ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
По настоящей заявке испрашивается приоритет временной заявки на патент США № 62/202772, поданной 7 августа 2015; временной заявки на патент США № 62/202775, поданной 7 августа 2015; временной заявки на патент США № 62/202779, поданной 7 августа 2015; временной заявки на патент США № 62/276801, поданной 8 января 2016; временной заявки на патент США № 62/265887, поданной 10 декабря 2015; временной заявки на патент США № 62/276796, поданной 8 января 2016 и временной заявки на патент США № 62/346414, поданной 6 июня 2016, каждая из которых полностью включена в настоящее описание путем ссылки.
КРАТКОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В рамках изобретения в некоторых вариантах осуществления раскрыты полипептиды, содержащие: вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92. В некоторых вариантах осуществления домен D1 SIRP-α дикого типа имеет последовательность согласно любой из SEQ ID NO: 1-10. В некоторых вариантах осуществления домен D1 SIRP-α содержит от одной до девяти дополнительных аминокислотных мутаций по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54 остатка 56, остатка 66 и остатка 92. В некоторых вариантах осуществления вариант D1 SIRP-α содержит аминокислотную последовательность
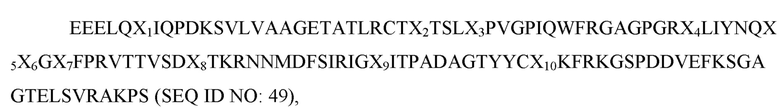
где X1 представляет собой V, L или I; X2 представляет собой A, I, V или L; X3 представляет собой I, F, S или T; X4 представляет собой E, V или L; X5 представляет собой K или R; X6 представляет собой E или Q; X7 представляет собой H, P или R; X8 представляет собой L, T, S или G; X9 представляет собой A, а X10 представляет собой V или I; и при этом вариант D1 SIRP-α имеет по меньшей мере две аминокислотные замены по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность согласно SEQ ID NO: 1.В некоторых вариантах осуществления вариант D1 SIRP-α имеет аминокислотную последовательность согласно любой из SEQ ID NO: 78-85. В некоторых вариантах осуществления вариант D1 SIRP-α содержит аминокислотную последовательность
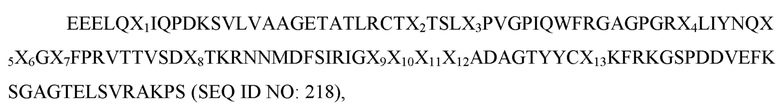
где X1 представляет собой V, L или I; X2 представляет собой A, V, L или I; X3 представляет собой I, S, T или F; X4 представляет собой E, L или V; X5 представляет собой K или R; X6 представляет собой E или Q; X7 представляет собой H, R или P; X8 представляет собой S, G, L или T;X9 представляет собой любую аминокислоту; X10 представляет собой любую аминокислоту; X11 представляет собой любую аминокислоту; X12 представляет собой любую аминокислоту, а X13 представляет собой V или I; и при этом вариант D1 SIRP-α имеет по меньшей мере две аминокислотные замены по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность согласно SEQ ID NO: 1. В некоторых вариантах осуществления X9 представляет собой A. В некоторых вариантах осуществления X9 представляет собой N. В некоторых вариантах осуществления X10 представляет собой I. В некоторых вариантах осуществления X9 представляет собой N, а X10 представляет собой P. В некоторых вариантах осуществления X9 представляет собой N, а X11 представляет собой любую аминокислоту, отличную от S, T или C. В некоторых вариантах осуществления X11 представляет собой T. В некоторых вариантах осуществления X11 представляет собой аминокислоту, отличную от T. В некоторых вариантах осуществления X12 представляет собой P. В некоторых вариантах осуществления X9 представляет собой N, а X12 представляет собой любую аминокислоту, отличную от P. В некоторых вариантах осуществления вариант D1 SIRP-α содержит аминокислотную последовательность
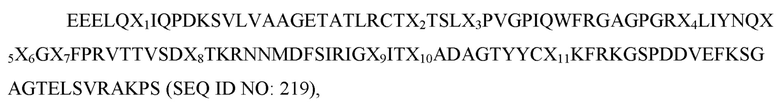
где X1 представляет собой V, L или I; X2 представляет собой A, V, L или I; X3 представляет собой I, S, T или F; X4 представляет собой E, L или V; X5 представляет собой K или R; X6 представляет собой E или Q; X7 представляет собой H, R или P; X8 представляет собой S, G, L или T; X9 представляет собой N; X10 представляет собой любую аминокислоту, отличную от P, а X11 представляет собой V или I; и при этом вариант D1 SIRP-α имеет по меньшей мере две аминокислотные замены по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность согласно SEQ ID NO: 1. В некоторых вариантах осуществления вариант D1 SIRP-α содержит аминокислотную последовательность
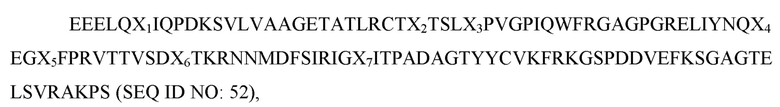
где X1 представляет собой V, L или I; X2 представляет собой A, I или L; X3 представляет собой I, T, S или F; X4 представляет собой K или R; X5 представляет собой H, P или R; X6 представляет собой L, T или G, а X7 представляет собой A; и при этом вариант D1 SIRP-α имеет по меньшей мере две аминокислотные замены по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность согласно SEQ ID NO: 1. В некоторых вариантах осуществления X1 представляет собой V или I, X2 представляет собой A или I, X3 представляет собой I или F, X4 представляет собой K или R, X5 представляет собой H или P, X6 представляет собой L или T, а X7 представляет собой A. В некоторых вариантах осуществления вариант D1 SIRP-α имеет по меньшей мере три аминокислотных замены по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность согласно SEQ ID NO: 1. В некоторых вариантах осуществления вариант D1 SIRP-α имеет по меньшей мере четыре аминокислотных замены по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность согласно SEQ ID NO: 1. В некоторых вариантах осуществления вариант D1 SIRP-α имеет по меньшей мере пять аминокислотных замен по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность согласно SEQ ID NO: 1. В некоторых вариантах осуществления вариант D1 SIRP-α имеет по меньшей мере шесть аминокислотных замен по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность согласно SEQ ID NO: 1. В некоторых вариантах осуществления вариант D1 SIRP-α имеет по меньшей мере семь аминокислотных замен по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность согласно SEQ ID NO: 1. В некоторых вариантах осуществления X1 представляет собой I. В некоторых вариантах осуществления X2 представляет собой I. В некоторых вариантах осуществления X3 представляет собой F. В некоторых вариантах осуществления X4 представляет собой R. В некоторых вариантах осуществления X5 представляет собой P. В некоторых вариантах осуществления X6 представляет собой T. В некоторых вариантах осуществления каждый из X1, X2, X3, X4, X5 и X6 не является аминокислотой дикого типа. В некоторых вариантах осуществления вариант D1 SIRP-α имеет аминокислотную последовательность согласно любой из SEQ ID NO: 81-85. В некоторых вариантах осуществления вариант D1 SIRP-α содержит аминокислотную последовательность
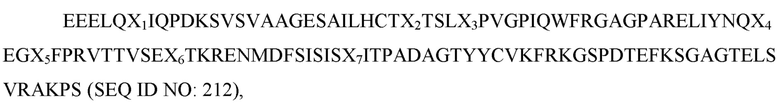
где X1 представляет собой V, L или I; X2 представляет собой V, I или L; X3 представляет собой I, T, S или F; X4 представляет собой K или R; X5 представляет собой H, P или R; X6 представляет собой S, T или G, а X7 представляет собой A; и при этом вариант D1 SIRP-α имеет по меньшей мере две аминокислотные замены по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность SEQ ID NO: 2. В некоторых вариантах осуществления полипептид связывается с CD47 человека с KD менее чем около 5×10 -9 M. В некоторых вариантах осуществления полипептид дополнительно содержит мономер домена Fc, связанный с N-концом или С-концом полипептида, причем мономер домена Fc представляет собой область Fc IgG1, IgG2 или IgG4 человека. В некоторых вариантах осуществления мономер домена Fc содержит по меньшей мере одну мутацию по сравнению с областью Fc дикого типа IgG1, IgG2 или IgG4 человека. В некоторых вариантах осуществления полипептид имеет аминокислотную последовательность любой из SEQ ID NO: 135, SEQ ID NO: 136 или SEQ ID NO: 137. В некоторых вариантах осуществления мономер домена Fc содержит (а) одну из следующих аминокислотных замен по сравнению с IgG1 человека дикого типа: T366W, T366S, L368A, Y407V, T366Y, T394W, F405W, Y349T, Y349E, Y349V, L351T, L351H, L351N, L351K, P353S, S354D, D356K, D356R, D356S, E357K, E357R, E357Q, S364A, T366E, L368T, L368Y, L368E, K370E, K370D, K370Q, K392E, K392D, T394N, P395N, P396T, V397T, V397Q, L398T, D399K, D399R, D399N, F405T, F405H, F405R, Y407T, Y407H, Y407I, K409E, K409D, K409T, или K409I; или (b) (i) мутацию N297A по сравнению с областью Fc IgG1 человека; (ii) мутацию L234A, L235A и G237A по сравнению с областью Fc IgG1 человека; (iii) мутацию L234A, L235A, G237A и N297A по сравнению с областью Fc IgG1 человека; (iv) мутацию N297A по сравнению с областью Fc IgG2 человека; (v) мутацию A330S и P331S по сравнению с областью Fc IgG2 человека; (vi) мутацию A330S, P331S и N297A по сравнению с областью Fc IgG2 человека; (vii) мутацию S228P, E233P, F234V, L235A и delG236 по сравнению с областью Fc IgG4 человека, или (viii) мутацию S228P, E233P, F234V, L235A, delG236 и N297A по сравнению с областью Fc IgG4 человека. В некоторых вариантах осуществления мономер домена Fc содержит (а) одну из следующих аминокислотных замен по сравнению с IgG1 человека дикого типа: T366W, T366S, L368A, Y407V, T366Y, T394W, F405W, Y349T, Y349E, Y349V, L351T, L351H, L351N, L351K, P353S, S354D, D356K, D356R, D356S, E357K, E357R, E357Q, S364A, T366E, L368T, L368Y, L368E, K370E, K370D, K370Q, K392E, K392D, T394N, P395N, P396T, V397T, V397Q, L398T, D399K, D399R, D399N, F405T, F405H, F405R, Y407T, Y407H, Y407I, K409E, K409D, K409T, или K409I; и (b) мономер домена Fc, дополнительно содержащий (i) мутацию N297A по сравнению с областью Fc IgG1 человека; (ii) мутацию L234A, L235A и G237A по сравнению с областью Fc IgG1 человека; (iii) мутацию L234A, L235A, G237A и N297A по сравнению с областью Fc IgG1 человека; (iv) мутацию N297A по сравнению с областью Fc IgG2 человека; (v) мутацию A330S и P331S по сравнению с областью Fc IgG2 человека; (vi) мутацию A330S, P331S и N297A по сравнению с областью Fc IgG2 человека; (vii) мутацию S228P, E233P, F234V, L235A и delG236 по сравнению с областью Fc IgG4 человека, или (viii) мутацию S228P, E233P, F234V, L235A, delG236 и N297A по сравнению с областью Fc IgG4 человека. В некоторых вариантах осуществления полипептид демонстрирует снижение фагоцитоза в анализе фагоцитоза по сравнению с полипептидом с областью Fc дикого типа IgG человека. В некоторых вариантах осуществления мономер домена Fc связан со вторым полипептидом, содержащим второй мономер домена Fc, с образованием димера домена Fc. В некоторых вариантах осуществления второй мономер домена Fc связан с дополнительным полипептидом. В некоторых вариантах осуществления дополнительный полипептид содержит вариабельный домен антитела. В некоторых вариантах осуществления вариабельный домен антитела нацелен на антиген, экспрессируемый в клетке. В некоторых вариантах осуществления клетка представляет собой раковую клетку. В некоторых вариантах осуществления вариабельный домен антитела нацелен на поверхностный белок клетки, вовлеченный в регуляцию иммунных клеток. В некоторых вариантах осуществления дополнительный полипептид содержит терапевтический белок. В некоторых вариантах осуществления терапевтический белок представляет собой цитокин, интерлейкин, антиген, стероид, противовоспалительный агент или иммуномодулирующий агент. В некоторых вариантах осуществления дополнительный полипептид содержит вариант D1 SIRP-α. В некоторых вариантах осуществления полипептид дополнительно содержит человеческий сывороточный альбумин (ЧСА) (SEQ ID NO: 12). В некоторых вариантах осуществления ЧСА содержит аминокислотную замену C34S или K573P по сравнению с SEQ ID NO: 12. В некоторых вариантах осуществления полипептид имеет аминокислотную последовательность согласно любой из SEQ ID NO: 152-159. В некоторых вариантах осуществления полипептид дополнительно содержит альбуминсвязывающий пептид. В некоторых вариантах осуществления альбуминсвязывающий пептид содержит аминокислотную последовательность DICLPRWGCLW (SEQ ID NO: 160). В некоторых вариантах осуществления полипептид дополнительно содержит полимер полиэтиленгликоль (ПЭГ). В некоторых вариантах осуществления полимер ПЭГ соединен с цистеиновой заменой в полипептиде.
В рамках изобретения в некоторых вариантах осуществления раскрыты полипептиды, содержащие: вариант D1 сигнального регуляторного белка α (SIRP-α), причем вариант D1 SIRP-α содержит аминокислотную последовательность
где X1 представляет собой E или G; X2 представляет собой L, I или V; X3 представляет собой V, L или I; X4 представляет собой S или F; X5 представляет собой L или S; X6 представляет собой S или T; X7 представляет собой A или V; X8 представляет собой I или T; X9 представляет собой H, R или L; X10 представляет собой A, V, I или L; X11 представляет собой I, T, S или F; X12 представляет собой A или G; X13 представляет собой E, V или L; X14 представляет собой K или R; X15 представляет собой E или Q; X16 представляет собой H, P или R; X17 представляет собой D или E; X18 представляет собой S, L, T или G; X19 представляет собой K или R; X20 представляет собой E или N; X21 представляет собой S или P; X22 представляет собой S или R; X23 представляет собой S или G; X24 представляет собой любую аминокислоту; X25 представляет собой любую аминокислоту; X26 представляет собой V или I; X27 представляет собой F, L или V; X28 представляет собой D или отсутствует; X29 представляет собой T или V; X30 представляет собой F или V, а X31 представляет собой A или G; и при этом вариант D1 SIRP-α имеет по меньшей мере две аминокислотных заменены по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность согласно любой из SEQ ID NO: 1-10; и вариант Fc, содержащий димер домена Fc, имеющий два мономера домена Fc, причем каждый мономер домена Fc независимо представляет собой (i) область Fc IgG1 человека, содержащую мутацию N297A; (ii) область Fc IgG1 человека, содержащую мутации L234A, L235A и G237A; (iii) область Fc IgG1 человека, содержащую мутации L234A, L235A, G237A и N297A; (iv) область Fc IgG2 человека, содержащую мутацию N297A; (v) область Fc IgG2 человека, содержащую мутации A330S и P331S; (vi) область Fc IgG2 человека, содержащую мутации A330S, P331S и N297A; (vii) область Fc IgG4 человека, содержащую мутации S228P, E233P, F234V, L235A и delG236; или (viii) область Fc IgG4 человека, содержащую мутации S228P, E233P, F234V, L235A, delG236 и N297A. В некоторых вариантах осуществления один из мономеров домена Fc в димере домена Fc содержит область Fc IgG1 человека, содержащую мутации L234A, L235A, G237A и N297A. В некоторых вариантах осуществления полипептид содержит аминокислотную последовательность согласно любой из SEQ ID NO: 98-104, 107-113, 116-122 или 135-137. В некоторых вариантах осуществления вариант Fc демонстрирует аблированное или уменьшенное связывание с рецептором Fcγ по сравнению с версией области Fc дикого типа IgG человека. В некоторых вариантах осуществления вариант Fc IgG1 или IgG2 демонстрирует аблированное или уменьшенное связывание с CD16a, CD32a, CD32b, CD32c и CD64 рецепторами Fcγ по сравнению с версией дикого типа области Fc IgG1 или IgG2 человека. В некоторых вариантах осуществления вариант Fc IgG4 демонстрирует аблированное или уменьшенное связывание с рецепторами Fcγ CD16a и CD32b по сравнению с версией области Fc дикого типа IgG4 человека. В некоторых вариантах осуществления вариант Fc IgG1 или IgG2 демонстрирует аблированное или уменьшенное связывание с C1q по сравнению с версией слитого Fc дикого типа и IgG1 или IgG2 человека. В некоторых вариантах осуществления вариант Fc связывается с рецептором Fcγ с KD более чем около 5×10-6 M.
В рамках изобретения в некоторых вариантах осуществления раскрыты полипептиды, содержащие вариант Fc, причем вариант Fc содержит димер домена Fc, имеющий два мономера домена Fc, при этом каждый мономер домена Fc независимо выбирают из (i) области Fc IgG1 человека, включающей в свой состав мутации L234A, L235A, G237A и N297A; (ii) области Fc IgG2 человека, включающей в свой состав мутации A330S, P331S и N297A; или (iii) области Fc IgG4 человека, содержащей мутации S228P, E233P, F234V, L235A, delG236 и N297A. В некоторых вариантах осуществления по меньшей мере один из мономеров домена Fc представляет собой область Fc IgG1 человека, состоящую из мутаций L234A, L235A, G237A и N297A. В некоторых вариантах осуществления по меньшей мере один из мономеров домена Fc представляет собой область Fc IgG2 человека состоящую из мутаций A330S, P331S и N297A. В некоторых вариантах осуществления вариант Fc демонстрирует аблированное или уменьшенное связывание с рецептором Fcγ по сравнению с версией области Fc дикого типа IgG человека. В некоторых вариантах осуществления вариант Fc демонстрирует аблированное или уменьшенное связывание с рецепторами Fcγ CD16a, CD32a, CD32b, CD32c и CD64 по сравнению с версией области Fc дикого типа IgG человека. В некоторых вариантах осуществления вариант Fc демонстрирует аблированное или уменьшенное связывание с C1q по сравнению с версией слитого Fc дикого типа и IgG человека. В некоторых вариантах осуществления по меньшей мере один из мономеров домена Fc представляет собой область Fc IgG4 человека, содержащую мутации S228P, E233P, F234V, L235A, delG236 и N297A. В некоторых вариантах осуществления вариант Fc демонстрирует аблированное или уменьшенное связывание с рецептором Fcγ по сравнению с областью Fc дикого типа IgG4 человека. В некоторых вариантах осуществления вариант Fc демонстрирует аблированное или уменьшенное связывание с рецепторами Fcγ CD16a и CD32b по сравнению с версией его области Fc дикого типа IgG4 человека. В некоторых вариантах осуществления вариант Fc связывается с рецептором Fcγ с KD более чем около 5×10-6 M. В некоторых вариантах осуществления полипептид дополнительно содержит CD47-связывающий полипептид. В некоторых вариантах осуществления вариант Fc демонстрирует аблированное или уменьшенное связывание с рецептором Fcγ по сравнению с версией области Fc дикого типа IgG человека. В некоторых вариантах осуществления CD47-связывающий полипептид не вызывает острой анемии у грызунов и приматов, отличных от человека. В некоторых вариантах осуществления CD47-связывающий полипептид не вызывает острой анемии у людей. В некоторых вариантах осуществления CD47-связывающий полипептид представляет собой полипептид сигнального регуляторного белка α (SIRP-α) или его фрагмент. В некоторых вариантах осуществления полипептид SIRP-α содержит вариант D1 SIRP-α, содержащий аминокислотную последовательность

где X1 представляет собой V или I; X2 представляет собой A или I; X3 представляет собой I или F; X4 представляет собой E или V; X5 представляет собой K или R; X6 представляет собой H или P; X7 представляет собой L или T; X8 представляет собой аминокислоту, отличную от N, а X9 представляет собой V или I. В некоторых вариантах осуществления полипептид SIRP-α содержит вариант D1 SIRP-α, где X1 представляет собой V или I; X2 представляет собой A или I; X3 представляет собой I или F; X4 представляет собой E; X5 представляет собой K или R; X6 представляет собой H или P; X7 представляет собой L или T; X8 представляет собой не N, а X9 представляет собой V.
В рамках изобретения в некоторых вариантах осуществления раскрыты полипептиды, содержащие: вариант D1 сигнального регуляторного белка α (SIRP-α), причем вариант D1 SIRP-α представляет собой неприродный высокоаффинный домен D1 SIRP-α, при этом вариант D1 SIRP-α связывается с CD47 человека с аффинностью, которая по меньшей мере в 10 раз больше, чем аффинность природного домена D1; и мономер домена Fc, причем мономер домена Fc связан со вторым полипептидом, содержащим второй мономер домена Fc, с образованием домена Fc, при этом домен Fc имеет аблированную или уменьшенную эффекторную функцию. В некоторых вариантах осуществления неприродный высокоаффинный домен D1 SIRP-α содержит аминокислотную мутацию в остатке 80.
В рамках изобретения в некоторых вариантах осуществления раскрыты полипептиды, содержащие вариант сигнального регуляторного белка α (SIRP-α) D1, причем вариант D1 SIRP-α связывает CD47 первого вида с KD менее 250 нМ; и при этом вариант D1 SIRP-α связывает CD47 второго вида с KD менее 250 нМ; и KD для CD47 первого вида и KD для CD47 второго вида отличается в 100 раз друг от друга; причем первый вид и второй вид выбраны из группы, состоящей из человека, грызуна и приматов, отличных от человека. В некоторых вариантах осуществления вариант D1 SIRP-α связывает CD47 по меньшей мере 3 различных видов. В некоторых вариантах осуществления примат, не являющийся человеком, представляет собой яванского макака.
В рамках изобретения в некоторых вариантах осуществления раскрыты полипептиды, содержащие: (a) домен D1 сигнального регуляторного белка α (SIRP-α), который связывает CD47 человека с KD менее 250 нМ; и (б) мономер домена Fc, связанный с N-концом или С-концом домена D1 SIRP-α, причем полипептид не вызывает острой анемии у грызунов и приматов, отличных от человека. В некоторых вариантах осуществления полипептид представляет собой неприродный вариант SIRP-α человека. В некоторых вариантах осуществления введение полипептида in vivo приводит к снижению гемоглобина менее чем на 50% в течение первой недели после введения. В некоторых вариантах осуществления введение полипептида людям приводит к снижению гемоглобина менее чем на 50% в течение первой недели после введения. В некоторых вариантах осуществления полипептид дополнительно содержит по меньшей мере один вариант Fc, причем вариант Fc выбирают из (i) области Fc IgG1 человека, включающей в свой состав мутации L234A, L235A, G237A и N297A; (ii) области Fc IgG2 человека, включающей в свой состав мутации A330S, P331S и N297A; или (iii) области Fc IgG4 человека, содержащей мутации S228P, E233P, F234V, L235A, delG236 и N297A. В некоторых вариантах осуществления вариант Fc представляет собой область Fc IgG1 человека, состоящую из мутаций L234A, L235A, G237A и N297A. В некоторых вариантах осуществления вариант Fc представляет собой область Fc IgG2 человека состоящую из мутаций A330S, P331S и N297A. В некоторых вариантах осуществления вариант Fc представляет собой область Fc IgG4 человека, содержащую мутации S228P, E233P, F234V, L235A, delG236 и N297A.
В рамках изобретения в некоторых вариантах осуществления раскрыты способы лечения индивидуума, имеющего болезнь или расстройство, причем способ включает введение пациенту описанного в рамках изобретения полипептида. В некоторых вариантах осуществления полипептид содержит вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92. В некоторых вариантах осуществления полипептид содержит вариант сигнального регуляторного белка α (SIRP-α) D1, причем вариант D1 SIRP-α содержит аминокислотную последовательность
где X1 представляет собой E или G; X2 представляет собой L, I или V; X3 представляет собой V, L или I; X4 представляет собой S или F; X5 представляет собой L или S; X6 представляет собой S или T; X7 представляет собой A или V; X8 представляет собой I или T; X9 представляет собой H, R или L; X10 представляет собой A, V, I или L; X11 представляет собой I, T, S или F; X12 представляет собой A или G; X13 представляет собой E, V или L; X14 представляет собой K или R; X15 представляет собой E или Q; X16 представляет собой H, P или R; X17 представляет собой D или E; X18 представляет собой S, L, T или G; X19 представляет собой K или R; X20 представляет собой E или N; X21 представляет собой S или P; X22 представляет собой S или R; X23 представляет собой S или G; X24 представляет собой любую аминокислоту; X25 представляет собой любую аминокислоту; X26 представляет собой V или I; X27 представляет собой F, L или V; X28 представляет собой D или отсутствует; X29 представляет собой T или V; X30 представляет собой F или V, а X31 представляет собой A или G; и при этом вариант D1 SIRP-α имеет по меньшей мере две аминокислотных заменены по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность согласно любой из SEQ ID NO: 1-10; и вариант Fc, содержащий димер домена Fc, имеющий два мономера домена Fc, причем каждый мономер домена Fc независимо представляет собой (i) область Fc IgG1 человека, содержащую мутацию N297A; (ii) область Fc IgG1 человека, содержащую мутации L234A, L235A и G237A; (iii) область Fc IgG1 человека, содержащую мутации L234A, L235A, G237A и N297A; (iv) область Fc IgG2 человека, содержащую мутацию N297A; (v) область Fc IgG2 человека, содержащую мутации A330S и P331S; (vi) область Fc IgG2 человека, содержащую мутации A330S, P331S и N297A; (vii) область Fc IgG4 человека, содержащую мутации S228P, E233P, F234V, L235A и delG236; или (viii) область Fc IgG4 человека, содержащую мутации S228P, E233P, F234V, L235A, delG236 и N297A. В некоторых вариантах осуществления полипептид содержит вариант Fc, причем вариант Fc содержит димер домена Fc, имеющий два мономера домена Fc, при этом каждый мономер домена Fc независимо выбирают из (i) области Fc IgG1 человека, включающей в свой состав мутации L234A, L235A, G237A и N297A; (ii) области Fc IgG2 человека, включающей в свой состав мутации A330S, P331S и N297A; или (iii) области Fc IgG4 человека, содержащей мутации S228P, E233P, F234V, L235A, delG236 и N297A. В некоторых вариантах осуществления полипептид содержит вариант D1 сигнального регуляторного белка α (SIRP-α), причем вариант D1 SIRP-α представляет собой неприродный высокоаффинный домен D1 SIRP-α, при этом вариант D1 SIRP-α связывается с CD47 человека с аффинностью, которая по меньшей мере в 10 раз больше, чем аффинность природного домена D1; и мономер домена Fc, причем мономер домена Fc связан со вторым полипептидом, содержащим второй мономер домена Fc, с образованием домена Fc, при этом домен Fc имеет аблированную или уменьшенную эффекторную функцию. В некоторых вариантах осуществления неприродный высокоаффинный домен D1 SIRP-α содержит аминокислотную мутацию в остатке 80. В некоторых вариантах осуществления полипептид содержит вариант сигнального регуляторного белка α (SIRP-α) D1, причем вариант D1 SIRP-α связывает CD47 первого вида с KD менее 250 нМ; и при этом вариант D1 SIRP-α связывает CD47 второго вида с KD менее 250 нМ; и KD для CD47 первого вида и KD для CD47 второго вида отличается в 100 раз друг от друга; причем первый вид и второй вид выбраны из группы, состоящей из человека, грызуна и приматов, отличных от человека. В некоторых вариантах осуществления полипептид содержит (a) домен сигнального регуляторного белка α (SIRP-α) D1, который связывает CD47 человека с KD менее 250 нМ; и (б) мономер домена Fc, связанный с N-концом или С-концом домена D1 SIRP-α, причем полипептид не вызывает острой анемии у грызунов и приматов, отличных от человека. В некоторых вариантах осуществления заболевание или расстройство представляет собой рак, аутоиммунное заболевание или воспалительное заболевание. В некоторых вариантах осуществления заболевание или расстройство представляет собой рак, а рак выбирают из солидной опухоли, гематологического рака, острого миелоидного лейкоза, хронического лимфоцитарного лейкоза, хронического миелоидного лейкоза, острого лимфобластного лейкоза, неходжкинской лимфомы, лимфомы Ходжкина, множественной миеломы, рака мочевого пузыря, рака поджелудочной железы, рака шейки матки, рака эндометрия, рака легкого, рака бронхов, рака печени, рака яичника, рака толстой кишки и прямой кишки, рака желудка, рака желудочно-кишечного тракта, рака желчного пузыря, желудочно-кишечной стромальной опухоли, рака щитовидной железы, рака головы и шеи, рака ротоглотки, рака пищевода, меланомы, немеланомного рака кожи, карциномы из клеток Меркеля, вирус-индуцированного рака, нейробластомы, рака молочной железы, рака предстательной железы, рака почки, почечно-клеточного рака, рака почечной лоханки, лейкоза, лимфомы, саркомы, глиомы, опухоли головного мозга и карциномы. В некоторых вариантах осуществления заболевание или расстройство представляет собой аутоиммунное заболевание или воспалительное заболевание, а аутоиммунное заболевание или воспалительное заболевание выбирают из рассеянного склероза, ревматоидного артрита, спондилоартропатии, системной красной волчанки, опосредованного антителом воспалительного или аутоиммунного заболевания, заболевания трансплантат против хозяина, сепсиса, диабетов, псориаза, атеросклероза, синдрома Шегрена, прогрессирующего системного склероза, склеродермии, острого коронарного синдрома, ишемической реперфузии, болезни Крона, эндометриоза, гломерулонефрита, миастении гравис, идиопатического легочного фиброза, астмы, острого респираторного дистресс-синдрома (ARDS), васкулита и воспалительного аутоиммунного миозита. В некоторых вариантах осуществления вариант D1 SIRP-α имеет последовательность согласно любой из SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159. В некоторых вариантах осуществления способ дополнительно включает введение по меньшей мере одного дополнительного агента. В некоторых вариантах осуществления по меньшей мере один дополнительный агент представляет собой антитело, опухолеассоциированный антиген или терапевтическое средство, отличное от антитела. В некоторых вариантах осуществления вводят по меньшей мере два дополнительных агента. В некоторых вариантах осуществления по меньшей мере два дополнительных агента включают два антитела. В некоторых вариантах осуществления по меньшей мере два дополнительных агента включают антитело и опухолеассоциированный антиген. В некоторых вариантах осуществления по меньшей мере один дополнительный агент представляет собой антитело. В некоторых вариантах осуществления антитело представляет собой изотип антитела IgG1 человека. В некоторых вариантах осуществления антитело представляет собой изотип антитела IgG2 человека. В некоторых вариантах осуществления антитело представляет собой изотип антитела IgG4 человека. В некоторых вариантах осуществления антитело выбирают из антитела против HER2, антитела против CD20, антитела против CD19, антитела против CS1, антитела против CD38, антитела против EGFR, антитела против PD1, антитела против OX40, антитела против PD-1, антитела против PD-L1, антитела против CD274, антитела против CTLA-4, антитела против CD137, антитела против 4-1BB, антитела против B7-H3, антитела против FZD7, антитела против CD27, антитела против CCR4, антитела против CD38, антитела против CSF1R, антитела против CSF, антитела против CD30, антитела против BAFF, антитела против VEGF или антитела против VEGFR2. В некоторых вариантах осуществления антитело выбирают из антитела против HER2, антитела против CD20, антитела против CD19, антитела против CS1, антитела против CD38, антитела против PD-1, антитела против RANKL или антитела против PD-L1. В некоторых вариантах осуществления по меньшей мере один дополнительный агент представляет собой по меньшей мере одно антитело, а антитело выбирают из цетуксимаба, нецитумумаба, пембролизумаба, ниволумаба, пидилизумаба, MEDI0680, MED16469, атезолизумаба, авелумаба, дурвалумаба, MEDI6383, RG7888, ипилимумаба, тремелимумаба, урелумаба, PF-05082566, эноблитузумаба, вантиктумаба, варлилумаба, могамалимузумаба, SAR650984, даратумумаба, трастузумаба, трастузумаб емтансина, пертузумаба, элотузумаба, ритуксимаба, офатумумаба, обинутузумаба, RG7155, FPA008, панитумумаба, брентуксимаб ведотина, MSB0010718C, белимумаба, бевацизумаба, деносумаба, панитумумаба, рамуцирумаба, нецитумумаба, ниволумаба, пембролизумаба, авелумаба, атезолизумаба, дувалумаба, MEDI0680, пидилизумаба или BMS-93659. В некоторых вариантах осуществления антитело представляет собой трастузумаб. В некоторых вариантах осуществления вариант D1 SIRP-α имеет последовательность согласно любой из SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159. В некоторых вариантах осуществления антитело представляет собой ритуксимаб. В некоторых вариантах осуществления вариант D1 SIRP-α имеет последовательность согласно любой из SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159. В некоторых вариантах осуществления антитело представляет собой цетуксимаб. В некоторых вариантах осуществления вариант D1 SIRP-α имеет последовательность согласно любой из SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159. В некоторых вариантах осуществления антитело представляет собой даратумумаб. В некоторых вариантах осуществления вариант D1 SIRP-α имеет последовательность согласно любой из SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159. В некоторых вариантах осуществления антитело представляет собой белимумаб. В некоторых вариантах осуществления вариант D1 SIRP-α имеет последовательность согласно любой из SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159. В некоторых вариантах осуществления антитело представляет собой бевацизумаб. В некоторых вариантах осуществления вариант D1 SIRP-α имеет последовательность согласно любой из SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159. В некоторых вариантах осуществления антитело представляет собой деносумаб. В некоторых вариантах осуществления вариант D1 SIRP-α имеет последовательность согласно любой из SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159. В некоторых вариантах осуществления антитело представляет собой пантимумаб. В некоторых вариантах осуществления вариант D1 SIRP-α имеет последовательность согласно любой из SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159. В некоторых вариантах осуществления антитело представляет собой рамуцирумаб. В некоторых вариантах осуществления вариант D1 SIRP-α имеет последовательность согласно любой из SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159. В некоторых вариантах осуществления антитело представляет собой нецитумумаб. В некоторых вариантах осуществления вариант D1 SIRP-α имеет последовательность согласно любой из SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159. В некоторых вариантах осуществления антитело представляет собой ниволумаб. В некоторых вариантах осуществления вариант D1 SIRP-α имеет последовательность согласно любой из SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159. В некоторых вариантах осуществления антитело представляет собой пембролизумаб. В некоторых вариантах осуществления вариант D1 SIRP-α имеет последовательность согласно любой из SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159. В некоторых вариантах осуществления антитело представляет собой авелумаб. В некоторых вариантах осуществления вариант D1 SIRP-α имеет последовательность согласно любой из SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159. В некоторых вариантах осуществления антитело представляет собой атезолизумаб. В некоторых вариантах осуществления вариант D1 SIRP-α имеет последовательность согласно любой из SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159. В некоторых вариантах осуществления антитело представляет собой дурвалумаб. В некоторых вариантах осуществления вариант D1 SIRP-α имеет последовательность согласно любой из SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159. В некоторых вариантах осуществления антитело представляет собой MEDI0680. В некоторых вариантах осуществления вариант D1 SIRP-α имеет последовательность согласно любой из SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159. В некоторых вариантах осуществления антитело представляет собой пидилизумаб. В некоторых вариантах осуществления вариант D1 SIRP-α имеет последовательность согласно любой из SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159. В некоторых вариантах осуществления антитело представляет собой BMS-93659. В некоторых вариантах осуществления вариант D1 SIRP-α имеет последовательность согласно любой из SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159. В некоторых вариантах осуществления по меньшей мере один дополнительный агент представляет собой опухолеассоциированный антиген, и опухолеассоциированный антиген вызывает иммунный ответ. В некоторых вариантах осуществления по меньшей мере один дополнительный агент представляет собой антитело, и антитело нацелено на комплекс HLA/пептид или ГКГС/пептид. В некоторых вариантах осуществления антитело нацелено на комплекс HLA/пептид или MHC/пептид, содержащий NY-ESO-1/LAGE1, SSX-2, семейство MAGE (MAGE-A3), gp100/pmel17, Melan-A/MART-1, gp75/TRP1, тирозиназу, TRP2, CEA, PSA, TAG-72, незрелый рецептор ламинина, MOK/RAGE-1, WT-1, Her2/neu, EphA3, SAP-1, BING-4, Ep-CAM, MUC1, PRAME, сурвивин, Mesothelin, BRCA1/2 (мутированный), CDK4, CML66, MART-2, p53 (мутированный), Ras (мутированный), β-катенин (мутированный), TGF-βRII (мутированный), HPV E6 или E7. В некоторых вариантах осуществления антитело представляет собой ESK1, RL1B, Pr20 или 3.2G1.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
В дальнейшем будут описаны новейшие детали изобретения, с подробностями в присоединенной формуле изобретения. Для лучшего понимания элементов и преимуществ настоящего изобретения рекомендуется изучить следующее подробное описание, в котором представлены иллюстративные варианты осуществления изобретения, использующие принципы изобретения, а также сопроводительные чертежи, на которых:
На фиг. 1 представлена иллюстрация конструкции SIRP-α, содержащей домен D1 SIRP-α или его вариант, присоединенный к первому мономеру Fc-домена, который образует домен Fc со вторым мономером домена Fc;
На фиг. 2 представлена иллюстрация конструкции SIRP-α, содержащей домен D1 SIRP-α или его вариант, присоединенный к первому мономеру домена Fc и вариабельный домен антитела, присоединенный ко второму мономеру домена Fc, причем первый мономер домена Fc и второй мономер домена Fc объединены с образованием домена Fc;
На фиг. 3 представлена иллюстрация конструкции SIRP-α, содержащей домен D1 SIRP-α или его вариант, присоединенный к первому мономеру домена Fc и терапевтический белок, присоединенный ко второму мономеру домена Fc, причем первый мономер домена Fc и второй мономер домена Fc объединены с образованием домена Fc;
На фиг. 4A представлена иллюстрация конструкции SIRP-α, содержащей домен D1 SIRP-α или его вариант, присоединенный к первому мономеру Fc-домена, имеющему мутацию выступа, который образует домен Fc со вторым мономером домена Fc, имеющим мутацию впадины; На фиг. 4B представлена иллюстрация конструкции SIRP-α, содержащей домен D1 SIRP-α или его вариант, присоединенный к первому мономеру Fc-домена, имеющему мутацию впадины, который образует домен Fc со вторым мономером домена Fc, имеющим мутацию выступа;
На фиг. 5A представлена иллюстрация конструкции SIRP-α, содержащей домен D1 SIRP-α или его вариант, присоединенный к мономеру домена Fc; На фиг. 5B представлена иллюстрация конструкции SIRP-α, которая представляет собой гомодимер конструкции, проиллюстрированной на фиг. 5A;
На фиг. 6 представлены данные связывания SPR для монофункциональных и бифункциональных конструкций SIRP-α, содержащих домен D1 SIRP-α;
На фиг. 7 представлен фагоцитоз опухолевых клеток DLD-1-GFP-люцифераза с помощью макрофагов, полученных из моноцитов человека, в присутствии различных концентраций полипептидных конструкций SIRP-α;
На фиг. 8 представлен фагоцитоз опухолевых клеток DLD-1-GFP-люцифераза с помощью макрофагов, полученных из моноцитов человека, в присутствии различных концентраций полипептидных конструкций SIRP-α;
На фиг. 9 представлен фагоцитоз опухолевых клеток DLD-1-GFP-люцифераза с помощью макрофагов, полученных из моноцитов человека, в присутствии различных концентраций полипептидных конструкций SIRP-α;
На фиг. 10 представлена стабильность периода полувыведения полипептидов SIRP-α в течение определенного периода времени;
На фиг. 11 представлены данные анализа гемагглютинации для конструкций полипептида SIRP-α;
На фиг. 12 представлены кривые выживания сингенных опухолевых моделей мыши, леченых конструкциями полипептида SIRP-α и анти-mPD-L1;
На фиг. 13 представлен анализ объема опухоли сингенных опухолевых моделей мыши, леченых полипептидными конструкциями SIRP-α, в комбинации с анти-mPD-L1;
На фиг. 14 представлено связывание различных концентраций комплемента C1q со слитыми SIRP-α-Fc;
На фиг. 15 представлен фагоцитоз клеток MM1R с помощью макрофагов, полученных из моноцитов человека, в присутствии различных концентраций полипептидных конструкций SIRP-α;
На фиг. 16 представлен фагоцитоз клеток MM1R с помощью макрофагов, полученных из моноцитов человека, в присутствии различных концентраций полипептидных конструкций SIRP-α;
На фиг. 17 представлен фагоцитоз клеток N87 с помощью макрофагов, полученных из моноцитов человека, в присутствии различных концентраций полипептидных конструкций SIRP-α;
На фиг. 18 представлен анализ молекулярной массы варианта D1 SIRP-α, имеющего мутацию P83V;
На фиг.19A представлен рост опухоли клеток лимфомы GFP-Luc-Raji человека в модели рака мыши NOD ТКИД гамма (NSG), леченой различными конструкциями SIRP-α с ритуксимабом или без него; На фиг. 19B представлена скатерограмма объема опухоли для опухолей, описанных на фиг. 19A; На фиг. 19C показаны значения гемоглобина леченых мышей, представленных на фиг. 19A; и
На фиг. 20 представлено количество эритроцитов, взятых у мышей, леченых либо конструкцией SIRP-α Fc IgG1 дикого типа, либо конструкцией варианта SIRP-α Fc IgG1.
ПОДРОБНОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Определения
Термин "около" или "приблизительно" означает пределы приемлемого диапазона ошибок для конкретного значения, определяемые обычным специалистом в данной области техники, что будет частично зависеть от того, как измеряется или определяется значение, то есть, ограничения измерительной системы. Например, термин "около" может означать значения в пределах 1 или более чем 1 стандартного отклонения, на практике в данной области техники. В альтернативном варианте, "около" может означать диапазон до 20%, до 10%, до 5% или до 1% от заданного значения. В альтернативном варианте, особенно в отношении биологических систем или процессов, этот термин может означать пределы порядка величины, предпочтительно в 5 раз и более предпочтительно в 2 раза от значения. В случаях, когда конкретные значения описаны в заявке и в формуле изобретения, если не указано иное, следует предполагать что термин "около" означает пределы приемлемого диапазона ошибок для конкретного значения.
Терминология, используемая в рамках изобретения, предназначена для описания определенных случаев и не предназначена для ограничения. В данном описании формы единственного числа включают также формы множественного числа, если контекст явно не указывает иного. Далее, в тех случаях, когда термины ʺвключающийʺ, ʺвключаетʺ, ʺимеющийʺ, ʺимеетʺ, ʺсʺ или их варианты использованы либо в подробном описании, либо в Формуле изобретения, такие термины предназначены для охвата тех же значений, что и термин ʺсодержитʺ.
Используемый в рамках изобретения термин «антитело» относится к интактным антителам; фрагментам антител, при условии, что они проявляют желаемую биологическую активность (например, связывание эпитопов); моноклональным антителам; поликлональным антителам; моноспецифическим антителам; мультиспецифическим антителам (например, биспецифическим антителам) и антителоподобным белкам.
Используемый в рамках изобретения термин «вариабельный домен антитела» относится к частям легкой и тяжелой цепей антитела, которые содержат аминокислотные последовательности областей, определяющих комплементарность (CDR, например, CDR L1, CDR L2, CDR L3, CDR H1, CDR H2 и CDR H3) и каркасных областей (FR).
Используемый в рамках изобретения термин «линкер» относится к соединению между двумя элементами, например, белковыми доменами. В некоторых вариантах осуществления линкер может представлять собой ковалентную связь или спейсер. Термин «спейсер» относится к фрагменту (например, полимеру полиэтиленгликоля (ПЭГ)) или аминокислотной последовательности (например, последовательности 1-200 аминокислот), встречающейся между двумя полипептидами или полипептидными доменами, который обеспечивает пространство или гибкость (или как пространство, так и гибкость) между двумя полипептидами или полипептидными доменами. В некоторых вариантах осуществления аминокислотный спейсер является частью первичной последовательности полипептида (например, соединен с разделенными полипептидами или полипептидными доменами через полипептидный скелет).
Используемый в рамках изобретения термин «терапевтически эффективное количество» относится к количеству полипептида или фармацевтической композиции, содержащей описанный в рамках изобретения полипептид, например, полипептид, имеющий домен D1 SIRP-α или его вариант, который является достаточным и эффективным для достижения желательного терапевтического эффекта при лечении пациента, имеющего заболевание, такое как рак, например, солидную опухоль или гематологический рак. В некоторых вариантах осуществления терапевтически эффективное количество полипептида будет предотвращать неблагоприятные побочные эффекты.
Используемый в рамках изобретения термин «фармацевтическая композиция» относится к лекарственной или фармацевтической композиции, которая содержит активный ингредиент, а также эксципиенты или разбавители (или как эксципиенты, так и разбавители), и позволяет вводить активный ингредиент подходящими способами введения. В некоторых вариантах осуществления раскрытые в рамках изобретения фармацевтические композиции содержат фармацевтически приемлемые компоненты, которые совместимы с полипептидом. В некоторых вариантах осуществления фармацевтическая композиция находится в форме таблетки или капсулы для перорального введения или в водной форме для внутривенного или подкожного введения, например, путем инъекции.
Используемые в рамках изобретения термины «индивидуум» и «пациент», используются взаимозаменяемо для обозначения позвоночного, например млекопитающего. Млекопитающие включают, но не ограничиваются ими, мышей, обезьян, людей, сельскохозяйственных животных, спортивных животных и домашних животных. Также охватываются ткани, клетки и их потомство биологического объекта, полученные in vivo или культивируемые in vitro. Ни один из этих терминов не предусматривает надзор медицинского работника.
Используемый в рамках изобретения термин «сродство» или «аффинность связывания» относится к прочности связывающего взаимодействия между двумя молекулами. Как правило, аффинность связывания относится к силе суммарного количества нековалентных взаимодействий между молекулой и ее связывающим партнером, таким как высокоаффинный вариант D1 SIRP-α и CD47. Если не указано иное, «аффинность связывания» относится к присущей молекуле аффинности связывания, отражающей взаимодействие между членами пары связывающихся компонентов при их соотношении 1:1. Аффинность связывания между двумя молекулами обычно описывается константой диссоциации (KD) или константой ассоциации (KA). Две молекулы с низкой аффинностью связывания друг с другом обычно связываются медленно, склонны легко диссоциировать и обладают большой KD. Две молекулы, которые имеют высокую аффинность друг к другу, обычно легко связываются, имеют тенденцию оставаться связанными дольше и проявляют небольшую KD. В некоторых вариантах осуществления KD двух взаимодействующих молекул определяют с использованием известных способов и методик, например, поверхностного плазмонного резонанса (SPR). KD может быть рассчитана как отношение koff/kon.
Используемый в рамках изобретения термин «KD менее чем» относится к значительно меньшему значению KD и увеличению аффинности связывания относительно указанного значения KD. Используемый в рамках изобретения термин «KD более чем» относится к значительно большему значению KD и уменьшающейся аффинности связывания относительно указанного значения KD.
Используемый в рамках изобретения термин «острая анемия» относится к уменьшению массы эритроцитов или гемоглобина на 30% в течение первых пяти суток после введения соединения или лечения.
I. Домен D1 сигнального регуляторного белка α (SIRP-α) и его варианты
В рамках изобретения, в некоторых вариантах осуществления, раскрыты полипептиды, содержащие: вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92.
В рамках изобретения, в некоторых вариантах осуществления также раскрыты полипептиды, содержащие вариант Fc, причем вариант Fc содержит димер домена Fc, имеющий два мономера домена Fc, при этом каждый мономер домена Fc независимо выбирают из (i) области Fc IgG1 человека, включающей в свой состав мутации L234A, L235A, G237A и N297A; (ii) области Fc IgG2 человека, включающей в свой состав мутации A330S, P331S и N297A; или (iii) области Fc IgG4 человека, содержащей мутации S228P, E233P, F234V, L235A, delG236 и N297A.
Сигнальный регуляторный белок α («SIRP-α» или «SIRP-альфа») представляет собой трансмембранный гликопротеин, принадлежащий к надсемейству Ig, который широко экспрессируется на мембране миелоидных клеток. SIRP-α взаимодействует с CD47 - белком, широко экспрессируемым во многих типах клеток в организме. Взаимодействие SIRP-α с CD47 предотвращает лизис «собственных» клеток, которые в противном случае могут быть распознаны иммунной системой. Было замечено, что высокий уровень экспрессии CD47 на опухолевых клетках может возникать при остром миелоидном лейкозе и некоторых солидных опухолях в качестве отрицательного прогностического фактора для выживания.
Нативный SIRP-α содержит 3 очень гомологичных иммуноглобулино (Ig)-подобных внеклеточных домена - D1, D2 и D3. Домен D1 SIRP-α («домен D1») относится к мембранному дистальному, внеклеточному домену SIRP-α и опосредует связывание SIRP-α с CD47. Используемый в рамках изобретения термин «полипептид SIRP-α» относится к любому полипептиду SIRP-α или его фрагменту, который способен связываться с CD47. Существует по меньшей мере десять вариантов SIRP-α человека дикого типа. В таблице 1 представлены аминокислотные последовательности доменов D1 из десяти встречающихся в природе вариантов домена D1 SIRP-α человека дикого типа (SEQ ID NO: 1-10). В некоторых вариантах осуществления полипептид SIRP-α содержит домен D1 SIRP-α. В некоторых вариантах осуществления полипептид SIRP-α содержит домен D1 дикого типа, такой как предоставленный в SEQ ID NO: 1-10. В некоторых вариантах осуществления полипептид SIRP-α содержит домен D2 или D3 (или оба домена D2 и D3) (Таблица 3) SIRP-α человека дикого типа.
Таблица 1. Последовательности доменов D1 SIRP-α дикого типа
Используемый в рамках изобретения термин «высокоаффинный вариант D1 SIRP-α», «высокоаффинный вариант SIRP-α» или «вариант D1 SIRP-α» относится к полипептиду, содержащему домен D1 SIRP-α или CD47-связывающую часть полипептида SIRP-α, который имеет более высокое сродство к CD47, чем SIRP-α дикого типа. Высокоаффинный вариант D1 SIRP-α содержит по меньшей мере одну замену, делецию или введение аминокислоты (или их комбинацию) по сравнению с SIRP-α дикого типа.
В некоторых вариантах осуществления высокоаффинные варианты D1 SIRP-α раскрытые в рамках изобретения, содержат домен D1 SIRP-α или его вариант. В некоторых вариантах осуществления высокоаффинный вариант D1 SIRP-α содержит одну или более аминокислотных замен, инсерций, вставок или делеций по сравнению с доменом D1 дикого типа, представленным в SEQ ID NO: 1-10. В таблице 2 приведены примеры аминокислотных замен в каждом варианте D1 SIRP-α (SEQ ID NO: 13-22). В некоторых вариантах осуществления полипептид домена D1 SIRP-α или высокоаффинный вариант D1 SIRP-α содержит фрагмент домена D1. В некоторых вариантах осуществления фрагмент полипептида SIRP-α или высокоаффинный вариант фрагмента SIRP-α содержит аминокислотную последовательность менее чем 10 аминокислот в длину, около 10 аминокислот в длину, около 20 аминокислот в длину, около 30 аминокислот в длину, около 40 аминокислот в длину, около 50 аминокислот в длину, около 60 аминокислот в длину, около 70 аминокислот в длину, около 80 аминокислот в длину, около 90 аминокислот в длину, около 100 аминокислот в длину или более чем около 100 аминокислот в длину. В некоторых вариантах осуществления фрагменты домена D1 SIRP-α сохраняют способность связываться с CD47.
В некоторых вариантах осуществления полипептид по раскрытию, содержащий высокоаффинный вариант D1 SIRP-α, связывается с более высокой аффинностью связывания с CD47, чем с доменом D1 SIRP-α дикого типа. В некоторых вариантах осуществления высокоаффинный вариант D1 SIRP-α связывается с CD47 человека с по меньшей мере 1-кратной (например, по меньшей мере 1,5-кратной, 2-кратной, 2,5-кратной, 3-кратной, 3,5-кратной, 4-кратной, 5-кратной или более чем 5-кратной) аффинностью по сравнению с аффинностью естественного домена D1. В некоторых вариантах осуществления высокоаффинный вариант D1 SIRP-α связывается с CD47 человека с по меньшей мере 1-кратной (например, по меньшей мере 10-кратной, 100-кратной, 1000-кратной или более чем 1000-кратной) аффинностью по сравнению с аффинностью естественного домена D1.
Используемый в рамках изобретения термин «оптимизированная аффинность» или «оптимизированная аффинность связывания» относится к оптимизированной прочности связывающего взаимодействия между описанным в рамках изобретения полипептидом, включая высокоаффинный вариант D1 SIRP-α и CD47. Например, в некоторых вариантах осуществления полипептид связывается в основном или с более высокой аффинностью с CD47 на раковых клетках и по существу не связывается или связывается с более низкой аффинностью с CD47 на не раковых клетках. В некоторых вариантах осуществления аффинность связывания между полипептидом и CD47 оптимизирована таким образом, что взаимодействие не вызывает клинически значимой токсичности или снижает токсичность по сравнению с вариантом, который связывается с максимальной аффинностью. В некоторых вариантах осуществления для достижения оптимизированной аффинности связывания между полипептидом, представленным в рамках изобретения и CD47, полипептид, содержащий высокоаффинный вариант D1 SIRP-α, развивается так, чтобы иметь более низкое связывание с CD47, чем максимально достижимое. В некоторых вариантах осуществления высокоаффинные варианты SIRP-α, раскрытые в рамках изобретения, перекрестно реагируют с CD47 грызуна, примата, отличного от человека (NHP) и человека.
Используемый в рамках изобретения термин «иммуногенность» относится к свойству белка (например, терапевтического белка), который вызывает иммунный ответ у хозяина, как если бы он был чужеродным антигеном. Иммуногенность белка может быть проанализирована in vitro различными способами, например, с помощью анализов пролиферации Т-клеток in vitro.
Используемый в рамках изобретения термин «минимальная иммуногенность» относится к иммуногенности белка (например, терапевтического белка), который был модифицирован, например, путем замены аминокислот, чтобы быть ниже (например, по меньшей мере на 10%, 25%, 50% или 100% ниже), чем иммуногенность перед введением аминокислотных замен (например, немодифицированный белок). В некоторых вариантах осуществления белок (например, терапевтический белок) модифицирован, чтобы иметь минимальную иммуногенность и не вызывает или вызывать очень маленький иммунный ответ хозяина, даже если он является чужеродным антигеном.
В некоторых вариантах осуществления высокоаффинный вариант D1 SIRP-α имеет минимальную иммуногенность. В некоторых вариантах осуществления полипептид SIRP-α согласно описанию, вводимый пациенту, имеет такую же аминокислотную последовательность, что и полипептид SIRP-α в биологическом образце пациента, за исключением аминокислотных изменений, которые увеличивают аффинность варианта D1 SIRP-α. В некоторых вариантах осуществления варианты полипептида, описанные в рамках изобретения, снижают риск побочных эффектов по сравнению с антителами против CD47 или SIRP-α дикого типа. В некоторых вариантах осуществления варианты полипептида, описанные в рамках изобретения, снижают риск анемии по сравнению с антителами против CD47 или SIRP-α дикого типа. В некоторых вариантах осуществления варианты полипептида, описанные в рамках изобретения, не вызывают острой анемии при исследованиях грызунов или приматов, отличных от человека (NHP).
В таблице 2 перечислены конкретные аминокислотные замены в высокоаффинном варианте D1 SIRP-α по сравнению с каждой последовательностью домена D1. В некоторых вариантах осуществления высокоаффинный вариант D1 SIRP-α содержит один или более (например, два, три, четыре, пять, шесть, семь, восемь, девять, десять, одиннадцать, двенадцать, тринадцать, четырнадцать или более) замен, перечисленных в таблице 2. В некоторых вариантах осуществления высокоаффинный вариант D1 SIRP-α содержит не более 14 аминокислотных замен по сравнению с доменом D1 дикого типа. В некоторых вариантах осуществления высокоаффинный вариант D1 SIRP-α содержит не более 10 аминокислотных замен по сравнению с доменом D1 дикого типа. В некоторых вариантах осуществления высокоаффинный вариант D1 SIRP-α содержит не более 7 аминокислотных замен по сравнению с доменом D1 дикого типа. В некоторых вариантах осуществления высокоаффинный вариант D1 SIRP-α согласно раскрытию имеет по меньшей мере 90% (например, по меньшей мере 92%, 95%, 97% или более 97%) идентичности аминокислотной последовательности с последовательностью домена D1 дикого типа.
В некоторых вариантах осуществления высокоаффинный вариант D1 SIRP-α представляет собой химерный высокоаффинный вариант D1 SIRP-α, который содержит часть двух или более доменов D1 дикого типа или их вариантов (например, часть одного домена D1 дикого типа или его вариант, и часть другого домена D1 дикого типа или его вариант). В некоторых вариантах осуществления вариант химерного высокоаффинного D1 SIRP-α содержит по меньшей мере две части (например, три, четыре, пять или более частей) доменов D1 дикого типа или их варианты, причем каждая из частей относится к разным диким типам домена D1. В некоторых вариантах осуществления химерный высокоаффинный вариант D1 SIRP-α дополнительно содержит одну или более аминокислотных замен, перечисленных в таблице 2.
Таблица 2. Аминокислотные замены в высокоаффинном варианте D1 SIRP-α
В некоторых вариантах осуществления полипептид содержит вариант D1 SIRP-α, имеющий последовательность:

где X1 представляет собой L, I или V; X2 представляет собой V, L или, I; X3 представляет собой A или V; X4 представляет собой A, I или L; X5 представляет собой I, T, S или F; X6 представляет собой E, V или L; X7 представляет собой K или R; X8 представляет собой E или Q; X9 представляет собой H, P или R; X10 представляет собой L, T или G; X11 представляет собой K или R; X12 представляет собой V или I; X13 представляет собой F, L или V; а X14 представляет собой F или V; и при этом вариант имеет по меньшей мере одну аминокислотную замену по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность SEQ ID NO: 1.
В некоторых вариантах осуществления полипептид содержит вариант D1 SIRP-α, имеющий последовательность:

где X1 представляет собой L, I или V; X2 представляет собой V, L или, I; X3 представляет собой A или V; X4 представляет собой A, I или L; X5 представляет собой I, T, S или F; X6 представляет собой E, V или L; X7 представляет собой K или R; X8 представляет собой E или Q; X9 представляет собой H, P или R; X10 представляет собой L, T или G; X11 представляет собой K или R; X12 представляет собой V или I; X13 представляет собой F, L или V; а X14 представляет собой F или V; и при этом вариант имеет по меньшей мере одну аминокислотную замену по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность SEQ ID NO: 4.
В некоторых вариантах осуществления полипептид содержит вариант D1 SIRP-α, имеющий последовательность:

где X1 представляет собой L, I или V; X2 представляет собой V, L или, I; X3 представляет собой A или V; X4 представляет собой A, I или L; X5 представляет собой I, T, S или F; X6 представляет собой E, V или L; X7 представляет собой K или R; X8 представляет собой E или Q; X9 представляет собой H, P или R; X10 представляет собой L, T или G; X11 представляет собой K или R; X12 представляет собой V или I; X13 представляет собой F, L или V; а X14 представляет собой F или V; и при этом вариант имеет по меньшей мере одну аминокислотную замену по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность SEQ ID NO: 5.
В некоторых вариантах осуществления полипептид содержит вариант D1 SIRP-α, имеющий последовательность:

где X1 представляет собой L, I или V; X2 представляет собой V, L или, I; X3 представляет собой A или V; X4 представляет собой A, I или L; X5 представляет собой I, T, S или F; X6 представляет собой E, V или L; X7 представляет собой K или R; X8 представляет собой E или Q; X9 представляет собой H, P или R; X10 представляет собой L, T или G; X11 представляет собой K или R; X12 представляет собой V или I; X13 представляет собой F, L или V; а X14 представляет собой F или V; и при этом вариант имеет по меньшей мере одну аминокислотную замену по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность SEQ ID NO: 6.
В некоторых вариантах осуществления полипептид содержит вариант D1 SIRP-α, имеющий последовательность:
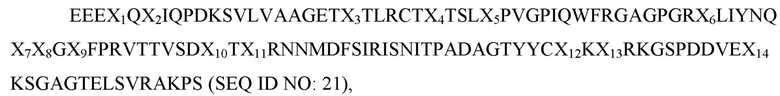
где X1 представляет собой L, I или V; X2 представляет собой V, L или, I; X3 представляет собой A или V; X4 представляет собой A, I или L; X5 представляет собой I, T, S или F; X6 представляет собой E, V или L; X7 представляет собой K или R; X8 представляет собой E или Q; X9 представляет собой H, P или R; X10 представляет собой L, T или G; X11 представляет собой K или R; X12 представляет собой V или I; X13 представляет собой F, L или V; а X14 представляет собой F или V; и при этом вариант имеет по меньшей мере одну аминокислотную замену по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность SEQ ID NO: 9.
В любом из вышеупомянутых вариантов осуществления полипептид содержит высокоаффинный вариант D1 SIRP-α, имеющий любую последовательность из SEQ ID NO: 13, 16-18 и 21, где X1 представляет собой L, I или V. В любом из вышеупомянутых вариантов осуществления X2 представляет собой V, L или I. В любом из вышеупомянутых вариантов осуществления X3 представляет собой A или V. В любом из вышеупомянутых вариантов осуществления X4 представляет собой A, I или L. В любом из вышеупомянутых вариантов осуществления X5 представляет собой I, T, S или F. В любом из вышеупомянутых вариантов осуществления X6 представляет собой E, V или L. В любом из вышеупомянутых вариантов осуществления X7 представляет собой K или R. В любом из вышеупомянутых вариантов осуществления X8 представляет собой E или Q. В любом из вышеупомянутых вариантов осуществления X9 представляет собой H, P или R. В любом из вышеупомянутых вариантов осуществления X10 представляет собой L, T или G. В любом из вышеупомянутых вариантов осуществления X11 представляет собой K или R. В любом из вышеупомянутых вариантов осуществления X12 представляет собой V или I. В любом из вышеупомянутых вариантов осуществления X13 представляет собой F, L, V. В любом из вышеупомянутых вариантов осуществления X14 представляет собой F или V. В некоторых вариантах осуществления полипептид по данному аспекту раскрытия содержит не более шести аминокислотных замен по сравнению с доменом D1 SIRP-α дикого типа, имеющим любую из последовательностей SEQ ID NO: 1, 4-6 и 9.
В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 10 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 1, 4-6 и 9. В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 100 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 1, 4-6 и 9. В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 1000 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 1, 4-6 и 9. В некоторых вариантах осуществления вариант D1 полипептида SIRP-α или его фрагмент связывается с CD47 с KD менее, чем 1×10-8 M, менее, чем 5×10-9 M, менее, чем 1×10-9 M, менее 5×10-10 M, менее, чем 1×10-10 M или менее, чем 1×10-11 M. В некоторых вариантах осуществления вариант D1 полипептида SIRP-α или его фрагмент связывается с CD47 с KD между около 500 нМ и 100 нМ, между около 100 нМ и 50 нМ, между около 50 нМ и 10 нМ, между около 10 нМ и 5 нМ, между около 5 нМ и 1 нМ, между около 1 нМ и 500 пМ, между около 500 пМ и 100 пМ, между около 100 пМ и 50 пМ или между около 50 пМ и 10 пМ.
В некоторых вариантах осуществления полипептид содержит вариант D1 SIRP-α, имеющий последовательность:

где X1 представляет собой L, I или V; X2 представляет собой V, L или, I; X3 представляет собой A или V; X4 представляет собой V, I или L; X5 представляет собой I, T, S или F; X6 представляет собой E, V или L; X7 представляет собой K или R; X8 представляет собой E или Q; X9 представляет собой H, P или R; X10 представляет собой S, T или G; X11 представляет собой K или R; X12 представляет собой V или I; X13 представляет собой F, L или V; а X14 представляет собой F или V; и при этом вариант имеет по меньшей мере одну аминокислотную замену по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность SEQ ID NO: 2.
В некоторых вариантах осуществления полипептид содержит вариант D1 SIRP-α, имеющий последовательность:

где X1 представляет собой L, I или V; X2 представляет собой V, L или, I; X3 представляет собой A или V; X4 представляет собой V, I или L; X5 представляет собой I, T, S или F; X6 представляет собой E, V или L; X7 представляет собой K или R; X8 представляет собой E или Q; X9 представляет собой H, P или R; X10 представляет собой S, T или G; X11 представляет собой K или R; X12 представляет собой V или I; X13 представляет собой F, L или V; а X14 представляет собой F или V; и при этом вариант имеет по меньшей мере одну аминокислотную замену по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность SEQ ID NO: 3.
В некоторых вариантах осуществления полипептид содержит вариант D1 SIRP-α, имеющий последовательность:

где X1 представляет собой L, I или V; X2 представляет собой V, L или, I; X3 представляет собой A или V; X4 представляет собой V, I или L; X5 представляет собой I, T, S или F; X6 представляет собой E, V или L; X7 представляет собой K или R; X8 представляет собой E или Q; X9 представляет собой H, P или R; X10 представляет собой S, T или G; X11 представляет собой K или R; X12 представляет собой V или I; X13 представляет собой F, L или V; а X14 представляет собой F или V; и при этом вариант имеет по меньшей мере одну аминокислотную замену по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность SEQ ID NO: 7.
В некоторых вариантах осуществления полипептид содержит вариант D1 SIRP-α, имеющий последовательность:

где X1 представляет собой L, I или V; X2 представляет собой V, L или, I; X3 представляет собой A или V; X4 представляет собой V, I или L; X5 представляет собой I, T, S или F; X6 представляет собой E, V или L; X7 представляет собой K или R; X8 представляет собой E или Q; X9 представляет собой H, P или R; X10 представляет собой S, T или G; X11 представляет собой K или R; X12 представляет собой V или I; X13 представляет собой F, L или V; а X14 представляет собой F или V; и при этом вариант имеет по меньшей мере одну аминокислотную замену по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность SEQ ID NO: 10.
В любом из вышеупомянутых вариантов осуществления в данном аспекте изобретения полипептид имеет последовательность любой из SEQ ID NO: 14, 15, 19 и 22, где X1 представляет собой L, I или V. В любом из вышеупомянутых вариантов осуществления X2 представляет собой V, L или I. В любом из вышеупомянутых вариантов осуществления X3 представляет собой A или V. В любом из вышеупомянутых вариантов осуществления X4 представляет собой V, I или L. В любом из вышеупомянутых вариантов осуществления X5 представляет собой I, T, S или F. В любом из вышеупомянутых вариантов осуществления X6 представляет собой E, V или L. В любом из вышеупомянутых вариантов осуществления X7 представляет собой K или R. В любом из вышеупомянутых вариантов осуществления X8 представляет собой E или Q. В любом из вышеупомянутых вариантов осуществления X9 представляет собой H, P или R. В любом из вышеупомянутых вариантов осуществления X10 представляет собой S, T или G. В любом из вышеупомянутых вариантов осуществления X11 представляет собой K или R. В любом из вышеупомянутых вариантов осуществления X12 представляет собой V или I. В любом из вышеупомянутых вариантов осуществления X13 представляет собой F, L или V. В любом из вышеупомянутых вариантов осуществления X14 представляет собой F или V. В некоторых вариантах осуществления полипептид по данному аспекту раскрытия содержит не более шести аминокислотных замен по сравнению с доменом D1 SIRP-α дикого типа, имеющим любую из последовательностей SEQ ID NO: 2, 3, 7 и 10
В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 10 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 2, 3, 7 и 10 В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 100 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 2, 3, 7 и 10 В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 1000 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 2, 3, 7 и 10 В некоторых вариантах осуществления вариант D1 полипептида SIRP-α или его фрагмент связывается с CD47 с KD менее чем 1×10-8 M, менее чем 5×10-9 M, менее чем 1×10-9 M, менее чем 5×10-10 M, менее чем 1×10-10 M или менее чем 1×10-11 M. В некоторых вариантах осуществления вариант D1 полипептида SIRP-α или его фрагмент связывается с CD47 с KD между около 500 нМ и 100 нМ, между около 100 нМ и 50 нМ, между около 50 нМ и 10 нМ, между около 10 нМ и 5 нМ, между около 5 нМ и 1 нМ, между около 1 нМ и 500 пМ, между около 500 пМ и 100 пМ, между около 100 пМ и 50 пМ или между около 50 пМ и 10 пМ.
В некоторых вариантах осуществления полипептид содержит вариант D1 SIRP-α, имеющий последовательность:
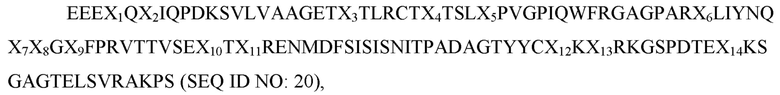
где X1 представляет собой L, I или V; X2 представляет собой V, L или, I; X3 представляет собой A или V; X4 представляет собой A, I или L; X5 представляет собой I, T, S или F; X6 представляет собой E, V или L; X7 представляет собой K или R; X8 представляет собой E или Q; X9 представляет собой H, P или R; X10 представляет собой S, T или G; X11 представляет собой K или R; X12 представляет собой V или I; X13 представляет собой F, L или V; а X14 представляет собой F или V; и при этом вариант имеет по меньшей мере одну аминокислотную замену по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность SEQ ID NO: 8.
В некоторых вариантах осуществления полипептид имеет последовательность SEQ ID NO: 20, где X1 представляет собой L, I или V. В любом из вышеупомянутых вариантов осуществления в данном аспекте раскрытия, X2 представляет собой V, L или I. В любом из вышеупомянутых вариантов осуществления X3 представляет собой A или V. В любом из вышеупомянутых вариантов осуществления X4 представляет собой A, I или L. В любом из вышеупомянутых вариантов осуществления X5 представляет собой I, T, S или F. В любом из вышеупомянутых вариантов осуществления X6 представляет собой E, V или L. В любом из вышеупомянутых вариантов осуществления X7 представляет собой K или R. В любом из вышеупомянутых вариантов осуществления X8 представляет собой E или Q. В любом из вышеупомянутых вариантов осуществления X9 представляет собой H, P или R. В любом из вышеупомянутых вариантов осуществления X10 представляет собой S, T или G. В любом из вышеупомянутых вариантов осуществления X11 представляет собой K или R. В любом из вышеупомянутых вариантов осуществления X12 представляет собой V или I. В любом из вышеупомянутых вариантов осуществления X13 представляет собой F, L или V. В любом из вышеупомянутых вариантов осуществления X14 представляет собой F или V. В некоторых вариантах осуществления полипептид по данному аспекту раскрытия содержит не более шести аминокислотных замен по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность SEQ ID NO: 8.
В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 10 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий последовательность SEQ ID NO: 8. В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 100 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий последовательность SEQ ID NO: 8. В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 1000 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий последовательность SEQ ID NO: 8. В некоторых вариантах осуществления вариант D1 полипептида SIRP-α или его фрагмент связывается с CD47 с KD менее чем 1×10-8 M, менее чем 5×10-9 M, менее чем 1×10-9 M, менее чем 5×10-10 M, менее чем 1×10-10 M или менее чем 1×10-11 M. В некоторых вариантах осуществления вариант D1 полипептида SIRP-α или его фрагмент связывается с CD47 с KD между около 500 нМ и 100 нМ, между около 100 нМ и 50 нМ, между около 50 нМ и 10 нМ, между около 10 нМ и 5 нМ, между около 5 нМ и 1 нМ, между около 1 нМ и 500 пМ, между около 500 пМ и 100 пМ, между около 100 пМ и 50 пМ или между около 50 пМ и 10 пМ.
В некоторых вариантах осуществления полипептид содержит вариант D1 SIRP-α, имеющий последовательность:
где X1 представляет собой E или G; X2 представляет собой L, I или V; X3 представляет собой V, L или, I; X4 представляет собой S или F; X5 представляет собой L или S; X6 представляет собой S или T; X7 представляет собой A или V; X8 представляет собой I или T; X9 представляет собой H или R; X10 представляет собой A, V, I или L; X11 представляет собой I, T, S или F; X12 представляет собой A или G; X13 представляет собой E, V или L; X14 представляет собой K или R; X15 представляет собой E или Q; X16 представляет собой H, P или R; X17 представляет собой D или E; X18 представляет собой S, L, T или G; X19 представляет собой K или R; X20 представляет собой E или D; X21 представляет собой S или P; X22 представляет собой S или R; X23 представляет собой S или G; X24 представляет собой V или I; X25 представляет собой F, L, V; X26 представляет собой D или отсутствует; X27 представляет собой T или V; X28 представляет собой F или V, а X29 представляет собой A или G; и при этом вариант имеет по меньшей мере одну аминокислотную замену по сравнению с доменом D1 SIRP-α дикого типа, имеющим любую из последовательностей SEQ ID NO: 1-10.
В любом из вышеупомянутых вариантов осуществления в данном аспекте раскрытия, X2 представляет собой L, I или V. В любом из вышеупомянутых вариантов осуществления X3 представляет собой V, L или I. В любом из вышеупомянутых вариантов осуществления X4 представляет собой S или F. В любом из вышеупомянутых вариантов осуществления X5 представляет собой L или S. В любом из вышеупомянутых вариантов осуществления X6 представляет собой S или T. В любом из вышеупомянутых вариантов осуществления X7 представляет собой A или V. В любом из вышеупомянутых вариантов осуществления X8 представляет собой I или T. В любом из вышеупомянутых вариантов осуществления X9 представляет собой H или R. В любом из вышеупомянутых вариантов осуществления X10 представляет собой A, V, I или L. В любом из вышеупомянутых вариантов осуществления X11 представляет собой I, T, S или F. В любом из вышеупомянутых вариантов осуществления X12 представляет собой A или G. В любом из вышеупомянутых вариантов осуществления X13 представляет собой E, V или L. В любом из вышеупомянутых вариантов осуществления X14 представляет собой K или R. В любом из вышеупомянутых вариантов осуществления X15 представляет собой E или Q. В любом из вышеупомянутых вариантов осуществления X16 представляет собой H, P или R. В любом из вышеупомянутых вариантов осуществления X17 представляет собой D или E. В любом из вышеупомянутых вариантов осуществления X18 представляет собой S, L, T или G. В любом из вышеупомянутых вариантов осуществления X19 представляет собой K или R. В любом из вышеупомянутых вариантов осуществления X20 представляет собой E или D. В любом из вышеупомянутых вариантов осуществления X21 представляет собой S или P. В любом из вышеупомянутых вариантов осуществления X22 представляет собой S или R. В любом из вышеупомянутых вариантов осуществления X23 представляет собой S или G. В любом из вышеупомянутых вариантов осуществления X24 представляет собой V или I. В любом из вышеупомянутых вариантов осуществления X25 представляет собой F, L, V. В любом из вышеупомянутых вариантов осуществления X26 представляет собой D или отсутствует. В любом из вышеупомянутых вариантов осуществления X27 представляет собой T или V. В любом из вышеупомянутых вариантов осуществления X28 представляет собой F или V. В любом из вышеупомянутых вариантов осуществления X29 представляет собой A или G. В некоторых вариантах осуществления полипептид по данному аспекту раскрытия содержит не более шести аминокислотных замен по сравнению с доменом D1 SIRP-α дикого типа, имеющим любую из последовательностей SEQ ID NO: 1-10.
В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 10 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 1-10. В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 100 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 1-10. В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 1000 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 1-10. В некоторых вариантах осуществления вариант D1 полипептида SIRP-α или его фрагмент связывается с CD47 с KD менее чем 1×10-8 M, менее чем 5×10-9 M, менее чем 1×10-9 M, менее чем 5×10-10 M, менее чем 1×10-10 M или менее чем 1×10-11 M. В некоторых вариантах осуществления вариант D1 полипептида SIRP-α или его фрагмент связывается с CD47 с KD между около 500 нМ и 100 нМ, между около 100 нМ и 50 нМ, между около 50 нМ и 10 нМ, между около 10 нМ и 5 нМ, между около 5 нМ и 1 нМ, между около 1 нМ и 500 пМ, между около 500 пМ и 100 пМ, между около 100 пМ и 50 пМ или между около 50 пМ и 10 пМ.
В некоторых вариантах осуществления полипептид раскрытия, содержащий высокоаффинный вариант D1 SIRP-α, дополнительно содержит домен D2, имеющий последовательность SEQ ID NO: 24, домен D3, имеющей последовательность SEQ ID NO: 25 или домен D2, имеющей последовательность SEQ ID NO: 24 и домен D3, имеющей последовательность SEQ ID NO: 25 SIRP-α человека дикого типа, как представлено в таблице 3. В некоторых вариантах осуществления высокоаффинный вариант D1 SIRP-α дополнительно содержит фрагмент или вариант домена D2, или фрагмент или вариант домена D3. В некоторых вариантах осуществления высокоаффинный вариант D1 SIRP-α дополнительно содержит фрагмент или вариант домена D2, и фрагмент или вариант домена D3. В некоторых вариантах осуществления высокоаффинный вариант D1 SIRP-α соединяется с доменом D2 или D3 посредством линкера. В некоторых вариантах осуществления высокоаффинный вариант D1 SIRP-α соединяется с доменом D2 и D3 посредством линкера.
Таблица 3. Аминокислотные последовательности доменов D2 и D3 SIRP-α
В некоторых вариантах осуществления полипептид раскрытия, содержащий высокоаффинный вариант D1 SIRP-α, присоединенный к мономеру домена Fc, альбумина сыворотки человека (ЧСА) или его варианта, сыворотко-связывающего белка или пептида или органической молекулы, например полимера (например, полимера ПЭГ), для улучшения фармакокинетических свойств полипептида, например увеличения периода полувыведения в сыворотке. В некоторых вариантах осуществления высокоаффинный вариант D1 SIRP-α присоединен к мономеру домена Fc, который не способен димеризоваться. В некоторых вариантах осуществления мономеры домена Fc, белки ЧСА, сывороточные связывающие белки или пептиды и органические молекулы, такие как PEG, служат для увеличения периода полувыведения в сыворотке описанных в рамках изобретения полипептидов. В некоторых вариантах осуществления полипептид раскрытия, содержащий высокоаффинный вариант D1 SIRP-α, не включает ни одну из последовательностей SEQ ID NO: 26-36 показанных в таблице 4.
Таблица 4.
В некоторых вариантах осуществления полипептиды и полипептидные конструкции, описанные в рамках изобретения, используют in vitro для анализа связывания, такого как иммунные анализы. Например, в некоторых вариантах осуществления описанные в рамках изобретения полипептиды и полипептидные конструкции используются в жидкой фазе или связаны с твердофазным носителем. В некоторых вариантах осуществления полипептиды, используемые для иммунологических анализов, детектируемо метят различными способами.
В некоторых вариантах осуществления полипептиды и полипептидные конструкции, описанные в рамках изобретения, связаны с различными носителями и используются для обнаружения присутствия специфических экспрессирующих антиген клеток. Примеры носителей включают стекло, полистирол, полипропилен, полиэтилен, декстран, найлон, амилазы, приро.дные и модифицированные целлюлозы, полиакриламиды, агарозы и магнетит. Характер носителя может быть либо растворимым, либо нерастворимым.
Известны различные метки и способы мечения. Примеры меток включают ферменты, радиоизотопы, флуоресцентные соединения, коллоидные металлы, хемилюминесцентные соединения и биолюминесцентные соединения. Доступны различные методики связывания меток с полипептидами, раскрытыми в рамках изобретения.
В некоторых вариантах осуществления полипептиды соединяют с низкомолекулярными гаптенами. Эти гаптены затем специфически обнаруживаются с помощью второй реакции. Например, в некоторых вариантах осуществления биотиновый гаптен используется с авидином или гаптенами динитрофенолом, пиридоксалом или флуоресцеином, обнаруживаются специфическими антителами против гаптена (например, антителами против динитрофенола, антипиридоксальными антителами и анти-флуоресцеиновыми антителами соответственно).
II. Высокоаффинные домены D1 SIRP-α с измененным гликозилированием
В рамках изобретения в некоторых вариантах осуществления раскрыты полипептиды, содержащие: вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92.
В рамках изобретения в некоторых вариантах осуществления также раскрыты полипептиды, содержащие вариант Fc, причем вариант Fc содержит димер домена Fc, имеющий два мономера домена Fc, при этом каждый мономер домена Fc независимо выбирают из (i) области Fc IgG1 человека, включающей в свой состав мутации L234A, L235A, G237A и N297A; (ii) области Fc IgG2 человека, включающей в свой состав мутации A330S, P331S и N297A; или (iii) области Fc IgG4 человека, содержащей мутации S228P, E233P, F234V, L235A, delG236 и N297A.
В некоторых вариантах осуществления полипептид в описанной в рамках изобретения композиции содержит высокоаффинный вариант D1 SIRP-α, который имеет уменьшенное или минимальное гликозилирование. Домен D1 каждого из десяти белков SIRP-α дикого типа человека (SEQ ID NO: 1-10 в таблице 1) содержит единственный потенциальный N-связанный сайт гликозилирования в аминокислоте N80 в последовательности N80ITP. Экспрессия домена D1 SIRP-α в клетках яичника китайского хомячка (CHO) приводит к появлению основной полосы 16 кДа (негликозилированной) и малой полосы с более высокой молекулярной массой, которая была удалена Endo Hf. Endo Hf представляет собой рекомбинантный слитый белок эндогликозидазы H и мальтозосвязывающего белка. Endo Hf расщепляет в пределах ядра хитобиозы высокоманнозные и некоторые гибридные олигосахариды из N-связанных гликопротеинов. Это означает, что пролин в аминокислотном положении 83 может снизить эффективность гликозилирования, что станет следствием белка с разной степенью гликозилирования и, следовательно, гетерогенности. Для разработки лекарственных средств гетерогенность может вызвать проблемы в процессе разработки. Поэтому для исследования возможности получения гомогенных негликозилированных форм высокоаффинных вариантов D1 SIPR-α в некоторых вариантах осуществления аминокислота N80 варианта D1 SIPR-α является мутированной на Ala. В некоторых вариантах осуществления для получения негликозилированного высокоаффинного варианта D1 SIRP-α аминокислоту N80 в высокоаффинном варианте D1 SIRP-α заменяют любой аминокислотой, включая любую естественную и неприродную аминокислоту, например, N80A и N80Q. В некоторых вариантах осуществления высокоаффинный вариант D1 SIRP-α содержит мутацию N80A и по меньшей мере 1 дополнительную мутацию (например, по меньшей мере 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных мутаций или более). В некоторых вариантах осуществления дополнительная мутация находится в сайте связывания CD47. В некоторых вариантах осуществления дополнительная мутация находится в гидрофобном ядре домена D1.
В некоторых вариантах осуществления полипептид в раскрытой в рамках изобретения композиции включает высокоаффинный вариант D1 SIRP-α, который имеет повышенное гликозилирование относительно домена D1 SIRP-α дикого типа. Другим вариантом повышения гомогенности конечного продукта является повышение эффективности гликозилирования в аминокислоте N80 и получение высокоаффинных вариантов D1 SIRP-α с повышенным гликозилированием относительно дикого типа. В некоторых вариантах осуществления аминокислота Р83 в последовательности NITP83 влияет на степень гликозилирования в аминокислоте N80. В некоторых вариантах осуществления изменение P83 на любую аминокислоту повышает эффективность гликозилирования в N80. В некоторых вариантах осуществления аминокислота P83 в высокоаффинном варианте D1 SIRP-α заменена любой аминокислотой, включая природные и неприродные аминокислоты, например, P83V, P83A, P83I и P83L. В некоторых вариантах осуществления полипептид по изобретению экспрессируется в клетке, которая оптимизирована не для гликозилированных белков, которые экспрессируются такой клеткой, например, путем генной инженерии клеточной линии (например, генетически модифицированных клеток-хозяев дрожжей или млекопитающих) или модификаций условия культивирования клеток, такие как добавление кифунзина или с использованием естественно негликозилирующего хозяина, такого как прокариот (E. coli и т.п.).
В таблице 5 перечислены конкретные аминокислотные замены в высокоаффинном варианте D1 SIRP-α по сравнению с каждой последовательностью варианта домена D1. В некоторых вариантах осуществления высокоаффинный вариант D1 SIRP-α содержит один или более (например, два, три, четыре, пять, шесть, семь, восемь, девять, десять, одиннадцать, двенадцать, тринадцать, четырнадцать или более) замен, перечисленных в таблице 5. В некоторых вариантах осуществления варианты D1 SIRP-α не являются гликозилированными или являются минимально гликозилированными. В некоторых вариантах осуществления варианты D1 SIRP-α являются полностью гликозилированными или почти полностью гликозилированными. В некоторых вариантах осуществления высокоаффинный вариант D1 SIRP-α содержит не более 14 аминокислотных замен по сравнению с доменом D1 дикого типа. В некоторых вариантах осуществления высокоаффинный вариант D1 SIRP-α содержит не более 10 аминокислотных замен по сравнению с доменом D1 дикого типа. В некоторых вариантах осуществления высокоаффинный вариант D1 SIRP-α содержит не более 7 аминокислотных замен по сравнению с доменом D1 дикого типа. В некоторых вариантах осуществления высокоаффинный вариант D1 SIRP-α согласно раскрытию имеет по меньшей мере 90% (например, по меньшей мере 92%, 95%, 97% или более 97%) идентичности аминокислотной последовательности с последовательностью домена D1 дикого типа.
В некоторых вариантах осуществления высокоаффинный вариант D1 SIRP-α представляет собой химерный высокоаффинный вариант D1 SIRP-α, который содержит часть двух или более доменов D1 дикого типа или их вариантов (например, часть одного домена D1 дикого типа или его вариант, и часть другого домена D1 дикого типа или его вариант). В некоторых вариантах осуществления вариант химерного высокоаффинного D1 SIRP-α содержит по меньшей мере две части (например, три, четыре, пять или более частей) доменов D1 дикого типа или их варианты, причем каждая из частей относится к разным диким типам домена D1. В некоторых вариантах осуществления химерный высокоаффинный вариант D1 SIRP-α дополнительно содержит одну или более аминокислотных замен, перечисленных в таблице 5.
Таблица 5. Аминокислотные замены в высокоаффинном варианте D1 SIRP-α
В некоторых вариантах осуществления полипептид содержит вариант D1 SIRP-α, имеющий последовательность:
где X1 представляет собой L, I или V; X2 представляет собой V, L или, I; X3 представляет собой A или V; X4 представляет собой A, I или L; X5 представляет собой I, T, S или F; X6 представляет собой E, V или L; X7 представляет собой K или R; X8 представляет собой E или Q; X9 представляет собой H, P или R; X10 представляет собой L, T или G; X11 представляет собой K или R; X12 представляет собой N, A, C, D, E, F, G, H, I, K, L, M, P, Q, R, S, T, V, W или Y; X13 представляет собой P, A, C, D, E, F, G, H, I, K, L, M, N, Q, R, S, T, V, W или Y; X14 представляет собой V или I; X15 представляет собой F, L или V, а X16 представляет собой F или V; и при этом вариант имеет по меньшей мере одну аминокислотную замену по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность SEQ ID NO: 1.
В некоторых вариантах осуществления полипептид содержит вариант D1 SIRP-α, имеющий последовательность:

где X1 представляет собой L, I или V; X2 представляет собой V, L или, I; X3 представляет собой A или V; X4 представляет собой A, I или L; X5 представляет собой I, T, S или F; X6 представляет собой E, V или L; X7 представляет собой K или R; X8 представляет собой E или Q; X9 представляет собой H, P или R; X10 представляет собой L, T или G; X11 представляет собой K или R; X12 представляет собой N, A, C, D, E, F, G, H, I, K, L, M, P, Q, R, S, T, V, W или Y; X13 представляет собой P, A, C, D, E, F, G, H, I, K, L, M, N, Q, R, S, T, V, W или Y; X14 представляет собой V или I; X15 представляет собой F, L или V, а X16 представляет собой F или V; и при этом вариант имеет по меньшей мере одну аминокислотную замену по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность SEQ ID NO: 4.
В некоторых вариантах осуществления полипептид содержит вариант D1 SIRP-α, имеющий последовательность:
где X1 представляет собой L, I или V; X2 представляет собой V, L или, I; X3 представляет собой A или V; X4 представляет собой A, I или L; X5 представляет собой I, T, S или F; X6 представляет собой E, V или L; X7 представляет собой K или R; X8 представляет собой E или Q; X9 представляет собой H, P или R; X10 представляет собой L, T или G; X11 представляет собой K или R; X12 представляет собой N, A, C, D, E, F, G, H, I, K, L, M, P, Q, R, S, T, V, W или Y; X13 представляет собой P, A, C, D, E, F, G, H, I, K, L, M, N, Q, R, S, T, V, W или Y; X14 представляет собой V или I; X15 представляет собой F, L или V, а X16 представляет собой F или V; и при этом вариант имеет по меньшей мере одну аминокислотную замену по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность SEQ ID NO: 5.
В некоторых вариантах осуществления полипептид содержит вариант D1 SIRP-α, имеющий последовательность:
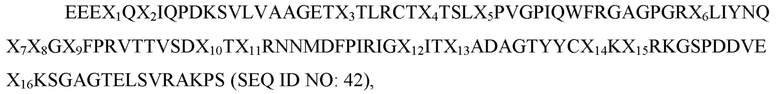
где X1 представляет собой L, I или V; X2 представляет собой V, L или, I; X3 представляет собой A или V; X4 представляет собой A, I или L; X5 представляет собой I, T, S или F; X6 представляет собой E, V или L; X7 представляет собой K или R; X8 представляет собой E или Q; X9 представляет собой H, P или R; X10 представляет собой L, T или G; X11 представляет собой K или R; X12 представляет собой N, A, C, D, E, F, G, H, I, K, L, M, P, Q, R, S, T, V, W или Y; X13 представляет собой P, A, C, D, E, F, G, H, I, K, L, M, N, Q, R, S, T, V, W или Y; X14 представляет собой V или I; X15 представляет собой F, L или V, а X16 представляет собой F или V; и при этом вариант имеет по меньшей мере одну аминокислотную замену по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность SEQ ID NO: 6.
В некоторых вариантах осуществления полипептид содержит вариант D1 SIRP-α, имеющий последовательность:

где X1 представляет собой L, I или V; X2 представляет собой V, L или, I; X3 представляет собой A или V; X4 представляет собой A, I или L; X5 представляет собой I, T, S или F; X6 представляет собой E, V или L; X7 представляет собой K или R; X8 представляет собой E или Q; X9 представляет собой H, P или R; X10 представляет собой L, T или G; X11 представляет собой K или R; X12 представляет собой N, A, C, D, E, F, G, H, I, K, L, M, P, Q, R, S, T, V, W или Y; X13 представляет собой P, A, C, D, E, F, G, H, I, K, L, M, N, Q, R, S, T, V, W или Y; X14 представляет собой V или I; X15 представляет собой F, L или V, а X16 представляет собой F или V; и при этом вариант имеет по меньшей мере одну аминокислотную замену по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность SEQ ID NO: 9.
В любом из вышеупомянутых вариантов осуществления, в данном аспекте раскрытия полипептид содержит вариант D1 SIRP-α, имеющий любую последовательность из SEQ ID NO: 37, 40-42 и 45, где X1 представляет собой L, I или V. В любом из вышеупомянутых вариантов осуществления X2 представляет собой V, L или I. В любом из вышеупомянутых вариантов осуществления X3 представляет собой A или V. В любом из вышеупомянутых вариантов осуществления X4 представляет собой A, I или L. В любом из вышеупомянутых вариантов осуществления X5 представляет собой I, T, S или F. В любом из вышеупомянутых вариантов осуществления X6 представляет собой E, V или L. В любом из вышеупомянутых вариантов осуществления X7 представляет собой K или R. В любом из вышеупомянутых вариантов осуществления X8 представляет собой E или Q. В любом из вышеупомянутых вариантов осуществления X9 представляет собой H, P или R. В любом из вышеупомянутых вариантов осуществления X10 представляет собой L, T или G. В любом из вышеупомянутых вариантов осуществления X11 представляет собой K или R. В любом из вышеупомянутых вариантов осуществления X12 представляет собой N, A, C, D, E, F, G, H, I, K, L, M, P, Q, R, S, T, V, W или Y. В любом из вышеупомянутых вариантов осуществления X13 представляет собой P, A, C, D, E, F, G, H, I, K, L, M, N, Q, R, S, T, V, W или Y. В любом из вышеупомянутых вариантов осуществления X14 представляет собой V или I. В любом из вышеупомянутых вариантов осуществления X15 представляет собой F, L, V. В любом из вышеупомянутых вариантов осуществления X16 представляет собой F или V.
В некоторых вариантах осуществления полипептид, предложенный в рамках изобретения, содержит не более десяти аминокислотных замен по сравнению с доменом D1 SIRP-α дикого типа, имеющим любую из последовательностей SEQ ID NO: 1, 4-6 и 9. В некоторых вариантах осуществления полипептид, предложенный в рамках изобретения, содержит не более семи аминокислотных замен по сравнению с доменом D1 SIRP-α дикого типа, имеющим любую из последовательностей SEQ ID NO: 1, 4-6 и 9.
В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 10 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 1, 4-6 и 9. В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 100 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 1, 4-6 и 9. В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 1000 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 1, 4-6 и 9. В некоторых вариантах осуществления вариант D1 полипептида SIRP-α или его фрагмент связывается с CD47 с KD менее чем 1×10-8 M, менее чем 5×10-9 M, менее чем 1×10-9 M, менее чем 5×10-10 M, менее чем 1×10-10 M или менее чем 1×10-11 M. В некоторых вариантах осуществления вариант D1 полипептида SIRP-α или его фрагмент связывается с CD47 с KD между около 500 нМ и 100 нМ, между около 100 нМ и 50 нМ, между около 50 нМ и 10 нМ, между около 10 нМ и 5 нМ, между около 5 нМ и 1 нМ, между около 1 нМ и 500 пМ, между около 500 пМ и 100 пМ, между около 100 пМ и 50 пМ или между около 50 пМ и 10 пМ.
В некоторых вариантах осуществления полипептид содержит вариант D1 SIRP-α, имеющий последовательность:

где X1 представляет собой L, I или V; X2 представляет собой V, L или, I; X3 представляет собой A или V; X4 представляет собой V, I или L; X5 представляет собой I, T, S или F; X6 представляет собой E, V или L; X7 представляет собой K или R; X8 представляет собой E или Q; X9 представляет собой H, P или R; X10 представляет собой S, T или G; X11 представляет собой K или R; X12 представляет собой N, A, C, D, E, F, G, H, I, K, L, M, P, Q, R, S, T, V, W или Y; X13 представляет собой P, A, C, D, E, F, G, H, I, K, L, M, N, Q, R, S, T, V, W или Y; X14 представляет собой V или I; X15 представляет собой F, L или V, а X16 представляет собой F или V; и при этом вариант имеет по меньшей мере одну аминокислотную замену по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность SEQ ID NO: 2.
В некоторых вариантах осуществления полипептид содержит вариант D1 SIRP-α, имеющий последовательность:

где X1 представляет собой L, I или V; X2 представляет собой V, L или, I; X3 представляет собой A или V; X4 представляет собой V, I или L; X5 представляет собой I, T, S или F; X6 представляет собой E, V или L; X7 представляет собой K или R; X8 представляет собой E или Q; X9 представляет собой H, P или R; X10 представляет собой S, T или G; X11 представляет собой K или R; X12 представляет собой N, A, C, D, E, F, G, H, I, K, L, M, P, Q, R, S, T, V, W или Y; X13 представляет собой P, A, C, D, E, F, G, H, I, K, L, M, N, Q, R, S, T, V, W или Y; X14 представляет собой V или I; X15 представляет собой F, L или V, а X16 представляет собой F или V; и при этом вариант имеет по меньшей мере одну аминокислотную замену по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность SEQ ID NO: 3.
В некоторых вариантах осуществления полипептид содержит вариант D1 SIRP-α, имеющий последовательность:

где X1 представляет собой L, I или V; X2 представляет собой V, L или, I; X3 представляет собой A или V; X4 представляет собой V, I или L; X5 представляет собой I, T, S или F; X6 представляет собой E, V или L; X7 представляет собой K или R; X8 представляет собой E или Q; X9 представляет собой H, P или R; X10 представляет собой S, T или G; X11 представляет собой K или R; X12 представляет собой N, A, C, D, E, F, G, H, I, K, L, M, P, Q, R, S, T, V, W или Y; X13 представляет собой P, A, C, D, E, F, G, H, I, K, L, M, N, Q, R, S, T, V, W или Y; X14 представляет собой V или I; X15 представляет собой F, L или V, а X16 представляет собой F или V; и при этом вариант имеет по меньшей мере одну аминокислотную замену по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность SEQ ID NO: 7.
В некоторых вариантах осуществления полипептид содержит вариант D1 SIRP-α, имеющий последовательность:

где X1 представляет собой L, I или V; X2 представляет собой V, L или, I; X3 представляет собой A или V; X4 представляет собой V, I или L; X5 представляет собой I, T, S или F; X6 представляет собой E, V или L; X7 представляет собой K или R; X8 представляет собой E или Q; X9 представляет собой H, P или R; X10 представляет собой S, T или G; X11 представляет собой K или R; X12 представляет собой N, A, C, D, E, F, G, H, I, K, L, M, P, Q, R, S, T, V, W или Y; X13 представляет собой P, A, C, D, E, F, G, H, I, K, L, M, N, Q, R, S, T, V, W или Y; X14 представляет собой V или I; X15 представляет собой F, L или V, а X16 представляет собой F или V; и при этом вариант имеет по меньшей мере одну аминокислотную замену по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность SEQ ID NO: 10.
В любом из вышеупомянутых вариантов осуществления, в данном аспекте раскрытия полипептид содержит вариант D1 SIRP-α, имеющий любую последовательность из SEQ ID NO: 38, 39, 43 и 46, где X1 представляет собой L, I или V. В любом из вышеупомянутых вариантов осуществления X2 представляет собой V, L или I. В любом из вышеупомянутых вариантов осуществления X3 представляет собой A или V. В любом из вышеупомянутых вариантов осуществления X4 представляет собой V, I или L. В любом из вышеупомянутых вариантов осуществления X5 представляет собой I, T, S или F. В любом из вышеупомянутых вариантов осуществления X6 представляет собой E, V или L. В любом из вышеупомянутых вариантов осуществления X7 представляет собой K или R. В любом из вышеупомянутых вариантов осуществления X8 представляет собой E или Q. В любом из вышеупомянутых вариантов осуществления X9 представляет собой H, P или R. В любом из вышеупомянутых вариантов осуществления X10 представляет собой S, T или G. В любом из вышеупомянутых вариантов осуществления X11 представляет собой K или R. В любом из вышеупомянутых вариантов осуществления X12 представляет собой N, A, C, D, E, F, G, H, I, K, L, M, P, Q, R, S, T, V, W или Y. В любом из вышеупомянутых вариантов осуществления X13 представляет собой P, A, C, D, E, F, G, H, I, K, L, M, N, Q, R, S, T, V, W или Y. В любом из вышеупомянутых вариантов осуществления X14 представляет собой V или I. В любом из вышеупомянутых вариантов осуществления X15 представляет собой F, L или V. В любом из вышеупомянутых вариантов осуществления X16 представляет собой F или V.
В некоторых вариантах осуществления полипептид содержит вариант D1 SIRP-α, имеющий не более десяти аминокислотных замен по сравнению с доменом D1 SIRP-α дикого типа, имеющим любую из последовательностей SEQ ID NO: 2, 3, 7 и 10 В некоторых вариантах осуществления полипептид содержит вариант D1 SIRP-α, имеющий не более семи аминокислотных замен по сравнению с доменом D1 SIRP-α дикого типа, имеющим любую из последовательностей SEQ ID NO: 2, 3, 7 и 10
В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 10 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 2, 3, 7 и 10 В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 100 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 2, 3, 7 и 10 В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 1000 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 2, 3, 7 и 10 В некоторых вариантах осуществления вариант D1 полипептида SIRP-α или его фрагмент связывается с CD47 с KD менее чем 1×10-8 M, менее чем 5×10-9 M, менее чем 1×10-9 M, менее чем 5×10-10 M, менее чем 1×10-10 M или менее чем 1×10-11 M. В некоторых вариантах осуществления вариант D1 полипептида SIRP-α или его фрагмент связывается с CD47 с KD между около 500 нМ и 100 нМ, между около 100 нМ и 50 нМ, между около 50 нМ и 10 нМ, между около 10 нМ и 5 нМ, между около 5 нМ и 1 нМ, между около 1 нМ и 500 пМ, между около 500 пМ и 100 пМ, между около 100 пМ и 50 пМ или между около 50 пМ и 10 пМ.
В некоторых вариантах осуществления полипептид содержит вариант D1 SIRP-α, имеющий последовательность:
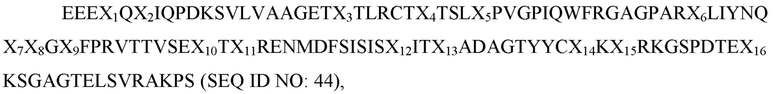
где X1 представляет собой L, I или V; X2 представляет собой V, L или, I; X3 представляет собой A или V; X4 представляет собой A, I или L; X5 представляет собой I, T, S или F; X6 представляет собой E, V или L; X7 представляет собой K или R; X8 представляет собой E или Q; X9 представляет собой H, P или R; X10 представляет собой S, T или G; X11 представляет собой K или R; X12 представляет собой N, A, C, D, E, F, G, H, I, K, L, M, P, Q, R, S, T, V, W или Y; X13 представляет собой P, A, C, D, E, F, G, H, I, K, L, M, N, Q, R, S, T, V, W или Y; X14 представляет собой V или I; X15 представляет собой F, L или V, а X16 представляет собой F или V; и при этом вариант имеет по меньшей мере одну аминокислотную замену по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность SEQ ID NO: 8.
В некоторых вариантах осуществления полипептид имеет последовательность SEQ ID NO: 44, где X1 представляет собой L, I или V. В любом из вышеупомянутых вариантов осуществления в данном аспекте раскрытия, X2 представляет собой V, L или I. В любом из вышеупомянутых вариантов осуществления X3 представляет собой A или V. В любом из вышеупомянутых вариантов осуществления X4 представляет собой A, I или L. В любом из вышеупомянутых вариантов осуществления X5 представляет собой I, T, S или F. В любом из вышеупомянутых вариантов осуществления X6 представляет собой E, V или L. В любом из вышеупомянутых вариантов осуществления X7 представляет собой K или R. В любом из вышеупомянутых вариантов осуществления X8 представляет собой E или Q. В любом из вышеупомянутых вариантов осуществления X9 представляет собой H, P или R. В любом из вышеупомянутых вариантов осуществления X10 представляет собой S, T или G. В любом из вышеупомянутых вариантов осуществления X11 представляет собой K или R. В любом из вышеупомянутых вариантов осуществления X12 представляет собой N, A, C, D, E, F, G, H, I, K, L, M, P, Q, R, S, T, V, W или Y. В любом из вышеупомянутых вариантов осуществления X13 представляет собой P, A, C, D, E, F, G, H, I, K, L, M, N, Q, R, S, T, V, W или Y. В любом из вышеупомянутых вариантов осуществления X14 представляет собой V или I. В любом из вышеупомянутых вариантов осуществления X15 представляет собой F, L или V. В любом из вышеупомянутых вариантов осуществления X16 представляет собой F или V.
В некоторых вариантах осуществления полипептид содержит вариант D1 SIRP-α, имеющий не более десяти аминокислотных замен по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность SEQ ID NO: 8. В некоторых вариантах осуществления полипептид содержит вариант D1 SIRP-α, имеющий не более семи аминокислотных замен по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность SEQ ID NO: 8.
В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 10 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий последовательность SEQ ID NO: 8. В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 100 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий последовательность SEQ ID NO: 8. В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 1000 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий последовательность SEQ ID NO: 8. В некоторых вариантах осуществления вариант D1 полипептида SIRP-α или его фрагмент связывается с CD47 с KD менее чем 1×10-8 M, менее чем 5×10-9 M, менее чем 1×10-9 M, менее чем 5×10-10 M, менее чем 1×10-10 M или менее чем 1×10-11 M. В некоторых вариантах осуществления вариант D1 полипептида SIRP-α или его фрагмент связывается с CD47 с KD между около 500 нМ и 100 нМ, между около 100 нМ и 50 нМ, между около 50 нМ и 10 нМ, между около 10 нМ и 5 нМ, между около 5 нМ и 1 нМ, между около 1 нМ и 500 пМ, между около 500 пМ и 100 пМ, между около 100 пМ и 50 пМ или между около 50 пМ и 10 пМ.
В другом аспекте в раскрытии предложен полипептид, содержащий вариант D1 SIRP-α, имеющий последовательность:
где X1 представляет собой E или G; X2 представляет собой L, I или V; X3 представляет собой V, L или I; X4 представляет собой S или F; X5 представляет собой L или S; X6 представляет собой S или T; X7 представляет собой A или V; X8 представляет собой I или T; X9 представляет собой H, R или L; X10 представляет собой A, V, I или L; X11 представляет собой I, T, S или F; X12 представляет собой A или G; X13 представляет собой E, V или L; X14 представляет собой K или R; X15 представляет собой E или Q; X16 представляет собой H, P или R; X17 представляет собой D или E; X18 представляет собой S, L, T или G; X19 представляет собой K или R; X20 представляет собой E или N; X21 представляет собой S или P; X22 представляет собой S или R; X23 представляет собой S или G; X24 представляет собой любую аминокислоту; X25 представляет собой любую аминокислоту; X26 представляет собой V или I; X27 представляет собой F, L, V; X28 представляет собой D или отсутствует; X29 представляет собой T или V; X30 представляет собой F или V, а X31 представляет собой A или G; и при этом вариант имеет по меньшей мере одну аминокислотную замену по сравнению с доменом D1 SIRP-α дикого типа, имеющим любую из последовательностей SEQ ID NO: 1-10.
В некоторых вариантах осуществления полипептид имеет последовательность SEQ ID NO: 47, где X1 представляет собой E или G. В любом из вышеупомянутых вариантов осуществления в данном аспекте раскрытия, X2 представляет собой L, I или V. В любом из вышеупомянутых вариантов осуществления X3 представляет собой V, L или I. В любом из вышеупомянутых вариантов осуществления X4 представляет собой S или F. В любом из вышеупомянутых вариантов осуществления X5 представляет собой L или S. В любом из вышеупомянутых вариантов осуществления X6 представляет собой S или T. В любом из вышеупомянутых вариантов осуществления X7 представляет собой A или V. В любом из вышеупомянутых вариантов осуществления X8 представляет собой I или T. В любом из вышеупомянутых вариантов осуществления X9 представляет собой H, R или L. В любом из вышеупомянутых вариантов осуществления X10 представляет собой A, V, I или L. В любом из вышеупомянутых вариантов осуществления X11 представляет собой I, T, S или F. В любом из вышеупомянутых вариантов осуществления X12 представляет собой A или G. В любом из вышеупомянутых вариантов осуществления X13 представляет собой E, V или L. В любом из вышеупомянутых вариантов осуществления X14 представляет собой K или R. В любом из вышеупомянутых вариантов осуществления X15 представляет собой E или Q. В любом из вышеупомянутых вариантов осуществления X16 представляет собой H, P или R. В любом из вышеупомянутых вариантов осуществления X17 представляет собой D или E. В любом из вышеупомянутых вариантов осуществления X18 представляет собой S, L, T или G. В любом из вышеупомянутых вариантов осуществления X19 представляет собой K или R. В любом из вышеупомянутых вариантов осуществления X20 представляет собой E или N. В любом из вышеупомянутых вариантов осуществления X21 представляет собой S или P. В любом из вышеупомянутых вариантов осуществления X22 представляет собой S или R. В любом из вышеупомянутых вариантов осуществления X23 представляет собой S или G. В любом из вышеупомянутых вариантов осуществления X24 представляет собой N, A, C, D, E, F, G, H, I, K, L, M, P, Q, R, S, T, V, W или Y. В любом из вышеупомянутых вариантов осуществления X25 представляет собой P, A, C, D, E, F, G, H, I, K, L, M, N, Q, R, S, T, V, W или Y. В любом из вышеупомянутых вариантов осуществления X26 представляет собой V или I. В любом из вышеупомянутых вариантов осуществления X27 представляет собой F, L, V. В любом из вышеупомянутых вариантов осуществления X28 представляет собой D или отсутствует. В любом из вышеупомянутых вариантов осуществления X29 представляет собой T или V. В любом из вышеупомянутых вариантов осуществления X30 представляет собой F или V. В любом из вышеупомянутых вариантов осуществления X31 представляет собой A или G.
В некоторых вариантах осуществления полипептид по данному аспекту раскрытия содержит не более десяти аминокислотных замен по сравнению с доменом D1 SIRP-α дикого типа, имеющим любую из последовательностей SEQ ID NO: 1-10. В некоторых вариантах осуществления полипептид по данному аспекту раскрытия содержит не более семи аминокислотных замен по сравнению с доменом D1 SIRP-α дикого типа, имеющим любую из последовательностей SEQ ID NO: 1-10.
В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 10 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 1-10. В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 100 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 1-10. В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 1000 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 1-10. В некоторых вариантах осуществления вариант D1 полипептида SIRP-α или его фрагмент связывается с CD47 с KD менее, чем 1×10-8 M, менее, чем 5×10-9 M, менее, чем 1×10-9 M, менее 5×10-10 M, менее, чем 1×10-10 M или менее, чем 1×10-11 M. В некоторых вариантах осуществления вариант D1 полипептида SIRP-α или его фрагмент связывается с CD47 с KD между около 500 нМ и 100 нМ, между около 100 нМ и 50 нМ, между около 50 нМ и 10 нМ, между около 10 нМ и 5 нМ, между около 5 нМ и 1 нМ, между около 1 нМ и 500 пМ, между около 500 пМ и 100 пМ, между около 100 пМ и 50 пМ или между около 50 пМ и 10 пМ.
В некоторых вариантах осуществления полипептид содержит вариант D1 SIRP-α, имеющий последовательность:

где X1 представляет собой V или I; X2 представляет собой L или S; X3 представляет собой T или S; X4 представляет собой T или I; X5 представляет собой R или H; X6 представляет собой A, V или I; X7 представляет собой I, R, Y, K или F; X8 представляет собой G или A; X9 представляет собой E или V; X10 представляет собой K или R; X11 представляет собой E, D или Q; X12 представляет собой H или P; X13 представляет собой D или E; X14 представляет собой S, L или T; X15 представляет собой N или E; X16 представляет собой R или S; X17 представляет собой G или S; X18 представляет собой N или A; X19 представляет собой V или I; X20 представляет собой S, I или M; X21 представляет собой P или отсутствует; X22 представляет собой D или P, а X23 представляет собой V или T, или его фрагмент.
В другом аспекте в раскрытии предложен полипептид, содержащий вариант D1 SIRP-α, имеющий последовательность:
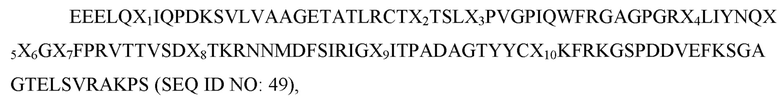
где X1 представляет собой V, L или I; X2 представляет собой A, I, V или L; X3 представляет собой I, F, S или T; X4 представляет собой E, V или L; X5 представляет собой K или R; X6 представляет собой E или Q; X7 представляет собой H, P или R; X8 представляет собой L, T, S или G; X9 представляет собой A, а X10 представляет собой V или I; и при этом вариант имеет по меньшей мере одну аминокислотную замену по сравнению с доменом D1 SIRP-α дикого типа, имеющим любую из последовательностей SEQ ID NO: 1.
В некоторых вариантах осуществления полипептид имеет последовательность SEQ ID NO: 49, где X1 представляет собой V, L или I. В любом из вышеупомянутых вариантов осуществления в данном аспекте раскрытия, X2 представляет собой A, I, V или L. В любом из вышеупомянутых вариантов осуществления X3 представляет собой I, F, S или T. В любом из вышеупомянутых вариантов осуществления X4 представляет собой E, V или L. В любом из вышеупомянутых вариантов осуществления X5 представляет собой K или R. В любом из вышеупомянутых вариантов осуществления X6 представляет собой E или Q. В любом из вышеупомянутых вариантов осуществления X7 представляет собой H, P или R. В любом из вышеупомянутых вариантов осуществления X8 представляет собой L, T, S или G. В любом из вышеупомянутых вариантов осуществления X9 представляет собой A. В любом из вышеупомянутых вариантов осуществления X10 представляет собой V или I.
В некоторых вариантах осуществления полипептид имеет высокоаффинный домен D1 SIRP-α, имеющий по меньшей мере 85% идентичности последовательности (например, по меньшей мере 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности) с SEQ ID NO: 49, где каждый из X1, X2, X3, X4, X5, X6, X7, X8, X9, и X10 не является аминокислотой дикого типа.
В некоторых вариантах осуществления полипептид по данному аспекту раскрытия содержит не более десяти аминокислотных замен по сравнению с доменом D1 SIRP-α дикого типа, имеющим любую из последовательностей SEQ ID NO: 1. В некоторых вариантах осуществления полипептид по данному аспекту раскрытия содержит не более семи аминокислотных замен по сравнению с доменом D1 SIRP-α дикого типа, имеющим любую из последовательностей SEQ ID NO: 1.
В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 10 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 1. В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 100 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 1. В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 1000 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 1. В некоторых вариантах осуществления вариант D1 полипептида SIRP-α или его фрагмент связывается с CD47 с KD менее чем 1×10-8 M, менее чем 5×10-9 M, менее чем 1×10-9 M, менее чем 5×10-10 M, менее чем 1×10-10 M или менее чем 1×10-11 M. В некоторых вариантах осуществления вариант D1 полипептида SIRP-α или его фрагмент связывается с CD47 с KD между около 500 нМ и 100 нМ, между около 100 нМ и 50 нМ, между около 50 нМ и 10 нМ, между около 10 нМ и 5 нМ, между около 5 нМ и 1 нМ, между около 1 нМ и 500 пМ, между около 500 пМ и 100 пМ, между около 100 пМ и 50 пМ или между около 50 пМ и 10 пМ.
В другом аспекте в раскрытии предложен полипептид, содержащий вариант D1 SIRP-α, имеющий последовательность:

где X1 представляет собой V или I; X2 представляет собой V или I; X3 представляет собой I или F; X4 представляет собой E или V; X5 представляет собой K или R; X6 представляет собой E или Q; X7 представляет собой H или P; X8 представляет собой S или T; X9 представляет собой N или A, а X10 представляет собой V или I; и при этом вариант имеет по меньшей мере одну аминокислотную замену по сравнению с доменом D1 SIRP-α дикого типа, имеющим любую из последовательностей SEQ ID NO: 2.
В некоторых вариантах осуществления полипептид имеет последовательность SEQ ID NO: 50, где X1 представляет собой V или I. В любом из вышеупомянутых вариантов осуществления в данном аспекте раскрытия, X2 представляет собой V или I. В любом из вышеупомянутых вариантов осуществления X3 представляет собой I или F. В любом из вышеупомянутых вариантов осуществления X4 представляет собой E или V. В любом из вышеупомянутых вариантов осуществления X5 представляет собой K или R. В любом из вышеупомянутых вариантов осуществления X6 представляет собой E или Q. В любом из вышеупомянутых вариантов осуществления X7 представляет собой H или P. В любом из вышеупомянутых вариантов осуществления X8 представляет собой S или R. В любом из вышеупомянутых вариантов осуществления X9 представляет собой N или A. В любом из вышеупомянутых вариантов осуществления X10 представляет собой V или I.
В некоторых вариантах осуществления полипептид имеет высокоаффинный домен D1 SIRP-α, имеющий по меньшей мере 85% идентичности последовательности (например, по меньшей мере 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности) с SEQ ID NO: 50, где каждый из X1, X2, X3, X4, X5, X6, X7, X8, X9, и X10 не является аминокислотой дикого типа.
В некоторых вариантах осуществления полипептид по данному аспекту раскрытия содержит не более десяти аминокислотных замен по сравнению с доменом D1 SIRP-α дикого типа, имеющим любую из последовательностей SEQ ID NO: 2. В некоторых вариантах осуществления полипептид по данному аспекту раскрытия содержит не более семи аминокислотных замен по сравнению с доменом D1 SIRP-α дикого типа, имеющим любую из последовательностей SEQ ID NO: 2.
В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 10 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 2. В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 100 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 2. В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 1000 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 2. В некоторых вариантах осуществления вариант полипептида D1 SIRP-α или его фрагмент связывается с CD47 с KD менее чем 1×10-8 M, менее чем 5×10-9 M, менее чем 1×10-9 M, менее чем 5×10-10 M, менее чем 1×10-10 M или менее чем 1×10-11 M. В некоторых вариантах осуществления вариант D1 полипептида SIRP-α или его фрагмент связывается с CD47 с KD между около 500 нМ и 100 нМ, между около 100 нМ и 50 нМ, между около 50 нМ и 10 нМ, между около 10 нМ и 5 нМ, между около 5 нМ и 1 нМ, между около 1 нМ и 500 пМ, между около 500 пМ и 100 пМ, между около 100 пМ и 50 пМ или между около 50 пМ и 10 пМ.
В другом аспекте в раскрытии предложен полипептид, содержащий вариант D1 SIRP-α, имеющий последовательность:

где X1 представляет собой V или I; X2 представляет собой A или I; X3 представляет собой I или F; X4 представляет собой E или V; X5 представляет собой K или R; X6 представляет собой H или P; X7 представляет собой L или T; X8 представляет собой аминокислоту, отличную от N, а X9 представляет собой V или I; и при этом вариант имеет по меньшей мере одну аминокислотную замену по сравнению с доменом D1 SIRP-α дикого типа, имеющим любую из последовательностей SEQ ID NO: 1.
В некоторых вариантах осуществления полипептид имеет последовательность SEQ ID NO: 51, где X1 представляет собой V или I. В любом из вышеупомянутых вариантов осуществления в данном аспекте раскрытия, X2 представляет собой A или I. В любом из вышеупомянутых вариантов осуществления X3 представляет собой I или F. В любом из вышеупомянутых вариантов осуществления X4 представляет собой E или V. В любом из вышеупомянутых вариантов осуществления X5 представляет собой K или R. В любом из вышеупомянутых вариантов осуществления X6 представляет собой H или P. В любом из вышеупомянутых вариантов осуществления X7 представляет собой L или T. В любом из вышеупомянутых вариантов осуществления X8 представляет собой N или A. В любом из вышеупомянутых вариантов осуществления X9 представляет собой V или I. В некоторых вариантах осуществления X4 представляет собой не V.
В некоторых вариантах осуществления полипептид имеет последовательность SEQ ID NO: 51, где X8 представляет собой A. В любом из вышеупомянутых вариантов осуществления в данном аспекте раскрытия, X8 представляет собой A, а X1 представляет собой V или I. В любом из вышеупомянутых вариантов осуществления в данном аспекте раскрытия, X8 представляет собой A, а X2 представляет собой A или I. В любом из вышеупомянутых вариантов осуществления X8 представляет собой A, а X3 представляет собой I или F. В любом из вышеупомянутых вариантов осуществления X8 представляет собой A, а X4 представляет собой E или V. В некоторых вариантах осуществления X4 представляет собой не V. В любом из вышеупомянутых вариантов осуществления X8 представляет собой A, а X5 представляет собой K или R. В любом из вышеупомянутых вариантов осуществления X8 представляет собой A, а X6 представляет собой H или P. В любом из вышеупомянутых вариантов осуществления X8 представляет собой A, а X7 представляет собой A или V. В любом из вышеупомянутых вариантов осуществления X8 представляет собой A, а X9 представляет собой V или I.
В некоторых вариантах осуществления полипептид имеет последовательность SEQ ID NO: 51, где X8 представляет собой A. В любом из вышеупомянутых вариантов осуществления в данном аспекте раскрытия, X8 представляет собой A, а X1 представляет собой I. В любом из вышеупомянутых вариантов осуществления в данном аспекте раскрытия, X8 представляет собой A, а X2 представляет собой I. В любом из вышеупомянутых вариантов осуществления X8 представляет собой A, а X3 представляет собой F. В любом из вышеупомянутых вариантов осуществления X8 представляет собой A, а X4 представляет собой V. В любом из вышеупомянутых вариантов осуществления X8 представляет собой A, а X5 представляет собой R. В любом из вышеупомянутых вариантов осуществления X8 представляет собой A, а X6 представляет собой P. В любом из вышеупомянутых вариантов осуществления X8 представляет собой A, а X7 представляет собой T. В любом из вышеупомянутых вариантов осуществления X8 представляет собой A, а X9 представляет собой I.
В некоторых вариантах осуществления полипептид имеет высокоаффинный домен D1 SIRP-α, имеющий по меньшей мере 85% идентичности последовательности (например, по меньшей мере 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности) с SEQ ID NO: 51, где каждый из X1, X2, X3, X4, X5, X6, X7, X8, и X9 не является аминокислотой дикого типа.
В некоторых вариантах осуществления полипептид по данному аспекту раскрытия содержит не более десяти аминокислотных замен по сравнению с доменом D1 SIRP-α дикого типа, имеющим любую из последовательностей SEQ ID NO: 1. В некоторых вариантах осуществления полипептид по данному аспекту раскрытия содержит не более семи аминокислотных замен по сравнению с доменом D1 SIRP-α дикого типа, имеющим любую из последовательностей SEQ ID NO: 1.
В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 10 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 1. В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 100 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 1. В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 1000 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 1. В некоторых вариантах осуществления вариант полипептида D1 SIRP-α или его фрагмент связывается с CD47 с KD менее чем 1×10-8 M, менее чем 5×10-9 M, менее чем 1×10-9 M, менее чем 5×10-10 M, менее чем 1×10-10 M или менее чем 1×10-11 M. В некоторых вариантах осуществления вариант D1 полипептида SIRP-α или его фрагмент связывается с CD47 с KD между около 500 нМ и 100 нМ, между около 100 нМ и 50 нМ, между около 50 нМ и 10 нМ, между около 10 нМ и 5 нМ, между около 5 нМ и 1 нМ, между около 1 нМ и 500 пМ, между около 500 пМ и 100 пМ, между около 100 пМ и 50 пМ или между около 50 пМ и 10 пМ.
В другом аспекте в раскрытии предложен полипептид, содержащий вариант D1 SIRP-α, имеющий последовательность:
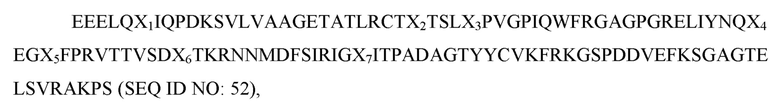
где X1 представляет собой V, L или I; X2 представляет собой A, I или L; X3 представляет собой I, T, S или F; X4 представляет собой K или R; X5 представляет собой H, P или R; X6 представляет собой L, T или G; X7 представляет собой N или A; и при этом вариант имеет по меньшей мере одну аминокислотную замену по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность согласно SEQ ID NO: 1.
В некоторых вариантах осуществления полипептид имеет последовательность SEQ ID NO: 52, где X1 представляет собой V, L или I. В любом из вышеупомянутых вариантов осуществления в данном аспекте раскрытия, X2 представляет собой A, I или L. В любом из вышеупомянутых вариантов осуществления X3 представляет собой I, T, S или F. В любом из вышеупомянутых вариантов осуществления X4 представляет собой K или R. В любом из вышеупомянутых вариантов осуществления X5 представляет собой H или P. В любом из вышеупомянутых вариантов осуществления X6 представляет собой L, T или G. В любом из вышеупомянутых вариантов осуществления X7 представляет собой N или A.
В некоторых вариантах осуществления полипептид имеет последовательность SEQ ID NO: 52, где X1 представляет собой V или I. В любом из вышеупомянутых вариантов осуществления в данном аспекте раскрытия, X2 представляет собой A или I. В любом из вышеупомянутых вариантов осуществления X3 представляет собой I или F. В любом из вышеупомянутых вариантов осуществления X4 представляет собой K или R. В любом из вышеупомянутых вариантов осуществления X5 представляет собой H или P. В любом из вышеупомянутых вариантов осуществления X6 представляет собой L или T. В любом из вышеупомянутых вариантов осуществления X7 представляет собой N или A.
В некоторых вариантах осуществления полипептид имеет последовательность SEQ ID NO: 52, где X7 представляет собой A. В любом из вышеупомянутых вариантов осуществления в данном аспекте раскрытия, X7 представляет собой A, а X1 представляет собой V или I. В любом из вышеупомянутых вариантов осуществления в данном аспекте раскрытия, X7 представляет собой A, а X2 представляет собой A или I. В любом из вышеупомянутых вариантов осуществления X7 представляет собой A, а X3 представляет собой I или F. В любом из вышеупомянутых вариантов осуществления X7 представляет собой A, а X4 представляет собой K или R. В любом из вышеупомянутых вариантов осуществления X7 представляет собой A, а X5 представляет собой H или P. В любом из вышеупомянутых вариантов осуществления X7 представляет собой A, а X6 представляет собой L или T.
В некоторых вариантах осуществления полипептид имеет последовательность SEQ ID NO: 52, где X7 представляет собой A. В любом из вышеупомянутых вариантов осуществления в данном аспекте раскрытия, X7 представляет собой A, а X1 представляет собой I. В любом из вышеупомянутых вариантов осуществления в данном аспекте раскрытия, X7 представляет собой A, а X2 представляет собой I. В любом из вышеупомянутых вариантов осуществления X7 представляет собой A, а X3 представляет собой F. В любом из вышеупомянутых вариантов осуществления X7 представляет собой A, а X4 представляет собой R. В любом из вышеупомянутых вариантов осуществления X7 представляет собой A, а X5 представляет собой P. В любом из вышеупомянутых вариантов осуществления X7 представляет собой A, а X6 представляет собой T.
В некоторых вариантах осуществления полипептид имеет высокоаффинный домен D1 SIRP-α, имеющий по меньшей мере 85% идентичности последовательности (например, по меньшей мере 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности) с SEQ ID NO: 52, где каждый из X1, X2, X3, X4, X5, X6, и X7 не является аминокислотой дикого типа.
В некоторых вариантах осуществления полипептид по данному аспекту раскрытия содержит не более десяти аминокислотных замен по сравнению с доменом D1 SIRP-α дикого типа, имеющим любую из последовательностей SEQ ID NO: 1. В некоторых вариантах осуществления полипептид по данному аспекту раскрытия содержит не более семи аминокислотных замен по сравнению с доменом D1 SIRP-α дикого типа, имеющим любую из последовательностей SEQ ID NO: 1.
В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 10 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 1. В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 100 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 1. В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 1000 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 1. В некоторых вариантах осуществления фрагменты включают полипептиды менее чем 10 аминокислот в длину, около 10 аминокислот в длину, около 20 аминокислот в длину, около 30 аминокислот в длину, около 40 аминокислот в длину, около 50 аминокислот в длину, около 60 аминокислот в длину, около 70 аминокислот в длину, около 80 аминокислот в длину, около 90 аминокислот в длину, около 100 аминокислот в длину или более чем около 100 аминокислот в длину. Фрагменты сохраняют возможность связываться с CD47. Предпочтительно варианты D1 полипептидов SIRP-α и их фрагменты связываются с CD47 с более высоким сродством, чем полипептид SIRP-α связывается с CD47. Например, в некоторых вариантах осуществления вариант D1 полипептида SIRP-α или его фрагмент связывается с CD47 с KD менее чем 1×10-8 M, менее чем 5×10-9 M, менее чем 1×10-9 M, менее чем 5×10-10 M, менее чем 1×10-10 M или менее чем 1×10-11 M. В некоторых вариантах осуществления вариант D1 полипептида SIRP-α или его фрагмент связывается с CD47 с KD между около 500 нМ и 100 нМ, между около 100 нМ и 50 нМ, между около 50 нМ и 10 нМ, между около 10 нМ и 5 нМ, между около 5 нМ и 1 нМ, между около 1 нМ и 500 пМ, между около 500 пМ и 100 пМ, между около 100 пМ и 50 пМ или между около 50 пМ и 10 пМ.
В другом аспекте в раскрытии предложен полипептид, содержащий вариант D1 SIRP-α, имеющий последовательность:

где X1 представляет собой V, L или I; X2 представляет собой V, I или L; X3 представляет собой I, T, S или F; X4 представляет собой K или R; X5 представляет собой H, P или R; X6 представляет собой S, T или G; X7 представляет собой A; и при этом вариант имеет по меньшей мере одну аминокислотную замену по сравнению с доменом D1 SIRP-α дикого типа, имеющим любую из последовательностей SEQ ID NO: 2.
В некоторых вариантах осуществления полипептид имеет последовательность SEQ ID NO: 212, где X1 представляет собой V, L или I. В любом из вышеупомянутых вариантов осуществления в данном аспекте раскрытия, X2 представляет собой V, I или L. В любом из вышеупомянутых вариантов осуществления X3 представляет собой I, T, S или F. В любом из вышеупомянутых вариантов осуществления X4 представляет собой K или R. В любом из вышеупомянутых вариантов осуществления X5 представляет собой H или P. В любом из вышеупомянутых вариантов осуществления X6 представляет собой S, T или G. В любом из вышеупомянутых вариантов осуществления X7 представляет собой A.
В некоторых вариантах осуществления полипептид имеет последовательность SEQ ID NO: 212, где X1 представляет собой V или I. В любом из вышеупомянутых вариантов осуществления в данном аспекте раскрытия, X2 представляет собой V или I. В любом из вышеупомянутых вариантов осуществления X3 представляет собой I или F. В любом из вышеупомянутых вариантов осуществления X4 представляет собой K или R. В любом из вышеупомянутых вариантов осуществления X5 представляет собой H или P. В любом из вышеупомянутых вариантов осуществления X6 представляет собой S или T. В любом из вышеупомянутых вариантов осуществления X7 представляет собой A.
В некоторых вариантах осуществления полипептид имеет последовательность SEQ ID NO: 212, где X7 представляет собой A. В любом из вышеупомянутых вариантов осуществления в данном аспекте раскрытия, X7 представляет собой A, а X1 представляет собой V или I. В любом из вышеупомянутых вариантов осуществления в данном аспекте раскрытия, X7 представляет собой A, а X2 представляет собой V или I. В любом из вышеупомянутых вариантов осуществления X7 представляет собой A, а X3 представляет собой I или F. В любом из вышеупомянутых вариантов осуществления X7 представляет собой A, а X4 представляет собой K или R. В любом из вышеупомянутых вариантов осуществления X7 представляет собой A, а X5 представляет собой H или P. В любом из вышеупомянутых вариантов осуществления X7 представляет собой A, а X6 представляет собой S или T.
В некоторых вариантах осуществления полипептид имеет последовательность SEQ ID NO: 212, где X7 представляет собой A. В любом из вышеупомянутых вариантов осуществления в данном аспекте раскрытия, X7 представляет собой A, а X1 представляет собой I. В любом из вышеупомянутых вариантов осуществления в данном аспекте раскрытия, X7 представляет собой A, а X2 представляет собой I. В любом из вышеупомянутых вариантов осуществления X7 представляет собой A, а X3 представляет собой F. В любом из вышеупомянутых вариантов осуществления X7 представляет собой A, а X4 представляет собой R. В любом из вышеупомянутых вариантов осуществления X7 представляет собой A, а X5 представляет собой P. В любом из вышеупомянутых вариантов осуществления X7 представляет собой A, а X6 представляет собой T.
В некоторых вариантах осуществления полипептид имеет высокоаффинный домен D1 SIRP-α, имеющий по меньшей мере 85% идентичности последовательности (например, по меньшей мере 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности) с SEQ ID NO: 212, где каждый из X1, X2, X3, X4, X5, X6, и X7 не является аминокислотой дикого типа.
В некоторых вариантах осуществления полипептид по данному аспекту раскрытия содержит не более десяти аминокислотных замен по сравнению с доменом D1 SIRP-α дикого типа, имеющим любую из последовательностей SEQ ID NO: 2. В некоторых вариантах осуществления полипептид по данному аспекту раскрытия содержит не более семи аминокислотных замен по сравнению с доменом D1 SIRP-α дикого типа, имеющим любую из последовательностей SEQ ID NO: 2.
В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 10 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 2. В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 100 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 2. В некоторых вариантах осуществления полипептид связывает CD47 с по меньшей мере в 1000 раз большей аффинностью связывания, чем домен D1 SIRP-α дикого типа, имеющий любую из последовательностей SEQ ID NO: 2. В некоторых вариантах осуществления фрагменты включают полипептиды менее чем 10 аминокислот в длину, около 10 аминокислот в длину, около 20 аминокислот в длину, около 30 аминокислот в длину, около 40 аминокислот в длину, около 50 аминокислот в длину, около 60 аминокислот в длину, около 70 аминокислот в длину, около 80 аминокислот в длину, около 90 аминокислот в длину, около 100 аминокислот в длину или более чем около 100 аминокислот в длину. Фрагменты сохраняют возможность связываться с CD47. Предпочтительно варианты D1 полипептидов SIRP-α и их фрагменты связываются с CD47 с более высоким сродством, чем полипептид SIRP-α связывается с CD47. Например, в некоторых вариантах осуществления вариант D1 полипептида SIRP-α или его фрагмент связывается с CD47 с KD менее чем 1×10-8 M, менее чем 5×10-9 M, менее чем 1×10-9 M, менее чем 5×10-10 M, менее чем 1×10-10 M или менее чем 1×10-11 M. В некоторых вариантах осуществления вариант D1 полипептида SIRP-α или его фрагмент связывается с CD47 с KD между около 500 нМ и 100 нМ, между около 100 нМ и 50 нМ, между около 50 нМ и 10 нМ, между около 10 нМ и 5 нМ, между около 5 нМ и 1 нМ, между около 1 нМ и 500 пМ, между около 500 пМ и 100 пМ, между около 100 пМ и 50 пМ или между около 50 пМ и 10 пМ.
В рамках изобретения, в некоторых вариантах осуществления описан полипептид, содержащий вариант D1 SIRP-α, имеющий последовательность согласно: 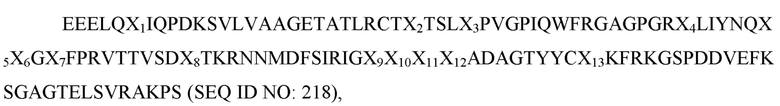
где X1 представляет собой V, L или I; X2 представляет собой A, V, L или I; X3 представляет собой I, S, T или F; X4 представляет собой E, L или V; X5 представляет собой K или R; X6 представляет собой E или Q; X7 представляет собой H, R или P; X8 представляет собой S, G, L или T;X9 представляет собой любую аминокислоту; X10 представляет собой любую аминокислоту; X11 представляет собой любую аминокислоту; X12 представляет собой любую аминокислоту, а X13 представляет собой V или I; и при этом вариант D1 SIRP-α имеет по меньшей мере две аминокислотные замены по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность согласно SEQ ID NO: 1.
В некоторых вариантах осуществления полипептид имеет последовательность SEQ ID NO: 212, X9 представляет собой A. В любом из вышеупомянутых вариантов осуществления в данном аспекте раскрытия, X9 представляет собой N. В любом из вышеупомянутых вариантов осуществления в данном аспекте раскрытия, X10 представляет собой I. В любом из вышеупомянутых вариантов осуществления в данном аспекте раскрытия, X9 представляет собой N, а X10 представляет собой P. В любом из вышеупомянутых вариантов осуществления в данном аспекте раскрытия, X9 представляет собой N, а X11 представляет собой аминокислоту, отличную от S, T или C. В любом из вышеупомянутых вариантов осуществления в данном аспекте раскрытия, X11 представляет собой T. В любом из вышеупомянутых вариантов осуществления в данном аспекте раскрытия, X11 представляет собой аминокислоту, отличную от T. В любом из вышеупомянутых вариантов осуществления в данном аспекте раскрытия, X12 представляет собой P. В любом из вышеупомянутых вариантов осуществления в данном аспекте раскрытия, X9 представляет собой N, а X12 представляет собой аминокислоту, отличную от P.
В рамках изобретения, в некоторых вариантах осуществления описан полипептид, содержащий вариант D1 SIRP-α, имеющий последовательность согласно:
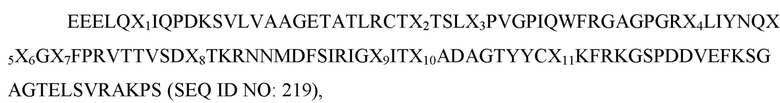
где X1 представляет собой V, L или I; X2 представляет собой A, V, L или I; X3 представляет собой I, S, T или F; X4 представляет собой E, L или V; X5 представляет собой K или R; X6 представляет собой E или Q; X7 представляет собой H, R или P; X8 представляет собой S, G, L или T; X9 представляет собой N; X10 представляет собой любую аминокислоту, отличную от P, а X11 представляет собой V или I; и при этом вариант D1 SIRP-α имеет по меньшей мере две аминокислотные замены по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность согласно SEQ ID NO: 1.
В другом аспекте изобретения раскрыты композиции, которые содержат вариант D1 полипептида SIRP-α, имеющий аминокислотную последовательность SEQ ID NO: 48 или его фрагмент. В некоторых вариантах осуществления вариантный D1 полипептида SIRP-α или его фрагмент связывается с CD47 с более высокой аффинностью по сравнению с аффинностью, с которой полипептид SIRP-α связывается с CD47. В некоторых вариантах осуществления вариант D1 полипептида SIRP-α связывается с CD47 с KD менее чем 1×10-8 М, или менее чем 1×10-9 М, менее чем 1×10-10 М или менее чем 1×10-11 М. В некоторых вариантах осуществления вышеупомянутые варианты D1 полипептида SIRP-α присоединены или слиты со вторым полипептидом. В некоторых вариантах осуществления второй полипептид включает, без ограничения, полипептид Fc, вариант Fc, полипептид ЧСА, пептид альбумина, полимер ПЭГ или фрагмент вышеуказанного.
Без ограничения вышеизложенного в некоторых вариантах осуществления вариант D1 полипептида SIRP-α выбран из любого из SEQ ID NO: 53-87 и 213, представленных в таблице 6.
Таблица 6. Варианты полипептидов SIRP-α
В некоторых вариантах осуществления полипептид содержит высокоаффинный домен D1 SIRP-α, имеющий по меньшей мере 85% идентичности последовательности (например, по меньшей мере 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности) с любым вариантом, приведенным в таблице 6.
В некоторых вариантах осуществления полипептид содержит высокоаффинный домен D1 SIRP-α, имеющий по меньшей мере 85% идентичности последовательности (например, по меньшей мере 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности) с SEQ ID NO: 80, 81 или 85 в таблице 6.
III. Варианты домена Fc и химерные конструкции
В рамках изобретения, в некоторых вариантах осуществления, раскрыты полипептиды, содержащие: вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92.
В рамках изобретения, в некоторых вариантах осуществления также раскрыты полипептиды, содержащие вариант Fc, причем вариант Fc содержит димер домена Fc, имеющий два мономера домена Fc, при этом каждый мономер домена Fc независимо выбирают из (i) области Fc IgG1 человека, включающей в свой состав мутации L234A, L235A, G237A и N297A; (ii) области Fc IgG2 человека, включающей в свой состав мутации A330S, P331S и N297A; или (iii) области Fc IgG4 человека, содержащей мутации S228P, E233P, F234V, L235A, delG236 и N297A.
Антитела, которые нацелены на клеточные поверхностные антигены, могут вызывать иммуностимулирующие и эффекторные функции, которые связаны с взаимодействием рецептора Fc (FcR) с иммунными клетками. Существует множество рецепторов Fc, которые специфичны для определенных классов антител, включая IgG (гамма-рецепторы), рецепторы IgE (эта), IgA (альфа-рецепторы) и IgM (мю-рецепторы). Связывание области Fc с рецепторами Fc на поверхностях клеток может инициировать ряд биологических реакций, включая фагоцитоз частиц, покрытых антителом (антителозависимый клеточно-опосредованный фагоцитоз или ADCP), клиренс иммунных комплексов, лизис клеток, покрытых антителами, посредством клеток-киллеров (антителозависимая клеточно-опосредованная цитотоксичность или ADCC) и высвобождение медиаторов воспаления, плацентарный перенос и контроль за продуцированием иммуноглобулина. Кроме того, связывание компонента C1 комплемента с антителами может активировать систему комплемента. Активация комплемента может быть важной для лизиса клеточных патогенов. Однако активация комплемента может также стимулировать воспалительный ответ и может также участвовать в аутоиммунной гиперчувствительности или других иммунологических расстройствах. Варианты областей Fc со сниженной или абляционной способностью связывать определенные рецепторы Fc полезны для разработки терапевтических антител и слитых Fc полипептидных конструкций, которые действуют путем нацеливания, активации или нейтрализации лигандных функций, не повреждая или не разрушая локальные клетки или ткани.
В некоторых вариантах осуществления полипептидная конструкция D1 SIRP-α содержит неприродный высокоаффинный вариант D1 SIRP-α, связанный с мономерным доменом Fc, который образует домен Fc, имеющий абляционную или уменьшенную эффекторную функцию.
В некоторых вариантах осуществления мономер домена Fc относится к полипептидной цепи, которая содержит второй и третий константные домены антител (например, CH2 и CH3). В некоторых вариантах осуществления мономер домена Fc также содержит шарнирный домен. В некоторых вариантах осуществления мономер домена Fc представляет собой изотип антитела иммуноглобулина, включая IgG, IgE, IgM, IgA и IgD. Кроме того, в некоторых вариантах осуществления мономер домена Fc представляет собой любой подтип IgG (например, IgG1, IgG2, IgG2a, IgG2b, IgG2c, IgG3 и IgG4). В некоторых вариантах осуществления мономеры домена Fc содержат до десяти изменений по сравнению с мономерной последовательностью домена Fc дикого типа (например, 1-10, 1-8, 1-6, 1-4 аминокислотных замен, добавок или вставок, делеций, или их комбинации), которые изменяют взаимодействие между доменом Fc и рецептором Fc.
Используемый в рамках изобретения термин «домен Fc» относится к димеру двух мономеров домена Fc. В домене Fc дикого типа два мономера домена Fc димеризуются взаимодействием между двумя константными доменами антитела CH3, а также одной или более дисульфидными связями, которые образуются между шарнирными доменами двух димеризованных мономеров домена Fc. В некоторых вариантах осуществления домен Fc мутирует с отсутствием эффекторных функций, например, «мертвый домен Fc». В некоторых вариантах осуществления каждый из мономеров домена Fc в домене Fc содержит аминокислотные замены в константной области антитела CH2 для уменьшения взаимодействия или связывания между доменом Fc и рецептором Fc, таким как рецептор Fcγ (FcγR), рецептор Fcα (FcαR) или Fcε (FcεR).
В некоторых вариантах осуществления высокоаффинный вариант D1 SIRP-α (например, любой из вариантов, описанных в таблицах 2, 5 и 6) слит с мономерным доменом Fc иммуноглобулина или фрагментом мономера домена Fc. В некоторых вариантах осуществления мономер домена Fc иммуноглобулина или фрагмент мономера домена Fc способен образовывать домен Fc с другим мономером домена Fc. В некоторых вариантах осуществления мономер домена Fc иммуноглобулина или фрагмент мономера домена Fc не способен образовывать домен Fc с другим мономером домена Fc. В некоторых вариантах осуществления мономер домена Fc или фрагмент домена Fc сливают с полипептидом раскрытия для увеличения периода полувыведения полипептида в сыворотке. В некоторых вариантах осуществления мономер домена Fc или фрагмент мономера домена Fc, слитого с полипептидом данного описания, димеризуется вторым мономером домена Fc с образованием домена Fc, который связывает рецептор Fc, или, альтернативно, мономер домена Fc связывается с рецептором Fc. В некоторых вариантах осуществления домен Fc или фрагмент домена Fc, слитый с полипептидом для увеличения периода полувыведения в сыворотке полипептида, не вызывает никакой реакции, связанной с иммунной системой.
В некоторых вариантах полипептид или конструкция SIRP-α, представленные в рамках изобретения, содержат домен D1 SIRP-α или его вариант, присоединенный к первому мономеру домена Fc и вариабельный домен антитела, присоединенный ко второму мономеру домена Fc, в котором первый и второй мономеры домена Fc объединяются для образования домена Fc (например, гетеродимерного домена Fc). Домен Fc представляет собой структуру белка, которая находится на С-конце иммуноглобулина. В домен Fc содержит два мономера домена Fc, которые димеризуются взаимодействием между константными доменами антитела CH3. Домен Fc дикого типа образует минимальную структуру, которая связывается с рецептором Fc, например FcγRI, FcγRIIa, FcγRIIb, FcγRIIIa, FcγRIIIb и FcγRIV.
Домен Fc не участвует непосредственно в связывании антитела с его мишенью, но может участвовать в различных эффекторных функциях, таких как участие антитела в антителозависимой клеточной токсичности. В некоторых вариантах осуществления домен Fc в полипептиде SIRP-α или конструкции раскрытия содержит аминокислотные замены, добавления или вставки, делеции или любые их комбинации, которые приводят к снижению эффекторной функции, такой как снижение антителозависимой клеточно-опосредованной цитотоксичности (ADCC), снижение комплементарнозависимого цитолиза (CDC), снижения антителозависимый клеточноопосредованного фагоцитоза (ADCP) или любой их комбинации. В некоторых вариантах осуществления полипептиды SIRP-α или конструкции данного описания характеризуются уменьшенным связыванием (например, минимальным связыванием или отсутствием связывания) с рецептором Fc человека и уменьшенным связыванием (например, минимальным связыванием или отсутствием связывания), к белку C1q комплемента. В некоторых вариантах осуществления конструкции SIRP-α описания характеризуются уменьшенным связыванием (например, минимальным связыванием или отсутствием связывания) с FcγRI, FcγRIIA, FcγRIIB, FcγRIIIB, FcγRIIIB человека или любыми их комбинациями и C1q. Чтобы изменить или уменьшить антителозависимую эффекторную функцию, такую, как ADCC, CDC, ADCP или любые их комбинации, в некоторых вариантах осуществления домены Fc в конструкциях SIRP-α раскрытия относятся к классу IgG и содержат одну или более аминокислотных замен в E233, L234, L235, G236, G237, D265, D270, N297, E318, K320, K322, A327, A330, P331 или P329 (нумерация в соответствии с индексом EU Kabat (Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991)).
В некоторых вариантах осуществления полипептидные конструкции, содержащие ненативную область Fc, описанную в рамках изобретения, демонстрируют восстановленное или абляционное связывание, по меньшей мере, с одним из рецепторов Fcγ CD16a, CD32a, CD32b, CD32c и CD64 по сравнению с полипептидной конструкцией, содержащей нативную область Fc. В некоторых случаях описанные в рамках изобретения полипептидные конструкции демонстрируют уменьшенное или аблированное связывание с CD16a, CD32a, CD32b, CD32c и CD64 рецепторов Fcγ.
CDC относится к форме цитотоксичности, в которой каскад комплемента активируется компонентом комплемента C1q, связывающимся с антителом Fc. В некоторых вариантах осуществления полипептидные конструкции, содержащие ненативную область Fc, описанную в рамках изобретения, демонстрирует, по меньшей мере 5%, 10%, 15%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или большее сокращения в связывании C1q по сравнению с полипептидной конструкцией, содержащей область Fc дикого типа. В некоторых случаях полипептидные конструкции, содержащие ненативную область Fc, описанную в рамках изобретения, демонстрируют уменьшенный CDC по сравнению с полипептидной конструкцией, содержащей область Fc дикого типа. В некоторых вариантах осуществления полипептидные конструкции, содержащие ненативную область Fc, описанную в рамках изобретения, демонстрирует, по меньшей мере 5%, 10%, 15%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или большее сокращения в связывании CDC по сравнению с полипептидной конструкцией, содержащей область Fc дикого типа. В некоторых случаях полипептидные конструкции, содержащие ненативный вариант Fc, описанный в рамках изобретения, демонстрируют незначительный CDC по сравнению с полипептидной конструкцией, содержащей область Fc дикого типа.
В некоторых вариантах осуществления варианты Fc в рамках изобретения минимально гликозилированы или имеют сниженное гликозилирование относительно последовательности дикого типа. В некоторых вариантах осуществления дегликозилирование осуществляют посредством мутации N297A или посредством мутации N297 с любой аминокислотой, которая не является N. В некоторых вариантах осуществления дегликозилирование осуществляют путем разрушения мотива N-Xaa1-Xaa2-Xaa3, где N=аспарагин; Xaa1=любая аминокислота, кроме P (пролина); Xaa2=T (треонин), S (серин) или C (цистеин), а Xaa3=любая аминокислота, кроме P (пролина). В одном варианте осуществления мотив N-Xaa1-Xaa2-Xaa3 относится к остаткам 297-300, указанным в соответствии с Kabat et al., 1991. В некоторых вариантах осуществления мутация в любом одном или более из N, Xaa1, Xaa2 или Xaa3 приводит к дегликозилированию варианта Fc.
В некоторых вариантах осуществления варианты константных областей IgG антитела (например, варианты Fc) обладают ограниченной способностью специфически связывать рецепторы Fcγ или имеют уменьшенную способность индуцировать фагоцитоз. В некоторых вариантах осуществления варианты константных областей IgG антитела (например, варианты Fc) обладают ограниченной способностью специфически связывать рецепторы Fcγ и имеют уменьшенную способность индуцировать фагоцитоз. Например, в некоторых вариантах осуществления домен Fc мутируется с отсутствием эффекторных функций, типичных для «мертвого» домена Fc. Например, в некоторых вариантах осуществления домен Fc содержит определенные аминокислотные замены, которые, как известно, минимизируют взаимодействие между доменом Fc и рецептором Fcγ. В некоторых вариантах осуществления мономер домена Fc представляет собой антитело IgG1 и содержит одну или более аминокислотных замен L234A, L235A, G237A и N297A (обозначенных в соответствии с системой нумерации EU Kabat et al., 1991). В некоторых вариантах осуществления одна или более дополнительных мутаций содержатся в таком варианте Fc IgG1. Неограничивающие примеры таких дополнительных мутаций для вариантов Fc IgG1 человека включают E318A и K322A. В некоторых случаях вариант Fc IgG1 человека имеет до 12, 11, 10, 9, 8, 7, 6, 5 или 4 или менее мутаций по сравнению с последовательностью IgG1 человека дикого типа. В некоторых вариантах осуществления одна или более дополнительных делеций содержатся в таком варианте Fc IgG1. Например, в некоторых вариантах осуществления С-концевой лизин константной области тяжелой цепи Fc IgG1, представленный в SEQ ID NO: 88 в Таблице 7 удаляется, например, для увеличения гомогенности полипептида, когда полипептид продуцируется в клетках бактерий или млекопитающих. В некоторых случаях вариант Fc IgG1 человека имеет до 12, 11, 10, 9, 8, 7, 6, 5 или 4 или менее делеций по сравнению с последовательностью IgG1 человека дикого типа. В некоторых вариантах осуществления вариант Fc IgG1 имеет последовательность согласно любой из SEQ ID NO: 135, SEQ ID NO: 136 или SEQ ID NO: 137.
В некоторых вариантах осуществления мономер домена Fc представляет собой антитело IgG2 или IgG4 и содержит аминокислотные замены A330S, P331S или обе A330S и P331S. Вышеупомянутые положения аминокислот определены согласно Kabat et al. (1991). Kabat-нумерация остатков может быть определена для данного антитела путем выравнивания в областях гомологии последовательности антитела со «стандартной» нумерацией Kabat. В некоторых вариантах осуществления вариант Fc содержит последовательность Fc IgG2 человека, содержащую одну или более аминокислотных замен A330S, P331S и N297A (обозначенных в соответствии с системой нумерации EU Kabat et al. (1991). В некоторых вариантах осуществления одна или более дополнительных мутаций содержатся в таких вариантах Fc IgG2. Неограничивающие примеры таких дополнительных мутаций для варианта Fc IgG2 человека включают V234A, G237A, P238S, V309L и H268A (обозначенные в соответствии с системой нумерации EU Kabat et al. (1991)). В некоторых случаях вариант Fc IgG2 человека имеет до 12, 11, 10, 9, 8, 7, 6, 5, 4, 3 или менее мутаций по сравнению с последовательностью IgG2 человека дикого типа. В некоторых вариантах осуществления одна или более дополнительных делеций содержатся в таком варианте Fc IgG2. Например, в некоторых вариантах осуществления С-концевой лизин константной области тяжелой цепи Fc IgG2, представленный в SEQ ID NO: 89 в Таблице 7 удаляется, например, для увеличения гомогенности полипептида, когда полипептид продуцируется в клетках бактерий или млекопитающих. В некоторых случаях вариант Fc IgG2 человека имеет до 12, 11, 10, 9, 8, 7, 6, 5 или 4 или менее делеций по сравнению с последовательностью IgG2 человека дикого типа.
Когда вариант Fc представляет собой вариант Fc IgG4, в некоторых вариантах осуществления такой вариант Fc содержит мутацию S228P (как указано в Kabat et al. (1991)). В некоторых случаях вариант Fc IgG2 человека имеет до 12, 11, 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1 мутацию(и) по сравнению с последовательностью IgG4 человека дикого типа.
В некоторых вариантах осуществления вариант Fc содержит по меньшей мере одну из мутаций L234A, L235A, G237A или N297A области Fc IgG1 или по меньшей мере одну из мутаций A330S, P331S или N297A области Fc IgG2. В некоторых вариантах осуществления вариант Fc содержит по меньшей мере две из мутаций L234A, L235A, G237A или N297A области Fc IgG1 или по меньшей мере две из мутаций A330S, P331S или N297A области Fc IgG2. В некоторых вариантах осуществления вариант Fc содержит по меньшей мере три из мутаций L234A, L235A, G237A или N297A области Fc IgG1 или состоит из мутаций A330S, P331S и N297A области Fc IgG2. В некоторых вариантах осуществления вариант Fc состоит из мутаций L234A, L235A, G237A и N297A.
В некоторых вариантах осуществления вариант Fc демонстрирует уменьшенное связывание с рецептором Fc пациента по сравнению с областью Fc IgG дикого типа человека. В некоторых вариантах осуществления вариант Fc демонстрирует аблированное связывание с рецептором Fc пациента по сравнению с областью Fc IgG дикого типа человека. В некоторых вариантах осуществления вариант Fc демонстрирует уменьшение фагоцитоза по сравнению с областью Fc IgG человека дикого типа. В некоторых вариантах осуществления вариант Fc демонстрирует аблированный фагоцитоз по сравнению с областью Fc IgG человека дикого типа.
SEQ ID NO: 88 или SEQ ID NO: 89 представляют собой аминокислотные последовательности Fc константных областей тяжелой цепи IgG1 и IgG2. В некоторых вариантах осуществления вариант Fc представляет собой любой вариант SEQ ID NO: 90-95, как показано в таблице 7.
Таблица 7. Аминокислотные последовательности вариантов Fc
"Антителозависимая клеточно-опосредованная цитотоксичность" также называемая в рамках изобретения как ADCC относится к форме цитотоксичности, при которой секретируемый Ig связывается с рецепторами Fc (FcR), присутствующими на некоторых цитотоксических клетках (например, клетки естественные киллеры (NK) и нейтрофилы), позволяя этим цитотоксическим эффекторным клеткам специфически связываться с антигенсодержащей клеткой-мишенью и впоследствии убивать клетку-мишень. Антителозависимый клеточно-опосредованный фагоцитоз, который также упоминается в рамках изобретения как ADCP, относится к форме цитотоксичности, в которой секретируемый Ig, связанный с рецепторами Fc (FcR), присутствующими на определенных фагоцитарных клетках (например, макрофагах), что позволяет этим фагоцитарным эффекторным клеткам специфично связывать с антигенсодержащей клеткой-мишенью и затем поглощать и переваривать клетку-мишень. Лиганд-специфичные высокоаффинные антитела IgG, направленные на поверхность клеток-мишеней, могут стимулировать цитотоксические или фагоцитарные клетки и могут быть использованы для такого убийства. В некоторых вариантах осуществления полипептидные конструкции, содержащие вариант Fc, описанный в рамках изобретения, демонстрируют уменьшенный ADCC или ADCP по сравнению с полипептидной конструкцией, содержащей область Fc дикого типа. В некоторых вариантах осуществления полипептидные конструкции, содержащие вариант Fc, описанный в рамках изобретения, демонстрирует, по меньшей мере 5%, 10%, 15%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или большее сокращения в связывании ADCC или ADCP по сравнению с полипептидной конструкцией, содержащей область Fc дикого типа. В некоторых вариантах осуществления полипептидные конструкции, содержащие вариант Fc, описанный в рамках изобретения, демонстрируют аблированную ADCC или ADCP по сравнению с полипептидной конструкцией, содержащей область Fc дикого типа.
Цитотоксичность, направленная на комплемент, которая также упоминается в рамках изобретения как CDC относится к форме цитотоксичности, в которой каскад комплемента активируется компонентом комплемента C1q, связывающимся с антителом Fc. В некоторых вариантах осуществления полипептидные конструкции, содержащие вариант Fc, описанный в рамках изобретения, демонстрирует, по меньшей мере 5%, 10%, 15%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или большее сокращения в связывании C1q по сравнению с полипептидной конструкцией, содержащей область Fc дикого типа. В некоторых случаях полипептидные конструкции, содержащие вариант Fc, описанный в рамках изобретения, демонстрируют незначительный CDC по сравнению с полипептидной конструкцией, содержащей область Fc дикого типа. В некоторых вариантах осуществления полипептидные конструкции, содержащие вариант Fc, описанный в рамках изобретения, демонстрирует, по меньшей мере 5%, 10%, 15%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или большее сокращения в связывании CDC по сравнению с полипептидной конструкцией, содержащей область Fc дикого типа. В некоторых случаях полипептидные конструкции, содержащие вариант Fc, описанный в рамках изобретения, демонстрируют незначительный CDC по сравнению с полипептидной конструкцией, содержащей область Fc дикого типа.
Варианты Fc в рамках изобретения включают те, которые проявляют сниженное связывание с рецептором Fcγ по сравнению с областью Fc IgG человека дикого типа. Например, в некоторых вариантах осуществления вариант Fc проявляет связывание с рецептором Fcγ, которое меньше связывания, проявляемого областью Fc IgG человека дикого типа, с рецептором Fcγ, как описано в примерах. В некоторых случаях вариант Fc имеет связывание с рецептором Fcγ уменьшенное на 10%, 20% 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98%, 99% или 100% (полностью аблированная эффекторная функция). В некоторых вариантах осуществления уменьшенное связывание относится к любому одному или более рецепторам Fcγ, например, CD16a, CD32a, CD32b, CD32c или CD64.
В некоторых случаях варианты Fc, описанные в рамках изобретения, демонстрируют снижение фагоцитоза по сравнению с областью Fc IgG человека дикого типа. Такие варианты Fc демонстрируют уменьшение фагоцитоза по сравнению с областью Fc IgG человека дикого типа, причем снижение активности фагоцитоза проявляется, например, на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98%, 99% или 100%. В некоторых случаях вариант Fc демонстрирует аблированный фагоцитоз по сравнению с областью Fc IgG человека дикого типа.
В некоторых вариантах осуществления варианты Fc, описанные в рамках изобретения, связаны с одним или более партнерами по слиянию. В некоторых случаях партнер по слиянию представляет собой терапевтический фрагмент. В некоторых случаях партнер по слиянию выбирают для обеспечения таргетинга экспрессированного белка, очистки, скрининга, отображения и т.п. В некоторых вариантах осуществления партнер по слиянию также влияет на степень связывания с рецепторами Fc или степень уменьшения фагоцитоза. Как описано в рамках изобретения, в некоторых вариантах осуществления, когда вариант Fc связан с партнером по слиянию, он образует полипептидную конструкцию, как описано ниже.
В некоторых вариантах осуществления партнеры по слиянию связаны с вариантной последовательностью Fc через линкерную последовательность. В некоторых вариантах осуществления линкерная последовательность обычно содержит небольшое количество аминокислот, такое как менее десяти аминокислот, хотя также используются более длинные линкеры. В некоторых случаях линкер имеет длину менее 10, 9, 8, 7, 6 или 5 аминокислот или короче. В некоторых случаях линкер имеет длину по меньшей мере 10, 11, 12, 13, 14, 15, 20, 25, 30 или 35 аминокислот или длинее. Необязательно, в некоторых вариантах осуществления используется расщепляемый линкер.
В некоторых вариантах осуществления партнер по слиянию представляет собой целевую или сигнальную последовательность, которая направляет белок варианта Fc и любые ассоциированные партнеры по слитию в желаемое местоположение в клетке или во внеклеточную среду. В некоторых вариантах осуществления некоторые сигнальные последовательности направляют белок, который либо секретируется в культуральную среду, либо в периплазматическое пространство, расположенное между внутренней и внешней мембраной клетки. В некоторых вариантах осуществления партнер по слиянию представляет собой последовательность, которая кодирует пептид или белок, который обеспечивает очистку или скрининг. Такие партнеры по слиянию включают, но не ограничиваются ими, полигистидиновые метки (His-метки) (например, His6 и His10) или другие метки для использования с системами иммобилизованной металлической аффинной хроматографии (IMAC) (например, аффинные колонки Ni+2), слитые GST, слитые MBP, стрептавидиновые метки (Strep-метки), целевую последовательность биотинилирования BSP бактериального фермента BirA и метки эпитопа, которые нацелены на антитела (например, метки c-myc, метки flag, и подобные).
В некоторых вариантах осуществления такие метки пригодны для очистки, для скрининга или того и другого. Например, в некоторых вариантах осуществления вариант Fc очищают с использованием His-метки, иммобилизуя его в аффинную колонку с Ni+2, а затем после очистки одну и ту же His-метку используют для иммобилизации антитела на пластине с покрытием Ni+2 для проведения ELISA или другого анализа связывания, как описано в рамках изобретения в другом месте. В некоторых вариантах осуществления партнер по слиянию позволяет использовать способ выбора для экранирования вариантов Fc, как описано в рамках изобретения.
Доступны различные партнеры по слиянию, которые позволяют использовать различные методы выбора. Например, путем слияния членов библиотеки вариантов Fc с белком гена III можно использовать фаговый дисплей. В некоторых вариантах осуществления партнеры по слиянию позволяют метить варианты Fc. Альтернативно, в некоторых вариантах осуществления партнер по слиянию связывается с определенной последовательностью на векторе экспрессии, позволяя партнеру по слиянию и связанному варианту Fc связываться ковалентно или нековалентно с нуклеиновой кислотой, которая их кодирует.
В некоторых вариантах осуществления, когда партнером по слиянию является терапевтический фрагмент, терапевтическая часть представляет собой, например, пептид, белок, антитело, миРНК или небольшую молекулу. Неограничивающие примеры терапевтических антител, которые связаны с вариантами Fc настоящего изобретения, включают, но не ограничиваются ими, антитела, которые распознают CD47. Неограничивающие примеры терапевтических полипептидов, которые связаны с вариантами Fc настоящего изобретения, включают, но не ограничиваются ими, CD47-связывающие полипептиды, включая полипептиды SIRP-α. В таких случаях связывающий СD47 полипептид присоединен или слит с вариантом Fc раскрытия. Примеры связывающих CD47 полипептидов включают, но не ограничиваются ими, антитела против CD47 или их фрагменты и лиганды CD47, такие как SIRP-α или его фрагмент. Дополнительные примеры связывающих CD47 полипептидов включают, но не ограничиваются ими, природные формы SIRP-α, а также их мутанты.
В некоторых вариантах осуществления, в рамках изобретения раскрыт полипептид, содержащий вариант Fc, причем вариант Fc содержит димер домена Fc, имеющий два мономера домена Fc, при этом каждый мономер домена Fc независимо выбирают из (i) области Fc IgG1 человека, включающей в свой состав мутации L234A, L235A, G237A и N297A; (ii) области Fc IgG2 человека, включающей в свой состав мутации A330S, P331S и N297A или (iii) области Fc IgG4 человека, содержащей мутации S228P, E233P, F234V, L235A, delG236 и N297A. В некоторых вариантах осуществления мономеры домена Fc являются идентичными (то есть гомодимер). В некоторых вариантах осуществления мономеры домена Fc являются различными (то есть гетеродимер). В некоторых вариантах осуществления по меньшей мере один из мономеров домена Fc в димере домена Fc представляет собой область Fc IgG1 человека, состоящую из мутаций L234A, L235A, G237A и N297A. В некоторых вариантах осуществления по меньшей мере один из мономеров домена Fc в димере домена Fc представляет собой область Fc IgG2 человека, состоящую из мутаций A330S, P331S и N297A. В некоторых вариантах осуществления вариант Fc демонстрирует аблированное или уменьшенное связывание с рецептором Fcγ по сравнению с версией области Fc дикого типа IgG человека. В некоторых вариантах осуществления вариант Fc демонстрирует аблированное или уменьшенное связывание с рецепторами Fcγ CD16a, CD32a, CD32b, CD32c и CD64 по сравнению с версией области Fc дикого типа IgG человека. В некоторых вариантах осуществления вариант Fc демонстрирует аблированное или уменьшенное связывание с C1q по сравнению с версией слитого Fc дикого типа и IgG человека. В некоторых вариантах осуществления по меньшей мере один из мономеров домена Fc в димере домена Fc представляет собой область Fc IgG4 человека, содержащую мутации S228P, E233P, F234V, L235A, delG236 и N297A. В некоторых вариантах осуществления вариант Fc демонстрирует аблированное или уменьшенное связывание с рецептором Fcγ по сравнению с областью Fc дикого типа IgG4 человека. В некоторых вариантах осуществления вариант Fc демонстрирует аблированное или уменьшенное связывание с рецепторами Fcγ CD16a и CD32b по сравнению с версией его области Fc дикого типа IgG4 человека. В некоторых вариантах осуществления вариант Fc связывается с рецептором Fcγ с KD более чем около 5×10-6 M.
В некоторых вариантах осуществления вариант Fc дополнительно содержит CD47-связывающий полипептид. В некоторых вариантах осуществления вариант Fc демонстрирует аблированное или уменьшенное связывание с рецептором Fcγ по сравнению с версией области Fc дикого типа IgG человека. В некоторых вариантах осуществления CD47-связывающий полипептид не вызывает острой анемии у грызунов и приматов, отличных от человека. В некоторых вариантах осуществления CD47-связывающий полипептид не вызывает острой анемии у людей.
В некоторых вариантах осуществления CD47-связывающий полипептид представляет собой полипептид сигнального регуляторного белка α (SIRP-α) или его фрагмент. В некоторых вариантах осуществления полипептид SIRP-α содержит вариант D1 SIRP-α, содержащий аминокислотную последовательность

