Изобретение относится к биотехнологии, в частности к генетической инженерии, и позволяет получить линию клеток HT1080 с искусственной хромосомой человека в геноме, которая включает два гена GFP с последовательностями CDT1 и GEMININ, позволяющею использовать для поиска ранее неизвестных генов, участвующих в процессах репликации и сегрегации хромосом человека, а также поиска лекарственных кандидатов, вызывающих хромосомную нестабильность.
Расхождение хромосом во время клеточного деления обеспечивается огромным количеством регуляторных элементов. Нарушение работы этих элементов приводит к хромосомной нестабильности и образованию клеток с набором хромосом не кратным гаплоидному (анеуплоидных клеток). Часто нарушение в работе клеточного деления и возникновение хромосомной нестабильности связано с мутациями в генах, регулирующих данные процессы. На сегодняшний день доказано участие приблизительно 400 генов в процессе трансмиссии хромосом человека, тогда как при исследовании дрожжей S. сerevisiae и S. pombe было обнаружено 692 гена, участвующих в данных процессах (Stirling, P. C., Bloom, M. S., Solanki-Patil, T., Smith, S., Sipahimalani, P., Li, Z., Hieter, P. (2011), The complete spectrum of yeast chromosome instability genes identifies candidate CIN cancer genes and functional roles for ASTRA complex components. PLoS Genetics, 7(4), e1002057. https://doi.org/10.1371/journal.pgen.1002057; Stirling, P. C., Crisp, M. J., Basrai, M. A., Tucker, C. M., Dunham, M. J., Spencer, F. A., & Hieter, P. 2012; Mutability and mutational spectrum of chromosome transmission fidelity genes. Chromosoma, 121(3), 263–275. https://doi.org/10.1007/s00412-011-0356-3).
Принимая во внимание, что митоз — эволюционно древний и консервативный процесс, можно предположить, что в геноме человека есть неописанные гены – регуляторы процессов сегрегации и репликации хромосом, родственные (ортологичные) генам дрожжей.
С другой стороны, явление хромосомной нестабильности характерно для злокачественных новообразований и может достигать порогового уровня, за которым прекращается деление клеток и рост опухоли (Janssen, A., Kops, G. J. P. L., & Medema, R. H. 2009, Elevating the frequency of chromosome mis-segregation as a strategy to kill tumor cells. Proceedings of the National Academy of Sciences of the United States of America, 106(45), 19108–19113 https://doi.org/10.1073/pnas.0904343106).
Эта особенность раковых клеток используется для лечения некоторых форм онкологических заболеваний. В основе данного лечения лежит медикаментозное увеличение хромосомной нестабильности. Мишенями для подобного рода противоопухолевых терапий служат продукты экспрессии генов, вовлеченные в процессы стабилизации расхождения хромосом.
Известна тест-система для определения уровня хромосомной нестабильности, которая представляет собой линию клеток человеческой фибросаркомы HT1080 с белок-кодирующими фрагментами GFP (Green Fluorescence Protein) в нативных хромосомах и дополнительно содержащую искусственную хромосому человека (Human Artificial Chromosome, HAC) с рекомбинантной плазмидной ДНК р264, которая включает белок-кодирующий фрагмент гена mCherry. (Kim, J.-H., Lee, H.-S., Lee, N. C. O., Goncharov, N. V., Kumeiko, V., Masumoto, H., … Larionov, V. (2016). Development of a novel HAC-based “gain of signal” quantitative assay for measuring chromosome instability (CIN) in cancer cells. Oncotarget, 7(12), 14841–14856. https://doi.org/10.18632/oncotarget.7854).
Принцип действия данной тест-системы основан на двухцветной флуоресцентной индикации, обусловленной регуляцией экспрессией mCherry (красное свечение) и GFP (зеленое свечение). Линия клеток человеческой фибросаркомы HT1080 в составе искусственной хромосомы человека содержит ген mCherry и shRNA (short hairpin RNA), подавляющую экспрессию GFP в нативных хромосомах. При возникновении хромосомной нестабильности клетки теряют искусственную хромосому человека, из-за чего прекращается подавление экспрессии GFP, и происходит изменение флуоресцентного свечения с красного на зеленый. По соотношению клеток, имеющих красное или зеленое свечение, можно рассчитать уровень хромосомной нестабильности.
Однако каждый из флуоресцентных белков (mCherry и GFP) отличается долгой естественной деградацией.
Таким образом, к недостаткам данного технического решения следует отнести низкий уровень точности и длительности определения хромосомной нестабильности. Это обусловлено длительной естественной деградацией флуоресцентных белков mCherry и GFP. Поэтому известное решение позволяет получить данные только через 14 дней эксперимента, что существенно снижает точность измерения хромосомной нестабильности. Потому что клетки в течение 14 дней проходят до 20 делений, а эффект от хромосомной нестабильности на 14 день отражает события, произошедшие в первом делении. Известное решение обладает низкой производительностью, так как для 20 клеточных делений необходимо использовать культуральных посуду большой площади, что невозможно реализовать в принятом для высокопроизводительного скрининга формате 96 или 384 луночных планшетов.
В качестве прототипа принята тест-система для определения уровня хромосомной нестабильности на основе линии клеток человеческой фибросаркомы HT1080, содержащей в геноме искусственную хромосому человека, которая включает фрагмент гена GFP (Lee, H. S., Lee, N. C. O., Grimes, B. R., Samoshkin, A., Kononenko, A. V., Bansal, R., … Larionov, V. (2013). A new assay for measuring chromosome instability (CIN) and identification of drugs that elevate CIN in cancer cells. BMC Cancer, 13, 1–12. https://doi.org/10.1186/1471-2407-13-252).
Принцип действия прототипа основан на флуоресцентной индикации, обусловленной экспрессией белка GFP. При возникновении хромосомной нестабильности клетки теряют искусственную хромосому человека и, следовательно, происходит потеря зеленой флуоресценции. По соотношению светящийся зеленым клеток и клеток, потерявших зеленое свечение, можно определить уровень хромосомной нестабильности.
К существенным недостаткам прототипа следует отнести:
- длительность определения хромосомной нестабильности за счет продолжительной естественной деградации флуоресцентных белков;
- низкую точность определения хромосомной нестабильности, обусловленную не способностью прототипа детектировать хромосомную нестабильность в момент ее возникновения.
Технической проблемой, поставленной перед данным изобретением, является повышение скорости и точности определения хромосомной нестабильности, за счет создания новой тест-системы для определения уровня хромосомной нестабильности.
Заявленная техническая проблема решается тем, что в известной тест-системе для определения уровня хромосомной нестабильности на основе линии клеток человеческой фибросаркомы HT1080, содержащей искусственную хромосому c геном GFP, согласно изобретению искусственная хромосома включает два гена GFP с последовательностями CDT1 и GEMININ, функционирующими в качестве индикаторов клеточного цикла.
Включение в искусственную хромосому, кодирующих последовательностей рекомбинантных флуоресцентных белков – индикаторов клеточного цикла – GFP-CDT1 и GFP-GEMINI, белковые продукты экспрессии которых представляют собой рекомбинантные варианты зеленого флуоресцентного белка, GFP-CDT1 и GFP-GEMENIN, функционирующие ограниченное время - в течении определенных периодов клеточного цикла. Это позволяет сократить время детекции хромосомной нестабильности и увеличить точность ее определение.
Наличие GFP-CDT1 и GFP-GEMENIN обеспечивает флуоресцентную индикацию всего клеточного цикла, при этом GFP-CDT1 функционирует только в G1 периоде клеточного цикла, а GFP-GEMENIN - в течение G2 и S периода. При возникновении хромосомной нестабильности происходит потеря искусственной хромосомы, что приводит к мгновенной потере флуоресцентного сигнала клетками HT1080. Вследствие чего, значительно сокращается длительность определения хромосомной нестабильности и увеличивается точность ее определения.
Использование искусственной хромосомы, содержащей кодирующие последовательности рекомбинантных флуоресцентных белков и индикаторов клеточного цикла CDT1 и GEMININ, позволяет сократить время получения данных о возникновении хромосомной нестабильности в 4,7 раз и получить квантованный сигнал сразу после потери искусственной хромосомы за счет короткого времени деградации рекомбинантных вариантов зеленого флуоресцентного белка, GFP-CDT1 и GFP-GEMENIN.
