ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к сайт-специфическим конъюгатам антитела к HER2 и лекарственного средства. Настоящее изобретение также относится к способам применения таких конъюгатов антитела-лекарственного средства для лечения рака.
УРОВЕНЬ ТЕХНИКИ
Представители семейства ErbB трансмембранных рецепторных тирозинкиназ являются важными медиаторами роста клеток, дифференцировки клеток, миграции клеток и апоптоза. Семейство рецепторов включает четыре представителя, включающих рецептор эпидермального фактора роста (EGFR или ErbB1), HER2 (ErbB2 или p185), HER3 (ErbB3) и HER4 (ErbB4 или tyro2).
Первоначально HER2 идентифицировали как продукт трансформирующего гена из нейробластом химически обработанных крыс. Оверэкспрессия HER2 была подтверждена как онкогенная, как in vitro (Di Fiore et al., 1987, Science 237(4811): 178-82; Hudziak et al., 1987, PNAS 84(20): 7159-63; Chazin et al., 1992, Oncogene 7(9): 1859-66), так и в моделях на животных (Guy et al., 1992, PNAS 89(22): 10578-82). Амплификация гена, кодирующего HER2, с последующей оверэкспрессией рецептора наблюдается при раке молочной железы и раке яичника и коррелирует с неблагоприятным прогнозом (Slamon et al., 1987, Science 235(4785): 177-82; Slamon et al., 1989, Science 244: 707-12; Anbazhagan et al., 1991, Annals Oncology 2(1): 47-53; Andrulis et al., 1998, J Clinical Oncology 16(4): 1340-9). Оверэкспрессию HER2 (часто, но не обязательно вследствие амплификации гена) также наблюдали в других типах опухолей, в том числе при раке желудка, эндометрия, немелкоклеточном раке легкого, толстой кишки, поджелудочной железы, мочевого пузыря, почки, предстательной железы и шейки матки (Scholl et al., 2001, Annals Oncology 12 (Suppl. 1): S81-7; Menard et al., 2001, Ann Oncol 12 (Suppl 1): S15-9; Martin et al., 2014, Future Oncology 10: 1469-86).
Герцептин® (трастузумаб) является гуманизированным моноклональным антителом, которое связывается с внеклеточным доменом HER2 (Carter et al. 1992, PNAS 89:4285-9 и патент США 5,821,337). Герцептин® получил разрешение на продажу от Управления по контролю качества пищевых продуктов и лекарственных препаратов 25 сентября 1998 года для лечения больных метастатическим раком молочной железы, при котором опухоли повышенно экспрессируют белок HER2. Хотя Герцептин® является прорывом в лечении пациентов со злокачественными опухолями молочной железы, оверэкспрессирующими HER2, которые ранее получали широкую противоопухолевую терапию, некоторые категории пациентов в данной группе не поддаются, плохо поддаются или становятся устойчивыми к лечению Герцептином®.
Кадсила® (трастузумаб-DM1 или T-DM1) является конъюгатом антитела-лекарственного средства, состоящим из трастузумаба, конъюгированного с майтанзиноидным средством DM1 через стабильный тиоэфирный линкер MCC (4-[N-малеимидометил]циклогексан-1-карбоксилат) (Lewis et al., 2008, Cancer Res. 68:9280-90; Krop et al., 2010, J Clin Oncol. 28:2698-2704; патент США 8,337,856). Кадсила® получил разрешение на продажу от Управления по контролю качества пищевых продуктов и лекарственных препаратов 22 февраля 2013 года для лечения HER2-положительного метастатического рака молочной железы у больных, которые ранее проходили лечение Герцептином® и таксановым лекарственным препаратом, и стали рефракторными к Герцептину®. Как отмечали в отношении Герцептина®, существуют категории пациентов в группе рака молочной железы с оверэкспрессией HER2, которые не поддаются успешной долговременной терапии Кадсилой®.
Таким образом, существует значительная клиническая потребность в разработке других HER2-направленных противоопухолевых терапий для пациентов с оверэкспрессирующими HER2 опухолями или другими заболеваниями, связанными с оверэкспрессией HER2, которые не поддаются, плохо поддаются или становятся резистентными к лечению Кадсилой® и/или Герцептином®.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В настоящем изобретении предложены сайт-специфические конъюгаты антитела к HER2-лекарственного средства (ADC) и их применение в лечении HER2-экспрессирующих форм рака. ADC обеспечивают направленную доставку терапевтических средств к раковым клеткам и дают возможность проведения более селективной терапии при одновременном уменьшении известных нецелевых токсических эффектов.
Сайт-специфический HER2 ADC согласно изобретению обычно имеет следующую формулу: Ab-(L-D), в которой Ab является антителом или его антигенсвязывающим фрагментом, который связывается с HER2; и L-D является молекулой линкера-лекарственного средства, где L является линкером, а D является лекарственным средством.
Антитело (Ab) в ADC согласно изобретению может быть любым HER2-связывающим антителом. В некоторых аспектах изобретения Ab связывается с тем же эпитопом на HER2, что и трастузумаб (Герцептин®). В других аспектах изобретения Ab имеет такую же тяжелую цепь и CDR-области легкой цепи, как и трастузумаб. В определенных аспектах изобретения Ab имеет такую же вариабельную область тяжелой цепи (VH) и такую же вариабельную область легкой цепи (VL), как и трастузумаб.
HER2 ADC настоящего изобретения конъюгированы с лекарственным средством сайт-специфическим образом. Для применения данного типа конъюгирования антитело должно быть дериватизировано с предоставлением либо реакционноспособного остатка цистеина, введенного с помощью методов генной инженерии в один или более специфических сайтов, либо ацил-донорного остатка глутамина (введенного с помощью методов генной инженерии в один или более специфических сайтов или в присоединенной пептидной метке). Такие модификации должны присутствовать в сайтах, которые не нарушают антигенсвязывающей способности антитела. В предпочтительных вариантах осуществления одна или более модификаций сделаны в константной области тяжелой и/или легкой цепей антитела.
В некоторых вариантах осуществления настоящего изобретения в сайт-специфических HER2 ADC могут применяться антитела, включающие CDR-области вариабельной области тяжелой цепи и CDR-области вариабельной области легкой цепи трастузумаба (VH CDR-области SEQ ID NO: 2-4 и VL CDR-области SEQ ID NO: 8-10), а также любую комбинацию константных областей тяжелой и легкой цепи, раскрытых в Таблице 1, при условии, что в том случае, когда константная область тяжелой цепи является SEQ ID NO: 5, то тогда константная область легкой цепи не является SEQ ID NO: 11. В таких вариантах осуществления константная область тяжелой цепи может быть выбрана из любого из SEQ ID NO: 17, 5, 13, 21, 23, 25, 27, 29, 31, 33, 35, 37 или 39, тогда как константная область легкой цепи может быть выбрана из любого из SEQ ID NO: 41, 11 или 43, при условии, что такая комбинация не является SEQ ID NO: 5 и SEQ ID NO: 11.
В определенном варианте осуществления антитело, применяемое для получения сайт-специфического HER2 ADC, включает VH домен с CDR-областями SEQ ID NO: 2-4 и VL домен с CDR-областями SEQ ID NO: 8-10, присоединенные к константной области тяжелой цепи SEQ ID NO: 17 и константной области легкой цепи SEQ ID NO: 41. В другом определенном варианте осуществления антитело, применяемое для получения сайт-специфического HER2 ADC, включает VH домен с CDR-областями SEQ ID NO: 2-4 и VL домен с CDR-областями SEQ ID NO: 8-10, присоединенные к константной области тяжелой цепи SEQ ID NO: 13 и константной области легкой цепи SEQ ID NO: 43.
В других вариантах осуществления в ADC согласно изобретению могут применяться антитела, включающие любую комбинацию тяжелых и легких цепей, раскрытых в Таблице 1, при условии, что в том случае, если тяжелая цепь является SEQ ID NO: 6, то тогда легкая цепь не является SEQ ID NO: 12. В таких вариантах осуществления тяжелая цепь может быть выбрана из любого из SEQ ID NO: 18, 6, 14, 22, 24, 26, 28, 30, 32, 34, 36, 38 или 40, тогда как легкая цепь может быть выбрана из любого из SEQ ID NO: 42, 12 или 44, при условии, что такая комбинация не является SEQ ID NO: 6 и SEQ ID NO: 12.
В определенном варианте осуществления в ADC согласно изобретению может применяться антитело, включающее тяжелую цепь SEQ ID NO: 18 и легкую цепь SEQ ID NO: 42. В другом определенном варианте осуществления в ADC согласно изобретению может применяться антитело, включающее тяжелую цепь SEQ ID NO: 14 и легкую цепь SEQ ID NO: 44.
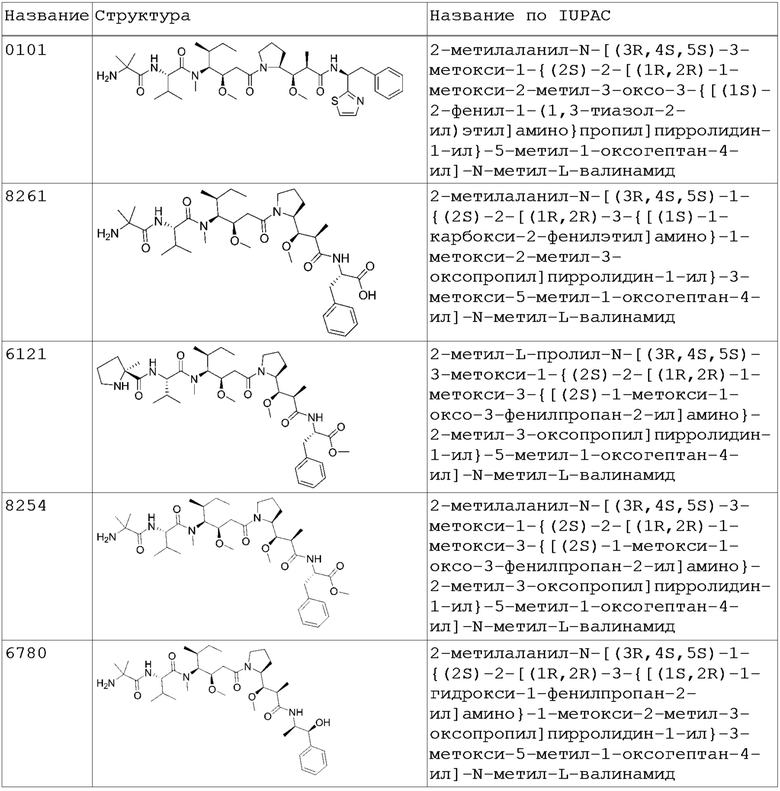
Любые сайт-специфические HER2 ADC, раскрытые в настоящей заявке, могут быть получены с лекарственным средством (D), которое является терапевтическим средством, применяемым для лечения рака. В определенном варианте осуществления терапевтическим средством является антимитотическое средство. В другом определенном варианте осуществления антимитотическим лекарственным компонентом в ADC конъюгатах согласно изобретению является ауристатин (например, 0101, 8261, 6121, 8254, 6780 и 0131). В более конкретном варианте осуществления ауристатин в качестве лекарственного компонента в ADC конъюгатах согласно изобретению представляет собой 2-метилаланил-N-[(3R,4S,5S)-3-метокси-1-{(2S)-2-[(1R,2R)-1-метокси-2-метил-3-оксо-3-{[(1S)-2-фенил-1-(1,3-тиазол-2-ил)этил]амино}пропил]пирролидин-1-ил}-5-метил-1-оксогептан-4-ил]-N-метил-L-валинамид (также известный как 0101). Предпочтительно лекарственный компонент ADC конъюгатов согласно изобретению обладает мембранной проницаемостью.
Любые сайт-специфические HER2 ADC, раскрытые в настоящей заявке, могут быть получены с линкером (L), который является расщепляемым или нерасщепляемым. Предпочтительно линкер является расщепляемым. Расщепляемые линкеры включают, без ограничения перечисленными, vc, AcLysvc и m(H20)c-vc. Более предпочтительно линкером является vc или AcLysvc.
В конкретном аспекте изобретения сайт-специфический HER2 ADC формулы Ab-(L-D) включает: (a) антитело, Ab, включающее тяжелую цепь SEQ ID NO: 18 и легкую цепь SEQ ID NO: 42; и (b) молекулу линкера-лекарственного средства, L-D, где L является линкером, а D является лекарственным средством, где линкером является vc, и где лекарственным средством является 0101.
В другом конкретном аспекте изобретения сайт-специфический HER2 ADC формулы Ab-(L-D) включает: (a) антитело, Ab, включающее тяжелую цепь SEQ ID NO: 14 и легкую цепь SEQ ID NO: 44; и (b) молекулу линкера-лекарственного средства, L-D, где L является линкером, а D является лекарственным средством, где линкером является AcLysvc, и где лекарственным средством является 0101.
Другой аспект изобретения включает способы создания, способы получения, способы синтеза, способы конъюгирования и способы очистки конъюгатов антитела-лекарственного средства, раскрытых в настоящей заявке, и промежуточных соединений для получения, синтеза и конъюгирования конъюгатов антитела-лекарственного средства, раскрытых в настоящей заявке.
Также предложены фармацевтические композиции, включающие сайт-специфический HER2 ADC, раскрытый в настоящей заявке, и фармацевтически приемлемый носитель.
Нуклеиновые кислоты, кодирующие часть антитела в сайт-специфических HER2 ADC, рассматриваются в изобретении. Дополнительные векторы и клетки-хозяева, включающие нуклеиновые кислоты, также рассматриваются в изобретении.
В настоящем изобретении также преложен способ применения сайт-специфических HER2 ADC конъюгатов в лечении HER2-экспрессирующих форм рака. HER2-экспрессирующий рак, который предполагается лечить сайт-специфическими HER2 ADC конъюгатами согласно изобретению, может экспрессировать HER2 на высоком, среднем или низком уровне. В некоторых вариантах осуществления рак, который предполагается лечить, является резистентным к лечению, рефракторным к лечению и/или рецидивирующим после лечения трастузумабом и/или трастузумабом эмтанзином (T-DM1), один из которых применяют отдельно или в комбинации с таксаном. Онкологические заболевания, которые предполагается лечить, включают, без ограничения перечисленными, рак молочной железы, рак яичника, рак легкого, рак желудка, рак пищевода, рак толстой и прямой кишки, уротелиальный рак, рак поджелудочной железы, рак слюнной железы и рак головного мозга или метастазы вышеуказанных форм рака. В более конкретном варианте осуществления рак молочной железы является эстроген-рецептор и прогестерон-рецептор негативным раком молочной железы или трижды негативным раком молочной железы (ТНРМЖ). В другом варианте осуществления рак легкого является немелкоклеточным раком легкого (НМРЛ).
Эти и другие аспекты изобретения будут ясны при рассмотрении заявки в целом.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На ФИГ. 1A-1B показан (A) ADC T(kK183C+K290C)-vc0101 и (B) ADC T(LCQ05+K222R)-AcLysvc0101. Каждый черный круг обозначает линкер/полезную нагрузку, которые конъюгированы с моноклональным антителом. Структура одного такого линкера/полезной нагрузки показана для каждого ADC. Подчеркнутый атом предоставлен аминокислотным остатком на антителе, через который происходит конъюгирование.
На ФИГ. 2A-2E показаны спектры выбранных ADC, полученные с помощью хроматографии гидрофобного взаимодействия (HIC) и показывающие изменения времени удерживания при конъюгировании полученных на основе трастузумаба антител с различными линкерами-полезными нагрузками.
На ФИГ. 3A-3B показаны кривые связывания ADC коъюгатов с HER2. (A) прямое связывание с HER2-положительными клетками BT474 и (B) конкурентное связывание с ФЭ-меченым трастузумабом с клетками BT474. Эти результаты указывают, что связывающие свойства антитела в таких ADC коъюгатах не были изменены в результате процесса конъюгирования.
На ФИГ. 4 показаны ADCC активности ADC, полученных на основе трастузумаба.
На ФИГ. 5 показаны данные исследования цитотоксичности in vitro (IC50), представленные в нМ концентрации полезной нагрузки для ряда полученных на основе трастузумаба ADC в отношении ряда клеточных линий с различными уровнями экспрессии HER2.
На ФИГ. 6 показаны данные исследования цитотоксичности in vitro (IC50), представленные в концентрации антитела нг/мл для ряда полученных на основе трастузумаба ADC в отношении ряда клеточных линий с различными уровнями экспрессии HER2.
На ФИГ. 7A-7I показана противоопухолевая активность девяти полученных на основе трастузумаба ADC в отношении ксенотрансплантатов N87, объем опухоли представлен на кривой в зависимости от времени. (A) T(kK183C+K290C)-vc0101; (B) T(kK183C)-vc0101; (C) T(K290C)-vc0101; (D) T(LCQ05+K222R)-AcLysvc0101; (E) T(K290C+K334C)-vc0101; (F) T(K334C+K392C)-vc0101; (G) T(N297Q+K222R)-AcLysvc0101; (H) T-vc0101; (I) T-DM1. Клетки рака желудка N87 экспрессируют высокие уровни HER2.
На ФИГ. 8A-8E показана противоопухолевая активность шести полученных на основе трастузумаба ADC в отношении ксенотрансплантатов HCC1954, объем опухоли представлен на кривой в зависимости от времени. (A) T(LCQ05+K222R)-AcLysvc0101; (B) T(K290C+K334C)-vc0101; (C) T(K334C+K392C)-vc0101; (D) T(N297Q+K222R)-AcLysvc0101; (E) T-DM1. Клетки рака молочной железы HCC1954 экспрессируют высокие уровни HER2.
На ФИГ. 9A-9G показана противоопухолевая активность семи полученных на основе трастузумаба ADC в отношении ксенотрансплантатов JIMT-1, объем опухоли представлен на кривой в зависимости от времени. (A) T(kK183C+K290C)-vc0101; (B) T(LCQ05+K222R)-AcLysvc0101; (C) T(K290C+K334C)-vc0101; (D) T(K334C+K392C)-vc0101; (E) T(N297Q+K222R)-AcLysvc0101; (F) T-vc0101; (G) T-DM1. Клетки рака молочной железы JIMT-1 экспрессируют средние/низкие уровни HER2.
На ФИГ. 10A-10D показана противоопухолевая активность пяти полученных на основе трастузумаба ADC в отношении ксенотрансплантатов MDA-MB-361 (DYT2), объем опухоли представлен на кривой в зависимости от времени. (A) T(LCQ05+K222R)-AcLysvc0101; (B) T(N297Q+K222R)-AcLysvc0101; (C) T-vc0101; (D) T-DM1. Клетки рака молочной железы MDA-MB-361 (DYT2) экспрессируют средние/низкие уровни HER2.
На ФИГ. 11A-11E показана противоопухолевая активность пяти полученных на основе трастузумаба ADC в отношении полученных от пациента ксенотрансплантатов PDX-144580, объем опухоли представлен на кривой в зависимости от времени. (A) T(kK183C+K290C)-vc0101; (B) T(LCQ05+K222R)-AcLysvc0101; (C) T(N297Q+K222R)-AcLysvc0101; (D) T-vc0101; (E) T-DM1. Полученные от пациента клетки PDX-144580 являются PDX-моделью ТНРМЖ.
На ФИГ. 12A-12D показана противоопухолевая активность четырех полученных на основе трастузумаба ADC в отношении полученных от пациента ксенотрансплантатов PDX-37622, объем опухоли представлен на кривой в зависимости от времени. (A) T(kK183C+K290C)-vc0101; (B) T(N297Q+K222R)-AcLysvc0101; (C) T(K297C+K334C)-vc0101; (D) T-DM1. Полученные от пациента клетки PDX-37622 являются PDX-моделью НМРЛ, экспрессирующей средние уровни HER2.
На ФИГ. 13A-13B показана иммуногистоцитохимия ксенотрансплантатов опухоли N87, обработанных (A) T-DM1 или (B) T-vc0101 и окрашенных на фосфогистон H3 и IgG антитело. Неспецифическую активность наблюдали с T-vc0101.
На ФИГ. 14 показаны данные исследования цитотоксичности in vitro (IC50), представленные в нМ концентрации полезной нагрузки и концентрации нг/мл антитела для ряда полученных на основе трастузумаба ADC и свободных полезных нагрузок в отношении клеток, сделанных устойчивыми к T-DM1 in vitro (N87-TM1 и N87-TM2), или исходных клеток, чувствительных к T-DM1 (клетки N87). Клетки рака желудка N87 экспрессируют высокий уровень HER2.
На ФИГ. 15A-15G показана противоопухолевая активность семи полученных на основе трастузумаба ADC в отношении T-DM1-чувствительных (клетки N87) и резистентных (N87-TM1 и N87-TM2) клеток рака желудка. (A) T-DM1; (B) T-mc8261; (C) T(297Q+K222R)-AcLysvc0101; (D) T(LCQ05+K222R)-AcLysvc0101; (E) T(K290C+K334C)-vc0101; (F) T(K334C+K392C)-vc0101; (G) T(kK183C+K290C)-vc0101.
На ФИГ. 16A-16B показаны Вестерн-блоты, на которых показана: (A) экспрессия белка MRP1, эффлюксного насоса, обеспечивающего лекарственную устойчивость, и (B) белка MDR1, эффлюксного насоса, обеспечивающего лекарственную устойчивость, на T-DM1-чувствительных (клетки N87) и резистентных (N87-TM1 и N87-TM2) клетках рака желудка.
На ФИГ. 17A-17B показана экспрессия HER2 и связывание с трастузумабом T-DM1-чувствительных (клетки N87) и резистентных (N87-TM1 и N87-TM2) клеток рака желудка. (A) Вестерн-блот, на котором показана экспрессия белка HER2, и (B) связывание трастузумаба с HER2 клеточной поверхности.
На ФИГ. 18A-18D показан анализ уровней экспрессии белка в T-DM1-чувствительных (клетки N87) и резистентных (N87-TM1 и N87-TM2) клетках рака желудка. (A) Изменения уровня экспрессии 523 белков; (B) Вестерн-блоты, на которых показана экспрессия белков IGF2R, LAMP1 и CTSB; (C) Вестерн-блот, на котором показана экспрессия белка CAV1; (D) ИГХ экспрессии белка CAV1 в опухолях, полученных in vivo при имплантации клеток N87 (левая панель) и клеток N87-TM2 (правая панель).
На ФИГ. 19A-19C показана чувствительность к трастузумабу и различным полученным на основе трастузумаба ADC конъюгатам опухолей, полученных in vivo при имплантации (A) T-DM1-чувствительных исходных клеток N87; (B) T-DM1-резистентных клеток N87-TM1; (C) T-DM1-резистентных клеток N87-TM2.
На ФИГ. 20A-20F показана чувствительность к трастузумабу и различным полученным на основе трастузумаба ADC конъюгатам опухолей, полученных in vivo при имплантации T-DM1-чувствительных исходных клеток N87 и T-DM1-резистентных клеток N87-TM2 или N87-TM1. (A) размер опухоли N87 наносили на кривую в зависимости от времени, в присутствии трастузумаба или двух полученных на основе трастузумаба ADC; (B) размер опухоли N87-TM2 наносили на кривую в зависимости от времени, в присутствии трастузумаба или двух полученных на основе трастузумаба ADC; (C) время до двухкратного увеличения размера опухоли из клеток N87 в присутствии трастузумаба или двух полученных на основе трастузумаба ADC; (D) время до двухкратного увеличения размера опухоли из клеток N87-TM2 в присутствии трастузумаба или двух полученных на основе трастузумаба ADC; (E) размер опухоли N87-TM2 наносили на кривую в зависимости от времени, в присутствии семи различных, полученных на основе трастузумаба, ADC; (F) размер опухоли N87-TM1 наносили на кривую в зависимости от времени, при этом полученный на основе трастузумаба ADC добавляли в день 14.
На ФИГ. 21A-21E показано создание и исследование T-DM1-резистентных клеток, полученных in vivo. (A) клетки рака желудка N87 были изначально чувствительны к T-DM1 при имплантации in vivo. (B) с течение времени имплантированные клетки N87 стали устойчивыми к T-DM1, но остались чувствительными к (C) T-vc0101, (D) T(N297Q+K222R)-AcLysvc0101 и (E) T(kK183+K290C)-vc0101.
На ФИГ. 22A-22D показана цитотоксичность in vitro четырех полученных на основе трастузумаба ADC в отношении T-DM1-резистентных клеток (N87-TDM), полученных in vivo, по сравнению с исходными T-DM1-чувствительными клетками N87, объем опухоли представлен на кривой в зависимости от времени. (A) T-DM1; (B) T(kK183+K290C)-vc0101; (C) T(LCQ05+K222R)-AcLysvc0101; (D) T(N297Q+K222R)-AcLysvc0101.
На ФИГ. 23A-23B показаны уровни экспрессии белка HER2 на T-DM1-резистентных клетках (N87-TDM1, у мышей 2, 17 и 18), полученных in vivo, по сравнению с исходными T-DM1-чувствительными клетками N87. (A) FACS анализ и (B) Вестерн-блот анализ. В экспрессии белка HER2 не наблюдали никакого значимого различия.
На ФИГ. 24A-24D показано, что устойчивость к T-DM1 в N87-TDM1 (мыши 2, 7 и 17) не обусловлена обеспечивающими лекарственную устойчивость эффлюксными насосами. (A) Вестерн-блот, на котором показана экспрессия белка MDR1. Цитотоксичность in vitro T-DM1-резистентных клеток (N87-TDM1) и исходных T-DM1-чувствительных клеток N87 в присутствии свободного лекарственного средства (B) 0101; (C) доксорубицина; (D) T-DM1.
На ФИГ. 25A-25B показаны профили зависимости концентрации от времени и фармакокинетика/токсикокинетика: (A) суммарного Ат и ADC на основе трастузумаба (T-vc0101) или сайт-специфического ADC T(kK183C+K290C) после введения дозы яванским макакам и (B) ADC аналита трастузумаба (T-vc0101) или различных сайт-специфических ADC после введения дозы яванским макакам.
На ФИГ. 26 показаны относительные значения времени удерживания хроматографии гидрофобного взаимодействия (HIC) в зависимости от экспозиции (AUC) у крыс. На оси X представлено Относительное время удерживания в HIC; тогда как на оси Y представлено фармакокинетическая нормализованная по дозе экспозиция у крыс ("площадь под кривой", AUC для антитела, от 0 до 336 часов, деленная на дозу лекарственного средства 10 мг/кг). Форма символа обозначает приблизительную нагрузку лекарственного средства (DAR): ромб=DAR 2; круг=DAR 4. Стрелка обозначает T(kK183C+K290C)-vc0101.
На ФИГ. 27 показано токсикологическое исследование с использованием обычного ADC конъюгата T-vc0101 и сайт-специфического ADC T(kK183C+K290C)-vc0101. T-vc0101 вызывал тяжелую нейтропению при дозе 5 мг/кг, тогда как T(kK183C+K290C)-vc0101 вызывал минимальное снижение количества нейтрофилов при дозе 9 мг/кг.
На ФИГ. 28A-28C показана кристаллическая структура (A) T(K290C+K334C)-vc0101; (B) T(K290C+K392C)-vc0101; и (C) T(K334C+K392C)-vc0101.
На ФИГ. 29 показана эффективность in vivo в ксенотрансплантатной модели с использованием клеточной линии N87. Все протестированные ADC показали эффективность при дозе 3 мг/кг.
На ФИГ. 30 показана противоопухолевая активность трастузумаба и двух полученных на основе трастузумаба ADC в отношении полученных от пациента ксенотрансплантатов PDX-GA0044, объем опухоли представлен на кривой в зависимости от времени. Животные получали растворитель (незакрашенные ромбы), трастузумаб (незакрашенные треугольники), T-DM1 (незакрашенные круги) или T(kK183C+K290C)-vc0101 (закрашенные белым круги и закрашенные квадраты). Полученные от пациента клетки PDX-GA0044 являются желудочной PDX моделью, экспрессирующей средние уровни HER2.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В настоящем изобретении предложены сайт-специфические конъюгаты антитела к HER2 и лекарственного средства (ADC), способы получения таких конъюгатов с применением антитела к HER2, линкеры и лекарственные полезные нагрузки, а также нуклеиновые кислоты, кодирующие антитела, применяемые в получении ADC конъюгатов. ADC конъюгаты изобретения могут применяться для получения и прозводства композиций, таких как лекарственные препараты, которые могут применяться в лечении HER2-экспрессирующих форм рака.
ADC конъюгаты состоят из антитела (иммуноглобулинового компонента), конъюгированного с лекарственным средством (полезной нагрузкой) с помощью линкера. Обычные стратегии конъюгирования ADC основаны на случайном конъюгировании лекарственной полезной нагрузки с антителом через лизины или цистеины, которые эндогенно присутствуют на тяжелой и/или легкой цепи антитела. Соответственно, такие ADC конъюгаты представляют собой гетерогенную смесь молекул, демонстрирующую различные отношения лекарственного средства:антитела (DAR). Напротив, ADC конъюгаты, раскрытые в настоящей заявке, являются сайт-специфическими ADC конъюгатами, в которых лекарственная полезная нагрузка конъюгирована с антителом по конкретным остаткам, введенным с помощью методов генной инженерии, на тяжелой и/или легкой цепи антитела. Таким образом, сайт-специфические ADC являются гомогенной популяцией ADC конъюгатов, состоящих из молекул с определенным отношением лекарственного средства:антитела (DAR). Таким образом, сайт-специфические ADC демонстрирует единообразную стехиометрию, приводящую к улучшению фармакокинетики, биораспределения и профилю безопасности конъюгата. ADC конъюгаты изобретения включают антитела изобретения, конъюгированные с одной или более молекулами линкера/полезной нагрузки.
В настоящем изобретении предложены конъюгаты антитела-лекарственного средства формулы Ab-(L-D), в которой: (a) Ab является антителом или его антигенсвязывающим фрагментом, которые связываются с HER2, и (b) L-D является молекулой линкера-лекарственного средства, где L является линкером, а D является лекарственным средством.
Также настоящее изобретение охватывает конъюгаты антитела-лекарственного средства формулы Ab-(L-D)p, в которой: (a) Ab является антителом или его антигенсвязывающим фрагментом, которые связываются с HER2, (b) L-D, является молекулой линкера-лекарственного средства, где L является линкером, а D является лекарственным средством, и (c) p является количеством молекул линкера/лекарственного средства, присоединенных к антителу. Для сайт-специфических ADC, p является целым числом вследствие гомогенной природы ADC. В некоторых вариантах осуществления p равно 4. В других вариантах осуществления p равно 3. В других вариантах осуществления p равно 2. В других вариантах осуществления p равно 1. В других вариантах осуществления p больше 4.
При использовании в настоящей заявке термин "HER2" относится к трансмембранному тирозинкиназному рецептору, который относится к семейству EGFR. HER2 также известен как ErbB2, p185 и CD340. Это семейство рецепторов включает четыре члена (EGFR/HER1, HER2, HER3 и HER4), функция которых заключается в стимуляции сигнальных путей факторов роста, таких как путь PI3K-AKT-mTOR. Амплификация и/или оверэкспрессия HER2 ассоциированы с различными злокачественными опухолями человека. Человеческий белок HER2 дикого типа описан, например, в публикациях Semba et al., 1985, PNAS 82:6497-6501 и Yamamoto et al., 1986, Nature 319: 230-4, а также под регистрационным номером Genbank X03363.
При использовании в настоящей заявке, термин "Антитело (Ab)" относится к молекуле иммуноглобулина, способной распознавать и связываться со специфической мишенью или антигеном, таким как полипептид, по меньшей мере через один участок распознавания антигена, расположенный в вариабельной области молекулы иммуноглобулина. Термин может охватывать любой тип антитела, в том числе, без ограничения перечисленными, моноклональные антитела, антигенсвязывающие фрагменты интактных антител, которые сохраняют способность специфично связываться с данным антигеном (т.е. Fab, Fab', F(ab')2, Fd, Fv, Fc и т.д.) и их мутантные формы.
Нативные или природные антитела и нативные иммуноглобулины, как правило, являются гетеротетрамерными гликопротеинами массой приблизительно 150000 дальтон, состоящими из двух идентичных легких (L) цепей и двух идентичных тяжелых (H) цепей. Каждая легкая цепь связана с тяжелой цепью одной ковалентной дисульфидной связью, при этом количество дисульфидных связей в тяжелых цепях иммуноглобулинов различных изотипов варьирует. Каждая тяжелая и легкая цепь также имеет расположенные с равными интервалами внутрицепочечные дисульфидные связи. Каждая тяжелая цепь имеет на одном конце вариабельный домен (VH), после которого следует ряд константных доменов. Каждая легкая цепь имеет вариабельный домен на одном конце (VL) и константный домен на другом своем конце; константный домен легкой цепи выровнен с первым константным доменом тяжелой цепи, и вариабельный домен легкой цепи выровнен с вариабельный доменом тяжелой цепи. Термин "вариабельный" относится к тому, что некоторые части вариабельных доменов широко отличаются по последовательности у различных антител.
Антитело, применяемое в настоящем изобретении, специфично связывается с HER2. В определенном варианте осуществления антитело HER2 связывается с тем же эпитопом на HER2, как и трастузумаб (Герцептин®). В более конкретном варианте осуществления антитело к HER2 имеет такие же вариабельные CDR-области, как и трастузумаб (Герцептин®). В еще более конкретном варианте осуществления антитело к HER2 имеет такие же вариабельные области (т.е. VH и VL), как и трастузумаб (Герцептин®).
При использовании в настоящей заявке термин "Линкер (L)" описывает прямое или косвенное соединение антитела с лекарственной полезной нагрузкой. Присоединение линкера к антителу может быть достигнуто множеством способов, например, через поверхностные лизины, восстановительное связывание с окисленными углеводами, остатками цистеина, освобожденными при восстановлении межцепочечных дисульфидных связей, реакционноспособные остатки цистеина, введенные с помощью методов генной инженерии в определенные сайты, и ацил-донорную глутаминсодержащую метку или эндогенный глутамин, сделанный реакционноспособным посредством инженерии полипептида в присутствии трансглутаминазы и амина. В настоящем изобретении применяются сайт-специфические способы соединения антитела с лекарственной полезной нагрузкой. В одном варианте осуществления конъюгирование проходит через остатки цистеина, которые были введены с помощью методов генной инженерии в константную область антитела. В другом варианте осуществления конъюгирование проходит через ацил-донорные остатки глутамина, которые были: a) добавлены в константную область антитела посредством пептидной метки, b) введены с помощью методов генной инженерии в константную область антитела или c) сделаны доступными/реакционноспособными в результате изменения окружающих остатков с помощью методов генной инженерии. Линкеры могут быть расщепляемыми (т.е. поддающимися расщеплению во внутриклеточных условиях) или нерасщепляемыми. В некоторых вариантах осуществления линкер является расщепляемым линкером.
При использовании в настоящей заявке термин "Лекарственное средство (D)" относится к любому терапевтическому средству, применимому в лечении рака. Лекарственное средство обладает биологической или поддающейся обнаружению активностью, например, цитотоксические средства, химиотерапевтические средства, цитостатические средства и иммуномодулирующие средства. В предпочтительных вариантах осуществления терапевтические средства оказывают цитотоксическое действие на опухоли, включающее истощение, элиминирование и/или уничтожение опухолевых клеток. Термины лекарственное средство, полезная нагрузка и лекарственная полезная нагрузка используются попеременно. В определенном варианте осуществления лекарственное средство является антимитотическим средством. В более конкретном варианте осуществления лекарственным средством является ауристатин. В еще более конкретном варианте осуществления лекарственным средством является 2-метилаланил-N-[(3R,4S,5S)-3-метокси-1-{(2S)-2-[(1R,2R)-1-метокси-2-метил-3-оксо-3-{[(1S)-2-фенил-1-(1,3-тиазол-2-ил)этил]амино}пропил]пирролидин-1-ил}-5-метил-1-оксогептан-4-ил]-N-метил-L-валинамид (также известный как 0101). В некоторых вариантах осуществления лекарственное средство предпочтительно обладает мембранной проницаемостью.
При использовании в настоящей заявке термин "L-D" относится к молекуле линкера-лекарственного средства, полученной в результате лекарственного средства связывания (D) с линкером (L).
Дополнительные научно-технические термины, используемые применительно к настоящему изобретению, если в настоящем документе не указано иное, должны иметь значения, которые обычно известны средним специалистам в данной области. Кроме того, если из контекста не следует иное, термины в единственном числе должны включать множества, и термины во множественном числе должны включать единственное число. Как правило, номенклатура, используемая в отношении, а также методики, культивирования клеток и тканей, молекулярной биологии, иммунологии, микробиологии, генетики, а также химии и гибридизации белков и нуклеиновых кислот, описанные в настоящей заявке, являются известными и широко используются в данной области.
I. Антитела к HER2
Для получения сайт-специфических HER2 ADC конъюгатов изобретения антитело может быть любым антителом, которое специфично связывается с внеклеточным доменом HER2. В одном варианте осуществления антитело, применяемое для получения ADC, связывается с тем же эпитопом HER2, как и трастузумаб, и/или конкурирует с трастузумабом за связывание HER2. В другом варианте осуществления антитело, применяемое для получения ADC, имеет такие же CDR-области вариабельной области тяжелой цепи и CDR-области вариабельной области легкой цепи, как и трастузумаб. В еще одном варианте осуществления антитело, применяемое для получения ADC, имеет такую же вариабельную область тяжелой цепи и вариабельную область легкой цепи, как и трастузумаб.
Термин "конкурирует" при использовании в настоящей заявке в отношении антитела, означает, что первое антитело или его антигенсвязывающий фрагмент, связывается с эпитопом по существу аналогично связыванию второго антитела или его антигенсвязывающего фрагмента, что результат связывания первого антитела с его когнатным эпитопом с возможностью обнаружения снижается в присутствии второго антитела по сравнению со связыванием первого антитела в отсутствие второго антитела. Альтернативный вариант, в котором связывание второго антитела с его эпитопом также с возможностью обнаружения снижается в присутствии первого антитела, может, но не должен обязательно иметь место. Таким образом, первое антитело может ингибировать связывание второго антитела с его эпитопом без того, что второе антитело ингибирует связывание первого антитела с его соответствующим эпитопом. Впрочем, когда каждое антитело с возможностью обнаружения ингибирует связывание другого антитела с его когнатным эпитопом или лигандом, в такой же, в большей или в меньшей степени, говорят, что антитела "перекрестно конкурируют" друг с другом за связывание своего соответствующего эпитопа(ов). Как конкурирующие, так и перекрестно конкурирующие антитела охвачены настоящим изобретением. Независимо от механизма, по которому такая конкуренция или перекрестная конкуренция проходит (например, стерическое затруднение, конформационное изменение или связывание с общим эпитопом или его частью), специалисту на основании описания, представленного в настоящей заявке, будет очевидно, что такие конкурирующие и/или перекрестно конкурирующие антитела включены в изобретение и могут применяться в способах, раскрытых в настоящей заявке.
Трастузумаб (торговое наименование Герцептин®) является гуманизированным моноклональным антителом, которое связывается с внеклеточным доменом HER2. Аминокислотные последовательности его вариабельных доменов раскрыты в патенте США 5,821,337 (VH - SEQ ID NO: 42, и VL - SEQ ID NO: 41 в патенте США 5,821,337), а также в Таблице 1 ниже (SEQ ID NO: 1 и 7, соответственно). Аминокислотные последовательности CDR-областей вариабельной области тяжелой цепи - SEQ ID NO: 2-4, тогда как аминокислотные последовательности CDR-областей легкой цепи - SEQ ID NO: 6-10 (Таблица 1 ниже). Аминокислотные последовательности полноразмерых тяжелой и легкой цепей - SEQ ID NO: 6 и 12, соответственно (Таблица 1 ниже).
T-DM1 (торговое наименование Кадсила®) является конъюгатом антитела-лекарственного средства, состоящим из трастузумаба, конъюгированного с майтанзиноидным средством DM1 через стабильный тиоэфирный линкер MCC (4-[N-малеимидометил]циклогексан-1-карбоксилат) (патент США 8,337,856). Иммуноглобулиновый компонент данного ADC идентичен трастузумабу. Конъюгирование полезной нагрузки с трастузумабом выполняют при использовании стандартных методов конъюгирования (не сайт-специфически), таким образом, что ADC представляет собой гетерогенную популяцию молекул с разными количествами DM1, конъюгированного с каждой из них. Полезная нагрузка DM1 ингибирует пролиферацию клеток, ингибируя формирование микротрубочек во время митоза посредством ингибирования полимеризации тубулина (Remillard et al., 1975, Science 189:1002-5). Кадсила® одобрена для лечения HER2-положительного метастатического рака молочной железы у больных, которые ранее проходили лечение Герцептином® и лекарственным средством из группы таксана, и стали рефракторными к Герцептину®. T-DM1, применяемый в экспериментах, описанных в Разделе Примеры, был получен в собственной лаборатории при использовании информации, имеющейся в общем доступе.
ADC конъюгаты настоящего изобретения конъюгированы с полезной нагрузкой сайт-специфическим способом. Для применения такого типа конъюгирования антитело должно быть дериватизировано с предоставлением реакционноспособного остатка цистеина, введенного с помощью методов генной инженерии в один или более специфических сайтов, ацил-донорной глутаминсодержащей метки или эндогенного глутамина, сделанного реакционноспособным в результате инженерии полипептида в присутствии трансглутаминазы и амина. Модификации аминокислот могут быть выполнены с помощью любого способа, известного в данной области, и многие такие способы известны и являются стандартными для специалиста. Например, но не в качестве ограничения, аминокислотные замены, делеции и вставки могут быть выполнены при использовании любой известной методики на основе ПЦР. Аминокислотные замены могут быть выполнены с помощью сайт-направленного мутагенеза (см., например, Zoller and Smith, 1982, Nucl. Acids Res. 10:6487-6500; и Kunkel, 1985, PNAS 82:488).
В таких применениях, когда требуется сохранение связывания антигена, подобные модификации следует вводить в участки, которые не нарушают антигенсвязывающей способности антитела. В предпочтительных вариантах осуществления одна или более модификаций сделаны в константной области тяжелой и/или легкой цепей.
При использовании в настоящей заявке термин "константная область" антитела относится к константной области легкой цепи антитела или константной области тяжелой цепи антитела, отдельно или в комбинации. Константные области антител, применяемых для получения ADC конъюгатов изобретения, могут быть получены из константных областей любого из IgA, IgD, IgE, IgG, IgM, или их любых изотипов, а также их субклассов и мутантных вариантов.
Константные домены не участвуют непосредственно в связывании антитела с антигеном, но проявляют различные эффекторные функции, такие как связывание Fc-рецептора (FcR), участие антитела в антителозависимой клеточной токсичности (ADCC), опсонизация, инициирование комплементзависимой цитотоксичности и дегрануляции тучных клеток. Как известно в уровне техники, термин "Fc-область" используется для определения C-концевой области тяжелой цепи иммуноглобулина. "Fc-область" может быть нативной последовательностью Fc-области или вариантом Fc-области. Хотя границы Fc-области тяжелой цепи иммуноглобулина моут изменяться, Fc-область тяжелой цепи IgG человека, как обычно определяют, идет от аминокислотного остатка в положении Cys226 или от Pro230, до C-конца. Нумерация остатков в Fc-области соответствует EU индексу Кэбата (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md., 1991). Fc-область иммуноглобулина обычно имеет две константных области, CH2 и CH3.
Существует две разных константных области легких цепей для применения в антителах, CLκ и CLλ. CLκ имеет известные полиморфные локусы CLκ-V/A45 и CLκ-L/V83 (при использовании системы нумерации Кэбата, как указано в Kabat et al. (1991, NIH Publication 91-3242, National Technical Information Service, Springfield, VA), таким образом, все положения каппа и лямбда пронумерованы согласно системе Кэбата), что допускает возможность существования полиморфизмов Km(1): CLκ-V45/L83; Km(1,2): CLκ-A45/L83; и Km(3): CLκ-A45/V83. Полипептиды, антитела и ADC конъюгаты изобретения могут иметь иммуноглобулиновые компоненты с любой из таких константных областей легкой цепи.
Для ясности, если не указано иное, аминокислотные остатки в константном домене тяжелой цепи человеческого IgG антитела пронумерованы согласно EU индексу согласно Edelman et al., 1969, Proc. Natl. Acad. Sci. USA 63(1): 78-85, как описано в Kabat et al., 1991, указанному в настоящей заявке как "EU индекс Кэбата". Как правило, Fc-домен включает приблизительно от аминокислотного остатка 236 до приблизительно 447 константного домена человеческого IgG1. Соответствие между C-нумерациями можно найти, например, в базе данных IGMT. Аминокислотные остатки константного домена легкой цепи пронумерованы согласно Kabat et al., 1991. Нумерация аминокислотных остатков константных доменов антител также показана в публикации международной заявки на патент WO 2013/093809. Единственным исключением для использования EU индекса Кэбата в константном домене тяжелой цепи IgG является остаток A114, описанный в примерах. A114 соответствует нумерации Кэбата, а соответствующий номер согласно EU индексу - 118. Это вызвано тем, что в первоначальной публикации сайт-специфического конъюгирования в этом сайте использовали нумерацию Кэбата, при этом данный сайт указали как A114C и с тех пор широко использовали в уровне техники как сайт "114". См. Junutula et al., Nature Biotechnology 26, 925-932 (2008). Для соответствия стандартному употреблению данного сайта в уровне техники в примерах используются "A114", "A114C", "C114" или "114C".
Нуклеиновые кислоты, кодирующие тяжелые и легкие цепи антител, применяемых для получения ADC конъюгатов изобретения, могут быть клонированы в вектор для экспрессии или размножения. Последовательность, кодирующая представляющее интерес антитело, можно поддерживаться в векторе в клетке-хозяине, и затем клетку-хозяина можно размножать и замораживать для последующего применения.
При использовании в настоящей заявке термин "вектор" относится к конструкции, которая способна к доставке и, предпочтительно, экспрессии одного или более генов или последовательностей, представляющих интерес, в клетке-хозяине. Примеры векторов включают, без ограничения перечисленными, вирусные векторы, "голые" ДНК или РНК векторы экспрессии, плазмидные, космидные или фаговые векторы, ДНК или РНК векторы экспрессии, ассоциированные с катионными конденсирующими агентами, ДНК или РНК векторы экспрессии, инкапсулированные в липосомах, а также некоторые эукариотические клетки, такие как клетки-продуценты.
При использовании в настоящей заявке термин "клетка-хозяин" включает отдельную клетку или культуру клеток, которая может быть или уже является реципиентом для вектора(ов), для включения полинуклеотидных вставок. Клетки-хозяева включают потомство одной клетки-хозяина, причем потомство может не быть полностью идентичным (по морфологии или по комплементу геномной ДНК) исходной родительской клетке вследствие природной, случайной или преднамеренной мутации. Клетка-хозяин включает клетки, трансфицированные in vivo нуклеиновыми кислотами или векторами согласно настоящему изобретению.
В Таблице 1 представлены аминокислотные (белковые) последовательности и соответствующие последовательности нуклеиновых кислот (ДНК) гуманизированных антител к HER2, применяемых в конструировании сайт-специфических ADC конъюгатов согласно изобретению. Показанные CDR-области определены согласно схеме нумерации Кэбата.
Тяжелые цепи и легкие цепи антител, показанные в Таблице 1, имеют вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL) трастузумаба. Константная область тяжелой цепи и константная область легкой цепи получены на основе трастузумаба и содержат одну или более модификаций, позволяющих применение сайт-специфического конъюгирования при получении ADC конъюгатов согласно изобретению.
Модификации в аминокислотных последовательностях в константной области антитела, позволяющие применять сайт-специфическое конъюгирование, подчеркнуты и выделены полужирным шрифтом. Обозначение антител, полученных на основе трастузумаба, включает T (от трастузумаба) и затем в круглых скобках положение аминокислотной модификации между однобуквенным кодом аминокислоты для остатка дикого типа и однобуквенным кодом аминокислоты для остатка, который теперь присутствует в данном положении в производном антителе. Двумя исключениями в данной номенклатуре являются "kK183C", которое обозначает, что положение 183 легкой (каппа) цепи была модифицировано с заменой лизина на цистеин, и "LCQ05", которое обозначает содержащую восемь аминокислотных остатков глутамина метку, которая была присоединена к C-концу константной области легкой цепи.
Одна из модификаций, показанных в Таблице 1, не используется для конъюгирования. Остаток в положении 222 тяжелой цепи (согласно EU индексу схемы нумерации Кэбата) может быть изменен для получения более гомогенного конъюгата антитела и полезной нагрузки, лучшего межмолекулярного поперечного сшивания между антителом и полезной нагрузкой и/или значительного уменьшения межцепочечного поперечного сшивания.
Таблица 1: Последовательности гуманизированных антител к HER2
VL трастузумаба
VL CDR1
VL CRD2
VL CDR3
Белок константной области тяжелой цепи
Белок тяжелой цепи
Белок константной области тяжелой цепи
Белок тяжелой цепи
Белок константной области тяжелой цепи
Белок тяжелой цепи
Белок константной области тяжелой цепи
Белок тяжелой цепи
Белок константной области тяжелой цепи
Белок тяжелой цепи
Белок константной области тяжелой цепи
Белок тяжелой цепи
Белок константной области тяжелой цепи
Белок тяжелой цепи
Белок константной области тяжелой цепи
Белок тяжелой цепи
Белок константной области тяжелой цепи
Белок тяжелой цепи
Белок константной области тяжелой цепи
Белок тяжелой цепи
Белок константной области тяжелой цепи
Белок тяжелой цепи
Белок константной области тяжелой цепи
Белок тяжелой цепи
Белок константной области тяжелой цепи
Белок тяжелой цепи
Белок константной области тяжелой цепи
Белок тяжелой цепи
Белок константной области легкой цепи
Белок легкой цепи
Белок константной области легкой цепи
Белок легкой цепи
ДНК константной области тяжелой цепи
ДНК тяжелой цепи
ДНК константной области тяжелой цепи
ДНК тяжелой цепи
ДНК константной области тяжелой цепи
ДНК тяжелой цепи
ДНК константной области тяжелой цепи
ДНК тяжелой цепи
ДНК константной области тяжелой цепи
ДНК тяжелой цепи
ДНК константной области тяжелой цепи
ДНК тяжелой цепи
ДНК константной области тяжелой цепи
ДНК тяжелой цепи
ДНК константной области тяжелой цепи
ДНК тяжелой цепи
ДНК константной области тяжелой цепи
ДНК тяжелой цепи
ДНК константной области тяжелой цепи
ДНК тяжелой цепи
ДНК константной области тяжелой цепи
ДНК тяжелой цепи
ДНК константной области тяжелой цепи
ДНК тяжелой цепи
ДНК константной области тяжелой цепи
ДНК тяжелой цепи
ДНК константной области тяжелой цепи
ДНК тяжелой цепи
ДНК константной области легкой цепи
ДНК легкой цепи
ДНК константной области легкой цепи
ДНК легкой цепи
В некоторых вариантах осуществления в ADC конъюгатах согласно изобретению могут применяться антитела, включающие CDR-области вариабельной области тяжелой цепи и CDR-области вариабельной области легкой цепи трастузумаба (VH CDR-области SEQ ID NO: 2-4 и VL CDR-области SEQ ID NO: 8-10), а также любую комбинацию константных областей тяжелой и легкой цепи, раскрытых в Таблице 1, при условии, что в том случае, когда константная область тяжелой цепи является SEQ ID NO: 5, то тогда константная область легкой цепи не является SEQ ID NO: 11 (вследствие того, что данная комбинация воссоздает трастузумаб дикого типа и, таким образом, не позволяет проводить сайт-специфическое конъюгирование). В таких вариантах осуществления константная область тяжелой цепи может быть выбрана из любого из SEQ ID NO: 17, 5, 13, 21, 23, 25, 27, 29, 31, 33, 35, 37 или 39, тогда как константная область легкой цепи может быть выбрана из любого из SEQ ID NO: 41, 11 или 43, при условии, что данная комбинация не является SEQ ID NO: 5 и SEQ ID NO: 11, как обсуждается выше.
В более конкретных вариантах осуществления в ADC конъюгатах согласно изобретению могут применяться антитела, включающие CDR-области вариабельной области тяжелой цепи и CDR-области вариабельной области легкой цепи трастузумаба (VH CDR-области SEQ ID NO: 2-4 и VL CDR-области SEQ ID NO: 8-10), и комбинацию константной области тяжелой и легкой цепи, выбранную из следующих:
(a) константная область тяжелой цепи SEQ ID NO: 17 и константная область легкой цепи SEQ ID NO: 41;
(b) константная область тяжелой цепи SEQ ID NO: 5 и константная область легкой цепи SEQ ID NO: 41;
(c) константная область тяжелой цепи SEQ ID NO: 17 и константная область легкой цепи SEQ ID NO: 11;
(d) константная область тяжелой цепи SEQ ID NO: 21 и константная область легкой цепи SEQ ID NO: 11;
(e) константная область тяжелой цепи SEQ ID NO: 23 и константная область легкой цепи SEQ ID NO: 11;
(f) константная область тяжелой цепи SEQ ID NO: 25 и константная область легкой цепи SEQ ID NO: 11;
(g) константная область тяжелой цепи SEQ ID NO: 27 и константная область легкой цепи SEQ ID NO: 11;
(h) константная область тяжелой цепи SEQ ID NO: 23 и константная область легкой цепи SEQ ID NO: 41;
(i) константная область тяжелой цепи SEQ ID NO: 25 и константная область легкой цепи SEQ ID NO: 41;
(j) константная область тяжелой цепи SEQ ID NO: 27 и константная область легкой цепи SEQ ID NO: 41;
(k) константная область тяжелой цепи SEQ ID NO: 29 и константная область легкой цепи SEQ ID NO: 11;
(l) константная область тяжелой цепи SEQ ID NO: 31 и константная область легкой цепи SEQ ID NO: 11;
(m) константная область тяжелой цепи SEQ ID NO: 33 и константная область легкой цепи SEQ ID NO: 43;
(n) константная область тяжелой цепи SEQ ID NO: 35 и константная область легкой цепи SEQ ID NO: 11;
(o) константная область тяжелой цепи SEQ ID NO: 37 и константная область легкой цепи SEQ ID NO: 11;
(p) константная область тяжелой цепи SEQ ID NO: 39 и константная область легкой цепи SEQ ID NO: 11; или
(q) константная область тяжелой цепи SEQ ID NO: 13 и константная область легкой цепи SEQ ID NO: 43.
В еще более конкретном варианте осуществления ADC согласно изобретению включает антитело с VH CDR-областями SEQ ID NO: 2-4 и VL CDR-областями SEQ ID NO: 8-10, а также константную область тяжелой цепи SEQ ID NO: 17 и константную область легкой цепи SEQ ID NO: 41.
В другом более конкретном варианте осуществления ADC согласно изобретению включает антитело с VH CDR-областями SEQ ID NO: 2-4 и VL CDR-областями SEQ ID NO: 8-10, а также константную область тяжелой цепи SEQ ID NO: 13 и константную область легкой цепи SEQ ID NO: 43.
