ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] Настоящая заявка испрашивает приоритет по предварительной заявке на патент США с серийным №61/910026, поданной 27 ноября 2013 года; по предварительной заявке на патент США с серийным №62/000908, поданной 20 мая 2014 года; и по предварительной заявке на патент США с серийным №62/009125, поданной 6 июня 2014 года, все из которых включены в данный документ в полном объеме посредством ссылки.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0002] В настоящей заявке содержится перечень последовательностей, который будет подан через EFS-Web и включен в данный документ в полном объеме посредством ссылки. Указанная копия ASCII, созданная в XX месяце 20ХХ года, называется XXXXXUS sequencelisting.txt, и ее размер составляет ХХХХХХХ байт.
УРОВЕНЬ ТЕХНИКИ
[0003] Большинство из коммерчески доступных в настоящее время терапевтических средств на основе антител представляют собой бивалентные моноспецифические антитела, оптимизированные и отобранные для высокоаффинного связывания и авидности, обеспечиваемой двумя антигенсвязывающими доменами. Чтобы повысить эффективность антител через реализацию Fc-зависимых механизмов клеточной цитотоксичности, было использовано дефукозилирование или улучшение связывающей способности FcgR посредством мутагенеза. Дефукозилированные антитела или антитела с улучшенной связывающей способностью FcgR все еще характеризуются неполной терапевтической эффективностью в клинических испытаниях, и для любого из этих антител все еще должен быть достигнут статус коммерчески доступного лекарственного вещества. Обычные бивалентные антитела, конъюгированные с токсинами (конъюгат антитела с лекарственным средством), являются более эффективными, но более широкое клиническое применение ограничено дозолимитирующей токсичностью.
[0004] В идеальном случае, терапевтические антитела должны обладать определенными минимальными характеристиками, включая специфичность по отношению к мишени, биостабильность, биодоступность и биораспределение после введения испытуемому пациенту, и достаточной аффинностью связывания с мишенью, а также высокой степенью заполненности мишени для оказания максимального антителозависимого терапевтического эффекта., как правило, терапевтические антитела являются моноспецифическими. Однако моноспецифическое направленное связывание не предусматривает связывания с другими эпитопами-мишенями, которые могут иметь отношение к сигнальным путям и патогенезу заболеваний, обеспечивающих устойчивость к лекарственным препаратам и механизм ускользания от их действия. Некоторые из современных терапевтических парадигм предусматривают применение комбинации двух терапевтических моноспецифических антител, направленно связывающихся с двумя разными эпитопами одного и того же антигена-мишени. Одним из таких примеров является применение комбинации трастузумаба и пертузумаба, оба из которых направленно связываются с рецепторным белком HER2, находящимся на поверхности некоторых раковых клеток, но, тем не менее, у некоторых пациентов данное заболевание прогрессирует, тогда как у других с более низкими уровнями экспрессии HER2-рецепторов (HER2 <3 + согласно тесту с герцептином) не отмечается никакого благоприятного терапевтического эффекта. Терапевтические антитела, направлено СВЯЗЫВАЮЩИЕСЯ с HER2, раскрыты в WO 2012/143523 для GenMab и WO 2009/154651 для Genentech. Антитела также описаны в WO 2009/068625 и WO 2009/068631.
[0005] В патентных заявках того же заявителя РСТ/СА2011/001238, поданной 4 ноября 2011 года, РСТ/СА2012/050780, поданной 2 ноября 2012 года, РСТ/СА2013/00471, поданной 10 мая 2013 года, и РСТ/СА2013/050358, поданной 8 мая 2013 года, описаны терапевтические антитела. Каждая заявка включена в данный документ в полном объеме посредством ссылки для всех целей.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В данном документе описаны бивалентные антигенсвязывающие конструкции, которые связывают HER2. Антигенсвязывающие конструкции содержат первую антигенсвязывающую полипептидную конструкцию, которая моновалентно и специфически связывается с антигеном HER2 (рецептор эпидермального фактора роста 2) ECD2 (внеклеточный домен 2), находящимся на HER2-экспрессирующих клетках, и вторую антигенсвязывающую полипептидную конструкцию, которая моновалентно и специфически связывается с антигеном HER2 ECD4 (внеклеточный домен 4), находящимся на HER2-экспрессирующих клетках, первый и второй линкерные полипептиды, причем первый линкерный полипептид функционально связан с первой антигенсвязывающей полипептидной конструкцией, а второй линкерный полипептид функционально связан со второй антигенсвязывающей полипептидной конструкцией;, при этом линкерные полипептиды способны к образованию ковалентной связи друг с другом, причем по меньшей мере одна ECD2- или ECD4-связывающая полипептидная конструкция представляет собой scFv. В определенных вариантах реализации изобретения ECD2-связывающая полипептидная конструкция представляет собой scFv, и ECD2-связывающая полипептидная конструкция представляет собой Fab. В определенных вариантах реализации изобретения ECD2-связывающая полипептидная конструкция представляет собой Fab, и ECD4-связывающая полипептидная конструкция представляет собой scFv. В некоторых вариантах реализации изобретения как ECD2-, так и ECD4-связывающие полипептидные конструкции представляют собой scFv. В некоторых вариантах реализации изобретения в составе антигенсвязывающих конструкций присутствует димерный Fc, содержащий последовательность CH3. В некоторых вариантах реализации изобретения Fc-домен представляет собой гетеродимер, характеризующийся одной или несколькими модификациями в последовательности СН3, которые способствуют образованию гетеродимера со стабильностью, сравнимой со стабильностью гомодимерного Fc-домена дикого типа. В некоторых вариантах реализации изобретения последовательность гетеродимерного CH3 имеет температуру плавления (Тпл) 68°С или выше. Также описаны нуклеиновые кислоты, кодирующие антигенсвязывающие конструкции, и веторы, а также клетки. Также описаны способы лечения заболевания, например, злокачественной опухоли, с использованием антигенсвязывающих конструкций, описанных в данном документе.
[0006] Также в настоящем изобретении предложена модифицированная конструкция пертузумаба, в которой содержатся мутации Y96A в VL-области и T30A/A49G/L70F в VH-области (пронумерованные согласно нумерации по Кабат). В одном варианте реализации изобретения модифицированная конструкция пертузумаба является моновалентной и характеризуется в 7-9 раз более высокой аффинностью к ECD2 HER2, чем пертузумаб. В определенных вариантах реализации изобретения модифицированная конструкция пертузумаба характеризуется форматом Fab/Fab, Fab/scFv или scFv/scFv.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0007] На Фигуре 1А представлена структура бипаратопного антитела в формате Fab-Fab. На Фигурах 1В-1Е представлена структура возможных вариантов бипаратопного антитела в формате scFv-Fab. На Фигуре 1В антигенсвязывающий домен 1 представляет собой scFv, слитый с Цепью А, тогда как антигенсвязывающий домен 2 представляет собой Fab, слитый с Цепью В. На Фигуре 1С антигенсвязывающий домен 1 представляет собой Fab, слитый с Цепью А, тогда как антигенсвязывающий домен 2 представляет собой scFv, слитый с Цепью В. На Фигуре 1D антигенсвязывающий домен 2 представляет собой Fab, слитый с Цепью А, тогда как антигенсвязывающий домен 1 представляет собой scFv, слитый с Цепью В. На Фигуре 1Е антигенсвязывающий домен 2 представляет собой scFv, слитый с Цепью А, тогда как антигенсвязывающий домен 1 представляет собой Fab, слитый с Цепью В. На Фигуре 1F оба ангигенсвязывающих домена представляют собой scFv.
[0008] На Фигуре 2 представлена характеристика экспрессии и очистки иллюстративных бипаратопных антител к HER2. На Фигуре 2А и Фигуре 2В представлена SEC-хроматограмма для антитела, очищенного с помощью хроматографии на белке А, и результаты экспрессии 10L, полученные с помощью электрофореза в полиакриламидном геле в невосстанавливающих условиях, а также результаты по очистке v5019. На Фигуре 2С представлены результаты экспрессии a25L, полученные с помощью электрофореза в полиакриламидном геле, а также результаты по очистке v10000.
[0009] На Фигуре 3 представлены результаты анализа с помощью СВЭЖХ-ЭХ иллюстративных бипаратопных антител к HER2, очищенных с помощью хроматографии на белке А и SEC. На Фигуре 3А показаны результаты для v5019, причем на верхней панели показаны результаты очистки, а на нижней панели показан то же самый результат с увеличенным масштабом для оси у. Полученные данные в краткой форме представлены ниже результатов, полученных с помощью СВЭЖХ-ЭХ. На Фигуре 3В показаны результаты для v10000.
[0010] На Фигуре 4 представлены результаты LCMS-анализа степени очистки гетеродимеров иллюстративных бипаратопных антител к HER2. На Фигуре 4А представлены результаты ЖХ-МС-анализа объединенных фракций v5019, выделенных с помощью ЭХ. На Фигуре 4В представлены результаты ЖХ-МС-анализа объединенных фракций v10000, выделенных с помощью хроматографии на белке А.
[0011] На Фигуре 5 представлены результаты анализа of a 25L-scale получение иллюстративного к бипаратопного антитела HER. На Фигуре 5А представлен профиль иллюстративного бипаратопного антитела к HER2, полученный с помощью электрофореза в полиакриламидном геле, после очистки с помощью MabSelect™ и HiTrap™ SP FF. На Фигуре 5В представлены результаты LCMS-анализа очищенного антитела.
[0012] На Фигуре 6 сравнивается способность иллюстративных бипаратопных антител к HER2 связываться с цельными клетками HER2+, характеризующимися разной плотностью HER2-рецепторов, по сравнению с контрольными антителами, что было измерено с помощью FACS. На Фигуре 6А и Фигуре 6Е представлено связывание с клетками SKOV3; на Фигуре 6В представлено связывание с клетками JIMT1; на Фигуре 6С и Фигуре 6F представлено связывание с клетками MCF7; на Фигуре 6D представлено связывание с клетками MDA-MB-231; и Фигуре 6G представлено связывание с клетками WI-38.
[0013] На Фигуре 7 представлена способность иллюстративных бипаратопных антител к HER2 ингибировать рост клеток HER2+. На Фигуре 7А и Фигуре 7D показано ингибирование роста клеток SKOV3; на Фигуре 7В показано ингибирование роста клеток ВТ-474; на Фигуре 1С показано ингибирование роста клеток SKBR3 и на Фигуре 7Е показано ингибирование роста клеток JIMT-1.
[0014] На Фигуре 8 представлены данные связывания, полученные с помощью ППР, относящиеся к паратопам иллюстративных бипаратопных антител к HER2. На Фигуре 8А проиллюстрированы значения KD (нМ) моновалентного антитела к HER2 (v1040: представляющего собой антигенсвязывающий домен, находящийся на СН-В иллюстративного бипаратопного антитела к HER2), для связывания с иммобилизированным Her2 ECD или димерным Her2-Fc. На Фигуре 8В проиллюстрированы значения KD (нМ) моновалентного антитела к HER2 (v4182; представляющего собой антигенсвязывающий домен, находящийся на СН-А иллюстративного бипаратопного антитела к HER2) для связывания с иммобилизированным Her2 ECD или димерным Her2-Fc.
[0015] На Фигуре 9 представлена способность иллюстративного бипаратопного антитела к HER2 интернализироваться в клетки HER2+. На Фигуре 9А представлена интернализация в клетки ВТ-474, тогда как как Фигуре 9B представлена интернализация в клетки JIMT-1.
[0016] На Фигуре 10 представлено поверхностное связывание и интернализация иллюстративных бипаратопных антител к HER2. На Фигуре 10А (v5019) представлен результат для клеток ВТ-474; на Фигуре 10В (v5019) и Фигуре 10F (v5019 и v10000) представлен результат для in JIMT1; на Фигуре 10С (v5019) и Фигуре 10Е (v5019 и v10000) представлен результат для клеток SKOV3 и на Фигуре 10D (v5019) представлен результат для клеток MCF7.
[0017] На Фигуре 11 представлена способность иллюстративного бипаратопного антитела к HER2 опосредовать АЗКЦ в клетках SKOV3. На Фигуре 11А показаны результаты анализа, проведенного с использованием соотношения эффекторных клеток к клеткам-мишеням 5:1; на Фигуре 11В показаны результаты анализа, проведенного с использованием соотношения эффекторных клеток к клеткам-мишеням 3:1; и на Фигуре 11С показаны результаты анализа, проведенного с использованием соотношения эффекторных клеток к клеткам-мишеням 1:1.
[0018] На Фигуре 12 представлена характеристика аффинности и кинетики связывания моновалентного антитела к HER2 (v630 и v4182) и иллюстративного бипаратопного антитела к HER2 (v5019) с рекомбинантным HER2 человека. На Фигуре 12А показаны результаты измерения ka (1/Мс). На Фигуре 12В показаны результаты измерения kd (1/с). На Фигуре 12С показаны результаты измерения KD (М).
[0019J На Фигуре 13 представлена характеристики аффинности и связывания иллюстративного бипаратопного антитела к HER2 с рекомбинантным HER2 человека для целога ряда уровней захвата антител. На Фигуре 13А представлены результаты измерения kd (1/с) для HER2 ECD, определенной для целога ряда уровней захвата антител для иллюстративного бипаратопного антитела к HER2 (v5019). На Фигуре 13В представлены результаты измерения kd (1/с) для HER2 ECD, определенной для целога ряда уровней захвата антител для моновалентного антитела к HER2 (v4182). На Фигуре 13С представлены результаты измерения kd (1/с) для HER2 ECD, определенной для целога ряда уровней захвата антител для моновалентного антитела к HER2 (v630).
[0020] На Фигуре 14 показано сравнение механизма связывания моноспецифического антитела к ECD4 HER2 (слева) и бипаратопного антитела Fab-scFv к ECD2x ECD4 HER2 (справа). Моноспецифическое антитело к ECD4 HER2 способно обеспечивать связывание одной молекулы антитела с двумя молекулами HER2; тогда как бипаратопное антитело к ECD2 х ECD4 HER2 способно обеспечивать связывание одного антитела с двумя молекулами HER2, а также 2 антитела с одной молекулой HER2 и их кобинации, что приводит к перекрестному сшиванию HER2-рецептора и формированию решетчатой структуры с последующими нисходящими биологическими эффектами, такими как интернализация и/или ингибирования роста, которые обозначены стрелками. ИЭК означает «иммунные эффекторные клетки». Четыре внеклеточные домена HER2 пронумерованы как 1, 2, 3 или 4, где 1=ECD1, 2=ECD2, 3=ECD3 и 4=ECD4.
[0021] На Фигуре 15 представлено влияние иллюстративного бипаратопного антитела к HER2 на фосфорилирование АКТ в клетках ВТ-474.
[0022] На Фигуре 16 представлено влияние иллюстративного бипаратопного антитела к HER2 на жизнеспособность кардиомиоцитов. На Фигуре 16А представлено влияние v5019 и соответствующего ADC конъюгата антитела с лекарственным средством v6363 на жизнеспособность кардиомиоцитов; на Фигуре 16В представлено влияние v5019, v7091 и v10000 и соответствующих ADC v6363, 7148, 10553 на жизнеспособность кардиомиоцитов и на Фигуре 16С представлено влияние v5019, v7091 и v10000 и соответствующих ADC v6363, 7148, 10553 на жизнеспособность кардиомиоцитов, предварительно обработанных доксорубицином.
[0023] На Фигуре 17 представлена способность иллюстративных ADC на основе бипаратопного антитела к HER2 ингибировать рост клеток HER2+. На Фигуре 17А показана способность конъюгата антитела с лекарственным средством v6363 ингибировать рост клеток JIMT1. На Фигуре 17В показана способность конъюгата антитела с лекарственным средством v6363 ингибировать рост клеток SKOV3. На Фигуре 17С показана способность конъюгата антитела с лекарственным средством v6363 ингибировать рост клеток MCF7. На Фигуре 17D показана способность конъюгата антитела с лекарственным средством v6363 ингибировать рост клеток MDA-MB-231. На Фигуре 17Е показана способность ADC v6363, v10553 и v1748 ингибировать рост клеток SKOV3. На Фигуре 17F показана способность ADC v6363, v10553 и v1748 ингибировать рост клеток JIMT-1. На Фигуре 17G показана способность ADC v6363, v10553 и v1748 ингибировать рост клеток NCI-N87.
[0024] На Фигуре 18 представлено влияние бипаратопного антитела к HER2 в модели ксенотрансплантации линии клеток рака яичников человека (SKOV3). На Фигуре 18А показано влияние антитела на средний объем опухоли. На Фигуре 18В показано влияние антитела на процент выживаемости животных.
[0025] На Фигуре 19 представлено влияние конъюгата ADC на основе бипаратопного антитела к HER2 (ADC) в модели ксенотрансплантации линии клеток рака яичников человека (SKOV3). На Фигуре 19А показано влияние антитела на средний объем опухоли. На Фигуре 19В показано влияние антитела на процент выживаемости животных.
[0026] На Фигуре 20 представлено влияние конъюгата ADC на основе бипаратопного антитела к HER2 (ADC) на средний объем опухоли в модели ксенотрансплантации первичной культуры клеток рака молочной железы человека (НВСх-13b).
[0027] На Фигуре 21 представлено влияние конъюгата ADC на основе бипаратопного антитела к HER2 (ADC) на средний объем опухоли в модели ксенотрансплантации первичной культуры клеток рака молочной железы человека (T226).
[0028] На Фигуре 22 представлено влияние конъюгата ADC на основе бипаратопного антитела к HER2 (ADC) на средний объем опухоли в модели ксенотрансплантации первичной культуры клеток рака молочной железы человека (НВСх-5).
[0029] На Фигуре 23 представлено влияние конъюгата ADC на основе бипаратопного антитела к HER2 (ADC) на опухоли, резистентные к лечению, направленному на подавление экспрессии HER2, в модели ксенотрансплантации линии клеток человека (SKOV3).
[0030] На Фигуре 24 представлено влияние конъюгата ADC на основе бипаратопного антитела к HER2 (ADC) на опухоли, резистентные к лечению, направленному на подавление экспрессии HER2, в модели ксенотрансплантации клеток первичной культуры человека (НВСх-13b).
[0031] На Фигуре 25 представлена термостабильность иллюстративных бипаратопных антител к HER2. На Фигуре 25А представлена термостабильность v5019. На Фигуре 25В представлена термостабильность v10000. На Фигуре 25С представлена термостабильность v7091.
[0032] На Фигуре 26 представлена термостабильность иллюстративных конъюгатов бипаратопных антител с лекарственными средствами. На Фигуре 26А представлена термостабильность v6363. На Фигуре 26В представлена термостабильность v10553. На Фигуре 26С представлена термостабильность v7148.
[0033] На Фигуре 27 представлена способность бипаратопных антител к HER2 опосредовать уничтожение клеток HER2+, путем АЗКЦ. Легенда, показанная на Фигуре 27С, применима к Фигуре 27А и Фигуре 27В. На Фигуре 27А представлена эта способность в клетках SKBR3; на Фигуре 27В представлена эта способность в клетках JIMT-1; на Фигуре 27С представлена эта способность в клетках MDA-MB-231; и на Фигуре 27D представлена эта способность в клетках WI-38.
[0034] На Фигуре 28 представлено влияние дефукозилирования на способность бипаратопных антител к HER2 опосредовать АЗКЦ. Легенда, показанная на Фигуре 28В. также применима к Фигуре 28А. На Фигуре 28А сравнивается способность дефукозилированного варианта v5019 опосредовать АЗКЦ со способностью Herceptin™ в клетках SKOV3. На Фигуре 28В сравнивается способность дефукозилированного варианта v5019 опосредовать АЗКЦ со способностью Herceptin™ в клетках MDA-MB-231. На Фигуре 28С сравнивается способность v10000 и дефукозилированного варианта v10000 опосредовать АЗКЦ со способностью Herceptin™ в клетках ZR-75-1.
[0035] На Фигуре 29 представлена способность v5019 ингибировать рост клеток ВТ-474 в присутствии или в отсутствие рост-стимулирующих лигандов.
[0036] На Фигуре 30 представлено влияние дефукозилированного варианта v5019 (v7187) на объем опухоли в модели ксенотрансплантации злокачественной опухоли молочной железы человека (НВСх13В).
[0037] На Фигуре 31 представлена способность бипаратопных антител к HER2 и ADC на основе бипаратопного антитела к HER2 связываться с HER2 + опухолевыми клетками. На Фигуре 31А сравнивается связывание v6363 с аналогом T-DM1, v6246, в клетках SKOV3. На Фигуре 31В сравнивается связывание v6363 с аналогом T-DM1, v6246, в клетках JIMT-1. На Фигуре 31С сравнивается связывание нескольких иллюстративных бипаратопных антител к HER2 и ADC на основе бипаратопного антитела к HER2 с контролями в клетках SKOV3. На Фигуре 31D сравнивается связывание нескольких иллюстративных бипаратопных антител к HER2 и ADC на основе бипаратопного антитела к HER2 с контролями в клетках JIMT-1.
[0038] На Фигуре 32 представлено Дозозависимое ингибирование опухолевого роста иллюстративным конъюгатом бипаратопного антитела к HER2 с лекарственным средством в модели ксенотрансплантации (ER-PR-негативной) опухоли пациента с уровнем экспрессии HER2 3+ (HBCx13b). На Фигуре 32А показано влияние v6363 на объем опухоли, тогда как как Фигуре 32В показано влияние на процент выживаемости.
[0039] На Фигуре 33 представлено влияние ADC на основе бипаратопного антитела к HER2 v6363 по сравнению со комбинациями, соответствующими стандарту лечения, в модели ксенотрансплантации PDX НВСх-13b, резистентных к трастузумабу. Фигура 33А представлено влияние лечения на объем опухоли, тогда как Фигуре 33В представлено влияние лечения на выживаемость.
[0040] На Фигуре 34 представлена эффективность ADC на основе бипаратопного антитела к HER2 в модели ксенотрансплантации опухоли, полученной из HER2 + клеток рака молочной железы, резистентных к трастузумабу (JIMT-1).
[0041] На Фигуре 35 представлена эффективность иллюстративных бипаратопных антител к HER2 in vivo в модели ксенотрансплантации опухоли, полученной из клеток рака яичников, чувствительных к трастузумабу (SKOV3). На Фигуре 35А представлено влияние лечения на объем опухоли, тогда как на Фигуре 35В представлено влияние лечения на выживаемость.
[0042] На Фигуре 36 представлена дозозависимая эффективность иллюстративных бипаратопных антител к HER2 in vivo в модели ксенотрансплантации опухоли, полученной из клеток рака яичников, чувствительных к трастузумабу (SKOV3).
[0043] На Фигуре 37 представлена способность бипаратопного антитела к HER2 и ADC на основе бипаратопного антитела к HER2 ингибировать рост клеточных линий, экспрессирующих HER2 и EGFR и/или HER3 на уровне 3+, 2+ или 1+. На Фигуре 37А представлена способность v10000 ингибировать рост выбранных клеточных линий. На Фигуре 37В представлена способность v10553 ингибировать рост выбранных клеточных линий.
[0044] На Фигуре 38 представлены общие результаты по способности v10000 и v10553 ингибировать рост ряда клеточных линий. Разделенные косой чертой значения (например, 1/2) обозначают противоречивые данные по уровню экспрессии рецептора erbb, которые сообщаются в литературе; ИГХ-значения для экспрессии Erbb были получены авторами изобретения или из литературы. Там, где не приведено никакого значения, количество рецепторов неизвестно и/или не сообщается. *Уровень экспрессии по ИГХ-анализу позволяет оценить данные экспрессии гена erBb2 (Crown BioSciences). Пронумерованные литературные источники описаны ниже.
[0045] На Фигуре 39 представлена способность v 10000 опосредовать уничтожение клеток HER2+, путем АЗКЦ,. На Фигуре 39А представлены результаты для клеток FaDu. На Фигуре 39В представлены результаты для клеток А549. На Фигуре 39С представлены результаты для клеток ВхРСЗ. На Фигуре 39D представлены результаты для клеток MiaPaca2.
[0046] На Фигуре 40 представлена способность бипаратопных антител к HER2 опосредовать уничтожение клеток HER2+, путем АЗКЦ. На Фигуре 40А представлены результаты для клеток А549. На Фигуре 40В представлены приводит к NCI-N87 клетки. Фигура 40С представлены результаты для клеток НСТ-116.
[0047] На Фигуре 41 представлено влияние формата бипаратопного антитела к HER2 на связывание с клеток HER2+. На Фигуре 41А представлено влияние формата на связывание с клетками ВТ-474. На Фигуре 41В представлено влияние формата на связывание с клетками JIMT-1. На Фигуре 41С представлено влияние формата на связывание с клетками MCF7. На Фигуре 41D представлено влияние формата на связывание с клетками MDA-MB-231.
[0048] На Фигуре 42 представлено влияние формата бипаратопного антитела к HER2 на интернализацию антитела в клетках HER2+. На Фигуре 42А представлено влияние на интернализацию в клетках ВТ-474. На Фигуре 42В представлено влияние на интернализацию в клетках JIMT-1. На Фигуре 42С представлено влияние на интернализацию в клетках MCF7.
[0049] На Фигуре 43 представлено влияние формата бипаратопного антитела к HER2 на способность опосредовать уничтожение клеток HER2+, путем АЗКЦ. На Фигуре 43А представлено влияние на клетки JIMT-1. На Фигуре 43В представлено влияние на клетки MCF7. На Фигуре 43С представлено влияние на клетки опухоли молочной железы HER2 0/1+MDA-MB-231.
[0050] На Фигуре 44 представлено влияние формата бипаратопного антитела к HER2 на способность антитела ингибировать рост HER2+опухолевых клеток ВТ-474 в присутствии или в отсутствие рост-стимулирующих лигандов.
[0051] На Фигуре 45 представлено влияние формата бипаратопного антитела к HER2 на способность антитела ингибировать рост клеток SKBR3.
[0052] На Фигуре 46 представлено влияние формата бипаратопного антитела к HER2 на способность антитела ингибировать рост HER2+ опухолевых клеток. На Фигуре 46А представлено ингибирование роста клеток SKOV3. На Фигуре 46В представлено ингибирование роста клеток JIMT-1. На Фигуре 46С представлено ингибирование роста клеток MCF7.
[0053] На Фигуре 47 представлены результаты сравнения характеристик связывания бипаратопных антител к HER2 различных форматов, что были измерены с помощью ППР. На Фигуре 47А представлен график и результаты линейного регрессионного анализа для kd (1/с) при разных уровнях захвата антител v6903 и v7091. Фигура 47В представлен график и результаты линейного регрессионного анализа для KD (М) при разных уровнях захвата антител v6903 и v7091.
[0054] Информация, представленная на Фигуре 38, взята из следующих литературных источников: 1. Labouret et al. 2012, Neoplasia 14:121-130; 2. Ghasemi et al. 2014, Oncogenesis doi:10.1038/oncsis.2014.31; 3. Gaborit et al. 2011 J Bio Chem, 286:1133-11345; 4. Kimura et al. 2006, Clin Cancer Res; 12:4925-4932; 5. Komoto et al. 2009, Cane Sci; 101:468-473; 6. Cretellaet al. 2014, Молекулярных Cancer 13:143-155; 7. Bunn et al. 2001, Clin Cancer Res; 7:3239-3250; 8. Lewis Phillips et al. 2013, Clin Cancer Res. 20:456-468; 9. McDonagh et al. 2012, 11:582-593; 10. Coldren et al. 2006, Mol Cancer Res:521-528; 11. Cavazzoni et al. 2012 Mol Cancer, 11:91-115; 12. Li et al. 2014, Mol Cancer Res, doi:10.1158/1541-7786.MCR-13-0396; 13. Chmielewski et al. 2004, Immunology, 173:7647-7653; 14. Kuwada et al. 2004, Int J Cancer, 109:291-301; 15. Fujimoto-Ouchi et al. 2007, Clin Chemother Pharmacol, 59:795-805; 16. Chavez-Bianco et al. 2004, BMC Cancer, 4:59; 17. Campiglio et al. 2004, J Cellular Physiology. 198:259-268; 18. Lehmann et al. 2011, J Clin Investigation, 121:2750-2767; 19. Collins et al. 2011. Annals Oncology, 23:1788-1795; 20. Takai et al. 2005, Cancer, 104:2701-2708; 21. Rusnack et al. 2007, Клетка Prolif, 40:580-594; 22. Ma et al. 2013, PLOS ONE, 8:e73261-e73261; 23. Meira et al. 2009, British J Cancer, 101:782-791; 24. Hayashi MP28-14 poster; 25. Wang et al. 2005 J Huazhong Univ Sci Technolog Med Sci. 25:326-8; 26. Makhja et al. 2010. J Clinc Oncolo 28:1215-1223.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0055] В данном документе описаны антигенсвязывающие конструкции, содержащие первую антигенсвязывающую полипептидную конструкцию, которая моновалентно и специфически связывается с антигеном HER2 (рецептор эпидермального фактора роста 2) ECD2 (внеклеточный домен 2), находящимся на HER2-экспрессирующих клетках, и вторую антигенсвязывающую полипептидную конструкцию, которая моновалентно и специфически связывается с антигеном HER2 ECD4 (внеклеточный домен 4), находящимся на HER2-экспрессирующих клетках, причем по меньшей мере одна ECD2- или ECD4-связывающая полипептидная конструкция представляет собой scFv. В определенных вариантах реализации изобретения ECD2-связывающая полипептидная конструкция представляет собой scFv, и ECD4-связывающая полипептидная конструкция представляет собой Fab. В определенных вариантах реализации изобретения ECD2-связывающая полипептидная конструкция представляет собой Fab, и ECD4-связывающая полипептидная конструкция представляет собой scFv. В некоторых вариантах реализации изобретения как ECD2-, так и ECD4-связывающие полипептидные конструкции представляют собой scFv. В некоторых вариантах реализации изобретения в составе антигенсвязывающих конструкций присутствует димерный Fc, содержащий последовательность СН3. В некоторых вариантах реализации изобретения Fc-домен представляет собой гетеродимер, характеризующийся одной или несколькими модификациями в последовательности СН3, которые способствуют образованию гетеродимера со стабильностью, сравнимой со стабильностью гомодимерного Fc-домена дикого типа. В некоторых вариантах реализации изобретения последовательность гетеродимерного CH3 имеет температуру плавления (Tra) 68°С или выше.
[0056] Антигенсвязывающие конструкции проявляют противоопухолевое действие in vitro, как, например, (i) способность ингибировать рост раковых клеток как в присутствии, так и в отсутствие стимуляции эпидермальным фактором роста или херегулином, (ii) способность к интернализации в раковые клетки и (iii) способность опосредовать антителозависимую клеточноопосредованную цитотоксичность (АЗКЦ). Это действие in vitro наблюдается как в случае использования свободных антигенсвязывающих конструкций (т.е. неконъюгированные с лекарственным веществом или токсином), так и в случае использования антигенсвязывающих конструкций, конъюгированных с майтансином, а также при различных уровнях экспрессии HER2 (1+, 2+ и 3+).
[0057] В данном документе показано, что формат (scFv/scFv, scFv/Fab или Fab/Fab) антигенсвязывающих конструкций имеет важное значение в определении его функционального профиля. В определенных вариантах реализации изобретения конструкции, обладающие специфичностью связывания по отношению к HER2, характеризуются повышенной способностью к интернализации HER2-экспрессирующими опухолевыми клетками по сравнению с эталонной бипаратопной антигенсвязывающей конструкцией, в которой как ECD2-, так и ECD4-связывающие полипептидные конструкции представляют собой Fab. Один вариант реалицазии, в котором как ECD2-, так и ECD4-связывающие полипептиды представляют собой scFv, интернализуются в большей степени опухолевыми клетками, экспрессирующими HER2 на уровне 1+, 2+ или 3+, чем обладающие эквивалентой аффинностью конструкции, которые характеризуются форматом Fab/scFv, что в свою очередь интернализуются более эффективно, чем обладающие эквивалентой аффинностью конструкции, которые характеризуются форматом Fab/Fab. Варианты реализации изобретения, которые быстро интернализуются, являются хорошими кандидатами для получения конъюгатов антитела с лекарственным средством, которые требуют интернализации опухолевой клеткой для обеспечения уничтожения клеток.
[0058] В определенных вариантах реализации изобретения антигенсвязывающие конструкции характеризуются повышенной способностью к уничтожению опухолевых клеток путем АЗКЦ, которые экспрессируют низкие уровни HER2, по сравнению с обладающими эквивалентой аффинностью конструкциями, которые характеризуются форматом Fab/Fab. В одном варианте реализации изобретения антигенсвязывающая конструкция, характеризующаяся форматом Fab/scFv, обладает большей активностью при уничтожении опухолевых клеток путем АЗКЦ, экспрессирующих низкие уровни HER2 (HER2 0-1+ или 1+), чем конструкция, обладающая специфичностью по отношению к HER2, характеризующаяся форматом Fab/Fab, которая в свою очередь обладает большей активностью, чем антигенсвязывающая конструкция, характеризующаяся форматом scFv/scFv.
[0059] В некоторых вариантах реализации изобретения антигенсвязывающие конструкции, обладающие специфичностью по отношению к HER2, гликозилированы.
[0060] В некоторых вариантах реализации изобретения конструкции, обладающие специфичностью связывания по отношению к HER2, дефукозилированы. В некоторых вариантах реализации конструкции, обладающие специфичностью связывания по отношению к HER2, конъюгированы с лекарственным веществом. В некоторых вариантах реализации изобретения конструкции, обладающие специфичностью связывания по отношению к HER2, конъюгированы с майтансином (DM1) через линкер SMCC.
[0061] Также в данном документе описаны способы лечения субъекта, имеющею HER2+опухоль путем введения субъекту антигенсвязывающей конструкции, обладающей специфичностью по отношению к HER2. В некоторых вариантах реализации изобретения уровень экспрессии HER2 на опухолевых клетках составляет 2+ или ниже. В некоторых вариантах реализации изобретения антигенсвязывающая конструкция конъюгирована с майтансином. В определенных вариантах реализации изобретения опухоль представляет собой рак поджелудочной железы, рак головы и шеи, рак ЖКТ, колоректальный рак, рак молочной железы, рак почек, рак шейки матки, рак яичников, рак эндометрия, рак матки, злокачественная меланома. рак глотки, рак ротовой полости или рак кожи. В некоторых вариантах реализации изобретения опухоль представляет собой (i) негативный по рецепторам эстрогенов (ER-), негативный по рецепторам прогестерона (PR-), резистентный к трастузумабу, резистентный к химиотерапии инвазивный протоковый рак молочной железы с уровнем экспрессии HER2 3+, (ii) резистентный к трастузумабу отечно-инфильтративный рак молочной железы (ER-, PR-) с уровнем экспрессии HER2 3+, (iii) инвазивный протоковый рак молочной железы (ER-, PR-) с уровнем экспрессии HER2 3+ или (iv) резистентный к трастузумабу и пертузумабу рак молочной железы с амплификацией гена HER2 и уровнем экспресии HER2 2+.
[0062] Также в настоящем изобретении предложены способы ингибирования роста опухолевых клеток или уничтожения опухолевых клеток путем введения антигенсвязывающих конструкций.
[0063] Также в настоящем изобретении предложена модифицированная конструкция пертузумаба, в которой содержатся мутации Y96A в VL-области и T30A/A49G/L70F в VH-области (согласно нумерации по Кабат). В одном варианте реализации изобретения модифицированная конструкция пертузумаба является моновалентной и характеризуется в 7-9 раз более высокой аффинностью к ECD2 HER2, чем пертузумаб. В определенных вариантах реализации изобретения модифицированная конструкция пертузумаба характеризуется форматом Fab/Fab, Fab/scFv или scFv/scFv.
Биспецифические антигенсвязывающие конструкции
[0064] В настоящем изобретении предложены биспецифические антигенсвязывающие конструкции, которые связывают HER2. Биспецифическая антигенсвязывающая конструкция включает две антигенсвязывающие полипептидные конструкции, причем каждая из них специфически связывается с разным эпитопом HER2. В некоторых вариантах реализации изобретения антигенсвязывающая конструкция получена из известного антитела или антигенсвязывающей конструкции., как описано более подробно ниже, антигенсвязывающие полипептидные конструкции могут представлять собой, в том числе, конструкции белковой природы, такие как Fab (антигенсвязывающий фрагмент), scFv (одноцепочечный Fv) и sdab (однодоменное антитело)., как правило, антигенсвязывающая конструкция включает Fc.
[0065] Термин «антигенсвязывающая конструкция» относится к любому средству, например, полипептиду или полипептидному комплексу, который способен связываться с антигеном. В некоторых аспектах антигенсвязывающая конструкция представляет собой полипептид, который специфически связывается с с представляющим итерес антигеном. Антигенсвязывающая конструкция может представлять собой мономер, димер, мультимер, белки, пептид или белки, или пептидный комплекс; антитело, фрагмент антитела или его антигенсвязывающий фрагмент; scFv и тому подобное. Антигенсвязывающая конструкция может представлять собой полипептидную конструкцию, которая является моноспецифической, биспецифической или мультиспецифической. В некоторых аспектах антигенсвязывающая конструкция может включать, например, один или более антигенсвязывающих компонентов (например, Fab или scFv), связанных с одним или более Fc. Дополнительные примеры антигенсвязывающих конструкций описаны ниже и приведены в примерах.
[0066] Подразумевается, что термин «биспецифический» охватывает любое средство, например, антигенсвязывающую конструкцию, которая имеет два антигенсвязывающих фрагмента (например, антигенсвязывающие полипептидные конструкции), причем каждый из них обладает уникальной специфичностью связывания. Например, первый антигенсвязывающий фрагмент связывается с эпитопом на первом антигене, и второй антигенсвязывающий фрагмент связывается с эпитопом на втором антигене. Используемый в данном документе термин «бипаратопное», относится к биспецифическому антителу, в котором первый антигенсвязывающий фрагмент и второй антигенсвязывающий фрагмент связываются с разными эпитопами на одном и том же антигене. Бипаратопное биспецифическое антитело может связываться с двумя эпитопами на одной и той же молекуле антигена или оно может связываться с эпитопами на двух разных молекулах антигена.
[0067] Моноспецифическая антигенсвязывающая конструкция относится к антигенсвязывающей конструкции с одной специфичностью связывания. Другими словами, оба антигенсвязывающих фрагмента связываются с одним и тем же эпитопом на одном и том же антигене. Примеры моноспецифических антигенсвязывающих конструкций включают трастузумаб, пертузумаб, которые связываются, например, с HER2.
[0068] Антигенсвязывающая конструкция может представлять собой антитело или его антигенсвязывающий участок. Используемый в данном документе термин «антитело» или «иммуноглобулин» относится к полипептиду, который, по существу, кодируется геном иммуноглобулина или генами иммуноглобулинов, или его фрагментам, которые специфически связываются и распознают анализируемое вещество (например, антиген). Общеизвестные гены иммуноглобулинов включают каппа-, лямбда-, альфа-, гамма-, дельта-, эпсилон- и мю-гены константной области, а также большое число генов вариабельной области иммуноглобулинов. Легкие цепи классифицируют либо на каппа, либо лямбда. «Класс» антитела или иммуноглобулина относится к типу константного домена или константной области, содержащихся в его тяжелой цепи. Выделяют пять основных классов антител: IgA, IgD, IgE, IgG и IgM, и некоторые из них могут дополнительно разделяться на подклассы (изотипы), например, IgGi, IgG2, IgG3, IgG4, IgAi и IgA2. Константные домены тяжелой цепи, которые соответствуют разным классам иммуноглобулинов, называются α, δ, ε, γ и μ, соответственно.
[0069] Структурная единица иллюстративного иммуноглобулина (антитела) состоит из двух пар полипептидных цепей, причем каждая пара имеет одну «легкую» (около 25 кДа) и одну «тяжелую» цепь (около 50-70 кДа). N-концевой домен каждой цепи определяет вариабельную область, состоящую из около 100-110 или более аминокислот главным образом отвечающих за распознавание антигена. Термины вариабельная область легкой цепи (VL) и вариабельная область тяжелой цепи (VH) относится к этим доменам легкой и тяжелой цепей, соответственно. Тяжелая цепь IgG1 состоит из VH-, СН1-, СН2- и СН3-доменов, соответственно, от N- к С-концу. Легкая цепь состоит из VL- и CL-доменов от N- к С-концу. Тяжелая цепь IgG1 содержит шарнир между СН1- и СН2-доменами. В определенных вариантах реализации изобретения иммуноглобулиновые конструкции содержат по меньшей мере один иммуноглобулиновый домен из IgG, IgM, IgA, IgD или IgE, соединенный с терапевтическим полипептидом. В некоторых вариантах реализации изобретения иммуноглобулиновый домен, встречающийся в антигенсвязывающей конструкции, представленной в настоящем изобретении, происходит из или получен из конструкции на основе иммуноглобулина, такой как диатело или нанотело. В определенных вариантах реализации изобретения иммуноглобулиновые конструкции, описанные в данном документе, содержат по меньшей мере один иммуноглобулиновый домен из тяжелой цепи антитела, как, например, верблюжьего антитела. В определенных вариантах реализации изобретения иммуноглобулиновые конструкции, представленные в настоящем изобретении, содержат по меньшей мере один иммуноглобулиновый домен из антитела млекопитающих, как, например, бычьего антитела, человеческого антитела, верблюжьего антитела, мышиного антитела или любого рекомбинантного антитела.
[0070] Используемый в данном документе термин «гипервариабельная область» или «HVR» относится к каждой из областей вариабельного домена антитела, которые имеют гипервариабельную последовательность и/или образуют структурно определенные петли («гипервариабельные петли»). Как правило, нативные четырехцепочечные антитела содержат шесть HVR; три в VH (H1, Н2, Н3) и три в V1 (L1, L2, L3). HVR, как правило, содержат аминокислотные остатки из гипервариабельных петель и/или из участков, определяющих комплементарность (CDR), последние характеризуются самой высокой вариабельностью последовательностей и/или вовлечены в распознавание антигена. За исключением CDR1 в VH, CDR, как правило, содержат аминокислотные остатки, которые формируют гипервариабельные петли. Гипервариабельные области (HVR) также называются «участками, определяющими комплементарность» (CDR), и эти термины используются в данном документе взаимозаменяемо в отношении участков вариабельной области, которые формируют антигенсвязывающие области. Эта определенная область была описана Kabat et al., U.S. Dept. of Health and Human Services, Sequenses of Proteins of Immunological Interest (1983) и Chothia et al., J Mol Biol 196:901-917 (1987), где определения подразумевают перекрывание подгрупп аминокислотных остатков при сравнении друг с другом. Несмотря на это, предполагается, что определение термина, использованного в данном документе для обозначения CDR антитела или его вариантов, охватывает применение любого из этих определений. Соответствующее аминокислотные остатки, которые входят в состав CDR, как определено в каждом из процитированных выше источников, изложены ниже в Таблице 1 для сравнения. Точное число остатков, которые входят в состав специфического CDR, будет зависеть от последовательности и размера CDR. Специалисты в данной области техники смогут стандартным способом определить, какие остатки содержатся в определенном CDR, если известна вариабельная область аминокислотной последовательности антитела.
[0071] Используемый в данном документе термин «одноцепочечный» относится к молекуле, содержащей аминокислотные мономеры, связанные с помощью пептидных связей с образованием линейной последовательности. В определенных вариантах реализации изобретения одна из антигенсвязывающих полипептидных конструкций представляет собой одноцепочечную Fab-молекулу, т.е. Fab-молекулу, в которой Fab легкой цепи и Fab тяжелой цепи соединяются с помощью пептидного линкера с образованием одной пептидной цепи. В таком конкретном варианте реализации изобретения С-конец Fab легкой цепи соединяется с N-концом Fab тяжелой цепи с образованием одноцепочечной Fab-молекулы. В определенных других вариантах реализации одна из антигенсвязывающих полипептидных конструкций представляет собой одноцепочечную Fv-молекулу (scFv)., как описано более подробно в данном документе, в scFv содержится вариабельный домен легкой цепи (VL), соединенный со стороны его С-конца с N-концевой областью вариабельного домена тяжелой цепи (VH) с помощью полипептидной цепи. В качестве альтернативы, scFv состоит из полипептидной цепи, в которой С-концевая область VH соединяется с N-концевой областью VL с помощью полипептидной цепи.
Антигенсвязывающая полипептидная конструкция
[0072] Биспецифическая антигенсвязывающая конструкция содержит две антигенсвязывающие полипептидные конструкции, каждая из которых связывается с определенным доменом или эпитопом HER2. В одном варианте реализации изобретения каждая антигенсвязывающая полипептидная конструкция связывается с внеклеточным доменом HER2, например, ECD2 или ECD4. Антигенсвязывающая полипептидная конструкция может представлять собой, например, Fab или scFv, в зависимости от применения.
[0073] Формат биспецифической антигенсвязывающей конструкции определяет функциональные характеристики биспецифической антигенсвязывающей конструкции. В одном варианте реализации изобретения биспецифическая антигенсвязывающая конструкция характеризуется форматом scFv-Fab (т.е. одна антигенсвязывающая полипептидная конструкция представляет собой scFv, и другая антигенсвязывающая полипептидная конструкция представляет собой Fab, которые также назваются форматом Fab-scFv). В другом варианте реализации изобретения биспецифическая антигенсвязывающая конструкция характеризуется форматом scFv-scFv (т.е. обе антигенсвязывающие полипептидные конструкции представляют собой scFv).
[0074] «Fab-фрагмент» (также называется антигенсвязывающим фрагментом) содержит константный домен (CL) легкой цепи и первый константный домен (СН1) тяжелой цепи наряду с вариабельными доменами VL и VH, находящимися в легкой и тяжелой цепях, соответственно. Вариабельные домены содержат петли, определяющие комплементарность (CDR, также называется гипервариабельной областью), которые вовлечены в процесс связывания антигена. Fab'-фрагменты отличаются от Fab-фрагментов по дополнительным нескольким остаткам на карбоксильном конце СН1-домена тяжелой цепи, включая один или более цистеинов из шарнирной области антитела.
[0075] «Одноцепочечный Fv» или «scFv» включает VH- и VL-домены антитела, причем эти домены содержатся в одной полипептидной цепи. В одном варианте реализации изобретения Fv-полипептид дополнительно содержит полипептидный линкер между VH- и VL-доменами, что обеспечивает формирование необходимой структуры scFv для связывания антигена. В отношении обзора, касающегося scFv, см. Pluckthun в The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds., Springer-Verlag, New York, pp. 269-315 (1994). ScFv-фрагменты антитела к HER2 описаны в WO 93/16185; в патенте США №5571894; и в патенте США №5587458.
[0076] Формат «однодоменных антител» или «sdAb» представляет собой отдельный иммуноглобулиновый домен. Sdab являются довольно стабильными и без труда экспрессируются как партнеры по слиянию с Fc-цепью антитела (Harmsen ММ, De Haard HJ (2007). "Properties, production и applications of camelid single- домен antibody fragments ". Appl. Microbiol Biotechnol. 77(1): 13-22).
Формат и функция антигенсвязывающих конструкций
[0077] В настоящем изобретении предложены бипаратопные конструкции, связывающие антиген HER2, имеющие две антигенсвязывающие полипептидные конструкции, первая из которых специфически связывается с ECD2 HER2, а вторая из которых специфически связывается с HER2 ECD4. Формат антигенсвязывающей конструкции является таким, что по меньшей мере один из первого или второго антигенсвязывающего полипептида представляют собой scFv. Формат антигенсвязывающей конструкции может представлять собой scFv-scFv или Fab-scFv, или scFv-Fab (первая антигенсвязывающая полипептидная конструкция-второй антигенсвязывающий полипептид, соответственно).
[0078] В определенных вариантах реализации изобретения антигенсвязывающие конструкции проявляют противоопухолевое действие in vitro, как, например, (i) способность ингибировать рост раковых клеток как в присутствии, так и в отсутствие стимуляции эпидермальным фактором роста или херегулином, (ii) способность к интернализации в раковые клетки (посредством связывания с HER2-антигеном и его интернализации) и (iii) способность опосредовать антителозависимую клеточноопосредованную цитотоксичность (АЗКЦ). Это действие in vitro наблюдается и в случае использования свободных антигенсвязывающих конструкций и антигенсвязывающих конструкций, конъюгированных с майтансином, и при различных уровнях экспрессии HER2 (1+, 2+ и 3+).
[0079] Примеры в данном документе показывают, что формат (scFv/scFv, scFv/Fab или Fab/Fab) антигенсвязывающих конструкций имеет важное значение в определении их функционального профиля. В определенных вариантах реализации изобретения конструкции, обладающие специфичностью связывания по отношению к HER2, характеризуются повышенной способностью к интернализации HER2-экспрессирующими опухолевыми клетками по сравнению с эталонной антигенсвязывающей конструкцией, в которой как ECD2-, так и ECD4-связывающие полипептидные конструкции представляют собой Fab. Подразумевается, что эффективность интернализации антигенсвязывающих конструкций, обладающих специфичностью по отношению к HER2, может быть дополнительно повышена путем увеличения аффинности одной или обеих антигенсвязывающих полипептидных конструкций по отношению к ECD2 или ECD4. Один вариант реалицазии, в котором как ECD2-, так и ECD4-связывающие полипептиды представляют собой scFv, интернализуются в большей степени опухолевыми клетками, экспрессирующими HER2 на уровне 1+, 2+ или 3+, чем обладающие эквивалентой аффинностью конструкции, которые характеризуются форматом Fab/scFv, что в свою очередь интернализуются более эффективно, чем обладающие эквивалентой аффинностью конструкции, которые характеризуются форматом Fab/Fab. Варианты реализации изобретения, которые быстро интернализуются, являются хорошими кандидатами для получения конъюгатов антитела с лекарственным средством, которые требуют интернализации опухолевой клеткой для обеспечения уничтожения клеток. В противоположность этому, в определенных вариантах реализации изобретения антигенсвязывающие конструкции, которые не так быстро интернализуются, характеризуются повышенной способностью к уничтожению опухолевых клеток путем АЗКЦ, которые экспрессируют низкие уровни HER2, по сравнению с конструкциями, обладающими эквивалентной аффинностью, которые характеризуются форматом Fab/Fab. В одном варианте реализации изобретения антигенсвязывающая конструкция, характеризующаяся форматом Fab/scFv, обладает большей активностью при уничтожении опухолевых клеток путем АЗКЦ, экспрессирующих низкие уровни HER2 (HER2 0-1+или1+), чем конструкция, обладающая специфичностью по отношению к HER2, характеризующаяся форматом Fab/Fab, которая в свою очередь обладает большей активностью, чем антигенсвязывающая конструкция, характеризующаяся форматом scFv/scFv. Повышенная способность некоторых вариантов реализации изобретения к уничтожению опухолевых клеток путем АЗКЦ может быть обусловлена 1) их повышенной способностью к авидному связыванию с клетками с низкой плотностью рецепторов HER2 и последующей кластеризации HER2-рецепторов на поверхности клетки-мишени, и тем, что они опосредуют downstream клеточно-опосредованное уничтожение клеток; и/или 2) их повышенной способностью оставаться на клеточной поверхности (а не вызывать интернализацию); следовательно, они более доступны для клеточно-опосредованного уничтожения клеток.
HER2
[0080] Антигенсвязывающие конструкции, описанные в данном документе, представляют собой антигенсвязывающие полипептидные конструкции, которые связываются с ECD2 и ECD4 HER2.
[0081] Термины «ErbB2» и «HER2» используются в данном документе взаимозаменяемо и относятся к белку HER2 человека, описанному, например, в Semba et al., PNAS (США) 82:6497-6501 (1985) и Yamamoto et al. Nature 319:230-234 (1986) (номер доступа в Genebank X03363). Термин «erbB2» и «neu» относится к гену, кодирующему белок ErbB2 человека. Термины p185 или p185neu относится к белковому продукту гена neu.
[0082] HER2 представляет собой рецептор HER. «HER-рецептор» представляет собой рецепторную тирозинкиназу, которая принадлежит к семейству рецепторов эпидермального фактора роста (HER) и включает рецепторы EGFR, HER2, HER3, а также HER4. HER-рецептор, как правило, содержит внеклеточный домен, который может связываться с HER-лигандом; липофильный трансмембранный домен; консервативный внутриклеточный тирозинкиназный домен; и карбокситерминальный сигнальный домен, содержащий несколько тирозиновых остатков, которые могут быть фосфорилированными. Под выражением «HER-лиганд» подразумевавется полипептид, который связывается и/или активирует HER-рецептор.
[0083] Внеклеточный (экто-) домен HER2 содержит четыре домена, Домен I (ECD1, аминокислотные остатки в диапазоне около 1-195), Домен II (ECD2, аминокислотные остатки в диапазоне около 196-319), Домен III (ECD3, аминокислотные остатки в диапазоне около 320-488) и Домен IV (ECD4, аминокислотные остатки в диапазоне около 489-630) (нумерация остатков без сигнального пептида). См. Garrett et al. Mol. Клетка. 11: 495-505 (2003), Cho et al. Nature All: 756-760 (2003), Franklin et al. Cancer Клетка 5:317-328 (2004), Tse et al. Cancer Treat Rev. 2012 Apr; 38(2): 133-42 (2012) или Plowman et al. Proc. Natl. Acad. Sci. 90:1746-1750 (1993).
[0084] Последовательность HER2 представлена ниже; границы ECD представляют собой Домен I: 1-165;
Домен II: 166-322; Домен III: 323-488; Домен IV: 489-607.

[0085] «Эпитоп 2С4» представляет собой область во внеклеточном домене HER2, с которым связывается антитело 2С4. Эпитоп 2С4 содержит остатки из домена II во внеклеточном домене HER2. 2С4 и пертузумаб связываются с внеклеточным доменом HER2 в месте соединения доменов I, II и III. Franklin et al. Cancer Клетка 5:317-328 (2004). Для скрининга антител, которые связываются с эпитопом 2С4, можно осуществить стандартный эпитоп-перекрестный конкурентный анализ, как, например, описанный в Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory, Ed Harlow and David Дорожка (1988). В качестве альтернативы, можно осуществить картирование эпитопов для оценки того, связывается ли антитело с эпитопом 2С4 HER2 с использованием способов, известных в данной области техники, и/или можно исследовать структуру антитело-HER2 (Franklin et al. Cancer Клетка 5:317-328 (2004)), чтобы определить с каким (какими) доменом (доменами) HER2 связывается антитело.
[0086] «Эпитоп 4D5» представляет собой область во внеклеточном домене HER2, с которым связываются антитело 4D5 (АТСС CRL 10463) и трастузумаб. Этот эпитоп близко расположен к трансмембранному домену HER2 и находится в составе Домена IV HER2. Для скрининга антител, которые связываются с эпитопом 4D5, можно осуществить стандартный эпитоп-перекрестный конкурентный анализ, как, например, описанный в Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory, Ed Harlow and David Дорожка (1988). В качестве альтернативы, можно осуществить картирование эпитопов для оценки того, связывается ли антитело с эпитопом 4D5 HER2 (например, с одним или более остатками в области от около остатка 529 до около остатка 625, включительно, см. ФИГ. 1 заявки на патент США №2006/0018899).
[0087] «Специфически связывается», «специфическое связывание» или «избирательное связывание» означает, что связывание является избирательным в отношении антигена и не относится к нежелательным или неспецифическим взаимодействиям. Способность антигенсвязывающей конструкции связываться со специфической антигенной детерминантой можно оценить либо с помощью твердофазного иммуноферментного анализа (ИФА), либо другими методами, известными специалисту в данной области техники, например, методом поверхностного плазмонного резонанса (ППР) (анализ осуществляется на приборе BIAcore) (Liljeblad et al., Glyco J 17, 323-329 (2000)) и с помощью традиционных анализов связывания (Heeley, Endocr Res 28, 217-229 (2002)). В одном варианте реализации изобретения степень связывания антигенсвязывающего фрагмента с неродственным белком составляет менее, чем около 10% от связывания антигенсвязывающей конструкции с антигеном, что оценивается, например, с помощью ППР. В определенных вариантах реализации изобретения антигенсвязывающая конструкция, которая связывается с антигеном, или антигенсвязывающая молекула, содержащая этот антигенсвязывающий фрагмент, имеет константу диссоциацию (КD)<1 мкМ, <100 нМ, <10 нМ, <1 нМ, <0,1 нМ, <0,01 нМ или <0,001 нМ (например, 10~8 М или меньше, например, от 10~8 М до 10''13 М, например, от 10''9 М до 10''13 М).
[0088] «Херегулин» (HRG) при использовании в данном документе относится к полипептиду, кодируеомому геном херегулина, как описано в в патенте США №5641869 или Marchionni et al, Nature, 362:312-318 (1993). Примеры херегулинов включают херегулин-α, херегулин-β1, херегулин-β2 и херегулин-β3 (Holmes et al., Science, 256:1205-1210 (1992); и патент США №5641869); фактор дифференцировки neu (NDF) (Peles et al. Клетка 69: 205-216 (1992)); фактор индукции синтеза ацетилхолиновых рецепторов (acetylcholine рецептор-inducing activity (ARIA)) (Falls et al. Клетка 72:801-815 (1993)); глиальные факторы роста (GGF) (Marchionni et al, Nature, 362:312-318 (1993)); фактор, полученный из сенсорных нейронов и мотонейронов (SMDF) (Но et al. J. Biol. Chem. 270:14523-14532 (1995)); γ-херегулин (Schaefer et al. Oncogene 15:1385-1394 (1997)). Термин охватывает биологически активные фрагменты и/или варианты аминокислотной последовательности, полученные из нативной последовательности полипептида HRG, как, например, фрагмент его EGF-подобного домена (например, HRGβ1177-244).
[0089] «Активация HER» или «активация HER2» относится к активации или фосфорилированию любого одного или более HER-рецепторов или HER2-рецепторов. В целом, активация HER приводит к передаче сигнала (которая, например, обусловлена фосфорилированием остатков тирозина в HER-рецепторе или субстратном полипептиде под воздействием внутриклеточного киназного домена HER-рецептора). Активация HER может быть опосредована связыванием лиганда HER с димером HER, содержащим целевой HER-рецептор. Связывание лиганда HER с димером HER может активировать киназный домен одного или нескольких HER-рецепторов в димере и тем самым приводить к фосфорилированию остатков тирозина в одном или нескольких HER-рецепторах и/или к фосфорилированию остатков тирозина в дополнительном субстратном полипептиде (субстратных полипептидах), как, например, внутриклеточные киназы Akt или МАРК.
Полученные антигенсвязывающие полипептидные конструкции
[0090] Антигенсвязывающие полипептидные конструкции могут быть получены из известных антител или связывающих доменов, обладающих специфичностью по отношению к HER2, независимо от типа домена. Примеры типов доменов включают Fab-фрагменты, scFv и Sdab. Кроме того, если антигенсвязывающие фрагменты известного антитела к HER2 или связывающий домен представляют собой Fab, Fab может быть превращен scFv. Аналогичным образом, если антигенсвязывающий фрагмент известного антитела к HER2 или связывающий домен представляют собой scFv. scFv может быть превращен в Fab. Способы превращения одних типов антигенсвязывающих доменов в другие известны в данной области техники (см., например, способы превращения scFv в Fab-формат, описанные, например, в Zhou et al. (2012) Mol Cancer Ther 11:1167-1476. Способы, описанные в данном документе, включены посредством ссылки).
[0091] Антигенсвязывающие конструкции, описанные в данном документе, могут быть получены из известных антител к HER2, которые связываются с ECD2 или ECD4., как описано в других местах в данном документе, антитела, которые связываются с HCD2 или ECD4, известны в данной области техники и включают, например, 2С4 или пертузумаб (который связывается с ECD2), 4D5 или трастузумаб (который связывается с ECD4). Другие антитела, которые связываются с ECD2 или ECD4 HER2, также были описаны в данной области техники, например, в WO 2011/147982 (Genmab A/S).
[0092] В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция антигенсвязывающей конструкции получена из антитела, которое блокирует на 50% или больше связывание антитела 4D5 или трастузумаба с ECD4 HER2. В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция антигенсвязывающей конструкции получена из антитела, которое блокирует на 50% или больше связывание антитела 2С4 или пертузумаба с ECD2 HER2. В некотором варианте реализации изобретения антигенсвязывающая конструкция получена из антитела, которое блокирует на 30% или больше связывание антитела 2С4 или пертузумаба с ECD2 HER2.
[0093] В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция получена из Fab-фрагмента трастузумаба или пертузумаба. В одном варианте реализации изобретения антигенсвязывающий полипептид получен из scFv.
[0094] В определенных вариантах реализации антигенсвязывающий полипептид получен из гуманизированных или рекомбинантных вариантов этих антител.
[0095] «Гуманизированные» формы отличных от человеческих антител (например, антител грызунов) представляют собой рекомбинантные антитела, которые содержат минимальную последовательность, полученную из отличного от человеческого иммуноглобулина. В большинстве случаев, гуманизированные антитела представляют собой иммуноглобулины человека (реципиентное антитело), в которых остатки из гипервариабельной области реципиентного антитела заменены на остатки из гипервариабельной области антител отличных от человека видов животных (донорское антитело), таких как мышь, крыса, кролик или нечеловекообразный примат, обладающих желательной специфичностью, аффинностью и связывающей способностью. В некоторых случаях, остатки каркасной области (FR) иммуноглобулина человека заменены на соответствующее остатки отличного от человеческого иммуноглобулина. Кроме того, гуманизированные антитела могут содержать остатки, которые не встречаются в реципиентном антителе или в донорском антителе. Эти модификации вносятся для дополнительного улучшения эффективности антитела. В целом, гуманизированное антитело, по существу, будет содержать все из по меньшей мере одного и, как правило, двух, вариабельных доменов, в которых все или, по существу, все из гипервариабельных петель соответствуют петлям отличного от человеческого иммуноглобулина, и все или, по существу, все из FR представляют собой FR последовательности иммуноглобулина человека. Гуманизированное антитело необязательно также будет содержать по меньшей мере часть константной области иммуноглобулина (Fc), как правило, иммуноглобулина человека. Для получения более подробной информации см. Jones et al., Nature 321:522-525 (1986); Riechmann et al., Nature 332:323-329 (1988); и Presta, Curr. Op. Struct. Biol. 2:593-596(1992).
[0096] Гуманизированные антитела к HER2 включают huMAb4D5-1, huMAb4D5-2, huMAb4D5-3, huMAb4D5-4, huMAb4D5-5, huMAb4D5-6, huMAb4D5-7 и huMAb4D5-8 или трастузумаб (HERCEPTIN® ), как представлено в Таблице 3 патента США №5821337, в явной форме включенного в данный документ посредством ссылки; гуманизированные 520С9 (WO 93/21319) и 20' гуманизированные антитела 2С4, как описано в публикации патента США №. 2006/0018899.
Созревание аффинности
[0097] В некоторых вариантах реализации изобретения антигенсвязывающая конструкция получена из известных антител, связывающихся с HER2, с использованием созревания аффинности.
[0098] В тех случаях, когда желательно повысить аффинность антигенсвязывающего полипептида по отношению к распознаваемому им антигену, могут быть использованы способы, известные в данной области техники, чтобы повысить аффинность антигенсвязывающего полипептида по отношению к его антигену. Примеры таких способов описаны в следующих источниках, Birtalan et al. (2008) JMB 311, 1518-1528; Gerstner et al. (2002) JMB 321, 851-862; Kelley et al. (1993) Biochem 32(27), 6828-6835; Li et al. (2010) JBC 285(6), 3865-3871 и Vajdos et al. (2002) JMB 320, 415-428.
[0099] Далее описан одиниллюстративный способ обеспечения созревания аффинности доменов, связывающих антиген HER2. Структуры комплекса трастузумаб/HER2 (PDB-код 1N8Z) и комплекса пертузумаб/HER2 (PDB-код 1S78) используются для моделирования. Для оценки внутренней динамической природы комплекса дикого типа в водной среде может быть использована молекулярная динамика (МД). Методы отсечения среднего поля и отсечения тупиковых ветвей наряду с гибкими скелетами молекул могут быть использованы для оптимизации и получения модельных структур для мутантных молекул, которые подлежат скринингу. После оценки упаковки оценивается целый ряд признаков, включая плотность контакта, оценку столкновений, гидрофобность и электростатику. Обобщенный метод Борна позволяет точно моделировать эффект среды растворителя и рассчитать различия в свободной энергии после замены с помощью мутации остатков в конкретных положениях в белке на альтернативные типы остатков. Плотность контакта и оценка столкновений служит мерой комплементарности, которая является крайне важным аспектом эффективной упаковки белка. Процедура скрининга предполагает использование статистических потенциалов, а также схемы анализа взаимодействий, основанные на энергии попарного взаимодействия остатков и расчетах величины энтропии. Мутации, которые, как известно из литературных источников, усиливают связывание с HER2 и их комбинации, в кратком виде представлены в следующих таблицах:


Fc-домен антигенсвязывающих конструкций.
[00100] В некоторых вариантах реализации изобретения антигенсвязывающие конструкции, описанные в данном документе, содержат Fc, например, димерный Fc.
[00101] Термин «Fc-домен» или «Fc-область» в данном документе используется для обозначения С-концевой области тяжелой цепи иммуноглобулина, которая содержит по меньшей мере часть константной области. Термин охватывает Fc-области с нативными последовательностями и вариантные Fc-области. Если в данном документе не указано иное, нумерация аминокислотных остатков в Fc-области или константной области осуществляется согласно системе нумерации EU, также называемой индекс EU, как описано в Kabat et al, Sequences of Белки of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD, 1991. Используемый в данном документе термин «Fc-полипептид» димерного Fc-домена относится к одному из двух полипептидов, формирующих димерный Fc-домен, т.е. полипептиду, содержащему С-концевые константные области тяжелой цепи иммуноглобулина, способные к стабильной самоассоциации. Например, Fc-полипептид димерного IgG Fc содержит последовательность константного домена IgG СН2 и IgG CH3.
[00102] Fc-домен содержит либо СН3-домен, либо СН3- и СН2-домен. СН3-домен содержит две последовательности CH3, одну из каждого из двух Fc-полипептидов димерного Fc-домена. The СН2-домена содержит две СН2 последовательности, одну из каждого из двух Fc-полипептидов димерного Fc-домена.
[00103] В некоторых аспектах Fc-домен содержит по меньшей мере одну или две последовательности CH3. В некоторых аспектах Fc-домен связан, с помощью одного или более линкеров, или другим способом, с первой антигенсвязывающей конструкцией и/или второй антигенсвязывающей конструкцией. В некоторых аспектах Fc-домен представляет собой Fc-домен антитела человека. В некоторых аспектах Fc-домен представляет собой Fc-домен IgG или IgG1 человека. В некоторых аспектах Fc-домен представляет собой гетеродимерный Fc-фрагмент. В некоторых аспектах Fc-домен содержит по меньшей мере одну или две последовательности СН2.
[00104] В некоторых аспектах В Fc-домене содержится одна или более модификаций по меньшей мере в одной из последовательностей CH3. В некоторых аспектах В Fc-домене содержится одна или более модификаций по меньшей мере в одной из последовательностей СН2. В некоторых аспектах Fc-домен состоит из одного полипептида. В некоторых аспектах Fc-домен сосоит из многих пептидов, например, двух полипептидов.
[00105] В некоторых аспектах Fc-домен представляет собой Fc-домен, описанный в патентных заявках РСТ/СА2011/001238, поданной 4 ноября 2011 года, или РСТ/СА2012/050780, поданной 2 ноября 2012 года, причем полное описание каждой из них включено в данный документ в полном объеме посредством ссылки для всех целей.
Модифицированные СН3-домены
[00106] В некоторых аспектах антигенсвязывающая конструкция, описанная в данном документе, содержит гетеродимерный Fc-фрагмент, содержащий модифицированный СН3-домен, который был асимметрично модифицирован. Гетеродимерный Fc-фрагмент может содержать два полипептида константных доменов тяжелой цепи: первый Fc-полипептид и второй Fc-полипептид, которые могут быть использованы взаимозаменяемо при условии, что Fc-домен содержит один первый Fc-полипептид и один второй Fc-полипептид. Как правило, первый Fc-полипептид содержит первую последовательность CH3, и второй Fc-полипептид содержит вторую последовательность CH3.
[00107] В результате введения одной или более асимметричных аминокислотных модификаций в две последовательности CH3, как правило, образуется гетеродимерный Fc-фрагмент, а не гомодимер, при этом происходит димеризация двух последовательностей CH3. Используемый в данном документе терин «асимметричные аминокислотные модификации» относится к любой модификации, при которой аминокислота в определенном положении в первой последовательности CH3 отличается от аминокислоты, находящейся во второй последовательности CH3 в том же самом положении, и первая, а вторая последовательность CH3 преимущественно соединяются с образованием гетеродимера, а не гомодимера. Эта гетеродимеризация может происходить в результате модификации только одной из двух аминокислот в одном и том же положении соответствующей аминокислоты в каждой последовательности; или модификации обеих аминокислот в каждой последовательности в одном и том же соответствующем положении в каждой первой и второй последовательности CH3. Первая и вторая последовательность CH3 гетеродимерного Fc-фрагмента могут содержать одну или более, чем одну асимметричную аминокислотную модификацию.
[00108] В Таблице А представлена аминокислотная последовательность Fc-домена IgG1 человека, соответствующая аминокислотам от 231 до 447 тяжелой цепи полноразмерного IgG1 человека. Последовательность CH3 содержит аминокислоты 341-447 тяжелой цепи полноразмерного IgG1 человека.
[00109] Как правило, Fc-домен может включать две непрерывные последовательности тяжелой цепи (А и В), которые способны к димеризации. В некоторых аспектах одна или обе последовательности Fc-домена включают одну или более мутаций, или модификаций в следующих положениях: L351, F405, Y407, Т366, К392, Т394, Т350, S400 и/или N390 согласно нумерации EU. В некоторых аспектах Fc-домен включает последовательность с аминокислотными заменами, показанную в Таблице X. В некоторых аспектах Fc-домен содержит мутации Варианта 1 А-В. В некоторых аспектах Fc-домен содержит мутации Варианта 2 А-В. В некоторых аспектах Fc-домен содержит мутации Варианта 3 А-В. В некоторых аспектах Fc-домен содержит мутации Варианта 4 А-В. В некоторых аспектах Fc-домен содержит мутации Варианта 5 А-В.
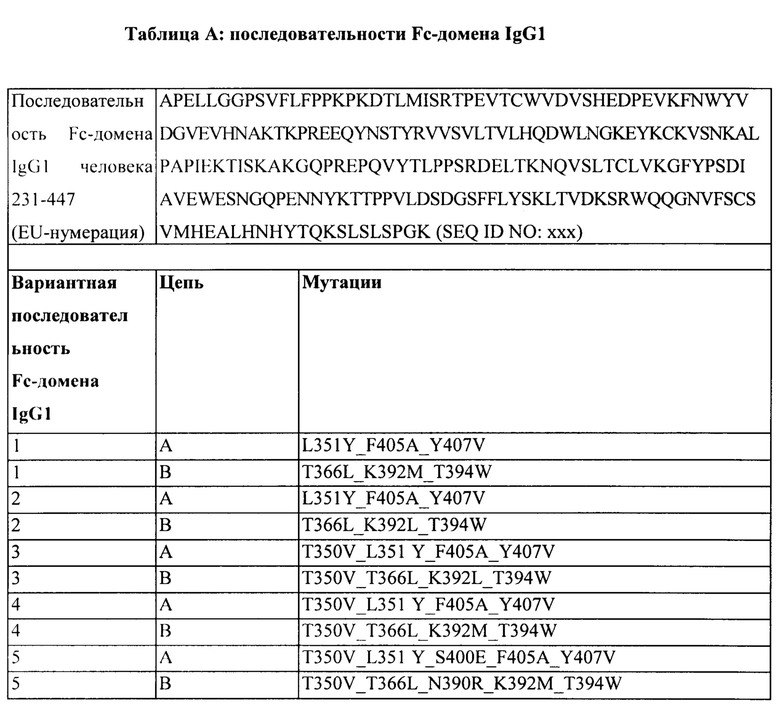
[00110] Первая и вторая последовательности CH3 могут содержать аминокислотные замены, которые описаны в данном документе, что касается аминокислот 231-447 тяжелой цепи полноразмерного IgG1 человека. В одном варианте реализации изобретения гетеродимерный Fc-фрагмент содержит модифицированный СН3-домен, причем в первой последовательности CH3 содержатся аминокислотные модификации в положениях F405 и Y407, и во второй последовательности CH3 содержатся аминокислотные модификации в положении Т394. В одном варианте реализации изобретения гетеродимерный Fc-фрагмент содержит модифицированный СН3-домен, причем в первой последовательности CH3 содержится одна или более аминокислотных модификаций, выбранных из L351Y, F405A и Y407V, и во второй последовательности CH3 содержится одна или более аминокислотных модификаций, выбранных из T366L, T366I, K392L, К392М и T394W.
[00111] В одном варианте реализации изобретения гетеродимерный Fc-фрагмент содержит модифицированный СН3-домен, причем в первой последовательности CH3 содержатся аминокислотные модификации в положениях L351, F405 и Y407, и во второй последовательности CH3 содержатся аминокислотные модификации в положениях Т366, К392 и Т394, и в одной из первой или второй последовательности CH3 дополнительно содержатся аминокислотные модификации в положении Q347, и в другой последовательности CH3 дополнительно содержится аминокислотная модификация в положении К360. В другом варианте реализации изобретения гетеродимерный Fc-фрагмент содержит модифицированный СН3-домен, причем в первой последовательности CH3 содержатся аминокислотные модификации в положениях L351, F405 и Y407, и во второй последовательности CH3 содержатся аминокислотные модификации в положении Т366, К392 и Т394, причем в одной из первой или второй последовательности CH3 дополнительно содержатся аминокислотные модификации в положении Q347, и в другой последовательности CH3 дополнительно содержится аминокислотная модификация в положении К360, и в одной или обеих из указанных последовательностей CH3 дополнительно содержится аминокислотная модификация T350V.
[00112] В одном варианте реализации изобретения гетеродимерный Fc-фрагмент содержит модифицированный СН3-домен, причем в первой последовательности CH3 содержатся аминокислотные модификации в положениях L351, F405 и Y407, и во второй последовательности CH3 содержатся аминокислотные модификации в положениях Т366, К392 и Т394, и в одной из указанных первой и второй последовательности CH3 дополнительно содержится аминокислотная модификация D399R или D399K, и причем в другой последовательности CH3 содержится одна или более модификаций из Т411Е, T411D, К409Е, K409D, К392Е и K392D. В другом варианте реализации изобретения гетеродимерный Fc-фрагмент содержит модифицированный СН3-домен, причем в первой последовательности CH3 содержатся аминокислотные модификации в положениях L351, F405 и Y407, и во второй последовательности CH3 содержатся аминокислотные модификации в положениях Т366, К392 и Т394, причем в одной из указанных первой и второй последовательностей CH3 дополнительно содержится аминокислотная модификация D399R или D399K, и в другой последовательности CH3 содержится одна или более модификаций из Т411Е, T411D, К409Е, K409D, К392Е и K392D, и в одной или обеих из указанных последовательностей CH3 дополнительно содержится аминокислотная модификация T350V.
[00113] В одном варианте реализации изобретения гетеродимерный Fc-фрагмент содержит модифицированный СН3-домен, причем в первой последовательности CH3 содержатся аминокислотные модификации в положениях L351, F405 и Y407, и во второй последовательности CH3 содержатся аминокислотные модификации в положениях Т366, К392 и Т394, причем в одной или обеих из указанных последовательностей CH3 дополнительно содержится аминокислотная модификация T350V.
[00114] В одном варианте реализации изобретения гетеродимерный Fc-фрагмент содержит модифицированный СН3-домен, содержащий следующие ниже аминокислотные модификации, где «А» представляет собой аминокислотную модификацию в первой последовательности CH3, и «В» представляет собой аминокислотную модификацию в второй последовательности CH3: A:L351Y_F405A_Y407V, B:T366L_K392M_T394W, A:L351Y_F405A_Y407V, B:T366L_K392L_T394W, A:T350V_L351 Y_F405A_Y407V, B:T350V_T366L_K392L_T394W, A:T350V_L351Y_F405A_Y407V, B:T350V_T366L_K392M_T394W, A:T350V_L351 Y_S400E_F405A_Y407V и/или B:T350V_T366L_N390R_K392M_T394W.
[00115] Одна или более асимметричных аминокислотных модификаций могут способствовать образованию гетеродимерного Fc-фрагмента, причем гетеродимерный СН3-домен обладает стабильностью, которая сравнима с гомодимерным СН3-доменом дикого типа. В одном варианте реализации изобретения одна или более асимметричных аминокислотных модификаций способствуют образованию гетеродимерного Fc-фрагмента, причем гетеродимерный Fc-фрагмент обладает стабильностью, которая сравнима с гомодимерным Fc-фрагментом дикого типа домен. В одном варианте реализации изобретения одна или более асимметричных аминокислотных модификаций способствуют образованию гетеродимерного Fc-фрагмента, причем гетеродимерный Fc-фрагмент обладает стабильностью, определяемой по температуре плавления (Тпл) в исследовании методом дифференциальной сканирующей калориметрии, и причем температура плавления находится в пределах 4°С по отношению к температуре, наблюдаемой для соответствующего симметричного гомодимерного Fc-домена дикого типа. В некоторых аспектах В Fc-домене содержится одна или более модификаций по меньшей мере в одной из последовательностей Снз, которые способствуют образованию гетеродимерного Fc-фрагмента со стабильностью, сравнимой со стабильностью гомодимерного Fc-домена дикого типа.
[00116] В одном варианте реализации изобретения стабильность СН3-домена может быть оценена путем измерения температуры плавления СН3-домена, например, с помощью дифференциальной сканирующей калориметрии (ДСК). Таким образом, в дополнительном варианте реализации изобретения СН3-домен имеет температуру плавления около 68°С или выше. В другом варианте реализации изобретения СН3-домен имеет температуру плавления около 70°С или выше. В другом варианте реализации изобретения СН3-домен имеет температуру плавления около 72°С или выше. В другом варианте реализации изобретения СН3-домен имеет температуру плавления около 73°С или выше. В другом варианте реализации изобретения СН3-домен имеет температуру плавления около 75°С или выше. В другом варианте реализации изобретения СН3-домен имеет температуру плавления около 78°С или выше. В некоторых аспектах температура плавления (Тпл) димеризованных СН3-доменов составляет около 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 77,5, 78, 79, 80, 81, 82, 83, 84 или 85°С, или выше.
[00117] В некоторых вариантах реализации изобретения гетеродимерный Fc-фрагмент, содержащий модифицированные последовательности CH3, может образоваться с показателем чистоты по меньшей мере около 75% по сравнению с гомодимерным Fc-доменом в экспресированном продукте. В другом варианте реализации изобретения гетеродимерный Fc-фрагмент образуется с показателем чистоты, большим чем около 80%. В другом варианте реализации изобретения гетеродимерный Fc-фрагмент образуется с показателем чистоты, большим, чем около 85%. В другом варианте реализации изобретения гетеродимерный Fc-фрагмент образуется с показателем чистоты, большим чем около 90%. В другом варианте реализации изобретения гетеродимерный Fc-фрагмент образуется с показателем чистоты, большим, чем около 95%. В другом варианте реализации изобретения гетеродимерный Fc-фрагмент образуется с показателем чистоты, большим чем около 97%. В некоторых аспектах Fc-домен представляет собой гетеродимер, вырабатываемый с чистотой более, чем около 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% при экспрессии. В некоторых аспектах Fc-домен представляет собой гетеродимер, вырабатываемый с чистотой более, чем около 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% при экспрессии на основе генетического материала одной клетки.
[00118] Дополнительные способы модификации мономерных Fc-полипептидов для обеспечения образования гетеродимерных Fc-доменов, описаны в публикации международной заявки на патент №WO 96/027011 (выступы-во-впадины), Gunasekaran et al. (Gunasekaran K. et al. (2010) J Biol Chem. 285, 19637-46, electrostatic design to achieve selective heterodimerization), в Davis et al. (Davis, JH. et al. (2010) Prot Eng Des Sel;23(4): 195-202, strand exchange engineered domain (SEED) technology) и в Labrijn et al. [Efficient generation of stable bispecific IgG1 by controlled Fab-arm exchange. Labrijn AF, Meesters JI, de Goeij BE, van den Bremer ET, Neijssen J, van Kampen MD, Strumane K, Verploegen S, Kundu A, Gramer MJ, van Berkel PH, van de Winkel JG, Schuurman J, Parren PW. Proc Natl Acad Sci США. 2013 Mar 26;110(13):5145-50.
СН2-домены
[00119] В некоторых вариантах реализации изобретения Fc-фрагмент антигенсвязывающей конструкции содержит СН2-домен. Одним примером СН2-домена Fc-фрагмента являются аминокислоты 231-340 последовательности, показанной в Таблице А. Несколько эффекторных функций опосредованы Fc-рецепторами (FcR), которые связываются с Fc-фрагментом антитела.
[00120) Термины «Fc-рецептор» и «FcR» используются для описания рецептора, который связывается с Fc-областью антитела. Например, FcR может представлять собой FcR человека с нативной последовательностью. Как правило, FcR - это рецептор, который связывается с антителом IgG (гамма-рецептор) и включает рецепторы подклассов FcγRI, FcγRII и FcγRIII, включая аллельные варианты и формы этих рецепторов, образовавшиеся в результате альтернативного сплайсинга. Рецепторы FcγRII включают FcγRIIA («активирующий рецептор») и FcγRIIB («ингибирующий рецептор»), которые имеют сходные аминокислотные последовательности, что отличаются главным образом по их цитоплазматическим доменам. Иммуноглобулины других изотипов также могут связываться с определенными FcR (см., например, Janeway et al., Immuno Biology: the immune system in health and disease (Elsevier Science Ltd., NY) (4th ed., 1999)). Активирующий рецептор FcγRIIA содержит иммунорецепторный тирозиновый ингибирующий мотив (ITAM) в его цитоплазматическом домене. Ингибирующий рецептор FcγRIIB содержит иммунорецепторный тирозиновый ингибирующий мотив (ITIM) в его цитоплазматическом домене (обзор содержится в  Annu. Rev. Immunol. 15:203-234 (1997)). Обзор no FcR содержится в Ravetch and Kinet, Annu. Rev. Immunol 9:457-92 (1991); Capel et al., Immunomethods 4:25-34 (1994); и de Haas et al., J. Lab. Clin. Med. 126:330-41 (1995). Другие FcR, включая те, что будут идентифицированы в будущем, охватываются термином «FcR» в данном документе. Термин также охватывает неонатальный рецептор, FcRn, который отвечает за передачу материнских IgG плоду (Guyer et al, J. Immunol. 117:587 (1976); and Kim et al, J. Immunol. 24:249 (1994)).
Annu. Rev. Immunol. 15:203-234 (1997)). Обзор no FcR содержится в Ravetch and Kinet, Annu. Rev. Immunol 9:457-92 (1991); Capel et al., Immunomethods 4:25-34 (1994); и de Haas et al., J. Lab. Clin. Med. 126:330-41 (1995). Другие FcR, включая те, что будут идентифицированы в будущем, охватываются термином «FcR» в данном документе. Термин также охватывает неонатальный рецептор, FcRn, который отвечает за передачу материнских IgG плоду (Guyer et al, J. Immunol. 117:587 (1976); and Kim et al, J. Immunol. 24:249 (1994)).
[00121] Модификации в СН2-домене могут влиять на связывание FcR с Fc. В данной области техники известен целый ряд аминокислотных модификациий в Fc-области по избирательному изменению аффинности Fc-домена по отношению к разным Fc-гамма-рецепторам. В некоторых аспектах в Fc-домене содержится одна или более модификаций, что способствуют избирательному связыванию Fc-гамма рецепторов.
[00122] Иллюстративные мутации, которые влияют на связывание FcR с Fc приведены ниже:
[00123] S298A/E333A/K334A, S298A/E333A/K334A/K326A (Lu Y, Vemes JM, Chiang N, et al. J Immunol Methods. 2011 Feb 28;365(1-2): 132-41);
[00124] F243L/R292P/Y300L/V305I/P396L, F243L/R292P/Y300L/L235V/P396L (Stavenhagen JB, Gorlatov S, Tuaillon N, et al. Cancer Res. 2007 Sep 15; 67(18):8882-90; Nordstrom JL, Gorlatov S, Zhang W, et al. Молочной железы Cancer Res. 2011 Nov 30; 13(6):R123);
[00125] F243L (Stewart R, Thorn G, Levens M, et al. Белок Eng Des Sel. 2011 Sep;24(9):671-8.), S298A/E333A/K334A (Shields RL, Namenuk AK, Hong K, et al. J Biol Chem. 2001 Mar 2; 276(9):6591-604);
[00126] S239D/I332E/A330L, S239D/I332E (Lazar GA, Dang W, Karki S, et al. Proc Natl Acad Sci США. 2006 Mar 14; 103(11):4005-10);
[00127] S239D/S267E, S267E/L328F (Chu SY, Vostiar I, Karki S, et al. Mol Immunol. 2008 Sep;45(15):3926-33);
[00128] S239D/D265S/S298A/I332E, S239E/S298A/K326A/A327H, G237F/S298A/A33 0L/I332E, S239D/I332E/S298A, S239D/K326E/A330L/I332E/S298A, G236A/S239D/D270L/I3 32E, S239E/S267E/H268D, L234F/S267E/N325L, G237F/V266L/S267D и другие мутации, приведенные в WO2011/120134 и WO2011/120135. включены в данный документ посредством ссылки. В Therapeutic Antibody Engineering (William R. Strohl and Lila M. Strohl, Woodhead Publishing series in Biomedicine No 11, ISBN 1 907568 37 9, Oct 2012) приведены мутации на странице 283.
[00129] В некоторых вариантах реализации изобретения антигенсвязывающая конструкция, описанная в данном документе, содержит антигенсвязывающую полипептидную конструкцию, которая связывается с антигеном; и димерный Fc-домен, который обладает улучшенными биофизическими свойствами, такими как стабильность и легкость изготовления по сравнению с антигенсвязывающей конструкцией, которая не содержит такой же самый димерный Fc-домен. В некоторых вариантах реализации изобретения СН2-домен содержит одну или более асимметричных аминокислотных модификаций. Иллюстративные асимметричные мутации описаны в международной заявке на патент №. РСТ/СА2014/050507.
Дополнительные модификации, улучшающие эффекторные функции.
[00130] В некоторых вариантах реализации изобретения антигенсвязывающая конструкция, описанная в данном документе, содержит модификации, улучшающие ее способность опосредовать эффекторную функцию. Такие модификации известны в данной области техники и включают дефукозилирование или созревание аффинности Fc-домена по отношению к активирующему рецептору, главным образом FCGR3a для АЗКЦ и по отношению к CIq для (КЗЦТ). В приведенной ниже Таблице В в краткой форме представлены различные дизайны, описанные в литературе, для реализации эффекторных функций.
[00131] Способы получения антигенсвязывающих конструкций с небольшим количеством остатков фукозы в сайте гликозилирования Fc-домена или без фукозы (Asn 297, EU-нумерация) без изменения аминокислотной последовательности хорошо известны в данной области техники. Технология GlymaX® (ProBioGen AG) основана на введения гена фермента, который изменяет направление клеточного пути биосинтеза фукозы в клетках, используемых для получения антигенсвязывающих конструкций. Это препятствует присоединению сахара «фукозы» к N-связанной углеводной части антитела в клетках, продуцирующих антигенсвязывающую конструкцию (von Horsten et al. (2010) Glycobiology. 2010 Dec; 20 (12): 1607-18. Другой подход к получению антигенсвязывающих конструкций с пониженными уровнями фукозилирования можно найти в патенте США 8409572, идея которого заключается в отборе клеточных линий для получения антигенсвязывающих конструкций по их способности обеспечивать более низкие уровни фукозилирования антигенсвязывающих конструкций. Антигенсвязывающие конструкции могут быть полностью дефукозилированными (что означает, что они содержат неопределяемое количество фукозы) или они могут быть частично дефукозилированными, что означает, что выделенное антитело содержит менее, чем 95%. менее, чем 85%, менее, чем 75%, менее, чем 65%, менее, чем 55%, менее, чем 45%, менее, чем 35%, менее, чем 25%, менее, чем 15% или менее, чем 5% от количества фукозы, которое обычно определяется в подобном антителе, получаемом с использованием системы экспрессии генов в клетках млекопитающих.
[00132] Таким образом, в одном варианте реализации изобретения конструкция, описанная в данном документе, может включать димерный Fc-домен, который содержит одну или более аминокислотных модификациий, как отмечается в Таблице В, которые способствуют улучшению эффекторных функций. В другом варианте реализации изобретения конструкция может быть дефукозилированной для улучшения эффекторных функций.