Изобретение иллюстрируется следующими чертежами:
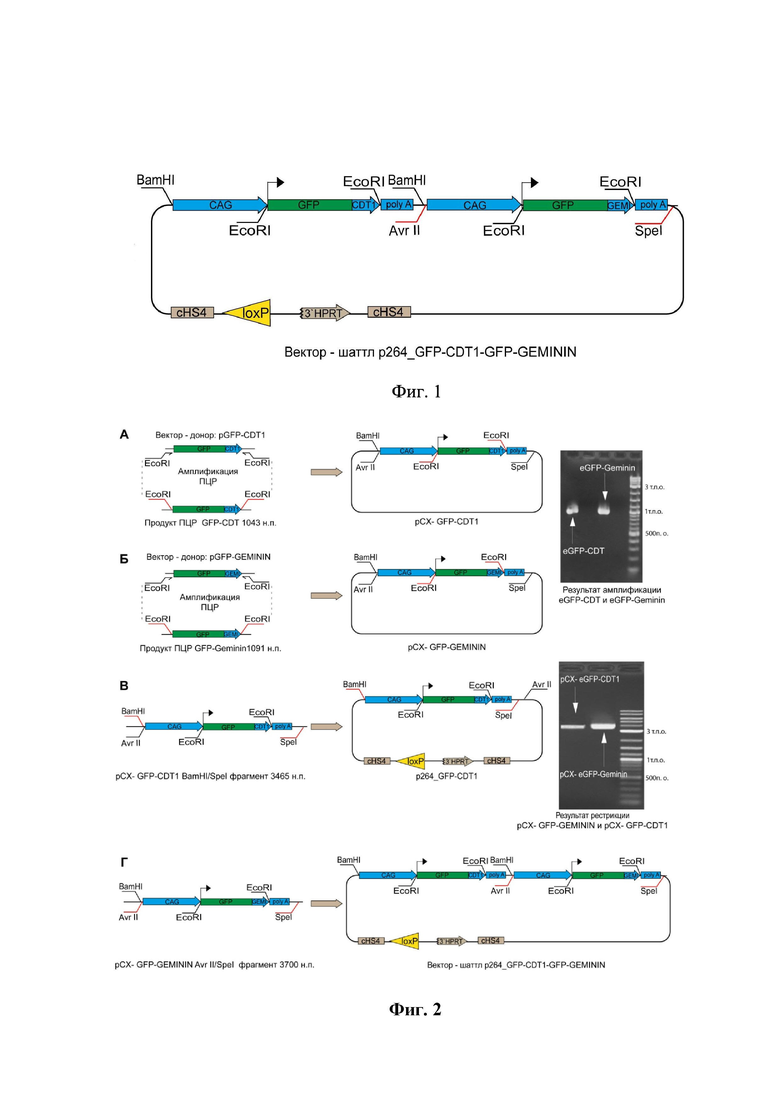
на фиг.1 приведена карта-схема плазмидной рекомбинантной ДНК p264-GFP-CDT1-GFP-GEMININ, и ее область, ответственная за экспрессию рекомбинантных флуоресцентных белков и индикаторов клеточного цикла CDT1 и GEMININ;
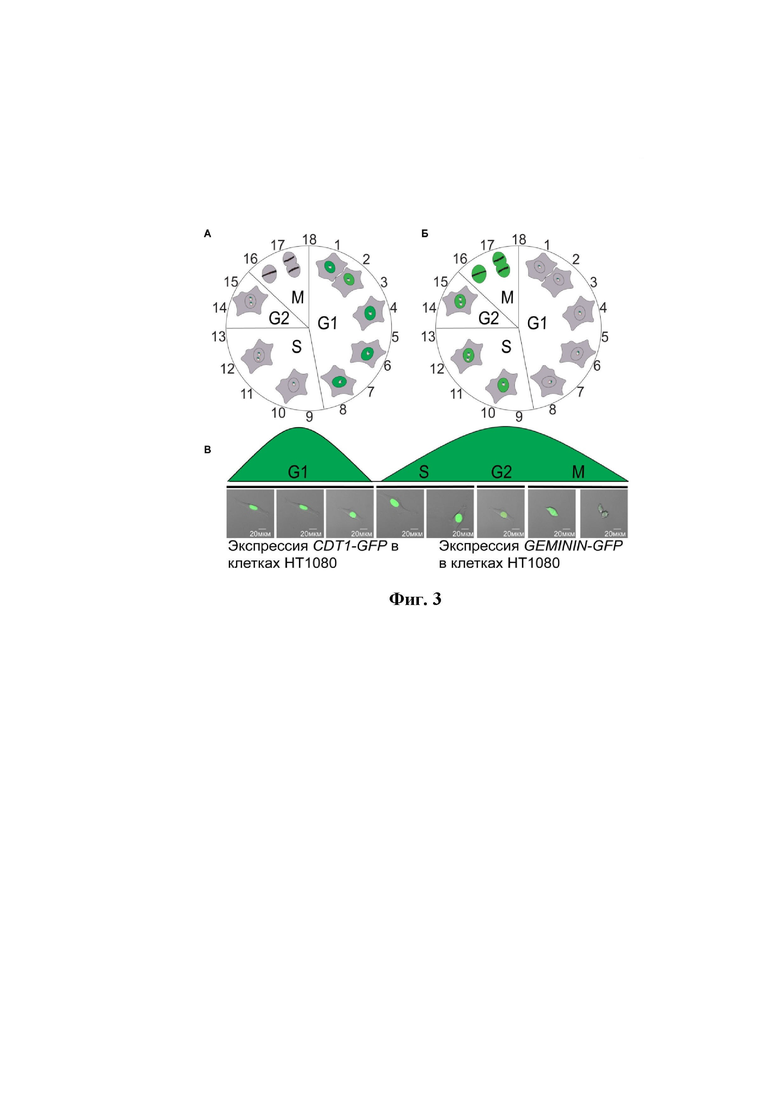
на фиг.2 показана сборка вектора p264, отличающегося наличием кодирующих последовательностей рекомбинантных флуоресцентных белков и индикаторов клеточного цикла CDT1 и GEMININ, где:
А – схема амплификации фрагмента, содержащего индикатор клеточного цикла GFP–CDT1 и его лигирования в плазмидный вектор pCX;
Б – схема амплификации фрагмента, содержащего индикатор клеточного цикла GFP–GEMININ, и его лигирования в плазмидный вектор pCX;
В – схема рестрикции и лигирования вектора pCX–GFP–CDT1 и вектора p264;
Г – схема лигирования вектора pCX–GFP–CDT1 и pCX–GFP–GEMININ;
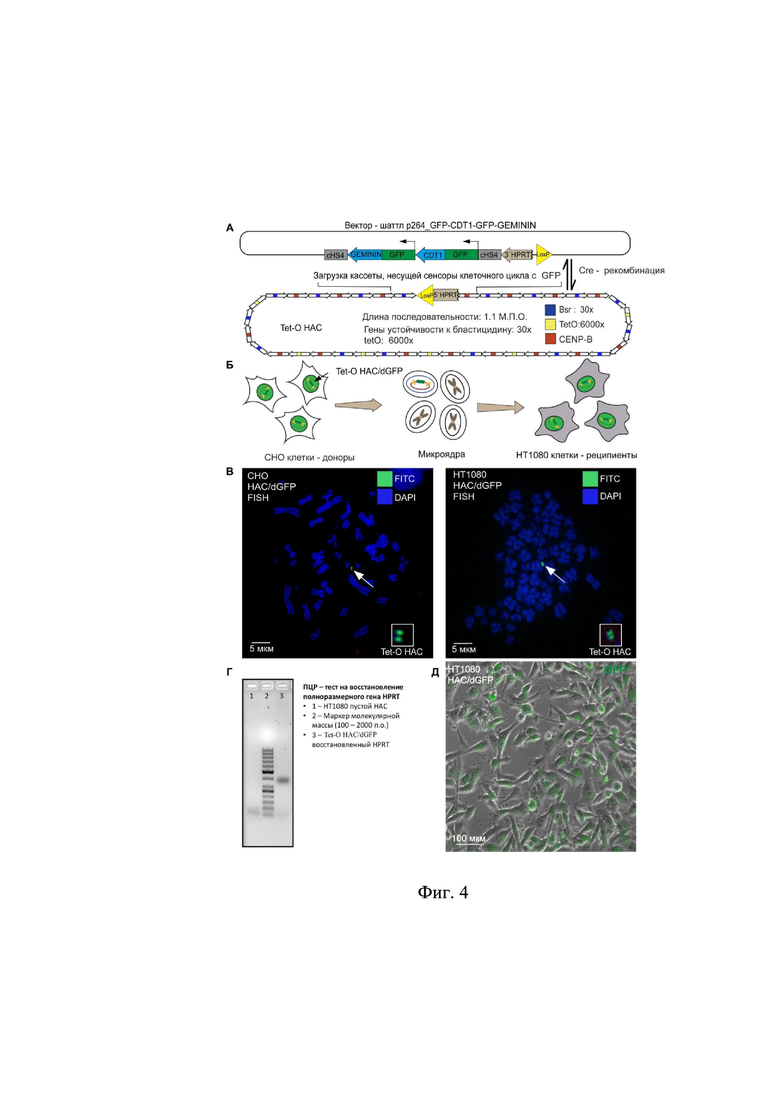
на фиг.3 представлена экспрессия индикаторов клеточного цикла в линии клеток человеческой фибросаркомы HT1080, где:
А – экспрессия вектора pCX–GFP–CDT1 в линии клеток человеческой фибросаркомы HT1080 в течение клеточного цикла;
Б – экспрессия pCX–GFP–GEMININ в линии клеток человеческой фибросаркомы HT1080 в течение клеточного цикла;
В – экспрессия шаттл-вектора p264-GFP–CDT1-GFP–GEMININ в линии клеток человеческой фибросаркомы HT1080 в течение клеточного цикла;
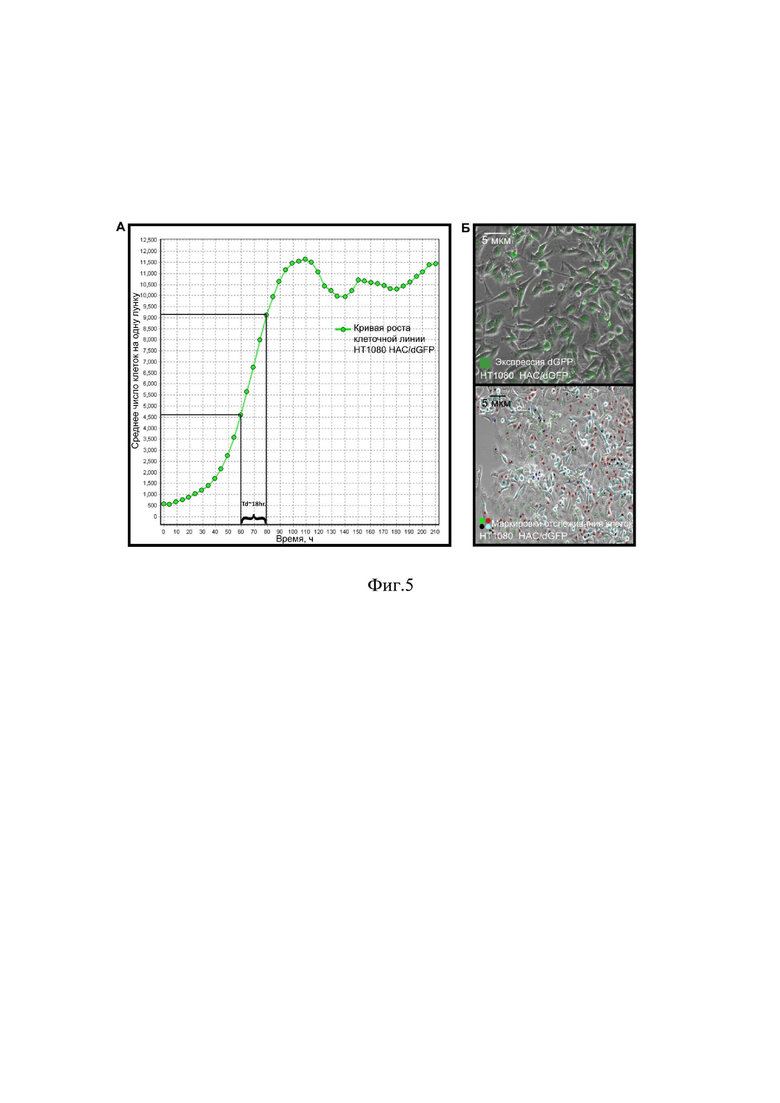
на фиг.4 представлена схема получения линии клеток человеческой фибросаркомы HT1080 с искусственной хромосомой человека с кодирующими последовательностями рекомбинантных флуоресцентных белков и индикаторов клеточного цикла CDT1 и GEMININ, где:
А – Cre опосредованная рекомбинация через сайт loxP, для помещения шаттл–вектора p264-GFP–CDT1-GFP–GEMININ в состав искусственной хромосомы в клетках яичника китайского хомячка CHO; карты искусственной хромосомы человека (Alphoid tetO – HAC) и вектора p264–GFP–CDT1-GFP–GEMININ;
Б – схема протокола переноса искусственной хромосомы из линии клеток яичника китайского хомячка CHO в линию клеток человеческой фибросаркомы HT1080;
В – флуоресцентная гибридизация in situ искусственной хромосомы в линии клеток человеческой фибросаркомы HT1080 и линии клеток яичника китайского хомячка CHO,
Г – ПЦР восстановленного в результате Cre-loxP рекомбинации полноразмерного гена,
Д - снимок популяции клеток HT1080-HAC/dGFP после проведения MMCT (microcell-mediated chromosome transfer).
на фиг.5 представлено определение продолжительности клеточного цикла для линии клеток человеческой фибросаркомы HT1080, содержащей искусственную хромосому человека, включающую два гена GFP с последовательностями CDT1 и GEMININ (HT1080-HAC/dGFP, методом высокоэффективной автоматизированной микроскопии на платформе Cell iQ, где:
А – продолжительность клеточного цикла (время удвоения клеточной популяции, перевод с англ. doubling time, Td), рассчитанное в логарифмической фазе кривой роста клеток HT1080-HAC/dGFP; один клеточный цикл занимает приблизительно 18 ч;
Б – изображения клеток HT1080-HAC/dGFP, экспрессирующих трансген dGFP (вверху) и клетки HT1080-HAC/dGFP с маркерами автоматической идентификации клеток в системе Cell iQ (внизу): распластанные клетки (красная маркировка), делящиеся клетки (голубая маркировка), мертвые клетки (черная маркировка), клеточный дебрис (зеленая маркировка);
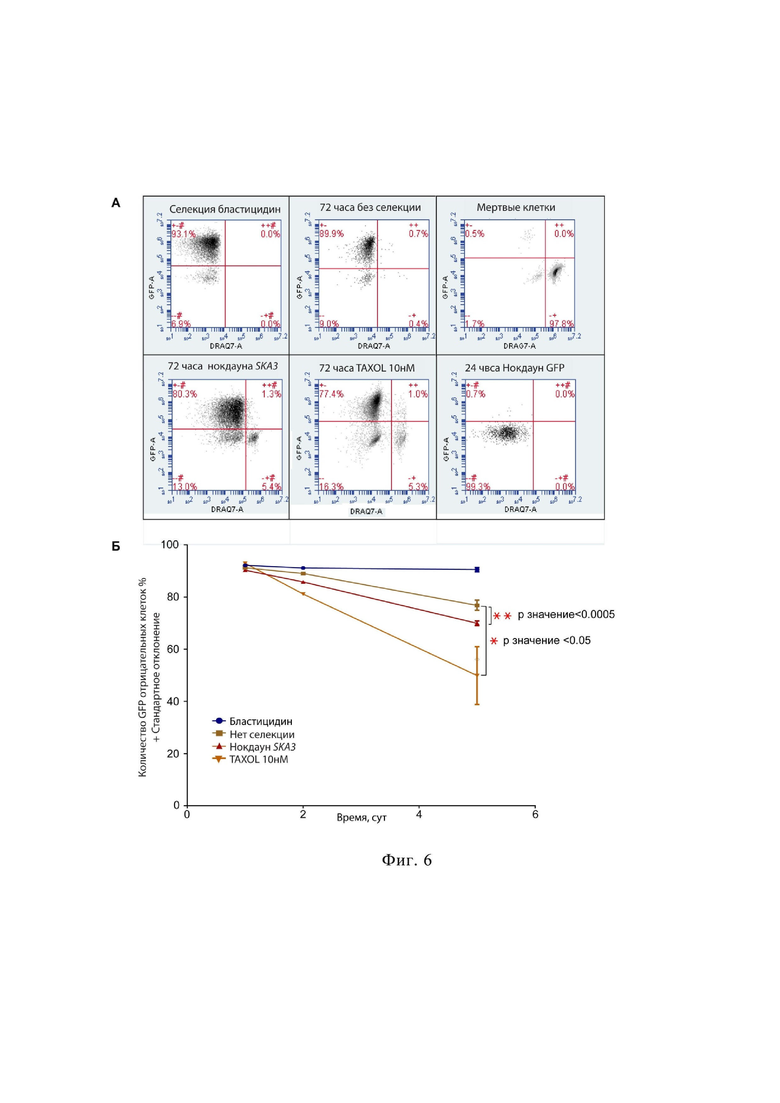
на фиг.6 приведена оценка эффективности работы тест-системы HT1080-HAC/dGFP методом проточной цитофлуориметрии, где
А – результаты проточной цитофлуориметрии клеток линии HT1080 HAC/dGFP через 72 ч после нокдауна SKA3, GFP и обработки таксолом;
Б – динамика потери сигнала GFP клетками линии HT1080 HAC/dGFP после нокдауна SKA3 и обработкой таксолом;
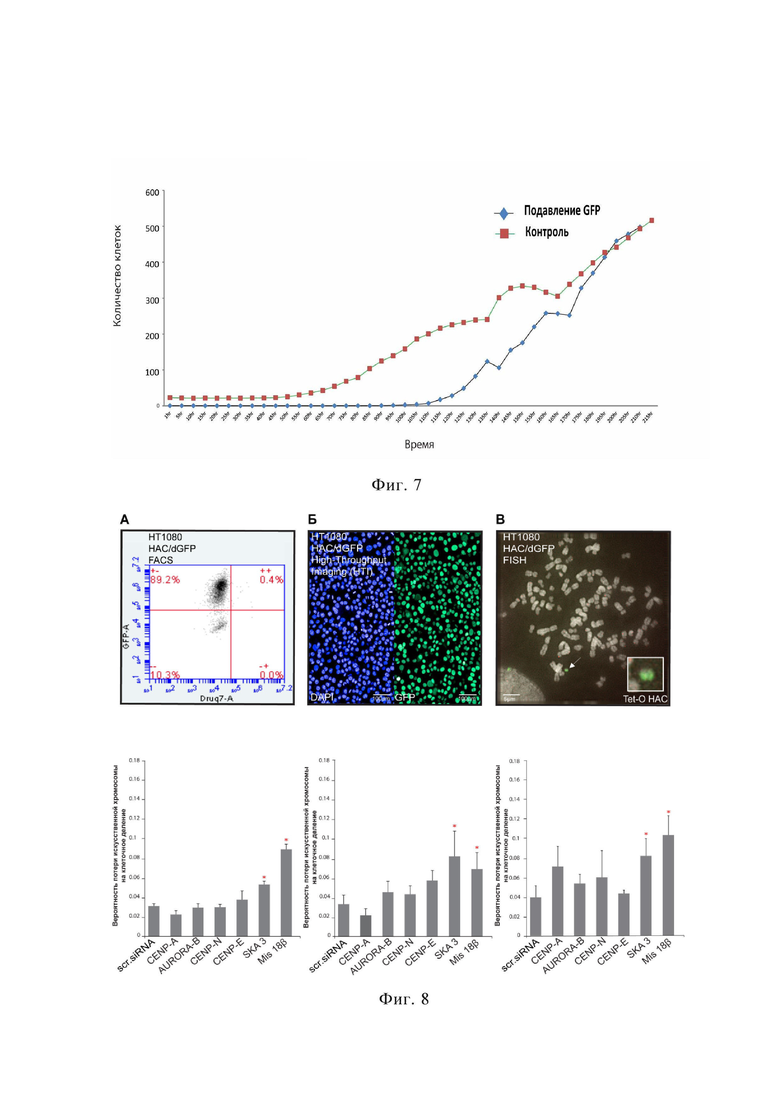
на фиг.7 приведено время восстановления флуоресцентного сигнала после миРНК-опосредованного ингибирования экспрессии GFP в клетках линии HT1080 HAC/dGFP;
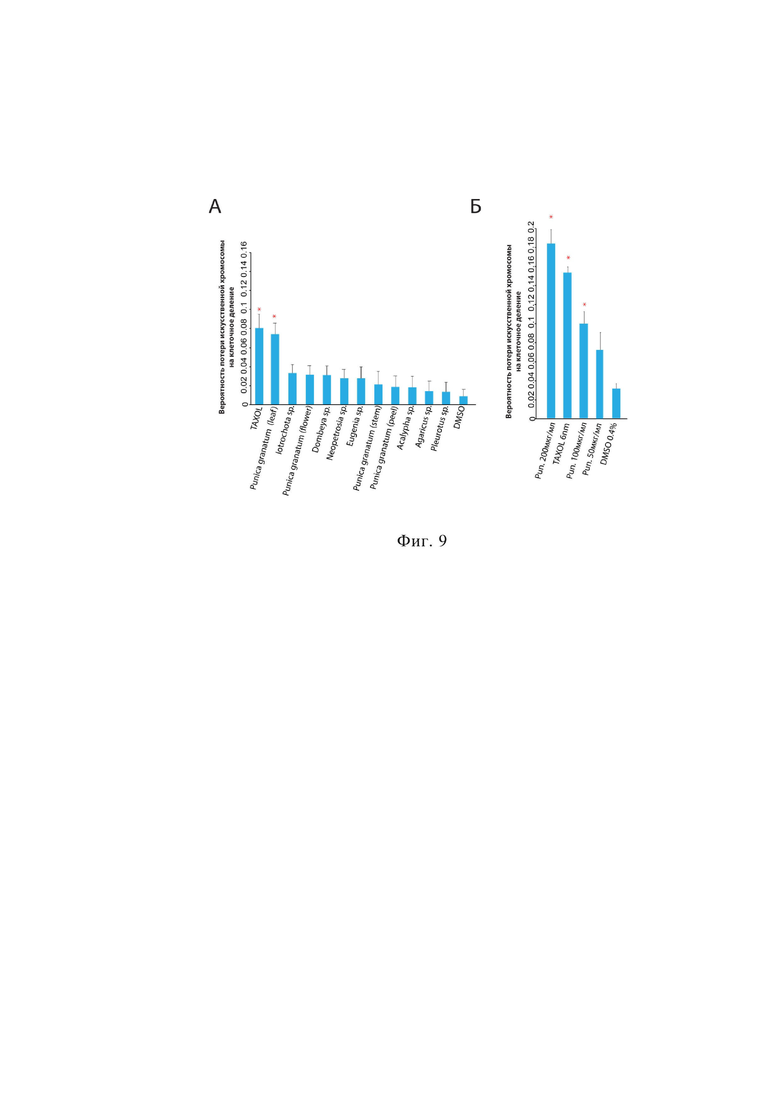
на фиг.8 продемонстрирована вероятность потери искусственной хромосомы клетками линии HT1080 HAC/dGFP после нокдауна выбранных генов, где:
A – вероятность потери искусственной хромосомы в клетках линии HT1080 HAC/dGFP спустя 72часа после нокаута контрольных генов методом проточной цитофлуориметрии;
Б - вероятность потери искусственной хромосомы в клетках HT1080 HAC/dGFP спустя 72часа после нокаута контрольных генов методом конфокальной микроскопии на микроскопе Yokogawa CV7000 Imaging;
В - вероятность потери искусственной хромосомы человека в клетках HT1080 HAC/dGFP спустя 72часа после нокаута контрольных генов методом FISH – гибридизации.
Звездочка указывает на статистическую значимость (р <0,005) по сравнению с отрицательным контролем;
на фиг. 9 представлена вероятность потери искусственной хромосомы в клетках линии HT1080 HAC/dGFP спустя 72часа после обработки клеток различными природными экстрактами, потенциально влияющими на развитие хромосомной нестабильности, где:
A - вероятность потери искусственной хромосомы клетками линии HT1080 HAC/dGFP спустя 72 часа под действием различных природных экстрактов;
Б - вероятность потери искусственной хромосомы клетками линии HT1080 HAC/dGFP спустя 72 часа под действием различных концентраций экстракта Punica granatum leaves.