В других вариантах осуществления в ADC конъюгатах согласно изобретению могут применяться антитела, включающие любую комбинацию тяжелых и легких цепей, раскрытых в Таблице 1, при условии, что в том случае, если тяжелая цепь является SEQ ID NO: 6, то тогда легкая цепь не является SEQ ID NO: 12 (вследствие того, что данная комбинация воссоздает трастузумаб дикого типа и, таким образом, не позволяет применять сайт-специфическое конъюгирование). В таких вариантах осуществления тяжелая цепь может быть выбрана из любого из SEQ ID NO: 18, 6, 14, 22, 24, 26, 28, 30, 32, 34, 36, 38 или 40, тогда как легкая цепь может быть выбрана из любого из SEQ ID NO: 42, 12 или 44, если данная комбинация не является SEQ ID NO: 6 и SEQ ID NO: 12, как обсуждается выше.
В более конкретных вариантах осуществления в ADC конъюгатах согласно изобретению могут применяться антитела, включающие комбинацию тяжелой цепи и легкой цепи, выбранную из следующих:
(a) тяжелая цепь SEQ ID NO: 18 и легкая цепь SEQ ID NO: 42;
(b) тяжелая цепь SEQ ID NO: 6 и легкая цепь SEQ ID NO: 42;
(c) тяжелая цепь SEQ ID NO: 18 и легкая цепь SEQ ID NO: 12;
(d) тяжелая цепь SEQ ID NO: 22 и легкая цепь SEQ ID NO: 12;
(e) тяжелая цепь SEQ ID NO: 24 и легкая цепь SEQ ID NO: 12;
(f) тяжелая цепь SEQ ID NO: 26 и легкая цепь SEQ ID NO: 12;
(g) тяжелая цепь SEQ ID NO: 28 и легкая цепь SEQ ID NO: 12;
(h) тяжелая цепь SEQ ID NO: 24 и легкая цепь SEQ ID NO: 42;
(i) тяжелая цепь SEQ ID NO: 26 и легкая цепь SEQ ID NO: 42;
(j) тяжелая цепь SEQ ID NO: 28 и легкая цепь SEQ ID NO: 42;
(k) тяжелая цепь SEQ ID NO: 30 и легкая цепь SEQ ID NO: 12;
(l) тяжелая цепь SEQ ID NO: 32 и легкая цепь SEQ ID NO: 12;
(m) тяжелая цепь SEQ ID NO: 34 и легкая цепь SEQ ID NO: 44;
(n) тяжелая цепь SEQ ID NO: 36 и легкая цепь SEQ ID NO: 12;
(o) тяжелая цепь SEQ ID NO: 38 и легкая цепь SEQ ID NO: 12;
(p) тяжелая цепь SEQ ID NO: 40 и легкая цепь SEQ ID NO: 12; или
(q) тяжелая цепь SEQ ID NO: 14 и легкая цепь SEQ ID NO: 44.
В еще более конкретном варианте осуществления ADC изобретения включает антитело с тяжелой цепью SEQ ID NO: 18 и легкой цепью SEQ ID NO: 42. Плазмиды, содержащие нуклеиновые кислоты, кодирующие тяжелую цепь SEQ ID NO: 18 и легкую цепь SEQ ID NO: 42, были депонированы в Американской коллекции типовых культур (ATCC), 10801 University Blvd., Manassas, VA 20110-2209, 17 ноября 2015 года с присвоением регистрационных номеров PTA-122672 и PTA-122673, соответственно. Депозиты были сделаны в соответствии с положениями Будапештского соглашения о Международном признании депонирования микроорганизмов в рамках патентной процедуры, а также нормативных актов на его основе (Будапештское соглашение). Это гарантирует сохранение жизнеспособной депонированной культуры в течение 30 лет с даты депонирования. Депозит будет доступен в ATCC в соответствии с Будапештским соглашением и по согласнованию между Pfizer Inc. и ATCC, которая гарантирует постоянную и неограниченную доступность потомства депонированной культуры неограниченному кругу лиц после выдачи соответствующего патента США или после публикации в открытом доступе любой заявки на патент США или иностранной заявки на патент, в зависимости от того, что наступит раньше, и гарантирует доступность потомства лицу, определенному комиссаром патентного ведомства США, имеющему право на него согласно 35 U.S.C., раздел 122, и регламенту комиссара в соответствии с этим (включая 37 C.F.R., раздел 1.14, с уделением особого внимания 886 OG 638).
Настоящий заявитель согласился с тем, что если культура депонированных материалов погибнет или будет потеряна или уничтожена при культивировании в подходящих условиях, материалы будут незамедлительно заменены при уведомлении другим таким же материалом. Доступность депонированного материала не следует расценивать как лицензию на практическое осуществление изобретения в нарушение прав, предоставляемых в соответствии с патентным законодательством любого государства.
В другом более конкретном варианте осуществления ADC изобретения включает антитело с тяжелой цепью SEQ ID NO: 14 и легкой цепью SEQ ID NO: 44.
В некоторых аспектах изобретения ADC изобретения включает антитело, имеющее тяжелую цепь и/или легкую цепь, включающую аминокислотную последовательность, которая по меньшей мере на 90%, 95%, 98% или 99% идентична любой из тяжелых или легких цепей, раскрытых выше. Остатки, которые были изменены, могут находиться в вариабельной области или в константной области антитела. В некоторых вариантах осуществления не больше 1, 2, 3, 4 или 5 остатков были изменены по сравнению с любой из тяжелых или легких цепей, раскрытых выше. В других вариантах осуществления измененные остатки не присутствуют ни в одной из CDR-области вариабельной области.
Термин "процент идентичности" (или "% идентичности") в отношении аминокислотных последовательностей означает количество остатков в двух последовательностях, которые являются однаковыми при выравнивании с максимальным соответствием. Существует множество различных алгоритмов, известных в уровне техники, которые могут применяться для измерения процента идентичности аминокислот (т.е. Средство поиска основного локального выравнивания или BLAST®). Если не определено иное, для конкретной программы или алгоритма используются параметры по умолчанию.
Для использования в получении ADC конъюгатов, антитела к HER2, описанные в настоящей заявке, могут быть по существу чистыми, т.е. по меньшей мере на 50% чистыми (т.е. не содеращими примесей), более предпочтительно по меньшей мере на 90% чистыми, более предпочтительно по меньшей мере на 95% чистыми, еще более предпочтительно по меньшей мере на 98% чистыми и наиболее предпочтительно по меньшей мере на 99% чистыми.
II. Лекарственные средства
Лекарственные средства, применимые при получении сайт-специфических HER2 ADC конъюгатов согласно изобретению, включают любое терапевтическое средство, применимое для лечения рака, в том числе, без ограничения перечисленными, цитотоксические средства, цитостатические средства, иммуномодулирующие средства и химиотерапевтические средства. Цитотоксическое действие относится к истощению, элиминированию и/или уничтожению клетки-мишени (то есть опухолевой клетки). Цитотоксическое средство относится к средству, которое оказывает цитотоксическое воздействие на клетку. Цитостатическое действие относится к ингибированию клеточной пролиферации. Цитостатическое средство относится к средству, которое оказывает цитостатическое воздействие на клетку, препятствуя тем самым росту и/или размножению специфической субпопуляции клеток (то есть опухолевых клеток). Иммуномодулирующее средство относится к средству, которое стимулирует иммунный ответ посредством продуцирования цитокинов и/или антител и/или модулирования функции T-клеток, в результате чего наблюдается ингибирование или уменьшение роста субпопуляции клеток (то есть опухолевых клеток), прямое или опосредованное, что позволяет другому средству быть более эффективным. Химиотерапевтическое средство относится к средству, которое является химическим соединением, применимым для лечения рака. Лекарственное средство также может быть производным лекарственного средства, где лекарственное средство было функционализировано для обеспечения конъюгирования с антителом согласно изобретению.
В некоторых вариантах осуществления лекарственное средство является лекарственным средством, обладающим мембранной проницаемостью. В таких вариантах осуществления полезная нагрузка (т.е. лекарственное средство) может оказывать неспецифическое воздействие, при котором клетки, окружающие клетку, которая первоначально интернализировала ADC, уничтожаются полезной нагрузкой. Это происходит при высвобождении полезной нагрузки от антитела (т.е. при расщеплении расщепляемого линкера) и проникновении через клеточную мембрану и, при диффузии, вызывает гибель окружающих клеток.
В соответствии с раскрытыми способами, лекарственные средства применяются для получения конъюгатов антитела-лекарственного средства формулы Ab-(L-D), в которой: (a) Ab является антителом, которое связывается с HER2; и (b) L-D является молекулой линкера-лекарственного средства, где L является линкером, а D является лекарственным средством.
Отношение лекарственного средства к антителу (DAR) или лекарственная нагрузка указывает количество молекул лекарственного средства (D), которые конъюгированы с каждой молекулой антитела. В конъюгатах антитела-лекарственного средства согласно настоящему изобретению применяется сайт-специфическое конъюгирование, в результате чего присутствует по существу гомогенная популяция ADC конъюгатов, имеющих одно DAR в композиции ADC конъюгатов. В некоторых вариантах осуществления DAR равно 1. В некоторых вариантах осуществления DAR равно 2. В других вариантах осуществления DAR равно 3. В других вариантах осуществления DAR равно 4. В других вариантах осуществления DAR больше 4.
Применение обычного конъюгирования (а не сайт-специфического конъюгирования) приводит к гетерогенной популяции различных ADC конъюгатов, каждый из которых имеет индивидуальное различное DAR. Композиции ADC конъюгатов, полученные таким образом, включают множество антител, причем каждое антитело конъюгировано с определенным количеством молекул лекарственного средства. Фактически композиции имеют среднее DAR. В T-DM1 (Кадсила®) применяется обычное конъюгирование по остаткам лизина, при этом среднее DAR составляет приблизительно 4, с широким распределением, которое включает ADC конъюгаты, нагруженные 0, 1, 2, 3, 4, 5, 6, 7 или 8 молекулами лекарственного средства (Kim et al., 2014, Bioconj Chem 25(7):1223-32).
Композиции, партии и/или лекарственные формы множества ADC конъюгатов могут характеризоваться средним DAR. DAR и среднее DAR могут быть определены с помощью различных стандартных способов, таких как УФ-спектроскопия, масс-спектроскопия, ИФА, радиометрические методы, хроматография гидрофобного взаимодействия (HIC), электрофорез и ВЭЖХ.
В аспектах изобретения HER2 ADC может иметь DAR, равное 1, DAR, равное 2, DAR, равное 3, DAR, равное 4, DAR, равное 5, DAR, равное 6, DAR, равное 7, DAR, равное 8, DAR, равное 9, DAR, равное 10, DAR, равное 11, DAR, равное 12, или DAR больше 12. В аспектах изобретения HER2 ADC может содержать одну молекулу лекарственного средства или 2 молекулы лекарственного средства, или 3 молекулы лекарственного средства, или 4 молекулы лекарственного средства, или 5 молекул лекарственного средства, или 6 молекул лекарственного средства, или 7 молекул лекарственного средства, или 8 молекул лекарственного средства, или 9 молекул лекарственного средства, или 10 молекул лекарственного средства, или 11 молекул лекарственного средства, или 12 молекул лекарственного средства или больше 12 молекул.
В аспектах изобретения HER2 ADC может иметь среднее DAR в пределах от приблизительно 2 до приблизительно 4 или среднее DAR в пределах от приблизительно 3 до приблизительно 5, или среднее DAR в пределах от приблизительно 4 до приблизительно 6, или среднее DAR в пределах от приблизительно 5 до приблизительно 7, или среднее DAR в пределах от приблизительно 6 до приблизительно 8, или среднее DAR в пределах от приблизительно 7 до приблизительно 9, или среднее DAR в пределах от приблизительно 8 до приблизительно 10, или среднее DAR в пределах от приблизительно 9 до приблизительно 11, или среднее DAR в пределах от приблизительно 10 до приблизительно 12 и т.д. В некоторых аспектах композиции, партии и/или лекарственные формы HER2 ADC конъюгатов могут иметь среднее DAR приблизительно 1 или среднее DAR приблизительно 2, среднее DAR приблизительно 3 или среднее DAR приблизительно 4, или среднее DAR приблизительно 5, или среднее DAR приблизительно 6, или среднее DAR приблизительно 7, или среднее DAR приблизительно 8, или среднее DAR приблизительно 9, или среднее DAR приблизительно 10, или среднее DAR приблизительно 11, или среднее DAR приблизительно 12 или среднее DAR больше 12. При использовании в предыдущих диапазонах среднего DAR, термин "приблизительно" означает +/-0,5%.
Композиция, партия и/или лекарственная форма HER2 ADC конъюгатов могут характеризоваться предпочтительным диапазоном среднего DAR, например, среднего DAR в пределах от приблизительно 3 до приблизительно 5, среднего DAR в пределах от приблизительно 3 до приблизительно 4 или среднего DAR в пределах от приблизительно 4 до приблизительно 5. Кроме того, композиция, партия и/или лекарственная форма HER2 ADC конъюгатов могут характеризоваться предпочтительным диапазоном среднего DAR, например, среднего DAR в пределах 3-5, среднего DAR в пределах 3-4 или среднего DAR в пределах 4-5.
В некоторых аспектах изобретения композиция, партия и/или лекарственная форма HER2 ADC конъюгатов могут характеризоваться средним DAR, равным приблизительно 1,0, или средним DAR, равным 1,0, или средним DAR, равным 1,1, или средним DAR, равным 1,2, или средним DAR, равным 1,3, или средним DAR, равным 1,4, или средним DAR, равным 1,5, или средним DAR, равным 1,6, или средним DAR, равным 1,7, или средним DAR, равным 1,8, или средним DAR, равным 1,9. В другом аспекте композиция, партия и/или лекарственная форма HER2 ADC конъюгатов могут характеризоваться средним DAR, равным приблизительно 2,0, или средним DAR, равным 2,0, или средним DAR, равным 2,1, или средним DAR, равным 2,2, или средним DAR, равным 2,3, или средним DAR, равным 2,4, или средним DAR, равным 2,5, или средним DAR, равным 2,6, или средним DAR, равным 2,7, или средним DAR, равным 2,8, или средним DAR, равным 2,9. В другом аспекте композиция, партия и/или лекарственная форма HER2 ADC конъюгатов могут характеризоваться средним DAR, равным приблизительно 3,0, или средним DAR, равным 3,0, или средним DAR, равным 3,1, или средним DAR, равным 3,2, или средним DAR, равным 3,3, или средним DAR, равным 3,4, или средним DAR, равным 3,5, или средним DAR, равным 3,6, или средним DAR, равным 3,7, или средним DAR, равным 3,8, или средним DAR, равным 3,9. В другом аспекте композиция, партия и/или лекарственная форма HER2 ADC конъюгатов могут характеризоваться средним DAR, равным приблизительно 4,0, или средним DAR, равным 4,0, или средним DAR, равным 4,1, или средним DAR, равным 4,2, или средним DAR, равным 4,3, или средним DAR, равным 4,4, или средним DAR, равным 4,5, или средним DAR, равным 4,6, или средним DAR, равным 4,7, или средним DAR, равным 4,8, или средним DAR, равным 4,9, или средним DAR, равным 5,0.
В другом аспекте композиция, партия и/или лекарственная форма HER2 ADC конъюгатов могут характеризоваться средним DAR, равным 12 или меньше, средним DAR, равным 11 или меньше, средним DAR, равным 10 или меньше, средним DAR, равным 9 или меньше, средним DAR, равным 8 или меньше, средним DAR, равным 7 или меньше, средним DAR, равным 6 или меньше, средним DAR, равным 5 или меньше, средним DAR, равным 4 или меньше, средним DAR, равным 3 или меньше, средним DAR, равным 2 или меньше, или средним DAR, равным 1 или меньше.
В других аспектах композиция, партия и/или лекарственная форма HER2 ADC конъюгатов могут характеризоваться средним DAR, равным 11,5 или меньше, средним DAR, равным 10,5 или меньше, средним DAR, равным 9,5 или меньше, средним DAR, равным 8,5 или меньше, средним DAR, равным 7,5 или меньше, средним DAR, равным 6,5 или меньше, средним DAR, равным 5,5 или меньше, средним DAR, равным 4,5 или меньше, средним DAR, равным 3,5 или меньше, средним DAR, равным 2,5 или меньше, средним DAR, равным 1,5 или меньше.
В некоторых аспектах настоящего изобретения способы обычного конъюгирования через остатки цистеина и условия очистки, раскрытые в настоящей заявке, дают композицию, партию и/или лекарственную форму HER2 ADC конъюгатов с оптимизированным средним DAR в пределах приблизительно 3-5, предпочтительно приблизительно 4.
В некоторых аспектах настоящего изобретения способы сайт-специфического конъюгирования через модифицированные остатки цистеина и условия очистки, раскрытые в настоящей заявке, дают композицию, партию и/или лекарственную форму HER2 ADC конъюгатов с оптимизированным средним DAR в пределах приблизительно 3-5, предпочтительно приблизительно 4.
В некоторых аспектах настоящего изобретения способы сайт-специфического конъюгирования посредством конъюгирования на основе трансглутаминазы и условия очистки, раскрытые в настоящей заявке, дают композицию, партию и/или лекарственную форму HER2 ADC конъюгатов с оптимизированным средним DAR в пределах приблизительно 1-3, предпочтительно приблизительно 2.
Также настоящее изобретение охватывает конъюгаты антитела-лекарственного средства формулы Ab-(L-D)p, в которой: (a) Ab является антителом или его антигенсвязывающим фрагментом, который связывается с HER2, (b) L-D является молекулой линкера-лекарственного средства, где L является линкером, а D является лекарственным средством, и (c) p является количеством молекул линкера/лекарственного средства, которые присоединены к антителу. В случае сайт-специфических ADC конъюгатов, p является целым числом вследствие гомогенной природы ADC. В некоторых вариантах осуществления p равно 4. В других вариантах осуществления p равно 3. В других вариантах осуществления p равно 2. В других вариантах осуществления p равно 1. В других вариантах осуществления p больше 4.
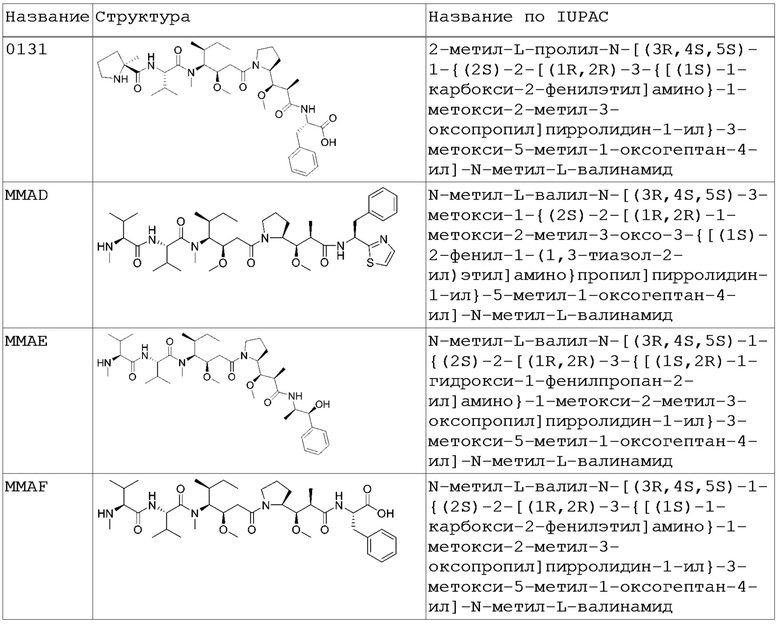
В одном варианте осуществления лекарственный компонент ADC конъюгатов согласно изобретению является антимитотическим лекарственным средством. В определенном варианте осуществления антимитотическим лекарственным средством является ауристатин (например, 0101, 8261, 6121, 8254, 6780 и 0131; см. Таблицу 2 ниже). В более конкретном варианте осуществления лекарственным средством из группы ауристатинов является 2-метилаланил-N-[(3R,4S,5S)-3-метокси-1-{(2S)-2-[(1R,2R)-1-метокси-2-метил-3-оксо-3-{[(1S)-2-фенил-1-(1,3-тиазол-2-ил)этил]амино}про-пил]пирролидин-1-ил}-5-метил-1-оксогептан-4-ил]-N-метил-L-валинамид (также известный как 0101).
Ауристатины ингибируют пролиферацию клеток, ингибируя формирование микротрубочек во время митоза посредством ингибирования полимеризации тубулина. В публикации международной заявки PCT WO 2013/072813, которая полностью включена посредством отсылки, раскрыты ауристатины, которые могут применяться в производстве ADC конъюгатов согласно изобретению, и предложены способы получения таких ауристатинов.
Таблица 2: Лекарственные средства
В некоторых аспектах изобретения цитотоксическое средство может быть получено с применением липосомы или биосовместимого полимера. Антитела к HER2, как описано в настоящей заявке, могут быть конъюгированы с биосовместимым полимером для увеличения полупериода существования в сыворотке и биоактивности, и/или увеличения периода полувыведения in vivo. Примеры биосовместимых полимеров включают водорастворимые полимеры, такие как полиэтиленгликоль (ПЭГ) или его производные, и цвиттер-ион-содержащие биосовместимые полимеры (например, фосфорилхолин-содержащий полимер).
III. Линкеры
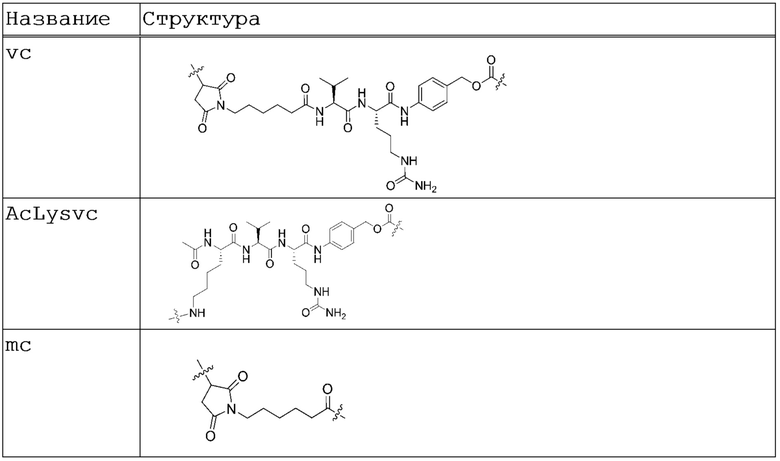
Сайт-специфические HER2 ADC конъюгаты согласно изобретению получают с использованием линкера для соединения или конъюгирования лекарственного средства с антителом к HER2. Линкер является бифункциональным соединением, которое может применяться для связывания лекарственного средства и антитела с получением конъюгата антитела-лекарственного средства (ADC). Такие конъюгаты обеспечивают селективную доставку лекарственных средств к опухолевым клеткам. Подходящие линкеры включают, например, расщепляемые и нерасщепляемые линкеры. Расщепляемый линкер обычно подвергается расщеплению во внутриклеточных условиях. Основные механизмы, по которым конъюгированное лекарственное средство отщепляется от антитела, включают гидролиз при кислотном pH лизосом (гидразоны, ацетали и цис-аконитат-подобные амиды), расщепление пептидов лизосомальными ферментами (катепсинами и другими лизосомальными ферментами) и восстановление дисульфидов. В результате такого разнообразия механизмов расщепления, механизмы соединения лекарственного средства с антителом также широко варьируют, при этом может применяться любой подходящий линкер.
Подходящие расщепляемые линкеры включают, без ограничения перечисленными, пептидный линкер, расщепляемый внутриклеточной протеазой, такой как лизосомальная протеаза или эндосомальная протеаза, такой как малеимидокапроил-валин-цитруллин-п-аминобензилоксикарбонил (vc), N~2~-ацетил-L-лизил-L-валил-L-цитруллин-п-аминобензилоксикарбонил-N,N'-диметиламиноэтил-CO-(AcLysvc) и m(H20)c-vc (Таблица 3, ниже). В определенных вариантах осуществления линкер является расщепляемым линкером, таким, что полезная нагрузка может оказывать неспецифическое действие сразу после расщепления линкера. Неспецифическое действие состоит в том, что при высвобождении обладающего мембранной проницаемостью лекарственного средства от антитела (т.е. при расщеплении расщепляемого линкера) и проникновении через клеточную мембрану и диффузии, лекарственное средство вызывает гибель клеток, окружающих клетку, которая первоначально интернализировала ADC.
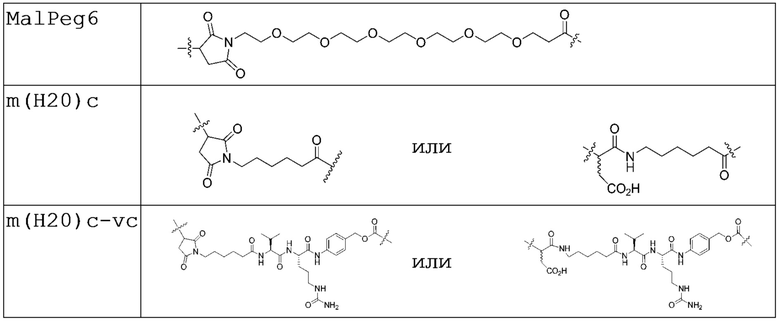
Подходящие нерасщепляемые линкеры включают, без ограничения перечисленными, малеимидокапроил (mc), малеимид-(полиэтиленгликоль)6 (MalPeg6), Mal-Peg2C2, Mal-Peg3C2 и m(H20)c (Таблица 3, ниже).
Другие подходящие линкеры включают линкеры, гидролизуемые при определенном pH или в диапазоне pH, такие как гидразоновый линкер. Дополнительные подходящие расщепляемые линкеры включают дисульфидные линкеры. Линкер может быть ковалентно связан с антителом в такой степени, что антитело должно расщепляться внутриклеточно для высвобождения лекарственного средства, например, mc линкер и т.п.
В определенных аспектах изобретения линкеры в сайт-специфических HER2 ADC конъюгатах согласно изобретению являются расщепляемыми и могут быть vc или AcLysvc.
Многие терапевтические средства (лекарственные средства), конъюгированные с антителами, обладают низкой, если вообще обладают, растворимостью в воде, и это может ограничивать лекарственную нагрузку на конъюгате вследствие агрегации конъюгата. Один из способов решения данной проблемы заключается в добавлении к линкеру солюбилизирующих групп. Могут примененяться конъюгаты, полученные с применением линкера, состоящего из ПЭГ и дипептида, в том числе такие, которые содержат ПЭГ-дикислоту, тиол-кислоту или малеимид-кислоту, присоединенные к антителу, дипептидный спейсер и амидную связь с амином аналога антрациклина или дуокармицина. Другим примером является конъюгат, полученный с ПЭГ-содержащим линкером, в котором дисульфид связан с цитотоксическим средством, а амид связан с антителом. Способы, которые включают ПЭГ-группы, могут быть полезными в преодолении агрегации и ограничений в лекарственной нагрузке.
Таблица 3: Линкеры
Линкеры соединены с моноклональным антителом с левой стороны молекулы и с лекарственным средством с правой стороны молекулы, как показано в Таблице 3.
IV. Способы получения сайт-специфических HER2 ADC конъюгатов
Также предложены способы получения конъюгатов антитела-лекарственного средства согласно настоящему изобретению. Например, способ получения сайт-специфического HER2 ADC, как раскрыто в настоящей заявке, может включать: (a) конъюгирование линкера с лекарственным средством; (b) конъюгирование молекулы линкера-лекарственного средства с антителом; и (c) очистку конъюгата антитела-лекарственного средства.
В HER2 ADC конъюгатах согласно настоящему изобретению применены сайт-специфические способы конъюгирования антитела к HER2 с лекарственной полезной нагрузкой.
В одном варианте осуществления сайт-специфическое конъюгирование осуществляют через один или более остатков цистеина, которые были введены с помощью методов генной инженерии в константную область антитела. Способы получения антител к HER2 для сайт-специфического конъюгирования через остатки цистеина могут быть осуществлены, как описано в публикации PCT WO 2013/093809, которая полностью включена посредством отсылки. Одно или более следующих положений (при использовании EU индекса нумерации Кэбата для константной области IgG1 и нумерации Кэбата для константной области каппа-цепи) могут быть изменены на цистеин и, таким образом, могут служить в качестве сайта для конъюгирования: a) на константной области тяжелой цепи, остатки 114, 246, 249, 265, 267, 270, 276, 278, 283, 290, 292, 293, 294, 300, 302, 303, 314, 315, 318, 320, 327, 332, 333, 334, 336, 345, 347, 354, 355, 358, 360, 362, 370, 373, 375, 376, 378, 380, 382, 386, 388, 390, 392, 393, 401, 404, 411, 413, 414, 416, 418, 419, 421, 428, 431, 432, 437, 438, 439, 443 и 444, и/или b) на константной области каппа-цепи, остатки 111, 149, 183, 188, 207 и 210.
В определенном варианте осуществления одно или более положений (при использовании EU индекса нумерации Кэбата), которые могут быть изменены на цистеин: a) на константной области тяжелой цепи, 290, 334, 392 и/или 443, и/или b) на константной области легкой цепи является 183 (нумерация Кэбата).
В более конкретном варианте осуществления положения 290 на константной области тяжелой цепи и положение 183 на константной области легкой цепи изменены на цистеин для конъюгирования.
В другом варианте осуществления сайт-специфическое конъюгирование осуществляют через один или более ацил-донорных остатков глутамина, которые были введены с помощью методов генной инженерии в константную область антитела. Способы получения антител к HER2 для сайт-специфического конъюгирования через остатки глутамина могут быть осуществлены, как описано в публикации PCT WO2012/059882, которая полностью включена посредством отсылки. Антитела могут быть модифицированы с помощью методов генной инженерии для экспрессии остатка глутамина, используемого для сайт-специфического конъюгирования, тремя различными способами.
Короткую пептидную метку, содержащую остаток глутамина, можно ввести в ряд различных положений легкой и/или тяжелой цепи (т.е. на N-конец, на C-конец, внутренне). В первом варианте осуществления короткая пептидная метка, содержащая остаток глутамина, может быть присоединена к C-концу тяжелой и/или легкой цепи. Одна или более следующих глутаминсодержащих меток может быть присоединена в качестве донора ацильной группы для конъюгирования лекарственного средства: GGLLQGPP (SEQ ID NO: 81), GGLLQGG (SEQ ID NO: 82), LLQGA (SEQ ID NO: 83), GGLLQGA (SEQ ID NO: 84), LLQG (SEQ ID NO: 85), LLQGPG (SEQ ID NO: 86), LLQGPA (SEQ ID NO: 87), LLQGP (SEQ ID NO: 88), LLQP (SEQ ID NO: 89), LLQPGK (SEQ ID NO: 90), LLQGAPGK (SEQ ID NO: 91), LLQGAPG (SEQ ID NO: 92), LLQGAP (SEQ ID NO: 93), LLQX1X2X3X4X5, где X1 является G или P, где X2 является A, G, P или отсутствует, где X3 является A, G, K, P или отсутствует, где X4 является G, K или отсутствует, и где X5 является K или отсутствует (SEQ ID NO: 94), или LLQX1X2X3X4X5, где X1 является любой природной аминокислотой, и где X2, X3, X4 и X5 являются любыми природными аминокислотами или отсутствуют (SEQ ID NO: 95).
В определенном варианте осуществления GGLLQGPP (ID NO: 81 SEQ) присоединен к C-концу легкой цепи.
Во втором варианте осуществления остаток на тяжелой и/или легкой цепи может быть изменен на остаток глутамина с помощью сайт-направленного мутагенеза. В определенном варианте осуществления остаток в положении 297 на тяжелой цепи (при использовании EU индекса нумерации Кэбата) может быть изменен на глутамин (Q) и, таким образом, может служить в качестве сайта для конъюгирования.
В третьем варианте осуществления остаток на тяжелой цепи или легкой цепи может быть изменен так, что это приводит к агликозилированию в данном положении, в результате чего один или более эндогенных глутаминов становятся доступными/реакционноспособными для конъюгирования. В определенном варианте осуществления остаток в положении 297 тяжелой цепи (при использовании EU индекса нумерации Кэбата) изменен на аланин (A). В таких случаях глутамин (Q) в положении 295 тяжелой цепи может использоваться в конъюгировании.
Оптимальные условия реакции для образования конъюгата можно эмпирически определить путем изменения переменных параметров реакции, таких как температура, pH, ввод молекулы линкера-полезной нагрузки и концентрация добавляемых реагентов. Условия, подходящие для конъюгирования других лекарственных средств, могут быть определены специалистами в данной области без излишнего экспериментирования. Сайт-специфическое конъюгирование через модифицированные остатки цистеина проиллюстрировано в Примере 5А ниже. Сайт-специфическое конъюгирование через остатки глутамина проиллюстрировано в Примере 5B ниже.
Чтобы дополнительно увеличить число молекул лекарственного средства на конъюгат антитела-лекарственного средства, лекарственное средство можно конъюгировать с полиэтиленгликолем (ПЭГ), включая линейные или разветвленные полимеры и мономеры полиэтиленгликоля. Мономер ПЭГ имеет формулу: -(CH2CH2O)-. Лекарственные средства и/или аналоги пептидов могут быть связаны с ПЭГ прямо или косвенно, т.е. через подходящие спейсерные группы, такие как сахара. Композиция ПЭГ-антитела-лекарственного средства также может включать дополнительные липофильные и/или гидрофильньные молекулы, способствующие стабильности лекарственного средства и доставке к целевому участку in vivo. Репрезентативные способы получения ПЭГ-содержащих композиций можно найти, например, в патентах США 6,461,603; 6,309,633 и 5,648,095.
После конъюгирования конъюгаты можно отделять и очищать от неконъюгированных реагентов и/или агрегированных форм конъюгатов с помощью стандартных методов. Это может включать такие процессы, как эксклюзионную хроматографию (SEC), ультрафильтрацию/диафильтрацию, ионообменную хроматографию (IEC), хроматофокусирование (CF), ВЭЖХ, FPLC или хроматографию на Sephacryl S-200. Разделение также может быть выполнено с помощью хроматографии гидрофобного взаимодействия (HIC). Подходящие среды для HIC включают хроматографическую среду Phenyl Sepharose 6 Fast Flow, хроматографическую среду Butyl Sepharose 4 Fast Flow, хроматографическую среду Octyl Sepharose 4 Fast Flow, хроматографическую среду Toyopearl Ether-650M, среду Macro-Prep methyl HIC или среду Macro-Prep t-Butyl HIC.
В Таблице 4, ниже, показаны HER2 ADC конъюгаты, использованные для получения данных в разделе Примеры, представленном в настоящей заявке. Сайт-специфические HER2 ADC конъюгаты, показанные в Таблице 4 (в строках 1-17), являются примерами сайт-специфических ADC конъюгатов согласно изобретению.
Для получения сайт-специфического HER2 ADC согласно изобретению, любое антитело к HER2, раскрытое в разделе I выше, может быть конъюгировано с применением сайт-специфических методик с любым лекарственным средством, раскрытым в разделе II выше, через любой линкер, раскрытый в разделе III выше. В предпочтительных вариантах осуществления линкер является расщепляемым (например, vc или AcLysvc). В других предпочтительных вариантах осуществления лекарственным средством является ауристатин (например, 0101).
В конкретном аспекте изобретения сайт-специфический HER2 ADC формулы Ab-(L-D) включает: (a) антитело, Ab, включающее тяжелую цепь SEQ ID NO: 18 и легкую цепь SEQ ID NO: 42; и (b) молекулу линкера-лекарственного средства, L-D, где L является линкером, а D является лекарственным средством, где линкером является vc, и где лекарственным средством является 0101. Схема такого ADC показана на ФИГ. 1A.
В другом конкретном аспекте изобретения сайт-специфический HER2 ADC формулы Ab-(L-D) включает: (a) антитело, Ab, включающее тяжелую цепь SEQ ID NO: 14 и легкую цепь SEQ ID NO: 44; и (b) молекулу линкера-лекарственного средства, L-D, где L является линкером, а D является лекарственным средством, где линкером является AcLysvc, и где лекарственным средством является 0101. Схема такого ADC показана на ФИГ. 1B.
В другом конкретном аспекте изобретения сайт-специфический HER2 ADC формулы Ab-(L-D) включает: (a) антитело, Ab, включающее тяжелую цепь SEQ ID NO: 24 и легкую цепь SEQ ID NO: 42; и (b) молекулу линкера-лекарственного средства, L-D, где L является линкером, а D является лекарственным средством, где линкером является vc, и где лекарственным средством является 0101.
В другом конкретном аспекте изобретения сайт-специфический HER2 ADC формулы Ab-(L-D) включает: (a) антитело, Ab, включающее тяжелую цепь SEQ ID NO: 26 и легкую цепь SEQ ID NO: 42; и (b) молекулу линкера-лекарственного средства, L-D, где L является линкером, а D является лекарственным средством, где линкером является vc, и где лекарственным средством является 0101.
В другом конкретном аспекте изобретения сайт-специфический HER2 ADC формулы Ab-(L-D) включает: (a) антитело, Ab, включающее тяжелую цепь SEQ ID NO: 28 и легкую цепь SEQ ID NO: 42; и (b) молекулу линкера-лекарственного средства, L-D, где L является линкером, а D является лекарственным средством, где линкером является vc, и где лекарственным средством является 0101.
В другом конкретном аспекте изобретения сайт-специфический HER2 ADC формулы Ab-(L-D) включает: (a) антитело, Ab, включающее тяжелую цепь SEQ ID NO: 30 и легкую цепь SEQ ID NO: 12; и (b) молекулу линкера-лекарственного средства, L-D, где L является линкером, а D является лекарственным средством, где линкером является vc, и где лекарственным средством является 0101.
В другом конкретном аспекте изобретения сайт-специфический HER2 ADC формулы Ab-(L-D) включает: (a) антитело, Ab, включающее тяжелую цепь SEQ ID NO: 32 и легкую цепь SEQ ID NO: 12; и (b) молекулу линкера-лекарственного средства, L-D, где L является линкером, а D является лекарственным средством, где линкером является vc, и где лекарственным средством является 0101.
В другом конкретном аспекте изобретения сайт-специфический HER2 ADC формулы Ab-(L-D) включает: (a) антитело, Ab, включающее тяжелую цепь SEQ ID NO: 34 и легкую цепь SEQ ID NO: 44; и (b) молекулу линкера-лекарственного средства, L-D, где L является линкером, а D является лекарственным средством, где линкером является AcLysvc, и где лекарственным средством является 0101.
В другом конкретном аспекте изобретения сайт-специфический HER2 ADC формулы Ab-(L-D) включает: (a) антитело, Ab, включающее тяжелую цепь SEQ ID NO: 36 и легкую цепь SEQ ID NO: 12; и (b) молекулу линкера-лекарственного средства, L-D, где L является линкером, а D является лекарственным средством, где линкером является AcLysvc, и где лекарственным средством является 0101.
В другом конкретном аспекте изобретения сайт-специфический HER2 ADC формулы Ab-(L-D) включает: (a) антитело, Ab, включающее тяжелую цепь SEQ ID NO: 38 и легкую цепь SEQ ID NO: 12; и (b) молекулу линкера-лекарственного средства, L-D, где L является линкером, а D является лекарственным средством, где линкером является vc, и где лекарственным средством является 0101.
В другом конкретном аспекте изобретения сайт-специфический HER2 ADC формулы Ab-(L-D) включает: (a) антитело, Ab, включающее тяжелую цепь SEQ ID NO: 40 и легкую цепь SEQ ID NO: 12; и (b) молекулу линкера-лекарственного средства, L-D, где L является линкером, а D является лекарственным средством, где линкером является vc, и где лекарственным средством является 0101.
Таблица 4: HER2 ADC конъюгаты
1 C=расщепляемый; N=нерасщепляемый
V. Применение сайт-специфических конъюгатов HER2-антитела-лекарственного средства
Конъюгаты антитела-лекарственного средства согласно настоящему изобретению могут применяться в терапевтических способах для лечения HER2-экспрессирующего рака. В некоторых аспектах изобретения предложен способ ингибирования роста или прогрессирования опухоли у субъекта, который имеет HER2-экспрессирующую опухоль, включающий введение нуждающемуся в этом субъекту эффективного количества композиции (т.е. фармацевтической композиции), содержащей один или более ADC конъюгатов, описанных в настоящей заявке. В других аспектах изобретения предложен способ ингибирования метастаза HER2-экспрессирующих раковых клеток у субъекта, включающий введение нуждающемуся в этом субъекту эффективного количества композиции (т.е. фармацевтической композиции), содержащей один или более ADC конъюгатов, описанных в настоящей заявке. В других аспектах изобретения предложен способ вызывания регрессии HER2-экспрессирующей опухоли у субъекта, включающий введение нуждающемуся в этом субъекту эффективного количества композиции (т.е. фармацевтической композиции), содержащей один или более ADC конъюгатов, описанных в настоящей заявке. В других аспектах изобретения предложена фармацевтическая композиция, включающая один или более ADC конъюгатов, описанных в настоящей заявке, для применения в способе, как описано выше. В других аспектах изобретения предложено применение одного или более ADC конъюгатов, как описано в настоящей заявке, или фармацевтической композиции, включающей ADC конъюгаты, как описано в настоящей заявке, в производстве лекарственного средства для применения в способах, описанных выше.
Требуемые результаты применения раскрытых терапевтических способов в целом являются поддающимися количественному определению показателями по сравнению с контрольным или исходным измерением. При использовании в настоящей заявке относительные термины, такие как "улучшает", "повышает" или "снижает" обозначают значения по отношению к контролю, такие как измерение у того же субъекта до начала лечения, описанного в настоящей заявке, или измерение у контрольного субъекта (или множества контрольных субъектов) без лечения, описанного в настоящей заявке. Репрезентативный контрольный субъект является субъектом, страдающим той же формой рака, что и субъект, подвергаемый лечению, имеет примерно такой же возраст, что и подвергаемый лечению субъект (чтобы гарантировать, что стадии нарушения у подвергаемого лечению субъекта и контрольного субъекта сопоставимы).
Изменения или улучшения ответа на терапию обычно являются статистически значимыми. При использовании в настоящей заявке термин "значимость" или "значимый" относятся к статистическому анализу вероятности присутствия неслучайной связи между двумя или более единицами. Для определения, является ли отношение "значимым" или имеет "значимость", статистическая обработка данных может включать "p-значение". P-значения ниже определенной пользователем пороговой точки считаются значимыми. P-значение, меньше или равное 0,1, меньше 0,05, меньше 0,01, меньше 0,005 или меньше 0,001 может считаться значимым.
V.A. Формы рака
ADC конъюгаты согласно настоящему изобретению могут применятся в лечении HER2-экспрессирующих форм рака. В одном варианте осуществления HER2-экспрессирующий рак является солидной опухолью. В более конкретном варианте осуществления HER2-экспрессирующие солидные опухоли включают, без ограничения перечисленными, рак молочной железы (например, эстроген и прогестерон-рецептор негативный рак молочной железы, трижды негативный рак молочной железы), рак яичника, рак легкого (например, немелкоклеточный рак легкого (в том числе аденокарциномы, плоскоклеточные карциномы и крупноклеточные карциномы) и мелкоклеточный рак легкого), рак желудка, рак пищевода, рак толстой и прямой кишки, уротелиальный рак (например, микропапиллярный уротелиальный рак и типичный уротелиальный рак), рак поджелудочной железы, рак слюнной железы (например, мукоэпидермоидные карциномы, аденокистозные карциномы и аденокарциному терминальных протоков) и рак головного мозга или метастазы вышеуказанных форм рака (т.е. метастаз HER2+ рака молочной железы в легких) (Martin et al., 2014, Future Oncol. 10(8): 1469-86).
В еще более конкретном варианте осуществления HER2-экспрессирующие солидные опухоли включают, без ограничения перечисленными, рак молочной железы, рак яичника, рак легкого и рак желудка.
В другом варианте осуществления рак молочной железы является эстроген-рецептор и прогестерон-рецептор негативным. В более конкретном варианте осуществления рак молочной железы является трижды негативным раком молочной железы (ТНРМЖ).
В другом варианте осуществления рак легкого является немелкоклеточным раком легкого (НМРЛ).
В одном аспекте изобретения ADC конъюгаты, раскрытые в настоящей заявке, могут применяться для лечения HER2-экспрессирующих форм рака, которые ранее подвергались лечению терапевтическим средством (т.е. терапии первого ряда).
В другом аспекте изобретения ADC конъюгаты, раскрытые в настоящей заявке, могут применяться для лечения HER2-экспрессирующих форм рака, которые являются резистентными к лечению, рефракторными к лечению и/или рецидивирующими после лечения другим терапевтическим средством (т.е. терапии второго ряда). В одном варианте осуществления первичным лечением являлся трастузумаб (трастузумаб или Герцептин®), отдельно или в комбинации с дополнительным терапевтическим средством (т.е. таксаном, таким как паклитаксел, доцетаксел, кабазитаксел и т.д.). В другом варианте осуществления первичным лечением являлся трастузумаб эмтанзин (T-DM1 или Кадсила®), отдельно или в комбинации с дополнительным терапевтическим средством (т.е. таксаном, таким как паклитаксел, доцетаксел, кабазитаксел и т.д.).
В другом аспекте изобретения ADC конъюгаты, раскрытые в настоящей заявке, могут применяться для лечения HER2-экспрессирующих форм рака, которые являются резистентными к лечению, рефракторными к лечению и/или рецидивирующими после лечения больше чем одним другим терапевтическим средством (т.е. в качестве терапии третьего ряда или терапии четвертого ряда и т.д.).
ADC конъюгаты согласно настоящему изобретению могут применяться для лечения форм рака, которые экспрессируют высокие уровни HER2 (т.е. ИГХ 3+), средние уровни HER2 (т.е. 2+ ИГХ или 2+/3+ ИГХ) или низкие уровни HER2 (т.е. ИГХ 1+, ИГХ 2+ или ИГХ 1+/2+) (см. раздел IVB в отношении способов обнаружения HER2). Это отличается от трастузумаба и T-DM1, так как они не эффективны в случае форм рака с низкой или средней экспрессией HER2 (Burris et al., 2011, J Clinical Oncology 29(4):398-405).
ADC конъюгаты согласно настоящему изобретению могут применяться для лечения форм рака, которые являются гомогенными по природе, когда большинство опухолевых клеток экспрессирует аналогичное количество HER2. В альтернативе ADC конъюгаты согласно настоящему изобретению могут применяться для лечения форм рака, которые являются гетерогенными по природе, когда присутствуют популяции разных опухолевых клеток, экспрессирующих разные уровни HER2.
V.B. Способы обнаружения HER2
Обсуждались аспекты относительно наилучшего способа оценки уровней экспрессии HER2 на опухоли, и были описаны клинические проявления (Sauter et al., 2009, J Clin Oncol. 27:1323-33; Wolff et al., 2007, J Clinical Oncology 25: 118-45; Wolff et al., 2013, J Clinical Oncology 31: 3997-4014). В настоящее время HER2 статус можно оценивать с помощью иммуногистохимии (ИГХ), флуоресцентной гибридизации in situ (FISH) и хромогенной гибридизации in situ (CISH).
ИГХ позволяет определять экспрессию белка HER2 на клеточной мембране. Результаты обычно выражают при использовании полуколичественной оценочной системы в баллах от 0+ (нет экспрессии) до 3+ (высокая экспрессия). Опухоли, которые показывают отсутствие (0+) или низкие уровни (1+) экспрессии, считаются HER2-негативными; напротив, опухоли, которые показывают высокий уровень (3+) экспрессии, следует считать HER2-позитивными. Данный метод экономически выгоден и легко доступен, но имеет недостатки, связанные с низкой чувствительностью и высокой вариабельностью результатов, получаемых разными исследователями (Gancberg et al., 2002, Breast Cancer Res Treat. 74: 113-20).
Существует четыре одобренных FDA коммерческих набора, доступных для обнаружения HER2 с помощью ИГХ: HercepTestTM (Dako Denmark A/S); Pathway (Ventana Medical Systems, Inc.); набор Insite HER2/NEU (Biogenex Laboratories, Inc.) и Bond Oracle HER2 IHC System (Leica Biosystems). Это высоко стандартизированные, полуколичественные анализы, в которых уровни экспрессии HER2 оценивают следующим образом: 0 (<20000 рецепторов на клетку, видимая экспрессия отсутствует), 1+ (~100000 рецепторов на клетку, частичное окрашивание мембраны, <10% клеток с оверэкспрессией HER-2), 2+ (~500000 рецепторов на клетку, легкое или умеренное полное окрашивание мембраны, >10% клеток с оверэкспрессией HER-2) и 3+ (~2000000 рецепторов на клетку, сильное полное окрашивание мембраны, >10% клеток с оверэкспрессией HER-2). Присутствие цитоплазматической экспрессии не учитывают.
Метод FISH позволяет обнаруживать амплификацию гена HER2 с помощью ДНК-зонда и является более специфичным и чувствительным, чем ИГХ (Owens et al., 2004, Clin Breast Cancer. 5:63-69; Press et al., 2005, Clin Cancer Res. 11: 6598-6607; Vogel et al., 2002, J Clinical Oncology 20(3): 719-726). FISH дает количественные результаты, представленные числом копий гена HER2 относительно центромеров хромосомы 17. Результаты приводят в виде отношения числа сигналов HER2 к сигналам центромерных областей хромосомы 17. Отношение меньше 1,8 считается находящимся в пределах нормы. Отношение 1,8-2,0 является неопределенным и требует дальнейшего исследования. Отношение больше 2,0 соответствует амплификации последовательностей гена HER2.
Существует четыре одобренных FDA коммерческих набора, доступных для обнаружения HER2 с помощью FISH: набор HER2 FISH Pharm DxTM (Dako Denmark A/S); набор Pathvysion HER2 DNA Probe (Abbott Molecular Inc.); Inform HER2/NEU и Inform HER2 Dual ISH DNA Probe Cocktail (Ventana Medical Systems, Inc.).
Другим методом оценки амплификации гена HER2 является CISH. Метод CISH похож на FISH, но в нем используют обычные реакции пероксидазы или щелочной фосфатазы, визуализируемые с помощью стандартного светлопольного микроскопа. Существует два одобренных FDA коммерческих набора, доступных для обнаружения HER2 с помощью CISH: набор HER2 CISH PharmDx (Dako Denmark A/S) и набор Spot-Light HER2 CISH (Life Technologies, Inc.).
Амплификация гена, обнаруженная с помощью FISH или CISH, и экспрессия белка, определенная с помощью ИГХ, обычно используются в качестве первичного анализа для оценки HER2-статуса. Между двумя указанными методами существует хорошая корреляция (Jacobs et al., 1999, J Clinical Oncology 17(7): 1974-82). Впрочем, в тех случаях, когда опухоль оценивают как неопределенную (т.е. ИГХ 2+ или отношение FISH/CISH 1,8-2,2 или среднее число копий гена HER2 четыре-шесть сигналов на ядро), стандартным методом является проверка опухоли с помощью альтернативного метода (Wolff et al., 2007, J Clinical Oncology 25: 118-45).
Таким образом, экспрессию HER2 считают высокой в опухолях с уровнем 3+, определенным с помощью иммуногистохимии (ИГХ), и/или отношением амплификации флуоресцентной гибридизации in situ (FISH) ≥2,0. Экспрессию HER2 считают средней в опухолях с уровнем 2+, определенным с помощью иммуногистохимии (ИГХ), и/или отношением амплификации флуоресцентной гибридизации in situ (FISH) <2,0. Экспрессию HER2 считают низкой в опухолях с уровнем 1+, определенным с помощью иммуногистохимии (ИГХ), и/или отношением амплификации флуоресцентной гибридизации in situ (FISH) <2,0.
В одном варианте осуществления уровни HER2 определяют с помощью ИГХ. В более конкретном варианте осуществления ИГХ проводят при использовании анализа Dako HercptestTM.
В другом варианте осуществления уровни HER2 определяют с помощью FISH. В более конкретном варианте осуществления FISH проводят при использовании анализа Dako HER2 FISH Pharm DxTM.
Репрезентативные образцы опухоли включают любой биологический или клинический образец, который содержит опухолевые клетки, например, образец ткани, биопсию, образец крови, образец плазмы, образец слюны, образец мочи и т.д.
VI. Лекарственные формы
В настоящем изобретении предложены фармацевтические композиции, включающие любые сайт-специфические конъюгаты HER2-антитела-лекарственного средства, раскрытые в настоящей заявке, и фармацевтически приемлемый носитель. Кроме того, композиции могут включать больше одного из сайт-специфических HER2 ADC конъюгатов, раскрытых в настоящей заявке.
Композиции, применяемые в настоящем изобретении, могут дополнительно включать фармацевтически приемлемые носители, вспомогательные вещества или стабилизаторы (Remington: The Science and practice of Pharmacy 21st Ed., 2005, Lippincott Williams and Wilkins, Ed. K. E. Hoover) в форме лиофилизированных лекарственных форм или водных растворов. Приемлемые носители, вспомогательные вещества или стабилизаторы нетоксичны для реципиентов в таких дозах и концентрациях, и могут включать буферы, такие как фосфат, цитрат, и другие органические кислоты; антиоксиданты, в том числе аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония, хлорид бензетония; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил или пропилпарабен; пирокатехин; резорцин; циклогексанол; 3-пентанол; и м-крезол); полипептиды с низкой молекулярной массой (меньше чем приблизительно 10 остатков); белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильньные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, в том числе глюкозу, маннозу или декстраны; хелатообразующие вещества, такие как ЭДТА; сахара, такие как сахарозу, маннит, трегалозу или сорбит; солеобразующие противоионы, такие как натрий; комплексы металлов (например, Zn-белковые комплексы); и/или неионогенные поверхностно-активные вещества, такие как TWEENTM, PLURONICSTM или полиэтиленгликоль (ПЭГ). "Фармацевтически приемлемая соль" при использовании в настоящей заявке относится к фармацевтически приемлемым органическим или неорганическим солям молекулы или макромолекулы. Фармацевтически приемлемые вспомогательные вещества также описаны в настоящей заявке.
Для введения могут применяться различные лекарственные формы одного или более сайт-специфических HER2 ADC конъюгатов, в том числе, без ограничения перечисленными, лекарственные формы, включающие один или более фармацевтически приемлемых вспомогательных веществ. Фармацевтически приемлемые вспомогательные вещества известны в уровне техники и являются относительно инертными веществами, которые облегчают введение фармакологически эффективного вещества. Например, вспомогательное вещество может придавать форму или консистенцию, или действовать как разбавитель. Подходящие вспомогательные вещества включают, без ограничения перечисленными, стабилизирующие вещества, смачивающие и эмульгирующие вещества, соли для различной осмолярности, инкапсулирующие вещества, буферы и вещества, усиливающие проникновение через кожу. Вспомогательные вещества, а также лекарственные формы для парентеральной и непарентеральной доставки лекарственного средства представлены в справочнике Remington, The Science and Practice of Pharmacy 20th Ed. Mack Publishing, 2000.
В некоторых аспектах изобретения такие средства изготовлены в форме для введения путем инъекции (например, внутрибрюшинно, внутривенно, подкожно, внутримышечно и т.д.). Таким образом, эти вещества могут быть объединены с фармацевтически приемлемыми растворителями, такими как солевой раствор, раствор Рингера, раствор декстрозы и т.п. Конкретная схема дозирования, т.е. доза, время введения и повтор, будет зависеть от конкретного субъекта и сведений о перенесенных им заболеваниях.
