ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
В данной заявке заявлен приоритет на основании временных заявок США №№60/823319, поданной 23 августа 2006, и 60/953910, поданной 3 августа 2007, которые включены сюда путем ссылки в полном объеме.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к нуклеиновым кислотам, содержащим нуклеотидные последовательности, соответствующие изолированным и очищенным последовательностям MAR, имеющим происхождение от человека и животных, отличных от человека, или основанные на них. Эти нуклеиновые кислоты в целом обладают активностями усиления транскрипции и/или продуцирования белка. Изобретение также относится к способам идентификации таких последовательностей и к системам, в которых они использованы, например, для высокого выхода при продуцировании белков.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Публикации и другие материалы, включая патенты, используемые здесь для иллюстрации изобретения и, в частности, для приведения аналогичных деталей, касающихся практики, включены здесь путем ссылки. Для удобства публикации, если они не указаны полностью в тексте, перечислены в алфавитном порядке в прилагаемой библиографии. EMBL номер по каталогу АС102666 и последовательности, фланкирующие EMBL, номер по каталогу ВН101870 и ВН101901, а также EMBL номера по каталогу (синонимы) 126658, 23119391, 22981746 также включены здесь путем ссылки в полном объеме.
В настоящее время модель организации эукариотических хромосом в домены хроматиновых петель примерно от 50 до 100 кб широко признана [Bodnar JW, Breyene Р, Van Montagu M and Gheyseu G, Razin SV]. Считают, что наружные концы этих петель соответствуют специфичным последовательностям ДНК, которые присоединены к ядерному матриксу, белковой сети, состоящей из РНП (рибонуклеопротеинов) и других негистоновых белков [Bode J, Benham С, Knopp А и Mielke С]. Хромосомные последовательности ДНК, которые присоединены к ядерному матриксу, называют SAR или MAR, соответственно, для каркасных (во время метафазы) или матриксных (интерфаза) областей присоединения (SAR от scaffold attachment region, и MAR от matrix attachment region). S/MAR, элементы MAR или последовательности MAR, или, для краткости, MAR представляют собой полиморфные области типичной длины 300-3000 п.о. Установлено, что в ядре млекопитающих находится примерно 100000 MAR [Bode J, Stengert-lber M, Kay V, Schlake Т и Dietz-Pfeilstetter A].
Считают, что посредством структурной и функциональной сегрегации хроматина в петлевые домены элементы MAR играют критическую роль в репликации и регуляции экспрессии генов, так чтобы облегчить структурную сборку и разборку центров транскрипции в ядре млекопитающих. Множество косвенных данных собрано в подтверждение этого мнения; например, в различных эукариотических геномах точки начала репликации ДНК были картированы внутри элементов MAR [Amati В и Gasser SM (1988), Amati В и Gasser SM (1990)]. MAR также почти всегда обнаруживают в некодирующих межгенных областях, внутри интронов [Girod PA, Zahn-Zabal M и Mermod N] или на границах транскрипционных единиц [Gasser SM и Laemmli UK; National Center for Biotechnology Information], где они могут связывать универсальные и/или тканеспецифические факторы транскрипции. В целом в трансгенных экспериментах на растениях и на линиях животных клеток элементы MAR успешно использованы для повышения экспрессии и стабильности трансгена [Allen GC, Spiker S, Thompson WF, Bode J, Schlake T, Rios-Ramirez M, Mielke C, Stengart M, Kay V и Klehr-Wirth D, Girod PA, Zahn-Zabal M и Mermod N]. Например, MAR использованы для повышения продуцирования различных рекомбинантных белков в клетках, релевантных для биотехнологии и терапевтических применений, таких как клетки СНО (яичника китайского хомячка) [Girod PA, Zahn-Zabal M и Mermod N, Kim JM, Kim JS, Park DH, Kang HS, Yoon J, Baek К и Yoon Y, Zahn-Zabal M, Kobr M, Girod PA, Imhof M, Chatellard P, de Jesus M, Wurm F и Mermod N] (Mermod et al., "Development of stable cell lines for production or regulated expression using matrix attachment regions," WO 02074969, а также публикация патента США 20030087342).
Функциональная активность MAR вероятнее связана с их структурными свойствами, чем с их первичной последовательностью ДНК. Действительно, MAR имеют высокое содержание А и Т [Boulikas Т (1993)], и наблюдаются некоторые конкретные конформационные и физико-химические свойства, такие как природная кривизна молекулы, узкая малая бороздка, высокий потенциал раскручивания/неспаривания или склонность к денатурации [Bode J, Schlake T, Rios-Ramirez М, Mielke С, Stengart М, Кау V и Klehr-Wirth D, Boulikas Т (1993), Boulikas T (1995)]. В действительности именно эти свойства использованы для идентификации MAR способом, называемым SMAR Scan. Кроме того, активность MAR может быть также опосредована ДНК-связывающими белками, такими как ферменты ремоделирования хроматина и/или факторы транскрипции, которые могут распознавать специфичные структурные признаки элементов MAR, такие как однонитевая и/или изогнутая ДНК [Bode J, Stengert-Iber М, Кау V, Schlake Т и Dietz-Pfeilstetter A]. He обнаружено отчетливого сайта связывания белка или консенсусной последовательности MAR [Boulikas T (1993)], что затрудняет предсказание MAR на основании геномных последовательностей.
Хотя некоторые функциональные и структурные свойства MAR описаны, их идентификация затруднительна, поскольку они имеют мало общего в отношении первичной структуры. Хотя элементы MAR могут быть функционально консервативными в эукариотических геномах, где это предположение подтверждается тем фактом, что MAR животных могут связываться с растительными ядерными каркасами и наоборот [Breyne P, Van Montagu М, Depicker А и Gheysen G, Mielke С, Kohwi Y, Kohwi-Shigematsu Т и Bode J], мало можно сказать о том, какой признак делает последовательность MAR последовательностью эффективного продуцирования белка. Также могут быть получены варьирующие результаты в зависимости от используемого анализа [Razin SV, Boulikas Т (1995), Кау V и Bode J]. С учетом огромного числа ожидаемых MAR в эукариотическом организме и количества последовательностей, опубликованных геномными проектами, были разработаны инструменты/программы для обнаружения структурных признаков последовательностей ДНК MAR (SMAR Scan I) или функциональных последовательностей, таких как сайты связывания для специфичных белков, которые действуют в качестве регуляторных белков или факторов транскрипции (SMAR Scan II) [временная патентная заявка США 60/953910, поданная 3 августа 2007, публикация патента США 20070178469 авторов Mermod et al.]. Такие программы были разработаны для идентификации новых потенциальных последовательностей MAR путем обнаружения кластеров признаков последовательностей ДНК, соответствующих изгибу ДНК, потенциалам глубины большой бороздки и ширины малой бороздки, а также сайтам связывания специфичных регуляторных белков транскрипции. Эти программы использованы для сканирования человеческого генома для идентификации предполагаемых последовательностей ДНК MAR, для некоторых из которых показано повышение экспрессии трансгена при встраивании в экспрессионную плазмиду, которая была трансфицирована в клетки СНО (Girod et al., "Identification of S/MAR from genomic sequences with bioinformatics and use to increase protein production in industrial and therapeutic processes," публикация патента США 20070178469 авторов Mermod et al.]. Это показало, что программы SMAR Scan могут эффективно идентифицировать человеческие генетические элементы, которые, в свою очередь, можно использовать для повышения синтеза белка. Хотя проведенные до сих пор функциональные скрининги были ограничены человеческим геномом, при широкомасштабной продукции интересующий белок часто экспрессируют в клетках млекопитающих, отличных от человека.
В человеческом геноме идентифицировано около шестнадцати сотен MAR с помощью SMAR Scan, и для шести из восьми было продемонстрировано, что они запускают усиленную экспрессию генов (таких как ген зеленого флуоресцентного белка (GFP), антитела и рецепторы) в клетках СНО при помещении их выше энхансера/промотора. Длина ДНК, которая, как показано, обладает эктопической активностью MAR, находится в интервале от 2,5 кб до 6 кб. Однако отсутствие структурной характеристики MAR до сих пор ограничивало получение "дизайна" MAR. Таким образом, существует необходимость в характеристике MAR, в частности функциональных и/или структурных областей MAR, чтобы дать возможность конструирования и дизайна MAR.
Проведенные до сих пор функциональные скрининги были ограничены человеческим геномом. Поскольку при широкомасштабной продукции интересующий белок часто экспрессируют в клетках млекопитающих, также существует необходимость в идентификации более эффективных встречающихся в природе MAR, которые усиливают транскрипцию и/или экспрессию генов и/или эффективных клеток-продуцентов белков в человеческих клетках и/или в клетках млекопитающих, отличных от человека.
В целом существует необходимость идентификации и/или получения MAR, обладающих преимущественными свойствами, например, путем идентификации дополнительных встречающихся в природе MAR, путем конструирования идентифицированных MAR и/или путем получения синтетических MAR. Преимущественные свойства проявляются, но не ограничены ими, в свойствах усиленной транскрипции и/или продуцирования белка/экспрессии гена; уменьшенной длины относительно встречающихся в природе MAR, что, таким образом, дает возможность более многостороннего применения в генной инженерии; специфичность к тканям, клеткам или органам и/или способность к индукции при добавлении внешнего стимулятора, такого как лекарство.
Для адресации одной или более чем одной из этих потребностей и других потребностей, которые станут очевидными на основании последующего описания, использовали несколько подходов, включая широкомасштабный биоинформационный анализ генома мыши для идентификации предполагаемых последовательностей ДНК MAR. Геном мыши анализировали, используя программное обеспечение для предсказания MAR SMAR Scan I. Вновь идентифицированные последовательности грызунов оценивали на их способность опосредовать улучшенное продуцирование рекомбинантных белков, представляющих фармацевтический интерес, из культивируемых клеток. Наконец, транскрипционную активность вновь идентифицированных MAR оценивали в трансфекционных анализах трансгена.
Кроме того, были исследованы MAR, такие как человеческая 1_68 MAR и мышиная MAR S4. Были идентифицированы блоки, в частности блоки, содержащие некоторые специфичные по структуре/последовательности MAR, и эти блоки использовали для конструирования MAR, обладающих преимущественными свойствами, путем, например, перегруппировки, делеции и/или дупликации последовательностей. Эти блоки также объединяли с другими элементами, например с синтетическими нуклеотидными последовательностями, содержащими некоторые сайты связывания, в частности сайты связывания факторов транскрипции (TFBS).
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На фиг.1 показан эффект различных MAR на продуцирование рекомбинантного зеленого флуоресцентного белка (GFP).
На фиг.2 показан эффект различных человеческих и мышиных MAR элементов на процент очень высоких продуцентов (% МЗ) в клетках СНО рекомбинантного зеленого флуоресцентного белка (GFP).
На фиг.3 показан эффект различных человеческих 1_68 и мышиных S4 MAR элементов на экспрессию рекомбинантного зеленого флуоресцентного белка (GFP).
На фиг.4 показан эффект мышиных элементов MAR на продуцирование рекомбинантных моноклональных антител.
На фиг.5 показано, что стабильные поликлональные популяции могут быть получены из популяции клеток СНО, трансфицированных векторами, направляющими экспрессию тяжелой и легкой цепи IgG без MAR (нет MAR) или с MAR S4, присоединенной в цис положении.
На фиг.6 (А) и (Б) показано, что стабильные индивидуальные клоны могут быть получены путем ограничения разведения популяции клеток СНО, трансфицированных векторами, направляющими экспрессию тяжелой и легкой цепи IgG без MAR (нет MAR) в (Б) или с MAR S4 и MAR 1_68, присоединенными в цис положении.
На фиг.7 (А) и (Б) показана экспрессия гена (GFP) без MAR (А) и с MAR (Б) со временем (2 недели и 26 недель).
На фиг.8 (А) и (Б) изображены признаки изгиба (А) и последовательности (Б) человеческой 1_68 MAR.
Фиг.9 (А)-(В): На (А) показаны различные конструкции MAR, полученные путем группировки идентифицированных областей и достигнутый прирост транскрипции; на (Б) показан паттерн изгиба конструкции MAR 6; на (В) приведены детали структурных параметров, таких как сайты связывания конструкции MAR 6.
На фиг.10 показан эффект различных конструкций MAR S4 на экспрессию рекомбинантного зеленого флуоресцентного белка (GFP), которая выявлена с помощью средней флуоресценции всей популяции (Avg Gmean М0).
На фиг.11 показаны различные конструкции MAR S4, выведенные на основании экспрессии рекомбинантного зеленого флуоресцентного белка (GFP), которая выявлена путем анализа средней флуоресценции всей популяции (Avg Gmean М0).
На фиг.12 показана карта потенциальных сайтов связывания факторов транскрипции человеческой 1_68 MAR, которые предсказаны программным обеспечением MATInspector.
Фиг.13 представляет собой карту плазмиды, использованной для тестирования на активность синтетических MAR, сконструированных в результате сборки АТ-богатой внутренней последовательности (MAR 1429-2880) и синтезированных химическим путем сайтов связывания ДНК для факторов транскрипции, помещенных выше промотора, и зеленого флуоресцентного белка (GFP).
Фиг.14 представляет собой иллюстрацию усиления транскрипции синтетическими MAR, сконструированными, как описано на фиг.13.
Фиг.15 представляет собой иллюстрацию усиления транскрипции синтетическими MAR, содержащими сайты связывания ДНК, подробно описанные в таблице 5.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Настоящее изобретение в одном воплощении направлено на экспрессионную систему для экспрессии на высоком уровне по меньшей мере одного гена, включающую:
промотор для оперативного сцепления с нуклеотидной последовательностью, кодирующей интересующий ген, и
по меньшей мере одну нуклеотидную последовательность MAR млекопитающего, отличного от человека, для усиления экспрессии указанного гена в клетке, трансформированной указанной экспрессионной системой,
где указанная нуклеотидная последовательность MAR млекопитающего, отличного от человека, повышает экспрессию указанного гена примерно в 2, примерно в 3, примерно в 4, примерно в 5, примерно в 6, примерно в 7, примерно в 8, примерно в 9, примерно в 10 раз или более после трансформации указанной клетки указанной конструкцией.
Указанная нуклеотидная последовательность MAR млекопитающего, отличного от человека, может включать, по существу состоять или состоять из:
(i) SEQ ID No. 3, SEQ ID No. 10 или ее функционального фрагмента; или
(ii) нуклеотидной последовательности, имеющей примерно 80%, примерно 90%, примерно 95% или примерно 98% идентичности последовательности с любой из последовательностей (i).
Изобретение также направлено на изолированную и очищенную молекулу нуклеиновой кислоты, содержащую, по существу состоящую или состоящую из:
(а) нуклеотидной последовательности SEQ ID No. 3 или SEQ ID No. 10 или ее функционального фрагмента; или
(б) нуклеотидной последовательности, имеющей примерно 80%, примерно 90%, примерно 95% или примерно 98% идентичности последовательности с последовательностью (а) и обладающей активностью MAR.