[00133] В данной области техники известны модификации Fc-домена, снижающие уровень связывания с FcγR и/или комплементом, и/или эффекторные функции. В последних публикациях описаны стратегии, которые были использованы для конструирования антител со сниженной или подавленной эффекторной активностью (см. Strohl, WR (2009), Curr Opin Biotech 20:685-691, и Strohl, WR, и Strohl LM, "Antibody Fc engineering for оптимальный antibody performance" в Therapeutic Antibody Engineering, Cambridge: Woodhead Publishing (2012), стр. 225-249). Эти стратегии включают снижение эффекторных функций посредством модификации гликозилирования, применения пептидных остовов IgG2/IgG4 или введения мутаций в гены, кодирующие шарнирную или СН2-области Fc-домена. Например, в публикации международной заявки на патент №. 2011/0212087 (Strohl), публикации международной заявки на патент №WO 2006/105338 (Xencor), публикации международной заявки на патент №2012/0225058 (Xencor), публикации международной заявки на патент №2012/0251531 (Genentech) и Strop et al ((2012) J. Mol. Biol. 420: 204-219) описаны специфические модификации, снижающие уровень связывания FcγR или комплемента с Fc-доменом.
[00134] Конкретные, неограничивающие примеры известных аминокислотных модификациий, снижающих уровень связывания FcγR или комплемента с Fc-доменом, включают модификациий, указаные в следующей таблице:

[00135] В одном варианте реализации изобретения Fc-домен содержит по меньшей мере одну аминокислотную модификацию, которая указана в приведенной выше таблице. В другом варианте реализации изобретения в Fc-домен содержит аминокислотную модификацию по меньшей мере одну из L234, L235 или D265. В другом варианте реализации изобретения Fc-домен содержит аминокислотную модификацию в положении L234, L235 и D265. В другом варианте реализации изобретения Fc-домен содержит аминокислотную модификацию L234A, L235A и D265S.
Линкеры и линкерные полипептиды
[00136] Каждая из антигенсвязывающих полипептидных конструкций антигенсвязывающей конструкции функционально связана с линкерным полипептидом, при этом линкерные полипептиды способны к образованию ковалентной связи друг с другом. Пространственная конформация антигенсвязывающей конструкции, содержащей первую и вторую антигенсвязывающие полипептидные конструкции с линкерными полипептидами, сходна с относительной пространственной конформацией паратопов F(ab’)2-фрагмента, полученных с помощью расщепления папаином, хотя в контексте биспецифических антигенсвязывающих конструкций, описанных в данном документе, две антигенсвязывающие полипептидные конструкции характеризуются форматом Fab-scFv или scFv-scFv.
[00137] Таким образом, линкерные полипептиды подбираются таким образом, чтобы они поддерживали относительную пространственную конформацию паратопов F(ab’)-фрагмента и были способны к образованию ковалентной связи, эквивалентной дисульфидной связи в коровом шарнире IgG. Подходящие линкерные полипептиды включают шарнирные области IgG, такие, как, например, шарнирные области из IgG1, IgG2 или IgG4. Также могут быть использованы модифицированные варианты этих иллюстративных линкеров. Например, в данной области техники известны модификации для улучшения стабильности шарнира IgG4 (см., например, Labrijn et al. (2009) Nature Biotechnology 27, 767-771).
[00138] В одном варианте реализации изобретения линкерные полипептиды функционально связаны с пептидным остовом, как описано в данном документе, например, Fc-доменом. В некоторых аспектах Fc-домен связан с одной или более антигенсвязывающих полипептидных конструкций с помощью одного или более линкеров. В некоторых аспектах Fc-домен связан с тяжелой цепью каждого антигенсвязывающего полипептида посредством линкера.
[00139] В других вариантах реализации изобретения линкерные полипептиды функционально связаны с пептидными остовами, отличными от Fc-домена. В данной области техники известен целый ряд альтернативных белковых или молекулярных доменов, и они могут быть использованы для образования характеризующихся избирательным взаимодействием пар из двух разных антигенсвязывающих полипептидов. Одним из примеров являются домены лейциновой застежки, такие как Fos и Jun, которые избирательно взаимодействуют друг с другом [S A Kostelny, М S Cole, and J Y Tso. Formation of a bispecific antibody by the use of leucine zippers. J Immunol 1992 148:1547-53; Bemd J. Wranik, Erin L. Christensen, Gabriele Schaefer, Janet K. Jackman, Andrew С Vendel, and Dan Eaton. LUZ-Y, a Novel Platform for the Mammalian Клетка Продукта ion of Полноразмерная IgG-bispecific AntibodiesJ. Biol. Chem. 2012 287: 43331-43339]. В качестве альтернативы, также могут быть использованы другие молекулярные пары, характеризующиеся избирательным взаимодействием, как, например, пара барназы и барстара [Deyev, S.М., Waibel, R., Lebedenko, Е. N., Schubiger, А. P. and Pluckthun, А. (2003). Design of multivalent complexes using the barnase*barstar module. Nat Biotechnol 21, 1486-1492], пары нитей ДНК [ZahidaN. Chaudri. Michael Bartlet-Jones, George Panayotou, Thomas Klonisch, Ivan M. Roitt, Torben Lund, Peter J. Delves, Dual specificity antibodies using a double-stranded oligonucleotide bridge, FEBS Letters, Volume 450, Issues 1-2, 30 April 1999, Pages 23-26], пары расщепленных флуоресцентных белков [Ulrich Brinkmann, Alexander Haas. Fluorescent antibody fusion белок, its продуктаюп and use, WO 2011135040 А1].
Константа диссоциации (KD) и максимальное связывание (Вмакс)
[00140] В некоторых вариантах реализации изобретения антигенсвязывающая конструкция описана по функциональным характеристикам, включая, в том числе константу диссоциации и максимальное связывание.
[00141] Подразумевается, что используемый в данном документе термин «константа диссоциации (KD)» относятся к равновесной константе диссоциации, характеризующей определенное взаимодействие лиганда с белком. В данном контексте взаимодействия лиганда с белком относятся в том числе к белок-белковым взаимодействиям или взаимодействиям антитела с антигеном. KD характеризует склонность двух белков (например, АВ) к обратимой диссоциации на меньшие компоненты (А+В) и определяется как соотношение скорости диссоциации, также называемой «скорость диссоциации (koff)», к скорости ассоциации или «on-rate (kon)». Таким образом, KD равна koff/kon и выражается как молярная концентрация (М). Из этого следует, что чем меньше KD, тем сильнее аффинность связывания. Таким образом, KD 1 мМ обозначает слабую аффинность связывания по сравнению с KD 1 нМ. Значения KD для антигенсвязывающих конструкций можно определить с использованием способов, устоявшихся в данной области техники. Один из способов определения KD антигенсвязывающей конструкции заключается в использовании поверхностного плазмонного резонанса (ППР), как правило, с использованием биосенсорной системы, как, например, система Biacore® . Изотермическая титрационная калориметрия (ИТК) является другим способом, который может быть использован для данного определения.
[00142] Характеристики связывания антигенсвязывающей конструкции можно определить с помощью различных методов. Один из них заключается в измерении уровня связывания с клетками-мишенями, экспрессирующими антиген, с помощью проточной цитометрии (FACS, флуоресцентно-активируемая клеточная сортировка). Как правило, в таком эксперименте клетки-мишени, экспрессирующие целевой антиген, инкубируют с антигенсвязывающими конструкциями в разных концентрациях, промывают, инкубируют со вторым средством для обнаружения антигенсвязывающей конструкции, промывают и анализируют в проточном цитометре для измерения медианы интенсивности флуоресценции (MFI), характеризующей силу обнаруживаемого сигнала на клетках, которая в свою очередь связана с числом антигенсвязывающих конструкций, связавшихся с клетками. Концентрацию антигенсвязывающей конструкции в зависимости от данных по MFI затем подставляют в уравнение связывания с насыщением для получения двух ключевых параметров связывания, Вмакс и кажущейся KD.
[00143] Кажущаяся KD или кажущаяся равновесная константа диссоциации представляет собой концентрацию антигенсвязывающей конструкции, при которой наблюдается полумаксимальное связывание с клетками. Очевидно, что чем меньше значение KD, тем меньшая концентрация антигенсвязывающей конструкции требуется для достижения максимального связывания с клетками и, таким образом, тем выше аффинность антигенсвязывающей конструкции. Кажущаяся KD зависит от условий эксперимента по связыванию клеток, как, например, разные уровни экспрессии рецептора на клетках и условия инкубация, и таким образом, кажущаяся KD, как правило, отличается от значений KD, определенных в бесклеточных молекулярных экспериментах, таких как ППР и ИТК. Однако, как правило, разные способы хорошо согласуются друг с другом.
[00144] Термин «Вмакс» или максимальное связывание, относится к максимальному уровню связывания антигенсвязывающих конструкций на клетках при насыщающих концентрациях антигенсвязывающей конструкции. Этот параметр может быть представлен в относительных единицах MFI для относительного сравнения или превращен в абсолютное значение, соответствующее количеству антигенсвязывающих конструкций, связавшихся с клеткой, с применением калибровочной кривой. В некоторых вариантах реализации изобретения антигенсвязывающие конструкции проявляют Вмакс, которое в 1,1, 1,2, 1,3, 1,4, 1,5, 1,6, 1,7, 1,8, 1,9 или 2,0 раза выше Вмакс эталонной антигенсвязывающей конструкции.
[00145] Для антигенсвязывающих конструкций, описанных в данном документе, наиболее точное разделение по Вмакс в сравнении с FSA происходит при насыщающих концентрациях и в том случае, когда Вмакс FSA не может больше увеличиваться. Статическая значимость снижается при ненасыщающих концентрациях. В одном варианте реализации изобретения увеличение Вмакс и KD антигенсвязывающей конструкции по сравнению с эталонной антигенсвязывающей конструкцией зависит от уровня экспрессии антигена-мишени на клетке-мишени.
[00146] В некоторых вариантах реализации изобретения представлена выделенная антигенсвязывающая конструкция, описанная в данном документе, причем указанная антигенсвязывающая конструкция характеризуется повышением Вмакс (максимальное связывание) при связывании с клеткой-мишенью, экспонирующей указанный антиген, по сравнению с соответствующей эталонной антигенсвязывающей конструкцией. В некоторых вариантах реализации изобретения указанное повышение Вмакс составляет по меньшей мере около 125% от Вмакс соответствующей эталонной антигенсвязывающей конструкции. В определенных вариантах реализации изобретения повышение Вмакс составляет по меньшей мере около 150% от Вмакс соответствующей эталонной антигенсвязывающей конструкции. В некоторых вариантах реализации изобретения повышение Вмакс составляет по меньшей мере около 200% от Вмакс соответствующей эталонной антигенсвязывающей конструкции. В некоторых вариантах реализации изобретения повышение Вмакс сотавляет более чем около 110 % от Вмакс соответствующей эталонной антигенсвязывающей конструкции.
Повышенные эффекторные функции
[00147] В одном варианте реализации изобретения биспецифическая антигенсвязывающая конструкция, описанная в данном документе, характеризуется повышенной эффекторной функцией по сравнению с каждой соответствующей моноспецифической бивалентной антигенсвязывающей конструкцией (т.е. по сравнению с моноспецифической бивалентной антигенсвязывающей конструкцией, которая связывается с ECD2, или моноспецифической бивалентной антигенсвязывающей конструкцией, которая связывается с ECD4) и/или по сравнению с комбинацией двух моноспецифических бивалентных антигенсвязывающих конструкций. «Эффекторные функции» антител относятся к биологическим активностям, свойственным Fc-домену (Fc-домен с нативной последовательностью или Fc-домен с вариантной аминокислотной последовательностью) антитела. Примеры эффекторных функций антител включают связывание Clq; комплементзависимая цитотоксичность (КЗЦ); связывание с Fc-рецептором; антителозависимая клеточноопосредованная цитотоксичность (АЗКЦ); антителозависимый клеточный фагоцитоз (АЗКФ); пониженная экспрессия рецепторов клеточной поверхности (например, В-клеточный рецептор; BCR) и т.д.
АЗКЦ
[00148] Таким образом, в одном варианте реализации изобретения биспецифическая антигенсвязывающая конструкция характеризуется форматом Fab-scFv и характеризуется более высокой активностью в анализе АЗКЦ, чем эталонная антигенсвязывающая конструкция, которая характеризуется форматом Fab-Fab, в клетках, экспрессирующих HER2 на уровне 1+.
[00149] В одном варианте реализации изобретения биспецифическая антигенсвязывающая конструкция характеризуется более выраженным максимальным клеточным лизисом в анализе АЗКЦ, чем эталонная антигенсвязывающая конструкция, которая представляет собой трастузумаб или его аналог. В одном варианте реализации изобретения биспецифическая антигенсвязывающая конструкция характеризуется форматом Fab-scFv и более выраженным максимальным клеточным лизисом в анализе АЗКЦ, чем эталонная антигенсвязывающая конструкция, которая представляет собой трастузумаб или его аналог, или комбинацию аналогов трастузумаба или пертузумаба. В одном варианте реализации изобретения биспецифическая антигенсвязывающая конструкция характеризуется форматом Fab-scFv и более выраженным максимальным клеточным лизисом в анализе АЗКЦ, чем эталонная антигенсвязывающая конструкция, которая представляет собой трастузумаб или его аналог, в клетках, экспрессирующих HER2 на уровне 1+ или больше. В одном варианте реализации изобретения биспецифическая антигенсвязывающая конструкция характеризуется форматом Fab-scFv и более высокой активностью в анализе АЗКЦ, чем эталонная антигенсвязывающая конструкция, которая представляет собой трастузумаб или его аналог, в клетках фенотипа HER2 2+/3+.
Интернализации
[00150] Биспецифические антигенсвязывающие конструкции, описанные в данном документе, интернализуются в клетки HER2+ посредством связывания с рецептором HER2. Таким образом, биспецифические антигенсвязывающие конструкции, описанные в данном документе, способны индуцировать интернализацию рецептора в клетки HER2+. В одном варианте реализации изобретения биспецифическая антигенсвязывающая конструкция характеризуется форматом Fab-scFv и индуцирует интернализацию HER2 в большей степени, чем эталонная антигенсвязывающая конструкция, которая характеризуется форматом Fab-Fab, в клетках, экспрессирующих HER2 на уровне 3+. В одном варианте реализации изобретения биспецифическая антигенсвязывающая конструкция характеризуется форматом Fab-scFv и индуцирует интернализацию HER2 в большей степени, чем эталонная антигенсвязывающая конструкция, которая характеризуется форматом Fab-Fab, в клетках, экспрессирующих HER2 на уровне 2+ или 3+. В одном варианте реализации изобретения биспецифическая антигенсвязывающая конструкция характеризуется форматом scFv-scFv и индуцирует интернализацию HER2 в большей степени, чем эталонная антигенсвязывающая конструкция, которая характеризуется форматом Fab-Fab, в клетках, экспрессирующих HER2 на уровне 1+, 2+ или 3+.
Клеточная цитотоксичность
[00151] Биспецифическая антигенсвязывающая конструкция может быть получена в виде ADC, как описано в других местах в данном документе, и может оказывать цитотоксическое действие на клетки. В одном варианте реализации изобретения ADC, содержащий биспецифическую антигенсвязывающую конструкцию, характеризуется более высокой активностью в анализе цитотоксичности и выживаемости клеток при использовании HER2+ клеток рака молочной железы, чем эталонная антигенсвязывающая конструкция, которая представляет собой трастузумаб или его аналог, или эталонная антигенсвязывающая конструкция, которая представляет собой комбинацию T-DM1 и пертузумаба, в клетках с уровнем экспрессии HER2 1+, 2+, 2+/3+ или 3+.
Повышенная способность к связыванию с FcγR
[00152] В некоторых вариантах реализации изобретения биспецифические антигенсвязывающие конструкции характеризуются более выраженной способностью к связыванию (Rmax) с одним или более FcγR. В одном варианте реализации изобретения биспецифическая антигенсвязывающая конструкция характеризуется повышением Rmax при связывании с одним или более FcγR по сравнению с эталонной антигенсвязывающей конструкцией, которая представляет собой v506 или v6246, имеющие гомодимераный Fc-фрагмент, в около 1,3-2 раза. В одном варианте реализации изобретения биспецифическая антигенсвязывающая конструкция характеризуется повышением Rmax при связывании с CD 16 FcγR в около 1,3-1,8 раза по сравнению с эталонной бивалентной антигенсвязывающей конструкцией. В одном варианте реализации изобретения биспецифическая антигенсвязывающая конструкция характеризуется повышением Rmax при связывании с CD32 FcγR в около 1,3-1,8 раза по сравнению с эталонной бивалентной антигенсвязывающей конструкцией. В одном варианте реализации изобретения биспецифическая антигенсвязывающая конструкция характеризуется повышением Rmax при связывании с CD64 FcγR в около 1,3-1,8 раза по сравнению с эталонной бивалентной антигенсвязывающей конструкцией.
Повышенная аффинность по отношению к FcγR
[00153] Биспецифические антигенсвязывающие конструкции, предложенные в настоящем изобретении, характеризуются повышенной аффинностью по отношению к FcγR по сравнению с эталонной антигенсвязывающей конструкцией, такой как трастузумаб. Повышение концентрации молекул Fc-фрагмента, возникающее в результате связывания, согласуется с повышением способности к АЗКЦ и/или другими механизмами уничтожения клеток иммунными эффекторами.
[00154] В некоторых вариантах реализации изобретения биспецифические антигенсвязывающие конструкции характеризуются повышенной аффинностью по отношению к одному или более FcγR. В одном варианте реализации изобретения, в котором биспецифическая антигенсвязывающая конструкция содержит антигенсвязывающий полипептид, который связывается с HER2, биспецифические антигенсвязывающие конструкции характеризуются повышенной аффинностью по отношению по меньшей мере к одному FcγR. В соответствии с этим вариантом реализации изобретения, биспецифическая антигенсвязывающая конструкция характеризуются повышенной аффинностью по отношению к CD32.
Связывание с FcRn и РК-параметры
[00155] В некоторых вариантах реализации изобретения антигенсвязывающие конструкции из описанных в данном документе способны связываться с FcRn. Как известно в данной области техники, связывание с FcRn приводит к рециркуляции подвергшегося эндоцитозу антитела из эндосомы назад в кровоток (Raghavan et al, 1996, Annu Rev Клетка Dev Biol 12:181-220; Ghetie et al, 2000, Annu Rev Immunol 18:739-766). Этот процесс в сочетании с уменьшением почечной фильтрации, обусловленным большим размером полноразмерной молекулы, приводит к более продолжительному периоду полувыведения антитела из сыворотки крови в пределах от одной до трех недель. Связывание Fc-домена с FcRn также играет ключевую роль в транспорте антитела.
Фармакокинетические параметры
[00156] В определенных вариантах реализации изобретения биспецифическая антигенсвязывающая конструкция, предложенная в настоящем изобретении, характеризуется фармакокинетическими (ФК) свойствами, сопоставимыми с коммерчески доступными терапевтическими антителами. В одном варианте реализации изобретения биспецифические антигенсвязывающие конструкции, описанные в данном документе, характеризуются ФК-свойствами, сходными со свойствами известных терапевтических антител, что касается концентрации в сыворотке крови, t1/2, периода полувыведения в бета-фазе и/или CL. В одном варианте реализации изобретения биспецифические антигенсвязывающие конструкции характеризуются стабильностью in vivo, сопоставимой или большей, чем стабильность указанной моноспецифической бивалентной антигенсвязывающей конструкции. Такие параметры стабильности in vivo включают концентрацию в сыворотке крови, t1/2, период полувыведения в бета-фазе и/или CL.
Исследование биспецифических антигенсвязывающих конструкций. Связывание с FcγR, FcRn и Clq
[00157] Эффекторные функции биспецифических антигенсвязывающих конструкций могут быть исследованы следующим образом. Для оценки АЗКФ, КЗЦ и/или АЗКЦ может быть проведена реакция цитотоксичности in vitro и/или in vivo. Например, для измерения уровня связывания с FcγR могут быть проведены анализы связывания с Fc-рецептором (FcR). Основные клетки, которые опосредуют АЗКЦ, NK-клетки. экспрессируют только FcγRIII, тогда как моноциты экспрессируют FcγRI, FcγRII и FcγRIII. Данные по экспрессии FcR на гематопоэтических клетках приведены в Таблице 3 на странице 464 в Ravetch and Kinet, Annu. Rev. Immunol 9:457-92 (1991). Например, in vitro анализ для оценки влияния представляющей интерес молекулы на АЗКЦ описан в патенте США №5500362 и 5821337. Подходящие эффекторные клетки для таких анализов включают мононуклеары периферической крови (МКПК) и натуральные киллеры (NK). В качестве альтернативы или в дополнение к этому, влияние представляющей интерес молекулы на АЗКЦ можно оценить in vivo, например, в экспериментальной модели на животных, что, например, описано в Clynes et al. PNAS (США) 95:652-656 (1998). Для определения того, способны ли биспецифические антигенсвязывающие конструкции связываться с Clq и, следовательно, активировать КЗЦ, также можно провести анализы свзывания с Clq. Для оценки активации комплемента, можно провеси анализ КЗЦ, например, как описано в Gazzano-Santoro et al., J. Immunol. Methods 202:163 (1996). Также могут быть определены связывание с FcRn. как. например, с помощью ППР, и ФК-параметры антител in vivo с использованием способов, хорошо известных в данной области техники.
Исследование антигенсвязывающих конструкций; связывание с HER2
[00158] Антигенсвязывающие конструкции или фармацевтические композиции, описанные в данном документе, исследуют in vitro, а затем in vivo, перед использованием на людях. Например, in vitro анализы, используемые для того, чтобы продемонстрировать пригодность соединения или фармацевтичекой композиции для использования в терапевтических и профилаткических целях, включают оценку влияния соеденения на линию клеток или образец ткани пациента. Влияние соеденения или композиции на линию клеток и/или образец ткани можно определить, используя способы, известные специалистам в данной области техники, включая, в том числе, анализы розеткообразования и анализы лизиса клеток. В соответствии с настоящим изобретением, анализы in vitro, которые могут быть использованы для определения наличия показаний к применению специфической антигенсвязывающей конструкции, включают анализы in vitro на культуре клеток или анализы in vitro на образце культивируемой ткани пациента, которые экспонируют или иным образом подвергают воздействию антигенсвязывающей конструкции, что позволяет оценить влияние такой антигенсвязывающей конструкции на образец ткани.
[00159] Антигенсвязывающие конструкции-кандидаты могут быть исследованы с использованием клеток, например, клеточных линий рака молочной железы, экспрессирующих HER2. В следующей ниже Таблице D приведено описание уровня экспрессии HER2 в нескольких репрезентативных линиях раковых клеток.
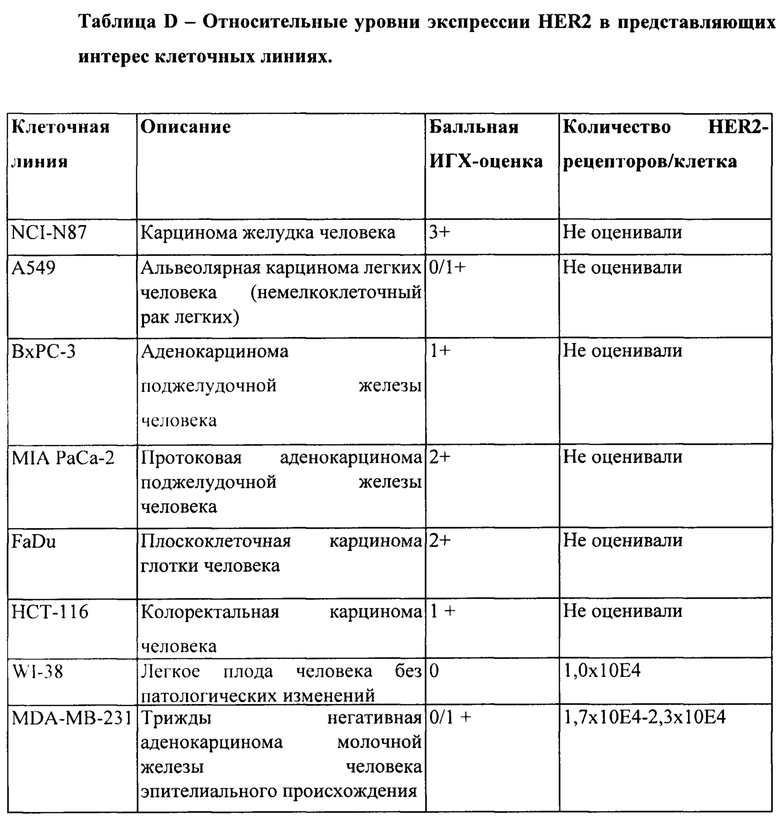
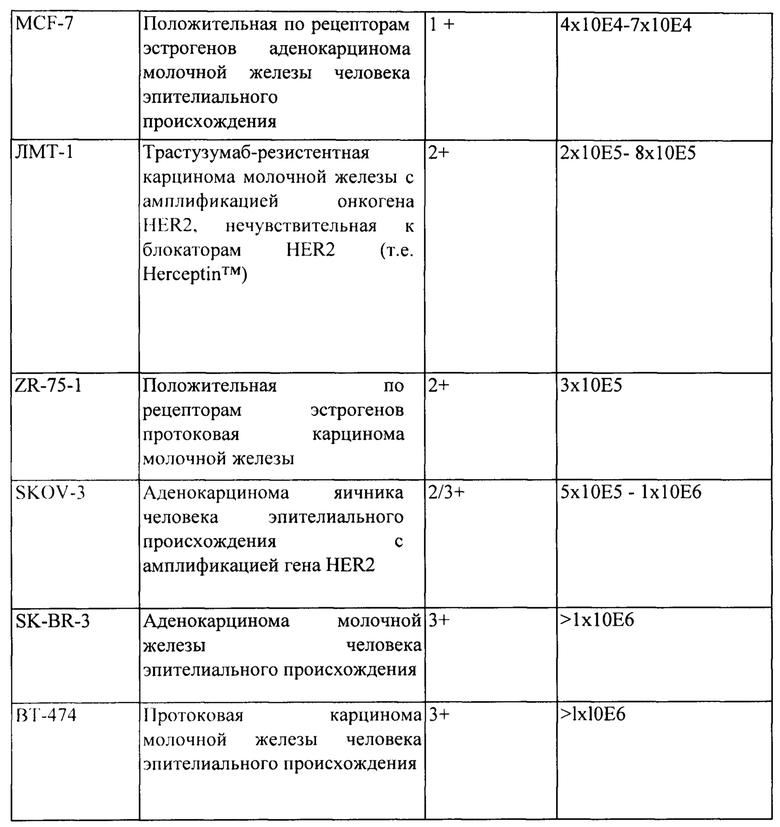
[00160] McDonagh et al Mol Cancer Ther. 2012 Mar; 11(3):582-93; Subik et al. (2010) Breast Cancer: Basic Clinical Research:4; 35-41; Carter et al. PNAS, 1994: 89; 4285-4289; Yarden 2000, HER2: Basic Research, Prognosis and Therару; Hendricks et al. Mol Cancer Ther 2013; 12:1816-28.
[00161] Как известно в данной области техники, для идентификации антигенсвязывающих конструкций, подходящих для использования в способах, описанных в данном документе, может быть использован целый ряд анализов. Эти анализы могут проводиться на раковых клетках, экспрессирующих HER2. Примеры подходящих раковых клеток преведены в Таблице А5. Примеры анализов, которые могут проводиться, описаны ниже.
[00162] Например, для идентификации ингибирующей рост антигенсвязывающей конструкции-кандидата, которая связывается с HER2, можно провести скрининг антител, которые ингибируют рост раковых клеток, что экспрессируют HER2. В одном варианте реализации изобретения предпочтительная антигенсвязывающая конструкция-кандидат способна ингибировать рост раковых клеток в культуре клеток на около 20-100% и предпочтительно на около 50-100% по сравнению с контрольной антигенсвязывающей конструкцией.
[00163] Для подбора антигенсвязывающией конструкции-кандидата, которая вызывает гибель клеток, можно оценить потерю целостности мембраны, например,по поглощению PI (фосфатидилинозитол), трипанового синего или 7AAD, по сравнению с контролем.
[00164] Для подбора антигенсвязывающией конструкции-кандидата, которая вызывает апоптоз, может быть использован анализ связывания аннексина. В дополнение к анализу связывания аннексина, также может быть использован анализ окрашивания ДНК.
[00165] В одном варианте реализации изобретения представляющая интерес антигенсвязывающая конструкция-кандидат может блокировать херегулин-зависимую ассоциацию ErbB2 и ErbB3 как в клетках MCF7, так и SK-BR-3, что установлено в эксперименте по коиммунопреципитации, по существу, более эффективно, чем моноклональное антитело 4D5 и предпочтительно, по существу, более эффективно, чем моноклональное антитело 7F3.
[00166] Для скрининга антигенсвязывающих конструкций, которые связываются с эпитопом на ErbB2, с которым связывается представляющее интерес антитело, можно провести стандартный эпитоп-перекрестный конкурентный анализ, как, например, описанный в Antibodies, A LaboratoryManual, Cold Spring Harbor Laboratory, Ed Harlow and David Lane (1988). В качестве альтернативы или в дополнение к этому, можно осуществить картирование эпитопов с помощью способов, известных в данной области техники.
[00167] Конкурентное связывание между антигенсвязывающими конструкциями можно определить путем анализа, в котором исследуемая антигенсвязывающая конструкция ингибирует или блокирует специфическое связывание эталонной антигенсвязывающей конструкцией с общим антигеном (см., например, Junghansetal, CancerRes. 50:1495, 1990; Fendlyetal. Cancer Research 50: 1550-1558; US 6,949,245). Исследуемая антигенсвязывающая конструкция конкурирует с эталонной антигенсвязывающей конструкцией, если взятая в избытке исследуемая антигенсвязывающая конструкция (например, по меньшей мере 2х, 5х, 10х, 20х или 100х) ингибирует или блокирует связывание эталонной антигенсвязывающей конструкцией, например, по меньшей мере на 50%, 60%, 70%, 75%, 80%, 85%, 90%, 95% или 99% по результатам анализа конкурентного связывания. Антигенсвязывающие конструкции, идентифицированные путем конкурентного анализа (конкурирующая антигенсвязывающая конструкция), включают антигенсвязывающие конструкции, связывающиеся с одним и тем же эпитопом, что и эталонная антигенсвязывающая конструкция, и антигенсвязывающие конструкции, связывающиеся с соседним эпитопом, расположенным достаточно проксимально к эпитопу, с которым связывается эталонная антигенсвязывающая конструкция, для обеспечения стерического несоответствия. Например, может быть идентифицирована вторая конкурирующая антигенсвязывающая конструкция, которая конкурирует с первой антигенсвязывающей конструкцией, описанной в данном документе, за связывание с HER2. В определенных случаях вторая конструкция может блокировать или ингибировать связывание первой конструкции, например, по меньшей мере на 50%. 60%, 70%, 75%, 80%, 85%, 90%, 95% или 99% по результатам анализа конкурентного связывания. В определенных случаях вторая конструкция может вытеснять первую конструкцию более чем на 50%, 60%, 70%, 75%, 80%, 85%, 90%, 95% или 99%.
[00168] В некоторых вариантах реализации изобретения анализируют функциональные свойства антигенсвязывающих конструкций, описанных в данном документе, in vivo, например, в экспериментальных моделях на животных. В некоторых вариантах реализации изобретения используются экспериментальные модели на животных, которые описаны в Таблице Е. В некоторых вариантах реализации изобретения антигенсвязывающие конструкции характеризуются повышенной эффективностью воздействия в экспериментальных моделях на животных по сравнению с эталонной антигенсвязывающей конструкцией.
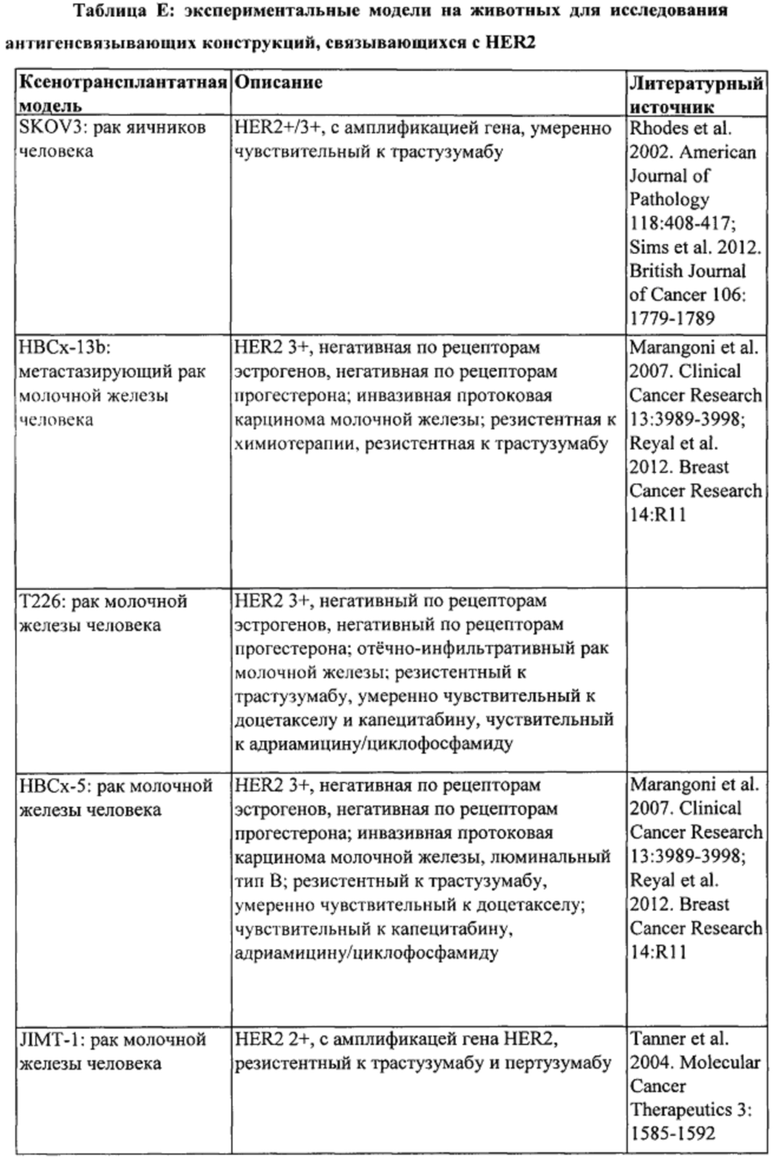
Эталонная антигенсвязывающая конструкция
[00169] В некоторых вариантах реализации изобретения функциональные характеристики биспецифических антигенсвязыващих конструкций, описанных в данном документе, сравнениваются с характеристиками эталонной антигенсвязывающей конструкции. Отличительные черты эталонной антигенсвязывающей конструкции зависят от определяемой функциональной характеристики или от устанавливаемого различия. Например, при сравнении функциональных характеристик иллюстративных биспецифическиех антигенсвязывающих конструкций эталонная антигенсвязывающая конструкция может представлять собой аналог трастузумаба, такой как, например, v506, или может представлять собой комбинацию антител, таких как трастузумаб и пертузумаб (v4184). В вариантах реализации изобретения, где сравнивается формат биспецифической антигенсвязывающей конструкции, эталонная антигенсвязывающая конструкция может представлять собой, например, бипаратопное антитело к HER2, где оба антигенсвязывающие фрагменты характеризуются форматом Fab-Fab (эталонная антигенсвязывающая конструкция определенного формата). Примеры последней конструкции включают v6902 и v6903.
Антигенсвязывающие конструкции и конъюгаты антитела с лекарственным средством (ADC)
[00170] В определенных вариантах реализации изобретения антигенсвязывающая конструкция конъюгирована с лекарственным средством, например, токсином, химиотерапевтическим средством, иммуномодулятором или радиоизотопом. Несколько способов получения ADC (конъюгат антитела с лекарственным средством или конъюгат антигенсвязывающей конструкции с лекарственным средством) известны в данной области техники и описаны, например, в патентах США №8624003 (способ синтеза в одном сосуде), 8163888 (одностадийный) и 5208020 (двухстадийный способ).
[00171] В некоторых вариантах реализации изобретения лекарственный препарат выбран из майтансина, ауристатина, калихимицин или их производных. В других вариантах реализации изобретения лекарственный препарат представляет собой майтансин, выбранный из DM1 и DM4. Дополнительные примеры описаны ниже.
[00172] В некоторых вариантах реализации изобретения лекарственный препарат конъюгирован с выделенной антигенсвязывающией конструкцией при помощи линкера SMCC (DM1) или линкера SPDB (DM4). Дополнительные примеры описаны ниже. Соотношение лекарственного препарата к антигенсвязывающему белку (DAR) может составлять, например, от 1,0 до 6,0, или от 3,0 до 5,0, или 3,5-4,2.
[00173] В некоторых вариантах реализации изобретения антигенсвязывающая конструкция конъюгирована с цитотоксическим средством. Используемый в данном документе термин «цитотоксическое средство» относится к веществу, которое ингибирует или подавляет функциональную активность клеток и/или вызывает разрушение клеток. Подразумевается, что термин охватывает радиоактивные изотопы (например, At211, I131, I 125, Y90, Re186, Re188, Sm153, Bi212, P32 и Lul77), хемотерапевтические средства и токсины, как, например, низкомолекулярные токсины, или обладающие ферментативной активностью токсины бактериального, грибного, растительного или животного происхождения, включая их фрагменты и/или варианты. Дополнительные примеры описаны ниже.
Лекарственные препараты
[00174] Неограничивающие примеры лекарственных препаратов или молекулярных грузов, используемых в различных вариантах реализации ADC, включают DM1 (майтансин, N2'-диацетил-N2'-(3-меркапто-1-оксопропил)- или N2'-диацетил-N2'-(3-меркапто-1-оксопропил)-майтансин), mc-MMAD(6-малеимидокапроил-монометилауристатин-D или N-метил-L-валил-N-[(1S,2R)-2-метокси-4-[(2S)-2-[(1R,2R)-l-метокси-2-метил-3-оксо-3-[[(1S)-2-фенил-1-(2-тиазолил)этил]амино]пропил]-1-пирролидинил]-1-[(1S)-1-метилпропил]-4-оксобутил]-N-метил-(9Cl)-L-валинамид), mc-MMAF (малеимидокапроил-монометилауристатин F или N-[6-(2,5-дигидро-2,5-диоксо-1Н-пиррол-1-ил)-1оксогексил]-N-метил-L-валил-L-валил-(3R,4S,5S)-3-метокси-5-метил-4-(метиламино)гептаноил-(αR,βR,2S)-β-метокси-α-метил 1-2-пирролидинпропаноил-L-фенилаланин) и mc-Val-Cit-PABA-MMAE (6-малеимидокапроил-ValcCit-(р-аминобензилоксикарбонил)-монометилауристатин Е или N-[[[4-[[N-[6-(2,5-дигидро-2,5-диоксо-1Н-пиррол-1-ил)-1-оксогексил]-L-валил-N5-(аминокарбонил)-L-орнитил]амино]фенил]метокси]карбонил]-N-метил-L-валил-N-[(1S,2R)-4-[(2S)-2-[(1R,2R)-3-[[(1R,2S)-2-гидрокси-1-метил-2-фенилеэтил]амино]-1-метокси-2-метил-3-оксопропил]-1-пирролидинил]-2-метокси-1-[(1S)-1-метилпропил]-4-оксобутил]-N-метил-L-валинамид). DM1 представляет собой производное ингибитора полимеризации тубулина - майтансина, тогда как MMAD, ММАЕ и MMAF представляют собой производные ауристатина.
Лекарственные фрагменты майтанзиноидов
[00175] Как указано выше, в некоторых вариантах реализации изобретения в качестве лекарственного препарата использован майтанзиноид. Иллюстративные майтанзиноиды включают DM1, DM3 (N2'-диацетил-N2'-(4-меркапто-1-оксопентил) майтансин) и DM4 (N2'-диацетил-N2'-(4-метил-4-меркапто-1-оксопентил)метилмайтансин) (см. US 20090202536).
[00176] Многие положения на соединениях майтансина, как известно, могут использоваться для формирования связи, в зависимости от типа связи. Например, для формирования сложноэфирной связи все положения, например, С-3 положение, в котором расположена гидроксильная группа, С-14 положение, модифицированное с помощью гидроксиметильной группы, С-15 положение, модифицированное с помощью гидроксильной группы, и С-20 положение, в котором расположена гидроксильная группа, являются подходящими.
[00177] Для получения ADC, описанных в данном документе, предусмотрено использование всех стереоизомеров лекарственных фрагментов майтанзиноидов, т.е. любая комбинация R- и S-конфигураций в хиральных атомах углерода D.
Ауристатины
[00178] В некоторых вариантах реализации изобретения лекарственным средством является ауристатин, как, например, ауристатин Е (также известный в данной области техники как производное доластатина-10), или его производные. Ауристатин может представлять собой, например, сложный эфир, образующийся при реакции между ауристатином Е и кетокислотой. Например, ауристатин Е может вступать в реакцию с пара-ацетилбензойной кислотой или бензоилвалериановой кислотой с образованием АЕВ и AEVB, соответственно. Другие обычные ауристатины включают AFP, MMAF и ММАЕ. Синтез и структура иллюстративных ауристатинов описаны в патентах США №. 6884869, 7098308, 7256257, 7423116, 7498298 и 7745394, каждый из которых включен в данный документ посредством ссылки в полном объеме и для всех целей.
Химиотерапевтические средства
[00179] В некоторых вариантах реализации изобретения антигенсвязывающая конструкция конъюгирована с хемотерапевтическим средством. Примеры, включают, в том числе, Цисплантин и Лапатиниб. Термин «хемотерапевтическое средство» относится к химическому соединению, которое можно использовать в лечении рака.
[00180] Примеры хемотерапевтических средств, включают алкилирующие агенты, такие как тиотепа и циклофосфамид (CYTOXAN™); алкилсульфонаты, такие как бусульфан, импросульфан и пипосульфан; азиридины, такие как бензодопа, карбоквон, метуредопа и уредопа; этиленимины и метиламеламины, включая альтретамин, триэтиленмеламин, триэтиленфосфорамид, триэтилентиофосфаорамид и триметилоломеламин; азотистые иприты, такие как хлорамбуцил, хлорнафазин, хлорфосфамид, эстрамустин, ифосфамид, мехлорэтамин, гидрохлорид оксида мехлорэтамина, мелфалан, новэмбихин, фенестерин, преднимустин, трофосфамид, урамустин; нитрозумочевины, такие как кармустин, хлорозотоцин, фотемустин, ломустин, нимустин, ранимустин; антибиотики, такие как аклациномицины, актиномицин, аутрамицин, азасерин, блеомицины, кактиномицин, калихеамицин, карабицин, карминомицин, карзинофилин, хромомицины, дактиномицин, даунорубицин. деторубицин, 6-диазо-5-оксо-L-норлейцин, доксорубицин, эпирубицин, эзорубицин, идарубицин, марцелломицин, митомицины, микофеноловая кислота, ногаламицин, оливомицины, пепломицин, потфиромицин, пуромицин, квеламицин, родорубицин, стрептонигрин, стрептозоцин, туберцидин, убенимекс, зиностатин, зорубицин; антиметаболиты, такие как метотрексат и 5-фторурацил (5-ФУ); аналоги фолиевой кислоты, такие как деноптерин, метотрексат, птероптерин, триметрексат; аналоги пурина, такие как флударабин, 6-меркаптопурин, тиамиприн, тиогуанин; аналоги пиримидина, такие как анцитабин, азацитидин, 6-азауридин, кармофур, цитарабин, дидеоксиуридин, доксифлуридин, эноцитабин, флоксуридин, 5-ФУ; андрогены, такие как калустерон, пропионат дромостанолона, эпитиостанол, мепитиостан, тестолактон; антиадреналины, такие как аминоглутетимид, митотан, трилостан; добавка для восполнения фолиевой кислоты, такая как фролиновая кислота; ацеглатон; гликозид альдофосфамида; аминолевулиновая кислота; амсакрин; бестрабуцил; бисантрен; эдатраксат; дефофамин; демеколцин; диазиквон; элфорнитин; ацетат эллиптиния; этоглуцид; нитрат галлия; гидроксимочевина; лентинан; лонидамин; мигогуазон; митоксантрон; мопидамол; нитракрин; пентостатин; фенамет; пирарубицин; подофиллиновая кислота; 2-этилгидразид; прокарбазин; PSK7; разоксан; сизофиран; спирогерманий; тенуазоновая кислота; триазиквон; 2,2',2'=-трихлоротриэтиламин; уретан; виндезин; дакарбазин; манномустин; митобронитол; митолактол; пипоброман; гацитозин; арабинозид («Ara-С»); циклофосфамид; тиотепа; таксаны, например, паклитаксел (TAXOL® , Bristol-Myers Squibb Oncology, Принстон, Нью-Джерси) и доксетаксел (TAXOTERE®, Rhone-Poulenc Rorer, Антони, Франция); хлорамбуцил; гемцитабин; 6-тиогуанин; меркаптопурин; метотрексат; аналоги платины, такие как цисплатин и карбоплатин; винбластин; платина; этопозид (VP-16); ифосфамид; митомицин С; митоксантрон; винкристин; винорелбин; навелбин; новантрон; тенипозид; дауномицин; аминоптерин; кселода; ибандронат; СРТ-11; ингибитор топоизомеразы RFS 2000; дифторометилорнитин (DMFO); ретиноевая кислота; эсперамицины; капецитабин; и фармацевтически приемлемые соли, кислоты и производные любого из вышеупомянутого. Также этим определением охватываются антигормональные средства, действие которых направлено на регуляцию или ингибирование действия горонов на опухоли, такие как антиэстрогены, включающие например, тамоксифен, ралоксифен, 4(5)-имидазолы, ингибирующис ароматазу, 4-гидрокситамоксифен, триоксифен, кеоксифен, LY117018, онапристон и торемифен (Fareston); и антиандрогены, такие как флутамид, нилутамид, бикалутамид, лейпролид и гозерелин; и фармацевтически приемлемые соли, кислоты и производные любого из вышеупомянутого.
Конъюгированные линкеры
[00181] В некоторых вариантах реализации изобретения лекарственное средство связано с антигенсвязывающей конструкцией, например, антителом, посредством линкера. Присодинение линкера к антителу может осуществляться различными способами, как, например, через поверхностные остатки лизина, восстановительное сочетание с окисленными углеводами и через высвобождение остатков цистеина путем восстановления межцепочечных дисульфидных связей. В данной области техники известны различные системы связи для образования ADC, включая гидразоновые, дисульфидные и пептидные связи.
[00182] Подходящие линкеры включают, например, расщепляемые и нерасщепляемые линкеры. Расщепляемый линкер, как правило, обладает чувствительностью к расщеплению при внутриклеточных условиях. Подходящие расщепляемые линкеры включают, например, пептидный линкер, расщепляемый внутриклеточной протеазой, как, например, лизосомальная протеаза или эндосомальная протеаза. В иллюстративных вариантах реализации изобретения, линкер может представлять собой дипептидный линкер, как, например, валин-цитрулиновый (val-cit), фенилаланин-лизиновый (phe-lys) линкер или линкер, представляющий собой малеимидокапроил-валин-цитруллин-п-аминобензилоксикарбонил (mc-Val-Cit-PABA). Другой линкер представляет собой сульфосукцинимидил-4-[N-малеимидометил]циклогексан-1-карбоксилат (SMCC). Конъюгирование с использованием сульфо-smcc происходит через малеимидную группу, которая вступает в реакцию с сульфгидрилами (тиолы, - SH), тогда как ее сульфо-NHS эфир реагирует с первичными аминами (которые встречаются в лизине и N-конце белков или пептидов). Еще один линкер представляет собой малеимидокапроил (МС). Другие подходящие линкеры включают линкеры, гидролизуемые при определенном рН или диапазоне рН, как, например, гидразоновый линкер. Дополнительные подходящие расщепляемые линкеры включают дисульфидные линкеры. Линкер может быть ковалентно связан с антителом в такой степени, что антитело неизбежно будет подвергаться внутриклеточной деградации для высвобождения лекарственного средства, например, МС-линкер и тому подобное.
Получение ADC
[00183] ADC может быть получен несколькими способами с применением реакций органической химии, условий и реактивов, известных специалистам в данной области техники, включая: (1) реакцию нуклеофильных групп или электрофильных групп антитела с бивалентным линкерным реагентом с образованием промежуточного продукта антитело-линкер, Ab-L, через ковалентную связь с последующей реакцией с активизированным лекарственным фрагментом D; и (2) реакцию нуклеофильных групп или электрофильных групп лекарственного фрагмента с линкерным реагентом с образованием промежуточного продукта лекарственное средство-линкер, D-L, через ковалентную связь с последующей реакцией с нуклеофильной группой или электрофильной группой антитела. Способы конъюгирования (1) и (2) могут быть использованы с различными антителами, лекартвенными фрагментами и линкерами для получения описанных в настоящем документе коньюгатов антитела с лекарственным средством.
[00184] Несколько конкретых примеров способов получения ADC известны в данной области техники и описаны в патентах США 8624003 (способ синтеза в одной реакции), 8163888 (одностадийный) и 5208020 (двухстадийный способ).
Способы получения антигенсвязывающих конструкций
[00185] Антигенсвязывающие конструкции, описанные в данном документе, могут быть получены с использованием методов рекомбинации и композиций, например, как описано в патенте США №4816567.
[00186] В одном варианте реализации изобретения предложена выделенная нуклеиновая кислота, кодирующая антигенсвязывающую конструкцию, описанную в данном документе. Такая нуклеиновая кислота может кодировать аминокислотную последовательность, содержащую VL-область, и/или аминокислотную последовательность, содержащую VH-область антигенсвязывающей конструкции (например, легкая и/или тяжелая цепи антигенсвязывающей конструкции). В дополнительном варианте реализации изобретения предложены один или более векторов (например, экспрессирующие векторы), содержащих такие нуклеиновые кислоты. В одном варианте реализации изобретения предложена нуклеиновая кислота в полицистронном векторе. В дополнительном варианте реализации изобретения предложена клетка-хозяин, содержащая такую нуклеиновую кислоту. В одном таком варианте реализации изобретения клетка-хозяин содержит (например, была трансформирована с использованием): (1) вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VL-область антигенсвязывающей конструкции, и аминокислотную последовательность, содержащую VH-область антигенсвязывающей полипептидной конструкции, или (2) первый вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VL-область антигенсвязывающей полипептидной конструкции, и второй вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VH-область антигенсвязывающей полипептидной конструкции. В одном варианте реализации изобретения клетка-хозяин представляет собой эукариотическую клетку, например, клетку яичника китайского хомячка (СНО) или человеческую эмбриональную клетку почек (НЕК) или лимфатическую клетку (например, Y0, NS0, Sp20 клетка). В одном варианте реализации изобретения предложен способ получения антигенсвязывающей конструкции, причем способ включает культивирование клетки-хазяина, содержащей нуклеиновую кислоту, кодирующую антигенсвязывающую конструкцию, которая представлены выше, в условиях, подходящих для экспрессии антигенсвязывающей конструкции, и необязательно выделение антигенсвязывающей конструкции из клетки-хозяина (или культуральной среды клетки-хазяина).
[00187] Для получения антигенсвязывающей конструкции методами рекомбинации нуклеиновую кислоту, которая кодирует антигенсвязывающую конструкцию, например, которая описана выше, выделяют и встравивают в один или более векторов для дальнейшего клонирования и/или экспрессии в клетке-хозяина. Такая нуклеиновая кислота может быть без труда выделена и секвенирована с использованием общепринятых методов (например, с использованием олигонуклеотидных зондов, которые способны специфически связываться с генами, кодирующими тяжелые и легкие цепи антигенсвязывающей конструкции).
[00188] Термин «очищенные по существу» относится к конструкции, описанной в данном документе, или ее вариантам, которые по существу или по сути могут не содержать компонентов, которые обычно сопутствуют или взаимодействуют с белком, который находится в естественном для него окружении, т.е. применение нативной клетки или клетки-хозяина в случае получения гетеромультимера методами рекомбинации, который в определенных вариантах реализации изобретения, по существу, не содержит клеточного материала, обеспечеивает получение препаратов белка, содержащих менее, чем около 30%, менее, чем около 25%, менее, чем около 20%, менее, чем около 15%, менее, чем около 2% или менее, чем около 1% (от сухого веса) белковых примесей. Когда гетеромультимеры или их варианты получают методами рекомбинации с использованием клеток-хозяев, в определенных вариантах реализации изобретения содержание белка составляет около 30%, около 25%, около 20%, около 15%, около 10%, около 5%, около 4%, около 3%, около 2% или около 1% или меньше в пересчете на сухой вес клеток. Когда гетеромультимеры или их варианты получают методами рекомбинации с использованием клеток-хозяев, в определенных вариантах реализации изобретения содержание белка в культурной среде составляет около 5 г/л, около 4 г/л, около 3 г/л, около 2 г/л, около 1 г/л, около 750 мг/л, около 500 мг/л, около 250 мг/л, около 100 мг/л, около 50 мг/л, около 10 мг/л или около 1 мг/л или меньше в пересчете на сухой вес клеток. В определенных вариантах реализации изобретения содержание «очищенного по существу» гетеромультимера, полученного способами, описанными в данном документе, составляет по меньшей мере около 30%, по меньшей мере около 35%, по меньшей мере около 40%, по меньшей мере около 45%, по меньшей мере около 50%, по меньшей мере около 55%, по меньшей мере около 60%, по меньшей мере около 65%, по меньшей мере около 70%, конкретно, содержание составляет по меньшей мере около 75%, 80%, 85%, и более конкретно, содержание составляет по меньшей мерео коло 90%, содержание составляет по меньшей мере около 95%, содержание составляет по меньшей мере около 99% или больше, что установлено соответствующими способами, такими как ДСН-ПААГ-анализ, ОФ-ВЭЖХ, ЭХ и капиллярный электрофорез.
[00189] Клетки-хозяева, подходящие для клонирования и экспрессии векторов, кодирующих антигенсвязывающие конструкции, включают прокариотические и эукариотические клетки, описанные в данном документе.
[00190] «Рекомбинантная клетка-хозяин» или «клетка-хозяин» относится к клетке, которая включает экзогенный полинуклеотид, независимо от способа, используемого для вставки, например, прямое поглощение, трансдукция, конъюгация с использованием f-фактора или другие способы, известные в данной области техники, для получения рекомбинантной клетки-хозяина. Экзогенный полинуклеотид может существовать как неинтегрированный вектор, например, плазмида, или в качестве альтернативы, может интегрироваться в геном хозяина.
[00191] Используемый в данном документе термин «эукариот», относится к организмам, относящимся к филогенетической группе Eucarya, как, например, животные (включая, в том числе, млекопитающих, насекомых, рептилий, птиц и т.д.), инфузории, растения (включая, в том числе, однодольное растение, двудольное растение, водоросли и т.д.), грибы, дрожжевые грибы, жгутиковые, микроспориды, простейшие и т.д.
[00192] Используемый в данном документе, термин «прокариот» относится к прокариотическим организмам. Например, организм, отличный от эукариотического, может относиться к филогенетической группе Eubacteria (включая, в том числе, Escherichia coli, Thermusthermophilus, Bacillusstearothermophilus, Pseudomonasfluorescens, Pseudomonasaeruginosa, Pseudomonasputida и т.д.) или филогенетической группе Archaea (включая, в том числе, Methanococcus jannaschii, Methanobacterium thermoautotrophicum, Halobacterium, как, например, Haloferax volcanii и вид NRC-1 Halobacterium, Archaeoglobus fulgidus, Pyrococcus furiosus, Pyrococcus horikoshii, Aeuropyrum pemix и т.д.).
[00193] Например, антигенсвязывающая конструкция может может быть получена в клетках бактерий, в частности, когда не требуется гликозилирование и эффекторная функция Fc-фрагмента. В отношении экспрессии фрагментов и полипептидов антигенсвязывающих конструкций в клетках бактерий, см., например, патенты США. №. 5648237, 5789199 и 5840523. (см. также Charlton, Methods in Molecular Biology, Vol. 248 (B.K.C. Lo, ed., Humana Press, Totowa, N.J., 2003), стр. 245-254, где описана экспрессия фрагментов антител в Е. coli). После экспрессии растворимую фракцию антигенсвязывающих конструкций можно выделить из суспензии бактериальных клеток и подвергнуть дополнительной очистке.
[00194] В дополнение к прокариотам подходящими клетками-хозяевами для клонирования или экспрессии векторов, кодирующих антигенсвязывающие конструкции, являются эукариотические микроорганизмы, такие как мицелиальные грибы или дрожжи, включая штаммы грибов и дрожжей, пути гликозилирования которых были «гуманизированы», что приводит к получению антигенсвязывающей конструкции с профилем гликозилирования, который частично или полностью соответствует человеческому. См. Gerngross, Nat. Biotech. 22:1409-1414 (2004) и Lietal, Nat. Biotech. 24:210-215 (2006).
[00195] Подходящие клетки-хозяева для экспрессии гликозилированных антигенсвязывающих конструкций также получают у многоклеточных организмов (беспозвоночных и позвоночных животных). Примеры клеток беспозвоночных включают растительные клетки и клетки насекомых. Было идентифицировано большое число штаммов бакуловирусов, которые могут быть использованы в сочетании с клетками насекомых, в частности, для трансфекции клеток Spodoptera frugiperda.
[00196] Культуры растительных клеток также могут быть использованы в качестве клеток-хозяев. См., например, патенты США №. 5959177, 6040498, 6420548, 7125978 и 6417.429 (в которых описана технология PLANTIBODIES™ для получения антигенсвязывающих конструкций в трансгенных растениях).
[00197] Клетки позвоночных животных также могут быть использованы в качестве клеток-хозяев. Например, могут быть использованы клеточные линии млекопитающих, которые адаптированы к росту в суспензии. К другим примерам подходящих клеточных линий млекопитающих в качестве клеток-хозяев относятся линия клеток CV1 почек обезяны, трансформированная SV40 (COS-7); линия человеческих эмбриональных клеток почек (293 или клетки 293, которые описаны, например, в Graham et al, J. Gen Virol. 36:59 (1977)); клетки почек новорожденного хомяка (ВНК); клетки Сертоли мыши (клетки ТМ4, которые описаны, например, в Mather, Biol. Reprod. 23:243-251 (1980)); клетки почек обезяны (CV1); клетки почек африканской зеленой мартышки (VERO-76); клетки карциномы шейки матки человека (HELA): клетки почек собаки (MDCK; клетки печени крысы линии buffalo (BRL 3А); клетки легких человека (W138); клетки печени человека (Hep G2); клетки опухоли молочной железы мыши (ММТ 060562); клетки TRI, которые описаны, например, в Mather et al., Annals N.Y. Acad. Sci. 383:44-68 (1982); клетки MRC 5; и клетки FS4. Другие подходящие клеточные линии млекопитающих в качестве клеток-хозяев включают клетки яичника китайского хомячка (СНО), включая клетки DHFR -СНО (Urlaub et al., Proc. Natl. Acad. Sci. США 77:4216 (1980)); и линии клеток миеломы, такие как Y0, NS0 и Sp2/0. В отношении обзора по определенным клеточным линиям клеток-хозяев млекопитающих, подходящих для получения антигенсвязывающих конструкций, см., например, Yazaki and Wu, Methods in Molecular Biology, Vol. 248 (B.K.C. Lo, ed., Humana Press, Totowa, N.J.), стр. 255-268 (2003).
[00198] В одном варианте реализации изобретения антигенсвязывающие конструкции, описанные в данном документе, получают в поддающихся стабильной трансфекции клетках млекопитающих по способу, который включает: трансфекцию по меньшей мере одной поддающейся стабильной трансфекции клетки млекопитающего нуклеиновой кислотой, кодирующей антигенсвязывающую конструкцию, в заранее определенном соотношении, и экспрессию нуклеиновой кислоты по меньшей мере в одной клетке млекопитающего. В некоторых вариантах реализации изобретения соотношение нуклеиновой кислоты заранее определяют в экспериментах по временной трансфекции для определения относительного соотношения вводимой нуклеиновой кислоты, которое бы приводило к получению наиболее высокого процентного содержания антигенсвязывающей конструкции в экспресированном продукте.
[00199] В некоторых вариантах реализации изобретения предложен способ получения антигенсвязывающей конструкции в поддающейся стабильной трансфекции клетке млекопитающего, который описана в данном документе, причем продукт экспрессии по меньшей мере одной поддающейся стабильной трансфекции клетки млекопитающего содержит большее количество желаемой гликозилированной антигенсвязывающей конструкции в процентах по сравнению с мономерными полипептидами тяжелой или легкой цепей или другими антителами.
[00200] В некоторых вариантах реализации изобретения предложен способ получения гликозилированой антигенсвязывающией конструкции в поддающихся стабильной трансфекции клетках млекопитающего, описанных в данном документе, причем указанный способ включает идентификацию и выделение желаемой гликозилированной антигенсвязывающей конструкции. В некоторых вариантах реализации изобретения указанная идентификация осуществляется одним или двумя способами из жидкостной хромотографии и масс-спектрометрии.
[00201] При необходимости антигенсвязывающие конструкции могут быть очищены или выделены после экспрессии. Белки могут быть выделены или очищены различными способами, известными специалистам в данной области техники. Стандартные способы очистки включают хроматографические методы, включая ионообменную хроматографию, хроматографию гидрофобного взаимодействия, аффинную хроматографию, эксклюзионную хроматографию или гель-фильтрацию и обращенно-фазовую хроматографию, которые осуществляются при атмосферном давлении или высоком давлением, используя системы, такие как ЖХБР и ВЭЖХ. Способы очистки также включают методы электрофореза, иммунологические методы, преципитацию, диализ и методы хроматофокусирования. Также можно использовать методы ультрафильтрации и диафильтрации в сочетании с концентрированием белка. Как хорошо известно в данной области техники, различные природные белки связываются с Fc-участком и антителами, и эти белки могут найти применение в настоящем изобретении для очистки антигенсвязывающих конструкций. Например, бактериальные белки А и G связываются с Fc-участком. Аналогичным образом, бактериальный белок L связывается с Fab-участком некоторых антител. Часто очистка может обеспечиваться определенным партнером по слиянию. Например, антитела могут быть очищены с использованием глутатионовой смолы, если для слияния используется GST, аффинной хроматографии с ионами Ni+2, если используется His-метка, или иммобилизации антител к flag, если используется flag-тэг. Для общего руководства по подходящим методам очистки, см., например, Protein Purification: Principles and Practice, 3rd Ed., Scopes, ППPinger-Verlag, NY, 1994, который включен в полном объеме посредством ссылки. Степень очистки обязательно будет зависеть от применения антигенсвязывающих конструкций. В некоторых случаях проводить очистку нет необходимости.
[00202] В определенных вариантах реализации антигенсвязывающие конструкции очищают с использованием анионообменной хроматографии, включая, в том числе, хроматографию на Q-сефарозе, ДЭАЭ-сефарозе, колонки на poros HQ, poros DEAF, Toyopearl Q, Toyopearl QAE, Toyopearl DEAE, Resource/Source Q и DEAE, Fractogel Q и DEAE.
[00203] В конкретных вариантах реализации белки, описанные в данном документе, очищают с использованием катионообменной хроматографии, включая, в том числе, колонки на SP-сефарозе, СМ-сефарозе, poros HS, poros CM, Toyopearl SP, Toyopearl CM, Resource/Source S и CM, Fractogel S и CM и их эквиваленты и аналоги.
[00204] В дополнение к этому, антигенсвязывающие конструкции, описанные в данном документе, могут быть получены путем химического синтеза с использованием методов, известных в данной области техники (например, см. Creighton, 1983, Proteins: Structures and Molecular Principles, W.H. Freeman & Co., N.Y and Hunkapiller et al, Nature, 310:105-111 (1984)). Например, полипептид, соответствующий фрагменту полипептида, может быть синтезирован с использованием синтезатора пептидов. Кроме того, при желании, неканонические аминокислоты или химические аминокислотные аналоги могут быть введены в виде замены или вставки в полипептидную последовательность. Неканонические аминокислот включают, в том числе, D- изомеры традиционных аминокислот, 2,4-диаминомасляную кислоту, альфа-аминоизомасляную кислоту, 4-аминомасляную кислоту, Abu, 2-аминомасляную кислоту, g-Abu, e-Ahx, 6-аминокапроновую кислоту, Aib, 2-аминоизомасляную кислоту, 3-аминопропионовую кислоту, орнитин, норлейцин, норвалин, гидроксипролин, саркозин, цитруллин, гомоцитруллин, цистеиновую кислоту, t-бутилглицин, t-бутилаланин, фенилглицин, циклогексилаланин, b-аланин, фторированные аминокислоты, сконструированные аминокислоты, такие как b-метиламинокислоты, Са-метиловые аминокислоты, Na-метиловые аминокислоты, а также, в целом, аналоги аминокислот. Кроме того, аминокислота может быть правовращающей (D) или левовращающей (L).
Посттрансляционные модификации:
[00205] В определенных вариантах реалицазии антигенсвязывающие конструкции, описанные в данном документе, дифференциально модифицируют на протяжении или по окончании трансляции.
[00206] Термин «модифицированный», используемый в данном документе, относится к любым изменениям, вносимым в тот или иной полипептид, как, например, изменения длины полипептида, аминокислотной последовательности, химической структуры, ко-трансляционная модификация или пост-трансляционная модификация полипептида. Формулировка термина «(модифицированный)» означает, что полипептиды, о которых идет речь, модифицируют необязательно, то есть рассматриваемые полипептиды могут быть модифицированными или немодифицированными.
[00207] Термин «посттрансляционная модификация» относится к любой модификации природной или искусственной аминокислоты, которая происходит в такой аминокислоте после ее встраивания в полипептидную цепь. В качестве примера данный термин охватывает котрансляционные модификации in vivo, котрансляционные модификации in vitro (как, например, бесклеточные трансляционные системы), посттрансляционные модификации in vivo и посттрансляционные модификации in vitro.
[00208] В некоторых вариантах реализации изобретения, модификация представляет собой по меньшей мере один из процессов: гликозилирования, ацетилирования, фосфорилирования, амидирования, дериватизации путем использования известных защитных/блокирующих групп, протеолитического расщепления и связи с молекулой антитела или антигенсвязывающей конструкцией, или другим клеточным лигандом. В некоторых вариантах реализации изобретения антигенсвязывающая конструкция химически модифицирована с помощью известных способов, включая, в том числе, специфическое химическое расщепление с помощью бромциана, трипсина, химотрипсина, папаина, протеазы V8, NaBH4; ацетилирования, формилирования, окисления, восстановления и метаболического синтеза в присутствии туникамицина.
[00209] Дополнительные посттрансляционные модификации антигенсвязывающих конструкций, описанных в данном документе, включают, например, образование N-связанных или О- связанных углеводных цепей, (процессинг N-концевых или С- концевых участков), присоединение химических фрагментов к аминокислотному остову, химическую модификацию N- связанных или О- связанных углеводных цепей и вставку или делецию N-концевого остатка метионина в результате экспрессии в прокариотической клетке-хозяине. Антигенсвязывающие конструкции, описанные в данном документе, модифицированы с использованием детектируемой метки, как, например, ферментная, флюоресцентная, изотопная или аффинная метка, для обеспечения детекции и выделения белка. В определенных вариантах реализации изобретения примеры подходящих ферментных меток, включают, пероксидазу хрена, щелочную фосфатазу, бета-галактозидазу или ацетилхолинэстеразу; примеры подходящих комплексов простетических групп, включают стрептавидин-биотин и авидин/биотин; примеры подходящих флуоресцентных веществ, включают, умбеллиферон, флюоресцеин, флуоресцинизотиоцианат, родамин, дихлортрициниламин флюоресцеин, дансилхлорид или фикоэритрин; пример люминисцентного вещества включает люминол; примеры биолюминесцентных веществ включают люциферазу, люциферин и экворин; и примеры подходящего радиактивного вещества включают йод, углерод, сера, тритий, индий, технеций, таллий, галлий, палладий, молибден, ксенон, фтор.
[00210] В конкретных вариантах реалицазии изобретения антигенсвязывающие конструкции, описанные в данном документе, присоединяют к макроциклическим хелатирующим агентам, которые ассоциируют с ионами радиоактивных металлов.
[00211] В некоторых вариантах реализации изобретения антигенсвязывающие конструкции, описанные в данном документе, модифицированы либо с использованием естественных процессов, таких как посттрансляционный процессинг, либо методами химической модификации, которые хорошо известны в данной области техники. В определенных вариантах реализации изобретения одни и те же типы модификаций могут встречаться с одинаковой или разной степенью в различных участках в конкретном полипептиде. В определенных вариантах реализации изобретения полипептиды из антигенсвязывающих конструкций, описанных в данном документе, являются разветвленными, например, в результате убиквитинирования, и в некоторых вариантах реалицазии изобретения, являются циклическими, с или без разветвления. Циклические, разветвленые и разветвленные циклические полипептиды образуются в результате посттрансляционных естественных процессов или с использованием методов синтеза. Модификации включают ацетилирование, ацилирование, АДФ-рибозилирование, амидирование, ковалентное присоединение флавина, ковалентное присоединение молекулы гема, ковалентное присоединение полинуклеотида или производного полинуклеотида, ковалентное присоединение липида или производного липида, ковалентное присоединение фосфотидилинозитола, поперечное сшивание, циклизацию, образование дисульфидной связи, деметилирование, образование ковалентных поперечных связей, образование цистина, образование пироглутамата, формилирование, гамма-карбоксилирование, глицирование, гликозилирование, образование GPI-якоря, гидроксилирование, йодирование, метилирование, миристоилирование, окисление, протеолитическую обработку, фосфорилирование, пренилирование, рацемизацию, селеноирование. сульфатирование, опосредованное тРНК добавление аминокислот к белкам, такое как аргинилирование и убиквитинизация. (См., к примеру, PROTEINS-STRUCTURE AND MOLECULAR PROPERTIES, 2nd Ed., Т.E. Creighton, W.H. Freeman and Company, New York (1993); POST-TRANSLATIONAL COVALENT MODIFICATION OF PROTEINS, В. С Johnson, Ed., Academic Press, New York, pgs. 1-12 (1983); Seifter et al., Meth. Enzymol. 182:626-646 (1990); Rattan et al., Ann. N.Y. Acad. Sci. 663:48-62(1992)).
[00212] В определенных вариантах реализации изобретения антигенсвязьпзающие конструкции, описанные в данном документе, присоединяются к твердым подложкам, которые особенно удобны для иммуноанализа или очистки полипептидов, с которыми связываются или которые связываются с, или ассоциирут с белками, описанными в данном документе. Такие твердые подложки включают, в том числе, стекло, целлюлозу, поли акрил амид, нейлон, полистирол, поливини лхлорид или полипропилен.
Фармацевтические композиции
[00213] Также в настоящем изобретении предложены фармацевтические композиции, содержащие антиген-связывающую конструкцию, описанную в данном документе. Фармацевтические композиции содержат конструкцию и фармацевтически приемлемый носитель.
[00214] Термин «фармацевтически приемлемый» означает одобренный регулирующим органом федерального правительства или правительства штата или перечислен в Фармакопеи США или других общепризнанных фармакопеях для использования на животных и конкретнее на людях. Термин «носитель» относится к разбавителю, адъюванту, вспомогательному веществу или растворителю, с которым терапевтическое средство применяется. Такие фармацевтические носители могут быть стерильными жидкостями, такими как вода и масла, включая нефтяные масла, масла животного, растительного или синтетического происхождения, такие как ореховое масло, соевое масло, минеральное масло, кунжутное масло и тому подобное. В некоторых аспектах носитель представляет собой исскуственный носитель, не встречающий в природе. Вода может использоваться как носитель, когда лекарственный препарат вводится внутривенно. Солевые растворы и водные растворы декстрозы и глицерина также могут быть использованы в качестве жидких носителей, особенно для инъекционных препаратов. Подходящие фармацевтические вспомогательные вещества включают крахмал, глюкозу, лактозу, сахарозу, желатин, солод, рис, муку, мел, силикатный гель, стеарат натрия, глицеролмоностеарат, тальк, хлорид натрия, сухое обезжиренное молоко, глицерол, пропилен, гликоль, воду, этанол и тому подобное. При желании композиция может также содержать незначительные количества увлажнителей, или эмульгаторов, или рН буферных веществ. Эти композиции могут быть в виде растворов, суспензий, эмульсии, таблеток, драже, капсул, порошков, препаратов длительного высвобождения и подобных им. Композиция может быть приготовлена в виде суппозитория с традиционным связующими веществами и носителями, такими как триглецириды. Композиция может быть приготовлена как суппозиторий с традиционными связующими и носителями, такими как триглицериды. Состав для перорального введения может включать стандартные носители, такие как фармацевтические сорта маннита, лактозы, крахмала, стеарата магния, натриевой соли сахарина, целлюлозы, карбоната магния и т.д. Примеры подходящих фармацевтических носителей описаны в «Remington's Pharmaceutical Sciences» by E.W. Martin. Такие композиции содержат терапевтически эффективное количество композиции, преимущественно в очищенном виде, вместе с подходящим количеством носителя для получения требуемой формы для введения пациенту. Препарат должен соответствовать способу введения.
[00215] В определенных вариантах реализации изобретения композицию, содержащую конструкцию, получают по стандартным методикам в виде фармацевтической композиции, адаптированной для внутривенного введения человеку. Как правило, композиции для внутривенного введения представляют собой растворы в стерильном изотоническом буфере на водной основе. При необходимости композиция также может содержать солюбилизирующее средство и местный анестетик, такой как лидокаин, для облегчения боли в участке инъекции. Как правило, ингредиенты поставляются либо по отдельности, либо смешанные друг с другом в виде единичной лекарственной формы, например, в виде сухого лиофилизированного порошка или безводного концентрата в герметичной емкости, такой как ампула или пакет-саше с указанием количества действующего вещества. Если композицию вводят путем инфузии ее можно распределить с использованием инфузионного флакона, содержащего стерильную воду фармацевтической степени чистоты или солевой раствор. Если композицию вводят путем инъекции, может предоставляться ампула со стерильной водой для инъекций или физиологическим раствором, с тем чтобы можно было смешать инградиенты до введения.
[00216] В определенных вариантах реализации изобретения композиции, описанные в данном документе, получают в форме нейтральных солей. Фармацевтически приемлемые соли включают соли, образующиеся с анионами, как, например, соли-производные соляной, фосфорной, уксусной, щавелевой, винной кислот и т.д., и соли, образующиеся с катионами, как, например, соли-производные натрия, калия, аммония, кальция, гидроксида железа, изопропиламина, триэтиламина, 2-этиламиноэтанола, гистидина, прокаина и т.д.
Способы лечения
[00217] В определенных вариантах реализации изобретения предложены способы лечения заболевания или нарушения, включающие введение субъекту, который нуждается в таком лечении, предотвращении или улучшении состояния, антигенсвязывающей конструкции, описанной в данном документе, в количестве, необходимом для эффективного лечения, предотвращения или улучшения состояния при заболевании или нарушении.
[00218] «Нарушение» относится к любому заболеванию, на течение которого благоприятное воздействие оказывает антигенсвязывающая конструкциая или способ, описанные в данном документе. Этот термин охватывает хронические и острые нарушения или заболевания, включая те патологические состояния, которые увеличивают предрасположенность млекопитающего к рассматриваемому нарушению. В некоторых вариантах реализации изобретения нарушение представляет собой рак, что описано более подробно ниже.
[00219] Термин «субъект» относится к животному, в некоторых вариантах реализации изобретения млекопитающему, который представляет собой объект лечения, изучения или эксперимента. Животное может подразумевать под собой человека, низшего примата, домашнее животное (например, собак, котов и тому подобное), сельскохозяйственное животное (например, коров, овец, свиней, коней и тому подобное) или лабораторных животных (например, крыс, мышей, морских свинок и тому подобное).
[00220] Термин «млекопитающее» используемый в данном документе, включает, но не ограничивается этим, людей, низших приматов, псовых, кошачьих, мышиных, бычьих, конячих и свиней.
[00221] «Лечение» относится к клиническим вмешательствам, стремясь переделать естественное течение болезни, отдельно или клеточно пройти лечение, и можно провести либо для профилактики либо в процессе течении болезни клинической патологии. Желательные эффекты лечения включают предотвращение частоты или возврата болезни, облегчение симптомов, уменьшение любого открытого или скрытого патологического последствия болезни, предотвращение метастазов, уменьшение скорости развития болезни, улучшение или временное облегчение стадии заболевания и ремиссию или улучшение прогноза. В некоторых вариантах реализации изобретения антигенсвязывающие конструкции, и способы, описанные в данном документе, используют для замедления развития болезни или нарушения. В одном варианте реализации изобретения антигенсвязывающие конструкции и способы описанные в данном документе оказывают эффект на регрессию опухоли. В одном варианте реализации изобретения антигенсвязывающие конструкции и способы описанные в данном документе оказывают эффект на ингибирование рост опухоли/рака.
[00222] Желательные эффекты лечения включают, но не ограничиваются этим, предупреждение частоты или возврат болезни, облегчение симптомов, уменьшение любого открытого или закрытого патологического последствия болезни, предотвращение метастазов, уменьшение скорости развития болезни, улучшение или временное облегчение стадии заболевания, улучшение выживаемости и ремиссия или улучшение прогноза. В некоторых вариантах реализации изобретения антигенсвязывающие конструкции, описанные в данном документе, используют для замедления развития болезни или медленного прогрессирования болезни.
[00223] Термин «эффективное количество» используемый в данном документе относится к тому количеству введенной конструкции, которая выполнит назначение перечисленного способа, например, облегчит до некоторой степени один или более симптомов заболевания, состояния или нарушения лечения. Количество композиции, описанное в данном документе, которая будет эффективна в лечении, ингибировать и предотвращать заболевание или нарушение, связанное с неправильной экспрессией и/или активностью терапевтического бежа, можно определить стандартными клиническими методами. В дополнение, in vitro анализы могут выборочно служить для помощи в определении оптимального диапазона доз. Точная доза используется в препарате, также в зависимости от способа введения, и серьезности заболевания или нарушения, и решения согласно заключению практикующего врача и обстоятельствах каждого пациента. Эффективные дозы экстраполированны из кривой зависимости дозаэффект, полученные от in vitro или тестовых систем экспериментальной модели на животном.
[00224] Антигенсвязывающую конструкцию вводят субъекту. Различные системы доставки известны и могут быть использованы для введения препарата антигенсвязывающей конструкции, описанной в данном документе, например, инкапсуляция в липосомах, микрочастицы, микрокапсулы, рекомбинантные клетки, способные к экспрессии соединения, рецепторно-опосредованный эндоцитоз (см., например, WuandWu, J. Biol. Chem. 262:4429-4432 (1987)), конструкция на нуклеиновой кислотекак часть ретровируса или другого вектора и т.д. Способы применения, включают но не ограничиваються ими, внутридермальный, внутримышечный, внутрибрюшинный, внутривенный, подкожный, интраназальный, эпидуральный и оральный путь. Компоненты или композиции можна вводить любым удобным способом применения, например, при помощи инфузии или болюсной инъекции, поглощение через эпителиальный или кожно-слизистую оболочку матки (например, слизистая оболочка полости рта, прямокишечная и слизистая оболочка кишки и т.д.) и может представлять собой введение вместе с другими биологически активными веществами. Введение может быть системным или местным. В добавление, в определенных вариантах реализации изобретения, желательно применять антигенсвязывающую конструкцию композиций, описанных в данном документе, в центральную нервную систему любым подходящим способом применения, включая интравентрикулярное и субарахноидальное введения, интравентрикулярное введение можно провести при помощи интравентрикулярного катетера, например, присоединить резервуар, такой как резервуар Оммайя. Ингаляционное введение также может применяться, например, с помощью ингалятора или распылителья и препарата с аэрозольным веществом.
[00225] В специфическом варианте реализации изобретения, желательно вводить антигенсвязывающие конструкции или композиции, описанных в данном документе, местно в облать нуждающийся в лечении; это можна достичь путем, например, и не в качестве ограничения, местной инфузии в течение операции, местного нанесения, например, в сочетании с раневой повязкой после операции, путем иньекции, с помощью катетера, с помощью суппозиторий или с помощью имплантанта, указанная имплантант состоит с пористого, не-пористого или гелеобразного материалов, включая мембраны, такие как силиконовые мембраны или нити. Предчтительно, когда введение белка, включая антигенсвязывающую конструкцию, описанной в данном документе, должно тщательно отбираться к использованию материала, к которому белок не является сорбентом.
[00226] В другом варианте реализации изобретения антигенсвязывающие конструкции или композиции могут поставляться в везикулу, в специальную липосому (см. Langer, Science 249:1527-1533 (1990); Treat et al., in Liposomes in the Therapy of Infectious Disease and Cancer, Lopez-Berestein and Fidler (eds.), Liss, New York, pp. 353-365 (1989); Lopez-Berestein, ibid., pp. 317-327; см., как правило, ibid.)
[00227] В еще другом варианте реализации изобретения, антигенсвязывающие конструкции или композия может поставляться в систему с контролируемым высвобождением. В одном варианте реализации изобретения накачивание разрешается использовать (см. Langer, supra; Sefton, CRCCrit. Ref Biomed. Eng. 14:201 (1987); Buchwald et al. Surgery 88:507 (1980); Saudek et al., N. Engl. J. Med. 321:574 (1989)). В другом варианте реализации изобретения полимерные материалы могут использоваться (см. Medical Applications of Controlled Release, Langer and Wise (eds.),CRC Pres., Boca Raton, Fla. (1974); Controlled Drug Bioavailability, Drug Product Design and Performance, Smolen and Ball (eds.), Wiley, New York (1984); Ranger and Peppas, J., Macromol. Sci. Rev. Macromol. Chem. 23:61 (1983); см. также Levy et al., Science 228:190 (1985); During et al., Ann. Neurol. 25:351 (1989); Howard et al., J. Neurosurg. 71:105 (1989)). В еще другом варианте реализации изобретения, система с контролируемым высвобождением может располагаться вблизи от мишени для терапевтического воздействия, например, головной мозг, таким образом требующий несравненно меньше, чем системная доза (см., например, Goodson, in Medical Applications of Controlled Release, vol. 2, pp. 115-138 (1984)).
[00228] В специфическом варианте реализации изобретения, содержащий нуклеиновую кислоту, кодирующую антигенсвязывающие конструкции, описанные в данном документе, нуклеиновая кислота может осуществлять in vivo обеспечивание экспрессии своим кодирующим белком, конструирование его в составе экспрессирующего вектора необходимой нуклеиновой кислоты и вводить его так, чтобы становился внутриклеточно, например, с помощью ретровирусного вектора (см. в патенте США №4980286) или при помощи прямого впрыскивания или с помощью облучения микрочастиц (например, генная пушка; Biolistic, Dupont) или нанесение липидов или рецепторов клеточной поверхности или трансфицирование веществ или путем введения их в связывании гомеобок-подобного пептида, который как известно вводит ядра (см. например, Joliot et al., Proc. Natl. Acad. Sci. USA 88:1864-1868 (1991)) и т.д. В качестве альтернативы, нуклеиновая кислота может быть внесена внутриклеточно и инкорпорирована в клетку-хозяина DNA для экспрессии, при помощи гомологичной рекомбинации.
[00229] В определенных вариантах реалицазии антигенсвязывающей конструкции, описанной в данном документе, введены как комбинации с анти ген связываю щи ми конструкциями с не-перекрыванием связывающих эпи топов-мишеней.
[00230] Количество антигенсвязывающей конструкции, которая будет эффективна в лечении, ингибировании и предотвращении заболевания или нарушения можно определить с помощью стандартных клинических методов. В добавление, in vitro анализы могут необязательно применяться для облегчения определения оптимальных диапазонов доз. Точные дозы применяются в препаратах, будучи также зависимы от способа введения и серьезности заболевания или нарушения и должны принимать решения согласно заключению практикующего врача и любых обстоятельствах пациента. Эффективные дозы экстраполированы из кривой зависимости от дозы полученные из in vitro или тестовых систем экспериментальной модели на животном.
[00231] Антигенсвязывающие конструкции, описанные в данном документе, могут вводиться одиночно или в комбинации с другими типами лечений (например, радиотерапии, химиотерапии, гормонотерапии, иммунотерапия и анти-опухолевых препаратов). Как правило, предпочтительным является введение видоспецифических продуктов или обладающих видоспецифической реактивностью продуктов (в случае с антителами), которые относятся к тому же виду что и пациент. Таким образом, в одном варианте реализации изобретения антигенсвязывающие конструкции человека, производные фрагментов, аналоги или нуклеиновые кислоты, введение пациенту для терапии или профилактики.
Способы лечения различных раковых заболеваний
[00232] В данном документе описаны способы лечения рака или опухоли с уровнем экспрессии HER2+ на субъекте, и способы ингибирования роста опухолевой клетки с уровнем экспрессии HER2+ или уничтожение опухолевой клетки с уровнем экспрессии HER2+ с использованием антигенсвязывающих конструкций, описанные в данном документе.
[00233] Под раком с уровнем экспрессии HER2+ подразумевавется рак, который экспрессирует такие HER2, которые антиген-связываающие конструкции, описанные в данном документе, в состоянии связываться с раком. Как известно в данной области техники, раки с уровнем экспрессии HER2+ экспрессируют HER2 при различных уровнях. Определить ErbB, например, ErbB2 (HER2) экспрессию в раке, различными диагностически/прогностическими анализами является доступным. В одном варианте реализации изобретения ErbB2 повышенная экспрессия может быть анализирована с помощью IHC, например, с использованием HERCEPTEST® (Dako). Заключенные в парафин срезы тканей из биопсии опухоли могут подвергаться ИГХ-анализу и привлекать ErbB2 белок окрашиванию интенсивным критерием следующим образом:
[00234] У менее чем 10% опухолевых клеток не наблюдается или наблюдается окрашивание мембраны с баллом 0.
[00235] У более чем 10% опухолевых клеток обнаружено слабое/едва заметное окрашивание мембраны с баллом 1+. Клетки окрашены только в части мембраны.
[00236] У более чем 10% опухолевых клеток наблюдается окрашивание мембраны от слабой до умеренной с баллом 2+.
[00237] У более чем 10% опухолевых клеток наблюдается окрашивание мембраны от умеренной до сильной с баллом 3+.
[00238] Эти опухоли с баллами 0 или 1+ для оценки сверхэкспрессирующих ErbB2 могут быть охарактеризованы как не сверхэкспрессирующие ErbB2, тогда как эти опухоли с баллами 2+ или 3+ могут быть охарактеризованы как сверхэкспрессирующие ErbB2.
[00239] В качестве альтернативы или дополнительно, флуоресцентная гибридизация in situ (FISH) анализов, таких как INFORM™ (реализованы с помощью Ventana. Ariz.) или PATHVISION™ (Vysis, Ill.) может осуществляться фиксированием в формалине, заливанием парафином ткани опухоли определяя степень (в случае необходимости) ErbB2 сверхэкспрессии в опухоль. В сравнении с IHC анализом, FISH анализ, который показывает HER2 амплификацию гена, по-видимому, находиться в соответствии лучше с восприимчивости пациента к лечению с HERCEPTIN® и на сегодняшний день считается оправданным, будучи предпочтительным анализом определенных пациентов, способных извлекать пользу от лечения HERCEPTIN® .
|00240] Таблица D описывает уровень экспрессии HER2 в нескольких типичных рака молочной железы и другого рака клеточных линий (Subik et al. (2010) Breast Cancer: Basic Clinical Research:4; 35-41; Prang et a. (2005) British Journal of Cancer Research:92; 342-349). Как, показано в таблице, клетки MCF-7 и MDA-MB-231 считаются клетками с низким уровнем экспрессии HER2; клетки JIMT-1 и ZR-75-1 считаются клетками со средним уровнем экспрессии HER2, и клетки SKBR3 и ВТ-474 считаются клетками с высоким уровнем экспрессии HER2. Клетки SKOV3 (рак яичников) считаются клетками со средним уровнем экспрессии HER2.
[00241] В данном документе описаны способы лечения субъекта, с уровенем экспрессии HER2+ в раке или опухоли, содержащие обеспечение субъекту эффективного количества фармацевтической композиции, содержащей антигенсвязывающую конструкцию, описанной в данном документе.
[00242] Также описаны в данном документе применения уровня экспрессии HER2 антигенсвязывающей конструкции, описанной в данном документе, для изготовления лекарственного средства для лечения рака или опухоли. Также в данном документе описаны уровни экспрессии HER2 антигенсвязывающих конструкций для использования в лечении рака или опухоли.
[00243] В специфических вариантах реализации изобретения, антигенсвязывающая конструкция представляет собой v10000, v7091, v5019 и v5020. В одном варианте реализации изобретения антигенсвязывающая конструкция представляет собой v10000. В некоторых вариантах реализации изобретения антигенсвязывающая конструкция конъюгирована с майтансином, (DM1). Если антигенсвязывающая конструкция конъюгирована с DM1, она интернализуется в опухолевые клетки, DM1 отщепляется внутриклеточно от конструкции и убивает опухолевые клетки.
[00244] В некоторых вариантах реализации изобретения субъект лечиться, имея рак поджелудочной железы, рак головы и шеи рак ЖКТ, колоректальный рак, рак молочной железы, рак почек, рак шейки матки, рак яичников, рак мозга, рак эндометрия, рак мочевого пузыря, немелкоклеточный рак легкого или эпидермально-полученный рак. В некоторых вариантах реализации изобретения опухоль метастазирует.
[00245] В целом, опухоль в субъекте лечиться экспрессией в среднем 10000 или больше копий уровней экспрессии HER2 на опухолевую клетку. В определенных вариантах реализации опухоли представляют собой уровни экспрессии HER2 0-1+, 1+, HER2 2+ or HER2 3+, которые определяют с помощью ИГХ. В некоторых вариантах реализации изобретения опухоль представляет собой ниже уровня экспрессии HER2 2+ или ниже уровня экспрессии HER2 1+.
[00246] В некоторых вариантах реализации изобретения опухоль субъекта лечиться с антигенсвязывающими конструкциями как рак молочной железы. В специфическом варианте реализации изобретения, рак молочной железы экспрессирует HER2 с уровня 2+ или ниже. В специфическом варианте реализации изобретения, рак молочной железы экспрессирует HER2 с 1+ уровеня или ниже. В некоторых вариантах реализации изобретения рак молочной железы экспрессирует рецептор к эстрогену (ЕР+) и/или рецепторы к прогестерону (ПР+). В некоторых вариантах реализации изобретения рак молочной железы представляет собой ER- и/или PR-. В некоторых вариантах реалицазии изобретения рак молочной железы имеет амплифицированный ген HER2. В некоторых вариантах реализации изобретения рак молочной железы представляет собой с уровнем экспрессии HER2 3+негативный по рецепторам эстрогенов (ER-), негативный по рецепторам прогестерона (PR-), резистентный к трастузумабу, резистентный к химиотерапии инвазивного-протокового рака молочной железы. В другом варианте реализации изобретения рак молочной железы является резистентным к трастузумабу, отечно-инфильтративному раку молочной железы (ER-, PR-) с уровнем экспресии HER2 3+. В другом варианте реализации изобретения рак молочной железы является инвазивным протоковым раком молочной железы (ER-, PR-) с уровнем экспресии HER2 3+. В другом варианте реализации изобретения рак молочной железы является резистентным к трастузумабу и пертузумабу раку молочной железы с амплификацией гена HER2 и уровнем экспресии HER2 2+. В некоторых вариантах реализации изобретения рак молочной железы является триадой негативов (ER-, PR- и ниже уровня экспрессии HER2).
[00247] В одном варианте реализации изобретения опухоль представляет собой аденокарцинома яичника эпителиального происхождения с уровнем экспрессии HER2 2/3+, имеющая амплификацию гена HER2.
[00248] В настоящем изобретении предложены способы лечения субъекта, имеющего опухоль HER2+, которая резистента или приобретает резистентность к другому стандартному лечению, включающие введение субъекту фармацевтической композиции, содержащей антигенсвязывающие конструкции, описанные в данном документе. В определенных вариантах реализации антигенсвязывающие конструкции, описанные в данном документе, предоставляются субъектам, которые не поддаются лечению современными методами, необязательно в комбинации с одним или более современными методами терапии, направленной на подавление экспрессии HER2. В некоторых вариантах реализации изобретения, действующие на сегодняшний день HER2 препараты, включают в том числе, к HER2 или анти-HER3 моноспецифические бивалентные антитела, трастузумаб, пертузумаб, T-DM1, биспецифическое HER2/HER3 scFv или их комбинации. В некоторых вариантах реализации изобретения рак резистентный к различным химиотерапевтическим средствам, такие как таксаны. В некоторых вариантах реалицазии изобретения рак резистентный к трастузумабу. В некотором варианте реализации изобретения рак резистентный к пертузумабу. В одном варианте реализации изобретения рак резистентный к TDM1 (трастузумаб конъюгирован с DM1). В некоторых вариантах реализации изобретения субъект предварительно проходил лечение с антитело к анти-HER2, такие как трастузумаб, пертузумаб или DM1. В некоторых вариантах реализации изобретения субъект предварительно не проходил лечение с антитело к HER2. В одном варианте реализации изобретения антигенсвязывающая конструкция обеспечивалась субъекту для лечения метастазированного рака, когда пациент имел улучшение на предыдущем лечении HER2.
[00249] В настоящем изобретении предложены способы лечения субъекта, имеющего опухоль HER2+, включающие предоставление эффективного количества фармацевтической композиции, содержащей антигенсвязывающую конструкцию, описанную в данном документе, вместе с дополнительным противоопухолевым средством.
Дополнительное противоопухолевое средство может представлять собой терапевтическое антитело, как отмечалось выше, или химиотерапевтическое средство. Химиотерапевтические средства, которве можно использовать в комбинации с антигенсвязывающими конструкциями по настоящему изобретению, включают цисплатин, карбоилатин. паклитаксел, альбумин-связанный паклитаксел, доцетаксел, гемцитабин, винорелбин, иринотекан, этопозид, винбластин, пеметрексед, 5-фтороурацил (в присутствии или в отсутствие фолиновой кислоты), капецитабин, карбоплатин, эпирубицин, оксалиплатин, фолфиринокс, абраксан и циклофосфамид.
[00250] В некоторых вариантах реализации изобретения опухоль представляет собой немелкоклеточный рак легких, и дополнительное средство представляет собой одно или более средств из цисплатина, карбоплатина, паклитаксела, альбумин-связанного паклитаксела, доцетаксела, гемцитабина, винорелбина, иринотекана, этопозида, винбластина или пеметрекседа. В вариантах реалицазии изобретения, опухоль представляет собой рак ЖКТ или желудка, и дополнительное средство представляет собой одно или более средств из 5-фторурацила (в присутствии или в отсутствие фолиновой кислоты), капецитабина, карбоплатина, цисплатина, доцетаксела, эпирубицина, иринотекана, оксалиплатина или паклитаксела. В других вариантах реалицазии изобретения опухоль представляет собой рак поджелудочной железы, и дополнительное средство представляет собой одно или более средств из гемцитабина, фолфиринокса, абраксана или 5-фтороурацила. В других вариантах реалицазии изобретения опухоль представляет собой эстроген- и/или прогестерон-позитивный рак молочной железы и дополнительное средство - один или более из комбинации (а) доксорубицина и эпирубицина, (b) комбинации паклитаксела и доцетаксела или (с) комбинация 5-фтороурацила, циклофосфамида и карбоплатина. В других вариантах реалицазии изобретения опухоль представляет собой рак головы и шеи, и дополнительное средство представляет собой одно или более средств из паклитаксела, карбоплатина, доксорубицина или цисплатина. В других вариантах реалицазии изобретения опухоль представляет собой рак яичников и дополнительное средство может представлять собой один или более цисплатина, карбоплатина или таксана, как, например, паклитаксела или доцетаксела.
[00251] Дополнительные средства можно вводить субъекту, проходящего лечение, одновременно с антигенсвязывающими конструкциями или последовательно.
[00252] Субъект, проходящий лечение с антигенсвязывающими конструкциями может представлять собой человека, низшего примата или другое млекопитающее, такое как мышь.
[00253] В некоторых вариантах реализации изобретения результат проведения обеспечивания эффективного количества антигенсвязывающей конструкции субъекту имеющего опухоль, является уменьшение объема опухоли, ингибирование роста опухоли, увеличение периода времени до начала прогрессии опухоли, продление выживаемости субъекта без признаков заболевания, уменьшение метастазов, увеличение выживаемости субъекта без прогрессирования заболевания или увеличение общей выживаемости субъекта или увеличение общей выживаемости групп субъектов получающих лечение.
[00254] Также в данном документе описаны способы уничтожения или ингибирования роста HER2- экспрессирующей опухолевой клетки, включающие воздействие на клетку антигенсвязывающей конструкцией, предложенной в настоящем изобретении.
[00255] В различных вариантах реалицазии изобретения опухолевая клетка может представлять собой клетку рака поджелудочной железы человека с уровнями экспрессии HER2 1+ или 2+, клетку карциномы легких человека с уровнем экспрессии HER2 3+, клетку бронхоальвеолярной карциномы человека европеоидной расы с уровнем экспрессии HER2 2+, клетку карциномы глотки человека, клетку плоскоклеточной карциномы языка человека с уровнем экспрессии HER2 2+, клетку плоскоклеточной карциномы глотки с уровнем экспрессии HER2 2+, клетку колоректальной карциномы человека с уровнем экспрессии HER2 1+ или 2+, клетку карциномы желудка человека с уровнем экспрессии HER2 3+, клетку протоковой карциномы молочной железы человека (ЕР+, позитивной по рецепторам эстрогена) с уровнем экспрессии HER2 1+, клетку карциномы молочной железы человека (ЕР+) с амплификацией гена HER2 и уровнем экспрессии HER2 2+/3+, клетку трижды негативной карциномы молочной железы человека с уровнем экспрессии HER2 0+/1+, клетку эндометриоидной карциномы человека с уровнем экспрессии HER2 2+, клетку метастатической злокачественной меланомы легких с уровнем экспрессии HER2 1+, клетку карциномы шейки матки человека с уровнем экспрессии HER2 1+, клетку почечно-клеточной карциномы человека с уровнем экспрессии Her2 1+ или клетку карциномы яичника человека с уровнем экспрессии HER2 1+.
[00256] В вариантах реалицазии изобретения, в котором антигенсвязывающие конструкции конъюгированы с DM1, опухолевая клетка может представлять собой клетку карциномы поджелудочной железы человека с уровнем экспрессии HER2 1+ и 2+ или 3+, клетку метастатической карциномы поджелудочной железы человека с уровнем экспрессии HER2 2+, клетку карциномы легких человека с уровнем экспрессии HER2 0+/1+, +3+, клетку бронхоальвеолярной карциномы человека европеоидной расы с уровнем экспрессии HER2 2+, клетку анапластической карциномы легких с уровнем экспрессии HER2 0+, клетку немелкоклеточной карциномы легких человека, клетку карциномы глотки человека, клетку плоскоклеточной карциномы языка человека с уровнем экспрессии HER2 2+, клетку плоскоклеточной карциномы глотки с уровнем экспрессии HER2 2+, клетку колоректальной карциномы человека с уровнем экспрессии HER2 1+ или 2+, клетку карциномы желудка человека с уровнем экспрессии HER2 0+, 1+ или 3+, клетку протоковой карциномы молочной железы человека (ER+, позитивной по рецепторам эстрогена) с уровнем экспрессии HER2 1+, клетку карциномы молочной железы человека (ER+) с амплификацией гена HER2 и уровнем экспрессии HER2 2+/3+, клетку трижды негативной карциномы молочной железы человека с уровнем экспрессии HER2 0+/1+, клетку протоковой карциномы молочной железы человека с уровнем экспрессии HER2 0+ (BasalB, Mesenchymal-liketriplenegative), клетку карциномы молочной железы (ER+) с уровнем экспрессии HER2 2+, клетку метастатической карциномы молочной железы человека с уровнем экспрессии HER2 0+ (трижды негативный рак (ER-) люминального типа А с амплификацией гена HER2), клетку мезодермальной опухоли матки человека (смешанная форма рака третьей стадии), клетку эндометриоидной карциномы человека с уровнем экспрессии HER2 2+, клетку эпидермоидный карциномы кожи человека с уровнем экспрессии HER2 1+, клетку метастатической злокачественной меланомы легких с уровнем экспрессии HER2 1+, клетку злокачественной меланомы с уровнем экспрессии HER2 1+, клетку эпидермоидной карциномы шейки матки человека, клетку карциномы мочевого пузыря человека с уровнем экспрессии HER2 1+, клетку карциномы шейки матки человека с уровнем экспрессии HER2 1+, клетку почечно-клеточной карциномы человека с уровнем экспрессии Her2 1+ или клетку карциномы яичника человека с уровнем экспрессии HER2 1+, 2+ или 3+.
[00257] В некоторых вариантах реалицазии изобретения опухолевая клетка может представлять собой одну или более следующие клеточные линии (показвано на Фигурах 37 и 38): клеточные линии опухоли поджелудочной железы ВхРС3, Capan-1, MiaPaca2; клеточные линии опухоли легких Calu-3, NCI-H322; клеточные линии опухоли головы и шеи Detroit 562, SCC-25, FaDu; клеточные лини колоректальной опухоли НТ29, SNU-C2B; клеточные линии опухоли желудка NCI-N87; клеточные линии опухоли молочной железы MCF-7, MDAMB175, MDAMB361, MDA-MB-231, ВТ-20, JIMT-1, SkBr3, ВТ-474; клеточные линии опухоли матки TOV-112D; клеточные линии опухоли кожи Malme-3М; клеточные линии опухоли шейки матки Caski, MS751; клеточные линии опухоли мочевого пузыря Т24, клеточные линии опухоли яичника CaOV3 и SKOV3.
[00258] В некоторых вариантах реалицазии изобретения, в которых антигенсвязывающие конструкции конъюгированы с DM1, опухолевая клетка может относиться к одной или более следующих клеточных линий (показаны на Фигурах 37 и 38): клеточные линии опухоли поджелудочной железы ВхРС3, Capan-1, MiaPaca2, SW 1990, Panc 1; клеточные линии опухоли легких А549, Calu-3, Calu-6, NCI-H2126, NCI-H322; клеточные линии опухоли головы и шеи Detroit 562, SCC-15, SCC-25, FaDu; клеточные линии колоректальной опухоли Colo201, DLD-1, НСТ116, НТ29, SNU-C2B; клеточные линии опухоли желудка SNU-1, SNU-16, NCI-N87; клеточные линии опухоли молочной железы SkBr3, MCF-7, MDAMB175, MDAMB361, MDA-MB-231, ZR-75-1, ВТ-20, ВТ549, ВТ-474, САМА-1, MDAMB453, JIMT-1, T47D; клеточные линий опухоли матки SK-UT-1, TOV-112D; клеточные линии опухоли кожи А431, Malme-3M, SKEMEL28; клеточные линии опухоли шейки матки Caski, MS751; клеточная линия опухоли мочевого пузыря Т24, клеточная линия опухоли почек ACHN; клеточные линии опухоли яичника CaOV3, Ovar-3 и SKOV3.
Наборы и изделия
[00259] Также в данном документе описаны наборы, содержащие одну или более антигенсвязывающих конструкций. Отдельные компоненты указанного набора упакованы в отдельные емкости и вместе с такими емкостями может прилагаться уведомление в форме, установленной правительственным учреждением, регулирующим производство, использование или сбыт фармацевтической или биологической продукции, в котором отражены разрешения этого учреждения для производства, использования или сбыта препарата. Набор может необязательно содержать инструкции или указания, в которых изложен способ применения или схема применения антигенсвязывающей конструкции.
[00260] Если один или более компонентов набора используются как растворы, например, водные растворы или стерильные водные растворы, сам контейнер может представлять собой ингалятор, шприц, пипетку, глазную капельницу или другие такие подобные приспособления, из которых раствор может вводиться субъекту или применяться и смешиваться с другими компонентами набора.
[00261] Компоненты набора могут также могут быть предоставлены в сухой или лиофилизированной форме, а набор может дополнительно содержать подходящий растворитель для восстановления лиофилизированных компонентов. Независимо от количества или типа емкостей наборы, описанные в данном документе, также могут содержать устройство, помогающее пациенту с введением композиции. Такое устройство может представлять собой ингалятор, дозатор для назального спрея, шприц, пипетку, пинцет, мерную ложку, глазную капельницу или подобные средства доставки, одобренные для применения в лечебных целях.
[00262] В другом аспекте, описанном в данном документе, используются изделия, содержащие материалы, используемые для лечения, предотвращения и/или диагностики нарушений, описанных выше. Изделия содержат контейнер и этикетку или инструкцию по применению препарата подходящую или имеющую отношение к контейнеру. Подходящие контейнеры, включают например, пробирки больших размеров, ампулы, шприцы, полиэтиленовые пакеты-сумки с инфузионным раствором и т.д. Контейнеры могут быть образованы с различных материалов, такие как стекло или пластик. Контейнер заключает в себе композицию, которая является сама по себе или сочетать в себе другую композицию эффективную для лечения, предотвращения и/или диагностики состояния и может иметь стерильный входное отверстие (например, контейнер может быть полиэтиленовый пакет-сумка с раствором для внутривенного введения или ампула, имеющая пробку, которая поддается прокалыванию иголкой для подкожной инъекции). По меньшей мере один активнодействующий агент в композиции является Т-клетка активирующая антигенсвязывающую конструкцию, описанную в данном документе. Этикетка или инструкция по применению препарата обозначает, какая композиция используется наиболее эффективного для лечения состояния. Кроме того, изделия могут содержать (а) первый контейнер с композицией, содержащей в данном документе, отличающаяся тем, что композиция содержит антигенсвязывающую конструкцию, описанную в данном документе; и (b) второй контейнер с композицией, содержащей в данном документе, отличающаяся тем, что композиция содержит дополнительно цитотоксическое или другое лечебное средство. Изделия в этом варианте реализации изобретения, описанные в данном документе, могут дополнительно содержать инструкцию по применению препарата указывающую, что композиции могут применяться для лечения того или иного состояния. В качестве альтернативы или дополнительно, изделия могут дополнительно содержать второй (или третий) контейнер, содержащий а фармацевтически приемлемый буфер, такое как бактериостатическая вода для инъекций (BWFT), физиологический раствор с фосфатным буфером, раствор Рингера и раствор Д-глюкозы. Возможно дополнительно включать остальные материалы, требуемые по рыночным и потребительским позициям, включая другие буферы, растворители, фильтры, иголки и шприцы.
Полипептиды и полинуклеотиды
[00263] Антигенсвязывающие конструкции, описанные в данном документе, содержат, по меньшей мере, один полипептид. Также описаны полинуклеотиды, кодирующие полипептиды, описанные в данном документе. Антигенсвязывающие конструкции являются, как правило, выделенными.
[00264] Используемый в данном документе, термин «выделенный» значит вещество (например, полипептид или полинуклеотид), которое было распознано и отделено и/или избавлено от компонента природной среды клеточной культуры. Примнсь компонентов этой натуральной среды являются материалы, которые сталкиваются с использованием диагностических или терапевтических целях для антигенсвязывающей конструкции, и могут включать ферменты, гормоны и другие белковоподобные или не- белковоподобные растворы. Термин «выделенный» также относится к средствам, которые были искусственно изготовлены, например, через посредство вмешательства человека
[00265] Термины «полипептид», «пептид» и «белок» используются равноправно в данном документе для обозначения полимерных аминокислотных остатков. Другими словами, представляет собой описание, направленное на полипептид, для употребления равным образом описания пептида и описания белка и наоборот. Термины относятся к встречающихся в природе аминокислотных полимеров, так и к аминокислотным полимерам в которых один или более аминокислотных остатков являются не естественно закодирована аминокислота. Используемые в данном документе термины охватывают аминокислотные цепи любой длины, включая непроцессированный белок, отличающаяся тем, что аминокислотные остатки соединены с помощью ковалентной пептидной связью.
[00266] Термин «аминокислота» относится к встречающийся в природе и не не встречающийся в природе аминокислот, а также аминокислотных аналогов и аминокислотных миметиков, которые функционируют в некоторой степени сходными с встречающимися в природе аминокислотами. Естественно закодированных аминокислот 20 общепринятых аминокислот (аланин, аргинин, аспарагин, аспарагиновая кислота, цистеин, глутамин, глютаминовая кислота, глицин, гистидин, изолейцин, лейцин, лизин, метионин, фенилоланин, пралин, серин, треонин, триптофан, тирозин и валин) и пиролизин и селеноцистеин. Аминокислотные аналоги относятся к компонентам, которые сходны основной химической структурой как встречающийся в природе аминокислота, т.е., карбон, который связывается с гидрогеном, карбоновой группой, аминогруппой и R-группой, такие как, гомосерин, норлейцин, метионина сульфоксид, метионина сульфониевый метил. Такие аналоги имеют модифицированные R-группы (такие, как норлейцин) или модифицированные пептидные цепи, но удерживают сходство основной химической структуры, как встречающаяся в природе аминокислота. Эталон аминокислоты включает, например, встречающаяся в природе протеогенная L-аминокислота; D-аминокислота, химически модифицированные аминокислоты, такие как варианты аминокислот и производные; встречающаяся в природе не протеогенные аминокислоты, такие как Р-аланин, орнитин и т.д.; и химически синтезированные компоненты, имеющие свойства, известных в данной области техники, характерных аминокислот. Примеры не встречающихся в природе аминокислот включают, но не ограничиваются этим, а-метил аминокислоты (например, а-метилаланин), D-аминокислоты, гистидин-подобные аминокислоты (например, 2-амино-гистидин, Р-гидрокси-гистидин, гомогистидин), аминокислоты, имеющие экстра-метилен в боковой цепи («гомо» аминокислоты) и аминокислоты в которых функциональная группа в боковой цепи карбоновая кислота заменяется на функциональную группу сульфоновую кислоту (например, цистеиновая кислота). Смешивание различными путями не естественных аминокислот, включая синтетически не нативных аминокислот, замещенных аминокислот, или одной или более D-аминокислот в белках, в настоящем изобретении может быть эффективно. D-аминокислота-содержащая пептиды и т.д., характеризуется повышенной стабильностью in vitro или in vivo по сравнению с L-аминокислотой-содержащей аналоги. Таким образом, конструкции пептидов и т.д., смешивание D-аминокислот может быть особенно удобным, если большая внутриклеточная стабильность является желаемой или требуемой. Конкретнее, D-пептиды и т.д., резистентны к эндогенным пептидазам и протеазам, в связи с этим обеспечивается улучшение биодоступности молекулы и увеличивается срок службы in vivo, когда требуются такие свойства. Дополнительно, D-пептиды и т.д., не могут прерабатываться эффективно для главного комплекса гистосовместимости класса II- рестриктции, представленные клетками Т-хелперами, и являются, таким образом, менее подходящими, чтобы вызывать гуморальный иммунный ответ в целом организме.
[00267] Аминокислоты могут называться, в данном документе, либо общеизвестным трехзначным основанным кодом, либо однобуквенным обозначением с помощью IUPAC-IUB Biochemical Nomenclature Commission. Нуклеотиды, аналогичным образом, могут называться общепринятым однобуквенным кодом.
[00268] Также включены в изобретение полинуклеотиды, кодирующие полипептиды антигенсвязывающих конструкций. Термин «полинуклеотид» или «нуклеотидная последовательность» подразумевает обозначение последующего отрезка из двух или более нуклеотидных молекул. Нуклеотидная последовательность может представлять собой геном, кДНК, РНК, полусинтетического или синтетического происхождения или любую их комбинацию.
[00269] Термин «нуклеиновая кислота» относится к дезоксирибонуклеотидам, дезоксирибонуклеозидам, рибонуклеозидам или рибонуклеотидам и их полимеры в либо одно- либо двухцепочечной форме. Не считая специальные ограничения, термин, охватывающий нуклеиновые кислоты, содержит известные аналоги из природных нуклеотидов, которые имеют похожие связующие способности, как эталонная нуклеиновая кислота, и метаболизирование в некоторой степени сходно с встречающимися в природе нуклеотидами. Не считая другие специальные ограничения, термин также относится к аналогам олигонуклеотидов, включая РНК (пептидонуклеиновые кислоты), аналоги ДНК, используемые в антисмысловой технологии (фосфотиоатах, фосфоамидатах, и тому подобное). Если не указано иное, особые последовательности нуклеиновой кислоты также косвенно охватывают их варианты с консервативными модификациями (включая в том числе, замещение вырожденного кодона) и комплементарные последовательности, также как ясно обозначают последовательность. Специфически, замещение вырожденного кодона может обеспечиваться произведением последовательности, в которой третье положение одного или более (или все) выбранных кодонов заменено с использованием смешанного основания и/или деоксиинозиновых остатков (Batzer et al, Nucleic Acid Res. 19:5081 (1991); Ohtsuka et al., J. Biol. Chem. 260:2605-2608 (1985); Rossolini et al., Mol. Cell. Probes 8:91-98 (1994)).
[00270] Термин «консервативно модифицированные варианты» применяют как к последовательностям аминокислот, так и нуклеиновых кислот. Что касается специфических последовательностей нуклеиновых кислот, то консервативно модифицированные варианты относятся к тем нуклеиновым кислотам, которые кодируют идентичные или в значительной степени идентичные аминокислотные последовательности, или, в случае если нуклеиновая кислота не кодирует аминокислотную последовательность, к последовательностям, идентичным в значительной степени. Из-за вырожденности генетического кода любой заданный белок кодируется значительным количеством функционально идентичных нуклеиновых кислот. Например, все кодоны GCA, GCC, GCG и GCU кодируют аминокислоту аланин. Таким образом, в каждом положении, в котором кодоном предусмотрен аланин, кодон может изменяться на любой из соответствующих кодонов без изменения кодируемого полипептида. Такие варианты нуклеиновых кислот являются «молчащими вариантами», которые представляют собой один вид модифицированных вариантов. Каждая последовательность нуклеиновых кислот в этом документе, которая кодирует полипептид, также включает любую возможную молчащую вариацию нуклеиновой кислоты. Специалисты в этой области техники признают, что каждый кодон в нуклеиновой кислоте (за исключением AUG, который, как правило, является единственным кодоном для метионина, и TGG, который, как правило, является единственным кодоном для триптофана) может быть модифицирован так, чтобы произвести функционально идентичную молекулу. Таким образом, в каждой описанной последовательности подразумевают каждую молчащую вариацию нуклеиновой кислоты, которая кодирует полипептид.
[00271] Что касается аминокислотных последовательностей, специалисту в данной области техники будет понятно, что отдельные замены, делении или вставки в последовательность нуклеиновой кислоты, пептида, полипептида или белка, которые приводят к изменению, вставке или делеции одной аминокислоты или небольшого количества в процентах аминокислот в кодируемой последовательности, относятся к «варианту с консервативными модификациями», причем указанное изменение приводит к делеции аминокислоты, вставке аминокислоты или замене аминокислоты на аминокислоту с подобными химическими свойствами. Таблицы с консервативными заменами, в которых приведены аминокислоты с подобными функциональными свойствами, известны специалисту в данной области техники. Такие варианты с консервативными модификациями включают, в том числе, полиморфные варианты, межвидовые гомологи и аллели, описанные в данном документе.
[00272] Таблицы консервативных замен, в которых приведены функционально близкие аминокислоты, хорошо известны специалисту в данной области техники. Далее представлены восемь групп включают аминокислоты, которые представляют собой консервативные замены друг для друга: 1) Аланин (А), Глицин (G); 2) Аспарагиновая кислота (D), Глютаминовая кислота (Е); 3) Аспарагин (N), Глютамин (Q); 4) Аргинин (R), Лизин (K); 5) Изолейцин (I), Лейцин (L), Метионин (М), Валин (V); 6) Фенилаланин (F), Тирозин (Y), Триптофан (W); 7) Серии (S), Треонин (Т); и [0139] 8) Цистеин (С), Метионин (М) (см., например, Creighton, Proteins: Structures and Molecular Properties (W H Freeman & Co.; 2nd edition (December 1993).
[00273] Термины «идентичный» или процентная «идентичность» в контексте двух или более последовательностей нуклеиновых кислот или полипептидных последовательностей, относятся к двум или более последовательностям, или подпоследовательностям, которые являются одинаковыми. Последовательности являются «по существу идентичными», если они содержат определенный процент аминокислотных остатков или нуклеотидов, которые являются одинаковыми (т.е. идентичными на около 60%, около 65%, около 70%, около 75%, около 80%, около 85%, около 90% или около 95% в пределах конкретной области) при сравнении и выравнивании на предмет максимального соответствия по окну сравнения или обозначенной области при оценке с использованием алгоритмов сравнения последовательности (или других алгоритмов, доступных специалистам в данной области техники) или путем выравнивания вручную и визуального наблюдения. Это определение также относится к последовательности, комплементарной исследуемой последовательности. Идентичность может распространяться на область, длина которой составляет по меньшей мере около 50 аминокислот или нуклеотидов, или на область, длина которой составляет 75-100 аминокислот или нуклеотидов в участке, или если не указано иное, на всю последовательность полинуклеотида или полипептида. Полинуклеотид, кодирующий полипептид по настоящему изобретению, включая гомологи, принадлежащие другим видам, отличным от человека, может быть получен способом, включающем стадии скриннинга библиотеки в жестких условиях гибридизации с использованием зонда, содержащего полинуклеотидную последовательность, описанную в данном документе, или ее фрагмент, и выделения полноразмерной кДНК и геномных клонов, содержащих указанную полинуклеотидную последовательность. Такие методы гибридизации хорошо известны специалистам в данной области.
[00274] Для сравнения последовательностей обычно одну последовательность рассматривают как эталонную, с которой сравнивают тестируемые последовательности. При использовании алгоритма для сравнения последовательностей тестируемую и исходную последовательность вводят в компьютер, при необходимости обозначают координаты подпоследовательностей и устанавливают параметры алгоритма для сравнения последовательностей. Можно использовать параметры программы по умолчанию или можно устанавливать альтернативные параметры. Алгоритм для сравнения последовательностей затем рассчитывает процент идентичности для тестируемых последовательностей относительно исходной последовательности на основе параметров программы.
[00275] Термин «окно сравнения», используемый в данном документе, включает ссылку на сегмент, представляющего собой какое-либо одно из множества смежных положений, выбираемых из группы, состоящей из от 20 до 600, обычно примерно от 50 до примерно 200, более часто примерно от 100 до примерно 150 положений, в которых последовательность можно сравнивать с исходной последовательностью из того же самого числа смежных положений после того, как две последовательности оптимально выровнены. Способы выравнивания последовательностей для сравнения хорошо известны в данной области техники. Оптимальное выравнивание последовательностей для сравнения можно проводить, например, с помощью алгоритма локальных гомологии Smith & Waterman, Adv. Appl. Math. 2:482 (1981), с помощью алгоритма выравнивания гомологии Needleman & Wunsch, J. Mol. Biol. 48:443 (1970), с помощью метода поиска сходства Pearson & Lipman, Proc. Nat'l. Acad. Sci. USA 85:2444 (1988), с помощью компьютерного обеспечения этих алгоритмов (GAP, BESTFIT, FASTA и TFASTA в пакете программ Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, WI) или с помощью выравнивания вручную и визуального просмотра (смотри, например, Current Protocols in Molecular Biology (Ausubel et al., eds. 1995 supplement).
[00276] Одним из примеров алгоритма, который пригоден для определения процента идентичности последовательностей и сходства последовательностей являются алгоритмы BLAST и BLAST 2.0, которые описаны в работах Altschul et al., Nuc. Acids Res. 25:3389-3402 (1977) и Altschul et al., J. Mol. Biol. 215:403-410 (1990), соответственно. Программное обеспечение для проведения анализов BLAST находится в открытом доступе в Национальном центре биотехнологической информации (the National Center for Biotechnology Information) (см. веб-сайт ncbi.nlm.nih.gov/). Параметры алгоритма BLAST W, Т и X определяют чувствительность и скорость выравнивания. В программе BLASTN (для нуклеотидных последовательностей) используются в качестве параметров по умолчанию длина слова (W), равная 11, ожидаемое значение (Е), равное 10, М=5, N=-4 и сравнение обеих нитей. Для аминокислотных последовательностей в программе BLASTP используются в качестве параметров по умолчанию длина слова (W), равная 3, ожидание (Е), равное 10, и матрицу подсчета баллов BLOSUM62 (см. Henikoff and Henikoff (1992) Proc. Natl. Acad. Sci. США 89:10915), выравнивание (В) 50, ожидаемое значение (Е), равное 10, М=5, N=-4 и сравнение обеих цепей Алгоритм BLAST, как правило, осуществляется с выключенным фильтром «низкая сложность».
[00277] Алгоритм BLAST также выполняет статистический анализ сходства между двумя последовательностями (см., например, Karlin & Altschul, Proc. Nat'l. Acad. Sci. USA 90:5873-5787 (1993)). Один показатель сходства, предоставляемый алгоритмом BLAST, представляет собой наименьшую сумму вероятностей (P(N)), которая является обозначением вероятности, с которой случайным образом будет возникать совпадение между двумя нуклеотидными или аминокислотными последовательностями. Например, нуклеиновую кислоту считают сходной с последовательностью сравнения, если наименьшая сумма вероятности при сравнении тестируемой нуклеиновой кислоты с нуклеиновой кислотой сравнения будет меньше чем примерно 0,2, более предпочтительно менее чем примерно 0,01 и наиболее предпочтительно менее чем примерно 0,001.
[00278] Фраза «избирательно (или специфически) гибридизуется с» относится к связыванию, образованию дуплексов или гибридизации молекулы только с определенной нуклеотидной последовательностью в жестких условиях гидридизации, когда эта последовательность содержится в сложной смеси (включая, в том числе, общую клеточную ДНК или РНК или библиотеку ДНК или РНК).
[00279] Фраза «жесткие условия гидридизации» относится к гибридизации последовательностей ДНК, РНК или других нуклеиновых кислот, или их комбинаций в условиях низкой ионной силы и высокой температуры, как известно в данной области техники. Как правило, под жесткими условиями гибридизации зонд будет гибридизироваться с его последовательностью-мишенью в сложной смеси нуклеиновых ксило (включая, в том числе, общую клеточную ДНК или РНК или библиотеку ДНК или РНК), но не будет гибридизироваться с другими последовательностями, содержащимися в сложной смеси. Жесткие условия зависят от последовательности и будут различными в различных обстоятельствах. Специфическая гибридизация более длинных последовательностей происходит при более высоких температурах. Исчерпывающее руководство по гибридизации нуклеиновых кислот представлено в Tijssen, Laboratory Techniques in Biochemistry and Molecular Biology - Hybridization with Nucleic Probes, "Overview of principles of hybridization and the strategy of nucleic acid assays" (1993).
[00280] Используемые в данном документе термины «конструировать, сконструированный, конструирование» включают любую манипуляцию с пептидным остовом или посттрансляционные модификации природного или рекомбинантного полипептида или иго фрагмента. Конструирование включает модификации аминокислотной последовательности, профиля гликозилирования или группы боковой цепи отдельной аминокислоты, а также комбинации этих подходов. Экспрессия и получение сконструированных белков осуществляется стандартными методами молекулярной биологии.
[00281] Под «выделенной молекулой нуклеиновой кислоты или полинуклеотидом» подразумевается молекула нуклеиновой кислоты, ДНК или РНК, которая была выделена из природной среды. Например, рекомбинатный полинуклеотид, кодирующий полипептид, содержащийся в векторе, считается выделенным. Дополнительные примеры выделенного полинуклеотида включают рекомбинантные полинуклеотиды, поддерживаемые в гетерологичных клетках-хозяевах или очищенные (частично или существенно) полинуклеотиды в растворе. Выделеный полинуклеотид включает полинуклеотидную молекулу, содержащуюся в клетках, которые обычно содержат полинуклеотидную молекулу, но полинуклеотидная молекула находится вне хромосомы или в таком положении в хромосоме, которое отличается от его естественного положения в хромосоме. Выделенные молекулы РНК включают РНК-транскрипты, полученные in vivo или in vitro, а также формы положительной и отрицательных цепей, двухцепочечные формы. Выделенные полинуклеотиды или нуклеиновые кислоты, описанные в настоящем изобретении дополнительно включают такие молекулы, полученные синтетически, например, посредством ПЦР или химического синтеза. Кроме того, полинуклеотид или нуклеиновая кислота в определенных вариантах реализации включают регуляторный элемент, такой как промотор, участок связывания рибосом или терминатор транскрипции.
[00282] Термин «полимеразная цепная реакция» или «ПЦР», как правило, относится к способу амплификации требуемой нуклеотидной последовательности in vitro, как описано, например, в патенте США №4683195. В общем случае метод ПЦР заключается в многократном повторении цикла реакций удлинения праймера с помощью олигонуклеотидных праймеров, способных предпочтительно гибридизироваться с матричной нуклеиновой кислотой.
[00283] Под нуклеиновой кислотой или полинуклеотидом, который имеет нуклеотидную последовательность, что по меньшей мере, например, на 95% «идентична» эталонной нуклеотидной последовательности по настоящему изобретению, подразумевается, что нуклеотидная последовательность полинуклеотида идентична эталонной последовательности, кроме того, что полинуклеотидная последовательность может включать вплоть до пяти точечных мутаций на каждые 100 нуклеотидов эталонной нуклеотидной последовательности. Другими словами, для получения полинуклеотида, имеющего нуклеотидную последовательность, по меньшей мере на 95% идентичную эталонной нуклеотидной последовательности, в эталонной последовательности могут быть удалены или замещены на другой нуклеотид вплоть до 5% нуклеотидов, или целый ряд нуклеотидов вплоть до 5% от общего числа нуклеотидов, содержащихся в эталонной последовательности, могут быть включены в эталонную последовательность. Эти изменения эталонной последовательности могут происходить в 5'- или 3'-концевых положениях эталонной нуклеотидной последовательности или в любом участке между этими концевыми положениями, диспергированные либо отдельно между остатками в эталонной последовательности или в составе одной или более непрерывных групп в пределах эталонной последовательности. На практике, для того чтобы определить, идентична ли любая определенная полинуклеотидная последовательность по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% нуклеотидной последовательности настоящего изобретения, можно использовать стандартные способы с использованием известных компьютерных программ, как, например, программ, которые рассматривались выше для полипептидов (например, ALIGN-2).
[00284] Считается, что производное или вариант полипептида характеризуется «гомологичностью» или является «гомологичным» пептиду, если аминокислотные последовательности производного или варианта по меньшей мере на 50% идентичны аминокислотной последовательности, состоящей из 100 аминокислот, из оригинального пептида. В определенных вариантах реализации изобретения производное или вариант полипептида характеризуется по меньшей мере 75% сходством либо с пептидом, либо фрагментом пептида, имеющим такое же число аминокислотных остатков, что и производное. В определенных вариантах реализации изобретения производное или вариант полипептида характеризуется сходством по меньшей мере на 85% либо с пептидом, либо фрагментом пептида, имеющим такое же число аминокислотных остатков, что и производное. В определенных вариантах реализации изобретения аминокислотная последовательность производного полипептида характеризуется сходством по меньшей мере на 90% либо с пептидом, либо фрагментом пептида, имеющим такое же число аминокислотных остатков, что и производное. В некоторых вариантах реализации изобретения аминокислотная последовательность производного характеризуется сходством по меньшей мере на 95% либо с пептидом, либо фрагментом пептида, имеющим такое же число аминокислотных остатков, что и производное. В определенных вариантах реализации изобретения производное или вариант полипептида характеризуется сходством по меньшей мере на 99% либо с пептидом, либо фрагментом пептида, имеющим такое же число аминокислотных остатков, что и производное.
[00285] Термин «модифицированный», используемый в данном документе, относится к любым изменениям, вносимым в тот или иной полипептид, как, например, изменения длины полипептида, аминокислотной последовательности, химической структуры, ко-трансляционная модификация или пост-трансляционная модификация полипептида. Формулировка термина «(модифицированный)» означает, что полипептиды, о которых идет речь, модифицируют необязательно, то есть рассматриваемые полипептиды могут быть модифицированными или немодифицированными.
[00286] В некоторых аспектах, антигенсвязывающая конструкция содержит аминокислотную последовательность, которая по меньшей мере на 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентична соответствующей эталонной аминокислотной последовательности или ее фрагменту, которые изложены в Таблице (Таблицах) или используются под номером(номерами) доступа, описанными в данном документе. В некоторых аспектах выделенная антигенсвязывающая конструкция содержит аминокислотную последовательность, кодируемую полинуклеотидом, который по меньшей мере на 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичен соответствующей эталонной нуклеотидной последовательности или ее фрагменту, которые изложены в Таблице(Таблицах) или используются под номером(номерами) доступа, описанными в данном документе.
[00287] Надо понимать, что настоящее изобретение не ограничивается никакими определенными протоколами; клеточными линиями, конструкциями и реагентами, описанными в данном документе, и по сути может изменяться. Надо также понимать, что терминология, использована в данном документе с целью описания только конкретных вариантов реализации изобретения, и не предназначена для ограничения объема настоящего изобретения.
[00288] Все публикации и патенты, упомянутые в данном документе, включены в данный документ посредством ссылки в целях описания и раскрытия, например, конструкций и методологий, которые описаны в публикациях, которые могут быть использованы в связи с описанием настоящего изобретения. Публикации, рассмотренные в данном документе, приведены исключительно для их раскрытия до даты подачи настоящей заявки. Ничего в данном документе не следует трактовать как допущение того, что авторы настоящего изобретения не имеют права датировать более ранним числом такое описание вследствие предшествующего изобретения или любой другой причины.
ЛИТЕРАТУРНЫЕ ИСТОЧНИКИ
[00289] Bowles JA, Wang SY, Link BK, Allan В, Beuerlein G, Campbell MA, Marquis D, Ondek B, Wooldridge JE, Smith BJ, Breitmeyer JB, Weiner GJ. Anti-CD20 monoclonal antibody with enhanced affinity for CD 16 activates NK cells at lower concentrations and more effectively than rituximab. Blood. 2006 Oct 15; 108(8): 2648-54. Epub 2006 Jul 6.
[00290] Desjarlais JR, Lazar GA. Modulation of antibody effector function. Exp Cell Res. 2011 May 15; 317(9): 1278-85.
[00291] Ferrara C, Grau S, Jager C, Sondermann P, Brunker P, Waldhauer I, Hennig M, Ruf A, Rufer AC, Stihle M, Umana P, Benz J. Unique carbohydrate-carbohydrate interactions are required for high affinity binding between FcgammaRIII and antibodies lacking core fucose. ProcNatl Acad Sci USA. 2011 Aug 2; 108(31): 12669-74.
[00292] Heider KH, Kiefer K, Zenz T, Volden M, Stilgenbauer S, Ostermann E, Baum A, Lamche H, Kiipcii Z, Jacobi A, Muller S, Hirt U, Adolf GR, Borges E. A novel Fc-engineered monoclonal antibody to CD37 with enhanced ADCC and high proapoptotic activity for treatment of B-cell malignancies. Blood. 2011 Oct 13; 118(15): 4159-68. Epub 2011 Jul 27. Blood. 2011 Oct 13; 118(15):4159-68. Epub 2011 Jul 27.
[00293] Lazar GA, Dang W, Karki S, Vafa O, Peng JS, Hyun L, Chan C, Chung HS, Eivazi A, Yoder SC, Vielmetter J, Carmichael DF, Hayes RJ, Dahiyat BI. Engineered antibody Fc variants with enhanced effector function. Proc Natl Acad Sci USA. 2006 Mar 14; 103(11): 4005-10. Epub 2006 Mar 6.
[00294] Lu Y, Vernes JM, Chiang N, Ou Q, Ding J, Adams C, Hong K, Truong ВТ. Ng D. Shen A. Nakamura G, Gong Q, Presta LG, Beresini M, Kelley B, Lowman H, Wong WL, Meng YG Identification of IgG(l) variants with increased affinity to FcγRIIIa and unaltered affinity to FcγRI and FcRn: comparison of soluble receptor-based and cell-based binding assays. J Immunol Methods. 2011 Feb 28; 365(1-2): 132-41. Epub 2010 Dec 23.
[00295] Mizushima T, Yagi H, Takemoto E, Shibata-Koyama M, Isoda Y, Iida S, Masuda K, Satoh M, Kato K. Structural basis for improved efficacy of therapeutic antibodies on defucosylation of their Fc glycans. Genes Cells. 2011 Nov; 16(11): 1071-1080.
[00296] Moore GL, Chen H, Karki S, Lazar GA. Engineered Fc variant antibodies with enhanced ability to recruit complement and mediate effector functions. MAbs. 2010 Mar-Apr; 2(2): 181-9.
[00297] Nordstrom JL, Gorlatov S, Zhang W, Yang Y, Huang L, Burke S, Li H, Ciccarone V, Zhang T, Stavenhagen J, Koenig S, Stewart SJ, Moore PA, Johnson S, Bonvini E. Anti-tumor activity and toxicokinetics analysis of MGAH22, an anti-HER2 monoclonal antibody with enhanced Fc-gamma receptor binding properties. Breast Cancer Res. 2011 Nov 30; 13(6): R123. [Epub ahead of print]
[00298] Richards JO, Karki S, Lazar GA, Chen H, Dang W, Desjarlais JR. Optimization of antibody binding to FcgammaRIIa enhances macrophage phagocytosis of tumor cells. Mol Cancer Ther. 2008 Aug; 7(8): 2517-27.
[00299] Schneider S, Zacharias M. Atomic resolution model of the antibody Fc interaction with the complement Clq component. Mol Immunol. 2012 May; 51(l): 66-72.
[00300] Shields RL, Namenuk AK, Hong K, Meng YG, Rae J, Briggs J, Xie D, Lai J, Stadlen A, Li B, Fox JA, Presta LG. High resolution mapping of the binding site on human IgG1 for Fc gamma RI, Fc gamma RII, Fc gamma RIII, and FcRn and design of IgG1 variants with improved binding to the Fc gamma R. J Biol Chem. 2001 Mar 2; 276(9): 6591-604,
[00301] Stavenhagen JB, Gorlatov S, Tuaillon N, Rankin CT, Li H, Burke S, Huang L, Vijh S, Johnson S, Bonvini E, Koenig S. Fc optimization of therapeutic antibodies enhances their ability to kill tumor cells in vitro and controls tumor expansion in vivo via low-affinity activating Fcgamma receptors. Cancer Res. 2007 Sep 15; 67(18): 8882-90.
[00302] Stewart R, Thorn G, Levens M, Guler-Gane G, Holgate R, Rudd PM, Webster C, Jermutus L, Lund J. A variant human IgG1-Fc mediates improved ADCC. Protein Eng Des Sel. 2011 Sep; 24(9): 671-8. Epub2011 May 18.
ПРИМЕРЫ
[00303] Ниже приведены примеры конкретных вариантов реализации для осуществления настоящего изобретения. Указанные примеры предложены только в иллюстративных целях и не предназначены для ограничения объема настоящего изобретения каким-либо образом. Были предприняты попытки обеспечить точность, что касается использованных числовых значений (например, количество, температуры и т.д.), но разумеется, что должны быть приняты во внимание некоторые экспериментальные ошибки и отклонения.
[00304] При практическом осуществлении настоящего изобретения будут использоваться, если не указано иное, традиционные способы химии белков, биохимии, методы рекомбинантной ДНК и фармакологии, известные специалистам. Такие методы подробно объясняются в литературе. См., например, Т.Е. Creighton, Proteins: Structures and Molecular Properties (W.H. Freeman and Company, 1993); A.L. Lehninger, Biochemistry (Worth Publishers, Inc., current addition); Sambrook, et al, Molecular Cloning: A Laboratory Manual (2nd Edition, 1989); Methods In Enzymology (S. Colowick and N. Kaplan eds., Academic Press, Inc.); Remington's Pharmaceutical Sciences, 18th Edition (Easton, Pennsylvania: Mack Publishing Company, 1990); Carey and Sundberg Advanced Organic Chemistry 3rd Ed. (Plenum Press) Vols A and B(1992).
Пример 1: получение иллюстративных биспецифических антител к HER2 и контролей
[00305] Были получены многие иллюстративные бипаратопные антитела к HER2 (или антигенсвязывающие конструкции) и контроли, как описано ниже. Антитела и контроли были получены в разных форматах, и изображения иллюстративных бипаратопных форматов показаны на Фигуре 1. Во всех форматах, показанных на Фигуре 1, одна цепь (Цепь А) гетеродимерного Fc-фрагмента показана черным цветом, а другая (Цепь В) показана серым цветом, тогда как один антигенсвязывающий домен (1) показан с помощью штриховки, а другой антигенсвязывающий домен (2) показан белым цветом.
(00306] На Фигуре 1А представлена структура бипаратопного антитела в формате Fab-Fab. На Фигурах 1В-1Е представлена структура возможных вариантов бипаратопного антитела в формате scFv-Fab. На Фигуре 1В, антигенсвязывающий домен 1 представляет собой scFv, слитый с Цепью А, тогда как антигенсвязывающий домен 2 представляет собой Fab, слитый с Цепью В. На Фигуре 1С, антигенсвязывающий домен 1 представляет собой Fab, слитый с Цепью А, тогда как антигенсвязывающий домен 2 представляет собой scFv, слитый с Цепью В. На Фигуре 1D, антигенсвязывающий домен 2 представляет собой Fab, слитый с Цепью А, тогда как антигенсвязывающий домен 1 представляет собой scFv, слитый с Цепью В. На Фигуре 1E, антигенсвязывающий домен 2 представляет собой scFv, слитый с Цепью А, тогда как антигенсвязывающий домен 1 представляет собой Fab, слитый с Цепью В. На Фигуре 1F, оба антигенсвязывающих домена представляют собой scFv.
[00307] Последовательности описанных ниже вариантов приведены в Таблице последовательностей, которая находится за Примерами. Области CDR были идентифицированы с использованием комбинации методов Кабат и Чотиа. Области могут несколько варьировать в зависимости от способа, используемого для идентификации.
Иллюстративные бипаратопные антитела к HER2
[00308] Были получены иллюстративные бипаратопные антитела к HER2, которые показаны в Таблице 1.