Звездочка указывает на статистическую значимость (р <0,005) по сравнению с отрицательным контролем;
В качестве тест-системы для определения уровня хромосомной нестабильности создавали линию клеток человеческой фибросаркомы HT1080, содержащую в геноме искусственную хромосому человека с рекомбинантной плазмидной ДНК на основе вектора p264 (фиг.1.). В состав вектора помещали кодирующие последовательности индикаторов клеточного цикла GFP-CDT1 и GFP-GEMININ, белковые продукты экспрессии которых позволяют получить рекомбинантные белки GFP-CDT1 и GFP-GEMENIN.
Экспрессия генов GFP–CDT1 и GFP–GEMININ в клетках человеческой фибросаркомы HT1080 продемонстрирована на фиг.3. Как видно из результатов, флуоресценция GFP имеет ядерную локализацию, а период флуоресценции соответствует изменению фаз клеточного цикла: G1 для GFP–CDT1 (фиг. 3Б) и G2, S и М для GFP–GEMININ (фиг. 3В). Продолжительность флуоресценции составила 8–9 ч для каждого из рекомбинантных белков. При этом в клетках экспрессирующих оба рекомбинантных белка происходит визуализация всего клеточного цикла (фиг. 3А). Данный результат демонстрирует корректность сборки рекомбинантных конструкций.
Наконец, кодирующие последовательности данных рекомбинантных белков помещали в состав искусственной хромосомы. На фиг. 4А представлена схема LoxP Cre рекомбинации шаттл-вектора и пустой искусственной хромосомы в линии клеток яичника китайского хомячка CHO. Как видно на фиг. 4А, последовательность искусственной хромосомы человека имеет половину гена HPRT (hypoxanthine–guanine phosphoribosyl transferase), полноразмерная последовательность которого, экспрессируясь, обеспечивает резистентность при культивировании линии клеток в селективной среде с содержанием гипоксантин – 5 мМ, аминоптерин – 20 мМ, тимидин – 0,8 мМ (hypoxanthine–aminopterin–thymidine, HAT). После рекомбинации LoxP (фиг.4А), происходило восстановление последовательности HPRT и шаттл–вектор был интегрирован в состав искусственной хромосомы человека (фиг. 4Г). Восстановление HPRT позволило провести селекцию клонов, содержащих последовательности интереса в составе искусственной хромосомы человека и получить необходимое количество клеток CHO для проведения завершающего этапа сборки системы – переноса полученной хромосомы человека в клеточную линию HT1080 с помощью протокола микроклеточно-опосредованного переноса хромосом (фиг 4Д) (MMCT).
На фиг. 4Б показана схема переноса искусственной хромосомы в клетки-реципиенты HT1080. Для подтверждения корректности переноса и наличия искусственной хромосомы в клетках донорах и реципиентах, CHO и HT1080 соответственно, проводили гибридизацию in situ препаратов метафазных пластинок (фиг. 4В).