Изобретение, кроме того, направлено на способ идентификации последовательностей MAR млекопитающего, отличного от человека, при котором:
- получают по меньшей мере одну молекулу нуклеиновой кислоты млекопитающего, отличного от человека, предпочтительно геном млекопитающего, отличного от человека, или его часть,
- подвергают указанную молекулу нуклеиновой кислоты методике сканирования на MAR последовательности, включающей:
- установление размера окна для молекул нуклеиновой кислоты, подлежащих оценке,
- выбор по меньшей мере 1 или по меньшей мере 2, предпочтительно 3, более предпочтительно 4 или более признаков, связанных с MAR,
- установление пороговых значений для последовательностей, проявляющих этот признак/эти признаки и
- выбор нуклеотидных последовательностей-кандидатов MAR, для которых превышены эти пороговые значения,
- подтверждение того, что указанная нуклеотидная последовательность MAR млекопитающего, отличного от человека, повышает экспрессию гена примерно в 2, примерно в 3, примерно в 4, примерно в 5, примерно в 6, примерно в 7, примерно в 8, примерно в 9, примерно в 10 или более раз после трансформации человеческой клетки и/или клетки млекопитающего, отличного от человека, экспрессионной системой, содержащей указанные нуклеотидные последовательности MAR млекопитающего, отличного от человека.
Признак может здесь представлять собой значение угла изгиба ДНК, которое умножают на значение окна с получением величины умножения между примерно 320 и 1320, как, например, примерно 420 и примерно 1220, примерно 520 и примерно 1120, примерно 620 и примерно 1020, примерно 720 и примерно 920; признак может здесь представлять собой значение глубины большой бороздки, которое умножают на значение окна с получением величины умножения между примерно 900 и примерно 4000, как, например, примерно 1200 и 3700, примерно 1500 и примерно 3400, примерно 1800 и примерно 3100, примерно 2100 и примерно 2800 и/или признак может здесь представлять собой значение ширины малой бороздки, которое умножают на значение окна с получением величины умножения между примерно 500 и примерно 2500, как, например, примерно 750 и примерно 2250, примерно 1000 и примерно 2000, примерно 1250 и 1750.
Изобретение также направлено на конструкции MAR, содержащие:
(а) (i) изолированную нуклеотидную последовательность, содержащую по меньшей мере часть концевого участка идентифицированной MAR, и
(ii) дополнительную изолированную нуклеотидную последовательность, содержащую примерно 10%, примерно 15%, примерно 20%, примерно 25%, примерно 30% или более указанной идентифицированной MAR или другой идентифицированной MAR; или
(б) (i) нуклеотидную последовательность, имеющую примерно 90%, примерно 95%, примерно 96%, примерно 97% примерно 98%, примерно 99% идентичности последовательности с нуклеотидной последовательностью (а) (i), и
(ii) нуклеотидную последовательность, имеющую примерно 90%, примерно 95%, примерно 96%, примерно 97%, примерно 98%, примерно 99% идентичности последовательности с нуклеотидной последовательностью (б) (i).
Другие конструкции MAR согласно изобретению включают:
области идентифицированной последовательности MAR или ее части в последовательном расположении, где порядок и/или ориентация отличается от таковой идентифицированной последовательности MAR.
Еще одни другие конструкции MAR согласно изобретению включают:
(а) внутреннюю нуклеотидную последовательность, содержащую
(i) по меньшей мере одну изолированную или синтетическую АТ-богатую область идентифицированной последовательности MAR или
(ii) по меньшей мере одну АТ-богатую область, имеющую по меньшей мере 80%, 85%, 90%, 95%, 98% или 99% идентичности последовательности с АТ-богатой областью (а) (i),
(б) нуклеотидную последовательность, содержащую
по меньшей мере один сайт связывания ДНК-белок, прилежащий к указанной нуклеотидной последовательности (а), где указанный сайт связывания представляет собой
(i) сайт связывания ДНК-белок дополнительной идентифицированной последовательности MAR,
(ii) сайт связывания ДНК-белок идентифицированной последовательности MAR (а), где указанный сайт связывания ДНК-белок в идентифицированной последовательности MAR расположен снаружи от внутренней нуклеотидной последовательности (а), либо
(iii) первый сайт связывания ДНК-белок присутствует во внутренней последовательности (а), но прилежит по меньшей мере к одному дополнительному сайту связывания ДНК-белок, где первый и по меньшей мере один из указанных дополнительных сайтов связывания ДНК-белок не прилежат к внутренней последовательности (а), либо
(iv) сайты связывания ДНК-белок последовательности, представляющей собой не MAR.
Изобретение также направлено на экспрессионные системы, содержащие любую из указанных конструкций MAR, набор, содержащий любую из указанных экспрессионных систем, и применение любой из конструкций MAR, экспрессионных систем, клеток, трансгенных животных, отличных от человека, наборов и/или способов, относящихся к изобретению, (1) для продуцирования белков, таких как антитела, распознающие патогенные белки человека или белки клеточной поверхности человека, и таких белков, как эритропоэтин, интерфероны или другие терапевтические или диагностические белки и/или (2) in vitro, in vivo генотерапии, клеточной терапии или регенерационной терапии тканей.
ПОДРОБНОЕ ОПИСАНИЕ РАЗЛИЧНЫХ И ПРЕДПОЧТИТЕЛЬНЫХ ВОПЛОЩЕНИЙ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к изолированным и очищенным последовательностям MAR из животных, отличных от человека, к способу идентификации этих последовательностей и к системе, в которой использованы такие последовательности для высокого выхода при продуцировании белков как в человеческих клетках, так и в не человеческих клетках, таких как клетки грызунов.
Изобретение также направлено на конструкции MAR, в частности усиленные конструкции MAR, на экспрессионные системы и наборы, в которых используют эти конструкции MAR, и на их применение при продуцировании, в частности при широкомасштабной продукции белков, а также в терапии.
Кроме того, изобретение направлено на способы продуцирования с высоким выходом белков, как в человеческих клетках, так и в клетках млекопитающих, отличных от человека, посредством MAR/конструкций MAR.
Если они не определены иначе, все технические и научные термины, используемые здесь, имеют такое же значение, как общепринято понимают специалисты в области техники, к которой принадлежит данное изобретение. Хотя методы и материалы, отличающиеся от описанных здесь, можно использовать в практике настоящего изобретения, примерные пригодные методы и материалы описаны ниже.
Экспрессионная кассета согласно настоящему изобретению представляет собой нуклеиновую кислоту, содержащую по меньшей мере один ген, а также элементы, необходимые для транскрипции этого гена.
Промотор согласно настоящему изобретению представляет собой регуляторную область ДНК, которая при локализации выше гена способствует транскрипции этого гена.
Экспрессия в клетке, например экспрессия в клетке млекопитающего, отличного от человека, относится в контексте настоящего изобретения к экспрессии in vitro и in vivo. Экспрессия in vitro включает, например, экспрессию в клеточной линии, такой как клеточная линия HeLa или клеточная линия СНО, и в клетках, используемых для генотерапии in vitro. Экспрессия in vivo включает экспрессию в трансгенном животном, отличном от человека, и экспрессию в человеческих клетках, используемых для генотерапии in vivo или для генотерапии in vitro после возвращения клеток в организм человека-реципиента генотерапии.
Клетка млекопитающего, такая как клетка млекопитающего, отличного от человека, согласно настоящему изобретению способна поддерживаться в условиях клеточной культуры. Не ограничивающим примером такого типа клеток являются клетки яичника китайского хомячка (СНО).
Конструкция MAR, элемент MAR, последовательность MAR, S/MAR или просто MAR согласно настоящему изобретению представляет собой нуклеотидную последовательность, имеющую одну или более чем одну (как, например, две, три или четыре) общую характеристику с встречающимися в природе "SAR" или "MAR" и обладающую по меньшей мере одним свойством, которое способствует экспрессии белка любого гена, на который влияет такая MAR. Конструкция MAR также обладает признаком изолированной и/или очищенной нуклеиновой кислоты с активностью MAR, в частности с активностью модулирования транскрипции, предпочтительно с энхансерной активностью, но также, например, с активностью стабилизации экспрессии и/или с другими активностями, которые также описаны под термином "усиленные конструкции MAR." Конструкции MAR могут быть определены на основе идентифицированных MAR, на которых они первично основаны: конструкция MAR S4, соответственно, представляет собой конструкцию MAR, большинство нуклеотидов которой (50% и более) основаны на MAR S4. Встречающиеся в природе SAR или MAR согласно широко признанной модели опосредуют заякоривание специфичных последовательностей ДНК в ядерном матриксе, образуя домены хроматиновых петель, которые распространяются вовне из гетерохроматиновых сердцевин. Хотя SAR или MAR не содержат какой-либо очевидной консенсусной или распознаваемой последовательности, их наиболее единообразным признаком оказывается общее высокое содержание А и Т и преобладание оснований С на одной нити. MAR в целом обладают склонностью к образованию изогнутых вторичных структур, которые могут быть склонны к разделению нитей. Несколько простых мотивов последовательности с высоким содержанием А и Т часто обнаруживают внутри SAR и/или MAR, но для большей части их функциональная значимость и потенциальный способ действия не выяснены. Они включают А-бокс, Т-бокс, мотивы раскручивания спирали ДНК, сайты связывания SATB1 (Н-бокс, А/T/С25) и консенсусные сайты топоизомеразы II для позвоночных или Drosophila.
Кандидат MAR или последовательность-кандидат MAR согласно настоящему изобретению представляет собой последовательность, имеющую одну или более чем одну общую характеристику, как, например, две, три или четыре общие характеристики, с природными SAR или MAR.
Идентифицированная MAR или идентифицированная последовательность MAR согласно настоящему изобретению представляет собой изолированную нуклеотидную последовательность и соответствует встречающейся в природе последовательности MAR в том, что она содержит все области ("блоки" или "элементы"), которые дают возможность полного усиления экспрессии белка/гена, соответствующие ее природному прототипу.
Блоки (также называемые здесь "области", "область ДНК', "участки", "домены") идентифицированной MAR все необходимы для возможности усиления экспрессии белка/гена, соответствующего способности встречающейся в природе MAR. Ни один из этих блоков сам по себе обычно не способен к достижению полной активности MAR. Некоторые из этих областей специфичны по последовательности, такие как обогащенные АТ-динуклеотидами области изгиба и области сайтов связывания факторов транскрипции (TFBS), описанные ниже. Другие "области" характеризуются их локализацией, например 5' и 3' концевые области идентифицированной последовательности MAR.
Обогащенная АТ/ТА-динуклеотидами область изгиба ДНК (здесь называемая "АТ-богатой областью") представляет собой область изгиба ДНК, содержащую высокое число А и Т, в форме динуклеотидов AT и ТА. В предпочтительном воплощении она содержит по меньшей мере 10% динуклеотида ТА и/или по меньшей мере 12% динуклеотида AT на отрезке из 100 непрерывных пар оснований, предпочтительно по меньшей мере 33% динуклеотида ТА и/или по меньшей мере 33% динуклеотида AT на отрезке из 100 непрерывных пар оснований (или на соответствующем более коротком отрезке, когда АТ-богатая область имеет более короткую длину), в то же время имея изогнутую вторичную структуру. Однако "АТ-богатые области" могут быть настолько короткими, как примерно 30 нуклеотидов или менее, но предпочтительно имеют длину примерно 50 нуклеотидов, примерно 75 нуклеотидов, примерно 100 нуклеотидов, примерно 150, примерно 200, примерно 250, примерно 300, примерно 350 или примерно 400 нуклеотидов или длиннее.
Как обсуждено ниже, АТ-богатую область можно отличить от соседней области, такой как область сайта связывания, с помощью, например, ее относительно высокого угла изгиба.
Некоторые сайты связывания также часто имеют относительно высокое содержание А и Т, как, например, сайт связывания SATB1 (Н-бокс, А/Т/С25) и консенсусные сайты топоизомеразы II для позвоночных или Drosophila. Однако область сайта связывания (блок), в частности область TFBS, которая содержит кластер сайтов связывания, можно легко отличить от областей, обогащенных динуклеотидами AT и ТА ("АТ-богатых областей"), от сайтов связывания с высоким содержанием А и Т путем сравнения паттерна изгиба этих областей. Например, для человеческой MAR 1_68 последний может иметь среднюю степень кривизны, превышающую примерно 3,8 или примерно 4,0, тогда как область TFBS может иметь среднюю степень кривизны ниже примерно 3,5 или примерно 3,3. Области идентифицированных MAR могут быть также подтверждены альтернативными способами, такими как, но не ограниченными ими, относительные температуры плавления, как описано здесь в другом месте. Однако такие значения видоспецифичны и, таким образом, могут варьироваться от вида к виду, и могут, например, быть более низкими. Следовательно, соответствующие области, обогащенные динуклеотидами AT и ТА, могут иметь более низкие степени кривизны, такие как от примерно 3,2 до примерно 3,4, или от примерно 3,4, до примерно 3,6, или от примерно 3,6 до примерно 3,8, и области TFBS могут иметь пропорционально более низкие степени кривизны, как, например, примерно 2,7, ниже примерно 2,9, ниже примерно 3,1, ниже примерно 3,3. В программе SMAR Scan II специалистом в данной области техники должны быть выбраны относительно более низкие размеры окна.
Концевая область идентифицированной MAR/последовательности MAR согласно настоящему изобретению включает по меньшей мере примерно 5%, примерно 6%, примерно 7%, примерно 8%, примерно 9% или примерно 10% идентифицированной MAR.
Сайт связывания или сайт связывания ДНК-белок представляет собой любую нуклеотидную последовательность, которая может связывать ДНК-связывающий белок. Сайты связывания для ДНК-связывающих белков типично представляют собой TFBS. TFBS представляет собой любую последовательность, которая может связывать фактор транскрипции. TFBS может иметь любое происхождение, как, например, но не ограниченное ими, от человека или мыши. TFBS могут быть также созданы генно-инженерным или синтетическим путем. Однако в некоторых воплощениях TFBS имеют прототип в последовательности MAR, такой как последовательность MAR того же организма, того же вида или того же рода. Однако TFBS могут принадлежать к последовательности MAR другого вида или другого рода. TFBS, которые не имеют известного в настоящее время прототипа в последовательности MAR, также находятся в пределах объема настоящего изобретения. Такие TFBS могут включать, но не ограничены ими, сайты связывания для USF1 (левый стимулирующий фактор 1) или белок с "цинковыми пальцами" CTCF. TFBS могут быть модифицированы путем 1, 2, 3, 4, 5 или большего количества замен, добавлений и/или делеций и могут быть полностью или частично синтезированы. Оптимизированные TFBS, которые представляют собой TFBS с оптимизированным сродством связывания для соответствующего ДНК-связывающего белка и которые часто не имеют известного природного прототипа, также находятся в пределах объема настоящего изобретения. Эти оптимизированные TFBS также могут быть созданы путем вышеописанных модификаций встречающихся в природе TFBS или синтетическим путем, в частности путем химического синтеза. В некоторых воплощениях изобретения сайт(ы) связывания или TFBS придают тканеспецифичность MAR посредством, например, связывания тканеспецифичными природными, полученными генно-инженерным путем или синтетическими регуляторными белками или другими природными, полученными генно-инженерным путем или синтетическими белками, которые, например, могут реагировать на специфичные лекарства и молекулы. Генная и/или клеточная терапия являются типичными случаями, в которых полезна тканеспецифичность, а также она полезна для способности MAR специфично реагировать на определенное лекарство, то есть способность к индукции этим лекарством. В первом случае, например, интересующий ген должен экспрессироваться только в специфичных органах или тканях, в последнем случае экспрессию можно запускать, например, только в ответ на определенное лекарство. Другими не ограничивающими примерами факторов транскрипции, для которых могут быть включены TFBS, являются, например, SATB1, NMP4, MEF2, S8, DLX1, FREAC7, BRN2, GATA 1/3, ТАТА, Bright, MSX, AP1, С/ЕВР, CREBP1, FOX, Freac7, HFH1, HNF3alpha, Nkx25, POU3F2, Pit1, TTF1, XFD1, AR, С/ЕВР гамма, Cdc5, FOXD3, HFH3, HNF3 бета, MRF2, Oct1, POU6F1, SRF, V$MTATA_B, XFD2, Bach2, CDP CR3, Cdx2, FOXJ2, HFL, HP1, Мус, PBX, Pax3, TEF, VBP, XFD3, Brn2, COMF1, Evil, FOXP3, GATA4, HFN1, Lhx3, NKX3A, POU1F1, Рах6 и/или TFIIA.
