ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
[1] Использование антител определенной специфичности в различных терапевтических целях постоянно растет. Для получения антител, пригодных для терапевтического использования у человека, использовали ряд способов. Они включают химерные и гуманизированные антитела, и полностью принадлежащие человеку антитела, выбранные из библиотек, например, библиотек фагового дисплея, или от трансгенных животных. Библиотеки иммуноглобулина, сконструированные в бактериофаге, могут происходить из продуцирующих антитела клеток от не получавших лечение или конкретным образом иммунизированных индивидуумов и могут в основном включать новые и разнообразные пары тяжелых и легких цепей иммуноглобулина человека. Несмотря на то, что эта стратегия не страдает от внутреннего ограничения репертуара, необходимо, чтобы определяющие комплементарность области (CDR) экспрессируемого фрагмента иммуноглобулина синтезировались и правильно сворачивались в бактериальных клетках. Однако многие антигенсвязывающие области трудно правильно собирать в виде слитого белка в бактериальных клетках. Кроме того, белок не подвергается нормальным посттрансляционным модификациям, характерным для эукариотических клеток. В результате, этот способ ограничен различным избирательным фильтром в отношении специфичности антител, которые можно получать. Альтернативно, полностью принадлежащие человеку антитела можно выделять из библиотек в эукариотических системах, например, дрожжевом дисплее, дисплее на основе ретровирусов или экспрессии в ДНК-вирусах, таких как поксвирусы. См., например, патент США № 7858559 и публикацию заявки на патент США № 2013-028892, которые полностью включены в настоящее описание посредством ссылки.
[2] Многими важными мишенями терапевтических антител являются интегральные мембранные белки (IMP), например, пронизывающие несколько раз мембрану белки (GPCR, ионные каналы и т.д.), которые трудно экспрессировать и очищать в конформационно интактном состоянии. Отсутствие правильно свернутых целевых белков в выделенном состоянии делает идентификацию и селекцию антител к этим мишеням затруднительной. Несмотря на то, что определенные IMP можно экспрессировать на поверхности клеток, например, клеток млекопитающих, применение целых клеток при обнаружении антител может вызывать проблемы, т.к. они представляют собой сложные антигенные смеси, экспрессия мишени может быть низкой, и потому что определенные наборы дисплеев, используемые для конструкции библиотек антител (например, библиотеки антител к вирусу осповакцины) могут неспецифически связываться с целыми клетками. Сохраняется потребность в новых способах экспрессии и предоставлении представляющих интерес IMP-мишеней в их нативной конформации в достаточной концентрации и с минимальной конкуренцией со стороны других клеточных белков для обеспечения возможности идентификации и селекции терапевтических антител и антителоподобных молекул из библиотек дисплея.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[3] Настоящее изобретение относится к композициям и способам экспрессии и дисплея выделенных интегральных мембранных белков (IMP) или их фрагментов в нативной конформации для применения в скрининге, селекции и идентификации антител или антителоподобных молекул, которые связываются с представляющим интерес IMP-мишенью.
[4] В некоторых вариантах осуществления изобретение относится к выделенному полинуклеотиду, который содержит: фрагмент первой нуклеиновой кислоты, кодирующий интегральный мембранный белок (IMP) или его фрагмент, где IMP или его фрагмент содержит по меньшей мере одну внемембранную область, по меньшей мере один трансмембранный домен и по меньшей мере одну внутримембранную область, и где участок фрагмента первой нуклеиновой кислоты, кодирующей по меньшей мере одну внутримембранную область располагается на 5'- или 3'-конце фрагмента первой нуклеиновой кислоты; и фрагмент второй нуклеиновой кислоты, кодирующей белок F13L вируса осповакцины или функциональный его фрагмент, где фрагмент второй нуклеиновой кислоты является слитым в рамке чтения с участком фрагмента первой нуклеиновой кислоты, кодирующей внутримембранную область IMP. В соответствии с эти вариантами осуществления инфицированная поксвирусом клетка, содержащая полинуклеотид, может экспрессировать слитый белок IMP-F13L в виде части внешней мембрана оболочки внеклеточного оболочечного вириона (EEV). В некоторых аспектах белок F13L или его функциональный фрагмент может содержать аминокислотную последовательность SEQ ID NO:1 или его функциональный фрагмент. В определенных аспектах IMP представляет собой пронизывающий несколько раз мембрану белок, содержащий по меньшей мере два, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять, по меньшей мере шесть или по меньшей мере семь трансмембранных домена. В определенных аспектах IMP представляет собой пронизывающий несколько раз мембрану белок, приведенный в таблице 1.
[5] В некоторых аспектах пронизывающий несколько раз мембрану IMP может содержать нечетное число трансмембранных доменов, 5'-конец фрагмента первой нуклеиновой кислоты может кодировать внемембранную область, и 3'-конец фрагмента первой нуклеиновой кислоты может кодировать внутримембранную область, слитую с 5'-концом фрагмента второй нуклеиновой кислоты. В некоторых аспектах фрагмент первой нуклеиновой кислоты этого типа может кодировать, например, сопряженный с G-белком рецептор (GPCR). В некоторых аспектах GPCR может представлять собой белок Фрайзлед-4 человека (FZD4) или его фрагмент, и полинуклеотид может кодировать полипептид, который содержит аминокислоты от 20 до 892 SEQ ID NO:2. В некоторых аспектах полипептид может дополнительно содержать сигнальный пептид, например, аминокислоты от 1 до 19 SEQ ID NO:2. В некоторых аспектах GPCR может представлять собой рецептор хемокинов CXC, например, CXCR4, или его фрагмент, и полинуклеотид может кодировать полипептид, который содержит аминокислотную последовательность SEQ ID NO:3.
[6] В некоторых аспектах пронизывающий несколько раз мембрану IMP может содержать четное число трансмембранных доменов, и 5'- и 3'-концы фрагмента первой нуклеиновой кислоты могут кодировать внутримембранные области. В некоторых аспектах фрагмент второй нуклеиновой кислоты может быть слитым с 3'-концом фрагмента первой нуклеиновой кислоты. В некоторых аспектах IMP может представлять собой, например, белок CD20 человека или его фрагмент, и полинуклеотид может кодировать полипептид, который содержит аминокислотную последовательность SEQ ID NO:4.
[7] В некоторых аспектах первый и второй фрагменты нуклеиновой кислоты полинуклеотида, предоставленного в настоящем описании, могут являться непосредственно слитыми. В некоторых аспектах полинуклеотид, как предоставлено в настоящем описании, может содержать третий фрагмент нуклеиновой кислоты, кодирующий гетерологичный пептид, например, линкерную последовательность, аминокислотную метку или пептидную или полипептидную последовательность, которая облегчает очистку, такую как гистидиновая метка. В некоторых аспектах полинуклеотид, как предоставлено в настоящем описании, можно функционально связывать с промотором поксвируса, например, промотор p7.5, T7 или H5.
[8] Изобретение дополнительно относится к слитому белку F13L, кодируемому полинуклеотидом, как предоставлено в настоящем описании. Изобретение дополнительно относится к геному поксвируса, например, геному вируса осповакцины, который содержит полинуклеотид, как предоставлено в настоящем описании. Изобретение дополнительно относится к рекомбинантному EEV вируса осповакцины, который содержит геном поксвируса, как предоставлено в настоящем описании.
[9] Изобретение дополнительно относится к способу получения рекомбинантного EEV вируса осповакцины, как предоставлено в настоящем описании, где способ включает инфицирование клетки-хозяина, чувствительной к инфекции вирусом осповакцины, вирусом осповакцины, содержащем геном поксвируса, как предоставлено в настоящем описании, и выделение EEV, высвобождаемого клеткой-хозяином.
[10] Изобретение дополнительно относится к способу дисплея интегрального мембранного белка (IMP) или его фрагмента в нативной конформации, где способ включает инфицирование клеток-хозяев, чувствительных к инфекции поксвирусом, рекомбинантным поксвирусом, который экспрессирует IMP или его фрагмент в виде белка, слитого с характерным для EEV поксвируса белком или его мембраносвязанным фрагментом, где EEV, продуцируемый инфицированной клеткой-хозяином, содержит слитый белок IMP как часть внешней мембраны оболочки EEV и выделение EEV, высвобождаемого клеткой-хозяином. В некоторых аспектах IMP или его фрагмент предоставляют на поверхности EEV в нативной конформации. В некоторых аспектах характерный для EEV белок может представлять собой белок A33R, белок A34R, белок A56R, белок B5R, белок A36R, F13L белок вируса осповакцины, любой его мембраносвязанный фрагмент или любое их сочетание.
[11] В некоторых аспектах характерный для EEV белок представляет собой F13L (SEQ ID NO:1) или его функциональный фрагмент. В некоторых аспектах IMP представляет собой пронизывающий несколько раз мембрану белок, который содержит по меньшей мере два, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять, по меньшей мере шесть или по меньшей мере семь трансмембранных доменов. В некоторых аспектах IMP может представлять собой сопряженный с G-белком рецептор (GPCR), например, FZD4 или CXCR4 человека, как описано выше, который содержит семь трансмембранных доменов, и белок F13L может являться слитым с C-конец IMP. В некоторых аспектах IMP или его фрагмент может содержать четное число трансмембранных доменов, например, CD20 человека, как описано выше, где N-конец и C-конец IMP или его фрагмент являются внутримембранными, и F13L может являться слитым с N-концом или C-концом IMP.
[12] В некоторых аспектах мембраносвязанный характерный для EEV белковый фрагмент может содержать или состоять из стебля, трансмембранного и внутримембранного доменов белка A56R вируса осповакцины, например, аминокислот от 108 до 314 SEQ ID NO:5. В некоторых аспектах участок IMP слитого белка A56R может содержать внеклеточный домен FZD4 человека, например, слитый белок может содержать аминокислоты от 20 до 370 SEQ ID NO:6, внеклеточный домен ErbB2 человека (Her2), например, слитый белок может содержать аминокислоты от 20 до 855 SEQ ID NO:7, или внеклеточный домен CD100 человека (семафорин 4D), например, слитый белок может содержать аминокислоты от 20 до 935 SEQ ID NO:8.
[13] В некоторых аспектах мембраносвязанный характерный для EEV белковый фрагмент может содержать или состоять из трансмембранного и внутримембранного доменов или стебля, трансмембранного и внутримембранного доменов белка B5R вируса осповакцины, например, аминокислот от 276 до 317 SEQ ID NO:9 или аминокислот от 238 до 317 SEQ ID NO:9, соответственно. В некоторых аспектах участок IMP слитого белка B5R может содержать внеклеточный домен FZD4 человека, например, слитый белок может содержать аминокислоты от 20 до 243 SEQ ID NO:10 или аминокислоты от 20 до 281 SEQ ID NO:11.
[14] Изобретение дополнительно относится к слитому белку, содержащему: аминокислоты от 20 до 892 SEQ ID NO:2; SEQ ID NO:3; SEQ ID NO:4; аминокислоты 20 до 370 SEQ ID NO:6; аминокислоты от 20 до 855 SEQ ID NO:7; аминокислоты от 20 до 935 SEQ ID NO:8; аминокислоты от 20 до 243 SEQ ID NO:10; аминокислоты от 20 до 281 SEQ ID NO:11, аминокислоты от 20 до 506 SEQ ID NO:16 или аминокислоты от 20 до 235 SEQ ID NO:17. Слитый белок, как предоставлено, при экспрессии рекомбинантным поксвирусом, например, вирусом осповакцины, может появляться на поверхности внеклеточного оболочечного вириона (EEV) поксвируса в нативной конформации. Также предоставлен рекомбинантный EEV поксвируса, содержащий слитый белок. Изобретение дополнительно относится к рекомбинантному EEV поксвируса, который содержит гетерологичный IMP или его фрагмент, слитый с характерным для EEV поксвируса белком или его мембраносвязанным фрагментом, где слитый белок находится во внешней мембране оболочки EEV, и где IMP или его фрагмент предоставляют на поверхности EEV его нативной конформации. В некоторых аспектах рекомбинантный EEV поксвируса представляет собой EEV вируса осповакцины.
[15] Изобретение дополнительно относится к способу селекции антител, которые связываются с пронизывающим несколько раз мембрану белком, где способ включает связывание рекомбинантного EEV, как предоставлено в настоящем описании, с твердой подложкой; предоставление библиотеки дисплея антител, где библиотека содержит пакеты дисплея, предоставляющие совокупность антигенсвязывающих доменов; приведение библиотека дисплея в контакт с EEV, таким образом, что пакеты дисплея, предоставляющие антигенсвязывающие домены, которые специфически связываются с IMP, экспрессируемым на EEV, могут связываться с ним; удаление несвязавшихся пакетов дисплея и выделение пакетов дисплея, которые предоставляют антигенсвязывающий домен, специфичный к IMP, экспрессируемому на EEV. В некоторых аспектах этого способа рекомбинантный EEV инактивируют перед связыванием с твердой подложкой, например, инкубацией с псораленом (триоксаленом, 4'-аминометилгидрохлоридом) в присутствии УФ-облучения. В некоторых аспектах этого способа рекомбинантный EEV связывают с твердой поверхностью с помощью реакции тозильных групп, связанных с поверхностью. В некоторых аспектах твердую поверхность может представлять собой активированные тозилом магнитные гранулы. В некоторых аспектах этого способа рекомбинантный EEV подвергают биотинилированию и связывают с твердой поверхностью, покрытый стрептавидином, например, покрытыми стрептавидином магнитными гранулами.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ/ФИГУР
[16] Фигура 1A-C: Схематичное описание интегральных мембранных белков (IMP) или их фрагмента, слитого с белками или их фрагментами, характерными для внеклеточного оболочечного вириона вируса осповакцины (EEV). Параллельные горизонтальные линии являются схематическим изображением наружной мембраны EEV. Фиг. 1A схематические изображения внеклеточного домена (ECD) IMP, слитого с фрагментом белка A56R осповакцины, который содержит трансмембранный домен и внутримембранный домен. Фиг. 1B схематическое отображение топологии характерного сопряженного с G-белком рецептора, слитого с характерным для EEV вируса осповакцины белком F13L. F13L связывают с внутренней стороной наружной мембраны EEV посредством пальмитоилирования. Фиг. 1C схематическое отображение топологии IMP с четным числом трансмембранных доменов, например, CD20, слитого с F13L.