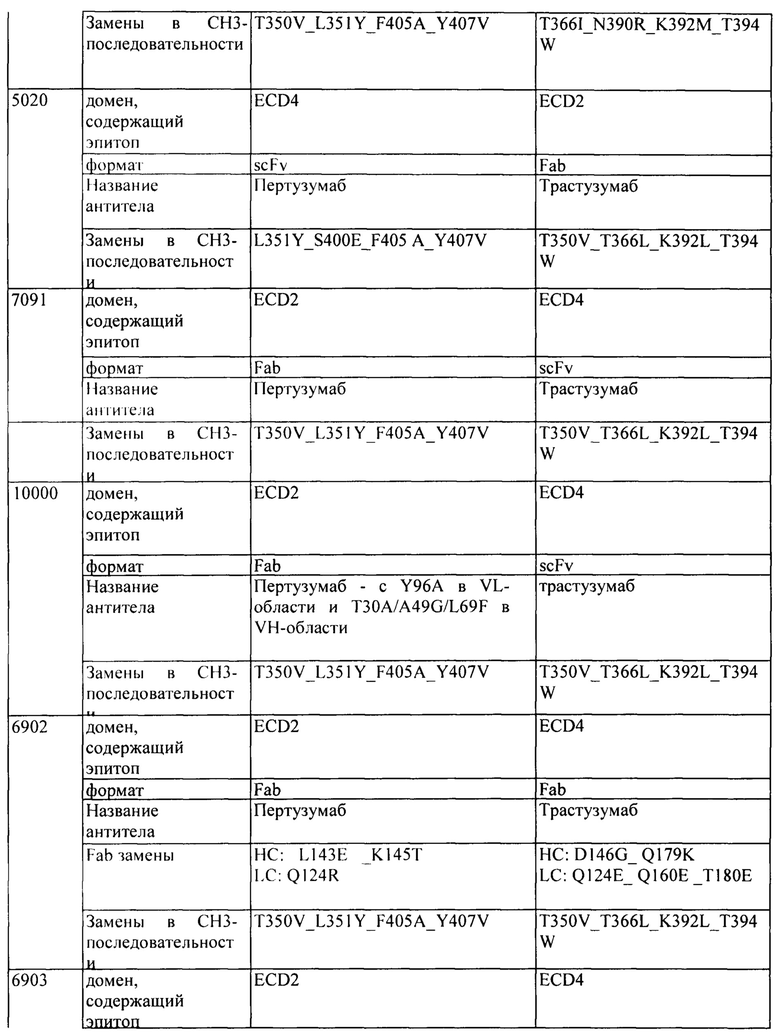
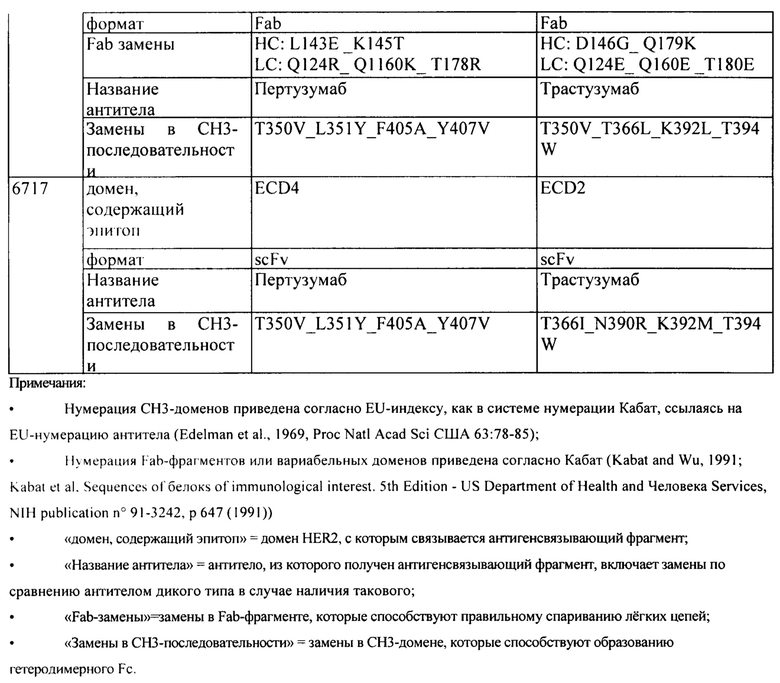
Иллюстративные моновалентные контрольные антитела к HER2
[00309] v1040: моновалентное антитело к HER2, причем HER2-связывающий домен представляет собой Fab-фрагмент на цепи А, полученный из трастузумабаа, и Fc-область представляет собой гетеродимер, имеющий мутации T350V_L351Y_F405A_Y407V в Цепи А, T350V_T366L_K392L_T394W в Цепи В, и при этом в шарнирной области Цепи В имеется мутация C226S; антигенсвязывающий домен связывается с доменом 4 HER2.
[00310] v630 - моновалентное антитело к HER2, причем HER2-связывающий домен представляет собой scFv на цепи А, полученный из трастузумабаа, и Fc-область представляет собой гетеродимер, имеющий мутации L351Y_S400E_F405A_Y407V в Цепи А, T366I_N390R_K392M_T394W в Цепи В; и при этом в шарнирной области имеется мутация C226S (EU-нумерация) в обеих цепях; антигенсвязывающий домен связывается с доменом 4 HER2.
[00311] v4182: моновалентное антитело к HER2, причем HER2-связывающий домен представляет собой Fab-фрагмент на цепи А, полученный из пертузумаба, и Fc-область представляет собой гетеродимер, имеющий мутации T350V_L351Y_F405A_Y407V в Цепи A, T350V_T366L_K392L_T394W в Цепи В; и при этом в шарнирной области Цепи В имеется мутация C226S; антигенсвязывающий домен связывается с доменом 2 HER2.
Иллюстративные моноспецифические бивалентные контрольные антитела к HER2 (полноразмерные антитела, FSA)
[00312] v506 представляет собой антитело к HER2 дикого типа собственного производства в клетках яичника китайского хомячка (СНО), используемое в качестве контроля. Оба HER2-связывающие домены получают из трастузумаба в Fab-формате, и Fc-домен представляет собой гомодимер дикого типа; антигенсвязывающий домен связывается с доменом 4 HER2. Это антитело также называется аналогом трастузумаба.
[00313] v792 представляет собой трастузумаб дикого типа с шарнирной областью IgG1, причем оба HER2-связывающие домены получают из трастузумаба в Fab-формате, и Fc-область представляет собой гетеродимер, имеющий мутации T350V_L351Y_F405A_Y407V в Цепи А и T350V_T366L_JC392L_T394W в Цепи В; антигенсвязывающий домен связывается с доменом 4 HER2. Это антитело также называется аналогом трастузумаба.
[00314] v4184 представляет собой бивалентное антитело к HER2, причем оба HER2-связывающие домены получают из пертузумаба в Fab-формате, и Fc-область представляет собой гетеродимер, имеющий мутации T350V_L351Y_F405A_Y407V в Цепи А и T350V_T366L_K392L_T394W в Цепи В. Антигенсвязывающий домен связывается с доменом 2 HER2. Это антитело также называется аналогом пертузумаба.
[00315] hIgG представляет собой коммерческое неспецифическое поликлональное контрольное антитело (Jackson ImmunoResearch, №009-000-003).
[00316] Эти антитела и контролей (отличные от IgG человека) клонировали экспрессировали следующим образом. Гены, кодирующие тяжелые и легкие цепи антител, были созданы искусственным путем посредством генного синтеза с использованием кодонов, оптимизированных для экспрессии в клетках человека/млекопитающих. Последовательность Fab-фрагмента трастузумаба была создана на основе известного антитела, связывавающегося с доменом 4 HER2/neu (Carter P. et al. (1992) Humаnization of an anti pi 85 HER2 антитело for человека cancer therapy. Proc Natl Acad Sci 89, 4285.) Fc-фрагмент был получен из изотипа IgG1. ScFv последовательность была создана на основе VH- и VL-доменов трастузумаба с использованием глицин-серинового линкера (Carter P. et al. (1992) Humаnization of an anti p1 85 her2 antibody for human cancer therapy. Proc Natl Acad Sci 89, 4285.). Последовательность Fab-фрагмента пертузумаба была создана на основе известного Ат, связывающегося с доменом 2 HER2/neu (Adams CW et al. (2006) Humаnization of а рекомбинантный monoclonal antibody to produce a therapeutic her dimerization inhibitor, Pertuzumab. Cancer Immunol Immunother. 2006; 55(6): 717-27).
[00317] Полученные в результате генные продукты субклонировали в экспрессирующий вектор РТТ5 млекопитающих (NRC-BRI, Канада), экспрессировали в клетках СНО (Durocher, Y., Perret, S. & Kamen, A. High-level and high-throughput recombinant protein production by transient transfection of suspension-growing СНО Cells. Nucleic acids research 30, e9 (2002)).
[00318] Клетки СНО трансфецировали в экспоненциальной фазе роста (от 1,5 до 2 миллионов клеток/мл) с использованием водного раствора 25 кДа полиэтиленимина в концентрации 1 г/мл (ПЭИ, polysciences) с соотношением ПЭИ-ДНК 2.5:1 (Raymond С et al. A simplified polyethylenimine-mediated transfection process for large-scale and high-throughput applications. Methods. 55(1):44-51 (2011)). Чтобы определить оптимальный диапазон концентраций для формирования гетеродимеров, трансфецировали ДНК в оптимальных соотношениях ДНК тяжелой цепи (НС-А), легкой цепи (LC) и тяжелой цепи В (НС-В), что обеспечивает формирование гетеродимеров (например, соотношения HC-A/HC-B/LC=30:30:40 (v5019). Трансфецированные клетки собирали через 5-6 суток, причем культуральную среду собирали после центрифугирования при 4000 об/мин и осветляли с использованием фильтра с размером пор 0,45 мкм.
[00319] Осветленную культуральную среду наносили на колонку MabSelect SuRe (GE Healthcare) с белком А и промывали с использованием буфера PBS при рН 7,2 в объеме, равном 10 объемам колонки. Антитела элюировали с использованием 10 объемов колонки цитратного буфера при рН 3,6, причем объединенные фракции содержали антитела, нейтрализованные с помощью TRIS при рН 11.
[00320] Элюат, содержащий белок А и антитела, дополнительно очищали с помощью гель-фильтрации (SEC). Для проведения гель-фильтрации 3,5 мг смеси антител концентрировали до 1,5 мл и наносили на колонку Sephadex 200 HiLoad 16/600 200 pg (GE Healthcare) через АКТА Express FPLC со скоростью потока 1 мл/мин. Буфер PBS при рН 7,4 использовали со скоростью потока 1 мл/мин. Фракции, соответствующие раствору очищенных антител, собирали, концентрировали до ~1 мг/мл.
[00321] Иллюстративные бипаратопные антитела к ECD2 HER2 × ECD4 с разными форматами молекул (например, v6717, scFv-scFv IgG1; v6903 и v6902 Fab-Fab IgG1; v5019, v7091 и v10000 Fab-scFv IgG1) клонировали, экспрессировали и очищали, как описано выше.
[00322] Для количественной оценки чистоты ратвора антител и для определения количества целевого гетеродимерного белка и возможного наличия гомодимеров и/или полуантител и/или примесей с неправильно спаренными легкими цепями, проводили ЖХ-МС-анализ интактных антител по массе. ЖХ-МС-анализ интактных антител по массе проводили, как описано в Примере 2, за исключением расчетов, используемых в DAR-анализе молекул ADC.
[00323] Данные показаны в Таблице 2. В таблице 2 показано, что в результате экспрессии и очистки этих бипаратопных антител содержание желаемого продукта в случае v6717 составляло 100%, в случае v6903 - 91% желаемого гетеродимерного продукта и в случае v6902 - 62% желаемого продукта. Числа в скобках обозначают количество, установленное по главному пику и побочному пику +81 Da. Этот побочный пик, как правило, характерен для вариантов, которые содержат С-концевые НА-теги (например, v6903 и v6902). В результате сложения главного и побочных пиков содержание гетеродимера составляет приблизительно 98% и 67% для v6903 и v6903. На основе высокого содержания гетеродимеров, было установлено, что v6903 является репрезентативным вариантом бипаратопного антитела к HER2 формата Fab-Fab для прямого сравнения с форматами scFv-scFv и Fab-scFv. v6903 был включен во все анализы сравнения форматов.

Пример 2: получение иллюстративных конъюгатов бипаратопных антител с лекарственными средствами (ADC)
[00324] Были получены следующие конъюгаты бипаратопных антител с лекарственными средствами (ADC на основе бипаратопного антитела к HER2). Были получены ADC из вариантов 5019, 7091, 10000 и 506. Эти ADC обозначают следующим образом:
v6363 (v5019, конъюгированный с DM1)
v7148 (v7091, конъюгированный с DM1)
v 10553 (v10000, конъюгированный с DM1)
v6246 (v506, конъюгированный с DM1, который является аналогом
T-DM1, трастузумаб-эмтанзин)
v6249 (IgG человека, конъюгированный с DM1)
[00325] ADC были получены посредством прямой конъюгации с майтансином. Антитела, очищенные с использованием белка А и SEC, как описано в Примере 1 (содержание >95%), были использованы для получения молекул ADC. ADC конъюгировали по способу, описанному в Kovtun YV, Audette СА, Ye Y, et al. Antibody-drug conjugates designed to eradicate опухоли with homogeneous and heterogeneous expression of the target antigen. Cancer Res 2006; 66: 3214-21. ADC имели среднее молярное соотношение, которое сотавляло 3,0 молекулы майтанзиноида на антитело, как было установлено методом ЖХ/МС и описано ниже.
[00326) Подробная информация по реактивам, использованным в реакции конъюгации ADC приведена далее: буфер для конъюгации 1:50 мМ фосфат калия/50 мМ хлорид натрия, рН 6,5, 2 мМ EDTA. Буфер для конъюгации 2: 50 мМ сукцинат натрия, рН 5,0. Буферная смесь для получения ADC: 20 мМ сукцинат натрия, 6% (вес/объем) трегалоза, 0,02% полисорбат 20, рН 5,0, диметилацетамид (ДМА); 10 мМ SMCC в ДМА (готовится перед конъюгацией), 10 мМ DM1-SH в ДМА (готовится перед конъюгацией), 1 мМ DTNB в PBS, 1 мМ цистеин в буфере, 20 мМ сукцинат натрия, рН 5,0. спектрофотометр УФ-видимого диапазона (Nano drop 100 от Fisher Scientific), колонки PD-10 (GE Healthcare).
[00327] ADC были получены следующим образом, исходный раствор антител наносили на колонку PD-10, которую предварительно уравновешивали 25 мл буфера для конъюгации 1, затем 0,5 мл буфера для конъюгации 1. Собирали элюат антител, и измеряли концентрацию при А280, и концентрацию доводили до 20 мг/мл. Готовили 10 мМ раствор SMCC-DM1 в ДМА. 7,5 молярных эквивалентов SMCC-DM1 в расчете на эквивалент антител добавляли в раствор антител, и ДМА добавляли до достижения конечного объема 10 об. %. Реакционную сместь перемешивали в течение короткого периода и инкубировали при комнатной температуре (КТ) в течение 2 часов. Вторую PD-10 колонку уравновешивали 25 мл буфера для конъюгации 1, и раствор антитела-MCC-DM1 добавляли на колонку с последующим добавлением 0,5 мл Буфера 1. Собирали элюат антитела-MCC-DM1, и измеряли поглощение раствора антител при А252 и А280. Рассчитывали концентрацию антитела-MCC-DMl (ð=1,45 мг-1 см-1 или 217500 М-1 см-1). ADC анализировали на колонке для эксклюзионной ВЭЖХ, используемой для анализа высокомолекулярных соединений (колонка TOSOH для эксклюзионной ВЭЖХ, G3000-SWXL, 7,8 мм × 30 см, Буфер, 100 мМ фосфат натрия, 300 мМ хлорид натрия, рН 7,0, скорость потока: 1 мл/мин).
[00328] Соотношение лекарственного средства к антителу (DAR) в ADC анализировали с помощью гидрофобной ВЭЖХ с использованием колонки Tosoh TSK гель Butyl-NPR (4,6 мМ × 3,5 мМ × 2,5 мм). Элюирование проводили при 1 мл/мин в градиенте 10-90% буфера В в течение 25 минут, затем 100% буфера В в течение 4 минут. Буфер А содержит 20 мМ фосфата натрия, 1,5 М сульфата аммония, рН 7,0. Буфер В содержит 20 мМ фосфата натрия, 25 об. % изопропанола, рН 7,0.
[00329] Соотношение лекарственного средства к антителу (DAR) в ADC определяли с помощью ЖХ-МС следующим способом. Антитела дегликозилировали с помощью пептид-N-гликозидазы F (PNGase F) перед нанесением на колонку для ЖХ-МС. Жидкостную хромотографию проводили на оборудовании Agilent 1100 Series HPLC в следующих условиях:
[00330] Скорость потока: 1 мл/мин разделяется после колонки до 100 мкл/мин до МС. Растворители: А=0,1% муравьиная кислота в ddH20, В=65% ацетонитрил, 25% THF, 9,9% ddH2O, 0,1% муравьиная кислота. Колонка: 2,1×30 мм PorosR2. Температура колонки: 80°С; растворитель также предварительно нагревают. Градиент: 20% В (0-3 мин), 20-90% В (3-6 мин), 90-20% В (6-7 мин), 20% В (7-9 мин).
[00331] Впоследствии проводили масс-спектрометрию (МС) на масс-спектрометре LTQ-Orbitrap XL в следующих условиях: ионизационный метод с использованием Ion Max Electrospray. Метод калибровки и настройки: 2 мг/мл раствора CsI впрыскивается со скоростью потока 10 мкл/мин. Орбитрэп настраивали на m/z 2211 с использованием параметра автоматической настройки (общий наблюдаемый диапазон ионов CsI: от 1690 до 2800). Напряжение на конусе: 40 В; цилиндрическая линза: 115 В; разрешение с преобразованием Фурье: 7500; диапазон сканирования m/z 400-4000; задержка сканирования: 1,5 мин. Профиль молекулярных масс данных был составлен с использованием программного обеспечения Thermo's Promass deconvolution. Средний DAR образца определяли в зависимости от DAR, наблюдаемом при каждом относительном пике (с использованием расчета: £ (DAR × интенсивность относительного пика)).
[00332] В Таблице 3 в краткой форме предстален средний DAR для Молекул ADC. Средний DAR для иллюстративного бипаратопного антитела к HER2 и контроля составлял приблизительно 3.
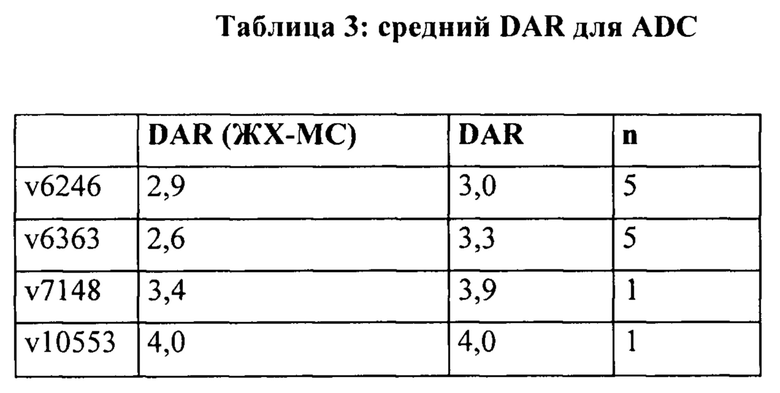
Пример 3: экспрессия и лабораторная очистка бипаратопного антитела к HER2
[00333] Бипаратопные антитела к HER2 (v5019, v7091 и v10000), описанные в Примере 1, экспрессировали в объеме 10 и/или 25 л и очищали с помощью хроматографии на белке А и эксклюзионной хроматографии (SEC) следующим образом.
[00334] Осветленную культуральную среду наносили на колонку MabSelect SuRe (GE Healthcare) с белком А и промывали с использованием 10 объемов колонки буфера PBS при рН 7,2. Антитела элюировали с использованием 10 объемов колонки нитратного буфера при рН 3,6, причем объединенные фракции содержали антитела, нейтрализованные с помощью Tris при рН 11.
[00335] Элюат, содержащий белок А и антитела, дополнительно очищали с помощью гель-фильтрации (SEC). Для проведения гель-фильтрации 3,5 мг смеси антител концентрировали до 1,5 мл и наносили на колонку Sephadex 200 HiLoad 16/600 200 pg (GE Healthcare) через АКТА Express FPLC со скоростью потока 1 мл/мин. Буфер PBS при рН 7,4 использовали со скоростью потока 1 мл/мин. Фракции, соответствующие раствору очищенных антител, собирали, концентрировали до ~1 мг/мл. Очищенные белки анализировали с помощью ЖХ-МС, как описано в Примере 2.
[00336] Результаты экспрессии в объеме 10 л и лабораторной очистки с использованием белка А и ЭХ показаны на Фигуре 2А и 2В. На Фигуре 2А показана SEC-хроматограмма v5019, очищенного с помощью белка А, и на Фигуре 2В показано изображение SDS-PAGE геля в восстанавливающих условиях, на котором сравнивается относительная чистота очищенной с использованием белка А объединенной фракции, а также фракции 15 и 19 и объединенные фракции 16-18, очищенные с помощью SEC. Эти результаты показывают экспрессию бипаратопного антитела к HER2 и то, что в результате очистки с использованием белка А и ЭХ был получен образец чистого белка. Дополнительная количественная оценка была проведена с использованием СВЭЖХ-ЭХ и ЖХ-МС-анализа и описана в Примере 4.
[00337] Результаты экспрессии в объеме 25 л и лабораторной очистки с использованием белка А показаны на Фигуре 2С. На Фигуре 2С показано изображение SDS-PAGE геля, на котором сравнивается относительная чистота v10000, очищенного с использованием белка А. На дорожке М представлен: белковый маркер молекулярной массы; на дорожке 1 представлен: v10000 в восстанавливающих условиях; на дорожке 2 представлен v10000 в неосстанавливающих условиях. На SDS-PAGE геле показано, что v10000 очищен и разганяется с точно предсказуемой молекулярной массой приблизительно 125 кДа в неосстанавливающих условиях. В восстанавливающих условиях визуализируются характерные для двух тяжелых цепей полоски, соответствующие тяжелой цепи СН-А (приблизительно 49 кДа) и тяжелой цепи СН-В (приблизительно 52.5 кДа); легкая цепь СН-А визуализируется и разганяется с точно предсказуемой массой приблизительно 23,5 кДа. Эти результаты показывают экспрессию бипаратопного антитела к HER2 и то, что в результате одностадийной очистки с использованием белка А получают образец чистого белка. Дополнительная количественная оценка была проведена с использованием СВЭЖХ-ЭХ и ЖХ-МС-анализа и описана в Примере 4.
Пример 4: анализ содержания бипаратопного антитела к HER2 с помощью СВЭЖХ-ЭХ и ЖХ-МС
[00338] Содержание и процент агрегировавших иллюстративных гетеромультимеров бипаратопных антител к HER2, очищенных с использованием белка А и ЭХ, определяли с помощью СВЭЖХ-ЭХ по описанному способу.
[00339] СВЭЖХ-ЭХ-анализ осуществляли с использованием колонки Waters BEH200 SEC с заданной температурой 30°С (2,5 мл, 4,6×150 мм, нержавеющая сталь, 1,7 мкм частицы) при 0,4 мл/мин. Время разгона составляло 7 мин, и общий объем на нанесение составлял 2,8 мл, при этом использовали подвижные буферы, состоящие из 25 мМ фосфата натрия, 150 мМ ацетата натрия, рН 7,1; и 150 мМ фосфати натрия, рН 6,4-7,1. Детекцию по абсорбции проводили при 190-400 нм и по флюоресценции с длиной волны возбуждения 280 нм и длиной волны излучения, регистрируемой при 300-360 нм. Интегрированные пики анализировали с помощью программного обеспечения Empower 3.
[00340] Результаты по СВЭЖХ-ЭХ для объединенных ЭХ-фракций v5019 показаны на Фигуре 3А. Эти результаты свидетельствуют о том, что в результате очистки с использованием хроматографии на основе белка А содержание иллюстративного бипаратопного антитела к HER2 составляло до >99%, причем содержание высокомолекулярных соединений составляло менее 1%.
[00341] Результаты по СВЭЖХ-ЭХ для объединенных фракций v10000, полученных с использованием Белка А, показаны на Фигуре 3В. Эти результаты свидетельствуют о том, что в результате очистки с использованием хроматографии на белке А содержание иллюстративного бипаратопного антитела к HER2 составляло до >96%, причем содержание высокомолекулярных соединений составляло менее 1%.
[00342] Содержание иллюстративных бипаратопных антител к HER2 определяли с использованием ЖХ-МС в стандартных условиях по способу, описанному в Примере 2. Результаты ЖХ-МС-анализа объединенных ЭХ-фракций v5019 показаны на Фигуре 4А. Эти данные показывают, что содержание гетеродимеров иллюстративных бипаратопных атител к HER2 составляет 100%. Результаты ЖХ-МС-анализа объединенных фракций v10000, полученных с использованием Белка А, показаны на Фигуре 4В. Эти данные показывают, что содержание гетеродимеров иллюстративных бипаратопных атител к HER2 составляет 98% после одностадийной очистки с использованием белка А.
[00343] Антитела, очищенные с помощью хроматографии на белке А, и/или белка А и ЭХ, были использованы для анализов, описанных в следующих Примерах.
Пример 5. экспрессия в больших объемах и оценка возможности производства бипаратопного антитела к HER2, очищенного с помощью хроматографии на белке А и катионообменной хроматографии
[00344] Иллюстративное бипаратопное антитела v5019 к HER2, описанное в Примере 1, экспрессировали в объеме 25 л и очищали следующим образом.
[00345] Антитело получали из супернатанта, затем использовали способ двустадийной очистки, который включал очистку с использованием Белка А (смола MabSelect™; GE Healthcare) с последующей катионообменной хроматографией (смола HiTrap™ SP FF; GE Healthcare) по описанному протоколу.
[00346] Клетки CHO-3E7 поддерживали в бессывороточной среде Freestyle СНО для экспрессии (Invitrogen, Карлсбад, Калифорния, США) в колбах Эрленмейера при 37°С с 5% CO2 (Coming Inc., Актон, Массачусетс) на орбитальном встряхивателе (VWR Scientific, Честер, Пенсильвания). За двое суток до трансфекция клетки высевали с соответствующей плотностью в 50 L емкости с объемом 25 л с использованием Wave Bioreactor System 20/50 (GE Healthcare Bio-Science Corp). В день трансфекции ДНК и ПЭИ (Polysciences, Эппельхейм, Германия) смешивали в оптимальном соотношении и добавляли к клеткам с использованием способа, описанного в Примере 1. Клеточные супернатанты, собранные на 6-е сутки, использовали для дополнительной очистки.
[00347] Культуральную жидкость центрифугировали и фильтровали перед нанесением на 30 мл смолы Mabselect™, упакованную в ХК26/20 (GE Healthcare, Уппсала. Швеция) при 10,0 мл/мин. После промывки и элюирования с использованием соответствующего буфера фракции собирали и нейтрализовали с использованием 1 М Tris-HCl, рН 9,0. Белок-мишень дополнительно очищали с использованием 20 мл смолы SP FF, упакованной в ХК16/20 (GE Healthcare, Уппсала, Швеция). Образец, очищенный с использованием MabSelect™, разводили 20 мМ NaAC, рН 5,5, для доведения проводимости до <5 мс/см, и 50 мМ лимонную кислоту (рН 3,0) добавляли для доведения значения рН образца до 5,5. Образец наносили при 1 мл/мин на смолу HiTrap™ SP FF (GE Healthcare) и промывали 20 мМ NaAC. Белок элюировали с использованием градиентного элюирования 0-100% 20 мМ NaAC, 1 М NaCl, рН 5,5, 10 объемов колонки при 1 мл/мин.
[00348] Очищенный белок анализировали с помощью SDS-PAGE, как описано в Примере 1, и ЖХ-МС на содержание гетеродимеров по способу, описанному в Примере 4. Результаты показаны на Фигуре 5А и 5В. На Фигуре 5А показаны результаты по SDS-PAGE v5019 после очистки с помощью MabSelect™ и HiTrap™ SP FF; на дорожке М представлен: белковый маркер молекулярной массы; на дорожке 1 представлен: v5019 в восстанавливающих условиях (3 мкг); на дорожке 2 представлен: v5019 в неосстанавливающих условиях (2,5 мкг). На SDS-PAGE геле показано, что v5019 характеризуется относительной чистотой после очистки с использованием MabSelect™ и HiTrap™ SP FF и в неосстанавливающих условиях разганяется с точно предсказуемой молекулярной массой приблизительно 125 кДа. В восстанавливающих условиях визуализируются характерные для двух тяжелых цепей полоски, соответствующие тяжелой цепи СН-А (приблизительно 49 кДа) и тяжелой цепи СН-В (приблизительно 52.5 кДа); легкая цепь СН-А визуализируется и разганяется с точно предсказуемой массой приблизительно 23,5 кДа.
[00349] Чтобы определить содержание гетеродимеров, осуществляли ЖХ-МС-анализ v5019, очищенного с использованием MabSelect™ и HiTrap™ SP FF, с использованием способа, описанного в Примере 4. Результаты ЖХ-МС-анализа показаны на Фигуре 5В. Эти результаты показывают, что в результате очистки v5019 с использованием MabSelect™ и HiTrap™ SP FF получают белок с содержанием гетеродимеров >99% и с незначительным (<1%) или неопределяемым количеством примесей гомодимеров или полуантител.
Пример 6: сравнение Вмакс бипаратопного антитела к HER2 по сравнению с Вмакс контролей в клеточных линиях, экспрессирующих HER2 на низком или высоком уровнях
[00350] Для оценки способности иллюстративного бипаратопного антитела к HER2 связываться с клетками, экспрессирующими различные уровни HER2, по сравнению с контролями проводили следующий эксперимент. Использовали следующие клеточные линии SKOV3 (HER2 2+/3+), JIMT-1 (HER2 2+), MDA-MB-231 (HER2 0/1+) и MCF7 (HER2 1+). Исследуемые бипаратопные антитела к HER2 включают v5019, v7091 и v 10000. Способность бипаратопных антител к HER2 связываться HER2-экспрессирующими (HER2+) клетками определяли, как описано ниже, при этом специально измеряли Вмакс и кажущуюся KD (равновесная константа диссоциации).
[00351] Связывание исследуемых антител с поверхностью клеток HER2+, определяли с помощью проточной цитометрии. Клетки промывали PBS ресуспендировали в DMEM при 1×105 клеток/100 мкл. 100 мкл клеточной суспензии добавляли в каждую микроцентрифужную пробирку, затем добавляли варианты антител в объеме 10 мкл/пробирку. Пробирки инкубировали в течение 2 ч при 4°С на встряхивателе. Микроцентрифужные пробирки центрифугировали в течение 2 мин при 2000 об/мин при комнатной температуре, и клеточные осадки промывали 500 мкл среды. Каждый клеточный осадок ресуспендировали в 100 мкл меченного флуорохромом вторичного антитела, разведенного в среде до 2 мкг/образец. Образцы затем инкубировали в течение 1 ч при 4°С на встряхивателе. После инкубации клетки центрифугировали в течение 2 мин при 2000 об/мин и промывали в среде. Клетки в ресуспендировали в 500 мкл среды, фильтровали в пробирку, содержащую 5 мкл пропидиум иодида (ПИ), и анализировали на проточном цитометре BD LSRII согласно инструкциям производителя. KD иллюстративного бипаратопного гетеродимерного антитела к HER2 и контрольных антител оценивали с помощью FACS, причем анализ данных и подбор аппроксимирующей кривой осуществляли в GraphPad Prism.
[00352] Результаты показаны на Фигурах 6A-6G. Эти результаты демонстрируют, что иллюстративные бипаратопные антитела к HER2 (v5019, v7091 и v10000) могут связываться с клетками HER2+, с приблизительно в 1,5 раза более высоким значением Вмакс по сравнению с FSA к HER2 (v506). Результаты, представленные на Фигурах 6A-6G, также показывают, что бипаратопные антитела к HER2 (v5019, v7091 и v10000) могут связываться с клетками HER2+, со сходным значнием Вмакс по сравнению с комбинацией двух FSA к HER2 (v506+v4184).
[00353] Результаты по связыванию для клеток HER2+ SKOV3, (HER2 2/3+) показаны на Фигурах 6А, 6Е и в Таблице 4, и в Таблице 5. Результаты, представленные на Фигуре 6А и в Таблица 4, показывают, что иллюстративное бипаратопное антитело к HER2 (v5019) характеризуется приблизительно в 1,5 раза более высоким значением Вмакс при связывании с клетками SKOV3 по сравнению с двумя разными FSA к HER2 (v506 или v4184). Эти результаты также показывают, что иллюстративное бипаратопное антитело к HER2 (v5019) характеризуется эквивалентым значением Вмакс по сравнению с комбинацией двух FSA к HER2 (v506+v4184). Кажущаяся KD v5019 для связывания с SKOV3 была приблизительно в 2-4 раза выше по сравнению либо с FSA к HER2 отдельно (v506 или v4184), либо с комбинацией двух FSA к HER2 (v506+v4184).
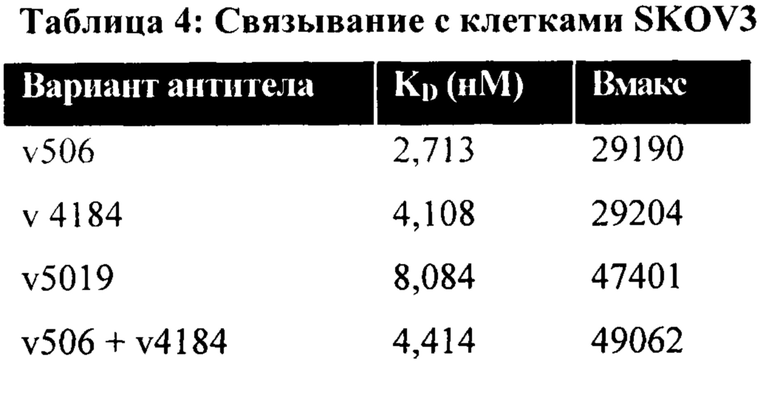
[00354] Результаты, представленные на Фигуре 6Е и в Таблице 5, показывают, что иллюстративные бипаратопные антитела к HER2 (v5019, 7091 и v10000) характеризуются приблизительно в 1,5-1,6 раза более высоким значением Вмакс при связывании с клетками SKOV3 по сравнению с двумя разными FSA к HER2 (v506 или v4184). Эти результаты также показывают, что иллюстративные бипаратопные антитела к HER2 (v5019, 7091 и v10000) характеризуются эквивалентым значением Вмакс по сравнению с комбинацией двух FSA к HER2 (v506+v4184). Кажущаяся KD v5019, v7091, v10000 и комбинации двух FSA к HER2 (v506+v4184) для связывания с SKOV3 была приблизительно в 2-3 раза выше по сравнению с любым FSA к HER2 отдельно (v506 или v4184).

[00355] Кривые связывания в клеточной линии JIMT-1 (HER2 2+) показаны на Фигуре 6В и в Таблице 6. Эти результаты показывают, что иллюстративное бипаратопное антитело к HER2 (v5019) характеризуется приблизительно в 1,5 раза более высоким значением Вмакс при связывании с клетками JIMT-1 по сравнению с FSA к HER2 (v506). Эти результаты также показывают, что иллюстративное бипаратопное антитело к HER2 (v5019) характеризуется эквивалентым значением Вмакс по сравнению с комбинацией двух FSA к HER2 (v506+v4184). Кажущаяся KD v5019 для связывания с JIMT-1 была приблизительно в 2 раза выше по сравнению с FSA к HER2 (v506) и была сходной (приблизительно в 1,2 раза выше) по сравнению с комбинацией двух FSA к HER2 (v506+v4184).

[00356] Кривые связывания в клеточной линии MCF7 (HER2 1+) показаны на Фигуре 6С, 6F и в Таблицах 7 и 8. Эти результаты показывают, что иллюстративные бипаратопные антитела к HER2 (v5019, 7091 и v10000) характеризуются приблизительно в 1,5 раза более высоким значением Вмакс при связывании с клетками MCF7 по сравнению с FSA к HER2 (v506). Результаты, представленные на Фигуре 6С, также показывают, что иллюстративное бипаратопное антитело к HER2 (v5019) характеризуется эквивалентам значением Вмакс по сравнению с комбинацией двух FSA к HER2 (v506+v4184). Кажущаяся KD v5019 для связывания с MCF7 была сходна с FSA к HER2 (v506) и комбинацией двух FSA к HER2 (v506+v4184).

Результаты, представленные на Фигуре 6F и в Таблице 8, показывают, что иллюстративные бипаратопные антитела к HER2 (v5019, v7091 и v 10000) характеризуются приблизительно в 1,6-1,7 раза более высоким значением Вмакс по сравнению с моноспецифическим FSA v506. Кажущаяся KD для v5019, v7091 и v10000 была сходна с FSA к HER2 (v506).

(00357] Кривые связывания в клеточной линии MDA-MB-231 (HER2 0/1+) показаны на Фигуре 6D и в Таблице 9. Эти результаты показывают, что иллюстративное бипаратопное антитело к HER2 (v5019) характеризуется приблизительно в 1,5 раза более высоким значением Вмакс при связывании с клетками MDA-MB-231 по сравнению с FSA к HER2 (v506). Эти результаты также показывают, что иллюстративное бипаратопное антитело к HER2 (v5019) характеризуется эквивалентам значением Вмакс по сравнению с комбинацией двух FSA к HER2 (v506+v4184). Кажущаяся KD v5019 для связывания с MDA-MB-231 была приблизительно в 2,4 раза ниже по сравнению с FSA к HER2 (v506) и была приблизительно в 1,7 раза выше по сравнению с комбинацией двух FSA к HER2 (v506+v4184).

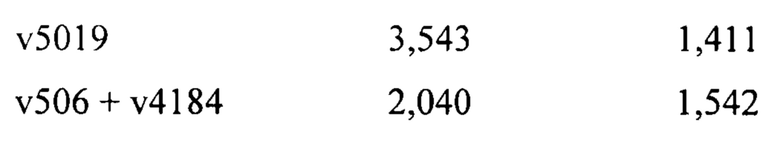
[00358] Кривые связывания в клеточной линии фибробластов легких WI-38 показаны на Фигуре 6G и в Таблице 10. Клеточная линия WI-38 представляет собой нормальный легочный эпителий, в клетках которого HER2 экспрессируется на базальном уровне (HER2 0+, ~10000 рецепторов/клетка) (Carter et al. 1992, PNAS, 89:4285-4289; Yarden 2000, HER2: Basic Research, Prognosis игару). Эти результаты показывают, что иллюстративные бипаратопные антитела к HER2 (v5019, v7091, v10000) характеризуются эквивалентной декорацией клеточной поверхности (Вмакс) при связывании с клетками WI-38 по сравнению с FSA к HER2 (v506); однако необходимо отметить, что в случае v506 оказалось, что связывание, не достигало насыщения и, таким образом, невозможно было определить KD. Кажущаяся KD среди иллюстративных бипаратопных антител к HER2 была эквивалентной.

[00359] Эти результаты показывают, что иллюстративное бипаратопное антитело к HER2 может связываться с опухолевыми клетками с уровнем экспрессии HER2 1+, 2+ и 3+ на уровне, который приблизительно 1,5-1,6 раза выше, чем моноспецифическое FSA к HER2, и что иллюстративные бипаратопные антитела к HER2 могут связываться с опухолевыми клетками с уровнем экспрессии HER2 1+, 2+ и 3+ на эквивалентном уровне по сравнению с комбинацией двух уникальных моноспецифических FSA к HER2 с разной специфичностью эпитопов. Эти результаты также показывают, что бипаратопные антитела к HER2 не характеризуются повышенным уровнем связывания (т.е. по сравнению с моноспецифическим антителом к HER2, v506) с базальными HER2-экспрессирующими клетками, которые экспрессируют приблизительно 10000 HER2-рецепторов/клетку или меньше, и что пороговая величина для повышенного уровня связывания бипаратопных антител к HER2 с клеточными поверхностями достигается, когда уровень HER2-рецепторов составляет приблизительно >10000 рецепторов/клетку. На основе этих данных следует ожидать, что иллюстративные бипаратопные антитела к HER2 будут характеризоваться повышенным уровнем связывания с клеточными поверхностями опухолевых клекок с уровнем экспрессии HER2 3+, 2+ и 1+, но не будут характеризоваться повышенным уровнем связывания с клеточными поверхностями неопухолевых клеток, которые экспрессируют HER2-рецепторы на базальном уровне приблизительно 10000 рецепторов или меньше.
Пример 7: способность бипаратопного антитела к HER2 ингибировать рост клеток HER2+
(00360] Изучали способность иллюстративного бипаратопного антитела к HER2 ингибировать рост клеток, экспрессирующих HER2 на уровне 3+и 2+. Эксперимент проводили на клеточных линиях ВТ-474, SKBr3, SKOV3 с уровнем экспрессии HER2 3+ и JIMT-1 с уровнем экспрессии HER2 2+. Исследовали бипаратопные антитела к HER2: v5019, v7091 и v10000. Изучали способность бипаратопных антител к HER2 ингибировать рост клеток ВТ-474 (конценрация антитела 200 нМ); SKOV3, SKBr3 и JIMT-1 (конценрация антитела 300 нМ), как описано ниже.
[00361] Исследуемые антитела разводили в среде и добавляли к клеткам в количестве 10 мкл/лунку в трех повторностях. Планшеты инкубировали в течение 3 суток при 37°С. Жизнеспособность клеток оценивали либо с использованием AlamarBlue™ (Biosource № dal1100), либо Celltiter-Glo® , и абсорбция измеряли согласно инструкциям производителя. Данные нормализовали по контрольным значениям группы без воздействия, и анализ осуществляли в GraphPad prism.
|00362) Результаты по ингибированию роста показаны на Фигуре 7А-Е. Результаты в кратком виде представлены в Таблицах 11А и 11В. Результаты, представленные на Фигурах 7А-В и в Таблице11А, свидетельствуют о том, что иллюстративное бипаратопное антитело к HER2 (v5019) способно ингибировать рост клеточных линий HER2+SKOV3 и ВТ-474. На Фигуре 10А показано, что бипаратопное антитело к HER2 опосредовало наиболее выраженное ингибирование роста SKOV3 по сравнению с FSA к HER2 (v506) и по сравнению с комбинацией двух антител FSA к HER2(v506+v4184).

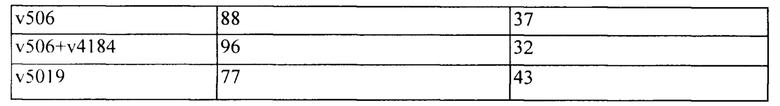
[00363] Результаты, представленные на Фигурах 7С-Е и в Таблице 11В, свидетельствуют о том, что иллюстративные бипаратопные антитела к HER2 (V5019, v7091 и v10000) могут ингибировать рост опухолевых клеток линии SKBR3 с уровнем экспрессии HER2 3+, SKOV3 с уровнем экспрессии HER2 2+/3+ и JIMT-1 с уровнем экспрессии HER2 2+. На Фигуре 1С показано, что бипаратопные антитела к HER2 v7091 и v10000 опосредовали наиболее выраженное ингибирование опухолевых клеток молочной железы SKBr3 с уровнем экспрессии HER2 3+. На Фигуре 7D показано, что бипаратопные антитела к HER2 (v7091 и v10000) опосредовали наиболее выраженное ингибирование роста клеток опухоли яичника SKOV3 с уровнем экспрессии HER2 3+. На Фигуре 7Е показано, что бипаратопные антитела к HER2 (v7091 и v10000) опосредовали наиболее выраженное ингибирование роста герцептин-резистентных опухолевых клеток JIMT-1 уровнем экспрессии HER2 2+. Во всех исследованных клеточных линиях иллюстративные бипаратопные антитела к HER2 (v7091 и v10000) опосредовали более выраженное ингибирование роста по сравнению с моноспецифическим антителом FSA к HER2 (v506).

[00364] Эти результаты показывают, что иллюстративные насыщающие концентрации бипаратопных антител к HER2 могут ингибировать рост опухолевых клеток молочной железы и яичника с уровнем экпрессии HER2 3+ и 2+, а также трастузумаб-резистентных опухолевых клеток с уровнем экпрессии HER2 2+ приблизительно на 20% больше, чем моноспецифическое антитело FSA к HER2.
Пример 8: избирательное связывание паратопов бипаратопных антител к HER2 с димерным HER2 по сравнению с HER2 ECD
[00365] Этот эксперимент осуществляли для того, чтобы определить способность отдельных паратопов иллюстративных бипаратопных антител к HER2 связываться с димерным HER2 и HER2 ECD в качестве имитирующего образца для оценки избирательного связывания с мембраносвязанным HER2 (HER2-Fc) и свободным HER2 ECD. Эксперимент проводили следующим образом.
[00366] Анализ с использованием поверхностного плазмонного резонанса (ППР): аффинность связывания моновалентных антител к HER2 (v1040 или v4182) с внеклеточным доменом (sHER-2, Ebioscience BMS362, содержащий аминокислоты 23 - 652 полноразмерного белка) и HER2-Fc (димерный слитый белок HER2-Fc, содержащий аминокислоты 1 - 652 внеклеточного домена; Sino Biological Inc., 10004-Н02Н) измеряли с помощью ППР с использованием системы Т200 от Biacore (GE Healthcare). Уровень связывания с HER2 ECD определяли с использованием следующего способа. HER2 ECD в 10 мМ Hepes рН 6,8 иммобилизировали на чипе СМ5 посредством связывания через аминогруппу до достижения уровня 44 RU (единицы ответа). Моновалентные антитела к HER2 пропускали по поверхности чипа с иммобилизированным HER2 в концентрациях в диапазоне 0,76-60 нМ. Уровень связывания с HER2-Fc определяли с использованием следующего способа. HER2-Fc в 10 мМ Hepes рН 6.8 иммобилизировали на чипе СМ5 посредством связывания через аминогруппу до достижения уровня 43 RU. Моновалентные антитела к HER2 пропускали по поверхности чипа с иммобилизированным HER2 в концентрациях в диапазоне 0,76-60 нМ. Разные концентрации антител анализировали на предмет связывания в трех повторностях. Равновесные константы связывания/диссоциации (KD) и кинетику (ка и kd) определяли с использованием способа оценки одноцикловой кинетики. Сенсограммы подвергали глобальной аппроксимации к 1:1 по модели связывания Ленгмюра. Все эксперименты проводили при комнатной температуре.
[00367] Результаты показаны на Фигуре 8А, Фигуре 8В, в Таблице 11С и Таблице 11D. В результатах, представленных на Фигуре 8А и в Таблице 11С, показаны полученные с помощью ППР данные по связыванию моновалентного антитела к HER2 (v1040; представляющего собой антигенсвязывающий домен, находящийся на СН-В иллюстративного бипаратопного антитела к HER2). На Фигуре 8А проиллюстрированы значения KD (нМ) для связывания v 1040 с иммобилизированным Her2 ECD или HER2-Fc и показано, что моновалентное антитело к HER2 имеет более низкую КDо для связывания с HER2-Fc по сравнению с HER2 ECD. В Таблице 11С показаны значения ka (1/М с) и kd (1/с) для моновалентного антитела к HER2 (OA) по сравнению с полноразмерным антителом к HER2 (FSA) при связывании с HER2 ECD и HER2-FC («HER2 mem»). Эти данные показывают сопоставимые скорости ассоциации (kа) и диссоциации (kd) OA и FSA для связывания с HER2 ECD и HER2-FC.
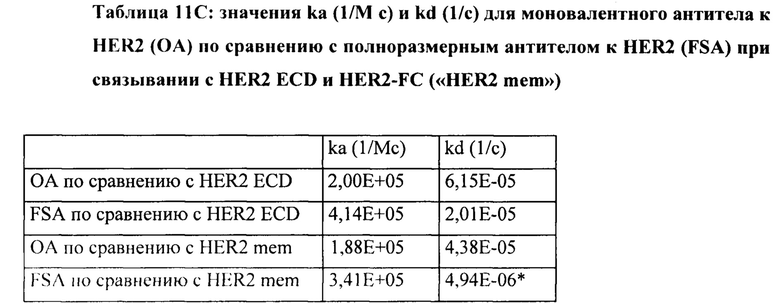
[00368] В результатах, представленных на Фигуре 8В и в Таблице 11D, показаны полученные с помощью ППР данные по связыванию моновалентного антитела к HER2 (v4182; представляющего собой антигенсвязывающий домен, находящийся на СН-А иллюстративного бипаратопного антитела к HER2). На Фигуре 8В проиллюстрированы значения KD (нМ) для связывания v4182 с иммобилизированным Her2 ECD или HER2-FC и показано, что моновалентное антитело к HER2 имеет более низкую KD для связывания с HER2-Fc по сравнению с HER2 ECD. В Таблице 11D показаны значения ka (1/М с) и kd (1/с) для моновалентного антитела к HER2 (OA) по сравнению с полноразмерным антителом к HER2 (FSA) при связывании с HER2 ECD и HER2-FC («HER2 mem»). Эти данные показывают сопоставимые скорости ассоциации (ka) и диссоциации (kd) OA и FSA для связывания с HER2 ECD и HER2-Fc.
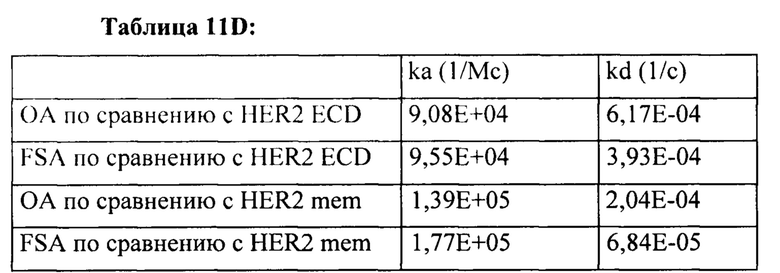
[00369] Эти данные показывают, что каждый из паратопов иллюстративного бипаратопного антитела к HER2 имеет более низкие значения KD для связывания с димерным HER2-антигеном, относящимся к мембраносвязанному HER2, по сравнению с HER2 ECD. На основе этих данных можно предположить, что иллюстративное антитело к HER2 будет обладать более высокой аффинностью связывания по отношению к мембраносвязанному HER2-антигену по сравнению со свободным HER2 ECD, который содержится в сыворотке крови больных пациентов, и может выступать в качестве ловушки для терапевтического антитела (Brodowicz Т, et al. Soluble HER-2/neu neutralizes biologic effects of anti-HER-2/neu antibody on breast cancer cells in vitro. Int J Cancer. 1997; 73:875-879). Например, исходные уровни HER2 HCD≤15 нг/мл; тогда как у пациентов с прогрессирующим заболеванием HER2 ECD ≥ 38 нг/мл.
Пример 9: нагрузка цельных клеток и интернализация бипаратопного антитела к HER2 в клетки HER2+
[00370] Этот эксперимент осуществляли для оценки способности иллюстративного бипаратопного антитела к HER2 к интернализации в клетки с уровнем экспрессии HER2 2+. Способ прямой интернализации осуществляли по протоколу, подробно описанному в Schmidt, М. et al. Kinetics of anti-car cinoembryoni с antigen antibody internalization: effects of affinity, bivalency, and stability. Cancer Immunol Immun другие (2008) 57:1879-1890. Более определенно, антитела подвергали прямому мечению с использованием набора AlexaFluor® 488 Protein Labeling Kit (Invitrogen, номер по каталогу А10235) согласно инструкциям производителя.
[00371] Для анализа интернализации в 12-луночные планшеты высевали 1×105 клеток/лунку и инкубировали на протяжении ночи при 37°С+5% CO2. На следующие сутки добавляли меченные антитела в концентрации 200 нМ в DMEM+10% FBS и инкубировали 24 часа при 37°С+5% CO2. Среду отбирали в условиях темноты, и лунки промывали 2×500 мкл PBS. Для сбора клеток добавляли буфер для диссоциации клеток (250 мкл) при 37°С. Клетки осаждали и ресуспендировали в 100 мкл DMEM+10% FBS в присутствии или в отсутствие антитела фракции IgG кролика к Alexa Fluor 488 (Molecular Probes, Al 1094) в концентрации 50 мкг/мл и инкубировали на льду в течение 30 минут. До анализа в образцы добавляли 300 мкл DMEM+10% FBS, содержащие 4 мкл фильтрованного пропидиум иодида. Образцы анализировали с использованием проточного питометра LSRII.
[00372] Способность иллюстративного бипаратопного антитела к HER2 к интернализации в клетки HER2+, показана на Фигуре 9А и Фигуре 9В. На Фигуре 9А показаны результаты по определению присутствия антитела на поверхности и внутри клеток в клетках ВТ-474 через 24 часа после инкубации с иллюстративным бипаратопным антителом к HER2 и контрольным FSA к HER2. Эти результаты показывают, что в результате инкубации с иллюстративным бипаратопным антителом к HER2 (v5019) степень интернализации антитела в клетки ВТ-474 повышается приблизительно в 2 раза по сравнению с контрольным FSA к HER2. На Фигуре 9 В показаны результаты по определению присутствия антитела на поверхности и внутри клеток в клетках JIMT-1 через 24 часа после инкубации с иллюстративным бипаратопным антителом к HER2 и контрольным FSA к HER2. Эти результаты показывают, что в результате инкубации с иллюстративным бипаратопным антителом к HER2 (v5019) степень интернализации антитела в клетки JIMT-1 повышается приблизительно в 2 раза по сравнению с контрольным FSA к HER2. Количество окрашенных поверхостных антигенов через 24 часа было сопоставимым для бипаратопного антитела к HER2 и FSA к HER2 как в клетках ВТ-474, так и JIMT-1.
[00373] В результатах, представленных на Фигуре 10A-F, показано сравнение обнаруживаемого количества антител, связавшихся с поверхностью цельной клетки через 2 часа при 4°С по сравнению с количеством антител, связавшихся с поверхностью после инкубации в течение 24 часов при 37°С; в дополнение к количеству интернализированных антител через 24 часа при 37°С. На Фигуре 10А показаны результаты для клеток ВТ-474 после инкубации с иллюстративным бипаратопным антителом к HER2 и контрольным FSA к HER2. Эти результаты показывают, что инкубация иллюстративного бипаратопного антитела к HER2 с клетками ВТ-474 в течение 24 часов приводит к снижению приблизительно на 15% количества антител, обнаруженных на поверхности цельной клетки. На Фигуре 10А также показано, что в результате инкубации с иллюстративным бипаратопным антителом к HER2 (v5019) степень интернализации антитела в клетки ВТ-474 повышается приблизительно в 2 раза по сравнению с контрольным FSA к HER2.
[00374] На Фигуре 10В показаны результаты для клеток JIMT-1 после инкубации с иллюстративным бипаратопным антителом к HER2 и контрольным FSA к HER2. На Фигуре 10 В представлены результаты повторного эксперимента, показанного на Фигуре 9В. с добавлением результатов по окрашиванию поверхности через 2 часа при 4°С. Эти результаты показывают, что инкубация иллюстративного бипаратопного антитела к HER2 с клетками JIMT-1 в течение 24 часов приводит к снижению приблизительно на 57% количества антител, обнаруженных на поверхности цельной клетки. На Фигуре 10В также показано, что инкубация с иллюстративным бипаратопным антителом к HER2 (v5019) приводит к более выраженной интернализации антител в клетки ВТ-474 через 24 после инкубации при 37°С, по сравнению с контрольным FSA к HER2.
[00375] На Фигуре 10С показаны результаты для клеток SKOV3 после инкубации с иллюстративным бипаратопным антителом к HER2. Эти результаты показывают, что инкубация иллюстративного бипаратопного антитела к HER2 с клетками SKOV3 в течение 24 часов приводит к снижению приблизительно на 32% количества антител, обнаруженных на поверхности цельной клетки.
[00376] На Фигуре 10D показаны результаты для клеток MCF7 после инкубации с иллюстративным бипаратопным антителом к HER2. Эти результаты показывают, что инкубация иллюстративного бипаратопного антитела к HER2 с клетками MCF7 в течение 24 часов приводит к снижению приблизительно на 45% количества антител, обнаруженных на поверхности цельной клетки.
[00377] На Фигуре 10Е показаны результаты для клеток SKOV3 после инкубации с иллюстративными бипаратопными антителами к HER2, v5019, v7091 и v10000. Эти результаты показывают, что в результате инкубации с иллюстративными бипаратопными антителами к HER2 степень интернализации антител в клетки SKOV3 повышается в 1,5-1,8 раза по сравнению с контрольным FSA к HER2. В результате инкубации с контрольным FSA к HER2 в течение 24 часов количество антител, обнаруженных на поверхности цельной клетки, снижалось в наибольшей степени (~77%).
[00378] На Фигуре 10F показаны результаты для клеток JIMT-1 после инкубации с иллюстративными бипаратопными антителами к HER2, v5019, v7091 и v10000. Эти результаты показывают, что в результате инкубации с иллюстративными бипаратопными антителами к HER2 степень интернализации атитела в клетки JIMT-1 повышается в 1,4-1,8 раз по сравнению с контрольным FSA к HER2. В результате инкубации с бипаратопными антителами к HER2 (v5019 и v10000) в течение 24 часов количество антител, обнаруженных на поверхности цельной клетки, снижалось в наибольшей степени (~64%).
[00379] Эти результаты показывают, что иллюстративные бипаратопные антитела к HER2 обладают лучшими свойствами в отношении интернализации в клетки HER2+, по сравнению с моноспецифическим FSA к HER2. Уменьшение количества антител на поверхности клеток, обнаруженное через 24 часа после инкубации при 37°С, показывает, что иллюстративное бипаратопное антитело к HER2 способно к уменьшению количества HER2-рецепторов клеточной поверхности после инкубации с клетками HER2+, и что уменьшение количества HER2 после инкубации наиболее выражено в опухолевых клетках с уровнем экспрессии HER2 2+.
Пример 10: клеточное окрашивание и локализация бипаратопного антитела к HER2 после инкубации с клетками HER2+, через 1,3 и 16 часов
[00380] Этот эксперимент осуществляли для анализа интернализации иллюстративного бипаратопного антитела к HER2 в клетки HER2+ JIMT-1 через разные интервалы времени и в качестве независимого способа по сравнению со способом, который приведен в Примере 9, для анализа нагрузки цельных клеток и интернализации.
[00381] Клетки JIMT-1 инкубировали с антителом (v506, v4184, v5019 или комбинацией v506 и v4184) в концентрации 200 нМ в бессывороточной DMEM, 37°С+5% СО2 в течение 1 ч, 3 ч и 16 ч. Клетки мягко промывали два раза теплым стерильным PBS (500 мл/лунку). Клетки фиксировали с использованием 250 мкл раствора 10% формалина/PBS в течение 10 мин при КТ. Фиксированные клетки промывали три раза PBS (500 мкл/лунку). пермеабилизировали с использованием PBS в объеме 250 мкл /лунку, содержащего 0,2%Triton Х-100 в течение 5 мин и промывали три раза с использованием PBS в объеме 500 мкл /лунку. Клетки блокировали с использованием PBS+5% козьей сывороти в объеме 500 мкл/лунку в течение 1 часа при КТ. Блокирующий буфер удаляли и клетки инкубировали вторичным антителом (конъюгированный с Alexa Fluor 488 AffiniPure Fab-фрагмента антитела козы к IgG человека (H+L); Jackson ImmunoResearch Laboritories, Inc.; 109-547-003) в объеме 300 мкл/лунку инкубировали в течение 1 часа при КТ. Клетки промывали три раза PBS в объеме 500 мкл/лунку, и покровные стекла, содержащие фиксированные клетки, затем монтировали на предметное стекло с использованием Prolong gold anti-fade с DAPI (Life Technologies; №P36931). Получали отдельные изображения под увеличением 60Х с использованием конфокального микроскопа Olympus FV1000.
[00382] Результаты свидетельствовали об интернализации иллюстративного бипаратопного антитела к HER2 (v5019) в клетки JIMT-1 через 3 часа и его локализации главным образом вблизи ядер. При сравнении изображений через 3 часа после инкубации была отмечена более выраженная интенсивность внутриклеточного окрашивания, ассоциированного с бипаратопным антителом к HER2, по сравнению с комбинацией двух FSA к HER2 (v506+v4184) и по сравнению с отдельным FSA к HER2 (v506 или v4184). Различия по локализации окрашивания антителами в клетках были выявлены, когда было проведено сравнение результатов по окрашиванию бипаратопным антителом к HER2 (V5019) с FSA к HER2 (v4184); причем в случае использования FSA к HER2 (v4184) было отмечено явно выраженное окрашивание плазматической мембраны через интервали времени 1, 3 и 16 ч. Количество поддающихся обнаружению антител уменьшалось через 16 ч, что касается FSA к HER2 (v506), комбинации двух FSA к HER2 (v506+v4184) и воздействий бипаратопным антителом к HER2 (данные не показаны).
[00383] Эти результаты показывают, что произошла нтернализация иллюстративного бипаратопного антитела v5019 к HER2 в клетки HER2+, и что интернализованное антитело можно было обнаружить через 3 ч после инкубации. Эти результаты согласуются с результатами, представленными в Примере 9, которые показывают, что иллюстративное бипаратопное антитело к HER2 может интернализироваться в больших количествах в клетки HER2+, по сравнению с FSA к HER2.
Пример 11: уничтожение клеток HER2+, с помощью АЗКЦ, опосредованной бипаратопным антителом к HER2 по сравнению с контролями
[00384] Этот эксперимент осуществляли для оценки способности иллюстративного бипаратопного антитела к HER2 опосредовать уничтожение клеток SKOV3 с помощью АЗКЦ (рак яичников, HER2 2+/3+).
[00385] Клетки-мишени предварительно инкубировали с исследуемыми антителами (10-кратно уменьшающиеся концентрации с 45 мкг/мл) в течение 30 мин с последующим добавлением эффекторных клеток, причем соотношение эффекторные клетки/клетки-мишени составляло 5:1, и инкубация продолжалась в течение 6 часов при 37°С+5% СО2. Образцы исследовали с использованием 8 концентраций, уменьшающихся 10-кратно с 45 мкг/мл. Высвобождение ЛДГ измеряли с использованием набора ЛДГ анализ kit.
[00386] Исследования дозозависимого эффекта осуществляли с использованием различных концентраций образцов, причем соотношение соотношение эффекторные клетки/клетки-мишени (Е/Т) составляло 5:1, 3:1 и 1:1. Значения полумаксимальной эффективной концентрации (ЕС50) анализировали с помощью сигмоидальной нелинейной регрессионной аппроксимации дозозависимого эффекта с использованием GraphPad prism.
[00387] Клетки поддерживали в полной среде Мак-Коя 5а при 37°С / 5% СО2 и регулярно субкультивировали в подходящей среде, дополненной 10% FBS, согласно протоколу от АТСС. Клетки с номером пассажа менее р10 были использованы для данных анализов. Перед использованием в данном анализе образцы разводили до достижения концентраций в пределах 0,3-300 нМ средой DMEM, не содержащей фенолового красного, дополненной 1% FBS и 1% пенициллина/стрептомицина.
[00388] Результаты по уничтожению позитивных по HER2+ клеток SKOV3 путем АЗКЦ при соотношении эффекторных клеток к клеткам-мишеням 5:1 показаны на Фигуре 11А и в Таблице 12. Эти результаты показывают, что иллюстративное бипаратопное антитело к HER2 (v5019) опосредовало наибольший в процентном отношении максимальная степень лизиса клеток-мишеней путем АЗКЦ по сравнению с FSA к HER2 (v792) и комбинацией двух разных FSA к HER2 (v792+v4184). Разница по максимальному лизису клеток, опосредованному иллюстративным бипаратопным антителом к HER2, была приблизительно в 1,6 раза больше по сравнению с FSA к HER2 и приблизительно в 1,2 раза больше по сравнению с комбинацией двух разных FSA к HER2 (v792+v4184).
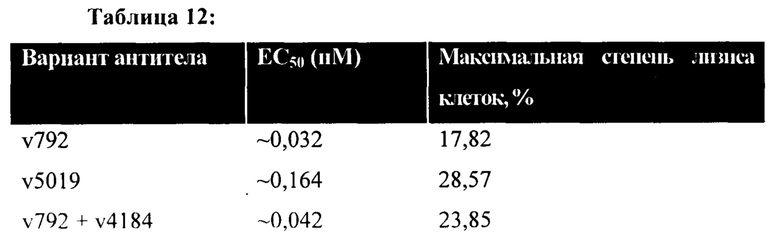
[00389] Результаты по уничтожению позитивных по HER2+клеток SKOV3 путем АЗКЦ при соотношении эффекторных клеток к клеткам-мишеням 3:1 показаны на Фигуре 11 В и в Таблице 13. Эти результаты показывают, что иллюстративное бипаратопное антитело к HER2 (v5019) опосредовало наибольший в процентном отношении максимальная степень лизиса клеток-мишеней путем АЗКЦ по сравнению с FSA к HER2 (v792) и комбинацией двух разных FSA к HER2 (v792+v4184). Разница по максимальному лизису клеток, опосредованному иллюстративным бипаратопным антителом к HER2, была приблизительно в 1,3 раза больше по сравнению с FSA к HER2 и приблизительно в 1,8 раза больше по сравнению с комбинацией двух разных FSA к HER2(v792+v4184).