Терапевтические лекарственные формы сайт-специфических HER2 ADC конъюгатов, применяемых в соответствии с настоящим изобретением, подготавливают для хранения путем смешивания ADC, обладающего требуемой степенью чистоты, с дополнительными фармацевтически приемлемыми носителями, вспомогательными веществами или стабилизаторами (Remington, The Science and Practice of Pharmacy 21st Ed. Mack Publishing, 2005), в форме лиофилизированных лекарственных форм или водных растворов. Приемлемые носители, вспомогательные вещества или стабилизаторы нетоксичны для реципиентов в применяемых дозах и концентрациях, и могут включать буферы, такие как фосфат, цитрат, и другие органические кислоты; соли, такие как хлорид натрия; антиоксиданты, в том числе аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония, хлорид бензетония; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил или пропилпарабен; пирокатехин; резорцин; циклогексанол; 3-пентанол; и м-крезол); полипептиды с низкой молекулярной массой (меньше чем приблизительно 10 остатков); белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильньные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, в том числе глюкозу, маннозу или декстрины; хелатообразующие вещества, такие как ЭДТА; сахара, такие как сахарозу, маннит, трегалозу или сорбит; солеобразующие противоионы, такие как натрий; комплексы металлов (например, Zn-белковые комплексы); и/или неионогенные поверхностно-активные вещества, такие как TWEENTM, PLURONICSTM или полиэтиленгликоль (ПЭГ).
Липосомы, содержащие сайт-специфические HER2 ADC конъюгаты, могут быть получены с помощью способов, известных в уровне техники, таких как описаннные в Eppstein, et al., 1985, PNAS 82:3688-92; Hwang, et al., 1908, PNAS 77:4030-4; и патентах США 4,485,045 и 4,544,545. Липосомы с увеличенной продолжительностью циркуляции раскрыты в патенте США 5,013,556. Особенно полезные липосомы могут быть получены методом обращенно-фазового выпаривания с липидной композицией, включающей фосфатидилхолин, холестерин и ПЭГилированный фосфатидилэтаноламин (ПЭГ-ФЭА). Липосомы экструдируют через фильтры с порами определенного размера, получая липосомы нужного диаметра.
Активные компоненты также могут быть заключены в микрокапсулах, изготовленных, например, с помощью методов коацервации или полимеризации на границе раздела фаз, например, гидроксиметилцеллюлозных или желатиновых микрокапсулах и полиметилметакрилатных микрокапсулах, соответственно, в коллоидных системах доставки лекарственных средств (например, липосомах, альбуминовых микросферах, микроэмульсиях, наночастицах и нанокапсулах) или в макроэмульсиях. Такие технологии раскрыты в Remington, The Science and Practice of Pharmacy 21st Ed. Mack Publishing, 2005.
Могут быть изготовлены препараты с замедленным высвобождением. Подходящие примеры препаратов с замедленным высвобождением включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащих антитело, где матрицы изготовлены в виде изделий, имеющих определенную форму, например, пленок или микрокапсул. Примеры матриц с замедленным высвобождением включают полиэфиры, гидрогели (например, поли-(2-гидроксиэтил-метакрилат), или поливиниловый спирт), полилактиды (патент США 3,773,919), сополимеры L-глутаминовой кислоты и 7-этил-L-глутамата, неразлагаемые сополимеры этилена-винилацетата, разлагаемые сополимеры молочной кислоты-гликолевой кислоты, такие как LUPRON DEPOTTM (микросферы для инъекций, состоящие из сополимера молочной кислоты-гликолевой кислоты и лейпролида ацетата), ацетат-изобутират сахарозы и поли-D-(-)-3-гидроксимасляную кислоту.
Лекарственные формы, применяемые для введения in vivo, должны быть стерильными. Это легко достигается, например, при фильтрации через стерилизующие фильтрующие мембраны. Терапевтические композиции сайт-специфических HER2 ADC конъюгатов обычно помещены в контейнер, имеющий стерильный порт доступа, например, пакет с раствором для внутривенного введения или флакон с пробкой, прокалываемой иглой для подкожной инъекции.
Подходящие поверхностно-активные вещества включают, в частности, неионогенные ПАВ, такие как полиоксиэтиленсорбитаны (например, TWEENTM 20, 40, 60, 80 или 85) и другие сорбитаны (например, SpanTM 20, 40, 60, 80 или 85). Композиции с поверхностно-активным веществом обычно включают 0,05-5% поверхностно-активного вещества и могут включать 0,1-2,5%. Следует понимать, что могут быть добавлены другие компоненты, например, маннит или другие фармацевтически приемлемые разбавители, при необходимости.
Подходящие эмульсии могут быть приготовлены при использовании коммерчески доступных жировых эмульсий, таких как INTRALIPIDTM, LIPOSYNTM, INFONUTROLTM, LIPOFUNDINTM и LIPIPHYSANTM. Действующее вещество может быть растворено в предварительно смешанной эмульсионной композиции или, в альтернативе, оно может быть растворено в масле (например, соевом масле, сафлоровом масле, хлопковом масле, кунжутном масле, кукурузном масле или миндальном масле) и эмульсии, полученной после смешивания с фосфолипидом (например, яичными фосфолипидами, соевыми фосфолипидами или соевым лецитином) и водой. Следует понимать, что могут быть добавлены другие компоненты, например глицерин или глюкоза, для регулирования тоничности эмульсии. Подходящие эмульсии, как правило, будут содержать 20% масла, например 5-20%. Жировая эмульсия может включать капли жира размером 0,1-1,0 мкм, в особенности 0,1-0,5 мкм, и иметь pH в пределах 5,5-8,0. Эмульсионные композиции могут быть получены при смешивании сайт-специфического HER2 ADC с INTRALIPIDTM или компонентами, входящими в его состав (соевым маслом, яичными фосфолипидами, глицерином и водой).
В изобретении также предложены наборы для применения в настоящих способах. Наборы согласно изобретению включают один или более контейнеров, включающих один или более сайт-специфических HER2 ADC конъюгатов, как описано в настоящей заявке, и инструкции по применении в соответствии с любым из способов изобретения, описанных в настоящей заявке. Обычно такие инструкции включают описание применения сайт-специфического HER2 ADC для вышеописанного терапевтического лечения.
Инструкции по применению сайт-специфических HER2 ADC конъюгатов, как описано в настоящей заявке, обычно включают информацию в отношении дозирования, схемы дозирования и пути введения для предполагаемого лечения. Контейнеры могут представлять собой стандартные дозы, упаковки с множеством доз (например, многодозовые упаковки) или субъединичные дозы. Инструкции, поставляемые с наборами изобретения, являются, как правило, письменными инструкциями на этикетке или вкладыше в упаковку (например, бумажным листом, включенный в набор), однако также допускаются машиночитаемые инструкции (например, инструкции на магнитном или оптическом носителе данных).
Наборы согласно настоящему изобретению находятся в подходящей упаковке. Подходящая упаковка включает, без ограничения перечисленными, флаконы, бутылки, банки, гибкую упаковку (например, герметичные майларовые или полиэтиленовые пакеты) и т.п. Также предусмотрены упаковки для применения в сочетании со специальным устройством, таким как инфузионное устройство, такое как мининасос. Набор может иметь стерильный порт доступа (например, контейнер может быть пакетом с раствором для внутривенного введения или флаконом с пробкой, прокалываемой иглой для подкожной инъекции). Контейнер также может иметь стерильный порт доступа (например, контейнер может быть пакетом с раствором для внутривенного введения или флаконом с пробкой, прокалываемой иглой для подкожной инъекции). По меньшей мере одним действующим веществом в композиции является сайт-специфический HER2 ADC. Контейнер может дополнительно включать второе фармацевтически активное вещество.
В наборах необязательно могут быть предоставлены дополнительные компоненты, такие как буферы и информация по интерпретации результатов. Обычно набор включает контейнер и этикетку или вкладыш(и) в упаковку на контейнере или связанные с контейнером.
VII. Доза и введение
Для применений in vivo, сайт-специфические HER2 ADC конъюгаты предоставляют или вводят в эффективной дозе. Фразы "эффективная доза" или "эффективное количество" при использовании в настоящей заявке относятся к количеству лекарственного средства, соединения или фармацевтической композиции, необходимому для достижения одного или более полезных или требуемых терапевтических результатов, непосредственно или опосредовано. Например, в случае введения имеющему раковую опухоль субъекту, эффективная доза включает количество, достаточное, чтобы вызывать противоопухолевую активность, включая цитолиз раковых клеток, ингибирование пролиферации раковых клеток, индукцию апоптоза раковых клеток, снижение антигенов раковых клеток, задержку роста опухоли и/или ингибирование метастаза. Уменьшение размеров опухоли является общепринятым клиническим суррогатным маркером эффективности. Другим общепринятым маркером эффективности является выживаемость без прогрессирования.
Эффективную дозу можно вводить в одно или более введений. Эффективная доза лекарственного средства, соединения или фармацевтической композиции может или не может быть достигнута в сочетании с другим лекарственным средством, соединением или фармацевтической композицией. Таким образом, эффективную дозу можно рассматривать в контексте применения одного или более терапевтических средств, при этом можно считать, что одно средство вводят в эффективном количестве, если вместе с одним или более другими средствами требуемый результат достигнут или может быть достигнут.
Сайт-специфические HER2 ADC конъюгаты могут вводить человеку любым подходящим путем. Специалистам в данной области должно быть известно, что примеры, описанные в настоящей заявке, не должны быть ограничивающими, а должны иллюстрировать доступные методики. Таким образом, в некоторых аспектах изобретения сайт-специфический HER2 ADC вводят человеку в соответствии с известными способами, такими как внутривенное введение, например, в виде болюса или непрерывной инфузии в течение некоторого периода времени, внутримышечным, внутрибрюшинным, внутриспинномозговым, внутричерепным, трансдермальным, подкожным, внутрисуставным, подъязычным, внутрисиновиальным, посредством инсуффляции, интратекальным, пероральным, ингаляционным или наружным путями. Введение может быть системным, например внутривенным введением, или локализованным. Доступные в продаже небулайзеры для жидких лекарственных форм, в том числе струйные небулайзеры и ультразвуковые небулайзеры, могут применяться для введения. Жидкие лекарственные формы можно распылять непосредственно, а лиофилизированный порошок можно распылять после восстановления. В альтернативе сайт-специфический HER2 ADC можно распылять при использовании фтороуглеводородной композиции и дозирующего ингалятора, или вдыхать в виде лиофилизированного и измельченного порошка.
В некоторых аспектах изобретения сайт-специфический HER2 ADC вводят посредством технологий сайт-специфической или направленной локальной доставки. Примеры технологий сайт-специфической или направленной локальной доставки включают различные имплантируемые депо-источники сайт-специфического HER2 ADC или катетеры локальной доставки, такие как инфузионные катетеры, постоянные катетеры или игольчатые катетеры, синтетические трансплантаты, адвентициальные манжетки, шунты и стенты или другие имплантируемые устройства, сайт-специфические носители, прямую инъекцию или прямое применение. См., например, публикацию международной заявки PCT WO 2000/53211 и патент США 5,981,568.
В рамках настоящего изобретения подходящая доза сайт-специфических HER2 ADC конъюгатов будет зависеть от конкретного применяемого ADC (или его композиций), типа и тяжести симптомов, подлежащих лечению, от того, вводят ли средство в терапевтических целях, от предыдущей терапии, истории болезни пациента и реакции на средство, скорости клиренса вводимого средства у пациента и решения лечащего врача. Медработник может вводить сайт-специфический HER2 ADC, пока не будет достигнута дозировка, которая обеспечивает требуемый результат, или более высокая. Доза и/или частота могут изменяться в течение курса лечения, но могут также оставаться постоянными. Эмпирические факторы, такие как полупериод существования, обычно будут способствовать определению дозы. Например, антитела, которые совместимы с человеческой иммунной системой, такие как гуманизированные антитела или полностью человеческие антитела, могут применяться для увеличения полупериода существования антитела и предотвращения атаки антитела иммунной системой реципиента. Частота введения может быть определена и скорректирована в течение курса терапии и обычно, но не обязательно, основана на лечении и/или супрессии, и/или уменьшении тяжести симптомов, например, ингибировании или задержке роста опухоли, и т.д. В альтернативе могут быть подходящими лекарственные формы с замедленным непрерывным высвобождением сайт-специфических HER2 ADC конъюгатов. Различные лекарственные формы и устройства для обеспечения замедленного высвобождения известны в уровне техники.
Обычно для введения сайт-специфического HER2 ADC, начальная возможная доза может составить приблизительно 2 мг/кг. В рамках настоящего изобретения типичная ежедневная доза может изменяться от приблизительно любого из 3 мкг/кг до 30 мкг/кг, до 300 мкг/кг, до 3 мг/кг, до 30 мг/кг, до 100 мг/кг или больше. Например, может использоваться доза приблизительно 1 мг/кг, приблизительно 2,5 мг/кг, приблизительно 5 мг/кг, приблизительно 10 мг/кг и приблизительно 25 мг/кг. В случае повторных введений в течение нескольких дней или больше, в зависимости от нарушения, лечение продолжают, пока не будет достигнута требуемая супрессия симптомов, или пока не будут достигнуты терапевтические уровни, достаточные, например, для ингибирования или задержки роста/прогрессирования опухоли или метастаза раковых клеток. Примерная схема дозирования включает введение начальной дозы приблизительно 2 мг/кг с последующей еженедельной поддерживающей дозой приблизительно 1 мг/кг сайт-специфического HER2 ADC или с последующей поддерживающей дозой приблизительно 1 мг/кг раз в две недели. Другие примерные схемы дозирования включают введение увеличиваемых доз (например, начальной дозы 1 мг/кг с постепенным увеличением до одной или больше более высоких доз каждую неделю или в течение более длительного периода времени). Другие схемы дозирования также могут применяться, в зависимости от структуры фармакокинетического спада, которая требуется медработнику. Например, в некоторых аспектах изобретения предусмотрено дозирование от одного до четырех раз в неделю. В других аспектах предусмотрено дозирование один раз в месяц или один раз в два месяца или один раз в три месяца, а также один раз в неделю, в две недели и в три недели. Эффективность такой терапии можно легко отслеживать с помощью стандартных методов и анализов. Схема дозирования (включая конкретный применяемый сайт-специфический HER2 ADC) может изменяться в течение времени.
VIII. Комбинированная терапия
В некоторых аспектах изобретения способы, описанные в настоящей заявке, дополнительно включают этап лечения субъекта с применением дополнительной формы терапии. В некоторых аспектах дополнительная форма терапии является дополнительной противоопухолевой терапией, включающей, без ограничения перечисленным, химиотерапию, радиацию, хирургию, гормональную терапию и/или дополнительную иммунотерапию.
Раскрытые сайт-специфические HER2 ADC конъюгаты могут быть введены в качестве первичного лечения или для лечения форм рака, которые не поддаются лечению с применением стандартной терапии. Кроме того, сайт-специфические HER2 ADC конъюгаты могут применяться в комбинации с другими терапиями (например, хирургическим удалением, радиацией, дополнительными противоопухолевыми препаратами и т.д.), что позволяет добиться аддитивных или потенцированных терапевтическим эффектов и/или снизить цитотоксичность некоторых противоопухолевых средств. Сайт-специфические HER2 ADC конъюгаты согласно изобретению можно совместно вводить или включать в лекарственную форму с дополнительными средствами, или изготавливать в лекарственной форме для последовательного введения с дополнительными средствами в любом порядке.
Сайт-специфические HER2 ADC конъюгаты согласно изобретению могут применятся в комбинации с другими терапевтическими средствами, включающими, без ограничения перечисленными, терапевтические антитела, ADC конъюгаты, иммуномодулирующие средства, цитотоксические средства и цитостатические средства. Цитотоксическое действие относится к истощению, элиминированию и/или уничтожению клеток-мишеней (т.е. опухолевых клеток). Цитотоксическое средство относится к средству, которое оказывает цитотоксическое и/или цитостатическое действие на клетку. Цитостатическое действие относится к ингибированию пролиферации клеток. Цитостатическое средство относится к средству, которое оказывает цитостатическое действие на клетку, вызывая ингибирование роста и/или размножения определенной субпопуляции клеток (т.е. опухолевых клеток). Иммуномодулирующее средство относится к средству, которое стимулирует иммунный ответ посредством продукции цитокинов и/или антител и/или модуляции функции Т-клеток, вызывая прямое или опосредованное ингибирование или снижение роста субпопуляции клеток (т.е. опухолевых клеток), что позволяет другому средству действовать с большей эффективностью.
В случае комбинированной терапии сайт-специфический HER2 ADC и/или одно или более дополнительных терапевтических средств вводят в течение любого периода времени, подходящего для проведения предполагаемой терапии. Таким образом, отдельные средства могут вводить по существу одновременно (т.е. в виде одной лекарственной формы или в течение минут или часов) или последовательно, в любом порядке. Например, отдельные средства могут вводить в пределах приблизительно 1 года друг после друга, например, в течение приблизительно 10, 8, 6, 4 или 2 месяцев, или в течение 4, 3, 2 или 1 недели (недель), или в течение приблизительно 5, 4, 3, 2 или 1 дня (дней).
Раскрытые комбинированные терапии могут вызывать синергический терапевтический эффект, т.е. эффект, превышающий величину их отдельных эффектов или терапевтических результатов. Например, синергический терапевтический эффект может быть эффектом, по меньшей мере приблизительно в два раза больше терапевтического эффекта, вызываемого одним средством, или суммы терапевтических эффектов, вызываемых отдельными средствами данной комбинации, или по меньшей мере приблизительно в пять раз больше, или по меньшей мере приблизительно в десять раз больше, или по меньшей мере приблизительно в двадцать раз больше, или по меньшей мере приблизительно в пятьдесят раз больше, или по меньшей мере приблизительно в сто раз больше. Синергический терапевтический эффект также может наблюдаться как увеличение терапевтического эффекта по меньшей мере на 10% по сравнению с терапевтическим эффектом, вызываемым одним средством, или с суммой терапевтических эффектов, вызываемых отдельными средствами данной комбинации, или по меньшей мере на 20%, или по меньшей мере на 30%, или по меньшей мере на 40%, или по меньшей мере на 50%, или по меньшей мере на 60%, или по меньшей мере на 70%, или по меньшей мере на 80%, или по меньшей мере на 90%, или по меньшей мере на 100% или больше. Синергический эффект также является эффектом, который позволяет снижать дозу терапевтических средств в случае их применения в комбинации.
ПРИМЕРЫ
Следующие примеры представлены исключительно в иллюстративных целях и не должны ограничивать объем настоящего изобретения никоим образом. Действительно, различные модификации изобретения, в дополнение к показанным и описанным в настоящей заявке, будут очевидны для специалистов в данной области из предшествующего описания и находятся в рамках объема прилагаемой формулы изобретения.
Пример 1: Получение антител на основе трастузумаба для сайт-специфического конъюгирования
A. В случае конъюгирования посредством цистеина
Способы получения производных трастузумаба для сайт-специфического конъюгирования через остатки цистеина в целом выполняли, как описано в публикации PCT WO 2013/093809 (которая полностью включена в настоящую заявку). Один или более остатков на легкой цепи (183 при использовании схемы нумерации Кэбата) или на тяжелой цепи (290, 334, 392 и/или 443 при использовании EU индекса схемы нумерации Кэбата) изменяли на остаток цистеина (C) с помощью сайт-направленного мутагенеза.
B. В случае конъюгирования посредством трансглутаминазы
Способы получения производных трастузумаба для сайт-специфического конъюгирования через остатки глутамина обычно выполняли, как описано в публикации PCT WO 2012/059882 (которая полностью включена в настоящую заявку). Трастузумаб подвергали генно-инженерной модификации с целью экспрессии остатка глутамина, применямого для конъюгирования тремя различными способами.
В первом способе метку из 8 аминокислотных остатков (LCQ05), содержащую остаток глутамина, присоединяли на C-конец легкой цепи (т.е. SEQ ID NO: 81).
Во втором способе остаток на тяжелой цепи (положение 297 при использовании EU индекса схемы нумерации Кэбата) изменяли с аспарагина (N) на остаток глутамина (Q) с помощью сайт-направленного мутагенеза.
В третьем способе остаток на тяжелой цепи (положение 297 при использовании EU индекса системы нумерации Кэбата) изменяли с аспарагина (N) на аланин (A). Это приводит к агликозилированию в положении 297 и доступному/реакционноспособному эндогенному глутамину в положении 295.
Кроме того, некоторые производные трастузумаба имеют изменение, которое не используется для конъюгирования. Остаток в положении 222 на тяжелой цепи (при использовании EU индекса схемы нумерации Кэбата) изменяли с лизина (K) на остаток аргинина (R). Замена K222R, как обнаружили, приводила к более гомогенному конъюгату антитела и полезной нагрузки, лучшему межмолекулярному сшиванию между антителом и полезной нагрузкой и/или значительному уменьшению межцепочечного сшивания с глутаминовой меткой на C-конце легкой цепи антитела.
Пример 2: Получение стабильно трансфицированных клеток, экспрессирующих антитела-производные трастузумаба
A. Цистеиновые мутанты
Для определения, что содержащие одиночную и двойную модификацию цистеина варианты антител-производных трастузумаба можно было стабильно экспрессировать в клетках и получать в крупном масштабе, клетки CHO трансфицировали ДНК, кодирующей девять вариантов антител-производных трастузумаба (T(ĸK183C), T(K290C), T(K334C), T(K392C), T(ĸK183C+K290C), T(ĸK183C+K392C), T(K290C+K334C), T(K334C+K392C) и T(K290C+K392C)), и стабильные высокопродуктивные пулы выделяли при помощи стандартных методик, известных в уровне техники. Для получения T(ĸK183C+K334C) с целью исследования конъюгирования, клетки HEK-293 (рег. номер ATCC CRL-1573) транзиентно совместно трансфицировали ДНК тяжелой и легкой цепи, кодирующими вариант антитела с двойной цистеиновой модификацией, при использовании стандартных методов. Двухколоночный процесс, т.е. аффинный захват на белке A с последующей колонкой TMAE, или трехколоночный процесс, т.е. аффинный захват на белке A с последующей колонкой TMAE, а затем колонкой CHA-TI, использовали для выделения указанных вариантов трастузумаба из концентрированного исходного материала пулов CHO. При использовании такого процесса очистки все препараты цистеин-модифицированных вариантов антител-производных трастузумаба содержали >97% представляющего интерес пика (POI), как определяли с помощью аналитической эксклюзионной хроматографии (Таблица 5). Эти результаты, показанные в Таблице 5, демонстрируют, что допустимые уровни агрегированных молекул высокой молекулярной массы (HMW) обнаруживали после элюции со смолы с белком A в случае всех десяти цистеиновых вариантов, полученных на основе трастузумаба, и что такие нежелательные HMW компоненты можно было удалить при помощи гель-фильтрации. Кроме того, данные продемонстрировали, что участок связывания белка A в константной области IgG1 человека не был изменен в результате присутствия модифицированных остатков цистеина.
Таблица 5: Получение цистеиновых вариантов антител, полученных на основе трастузумаба
(ProA)
(%POI)
(конечный)
ND=не определяли
Пример 3: целостность трастузумаба производные антитела
Молекулярную оценку модифицированных цистеиновых и трансглутаминазных вариантов проводили для оценки ключевых биофизических свойств по сравнению с антителом дикого типа (трастузумабом), чтобы гарантировать, что варианты будут пригодными для стандартного процесса производства антител.
A. Цистеиновые мутанты
Для определения целостности очищенных препаратов модифицированных цистеиновых вариантов антител, полученных путем стабильной CHO экспрессии, процентную чистоту пиков вычисляли при помощи невосстанавливающего капиллярного гель-электрофореза (Caliper LabChip GXII: Perkin Elmer Waltham, MA). Результаты показывают, что модифицированные цистеиновые варианты антител T(ĸK183C+K290C) и T(K290C+K334C), содержащие низкие уровни фрагментов и компонентов высокой молекулярной массы (HMMS), подобных антителу дикого типа (трастузумабу). Для сравнения, T(K334C+K392C) содержал высокие уровни пиков фрагментированного антитела по сравнению с другими исследованными вариантами с двойной цистеиновой модификацией (Таблица 6). Эти результаты позволяют предположить, что определенные комбинации модифицированных цистеинов могут оказывать влияние на целостность антитела, предназначенного для сайт-специфического конъюгирования.
Таблица 6: Процентная чистота пиков, вычисленная из электрофореграммы в невосстанавливающих условиях
Пример 4: Получение лекарственных соединений (полезной нагрузки)
Лекарственные соединения группы ауристатина, 0101, 0131, 8261, 6121, 8254 и 6780, были получены согласно способам, описанным в публикации PCT WO 2013/072813 (которая полностью включена в настоящую заявку). В опубликованной заявке ауристатиновые соединения обозначены согласно системе нумерации, показанной в Таблице 7.
Таблица 7
Согласно публикации PCT WO 2013/072813 лекарственное соединение 0101 было получено согласно следующей методике.
Стадия 1. Синтез N-[(9H-флуорен-9-илметокси)карбонил]-2-метилаланил-N-[(3R,4S,5S)-3-метокси-1-{(2S)-2-[(1R,2R)-1-метокси-2-метил-3-оксо-3-{[(1S)-2-фенил-1-(1,3-тиазол-2-ил)этил]амино}пропил]пирролидин-1-ил}-5-метил-1-оксогептан-4-ил]-N-метил-L-валинамида (#53). Согласно общей методике D, из #32 (2,05 г, 2,83 ммоль, 1 экв.) в дихлорметане (20 мл, 0,1 М) и N,N-диметилформамиде (3 мл), амина #19 (2,5 г, 3,4 ммоль, 1,2 экв.), HATU (1,29 г, 3,38 ммоль, 1,2 экв.) и триэтиламина (1,57 мл, 11,3 ммоль, 4 экв.) синтезировали неочищенный требуемый материал, который очищали с помощью хроматографии на силикагеле (градиент: от 0% до 55% ацетона в гептане), получив #53 (2,42 г, 74%) в виде твердого вещества. ЖХ-МС: m/z 965,7 [M+H+], 987,6 [M+Na+], время удерживания=1,04 минуты; ВЭЖХ (метод A): m/z 965,4 [M+H+], время удерживания=11,344 минут (чистота >97%); 1H-ЯМР (400 МГц, ДМСО-d6), предположительно является смесью ротамеров, характеристические сигналы: δ 7,86-7,91 (м, 2H), [7,77 (д, J=3,3 Гц) и 7,79 (д, J=3,2 Гц), всего 1H], 7,67-7,74 (м, 2H), [7,63 (д, J=3,2 Гц) и 7.65 (д, J=3,2 Гц), всего 1H], 7,38-7,44 (м, 2H), 7,30-7,36 (м, 2H), 7,11-7,30 (м, 5H), [5,39 (ддд, J=11,4, 8,4, 4,1 Гц) и 5,52 (ддд, J=11,7, 8,8, 4,2 Гц), всего 1H], [4,49 (дд, J=8,6, 7,6 Гц) и 4,59 (дд, J=8,6, 6,8 Гц), всего 1H], 3,13, 3,17, 3,18 и 3,24 (4 с, всего 6H), 2,90 и 3,00 (2 шс, всего 3H), 1,31 и 1,36 (2 шс, всего 6H), [1,05 (д, J=6,7 Гц) и 1,09 (д, J=6,7 Гц), всего 3H].
Стадия 2. Синтез 2-метилаланил-N-[(3R,4S,5S)-3-метокси-1-{(2S)-2-[(1R,2R)-1-метокси-2-метил-3-оксо-3-{[(1S)-2-фенил-1-(1,3-тиазол-2-ил)этил]амино}пропил]пирролидин-1-ил}-5-метил-1-оксогептан-4-ил]-N-метил-L-валинамида (#54 или 0101). Согласно общей методике A, из #53 (701 мг, 0,726 ммоль) в дихлорметане (10 мл, 0,07 М) синтезировали неочищенный требуемый материал, который очищали с помощью хроматографии на силикагеле (градиент: от 0% до 10% метанола в дихлорметане). Остаток разбавляли диэтиловым эфиром и гептаном и выпаривали в вакууме с получением #54 (или 0101) (406 мг, 75%) в виде белого твердого вещества. ЖХ-МС: m/z 743,6 [M+H+], время удерживания=0,70 минуты; ВЭЖХ (метод A): m/z 743,4 [M+H+], время удерживания=6,903 минут, (чистота >97%); 1H-ЯМР (400 МГц, ДМСО-d6), предположительно является смесью ротамеров, характеристические сигналы: δ [8,64 (шд, J=8,5 Гц) и 8,86 (шд, J=8,7 Гц), всего 1H], [8.04 (шд, J=9.3 Гц) и 8.08 (шд, J=9.3 Гц), всего 1H], [7,77 (д, J=3,3 Гц) и 7,80 (д, J=3,2 Гц), всего 1H], [7,63 (д, J=3,3 Гц) и 7,66 (д, J=3,2 Гц), всего 1H], 7,13-7,31 (м, 5H), [5,39 (ддд, J=11, 8,5, 4 Гц) и 5,53 (ддд, J=12, 9, 4 Гц), всего 1H], [4,49 (дд, J=9, 8 Гц) и 4,60 (дд, J=9, 7 Гц), всего 1H], 3,16, 3,20, 3,21 и 3,25 (4 с, всего 6H), 2,93 и 3,02 (2 шс, всего 3H), 1,21 (с, 3H), 1,13 и 1,13 (2 с, всего 3H), [1,05 (д, J=6,7 Гц) и 1,10 (д, J=6,7 Гц), всего 3H], 0,73-0,80 (м, 3H).
Лекарственные соединения MMAD, MMAE и MMAF получали в собственной лаборатории согласно способам, раскрытым в публикации PCT WO 2013/072813.
Лекарственное соединение DM1 получали в собственной лаборатории из приобретенного майтанзинола с применением методик, описанных в патенте США 5,208,020.
Пример 5: Биоконъюгирование антител, полученных на основе трастузумаба
Полученные на основе трастузумаба антитела согласно настоящему изобретению конъюгировали с полезной нагрузкой через линкеры, с получением ADC конъюгатов. Применяли либо способ сайт-специфического конъюгирования (т.е. через определенные остатки цистеина или определенные остатки глутамина), либо обычного конъюгирования.
A. Цистеин сайт-специфические
ADC конъюгаты в Таблице 8 конъюгировали с применением цистеин сайт-специфических способов, описанных ниже.
Таблица 8
T(K290C)-vc0101
T(K334C)-vc0101
T(K392C)-vc0101
T(kK183C+K290C)-vc0101
T(kK183C+K392C)-vc0101
T(K290C+K334C)-vc0101
T(K290C+K392C)-vc0101
T(K334C+K392C)-vc0101
500 мМ раствор гидрохлорида трис(2-карбоксиэтил)фосфина (TCEP) (50-100 молярных эквивалентов) добавляли к антителу (5 мг), при этом конечная концентрация антитела составила 5-15 мг/мл в PBS, содержащем 20 мМ ЭДТА. Реакцию оставляли при 37°C на 2,5 часа, после чего антитело подвергали замене буфера на PBS, содержащий 5 мМ ЭДТА, при использовании гель-фильтрационной колонки (обессоливающая колонка PD-10, GE Healthcare). Полученное в результате антитело (5-10 мг/мл) в PBS, содержащем 5 мМ ЭДТА, обрабатывали недавно приготовленным 50 мМ раствором DHA в 1:1 PBS/EtOH (конечная концентрация DHA=1-4 мМ) и оставляли при 4°C на ночь.
Смесь антитела/DHA подвергали замене буфера на PBS, содержащий 5 мМ ЭДТА (pH уравновешивающего буфера доводили до ~7,0 фосфорной кислотой), и концентрировали при использовании центрифужного концентрирующего устройства с отсечением по молекулярной массе 50 кДа. Полученное в результате антитело в PBS (концентрация антитела ~5-10 мг/мл), содержащем 5 мМ ЭДТА, обрабатывали 5-7 молярными эквивалентами 10 мМ малеимидной полезной нагрузки в DMA. После выдерживания в течение 1,5-2,5 часов материал подвергали замене буфера (PD-10). Очистку с помощью SEC проводили (при необходимости) для удаления агрегированного материала и оставшейся свободной полезной нагрузки.
B. Трансглутаминаза сайт-специфические
ADCs Таблицы 9 спрягались через трансглутаминаза сайт-специфические методы, описанные ниже.
Таблица 9
T (LCQ05+K222R)-AcLysvc0101
T (N297Q+K222R)-AcLysvc0101
T (N297A+K222R-LCQ05)-AcLysvc0101
В реакции трансамидирования глутамин на антителе действовал в качестве донора ацила, а аминсодержащее соединение действовало в качестве акцептора ацила (донора амина). Очищенное антитело к HER2 в концентрации 33 мкМ инкубировали с 10-25 М избытком акцептора ацила, в пределах 33-83,3 мкМ AcLysvc-0101, в присутствии 2% (в/об) трансглутаминазы Streptoverticillium mobaraense (ACTIVATM, Ajinomoto, Japan) в 150 мМ хлорида натрия и Трис HCl буфере при pH 7,5-8, с 0,31 мМ восстановленного глутатион, если не указано. Условия реакции корректировали в соответствии с индивидуальными донорами ацила, причем с T(LCQ05+K222R) использовали 10M избыток акцептора ацила при pH 8,0 без восстановленного глутатиона, с T(N297Q+K222R) и T(N297Q) использовали 20M избыток акцептора ацила при pH 7,5 и с T(N297A+K222R+LCQ05) использовали 25M избыток акцептора ацила при pH 7,5. После инкубирования при 37°C в течение 16-20 часов, антитело очищали на смоле MabSelect SuReO или Butyl Sepharose High Performance (GE Healthcare, Piscataway, NJ) при использовании стандартных хроматографических методов, известных специалистам в данной области, таких как коммерческая афинная хроматография и хроматография гидрофобного взаимодействия, предоставляемая GE Healthcare.
C. Стандартное конъюгирование
ADC конъюгаты в Таблицах 10 и 11 конъюгировали с помощью стандартных методов конъюгирования, описанных ниже.
Таблица 10
T-mc8261
T-MalPeg8261
T-mc6121
T-MalPeg6121
T-MalPegMMAD
T-vc0101
T-vc8261
T-vc8254
T-vc6780
T-vc0131
T-vcMMAE
Таблица 11
T-m(H20)cvc0101
Антитело диализировали в фосфатно-солевом буфере Дульбекко (DPBS, Lonza). Диализированное антитело разводили до 15 мг/мл в PBS, содержащем 5 мМ 2,2',2",2"'-(1,2-этандиилдинитрило)-тетрауксусной кислоты (ЭДТА), pH 7. Полученное в результате антитело обрабатывали 2-3 эквивалентами гидрохлорида трис(2-карбоксиэтил)фосфина (TCEP, 5 мМ в дистиллированной воде) и оставляли при 37°C на 1-2 часа. После охлаждения до комнатной температуры добавляли диметилацетамид (DMA) до получения 10% (об/об) органической фазы от общего объема. Смесь обрабатывали 8-10 эквивалентами соответствующего линкера-полезной нагрузки в виде 10 мМ стокового раствора в DMA. Реакцию оставляли на 1-2 часа при комнатной температуре и затем меняли буфер на DPBS (pH 7,4) при использовании колонок для замены буфера GE Healthcare Sephadex G-25 M согласно инструкциям производителя.
Материал, который должен был остаться с замкнутым кольцом (ADC конъюгаты в Таблице 10), очищали с помощью гель-фильтрационной хроматографии (SEC) при использовании системы GE AKTA Explorer с колонкой GE Superdex200 и PBS (pH 7,4) в качестве элюента. Полученные образцы концентрировали до ~5 мг/мл белка, стерилизовали фильтрацией и проверяли на нагрузку при использовании условий масс-спектроскопии, описанных ниже.
Материал, используемый для гидролиза сукцинимидного кольца (ADC конъюгаты Таблицы 11), подвергали замене буфера на 50 мМ боратный буфер (pH 9,2) при использовании ультрафильтрационного устройства (с отсечением по молекулярной массе 50 кДа). Полученный в результате раствор нагревали до 45°C в течение 48 ч. Затем раствор охлаждали, меняли буфер на PBS и очищали с помощью SEC (как описано ниже) для удаления агрегированного материала. Полученные образцы концентрировали до ~5 мг/мл белка, стерилизовали фильтрацией и проверяли на нагрузку при использовании условий масс-спектроскопии, описанных ниже.
D. Конъюгирование T-DM1
Конъюгат трастузумаба-майтанзиноида (T-DM1) структурно подобен трастузумабу эмтанзину (Кадсила®). T-DM1 состоит из антитела трастузумаба, ковалентно связанного с майтанзиноидом DM1 через бифункциональный линкер сульфосукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (сульфо-SMCC). Сульфо-SMCC сначала конъюгировали со свободными аминами на антителе в течение одного часа при 25°C в 50 мМ фосфате калия, 2 мМ ЭДТА, pH 6,8, при стехиометрии реакции 10:1, и затем несвязанный линкер удаляли при обессоливании из конъюгированного антитела. Этот промежуточный продукт антитела-MCC затем конъюгировали с DM1 сульфидом на свободном малеимидном конце MCC линкера антитела в течение ночи при 25°C в 50 мМ фосфате калия, 50 мМ NaCl, 2 мМ ЭДТА, pH 6,8, при стехиометрии реакции 10:1. Оставшийся непрореагировавший малеимид затем кэпировали L-цистеином, после чего ADC фракционировали через колонку Superdex200 для удаления немономерных компонентов (Chari et al., 1992, Cancer Res 52:127-31).
Пример 6: Очистка ADC конъюгатов
ADC конъюгаты обычно очищали и исследовали с помощью гель-фильтрационной хроматографии (SEC), как описано ниже. Нагрузку лекарственного средства в предполагаемом сайте конъюгирования определяли при использовании множества методов, в том числе масс-спектрометрии (МС), обращенно-фазовой ВЭЖХ и хроматографии гидрофобного взаимодействия (HIC), как более подробно описано ниже. Комбинация трех указанных аналитических методов обеспечивает различные способы подтверждения и количественного определения присутствия полезной нагрузки на антителе, что позволяет точно определять DAR для каждого конъюгата.
A. Препаративная SEC
ADC конъюгаты обычно очищали с помощью SEC хроматографии при использовании колонки Waters Superdex200 10/300GL на системе Akta Explorer FPLC, для удаления агрегатов белка и удаления следов линкера-полезной нагрузки, оставшихся в реакционной смеси. В некоторых случаях ADC конъюгаты не содержали агрегатов и низкомолекулярных компонентов перед SEC очисткой и поэтому не были подвергнуты препаративной SEC. В качестве элюента использовали PBS при скорости потока 1 мл/мин. В таких условиях агрегированный материал (элюирующийся приблизительно через 10 минут при комнатной температуре) легко отделяли от неагрегированного материала (элюирующегося приблизительно через 15 минут при комнатной температуре). Гидрофобные комбинации линкера-полезной нагрузки часто приводили к "правому сдвигу" пиков SEC. Без ограничения какой-либо конкретной теорией, такой сдвиг пиков SEC может быть вызван гидрофобными взаимодействиями линкера-полезной нагрузки с неподвижной фазой. В некоторых случаях правый сдвиг позволял частично отделять конъюгированный белок от неконъюгированного белка.
B. Аналитическая SEC
Аналитическую SEC проводили на системе ВЭЖХ Agilent 1100 при использовании PBS в качестве элюента для оценки чистоты и мономерного статуса ADC конъюгатов. Элюент контролировали при 220 и 280 нм. В случае, когда колонкой являлась колонка TSKGel G3000SW (7,8×300 мм, номер по каталогу R874803P), в качестве подвижной фазы использовали PBS со скоростью потока 0,9 мл/мин в течение 30 минут. В случае, когда колонкой являлась колонка BiosepSEC3000 (7,8×300 мм), в качестве подвижной фазы использовали PBS со скоростью потока 1,0 мл/мин в течение 25 минут.
Пример 7: Анализ ADC конъюгатов
A. Масс-спектроскопия (МС)
Образцы подготавливали к ЖХ-МС анализу, объединяя приблизительно 20 мкл образца (приблизительно 1 мг/мл ADC в PBS) с 20 мкл 20 мМ дитиотреитол (ДТТ). Смесь оставляли при комнатной температуре на 5 минут, после чего образцы вводили в систему ВЭЖХ Agilent 110, оснащенную колонкой Agilent Poroshell 300SB-C8 (2,1×75 мм). Температуру системы устанавливали на 60°C. Использовали 5-минутный градиент с 20% до 45% ацетонитрила в воде (с 0,1% муравьиной кислоты в качестве модификатора). Элюент контролировали с помощью УФ (220 нм) и масс-спектрометра Waters Micromass ZQ (ЭРИ ионизация; напряжение на конусе: 20 В; Темп. источника: 120°C; Темп. десольватации: 350°C). Необработанный спектр, содержащий многозарядные ионы, подвергали деконволюции при использовании MaxEnt1 в пакете программ MassLynx 4.1 согласно инструкциям производителя.
B. Определение нагрузки на антителе с помощью МС
Общее количество полезной нагрузки на антителе для получения ADC называют Отношение антитела-лекарственного средства или DAR. DAR вычисляли для каждого из полученных ADC конъюгатов (Таблица 12).
Спектры для полного окна элюции (обычно 5 минут) объединяли в один суммированный спектр (т.е. масс-спектр, который представляет МС всего образца). Результаты МС для образцов ADC сравнивали непосредственно с соответствующим МС идентичного контрольного антитела без нагрузки. Это позволило идентифицировать пики нагруженной/ненагруженной тяжелой цепи (HC) и пики нагруженной/ненагруженной легкой цепи (LC). Отношение различных пиков можно использовать для установления нагрузки с помощью уравнения, приведенного ниже (Уравнение 1). Вычисления основаны на предположении, что нагруженные и ненагруженные цепи ионизируются одинаково, что определили как в целом действительное предположение.
Следующее вычисление выполняли, чтобы установить DAR:
Уравнение 1:
Нагрузка=2*[LC1/(LC1+LC0)]+2*[HC1/(HC0+HC1+HC2)]+4*[HC2/(HC0+HC1+HC2)]
Где указанные переменные представляют собой относительную интенсивность: LC0=ненагруженной легкой цепи, LC1=одиночно нагруженной легкой цепи, HC0=ненагруженной тяжелой цепи, HC1=одиночно нагруженной тяжелой цепи и HC2=дважды нагруженной тяжелой цепи. Среднему специалисту в данной области будет известно, что изобретение охватывает расширенный вариант данного вычисления, охватывающий молекулы с более высокой нагрузкой, такие как LC2, LC3, HC3, HC4, HC5 и т.п.
Уравнение 2, представленое ниже, применяется для оценки величины нагрузки на немодифицированных остатках цистеина. В случае модифицированных Fc мутантов, нагрузка на легкой цепи (LC), как предполагали, по определению, является неспецифичной нагрузкой. Кроме того, предполагали, что нагрузка только на LC была результатом случайного восстановления HC-LC дисульфидного мостика (т.е. антитело было "перевосстановлено"). Учитывая, что в реакциях конъюгирования использовали большой избыток малеимидного электрофила (обычно приблизительно 5 эквивалентов для одиночных мутантов и 10 эквивалентов для двойных мутантов), предполагали, что любая неспецифичная нагрузка на легкой цепи сопровождалась соответствующим количеством неспецифичной нагрузки на тяжелой цепи (т.е. другой "половине" расщепленного HC-LC дисульфида). С учетом таких предположений, следующее уравнение (Уравнение 2) использовали для оценки величины неспецифичной нагрузки на белке:
Уравнение 2:
Неспецифичная нагрузка=4*[LC1/(LC1+LC0)]
Где обозначенные переменные представляют собой относительную интенсивность: LC0=ненагруженной легкой цепи, LC1=одиночно нагруженной легкой цепи.
Таблица 12: Отношение антитела-лекарственного средства (DAR) ADC конъюгатов
C. Протеолиз с использовнием FabRICATOR® для определения сайта нагрузки
В случае цистеин-мутантных ADC конъюгатов любое неспецифичное связывание электрофильной полезной нагрузки на антителе, как предполагают, проходит по "межцепочечным", также называемым "внутренними", остаткам цистеина (т.е. таким остаткам, которые, как правило, являются частью HC-HC или HC-LC дисульфидных мостиков). Для того чтобы отличить связывание электрофила на модифицированных цистеинах в Fc-домене от связывания на внутренних остатках цистеина (в ином случае, как правило, образующих S-S связи между HC-HC или HC-LC), конъюгаты обрабатывали протеазой, которая, как известно, расщепляет антитело между Fab-доменами и Fc-доменом. Одной из таких протеаз является цистеиновая протеаза IdeS, выпускаемая как "FabRICATOR®" фирмой Genovis, и описанная в публикации von Pawel-Rammingen et al., 2002, EMBO J. 21:1607.
Коротко, согласно предложенным производителем условиям, ADC обрабатывали протеазой FabRICATOR® и инкубировали образец при 37°C в течение 30 минут. Образцы подготавливали для ЖХ-МС анализа, объединяя приблизительно 20 мкл образца (приблизительно 1 мг/мл в PBS) с 20 мкл 20 мМ дитиотреитола (ДТТ), и оставляли смесь при комнатной температуре на 5 минут. Такая обработка человеческого IgG1 приводила к образованию трех фрагментов антитела размером в пределах от приблизительно 23-26 кДа: LC фрагмент, включающий внутренний цистеин, который, как правило, образует LC-HC дисульфидную связь; N-концевой HC фрагмент, включающий три внутренних цистеина (где один, как правило, образует LC-HC дисульфидную связь, а другие два цистеина, присутствующие в шарнирной области антитела, как правило, образуют HC-HC дисульфидные связи между двумя тяжелыми цепями антитела); и C-концевой HC фрагмент, который не содержит реакционноспособных цистеинов кроме цистеинов, введенных в результате мутации в конструкции, раскрытые в настоящей заявке. Образцы анализировали с помощью МС, как описано выше. Вычисления нагрузки выполняли так же, как описано ранее (выше), для количественного определения нагрузки на LC, N-концевой области HC и C-концевой области HC. Нагрузку на C-концевой области HC считают "специфичной" нагрузкой, тогда как нагрузку на LC и N-концевой области HC считают "неспецифичной" нагрузкой.
Для перекрестной проверки вычислений нагрузки, подгруппу ADC конъюгатов также оценивали на нагрузку с использованием альтернативных способов (способов, основанных на обращенно-фазовой высокоэффективной жидкостной хроматографии [офВЭЖХ] и хроматографии гидрофобного взаимодействия [HIC]), как более подробно описано в разделах ниже.
D. Анализ с помощью обращенно-фазовой ВЭЖХ
Образцы для анализа методом обращенно-фазовой ВЭЖХ подготавливали, объединяя приблизительно 20 мкл образца (приблизительно 1 мг/мл в PBS) с 20 мкл 20 мМ дитиотреитола (ДТТ). Смесь оставляли при комнатной температуре на 5 минут, после чего образцы вводили в систему ВЭЖХ Agilent 1100, оснащенную колонкой Agilent Poroshell 300SB-C8 (2,1×75 мм). Температуру системы устанавливали на 60°C и контролировали элюент с помощью УФ (220 нм и 280 нм). Использовали 20-минутный градиент с 20% до 45% ацетонитрила в воде (с 0,1% ТФУ в качестве модификатора): T=0 мин: 25% ацетонитрила; T=2 мин: 25% ацетонитрила; T=19 мин: 45% ацетонитрила; и T=20 мин: 25% ацетонитрила. При использовании таких условий, HC и LC антитела разделяли до нулевой линии. Результаты данного анализа указывают, что LC остается по существу немодифицированной (за исключением антител, содержащих T(kK183C) и T(LCQ05)), тогда как HC модифицирована (данные не показаны).
E. Хроматография гидрофобного взаимодействия (HIC)
Соединения подготавливали к HIC анализу путем разведения образцов PBS до приблизительно 1 мг/мл. Образцы исследовали при автоматическом вводе 15 мкл в систему ВЭЖХ Agilent 1200 с колонкой TSK-GEL Butyl NPR (4,6×3,5 мм, размер пор 2,5 мкм; Tosoh Biosciences #14947). Система включает автодозатор с термостатом, нагреватель колонки и УФ-детектор.
Метод градиента использовали следующим образом:
Подвижная фаза A: 1,5М сульфата аммония, 50 мМ двухосновного фосфата калия, (pH 7); Подвижная фаза B: 20% изопропилового спирта, 50 мМ двухосновного фосфата калия (pH 7); T=0 мин: 100% A; T=12 мин: 0% A.
Значения времени удерживания показаны в Таблице 13. Выбранные спектры показаны на ФИГ. 2A-2E. ADC конъюгаты, в которых применяется сайт-специфическое конъюгирование (T(kK183C+K290C)-vc0101, T(K334C+K392C)-vc0101 и T(LCQ05+K222R)-AcLysvc0101) (ФИГ. 1A-1C), показали, главным образом, один пик, тогда как ADC конъюгаты, в которых применяется обычное конъюгирование (T-vc0101 и T-DM1) (ФИГ. 2D-2E), показали смесь переменно нагруженных конъюгатов.
Таблица 13: Значения времени удерживания ADC при анализе с помощью хроматографии гидрофобного взаимодействия (HIC)
RT=время удерживания (в мин) на HIC
RRT=среднее относительное время удерживания, вычисленное по RT ADC конъюгата, деленное на RT референсного некоъюгированного трастузумаба дикого типа, имеющего типичное время удерживания 5,0-5,2 мин
F. Термостабильность
Дифференциальную сканирующую калориметрию (ДСК) использовали для определения термической стабильности цистеин- и трансглутаминаза-модифицированных вариантов антител и соответствующих сайт-специфических Aur-06380101 конъюгатов. Для данного анализа образцы, приготовленные в PBS-CMF, pH 7,2, вводили в кювету для образцов MicroCal VP-Capillary DSC с помощью автодозатора (GE Healthcare Bio-Sciences, Piscataway, NJ), уравновешивали в течение 5 минут при 10°C и затем сканировали до 110°C со скоростью 100°C в час. Был выбран период фильтрации 16 секунд. Исходные данные корректировали по нулевой линии и нормализовали концентрацию белка. Программу Origin Software 7.0 (OriginLab Corporation, Northampton, MA) использовали для выравнивания данных согласно модели MN2State с соответствующим количеством переходов.
Все варианты антител с одиночной и двойной цистеиновой модификацией, а также антитело с LCQ05 ацил-донорной глутамин-содержащей меткой показало превосходную термическую стабильность, как было определено первым переходом в расплав (Tm1) >65°C (Таблица 14).
Также оценивали моноклональные антитела, полученные на основе трастузумаба, конъюгированные с 0101 при использовании методов сайт-специфического конъюгирования, и которые также показали исключительную термическую стабильность (Таблица 15). Однако Tm1 для T(K392C+L443C)-vc0101 ADC подверглась наиболее сильному воздействию при конъюгировании полезной нагрузки, так как она составила -4,35°C по сравнению с неконъюгированным антителом.
В совокупности эти результаты продемонстрировали, что цистеин-модифицированные варианты антител и варианты антител с ацил-донорной глутамин-содержащей меткой являлись термически стабильными, и что сайт-специфическое конъюгирование 0101 через vc-линкер давало конъюгаты с превосходной термической стабильностью. Кроме того, более низкая термическая стабильность, наблюдаемая для T(K392C+L443C)-vc0101 по сравнению с неконъюгированным антителом, указывает, что конъюгирование 0101 через vc-линкер с некоторыми комбинациями модифицированных остатков цистеина может влиять на стабильность ADC.
Таблица 14: Термическая стабильность модифицированных вариантов на основе трастузумаба
Таблица 15: Термическая стабильность сайт-специфических конъюгатов, конъюгированных с ауристатином 0101
Пример 8: Связывание ADC с HER2
A. Прямое связывание
Клетки BT474 (HTB-20) трипсинизировали, центрифугировали и ресуспендировали в новой среде. Затем клетки инкубировали с серийными разведениями ADC конъюгатов или неконъюгированного трастузумаба с начальной концентрацией 1 мкг/мл в течение одного часа при 4°C. Затем клетки два раза промывали охлажденным во льду PBS и инкубировали с вторичным Alexafluor-488 антителом против антител человека (кат# A-11013, Life technologies) в течение 30 мин. Затем клетки два раза промывали, после чего ресуспендировали в PBS. Среднюю интенсивность флуоресценции считывали при помощи проточного цитометра Accuri (BD Biosciences San Jose, CA).
Таблица 16: Связываниие ADC с HER2
EC50=концентрация антитела или ADC, которая дает полумаксимальное связывание.
Как показано на ФИГ. 3A и в Таблице 16, ADC конъюгаты T(LCQ05+K222R)-AcLysvc0101, T(N297Q+K222R)-AcLysvc0101, T(kK183C+K290C)-vc0101, T(kK183C+K392C)-vc0101, T(K290C+K392C)-vc0101 обладали подобными аффинностями связывания, как T-DM1 и трастузумаб, при прямом связывании. Это указывает, что модификации в антителе в ADC конъюгатах согласно настоящему изобретению и добавление линкера-полезной нагрузки не оказывали значимого влияния на связывание.
B. Конкурентное связывание согласно FACS
Клетки BT474 трипсинизировали, центрифугировали и ресуспендировили в новой среде. Затем клетки нкубировали в течение одного часа при 4°C с серийными разведениями ADC конъюгатов или неконъюгированного трастузумаба, объединенного с 1 мкг/мл трастузумаба-ФЭ (1:1 ФЭ-меченый трастузумаб, специально синтезированный в eBiosciences (San Diego, CA)). Затем клетки два раза промывали, после чего ресуспендировали в PBS. Среднюю интенсивность флуоресценции считывали при помощи проточного цитометра Accuri (BD Biosciences San Jose, CA).
Как показано на ФИГ. 3B, ADC конъюгаты T(LCQ05+K222R)-AcLysvc0101, T(N297Q+K222R)-AcLysvc0101, T(kK183C+K290C)-vc0101, T(kK183C+K392C)-vc0101, T(K290C+K392C)-vc0101 обладали подобными аффинностями связывания, как T-DM1 и трастузумаб, при конкурентном связывании с ФЭ-меченым трастузумабом. Это указывает, что модификации в антителе в ADC конъюгатах согласно настоящему изобретению и добавление линкера-полезной нагрузки не оказывало значимого влияния на связывание.
Пример 9: Связывание ADC с человеческим FcRn
На данный момент считается, что FcRn взаимодействует с IgG независимо от субтипа pH-зависимым образом и защищает антитело от деградации, препятствуя его попаданию в лизосомальный компартемент, где антитело расщепляется. Таким образом, соображение по выбору положений для введения реакционноспособных цистеинов в IgG1-Fc область дикого типа состояло в том, чтобы избежать изменения связывающих свойств FcRn и полупериода существования антитела, включающего модифицированный цистеин.