где X1 представляет собой V или I; X2 представляет собой A или I; X3 представляет собой I или F; X4 представляет собой E или V; X5 представляет собой K или R; X6 представляет собой H или P; X7 представляет собой L или T; X8 представляет собой аминокислоту, отличную от N, а X9 представляет собой V или I. В некоторых вариантах осуществления полипептид SIRP-α содержит вариант D1 SIRP-α, где X1 представляет собой V или I; X2 представляет собой A или I; X3 представляет собой I или F; X4 представляет собой E; X5 представляет собой K или R; X6 представляет собой H или P; X7 представляет собой L или T; X8 представляет собой не N, а X9 представляет собой V.
В рамках изобретения, в некоторых вариантах осуществления раскрыты полипептиды, содержащие: вариант D1 SIRP-α, причем вариант D1 SIRP-α представляет собой неприродный высокоаффинный домен D1 SIRP-α, при этом вариант D1 SIRP-α связывается с CD47 человека с аффинностью, которая по меньшей мере в 10 раз больше, чем аффинность природного домена D1; и мономер домена Fc, причем мономер домена Fc связан со вторым полипептидом, содержащим второй мономер домена Fc, с образованием домена Fc, при этом домен Fc имеет аблированную или уменьшенную эффекторную функцию. В некоторых вариантах осуществления неприродный высокоаффинный домен D1 SIRP-α содержит аминокислотную мутацию в остатке 80.
В некоторых вариантах осуществления в рамках изобретения раскрыт вариант D1 SIRP-α, причем вариант D1 SIRP-α связывает CD47 первого вида с KD менее 250 нМ; и при этом вариант D1 SIRP-α связывает CD47 второго вида с KD менее 250 нМ; и KD для CD47 первого вида и KD для CD47 второго вида отличается в 100 раз друг от друга; причем первый вид и второй вид выбраны из группы, состоящей из человека, грызуна и приматов, отличных от человека. В некоторых вариантах осуществления вариант D1 SIRP-α связывает CD47 по меньшей мере 3 различных видов. В некоторых вариантах осуществления примат, не являющийся человеком, представляет собой яванского макака.
В некоторых вариантах осуществления в рамках изобретения раскрыт полипептид, содержащий: (a) домен D1 SIRP-α, который связывает CD47 человека с KD менее 250 нМ; и (б) мономер домена Fc, связанный с N-концом или С-концом домена D1 SIRP-α, причем полипептид не вызывает острой анемии у грызунов и приматов, отличных от человека. В некоторых вариантах осуществления полипептид представляет собой неприродный вариант SIRP-α человека. В некоторых вариантах осуществления введение полипептида in vivo приводит к снижению гемоглобина менее чем на 50% в течение первой недели после введения. В некоторых вариантах осуществления введение полипептида людям приводит к снижению гемоглобина менее чем на 50% в течение первой недели после введения. В некоторых вариантах осуществления полипептид дополнительно содержит по меньшей мере один вариант Fc, причем вариант Fc, содержит мономер домена Fc, который выбирают из (i) области Fc IgG1 человека, включающей в свой состав мутации L234A, L235A, G237A и N297A; (ii) области Fc IgG2 человека, включающей в свой состав мутации A330S, P331S и N297A; или (iii) области Fc IgG4 человека, содержащей мутации S228P, E233P, F234V, L235A, delG236 и N297A. В некоторых вариантах осуществления мономер домена Fc представляет собой область Fc IgG1 человека, состоящую из мутаций L234A, L235A, G237A и N297A. В некоторых вариантах осуществления мономер домена Fc представляет собой область Fc IgG2 человека состоящую из мутаций A330S, P331S и N297A.
Конструкции SIRP-α раскрытия содержат домен SIRP-α или его вариант, который своим C-концом присоединен к N-концу мономера домена Fc посредством линкера с использованием обычных генетических или химических средств, например, химической конъюгации. В некоторых вариантах осуществления линкер (например, спейсер) вставлен между полипептидом и мономером домена Fc. В некоторых вариантах осуществления полипептид раскрытия, включая высокоаффинный вариант D1 SIRP-α, слит с мономером домена Fc, который не способен образовывать димер. В некоторых вариантах осуществления полипептид изобретения слит с мономером домена Fc, который способен образовывать димер, например, гетеродимер, с другим мономером домена Fc. В некоторых вариантах осуществления полипептид по изобретению слит с мономером домена Fc, и этот слитый белок образует гомодимер. В некоторых вариантах осуществления полипептид изобретения слит с первым мономером домена Fc, а другой белок или пептид (например, вариабельная область антитела) слит со вторым мономером домена Fc. В некоторых вариантах осуществления домен или вариант D1 SIRP-α соединен с первым мономером домена Fc и терапевтическим белком (например, цитокином, интерлейкином, антигеном, стероидом, противовоспалительным агентом или иммуномодулирующим агентом) присоединен ко второму мономеру домена Fc. В некоторых вариантах осуществления первый и второй мономеры домена Fc образуют гетеродимер.
Без ограничения вышеизложенного в некоторых вариантах осуществления вариант D1 полипептида SIRP-α (например, любой из вариантов, описанных в таблицах 2, 5 и 6) слит с полипептидом Fc или вариантом Fc полипептида, таким как мономер домена Fc. Примеры полипептидов, включая вариант D1 полипептида SIRP-α и вариант слитого Fc полипептида, включают, но не ограничиваются ими, SEQ ID NO: 96-137, 214 и 216 представлены в таблице 8.
Таблица 8. Полипептиды содержащие варианты D1 SIRP-α слитый с вариантами Fc
В некоторых вариантах осуществления полипептид содержит высокоаффинный домен D1 SIRP-α, имеющий по меньшей мере 85% идентичности последовательности (например, по меньшей мере 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности) с любым вариантом, приведенным в таблице 8.
В некоторых вариантах осуществления полипептид содержит высокоаффинный домен D1 SIRP-α, имеющий по меньшей мере 85% идентичности последовательности (например, по меньшей мере 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности) с SEQ ID NO: 98-104, 107-113, 116-122 или 135-137 в таблице 8.
В некоторых вариантах осуществления полипептид содержит (a) вариант сигнального регуляторного белка α (SIRP-α) D1, причем вариант D1 SIRP-α содержит аминокислотную последовательность
где X1 представляет собой E или G; X2 представляет собой L, I или V; X3 представляет собой V, L или I; X4 представляет собой S или F; X5 представляет собой L или S; X6 представляет собой S или T; X7 представляет собой A или V; X8 представляет собой I или T; X9 представляет собой H, R или L; X10 представляет собой A, V, I или L; X11 представляет собой I, T, S или F; X12 представляет собой A или G; X13 представляет собой E, V или L; X14 представляет собой K или R; X15 представляет собой E или Q; X16 представляет собой H, P или R; X17 представляет собой D или E; X18 представляет собой S, L, T или G; X19 представляет собой K или R; X20 представляет собой E или N; X21 представляет собой S или P; X22 представляет собой S или R; X23 представляет собой S или G; X24 представляет собой любую аминокислоту; X25 представляет собой любую аминокислоту; X26 представляет собой V или I; X27 представляет собой F, L или V; X28 представляет собой D или отсутствует; X29 представляет собой T или V; X30 представляет собой F или V, а X31 представляет собой A или G; и при этом вариант D1 SIRP-α имеет по меньшей мере две аминокислотных заменены по сравнению с доменом D1 SIRP-α дикого типа, имеющим последовательность согласно любой из SEQ ID NO: 1-10; и (b) вариант Fc, содержащий димер домена Fc, имеющий два мономера домена Fc, причем каждый мономер домена Fc независимо представляет собой (i) область Fc IgG1 человека, содержащую мутацию N297A; (ii) область Fc IgG1 человека, содержащую мутации L234A, L235A и G237A; (iii) область Fc IgG1 человека, содержащую мутации L234A, L235A, G237A и N297A; (iv) область Fc IgG2 человека, содержащую мутацию N297A; (v) область Fc IgG2 человека, содержащую мутации A330S и P331S; (vi) область Fc IgG2 человека, содержащую мутации A330S, P331S и N297A; (vii) область Fc IgG4 человека, содержащую мутации S228P, E233P, F234V, L235A и delG236; или (viii) область Fc IgG4 человека, содержащую мутации S228P, E233P, F234V, L235A, delG236 и N297A.
В некоторых вариантах осуществления полипептид содержит вариант D1 SIRP-α, в котором вариант D1 SIRP-α содержит аминокислотную последовательность в соответствии с SEQ ID NO: 47; вариант Fc, содержащий димер домена Fc, имеющий два мономера домена Fc, причем один из мономеров домена Fc в димере домена Fc содержит область Fc IgG1 человека, содержащую мутации L234A, L235A, G237A и N297A.
Димеризация мономеров домена Fc
В некоторых вариантах осуществления вариант полипептида D1 SIRP-α (например, любой из вариантов, описанных в таблицах 2, 5 и 6) слит с первым мономером домена Fc либо на N-конце, либо на С-конце. В некоторых вариантах осуществления первый мономер домена Fc не способен образовывать домен Fc или димер. В некоторых вариантах осуществления первый мономер домена Fc объединяется со вторым мономером домена Fc с образованием домена Fc или димера. В некоторых вариантах осуществления первый и второй мономеры домена Fc содержат аминокислотные замены, которые способствуют гетеродимеризации между первым и вторым мономерами домена.
В некоторых вариантах осуществления каждый из двух мономеров домена Fc в домене Fc включает аминокислотные замены, которые способствуют гетеродимеризации двух мономеров. В некоторых вариантах осуществления конструкция SIRP-α формируется, например, из первой субъединицы, содержащей вариант D1 полипептида SIRP-α, слитый с первым мономером домена Fc, и второй субъединицы, содержащей второй мономер домена Fc (например, без SIRP -α D1 или любой другой полипептид). В некоторых вариантах осуществления конструкция имеет один вариант D1 полипептида SIRP-α, связанный с доменом Fc (например, одноплечевой). В некоторых вариантах осуществления конструкция имеет два варианта D1 полипептида SIRP-α, связанные с доменом Fc (например, двуплечевой). В некоторых вариантах осуществления вариант D1 SIRP-α, имеющий KD около 500 нМ, особенно полезен в двуплечевой конструкции. В некоторых вариантах осуществления вариант D1 SIRP-α, имеющий KD около 50 нМ, особенно полезен в двуплечевой конструкции. В некоторых вариантах осуществления вариант D1 SIRP-α, имеющий KD около 5 нМ, полезен в двуплечевой конструкции и одноплечевой конструкции. В некоторых вариантах осуществления вариант D1 SIRP-α, имеющий KD около 500 нМ, полезен в двуплечевой конструкции и одноплечевой конструкции. В некоторых вариантах осуществления вариант D1 SIRP-α, имеющий KD около 100 нМ, полезен в двуплечевой конструкции и одноплечевой конструкции. В некоторых вариантах осуществления вариант D1 SIRP-α, имеющий KD около 50 нМ, полезен в двуплечевой конструкции и одноплечевой конструкции. В некоторых вариантах осуществления вариант D1 SIRP-α, имеющий KD около 10 нМ, полезен в двуплечевой конструкции и одноплечевой конструкции.
В некоторых вариантах осуществления гетеродимеризация мономеров доменов Fc обеспечивается внедрением различных, но совместимых замен в двух мономерах домена Fc, таких как пары остатков «выступ во впадину» и пары заряженных остатков. Взаимодействие выступ и впадина благоприятствует образованию гетеродимеров, тогда как взаимодействия выступ-выступ и впадина-впадина препятствуют формированию гомодимера из-за стерического столкновения и делеции благоприятных взаимодействий. Впадина относится к пустоте, которая создается, когда исходная аминокислота в белке заменена другой аминокислотой, имеющей меньший объем боковой цепи. Выступ относится к выпячиванию, которое создается, когда исходная аминокислота в белке заменена другой аминокислотой, имеющей больший объем боковой цепи. Например, в некоторых вариантах осуществления заменяемая аминокислота находится в константном домене СН3 мономера домена Fc и участвует в димеризации двух мономеров домена Fc. В некоторых вариантах осуществления отверстие в одном константном домене антитела СН3 создано для размещения выступа в другом константном домене СН3, так что аминокислоты с выступом и впадиной действуют так, чтобы способствовать или облегчать гетеродимеризацию двух мономеров домена Fc. В некоторых вариантах осуществления впадина в одном константном домене антитела СН3 создана для лучшего размещения исходной аминокислоты в другом константном домене СН3. В некоторых вариантах осуществления выступ в одном константном домене антитела СН3 создан для образования дополнительных взаимодействий с исходными аминокислотами в другом константном домене СН3.
В некоторых вариантах осуществления впадина создана путем замены аминокислот, имеющих более крупные боковые цепи, такие как тирозин или триптофан, на аминокислоты, имеющие меньшие боковые цепи, такие как аланин, валин или треонин, например, мутация Y407V в константном домене СН3. Аналогичным образом, в некоторых вариантах осуществления выступ создан путем замены аминокислот с меньшими боковыми цепями аминокислотами, имеющими более крупные боковые цепи, например, мутация T366W в константном домене антитела CH3. В некоторых вариантах осуществления один мономер домена Fc содержит мутацию T366W с выступом, а другой мономер домена Fc содержит мутации впадины T366S, L358A и Y407V. В некоторых вариантах осуществления полипептид раскрытия, включая высокоаффинный вариант D1 SIRP-α, слит с мономером домена Fc, содержащим мутацию выступа T366W для ограничения нежелательного образования гомодимера выступ-выступ. Примеры пар аминокислот в форме «выступ во впадину» включены, без ограничения, в таблицу 9, а примеры вариантов Fc «выступ во впадину» и слитых SIRP-α-Fc приведены в таблице 10.
Таблица 9. Пары аминокислот «выступ во впадину»
Y407T
Y407A
Таблица 10. Примеры вариантов Fc и слитых SIRP-α - Fc
В дополнение к стратегии с выступ во впадину, в некоторых вариантах осуществления электростатическое взаимодействие также используется для управления димеризацией мономеров домена Fc. Электростатическое взаимодействие относится к использованию благоприятных электростатических взаимодействий между противоположно заряженными аминокислотами в пептидах, белковых доменах и белках для управления образованием высших упорядоченных белковых молекул. В частности, для контроля димеризации мономеров доменов Fc с использованием электростатического взаимодействия один или более аминокислотных остатков, которые составляют границу CH3-CH3, заменяются положительно или отрицательно заряженными аминокислотными остатками, так что взаимодействие становится электростатически благоприятным или неблагоприятным в зависимости от введенных специфических заряженных аминокислот. В некоторых вариантах осуществления положительно заряженная аминокислота на границе раздела, такая как лизин, аргинин или гистидин, заменена отрицательно заряженной аминокислотой, такой как аспарагиновая кислота или глутаминовая кислота. В некоторых вариантах осуществления отрицательно заряженная аминокислота на границе взаимодействия заменена положительно заряженной аминокислотой. В некоторых вариантах осуществления заряженные аминокислоты вводят в один из взаимодействующих константных доменов СН3-антитела или обоих. В некоторых вариантах осуществления введение заряженных аминокислот во взаимодействующие константные домены антитела СН3 двух мономеров домена Fc способствует селективному образованию гетеродимеров мономеров домена Fc, контролируемым эффектами электростатического взаимодействия, возникающими в результате взаимодействия между заряженными аминокислотами. Примеры электростатических взаимодействий аминокислотных пар включены, без ограничения, в Таблицу 11.
Таблица 11. Электростатические взаимодействия аминокислотных пар
K392D
K409D
K439E
D356K
D399K
Доступны другие способы, используемые для контроля гетеродимеризации мономеров домена Fc, особенно в контексте конструирования биспецифического антитела.
В некоторых вариантах осуществления первый мономер домена Fc и второй мономер домена Fc каждый содержит одну или более следующих аминокислотных замен: T366W, T366S, L368A, Y407V, T366Y, T394W, F405W, Y349T, Y349E, Y349V, L351T, L351H, L351N, L351K, P353S, S354D, D356K, D356R, D356S, E357K, E357R, E357Q, S364A, T366E, L368T, L368Y, L368E, K370E, K370D, K370Q, K392E, K392D, T394N, P395N, P396T, V397T, V397Q, L398T, D399K, D399R, D399N, F405T, F405H, F405R, Y407T, Y407H, Y407I, K409E, K409D, K409T и K409I; относительно последовательности IgG1 человека.
В некоторых вариантах осуществления мономер домена Fc содержит (а) одну из следующих аминокислотных замен по сравнению с IgG1 человека дикого типа: T366W, T366S, L368A, Y407V, T366Y, T394W, F405W, Y349T, Y349E, Y349V, L351T, L351H, L351N, L351K, P353S, S354D, D356K, D356R, D356S, E357K, E357R, E357Q, S364A, T366E, L368T, L368Y, L368E, K370E, K370D, K370Q, K392E, K392D, T394N, P395N, P396T, V397T, V397Q, L398T, D399K, D399R, D399N, F405T, F405H, F405R, Y407T, Y407H, Y407I, K409E, K409D, K409T, или K409I; или (b) (i) мутацию N297A по сравнению с областью Fc IgG1 человека; (ii) мутацию L234A, L235A и G237A по сравнению с областью Fc IgG1 человека; (iii) мутацию L234A, L235A, G237A и N297A по сравнению с областью Fc IgG1 человека; (iv) мутацию N297A по сравнению с областью Fc IgG2 человека; (v) мутацию A330S и P331S по сравнению с областью Fc IgG2 человека; (vi) мутацию A330S, P331S и N297A по сравнению с областью Fc IgG2 человека; (vii) мутацию S228P, E233P, F234V, L235A и delG236 по сравнению с областью Fc IgG4 человека, или (viii) мутацию S228P, E233P, F234V, L235A, delG236 и N297A по сравнению с областью Fc IgG4 человека. В некоторых вариантах осуществления мономер домена Fc содержит (а) одну из следующих аминокислотных замен по сравнению с IgG1 человека дикого типа: T366W, T366S, L368A, Y407V, T366Y, T394W, F405W, Y349T, Y349E, Y349V, L351T, L351H, L351N, L351K, P353S, S354D, D356K, D356R, D356S, E357K, E357R, E357Q, S364A, T366E, L368T, L368Y, L368E, K370E, K370D, K370Q, K392E, K392D, T394N, P395N, P396T, V397T, V397Q, L398T, D399K, D399R, D399N, F405T, F405H, F405R, Y407T, Y407H, Y407I, K409E, K409D, K409T, или K409I; и (b) дополнительно содержащий (i) мутацию N297A по сравнению с областью Fc IgG1 человека; (ii) мутацию L234A, L235A и G237A по сравнению с областью Fc IgG1 человека; (iii) мутацию L234A, L235A, G237A и N297A по сравнению с областью Fc IgG1 человека; (iv) мутацию N297A по сравнению с областью Fc IgG2 человека; (v) мутацию A330S и P331S по сравнению с областью Fc IgG2 человека; (vi) мутацию A330S, P331S и N297A по сравнению с областью Fc IgG2 человека; (vii) мутацию S228P, E233P, F234V, L235A и delG236 по сравнению с областью Fc IgG4 человека, или (viii) мутацию S228P, E233P, F234V, L235A, delG236 и N297A по сравнению с областью Fc IgG4 человека.
В некоторых вариантах осуществления первый и второй мономеры домена Fc содержат различные аминокислотные замены. В некоторых вариантах осуществления первый мономер домена Fc содержит T366W. В некоторых вариантах осуществления второй мономер домена Fc содержит T366S, L368A и Y407V. В некоторых вариантах осуществления первый мономер домена Fc содержит D399K. В некоторых вариантах осуществления второй мономер домена Fc содержит K409D.
IV. Сывороточный альбумин
В рамках изобретения, в некоторых вариантах осуществления, раскрыты полипептиды, содержащие: вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92.
В рамках изобретения, в некоторых вариантах осуществления также раскрыты полипептиды, содержащие вариант Fc, причем вариант Fc содержит димер домена Fc, имеющий два мономера домена Fc, при этом каждый мономер домена Fc независимо выбирают из (i) области Fc IgG1 человека, включающей в свой состав мутации L234A, L235A, G237A и N297A; (ii) области Fc IgG2 человека, включающей в свой состав мутации A330S, P331S и N297A; или (iii) области Fc IgG4 человека, содержащей мутации S228P, E233P, F234V, L235A, delG236 и N297A.
Слияние с сывороточными альбуминами может улучшить фармакокинетику белковых фармацевтических препаратов, и в некоторых вариантах осуществления полипептиды раскрытия, включая описанный в рамках изобретения высокоаффинный вариант D1 SIRP-α, соединены с сывороточным альбумином.
Сывороточный альбумин представляет собой глобулярный белок, который в изобилии содержится в крови у млекопитающих. Сывороточный альбумин продуцируется в печени и может составлять около половины белков сыворотки крови. Он мономерный и растворим в крови. Некоторые из наиболее важных функций сывороточного альбумина включают транспортировку гормонов, жирных кислот и других белков в организме, буферизацию рН и поддержание осмотического давления, необходимого для правильного распределения жидкостей организма между кровеносными сосудами и тканями организма. В предпочтительных вариантах осуществления сывороточный альбумин представляет собой сывороточный альбумин человека (ЧСА). В некоторых вариантах осуществления ЧСА соединяют с C-концом полипептида раскрытия для увеличения периода полужизни в сыворотке полипептида. В некоторых вариантах осуществления N-конец ЧСА соединяют с C-концом полипептида по данному описанию. В некоторых вариантах осуществления ЧСА соединяют либо непосредственно, либо через линкер с C-концом полипептида. В некоторых вариантах осуществления ЧСА соединяют либо непосредственно, либо через линкер с N-концом полипептида.
В некоторых вариантах осуществления сывороточный альбумин человека содержит последовательность аминокислот (ак) 25-609 UniProt ID NO: P02768 (SEQ ID NO: 12), как показано в таблице 12. В некоторых вариантах осуществления ЧСА, присоединенный к высокоаффинному варианту D1 SIRP-α (например, любой вариант D1 SIRP-α, описанный в таблицах 2, 5 и 6), содержит аминокислоты 25-609 (SEQ ID NO: 12) последовательности UniProt ID NO: P02768. В некоторых вариантах осуществления ЧСА содержит замены C34S или K573P по сравнению с SEQ ID NO:12. В некоторых вариантах осуществления ЧСА содержит замены C34S и K573P по сравнению с SEQ ID NO:12.
Таблица 12. Последовательность ЧСА
В некоторых вариантах осуществления сывороточный альбумин слит генетически с полипептидом данного изобретения или присоединен к полипептиду посредством химических средств, например химической конъюгации. В некоторых вариантах осуществления спейсер вставляется между полипептидом и ЧСА. Некоторые примеры спейсеров подробно описаны в рамках изобретения в другом месте. В некоторых вариантах осуществления спейсер представляет собой A или AAAL. В некоторых вариантах осуществления слияние ЧСА в полипептиде по данному описанию приводит к длительному удержанию полипептида, а также к увеличению периода полужизни.
Полипептиды, содержащие полипептид D1 SIRP-α и слитый ЧСА, включают, но не ограничиваются ими, SEQ ID NO: 150-159 приведенные в таблице 13.
Таблица 13. Полипептиды, содержащие варианты SIRP-α слитые с ЧСА
В некоторых вариантах осуществления полипептид содержит высокоаффинный домен D1 SIRP-α, имеющий по меньшей мере 85% идентичности последовательности (например, по меньшей мере 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности) с любым вариантом, приведенным в таблице 13.
В некоторых вариантах осуществления полипептид содержит высокоаффинный домен D1 SIRP-α, имеющий по меньшей мере 85% идентичности последовательности (например, по меньшей мере 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности) с SEQ ID NO: 154, 155 и 159 в таблице 13.
V. Альбумин-связывающий пептид
В рамках изобретения, в некоторых вариантах осуществления, раскрыты полипептиды, содержащие: вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92.В рамках изобретения, в некоторых вариантах осуществления также раскрыты полипептиды, содержащие вариант Fc, причем вариант Fc содержит димер домена Fc, имеющий два мономера домена Fc, при этом каждый мономер домена Fc независимо выбирают из (i) области Fc IgG1 человека, включающей в свой состав мутации L234A, L235A, G237A и N297A; (ii) области Fc IgG2 человека, включающей в свой состав мутации A330S, P331S и N297A; или (iii) области Fc IgG4 человека, содержащей мутации S228P, E233P, F234V, L235A, delG236 и N297A.
Связывание с белками сыворотки может улучшить фармакокинетику белковых фармацевтических препаратов, и, в частности, в некоторых вариантах осуществления описанные в рамках изобретения полипептиды слиты с белок-связывающими пептидами или белками.
Используемый в рамках изобретения термин «альбуминсвязывающий пептид» относится к аминокислотной последовательности, содержащей около 12-16 аминокислот, которая имеет сродство и функцию для связывания белка альбумина в сыворотке. В некоторых вариантах осуществления альбуминсвязывающий пептид происходит от человека, мыши или крысы.
В некоторых вариантах осуществления полипептид раскрытия, включающий высокоаффинный вариант D1 SIRP-α (например, любой вариант, представленный в таблицах 2, 5 и 6), слит с альбуминсвязывающим пептидом, который проявляет связывающую активность с сывороточным альбумином для увеличения периода полувыведения полипептида. Доступны различные альбуминсвязывающие пептиды, которые могут быть использованы в описанных в рамках изобретения способах и композициях. В некоторых вариантах осуществления пептид, альбуминсвязывающий, содержит последовательность DICLPRWGCLW (SEQ ID NO: 160). В некоторых вариантах осуществления альбуминсвязывающийся пептид слит генетически с полипептидом данного изобретения или прикрепляется к полипептиду с помощью химических средств, например химической конъюгации.
В некоторых вариантах осуществления линкер (например, спейсер) вставлен между полипептидом и альбуминсвязывающим пептидом, чтобы обеспечить дополнительную структурную и пространственную гибкость слитого белка. Конкретные линкеры (например, спейсер) и их аминокислотные последовательности подробно описаны в рамках изобретения далее. В некоторых вариантах осуществления альбуминсвязывающий пептид слит с N- или С-концом полипептида по данному описанию. В одном примере N-конец альбуминсвязывающего пептида непосредственно слит с С-концом полипептида раскрытия через пептидную связь. В другом примере C-конец альбуминсвязывающего пептида непосредственно слит с N-концом полипептида раскрытия через пептидную связь. В некоторых вариантах осуществления слияние альбуминсвязывающегося пептида с полипептидом раскрытия приводит к длительному удержанию полипептида посредством его связывания с сывороточным альбумином.
VI. Полимер полиэтиленгликоля (ПЭГ)
В рамках изобретения, в некоторых вариантах осуществления, раскрыты полипептиды, содержащие: вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92.
В рамках изобретения, в некоторых вариантах осуществления также раскрыты полипептиды, содержащие вариант Fc, причем вариант Fc содержит димер домена Fc, имеющий два мономера домена Fc, при этом каждый мономер домена Fc независимо выбирают из (i) области Fc IgG1 человека, включающей в свой состав мутации L234A, L235A, G237A и N297A; (ii) области Fc IgG2 человека, включающей в свой состав мутации A330S, P331S и N297A; или (iii) области Fc IgG4 человека, содержащей мутации S228P, E233P, F234V, L235A, delG236 и N297A.
В некоторых вариантах осуществления полипептид, содержащий высокоаффинный домен D1 SIRP-α (например, любой вариант, представленный в таблицах 2, 5 и 6), слит с полимером (например, полиэтиленгликолем, ПЭГ). В некоторых вариантах осуществления присоединение полимера к белковому препарату «маскирует» белковое фармацевтическое средство от иммунной системы хозяина. Кроме того, в некоторых вариантах осуществления некоторые полимеры, такие как гидрофильные полимеры, обеспечивают растворимость в воде гидрофобных белков и лекарственных средств. Например, в некоторых вариантах осуществления такие полимеры включают ПЭГ, полисиалевую кислотную цепь и молекулы цепи ПАС. В некоторых вариантах осуществления полимер, такой как ПЭГ, ковалентно присоединен к замене или добавлению цистеина в полипептиде. В некоторых вариантах замещение цистеина в полипептиде представляет собой I7C, A16C, S20C, T20C, A45C, G45C, G79C, S79C или A84C, относительно последовательности любой из последовательностей, представленных в таблицах 2, 5 и 6. В некоторых вариантах осуществления добавление остатка цистеина в полипептиде вводят с использованием пептидного синтеза, генетической модификации, молекулярного клонирования или любых их комбинаций. В некоторых вариантах осуществления полимер, например ПЭГ, присоединен к остатку цистеина с использованием конъюгации цистеин-малеимид. В некоторых вариантах осуществления полимер, такой как ПЭГ, ковалентно присоединен к полипептиду, содержащему высокоаффинный вариант D1 SIRP-α или на N-, или С-конце, либо во внутреннем положении, используя обычные химические способы, такие как химическое конъюгатирование.
VII. Биспецифическая конструкция
В рамках изобретения, в некоторых вариантах осуществления, раскрыты полипептиды, содержащие: вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92.
В рамках изобретения, в некоторых вариантах осуществления также раскрыты полипептиды, содержащие вариант Fc, причем вариант Fc содержит димер домена Fc, имеющий два мономера домена Fc, при этом каждый мономер домена Fc независимо выбирают из (i) области Fc IgG1 человека, включающей в свой состав мутации L234A, L235A, G237A и N297A; (ii) области Fc IgG2 человека, включающей в свой состав мутации A330S, P331S и N297A; или (iii) области Fc IgG4 человека, содержащей мутации S228P, E233P, F234V, L235A, delG236 и N297A.
В некоторых вариантах осуществления полипептид, имеющий высокоаффинный вариант D1 SIRP-α (например, любой из вариантов, представленных в таблицах 2, 5 и 6), содержит биспецифическую конструкцию. Биспецифическая конструкция относится к конструкции, которая имеет два домена, взаимодействующих с объектом. В некоторых вариантах осуществления биспецифическая конструкция содержит домен Fc и два домена, взаимодействующие с мишенями: (1) домен D1 SIRP-α или его вариант (например, любой из вариантов, представленных в таблицах 2, 5 и 6) и (2) вариабельный домен антитела. В некоторых вариантах осуществления биспецифическая конструкция содержит первый полипептид и второй полипептид. В некоторых вариантах осуществления первый полипептид имеет формулу A-L-B, где A включает домен D1 SIRP-α или его вариант, L прелставляет собой линкер, а B включает первый мономер домена Fc. В некоторых вариантах осуществления второй полипептид имеет формулу A'-L'-B', где A' включает вариабельный домен антитела, L' представляет собой линкер; а B 'включает второй мономер домена Fc. В некоторых вариантах осуществления ориентация первого и второго полипептидов представляет собой B-L-A и B'-L'-A', соответственно. В некоторых вариантах осуществления первый и второй мономеры домена Fc объединяются для образования домена Fc в биспецифической конструкции. В некоторых вариантах осуществления биспецифическая конструкция представляет собой любые изотипы антитела иммуноглобулина (например, IgG, IgE, IgM, IgA и IgD). Вариант домена D1 SIRP-α включает домен D1 SIRP-α человека дикого типа и одну или более аминокислотных замен относительно доменов D1 дикого типа (например, любой вариант D1 SIRP-α, как описано в таблицах 2, 5 и 6). В некоторых вариантах осуществления вариант D1 SIRP-α связывается с более высокой аффинностью связывания с CD47, чем домен D1 SIRP-α человека дикого типа. В некоторых вариантах осуществления вариабельный домен антитела в биспецифической конструкции нацелен на клеточный антиген (например, клеточный антиген в раковой клетке).
Вариабельный домен антитела относится к частям легкой и тяжелой цепей антитела, которые содержат аминокислотные последовательности областей, определяющих комплементарность (CDR, например, CDR L1, CDR L2, CDR L3, CDR H1, CDR H2 и CDR H3) и каркасных областей (FR). Вариабельный домен антитела может придать антителу способность связываться с конкретными антигенами. Могут быть созданы многие различные молекулы вариабельного домена антитела. В некоторых вариантах осуществления используемые молекулы вариабельного домена антитела включают, но не ограничиваются ими, одноцепочечные Fv.
В некоторых вариантах осуществления вариабельный домен антитела в биспецифической конструкции нацелен на клеточный антиген (например, клеточный антиген в раковой клетке или иммунную клетку). Некоторые белки экспрессируются на более высоких уровнях в раковых клетках, чем в нераковых клетках. Например, раковый антиген представляет собой белок, который предпочтительно экспрессируется раковыми клетками (например, он экспрессируется на более высоких уровнях на раковых клетках, чем на нераковых клетках), а в некоторых случаях он экспрессируется исключительно раковыми клетками. В некоторых вариантах осуществления белки, например, белки, экспрессируемые раковыми клетками, которые нацелены на вариабельный домен антитела, образующий домен Fc с высокоаффинным доменом SIRP-α-или его вариантом, включают, но не ограничиваются ими: 5T4, AGS-16, ALK1, ANG-2, B7-H3, B7-H4, c-fms, c-Met, CA6, CD123, CD19, CD20, CD22, EpCAM, CD30, CD32b, CD33, CD37, CD38, CD40, CD52, CD70, CD74, CD79b, CD98, CEA, CEACAM5, CLDN18.2, CLDN6, CS1, CXCR4, DLL-4, EGFR, EGP-1, ENPP3, EphA3, ETBR, FGFR2, фибронектин, FR-альфа, GCC, GD2, глипикан-3, GPNMB, HER-2, HER3, HLA-DR, ICAM-1, IGF-1R, ИЛ-3R, LIV-1, мезотелин, MUC16, MUC1, NaPi2b, нектин-4, Notch 2, Notch 1, PD-L1, PD-L2, PDGFR-a, PS, PSMA, SLTRK6, STEAP1, TEM1, VEGFR, CD25, CD27L, DKK-1 или CSF-1R. В некоторых вариантах осуществления вариабельный домен антитела в биспецифической конструкции не предназначен для связывания белка человека.
В некоторых вариантах осуществления каждый из первого и второго мономеров домена Fc в домене Fc биспецифической конструкции содержит одну или более аминокислотных замен, которые способствуют гетеродимеризации первого и второго мономеров домена Fc. Способы промотирования гетеродимеризации мономеров доменов Fc подробно описаны далее в рамках изобретения, см., например, стратегию «выступ во впадину» и стратегию электростатического взаимодействия.
В некоторых вариантах осуществления домен Fc биспецифической конструкции мутирован с отсутствием одной или более эффекторных функций, типичных для «мертвого домена Fc». В некоторых вариантах осуществления домен Fc биспецифической конструкции представляет собой антитело IgG1 и содержит аминокислотные замены L14A, L15A и G17A по отношению к последовательности SEQ ID NO: 161 (таблица 14) для уменьшения взаимодействия или связывания между Fc и рецептор Fcγ. В некоторых вариантах осуществления мономер домена Fc представляет собой антитело IgG1 и содержит одну или более аминокислотных замен L234A, L235A, G237A и N297A (обозначенных в соответствии с системой нумерации EU Kabat et al., 1991). В некоторых вариантах осуществления варианты Fc, описанные в рамках изобретения, являются минимально гликозилированными или имеют уменьшенное гликозилирование. В некоторых вариантах осуществления дегликозилирование осуществляют посредством мутации N297A или посредством мутации N297 с любой аминокислотой, которая не является N ((обозначенных в соответствии с системой нумерации EU Kabat et al. (1991)). В некоторых вариантах осуществления биспецифическая конструкция сконструирована таким образом, что она имеет преимущественное связывание с белками (например, рецепторами, такими как рецепторы Fc), экспрессируемыми различными типами клеток. Исследования показали, что аминокислотные замены в шарнире, константных доменах (например, константных доменах CH2 и CH3) или шарнирные и константные домены антитела могут эффективно изменять аффинности связывания антитела по отношению к специфическим рецепторам (например, рецепторам Fc), экспрессированным на различных типах клеток (например, регуляторные Т-клетки и эффекторные Т-клетки). IgG2, имеющий аминокислотные замены A111S и P112S (относительно SEQ ID NO: 162, таблица 14), демонстрирует значительно уменьшенное связывание с FcγRIIIa 131 H по сравнению с IgG2 дикого типа. В некоторых вариантах осуществления варианты Fc, описанные в рамках изобретения, являются минимально гликозилированными или имеют уменьшенное гликозилирование. В некоторых вариантах осуществления дегликозилирование осуществляют посредством мутации N297A или посредством мутации N297 с любой аминокислотой, которая не является N ((обозначенных в соответствии с системой нумерации EU Kabat et al. (1991)). В некоторых вариантах осуществления биспецифическая конструкция содержит домен Fc подкласса IgG2 или IgG4. В некоторых вариантах осуществления биспецифическая конструкция, содержащая домен Fc подкласса IgG2, содержит аминокислотные замены A111S и P112S относительно SEQ ID NO: 162 (таблица 14). В некоторых вариантах осуществления вариант Fc содержит последовательность Fc IgG2 человека, содержащую одну или более аминокислотных замен A330S, P331S и N297A (обозначенных в соответствии с системой нумерации EU Kabat et al. (1991)).
Таблица 14. Аминокислотные последовательности IgG
Пример конструкции SIRP-α, содержащей домен D1 SIRP-α или его вариант, присоединенный к первому мономерному домену Fc посредством линкера и второго мономера домена Fc, в котором первый и второй мономеры домена Fc объединяются, чтобы сформировать область Fc, представленную на фиг. 1. В некоторых вариантах осуществления не существует вариабельного домена белка или антитела, присоединенного ко второму мономеру Fc. В некоторых вариантах осуществления конструкция SIRP-α содержит домен D1 SIRP-α или его вариант, присоединенный к первому мономеру домена Fc посредством линкера и вариабельного домена антитела, присоединенного ко второму мономеру домена Fc посредством линкера, в который первый и второй мономеры домена Fc объединяются, чтобы сформировать домен Fc (как показано на фиг. 2). В некоторых вариантах осуществления конструкция SIRP-α содержит домен D1 SIRP-α или его вариант, присоединенный к первому мономеру домена Fc посредством линкера и терапевтического белка (например, цитокина, интерлейкина, антигена, стероида, противовоспалительного агента или иммуномодулирующего агента), присоединенный ко второму мономерному домену Fc посредством линкера, в котором первый и второй мономеры домена Fc объединяются для образования домена Fc (как показано на фиг. 3). В некоторых вариантах осуществления каждый из двух мономеров домена Fc в домене Fc конструкций SIRP-α, описанных ранее (например, конструкции SIRP-α, как показано на фиг. 1-3), включает аминокислотные замены, которые способствуют гетеродимеризации двух мономеров. Различные стратегии (например, стратегия «выступ во впадину», стратегия электростатического взаимодействия) и аминокислотные замены домена Fc, которые способствуют гетеродимеризации двух мономеров домена Fc, подробно описаны в рамках изобретения. Например, на фиг. 4A проиллюстрирована конструкция SIRP-α, имеющая домен D1 SIRP-α или его вариант, присоединенный к домену Fc мономера, содержащий мутации выступа, например, T366W, чтобы ограничить образование нежелательного образования гомодимера выступ-выступ. На фиг. 4B проиллюстрирована конструкция SIRP-α, имеющая домен D1 SIRP-α или его вариант, присоединенный к мономерному домену Fc, включая мутации впадины, например, T366S, L358A и Y407V. В некоторых вариантах осуществления аналогичные стратегии гетеродимеризации домена Fc применяются к доменам Fc в конструкциях, описанных на фиг. 2 и 3. В некоторых вариантах осуществления конструкция SIRP-α содержит слитый белок домена D1 SIRP-α или его вариант, присоединенный к мономеру домена Fc (как показано на фиг. 5A). В некоторых вариантах осуществления этот слитый белок образует гомодимер (как показано на фиг. 5B).
Варианты Fc раскрытия, связанные с партнером по слиянию, предпочтительно проявляют уменьшенное или аблированное связывание, по меньшей мере, с одним из рецепторов Fcγ CD16a, CD32a, CD32b, CD32c и CD64 по сравнению с аналогичной полипептидной конструкцией, содержащей нативную область Fc антитела или область Fc антитела дикого типа (немутированную). В некоторых случаях описанный в рамках изобретения вариант Fc или партнера по слиянию демонстрируют уменьшенное или аблированное связывание с Fcγ рецепторами CD16a, CD32a, CD32b, CD32c и CD64.
В некоторых вариантах осуществления варианты Fc описания, связанные с партнером по слиянию, демонстрируют восстановленное связывание с компонентом C1q и CDC комплемента по сравнению с аналогичной полипептидной конструкцией, содержащей природную или не мутированную область Fc. В некоторых вариантах осуществления вариант Fc демонстрирует, по меньшей мере 5%, 10%, 15%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или большее сокращения в связывании C1q по сравнению с полипептидной конструкцией, содержащей область Fc дикого типа. В некоторых случаях вариант Fc демонстрирует уменьшенный CDC по сравнению с полипептидной конструкцией, содержащей природную или немутантную область Fc. В некоторых вариантах осуществления вариант Fc демонстрирует, по меньшей мере 5%, 10%, 15%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или большее сокращения в связывании CDC по сравнению с полипептидной конструкцией, содержащей область Fc дикого типа.
VIII. Линкеры
В рамках изобретения, в некоторых вариантах осуществления, раскрыты полипептиды, содержащие: вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92.
В рамках изобретения, в некоторых вариантах осуществления также раскрыты полипептиды, содержащие вариант Fc, причем вариант Fc содержит димер домена Fc, имеющий два мономера домена Fc, при этом каждый мономер домена Fc независимо выбирают из (i) области Fc IgG1 человека, включающей в свой состав мутации L234A, L235A, G237A и N297A; (ii) области Fc IgG2 человека, включающей в свой состав мутации A330S, P331S и N297A; или (iii) области Fc IgG4 человека, содержащей мутации S228P, E233P, F234V, L235A, delG236 и N297A.
В настоящем описании линкер используется для описания связи или соединения между полипептидами или белковыми доменами или связанными с ними белковыми фрагментами. В некоторых вариантах осуществления линкер представляет собой соединение или связь между мономерным доменом Fc, пептидом, связывающим альбумин, или ЧСА, и с высокоаффинным вариантом D1 SIRP-α. В некоторых вариантах осуществления линкер соединяет С-конец варианта D1 SIRP-α и N-конец мономера домена Fc, пептида, связывающего альбумин, или ЧСА, так что два полипептида соединены друг с другом в тандемные серии.
В некоторых вариантах осуществления линкер представляет собой простую ковалентную связь, например, пептидную связь, синтетический полимер, такой как полимер полиэтиленгликоля (ПЭГ), или любую связь, образованную из химической реакции, например, химическое сопряжение. Когда линкером является пептидная связь, в некоторых вариантах осуществления группа карбоновой кислоты на С-конце одного домена белка реагирует с аминогруппой на N-конце другого белкового домена в реакции конденсации с образованием пептидной связи. В некоторых вариантах осуществления пептидная связь образуется из синтетических средств посредством обычной реакции органической химии или путем естественного получения из клетки-хозяина, где молекула нуклеиновой кислоты, кодирующая последовательности ДНК обоих белков (например, мономер домена Fc и высокоаффинный вариант D1 SIRP-α) в тандемных сериях могут быть непосредственно транскрибированы и переведены в смежный полипептид, кодирующий оба белка с помощью необходимых молекулярных механизмов (например, ДНК-полимеразы и рибосомы) в клетке-хозяине.
Когда линкер представляет собой синтетический полимер (например, полимер ПЭГ), в некоторых вариантах осуществления полимер функционализирован реакционно-способными химическими функциональными группами на каждом конце для взаимодействия с концевыми аминокислотами на соединительных концах двух белков.
Когда линкером (за исключением упомянутой выше пептидной связи) является химическая реакция, в некоторых вариантах осуществления химические функциональные группы (например, амин, карбоновая кислота, сложный эфир, азид или другие функциональные группы) присоединены синтетически к С-концу одного белка и N-конца другого белка, соответственно. В некоторых вариантах осуществления две функциональные группы затем реагируют с помощью синтетической химии, чтобы образовать химическую связь, тем самым соединяя два белка вместе.
Спейсеры
В настоящем описании в некоторых вариантах осуществления линкер между мономером домена Fc, альбуминсвязывающим пептидом или ЧСА и полипептидом раскрытия представляет собой аминокислотный спейсер, содержащий около 1-200 аминокислот. Подходящие пептидные спейсеры включают пептидные линкеры, содержащие гибкие аминокислотные остатки, такие как глицин и серин. Примеры линкерных последовательностей приведены в таблице 15. В некоторых вариантах осуществления спейсер содержит мотивы, например, множественные или повторяющиеся мотивы, GS, GG, GGS, GGG, GGGGS (SEQ ID NO: 163), GGSG (SEQ ID NO: 164) или SGGG (SEQ ID NO: 165). В некоторых вариантах осуществления спейсер содержит от 2 до 12 аминокислот, включая мотивы GS, например, GS, GSGS (SEQ ID NO: 166), GSGSGS (SEQ ID NO: 167), GSGSGSGS (SEQ ID NO: 168), GSGSGSGSGS (SEQ ID NO: 169) или GSGSGSGSGSGS (SEQ ID NO: 170). В некоторых вариантах осуществления спейсер содержит от 3 до 12 аминокислот, включая мотивы GGS, например, GGS, GGSGGS (SEQ ID NO: 171), GGSGGSGGS (SEQ ID NO: 172) и GGSGGSGGSGGS (SEQ ID NO: 173). В некоторых вариантах осуществления спейсер содержит от 4 до 12 аминокислот, включая мотивы GGSG (SEQ ID NO: 164), например, GGSG (SEQ ID NO: 164), GGSGGGSG (SEQ ID NO: 174) или GGSGGGSGGGSG (SEQ ID NO: 175). В некоторых вариантах осуществления спейсер содержит мотивы GGGGS (SEQ ID NO: 163), например, GGGGSGGGGSGGGGS (SEQ ID NO: 176). В некоторых вариантах осуществления спейсер содержит аминокислоты, отличные от глицина и серина, напрмер, AAS (SEQ ID NO: 177), AAAL (SEQ ID NO: 178), AAAK (SEQ ID NO: 179), AAAR (SEQ ID NO: 180), EGKSSGSGSESKST (SEQ ID NO: 181), GSAGSAAGSGEF (SEQ ID NO: 182), AEAAAKEAAAKA (SEQ ID NO: 183), KESGSVSSEQLAQFRSLD (SEQ ID NO: 184), GGGGAGGGG (SEQ ID NO: 185), GENLYFQSGG (SEQ ID NO: 186), SACYCELS (SEQ ID NO: 187), RSIAT (SEQ ID NO: 188), RPACKIPNDLKQKVMNH (SEQ ID NO: 189), GGSAGGSGSGSSGGSSGASGTGTAGGTGSGSGTGSG (SEQ ID NO: 190), AAANSSIDLISVPVDSR (SEQ ID NO: 191) или GGSGGGSEGGGSEGGGSEGGGSEGGGSEGGGSGGGS (SEQ ID NO: 192).
В некоторых вариантах осуществления спейсер содержит мотивы, например, множественные или повторяющиеся мотивы EAAAK (SEQ ID NO: 193). В некоторых вариантах осуществления спейсер содержит мотивы, например, множественные или повторяющиеся мотивы, богатых пролином последовательностей, таких как (XP)n, в которых X представляет собой любую аминокислоту (например, A, K или E) и n равен 1-5, и PAPAP(SEQ ID NO: 194).
Таблица 15. Последовательности линкера
В некоторых вариантах осуществления длина пептидного спейсера и используемых аминокислот регулируется в зависимости от двух вовлеченных белков и степени гибкости, требуемой в конечном полипептиде слияния белка. В некоторых вариантах осуществления длина спейсера регулируется для обеспечения правильного сгибания белка и предотвращения образования агрегатов. В некоторых вариантах осуществления спейсер, такой как спейсер между ЧСА и полипептидом, описанным в рамках изобретения, представляет собой A или AAAL (SEQ ID NO: 178).
IX. Векторы, клетки-хозяева и получение белков
В рамках изобретения, в некоторых вариантах осуществления, раскрыты полипептиды, содержащие: вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92.
В рамках изобретения, в некоторых вариантах осуществления также раскрыты полипептиды, содержащие вариант Fc, причем вариант Fc содержит димер домена Fc, имеющий два мономера домена Fc, при этом каждый мономер домена Fc независимо выбирают из (i) области Fc IgG1 человека, включающей в свой состав мутации L234A, L235A, G237A и N297A; (ii) области Fc IgG2 человека, включающей в свой состав мутации A330S, P331S и N297A; или (iii) области Fc IgG4 человека, содержащей мутации S228P, E233P, F234V, L235A, delG236 и N297A.
В некоторых вариантах осуществления полипептиды раскрытия получают из клетки-хозяина. Клетка-хозяин относится к носителю, который содержит необходимые клеточные компоненты, например, органеллы, необходимые для экспрессии полипептидов и гибридных полипептидов, описанных в рамках изобретения, из их соответствующих нуклеиновых кислот. В некоторых вариантах осуществления нуклеиновые кислоты включены в векторы нуклеиновой кислоты, вводимые в клетку-хозяина путем трансформации, трансфекции, электропорации, осаждения фосфатом кальция, прямой микроинъекции, инфекции и т.д. В некоторых вариантах осуществления выбор векторов нуклеиновых кислот зависит от используемой клетки-хозяина. В некоторых вариантах осуществления клетки-хозяева имеют либо прокариотическое (например, бактерии), либо эукариотическое (например, клетки млекопитающих) происхождение.
В некоторых вариантах осуществления полипептид, например полипептидная конструкция, содержащая вариант D1 SIRP-α (например, любой вариант, представленный в таблицах 2, 5 и 6), и партнер слияния, такой как вариант Fc, ЧСА и альбуминсвязывающий пептид получают путем культивирования клетки-хозяина, трансформированной нуклеиновой кислотой, предпочтительно экспрессионным вектором, содержащей нуклеиновую кислоту, кодирующую полипептидную конструкцию (например, вариант Fc, линкер и партнер слияния) в соответствующих условиях, чтобы индуцировать или вызывать экспрессию полипептидной конструкции. В некоторых вариантах осуществления условия, подходящие для экспрессии, варьируют в зависимости от вектора экспрессии и выбранной клетки-хозяина. В некоторых вариантах осуществления используется большое количество подходящих клеток-хозяев, включая, но не ограничиваясь ими, клетки млекопитающих, бактерии, клетки насекомых и дрожжи. Например, различные линии клеток, которые находят применение в настоящем раскрытии, описаны в каталоге клеточной линии ATCC®, доступном в Американской коллекции типовых культур. В некоторых вариантах осуществления варианты Fc данного раскрытия экспрессируются в клетке, которая оптимизирована не для гликозилированных белков, которые экспрессируются такой клеткой, либо путем генной инженерии клеточной линии, либо с изменением условий культивирования клеток, таких как добавление кифунзина или с использованием естественно негликозилирующих хозяев, таких как прокариоты (E.coli и т. д.), и в некоторых случаях модификация последовательности гликозилирования в Fc не требуется.
Создание вектора нуклеиновой кислоты и клеток-хозяев
Последовательность нуклеиновой кислоты, кодирующая аминокислотную последовательность полипептида по изобретению, может быть получена различными способами. Эти способы включают, но не ограничиваются ими, олигонуклеотид-опосредованный (или сайт-направленный) мутагенез и мутагенез ПЦР. В некоторых вариантах осуществления молекула нуклеиновой кислоты, кодирующая полипептид, раскрывается с использованием стандартных методик, например, генного синтеза. Альтернативно, молекула нуклеиновой кислоты, кодирующая домен D1 SIRP-α дикого типа, мутируется с включением конкретных аминокислотных замен с использованием стандартных методик, например мутагенеза QuikChangeTM. В некоторых случаях молекулы нуклеиновых кислот синтезируются с использованием синтезатора нуклеотидов или методик ПЦР.
В некоторых вариантах осуществления нуклеиновые кислоты, которые кодируют полипептидную конструкцию, например, полипептидную конструкцию, содержащую вариант D1 SIRP-α (например, любой вариант, представленный в таблицах 2, 5 и 6), и партнера по слиянию, такого как вариант Fc, ЧСА и альбуминсвязывающий пептид, включены в экспрессирующий вектор, чтобы экспрессировать белок. Для экспрессии белка можно использовать множество векторов экспрессии. Экспрессионные векторы могут включать самореплицирующиеся, внехромосомные векторы или векторы, которые интегрируются в геном хозяина. Вектор может также содержать различные компоненты или элементы. Например, в некоторых вариантах осуществления векторные компоненты включают, но не ограничиваются ими, транскрипционные и трансляционные регуляторные последовательности, такие как промоторная последовательность, сайт связывания рибосом, сигнальная последовательность, последовательности начала и остановки транскрипции, последовательности начала и остановки трансляции, 3 'и 5' нетранслируемых областей (UTR), и энхансерные или активаторные последовательности; точки начала репликации; ген маркера селекции; и последовательность нуклеиновой кислоты, кодирующую представляющий интерес полипептид, и последовательность терминации транскрипции. В некоторых вариантах осуществления векторы экспрессии содержат белок, функционально связанный с контрольными или регуляторными последовательностями, выбираемыми маркерами, любыми партнерами по слиянию, дополнительными элементами или любыми их комбинациями. Термин нуклеиновая кислота является "функционально связанной", означает, что нуклеиновая кислота вступает в функциональное взаимоотношение с другой последовательностью нуклеиновой кислоты. Как правило, эти векторы экспрессии включают транскрипционную и трансляционную регуляторную нуклеиновую кислоту, функционально связанную с нуклеиновой кислотой, кодирующей вариант Fc, и обычно подходят для клетки-хозяина, используемой для экспрессии белка. Ген или маркер селекции, такой как, но не ограничиваясь этим, ген устойчивости к антибиотику или ген флуоресцентного белка, можно использовать для отбора клеток-хозяев, содержащих вектор экспрессии, например, путем экспрессии антибиотика или флуоресценции. Доступны различные селективные гены.
В некоторых вариантах осуществления компоненты или элементы вектора оптимизированы таким образом, что векторы экспрессии совместимы с типом клетки-хозяина. Экспрессионные векторы, которые находят применение в настоящем описании, включают, но не ограничиваются ими, те, которые обеспечивают экспрессию белка в клетках млекопитающих, бактериях, клетках насекомых, дрожжах и в системах in vitro.
В некоторых вариантах осуществления клетки млекопитающих используют в качестве клеток-хозяев для получения полипептидов раскрытия. Примеры типов клеток млекопитающих включают, но не ограничиваются ими, клетку почки эмбриона человека (HEK) (например, HEK293, HEK 293F), клетки яичника китайского хомячка (CHO), HeLa, COS, PC3, Vero, MC3T3, NS0, Sp2/0, VERY, BHK, MDCK, W138, BT483, Hs578T, HTB2, BT20, T47D, NS0 (клеточная линия миеломы мыши, которая не эндогенно продуцирует любые цепи иммуноглобулина), CRL7O3O и HsS78Bst. В некоторых вариантах осуществления клетки E. coli используют в качестве клеток-хозяев для получения полипептидов раскрытия. Примеры штаммов E.coli включают, но не ограничиваются ими, E. coli 294 (ATCC® 31,446), E. coli λ 1776 (ATCC® 31,537, E. coli BL21 (DE3) (ATCC® BAA-1025) и E. coli RV308 (ATCC® 31,608).
Различные клетки-хозяева обладают характерными и специфическими механизмами посттрансляционного процессинга и модификации белков и продуктов генов. В некоторых вариантах осуществления подходящие клеточные линии или хост-системы выбраны для обеспечения правильной модификации и обработки экспрессируемого полипептида. После того как векторы вводятся в клетки-хозяева для производства белка, клетки-хозяева культивируют на обычных питательных средах, модифицированных по мере необходимости для индуцирования промоторов, отбора трансформантов или амплификации генов, кодирующих желаемые последовательности.
В некоторых вариантах осуществления полипептидная конструкция, например, полипептидная конструкция, содержащая вариант D1 SIRP-α (например, любой вариант, представленный в таблицах 2, 5 и 6), и партнер слияния, такой как вариант Fc, ЧСА и альбуминсвязывающий пептид, экспрессируются в системах экспрессии млекопитающих, включая системы, в которых экспрессирующие конструкции вводят в клетки млекопитающих с использованием вируса, такого как ретровирус или аденовирус. В некоторых вариантах осуществления используются клетки человека, мыши, крысы, хомячка или приматов. Подходящие клетки также включают известные исследовательские клетки, включая, но не ограничиваясь ими, клетки Jurkat T, NIH3T3, CHO, COS и 293. Альтернативно, в некоторых вариантах осуществления белки экспрессируются в бактериальных клетках. Бактериальные системы экспрессии хорошо известны в данной области техники и включают Escherichia coli (E. coli), Bacillus subtilis, Streptococcus cremoris и Streptococcus lividans. В некоторых случаях полипептидные конструкции, содержащие варианты Fc, продуцируются в клетках насекомых, таких как, но не ограничиваясь ими, клетки Sf9 и Sf21 или дрожжевые клетки, такие как, но не ограничиваясь ими, организмы из родов Saccharomyces, Pichia, Kluyveromyces, Hansenula и Yarrowia. В некоторых случаях полипептидные конструкции, содержащие варианты Fc, экспрессируются in vitro с использованием бесклеточных систем трансляции. Системы трансляции in vitro, полученные как из прокариотических (например, E. coli), так и эукариотических клеток (например, ростков пшеницы, кроличьих ретикулоцитов), доступны и, в некоторых вариантах осуществления, выбираются на основе уровней экспрессии и функциональных свойств представляющего интерес белка. Например, как оценивают специалисты в данной области техники, трансляция in vitro необходима для некоторых технологий дисплея, например, рибосомного дисплея. Кроме того, в некоторых вариантах осуществления варианты Fc получают способами химического синтеза, такими как, но не ограничиваясь ими, жидкофазный пептидный синтез и твердофазный пептидный синтез. В случае транскрипции in vitro с использованием негликозилирующей системы, такой как бактериальные экстракты, Fc не будет гликозилирован даже в присутствии природного сайта гликозилирования, и поэтому инактивация Fc будет получена эквивалентно.
В некоторых вариантах осуществления полипептидная конструкция включает неприродные аминокислоты, аминокислотные аналоги, аминокислотные миметики или любые их комбинации, которые функционируют аналогично природным аминокислотам. Природными аминокислотами обычно называют 20 общепринятых аминокислот (аланин, аргинин, аспарагин, аспарагиновая кислота, цистеин, глутамин, глютаминовая кислота, глицин, гистидин, изолейцин, лейцин, лизин, метионин, фенилаланин, пролин, серин, треонин, триптофан, тирозин и валин) и пиролизин и селеноцистеин. Аминокислотные аналоги относятся к соединениям, которые имеют основную химическую структуру, идентичную с аминокислотой, встречающейся в природе, т.е., углерод, который связан с водородом, карбоксильная группа, амино группа, и R группа, например, гомосерин, норлейцин, метионилсульфоксид, метионинметилсульфоний. В некоторых вариантах осуществления такие аналоги могут иметь модифицированные R группы (например, норлейцин) или модифицированные пептидные остовы, но сохраняют такую же основную химическую структуру, что и аминокислота, встречающаяся в природе.
Получение, восстановление и очистка белка
В некоторых вариантах осуществления клетки-хозяева, используемые для получения полипептидов раскрытия, растут на средах, подходящих для культивирования выбранных клеток-хозяев. Примеры подходящих сред для клеток-хозяев млекопитающих включают минимальную основную среду (MEM), среду Игла, модифицированную по Дульбекко (DMEM), экспрессионную среду Expi293™, DMEM с добавленной фетальной бычьей сывороткой (ФБС) и RPMI-1640. Примеры подходящей среды для бактериальных клеток-хозяев включают бульон Luria (LB) плюс необходимые добавки, такие как селективный агент, например, ампициллин. В некоторых вариантах осуществления клетки-хозяева культивируют при подходящих температурах, например, от около 20°С до около 39°С, например, от около 25°С до около 37°С, предпочтительно, 37°С, и уровней СО2, таких как около 5%-10%. В некоторых вариантах осуществления рН среды составляет от около 6,8 до 7,4, например, рН 7,0, в зависимости, главным образом, от организма-хозяина. Если в экспрессирующем векторе используют индуцибельный промотор, экспрессию белка можно индуцировать в условиях, подходящих для активации промотора.
В некоторых вариантах осуществления восстановление белка включает разрушение клетки-хозяина, например, путем осмотического шока, ультразвука или лизиса. Как только клетки разрушаются, клеточный дебрис удаляют центрифугированием или фильтрацией. Затем белки могут быть дополнительно очищены. В некоторых вариантах осуществления полипептид изобретения очищают различными способами очистки белков, например, хроматографией (например, ионообменной хроматографией, аффинной хроматографией и хроматографией с исключением по размеру), центрифугированием, дифференциальной растворимостью или любой другой стандартной методикой очистки белков. Например, в некоторых вариантах осуществления белок выделяют и очищают путем соответствующего отбора и объединения аффинных колонок, таких как колонка с белком А (например, хроматография POROS-белок А) с хроматографическими колонками (например, катионообменная хроматография POROS HS-50), фильтрация, ультрафильтрация, процедуры деминерализации и диализа. В некоторых вариантах осуществления полипептид конъюгирован с маркерными последовательностями, такими как пептид для облегчения очистки. Примером маркерной аминокислотной последовательности является гексагистидиновый пептид (His6-tag), который может связываться с никель-функционализованной аффинной колонкой с микромолярной аффинностью. В качестве альтернативы можно использовать метку гемагглютинина «HA», которая соответствует эпитопу, полученному из белка гемагглютинина гриппа.
В некоторых вариантах осуществления полипептиды раскрытия, например полипептидная конструкция, содержащая вариант D1 SIRP-α (например, любой вариант, представленный в таблицах 2, 5 и 6), и партнер по слиянию, такой как вариант Fc, ЧСА и альбуминсвязывающий пептид, продуцируются клетками пациента (например, человека), например, в контексте генной терапии, путем введения вектора, такого как вирусный вектор (например, ретровирусного вектора, аденовирусного вектора, поксвирусного вектора (например, вектор вируса коровьей оспы, такой как модифицированная вакцинация Анкара (MVA)), аденоассоциированный вирусный вектор и альфавирусный вектор), содержащий молекулу нуклеиновой кислоты, кодирующую полипептид раскрытия. Вектор, когда он находится внутри клетки пациента (например, путем трансформации, трансфекции, электропорации, осаждения фосфатом кальция, прямой микроинъекции, инфекции и т.д.) может быть использован для экспрессии описанного в рамках изобретения полипептида. В некоторых случаях полипептид выделяют из клетки. В некоторых вариантах осуществления, если лечение заболевания или расстройства является желаемым результатом, никаких дополнительных действий не требуется. В некоторых вариантах осуществления, если требуется сбор белка, кровь собирается от пациента и белок, очищают от крови различными способами.
X. Фармацевтические композиции и препараты
В рамках изобретения, в некоторых вариантах осуществления, раскрыты полипептиды, содержащие: вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92.
В рамках изобретения, в некоторых вариантах осуществления также раскрыты полипептиды, содержащие вариант Fc, причем вариант Fc содержит димер домена Fc, имеющий два мономера домена Fc, при этом каждый мономер домена Fc независимо выбирают из (i) области Fc IgG1 человека, включающей в свой состав мутации L234A, L235A, G237A и N297A; (ii) области Fc IgG2 человека, включающей в свой состав мутации A330S, P331S и N297A; или (iii) области Fc IgG4 человека, содержащей мутации S228P, E233P, F234V, L235A, delG236 и N297A.
Раскрытие изобретения включает фармацевтические композиции, которые содержат описанные в рамках изобретения полипептиды, такие как полипептиды, имеющие высокоаффинный вариант D1 SIRP-α. В некоторых вариантах осуществления фармацевтическая композиция изобретения содержит полипептид раскрытия в качестве терапевтического белка. В некоторых вариантах осуществления фармацевтическая композиция раскрытия, содержащая описанный в рамках изобретения полипептид, используется в комбинации с другими агентами или композициями (например, терапевтическими агентами, биологическими веществами, низкомолекулярными веществами или любыми их комбинациями) в терапии. В некоторых вариантах осуществления один или более дополнительных терапевтически активных агентов, таких как, например, низкомолекулярные вещества, химическое соединение или биологическое соединение, такое как полинуклеотиды и полипептиды, включая, но не ограничиваясь ими, миРНК, короткие полипептиды и антитела с терапевтической активностью, являются необязательными для в фармацевтических композиций полипептидов, описанных в рамках изобретения. В некоторых вариантах осуществления композиции полипептидных конструкций, описанные в рамках изобретения, готовят для хранения путем смешивания описанной в рамках изобретения полипептидной конструкции с требуемой степенью чистоты с необязательными фармацевтически приемлемыми носителями, эксципиентами или стабилизаторами в форме лиофилизированных составов или водных растворов. В некоторых вариантах осуществления фармацевтическая композиция раскрытия содержит молекулу нуклеиновой кислоты (ДНК или РНК, например, мРНК), кодирующую полипептид раскрытия, или вектор, содержащий такую молекулу нуклеиновой кислоты.
Приемлемые носители, эксципиенты или стабилизаторы в фармацевтической композиции предпочтительно являются нетоксичными для реципиентов при вводимых дозах и концентрациях. В некоторых вариантах осуществления приемлемые носители, эксципиенты и стабилизаторы включают буферы, такие как фосфатный, цитратный, HEPES, TAE и другие органические кислоты; антиоксиданты, такие как аскорбиновая кислота и метионин; консерванты (например, гексаметонийхлорид, хлорид октадецилдиметилбензиламмония, хлорид бензалкония, бензетонийхлорид, фенол, бутилбензиловый спирт, алкилпарабены, такие как метил или пропилпарабен, катехол, резорцин, циклогексанол, 3-пентанол и м-крезол); низкомолекулярные (например, менее 10 остатков) полипептиды; белки, такие как человеческий сывороточный альбумин, желатин, декстран и иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, гистидин и лизин; моносахариды, дисахариды и другие углеводы, такие как глюкоза, манноза, сахароза и сорбит; хелатирующие агенты, такие как сахара ЭДТК, такие как сахароза, маннит, трегалоза или сорбит; подсластители и другие ароматизаторы; наполнители, такие как микрокристаллическая целлюлоза, лактоза, кукурузный и другие крахмалы; связывающие агенты; добавки; красители; солеобразующие противоионы, такие как натрий; комплексы металлов (например, Zn-белковые комплексы); неионные поверхностно-активные вещества, такие как TWEEN ™, PLURONICS TM, и полиэтиленгликоль (ПЭГ), или любых их комбинации.
В некоторых вариантах осуществления фармацевтические композиции, которые содержат полипептиды, описанные в рамках изобретения, находятся в водорастворимой форме, такой как в виде фармацевтически приемлемых солей, которые, как предполагается, содержат кислотную и основную аддитивные соли. Термин «фармацевтически приемлемая кислотно-аддитивная соль» относится к тем солям, которые сохраняют биологическую эффективность свободных оснований и которые в противном случае нежелательны, образуются с неорганическими кислотами, такими как хлористоводородная кислота, бромистоводородная кислота, серная кислота, азотная кислота, фосфорная кислота и подобные, и органическими кислотами, такими как уксусная кислота, пропионовая кислота, гликолевая кислота, пировиноградная кислота, щавелевая кислота, малеиновая кислота, малоновая кислота, янтарная кислота, фумаровая кислота, винная кислота, лимонная кислота, бензойная кислота, коричная кислота, миндальная кислота, метансульфоновая кислота, этансульфоновая кислота, п-толуолсульфоновая кислота, салициловая кислота и тому подобное. Термин «фармацевтически приемлемые основно-аддитивные соли» включает соли, являющиеся производными неорганических оснований, таких как натриевые, калиевые, литиевые, аммониевые, кальциевые, магниевые, железные, цинковые, медные, марганцевые, алюминиевые соли и т.д. Особенно предпочтительными являются соли аммония, калия, натрия, кальция и магния. Соли, производные от фармацевтически приемлемых органических нетоксических оснований, включают соли первичных, вторичных и третичных аминов, замещенных аминов, включающих встречающиеся в природе замещенные амины, циклические амины и основные ионообменные смолы, такие как изопропиламин, триметиламин, диэтиламин, триэтиламин, трипропиламин и этаноламин. Композиции, предназначенные для применения in vivo, предпочтительно являются стерильными. Это может быть достигнуто путем фильтрации через стерильные фильтрующие мембраны или другими способами.
В некоторых вариантах осуществления фармацевтические композиции по изобретению вводят парентерально в форме инъекционной композиции. В некоторых вариантах осуществления фармацевтические композиции для инъекций получают с использованием стерильного раствора или любой фармацевтически приемлемой жидкости в качестве носителя. Фармацевтически приемлемые носители включают, но не ограничиваются ими, стерильную воду, физиологический раствор и среду для культивирования клеток (например, среду Игла, модифицированную по Дульбекко (DMEM), α-модифицированную среду Игла (α-MEM) и среду F-12). Доступны различные методы составления.
В некоторых вариантах осуществления описанные в рамках изобретения полипептиды представлены в виде иммунолипосом. Липосома представляет собой небольшую везикулу, содержащую различные типы липидов, фосфолипиды или поверхностно-активные вещества, которые полезны для доставки терапевтического агента млекопитающему. Липосомы, содержащие антитело или слитый Fc, могут быть получены различными способами, известными в данной области техники. В некоторых вариантах осуществления компоненты липосомы организованы в виде двухслойного состава, схожего с распределением липидов в биологических мембранах. В некоторых вариантах осуществления липосомы получают при помощи способа обращенно-фазового выпаривания с применением композиции липидов, содержащей фосфатидилхолин, холестерин и производные фосфатидилэтаноламина с ПЭГ (ПЭГ-PE). В некоторых вариантах осуществления липосомы экструдируют через фильтры с определенным размером пор с получением липосом, имеющих целевой диаметр. В некоторых вариантах осуществления химиотерапевтический агент или другой терапевтически активный агент необязательно содержится в липосоме.
В некоторых вариантах осуществления описанные в рамках изобретения полипептидные конструкции и другие терапевтически активные агенты захватываются в микрокапсулы, полученные способами, включая, но не ограничиваясь ими, способы коацервации, межфазную полимеризацию (например, с использованием гидроксиметилцеллюлозы или желатиновых микрокапсул или поли-(метилметакрилатных) микрокапсул), системы доставки коллоидных лекарственных средств (например, липосомы, микросферы альбумина, микроэмульсии, наночастицы и нанокапсулы) и макроэмульсии.
В некоторых вариантах осуществления готовят препараты с замедленным высвобождением. Подходящие примеры препаратов с пролонгированным высвобождением включают полупроницаемые матрицы из твердых гидрофобных полимеров, причем матрицы находятся в форме формованных изделий, например, пленок или микрокапсул. Примеры матриц с замедленным высвобождением включают сложные полиэфиры, гидрогели (например, поли (2-гидроксиэтилметакрилат) или поливиниловый спирт), полилактиды, сополимеры L-глутаминовой кислоты и гамма-этил-L-глутамата, нерастворимый этилен-винилацетат, разлагаемые сополимеры молочной кислоты и гликолевой кислоты, такие как LUPRON DEPOT ™ (которые представляют собой инъецируемые микросферы, состоящие из сополимера молочной кислоты и гликолевой кислоты и ацетата лейпролида), поли-D -(-)-3-гидроксимасляной кислоты и ProLease® (коммерчески доступный от Alkermes), который представляет собой систему доставки на основе микросфер, состоящую из желаемой биоактивной молекулы, включенной в матрицу поли-DL-лактид-со-гликолида (PLG). Некоторые препараты с замедленным высвобождением позволяют высвобождать молекулы в течение нескольких месяцев, например, от одного до шести месяцев, тогда как другие композиции высвобождают фармацевтические композиции раскрытия для более коротких периодов времени, например, от суток до недель.
В некоторых вариантах осуществления концентрация полипептида, описанного в рамках изобретения в фармацевтической композиции, варьирует от около 0,1 до 100 мас.%. В некоторых случаях концентрация описанного в рамках изобретения полипептида находится в диапазоне от 0,003 до 1,0 моля. В некоторых случаях концентрация полипептида в фармацевтической композиции варьирует от около 5 мг/мл до около 50 мг/мл (например, от около 10 до около 40 мг/мл или от около 20 мг/мл до около 30 мг/мл). В некоторых вариантах осуществления для лечения пациента вводят терапевтически эффективную дозу описанного в рамках изобретения полипептида. Термин «терапевтически эффективная доза» относится к дозе, которая производит эффекты, для которых она вводится. Точная доза будет зависеть от цели лечения. В некоторых вариантах осуществления дозы составляют от 0,01 до 100 мг/кг массы тела или выше, например, 0,1, 1, 5, 10, 15, 20, 25, 30, 35, 40, 45 или 50 мг/кг массы тела. В некоторых вариантах осуществления необходимы корректировки для деградации полипептидной конструкции, системной и локализованной доставки и скорости синтеза новой протеазы, а также возраста, массы тела, общего состояния здоровья, пола, питания, времени введения, взаимодействия лекарственного средства и тяжести состояния.
В некоторых вариантах осуществления композиции и препараты, описанные в рамках изобретения, вводят пациенту, нуждающемуся в этом. В некоторых вариантах осуществления такое введение осуществляют in vivo. В некоторых вариантах осуществления такое введение осуществляют ex vivo. В некоторых вариантах осуществления введение фармацевтической композиции, содержащей описанный в рамках изобретения полипептид, осуществляют различными способами, включая, но не ограничиваясь ими, пероральное, подкожное, внутривенное, интраназальное, внутрикостное, трансдермальное, местное (например, гели, мази, лосьоны, кремы и т.д.), внутрибрюшинное, внутримышечное, внутрилегочное (например, ингаляционная технология AERx®, коммерчески доступная от системы легочной доставки Aradigm или Inhance ™, коммерчески доступная от Inhale Therapeutics), вагинальное, парентеральное, ректальное или внутриглазное. В некоторых вариантах осуществления фармацевтическая композиция создана соответственно в зависимости от способа введения.
В некоторых вариантах осуществления фармацевтическая композиция для генной терапии находится в приемлемом разбавителе или включает матрицу с замедленным высвобождением, в которую внедрен носитель доставки гена. В некоторых вариантах осуществления векторы, используемые в качестве носителей доставки гена in vivo, включают, но не ограничиваются ими, ретровирусные векторы, аденовирусные векторы, поксвирусные векторы (например, векторы на основе вируса осповакцины, такие как модифицированная вакцинация Анкара), адено-ассоциированные вирусные векторы и альфавирусные векторы.
XI. Пути введения, дозировка и способ применения
В рамках изобретения, в некоторых вариантах осуществления, раскрыты полипептиды, содержащие: вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92.
В рамках изобретения, в некоторых вариантах осуществления также раскрыты полипептиды, содержащие вариант Fc, причем вариант Fc содержит димер домена Fc, имеющий два мономера домена Fc, при этом каждый мономер домена Fc независимо выбирают из (i) области Fc IgG1 человека, включающей в свой состав мутации L234A, L235A, G237A и N297A; (ii) области Fc IgG2 человека, включающей в свой состав мутации A330S, P331S и N297A; или (iii) области Fc IgG4 человека, содержащей мутации S228P, E233P, F234V, L235A, delG236 и N297A.
В некоторых вариантах осуществления фармацевтические композиции, которые содержат полипептиды раскрытия в качестве терапевтических белков, составлены для, например, внутривенного введения, парентерального введения, подкожного введения, внутримышечного введения, внутриартериального введения, интратекального введения или внутрибрюшинного введения. В некоторых вариантах осуществления фармацевтическая композиция составлена или введена путем перорального, назального, спреевого, аэрозольного, ректального или вагинального введения. Для инъекционных препаратов доступны различные эффективные фармацевтические носители.
В некоторых вариантах осуществления фармацевтическая композиция, которая содержит молекулу нуклеиновой кислоты, кодирующую полипептид раскрытия, или вектор, содержащий такую молекулу нуклеиновой кислоты, вводят путем доставки гена. Доступны различные способы доставки генов. В некоторых вариантах осуществления векторы, используемые для доставки и экспрессии гена in vivo, включают, но не ограничиваются ими, ретровирусные векторы, аденовирусные векторы, поксвирусные векторы (например, векторы на основе вируса осповакцины, такие как модифицированная вакцинация Анкара (MVA)), адено-ассоциированные вирусные векторы и альфавирусные векторы. В некоторых вариантах осуществления молекулы мРНК, кодирующие полипептиды раскрытия, вводят непосредственно пациенту.
Дозировка фармацевтических композиций раскрытия зависит от факторов, включая путь введения, заболевание, подлежащее лечению, и физические характеристики, например, возраст, массу, общее состояние здоровья пациента. В некоторых вариантах осуществления количество полипептида, содержащегося в разовой дозе, представляет собой количество, которое эффективно предотвращает, задерживает или лечит болезнь, не вызывая значительную токсичность. В некоторых вариантах осуществления фармацевтическая композиция по изобретению содержит дозу полипептида по изобретению в диапазоне от 0,01 до 500 мг/кг (например, 0,01, 0,1, 0,2, 0,3, 0,4, 0,5, 1, 2, 3, 4, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 100, 150, 200, 250, 300, 350, 400, 450 или 500 мг/кг) и в более конкретном варианте осуществления от около 0,1 до около 50 мг/кг и в более конкретном варианте осуществления от около 1 до около 30 мг/кг. В некоторых вариантах осуществления доза адаптируется врачом в соответствии с масштабом заболевания и различными параметрами пациента.
В некоторых вариантах осуществления токсичность терапевтических агентов и полипептидов, описанных в рамках изобретения, определяется стандартными фармацевтическими процедурами в культурах клеток или экспериментальных животных, например, путем определения LD50 (доза, летальная для 50% популяции) или LD100 (доза, летальная для 100% популяции). В некоторых вариантах осуществления данные, полученные из этих анализов клеточной культуры и исследований на животных, используются для формулирования диапазона доз, которые не токсичны для применения человеком. Доза описанных в рамках изобретения белков предпочтительно находится в пределах диапазона циркулирующих концентраций, которые включают эффективную дозу с небольшой токсичностью или без нее. В некоторых вариантах осуществления дозировка варьирует в пределах этого диапазона в зависимости от используемой лекарственной формы и используемого пути введения. В некоторых вариантах осуществления точная рецептура, способ введения и дозировка выбирается конкретным врачом с учетом состояния пациента.
В некоторых вариантах осуществления фармацевтические композиции вводят способом, совместимым с лекарственным препаратом, и в таком количестве, которое является терапевтически эффективным и приводит к улучшению или устранению симптомов заболевания или расстройства. В некоторых вариантах осуществления фармацевтические композиции вводят посредством множественных дозированных форм, например, внутривенных лекарственных форм, подкожных дозированных форм и пероральных лекарственных форм (например, проглатываемых растворов, капсул для высвобождения лекарственного средства). Обычно терапевтические белки дозируют в количестве 0,1-100 мг/кг, например, 1-50 мг/кг. В некоторых вариантах осуществления фармацевтические композиции, которые содержат полипептид раскрытия, вводят пациенту, нуждающемуся в этом, например, один или более раз (например, 1-10 раз или более) ежесуточно, еженедельно, ежемесячно, раз в два года, ежегодно или в зависимости от медицинских нужд. Дозировки могут быть предусмотрены в виде одного или множественных доз. В некоторых вариантах осуществления время между введениями уменьшается, поскольку состояние здоровья улучшается, или увеличивается по мере ухудшения здоровья пациента.
XII. Способы лечения
В рамках изобретения, в некоторых вариантах осуществления, раскрыты полипептиды, содержащие: вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92.
В рамках изобретения, в некоторых вариантах осуществления также раскрыты полипептиды, содержащие вариант Fc, причем вариант Fc содержит димер домена Fc, имеющий два мономера домена Fc, при этом каждый мономер домена Fc независимо выбирают из (i) области Fc IgG1 человека, включающей в свой состав мутации L234A, L235A, G237A и N297A; (ii) области Fc IgG2 человека, включающей в свой состав мутации A330S, P331S и N297A; или (iii) области Fc IgG4 человека, содержащей мутации S228P, E233P, F234V, L235A, delG236 и N297A.
Далее, в некоторых вариантах осуществления в рамках изобретения раскрыты способы лечения, включающие введение полипептидов, содержащих вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92.
В некоторых вариантах осуществления настоящее изобретение относится к фармацевтическим композициям и способам лечения, которые используются для лечения пациентов, страдающих заболеваниями и расстройствами, связанными с активностью SIRP-α или CD47, такими как рак и иммунологические заболевания (например, аутоиммунные заболевания и воспалительные заболевания). В некоторых вариантах осуществления описанные в рамках изобретения полипептиды вводят пациенту в способе увеличения фагоцитоза клетки-мишени (например, раковой клетки) у пациента. В некоторых вариантах осуществления полипептиды вводят пациенту в способе уничтожения раковых клеток у пациента. В некоторых вариантах осуществления полипептиды вводят пациенту в способе устранения регуляторных Т-клеток у пациента. В некоторых вариантах осуществления описанные в рамках изобретения полипептиды вводят пациенту в способе увеличения приживления гемопоэтических стволовых клеток у пациента, причем способ включает модулирование взаимодействия между SIRP-α и CD47 у пациента. В некоторых вариантах осуществления описанные в рамках изобретения полипептиды вводят пациенту в способе изменения иммунного ответа (например, подавления иммунного ответа) у пациента. В некоторых вариантах осуществления вышеупомянутые способы сочетаются с другими способами лечения заболевания. В некоторых вариантах осуществления, в рамках изобретения раскрыты комбинации полипептида (например, варианта D1 SIRP-α) и второго терапевтического агента. В некоторых вариантах осуществления комбинация содержит полипептид (например, вариант D1 SIRP-α) и второй терапевтический агент, причем второй терапевтический агент представляет собой антитело. В некоторых вариантах осуществления комбинация содержит вариант D1 SIRP-α, содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92, и антитело. В некоторых вариантах осуществления комбинация содержит полипептид, имеющий последовательность согласно любой из SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159, и антитело.
В некоторых вариантах осуществления вышеупомянутые способы используются со стратегиями лечения заболевания, при которых введение полипептида является терапевтическим вариантом. Неограничивающие примеры вышеизложенного включают применение антитела или фрагмента белка. Например, в некоторых вариантах осуществления антитело или фрагмент белка вводят в комбинации с вариантами Fc полипептидов, описанными в рамках изобретения. В некоторых вариантах осуществления описанные в рамках изобретения полипептидные конструкции применяются для улучшения фагоцитоза других агентов.
Способы лечения включают введение пациенту, имеющему заболевание (например, рак) (i) полипептида, содержащего вариант D1 SIRP-α и необязательно (ii) антитела. В некоторых вариантах осуществления перед лечением заболевания (например, рака) у пациента аминокислотную последовательность(и) SIRP-α у пациента определяют, например, из каждого из двух аллелей, кодирующих ген SIRP-α. В данном способе лечения сначала определяют аминокислотную последовательность(и) полипептидов SIRP-α в биологическом образце от пациента. Затем пациенту вводят терапевтически эффективное количество полипептида по данному изобретению. В некоторых вариантах осуществления высокоаффинный вариант D1 SIRP-α имеет такую же аминокислотную последовательность, что и полипептиды SIRP-α в биологическом образце пациента, за исключением введения аминокислотных изменений, которые увеличивают аффинность полипептида SIRP-α-к CD47. Высокоаффиный вариант D1 SIRP-α в полипептиде предпочтительно имеет минимальную иммуногенность у пациента после введения полипептида.
В некоторых вариантах осуществления антитело вводят в дополнение к описанным в рамках изобретения полипептидам. В некоторых вариантах осуществления антитело вводят совместно с полипептидом. В некоторых вариантах осуществления антитело вводят одновременно, например, в фармацевтической композиции, имеющей как полипептид, так и антитело. Альтернативно, антитело вводят либо до, либо после введения полипептида. В некоторых вариантах осуществления полипептид и антитело вводят по существу одновременно (например, в течение одной недели, 6, 5, 4, 3, 2, 1 суток, 12, 6, 3, 2, 1 часов друг от друга или по существу одновременно), с последующим введением антитела отдельно. В некоторых вариантах осуществления сначала вводят антитело с последующим введением полипептида и антитела по существу одновременно (то есть в течение одной недели, 6, 5, 4, 3, 2, 1 суток, 12, 6, 3, 2, 1 часов друг от друга или по существу одновременно).
Антитело, вводимое совместно или представленное в композиции или способе, описанном в рамках изобретения, относится к антителу, которое нацелено на клетку, такую как раковая клетка или клетка иммунной системы, такая как Т-клетка (например, регуляторная Т- клетка). Антитело может представлять собой любые изотипы антител иммуноглобулинов, например, IgG, IgE, IgM, IgA или IgD. В некоторых вариантах осуществления антитело представляет собой изотип антитела IgG1 человека. В некоторых вариантах осуществления антитело представляет собой изотип антитела IgG2 человека. В некоторых вариантах осуществления антитело представляет собой изотип антитела IgG4 человека.
Термин "антитело" в настоящем документе применяется в самом широком смысле и включает различные структуры антител, включая моноклональные антитела, поликлональные антитела, полиспецифические антитела (например, биспецифические антитела) и фрагменты антител, при условии, что они проявляют желательную антигенсвязывающую активность, но не ограничивается ими. «Фрагменты антитела» включают часть интактного антитела, предпочтительно антигенсвязывающий или вариабельный участок интактного антитела. Примеры фрагментов антител включают Fab, Fab', F(аb')2 и Fv-фрагменты; диатела; линейные антитела; одноцепочечные антитела и полиспецифические антитела. «Моноклональное антитело» относится к антителу, полученному из популяции практически однородных антител, например, отдельные антитела, составляющие популяцию, являются идентичными, за исключением возможных естественных мутаций, которые могут присутствовать в незначительных количествах. Моноклональные антитела могут являться высокоспецифичными и направлены против единственного антигенного сайта (например, эпитопа или ракового антигена). В отличие от препаратов поликлональных антител, которые, как правило, включают различные антитела против различных детерминант (эпитопов), каждое моноклональное антитело направлено против одной детерминанты на антигена. Модификатор "моноклональное" указывает на то, что антитело получают по существу из однородной популяции антител; его не следует интерпретировать как требование относительно продукции антитела с помощью какого-либо конкретного способа. В некоторых вариантах осуществления антитело в композиции настоящего изобретения вызывает антителозависимый клеточный фагоцитоз (ADCP) или антителозависимую клеточную цитотоксичность (АЗКЦ). Неограничивающие примеры заболеваний, которые лечат с использованием таких стратегий, включают раковые заболевания, такие как гематологические раковые заболевания, например, лейкозы (например, острый миелоидный лейкоз); иммунные нарушения (например, для улучшения ослабленного или уменьшенного иммунного ответа пациента или поочередно для ограничения чрезмерного активного ответа пациента); и патогенных инфекций.
В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение описанного в рамках изобретения полипептида (например, варианта D1 SIRP-a) и антитела, которое нацелено на раковый антиген. В некоторых вариантах осуществления антиген рака, на который нацелено антитело или антителоподобный белок, подвергают воздействию пептидов, полученных из внутриклеточных опухолеассоциированных антигенов (TAA), в комплексе с молекулами класса I лейкоцитарного антигена человека (HLA) на поверхности (также известный как ГКГС/ пептидный комплекс). Неограничивающие примеры таких раковых антигенов, например, пептиды в комплексе с молекулами HLA, расположенные на поверхности раковых клеток, на которые нацелены антитело или антителоподобные белки в композиции раскрытия, включают: NY-ESO-1/LAGE1, SSX-2, семейство MAGE (MAGE-A3), gp100/pmel17, Melan-A/MART-1, gp75/TRP1, тирозиназу, TRP2, CEA, PSA, TAG-72, незрелый рецептор ламинина, MOK/RAGE-1, WT-1, Her2/neu, EphA3, SAP-1, BING-4, Ep-CAM, MUC1, PRAME, сурвивин, Mesothelin, BRCA1/2 (мутированный), CDK4, CML66, MART-2, p53 (мутированный), Ras (мутированный), β-катенин (мутированный), TGF-βRII (мутированный), HPV E6, E7. Примеры таких антител включают ESK1 (WT-1), RL1B (Her2-E75), Pr20 (PRAME) и 3.2G1 (hCGβ).
В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение полипептида, содержащего вариант D1 SIRP-α, содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92, и антитела, которое связывает раковый антиген. В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение полипептида, содержащего вариант D1 SIRP-α, содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92, и антитела, которое связывает раковый NY-ESO-1/LAGE1, SSX-2, семейство MAGE (MAGE-A3), gp100/pmel17, Melan-A/MART-1, gp75/TRP1, тирозиназу, TRP2, CEA, PSA, TAG-72, незрелый рецептор ламинина, MOK/RAGE-1, WT-1, Her2/neu, EphA3, SAP-1, BING-4, Ep-CAM, MUC1, PRAME, сурвивин, Mesothelin, BRCA1/2 (мутированный), CDK4, CML66, MART-2, p53 (мутированный), Ras (мутированный), β-катенин (мутированный), TGF-βRII (мутированный), HPV E6, E7. В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение полипептида, содержащего вариант D1 SIRP-α, содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92, и антитело, которое связывает раковый антиген, причем антитело представляет собой ESK1 (WT-1), RL1B (Her2-E75), Pr20 (PRAME) и 3.2G1 (hCGβ).
В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение полипептида, имеющего последовательность в соответствии с SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159, и антитела, которое нацелено на раковый антиген. В некоторых вариантах осуществления, раскрытые в рамках изобретения способы включают введение полипептида, имеющего последовательность в соответствии с любой из SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159; и антитела, которое связывает NY-ESO-1/LAGE1, SSX-2, семейство MAGE (MAGE-A3), gp100/pmel17, Melan-A/MART-1, gp75/TRP1, тирозиназу, TRP2, CEA, PSA, TAG-72, незрелый рецептор ламинина, MOK/RAGE-1, WT-1, Her2/neu, EphA3, SAP-1, BING-4, Ep-CAM, MUC1, PRAME, сурвивин, Mesothelin, BRCA1/2 (мутированный), CDK4, CML66, MART-2, p53 (мутированный), Ras (мутированный), β-катенин (мутированный), TGF-βRII (мутированный), HPV E6, E7. В некоторых вариантах осуществления, раскрытые в рамках изобретения способы включают введение полипептида, имеющего последовательность в соответствии с любой из SEQ ID NO: SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159, и антитела, причем антитело представляет собой ESK1 (WT-1), RL1B (Her2-E75), Pr20 (PRAME) и 3.2G1 (hCGβ).
В некоторых вариантах осуществления антитело направлено на раковые клетки, например, путем связывания с белками, экспрессируемыми раковыми клетками. Некоторые белки экспрессируются на более высоких уровнях в раковых клетках, чем в нераковых клетках. Например, раковый антиген представляет собой белок, который предпочтительно экспрессируется раковыми клетками (например, он экспрессируется на более высоких уровнях на раковых клетках, чем на нераковых клетках), а в некоторых случаях он экспрессируется исключительно раковыми клетками. Неограничивающие примеры белков, например, белков, экспрессируемых раковыми клетками, на которые нацелено антитело в композициях раскрытия, включают: 4-1BB, 5T4, AGS-16, ALK1, ANG-2, B7-H3, B7-H4, c-fms, c-Met, CA6, CCR4, CD123, CD19, CD20, CD22, CD27, EpCAM, CD30, CD32b, CD33, CD37, CD38, CD40, CD52, CD70, CD74, CD79b, CD98, CEA, CEACAM5, CLDN18.2, CLDN6, CS1, CTLA-4, CXCR4, DLL-4, EGFR, EGP-1, ENPP3, EphA3, ETBR, FGFR2, фибронектин, FR-альфа, рецептор Frizzled, GCC, GD2, глипикан-3, GPNMB, HER-2, HER3, HLA-DR, ICAM-1, IGF-1 R, ИЛ-3R, LAG-3, LIV-1, мезотелин, MUC16, MUC1, NaPi2b, нектин-4, Notch 2, Notch 1, OX40, PD-1, PD-L1, PD-L2, PDGFR-α, PS, PSMA, SLTRK6, STEAP1, TEM1, VEGFR, CD25, CD27L, DKK-1, CSF-1 R или любых их комбинации. В некоторых вариантах осуществления описанные в рамках изобретения полипептиды вводят в комбинации с ингибитором контрольных точек, таким как ингибитор антитела CTLA-4 (например, ипилимумаб, тремелимумаб), PD-1 (например, ниволумаб, пидилизумаб, MK3475, также известный как пембролизумаб, BMS936559 и MPDL3280A) и LAG-3 (например, BMS986016).
В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение полипептида, содержащего вариант D1 SIRP-α, содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92, и антитела, которое связывает белок, экспрессируемый раковой клеткой. В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение полипептида, содержащего вариант D1 SIRP-α, содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92, и антитела, которое связывает 4-1BB, 5T4, AGS-16, ALK1, ANG-2, B7-H3, B7-H4, c-fms, c-Met, CA6, CCR4, CD123, CD19, CD20, CD22, CD27, EpCAM, CD30, CD32b, CD33, CD37, CD38, CD40, CD52, CD70, CD74, CD79b, CD98, CEA, CEACAM5, CLDN18.2, CLDN6, CS1, CTLA-4, CXCR4, DLL-4, EGFR, EGP-1, ENPP3, EphA3, ETBR, FGFR2, фибронектин, FR-альфа, рецептор Frizzled, GCC, GD2, глипикан-3, GPNMB, HER-2, HER3, HLA-DR, ICAM-1, IGF-1 R, ИЛ-3R, LIV-1, мезотелин, MUC16, MUC1, NaPi2b, Nectin-4, Notch 2, Notch 1, OX40, PD-1, PD-L1, PD-L2, PDGFR-α, PS, PSMA, SLTRK6, STEAP1, TEM1, VEGFR, CD25, CD27L, DKK-1, CSF-1 R или любую их комбинацию. В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение полипептида, содержащего вариант D1 SIRP-α, содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92, и антитела, причем антитело представляет собой антитело-ингибитор CTLA-4 (например, ипилимумаб, тремелимумаб), PD-1 (например, ниволумаб, пидилизумаб, MK3475 также известный как пембролизумаб, BMS936559 и MPDL3280A) или LAG-3 (например, BMS986016).
В некоторых вариантах осуществления, раскрытые в рамках изобретения способы включают введение полипептида, имеющего последовательность в соответствии с любой из SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159, и антитела, которое нацелено на белок, экспрессируемый раковой клеткой. В некоторых вариантах осуществления, раскрытые в рамках изобретения способы включают введение полипептида, имеющего последовательность в соответствии с любой из SEQ ID NO: SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159, и антитела, которое связывает 4-1BB, 5T4, AGS-16, ALK1, ANG-2, B7-H3, B7-H4, c-fms, c-Met, CA6, CCR4, CD123, CD19, CD20, CD22, CD27, EpCAM, CD30, CD32b, CD33, CD37, CD38, CD40, CD52, CD70, CD74, CD79b, CD98, CEA, CEACAM5, CLDN18.2, CLDN6, CS1, CTLA-4, CXCR4, DLL-4, EGFR, EGP-1, ENPP3, EphA3, ETBR, FGFR2, фибронектин, FR-альфа, рецептор Frizzled, GCC, GD2, глипикан-3, GPNMB, HER-2, HER3, HLA-DR, ICAM-1, IGF-1 R, ИЛ-3R, LAG-3, LIV-1, мезотелин, MUC16, MUC1, NaPi2b, нектин-4, Notch 2, Notch 1, OX40, PD-1, PD-L1, PD-L2, PDGFR-α, PS, PSMA, SLTRK6, STEAP1, TEM1, VEGFR, CD25, CD27L, DKK-1, CSF-1 R или любые их комбинации.В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение полипептида, имеющего последовательность в соответствии с любой из SEQ ID NO: SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159, и антитела, причем антитело представляет собой ингибитор CTLA-4 (например, ипилимумаб, тремелимумаб), PD-1 (например, ниволумаб, пидилизумаб, MK3475 также известный как пембролизумаб, BMS936559 и MPDL3280A) или LAG-3 (например, BMS986016).
В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение описанного в рамках изобретения полипептида (например, варианта D1 SIRP-α) и иммуноонкологического антитела. В некоторых вариантах осуществления антитела, которые используются в композициях раскрытия, включают, но не ограничиваются ими: цетуксимаб, нецитумумаб, пембролизумаб, ниволумаб, пидилизумаб, MEDI0680, MED16469, атезолизумаб, авелумаб, дурвалумаб, MEDI6383, RG7888, ипилимумаб, тремелимумаб, урелумаб, PF-05082566, еноблитузумаб, вантицитумаб, варлилумаб, могамализумаб, SAR650984, даратумумаб, трастузумаб, трастузумаб эмтанзин, пертузумаб, элотузумаб, ритуксимаб, офатумумаб, обинутузумаб, RG7155, FPA008, панитумумаб, брентуксимаб ведотин, MSB0010718C, белимумаб, бевацизумаб, деносумаб, панитумумаб, рамуцирумаб, нецитумумаб, ниволумаб, пембролищумаб, авелумаб, атезолизумаб, дурвалумаб, MEDI0680, пидилизумаб или BMS-93659, антитело против HER2, антитело против CD20, антитело против CD19, антитело против CS1, антитело против CD38, антитело против EGFR, антитело против PD1, антитело против RANKL, антитело против OX40, антитело против PD-1, антитело против PD-L1, антитело против CD274, антитело против CTLA-4, антитело против CD137, антитело против 4-1BB, антитело против B7-H3, антитело против FZD7, антитело против CD27, антитело против CCR4, антитело против CD38, антитело против CSF1R, антитело против CSF, антитело против CD30, антитело против BAFF, антитело против VEGF или антитело против VEGFR2. В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение полипептида, содержащего вариант D1 SIRP-α, содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92, и антитело, которое связывает раковый антиген, причем антитело представляет собой антитело против HER2, антитело против CD20, антитело против CD19, антитело против CS1, антитело против CD38, антитело против EGFR, антитело против PD1, антитело против RANKL, антитело против OX40, антитело против PD-1, антитело против PD-L1, антитело против CD274, антитело против CTLA-4, антитело против CD137, антитело против 4-1BB, антитело против B7-H3, антитело против FZD7, антитело против CD27, антитело против CCR4, антитело против CD38, антитело против CSF1R, антитело против CSF, антитело против CD30, антитело против BAFF, антитело против VEGF или антитело против VEGFR2. В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение полипептида, содержащего вариант D1 SIRP-α, содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92, и антитела, причем антитело представляет собой цетуксимаб, нецитумумаб, пембролизумаб, ниволумаб, пидилизумаб, MEDI0680, MED16469, атезолизумаб, авелумаб, дурвалумаб, MEDI6383, RG7888, ипилимумаб, тремелимумаб, урелумаб, PF-05082566, еноблитузумаб, вантицитумаб, варлилумаб, могамализумаб, SAR650984, даратумумаб, трастузумаб, трастузумаб эмтанзин, пертузумаб, элотузумаб, ритуксимаб, офатумумаб, обинутузумаб, RG7155, FPA008, панитумумаб, брентуксимаб ведотин, MSB0010718C, белимумаб, бевацизумаб, деносумаб, панитумумаб, рамуцирумаб, нецитумумаб, ниволумаб, пембролищумаб, авелумаб, атезолизумаб, дурвалумаб, MEDI0680, пидилизумаб или BMS-93659.
В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение полипептида, содержащего вариант D1 SIRP-α, содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92, и антитела, причем антитело представляет собой трастузумаб.
В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение полипептида, содержащего вариант D1 SIRP-α, содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92, и антитела, причем антитело представляет собой ритуксимаб.
В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение полипептида, содержащего вариант D1 SIRP-α, содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92, и антитела, причем антитело представляет собой цетуксимаб.
В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение полипептида, содержащего вариант D1 SIRP-α, содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92, и антитела, причем антитело представляет собой даратумумаб.
В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение полипептида, содержащего вариант D1 SIRP-α, содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92, и антитела, причем антитело представляет собой белимумаб.
В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение полипептида, содержащего вариант D1 SIRP-α, содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92, и антитела, причем антитело представляет собой бевацизумаб.
В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение полипептида, содержащего вариант D1 SIRP-α, содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92, и антитела, причем антитело представляет собой деносумаб.
В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение полипептида, содержащего вариант D1 SIRP-α, содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92, и антитела, причем антитело представляет собой пантимумаб.
В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение полипептида, содержащего вариант D1 SIRP-α, содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92, и антитела, причем антитело представляет собой рамуцирумаб.
В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение полипептида, содержащего вариант D1 SIRP-α, содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92, и антитела, причем антитело представляет собой нецитумумаб.
В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение полипептида, содержащего вариант D1 SIRP-α, содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92, и антитела, причем антитело представляет собой ниволумаб.
В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение полипептида, содержащего вариант D1 SIRP-α, содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92, и антитела, причем антитело представляет собой пембролизумаб.
В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение полипептида, содержащего вариант D1 SIRP-α, содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92, и антитела, причем антитело представляет собой авелумаб.
В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение полипептида, содержащего вариант D1 SIRP-α, содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92, и антитела, причем антитело представляет собой атезолизумаб.
В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение полипептида, содержащего вариант D1 SIRP-α, содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92, и антитела, причем антитело представляет собой дурвалумаб.
В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение полипептида, содержащего вариант D1 SIRP-α, содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92, и антитела, причем антитело представляет собой MEDI0680.
В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение полипептида, содержащего вариант D1 SIRP-α, содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92, и антитела, причем антитело представляет собой пидилизумаб.
В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение полипептида, содержащего вариант D1 SIRP-α, содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92, и антитела, причем антитело представляет собой BMS-93659.
В некоторых вариантах осуществления, раскрытые в рамках изобретения способы включают введение полипептида, имеющего последовательность в соответствии с любой из SEQ ID NO: SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159, и антитела, причем антитело выбирают из антитела против HER2, антитела против CD20, антитела против CD19, антитела против CS1, антитела против CD38, антитела против EGFR, антитела против PD1, антитела против OX40, антитела против PD-1, антитела против RANKL, антитела против CD274, антитела против CTLA-4, антитела против CD137, антитела против 4-1BB, антитела против B7-H3, антитела против FZD7, антитела против CD27, антитела против CCR4, антитела против CD38, антитела против CSF1R, антитела против CSF, антитела против CD30, антитела против BAFF, антитела против VEGF или антитела против VEGFR2. В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение полипептида, имеющего последовательность в соответствии с SEQ ID NO: SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159, и антитела, причем антитело выбирают из цетуксимаба, нецитумумаба, пембролизумаба, ниволумаба, пидилизумаба, MEDI0680, MED16469, атезолизумаба, авелумаба, дурвалумаба, MEDI6383, RG7888, ипилимумаба, тремелимумаба, урелумаба, PF-05082566, эноблитузумаба, вантиктумаба, варлилумаба, могамалимузумаба, SAR650984, даратумумаба, трастузумаба, трастузумаб емтансина, пертузумаба, элотузумаба, ритуксимаба, офатумумаба, обинутузумаба, RG7155, FPA008, панитумумаба, брентуксимаб ведотина, MSB0010718C, белимумаба, бевацизумаба, деносумаба, панитумумаба, рамуцирумаба, нецитумумаба, ниволумаба, пембролизумаба, авелумаба, атезолизумаба, дувалумаба, MEDI0680, пидилизумаба или BMS-93659.
В некоторых вариантах осуществления, раскрытые в рамках изобретения способы включают введение полипептида, имеющего последовательность в соответствии с любой из SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159, и антитела, причем антитело представляет собой трастузумаб.
В некоторых вариантах осуществления, раскрытые в рамках изобретения способы включают введение полипептида, имеющего последовательность в соответствии с любой из SEQ ID NO: 7, SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159, и антитела, причем антитело представляет собой ритуксимаб.
В некоторых вариантах осуществления, раскрытые в рамках изобретения способы включают введение полипептида, имеющего последовательность в соответствии с любой из SEQ ID NO: SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159, и антитела, причем антитело представляет собой цетуксимаб.
В некоторых вариантах осуществления, раскрытые в рамках изобретения способы включают введение полипептида, имеющего последовательность в соответствии с любой из SEQ ID NO: SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159, и антитела, причем антитело представляет собой даратумумаб.
В некоторых вариантах осуществления, раскрытые в рамках изобретения способы включают введение полипептида, имеющего последовательность в соответствии с любой из SEQ ID NO: SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159, и антитела, причем антитело представляет собой белимумаб.
В некоторых вариантах осуществления, раскрытые в рамках изобретения способы включают введение полипептида, имеющего последовательность в соответствии с любой из SEQ ID NO: SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159, и антитела, причем антитело представляет собой бевацизумаб.
В некоторых вариантах осуществления, раскрытые в рамках изобретения способы включают введение полипептида, имеющего последовательность в соответствии с любой из SEQ ID NO: SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159, и антитела, причем антитело представляет собой деносумаб.
В некоторых вариантах осуществления, раскрытые в рамках изобретения способы включают введение полипептида, имеющего последовательность в соответствии с любой из SEQ ID NO: SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159, и антитела, причем антитело представляет собой пантимумаб.
В некоторых вариантах осуществления, раскрытые в рамках изобретения способы включают введение полипептида, имеющего последовательность в соответствии с любой из SEQ ID NO: SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159, и антитела, причем антитело представляет собой рамуцирумаб.
В некоторых вариантах осуществления, раскрытые в рамках изобретения способы включают введение полипептида, имеющего последовательность в соответствии с любой из SEQ ID NO: SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159, и антитела, причем антитело представляет собой нецитумумаб.
В некоторых вариантах осуществления, раскрытые в рамках изобретения способы включают введение полипептида, имеющего последовательность в соответствии с любой из SEQ ID NO: SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159, и антитела, причем антитело представляет собой ниволумаб.
В некоторых вариантах осуществления, раскрытые в рамках изобретения способы включают введение полипептида, имеющего последовательность в соответствии с любой из SEQ ID NO: SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159, и антитела, причем антитело представляет собой пембролизумаб.
В некоторых вариантах осуществления, раскрытые в рамках изобретения способы включают введение полипептида, имеющего последовательность в соответствии с любой из SEQ ID NO: SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159, и антитела, причем антитело представляет собой авелумаб.
В некоторых вариантах осуществления, раскрытые в рамках изобретения способы включают введение полипептида, имеющего последовательность в соответствии с любой из SEQ ID NO: SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159, и антитела, причем антитело представляет собой атезолизумаб.
В некоторых вариантах осуществления, раскрытые в рамках изобретения способы включают введение полипептида, имеющего последовательность в соответствии с любой из SEQ ID NO: SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159, и антитела, причем антитело представляет собой дурвалумаб.
В некоторых вариантах осуществления, раскрытые в рамках изобретения способы включают введение полипептида, имеющего последовательность в соответствии с любой из SEQ ID NO: SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159, и антитела, причем антитело представляет собой MEDI0680.
В некоторых вариантах осуществления, раскрытые в рамках изобретения способы включают введение полипептида, имеющего последовательность в соответствии с любой из SEQ ID NO: SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159, и антитела, причем антитело представляет собой пидилизумаб.
В некоторых вариантах осуществления, раскрытые в рамках изобретения способы включают введение полипептида, имеющего последовательность в соответствии с любой из SEQ ID NO: SEQ ID NO: 78-85, 98-104, 107-113, 116-122, 135-137 или 152-159, и антитела, причем антитело представляет собой BMS-93659.
В некоторых вариантах осуществления описанные в рамках изобретения полипептиды усиливают противоопухолевую активность ритуксимаба. В некоторых вариантах осуществления описанные в рамках изобретения полипептиды усиливают противоопухолевую активность ритуксимаба в модели ксенотрансплантата Raji-NSG. В некоторых вариантах осуществления описанные в рамках изобретения полипептиды усиливают ритуксимаб-опосредованное истощение B-клеток у приматов, отличных от человека (NHP).
В некоторых вариантах осуществления полипептиды и фармацевтические композиции раскрытия используются в различных способах лечения рака. Раковые опухоли, пригодные для лечения в соответствии с раскрытием, включают, но не ограничиваются ими солидную опухоль, гематологический рак, острый миелоидный лейкоз, хронический лимфоцитарный лейкоз, хронический миелоидный лейкоз, острый лимфобластный лейкоз, неходжкинскую лимфому, лимфому Ходжкина, множественную миелому, рак мочевого пузыря, рак поджелудочной железы, рак шейки матки, рак эндометрия, рак легкого, рак бронхов, рак печени, рак яичника, рак толстой кишки и прямой кишки, рак желудка, рак желудочно-кишечного тракта, рак желчного пузыря, желудочно-кишечную стромальную опухоль, рак щитовидной железы, рак головы и шеи, рак ротоглотки, рак пищевода, меланому, немеланомный рак кожи, карциному из клеток Меркеля, вирус-индуцированный рак, нейробластому, рак молочной железы, рак предстательной железы, рак почки, почечно-клеточный рак, рак почечной лоханки, лейкоз, лимфому, саркому, глиому, опухоль головного мозга и карциному. В некоторых вариантах осуществления раковые состояния, поддающиеся лечению в соответствии с описанием, включают метастатические раковые заболевания. В некоторых вариантах осуществления рак, поддающийся лечению согласно раскрытию, представляет собой солидную опухоль или гематологический рак.
В некоторых вариантах осуществления антитело направлено на клетки иммунной системы, такие как Т-клетки, например регуляторные Т-клетки, путем связывания с белками, экспрессируемыми клетками иммунной системы. В некоторых вариантах осуществления раскрытые в рамках изобретения способы включают введение описанного в рамках изобретения полипептида (например, варианта D1 SIRP-α) и антитела, которое нацелено на клетки иммунной системы. Примеры белков, экспрессируемых клетками иммунной системы, включают, но не ограничиваются ими, 41BB, CD40, CD40L, CD163, CD206, CTLA4, PD1, TIM-3, BTLA, VISTA, LAG-3, CD28, OX40, GITR, CD137, CD27, HVEM, CCR4, CD25, CD103, KIrg1, Nrp1, CD278, Gpr83, TIGIT, CD154, CD160 и PD1H. В некоторых вариантах осуществления антитело сконструировано таким образом, что оно имеет преимущественное связывание с белками (например, рецепторами), экспрессируемыми Т-клетками (например, регуляторными Т-клетками) по сравнению с другими клетками иммунной системы. В некоторых вариантах осуществления антитело в композиции раскрытия содержит домен Fc подкласса IgG1, IgG2 или IgG4.
В некоторых вариантах осуществления способы раскрытия включают изменение иммунного ответа у пациента. Способы включают введение пациенту полипептида, содержащего высокоаффинный вариант D1 SIRP-α и антитела, тем самым изменяя иммунный ответ у пациента. В некоторых вариантах осуществления изменение иммунного ответа включает подавление иммунного ответа.
В некоторых вариантах осуществления полипептиды и фармацевтические композиции раскрытия используются в различных способах лечения для лечения иммунологических заболеваний. Аутоиммунные заболевания и воспалительные заболевания, поддающиеся лечению согласно раскрытию, включают, но не ограничиваются ими, аутоиммунное заболевание или воспалительное заболевание, а аутоиммунное заболевание или воспалительное заболевание выбирают из рассеянного склероза, ревматоидного артрита, спондилоартропатии, системной красной волчанки, опосредованного антителом воспалительного или аутоиммунного заболевания, заболевания трансплантат против хозяина, сепсиса, диабетов, псориаза, атеросклероза, синдрома Шегрена, прогрессирующего системного склероза, склеродермии, острого коронарного синдрома, ишемической реперфузии, болезни Крона, эндометриоза, гломерулонефрита, миастении гравис, идиопатического легочного фиброза, астмы, острого респираторного дистресс-синдрома (ARDS), васкулита и воспалительного аутоиммунного миозита.
В некоторых вариантах осуществления доставка полипептида в клетку включает контактирование клетки с одной или более описанными в рамках изобретения композициями.
Эффективные дозы для таких вариантов лечения варьируют в зависимости от множества различных факторов, включая способы введения, целевой сайт, физиологическое состояние пациента, будь то человек или животное, другие назначенные лекарства и является ли лечение профилактическим или терапевтическим. В некоторых вариантах осуществления пациент является человеком, но млекопитающие, отличные от человека также подвергаются лечению, например, животные-компаньоны, такие как собаки, кошки, лошади и т.д., лабораторные млекопитающие, такие как кролики, мыши, крысы и т.д. и тому подобное. В некоторых вариантах осуществления дозы лечения титруются для оптимизации безопасности и эффективности.
В некоторых вариантах осуществления терапевтическая доза составляет от около 0,0001 до 100 мг/кг и более обычно от 0,01 до 30 мг/кг массы тела хозяина. В некоторых вариантах осуществления, например, дозы составляют 1 мг/кг массы тела или 30 мг/кг массы тела или в пределах 1-30 мг/кг. В некоторых вариантах осуществления примерный режим лечения включает введение один раз в неделю или один раз в две недели или один раз в месяц или один раз каждые 3-6 месяцев. В некоторых вариантах осуществления терапевтические агенты и полипептидные конструкции, описанные в рамках изобретения, вводятся несколько раз. В некоторых вариантах осуществления интервал между разовыми дозами являются еженедельным, ежемесячным или ежегодным. В некоторых вариантах осуществления интервалы также являются нерегулярными, как указано путем измерения уровней крови в терапевтических целях у пациента. Альтернативно, терапевтические агенты или полипептидные конструкции, описанные в рамках изобретения, вводят в виде композиции с замедленным высвобождением, и в этом случае возможно более частое введение. В некоторых вариантах осуществления доза и частота варьируются в зависимости от периода полувыведения полипептида у пациента.
В профилактических применениях в некоторых вариантах осуществления относительно низкая доза вводится с относительно небольшими интервалами в течение длительного периода времени. В некоторых вариантах осуществления пациенты продолжают получать лечение всю оставшуюся жизнь. В других терапевтических применениях требуется относительно высокая доза с относительно короткими интервалами, пока прогрессия заболевания не будет уменьшена или не прекращена, и предпочтительно до тех пор, пока пациент не продемонстрирует частичное или полное улучшение симптомов заболевания. После этого в некоторых вариантах осуществления пациенту вводят профилактический режим.
Используемые в рамках изобретения термины «лечение», «лечить» и т.п. относятся к введению агента или проведению процедуры для целей получения эффекта. В некоторых вариантах осуществления эффект является профилактическим с точки зрения полного или частичного предотвращения заболевания или его симптома. В некоторых вариантах осуществления эффект является терапевтическим с точки зрения воздействия на частичное или полное излечение от заболевания или симптомов заболевания.
XIII. Наборы
В рамках изобретения, в некоторых вариантах осуществления, раскрыты полипептиды, содержащие: вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий домен D1 SIRP-α или его фрагмент, имеющий аминокислотную мутацию в остатке 80 по сравнению с доменом D1 SIRP-α дикого типа; и по меньшей мере одну дополнительную аминокислотную мутацию по сравнению с доменом D1 SIRP-α дикого типа в остатке, выбранном из группы, состоящей из: остатка 6, остатка 27, остатка 31, остатка 47, остатка 53, остатка 54, остатка 56, остатка 66 и остатка 92.
В рамках изобретения, в некоторых вариантах осуществления также раскрыты полипептиды, содержащие вариант Fc, причем вариант Fc содержит димер домена Fc, имеющий два мономера домена Fc, при этом каждый мономер домена Fc независимо выбирают из (i) области Fc IgG1 человека, включающей в свой состав мутации L234A, L235A, G237A и N297A; (ii) области Fc IgG2 человека, включающей в свой состав мутации A330S, P331S и N297A; или (iii) области Fc IgG4 человека, содержащей мутации S228P, E233P, F234V, L235A, delG236 и N297A.
Также представлены наборы, которые содержат полипептиды, описанные в рамках изобретения, и инструкции по их применению. Необязательно, наборы могут дополнительно содержать по меньшей мере один дополнительный реагент. В качестве неограничивающего примера химиотерапевтический агент или противоопухолевое антитело могут служить в качестве по меньшей мере одного дополнительного агента. В некоторых вариантах осуществления наборы содержат этикетку, указывающую предполагаемое использование содержимого набора. Термин «этикетка» означает любую запись или записанный материал, поставляемый на комплекте или с комплектом, или который в противном случае прилагается к комплекту.
В некоторых вариантах осуществления набор содержит (i) полипептид, содержащий высокоаффинный вариант D1 SIRP-α; необязательно (ii) антитело; и (iii) инструкции по введению (i) и (ii) (если предоставлено) пациенту, имеющему заболевание. В некоторых вариантах осуществления наборы содержит (i) полипептид, содержащий высокоаффинный вариант D1 SIRP-α; и (ii) инструкции для введения (i) с антителом, например антителом, которое не предусмотрено в наборе, пациенту, имеющему заболевание. В некоторых вариантах осуществления наборы содержат (i) антитело; и (ii) инструкции для введения (i) с полипептидом, содержащим высокоаффинный вариант D1 SIRP-α пациенту, имеющему заболевание.
В некоторых вариантах осуществления наборы используются для лечения пациента, имеющего рак, такой как солидная опухоль, гематологический рак, острый миелоидный лейкоз, хронический лимфоцитарный лейкоз, хронический миелоидный лейкоз, острый лимфобластный лейкоз, неходжкинская лимфома, лимфома Ходжкина, множественная миелома, рак мочевого пузыря, рак поджелудочной железы, рак шейки матки, рак эндометрия, рак легкого, рак бронхов, рак печени, рак яичника, рак толстой кишки и прямой кишки, рак желудка, рак желудочно-кишечного тракта, рак желчного пузыря, желудочно-кишечная стромальная опухоль, рак щитовидной железы, рак головы и шеи, рак ротоглотки, рак пищевода, меланома, немеланомный рак кожи, карцинома из клеток Меркеля, вирус-индуцированный рак, нейробластома, рак молочной железы, рак предстательной железы, рак почки, почечно-клеточный рак, рак почечной лоханки, лейкоз, лимфома, саркома, глиома, опухоль головного мозга и карцинома, или их комбинацию. В некоторых вариантах осуществления наборы используются для лечения пациента, имеющего солидную опухоль или гематологический рак.
В некоторых вариантах осуществления наборы используются для лечения пациента, обладающего иммунологическими заболеваниями. В некоторых вариантах осуществления иммунологическое заболевание представляет собой аутоиммунное заболевание или воспалительное заболевание, такое как рассеянный склероз, ревматоидный артрит, спондилоартропатию, системная красная волчанка, опосредованное антителом воспалительное или аутоиммунное заболевание, заболевание трансплантат против хозяина, сепсис, диабеты, псориаз, атеросклероз, синдром Шегрена, прогрессирующий системный склероз, склеродермия, острый коронарный синдром, ишемическая реперфузия, болезнь Крона, эндометриоз, гломерулонефрит, миастения гравис, идиопатический легочный фиброз, астму, острый респираторный дистресс-синдром (ARDS), васкулит и воспалительный аутоиммунный миозит.
ПРИМЕРЫ
Пример 1 - Полипептиды варианта D1 SIRP-α
Получение полипептидов раскрытия
Полипептид раскрытия, содержащий высокоаффинный вариант D1 SIRP-α, создавали с использованием обычных методик молекулярного клонирования и экспрессии белка. Возможные аминокислотные замены в варианте D1 SIRP-α по сравнению с доменом D1 SIRP-α дикого типа приведены в таблицах 2 и 5. Молекулу нуклеиновой кислоты, кодирующую полипептид раскрытия, клонируют в вектор, оптимизированный для экспрессии в клетках бактерий или млекопитающих с использованием хорошо известных методик молекулярной биологии. После индукции экспрессии белка клетки собирают и экспрессируемые полипептиды очищают от супернатанта клеточной культуры с использованием аффинной колоночной хроматографии. Затем очищенные полипептиды анализируют с помощью SDS-PAGE с последующим окрашиванием Кумасси синим (Coomassie Blue), чтобы подтвердить присутствие белковых бэндов ожидаемого размера.
Очищенные полипептиды оценивали на связывание с CD47 с использованием доступных методик в данной области техники, таких как фаговый дисплей, дрожжевой дисплей, поверхностный плазмонный резонанс (SPR), сцинтилляционный анализ сближения, твердофазный иммуноферментный анализ (ELISA), иммуноанализ ORIGEN (IGEN), гашение флуоресценции, перенос флуоресценции или любой подходящий биоанализ. Искомые полипептиды связываются с более высоким сродством с CD47, например CD47 человека, чем SIRP-α дикого типа.
Аффинность связывания варианта D1 полипептидов SIRP-α
В серии экспериментов полипептиды с доменом D1 SIRP-α дикого типа и высокоаффинные варианты D1 SIRP-α были получены с использованием обычных методик молекулярного клонирования и экспрессии белка. Связывание с CD47 человека определяли с использованием SPR следующим образом: вкратце, связывание CD47 человека (системы R и D, каталожный номер 4670-CD или собственный продукт, произведенный как мономерный внеклеточный домен - ECD) с SIRP-α дикого типа и вариантами D1 полипептида SIRP-α были проанализированы на приборе Biacore T100 (GE Healthcare) или Proteon XPR36 (Bio-rad, Hercules, CA) с использованием забуференного фосфатом физиологического раствора (PBS, pH 7,4), дополненного 0,01% Tween-20 (PBST) в качестве подвижного буфера. 200-1000 RU лиганда иммобилизировали в 10 мМ буфере для ацетата натрия (рН 4,5) на чипе-сенсоре Biacore CM4 или чипе Proteon GLC стандартным аминным соединением согласно рекомендациям производителя. Несколько концентраций аналита (или варианта D1 полипептидов SIRP-α), например, ранжированных по значению KD от по меньшей мере 0,1х до 10х, инъецировали в течение двух минут со скоростью потока 100 мкл/мин с последующим десятиминутным временем диссоциации. После каждой инъекции аналита поверхность восстанавливали с использованием смеси элюирующего буфера Pierce IgG 2:1 (Life Technologies, каталожный номер 21004) и 4 М NaCl, вводимой в течение 30 секунд. Полное восстановление поверхности было подтверждено базовым анализом и инъекцией того же аналита в начале и в конце эксперимента. Все сенсограммы были дважды откалиброваны, с использованием базовой поверхности и инъекции буфера и установлены до 1:1 Ленгмюра. Аналит был преимущественно мономерным, либо CD47 ECD, либо SIRP-α без Fc. Лиганд на чипе может быть мономерным или слитым Fc. Данные связывания приведены в таблице 16. Все анализы SPR проводили при 25 °C.
Таблица 16. Вариант полипептида SIRP-α и связанные значения KD
Также было определено, что наличие остатка глутамата или аспартата в положении 54 улучшает связывание вариантов D1 полипептидов SIRP-α с CD47 мыши. В качестве неограничивающего примера варианты D1 полипептидов SIRP-α, указанные в таблице 17 ниже, демонстрируют высокую аффинность с CD47 мыши. Аффинность связывания с CD47 человека нескольких вариантов D1 полипептидов SIRP-α сравнивали с аффинностью связывания с CD47 мыши, с использованием SPR, как описано ранее, при этом белок CD47 мыши использовался вместо CD47 человека, где это необходимо. Результаты представлены в таблице 18.
Таблица 17. Последовательности вариантов полипептида SIRP-α, имеющие улучшенное связывание с CD47 мыши
Таблица 18. Связывание вариантов полипептидов SIRP-α с CD47 человека и мыши
Также было определено, что мутация N80A, которая может минимизировать или аннулировать частичное гликозилирование, присутствующую в некоторых вариантах D1 полипептидов SIRP-α, дает функциональное преимущество увеличения гомогенности, связанной с вариантами D1 полипептидов SIRP-α, содержащих такую мутацию. Когда варианты полипептидов SIRP-α экспрессируются в E.coli, гликозилирование N80 не произойдет из-за отсутствия системы гликозилирования в E.coli по сравнению с системой млекопитающих. В таблице 19 показано, что эффективное связывание между вариантом D1 полипептида SIRP-α, продуцируемым в E.coli и CD47 человека, все еще может иметь место, тем самым демонстрируя, что дегликозилирование не влияет на аффинность связывания, с которой варианты D1 SIRP-α все еще могут связываться с CD47. В дополнение к мутации N80A дегликозилирование может быть осуществлено путем мутации N80 до любой аминокислоты, которая не является N или путем нарушения мотива N-Xaa1-Xaa2 где N=аспарагин; Xaa1=любая аминокислота, кроме P (пролина); Xaa2=T (треонин), S (серин) или C (цистеин), причем мотив относится к остаткам 80-82 варианта D1 полипептида SIRP-α. Путем мутации P83 на валин или другой остаток, который не является P, может возникать повышенное гликозилирование при N80 и могут быть получены гомогенно гликозилированные варианты D1 полипептидов SIRP-α.
Аминокислота Р83 может также влиять на степень гликозилирования. Изменение P83 на любую аминокислоту может повысить эффективность гликозилирования в N80. Вариант D1 SIRP-α, имеющий валин (V) в положении 83 (SEQ ID NO: 213) экспрессировался в клетках млекопитающих HEK293FS. Размер экспрессированного белка сравнивали с вариантом D1 SIRP-α, имеющим аминокислотный остаток дикого типа (например, пролин, P) в положении 83 (SEQ ID NO: 71). Анализ молекулярной массы экспрессированного белка на белковом геле (фиг. 18) продемонстрировал, что вариант, имеющий мутацию P83V (SEQ ID NO: 213, дорожка 2) имеет более высокую молекулярную массу (например, ~22 кДа) по сравнению с вариантом, который не имеет мутацию в положении 83 (дорожка 1). Как показано на фиг. 18, когда остаток 83 мутирован на Val, вариант полипептида SIRP-α, экспрессированный в клетке-хозяине млекопитающего, представляет собой прежде всего молекулу с более высокой молекулярной массой (~22 кДа), указывающей на то, что эффективность для гликозилирования в N80, может быть увеличена.
Таблица 19. Репрезентативные данные связывания для полипептидных последовательностей SIRP-α, имеющих различные профили гликозилирования
Пример 2. Создание однополярных и биспецифических полипептидов SIRP-α.
Способность конструкций, содержащих гетеродимеры (i) слитого белка SIRP-α-Fc и (ii) мономера домена Fc, слитого с полипептидом, таким как антигенсвязывающий домен, для связывания как CD47, так и антигена, например EGFR, была определена при помощи SPR, как описано выше в данном примере. Слитые белки Fc для образования гетеродимеров приведены в таблице 20. Три монофункциональных (например, связывание одной мишени) слитых SIRP-α - Fc были протестированы. Эти слитые белки изображены как A, B, C на фиг. 6A. Первый монофункциональный слитый SIRP-α - Fc (ʺAʺ) представлял собой гомодимер SEQ ID NO: 136. Второй и третий монофункциональные слитые SIRP-α - Fc представляли собой гетеродимеры (i) слитый SIRP-α - Fc и (ii) мономер домена Fc без присоединенного к нему дополнительного полипептида. Они были созданы с использованием инженерных стратегий мутации Knob & Hole («выступ и впадина»), изображенных на фиг. 4A и 4B. Один монофункциональный слитый SIRP-α - Fc (ʺBʺ) был образован от гетеродимеризации SEQ ID NO: 139 (вариант Fc) и SEQ ID NO: 142 (слитый SIRP-α - Fc). Другой монофункциональный слитый SIRP-α - Fc (ʺCʺ) был образован от гетеродимеризации SEQ ID NO: 139 (вариант Fc) и SEQ ID NO: 138 (слитый SIRP-α - Fc). Бифункциональный (например, связывающий две мишени) слитый SIRP-α - Fc (ʺDʺ) был образован от гетеродимеризации SEQ ID NO: 127 (слитый SIRP-α Fc) и SEQ ID NO: 144 (антигенсвязывающий участок Erbitux, связанный с вариантом Fc). SEQ ID NO: 220 представляет собой легкую цепь антитела Erbitux.
Таблица 20. Аминокислотные последовательности слитых белков Fc для формирования гетеродимеров
Вкратце, CD47 иммобилизуют на чипе Proteon GLC при помощи химии аминов, как описано выше. В первой инъекции аналиты (например, A, B, C, D и Erbitux) вводили при 30 мкл/мин в PBST в течение 60 с при 100 нМ, а связывание с поверхностью CD47 было определено при помощи SPR. Через секунду вводили инъекцию 100 нМ EGFR-ECD (внеклеточный домен рецептора эпидермального фактора роста, продуцируемый в клетках HEK293), и измеряли связывание EGFR-ECD со связанными с CD47 аналитами. Erbitux не связывает CD47 на чипе, и поэтому он не смог связать EGFR во второй инъекции, как показано кривой, обозначенной «Erbitux» на фиг. 6B и показано на фиг. 6A. Слитые SIRP-α - Fc (например, A, B и C) связывают CD47, но не связывают EGFR во второй инъекции, как показано на кривых ʺA,ʺ ʺB,ʺ и ʺCʺ представленных на фиг. 6B и показано на фиг. 6A. Мономерные белки или белки с одним доменом D1 SIRP-α (например, B и C) с более высоким резонансом, чем димерный белок (например, A) из-за большего количества молекул, связанных с одними и теми же сайтами CD47, доступными на чипе, как показанные кривыми с надписью «B» и «C», указывающие на связывание с иммобилизованным CD47 и незначительное связывание с EGFR-ECD (например, монофункциональность). Гетеродимерный SIRP-α-Erbitux-Fc связал CD47 на чипе и также смог связать EGFR-ECD во второй инъекции, как показано кривой, обозначенной «D» на фиг. 6B, что указывает на связывание с иммобилизованным CD47 и связывание EGFR-ECD (например, бифункциональность).
Пример 3. Тестирование полипептидов с высокой аффинностью с CD47 у мышей
Генетически сконструированные мышиные модели различных видов рака, например солидная опухоль и гематологический рак, используются для проверки связывания полипептидов раскрытия с CD47. Полипептид раскрытия вводят мыши, которая в дальнейшем подвергается расчленению для обнаружения присутствия комплекса полипептида и CD47. При обнаружении используют антитела, специфичные к SIRP-α или CD47.
Пример 4. Тестирование полипептидов на иммуногенность.
Полипептиды, включая высокоаффинный вариант D1 SIRP-α, испытывают в анализах иммуногенности. Полипептиды тестируются как in silico, так и in vitro в анализах пролиферации Т-клеток, некоторые из которых являются коммерчески доступными. Полипептиды, которые вызывают минимальную реакцию иммуногенности в анализе пролиферации Т-клеток in vitro и демонстрируют более высокую аффинность связывания с CD47, чем SIRP-α дикого типа, выбраны для дальнейшей разработки.
Пример 5. Тестирование полипептидов на токсичность in vivo
Различные полипептиды, включая различные высокоаффинные варианты D1 SIRP-α, которые проявляют различную степень повышенной аффинности связывания с CD47 по сравнению с SIRP-α дикого типа, вводят в модель рака животного (например, модель рака мыши) для анализа влияния различных аффинностей связывания на токсичность в организме. Примат, отличный от человека (NHP) также может быть использован для тестирования высокоаффинных вариантов D1 SIRP-α, так как уровень кросс реактивности для CD47 примата, отличного от человека (NHP) и CD47 мыши может быть различным.
Пример 6 - Связывание рецептора Fcγ вариантами Fc
В дополнение к их способности модулировать целевую функцию терапевтические моноклональные антитела и Fc, содержащие слитые белки, также способны вызывать два первичных иммунных эффекторных механизма: антителозависимую клеточную цитотоксичность (ADCC) и комплементзависимую цитотоксичность (CDC). ADCC опосредованную связыванием области Fc с активацией рецепторов Fcγ и полипептидные конструкции, содержащие варианты Fc, описанные в рамках изобретения, тестируют на связывание рецептора Fcγ. Как показано в таблице 21 ниже, полипептидные конструкции продемонстрировали снижение связывания с одним или более рецепторами Fcγ по сравнению с соответствующим Fc IgG дикого типа. В отношении к IgG1 мутации L234A, L235A, G237A и N297A Fc IgG1 привели к серьезной потере связывания с рецепторами Fcγ CD16a, CD32a, CD32b, CD32c и CD64 по сравнению с IgG1 дикого типа или конструкции, в которой отсутствует одна или более из этих мутаций. Соответственно, мутации L234A, L235A, G237A (например, IgG1 AAA) наряду с агликозилированием или мутацией дегликозилирования N297A приводят к полной потере связывания с изученными рецепторами Fcγ. Поскольку известно, что связывание рецептора Fcγ играет важную роль в фагоцитозе, мутации L234A, L235A, G237A и N297A могут приводить к уменьшению фагоцитоза конструкции, содержащей вариант Fc.
В данном примере использовались следующие материалы и методы. Связывание рецепторов Fcγ человека RI (CD64), RIIA (CD32a), RIIB/C (CD32b/c) и RIIIA (CD16a) (R & D Systems, каталожные номера 1257-FC-050, 1330-CD-050, 1875-CD-050 и 4325-FC-050 соответственно) к вариантам Fc были проанализированы на приборе ProteOn XPR36 (Bio-Rad, Hercules, CA) используя забуференный фосфатом физиологический раствор (PBS, pH 7,4) дополненный 0,01% Tween-20 в качестве подвижного буфера. Примерно 400 единиц резонанса (RU) минимально биотинилированных конструкций Fc были иммобилизованы на проточных ячейках сенсорного чипа NLC (Bio-rad, Hercules, CA) путем взаимодействия авидин-нейтравидин. Биотинилирование проводили в соответствии с инструкциями изготовителя, используя Pierce EZ-Link Sulfo-NHS-LC-LC-Biotin и эквимолярное соотношение линкер:белок. Аналиты (hFcγR) были введены в кинетическом «одностадийном» режиме при номинальных концентрациях 0, 61, 185, 555, 1666 и 5000 нМ. Время ассоциации и диссоциации наблюдали на 90 с и 600 с соответственно. После каждой инъекции поверхность восстанавливали с использованием смеси элюирующего буфера Pierce IgG 2:1 об./об. (Life Technologies, каталожный номер 21004) и 4 М NaCl. Полное восстановление поверхности было подтверждено инъекцией вариантов Fc в начале и в конце эксперимента. Данные биосенсора были дважды откалиброваны, путем вычитания данных интерпота (не содержащих иммобилизованного белка) из данных о пятне реакции (иммобилизованный белок), а затем вычитания реакции буферной «заготовки» аналита с помощью инъекции аналита. Дважды откалиброванные данные были подобраны для равновесного анализа с использованием простой изотермы связывания. KD,app. Для молекул Fc с сильным связыванием с hFcγRI данные также были глобально сопоставимы с простой моделью Лэнгмюра и значение KD,app было рассчитано из соотношения кажущихся констант кинетической скорости (KDapp=kd,app/ka,app).
Как показано в таблице 21, мутации A330S, P331S и N297A области Fc IgG2 приводили к серьезной потере связывания с рецепторами Fcγ CD16a, CD32a, CD32b, CD32c и CD64 по сравнению с IgG дикого типа или конструкцией без этих мутаций. Соответственно, мутации A330S и P331S вместе с агликозилированием или дегликозилирующей мутацией N297A приводили к полной потере связывания с изученными рецепторами Fcγ. Поскольку известно, что связывание рецептора Fcγ играет важную роль в фагоцитозе, предполагается, что мутации A330S, P331S и N297A приводят к уменьшению фагоцитоза варианта Fc. Также представлены данные связывания для IgG4 и различные мутации.
Таблица 21. Данные связывания (KD) для связывания рецептора Fcγ с вариантами Fc.
Отсутствие связывания представлено ʺ-ʺ.
Пример 7 - Определение связывания C1q вариантов Fc
Комплементзависимая цитотоксичность (CDC) опосредуется белком комплемента C1q и активацией каскада комплемента. Связывание различных концентраций комплемента C1q с различными конструкциями Fc SIRP-α определяли с помощью твердофазного иммуноферментного анализа (ELISA). Слитые Fc SIRP-α готовили в 5 мкг/мл в PBS pH 7,4 и использовали для двойного покрытия лунок Nunc Immulon 4HBX ELISA 96-луночных планшетов (используя 50 мкл/лунка) в течение ночи при 4°C. На следующие сутки планшеты промывали пять раз при помощи промывочного буфера (PBS и 0,05% Tween-20) и инкубировали с 200 мкл/лунка блокирующего буфера (PBS и 0,5% БСА) в течение 1 часа при комнатной температуре. Планшеты промывали пять раз и инкубировали в течение 2 часов при комнатной температуре с 0, 0,13, 0,41, 1,23, 3,7, 11,1, 33,3, 100 мкг/мл C1q в буфере для анализа (PBS, 0,5% БСА, 0,05% Tween-20, 0,25% CHAPS, 5 мМ ЭДТА и 0,35% NaCl). Планшеты промывали и инкубировали в течение 1 часа с 50 мкл/лунка HRP конъюгированным овечьим-анти-человеческим-C1q при 2,0 мкг/мл в аналитическом буфере. Планшеты промывали пять раз и инкубировали в течение ~10 минут с TMB (1-Step Ultra TMB-ELISA, Thermo Sci. кат. № 34028). Наконец, добавляли 50 мкл/лунка Pierce/Thermo Sci. стоп-раствора (0,16 M серная кислота, кат. № N600) и планшеты считывали с поглощением 450 нм с эмиссией при 570 нм. Планшеты без слитого Fc SIRP-α, запускали для контроля за неспецифическим обнаружения связыванием C1q или HRP-конъюгированного антитела с планшетом. Лунки без C1q, запускались для контроля неспецифического связывания конъюгированного с HRP-антитела со слитым Fc SIRP-α-или с планшетом.
Как показано на фиг. 14, оба IgG1 дикого типа (SEQ ID NO: 123) и IgG2 дикого типа (SEQ ID NO: 126) связывают C1q в зависимости от дозы. И наоборот, варианты IgG1 IgG1_AAA (SEQ ID NO: 124); IgG1_N297A (SEQ ID NO: 125) и IgG1_AAA_N297A (SEQ ID NO: 96) продемонстрировали значительно сниженную и минимально обнаруживаемую активность связывания C1q. Аналогично, варианты IgG2 IgG2_A330S, P331S (SEQ ID NO: 127); IgG2_N297A (SEQ ID NO: 128) и IgG2_N297A, A330S, P331S (SEQ ID NO: 129) также продемонстрировали значительно уменьшенную и минимально определяемую активность связывания C1q. Эта уменьшенная и минимально обнаружимая C1q-связывающая активность вариантов IgG1 и IgG2 была сопоставима с IgG4 дикого типа (SEQ ID NO: 130), который не связывает C1q.
Пример 8 - Получение вариантов Fc и Fc дикого типа
Используя способы, описанные в рамках изобретения и в соответствии с вариантами осуществления раскрытия, были получены полипептиды Fc дикого типа и варианты Fc в таблице 7.
Пример 9 - Получение варианта полипептидов SIRP-α и варианта Fc
Используя способы, описанные в рамках изобретения и в соответствии с вариантами осуществления раскрытия, были получены следующие варианты полипептидов варианта D1 SIRP-α и варианта Fc, представленные в таблице 22 ниже. Связывание с CD47 человека определяли по методикам, описанным в примере 1.
Таблица 22. Аффинность связывания CD47 полипептидов вариантов SIRP-α вариантов Fc.
Пример 10 - Фагоцитоз вариантов Fc SIRP-α
Для получения количественных измерений фагоцитоза был использован анализ фагоцитоза, в котором первичные человеческие макрофаги и меченые GFP+или CFSE опухолевые клетки были совместно культивированы с конструкциями полипептида Fc, описанными в рамках изобретения. Использовались следующие материалы и методы:
Культура опухолевых клеточных линий
Клетки DLD-1-GFP-люцифераза, MM1R и N87 поддерживали в культуральной среде, содержащей RPMI (Gibco) дополненной 10% инактивированной теплом фетальной бычьей сыворотки (Gibco), 1% пенициллина/стрептомицина (Gibco) и 1% Glutamax (Gibco). Клетки DLD-1-GFP-люцифераза и N87 выращивали в виде адгезивных монослоев, а клетки MM1R выращивали в суспензии.
Получение и культивирование макрофагов, полученных из моноцитов человека
Лейкоцитарный слой сгустка цельной крови разбавляли 1:2 фосфатным буферным раствором (PBS, Gibco). Разведенную кровь разделяли на две пробирки и помещали сверху на 20 мл Ficoll-Paque Plus (GE Healthcare). Пробирки центрифугировали в течение 30 минут при 400 × g. Мононуклеарные клетки периферической крови (РВМС) собирали из интерфейса, дважды промывали добавлением 40 мл PBS, центрифугировали в течение 10 минут при 100 × g и ресуспендировали в буфере FACS (PBS с 0,5% бычьим сывороточным альбумином (Gibco)). CD14+моноциты очищали путем негативного отбора с использованием колонок Monocyte Isolation Kit II (Miltenyi Biotec) и LS (Miltenyi Biotec) в соответствии с протоколом производителя. CD14+моноциты высевали в 15 см чашки Петри для культивирования тканей (Corning) в количестве 10 миллионов клеток на чашку в 25 мл среды дифференцировки, состоящей из IMDM (Gibco), дополненной 10% AB-сывороткой человека (Corning), 1% пенициллином/стрептомицином и 1% Glutamax. Клетки культивировали в течение 7-10 суток.
Анализы фагоцитоза in vitro
Клетки DLD-1-GFP-люцифераза и N87 отделяли от культуральных планшетов, дважды промывая 20 мл PBS и инкубировали в 10 мл TrypLE Select (Gibco) в течение 10 минут при 37°C. Клетки удаляли с помощью скребка клеток (Corning), центрифугировали, промывали в PBS и ресуспендировали в IMDM. Клетки MM1R и N87 маркировали комплектом Celltrace CFSE Cell Proliferation kit (Thermo Fisher) в соответствии с инструкциями производителя и ресуспендировали в IMDM. Макрофаги отделяли от культуральных планшетов, дважды промывая 20 мл PBS и инкубировали в 10 мл TrypLE Select в течение 20 минут при 37°C. Клетки удаляли с помощью скребка клеток (Corning), промывали в PBS и ресуспендировали в IMDM.
Анализы фагоцитоза были проведены в 96-луночных планшетах с U-образным дном ультранизким содержанием (Corning) содержащих 100000 клеток DLD-1-GFP-люцифераза, MM1R или N87, пятикратные серийные разведения вариантов Fc SIRP-α от 1000 нМ до 64 пМ, и цетуксимумаб (Absolute Antibody), даратумумаб или контрольное антитело того же изотипа (Southern Biotech) в количестве 1 мкг/мл. Планшеты предварительно инкубировали 30 минут при 37°С во влажном инкубаторе с 5% диоксидом углерода, затем добавляли 50000 макрофагов. Плиты инкубировали два часа при 37°С во влажном инкубаторе с 5% диоксидом углерода. Клетки осаждали центрифугированием в течение пяти минут при 400 × g и промывали в 250 мкл буфера FACS. Макрофаги помещали на лед в течение 15 минут в 50 мкл буфера FACS, содержащего 10 мкл реагента для блокирования FcR человека (Miltenyi Biotec), 0,5 мкл анти-CD33 BV421 (Biolegend) и 0,5 мкл анти-CD206 APC-Cy7 (Biolegend). Клетки промывали в 200 мкл буфера FACS, промывали в 250 мкл PBS и помещали на лед в течение 30 минут в 50 мкл красителя Fixable Viability eFluor 506 (ebioscience), разбавленного 1:1000 в PBS. Клетки дважды промывали в 250 мкл буфера FACS и помещали на 30 минут на лед в 75 мкл Cytofix (BD Biosciences). Клетки промывали в 175 мкл буфера FACS и ресуспендировали в 75 мкл буфера FACS. Клетки анализировали на FACS Canto II (BD Biosciences) с последующим анализом данных Flowjo 10.7 (Treestar). Мертвые клетки были исключены из строя на отрицательной популяции e506. Макрофаги, которые имели фагоцитированные опухолевые клетки, были идентифицированы как клетки, позитивные в отношении CD33, CD206 и GFP или CFSE. Пять полипептидных конструкций, содержащих варианты домена D1 SIRP-α, слитые с соответствующими вариантами Fc, были протестированы на фагоцитоз in vitro:


Результаты
На фиг. 7 представлен фагоцитоз опухолевых клеток DLD-1-GFP-люцифераза с помощью макрофагов, полученных из моноцитов человека, в присутствии SEQ ID NO: 105 (вариант Fc IgG2_A330S, P331S, N297A) и SEQ ID NO: 127 (вариант Fc IgG2_A330S, P331S). В частности, на фиг. 7 показано, что SEQ ID NO: 105 (вариант Fc IgG2_A330S, P331S, N297A) (в присутствии или отсутствии контрольного антитела IgG1,k) имеет аблированный фагоцитоз в анализе фагоцитоза в качестве единственного агента, тогда как он способен увеличивать фагоцитоз цетуксимаба (CTX) (SEQ ID NO: 105+CTX). Напротив, полипептид с вариантом Fc IgG2_A330S, P331S (SEQ ID NO: 127+IgG1,k) имеет измеримую активность фагоцитоза в качестве единственного агента. Процент макрофагов, которые фагоцитировали опухолевые клетки и являлись GFP+ обозначены на оси y (фиг. 7). Концентрация сайтов связывания CD47 с добавлением SEQ ID NO: 105 и SEQ ID NO: 127 указана на оси х. Клетки DLD-1-GFP-люцифераза и макрофаги инкубировали с указанными концентрациями SEQ ID NO: 105, SEQ ID NO: 107 и CTX (1 мкг/мл) и контрольным антителом (IgG1, k). Клетки также инкубировали с PBS и цетуксимабом (линия с маркировкой PBS+CTX) или PBS и контрольным антителом того же изотипа (линия с маркировкой PBS+IgG1k).
На фиг. 8 представлен фагоцитоз опухолевых клеток DLD-1-GFP-люцифераза с помощью макрофагов, полученных из моноцитов человека, в присутствии SEQ ID NO: 96 (вариант Fc IgG1 L234A, L235A, G237A, N297A и SEQ ID NO: 124 (вариант Fc IgG1 L234A, L235A, G237A). В частности, на фиг. 8 показано, что вариант Fc IgG1 L234A, L235A, G237A, N297A (SEQ ID NO: 96) и вариант Fc IgG1 L234A, L235A, G237A (SEQ ID NO: 124) имеют аблированный фагоцитоз в анализе фагоцитоза в качестве отдельных агентов. Они представлены линиями, помеченными SEQ ID NO 96+IgG1, k и SEQ ID NO: 124+IgG1,k соответственно. Интересно, что оба полипептида SEQ ID NO: 96 и SEQ ID NO: 124 увеличивает фагоцитоз опухолеспецифического антитела - CTX. Как показано на фиг. 8, процент макрофагов, которые фагоцитировали опухолевые клетки и являлись GFP+ обозначены на оси y. Концентрация сайтов связывания CD47 с добавлением SEQ ID NO: 96 и SEQ ID NO: 124 указана на оси х. Клетки DLD-1-GFP-люцифераза и макрофаги инкубировали с CTX в концентрации 1 мкг/мл и указанными концентрациями SEQ ID NO: 96 (линия, обозначенная SEQ ID NO: 96+CTX) или SEQ ID NO: 124 (линия, обозначенная SEQ ID NO: 124+CTX). Для выявления неспецифических эффектов цетуксимаба при фагоцитозе клетки инкубировали с контрольным антителом того же изотипа, что и цетуксимаб, и указанными концентрациями SEQ ID NO: 96 (линия, обозначенная SEQ ID NO: 96+IgG1,k) или SEQ ID NO: 124 (линия, обозначенная SEQ ID NO: 124+IgG1,k). Клетки также инкубировали с PBS и цетуксимабом (линия с маркировкой PBS+CTX) или PBS и контрольным антителом того же изотипа (линия с маркировкой PBS+IgG1k).
На фиг. 9 представлен фагоцитоз опухолевых клеток DLD-1-GFP-люцифераза с помощью макрофагов, полученных из моноцитов человека, в присутствии SEQ ID NO: 134 (вариант Fc IgG4_S228P). В частности, на фиг. 9 представлено, что конструкция SEQ ID NO: 134 обладает значительной фагоцитозной активностью в качестве единственного агента в фагоцитозе in vitro. Как показано на фиг. 9, процент макрофагов, которые фагоцитировали опухолевые клетки и являлись GFP+ обозначены на оси y. Концентрация сайтов связывания CD47 с добавлением SEQ ID NO: 134 указана на оси х. Клетки DLD-1-GFP-люцифераза и макрофаги инкубировали с указанными концентрациями SEQ ID NO: 134 (линия, обозначенная SEQ ID NO: 134+среднее). Клетки также инкубировали с контрольным антителом (IgG1, k, черный квадрат).
Пример 11 - Получение варианта полипептидов SIRP-α и варианта ЧСА
Кроме того, используя способы, описанные в рамках изобретения и в соответствии с вариантами осуществления раскрытия, вариант D1 полипептида SIRP-α экспрессируется путем слияния с полипептидами ЧСА, как показано в таблице 23 ниже. Связывание с CD47 человека определяли по методикам, описанным в примере 1.
Таблица 23. Аффинность связывания CD47 полипептидов вариантов SIRP-α вариантов ЧСА.
Пример 12 - Увеличенный период полужизни, связанный с вариантами полипептидов SIRP-α
Как показано в таблице 24 и фиг. 10, варианты D1 полипептидов SIRP-α, содержащие слитые Fc и ЧСА могут иметь увеличенный период полужизни по сравнению с вариантом D1 SIRP-α одельно. Например, вариант D1 полипептида SIRP-α, слитый с Fc, представленный SEQ ID NO: 104 и вариант D1 полипептида SIRP-α слитый с ЧСА, представленный в SEQ ID NO: 159 имеют увеличенный период полужизни по сравнению с вариантом D1 полипептида SIRP-α который не слит с Fc или ЧСА, как представлено в SEQ ID NO: 85. Увеличение полужизни можно объяснить способностью вариантов D1 полипептидов SIRP-α, которые слиты с Fc и ЧСА для связывания с FcRn, что может быть связано с длительным циклом.
Таблица 24. Измерения продолжительности жизни для однократных доз, с вариантами полипептидов SIRP-α
Методологии, используемые для данного примера, следующие. Кратко, самцы мыши CD-1 массой около 25 грамм были получены от Harlan Labs и использовались для однократного исследования PK соединений, представленных SEQ ID NO: 104, SEQ ID NO: 159 и SEQ ID NO: 85. Каждое соединение было приготовлено в рабочей дозе 5 мг/мл. Объем дозы корректировали в зависимости от массы каждой мыши, гарантируя, что каждая мышь получала дозу 1, 3 и 10 мг/кг. Соединения вводили внутривенно через хвостовую вену мыши. Трем мышам вводили для каждой точки времени на каждом уровне дозы для каждого соединения. После дозирования у мышей поводили заборы крови в следующие 8 временных точек: 0,25, 1, 4, 8, 24, 48, 72 и 120 часов. 500 мкл цельной крови собирали в микро пробирки с помощью глазничного иссечения. Образцы цельной крови выдерживали в течение 30 минут, чтобы обеспечить отделение сыворотки. Образцы затем центрифугировали в течение 10 мин при 4 °С при RCF, равном 1000. Затем сыворотку переносили в 0,5 мл пробирки в течение 40 минут обработки и хранили замороженную до анализа.
Данные для SEQ ID NO: 104 были получены с использованием протокола ELISA Fc человека. Кратко, 96-луночные планшеты Immulon 4HBX ELISA были покрыты (Thermo Scientific кат. № 3855) 2 мкг/мл, 100 мкл/лунка очищенного CD47 в течение ночи при комнатной температуре в 1x буфер для хранения антигена (ImmunoChemistry Technologies, кат. № 6248). Лунки промывали 5 раз 200-300 мкл/лунка 1x TBST (Трис-буферный раствор+0,05% Tween-20) (Thermo Scientific 20x, кат. № 28360). Лунки блокировали 200 мкл/лунку 7,5% БСА в PBS (GIBCO, кат. № 15260-037) в течение 1-2 часов. Лунки промывали 5 раз 200-300 мкл/лунка 1x TBST. Добавляли 50 мкл/лунка стандартной кривой, контроли качества (QC) и неизвестные образцы, разбавленные нормальной сыворотки мыши CD1, разбавленной 1:4 в TBS. Стандартную кривую, QC и неизвестные образцы инкубировали при комнатной температуре в течение 1 часа. Концентрации для стандартной кривой были следующими: 0,2500 мкг/мл; 0,1250 мкг/мл; 0,0625 мкг/мл; 0,0313 мкг/мл; 0,0156 мкг/мл; 0,0078 мкг/мл; 0,0039 мкг/мл; 0,0020 мкг/мл; 0,0010 мкг/мл; 0,0005 мкг/мл; 0,00025 мкг/мл и 0,00000 мкг/мл. Контроли качества (КК) были заморожены и аликвотированы, и белок стандартной кривой в «высоких», «средних» и «низких» концентрациях на линейной кривой стандартной кривой, которые служили контролем, чтобы гарантировать, что анализ работает хорошо, следующим образом: QC высокий=0,125 мкг/мл; QC средний=0,016 мкг/мл и QC низкий=0,004 мкг/мл.
Затем, лунки промывали 5 раз 200-300 мкл/лунка 1x TBST. 50 мкл/лунка 0,25 мкг/мл Abbexa козьего анти-человеческого IgG Fc поликлонального антитела (11,6 мг/мл исходный раствор, Abbexa кат. № abx023511) разведенный в 1x TBST+1% БСА добавляли и инкубировали в течение 1 часа при комнатной температуре. Планшеты промывали 5 раз 200-300 мкл/лунка 1x TBST. 50 мкл/лунка 0,125 мкг/мл ZyMax/Invitrogen кроличьего анти-козьего конъюгата IgG - HRP (Thermo Scientific, кат. № 81-1620), разведенного в TBST+1% БСА добавляли и инкубировали в течение 1 часа при комнатной температуре. Лунки промывали 6 раз 200-300 мкл/лунка 1x TBST. Следующие этапы и реагенты использовали при комнатной температуре: 0 мкл/лунка 1-стадийный Ultra TMB - ELISA (Thermo Scientific ка № 34028) комнатной температуры добавляли и инкубировали 2-5 минут при комнатной температуре до тех пор, пока окрашивание не было достаточным. 50 мкл/лунка Stop Solution (0,16 M серной кислоты, Thermo Scientific кат. № N600) комнатной температуры добавляли сразу и хорошо перемешивали. Планшеты сразу считывали на спектрофотометре при OD, равной 450 и OD, равной 570. Считывание при OD 570 было фоновым считыванием, которое было вычтено из считывания при OD 450. Используя программное обеспечение, например Molecular Devices SoftMax Pro или Graph Pad Prism, стандартные значения кривой были построены с использованием кривой с 4 параметрами, а концентрации неизвестных образцов были интерполированы со стандартной кривой с использованием программного обеспечения.
Данные для SEQ ID NO: 85 был получен с использованием протокола ELISA His Tag. 96-луночные планшеты Immulon 4HBX ELISA были покрыты (Thermo Scientific кат. № 3855) 2 мкг/мл, 100 мкл/лунка очищенного CD47 в течение ночи при комнатной температуре в 1x буфер для хранения антигена (ImmunoChemistry Technologies, кат. № 6248). Лунки промывали 5 раз 200-300 мкл/лунка 1x TBST (Трис-буферный раствор+0,05% Tween-20) (Thermo Scientific 20x, кат. № 28360). Лунки блокировали 200 мкл/лунку 7,5% БСА в PBS (GIBCO, кат. № 15260-037) в течение 1-2 часов. Лунки промывали 5 раз 200-300 мкл/лунка 1x TBST. Добавляли 50 мкл/лунка стандартной кривой, контроли качества (QC) и неизвестные образцы, разбавленные нормальной сыворотки мыши CD1, разбавленной 1:4 в TBS. Стандартную кривую, QC и неизвестные образцы инкубировали при комнатной температуре в течение 1 часа. Концентрации стандартной кривой были следующими: 0,12500 мкг/мл; 0,06250 мкг/мл; 0,03125 мкг/мл; 0,01563 мкг/мл; 0,00781 мкг/мл; 0,00391 мкг/мл; 0,00195 мкг/мл; 0,00098 мкг/мл и 0,00000 мкг/мл. Контроли качества (КК) были заморожены и аликвотированы, и белок стандартной кривой в «высоких», «средних» и «низких» концентрациях на линейной кривой стандартной кривой, которые служили контролем, чтобы гарантировать, что анализ работает хорошо, следующим образом: QC высокий=0,02 мкг/мл; QC средний=0,01 мкг/мл и QC низкий=0,005 мкг/мл.
После этого лунки промывали 5 раз 200-300 мкл/лунка 1x TBST. 50 мкл/лунка 0,2 мкг/мл Abcam кроличьего анти-6x His Tag -HRP конъюгата поликлонального антитела (1 мг/мл стокового раствора, abcam кат. № ab1187) разведенного в TBST+1% БСА добавляли и инкубировали в течение 1 часа при комнатной температуре. Планшеты промывали 6 раз 200-300 мкл/лунка 1x TBST. После этого, следующие этапы и агенты использовали при комнатной температуре: 50 мкл/лунка 1-стадийный Ultra TMB - ELISA (Thermo Scientific ка № 34028) комнатной температуры добавляли и инкубировали 3-5 минут при комнатной температуре до тех пор, пока окрашивание не было достаточным. 50 мкл/лунка Stop Solution (0,16 M серной кислоты, Thermo Scientific кат. № N600) комнатной температуры добавляли сразу и хорошо перемешивали. Планшеты сразу считывали на спектрофотометре при OD, равной 450 и OD, равной 570. Считывание при OD 570 было фоновым считыванием, которое было вычтено из считывания при OD 450. Используя программное обеспечение, такое как Molecular Devices SoftMax Pro или Graph Pad Prism, стандартные значения кривой были построены с использованием кривой с 4 параметрами, а концентрации неизвестных образцов были интерполированы со стандартной кривой с использованием программного обеспечения.
Данные для SEQ ID NO: 159 были получены с использованием протокола ELISA ЧСА человека. 96-луночные планшеты Immulon 4HBX ELISA были покрыты (Thermo Scientific кат. № 3855) 2 мкг/мл, 100 мкл/лунка очищенного CD47 в течение ночи при комнатной температуре в 1x буфер для хранения антигена (ImmunoChemistry Technologies, кат. № 6248). Лунки промывали 5 раз 200-300 мкл/лунка 1x TBST (Трис-буферный раствор+0,05% Tween-20) (Thermo Scientific 20x, кат. № 28360). Лунки блокировали 200 мкл/лунка блокирующим буфером Li-Cor Odyssey (TBS) (Li-Cor, кат. № 927-50000) в течение 2 часов, а блокирующие буферы, содержащие альбумин, не использовались. Лунки промывали 5 раз 200-300 мкл/лунка 1x TBST. Добавляли 50 мкл/лунка стандартной кривой, контроли качества (QC) и неизвестные образцы, разбавленные нормальной сыворотки мыши CD1, разбавленной 1:4 в TBS. Стандартную кривую, QC и неизвестные образцы инкубировали при комнатной температуре в течение 1 часа.
Концентрации стандартной кривой были следующими: 3,20 мкг/мл; 1,60 мкг/мл; 0,80 мкг/мл; 0,40 мкг/мл; 0,20 мкг/мл; 0,10 мкг/мл; 0,05 мкг/мл; 0,025 мкг/мл и 0,00 мкг/мл. Контроли качества (КК) были заморожены и аликвотированы, и белок стандартной кривой в «высоких», «средних» и «низких» концентрациях на линейной кривой стандартной кривой, которые служили контролем, чтобы гарантировать, что анализ работает хорошо, следующим образом: QC высокий=0,6 мкг/мл; QC средний=0,3 мкг/мл; QC низкий=0,15 мкг/мл и QC низкий=0,01 мкг/мл.
После этого лунки промывали 5 раз 200-300 мкл/лунка 1x TBST. 50 мкл/лунка 1 мкг/мл Thermo Scientific/Pierce кроличьего анти-HSA-HRP конъюгата (Thermo Scientific, кат. № PA1-26887), разведенного в 1x TBST добавляли и инкубировали в течение 1 часа при комнатной температуре. Планшеты промывали 6 раз 200-300 мкл/лунка 1x TBST. После этого, следующие этапы и реагенты использовали при комнатной температуре: 50 мкл/лунка 1-стадийный Ultra TMB - ELISA (Thermo Scientific ка № 34028) комнатной температуры добавляли и инкубировали 3-5 минут при комнатной температуре до тех пор, пока окрашивание не было достаточным. 50 мкл/лунка TMB Stop Solution (0,16 M раствор серной кислоты, Thermo Scientific кат. № N600) комнатной температуры добавляли и хорошо перемешивали. Планшеты сразу считывали на спектрофотометре при OD, равной 450 и OD, равной 570. Считывание при OD 570 было фоновым считыванием, которое было вычтено из считывания при OD 450. Используя программное обеспечение, такое как Molecular Devices SoftMax Pro или Graph Pad Prism, стандартные значения кривой были построены с использованием кривой с 4 параметрами, а концентрации неизвестных образцов были интерполированы со стандартной кривой с использованием программного обеспечения.
Пример 13 - Уменьшенная гемагглютинация, продемонстрированная вариантами полипептида SIRP-α
Как показано на фиг. 11, вариант D1 полипептидов SIRP-α демонстрирует снижение или аблацию гемагглютинации. В частности, когда происходит гемагглютинация, вместо красной точки присутствует рассеянная красная окраска, как показано для положительного контроля B6H12. Для вариантов D1 полипептидов SIRP-α, проверенных на фиг. 11, произошло уменьшение или абляция гемагглютинации.
Методики, используемые для данного примера, были следующими: лейкоцитарный слой сгустка цельной крови человека был получен из центра крови Стэнфордского университета и разбавлен 1:2 фосфатным буферным раствором (PBS, Gibco). Разведенную кровь разделяли на две пробирки и помещали сверху на 20 мл Ficoll-Paque Plus (GE Healthcare). Пробирки центрифугировали в течение 30 минут при 400 × g. Супернатанты удаляли, а гранулы эритроцитов дважды промывали добавлением 30 мл PBS и центрифугированием при 3500 об/мин. Затем анализ гемагглютинации проводили следующим образом: эритроциты человека разводили в PBS и переносили в 96-луночные полистирольные планшеты (Corning) с 4 миллионами клеток на лунку в объеме 75 мкл. Пятикратные серийные разведения указанных белков добавляли в лунки в объеме 75 мкл PBS с конечной концентрацией от 1000 нМ до 0,488 нМ. В качестве негативного контроля PBS отдельно был добавлен в один ряд лунок. Эритроциты оседают на дно лунки, образуя небольшой и четко определенный осадок. В качестве позитивного контроля клетки обрабатывали антителом B6H12 против CD47 (ebioscience). Это антитело вызывало гемагглютинацию в концентрациях от 8 до 63 нМ, что указывалось образованием крупного и диффузного клеточного осадка. Среди тестируемых конструкций полипептиды на основе IgG2 (SEQ ID NO: 109 и SEQ ID NO: 113) вызывали небольшую гемагглютинацию при 4 и 8 нМ. Гемагглютинации не наблюдалось для всех других полипептидных конструкций (на основе IgG1 и HSA).
Пример 14 - Противоопухолевая активность SEQ ID NO: 211 в сингенной модели опухоли
Использовали мышей C57BL/6 (самки животных в возрасте 7-10-недель), полученных из лаборатории Charles River. Линию клеток аденокарциномы толстой кишки мыши MC38 извлекали из замороженных запасов и выращивали в RPMI 1640, содержащей 10% фетальной бычьей сыворотки, пенициллина-стрептомицина и L-глутамина. Клетки центрифугировали и ресуспендировали в концентрации 2E+07 клеток/мл в бессывороточной среде без добавок. На -7 сутки (т.е. за 7 суток до прогнозируемых промежуточных суток) мышам имплантировали путем подкожной инъекции в левый бок 100 мкл (2,0 × 106 клеток) на мышь свежеприготовленного МС38 в забуференном фосфатом физиологическом растворе (PBS). Когда опухоли достигли среднего объема приблизительно 50 мм3, пятьдесят животных с установленными опухолями и умеренными массами тела были рандомизированы в 5 групп лечения (группа 1-5, n=10 мышей в каждой). Начиная с 1-х суток мышей групп 1-5 обрабатывали носителем (PBS), антителом против mPD-L1 (клон 10F.9G2, 200 мкг), SEQ ID NO: 211 (200 мкг), антителом против mPD-L1 (200 мкг)+SEQ ID NO: 211 (100 мкг) или антителом против mPD-L1 (200 мкг)+SEQ ID NO: 211 (200 мкг), соответственно. Дозы вводили посредством внутрибрюшинной (IP) инъекции 0,05 мл/мышь на сутки 1, 4 и 7.
SEQ ID NO: 211 была создана путем генетического слияния SED ID NO: 206 с мономером домена Fc. SEQ ID NO: 206 содержит мутации, которые, как показано улучшают связывание с CD47 мыши. Данные связывания представлены в таблице 18.