[00390] Результаты по уничтожению позитивных по HER2+ клеток SKOV3 путем АЗКЦ при соотношении эффекторных клеток к клеткам-мишеням 1:1 показаны на Фигуре 11С и в Таблице 14. Эти результаты показывают, что иллюстративное бипаратопное антитело к HER2 (V5019) опосредовало наибольший в процентном отношении максимальная степень лизиса клеток-мишеней путем АЗКЦ по сравнению с FSA к HER2 (v792) и комбинацией двух разных FSA к HER2 (v792+v4184). Разница по максимальному лизису клеток, опосредованному иллюстративным бипаратопным антителом к HER2, была приблизительно в 1,8 раза больше по сравнению с FSA к HER2 и приблизительно 1,13 раза больше по сравнению с комбинацией двух разных FSA к HER2 (v792+v4184).
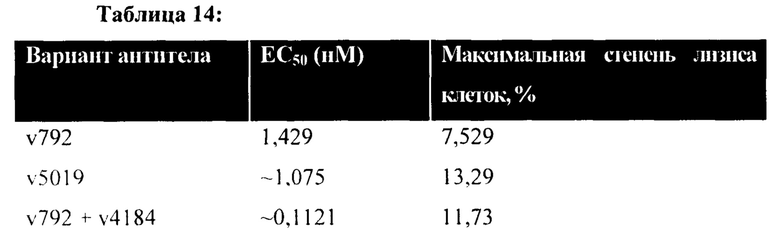
[00391] Результаты, представленные на Фигуре 11 и Таблицах 12-14, показывают, что иллюстративное бипаратопное антитело к HER2 опосредует наиболее выраженное уничтожение клеток SKOV3 путем АЗКЦ при разных соотношениях Е:Т по сравнению с FSA к HER2 и комбинацией двух FSA к HER2. Следует полагать, что повышенная способность к АЗКЦ, опосредованная бипаратопным антителом к HER2, будет наблюдаться у больных пациентов, позитивных по HER2+, у которых отмечается изменяющееся и/или сниженное количество циркулирующих эффекторных клеток после химиотерапии (Suzuki Е. et al. Clin Cancer Res 2007; 13:1875-1882). Результаты наблюдений, представленные на Фигуре 11, согласуются со всеми данными по Вмакс при связывании, представленными в Примере 6, в которых показано приблизительное повышение в 1,5 раза уровня связывания иллюстративного бипаратопного антитела к HER2 с клетками с по сравнению с FSA к HER2.
Пример 12: Способность иллюстративного антитела к HER2 связываться с HER2 ECD
[00392] ПРR-анализ использовали для оценки механизма, посредством которого Иллюстративное бипаратопное антитело к HER2 связывается с HER2 ECD; в частности, чтобы установить, оба ли паратопа одной молекулы бипаратопного антитела могут связываться с одним HER2 ECD (цис-связывание; соотношение антител к молекулам HER2 составляет 1:1) или каждый ли паратоп одного бипаратопного антитела может связываться с двумя разными HER2 ECD (транс-связывание; соотношение антител к молекулам HER2 составляет 1:2). Графическое представление цис- по сравнению с транс-связыванием проиллюстрировано на Фигуре 14. Корреляция между снижением (замедлением) скорости диссоциации с увеличением уровней захвата антител (количество молекул на поверхности) является показателем транс-связывания (т.е. одна молекула антитела связывается с двумя молекулами HER2.
[00393] Аффинность и кинетику связывания иллюстративного бипаратопного антитела к H1-R2 (v5019) с рекомбинантным HER2 человека оценивали и сравнивали с этими параметрами моновалентных антител к HER2 (v630 или v4182; содержащие отдельные паратопы v5019) с помощью ППР с использованием системы Т200 от Biacore (GE Healthcare). От 2000 до 4000 RU антитела к Fc-фрагменту человека, которое наносили в концентрациях от 5 до 10 мкг/мл, иммобилизировали на чипе СМ5 с использованием стандартного присоединения через аминогруппу. Осуществляли захват моновалентного антитела к HER2 (v630 или v4182) и иллюстративного бипаратопного антитела к HER2 (v5019) на антитело к Fc-фрагменту человка (наносили в концентрации от 0,08 до 8 мкг/мл в PBST в течение 1 мин при 10 мкл/мин) с уровнями ответа в диапазоне 350 - 15 RU. Рекомбинантный HER2 человека разводили в PBST и наносили с исходной концентрацией либо 120 нМ, 200 нМ, либо 300 нМ с 3-кратными разведениями и наносили со скоростью потока 50 мкл/мин в течение 3 минут с последующей диссоциацией еще в течение 30 минут по окончанию последнего этапа нанесения. Разведенные образцы HER2 анализировали в двух повторностях. Сенсограммы подвергали глобальной аппроксимации к 1: 1 по модели связывания Ленгмюра. Все эксперименты проводили при 25°С.
[00394] Результаты показаны на Фигуре 12 и Фигуре 13.
[00395] В результатах, представленных на Фигуре 12А, показана ка (1/Мс) моновалентного антитела к HER2 (v630 и v4182) и иллюстративного бипаратопного антитела к HER2 (v5019) для связывания с рекомбинантным HER2 человека для целого ряда разных концентраций антител, нанесение и захват которых производились на поверхности чипа. Эти результаты показывают, что ka v630, v4182 и v5019 не изменяется при разных уровнях захвата антител.
[00396] В результатах, представленных на Фигуре 12В, показана kd (1/с) моновалентного антитела к HER2 (v630 и v4182) и иллюстративного бипаратопного антитела к HER2 (V5019) для связывания с рекомбинантным HER2 человека для целого ряда разных концентраций антител, нанесение и захват которых производились на поверхности чипа. Эти результаты показывают, что уменьшалась только kd иллюстративного бипаратопного антитела к HER2 (v5019) с увеличением уровней захвата антител.
[00397] В результатах, представленных на Фигуре 12С, показана KD (М) моновалентного антитела к HER2 (v630 и v4182) и иллюстративного бипаратопного антитела к HER2 (v5019) для связывания с рекомбинантным HER2 человека для целого ряда разных концентраций антител, нанесение и захват которых производились на поверхности чипа. Эти результаты показывают, что уменьшалась только kd иллюстративною бипаратопного антитела к HER2 (v5019) с увеличеникм уровней захвата антител. Этот результат коррелировал с уменьшением значений kd, показанных на Фигуре 15 В.
[00398] В результатах, представленных на Фигуре 13А, показана kd (1/с) иллюстративного бипаратопного антитела к HER2 (v5019) для связывания с рекомбинантным HER2 человека для целога ряда уровней захвата антител. Эти результаты показывают, что значения kd обратно пропорциональны более высоким RU антитела, захваченного на поверхности чипа (т.е. более низкие скорости диссоциации при более высоких уровнях захвата). Результаты свидетельствуют о том, что иллюстративное бипаратопное антитело к HER2 (v5019) способно связываться с ECD2 HER2 и HER2 ECD4 на двух отдельных молекулах HER2 (т.е. транс-связывание), о чем свидетельствует снижение скорости диссоциации при более высоких уровнях захвата. Эти данные подтверждаются данными похожего эксперимента, которые представлены на Фигуре 47 и рассматриваются в Примере 43, в котором было продемонстрировано цис-связывание (соотношение антитела к HER2 1:1) бивалентного моноспецифического FSA к HER2 (v506), причем значения kd (1/с) и KD (М) оставались неизменными с увеличением уровней захвата антител, что и ожидается для этой молекулы.
[00399] В результатах, представленных на Фигуре 13В, показана kd (1/с) моновалентного антитела к HER2 (v4182) для связывания с рекомбинантным HER2 человека для целого ряда уровней захвата антител. Эти результаты показывают, что значения kd не изменялись для целого ряда антител с разными RU, захвачнных на поверхности чипа. Эти результаты показывают, что моновалентное антитело к HER2 (v4182) связывается моновалентно в соотношении 1:1 (цис-связывание).
[00400] В результатах, представленных на Фигуре 13С, показана kd (1/с) моновалентного антитела к HER2 (v630) для связывания с рекомбинантным HER2 человека для целога ряда уровней захвата антител. Эти результаты показывают, что значения kd не изменялись для целого ряда антител с разными RU, захвачнных на поверхности чипа. Эти результаты показывают, что моновалентное антитело к HER2 (v630) связывается моновалентно в соотношении 1:1 (цис-связывание). Эти данные подтверждаются данными эксперимента, которые представлены на Фигуре 47 и рассматриваются в Примере 43Х, в котором не показано никаких изменений kd (1/с) бивалентного моноспецифического FSA к HER2 (v506).
[00401] [0098] Результаты, представленные на Фигурах 12 и Фигуре 13, свидетельствуют о том, что иллюстративное бипаратопное антитело к HER2 (v5019) способно к одновременному транс-связыванию с двумя молекулами HER2 (соотношение антитела к HER2 составляет 1:2). Транс-механизм связывания, выявленный с помощью ППР, согласуется с данными по связыванию с более высоким насыщением клеточной поверхности (Вмакс), представленными в Примере 6, в комбинации с данными по интернализации, представленными в Примерах 9 и 10.
Пример 13: эффект инкубации с иллюстративным бипаратопным антителом к HER2 на фосфорилирование АКТ в клетках ВТ-474
[00402] Способность иллюстративного бипаратопного антитела к HER2 к уменьшению передачи сигнала, опосредованной рАКТ, исследовали в клетках ВТ-474 с использованием набора АКТ Colorimetric In-Cell ELISA Kit (Thermo Scientifiic; номер по каталогу 62215) согласно инструкциям производителя со следующими модификациями. Клетки высевали в концентрации 5×103/лунку и инкубировали 24 ч при 37°С+5% СО2. Клетки инкубировали в присутствии 100 нМ антитела в течение 30 мин с последующей инкубацией в течение 15 мин с rhHRG-β1. Клетки промывали, фиксировали и пермеабилизировали согласно указанным инструкциям. Добавляли вторичные антитела (1:5000; Jackson ImmunoReasearch, меченное пероксидазой хрена антитело осла к IgG мыши, JIR, Cat№715-036-150, меченное пероксидазой хрена антитело осла к IgG кролика, JIR, Cat№711-036-452), и анализ производился согласно инструкциям производителя.
[00403] Результаты, представленные на Фигуре 15, показывают, что инкубация с иллюстративным бипаратопном антителом к HER2 опосредовала снижение уровней p-Akt приблизительно в 1,2 раза в присутствии HRGβ1 по сравнению с контрольным IgG человека (КОНТР.). Комбинация двух FSA к HER2 (v506+v4184) опосредовала наибольшее снижение уровней p-Akt в присутствии HRGβ1, которое было приблизительно в 1,5 раза меньше по сравнению с контрольным IgG человека. Умеренное снижение уровней p-Akt было выявлено в случае с иллюстративным бипаратопным антителом к HER2 в отсутствие лиганда (HRGβ1) по сравнению с контрольным антителом IgG человека.
[00404] Эти данные показывают, что иллюстративное бипаратопное антитело к HER2 может блокировать лиганд-активируемую передачу сигнала в клетки HER2+.
Пример 14: эффект бипаратопного антитела к HER2 на жизнеспособность кардиомиоцитов
[00405] Эффект иллюстративных бипаратопных антител к HER2 и ADC на жизнеспособность кардиомиоцитов оценивали для заблаговременного выявления потенциальных кардиотоксических эффектов.
[00406] Кардиомиоциты iCell (Cellular Dynamics International, CMC-100-010), которые экспрессируют HER2-рецептор на базальном уровне, выращивали согласно инструкциям производителя и использовали в качестве клеток-мишеней для оценки жизнеспособности кардиомиоцитов после воздействия антитела. Анализ осуществляли следующим образом. Клетки высевали в 96-луночные планшеты (15000 клеток/лунку) и поддерживали в течение 48 ч. Культуральную среду заменяли на поддерживающую среду и клетки поддерживали в течение 72 ч. Для оценки влияния антителоиндуцированной кардиотоксичности клетки подвергали воздействию вариантов антител в концентрациях 10 и 100 нМ в течение 72 ч отдельно или в различных комбинациях. Для оценки влияния антрациклин-индуцированной кардиотоксичности (отдельно или в комбинации с иллюстративными бипаратопными антителами к HER2). клетки подвергали воздействию доксорубицина в концентрации 3 мкМ (~IC20) в течение 1 часа с последующим воздействием вариантов антител в течение 72 ч в концентрациях 10 и 100 нМ отдельно или в различных комбинациях. Жизнеспособность клеток оценивали путем количественного определения клеточных уровней АТФ с использованием набора CellTiter-Glo® Luminescent Cell Viability Assay (Promega, G7570) и/или сульфородамина (Sigma 230162-5G) согласно инструкциям производителя.
[00407] Результаты показаны на Фигуре 16А-С. Результаты, представленные на Фигуре 16А, показывают, что инкубация кардиомиоцитов с терапевтически значимыми концентрациями иллюстративного бипаратопного антитела к HER2 (v5019) и иллюстративного ADC на основе бипаратопного антитела к HER2 (V6363) не влияла на жизнеспособность кардиомиоцитов по сравнению с контрольной группой без воздействия («Имитирующий образец»).
[00408] Результаты, представленные на Фигуре 16 В, показывают, что инкубация кардиомиоцитов с терапевтически значимыми концентрациями иллюстративных бипаратопных антител к HER2 (v5019, v7091 и v10000) и иллюстративных ADC на основе бипаратопного антитела к HER2 (v6363, v7148 и v10553) не оказывала никакого влияния на жизнеспособность кардиомиоцитов по сравнению с контрольной группой без воздействия («Имитирующий образец»). На основе результатов, представленных на Фигуре 16A и 16В. предполагается, что иллюстративные бипаратопные антитела к HER2 и иллюстративные ADC на основе бипаратопного антитела к HER2 не должны индуцировать кардиомиопатию, например, посредством нарушения функций митохондрий, что описано для других антител, специфически связывающихся с HER2, (Grazette L.P. et al. Ингибирование of ErbB2 Causes Mitochondrial Dysfunction in Cardiomyocytes; Journal of the American College of Cardiology: 2004; 44:11).
[00409] Результаты, представленные на Фигуре 16С показывают, что предобработка кардиомиоцитов доксорубицином с последующей инкубацией с терапевтически значимыми концентрациями иллюстративных бипаратопных антител к HER2 (v5019, v7091 и v10000) и иллюстративных ADC на основе бипаратопного антитела к HER2 (v6363, v7148 и v10553) не оказывала никакого влияния на жизнеспособность кардиомиоцитов по сравнению с контрольной группой без воздействия+доксорубицин («Имитирующий образец + Докс.»). На основе результатов, представленных на Фигуре 16С, предполагается, что иллюстративные бипаратопные антитела к HER2 и иллюстративные ADC на основе бипаратопного антитела к HER2 не должны приводить к повышению риска дисфункции сердца у пациентов, получающих сопутствущее лечение антрациклином (Seidman A, Hudis С, Pierri МК, et al. Cardiac dysfunction in the trastuzumab clinical trials experience. J Clin Oncol (2002) 20:1215-1221).
[00410] На Фигурах 16A-C показано, что инкубация кардиомиоцитов с бипаратопными антителами к HER2 и ADC оказывала эквивалентное воздействие по сравнению с моноспецифическим антителом FSA к HER2 (v506), комбинацией FSA к HER2 (v506+v4184) и ADC (v6246) при воздействии либо отдельно, либо в комбинации доксорубицином. На основе этих результатов, предполагается, что иллюстративные бипаратопные антитела к HER2 и ADC не будут оказывать более выраженное кардиотоксическое воздействие по сравнению с моноспецифическим FSA к HER2, трастузумабом или ADC, T-DM1.
Пример 15: цитотоксическое воздействие иллюстративных ADC на основе бипаратопного антитела к HER2+ на клетки, позитивные по HER2
[00411] Оценивали способность иллюстративных ADC на основе бипаратопного антитела к HER2 (v6363, v7148 и v10553) опосредовать клеточную цитотоксичность в клетках, позитивных по HER2+. В некоторых случаях в качестве контроля использовали IgG человека, конъюгированный с DM1 (v6249). Эксперимент проводили на позитивных по HER2+ клеточных линиях опухоли молочной железы JIMT-1, MCF7, MDA-MB-231, позитивной по HER2+ клеточной линии опухоли яичника SKOV3 и позитивной по HER2+ клеточной линии опухоли желудка NCI-N87. Оценивали цитотоксическое воздействие иллюстративных ADC на основе бипаратопного антитела к HER2 на клетки, позитивные по HER2+, и сравнивали с ADC на основе моноспецифического FSA к HER2 (v6246) и ADC на основе FSA к HER2+контрольные FSA к HER2 (v6246+v4184). Способ осуществляли, как описано в Примере 7, со следующими модификациями. ADC к HER2 инкубировали с клетками-мишенями SKOV3 и JIMT-1 (Фигура 17А и В) в течение 24 часов, клетки промывали, среду заменяли, и выживаемость клеток оценивали через 5 суток после инкубации при 37°С. ADC к HER2 инкубировали с клетками-мишенями MCF7 и MDA-MB-231 в течение 6 ч (Фигура 17С и D), клетки промывали, среду заменяли, и выживаемость клеток оценивали через 5 суток после инкубации при 37°С. На Фигуре 17E-G ADC к HER2 непрерывно инкубировали с клетками-мишенями SKOV3, JIMT-1, NCI-N87 в течение 5 суток. Жизнеспособность клеток оценивали, как описано в Примере 7, либо с использованием AlamarBlue™ (Фигуры 17A-D), либо Celltiter-Glo® (Фигуры 17E-G).
[00412] Результаты показаны на Фигуре 17A-G, и данные в краткой форме предствалены в Таблицах 15 и 16.
[00413] Результаты, представленные на Фигуре 17А и в Таблице 15 и 16 показывают, что иллюстративные ADC на основе бипаратопного антитела к HER2 (v6363) оказывает более выраженное цитотоксическое воздействие на JIMT-1 по сравнению с ADC на основе FSA к HER2 (v6246) и комбинацией к ADC на основе FSA к HER2+FSA к HER2 (v6246+v4184). Иллюстративный ADC на основе бипаратопного антитела к HER2 обладал лучшей ЕС50, которая была приблизительно в 13 раз ниже по сравнению с контрольным ADC на основе FSA к HER2.
[00414] Результаты, представленные на Фигуре 17В и в Таблице 15, показывают, что иллюстративный ADC на основе бипаратопного антитела к HER2 (v6363) оказывает более выраженное цитотоксическое воздействие на SKOV3 по сравнению с ADC на основе FSA к HER2 (v6246) и комбинацией к ADC на основе FSA к HER2+FSA к HER2 (v6246+v4184). Иллюстративный ADC на основе бипаратопного антитела к HER2 обладал лучшей ЕС50, которая была приблизительно в 5 раз ниже по сравнению с контрольным ADC на основе FSA к HER2.
[00415] Результаты, представленные на Фигуре 17С и в Таблице 15, показывают, что иллюстративный ADC на основе бипаратопного антитела к HER2 (v6363) оказывает более выраженное цитотоксическое воздействие на MCF7 по сравнению с ADC на основе FSA к HER2 (v6246) и комбинацией к ADC на основе FSA к HER2+FSA к HER2 (v6246+v4184). Иллюстративный ADC на основе бипаратопного антитела к HER2 обладал лучшей ЕС50, которая была приблизительно в 2 раза ниже по сравнению с контрольным ADC на основе FSA к HER2.
[00416] Результаты, представленные на Фигуре 17D и в Таблице 15, показывают, что иллюстративный ADC на основе бипаратопного антитела к HER2 (v6363) оказывает более выраженное цитотоксическое воздействие на MDA-MB-231 по сравнению с ADC на основе FSA к HER2 (v6246) и комбинацией к ADC на основе FSA к HER2+FSA к HER2 (v6246+v4184). Иллюстративный ADC на основе бипаратопного антитела к HER2 обладал лучшей ЕС50, которая была приблизительно в 2 раза ниже по сравнению с контрольным ADC на основе FSA к HER2.

[00417] Результаты, представленные на Фигуре 17Е и в Таблице 16, показывают, что иллюстративные ADC на основе бипаратопного антитела к HER2 (v6363, v7148 и v10553) оказывают более выраженное цитотоксическое воздействие на клетки опухоли яичника SKOV3 по сравнению с ADC на основе FSA к HER2 (v6246). Иллюстративные ADC на основе бипаратопного антитела к HER2 обладали лучшими значениями ЕС50, которые были приблизительно в 2-7 раз ниже по сравнению с контрольным ADC на основе FSA к HER2.
[00418] Результаты, представленные на Фигуре 17F и в Таблице 16, показывают, что иллюстративные ADC на основе бипаратопного антитела к HER2 (v6363, v7148 и v10553) оказывают более выраженное цитотоксическое воздействие на клетки опухоли молочной железы JIMT-1 по сравнению с ADC на основе FSA к HER2 (v6246). Иллюстративные ADC на основе бипаратопного антитела к HER2 обладали лучшими значениями EC50, которые были приблизительно в 6-9- раз ниже по сравнению с контрольным ADC на основе FSA к HER2.
[00419] Результаты, представленные на Фигуре 17G и в Таблице 16, показывают, что иллюстративные ADC на основе бипаратопного антитела к HER2 (v6363, v7148 и v10553) оказывают цитотоксическое воздействие на клетки опухоли желудка NCI-N87. Иллюстративные ADC на основе бипаратопного антитела к HER2 обладали приблизительно эквивалентными значениями ЕС50 по сравнению с контрольным ADC на основе FSA к HER2.
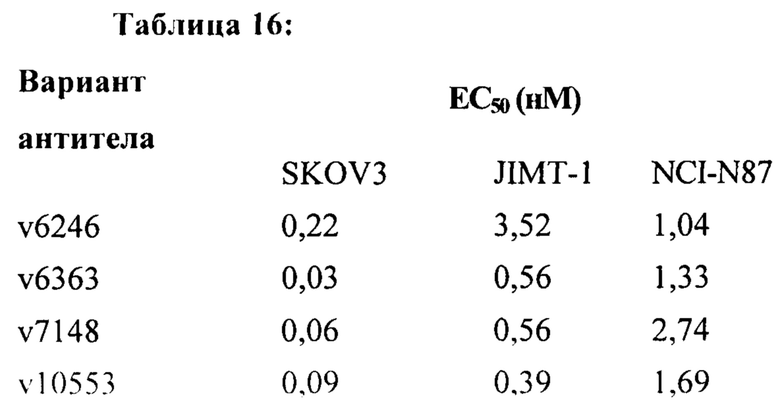
Эти результаты показывают, что иллюстративные ADC на основе бипаратопного антитела к HER2 (v6363, v7148 и v10553) оказывают более выраженное цитотоксическое воздействие по сравнению с контрольным ADC на основе FSA к HER на клетки опухоли молочной железы с уровнем экспрессии HER2 3+, 2+ и 1+. Эти результаты также показывают, что иллюстративные ADC на основе бипаратопного антитела к HER2 (v6363, v7148 и v10553) оказывают цитотоксическое воздействие на клетки опухоли желудка с уровнем экспрессии HER2 2/3+. Эти результаты согласуются с результатами по интернализации, представленными в Примере 9.
Пример 16: эффект бипаратопного антитела к HER2 в модели ксенотрансплантации клеток рака яичников человека
[00420] Разработанную модель на основе ксенотрансплантации клеток рака яичников человека SKOV3 использовали для оценки противоопухолевой эффективности иллюстративного бипаратопного антитела к HER2.
[00421] Самкам безтимусных мышей инокулировали опухоль путем подкожной трансплантации фрагмента опухоли объемом 1 мм3. Проводили мониторинг роста опухолей до тех пор, пока они не достигали среднего объема 220 мм3; животных затем рандомизировали на 3 группы воздействия: контрольный IgG, FSA к HER2 (v506) и бипаратопное антитело к HER2 (v5019).
[00422] Пятнадцать животных были включены в состав каждой группы. Далее представлена схема введения для каждой группы:
[00423] А) контрольный IgG вводили внутривенно с использованием нагрузочной дозы 30 мг/кг на 1-е сутки исследования, затем с использованием поддерживающих доз 20 мг/кг два раза в неделю до 39-х суток исследования.
[00424] В) FSA к HER2 (v506) вводили внутривенно с использованием нагрузочной дозы 15 мг/кг на 1-е сутки исследования, затем с использованием поддерживающих доз 10 мг/кг два раза в неделю до 18-х суток исследования. С 22-х по 39-е сутки 5 мг/кг FSA к HER2 вводили внутривенно два раза в неделю. FSA к HER2 (V4184) вводили одновременно в дозе 5 мг/кг внутрибрюшинно два раза в неделю.
[00425] С) бипаратопное антитело к HER2 вводили внутривенно с использованием нагрузочной дозы 15 мг/кг на 1-е сутки исследования, затем с использованием поддерживающих доз 10 мг/кг два раза в неделю до 39-х суток исследования.
[00426] Объем опухоли измеряли два раза в неделю в ходе исследования, число респондеров и медиану выживаемости оценивали на 22-е сутки. Результаты показаны на Фигуре 18 и в Таблице 17.
|00427] Бипаратопное антитело к HER2 и FSA к HER2 обеспечивали более выраженное ингибирование роста опухоли по сравнению с контрольным IgG. Бипаратопное антитело к HER2 индуцировало более выраженное ингибирование роста опухоли по сравнению с FSA к HER2 комбинация (Фигура 18А). Использование бипаратопного антитела к HER2 было ассоциировано с повышением числа поддающихся лечению опухолей по сравнению с FSA к HER2 v506 на 22-е сутки (11 и 5, соответственно) (Таблица 17). Иллюстративное бипаратопное антитело к HER2 и FSA к HER2 обеспечивали повышение медианы выживаемости по сравнению с контрольным IgG. Бипаратопное антитело к HER2 обеспечивало повышение медианы выживаемости (61 суток) по сравнению с FSA к HER2 (36 суток) (Фигура 18В и Таблица 17). На 22-е сутки исследования добавляли второй FSA к HER2 (v4184) в комбинации с FSA к HER2 (v506). Комбинация двух FSA к HER2 индуцировала дополнительное ингибирование роста опухоли по сравнению с FSA к HER2 (v506), использованным отдельно.

Пример 17: эффект ADC на основе бипаратопного антитела к HER2 (ADC) в модели ксенотрансплантации клеточной линии рака яичников человека
[00428] Разработанную модель на основе ксенотрансплантации клеток рака яичников человека SKOV3 использовали для оценки противоопухолевой эффективности иллюстративного бипаратопного антитела к HER2, конъюгированного с DM1 (v6363).
[00429] Самкам безтимусных мышей инокулировали опухоль путем подкожной трансплантации фрагмента опухоли объемом 1 мм3. Проводили мониторинг роста опухолей до тех пор. пока они не достигали среднего объема 220 мм3; животных затем рандомизировали на 3 группы воздействия: контрольный IgG, ADC на основе FSA к HER2 и ADC на основе бипаратопного антитела к HER2.
[00430] Пятнадцать животных были включены в состав каждой группы. Далее представлена схема введения для каждой группы:
[00431] А) контрольный IgG вводили внутривенно с использованием нагрузочной дозы 30 мг/кг на 1-е сутки исследования, затем с использованием поддерживающих доз 20 мг/кг два раза в неделю до 39-х суток исследования.
[00432] В) ADC на основе FSA к HER2 (v6246) вводили внутривенно с использованием нагрузочной дозы 10 мг/кг на 1-е сутки исследования, затем с использованием поддерживающей дозы 5 мг/кг на 15-е и 29-е сутки.
[00433] С) бипаратопное антитело к HER2-ADC (v6363) вводили внутривенно с использованием нагрузочной дозы 10 мг/кг на 1-е сутки исследования, затем с использованием поддерживающей дозы 5 мг/кг на 15-е и 29-е сутки.
[00434] Объем опухоли измеряли в ходе исследования, а число респондеров, а также медиану выживаемости оценивали на 22-е сутки. Результаты показаны на Фигуре 19. Обобщающие результаты показаны в Таблице 18.
[00435] ADC на основе бипаратопного антитела к HER2 и ADC на основе FSA к HER2 обеспечивали более выраженное ингибирование роста опухоли, чем контрольный IgG (Фигура 19А и Таблица 18). ADC на основе бипаратопного антитела к HER2 ингибировали рост опухоли в большей степени, чем ADC на основе FSA к HER2. Использование группы ADC на основе бипаратопного антитела к HER2 было ассоциировано с повышением числа поддающихся лечению опухолей по сравнению с ADC на основе FSA к HER2 (11 и 9, соответственно). В группах ADC на основе бипаратопного антитела к HER2 и ADC на основе FSA к HER2 была отмечена более высокая выживаемость по сравнению с контрольным IgG (Фигура 19 В и Таблица 18). В группе бипаратопного антитела к HER2 было отмечена повышение медианы выживаемости на уровне 61 суток по сравнению с ADC на основе FSA к HER2, для которого медиана выживаемости составляла 36 суток (Фигура 19В и Таблица 18).


Пример 18: эффект ADC на основе бипаратопного антитела к HER2 (ADC) в модели ксенотрансплантации первичной культуры опухолевых клеток (НВСх-13b)
[00436] Модель ксенотрансплантации резистентной к трастузумабу злокачественной опухоли молочной железы пациента, НВСх-13В, использовали для оценки противоопухолевой эффективности иллюстративного бипаратопного антитела к HER2, конъюгированного с DM1.
[00437] Самкам безтимусных мышей инокулировали опухоль путем подкожной трансплантации фрагмента опухоли объемом 20 мм3. Проводили мониторинг роста опухолей до чех пор, пока они не достигали среднего объема 100 мм3; животных затем рандомизировали на 3 группы воздействия: FSA к HER2 (v506), ADC на основе FSA к HER2 (v6246) и ADC на основе бипаратопного антитела к HER2 (v6363). Семь животных были включены в состав каждой группы. Далее представлена схема введения для каждой группы:
[00438] A) FSA к HER2 вводили внутривенно с использованием нагрузочной дозы 15 мг/кг на 1-е сутки исследования, и поддерживающие дозы 10 мг/кг вводили на 4-е. 8-е, 11-е. 15-е, 18-е, 22-е и 25-е сутки исследования.
[00439] В) ADC на основе FSA к HER2 вводили внутривенно с использованием нагрузочной дозы 10 мг/кг на 1-е сутки исследования, затем с использованием поддерживающей дозы 5 мг/кг на 22-е сутки.
[00440] С) бипаратопное антитело к HER2-ADC вводили внутривенно с использованием нагрузочной дозы 10 мг/кг на 1-е сутки исследования, затем с использованием поддерживающей дозы 5 мг/кг на 22-е сутки.
[00441] Объем опухоли измеряли в ходе исследования, и такие параметры, как средний объем опухоли, полный ответ и отсутствие остаточных явлений болезни, оценивали на 50-е сутки. Результаты показаны на Фигуре 20. Обобщающие результаты показаны в Таблице 19.
[00442] ADC на основе бипаратопного антитела к HER2 и ADC на основе FSA к HER2 обеспечивали более выраженное ингибирование роста опухоли по сравнению с FSA к HER2 (v506). ADC на основе бипаратопного антитела к HER2 обеспечивал более выраженное ингибирование роста опухоли, чем ADC на основе FSA к HER2. Использование группы ADC на основе бипаратопного антитела к HER2 по сравнению с группой ADC на основе FSA к HER2 было ассоциировано с повышением числа опухолей, характеризующихся полными ответами (уменьшение больше, чем на 10% относительно исходных значений), 7 и 4, соответственно, и характеризующихся отсутствием остаточных явлений болезни, 5 и 2, соответственно.
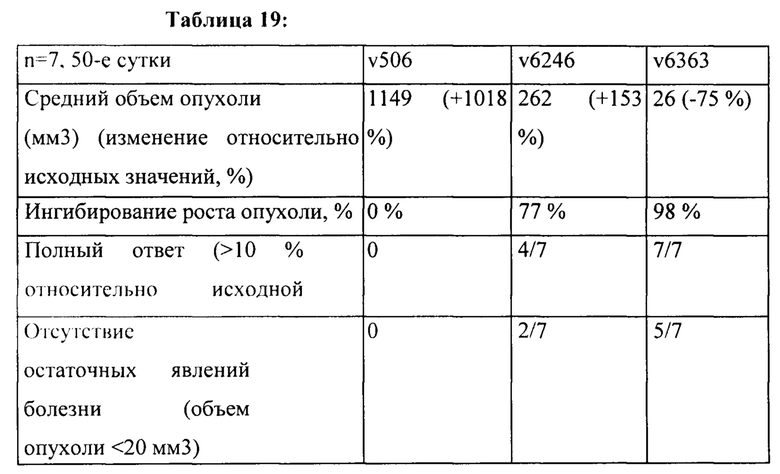
Пример 19: эффект ADC на основе бипаратопного антитела к HER2 (ADQ в модели ксенотрансплантации первичной культуры опухолевых клеток (Т226)
[00443] Модель ксенотрансплантации резистентной к трастузумабу злокачественной опухоли молочной железы пациента, Т226, использовали для оценки противоопухолевой эффективности иллюстративного ADC на основе бипаратопного антитела к HER2.
[00444] Самкам безтимусных мышей инокулировали опухоль путем подкожной трансплантации фрагмента опухоли объемом 20 мм3. Проводили мониторинг роста опухолей до тех пор, пока они не достигали среднего объема 100 мм3; животных затем рандомизировали на 4 группы воздействия: контрольный IgG (n=15), FSA к HER2 (v506; n=15), ADC на основе FSA к HER2 (v6246; n=16) и конъюгат ADC на основе бипаратопного антитела к HER2 (v6363; n=16). Далее представлена схема введения для каждой группы:
[00445] А) контрольный IgG вводили внутривенно с использованием нагрузочной дозы 15 мг/кг на 1-е сутки исследования, и поддерживающие дозы 10 мг/кг вводили на 4-е, 8-е, 11-е, 15-е, 18-е, 22-е и 25-е сутки исследования
[00446] В) FSA к HER2 вводили внутривенно с использованием нагрузочной дозы 15 мг/кг на 1-е сутки исследования, и поддерживающие дозы 10 мг/кг вводили на 4-е, 8-е, 11-е, 15-е, 18-е, 22-е и 25-е сутки исследования
[00447] С) ADC на основе FSA к HER2 вводили внутривенно в дозе 5 мг/кг на 1-е и 15-е сутки исследования
[00448] D) конъюгат ADC на основе бипаратопного антитела к HER2 вводили внутривенно в дозе 5 мг/кг на 1-е и 15-е сутки исследования.
[00449] Объем опухоли измеряли измеряли в ходе исследования, и такие параметры, как средний объем опухоли и полный ответ, оценивали на 31-е сутки. Результаты показаны на Фигуре 21. Обобщающие результаты показаны в Таблице 20.
[00450] ADC на основе бипаратопного антитела к HER2 и ADC на основе FSA к HER2 обеспечивали более выраженное ингибирование роста опухоли по сравнению с FSA к HER2 (v506) и контрольным IgG. Иллюстративный ADC на основе бипаратопного антитела к HER2 индуцировал эквивалентное ингибирование роста опухоли и полную регрессию относительно исходного уровня по сравнению с ADC на основе FSA к HER2 (Фигура 21 и Таблица 20) в этой модели.

Пример 20: Эффект ADC на основе бипаратопного антитела к HER2 (ADC) в модели ксенотрансплантации первичной культуры опухолевых клеток (НВСх-5)
[00451] Модель ксенотрансплантации резистентной к трастузумабу злокачественной опухоли молочной железы пациента, НВСх-5 (инвазивный протоковый рак молочной железы, тип люминальный В), использовали для оценки противоопухолевой эффективности иллюстративного ADC на основе бипаратопного антитела к HER2.
[00452] Самкам безтимусных мышей инокулировали опухоль путем подкожной трансплантации фрагмента опухоли объемом 20 мм3. Проводили мониторинг роста опухолей до тех пор, пока они не достигали среднего объема 100 мм3; животных затем рандомизировали на 4 группы воздействия: контрольный IgG (n=15), FSA к HER2 (v506; n=15), ADC на основе FSA к HER2 (v6246; n=16) и ADC на основе бипаратопного антитела к HER2 (v6363; n=16). Далее представлена схема введения для каждой группы:
[00453] [А) контрольный IgG вводили внутривенно с использованием нагрузочной дозы 15 мг/кг на 1-е сутки исследования, и поддерживающие дозы 10 мг/кг вводили на 4-е, 8-е, 11-е, 15-е, 18-е, 22-е и 25-е сутки исследования
[00454] В) FSA к HER2 вводили внутривенно с использованием нагрузочной дозы 15 мг/кг на 1-е сутки исследования, и поддерживающие дозы 10 мг/кг вводили на 4-е, 8-е, 11-е, 15-е, 18-е, 22-е и 25-е сутки исследования
[00455] С) ADC на основе FSA к HER2 вводили внутривенно в дозе 10 мг/кг на 1-е и 15-е, 22-е, 29-е, 36-е сутки исследования
[00456] D) ADC на основе бипаратопного антитела к HER2 вводили внутривенно в дозе 10 мг/кг на 1-е и 15-е сутки исследования, 22, 29, 36.
[00457] Объем опухоли измеряли измеряли в ходе исследования, и такие параметры, как средний объем опухоли, соотношение Т/С, число респондеров, полный ответ и отсутствие остаточных явлений болезни, оценивали на 43-е сутки. Результаты показаны на Фигуре 22. Обобщающие результаты показаны в Таблице 21.
[00458] ADC на основе бипаратопного антитела к HER2 и ADC на основе FSA к HER2 обеспечивали более выраженное ингибирование роста опухоли по сравнению с FSA к HER2 (v506) и контрольным IgG. Иллюстративный ADC на основе бипаратопного антитела к HER2 индуцировал эквивалентное ингибирование роста опухоли и обесчпечивал повышение числа респондеров по сравнению с ADC на основе FSA к HER2 (Фигура 22 и Таблица 21) в модели ксенотрансплантации клеток HBCx-5 резистентной к трастузумабу злокачественной опухоли молочной железы.

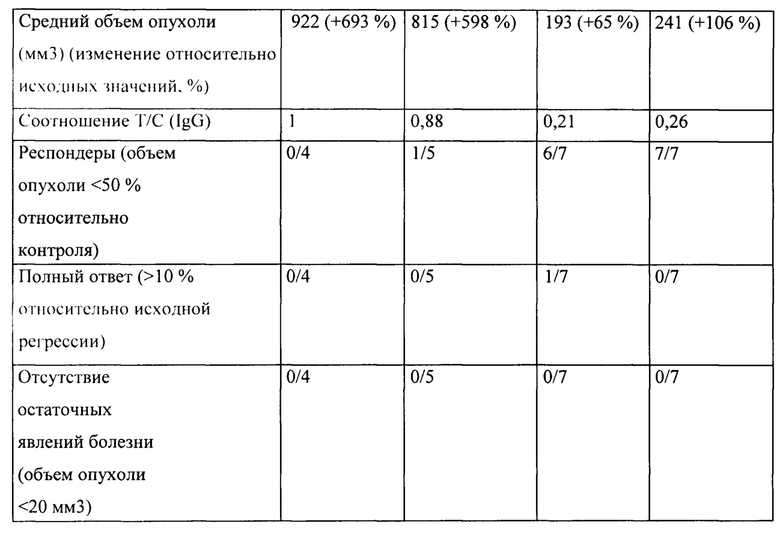
Пример 21: эффект ADC на основе бипаратопного антитела к HER2 (ADC) на опухоли, резистентные к лечению, направленному на подавление экспрессии HER2, в модели ксенотрансплантации линии клеток человека (SKOV3)
[00459] Разработанную модель на основе ксенотрансплантации клеток рака яичников человека SKOV3, описанную в Примере 17, использовали для оценки противоопухолевой эффективности иллюстративного ADC на основе бипаратопного антитела к HER2 на опухолях, рзистентных к лечению, направленному на подавление экспрессии HER2.
[00460] Способы осуществляли, как описано в Примере 17 со следующими модификациями. Когорте животных внутривенно вводили антитело к HER2 в дозе 15 мг/кг на 1-е сутки исследования и в дозе 10 мг/кг на 4-е, 8-е, 15-е сутки; однако данное воздействие не продемонстрировало эффективного ответа на 15-е сутки реализации этой модели. Эту группу воздействия затем перевели на воздействие с использованием конъюгата иллюстративного бипаратопного антитела к HER2 с лекарственным средством (v6363), который вводили в дозе 5 мг/кг на 9-е и 27-е сутки исследования и 15 мг/кг на 34-е, 41-е и 48-е сутки исследования.
[004611 Объем опухоли измеряли два раза в неделю в ходе эксперимента.
[00462] Результаты показаны на Фигуре 23 и свидетельствуют о том, что в группе, которая получала иллюстративный ADC на основе бипаратопного антитела к HER2, (v6363) была отмечена регрессия опухоли до значений среднего объема опухоли, которые были меньше значений исходного среднего объема 220 мм3.
Пример 22: эффект ADC на основе бипаратопного антитела к HER2 (ADC) на опухоли, резистентные к лечению, направленному на подавление экспрессии HER2, в модели ксенотрансплантации первичной культуры опухолевых клеток человека (НВСх-13b)
[00463] Модель ксенотрансплантации резистентной к трастузумабу злокачественной опухоли молочной железы пациента, НВСх-13В, использовали для оценки противоопухолевой эффективности иллюстративного бипаратопного антитела к HER2. конъюгированного с DM1.
[00464] Способы осуществляли, как описано в Примере 18 со следующими модификациями. Когорте животных вводили внутривенно биспецифическое антитело, направленно связывающееся с антигенами семейства ErbB, в дозе 15 мг/кг на 1-е сутки исследования и в дозе 10 мг/кг на 4-е, 8-е, 15-е, 18-е, 22-е и 25-е сутки; однако данное воздействие не продемонстрировало эффективного ответа. Эту группу воздействия затем перевели на воздействие с использованием конъюгата иллюстративного бипаратопного антитела к HER2 с лекарственным средством (v6363), который вводили в дозе 10 мг/кг на 31-е, 52-е сутки и 5 мг/кг на 45-е сутки. Объем опухоли измеряли на протяжении данного исследования.
[00465] Результаты показаны на Фигуре 24. Эти результаты показывают, что иллюстративный ADC на основе бипаратопного антитела к HER2 (v6363) предотвращал прогрессию опухоли. Начиная с введения первой дозы и до 57-х суток объем опухоли в группе, получавшей v6363, повышался менее, чем на 2%, тогда как, за тот же самый итервал времени в группе, получавшей v506, он увеличивался более, чем на 110%.
Пример 23: анализ содержания фукозы в иллюстративном бипаратопном антителе к HER2
[00466] Анализ содержания гликопептидов осуществляли для количественной оценки содержания фукозы в N-связанных гликанах иллюстративных бипаратопных антител к HER2 (v5019, v7091 и v10000).
[00467] Анализ содержания гликопептидов осуществляли следующим образом. Образцы антител подвергали восстанавлению с использованием 10 мМ DTT при 56°С в течение 1 ч и алкилированию с использованием 55 мМ иодоацетамида при КТ в течение 1 ч, и ращеплению в растворе, содержащем трипсин в 50 мМ гидрокарбоната аммония на протяжении ночи при 37°С. Триптические гидролизаты анализировали с помощью nanoLC-MS/MS на QTof-Ultima. Для идентификации белковых последовательностей осуществляли поиск базе данных в NCBI с использованием Mascot, MaxEnt3 (MassLynx) использовали для деконволюции ионов гликопептидов и для количественной оценки разных гликоформ.
[00468] Обобщенные результаты анализа гликопептидов представлены в Таблице 22. N-связанные гликаны иллюстративных бипаратопных антител к HER2 (v5019, v7091 и v10000) фукозилированы приблизительно на 90% (10% N-связанных гликанов без фукозы). N-связанные гликаны моноспецифического FSA к HER2 (v506) фукозилированы приблизительно на 96% (4% N-связанных гликанов без фукозы), и Herceptin® фукозилирован приблизительно на 87% (4% N-связанных гликанов без фукозы).
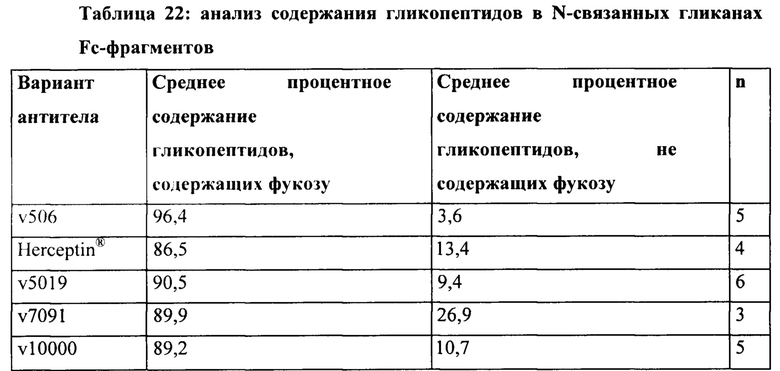
[00469] Эти результаты показывают, что содержание фукозы в бипаратопных антителах к HER2 (с гетеродимерным Fc-фрагментом), временно экспрессированных в клетках СНО, приблизительно на 3% выше в N-гликане по сравнению с коммерческим Herceptin® . Гомодимерное FSA к HER2 (v506), временно экспрессированное в клетках СНО, характеризуется наиболее высоким содержанием фукозы приблизительно 96%.
Пример 24: термостабильность иллюстративного бипаратопного антитела к HER2
[00470] Термостабильность иллюстративных бипаратопных антител к HER2 (v5019, v7091 и v10000) и ADC (v6363, v7148 и VL0533) оценивали с помощью ДСК, как описано ниже.
[00471] ДСК осуществляли в MicroCal™ VP-Capillary DSC (GE Healthcare) с использованием образца очищенных белков (бипаратопные антитела к HER2 и ADC на основе бипаратопного антитела к HER2) в концентрации, доведенной до около 0,3 мг/мл в PBS. Образец сканировали от 20 до 100°С со скоростью 60°С/ч, с низкой обратной связью, фильтром 8 с, 5 мин preTstat и давлением азота 70 фунтов/кв. дюйм. Полученную в результате термограмму анализировали с использованием программного обеспечения Origin 7.
[00472] Результаты по термостабильности иллюстративных бипаратопных антител к HER2 (v5019, v7091 и v10000) показаны на Фигуре 25А-С. На Фигуре 25А показана термограмма для v5019; Тпл Fc-фрагмента и цепи А Fab-фрагмента составляют 75° Цельсия, и Тпл цепи В scFv 5019 составляет 69° Цельсия. На Фигуре 25В показана термограмма для v10000; Тпл Fc-фрагмента СН3-домена составляет 82° Цельсия, Тпл цепи А Fab-фрагмента составляет 76,5° Цельсия, и Тпл цепи В scFv 5019 составляет 69,5° Цельсия. На Фигуре 25С показана термограмма для v7091; Тпл Fc-фрагмента СН3-домена составляет 82° Цельсия, Тпл цепи А Fab-фрагмента составляет 76,7° Цельсия, и Тпл цепи В scFv составляет 69,5° Цельсия.
[00473] Результаты по термостабильности иллюстративных бипаратопных ADC, специфичных к HER2. (v6363, v7148 и V10533) показаны на Фигуре 26А-С. На Фигуре 26А показана термограмма для v6363; Тпл Fc-фрагмента составляет 75° Цельсия и Тпл цепи А Fab-фрагмента и Fc-фрагмента СН3-домена составляют 75° Цельсия. Тпл цепи В scFv 6363 составляет 69° Цельсия. На Фигуре 26В показана термограмма для v10553; Тпл Fc-фрагмента СН3-домена составляет 83° Цельсия, Тпл цепи А Fab-фрагмента составляет 75,7° Цельсия, и Тпл цепи В scFv составляет 66,2° Цельсия. На Фигуре 26С показана термограмма для v7148; Тпл Fc-фрагмента СН3-домена составляет 82,6° Цельсия, Тпл цепи А Fab-фрагмента составляет 74,8° Цельсия, и Тпл цепи В scFv составляет 66,6° Цельсия.
[00474] Иллюстративные бипаратопные антитела и ADC обладают термостабильностью, сопоставимой с термостабильностью IgG дикого типа.
Пример 25: способность иллюстративного бипаратопного антитела к HER2 вызывать уничтожение опухолевых клеток молочной железы, экспрессирующих различные уровни HER2, путем АЗКЦ
[00475] Изучали способность иллюстративного бипаратопного антитела (v5019) вызывать дозозависимое уничтожение путем АЗКЦ HER2-позитивных (3+, 2+) и HER-2-экспрессирующих (0/1+, трижды негативный рак) клеток линий рака молочной железы. Эксперименты по оценке АЗКЦ осуществляли, как описано в Примере 11, за исключением того, что соотношение эффекторных NK-клеток к клеткам-мишеням оставалось неизменным на уровне 5:1.
[00476] Результаты по АЗКЦ показаны на Фигуре 27 и в Таблице 23. Результаты, представленные на Фигуре 27А-С, показывают, что иллюстративное бипаратопное антитело (v5019) вызывает приблизительно в 1,2-1,3 раза больший максимальный лизис HER-2-позитивных (3+, 2+) и HER2-экспрессирующих (0/1+) клеток рака молочной железы по сравнению с Herceptin® . Эти результаты также показывают, что v5019 (90% N-гликанов с фукозой) более эффективно опосредует АЗКЦ HER2-позитивных (3+, 2+) и HER2-экспрессирующих (0/1+) клеток рака молочной железы несмотря на то, что в его N-гликане содержание фукозы приблизительно на 4% выше (что приводит к более низкой аффинности связывания с CD 16 на NK-клетках) по сравнению с Herceptin® (86% N-гликанов с фукозой; Пример 23). Более выраженное уничтожение клеток-мишеней, вызванное v5019, предположительно обусловлено повышенной декорацией опухолевых клеток, как описано в Примере 6.
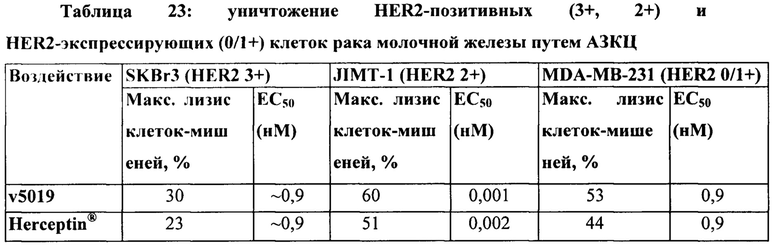
[00477] Результаты по АЗКЦ, представленные на Фигуре 27D показывают, что иллюстративные бипаратопные антитела (v7091 и v10000) вызывают подобный максимальная степень лизиса клеток по сравнению с Herceptin® в линии клеток WI-38, экспрессирующих HER2 на базальном уровне. Результаты по АЗКЦ подтверждают данные по связыванию с клетками (Пример 6), что показывает, что пороговая величина для повышенного уровня связывания АЗКЦ достигается, когда количство HER2-рецепторов превышает 10000 HER2/клетку. На основе этих данных можно предположить, что иллюстративные бипаратопные антитела к HER2 будут характеризоваться повышенным уровнем связывания с клеточными поверхностями и уничтожения путем АЗКЦ опухолевых клеток с уровнем экспрессии 3+, 2+ и 1+, но не будут характеризоваться повышенным уровнем связывания с клеточными поверхностями и уничтожения путем АЗКЦ неопухолевых клеток, которые экспрессируют HER2-рецепторы на базальном уровне приблизительно 10000 рецепторов или меньше.
Пример 26: эффект дефукозилированных антител на АЗКЦ
[00478] Изучали способность дефукозилированных иллюстративных бипаратопных антител (v5019-afuco, 10000-afuco) вызывать дозозависимое уничтожение путем АЗКЦ HER2-позитивных (2/3+, 2+) и HER2-экспрессирующих (0/1+, трижды негативный фенотип) клеток линий рака молочной железы. Эксперименты по оценке АЗКЦ осуществляли, как описано в Примере 11, на клетках SKOV3, клетках MDA-MB-231 и клетках ZR75-1 за исключением того, что использовали постоянное соотношение эффекторных NK-клеток или эффекторных МКПК к клеткам-мишеням (Е:Т) на уровне 5:1. Дефукозилированные иллюстративные бипаратопные антитела получали путем временной экспрессии в клетках СНО, как описано в Примере 1, с использованием временно экспрессированного фермента RMD, как описано в von Horsten et al. 2010 Glycobiology 20:1607-1618. Содержание фукозы в v5019-afuco и v10000-afuco было измерено, как описано в Примере 23, и было установлено, что оно составляет менее <2% (данные не показаны). Данные, полученные с использованием Эффекторных NK-клеток показаны на Фигуре 28А-В, тогда как данные, полученные с использованием МКПК показаны на Фигуре 28С.
[00479] На Фигуре 28А, Фигуре 28В и в Таблице 24 показано, что дефукозилированный v5019 (v5019-afuco) вызывает приблизительно в 1,5-1,7 раза больший максимальный лизис HER 2/3+ и HER2-экспрессирующих (0/1+) клеток рака молочной железы путем АЗКЦ, чем Herceptin®.
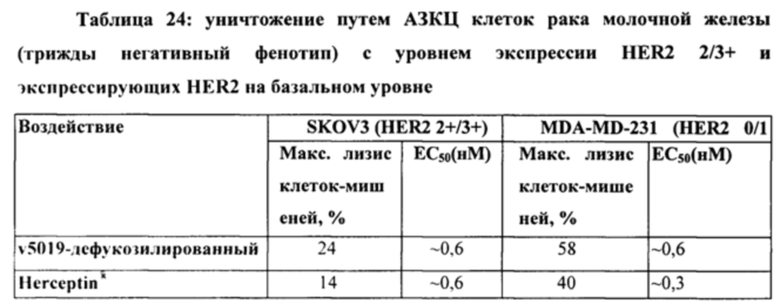
[00480] Результаты, представленные на Фигуре 28С и в Таблице 25 показывают, что v10000 вызывает приблизительно в 1,3 раза больший максимальная степень лизиса клеток рака молочной железы ZR-75-1 (HER2 2+) путем АЗКЦ, чем Herceptin® , и v10000-afuco вызывает приблизительно в 1,5 раза больший максимальная степень лизиса клеток, чем Herceptin® .
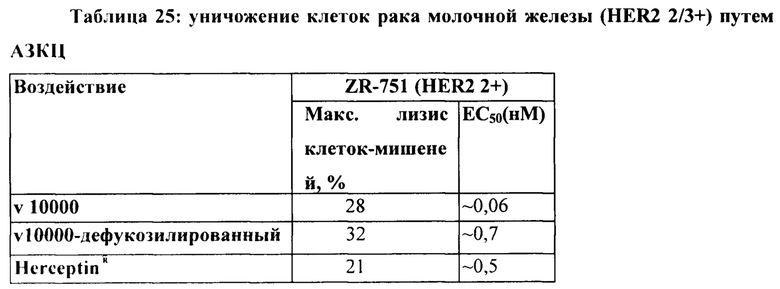
[00481] Результаты по АЗКЦ показывают, что иллюстративные дефукозилированные бипаратопные антитела (v5019-afuco, v10000-afuco) вызывают приблизительно на 15-25% больший максимальная степень лизиса клеток по сравнению с фукозилированными антителами (v5019, Пример 25, v10000), причем в качестве эталона для сравнения используется Herceptin® . Эти результаты показывают, что уменьшение содержания фукозы в N-гликане Fc-домена приводит к повышению максимального лизиса клеток путем АЗКЦ.
Пример 27: способность иллюстративного бипаратопного антитела к HER2 ингибировать рост клеток рака молочной железы (HER2 3+) в присутствии экзогенных рост-стимулирующих лигандов (EGF и HRG)
[00482] Изучали способность 5019 ингибировать рост клеток рака молочной железы (HER2 3+) в присутствии экзогенных рост-стимулирующих лигандов (EGF и HRG).
[00483] Исследуемые антитела и экзогенный лиганд (10 нг/мл HRG или 50 нг/мл EGF) добавляли к клеткам-мишеням ВТ-474 (HER2 3+) в трех повторностях и инкубировали в течение 5 суток при 37°С. Жизнеспособность клеток оценивали с использованием AlamarBlue™ (37°С в течение 2 ч), абсорбцию измеряли при 530/ 580 нм. Данные нормализовали по контрольной группе без воздействия, и анализ осуществляли с использованием GraphPad Prism.
(00484] Результаты показаны на Фигуре 29 и в Таблице 26. Результаты показывают, что иллюстративное бипаратопное антитело v5019 ингибирует рост клеток рака молочной железы (HER2 3+) в отсутствие рост-стимулирующего лиганда (ингибирование на 70%), а также в присутствии EGF (ингибирование на 40%) or HRG (ингибирование на ~10%). Моноспецифическое FSA к HER2(v506) не блокирует EGF-или HRG-индуцированный рост опухолевых клеток через другие erbB-рецепторы EGFR и HER3. v5019 превосходит v506 по ингибированию HER2- и лиганд-зависимой димеризации и роста через другие дополнительные erbB-рецепторы.
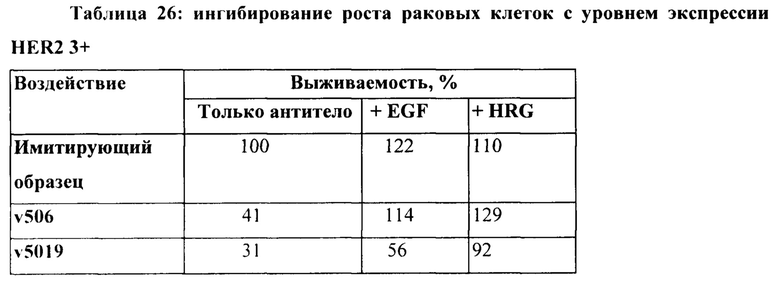
[00485] Эти результаты показывают, что иллюстративное бипаратопное антитело способно к уменьшению лиганд-зависимого роста клеток HER2+, предположительно посредством связывания плеча Fab-фрагмента цепи А антитела к ECD2 и последующей блокировки лиганд-стимулированной гомо- и гетеродимеризации рецепторов и передачи сигнала, опосредованной erbB.
Пример 28: эффект бипаратопного антитела к HER2 в модели ксенотрансплантации клеток трастузумаб-резистентного и резистентного к химиотерапии метастатического рака молочной железы (HER2 3+) пациента (PDX) на примере инвазивной протоковой карциномы молочной железы
[00486] Модель ксенотрансплантации опухоли пациента, на основе использования клеток НВСх-13В инвазивного протокового рака молочной железы человека (ER-PR-негативный фенотип) с уровнем экспрессии HER2 3+, использовали для оценки противоопухолевой эффективности иллюстративного бипаратопного антитела к HER2, v7187. v7187 представляет собой дефукозилированный вариант v5019.
Модельный объект обладает резистентностью к монопрепарату трастузумабу, комбинации трастузумаба и пертузумаба (см. пример 31), капецитабину, доцетакселу и адриамицину/циклофосфамиду.
[00487] Самкам безтимусных мышей подкожно инокулировали фрагмент опухоли объемом 20 мм3. Проводили мониторинг роста опухолей до тех пор, пока их средний объем не достигал 140 мм3. Животных затем рандомизировали на на 2 группы воздействия: контроль растворителем и v7187, причем в каждая группа включала восемь животных. Далее представлена схема внутривенного введения. Растворитель вводили внутривенно в дозе 5 мл/кг в виде буферной смеси два раза в неделю до 43-х суток исследования. v7187 вводили внутривенно в дозе 10 мг/кг два раза в неделю до 43-х суток исследования. Объем опухоли измеряли в ходе исследования, и другие параметры оценивали на 43-е сутки, как показано в Таблице 27.
[00488) Результаты показаны на Фигуре 30 и в Таблице 27. Результаты показывают, что опухоли, на которые воздействовали растворителем, характеризовались постоянной прогрессией и превышали объем 1600 мм3 на 43-е сутки исследования. У мышей, которым вводили v7187, было отмечено более выраженное ингибирование роста опухоли (Т/С - 0,44), причем средний объем опухоли на 43-е сутки составлял 740 мм3. v7187 индуцировал ответы у 5/8 опухолей, причем одна опухоль характеризовалась полной регрессией с отсутствием остаточных явлений болезни на 43-и сутки исследования. У животных, которым вводили v7187 отмечалась более высокая частота ответа, причем 5/8 опухолей отвечали на терапиюпо сравнению с 0/8 у мышей, которым вводили растворитель. В дополнение к этому, воздействие с использованием v7187 в значительной степени задерживало прогрессию опухоли по сравнению с растворителем, причем время удвоения составляло 19 и 11 суток, соответственно.
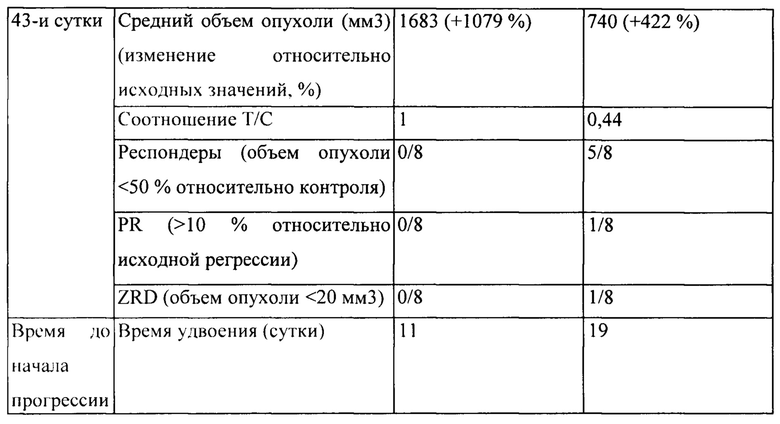
[00489] Эти данные показывают, что иллюстративное бипаратопное антитело к HER2 (v7187) является эффективным в модели ксенотрансплантации злокачественной метастатической опухоли молочной железы, резистентной к трастузумабу и пертузумабу, с уровнем экспресси HER2 3+. Воздействие v7187 обеспечивает повышение частоты ответа и может в значительной степени уменьшать прогрессию злокачественных опухолей молочной железы с уровнем экспресси HER2 3+, резистентных к стандартному лечению.
Пример 29: оценка связывания ADC, специфичных к HER2, с опухолевыми клетками HER2+
[00490] Способность иллюстративных бипаратопных ADC, специфичных к HER2, к связыванию и насыщению поверхностей клеток HER2-позитивных (3+, 2+) опухолей молочной железы и опухолей яичника анализировали с помощью FACS, как описано в Примере 6.
[00491J Данные показаны на Фигуре 31. На Фигуре 31А показаны результаты по связыванию v6363 с опухолевыми клетками линии SKOV3 с приблизительно в 2,0 раза большим Вмакс (MFI), чем у T-DM1 (v6246) при насыщающих концентрациях. На Фигуре 31 В показано, что v6363 связывается с опухолевыми клетками линии JIMT-1 с приблизительно в 1,6 раза большим Вмакс (MFI), чем у T-DM1 (v6246) при насыщающих концентрациях. Эти данные показывают, что v6363 (ADC) обладает подобными свойствами связывания с опухолевыми клетками с повышенным уровнем связывания с клеточными поверхностями по сравнению с исходным неконъюгированным антителом v5019 (Пример 6). Конъюгация v5019 с SMCC-DM1 (v6363) не влияет на антигенсвязывающие свойства данного антитела.
[00492] FACS-анализ связывания повторяли с целью обеспечение прямого сравнения с иллюстративными бипаратопными антителами (v5019, v7091 и v10000) и ADC (v6363. v7148 и v10553). Данные показаны на Фигуре 31С и Фигуре 31D. Иллюстративные бипаратопные ADC, специфичные к HER2 (v6363, v7148 и v10553), характеризуюся эквивалентным уровнем насыщения клеточной поверхности (Вмакс) по сравнению с немеченными бипаратопными антителами (v5019, v7091 и v10000).
[00493] Эти данные показывают, что конъюгация иллюстративных бипаратопных антител (v5019, v7091 и v10000) с SMCC-DM1 не влияет на связывающие свойства. Уровень связывания иллюстративных ADC на основе бипаратопного антитела к HER2 (v6363, v7148 и v10553) с клеточными поверхностями приблизительно в 1,5 раза (или больше) выше по сравнению с моноспецифическим ADC, специфичным к HER2 (v6246, T-DM1).
Пример 30: дозозависимое ингибирование роста опухоли иллюстративным ADC на основе бипаратопного антитела к HER2 в модели ксенотрансплантации (ER-PR-негативной) опухоли пациента с уровнем экспрессии HER2 3+
[00494] Для оценки противоопухолевой эффективности иллюстративного бипаратопного ADC к HER2, v6363, использовали модель ксенотрансплантации опухоли пациента на основе использования клеток НВСх-13В инвазивного протокового рака молочной железы человека (ER-PR-негативный фенотип) с уровнем экспрессии HER2 3+. Модельный объект обладает резистентностью к монопрепарату трастузумабу, комбинации трастузумаба и пертузумаба (см. пример 31), капецитабину, доцетакселу и адриамицину/циклофосфамиду.
[00495] Самкам безтимусных мышей инокулировали опухоль путем подкожной трансплантации фрагмента опухоли объемом 20 мм3. Проводили мониторинг роста опухолей до тех пор. пока они не достигали среднего объема 160 мм3; животных затем рандомизировали на 5 групп воздействия: неспецифический контрольный IgG человека и 4 повышающиеся дозы v6363. 8-10 животных были включены в состав каждой группы. Далее представлена схема введения для каждой группы. Контрольный IgG вводили внутривенно в дозе 10 мг/кг два раза в неделю до 29-х суток исследования. v6363 вводили внутривенно в дозе 0,3, 1, 3 или 10 мг/кг на 1-е, 15-е и 29-е сутки исследования. В ходе исследования оценивали объем опухоли и параметры, которые указаны в Таблице 29.
[00496) Результаты показаны на Фигуре 32 и в Таблице 28. Эти результаты показывают, что иллюстративный ADC на основе бипаратопного антитела к HER2 (v6363) опосредовало дозозависимое ингибирование роста опухоли в PDX-модели трастузумаб-резистентной опухоли на основе клеток НВСх-13b (Фигура 32А). В дополнение к этому, v6363 улучшал общую выживаемость в зависимости от дозы, причем медиана продолжительности выживаемости составляла более, чем 63 суток в случае использования доз 3 мг/кг и 10 мг/кг по сравнению с 43 сутками в случае использования контрольного IgG (Фигура 32 В и Таблица 28). Использования дозы 3 мг/кг было ассоциировано с повышением частоты ответа (5/10) по сравнению с контролем (0/8). У всех мышей, которым вводили v6363 в дозе 10 мг/кг, не только отмечался ответ на терапию (9/9), но также предотвращение прогрессии опухоли. Более того, большинство опухолей характеризовались объективными частичными ответами (7/9) и в конце исследования у многих мышей отсутствовали остаточные явления болезни (6/9). v6363 хорошо переносился во всех дозах, причем не наблюдалось никаких нежелательных явлений и потери массы тела.


[00497] Эти данные показывают, что иллюстративный ADC на основе бипаратопного антитела к HER2 (v6363) является эффективным в модели ксенотрансплантации злокачественной метастатической опухоли молочной железы, резистентной к трастузумабу и пертузумабу, с уровнем экспресси HER2 3+. Воздействие v6363 ассоциировано с высокой частотой ответа, в значительной степени уменьшает прогрессию опухоли и способствует увеличению выживаемости при злокачественных опухолях молочной железы с уровнем экспресси HER2 3+, резистентных к стандартному лечению.
Пример 31: ADC на основе бипаратопного антитела к HER2 в сравнении с комбинациями, соответствующими стандарту лечения, при PDX трастузумаб-резистентной опухоли на основе НВСх-13b
[00498] Оценивали эффективность v6363 в модели ксенотрансплантации (НВСх-13b) ER-PR-негативной трастузумаб-резистентной злокачественной опухоли молочной железы пациента с уровнем экспрессии HER2 3+ и сравнивали с комбинацией: Herceptin™ + Perjeta™; и Herceptin™ + Доцетаксел.
[00499] Самкам безтимусных мышей инокулировали опухоль путем подкожной трансплантации фрагмента опухоли объемом 20 мм3. Проводили мониторинг роста опухолей до тех пор, пока они не достигали среднего объема 100 мм3; животных затем рандомизировали на 4 группы воздействия (8-10 животных/группу): неспецифический контрольный IgG человека, Herceptin™ + Доцетаксел, Herceptin™ + Perjeta™ и v6363. Далее представлена схема введения для каждой группы. Контрольный IgG вводили внутривенно в дозе 10 мг/кг два раза в неделю до 29-х суток исследования. Комбинацию Herceptin™ + Доцетаксел вводили внутривенно в дозе 10 мг/кг два раза в неделю до 29-х суток исследования, и Доцетаксел вводили внутрибрюшинно в дозе 20 мг/кг на 1-е и 22-е сутки исследования. Комбинацию Herceptin™ + Perjeta™ вводили внутривенно в дозе 5 мг/кг два раза в неделю до 29-х суток исследования и Perjeta вводили внутривенно в дозе 5 мг/кг два раза в неделю до 29-х суток исследования. Herceptin™ и Perjeta™ вводили одновременно. v6363 вводили внутривенно в дозе 10 мг/кг на 1-е, 15-е и 29-е сутки исследования.
[00500] Результаты показаны на Фигуре 33 и в Таблице 29. На Фигуре 33А показано изменение объема опухоли с течением времени и на Фигуре 33В показан график выживаемости. Эти результаты показывают, что комбинация Herceptin™ + Perjeta™ не обеспечивала никакого ингибирования роста опухоли по сравнению с контрольным IgG, и объем опухоли превышал 1800 мм3 на 39-е сутки. Комбинация Herceptin™ + Доцетаксел незначительно уменьшала рост опухоли, но не увеличивала медиану выживаемости до 53 суток по сравнению с 43 сутками в случае использования контрольного IgG. v6363 обеспечивал значительное ингибирование роста опухоли (Т/С - 0,04), причем отмечался ответ всех опухолей на терапию и 7/10 опухолей подвергались полной регрессии (отсутствие остаточных явлений болезни). v6363 в значительной степени увеличивал выживаемость по сравнению с обоими комбинированными препаратами. Препараты оказывали незначительное влияние на массу тела животных из всех когорт.
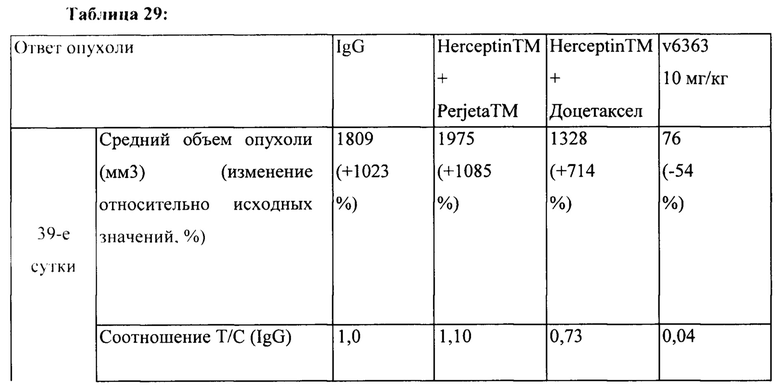

[00501] Эти результаты показывают, что в этой модели ксенотрансплантации иллюстративный ADC на основе бипаратопного антитела к HER2 (v6363) превосходит комбинации, соответствующие стандарту лечения, по всем исследованным параметрам.
Пример 32: эффективность ADC на основе бипаратопного антитела к HER2 в модели ксенотрансплантации опухоли на основе клеток трастузумаб-резистентного рака молочной железы с уровнем экпрессии HER2+
[00502] Оценивали эффективность v6363 в модели ксенотрансплантации опухоли на основе клеток (JIMT-1, HER2 2+) трастузумаб-резистентного рака молочной железы с уровнем экспрессии HER2 3+ (Tanner et al. 2004. Molecular Cancer Therapeutics 3: 1585-1592).
[00503] Самкам мышей линии RAG2 инокулировали опухоль подкожно. Проводили мониторинг роста опухолей до тех пор, пока они не достигали среднего объема 115 мм3; животных затем рандомизировали на на 2 группы воздействия: трастузумаб (n=10) и v6363. Далее представлена схема введения для каждой группы. Трастузумаб вводили внутривенно в дозе 15 мг/кг на 1-е сутки исследования и 10 мг/кг два раза в неделю до 26-х суток исследования. v6363 вводили внутривенно в дозе 5 мг/кг на 1-е и 15-е сутки исследования и в дозе 10 мг/кг на 23-и и 30-е сутки, а также 9 мг/кг на 37-е и 44-е сутки.
[00504] Результаты показаны на Фигуре 34 и в Таблице 30. Эти результаты показывают, что v6363 в значительной степени ингибировал рост опухоли (Т/С - 0,74) по сравнению с трастузумабом на 36-е сутки исследования. Воздействие v6363 и трастузумаба не оказывало значительного влияния на изменение массы тела. Экспозиция v6363 в сыворотке крови составляла 17,9 мкг/мл через 7 суток после введения первой дозы 10 мг/кг.
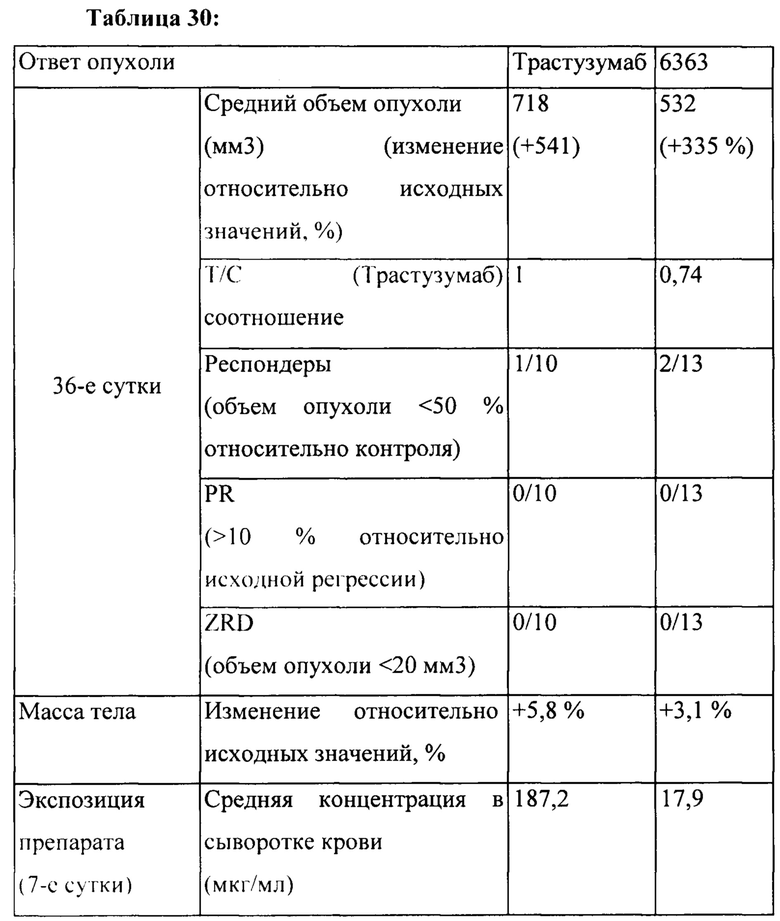
[00505] Эти результаты показывают, что иллюстративный ADC на основе бипаратопного антитела к HER2 (v6363) является эффективным в отношении лечения трастузумаб-резистентного рака молочной железы и его потенциально можно использовать в лечении злокачественных опухолей молочной железы, которые резистентны к современным стандартам лечения.
Пример 33: связывание FcγR с гетеродимерным Fc-фрагментом бипаратопных антител к HER2 и ADC на основе бипаратопного антитела к HER2
[00506] Оценивали связывание бипаратопного антитела к HER2 (v5019, v7019 v10000) и ADC (v6363, v7148 и v10553), содержащего гетеродимерный Fc-фрагмент, с FcγR человека и сравнивали с FSA к HER2 (v506) и ADC (v6246), содержащим гомодимераный Fc-фрагмент.
[00507] Аффинность FcγR по отношению к Fc-области антитела определяли с помощью ППР с использованием ProteOn XPR36 (BIO-RAD). HER2 иммобилизировали (3000 RU) на чипе СМ5 путем стандартного присоединения через аминогруппу. Антитела улавливались с помощью антигенов на поверхности HER2. Очищенный FcγR наносили в различных концентрациях (20-30 мкл/мин) в течение 2 минут с последующей диссоциацией в течение 4 минут. Сенсограммы подвергали глобальной аппроксимации к 1: 1 по модели связывания Ленгмюра. Эксперименты проводили при 25°С.
[00508] Результаты показаны в Таблице 31. Иллюстративные гетеродимерные бипаратопные антитела к HER2 и ADC связывались с CD16aF, CD16aV158, CD32aH, CD32aR131, CD32bY163 и CD64A с сопоставимыми уровнями аффинности. Конъюгация антитела с SMCC-DM1 не оказывает негативного влияния на связывание с FcγR. Гетеродимерные бипаратопные антитела к HER2 обладают приблизительно в 1,3-2 раза более высокой аффинность по отношению к CD16aF, CD32aR131, CD32aH по сравнению с гомодимерным FSA к HER2 (v506) и ADC (v6246). Эти результаты показывают что гетеродимерные бипаратопные антитела к HER2 и ADC связываются разными полиморфными формами FcγR на иммунных эффекторных клетках с подобной или большей аффинностью, чем гомодимерный IgG1 дикого типа.
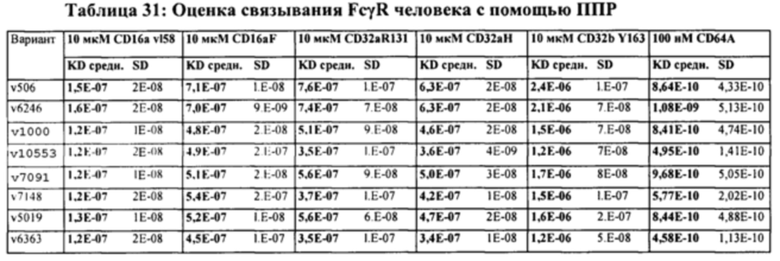
Пример 34: эффективность иллюстративных бипаратопных антител к HER2 в in vivo модели ксенотрансплантации опухоли, полученной из клеток рака яичников, чувствительных к трастузумабу
[00509] Разработанную модель ксенотрансплантации клеток рака яичников человека SKOV3, описанную в Примере 17, использовали для оценки противоопухолевой эффективности иллюстративных бипаратопных антител к HER2, v5019, v7091 и v10000.
[00510] Самкам безтимусных мышей подкожно инокулировали суспензию опухолевых клеток в количестве 325000 клеток в HBSS в левый бок. Проводили мониторинг роста опухолей до тех пор, пока они не достигали среднего объема 190 мм3, а мышей поочередно рандомизировали на 4 группы воздействия: неспецифический контрольный IgG человека, v5019, v7091 и v10000. Далее представлена схема введения для каждой группы. Неспецифический IgG человека вводили внутривенно в дозе 10 мг/кг, начиная с 1-х суток исследования, два раза в неделю до 26-х суток исследования. V5019, v7091 и v10000 вводили внутривенно в дозе 3 мг/кг, начиная с 1-х суток исследования, два раза в неделю до 26-х суток исследования. Объем опухоли измеряли в ходе исследования, и параметры, приведенные в Таблице 32, оценивались на 29-е сутки.
[00511] Данные представлены на Фигуре 35А (рост опухоли), Фигуре 35В (график выживаемости) и в Таблице 32 и показывают, что воздействие v5019, v7091 и v10000 приводило к сопоставимому ингибированию роста опухоли (Т/С: 0,53-0.71), количеству поддающихся лечению опухолей, времени до начала прогрессии и выживаемости на 29-сутки исследования по сравнению с контрольным IgG. Концентрация v5019. v7091 и v10000 в сыворотке крови была сходной (31-41 микрограмм/мл) на 7-сутки исследования.

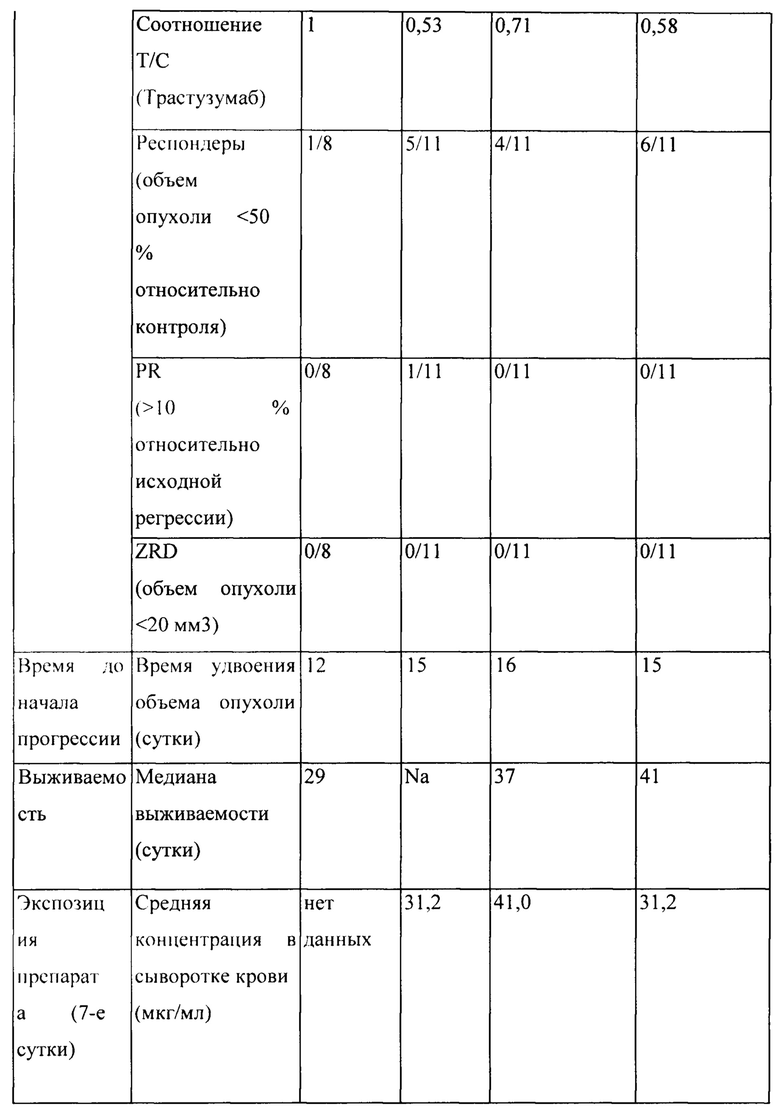
[00512] Эти результаты показывают, что иллюстративные бипаратопные антитела к HER2, v5019, v7091 и v10000) потенциально можно использовать в лечении умеренно чувствительных к злокачественных опухолей яичников с избыточной экспрессией HER2.
Пример 35: иллюстративные бипаратопные антитела к HER2 дозозависимо ингибируют рост опухоли в модели ксенотрансплантации чувствительных к трастузумабу клеток рака яичников
[00513] Разработанную модель на основе ксенотрансплантации клеток рака яичников человека SKOV3, описанную в Примере 17, использовали для оценки дозозависимой эффективности иллюстративного бипаратопного антитела к HER2, v10000.
[00514] Самкам безтимусных мышей подкожно инокулировали суспензию опухолевых клеток в количестве 325000 клеток в HBSS в левый бок. Проводили мониторинг роста опухолей до тех пор, пока они не достигали среднего объема 190 мм3 а мышей поочередно рандомизировали на 6 групп воздействия: неспецифический контрольный IgG человека и v10000 в 5 повышающихся дозах. 9-13 животных были включены в состав каждой группы. Далее представлена схема введения для каждой группы. Контрольный IgG вводили внутривенно в дозе 10 мг/кг два раза в неделю до 26-х суток исследования. v10000 вводили внутривенно в дозе 0,1, 0,3, 1, 3 или 10 мг/кг два раза в неделю.
[00515] Данные представлены на Фигуре 36 и в Таблице 33 и показывают, что воздействие v10000 индуцирует дозозависимое ингибирование роста опухоли (Т/С: 0.28-0.73) по сравнению с контрольным IgG. В дополнение к этому, воздействие v10000 было ассоциировано с дозозависимым увеличением числа опухолей, поддающихся лечению (7/9 при 10 мг/кг и 3/11 при 0,1 мг/кг), увеличением периода времени (24 суток при 10 мг/кг и 12 суток при 0,1 мг/кг) до начала прогрессии на 29-сутки исследования. Изменение концентрации v10000 в сыворотке крови на 7-е сутки носило дозозависимый характер, и концентрация повышалась от 0,46 микрограмм/мл при использовании дозы 0,1 мг/кг до 79,3 микрограмм/мл мл при использовании дозы 10 мг/кг.