Сайт связывания, такой как TFBS, указан как прилежащий к внутренней нуклеотидной последовательности, если внутренняя нуклеотидная последовательность и сайт связывания разделены не более чем примерно 200, предпочтительно не более чем примерно 100 нуклеотидами, даже более предпочтительно не более чем примерно 50 нуклеотидами, даже более предпочтительно не более чем примерно 25, не более чем примерно 15, не более чем примерно 5 или не разделены нуклеотидами. В предпочтительном воплощении сайты связывания, в частности TFBS, сами содержат короткие линкеры или адаптеры вплоть до 25 нуклеотидов с каждой стороны TFBS. В еще более предпочтительном воплощении TFBS составляют часть олигомера вплоть до примерно 50 нуклеотидов, вплоть до примерно 40 нуклеотидов или вплоть до примерно 30 нуклеотидов. Серия сайтов связывания, таких как TFBS, в соответствии с настоящим изобретением представляет собой ряд TFBS, расположенных в последовательности друг за другом. Серия TFBS указана как прилежащая к внутренней нуклеотидной последовательности, если TFBS из этой серии, который является ближайшим к внутренней последовательности, находится на расстоянии, указанном выше. Сайт связывания указан как фланкирующий "АТ-богатую область", если этот сайт связывания представляет собой сайт связывания, который является частью внутренней нуклеотидной последовательности и имеет прототип в идентичной локализации в природной MAR.
Сайт связывания может быть модифицирован путем 1, 2, 3, 4, 5 или большего количества замен, добавлений и/или делеций. Предпочтительно эти замены, добавления и/или делеции включают таким образом, чтобы сайт связывания совпадал с консенсусной последовательностью соответствующего сайта связывания.
Разнообразные усиленные конструкции MAR составляют часть настоящего изобретения и обладают свойствами, которые состоят в усилении по сравнению с природными и/или идентифицированными MAR, на которых может быть основана конструкция MAR согласно настоящему изобретению, в частности с природными MAR, на которых основана внутренняя нуклеиново-кислотная последовательность. Такие свойства включают, но не ограничены ими, уменьшенную длину относительно полноразмерной природной и/или идентифицированной MAR, усиление экспрессии/транскрипции гена, усиление стабильности экспрессии, тканеспецифичность, способность к индукции или их комбинации. Соответственно, конструкция MAR, которая является усиленной, может, например, содержать менее чем примерно 90%, предпочтительно менее чем примерно 80%, даже более предпочтительно менее чем примерно 70%, менее чем примерно 60% или менее чем примерно 50% от числа нуклеотидов идентифицированной последовательности MAR. Конструкция MAR может усиливать экспрессию гена и/или транскрипцию гена после трансформации соответствующей клетки указанной конструкцией. Если в контексте настоящего изобретения ссылаются на конструкции МАР/(нуклеотидные) последовательности MAR, которые "усиливают экспрессию", обладают "активностью усиления экспрессии гена", "усиливают экспрессию белка" или тому подобное, это "усиление" представляет собой усиление относительно экспрессии, например, гена, экспрессируемого в эквивалентных условиях во всем остальном, но в отсутствие такой последовательности. Усиление может, например, быть примерно 2, примерно 3, примерно 4, примерно 5, примерно 6, примерно 7, примерно 8, примерно 9, примерно 10-кратным или примерно 15-кратным, примерно 20-кратным или примерно 25-кратным или выше.
Конструкция MAR может также увеличивать средний процент очень высокопродуцирующих клеток примерно в 5 раз, примерно в 10 раз, примерно в 15 раз или более. Таким образом, помимо более высокой средней экспрессии гена, повышение процента очень высокоэкспрессирующих клеток, а также встречаемости стабильных ("устойчивых") колоний (примерно 100%, примерно 200%, примерно 300% или примерно 400% или большее повышение и/или более низкая вариабельность экспрессии (снижение cv (коэффициента вариации) примерно на 30%, примерно на 40%, примерно на 50% или более)) находится в пределах объема настоящего изобретения.
Конструкция MAR или подобная конструкция может "усиливать стабильность экспрессии". Это "усиление" является относительным к экспрессии, например, гена, экспрессируемого в эквивалентных условиях во всем остальном, но в отсутствие такой конструкции MAR/последовательности MAR. Усиление стабильности может, например, поддерживать 100% усиление вплоть до примерно 5, 10, 20, 25, 30, 35, 40, 45 или 50 недель. Конструкция MAR может быть специфична, например, для мышц, печени, центральной нервной системы или других тканей и/или может индуцироваться при введении вещества, такого как антибиотики, гормоны и/или промежуточные соединения метаболизма.
Конструкция MAR/последовательность MAR может быть встроена предпочтительно выше промоторной области, которая относится к интересующему гену, либо может быть оперативно сцеплена с ним. Однако в некоторых воплощениях предпочтительно, чтобы конструкция MAR была локализована как выше, так и ниже или непосредственно ниже интересующего гена/нуклеиново-кислотной последовательности. Другие множественные конфигурации MAR как в цис, так и/или в транс положении также находятся в пределах объема настоящего изобретения.
Конструкция MAR или область MAR указана как основанная, например, на идентифицированной MAR или на области идентифицированной MAR, если она обладает одной или более чем одной общей характеристикой (как, например, двумя, тремя или четырьмя) общими характеристиками с природными "SAR" или "MAR" или их соответствующей областью и обладает по меньшей мере одним свойством, которое способствует экспрессии любого гена, на который влияет такая MAR. Эти конструкции MAR или области MAR, как правило, обладают "существенной идентичностью" с идентифицированными MAR, на которых они основаны, в соответствии с определением этого термина, приведенным здесь. Несмотря на эти и/или другие модификации их нуклеотидной последовательности, они должны сохранять по меньшей мере одну функцию/характеристику лежащих в основе идентифицированных MAR.
Настоящее изобретение также направлено на применения конструкций MAR, включая усиленные конструкции MAR. В этих применениях конструкцию MAR можно также объединять с одним или более чем одним инструментом эпигенетической регуляции гена, представляющим собой не MAR, как, например, но не ограничиваясь ими, модификаторы гистонов, такие как деацетилаза гистонов (HDAC), другие элементы ДНК, такие как регуляторные области локуса (LCR), инсуляторы, такие как cHS4, или антирепрессорные элементы (например, стабилизаторные и антирепрессорные элементы (элементы STAR или UCOE) или горячие точки (Kwaks THJ и Otte АР)).
Синтетические при использовании в контексте MAR/конструкций MAR относятся к MAR, в дизайн которых вовлечено больше действий, чем простая перегруппировка, дупликация и/или делеция последовательностей/областей или частичных областей идентифицированных MAR или основанных на них MAR. В частности, синтетические MAR/конструкции MAR, как правило, содержат одну или более чем одну, предпочтительно одну область идентифицированной MAR, которая, однако, в некоторых воплощениях может быть синтезирована или модифицирована, а также специально сконструированные, хорошо охарактеризованные элементы, такие как один или серия TFBS, которые в предпочтительном воплощении получены синтетическим путем. Эти элементы дизайна во многих воплощениях являются относительно короткими, в частности их длина, как правило, составляет не более чем примерно 300 п.о., предпочтительно не более чем примерно 100, примерно 50, примерно 40, примерно 30, примерно 20 или примерно 10 п.о. Эти элементы в некоторых воплощениях могут быть мультимеризованы.
MAR млекопитающего, отличного от человека, согласно настоящему изобретению представляет собой MAR/последовательность MAR, которая, по меньшей мере частично, определена геномом или частями генома организма млекопитающего, отличного от человека. Она включает, например, MAR/последовательности MAR, идентифицированные посредством анализа генома грызунов, такого как, но не ограниченного им, геном мыши.
Вектор согласно настоящему изобретению представляет собой молекулу нуклеиновой кислоты, способную транспортировать другую молекулу нуклеиновой кислоты, с которой она сцеплена. Например, плазмида является типом вектора, ретровирус или лентивирус является другим типом вектора.
Трансфекция согласно настоящему изобретению представляет собой введение нуклеиновой кислоты в эукариотическую клетку-реципиент, как, например, но не ограничиваясь этим, путем электропорации, липофекции, посредством вирусного вектора или химическими способами.
Трансформация, как используют здесь, относится к модификации эукариотической клетки путем добавления нуклеиновой кислоты. Например, трансформация клетки может включать трансфекцию клетки нуклеиновой кислотой, как, например, путем введения векторной ДНК посредством электропорации. Однако во многих воплощениях изобретения путь введения усиленных MAR по настоящему изобретению в клетку не ограничен каким-либо конкретным способом.
Транскрипция означает синтез РНК с ДНК-матрицы.
Цис относится к положению двух или более чем двух элементов (таких как хроматиновые элементы) на одной и той же молекуле нуклеиновой кислоты, такой как, но не ограниченной ими, один и тот же вектор или хромосома.
Транс относится к положению двух или более чем двух элементов (таких как хроматиновые элементы) на двух или более чем двух молекулах нуклеиновой кислоты, таких как, но не ограниченных ими, два или более чем два вектора или две или более чем две хромосомы.
Последовательность указана как действующая в цис и/или в транс положении, например, на ген, когда она проявляет свою активность из цис/транс положения.
Окно согласно настоящему изобретению описывает число пар оснований, оцениваемых на MAR, например, во время методики SMAR Scan. Это число обычно составляет примерно 50 п.о., примерно 100 п.о., примерно 200 п.о., примерно 300 п.о. Однако окна 400, 500, 600 или более п.о. также находятся в пределах объема настоящего изобретения.
Нуклеотидная последовательность или ее фрагмент обладает существенной идентичностью с другими, если они при оптимальном выравнивании (с соответствующими нуклеотидными инсерциями или делециями) с другой нуклеотидной последовательностью (или ее комплементарной нитью) существует идентичность нуклеотидной последовательности по меньшей мере примерно на 60% оснований нуклеотидов, обычно по меньшей мере примерно на 70%, более типично по меньшей мере примерно на 80%, предпочтительно по меньшей мере примерно на 90% и более предпочтительно по меньшей мере примерно на 95-98% оснований нуклеотидов.
Идентичность означает степень родственности последовательности между двумя нуклеотидными последовательностями, которую определяют по идентичности совпадения между двумя нитями таких последовательностей, как, например, полная и полноразмерная последовательность. Идентичность можно легко вычислить. Хотя существует ряд способов измерения идентичности между двумя нуклеотидными последовательностями, термин "идентичность" хорошо известен специалистам в данной области техники (Computational Molecular Biology, Lesk, A. M., ed., Oxford University Press, New York, 1988; Biocomputing: Informatics and Genome Projects, Smith, D. W., ed., Academic Press, New York, 1993; Computer Analysis of Sequence Data, Part I, Griffin, A. M., and Griffin, H. G., eds., Humana Press, New Jersey, 1994; Sequence Analysis in Molecular Biology, von Heinje, G., Academic Press, 1987; и Sequence Analysis Primer, Gribskov, M. и Devereux, J., eds., M Stockton Press, New York, 1991). Способы, обычно применяемые для определения идентичности между двумя последовательностями, включают, но не ограничены ими, способы, описанные в Guide to Huge Computers, Martin J. Bishop, ed., Academic Press, San Diego, 1994, и Carillo, H., и Lipman, D., SIAM J Applied Math. 48: 1073 (1988). Предпочтительные способы определения идентичности предназначены для того, чтобы давать самое большое совпадение между двумя тестируемыми последовательностями. Такие способы закодированы в компьютерных программах. Предпочтительные компьютерные программные способы определения идентичности между двумя последовательностями включают, но не ограничены ими, пакет программ GCG (Genetics Computer Group, Madison Wis.) (Devereux, J., et al., Nucleic Acids Research 12(1). 387 (1984)), BLASTP, BLASTN, FASTA (Altschul et al. (1990); Altschul et al. (1997)). Хорошо известный алгоритм Smith Waterman можно также использовать для определения идентичности.
В качестве иллюстрации, под нуклеиновой кислотой, содержащей нуклеотидную последовательность, имеющую по меньшей мере, например, 95% "идентичность" со сравнительной нуклеотидной последовательностью, подразумевают, что нуклеотидная последовательность этой нуклеиновой кислоты идентична сравнительной последовательности за исключением того, что эта нуклеотидная последовательность может включать вплоть до пяти точечных мутаций на каждые 100 нуклеотидов сравнительной нуклеотидной последовательности. Иными словами, для получения нуклеотидной последовательности, имеющей по меньшей мере 95% нуклеотидов, идентичных сравнительной нуклеотидной последовательности, вплоть до 5% нуклеотидов в сравнительной последовательности могут быть делегированы или заменены другим нуклеотидом, или число нуклеотидов вплоть до 5% общего числа нуклеотидов в сравнительной последовательности может быть встроено в сравнительную последовательность. Эти мутации сравнительной последовательности могут встречаться в 5' или 3' концевых положениях сравнительной нуклеотидной последовательности или где-либо между этими концевыми положениями, чередуясь либо индивидуально в сравнительной последовательности, либо в одной или более чем одной непрерывной группе в пределах сравнительной последовательности.
Функциональные фрагменты нуклеотидных последовательностей также составляют часть настоящего изобретения. Фрагменты считают функциональными настолько, насколько они сохраняют желаемую функцию встречающихся в природе прототипов последовательностей, в частности повышение экспрессии гена, на который они влияют. Фрагмент MAR или области MAR все еще считают функциональным фрагментом, если его делеция уменьшает активность усиления транскрипции MAR/области, но не прерывает ее. "Полностью функциональный фрагмент" представляет собой фрагмент, в котором какое-либо уменьшение активности, если его вообще наблюдают, невозможно подтвердить статистически, когда этот фрагмент используют без других последовательностей MAR. В объем настоящего изобретения также включены функциональные фрагменты, имеющие существенную идентичность в соответствии с определением, предложенным здесь, например, с природной MAR, идентифицированной MAR, областью MAR или фрагментом любой из них.