Клинические наблюдения и масса тела контролировались на протяжении всего исследования до 42-х суток. Размеры опухолей измеряли два раза в неделю, а при завершении исследования минимальные (ширина, W и высота, H) и максимальные (длина, L) перпендикулярные размеры измеряли с использованием микрометра (Mitutoyo, Аврора, штат Иллинойс). Объем опухоли (мм3) был рассчитан по формуле для объема эллипсоидной сферы (L x W x H/2). Изучаемые животные подвергались гуманной жертве во время исследования, когда объемы опухолей у отдельных животных превышали (или приближались) 2,500 мм3. Количество животных, оставшихся в исследовании до 42 суток, использовалось для анализа выживаемости.
Опухоли выросли в разной степени во всех пяти группах. Среди мышей, которым вводили носитель или SEQ ID NO: 211 (200 мкг) (Группы 1 и 3, соответственно), жертвы начались во время 4-ой недели (с 25 суток), и все животные в этих группах были мертвы к концу 5-й недели (35-е сутки). Среди мышей, получавших антитело против mPD-L1 (отдельно или в комбинации с SEQ ID NO: 211, группы 2, 4 и 5), жертвы начались в течение 5-й недели (с 29-х суток или 32-х суток), но подмножество (40-70%) этих животных сохранились до запланированного окончания исследования (42-е сутки). На фиг. 12 показаны кривые выживаемости для каждой группы лечения в течение периода исследования. В численном виде группа лечения антитело против mPD-L1 и 200 мкг SEQ ID NO: 211 имела наибольшее количество выживших животных, за ней следовала группа лечения антитело против mPD-L1 и 100 мкг SEQ ID NO: 211 и группа антитело против mPD-L1 отдельно, с 7 из 10 (70%), 5 из 10 (50%) и 4 из 10 (40%) оставшихся мышей на 42 сутки, соответственно (таблица 25). Медианная выживаемость составляла 29 и 30,5 суток соответственно для плацебо (группа 1) и SEQ ID NO: 211 отдельно (группа 3). Медианная выживаемость увеличилась до 42 суток для антитела против mPD-L1 (группа 2) отдельно и антитела против PD-L1 и 100 мкг SEQ ID NO: 211 (группа 4). Медианная выживаемости для антитела против mPD-L1 и 200 мкг SEQ ID NO: 211 (Группа 5) не определялась, поскольку более 50% животных оставалось в конце исследования (42-е сутки).
Таблица 25. Данные о выживании животных.
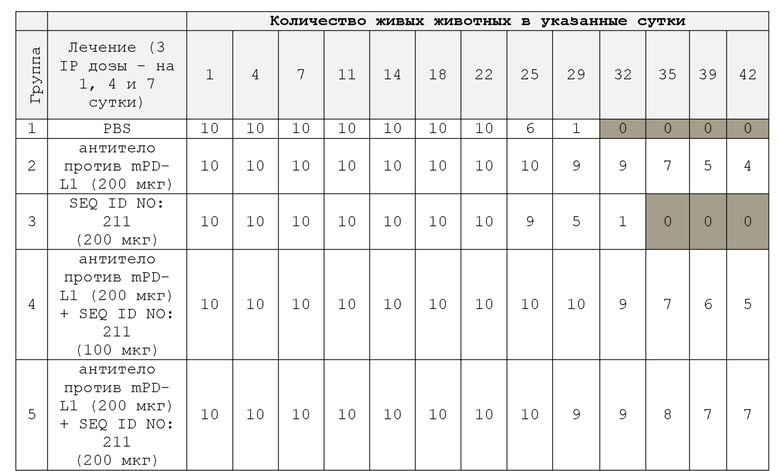
Опухоли проявляли быстрый рост в группе, леченой плацебо, что указывает на продолжающийся рост опухоли в отсутствие эффективного лечения. Дозирование 200 мкг SEQ ID NO: 211 (группа 3) обеспечило значительное ослабление роста опухоли только в промежуточные моменты времени (7-е и 14-е сутки, как для исходного, так и нормализованного объема опухоли) по сравнению с дозированием с носителем. Дозирование 200 мкг антитела против mPD-L1, отдельно или в комбинации с SEQ ID NO: 211 (группы 2, 4 и 5), обеспечило значительном ослаблении роста опухоли с 4 или 7 суток (для исходного или нормализованного объема опухоли, соответственно) по сравнению с дозированием с носителем (фиг. 13 и таблица 26). Добавление SEQ ID NO: 211 к режиму антитела против mPD-L1 (группа 2 против 4 или группа 2 против 5) приводило к дополнительному ингибированию роста опухоли только по сравнению с антителом против mPD-L1 отдельно. Объемы опухоли на 22 сутки, включая ингибирование роста опухоли (% TGI), приведены в таблице 26. 22 сутки использовали для сравнения, потому что эти сутки являются последним моментом, когда все животные все еще были живы. Ингибирование роста опухоли (% TGI) на 22 сутки по сравнению с первыми сутками составляло 83%, 81% и 77% для группы антитело mPD-L1 и 200 мкг SEQ ID NO: 211, группы антитело mPD-L1 и 100 мкг SEQ ID NO: 211 и группы антитело против mPD-L1 отдельно, соответственно (таблица 26).
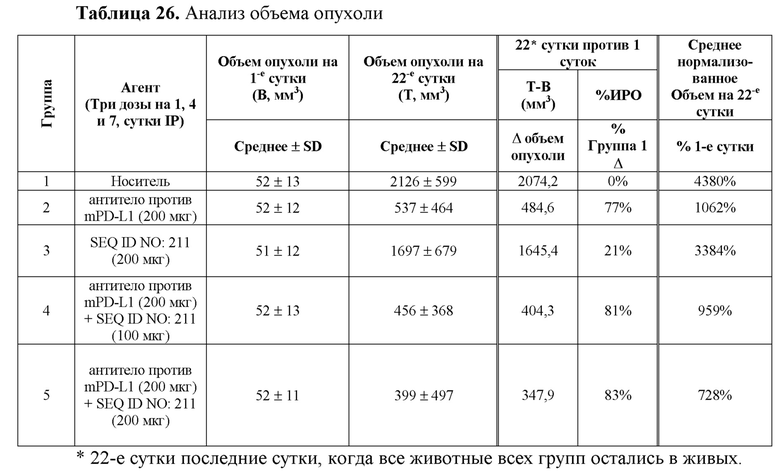
Пример 15 - Оптимизация комбинированной терапии для лечения рака
Полипептиды, включая высокоаффинный вариант D1 SIRP-α, вводят совместно с ингибиторами контрольных точек для лечения мышиных моделей различных видов рака, например, солидной опухоли и гематологического рака. Рак может быть распознан иммунной системой, и при некоторых обстоятельствах иммунная система может участвовать в устранении опухолей. Блокада ко-ингибирующих молекул, таких как CTLA-4, PD-1 и LAG-3, может быть вовлечена в усиление ответов Т-клеток против опухолей. Полипептиды, описанные в рамках изобретения, вводят в комбинации с ингибитором контрольных точек, таким как ингибитор антител CTLA-4 (например, ипилимумаб, тремелимумаб), PD-1 (ниволумаб, пидилизумаб, MK3475, также известный как пембролизумаб, BMS936559 и MPDL3280A) и LAG-3 (например, BMS986016).
Установленные опухоли A20 у мышей BALB/c (например, модели лимфомы) обрабатывают ингибитором антител CTLA-4 и высокоаффинным вариантом D1 SIRP-α, слитым с вариантом Fc IgG, представленным в рамках изобретения (например, конструкции SIRP-α). Начиная с 1-х суток, мышей лечили плацебо (PBS), треммулимумабом (200 мкг)+конструкцией SIRP-α (100 мкг) или тиремимумабом (200 мкг)+конструкцией SIRP-α (200 мкг). Дозы вводили путем внутрибрюшинной (IP) инъекции 0,05 мл/мышь на 1, 4 и 7 сутки. Ответ опухоли на комбинированную терапию определяли ежесуточно, измеряя объем опухоли. Если на 4-е сутки объем опухоли у мышей, получавших комбинированную терапию, не демонстрировал значительного улучшения, тремлимумаб заменяли ипилимумабом. Аналогично, если на 7-е сутки объем опухоли у мышей, получавших комбинированную терапию, не демонстрировал значительного улучшения, тремлимумаб заменяли ипилимумабом. Ожидается, что в то время как тремелимумаб и ипилимумаб нацелены на один и тот же контрольный белок, они обладают различной терапевтической эффективностью и синергическими эффектами с конструкцией SIRP-α из-за их различных областей Fc. Тремелимумаб представляет собой антитело IgG2, которое может быть более эффективным при фиксации комплемента, в то время как ипилимуммаб представляет собой антитело IgG1, которое может быть полезным для предотвращения элиминации активированных Т-клеток.
Пример 16 - Способ лечения рака, экспрессирующего эпителиальный маркер
Конструкции полипептида SIRP-α, такие как высокоаффинный вариант D1 SIRP-α (например, любой вариант представленный в таблицах 2, 5 и 6) вариант Fc, слитый с IgG, вводили для лечения рака, экспрессирующего маркер эпителиальных клеток. Увеличенный фагоцитоз, возникающий в результате блокады передачи CD47, например, с помощью конструкции D1 SIRP-α, может зависеть от наличия макрофагов. Поэтому введение полипептидной конструкции D1 SIRP-α в комбинации с антителом, нацеленным на эпителиальный маркер, экспрессируемый в раковой клетке или на ней, используемый для лечения рака, снижает риск побочных эффектов, например, фагоцитоз эпителиальных клеток, из-за низкого содержания макрофагов на периферии кожи.
Мышиной модели рака, экспрессирующего эпителиальный маркер, например, EGFR или EpCAM, вводят конструкцию SIRP-α в сочетании с антителом, которое нацелено на эпителиальный маркер, например, антитело против EGFR или антитело против EpCAM. Антитела, нацеленные на эпителиальные маркеры, могут распознавать как клетки рака, так и не клетки, не являющиеся клетками рака, например, нераковые клетки на периферии кожи. Однако ожидается, что нераковые клетки на периферии кожи не будут подвержены фагоцитозу из-за низкого содержания макрофагов вблизи кожи.
Пример 17 - Фагоцитоз за счет слитого одноплечевого SIRP-α - Fc
Для получения количественных измерений фагоцитоза, индуцированных слитым SIRP-α-Fc, имеющими одну молекулу SIRP-α (например, молекулу одного плеча) (изображенного на фиг. 1, 4A и 4B), анализ фагоцитоза с различными типами клеток MM1R и N87 проводили с использованием способов, описанных в примере 8.
Шесть конструкций с одним плечом были протестированы на фагоцитоз in vitro. Эти конструкции с одним плечом создавали с использованием стратегий выступов и впадин. Гомодимер слитого SIRP-α Fc SEQ ID NO: 136 был использован в качестве двухплечевого сравнения (контроль). Первый слитый одноплечевой SIRP-α Fc (например, A) был образован из гетеродимера SEQ ID NO: 139 (вариант Fc) и SEQ ID NO: 138 (слитый SIRP-α - Fc). Второй слитый одноплечевой SIRP-α Fc (например, B) был образован из гетеродимера SEQ ID NO: 141 (вариант Fc) и SEQ ID NO: 140 (слитый SIRP-α - Fc). Третий слитый одноплечевой SIRP-α Fc (например, C) был образован из гетеродимера SEQ ID NO: 139 (вариант Fc) и SEQ ID NO: 142 (слитый SIRP-α - Fc). Четвертый слитый одноплечевой SIRP-α Fc (например, D) был образован из гетеродимера SEQ ID NO: 141 (вариант Fc) и SEQ ID NO: 143 (слитый SIRP-α - Fc). Пятый слитый одноплечевой SIRP-α Fc (например, E) был образован из гетеродимера SEQ ID NO: 147 (вариант Fc) и SEQ ID NO: 146 (слитый SIRP-α - Fc). Шестой слитый одноплечевой SIRP-α Fc (например, F) был образован из гетеродимера SEQ ID NO: 149 (вариант Fc) и SEQ ID NO: 148 (слитый SIRP-α - Fc). Аффинность связывания CD47 (KD) одноплечевого SIRP-α при испытании в виде мономера являлась следующей: ~10 пМ (A,B), ~100 пМ (C, D) и ~5 нМ (E, F). Последовательности представлены в таблице 27 ниже.
Таблица 27. Аминокислотные последовательности слитых SIRPα - Fc для создания гетеродимеров
На фиг. 15-17 представлен фагоцитоз линии множественной миеломы 1R (MM1R) и линии карциномы желудка N87 с помощью неполяризованных макрофагов, полученных из моноцитов человека. ʺ+ʺ и ʺ-ʺ обозначает добавление или отсутствие даратумумаба (Dara) соответственно на фиг. 15-16. На фиг. 17, ʺ+ʺ и ʺ-ʺ означает добавление и отсутствие герцептина/трастузумаба (Her) соответственно.
На фиг. 15 представлена конструкция A в присутствии контрольного антитела (IgG1, k), например, ʺA-ʺ имеет аблированный фагоцитоз в качестве единственного агента, тогда как он способен увеличивать фагоцитоз даратумумаба (Dara), например, ʺA+ʺ. Аналогично, конструкция B в присутствии контрольного антитела (IgG1, k), например, ʺB-ʺ имеет аблированный фагоцитоз в качестве единственного агента, тогда как он способен увеличивать фагоцитоз даратумумаба (Dara), например, ʺB+ʺ. Процент макрофагов, которые фагоцитировали MM1R и являлись CFSE+ обозначены на оси y. Концентрация сайтов связывания CD47 от добавления конструкции A, B или контрольной конструкции указана на оси x. Уровни фагоцитоза сравнимы с контрольной конструкцией (которая представляет собой двуплечевой SIRP-α). Уровень фагоцитоза, возникающий в результате инкубации с антителом против CD47, например, B6H12 (100 нМ), сравним с инкубацией с Dara, например, PBS +. Как показано, слитые одноплечевой SIRP-α-Fc могут увеличить фагоцитоз Dara по сравнению со слитым двуплечевой SIRP-α-Fc.
На фиг. 16 представлена конструкция C в присутствии контрольного антитела (IgG1, k), например, ʺC-ʺ имеет аблированный фагоцитоз в анализе фагоцитоза в качестве единственного агента, тогда как он способен увеличивать фагоцитоз даратумумаба (Dara), например, ʺC+ʺ. Аналогично, конструкция D в присутствии контрольного антитела (IgG1, k), например, ʺD-ʺ имеет аблированный фагоцитоз в анализе фагоцитоза в качестве единственного агента, тогда как он способен увеличивать фагоцитоз даратумумаба (Dara), например, ʺD+ʺ. Процент макрофагов, которые фагоцитировали MM1R и являлись CFSE+ обозначены на оси y. Концентрация сайтов связывания CD47 от добавления конструкции C, D или контрольной конструкции указана на оси x. Уровни фагоцитоза сравнимы с контрольной конструкцией (которая представляет собой двуплечевой SIRP-α). Уровень фагоцитоза, возникающий в результате инкубации с антителом против CD47, например, B6H12 (100 нМ), сравним с инкубацией с Dara, например, PBS +. Как показано, слитые одноплечевой SIRP-α-Fc могут увеличить фагоцитоз Dara по сравнению со слитым двуплечевой SIRP-α-Fc. Как показано, слитые одноплечевой SIRP-α-Fc могут увеличить фагоцитоз Dara по сравнению со слитым двуплечевой SIRP-α-Fc.
На фиг. 17 представлен фагоцитоз в присутствии низкоаффинных одноплечевых конструкций SIRP-α (E, F), выполненный аналогично вышеприведенным примерам. Как показано, эти низкоаффинные одноплечевые конструкции SIRP-α (E, F) в сочетании с герцептином продемонстрировали сопоставимый фагоцитоз клеток N87 по сравнению с герцептином отдельно (PBS+). Следовательно, аффинность 5 нМ с CD47 недостаточно для слитых одноплечевой SIRP-α-Fc для усиления дальнейшего фагоцитоза in vitro в сочетании с герцептином.
Пример 17 - Кросс-реактивность высокоаффинных вариантов D1 SIRP-α
Полипептиды высокоаффинных вариантов D1 SIRP-α были получены, как описано выше. Связывание с CD47 человека, мыши и крысы определяли с использованием SPR, измеренное прибором Biacore T100 (GE Healthcare) и Proteon XPR36 (Bio-rad, Hercules, CA), как описано в примере 1. SEQ ID NO: 215 представляет собой сконструированный вариант D1 SIRP-α, который не связывается с CD47 человека, мыши или крысы и использовался в качестве негативного контроля.
Таблица 28. Репрезентативная кросс-видовая аффинность связывания CD47 для высокоаффинных вариантов SIRP-α