В качестве маркера использовали зонд PNA, меченый FITC, на последовательность tetO в составе искусственной хромосомы, данная последовательность уникальна по отношению к остальным последовательностям нативных хромосом линии клеток человеческой фибросаркомы HT1080. На фиг. 4В слева видно, что клетки-реципиенты имеют четкий сигнал зеленой флуоресценции (FITC) исходящий от комплекса tetO в последовательности искусственной хромосомы и флуоресцентной метки PNA–FITC. После осуществления переноса из линии клеток CHO в линию клеток HT1080, в них также наблюдался сигнал от последовательности tetO, которая является свидетельством наличия искусственной хромосомы в геноме линии клеток человеческой фибросаркомы HT1080 (фиг. 4В). После проведения MMCT клетки линии HT1080 подвергали селекции в присутствии селективного антибиотика бластицидина. Гены устойчивости к бластицидину были локализованы в искусственной хромосоме человека (фиг 4А) и позволяли провести селекцию клеток с искусственной хромосомой человека. После селекции выбирали клон линии клеток HT1080-HAC-dGFP с наиболее стабильной экспрессией GFP и использовали его для дальнейших экспериментов в качестве тест-системы для определения уровня хромосомной нестабильности.
Линия клеток (созданная тест-система) HT1080-HAC/dGFP характеризуется следующими признаками:
1) культурально-морфологические признаки:
Клетки представляют собой адгезивную линию, имеют длинные отростки, составляющие 2/3 от диаметра ядра, скорость деления клеток составляет 18 часов. Данная линия клеток активно пролиферирует на таких средах, как Среда Игла модифицированной Дульбекко (DMEM), c 10% содержанием бычьей сыворотки Fetal Bovine Serum (FBS) и F12 в присутствии 10% FBS, 1% P/S. Для культивирования и поддержания стабильности искусственной хромосомы необходимо присутствие 10мкг/мл бластицидина. Наиболее оптимальной для роста клеток является температура 37ºС, 5%СО2 и 100% влажность. Для культивирования необходимо использовать культуральную посуду с поверхностью Nunclon ρ, покрытую поли-D-лизином или коллагеном I.
2) физико-биологические признаки:
Клетки содержат в геноме искусственную хромосому, содержащую кодирующие последовательности рекомбинантных флуоресцентных белков и индикаторов клеточного цикла, - GFP–CDT1 и GFP–GEMININ. Благодаря наличию данных последовательностей клетки имеют флуоресцентное свечение от зеленого флуоресцентного белка (GFP), локализованного в ядре и обладают резистентностью к бластицидину. Линия клеток не патогенна и нетоксична. Для хранения используют раствор: 10% культуральной среды DMEM; 80% FBS; 10% DMSO и хранят при температуре -196°С.
Полученную тест-систему HT1080-HAC/dGFP использовали в дальнейших экспериментах.
В экспериментах по определению уровня хромосомной нестабильности, клетки линии HT1080-HAC/dGFP, подвергшиеся обработке факторами, вызывающими хромосомную нестабильность, проходят несколько делений. Определение вероятности потери искусственной хромосомы человека на одно клеточное деление (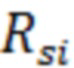 ) вычисляли по формуле:
) вычисляли по формуле:
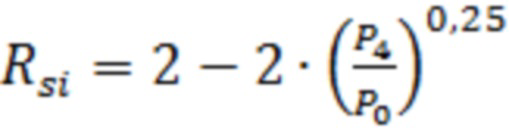 , где
, где
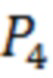 - количество GFP-положительных (флуоресцирующих зеленым цветом) клеток;
- количество GFP-положительных (флуоресцирующих зеленым цветом) клеток;
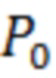 - количество GFP-отрицательных (без зеленой флуоресценции) клеток.
- количество GFP-отрицательных (без зеленой флуоресценции) клеток.
По фактическому значению вероятности потери искусственной хромосомы на одно клеточное деление определяют количественный уровень хромосомной нестабильности.
Для определения количества делений, которые клетки линии HT1080-HAC/dGFP проходят в течение эксперимента проводили анализ скорости клеточного цикла (фиг 5). Кривую роста строили посредством прижизненного мониторинга GFP-позитивных клеток методом высокоэффективной автоматизированной микроскопии Cell IQ (фиг 5А).
Используя данные анализа продолжительности клеточного цикла линии клеток HT1080-HAC/dGFP, разрабатывали модель вычисления вероятности потери искусственной хромосомы под действием факторов, вызывающих хромосомную нестабильность.
Экспрессия GFP с индикаторами клеточного цикла CDT1 и GEMININ позволяет детектировать потерю искусственной хромосомы непосредственно после развития хромосомной нестабильности. При действии на клеточную популяцию химических веществ, вызывающих хромосомную нестабильность, или подавления экспрессии важных для процесса сегрегации хромосом генов, происходит нарушение хромосомной трансмиссии и потеря флуоресценции утратившими искусственную хромосому человека клетками. По соотношению GFP позитивных и GFP негативных клеток в популяции, можно определять уровень хромосомной нестабильности.
Изобретение осуществляется следующим образом.
Для сборки заявленной тест-системы была сконструированна рекомбинантная плазмидная ДНК, для этого при помощи метода ПЦР амплифицируют последовательность индикаторов клеточного цикла CDT1 и GEMENIN вместе с белок-кодирующими районами зеленых флуоресцентных белков (GFPs). При проведении ПЦР используют праймеры, имеющие в своем составе последовательности сайтов рестрикции EcoRI, в частности:
- праймер 1: EcoRI-GFP-FWD 5'-AGAATTCGCCACCATGGTGAGCA-3′
- праймер 2: EcoRI-GFP-hCDT1-REV 5'-TGAATTCTTAGATGGTGTCCTGGTCCT-3′
- праймер 3: EcoRI-GFP-hGEMININ-REV 5'-TGAATTCTTACAGCGCCTTTCTCCG-3′
В качестве матрицы для ПЦР используют векторы pCX–GFP–CDT1 и pCX–GFP–GEMININ. Схема амплификации и электрофорез продуктов ПЦР представлены на фиг. 2 (A и Б), длины ампликонов составили 1043 и 1091 н.п. для CDT1 и GEMININ, соответственно. Далее последовательности помещают под контроль плазмидного вектора pCX, который служит источником промоторной области. Для этого продукты амплификации рестрицируют по сайтам EcoRI и лигируют в вектор pCX, также имеющий липкие концы после рестрикции по EcoRI (фиг. 2A). На первом этапе сборки получают 2 вектора: pCX–GFP–CDT1 длиной 5813 н.п. и вектор pCX–GFP–GEMININ размером 5861 н.п. Каждая пара индикаторов клеточного цикла и зеленых флуоресцентных белков находится в отдельной рамке считывания под контролем промоторов CV40.
На втором этапе полученные векторы линеаризируют посредством рестрикции по сайтам BamHI/SpeI для вектора pCX–CDT1-GFP и AvaI/SpeI для вектора pCX–GFP–GEMININ. На электрофореграмме (фиг. 2Б) видно, что длина продуктов рестрикции соответствует расчетной и составляет 5813 н.п. и 5861 н.п. для pCX–GFP–CDT1 и pCX–GFP–GEMININ соответственно. Далее проводили последовательное лигирование продуктов рестрикции и «пустого» шаттл–вектора p264 (фиг 2В, Г), сначала выполняют лигирование pCX–GFP–CDT1 и шаттл–вектора (фиг 2В), а после чего к получившейся конструкции лигируют pCX–GFP–GEMININ (фиг. 2Г). Клонирование рекомбинантной плазмидной ДНК осуществляют с использованием штамма DH10β E. coli.
Таким образом, на последнем этапе сборки получали вектор p264, отличающийся наличием кодирующих последовательностей рекомбинантных флуоресцентных белков и индикаторов клеточного цикла, - GFP–CDT1 и GFP–GEMININ. Открытые рамки считывания GFP–CDT1 и GFP–GEMININ находятся под контролем независимых промоторов, подходящих для экспрессии в клетках человека. Также в последовательности данного вектора присутствует сайт рекомбинации LoxP, который используют для рекомбинантного встраивания созданного вектора в состав искусственной хромосомы человека в клетках яичника китайского хомячка CHO на этапе загрузки шаттл-вектора.
Для сравнения нуклеотидной последовательности полученного фрагмента с последовательностью целевого гена его секвенируют с использованием смеси флуоресцентно-меченных нуклеотидов BigDye™ Terminator v3.1 Cycle Sequencing Kit по методике производителя (Applied Biosystems). Анализ последовательностей показывает отсутствие замен в рамках считывания индикаторов клеточного цикла CDT1 и GEMININ, а также связанных с ними белков GFP.
Поскольку GFP связан с каждым из индикаторов клеточного цикла (CDT1 и GEMININ), то флуоресцентный сигнал детектируется только в период, когда индикатор клеточного цикла находится в нуклеоплазме, согласно сигналу ядерной локализации.