Анализ BIAcore® проводили с целью определения аффинности в равновесном состоянии (KD) для полученных на основе трастузумаба моноклональных антител и их соответствующих ADC конъюгатов при связывании с человеческим FcRn. Технология BIAcore® использует изменения показателя преломления в поверхностном слое сенсора при связывании моноклональных антител на основе трастузумаба или их соответствующих ADC конъюгатов с человеческим белком FcRn, иммобилизированным на поверхности сенсора. Связывание обнаруживали с помощью поверхностного плазмонного резонанса (SPR) при преломлении луча лазера в поверхностном слое. Человеческий FcRn специфично биотинилировали через введенную Avi-метку при использовании реактива BirA (номер по каталогу: BIRA500, Avidity, LLC, Aurora, Colorado) и иммобилизировали на стрептавидиновом (SA) сенсоре для обеспечения одинаковой ориентации белка FcRn на сенсоре. Затем над поверхностью чипа пропускали различные концентрации моноклональных антител на основе трастузумаба или их соответствующих ADC конъюгатов в 20 мМ MES (2-(N-морфолино)этансульфоновой кислоты, pH 6,0, с 150 мМ NaCl, 3 мМ ЭДТА (этилендиаминтетрауксусная кислота), 0,5% ПАВ P20 (MES-EP). Поверхность регенерировали при использовании HBS-EP+0,05% ПАВ P20 (GE Healthcare, Piscataway, NJ), pH 7,4, между циклами ввода образцов. Аффинности связывания в равновесном состоянии моноклональных антител на основе трастузумаба или их соответствующих ADC конъюгатов определяли и сравнивали с антителом дикого типа (трастузумабом, который не содержит никаких цистеиновых мутаций в Fc-области IgG1, не имеет никакой ТГаза-модифицированной метки или сайт-специфического конъюгирования полезной нагрузки).
Эти данные продемонстрировали, что включение модифицированных остатков цистеина в Fc-область IgG в указанных положениях согласно изобретению не изменяла аффинность к FcRn (Таблица 17).
Таблица 17: Аффинность в равновесном состоянии сайт-специфических конъюгатов при связывании с FcRn человека
ND=не определяли
Пример 10: Связывание ADC с Fcγ-рецепторами
Связывание ADC конъюгатов с применением сайт-специфического конъюгирования с рецепторами Fc-γ человека оценивали с целью исследования, вызывает ли конъюгирование с полезной нагрузкой изменение связывания, что может влиять на функциональные свойства антитела, такие как антителозависимую клеточно-опосредованную цитотоксичность (ADCC). FcγIIIa (CD16) экспрессируется на NK-клетках и макрофагах, и одновременное связывание этого рецептора с клетками, экспрессирующими мишень, при связывании с антителом вызывает ADCC. Анализ BIAcore® использовали для исследования связывания моноклональных антител, полученных на основе трастузумаба, и их соответствующих ADC конъюгатов с Fc-γ рецепторами IIa (CD32a), IIb (CD32b), IIIa (CD16) и FcγRI (CD64).
Для этого анализа, основанного на явлении поверхностного плазмонного резонанса (SPR), внеклеточный домен рекомбинантного белка человеческого рецептора эпидермального фактора роста 2 (Her2/neu) (Sino Biological Inc., Beijing, P.R. China) иммобилизировали на чипе CM5 (GE Healthcare, Piscataway, NJ) и захватывали ~300-400 единиц ответа (RU) моноклонального антитела на основе трастузумаба или его соответствующего ADC. T-DM1 включали в данный анализ в качестве положительного контроля, так как было показано, что он сохраняет связывающие свойства после конъюгирования с Fcγ-рецепторами на уровне, сопоставимом с неконъюгированным антителом трастузумабом. Затем различные концентрации Fcγ-рецепторов FcγIIa (CD32a), FcγIIb (CD32b), FcγIIIa (CD16a) и FcγRI (CD64) пропускали над поверхностью и определяли связывание.
FcγR IIa, IIb и IIIa показали высокие скорости ассоциации/диссоциации, и поэтому сенсограммы выравнивали при использовании модели равновесного состояния с получением значений Kd. FcγRI показал более низкие скорости ассоциации/диссоциации, таким образом, данные выравнивали при использовании кинетической модели с получением значений Kd.
Конъюгирование полезной нагрузки в положениях модифицированных цистеинов 290 и 334 показало умеренную потерю аффинности к FcγR, в особенности к CD16a, CD32a и CD64, по сравнению с соответствующими неконъюгированными антителами-аналогами и T-DM1 (Таблица 18). Впрочем, одновременное конъюгирование на сайтах 290, 334 и 392 приводило к существенной потере аффинности к CD16a, CD32a и CD32b, но не к CD64, как наблюдали в случае с T(K290C+K334C)-vc0101 и T(K334C+K392C)-vc0101 (Таблица 18). Примечательно, что T(ĸK183C+K290C)-vc0101 демонстрировал сопоставимое связывание со всеми FcγR, которые оценивали в данном исследовании, несмотря на присутствие лекарственной полезной нагрузки в положении K29°C (Таблица 18). Как ожидали, трансглутаминаза-опосредованно конъюгированный T(N297Q+K222R)-AcLysvc0101 не связывался ни с одним из оцениваемых Fcγ-рецепторов, поскольку присутствие в таком положении ацил-донорной глутамин-содержащей метки приводит к устранению N-связанного гликозилирования. С другой стороны, T(LCQ05+K222R)-AcLysvc0101 сохранял полное связывание с Fcγ-рецепторами, поскольку глутамин-содержащую метку вводили в константную область легкой каппа цепи человека.
В совокупности, эти результаты указывают, что положение конъюгированной полезной нагрузки может влиять на связывание ADC с FcγR и может влиять на функциональность антитела в конъюгате.
Таблица 18: Аффинность связывания сайт-специфических конъюгатов для связывания Fcγ-рецепторов с CD16a, CD32a, CD32b и CD64
[мкМ]
(CD32a)
[мкМ]
(CD32b)
[мкМ]
[нМ]
NB=не определяли, NB=нет связывания
Пример 11: ADCC активности
В анализах ADCC, Her2-экспрессирующие линии клеток BT474 и SKBR3 использовали в качестве клеток-мишеней, тогда как клетки NK-92 (линия интерлейкин-2-зависимых NK-клеток, полученных из мононуклеарных клеток периферической крови 50-летнего белого мужчины в компании Conkwest) или мононуклеары периферической крови человека (МКПК), выделенные из недавно забранной крови здорового донора (#179), использовали в качестве эффекторных клеток.
Клетки-мишени (BT474 или SKBR3) в количестве 1×104 клеток/100 мкл/лунка помещали в 96-луночный планшет и культивировали в течение ночи в среде RPMI1640 при 37°C/5% CO2. На следующий день среду удаляли и заменяли 60 мкл буфера для анализа (среда RPMI1640, содержащая 10 мМ HEPES), 20 мкл 1 мкг/мл антитела или ADC, после чего добавляли 20 мкл суспензии 1×105 (для SKBR3) или 5×105 (для BT474) МКПК или 2,5×105 клеток NK92 обеих клеточных линий в каждую лунку, с получением отношения эффектора к мишени 50:1 для BT474 или 25:1 для SKBR3 в случае МКПК, 10:1 в случае NK92. Все образцы исследовали в трех повторностях.
Планшеты инкубировали при 37°C/5% CO2 в течение 6 часов и затем уравновешивали до комнатной температуры. Высвобождение ЛДГ при лизисе клеток измеряли при использовании реактива CytoTox-OneTM при длине волны возбуждения 560 нм и длине волны эмиссии 590 нм. В качестве положительного контроля добавляли 8 мкл Triton для получения максимального высвобождения ЛДГ в контрольных лунках. Специфическую цитотоксичность, показанную на ФИГ. 4, вычисляли при использовании следующей формулы:

"Экспериментальный" соответствует сигналу, измеренному при одном из условий, описанных выше.
"Эффектор спонтанный" соответствует сигналу, измеренному в присутствии только МКПК.
"Мишень спонтанный" соответствует сигналу, измеренному в присутствии только клеток-мишеней.
"Мишень максимальный" соответствует сигналу, измеренному в присутствии только лизированных детергентом клеток-мишеней.
На ФИГ. 4 показаны ADCC активности, полученные для трастузумаба, T-DM1 и vc0101 ADC конъюгатов. Данные соотвестсвуют ADCC активностям, описанным для Трастузумаба и T-DM1. Поскольку мутация N297Q прсутствует на сайте гликозилирования, предполагали, что T(N297Q+K222R)-AcLysvc0101 не будет обладать ADCC активностью, что было также подтверждено в анализах. В случае ADC конъюгатов с одиночной мутацией (K183C, K290C, K334C, K392C, включающих LCQ05) ADCC активность сохранялась. Неожиданно, ADC конъюгаты с двойной мутацией (K183C+K290C, K183C+K392C, K183C+K334C K290C+K392C, K290C+K334C, K334C+K392C) сохраняли ADCC активность во всех случаях кроме двух ADC конъюгатов с двойной мутацией, связанных с сайтом K334C (K290C+K334C и K334C+K392C).
Пример 12: Исследования цитотоксичности in vitro
Конъюгаты антитела-лекарственного средства получали, как указано в Примере 3. Клетки сеяли в 96-луночные планшеты при низкой плотности, затем на следующий день обрабатывали ADC конъюгатами и неконъюгированными полезными нагрузками в 3-кратных серийных разведениях при 10 концентрациях в двойной повторности. Клетки инкубировали в течение 4 дней при 37°C/5% CO2 в увлажняемой камере. Материал с планшетов собирали при инкубировании с раствором CellTiter® 96 AQueous One MTS (Promega, Madison, WI) в течение 1,5 часов и измеряли оптическую плотность на планшетном спектрофотометре Victor (PerkinElmer, Waltham, MA) при длине волны 490 нм. Значения IC50 вычисляли при использовании четырехпараметрической логистической модели с XLfit (IDBS, Bridgewater, NJ) и представляли в концентрации полезной нагрузки в нМ на Фиг. 5 и концентрации антитела в нг/мл на ФИГ. 6. Представлены значения IC50 +/- стандартное отклонение с количеством независимых определений в круглых скобках.
ADC конъюгаты, содержащие линкер-полезную нагрузку vc-0101 или AcLysv-0101, обладали высокой активностью против моделей Her2-позитивных клеток и селективностью против Her2-негативных клеток по сравнению с референсным ADC, T-DM1 (Кадсила).
ADC конъюгаты, синтезированные с применением сайт-специфического конъюгирования с трастузумабом, показали высокую активность и селективность против моделей Her2-клеток. В частности, несколько трастузумаб-vc0101 ADC конъюгатов являлись более активными, чем T-DM1, в моделях клеток со средней или низкой экспрессией Her2. Например, IC50 цитотоксичности in vitro для T(kK183C+K290C)-vc0101 на клетках MDA-MB-175-VII (с 1+ экспрессией Her2) составляет 351 нг/мл, по сравнению с 3626 нг/мл для T-DM1 (в ~10 раз ниже). Для клеток с 2++ уровнем экспрессии Her2, таких как клетки MDA-MB-361-DYT2 и MDA-MB-453, IC50 для T(kK183C+K290C)-vc0101 составила 12-20 нг/мл, по сравнению с 38-40 нг/мл для T-DM1.
Пример 13: Ксенотрансплантатные модели
Полученные на основе трастузумаба ADC конъюгаты согласно изобретению тестировали в ксенотрансплантатной модели рака желудка N87, ксенотрансплантатной модели рака легкого 37622 и различных ксенотрансплантатных моделях рака молочной железы (т.е. моделях HCC 1954, JIMT-1, MDA-MB-361 (DYT2) и 144580 (PDX)). В случае каждой модели, описанной ниже, первую дозу вводили в День 1. Опухоли измеряли по меньшей мере один раз в неделю, и их объем вычисляли согласно формуле: объем опухоли (мм3)=0,5×(ширина опухоли2)(длина опухоли). Средние объемы опухоли (±S.E.M.) для каждой контрольной группы вычисляли при включении в группу максимум 8-10 животных и минимум 6-8 животных.
A. Ксенотрансплантаты рака желудка N87
Действие полученных на основе трастузумаба ADC конъюгатов у иммунодефицитных мышей исследовали в отношении роста ксенотрансплантатов человеческих опухолей in vivo, которые были получены из клеточной линии N87 (ATCC CRL-5822), которая имеет высокий уровень экспрессии HER2. Для получения ксенотрансплантатов самкам голых мышей (Nu/Nu, Charles River Lab, Wilmington, MA) подкожно имплантировали 7,5×106 клеток N87 в 50% Матригеле (BD Biosciences). Когда опухоли достигли объема 250-450 мм3, стадию опухоли определяли, чтобы гарантировать однородность опухолевой массы в разных группах лечения. Животным в модели рака желудка N87 4 раза внутривенно, с интервалом 4 дня (Q4d×4) вводили растворитель PBS, трастузумаб ADC конъюгаты (в дозе 0,3, 1 и 3 мг/кг) или T-DM1 (1, 3 и 10 мг/кг) (ФИГ. 7).
Данные демонстрируют, что полученные на основе трастузумаба ADC конъюгаты ингибировали рост ксенотрансплантатов рака желудка N87 дозо-зависимым образом (ФИГ. 7A-7H).
Как показано на ФИГ. 7I, T-DM1 ингибировал рост опухолей при дозе 1 и 3 мг/кг и вызывал полную регрессию опухолей при дозе 10 мг/кг. При этом T(kK183C+K290C)-vc0101 обеспечивал полную регрессию при дозе 1 и 3 мг/кг и частичную регрессию при дозе 0,3 мг/кг (ФИГ. 7A). Данные показывают, что T(kK183C+K290C)-vc0101 значительно более активный (в ~10 раз), чем T-DM1 в данной модели.
Аналогичная эффективность in vivo ADC конъюгатов с DAR4 (ФИГ. 6E, 6F и 6G) была получена по сравнению с 183+290 (ФИГ. 7A). Кроме того, оценивали одиночные мутанты, которые являются DAR2 ADC конъюгатами (ФИГ. 7B, 7C и 7D). В целом такие ADC конъюгаты менее эффективны по сравнению с DAR4 ADC конъюгатами, но более эффективны, чем T-DM1. Среди DAR2 ADC конъюгатов LCQ05 представляется наиболее активным ADC на основе данных эффективности in vivo.
B. Ксенотрансплантаты рака молочной железы HCC1954
HCC1954 (ATCC# CRL-2338) является линией клеток рака молочной железы с высокой экспрессией HER2. Для получения ксенотрансплантатов самкам мышей SHO (Charles River, Wilmington, MA) подкожно имплантировали 5×106 клеток HCC1954 в 50% Матригеле (BD Biosciences). Когда опухоли достигали объема 200-250 мм3, определяли стадию опухолей, чтобы гарантировать однородность опухолевой массы в различных группах лечения. В модели рака молочной железы HCC1954 животным внутривенно вводили Q4d×4 растворитель PBS, трастузумаб-производные ADC конъюгаты и ADC отрицательного контроля (ФИГ. 8A-8E).
Данные демонстрируют, что трастузумаб ADC конъюгаты ингибировали рост ксенотрансплантатов рака молочной железы HCC1954 дозо-зависимым образом. При сравнении в дозе 1 мг/кг, vc0101 конъюгаты являлись более эффективными, чем T-DM1. При сравнении в дозе 0,3 мг/кг, DAR4 нагруженные ADC конъюгаты (ФИГ. 8B, 8C и 8D) являлись более эффективными, чем DAR2 нагруженный ADC (ФИГ. 8A). Кроме того, ADC отрицательного контроля в дозе 1 мг/кг оказывал лишь незначительное влияние на рост опухоли по сравнению с контролем растворителем (ФИГ. 8D). Однако T(N297Q+K222R)-AcLysvc0101 вызывал полную регрессию опухолей, что указывает на специфичность к мишени.
C. Ксенотрансплантаты рака молочной железы JIMT-1
JIMT-1 - линия клеток рака молочной железы, экспрессирующая средние/низкие уровни Her2 и изначально устойчивая к трастузумабу. Для получения ксенотрансплантатов самкам голых (Nu/Nu) мышей подкожно имплантировали 5×106 клеток JIMT-1 (DSMZ# ACC-589) в 50% Матригеле (BD Biosciences). Когда опухоли достигали объема 200-250 мм3, определяли стадию опухолей, чтобы гарантировать однородность опухолевой массы в различных группах лечения. В модели рака молочной железы JIMT-1 животным внутривенно вводили Q4dx4 растворитель PBS, T-DM1 (ФИГ. 9G), трастузумаб-производные ADC конъюгаты с применением сайт-специфического конъюгирования (ФИГ. 9A-9E), трастузумаб-производный ADC с применением обычного конъюгирования (ФИГ. 9F) и huNeg-8.8 ADC отрицательного контроля.
Данные демонстрируют, что все исследованные vc0101 конъюгаты вызывали уменьшение опухолей дозо-зависимым образом. Эти ADC конъюгаты могут вызывать регрессию опухоли в дозе 1 мг/кг. Однако T-DM1 неактивен в данной модели со средней/низкой экспрессией Her2 даже в дозе 6 мг/кг.
D. Ксенотрансплантаты рака молочной железы MDA-MB-361 (DYT2)
MDA-MB-361 (DYT2) является линией клеток рака молочной железы, экспрессирующей средние/низкие уровни Her2. Для получения ксенотрансплантатов самок голых (Nu/Nu) мышей облучали при мощности дозы 100 сГр/мин в течение 4 минут и через три дня подкожно имплантировали 1,0×107 клеток MDA-MB-361 (DYT2) (ATCC# HTB-27) в 50% Матригеле (BD Biosciences). Когда опухоли достигали объема 300-400 мм3, определяли стадию опухоли, чтобы гарантировать однородность опухолевой массы в различных группах лечения. В модели рака молочной железы DYT2 животным внутривенно вводили Q4dx4 растворителя PBS, трастузумаб-производных ADC конъюгатов с использованием сайт-специфического и обычного конъюгирования, T-DM1 и ADC отрицательного контроля (ФИГ. 10A-10D).
Данные демонстрируют, что трастузумаб ADC конъюгаты ингибировали рост ксенотрансплантатов рака молочной железы DYT2 дозо-зависимым образом. Хотя DYT2 является линией клеток со средними/низкими уровнями экспрессии Her2, она более чувствительна к ингибиторам микротрубочек, чем другие линии клеток с низкой/средней экспрессией Her2.
E. Полученные от пациента ксенотрансплантаты рака молочной железы 144580
Действие полученных на основе трастузумаба ADC конъюгатов исследовали у иммунодефицитных мышей на рост in vivo ксенотрансплантатов человеческих опухолей, которые были получены из фрагментов недавно удаленных опухолей молочной железы 144580, полученных в соответствии с надлежащими процедурами согласия. Анализ опухоли 144580 при взятии новой биопсии показал трижды негативную (ER-, PR- и HER2-) опухоль молочной железы. Полученные от пациента ксенотрансплантаты рака молочной железы 144580 подкожно перевивали in vivo в виде фрагментов от животного животному у самок голых (Nu/Nu) мышей. Когда опухоли достигали объема 150-300 мм3, определяли их стадию, чтобы гарантировать однородность размера опухолей в различных группах лечения. В модели рака молочной железы 144580 животным внутривенно вводили четыре дозы раз в четыре дня (Q4d×4) растворителя PBS, трастузумаб ADC конъюгатов с использованием сайт-специфического конъюгирования, трастузумаб-производного ADC с использованием обычного конъюгирования и ADC отрицательного контроля (ФИГ. 11A-11E).
В этой HER2-(по клиническому определению) PDX модели, T-DM1 не был эффективен ни в одной из протестированных доз (1, 5, 3 и 6 мг/кг) (ФИГ. 10E). В случае DAR4 vc0101 ADC конъюгатов (ФИГ. 11А, 11C и 11D), доза 3 мг/кг могла вызывать регрессию опухоли (даже в дозе 1 мг/кг на ФИГ. 11C). DAR2 VC0101 ADC (ФИГ. 11B) оказался менее эффективным, чем DAR4 ADC конъюгаты при дозе 3 мг/кг. Однако DAR 2 vc0101 ADC эффективен при дозе 6 мг/кг в отличие от T-DM1.
F. Полученный от пациента ксенотрансплантат немелкоклеточного рака легкого 37622
Несколько ADC конъюгатов тестировали в модели с полученным от пациента ксенотрансплантатом немелкоклеточного рака легкого 37622, полученного в соответствии с надлежащими процедурами согласия. Полученные от пациента ксенотрансплантаты 37622 подкожно перевивали in vivo в виде фрагментов от животного животному у самок голых (Nu/Nu) мышей. Когда опухоли достигали объема 150-300 мм3, определяли их стадию, чтобы гарантировать однородность размера опухолей в различных группах лечения. В модели PDX 37622 животным внутривенно вводили четыре дозы раз в четыре дня (Q4dx4) растворителя PBS, трастузумаб-производных ADC конъюгатов с использованием сайт-специфического конъюгирования, T-DM1 и ADC отрицательного контроля (ФИГ. 12A-12D).
Профили экспрессии Her2 определяли с помощью модифицированного теста Hercept и классифицировали как 2+ с более высокой гетерогенностью, чем наблюдаемой в клеточных линиях. ADC конъюгаты, конъюгированные с vc0101 в качестве линкера-полезной нагрузки (ФИГ. 12A-12C), были эффективными в дозах 1 и 3 мг/кг, вызывающих регрессию опухоли. Однако T-DM1 показали лишь небольшую терапевтическую эффективность при дозе 10 мг/кг (ФИГ. 12D). По-видимому, vc0101 ADC конъюгаты в 10 раз более активны, чем T-DM1, при сравнении результатов в случае дозы 10 мг/кг для T-DM1 и 1 мг/кг для vc0101 ADC конъюгатов. Вероятно, что неспецифическое действие важно для эффективности в отношении гетерогенной опухоли.
G. Полученный от пациента ксенотрансплантат рака желудка GA0044
Трастузумаб и ADC конъюгаты против HER2 тестировали в полученной от пациента ксенотрансплантатной модели рака желудка (GA0044), полученной в соответствии с надлежащими процедурами согласия. Полученные от пациента ксенотрансплантаты GA0044 подкожно перевивали in vivo в виде фрагментов от животного животному у самок голых (Nu/Nu) мышей. Когда опухоли достигали объема 150-300 мм3, определяли их стадию, чтобы гарантировать однородность размера опухолей в различных группах лечения. В PDX модели GA0044 животным внутривенно вводили четыре дозы раз в четыре дня (Q4dx4) растворителя PBS, трастузумаба, T-DM1 или трастузумаб-производного ADC с использованием сайт-специфического конъюгирования с vc0101 (ФИГ. 30).
Профили экспрессии HER2 в GA0044 определяли с помощью модифицированного теста Hercept и классифицировали как 2+ с гетерогенным распределением. ADC, конъюгированный с vc0101 в качестве полезной нагрузки (а именно T(kK183C+K290C)-vc0101), был эффективен и приводил к полной регрессии опухоли в дозах 1 и 3 мг/кг. Трастузумаб и T-DM1 не показали какого-либо значимого различия в росте опухоли по сравнению с опухолями, обработанными растворителем. Вероятно, что неспецифическое действие важно для эффективности в отношении данной опухоли с гетерогенной экспрессией мишени (т.е. HER2).
H. Демонстрация неспецифического действия T-vc0101 ADC на ксенотрансплантате рака желудка N87
Высвобождаемый метаболит T-DM1 ADC, как было показано, являлся лизином-кэпированным линкером-полезной нагрузкой mcc-DM1 (т.е. Lys-mcc-DM1), который является соединением, не способным проникать через мембрану (Kovtun et al., 2006, Cancer Res 66:3214-21; Xie et al., 2004, J Pharmacol Exp Ther 310:844). При этом высвобождаемым из T-vc0101 ADC метаболитом является ауристатин 0101, соединение с более высокой мембранной проницаемостью, чем Lys-mcc-DM1. Способность высвобождаемой из ADC полезной нагрузки уничтожать соседние клетки известна как неспецифическое действие. Вследствие высвобождения мембранопроникающей полезной нагрузки, T-vc0101 способен вызывать сильное неспецифическое действие, а T-DM1 нет. На ФИГ. 13 показана иммуногистоцитохимия ксенотрансплантатных опухолей из линии клеток N87, которые обрабатывали однократной дозой 6 мг/кг T-DM1 (ФИГ. 13A) или 3 мг/кг T-vc0101 (ФИГ. 13B), а затем собирали и фиксировали формалином через 96 часов. Срезы опухолей окрашивали на IgG человека для обнаружения ADC, связанного с опухолевыми клетками, и фосфогистон H3 (pHH3), для обнаружения митотических клеток в качестве наблюдения предполагаемого механизма действия для полезных нагрузок обоих ADC конъюгатов.
ADC обнаруживали по периферии опухолей в обоих случаях. В опухолях, обработанных T-DM1 (ФИГ. 13A), большинство pHH3-положительных опухолевых клеток располагалось около ADC. Однако в опухолях, обработанных T-vc0101 (ФИГ. 13B), большинство pHH3-положительных опухолевых клеток выходило за пределы локализации ADC (черными стрелками указаны несколько примеров), и находилось внутри опухоли. Эти данные позволяют предположить, что ADC с расщепляемым линкером и мембранопроникающей полезной нагрузкой может вызывать сильное неспецифическое действие in vivo.
Пример 14: Модели резистентности к T-DM1 in vitro
A. Получение T-DM1 резистентных клеток in vitro
Клетки N87 пересевали в две отдельных колбы и каждую колбу идентично обрабатывали в соответствии с протоколом получения резистентности для получения биологических дубликатов. Клетки подвергали пяти циклам обработки конъюгатом T-DM1 приблизительно при IC80 концентрациях (концентрация полезной нагрузки 10 нМ) в течение 3 дней, с последующим восстановлением в течение приблизительно 4-11 дней без обработки. После пяти циклов при 10 нМ конъюгата T-DM1, клетки подвергали шести дополнительным циклам обработки 100 нМ T-DM1 аналогичным образом. Процедура была предназначена для моделирования долговременного, мультицикличного (обработка/перерыв) дозирования в максимально переносимых дозах, обычно используемых для цитотоксических средств в клинике, с последующим периодом восстановления. Исходные клетки, полученные из N87, обозначены как N87, и клетки, которые подвергали длительному воздействию T-DM1, обозначены как N87-ТМ. Умеренная или высокая устойчивость к лекарственному средству развивалась в течение 4 месяцев для клеток N87-ТМ. Слективное давление лекарственного средства снимали через ~3-4 месяца циклов обработки, когда уровень устойчивости уже не увеличивался после продолжения воздействия лекарственного средства. Реакции и фенотипы оставались стабильными в культивируемых линиях клеток в течение приблизительно 3-6 месяцев после этого. Затем иногда наблюдали снижение величины фенотипа устойчивости при измерении с помощью анализов цитотоксичности, и в этом случае криоконсервированные резистентные к Т-DM1 клетки ранних пассажей размораживали для дополнительных исследований. Все описанные исследования проводили после снятия селективного пресса T-DM1 по меньшей мере на 2-8 недель, чтобы обеспечить стабилизацию клеток. Данные собирали с различными размороженными криоконсервированными популяциями, полученными при одном отборе, за приблизительно 1-2 года после разработки модели, чтобы гарантировать согласованность результатов. Линию клеток рака желудка N87 отбирали по резистентности к конъюгату антитела-лекарственного средства, трастузумабу-майтанзиноиду (T-DM1), в циклах обработки в дозах, которые составляли приблизительно IC80 (~10 нМ концентрацию полезной нагрузки) для соответствующей клеточной линии. Исходные клетки N87 были изначально чувствительными к конъюгату (IC50=1,7 нМ концентрация полезной нагрузки; концентрация антитела 62 нг/мл) (ФИГ. 14). Две популяции исходных клеток N87 подвергали циклам обработки, и после цикличного воздействия в течение всего лишь приблизительно четырех месяцев 100 нМ T-DM1 две указанных популяции (далее называемых N87-TM-1 и N87-TM-2) стали рефракторными к ADC в 114 и 146 раз, соответственно, по сравнению с исходными клетками (ФИГ. 14 и ФИГ. 15A). Примечательно, что наблюдалась минимальная перекрестная резистентность (~2,2-2,5×) к соответствующему неконъюгированному свободному майтанзиноидному лекарственному средству, DM1 (ФИГ. 14).
B. Исследования цитотоксичности
ADC конъюгаты были получены, как указано в Примере 3. Неконъюгированный аналог майтанзина (DM1) и аналоги ауристатина были получены в Pfizer Worldwide Medicinal Chemistry (Groton, CT). Другие химиотерапевтические средства стандарта лечения приобретали в Sigma (St. Louis, MO). Клетки сеяли в 96-луночные планшеты при низкой плотности, затем на следующий день обрабатывали ADC конъюгатами и неконъюгированными полезными нагрузками в 3-кратных серийных разведениях при 10 концентрациях в двойной повторности. Клетки инкубировали в течение 4 дней при 37°C/5% CO2 в увлажняемой камере. Материалы с планшетов собирали при инкубировании с раствором CellTiter® 96 AQueous One MTS (Promega, Madison, WI) в течение 1,5 часов и оптическое поглощение измеряли на планшетном спектрофотометре Victor (PerkinElmer, Waltham, MA) при длине волны 490 нм. Значения IC50 вычисляли при использовании четырехпараметрической логистической модели с XLfit (IDBS, Bridgewater, NJ).
Определяли профиль перекрестной резистентности к другим трастузумаб-производным ADC конъюгатам. Наблюдали значимую перекрестную резистентность ко многим полученным на основе трастузумаба ADC конъюгатам, состоящим из нерасщепляемых линкеров и доставляемых полезных нагрузок с антитубулиновым механизмом действия (ФИГ. 14). Например, в клетках N87-ТМ в сравнении с исходными клетками N87 наблюдали >330- и >272-кратное снижение активности с T-mc8261 (ФИГ. 14 и ФИГ. 15B) и T-MalPeg8261 (ФИГ. 14), которые представляют собой полезные нагрузки на основе ауристатина, связанные с трастузумабом через нерасщепляемые малеимидокапроил или Mal-PEG линкеры, соответственно. Более чем 235-кратную резистентность наблюдали в клетках N87-ТМ против T-mcMalPegMMAD, другого трастузумаб-ADC, с другим нерасщепляемым линкером, доставляющим монометил-доластатин (MMAD) (ФИГ. 14).
Примечательно то, что наблюдали, что линия клеток N87-ТМ сохраняла чувствительность к полезным нагрузкам в случае доставки через расщепляемый линкер, даже несмотря на то, что такие лекарственные средства функционально ингибируют подобные мишени (т.е. деполимеризация микротрубочек). Примеры ADC конъюгатов, которые преодолевают устойчивость, включают, без ограничения перечисленными, T(N297Q+K222R)-AcLysvc0101 (ФИГ. 14 и ФИГ. 15C), T(LCQ05+K222R)-AcLysvc0101 (ФИГ. 14 и ФИГ. 15D), T(K290C+K334C)-vc0101 (ФИГ. 10 и ФИГ. 11E), T(K334C+K392C)-vc0101 (ФИГ. 14 и ФИГ. 15F) и T(kK183C+K290C)-vc0101 (ФИГ. 14 и ФИГ. 15G). Они представляют собой ADC конъюгаты на основе трастузумаба, доставляющие аналог ауристатина 0101, но высвобождают полезные нагрузки внутриклеточно, при протеолитическом расщеплении vc линкера.
Для определения, обладают ли такие ADC-резистентные раковые клетки широкой устойчивостью к другим терапиям, модели с клетками N87-TM обрабатывали группой химиотерапевтических средств стандарта лечения, имеющих разные механизмы действия. В целом, низкомолекулярные ингибиторы микротрубочек и функции ДНК сохраняли эффективность против резистентных линий клеток N87-TM (ФИГ. 14). Хотя такие клетки приобрели резистентность к ADC, доставляющему аналог деполимеризующего микротрубочки средства, майтанзина, минимальную перекрестную резистентность или ее отсутствие наблюдали к нескольким деполимеризующим или полимеризующим тубулин средствам. Аналогичным образом, обе клеточные линии сохраняли чувствительность к средствам, которые нарушали функцию ДНК, в том числе к ингибиторам топоизомеразы, антиметаболитам и алкилирующим/сшивающим средствам. В целом, клетки N87-TM не были рефракторными к широкому спектру цитотоксических средств, за исключением характерных нарушений развития или клеточного цикла, которые могут имитировать резистентность к лекарственным средствам.
Обе популяции N87-ТМ также сохраняли чувствительность к соответствующим неконъюгированным лекарственным средствам (т.е. DM1 и 0101; ФИГ. 14). Следовательно, клетки N87-ТМ, сделанные рефракторными к конъюгату трастузумаба-майтанзиноида, демонстрировали перекрестную резистентность к другим ADC конъюгатам на основе ингибиторов микротрубочек, при доставке посредством нерасщепляемых линкеров, но оставались чувствительными к неконъюгированным ингибиторам микротрубочек и другим химиотерапевтическим средствам.
Для определения молекулярного механизма устойчивости к T-DM1 в клетках N87-ТМ, определяли уровни экспрессии белков MDR1 и MRP1, эффлюксных насосов, обеспечивающих лекарственную резистентность. Это объяснялось тем, что низкомолекулярные ингибиторы тубулина являются известными субстратами эффлюксных насосов лекарственных средств MDR1 и MRP1 (Thomas and Coley, 2003, Cancer Control 10(2):159-165). Определяли уровни экспрессии этих двух белков в суммарных клеточных лизатах исходных N87 и N87-ТМ резистентных клеток (ФИГ. 16). Иммуноблот-анализ показал, что резистентные клетки N87-ТМ имеют незначительно повышенную экспрессию белков MRP1 (ФИГ. 16A) или MDR1 (ФИГ. 16B). В совокупности эти данные, в сочетании с отсутствием перекрестной резистентности к известным субстратам эффлюксных насосов лекарственных средств (например, паклитакселу, доксорубицину) в клетках N87-ТМ, дают основание предполагать, что оверэкспрессия эффлюксных насосов лекарственных средств не является молекулярным механизмом резистентности к T-DM1 в клетках N87-ТМ.
Поскольку механизм действия ADC конъюгатов требует связывания со специфическим антигеном, истощение антигена или пониженное связывание антитела могут объяснять резистентность к T-DM1 в клетках N87-ТМ. Для определения, был ли антиген T-DM1 значимо истощен в клетках N87-ТМ, сравнивали уровни экспрессии белка HER2 в суммарных клеточных лизатах исходных N87 и N87-ТМ резистентных клеток (ФИГ. 17A). Иммуноблот-анализ показал, что в клетках N87-ТМ уровень экспрессии белка HER2 не был заметно понижен по сравнению с исходными клетками N87.
Определяли количество антитела, связанного с HER2 антигенами клеточной поверхности клеток N87-ТМ. В исследовании связывания с клеточной поверхностью при использовании сортировки флуоресцентно активированных клеток, клетки N87-ТМ действительно показали ~50% уменьшение связывания трастузумаба с антигенами клеточной поверхности (ФИГ. 17B). Поскольку клетки N87 имеют высокую экспрессию белка HER2 среди линий раковых клеток (Fujimoto-Ouchi et al., 2007, Cancer Chemother Pharmacol 59(6):795-805), ~50% уменьшение связывания антитела с HER2 в этих клетках, вероятно, не представляет ведущий механизм устойчивости к T-DM1 в клетках N87-ТМ. Доказательством, подтверждающим такую интерпретацию, служит то, что резистентные клетки N87-ТМ остаются чувствительными к другим HER2-связывающим, полученным на основе трастузумаба ADC конъюгатам с другими линкерами и полезными нагрузками (ФИГ. 14).
Для определения потенциальных механизмов резистентности к T-DM1 в объективном методе, модели с исходными N87 и N87-ТМ резистентными клетками подвергали протеомному анализу для глобального определения изменений уровней экспрессии мембранных белков, которые могут отвечать за устойчивость T-DM1. Наблюдали значительные изменения уровня экспрессии 523 белков в обеих моделях клеточных линий (ФИГ. 18A). Для подтверждения выбора таких предполагаемых белковых изменений, проводили иммуноблоттинг лизатов клеток N87 и N87-ТМ для анализа белков с продполагаемой пониженной экспрессией (IGF2R, LAMP1, CTSB) (ФИГ. 18B) и повышенной экспрессией (CAV1) (ФИГ. 18C) в клетках N87-ТМ по сравнению с клетками N87. Опухоли in vivo были получены при подкожной имплантации клеток N87 и N87-TM-2 мышам NSG для оценки, повторяют ли белковые изменения, наблюдаемые in vivo, результаты, отмеченные in vitro. Опухоли N87-TM-2 сохраняли оверэкспрессию белка CAV1 по сравнению с опухолями N87 (ФИГ. 18D). Хотя ожидали окрашивание CAV1 в строме мышей в обеих моделях, эпителиальное окрашивание CAV1 наблюдали только в модели N87-TM-2.
C. Исследования эффективности in vivo
Для определения, повторяется ли in vivo устойчивость, наблюдаемая в клеточной культуре, исходные клетки N87 и клетки N87-TM-2 размножали и вводили в бока самок мышей NOD scid gamma (NSG) с иммунодефицитом (NOD.Cg-Prkdcscid Il2rgtm1Wjl/SzJ), полученных из Jackson Laboratory (Bar Harbor, ME). Мышам в правый бок подкожно вводили суспензии клеток N87 или N87-ТМ (7,5×106 клеток на одну инъекции с 50% Матригелем). Мышей рандомизировано распределяли в исследуемые группы, когда опухоли достигали ~0,3 г (~250 мм3). Конъюгат T-DM1 или растворитель вводили внутривенно в солевом растворе в день 0 и повторно, с введением в общей сложности четырех доз, с интервалом в четыре дня (Q4D×4). Опухоли измеряли еженедельно и вычисляли массу как объем=(ширина×ширина×длина)/2. Проводили анализ времени до наступления события (двухкратное увеличение опухоли) и оценивали значимость с использованием логарифмического рангового критерия (Мантела-Кокса). В этих исследованиях у мышей ни в одной из групп лечения не наблюдали потери веса.
Мышей обрабатывали следующими средствами: (1) контроль растворителем PBS, (2) антитело трастузумаб в дозе 13 мг/кг, затем 4,5 мг/кг; (3) T-DM1 в дозе 6 мг/кг; (4) T-DM1 в дозе 10 мг/кг; (5) T-DM1 в дозе 10 мг/кг, затем T(N297Q+K222R)-AcLysvc0101 в дозе 3 мг/кг; (6) T(N297Q+K222R)-AcLysvc0101 в дозе 3 мг/кг. Размеры опухолей контролировали, и результаты показаны на Фигуре 20. Опухоли N87 (ФИГ. 19 и ФИГ. 20A) и N87-TM-2 (ФИГ. 19 и ФИГ. 20B) показали профиль эффективности ADC, аналогичный наблюдаемому в анализах цитотоксичности in vitro (ФИГ. 19 и 20B), где устойчивые к лекарственному средству клетки N87-ТМ были рефракторными к T-DM1, но при этом отвечали на полученные на основе трастузумаба ADC конъюгаты с расщепляемыми линкерами. Фактически, опухоли, которые были рефракторными к T-DM1 и выросли до массы приблизительно 1 грамм, были переведены на терапию T(N297Q+K222R)-AcLysvc0101 и эффективно регрессировали (ФИГ. 20B). В анализе времени до наступления события в этом исследовании, T-DM1 в дозах 6 и 10 мг/кг предотвращал двухкратное увеличение опухоли у >50% мышей в течение по меньшей мере 60 дней в модели N87, однако T-DM1 не был так же эффективен в модели N87-TM-2 (ФИГ. 20C и 20D). T(N297Q+K222R)-AcLysvc0101, вводимый в дозе 3 мг/кг, предотвращал двухкратное увеличение обеих опухолей N87 и N87-ТМ у мышей в течение всего исследования (~80 дней) (ФИГ. 20C и 20D).
В другом исследовании все связанные расщепляемыми линкерами ADC конъюгаты, которые преодолевали устойчивость к T-DM1 in vitro, оставались эффективными в данной модели опухоли N87-TM2, которая не отвечала на T-DM1 (ФИГ. 19 и ФИГ. 20E).
После этого оценивали, был ли способен T(kK183+K290C)-vc0101 ADC ингибировать рост опухолей, которые были рефракторными к TDM1. Опухоли N87-ТМ, обработанные растворителем или T-DM1, росли в ходе такой обработки, однако опухоли, переведенные на терапию T(kK183C+K290C)-vc0101 в день 14, сразу регрессировали (ФИГ. 20F).
Пример 15: In vivo T-DM1-резистентные модели
A. Получение резистентных к T-DM1 клеток in vivo
Все исследования на животных были одобрены Институциональным комитетом по уходу за животными и их использованию исследовательского центра компании Pfizer в Перл-Ривер согласно установленным рекомендациям. Для получения ксенотрансплантатов самкам голых (Nu/Nu) мышей подкожно имплантировали 7,5×106 клеток N87 в 50% Матригеле (BD Biosciences). Когда средний объем опухоли достиг ~300 мм3, животных рандомизировано распределяли в две группы: 1) контроль растворителем (n=10) и 2) введение T-DM1 (n=20). T-DM1 ADC (6,5 мг/кг) или растворитель (PBS) вводили внутривенно в солевом растворе в день 0, после чего животным раз в неделю вводили дозу 6,5 мг/кг в течение периода продолжительностью до 30 недель. Опухоли измеряли два или один раз в неделю и вычисляли массу как объем=(ширина×ширина×длина)/2. В этих исследованиях у мышей ни в одной из групп лечения не наблюдали потери веса.
Животных считали рефракторными или рецидивирующими при обработке T-DM1, когда объем отдельных опухолей достигал ~600 мм3 (двухкратное увеличение от исходного размера опухоли при рандомизации). По сравнению с контрольной группой большинство опухолей первоначально отвечали на обработку T-DM1, как показано на ФИГ. 21A. В частности, 17 из 20 мышей отвечали на первичную обработку T-DM1, однако значимое количество опухолей (13 из 20) рецидивировали при обработке T-DM1. С течением времени имплантированные опухолевые клетки N87 становились резистентными к T-DM1 (ФИГ. 21B). Три опухоли, которые первоначально не отвечали на лечение T-DM1, собирали для определения экспрессии Her2 с помощью ИГХ, показавшей отсутствие изменений экспрессии HER2. Остальные 10 рецидивировавших опухолей описаны ниже.
Четыре опухоли, которые первоначально отвечали на обработку T-DM1 и затем рецидивировали, были переведены на обработку T-vc0101 раз в неделю в дозе 2,6 мг/кг в день 77 (мыши 1 и 16), 91 (мышь 19), 140 (мышь 6). Как показано на ФИГ. 19C, T-DM1-резистентные опухоли, полученные in vivo, отвечали на T-vc0101, что указывает на то, что полученные T-DM1-резистентные опухоли чувствительны к обработке vc0101 ADC.
Еще три опухоли первоначально отвечали на обработку T-DM1 и затем рецидивировали, после чего были переведены на обработку T (N297Q+K222R)-AcLysvc0101 раз в неделю в дозе 2,6 мг/кг в день 110 (мыши 4, 13 и 18). Как показано на ФИГ. 21D, T-DM1-резистентные опухоли, полученные in vivo, также отвечали на T(N297Q+K222R)-AcLysvc0101. Последующий эксперимент проводили для оценки T(kK183C+K290C)-vc0101, при этом были получены подобные результаты, что указывает на то, что T-DM1-резистентные опухоли, полученные in vivo, были чувствительны к обработке T(kK183C+K290C)-vc0101, как показано на ФИГ. 21E.
В заключение следует отметить, что все рефракторные к T-DM1 опухоли, подвергнутые последующей обработке, были чувствительны к vc0101 ADC (7 из 7), что указывает на то, что in vivo резистентные к T-DM1 опухоли можно лечить расщепляемыми vc0101 конъюгатами.
Дополнительные три опухоли (мышь 7, 17 и 2, как показано на ФИГ. 21B) первоначально отвечали на T-DM1 и затем рецидивировали, после чего были удалены для исследования in vitro. После 2-5 месяцев культивирования изъятых опухолей in vitro эти клетки оценивали на резистентность к T-DM1 и исследовали in vitro (см. Разделы B и C данного Примера ниже).
B. Исследования цитотоксичности
Клетки, рецидивировавшие при обработке T-DM1 и культивируемые in vitro (как описано в Разделе A данного Примера), сеяли в 96-луночные планшеты и на следующий день вводили 4-кратные серийные разведения ADC конъюгатов или неконъюгированных полезных нагрузок. Клетки инкубировали в течение 96 часов при 37°C/5% CO2 в увлажняемой камере. В планшеты добавляли раствор CellTiter Glo (Promega, Madison, WI) и измеряли оптическую плотность на планшетном спектрофотометре Victor (PerkinElmer, Waltham, MA) при длине волны 490 нм. Значения IC50 вычисляли при использовании четырехпараметрической логистической модели с XLfit (IDBS, Bridgewater, NJ).
Результаты скрининга цитотоксичности представлены в Таблицах 19 и 20. Клетки были резистентными к T-DM1 (ФИГ. 22A) по сравнению с исходным, но чувствительными к расщепляемым vc0101 конъюгатам T-vc0101 (данные не показаны), T(kK183C+K290C)-vc0101 (ФИГ. 22B), T(LCQ05+K222R)-AcLysvc0101 (ФИГ. 22C) и T(N297Q+K222R)-AcLysvc0101 (ФИГ. 22D) (Таблица 19). Резистентные к T-DM1 клетки неожиданно оказались чувствительными к исходной полезной нагрузке DM1, а также к полезной нагрузке 0101 (Таблица 20).
Таблица 19: Чувствительность резистентных клеток к ADC конъюгатам
Мышь #7
Мышь #17
Мышь #2
Значения IC50 показаны для каждой из клеточных линий
Таблица 20: Чувствительность резистентных клеточных линий к свободной полезной нагрузке
C. Экспрессия Her2 согласно FACS и Вестерн-блоттингу
Экспрессию Her2 исследовали на клетках, рецидивировавших при обработке T-DM1 и культивируемых in vitro (как описано в Разделе A данного Примера). Для анализа FACS клетки трипсинизировали, центрифугировали и ресуспендировили в новой среде. Затем клетки инкубировали в течение одного часа при 4°C с 5 мкг/мл Трастузумаба-ФЭ (меченый 1:1 ФЭ трастузумаб, специально синтезированный в eBiosciences (San Diego, CA)). Затем клетки два раза промывали и ресуспендировали в PBS. Среднюю интенсивность флуоресценции регистрировали при использовании проточного цитометра Accuri (BD Biosciences, San Jose, CA).
Для Вестерн-блоттинга клетки лизировали при использовании лизирующего буфера RIPA (с ингибиторами протеаз и ингибитором фосфатазы) на льду в течение 15 минут, затем обрабатывали на вортексе и центрифугировали на максимальной скорости в микроцентрифуге при 4°C. Супернатант собирали и добавляли 4× буфер для образцов и восстанавливающий агент с нормализацией образцов по общему содержанию белка в каждом образце. Образцы разделяли в 4-12% бис-трис геле и переносили на нитроцеллюлозную мембрану. Мембраны блокировали в течение часа и инкубировали с антителом к HER2 (Cell Signalling, 1:1000) в течение ночи при 4°C. Затем мембраны 3 раза промывали в 1× TBST и инкубировали с HRP-антителом против антител мыши (Cell Signalling, 1:5000) в течение 1 часа, 3 раза промывали и проявляли.
Уровни экспрессии HER2 в T-DM1 рецидивировавших опухолях были аналогичны показателям в контрольных опухолях (без обработки T-DM1) согласно анализу FACS (ФИГ. 23A) и Вестерн-блоттингу (ФИГ. 23B).
D. Резистентность к T-DM1 не обусловлена экспрессией эффлюксных насосов лекарственных средств
Клеточные линии не экспрессируют MDR1 согласно Вестерн-блоттингу (ФИГ. 24A), при этом клетки не резистентны к субстрату MDR-1, свободному лекарственному средству 0101 (ФИГ. 24B). Не наблюдали резистентности к доксорубицину (ФИГ. 24C), что указывает на то, что механизм резистентности не опосредован MRP1. Однако клетки все же являются резистентными к свободному DM1 (ФИГ. 24D).
Пример 16: Фармакокинетика (ФК)
Экспозицию обычных или сайт-специфических vc0101 конъюгатов антитела-лекарственного средства определяли после в/в болюсного введения дозы 5 или 6 мг/кг яванским макакам. Концентрации суммарного антитела (суммарное Ат; измерение конъюгированного мАт и неконъюгированного мАт), и ADC (мАт, который конъюгирован по меньшей мере с одной молекулой лекарственного средства) измеряли с помощью анализов связывания лиганда (LBA). ADC был получен с использованием vc0101 во всех случаях, за исключением T(LCQ05), в котором использовали AcLysvc0101. Обычное конъюгирование (не сайт-специфическое конъюгирование) использовали для получения ADC из трастузумаба.
Профили зависимости концентрации от времени и фармакокинетика/токсикокинетика суммарного Ат и трастузумаб-ADC (T-vc0101) (5 мг/кг) или T(kK183C+K290C) сайт-специфического ADC (6 мг/кг) после введения дозы яванским макакам (ФИГ. 25A и Таблица 21). Воздействие T(kK183C+K290C) сайт-специфического ADC характеризовалось повышенной экспозицией и стабильностью по сравнению с обычным конъюгатом.
Профили зависимости концентрации от времени и фармакокинетика/токсикокинетика ADC аналита трастузумаба (T-vc0101) (5 мг/кг) или T(kK183C+K290C), T(LCQ05), T(K334C+K392C), T (K290C+K334C), T(K290C+K392C) и T(kK183C+K392C) сайт-специфического ADC (6 мг/кг) после введения дозы яванским макакам (ФИГ. 25B и Таблица 21). Экспозиция нескольких сайт-специфических ADC (T(LCQ05), T(kK183C+K290C), T(K290C+K392C) и T(kK183C+K392C)) более высокая по сравнению с трастузумабом-ADC с использованием обычного конъюгирования. Однако экспозиция двух других сайт-специфических ADC (T(K290C+K334C) и T(K334C+K392C)) не превышала экспозицию трастузумаба-ADC, что указывает на то, что не все сайт-специфические ADC конъюгаты будут обладать более высокими фармакокинетическими свойствами, чем трастузумаб-ADC, полученный при использовании обычного конъюгирования.
Таблица 21: Фармакокинетика
Пример 17: Значения относительного удерживания в хроматографии гидрофобного взаимодействия в зависимости от экспозиции (AUC) у крыс.
Гидрофобность является физическим свойством белка, которое можно оценивать с помощью хроматографии гидрофобного взаимодействия (HIC), при этом время удерживания образцов белка отличается в зависимости от их относительной гидрофобности. ADC конъюгаты можно сравнивать с их соответствующим антителом путем вычисления относительного времени удерживания (RRT), которое является отношением времени удерживания в HIC для ADC, деленного на время удерживания в HIC соответствующего антитела. Сильно гидрофобные ADC конъюгаты имеют большее RRT, при этом возможно, что такие ADC конъюгаты также могут иметь более высокую фармакокинетическую лабильность, в частности меньшую площадь под кривой (AUC или экспозицию). Когда HIC значения ADC конъюгатов с мутациями различных сайтов сравнивали с их измеренной AUC у крыс, наблюдали распределение, показанное на ФИГ. 26.
ADC конъюгаты с RRT≥1,9 показали меньшие значения AUC, тогда как ADC конъюгаты с меньшим RRT обычно имели большую AUC, хотя отношение не было прямым. ADC T(kK183C+K290C)-vc0101, как наблюдали, имел относительно большее RRT (среднее значение 1,77), и поэтому ожидали, что он будет иметь относительно меньшую AUC. Неожиданно, наблюдаемая AUC была относительно высокой, следовательно, не возможно однозначно прогнозировать экспозицию такого ADC на основе данных гидрофобности.
Пример 18: Исследования токсичности
В двух независимых поисковых исследованиях токсичности в общей сложности десять самок и самцов яванских макаков распределяли в 5 групп дозирования (1/пол/доза) и в/в вводили дозы раз в 3 недели (дни исследования 1, 22 и 43). В день исследования 46 (через 3 дня после 3-го введения дозы) животных усыпляли и забирали указанные в протоколе образцы крови и ткани. Клинические наблюдения, клиническую лабораторную диагностику, макроскопические и микроскопические лабораторные исследования проводили прижизненно и при вскрытии. Для исследования анатомической патологии степень выраженности нарушений, обнаруженных в гистопатологических исследованиях, регистрировали на субъективной, относительной, специфической для исследования основе.
В поисковых исследованиях токсичности у яванских макаков при дозе 3 и 5 мг/кг, T-vc0101 вызывал транзиторную, но заметную (390/мкл) или тяжелую (40/мкл до не поддающейся обнаружению) нейтропению в День 11 после первой дозы. В то же время, при дозе 9 мг/кг, ни у одного из яванских макаков, получавших T(kK183C+K290C)-vc0101, не наблюдали или наблюдали минимальную нейтропению, причем количество нейтрофилов значимо превышало 500/мкл в любой момент тестирования (ФИГ. 27). Фактически, получавшие T(kK183C+K290C)-vc0101 животные показали среднее количество нейтрофилов (>1000 в мкл) в день 11 и 14 по сравнению с контролем растворителем.
При микроскопическом исследовании в костном мозге при дозе 3 и 5 мг/кг, яванские макаки, получавшие T-vc0101, имели связанное с лекарственным соединением повышение М/Э отношения. Повышенное миелоидноэритроидное (М/Э) отношение было обусловлено снижением эритроидных клеток-предшественников в сочетании с увеличением первичных зрелых гранулоцитов. В то же время, при дозе 6 и 9 мг/кг только самец, получавший T(kK183C+K290C)-vc0101 в 6 мг/кг/доза, имел минимально или умеренно повышенную насыщенность ткани зрелыми гранулоцитами (данные не показаны).
Таким образом, гемологические и микроскопические данные четко показали, что ADC конъюгат на основе технологии сайт-специфической мутации, T(kK183C+K290C)-vc010, заметно снижал вызванную T-vc010 миелотоксичность и нейтропению.
Пример 19: Кристаллическая структура ADC
Кристаллические структуры были получены для T(K290C+K334C)-vc0101, T(K290C+K392C)-vc0101 и T(K334C+K392C)-vc0101. Эти конкретные ADC конъюгаты были выбраны для кристаллографии, так как конъюгирование с двойными цистеиновыми вариантами K290C+K334C и K334C+K392C, но не с K290C+K392C, устраняло ADCC активность.
Конъюгированные Fc-области подготавливали к кристаллографии путем расщепления ADC конъюгатов папаином. Кристаллы одной и той же морфологии были получены для трех конъюгированных IgG1-Fc областей при использовании одинаковых условий: 100 мМ цитрата Na, pH 5,0,+100 мМ MgCl2+15% ПЭГ-4000.
Структуры человеческого IgG1-Fc дикого типа, депонированные в PDB, являются относительно подобными, что демонстрирует то, что CH2-CH2 домены контактируют друг с другом через Asn297-связанные гликаны (углеводные или гликановые антенны), и что CH3-CH3 домены формируют стабильный интерфейс, который является относительно константным в различных структурах. Структуры Fc существуют в "закрытой" или "открытой" конформации, причем структура дегликозилированных Fc принимает структуру "открытой" конформации, что демонстрирует то, что гликановые антенны удерживают вместе CH2 области. Кроме того, опубликованная структура неконъюгированного Phe241Ala-IgG1 Fc мутанта (Yu et al., "Engineering Hydrophobic Protein-Carbohydrate interactions to fine-tune monoclonal antibodies". JACS 2013) показывает один частично разупорядоченный CH2 домен, поскольку такая мутация приводит к дестабилизации CH2-гликанового интерфейса и CH2-CH2 интерфейса, так как ароматический остаток Phe не может стабилизировать углевод.