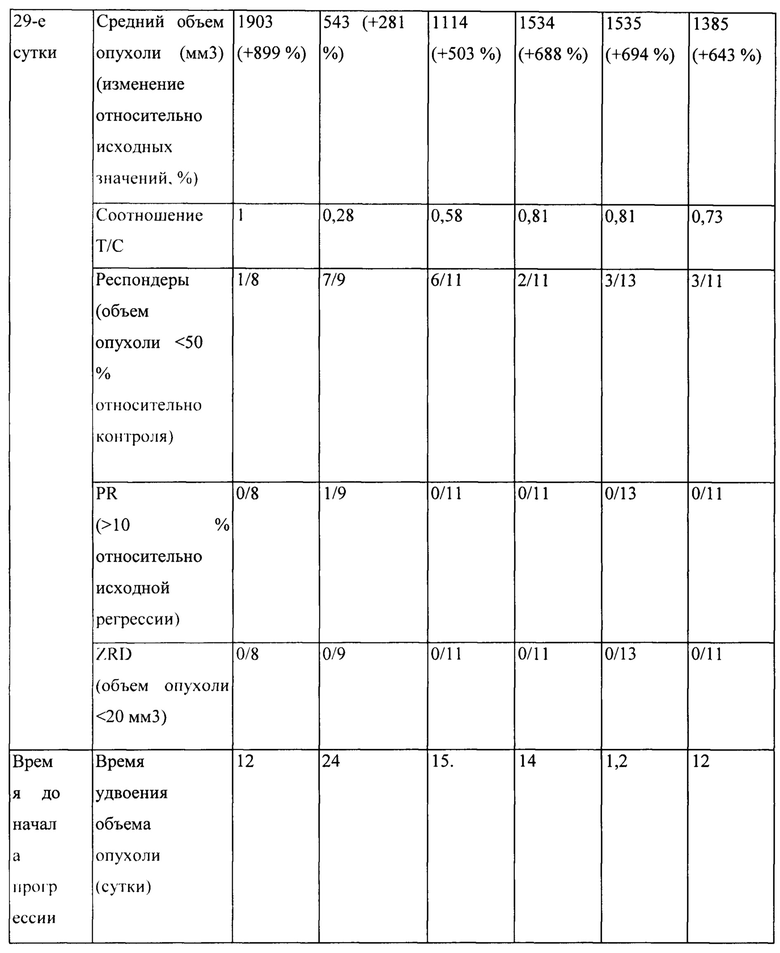
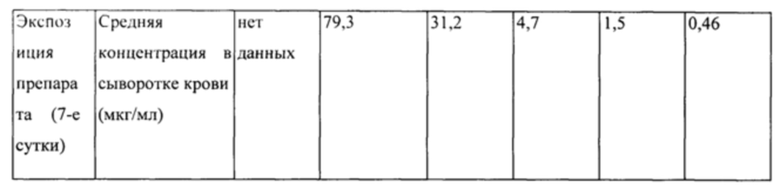
[00516] Эти результаты показывают, что иллюстративное бипаратопное антитело к HER2, v10000. ингибирует прогрессию опухоли в зависимости от дозы.
Пример 36: способность бипаратопного антитела к HER2 и ADC на основе бипаратопного антитела к HER2 ингибировать рост клеточных линий, экспрессирующих HER2 и EGFR и/или HER3 на уровне 3+, 2+ или 1+
[00517] Следующий эксперимент осуществляли для оценки способности иллюстративного бипаратопного антитела к HER2 (v10000) и соответствующего ADC (v10553) на основе бипаратопного антитела, специфичного к HER2, ингибировать рост целого ряда линий клеток рака молочной железы, колоректального рака, рака желудка, легких, кожи, яичников, поджелудочной железы, головы и шеи, матки и мочевого пузыря, которые экспрессируют HER2 и EGFR и/или HER3 на уровне 3+, 2+, 1+ или 0+, который определяют с помощью ИГХ.
[00518] Эксперимент осуществляли следующим образом. Оптимальную плотность посева для каждой клеточной линии определяли отдельно для установления плотности посева, которая бы обеспечивала приблизительно 60-90% конфлуентность после культивирования в течение 72 часов. Клетки каждой линии высевали с оптимальной плотностью посева в соответствующей ростовой среде на клеточную линию в 96-луночные планшеты и инкубировали в течение 24 ч при 36°С и 5% СО2. Антитела добавляли в трех концентрациях (v10000 при 300, 30 и 0,3 нМ; v10553 при 300, 1, 0,1 нМ) наряду с позитивным контролем и контролем растворителем. Комбинация химиотерапевтических лекарственных средств позитивного контроля состояла из 5-ФУ (5-фтороурацила), паклитаксела, цисплатина, этопозида (25 микроМ), контроль растворителем состоял из PBS. Препараты антител и контроли инкубировали с клетками в течение 72 ч в инкубаторе для культивирования клеток при 36°С и 5% СО2. Планшеты центрифугировали при 1200 об/мин в течение 10 мин, и культуральную среду полностью удаляли путем отсасывания. Среду RPMI SFM (200 микролитров) и MTS (20 микролитров) добавляли в каждую лунку и инкубировали при 36°С и 5% СО2 в течение 3 ч. Оптическую плотность измеряли при 490 нм, и ингибирование роста в процентах определяли по сравнению с контролем растворителем.
[00519] Результаты показаны на Фигуре 37, и все итоговые результаты испытаний показаны на Фигуре 38. На Фигуре 37А показаны результаты по ингибированию роста для v10000. Эти результаты показывают, что v10000 может ингибировать рост клеток линий рака молочной железы, колоректального рака, рака желудка, легких, кожи, яичников, поджелудочной железы, головы и шеи, матки и эндометрия, которые экспрессируют HER2 и коэкспрессируют EGFR и/или HER3 на уровне 3+, 2+, 1+ или 0+. Данные по активности v10000 и v10553 при 300 нМ в краткой форме представлены на Фигуре 38, где «+» обозначены клеточные линии, для которых показано снижение жизнеспособности клеток при 300 нМ, которая составляла >5% относительно контроля растворителем, и «-» обозначена жизнеспособность <5% относительно контроля растворителем.
[00520] На Фигуре 37В показанi результаты по ингибированию роста v10553. Эти результаты показывают, что v10553 может ингибировать рост клеток линий рака молочной железы, колоректального рака, рака желудка, легких, кожи, яичников, поджелудочной железы, головы и шеи, матки и мочевого пузыря, которые экспрессируют HER2 и коэкспрессируют EGFR и/или HER3 на уровне 3+, 2+, 1+ или 0+ (см. также Фигуру 38). Результаты, представленные на графике на Фигуре 37В, относятся к клеточным линиям, у которых отмечалось минимальное дозозависимое ингибирование роста при 300 и 1 нМ, и у которых ингибирование роста при 1 нМ равно или больше, чем 5% (Фигура 37В).
[00521] Эти результаты показывают, что иллюстративное бипаратопное антитело v10000 и ADC v10553 могут ингибировать рост опухолевых клеток по результатам гистологических исследований тканей рака молочной железы, колоректального рака, рака желудка, легких, кожи, яичников, поджелудочной железы, головы и шеи, матки и мочевого пузыря, которые экспрессируют HER2 на уровне 3+, 2/3+, 2+, 1+ и 0/1+ и которые коэкспрессируют EGFR и/или HER3 на уровне 2+, 1+.
Пример 37: способность бипаратопных антител к HER2 опосредовать уничтожение раковых клеток с уровнем экспрессии HER2 2+, 1+ и 0/1+ путем АЗКЦ
[00522] Следующий эксперимент осуществляли, чтобы определить способность бипаратопных антител к HER2 опосредовать уничтожение путем АЗКЦ опухолевых клеток, которые экспрессируют HER2 на уровне 2+, 1+ и/или 0/1+, и которые коэкспрессируют EGFR и/или HER3 на уровне 2+ или 1+. В качестве бипаратопных антител к HER2 исследовали 5019, 10000 и 10154 (дефукозилированный вариант v10000), в качестве контролей использовали Herceptin™ и v506.
[00523] Эксперимент по АЗКЦ осуществляли, как описано в Примере 11 и Примере 25 причем соотношение Е/Т составляло 5:1 и в качестве эффекторных клеток были использованы NK-92 (Фигура 39), и как описано в Примере 26, с Е/Т 30:1 и МКПК в качестве эффекторных клеток.
[00524] Результаты показаны на Фигуре 39 (эффекторные клетки NK-92) и Фигуре 40 (эффекторные клетки МКПК). На Фигуре 39А показаны результаты по АЗКЦ для клеточной линии опухоли головы и шеи с уровнем экспрессии HER2 2+ (гипофарингеальная карцинома), FaDu, при этом бипаратопное антитело к HER2 вызывает максимальная степень лизиса клеток приблизительно на уровне 15%. На Фигуре 39С показаны результаты по АЗКЦ для клеточной линии ВхРСЗ опухоли поджелудочной железы с уровнем экспрессии HER2 1+, и на Фигуре 39D показаны результаты для клеточной линии MiaPaca2 опухоли поджелудочной железы с уровнем экспрессии HER2 2+. На Фигуре 39В показаны результаты по АЗКЦ для клеточной линии А549 НМРЛ (немелкоклеточный рак легкого) с уровнем экспрессии HER2 0/1+HER2 0/1+. В отношении линий опухолевых клеток ВхРСЗ, MiaPaca2 и А549, v10000 опосредовал максимальный лизис опухолевых клеток приблизительно на уровне 5%.
[00525] На Фигуре 40 показаны результаты по АЗКЦ для клеток А549, NCI-N87 и НСТ-116, где МКПК были использованы в качестве эффекторных клеток. На Фигуре 40А показаны результаты по АЗКЦ для линии опухолевых клеток А549 НМРЛ с уровнем экспрессии HER2 0/1+, при этом v10000 вызывал максимальная степень лизиса клеток на уровне ~28%, и это было сравнимым с Herceptin™, в N-связанном гликане которого содержится фукоза в эквивалентном количестве. Иллюстративное дефукозилированное на 100% (0% фукозы) бипаратопное антитело v10154 характеризуется повышенным максимальным лизисом клеток (40% максимальная степень лизиса клеток) и повышенной активностью по сравнению с v10000 и Герцептином, в N-связанном гликане которого содержится приблизительно 88% фукозы.
[00526] На Фигуре 40В показаны результаты по АЗКЦ для клеточной линии NCI-N87 опухоли желудка с уровнем экспрессии HER2 3+. На Фигуре 40В показано, что иллюстративное бипаратопное антитело v5019 (фукозилированное приблизительно на 88%) опосредует максимальная степень лизиса клеток на уровне приблизительно 23% и характеризуется более низким значением ЕС50 по сравнению с трастузумабом v506 (фукозилирован приблизительно на 98%).
[00527] На Фигуре 40С показаны результаты по АЗКЦ для клеточной линии НСТ-116 колоректальной опухоли с уровнем экспрессии HER2 1+. На Фигуре 40С показано, что иллюстративное бипаратопное антитело v5019 (фукозилированное приблизительно на 88%) опосредует максимальная степень лизиса клеток на уровне приблизительно 25% и обладает большей активностью по сравнению с трастузумабом v506 (фукозилирован приблизительно на 98%).
[00528] Эти результаты показывают, что иллюстративные бипаратопные антитела к HER2 могут вызывать уничтожение путем АЗКЦ по результатам гистологических исследований тканей опухолей рака и шеи, желудка, НМРЛ и поджелудочной железы. Для реализации АЗКЦ в присутствии NK-92 клеток в качестве эффекторных клеток очевидно потребовалась экспрессия рецептора HER2 на уровне 2+ (т.е. 2+ или больше) для обеспечение более высокого процента (>5%) максимального лизиса клеток. Однако если в качестве эффекторных клеток были использованы клетки МКПК, были достигнуты более высокие уровни максимального лизиса клеток (>5% и вплоть до 28% или 40%; v10000 и v10154, соответственно), и это зависело от плотности НЕR2-рецепторов, поскольку при плотности HER2-рецепторов на уровне 0/1+, 1+и 3+ отмечалась АЗКЦ>5%.
Пример 38: аффинность и кинетика связывания с HER2, измеренные с помощью ППР
[00529] Как указано в Примере 1, были сконструированы бипаратопные антитела к HER2, имеющие разные форматы антигенсвязывающих фрагментов, которые описаны в Таблице 1. Указанные форматы включали формат scFv-scFv (v6717), формат Fab-Fab (v6902 и v6903) наряду с форматом Fab-scFv (v5019, v7091 и v10000). Следующий эксперимент осуществляли для сравнения аффинности и кинетики связывания с HER2 этих иллюстративных бипаратопных антител к HER2 в указанных форматах.
[00530] Аффинность и кинетику связывания с HER2 ECD мыши (Sino Biological 50714-М08Н) измеряли посредством одноцикловой кинетики с использованием системы Т200 SPR от Biacore (GE Healthcare). В пределах 2000-4000 RU антитела к Fc-фрагменту человека иммобилизировали на чипе СМ5 с использованием стандартного присоединения через аминогруппу. Захват 5019 на поверхности Fc-фрагмента к IgG человека происходил при 50 RU. Рекомбинантный HER2 ECD (1.8-120 nM) наносили при 50 мкл/мин в течение 3 минут с последующей диссоциацией в течение 30 минут вслед за последним нанесением. Разведенные образцы HER2 анализировали в двух повторностях. Сенсорграммы апроксимировали к модели Ленгмюра для связывания 1:1. Все эксперименты проводили при комнатной температуре, 25°С.
[00531] Результаты в Таблице 34 показывают, что бипаратопные антитела Fab-scFv (v5019 и v7091), варианты Fab-Fab (v6902 и v6903) и вариант scFv-scFv (v6717) обладают сопоставимой аффинностью связывания (1-4 нМ). Вариант Fab-scFv v10000 обладал более высокой аффинностью связывания (более низкой KD) приблизительно 0,6 нМ. Моноспецифическое антитело к HER2 ECD4 (v506) и антитело к ECD2 HER2 (v4184) были включены в анализ в качестве контролей. Эти результаты свидетельствуют о том, что молекулярные форматы, включая v6717, v6902, v6903, v5019 и/или v7091, характеризуются эквивалентной аффинностью связывания, и таким образом, можно предположить, что функциональные различия этих антител могут быть обусловлены различиями в формате.
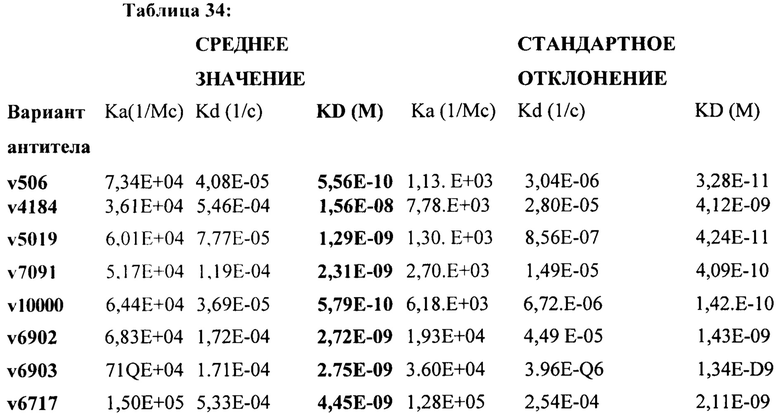
Пример 39: эффект формата бипаратопного антитела к HER2 на связывание с опухолевыми клетками с уровнем экспрессии HER2+
[00532] Следующий эксперимент осуществляли для сравнения свойств связывания иллюстративных бипаратопных антител к ECD2 HER2 × ECD4 с цельными клетками (Вмакс и кажущаяся KD), которые имеют разный формат молекул (например, v6717, scFv-scFv IgG1; v6903 и v6902 Fab-Fab IgG1; v5019, v7091 и v10000 Fab-scFv IgG1).
[00533] Эксперимент осуществляли, как описано в Примере 6. Результаты показаны на Фигуре 41 и в Таблицах 35-38. На Фигуре 41А и в Таблице 35 показаны результаты FACS-анализа по связыванию иллюстративных бипаратопных антител с клетками линии опухоли молочной железы ВТ474 с уровнем экспрессии HER2 3+. Результаты показывают, что все антитела к HER2 имеют более высокое значение Вмакс (в 1,5-1,7 раза выше) по сравнению с моноспецифическим бивалентным антителом v506 к HER2. Значение Вмакс форматов Fab-scFv (v5019, v7091 и v10000) и Fab-Fab (v6903) повышалось приблизительно в 1,7 раза, а значение Вмакс формата scFv-scFv (v6717) повышалось в 1,5 раза по сравнению с v506. При использовании эквимолярной комбинации FSA v506 и v4184 Вмакс повышалась в 1,7 раза. Кажущаяся KD иллюстративных бипаратопных антител к HER2 была приблизительно в 2-3 раза выше по сравнению с моноспецифическим v506.
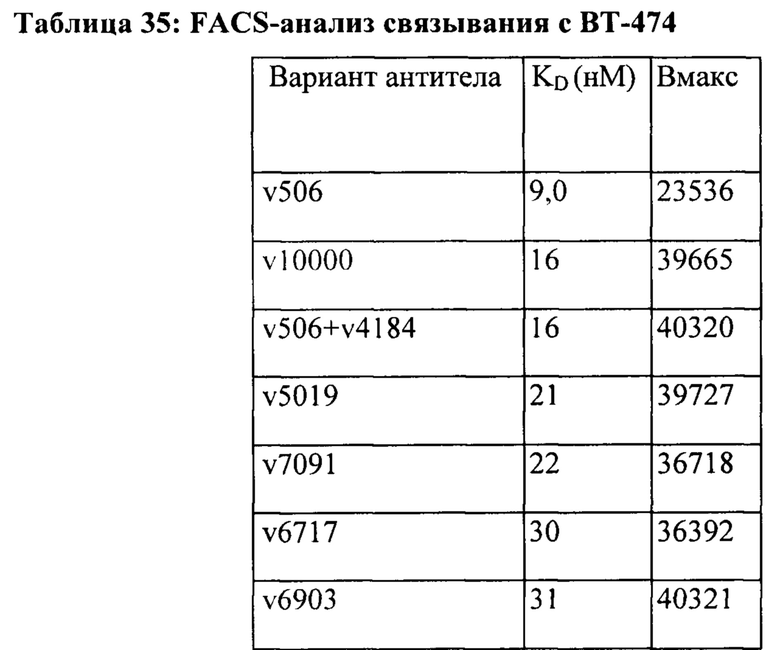
[00534] На Фигуре 41В и в Таблице 36 показаны результаты FACS-анализа по связыванию с клетками линии опухоли молочной железы JIMT-1 с уровнем экспрессии HER2 2+. Результаты показывают, что все антитела к HER2 имеют более высокое значение Вмакс (в 1,5-1,8 раза выше) по сравнению с моноспецифическим бивалентным антителом v506 к HER2. Значение Вмакс форматов Fab-scFv (v7091 и v10000) и Fab-Fab (v6903) повышалось приблизительно 1,7 раза, значение Вмакс формата scFv-scFv (v6717) повышалось в 1,5 раза и Fab-scFv (v5019) и значение Вмакс комбинации FSA (v506 + v4184) повышалось в 1.8 раза по сравнению с v506. Кажущаяся KD иллюстративных бипаратопных антител Fab-scFv к HER2 была приблизительно в 2-4 раза выше по сравнению с моноспецифическим v506; тогда как kD Fab-Fab (v6903) и scFv-scFv (v6717) была приблизительно в 8 раз выше по сравнению с v506.
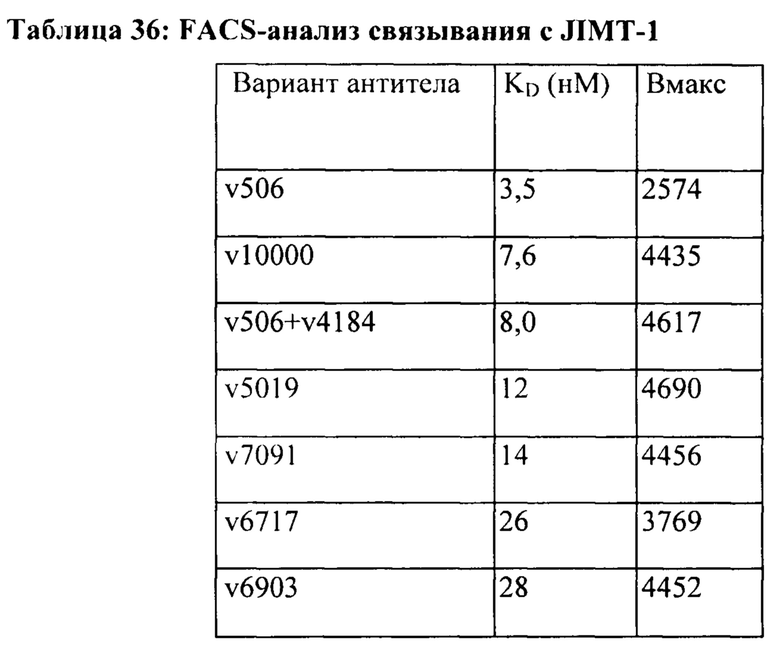
[00535] На Фигуре 41С и в Таблице 37 показаны результаты FACS-анализа по связыванию иллюстративных бипаратопных антител с клетками линии опухоли молочной железы MCF7 с уровнем экспрессии HER2 1+. Результаты показывают, что значение Вмакс антитела v10000 к HER2 и комбинации FSA (v506+v4184) было в 1,6 раза выше по сравнению с моноспецифическим бивалентным антителом v506 к HER2. Значение Вмакс Fab-scFv (v5019, v7091) повышалось приблизительно в 1,4 раза; формата scFv-scFv (v6717) - в 1,3 раза и формата Fab-Fab (v6903) - в 1,2 раза по сравнению с v506. Кажущаяся KD иллюстративного бипаратопного антитела к HER2 Fab-scFv, Fab-Fab (v6903) и комбинации FSA (v506+v4184) была приблизительно в 2-3 раза ниже по сравнению с v506; тогда как kD scFv-scFv (v6717) была приблизительно в 3 раза выше по сравнению с v506.
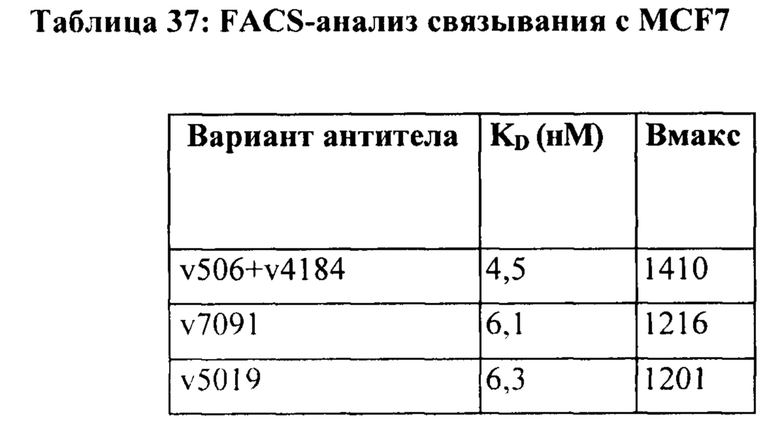

[00536] На Фигуре 41D и в Таблице 38 показаны результаты FACS-анализа по связыванию иллюстративных бипаратопных антител с клетками линии опухоли молочной железы MDA-MD-231 с уровнем экспрессии HER2 0/1+. Результаты показывают, что значение Вмакс иллюстративных бипаратопных антител к HER2 повышалось приблизительно в 1,3-1,4 раза по сравнению с моноспецифическим бивалентным антителом v506 к HER2. Значение Вмакс комбинации FSA (v506+v4184) повышалось в 1,7 раза. Кажущаяся KD иллюстративных бипаратопных антител Fab-scFv к HER2 (v5019, v7091, v10000) и комбинации FSA (v506+v4184) была приблизительно эквивалентной по сравнению с v506; тогда как KD Fab-Fab (v6903) и scFv-scFv (v6717) была приблизительно в 4 и 16 раза выше, соответственно, по сравнению с v506.
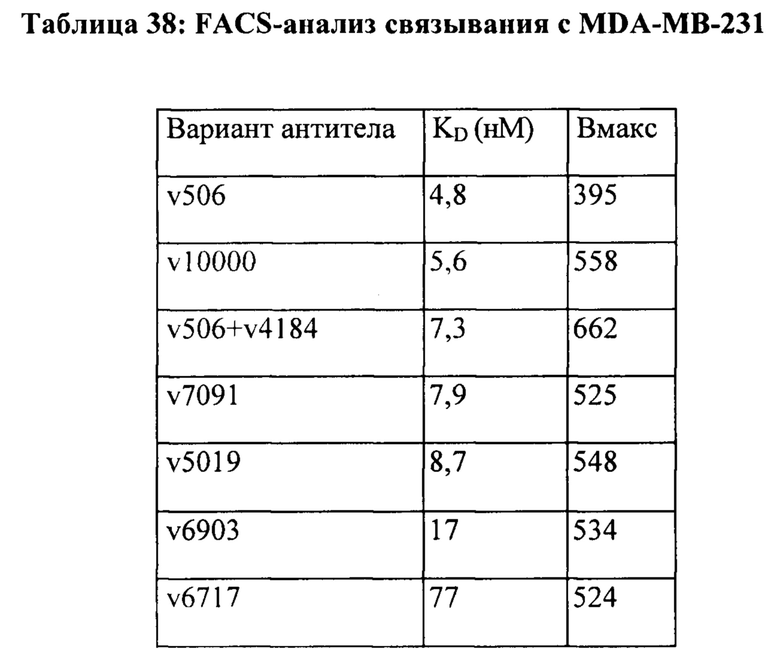
[00537] Результаты по связыванию с опухолевыми клетками показывают, что значение Вмакс бипаратопных антител к HER2 с разными форматами молекул выше при связывании с опухолевыми клетами с уровнем экспрессии HER2 3+, 2+, 1+ и 0/1+ по сравнению с бивалентным моноспецифическим антителом к HER2. Из разных бипаратопных антител к HER2 формат scFv-scFv обладает самым низким значением Вмакс по сравнению с v506 для опухолевых клеток с уровнем экспрессии HER2 3+, 2+, 1+ и 0/1+. Эти результаты также показывают, что KD форматов scFv-scFv и Fab-Fab повышается в наибольшей степени при связывании с опухолевыми клетками с уровнем экспрессии HER2 3+, 2+, 1+ и 0/1+ по сравнению с моноспецифическим v506 (повышение в 3-16 раз) и форматами бипаратопных антител Fab-scFv (повышение приблизительно в 2 раза или больше). Увеличение KD является признаком уменьшения авидности связывания и предполагает, что разные бипаратопные форматы характеризуются уникальными механизмамии связывания с HER2 на клеточной поверхности.
Пример 40: эффект формата бипаратопного антитела к HER2 на интернализацию клеток HER2+
[00538] Следующий эксперимент осуществляли для сравнения способности иллюстративных бипаратопных антител к ECD2 HER2 × ECD4, которые имеют разный формат молекул (например, v6717, scFv-scFv IgG1; v6903 и v6902 Fab-Fab IgG1: v5019, v7091 и v10000 Fab-scFv IgG1), к интернализации в клетки HER2+, экспрессирующие HER2 при различных уровнях.
[00539] Эксперимент осуществляли, как подробно описано в Примере 9. Результаты показаны на Фигуре 42 и Таблицах 39-41. На Фигуре 42А и в Таблице 39 показаны результаты по интернализации в клетки ВТ-474 с уровнем экспрессии HER2 3+. Эти результаты показывают, что внутриклеточное количество антител формата Fab-scFv (v10000) и комбинации FSA (v506+v4184) в 2,2 раза выше по сравнению с моноспецифическим антителом v506 к HER2. Внутриклеточное количество антител формата scFv-scFv (v6717) в 1,9 раза выше; антител форматов Fab-scFv (v5019 и v7091) в 1,5-1,7 раза выше; и количество накопленных внутри клетки антител форматов Fab-Fab (v6902 и v6903) в 1,2-1,3 раза выше по сравнению с v506.
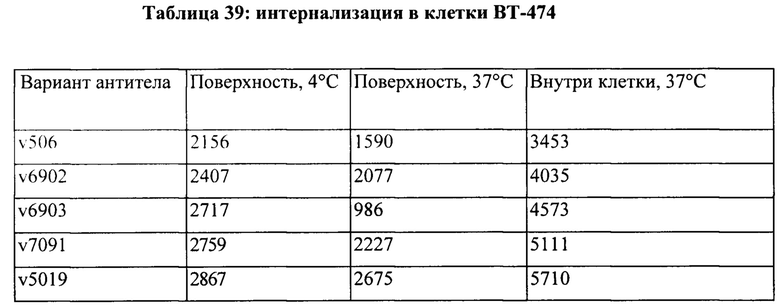

[00540] На Фигуре 42В и в Таблице 40 показаны результаты по интернализации в клетки JIMT-1 с уровнем экспрессии HER2 2+. Эти результаты показывают, что внутриклеточное количество антител формата Fab-scFv (v10000) и комбинации FSA (v506+v4184), соответственно, в 1,8 и 1,9 раза выше по сравнению с моноспецифическим антителом v506 к HER2. Внутриклеточное количество антител форматов scFv-scFv (v6717) и Fab-scFv (v5019) в 1,4 раза выше; и количество накопленных внутри клетки антител форматов Fab-scFv (v7091) и Fab-Fab (v6902 и v6903) в 1,2 раза выше по сравнению с v506.
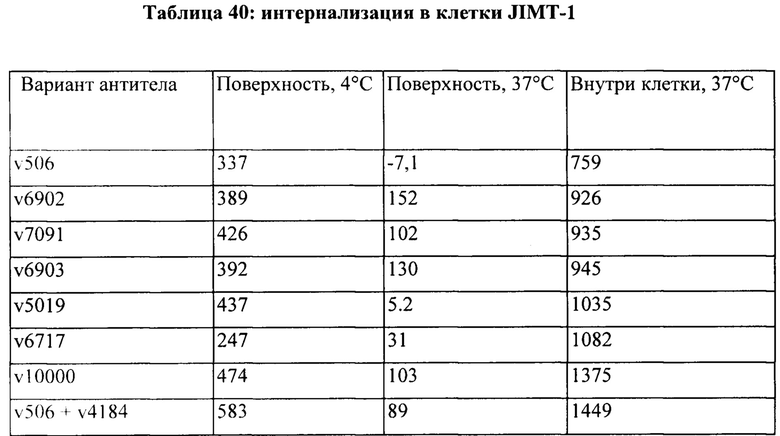
[00541] Фигура 42С и в Таблице 41 показаны результаты по интернализации клетки MCF7 с уровнем экспрессии HER2 1+. Эти результаты показывают, что внутриклеточное количество антител формата scFv-scFv и форматов Fab-scFv в 3,0 и 2,8 раза выше в сравнении с моноспецифическим антителом v506 к HER2. Внутриклеточное количество антител формата Fab-scFv (v10000) и комбинации FSA (v506+v4184) приблизительно в 2,0 раза выше; количество накопленных внутри клетки антител форматов Fab-scFv (v7091) и Fab-Fab (v6903) в 1,8 раза выше по сравнению с v506.

[00542] Эти результаты показывают, что бипаратопные антитела к HER2 с разными форматами молекул характеризуются уникальной степенью интернализации в опухолевые клетки с уровнем экспрессии HER2 3+, 2+ и 1+, которая изменяется в зависимости от структуры и формата антигенсвязывающих доменов. В целом, комбинация моноспецифических FSA v506 и v4184, Fab-scFv (v10000, v7091 и v5019) и бипаратопных форматов scFv-scFv (v6717) характеризуется более высокими значениями степени интернализации в опухолевые клетки с уровнем экспрессии HER2 3+, 2+ и 1+. В то же время бипаратопные форматы Fab-Fab (v6902 и v6903) характеризуется самыми низкими значениями степени интернализации в опухолевые клетки с уровнем экспрессии HER2 3+, 2+ и 1+. Эти данные предполагают, что формат молекулы и геометрическое расположение антигенсвязывающих доменов оказывает влияние на способность бипаратопных антител к поперечной сшивке HER2-рецепторов и впоследствии к интернализации в позитивные по HER2 опухолевые клетки. Бипаратопный формат Fab-Fab, характеризующийся наибольшим расстоянием между двумя антигенсвязывающими доменами, обеспечивал наиболее низкую степень интернализации, тогда как форматы Fab-scFv и scFv-scFv, характеризующиеся меньшими расстояниями между антигенсвязывающими доменами, обеспечивали большую степень интернализации в клетки HER2+. Это согласуется с корреляцией активности с меньшей длиной линкера, как описано в Jost et al. 2013, Structure 21, 1979-1991).
Пример 41: эффект формата бипаратопного антитела к HER2 на уничтожение клеток HER2+, путем АЗКЦ
[00543] Следующий эксперимент осуществляли для сравнения способности иллюстративных бипаратопных антител к ECD2 HER2 × ECD4, которые имеют разный формат молекул (например, v6717, scFv-scFv IgG1; v6903 и v6902 Fab-Fab IgG1; v5019, v7091 и v10000 Fab-scFv IgG1), опосредовать уничтожение клеток HER2+, путем АЗКЦ, экспрессирующих HER2 при различных уровнях.
[00544] Для количественной оценки содержания фукозы в N-связанном гликопептиде до осуществления АЗКЦ-анализа осуществляли анализ содержания гликопептидов на образцах антител. Способ осуществляли, как описано в Примере 23. Результаты показаны в Таблице 42; данные показывают, что в N-связанном гликане иллюстративных бипаратопных вариантов v5019, v6717, v6903 содержится эквивалентное количество фукозы (91-93%). Образцы антител с эквивалентными уровнями фукозы в N-гликане были отобраны для проведения АЗКЦ-анализа, чтобы нормализировать содержание фукозы при интерпретации результатов АЗКЦ-анализа.

[00545] Эксперимент по АЗКЦ осуществляли, как описано в Примере 11, причем соотношение Е/Т составляло 5:1 и в качестве эффекторных клеток были использованы NK-92. Результаты по АЗКЦ показаны на Фигуре 43 и Таблицах 43-45. На Фигуре 43А и в Таблице 43 показаны результаты по уничтожению опухолевых клеток молочной железы JIMT-1 с уровнем экспрессии HER2 2+ путем АЗКЦ. Эти данные показывают, что v5019, v6717 и v6903 вызывают максимальную степень лизиса клеток на похожем уровне, и что антитела в формате scFv-scFv (v6717) обладают меньшей активностью по сравнению с v5019 и v6903, если в качестве клеток-мишеней используются опухолевые клетки с уровнем экспрессии HER2 2+.
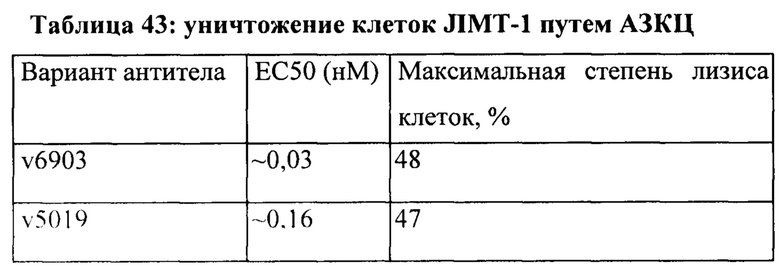

[00546] На Фигуре 43В и в Таблице 44 показаны результаты по уничтожению опухолевых клеток молочной железы MCF7 с уровнем экспрессии HER2 1+ путем АЗКЦ. Эти данные показывают, что v5019 и v6717 обуславливают немного более высокую максимальную степень лизиса клеток (27-30%) по сравнению с v6903 (24%). Эти данные также показывают, что v6717 обладает меньшей активностью, за ним следует v6903 и v5019, которые имеют более низкие значения ЕС50.
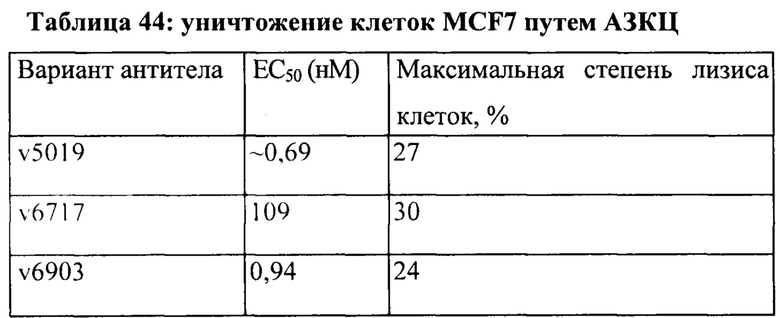
[00547] На Фигуре 43С и в Таблице 45 показаны результаты по по уничтожению опухолевых клеток молочной железы MDA-MB-231 с уровнем экспрессии HER2 0/1+ путем АЗКЦ. Эти данные показывают, что v5019 обуславливает немного более высокую максимальную степень лизиса клеток (77%) по сравнению с v6903 (62%) и v6717 (63%). Эти данные также показывают, что v6717 обладает меньшей активностью, за ним следуют v6903 и v5019, которые имеют более низкие значения ЕС50.
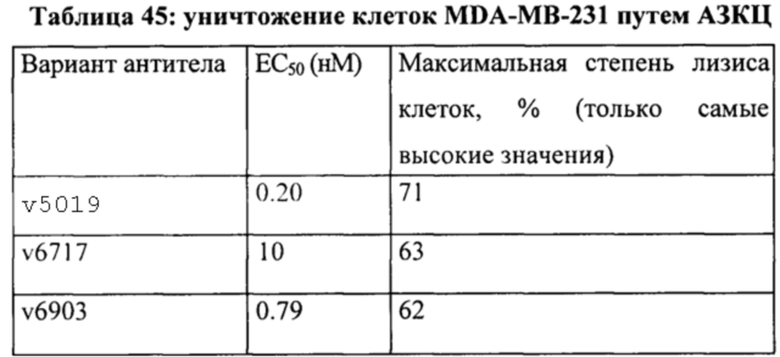
[00548] Эти данные показывают, что иллюстративные бипаратопное антитела к ECD2 HER2 × ECD4 вызывают максимальную степень лизиса опухолевых клеток с уровнем экспрессии HER2 2+ и 1+ на похожем уровне путем АЗКЦ. Несмотря на похожую максимальную степень лизиса клеток, эти данные также показывают, что молекулы разных форматов характеризуются уникальной АЗКЦ-активностью. ScFv-scFv обладал наименее выраженной активностью (наибольшие значения ЕС50) в клетках с уровнем экспрессии HER2 2+ и HER2 1+. Различная активность трех форматов наблюдалась в данных по АЗКЦ, направленной против клеток HER2 1+, где значения ЕС50 для располагаются в порядке v6717>v6903>v5019. Эти данные согласуются с данными наблюдений, которые представлены в Примере 40 (FACS-анализ связывания), где наблюдалось увеличение KD (уменьшение аффинности) при использовании форматов Fab-Fab и scFv-scFv.
Пример 42: эффект формата бипаратопного антитела к HER2 на рост опухолевых клеток HER2+
[00549] Следующий эксперимент осуществляли для сравнения эффекта формата бипаратопного антитела к HER2 на рост опухолевых клеток с уровнем экспрессии HER2 3+, 2+ и 1+, либо базальный рост, либо лиганд-стимулированный. Базальный рост оценивали, как описано в Примере 15, тогда как лиганд-стимулированный рост оценивали, как описано в Примере 27. В обоих типах экспериментов рост опухолевых клеток оценивали как % их выживаемости под воздействием контроля.
[00550] На Фигуре 44 и в Таблице 46 показан эффект иллюстративных бипаратопных антител к ECD2 HER2 × ECD4 на рост клеток рака молочной железы (HER2 3+) (ВТ-474) в присутствии экзогенных рост-стимулирующих лигандов (EGF и HRG). В отсутствие EGF или HRG бипаратопные антитела к HER2 были способны ингибировать рост клеток ВТ-474, причем процент выживаемости в зависимости от каждой группы воздействия располагался в следующем порядке: v6903<v506+v4184<506<v7091<v5019<v10000<v6717. В присутствии HRG ингибирование роста по сравнению с контрольным имитирующим образцом достигалось только при использовании комбинации FSA, состоящей из v506+v4184. В присутствии EGF достигалось ингибирование роста по сравнению с контрольным имитирующим образцом, причем каждая группа воздействия по показателю выживаемости (%) располагалась следующим образом: v6903<v506+v4184<709K v10000<5019.
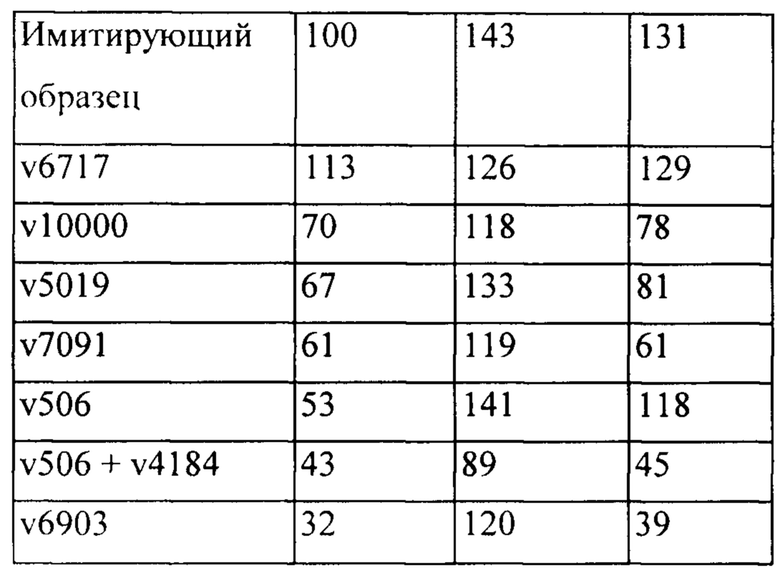
[00551] На Фигуре 45 показан дозозависимый эффект форматов бипаратопного антитела к HER2 на ингибирование роста клеток линии SKBr3 с уровнем экспрессии HER2 3+. Данные согласуются с результатами, представленными на Фигуре 44, где активность/эффективность бипаратопных форматов располагаются в следующем порядке Fab-Fab>Fab-scFv>scFv-scFv в опухолевых клетках HER2 3+.
[00552] Эффект форматов бипаратопных антител к HER2 на выживаемость клеток HER2+, показан на Фигуре 46, причем на Фигуре 46А показан результат, полученный на чувствительных к трастузумабу клетках линии SKOV3 с уровнем экспрессии HER2 2+/3+ при 300 нМ; на Фигуре 46В показан результат, полученный на (трастузумаб-резистентных) клетках JIMT-1 с уровнем экспрессии HER2 2+ при 300 нМ и на Фигуре 46С показан результат, полученный на клетках линии MCF7 с уровнем экспрессии HER2 1+ при 300 нМ. При использовании клеток линии SKOV3 наблюдались небольшие различия среди бипаратопных форматов по степени ингибирования роста, и никакого ингибирования роста не наблюдалось при использовании любого из исследуемых антител на клетках JIMT-1 и MCF7.
[00553] Данные на Фигуре 44 и Фигуре 45 показывают, что бипаратопные антитела к ECD2 HER2 × ECD4 с форматами Fab-scFv and Fab-Fab (v5019, v7091, v10000, v6903) способны ингибировать рост опухолевых клеток с уровнем экспрессии HER2 3+ в отсутствие и в присутствии EGF или HRG. В клеточных линиях ВТ-474 и SKBR3 с уровнем экспрессии HER2 3+ антитела располагались в порядке увеличения степени ингибирования роста по сравнению с контрольным имитирующим образцом следующим образом, где v506+v4184>v6903>v7091>v10000>v5019>v506>v6717. Расстояние между антигенсвязывающими доменами (Fab-Fab>Fab-scFv>scFv-scFv) коррелирует с порядком расположения антител по степени ингибирования роста в опухолевых клетках HER2 3+. На основе данных, полученных на чувствительных к трастузумабу опухолевых клетках ВТ-474 и SKBr3, можно предположить, что в разница в ингибировании роста среди форматов значительна на уровне экспрессии HER2 3+, но менее значительна на уровне экспрессии HER2 2+ или HER2 1+.
Пример 43: оценка аффинности и кинетики связывания с HER2 при различных уровнях захвата антител
[00554] Следующий эксперимент осуществляли для сравнения кинетики связывания с HER2 (kd, скорость диссоциации) иллюстративных бипаратопных антител к ECD2 HER2 × ECD4 при захвате при различных плотностях рецепторов на поверхности с помощью ППР. Корреляция между снижением (замедлением) скорости диссоциации с увеличением уровней захвата антител (плотность рецепторов на поверхности) является показателем транс-связывания (т.е. одна молекула антитела связывается с двумя молекулами HER2, как описано в Примере 12). В этом эксперименте формат Fab-Fab (v6903) сравнивали с форматом Fab-scFv (v7091), чтобы определить потенциальную разницу в транс-связывании среди вариантов. Из-за большего пространственного расстояния между антигенсвязывающими доменами, выдвигается гипотеза, согласно которой формат Fab-Fab может быть способен к цис-связыванию (вовлечение ECD 2 и 4 на одной молекуле HER2); в то же время Fab-scFv не способен к цис-связыванию из-за более короткого расстояния между его антигенсвязывающими доменами. Моноспецифическое антитело к HER2 v506 было включено в качестве контроля.
[00555] Эксперимент осуществляли с помощью ППР, как описано в Примере 12. Данные показаны на Фигуре 47. На Фигуре 47А показан график и результаты линейного регрессионного анализа для kd (1/с) при разных уровнях захвата антител при использовании v6903 и v7091. При использовании как v7091, так и v6093 наблюдалась тенденция к уменьшению скорости диссоциации с увеличением уровней захвата антител на поверхности; однако корреляция статистически значима лишь при использовании варианта Fab-scFv (v7091; Р-значение = 0,023), но не формата Fab-Fab (v6093; Р-значение = 0,053). Скорость диссоциации не изменялась при различных уровнях захвата антител для моноспецифического контрольного антитела v506 к HER2.
[00556] На Фигуре 47В показан график и результаты линейного регрессионного анализа для kD (М) при разных уровнях захвата антител при использовании v6903 и v7091. Подобно сравнению скорости диссоциации при использовании как v7091, так и v6093 наблюдалась тенденция к увеличению аффинности (более низкие значение KD) с увеличением уровней захвата антител на поверхности. Однако корреляция статистически значима лишь при использовании варианта Fab-scFv (v7091; Р-значение = 0,04), но не формата Fab-Fab (v6093; Р-значение = 0,51). KD не изменялась при различных уровнях захвата антител для моноспецифического контрольного антитела v506 к HER2. Данные на Фигуре 47 показывают, что при использовании форматов Fab-Fab и Fab-scFv бипаратопных антител к HER2 наблюдалась тенденция к меньшению скорости диссоциации с увеличением уровней захвата антител на поверхности; эти тенденции являются уникальными по сравнению с моноспецифическим антителом к Her2.
Пример 44: повышение аффинности и стабильности Fab-фрагмента пертузумаба генно-инженерным путем
[00557] Как указано в Таблице 1, один вариант (v10000) содержит мутации в Fab-фрагментс пертузумаба. Этот Fab-фрагмент был получен в результате повышения аффинности и стабильности генно-инженерным путем in silico, что оценивали экспериментальным путем в качестве моновалентных одноплечих антител (ОАА).
[00558] Вариант 9996: моновалентное антитело к HER2, причем НЕР2-связывающий домен представляет собой Fab-фрагмент полученный из цепи А пертузумаба, причем в VL-области содержится Y96A, и в VH-области содержится T30A/A49G/L69F (нумерация по Кабат), и Fc-область представляет собой гетеродимер, имеющий мутации T350V_L351Y_F405A_Y407V (EU-нумерация) в Цепи А, T350V_T366L_K392L_T394W (EU-нумерация) в Цепи В, и при этом в шарнирной области Цепи В имеется мутация C226S; антигенсвязывающий домен связывается с доменом 4 HER2.
[00559] Вариант 10014: моновалентное антитело к HER2, причем HER2-связывающий домен представляет собой Fab-фрагмент полученный из цепи А пертузумаба, причем в VL-области содержится Y96A, и VH-области содержится Т30А (нумерация по Кабат), и Fc-область представляет собой гетеродимер, имеющий мутации T350V_L351Y_F405A_Y407V (EU-нумерация) в Цепи А, T350V_T366L_K392L_T394W (EU-нумерация) в Цепи В, и при этом в шарнирной области Цепи В имеется мутация C226S; антигенсвязывающий домен связывается с доменом 4 HER2.
[00560] Вариант 10013: моновалентное антитело к HER2, причем HER2-связывающий домен представляет собой Fab-фрагмет, полученный из цепи А пертузумаба дикого типа, и Fc-область представляет собой гетеродимер, имеющий мутации T350V_L351Y_F405A_Y407V (EU-нумерация) в Цепи А, T350V_T366L_K392L_T394W (EU-нумерация) в Цепи В, и при этом в шарнирной области Цепи В имеется мутация C226S; антигенсвязывающий домен связывается с доменом 4 HER2.
[00561] Следующие эксперименты проводили для сравнения аффинности и стабильности связывания HER2 полученных генно-инженерным путем вариантов пертузумаба.
[00562] Варианты ОАА клонировали и экспрессировали, как описано в Примере 1.
[00563] ОАА очищали с помощью хроматографии на белке А и эксклюзионной хроматографии, как описано в Примере 1.
[00564] Содержание гетеродимеров (т.е., количество ОАА с гетеродимерным Fc-фрагментом) оценивали путем анализа высокоэффективной экспрессии белка в невосстанавливающих условиях с использованием Caliper LabCH1p GXII (Perkin Elmer №760499). Эксперименты проводили согласно руководству НТ Protein Express LabChip User Guide version2 LabChip GXII User Manual со следующими модификациями. Образцы гетеродимерных антител в концентрации либо 2 мкл, либо 5 мкл (диапазон концентраций 5-2000 нг/мкл) добавляли в отдельные лунки в 96-луночные планшеты (BioRad №HSP9601) наряду с 7 мкл буфера НТ Protein Express Sample Buffer (Perkin Elmer №760328). Образцы гетеродимерных антител затем подвергали денатурации при 70°С в течение 15 минут. Прибор LabChip функционирует с использованием чипа НТ Protein Express Chip (Perkin Elmer №760499) и условий анализа Ab-200. После использования чип промывали водой MilliQ и хранили при 4°С.
[00565] Стабильность образцов оценивали путем измерения температуры плавления или Тпл, определенной с помощью ДСК согласно протоколу, показанному в Примере 24. Измерения с помощью ДСК проводили до и после ЭХ-очистки.
[00566] Аффинность образцов по отношению к HER2 ECD измеряли с помощью ППР по протоколу из примера 12. Измерения с помощью ППР проводили до и после ЭХ-очистки. Как приведено в Таблице 47А и 47В, мутации в вариабельном домене обеспечивают повышение аффинности HER2 Fab-фрагмента по отношению к по сравнению с молекулой пертузумаба дикого типа с сохранением WT стабильности, присущей молекуле дикого типа. (1Содержание, определенное с помощью Caliper LabChip; 2KD(дикий тип)/RD(с мутацией)
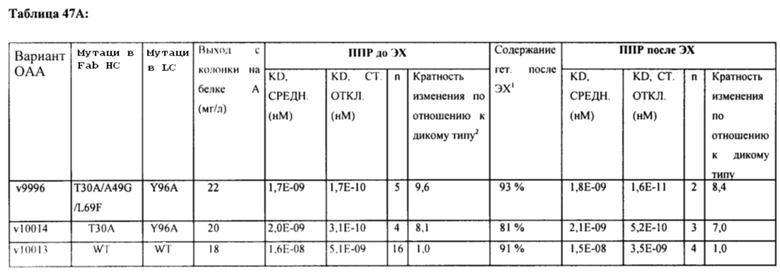
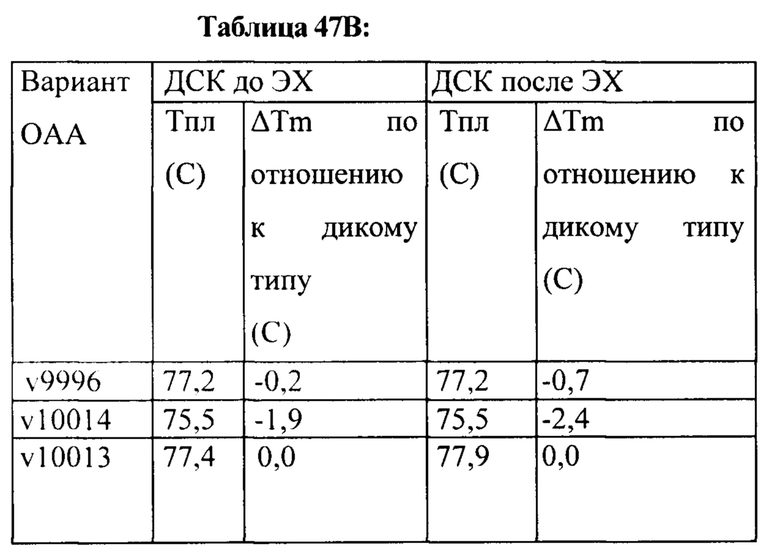
[00567] Реактивы, использованные в примерах, как правило, коммерчески доступны или могут быть получены с использованием коммерчески доступного оборудования, способов или реактивов, известных в данной области техники. Вышеизложенные примеры иллюстрируют различные аспекты, описанные в данном документе, и практическое воплощение способов, описанных в данном документе. Указанные примеры не предназначены для предоставления исчерпывающего описания многих разных вариантов реализации настоящего изобретения. Таким образом, несмотря на то, что вышеизложенное изобретение, которое было довольно подробно описано с иллюстративной целью, и примеры, описанные для ясности толкования, специалистам в данной области техники совершенно понятно, что многие изменения и модификации могут быть внесены, не отклоняясь от сущности и объема прилагаемой формулы изобретения.
[00568] Все публикации, патенты и патентные заявки, упомянутые в данном описании, включены в данное описание посредством ссылки в той же степени, как если бы каждая отдельная публикация, патент или патентная заявка были определенным образом и отдельно включены в данный документ посредством ссылки.
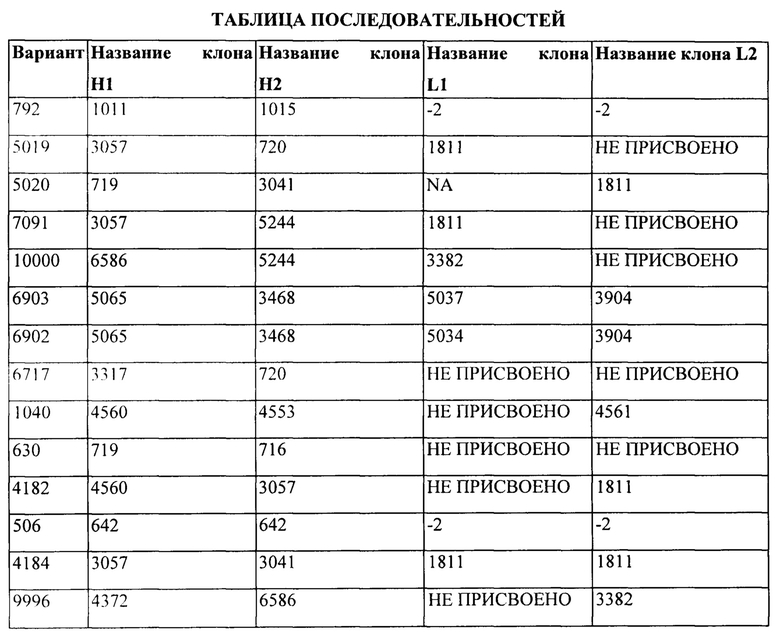