[17] Фигура 2: Иллюстрация встраивания слитых белков CD20-F13L и CD20 ECD-A56R в частицы EEV вируса осповакцины.
[18] Фигура 3A: Иллюстрация предпочтительного встраивания слитого белка CD20-F13L по сравнению с немеченым CD20 в частицы EEV вируса осповакцины.
[19] Фигура 3B: Иллюстрация предпочтительного встраивания слитого белка FZD4-F13L по сравнению с немеченым (неслитым) FZD4 в частицы EEV вируса осповакцины.
[20] Фигура 4: Встраивания дополнительных слияний белок IMP-EEV в EEV вируса осповакцины. "CD20" представляет собой слитый белок CD20-F13L, "CXCR4" представляет собой слитый белок CXCR4-F13L, "Her2" представляет собой слитый белок Her2 ECD-A56R, и "CD100" представляет собой слитый белок CD100 ECD-A56R.
[21] Фигура 5: Схема анализа скрининга библиотеки дисплея антител на пакеты дисплея, которые связываются с представляющим интерес IMP, экспрессируемым на EEV вируса осповакцины.
[22] Фигура 6A: Связывание EEV вируса осповакцины, экспрессирующего антитело к HER-2, с EEV вируса осповакцины, экспрессирующего HER2 ECD в виде слияния с белком A56R вируса осповакцины, связанного тозильными группами с магнитными гранулами.
[23] Фигура 6B: Связывание EEV вируса осповакцины, экспрессирующего антитело к FZD, с EEV вируса осповакцины, экспрессирующего FZD4 в виде слияния с белком F13L вируса осповакцины, связанного тозильными группами с магнитными гранулами.
[24] Фигура 6C: Связывание EEV вируса осповакцины, экспрессирующего антитело к CXCR4, с EEV вируса осповакцины, экспрессирующего CXCR4 в виде слияния с белком F13L вируса осповакцины, связанного тозильными группами с магнитными гранулами.
[25] Фигура 6D: Связывание EEV вируса осповакцины, экспрессирующего антитело к CD100 ("sema"), с EEV вируса осповакцины, экспрессирующим ECD CD100 в виде слияния с белком A56R вируса осповакцины, связанного тозильными группами с магнитными гранулами.
[26] Фигура 7: Сканирования FACS, демонстрирующие обогащения антителами к FZD4 после пэннинга на активированных экспрессирующих FZD-ECD-A45R EEV, связанных тозильными группами с магнитными гранулами, после 3 (Rd3), 4 (Rd4) и 5 (Rd5) циклов пэннинга. В верхнем ряду представлены инфицированные вирусом, экспрессирующим антитело, клетки, окрашенные 10 мкг/мл FZD-His с последующим окрашиванием антителом к His-Dyelight650 и антителом к Fab-FITC. В нижнем ряду представлены инфицированные вирусом, экспрессирующим антитело, клетки, окрашенные 10 мкг/мл CD100-His (отрицательный контроль) с последующим окрашиванием антителом к His-Dyelight650 и антителом к Fab-FITC.
[27] Фигура 8: Встраивание двух различных слияний белков (слияние HA-A56R и слияние FZD4-F13L) в EEV вируса осповакцины. EEV, экспрессирующий только слияние HA-A56R, только слияние FZD4-F13L или оба слитых белка, тестировали на связывание с покрытиями антителами к FZD4 гранулами или покрытыми антителами к HA гранулами.
[28] Фигура 9: Конкретное спасение экспрессирующим антитело к CXCR4 EEV магнитными гранулами, покрытыми EEV, экспрессирующим слияние HA-A56R и слияние CXCR4-F13L. Антиген-EEV связывали с покрытыми антителами к HA гранулами.
[29] Фигура 10: Связывание биотинилированного EEV вируса осповакцины, экспрессирующего сконструированные слитые белки, покрытыми стрептавидином магнитными гранулами.
ПОДРОБНОЕ ОПИСАНИЕ
[30] Настоящее изобретение относится к способам и композициям для экспрессии и дисплея интегральных мембранных белков (IMP), например, пронизывающими несколько раз мембрану (IMP), в конформационно интактном или нативном состоянии на поверхности внеклеточных оболочечных вирионных частиц, (EEV) поксвируса, например, вируса осповакцины, в виде слияния с сегментом полипептида характерного для EEV мембраносвязанного белка, например, F13L.
Определения
[31] Термин в единственном числе относится к одному или более объектов; например, понимают, что "связывающая молекула" представляет собой одну или более связывающих молекул. Таким образом, термины "один или более" и "по меньшей мере один" можно использовать взаимозаменяемо в настоящем описании.
[32] Кроме того, "и/или", при использовании в настоящем описании, следует рассматривать как конкретное описание каждого из двух конкретных признаков или компонентов с другим или без него. Таким образом, термин и/или", как используют в такой фразе, как "A и/или B" в настоящем описании, предназначен включать "A и B", "A или B", "A" (отдельно) и "B" (отдельно). Подобным образом, термин "и/или" как используют в такой фразе, как "A, B и/или C" предназначен включать каждый из следующих вариантов осуществления: A, B и C; A, B или C; A или C; A или B; B или C; A и C; A и B; B и C; A (отдельно); B (отдельно) и C (отдельно).
[33] Если не определено иное, технические и научные термины, используемые в настоящем описании, имеют такое же значение, как общепринято понимает специалист в данной области, к которой принадлежит изобретение. Например, в Concise Dictionary of Biomedicine and Molecular Biology, Juo, Pei-Show, 2nd ed., 2002, CRC Press; The Dictionary of Cell and Molecular Biology, 3rd ed., 1999, Academic Press; and the Oxford Dictionary Of Biochemistry And Molecular Biology, Revised, 2000, Oxford University Press, специалист может найти общий словарь многих терминов, используемых в этом описании.
[34] Единицы, префиксы и символы обозначаются в их принятой форме в Международной системе единиц (СИ) (Système International de Unites (SI)). Числовые диапазоны включают числа, определяющие диапазон. Если не указано иное, аминокислотные последовательности записаны слева направо в ориентации от N-конца к С-концу. Заголовки, предоставленные в настоящем описании, не являются ограничениями различных аспектов или аспектов изобретения, которые могут быть приведены посредством ссылки на описание в целом. Таким образом, термины, определения которых приведены непосредственно ниже, более полно определяют посредством ссылки на описание в его полном объеме.
[35] В рамках изобретения, термин "неприродное" вещество, композиция, соединение и/или любая комбинация веществ, композиций или соединений, или любой их грамматический вариант, является условным термином, который явно исключает, но только исключает, такие формы вещества, композиции, соединения и/или любую комбинацию веществ, композиций или соединений, которые хорошо понятны специалистам в данной области как "природные" или которые определены или интерпретированы судьей или административным или юридическим органом как "природные" или в любой момент времени могут быть определены или интерпретированы так.
[36] В рамках изобретения, термин "полипептид" предназначен включать один "полипептид", а также несколько "полипептидов" и относится к молекуле, состоящей из линейных мономеров (аминокислот), связанных амидными связями (также известным как пептидные связи). Термин "полипептид" относится к любой цепи или цепям двух или более аминокислот, и не относится к конкретной длине продукта. Таким образом, пептиды, дипептиды, трипептиды, олигопептид, "белок", "аминокислотная цепь" или любой другой термин, используемой для обозначения цепи или цепей двух или более аминокислот, входит в определение "полипептида", и термин "полипептид" можно использовать вместо него или взаимозаменяемо с любым из этих терминов. Термин "полипептид" предназначен включать продукты постэкспрессионных модификаций полипептида, включая без ограничения гликозилирование, ацетилирование, фосфорилирование, амидирование и дериватизация известными защитными/блокирующими группами, протеолитическое расщепление или модификацию неприродными аминокислотами. Полипептид можно получать из биологического источника или получать рекомбинантной технологией, но он необязательно транслируется из сконструированной последовательности нуклеиновой кислоты. Его можно получать любым способом, включая путем химического синтеза.
[37] Размер полипептида, как описано в настоящем описании, может составлять приблизительно 3 или более, 5 или более, 10 или более, 20 или более, 25 или более, 50 или более, 75 или более, 100 или более, 200 или более, 500 или более, 1000 или более, или 2000 или более аминокислот. Полипептиды могут иметь определенную трехмерную структуру, хотя они необязательно имеют такую структуру. Полипептиды с определенной трехмерной структурой обозначают как свернутые и полипептиды, которые не обладают трехмерной структурой, а наоборот могут принимать большое число различных конформаций, обозначают как несвернутые. В рамках изобретения, термин гликопротеин относится к белку, связанному по меньшей мере с одной углеводной группой, которая является связанной с белком через содержащую кислород или содержащую азот боковую цепь аминокислоты, например, серин или аспарагин.
[38] Под "выделенным" полипептидом или его фрагментом, вариантом или производным подразумевают полипептид, который не находится в своем природном окружении. Не требуется конкретного уровня очистки. Например, выделенный полипептид можно удалять из его нативной или природной среды. Рекомбинантно продуцируемые полипептиды и белки, экспрессируемые в клетках-хозяевах считают выделенными, как описано в настоящем описании, как нативные или рекомбинантные полипептиды, которые отделяли, фракционировали или частично или в значительной степени очищали любым подходящим способом.
[39] В рамках изобретения, термин "неприродный" полипептид или любые его грамматические варианты является условным термином, который явно исключает, но только исключает, такие формы полипептида, которые хорошо понятны специалистам в данной области как "природные" или которые определены или интерпретированы судьей или административным или юридическим органом как "природные" или в любой момент времени могут быть определены или интерпретированы так.
[40] Другие полипептиды, описываемые в настоящем описании, представляют собой фрагменты, производные, аналоги или варианты указанных выше полипептидов и любого их сочетания. Термины "фрагмент", "вариант", "производное" и "аналог", как описано в настоящем описании, включают любые полипептиды, которые сохраняют по меньшей мере некоторые из свойств соответствующего нативного антитела или полипептида, например, специфическое связывание с антигеном. Фрагменты полипептидов включают, например, протеолитические фрагменты, а также фрагменты делеции, в дополнение к конкретным фрагментам антител, описываемы в другом месте в настоящем описании. Варианты, например, полипептида включают фрагменты, как описано выше, а также полипептиды с измененными аминокислотными последовательностями вследствие замены, делеций или вставок аминокислот. В некоторых аспектах варианты могут являться неприродными. Неприродные варианты можно получать с использованием известных в данной области способов мутагенеза. Полипептиды-варианты могут содержать консервативные или неконсервативные замены, делеции или добавления аминокислот. Производные представляют собой полипептиды, которые изменяли таким образом, чтобы они обладали дополнительными признаками, не встречающимися в исходном полипептиде. Примеры включают слитые белки. Полипептиды-варианты также могут быть обозначены в настоящем описании как "аналоги полипептидов". В рамках изобретения "производное" полипептида также может относиться к отдельному полипептиду, содержащему одну или более аминокислот химически дериватизированных с помощью реакции функциональной группы боковой цепи. Также в определение "производные" входят такие пептиды, которые содержат одно или более производных двадцати стандартных аминокислот. Например, 4-гидроксипролин можно заменять пролином; 5-гидроксилизин можно заменять лизином; 3-метилгистидин можно заменять гистидином; гомосерин можно заменять серином и орнитин можно заменять лизином.
[41] "Консервативная замена аминокислот" представляет собой замену, при которой одну аминокислоту заменяют другой аминокислотой с аналогичной боковой цепью. Семейства аминокислот, содержащих аналогичные боковые цепи, определены в данной области, включая основные боковые цепи (например, лизин, аргинин, гистидин), кислые боковые цепи (например, аспарагиновая кислота, глутаминовая кислота), незаряженные полярные боковые цепи (например, аспарагин, глутамин, серин, треонин, тирозин, цистеин), неполярные боковые цепи (например, глицин, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), бета-разветвленные боковые цепи (например, треонин, валин, изолейцин) и ароматические боковые цепи (например, тирозин, фенилаланин, триптофан, гистидин). Например, замена фенилаланина тирозином является консервативной заменой. В некоторых вариантах осуществления консервативные замены в последовательностях полипептидов и антител по настоящему изобретению не подавляют связывание полипептида или антитела, содержащего аминокислотную последовательность, с антигеном, с которым связывается связывающая молекула. Способы идентификации консервативных замен нуклеотида и аминокислоты, которые не устраняют связывание антигена, хорошо известны в данной области (см., например, Brummell et al., Biochem. 32:1180-1 187 (1993); Kobayashi et al., Protein Eng. 12(10):879-884 (1999); and Burks et al., Proc. Natl. Acad. Sci. USA 94:.412-417 (1997)).
[42] В рамках изобретения, термин "интегральный мембранный белок" или "IMP" относится к белку или полипептиду, который прикреплен к биологической мембране. Одним из примеров IMP является трансмембранный белок, которые проходят через липидный бислой биологической мембраны один или более раз. Пронизывающие мембрану один раз мембранные белки пересекают через мембрану только один раз, тогда как пронизывающие несколько раз мембрану мембранные белки хаотично располагаются, пересекая мембрану несколько раз. I тип пронизывающие мембрану один раз белки располагаются так, что их N-конец находится на внешней стороне мембраны или "внемембранный", и их C-конец противоположной стороне мембраны или "внутримембранный." N-конец пронизывающих мембрану один раз белков II типа располагается на внутримембранной стороне. Пронизывающие несколько раз мембрану трансмембранные белки проходят через мембрану два или более раз и могут характеризоваться рядом различных топологий. N-конец и C-конец таких белков с четным числом трансмембранных доменов располагаются на одной и той же стороне мембраны. Одним из примеров такого белка является CD20, который экспрессируется B-клетками. N- и C-концы белков с нечетным числом трансмембранных доменов располагаются на противоположных сторонах мембраны. Примеры включают сопряженные с G-белком рецепторы, которые, как правило, содержат 7 трансмембранных доменов, с N-концом на внемембранной стороне и C-конец на внутримембранной стороне. Определенные IMP не содержат трансмембранные домены, а вместо этого и заякореваются в мембране, например, посредством липида, такого как гликозилфосфатидилинозитол или пальмитоильной группы. IMP обладают огромных числом биологических функций, включая, но, не ограничиваясь ими, транспортеры, линкеры, каналы, рецепторы, ферменты, преобразование энергии или адгезия клеток.