"CH2 домен" Fc-области IgG человека (также называемый "Cγ2" доменом) обычно располагается от аминокислоты 231 до приблизительно аминокислоты 340. CH2 домен уникален тем, что он непосредственно не спарен с другим доменом. Напротив, две N-связанных разветвленных углеводных цепи расположены между двумя CH2 доменами интактной нативной молекулы IgG. Предполагали, что углевод способен заменять междоменное спаривание и может способствовать стабилизации CH2 домена (Burton et al., 1985, Molec. Immunol. 22: 161-206).
"CH3 домен" включает фрагмент, расположенный на C-конце относительно CH2 домена в Fc-области (т.е. от аминокислотного остатка 341 до приблизительно аминокислотного остатка 447 в IgG).
Установленные структуры Fc-областей T(K290C+K334C)-vc0101 и T(K290C+K392C)-vc0101 являлись сходными, что демонстрирует то, что Fc-димер содержал один CH2 и оба CH3, которые были высокоупорядоченными (как в Fc дикого типа). Однако они также содержат разупорядоченный CH2 с присоединенным гликаном (ФИГ. 28A и ФИГ. 28B). Более высокую степень дестабилизации одного CH2 домена связывали с непосредственной близостью сайтов конъюгирования с гликановыми антеннами. С учетом геометрии полезной нагрузки 0101, конъюгирование на любом из сайтов K290, K334, K392 может нарушать общую траекторию гликана с его удалением от поверхности CH2, вызывая дестабилизацию гликана и структуры самого CH2, и, как следствие, CH2-CH2 интерфейса (ФИГ. 28C). Более высокая степень гетерогенности доступна для таких 0101 сайт-специфически конъюгированных двойных цистеиновых Fc-вариантов по сравнению с Fc WT, Phe241Ala-Fc или дегликозилированным Fc. Когда положения модифицированных цистеиновых вариантов картировали на структуре Fc WT в комплексе с FcγR IIb типа, было показано, что конъюгирование по C334 могло напрямую препятствовать связыванию с FcγRIIb (ФИГ. 28C). Такая гетерогенность в расположении CH2, вызванная мутацией или конъюгированием, могла приводить к значимому уменьшению связывания с FcRIIb. Поэтому такие результаты позволяют предположить, что гетерогенность конформации или конъюгирование 0101 с некоторыми комбинациями модифицированных цистеинов в IgG1-Fc могли по отдельности или вместе влиять на ADCC активность двойных цистеиновых вариантов содержащих сайт K334C.
Пример 20: Разные сайты конъюгирования приводят к разным свойствам ADC
A. Общая методика синтеза cys-мутантных ADC конъюгатов
Раствор трастузумаба, включающего один или более модифицированных цистеиновых остатков (как показано в Таблице 22), приготавливали в 50 мМ фосфатном буфере, pH 7,4. PBS, ЭДТА (0,5 М сток) и TCEP (0,5 М сток) добавляли таким образом, чтобы конечная концентрация белка составила 10 мг/мл, конечная концентрация ЭДТА составила 20 мМ, и конечная концентрация TCEP составила приблизительно 6,6 мМ (100 молярных экв.). Реакцию оставляли при кт на 48 ч, затем меняли буфер на PBS при использовании колонок GE PD-10 Sephadex G25 согласно инструкции производителя. Полученный раствор обрабатывали приблизительно 50 эквивалентами дегидроаскорбата (50 мМ сток в EtOH/воде 1:1). Антитело оставляли на ночь при 4°C и затем меняли буфер на PBS при использовании колонок GE PD-10 Sephadex G25 согласно инструкции производителя. Варианты вышеуказанной методики с небольшими изменениями применяли для некоторых мутантов.
Антитело, полученное таким образом, разбавляли до ~2,5 мг/мл в PBS, содержащем 10% DMA (об/об), и обрабатывали vc0101 (10 молярных экв.) в виде 10 мМ стокового раствора в DMA. Через 2 ч при кт, смесь подвергали замене буфера на PBS (как указано выше) и очищали с помощью гель-фильтрационной хроматографии на колонке Superdex200. Мономерные фракции концентрировали и стерилизовали фильтрацией с получением конечного ADC. См. Таблицу 22 ниже по поводу показателей продукта.
Таблица 22: Описание свойств ADC
(моль/моль)
B. Общие аналитические методы для примеров конъюгирования
ЖХ-МС: Колонка=Waters BEH300-C4, 2,1×100 мм (P/N=186004496); Прибор=Acquity UPLC с масс-спектрометрическим детектором SQD2; Скорость потока=0,7 мл/мин; Температура=80°C; Буфер A=вода+0,1% муравьиной кислоты; Буфер B=ацетонитрил+0,1% муравьиной кислоты. Градиент: с 3% B до 95% B за 2 минуты, выдержка при 95% B в течение 0,75 мин, затем повторное уравновешивание при 3% B. Образец восстанавливали TCEP или ДТТ непосредственно перед вводом. Элюат контролировали с помощью ЖХ-МС (400-2000 дальтон), и деконволюцию белкового пика выполняли при использовании MaxEnt1. DAR указана как средневесовая нагрузка.
SEC: Колонка: Superdex200 (5/150 GL); Подвижная фаза: Фосфатно-солевой буферный раствор, содержащий 2% ацетонитрила, pH 7,4; Скорость потока=0,25 мл/мин; Температура=окружающая; Прибор: Agilent 1100 HPLC.
HIC: Колонка: TSKGel Butyl NPR, 4,6 мм × 3,5 см (P/N=S0557-835); Буфер A=1,5 М сульфат аммония, содержащий 10 мМ фосфат, pH 7; Буфер B=10 мМ фосфат, pH 7,+20% изопропилового спирта; Скорость потока=0,8 мл/мин; Температура=окружающая; Градиент=0% B до 100% B за 12 минут, выдержка при 100% B в течение 2 минут, затем повторное уравновешивание при 100% A; Прибор: Agilent 1100 HPLC.
C. Определение гидрофобности сайт-специфических vc0101 конъюгатов
ADC конъюгаты Таблицы 22 оценивали с помощью хроматографии гидрофобного взаимодействия (метод выше) для определения относительной гидрофобности различных конъюгатов. Гидрофобность ADC, как было описано, коррелировала с общей экспозицией антитела.
Конъюгаты по сайтам 334, 375 и 392 демонстрировали наименьшее изменение времени удерживания по сравнению с немодифицированным антителом, тогда как конъюгаты по сайтам 421, 443 и 347 показали наибольшее изменение времени удерживания. Относительную гидрофобность каждого ADC вычисляли путем деления времени удерживания ADC на время удерживания немодифицированного антитела, с получением, таким образом, "относительного времени удерживания" или "RRT". RRT ~1 указывает, что ADC обладает приблизительно такой же гидрофобностью, что и немодифицированное антитело. RRT для каждого ADC показано в Таблице 22.
D. Стабильность сайт-специфических vc0101 ADC конъюгатов в плазме
Образцы ADC (~1,5 мг/мл) разводили в мышиной, крысиной или человеческой плазме с получением конечного раствора 50 мкг/мл ADC в плазме. Образцы инкубировали при 37°C в атмосфере 5% CO2, и отбирали аликвоты в трех моментах времени (0, 24 ч и 72 ч). В каждый момент времени образцы ADC после инкубирования с плазмой (25 мкл) дегликозилировали IgG0 при 37°C в течение 1 ч. После дегликозилирования добавляли захватывающее антитело (биотинилированный фрагмент Fcγ IgG1 козы против антител человека, специфичное при 1 мг/мл, в случае плазмы мыши и крысы или биотинилированное антитело против трастузумаба при 1 мг/мл в случае человеческой плазмы) и нагревали смесь при 37°C в течение 1 ч с последующим мягким встряхиванием при комнатной температуре в течение второго часа. Магнитные гранулы Dynabead MyOne Streptavidin T1 добавляли к образцам и инкубировали при комнатной температуре в течение 1 ч с мягким встряхиванием. Затем планшет с образцами промывали 200 мкл PBS+0,05% Tween-20, 200 мкл PBS и водой для ВЭЖХ. Связанный ADC элюировали 55 мкл 2% муравьиной кислоты (МК) (об/об). Аликвоту объемом 50 мкл каждого образца переносили в новый планшет, после чего добавляли еще 5 мкл 200 мМ TCEP.
Анализ интактного белка проводили с помощью масс-спектрометра Xevo G2 Q-TOF, сопряженного с nanoAcquity UPLC (Waters), при использовании колонки BEH300 C4, 1,7 мкм, 0,3×100 мм iKey. Подвижная фаза A (MPA) состояла из 0,1% МК в воде (об/об), и подвижная фаза B (MPB) состояла из 0,1% МК в ацетонитриле (об/об). Хроматографическое разделение выполняли при скорости потока 0,3 мкл/мин с использованием линейного градиента MPB с 5% до 90% за 7 мин. Температуру ЖХ-колонки устанавливали на 85°C. Сбор данных проводили с помощью программы MassLynx версии 4.1. Диапазон регистрации массы составлял от 700 дальтонов до 2400 дальтонов. Анализ данных, включая деконволюцию, проводили при использовании Biopharmalynx версии 1.33.
Присоединение нагрузки и раскрытие сукцинимидного кольца (пик +18 дальтон) контролировали во времени. Данные по присоединению нагрузки представлены как %DAR потеря по сравнению с DAR 0 ч. Данные по раскрытию кольца представлены как % молекул с раскрытым кольцом в сравнении с общим количеством молекул, присутствующих через 72 ч. Несколько сайт-мутантов давали очень стабильные ADC конъюгаты (334C, 421C и 443C), тогда как некоторые сайты теряли значительное количество линкера-полезной нагрузки (38°C и 114C). Скорость раскрытия кольца значительно отличалась в зависимости от сайта. Несколько сайтов, таких как 392C, 183C и 334C, приводили к очень небольшому раскрытию кольца, тогда как другие сайты, такие как 421C, 388C и 347C, приводили к быстрому и спонтанному раскрытию кольца.
Сайты, которые приводят к быстрому и спонтанному раскрытию кольца, могут быть полезными для получения конъюгатов, которые обладают уменьшенной гидрофобностью и/или увеличенной ФК экспозицией. Такое обнаружение находится в противоречии с преобладающим пониманием того, что стабильность кольца коррелирует со стабильностью в плазме. В некоторых аспектах, таким образом, конъюгирование по одному или более сайтам 421C, 388C и 347C может обеспечивать особое преимущество при использовании линкера-полезной нагрузки с высокой гидрофобностью. В некоторых аспектах высокая гидрофобность является значением относительного времени удерживания (RRT) (согласно измерению с помощью HIC) 1,5 или больше. В некоторых аспектах высокая гидрофобность является значением RRT 1,7 или больше. В некоторых аспектах высокая гидрофобность является значением RRT 1,8 или больше. В некоторых аспектах высокая гидрофобность является значением RRT 1,9 или больше. В некоторых аспектах высокая гидрофобность является значением RRT 2,0 или больше.
Таблица 23: Стабильность различных ADC конъюгатов в плазме
E. Стабильность сайт-специфических vc0101 конъюгатов в глутатионе
Образцы ADC разводили в водном глутатионе с получением конечной концентрации GSH 0,5 мМ и конечной концентрации белка ~0,1 мг/мл в фосфатном буфере, pH 7,4. Затем образцы инкубировали при 37°C и отбирали аликвоты в трех моментах времени для определения DAR (T-0, T-3 день, T-6 день). Аликвоту каждого момента времени обрабатывали TCEP и анализировали с помощью ЖХ-МС согласно методу, описанному в Примере 20 A.
Присоединение нагрузки и раскрытие сукцинимидного кольца (пик +18 дальтон) контролировали во времени. Данные по присоединению нагрузки представлены как %DAR потеря по сравнению с DAR 0 ч (Таблица 24). Данные по раскрытию кольца представлены как % молекул с раскрытым кольцом в сравнении с общим количеством молекул, присутствующих через 72 ч. Несколько сайт-мутантов давали очень стабильные ADC конъюгаты (334C, 421C и 443C), тогда как некоторые сайты теряли значительное количество линкера-полезной нагрузки (38°C и 114C). Скорость раскрытия кольца значительно отличалась в зависимости от сайта. Несколько сайтов, таких как 392C, 183C и 334C, приводили к очень небольшому раскрытию кольца, тогда как другие сайты, такие как 421C, 388C и 347C, приводили к быстрому и спонтанному раскрытию кольца. Результаты данного анализа достаточно хорошо коррелируют с результатами стабильности в плазме (Пример 20 D), что позволяет предположить, что тиол-опосредованное деконъюгирование является основным путем потери полезной нагрузки в плазме. В совокупности эти результаты указывают, что определенные сайты, такие как 334, 443, 290 и 392, могут быть особенно полезными для конъюгирования линкеров-полезных нагрузок, которые могут быть легко потеряны при тиол-опосредованном деконъюгировании. Такие линкеры-полезные нагрузки включают линкеры-полезные нагрузки, в которых используются обычные mc и vc связи (например, vc-101, vc-MMAE, mc-MMAF и т.д.).
Таблица 24: Стабильность различных vc0101 сайт-специфических конъюгатов в глутатионе
F. Фармакокинетическая оценка отобранных сайт-специфических vc0101 конъюгатов на мышах
Не несущие опухоли самки бестимусных Nu/Nu (голых) мышей (возрастом 6-8 недель) были получены из Charles River Laboratories. Все процедуры с использованием мышей были одобрены Институциональным комитетом по уходу за животными и их использованию согласно установленным рекомендациям. Мышам (n=3 или 4) вводили однократную внутривенную дозу ADC 3 мг/кг в расчете на иммуноглобулиновый компонент. Образцы крови забирали у каждой мыши из хвостовой вены через 0,083, 6, 24, 48, 96, 168 и 336 часов после введения дозы. Суммарные концентрации антител (Tab) и концентрации ADC определяли с помощью LBA, где антитело овцы против IgG человека использовали для захвата, антитело козы против IgG человека использовали для детектирования Tab или антитело против полезной нагрузки использовали для детектирования ADC. Данные по концентрации в плазе у каждого животного анализировали при использовании Watson LIMS версии 7.4 (Thermo). Экспозиция изменилась в зависимости от сайта. ADC конъюгаты, полученные из 29°C и 443C мутантов, показали наименьшую экспозицию, тогда как ADC конъюгаты, полученные с сайтами 183C и 392C, показали наибольшую экспозицию. Для многих применений могут быть предпочтительны сайты с высокой экспозицией, поскольку это приведет к увеличенной продолжительности действия терапевтического средства. Однако в случае некоторых применений, может быть предпочтительным использование конъюгата с более низкой экспозицией (такого как 29°C и 443C). В частности, применения, в которых требуется более низкая экспозиция (т.е. ниже ФК), могут включать, без ограничения, применение в головном мозге, центральной нервной системе и глазах. Показания включают рак, в особенности головного мозга, центральной нервной системы и/или глаза.
Таблица 25: ФК экспозиция различных сайт-специфических vc0101 ADC конъюгатов
G. Расщепление сайт-специфических vc0101 конъюгатов катепсином
Катепсин B активировали при использовании 6 мМ дитиотреитола (ДТТ) в 150 мМ ацетате натрия, pH 5,2, в течение 15 минут при 37°C. Затем 50 нг активированного катепсина-B смешивали с 20 мкл 1 мг/мл ADC при конечной концентрации 2 мМ ДТТ, 50 мМ ацетата натрия, pH 5,2. Реакции останавливали при использовании 10 мкМ ингибитора цистеиновых протеаз E-64 в 250 мМ боратном буфере, pH 8,5, с последующим инкубирование при 37°C в течение 20 мин, 1 ч, 2 ч и 4 ч. После анализа образцы восстанавливали при использовании TCEP и анализировали с помощью ЖХ/МС при использовании условий, описанных в Примере 21 A. Данные показали, что скорость расщепления линкера сильно зависит от сайта конъюгирования. Определенные сайты расщепляются очень быстро, например, 443C, 388C и 290C, тогда как другие сайты расщепляются очень медленно, например, 334C, 375C и 392C. В некоторых аспектах может быть предпочтительно проводить конъюгирование на сайтах, которые обеспечивают замедление расщепления. В других аспектах предпочтительно быстрое расщепление. Например, может быть предпочтительным быстрое высвобождение полезной нагрузки, чтобы сократить время, проведенное в эндосоме. В других аспектах быстрое расщепление полезной нагрузки может с преимуществом обеспечивать проникновение полезной нагрузки, когда конъюгированная молекула может не позволить сделать это, например, в некоторых солидных опухолях. В других аспектах быстрое расщепление может обеспечивать доставку полезной нагрузки к соседним клеткам, которые не экспрессируют антиген, распознаваемый антителом, что дает возможность, например, проводить терапию гетерогенной опухоли.
Таблица 26: Кинетика расщепления линкеров в различных сайт-специфических vc0101 ADC конъюгатах
линкера за 20 мин
линкера за 1 ч
линкера за 2 ч
линкера за 4 ч
H. Термическая стабильность сайт-специфических vc0101 конъюгатов
ADC разводили до 0,2 мг/мл в PBS (pH 7,4), содержащем 10 мМ ЭДТА. ADC конъюгаты помещали в изолированный сосуд и нагревали до 45°C. Аликвоту (10 мкл) отбирали с интервалом в 1 неделю для оценки уровня высокомолекулярных соединений (HMWS) и низкомолекулярных соединений (LMWS), которые образовывались с течением времени, с помощью гель-фильтрационной хроматографии (SEC). Условия SEC представлены в Примере 21 A. При таких условиях мономер элюировался приблизительно через 3,6 минуты. Любой белковый материал, элюирующийся слева от пика мономера, считали HMWS, и любой белковый материал, элюирующийся справа от пика мономера, считали LMWS. Результаты показаны в Таблице 27 ниже. Отдельные ADC конъюгаты показали превосходную термическую стабильность, например, 183C, 375C и 334C, тогда как другие ADC конъюгаты показали значимое разложение, например, 443C и 392C+443C.
Таблица 27: Термическая стабильность различных сайт-специфических vc0101 ADC конъюгатов
I. Эффективность различных vc0101 сайт-мутантов
Исследования эффективности in vivo конъюгатов антитела-лекарственного средства проводили в мишень-экспрессирующей ксенотрансплантатной модели с использованием клеточной линии N87. Приблизительно 7,5 миллионов опухолевых клеток в 50% матригеле подкожно имплантировали голым мышам возрастом 6-8 недель, пока размер опухоли не достигал 250-350 мм3. Лекарственное средство вводили путем болюсной инъекции в хвостовую вену. Животным вводили 10, 3 или 1 мг/кг конъюгата антитела-лекарственного средства в общей сложности четыре раза, один раз в 4 дня (в дни 1, 5, 9 и 13). Всех подопытных животных еженедельно контролировали на изменения массы тела. Объем опухоли измеряли два раза в неделю в течение первых 50 дней, а затем один раз в неделю, с помощью измерительного устройства и вычисляли согласно следующей формуле: Объем опухоли=(длина×ширина2)/2. Животных гуманно умерщвляли, до того как объемы их опухоли достигали 2500 мм3. Размер опухоли, как обычно наблюдали, уменьшался после первой недели лечения. Животных непрерывно контролировали на возобновление роста опухоли после прекращения лечения (до 100 дней после лечения). Данные группы, получавшей дозу 3 мг/кг, показаны на ФИГ. 29. ADC конъюгаты, полученные из 388C и 347C мутантов, показали немного более низкую активность, чем ADC конъюгаты из 334C, 183C, 392C и 443C мутантов.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Pfizer Inc.
Ma, Dangshe
<120> САЙТ-СПЕЦИФИЧЕСКИЕ КОНЪЮГАТЫ АНТИТЕЛА К HER2 И ЛЕКАРСТВЕННОГО СРЕДСТВА
<130> PC72091
<150> US 62/260,854
<151> 2015-11-30
<150> US 62/289,744
<151> 2016-02-01
<150> US 62/409,105
<151> 2016-10-17
<150> US 62/289,727
<151> 2016-02-01
<160> 95
<170> PatentIn version 3.5
<210> 1
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 1
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 2
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 2
Asp Thr Tyr Ile His
1 5
<210> 3