Вариант CDR-L3 пертузумаба: QQYYIPAT
Клон 3382, вариант 10000 (SEQ ID NO: 347)
Вариант CDR-H1 пертузумаба: GFTFADYT
Клон 6586. вариант 10000 (SEQ ID NO: 348)
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ЗАЙМВОРКС ИНК.
<120> БИСПЕЦИФИЧЕСКИЕ АНТИГЕНСВЯЗЫВАЮЩИЕ КОНСТРУКЦИИ ПРОТИВ HER2
<130> 30712-28087/PCT
<140> PCT/CA2014/051140
<141> 2014-11-27
<150> 62/009,125
<151> 2014-06-06
<150> 62/000,908
<151> 2014-05-20
<150> 61/910,026
<151> 2013-11-27
<160> 350
<170> PatentIn version 3.5
<210> 1
<211> 450
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 1
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 2
<211> 1350
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 2
gaggtgcagc tggtggaaag cggaggagga ctggtgcagc caggaggatc tctgcgactg 60
agttgcgccg cttcaggatt caacatcaag gacacctaca ttcactgggt gcgacaggct 120
ccaggaaaag gactggagtg ggtggctcga atctatccca ctaatggata cacccggtat 180
gccgactccg tgaaggggag gtttactatt agcgccgata catccaaaaa cactgcttac 240
ctgcagatga acagcctgcg agccgaagat accgctgtgt actattgcag tcgatgggga 300
ggagacggat tctacgctat ggattattgg ggacagggga ccctggtgac agtgagctcc 360
gcctctacca agggccccag tgtgtttccc ctggctcctt ctagtaaatc cacctctgga 420
gggacagccg ctctgggatg tctggtgaag gactatttcc ccgagcctgt gaccgtgagt 480
tggaactcag gcgccctgac aagcggagtg cacacttttc ctgctgtgct gcagtcaagc 540
gggctgtact ccctgtcctc tgtggtgaca gtgccaagtt caagcctggg cacacagact 600
tatatctgca acgtgaatca taagccctca aatacaaaag tggacaagaa agtggagccc 660
aagagctgtg ataagaccca cacctgccct ccctgtccag ctccagaact gctgggagga 720
cctagcgtgt tcctgtttcc ccctaagcca aaagacactc tgatgatttc caggactccc 780
gaggtgacct gcgtggtggt ggacgtgtct cacgaggacc ccgaagtgaa gttcaactgg 840
tacgtggatg gcgtggaagt gcataatgct aagacaaaac caagagagga acagtacaac 900
tccacttatc gcgtcgtgag cgtgctgacc gtgctgcacc aggactggct gaacgggaag 960
gagtataagt gcaaagtcag taataaggcc ctgcctgctc caatcgaaaa aaccatctct 1020
aaggccaaag gccagccaag ggagccccag gtgtacacac tgccacccag cagagacgaa 1080
ctgaccaaga accaggtgtc cctgacatgt ctggtgaaag gcttctatcc tagtgatatt 1140
gctgtggagt gggaatcaaa tggacagcca gagaacaatt acaagaccac acctccagtg 1200
ctggacagcg atggcagctt cttcctgtat tccaagctga cagtggataa atctcgatgg 1260
cagcagggga acgtgtttag ttgttcagtg atgcatgaag ccctgcacaa tcattacact 1320
cagaagagcc tgtccctgtc tcccggcaaa 1350
<210> 3
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 3
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 4
<211> 360
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 4
gaggtgcagc tggtggaaag cggaggagga ctggtgcagc caggaggatc tctgcgactg 60
agttgcgccg cttcaggatt caacatcaag gacacctaca ttcactgggt gcgacaggct 120
ccaggaaaag gactggagtg ggtggctcga atctatccca ctaatggata cacccggtat 180
gccgactccg tgaaggggag gtttactatt agcgccgata catccaaaaa cactgcttac 240
ctgcagatga acagcctgcg agccgaagat accgctgtgt actattgcag tcgatgggga 300
ggagacggat tctacgctat ggattattgg ggacagggga ccctggtgac agtgagctcc 360
<210> 5
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 5
Gly Phe Asn Ile Lys Asp Thr Tyr
1 5
<210> 6
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 6
ggattcaaca tcaaggacac ctac 24
<210> 7
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 7
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr
1 5 10
<210> 8
<211> 39
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 8
agtcgatggg gaggagacgg attctacgct atggattat 39
<210> 9
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 9
Ile Tyr Pro Thr Asn Gly Tyr Thr
1 5
<210> 10
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 10
atctatccca ctaatggata cacc 24
<210> 11
<211> 98
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 11
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val
<210> 12
<211> 294
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 12
gcctctacca agggccccag tgtgtttccc ctggctcctt ctagtaaatc cacctctgga 60
gggacagccg ctctgggatg tctggtgaag gactatttcc ccgagcctgt gaccgtgagt 120
tggaactcag gcgccctgac aagcggagtg cacacttttc ctgctgtgct gcagtcaagc 180
gggctgtact ccctgtcctc tgtggtgaca gtgccaagtt caagcctggg cacacagact 240
tatatctgca acgtgaatca taagccctca aatacaaaag tggacaagaa agtg 294
<210> 13
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 13
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
100 105 110
<210> 14
<211> 330
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 14
gctccagaac tgctgggagg acctagcgtg ttcctgtttc cccctaagcc aaaagacact 60
ctgatgattt ccaggactcc cgaggtgacc tgcgtggtgg tggacgtgtc tcacgaggac 120
cccgaagtga agttcaactg gtacgtggat ggcgtggaag tgcataatgc taagacaaaa 180
ccaagagagg aacagtacaa ctccacttat cgcgtcgtga gcgtgctgac cgtgctgcac 240
caggactggc tgaacgggaa ggagtataag tgcaaagtca gtaataaggc cctgcctgct 300
ccaatcgaaa aaaccatctc taaggccaaa 330
<210> 15
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 15
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
65 70 75 80
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 16
<211> 318
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 16
ggccagccaa gggagcccca ggtgtacaca ctgccaccca gcagagacga actgaccaag 60
aaccaggtgt ccctgacatg tctggtgaaa ggcttctatc ctagtgatat tgctgtggag 120
tgggaatcaa atggacagcc agagaacaat tacaagacca cacctccagt gctggacagc 180
gatggcagct tcttcctgta ttccaagctg acagtggata aatctcgatg gcagcagggg 240
aacgtgttta gttgttcagt gatgcatgaa gccctgcaca atcattacac tcagaagagc 300
ctgtccctgt ctcccggc 318
<210> 17
<211> 448
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 17
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Asp Tyr
20 25 30
Thr Met Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asp Val Asn Pro Asn Ser Gly Gly Ser Ile Tyr Asn Gln Arg Phe
50 55 60
Lys Gly Arg Phe Thr Leu Ser Val Asp Arg Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Leu Gly Pro Ser Phe Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Lys Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Val
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Leu
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Leu Thr Trp Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 18
<211> 1344
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 18
gaagtgcagc tggtcgaatc tggaggagga ctggtgcagc caggagggtc cctgcgcctg 60
tcttgcgccg ctagtggctt cacttttacc gactacacca tggattgggt gcgacaggca 120
cctggaaagg gcctggagtg ggtcgccgat gtgaacccaa atagcggagg ctccatctac 180
aaccagcggt tcaagggccg gttcaccctg tcagtggacc ggagcaaaaa caccctgtat 240
ctgcagatga atagcctgcg agccgaagat actgctgtgt actattgcgc ccggaatctg 300
gggccctcct tctactttga ctattggggg cagggaactc tggtcaccgt gagctccgcc 360
tccaccaagg gaccttctgt gttcccactg gctccctcta gtaaatccac atctggggga 420
actgcagccc tgggctgtct ggtgaagggc tacttcccag agcccgtcac agtgtcttgg 480
aacagtggcg ctctgacttc tggggtccac acctttcctg cagtgctgaa gtcaagcggg 540
ctgtacagcc tgtcctctgt ggtcaccgtg ccaagttcaa gcctgggaac acagacttat 600
atctgcaacg tgaatcacaa gccatccaat acaaaagtcg acaagaaagt ggaacccaag 660
tcttgtgata aaacccatac atgcccccct tgtcctgcac cagagctgct gggaggacca 720
agcgtgttcc tgtttccacc caagcctaaa gatacactga tgattagtag gaccccagaa 780
gtcacatgcg tggtcgtgga cgtgagccac gaggaccccg aagtcaagtt taactggtac 840
gtggacggcg tcgaggtgca taatgccaag actaaaccca gggaggaaca gtacaacagt 900
acctatcgcg tcgtgtcagt cctgacagtg ctgcatcagg attggctgaa cgggaaagag 960
tataagtgca aagtgagcaa taaggctctg cccgcaccta tcgagaaaac aatttccaag 1020
gcaaaaggac agcctagaga accacaggtg tacgtgctgc ctccatcaag ggatgagctg 1080
acaaagaacc aggtcagcct gctgtgtctg gtgaaaggat tctatccctc tgacattgct 1140
gtggagtggg aaagtaatgg ccagcctgag aacaattacc tgacctggcc ccctgtgctg 1200
gactcagatg gcagcttctt tctgtatagc aagctgaccg tcgacaaatc ccggtggcag 1260
caggggaatg tgtttagttg ttcagtcatg cacgaggcac tgcacaacca ttacacccag 1320
aagtcactgt cactgtcacc aggg 1344
<210> 19
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 19
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Asp Tyr
20 25 30
Thr Met Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asp Val Asn Pro Asn Ser Gly Gly Ser Ile Tyr Asn Gln Arg Phe
50 55 60
Lys Gly Arg Phe Thr Leu Ser Val Asp Arg Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Leu Gly Pro Ser Phe Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 20
<211> 357
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 20
gaagtgcagc tggtcgaatc tggaggagga ctggtgcagc caggagggtc cctgcgcctg 60
tcttgcgccg ctagtggctt cacttttacc gactacacca tggattgggt gcgacaggca 120
cctggaaagg gcctggagtg ggtcgccgat gtgaacccaa atagcggagg ctccatctac 180
aaccagcggt tcaagggccg gttcaccctg tcagtggacc ggagcaaaaa caccctgtat 240
ctgcagatga atagcctgcg agccgaagat actgctgtgt actattgcgc ccggaatctg 300
gggccctcct tctactttga ctattggggg cagggaactc tggtcaccgt gagctcc 357
<210> 21
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 21
Gly Phe Thr Phe Thr Asp Tyr Thr
1 5
<210> 22
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 22
ggcttcactt ttaccgacta cacc 24
<210> 23
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 23
Ala Arg Asn Leu Gly Pro Ser Phe Tyr Phe Asp Tyr
1 5 10
<210> 24
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 24
gcccggaatc tggggccctc cttctacttt gactat 36
<210> 25
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 25
Val Asn Pro Asn Ser Gly Gly Ser
1 5
<210> 26
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 26
gtgaacccaa atagcggagg ctcc 24
<210> 27
<211> 98
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 27
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Gly Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Lys Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val
<210> 28
<211> 294
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 28
gcctccacca agggaccttc tgtgttccca ctggctccct ctagtaaatc cacatctggg 60
ggaactgcag ccctgggctg tctggtgaag ggctacttcc cagagcccgt cacagtgtct 120
tggaacagtg gcgctctgac ttctggggtc cacacctttc ctgcagtgct gaagtcaagc 180
gggctgtaca gcctgtcctc tgtggtcacc gtgccaagtt caagcctggg aacacagact 240
tatatctgca acgtgaatca caagccatcc aatacaaaag tcgacaagaa agtg 294
<210> 29
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 29
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
100 105 110
<210> 30
<211> 330
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 30
gcaccagagc tgctgggagg accaagcgtg ttcctgtttc cacccaagcc taaagataca 60
ctgatgatta gtaggacccc agaagtcaca tgcgtggtcg tggacgtgag ccacgaggac 120
cccgaagtca agtttaactg gtacgtggac ggcgtcgagg tgcataatgc caagactaaa 180
cccagggagg aacagtacaa cagtacctat cgcgtcgtgt cagtcctgac agtgctgcat 240
caggattggc tgaacgggaa agagtataag tgcaaagtga gcaataaggc tctgcccgca 300
cctatcgaga aaacaatttc caaggcaaaa 330
<210> 31
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 31
Gly Gln Pro Arg Glu Pro Gln Val Tyr Val Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Thr Lys Asn Gln Val Ser Leu Leu Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Leu Thr Trp Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
65 70 75 80
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 32
<211> 318
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 32
ggacagccta gagaaccaca ggtgtacgtg ctgcctccat caagggatga gctgacaaag 60
aaccaggtca gcctgctgtg tctggtgaaa ggattctatc cctctgacat tgctgtggag 120
tgggaaagta atggccagcc tgagaacaat tacctgacct ggccccctgt gctggactca 180
gatggcagct tctttctgta tagcaagctg accgtcgaca aatcccggtg gcagcagggg 240
aatgtgttta gttgttcagt catgcacgag gcactgcaca accattacac ccagaagtca 300
ctgtcactgt caccaggg 318
<210> 33
<211> 214
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 33
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Ile Gly
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Tyr Ile Tyr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 34
<211> 642
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 34
gatattcaga tgacccagtc cccaagctcc ctgagtgcct cagtgggcga ccgagtcacc 60
atcacatgca aggcttccca ggatgtgtct attggagtcg catggtacca gcagaagcca 120
ggcaaagcac ccaagctgct gatctatagc gcctcctacc ggtataccgg cgtgccctct 180
agattctctg gcagtgggtc aggaacagac tttactctga ccatctctag tctgcagcct 240
gaggatttcg ctacctacta ttgccagcag tactatatct acccatatac ctttggccag 300
gggacaaaag tggagatcaa gaggactgtg gccgctccct ccgtcttcat ttttccccct 360
tctgacgaac agctgaaaag tggcacagcc agcgtggtct gtctgctgaa caatttctac 420
cctcgcgaag ccaaagtgca gtggaaggtc gataacgctc tgcagagcgg caacagccag 480
gagtctgtga ctgaacagga cagtaaagat tcaacctata gcctgtcaag cacactgact 540
ctgagcaagg cagactacga gaagcacaaa gtgtatgcct gcgaagtcac acatcagggg 600
ctgtcctctc ctgtgactaa gagctttaac agaggagagt gt 642
<210> 35
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 35
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Ile Gly
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Tyr Ile Tyr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 36
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 36
gatattcaga tgacccagtc cccaagctcc ctgagtgcct cagtgggcga ccgagtcacc 60
atcacatgca aggcttccca ggatgtgtct attggagtcg catggtacca gcagaagcca 120
ggcaaagcac ccaagctgct gatctatagc gcctcctacc ggtataccgg cgtgccctct 180
agattctctg gcagtgggtc aggaacagac tttactctga ccatctctag tctgcagcct 240
gaggatttcg ctacctacta ttgccagcag tactatatct acccatatac ctttggccag 300
gggacaaaag tggagatcaa g 321
<210> 37
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 37
Gln Asp Val Ser Ile Gly
1 5
<210> 38
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 38
caggatgtgt ctattgga 18
<210> 39
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 39
Gln Gln Tyr Tyr Ile Tyr Pro Tyr Thr
1 5
<210> 40
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 40
cagcagtact atatctaccc atatacc 27
<210> 41
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 41
Ser Ala Ser
1
<210> 42
<211> 9
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 42
agcgcctcc 9
<210> 43
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 43
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 44
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 44
aggactgtgg ccgctccctc cgtcttcatt tttccccctt ctgacgaaca gctgaaaagt 60
ggcacagcca gcgtggtctg tctgctgaac aatttctacc ctcgcgaagc caaagtgcag 120
tggaaggtcg ataacgctct gcagagcggc aacagccagg agtctgtgac tgaacaggac 180
agtaaagatt caacctatag cctgtcaagc acactgactc tgagcaaggc agactacgag 240
aagcacaaag tgtatgcctg cgaagtcaca catcaggggc tgtcctctcc tgtgactaag 300
agctttaaca gaggagagtg t 321
<210> 45
<211> 222
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 45
Asp Tyr Lys Asp Asp Asp Asp Lys Asp Ile Gln Met Thr Gln Ser Pro
1 5 10 15
Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg
20 25 30
Ala Ser Gln Asp Val Asn Thr Ala Val Ala Trp Tyr Gln Gln Lys Pro
35 40 45
Gly Lys Ala Pro Lys Leu Leu Ile Tyr Ser Ala Ser Phe Leu Tyr Ser
50 55 60
Gly Val Pro Ser Arg Phe Ser Gly Ser Arg Ser Gly Thr Asp Phe Thr
65 70 75 80
Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
85 90 95
Gln Gln His Tyr Thr Thr Pro Pro Thr Phe Gly Gln Gly Thr Lys Val
100 105 110
Glu Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro
115 120 125
Ser Asp Glu Arg Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu
130 135 140
Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn
145 150 155 160
Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser
165 170 175
Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala
180 185 190
Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly
195 200 205
Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215 220
<210> 46
<211> 666
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 46
gactacaaag acgacgatga caaagatatc cagatgaccc agtcccctag ctccctgtcc 60
gcttctgtgg gcgatagggt cactattacc tgccgcgcat ctcaggacgt gaacaccgca 120
gtcgcctggt accagcagaa gcctgggaaa gctccaaagc tgctgatcta cagtgcatca 180
ttcctgtatt caggagtgcc cagccggttt agcggcagca gatctggcac cgatttcaca 240
ctgactattt ctagtctgca gcctgaggac tttgccacat actattgcca gcagcactat 300
accacacccc ctactttcgg ccaggggacc aaagtggaga tcaagcgaac tgtggccgct 360
ccaagtgtct tcatttttcc acccagcgat gaaagactga agtccggcac agcttctgtg 420
gtctgtctgc tgaacaattt ttaccccaga gaggccaaag tgcagtggaa ggtcgacaac 480
gctctgcaga gtggcaacag ccaggagagc gtgacagaac aggattccaa agactctact 540
tatagtctgt caagcaccct gacactgagc aaggcagact acgaaaagca taaagtgtat 600
gcctgtgagg tcacacatca ggggctgtca tcaccagtca ccaaatcatt caatcggggg 660
gagtgc 666
<210> 47
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 47
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 48
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 48
gatatccaga tgacccagtc ccctagctcc ctgtccgctt ctgtgggcga tagggtcact 60
attacctgcc gcgcatctca ggacgtgaac accgcagtcg cctggtacca gcagaagcct 120
gggaaagctc caaagctgct gatctacagt gcatcattcc tgtattcagg agtgcccagc 180
cggtttagcg gcagcagatc tggcaccgat ttcacactga ctatttctag tctgcagcct 240
gaggactttg ccacatacta ttgccagcag cactatacca caccccctac tttcggccag 300
gggaccaaag tggagatcaa g 321
<210> 49
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 49
Gln Asp Val Asn Thr Ala
1 5
<210> 50
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 50
caggacgtga acaccgca 18
<210> 51
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 51
Gln Gln His Tyr Thr Thr Pro Pro Thr
1 5
<210> 52
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 52
cagcagcact ataccacacc ccctact 27
<210> 53
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 53
Ser Ala Ser
1
<210> 54
<211> 9
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 54
agtgcatca 9
<210> 55
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 55
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Arg Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 56
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 56
cgaactgtgg ccgctccaag tgtcttcatt tttccaccca gcgatgaaag actgaagtcc 60
ggcacagctt ctgtggtctg tctgctgaac aatttttacc ccagagaggc caaagtgcag 120
tggaaggtcg acaacgctct gcagagtggc aacagccagg agagcgtgac agaacaggat 180
tccaaagact ctacttatag tctgtcaagc accctgacac tgagcaaggc agactacgaa 240
aagcataaag tgtatgcctg tgaggtcaca catcaggggc tgtcatcacc agtcaccaaa 300
tcattcaatc ggggggagtg c 321
<210> 57
<211> 222
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 57
Asp Tyr Lys Asp Asp Asp Asp Lys Asp Ile Gln Met Thr Gln Ser Pro
1 5 10 15
Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg
20 25 30
Ala Ser Gln Asp Val Asn Thr Ala Val Ala Trp Tyr Gln Gln Lys Pro
35 40 45
Gly Lys Ala Pro Lys Leu Leu Ile Tyr Ser Ala Ser Phe Leu Tyr Ser
50 55 60
Gly Val Pro Ser Arg Phe Ser Gly Ser Arg Ser Gly Thr Asp Phe Thr
65 70 75 80
Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
85 90 95
Gln Gln His Tyr Thr Thr Pro Pro Thr Phe Gly Gln Gly Thr Lys Val
100 105 110
Glu Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro
115 120 125
Ser Asp Glu Arg Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu
130 135 140
Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn
145 150 155 160
Ala Leu Gln Ser Gly Asn Ser Lys Glu Ser Val Thr Glu Gln Asp Ser
165 170 175
Lys Asp Ser Thr Tyr Ser Leu Ser Ser Arg Leu Thr Leu Ser Lys Ala
180 185 190
Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly
195 200 205
Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215 220
<210> 58
<211> 666
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 58
gactacaaag acgacgatga caaagatatc cagatgaccc agtcccctag ctccctgtcc 60
gcttctgtgg gcgatagggt cactattacc tgccgcgcat ctcaggacgt gaacaccgca 120
gtcgcctggt accagcagaa gcctgggaaa gctccaaagc tgctgatcta cagtgcatca 180
ttcctgtatt caggagtgcc cagccggttt agcggcagca gatctggcac cgatttcaca 240
ctgactattt ctagtctgca gcctgaggac tttgccacat actattgcca gcagcactat 300
accacacccc ctactttcgg ccaggggacc aaagtggaga tcaagcgaac tgtggccgct 360
ccaagtgtct tcatttttcc acccagcgat gaaagactga agtccggcac agcttctgtg 420
gtctgtctgc tgaacaattt ttaccccaga gaggccaaag tgcagtggaa ggtcgacaac 480
gctctgcaga gtggcaacag caaggagagc gtgacagaac aggattccaa agactctact 540
tatagtctgt caagcagact gacactgagc aaggcagact acgaaaagca taaagtgtat 600
gcctgtgagg tcacacatca ggggctgtca tcaccagtca ccaaatcatt caatcggggg 660
gagtgc 666
<210> 59
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 59
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 60
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 60
gatatccaga tgacccagtc ccctagctcc ctgtccgctt ctgtgggcga tagggtcact 60
attacctgcc gcgcatctca ggacgtgaac accgcagtcg cctggtacca gcagaagcct 120
gggaaagctc caaagctgct gatctacagt gcatcattcc tgtattcagg agtgcccagc 180
cggtttagcg gcagcagatc tggcaccgat ttcacactga ctatttctag tctgcagcct 240
gaggactttg ccacatacta ttgccagcag cactatacca caccccctac tttcggccag 300
gggaccaaag tggagatcaa g 321
<210> 61
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 61
Gln Asp Val Asn Thr Ala
1 5
<210> 62
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 62
caggacgtga acaccgca 18
<210> 63
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 63
Gln Gln His Tyr Thr Thr Pro Pro Thr
1 5
<210> 64
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 64
cagcagcact ataccacacc ccctact 27
<210> 65
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 65
Ser Ala Ser
1
<210> 66
<211> 9
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 66
agtgcatca 9
<210> 67
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 67
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Arg Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Lys Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Arg Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 68
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 68
cgaactgtgg ccgctccaag tgtcttcatt tttccaccca gcgatgaaag actgaagtcc 60
ggcacagctt ctgtggtctg tctgctgaac aatttttacc ccagagaggc caaagtgcag 120
tggaaggtcg acaacgctct gcagagtggc aacagcaagg agagcgtgac agaacaggat 180
tccaaagact ctacttatag tctgtcaagc agactgacac tgagcaaggc agactacgaa 240
aagcataaag tgtatgcctg tgaggtcaca catcaggggc tgtcatcacc agtcaccaaa 300
tcattcaatc ggggggagtg c 321
<210> 69
<211> 214
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 69
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Ile Gly
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Tyr Ile Tyr Pro Ala
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 70
<211> 642
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 70
gatattcaga tgacccagtc cccaagctcc ctgagtgcct cagtgggcga ccgagtcacc 60
atcacatgca aggcttccca ggatgtgtct attggagtcg catggtacca gcagaagcca 120
ggcaaagcac ccaagctgct gatctatagc gcctcctacc ggtataccgg cgtgccctct 180
agattctctg gcagtgggtc aggaacagac tttactctga ccatctctag tctgcagcct 240
gaggatttcg ctacctacta ttgccagcag tactatatct acccagccac ctttggccag 300
gggacaaaag tggagatcaa gaggactgtg gccgctccct ccgtcttcat ttttccccct 360
tctgacgaac agctgaaaag tggcacagcc agcgtggtct gtctgctgaa caatttctac 420
cctcgcgaag ccaaagtgca gtggaaggtc gataacgctc tgcagagcgg caacagccag 480
gagtctgtga ctgaacagga cagtaaagat tcaacctata gcctgtcaag cacactgact 540
ctgagcaagg cagactacga gaagcacaaa gtgtatgcct gcgaagtcac acatcagggg 600
ctgtcctctc ctgtgactaa gagctttaac agaggagagt gt 642
<210> 71
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 71
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Ile Gly
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Tyr Ile Tyr Pro Ala
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 72
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 72
gatattcaga tgacccagtc cccaagctcc ctgagtgcct cagtgggcga ccgagtcacc 60
atcacatgca aggcttccca ggatgtgtct attggagtcg catggtacca gcagaagcca 120
ggcaaagcac ccaagctgct gatctatagc gcctcctacc ggtataccgg cgtgccctct 180
agattctctg gcagtgggtc aggaacagac tttactctga ccatctctag tctgcagcct 240
gaggatttcg ctacctacta ttgccagcag tactatatct acccagccac ctttggccag 300
gggacaaaag tggagatcaa g 321
<210> 73
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 73
Gln Asp Val Ser Ile Gly
1 5
<210> 74
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 74
caggatgtgt ctattgga 18
<210> 75
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 75
Gln Gln Tyr Tyr Ile Tyr Pro Ala Thr
1 5
<210> 76
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 76
cagcagtact atatctaccc agccacc 27
<210> 77
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 77
Ser Ala Ser
1
<210> 78
<211> 9
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 78
agcgcctcc 9
<210> 79
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 79
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 80
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 80
aggactgtgg ccgctccctc cgtcttcatt tttccccctt ctgacgaaca gctgaaaagt 60
ggcacagcca gcgtggtctg tctgctgaac aatttctacc ctcgcgaagc caaagtgcag 120
tggaaggtcg ataacgctct gcagagcggc aacagccagg agtctgtgac tgaacaggac 180
agtaaagatt caacctatag cctgtcaagc acactgactc tgagcaaggc agactacgag 240
aagcacaaag tgtatgcctg cgaagtcaca catcaggggc tgtcctctcc tgtgactaag 300
agctttaaca gaggagagtg t 321
<210> 81
<211> 449
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 81
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Glu Val Thr Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Val Tyr Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Ala Leu Val Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly
<210> 82
<211> 1347
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 82
gaggtgcagc tggtcgaaag cggaggagga ctggtgcagc caggagggtc actgcgactg 60
agctgcgcag cttccggctt caacatcaag gacacctaca ttcactgggt ccgccaggct 120
cctggaaaag gcctggagtg ggtggcacga atctatccaa ctaatggata cacccggtat 180
gccgactccg tgaagggccg gttcaccatt tctgcagata caagtaaaaa cactgcctac 240
ctgcagatga acagcctgcg agccgaagat acagccgtgt actattgcag ccgatgggga 300
ggcgacggct tctacgctat ggattattgg gggcagggaa ccctggtcac agtgagctcc 360
gcatcaacaa aggggcctag cgtgtttcca ctggccccct ctagtaaatc cacctctggg 420
ggaacagcag ccctgggatg tgaggtgacc gactacttcc cagagcccgt cactgtgagc 480
tggaactccg gcgccctgac atctggggtc catacttttc ctgctgtgct gcagtcaagc 540
ggcctgtaca gcctgtcctc tgtggtcact gtgccaagtt caagcctggg gactcagacc 600
tatatctgca acgtgaatca caagccatcc aataccaaag tcgacaagaa agtggaaccc 660
aagtcttgtg ataaaacaca tacttgcccc ccttgtcctg caccagagct gctgggagga 720
ccaagcgtgt tcctgtttcc acccaagcct aaagacaccc tgatgattag taggactcca 780
gaagtcacct gcgtggtcgt ggacgtgagc cacgaggacc ccgaagtcaa gttcaactgg 840
tacgtggatg gcgtcgaggt gcataatgcc aagacaaaac ccagggagga acagtacaac 900
tccacttatc gcgtcgtgtc tgtcctgacc gtgctgcacc aggactggct gaacggcaag 960
gagtataagt gcaaagtgag caataaggct ctgcccgcac ctatcgagaa aacaatttcc 1020
aaggctaaag ggcagcctag agaaccacag gtgtacgtgt accctccatc tagggacgag 1080
ctgaccaaga accaggtcag tctgacatgt ctggtgaaag ggttctatcc cagcgatatc 1140
gcagtggagt gggaatccaa tggacagcct gagaacaatt acaagaccac accccctgtg 1200
ctggactctg atggaagttt cgccctggtg agtaagctga ccgtcgataa atcacggtgg 1260
cagcagggca acgtgttcag ctgttcagtg atgcacgaag cactgcacaa ccactacacc 1320
cagaaaagcc tgtccctgtc ccccggc 1347
<210> 83
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 83
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 84
<211> 360
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 84
gaggtgcagc tggtcgaaag cggaggagga ctggtgcagc caggagggtc actgcgactg 60
agctgcgcag cttccggctt caacatcaag gacacctaca ttcactgggt ccgccaggct 120
cctggaaaag gcctggagtg ggtggcacga atctatccaa ctaatggata cacccggtat 180
gccgactccg tgaagggccg gttcaccatt tctgcagata caagtaaaaa cactgcctac 240
ctgcagatga acagcctgcg agccgaagat acagccgtgt actattgcag ccgatgggga 300
ggcgacggct tctacgctat ggattattgg gggcagggaa ccctggtcac agtgagctcc 360
<210> 85
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 85
Gly Phe Asn Ile Lys Asp Thr Tyr
1 5
<210> 86
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 86
ggcttcaaca tcaaggacac ctac 24
<210> 87
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 87
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr
1 5 10
<210> 88
<211> 39
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 88
agccgatggg gaggcgacgg cttctacgct atggattat 39
<210> 89
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 89
Ile Tyr Pro Thr Asn Gly Tyr Thr
1 5
<210> 90
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 90
atctatccaa ctaatggata cacc 24
<210> 91
<211> 98
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 91
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Glu Val Thr Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val
<210> 92
<211> 294
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 92
gcatcaacaa aggggcctag cgtgtttcca ctggccccct ctagtaaatc cacctctggg 60
ggaacagcag ccctgggatg tgaggtgacc gactacttcc cagagcccgt cactgtgagc 120
tggaactccg gcgccctgac atctggggtc catacttttc ctgctgtgct gcagtcaagc 180
ggcctgtaca gcctgtcctc tgtggtcact gtgccaagtt caagcctggg gactcagacc 240
tatatctgca acgtgaatca caagccatcc aataccaaag tcgacaagaa agtg 294
<210> 93
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 93
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
100 105 110
<210> 94
<211> 330
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 94
gcaccagagc tgctgggagg accaagcgtg ttcctgtttc cacccaagcc taaagacacc 60
ctgatgatta gtaggactcc agaagtcacc tgcgtggtcg tggacgtgag ccacgaggac 120
cccgaagtca agttcaactg gtacgtggat ggcgtcgagg tgcataatgc caagacaaaa 180
cccagggagg aacagtacaa ctccacttat cgcgtcgtgt ctgtcctgac cgtgctgcac 240
caggactggc tgaacggcaa ggagtataag tgcaaagtga gcaataaggc tctgcccgca 300
cctatcgaga aaacaatttc caaggctaaa 330
<210> 95
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 95
Gly Gln Pro Arg Glu Pro Gln Val Tyr Val Tyr Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Ala Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
65 70 75 80
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 96
<211> 318
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 96
gggcagccta gagaaccaca ggtgtacgtg taccctccat ctagggacga gctgaccaag 60
aaccaggtca gtctgacatg tctggtgaaa gggttctatc ccagcgatat cgcagtggag 120
tgggaatcca atggacagcc tgagaacaat tacaagacca caccccctgt gctggactct 180
gatggaagtt tcgccctggt gagtaagctg accgtcgata aatcacggtg gcagcagggc 240
aacgtgttca gctgttcagt gatgcacgaa gcactgcaca accactacac ccagaaaagc 300
ctgtccctgt cccccggc 318
<210> 97
<211> 448
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 97
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ala Asp Tyr
20 25 30
Thr Met Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Asp Val Asn Pro Asn Ser Gly Gly Ser Ile Tyr Asn Gln Arg Phe
50 55 60
Lys Gly Arg Phe Thr Phe Ser Val Asp Arg Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Leu Gly Pro Ser Phe Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Val
340 345 350
Tyr Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Ala Leu Val Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 98
<211> 1344
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 98
gaggtgcagc tggtggaatc aggagggggc ctggtgcagc ccggagggtc tctgcgactg 60
tcatgtgccg cttctgggtt cactttcgca gactacacaa tggattgggt gcgacaggcc 120
cccggaaagg gactggagtg ggtgggcgat gtcaacccta attctggcgg gagtatctac 180
aaccagcggt tcaaggggag attcactttt tcagtggaca gaagcaaaaa caccctgtat 240
ctgcagatga acagcctgag ggccgaagat accgctgtct actattgcgc tcgcaatctg 300
ggccccagtt tctactttga ctattggggg cagggaaccc tggtgacagt cagctccgct 360
agcactaagg ggccttccgt gtttccactg gctccctcta gtaaatccac ctctggaggc 420
acagctgcac tgggatgtct ggtgaaggat tacttccctg aaccagtcac agtgagttgg 480
aactcagggg ctctgacaag tggagtccat acttttcccg cagtgctgca gtcaagcgga 540
ctgtactccc tgtcctctgt ggtcaccgtg cctagttcaa gcctgggcac ccagacatat 600
atctgcaacg tgaatcacaa gccatcaaat acaaaagtcg acaagaaagt ggagcccaag 660
agctgtgata aaactcatac ctgcccacct tgtccggcgc cagaactgct gggaggacca 720
agcgtgttcc tgtttccacc caagcctaaa gacaccctga tgatttcccg gactcctgag 780
gtcacctgcg tggtcgtgga cgtgtctcac gaggaccccg aagtcaagtt caactggtac 840
gtggatggcg tcgaagtgca taatgccaag accaaacccc gggaggaaca gtacaactct 900
acctatagag tcgtgagtgt cctgacagtg ctgcaccagg actggctgaa tgggaaggag 960
tataagtgta aagtgagcaa caaagccctg cccgccccaa tcgaaaaaac aatctctaaa 1020
gcaaaaggac agcctcgcga accacaggtc tacgtctacc ccccatcaag agatgaactg 1080
acaaaaaatc aggtctctct gacatgcctg gtcaaaggat tctacccttc cgacatcgcc 1140
gtggagtggg aaagtaacgg ccagcccgag aacaattaca agaccacacc ccctgtcctg 1200
gactctgatg ggagtttcgc tctggtgtca aagctgaccg tcgataaaag ccggtggcag 1260
cagggcaatg tgtttagctg ctccgtcatg cacgaagccc tgcacaatca ctacacacag 1320
aagtccctga gcctgagccc tggc 1344
<210> 99
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 99
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ala Asp Tyr
20 25 30
Thr Met Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Asp Val Asn Pro Asn Ser Gly Gly Ser Ile Tyr Asn Gln Arg Phe
50 55 60
Lys Gly Arg Phe Thr Phe Ser Val Asp Arg Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Leu Gly Pro Ser Phe Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 100
<211> 357
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 100
gaggtgcagc tggtggaatc aggagggggc ctggtgcagc ccggagggtc tctgcgactg 60
tcatgtgccg cttctgggtt cactttcgca gactacacaa tggattgggt gcgacaggcc 120
cccggaaagg gactggagtg ggtgggcgat gtcaacccta attctggcgg gagtatctac 180
aaccagcggt tcaaggggag attcactttt tcagtggaca gaagcaaaaa caccctgtat 240
ctgcagatga acagcctgag ggccgaagat accgctgtct actattgcgc tcgcaatctg 300
ggccccagtt tctactttga ctattggggg cagggaaccc tggtgacagt cagctcc 357
<210> 101
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 101
Gly Phe Thr Phe Ala Asp Tyr Thr
1 5
<210> 102
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 102
gggttcactt tcgcagacta caca 24
<210> 103
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 103
Ala Arg Asn Leu Gly Pro Ser Phe Tyr Phe Asp Tyr
1 5 10
<210> 104
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 104
gctcgcaatc tgggccccag tttctacttt gactat 36
<210> 105
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 105
Val Asn Pro Asn Ser Gly Gly Ser
1 5
<210> 106
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 106
gtcaacccta attctggcgg gagt 24
<210> 107
<211> 98
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 107
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val
<210> 108
<211> 294
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 108
gctagcacta aggggccttc cgtgtttcca ctggctccct ctagtaaatc cacctctgga 60
ggcacagctg cactgggatg tctggtgaag gattacttcc ctgaaccagt cacagtgagt 120
tggaactcag gggctctgac aagtggagtc catacttttc ccgcagtgct gcagtcaagc 180
ggactgtact ccctgtcctc tgtggtcacc gtgcctagtt caagcctggg cacccagaca 240
tatatctgca acgtgaatca caagccatca aatacaaaag tcgacaagaa agtg 294
<210> 109
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 109
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
100 105 110
<210> 110
<211> 330
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 110
gcgccagaac tgctgggagg accaagcgtg ttcctgtttc cacccaagcc taaagacacc 60
ctgatgattt cccggactcc tgaggtcacc tgcgtggtcg tggacgtgtc tcacgaggac 120
cccgaagtca agttcaactg gtacgtggat ggcgtcgaag tgcataatgc caagaccaaa 180
ccccgggagg aacagtacaa ctctacctat agagtcgtga gtgtcctgac agtgctgcac 240
caggactggc tgaatgggaa ggagtataag tgtaaagtga gcaacaaagc cctgcccgcc 300
ccaatcgaaa aaacaatctc taaagcaaaa 330
<210> 111
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 111
Gly Gln Pro Arg Glu Pro Gln Val Tyr Val Tyr Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Ala Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
65 70 75 80
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 112
<211> 318
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 112
ggacagcctc gcgaaccaca ggtctacgtc taccccccat caagagatga actgacaaaa 60
aatcaggtct ctctgacatg cctggtcaaa ggattctacc cttccgacat cgccgtggag 120
tgggaaagta acggccagcc cgagaacaat tacaagacca caccccctgt cctggactct 180
gatgggagtt tcgctctggt gtcaaagctg accgtcgata aaagccggtg gcagcagggc 240
aatgtgttta gctgctccgt catgcacgaa gccctgcaca atcactacac acagaagtcc 300
ctgagcctga gccctggc 318
<210> 113
<211> 226
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 113
Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Thr Gly Ser Asp Ile Gln Met
1 5 10 15
Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr
20 25 30
Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Ile Gly Val Ala Trp Tyr
35 40 45
Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Ser Ala Ser
50 55 60
Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly
65 70 75 80
Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala
85 90 95
Thr Tyr Tyr Cys Gln Gln Tyr Tyr Ile Tyr Pro Tyr Thr Phe Gly Gln
100 105 110
Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe
115 120 125
Ile Phe Pro Pro Ser Asp Glu Glu Leu Lys Ser Gly Thr Ala Ser Val
130 135 140
Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp
145 150 155 160
Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Glu Glu Ser Val Thr
165 170 175
Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Glu
180 185 190
Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val
195 200 205
Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly
210 215 220
Glu Cys
225
<210> 114
<211> 678
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 114
tatccctacg atgtgcctga ctacgctact ggctccgata tccagatgac ccagtctcca 60
agctccctga gtgcatcagt gggggaccga gtcaccatca catgcaaggc ttcccaggat 120
gtgtctattg gagtcgcatg gtaccagcag aagccaggca aagcacccaa gctgctgatc 180
tacagcgcct cctaccggta tactggggtg ccttccagat tctctggcag tgggtcagga 240
accgacttta ctctgaccat ctctagtctg cagcccgagg atttcgccac ctactattgc 300
cagcagtact atatctaccc ttataccttt ggccagggga caaaagtgga gatcaagagg 360
acagtggccg ctccaagtgt cttcattttt cccccttccg acgaagagct gaaaagtgga 420
actgcttcag tggtctgtct gctgaacaat ttctaccccc gcgaagccaa agtgcagtgg 480
aaggtcgata acgctctgca gagcggcaat tccgaggagt ctgtgacaga acaggacagt 540
aaagattcaa cttatagcct gtcaagcaca ctggagctgt ctaaggcaga ctacgagaag 600
cacaaagtgt atgcctgcga agtcacccat caggggctgt cctctcccgt gacaaagagc 660
tttaacagag gagagtgt 678
<210> 115
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 115
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Ile Gly
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Tyr Ile Tyr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 116
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 116
gatatccaga tgacccagtc tccaagctcc ctgagtgcat cagtggggga ccgagtcacc 60
atcacatgca aggcttccca ggatgtgtct attggagtcg catggtacca gcagaagcca 120
ggcaaagcac ccaagctgct gatctacagc gcctcctacc ggtatactgg ggtgccttcc 180
agattctctg gcagtgggtc aggaaccgac tttactctga ccatctctag tctgcagccc 240
gaggatttcg ccacctacta ttgccagcag tactatatct acccttatac ctttggccag 300
gggacaaaag tggagatcaa g 321
<210> 117
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 117
Gln Asp Val Ser Ile Gly
1 5
<210> 118
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 118
caggatgtgt ctattgga 18
<210> 119
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 119
Gln Gln Tyr Tyr Ile Tyr Pro Tyr Thr
1 5
<210> 120
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 120
cagcagtact atatctaccc ttatacc 27
<210> 121
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 121
Ser Ala Ser
1
<210> 122
<211> 9
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 122
agcgcctcc 9
<210> 123
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 123
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Glu Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Glu Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Glu Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 124
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 124
aggacagtgg ccgctccaag tgtcttcatt tttccccctt ccgacgaaga gctgaaaagt 60
ggaactgctt cagtggtctg tctgctgaac aatttctacc cccgcgaagc caaagtgcag 120
tggaaggtcg ataacgctct gcagagcggc aattccgagg agtctgtgac agaacaggac 180
agtaaagatt caacttatag cctgtcaagc acactggagc tgtctaaggc agactacgag 240
aagcacaaag tgtatgcctg cgaagtcacc catcaggggc tgtcctctcc cgtgacaaag 300
agctttaaca gaggagagtg t 321
<210> 125
<211> 450
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 125
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Val Tyr Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Ala Leu Val Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 126
<211> 1350
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 126
gaagtccagc tggtcgaaag cggaggagga ctggtgcagc caggagggtc tctgcgactg 60
agttgcgccg cttcaggctt caacatcaag gacacctaca ttcactgggt gcgccaggct 120
cctggaaaag gcctggagtg ggtggcacga atctatccaa ctaatggata cacccggtat 180
gcagacagcg tgaagggccg gttcaccatt agcgcagata catccaaaaa cactgcctac 240
ctgcagatga acagcctgcg agccgaagat actgctgtgt actattgcag tcggtgggga 300
ggcgacggct tctacgctat ggattattgg gggcagggaa ccctggtcac agtgagctcc 360
gcatctacaa aggggcctag tgtgtttcca ctggccccct ctagtaaatc cacctctggg 420
ggaacagcag ccctgggatg tctggtgaag gactatttcc cagagcccgt cactgtgagt 480
tggaactcag gcgccctgac atccggggtc catacttttc ctgctgtgct gcagtcaagc 540
ggcctgtact ctctgtcctc tgtggtcacc gtgccaagtt caagcctggg gactcagacc 600
tatatctgca acgtgaatca caagccaagc aatacaaaag tcgacaagaa agtggaaccc 660
aagagctgtg ataaaacaca tacttgcccc ccttgtcctg caccagagct gctgggagga 720
ccatccgtgt tcctgtttcc acccaagcct aaagacaccc tgatgatttc caggactcca 780
gaagtcacct gcgtggtcgt ggacgtgtct cacgaggacc ccgaagtcaa gttcaactgg 840
tacgtggatg gcgtcgaggt gcataatgcc aagacaaaac ccagggagga acagtacaac 900
tcaacttatc gcgtcgtgag cgtcctgacc gtgctgcacc aggactggct gaacggcaag 960
gagtataagt gcaaagtgag caataaggct ctgcccgcac ctatcgagaa aaccattagc 1020
aaggccaaag ggcagcctag agaaccacag gtctacgtgt atcctccaag cagggacgag 1080
ctgaccaaga accaggtctc cctgacatgt ctggtgaaag ggttttaccc cagtgatatc 1140
gctgtggagt gggaatcaaa tggacagcct gaaaacaatt ataagaccac accccctgtg 1200
ctggacagcg atggcagctt cgctctggtc tccaagctga ctgtggataa atctcggtgg 1260
cagcagggca acgtctttag ttgttcagtg atgcatgagg cactgcacaa tcattacacc 1320
cagaagagcc tgtccctgtc tcccggcaaa 1350
<210> 127
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 127
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 128
<211> 360
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 128
gaagtccagc tggtcgaaag cggaggagga ctggtgcagc caggagggtc tctgcgactg 60
agttgcgccg cttcaggctt caacatcaag gacacctaca ttcactgggt gcgccaggct 120
cctggaaaag gcctggagtg ggtggcacga atctatccaa ctaatggata cacccggtat 180
gcagacagcg tgaagggccg gttcaccatt agcgcagata catccaaaaa cactgcctac 240
ctgcagatga acagcctgcg agccgaagat actgctgtgt actattgcag tcggtgggga 300
ggcgacggct tctacgctat ggattattgg gggcagggaa ccctggtcac agtgagctcc 360
<210> 129
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 129
Gly Phe Asn Ile Lys Asp Thr Tyr
1 5
<210> 130
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 130
ggcttcaaca tcaaggacac ctac 24
<210> 131
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 131
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr
1 5 10
<210> 132
<211> 39
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 132
agtcggtggg gaggcgacgg cttctacgct atggattat 39
<210> 133
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 133
Ile Tyr Pro Thr Asn Gly Tyr Thr
1 5
<210> 134
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 134
atctatccaa ctaatggata cacc 24
<210> 135
<211> 98
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 135
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val
<210> 136
<211> 294
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 136
gcatctacaa aggggcctag tgtgtttcca ctggccccct ctagtaaatc cacctctggg 60
ggaacagcag ccctgggatg tctggtgaag gactatttcc cagagcccgt cactgtgagt 120
tggaactcag gcgccctgac atccggggtc catacttttc ctgctgtgct gcagtcaagc 180
ggcctgtact ctctgtcctc tgtggtcacc gtgccaagtt caagcctggg gactcagacc 240
tatatctgca acgtgaatca caagccaagc aatacaaaag tcgacaagaa agtg 294
<210> 137
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 137
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
100 105 110
<210> 138
<211> 330
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 138
gcaccagagc tgctgggagg accatccgtg ttcctgtttc cacccaagcc taaagacacc 60
ctgatgattt ccaggactcc agaagtcacc tgcgtggtcg tggacgtgtc tcacgaggac 120
cccgaagtca agttcaactg gtacgtggat ggcgtcgagg tgcataatgc caagacaaaa 180
cccagggagg aacagtacaa ctcaacttat cgcgtcgtga gcgtcctgac cgtgctgcac 240
caggactggc tgaacggcaa ggagtataag tgcaaagtga gcaataaggc tctgcccgca 300
cctatcgaga aaaccattag caaggccaaa 330
<210> 139
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 139
Gly Gln Pro Arg Glu Pro Gln Val Tyr Val Tyr Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Ala Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
65 70 75 80
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 140
<211> 318
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 140
gggcagccta gagaaccaca ggtctacgtg tatcctccaa gcagggacga gctgaccaag 60
aaccaggtct ccctgacatg tctggtgaaa gggttttacc ccagtgatat cgctgtggag 120
tgggaatcaa atggacagcc tgaaaacaat tataagacca caccccctgt gctggacagc 180
gatggcagct tcgctctggt ctccaagctg actgtggata aatctcggtg gcagcagggc 240
aacgtcttta gttgttcagt gatgcatgag gcactgcaca atcattacac ccagaagagc 300
ctgtccctgt ctcccggc 318
<210> 141
<211> 232
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 141
Glu Pro Lys Ser Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
1 5 10 15
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
20 25 30
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
35 40 45
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
50 55 60
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
65 70 75 80
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
85 90 95
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
100 105 110
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
115 120 125
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
130 135 140
Lys Asn Gln Val Ser Leu Ile Cys Leu Val Lys Gly Phe Tyr Pro Ser
145 150 155 160
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Arg Tyr
165 170 175
Met Thr Trp Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
180 185 190
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
195 200 205
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
210 215 220
Ser Leu Ser Leu Ser Pro Gly Lys
225 230
<210> 142
<211> 696
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 142
gagcccaaga gcagcgataa gacccacacc tgccctccct gtccagctcc agaactgctg 60
ggaggaccta gcgtgttcct gtttccccct aagccaaaag acactctgat gatttccagg 120
actcccgagg tgacctgcgt ggtggtggac gtgtctcacg aggaccccga agtgaagttc 180
aactggtacg tggatggcgt ggaagtgcat aatgctaaga caaaaccaag agaggaacag 240
tacaactcca cttatcgcgt cgtgagcgtg ctgaccgtgc tgcaccagga ctggctgaac 300
gggaaggagt ataagtgcaa agtcagtaat aaggccctgc ctgctccaat cgaaaaaacc 360
atctctaagg ccaaaggcca gccaagggag ccccaggtgt acacactgcc acccagcaga 420
gacgaactga ccaagaacca ggtgtccctg atctgtctgg tgaaaggctt ctatcctagt 480
gatattgctg tggagtggga atcaaatgga cagccagaga acagatacat gacctggcct 540
ccagtgctgg acagcgatgg cagcttcttc ctgtattcca agctgacagt ggataaatct 600
cgatggcagc aggggaacgt gtttagttgt tcagtgatgc atgaagccct gcacaatcat 660
tacactcaga agagcctgtc cctgtctccc ggcaaa 696
<210> 143
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 143
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
100 105 110
<210> 144
<211> 330
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 144
gctccagaac tgctgggagg acctagcgtg ttcctgtttc cccctaagcc aaaagacact 60
ctgatgattt ccaggactcc cgaggtgacc tgcgtggtgg tggacgtgtc tcacgaggac 120
cccgaagtga agttcaactg gtacgtggat ggcgtggaag tgcataatgc taagacaaaa 180
ccaagagagg aacagtacaa ctccacttat cgcgtcgtga gcgtgctgac cgtgctgcac 240
caggactggc tgaacgggaa ggagtataag tgcaaagtca gtaataaggc cctgcctgct 300
ccaatcgaaa aaaccatctc taaggccaaa 330
<210> 145
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 145
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Thr Lys Asn Gln Val Ser Leu Ile Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Arg Tyr Met Thr Trp Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
65 70 75 80
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 146
<211> 318
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 146
ggccagccaa gggagcccca ggtgtacaca ctgccaccca gcagagacga actgaccaag 60
aaccaggtgt ccctgatctg tctggtgaaa ggcttctatc ctagtgatat tgctgtggag 120
tgggaatcaa atggacagcc agagaacaga tacatgacct ggcctccagt gctggacagc 180
gatggcagct tcttcctgta ttccaagctg acagtggata aatctcgatg gcagcagggg 240
aacgtgttta gttgttcagt gatgcatgaa gccctgcaca atcattacac tcagaagagc 300
ctgtccctgt ctcccggc 318
<210> 147
<211> 481
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 147
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Gly Gly Ser Gly Gly
100 105 110
Gly Ser Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Ser Gly Glu
115 120 125
Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser
130 135 140
Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr Tyr
145 150 155 160
Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala
165 170 175
Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val Lys
180 185 190
Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr Leu
195 200 205
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ser
210 215 220
Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln Gly
225 230 235 240
Thr Leu Val Thr Val Ser Ser Ala Ala Glu Pro Lys Ser Ser Asp Lys
245 250 255
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
260 265 270
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
275 280 285
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
290 295 300
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
305 310 315 320
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
325 330 335
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
340 345 350
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
355 360 365
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
370 375 380
Tyr Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
385 390 395 400
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
405 410 415
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
420 425 430
Asp Glu Asp Gly Ser Phe Ala Leu Val Ser Lys Leu Thr Val Asp Lys
435 440 445
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
450 455 460
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
465 470 475 480
Lys
<210> 148
<211> 1443
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 148
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagagtcacc 60
atcacttgcc gggcaagtca ggacgttaac accgctgtag cttggtatca gcagaaacca 120
gggaaagccc ctaagctcct gatctattct gcatcctttt tgtacagtgg ggtcccatca 180
aggttcagtg gcagtcgatc tgggacagat ttcactctca ccatcagcag tctgcaacct 240
gaagattttg caacttacta ctgtcaacag cattacacta ccccacccac tttcggccaa 300
gggaccaaag tggagatcaa aggtggttct ggtggtggtt ctggtggtgg ttctggtggt 360
ggttctggtg gtggttctgg tgaagtgcag ctggtggagt ctgggggagg cttggtacag 420
cctggcgggt ccctgagact ctcctgtgca gcctctggat tcaacattaa agatacttat 480
atccactggg tccggcaagc tccagggaag ggcctggagt gggtcgcacg tatttatccc 540
acaaatggtt acacacggta tgcggactct gtgaagggcc gattcaccat ctccgcagac 600
acttccaaga acaccgcgta tctgcaaatg aacagtctga gagctgagga cacggccgtt 660
tattactgtt caagatgggg cggagacggt ttctacgcta tggactactg gggccaaggg 720
accctggtca ccgtctcctc agccgccgag cccaagagca gcgataagac ccacacctgc 780
cctccctgtc cagctccaga actgctggga ggacctagcg tgttcctgtt tccccctaag 840
ccaaaagaca ctctgatgat ttccaggact cccgaggtga cctgcgtggt ggtggacgtg 900
tctcacgagg accccgaagt gaagttcaac tggtacgtgg atggcgtgga agtgcataat 960
gctaagacaa aaccaagaga ggaacagtac aactccactt atcgcgtcgt gagcgtgctg 1020
accgtgctgc accaggactg gctgaacggg aaggagtata agtgcaaagt cagtaataag 1080
gccctgcctg ctccaatcga aaaaaccatc tctaaggcca aaggccagcc aagggagccc 1140
caggtgtaca catacccacc cagcagagac gaactgacca agaaccaggt gtccctgaca 1200
tgtctggtga aaggcttcta tcctagtgat attgctgtgg agtgggaatc aaatggacag 1260
ccagagaaca attacaagac cacacctcca gtgctggacg aggatggcag cttcgccctg 1320
gtgtccaagc tgacagtgga taaatctcga tggcagcagg ggaacgtgtt tagttgttca 1380
gtgatgcatg aagccctgca caatcattac actcagaaga gcctgtccct gtctcccggc 1440
aaa 1443
<210> 149
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 149
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 150
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 150
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagagtcacc 60
atcacttgcc gggcaagtca ggacgttaac accgctgtag cttggtatca gcagaaacca 120
gggaaagccc ctaagctcct gatctattct gcatcctttt tgtacagtgg ggtcccatca 180
aggttcagtg gcagtcgatc tgggacagat ttcactctca ccatcagcag tctgcaacct 240
gaagattttg caacttacta ctgtcaacag cattacacta ccccacccac tttcggccaa 300
gggaccaaag tggagatcaa a 321
<210> 151
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 151
Gln Asp Val Asn Thr Ala
1 5
<210> 152
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 152
caggacgtta acaccgct 18
<210> 153
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 153
Gln Gln His Tyr Thr Thr Pro Pro Thr
1 5
<210> 154
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 154
caacagcatt acactacccc acccact 27
<210> 155
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 155
Ser Ala Ser
1
<210> 156
<211> 9
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 156
tctgcatcc 9
<210> 157
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 157
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 158
<211> 360
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 158
gaagtgcagc tggtggagtc tgggggaggc ttggtacagc ctggcgggtc cctgagactc 60
tcctgtgcag cctctggatt caacattaaa gatacttata tccactgggt ccggcaagct 120
ccagggaagg gcctggagtg ggtcgcacgt atttatccca caaatggtta cacacggtat 180
gcggactctg tgaagggccg attcaccatc tccgcagaca cttccaagaa caccgcgtat 240
ctgcaaatga acagtctgag agctgaggac acggccgttt attactgttc aagatggggc 300
ggagacggtt tctacgctat ggactactgg ggccaaggga ccctggtcac cgtctcctca 360
<210> 159
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 159
Gly Phe Asn Ile Lys Asp Thr Tyr
1 5
<210> 160
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 160
ggattcaaca ttaaagatac ttat 24
<210> 161
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 161
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr
1 5 10
<210> 162
<211> 39
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 162
tcaagatggg gcggagacgg tttctacgct atggactac 39
<210> 163
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 163
Ile Tyr Pro Thr Asn Gly Tyr Thr
1 5
<210> 164
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 164
atttatccca caaatggtta caca 24
<210> 165
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 165
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
100 105 110
<210> 166
<211> 330
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 166
gctccagaac tgctgggagg acctagcgtg ttcctgtttc cccctaagcc aaaagacact 60
ctgatgattt ccaggactcc cgaggtgacc tgcgtggtgg tggacgtgtc tcacgaggac 120
cccgaagtga agttcaactg gtacgtggat ggcgtggaag tgcataatgc taagacaaaa 180
ccaagagagg aacagtacaa ctccacttat cgcgtcgtga gcgtgctgac cgtgctgcac 240
caggactggc tgaacgggaa ggagtataag tgcaaagtca gtaataaggc cctgcctgct 300
ccaatcgaaa aaaccatctc taaggccaaa 330
<210> 167
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 167
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Tyr Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Glu Asp Gly Ser Phe
50 55 60
Ala Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
65 70 75 80
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 168
<211> 318
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 168
ggccagccaa gggagcccca ggtgtacaca tacccaccca gcagagacga actgaccaag 60
aaccaggtgt ccctgacatg tctggtgaaa ggcttctatc ctagtgatat tgctgtggag 120
tgggaatcaa atggacagcc agagaacaat tacaagacca cacctccagt gctggacgag 180
gatggcagct tcgccctggt gtccaagctg acagtggata aatctcgatg gcagcagggg 240
aacgtgttta gttgttcagt gatgcatgaa gccctgcaca atcattacac tcagaagagc 300
ctgtccctgt ctcccggc 318
<210> 169
<211> 481
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 169
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Gly Gly Ser Gly Gly
100 105 110
Gly Ser Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Ser Gly Glu
115 120 125
Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser
130 135 140
Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr Tyr
145 150 155 160
Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala
165 170 175
Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val Lys
180 185 190
Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr Leu
195 200 205
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ser
210 215 220
Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln Gly
225 230 235 240
Thr Leu Val Thr Val Ser Ser Ala Ala Glu Pro Lys Ser Ser Asp Lys
245 250 255
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
260 265 270
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
275 280 285
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
290 295 300
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
305 310 315 320
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
325 330 335
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
340 345 350
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
355 360 365
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
370 375 380
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Ile
385 390 395 400
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
405 410 415
Ser Asn Gly Gln Pro Glu Asn Arg Tyr Met Thr Trp Pro Pro Val Leu
420 425 430
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
435 440 445
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
450 455 460
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
465 470 475 480
Lys
<210> 170
<211> 1443
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 170
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagagtcacc 60
atcacttgcc gggcaagtca ggacgttaac accgctgtag cttggtatca gcagaaacca 120
gggaaagccc ctaagctcct gatctattct gcatcctttt tgtacagtgg ggtcccatca 180
aggttcagtg gcagtcgatc tgggacagat ttcactctca ccatcagcag tctgcaacct 240
gaagattttg caacttacta ctgtcaacag cattacacta ccccacccac tttcggccaa 300
gggaccaaag tggagatcaa aggtggttct ggtggtggtt ctggtggtgg ttctggtggt 360
ggttctggtg gtggttctgg tgaagtgcag ctggtggagt ctgggggagg cttggtacag 420
cctggcgggt ccctgagact ctcctgtgca gcctctggat tcaacattaa agatacttat 480
atccactggg tccggcaagc tccagggaag ggcctggagt gggtcgcacg tatttatccc 540
acaaatggtt acacacggta tgcggactct gtgaagggcc gattcaccat ctccgcagac 600
acttccaaga acaccgcgta tctgcaaatg aacagtctga gagctgagga cacggccgtt 660
tattactgtt caagatgggg cggagacggt ttctacgcta tggactactg gggccaaggg 720
accctggtca ccgtctcctc agccgccgag cccaagagca gcgataagac ccacacctgc 780
cctccctgtc cagctccaga actgctggga ggacctagcg tgttcctgtt tccccctaag 840
ccaaaagaca ctctgatgat ttccaggact cccgaggtga cctgcgtggt ggtggacgtg 900
tctcacgagg accccgaagt gaagttcaac tggtacgtgg atggcgtgga agtgcataat 960
gctaagacaa aaccaagaga ggaacagtac aactccactt atcgcgtcgt gagcgtgctg 1020
accgtgctgc accaggactg gctgaacggg aaggagtata agtgcaaagt cagtaataag 1080
gccctgcctg ctccaatcga aaaaaccatc tctaaggcca aaggccagcc aagggagccc 1140
caggtgtaca cactgccacc cagcagagac gaactgacca agaaccaggt gtccctgatc 1200
tgtctggtga aaggcttcta tcctagtgat attgctgtgg agtgggaatc aaatggacag 1260
ccagagaaca gatacatgac ctggcctcca gtgctggaca gcgatggcag cttcttcctg 1320
tattccaagc tgacagtgga taaatctcga tggcagcagg ggaacgtgtt tagttgttca 1380
gtgatgcatg aagccctgca caatcattac actcagaaga gcctgtccct gtctcccggc 1440
aaa 1443
<210> 171
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 171
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 172
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 172
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagagtcacc 60
atcacttgcc gggcaagtca ggacgttaac accgctgtag cttggtatca gcagaaacca 120
gggaaagccc ctaagctcct gatctattct gcatcctttt tgtacagtgg ggtcccatca 180
aggttcagtg gcagtcgatc tgggacagat ttcactctca ccatcagcag tctgcaacct 240
gaagattttg caacttacta ctgtcaacag cattacacta ccccacccac tttcggccaa 300
gggaccaaag tggagatcaa a 321
<210> 173
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 173
Gln Asp Val Asn Thr Ala
1 5
<210> 174
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 174
caggacgtta acaccgct 18
<210> 175
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 175
Gln Gln His Tyr Thr Thr Pro Pro Thr
1 5
<210> 176
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 176
caacagcatt acactacccc acccact 27
<210> 177
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 177
Ser Ala Ser
1
<210> 178
<211> 9
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 178
tctgcatcc 9
<210> 179
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 179
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 180
<211> 360
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 180
gaagtgcagc tggtggagtc tgggggaggc ttggtacagc ctggcgggtc cctgagactc 60
tcctgtgcag cctctggatt caacattaaa gatacttata tccactgggt ccggcaagct 120
ccagggaagg gcctggagtg ggtcgcacgt atttatccca caaatggtta cacacggtat 180
gcggactctg tgaagggccg attcaccatc tccgcagaca cttccaagaa caccgcgtat 240
ctgcaaatga acagtctgag agctgaggac acggccgttt attactgttc aagatggggc 300
ggagacggtt tctacgctat ggactactgg ggccaaggga ccctggtcac cgtctcctca 360
<210> 181
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 181
Gly Phe Asn Ile Lys Asp Thr Tyr
1 5
<210> 182
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 182
ggattcaaca ttaaagatac ttat 24
<210> 183
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 183
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr
1 5 10
<210> 184
<211> 39
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 184
tcaagatggg gcggagacgg tttctacgct atggactac 39
<210> 185
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 185
Ile Tyr Pro Thr Asn Gly Tyr Thr
1 5
<210> 186
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 186
atttatccca caaatggtta caca 24
<210> 187
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 187
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
100 105 110
<210> 188
<211> 330
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 188
gctccagaac tgctgggagg acctagcgtg ttcctgtttc cccctaagcc aaaagacact 60
ctgatgattt ccaggactcc cgaggtgacc tgcgtggtgg tggacgtgtc tcacgaggac 120
cccgaagtga agttcaactg gtacgtggat ggcgtggaag tgcataatgc taagacaaaa 180
ccaagagagg aacagtacaa ctccacttat cgcgtcgtga gcgtgctgac cgtgctgcac 240
caggactggc tgaacgggaa ggagtataag tgcaaagtca gtaataaggc cctgcctgct 300
ccaatcgaaa aaaccatctc taaggccaaa 330
<210> 189
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 189
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Thr Lys Asn Gln Val Ser Leu Ile Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Arg Tyr Met Thr Trp Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
65 70 75 80
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 190
<211> 318
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 190
ggccagccaa gggagcccca ggtgtacaca ctgccaccca gcagagacga actgaccaag 60
aaccaggtgt ccctgatctg tctggtgaaa ggcttctatc ctagtgatat tgctgtggag 120
tgggaatcaa atggacagcc agagaacaga tacatgacct ggcctccagt gctggacagc 180
gatggcagct tcttcctgta ttccaagctg acagtggata aatctcgatg gcagcagggg 240
aacgtgttta gttgttcagt gatgcatgaa gccctgcaca atcattacac tcagaagagc 300
ctgtccctgt ctcccggc 318
<210> 191
<211> 214
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 191
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 192
<211> 642
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 192
gatattcaga tgacccagtc ccctagctcc ctgtccgctt ctgtgggcga cagggtcact 60
atcacctgcc gcgcatctca ggatgtgaac accgcagtcg cctggtacca gcagaagcct 120
gggaaagctc caaagctgct gatctacagt gcatcattcc tgtattcagg agtgcccagc 180
cggtttagcg gcagcagatc tggcaccgac ttcacactga ctatctctag tctgcagcct 240
gaggattttg ccacatacta ttgccagcag cactatacca caccccctac tttcggccag 300
gggaccaaag tggagatcaa gcgaactgtg gccgctccaa gtgtcttcat ttttccaccc 360
agcgacgaac agctgaaatc cggcacagct tctgtggtct gtctgctgaa caacttctac 420
cccagagagg ccaaagtgca gtggaaggtc gataacgctc tgcagagtgg caacagccag 480
gagagcgtga cagaacagga ctccaaagat tctacttata gtctgtcaag caccctgaca 540
ctgagcaagg cagactacga aaagcataaa gtgtatgcct gtgaggtgac ccatcagggg 600
ctgtcttctc ccgtgaccaa gtctttcaac cgaggcgaat gt 642
<210> 193
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 193
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 194
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 194
gatattcaga tgacccagtc ccctagctcc ctgtccgctt ctgtgggcga cagggtcact 60
atcacctgcc gcgcatctca ggatgtgaac accgcagtcg cctggtacca gcagaagcct 120
gggaaagctc caaagctgct gatctacagt gcatcattcc tgtattcagg agtgcccagc 180
cggtttagcg gcagcagatc tggcaccgac ttcacactga ctatctctag tctgcagcct 240
gaggattttg ccacatacta ttgccagcag cactatacca caccccctac tttcggccag 300
gggaccaaag tggagatcaa g 321
<210> 195
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 195
Gln Asp Val Asn Thr Ala
1 5
<210> 196
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 196
caggatgtga acaccgca 18
<210> 197
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 197
Gln Gln His Tyr Thr Thr Pro Pro Thr
1 5
<210> 198
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 198
cagcagcact ataccacacc ccctact 27
<210> 199
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 199
Ser Ala Ser
1
<210> 200
<211> 9
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 200
agtgcatca 9
<210> 201
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 201
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 202
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 202
cgaactgtgg ccgctccaag tgtcttcatt tttccaccca gcgacgaaca gctgaaatcc 60
ggcacagctt ctgtggtctg tctgctgaac aacttctacc ccagagaggc caaagtgcag 120
tggaaggtcg ataacgctct gcagagtggc aacagccagg agagcgtgac agaacaggac 180
tccaaagatt ctacttatag tctgtcaagc accctgacac tgagcaaggc agactacgaa 240
aagcataaag tgtatgcctg tgaggtgacc catcaggggc tgtcttctcc cgtgaccaag 300
tctttcaacc gaggcgaatg t 321
<210> 203
<211> 448
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 203
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Asp Tyr
20 25 30
Thr Met Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asp Val Asn Pro Asn Ser Gly Gly Ser Ile Tyr Asn Gln Arg Phe
50 55 60
Lys Gly Arg Phe Thr Leu Ser Val Asp Arg Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Leu Gly Pro Ser Phe Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Val
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Leu
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Leu Thr Trp Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 204
<211> 1344
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 204
gaagtgcagc tggtcgaatc tggaggagga ctggtgcagc caggagggtc cctgcgcctg 60
tcttgcgccg ctagtggctt cacttttacc gactacacca tggattgggt gcgacaggca 120
cctggaaagg gcctggagtg ggtcgccgat gtgaacccaa atagcggagg ctccatctac 180
aaccagcggt tcaagggccg gttcaccctg tcagtggacc ggagcaaaaa caccctgtat 240
ctgcagatga atagcctgcg agccgaagat actgctgtgt actattgcgc ccggaatctg 300
gggccctcct tctactttga ctattggggg cagggaactc tggtcaccgt gagctccgcc 360
tccaccaagg gaccttctgt gttcccactg gctccctcta gtaaatccac atctggggga 420
actgcagccc tgggctgtct ggtgaaggac tacttcccag agcccgtcac agtgtcttgg 480
aacagtggcg ctctgacttc tggggtccac acctttcctg cagtgctgca gtcaagcggg 540
ctgtacagcc tgtcctctgt ggtcaccgtg ccaagttcaa gcctgggaac acagacttat 600
atctgcaacg tgaatcacaa gccatccaat acaaaagtcg acaagaaagt ggaacccaag 660
tcttgtgata aaacccatac atgcccccct tgtcctgcac cagagctgct gggaggacca 720
agcgtgttcc tgtttccacc caagcctaaa gatacactga tgattagtag gaccccagaa 780
gtcacatgcg tggtcgtgga cgtgagccac gaggaccccg aagtcaagtt taactggtac 840
gtggacggcg tcgaggtgca taatgccaag actaaaccca gggaggaaca gtacaacagt 900
acctatcgcg tcgtgtcagt cctgacagtg ctgcatcagg attggctgaa cgggaaagag 960
tataagtgca aagtgagcaa taaggctctg cccgcaccta tcgagaaaac aatttccaag 1020
gcaaaaggac agcctagaga accacaggtg tacgtgctgc ctccatcaag ggatgagctg 1080
acaaagaacc aggtcagcct gctgtgtctg gtgaaaggat tctatccctc tgacattgct 1140
gtggagtggg aaagtaatgg ccagcctgag aacaattacc tgacctggcc ccctgtgctg 1200
gactcagatg gcagcttctt tctgtatagc aagctgaccg tcgacaaatc ccggtggcag 1260
caggggaatg tgtttagttg ttcagtcatg cacgaggcac tgcacaacca ttacacccag 1320
aagtcactgt cactgtcacc aggg 1344
<210> 205
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 205
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Asp Tyr
20 25 30
Thr Met Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asp Val Asn Pro Asn Ser Gly Gly Ser Ile Tyr Asn Gln Arg Phe
50 55 60
Lys Gly Arg Phe Thr Leu Ser Val Asp Arg Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Leu Gly Pro Ser Phe Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 206
<211> 357
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 206
gaagtgcagc tggtcgaatc tggaggagga ctggtgcagc caggagggtc cctgcgcctg 60
tcttgcgccg ctagtggctt cacttttacc gactacacca tggattgggt gcgacaggca 120
cctggaaagg gcctggagtg ggtcgccgat gtgaacccaa atagcggagg ctccatctac 180
aaccagcggt tcaagggccg gttcaccctg tcagtggacc ggagcaaaaa caccctgtat 240
ctgcagatga atagcctgcg agccgaagat actgctgtgt actattgcgc ccggaatctg 300
gggccctcct tctactttga ctattggggg cagggaactc tggtcaccgt gagctcc 357
<210> 207
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 207
Gly Phe Thr Phe Thr Asp Tyr Thr
1 5
<210> 208
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 208
ggcttcactt ttaccgacta cacc 24
<210> 209
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 209
Ala Arg Asn Leu Gly Pro Ser Phe Tyr Phe Asp Tyr
1 5 10
<210> 210
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 210
gcccggaatc tggggccctc cttctacttt gactat 36
<210> 211
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 211
Val Asn Pro Asn Ser Gly Gly Ser
1 5
<210> 212
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 212
gtgaacccaa atagcggagg ctcc 24
<210> 213
<211> 98
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 213
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val
<210> 214
<211> 294
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 214
gcctccacca agggaccttc tgtgttccca ctggctccct ctagtaaatc cacatctggg 60
ggaactgcag ccctgggctg tctggtgaag gactacttcc cagagcccgt cacagtgtct 120
tggaacagtg gcgctctgac ttctggggtc cacacctttc ctgcagtgct gcagtcaagc 180
gggctgtaca gcctgtcctc tgtggtcacc gtgccaagtt caagcctggg aacacagact 240
tatatctgca acgtgaatca caagccatcc aatacaaaag tcgacaagaa agtg 294
<210> 215
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 215
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
100 105 110
<210> 216
<211> 330
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 216
gcaccagagc tgctgggagg accaagcgtg ttcctgtttc cacccaagcc taaagataca 60
ctgatgatta gtaggacccc agaagtcaca tgcgtggtcg tggacgtgag ccacgaggac 120
cccgaagtca agtttaactg gtacgtggac ggcgtcgagg tgcataatgc caagactaaa 180
cccagggagg aacagtacaa cagtacctat cgcgtcgtgt cagtcctgac agtgctgcat 240
caggattggc tgaacgggaa agagtataag tgcaaagtga gcaataaggc tctgcccgca 300
cctatcgaga aaacaatttc caaggcaaaa 330
<210> 217
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 217
Gly Gln Pro Arg Glu Pro Gln Val Tyr Val Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Thr Lys Asn Gln Val Ser Leu Leu Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Leu Thr Trp Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
65 70 75 80
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 218
<211> 318
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 218
ggacagccta gagaaccaca ggtgtacgtg ctgcctccat caagggatga gctgacaaag 60
aaccaggtca gcctgctgtg tctggtgaaa ggattctatc cctctgacat tgctgtggag 120
tgggaaagta atggccagcc tgagaacaat tacctgacct ggccccctgt gctggactca 180
gatggcagct tctttctgta tagcaagctg accgtcgaca aatcccggtg gcagcagggg 240
aatgtgttta gttgttcagt catgcacgag gcactgcaca accattacac ccagaagtca 300
ctgtcactgt caccaggg 318
<210> 219
<211> 448
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 219
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Asp Tyr
20 25 30
Thr Met Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asp Val Asn Pro Asn Ser Gly Gly Ser Ile Tyr Asn Gln Arg Phe
50 55 60
Lys Gly Arg Phe Thr Leu Ser Val Asp Arg Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Leu Gly Pro Ser Phe Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Val
340 345 350
Tyr Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Ala Leu Val Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 220
<211> 1344
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 220
gaagtgcagc tggtcgaatc tggaggagga ctggtgcagc caggagggtc cctgcgcctg 60
tcttgcgccg ctagtggctt cacttttacc gactacacca tggattgggt gcgacaggca 120
cctggaaagg gcctggagtg ggtcgccgat gtgaacccaa atagcggagg ctccatctac 180
aaccagcggt tcaagggccg gttcaccctg tcagtggacc ggagcaaaaa caccctgtat 240
ctgcagatga atagcctgcg agccgaagat actgctgtgt actattgcgc ccggaatctg 300
gggccctcct tctactttga ctattggggg cagggaactc tggtcaccgt gagctccgcc 360
tccaccaagg gaccttctgt gttcccactg gctccctcta gtaaatccac atctggggga 420
actgcagccc tgggctgtct ggtgaaggac tacttcccag agcccgtcac agtgtcttgg 480
aacagtggcg ctctgacttc tggggtccac acctttcctg cagtgctgca gtcaagcggg 540
ctgtacagcc tgtcctctgt ggtcaccgtg ccaagttcaa gcctgggaac acagacttat 600
atctgcaacg tgaatcacaa gccatccaat acaaaagtcg acaagaaagt ggaacccaag 660
tcttgtgata aaacccatac atgcccccct tgtcctgcac cagagctgct gggaggacca 720
agcgtgttcc tgtttccacc caagcctaaa gatacactga tgattagtag gaccccagaa 780
gtcacatgcg tggtcgtgga cgtgagccac gaggaccccg aagtcaagtt taactggtac 840
gtggacggcg tcgaggtgca taatgccaag actaaaccca gggaggaaca gtacaacagt 900
acctatcgcg tcgtgtcagt cctgacagtg ctgcatcagg attggctgaa cgggaaagag 960
tataagtgca aagtgagcaa taaggctctg cccgcaccta tcgagaaaac aatttccaag 1020
gcaaaaggac agcctagaga accacaggtg tacgtgtatc ctccatcaag ggatgagctg 1080
acaaagaacc aggtcagcct gacttgtctg gtgaaaggat tctatccctc tgacattgct 1140
gtggagtggg aaagtaatgg ccagcctgag aacaattaca agaccacacc ccctgtgctg 1200
gactcagatg gcagcttcgc gctggtgagc aagctgaccg tcgacaaatc ccggtggcag 1260
caggggaatg tgtttagttg ttcagtcatg cacgaggcac tgcacaacca ttacacccag 1320
aagtcactgt cactgtcacc aggg 1344
<210> 221
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 221
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Asp Tyr
20 25 30
Thr Met Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asp Val Asn Pro Asn Ser Gly Gly Ser Ile Tyr Asn Gln Arg Phe
50 55 60
Lys Gly Arg Phe Thr Leu Ser Val Asp Arg Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Leu Gly Pro Ser Phe Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 222
<211> 357
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 222
gaagtgcagc tggtcgaatc tggaggagga ctggtgcagc caggagggtc cctgcgcctg 60
tcttgcgccg ctagtggctt cacttttacc gactacacca tggattgggt gcgacaggca 120
cctggaaagg gcctggagtg ggtcgccgat gtgaacccaa atagcggagg ctccatctac 180
aaccagcggt tcaagggccg gttcaccctg tcagtggacc ggagcaaaaa caccctgtat 240
ctgcagatga atagcctgcg agccgaagat actgctgtgt actattgcgc ccggaatctg 300
gggccctcct tctactttga ctattggggg cagggaactc tggtcaccgt gagctcc 357
<210> 223
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 223
Gly Phe Thr Phe Thr Asp Tyr Thr
1 5
<210> 224
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 224
ggcttcactt ttaccgacta cacc 24
<210> 225
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 225
Ala Arg Asn Leu Gly Pro Ser Phe Tyr Phe Asp Tyr
1 5 10
<210> 226
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 226
gcccggaatc tggggccctc cttctacttt gactat 36
<210> 227
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 227
Val Asn Pro Asn Ser Gly Gly Ser
1 5
<210> 228
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 228
gtgaacccaa atagcggagg ctcc 24
<210> 229
<211> 98
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 229
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val
<210> 230
<211> 294
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 230
gcctccacca agggaccttc tgtgttccca ctggctccct ctagtaaatc cacatctggg 60
ggaactgcag ccctgggctg tctggtgaag gactacttcc cagagcccgt cacagtgtct 120
tggaacagtg gcgctctgac ttctggggtc cacacctttc ctgcagtgct gcagtcaagc 180
gggctgtaca gcctgtcctc tgtggtcacc gtgccaagtt caagcctggg aacacagact 240
tatatctgca acgtgaatca caagccatcc aatacaaaag tcgacaagaa agtg 294
<210> 231
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 231
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
100 105 110
<210> 232
<211> 330
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 232
gcaccagagc tgctgggagg accaagcgtg ttcctgtttc cacccaagcc taaagataca 60
ctgatgatta gtaggacccc agaagtcaca tgcgtggtcg tggacgtgag ccacgaggac 120
cccgaagtca agtttaactg gtacgtggac ggcgtcgagg tgcataatgc caagactaaa 180
cccagggagg aacagtacaa cagtacctat cgcgtcgtgt cagtcctgac agtgctgcat 240
caggattggc tgaacgggaa agagtataag tgcaaagtga gcaataaggc tctgcccgca 300
cctatcgaga aaacaatttc caaggcaaaa 330
<210> 233
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 233
Gly Gln Pro Arg Glu Pro Gln Val Tyr Val Tyr Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Ala Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
65 70 75 80
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 234
<211> 318
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 234
ggacagccta gagaaccaca ggtgtacgtg tatcctccat caagggatga gctgacaaag 60
aaccaggtca gcctgacttg tctggtgaaa ggattctatc cctctgacat tgctgtggag 120
tgggaaagta atggccagcc tgagaacaat tacaagacca caccccctgt gctggactca 180
gatggcagct tcgcgctggt gagcaagctg accgtcgaca aatcccggtg gcagcagggg 240
aatgtgttta gttgttcagt catgcacgag gcactgcaca accattacac ccagaagtca 300
ctgtcactgt caccaggg 318
<210> 235
<211> 450
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 235
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Val Tyr Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Ala Leu Val Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 236
<211> 1350
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 236
gaggtgcagc tggtggaaag cggaggagga ctggtgcagc caggaggatc tctgcgactg 60
agttgcgccg cttcaggatt caacatcaag gacacctaca ttcactgggt gcgacaggct 120
ccaggaaaag gactggagtg ggtggctcga atctatccca ctaatggata cacccggtat 180
gccgactccg tgaaggggag gtttactatt agcgccgata catccaaaaa cactgcttac 240
ctgcagatga acagcctgcg agccgaagat accgctgtgt actattgcag tcgatgggga 300
ggagacggat tctacgctat ggattattgg ggacagggga ccctggtgac agtgagctcc 360
gcctctacca agggccccag tgtgtttccc ctggctcctt ctagtaaatc cacctctgga 420
gggacagccg ctctgggatg tctggtgaag gactatttcc ccgagcctgt gaccgtgagt 480
tggaactcag gcgccctgac aagcggagtg cacacttttc ctgctgtgct gcagtcaagc 540
gggctgtact ccctgtcctc tgtggtgaca gtgccaagtt caagcctggg cacacagact 600
tatatctgca acgtgaatca taagccctca aatacaaaag tggacaagaa agtggagccc 660
aagagctgtg ataagaccca cacctgccct ccctgtccag ctccagaact gctgggagga 720
cctagcgtgt tcctgtttcc ccctaagcca aaagacactc tgatgatttc caggactccc 780
gaggtgacct gcgtggtggt ggacgtgtct cacgaggacc ccgaagtgaa gttcaactgg 840
tacgtggatg gcgtggaagt gcataatgct aagacaaaac caagagagga acagtacaac 900
tccacttatc gcgtcgtgag cgtgctgacc gtgctgcacc aggactggct gaacgggaag 960
gagtataagt gcaaagtcag taataaggcc ctgcctgctc caatcgaaaa aaccatctct 1020
aaggccaaag gccagccaag ggagccccag gtgtacgtgt acccacccag cagagacgaa 1080
ctgaccaaga accaggtgtc cctgacatgt ctggtgaaag gcttctatcc tagtgatatt 1140
gctgtggagt gggaatcaaa tggacagcca gagaacaatt acaagaccac acctccagtg 1200
ctggacagcg atggcagctt cgccctggtg tccaagctga cagtggataa atctcgatgg 1260
cagcagggga acgtgtttag ttgttcagtg atgcatgaag ccctgcacaa tcattacact 1320
cagaagagcc tgtccctgtc tcccggcaaa 1350
<210> 237
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 237
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 238
<211> 360
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 238
gaggtgcagc tggtggaaag cggaggagga ctggtgcagc caggaggatc tctgcgactg 60
agttgcgccg cttcaggatt caacatcaag gacacctaca ttcactgggt gcgacaggct 120
ccaggaaaag gactggagtg ggtggctcga atctatccca ctaatggata cacccggtat 180
gccgactccg tgaaggggag gtttactatt agcgccgata catccaaaaa cactgcttac 240
ctgcagatga acagcctgcg agccgaagat accgctgtgt actattgcag tcgatgggga 300
ggagacggat tctacgctat ggattattgg ggacagggga ccctggtgac agtgagctcc 360
<210> 239
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 239
Gly Phe Asn Ile Lys Asp Thr Tyr
1 5
<210> 240
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 240
ggattcaaca tcaaggacac ctac 24
<210> 241
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 241
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr
1 5 10
<210> 242
<211> 39
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 242
agtcgatggg gaggagacgg attctacgct atggattat 39
<210> 243
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 243
Ile Tyr Pro Thr Asn Gly Tyr Thr
1 5
<210> 244
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 244
atctatccca ctaatggata cacc 24
<210> 245
<211> 98
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 245
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val
<210> 246
<211> 294
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 246
gcctctacca agggccccag tgtgtttccc ctggctcctt ctagtaaatc cacctctgga 60
gggacagccg ctctgggatg tctggtgaag gactatttcc ccgagcctgt gaccgtgagt 120
tggaactcag gcgccctgac aagcggagtg cacacttttc ctgctgtgct gcagtcaagc 180
gggctgtact ccctgtcctc tgtggtgaca gtgccaagtt caagcctggg cacacagact 240
tatatctgca acgtgaatca taagccctca aatacaaaag tggacaagaa agtg 294
<210> 247
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 247
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
100 105 110
<210> 248
<211> 330
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 248
gctccagaac tgctgggagg acctagcgtg ttcctgtttc cccctaagcc aaaagacact 60
ctgatgattt ccaggactcc cgaggtgacc tgcgtggtgg tggacgtgtc tcacgaggac 120
cccgaagtga agttcaactg gtacgtggat ggcgtggaag tgcataatgc taagacaaaa 180
ccaagagagg aacagtacaa ctccacttat cgcgtcgtga gcgtgctgac cgtgctgcac 240
caggactggc tgaacgggaa ggagtataag tgcaaagtca gtaataaggc cctgcctgct 300
ccaatcgaaa aaaccatctc taaggccaaa 330
<210> 249
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 249
Gly Gln Pro Arg Glu Pro Gln Val Tyr Val Tyr Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Ala Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
65 70 75 80
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 250
<211> 318
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 250
ggccagccaa gggagcccca ggtgtacgtg tacccaccca gcagagacga actgaccaag 60
aaccaggtgt ccctgacatg tctggtgaaa ggcttctatc ctagtgatat tgctgtggag 120
tgggaatcaa atggacagcc agagaacaat tacaagacca cacctccagt gctggacagc 180
gatggcagct tcgccctggt gtccaagctg acagtggata aatctcgatg gcagcagggg 240
aacgtgttta gttgttcagt gatgcatgaa gccctgcaca atcattacac tcagaagagc 300
ctgtccctgt ctcccggc 318
<210> 251
<211> 232
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 251
Glu Pro Lys Ser Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
1 5 10 15
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
20 25 30
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
35 40 45
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
50 55 60
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
65 70 75 80
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
85 90 95
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
100 105 110
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
115 120 125
Arg Glu Pro Gln Val Tyr Val Leu Pro Pro Ser Arg Asp Glu Leu Thr
130 135 140
Lys Asn Gln Val Ser Leu Leu Cys Leu Val Lys Gly Phe Tyr Pro Ser
145 150 155 160
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
165 170 175
Leu Thr Trp Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
180 185 190
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
195 200 205
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
210 215 220
Ser Leu Ser Leu Ser Pro Gly Lys
225 230
<210> 252
<211> 696
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 252
gaacctaaaa gcagcgacaa gacccacaca tgcccccctt gtccagctcc agaactgctg 60
ggaggaccaa gcgtgttcct gtttccaccc aagcccaaag atacactgat gatcagccga 120
actcccgagg tcacctgcgt ggtcgtggac gtgtcccacg aggaccccga agtcaagttc 180
aactggtacg tggacggcgt cgaagtgcat aatgcaaaga ctaaaccacg ggaggaacag 240
tacaactcta catatagagt cgtgagtgtc ctgactgtgc tgcatcagga ttggctgaac 300
ggcaaagagt ataagtgcaa agtgtctaat aaggccctgc ctgctccaat cgagaaaact 360
attagtaagg caaaagggca gcccagggaa cctcaggtct acgtgctgcc tccaagtcgc 420
gacgagctga ccaagaacca ggtctcactg ctgtgtctgg tgaaaggatt ctatccttcc 480
gatattgccg tggagtggga atctaatggc cagccagaga acaattacct gacctggccc 540
cctgtgctgg acagcgatgg gtccttcttt ctgtattcaa agctgacagt ggacaaaagc 600
agatggcagc agggaaacgt ctttagctgt tccgtgatgc acgaagccct gcacaatcat 660
tacacccaga agtctctgag tctgtcacct ggcaaa 696
<210> 253
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 253
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
100 105 110
<210> 254
<211> 330
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 254
gctccagaac tgctgggagg accaagcgtg ttcctgtttc cacccaagcc caaagataca 60
ctgatgatca gccgaactcc cgaggtcacc tgcgtggtcg tggacgtgtc ccacgaggac 120
cccgaagtca agttcaactg gtacgtggac ggcgtcgaag tgcataatgc aaagactaaa 180
ccacgggagg aacagtacaa ctctacatat agagtcgtga gtgtcctgac tgtgctgcat 240
caggattggc tgaacggcaa agagtataag tgcaaagtgt ctaataaggc cctgcctgct 300
ccaatcgaga aaactattag taaggcaaaa 330
<210> 255
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 255
Gly Gln Pro Arg Glu Pro Gln Val Tyr Val Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Thr Lys Asn Gln Val Ser Leu Leu Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Leu Thr Trp Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
65 70 75 80
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 256
<211> 318
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 256
gggcagccca gggaacctca ggtctacgtg ctgcctccaa gtcgcgacga gctgaccaag 60
aaccaggtct cactgctgtg tctggtgaaa ggattctatc cttccgatat tgccgtggag 120
tgggaatcta atggccagcc agagaacaat tacctgacct ggccccctgt gctggacagc 180
gatgggtcct tctttctgta ttcaaagctg acagtggaca aaagcagatg gcagcaggga 240
aacgtcttta gctgttccgt gatgcacgaa gccctgcaca atcattacac ccagaagtct 300
ctgagtctgt cacctggc 318
<210> 257
<211> 475
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 257
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Ile Gly
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Tyr Ile Tyr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu
115 120 125
Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys
130 135 140
Ala Ala Ser Gly Phe Thr Phe Thr Asp Tyr Thr Met Asp Trp Val Arg
145 150 155 160
Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Asp Val Asn Pro Asn
165 170 175
Ser Gly Gly Ser Ile Tyr Asn Gln Arg Phe Lys Gly Arg Phe Thr Leu
180 185 190
Ser Val Asp Arg Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu
195 200 205
Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asn Leu Gly Pro
210 215 220
Ser Phe Tyr Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser
225 230 235 240
Ser Ala Ala Glu Pro Lys Ser Ser Asp Lys Thr His Thr Cys Pro Pro
245 250 255
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
260 265 270
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
275 280 285
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
290 295 300
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
305 310 315 320
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
325 330 335
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
340 345 350
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
355 360 365
Gly Gln Pro Arg Glu Pro Gln Val Tyr Val Tyr Pro Pro Ser Arg Asp
370 375 380
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
385 390 395 400
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
405 410 415
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
420 425 430
Ala Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
435 440 445
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
450 455 460
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
465 470 475
<210> 258
<211> 1425
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 258
gacattcaga tgacccagag ccctagctcc ctgagtgcct cagtcgggga cagggtgact 60
atcacctgca aggcttcaca ggatgtcagc attggcgtgg catggtacca gcagaagcca 120
gggaaagcac ccaagctgct gatctatagc gcctcctaca ggtatacagg cgtgccatcc 180
cgcttctctg gcagtgggtc aggaactgac tttacactga ctatttctag tctgcagccc 240
gaagatttcg ccacatacta ttgccagcag tactatatct acccttatac ttttggccag 300
gggaccaaag tggagattaa gggcggagga ggctccggag gaggagggtc tggaggagga 360
ggaagtgagg tccagctggt ggaatctgga ggaggactgg tgcagccagg agggtccctg 420
aggctgtctt gtgccgctag tggcttcacc tttacagact acacaatgga ttgggtgcgc 480
caggcaccag gaaagggact ggaatgggtc gctgatgtga accctaatag cggaggctcc 540
atctacaacc agcggttcaa aggacggttc accctgtcag tggaccggag caagaacacc 600
ctgtatctgc agatgaacag cctgagagcc gaggatactg ctgtgtacta ttgcgccagg 660
aatctgggcc caagcttcta ctttgactat tgggggcagg gaacactggt cactgtgtca 720
agcgcagccg aacccaaatc ctctgataag actcacacct gcccaccttg tccagctcca 780
gagctgctgg gaggacctag cgtgttcctg tttccaccca agccaaaaga cactctgatg 840
atttctagaa cccctgaagt gacatgtgtg gtcgtggacg tcagtcacga ggaccccgaa 900
gtcaaattca actggtacgt ggatggcgtc gaggtgcata atgccaagac caaaccccga 960
gaggaacagt acaactcaac ctatcgggtc gtgagcgtcc tgacagtgct gcatcaggac 1020
tggctgaacg gcaaggagta taagtgcaaa gtgagcaaca aggctctgcc tgcaccaatc 1080
gagaagacca tttccaaggc taaagggcag ccccgcgaac ctcaggtcta cgtgtatcct 1140
ccaagccgag atgagctgac aaaaaaccag gtctccctga cttgtctggt gaagggattt 1200
tacccaagtg acatcgcagt ggagtgggaa tcaaatggcc agcccgaaaa caattataag 1260
accacacccc ctgtgctgga ctctgatggg agtttcgcac tggtctccaa actgaccgtg 1320
gacaagtctc ggtggcagca gggaaacgtc tttagctgtt ccgtgatgca cgaggccctg 1380
cacaatcatt acacacagaa atctctgagt ctgtcacctg gcaag 1425
<210> 259
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 259
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Ile Gly
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Tyr Ile Tyr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 260
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 260
gacattcaga tgacccagag ccctagctcc ctgagtgcct cagtcgggga cagggtgact 60
atcacctgca aggcttcaca ggatgtcagc attggcgtgg catggtacca gcagaagcca 120
gggaaagcac ccaagctgct gatctatagc gcctcctaca ggtatacagg cgtgccatcc 180
cgcttctctg gcagtgggtc aggaactgac tttacactga ctatttctag tctgcagccc 240
gaagatttcg ccacatacta ttgccagcag tactatatct acccttatac ttttggccag 300
gggaccaaag tggagattaa g 321
<210> 261
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 261
Gln Asp Val Ser Ile Gly
1 5
<210> 262
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 262
caggatgtca gcattggc 18
<210> 263
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 263
Gln Gln Tyr Tyr Ile Tyr Pro Tyr Thr
1 5
<210> 264
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 264
cagcagtact atatctaccc ttatact 27
<210> 265
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 265
Ser Ala Ser
1
<210> 266
<211> 9
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 266
agcgcctcc 9
<210> 267
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 267
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Asp Tyr
20 25 30
Thr Met Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asp Val Asn Pro Asn Ser Gly Gly Ser Ile Tyr Asn Gln Arg Phe
50 55 60
Lys Gly Arg Phe Thr Leu Ser Val Asp Arg Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Leu Gly Pro Ser Phe Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 268
<211> 357
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 268
gaggtccagc tggtggaatc tggaggagga ctggtgcagc caggagggtc cctgaggctg 60
tcttgtgccg ctagtggctt cacctttaca gactacacaa tggattgggt gcgccaggca 120
ccaggaaagg gactggaatg ggtcgctgat gtgaacccta atagcggagg ctccatctac 180
aaccagcggt tcaaaggacg gttcaccctg tcagtggacc ggagcaagaa caccctgtat 240
ctgcagatga acagcctgag agccgaggat actgctgtgt actattgcgc caggaatctg 300
ggcccaagct tctactttga ctattggggg cagggaacac tggtcactgt gtcaagc 357
<210> 269
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 269
Gly Phe Thr Phe Thr Asp Tyr Thr
1 5
<210> 270
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 270
ggcttcacct ttacagacta caca 24
<210> 271
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 271
Ala Arg Asn Leu Gly Pro Ser Phe Tyr Phe Asp Tyr
1 5 10
<210> 272
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 272
gccaggaatc tgggcccaag cttctacttt gactat 36
<210> 273
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 273
Val Asn Pro Asn Ser Gly Gly Ser
1 5
<210> 274
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 274
gtgaacccta atagcggagg ctcc 24
<210> 275
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 275
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
100 105 110
<210> 276
<211> 330
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 276
gctccagagc tgctgggagg acctagcgtg ttcctgtttc cacccaagcc aaaagacact 60
ctgatgattt ctagaacccc tgaagtgaca tgtgtggtcg tggacgtcag tcacgaggac 120
cccgaagtca aattcaactg gtacgtggat ggcgtcgagg tgcataatgc caagaccaaa 180
ccccgagagg aacagtacaa ctcaacctat cgggtcgtga gcgtcctgac agtgctgcat 240
caggactggc tgaacggcaa ggagtataag tgcaaagtga gcaacaaggc tctgcctgca 300
ccaatcgaga agaccatttc caaggctaaa 330
<210> 277
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 277
Gly Gln Pro Arg Glu Pro Gln Val Tyr Val Tyr Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Ala Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
65 70 75 80
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 278
<211> 318
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 278
gggcagcccc gcgaacctca ggtctacgtg tatcctccaa gccgagatga gctgacaaaa 60
aaccaggtct ccctgacttg tctggtgaag ggattttacc caagtgacat cgcagtggag 120
tgggaatcaa atggccagcc cgaaaacaat tataagacca caccccctgt gctggactct 180
gatgggagtt tcgcactggt ctccaaactg accgtggaca agtctcggtg gcagcaggga 240
aacgtcttta gctgttccgt gatgcacgag gccctgcaca atcattacac acagaaatct 300
ctgagtctgt cacctggc 318
<210> 279
<211> 450
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 279
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Val Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu
355 360 365
Leu Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Leu Thr Trp Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 280
<211> 1350
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 280
gaggtgcagc tggtggaaag cggaggagga ctggtgcagc caggaggatc tctgcgactg 60
agttgcgccg cttcaggatt caacatcaag gacacctaca ttcactgggt gcgacaggct 120
ccaggaaaag gactggagtg ggtggctcga atctatccca ctaatggata cacccggtat 180
gccgactccg tgaaggggag gtttactatt agcgccgata catccaaaaa cactgcttac 240
ctgcagatga acagcctgcg agccgaagat accgctgtgt actattgcag tcgatgggga 300
ggagacggat tctacgctat ggattattgg ggacagggga ccctggtgac agtgagctcc 360
gcctctacca agggccccag tgtgtttccc ctggctcctt ctagtaaatc cacctctgga 420
gggacagccg ctctgggatg tctggtgaag gactatttcc ccgagcctgt gaccgtgagt 480
tggaactcag gcgccctgac aagcggagtg cacacttttc ctgctgtgct gcagtcaagc 540
gggctgtact ccctgtcctc tgtggtgaca gtgccaagtt caagcctggg cacacagact 600
tatatctgca acgtgaatca taagccctca aatacaaaag tggacaagaa agtggagccc 660
aagagctgtg ataagaccca cacctgccct ccctgtccag ctccagaact gctgggagga 720
cctagcgtgt tcctgtttcc ccctaagcca aaagacactc tgatgatttc caggactccc 780
gaggtgacct gcgtggtggt ggacgtgtct cacgaggacc ccgaagtgaa gttcaactgg 840
tacgtggatg gcgtggaagt gcataatgct aagacaaaac caagagagga acagtacaac 900
tccacttatc gcgtcgtgag cgtgctgacc gtgctgcacc aggactggct gaacgggaag 960
gagtataagt gcaaagtcag taataaggcc ctgcctgctc caatcgaaaa aaccatctct 1020
aaggccaaag gccagccaag ggagccccag gtgtacgtgc tgccacccag cagagacgaa 1080
ctgaccaaga accaggtgtc cctgctgtgt ctggtgaaag gcttctatcc tagtgatatt 1140
gctgtggagt gggaatcaaa tggacagcca gagaacaatt acctgacctg gcctccagtg 1200
ctggacagcg atggcagctt cttcctgtat tccaagctga cagtggataa atctcgatgg 1260
cagcagggga acgtgtttag ttgttcagtg atgcatgaag ccctgcacaa tcattacact 1320
cagaagagcc tgtccctgtc tcccggcaaa 1350
<210> 281
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 281
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 282
<211> 360
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 282
gaggtgcagc tggtggaaag cggaggagga ctggtgcagc caggaggatc tctgcgactg 60
agttgcgccg cttcaggatt caacatcaag gacacctaca ttcactgggt gcgacaggct 120
ccaggaaaag gactggagtg ggtggctcga atctatccca ctaatggata cacccggtat 180
gccgactccg tgaaggggag gtttactatt agcgccgata catccaaaaa cactgcttac 240
ctgcagatga acagcctgcg agccgaagat accgctgtgt actattgcag tcgatgggga 300
ggagacggat tctacgctat ggattattgg ggacagggga ccctggtgac agtgagctcc 360
<210> 283
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 283
Gly Phe Asn Ile Lys Asp Thr Tyr
1 5
<210> 284
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 284
ggattcaaca tcaaggacac ctac 24
<210> 285
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 285
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr
1 5 10
<210> 286
<211> 39
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 286
agtcgatggg gaggagacgg attctacgct atggattat 39
<210> 287
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 287
Ile Tyr Pro Thr Asn Gly Tyr Thr
1 5
<210> 288
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 288
atctatccca ctaatggata cacc 24
<210> 289
<211> 98
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 289
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val
<210> 290
<211> 294
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 290
gcctctacca agggccccag tgtgtttccc ctggctcctt ctagtaaatc cacctctgga 60
gggacagccg ctctgggatg tctggtgaag gactatttcc ccgagcctgt gaccgtgagt 120
tggaactcag gcgccctgac aagcggagtg cacacttttc ctgctgtgct gcagtcaagc 180
gggctgtact ccctgtcctc tgtggtgaca gtgccaagtt caagcctggg cacacagact 240
tatatctgca acgtgaatca taagccctca aatacaaaag tggacaagaa agtg 294
<210> 291
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 291
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
100 105 110
<210> 292
<211> 330
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 292
gctccagaac tgctgggagg acctagcgtg ttcctgtttc cccctaagcc aaaagacact 60
ctgatgattt ccaggactcc cgaggtgacc tgcgtggtgg tggacgtgtc tcacgaggac 120
cccgaagtga agttcaactg gtacgtggat ggcgtggaag tgcataatgc taagacaaaa 180
ccaagagagg aacagtacaa ctccacttat cgcgtcgtga gcgtgctgac cgtgctgcac 240
caggactggc tgaacgggaa ggagtataag tgcaaagtca gtaataaggc cctgcctgct 300
ccaatcgaaa aaaccatctc taaggccaaa 330
<210> 293
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 293
Gly Gln Pro Arg Glu Pro Gln Val Tyr Val Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Thr Lys Asn Gln Val Ser Leu Leu Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Leu Thr Trp Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
65 70 75 80
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 294
<211> 318
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 294
ggccagccaa gggagcccca ggtgtacgtg ctgccaccca gcagagacga actgaccaag 60
aaccaggtgt ccctgctgtg tctggtgaaa ggcttctatc ctagtgatat tgctgtggag 120
tgggaatcaa atggacagcc agagaacaat tacctgacct ggcctccagt gctggacagc 180
gatggcagct tcttcctgta ttccaagctg acagtggata aatctcgatg gcagcagggg 240
aacgtgttta gttgttcagt gatgcatgaa gccctgcaca atcattacac tcagaagagc 300
ctgtccctgt ctcccggc 318
<210> 295
<211> 480
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 295
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Gly Gly Ser Gly Gly
100 105 110
Gly Ser Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Ser Gly Glu
115 120 125
Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser
130 135 140
Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr Tyr
145 150 155 160
Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala
165 170 175
Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val Lys
180 185 190
Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr Leu
195 200 205
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ser
210 215 220
Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln Gly
225 230 235 240
Thr Leu Val Thr Val Ser Ser Ala Ala Glu Pro Lys Ser Ser Asp Lys
245 250 255
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
260 265 270
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
275 280 285
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
290 295 300
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
305 310 315 320
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
325 330 335
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
340 345 350
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
355 360 365
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Val
370 375 380
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Leu
385 390 395 400
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
405 410 415
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Leu Thr Trp Pro Pro Val Leu
420 425 430
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
435 440 445
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
450 455 460
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
465 470 475 480
<210> 296
<211> 1440
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 296
gacattcaga tgacacagag ccccagctcc ctgagtgctt cagtcggcga cagggtgact 60
atcacctgcc gcgcatccca ggatgtcaac accgctgtgg catggtacca gcagaagcct 120
ggaaaagccc caaagctgct gatctacagc gcttccttcc tgtattctgg cgtgccaagt 180
cggttttctg gaagtagatc aggcactgac ttcacactga ctatctctag tctgcagccc 240
gaagattttg ccacctacta ttgccagcag cactatacca caccccctac attcggacag 300
ggcactaaag tggagattaa gggcgggtca ggcggaggga gcggaggagg gtccggagga 360
gggtctggag gagggagtgg agaggtccag ctggtggaat ctggaggagg actggtgcag 420
cctggaggct cactgcgact gagctgtgcc gcttccggct ttaacatcaa agacacatac 480
attcattggg tcaggcaggc accagggaag ggactggaat gggtggcccg catctatccc 540
acaaatgggt acactcgata tgccgacagc gtgaaaggac ggtttaccat ttctgctgat 600
accagtaaga acacagcata cctgcagatg aacagcctgc gcgcagagga tacagccgtg 660
tactattgca gtcgatgggg gggagacggc ttctacgcca tggattattg gggccagggg 720
actctggtca ccgtgtcaag cgcagccgaa cctaaatcct ctgacaagac ccacacatgc 780
ccaccctgtc ctgctccaga gctgctggga ggaccatccg tgttcctgtt tcctccaaag 840
cctaaagata cactgatgat tagccgcact cccgaagtca cctgtgtggt cgtggacgtg 900
tcccacgagg accccgaagt caagttcaac tggtacgtgg acggcgtcga ggtgcataat 960
gccaagacta aaccaagaga ggaacagtac aattcaacct atagggtcgt gagcgtcctg 1020
acagtgctgc atcaggattg gctgaacggc aaggagtata agtgcaaagt gtctaacaag 1080
gccctgcccg ctcctatcga gaagactatt agcaaggcaa aagggcagcc acgggaaccc 1140
caggtctacg tgctgccccc tagcagagac gagctgacca aaaaccaggt ctccctgctg 1200
tgtctggtga agggctttta tcctagtgat atcgctgtgg agtgggaatc aaatgggcag 1260
ccagaaaaca attacctgac atggccaccc gtgctggaca gcgatgggtc cttctttctg 1320
tattccaaac tgactgtgga caagtctaga tggcagcagg gaaacgtctt cagctgttcc 1380
gtgatgcacg aggccctgca caatcattac acccagaagt ctctgagtct gtcacccggc 1440
<210> 297
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 297
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 298
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 298
gacattcaga tgacacagag ccccagctcc ctgagtgctt cagtcggcga cagggtgact 60
atcacctgcc gcgcatccca ggatgtcaac accgctgtgg catggtacca gcagaagcct 120
ggaaaagccc caaagctgct gatctacagc gcttccttcc tgtattctgg cgtgccaagt 180
cggttttctg gaagtagatc aggcactgac ttcacactga ctatctctag tctgcagccc 240
gaagattttg ccacctacta ttgccagcag cactatacca caccccctac attcggacag 300
ggcactaaag tggagattaa g 321
<210> 299
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 299
Gln Asp Val Asn Thr Ala
1 5
<210> 300
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 300
caggatgtca acaccgct 18
<210> 301
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 301
Gln Gln His Tyr Thr Thr Pro Pro Thr
1 5
<210> 302
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 302
cagcagcact ataccacacc ccctaca 27
<210> 303
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 303
Ser Ala Ser
1
<210> 304
<211> 9
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 304
agcgcttcc 9
<210> 305
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 305
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 306
<211> 360
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 306
gaggtccagc tggtggaatc tggaggagga ctggtgcagc ctggaggctc actgcgactg 60
agctgtgccg cttccggctt taacatcaaa gacacataca ttcattgggt caggcaggca 120
ccagggaagg gactggaatg ggtggcccgc atctatccca caaatgggta cactcgatat 180
gccgacagcg tgaaaggacg gtttaccatt tctgctgata ccagtaagaa cacagcatac 240
ctgcagatga acagcctgcg cgcagaggat acagccgtgt actattgcag tcgatggggg 300
ggagacggct tctacgccat ggattattgg ggccagggga ctctggtcac cgtgtcaagc 360
<210> 307
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 307
Gly Phe Asn Ile Lys Asp Thr Tyr
1 5
<210> 308
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 308
ggctttaaca tcaaagacac atac 24
<210> 309
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 309
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr
1 5 10
<210> 310
<211> 39
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 310
agtcgatggg ggggagacgg cttctacgcc atggattat 39
<210> 311
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 311
Ile Tyr Pro Thr Asn Gly Tyr Thr
1 5
<210> 312
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 312
atctatccca caaatgggta cact 24
<210> 313
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 313
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
100 105 110
<210> 314
<211> 330
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 314
gctccagagc tgctgggagg accatccgtg ttcctgtttc ctccaaagcc taaagataca 60
ctgatgatta gccgcactcc cgaagtcacc tgtgtggtcg tggacgtgtc ccacgaggac 120
cccgaagtca agttcaactg gtacgtggac ggcgtcgagg tgcataatgc caagactaaa 180
ccaagagagg aacagtacaa ttcaacctat agggtcgtga gcgtcctgac agtgctgcat 240
caggattggc tgaacggcaa ggagtataag tgcaaagtgt ctaacaaggc cctgcccgct 300
cctatcgaga agactattag caaggcaaaa 330
<210> 315
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 315
Gly Gln Pro Arg Glu Pro Gln Val Tyr Val Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Thr Lys Asn Gln Val Ser Leu Leu Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Leu Thr Trp Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
65 70 75 80
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 316
<211> 318
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 316
gggcagccac gggaacccca ggtctacgtg ctgcccccta gcagagacga gctgaccaaa 60
aaccaggtct ccctgctgtg tctggtgaag ggcttttatc ctagtgatat cgctgtggag 120
tgggaatcaa atgggcagcc agaaaacaat tacctgacat ggccacccgt gctggacagc 180
gatgggtcct tctttctgta ttccaaactg actgtggaca agtctagatg gcagcaggga 240
aacgtcttca gctgttccgt gatgcacgag gccctgcaca atcattacac ccagaagtct 300
ctgagtctgt cacccggc 318
<210> 317
<211> 214
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 317
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 318
<211> 642
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 318
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagagtcacc 60
atcacttgcc gggcaagtca ggacgttaac accgctgtag cttggtatca gcagaaacca 120
gggaaagccc ctaagctcct gatctattct gcatcctttt tgtacagtgg ggtcccatca 180
aggttcagtg gcagtcgatc tgggacagat ttcactctca ccatcagcag tctgcaacct 240
gaagattttg caacttacta ctgtcaacag cattacacta ccccacccac tttcggccaa 300
gggaccaaag tggagatcaa acgaactgtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccaa 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gt 642
<210> 319
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 319
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 320
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 320
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagagtcacc 60
atcacttgcc gggcaagtca ggacgttaac accgctgtag cttggtatca gcagaaacca 120
gggaaagccc ctaagctcct gatctattct gcatcctttt tgtacagtgg ggtcccatca 180
aggttcagtg gcagtcgatc tgggacagat ttcactctca ccatcagcag tctgcaacct 240
gaagattttg caacttacta ctgtcaacag cattacacta ccccacccac tttcggccaa 300
gggaccaaag tggagatcaa a 321
<210> 321
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 321
Gln Asp Val Asn Thr Ala
1 5
<210> 322
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 322
caggacgtta acaccgct 18
<210> 323
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 323
Gln Gln His Tyr Thr Thr Pro Pro Thr
1 5
<210> 324
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 324
caacagcatt acactacccc acccact 27
<210> 325
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 325
Ser Ala Ser
1
<210> 326
<211> 9
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 326
tctgcatcc 9
<210> 327
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 327
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 328
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 328
cgaactgtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccaag agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagctcgcc cgtcacaaag 300
agcttcaaca ggggagagtg t 321
<210> 329
<211> 231
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 329
Glu Pro Lys Ser Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
1 5 10 15
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
20 25 30
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
35 40 45
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
50 55 60
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
65 70 75 80
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
85 90 95
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
100 105 110
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
115 120 125
Arg Glu Pro Gln Val Tyr Val Leu Pro Pro Ser Arg Asp Glu Leu Thr
130 135 140
Lys Asn Gln Val Ser Leu Leu Cys Leu Val Lys Gly Phe Tyr Pro Ser
145 150 155 160
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
165 170 175
Leu Thr Trp Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
180 185 190
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
195 200 205
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
210 215 220
Ser Leu Ser Leu Ser Pro Gly
225 230
<210> 330
<211> 693
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 330
gaacctaaat ccagcgacaa gacccacaca tgcccccctt gtccagctcc agaactgctg 60
ggaggaccaa gcgtgttcct gtttccaccc aagcccaaag atacactgat gatcagccga 120
actcccgagg tcacctgcgt ggtcgtggac gtgtcccacg aggaccccga agtcaagttc 180
aactggtacg tggacggcgt cgaagtgcat aatgcaaaga ctaaaccacg ggaggaacag 240
tacaactcta catatagagt cgtgagtgtc ctgactgtgc tgcatcagga ttggctgaac 300
ggcaaagagt ataagtgcaa agtgtctaat aaggccctgc ctgctccaat cgagaaaact 360
attagtaagg caaaagggca gcccagggaa cctcaggtct acgtgctgcc tccaagtcgc 420
gacgagctga ccaagaacca ggtctcactg ctgtgtctgg tgaaaggatt ctatccttcc 480
gatattgccg tggagtggga atctaatggc cagccagaga acaattacct gacctggccc 540
cctgtgctgg acagcgatgg gtccttcttt ctgtattcaa agctgacagt ggacaaaagc 600
agatggcagc agggaaacgt ctttagctgt tccgtgatgc acgaagccct gcacaatcat 660
tacacccaga agtctctgag tctgtcacct ggc 693
<210> 331
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 331
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
100 105 110
<210> 332
<211> 330
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 332
gctccagaac tgctgggagg accaagcgtg ttcctgtttc cacccaagcc caaagataca 60
ctgatgatca gccgaactcc cgaggtcacc tgcgtggtcg tggacgtgtc ccacgaggac 120
cccgaagtca agttcaactg gtacgtggac ggcgtcgaag tgcataatgc aaagactaaa 180
ccacgggagg aacagtacaa ctctacatat agagtcgtga gtgtcctgac tgtgctgcat 240
caggattggc tgaacggcaa agagtataag tgcaaagtgt ctaataaggc cctgcctgct 300
ccaatcgaga aaactattag taaggcaaaa 330
<210> 333
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 333
Gly Gln Pro Arg Glu Pro Gln Val Tyr Val Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Thr Lys Asn Gln Val Ser Leu Leu Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Leu Thr Trp Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
65 70 75 80
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 334
<211> 318
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 334
gggcagccca gggaacctca ggtctacgtg ctgcctccaa gtcgcgacga gctgaccaag 60
aaccaggtct cactgctgtg tctggtgaaa ggattctatc cttccgatat tgccgtggag 120
tgggaatcta atggccagcc agagaacaat tacctgacct ggccccctgt gctggacagc 180
gatgggtcct tctttctgta ttcaaagctg acagtggaca aaagcagatg gcagcaggga 240
aacgtcttta gctgttccgt gatgcacgaa gccctgcaca atcattacac ccagaagtct 300
ctgagtctgt cacctggc 318
<210> 335
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 335
Val Asn Pro Asn Ser Gly Gly Ser
1 5
<210> 336
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 336
Ala Arg Asn Leu Gly Pro Ser Phe Tyr Phe Asp Tyr
1 5 10
<210> 337
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 337
Gly Phe Thr Phe Thr Asp Tyr Thr
1 5
<210> 338
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 338
Ser Ala Ser
1
<210> 339
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 339
Gln Gln Tyr Tyr Ile Tyr Pro Tyr Thr
1 5
<210> 340
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 340
Gln Asp Val Ser Ile Gly
1 5
<210> 341
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 341
Ile Tyr Pro Thr Asn Gly Tyr Thr
1 5
<210> 342
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 342
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr
1 5 10
<210> 343
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 343
Gly Phe Asn Ile Lys Asp Thr Tyr
1 5
<210> 344
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 344
Ser Ala Ser
1
<210> 345
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 345
Gln Gln His Tyr Thr Thr Pro Pro Thr
1 5
<210> 346
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 346
Gln Asp Val Asn Thr Ala
1 5
<210> 347
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 347
Gln Gln Tyr Tyr Ile Tyr Pro Ala Thr
1 5
<210> 348
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 348
Gly Phe Thr Phe Ala Asp Tyr Thr
1 5
<210> 349
<211> 607
<212> PRT
<213> Homo sapiens
<400> 349
Thr Gln Val Cys Thr Gly Thr Asp Met Lys Leu Arg Leu Pro Ala Ser
1 5 10 15
Pro Glu Thr His Leu Asp Met Leu Arg His Leu Tyr Gln Gly Cys Gln
20 25 30
Val Val Gln Gly Asn Leu Glu Leu Thr Tyr Leu Pro Thr Asn Ala Ser
35 40 45
Leu Ser Phe Leu Gln Asp Ile Gln Glu Val Gln Gly Tyr Val Leu Ile
50 55 60
Ala His Asn Gln Val Arg Gln Val Pro Leu Gln Arg Leu Arg Ile Val
65 70 75 80
Arg Gly Thr Gln Leu Phe Glu Asp Asn Tyr Ala Leu Ala Val Leu Asp
85 90 95
Asn Gly Asp Pro Leu Asn Asn Thr Thr Pro Val Thr Gly Ala Ser Pro
100 105 110
Gly Gly Leu Arg Glu Leu Gln Leu Arg Ser Leu Thr Glu Ile Leu Lys
115 120 125
Gly Gly Val Leu Ile Gln Arg Asn Pro Gln Leu Cys Tyr Gln Asp Thr
130 135 140
Ile Leu Trp Lys Asp Ile Phe His Lys Asn Asn Gln Leu Ala Leu Thr
145 150 155 160
Leu Ile Asp Thr Asn Arg Ser Arg Ala Cys His Pro Cys Ser Pro Met
165 170 175
Cys Lys Gly Ser Arg Cys Trp Gly Glu Ser Ser Glu Asp Cys Gln Ser
180 185 190
Leu Thr Arg Thr Val Cys Ala Gly Gly Cys Ala Arg Cys Lys Gly Pro
195 200 205
Leu Pro Thr Asp Cys Cys His Glu Gln Cys Ala Ala Gly Cys Thr Gly
210 215 220
Pro Lys His Ser Asp Cys Leu Ala Cys Leu His Phe Asn His Ser Gly
225 230 235 240
Ile Cys Glu Leu His Cys Pro Ala Leu Val Thr Tyr Asn Thr Asp Thr
245 250 255
Phe Glu Ser Met Pro Asn Pro Glu Gly Arg Tyr Thr Phe Gly Ala Ser
260 265 270
Cys Val Thr Ala Cys Pro Tyr Asn Tyr Leu Ser Thr Asp Val Gly Ser
275 280 285
Cys Thr Leu Val Cys Pro Leu His Asn Gln Glu Val Thr Ala Glu Asp
290 295 300
Gly Thr Gln Arg Cys Glu Lys Cys Ser Lys Pro Cys Ala Arg Val Cys
305 310 315 320
Tyr Gly Leu Gly Met Glu His Leu Arg Glu Val Arg Ala Val Thr Ser
325 330 335
Ala Asn Ile Gln Glu Phe Ala Gly Cys Lys Lys Ile Phe Gly Ser Leu
340 345 350
Ala Phe Leu Pro Glu Ser Phe Asp Gly Asp Pro Ala Ser Asn Thr Ala
355 360 365
Pro Leu Gln Pro Glu Gln Leu Gln Val Phe Glu Thr Leu Glu Glu Ile
370 375 380
Thr Gly Tyr Leu Tyr Ile Ser Ala Trp Pro Asp Ser Leu Pro Asp Leu
385 390 395 400
Ser Val Phe Gln Asn Leu Gln Val Ile Arg Gly Arg Ile Leu His Asn
405 410 415
Gly Ala Tyr Ser Leu Thr Leu Gln Gly Leu Gly Ile Ser Trp Leu Gly
420 425 430
Leu Arg Ser Leu Arg Glu Leu Gly Ser Gly Leu Ala Leu Ile His His
435 440 445
Asn Thr His Leu Cys Phe Val His Thr Val Pro Trp Asp Gln Leu Phe
450 455 460
Arg Asn Pro His Gln Ala Leu Leu His Thr Ala Asn Arg Pro Glu Asp
465 470 475 480
Glu Cys Val Gly Glu Gly Leu Ala Cys His Gln Leu Cys Ala Arg Gly
485 490 495
His Cys Trp Gly Pro Gly Pro Thr Gln Cys Val Asn Cys Ser Gln Phe
500 505 510
Leu Arg Gly Gln Glu Cys Val Glu Glu Cys Arg Val Leu Gln Gly Leu
515 520 525
Pro Arg Glu Tyr Val Asn Ala Arg His Cys Leu Pro Cys His Pro Glu
530 535 540
Cys Gln Pro Gln Asn Gly Ser Val Thr Cys Phe Gly Pro Glu Ala Asp
545 550 555 560
Gln Cys Val Ala Cys Ala His Tyr Lys Asp Pro Pro Phe Cys Val Ala
565 570 575
Arg Cys Pro Ser Gly Val Lys Pro Asp Leu Ser Tyr Met Pro Ile Trp
580 585 590
Lys Phe Pro Asp Glu Glu Gly Ala Cys Gln Pro Cys Pro Ile Asn
595 600 605
<210> 350
<211> 217
<212> PRT
<213> Homo sapiens
<400> 350
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly Lys
210 215
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| БИСПЕЦИФИЧЕСКИЕ АНТИГЕНСВЯЗЫВАЮЩИЕ ПОЛИПЕПТИДЫ | 2015 |
|
RU2723940C2 |
| АНТИ-PVRIG АНТИТЕЛА И СПОСОБЫ ПРИМЕНЕНИЯ | 2016 |
|
RU2732042C2 |
| АНТИ-PVRIG АНТИТЕЛА И СПОСОБЫ ПРИМЕНЕНИЯ | 2016 |
|
RU2827926C2 |
| АНТИТЕЛА ПРОТИВ PD-L1 И ИХ ВАРИАНТЫ | 2017 |
|
RU2770590C2 |
| Антитела против CXCR2 и их применение | 2019 |
|
RU2807067C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ И ПУТИ ИХ ПРИМЕНЕНИЯ | 2020 |
|
RU2841244C2 |
| Биспецифическое антитело анти-HER2/анти-4-1BB и его применение | 2020 |
|
RU2831838C2 |
| ВВЕДЕНИЕ СКОНСТРУИРОВАННЫХ Т-КЛЕТОК ДЛЯ ЛЕЧЕНИЯ РАКА ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ | 2017 |
|
RU2757308C2 |
| ГИБРИДНЫЕ БЕЛКИ ВАРИАНТА sPD-1—FC | 2019 |
|
RU2785993C2 |
| HER-2-НАПРАВЛЕННЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СОДЕРЖАЩИЕ 4-1BBL | 2019 |
|
RU2815451C2 |
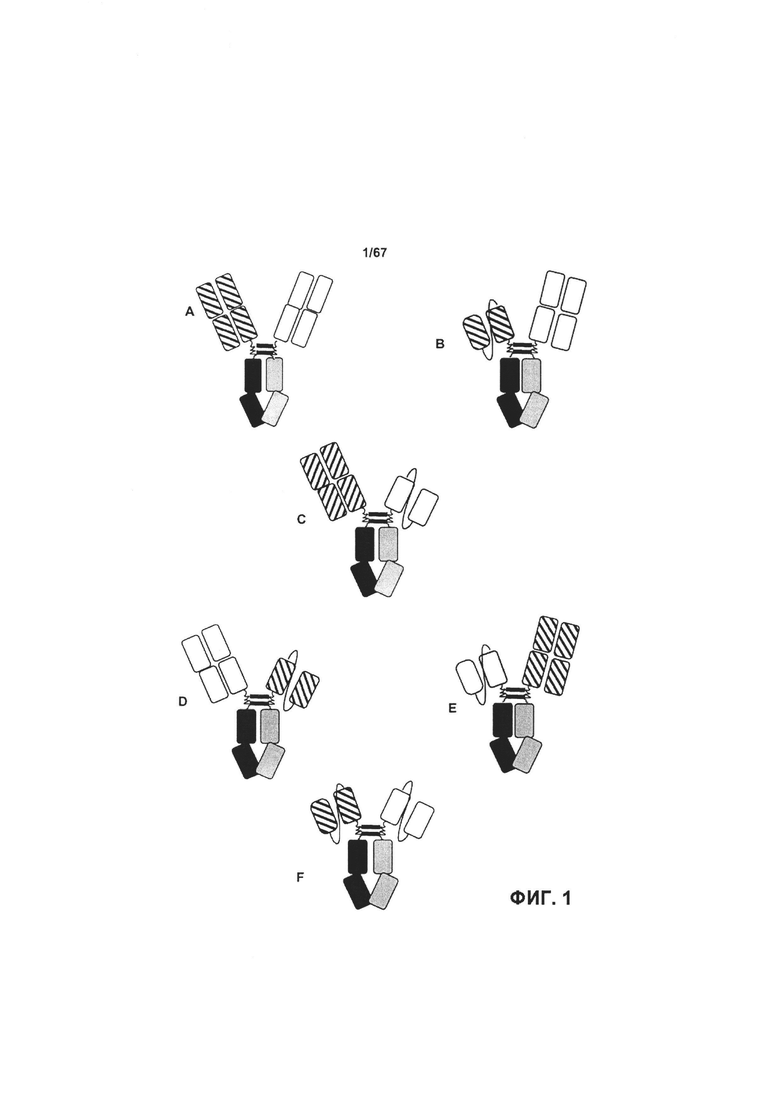
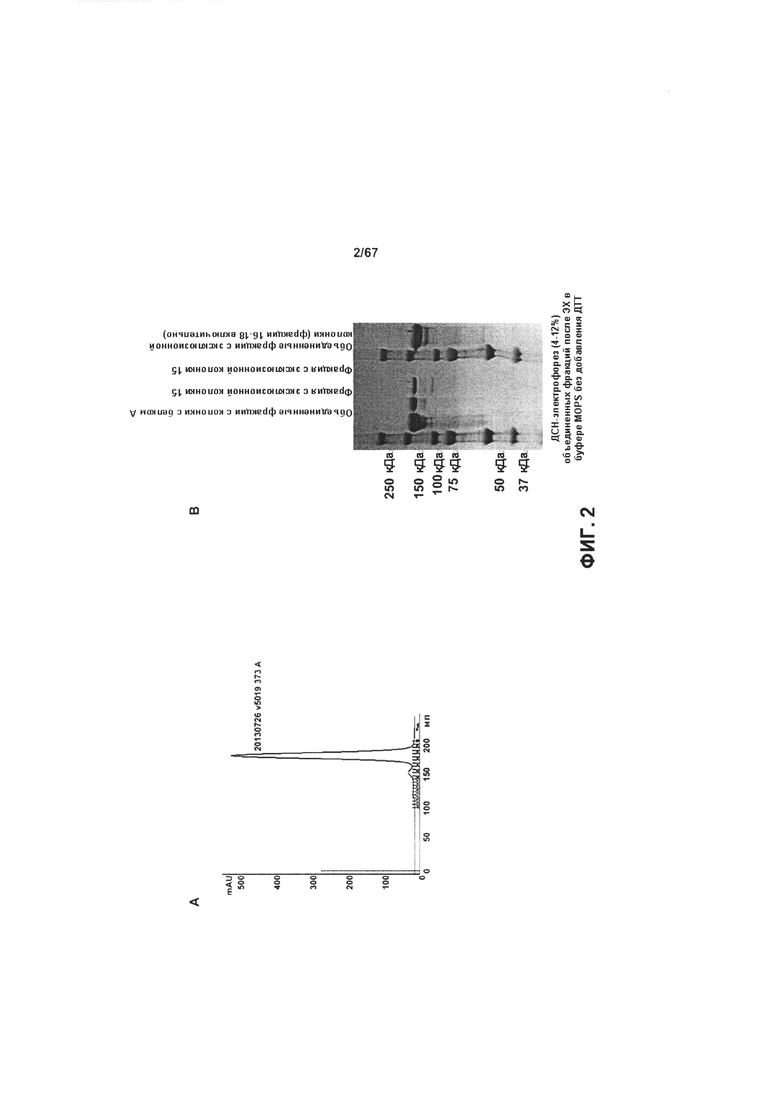
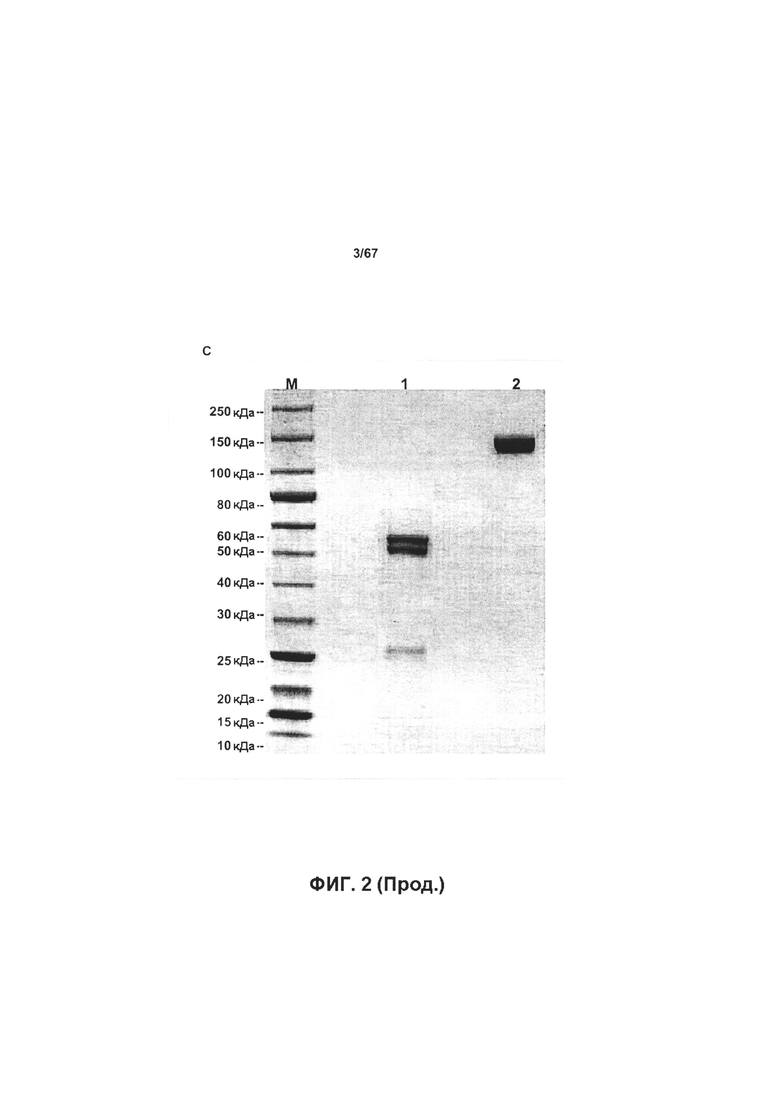
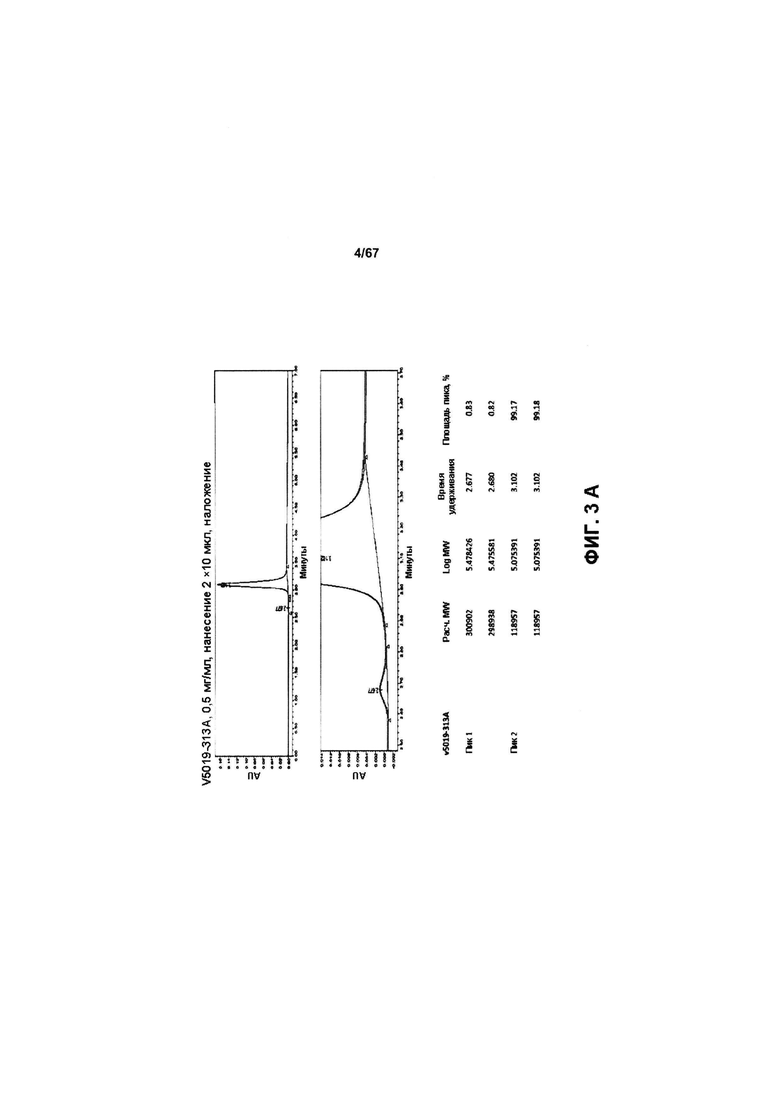