Как более подробно описано здесь ниже, в некоторых воплощениях их блоки или части перегруппируют, дуплицируют и/или подвергают делетированию. Как должно быть понятно специалистам в данной области техники, такая перегруппировка и/или дупликация областей может создать, например, новые сайты рестрикции, которые, в свою очередь, могут привести к новому рестрикционному паттерну конструкций, созданных таким образом, и могут привести к корректировкам длины их последовательностей. Эти корректировки могут действовать, но не ограничены ими, на 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 10-15, 15-20, 20-25, 25-30, 30-35, 35-40 нуклеотидов. Эти корректировки, а также другие модификации находятся в пределах объема настоящего изобретения. Последовательности перестроенных MAR, в частности, перегруппированных и/или дуплицированных MAR, которые обладают существенной идентичностью в соответствии с определением, предложенным здесь, с каждым из их соответствующих элементов (или областей/блоков) и/или фрагментов находятся в пределах объема настоящего изобретения.
Последовательности MAR могут быть перенесены из клеток растений в клетки млекопитающих или наоборот и сохраняют активность присоединения к ядерному матриксу в гетерологичных клетках-хозяевах [Breyne P, Van Montagu M, Depicker А и Gheysen G, Mielke С, Kohwi Y, Kohwi-Shigematsu T and Bode J]. Учитывая эту консервативность функций MAR для всех высших эукариот, следует ожидать, что последовательность MAR из одного рода должна работать как у рода, из которого она имеет происхождение, так и у другого рода.
Тем не менее, в связи с тем, что последовательности MAR, имеющие происхождение из грызунов, могут иметь некоторое преимущество для продуцирования рекомбинантных белков, весь геном мыши был подвергнут скринингу на идентификацию последовательностей-кандидатов MAR, используя компьютерную программу SMAR Scan I, которая, как описано ниже, обнаруживает структурные признаки последовательностей ДНК (например, изгиб ДНК).
Как обсуждено ниже, было неожиданно обнаружено, что не человеческие последовательности MAR, в частности из грызунов (здесь мыши), более эффективны в отношении усиления экспрессии, например, в клетках СНО, а также в человеческих клетках, таких как клетки HeLA. Даже более неожиданно было обнаружено, что некоторые не человеческие последовательности MAR работают существенно лучше как в не человеческих клетках, например в клетках СНО, так и в человеческих клетках, например в клетках HeLa, чем человеческие последовательности MAR.
Для некоторых из идентифицированных новых последовательностей ДНК S/MAR мышиного происхождения было возможно показать повышение экспрессии трансгена, таким образом, предоставив доказательство того, что программа SMAR Scan I, разработанная для человеческих последовательностей MAR и протестированная на них, является эффективным инструментом для идентификации элементов S/MAR, имеющих происхождение из множества геномов, например мышиного, в дополнение к человеческому. Однако было важно обнаружить, что более эффективные элементы MAR можно идентифицировать путем скрининга генома грызунов (например, мыши), чем путем скрининга человеческого генома. В частности, в изобретении установлено, что высокоактивные элементы S/MAR из мышиного генома можно использовать для повышения продуцирования рекомбинантных белков, таких как рекомбинантные белки, имеющие применение в фармацевтике, в ряде клеток, в частности мышиных и человеческих клеток. Было показано, что мышиная S/MAR S4 является наиболее эффективной из вновь выделенных мышиных MAR и из ранее клонированных человеческих MAR. Изобретение, таким образом, направлено на не человеческие MAR, обладающие усиленным продуцированием белка, и/или на MAR, усиливающие стабильность экспрессии белка со временем.
SMAR Scan I представляет собой программный инструмент, который идентифицирует последовательности-кандидаты MAR на основе структурных и физико-химических признаков этих последовательностей. Подробное обсуждение этого метода приведено в других местах (публикация патента США 20070178469 авторов Mermod et al.). По существу "SMAR Scan" описывает биоинформационные инструменты, содержащие алгоритмы, которые распознают профили, основанные на динуклеотидных массовых матрицах, для компьютерного вычисления теоретических значений для конформационных и физико-химических свойств ДНК. Предпочтительно SMAR Scan оценивает признаки последовательности ДНК, соответствующие потенциалам изгиба ДНК, глубины большой бороздки и ширины малой бороздки, температурам плавления в широком ряде комбинаций, используя окна сканирования вариабельных размеров. Для каждого признака нужно установить значение отсечения или пороговое значение. Программа выдает совпадение каждый раз, когда вычисленный балл данной области находится выше установленного значения отсечения/порогового значения.
Для управления этими совпадениями доступны два режима вывода данных, первый (названный "по профилю") просто выдает все совпадающие положения на исследуемой последовательности и их соответствующие значения для различных выбранных критериев. Второй режим (названный "непрерывные совпадения") выдает только положения нескольких непрерывных совпадений и их соответствующую последовательность. Для данного режима минимальное число непрерывных совпадений является другим значением отсечения/пороговым значением, которое можно установить, опять же, с настраиваемым размером окна. Для настройки значений отсечения/пороговых значений по умолчанию, например для четырех теоретических структурных критериев, можно использовать экспериментально подтвержденные MAR, например, из SMARt DB. Таким образом, например, все человеческие последовательности MAR из базы данных были изъяты и проанализированы с помощью SMAR Scan, используя режим "поиска по профилю" с четырьмя критериями и без установленного значения отсечения/порогового значения. Это дало возможность установления каждой функции для каждого положения последовательностей. Затем распределение по каждому критерию вычисляли на компьютере в соответствии с этими данными (см. фиг.1 и 3 публикации патента США 20070178469 авторов Mermod et al.).
Хотя использование технологии SMAR Scan предпочтительно для идентификации последовательностей MAR, специалистам в данной области техники должно быть понятно, что другие биоинформационные инструменты, которые позволяют идентифицировать мотивы S/MAR с подобной или даже несколько более низкой избирательностью, можно использовать в контексте настоящего изобретения. Предпочтительно такие инструменты могут быть установлены таким образом, что только те признаки, связанные с MAR, которые проявляют эти признаки выше определенного значения, которое представляет собой установленное пороговое значение или значение отсечения, дают положительный результат или могут быть установлены для его получения. Многие биоинформационные инструменты, используемые для идентификации MAR, были, однако, разработаны для идентификации активности связывания с матриксом. Эта активность необязательно коррелирует со способностью к повышению экспрессии гена [Phi-Van, L. & Stratling, W.H.].
Программа SMAR Scan 1 разработана для идентификации человеческих MAR. Следовательно, она была разработана с использованием структурных данных, собранных на основании известных человеческих MAR. Настроенная на человека программа SMAR Scan I была использована в контексте настоящего изобретения для оценки мышиного генома на последовательности MAR. Однако различия в составе оснований мышиного и человеческого геномов не давали возможности использовать программу SMAR Scan с установками, определенными ранее для сканирования человеческого генома (публикация патента США 20070178469 авторов Mermod et al.). Таким образом, отдельный размер окна и пороговые значения структурных параметров следовало определять путем проб и ошибок до тех пор, пока эта программа не дала бы возможность идентификации оперируемой коллекции мышиных последовательностей-кандидатов MAR. Некоторые из них при тестировании оказались "сверх MAR-последовательностями", которые представляют собой последовательности MAR, позволяющие существенно повысить продуцирование белка, при помещении, например, на вектор с геном, кодирующим соответствующий белок, и введении в клеточную линию грызунов.
Мышиная MAR S4 и мышиная MAR S46 являются примерами последовательностей MAR, которые находятся в пределах объема настоящего изобретения. Эти последовательности MAR в изолированном виде представлены в прилагаемом перечне последовательностей как SEQ ID No. 3 и SEQ ID No. 10. Однако, как должно быть понятно специалистам в данной области техники, инсерции, делеции, замены пар оснований, в частности их фрагментов и других не человеческих MAR, которые сами по себе могут содержать инсерции, делеции, замены пар оснований, находятся в пределах объема настоящего изобретения настолько, насколько они сохраняют желаемую функцию последовательностей дикого типа, в частности повышения экспрессии гена, на который они влияют. Например, инсерция, которая уменьшает активность усиления транскрипции/экспрессии гена последовательностью MAR, но не прерывает ее, считают по существу не препятствующей желаемой функции MAR, здесь усилению экспрессии гена. Подобным образом, фрагмент, например, идентифицированной MAR все еще считают функциональным фрагментом, если он обладает несколько сниженной активностью усиления транскрипции относительно идентифицированной MAR, но полностью не утрачивает активность усиления транскрипции. "Полностью функциональный фрагмент" представляет собой фрагмент, в котором какое-либо уменьшение активности, если его вообще наблюдают, невозможно подтвердить статистически. Как подробно описано здесь в другом месте, в объем настоящего изобретения также включены последовательности, обладающие "существенной идентичностью" с нуклеотидной последовательностью встречающейся в природе MAR или ее фрагментом.
БЛОЧНОЕ СТРОЕНИЕ MAR
Идентифицированные MAR были проанализированы, чтобы определить, содержат ли они блоки (или области), в частности блоки со специфичной последовательностью, которые можно было бы использовать в генной инженерии идентифицированных MAR или при получении синтетических MAR, включая MAR, содержащие синтезированные области. В действительности можно было подтвердить несколько специфичных по последовательности блоков идентифицированных MAR. Неожиданно было обнаружено, что перегруппировка и/или полная или частичная дупликация и даже делеция некоторых их блоков или участков приводит в результате к усиленным MAR, как описано выше.
Человеческая 1_68 MAR и S4 MAR из мыши служат в качестве модели для получения конструкций MAR путем перегруппировки, делеции и/или дупликации областей. Однако, как должно быть легко понятно специалистам в данной области техники, настоящее изобретение направлено на манипуляции с любыми идентифицированными MAR и конструкциями MAR, полученными в результате из них. Соответствующие корректировки, которые могут быть необходимы для приспособления других MAR, включая MAR другого происхождения, хорошо соответствуют компетенции специалистов в данной области техники. Примеры включают, но не ограничены ими, эукариотические организмы, предпочтительно млекопитающих, в частности модельные организмы, такие как мышь, и виды, важные для экономики, такие как крупный рогатый скот, свиньи, овцы, а также людей.
Блочное строение человеческих MAR
Человеческая 1_68 MAR служила в качестве модели для получения конструкций MAR путем перегруппировки и/или дупликации областей. Используя блоки, подтвержденные, как описано ниже, или их части, были получены конструкции MAR на основе идентифицированных MAR, таких как человеческая 1_68 MAR. Конструкции MAR были, в частности, получены путем перегруппировки и/или дупликации областей (блоков) или их частей.
Пример 1_68 MAR показывает, что блоки (также называемые здесь областями или элементами) идентифицированной MAR были все необходимы, чтобы дать возможность усиления экспрессии гена до способности природной MAR. Ни один из идентифицированных блоков сам по себе не был способен к достижению полной активности MAR. Неожиданно было обнаружено, что перегруппировка и полная или частичная дупликация некоторых блоков приводит в результате к дополнительному усилению экспрессии гена.
Было идентифицировано несколько не избыточных специфичных по последовательности блоков (областей). Эти блоки кооперированы по влиянию на локальную структуру хроматина. Организация MAR некоторым образом параллельно регулирует транскрипцию многоклеточного организма: разнообразная коллекция блоков, которые распределены вплоть до нескольких килобаз от сайта инициации, все вместе определяют, где будет происходить инициация транскрипции.
Идентифицированные блоки со специфичной последовательностью находились в конкретных (1) областях с высоким содержанием А и Т, таких как симметричные А-Т-богатые области (чередующиеся А и Т), в частности "АТ-богатые области", и (2) областях, обогащенных сайтами связывания, в частности, но не ограничиваясь ими, TFBS, разделенных А-Т-богатыми областями.
Сообщили, что изогнутую ДНК с высоким содержанием А и Т обычно обнаруживают в промоторных областях, MAR и репликаторах [Aladjem и Fanning 2004]. Ранее считали, что последовательности с высоким содержанием А и Т ("симметричные" последовательности, как описано выше, а также "асимметричные" последовательности, которые представляют собой последовательности, имеющие главным образом А на одной нити и главным образом Т на другой), в основном, облегчают раскрытие дуплекса. Однако эти области могут обладать широким рядом функций. Например, последовательности с высоким содержанием А и Т в репликаторе ламина В2 связывают комплекс распознавания точки начала репликации (ORC) [Abdurashidova, Danailov et al. 2003; Stefanovic, Stanojcic et al. 2003] и могут облегчить нагрузку геликазы Mcm4/6/7 и раскручивание дуплексной ДНК in vitro [You, Ishimi et al. 2003]. Архитектурные роли самостоятельно изогнутых ДНК с высоким содержанием А и Т также рассмотрены. "ДНК-связывающие мотивы АТ-крючка" ORC4 делящихся дрожжей, которые напоминают мотивы белка HMG-I/Y из группы высокомобильных белков, могут обладать такой архитектурной ролью [Strick и Laemmli 1995; Bell 2002]. Может также иметь место опосредованный белком изгиб, аналогичный опосредованному HMG-I/Y изгибу ДНК, который способствует рекомбинации V(D)J, a также сборке и стабилизации транскрипционных комплексов при энхансерах и промоторах у эукариот [Levine and Tjian 2003]. Не все области, которые имеют высокое содержание А и Т, соответствуют изогнутой ДНК. Однако те ДНК, которые являются изогнутыми, могут действовать в качестве 'магнита гистонов' для привлечения гистонов с образованием нуклеосом вокруг изогнутой ДНК, оставляя соседние области свободными, чтобы действовать в качестве подложки для белков пререпликации/транскрипции.
Как описано выше, MAR также содержат сайты связывания для других белков, в частности, в "областях, обогащенных сайтами связывания" или просто в "областях сайтов связывания" (см. (2) выше). Эти другие белки могут включать, но не ограничены ими, белок связывания элемента, раскручивающего ДНК (DUE-B), и факторы транскрипции, такие как белки Нох, SATB1, СЕВР и т.д., которые обнаружены в 1_68 MAR. Мутационный анализ показывает, что эти сайты связывания вносят вклад в функцию MAR.
Человеческая 1_68 MAR может быть усовершенствована путем обращения ее ориентации и путем удаления изогнутой ДНК с увеличением размера области сайтов связывания факторов транскрипции выше промоторной области. Как можно видеть на фиг.9, ряд этих перестроенных MAR (например, конструкция 6) значительно увеличивает транскрипцию относительно конструкции без MAR (в 10 раз) и даже относительно конструкции, включающей природную MAR (конструкции 1 и 16; примерно в 2 раза). Представленные данные также являются сильным показателем того, что сам по себе дистальный элемент регуляции транскрипции ограничивает инициацию транскрипции в нижележащем хроматине. Фрагмент 223 п.о., локализованный при 3'-конце области, показанный в виде прямо заштрихованного прямоугольника, в природной MAR полностью сохраняет активность данной области в конструкции 7 по сравнению с конструкцией 11. Это позволяет предположить, что данный важный участок должен в данном случае кооперативно взаимодействовать с областью изгиба и с 5'-концом остальной части (нуклеотиды 1-1425) элемента в конструкции 6. Было обнаружено, что два сайта HMG-I/Y локализованы возле этого конца. Конструкция 2 показывает, что соединение двух идентифицированных последовательностей MAR вместе также повышает экспрессию.