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 3
Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 4
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 4
Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr
1 5 10
<210> 5
<211> 329
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 5
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly
325
<210> 6
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 6
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly
<210> 7
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 7
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 8
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 8
Arg Ala Ser Gln Asp Val Asn Thr Ala Val Ala
1 5 10
<210> 9
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 9
Ser Ala Ser Phe Leu Tyr Ser
1 5
<210> 10
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 10
Gln Gln His Tyr Thr Thr Pro Pro Thr
1 5
<210> 11
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 11
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 12
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 12
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 13
<211> 330
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 13
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Arg Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 14
<211> 450
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 14
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Arg Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 15
<211> 329
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 15
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Cys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly
325
<210> 16
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 16
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Cys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly
<210> 17
<211> 329
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 17
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Cys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly
325
<210> 18
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 18
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Cys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly
<210> 19
<211> 330
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 19
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 20
<211> 450
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 20
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Ala Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 21
<211> 330
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 21
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Gln Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 22
<211> 450
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 22
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gln Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 23
<211> 329
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 23
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Cys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly
325
<210> 24
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 24
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Cys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly
<210> 25
<211> 329
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 25
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Cys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly
325
<210> 26
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 26
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Cys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly
<210> 27
<211> 329
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 27
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Cys Ser Pro Gly
325
<210> 28
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 28
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Cys Ser Pro
435 440 445
Gly
<210> 29
<211> 329
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 29
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Cys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Cys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly
325
<210> 30
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 30
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Cys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Cys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly
<210> 31
<211> 329
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 31
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Cys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Cys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly
325
<210> 32
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 32
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Cys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Cys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly
<210> 33
<211> 330
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 33
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Arg Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 34
<211> 450
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 34
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Arg Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Ala Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 35
<211> 330
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 35
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Arg Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Gln Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 36
<211> 450
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 36
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Arg Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gln Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 37
<211> 329
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 37
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Cys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Cys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly
325
<210> 38
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 38
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Cys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Cys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly
<210> 39
<211> 329
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 39
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Cys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Cys Ser Pro Gly
325
<210> 40
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 40
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Cys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Cys Ser Pro
435 440 445
Gly
<210> 41
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 41
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Cys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 42
<211> 213
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 42
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Cys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 43
<211> 115
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 43
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Leu Leu Gln
100 105 110
Gly Pro Pro
115
<210> 44
<211> 222
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 44
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys Gly Gly Leu Leu Gln Gly Pro Pro
210 215 220
<210> 45
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 45
gaggtgcagc tggtggaatc cggcggaggc ctggtccagc ctggcggatc tctgcggctg 60
tcttgcgccg cctccggctt caacatcaag gacacctaca tccactgggt ccgacaggca 120
cctggcaagg gactggaatg ggtggcccgg atctacccca ccaacggcta caccagatac 180
gccgactccg tgaagggccg gttcaccatc tccgccgaca cctccaagaa caccgcctac 240
ctgcagatga actccctgcg ggccgaggac accgccgtgt actactgctc cagatgggga 300
ggcgacggct tctacgccat ggactactgg ggccagggca ccctggtcac cgtgtctagc 360
<210> 46
<211> 1347
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 46
gaggtgcagc tggtggaatc cggcggaggc ctggtccagc ctggcggatc tctgcggctg 60
tcttgcgccg cctccggctt caacatcaag gacacctaca tccactgggt ccgacaggca 120
cctggcaagg gactggaatg ggtggcccgg atctacccca ccaacggcta caccagatac 180
gccgactccg tgaagggccg gttcaccatc tccgccgaca cctccaagaa caccgcctac 240
ctgcagatga actccctgcg ggccgaggac accgccgtgt actactgctc cagatgggga 300
ggcgacggct tctacgccat ggactactgg ggccagggca ccctggtcac cgtgtctagc 360
gcgtcgacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 420
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 480
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 540
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 600
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 660
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 720
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 780
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 840
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 900
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 960
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 1020
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 1080
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1140
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 1200
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 1260
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1320
cagaagagcc tctccctgtc cccgggt 1347
<210> 47
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 47
gacatccaga tgacccagtc cccctccagc ctgtccgcct ctgtgggcga cagagtgacc 60
atcacctgtc gggcctccca ggacgtgaac accgccgtgg cctggtatca gcagaagccc 120
ggcaaggccc ccaagctgct gatctactcc gcctccttcc tgtactccgg cgtgccctcc 180
cggttctccg gctccagatc tggcaccgac tttaccctga ccatctccag cctgcagccc 240
gaggacttcg ccacctacta ctgccagcag cactacacca ccccccccac ctttggccag 300
ggcaccaagg tggaaatcaa g 321
<210> 48
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 48
gacatccaga tgacccagtc cccctccagc ctgtccgcct ctgtgggcga cagagtgacc 60
atcacctgtc gggcctccca ggacgtgaac accgccgtgg cctggtatca gcagaagccc 120
ggcaaggccc ccaagctgct gatctactcc gcctccttcc tgtactccgg cgtgccctcc 180
cggttctccg gctccagatc tggcaccgac tttaccctga ccatctccag cctgcagccc 240
gaggacttcg ccacctacta ctgccagcag cactacacca ccccccccac ctttggccag 300
ggcaccaagg tggaaatcaa gcggaccgtg gccgctccct ccgtgttcat cttcccaccc 360
tccgacgagc agctgaagtc cggcaccgcc tccgtcgtgt gcctgctgaa caacttctac 420
ccccgcgagg ccaaggtgca gtggaaggtg gacaacgccc tgcagtccgg caactcccag 480
gaatccgtca ccgagcagga ctccaaggac agcacctact ccctgtcctc caccctgacc 540
ctgtccaagg ccgactacga gaagcacaag gtgtacgcct gcgaagtgac ccaccagggc 600
ctgtccagcc ccgtgaccaa gtccttcaac cggggcgagt gc 642
<210> 49
<211> 990
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 49
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtagtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg accgtactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 720
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggaaaa 990
<210> 50
<211> 1350
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 50
gaggtgcagc tggtggagtc cggcggcggc ctggttcagc ccggcggatc actgaggctc 60
tcctgtgccg ccagcggctt caacatcaag gacacataca tccactgggt tcgccaggct 120
cctggcaagg gactggagtg ggtcgctagg atctacccca ccaatgggta caccaggtac 180
gccgactccg tgaaggggcg gttcacaatc tcagccgata ctagcaaaaa tacagcctac 240
ttgcagatga actccctgag agcagaggat accgccgtgt actattgctc tcgctggggc 300
ggcgacggct tctacgctat ggattattgg ggccagggaa ccttggtcac cgtctcctca 360
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 420
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 480
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 540
ggactctact ccctcagcag cgtagtgacc gtgccctcca gcagcttggg cacccagacc 600
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 660
aaatcttgtg accgtactca cacatgccca ccgtgcccag cacctgaact cctgggggga 720
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 780
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 840
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 900
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 960
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 1020
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 1080
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1140
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 1200
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 1260
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1320
cagaagagcc tctccctgtc tccgggaaaa 1350
<210> 51
<211> 987
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 51
gcgtcgacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccatgcccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 720
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc cccgggt 987
<210> 52
<211> 1347
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 52
gaggtgcagc tggtggaatc cggcggaggc ctggtccagc ctggcggatc tctgcggctg 60
tcttgcgccg cctccggctt caacatcaag gacacctaca tccactgggt ccgacaggca 120
cctggcaagg gactggaatg ggtggcccgg atctacccca ccaacggcta caccagatac 180
gccgactccg tgaagggccg gttcaccatc tccgccgaca cctccaagaa caccgcctac 240
ctgcagatga actccctgcg ggccgaggac accgccgtgt actactgctc cagatgggga 300
ggcgacggct tctacgccat ggactactgg ggccagggca ccctggtcac cgtgtctagc 360
gcgtcgacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 420
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 480
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 540
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 600
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 660
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 720
ccgtcagtct tcctcttccc cccatgcccc aaggacaccc tcatgatctc ccggacccct 780
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 840
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 900
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 960
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 1020
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 1080
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1140
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 1200
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 1260
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1320
cagaagagcc tctccctgtc cccgggt 1347
<210> 53
<211> 987
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 53
gcgtcgacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacatgcc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 720
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc cccgggt 987
<210> 54
<211> 1347
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 54
gaggtgcagc tggtggaatc cggcggaggc ctggtccagc ctggcggatc tctgcggctg 60
tcttgcgccg cctccggctt caacatcaag gacacctaca tccactgggt ccgacaggca 120
cctggcaagg gactggaatg ggtggcccgg atctacccca ccaacggcta caccagatac 180
gccgactccg tgaagggccg gttcaccatc tccgccgaca cctccaagaa caccgcctac 240
ctgcagatga actccctgcg ggccgaggac accgccgtgt actactgctc cagatgggga 300
ggcgacggct tctacgccat ggactactgg ggccagggca ccctggtcac cgtgtctagc 360
gcgtcgacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 420
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 480
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 540
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 600
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 660
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 720
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 780
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 840
tacgtggacg gcgtggaggt gcataatgcc aagacatgcc cgcgggagga gcagtacaac 900
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 960
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 1020
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 1080
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1140
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 1200
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 1260
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1320
cagaagagcc tctccctgtc cccgggt 1347
<210> 55
<211> 990
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 55
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtagtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacgcc 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 720
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggaaaa 990
<210> 56
<211> 1350
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 56
gaggtgcagc tggtggagtc cggcggcggc ctggttcagc ccggcggatc actgaggctc 60
tcctgtgccg ccagcggctt caacatcaag gacacataca tccactgggt tcgccaggct 120
cctggcaagg gactggagtg ggtcgctagg atctacccca ccaatgggta caccaggtac 180
gccgactccg tgaaggggcg gttcacaatc tcagccgata ctagcaaaaa tacagcctac 240
ttgcagatga actccctgag agcagaggat accgccgtgt actattgctc tcgctggggc 300
ggcgacggct tctacgctat ggattattgg ggccagggaa ccttggtcac cgtctcctca 360
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 420
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 480
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 540
ggactctact ccctcagcag cgtagtgacc gtgccctcca gcagcttggg cacccagacc 600
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 660
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 720
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 780
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 840
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacgcc 900
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 960
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 1020
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 1080
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1140
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 1200
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 1260
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1320
cagaagagcc tctccctgtc tccgggaaaa 1350
<210> 57
<211> 1055
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 57
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtagtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtaccaa 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 720
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggaaaa gccgccagcg gcttcaacat caaggacaca 1020
tacatccact gggttcgcca ggctcctggc aaggg 1055
<210> 58
<211> 1415
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 58
gaggtgcagc tggtggagtc cggcggcggc ctggttcagc ccggcggatc actgaggctc 60
tcctgtgccg ccagcggctt caacatcaag gacacataca tccactgggt tcgccaggct 120
cctggcaagg gactggagtg ggtcgctagg atctacccca ccaatgggta caccaggtac 180
gccgactccg tgaaggggcg gttcacaatc tcagccgata ctagcaaaaa tacagcctac 240
ttgcagatga actccctgag agcagaggat accgccgtgt actattgctc tcgctggggc 300