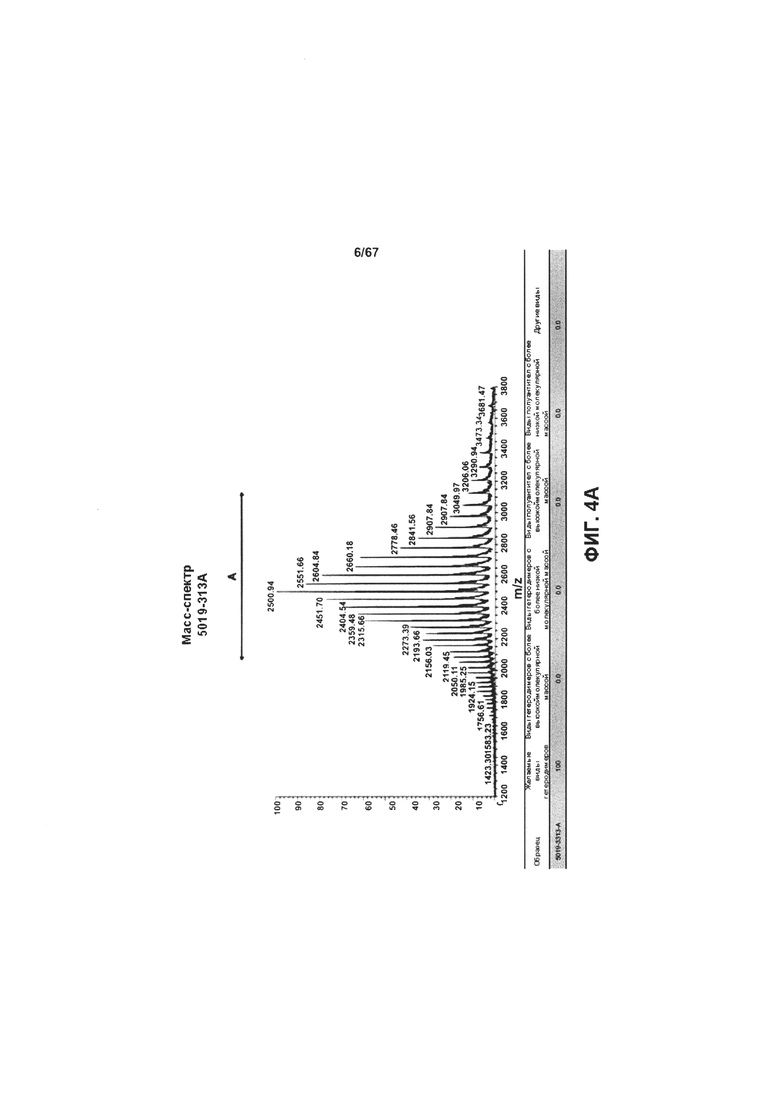
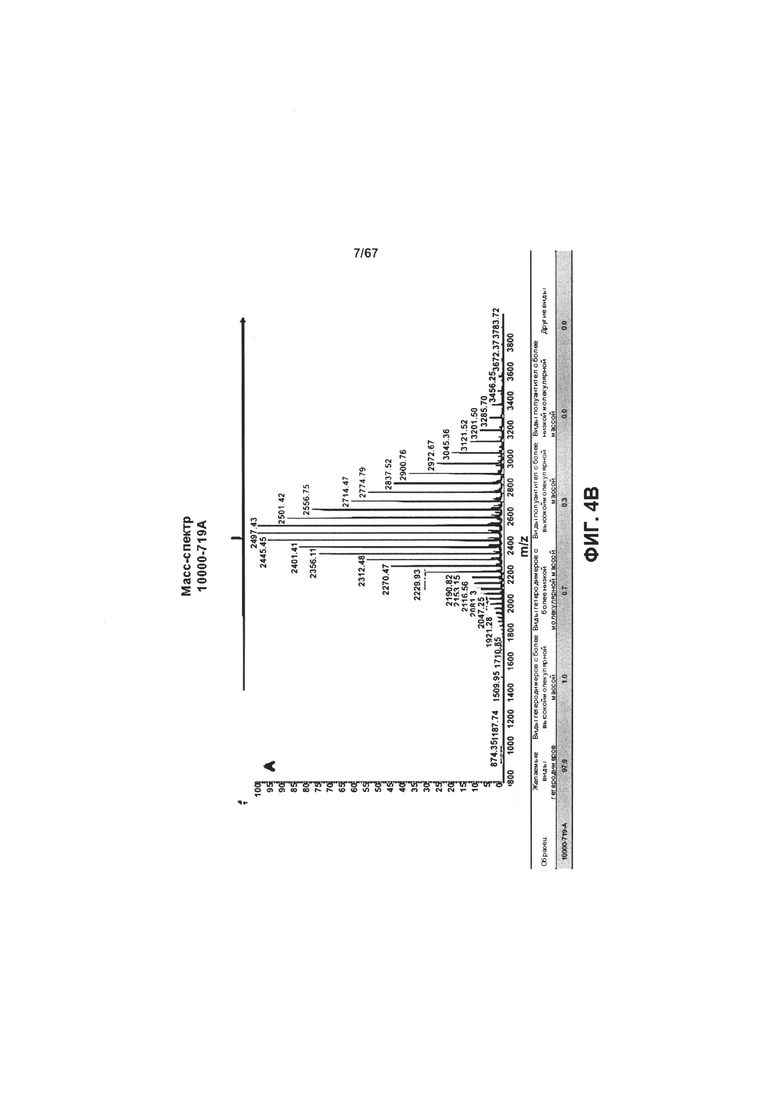
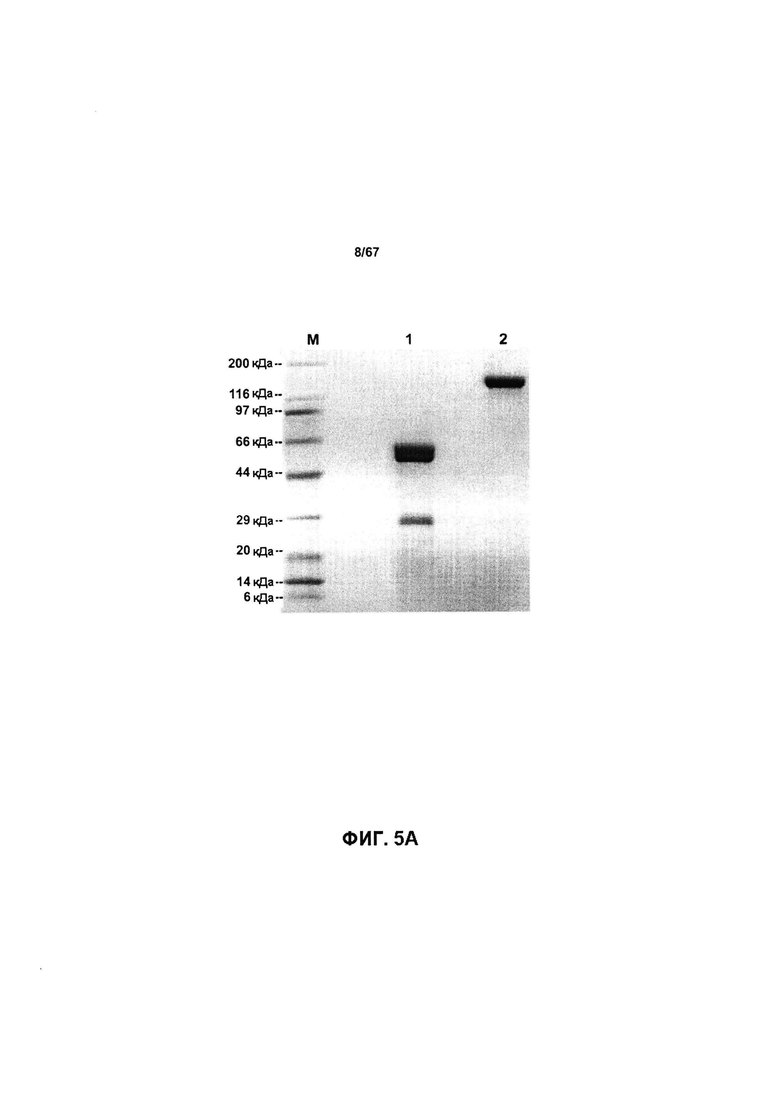
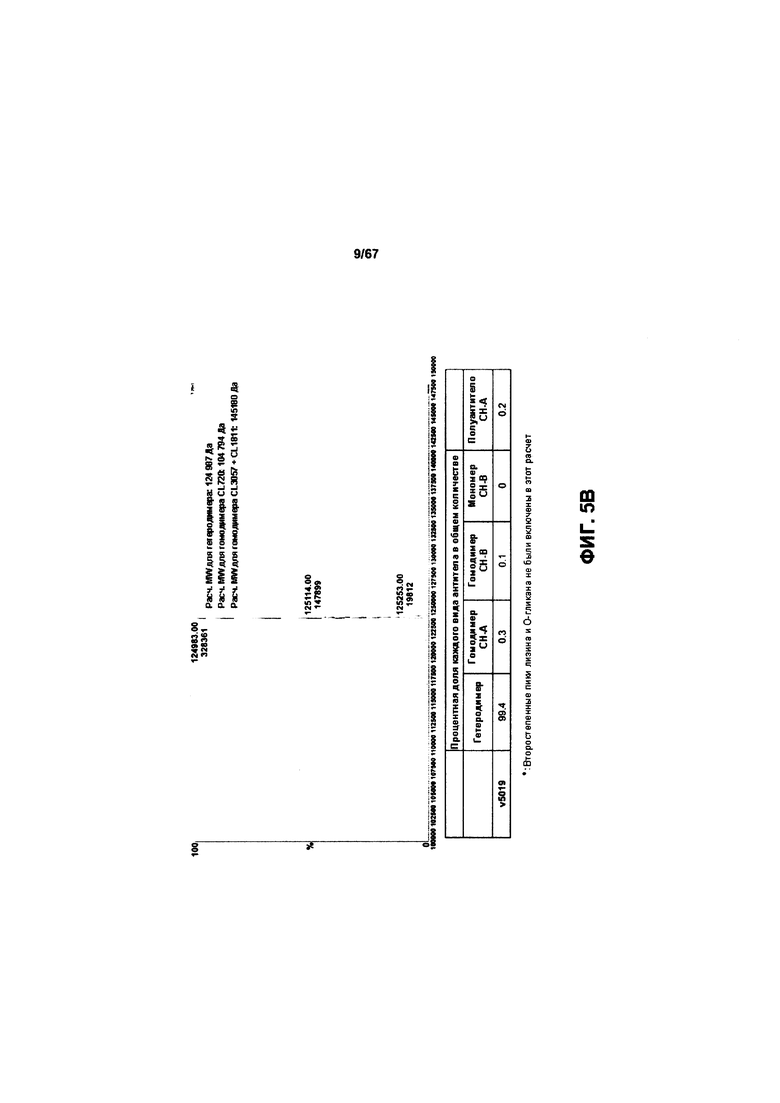
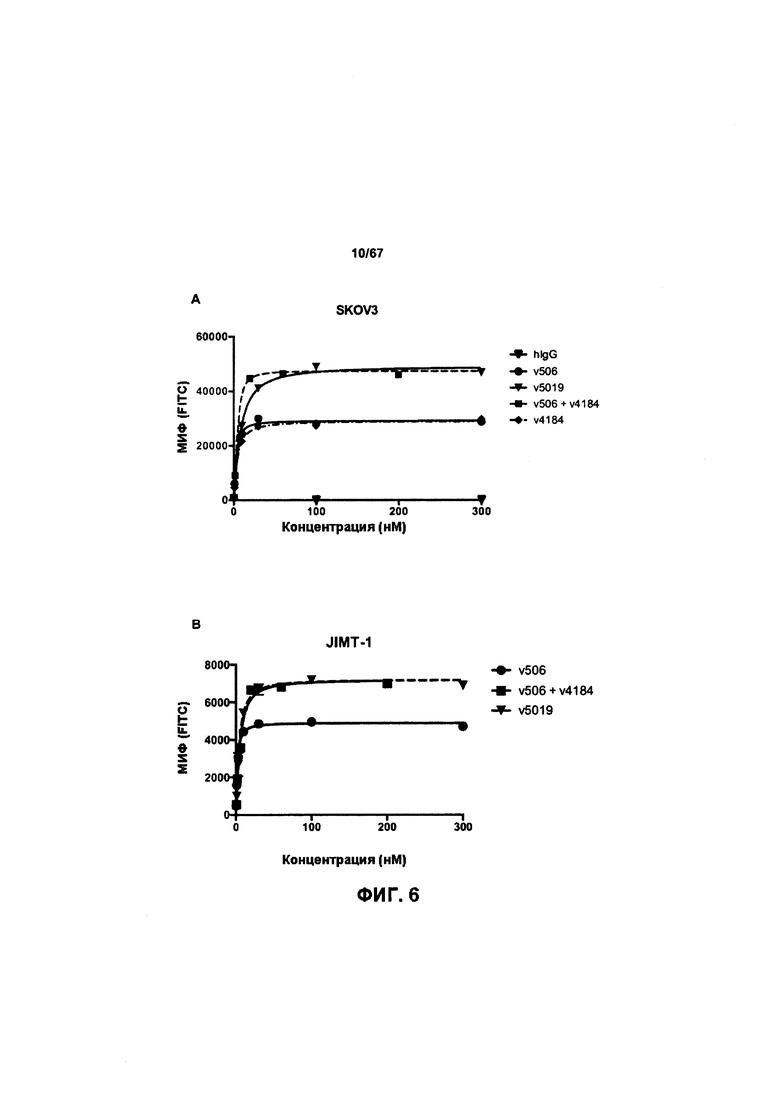
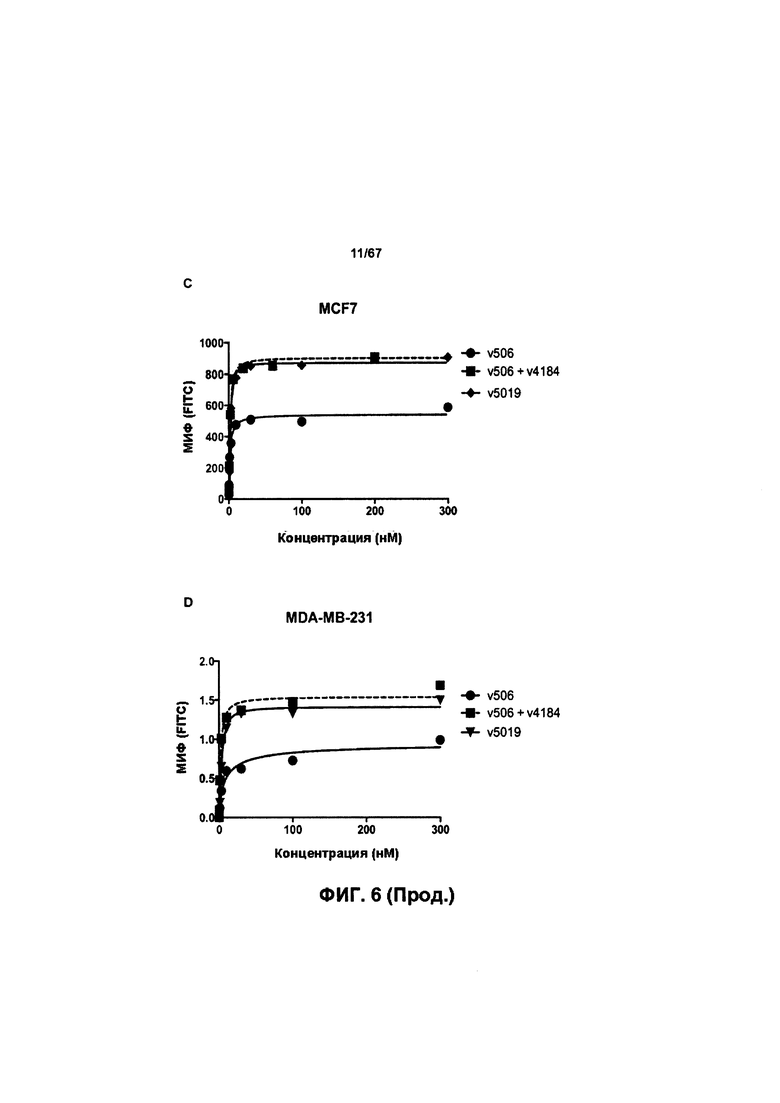
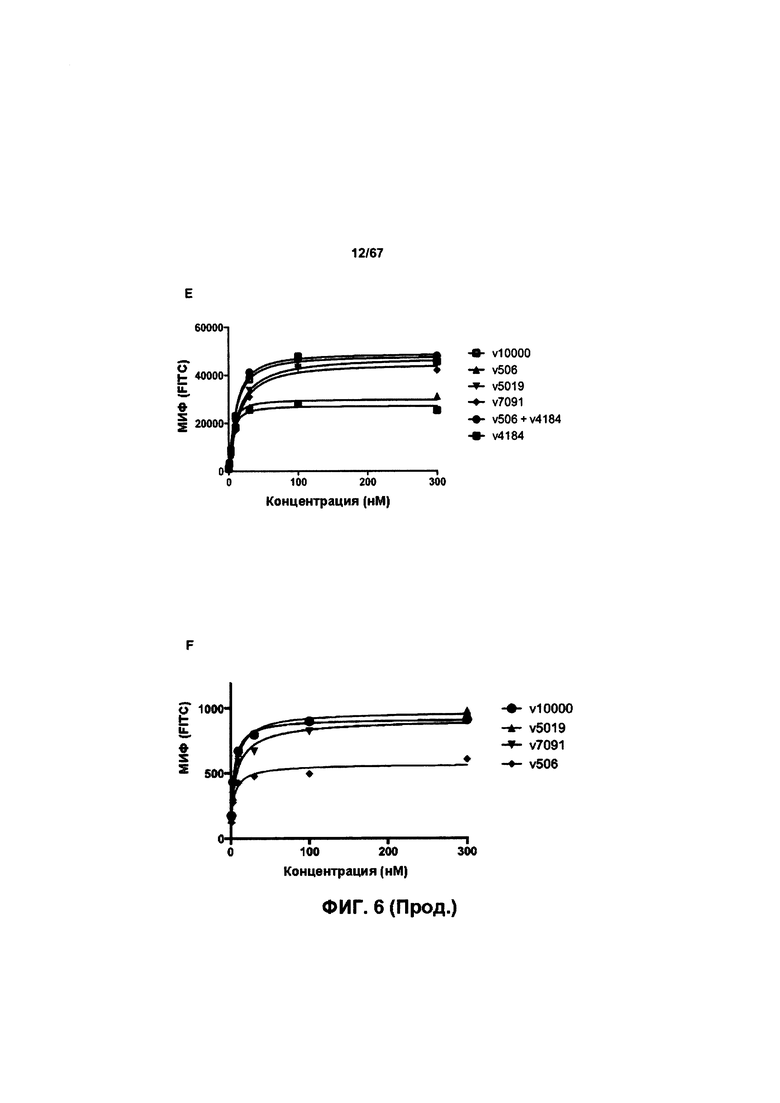
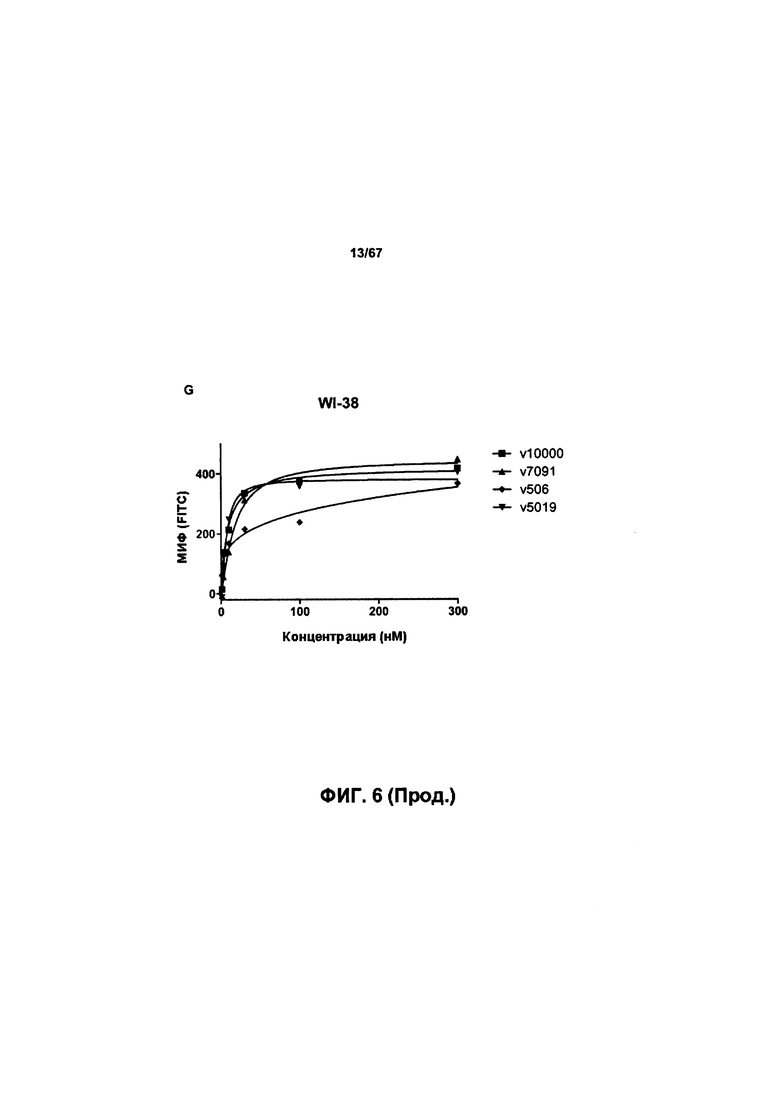
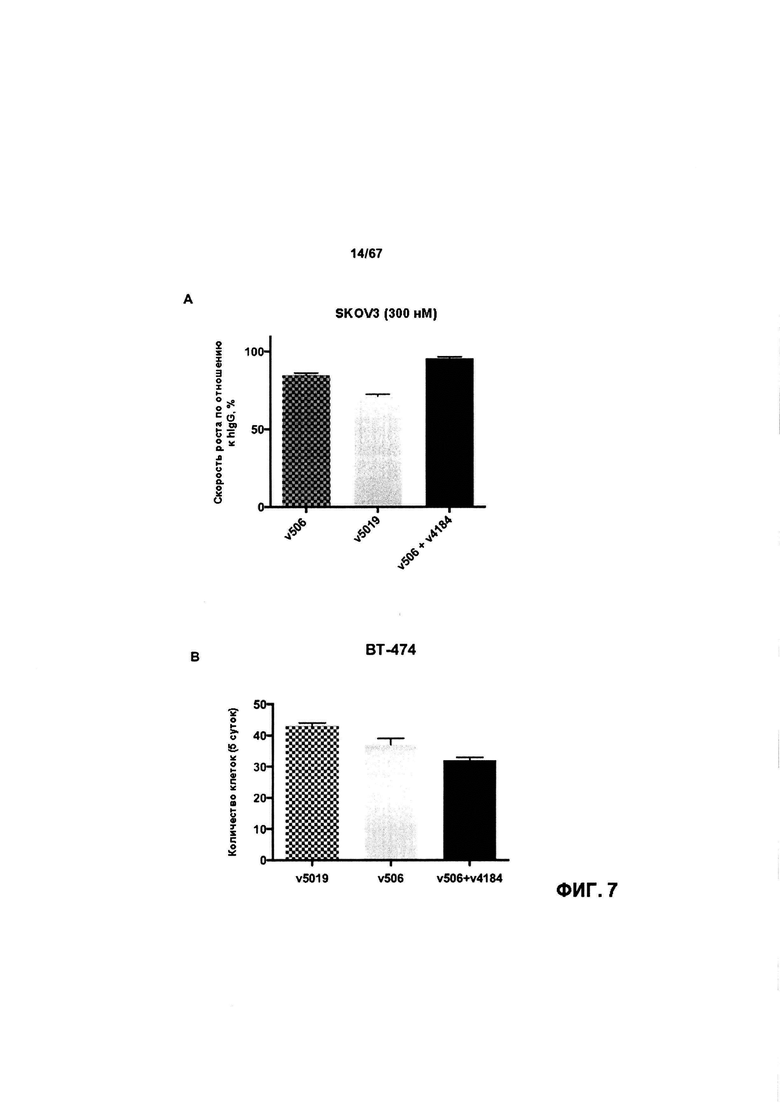
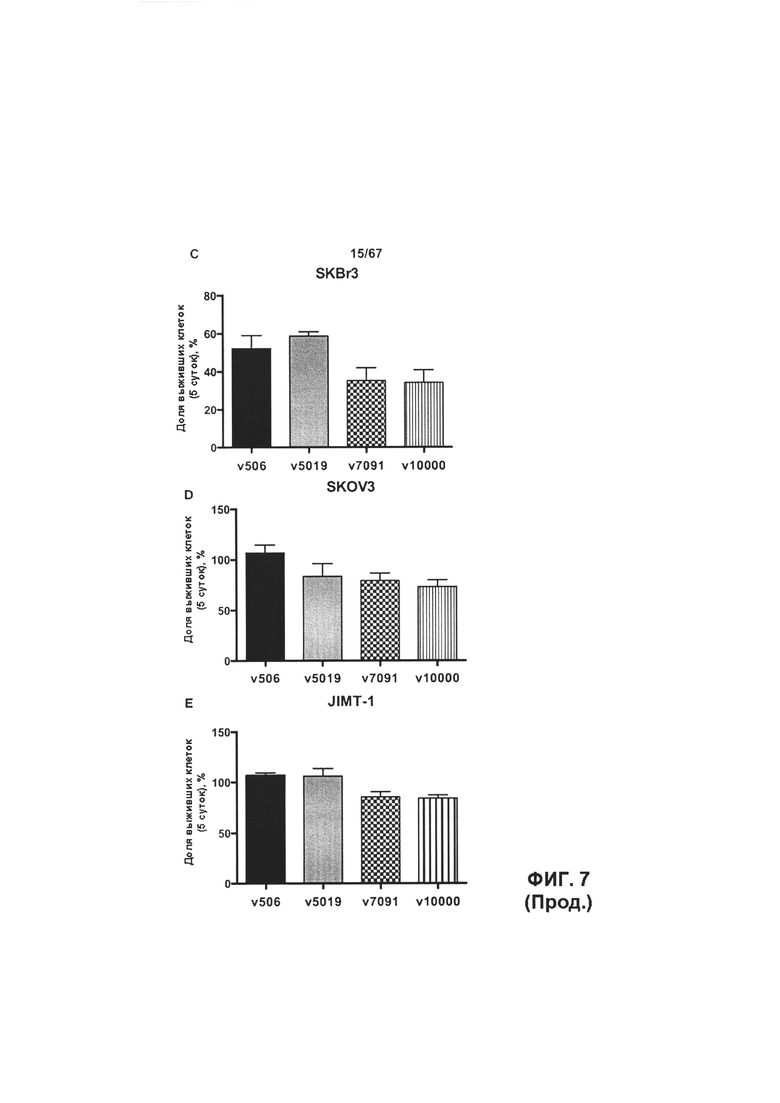
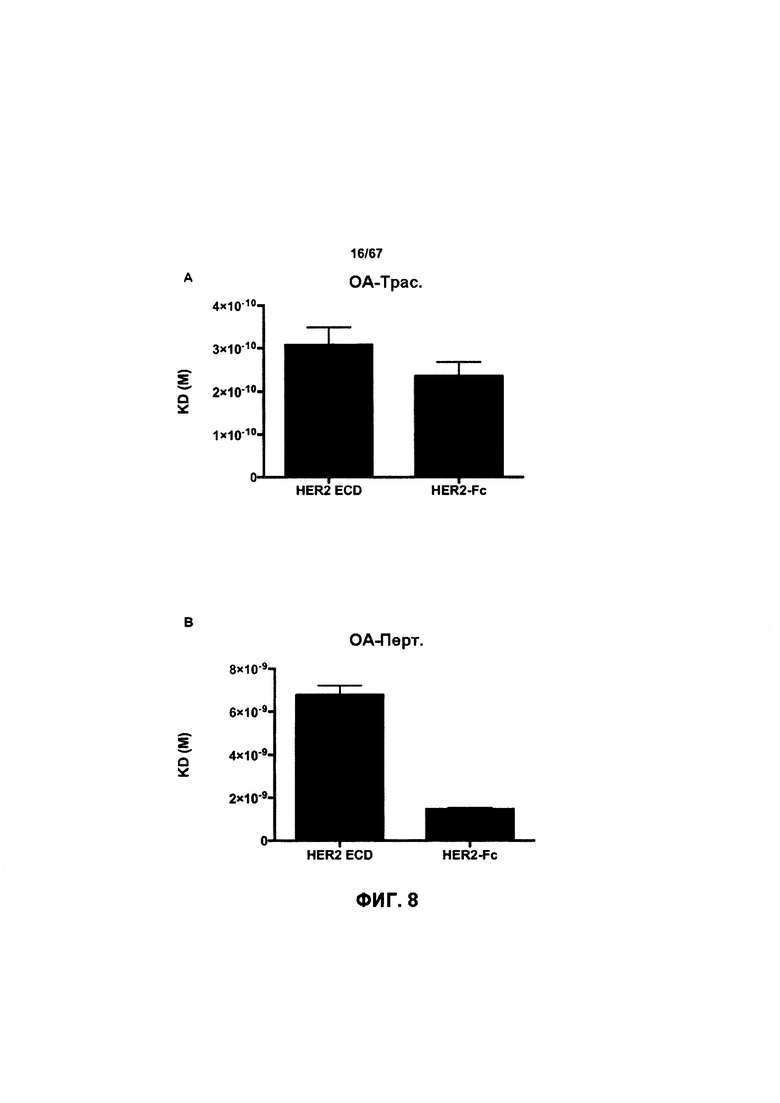
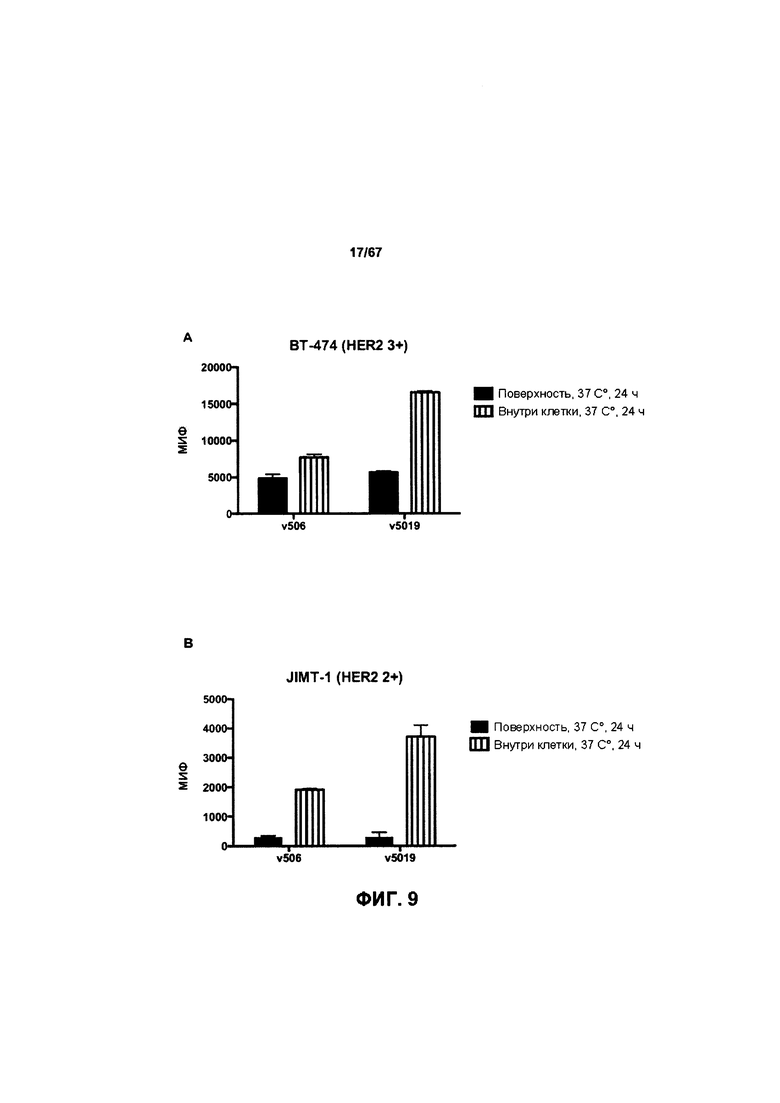
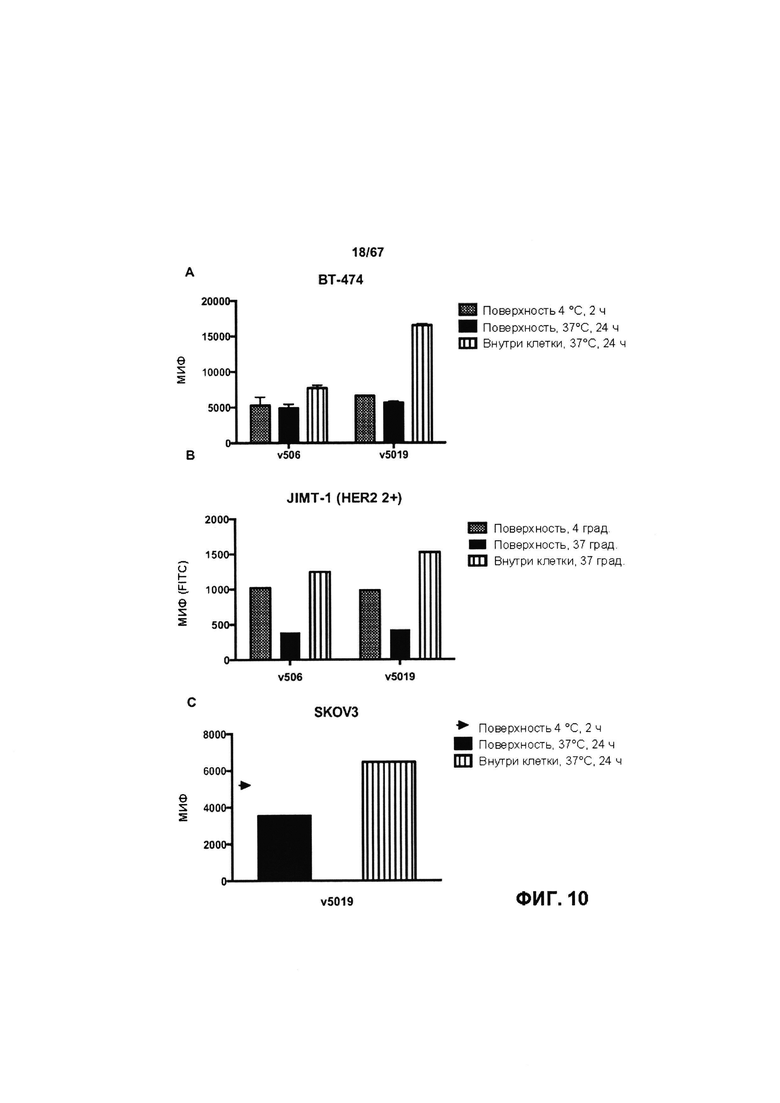
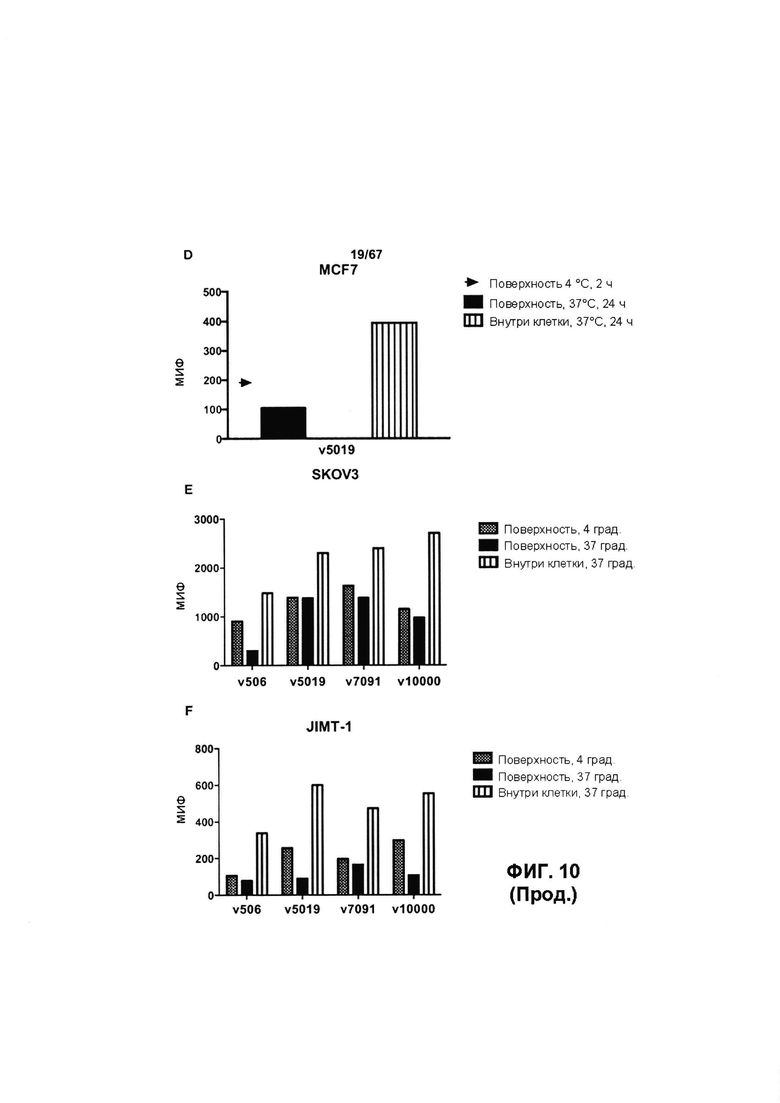
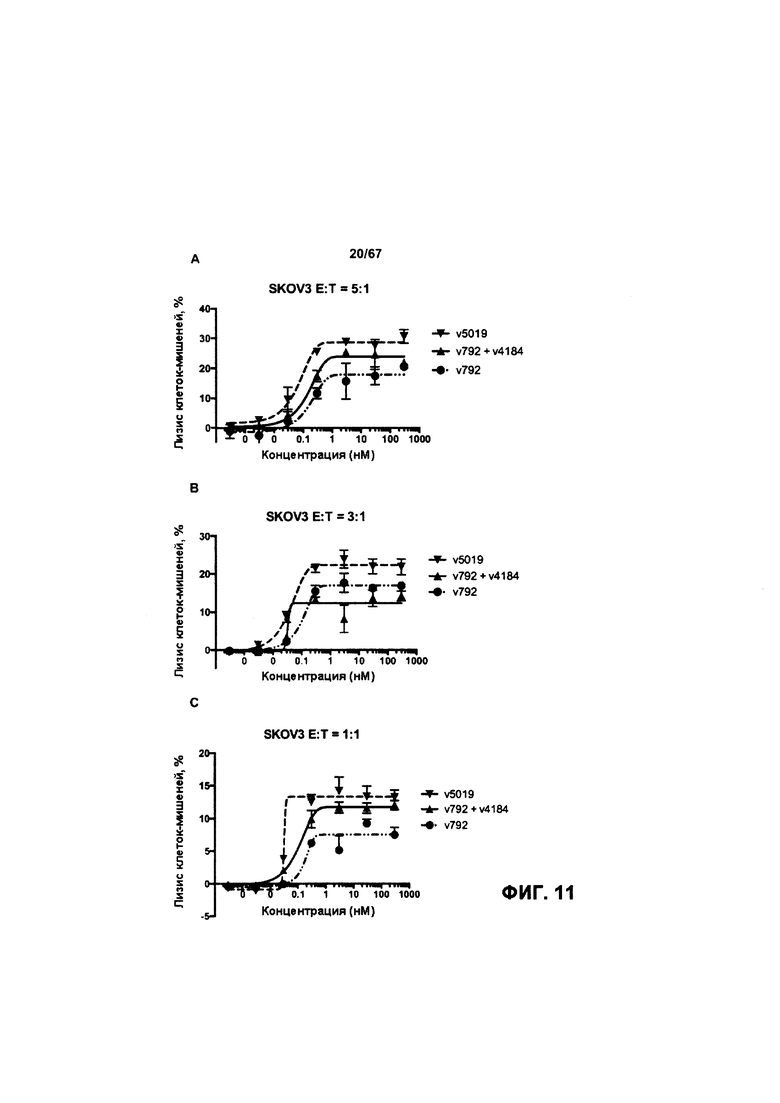
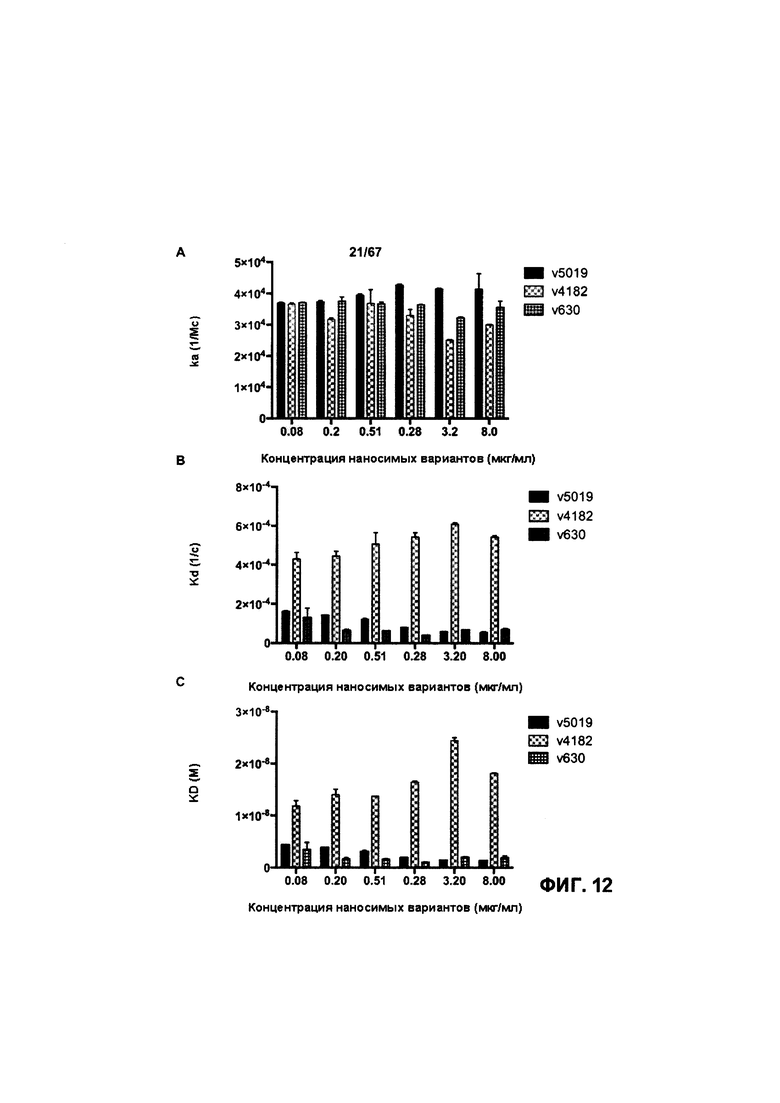
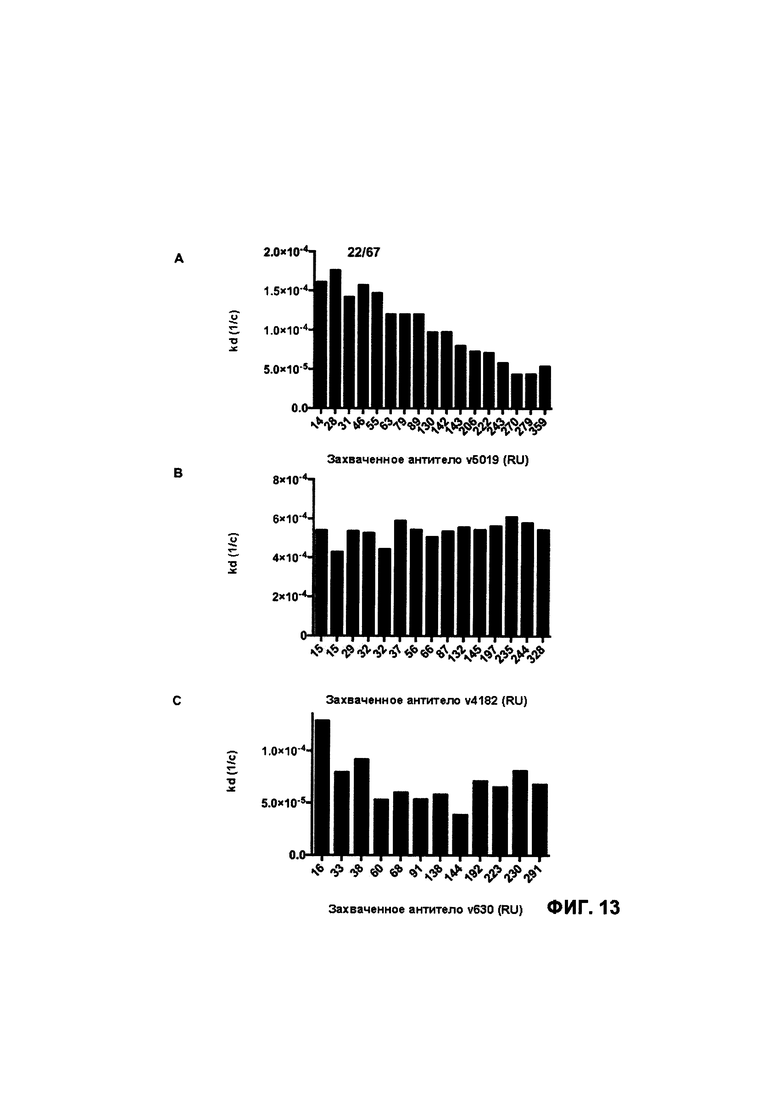
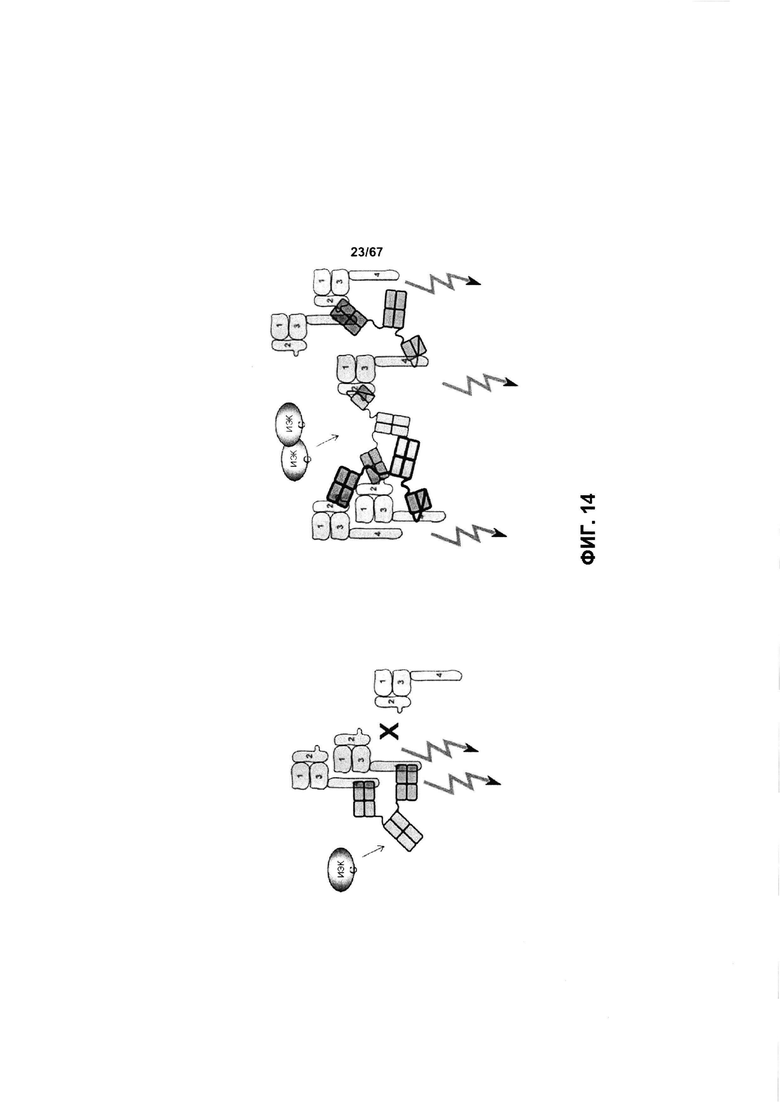
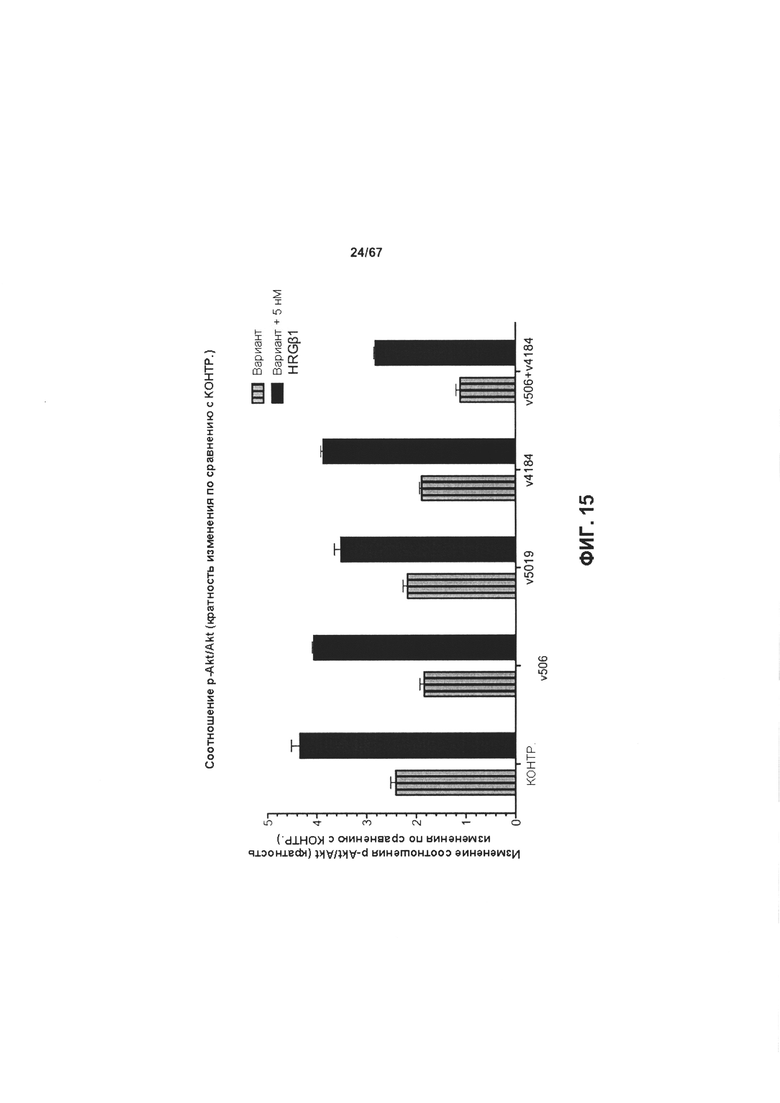
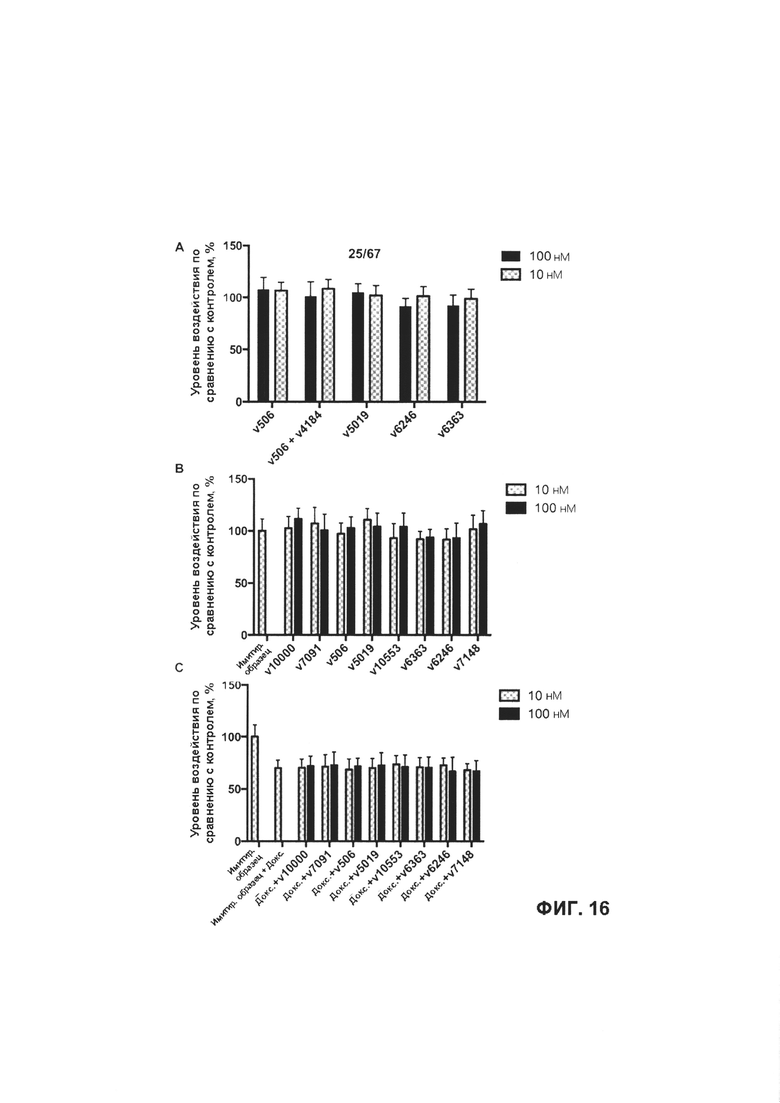
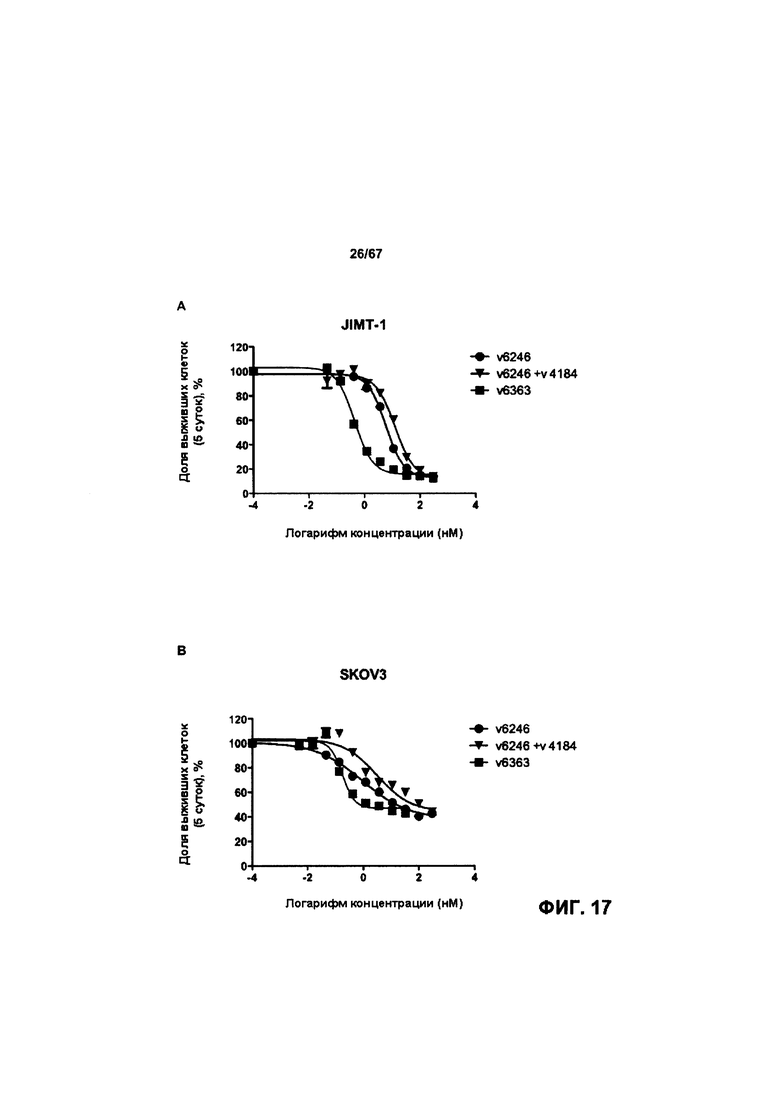
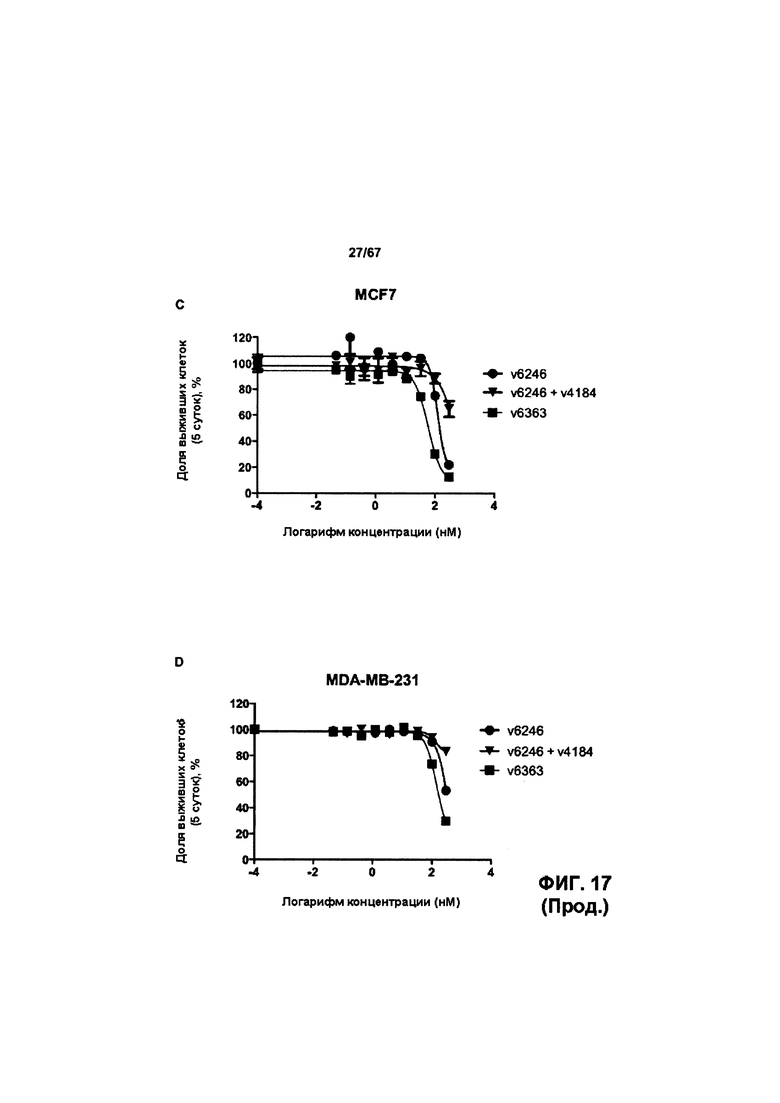
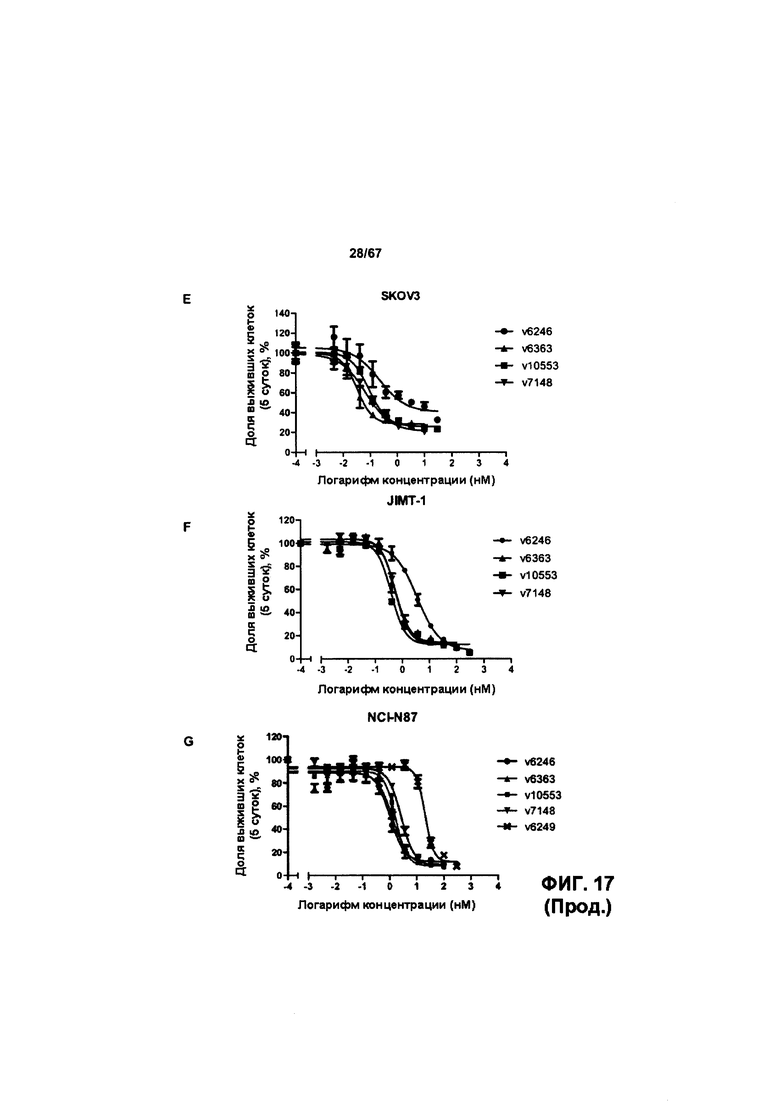
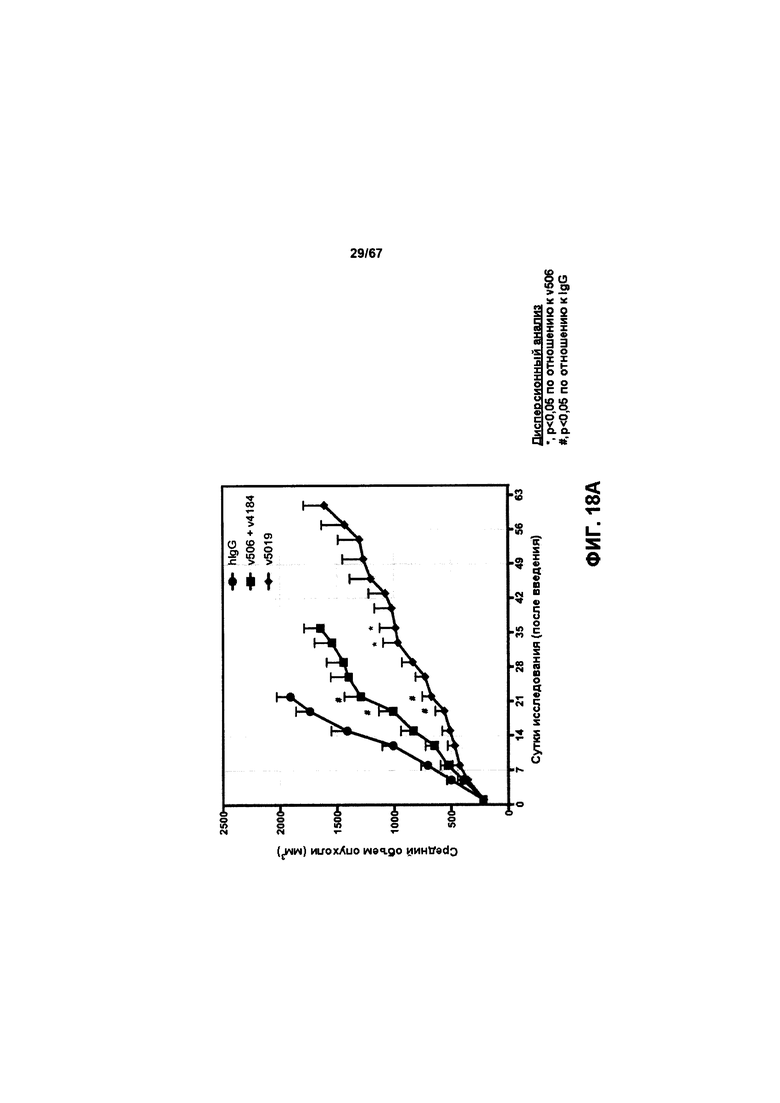
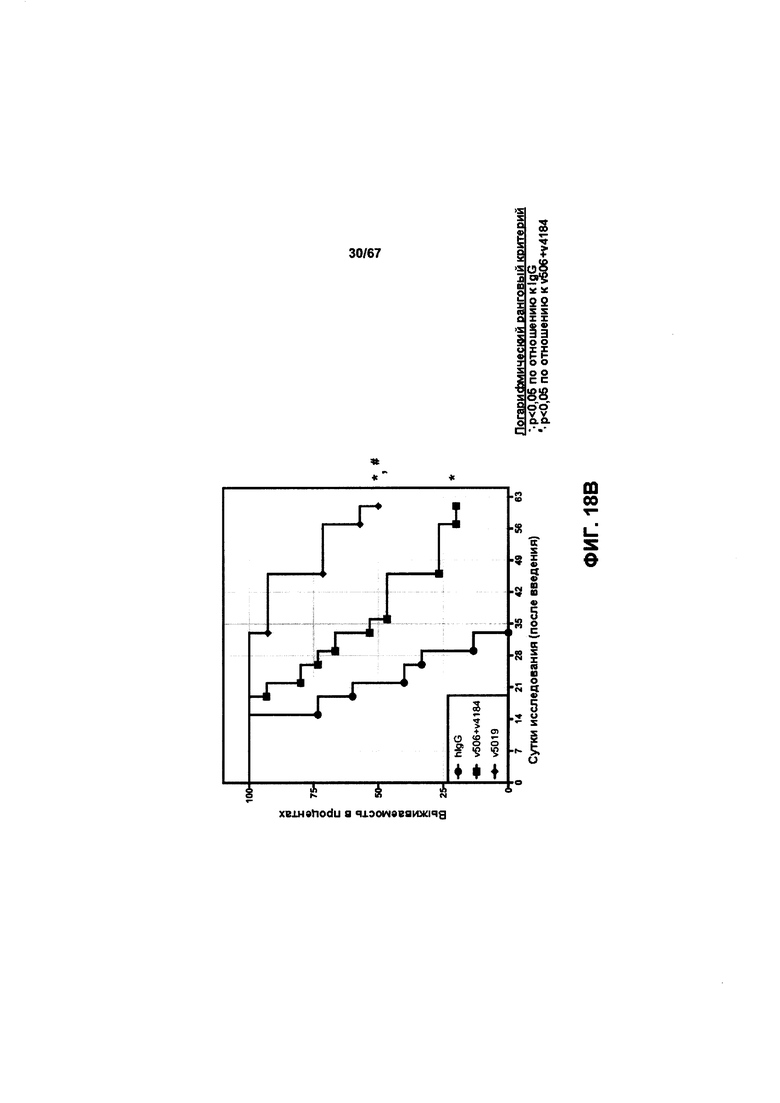
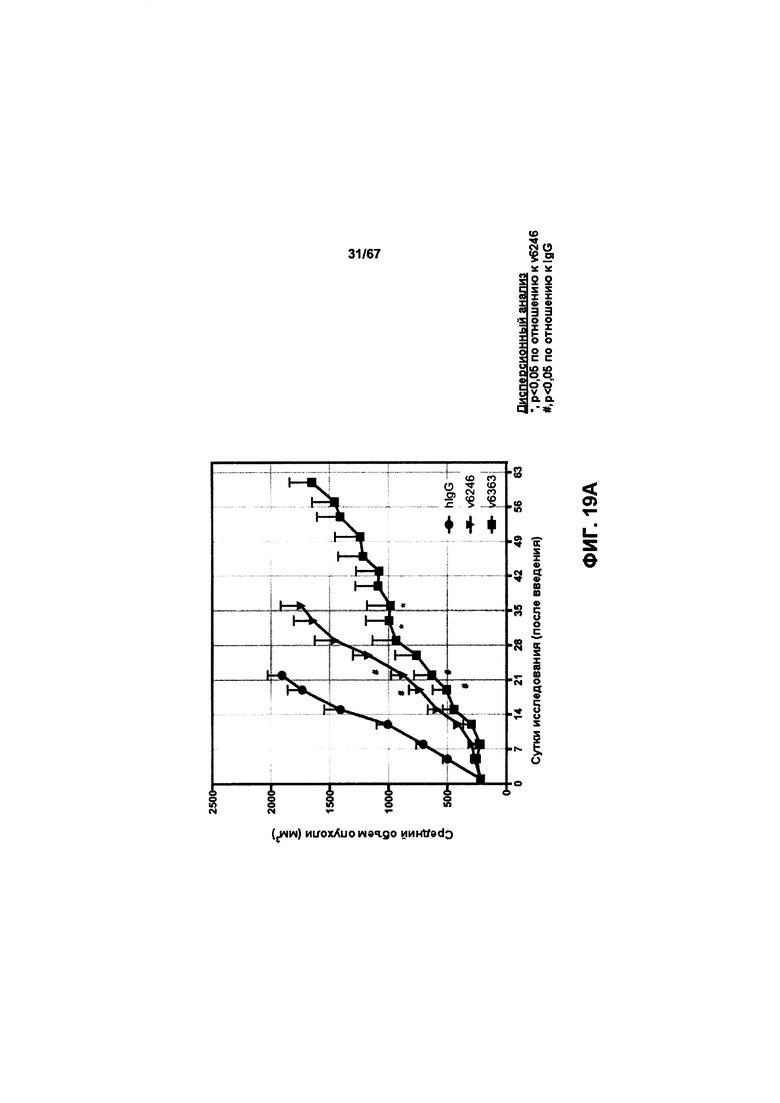
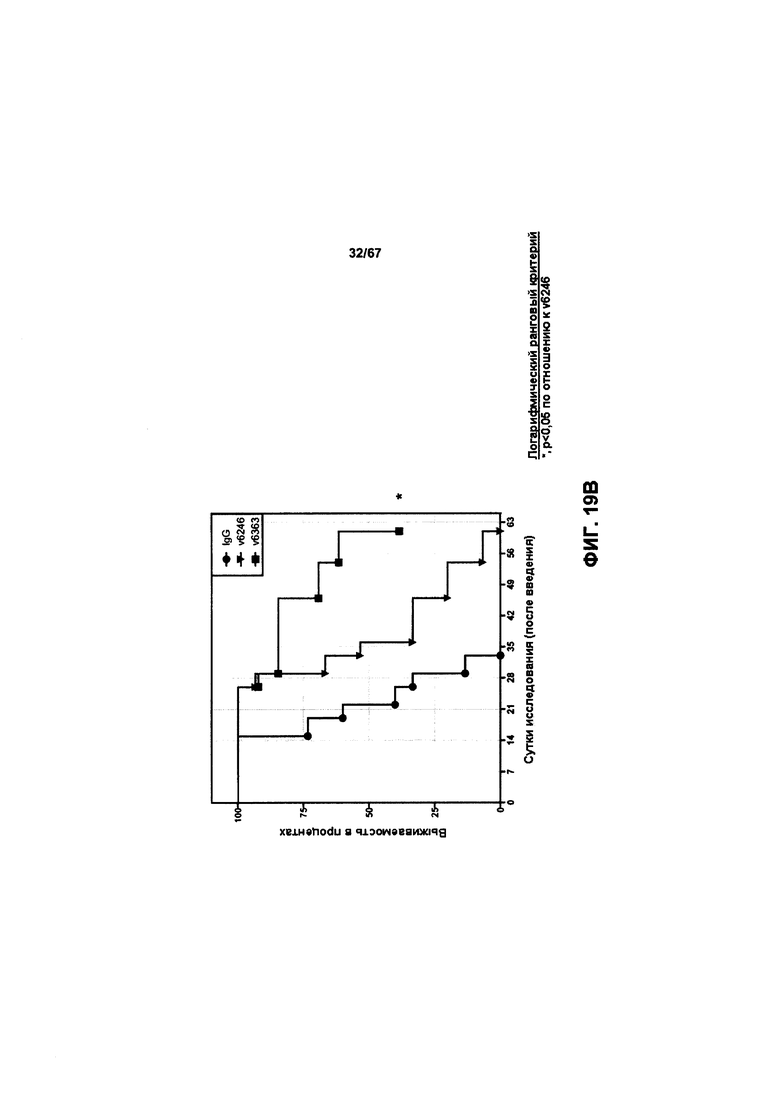
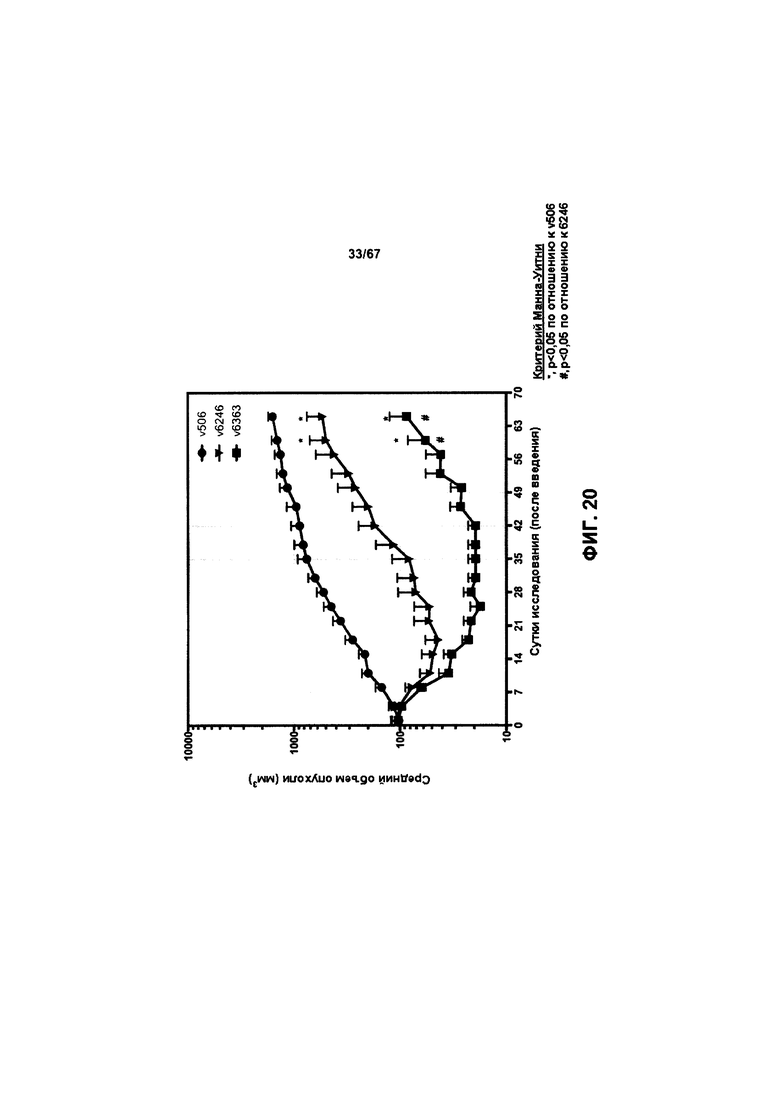
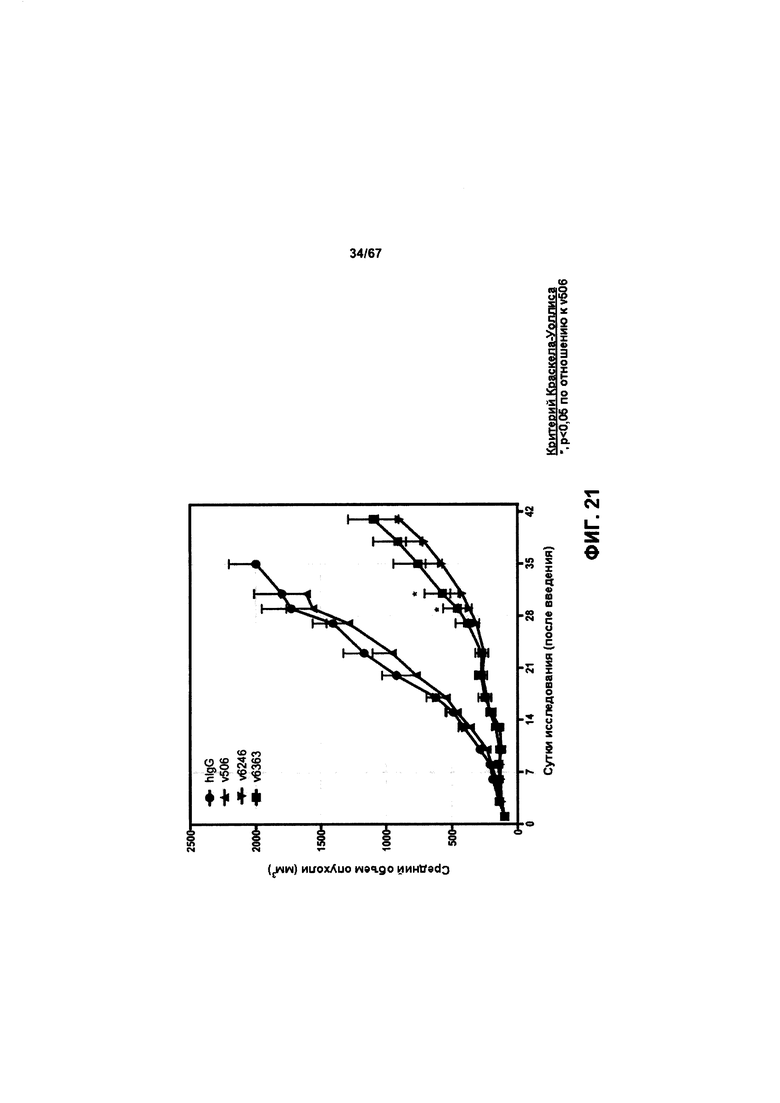
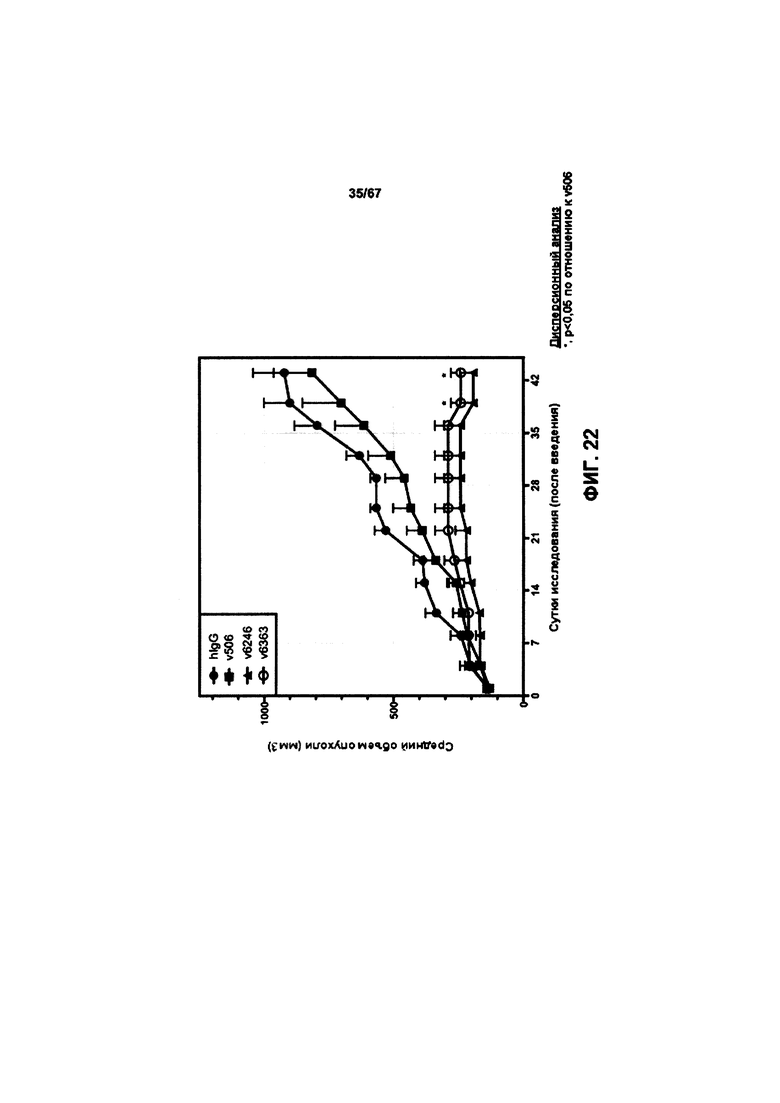
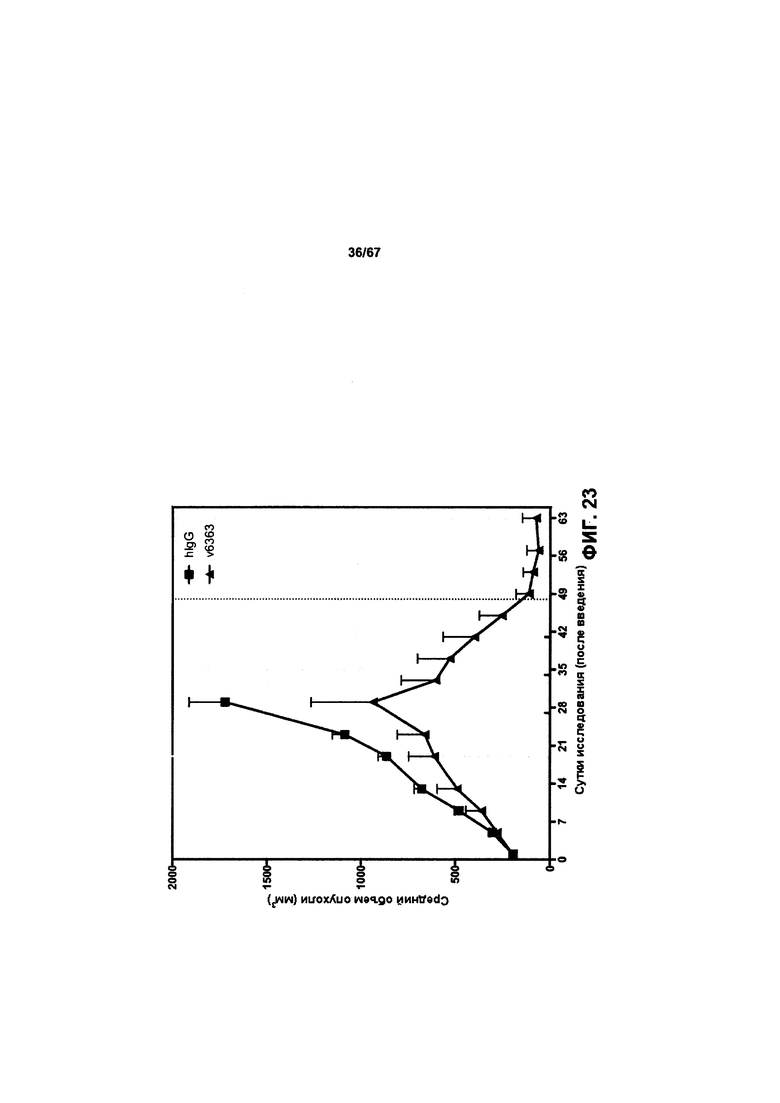
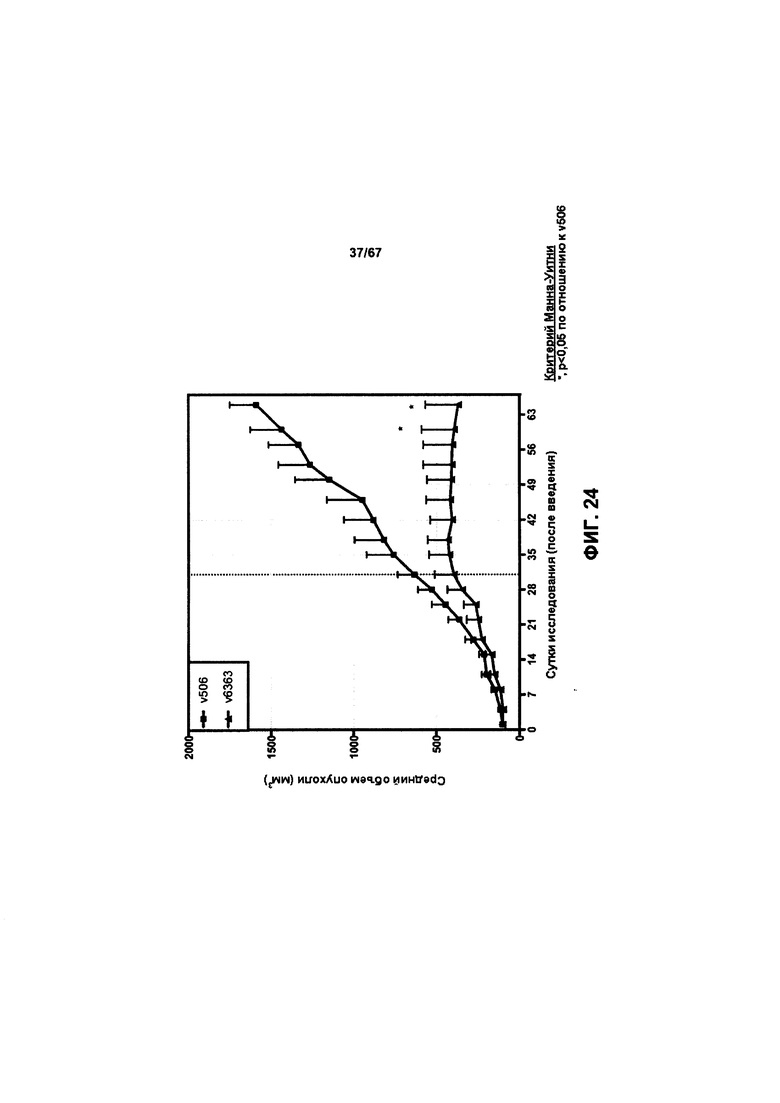
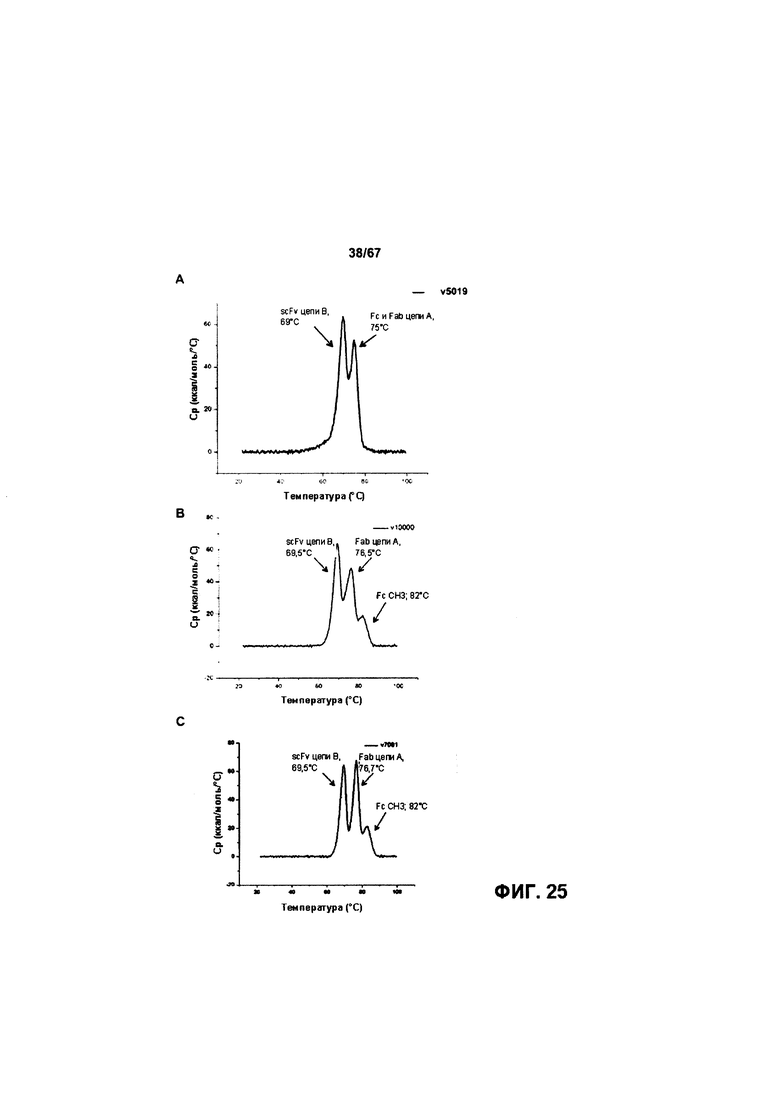
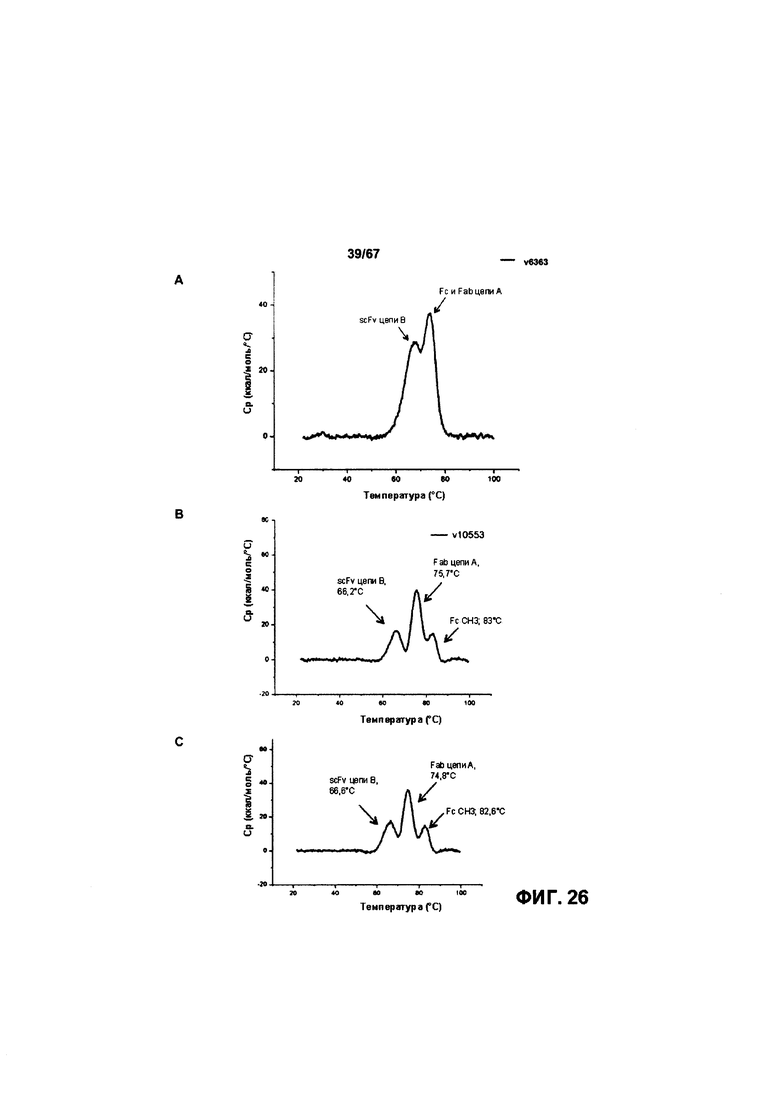
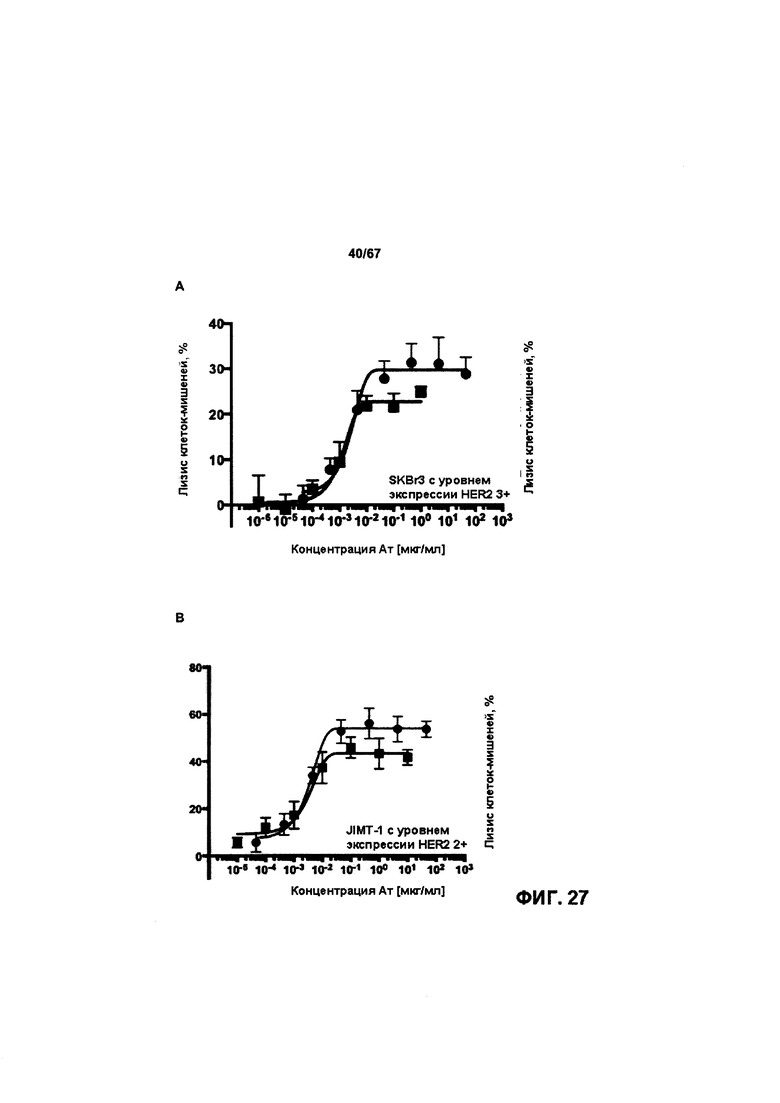
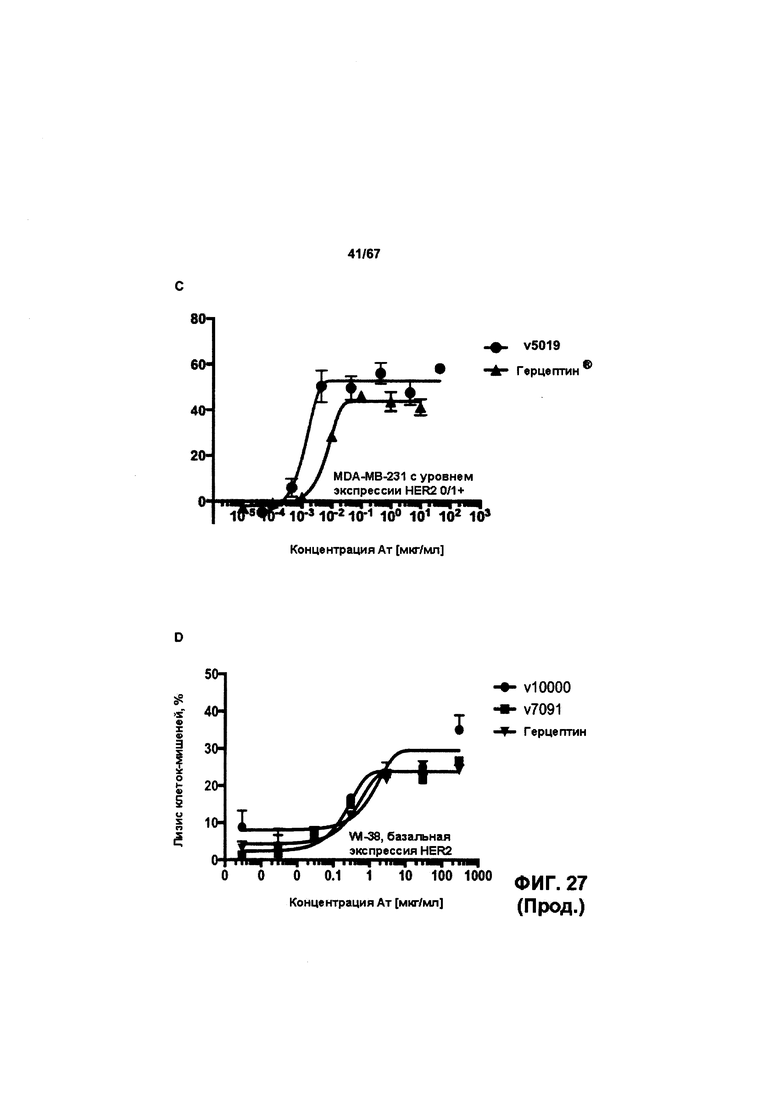
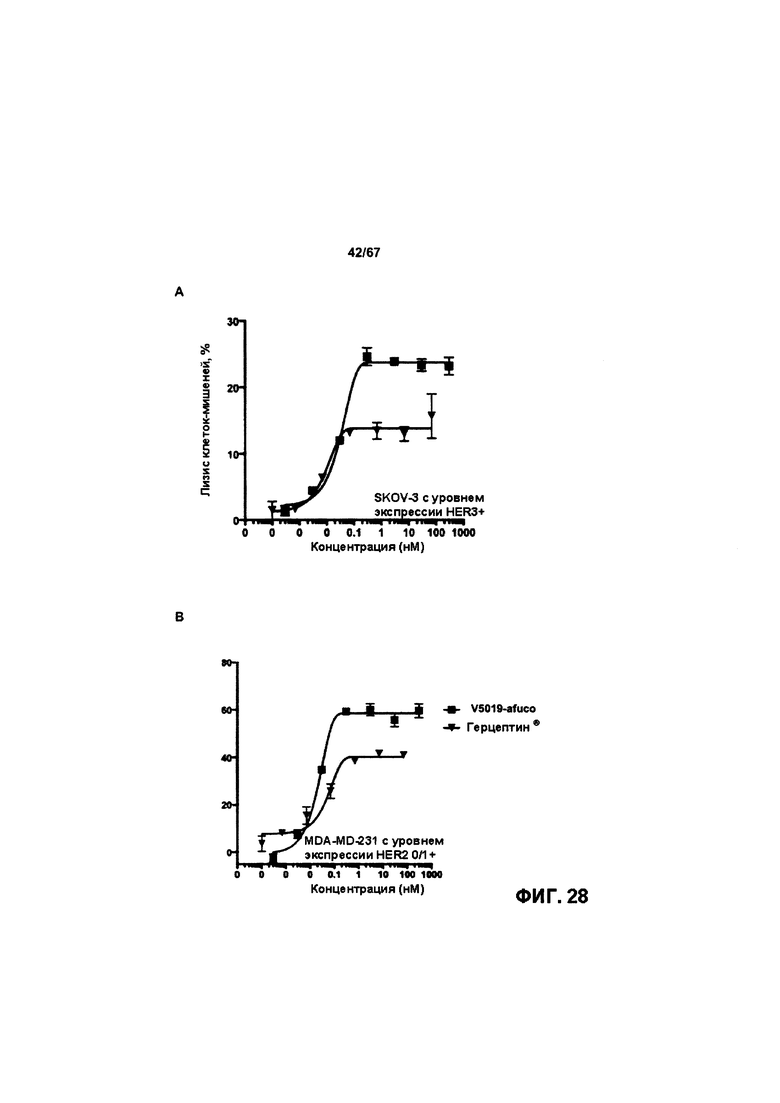
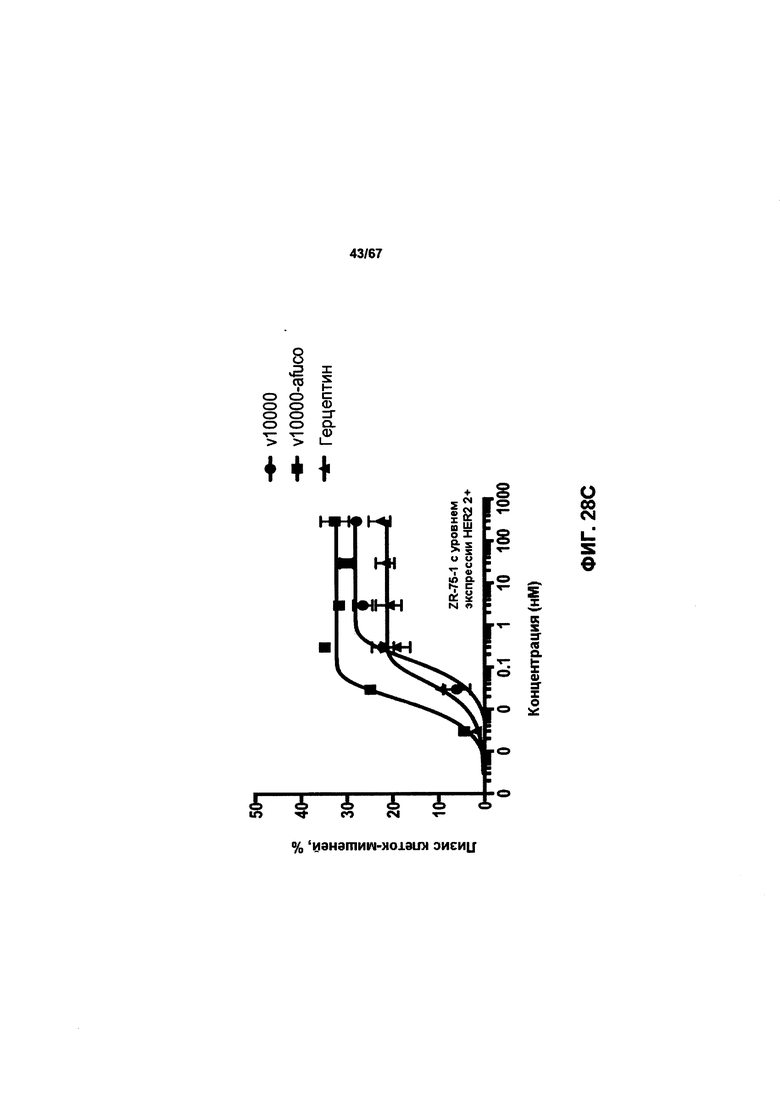
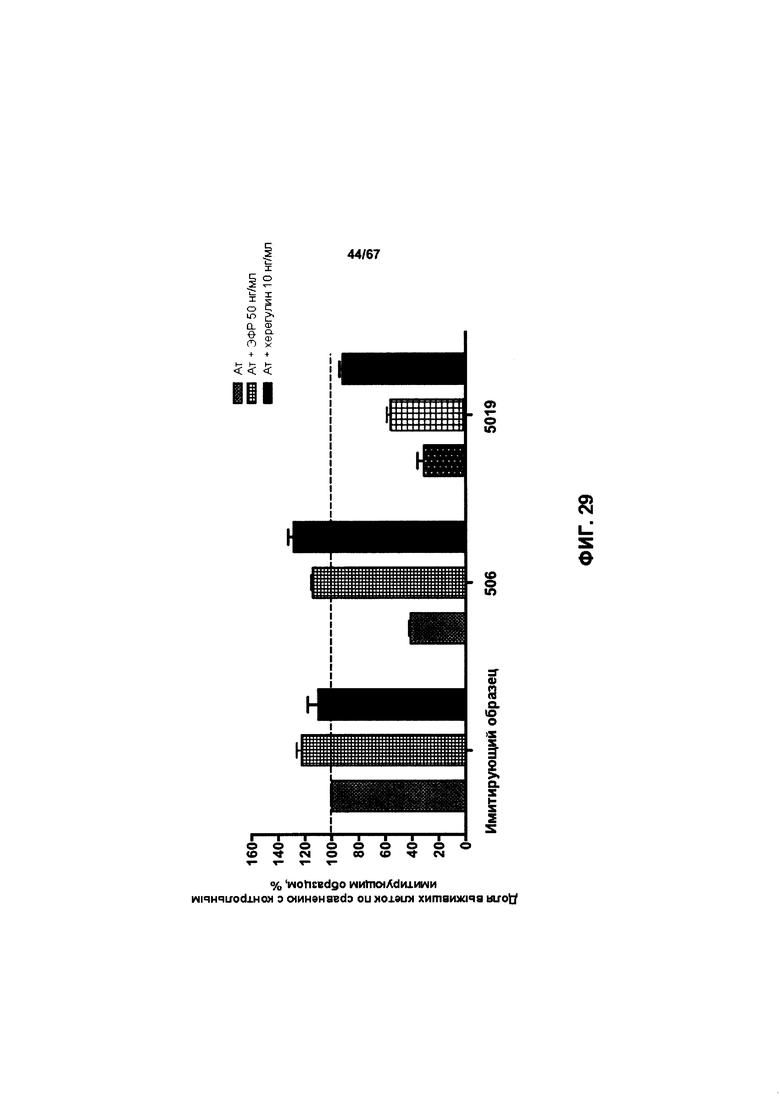
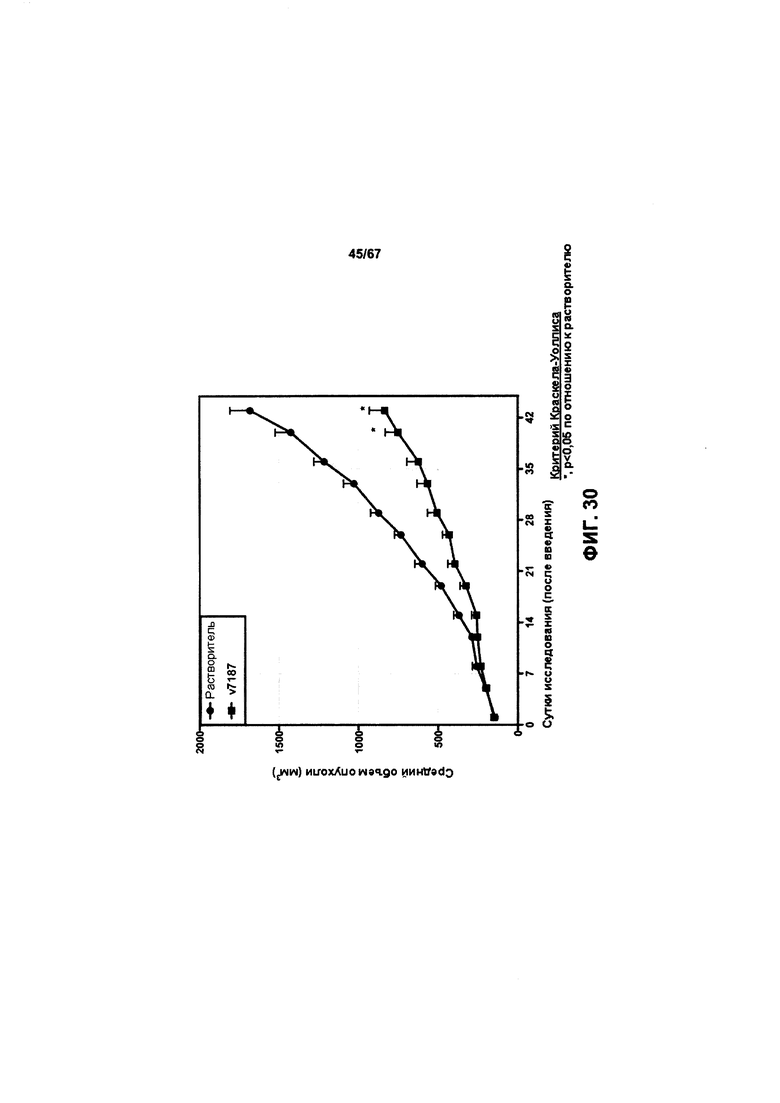
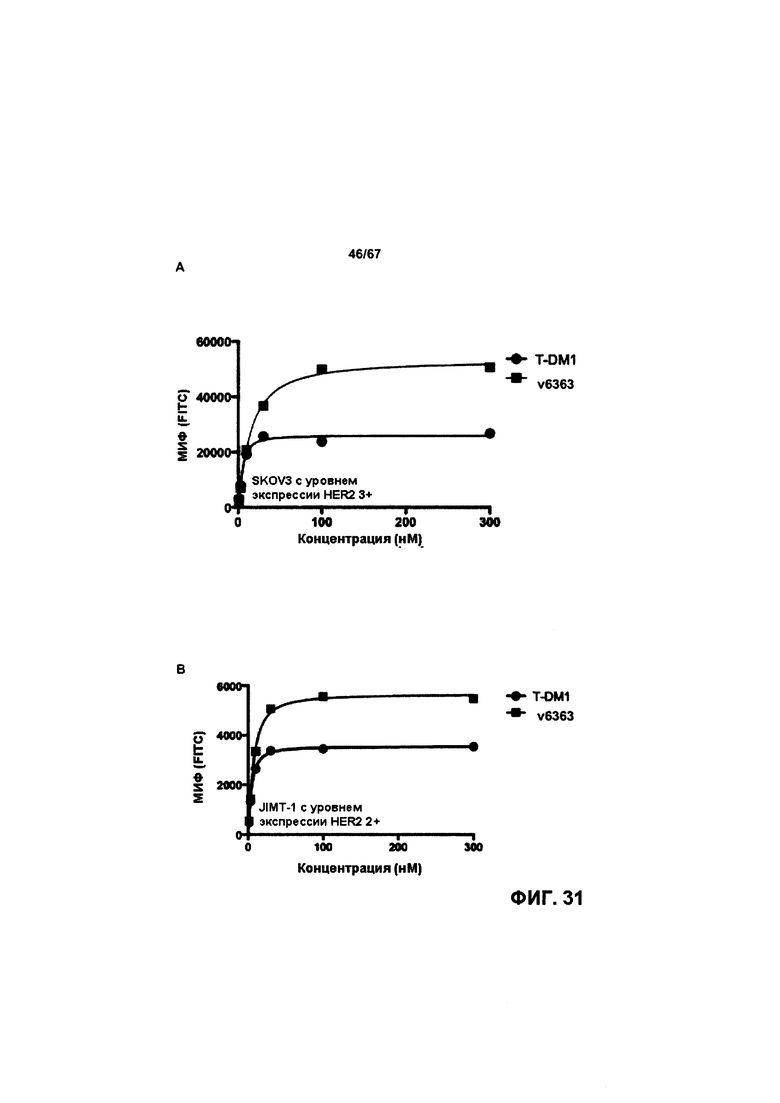
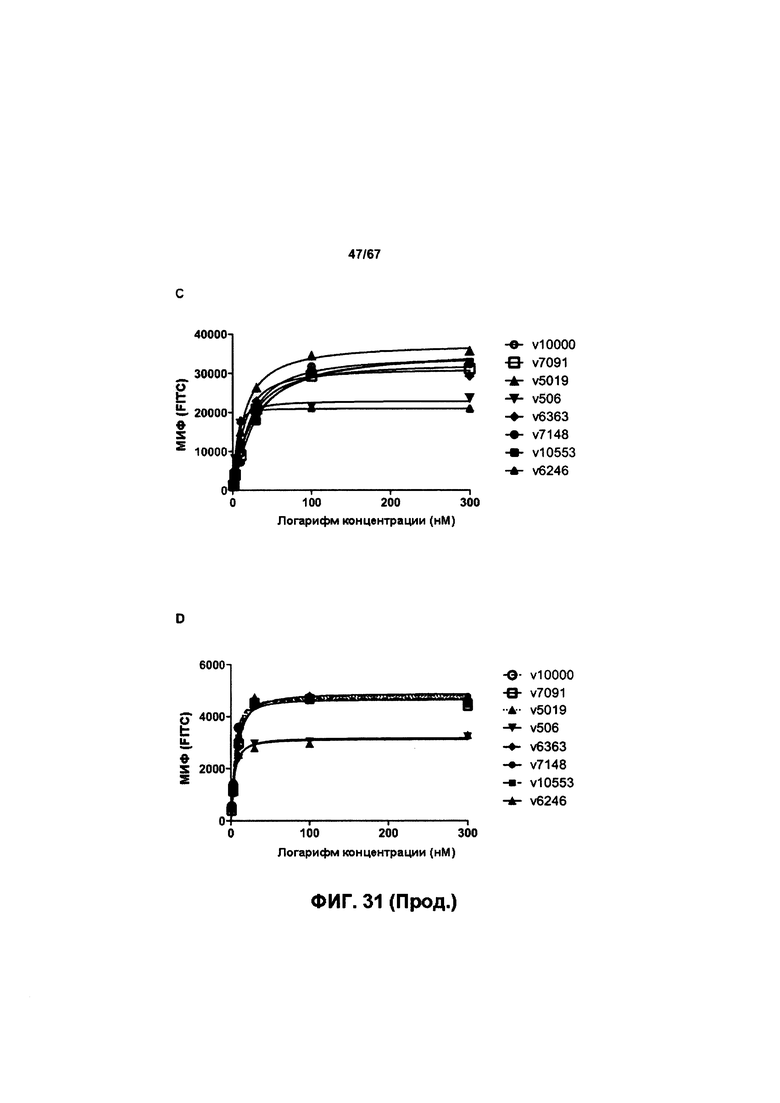