Пример 18 - Противоопухолевая активность высокоаффинных конструкций SIRP-α в опухолевой модели ксенотрансплантата мыши
Иммунодефицитные мыши NOD ТКИД гамма (NSG) (NOD.Cg-Prkdcscid Il2rgtm1Wjl/SzJ; 50 самок, плюс резервные) были приобретены в возрасте животных от 6 до 10 недель. Клетки линии клеток лимфомы человека GFP-Luc-Raji выращивали на RPMI 1640, содержащей 10% фетальной бычьей сыворотки, пенициллина, стрептомицина и L-глутамина. Затем клетки центрифугировали и повторно суспендировали в концентрации 1,0 × 107 клеток/мл в бессывороточной среде без добавок и объединяли 1:1 с Matrigel ™ (Trevigen, Гейтерсберг, штат Мэриленд). На -11 сутки (т.е. за 11 суток до прогнозируемых промежуточных суток) мышам имплантировали путем подкожной инъекции в левый бок 200 мкл (1,0 × 106 клеток) на мышь свежеприготовленного раствора GFP-Luc-Raji:Matrigel. Когда опухоли достигли среднего объема приблизительно 55 мм3, пятьдесят животных с установленными опухолями и умеренными массами тела были рандомизированы в 5 групп лечения (группа 1-5, n=10 мышей в каждой). Начиная с 1-х суток, Группы 1-5 лечили (1) SEQ ID NO: 215 [10 мг/кг (мг/кг массы тела), 3x/неделя]; (2) SEQ ID NO: 104 (10 мг/кг массы тела, 3x/неделя); (3) ритуксимаб (5 мг/кг массы тела, 2x/неделя)+SEQ ID NO: 100 (10 мг/кг массы тела, 3x/неделя); (4) ритуксимаб (5 мг/кг массы тела, 2x/неделя)+SEQ ID NO: 104 (10 мг/кг массы тела, 3x/неделя) или (5) ритуксимаб (5 мг/кг массы тела, 2x/неделя)+SEQ ID NO: 215 (10 мг/кг массы тела, 3x/неделя), соответственно. Дозы вводили путем внутрибрюшинной (IP) инъекции 0,05 мл/мышь. Для всех животных дозы вводили начиная со стадии постановки и продолжали в общей сложности 31 сутки (1-31 сутки).
Клинические наблюдения регистрировались два раза в сутки (утром и вечером). Дополнительные наблюдения регистрировались, когда наблюдались. Массу тела измеряли три раза в неделю с использованием электронных весов (Ohaus SCOUT® PRO). Размеры опухолей измеряли три раза в неделю, и при завершении исследования с использованием микрометра (Mitutoyo, Аврора, штат Иллинойс) для измерения перпендикулярных минимальных (ширина, W и высота, H) и максимальных (длина, L) размеров. Объем опухоли (мм3) был рассчитан по формуле для объема эллипсоидной сферы (L x W x H/2). Образцы крови были взяты от 20 животных на 1-е сутки (базовый уровень, до группового назначения) и от всех животных на 8-е сутки (неделя 1) и на 31-е сутки (при окончании). Образцы крови были отправлены на полный анализ крови (CBC) в соответствующие сутки забора.
Конструкция SIRP-α SEQ ID NO: 215 не имеет измеримого связывания с CD47 (см. таблицу 28). Опухоли в дозовой группе SEQ ID NO: 215 (группа 1) росли линейно до 31 суток (фиг. 19A), аналогичные опухолям, наблюдаемым в группе PBS с той же моделью (данные не представлены). Это наблюдение демонстрирует постоянный рост опухоли при отсутствии эффективного лечения.
Сравнение между группами 1 и 5 (SEQ ID NO: 215 с ритуксимабом или без него) и между группами 2 и 4 (SEQ ID NO: 104 с ритуксимабом или без него) демонстрирует, что комбинированные лечения обеспечили значительное уменьшение объема опухоли в виде исходных значений (с 9-х суток) и нормализованных значений (с 7-х суток). К 16-м суткам большинство мышей группы 3 (SEQ ID NO: 100+ритуксимаб) и группы 4 (SEQ ID NO: 104+ритуксимаб) больше не содержали обнаруживаемых опухолей; эти два комбинированных лечения продемонстрировали сходную эффективность. Напротив, рост опухоли, по-видимому, восстанавливался у животных группы 5 (SEQ ID NO: 215+ритуксимаб) с 18-х суток. Объемы опухолей всех пяти групп за исследуемый период (среднее +/- SEM и отдельные графики рассеяния) представлены на фиг. 19A и фиг. 19B, соответственно.
Показатели полной крови (CBC) (эритроциты, гемоглобин, гематокрит, тромбоциты и т.д.) измеряли перед дозированием (1-е сутки), через 1 неделю после введения дозы (9-е сутки) и через 4 недели после введения дозы (31-е сутки). Параметры не сильно отличались на первой или четвертой неделе среди пяти групп. Значения гемоглобина (HGB) представлены на фиг. 19C. Данные результаты демонстрируют, что высокоаффинные конструкции SIRP-α могут эффективно ослаблять рост опухоли и синергизировать с ритуксимабом в модели рака in vivo мыши. Кроме того, в отличие от лечений на основе антител против CD47, никаких острых эпизодов анемии не наблюдалось ни в одной из тестовых групп, леченых высокоаффинными конструкциями SIRP-α.
Пример 19: Варианты Fc конструкций SIRP-α демонстрируют уменьшенную токсичность в отношении красных кровяных телец
Потеря красных кровяных телец является проблемой при нацеливании на CD47. Чтобы исследовать влияние варианта Fc конструкции SIRP-α на токсичность в отношении красных кровяных телец, мышей лечили высокоаффинным вариантом конструкции SIRP-α, содержащим либо конструкцию Fc IgG1 дикого типа (SEQ ID NO: 216), либо вариант Fc конструкции IgG1 (SEQ ID NO: 96) с мутациями IgG1 L234A, L235A, G237A и N297A (IgG1_AAA_N297A). Мышей разделяли на пять групп по шесть и лечили на 1 и 7 сутки (см. сплошные стрелки на фиг. 20) или: (1) PBS; (2) 10 мг/кг SEQ ID NO: 216 (Fc IgG1 дикого типа); (3) 30 мг/кг SEQ ID NO: 216; (4) 10 мг/кг SEQ ID NO: 96 (IgG1_AAA_N297A), или (5) 30 мг/кг SEQ ID NO: 96. Измерения уровня полной крови (CBC) были взяты у всех животных на 7-е сутки и для трех из шести животных на 1-е сутки. Заборы крови (см фиг. 20) осуществляли между тремя мышами из каждой группы, чтобы не превышать количество крови, которое было разрешено в неделю. Как показано на фиг. 20, лечение конструкцией варианта D1 SIRP-α, содержащей IgG1 дикого типа, приводило к дозозависимому снижению количества эритроцитов. Напротив, лечение конструкцией варианта D1 SIRP-α, содержащей IgG1_AAA_N297A, приводило к тому, что количество эритроцитов было похоже на контрольную группу, леченую PBS.
Хотя в настоящем документе показаны и описаны предпочтительные варианты осуществления настоящего изобретения, для специалистов в данной области будет очевидно, что такие варианты осуществления предоставлены только в качестве примера. Специалистам в данной области понятны многочисленные варианты, изменения и замещения, без отклонения от идеи настоящего изобретения. Следует понимать, что различные альтернативы вариантов реализации настоящего изобретения, описанных в настоящем изобретении, могут быть использованы при практическом осуществлении настоящего изобретения. Подразумевается, что следующая формула изобретения определяет рамки настоящего изобретения, и таким образом охватываются способы и структуры в этой формуле, а также их эквиваленты.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ALEXO THERAPEUTICS INC.
<120> КОНСТРУКЦИИ, ИМЕЮЩИЕ SIRP-АЛЬФА ДОМЕН ИЛИ ЕГО ВАРИАНТ
<130> 75797-20004.40
<140> PCT/US2016/045914
<141> 2016-08-05
<150> 62/346,414
<151> 2016-06-06
<150> 62/276,801
<151> 2016-01-08
<150> 62/276,796
<151> 2016-01-08
<150> 62/265,887
<151> 2015-12-10
<150> 62/202,772
<151> 2015-08-07
<150> 62/202,775
<151> 2015-08-07
<150> 62/202,779
<151> 2015-08-07
<160> 223
<170> PatentIn версии 3.5
<210> 1
<211> 119
<212> Белок
<213> Homo sapiens
<400> 1
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Ile Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 2
<211> 118
<212> Белок
<213> Homo sapiens
<400> 2
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Val Thr Ser Leu Ile Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Ser Thr Lys Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 3
<211> 118
<212> Белок
<213> Homo sapiens
<400> 3
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu Leu Cys Thr Val Thr Ser Leu Ile Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Ser Thr Lys Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 4
<211> 119
<212> Белок
<213> Homo sapiens
<400> 4
Glu Glu Gly Leu Gln Val Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Ala Thr Ser Leu Ile Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 5
<211> 119
<212> Белок
<213> Homo sapiens
<400> 5
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Phe Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Ile Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 6
<211> 119
<212> Белок
<213> Homo sapiens
<400> 6
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Ile Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Pro Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 7
<211> 118
<212> Белок
<213> Homo sapiens
<400> 7
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Val Thr Ser Leu Ile Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Ser Thr Lys Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Gly Lys Pro Ser
115
<210> 8
<211> 118
<212> Белок
<213> Homo sapiens
<400> 8
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Ile Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Ser Thr Lys Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 9
<211> 119
<212> Белок
<213> Homo sapiens
<400> 9
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Ile Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 10
<211> 118
<212> Белок
<213> Homo sapiens
<400> 10
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Val Thr Ser Leu Ile Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Ser Thr Lys Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 11
<211> 119
<212> Белок
<213> Homo sapiens
<220>
<221> MOD_RES
<222> (3)..(3)
<223> E или G
<220>
<221> MOD_RES
<222> (12)..(12)
<223> S или F
<220>
<221> MOD_RES
<222> (14)..(14)
<223> L или S
<220>
<221> MOD_RES
<222> (20)..(20)
<223> T или S
<220>
<221> MOD_RES
<222> (22)..(22)
<223> T или I
<220>
<221> MOD_RES
<222> (24)..(24)
<223> R или H или L
<220>
<221> MOD_RES
<222> (27)..(27)
<223> A или V
<220>
<221> MOD_RES
<222> (45)..(45)
<223> G или A
<220>
<221> MOD_RES
<222> (65)..(65)
<223> D или E
<220>
<221> MOD_RES
<222> (66)..(66)
<223> L или S
<220>
<221> MOD_RES
<222> (70)..(70)
<223> N или E, или D
<220>
<221> MOD_RES
<222> (75)..(75)
<223> S или P
<220>
<221> MOD_RES
<222> (77)..(77)
<223> R или S
<220>
<221> MOD_RES
<222> (79)..(79)
<223> G или S
<220>
<221> MISC_FEATURE
<222> (99)..(99)
<223> Может как присутствовать, так и отсутствовать
<220>
<221> MOD_RES
<222> (100)..(100)
<223> D или P
<220>
<221> MOD_RES
<222> (102)..(102)
<223> V или T
<220>
<221> MOD_RES
<222> (116)..(116)
<223> A или G
<400> 11
Glu Glu Xaa Leu Gln Val Ile Gln Pro Asp Lys Xaa Val Xaa Val Ala
1 5 10 15
Ala Gly Glu Xaa Ala Xaa Leu Xaa Cys Thr Xaa Thr Ser Leu Ile Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Xaa Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Xaa Xaa Thr Lys Arg Xaa Asn Met Asp Phe Xaa Ile Xaa Ile Xaa Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Xaa Asp Xaa Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Xaa Lys Pro Ser
115
<210> 12
<211> 585
<212> Белок
<213> Homo sapiens
<400> 12
Asp Ala His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu
1 5 10 15
Glu Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln
20 25 30
Gln Cys Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu
35 40 45
Phe Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys
50 55 60
Ser Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu
65 70 75 80
Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro
85 90 95
Glu Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu
100 105 110
Pro Arg Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His
115 120 125
Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg
130 135 140
Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg
145 150 155 160
Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala
165 170 175
Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser
180 185 190
Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu
195 200 205
Arg Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro
210 215 220
Lys Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys
225 230 235 240
Val His Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp
245 250 255
Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser
260 265 270
Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His
275 280 285
Cys Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser
290 295 300
Leu Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala
305 310 315 320
Glu Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg
325 330 335
Arg His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr
340 345 350
Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu
355 360 365
Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro
370 375 380
Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu
385 390 395 400
Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro
405 410 415
Gln Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys
420 425 430
Val Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys
435 440 445
Ala Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His
450 455 460
Glu Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser
465 470 475 480
Leu Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr
485 490 495
Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp
500 505 510
Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala
515 520 525
Leu Val Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu
530 535 540
Lys Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys
545 550 555 560
Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val
565 570 575
Ala Ala Ser Gln Ala Ala Leu Gly Leu
580 585
<210> 13
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<220>
<221> MOD_RES
<222> (4)..(4)
<223> L или I, или V
<220>
<221> MOD_RES
<222> (6)..(6)
<223> V или L, или I
<220>
<221> MOD_RES
<222> (21)..(21)
<223> A или V
<220>
<221> MOD_RES
<222> (27)..(27)
<223> A или I, или L
<220>
<221> MOD_RES
<222> (31)..(31)
<223> I или T, или S, или F
<220>
<221> MOD_RES
<222> (47)..(47)
<223> E или V, или L
<220>
<221> MOD_RES
<222> (53)..(53)
<223> K или R
<220>
<221> MOD_RES
<222> (54)..(54)
<223> E или Q
<220>
<221> MOD_RES
<222> (56)..(56)
<223> H или P, или R
<220>
<221> MOD_RES
<222> (66)..(66)
<223> L или T, или G
<220>
<221> MOD_RES
<222> (68)..(68)
<223> K или R
<220>
<221> MOD_RES
<222> (92)..(92)
<223> V или I
<220>
<221> MOD_RES
<222> (94)..(94)
<223> F или L, или V
<220>
<221> MOD_RES
<222> (104)..(104)
<223> F или V
<400> 13
Glu Glu Glu Xaa Gln Xaa Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Xaa Thr Leu Arg Cys Thr Xaa Thr Ser Leu Xaa Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Xaa Leu
35 40 45
Ile Tyr Asn Gln Xaa Xaa Gly Xaa Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Xaa Thr Xaa Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Xaa Lys Xaa Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Xaa Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 14
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<220>
<221> MOD_RES
<222> (4)..(4)
<223> L или I, или V
<220>
<221> MOD_RES
<222> (6)..(6)
<223> V или L, или I
<220>
<221> MOD_RES
<222> (21)..(21)
<223> A или V
<220>
<221> MOD_RES
<222> (27)..(27)
<223> V или I, или L
<220>
<221> MOD_RES
<222> (31)..(31)
<223> I или T, или S, или F
<220>
<221> MOD_RES
<222> (47)..(47)
<223> E или V, или L
<220>
<221> MOD_RES
<222> (53)..(53)
<223> K или R
<220>
<221> MOD_RES
<222> (54)..(54)
<223> E или Q
<220>
<221> MOD_RES
<222> (56)..(56)
<223> H или P, или R
<220>
<221> MOD_RES
<222> (66)..(66)
<223> S или T, или G
<220>
<221> MOD_RES
<222> (68)..(68)
<223> K или R
<220>
<221> MOD_RES
<222> (92)..(92)
<223> V или I
<220>
<221> MOD_RES
<222> (94)..(94)
<223> F или L, или V
<220>
<221> MOD_RES
<222> (103)..(103)
<223> F или V
<400> 14
Glu Glu Glu Xaa Gln Xaa Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Xaa Ile Leu His Cys Thr Xaa Thr Ser Leu Xaa Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Xaa Leu
35 40 45
Ile Tyr Asn Gln Xaa Xaa Gly Xaa Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Xaa Thr Xaa Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Xaa Lys Xaa Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Xaa Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 15
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<220>
<221> MOD_RES
<222> (4)..(4)
<223> L или I, или V
<220>
<221> MOD_RES
<222> (6)..(6)
<223> V или L, или I
<220>
<221> MOD_RES
<222> (21)..(21)
<223> A или V
<220>
<221> MOD_RES
<222> (27)..(27)
<223> V или I, или L
<220>
<221> MOD_RES
<222> (31)..(31)
<223> I или T, или S, или F
<220>
<221> MOD_RES
<222> (47)..(47)
<223> E или V, или L
<220>
<221> MOD_RES
<222> (53)..(53)
<223> K или R
<220>
<221> MOD_RES
<222> (54)..(54)
<223> E или Q
<220>
<221> MOD_RES
<222> (56)..(56)
<223> H или P, или R
<220>
<221> MOD_RES
<222> (66)..(66)
<223> S или T, или G
<220>
<221> MOD_RES
<222> (68)..(68)
<223> K или R
<220>
<221> MOD_RES
<222> (92)..(92)
<223> V или I
<220>
<221> MOD_RES
<222> (94)..(94)
<223> F или L, или V
<220>
<221> MOD_RES
<222> (103)..(103)
<223> F или V
<400> 15
Glu Glu Glu Xaa Gln Xaa Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Xaa Ile Leu Leu Cys Thr Xaa Thr Ser Leu Xaa Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Xaa Leu
35 40 45
Ile Tyr Asn Gln Xaa Xaa Gly Xaa Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Xaa Thr Xaa Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Xaa Lys Xaa Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Xaa Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 16
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<220>
<221> MOD_RES
<222> (4)..(4)
<223> L или I, или V
<220>
<221> MOD_RES
<222> (6)..(6)
<223> V или L, или I
<220>
<221> MOD_RES
<222> (21)..(21)
<223> A или V
<220>
<221> MOD_RES
<222> (27)..(27)
<223> A или I, или L
<220>
<221> MOD_RES
<222> (31)..(31)
<223> I или T, или S, или F
<220>
<221> MOD_RES
<222> (47)..(47)
<223> E или V, или L
<220>
<221> MOD_RES
<222> (53)..(53)
<223> K или R
<220>
<221> MOD_RES
<222> (54)..(54)
<223> E или Q
<220>
<221> MOD_RES
<222> (56)..(56)
<223> H или P, или R
<220>
<221> MOD_RES
<222> (66)..(66)
<223> L или T, или G
<220>
<221> MOD_RES
<222> (68)..(68)
<223> K или R
<220>
<221> MOD_RES
<222> (92)..(92)
<223> V или I
<220>
<221> MOD_RES
<222> (94)..(94)
<223> F или L, или V
<220>
<221> MOD_RES
<222> (104)..(104)
<223> F или V
<400> 16
Glu Glu Gly Xaa Gln Xaa Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Xaa Ile Leu His Cys Thr Xaa Thr Ser Leu Xaa Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Xaa Leu
35 40 45
Ile Tyr Asn Gln Xaa Xaa Gly Xaa Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Xaa Thr Xaa Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Xaa Lys Xaa Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Xaa Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 17
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<220>
<221> MOD_RES
<222> (4)..(4)
<223> L или I, или V
<220>
<221> MOD_RES
<222> (6)..(6)
<223> V или L, или I
<220>
<221> MOD_RES
<222> (21)..(21)
<223> A или V
<220>
<221> MOD_RES
<222> (27)..(27)
<223> A или I, или L
<220>
<221> MOD_RES
<222> (31)..(31)
<223> I или T, или S, или F
<220>
<221> MOD_RES
<222> (47)..(47)
<223> E или V, или L
<220>
<221> MOD_RES
<222> (53)..(53)
<223> K или R
<220>
<221> MOD_RES
<222> (54)..(54)
<223> E или Q
<220>
<221> MOD_RES
<222> (56)..(56)
<223> H или P, или R
<220>
<221> MOD_RES
<222> (66)..(66)
<223> L или T, или G
<220>
<221> MOD_RES
<222> (68)..(68)
<223> K или R
<220>
<221> MOD_RES
<222> (92)..(92)
<223> V или I
<220>
<221> MOD_RES
<222> (94)..(94)
<223> F или L, или V
<220>
<221> MOD_RES
<222> (104)..(104)
<223> F или V
<400> 17
Glu Glu Glu Xaa Gln Xaa Ile Gln Pro Asp Lys Phe Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Xaa Thr Leu Arg Cys Thr Xaa Thr Ser Leu Xaa Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Xaa Leu
35 40 45
Ile Tyr Asn Gln Xaa Xaa Gly Xaa Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Xaa Thr Xaa Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Xaa Lys Xaa Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Xaa Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 18
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<220>
<221> MOD_RES
<222> (4)..(4)
<223> L или I, или V
<220>
<221> MOD_RES
<222> (6)..(6)
<223> V или L, или I
<220>
<221> MOD_RES
<222> (21)..(21)
<223> A или V
<220>
<221> MOD_RES
<222> (27)..(27)
<223> A или I, или L
<220>
<221> MOD_RES
<222> (31)..(31)
<223> I или T, или S, или F
<220>
<221> MOD_RES
<222> (47)..(47)
<223> E или V, или L
<220>
<221> MOD_RES
<222> (53)..(53)
<223> K или R
<220>
<221> MOD_RES
<222> (54)..(54)
<223> E или Q
<220>
<221> MOD_RES
<222> (56)..(56)
<223> H или P, или R
<220>
<221> MOD_RES
<222> (66)..(66)
<223> L или T, или G
<220>
<221> MOD_RES
<222> (68)..(68)
<223> K или R
<220>
<221> MOD_RES
<222> (92)..(92)
<223> V или I
<220>
<221> MOD_RES
<222> (94)..(94)
<223> F или L, или V
<220>
<221> MOD_RES
<222> (104)..(104)
<223> F или V
<400> 18
Glu Glu Glu Xaa Gln Xaa Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Xaa Thr Leu Arg Cys Thr Xaa Thr Ser Leu Xaa Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Xaa Leu
35 40 45
Ile Tyr Asn Gln Xaa Xaa Gly Xaa Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Xaa Thr Xaa Arg Asn Asn Met Asp Phe Pro Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Xaa Lys Xaa Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Xaa Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 19
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<220>
<221> MOD_RES
<222> (4)..(4)
<223> L или I, или V
<220>
<221> MOD_RES
<222> (6)..(6)
<223> V или L, или I
<220>
<221> MOD_RES
<222> (21)..(21)
<223> A или V
<220>
<221> MOD_RES
<222> (27)..(27)
<223> V или I, или L
<220>
<221> MOD_RES
<222> (31)..(31)
<223> I или T, или S, или F
<220>
<221> MOD_RES
<222> (47)..(47)
<223> E или V, или L
<220>
<221> MOD_RES
<222> (53)..(53)
<223> K или R
<220>
<221> MOD_RES
<222> (54)..(54)
<223> E или Q
<220>
<221> MOD_RES
<222> (56)..(56)
<223> H или P, или R
<220>
<221> MOD_RES
<222> (66)..(66)
<223> S или T, или G
<220>
<221> MOD_RES
<222> (68)..(68)
<223> K или R
<220>
<221> MOD_RES
<222> (92)..(92)
<223> V или I
<220>
<221> MOD_RES
<222> (94)..(94)
<223> F или L, или V
<220>
<221> MOD_RES
<222> (103)..(103)
<223> F или V
<400> 19
Glu Glu Glu Xaa Gln Xaa Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Xaa Ile Leu His Cys Thr Xaa Thr Ser Leu Xaa Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Xaa Leu
35 40 45
Ile Tyr Asn Gln Xaa Xaa Gly Xaa Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Xaa Thr Xaa Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Xaa Lys Xaa Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Xaa Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Gly Lys Pro Ser
115
<210> 20
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<220>
<221> MOD_RES
<222> (4)..(4)
<223> L или I, или V
<220>
<221> MOD_RES
<222> (6)..(6)
<223> V или L, или I
<220>
<221> MOD_RES
<222> (21)..(21)
<223> A или V
<220>
<221> MOD_RES
<222> (27)..(27)
<223> A или I, или L
<220>
<221> MOD_RES
<222> (31)..(31)
<223> I или T, или S, или F
<220>
<221> MOD_RES
<222> (47)..(47)
<223> E или V, или L
<220>
<221> MOD_RES
<222> (53)..(53)
<223> K или R
<220>
<221> MOD_RES
<222> (54)..(54)
<223> E или Q
<220>
<221> MOD_RES
<222> (56)..(56)
<223> H или P, или R
<220>
<221> MOD_RES
<222> (66)..(66)
<223> S или T, или G
<220>
<221> MOD_RES
<222> (68)..(68)
<223> K или R
<220>
<221> MOD_RES
<222> (92)..(92)
<223> V или I
<220>
<221> MOD_RES
<222> (94)..(94)
<223> F или L, или V
<220>
<221> MOD_RES
<222> (103)..(103)
<223> F или V
<400> 20
Glu Glu Glu Xaa Gln Xaa Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Xaa Thr Leu Arg Cys Thr Xaa Thr Ser Leu Xaa Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Xaa Leu
35 40 45
Ile Tyr Asn Gln Xaa Xaa Gly Xaa Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Xaa Thr Xaa Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Xaa Lys Xaa Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Xaa Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 21
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<220>
<221> MOD_RES
<222> (4)..(4)
<223> L или I, или V
<220>
<221> MOD_RES
<222> (6)..(6)
<223> V или L, или I
<220>
<221> MOD_RES
<222> (21)..(21)
<223> A или V
<220>
<221> MOD_RES
<222> (27)..(27)
<223> A или I, или L
<220>
<221> MOD_RES
<222> (31)..(31)
<223> I или T, или S, или F
<220>
<221> MOD_RES
<222> (47)..(47)
<223> E или V, или L
<220>
<221> MOD_RES
<222> (53)..(53)
<223> K или R
<220>
<221> MOD_RES
<222> (54)..(54)
<223> E или Q
<220>
<221> MOD_RES
<222> (56)..(56)
<223> H или P, или R
<220>
<221> MOD_RES
<222> (66)..(66)
<223> L или T, или G
<220>
<221> MOD_RES
<222> (68)..(68)
<223> K или R
<220>
<221> MOD_RES
<222> (92)..(92)
<223> V или I
<220>
<221> MOD_RES
<222> (94)..(94)
<223> F или L, или V
<220>
<221> MOD_RES
<222> (104)..(104)
<223> F или V
<400> 21
Glu Glu Glu Xaa Gln Xaa Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Xaa Thr Leu Arg Cys Thr Xaa Thr Ser Leu Xaa Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Xaa Leu
35 40 45
Ile Tyr Asn Gln Xaa Xaa Gly Xaa Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Xaa Thr Xaa Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Xaa Lys Xaa Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Xaa Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 22
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<220>
<221> MOD_RES
<222> (4)..(4)
<223> L или I, или V
<220>
<221> MOD_RES
<222> (6)..(6)
<223> V или L, или I
<220>
<221> MOD_RES
<222> (21)..(21)
<223> A или V
<220>
<221> MOD_RES
<222> (27)..(27)
<223> V или I, или L
<220>
<221> MOD_RES
<222> (31)..(31)
<223> I или T, или S, или F
<220>
<221> MOD_RES
<222> (47)..(47)
<223> E или V, или L
<220>
<221> MOD_RES
<222> (53)..(53)
<223> K или R
<220>
<221> MOD_RES
<222> (54)..(54)
<223> E или Q
<220>
<221> MOD_RES
<222> (56)..(56)
<223> H или P, или R
<220>
<221> MOD_RES
<222> (66)..(66)
<223> S или T, или G
<220>
<221> MOD_RES
<222> (68)..(68)
<223> K или R
<220>
<221> MOD_RES
<222> (92)..(92)
<223> V или I
<220>
<221> MOD_RES
<222> (94)..(94)
<223> F или L, или V
<220>
<221> MOD_RES
<222> (103)..(103)
<223> F или V
<400> 22
Glu Glu Glu Xaa Gln Xaa Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Xaa Ile Leu His Cys Thr Xaa Thr Ser Leu Xaa Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Xaa Leu
35 40 45
Ile Tyr Asn Gln Xaa Xaa Gly Xaa Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Xaa Thr Xaa Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Xaa Lys Xaa Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Xaa Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 23
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<220>
<221> MOD_RES
<222> (3)..(3)
<223> E или G
<220>
<221> MOD_RES
<222> (4)..(4)
<223> L или I, или V
<220>
<221> MOD_RES
<222> (6)..(6)
<223> V или L, или I
<220>
<221> MOD_RES
<222> (12)..(12)
<223> S или F
<220>
<221> MOD_RES
<222> (14)..(14)
<223> L или S
<220>
<221> MOD_RES
<222> (20)..(20)
<223> S или T
<220>
<221> MOD_RES
<222> (21)..(21)
<223> A или V
<220>
<221> MOD_RES
<222> (22)..(22)
<223> I или T
<220>
<221> MOD_RES
<222> (24)..(24)
<223> H или R
<220>
<221> MOD_RES
<222> (27)..(27)
<223> A или V, или I, или L
<220>
<221> MOD_RES
<222> (31)..(31)
<223> I или T, или S, или F
<220>
<221> MOD_RES
<222> (45)..(45)
<223> A или G
<220>
<221> MOD_RES
<222> (47)..(47)
<223> E или V, или L
<220>
<221> MOD_RES
<222> (53)..(53)
<223> K или R
<220>
<221> MOD_RES
<222> (54)..(54)
<223> E или Q
<220>
<221> MOD_RES
<222> (56)..(56)
<223> H или P, или R
<220>
<221> MOD_RES
<222> (65)..(65)
<223> D или E
<220>
<221> MOD_RES
<222> (66)..(66)
<223> S или L, или T, или G
<220>
<221> MOD_RES
<222> (68)..(68)
<223> K или R
<220>
<221> MOD_RES
<222> (70)..(70)
<223> E или D
<220>
<221> MOD_RES
<222> (75)..(75)
<223> S или P
<220>
<221> MOD_RES
<222> (77)..(77)
<223> S или R
<220>
<221> MOD_RES
<222> (79)..(79)
<223> S или G
<220>
<221> MOD_RES
<222> (92)..(92)
<223> V или I
<220>
<221> MOD_RES
<222> (94)..(94)
<223> F или L, или V
<220>
<221> MOD_RES
<222> (101)..(101)
<223> Может как присутствовать, так и отсутствовать
<220>
<221> MOD_RES
<222> (102)..(102)
<223> T или V
<220>
<221> MOD_RES
<222> (104)..(104)
<223> F или V
<220>
<221> MOD_RES
<222> (116)..(116)
<223> A или G
<400> 23
Glu Glu Xaa Xaa Gln Xaa Ile Gln Pro Asp Lys Xaa Val Xaa Val Ala
1 5 10 15
Ala Gly Glu Xaa Xaa Xaa Leu Xaa Cys Thr Xaa Thr Ser Leu Xaa Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Xaa Arg Xaa Leu
35 40 45
Ile Tyr Asn Gln Xaa Xaa Gly Xaa Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Xaa Xaa Thr Xaa Arg Xaa Asn Met Asp Phe Xaa Ile Xaa Ile Xaa Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Xaa Lys Xaa Arg Lys
85 90 95
Gly Ser Pro Asp Asp Xaa Glu Xaa Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Xaa Lys Pro Ser
115
<210> 24
<211> 102
<212> Белок
<213> Homo sapiens
<400> 24
Ala Pro Val Val Ser Gly Pro Ala Ala Arg Ala Thr Pro Gln His Thr
1 5 10 15
Val Ser Phe Thr Cys Glu Ser His Gly Phe Ser Pro Arg Asp Ile Thr
20 25 30
Leu Lys Trp Phe Lys Asn Gly Asn Glu Leu Ser Asp Phe Gln Thr Asn
35 40 45
Val Asp Pro Val Gly Glu Ser Val Ser Tyr Ser Ile His Ser Thr Ala
50 55 60
Lys Val Val Leu Thr Arg Glu Asp Val His Ser Gln Val Ile Cys Glu
65 70 75 80
Val Ala His Val Thr Leu Gln Gly Asp Pro Leu Arg Gly Thr Ala Asn
85 90 95
Leu Ser Glu Thr Ile Arg
100
<210> 25
<211> 99
<212> Белок
<213> Homo sapiens
<400> 25
Val Pro Pro Thr Leu Glu Val Thr Gln Gln Pro Val Arg Ala Glu Asn
1 5 10 15
Gln Val Asn Val Thr Cys Gln Val Arg Lys Phe Tyr Pro Gln Arg Leu
20 25 30
Gln Leu Thr Trp Leu Glu Asn Gly Asn Val Ser Arg Thr Glu Thr Ala
35 40 45
Ser Thr Val Thr Glu Asn Lys Asp Gly Thr Tyr Asn Trp Met Ser Trp
50 55 60
Leu Leu Val Asn Val Ser Ala His Arg Asp Asp Val Lys Leu Thr Cys
65 70 75 80
Gln Val Glu His Asp Gly Gln Pro Ala Val Ser Lys Ser His Asp Leu
85 90 95
Lys Val Ser
<210> 26
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 26
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Ile Thr Ser Leu Ile Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly His Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Thr Thr Arg Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Val Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 27
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 27
Glu Glu Glu Val Gln Val Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Leu Thr Ser Leu Ile Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly His Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Gly Thr Arg Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 28
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 28
Glu Glu Glu Val Gln Ile Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Val Ile Leu His Cys Thr Ile Thr Ser Leu Thr Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Leu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Thr Thr Arg Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Leu Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 29
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 29
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Ile Thr Ser Leu Ser Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Gly Thr Lys Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Leu Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 30
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 30
Glu Glu Glu Ile Gln Val Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Val Ile Ile His Cys Thr Val Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Arg Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Gly Thr Lys Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Val Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Val Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 31
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 31
Glu Glu Glu Val Gln Ile Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ile Ile Leu His Cys Thr Val Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Arg Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Gly Thr Arg Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Leu Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 32
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 32
Glu Glu Glu Val Gln Leu Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Val Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Gly Thr Lys Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Val Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 33
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 33
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 34
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 34
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Leu Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Thr Thr Lys Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 35
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 35
Glu Glu Glu Val Gln Ile Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Val Leu
35 40 45
Ile Tyr Asn Gln Lys Gln Gly Pro Phe Pro Arg Val Thr Thr Ile Ser
50 55 60
Glu Thr Thr Arg Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 36
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 36
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Ile Thr Ser Leu Thr Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Gly Thr Arg Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Val Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 37
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<220>
<221> MOD_RES
<222> (4)..(4)
<223> L или I, или V
<220>
<221> MOD_RES
<222> (6)..(6)
<223> V или L, или I
<220>
<221> MOD_RES
<222> (21)..(21)
<223> A или V
<220>
<221> MOD_RES
<222> (27)..(27)
<223> A или I, или L
<220>
<221> MOD_RES
<222> (31)..(31)
<223> I или T, или S, или F
<220>
<221> MOD_RES
<222> (47)..(47)
<223> E или V, или L
<220>
<221> MOD_RES
<222> (53)..(53)
<223> K или R
<220>
<221> MOD_RES
<222> (54)..(54)
<223> E или Q
<220>
<221> MOD_RES
<222> (56)..(56)
<223> H или P, или R
<220>
<221> MOD_RES
<222> (66)..(66)
<223> L или T, или G
<220>
<221> MOD_RES
<222> (68)..(68)
<223> K или R
<220>
<221> MOD_RES
<222> (80)..(80)
<223> N или A, или C, или D, или E, или F, или G, или
H, или I, или K, или L, или M, или P, или Q, или
R, или S, или T, или V, или W, или Y
<220>
<221> MOD_RES
<222> (83)..(83)
<223> P или A, или C, или D, или E, или F, или G, или
H, или I, или K, или L, или M, или N, или Q, или
R, или S, или T, или V, или W, или Y
<220>
<221> MOD_RES
<222> (92)..(92)
<223> V или I
<220>
<221> MOD_RES
<222> (94)..(94)
<223> F или L, или V
<220>
<221> MOD_RES
<222> (104)..(104)
<223> F или V
<400> 37
Glu Glu Glu Xaa Gln Xaa Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Xaa Thr Leu Arg Cys Thr Xaa Thr Ser Leu Xaa Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Xaa Leu
35 40 45
Ile Tyr Asn Gln Xaa Xaa Gly Xaa Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Xaa Thr Xaa Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Xaa
65 70 75 80
Ile Thr Xaa Ala Asp Ala Gly Thr Tyr Tyr Cys Xaa Lys Xaa Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Xaa Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 38
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<220>
<221> MOD_RES
<222> (4)..(4)
<223> L или I, или V
<220>
<221> MOD_RES
<222> (6)..(6)
<223> V или L, или I
<220>
<221> MOD_RES
<222> (21)..(21)
<223> A или V
<220>
<221> MOD_RES
<222> (27)..(27)
<223> V или I, или L
<220>
<221> MOD_RES
<222> (31)..(31)
<223> I или T, или S, или F
<220>
<221> MOD_RES
<222> (47)..(47)
<223> E или V, или L
<220>
<221> MOD_RES
<222> (53)..(53)
<223> K или R
<220>
<221> MOD_RES
<222> (54)..(54)
<223> E или Q
<220>
<221> MOD_RES
<222> (56)..(56)
<223> H или P, или R
<220>
<221> MOD_RES
<222> (66)..(66)
<223> S или T, или G
<220>
<221> MOD_RES
<222> (68)..(68)
<223> K или R
<220>
<221> MOD_RES
<222> (80)..(80)
<223> N или A, или C, или D, или E, или F, или G, или
H, или I, или K, или L, или M, или P, или Q, или
R, или S, или T, или V, или W, или Y
<220>
<221> MOD_RES
<222> (83)..(83)
<223> P или A, или C, или D, или E, или F, или G, или
H, или I, или K, или L, или M, или N, или Q, или
R, или S, или T, или V, или W, или Y
<220>
<221> MOD_RES
<222> (92)..(92)
<223> V или I
<220>
<221> MOD_RES
<222> (94)..(94)
<223> F или L, или V
<220>
<221> MOD_RES
<222> (103)..(103)
<223> F или V
<400> 38
Glu Glu Glu Xaa Gln Xaa Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Xaa Ile Leu His Cys Thr Xaa Thr Ser Leu Xaa Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Xaa Leu
35 40 45
Ile Tyr Asn Gln Xaa Xaa Gly Xaa Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Xaa Thr Xaa Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Xaa
65 70 75 80
Ile Thr Xaa Ala Asp Ala Gly Thr Tyr Tyr Cys Xaa Lys Xaa Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Xaa Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 39
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<220>
<221> MOD_RES
<222> (4)..(4)
<223> L или I, или V
<220>
<221> MOD_RES
<222> (6)..(6)
<223> V или L, или I
<220>
<221> MOD_RES
<222> (21)..(21)
<223> A или V
<220>
<221> MOD_RES
<222> (27)..(27)
<223> V или I, или L
<220>
<221> MOD_RES
<222> (31)..(31)
<223> I или T, или S, или F
<220>
<221> MOD_RES
<222> (47)..(47)
<223> E или V, или L
<220>
<221> MOD_RES
<222> (53)..(53)
<223> K или R
<220>
<221> MOD_RES
<222> (54)..(54)
<223> E или Q
<220>
<221> MOD_RES
<222> (56)..(56)
<223> H или P, или R
<220>
<221> MOD_RES
<222> (66)..(66)
<223> S или T, или G
<220>
<221> MOD_RES
<222> (68)..(68)
<223> K или R
<220>
<221> MOD_RES
<222> (80)..(80)
<223> N или A, или C, или D, или E, или F, или G, или
H, или I, или K, или L, или M, или P, или Q, или
R, или S, или T, или V, или W, или Y
<220>
<221> MOD_RES
<222> (83)..(83)
<223> P или A, или C, или D, или E, или F, или G, или
H, или I, или K, или L, или M, или N, или Q, или
R, или S, или T, или V, или W, или Y
<220>
<221> MOD_RES
<222> (92)..(92)
<223> V или I
<220>
<221> MOD_RES
<222> (94)..(94)
<223> F или L, или V
<220>
<221> MOD_RES
<222> (103)..(103)
<223> F или V
<400> 39
Glu Glu Glu Xaa Gln Xaa Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Xaa Ile Leu Leu Cys Thr Xaa Thr Ser Leu Xaa Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Xaa Leu
35 40 45
Ile Tyr Asn Gln Xaa Xaa Gly Xaa Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Xaa Thr Xaa Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Xaa
65 70 75 80
Ile Thr Xaa Ala Asp Ala Gly Thr Tyr Tyr Cys Xaa Lys Xaa Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Xaa Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 40
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<220>
<221> MOD_RES
<222> (4)..(4)
<223> L или I, или V
<220>
<221> MOD_RES
<222> (6)..(6)
<223> V или L, или I
<220>
<221> MOD_RES
<222> (21)..(21)
<223> A или V
<220>
<221> MOD_RES
<222> (27)..(27)
<223> A или I, или L
<220>
<221> MOD_RES
<222> (31)..(31)
<223> I или T, или S, или F
<220>
<221> MOD_RES
<222> (47)..(47)
<223> E или V, или L
<220>
<221> MOD_RES
<222> (53)..(53)
<223> K или R
<220>
<221> MOD_RES
<222> (54)..(54)
<223> E или Q
<220>
<221> MOD_RES
<222> (56)..(56)
<223> H или P, или R
<220>
<221> MOD_RES
<222> (66)..(66)
<223> L или T, или G
<220>
<221> MOD_RES
<222> (68)..(68)
<223> K или R
<220>
<221> MOD_RES
<222> (80)..(80)
<223> N или A, или C, или D, или E, или F, или G, или
H, или I, или K, или L, или M, или P, или Q, или
R, или S, или T, или V, или W, или Y
<220>
<221> MOD_RES
<222> (83)..(83)
<223> P или A, или C, или D, или E, или F, или G, или
H, или I, или K, или L, или M, или N, или Q, или
R, или S, или T, или V, или W, или Y
<220>
<221> MOD_RES
<222> (92)..(92)
<223> V или I
<220>
<221> MOD_RES
<222> (94)..(94)
<223> F или L, или V
<220>
<221> MOD_RES
<222> (104)..(104)
<223> F или V
<400> 40
Glu Glu Gly Xaa Gln Xaa Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Xaa Ile Leu His Cys Thr Xaa Thr Ser Leu Xaa Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Xaa Leu
35 40 45
Ile Tyr Asn Gln Xaa Xaa Gly Xaa Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Xaa Thr Xaa Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Xaa
65 70 75 80
Ile Thr Xaa Ala Asp Ala Gly Thr Tyr Tyr Cys Xaa Lys Xaa Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Xaa Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 41
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<220>
<221> MOD_RES
<222> (4)..(4)
<223> L или I, или V
<220>
<221> MOD_RES
<222> (6)..(6)
<223> V или L, или I
<220>
<221> MOD_RES
<222> (21)..(21)
<223> A или V
<220>
<221> MOD_RES
<222> (27)..(27)
<223> A или I, или L
<220>
<221> MOD_RES
<222> (31)..(31)
<223> I или T, или S, или F
<220>
<221> MOD_RES
<222> (47)..(47)
<223> E или V, или L
<220>
<221> MOD_RES
<222> (53)..(53)
<223> K или R
<220>
<221> MOD_RES
<222> (54)..(54)
<223> E или Q
<220>
<221> MOD_RES
<222> (56)..(56)
<223> H или P, или R
<220>
<221> MOD_RES
<222> (66)..(66)
<223> L или T, или G
<220>
<221> MOD_RES
<222> (68)..(68)
<223> K или R
<220>
<221> MOD_RES
<222> (80)..(80)
<223> N или A, или C, или D, или E, или F, или G, или
H, или I, или K, или L, или M, или P, или Q, или
R, или S, или T, или V, или W, или Y
<220>
<221> MOD_RES
<222> (83)..(83)
<223> P или A, или C, или D, или E, или F, или G, или
H, или I, или K, или L, или M, или N, или Q, или
R, или S, или T, или V, или W, или Y
<220>
<221> MOD_RES
<222> (92)..(92)
<223> V или I
<220>
<221> MOD_RES
<222> (94)..(94)
<223> F или L, или V
<220>
<221> MOD_RES
<222> (104)..(104)
<223> F или V
<400> 41
Glu Glu Glu Xaa Gln Xaa Ile Gln Pro Asp Lys Phe Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Xaa Thr Leu Arg Cys Thr Xaa Thr Ser Leu Xaa Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Xaa Leu
35 40 45
Ile Tyr Asn Gln Xaa Xaa Gly Xaa Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Xaa Thr Xaa Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Xaa
65 70 75 80
Ile Thr Xaa Ala Asp Ala Gly Thr Tyr Tyr Cys Xaa Lys Xaa Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Xaa Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 42
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<220>
<221> MOD_RES
<222> (4)..(4)
<223> L или I, или V
<220>
<221> MOD_RES
<222> (6)..(6)
<223> V или L, или I
<220>
<221> MOD_RES
<222> (21)..(21)
<223> A или V
<220>
<221> MOD_RES
<222> (27)..(27)
<223> A или I, или L
<220>
<221> MOD_RES
<222> (31)..(31)
<223> I или T, или S, или F
<220>
<221> MOD_RES
<222> (47)..(47)
<223> E или V, или L
<220>
<221> MOD_RES
<222> (53)..(53)
<223> K или R
<220>
<221> MOD_RES
<222> (54)..(54)
<223> E или Q
<220>
<221> MOD_RES
<222> (56)..(56)
<223> H или P, или R
<220>
<221> MOD_RES
<222> (66)..(66)
<223> L или T, или G
<220>
<221> MOD_RES
<222> (68)..(68)
<223> K или R
<220>
<221> MOD_RES
<222> (80)..(80)
<223> N или A, или C, или D, или E, или F, или G, или
H, или I, или K, или L, или M, или P, или Q, или
R, или S, или T, или V, или W, или Y
<220>
<221> MOD_RES
<222> (83)..(83)
<223> P или A, или C, или D, или E, или F, или G, или
H, или I, или K, или L, или M, или N, или Q, или
R, или S, или T, или V, или W, или Y
<220>
<221> MOD_RES
<222> (92)..(92)
<223> V или I
<220>
<221> MOD_RES
<222> (94)..(94)
<223> F или L, или V
<220>
<221> MOD_RES
<222> (104)..(104)
<223> F или V
<400> 42
Glu Glu Glu Xaa Gln Xaa Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Xaa Thr Leu Arg Cys Thr Xaa Thr Ser Leu Xaa Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Xaa Leu
35 40 45
Ile Tyr Asn Gln Xaa Xaa Gly Xaa Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Xaa Thr Xaa Arg Asn Asn Met Asp Phe Pro Ile Arg Ile Gly Xaa
65 70 75 80
Ile Thr Xaa Ala Asp Ala Gly Thr Tyr Tyr Cys Xaa Lys Xaa Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Xaa Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 43
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<220>
<221> MOD_RES
<222> (4)..(4)
<223> L или I, или V
<220>
<221> MOD_RES
<222> (6)..(6)
<223> V или L, или I
<220>
<221> MOD_RES
<222> (21)..(21)
<223> A или V
<220>
<221> MOD_RES
<222> (27)..(27)
<223> V или I, или L
<220>
<221> MOD_RES
<222> (31)..(31)
<223> I или T, или S, или F
<220>
<221> MOD_RES
<222> (47)..(47)
<223> E или V, или L
<220>
<221> MOD_RES
<222> (53)..(53)
<223> K или R
<220>
<221> MOD_RES
<222> (54)..(54)
<223> E или Q
<220>
<221> MOD_RES
<222> (56)..(56)
<223> H или P, или R
<220>
<221> MOD_RES
<222> (66)..(66)
<223> S или T, или G
<220>
<221> MOD_RES
<222> (68)..(68)
<223> K или R
<220>
<221> MOD_RES
<222> (80)..(80)
<223> N или A, или C, или D, или E, или F, или G, или
H, или I, или K, или L, или M, или P, или Q, или
R, или S, или T, или V, или W, или Y
<220>
<221> MOD_RES
<222> (83)..(83)
<223> P или A, или C, или D, или E, или F, или G, или
H, или I, или K, или L, или M, или N, или Q, или
R, или S, или T, или V, или W, или Y
<220>
<221> MOD_RES
<222> (92)..(92)
<223> V или I
<220>
<221> MOD_RES
<222> (94)..(94)
<223> F или L, или V
<220>
<221> MOD_RES
<222> (103)..(103)
<223> F или V
<400> 43
Glu Glu Glu Xaa Gln Xaa Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Xaa Ile Leu His Cys Thr Xaa Thr Ser Leu Xaa Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Xaa Leu
35 40 45
Ile Tyr Asn Gln Xaa Xaa Gly Xaa Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Xaa Thr Xaa Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Xaa
65 70 75 80
Ile Thr Xaa Ala Asp Ala Gly Thr Tyr Tyr Cys Xaa Lys Xaa Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Xaa Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Gly Lys Pro Ser
115
<210> 44
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<220>
<221> MOD_RES
<222> (4)..(4)
<223> L или I, или V
<220>
<221> MOD_RES
<222> (6)..(6)
<223> V или L, или I
<220>
<221> MOD_RES
<222> (21)..(21)
<223> A или V
<220>
<221> MOD_RES
<222> (27)..(27)
<223> A или I, или L
<220>
<221> MOD_RES
<222> (31)..(31)
<223> I или T, или S, или F
<220>
<221> MOD_RES
<222> (47)..(47)
<223> E или V, или L
<220>
<221> MOD_RES
<222> (53)..(53)
<223> K или R
<220>
<221> MOD_RES
<222> (54)..(54)
<223> E или Q
<220>
<221> MOD_RES
<222> (56)..(56)
<223> H или P, или R
<220>
<221> MOD_RES
<222> (66)..(66)
<223> S или T, или G
<220>
<221> MOD_RES
<222> (68)..(68)
<223> K или R
<220>
<221> MOD_RES
<222> (80)..(80)
<223> N или A, или C, или D, или E, или F, или G, или
H, или I, или K, или L, или M, или P, или Q, или
R, или S, или T, или V, или W, или Y
<220>
<221> MOD_RES
<222> (83)..(83)
<223> P или A, или C, или D, или E, или F, или G, или
H, или I, или K, или L, или M, или N, или Q, или
R, или S, или T, или V, или W, или Y
<220>
<221> MOD_RES
<222> (92)..(92)
<223> V или I
<220>
<221> MOD_RES
<222> (94)..(94)
<223> F или L, или V
<220>
<221> MOD_RES
<222> (103)..(103)
<223> F или V
<400> 44
Glu Glu Glu Xaa Gln Xaa Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Xaa Thr Leu Arg Cys Thr Xaa Thr Ser Leu Xaa Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Xaa Leu
35 40 45
Ile Tyr Asn Gln Xaa Xaa Gly Xaa Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Xaa Thr Xaa Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Xaa
65 70 75 80
Ile Thr Xaa Ala Asp Ala Gly Thr Tyr Tyr Cys Xaa Lys Xaa Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Xaa Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 45
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<220>
<221> MOD_RES
<222> (4)..(4)
<223> L или I, или V
<220>
<221> MOD_RES
<222> (6)..(6)
<223> V или L, или I
<220>
<221> MOD_RES
<222> (21)..(21)
<223> A или V
<220>
<221> MOD_RES
<222> (27)..(27)
<223> A или I, или L
<220>
<221> MOD_RES
<222> (31)..(31)
<223> I или T, или S, или F
<220>
<221> MOD_RES
<222> (47)..(47)
<223> E или V, или L
<220>
<221> MOD_RES
<222> (53)..(53)
<223> K или R
<220>
<221> MOD_RES
<222> (54)..(54)
<223> E или Q
<220>
<221> MOD_RES
<222> (56)..(56)
<223> H или P, или R
<220>
<221> MOD_RES
<222> (66)..(66)
<223> L или T, или G
<220>
<221> MOD_RES
<222> (68)..(68)
<223> K или R
<220>
<221> MOD_RES
<222> (80)..(80)
<223> N или A, или C, или D, или E, или F, или G, или
H, или I, или K, или L, или M, или P, или Q, или
R, или S, или T, или V, или W, или Y
<220>
<221> MOD_RES
<222> (83)..(83)
<223> P или A, или C, или D, или E, или F, или G, или
H, или I, или K, или L, или M, или N, или Q, или
R, или S, или T, или V, или W, или Y
<220>
<221> MOD_RES
<222> (92)..(92)
<223> V или I
<220>
<221> MOD_RES
<222> (94)..(94)
<223> F или L, или V
<220>
<221> MOD_RES
<222> (104)..(104)
<223> F или V
<400> 45
Glu Glu Glu Xaa Gln Xaa Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Xaa Thr Leu Arg Cys Thr Xaa Thr Ser Leu Xaa Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Xaa Leu
35 40 45
Ile Tyr Asn Gln Xaa Xaa Gly Xaa Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Xaa Thr Xaa Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Ser Xaa
65 70 75 80
Ile Thr Xaa Ala Asp Ala Gly Thr Tyr Tyr Cys Xaa Lys Xaa Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Xaa Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 46
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<220>
<221> MOD_RES
<222> (4)..(4)
<223> L или I, или V
<220>
<221> MOD_RES
<222> (6)..(6)
<223> V или L, или I
<220>
<221> MOD_RES
<222> (21)..(21)
<223> A или V
<220>
<221> MOD_RES
<222> (27)..(27)
<223> V или I, или L
<220>
<221> MOD_RES
<222> (31)..(31)
<223> I или T, или S, или F
<220>
<221> MOD_RES
<222> (47)..(47)
<223> E или V, или L
<220>
<221> MOD_RES
<222> (53)..(53)
<223> K или R
<220>
<221> MOD_RES
<222> (54)..(54)
<223> E или Q
<220>
<221> MOD_RES
<222> (56)..(56)
<223> H или P, или R
<220>
<221> MOD_RES
<222> (66)..(66)
<223> S или T, или G
<220>
<221> MOD_RES
<222> (68)..(68)
<223> K или R
<220>
<221> MOD_RES
<222> (80)..(80)
<223> N или A, или C, или D, или E, или F, или G, или
H, или I, или K, или L, или M, или P, или Q, или
R, или S, или T, или V, или W, или Y
<220>
<221> MOD_RES
<222> (83)..(83)
<223> P или A, или C, или D, или E, или F, или G, или
H, или I, или K, или L, или M, или N, или Q, или
R, или S, или T, или V, или W, или Y
<220>
<221> MOD_RES
<222> (92)..(92)
<223> V или I
<220>
<221> MOD_RES
<222> (94)..(94)
<223> F или L, или V
<220>
<221> MOD_RES
<222> (103)..(103)
<223> F или V
<400> 46
Glu Glu Glu Xaa Gln Xaa Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Xaa Ile Leu His Cys Thr Xaa Thr Ser Leu Xaa Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Xaa Leu
35 40 45
Ile Tyr Asn Gln Xaa Xaa Gly Xaa Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Xaa Thr Xaa Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Xaa
65 70 75 80
Ile Thr Xaa Ala Asp Ala Gly Thr Tyr Tyr Cys Xaa Lys Xaa Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Xaa Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 47
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<220>
<221> MOD_RES
<222> (3)..(3)
<223> E или G
<220>
<221> MOD_RES
<222> (4)..(4)
<223> L или I, или V
<220>
<221> MOD_RES
<222> (6)..(6)
<223> V или L, или I
<220>
<221> MOD_RES
<222> (12)..(12)
<223> S или F
<220>
<221> MOD_RES
<222> (14)..(14)
<223> L или S
<220>
<221> MOD_RES
<222> (20)..(20)
<223> S или T
<220>
<221> MOD_RES
<222> (21)..(21)
<223> A или V
<220>
<221> MOD_RES
<222> (22)..(22)
<223> I или T
<220>
<221> MOD_RES
<222> (24)..(24)
<223> H или R, или L
<220>
<221> MOD_RES
<222> (27)..(27)
<223> A или V, или I, или L
<220>
<221> MOD_RES
<222> (31)..(31)
<223> I или T, или S, или F
<220>
<221> MOD_RES
<222> (45)..(45)
<223> A или G
<220>
<221> MOD_RES
<222> (47)..(47)
<223> E или V, или L
<220>
<221> MOD_RES
<222> (53)..(53)
<223> K или R
<220>
<221> MOD_RES
<222> (54)..(54)
<223> E или Q
<220>
<221> MOD_RES
<222> (56)..(56)
<223> H или P, или R
<220>
<221> MOD_RES
<222> (65)..(65)
<223> D или E
<220>
<221> MOD_RES
<222> (66)..(66)
<223> S или L или T, или G
<220>
<221> MOD_RES
<222> (68)..(68)
<223> K или R
<220>
<221> MOD_RES
<222> (70)..(70)
<223> E или N
<220>
<221> MOD_RES
<222> (75)..(75)
<223> S или P
<220>
<221> MOD_RES
<222> (77)..(77)
<223> S или R
<220>
<221> MOD_RES
<222> (79)..(79)
<223> S или G
<220>
<221> MOD_RES
<222> (80)..(80)
<223> Любая аминокислота
<220>
<221> MOD_RES
<222> (83)..(83)
<223> Любая аминокислота
<220>
<221> MOD_RES
<222> (92)..(92)
<223> V или I
<220>
<221> MOD_RES
<222> (94)..(94)
<223> F или L, или V
<220>
<221> MOD_RES
<222> (101)..(101)
<223> Может как присутствовать, так и отсутствовать
<220>
<221> MOD_RES
<222> (102)..(102)
<223> T или V
<220>
<221> MOD_RES
<222> (104)..(104)
<223> F или V
<220>
<221> MOD_RES
<222> (116)..(116)
<223> A или G
<400> 47
Glu Glu Xaa Xaa Gln Xaa Ile Gln Pro Asp Lys Xaa Val Xaa Val Ala
1 5 10 15
Ala Gly Glu Xaa Xaa Xaa Leu Xaa Cys Thr Xaa Thr Ser Leu Xaa Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Xaa Arg Xaa Leu
35 40 45
Ile Tyr Asn Gln Xaa Xaa Gly Xaa Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Xaa Xaa Thr Xaa Arg Xaa Asn Met Asp Phe Xaa Ile Xaa Ile Xaa Xaa
65 70 75 80
Ile Thr Xaa Ala Asp Ala Gly Thr Tyr Tyr Cys Xaa Lys Xaa Arg Lys
85 90 95
Gly Ser Pro Asp Asp Xaa Glu Xaa Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Xaa Lys Pro Ser
115
<210> 48
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<220>
<221> MOD_RES
<222> (6)..(6)
<223> V или I
<220>
<221> MOD_RES
<222> (14)..(14)
<223> L или S
<220>
<221> MOD_RES
<222> (20)..(20)
<223> T или S
<220>
<221> MOD_RES
<222> (22)..(22)
<223> T или I
<220>
<221> MOD_RES
<222> (24)..(24)
<223> R или H
<220>
<221> MOD_RES
<222> (27)..(27)
<223> A или V, или I
<220>
<221> MOD_RES
<222> (31)..(31)
<223> I или R, или Y, или K, или F
<220>
<221> MOD_RES
<222> (45)..(45)
<223> G или A
<220>
<221> MOD_RES
<222> (47)..(47)
<223> E или V
<220>
<221> MOD_RES
<222> (53)..(53)
<223> K или R
<220>
<221> MOD_RES
<222> (54)..(54)
<223> E или D, или Q
<220>
<221> MOD_RES
<222> (56)..(56)
<223> H или P
<220>
<221> MOD_RES
<222> (65)..(65)
<223> D или E
<220>
<221> MOD_RES
<222> (66)..(66)
<223> S или L, или T
<220>
<221> MOD_RES
<222> (70)..(70)
<223> N или E
<220>
<221> MOD_RES
<222> (77)..(77)
<223> R или S
<220>
<221> MOD_RES
<222> (79)..(79)
<223> G или S
<220>
<221> MOD_RES
<222> (80)..(80)
<223> N или A
<220>
<221> MOD_RES
<222> (92)..(92)
<223> V или I
<220>
<221> MOD_RES
<222> (98)..(98)
<223> S или I, или M
<220>
<221> MOD_RES
<222> (99)..(99)
<223> Может как присутствовать, так и отсутствовать
<220>
<221> MOD_RES
<222> (100)..(100)
<223> D или P
<220>
<221> MOD_RES
<222> (102)..(102)
<223> V или T
<400> 48
Glu Glu Glu Leu Gln Xaa Ile Gln Pro Asp Lys Ser Val Xaa Val Ala
1 5 10 15
Ala Gly Glu Xaa Ala Xaa Leu Xaa Cys Thr Xaa Thr Ser Leu Xaa Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Xaa Arg Xaa Leu
35 40 45
Ile Tyr Asn Gln Xaa Xaa Gly Xaa Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Xaa Xaa Thr Lys Arg Xaa Asn Met Asp Phe Ser Ile Xaa Ile Xaa Xaa
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Xaa Lys Phe Arg Lys
85 90 95
Gly Xaa Pro Xaa Asp Xaa Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 49
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<220>
<221> MOD_RES
<222> (6)..(6)
<223> V или I, или L
<220>
<221> MOD_RES
<222> (27)..(27)
<223> A или I, или V, или L
<220>
<221> MOD_RES
<222> (31)..(31)
<223> I или F, или S, или T
<220>
<221> MOD_RES
<222> (47)..(47)
<223> E или V, или L
<220>
<221> MOD_RES
<222> (53)..(53)
<223> K или R
<220>
<221> MOD_RES
<222> (54)..(54)
<223> E или Q
<220>
<221> MOD_RES
<222> (56)..(56)
<223> H или P, или R
<220>
<221> MOD_RES
<222> (66)..(66)
<223> L или T, или S, или G
<220>
<221> MOD_RES
<222> (80)..(80)
<223> A
<220>
<221> MOD_RES
<222> (92)..(92)
<223> V или I
<400> 49
Glu Glu Glu Leu Gln Xaa Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Xaa Thr Ser Leu Xaa Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Xaa Leu
35 40 45
Ile Tyr Asn Gln Xaa Xaa Gly Xaa Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Xaa Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Xaa
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Xaa Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 50
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<220>
<221> MOD_RES
<222> (6)..(6)
<223> V или I
<220>
<221> MOD_RES
<222> (27)..(27)
<223> V или I
<220>
<221> MOD_RES
<222> (31)..(31)
<223> I или F
<220>
<221> MOD_RES
<222> (47)..(47)
<223> E или V
<220>
<221> MOD_RES
<222> (53)..(53)
<223> K или R
<220>
<221> MOD_RES
<222> (54)..(54)
<223> E или Q
<220>
<221> MOD_RES
<222> (56)..(56)
<223> H или P
<220>
<221> MOD_RES
<222> (66)..(66)
<223> S или T
<220>
<221> MOD_RES
<222> (80)..(80)
<223> N или A
<220>
<221> MOD_RES
<222> (92)..(92)
<223> V или I
<400> 50
Glu Glu Glu Leu Gln Xaa Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Xaa Thr Ser Leu Xaa Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Xaa Leu
35 40 45
Ile Tyr Asn Gln Xaa Xaa Gly Xaa Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Xaa Thr Lys Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Xaa
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Xaa Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 51
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<220>
<221> MOD_RES
<222> (6)..(6)
<223> V или I
<220>
<221> MOD_RES
<222> (27)..(27)
<223> A или I
<220>
<221> MOD_RES
<222> (31)..(31)
<223> I или F
<220>
<221> MOD_RES
<222> (47)..(47)
<223> E или V
<220>
<221> MOD_RES
<222> (53)..(53)
<223> K или R
<220>
<221> MOD_RES
<222> (56)..(56)
<223> H или P
<220>
<221> MOD_RES
<222> (66)..(66)
<223> L или T
<220>
<221> MOD_RES
<222> (80)..(80)
<223> Любая аминокислот, отличная от N
<220>
<221> MOD_RES
<222> (92)..(92)
<223> V или I
<400> 51
Glu Glu Glu Leu Gln Xaa Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Xaa Thr Ser Leu Xaa Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Xaa Leu
35 40 45
Ile Tyr Asn Gln Xaa Glu Gly Xaa Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Xaa Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Xaa
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Xaa Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 52
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<220>
<221> MOD_RES
<222> (6)..(6)
<223> V или L, или I
<220>
<221> MOD_RES
<222> (27)..(27)
<223> A или I, или L
<220>
<221> MOD_RES
<222> (31)..(31)
<223> I или T, или S, или F
<220>
<221> MOD_RES
<222> (53)..(53)
<223> K или R
<220>
<221> MOD_RES
<222> (56)..(56)
<223> H или P, или R
<220>
<221> MOD_RES
<222> (66)..(66)
<223> L или T, или G
<220>
<221> MOD_RES
<222> (80)..(80)
<223> N или A
<400> 52
Glu Glu Glu Leu Gln Xaa Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Xaa Thr Ser Leu Xaa Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Xaa Glu Gly Xaa Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Xaa Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Xaa
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 53
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 53
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Thr Thr Lys Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 54
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 54
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Val Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Ser Thr Lys Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 55
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 55
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Thr Thr Lys Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 56
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 56
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Val Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Thr Thr Lys Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 57
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 57
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Ile Thr Ser Leu Ile Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Thr Thr Lys Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 58
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 58
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Thr Thr Lys Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 59
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 59
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Val Leu
35 40 45
Ile Tyr Asn Gln Lys Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Thr Thr Lys Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 60
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 60
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Thr Thr Lys Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 61
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 61
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly His Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Thr Thr Lys Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 62
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 62
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Ser Thr Lys Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 63
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 63
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Thr Thr Lys Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 64
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 64
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Val Thr Ser Leu Ile Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Ser Thr Lys Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 65
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 65
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Val Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Ser Thr Lys Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 66
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 66
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Ser Thr Lys Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 67
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 67
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Thr Thr Lys Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 68
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 68
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Ser Thr Lys Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 69
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 69
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Ile Thr Ser Leu Ile Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Ser Thr Lys Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 70
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 70
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Thr Thr Lys Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 71
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 71
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 72
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 72
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 73
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 73
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 74
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 74
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 75
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 75
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 76
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 76
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 77
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 77
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 78
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 78
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 79
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 79
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 80
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 80
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 81
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 81
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 82
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 82
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 83
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 83
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 84
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 84
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 85
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 85
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 86
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 86
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 87
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 87
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Ile Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 88
<211> 232
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 88
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
1 5 10 15
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
20 25 30
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
35 40 45
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
50 55 60
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
65 70 75 80
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
85 90 95
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
100 105 110
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
115 120 125
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
130 135 140
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
145 150 155 160
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
165 170 175
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
180 185 190
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
195 200 205
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
210 215 220
Ser Leu Ser Leu Ser Pro Gly Lys
225 230
<210> 89
<211> 325
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 89
Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser
1 5 10 15
Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe
20 25 30
Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly
35 40 45
Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu
50 55 60
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr
65 70 75 80
Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Thr
85 90 95
Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro Pro
100 105 110
Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
115 120 125
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
130 135 140
Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val
145 150 155 160
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser
165 170 175
Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp Leu
180 185 190
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ala
195 200 205
Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu Pro
210 215 220
Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln
225 230 235 240
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
245 250 255
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
260 265 270
Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
275 280 285
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
290 295 300
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
305 310 315 320
Leu Ser Pro Gly Lys
325
<210> 90
<211> 227
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 90
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
1 5 10 15
Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Ala Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
130 135 140
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys
225
<210> 91
<211> 226
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 91
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
1 5 10 15
Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Ala Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
130 135 140
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly
225
<210> 92
<211> 223
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 92
Val Glu Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val
1 5 10 15
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
20 25 30
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
35 40 45
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
50 55 60
Thr Lys Pro Arg Glu Glu Gln Phe Ala Ser Thr Phe Arg Val Val Ser
65 70 75 80
Val Leu Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
85 90 95
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
100 105 110
Ser Lys Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
115 120 125
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
130 135 140
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
145 150 155 160
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser
165 170 175
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
180 185 190
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
195 200 205
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
210 215 220
<210> 93
<211> 222
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 93
Val Glu Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val
1 5 10 15
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
20 25 30
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
35 40 45
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
50 55 60
Thr Lys Pro Arg Glu Glu Gln Phe Ala Ser Thr Phe Arg Val Val Ser
65 70 75 80
Val Leu Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
85 90 95
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
100 105 110
Ser Lys Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
115 120 125
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
130 135 140
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
145 150 155 160
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser
165 170 175
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
180 185 190
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
195 200 205
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
210 215 220
<210> 94
<211> 228
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 94
Glu Arg Lys Ser Ser Val Glu Cys Pro Pro Cys Pro Ala Pro Pro Val
1 5 10 15
Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
20 25 30
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
35 40 45
His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu
50 55 60
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Ala Ser Thr
65 70 75 80
Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp Leu Asn
85 90 95
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser
100 105 110
Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu Pro Gln
115 120 125
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
130 135 140
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
145 150 155 160
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
165 170 175
Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
180 185 190
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
195 200 205
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
210 215 220
Ser Pro Gly Lys
225
<210> 95
<211> 227
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 95
Glu Arg Lys Ser Ser Val Glu Cys Pro Pro Cys Pro Ala Pro Pro Val
1 5 10 15
Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
20 25 30
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
35 40 45
His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu
50 55 60
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Ala Ser Thr
65 70 75 80
Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp Leu Asn
85 90 95
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser
100 105 110
Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu Pro Gln
115 120 125
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
130 135 140
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
145 150 155 160
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
165 170 175
Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
180 185 190
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
195 200 205
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
210 215 220
Ser Pro Gly
225
<210> 96
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 96
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Lys Thr His Thr Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 97
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 97
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Lys Thr His Thr Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 98
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 98
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Lys Thr His Thr Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 99
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 99
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Lys Thr His Thr Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 100
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 100
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Lys Thr His Thr Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 101
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 101
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Lys Thr His Thr Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 102
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 102
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Lys Thr His Thr Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 103
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 103
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Lys Thr His Thr Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 104
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 104
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Lys Thr His Thr Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 105
<211> 342
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 105
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Val Glu Cys Pro Pro Cys Pro Ala Pro
115 120 125
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
130 135 140
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
145 150 155 160
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
165 170 175
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Ala
180 185 190
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
195 200 205
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
210 215 220
Ser Ser Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
225 230 235 240
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
245 250 255
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
260 265 270
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
275 280 285
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
290 295 300
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
305 310 315 320
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
325 330 335
Ser Leu Ser Pro Gly Lys
340
<210> 106
<211> 342
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 106
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Val Glu Cys Pro Pro Cys Pro Ala Pro
115 120 125
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
130 135 140
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
145 150 155 160
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
165 170 175
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Ala
180 185 190
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
195 200 205
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
210 215 220
Ser Ser Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
225 230 235 240
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
245 250 255
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
260 265 270
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
275 280 285
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
290 295 300
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
305 310 315 320
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
325 330 335
Ser Leu Ser Pro Gly Lys
340
<210> 107
<211> 342
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 107
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Val Glu Cys Pro Pro Cys Pro Ala Pro
115 120 125
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
130 135 140
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
145 150 155 160
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
165 170 175
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Ala
180 185 190
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
195 200 205
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
210 215 220
Ser Ser Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
225 230 235 240
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
245 250 255
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
260 265 270
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
275 280 285
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
290 295 300
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
305 310 315 320
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
325 330 335
Ser Leu Ser Pro Gly Lys
340
<210> 108
<211> 342
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 108
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Val Glu Cys Pro Pro Cys Pro Ala Pro
115 120 125
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
130 135 140
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
145 150 155 160
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
165 170 175
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Ala
180 185 190
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
195 200 205
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
210 215 220
Ser Ser Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
225 230 235 240
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
245 250 255
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
260 265 270
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
275 280 285
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
290 295 300
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
305 310 315 320
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
325 330 335
Ser Leu Ser Pro Gly Lys
340
<210> 109
<211> 342
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 109
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Val Glu Cys Pro Pro Cys Pro Ala Pro
115 120 125
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
130 135 140
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
145 150 155 160
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
165 170 175
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Ala
180 185 190
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
195 200 205
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
210 215 220
Ser Ser Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
225 230 235 240
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
245 250 255
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
260 265 270
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
275 280 285
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
290 295 300
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
305 310 315 320
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
325 330 335
Ser Leu Ser Pro Gly Lys
340
<210> 110
<211> 342
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 110
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Val Glu Cys Pro Pro Cys Pro Ala Pro
115 120 125
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
130 135 140
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
145 150 155 160
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
165 170 175
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Ala
180 185 190
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
195 200 205
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
210 215 220
Ser Ser Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
225 230 235 240
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
245 250 255
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
260 265 270
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
275 280 285
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
290 295 300
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
305 310 315 320
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
325 330 335
Ser Leu Ser Pro Gly Lys
340
<210> 111
<211> 342
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 111
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Val Glu Cys Pro Pro Cys Pro Ala Pro
115 120 125
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
130 135 140
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
145 150 155 160
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
165 170 175
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Ala
180 185 190
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
195 200 205
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
210 215 220
Ser Ser Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
225 230 235 240
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
245 250 255
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
260 265 270
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
275 280 285
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
290 295 300
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
305 310 315 320
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
325 330 335
Ser Leu Ser Pro Gly Lys
340
<210> 112
<211> 342
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 112
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Val Glu Cys Pro Pro Cys Pro Ala Pro
115 120 125
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
130 135 140
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
145 150 155 160
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
165 170 175
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Ala
180 185 190
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
195 200 205
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
210 215 220
Ser Ser Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
225 230 235 240
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
245 250 255
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
260 265 270
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
275 280 285
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
290 295 300
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
305 310 315 320
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
325 330 335
Ser Leu Ser Pro Gly Lys
340
<210> 113
<211> 342
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 113
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Val Glu Cys Pro Pro Cys Pro Ala Pro
115 120 125
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
130 135 140
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
145 150 155 160
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
165 170 175
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Ala
180 185 190
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
195 200 205
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
210 215 220
Ser Ser Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
225 230 235 240
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
245 250 255
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
260 265 270
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
275 280 285
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
290 295 300
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
305 310 315 320
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
325 330 335
Ser Leu Ser Pro Gly Lys
340
<210> 114
<211> 347
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 114
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Glu Arg Lys Ser Ser Val Glu Cys Pro
115 120 125
Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro
130 135 140
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
145 150 155 160
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn
165 170 175
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
180 185 190
Glu Glu Gln Phe Ala Ser Thr Phe Arg Val Val Ser Val Leu Thr Val
195 200 205
Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
210 215 220
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Thr Lys
225 230 235 240
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
245 250 255
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
260 265 270
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
275 280 285
Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe
290 295 300
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
305 310 315 320
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
325 330 335
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 115
<211> 347
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 115
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Glu Arg Lys Ser Ser Val Glu Cys Pro
115 120 125
Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro
130 135 140
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
145 150 155 160
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn
165 170 175
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
180 185 190
Glu Glu Gln Phe Ala Ser Thr Phe Arg Val Val Ser Val Leu Thr Val
195 200 205
Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
210 215 220
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Thr Lys
225 230 235 240
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
245 250 255
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
260 265 270
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
275 280 285
Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe
290 295 300
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
305 310 315 320
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
325 330 335
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 116
<211> 347
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 116
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Glu Arg Lys Ser Ser Val Glu Cys Pro
115 120 125
Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro
130 135 140
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
145 150 155 160
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn
165 170 175
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
180 185 190
Glu Glu Gln Phe Ala Ser Thr Phe Arg Val Val Ser Val Leu Thr Val
195 200 205
Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
210 215 220
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Thr Lys
225 230 235 240
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
245 250 255
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
260 265 270
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
275 280 285
Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe
290 295 300
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
305 310 315 320
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
325 330 335
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 117
<211> 347
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 117
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Glu Arg Lys Ser Ser Val Glu Cys Pro
115 120 125
Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro
130 135 140
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
145 150 155 160
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn
165 170 175
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
180 185 190
Glu Glu Gln Phe Ala Ser Thr Phe Arg Val Val Ser Val Leu Thr Val
195 200 205
Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
210 215 220
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Thr Lys
225 230 235 240
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
245 250 255
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
260 265 270
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
275 280 285
Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe
290 295 300
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
305 310 315 320
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
325 330 335
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 118
<211> 347
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 118
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Glu Arg Lys Ser Ser Val Glu Cys Pro
115 120 125
Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro
130 135 140
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
145 150 155 160
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn
165 170 175
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
180 185 190
Glu Glu Gln Phe Ala Ser Thr Phe Arg Val Val Ser Val Leu Thr Val
195 200 205
Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
210 215 220
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Thr Lys
225 230 235 240
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
245 250 255
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
260 265 270
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
275 280 285
Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe
290 295 300
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
305 310 315 320
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
325 330 335
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 119
<211> 347
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 119
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Glu Arg Lys Ser Ser Val Glu Cys Pro
115 120 125
Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro
130 135 140
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
145 150 155 160
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn
165 170 175
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
180 185 190
Glu Glu Gln Phe Ala Ser Thr Phe Arg Val Val Ser Val Leu Thr Val
195 200 205
Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
210 215 220
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Thr Lys
225 230 235 240
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
245 250 255
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
260 265 270
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
275 280 285
Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe
290 295 300
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
305 310 315 320
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
325 330 335
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 120
<211> 347
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 120
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Glu Arg Lys Ser Ser Val Glu Cys Pro
115 120 125
Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro
130 135 140
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
145 150 155 160
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn
165 170 175
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
180 185 190
Glu Glu Gln Phe Ala Ser Thr Phe Arg Val Val Ser Val Leu Thr Val
195 200 205
Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
210 215 220
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Thr Lys
225 230 235 240
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
245 250 255
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
260 265 270
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
275 280 285
Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe
290 295 300
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
305 310 315 320
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
325 330 335
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 121
<211> 347
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 121
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Glu Arg Lys Ser Ser Val Glu Cys Pro
115 120 125
Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro
130 135 140
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
145 150 155 160
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn
165 170 175
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
180 185 190
Glu Glu Gln Phe Ala Ser Thr Phe Arg Val Val Ser Val Leu Thr Val
195 200 205
Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
210 215 220
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Thr Lys
225 230 235 240
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
245 250 255
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
260 265 270
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
275 280 285
Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe
290 295 300
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
305 310 315 320
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
325 330 335
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 122
<211> 347
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 122
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Glu Arg Lys Ser Ser Val Glu Cys Pro
115 120 125
Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro
130 135 140
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
145 150 155 160
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn
165 170 175
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
180 185 190
Glu Glu Gln Phe Ala Ser Thr Phe Arg Val Val Ser Val Leu Thr Val
195 200 205
Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
210 215 220
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Thr Lys
225 230 235 240
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
245 250 255
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
260 265 270
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
275 280 285
Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe
290 295 300
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
305 310 315 320
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
325 330 335
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 123
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 123
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Lys Thr His Thr Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 124
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 124
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Lys Thr His Thr Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 125
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 125
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Lys Thr His Thr Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 126
<211> 347
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 126
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Glu Arg Lys Cys Cys Val Glu Cys Pro
115 120 125
Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro
130 135 140
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
145 150 155 160
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn
165 170 175
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
180 185 190
Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val
195 200 205
Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
210 215 220
Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys
225 230 235 240
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
245 250 255
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
260 265 270
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
275 280 285
Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe
290 295 300
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
305 310 315 320
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
325 330 335
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 127
<211> 347
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 127
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Glu Arg Lys Cys Cys Val Glu Cys Pro
115 120 125
Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro
130 135 140
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
145 150 155 160
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn
165 170 175
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
180 185 190
Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val
195 200 205
Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
210 215 220
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Thr Lys
225 230 235 240
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
245 250 255
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
260 265 270
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
275 280 285
Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe
290 295 300
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
305 310 315 320
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
325 330 335
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 128
<211> 347
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 128
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Glu Arg Lys Cys Cys Val Glu Cys Pro
115 120 125
Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro
130 135 140
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
145 150 155 160
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn
165 170 175
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
180 185 190
Glu Glu Gln Phe Ala Ser Thr Phe Arg Val Val Ser Val Leu Thr Val
195 200 205
Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
210 215 220
Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys
225 230 235 240
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
245 250 255
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
260 265 270
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
275 280 285
Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe
290 295 300
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
305 310 315 320
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
325 330 335
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 129
<211> 347
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 129
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Glu Arg Lys Cys Cys Val Glu Cys Pro
115 120 125
Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro
130 135 140
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
145 150 155 160
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn
165 170 175
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
180 185 190
Glu Glu Gln Phe Ala Ser Thr Phe Arg Val Val Ser Val Leu Thr Val
195 200 205
Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
210 215 220
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Thr Lys
225 230 235 240
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
245 250 255
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
260 265 270
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
275 280 285
Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe
290 295 300
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
305 310 315 320
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
325 330 335
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 130
<211> 347
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 130
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Val Thr Ser Leu Ile Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Ser Thr Lys Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser
115 120 125
Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
130 135 140
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
145 150 155 160
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
165 170 175
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
180 185 190
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
195 200 205
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
210 215 220
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
225 230 235 240
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
245 250 255
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
260 265 270
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
275 280 285
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
290 295 300
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
305 310 315 320
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
325 330 335
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
340 345
<210> 131
<211> 348
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 131
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Glu Ser Lys Tyr Gly Pro Pro Cys Pro
115 120 125
Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe
130 135 140
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
145 150 155 160
Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe
165 170 175
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
180 185 190
Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
195 200 205
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
210 215 220
Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala
225 230 235 240
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln
245 250 255
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
260 265 270
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
275 280 285
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
290 295 300
Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu
305 310 315 320
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
325 330 335
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
340 345
<210> 132
<211> 348
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 132
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Glu Ser Lys Tyr Gly Pro Pro Cys Pro
115 120 125
Pro Cys Pro Ala Pro Glu Phe Glu Gly Gly Pro Ser Val Phe Leu Phe
130 135 140
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
145 150 155 160
Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe
165 170 175
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
180 185 190
Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
195 200 205
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
210 215 220
Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala
225 230 235 240
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln
245 250 255
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
260 265 270
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
275 280 285
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
290 295 300
Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu
305 310 315 320
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
325 330 335
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
340 345
<210> 133
<211> 347
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 133
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Glu Ser Lys Tyr Gly Pro Pro Cys Pro
115 120 125
Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro
130 135 140
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
145 150 155 160
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
165 170 175
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
180 185 190
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
195 200 205
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
210 215 220
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
225 230 235 240
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
245 250 255
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
260 265 270
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
275 280 285
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
290 295 300
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
305 310 315 320
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
325 330 335
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
340 345
<210> 134
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 134
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Ala Ala Ala Pro Pro Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 135
<211> 345
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 135
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Lys Thr His Thr Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Pro Gly
340 345
<210> 136
<211> 345
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 136
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Lys Thr His Thr Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Pro Gly
340 345
<210> 137
<211> 345
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 137
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Lys Thr His Thr Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Pro Gly
340 345
<210> 138
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 138
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Lys Thr His Thr Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 139
<211> 227
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 139
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
1 5 10 15
Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Ala Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
130 135 140
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys
225
<210> 140
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 140
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Lys Thr His Thr Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 141
<211> 227
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 141
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
1 5 10 15
Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Ala Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
130 135 140
Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys
225
<210> 142
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 142
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Lys Thr His Thr Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 143
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 143
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Lys Thr His Thr Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 144
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 144
Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val Gln Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Asn Tyr
20 25 30
Gly Val His Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Ser Gly Gly Asn Thr Asp Tyr Asn Thr Pro Phe Thr
50 55 60
Ser Arg Leu Ser Ile Asn Lys Asp Asn Ser Lys Ser Gln Val Phe Phe
65 70 75 80
Lys Met Asn Ser Leu Gln Ser Asn Asp Thr Ala Ile Tyr Tyr Cys Ala
85 90 95
Arg Ala Leu Thr Tyr Tyr Asp Tyr Glu Phe Ala Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Arg Lys
210 215 220
Thr His Thr Cys Pro Arg Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Ala Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 145
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 145
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Glu Lys Thr His Thr Cys Pro Glu Cys
115 120 125
Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Glu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 146
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 146
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Lys Thr His Thr Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 147
<211> 227
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 147
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
1 5 10 15
Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
130 135 140
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys
225
<210> 148
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 148
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Lys Thr His Thr Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 149
<211> 227
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 149
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
1 5 10 15
Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
130 135 140
Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys
225
<210> 150
<211> 704
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 150
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Ala His Lys Ser Glu Val Ala His
115 120 125
Arg Phe Lys Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu Val Leu Ile
130 135 140
Ala Phe Ala Gln Tyr Leu Gln Gln Ser Pro Phe Glu Asp His Val Lys
145 150 155 160
Leu Val Asn Glu Val Thr Glu Phe Ala Lys Thr Cys Val Ala Asp Glu
165 170 175
Ser Ala Glu Asn Cys Asp Lys Ser Leu His Thr Leu Phe Gly Asp Lys
180 185 190
Leu Cys Thr Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala Asp
195 200 205
Cys Cys Ala Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln His
210 215 220
Lys Asp Asp Asn Pro Asn Leu Pro Arg Leu Val Arg Pro Glu Val Asp
225 230 235 240
Val Met Cys Thr Ala Phe His Asp Asn Glu Glu Thr Phe Leu Lys Lys
245 250 255
Tyr Leu Tyr Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro Glu
260 265 270
Leu Leu Phe Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys Cys
275 280 285
Gln Ala Ala Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu Leu
290 295 300
Arg Asp Glu Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys Ala
305 310 315 320
Ser Leu Gln Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala Val Ala
325 330 335
Arg Leu Ser Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu Val Ser Lys
340 345 350
Leu Val Thr Asp Leu Thr Lys Val His Thr Glu Cys Cys His Gly Asp
355 360 365
Leu Leu Glu Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile Cys
370 375 380
Glu Asn Gln Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu Lys
385 390 395 400
Pro Leu Leu Glu Lys Ser His Cys Ile Ala Glu Val Glu Asn Asp Glu
405 410 415
Met Pro Ala Asp Leu Pro Ser Leu Ala Ala Asp Phe Val Glu Ser Lys
420 425 430
Asp Val Cys Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly Met
435 440 445
Phe Leu Tyr Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser Val Val Leu
450 455 460
Leu Leu Arg Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys Cys
465 470 475 480
Ala Ala Ala Asp Pro His Glu Cys Tyr Ala Lys Val Phe Asp Glu Phe
485 490 495
Lys Pro Leu Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys Glu
500 505 510
Leu Phe Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu Val
515 520 525
Arg Tyr Thr Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu Val Glu
530 535 540
Val Ser Arg Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys His Pro
545 550 555 560
Glu Ala Lys Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser Val Val Leu
565 570 575
Asn Gln Leu Cys Val Leu His Glu Lys Thr Pro Val Ser Asp Arg Val
580 585 590
Thr Lys Cys Cys Thr Glu Ser Leu Val Asn Arg Arg Pro Cys Phe Ser
595 600 605
Ala Leu Glu Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala Glu
610 615 620
Thr Phe Thr Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu Arg
625 630 635 640
Gln Ile Lys Lys Gln Thr Ala Leu Val Glu Leu Val Lys His Lys Pro
645 650 655
Lys Ala Thr Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe Ala Ala
660 665 670
Phe Val Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe Ala
675 680 685
Glu Glu Gly Lys Lys Leu Val Ala Ala Ser Gln Ala Ala Leu Gly Leu
690 695 700
<210> 151
<211> 704
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 151
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Ala His Lys Ser Glu Val Ala His
115 120 125
Arg Phe Lys Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu Val Leu Ile
130 135 140
Ala Phe Ala Gln Tyr Leu Gln Gln Ser Pro Phe Glu Asp His Val Lys
145 150 155 160
Leu Val Asn Glu Val Thr Glu Phe Ala Lys Thr Cys Val Ala Asp Glu
165 170 175
Ser Ala Glu Asn Cys Asp Lys Ser Leu His Thr Leu Phe Gly Asp Lys
180 185 190
Leu Cys Thr Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala Asp
195 200 205
Cys Cys Ala Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln His
210 215 220
Lys Asp Asp Asn Pro Asn Leu Pro Arg Leu Val Arg Pro Glu Val Asp
225 230 235 240
Val Met Cys Thr Ala Phe His Asp Asn Glu Glu Thr Phe Leu Lys Lys
245 250 255
Tyr Leu Tyr Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro Glu
260 265 270
Leu Leu Phe Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys Cys
275 280 285
Gln Ala Ala Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu Leu
290 295 300
Arg Asp Glu Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys Ala
305 310 315 320
Ser Leu Gln Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala Val Ala
325 330 335
Arg Leu Ser Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu Val Ser Lys
340 345 350
Leu Val Thr Asp Leu Thr Lys Val His Thr Glu Cys Cys His Gly Asp
355 360 365
Leu Leu Glu Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile Cys
370 375 380
Glu Asn Gln Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu Lys
385 390 395 400
Pro Leu Leu Glu Lys Ser His Cys Ile Ala Glu Val Glu Asn Asp Glu
405 410 415
Met Pro Ala Asp Leu Pro Ser Leu Ala Ala Asp Phe Val Glu Ser Lys
420 425 430
Asp Val Cys Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly Met
435 440 445
Phe Leu Tyr Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser Val Val Leu
450 455 460
Leu Leu Arg Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys Cys
465 470 475 480
Ala Ala Ala Asp Pro His Glu Cys Tyr Ala Lys Val Phe Asp Glu Phe
485 490 495
Lys Pro Leu Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys Glu
500 505 510
Leu Phe Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu Val
515 520 525
Arg Tyr Thr Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu Val Glu
530 535 540
Val Ser Arg Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys His Pro
545 550 555 560
Glu Ala Lys Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser Val Val Leu
565 570 575
Asn Gln Leu Cys Val Leu His Glu Lys Thr Pro Val Ser Asp Arg Val
580 585 590
Thr Lys Cys Cys Thr Glu Ser Leu Val Asn Arg Arg Pro Cys Phe Ser
595 600 605
Ala Leu Glu Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala Glu
610 615 620
Thr Phe Thr Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu Arg
625 630 635 640
Gln Ile Lys Lys Gln Thr Ala Leu Val Glu Leu Val Lys His Lys Pro
645 650 655
Lys Ala Thr Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe Ala Ala
660 665 670
Phe Val Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe Ala
675 680 685
Glu Glu Gly Lys Lys Leu Val Ala Ala Ser Gln Ala Ala Leu Gly Leu
690 695 700
<210> 152
<211> 704
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 152
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Ala His Lys Ser Glu Val Ala His
115 120 125
Arg Phe Lys Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu Val Leu Ile
130 135 140
Ala Phe Ala Gln Tyr Leu Gln Gln Ser Pro Phe Glu Asp His Val Lys
145 150 155 160
Leu Val Asn Glu Val Thr Glu Phe Ala Lys Thr Cys Val Ala Asp Glu
165 170 175
Ser Ala Glu Asn Cys Asp Lys Ser Leu His Thr Leu Phe Gly Asp Lys
180 185 190
Leu Cys Thr Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala Asp
195 200 205
Cys Cys Ala Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln His
210 215 220
Lys Asp Asp Asn Pro Asn Leu Pro Arg Leu Val Arg Pro Glu Val Asp
225 230 235 240
Val Met Cys Thr Ala Phe His Asp Asn Glu Glu Thr Phe Leu Lys Lys
245 250 255
Tyr Leu Tyr Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro Glu
260 265 270
Leu Leu Phe Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys Cys
275 280 285
Gln Ala Ala Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu Leu
290 295 300
Arg Asp Glu Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys Ala
305 310 315 320
Ser Leu Gln Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala Val Ala
325 330 335
Arg Leu Ser Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu Val Ser Lys
340 345 350
Leu Val Thr Asp Leu Thr Lys Val His Thr Glu Cys Cys His Gly Asp
355 360 365
Leu Leu Glu Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile Cys
370 375 380
Glu Asn Gln Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu Lys
385 390 395 400
Pro Leu Leu Glu Lys Ser His Cys Ile Ala Glu Val Glu Asn Asp Glu
405 410 415
Met Pro Ala Asp Leu Pro Ser Leu Ala Ala Asp Phe Val Glu Ser Lys
420 425 430
Asp Val Cys Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly Met
435 440 445
Phe Leu Tyr Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser Val Val Leu
450 455 460
Leu Leu Arg Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys Cys
465 470 475 480
Ala Ala Ala Asp Pro His Glu Cys Tyr Ala Lys Val Phe Asp Glu Phe
485 490 495
Lys Pro Leu Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys Glu
500 505 510
Leu Phe Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu Val
515 520 525
Arg Tyr Thr Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu Val Glu
530 535 540
Val Ser Arg Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys His Pro
545 550 555 560
Glu Ala Lys Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser Val Val Leu
565 570 575
Asn Gln Leu Cys Val Leu His Glu Lys Thr Pro Val Ser Asp Arg Val
580 585 590
Thr Lys Cys Cys Thr Glu Ser Leu Val Asn Arg Arg Pro Cys Phe Ser
595 600 605
Ala Leu Glu Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala Glu
610 615 620
Thr Phe Thr Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu Arg
625 630 635 640
Gln Ile Lys Lys Gln Thr Ala Leu Val Glu Leu Val Lys His Lys Pro
645 650 655
Lys Ala Thr Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe Ala Ala
660 665 670
Phe Val Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe Ala
675 680 685
Glu Glu Gly Lys Lys Leu Val Ala Ala Ser Gln Ala Ala Leu Gly Leu
690 695 700
<210> 153
<211> 704
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 153
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Ala His Lys Ser Glu Val Ala His
115 120 125
Arg Phe Lys Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu Val Leu Ile
130 135 140
Ala Phe Ala Gln Tyr Leu Gln Gln Ser Pro Phe Glu Asp His Val Lys
145 150 155 160
Leu Val Asn Glu Val Thr Glu Phe Ala Lys Thr Cys Val Ala Asp Glu
165 170 175
Ser Ala Glu Asn Cys Asp Lys Ser Leu His Thr Leu Phe Gly Asp Lys
180 185 190
Leu Cys Thr Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala Asp
195 200 205
Cys Cys Ala Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln His
210 215 220
Lys Asp Asp Asn Pro Asn Leu Pro Arg Leu Val Arg Pro Glu Val Asp
225 230 235 240
Val Met Cys Thr Ala Phe His Asp Asn Glu Glu Thr Phe Leu Lys Lys
245 250 255
Tyr Leu Tyr Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro Glu
260 265 270
Leu Leu Phe Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys Cys
275 280 285
Gln Ala Ala Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu Leu
290 295 300
Arg Asp Glu Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys Ala
305 310 315 320
Ser Leu Gln Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala Val Ala
325 330 335
Arg Leu Ser Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu Val Ser Lys
340 345 350
Leu Val Thr Asp Leu Thr Lys Val His Thr Glu Cys Cys His Gly Asp
355 360 365
Leu Leu Glu Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile Cys
370 375 380
Glu Asn Gln Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu Lys
385 390 395 400
Pro Leu Leu Glu Lys Ser His Cys Ile Ala Glu Val Glu Asn Asp Glu
405 410 415
Met Pro Ala Asp Leu Pro Ser Leu Ala Ala Asp Phe Val Glu Ser Lys
420 425 430
Asp Val Cys Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly Met
435 440 445
Phe Leu Tyr Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser Val Val Leu
450 455 460
Leu Leu Arg Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys Cys
465 470 475 480
Ala Ala Ala Asp Pro His Glu Cys Tyr Ala Lys Val Phe Asp Glu Phe
485 490 495
Lys Pro Leu Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys Glu
500 505 510
Leu Phe Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu Val
515 520 525
Arg Tyr Thr Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu Val Glu
530 535 540
Val Ser Arg Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys His Pro
545 550 555 560
Glu Ala Lys Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser Val Val Leu
565 570 575
Asn Gln Leu Cys Val Leu His Glu Lys Thr Pro Val Ser Asp Arg Val
580 585 590
Thr Lys Cys Cys Thr Glu Ser Leu Val Asn Arg Arg Pro Cys Phe Ser
595 600 605
Ala Leu Glu Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala Glu
610 615 620
Thr Phe Thr Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu Arg
625 630 635 640
Gln Ile Lys Lys Gln Thr Ala Leu Val Glu Leu Val Lys His Lys Pro
645 650 655
Lys Ala Thr Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe Ala Ala
660 665 670
Phe Val Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe Ala
675 680 685
Glu Glu Gly Lys Lys Leu Val Ala Ala Ser Gln Ala Ala Leu Gly Leu
690 695 700
<210> 154
<211> 704
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 154
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Ala His Lys Ser Glu Val Ala His
115 120 125
Arg Phe Lys Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu Val Leu Ile
130 135 140
Ala Phe Ala Gln Tyr Leu Gln Gln Ser Pro Phe Glu Asp His Val Lys
145 150 155 160
Leu Val Asn Glu Val Thr Glu Phe Ala Lys Thr Cys Val Ala Asp Glu
165 170 175
Ser Ala Glu Asn Cys Asp Lys Ser Leu His Thr Leu Phe Gly Asp Lys
180 185 190
Leu Cys Thr Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala Asp
195 200 205
Cys Cys Ala Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln His
210 215 220
Lys Asp Asp Asn Pro Asn Leu Pro Arg Leu Val Arg Pro Glu Val Asp
225 230 235 240
Val Met Cys Thr Ala Phe His Asp Asn Glu Glu Thr Phe Leu Lys Lys
245 250 255
Tyr Leu Tyr Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro Glu
260 265 270
Leu Leu Phe Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys Cys
275 280 285
Gln Ala Ala Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu Leu
290 295 300
Arg Asp Glu Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys Ala
305 310 315 320
Ser Leu Gln Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala Val Ala
325 330 335
Arg Leu Ser Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu Val Ser Lys
340 345 350
Leu Val Thr Asp Leu Thr Lys Val His Thr Glu Cys Cys His Gly Asp
355 360 365
Leu Leu Glu Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile Cys
370 375 380
Glu Asn Gln Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu Lys
385 390 395 400
Pro Leu Leu Glu Lys Ser His Cys Ile Ala Glu Val Glu Asn Asp Glu
405 410 415
Met Pro Ala Asp Leu Pro Ser Leu Ala Ala Asp Phe Val Glu Ser Lys
420 425 430
Asp Val Cys Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly Met
435 440 445
Phe Leu Tyr Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser Val Val Leu
450 455 460
Leu Leu Arg Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys Cys
465 470 475 480
Ala Ala Ala Asp Pro His Glu Cys Tyr Ala Lys Val Phe Asp Glu Phe
485 490 495
Lys Pro Leu Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys Glu
500 505 510
Leu Phe Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu Val
515 520 525
Arg Tyr Thr Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu Val Glu
530 535 540
Val Ser Arg Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys His Pro
545 550 555 560
Glu Ala Lys Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser Val Val Leu
565 570 575
Asn Gln Leu Cys Val Leu His Glu Lys Thr Pro Val Ser Asp Arg Val
580 585 590
Thr Lys Cys Cys Thr Glu Ser Leu Val Asn Arg Arg Pro Cys Phe Ser
595 600 605
Ala Leu Glu Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala Glu
610 615 620
Thr Phe Thr Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu Arg
625 630 635 640
Gln Ile Lys Lys Gln Thr Ala Leu Val Glu Leu Val Lys His Lys Pro
645 650 655
Lys Ala Thr Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe Ala Ala
660 665 670
Phe Val Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe Ala
675 680 685
Glu Glu Gly Lys Lys Leu Val Ala Ala Ser Gln Ala Ala Leu Gly Leu
690 695 700
<210> 155
<211> 704
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 155
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Ala His Lys Ser Glu Val Ala His
115 120 125
Arg Phe Lys Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu Val Leu Ile
130 135 140
Ala Phe Ala Gln Tyr Leu Gln Gln Ser Pro Phe Glu Asp His Val Lys
145 150 155 160
Leu Val Asn Glu Val Thr Glu Phe Ala Lys Thr Cys Val Ala Asp Glu
165 170 175
Ser Ala Glu Asn Cys Asp Lys Ser Leu His Thr Leu Phe Gly Asp Lys
180 185 190
Leu Cys Thr Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala Asp
195 200 205
Cys Cys Ala Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln His
210 215 220
Lys Asp Asp Asn Pro Asn Leu Pro Arg Leu Val Arg Pro Glu Val Asp
225 230 235 240
Val Met Cys Thr Ala Phe His Asp Asn Glu Glu Thr Phe Leu Lys Lys
245 250 255
Tyr Leu Tyr Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro Glu
260 265 270
Leu Leu Phe Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys Cys
275 280 285
Gln Ala Ala Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu Leu
290 295 300
Arg Asp Glu Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys Ala
305 310 315 320
Ser Leu Gln Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala Val Ala
325 330 335
Arg Leu Ser Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu Val Ser Lys
340 345 350
Leu Val Thr Asp Leu Thr Lys Val His Thr Glu Cys Cys His Gly Asp
355 360 365
Leu Leu Glu Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile Cys
370 375 380
Glu Asn Gln Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu Lys
385 390 395 400
Pro Leu Leu Glu Lys Ser His Cys Ile Ala Glu Val Glu Asn Asp Glu
405 410 415
Met Pro Ala Asp Leu Pro Ser Leu Ala Ala Asp Phe Val Glu Ser Lys
420 425 430
Asp Val Cys Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly Met
435 440 445
Phe Leu Tyr Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser Val Val Leu
450 455 460
Leu Leu Arg Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys Cys
465 470 475 480
Ala Ala Ala Asp Pro His Glu Cys Tyr Ala Lys Val Phe Asp Glu Phe
485 490 495
Lys Pro Leu Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys Glu
500 505 510
Leu Phe Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu Val
515 520 525
Arg Tyr Thr Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu Val Glu
530 535 540
Val Ser Arg Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys His Pro
545 550 555 560
Glu Ala Lys Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser Val Val Leu
565 570 575
Asn Gln Leu Cys Val Leu His Glu Lys Thr Pro Val Ser Asp Arg Val
580 585 590
Thr Lys Cys Cys Thr Glu Ser Leu Val Asn Arg Arg Pro Cys Phe Ser
595 600 605
Ala Leu Glu Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala Glu
610 615 620
Thr Phe Thr Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu Arg
625 630 635 640
Gln Ile Lys Lys Gln Thr Ala Leu Val Glu Leu Val Lys His Lys Pro
645 650 655
Lys Ala Thr Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe Ala Ala
660 665 670
Phe Val Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe Ala
675 680 685
Glu Glu Gly Lys Lys Leu Val Ala Ala Ser Gln Ala Ala Leu Gly Leu
690 695 700
<210> 156
<211> 704
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 156
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Ala His Lys Ser Glu Val Ala His
115 120 125
Arg Phe Lys Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu Val Leu Ile
130 135 140
Ala Phe Ala Gln Tyr Leu Gln Gln Ser Pro Phe Glu Asp His Val Lys
145 150 155 160
Leu Val Asn Glu Val Thr Glu Phe Ala Lys Thr Cys Val Ala Asp Glu
165 170 175
Ser Ala Glu Asn Cys Asp Lys Ser Leu His Thr Leu Phe Gly Asp Lys
180 185 190
Leu Cys Thr Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala Asp
195 200 205
Cys Cys Ala Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln His
210 215 220
Lys Asp Asp Asn Pro Asn Leu Pro Arg Leu Val Arg Pro Glu Val Asp
225 230 235 240
Val Met Cys Thr Ala Phe His Asp Asn Glu Glu Thr Phe Leu Lys Lys
245 250 255
Tyr Leu Tyr Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro Glu
260 265 270
Leu Leu Phe Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys Cys
275 280 285
Gln Ala Ala Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu Leu
290 295 300
Arg Asp Glu Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys Ala
305 310 315 320
Ser Leu Gln Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala Val Ala
325 330 335
Arg Leu Ser Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu Val Ser Lys
340 345 350
Leu Val Thr Asp Leu Thr Lys Val His Thr Glu Cys Cys His Gly Asp
355 360 365
Leu Leu Glu Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile Cys
370 375 380
Glu Asn Gln Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu Lys
385 390 395 400
Pro Leu Leu Glu Lys Ser His Cys Ile Ala Glu Val Glu Asn Asp Glu
405 410 415
Met Pro Ala Asp Leu Pro Ser Leu Ala Ala Asp Phe Val Glu Ser Lys
420 425 430
Asp Val Cys Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly Met
435 440 445
Phe Leu Tyr Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser Val Val Leu
450 455 460
Leu Leu Arg Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys Cys
465 470 475 480
Ala Ala Ala Asp Pro His Glu Cys Tyr Ala Lys Val Phe Asp Glu Phe
485 490 495
Lys Pro Leu Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys Glu
500 505 510
Leu Phe Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu Val
515 520 525
Arg Tyr Thr Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu Val Glu
530 535 540
Val Ser Arg Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys His Pro
545 550 555 560
Glu Ala Lys Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser Val Val Leu
565 570 575
Asn Gln Leu Cys Val Leu His Glu Lys Thr Pro Val Ser Asp Arg Val
580 585 590
Thr Lys Cys Cys Thr Glu Ser Leu Val Asn Arg Arg Pro Cys Phe Ser
595 600 605
Ala Leu Glu Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala Glu
610 615 620
Thr Phe Thr Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu Arg
625 630 635 640
Gln Ile Lys Lys Gln Thr Ala Leu Val Glu Leu Val Lys His Lys Pro
645 650 655
Lys Ala Thr Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe Ala Ala
660 665 670
Phe Val Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe Ala
675 680 685
Glu Glu Gly Lys Lys Leu Val Ala Ala Ser Gln Ala Ala Leu Gly Leu
690 695 700
<210> 157
<211> 704
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 157
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Ala His Lys Ser Glu Val Ala His
115 120 125
Arg Phe Lys Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu Val Leu Ile
130 135 140
Ala Phe Ala Gln Tyr Leu Gln Gln Ser Pro Phe Glu Asp His Val Lys
145 150 155 160
Leu Val Asn Glu Val Thr Glu Phe Ala Lys Thr Cys Val Ala Asp Glu
165 170 175
Ser Ala Glu Asn Cys Asp Lys Ser Leu His Thr Leu Phe Gly Asp Lys
180 185 190
Leu Cys Thr Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala Asp
195 200 205
Cys Cys Ala Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln His
210 215 220
Lys Asp Asp Asn Pro Asn Leu Pro Arg Leu Val Arg Pro Glu Val Asp
225 230 235 240
Val Met Cys Thr Ala Phe His Asp Asn Glu Glu Thr Phe Leu Lys Lys
245 250 255
Tyr Leu Tyr Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro Glu
260 265 270
Leu Leu Phe Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys Cys
275 280 285
Gln Ala Ala Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu Leu
290 295 300
Arg Asp Glu Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys Ala
305 310 315 320
Ser Leu Gln Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala Val Ala
325 330 335
Arg Leu Ser Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu Val Ser Lys
340 345 350
Leu Val Thr Asp Leu Thr Lys Val His Thr Glu Cys Cys His Gly Asp
355 360 365
Leu Leu Glu Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile Cys
370 375 380
Glu Asn Gln Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu Lys
385 390 395 400
Pro Leu Leu Glu Lys Ser His Cys Ile Ala Glu Val Glu Asn Asp Glu
405 410 415
Met Pro Ala Asp Leu Pro Ser Leu Ala Ala Asp Phe Val Glu Ser Lys
420 425 430
Asp Val Cys Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly Met
435 440 445
Phe Leu Tyr Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser Val Val Leu
450 455 460
Leu Leu Arg Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys Cys
465 470 475 480
Ala Ala Ala Asp Pro His Glu Cys Tyr Ala Lys Val Phe Asp Glu Phe
485 490 495
Lys Pro Leu Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys Glu
500 505 510
Leu Phe Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu Val
515 520 525
Arg Tyr Thr Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu Val Glu
530 535 540
Val Ser Arg Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys His Pro
545 550 555 560
Glu Ala Lys Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser Val Val Leu
565 570 575
Asn Gln Leu Cys Val Leu His Glu Lys Thr Pro Val Ser Asp Arg Val
580 585 590
Thr Lys Cys Cys Thr Glu Ser Leu Val Asn Arg Arg Pro Cys Phe Ser
595 600 605
Ala Leu Glu Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala Glu
610 615 620
Thr Phe Thr Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu Arg
625 630 635 640
Gln Ile Lys Lys Gln Thr Ala Leu Val Glu Leu Val Lys His Lys Pro
645 650 655
Lys Ala Thr Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe Ala Ala
660 665 670
Phe Val Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe Ala
675 680 685
Glu Glu Gly Lys Lys Leu Val Ala Ala Ser Gln Ala Ala Leu Gly Leu
690 695 700
<210> 158
<211> 704
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 158
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Ala His Lys Ser Glu Val Ala His
115 120 125
Arg Phe Lys Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu Val Leu Ile
130 135 140
Ala Phe Ala Gln Tyr Leu Gln Gln Ser Pro Phe Glu Asp His Val Lys
145 150 155 160
Leu Val Asn Glu Val Thr Glu Phe Ala Lys Thr Cys Val Ala Asp Glu
165 170 175
Ser Ala Glu Asn Cys Asp Lys Ser Leu His Thr Leu Phe Gly Asp Lys
180 185 190
Leu Cys Thr Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala Asp
195 200 205
Cys Cys Ala Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln His
210 215 220
Lys Asp Asp Asn Pro Asn Leu Pro Arg Leu Val Arg Pro Glu Val Asp
225 230 235 240
Val Met Cys Thr Ala Phe His Asp Asn Glu Glu Thr Phe Leu Lys Lys
245 250 255
Tyr Leu Tyr Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro Glu
260 265 270
Leu Leu Phe Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys Cys
275 280 285
Gln Ala Ala Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu Leu
290 295 300
Arg Asp Glu Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys Ala
305 310 315 320
Ser Leu Gln Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala Val Ala
325 330 335
Arg Leu Ser Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu Val Ser Lys
340 345 350
Leu Val Thr Asp Leu Thr Lys Val His Thr Glu Cys Cys His Gly Asp
355 360 365
Leu Leu Glu Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile Cys
370 375 380
Glu Asn Gln Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu Lys
385 390 395 400
Pro Leu Leu Glu Lys Ser His Cys Ile Ala Glu Val Glu Asn Asp Glu
405 410 415
Met Pro Ala Asp Leu Pro Ser Leu Ala Ala Asp Phe Val Glu Ser Lys
420 425 430
Asp Val Cys Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly Met
435 440 445
Phe Leu Tyr Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser Val Val Leu
450 455 460
Leu Leu Arg Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys Cys
465 470 475 480
Ala Ala Ala Asp Pro His Glu Cys Tyr Ala Lys Val Phe Asp Glu Phe
485 490 495
Lys Pro Leu Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys Glu
500 505 510
Leu Phe Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu Val
515 520 525
Arg Tyr Thr Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu Val Glu
530 535 540
Val Ser Arg Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys His Pro
545 550 555 560
Glu Ala Lys Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser Val Val Leu
565 570 575
Asn Gln Leu Cys Val Leu His Glu Lys Thr Pro Val Ser Asp Arg Val
580 585 590
Thr Lys Cys Cys Thr Glu Ser Leu Val Asn Arg Arg Pro Cys Phe Ser
595 600 605
Ala Leu Glu Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala Glu
610 615 620
Thr Phe Thr Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu Arg
625 630 635 640
Gln Ile Lys Lys Gln Thr Ala Leu Val Glu Leu Val Lys His Lys Pro
645 650 655
Lys Ala Thr Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe Ala Ala
660 665 670
Phe Val Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe Ala
675 680 685
Glu Glu Gly Lys Lys Leu Val Ala Ala Ser Gln Ala Ala Leu Gly Leu
690 695 700
<210> 159
<211> 704
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 159
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Ala His Lys Ser Glu Val Ala His
115 120 125
Arg Phe Lys Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu Val Leu Ile
130 135 140
Ala Phe Ala Gln Tyr Leu Gln Gln Ser Pro Phe Glu Asp His Val Lys
145 150 155 160
Leu Val Asn Glu Val Thr Glu Phe Ala Lys Thr Cys Val Ala Asp Glu
165 170 175
Ser Ala Glu Asn Cys Asp Lys Ser Leu His Thr Leu Phe Gly Asp Lys
180 185 190
Leu Cys Thr Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala Asp
195 200 205
Cys Cys Ala Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln His
210 215 220
Lys Asp Asp Asn Pro Asn Leu Pro Arg Leu Val Arg Pro Glu Val Asp
225 230 235 240
Val Met Cys Thr Ala Phe His Asp Asn Glu Glu Thr Phe Leu Lys Lys
245 250 255
Tyr Leu Tyr Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro Glu
260 265 270
Leu Leu Phe Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys Cys
275 280 285
Gln Ala Ala Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu Leu
290 295 300
Arg Asp Glu Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys Ala
305 310 315 320
Ser Leu Gln Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala Val Ala
325 330 335
Arg Leu Ser Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu Val Ser Lys
340 345 350
Leu Val Thr Asp Leu Thr Lys Val His Thr Glu Cys Cys His Gly Asp
355 360 365
Leu Leu Glu Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile Cys
370 375 380
Glu Asn Gln Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu Lys
385 390 395 400
Pro Leu Leu Glu Lys Ser His Cys Ile Ala Glu Val Glu Asn Asp Glu
405 410 415
Met Pro Ala Asp Leu Pro Ser Leu Ala Ala Asp Phe Val Glu Ser Lys
420 425 430
Asp Val Cys Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly Met
435 440 445
Phe Leu Tyr Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser Val Val Leu
450 455 460
Leu Leu Arg Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys Cys
465 470 475 480
Ala Ala Ala Asp Pro His Glu Cys Tyr Ala Lys Val Phe Asp Glu Phe
485 490 495
Lys Pro Leu Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys Glu
500 505 510
Leu Phe Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu Val
515 520 525
Arg Tyr Thr Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu Val Glu
530 535 540
Val Ser Arg Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys His Pro
545 550 555 560
Glu Ala Lys Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser Val Val Leu
565 570 575
Asn Gln Leu Cys Val Leu His Glu Lys Thr Pro Val Ser Asp Arg Val
580 585 590
Thr Lys Cys Cys Thr Glu Ser Leu Val Asn Arg Arg Pro Cys Phe Ser
595 600 605
Ala Leu Glu Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala Glu
610 615 620
Thr Phe Thr Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu Arg
625 630 635 640
Gln Ile Lys Lys Gln Thr Ala Leu Val Glu Leu Val Lys His Lys Pro
645 650 655
Lys Ala Thr Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe Ala Ala
660 665 670
Phe Val Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe Ala
675 680 685
Glu Glu Gly Lys Lys Leu Val Ala Ala Ser Gln Ala Ala Leu Gly Leu
690 695 700
<210> 160
<211> 11
<212> Белок
<213> Неизвестно
<220>
<223> Описание неизвестного: альбуминсвязывающий
пептид
<400> 160
Asp Ile Cys Leu Pro Arg Trp Gly Cys Leu Trp
1 5 10
<210> 161
<211> 226
<212> Белок
<213> Homo sapiens
<400> 161
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
130 135 140
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly
225
<210> 162
<211> 228
<212> Белок
<213> Homo sapiens
<400> 162
Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro Pro Val
1 5 10 15
Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
20 25 30
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
35 40 45
His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu
50 55 60
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr
65 70 75 80
Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp Leu Asn
85 90 95
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ala Pro
100 105 110
Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu Pro Gln
115 120 125
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
130 135 140
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
145 150 155 160
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
165 170 175
Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
180 185 190
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
195 200 205
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
210 215 220
Ser Pro Gly Lys
225
<210> 163
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственнаой последовательности: Синтетический
пептид
<400> 163
Gly Gly Gly Gly Ser
1 5
<210> 164
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственнаой последовательности: Синтетический
пептид
<400> 164
Gly Gly Ser Gly
1
<210> 165
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственнаой последовательности: Синтетический
пептид
<400> 165
Ser Gly Gly Gly
1
<210> 166
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственнаой последовательности: Синтетический
пептид
<400> 166
Gly Ser Gly Ser
1
<210> 167
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственнаой последовательности: Синтетический
пептид
<400> 167
Gly Ser Gly Ser Gly Ser
1 5
<210> 168
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственнаой последовательности: Синтетический
пептид
<400> 168
Gly Ser Gly Ser Gly Ser Gly Ser
1 5
<210> 169
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственнаой последовательности: Синтетический
пептид
<400> 169
Gly Ser Gly Ser Gly Ser Gly Ser Gly Ser
1 5 10
<210> 170
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственнаой последовательности: Синтетический
пептид
<400> 170
Gly Ser Gly Ser Gly Ser Gly Ser Gly Ser Gly Ser
1 5 10
<210> 171
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственнаой последовательности: Синтетический
пептид
<400> 171
Gly Gly Ser Gly Gly Ser
1 5
<210> 172
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственнаой последовательности: Синтетический
пептид
<400> 172
Gly Gly Ser Gly Gly Ser Gly Gly Ser
1 5
<210> 173
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственнаой последовательности: Синтетический
пептид
<400> 173
Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser
1 5 10
<210> 174
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственнаой последовательности: Синтетический
пептид
<400> 174
Gly Gly Ser Gly Gly Gly Ser Gly
1 5
<210> 175
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственнаой последовательности: Синтетический
пептид
<400> 175
Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Ser Gly
1 5 10
<210> 176
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственнаой последовательности: Синтетический
пептид
<400> 176
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 177
<211> 3
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственнаой последовательности: Синтетический
пептид
<400> 177
Ala Ala Ser
1
<210> 178
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственнаой последовательности: Синтетический
пептид
<400> 178
Ala Ala Ala Leu
1
<210> 179
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственнаой последовательности: Синтетический
пептид
<400> 179
Ala Ala Ala Lys
1
<210> 180
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственнаой последовательности: Синтетический
пептид
<400> 180
Ala Ala Ala Arg
1
<210> 181
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственнаой последовательности: Синтетический
пептид
<400> 181
Glu Gly Lys Ser Ser Gly Ser Gly Ser Glu Ser Lys Ser Thr
1 5 10
<210> 182
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственнаой последовательности: Синтетический
пептид
<400> 182
Gly Ser Ala Gly Ser Ala Ala Gly Ser Gly Glu Phe
1 5 10
<210> 183
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственнаой последовательности: Синтетический
пептид
<400> 183
Ala Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys Ala
1 5 10
<210> 184
<211> 18
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственнаой последовательности: Синтетический
пептид
<400> 184
Lys Glu Ser Gly Ser Val Ser Ser Glu Gln Leu Ala Gln Phe Arg Ser
1 5 10 15
Leu Asp
<210> 185
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственнаой последовательности: Синтетический
пептид
<400> 185
Gly Gly Gly Gly Ala Gly Gly Gly Gly
1 5
<210> 186
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственнаой последовательности: Синтетический
пептид
<400> 186
Gly Glu Asn Leu Tyr Phe Gln Ser Gly Gly
1 5 10
<210> 187
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственнаой последовательности: Синтетический
пептид
<400> 187
Ser Ala Cys Tyr Cys Glu Leu Ser
1 5
<210> 188
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственнаой последовательности: Синтетический
пептид
<400> 188
Arg Ser Ile Ala Thr
1 5
<210> 189
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственнаой последовательности: Синтетический
пептид
<400> 189
Arg Pro Ala Cys Lys Ile Pro Asn Asp Leu Lys Gln Lys Val Met Asn
1 5 10 15
His
<210> 190
<211> 36
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 190
Gly Gly Ser Ala Gly Gly Ser Gly Ser Gly Ser Ser Gly Gly Ser Ser
1 5 10 15
Gly Ala Ser Gly Thr Gly Thr Ala Gly Gly Thr Gly Ser Gly Ser Gly
20 25 30
Thr Gly Ser Gly
35
<210> 191
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственнаой последовательности: Синтетический
пептид
<400> 191
Ala Ala Ala Asn Ser Ser Ile Asp Leu Ile Ser Val Pro Val Asp Ser
1 5 10 15
Arg
<210> 192
<211> 36
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 192
Gly Gly Ser Gly Gly Gly Ser Glu Gly Gly Gly Ser Glu Gly Gly Gly
1 5 10 15
Ser Glu Gly Gly Gly Ser Glu Gly Gly Gly Ser Glu Gly Gly Gly Ser
20 25 30
Gly Gly Gly Ser
35
<210> 193
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственнаой последовательности: Синтетический
пептид
<400> 193
Glu Ala Ala Ala Lys
1 5
<210> 194
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственнаой последовательности: Синтетический
пептид
<400> 194
Pro Ala Pro Ala Pro
1 5
<210> 195
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 195
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Met Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 196
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 196
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Lys Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 197
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 197
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Arg Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 198
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 198
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Tyr Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 199
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 199
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Asp Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 200
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 200
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ile Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 201
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 201
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Met Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 202
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 202
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 203
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 203
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Ser Glu Pro Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu
100 105 110
Leu Ser Val Arg Ala Lys Pro Ser
115 120
<210> 204
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 204
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Arg Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Asp Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 205
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 205
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Arg Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ile Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 206
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 206
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Arg Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Asp Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ile Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 207
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 207
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Tyr Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Asp Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 208
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 208
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Tyr Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ile Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 209
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 209
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Tyr Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Asp Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ile Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 210
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 210
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Asp Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ile Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 211
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 211
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Arg Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Asp Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ile Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Lys Thr His Thr Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 212
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<220>
<221> MOD_RES
<222> (6)..(6)
<223> V или L, или I
<220>
<221> MOD_RES
<222> (27)..(27)
<223> V или I, или L
<220>
<221> MOD_RES
<222> (31)..(31)
<223> I или T, или S, или F
<220>
<221> MOD_RES
<222> (53)..(53)
<223> K или R
<220>
<221> MOD_RES
<222> (56)..(56)
<223> H или P, или R
<220>
<221> MOD_RES
<222> (66)..(66)
<223> S или T, или G
<220>
<221> MOD_RES
<222> (80)..(80)
<223> A
<400> 212
Glu Glu Glu Leu Gln Xaa Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Xaa Thr Ser Leu Xaa Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Xaa Glu Gly Xaa Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Xaa Thr Lys Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Xaa
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 213
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 213
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Val Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 214
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 214
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Glu Arg Lys Ser Ser Val Glu Cys Pro
115 120 125
Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro
130 135 140
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
145 150 155 160
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn
165 170 175
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
180 185 190
Glu Glu Gln Phe Ala Ser Thr Phe Arg Val Val Ser Val Leu Thr Val
195 200 205
Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
210 215 220
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Thr Lys
225 230 235 240
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
245 250 255
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
260 265 270
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
275 280 285
Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe
290 295 300
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
305 310 315 320
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
325 330 335
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
340 345
<210> 215
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 215
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Ile Pro
20 25 30
Arg Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Arg Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Lys Thr His Thr Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 216
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 216
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu
35 40 45
Ile Tyr Asn Gln Arg Gln Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Ile Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Asp Lys Thr His Thr Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 217
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 217
Glu Glu Glu Leu Gln Ile Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ile Thr Ser Leu Phe Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Arg Glu Gly Pro Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Thr Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Ala
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Glu Lys Thr His Thr Cys Pro Glu Cys
115 120 125
Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Glu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345
<210> 218
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<220>
<221> MOD_RES
<222> (6)..(6)
<223> V или L, или I
<220>
<221> MOD_RES
<222> (27)..(27)
<223> A или V или L, или I
<220>
<221> MOD_RES
<222> (31)..(31)
<223> I или S или T или F
<220>
<221> MOD_RES
<222> (47)..(47)
<223> E или L или V
<220>
<221> MOD_RES
<222> (53)..(53)
<223> K или R
<220>
<221> MOD_RES
<222> (54)..(54)
<223> E или Q
<220>
<221> MOD_RES
<222> (56)..(56)
<223> H или R или P
<220>
<221> MOD_RES
<222> (66)..(66)
<223> S или G, или L, или T
<220>
<221> MOD_RES
<222> (80)..(80)
<223> Любая аминокислота
<220>
<221> MOD_RES
<222> (81)..(81)
<223> Любая аминокислота
<220>
<221> MOD_RES
<222> (82)..(82)
<223> Любая аминокислота
<220>
<221> MOD_RES
<222> (83)..(83)
<223> Любая аминокислота
<220>
<221> MOD_RES
<222> (92)..(92)
<223> V или I
<400> 218
Glu Glu Glu Leu Gln Xaa Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Xaa Thr Ser Leu Xaa Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Xaa Leu
35 40 45
Ile Tyr Asn Gln Xaa Xaa Gly Xaa Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Xaa Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Xaa
65 70 75 80
Xaa Xaa Xaa Ala Asp Ala Gly Thr Tyr Tyr Cys Xaa Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 219
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<220>
<221> MOD_RES
<222> (6)..(6)
<223> V или L, или I
<220>
<221> MOD_RES
<222> (27)..(27)
<223> A или V, или L, или I
<220>
<221> MOD_RES
<222> (31)..(31)
<223> I или S, или T, или F
<220>
<221> MOD_RES
<222> (47)..(47)
<223> E или L, или V
<220>
<221> MOD_RES
<222> (53)..(53)
<223> K или R
<220>
<221> MOD_RES
<222> (54)..(54)
<223> E или Q
<220>
<221> MOD_RES
<222> (56)..(56)
<223> H или R, или P
<220>
<221> MOD_RES
<222> (66)..(66)
<223> S или G, или L, или T
<220>
<221> MOD_RES
<222> (83)..(83)
<223> Любая аминокислота, отличная от P
<220>
<221> MOD_RES
<222> (92)..(92)
<223> V или I
<400> 219
Glu Glu Glu Leu Gln Xaa Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Xaa Thr Ser Leu Xaa Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Xaa Leu
35 40 45
Ile Tyr Asn Gln Xaa Xaa Gly Xaa Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Xaa Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Xaa Ala Asp Ala Gly Thr Tyr Tyr Cys Xaa Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser
115
<210> 220
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 220
Asp Ile Leu Leu Thr Gln Ser Pro Val Ile Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Val Ser Phe Ser Cys Arg Ala Ser Gln Ser Ile Gly Thr Asn
20 25 30
Ile His Trp Tyr Gln Gln Arg Thr Asn Gly Ser Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Glu Ser Ile Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile Asn Ser Val Glu Ser
65 70 75 80
Glu Asp Ile Ala Asp Tyr Tyr Cys Gln Gln Asn Asn Asn Trp Pro Thr
85 90 95
Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 221
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<220>
<221> MOD_RES
<222> (6)..(6)
<223> V или L, или I
<220>
<221> MOD_RES
<222> (27)..(27)
<223> V или I, или L
<220>
<221> MOD_RES
<222> (31)..(31)
<223> I или T, или S, или F
<220>
<221> MOD_RES
<222> (53)..(53)
<223> K или R
<220>
<221> MOD_RES
<222> (56)..(56)
<223> H или P, или R
<220>
<221> MOD_RES
<222> (66)..(66)
<223> S или T, или G
<220>
<221> MOD_RES
<222> (80)..(80)
<223> N или A
<400> 221
Glu Glu Glu Leu Gln Xaa Ile Gln Pro Asp Lys Ser Val Ser Val Ala
1 5 10 15
Ala Gly Glu Ser Ala Ile Leu His Cys Thr Xaa Thr Ser Leu Xaa Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Ala Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Xaa Glu Gly Xaa Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Glu Xaa Thr Lys Arg Glu Asn Met Asp Phe Ser Ile Ser Ile Ser Xaa
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Thr Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser
100 105 110
Val Arg Ala Lys Pro Ser
115
<210> 222
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетическая
метка 6xHis
<400> 222
His His His His His His
1 5
<210> 223
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетическая
метка 10xHis
<400> 223
His His His His His His His His His His
1 5 10
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА ПРОТИВ СИГНАЛ-РЕГУЛЯТОРНОГО БЕЛКА АЛЬФА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2771964C2 |
| АНТИ-TIGIT АНТИТЕЛА И СПОСОБЫ ПРИМЕНЕНИЯ | 2016 |
|
RU2732591C2 |
| АНТИ-TIGIT АНТИТЕЛА И СПОСОБЫ ПРИМЕНЕНИЯ | 2016 |
|
RU2817838C2 |
| РЕКОМБИНАНТНЫЕ МОДИФИЦИРОВАННЫЕ ФАКТОРЫ РОСТА ФИБРОБЛАСТОВ И ИХ ТЕРАПЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2811435C2 |
| CD8A-СВЯЗЫВАЮЩИЕ ДОМЕНЫ ТИПА III ФИБРОНЕКТИНА | 2017 |
|
RU2759952C2 |
| АНТИТЕЛО, СПЕЦИФИЧНО СВЯЗЫВАЮЩЕЕСЯ С ICAM-1, И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2789757C2 |
| АНТИ-PD-1 АНТИТЕЛА | 2019 |
|
RU2788095C2 |
| АНТИ-PVRIG АНТИТЕЛА И СПОСОБЫ ПРИМЕНЕНИЯ | 2016 |
|
RU2732042C2 |
| ВАРИАНТЫ IgG-FC ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2018 |
|
RU2814952C2 |
| АНТИ-PVRIG АНТИТЕЛА И СПОСОБЫ ПРИМЕНЕНИЯ | 2016 |
|
RU2827926C2 |
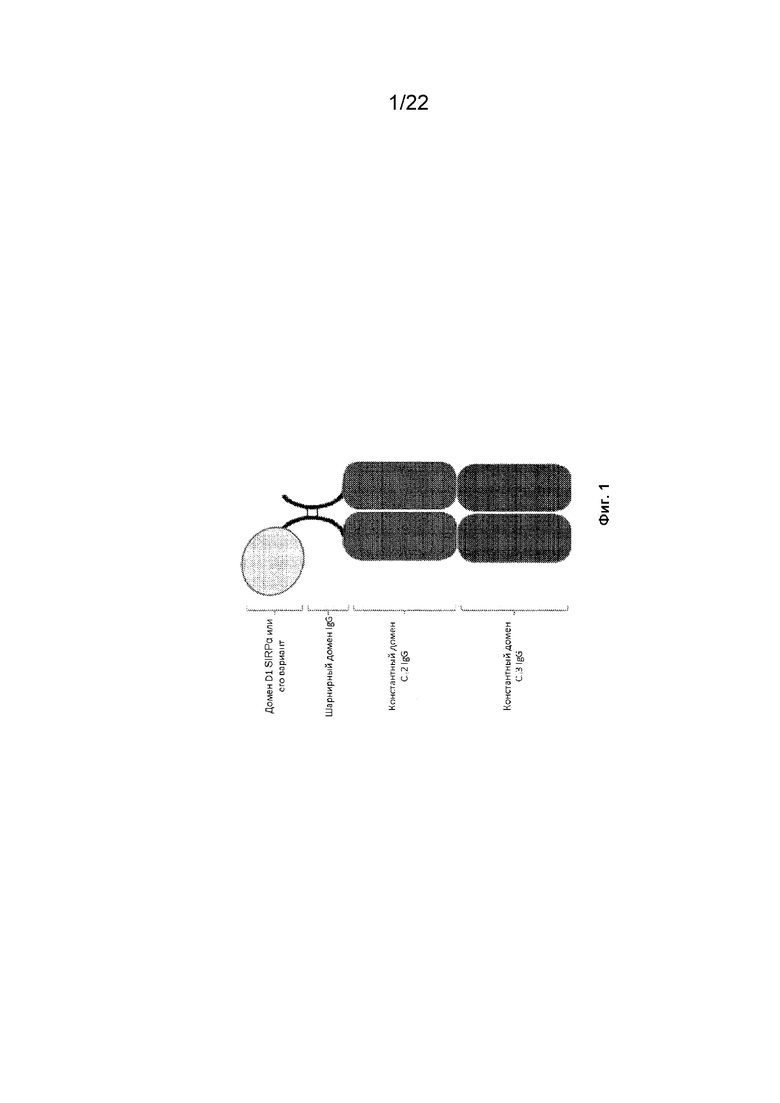
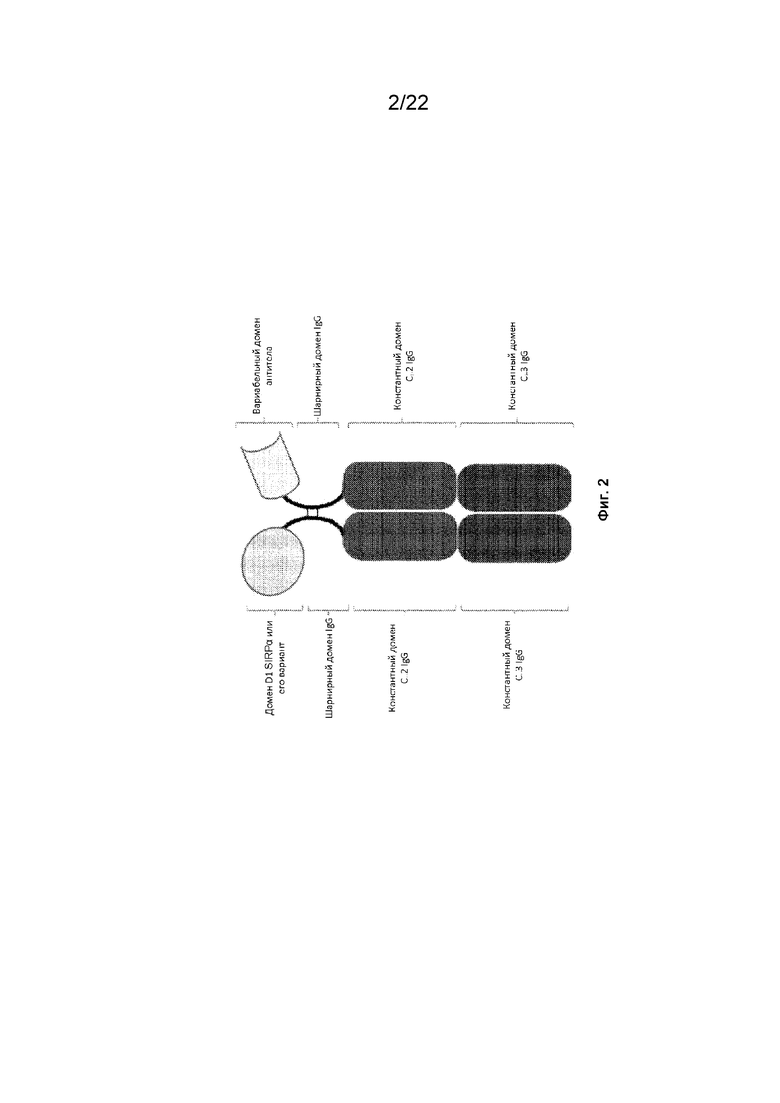