[43] Термин "полинуклеотид" предназначен включать одну нуклеиновую кислоту, а также несколько нуклеиновых кислот, и относится к выделенной молекуле нуклеиновой кислоты или конструкции, например, информационной РНК (иРНК), кДНК или плазмидной ДНК (пДНК). Полинуклеотид может содержать общепринятую фосфодиэфирную связь или необщепринятую связь (например, амидную связь, такую как связь, встречающуюся в пептидных нуклеиновых кислотах (ПНК)). Термины "нуклеиновая кислота" или "последовательность нуклеиновой кислоты" относятся к любому одному или более сегментам нуклеиновой кислоты, например, фрагментам ДНК или РНК, присутствующим в полинуклеотиде.
[44] Под "выделенной" нуклеиновой кислотой или полинуклеотидом понимают любую форму нуклеиновой кислоты или полинуклеотида, которую отделяют от его природного окружения. Например, считают, что очищенный в геле полинуклеотид или рекомбинантный полинуклеотид, кодирующий полипептид, содержащийся в векторе, является "выделенным". Также, считают, что полинуклеотидный сегмент, например, продукт ПЦР, который сконструировали так, чтобы он содержал участки рестрикции для клонирования, является "выделенным". Дополнительные примеры выделенного полинуклеотида включают рекомбинантные полинуклеотиды, содержащиеся в гетерологичных клетках-хозяевах или очищенные (частично или по существу) полинуклеотиды в ненативном растворе, таком как буфер или солевой раствор. Выделенные молекулы РНК включают транскрипты РНК in vivo или in vitro полинуклеотидов, где транскрипт не является транскриптом, встречающимся в природе. Выделенные полинуклеотиды или нуклеиновые кислоты дополнительно включают такие молекулы, получаемые синтетически. Кроме того, полинуклеотид или нуклеиновая кислота может представлять собой или может содержать регуляторный элемент, такой как промотор, участок связывания рибосомы или терминатор транскрипции.
[45] В рамках изобретения, "неприродный" полинуклеотид или любые его грамматические варианты является условным определением, которое явно исключает, но только исключает, такие формы полинуклеотида, которые хорошо понятны специалистам в данной области как "природные" или которые определены или интерпретированы судьей или административным или юридическим органом как "природные" или в любой момент времени могут быть определены или интерпретированы таким образом.
[46] В рамках изобретения, "кодирующая область" представляет собой участок нуклеиновой кислоты, который состоит из кодонов, транслируемых в аминокислоты. Несмотря на то, что "стоп-кодон" (TAG, TGA или TAA) не транслируется в аминокислоту, можно считать, что он является частью кодирующей области, но любые фланкирующие последовательности, например, промоторы, участки связывания рибосомы, терминаторы трансляции, интроны и т.п., не являются частью кодирующей области. Две или более кодирующие области могут присутствовать в одной полинуклеотидной конструкции, например, в одном векторе, или в отдельных полинуклеотидных конструкциях, например, в отдельных (различных) векторах. Кроме того, любой вектор может содержать одну кодирующую область или может содержать две или более кодирующие области, например, один вектор может раздельно кодировать вариабельную область тяжелой цепи иммуноглобулина и вариабельную область легкой цепи иммуноглобулина. Кроме того, вектор, полинуклеотид или нуклеиновая кислота может включать гетерологичные кодирующие области, слитые или не слитые с другой кодирующей областью. Гетерологичные кодирующие области включают без ограничения области, кодирующие специализированные элементы или мотивы, такие как секреторный сигнальный пептид или гетерологичный функциональный домен.
[47] В некоторых вариантах осуществления полинуклеотид или нуклеиновая кислота представляет собой ДНК. В случае ДНК полинуклеотид, содержащий нуклеиновую кислоту, кодирующую полипептид, обычно может содержать промотор и/или другие регулирующие транскрипцию или трансляцию элементы, функционально связанные с одной или более кодирующими областями. Функциональная связь является такой, когда кодирующая область продукта гена, например, полипептида, находится в связи с одной или более регуляторных последовательностей, таким образом, чтобы экспрессия продукта гена находилась под влиянием или контролем регуляторной последовательности (ей). Два фрагмента ДНК (такие как кодирующая полипептид область и промотор, связанный с ней) являются "функционально связанными", если индукция функции промотора приводит к транскрипции иРНК, кодирующей желаемый продукт гена, и если природа связи между двум фрагментами ДНК не влияет на способность регулирующих экспрессию последовательностей направлять экспрессию продукта гена или влить на способность ДНК-матрицы транскрибироваться. Таким образом, область промотора может являться функционально связанной с нуклеиновой кислотой, кодирующей полипептид, если промотор был способен влиять на транскрипцию такой нуклеиновой кислоты. Промотор может представлять собой клеткоспецифический промотор, которые направляет основную часть транскрипции ДНК в определенных клетках. Другие элементы контроля транскрипции, наряду с промотором, например, энхансеры, операторы, репрессоры и сигналы терминации транскрипции, можно функционально связывать с полинуклеотидом для направления клеткоспецифической транскрипции.
[48] Специалистам в данной области известен ряд областей контроля транскрипции. Они включают без ограничения области контроля транскрипции, которые функционируют в клетках позвоночных, такие как, но, не ограничиваясь ими, промоторные и энхансерные сегменты из цитомегаловирусов (предранний промотор в сочетании с интроном-A), вирус обезьян 40 (ранний промотор) и ретровирусы (такие как вирус саркомы Рауса). Другие области контроля транскрипции включают области контроля транскрипции, получаемые из генов позвоночных, такие как актин, белок теплового шока, бычий гормон роста и β-глобин кролика, а также другие последовательности, способные регулировать экспрессию гена в эукариотических клетках. Дополнительные подходящие области контроля транскрипции включают тканеспецифические промоторы и энхансеры, а также лимфокин-индуцибельные промоторы (например, промоторы, индуцируемые интерферонами или интерлейкинами).
[49] В качестве областей контроля транскрипции также можно использовать промоторы поксвирусов (например. p7.5 или H5) или промотор бактериофага T7. При использовании промотор T7 можно использовать индуцибельную экспрессирующуюу систему вируса осповакцины. Экспрессирующая система вируса осповакцины может включать, но не ограничена ими, первый рекомбинантный вирус осповакцины, который кодирует полную кодирующую область гена 1 бактериофага T7 для РНК-полимераза T7, и второй рекомбинантный вирус осповакцины, который кодирует представляющий интерес ген, фланкированный промотором T7 и регуляторными элементами терминации. Двойная инфекция эукариотических клеток обоими рекомбинантными вирусами осповакцины приводит к синтезу РНК-полимеразы T7 и экспрессии представляющего интерес гена под контролем промотора T7.
[50] Аналогично, специалистам в данной области известен ряд элементов контроля трансляции. Они включают, но не ограничиваются ими, участки связывания рибосомы, кодоны инициация трансляции и терминации, и элементы, получаемые из пикорнавирусов (в частности внутренний участок связывания рибосомы или IRES, также обозначаемый как последовательность CITE).
[51] В других вариантах осуществления полинуклеотид может представлять собой РНК, например, в форме информационной РНК (иРНК), транспортной РНК или рибосомной РНК.
[52] Полинуклеотид и нуклеиновая кислота, кодирующие области, можно связывать с дополнительными кодирующими областями, которые кодируют секреторные или сигнальные пептиды, которые направляют секрецию полипептида, кодируемого полинуклеотидом, как описано в настоящем описании. В соответствии с сигнальной гипотезой белки, секретируемые клетками млекопитающих, содержат последовательность сигнального пептида или секреторную лидерную последовательность, которая отщепляется от зрелого белка после начала экспорта растущей белковой цепи через шероховатый эндоплазматический ретикулум. Специалистам в данной области известно, что полипептиды, секретируемые клетками позвоночных, могут содержать сигнальный пептид, слитый с N-концом полипептида, который отщепляется от полного или "полноразмерного" полипептида с образованием секретируемой или "зрелой" формы полипептида. В некоторых вариантах осуществления используют нативный сигнальный пептид, например, сигнальный пептид тяжелой цепи или легкой цепи иммуноглобулина, или функциональное производное такой последовательности, которое сохраняет способность направлять секрецию полипептида, который является функционально связанным с ней. Альтернативно, можно использовать гетерологичный сигнальный пептид млекопитающих или его функциональное производное. Например, лидерную последовательность дикого типа можно заменять лидерной последовательностью тканевого активатора плазминогена человека (TPA) или β-глюкуронидазы мыши.
[53] В рамках изобретения, "библиотека" представляет собой характерный род полинуклеотидов, например, группу полинуклеотидов, связанную, например, их происхождением от одного вида животного, типа ткани, органа или типа клеток, где библиотека в совокупности содержит по меньшей мере два разных вида в данном роде полинуклеотидов. Библиотека полинуклеотидов может включать, например, по меньшей мере два, по меньшей мере 5, по меньшей мере 10, 100, 103, 104, 105, 106, 107, 108 или 109 различных видов в пределах данного рода n полинуклеотидов. В некоторых аспектах библиотека полинуклеотидов, как предоставлено в настоящем описании, может кодировать совокупность полипептидов, которые содержат представляющий интерес полипептид. В некоторых аспектах библиотека полинуклеотидов, как предоставлено в настоящем описании, может кодировать совокупность полипептидов субъединиц иммуноглобулина, например, полипептиды субъединицы тяжелой цепи или полипептиды субъединицы легкой цепи. В этом контексте, "библиотека", как предоставлено в настоящем описании, содержит полинуклеотиды общего рода, причем родом являются полинуклеотиды, кодирующие полипептиды субъединиц иммуноглобулина определенного типа и класса, например, библиотека может кодировать тяжелые γ-1, γ-2, γ-3, γ-4, α-1, α-2, ε- или δ-цепи человека или легкую κ- или λ-цепь человека. Несмотря на то, что каждый представитель любой библиотеки, сконструированной способами, предоставленными в настоящем описании, может кодировать одну и ту же константную область тяжелой или легкой цепи и/или заякоренный в мембране домен, библиотека может совокупно содержать по меньшей мере два, по меньшей мере 5, или по меньшей мере 10, 100, 103, 104, 105, 106, 107, 108 или 109 различных вариабельных областей, связанных с общей константной областью.
[54] В других вариантах осуществления библиотека может содержать совокупность одноцепочечных фрагментов иммуноглобулина, которые содержат вариабельную область, такую как вариабельная область легкой цепи или вариабельная область тяжелой цепи, и/или вариабельную область легкой цепи и вариабельную область тяжелой цепи, например, фрагмент scFv.
[55] В рамках изобретения, "библиотека дисплея" представляет собой библиотеку полинуклеотидов, каждый из которых введен в "пакет дисплея", который экспрессирует на своей поверхности полипептид, кодируемый полинуклеотидом библиотеки. Библиотека дисплея антител, например, может содержать совокупность пакетов дисплея, каждый из которых предоставляет антигенсвязывающий домен антитела на своей поверхности. Когда допускают взаимодействие библиотеки дисплея с представляющим интерес антигеном, например, иммобилизованным на твердой поверхности, то те пакеты дисплея, которые связывают антиген, можно выделять из оставшейся части библиотеки и восстанавливать. Затем можно выделять полинуклеотид, кодирующий антигенсвязывающий домен, предоставленный на поверхности пакета дисплея. Библиотеки дисплея включают, без ограничения библиотеки фагового дисплея в бактериях или библиотеки в эукариотических системах, например, дрожжевой дисплей, дисплей на основе ретровируса, или экспрессию в ДНК-вирусах, таких как поксвирусы. См., например, патент США № 7858559 и публикацию заявки на патент США № 2013-028892, которые полностью включены в настоящее описание посредством ссылки. В некоторых аспектах антитело библиотеку дисплея можно получать в поксвирусе, например, векторе на основе вируса осповакцины, в виде слитых белков с характерным для EEV белком, таким образом, что "пакеты дисплея" представляют собой частицы EEV. См. публикацию заявки на патент США № 2013-028892.
[56] Такие библиотеки дисплея можно подвергать скринингу против слитых белков IMP, предоставленных на поверхности EEV, как предоставлено в настоящем описании.
[57] Под "клетка-реципиентом" или "клеткой-хозяином", или "клеткой" подразумевают клетку или популяцию клеток, в которых рекомбинантный белок можно экспрессировать, вирус можно выращивать, или библиотеки полинуклеотидов, как предоставлено в настоящем описании, можно конструировать и/или расширять. Клетка-хозяин, как предоставлено в настоящем описании, как правило, представляет собой эукариотическую клетку или линию клеток, например, клетку позвоночного, млекопитающего, грызуна, мыши, примата или человека, или линию клеток. Под "популяцией клеток-хозяев" подразумевают группу культивируемых клеток, из которых, как предоставлено в настоящем описании, можно конструировать, выращивать и/или экспрессировать "библиотеку". Любая клетка-хозяин, которая является чувствительной к инфекции вирусом осповакцины, является подходящей для способов, предоставленных настоящим описанием. Клетки-хозяева для применения в способах, предоставленных в настоящем описании, могут быть прикрепленными, например, клетками-хозяевами, которые растут прикрепленными к твердому субстрату, или, альтернативно, клетки-хозяева могут находиться в суспензии.