Блочное строение мышиных MAR и уменьшение размера
Несколько MAR было сконструировано на основе S4 MAR (таблица 3) и охарактеризовано (фиг.10). Как можно видеть на фиг.10, внутренняя делеция фрагмента длиной более чем 1600 п.о. не приводит к значительной потере активности MAR (S4-1-703_2328-5457). Однако делеция близкого к промотору фрагмента 795 п.о. или замена этой последовательности фрагментом гена люциферазы подобной длины (S4_1-4661; S4_1-4661-Luc5489) вызывала полную утрату этой активности.
Блоки, неспецифичные по последовательности: Активность 3' концевых последовательностей MAR
Эксперимент с человеческой 1_68 MAR (фиг.9) уже показал значимость области сайтов связывания 3' HoxF и SATB1 человеческой 1-68 MAR. Значимость этой области была дополнительно продемонстрирована экспериментами с мышиной MAR S4, представленными на фиг.10. Как показано на фиг.11, для дополнительного анализа активности 3' концевых последовательностей MAR S4, этот участок MAR был дополнительно рассечен путем удаления или дупликации его областей. На фиг.11 также показан эффект различных производных MAR S4 на экспрессию гена. Интересно, что одно такое производное, имеющее укороченный 3' конец (4658-5054 по сравнению с 4658-5457 исходной MAR S4), проявляло в среднем несколько более высокую экспрессию трансгена по сравнению с более длинной исходной последовательностью MAR S4 (104% по сравнению со 100%). Это указывает на возможность получения более эффективных, а также более коротких производных элементов MAR.
Таким образом, настоящее изобретение включает конструкции MAR с высокой активностью, которые являются значительно более короткими по длине, чем их природные прототипы, что, таким образом, позволяет получить конструкции более удобного размера, например, для конструирования и переноса вектора.
В частности, конструкции MAR, содержащие менее чем примерно 90%, предпочтительно менее чем примерно 80%, даже более предпочтительно менее чем примерно 70%, менее чем примерно 60% или менее чем примерно 50% числа нуклеотидов идентифицированной последовательности MAR, находятся в пределах объема настоящего изобретения. Эти конструкции предпочтительно содержат 3' концевую область идентифицированной MAR, даже более предпочтительно по меньшей мере примерно 5%, примерно 6%, примерно 7%, примерно 8%, примерно 9% или примерно 10% 3' концевой области идентифицированной MAR/последовательности MAR. Однако конструкции MAR, которые содержат 5' концевую область идентифицированной MAR, также находятся в пределах объема настоящего изобретения.
СИНТЕТИЧЕСКИЕ MAR
Перестройка человеческой 1_68 MAR показала, что фрагмент 223 п.о. Нох-богатой области, локализованный при 3' конце прямо заштрихованного участка изолированной MAR, сохраняет в некоторых воплощениях активность полноразмерной области. Это позволяет предположить, что этот участок в некоторых воплощениях изобретения может быть важен при кооперировании с другими элементами. На фиг.12 показан порядок потенциальных сайтов связывания факторов транскрипции MAR 1_68, которые предсказаны программой MATInspector. Положение сайтов связывания С/ЕВР, NMP4, FAST1, SATB1 и HoxF показано в качестве примеров, иллюстрируя их большое количество в 5' (прямо заштрихованной) фланкирующей последовательности.
Обнаружение возможного кооперирования между АТ-богатой областью изгиба ДНК и сайтами связывания факторов транскрипции в человеческой MAR 1_68 привело в результате к конструированию MAR/конструкций MAR, содержащих АТ-богатую область MAR 1-68, прилежащую к одному или нескольким сайтам связывания факторов транскрипции. На фиг.13 изображена карта плазмиды, использованной для тестирования на активность синтетических MAR, сконструированных в результате сборки внутренней последовательности (MAR 1429-2880), содержащей как АТ-богатую область, так и TFBS идентифицированной MAR на каждом конце АТ-богатой области, а также химически синтезированные сайты связывания ДНК-связывающих белков для факторов транскрипции, помещенные выше промотора для зеленого флуоресцентного белка (GFP). На фиг.13 показано, в частности, что сайты связывания факторов транскрипции были встроены между АТ-богатым доменом и промотором SV40, направляющим экспрессию трансгена GFP, имитируя ситуацию, представленную на фиг.9, где участки MAR, содержащие сайты связывания, расположены между промотором и областью изгиба ДНК в наиболее выгодных положениях (конструкция 6). В таблице 4 показана последовательность ДНК синтезированных химическим путем олигонуклеотидов, которые были использованы.
Сайты связывания для факторов транскрипции С/ЕВР, NMP4, FAST1, SATB1 и HoxF (также названного Gsh) были идентифицированы на основании последовательности MAR 1-68 (фиг.12). Эти сайты связывания, как они встречаются в MAR 1-68, были использованы без изменения (FAST1, С/ЕВР, HOXF/Gsh), либо они были скорректированы в том случае, когда они имели одно или более чем одно несовпадение по сравнению с консенсусной (то есть правильной) последовательностью (HoxF, SATB1, NMP4).
Как можно видеть из фиг.14, добавление здесь синтетических сайтов связывания обеспечивает почти во всех случаях некоторое, а в некоторых случаях значительное усиление транскрипции по сравнению с внутренней последовательностью MAR, содержащей АТ-богатую область. С/ЕВР и Нох или Gsh2 были наиболее активными, причем следующими по активности были SATB1 и Fast1, тогда как один сайт NMP4 не обладал каким-либо обнаружимым эффектом.
На фиг.14 показан неожиданный результат, что вставка внутренней последовательности, здесь MAR 1429-2880, основанной на MAR 1_68, которую фланкируют сайты связывания идентифицированной MAR на основе АТ-богатой области, не дает значительного улучшения экспрессии, но конструкция MAR, дополнительно содержащая один или более чем один сайт связывания, в частности, при встраивании ниже АТ-богатой внутренней последовательности, но выше промотора, приводит в результате к значительному усилению экспрессии/продуцирования белка геном под контролем этого промотора (здесь идентифицированной как % клеток М3).
Хотя в предпочтительных воплощениях дополнительные сайты связывания находятся ниже АТ-богатой внутренней последовательности, но выше промотора, другие конфигурации, такие как, но не ограниченные ими, расположение выше АТ-богатой области, внутри АТ-богатой области, по соседству с АТ-богатой областью внутренней последовательности или ниже гена, также находятся в пределах объема настоящего изобретения.
В предпочтительном воплощении рассмотрены некоторые комбинации сайтов связывания белков, либо синтетических, либо изолированных, как, например, комбинации двух различных сайтов связывания белков, комбинации трех различных сайтов связывания белков, комбинации четырех, пяти, шести, семи, восьми, девяти, десяти или большего числа различных сайтов связывания белков. Эти комбинации могут быть мультимеризованы, полностью или частично. В предпочтительном воплощении комбинация включает Hox/Gsh и SATB1. Вставка этих комбинаций или мультимеризованных комбинаций, например, между внутренней последовательностью и подходящим промотором, может повысить встречаемость высокоэкспрессирующих клонов примерно в два раза или более, как, например, но не ограничиваясь этим, примерно в три, в четыре, в пять, в шесть, в семь, в восемь, в девять раз или более, предпочтительно примерно в 10 раз или более, даже более предпочтительно примерно в 11, 12, 13, 14, 15, 16, 17, 18, 19 раз или более или примерно в 20 или даже примерно в 25 или примерно в 30 раз или более относительно встречаемости высокоэкспрессирующих клонов, когда векторы, не содержащие конструкцию MAR/последовательность MAR, используют в условиях, эквивалентных во всем остальном.
Подводя итог, конструкции MAR могут быть собраны из строительных блоков. Эти строительные блоки могут включать или основываться на областях, таких как области специфичной последовательности идентифицированных MAR или их частей, синтетические строительные блоки (включая модификации для оптимизации их функциональности), такие как серии химически синтезированных сайтов связывания факторов транскрипции (TFBS), строительные блоки, имеющие происхождение или основанные на последовательностях, представляющих собой не MAR, или строительные блоки, имеющие происхождение или основанные на последовательностях MAR из другого вида или рода. В предпочтительном воплощении такие MAR содержат АТ-богатые области, связанные с областями TFBS или комбинациями специфичных сайтов ДНК-связывающих белков для факторов транскрипции, таких как показаны в таблице 5.
Специалистам в данной области техники должно быть понятно, что эти принципы не ограничены конкретными последовательностями или сайтами связывания, раскрытыми здесь, и что другие производные, гомологи или комбинации последовательностей также находятся в пределах объема настоящего изобретения.
Как упомянуто выше, конструкции MAR, экспрессионные системы и/или наборы по изобретению можно применять для продуцирования белков. Здесь конструкция MAR может быть включена в вектор, содержащий ген для интересующего белка, например инсулина, под контролем промотора. Этот вектор вводят в клетку, и клетки выращивают. Затем масштаб этого процесса увеличивают до широкомасштабного серийного производства инсулина. Высокую продукцию инсулина, например в 3-5 раз выше, чем без конструкции MAR, можно поддерживать в течение трех недель.
Как упомянуто выше, конструкции MAR, экспрессионные системы и/или наборы по изобретению можно применять для in vitro и/или in vivo генотерапии, а также в клеточной и тканевой заместительной терапии. Например, при генотерапии in vitro конструкцию MAR можно включать в вектор, содержащий ген, дефектный у пациента, нуждающегося в генотерапии in vitro, под контролем промотора. Затем конструкцию MAR вводят в клетки, такие как клетки костного мозга пациента. После трансформации конструкцией MAR эти клетки костного мозга вводят пациенту, и экспрессия интересующего гена может превышать уровень экспрессии без конструкции MAR в 5 раз. Таким образом, может быть экспрессировано эффективное количество белка.
При генотерапии in vivo вектор, содержащий конструкцию MAR, можно непосредственно вводить в клетки пациента, нуждающегося в этом, например, путем инъекции.
Подобным образом экспрессионные системы по настоящему изобретению можно вводить в стволовую клетку для приживления для регенерации ткани или, например, нейронной клеточной терапии для нейродегенеративных заболеваний. Не ограничивающими примерами стволовых клеток, которые можно применять в данном воплощении изобретения, являются гемопоэтические стволовые клетки (HSC) и мезенхимальные стволовые клетки (MSC), полученные из ткани костного мозга индивидуума в любом возрасте или из пуповинной крови новорожденного. Стволовые клетки трансфицируют экспрессионной системой согласно настоящему изобретению, и успешные трансформанты можно трансплантировать или обратно ввести пациенту, нуждающемуся в клеточной терапии или тканевой регенерационной терапии. Несколько способов доступно для получения трансформированных стволовых клеток, например, Nucleofection® (Cell Line Solution V (VCA-1003), Amaxa GmbH, Germany).
Трансгенные животные, которые могут продуцировать широкое разнообразие белков, включая антитела, которые связываются с человеческими антигенами, могут быть получены известными способами (например, но не ограничиваясь ими, патенты США №№5770428, 5569825, 5545806, 5625126, 5625825, 5633425, 5661016 и 5789650, опубликованные Lonberg et al.). Экспрессионные системы и конструкции MAR можно применять при продуцировании белков посредством, например, трансгенных коров, овец, коз или свиней, типично путем секреции белка в биологическую жидкость (например, молоко). См., например, патент США №5750172 авторов Meade et al. См. также получение трансгенных животных в патенте США №6518482 авторов Lubon et al.
ПРИМЕРЫ
Далее изобретение описано в приведенных ниже примерах, которые не ограничивают объем изобретения, изложенный здесь в формуле изобретения, кратком изложении сущности изобретения или в другом месте. Материалы, методы и примеры являются только иллюстративными и не предназначены для ограничения. С помощью приведенного здесь руководства специалист в данной области техники сможет осуществить модификации, дополнения и усовершенствования всего того, что находится в пределах объема настоящего изобретения.
Предсказание мышиного генома на S/MAR: SMAR Scan I
Все мышиные хромосомные последовательности, соответствующие мышиному набору хромосом NCBI m34, компилировали и анализировали с помощью программы SMAR Scan I. Были проведены скрининги низкой и высокой строгости с использованием либо порога для критерия изгиба ДНК 3,6 градуса и минимального размера окна 300 п.о., либо порога 4,2 градуса и минимального размера окна 100 п.о. соответственно.
Анализ низкой строгости с помощью SMAR Scan I всего мышиного генома дал в сумме 1496 предполагаемых S/MAR (кандидатов MAR), представляющих в сумме 622410 п.о. (0,024% от всего мышиного генома). В таблице 1 показаны для каждой хромосомы: ее размер, ее число генов, ее число предсказанных MAR (кандидатов MAR), ее плотности MAR на ген и среднее расстояние в кб между S/MAR. В этой таблице выявлено, что существуют различные плотности генов на предсказанные S/MAR (кандидаты MAR) на различных хромосомах (при стандартном отклонении, представляющем примерно 50% среднего плотности генов на MAR). Кратности различия между более высокой и более низкой плотностью генов на MAR составляет 6 без учета хромосомы Y, которая крайне богата предсказанными MAR (кандидатами MAR) относительно ее размера и ее числа генов, показывая сильное и неожиданное смещение в распределении этих MAR. В таблице 1 также показано, что среднее расстояние между S/MAR (кб на S/MAR) является вариабельным (стандартное отклонение составляет 38% от среднего в кб на S/MAR, и кратность различия между более высокой и более низкой плотностью генов на S/MAR составляет 8,3). Хромосомы 10, 11, X и Y вносят значительный вклад в высокое стандартное отклонение этих плотностей.
SMAR Scan I исходно настроена на человеческие последовательности и, следовательно, дает малое количество MAR с мышиными геномными последовательностями при использовании наиболее строгих параметров: поэтому значения отсечения по умолчанию были отрегулированы для скрининга высокой строгости (порог 4,2 градуса для критерия изгиба ДНК) до минимального размера непрерывных совпадений, которые следует учитывать как MAR, используя окно 100 п.о. вместо 300 п.о. Анализ мышиного генома с помощью SMAR Scan I предсказал 49 "сверх" MAR со значением >4,2 градуса для критерия изгиба ДНК.
Число генов на хромосому соответствует мышиному набору хромосом NCBI m34 (Национальный центр Биотехнологической информации). Размеры хромосом представляют собой сумму длин соответствующих эталонных последовательностей мыши.
Использование вновь идентифицированных мышиных MAR для повышения продуцирования рекомбинантных белков
Пять элементов MAR были отобраны из предполагаемых MAR (кандидатов MAR), полученных при скрининге высокой строгости полноразмерного генома мыши с помощью SMAR Scan. Они были клонированы в плазмидных векторах из бактериальных искусственных хромосом геномной ДНК мыши, приобретенных в Институте детской больницы Окленда (CHORI, http://bacpac.chori.org/).