ggcgacggct tctacgctat ggattattgg ggccagggaa ccttggtcac cgtctcctca 360
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 420
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 480
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 540
ggactctact ccctcagcag cgtagtgacc gtgccctcca gcagcttggg cacccagacc 600
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 660
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 720
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 780
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 840
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtaccaa 900
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 960
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 1020
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 1080
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1140
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 1200
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 1260
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1320
cagaagagcc tctccctgtc tccgggaaaa gccgccagcg gcttcaacat caaggacaca 1380
tacatccact gggttcgcca ggctcctggc aaggg 1415
<210> 59
<211> 987
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 59
gcgtcgacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagtg caccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 720
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc cccgggt 987
<210> 60
<211> 1347
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 60
gaggtgcagc tggtggaatc cggcggaggc ctggtccagc ctggcggatc tctgcggctg 60
tcttgcgccg cctccggctt caacatcaag gacacctaca tccactgggt ccgacaggca 120
cctggcaagg gactggaatg ggtggcccgg atctacccca ccaacggcta caccagatac 180
gccgactccg tgaagggccg gttcaccatc tccgccgaca cctccaagaa caccgcctac 240
ctgcagatga actccctgcg ggccgaggac accgccgtgt actactgctc cagatgggga 300
ggcgacggct tctacgccat ggactactgg ggccagggca ccctggtcac cgtgtctagc 360
gcgtcgacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 420
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 480
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 540
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 600
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 660
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 720
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 780
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 840
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 900
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 960
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagtg caccatctcc 1020
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 1080
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1140
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 1200
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 1260
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1320
cagaagagcc tctccctgtc cccgggt 1347
<210> 61
<211> 987
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 61
gcgtcgacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 720
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact actgcaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc cccgggt 987
<210> 62
<211> 1347
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 62
gaggtgcagc tggtggaatc cggcggaggc ctggtccagc ctggcggatc tctgcggctg 60
tcttgcgccg cctccggctt caacatcaag gacacctaca tccactgggt ccgacaggca 120
cctggcaagg gactggaatg ggtggcccgg atctacccca ccaacggcta caccagatac 180
gccgactccg tgaagggccg gttcaccatc tccgccgaca cctccaagaa caccgcctac 240
ctgcagatga actccctgcg ggccgaggac accgccgtgt actactgctc cagatgggga 300
ggcgacggct tctacgccat ggactactgg ggccagggca ccttggtcac cgtgtctagc 360
gcgtcgacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 420
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 480
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 540
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 600
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 660
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 720
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 780
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 840
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 900
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 960
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 1020
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 1080
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1140
gccgtggagt gggagagcaa tgggcagccg gagaacaact actgcaccac gcctcccgtg 1200
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 1260
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1320
cagaagagcc tctccctgtc cccgggt 1347
<210> 63
<211> 987
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 63
gcgtcgacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 720
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctcctgctc cccgggt 987
<210> 64
<211> 1347
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 64
gaggtgcagc tggtggaatc cggcggaggc ctggtccagc ctggcggatc tctgcggctg 60
tcttgcgccg cctccggctt caacatcaag gacacctaca tccactgggt ccgacaggca 120
cctggcaagg gactggaatg ggtggcccgg atctacccca ccaacggcta caccagatac 180
gccgactccg tgaagggccg gttcaccatc tccgccgaca cctccaagaa caccgcctac 240
ctgcagatga actccctgcg ggccgaggac accgccgtgt actactgctc cagatgggga 300
ggcgacggct tctacgccat ggactactgg ggccagggca ccctggtcac cgtgtctagc 360
gcgtcgacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 420
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 480
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 540
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 600
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 660
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 720
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 780
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 840
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 900
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 960
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 1020
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 1080
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1140
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 1200
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 1260
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1320
cagaagagcc tctcctgctc cccgggt 1347
<210> 65
<211> 987
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 65
gcgtcgacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacatgcc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagtg caccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 720
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc cccgggt 987
<210> 66
<211> 1347
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 66
gaggtgcagc tggtggaatc cggcggaggc ctggtccagc ctggcggatc tctgcggctg 60
tcttgcgccg cctccggctt caacatcaag gacacctaca tccactgggt ccgacaggca 120
cctggcaagg gactggaatg ggtggcccgg atctacccca ccaacggcta caccagatac 180
gccgactccg tgaagggccg gttcaccatc tccgccgaca cctccaagaa caccgcctac 240
ctgcagatga actccctgcg ggccgaggac accgccgtgt actactgctc cagatgggga 300
ggcgacggct tctacgccat ggactactgg ggccagggca ccctggtcac cgtgtctagc 360
gcgtcgacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 420
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 480
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 540
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 600
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 660
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 720
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 780
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 840
tacgtggacg gcgtggaggt gcataatgcc aagacatgcc cgcgggagga gcagtacaac 900
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 960
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagtg caccatctcc 1020
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 1080
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1140
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 1200
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 1260
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1320
cagaagagcc tctccctgtc cccgggt 1347
<210> 67
<211> 987
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 67
gcgtcgacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacatgcc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 720
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact actgcaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc cccgggt 987
<210> 68
<211> 1347
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 68
gaggtgcagc tggtggaatc cggcggaggc ctggtccagc ctggcggatc tctgcggctg 60
tcttgcgccg cctccggctt caacatcaag gacacctaca tccactgggt ccgacaggca 120
cctggcaagg gactggaatg ggtggcccgg atctacccca ccaacggcta caccagatac 180
gccgactccg tgaagggccg gttcaccatc tccgccgaca cctccaagaa caccgcctac 240
ctgcagatga actccctgcg ggccgaggac accgccgtgt actactgctc cagatgggga 300
ggcgacggct tctacgccat ggactactgg ggccagggca ccctggtcac cgtgtctagc 360
gcgtcgacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 420
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 480
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 540
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 600
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 660
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 720
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 780
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 840
tacgtggacg gcgtggaggt gcataatgcc aagacatgcc cgcgggagga gcagtacaac 900
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 960
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 1020
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 1080
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1140
gccgtggagt gggagagcaa tgggcagccg gagaacaact actgcaccac gcctcccgtg 1200
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 1260
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1320
cagaagagcc tctccctgtc cccgggt 1347
<210> 69
<211> 990
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 69
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtagtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg accgtactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacgcc 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 720
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggaaaa 990
<210> 70
<211> 1350
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 70
gaggtgcagc tggtggagtc cggcggcggc ctggttcagc ccggcggatc actgaggctc 60
tcctgtgccg ccagcggctt caacatcaag gacacataca tccactgggt tcgccaggct 120
cctggcaagg gactggagtg ggtcgctagg atctacccca ccaatgggta caccaggtac 180
gccgactccg tgaaggggcg gttcacaatc tcagccgata ctagcaaaaa tacagcctac 240
ttgcagatga actccctgag agcagaggat accgccgtgt actattgctc tcgctggggc 300
ggcgacggct tctacgctat ggattattgg ggccagggaa ccttggtcac cgtctcctca 360
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 420
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 480
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 540
ggactctact ccctcagcag cgtagtgacc gtgccctcca gcagcttggg cacccagacc 600
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 660
aaatcttgtg accgtactca cacatgccca ccgtgcccag cacctgaact cctgggggga 720
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 780
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 840
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacgcc 900
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 960
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 1020
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 1080
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1140
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 1200
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 1260
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1320
cagaagagcc tctccctgtc tccgggaaaa 1350
<210> 71
<211> 1055
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 71
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtagtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg accgtactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtaccaa 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 720
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggaaaa gccgccagcg gcttcaacat caaggacaca 1020
tacatccact gggttcgcca ggctcctggc aaggg 1055
<210> 72
<211> 1415
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 72
gaggtgcagc tggtggagtc cggcggcggc ctggttcagc ccggcggatc actgaggctc 60
tcctgtgccg ccagcggctt caacatcaag gacacataca tccactgggt tcgccaggct 120
cctggcaagg gactggagtg ggtcgctagg atctacccca ccaatgggta caccaggtac 180
gccgactccg tgaaggggcg gttcacaatc tcagccgata ctagcaaaaa tacagcctac 240
ttgcagatga actccctgag agcagaggat accgccgtgt actattgctc tcgctggggc 300
ggcgacggct tctacgctat ggattattgg ggccagggaa ccttggtcac cgtctcctca 360
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 420
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 480
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 540
ggactctact ccctcagcag cgtagtgacc gtgccctcca gcagcttggg cacccagacc 600
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 660
aaatcttgtg accgtactca cacatgccca ccgtgcccag cacctgaact cctgggggga 720
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 780
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 840
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtaccaa 900
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 960
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 1020
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 1080
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1140
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 1200
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 1260
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1320
cagaagagcc tctccctgtc tccgggaaaa gccgccagcg gcttcaacat caaggacaca 1380
tacatccact gggttcgcca ggctcctggc aaggg 1415
<210> 73
<211> 987
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 73
gcgtcgacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagtg caccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 720
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact actgcaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc cccgggt 987
<210> 74
<211> 1347
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 74
gaggtgcagc tggtggaatc cggcggaggc ctggtccagc ctggcggatc tctgcggctg 60
tcttgcgccg cctccggctt caacatcaag gacacctaca tccactgggt ccgacaggca 120
cctggcaagg gactggaatg ggtggcccgg atctacccca ccaacggcta caccagatac 180
gccgactccg tgaagggccg gttcaccatc tccgccgaca cctccaagaa caccgcctac 240
ctgcagatga actccctgcg ggccgaggac accgccgtgt actactgctc cagatgggga 300
ggcgacggct tctacgccat ggactactgg ggccagggca ccctggtcac cgtgtctagc 360
gcgtcgacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 420
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 480
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 540
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 600
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 660
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 720
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 780
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 840
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 900
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 960
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagtg caccatctcc 1020
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 1080
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1140
gccgtggagt gggagagcaa tgggcagccg gagaacaact actgcaccac gcctcccgtg 1200
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 1260
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1320
cagaagagcc tctccctgtc cccgggt 1347
<210> 75
<211> 987
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 75
gcgtcgacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 720
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact actgcaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctcctgctc cccgggt 987
<210> 76
<211> 1347
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 76
gaggtgcagc tggtggaatc cggcggaggc ctggtccagc ctggcggatc tctgcggctg 60
tcttgcgccg cctccggctt caacatcaag gacacctaca tccactgggt ccgacaggca 120
cctggcaagg gactggaatg ggtggcccgg atctacccca ccaacggcta caccagatac 180
gccgactccg tgaagggccg gttcaccatc tccgccgaca cctccaagaa caccgcctac 240
ctgcagatga actccctgcg ggccgaggac accgccgtgt actactgctc cagatgggga 300
ggcgacggct tctacgccat ggactactgg ggccagggca ccctggtcac cgtgtctagc 360
gcgtcgacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 420
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 480
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 540
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 600
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 660
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 720
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 780
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 840
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 900
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 960
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 1020
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 1080
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1140
gccgtggagt gggagagcaa tgggcagccg gagaacaact actgcaccac gcctcccgtg 1200
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 1260
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1320
cagaagagcc tctcctgctc cccgggt 1347
<210> 77
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 77
cggaccgtgg ccgctccctc cgtgttcatc ttcccaccct ccgacgagca gctgaagtcc 60
ggcaccgcct ccgtcgtgtg cctgctgaac aacttctacc cccgcgaggc caaggtgcag 120
tggaaggtgg acaacgccct gcagtccggc aactcccagg aatccgtcac cgagcaggac 180
tccaaggaca gcacctactc cctgtcctcc accctgaccc tgtcctgcgc cgactacgag 240
aagcacaagg tgtacgcctg cgaagtgacc caccagggcc tgtccagccc cgtgaccaag 300
tccttcaacc ggggcgagtg c 321
<210> 78
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 78
gacatccaga tgacccagtc cccctccagc ctgtccgcct ctgtgggcga cagagtgacc 60
atcacctgtc gggcctccca ggacgtgaac accgccgtgg cctggtatca gcagaagccc 120
ggcaaggccc ccaagctgct gatctactcc gcctccttcc tgtactccgg cgtgccctcc 180
cggttctccg gctccagatc tggcaccgac tttaccctga ccatctccag cctgcagccc 240
gaggacttcg ccacctacta ctgccagcag cactacacca ccccccccac ctttggccag 300
ggcaccaagg tggaaatcaa gcggaccgtg gccgctccct ccgtgttcat cttcccaccc 360
tccgacgagc agctgaagtc cggcaccgcc tccgtcgtgt gcctgctgaa caacttctac 420