[58] Клетки-хозяева, как предоставлено в настоящем описании, могут содержать конститутивный секреторный путь, где белки, например, представляющие интерес белки, экспрессируемые клеткой или библиотекой, секретируются из внутреннего пространства клетки для экспрессии на клетке или поверхности мембраны вируса или для полной секреции в виде растворимых полипептидов. В некоторых аспектах представляющие интерес белки, экспрессируемые на биологической мембране или в биологической мембране, например, IMP, экспрессируется на поверхности вируса в оболочке, продуцируемого клеткой-хозяином, например, внеклеточным вирусом в оболочке осповакцины или EEV. IMP могут следовать по тому же пути, как полностью секретируемые формы или белки, проходящие через просвет ER, за исключением того, что они могут удерживаться в мембране ER в результате присутствия одного или более стоп-сигналов транспорта или "трансмембранных доменов". Трансмембранные домены представляют собой гидрофобные участки приблизительно 20 аминокислот, которые принимают конформацию альфа-спирали, так только они пересекают мембрану. Погруженные в мембрану белки заякориваются в фосфолипидном бислое плазматической мембраны. В трансмембранных формах представляющих интерес полипептидов, например, заякоренных в мембране полипептидов тяжелой цепи иммуноглобулина, как правило, используются сигнальные пептиды V-конца как и секретируемый полностью формы.
[59] Сигнальные пептиды, трансмембранные домены и цитозольные или "внутримембранные" домены известны для широкого спектра связанных с мембранной и/или полностью секретируемых белков.
[60] Подходящие трансмембранные домены включают, но не ограничиваются ими, домен TM характерного для EEV вируса осповакцины HA белка A56R, или характерных для EEV вируса осповакцины трансмембранных белков A33R, A34R, A36R или B5R. См., например, публикацию заявки на патент США № 2013/0288927, опубликованную 31 октября 2013 года и полностью включенную в настоящее описание посредством ссылки. В некоторых аспектах характерный для EEV белок можно закреплять на внутренней поверхности оболочки вируса посредством пальмитоильной группы, например, белок F13L вируса осповакцины, описываемый более подробно в другом месте в настоящем описании.
[61] В рамках изобретения, термин "связывающая молекула" в широком смысле относится к молекуле, которая специфически связывается с рецептором, например, эпитопом или антигенной детерминантой. Как описано далее в настоящем описании, связывающая молекула может содержать один или более "антигенсвязывающие домены", описываемые в настоящем описании. Неограничивающий пример связывающей молекулы представляет собой антитело или его фрагмент, который сохраняет антигенспецифическое связывание.
[62] Термины "связывающий домен" и "антигенсвязывающий домен" в настоящем описании используют взаимозаменяемо, и он обозначает область связывающей молекулы, которая является обязательной и достаточной для специфического связывания с эпитопом. Например, считают, что "Fv", вариабельная область тяжелой цепи и вариабельная область легкой цепи антитела, в виде двух отдельных субъединиц полипептида или в виде одной цепи, является "связывающим доменом".
[63] Другие антигенсвязывающие домены включают без ограничения вариабельную область тяжелой цепи (VHH) антитела, получаемого от видов верблюжьих, или шесть определяющих комплементарность областей (CDR) иммуноглобулина, экспрессируемых в каркасе фибронектина.
[64] В настоящем описании термины "антитело" и "иммуноглобулин" можно использовать взаимозаменяемо. Антитело (или его фрагмент, вариант или производное, как описано в настоящем описании) содержит по меньшей мере вариабельную область тяжелой цепи (например, для видов верблюжьих) или по меньшей мере вариабельные области тяжелой цепи и легкой цепи. Основные структуры иммуноглобулина в системах позвоночных относительно хорошо изучены. См., например, Harlow et al., Antibodies: A Laboratory Manual, (Cold Spring Harbor Laboratory Press, 2nd ed. 1988). Если не указано иное, термин "антитело" включает все в диапазоне от небольшого антигенсвязывающего фрагмента антитела до полноразмерного антитела, например, антитело IgG, которое содержит две полные тяжелые цепи и две полные легкие цепи.
[65] Термин "иммуноглобулин" включает обширные классы полипептидов, которые можно различать на биохимическом уровне. Специалистам в данной области понятно, что тяжелые цепи классифицируют как гамма, мю, альфа, дельта или эпсилон (γ, μ, α, δ, ε) с некоторыми подклассами среди них (например, γ1-γ4 или α1-α2)). Именно природа этой цепи определяет "класс" антитела как IgG, IgM, IgA IgG или IgE, соответственно. Подклассы иммуноглобулина (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1, IgA2 и т.д. хорошо охарактеризованы, и известно, что они определяют функциональную специализацию.
[66] Легкие цепи классифицируют как каппа или лямбда (κ, λ). Каждый класс тяжелой цепи может быть связан с легкой каппа- или лямбда-цепью. В основном, легкие и тяжелые цепи ковалентно связаны друг с другом, и "хвостовые" участки двух тяжелых цепей связаны друг с другом ковалентными дисульфидными связями или нековалентными связями, когда иммуноглобулины получают посредством гибридом, B-клеток или генетически сконструированных клеток-хозяев. В тяжелой цепи, аминокислотные последовательности идут от N-конца к раздвоенным концам конфигурации Y к C-концу в нижней части каждой цепи. Основная структура определенных антител, например, антител IgG, включает две субъединицы тяжелых цепей и две субъединицы легких цепей, ковалентно связанных через дисульфидные связи с образованием "Y"-образной структуры, также обозначаемой в настоящем описании как структура "H2L2".
[67] Термин "эпитоп" включает любую молекулярную детерминанту, способную специфически связываться с антителом. В некоторых аспектах эпитоп может включать химически активные группы на поверхности молекул, такие как аминокислоты, боковые цепи сахаров, фосфорил или сульфонил, и в некоторых аспектах может обладать характеристиками трехмерной структуры и/или конкретными характеристиками заряда. Эпитоп представляет собой область мишени, с которой связывается антитело.
[68] Термин "мишень" используют в самом широком смысле обозначения веществ, которые может связывать связывающая молекула. Мишенью может являться, например, полипептид, нуклеиновая кислота, углевод, липид или другая молекула. Кроме того, "мишенью" может являться, например, клетка, орган, или организм, которые содержит эпитоп, который может связывать связывающая молекула.
[69] Легкую и тяжелую цепи делят на области структурной и функциональной гомологии. Термины "контрактная" и "вариабельная" используют в отношении функций. В связи с этим следует понимать, что вариабельные области (которые могут называться "вариабельными доменами" взаимозаменяемо в настоящем описании) вариабельных участков легкой (VL) цепи и вариабельных участков (VH) цепи определяют распознавание антигена и специфичность. В противоположность этому, константные домены легкой цепи (CL) и тяжелой цепи (например, CH1, CH2 или CH3) обеспечивают биологические свойства, такие как секреция, перенос через плаценту, связывание Fc-рецептора, связывание комплемента и т.п. Согласно правилу нумерация доменов константной области возрастает по мере того как они отдаляются от антигенсвязывающего участка или N-конца антителаав. N-концевая часть представляет собой вариабельную область, и в C-концевой части находится константная область; домены CH3 (или CH4 в случае IgM) и CL располагаются на C-конце тяжелых и легких цепей, соответственно.
[70] Шесть "определяющих комплементарность областей" или "CDR", присутствующих в антигенсвязывающем домене антитела, представляют собой короткие непрерывные последовательности аминокислот, которые располагаются в определенном порядке с образованием антигенсвязывающего домна, в результате чего антитело приобретает свою трехмерную конфигурацию в водной среде. Оставшаяся часть аминокислот в антигенсвязывающем домене, обозначаемая как "каркасные" области, обладает меньшей внутримолекулярной вариабельностью. Каркасные области в значительной степени принимают комформацию β-листа, и CDR образуют петли, которые соединяются и в некоторых случаях образуют часть структуры β-листа. Таким образом, каркасные области функционируют с образованием каркаса, который обеспечивает расположение CDR в правильной ориентации в результате межцепочечных, нековалентных взаимодействий. Антигенсвязывающий домен, образованный располагающимися в определенном порядке CDR, определяют поверхность, комплементарную эпитопу на иммунореактивного антигена. Такая комплементарная поверхность способствует нековалентному связыванию антитела с его когнатным эпитопом. Специалист в данной области может легко идентифицировать аминокислоты, которые составляют CDR и каркасные области, соответственно, для любой данной вариабельной области тяжелой или легкой цепи, т.к. их можно идентифицировать различными способами (см., "Sequences of Proteins of Immunological Interest," Kabat, E., et al., U.S. Department of Health and Human Services, (1983); и Chothia and Lesk, J. Mol. Biol., 196:901-917 (1987), которые полностью включены в настоящее описание посредством ссылки).
[71] В случае, когда существует два или более определения термина, который используют и/или который принят в данной области, определение термина, в рамках изобретения, предназначено включать все такие значения, если конкретно не указано иное. Конкурентным примером является использование термина "определяющая комплементарность область" ("CDR") для описания непрерывных антигенсвязывающих участков, встречающихся в вариабельной области тяжелых и легких цепей полипептидов. Эти конкретные области описаны, например, Kabat et al., U.S. Dept. of Health and Human Services, "Sequences of Proteins of Immunological Interest" (1983) и Chothia et al., J. Mol. Biol. 196:901-917 (1987), которые включены в настоящее описание посредством ссылки. Для идентификации сегментов вариабельной области, содержащих CDR, также можно анализировать вариабельные домены иммуноглобулина, например, с использованием информационной системы IMGT (www://imgt.cines.fr/) (IMGTВ®/V-Quest). (См., например, Brochet et al., Nucl. Acids Res., 36:W503-508, 2008).
[72] Kabat et al. также установили систему нумерации для последовательности вариабельных доменов, которую можно применять к любому антителу. Специалист в данной области может однозначно применять эту систему "нумерации по Kabat" к любой последовательности вариабельного домена, без учета каких-либо экспериментальных данных за пределами самой последовательности. В рамках изобретения, "нумерация по Kabat" относится к системе нумерации, указанной Kabat et al., U.S. Dept. of Health and Human Services, "Sequence of Proteins of Immunological Interest" (1983). Однако если использование системы нумерации по Kabat явно не указано, то используют последовательную нумерацию для всех аминокислотных последовательностей в этом описании.
[73] Связывающие молекулы, например, антитела или их антигенсвязывающие фрагменты, варианты или производные включают, но не ограничиваются ими, поликлональные, моноклональные антитела, антитела человека, гуманизированные или химерные антитела, одноцепочечные антитела, связывающие эпитоп фрагменты, например, Fab, Fab' и F(ab')2, Fd, Fvs, одноцепочечный Fvs (scFv), одноцепочечные антитела, связанный дисульфидными связями Fvs (sdFv), однодоменные антитела, такие как антитела VHH верблюжьих, фрагменты, содержащие домен VL или домен VH, фрагменты, получаемые посредством библиотеки экспрессии Fab. Молекулы scFv известны в данной области и описаны, например, в патенте США 5892019. Молекулы иммуноглобулинов или антител, которые охватывает настоящее описание, могут быть любого типа (например, IgG, IgE, IgM, IgD, IgA и IgY), класса (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2) или подкласса молекулы иммуноглобулина. Также предусматривают изотипы иммуноглобулина - новый антигенный рецептор (IgNAR), которые являются бивалентными и содержат одну цепь, которая содержит вариабельный домен IgNAR (VNAR). (См., Walsh et al., Virology 411:132-141, 2011).
[74] Под "специфически связывается", как правило, понимают, что связывающая молекула, например, антитело или его фрагмент, вариант или производное связывается с эпитопом через свой антигенсвязывающий домен, и что связывание подразумевает некоторую комплементарность между антигенсвязывающим доменом и эпитопом. Согласно этому определению считают, что связывающая молекула "специфически связывается" с эпитопом, кода она связывается с этим эпитопом через свой антигенсвязывающий домен легче, чем, если бы она связывалась со случайным неродственным эпитопом. Термин "специфичность" используют в настоящем описании для качественного описания относительной аффинности, с которой определенная связывающая молекула связывается с определенным эпитопом. Например, можно считать, что связывающая молекула "A" обладает более высокой специфичностью к данному эпитопу, чем связывающая молекула "B", или можно считать, что связывающая молекула "A" связывается с эпитопом "C" с более высокой специфичностью по сравнению со специфичностью, которой она обладает в отношении родственного эпитопа "D".
[75] В рамках изобретения, термин "аффинность" относится к мере силы связывания отдельного эпитопа с одним или более антигенсвязывающих доменов, например, молекулы иммуноглобулина. Cм., например, Harlow et al., Antibodies: A Laboratory Manual, (Cold Spring Harbor Laboratory Press, 2nd ed. 1988) страницы 27-28. В рамках изобретения, термин "авидность" относится к общей стабильности комплекса между популяцией антигенсвязывающих доменов и антигена. См., например, Harlow страницы 29-34. Авидность относится к аффинности отдельных антигенсвязывающих доменов в популяции с конкретными эпитопами, а также валентностям иммуноглобулинов и антигена. Например, взаимодействие между бивалентным моноклональным антителом и антигеном с структурой эпитопа с высоким содержанием повторяющихся элементов, таким как полимер, будет взаимодействием высокой авидности. Взаимодействие между бивалентным моноклональным антителом с рецептором, содержащимся в высокой плотности на поверхности клеток, также будет высокой авидности.
[76] В рамках изобретения, термин "субъединица тяжелой цепи" или "домен тяжелой цепи" включает аминокислотные последовательности, получаемые из тяжелой цепи иммуноглобулина, связывающей молекулы, например, антитело, содержащее субъединицу тяжелой цепи может содержать по меньшей мере один из: домена VH, домена CH1, шарнирного домена (например, верхней, средней и/или нижней шарнирной области), домена CH2, домена CH3, домена CH4 или его варианта или фрагмента.
[77] В рамках изобретения, термин "субъединица легкой цепи" или "домен легкой цепи" включает аминокислотные последовательности, получаемые из легкой цепи иммуноглобулина. Субъединица легкой цепи содержит по меньшей мере один домен из домена VL или CL (например, Cκ или Cλ).
[78] Связывающие молекулы, например, антитела или их антигенсвязывающие фрагменты, варианты или производные можно описывать или определять в отношении эпитоп(ов) или участка(ов) антигена, который они распознают или специфически связывают. Участком антигена-мишени, который специфически взаимодействует с антигенсвязывающим доменом антитела, является "эпитоп" или "антигенная детерминанта". Антиген-мишень может содержать один эпитоп или по меньшей мере два эпитопа и может содержать любое число эпитопов, в зависимости от размера, конформации и типа антигена.