Экспрессия генов GFP–CDT1 и GFP–GEMININ в клетках человеческой фибросаркомы HT1080 продемонстрирована на фиг.3. Как видно из результатов, флуоресценция GFP имеет ядерную локализацию, а период флуоресценции соответствует изменению фаз клеточного цикла: G1 для GFP–CDT1 (фиг 3Б) и G2, S и М для GFP–GEMININ (фиг. 3В). Продолжительность флуоресценции составила 8–9 ч для каждого из рекомбинантных белков. При этом в клетках экспрессирующих оба рекомбинантных белка происходит визуализация всего клеточного цикла (фиг. 3А). Данный результат демонстрирует корректность сборки рекомбинантных конструкций.
Наконец, кодирующие последовательности данных рекомбинантных белков помещали в состав искусственной хромосомы. На фиг. 4А представлена схема LoxPCre рекомбинации шаттл-вектора и пустой искусственной хромосомы в линии клеток яичника китайского хомячка CHO. Как видно на фиг. 4А, последовательность искусственной хромосомы человека имеет половину гена HPRT (hypoxanthine–guaninephosphoribosyltransferase), полноразмерная последовательность которого, экспрессируясь, обеспечивает резистентность при культивировании линии клеток в селективной среде с содержанием гипоксантин – 5 мМ, аминоптерин – 20 мМ, тимидин – 0,8 мМ (hypoxanthine–aminopterin–thymidine, HAT). После рекомбинации LoxP (фиг.4А), происходило восстановление последовательности HPRT и шаттл–вектор был интегрирован в состав искусственной хромосомы человека (фиг. 4Г). Восстановление HPRT позволило провести селекцию клонов, содержащих последовательности интереса в составе искусственной хромосомы человека и получить необходимое количество клеток CHO для проведения завершающего этапа сборки системы – переноса полученной хромосомы человека в клеточную линию HT1080 с помощью протокола микроклеточно-опосредованного переноса хромосом (фиг 4Д) (MMCT).
На фиг. 4Б показана схема переноса искусственной хромосомы в клетки-реципиенты HT1080. Для подтверждения корректности переноса и наличия искусственной хромосомы в клетках донорах и реципиентах, CHO и HT1080 соответственно, проводили гибридизацию in situ препаратов метафазных пластинок (фиг. 4В).
В качестве маркера использовали зонд PNA, меченый FITC, на последовательность tetO в составе искусственной хромосомы, данная последовательность уникальна по отношению к остальным последовательностям нативных хромосом линии клеток человеческой фибросаркомы HT1080. На фиг. 4В слева видно, что клетки-реципиенты имеют четкий сигнал зеленой флуоресценции (FITC) исходящий от комплекса tetO в последовательности искусственной хромосомы и флуоресцентной метки PNA–FITC. После осуществления переноса из линии клеток CHO в линию клеток HT1080, в них также наблюдался сигнал от последовательности tetO, которая является свидетельством наличия искусственной хромосомы в геноме линии клеток человеческой фибросаркомы HT1080 (фиг. 4В). После проведения MMCT клетки линии HT1080 подвергали селекции в присутствии селективного антибиотика бластицидина. Гены устойчивости к бластицидину были локализованы в искусственной хромосоме человека (фиг 4А) и позволяли провести селекцию клеток с искусственной хромосомой человека. После селекции выбирали клон линии клеток HT1080-HAC-dGFP с наиболее стабильной экспрессией GFP и использовали его для дальнейших экспериментов в качестве тест-системы для определения уровня хромосомной нестабильности.
Заявленный технический результат повышение скорости и точности определения хромосомной нестабильности обеспечивается также новыми техническим решениями - способами использования разработанной тест–системы, в частности, способом поиска генов, контролирующих трансмиссию хромосом, и способом скрининга противоопухолевых лекарственных кандидатов, вызывающих хромосомную нестабильность.
В настоящее время для поиска генов, контролирующих митотическую трансмиссию хромосом, используют два классических метода: тест на образование микроядер и флуоресцентную гибридизацию in situ (Kirsch-Vders M. Towards a validation of the micronucleus test // Mutat.Res.1997.Vol.392,№1-2.P. 1-4.).
Данные методы не позволяют проводить массовый поиск генов, вызывающих хромосомную нестабильность.
Для поиска генов, вызывающих хромосомную нестабильность заявителем использована заявленная тест-система.
Заявленный технический результат достигается способом осуществления поиска генов, контролирующих трансмиссию хромосом, который ведут следующим образом:
осуществляют посев линии клеток человеческой фибросаркомы HT1080, содержащей искусственную хромосому человека, включающую два гена GFP с последовательностями CDT1 и GEMININ, функционирующими в качестве индикаторов клеточного цикла в сосуды для культивирования клеток в питательную среду;
проводят трансфекцию линии клеток препаратами антисмысловых олигонуклеотидов, способными подавлять экспрессию генов, выбранных для анализа их вовлеченности в контроль трансмиссии хромосом;
культивируют клетки до периода прохождения не менее одного клеточного цикла;
регистрируют флуоресцентный сигнал индикаторов клеточного цикла, определяя вовлеченность гена интереса в механизм трансмиссии хромосом по эффекту воздействия препарата антисмысловых олигонуклеотидов, устанавливая долю нефлуоресцирующих клеток, утративших искусственную хромосому.
Применение заявленной тест-системы, содержащей искусственную хромосому с генами рекомбинантных флуоресцентных белков с последовательностями CDT1 и GEMININ, обеспечивает быструю и точную оценку потери искусственной хромосомы по утрате частью клеточной популяции флуоресцентного сигнала индикаторов клеточного цикла.
Трансфекция линии клеток антисмысловыми олигонуклеотидами обеспечивает подавление экспрессии генов, выбранных для анализа их вовлеченности в контроль трансмиссии хромосом, результатом которой станет утрата искусственной хромосомы и быстрая потеря флуоресценции в случае, если анализируемый ген действительно вовлечен в процессы контроля трансмиссии хромосом.
Регистрация потери искусственной хромосомы может осуществляться в течение одного клеточного цикла (18 часов), что обеспечивает значительное сокращение и точность идентификации новых генов хромосомной нестабильности.
Кроме того, регистрация потери флуоресцентного сигнала индикаторов клеточного цикла позволяет точно определить долю клеток, утративших искусственную хромосому после подавления экспрессии анализируемого гена, и позволяет идентифицировать его вовлеченность в механизм трансмиссии хромосом и точно определить уровень хромосомной нестабильности.
Пример 1
Для поиска генов хромосомной нестабильности были взяты гены, нокдаун которых может приводить к сбоям при расхождении хромосом во время митоза.
Данные гены были использованы в качестве положительных контролей, среди них: CNP-A, Aurora B, CNP-N, SKA-3, Mis18.
За день до трансфекции малых интерферирующих РНК (миРНК) клетки линии HT1080 HAC/dGFP рассаживают в количестве 12500 на лунку 24 луночного планшета в полной среде DMEM, содержащей 10 мкг/мл бластицидина. На следующий день проводят трансфекциию с помощью трасфецирующего реагента «Lipofectamine RNAiMAX™» Transfection Reagent (Thermo Fisher Scientific).
В реакционной смеси используют по 17 мкл интерферирующей РНК, 3 мкл липофектамина, которые по отдельности растворяют в 50 мкл среды Opti-Mem, полученную смесь инкубируют 5 минут при комнатной температуре. После 5-ти минутной инкубации, смешивают разбавленную РНК и разбавленный липофектамин. Аккуратно перемешивают и оставляют инкубироваться при комнатной температуре на 20-30 минут. Для трансфекции клетки промывают 1X PBS, и оставляют в 2 мл среды Opti-Mem. После чего к клеткам линии HT1080 HAC/dGFP по каплям приливают по 100 мкл трансфекционой смеси и инкубируют 12 часов при 37ºC в 5% CO2. После трансфекции заменяют среду на новую DMEM без бластицидина и культивируют 72 часа (время, необходимое клеткам для прохождения 4-х делений), после чего анализируют полученные результаты.
На фиг. 8 можно наблюдать вероятность потери искусственной хромосомы человека клетками линии HT1080 HAC/dGFP после нокдауна выбранных генов.
По результатам сравнения с контролем можно сделать вывод, что наиболее достоверный эффект на развитие хромосомной нестабильности показало подавление экспрессии генов SKA-3 и Mis 18, что подтверждается в ранее опубликованной работе (Brownlow, N., Pike, T., Zicha, D., Collinson, L., & Parker, P. J. (2014). Mitotic catenation is monitored and resolved by a PKCepsilon-regulated pathway. Nature Communications, 5, 5685. https://doi.org/10.1038/ncomms6685;
Liskovykh, M., Lee, N. C., Larionov, V., & Kouprina, N. (2016). Moving toward a higher efficiency of microcell-mediated chromosome transfer. Molecular Therapy. Methods & Clinical Development, 3, 16043. https://doi.org/10.1038/mtm.2016.43)
Полученные данные доказывают возможность заявляемого технического решения ускоренно и точно (через 72 часа) получать данные о развитии хромосомной нестабильности, при проведении поиска новых генов, участвующих в процессе управления митозом.
Поставленная перед изобретениями проблема решается так же способом скрининга противоопухолевых лекарственных кандидатов, вызывающих хромосомную нестабильность.