Эти вновь идентифицированные мышиные MAR были названы S4, S8, S15, S32 и S46 (соответственно порядку идентификации SMAR Scan I, "сверх" MAR S1 - S49). Человеческие MAR 1_3, 1_6, 1_9, 1_42,1_68, 3_S5 и X_S29 идентифицированы ранее, причем MAR 1_68 и X_S29 являются наиболее эффективными элементами человека (Mermod et al., "High efficiency gene transfer and expression in mammalian cells by a multiple transfection procedure of MAR sequences," WO 2005/040377, см. также публикацию патента США 20070178469 авторов Mermod et al.). Эти MAR были встроены в контрольный вектор pGEGFP выше промотора и энхансера SV40, направляющих экспрессию зеленого флуоресцентного белка, и этими плазмидами были трансфицированы культивируемые клетки СНО, как описано ранее [Girod PA, Zahn-Zabel M и Mermod N]. Затем экспрессию трансгена анализировали в общей популяции стабильно трансфицированных клеток, используя аппарат для флуоресцентного клеточного сортинга (FACS). На фиг.1 показан эффект различных S/MAR на продуцирование рекомбинантного зеленого флуоресцентного белка (GFP). Показаны популяции клеток СНО, трансфицированные экспрессионным вектором GFP pGEGFP, содержащим или не содержащим MAR, как показано с помощью клеточного сортера с возбуждением флуоресценции (FACS®), и типичные профили. На данной фигуре показаны только наиболее эффективные человеческие MAR 1_68 и X_S29. Профили выявляют подсчет числа клеток как функцию уровней флуоресценции GFP. Горизонтальными полосами представлены субпопуляции клеток М1, М2 и М3 со значением относительных световых единиц флуоресценции менее чем 2 (М1) или более чем 102 (М2) или 103 (М3).
Как можно видеть из фиг.1, все вновь идентифицированные мышиные MAR повышали экспрессию трансгена значительно выше уровня экспрессии, направляемой только GFP без MAR, причем "сверх" мышиная MAR S4 является наиболее эффективной из всех представленных MAR.
Клетки СНО были совместно трансфицированы плазмидой селекции на антибиотике и репортерной конструкцией pGEGFP или производными pGEGFP, содержащими либо человеческие MAR 1_68 и X_S29, либо указанные мышиные S4, S8, S15, S32 или S46 MAR. Поликлональная популяция стабильно трансфицированных клеток была отобрана по устойчивости к антибиотику в течение двух недель и протестирована на флуоресценцию GFP с помощью анализа FACS, как изображено на фиг.1. В таблице представлено среднее значение флуоресценции, его коэффициент вариации (CV) и процент клеток, показывающих значение относительных световых единиц флуоресценции менее чем 2 (М1) или значения относительных световых единиц флуоресценции более чем 102 (М2) или 103 (М3). Эти результаты представляют собой средние значения, и стандартная ошибка среднего (СОС) была получена из трех независимых экспериментов.
Транскрипционную активность наиболее эффективных человеческих MAR 1_68 и X_S29 сравнивали с активностями, полученными с вновь идентифицированными мышиными MAR. Пять мышиных MAR были первоначально протестированы с помощью экспрессионных анализов GFP, и было обнаружено, что все они повышают экспрессию GFP до различных уровней. Мышиные MAR S15 и S32 по меньшей мере относительно обладают транскрипционной активностью MAR (~ 2-кратное повышение по сравнению с одним GFP), S8 и S46 показали среднюю активность (3-4-кратное повышение) и MAR S4 проявляла очень высокую транскрипционную активность (7-кратное повышение). Кроме того, мышиная MAR S4 является наиболее эффективной из всех MAR, протестированных в данном исследовании. Сравнение между транскрипционной активностью человеческой MAR 1-68 и мышиной MAR S4 выявило 50% повышение средней флуоресценции всей популяции (Gmean М0) и клеток с высоким продуцированием GFP (M2), тогда как процент клеток с очень высоким продуцированием GFP (М3) был на 175% выше с мышиной MAR S4. Однородность всей популяции в отношении флуоресценции GFP (CV М0) всегда была на 1-2% ниже с мышиной MAR S4, что является преимуществом, поскольку указывает на более высокую стабильность продуктивности клеток.
После этого первого раунда клонирования было решено определить, можно ли систематически получать высокоактивные элементы MAR из мышиного генома. Таким образом, две дополнительные мышиные MAR (S6 и S10) были клонированы и охарактеризованы. Эти новые мышиные MAR были встроены в контрольный вектор pGEGFP и проанализированы с помощью FACS, как описано выше. Мышиная MAR S10 оказалась также более эффективной, чем лучшие человеческие MAR по всем различным параметрам, анализируемым с помощью FACS, и обладает почти такой же транскрипционной активностью, как MAR S4, по повышению общей экспрессии.
Для оценки очень высоких продуцентов процент клеток М3 нормализовали по результату, полученному для человеческой MAR 1_68. Результат представлен на фиг.2. На фиг.2 показан эффект различных человеческих и мышиных элементов S/MAR на процент очень высоких продуцентов (% М3) рекомбинантного зеленого флуоресцентного белка (GFP). Популяции клеток СНО, трансфицированных экспрессионным вектором GFP, содержащим или не содержащим элемент MAR, как указано, были проанализированы с помощью клеточного сортера с возбуждением флуоресценции (FACS®). Процент очень высоких продуцентов был нормализован по полученному результату с лучшей человеческой MAR для данного критерия, MAR 1_68, значение которого было принято за 100.
Мышиные MAR S10 и S4 давали в среднем 80% и 180% более высокопродуцирующих клеток, чем человеческая MAR 1_68, соответственно. В целом на основании сравнения 7 мышиных MAR и 7 человеческих MAR был сделан вывод, что более высокая экспрессия была достигнута из клеток СНО с использованием MAR грызунов.
Оценка эффективности вновь идентифицированных мышиных MAR в различных типах клеток
Эффективность S4 MAR оценивали в клетках СНО. Кроме того, экспрессионными векторами EGFP, содержащими либо человеческую MAR 1-68, либо мышиную MAR S4, либо не содержащими MAR, были стабильно трансфицированы человеческие клетки HeLa и проанализирована флуоресценция EGFP. На фиг.3 показан эффект различных человеческих 1_68 и мышиных элементов S4 MAR на экспрессию рекомбинантного зеленого флуоресцентного белка (GFP). Популяции клеток HeLa были трансфицированы и проанализированы, как описано для таблицы 2. При сравнении эффективности S4 и 1-68 MAR в клетках HeLa было обнаружено, что S4 превосходит 1_68 в нескольких отношениях: S4 дает более высокую среднюю флуоресценцию GFP (среднее Gmean М0), а также большее количество клеток в среде и более высокий уровень экспрессии (М1 и М2 соответственно), а также более низкую вариабельность экспрессии (среднее CV М0). Не было обнаружено клеток с очень высоким диапазоном экспрессии (М3) с использованием клеток HeLa.
Усиленная экспрессия моноклональных антител с использованием мышиных MAR
Чтобы определить, можно ли использовать мышиные MAR, в частности наиболее эффективные, для увеличения продуцирования белков для фармацевтических применений, их встраивали в векторы pMZ37 и pMZ59, кодирующие тяжелые и легкие цепи иммуноглобулина, распознающего резус-D [Miescher S, Zahn-Zabal M, De Jesus, M, Moudry, R, Fisch, I, Vogel, M, Kobr, M, Imboden, MA, Kragten, E, Bichler, J, Mermod, N, Stadler, ВС, Amstutz, H., Wurm, F]. Этими плазмидами трансфицировали клетки СНО, отбор и анализы на иммуноглобулин проводили, как описано ранее [Girod PA, Zahn-Zabal M и Mermod N]. На фиг.4 показан эффект элементов S/MAR на продуцирование рекомбинантных моноклональных антител. Так, клетки СНО трансфицировали вышеупомянутыми векторами, направляющими экспрессию тяжелых и легких цепей IgG без MAR (нет MAR) или с MAR S4, присоединенной в цис положении. Титры IgG измеряли в надосадочных жидкостях через 24, 48 и 72 часа. Кроме того, как изображено на фиг.5, были получены стабильные клоны из популяции клеток СНО, трансфицированных вышеупомянутыми векторами, направляющими экспрессию тяжелых и легких цепей IgG без MAR (нет MAR) или с MAR S4, присоединенной в цис положении. После отбора титры секретируемых IgG измеряли в среде, и удельную продуктивность оценивали путем подсчета клеток. На фиг.6 (А) показаны результаты, полученные после создания стабильных индивидуальных клонов путем ограничения разведения из популяции клеток СНО, трансфицированных векторами, направляющими экспрессию тяжелых и легких цепей IgG без MAR (нет MAR) или с MAR S4, присоединенной в цис положении. После отбора титры секретируемых IgG измеряли в среде и удельную продуктивность оценивали путем подсчета клеток. Также включены сравнительные результаты, полученные с MAR 1_68, а также на (Б) результаты, полученные с клонами, не содержащими MAR. Результаты, полученные и изображенные на фиг.3-6, показывают, что вновь идентифицированные мышиные MAR, в частности MAR S4, можно использовать для усиления продуцирования фармацевтических белков, таких как моноклональные антитела, в транзитных трансфектантах (фиг.4) и в стабильных трансфектантах (фиг.5 и 6). Стабильные клоны с удельными продуктивностями около или выше 5 пг/клетка/сутки (пкс) могут быть легко идентифицированы на основании анализа нескольких клонов-кандидатов при использовании MAR S4 (фиг.6(А)). Действительно, средняя продуктивность 21 лучшего клона с MAR S4 или без MAR составляла 7,28±0,78 пкс (фиг.6(А)) и 2,61±1,09 пкс соответственно. Эти результаты противоположны уровням титров, полученных с известной MAR куриного лизоцима (менее чем 1,5 мг/л) или без MAR (менее чем 0,5 мг/л). В частности, эти результаты указывают на то, что вновь идентифицированные мышиные MAR можно использовать для усиления продуцирования белков фармацевтического применения, таких как, но не ограничиваясь ими, моноклональные антитела, что придает мышиным MAR, таким как MAR S4, особый интерес для продуцирования рекомбинантных белков.
Стабильность экспрессии с человеческой MAR 1_68
MAR 1_68 использовали, чтобы продемонстрировать, что экспрессия генов, которые продуцируются клонами, не содержащими MAR, постепенно прекращается, эквивалентные клоны, содержащие MAR, не только сохраняют высокий уровень экспрессии со временем, но молчащие клоны восстанавливают экспрессию.
На фиг.7 показана совместная трансфекция экспрессионной плазмиды pEGFP, содержащей MAR 1-68, в клетки СНО с геном устойчивости к антибиотику G418, и стабильно экспрессирующие клетки были отобраны в присутствии G418 в течение трех недель, как описано в Girod et al., 2005. Клеточные клоны были получены путем ограничения разведения, и 9 индивидуальных клонов анализировали на флуоресценцию GFP. Типичный клон, экспрессирующий GFP, был отобран из каждой из двух популяций для дальнейшего анализа, и его культивировали далее вплоть до 26 недель в присутствии или в отсутствие селекции антибиотиком. Профили уровней флуоресценции GFP (ось х) и числовых значений счета клеток на оси у после двух недель культивирования представлены слева, тогда как профили справа были получены от клеток, культивируемых в течение 26 недель. Как можно видеть, клон, в котором отсутствует MAR, показывает сниженный уровень флуоресценции GFP в отсутствие антибиотика через 26 недель относительно уровня через две недели, тогда как клон, содержащий MAR, может сохранять уровень флуоресценции GFP на неделе 26 с селекцией или без селекции антибиотиком, что делает экспрессионные системы, содержащие MAR, полезными для стабильной экспрессии интересующего гена.
Блочное строение MAR и релевантность для усиления экспрессии генов
Структурный анализ MAR выявил области/модули последовательности ДНК, каждая из которых вносит вклад в усиленную экспрессию генов. На фиг.8 изображены результаты, полученные с помощью структурного анализа 1_68 MAR. На фиг.8 (А) показано, что центральная АТ-богатая область определяет изгиб ДНК в локусе MAR 1_68. На фиг.8 (Б) показано, что эта АТ-богатая область окружена областями, обогащенными сайтами связывания факторов транскрипции, как идентифицировано с помощью Matlnspector (Cartharius, Frech et al. 2005). Точно 729 потенциальных TFBS были обнаружены с помощью Matlnspector вдоль последовательности MAR. В нижней части фиг.8 (Б) показаны атрибуты кодирования идентифицированных областей.
На фиг.9 (А) показана 1_68 MAR, слева показаны различные MAR, в которые включены области или части 1_68 MAR и изменен порядок и/или ориентация ее областей или частей и/или дуплицированы такие ее области или части. Справа показана степень увеличения транскрипции, достигнутая посредством конструкций 1 -16, а также увеличение транскрипции, достигнутое с MAR 1_68 или без MAR. Все представленные последовательности MAR были встроены выше промотора, направляющего экспрессию гена-маркера eGFP. Стрелками показана ориентация областей или их фрагментов относительно последовательности MAR дикого типа, изображенной на фиг.8. Последовательности, окружающие АТ-богатую область, показаны в виде обратно заштрихованного прямоугольника со стрелкой (слева) и прямо заштрихованного прямоугольника (не в масштабе; справа). Изогнутая область показана в виде перекрестно заштрихованного прямоугольника.
На фиг.9 (Б) показан паттерн изгиба MAR, который соответствует конструкции 6 на фиг.9А. Этот паттерн изгиба был определен с помощью SMAR Scan I.
На фиг.9 (В) показаны результаты анализа Matlnspector [Cartharius, Frech et al. 2005]. Потенциальные сайты связывания факторов транскрипции (TFBS) были идентифицированы с помощью Matlnspector [Cartharius, Frech et al. 2005]. 731 потенциальный сайт обнаружен с помощью Matlnspector вдоль последовательности MAR. Снизу на фиг.9 (В) показана конструкция 6 с использованием кодирования, соответствующего фиг.8 (Б) и Фиг.9 (А). Кодирование нижней части этой фигуры соответствует показанному и обсужденному на фиг.9 (А).
Эксперименты, изображенные на фиг.9, показывают, что ни одна из областей сама по себе не проявляет полной активности MAR. Например, для усиления транскрипции ДНК, являющееся результатом встречающейся в природе человеческой 1_68 MAR, полностью необходимы три различных последовательности (фиг.8): сегмент 1189 п.о., который содержит сайты связывания для множества факторов транскрипции (то есть СЕВР) (фиг.9А сверху, показан в виде обратно заштрихованного прямоугольника со стрелкой), собственно изогнутая ДНК, которая продиктована 763 п.о. симметричной АТ-богатой областью (чередующиеся А и Т) (фиг.9А сверху, перекрестно заштрихованный прямоугольник), и дополнительный сегмент 1648 п.о., который включает множество сайтов связывания HoxF и SATBI (Фиг.9А сверху, показан в виде прямо заштрихованного прямоугольника со стрелкой).
На фиг.9 показано, что усовершенствование человеческой 1_68 MAR путем удаления изогнутой ДНК с увеличением размера области сайтов связывания факторов транскрипции выше промоторной области. Для достижения этого увеличения области сайта связывания факторов транскрипции (TFBS), здесь Нох-богатой области (SEQ ID No. 19) (здесь далее прямо заштрихованный прямоугольник со стрелкой), прилежащей к АТ-богатой (SEQ ID. No. 18) области, присоединяли к СЕВР-богатой области (SEQ ID No. 17) (здесь далее также обратно заштрихованная область (фиг.9)). Сравнение активности усиления транскрипции различных полученных в результате конструкций MAR, которые изображены справа на фиг.9А, показывает, что ориентация прямо заштрихованной области со стрелкой была важна для увеличения транскрипции (сравнение конструкций 5 и 6). Представленные данные также являются сильным показателем того, что дистальный элемент транскрипционной регуляции сам по себе ограничивает инициацию транскрипции в хроматине, расположенном ниже. С учетом того, что фрагмент 223 п.о. (SEQ ID No. 20), расположенный при 3'-конце прямо заштрихованной области со стрелкой, сохраняет полную активность этой области в конструкции 7, предполагают, что этот важный участок должен в данном случае кооперировать с областью изгиба и 5'-концом остальной части (нуклеотиды 1-1425) элемента в конструкции 6. Было обнаружено, что два сайта HMG-I/Y локализованы возле этого конца.