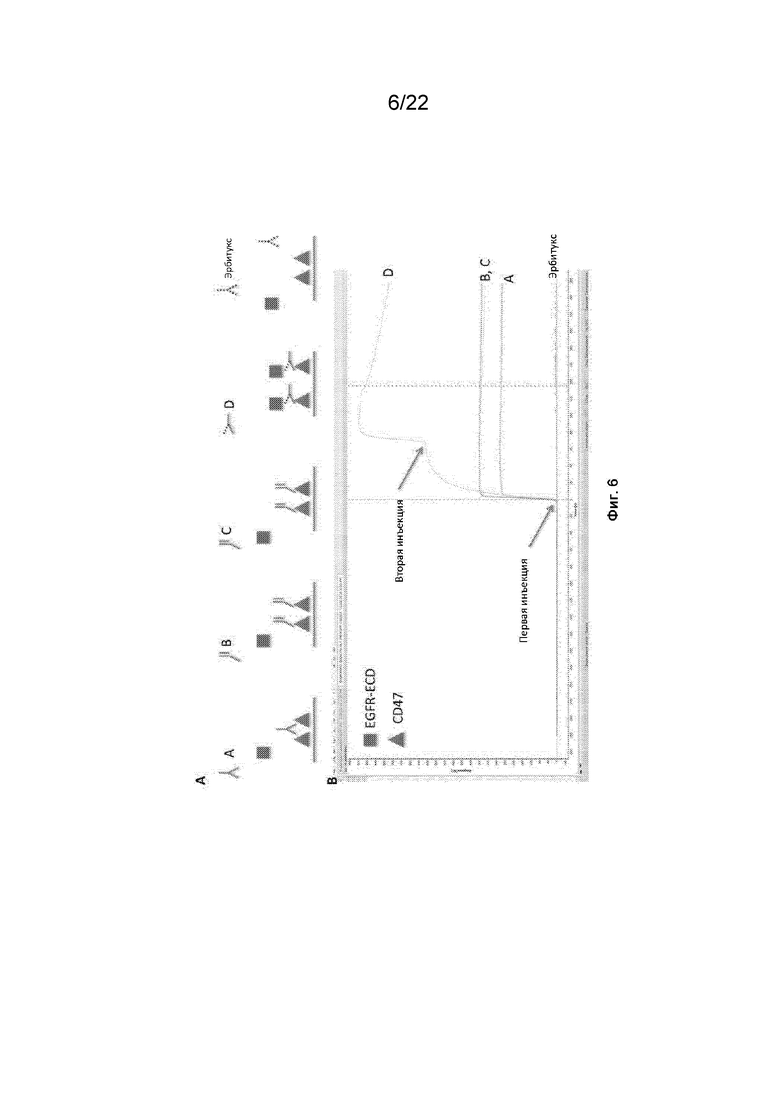
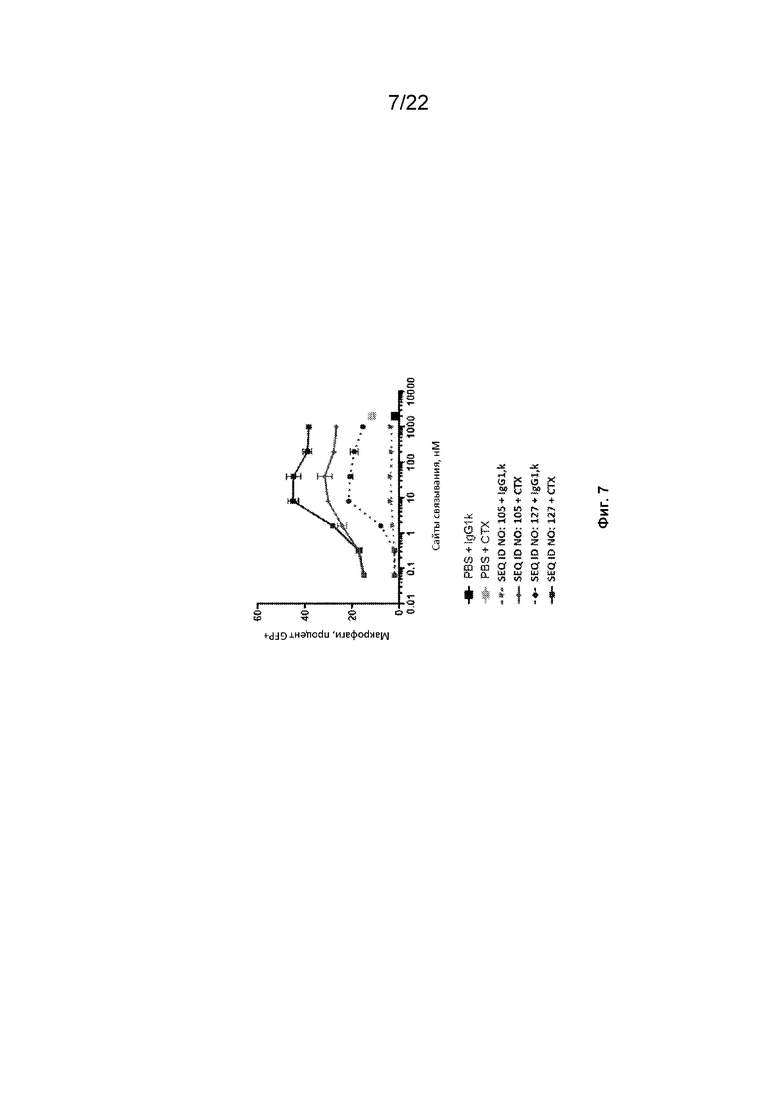
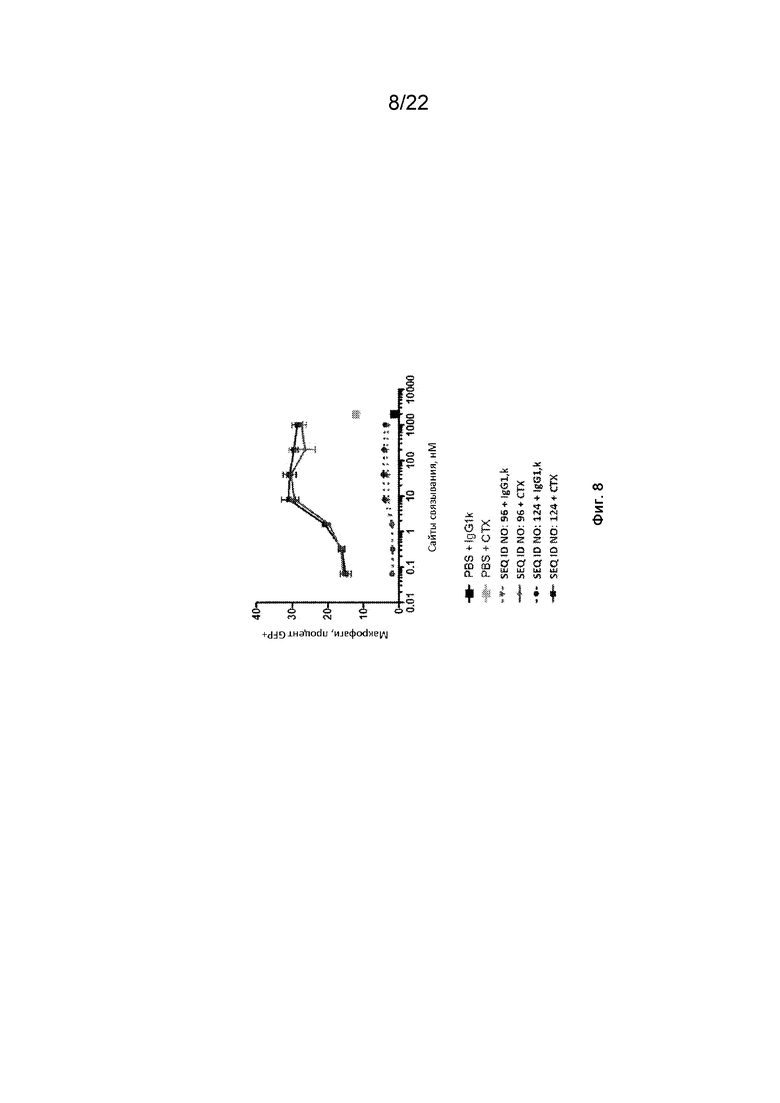
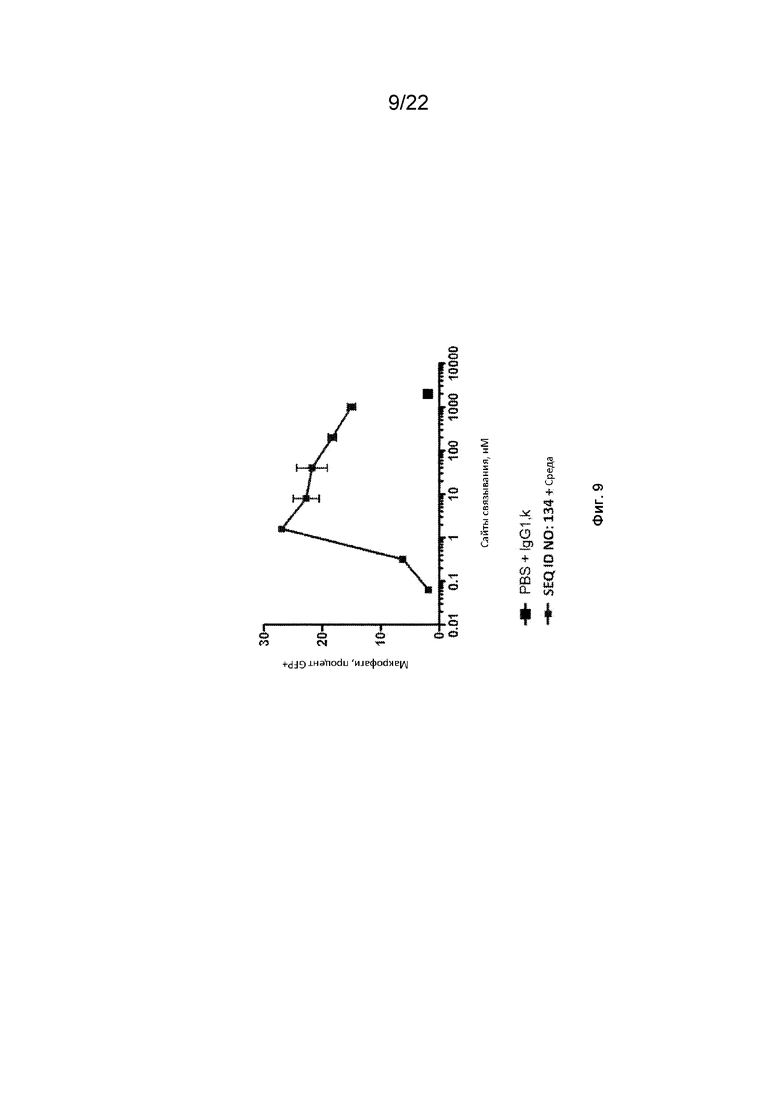


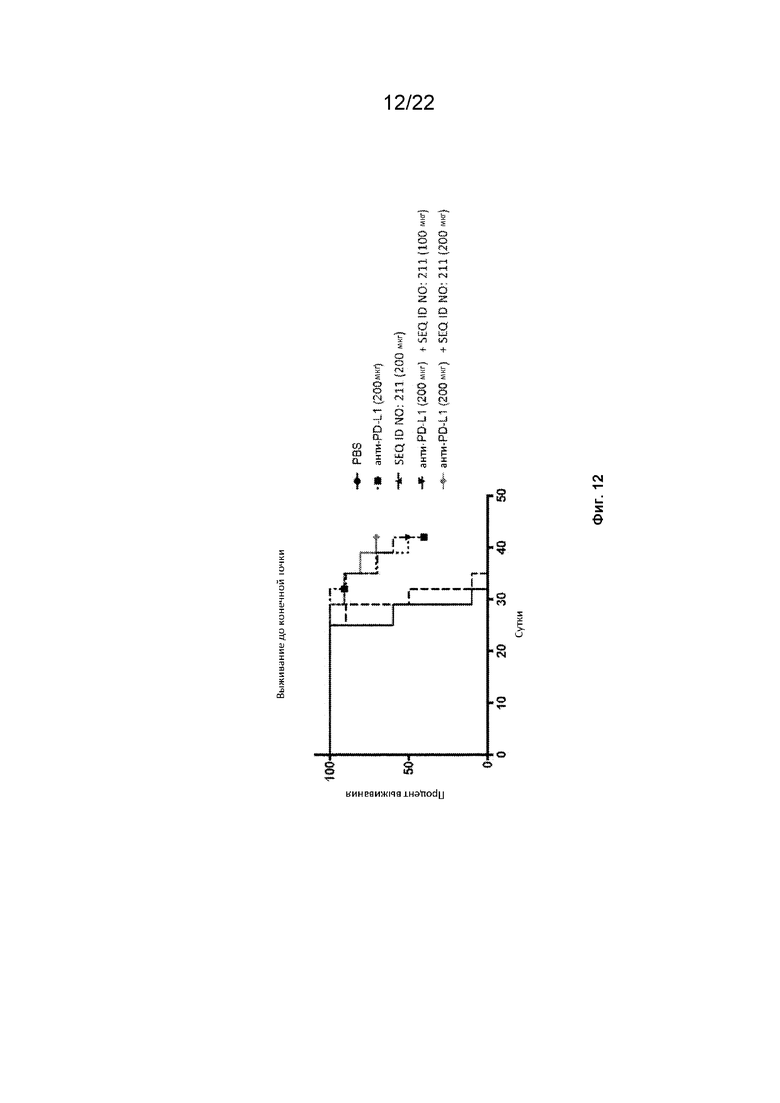






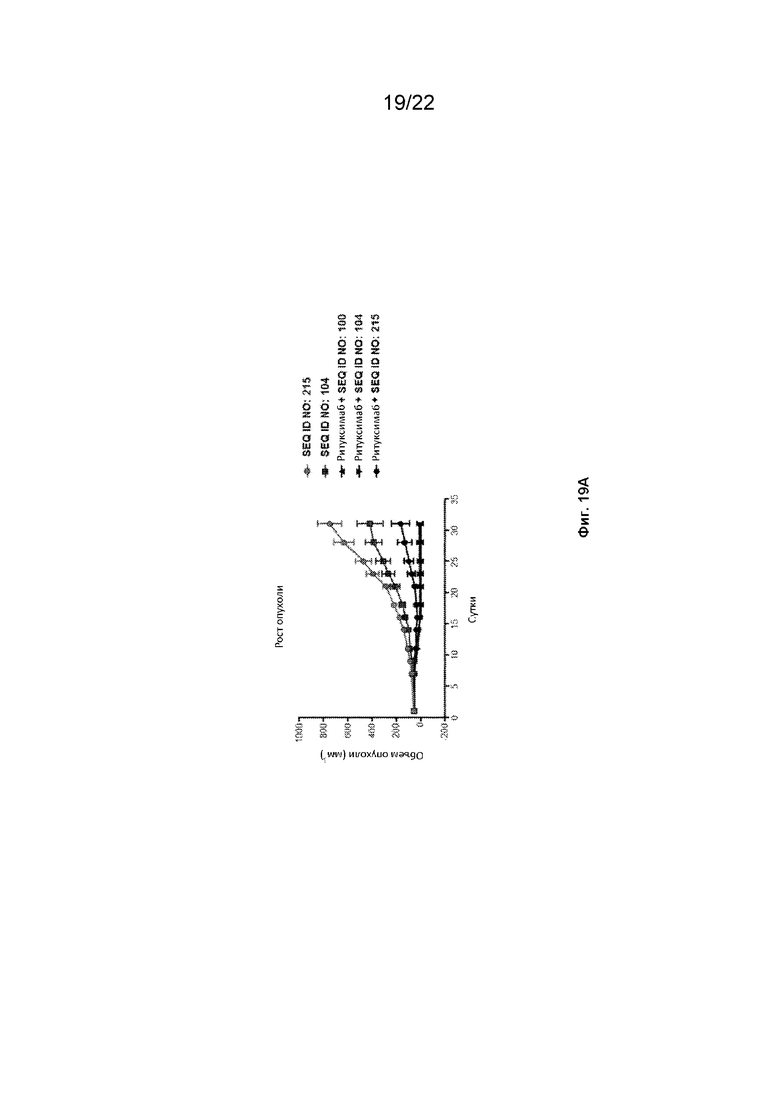



Предложенная группа изобретений относится к области медицины. Предложены полипептиды для лечения заболевания, связанного с активностью SIRPα или CD47, нуклеиновые кислоты, кодирующие полипептиды, векторы и клетка-хозяин, содержащие нуклеиновую кислоту. Предложены способы получения полипептида для лечения заболевания, связанного с активностью SIRPα или CD47, включающие культивирование клетки-хозяина и способы лечения рака у субъекта. Предложены димеры для лечения заболевания, связанного с активностью SIRPα или CD47, содержащие полипептид. Предложены фармацевтические композиции для лечения заболевания, связанного с активностью SIRPα или CD47, содержащие полипептид или димер и фармацевтически приемлемый эксципиент. Предложенная группа изобретений обеспечивает эффективные средства и методы лечения рака. 45 н. и 28 з.п. ф-лы, 28 табл., 20 пр., 20 ил.
1. Полипептид для лечения заболевания, связанного с активностью SIRPα или CD47, включающий:
(а) вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий аминокислотную последовательность, указанную в любой из SEQ ID NO: 78-85; и
(b) вариант Fc, причем вариант Fc представляет собой:
(i) Fc-область человеческого IgG1, содержащую мутации L234A, L235A, G237A и N297A, при этом нумерация соответствует индексу EU Kabat;
(ii) Fc-область человеческого IgG2, содержащую мутации A330S, P331S и N297A, при этом нумерация соответствует индексу EU Kabat;
(iii) Fc-область человеческого IgG4, содержащую мутации S228P, E233P, F234V, L235A и delG236, при этом нумерация соответствует индексу EU Kabat; или
(iv) Fc-область человеческого IgG4, содержащую мутации S228P, E233P, F234V, L235A, delG236 и N297A, при этом нумерация соответствует индексу EU Kabat.
2. Полипептид по п.1, в котором вариант Fc представляет собой Fc-область человеческого IgG1, содержащую мутации L234A, L235A, G237A и N297A.
3. Полипептид по п.1, в котором вариант Fc демонстрирует аблированное или пониженное связывание с рецептором Fcγпо сравнению с версией дикого типа области Fc человеческого IgG, причем необязательно рецептор Fcγ представляет собой CD16a, CD32a, CD32b, CD32c и CD64.
4. Полипептид по п.1, в котором вариант Fc демонстрирует уменьшение или абляцию фагоцитоза в анализе на фагоцитоз по сравнению с версией варианта Fc дикого типа.
5. Полипептид по п.1, в котором вариант D1 SIRP-α связывает CD47 с KD, составляющим: (а) 1 х 10-8 М или меньше, (b) 1 х 10-9 М или меньше или (с) 1 х 10-10 М или меньше.
6. Полипептид по п.1, причем полипептид содержит последовательность, выбранную из группы, состоящей из: SEQ ID NO: 98-104, 107-113, 116-122, 133 и 135-136.
7. Нуклеиновая кислота, кодирующая полипептид по п.1.
8. Вектор, содержащий нуклеиновую кислоту по п.7.
9. Клетка-хозяин, содержащая нуклеиновую кислоту по п.8.
10. Способ получения полипептида для лечения заболевания, связанного с активностью SIRPα или CD47, включающий культивирование клетки-хозяина по п.9 в подходящих условиях для того, чтобы вызвать экспрессию полипептида, и выделение полипептида.
11. Димер для лечения заболевания, связанного с активностью SIRPα или CD47, содержащий полипептид по п.1.
12. Димер по п.11, где димер представляет собой гомодимер.
13. Фармацевтическая композиция для лечения заболевания, связанного с активностью SIRPα или CD47, содержащая полипептид по любому из пп. 1-6 и фармацевтически приемлемый носитель.
14. Фармацевтическая композиция для лечения заболевания, связанного с активностью SIRPα или CD47, содержащая димер по п.11 или 12 и фармацевтически приемлемый носитель.
15. Способ лечения рака у субъекта, включающий введение субъекту эффективного количества фармацевтической композиции по п.13 или 14.
16. Способ по п.15, в котором рак представляет собой рак с солидной опухолью, гематологический рак, острый миелоидный лейкоз, хронический лимфоцитарный лейкоз, хронический миелоидный лейкоз, острый лимфобластный лейкоз, неходжкинскую лимфому, лимфому Ходжкина, множественную миелому, рак мочевого пузыря, рак поджелудочной железы, рак шейки матки, рак эндометрия, рак легкого, рак бронхов, рак печени, рак яичника, рак толстой кишки и прямой кишки, рак желудка, рак желудочно-кишечного тракта, рак желчного пузыря, желудочно-кишечную стромальную опухоль, рак щитовидной железы, рак головы и шеи, рак ротоглотки, рак пищевода, меланому, немеланомный рак кожи, карциному из клеток Меркеля, вирус-индуцированный рак, нейробластому, рак молочной железы, рак предстательной железы, рак почки, почечно-клеточный рак, рак почечной лоханки, лейкоз, лимфому, саркому, глиому, опухоль головного мозга и карциному.
17. Способ по п.15, дополнительно включающий введение терапевтического антитела.
18. Способ по п.17, в котором терапевтическое антитело связывается с 4-1BB, 5T4, ALK1, ANG-2, B7-H3, B7-H4, c-fms, c-Met, CA6, CCR4, CD123, CD19, CD20, CD22, CD27, EpCAM, CD30, CD32b, CD33, CD37, CD38, CD40, CD52, CD70, CD74, CD79b, CD98, CEA, CEACAM5, CLDN18.2, CLDN6, CS1, CTLA-4, CXCR4, DLL-4, EGFR, EGP-1, ENPP3, EphA3, ETBR, FGFR2, фибронектином, FR-альфа, Frizzled рецептором, GCC, GD2, глипиканом-3, GPNMB, HER2, HER3, HLA-DR, ICAM-1, IGF-1R, IL-3R, LAG-3, LIV-1, мезотелином, MUC16, MUC1, NaPi2b, нектином-4, Notch 2, Notch 1, OX-40, PD-1, PD-L1, PD-L2, PDGFR-α, PS, PSMA, SLTRK6, STEAP1, TEM1, VEGFR, CD25, DKK-1 или CSF-1R.
19. Способ по п.15, в котором терапевтическое антитело представляет собой цетуксимаб, нецитумумаб, пембролизумаб, ниволумаб, пидилизумаб, MEDI0680, атезолизумаб, авелумаб, дурвалумаб, MEDI6383, MEDI6469, RG7888, ипилимумаб, тремелимумаб, урелумаб, PF-05082566, эноблитузумаб, вантиктумаб, варлилумаб, могамулизумаб, SAR650984, даратумумаб, трастузумаб, трастузумаб эмтансин, пертузумаб, элотузумаб, ритуксимаб, офатумумаб, обинутузумаб, RG7155, FPA008, панитумумаб или брентуксимаб ведотин.
20. Полипептид для лечения заболевания, связанного с активностью SIRPα или CD47, включающий:
(a) вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий SEQ ID NO: 85; и
(b) Fc-область человеческого IgG1, содержащую мутации L234A, L235A, G237A и N297A, где нумерация соответствует индексу EU,
где С-конец варианта D1 SIRP-α связан с N-концом области Fc человеческого IgG1.
21. Нуклеиновая кислота, кодирующая полипептид, который содержит
(a) вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий SEQ ID NO: 85; и
(b) Fc-область человеческого IgG1, содержащую мутации L234A, L235A, G237A и N297A, где нумерация соответствует индексу EU,
где С-конец варианта D1 SIRP-α связан с N-концом области Fc человеческого IgG1.
22. Вектор, содержащий нуклеиновую кислоту по п.21.
23. Клетка-хозяин, содержащая нуклеиновую кислоту по п.21 или вектор по п.22.
24. Способ получения полипептида для лечения заболевания, связанного с активностью SIRPα или CD47, включающий культивирование клетки-хозяина по п.23 в условиях, подходящих для того, чтобы вызвать экспрессию полипептида, и выделение полипептида.
25. Фармацевтическая композиция для лечения заболевания, связанного с активностью SIRPα или CD47, содержащая:
полипептид, который содержит (а) вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий SEQ ID NO: 85, и (b) область Fc человеческого IgG1, содержащую мутации L234A, L235A, G237A и N297A, где нумерация соответствует индексу EU, где С-конец варианта D1 SIRP-α связан с N-концом области Fc человеческого IgG1; и
фармацевтически приемлемый эксципиент.
26. Фармацевтическая композиция для лечения заболевания, связанного с активностью SIRPα или CD47, содержащая
(а) димер, содержащий полипептид, который содержит
(i) вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий SEQ ID NO: 85; и
(ii) Fc-область человеческого IgG1, содержащую мутации L234A, L235A, G237A и N297A, где нумерация соответствует индексу EU,
где С-конец варианта D1 SIRP-α связан с N-концом области Fc человеческого IgG1, и
(b) фармацевтически приемлемый эксципиент.
27. Фармацевтическая композиция по п.26, где каждый мономер в димере содержит полипептид, который содержит
(i) вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий SEQ ID NO: 85; и
(ii) Fc-область человеческого IgG1, содержащую мутации L234A, L235A, G237A и N297A, где нумерация соответствует индексу EU,
где С-конец варианта D1 SIRP-α связан с N-концом области Fc человеческого IgG1.
28. Способ лечения индивидуума, страдающего раком, включающий введение индивидууму фармацевтической композиции, содержащей полипептид и фармацевтически приемлемый эксципиент, где полипептид содержит
(a) вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий SEQ ID NO: 85; и
(b) Fc-область человеческого IgG1, содержащую мутации L234A, L235A, G237A и N297A, где нумерация соответствует индексу EU,
где С-конец варианта D1 SIRP-α связан с N-концом области Fc человеческого IgG1.
29. Способ по п.28, дополнительно включающий введение индивидууму по меньшей мере одного дополнительного агента, где по меньшей мере один дополнительный агент представляет собой антитело, ассоциированный с опухолью антиген или терапевтическое средство, не являющееся антителом.
30. Способ лечения индивидуума, страдающего раком, включающий введение индивидууму фармацевтической композиции, содержащей димер и фармацевтически приемлемый эксципиент, где димер содержит полипептид, который содержит:
(a) вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий SEQ ID NO: 85; и
(b) Fc-область человеческого IgG1, содержащую мутации L234A, L235A, G237A и N297A, где нумерация соответствует индексу EU,
где С-конец варианта D1 SIRP-α связан с N-концом области Fc человеческого IgG1.
31. Способ по п.30, где каждый мономер в димере содержит полипептид, который содержит:
(a) вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий SEQ ID NO: 85; и
(b) Fc-область человеческого IgG1, содержащую мутации L234A, L235A, G237A и N297A, где нумерация соответствует индексу EU,
где С-конец варианта D1 SIRP-α связан с N-концом области Fc человеческого IgG1.
32. Способ по п.30, дополнительно включающий введение индивидууму по меньшей мере одного дополнительного агента, где по меньшей мере один дополнительный агент представляет собой антитело, ассоциированный с опухолью антиген или терапевтическое средство, не являющееся антителом.
33. Димер для лечения заболевания, связанного с активностью SIRPα или CD47, содержащий полипептид, который содержит аминокислотную последовательность, представленную в SEQ ID NO: 136.
34. Димер по п.33, где каждый мономер в димере содержит полипептид, который содержит аминокислотную последовательность, представленную в SEQ ID NO: 136.
35. Нуклеиновая кислота, кодирующая полипептид, который содержит аминокислотную последовательность, представленную в SEQ ID NO: 136.
36. Вектор, содержащий нуклеиновую кислоту по п.35.
37. Клетка-хозяин, содержащая нуклеиновую кислоту по п.35 или вектор по п.36.
38. Способ получения полипептида для лечения заболевания, связанного с активностью SIRPα или CD47, включающий культивирование клетки-хозяина по п.37 в условиях, подходящих для того, чтобы вызвать экспрессию полипептида, и выделение полипептида.
39. Фармацевтическая композиция для лечения заболевания, связанного с активностью SIRPα или CD47, содержащая полипептид, который содержит аминокислотную последовательность, представленную в SEQ ID NO: 136, и фармацевтически приемлемый эксципиент.
40. Фармацевтическая композиция для лечения заболевания, связанного с активностью SIRPα или CD47, содержащая (а) димер, который содержит полипептид, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 136, и (b) фармацевтически приемлемый эксципиент.
41. Фармацевтическая композиция по п.40, в которой каждый мономер в димере содержит полипептид, который содержит аминокислотную последовательность, представленную в SEQ ID NO: 136.
42. Способ лечения индивидуума, страдающего раком, включающий введение индивидууму фармацевтической композиции, содержащей полипептид, который содержит аминокислотную последовательность, представленную в SEQ ID NO: 136, и фармацевтически приемлемый эксципиент.
43. Способ по п.42, дополнительно включающий введение индивидууму по меньшей мере одного дополнительного агента, где по меньшей мере один дополнительный агент представляет собой антитело, ассоциированный с опухолью антиген или терапевтическое средство, не являющееся антителом.
44. Способ лечения индивидуума, страдающего раком, включающий введение индивидууму фармацевтической композиции, содержащей димер и фармацевтически приемлемый эксципиент, где димер содержит полипептид, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 136.
45. Способ по п.44, где каждый мономер в димере содержит полипептид, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 136.
46. Способ по п.44, дополнительно включающий введение индивидууму по меньшей мере одного дополнительного агента, где по меньшей мере один дополнительный агент представляет собой антитело, ассоциированный с опухолью антиген или терапевтическое средство, не являющееся антителом.
47. Полипептид для лечения заболевания, связанного с активностью SIRPα или CD47, включающий:
(a) вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий SEQ ID NO: 81; и
(b) Fc-область человеческого IgG1, содержащую мутации L234A, L235A, G237A и N297A, где нумерация соответствует индексу EU,
где С-конец варианта D1 SIRP-α связан с N-концом области Fc человеческого IgG1.
48. Нуклеиновая кислота, кодирующая полипептид, который содержит
(a) вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий SEQ ID NO: 81; и
(b) Fc-область человеческого IgG1, содержащую мутации L234A, L235A, G237A и N297A, где нумерация соответствует индексу EU,
где С-конец варианта D1 SIRP-α связан с N-концом области Fc человеческого IgG1.
49. Вектор, содержащий нуклеиновую кислоту по п.48.
50. Клетка-хозяин, содержащая нуклеиновую кислоту по п.48 или вектор по п.49.
51. Способ получения полипептида для лечения заболевания, связанного с активностью SIRPα или CD47, включающий культивирование клетки-хозяина по п.50 в условиях, подходящих для того, чтобы вызвать экспрессию полипептида, и выделение полипептида.
52. Фармацевтическая композиция для лечения заболевания, связанного с активностью SIRPα или CD47, содержащая:
полипептид, который содержит (а) вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий SEQ ID NO: 81; и (b) Fc-область человеческого IgG1, содержащую мутации L234A, L235A, G237A и N297A, где нумерация соответствует индексу EU, где С-конец варианта D1 SIRP-α связан с N-концом человеческая область Fc IgG1 и
фармацевтически приемлемый эксципиент.
53. Фармацевтическая композиция для лечения заболевания, связанного с активностью SIRPα или CD47, содержащая
(а) димер, содержащий полипептид, который содержит
(i) вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий SEQ ID NO: 81; и
(ii) Fc-область человеческого IgG1, содержащую мутации L234A, L235A, G237A и N297A, где нумерация соответствует индексу EU,
где С-конец варианта D1 SIRP-α связан с N-концом области Fc человеческого IgG1, и
(b) фармацевтически приемлемый эксципиент.
54. Фармацевтическая композиция по п.53, где каждый мономер в димере содержит полипептид, который содержит
(i) вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий SEQ ID NO: 81; и
(ii) Fc-область человеческого IgG1, содержащую мутации L234A, L235A, G237A и N297A, где нумерация соответствует индексу EU,
где С-конец варианта D1 SIRP-α связан с N-концом области Fc человеческого IgG1.
55. Способ лечения индивидуума, страдающего раком, включающий введение индивидууму фармацевтической композиции, содержащей полипептид и фармацевтический эксципиент, где полипептид содержит
(a) вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий SEQ ID NO: 81; и
(b) Fc-область человеческого IgG1, содержащую мутации L234A, L235A, G237A и N297A, где нумерация соответствует индексу EU,
где С-конец варианта D1 SIRP-α связан с N-концом области Fc человеческого IgG1.
56. Способ по п.55, дополнительно включающий введение индивидууму по меньшей мере одного дополнительного агента, где по меньшей мере один дополнительный агент представляет собой антитело, ассоциированный с опухолью антиген или терапевтическое средство, не являющееся антителом.
57. Способ лечения индивидуума, страдающего раком, включающий введение индивидууму фармацевтической композиции, содержащей димер и фармацевтически приемлемый эксципиент, где димер содержит полипептид, который содержит
(a) вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий SEQ ID NO: 81; и
(b) Fc-область человеческого IgG1, содержащую мутации L234A, L235A, G237A и N297A, где нумерация соответствует индексу EU,
где С-конец варианта D1 SIRP-α связан с N-концом области Fc человеческого IgG1.
58. Способ по п.57, где каждый мономер в димере содержит полипептид, содержащий:
(a) вариант D1 сигнального регуляторного белка α (SIRP-α), содержащий SEQ ID NO: 81; и
(b) Fc-область человеческого IgG1, содержащую мутации L234A, L235A, G237A и N297A, где нумерация соответствует индексу EU, где С-конец варианта D1 SIRP-α связан с N-концом человека IgG1 Fc регион.
59. Способ по п.57, дополнительно включающий введение индивидууму по меньшей мере одного дополнительного агента, где по меньшей мере один дополнительный агент представляет собой антитело, ассоциированный с опухолью антиген или терапевтическое средство, не являющееся антителом.
60. Димер для лечения заболевания, связанного с активностью SIRPα или CD47, содержащий полипептид, который содержит аминокислотную последовательность, представленную в SEQ ID NO: 135.
61. Димер по п.60, где каждый мономер в димере содержит полипептид, который содержит аминокислотную последовательность, представленную в SEQ ID NO: 135.
62. Нуклеиновая кислота, кодирующая полипептид, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 135.
63. Вектор, содержащий нуклеиновую кислоту по п.62.
64. Клетка-хозяин, содержащая нуклеиновую кислоту по п.62 или вектор по п.63.
65. Способ получения полипептида для лечения заболевания, связанного с активностью SIRPα или CD47, включающий культивирование клетки-хозяина по п.64 в условиях, подходящих для того, чтобы вызвать экспрессию полипептида, и выделение полипептида.
66. Фармацевтическая композиция для лечения заболевания, связанного с активностью SIRPα или CD47, содержащая полипептид, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 135, и фармацевтически приемлемый эксципиент.
67. Фармацевтическая композиция для лечения заболевания, связанного с активностью SIRPα или CD47, содержащая (а) димер, который содержит полипептид, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 135, и (b) фармацевтически приемлемый эксципиент.
68. Фармацевтическая композиция по п.67, где каждый мономер в димере содержит полипептид, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 135.
69. Способ лечения индивидуума, страдающего раком, включающий введение индивидууму фармацевтической композиции, содержащей полипептид, который содержит аминокислотную последовательность SEQ ID NO: 135, и фармацевтически приемлемый эксципиент.
70. Способ по п.69, дополнительно включающий введение индивидууму по меньшей мере одного дополнительного агента, где по меньшей мере один дополнительный агент представляет собой антитело, ассоциированный с опухолью антиген или терапевтическое средство, не являющееся антителом.
71. Способ лечения индивидуума, страдающего раком, включающий введение индивидууму фармацевтической композиции, содержащей димер и фармацевтически приемлемый эксципиент, где димер содержит полипептид, содержащий аминокислотную последовательность SEQ ID NO: 135.
72. Способ по п.71, в котором каждый мономер в димере содержит аминокислотную последовательность, представленную в SEQ ID NO: 135.
73. Способ по п.71, дополнительно включающий введение индивидууму по меньшей мере одного дополнительного агента, где по меньшей мере один дополнительный агент представляет собой антитело, ассоциированный с опухолью антиген или терапевтическое средство, не являющееся антителом.
| LEE W.Y | |||
| et al | |||
| Способ очищения сернокислого глинозема от железа | 1920 |
|
SU47A1 |
| J Immunol | |||
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| WO 2013109752 A1, 25.07.2013 | |||
| WO 2010070047 A1, 24.06.2010 | |||
| WO 2012142515 A2, 18.10.2012 | |||
| BORROK M.J | |||
| et al | |||
| Revisiting the Role of Glycosylation in the Structure of Human IgG Fc | |||
| ACS Chem Biol | |||
| Изложница с суживающимся книзу сечением и с вертикально перемещающимся днищем | 1924 |
|
SU2012A1 |