На сегодняшний день для поиска противоопухолевых фармакологических кандидатов применяются те же методы, что и для поиска новых генов хромосомой нестабильности, а именно флуоресцентная гибридизация in situ и тест на определение микроядер (Kirsch-Vders M. Towards a validation of the micronucleus test // Mutat.Res.1997.Vol.392,№1-2.P. 1-4.). Однако данные методы не подходят для массового количественного скрининга лекарственных препаратов, вызывающих хромосомную нестабильность. В литературных данных заявителем не обнаружены сведения об использовании новой заявленной тест-системы для скрининга лекарственных препаратов.
Заявленный технический результат повышение скорости и точности определения хромосомной нестабильности, также достигается способом скрининга противоопухолевых лекарственных кандидатов, вызывающих хромосомную нестабильность за счет использования новой тест-системы для определения уровня хромосомной нестабильности.
Способ скрининга включает следующие этапы:
осуществляют посев линии клеток человеческой фибросаркомы HT1080, содержащую искусственную хромосому человека включающую два гена GFP с последовательностями CDT1 и GEMININ, функционирующими в качестве индикаторов клеточного цикла в сосуды для культивирования клеток в питательную среду;
добавляют лекарственные кандидаты, предназначенные для проведения скрининга с целью идентификации противоопухолевой активности и способности вызывать хромосомную нестабильность;
культивируют клетки до периода прохождения не менее одного клеточного цикла;
регистрируют флуоресцентный сигнал индикаторов клеточного цикла, определяя хромосомную нестабильность и эффект воздействия анализируемого препарата по доле нефлуоресцирующих клеток, утративших искусственную хромосому.
Применение линии клеток HT1080, содержащей искусственную хромосому с генами рекомбинантных флуоресцентных белков с последовательностями CDT1 и GEMININ, обеспечивает быструю и точную оценку потери искусственной хромосомы по утрате частью клеточной популяции флуоресцентного сигнала индикаторов клеточного цикла.
Добавление к линии клеток лекарственных кандидатов, предназначенных для проведения скрининга с целью идентификации противоопухолевой активности и способности вызывать хромосомную нестабильность, позволяет выявить новые препараты-кандидаты, обладающие противоопухолевой активностью, обеспеченной развитием запредельного уровня хромосомной нестабильности, приводящего к гибели опухолевых клеток. В результате чего, может быть расширен ассортимент эффективных противоопухолевых лекарственных средств.
Проведение культивирования не менее одного клеточного цикла, позволяет точно и в короткий срок идентифицировать клетки, утратившие искусственную хромосому после деления, что является следствием воздействия препарата-кандидата. При этом в качестве контрольного лекарственного средства может быть применен известный противоопухолевый препарат таксол (паклитаксел), внесенный Всемирной организацией здравоохранения в список наиважнейших лекарств.
Регистрация флуоресцентного сигнала индикаторов клеточного цикла, позволяет в короткий срок с высокой точностью определить уровень хромосомной нестабильности и выявить эффект воздействия анализируемого препарата по доле нефлуоресцирующих клеток, утративших искусственную хромосому.
Способ осуществляют следующим образом.
Пример 2
Для проведения скрининга лекарственных кандидатов, потенциально вызывающих хромосомную нестабильность, использовали заявляемую тест-систему.
В качестве лекарственных кандидатов используют уникальную коллекцию экстрактов с целью идентификации веществ, вызывающих хромосомную нестабильность. Данная коллекция экстрактов была любезно предоставлена Александром Каганским, заведующим Центра геномной и регенеративной медицины Школы биомедицины Дальневосточного федерального университета.
В данном случае вместо трансфекции интерферирующих РНК, линию клеток HT1080 HAC/dGFP культивировали в присутствии различных концентраций экстрактов (фиг 9).
На первом этапе были подобраны эффективные концентрации экстрактов и лекарственных кандидатов. Выполнен тест по определению жизнеспособности клеток под воздействием данных веществ.
В качестве положительного контроля используют блокатор полимеризации микротрубочек – таксол (фиг. 9). Был проведен поиск лекарственных кандидатов и дана количественная оценка эффекта на развитие хромосомной нестабильности и изменения в жизнеспособности клеток.
Для проведения экспериментов использовали экстракты из тканей и органов следующих организмов: Punica sp.; Iotrochota sp.; Dombeya sp.; Neopetrosia sp.; Eugenia sp.; Acalypha sp; Agaricus sp; Pleurotus sp., для которых показана противоопухолевая активность, а значит они являются перспективными для обнаружения новых веществ, вызывающих хромосомную нестабильность. В качестве контрольного вещества используют известный ингибитор полимеризации микротрубочек Taxol, который приводит хромосомной нестабильность (Hee-Sheung Lee, Nicholas CO Lee, Brenda R Grimes, Alexander Samoshkin, Artem V Kononenko, Ruchi Bansal, Hiroshi Masumoto, William C Earnshaw, Natalay Kouprina and Vladimir Larionov A new assay for measuring chromosome instability (CIN) and identification of drugs that elevate CIN in cancer cells Lee et al. BMC Cancer 2013, 13:252).
Экстракты готовят по методике, опубликованной в статье (Rummun, N., Somanah, J., Ramsaha, S., Bahorun, T., & Neergheen-Bhujun, V. S. (2013). Bioactivity of Nonedible Parts of Punica granatum L.: A Potential Source of Functional Ingredients. International journal of food science, 2013, 602312. https://doi.org/10.1155/2013/602312).
За день до обработки экстрактами или веществами-кандидатами клетки линии HT1080 HAC/dGFP рассаживают в количестве 12500 клеток на лунку 24 луночного планшета в полной среде DMEM, содержащей 10мкг/мл бластицидина.
На следующий день проводят обработку экстрактами или веществами-кандидатами. Для этого разные концентрации экстрактов или веществ-кандидатов растворяют в 500 микролитрах среды DMEM и приливают к клеткам инкубируют 12 часов при 37ºC в 5% CO2.
После 24 часов инкубации заменяют среду на новую порцию DMEM без бластицидина и культивируют 72 часа (время, необходимое клеткам для прохождения 4-х делений), после чего анализируют полученные данные.
Для подготовки клеток лини HT1080 HAC/dGFP к анализу из шести луночных планшетов аспирируют культуральную среду, клетки промывают PBS (37ºC) и добавляют 300 мкл раствора трипсина 0.25%/PBS.
Клетки линии HT1080 HAC/dGFP инкубируют 5 минут при 37ºС, после добавляют новую среду и ресуспендируют открепившиеся клетки до одноклеточной суспензии. Клетки линии HT1080 HAC/dGFP центрифугируют при 120g 5 минут. Убирали надосадочную, осадок ресуспендируют в PBS.
Репортерным сигналом служил зеленый флуоресцентный белок GFP с максимумом возбуждения при длине волны 488 нм, и DRAQ7™ (Biolegend, USA) с максимумом возбуждения при 695nm. GFP и DRAQ7™ регистрируют разными каналами. DRAQ7™ проникает только в клетки с поврежденной мембраной и окрашивает их ДНК (синяя флуоресценция). DRAQ7™ используют в концентрации 0,02 мМ с последующей инкубацией в течение 10 минут при температуре 22°С. Результаты подсчитывают на проточном цитометре.
Для каждого образца было взято 20 тысяч клеток. Популяцию оценивают в плоскости двухпараметрической гистограммы, на которой можно наблюдать соотношения GFP-положительных, GFP-отрицательных и DRAQ7™ -положительных клеток.
На фиг 9 приведены результаты проведения скрининга фармакологических кандидатов, вызывающих хромосомную нестабильность.
По результатам скрининга наиболее значимый эффект на развитие хромосомной нестабильности был продемонстрирован экстрактом из листьев Punica granatum в концентрации 200 мкг/мл.
Полученные данные доказывают возможность заявляемого технического решения ускоренно (через 72 часа) получать данные о развитии хромосомной нестабильности, при проведении поиска новых лекарственных веществ, вызывающих хромосомную нестабильность.
Как следует из проведенных экспериментов и результатов, представленных на фиг. 1-9, созданная тест-система HT1080 HAC/dGFP позволяет в 4,7 раза сократить время и получить точные данные об уровне хромосомной нестабильности под влиянием различных факторов: неидентифицированных ранее генов, участвующих в процессах репликации и сегрегации хромосом человека, а также лекарственных кандидатов, потенциально вызывающих хромосомную нестабильность и обладающих противоопухолевой активностью.
Пример 3
В настоящем примере показана возможность одновременного использования разработанной тест-системы для поиска новых генов и фармакологических кандидатов способных вызывать хромосомную нестабильность.
Для этого воздействуют на клетки линии фибросаркомы человека HT1080 ингибитором деполимеризации микротрубочек – цитотоксического соединения таксол, (Weaver, 2014); и на нокдауном SKA3 (фиг 6A) – одного из ключевых генов, участвующих в процессе формирования кинетохора. Таксол и миРНК против SKA3 часто применяют в качестве положительных контролей в опытах для изучения митотического цитоскелета и хромосомной трансмиссии (Park et al., 2011; Lee et al., 2013).