Блочное строение мышиных MAR и уменьшение размера
На основании открытий для человеческой 1_68 MAR, S4 MAR была также проанализирована на блоки, в частности, ответственные за ее транскрипционную активность. Этот анализ также проводили с целью уменьшения размера S4 MAR, которая является относительно длинной. Таким образом, сконструировали несколько MAR из S4 MAR (таблица 3) и охарактеризовали (фиг.10). На фиг.10 слева показана специфичная конструкция MAR S4, а справа эффект различных конструкций MAR S4 на экспрессию рекомбинантного зеленого флуоресцентного белка (GFP), которая выявлена путем анализа средней флуоресценции всей популяции (Avg Gmean М0). Популяции клеток СНО, трансфицированных экспрессионным вектором GFP, содержащим или не содержащим конструкцию MAR, как указано, анализировали с помощью проточной цитометрии на цитометре FACScalibur (Becton Dickinson). Среднюю флуоресценцию всей популяции нормализовали по полученной с человеческой MAR 1_68, значение которой было принято за 100, поскольку GFP показывает экспрессию в отсутствие MAR. Другие конструкции MAR названы в соответствии с их содержанием оснований по сравнению с полноразмерной 1547 п.о. S4 MAR (см. таблицу 3). Заштрихованный прямоугольник показывает АТ-богатую область изгиба MAR S4. S_41-4662-Luc5489 обозначает конструкцию, где концевые (3') 795 пар оснований были удалены и заменены частью гена люциферазы (черный прямоугольник). Интересно, что, как видно на фиг.10, было обнаружено, что 1624-п.о. фрагмент EcoRI может быть делегирован из S4 MAR (S4-1-703_2328-5457) без значительной потери ее активности MAR. Однако делеция близкого к промотору 795-п.о. фрагмента или замена этой последовательности фрагментом гена люциферазы подобной длины (S4_1-4661; S4_1-4661-Luc5489) вызывала полную утрату этой активности. Это указывает на то, что некоторые варианты мышиной S4 MAR могут проявлять высокую активность, в то же время имея более короткую длину, что, таким образом, делает их размер более удобным, например, для конструирования и переноса вектора.

Активность 3' концевых последовательностей MAR
Для дальнейшего анализа активности 3' концевых последовательностей MAR S4 этот участок MAR был дополнительно рассечен путем удаления или дупликации его частей. На фиг.11 показан эффект различных производных MAR S4 на экспрессию рекомбинантного зеленого флуоресцентного белка (GFP), которая выявлена путем анализа средней флуоресценции всей популяции (Avg Gmean М0). Популяции клеток СНО получали и анализировали, как описано выше. Интересно, что одно такое производное, имеющее укороченный 3' конец (4658-5054 против 4658-5457 исходной MAR S4), проявляло в среднем несколько более высокую экспрессию трансгена по сравнению с более длинной исходной последовательностью MAR S4 (104% против 100%). Это указывает на возможность получения как более эффективных, так и более коротких производных элементов MAR.
СИНТЕТИЧЕСКИЕ MAR
На фиг.12 показана карта потенциальных сайтов связывания факторов транскрипции [1_68 MAR], которые предсказаны программой MATInspector. Положение сайтов связывания С/ЕВР, NMP4, FAST1, SATB1 и HoxF (также называемого Gsh) показано в качестве примеров, иллюстрируя обогащение ими в 5' прямо заштрихованной фланкирующей последовательности. Эти сайты связывания, как они встречаются в MAR 1-68, использовали без изменения (FAST1, С/ЕВР, HOXF/Gsh), либо они были скорректированы в случае, если они имели одно или два несовпадения по сравнению с консенсусной (то есть правильной) последовательностью (HoxF, SatB1,NMP4).
Обнаружение возможного кооперирования между АТ-богатой областью изгиба ДНК и сайтами связывания факторов транскрипции в человеческой MAR 1_68 способствовало конструированию синтетических MAR, содержащих АТ-богатую область MAR 1-68, прилежащую к одному или нескольким сайтам связывания факторов транскрипции. На фиг.13 изображена карта плазмиды, используемой для тестирования активности синтетических MAR, сконструированных в результате сборки внутренней последовательности, содержащей АТ-богатую область (MAR 1429-2880) и химического синтеза сайтов связывания ДНК-связывающих белков для факторов транскрипции, помещенных выше промотора и зеленого флуоресцентного белка (GFP). На фиг.13 показано, что сайты связывания факторов транскрипции были встроены между АТ-богатой внутренней последовательностью и промотором SV40, направляющим экспрессию трансгена в GFP, имитируя ситуацию, находящуюся на фиг.9, где участки MAR, содержащие сайты связывания, расположены между промотором и областью изгиба ДНК в наиболее выгодных положениях. В таблице 4 показана последовательность ДНК химически синтезированных олигонуклеотидов, которые были использованы.

На фиг.14 показано усиление транскрипции синтетическими MAR, 15 сконструированными, как описано на фиг.13. Встроенные элементы содержат 1 или несколько сайтов связывания ДНК-связывающих белков в дополнение к внутренней последовательности, как указано. Трансфекция плазмидами, содержащими один или несколько сайтов связывания в дополнение к внутренней последовательности, содержащей АТ-богатую область (АТ-богатую внутреннюю последовательность), показала, что включение сайтов связывания повышало усиление транскрипции по сравнению с одной АТ-богатой внутренней областью и что С/ЕВР и Нох или Gsh2 были наиболее активными, за которыми следуют SATB1 и Fasti, тогда как один сайт NMP4 не обладал обнаружимым эффектом.