[79] В рамках изобретения, термины "связанный", "слитый" или "слияние", или другие грамматические эквиваленты можно использовать взаимозаменяемо. Эти термины относятся к объединению друг с другом двух других элементов или компонентов любыми способами, включая химическую конъюгацию или рекомбинантные способы. "Слияние в рамке считывания" относится к объединению двух или более открытых рамок считывания (ORF) полинуклеотида с образованием непрерывной более длинной ORF, таким образом, чтобы поддерживать трансляционную рамка считывания исходных ORF. Таким образом, рекомбинантный слитый белок представляет собой отдельный белок, содержащий два или более сегментов, которые соответствуют полипептидам, кодируемым исходными ORF (сегменты которых, как правило, не являются объединенными таким образом в природе). Несмотря на то, что рамка считывания создает таким образом непрерывные на всем протяжении слитые сегменты, сегменты можно физически или пространственно разделять, например, линкерной последовательностью в рамке считывания. Например, полинуклеотиды, кодирующие IMP и характерный для EEV вируса осповакцины белок, могут являться слитыми в рамке считывания, но быть разделенными полинуклеотидом, кодирующим линкер или спейсер, при условии, что "слитые" открытые рамки считывания котранслируются в виде части непрерывного полипептида.
[80] В рамках изобретения, термин "гемагглютининовая метка" или "метка HA" представляет собой белок, получаемый из поверхностного гликопротеина гемагглютинина вируса гриппа человека (HA), соответствующего аминокислотам 98-106. Метку HA метка широко используют в качестве основной эпитопной метки в экспрессирующих векторах. Рекомбинантные белки можно конструировать с возможность экспрессии метки HA, которая, как полагают, не нарушает биологическую активность или биораспределение рекомбинантного белка. Такая метка облегчает детекцию, выделение и очистку представляющего интерес белка.
[81] В отношении полипептидов "линейная последовательность" или "последовательность" представляет собой порядок аминокислот в полипептиде от амино- или N-конца к карбоксильному или C-концу, в котором аминокислоты, которые располагаются по соседству друг с другом в последовательности, являются смежными в первичной структуре полипептида.
[82] Участок полипептида, который является "аминоконцевой" или "N-концевой" по отношению к другому участку полипептиду, представляет собой такой участок, который выступает раньше в последовательной полипептидной цепи. Подобным образом участок полипептида, который является "карбокси-концевым" или "C-концевым" по отношению к другому участку полипептида, представляет собой участок, который выступает позже в последовательной полипептидной цепи.
[83] В рамках изобретения, термин "экспрессия" относится к процессу, посредством которого ген продуцирует биохимическое вещество, например, полипептид. Процесс включает любое проявление функционального присутствия гена в клетке, включая без ограничения, нокдаун гена, а также транзиторную экспрессию и стабильную экспрессию. Он включает, без ограничения транскрипцию гена в информационную РНК (иРНК) и трансляцию такой иРНК в полипептид(ы). Если конечный желаемый продукт является биохимическим веществом, экспрессия включает создание такого биохимического вещества и любых предшественников. Экспрессия гена приводит к образованию "продукта гена". В рамках изобретения, продукт гена может представлять собой нуклеиновую кислоту, например, информационную РНК, продуцируемую транскрипцией гена, или полипептид, который транслируется из транскрипта. Продукты генов, описываемые в настоящем описании, дополнительно включают нуклеиновые кислоты с посттранскрипционными модификациями, например, полиаденилированием, или полипептиды с посттрансляционными модификациями, например, метилированием, гликозилированием, добавлением липидов, ассоциацией с другими белковыми субъединицами, протеолитическим расщеплением и т.п.
[84] Термин "эукариот" или "эукариотический организм" предназначен включать все организмы в царстве животных, растений и простейших, включая одноклеточных, грибы, дрожжи, зеленые водоросли, одноклеточные растения, многоклеточные растения и всех животных, как позвоночных, так и не позвоночных. Термин не включает бактерии или вирусы. "Эукариотическая клетка" предназначена включать отдельную "эукариотическую клетку", а также совокупность "эукариотических клеток", и включает клетки, получаемые от эукариот.
[85] Термин "позвоночное" предназначен включать отдельное "позвоночное", а также совокупность "позвоночных", и включает млекопитающих и птиц, а также рыб, пресмыкающихся и амфибий.
[86] Термин "млекопитающее" предназначен включать отдельное "млекопитающее" и совокупность "млекопитающих", и включает, но не ограничивается ими, людей; приматов, таких как человекообразные обезьяны, нечеловекообразные обезьяны, орангутанги и шимпанзе; собачьих, таких как собаки и волки; кошачьих, таких как кошки, львы и тигры; лошадиных, таких как лошади, ослы и зебры, животных, служащих для получения продуктов питания, таких как коровы, свиньи и овцы; копытных, таких как олень и жирафы; грызунов, таких как мыши, крысы, хомяки и морские свинки; и медведей. В некоторых аспектах млекопитающее представляет собой являющегося человеком индивидуума.
[87] Термины "тканевая культура" или "культура клеток", или "культура" или "культивирование" относится к поддержанию или росту растительной или животной ткани или клеток in vitro в условиях, которые обеспечивают сохранение архитектуры клеток, сохранение функции клеток, дальнейшую дифференцировку или все три пункта. "Клетки первичных тканей" представляют собой клетки, получаемые непосредственно из ткани, т.е. популяцию клеток одного и того же вида, выполняющего ту же функцию в организме. Обработка таких клеток тканей протеолитическим ферментом трипсином, например, приводит к их диссоциации на отдельные клетки первичной ткани, которые растут или сохраняют клеточную архитектуру при высевании на планшеты для культивирования. Культуры клеток, получаемые в результате размножения первичных клеток в тканевой культуре, называют "культурами вторичных клеток". Большая часть вторичных клеток делится конечное число раз, а затем погибает. Однако некоторые вторичные клетки, могут проходить через такие "кризисные периоды", после которых они способны бесконечно размножаться с образованием стабильной "линии клеток". Жидкую среду, в которой клетки культивируют, обозначают в настоящем описании как "среду для культивирования" или "среды для культивирования". Среда для культивирования, в которую секретируются желаемые молекулы, например, вирусы или белки, например, молекулы иммуноглобулина во время культивирования клеток, может быть обозначена как "кондиционированная среда".
[88] В рамках изобретения, термин "идентифицировать" относится к способам, в которых желаемую молекулу, например, полинуклеотид, кодирующий представляющий интерес белок с желаемыми характеристиками или функцией, отличают от совокупности или библиотеки таких молекул. Способы идентификации включают "селекцию" и "скрининг" или "пэннинг". В рамках изобретения, способы "селекции" представляет собой способы, в которых желаемые молекулы можно непосредственно выделять из библиотеки, например, посредством устойчивости к лекарственному средству. В рамках изобретения, способы "скрининга" или "пэннинга" представляют собой способы, в которых объединения образцов, содержащие желаемые молекулы, подвергают анализу, в котором можно детектировать желаемую молекулу. Аликвоты объединений образцов, в которых детектируют молекулу, затем разделяют на несколько меньших объединений, которые аналогично анализируют до тех пор, пока не получают совокупность, которая сильно обогащена желаемой молекулой.
Векторы на основе поксвирусов, например, EEV вируса осповакцины
[89] Слитые белки IMP, как предоставлено в настоящем описании, получают в векторах на основе поксвируса, например, векторах на основе вируса осповакцины. Термин "поксвирус" включает любого представителя семейства Poxviridae. См., например, B. Moss in: Virology, 2d Edition, B. N. Fields, D. M. Knipe et al., Eds., Raven Press, p. 2080 (1990). Род ортопоксвирусов включает, например, вирус осповакцины, вирус натуральной оспы (вирус, который взывает оспу) и поксвирус енотов. Вирус осповакцины является прототипом ортопоксвируса, и был разработан и хорошо охарактеризован как вектор для экспрессии гетерологичных белков.
[90] В этих вариантах осуществления где векторы на основе поксвируса, в частности векторы на основе вируса осповакцины, используют для экспрессии слитых белков IMP, как предоставлено в настоящем описании, можно использовать любой подходящий вектор на основе поксвируса. В некоторых аспектах локализация гена, кодирующего слитый белок IMP, может находиться в области вектора, который не является необходимыми для роста и репликации вируса, таким образом, что получают инфекционные вирусы. Хотя ряд не являющихся необходимыми областей генома вируса осповакцины был охарактеризован, наиболее широко используемым локусом для вставки чужеродных генов является локус тимидинкиназы, располагаемый во фрагменте J HindIII в геноме. В определенных векторах на основе вируса осповакцины локус tk был сконструирован таким образом, чтобы содержать один или два распознавания рестрикционных ферментов, что позволяет удобно использовать способ получения рекомбинантного вируса тримолекулярной рекомбинации, как описано в настоящем описании в другом месте.
[91] Полинуклеотиды, кодирующие слитые белки IMP, как предоставлено в настоящем описании, можно встраивать в векторы на основе поксвируса, в частности, векторы на основе вируса осповакцины, в функциональной связи с областью транскрипционного контроля, которая функционирует в цитоплазме инфицированной поксвирусом клетки.
[92] Области транскрипционного контроля поксвируса содержат промотор и сигнал терминации транскрипции. Экспрессия гена в поксвирусах временно регулируется, и промоторы для ранних, промежуточных и поздних генов обладают различными структурами. Определенные гены поксвирусов экспрессированы конститутивно, и промоторы для этих "ранних-поздних" генов несут гибридные структуры. Также разработаны синтетические ранние-поздние промоторы. Подходящие промоторы поксвируса для экспрессии слитых белков IMP, как предоставлено в настоящем описании, включают, но не ограничиваются ими, поздние промоторы, такие как промотор 7,5 кДа, промотор MIL, промотор 37 кДа, промотор 11 кДа, промотор 11L, промотор 12L, промотор 13L, промотор 15L, промотор 17L, промотор 28 кДа, промотор H1L, промотор H3L, промотор H5L, промотор H6L, промотор H8L, промотор D11L, промотор D12L, промотор D13L, промотор A1L, промотор A2L, промотор A3L и промотор P4b. См., например, Moss B., "Poxviridae и their Replication" IN Virology, 2d Edition, B. N. Fields, D. M. Knipe et al., Eds., Raven Press, p. 2090 (1990).
[93] Подходящие векторы на основе поксвируса включают вирус осповакцины дикого типа, например, штамм Western Reserve или WR, или аттенуированный вирус осповакцины, например, модифицированную осповакцину Ankara (MVA) (Mayr A. et al., Infection 3:6-14 (1975)).
[94] Во время своего цикла репликации поксвирус, например, вирус осповакцины продуцирует четыре инфекционный формы, которые отличаются по своей структуре мембран: внутриклеточный зрелый вирион (IMV), внутриклеточный оболочечный вирион (IEV), связанный с клеткой оболочечный вирион (CEV) и внеклеточный оболочечный вирион (EEV). Преобладает мнение, что IMV имеют единственную липопротеиновую мембрану, тогда как CEV и EEV оба окружены двумя мембранными слоями, а IEV имеет три оболочки. EEV выделяется из плазматической мембраны клетки-хозяина, и мембрана EEV происходит из транс-Гольджи.
[95] После инфекции вирус теряет свою мембрану (ы), и ядро ДНК/белковая сердцевиина транспортируется по микротрубочкам в клетку. Белки, кодируемые ранней иРНК осповакцины ("ранняя" определяют как репликация до ДНК), приводят к сбрасыванию оболочки сердцевины вируса осповакцины и последующей репликация ДНК. Эта репликация происходит в так называемых "фабриках вирусов", которые располагаются в основном на вершине ER. На фабрике вирусов незрелые вирионы (IV) подвергаются сборке и процессингу с образованием IMV (внутриклеточного зрелого вируса). IMV содержит мембрану, которая происходит из ER. Большая часть IMV высвобождается из клетки а результате лизиса клетки. Some IMV транспортируются по микротрубочкам в участки окутывания мембранами сети транс-Гольджи или ранних эндосом. Окутывание частиц IMV двойной мембраной приводит к форме вируса осповакцины, называемой IEV (внутриклеточный заключенный в оболочку вирус). Затем IEV транспортируются на поверхность клетки по микротрубочкам. Внешняя мембрана IEV сливается с плазматической мембраной в предоставление CEV (связанного с клеткой оболочечного вируса) на поверхности клетки. Полимеризация актина из клетки-хозяина может способствовать тому, что CEV инфицирует соседние клетки, или вирус может высвобождаться как EEV. См., например, Kim L. Roberts and Geoffrey L. Smith. Trends in Microbiology 16(10):472-479 (2008); Geoffrey L. Smith et al., Journal of General Virology 83:2915-2931 (2002).
[96] Сообщается по меньшей мере о шести кодируемых вирусом белков как компонентов мембраны оболочки EEV. Из них четыре белка (A33R, A34R, A56R и B5R) являются гликопротеинами, один (A36R) представляет собой негликозилированный трансмембранный белок, и один (F13L) представляет собой пальмитоилированный периферический мембранный белок. См., например, Lorenzo et al., Journal of Virology 74(22):10535 (2000). Во время инфекции эти белки локализуются в комплексе Гольджи, где они встраиваются в патогенный вирус, который затем транспортируется и высвобождается во внеклеточную среду. Как предоставлено в настоящем описании, слитые белки IMP являются направленными на мембрану EEV и экспрессируются на ней в виде слитого белка с характерным для EEV белком, например, F13L или A56R.
[97] Белок F13L связан с внутренней поверхностью наружной мембраны EEV в результате пальмитоилирования цистеинов 185 и 186. Smith Trends in Microbiol. 16:472-479 (2008). Вирусы осповакцины, в которых удаляют ген, кодирующий F13L, образует очень маленькие бляшки, и количество продуцируемых EEV значительно сокращается.