ccccgcgagg ccaaggtgca gtggaaggtg gacaacgccc tgcagtccgg caactcccag 480
gaatccgtca ccgagcagga ctccaaggac agcacctact ccctgtcctc caccctgacc 540
ctgtcctgcg ccgactacga gaagcacaag gtgtacgcct gcgaagtgac ccaccagggc 600
ctgtccagcc ccgtgaccaa gtccttcaac cggggcgagt gc 642
<210> 79
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 79
cgtacggtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagctcgcc cgtcacaaag 300
agcttcaaca ggggagagtg t 321
<210> 80
<211> 666
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеотидная последовательность
<400> 80
gatatccaga tgacacagtc cccctccagc ctctccgcta gtgtcggaga tagagtgaca 60
attacatgtc gggcaagcca ggacgtcaat accgccgtgg cctggtatca gcagaagcca 120
ggaaaggccc caaaactcct gatctactcc gcctccttcc tgtactcagg ggtcccttca 180
cgcttctccg gttcccggag cggcaccgac ttcactctga ctatctcaag cttgcagccc 240
gaggacttcg ccacatacta ttgccagcag cactatacca ccccccctac cttcggtcag 300
ggaactaagg tggaaattaa acgtacggtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gtggtggcct gcttcagggc 660
ccacca 666
<210> 81
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 81
Gly Gly Leu Leu Gln Gly Pro Pro
1 5
<210> 82
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 82
Gly Gly Leu Leu Gln Gly Gly
1 5
<210> 83
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 83
Leu Leu Gln Gly Ala
1 5
<210> 84
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 84
Gly Gly Leu Leu Gln Gly Ala
1 5
<210> 85
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 85
Leu Leu Gln Gly
1
<210> 86
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 86
Leu Leu Gln Gly Pro Gly
1 5
<210> 87
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 87
Leu Leu Gln Gly Pro Ala
1 5
<210> 88
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 88
Leu Leu Gln Gly Pro
1 5
<210> 89
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 89
Leu Leu Gln Pro
1
<210> 90
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 90
Leu Leu Gln Pro Gly Lys
1 5
<210> 91
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 91
Leu Leu Gln Gly Ala Pro Gly Lys
1 5
<210> 92
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 92
Leu Leu Gln Gly Ala Pro Gly
1 5
<210> 93
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<400> 93
Leu Leu Gln Gly Ala Pro
1 5
<210> 94
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<220>
<221> ПРОЧИЕ_ПРИЗНАКИ
<222> (4)..(4)
<223> Xaa является G или P
<220>
<221> ПРОЧИЕ_ПРИЗНАКИ
<222> (5)..(5)
<223> Xaa является A, G, P или отсутствует
<220>
<221> ПРОЧИЕ_ПРИЗНАКИ
<222> (6)..(6)
<223> Xaa является A, G, K, P или отсутствует
<220>
<221> ПРОЧИЕ_ПРИЗНАКИ
<222> (7)..(7)
<223> Xaa является G, K или отсутствует
<220>
<221> ПРОЧИЕ_ПРИЗНАКИ
<222> (8)..(8)
<223> Xaa является K или отсутствует
<400> 94
Leu Leu Gln Xaa Xaa Xaa Xaa Xaa
1 5
<210> 95
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность синтетического пептида
<220>
<221> ПРОЧИЕ_ПРИЗНАКИ
<222> (4)..(4)
<223> Xaa является любой природной аминокислотой
<220>
<221> ПРОЧИЕ_ПРИЗНАКИ
<222> (5)..(5)
<223> Xaa является любой природной аминокислотой или отсутствует
<220>
<221> ПРОЧИЕ_ПРИЗНАКИ
<222> (6)..(6)
<223> Xaa является любой природной аминокислотой или отсутствует
<220>
<221> ПРОЧИЕ_ПРИЗНАКИ
<222> (7)..(7)
<223> Xaa является любой природной аминокислотой или отсутствует
<220>
<221> ПРОЧИЕ_ПРИЗНАКИ
<222> (8)..(8)
<223> Xaa является любой природной аминокислотой или отсутствует
<400> 95
Leu Leu Gln Xaa Xaa Xaa Xaa Xaa
1 5
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА И ФРАГМЕНТЫ АНТИТЕЛ ДЛЯ САЙТ-СПЕЦИФИЧЕСКОГО КОНЪЮГИРОВАНИЯ | 2016 |
|
RU2757815C2 |
| АНТИТЕЛА ПРОТИВ EDB И КОНЪЮГАТЫ АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО | 2017 |
|
RU2758632C2 |
| КОНЪЮГАТЫ АНТИ-HER2 БИПАРАТОПНЫХ АНТИТЕЛ-ЛЕКАРСТВЕННЫХ ВЕЩЕСТВ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2806049C2 |
| КОНЪЮГАТЫ АНТИТЕЛА И ЛЕКАРСТВЕННОГО СРЕДСТВА, СОДЕРЖАЩИЕ МОДУЛЯТОР СПЛАЙСИНГА НА ОСНОВЕ ГЕРБОКСИДИЕНА, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2820607C2 |
| АМАТОКСИНОВЫЕ КОНЪЮГАТЫ АНТИТЕЛА С ЛЕКАРСТВЕННЫМ СРЕДСТВОМ И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2826004C2 |
| КОНЪЮГАТЫ СВЯЗУЮЩЕГО И АКТИВНОГО ВЕЩЕСТВА (ADC), ИМЕЮЩИЕ ФЕРМЕНТАТИВНО РАСЩЕПЛЯЕМЫЕ ГРУППЫ | 2017 |
|
RU2761390C2 |
| КОНЪЮГАТ АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО, ПРОМЕЖУТОЧНОЕ СОЕДИНЕНИЕ ДЛЯ ЕГО ПОЛУЧЕНИЯ, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2800137C1 |
| БИСПЕЦИФИЧЕСКИЕ АНТИГЕНСВЯЗЫВАЮЩИЕ КОНСТРУКЦИИ ПРОТИВ HER2 | 2014 |
|
RU2737882C2 |
| САЙТ-СПЕЦИФИЧЕСКАЯ КОНЪЮГАЦИЯ АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО ПОСРЕДСТВОМ ГЛИКОИНЖЕНЕРИИ | 2014 |
|
RU2831409C2 |
| СПОСОБ ПОЛУЧЕНИЯ КОНЪЮГАТА АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО НА ОСНОВЕ ЭРИБУЛИНА | 2020 |
|
RU2840013C1 |
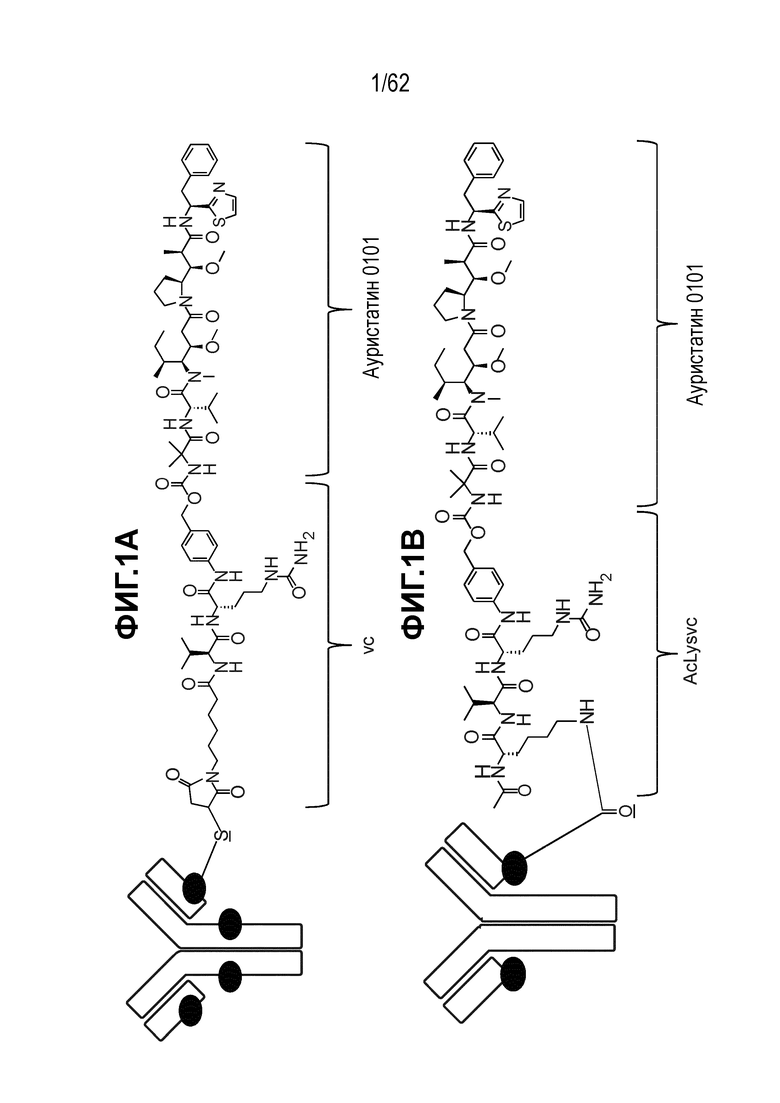
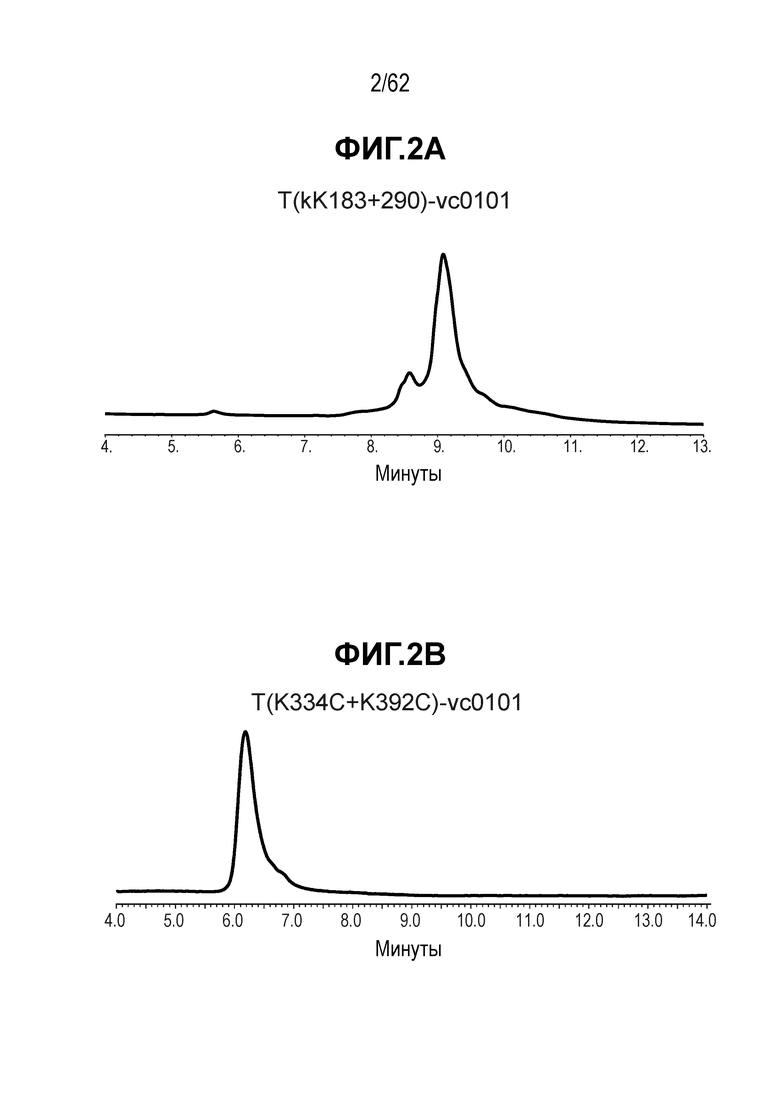
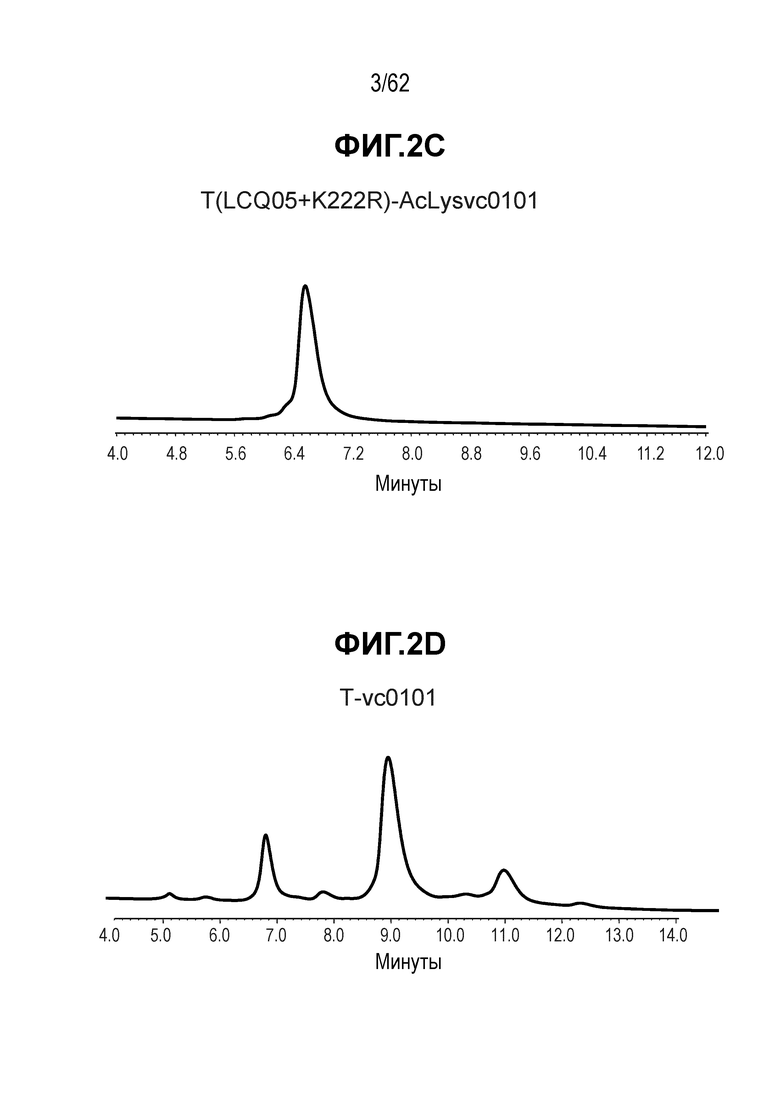
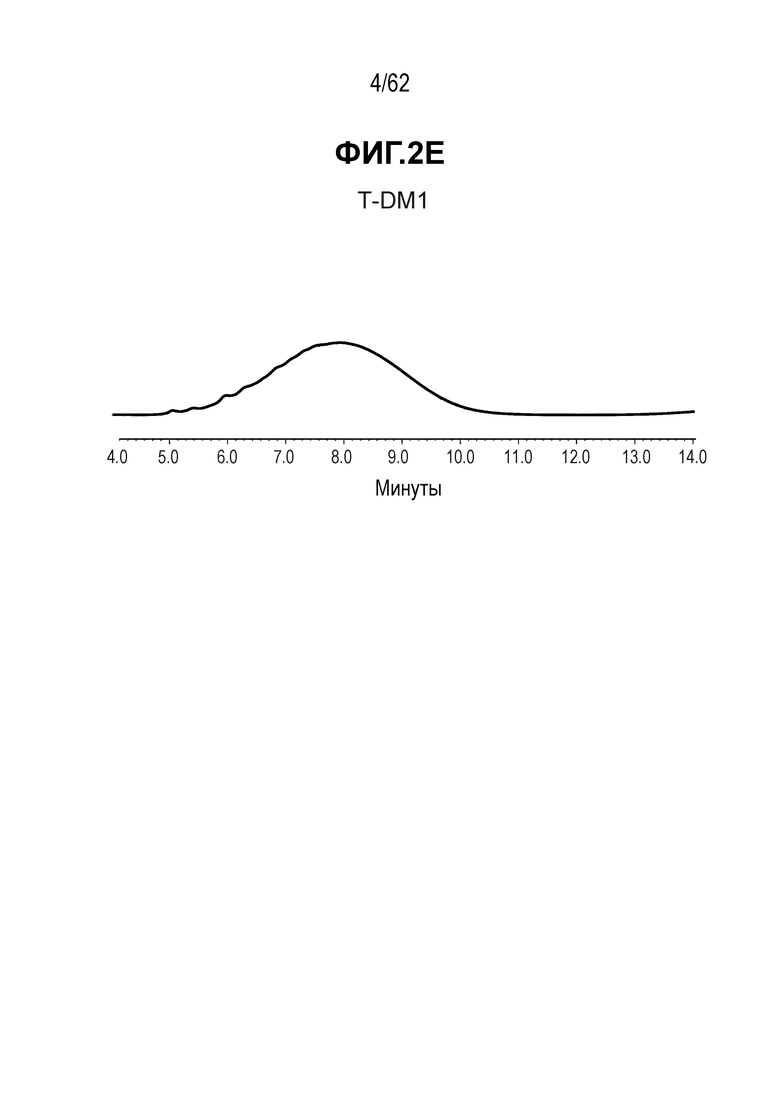
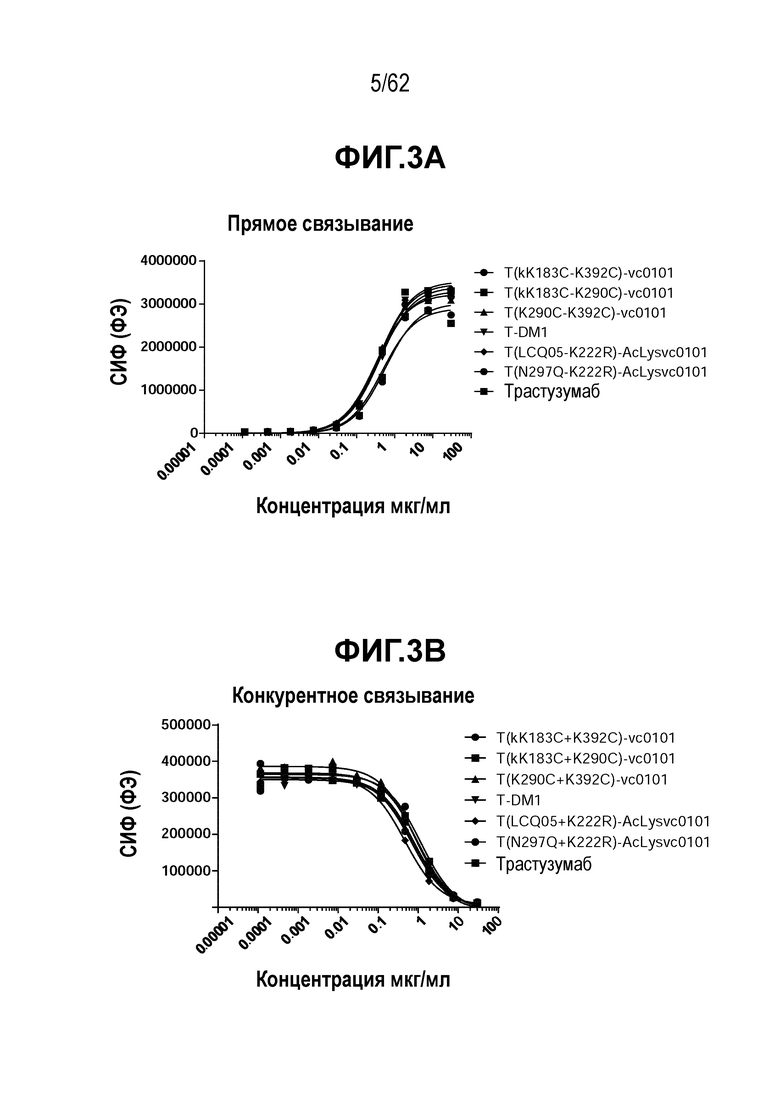
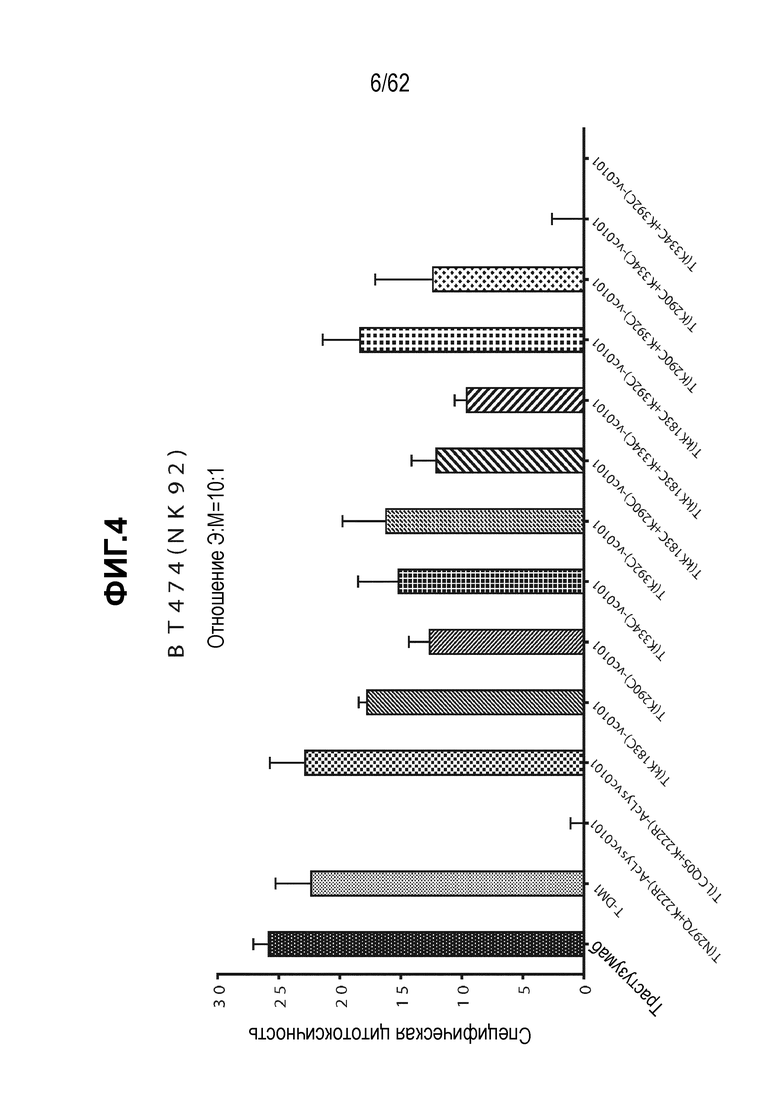

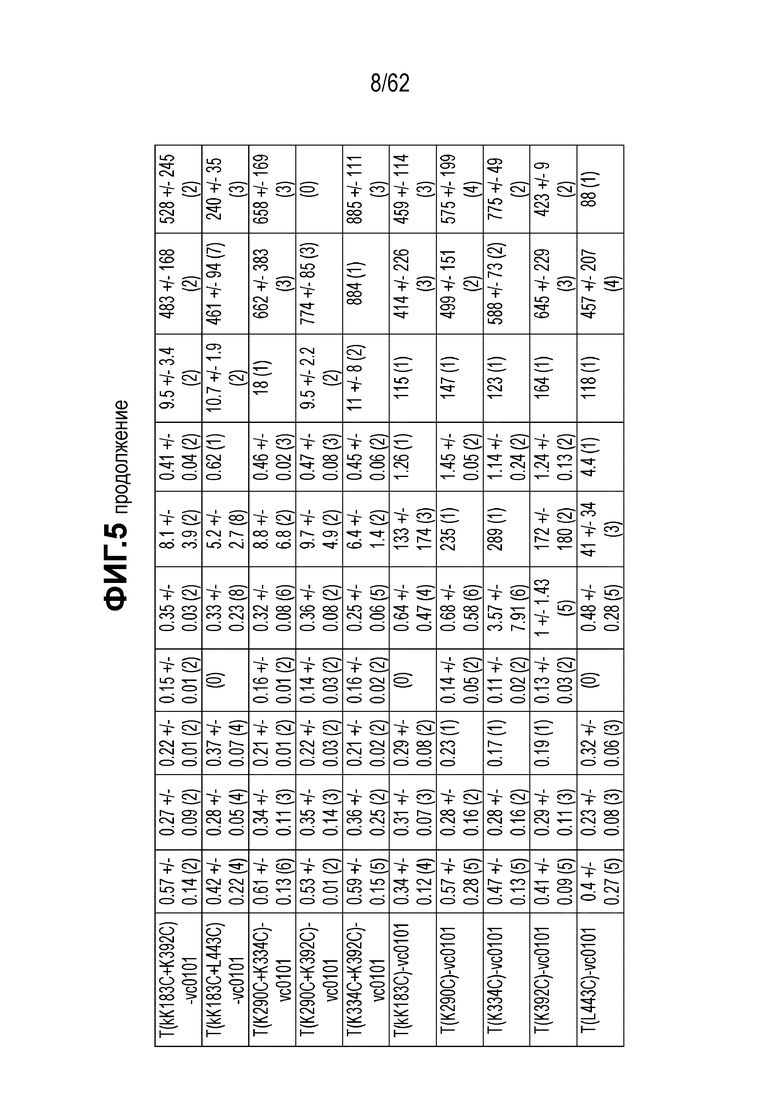
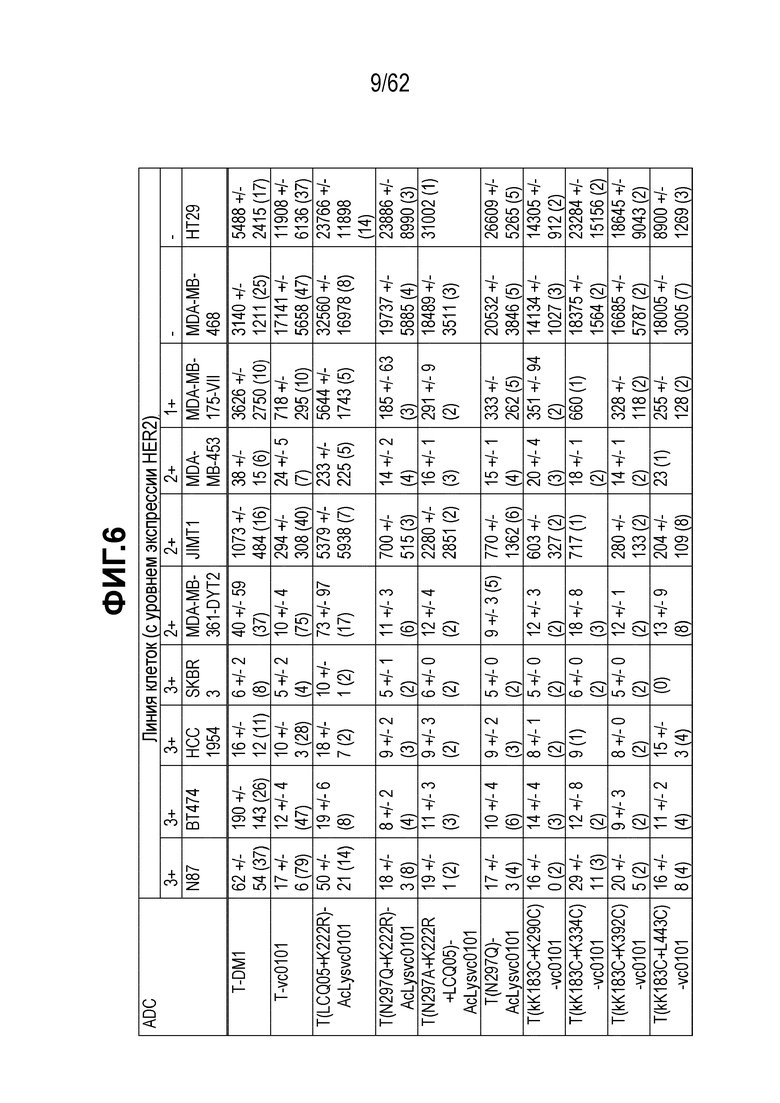

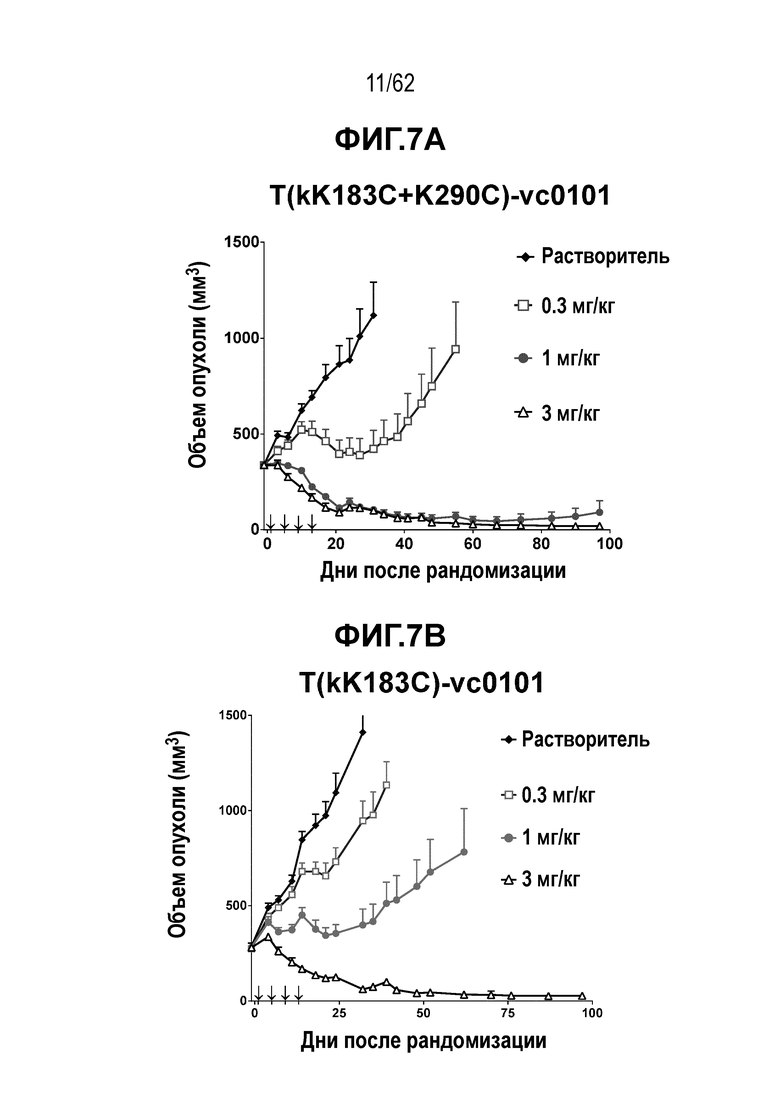
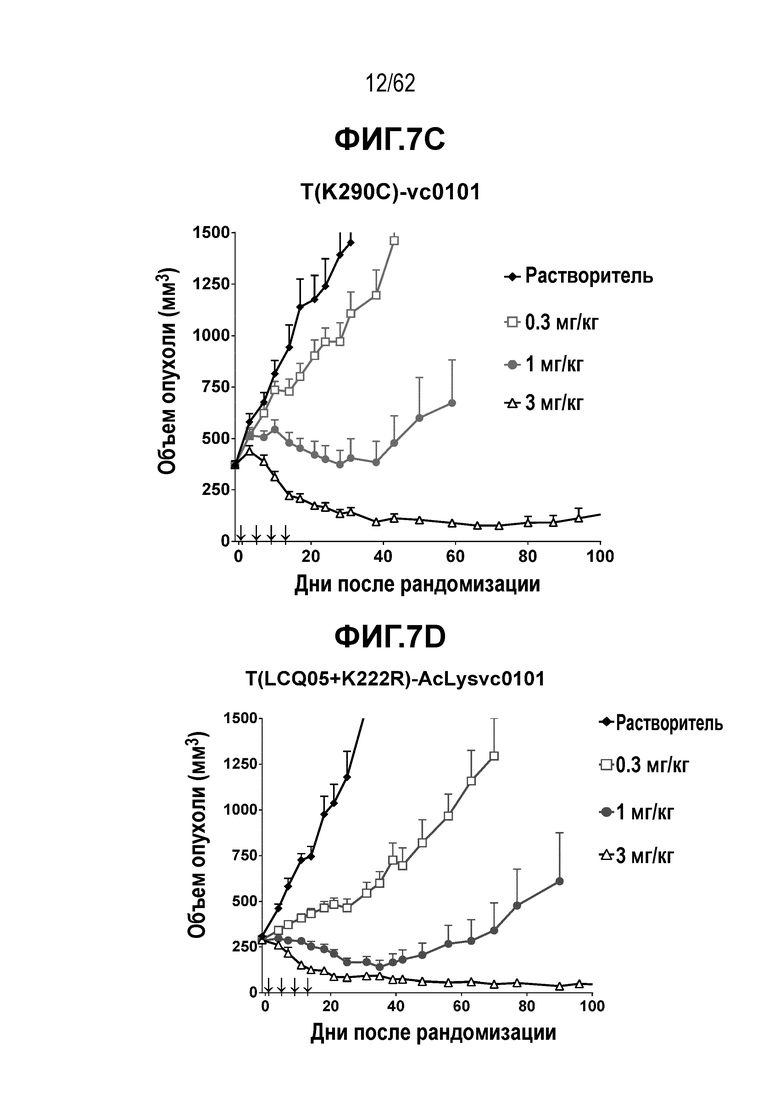
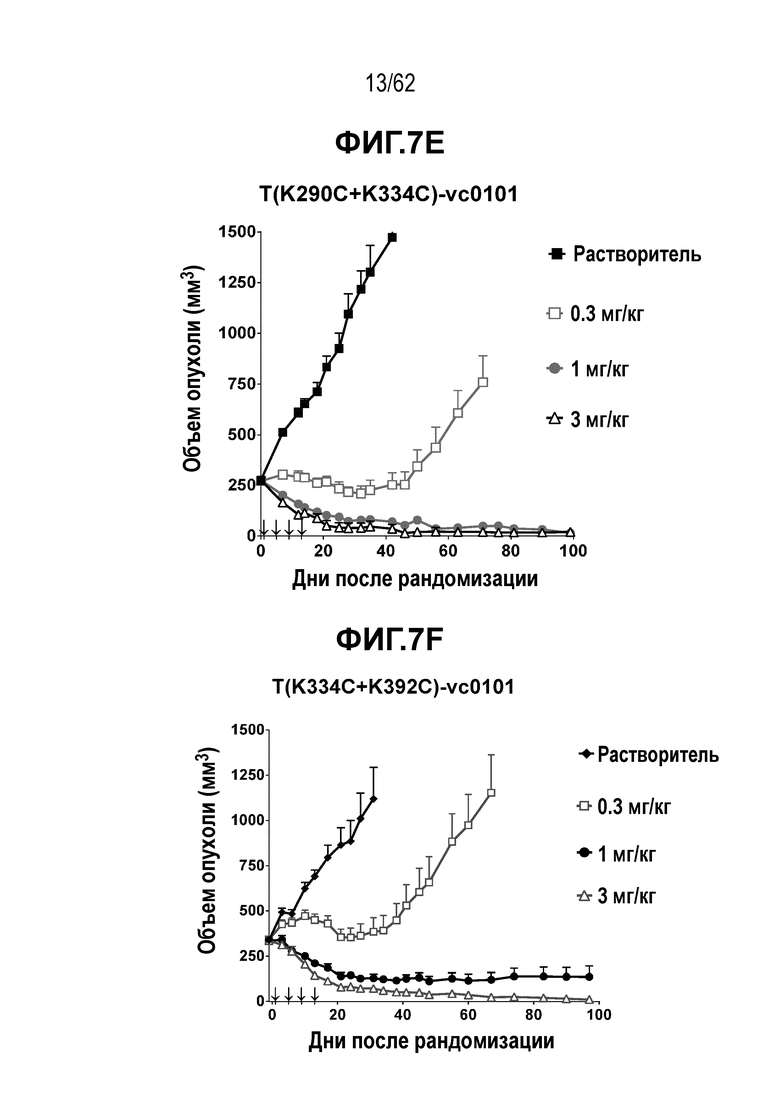
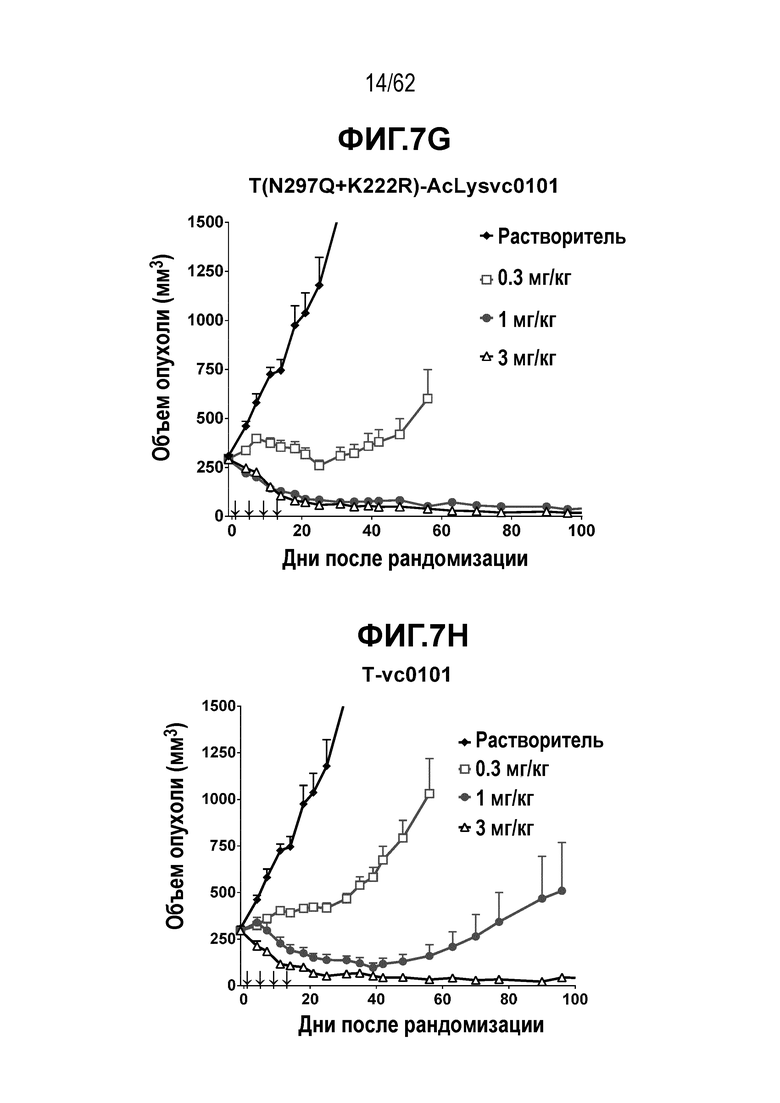
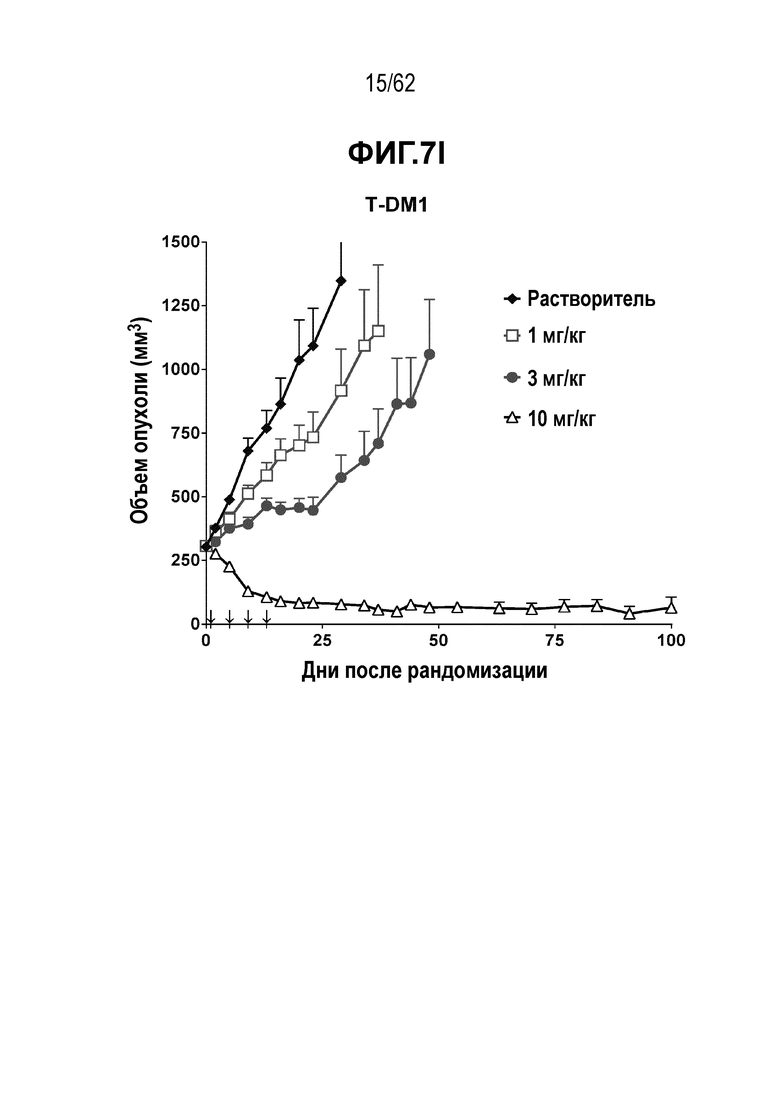
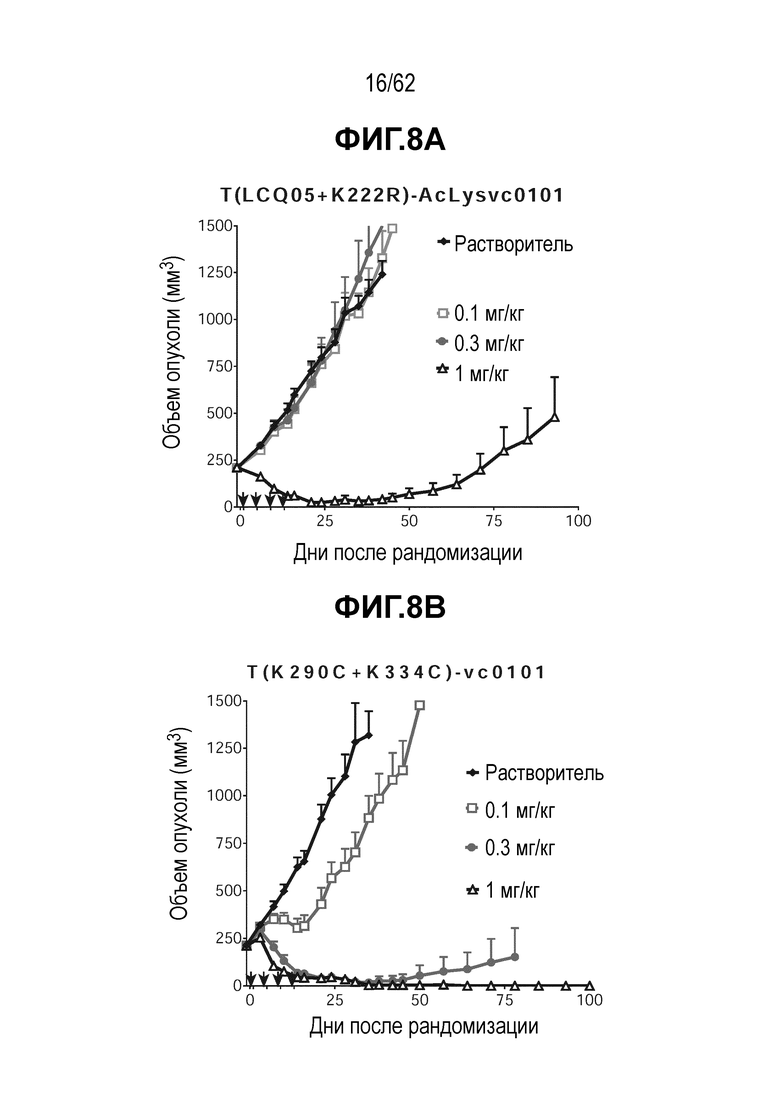
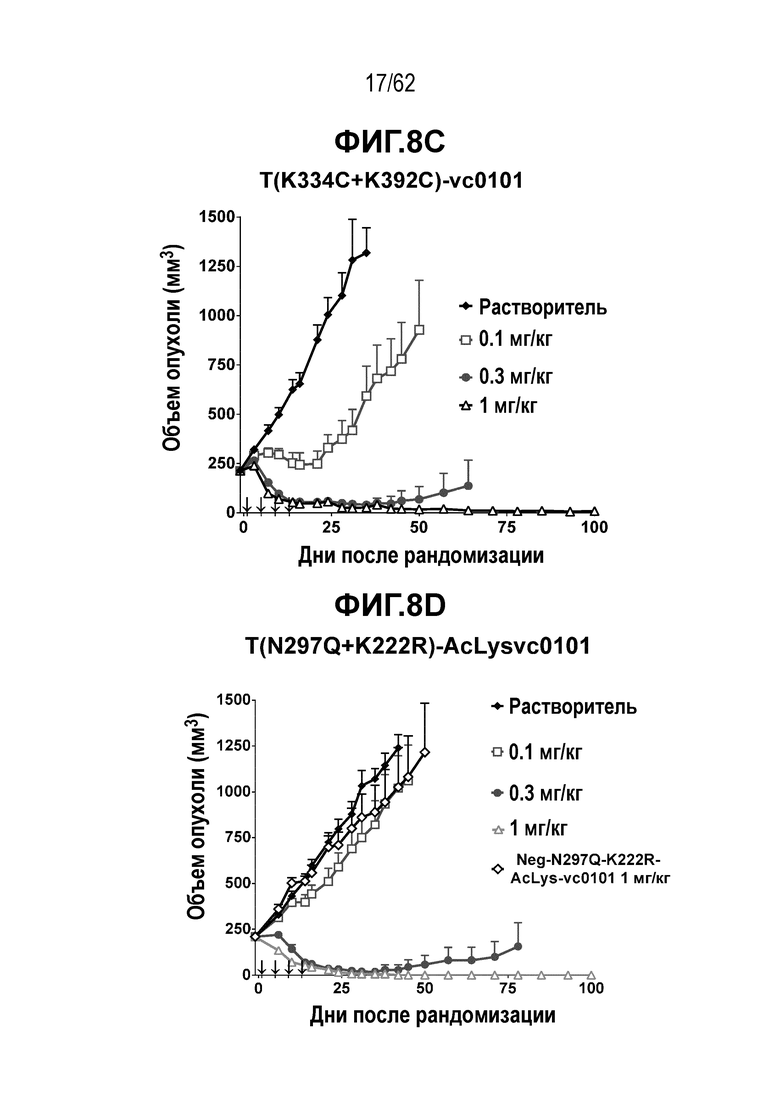
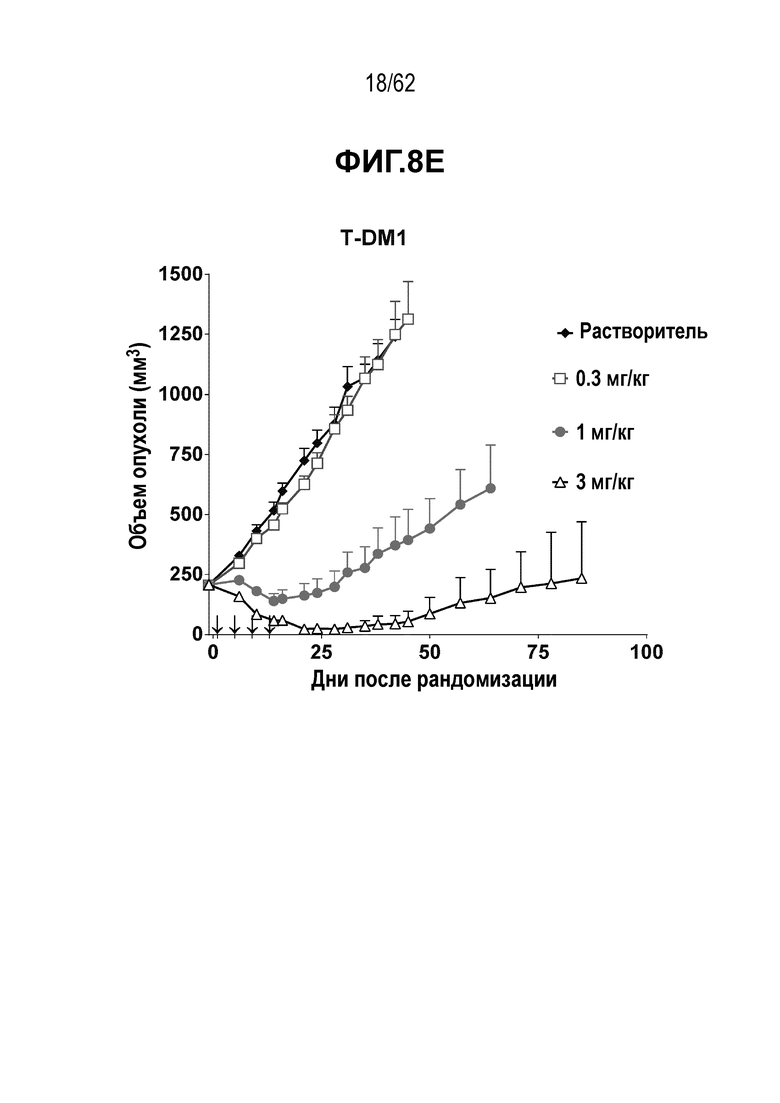
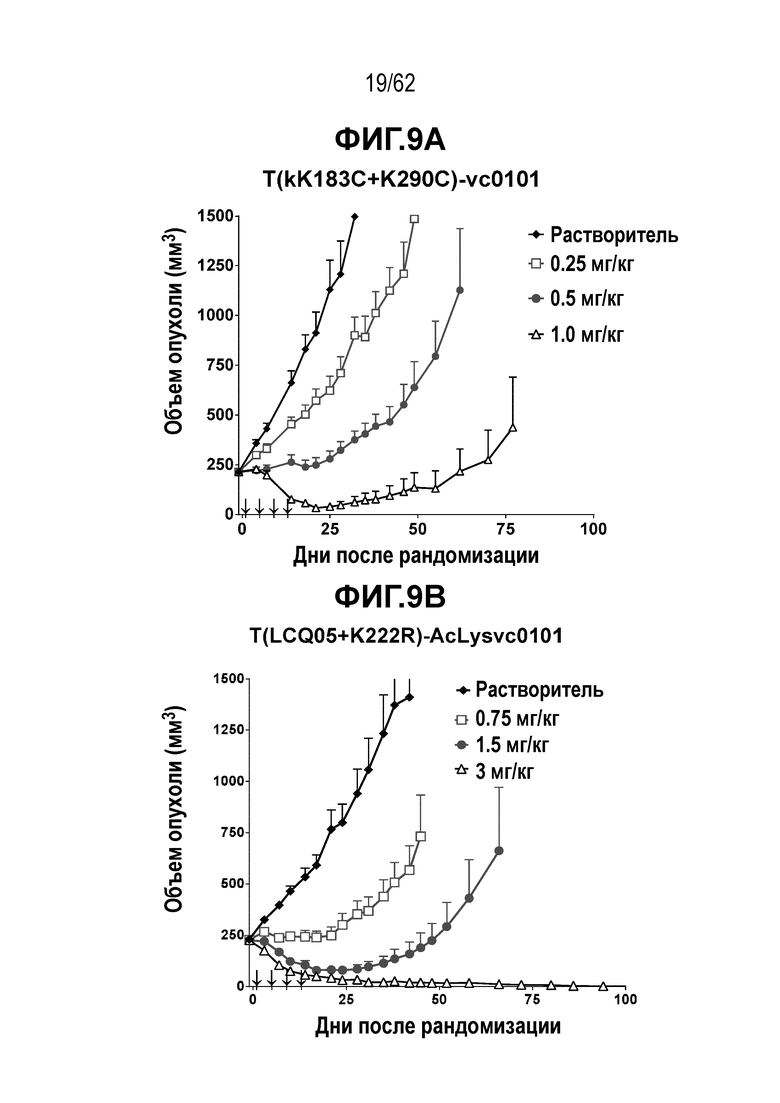
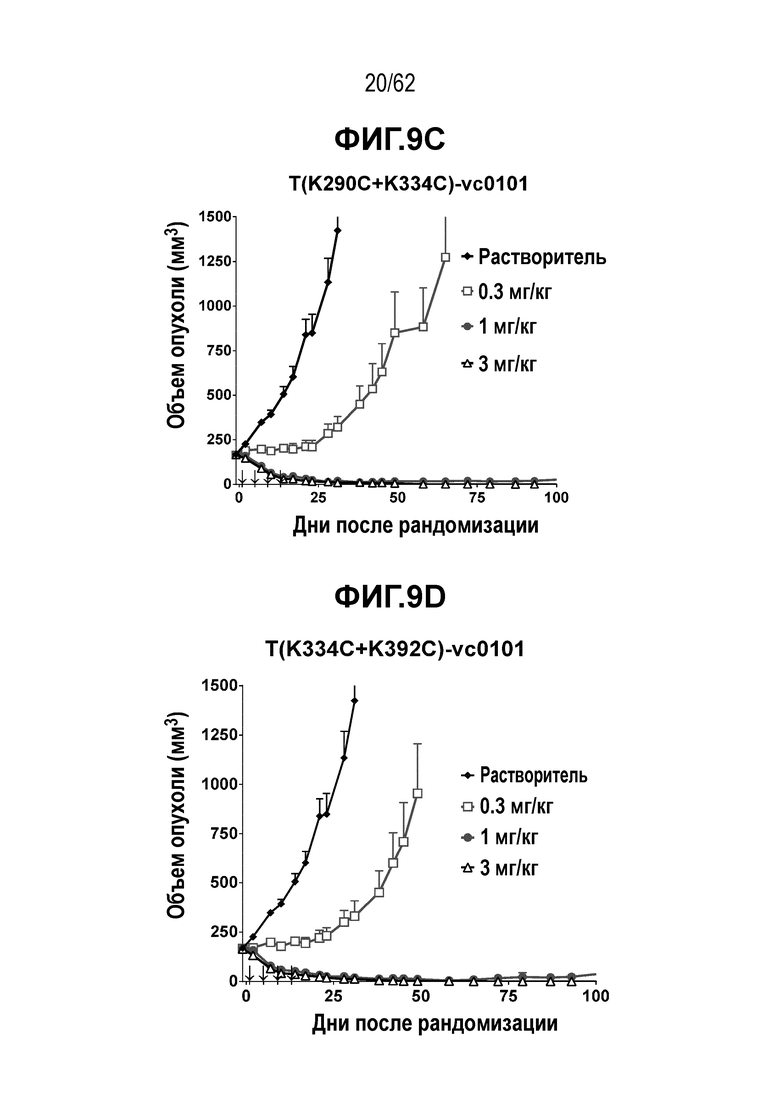
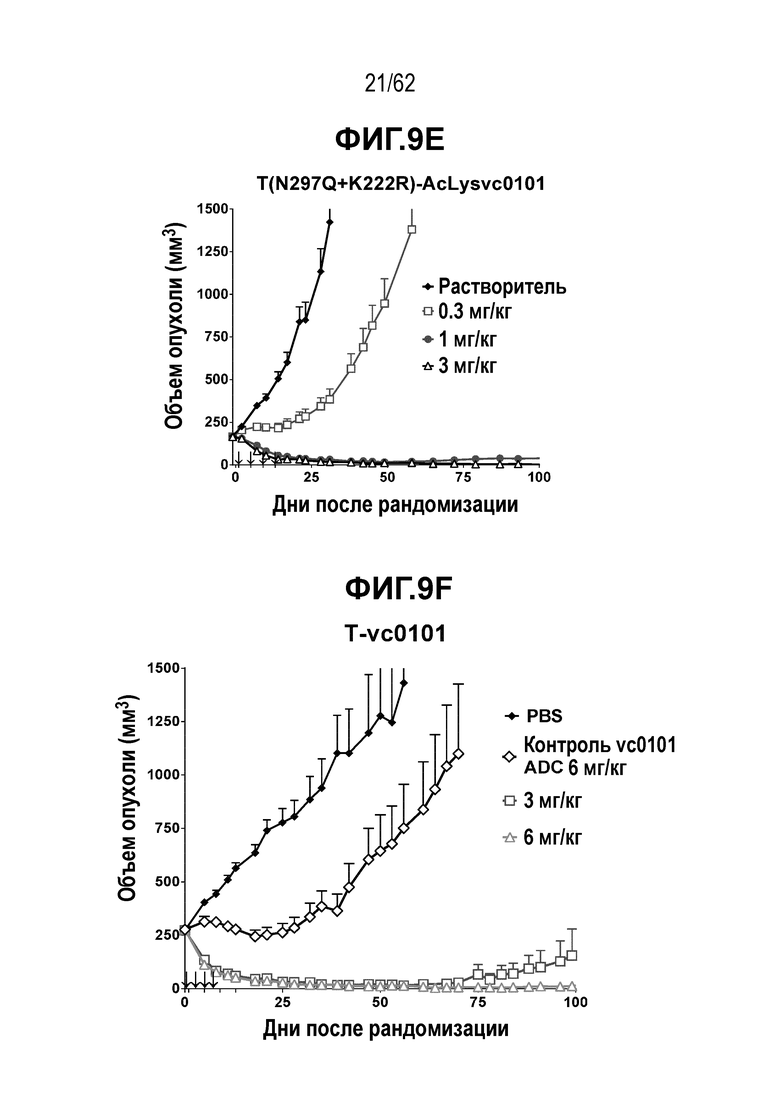
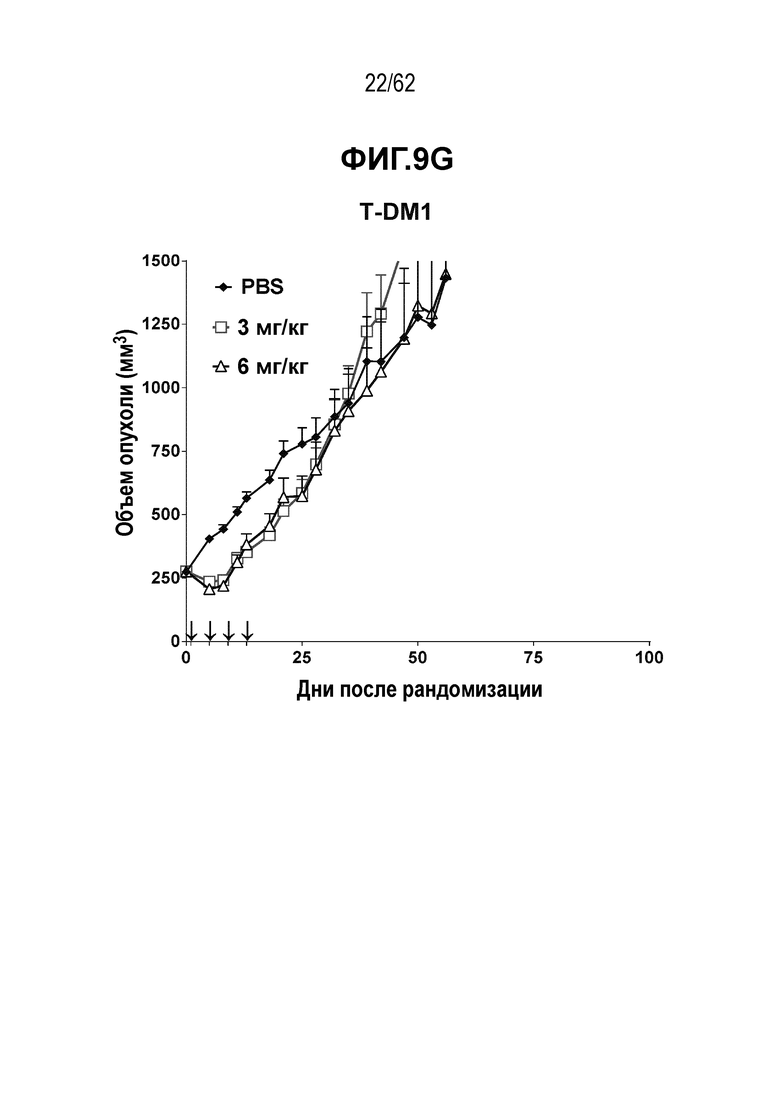
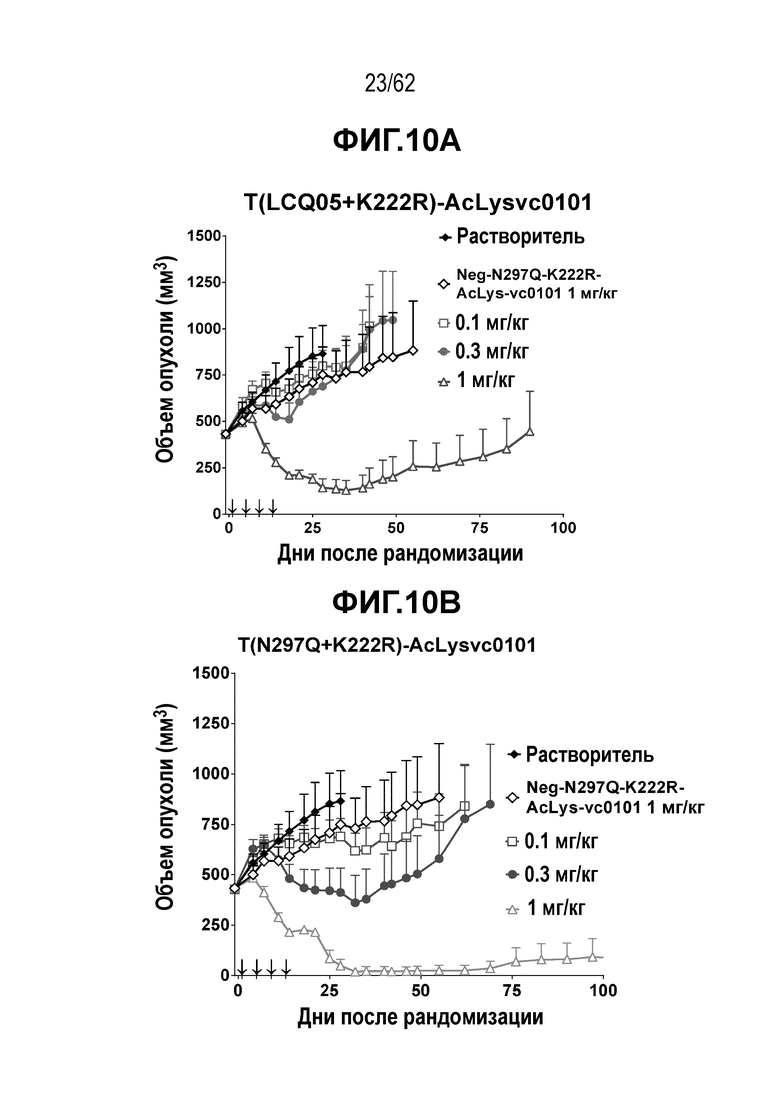
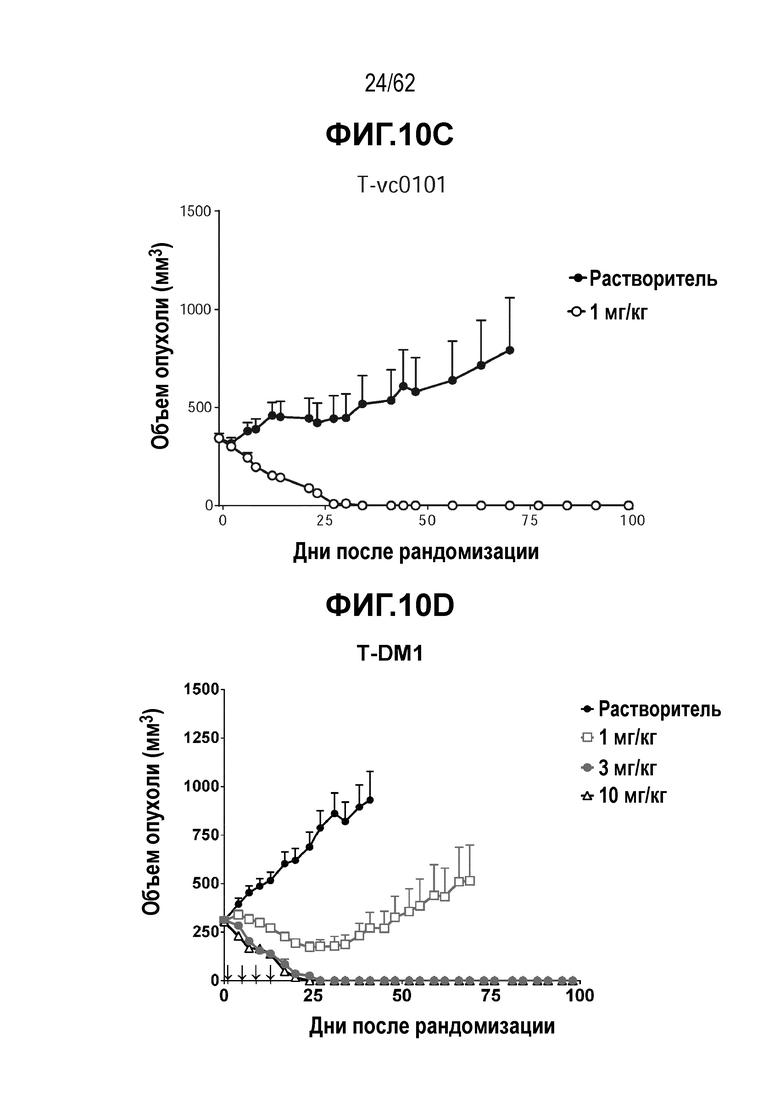
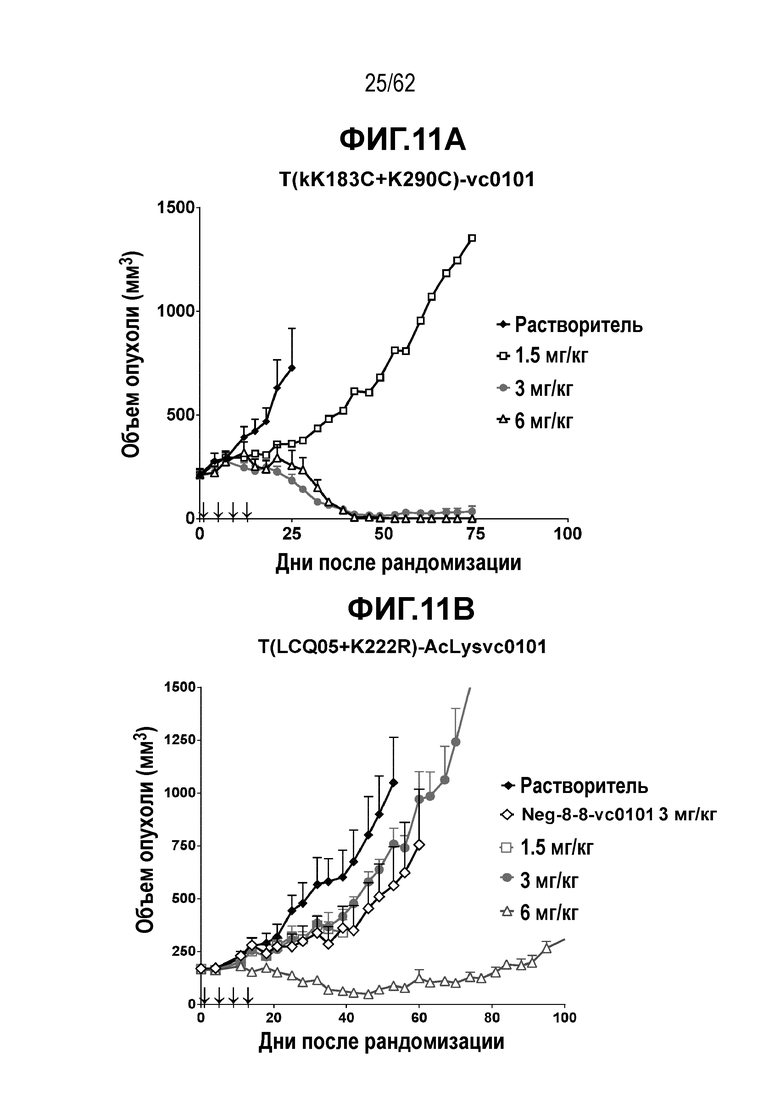
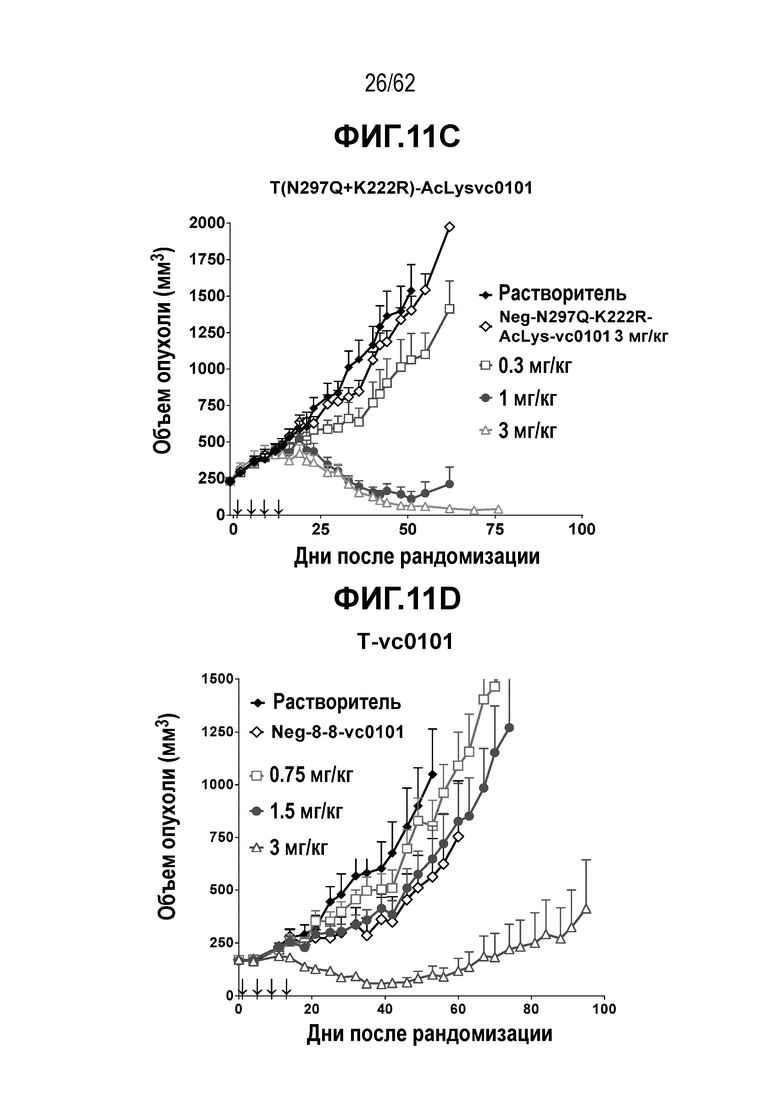
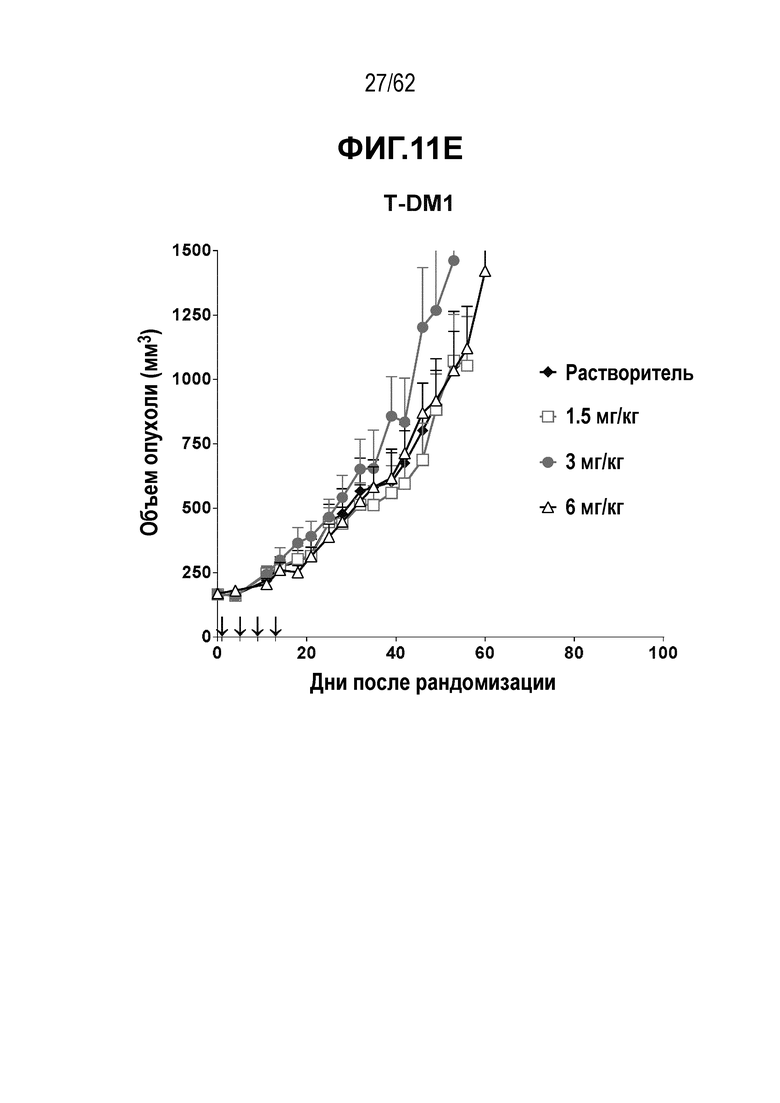
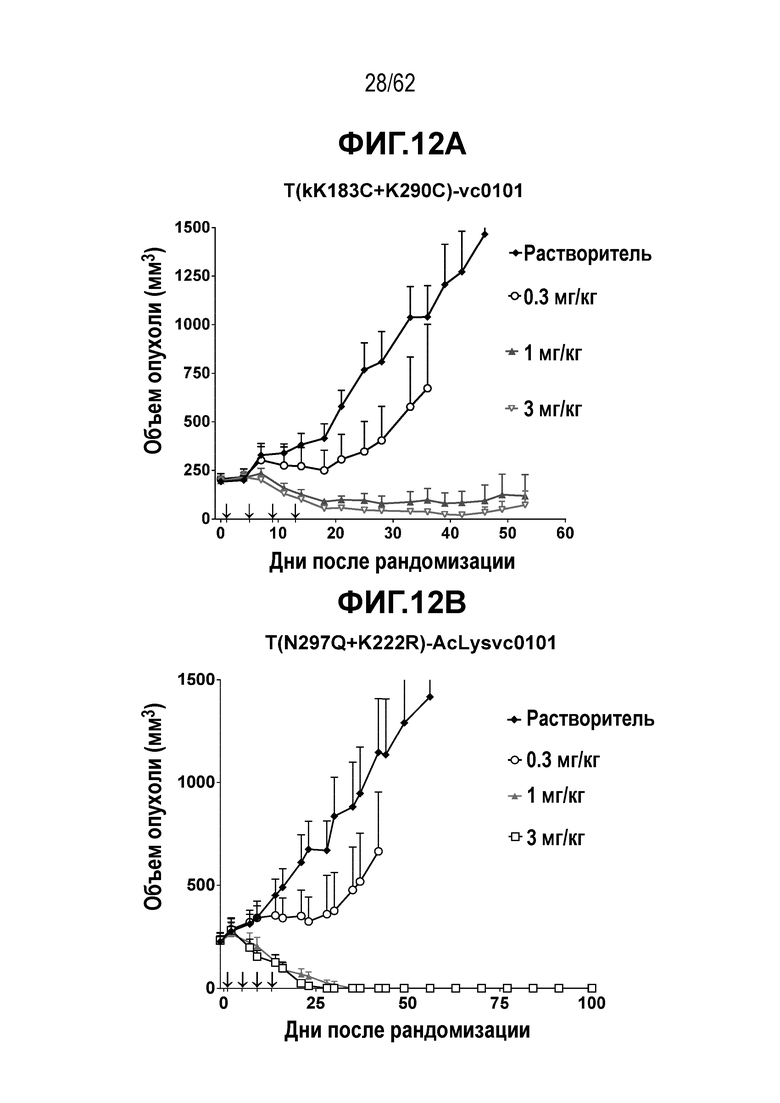
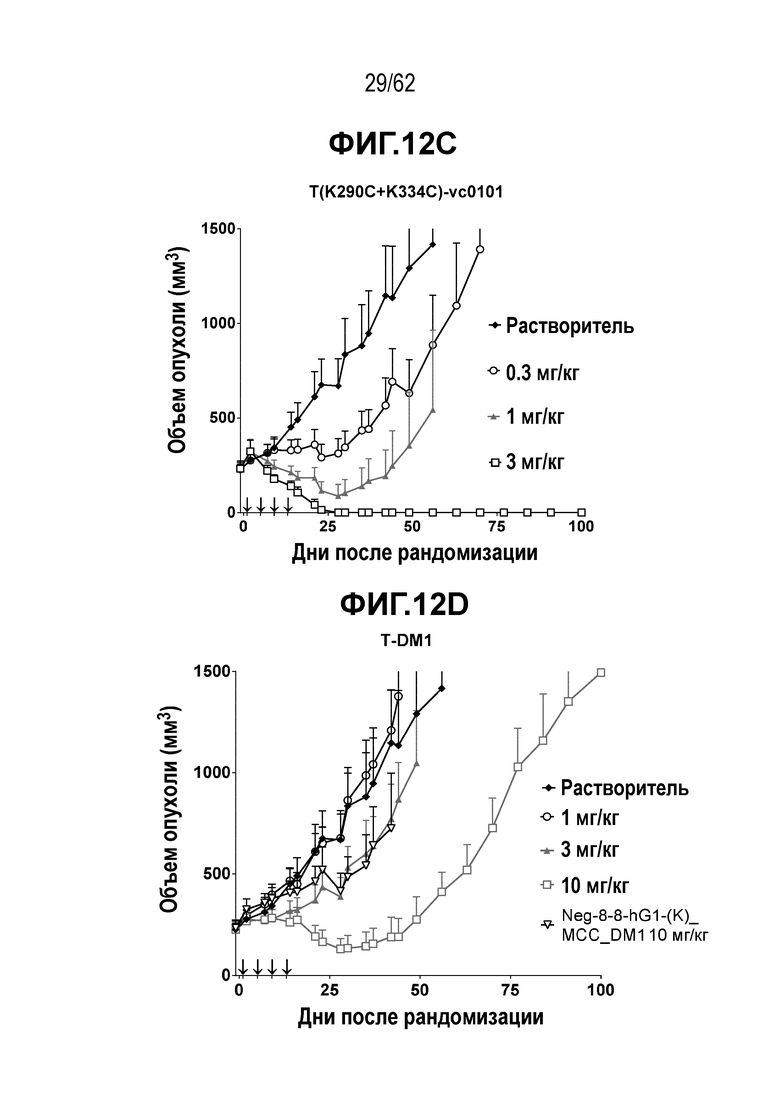
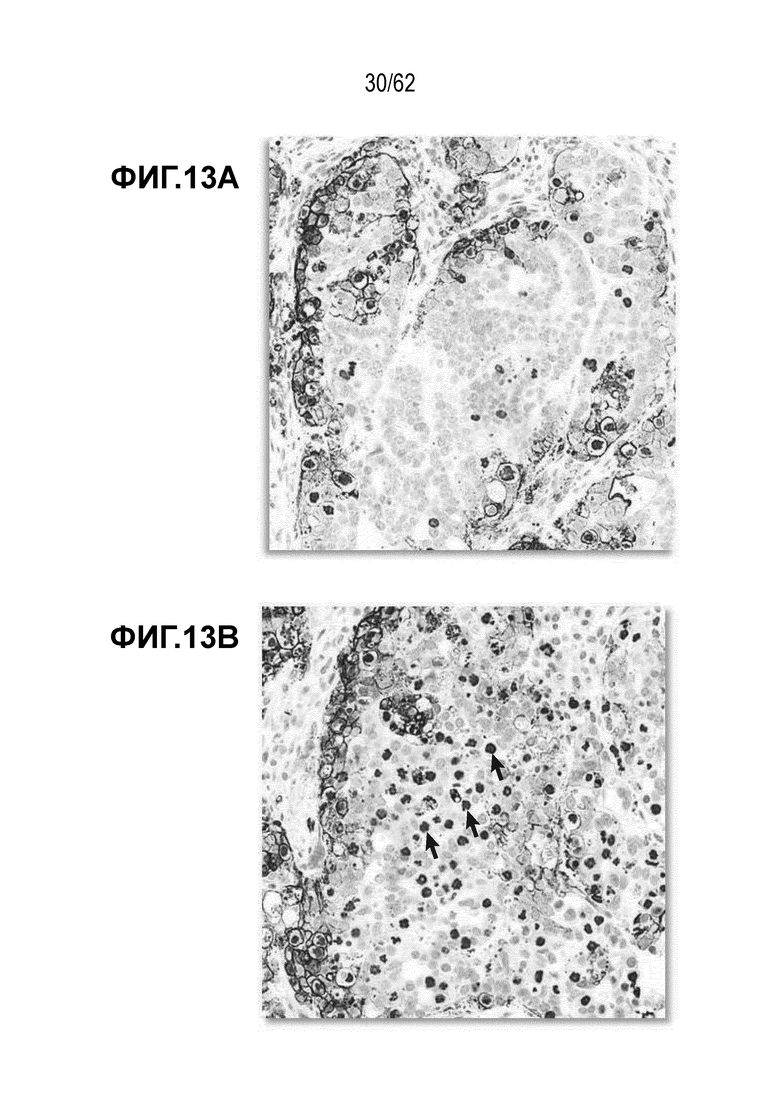
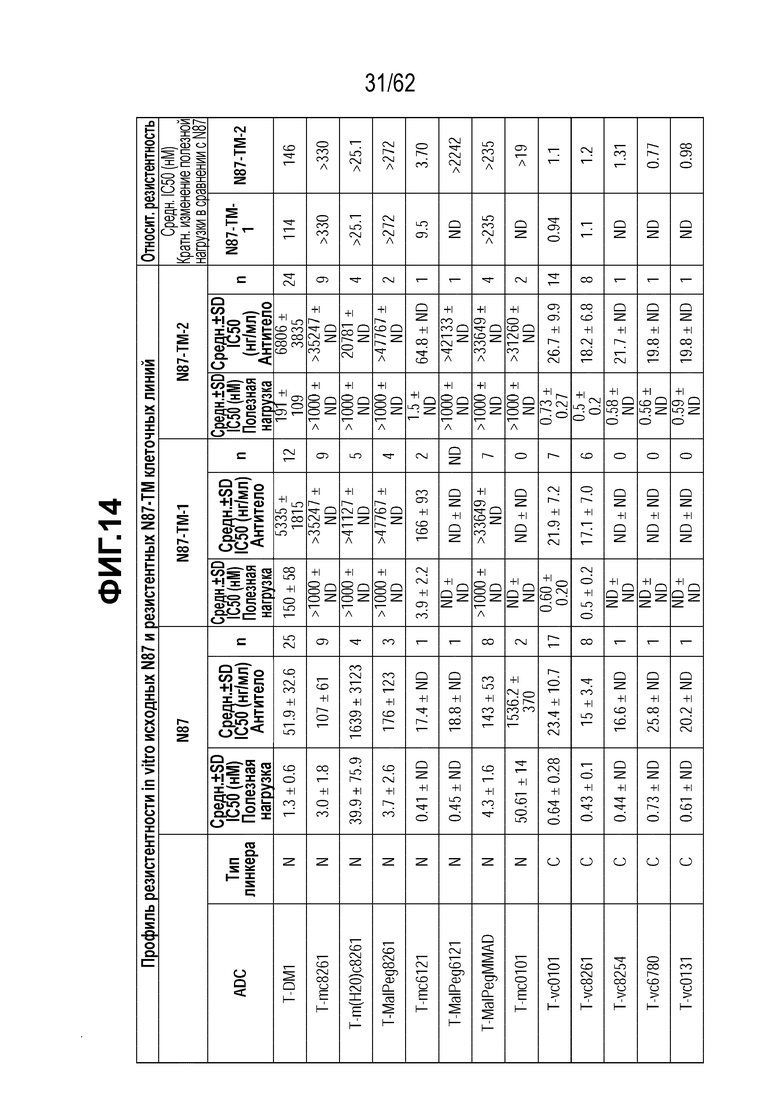
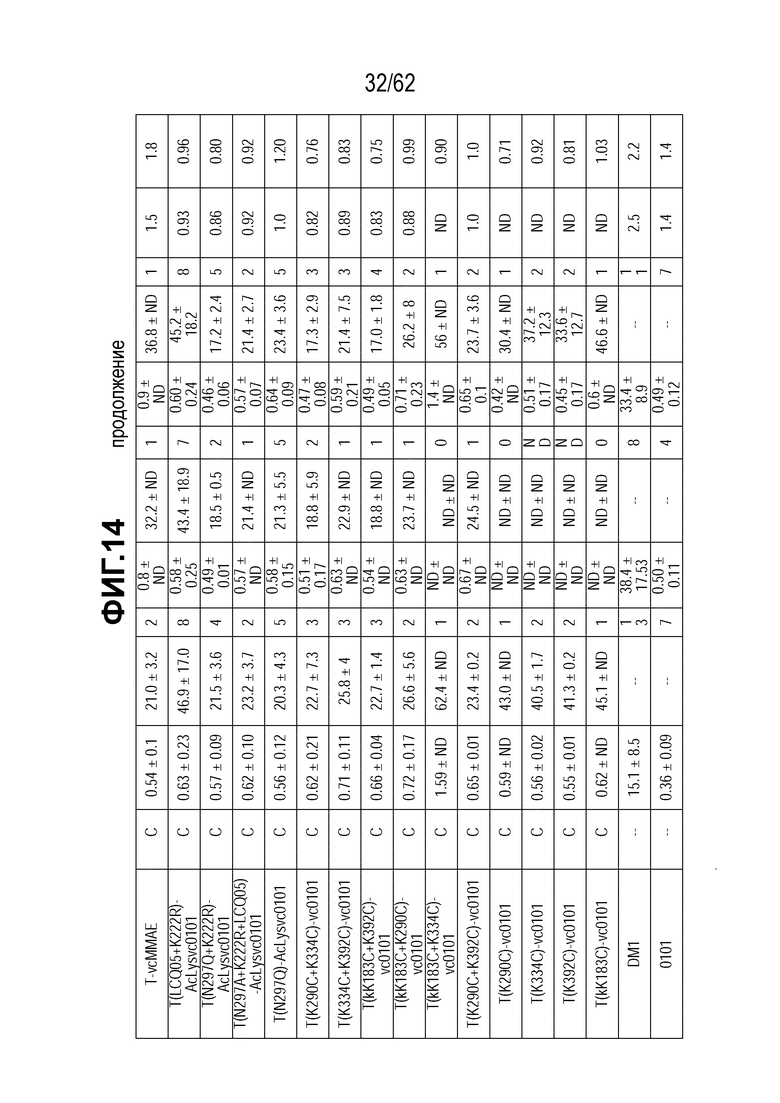
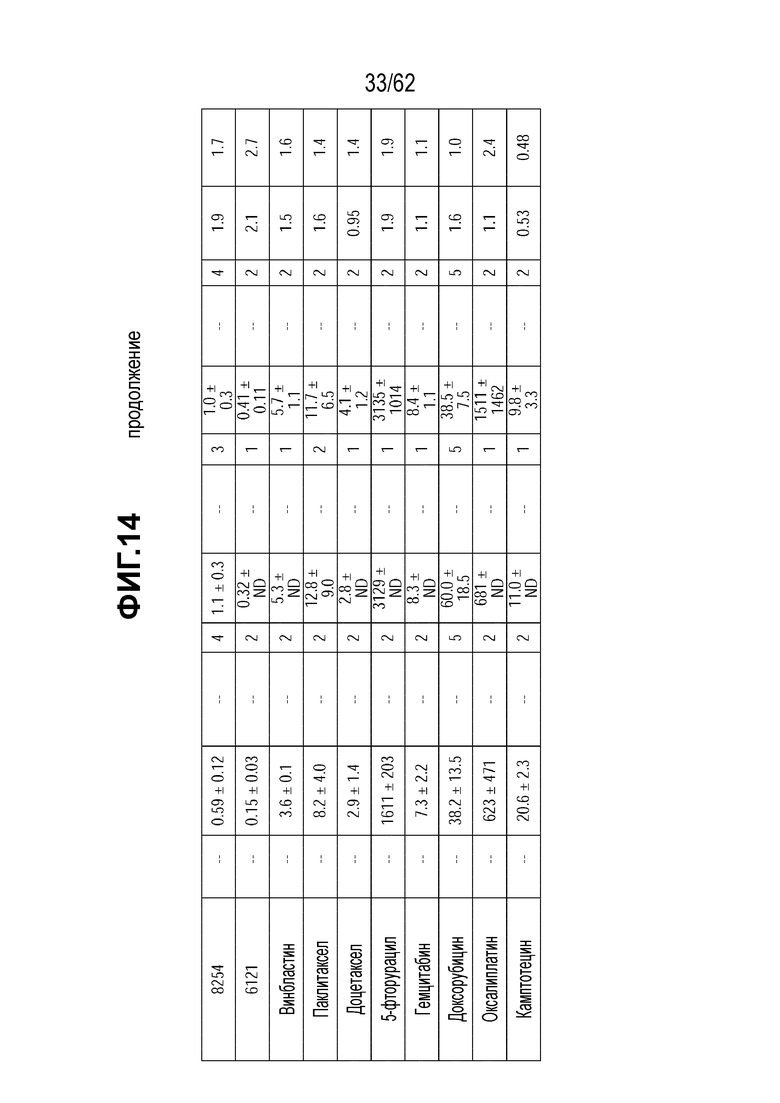
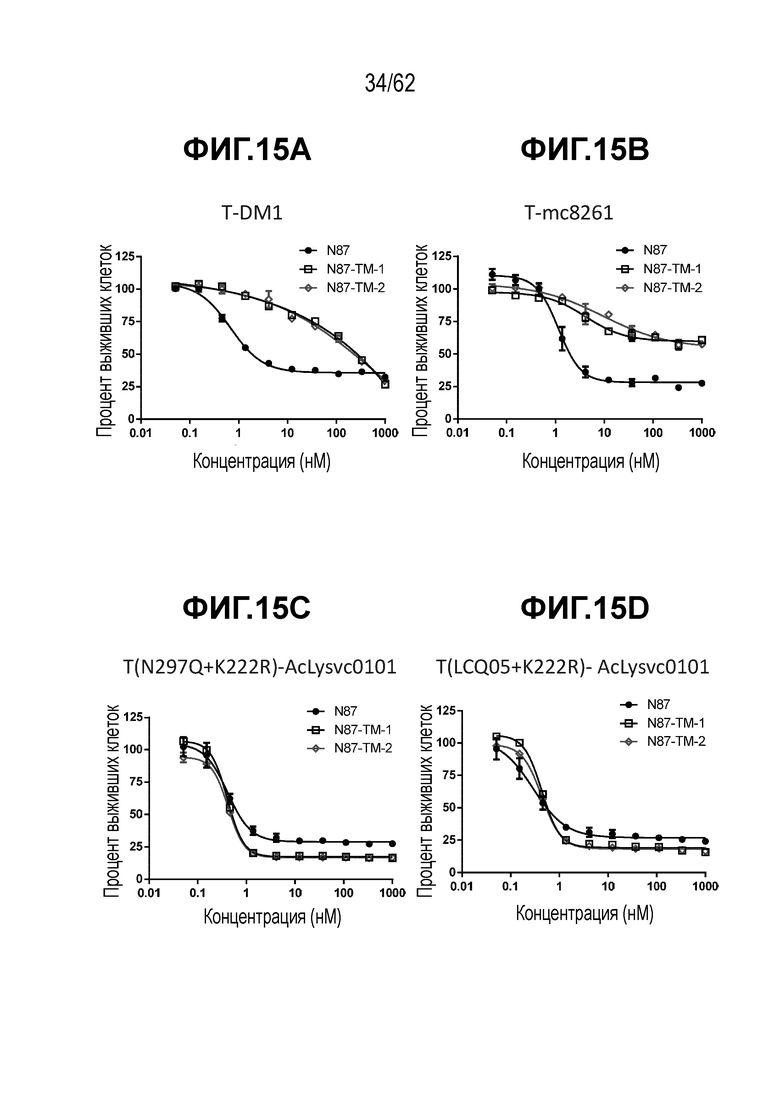
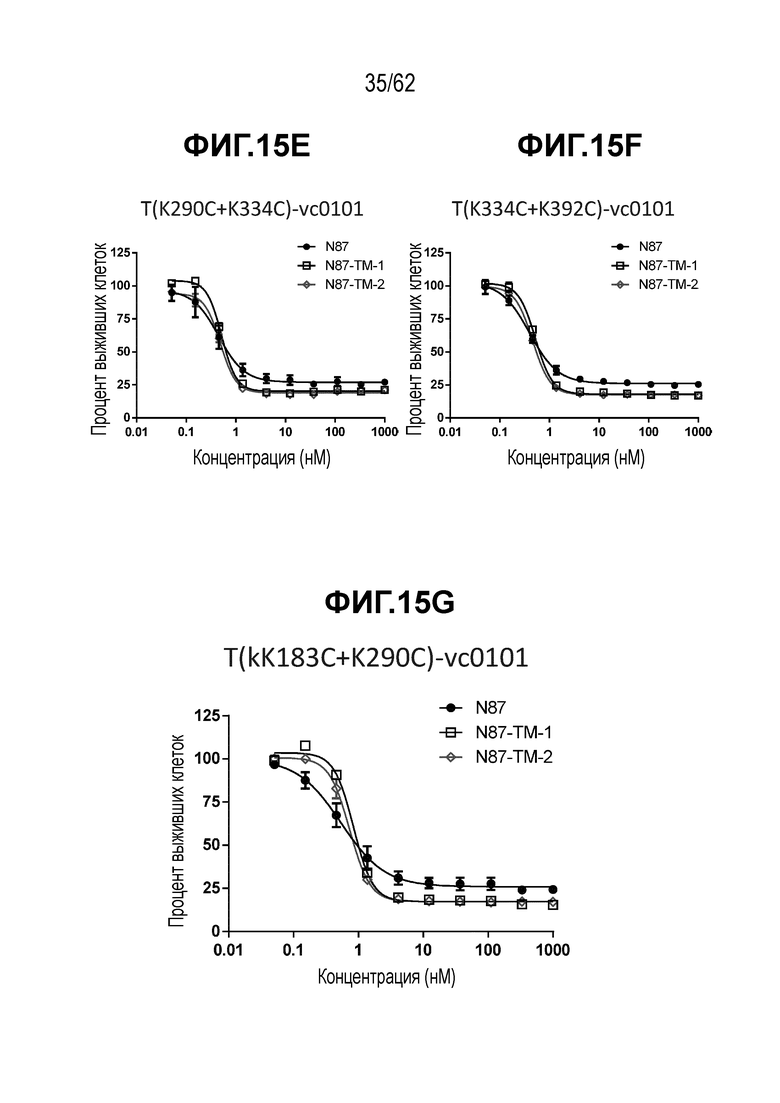
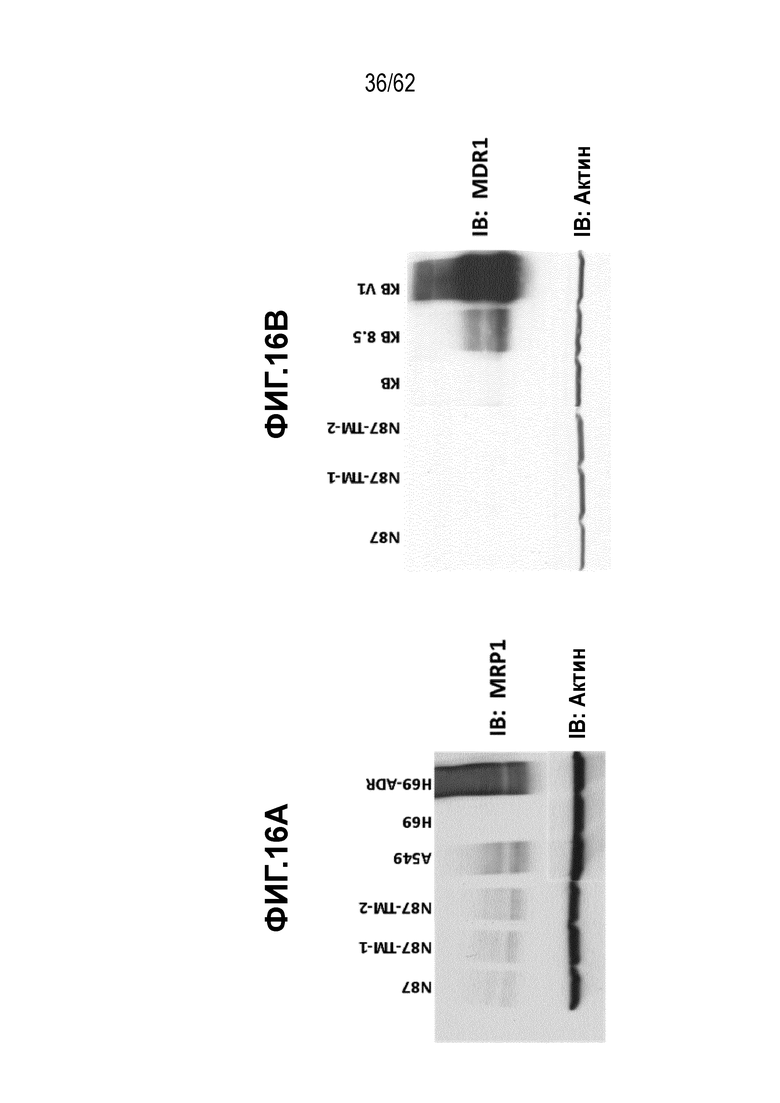
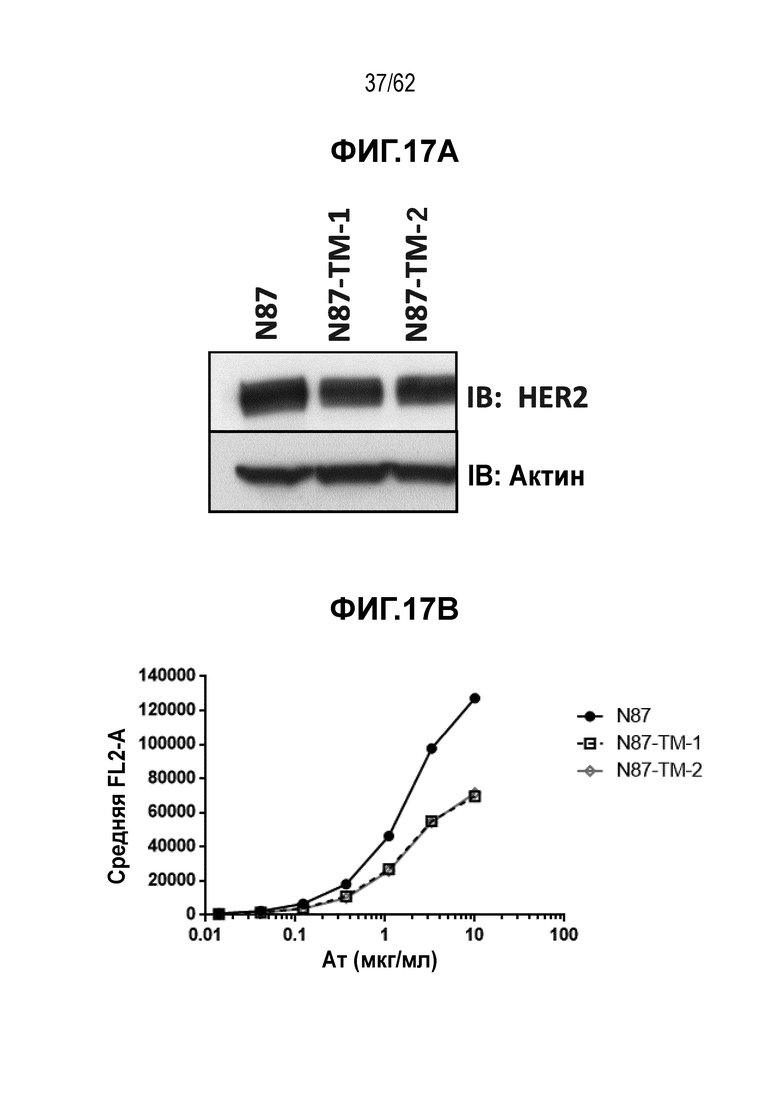
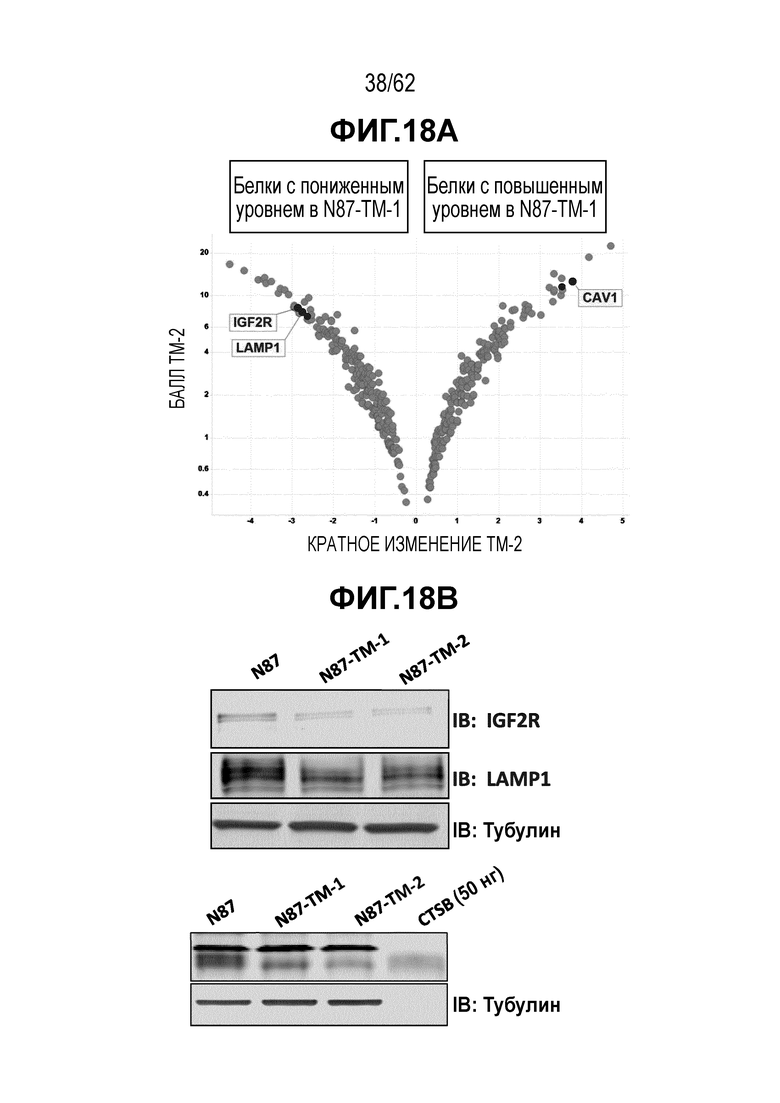
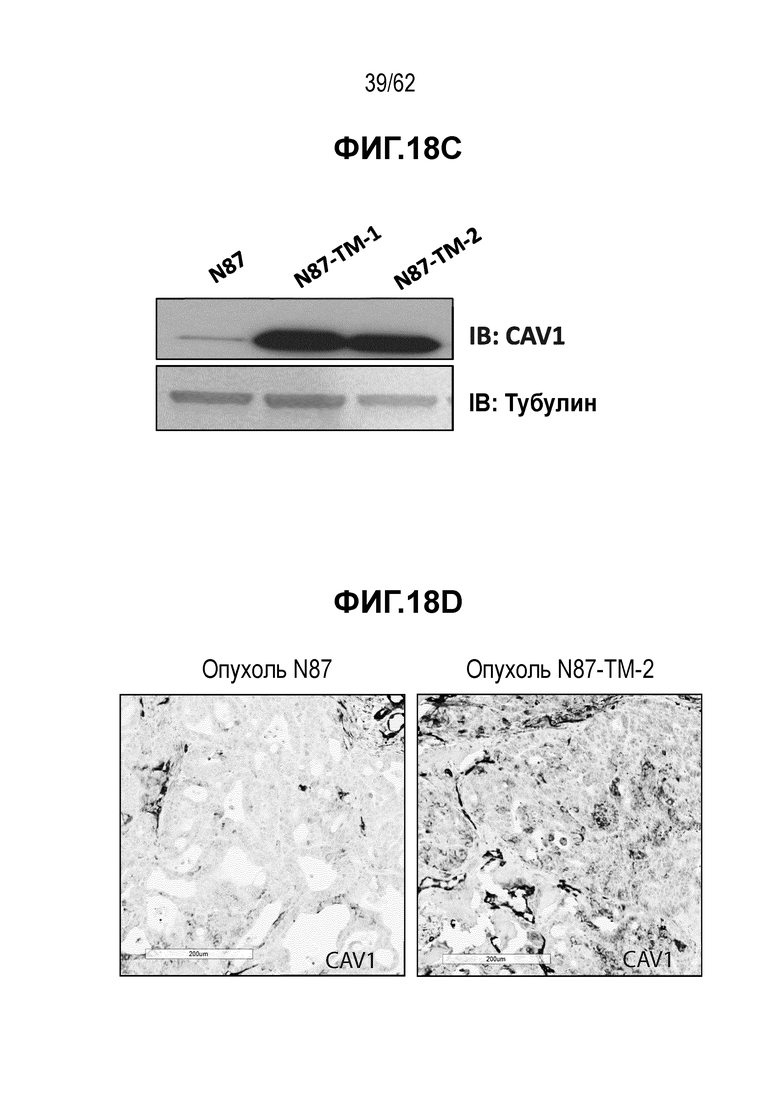
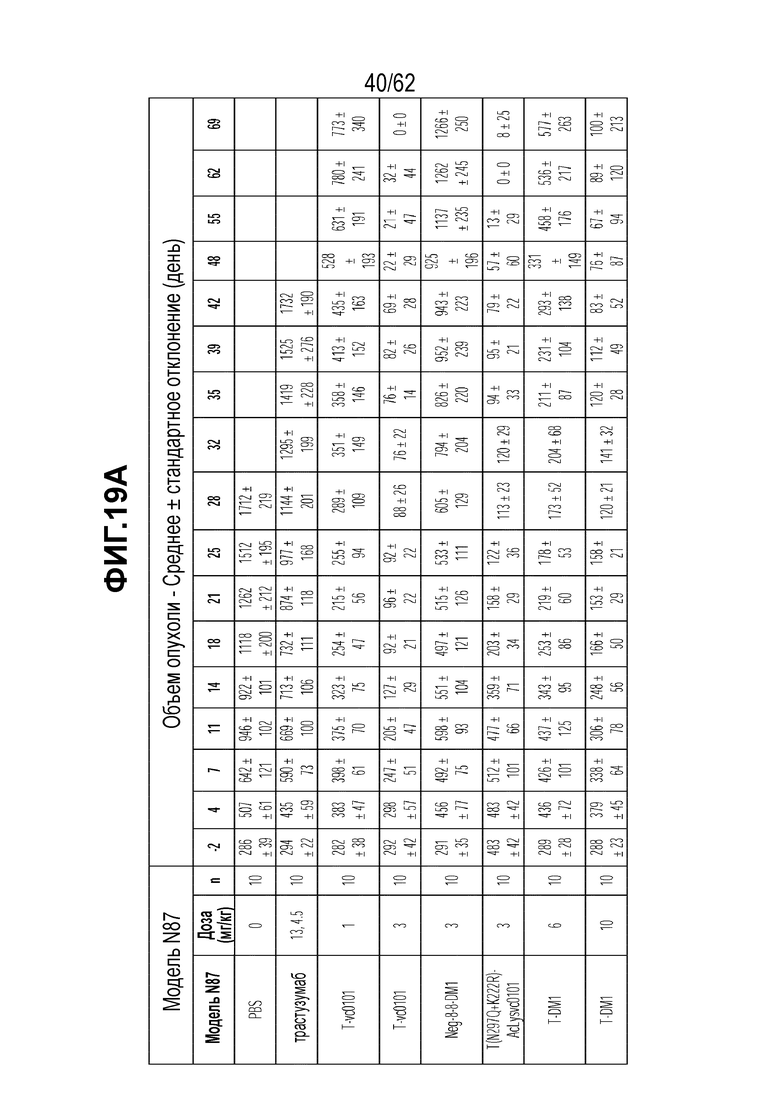
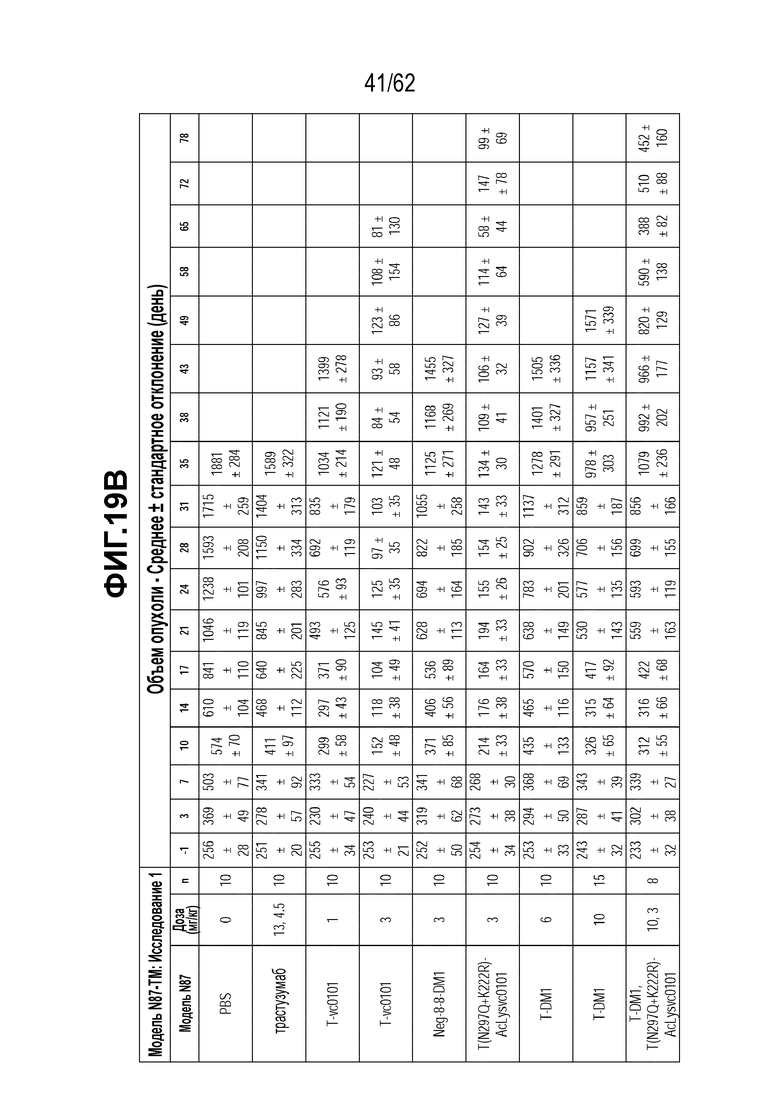
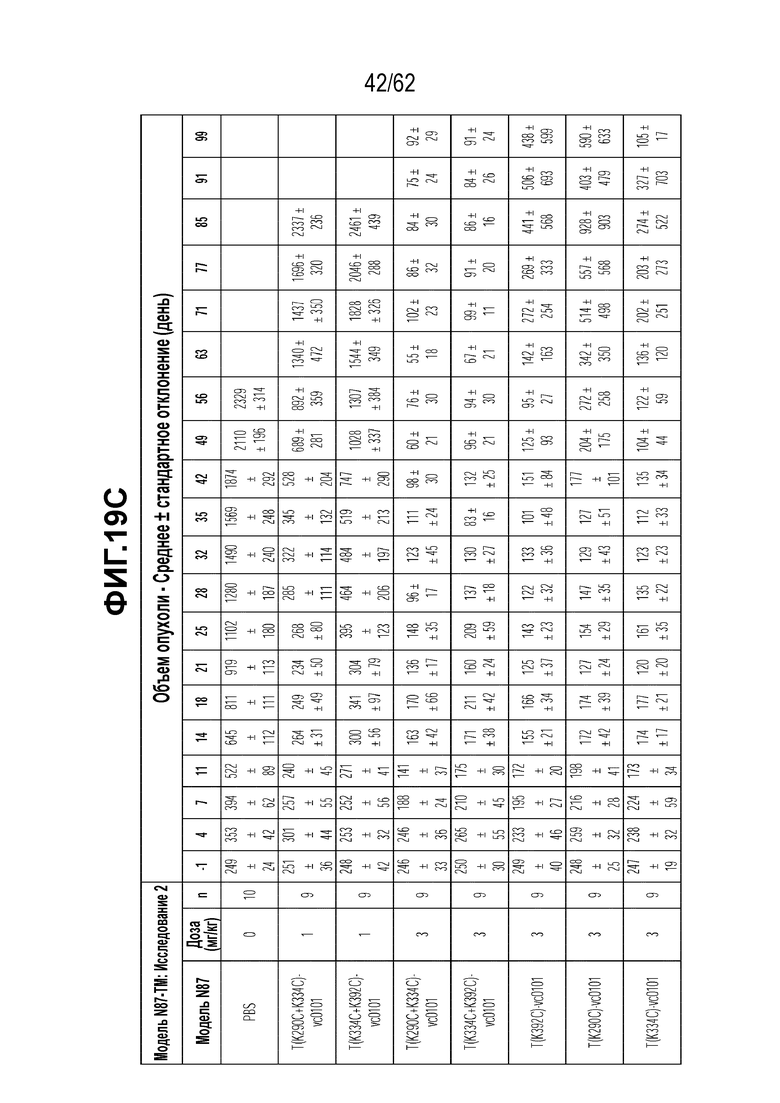
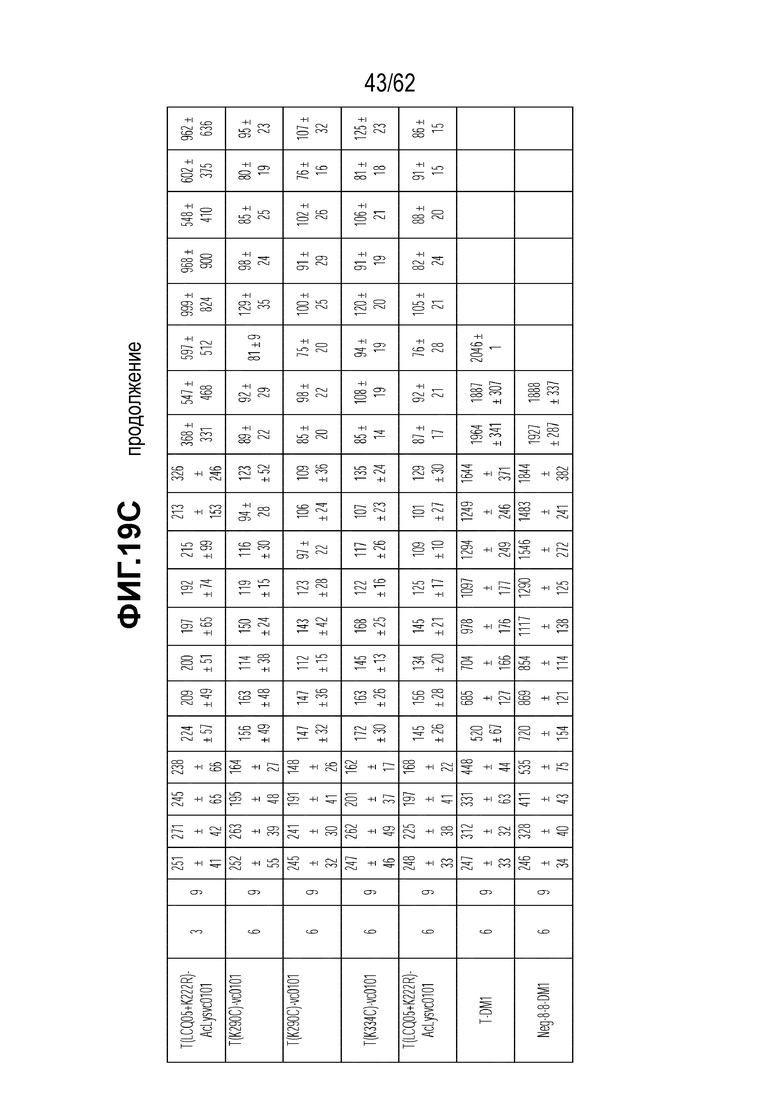
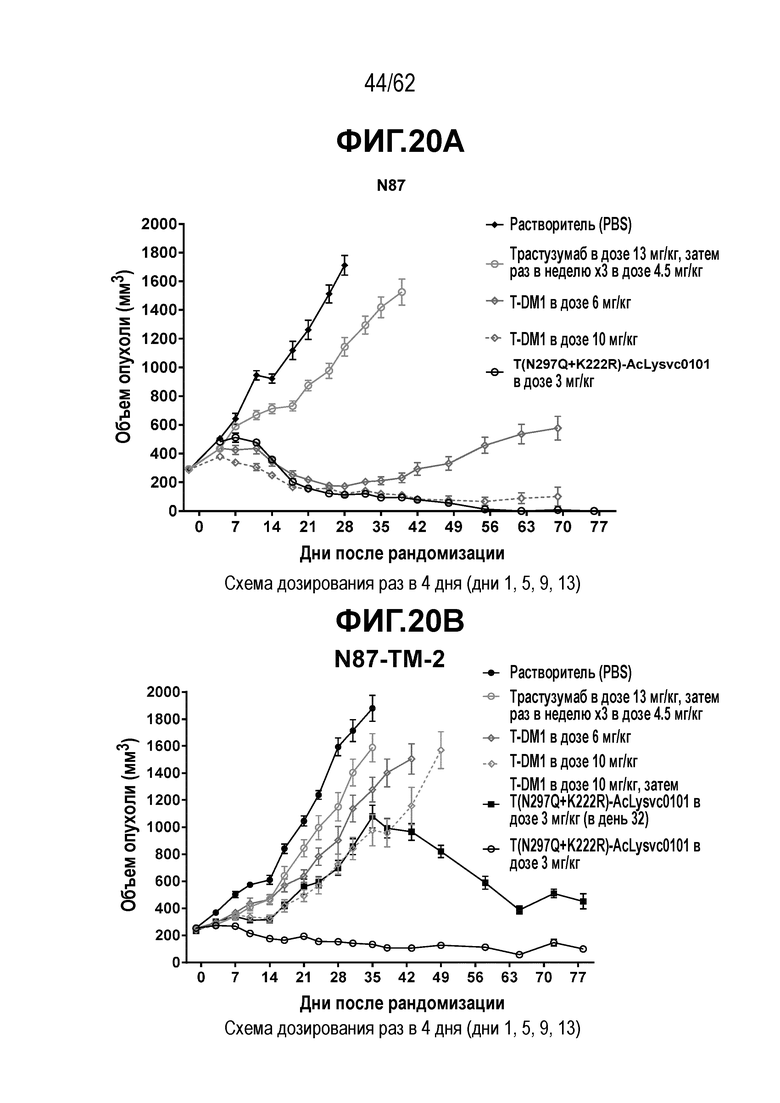
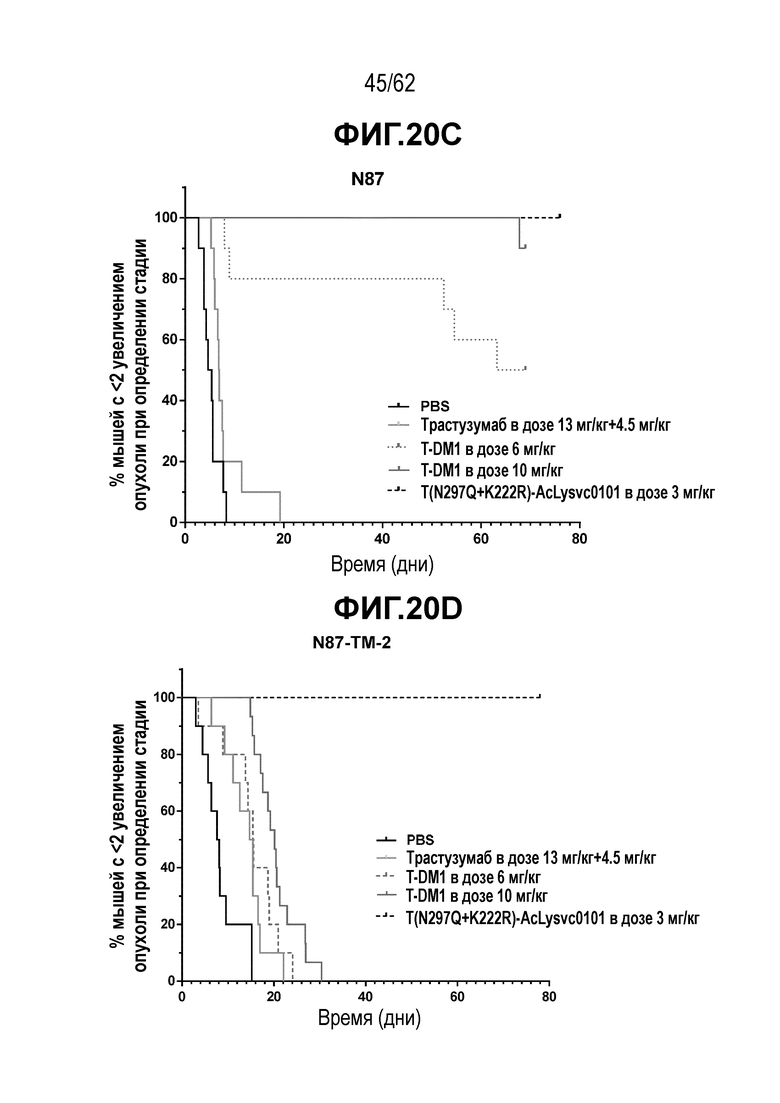
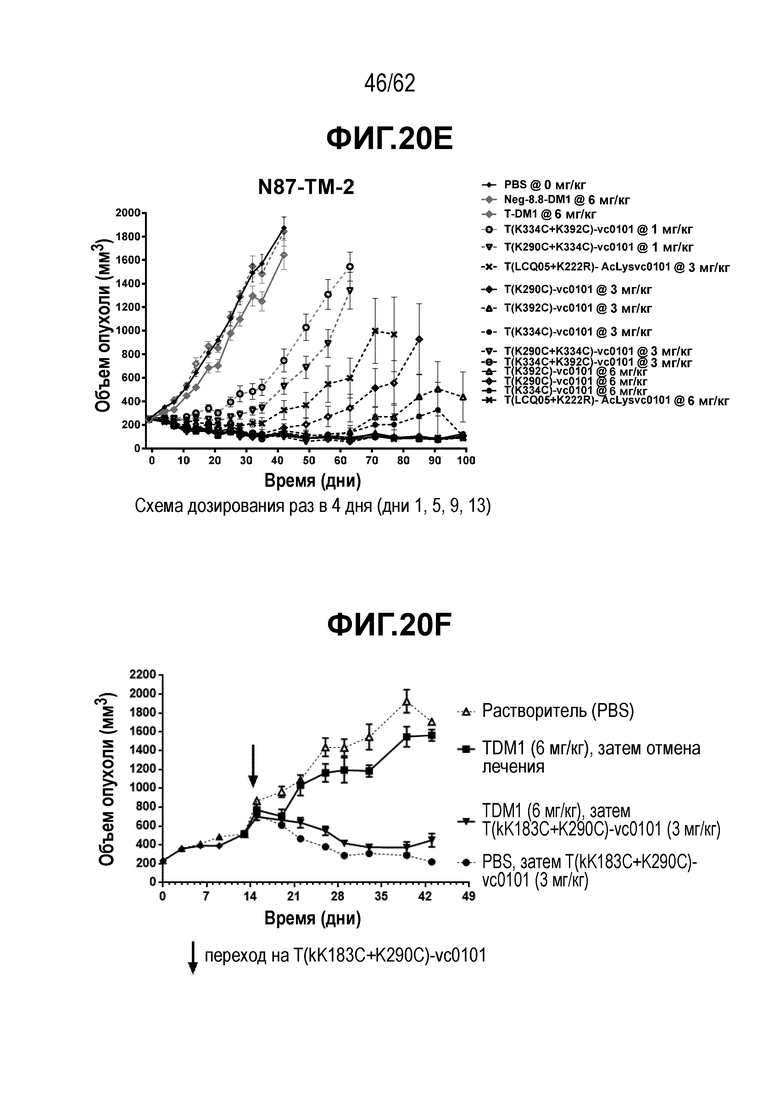
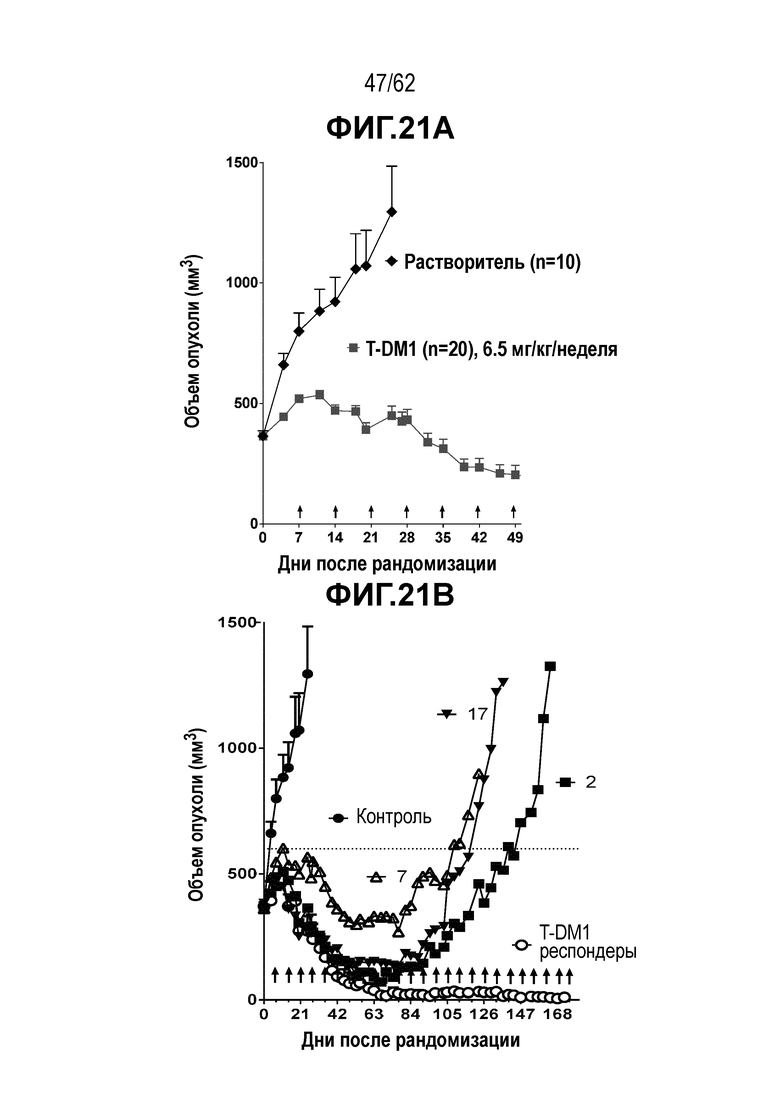
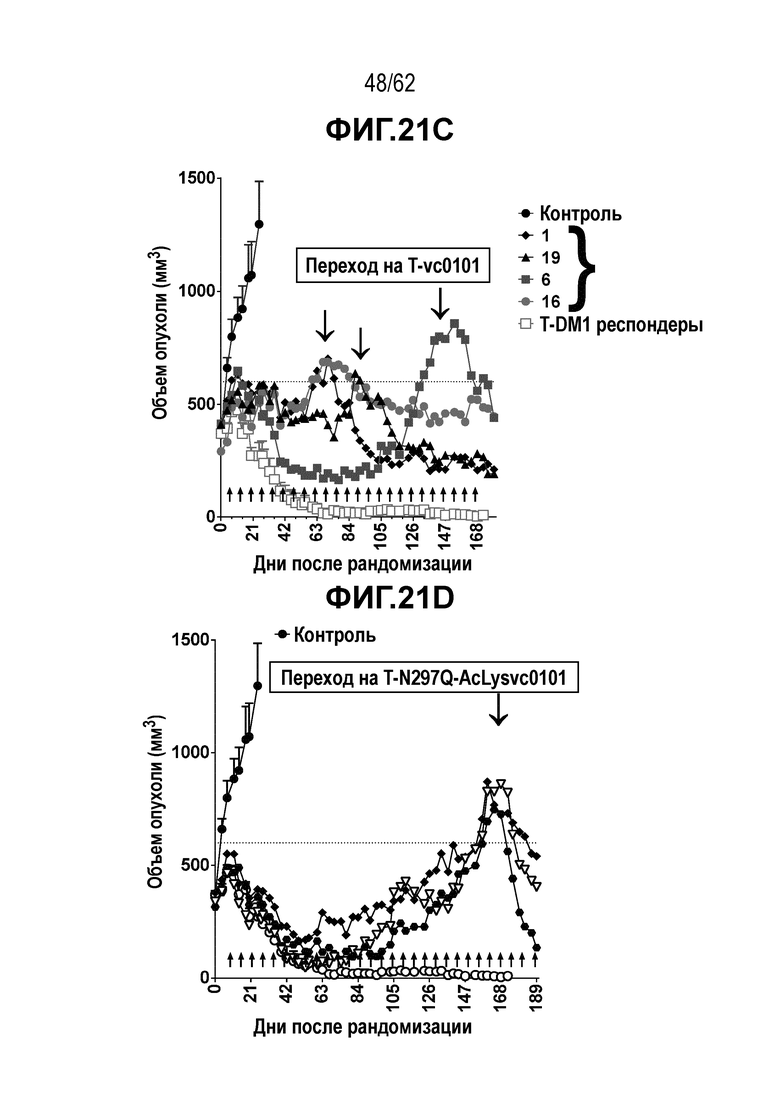
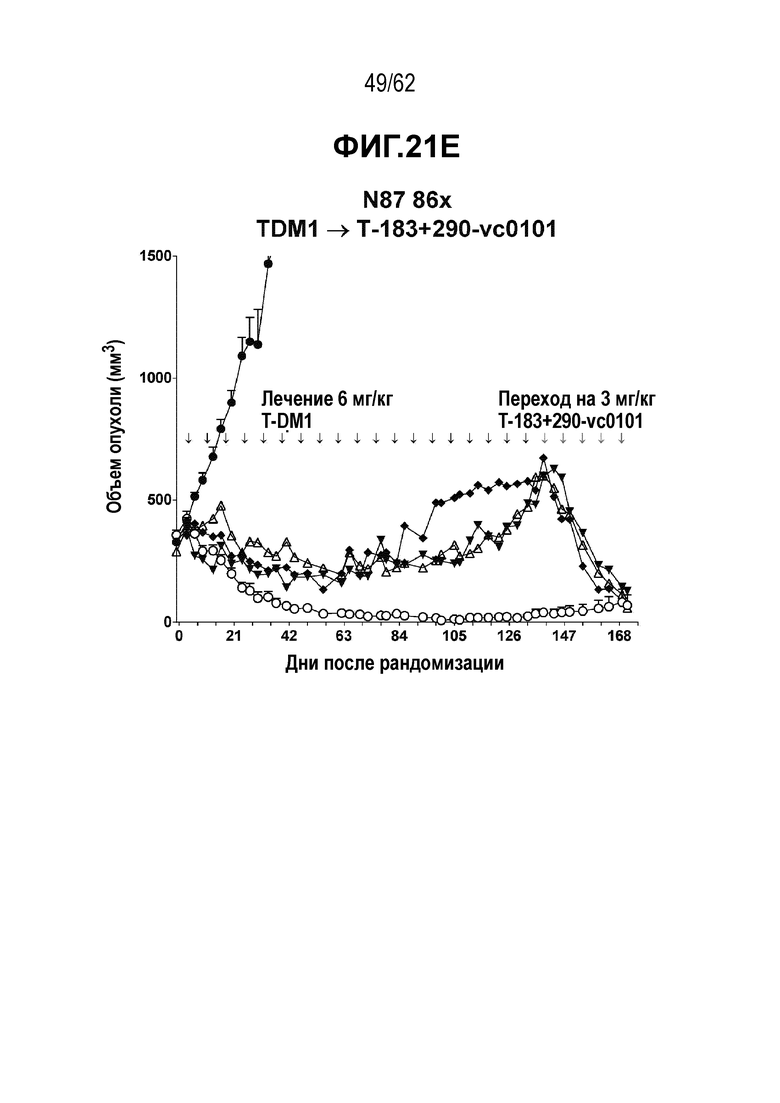
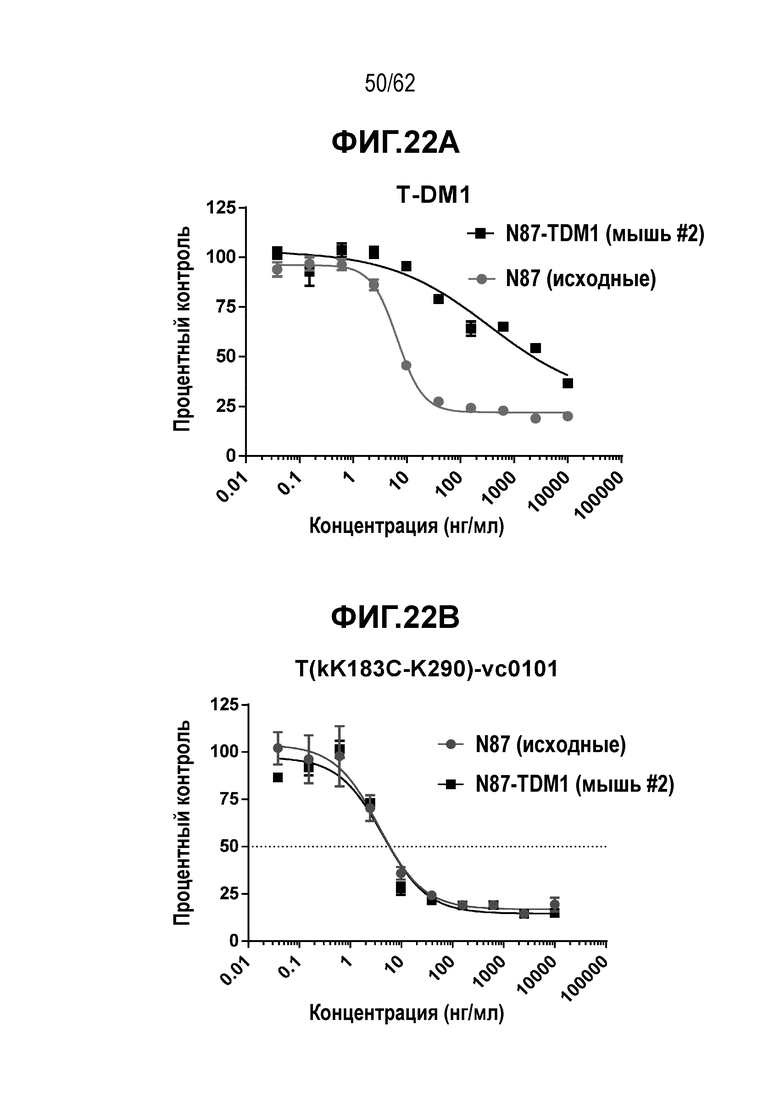
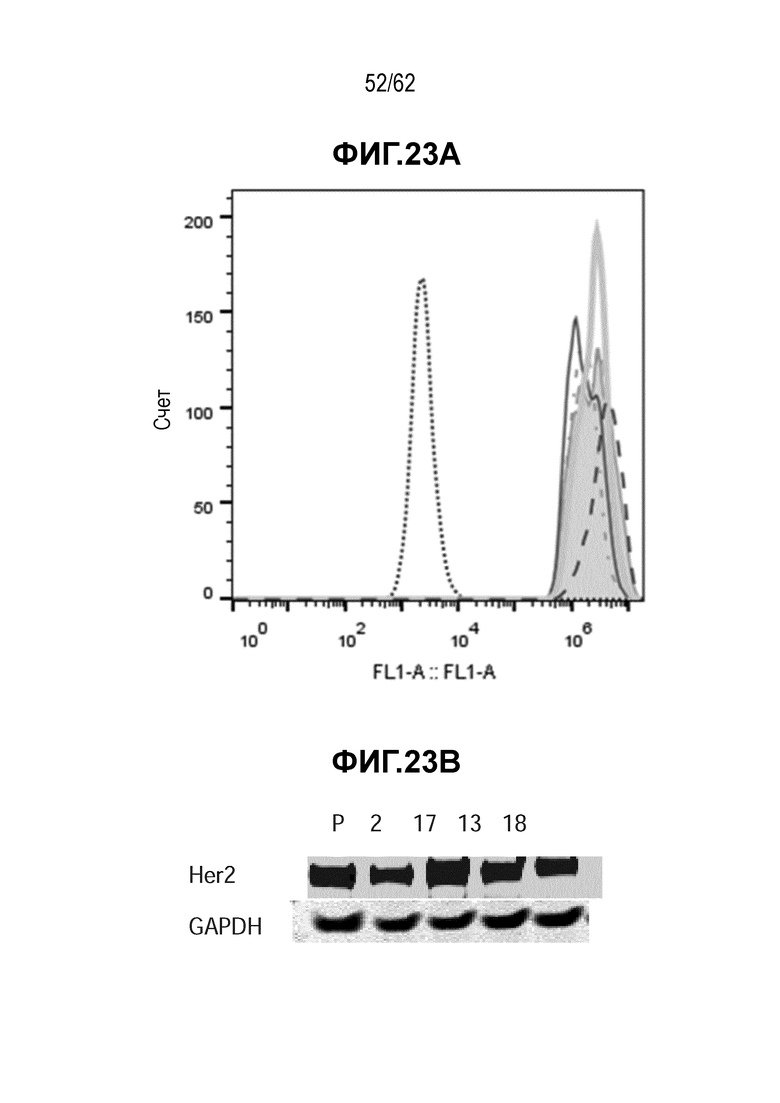
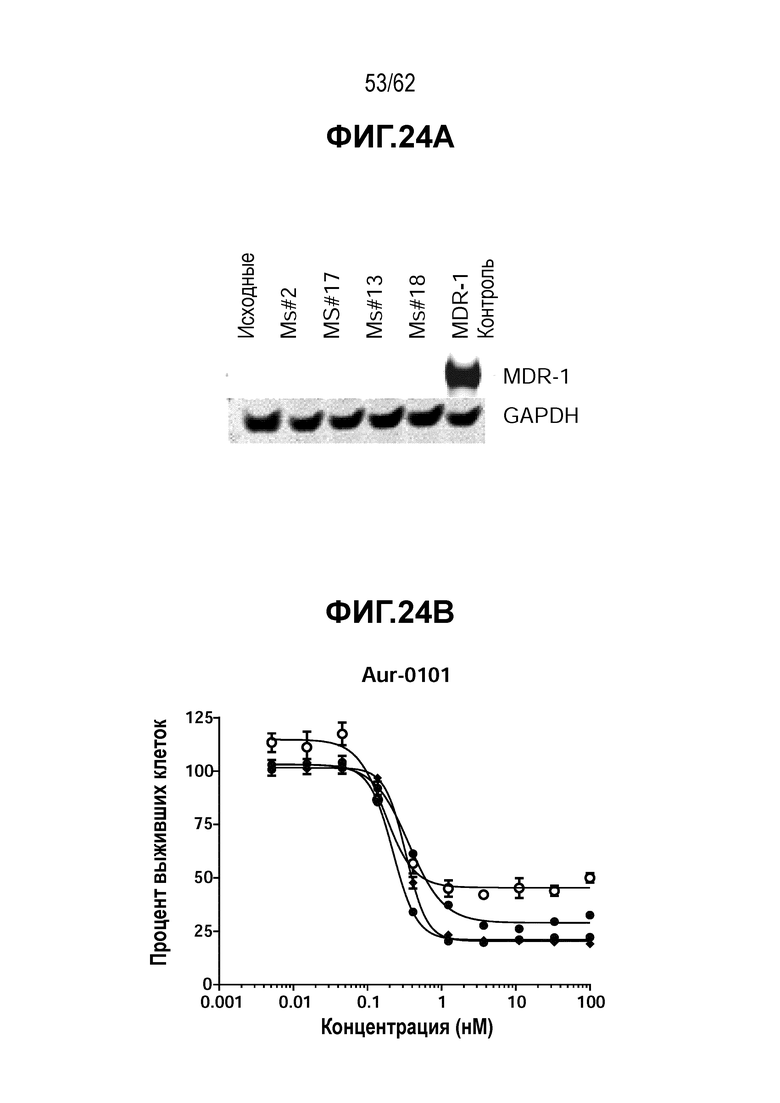
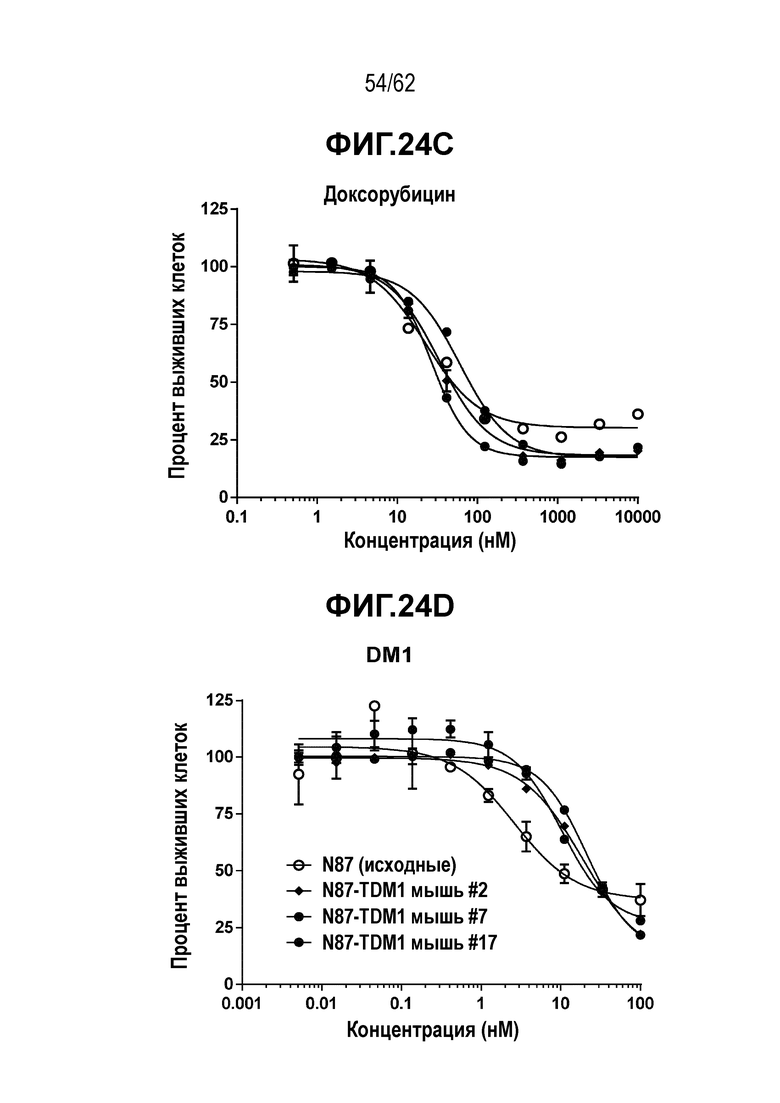
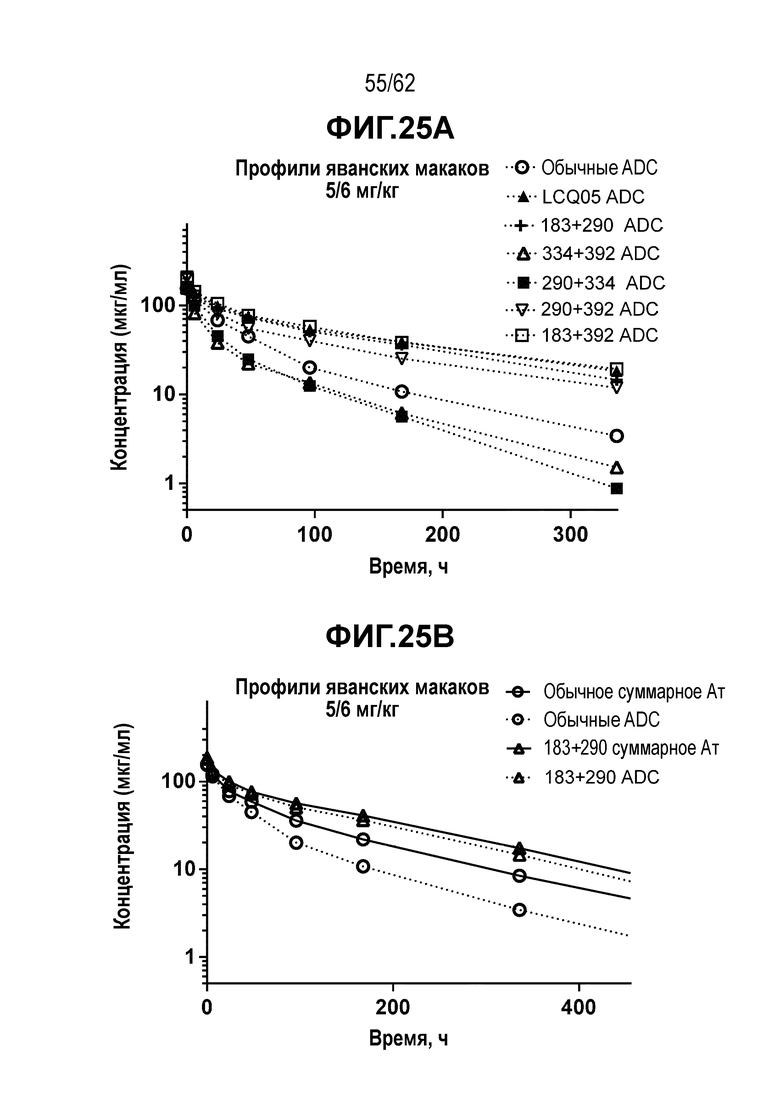
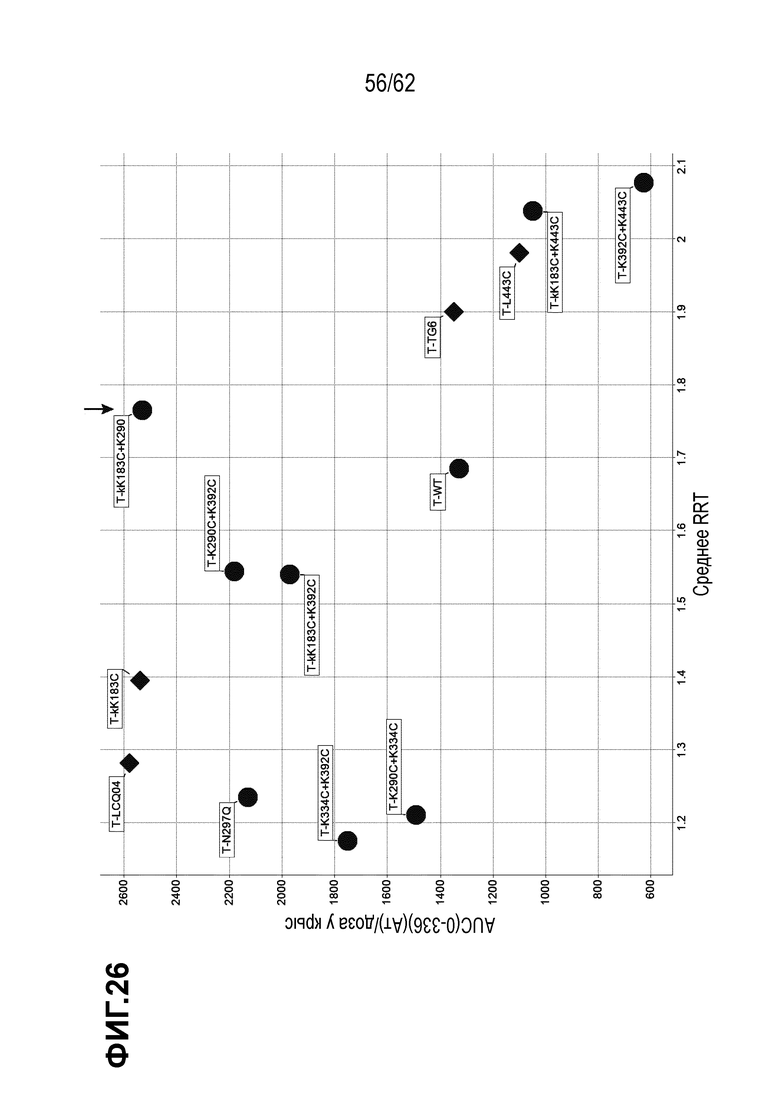
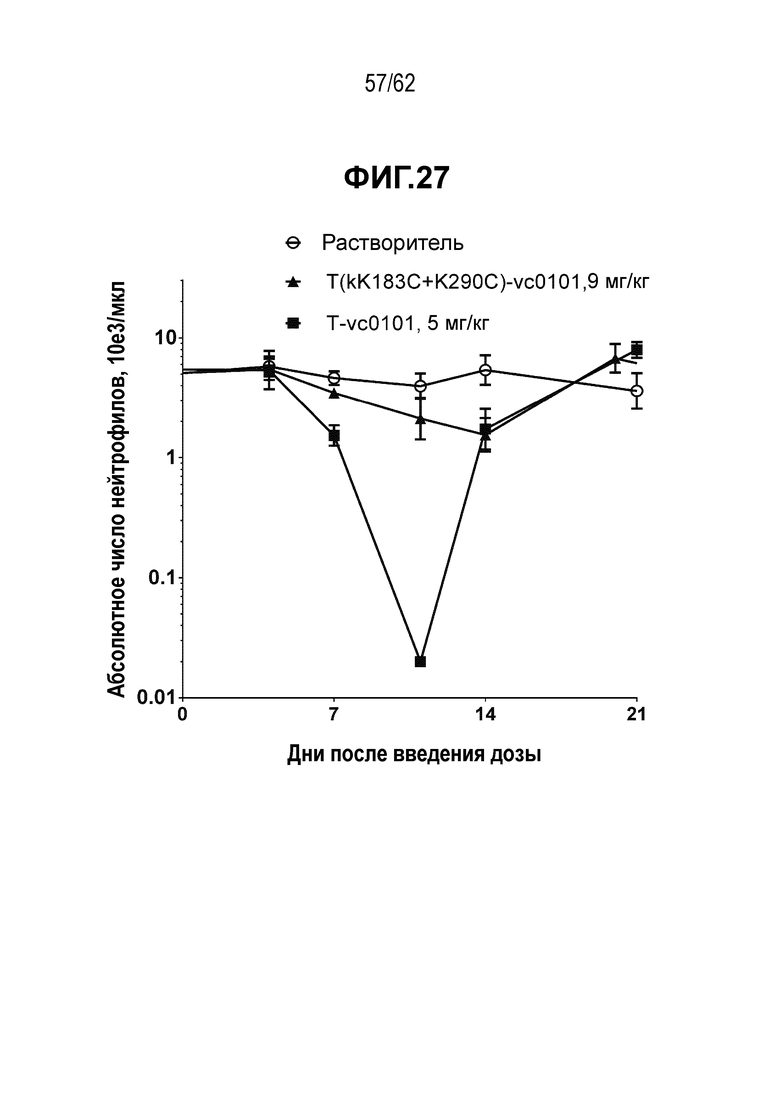
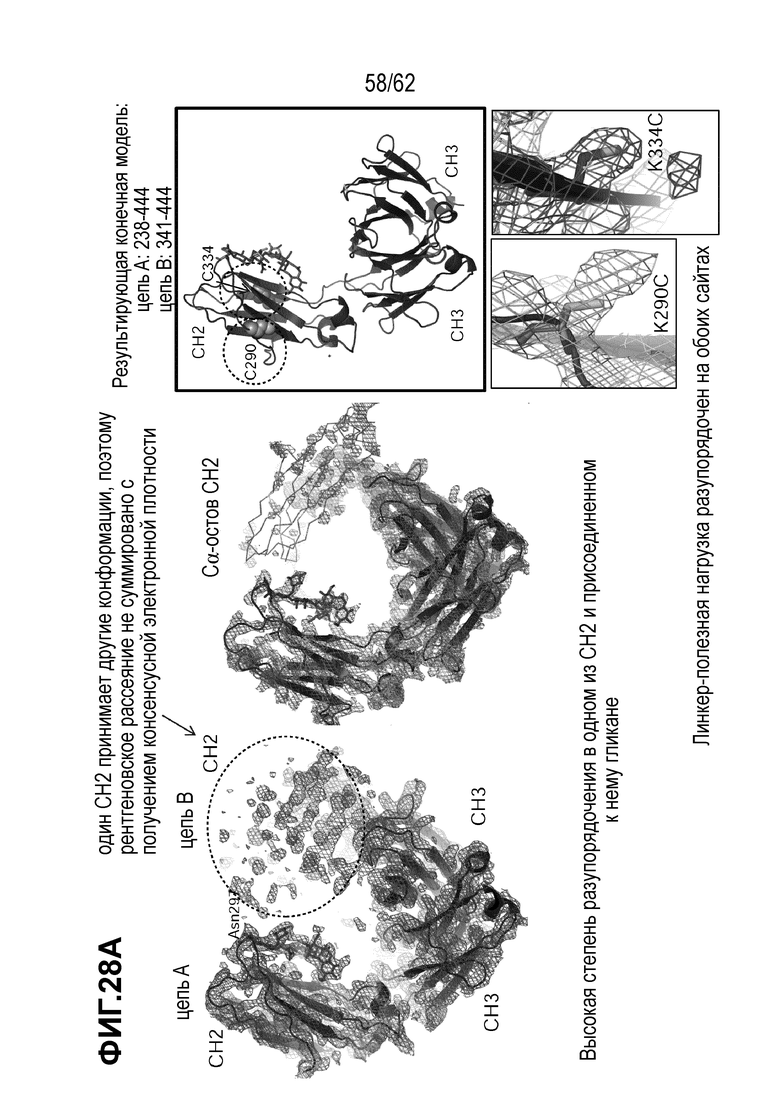
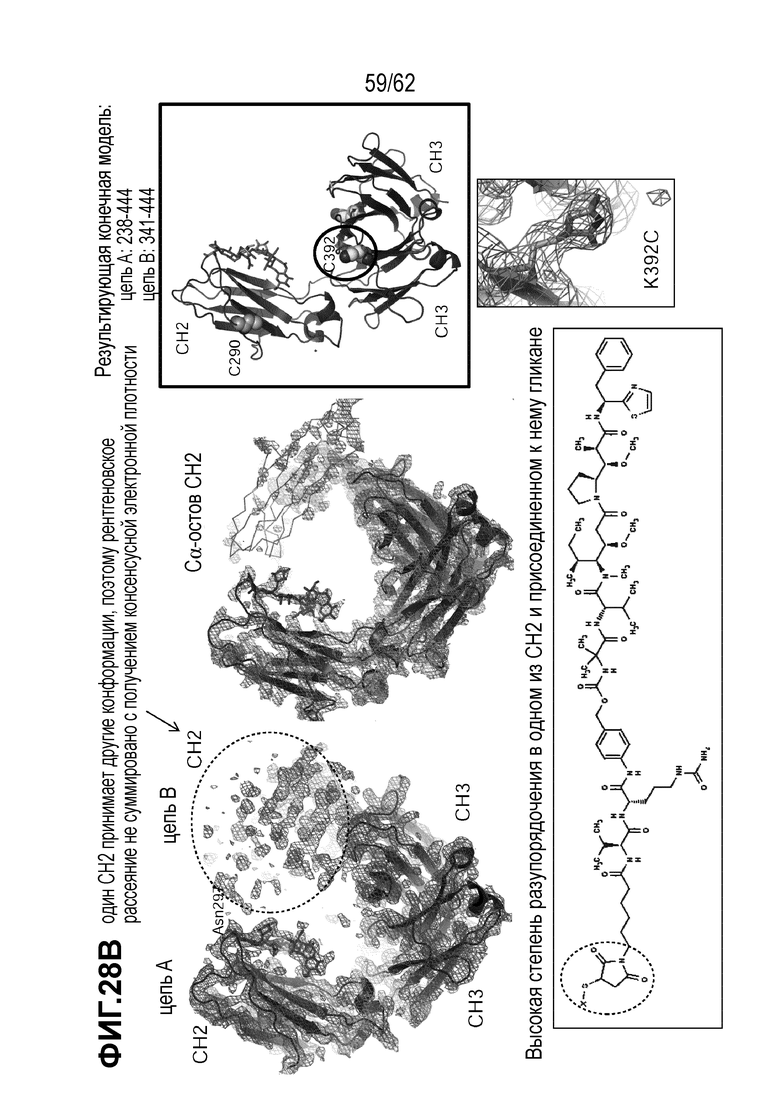
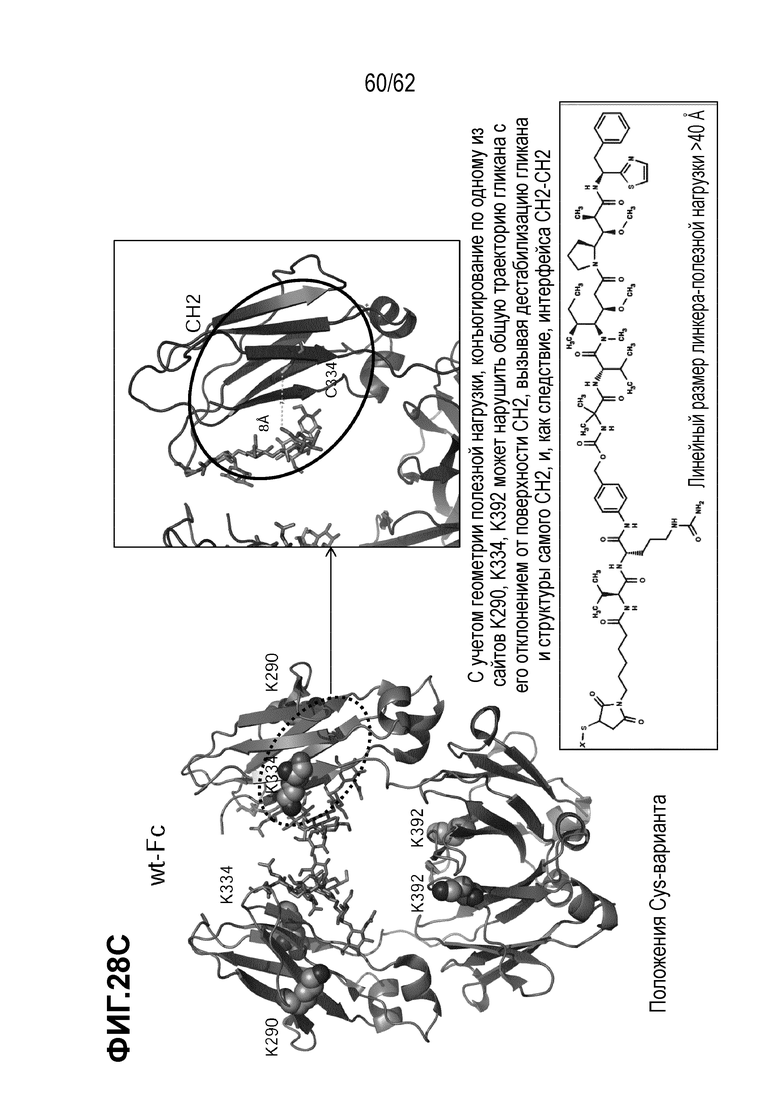
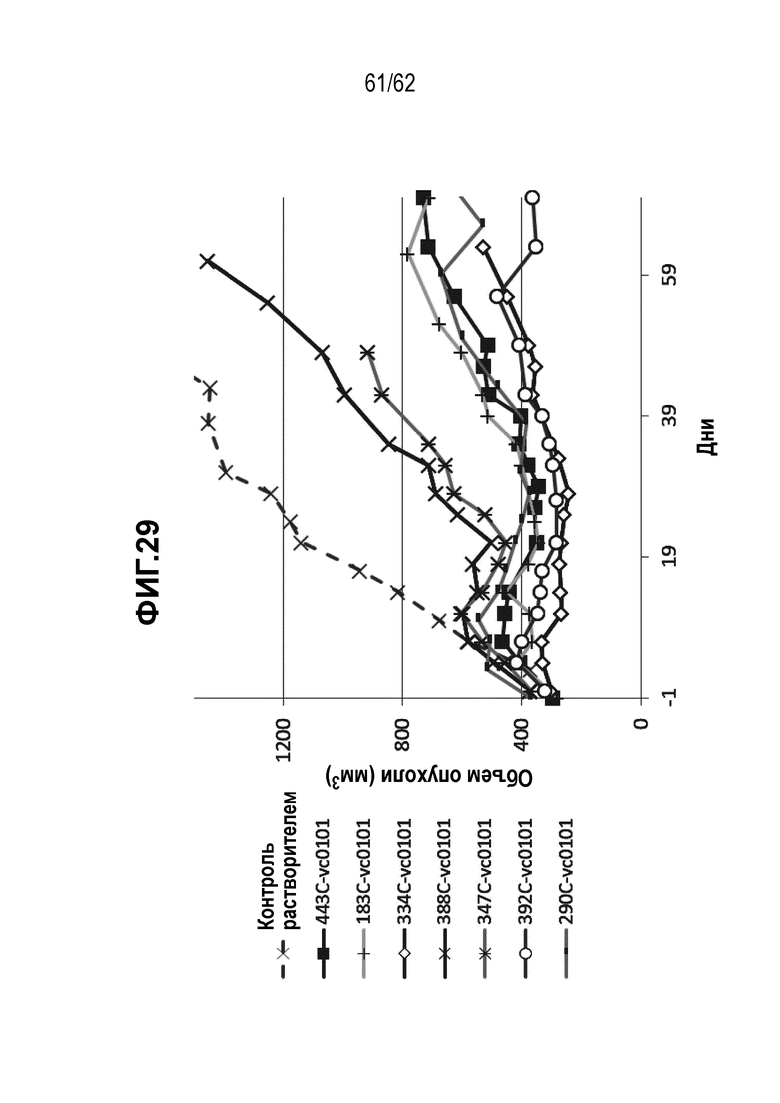
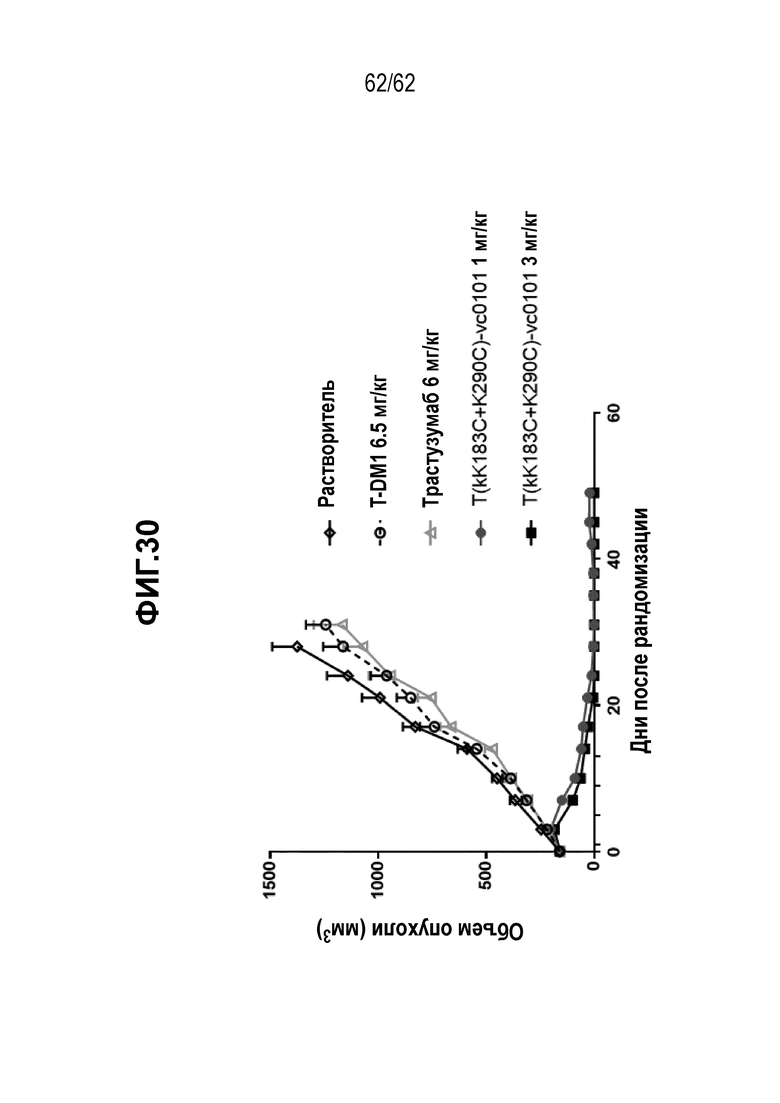
Изобретение относится к области биохимии, в частности к конъюгату антитело-лекарственное средство для лечения злокачественного новообразования, экспрессирующего Her2. Также раскрыты способ получения указанного конъюгата, способ лечения злокачественной опухоли с помощью указанного конъюгата и применение указанного конъюгата для получения лекарственного средства для лечения злокачественной опухоли, экспрессирующей HER2, у пациента. Раскрыта композиция, содержащая указанный конъюгат, нуклеиновая кислота, кодирующая антитело, используемое в получении указанного конъюгата, вектор экспрессии и клетка-хозяин, содержащие указанную нуклеиновой кислоту. Изобретение позволяет эффективно лечить злокачественное новообразование, экспрессирующее Her2. 10 н. и 41 з.п. ф-лы, 27 табл., 30 ил., 20 пр.
1. Конъюгат антитело-лекарственное средство для лечения злокачественного новообразования, экспрессирующего Her2, имеющий формулу:
Ab-(L-D),
где:
(а) Ab является антителом, которое связывается с HER2 и содержит
(1) вариабельную область тяжелой цепи, содержащую CDR, содержащие SEQ ID NO:2, 3 и 4;
(2) константную область тяжелой цепи SEQ ID NO:17;
(3) вариабельную область легкой цепи, содержащую CDR, содержащие SEQ ID NO:8, 9 и 10;
(4) константную область легкой цепи SEQ ID NO:41; и
(b) L-D представляет собой молекулу линкер-лекарственное средство, где L представляет собой линкер и D представляет собой лекарственное средство, и
где молекула линкер-лекарственное средство конъюгировано с антителом (i) на константной области тяжелой цепи через цистеин, соответствующий положению 173 последовательности SEQ ID NO:17, или (ii) на константной области легкой цепи через цистеин, соответствующий положению 76 последовательности SEQ ID NO:41.
2. Конъюгат антитело-лекарственное средство по п.1, где Ab содержит полипептид тяжелой цепи, кодируемый нуклеиновой кислотой с номером доступа в ATCC PTA-122672, и полипептид легкой цепи, кодируемый нуклеиновой кислотой с номером доступа ATCC PTA-122673.
3. Конъюгат антитело-лекарственное средство по п.1 или 2, где:
(a) тяжелая цепь содержит SEQ ID NO:18; и
(b) легкая цепь содержит SEQ ID NO:42.
4. Конъюгат антитело-лекарственное средство по любому из пп.1-3, где линкер выбран из группы, состоящей из малеимидокапроил-валин-цитруллин-p-аминобензилоксикарбонила (vc), AcLysvc, mc, MalPeg6, m(H20)c и m(H20)cvc.
5. Конъюгат антитело-лекарственное средство по п.4, где линкер является расщепляемым.
6. Конъюгат антитело-лекарственное средство по п.5, где линкером является малеимидокапроил-валин-цитруллин-p-аминобензилоксикарбонил (vc).
7. Конъюгат антитело-лекарственное средство по любому из пп.1-6, где лекарственное средство обладает мембранной проницаемостью.
8. Конъюгат антитело-лекарственное средство по любому из пп.1-7, где лекарственным средством является ауристатин.
9. Конъюгат антитело-лекарственное средство по п.8, где ауристатин является 2-метилаланил-N-[(3R,4S,5S)-3-метокси-1-{(2S)-2-[(1R,2R)-1-метокси-2-метил-3-оксо-3-{[(1S)-2-фенил-1-(1,3-тиазол-2-ил)этил]амино}пропил]пирролидин-1-ил}-5-метил-1-оксогептан-4-ил]-N-метил-L-валинамидом или его фармацевтически приемлемой солью или сольватом.
10. Конъюгат антитело-лекарственное средство для лечения злокачественной опухоли, эксперссирующей Her2, имеющий формулу:
Ab-(L-D),
где:
(a) Ab является антителом, которое связывается с HER2 и содержит тяжелую цепь, содержащую SEQ ID NO:18, и легкую цепь, содержащую SEQ ID NO:42; и
(b) L-D является молекулой линкера-лекарственного средства, где L является линкером малеимидокапроил-валин-цитруллин-p-аминобензилоксикарбонил (vc) и D является ауристатином 2-метилаланил-N-[(3R,4S,5S)-3-метокси-1-{(2S)-2-[(1R,2R)-1-метокси-2-метил-3-оксо-3-{[(1S)-2-фенил-1-(1,3-тиазол-2-ил)этил]амино}пропил]пирролидин-1-ил}-5-метил-1-оксогептан-4-ил]-N-метил-L-валинамидом или его фармацевтически приемлемой солью или сольватом, и
где молекула линкер-лекарственное средство конъюгировано с антителом (i) на тяжелой цепи через цистеин, соответствующий положению 293 последовательности SEQ ID NO:18, или (ii) на легкой цепи через цистеин, соответствующий положению 183 последовательности SEQ ID NO:42.
11. Фармацевтическая композиция для лечения злокачественной опухоли, экспрессирующей Her2, содержащая эффективное количество конъюгата антитело-лекарственное средство по любому из пп.1-9 и фармацевтически приемлемый носитель.
12. Композиция для лечения злокачественной опухоли, экспрессирующей Her2, содержащая эффективное количество конъюгата антитело-лекарственное средство по любому из пп.1-10 и, необязательно, фармацевтический носитель, где композиция имеет DAR, равное 4.
13. Нуклеиновая кислота, кодирующая тяжелую цепь или легкую цепь антитела, которое связывается с HER2 и содержит:
(1) вариабельную область тяжелой цепи, содержащую три CDR, содержащие SEQ ID NO:2, 3 и 4;
(2) константную область тяжелой цепи SEQ ID NO:17;
(3) вариабельную область легкой цепи, содержащую три CDR, содержащие SEQ ID NO:8, 9 и 10; и
(4) константную область легкой цепи SEQ ID NO:41.
14. Нуклеиновая кислота по п.13, содержащая нуклеотидную последовательность тяжелой цепи SEQ ID NO:54 или нуклеотидную последовательность SEQ ID NO:78.
15. Вектор экспрессии, содержащий нуклеиновую кислоту по любому из пп.13 или 14.
16. Клетка-хозяин для экспрессии антитела, которое связывается c HER2, содержащая нуклеиновую кислоту по любому из пп.13 или 14.
17. Способ получения конъюгата антитело-лекарственное средство по любому из пп.1-12, включающий:
(a) присоединение линкера к лекарственному средству;
(b) конъюгирование молекулы линкера-лекарственного средства с антителом; и
(c) очистку конъюгата антитело-лекарственное средство.
18. Способ по п.17, где конъюгирование является сайт-специфическим и имеет модифицированные цистеиновые остатки в положении 183 легкой цепи согласно нумерации Кэбата и в положении 290 тяжелой цепи согласно нумерации EU Кэбата антитела.
19. Способ лечения злокачественной опухоли, экспрессирующей HER2, включающий введение терапевтически эффективного количества композиции, включающей конъюгат антитело-лекарственное средство по любому из пп.1-11, пациенту.
20. Способ по п.19, где злокачественной опухолью является солидная опухоль.
21. Способ по п.20, где солидная опухоль выбрана из группы, состоящей из рака молочной железы, рака яичника, рака легкого, рака желудка, рака пищевода, рака толстой и прямой кишки, уротелиального рака, рака поджелудочной железы, рака слюнной железы и рака головного мозга.
22. Способ по п.21, где солидная опухоль выбрана из группы, состоящей из рака молочной железы, рака яичника, рака легкого и рака желудка.
23. Способ по п.22, где рак молочной железы является эстроген- и прогестерон-рецептор негативным или трижды негативным раком молочной железы (ТНРМЖ).
24. Способ по п.22, где рак легкого является немелкоклеточным раком (НМРЛ).
25. Способ по любому из пп.19-24, где пациент ранее получал лечение трастузумабом и/или трастузумабом эмтанзином, каждым отдельно или в комбинации с другим терапевтическим средством.
26. Способ по п.25, где терапевтическим средством является таксан.
27. Способ по п.25, где рак является резистентным к трастузумабу и/или трастузумабу эмтанзину, рефракторным к и/или рецидивирующим после лечения трастузумабом и/или трастузумабом эмтанзином, каждым отдельно или в комбинации с другим терапевтическим средством.
28. Способ по п.27, где терапевтическим средством является таксан.
29. Способ по любому из пп.19-28, где злокачественная опухоль экспрессирует HER2 на высоком уровне.
30. Способ по п.29, где злокачественная опухоль экспрессирует HER2 на уровне 3+, как определено с помощью иммуногистохимии (ИГХ), и/или по отношению амплификации при флуоресцентной гибридизации in situ (FISH) ≥2,0.
31. Способ по любому из пп.19-28, где злокачественная опухоль экспрессирует HER2 на среднем уровне.
32. Способ по п.31, где злокачественная опухоль экспрессирует HER2 на уровне 2+, как определено с помощью иммуногистохимии (ИГХ), и/или по отношению амплификации при флуоресцентной гибридизации in situ (FISH) <2,0.
33. Способ по любому из пп.19-28, где злокачественная опухоль экспрессирует HER2 на низком уровне.
34. Способ по п.33, где злокачественная опухоль экспрессирует HER2 на уровне 1+, как определено с помощью иммуногистохимии (ИГХ), и/или по отношению амплификации при флуоресцентной гибридизации in situ (FISH) <2,0.
35. Способ по любому из пп.30, 32 и 34, где ИГХ проводят при использовании анализа Dako HercptestTM и FISH проводят при использовании анализа Dako HER2 FISH Pharm DxTM.
36. Применение конъюгата антитело-лекарственное средство по любому из пп.1-11 для получения лекарственного средства для лечения злокачественной опухоли, экспрессирующей HER2, у пациента.
37. Применение по п.36, где злокачественной опухолью является солидная опухоль.
38. Применение по п.37, где солидная опухоль выбрана из группы, состоящей из рака молочной железы, рака яичника, рака легкого, рака желудка, рака пищевода, рака толстой и прямой кишки, уротелиального рака, рака поджелудочной железы, рака слюнной железы и рака головного мозга.
39. Применение по п.38, где солидная опухоль выбрана из группы, состоящей из рака молочной железы, рака яичника, рака легкого и рака желудка.
40. Применение по п.39, где рак молочной железы является эстроген- и прогестерон-рецептор негативным или трижды негативным раком молочной железы (ТНРМЖ).
41. Применение по п.39, где рак легкого является немелкоклеточным раком (НМРЛ).
42. Применение по любому из пп.37-41, где пациент ранее проходил лечение трастузумабом и/или трастузумабом эмтанзином, один из которых применяли отдельно или в комбинации с другим терапевтическим средством.
43. Применение по п.42, где терапевтическим средством является таксан.
44. Применение по п.42, где рак является резистентным к, рефракторным к и/или рецидивирующим после лечения трастузумабом и/или трастузумабом эмтанзином, один из которых применяют отдельно или в комбинации с другим терапевтическим средством.
45. Применение по п.44, где терапевтическим средством является таксан.
46. Применение по любому из пп.37-45, где злокачественная опухоль экспрессирует HER2 на высоком уровне.
47. Применение по п.46, где злокачественная опухоль экспрессирует HER2 на уровне 3+, как определено с помощью иммуногистохимии (ИГХ), и/или по отношению амплификации при флуоресцентной гибридизации in situ (FISH) ≥2,0.
48. Применение по любому из пп.37-45, где злокачественная опухоль экспрессирует HER2 на среднем уровне.
49. Применение по п.48, где злокачественная опухоль экспрессирует HER2 на уровне 2+, как определено с помощью иммуногистохимии (ИГХ), и/или по отношению амплификации при флуоресцентной гибридизации in situ (FISH) <2,0.
50. Применение по любому из пп.37-50, где злокачественная опухоль экспрессирует HER2 на низком уровне.
51. Применение по п.50, где злокачественная опухоль экспрессирует HER2 на уровне 1+, как определено с помощью иммуногистохимии (ИГХ), и/или по отношению амплификации при флуоресцентной гибридизации in situ (FISH) <2,0.
52. Применение по любому из пп.47, 49 или 51, где ИГХ проводят с помощью анализа Dako HercptestTM и FISH проводят с помощью анализа Dako HER2 FISH Pharm DxTM.
| 2005 |
|
RU2404810C2 | |
| WO 2012075581 A1, 14.06.2012 | |||
| АНТИ-CD79b АНТИТЕЛА И ИММУНОКОНЪЮГАТЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2009 |
|
RU2553566C2 |
| WO 2014022846 A1, 06.02.2014. | |||