Различные смеси активных сайтов связывания также тестировали, чтобы определить их синергические эффекты, которые можно наблюдать. Для этого различные комбинации олигонуклеотидов, содержащих сайты связывания для различных факторов транскрипции, смешивали в реакциях лигирования ДНК, и точный порядок и расположение сайтов связывания определяли путем секвенирования ДНК. Полученные комбинации показаны в таблице 5:
Полученные в результате плазмиды тестировали путем трансфекции, как ранее. На фиг.15 показано усиление транскрипции синтетическими MAR, сконструированными с комбинациями сайтов связывания ДНК-связывающих белков, показанными в таблице 5. Наиболее активные комбинации показаны знаком звездочки, и указана встречаемость сайтов HoxF/Gsh2 или SatB1. Результаты, представленные на фиг.15, показывают, что активность синтетических MAR в данном случае не зависит от числа встроенных сайтов связывания, но что конкретные комбинации сайтов связывания проявляют высокие активности усиления, тогда как у других отсутствует активность, либо они даже подавляют экспрессию гена. Конструкции с более высокими активностями содержали в данном случае комбинации белков Hox/Gsh2 и SATB1, и наиболее активная конструкция исключительно состояла из этих элементов. Вставка этой синтетической MAR повышала встречаемость высокоэкспрессирующих клонов примерно в 10 раз по сравнению с контрольным вектором pEGFP, в котором отсутствовала какая-либо последовательность MAR.






























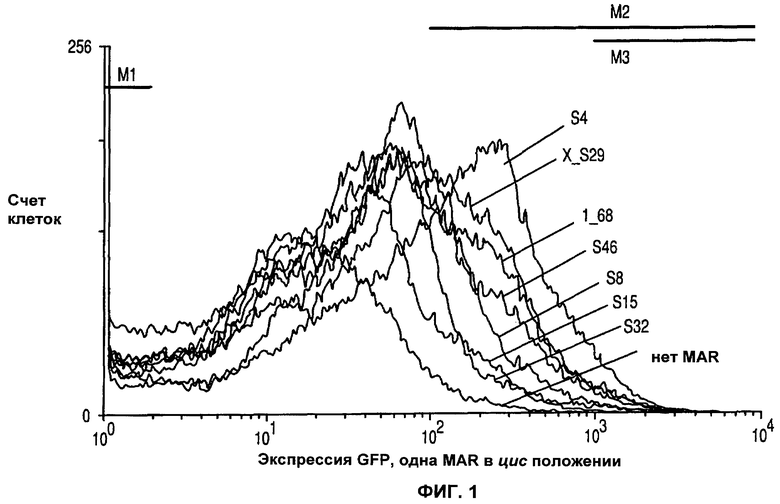
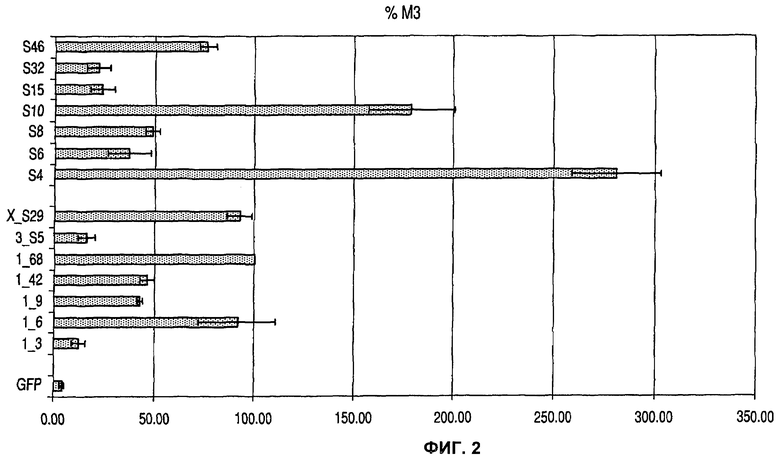
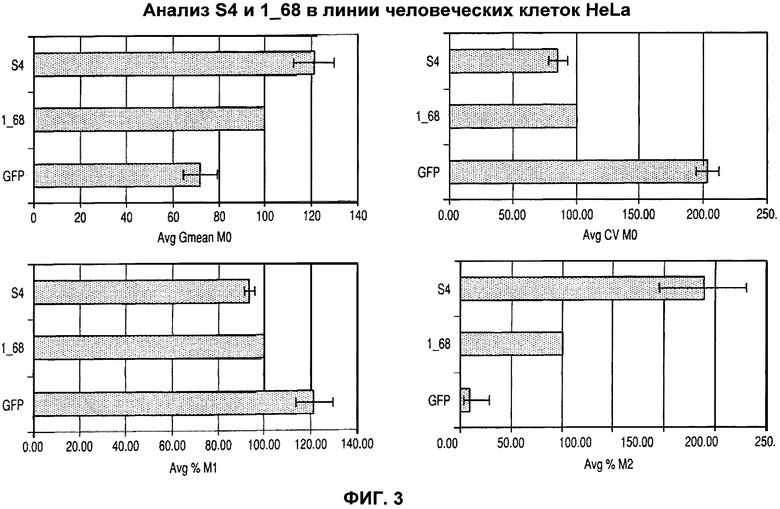
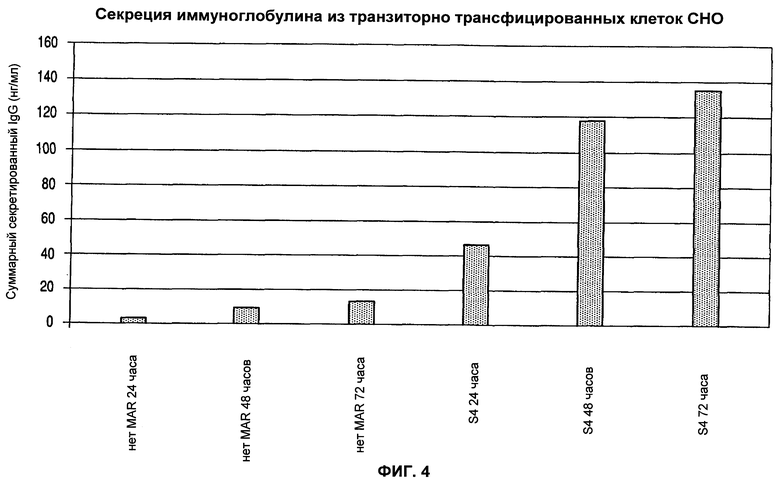
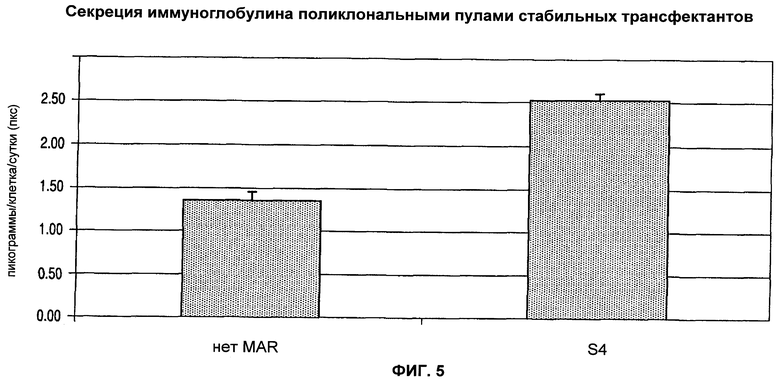
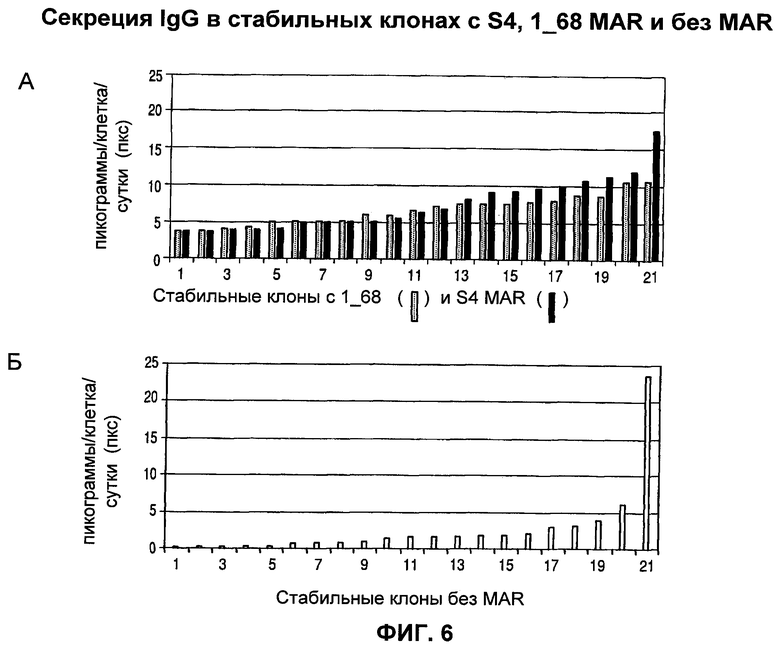
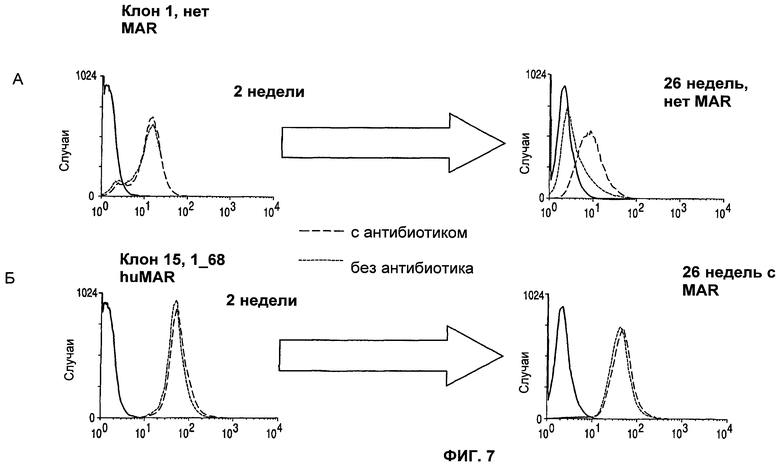
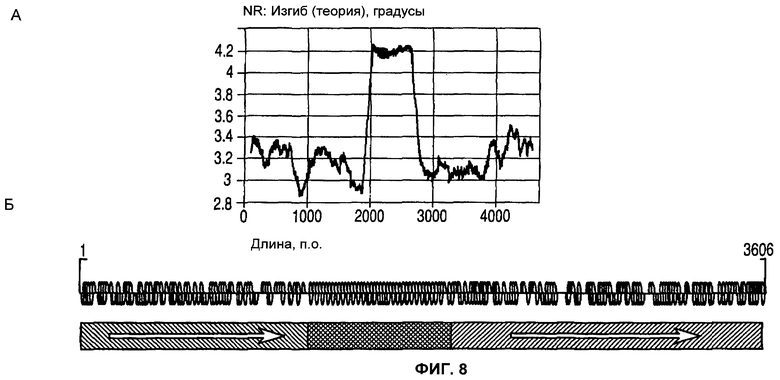
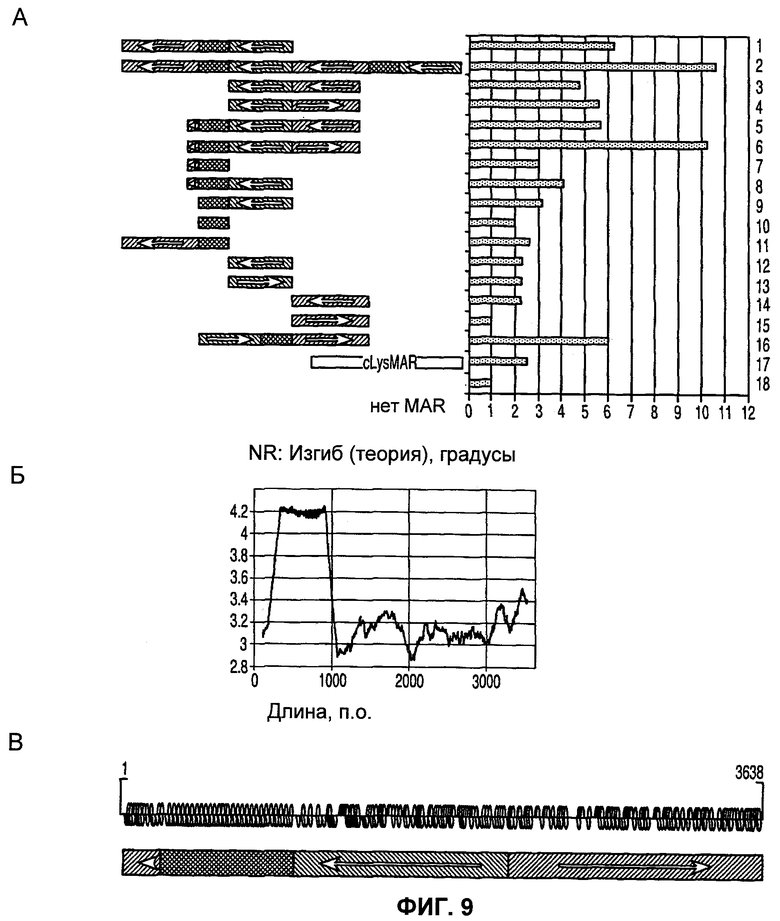
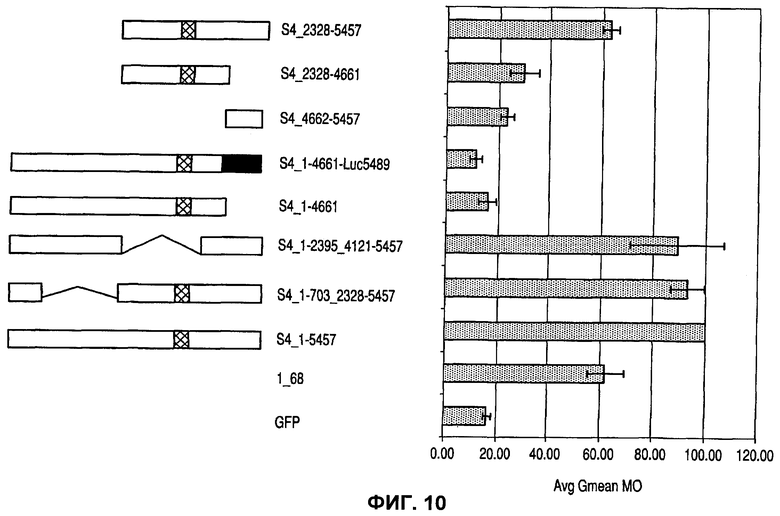
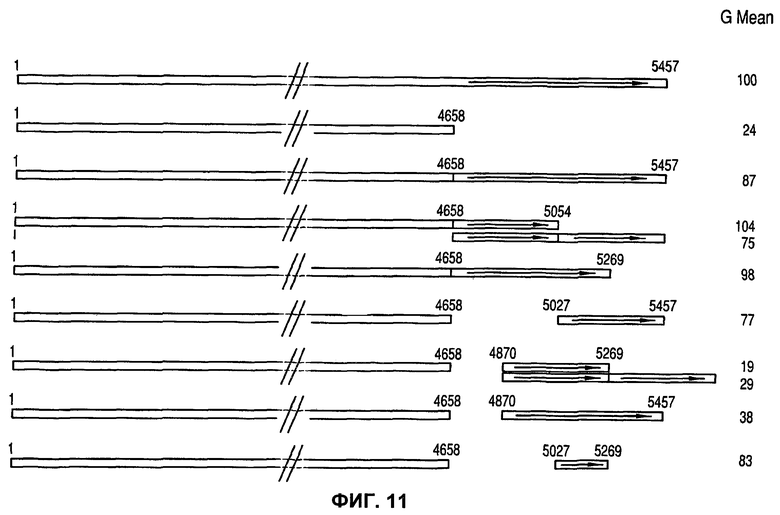
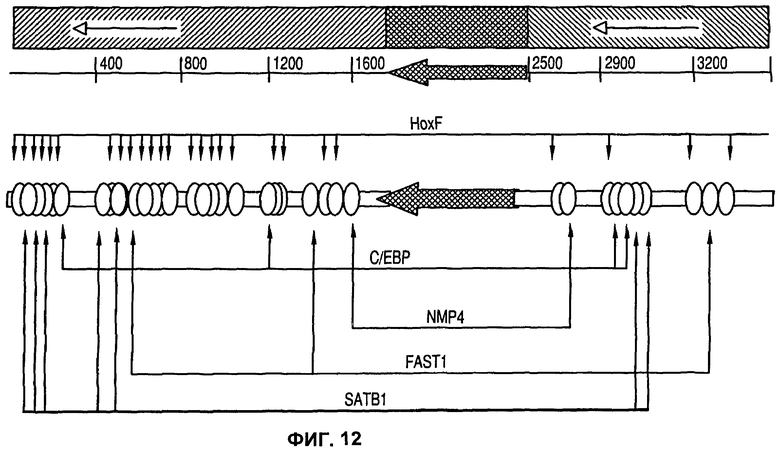
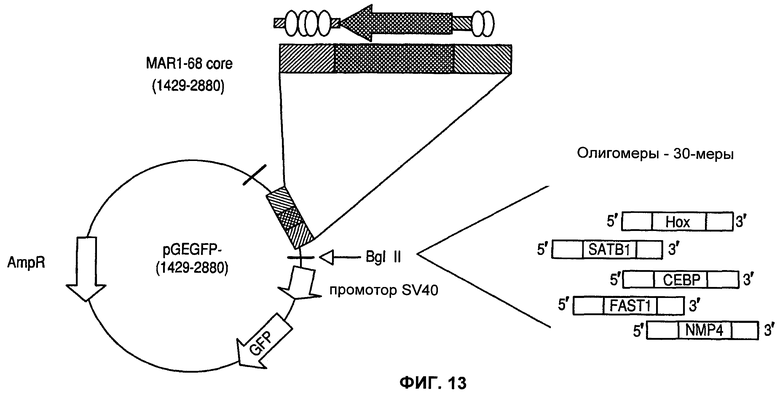
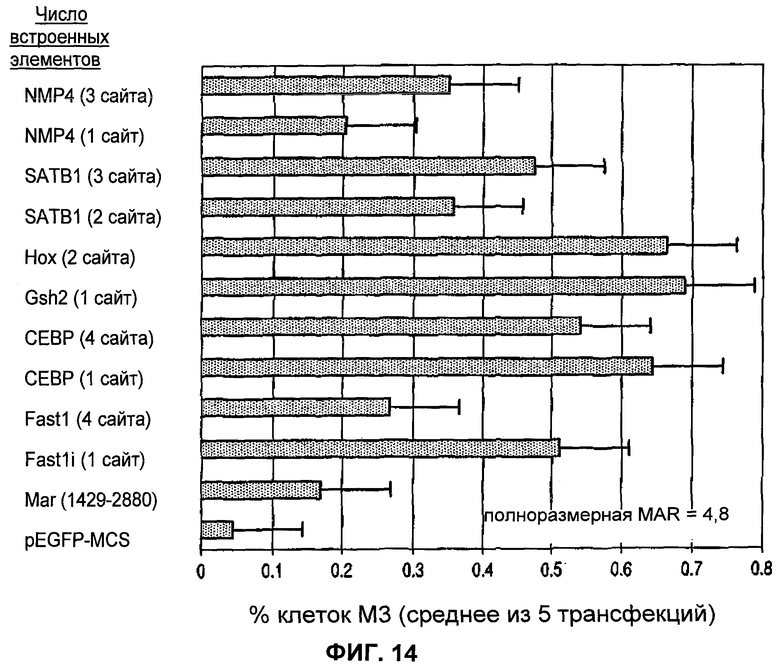
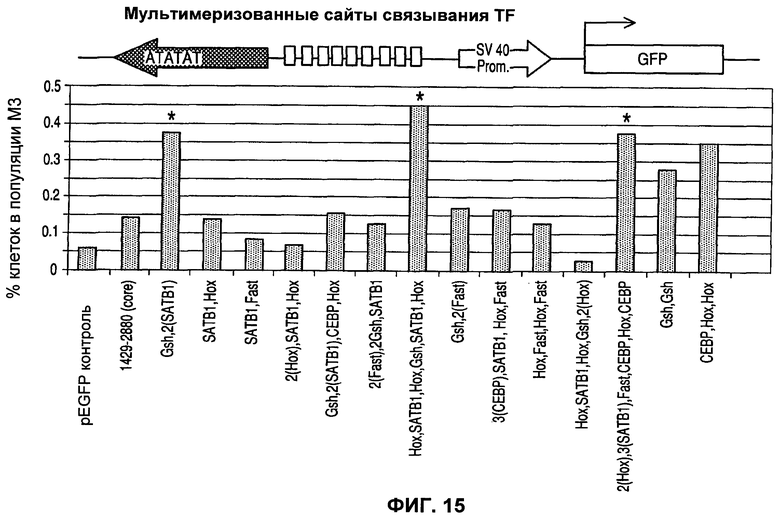
Изобретение относится к области молекулярной биологии. Предложены экспрессионная система для экспрессии высокого уровня гена, включающая промотор и по меньшей мере одну последовательность MAR; изолированная и очищенная молекула нуклеиновой кислоты, представляющая собой последовательность MAR; клетка; трансгенное животное; набор для экспрессии высокого уровня, а также применение указанной экспрессионной системы для повышенного продуцирования белка. Изобретение может быть использовано для получения широкого круга белков, включая антитела и интерфероны, посредством трансгенных животных и клеточных культур. 7 н.п. ф-лы, 15 ил., 5 табл., 1 пр.
1. Экспрессионная система для повышения уровня экспрессии по меньшей мере одного гена, включающая:
промотор для оперативного сцепления с нуклеотидной последовательностью, кодирующей интересующий ген, и
по меньшей мере одну нуклеотидную последовательность MAR млекопитающего, отличного от человека, для усиления экспрессии указанного гена в клетке, трансформированной указанной экспрессионной системой,
где указанная нуклеотидная последовательность MAR выбрана из группы, включающей в себя последовательности SEQ ID No.3, SEQ ID No.4, SEQ ID No.5, SEQ ID No.6, SEQ ID No.7, SEQ ID No.8, SEQ ID No.9, SEQ ID No.10.
2. Способ усиленного продуцирования белка в клетке, при котором
- получают клетку человека или млекопитающего, отличного от человека,
- вводят экспрессионную систему по п.1 в указанную клетку таким образом, что экспрессия гена повышается.
3. Изолированная и очищенная молекула нуклеиновой кислоты, обеспечивающая повышение уровня экспрессии по меньшей мере одного гена, содержащая нуклеотидную последовательность, выбранную из группы, включающей в себя последовательности SEQ ID No.3, SEQ ID No.4, SEQ ID No.5, SEQ ID No.6, SEQ ID No.7, SEQ ID No.8, SEQ ID No.9, SEQ ID No.10.
4. Клетка для экспрессии высокого уровня по меньшей мере одного гена, содержащая экспрессионную систему по п.1.
5. Трансгенное животное, отличное от человека, для экспрессии высокого уровня по меньшей мере одного гена, содержащее экспрессионную систему по п.1.
6. Набор для экспрессии высокого уровня по меньшей мере одного гена, содержащий:
экспрессионную систему по п.1 и
инструкции по применению указанной экспрессионной системы.
7. Применение экспрессионной системы по п.1 при продуцировании белков, таких как антитела, распознающие патогенные белки человека или белки клеточной поверхности человека, эритропоэтин, интерфероны.
| WO 2005040377, 06.05.2005 | |||
| Бесколесный шариковый ход для железнодорожных вагонов | 1917 |
|
SU97A1 |
| LUTZKO С ЕТ AL | |||
| Lentivirus Vectors Incorporating the Immunoglobulin Heavy Chain Enhancer and Matrix Attachment Regions Provide Position-Independent Expression in В Lymphocytes | |||
| Journal of virology, 2003, v.77, n 13, p.7341-7351 | |||
| AGARWAL M | |||
| et al | |||
| Scaffold attachment region-mediated enhancement of | |||