[98] Аминокислотная последовательность белка F13L из штамма WR вируса осповакцины предоставлена как SEQ ID NO:1. Два пальмитоилированных остатка цистеина (аминокислоты 85 и 86 SEQ ID NO:1) подчеркнуты. Вследствие того, что F13L не проходит через мембрану, он не содержит трансмембранный домен или сигнальный пептид.
F13L (SEQ ID NO:1)
MWPFASVPAGAKCRLVETLPENMDFRSDHLTTFECFNEIITLAKKYIYIASFCCNPLSTTRGALIFDKLKEASEKGIKIIVLLDERGKRNLGELQSHCPDINFITVNIDKKNNVGLLLGCFWVSDDERCYVGNASFTGGSIHTIKTLGVYSDYPPLATDLRRRFDTFKAFNSAKNSWLNLCSAACCLPVSTAYHIKNPIGGVFFTDSPEHLLGYSRDLDTDVVIDKLKSAKTSIDIEHLAIVPTTRVDGNSYYWPDIYNSIIEAAINRGVKIRLLVGNWDKNDVYSMATARSLDALCVQNDLSVKVFTIQNNTKLLIVDDEYVHITSANFDGTHYQNHGFVSFNSIDKQLVSEAKKIFERDWVSSHSKSLKI
[99] Белок A56R представляет собой гемагглютинин вируса осповакцины и является стандартным интегральным мембранным белком I типа, содержащим внеклеточный ("внемембранный") домен на N-конце, один трансмембранный домен и цитоплазматический ("внутримембранный") домен. A56R содержит сигнальный пептид приблизительно 33 аминокислоты на N-конце, Ig-подобный домен, простирающийся приблизительно от аминокислоты 34 приблизительно до аминокислоты 103, стеблевую область, простирающуюся приблизительно от аминокислоты 121 приблизительно до аминокислоты 275, трансмембранный домен, простирающийся приблизительно от аминокислоты 276 приблизительно до аминокислоты 303, и цитоплазматический ("внутримембранный") домен, простирающийся приблизительно от аминокислоты 304 до аминокислоты 314. См. DeHaven et al., J. Gen Virol. 92:1971-1980 (2011). A56R предоставлен как SEQ ID NO:5.
A56R (SEQ ID NO:5)
MTRLPILLLLISLVYATPFPQTSKKIGDDATLSCNRNNTNDYVVMSAWYKEPNSIILLAAKSDVLYFDNYTKDKISYDSPYDDLVTTITIKSLTARDAGTYVCAFFMTSTTNDTDKVDYEEYSTELIVNTDSESTIDIILSGSTHSPETSSKKPDYIDNSNCSSVFEIATPEPITDNVEDHTDTVTYTSDSINTVSASSGESTTDETPEPITDKEDHTVTDTVSYTTVSTSSGIVTTKSTTDDADLYDTYNDNDTVPPTTVGGSTTSISNYKTKDFVEIFGITALIILSAVAIFCITYYIYNKRSRKYKTENKV
[0100] Слитые белки IMP, как предоставлено в настоящем описании, можно экспрессировать в любом подходящем вирусе осповакцины. В некоторых вариантах осуществления ДНК, кодирующую слитый белок EEV можно вводить в область генома вируса осповакцины, которая не является существенной для роста и репликации вектора так, чтобы получать патогенные вирусы. Несмотря на то, что ряд несущественных областей генома вируса осповакцины был охарактеризован, наиболее широко используемым локусом для введения чужеродных генов является локус тимидинкиназы, расположенный во фрагменте J HindIII в геноме. Слитые белки IMP, как предоставлено в настоящем описании, можно вводить в векторы на основе вируса осповакцины в функциональной связи с областью транскрипционного контроля, которая функционирует в цитоплазме инфицированной поксвирусом клетки.
[0101] Подходящие промоторы для применения в способах, описываемых в настоящем описании, включают без ограничения ранний/поздний промотор 7,5 кДа или ранний/поздний промотор H5 (или их варианты).
Способ тримолекулярной рекомбинации
[0102] Тримолекулярная рекомбинация, как описано в публикация PCT № WO 00/028016 на имя Zauderer и в патенте США № 7858559, является высокоэффективным образующим высокие титры способом экспрессии представляющих интерес белков и/или получения библиотек в вирусе осповакцины. Способ тримолекулярной рекомбинации позволяет получать рекомбинантные вирусы с эффективностью по меньшей мере 90%, и титры по меньшей мере с величиной на 2 порядка выше, чем титры, получаемые прямым лигированием.
[0103] В некоторых аспектах слитые белки IMP для экспрессии в вирусе осповакцины и прдоставлении на EEV, как описано в настоящем описании, можно конструировать в векторах на основе поксвируса, например, векторах на основе вируса осповакцины посредством тримолекулярной рекомбинации.
[0104] В некоторых вариантах осуществления предоставлена переносящая плазмида для слитых белков IMP для экспрессии в EEV, которая содержит фланкирующие полинуклеотид области в гене Tk вируса осповакцины, промотор H5 вируса осповакцины и участки рестрикции NcoI и BsiWI для введения кодирующих областей для желаемых слитых белков.
Интегральные мембранные белки
[0105] Изобретение относится к способу экспрессии интегральных мембранных белков (IMP) в конформационно интактном состоянии, который приближает нативную конформацию белка, как это происходило бы в клетке, в которой белок экспрессируется естественным образом. По изобретению IMP экспрессируются в виде белков, слитых с белками поксвируса, которые экспрессируются на поксвирусе, например, EEV вируса осповакцины. Слитые белки IMP, как предоставлено в настоящем описании, в случае экспрессии и дисплея на поверхности EEV, являются пригодными в качестве антигенов-мишеней для скрининга библиотек связывающих молекул, например, библиотек дисплея антитела.
[0106] Любой IMP можно конструировать в виде слитого белка способами, предоставленными в настоящем описании. В некоторых аспектах IMP является мишенью для иммунотерапии. В некоторых аспектах IMP представляет собой пронизывающий несколько раз мембрану IMP, такой как CD20 или сопряженный с G-белком рецептор (GPCR). Подходящие пронизывающие несколько раз мембрану IMP человека для применения в конструкции слитых белков IMP, как предоставлено в настоящем описании, включают, без ограничения белки, перечисленные в таблице 1.
Таблица 1: Иллюстративные пронизывающие несколько раз интегральную мембрану белки человека
[0107] В некоторых аспектах пронизывающий несколько раз мембрану IMP представляет собой GPCR, например, FZD4 или CXCR4. В некоторых аспектах пронизывающий несколько раз мембрану IMP представляет собой CD20.
Полинуклеотиды, кодирующие слитые белки IMP для экспрессии на EEV поксвируса
[0108] Настоящее изобретение относится к выделенному полинуклеотиду для экспрессии интегрального мембранного белка или его фрагмента в конформационно интактной форме п отношению к биологической мембране, в виде слияния с белком или его фрагментом, характерным для EEV вируса осповакцины. Под "конформационно интактной" подразумевают то, что белок появляется или предоставлен в дисплее нативной или близкой к нативной конформации по отношению у биологическому мембране из липидного бислоя именно так, как белок он появляется в своем нативном состоянии.
[0109] В одном из аспектов изобретение относится к выделенному полинуклеотиду, который содержит фрагмент первой нуклеиновой кислоты, кодирующий интегральный мембранный белок (IMP) или его фрагмент, например, пронизывающий несколько раз мембрану IMP, где IMP или его фрагмент содержит по меньшей мере одну внемембранную область, по меньшей мере один трансмембранный домен и по меньшей мере одну внутримембранную область, и где участок фрагмента первой нуклеиновой кислоты, кодирующий по меньшей мере одну внутримембранную область, располагается на 5'- или 3'-конце фрагмента первой нуклеиновой кислоты; и фрагмент второй нуклеиновой кислоты, кодирующий белок F13L вируса осповакцины (SEQ ID NO:1) или его функциональный фрагмент, где фрагмент второй нуклеиновой кислоты является слитым в рамке считывания с участком фрагмента первой нуклеиновой кислоты, кодирующим внутримембранную область IMP. Фрагмент первой нуклеиновой кислоты и фрагмент второй нуклеиновой кислоты в некоторых случаях может быть разделен нуклеиновой кислотой, кодирующей линкер или другой спейсер. Полинуклеотид может дополнительно содержать промотор поксвируса, функционально связанный с первым и вторым фрагментом нуклеиновой кислоты, обеспечивающим экспрессию полинуклеотида в цитоплазме инфицированной поксвирусом клетки. По этому аспекту инфицированная поксвирусом клетка, которая содержит полинуклеотид, может экспрессировать слитый белок IMP-F13L как часть внешней мембраны оболочки внеклеточного оболочечного вириона (EEV). Схематические изображения, демонстрирующие экспрессию IMP в виде слияния с F13L, приведены на фиг. 1B и фиг. 1C.
[0110] В некоторых аспектах IMP или его фрагмент может представлять собой пронизывающий несколько раз мембрану белок, содержащий по меньшей мере два, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять, по меньшей мере шесть, по меньшей мере семь, по меньшей мере восемь, по меньшей мере девять, по меньшей мере десять, по меньшей мере одиннадцать, по меньшей мере двенадцать или даже больше трансмембранных (TM) доменов, таких как домены, перечисленные в таблице 1.
[0111] В случае, когда IMP содержит нечетное число доменов TM, один конец IMP N-конец или C-конец естественным образом располагается на внемембранной стороне биологической мембраны, а другой конец IMP располагается на внутримембранной стороне IMP. Поскольку белок F13L является полностью внутренним по отношению к внешней мембране EEV поксвируса, конец IMP, N-конец или C-конец, который находится в мембране, может являться слитым с F13L. Таким образом, для IMP, такого как характерный 7-доменный TM GPCR, в котором N-конец белка является внемембранным, а C-конец является внутримембранным, N-конец F13L можно сливать с C-концом GPCR, как показано на фиг. 1B. Таким образом, предоставлен полинуклеотид, как указано выше, в котором фрагмент первой нуклеиновой кислоты кодирует IMP с нечетным числом трансмембранных доменов, где 5'-конец фрагмента первой нуклеиновой кислоты кодирует внемембранную область, и 3'-конец фрагмента первой нуклеиновой кислоты кодирует внутримембранную область IMP, где последний является слитым с 5'-концом фрагмента нуклеиновой кислоты, кодирующей F13L или его фрагмент.
[0112] В иллюстративном полинуклеотиде такого типа, первый полинуклеотид может кодировать белок Фрайзлед 4 человека (FZD4) или его фрагмент, мишень для иммунотерапии определенных злокачественных опухолей человека, слитый с N-концом F13L. Таким образом, предоставлен полинуклеотид, который кодирует слитый белок FZD4-F13L. Иллюстративный полинуклеотид по этому аспекту кодирует зрелый слитый белок, аминокислоты от 20 до 892 SEQ ID NO:2, как показано ниже. Полинуклеотид может дополнительно кодировать сигнальный пептид, например, сигнальный пептид FZD4, аминокислоты от 1 до 19 SEQ ID NO:2.
FZD(FL)-F13L (SEQ ID NO:2)
MGWSCIILFLVATATGAHS FGDEEERRCDPIRISMCQNLGYNVTKMPNLVGHELQTDAELQLTTFTPLIQYGCSSQLQFFLCSVYVPMCTEKINIPIGPCGGMCLSVKRRCEPVLKEFGFAWPESLNCSKFPPQNDHNHMCMEGPGDEEVPLPHKTPIQPGEECHSVGTNSDQYIWVKRSLNCVLKCGYDAGLYSRSAKEFTDIWMAVWASLCFISTAFTVLTFLIDSSRFSYPERPIIFLSMCYNIYSIAYIVRLTVGRERISCDFEEAAEPVLIQEGLKNTGCAIIFLLMYFFGMASSIWWVILTLTWFLAAGLKWGHEAIEMHSSYFHIAAWAIPAVKTIVILIMRLVDADELTGLCYVGNQNLDALTGFVVAPLFTYLVIGTLFIAAGLVALFKIRSNLQKDGTKTDKLERLMVKIGVFSVLYTVPATCVIACYFYEISNWALFRYSADDSNMAVEMLKIFMSLLVGITSGMWIWSAKTLHTWQKCSNRLVNSGKVKREKRGNGWVKPGKGSETVV MWPFASVPAGAKCRLVETLPENMDFRSDHLTTFECFNEIITLAKKYIYIASFCCNPLSTTRGALIFDKLKEASEKGIKIIVLLDERGKRNLGELQSHCPDINFITVNIDKKNNVGLLLGCFWVSDDERCYVGNASFTGGSIHTIKTLGVYSDYPPLATDLRRRFDTFKAFNSAKNSWLNLCSAACCLPVSTAYHIKNPIGGVFFTDSPEHLLGYSRDLDTDVVIDKLKSAKTSIDIEHLAIVPTTRVDGNSYYWPDIYNSIIEAAINRGVKIRLLVGNWDKNDVYSMATARSLDALCVQNDLSVKVFTIQNNTKLLIVDDEYVHITSANFDGTHYQNHGFVSFNSIDKQLVSEAKKIFERDWVSSHSKSLKI
Одинарное подчеркивание - лидерный пептид (аминокислоты 1-19)
Жирный шрифт - Fzd4 человека (аминокислоты 20-520)
Курсив - F13L (аминокислоты 521-892)
[0113] В другом иллюстративном полинуклеотиде этого типа первый полинуклеотид может кодировать рецептор хемокинов A CXC или его фрагмент, слитый с N-концом F13L. рецепторы хемокинов CXC также являются мишенями для иммунотерапии определенных злокачественных опухолей человека. Иллюстративный рецептор хемокинов CXC представляет собой CXCR4 или его фрагмент. Таким образом, предоставлен полинуклеотид, который кодирует рецептор хемокинов CXC-F13L слитый белок, например, слитый белок CXCR4-F13L. Иллюстративный полинуклеотид по этому аспекту кодирует SEQ ID NO:3, как показано ниже.