За день до трансфекции малых интерферирующих РНК (миРНК) клетки линии HT1080 HAC/dGFP рассаживали в количестве 12 500 на лунку 24 луночного планшета в полной среде DMEM, содержащей 10мкг/мл бластицидина. На следующий день, с для половины лунок планшета проводили трансфекциию с помощью трансфецирующего реагента «Lipofectamine RNAiMAX™» Transfection Reagent (Thermo Fisher Scientific).
В реакционной смеси используют по 17 мкл интерферирующей РНК, 3 мкл липофектамина, которые по отдельности растворяли в 50 мкл среды Opti-Mem, полеченную смесь инкубируют 5 минут при комнатной температуре. После 5-ти минутной инкубации, смешивают разбавленную РНК и разбавленный липофектамин. Аккуратно перемешивают и оставляют инкубироваться при комнатной температуре на 20-30 минут. Для трансфекции клетки промывали 1X PBS, и оставляют в 2 мл среды Opti-Mem. После чего к клеткам линии HT1080 HAC/dGFP по каплям приливают по 100 мкл трансфекционой смеси и инкубируют 12 часов при 37ºC в 5% CO2.
После трансфекции заменяют среду на новую DMEM без бластицидина и культивируют 72 часа (время, необходимое клеткам для прохождения 4-х делений), после чего анализируют полученные результаты. В качестве отрицательного контроля используют клетки линии клеток HT1080 HAC/dGFP без селекции.
Вторую половину планшета используют для анализа воздействия таксола на клетки линии HT1080 HAC/dGFP. Для этого на следующий день после посадки клеток проводят обработку таксолом в концентрации 10 нМ и инкубируют 24 часа при 37ºC в 5% CO2. После 24 часов инкубации заменяют среду на новую порцию DMEM и культивируют 72 часа (время, необходимое клеткам для прохождения 4-х делений), после чего анализируют полученные данные.
Для подготовки клеток лини HT1080 HAC/dGFP к анализу асперируют культуральную среду, клетки промывают PBS (37ºC) и добавляют 100 мкл раствора трипсина 0.25%/PBS.
Клетки линии HT1080 HAC/dGFP инкубируют 5 минут при 37ºС, после добавляют новую среду и ресуспендируют открепившиеся клетки до одноклеточной суспензии. Клетки линии HT1080 HAC/dGFP центрифугируют при 120g 5 минут. Убирают надосадочную жидкость, осадок ресуспендируют в PBS и анализируют.
Репортерным сигналом служит зеленый флуоресцентный белок GFP с максимумом возбуждения при длине волны 488 нм, и DRAQ7™ (Biolegend, USA) с максимумом возбуждения при 695nm. GFP и DRAQ7™ регистрируют разными каналами. DRAQ7™ проникает только в клетки с поврежденной мембраной и окрашивает их ДНК (синяя флуоресценция). DRAQ7™ используют в концентрации 0,02 мМ с последующей инкубацией в течение 10 минут при температуре 22°С. Результаты подсчитывают на проточном цитометре.
Для каждого образца было взято 20 тысяч клеток. Популяцию оценивают в плоскости двухпараметрической гистограммы, на которой можно наблюдать соотношения GFP-положительных, GFP-отрицательных и DRAQ7™ -положительных клеток.
Фиг. 6 отражает результаты, полученные методом проточной цитофлуориметрии. Разница между количеством клеток, потерявших сигнал в отрицательном контроле и в эксперименте, является необходимым результатом для измерения вероятности возникновения хромосомной нестабильности под воздействием лекарственных веществ-кандидатов или нокдауна генов-регуляторов хромосомной трансмиссии. Для того, чтобы исключить из расчета мертвые клетки, используют витальный краситель DRAQ7™, количество клеток положительных по данному красителю можно наблюдать во всех нижних левых квадратах каждого плота.
На фиг. 7 представлен результат эксперимента по анализу динамики клеточной популяции после нокдауна GFP с помощью пула интерферирующих РНК. В эксперименте используют систему высокоэффективной автоматизированной микроскопии для слежения за клеточной популяцией в режиме реального времени (Cell iQ). При этом клетки линии HT1080 HAC/dGFP культивируют в течение 215 ч (до достижения конфлюентности) и детектируют изменение количества GFP положительных клеток в популяции. Как видно на фиг. 7, после трансфекции наблюдается стабильный рост клеточной популяции, при этом клетки, подвергшиеся трансфекции, сначала не флуоресцируют, а затем восстанавливают экспрессию GFP через 150 ч. Данный опыт доказывает способность тест-системы HT1080 HAC/dGFP мгновенно отвечать на остановку экспрессии GFP. Таким образом, линия клеток HT1080 HAC/dGFP позволяет детектировать потерю искусственной хромосомы непосредственно после развития хромосомной нестабильности, как после нокдауна гена, так и после воздействия вещества вызывающего хромосомную нестабильность.
Заявленные технические решения позволяют быстро и точно определять уровень хромосомной нестабильности при воздействии существующих лекарственных препаратов, а также обеспечивают поиск новых противоопухолевых веществ и генов, вызывающих хромосомную нестабильность.
Изобретение относится к биотехнологии, в частности к генетической инженерии, и позволяет получить линию клеток млекопитающих с искусственной хромосомой человека, которую можно использовать для обнаружения ранее неидентифицированных генов, участвующих в процессах репликации и сегрегации хромосом человека, а также для поиска лекарственных кандидатов, потенциально вызывающих хромосомную нестабильность. Заявленная тест-система для определения уровня хромосомной нестабильности на основе линии клеток млекопитающих содержит искусственную хромосому с генами рекомбинантных флуоресцентных белков с последовательностями CDT1 и GEMININ, функционирующих в качестве индикаторов клеточного цикла. Использование предлагаемой тест-системы позволяет выполнить быстрый скрининг противоопухолевых лекарственных кандидатов, вызывающих хромосомную нестабильность, а также поиск генов, контролирующих трансмиссию хромосом. 3 н.п. ф-лы, 9 ил., 3 пр.
1. Тест-система для определения уровня хромосомной нестабильности на основе линии клеток человеческой фибросаркомы HT1080, содержащей искусственную хромосому c геном GFP, отличающаяся тем, что искусственная хромосома включает два гена GFP с последовательностями CDT1 и GEMININ, функционирующими в качестве индикаторов клеточного цикла.
2. Способ скрининга противоопухолевых лекарственных кандидатов, вызывающих хромосомную нестабильность, заключающийся в следующем:
осуществляют посев линии клеток человеческой фибросаркомы HT1080, содержащей искусственную хромосому человека, включающую два гена GFP с последовательностями CDT1 и GEMININ, функционирующими в качестве индикаторов клеточного цикла, в сосуды для культивирования клеток в питательную среду;
добавляют препараты, предназначенные для проведения скрининга с целью идентификации противоопухолевой активности и способности вызывать хромосомную нестабильность;
культивируют клетки до периода прохождения не менее одного клеточного цикла;
регистрируют флуоресцентный сигнал индикаторов клеточного цикла, определяя хромосомную нестабильность и эффект воздействия анализируемого препарата по доле нефлуоресцирующих клеток, утративших искусственную хромосому.
3. Способ поиска генов, контролирующих трансмиссию хромосом, заключающийся в следующем:
осуществляют посев линии клеток человеческой фибросаркомы HT1080, содержащей искусственную хромосому человека, включающую два гена GFP с последовательностями CDT1 и GEMININ, функционирующими в качестве индикаторов клеточного цикла, в сосуды для культивирования клеток в питательную среду;
проводят трансфекцию линии клеток препаратами антисмысловых олигонуклеотидов, способными подавлять экспрессию генов, выбранных для анализа их вовлеченности в контроль трансмиссии хромосом;
культивируют клетки до периода прохождения не менее одного клеточного цикла;
регистрируют флуоресцентный сигнал индикаторов клеточного цикла, определяя вовлеченность гена интереса в механизм трансмиссии хромосом по эффекту воздействия препарата антисмысловых олигонуклеотидов, устанавливая долю нефлуоресцирующих клеток, утративших искусственную хромосому.
| Xouri G | |||
| et al., Cdt1 and geminin are down‐regulated upon cell cycle exit and are over‐expressed in cancer‐derived cell lines, European Journal of Biochemistry, 2004, Т | |||
| Искроудержатель для паровозов | 1920 |
|
SU271A1 |
| Устройство для электрической сигнализации | 1918 |
|
SU16A1 |
| АЭРОСАНИ | 1922 |
|
SU3368A1 |
| Petropoulou C | |||
| et al., Cdt1 and Geminin in cancer: markers or triggers of malignant transformation, Front Biosci | |||
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| - Т | |||
| Насос | 1917 |
|
SU13A1 |
| - No | |||
| ПРИСПОСОБЛЕНИЕ ДЛЯ РАБОТ НАД ПРЕПАРАТАМИ ПОД МИКРОСКОПОМ | 1926 |
|
SU4485A1 |
| Экономайзер | 0 |
|
SU94A1 |

