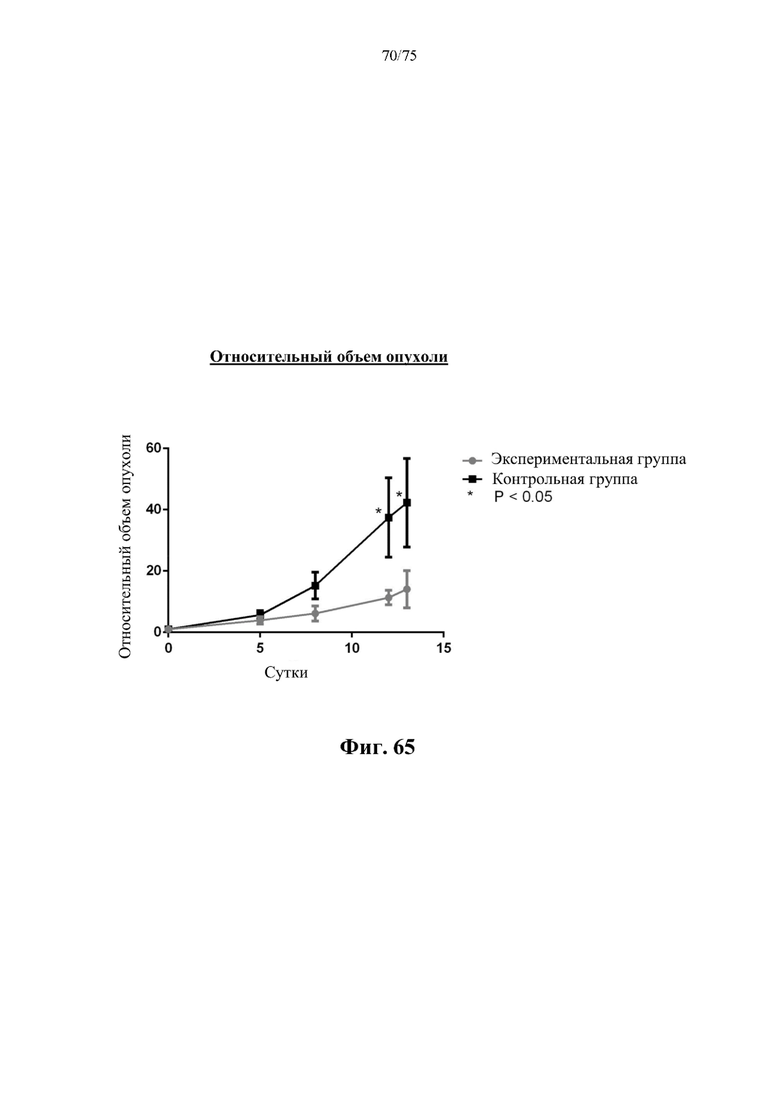
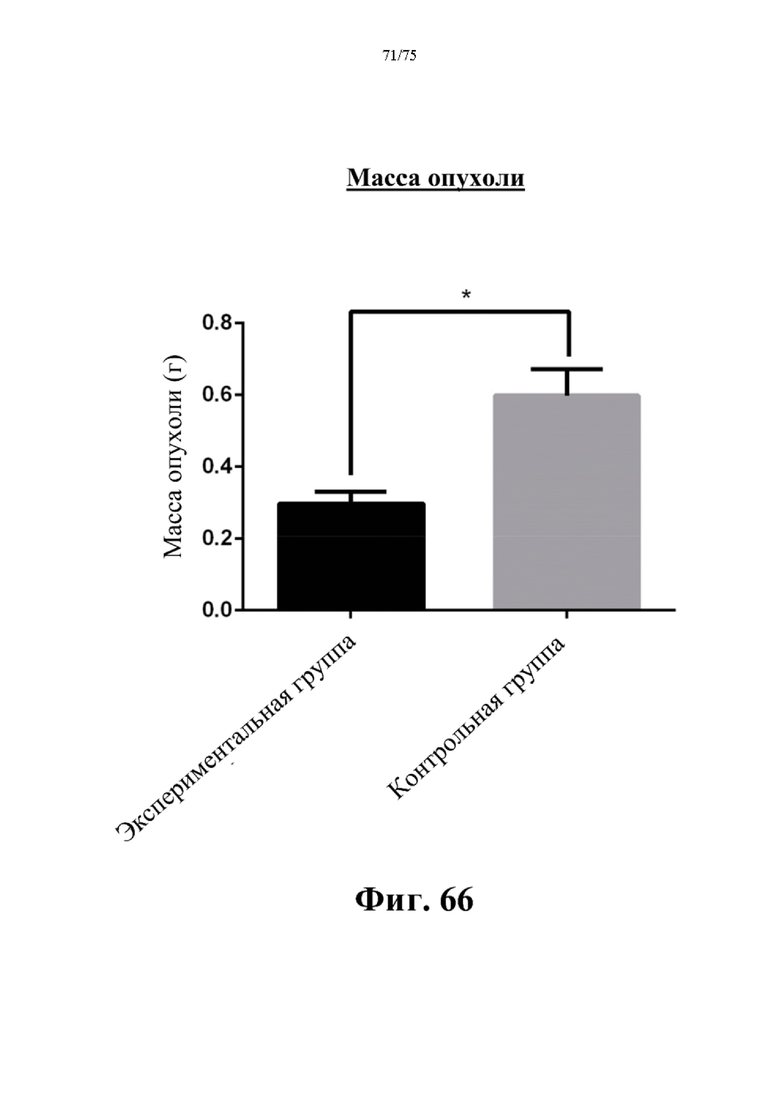

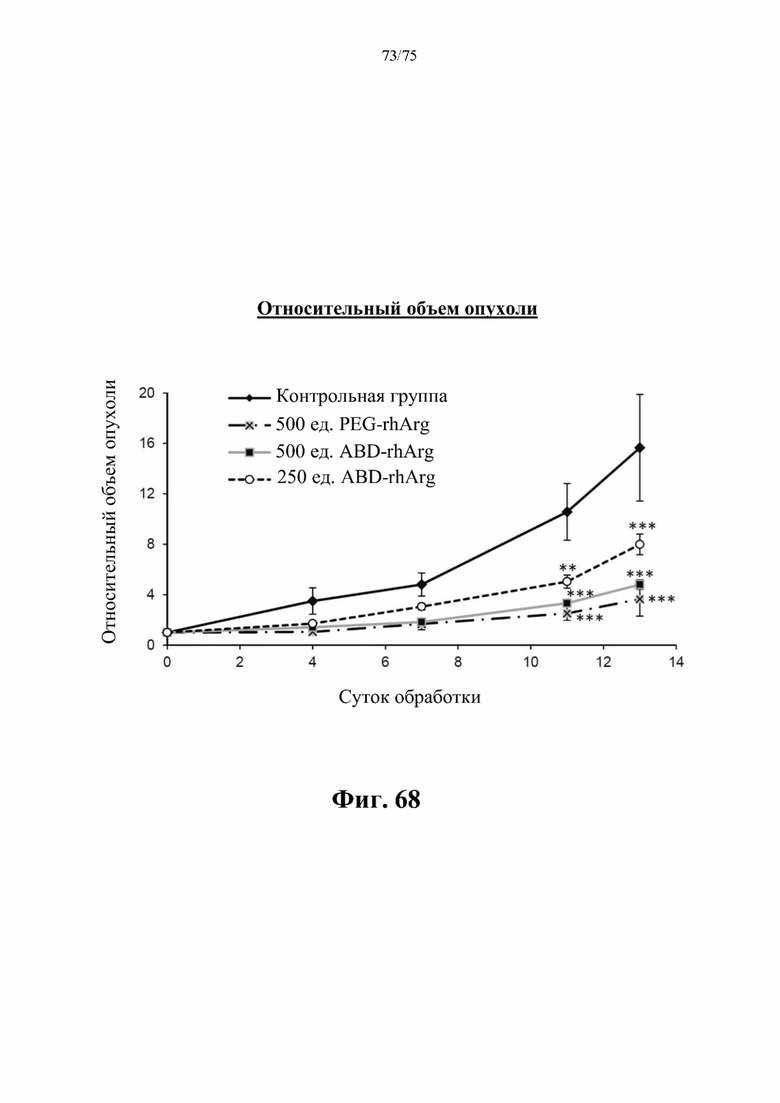
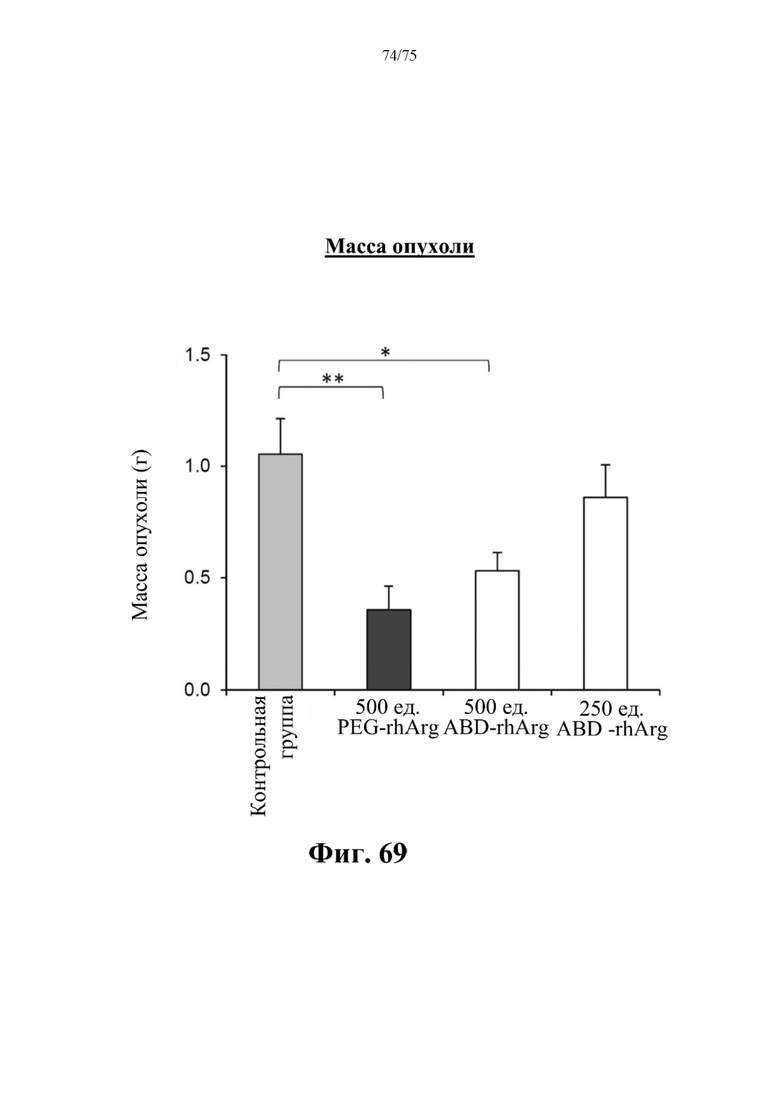
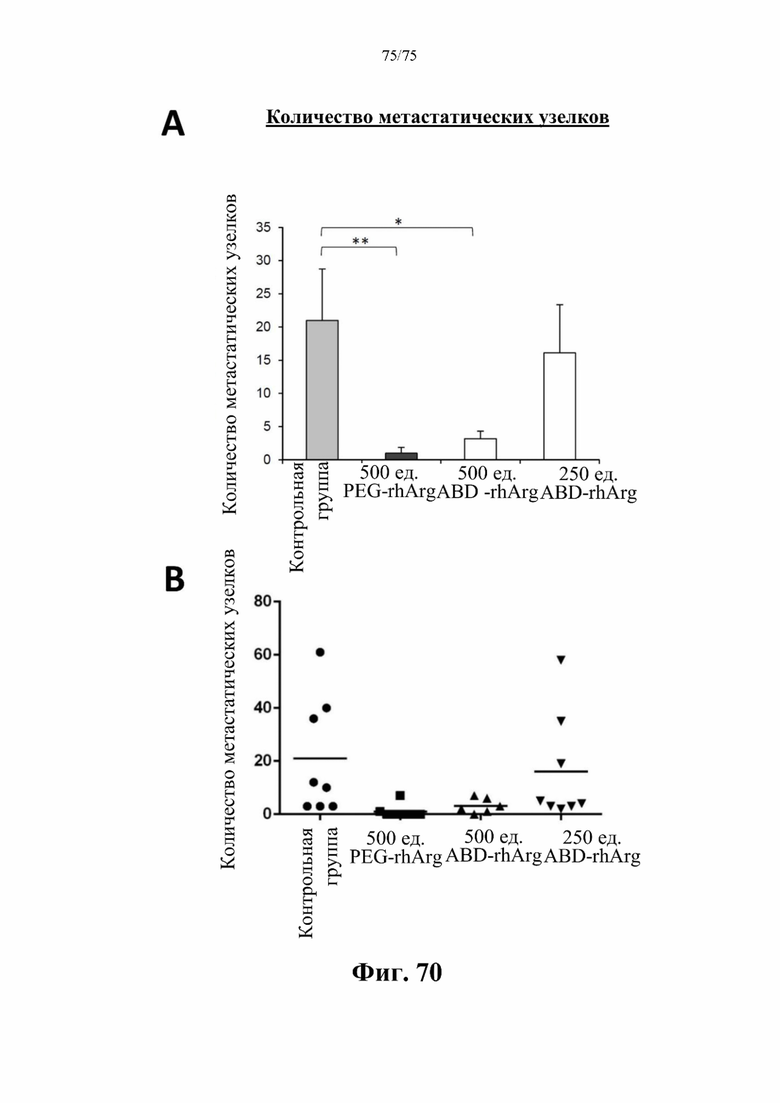
Изобретение относится к области биотехнологии, конкретно к терапевтическому применению истощающего запасы аргинина средства, и может быть использовано в медицине для лечения по меньшей мере одного заболевания или патологического состояния, при котором истощение запасов аргинина приводит к терапевтическому эффекту. Предложено применение истощающего запасы аргинина средства, представляющего собой катаболизирующий аргинин фермент, выбранный из белка аргиназы, аргининдезиминазы или аргининдекарбоксилазы. Изобретение обеспечивает эффективное лечение ожирения, а также метаболического нарушения, выбранного из непереносимости глюкозы, гипергликемии, сахарного диабета, стеатоза печени, стеатоза почек, стеатоза поджелудочной железы, стеатоза сердца и инсулинорезистентности, и связанного с ним осложнения, которое представляет собой одно или несколько патологических состояний, выбранных из диабетической нефропатии, диабетической васкулопатии, диабетической невропатии, гиперхолестеринемии, дислипидемии, гипертриглицеридемии, гиперлептинемии, гиперинсулинемии, стеатоза, стеатогепатита, гипертонии, сердечно-сосудистого заболевания и побеления бурого жира. 12 з.п. ф-лы, 70 ил., 19 табл., 43 пр.