CXCR4-F13L (SEQ ID NO:3)
MAIPLPLLQIYTSDNYTEEMGSGDYDSMKEPCFREENANFNKIFLPTIYSIIFLTGIVGNGLVILVMGYQKKLRSMTDKYRLHLSVADLLFVITLPFWAVDAVANWYFGNFLCKAVHVIYTVNLYSSVLILAFISLDRYLAIVHATNSQRPRKLLAEKVVYVGVWIPALLLTIPDFIFANVSEADDRYICDRFYPNDLWVVVFQFQHIMVGLILPGIVILSCYCIIISKLSHSKGHQKRKALKTTVILILAFFACWLPYYIGISIDSFILLEIIKQGCEFENTVHKWISITEALAFFHCCLNPILYAFLGAKFKTSAQHALTSVSRGSSLKILSKGKRGGHSSVSTESESSSFHSS MWPFASVPAGAKCRLVETLPENMDFRSDHLTTFECFNEIITLAKKYIYIASFCCNPLSTTRGALIFDKLKEASEKGIKIIVLLDERGKRNLGELQSHCPDINFITVNIDKKNNVGLLLGCFWVSDDERCYVGNASFTGGSIHTIKTLGVYSDYPPLATDLRRRFDTFKAFNSAKNSWLNLCSAACCLPVSTAYHIKNPIGGVFFTDSPEHLLGYSRDLDTDVVIDKLKSAKTSIDIEHLAIVPTTRVDGNSYYWPDIYNSIIEAAINRGVKIRLLVGNWDKNDVYSMATARSLDALCVQNDLSVKVFTIQNNTKLLIVDDEYVHITSANFDGTHYQNHGFVSFNSIDKQLVSEAKKIFERDWVSSHSKSLKI
Жирный шрифт - CXCR4 человека (аминокислоты 1-356)
Курсив - F13L (аминокислоты 357-728)
[0114] Как будет понятно специалисту в данной области, пронизывающий несколько раз мембрану белок с четным числом трансмембранных доменов водят в биологическую мембрану таким образом, что such его N-конец и его C-конец находятся на одной и той же стороне мембраны, на внемембранной стороне мембраны или на внутримембранной стороне мембраны. Вследствие того, что белок F13L располагается полностью на внутримембранной стороне EEV поксвируса, для формирования слитого белка IMP-F13L, соответствующим образом погруженного в мембрану, необходимым является, чтобы меньшей мере один из N-конца или C-конца IMP или его фрагмента являлся внутренним по отношению к мембране. Когда IMP имеет четное число доменов TM, и ба домена расположены внутри, белок F13L можно сливать с N-концом IMP или с C-концом IMP. Если полноразмерный IMP располагается таким образом, что N- и C-конец являются внемембранными, фрагмент IMP, содержащий нечетное число доменов TM, можно сливать с F13L.
[0115] Таким образом, изобретение относится к полинуклеотиду, как описано выше, кодирующему IMP с четным числом трансмембранных доменов, где 5'- и 3'-концы фрагмента первой нуклеиновой кислоты кодируют внутримембранные области. В некоторых аспектах 3'-конец фрагмента нуклеиновой кислоты, кодирующей F13L, может являться слитым с 5'-концом фрагмента нуклеиновой кислоты, кодирующего IMP, в некоторых аспектах 5'-конец фрагмента нуклеиновой кислоты, кодирующий F13L, может являться слитым с 3'-концом фрагмента нуклеиновой кислоты, кодирующего IMP.
[0116] Иллюстративный IMP этого типа представляет собой CD20 человека, 4-доменный TM IMP, экспрессируемый на B-клетках человека, который является мишенью для иммунотерапии B-клеточных лейкозов, лимфом и миелом. Схематическое изображение слитого белка CD20-F13L, в котором C-конец CD20 является слитым с N-концом F13L, продемонстрирован на фиг. 1C. Таким образом, предоставлен полинуклеотид, который кодирует слитый белок CD20-F13L. Иллюстративный полинуклеотид по этому аспекту кодирует SEQ ID NO:4, как показано ниже.
CD20-F13L (Seq ID NO: 4)
MATPRNSVNGTFPAEPMKGPIAMQSGPKPLFRRMSSLVGPTQSFFMRESKTLGAVQIMNGLFHIALGGLLMIPAGIYAPICVTVWYPLWGGIMYIISGSLLAATEKNSRKCLVKGKMIMNSLSLFAAISGMILSIMDILNIKISHFLKMESLNFIRAHTPYINIYNCEPANPSEKNSPSTQYCYSIQSLFLGILSVMLIFAFFQELVIAGIVENEWKRTCSRPKSNIVLLSAEEKKEQTIEIKEEVVGLTETSSQPKNEEDIEIIPIQEEEEEETETNFPEPPQDQESSPIENDSSP MWPFASVPAGAKCRLVETLPENMDFRSDHLTTFECFNEIITLAKKYIYIASFCCNPLSTTRGALIFDKLKEASEKGIKIIVLLDERGKRNLGELQSHCPDINFITVNIDKKNNVGLLLGCFWVSDDERCYVGNASFTGGSIHTIKTLGVYSDYPPLATDLRRRFDTFKAFNSAKNSWLNLCSAACCLPVSTAYHIKNPIGGVFFTDSPEHLLGYSRDLDTDVVIDKLKSAKTSIDIEHLAIVPTTRVDGNSYYWPDIYNSIIEAAINRGVKIRLLVGNWDKNDVYSMATARSLDALCVQNDLSVKVFTIQNNTKLLIVDDEYVHITSANFDGTHYQNHGFVSFNSIDKQLVSEAKKIFERDWVSSHSKSLKI
Жирный шрифт - CD20 человека (MS4A1)(аминокислоты 1-298)
Курсив - F13L (аминокислоты 299-669)
[0117] В полинуклеотидах, как предоставлено выше, первый и второй фрагменты нуклеиновой кислоты могут являться непосредственно слитыми, или, альтернативно, они могут быть разделены фрагментом нуклеиновой кислоты, кодирующим линкер или спейсер, или другой полипептидный фрагмент. В некоторых аспектах полинуклеотид, как предоставлено выше, может дополнительно содержать третий фрагмент нуклеиновой кислоты, кодирующий гетерологичный пептид/ полипептид, между первым и вторым фрагментами нуклеиновой кислоты или с одной стороны. Гетерологичный пептид может представлять собой, например, линкерную последовательность, аминокислотную метку или пептидную или полипептидную последовательность, которая облегчает очистку. В некоторых аспектах гетерологичный пептид представляет собой 6-гистидиновую метку, слитую, например, с C-концом слитого белка.
[0118] В некоторых аспектах полинуклеотид, как предоставлено в настоящем описании, является функционально связанным с промотором поксвируса. Подходящие промоторы описаны в другом месте в настоящем описании. В некоторых аспектах промотор представляет собой промотор p7.5 поксвируса или промотор H5 поксвируса.
[0119] Полинуклеотид, как предоставлено в настоящем описании, может являться геномом или быть частью генома поксвируса, где геном поксвируса, при введении в клетку-хозяина с подходящей чувствителью, может продуцировать инфекционные EEV, которые предоставляют слитый белок IMP-F13L на своей поверхности. В некоторых аспектах геному поксвируса представляет собой геном вируса осповакцины, например, геном вируса осповакцины WR или геном MVA. Геном поксвируса, содержащий полинуклеотид, как описано, можно получать стандартными молекулярными биологическими способами и используемыми в вирусологии способами, например, с использованием тримолекулярной рекомбинации, как описано в настоящем описании. Геном поксвируса, как предоставлено в настоящем описании, можно вводить в чувствительные клетки в виде части рекомбинантного поксвируса или в виде голой ДНК с последующим введением подходящих хелперных вирусов, например, вируса куриной оспы. Изобретение дополнительно относится к рекомбинантному поксвирусу, например, рекомбинантному вирусу осповакцины, содержащему предоставляемый геном поксвируса.
Слитые белки IMP-EEV, рекомбинантные EEV поксвируса и способы получения
[0120] Настоящее изобретение дополнительно относится к слитому белку IMP-F13L, такому как слитые белки, кодируемые полинуклеотидами, описанными выше. Кроме того, слитый белок IMP-F13L можно экспрессировать на поверхности рекомбинантного EEV поксвируса, например, рекомбинантного EEV вируса осповакцины. Изобретение относится к рекомбинантному EEV поксвируса, например, рекомбинантному EEV вируса осповакцины, содержащему предоставленный слитый белок. Рекомбинантный EEV поксвируса можно получать способом, который включает инфицирование клетки-хозяина, чувствительной к инфекции вирусом осповакцины, вирусом осповакцины, содержащим геном поксвируса, как предоставлено выше, и выделение EEV, высвобождающихся из инфицированной клетки-хозяина. Таким образом, предоставлен слитый белок IMP-F13L, кодируемый полинуклеотидом, как описано выше.
[0121] Кроме того изобретение относится к слитым белкам, содержащим IMP или его фрагмент, который может представлять собой пронизывающий несколько раз мембрану IMP и пронизывающий один раз IMP, или даже только внеклеточный домен (ECD) IMP, слитый с белком поксвируса, например, белком вируса осповакцины, характерным для EEV, таким как F13L, A56R, B5R, 33R, A34R или A36R, "слитый белок IMP-EEV". Иллюстративные слитые белки ECD описаны ниже. Слитый белок IMP-EEV, как предоставлено в настоящем описании, может предоставлять IMP, например, пронизывающий несколько раз мембрану IMP, пронизывающий один раз IMP или ECD IMP, в конформационно интактной форме на поверхности EEV поксвируса. Для применения в скрининге библиотек дисплея антител для антигенсвязывающих доменов, которые специфически связываются с мишенью IMP, предоставление IMP на поверхности EEV поксвируса обеспечивает много преимуществ над предоставлением IMP на поверхности рекомбинантных клеток, например, клеток CHO, как это широко распространено. Например, IMP можно экспрессировать с более высокой плотностью на EEV, чем на клетках. Кроме того, EEV экспрессирует только приблизительно шесть различных белков поксвируса на их поверхности (например, F13L, A56R, B5R, 33R, A34R и A36R), в отличие от нескольких тысяч, которые можно экспрессировать на поверхности клеток. Наконец, инактивированный EEV, экспрессирующий слитые белки IMP-F13L, как предоставлено в настоящем описании, можно связывать с твердыми подложками, что обеспечивает удобство при скрининге библиотек.
[0122] Таким образом, настоящее изобретение относится к способу дисплея интегрального мембранного белка (IMP) или его фрагмента в нативной конформации для использования, например, в скрининге библиотеках дисплея антител на антигенсвязывающие домены, специфичные к IMP. Способ включает: инфицирование клеток-хозяев, чувствительных к инфекции поксвирусом, рекомбинантным поксвирусом, который экспрессирует IMP или его фрагмент в виде слитого белка с характерным для EEV поксвируса белком или его мембраносвязанным фрагментом, где EEV продуцируемый инфицированной клеткой-хозяином, содержит IMP в виде части внешней мембраны оболочки EEV, и выделение EEV, высвобождаемого клеткой-хозяином. В некоторых аспектах характерный для EEV белок или его фрагмент может представлять собой белок вируса осповакцины A33R, белок A34R, белок A56R, белок B5R, белок A36R, F13L белок, любой его мембраносвязанный фрагмент или любое их сочетание.
[0123] В некоторых аспектах характерный для EEV белок представляет собой F13L (SEQ ID NO:1) или его функциональный фрагмент, и слитый белок можно экспрессировать в виде полинуклеотида, как предоставлено выше, например, где IMP представляет собой пронизывающий несколько раз мембрану белок, содержащий по меньшей мере два, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять, по меньшей мере шесть или по меньшей мере семь трансмембранных доменов.
[0124] В некоторых аспектах мембраносвязанный характерный для EEV белковый фрагмент содержит стебель, трансмембранный и внутримембранный домены белка A56R вируса осповакцины, где фрагмент содержит, состоит или состоит по существу из аминокислот от 108 до 314 SEQ ID NO:5. Один из нескольких иллюстративных партнеров по слиянию включает ECD FZD4 человека, показанную жирным шрифтом в SEQ ID NO:6 ниже. В соответствии с этим иллюстративным аспектом изобретение относится к способу дисплея конформационно интактного фрагмента FZD4 человека на поверхности EEV поксвируса, инфицирование клеток-хозяев, чувствительных к инфекции поксвируса, рекомбинантным поксвирусом, кодирующим слитый белок, содержащий аминокислоты от 20 до 370 SEQ ID NO:6. В некоторых аспектах слитый белок может дополнительно содержать сигнальный пептид, например, аминокислоты от 1 до 19 SEQ ID NO:6.
FZD-ECD-A56R (Seq ID NO: 6)
MGWSCIILFLVATATGAHS FGDEEERRCDPIRISMCQNLGYNVTKMPNLVGHELQTDAELQLTTFTPLIQYGCSSQLQFFLCSVYVPMCTEKINIPIGPCGGMCLSVKRRCEPVLKEFGFAWPESLNCSKFPPQNDHNHMCMEGPGDEEVPLPHKTPIQPGEE TSTTNDTDKVDYEEYSTELIVNTDSESTIDIILSGSTHSPETSSKKPDYIDNSNCSSVFEIATPEPITDNVEDHTDTVTYTSDSINTVSASSGESTTDETPEPITDKEDHTVTDTVSYTTVSTSSGIVTTKSTTDDADLYDTYNDNDTVPPTTVGGSTTSISNYKTKDFVEIFGITALIILSAVAIFCITYYIYNKRSRKYKTENKV.
Одинарное подчеркивание - лидерный пептид (аминокислоты 1-19)
Жирный шрифт - Внеклеточный домен FZD4 человека (аминокислоты 20-163)
Курсив - стебель, трансмембранный и внутримембранный A56R (аминокислоты от 164 до 370)
[0125] Другой иллюстративный партнер по слиянию включает ECD ErbB2 (Her2) человека, выделенный жирным шрифтом в SEQ ID NO:7 ниже. Согласно этому иллюстративному аспекту изобретение относится к способу дисплея конформационно интактного фрагмента Her2 человека на поверхности EEV поксвируса, включающему инфицирование клеток-хозяев, чувствительных к инфекции поксвируса, рекомбинантным поксвирусом, кодирующим слитый белок, содержащий аминокислоты от 20 до 855 SEQ ID NO:7. В некоторых аспектах слитый белок может дополнительно содержать сигнальный пептид, например, аминокислоты от 1 до 19 SEQ ID NO:7.