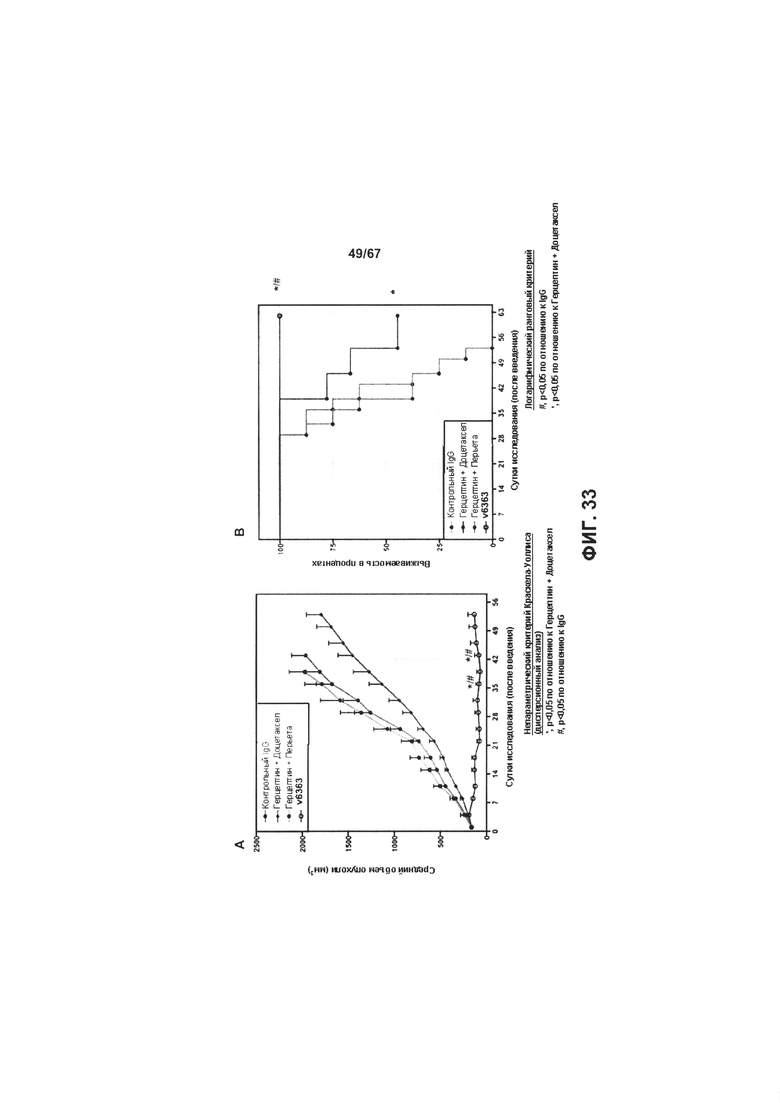
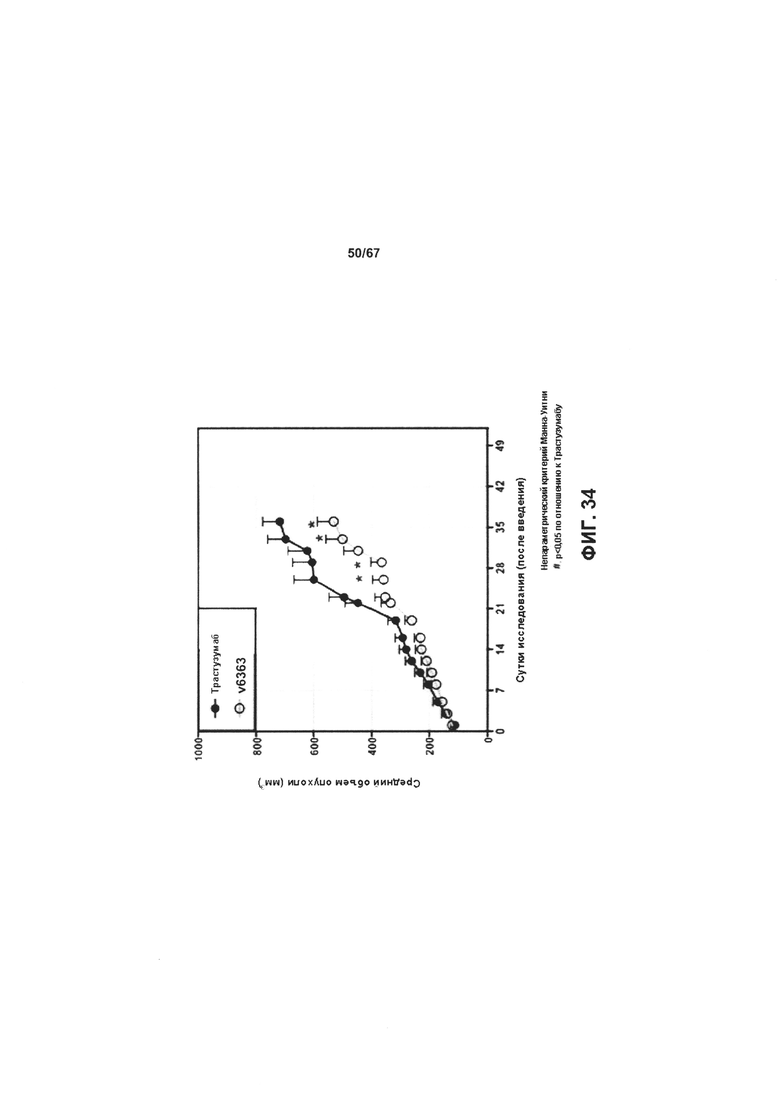
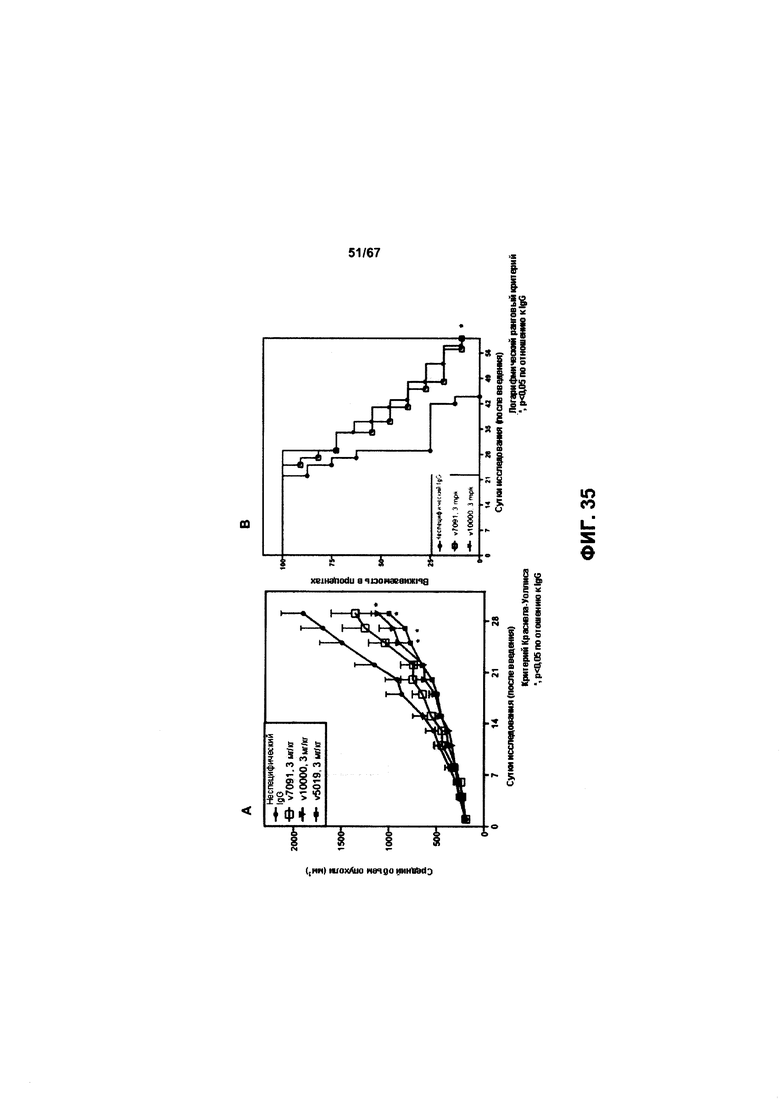
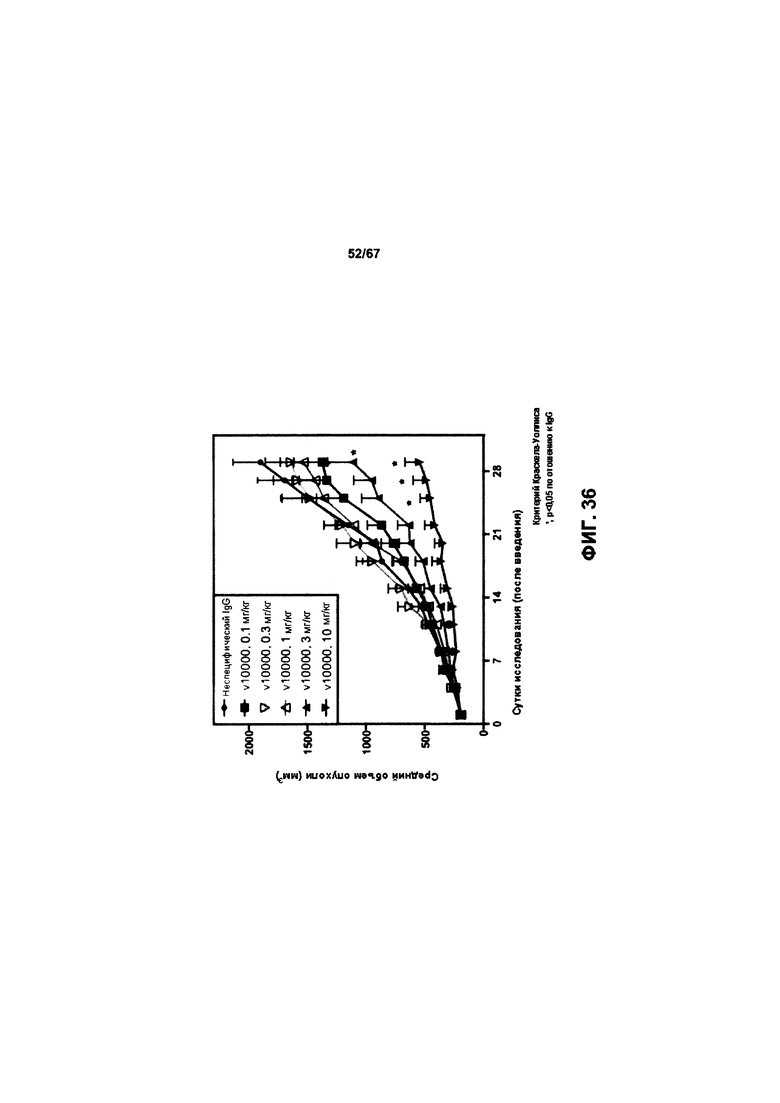
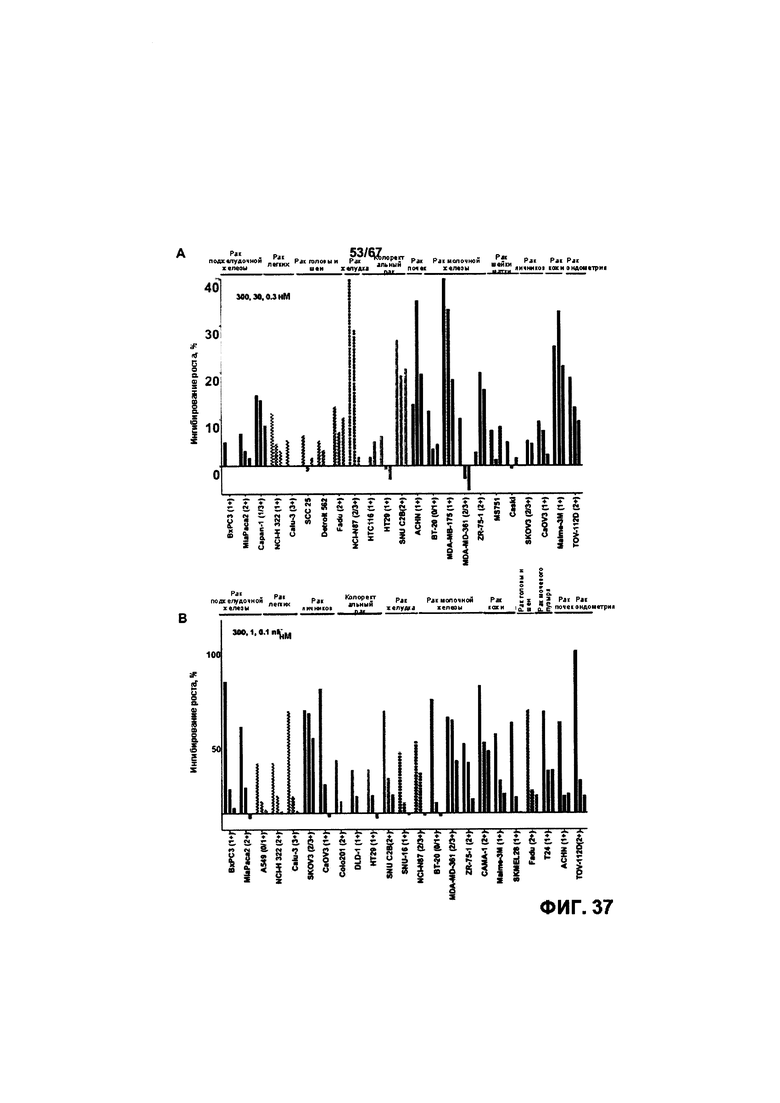
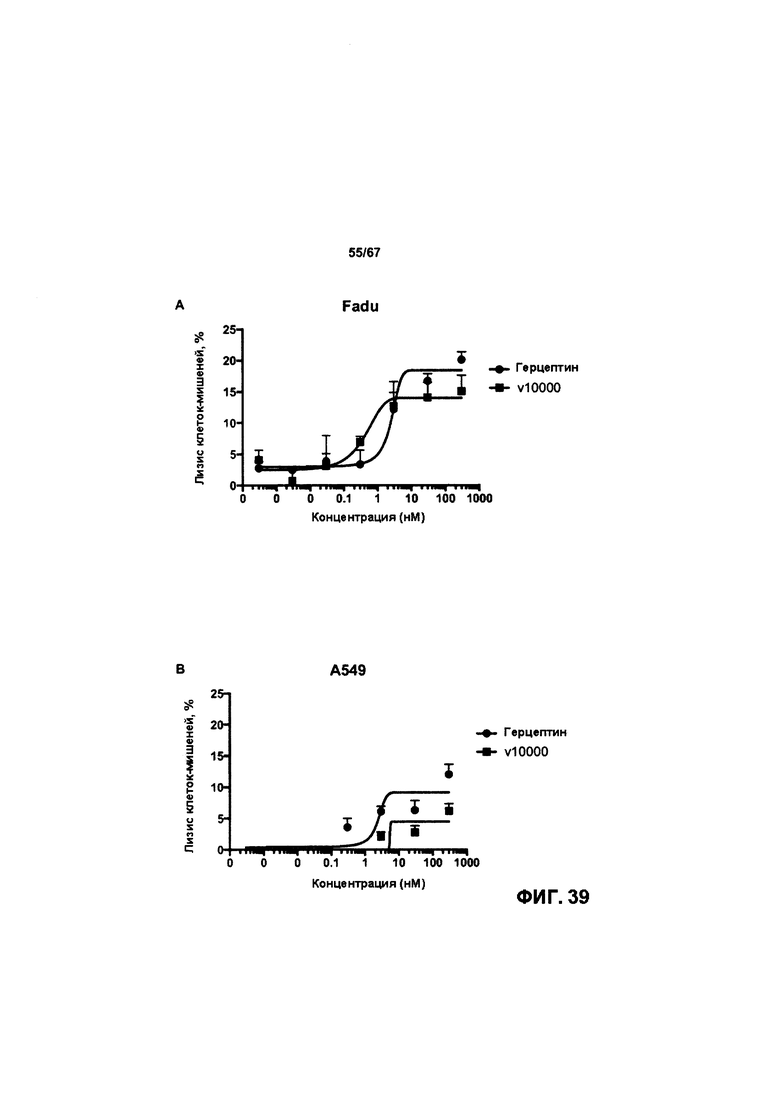

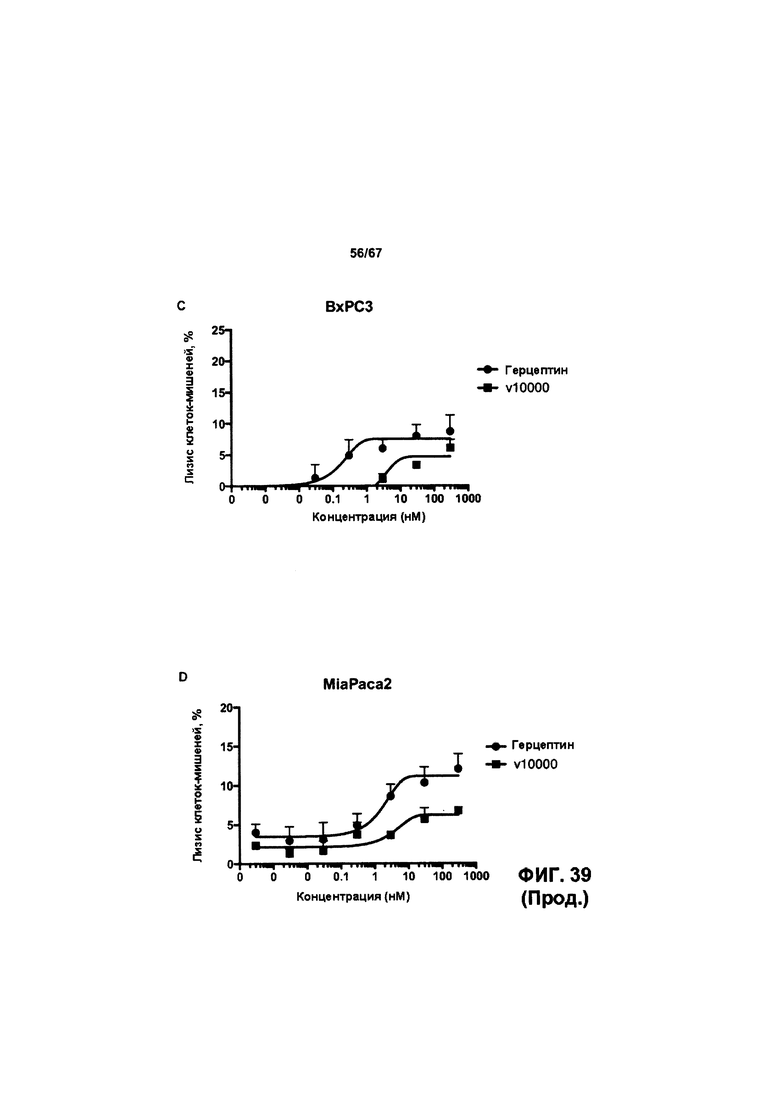
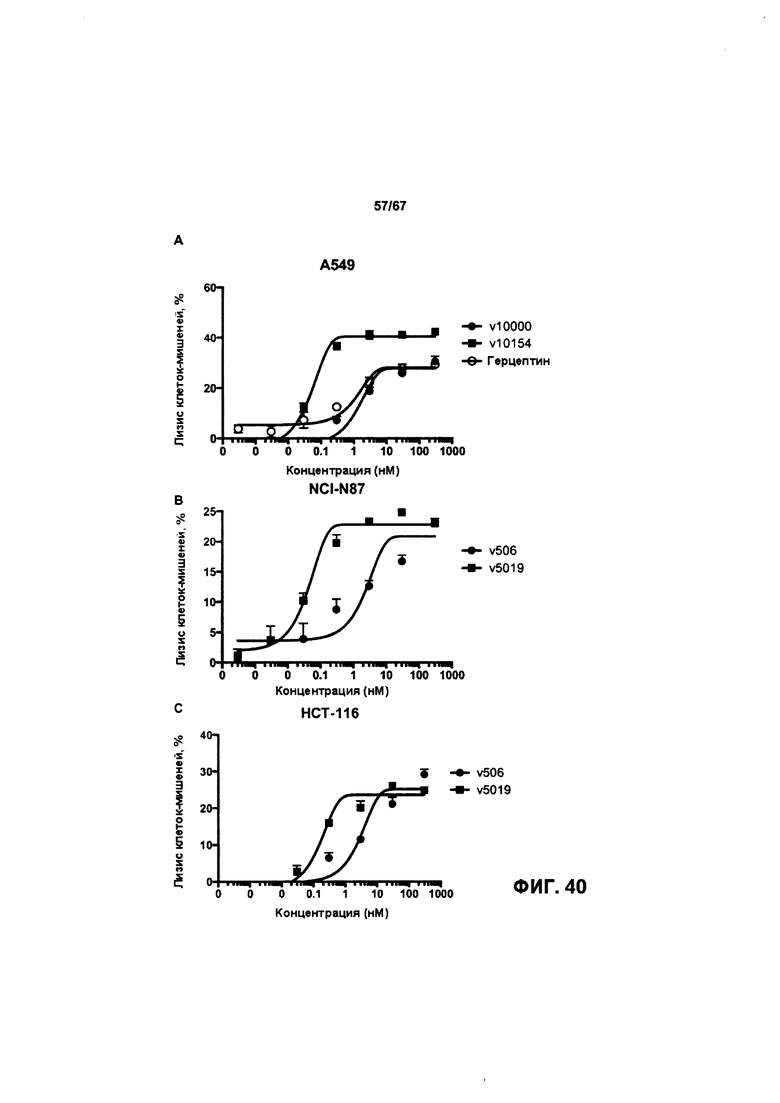
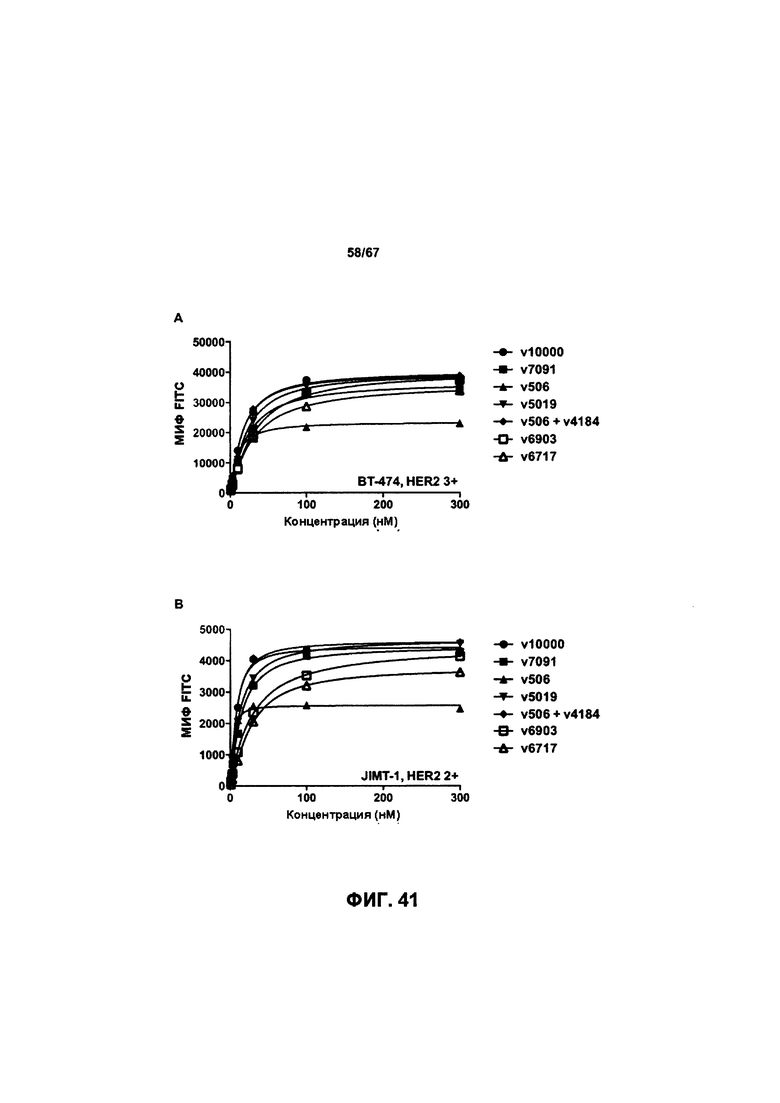
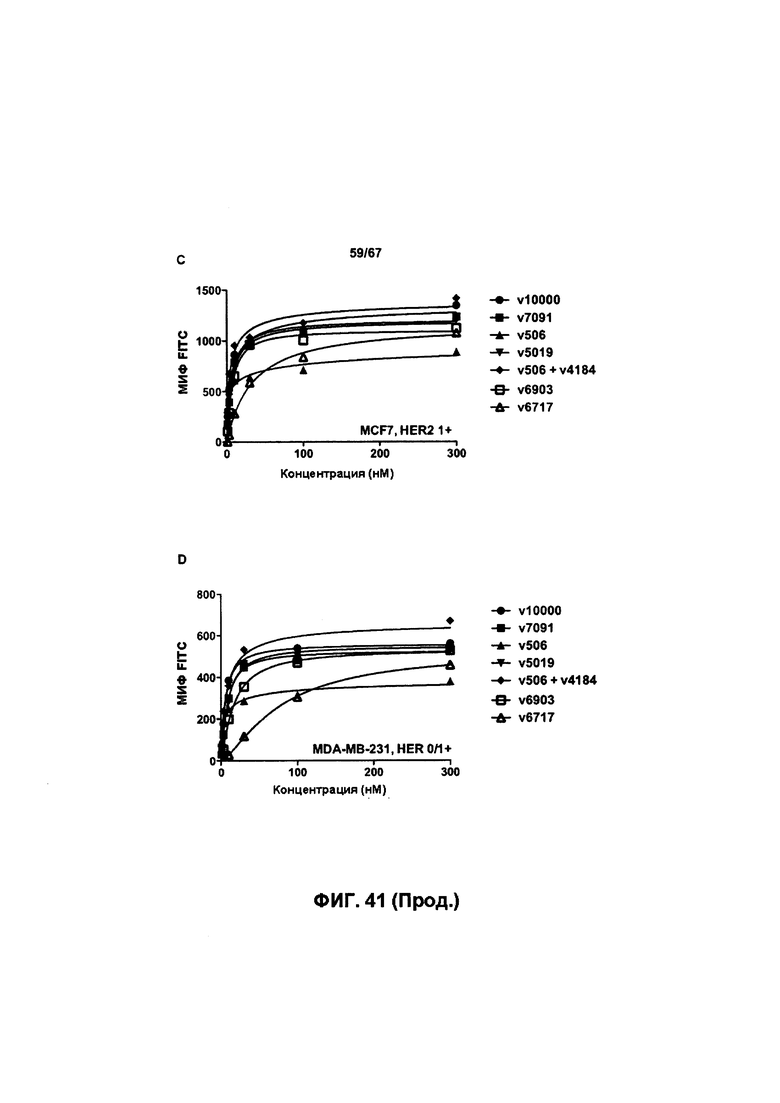
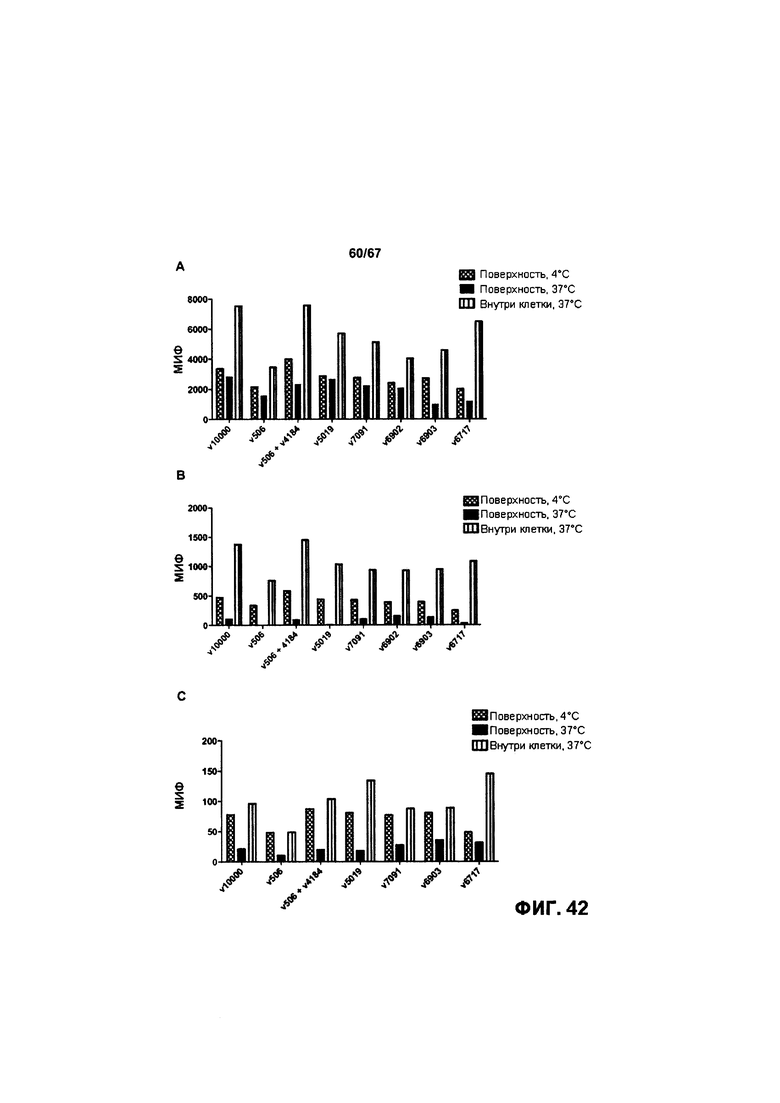
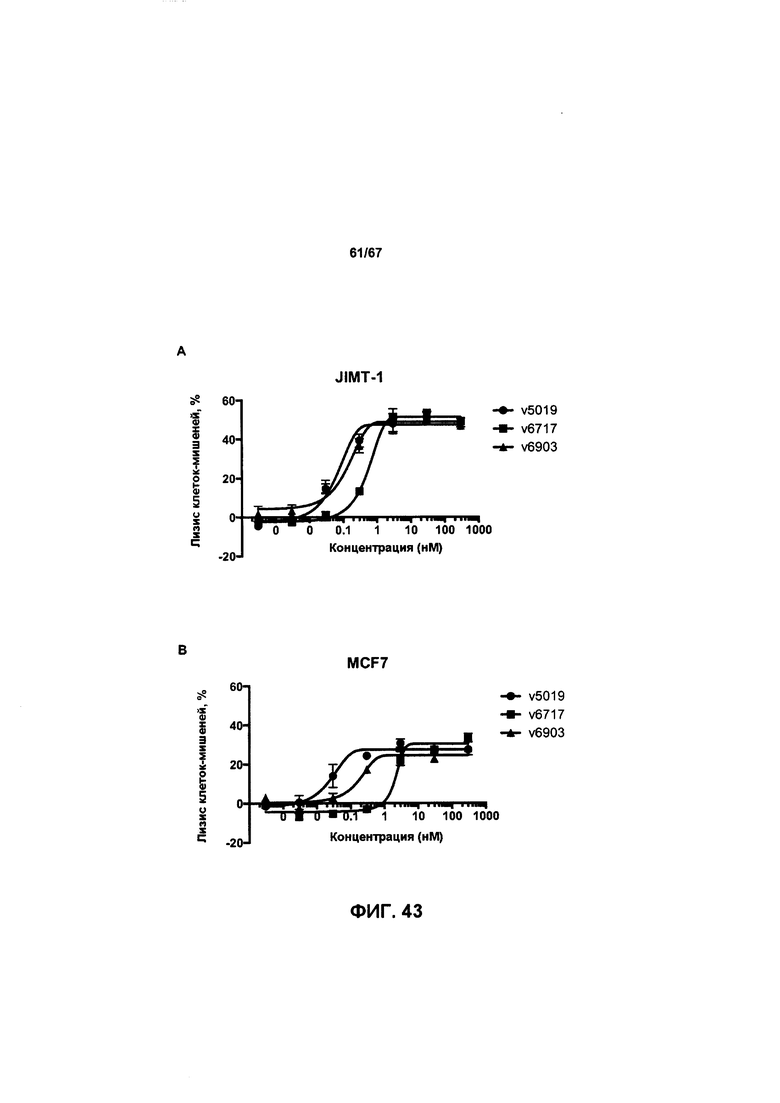
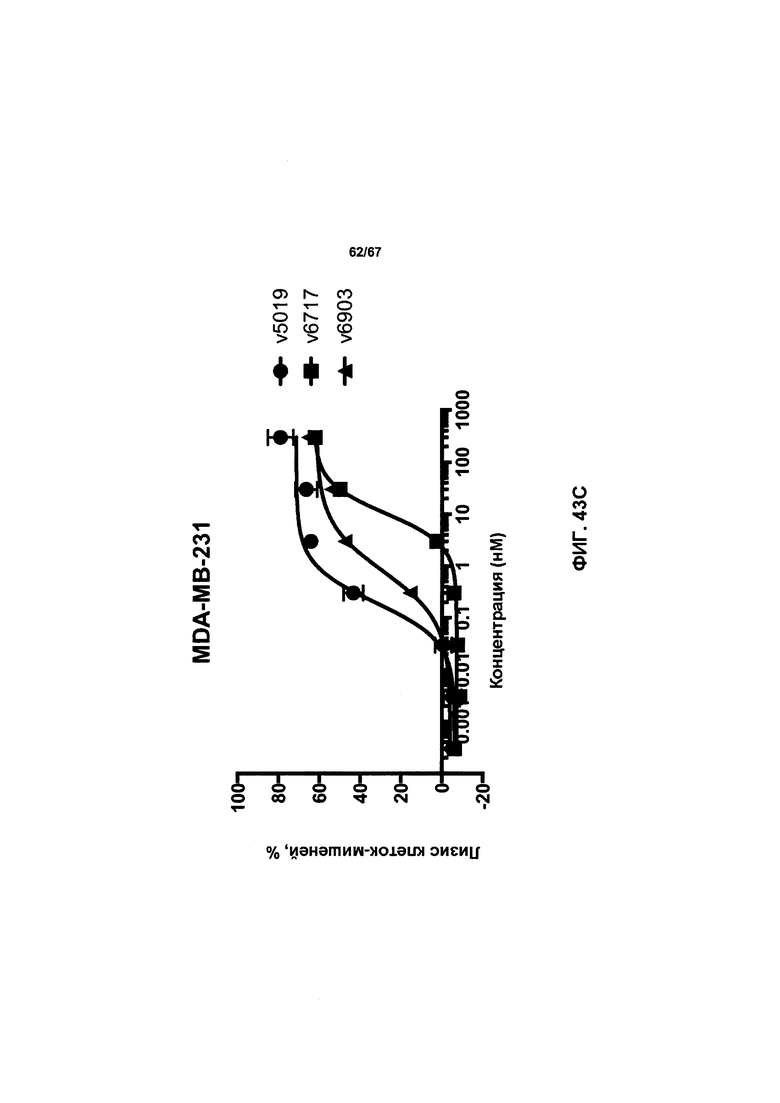
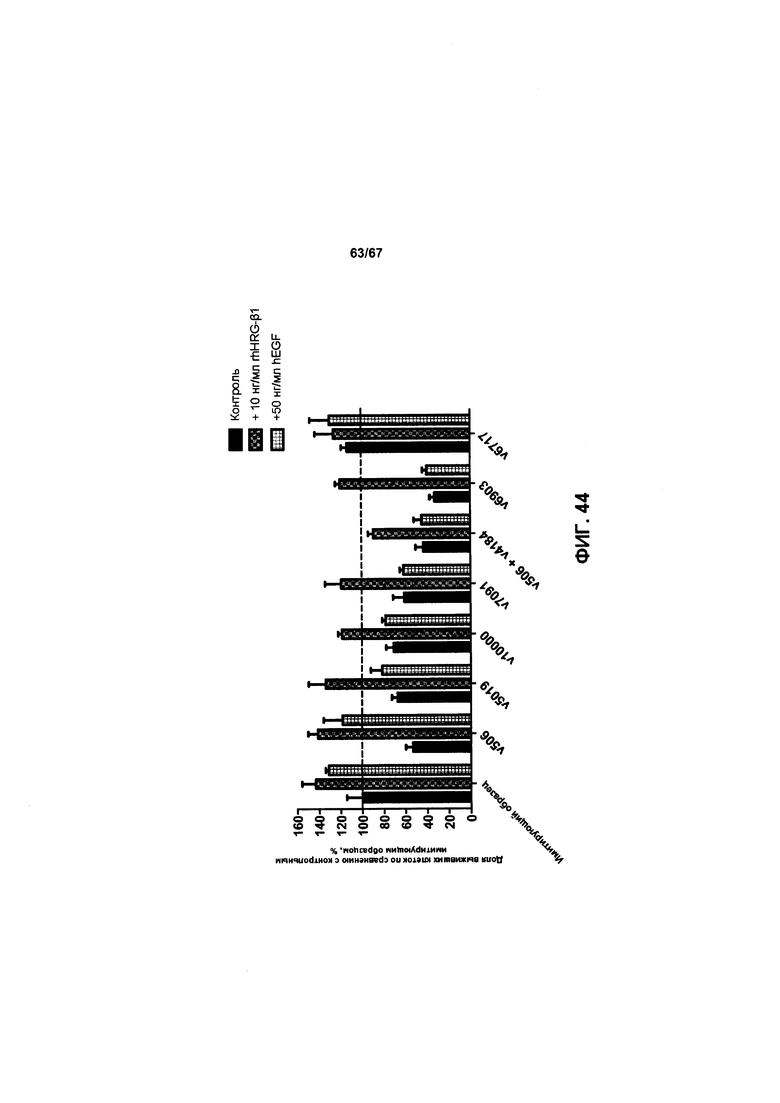
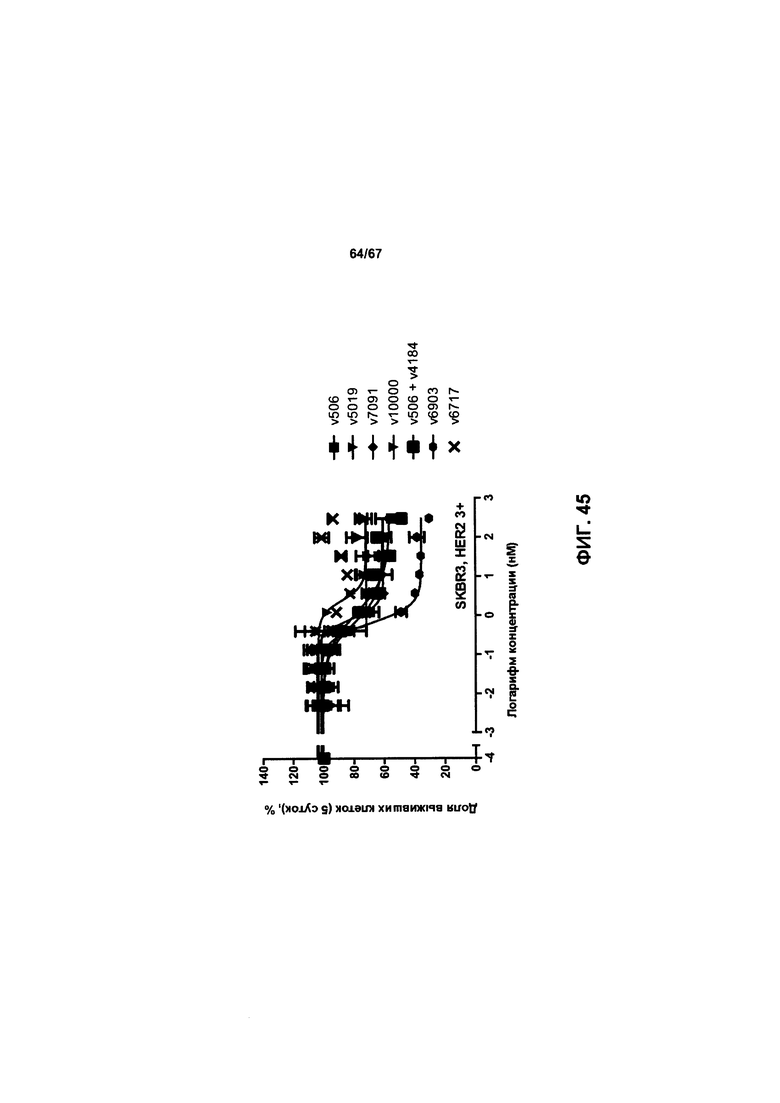
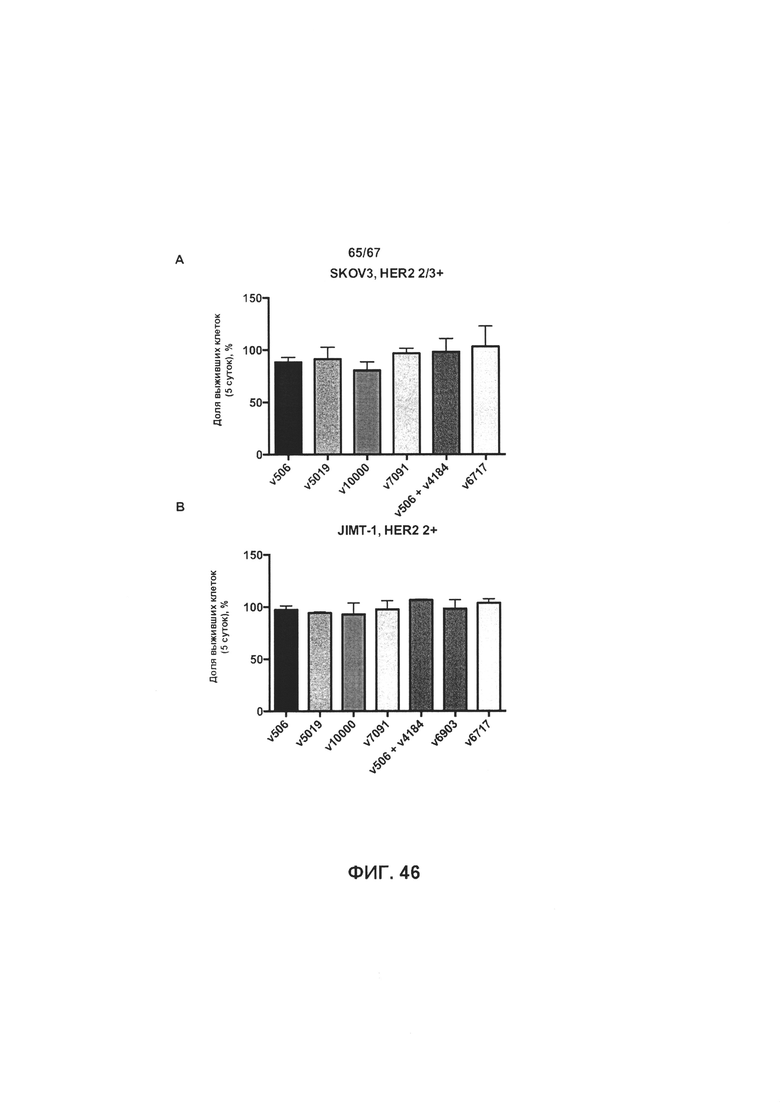
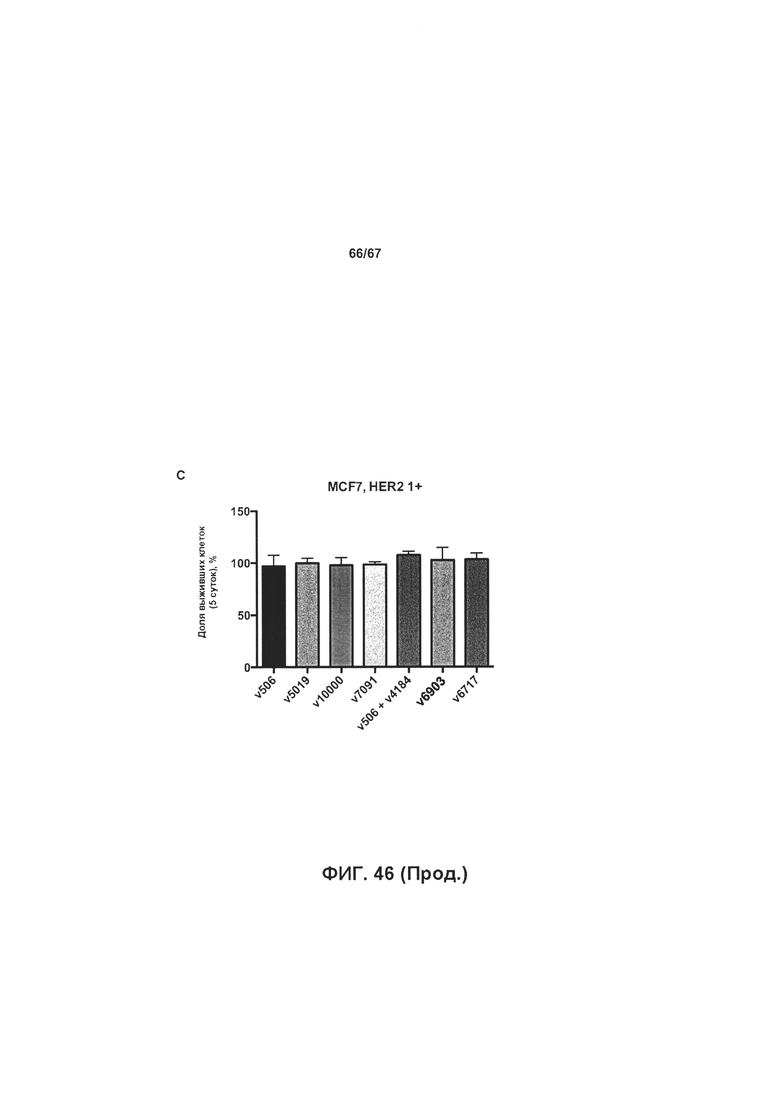
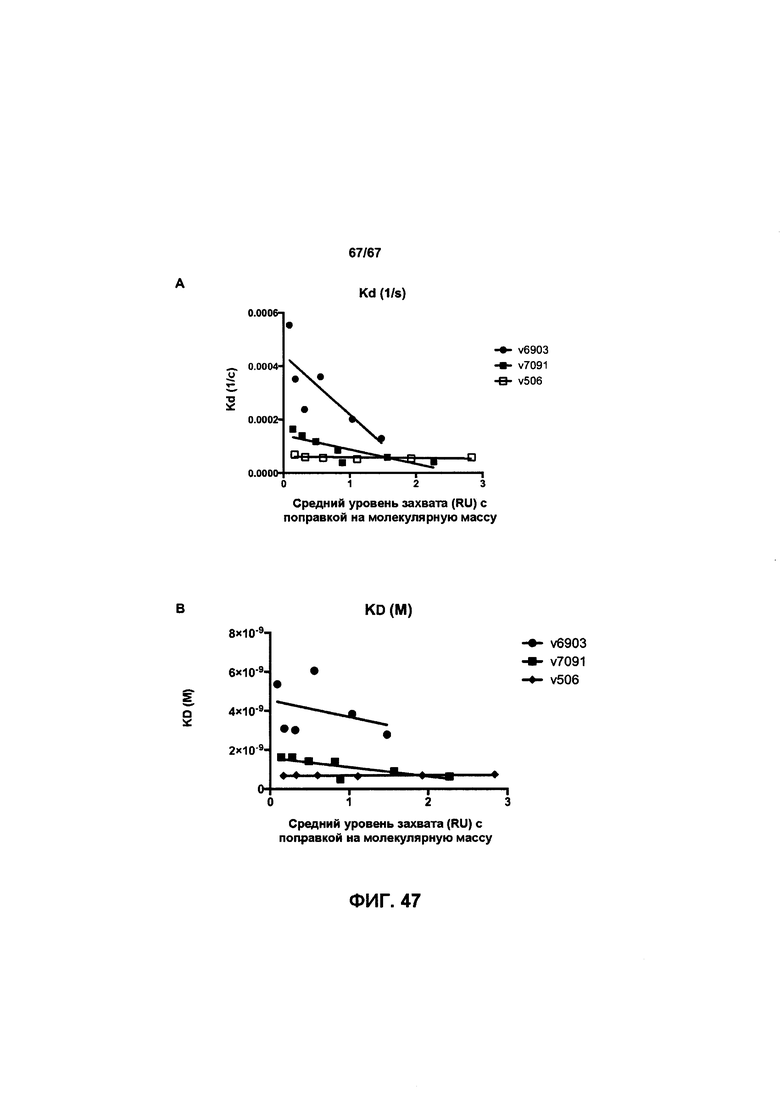
Изобретение относится к области биотехнологии. Описана группа изобретений, включающая антигенсвязывающую конструкцию, которая специфически связывается с HER2, фармацевтическую композицию для уничтожения или ингибирования роста HER2-экспрессирующей (HER2+) опухолевой клетки, фармацевтическую композицию для использования в лечении HER2-экспрессирующей (HER2+) опухоли, фармацевтическую композицию для использования в лечении рака, способ лечения субъекта, имеющего HER2-экспрессирующую (HER2+) опухоль, способ обнаружения или измерения количества HER2 в образце, способ подавления, снижения или блокировки передачи сигнала, опосредованного HER2, в клетке, способ уничтожения или ингибирования роста HER2-экспрессирующей опухолевой клетки, способ получения вышеуказанной конструкции, выделенный полинуклеотид (варианты), вектор (варианты), выделенную клетку для экспрессии антигенсвязывающей конструкции и набор для лечения рака. Изобретение расширяет арсенал средств, специфически связывающихся с HER2. 21 н. и 53 з.п. ф-лы, 54 табл., 44 пр., 47 ил.
1. Антигенсвязывающая конструкция, которая специфически связывается с HER2, содержащая:
первую антигенсвязывающую полипептидную конструкцию, которая моновалентно и специфически связывает ECD2 (внеклеточный домен 2) антигена HER2 (человеческий рецептор эпидермального фактора роста 2), расположенный на HER2-экспрессирующей клетке, причем первая антигенсвязывающая полипептидная конструкция содержит шесть последовательностей CDR, содержащих аминокислотные последовательности шести последовательностей CDR SEQ ID NO: 335, SEQ ID NO:336, SEQ ID NO:337, SEQ ID NO: 338, SEQ ID NO:339 и SEQ ID NO:340; или SEQ ID NO:335, SEQ ID NO:336, SEQ ID NO:348, SEQ ID NO:338, SEQ ID NO:347 и SEQ ID NO:340;
вторую антигенсвязывающую полипептидную конструкцию, которая моновалентно и специфически связывает ECD4 (внеклеточный домен 4) антигена HER2, расположенный на HER2-экспрессирующей клетке, причем вторая антигенсвязывающая полипептидная конструкция содержит шесть последовательностей CDR, содержащих аминокислотные последовательности шести последовательностей CDR SEQ ID NO: 341, SEQ ID NO:342, SEQ ID NO:343, SEQ ID NO: 344, SEQ ID NO:345 и SEQ ID NO:346;
первый и второй линкерные полипептиды, причем первый линкерный полипептид функционально связан с первой антигенсвязывающей полипептидной конструкцией, а второй линкерный полипептид функционально связан со второй антигенсвязывающей полипептидной конструкцией;
при этом линкерные полипептиды способны к образованию ковалентной связи друг с другом,
и причем (i) первая антигенсвязывающая полипептидная конструкция представляет собой scFv, а вторая антигенсвязывающая полипептидная конструкция представляет собой Fab; или (ii) первая антигенсвязывающая полипептидная конструкция представляет собой Fab, а вторая антигенсвязывающая полипептидная конструкция представляет собой scFv.
2. Антигенсвязывающая конструкция, которая специфически связывается с HER2, содержащая:
первую антигенсвязывающую полипептидную конструкцию, которая моновалентно и специфически связывает ECD2 антигена HER2, расположенный на HER2-экспрессирующей клетке, причем первая антигенсвязывающая полипептидная конструкция содержит шесть последовательностей CDR, содержащих аминокислотные последовательности, которые по меньшей мере на 90% идентичны шести последовательностям CDR SEQ ID NO: 335, SEQ ID NO:336, SEQ ID NO:337, SEQ ID NO: 338, SEQ ID NO:339 и SEQ ID NO:340; или SEQ ID NO:335, SEQ ID NO:336, SEQ ID NO:348, SEQ ID NO:338, SEQ ID NO:347 и SEQ ID NO:340;
вторую антигенсвязывающую полипептидную конструкцию, которая моновалентно и специфически связывает ECD4 антигена HER2, расположенный на HER2-экспрессирующих клетках, причем вторая антигенсвязывающая полипептидная конструкция содержит шесть последовательностей CDR, содержащих аминокислотные последовательности, которые по меньшей мере на 90% идентичны шести последовательностям CDR SEQ ID NO: 341, SEQ ID NO:342, SEQ ID NO:343, SEQ ID NO: 344, SEQ ID NO:345 и SEQ ID NO:346;
первый и второй линкерные полипептиды, причем первый линкерный полипептид функционально связан с первой антигенсвязывающей полипептидной конструкцией, а второй линкерный полипептид функционально связан со второй антигенсвязывающей полипептидной конструкцией;
при этом линкерные полипептиды способны к образованию ковалентной связи друг с другом,
и причем (i) первая антигенсвязывающая полипептидная конструкция представляет собой scFv, а вторая антигенсвязывающая полипептидная конструкция представляет собой Fab; или (ii) первая антигенсвязывающая полипептидная конструкция представляет собой Fab, а вторая антигенсвязывающая полипептидная конструкция представляет собой scFv.
3. Антигенсвязывающая конструкция, которая специфически связывается с HER2, содержащая:
первую антигенсвязывающую полипептидную конструкцию, которая моновалентно и специфически связывает ECD2 антигена HER2, расположенный на HER2-экспрессирующей клетке, причем первая антигенсвязывающая полипептидная конструкция выбрана из
i. полипептидной конструкции содержащей первый полипептид VH1 вариабельной области тяжёлой цепи, содержащий последовательность, представленную в SEQ ID NO:99, и первый полипептид VL1 вариабельной области легкой цепи, содержащий последовательность, представленную в SEQ ID NO:71; или
ii. полипептидной конструкции содержащей первый полипептид VH1 вариабельной области тяжёлой цепи, содержащий последовательность, которая по меньшей мере на 90% идентична SEQ ID NO:99, и первый полипептид VL1 вариабельной области легкой цепи, содержащий последовательность, которая по меньшей мере на 90% идентична SEQ ID NO:71;
вторую антигенсвязывающую полипептидную конструкцию, которая моновалентно и специфически связывает ECD4 антигена HER2, расположенный на HER2-экспрессирующей клетке, причем вторая антигенсвязывающая полипептидная конструкция выбрана из
iii. полипептидной конструкции содержащей второй полипептид VH2 вариабельной области тяжёлой цепи, содержащий последовательность, представленную в SEQ ID NO:305, и второй полипептид VL2 вариабельной области легкой цепи, содержащий последовательность, представленную в SEQ ID NO:297; или
iv. полипептидной конструкции содержащей второй полипептид VH2 вариабельной области тяжёлой цепи, содержащий последовательность, которая по меньшей мере на 90% идентична SEQ ID NO:305, и второй полипептид VL2 вариабельной области легкой цепи, содержащий последовательность, которая по меньшей мере на 90% идентична SEQ ID NO:297;
первый и второй линкерные полипептиды, причем первый линкерный полипептид функционально связан с первой антигенсвязывающей полипептидной конструкцией, а второй линкерный полипептид функционально связан со второй антигенсвязывающей полипептидной конструкцией;
при этом линкерные полипептиды способны к образованию ковалентной связи друг с другом,
и причем (i) первая антигенсвязывающая полипептидная конструкция представляет собой scFv, а вторая антигенсвязывающая полипептидная конструкция представляет собой Fab; или (ii) первая антигенсвязывающая полипептидная конструкция представляет собой Fab, а вторая антигенсвязывающая полипептидная конструкция представляет собой scFv.
4. Антигенсвязывающая конструкция, которая специфически связывается с HER2, содержащая:
первую антигенсвязывающую полипептидную конструкцию, которая моновалентно и специфически связывает ECD2 антигена HER2, расположенный на HER2-экспрессирующей клетке, причем первая антигенсвязывающая полипептидная конструкция выбрана из
i. полипептидной конструкции содержащей первую тяжелую цепь H1, содержащую последовательность, представленную в SEQ ID NO:97, и первую легкую цепь L1, содержащую последовательность, представленную в SEQ ID NO: 69; или
ii. полипептидной конструкции содержащей первую тяжелую цепь H1, содержащую последовательность, которая по меньшей мере на 90% идентична SEQ ID NO:97, и первую легкую цепь L1, содержащую последовательность, которая по меньшей мере на 90% идентична SEQ ID NO: 69;
вторую антигенсвязывающую полипептидную конструкцию, которая моновалентно и специфически связывает ECD4 антигена HER2, расположенный на HER2-экспрессирующей клетке, причем вторая антигенсвязывающая полипептидная конструкция выбрана из
iii. полипептидной конструкции содержащей последовательность, представленную в SEQ ID NO:295; или
iv. полипептидной конструкции содержащей последовательность, которая по меньшей мере на 90% идентична SEQ ID NO:295;
первый и второй линкерные полипептиды, причем первый линкерный полипептид функционально связан с первой антигенсвязывающей полипептидной конструкцией, а второй линкерный полипептид функционально связан со второй антигенсвязывающей полипептидной конструкцией;
при этом линкерные полипептиды способны к образованию ковалентной связи друг с другом,
и причем (i) первая антигенсвязывающая полипептидная конструкция представляет собой scFv, а вторая антигенсвязывающая полипептидная конструкция представляет собой Fab; или (ii) первая антигенсвязывающая полипептидная конструкция представляет собой Fab, а вторая антигенсвязывающая полипептидная конструкция представляет собой scFv.
5. Антигенсвязывающая конструкция по любому из пп. 1-4, отличающаяся тем, что каждый первый и второй линкерный полипептид содержат полипептид шарнирной области иммуноглобулина, выбранный из шарнирной области IgG1, IgG2 или IgG4.
6. Антигенсвязывающая конструкция по любому из пп. 1-5, отличающаяся тем, что первый и/или второй линкерные полипептиды функционально связаны с пептидным остовом, необязательно Fc.
7. Антигенсвязывающая конструкция по любому из пп. 1-5, отличающаяся тем, что первый и второй линкерные полипептиды функционально связаны с димерным Fc-доменом, содержащим первый и второй Fc-полипептиды, причем каждый из них содержит последовательность CH3, причем первый Fc-полипептид функционально связан с первым линкерным полипептидом, а второй линкерный полипептид функционально связан со вторым линкерным полипептидом.
8. Антигенсвязывающая конструкция по п. 3, отличающаяся тем, что
i. первая антигенсвязывающая полипептидная конструкция представляет собой Fab, а вторая антигенсвязывающая полипептидная конструкция представляет собой scFv, и первая антигенсвязывающая полипептидная конструкция Fab содержит
a. первый полипептид VH1 вариабельной области тяжёлой цепи, содержащий VH ECD2-связывающей полипептидной конструкции v10000 (SEQ ID NO:99) и
b. первый полипептид VL1 вариабельной области легкой цепи, содержащий VL ECD2-связывающей полипептидной конструкции v10000 (SEQ ID NO:71);
а вторая антигенсвязывающая полипептидная конструкция scFv содержит
(a) второй полипептид VH2 вариабельной области тяжёлой цепи, содержащий VH ECD4-связывающей полипептидной конструкции v10000 (SEQ ID NO:305), и
(b) второй полипептид VL2 вариабельной области легкой цепи, содержащий VL ECD4-связывающей полипептидной конструкции v10000 (SEQ ID NO:297); или
ii. первая антигенсвязывающая полипептидная конструкция представляет собой scFv, а вторая антигенсвязывающая полипептидная конструкция представляет собой Fab, и первая антигенсвязывающая полипептидная конструкция scFv содержит
(a) первый полипептид VH1 вариабельной области тяжёлой цепи, содержащий VH ECD2-связывающей полипептидной конструкции v10000 (SEQ ID NO:99), и
(b) первый полипептид VL1 вариабельной области легкой цепи, содержащий VL ECD2-связывающей полипептидной конструкции v10000 (SEQ ID NO:71),
а вторая антигенсвязывающая полипептидная конструкция Fab содержит
(a) второй полипептид VH2 вариабельной области тяжёлой цепи, содержащий VH ECD4-связывающей полипептидной конструкции v10000 (SEQ ID NO:305), и
(b) второй полипептид VL2 вариабельной области легкой цепи, содержащий VL ECD4-связывающей полипептидной конструкции v10000 (SEQ ID NO:297).
9. Антигенсвязывающая конструкция по любому из пп. 1-5, отличающаяся тем, что первая антигенсвязывающая полипептидная конструкция: (i) блокирует на 50 % или больше связывание пертузумаба с ECD2, и/или (ii) второй антигенсвязывающий полипептид блокирует на 50 % или больше связывание трастузумаба с ECD4.
10. Антигенсвязывающая конструкция по любому из пп. 1-5, отличающаяся тем, что первая антигенсвязывающая полипептидная конструкция представляет собой Fab, а вторая антигенсвязывающая полипептидная конструкция представляет собой scFv, и причем указанная антигенсвязывающая конструкция
(i) индуцирует повышенную степень интернализации рецептора в клетках HER2 3+ и/или
(ii) демонстрирует повышенную активность против клеток HER2 1+ в анализе АЗКЦ (антителозависимая клеточно-опосредованная цитотоксичность),
по сравнению с эталонной бипаратопной антигенсвязывающей конструкцией, имеющей два Fab.
11. Конструкция, которая специфически связывается с HER2, содержащая антигенсвязывающую полипептидную конструкцию, которая моновалентно и специфически связывается с ECD2 HER2, причем антигенсвязывающая полипептидная конструкция выбрана из:
(i) антигенсвязывающей полипептидной конструкции содержащей VH и VL, при этом VH содержит три последовательности CDR, содержащие аминокислотные последовательности трех последовательностей VH CDR SEQ ID NO:335, SEQ ID NO:336 и SEQ ID NO:348, и VL содержит три последовательности CDR, содержащие аминокислотные последовательности трех последовательностей VL CDR SEQ ID NO:338, SEQ ID NO:347 и SEQ ID NO:340; или
(ii) антигенсвязывающей полипептидной конструкции содержащей VH и VL, при этом VH содержит три последовательности CDR, которые по меньшей мере на 90% идентичны аминокислотным последовательностям трех последовательностей VH CDR SEQ ID NO:335, SEQ ID NO:336 и SEQ ID NO:348, и VL содержит три последовательности CDR, которые по меньшей мере на 90% идентичны аминокислотным последовательностям трех последовательностей VL CDR SEQ ID NO:338, SEQ ID NO:347 и SEQ ID NO:340.
12. Конструкция по п. 11, отличающаяся тем, что указанная конструкция является моновалентной, бивалентной или мультивалентной.
13. Конструкция по п. 11, отличающаяся тем, что указанная конструкция является (i) моновалентной и содержит Fab или scFv, (ii) является бивалентной и содержит Fab и scFv или (iii) является бивалентной и содержит два scFv.
14. Конструкция по п. 11, отличающаяся тем, что указанная конструкция является моновалентной и обладает аффинностью, повышенной в 7-9 раз, по отношению к ECD2 HER2 по сравнению с пертузумабом.
15. Конструкция по п. 11, содержащая димерный Fc-домен, связанный с антигенсвязывающей полипептидной конструкцией при помощи линкера или без него.
16. Конструкция по п. 11, отличающаяся тем, что указанная конструкция содержит первую и вторую антигенсвязывающие полипептидные конструкции и первый и второй Fc-полипептиды, причем каждый из них содержит последовательность CH3, причем первый Fc-полипептид функционально связан с первой антигенсвязывающей полипептидной конструкцией при помощи первого линкера или без него, и второй мономерный Fc-полипептид функционально связан со второй антигенсвязывающей полипептидной конструкцией с помощью второго линкера или без него.
17. Конструкция по п. 16, содержащая полипептидный линкер.
18. Конструкция по п. 17, отличающаяся тем, что указанный линкер содержит шарнирную область IgG1.
19. Конструкция по п. 16, отличающаяся тем, что Fc-домен представляет собой Fc-домен антитела человека.
20. Конструкция по п. 16, отличающаяся тем, что Fc-домен представляет собой Fc-домен IgG1 человека.
21. Конструкция по любому из пп. 1-5, 11, 17-20, содержащая гетеродимерный Fc-фрагмент, отличающаяся тем, что температура плавления (Тпл) димеризованных СН3-доменов составляет около 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 77,5, 78, 79, 80, 81, 82, 83, 84 или 85°C или выше.
22. Конструкция по любому из пп. 1-5, 11, 17-20, содержащая гетеродимерный Fc-фрагмент, вырабатываемый с чистотой, более чем около 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99 % при экспрессии.
23. Конструкция по любому из пп. 1-5, 11, 17-20, содержащая гетеродимерный Fc-фрагмент, вырабатываемый с чистотой, более чем около 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99 % при экспрессии на основе генетического материала одной клетки.
24. Конструкция по любому из пп. 1-5, 11, 17-20, содержащая гетеродимерный Fc-фрагмент, содержащий одну или более модификаций по меньшей мере в одной из последовательностей CH3.
25. Конструкция по любому из пп. 1-5, 11, 17-20, содержащая гетеродимерный Fc-фрагмент, содержащий одну или более модификаций по меньшей мере в одной из последовательностей CH3, которые способствуют образованию гетеродимера со стабильностью, сравнимой со стабильностью гомодимерного Fc-домена дикого типа.
26. Конструкция по любому из пп. 1-5, 11, 17-20, содержащая
i. гетеродимерный Fc-фрагмент IgG1, имеющий модификации
T350V_L351Y_F405A_Y407V в первом Fc-полипептиде и модификации T366I_N390R_K392M_T394W во втором Fc-полипептиде; или
ii. гетеродимерный Fc-фрагмент IgG1, имеющий модификации L351Y_S400E_F405A_Y407V в первом Fc-полипептиде и модификации T350V_T366L_K392L_T394W во втором Fc-полипептиде,
iii. гетеродимерный Fc-фрагмент IgG1, имеющий модификации L351Y_F405A_Y407V в первом Fc-полипептиде и модификации T366L_K392M_T394W во втором полипептиде;
iv. гетеродимерный Fc-фрагмент IgG1, имеющий модификации L351Y_F405A_Y407V в первом Fc-полипептиде и модификации T366L_K392L_T394W во втором Fc-полипептиде;
v. гетеродимерный Fc-фрагмент IgG1, имеющий модификации T350V_L351Y_F405A_Y407V в первом Fc-полипептиде и модификации T350V_T366L_K392L_T394W во втором Fc-полипептиде;
vi. гетеродимерный Fc-фрагмент IgG1, имеющий модификации T350V_L351Y_F405A_Y407V в первом Fc-полипептиде и модификации T350V_T366L_K392M_T394W во втором Fc-полипептиде; или
vii. гетеродимерный Fc-фрагмент IgG1, имеющий модификации T350V_L351Y_S400E_F405A_Y407V в первом Fc-полипептиде и модификации T350V_T366L_N390R_K392M_T394W во втором Fc-полипептиде,
согласно EU-нумерации по сравнению с гомодимерным Fc-фрагментом дикого типа.
27. Конструкция по любому из пп. 1-5, 11, 17-20, содержащая гетеродимерный Fc-фрагмент, содержащий по меньшей мере один CH2-домен.
28. Конструкция по п. 27, отличающаяся тем, что CH2-домен(ы) гетеродимерного Fc-фрагмента содержит одну или более модификаций.
29. Конструкция по любому из пп. 1-5, 11, 17-20, 28, содержащая гетеродимерный Fc-фрагмент, содержащий одну или более модификаций, что способствуют избирательному связыванию Fc-гамма рецепторов.
30. Конструкция по любому из пп. 1-5, 11, 17-20, 28, отличающаяся тем, что указанная конструкция гликозилирована.
31. Конструкция по любому из пп. 1-5, 11, 17-20, 28, отличающаяся тем, что указанная конструкция конъюгирована с лекарственным средством.
32. Конструкция по п. 31, отличающаяся тем, что лекарственное средство представляет собой майтансин (DM1).
33. Конструкция по п. 31, отличающаяся тем, что указанная конструкция конъюгирована с DM1 при помощи линкера SMCC.
34. Фармацевтическая композиция для уничтожения или ингибирования роста HER2-экспрессирующей (HER2+) опухолевой клетки, содержащая эффективное количество конструкции по любому из пп. 1-33 и фармацевтический носитель.
35. Фармацевтическая композиция по п. 34, отличающаяся тем, что фармацевтический носитель содержит буфер, антиоксидант, молекулу с низкой молекулярной массой, лекарственное средство, белок, аминокислоту, углевод, липид, хелатирующий агент, стабилизатор или вспомогательное вещество.
36. Фармацевтическая композиция для использования в лечении HER2-экспрессирующей (HER2+) опухоли, содержащая эффективное количество конструкции по любому из пп. 1-33.
37. Фармацевтическая композиция для использования в лечении рака, содержащая эффективное количество конструкции по любому из пп. 1-33.
38. Способ лечения субъекта, имеющего HER2-экспрессирующую (HER2+) опухоль, включающий введение субъекту эффективного количества конструкции по любому из пп. 1-33 или фармацевтической композиции по любому из пп. 34-37.
39. Способ по п. 38, отличающийся тем, что результатом лечения является уменьшение объема опухоли, ингибирование роста опухоли, увеличение периода времени до начала прогрессии опухоли, продление выживаемости субъекта без признаков заболевания, уменьшение метастазов, увеличение выживаемости субъекта без прогрессирования заболевания или увеличение общей выживаемости субъекта.
40. Способ по п. 38 или 39, отличающийся тем, что опухоль представляет собой рак поджелудочной железы, рак головы и шеи, рак ЖКТ, колоректальный рак, рак молочной железы, рак почек, рак шейки матки, рак яичников, рак эндометрия, рак матки, злокачественная меланома, рак глотки, рак ротовой полости или рак кожи.
41. Способ по пп. 38-39, отличающийся тем, что опухоль содержит клетки, которые экспрессируют в среднем 10000 или больше копий HER2 на опухолевую клетку.
42. Способ по пп. 38-39, отличающийся тем, что опухоль представляет собой HER2 1+, HER2 2+ или HER2 3+, что определяют с помощью иммуногистохимического исследования (ИГХ).
43. Способ по п. 42, отличающийся тем, что опухоль экспрессирует HER2 на уровне 2+ или ниже, который определяют с помощью ИГХ.
44. Способ по п. 38, отличающийся тем, что HER2+ опухоль представляет собой рак яичников с уровнем экспрессии HER2 2+/3+, который определяют с помощью ИГХ, и амплификацией гена HER2, и умеренно чувствительна к трастузумабу.
45. Способ по п. 38, отличающийся тем, что HER2+ опухоль представляет собой рак молочной железы с уровнем экспрессии HER2 2+ или ниже, что определяют с помощью иммуногистохимического исследования (ИГХ).
46. Способ по п. 38, отличающийся тем, что HER2+ опухоль представляет собой (i) негативный по рецепторам эстрогенов (ER-), негативный по рецепторам прогестерона (PR-), резистентный к трастузумабу, резистентный к химиотерапии инвазивный протоковый рак молочной железы с уровнем экспресии HER2 3+, (ii) резистентный к трастузумабу отёчно-инфильтративный рак молочной железы ER-, PR-, с уровнем экспресии HER2 3+, (iii) инвазивный протоковый рак молочной железы ER-, PR- с уровнем экспресии HER2 3+ или (iv) резистентный к трастузумабу и пертузумабу рак молочной железы c амплификацией гена HER2 и уровнем экспресии HER2 2+.
47. Способ по пп. 38-39, 43-46, отличающийся тем, что субъект ранее не получал лечение с использованием антитела к HER2.
48. Способ по пп. 38-39, 43-46, отличающийся тем, что опухоль резистентна или устойчива к пертузумабу, трастузумабу и/или TDM1.
49. Способ по любому из пп. 38-39, 43-46, отличающийся тем, что субъект ранее получал лечение с использованием пертузумаба, трастузумаба и/или TDM1.
50. Способ по пп. 38-39, 43-46, отличающийся тем, что введение осуществляют путем инъекции или инфузии.
51. Способ по пп. 38-39, 43-46, дополнительно включающий введение субъекту дополнительного средства, необязательно химиотерапевтического средства.
52. Способ по п. 51, отличающийся тем, что
i. опухоль представляет собой немелкоклеточный рак легких, и дополнительное средство представляет собой одно или более из цисплатина, карбоплатина, паклитаксела, альбумин-связанного паклитаксела, доцетаксела, гемцитабина, винорелбина, иринотекана, этопозида, винбластина или пеметрекседа;
ii. опухоль представляет собой рак ЖКТ или желудка, и дополнительное средство представляет собой одно или более из 5-фторурацила (в присутствии или в отсутствие фолиновой кислоты), капецитабина, карбоплатина, цисплатина, доцетаксела, эпирубицина, иринотекана, оксалиплатина или паклитаксела;
iii. опухоль представляет собой рак поджелудочной железы, и дополнительное средство представляет собой одно или более из гемцитабина, фолфиринокса, абраксана или 5-фторурацила;
iv. опухоль представляет собой эстроген- и/или прогестерон-позитивный рак молочной железы, и дополнительное средство представляет собой одно или более из комбинации (a) доксорубицина и эпирубицина, (b) комбинации паклитаксела и доцетаксела или (c) комбинации фторурацила, циклофосфамида и карбоплатина;
v. опухоль представляет собой рак головы и шеи, и дополнительное средство представляет собой одно или более из паклитаксела, карбоплатина, доксорубицина или цисплатина;
vi. опухоль представляет собой рак яичников, и дополнительное средство представляет собой одно или более из цисплатина, карбоплатина или таксана, такого как паклитаксел или доцетаксел.
53. Способ по пп.38-39, 43-46, 52, отличающийся тем, что субъект представляет собой человека.
54. Способ обнаружения или измерения количества HER2 в образце, включающий воздействие на образец антигенсвязывающей конструкцией по пп. 1-33 и обнаружение или измерение количества связанного комплекса.
55. Способ подавления, снижения или блокировки передачи сигнала, опосредованного HER2, в клетке, включающий введение эффективного количества антигенсвязывающей конструкции по пп. 1-33 в клетку.
56. Способ уничтожения или ингибирования роста HER2-экспрессирующей опухолевой клетки, включающий воздействие на клетку антигенсвязывающей конструкцией по пп. 1-33.
57. Способ по п. 56, отличающийся тем, что опухолевая клетка представляет собой клетку карциномы поджелудочной железы человека с уровнем экспрессии HER2 1+ или 2+, клетку карциномы легких человека с уровнем экспрессии HER2 3+, клетку бронхоальвеолярной карциномы человека европеоидной расы с уровнем экспрессии HER2 2+, клетку карциномы глотки человека, клетку плоскоклеточной карциномы языка человека с уровнем экспрессии HER2 2+, клетку плоскоклеточной карциномы глотки с уровнем экспрессии HER2 2+, клетку колоректальной карциномы человека с уровнем экспрессии HER2 1+ или 2+, клетку карциномы желудка человека с уровнем экспрессии HER2 3+, клетку протоковой карциномы молочной железы человека ER+ (позитивной по рецепторам эстрогена) с уровнем экспрессии HER2 1+, клетку карциномы молочной железы человека ER+ с амплификацией гена HER2 и уровнем экспрессии HER2 2+/3+, клетку трижды негативной карциномы молочной железы человека с уровнем экспрессии HER2 0+/1+, клетку эндометриоидной карциномы человека с уровнем экспрессии HER2 2+, клетку метастатической злокачественной меланомы легких с уровнем экспрессии HER2 1+, клетку карциномы шейки матки человека с уровнем экспрессии HER2 1+, клетку почечно-клеточной карциномы человека с уровнем экспрессии HER2 1+ или клетку карциномы яичника человека с уровнем экспрессии HER2 1+.
58. Способ по п. 56, отличающийся тем, что опухолевая клетка представляет собой клетку карциномы поджелудочной железы человека с уровнем экспрессии HER2 1+ и 2+ или 3+, клетку метастатической карциномы поджелудочной железы с уровнем экспрессии HER2 2+, клетку карциномы легких человека с уровнем экспрессии HER2 0+/1+, +3+, клетку бронхоальвеолярной карциномы человека европеоидной расы с уровнем экспрессии HER2 2+, клетку анапластической карциномы легких с уровнем экспрессии HER2 0+, клетку немелкоклеточной карциномы легких человека, клетку карциномы глотки человека, клетку плоскоклеточной карциномы языка человека с уровнем экспрессии HER2 2+, клетку плоскоклеточной карциномы глотки с уровнем экспрессии HER2 2+, клетку колоректальной карциномы человека с уровнем экспрессии HER2 1+ или 2+, клетку карциномы желудка человека с уровнем экспрессии HER2 0+, 1+ или 3+, клетку протоковой карциномы молочной железы человека ER+ (позитивной по рецепторам эстрогена) с уровнем экспрессии HER2 1+, клетку карциномы молочной железы человека ER+ с амплификацией гена HER2 и уровнем экспрессии HER2 2+/3+, клетку трижды негативной карциномы молочной железы человека с уровнем экспрессии HER2 0+/1+, клетку протоковой карциномы молочной железы человека с уровнем экспрессии HER2 0+ (базального типа B, мезенхимально-подобная, трижды негативная), клетку карциномы молочной железы ER+ с уровнем экспрессии HER2 2+, клетку метастатической карциномы молочной железы человека с уровнем экспрессии HER2 0+ (трижды негативный рак ER- люминального типа А с амплификацией гена HER2), клетку мезодермальной опухоли матки человека (смешанная форма рака третьей стадии), клетку эндометриоидной карциномы человека с уровнем экспрессии HER2 2+, клетку эпидермоидной карциномы кожи человека с уровнем экспрессии HER2 1+, клетку метастатической злокачественной меланомы легких с уровнем экспрессии HER2 1+, клетку злокачественной меланомы с уровнем экспрессии HER2 1+, клетку эпидермоидной карциномы шейки матки человека, клетку карциномы мочевого пузыря человека с уровнем экспрессии HER2 1+, клетку карциномы шейки матки человека с уровнем экспрессии HER2 1+, клетку почечно-клеточной карциномы человека с уровнем экспрессии HER2 1+ или клетку карциномы яичника человека с уровнем экспрессии HER2 1+, 2+ или 3+, и отличающийся тем, что антигенсвязывающая конструкция конъюгирована с майтансином (DM1).
59. Способ по п. 56, отличающийся тем, что опухолевая клетка представляет собой клетку, выбранную из клеточных линий опухоли поджелудочной железы ВхРСЗ, Capan-1, MiaPaca2; клеточных линий опухоли легких Calu-3, NCI-H322; клеточных линий опухоли головы и шеи Detroit 562, SCC-25, FaDu; клеточных линий колоректальной опухоли HT29,SNU-C2B; клеточной линии опухоли желудка NCI-N87; клеточных линий опухоли молочной железы MCF-7, MDAMB175, MDAMB361, MDA-MB-231, BT-20, JIMT-1, SkBr3, BT-474; клеточной линии опухоли матки TOV-112D; клеточной линии опухоли кожи Malme-3M; клеточных линий опухоли шейки матки Caski, MS751; клеточной линии опухоли мочевого пузыря T24, клеточных линий опухоли яичника CaOV3 и SKOV3.
60. Способ по п. 56, отличающийся тем, что опухолевая клетка представляет собой клетку, выбранную из клеточных линий опухоли поджелудочной железы ВхРСЗ, Capan-1, MiaPaca2, SW 1990, Panс 1; клеточных линий опухоли легких A549, Calu-3, Calu-6, NCI-H2126, NCI-H322; клеточных линий опухоли головы и шеи Detroit 562, SCC-15, SCC-25, FaDu; клеточных линий колоректальной опухоли Colo201, DLD-1, HCT116, HT29, SNU-C2B; клеточных линий опухоли желудка SNU-1, SNU-16, NCI-N87; клеточных линий опухоли молочной железы SkBr3, MCF-7, MDAMB175, MDAMB361, MDA-MB-231, ZR-75-1, BT-20, BT549, BT-474, CAMA-1, MDAMB453, JIMT-1, T47D; клеточных линий опухоли матки SK-UT-1, TOV-112D; клеточных линий опухоли кожи A431, Malme-3M, SKEMEL28; клеточных линий опухоли шейки матки Caski, MS751; клеточной линии опухоли мочевого пузыря T24, клеточной линии опухоли почек ACHN; клеточных линий опухоли яичника CaOV3, Ovar-3 и SKOV3, и отличающийся тем, что антигенсвязывающая конструкция конъюгирована с майтансином.
61. Способ по п. 56, отличающийся тем, что опухолевую клетку выбирают из клетки рака яичников с уровнем экспрессии HER2 2/3+ и амплификацией гена HER2, клетки трижды негативного рака молочной железы с уровнем экспрессии HER2 0+/1+; из клетки рака молочной железы ER+ с уровнем экспрессии HER2 1+; клетки резистентного к трастузумабу рака молочной железы с уровнем экспрессии HER2 2+; клетки рака молочной железы ER+ с уровнем экспрессии HER2+; или клетки рака молочной железы с уровнем экспрессии HER2 3+.
62. Способ получения конструкции по пп. 1-10, включающий культивирование клетки-хозяина в условиях, подходящих для экспрессии антигенсвязывающей конструкции, отличающийся тем, что указанная клетка-хозяин содержит полинуклеотид, кодирующий антигенсвязывающую конструкцию по п. 1-10, и выделение указанной конструкции.
63. Способ получения конструкции по пп. 11-33, включающий культивирование клетки-хозяина в условиях, подходящих для экспрессии антигенсвязывающей конструкции, отличающийся тем что, указанная клетка-хозяин содержит полинуклеотид, кодирующий антигенсвязывающую конструкцию по пп. 11-33, и выделение указанной конструкции.
64. Выделенный полинуклеотид, содержащий по меньшей мере одну нуклеотидную последовательность, которая кодирует по меньшей мере один полипептид антигенсвязывающей конструкции по любому из пп. 1-10.
65. Полинуклеотид по п. 64, отличающийся тем, что указанный полинуклеотид представляет собой кДНК.
66. Выделенный полинуклеотид, содержащий по меньшей мере одну нуклеотидную последовательность, которая кодирует по меньшей мере один полипептид конструкции по любому из пп. 11-33.
67. Полинуклеотид по п. 66, отличающийся тем, что указанный полинуклеотид представляет собой кДНК.
68. Полинуклеотид, кодирующий антигенсвязывающую конструкцию, которая специфически связывается с HER2, в которой один HER2-связывающий домен представляет собой Fab-фрагмент на цепи А, полученный из пертузумаба с Y96A в VL-области и T30A/A49G/L69F в VH-области, а другой HER2-связывающий домен представляет собой scFv-фрагмент на цепи В, полученный из трастузумаба, и Fc-область представляет собой гетеродимер, имеющий мутации T350V_L351Y_F405A_Y405V в цепи A, и мутации T350V_T366L_K392L_T394W в цепи B.
69. Вектор, содержащий один или более полинуклеотидов по пп. 64-68, который кодирует по меньшей мере один полипептид антигенсвязывающей конструкции по пп. 1-10.
70. Вектор, содержащий один или более полинуклеотидов по п. 69, который кодирует по меньшей мере один полипептид антигенсвязывающей конструкции по пп. 1-10, выбранный из группы, состоящей из плазмиды, вирусного вектора, неэписомального вектора млекопитающего, экспрессирующего вектора и рекомбинантного экспрессирующего вектора.
71. Выделенная клетка для экспрессии антигенсвязывающей конструкции по любому из пп. 1-10, содержащая полинуклеотид по пп. 64-68, или вектор по п. 69 или 70.
72. Выделенная клетка по п. 71, представляющая собой гибридому, клетку яичников китайского хомячка (CHO) или клетку HEK293.
73. Набор для лечения рака, содержащий конструкцию по любому из пп. 1-33 и инструкции к применению.
74. Конструкция по п. 30, отличающаяся тем, что указанная конструкция является дефукозилированной.
| WO 2012143523 A1 26.10.2012 | |||
| WO 2013063702 A1 10.05.2013 | |||
| WO 2013166604 A1 14.11.2013 | |||
| RU 2420537 C2 10.06.2011. |