1. Применение истощающего запасы аргинина средства для лечения по меньшей мере одного заболевания или патологического состояния, при котором истощение запасов аргинина приводит к терапевтическому эффекту, выбранного из группы, состоящей из ожирения, метаболического нарушения и связанного с ним осложнения у нуждающегося в том субъекта, при этом истощающее запасы аргинина средство представляет собой катаболизирующий аргинин фермент, выбранный из группы, состоящей из белка аргиназы, белка аргининдезиминазы и белка аргининдекарбоксилазы; метаболическое нарушение выбрано из группы, состоящей из непереносимости глюкозы, гипергликемии, сахарного диабета, стеатоза печени, стеатоза почек, стеатоза поджелудочной железы, стеатоза сердца и инсулинорезистентности; и связанное с ним осложнение представляет собой одно или несколько патологических состояний, выбранных из группы, состоящей из диабетической нефропатии, диабетической васкулопатии, диабетической невропатии, гиперхолестеринемии, дислипидемии, гипертриглицеридемии, гиперлептинемии, гиперинсулинемии, стеатоза, стеатогепатита, гипертонии, сердечно-сосудистого заболевания и побеления бурого жира.
2. Применение по п. 1, при котором метаболическое нарушение представляет собой инсулинорезистентность.
3. Применение по п. 1, при котором лечение ожирения предусматривает по меньшей мере одно из предупреждения увеличения жировой массы и уменьшения жировой массы.
4. Применение по п. 1, при котором метаболическое нарушение выбрано из группы, состоящей из стеатоза печени, стеатоза почек, стеатоза поджелудочной железы и стеатоза сердца.
5. Применение по п. 1, при котором концентрация аргинина в сыворотке субъекта поддерживается на уровне ниже 50 мкМ.
6. Применение по п. 1, при котором белок аргиназы, белок аргининдезиминазы или белок аргининдекарбоксилазы дополнительно содержит одну или несколько полиэтиленгликолевых (PEG) групп.
7. Применение по п. 6, при котором белок аргиназы содержит полипептид, имеющий последовательность под SEQ ID NO: 101, SEQ ID NO: 102, SEQ ID NO: 103 или SEQ ID NO: 104.
8. Применение по п. 1, при котором белок аргиназы содержит полипептид, имеющий по меньшей мере 95% гомологии последовательности с последовательностью под SEQ ID NO: 69, SEQ ID NO: 70, SEQ ID NO: 71 или SEQ ID NO: 72.
9. Применение по п. 1, при котором белок аргиназы, белок аргининдезиминазы или белок аргининдекарбоксилазы дополнительно содержит альбуминсвязывающий домен (ABD), или сывороточный альбумин человека, или Fc-домен IgG человека.
10. Применение по п. 9, причем ABD содержит полипептид, имеющий по меньшей мере 93% гомологии последовательности с последовательностью под SEQ ID NO: 66, SEQ ID NO: 67 или SEQ ID NO: 68.
11. Применение по п. 9, при котором катаболизирующий аргинин фермент представляет собой слитый белок, содержащий полипептид ABD и полипептид аргиназы, полипептид ABD и полипептид аргининдезиминазы или полипептид ABD и полипептид аргининдекарбоксилазы.
12. Применение по п. 11, при котором катаболизирующий аргинин фермент представляет собой слитый белок, содержащий полипептид ABD и полипептид аргиназы.
13. Применение по п. 12, при котором катаболизирующий аргинин фермент содержит полипептид, имеющий по меньшей мере 98% гомологии последовательности с последовательностью под SEQ ID NO: 49, SEQ ID NO: 50, SEQ ID NO: 51, SEQ ID NO: 52, SEQ ID NO: 53, SEQ ID NO: 54, SEQ ID NO: 55, SEQ ID NO: 56, SEQ ID NO: 75 или SEQ ID NO: 76.
| WO 2005027978 A2, 31.03.2005 | |||
| US 5196195 A, 23.03.1993 | |||
| Демпфер крутильных колебаний | 1979 |
|
SU956864A1 |
| US 2005244398 A1, 03.11.2005 | |||
| EA 201500826 A1, 30.12.2015 | |||
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ЛЕЧЕНИЯ ОПУХОЛЕЙ, ПРЕЗЕНТИРУЮЩИХ АНТИГЕНЫ СУРВИВИНА | 2006 |
|
RU2411040C2 |