Her2-A56R (SEQ ID NO:7)
MGWSCIILFLVATATGAHS STQVCTGTDMKLRLPASPETHLDMLRHLYQGCQVVQGNLELTYLPTNASLSFLQDIQEVQGYVLIAHNQVRQVPLQRLRIVRGTQLFEDNYALAVLDNGDPLNNTTPVTGASPGGLRELQLRSLTEILKGGVLIQRNPQLCYQDTILWKDIFHKNNQLALTLIDTNRSRACHPCSPMCKGSRCWGESSEDCQSLTRTVCAGGCARCKGPLPTDCCHEQCAAGCTGPKHSDCLACLHFNHSGICELHCPALVTYNTDTFESMPNPEGRYTFGASCVTACPYNYLSTDVGSCTLVCPLHNQEVTAEDGTQRCEKCSKPCARVCYGLGMEHLREVRAVTSANIQEFAGCKKIFGSLAFLPESFDGDPASNTAPLQPEQLQVFETLEEITGYLYISAWPDSLPDLSVFQNLQVIRGRILHNGAYSLTLQGLGISWLGLRSLRELGSGLALIHHNTHLCFVHTVPWDQLFRNPHQALLHTANRPEDECVGEGLACHQLCARGHCWGPGPTQCVNCSQFLRGQECVEECRVLQGLPREYVNARHCLPCHPECQPQNGSVTCFGPEADQCVACAHYKDPPFCVARCPSGVKPDLSYMPIWKFPDEEGACQPCPINCTHSCVDLDDKGCPAEQRASP TSTTNDTDKVDYEEYSTELIVNTDSESTIDIILSGSTHSPETSSKKPDYIDNSNCSSVFEIATPEPITDNVEDHTDTVTYTSDSINTVSASSGESTTDETPEPITDKEDHTVTDTVSYTTVSTSSGIVTTKSTTDDADLYDTYNDNDTVPPTTVGGSTTSISNYKTKDFVEIFGITALIILSAVAIFCITYYIYNKRSRKYKTENKV.
Одинарное подчеркивание - лидерный пептид (аминокислоты 1-19)
Жирный шрифт - внеклеточный домен ERBB2 (HER2) человека (аминокислоты 20-648)
Курсив - стебель, трансмембранный и внутримембранный A56R (аминокислоты от 649 до 855)
[0126] Другой иллюстративный партнер по слиянию включает ECDCD100 человека (семафорин 4D), выделенный жирным шрифтом в SEQ ID NO:8 ниже. Согласно этому иллюстративному аспекту изобретение относится к способу дисплея конформационно интактного фрагмента CD100 человека на поверхности EEV поксвируса, включающему инфицирование клеток-хозяев, чувствительных к инфекции поксвируса, рекомбинантным поксвирусом, кодирующим слитый белок, содержащий аминокислоты от 20 до 935 SEQ ID NO:8. В некоторых аспектах слитый белок может дополнительно содержать сигнальный пептид, например, аминокислоты от 1 до 19 SEQ ID NO:8.
CD100-A56R (SEQ ID NO:8)
MGWSCIILFLVATATGAHS FAPIPRITWEHREVHLVQFHEPDIYNYSALLLSEDKDTLYIGAREAVFAVNALNISEKQHEVYWKVSEDKKAKCAEKGKSKQTECLNYIRVLQPLSATSLYVCGTNAFQPACDHLNLTSFKFLGKNEDGKGRCPFDPAHSYTSVMVDGELYSGTSYNFLGSEPIISRNSSHSPLRTEYAIPWLNEPSFVFADVIRKSPDSPDGEDDRVYFFFTEVSVEYEFVFRVLIPRIARVCKGDQGGLRTLQKKWTSFLKARLICSRPDSGLVFNVLRDVFVLRSPGLKVPVFYALFTPQLNNVGLSAVCAYNLSTAEEVFSHGKYMQSTTVEQSHTKWVRYNGPVPKPRPGACIDSEARAANYTSSLNLPDKTLQFVKDHPLMDDSVTPIDNRPRLIKKDVNYTQIVVDRTQALDGTVYDVMFVSTDRGALHKAISLEHAVHIIEETQLFQDFEPVQTLLLSSKKGNRFVYAGSNSGVVQAPLAFCGKHGTCEDCVLARDPYCAWSPPTATCVALHQTESPSRGLIQEMSGDASVCPDKSKGSYRQHFFKHGGTAELKCSQKSNLARVFWKFQNGVLKAESPKYGLMGRKNLLIFNLSEGDSGVYQCLSEERVKNKTVFQVVAKHVLEVKVVPKPVVAPTLSVVQTEGSRIATKVLVASTQGSSPPTPAVQATSSGAITLPPKPAPTGTSCEPKIVINTVPQLHSEKTMYLKSSD TSTTNDTDKVDYEEYSTELIVNTDSESTIDIILSGSTHSPETSSKKPDYIDNSNCSSVFEIATPEPITDNVEDHTDTVTYTSDSINTVSASSGESTTDETPEPITDKEDHTVTDTVSYTTVSTSSGIVTTKSTTDDADLYDTYNDNDTVPPTTVGGSTTSISNYKTKDFVEIFGITALIILSAVAIFCITYYIYNKRSRKYKTENKV.
Одинарное подчеркивание - лидерный пептид (аминокислоты 1-19)
Жирный шрифт - внеклеточный домен CD100 человека (аминокислоты 20-728)
Курсив - стебель, трансмембранный и внутримембранный A56R (аминокислоты от 729 до 935)
[0127] В некоторых аспектах мембраносвязанный характерный для EEV белковый фрагмент содержит трансмембранный и внутримембранный домены белка B5R вируса осповакцины, где фрагмент содержит, состоит или состоит по существу из аминокислот от 276 до 317 SEQ ID NO:9. В некоторых аспектах мембраносвязанный характерный для EEV белковый фрагмент содержит стеблевй, трансмембранный и внутримембранный домены белка B5R вируса осповакцины, где фрагмент содержит, состоит или состоит по существу из аминокислот от 238 до 317 SEQ ID NO:9.
SEQ ID NO:9: белок WR B5R
MKTISVVTLLCVLPAVVYSTCTVPTMNNAKLTSTETSFNDKQKVTFTCDQGYHSSDPNAVCETDKWKYENPCKKMCTVSDYISELYNKPLYEVNSTMTLSCNGETKYFRCEEKNGNTSWNDTVTCPNAECQPLQLEHGSCQPVKEKYSFGEYMTINCDVGYEVIGASYISCTANSWNVIPSCQQKCDMPSLSNGLISGSTFSIGGVIHLSCKSGFTLTGSPSSTCIDGKWNPVLPICVRTNEEFDPVDDGPDDETDLSKLSKDVVQYEQEIESLEATYHIIIVALTIMGVIFLISVIVLVCSCDKNNDQYKFHKLLP
[0128] В определенных иллюстративных аспектах IMP партнер по слиянию для фрагмента B5R содержит внеклеточный домен FZD4 человека, выделенный жирным шрифтом в SEQ ID NO:10 и SEQ ID NO:11 ниже. Согласно этому иллюстративному аспекту изобретение относится к способу дисплея конформационно интактного фрагмента FZD4 человека на поверхности EEV поксвируса, включающему инфицирование клеток-хозяев, чувствительных к инфекции поксвируса, рекомбинантным поксвирусом, кодирующим слитый белок, содержащий аминокислоты от 20 до 243 SEQ ID NO:10 или аминокислоты от 20 до 281 SEQ ID NO:11. В некоторых аспектах слитый белок может дополнительно содержать сигнальный пептид, например, аминокислоты от 1 до 19 SEQ ID NO:10.
FZD-B5R (short) (SEQ ID NO:10)
MGWSCIILFLVATATGAYA FGDEEERRCDPIRISMCQNLGYNVTKMPNLVGHELQTDAELQLTTFTPLIQYGCSSQLQFFLCSVYVPMCTEKINIPIGPCGGMCLSVKRRCEPVLKEFGFAWPESLNCSKFPPQNDHNHMCMEGPGDEEVPLPHKTPIQPGEECHSVGTNSDQYIWVKRSLNCVLKCGYDAGLYSRSAKEC ATYHIIIVALTIMGVIFLISVIVLVCSCDKNNDQYKFHKLLP.
Одинарное подчеркивание - лидерный пептид (аминокислоты 1-19)
Жирный шрифт - внеклеточный домен FZD4 человека (аминокислоты 20-200)
Курсив - B5R TM и цитоплазматический хвост (аминокислоты 201-243)
FZD-B5R (полный) (SEQ ID NO:11)
MGWSCIILFLVATATGAYA FGDEEERRCDPIRISMCQNLGYNVTKMPNLVGHELQTDAELQLTTFTPLIQYGCSSQLQFFLCSVYVPMCTEKINIPIGPCGGMCLSVKRRCEPVLKEFGFAWPESLNCSKFPPQNDHNHMCMEGPGDEEVPLPHKTPIQPGEECHSVGTNSDQYIWVKRSLNCVLKCGYDAGLYSRSAKE YVRTNEEFDPVDDGPDDETDLSKLSKDVVQYEQEIESLEATYHIIIVALTIMGVIFLISVIVLVCSCDKNNDQYKFHKLLP.
Одинарное подчеркивание - лидерный пептид (аминокислоты 1-19)
Жирный шрифт - Внеклеточный домен FZD4 человека (аминокислоты 20-200)
Курсив - стебель, TM и цитоплазматический хвост B5R (аминокислоты 201-281)
[0129] Изобретение дополнительно относится к слитому белку, содержащему: аминокислоты от 20 до 892 SEQ ID NO:2; SEQ ID NO:3; SEQ ID NO:4; аминокислоты от 20 до 370 SEQ ID NO:6; аминокислоты от 20 до 855 SEQ ID NO:7; аминокислоты от 20 до 935 SEQ ID NO:8; аминокислоты от 20 до 243 SEQ ID NO:10, или аминокислоты от 20 до 281 SEQ ID NO:11, любое их сочетание, любому его фрагменту или любому его варианта, где слитый белок при экспрессии рекомбинантным поксвирусом появляется на поверхности внеклеточного оболочечного вириона (EEV) поксвируса в нативной конформации.
[0130] Также предоставлен рекомбинантный EEV поксвируса, содержащий любой EEV слитый белок, как предоставлено в настоящем описании.
Способ селекции антител
[0131] Настоящее изобретение дополнительно относится к способу выбора связывающих молекул, например, антител, антигенсвязывающих фрагментов антител или антителоподобных связывающих молекул, которые связываются с представляющим интерес пронизывающим несколько раз мембрану белком. Способ включает связывание рекомбинантного EEV, как предоставлено в настоящем описании, с твердой подложкой, где рекомбинантный EEV может предоставлять пронизывающими пронизывающий несколько раз мембрану белок на своей поверхности; предоставление библиотеки дисплея, например, библиотеки дисплея антител, где библиотека содержит пакеты дисплея, предоставляющих совокупность антигенасвязывающих доменов; приведение библиотеки дисплея в контакт с EEV, таким образом, что такие пакеты дисплея, предоставляющие антигенсвязывающие домены, которые специфически связываются с IMP, экспрессируемым на EEV, могут его связывать; удаление несвязанных пакетов дисплея и выделение пакетов дисплея, которые предоставляют антигенсвязывающий домен, специфический к IMP, экспрессируемому на EEV.
[0132] Любая библиотека дисплея, содержащая совокупность связывающих доменов, например, антител, антителоподобных молекул или других связывающих молекул, является подходящей для этого способа. Например, библиотека дисплея может представлять собой библиотеку фагового дисплея, библиотеку дрожжевого дисплея или библиотеку, сконструированную в векторе на основе вируса осповакцины, как описано в другом месте в настоящем описании.
[0133] В некоторых аспектах рекомбинантный EEV можно инактивировать перед связыванием с твердой подложкой. Например, EEV можно инактивировать инкубацией с псораленом (триоксаленом, 4'-аминометилгидрохлоридом) в присутствии УФ-облучения.
[0134] Можно использовать любую подходящую твердую подложку. В рамках изобретения, "твердая подложка" представляет собой любую подложку, способную связывать EEV, которая может находиться в любой из различных форм, как известно в данной области. Хорошо известные подложки включают пластик, стекло, полистирол, полипропилен, полиэтилен, декстран, нейлон, амилазы, природную и модифицированные целлюлозы, полиакриламиды, габбро и магнетит для культивирования тканей. Природа носителя может являться растворимой до некоторой степени или нерастворимой для целей настоящего изобретения. Материал подложки может иметь практически любую структурную конфигурацию при условии, что связанный EEV способен связываться с предоставляемой связывающей молекулой, такой как антитело. Таким образом, конфигурация подложки может быть сферической как в грануле или цилиндрической как на внутренней поверхности тестовой пробирки или внешней поверхности палочки. Альтернативно, поверхность может быть плоской, такой как лист, тест-полоска и т.д. Характерные подложки включат гранулы, например, магнитные полистироловые гранулы, такие как DYNABEADS®, которые можно удалять из суспензии посредством магнита. Конфигурация подложки может включать пробирку, гранулу, микрогранулу, лунку, планшет, планшет для культивирования тканей, чашку Петри, микропланшет, планшет для микротитрования, колбу, карандаш, полоску, флакон, лопасть мешалки и т.д., и т.д. Твердая подложка может быть магнитной или немагнитной. Специалистам в данной области хорошо известны многие другие подходящие носители для связывания EEV, как предоставлено в настоящем описании, или они могут легко их устанавливать. В некоторых аспектах EEV, как предоставлено в настоящем описании, можно связывать с твердой подложкой с помощью реакции, например, с тозильными группами, эпоксигруппами, карбоксильными группами или аминогруппами, связанными с поверхностью. Например, EEV можно связывать с поверхностью активируемых тозилом магнитных гранул, например, активируемых тозилом гранул MYONE™. Альтернативно, EEV можно подвергать биотинилированию и связывать с твердой поверхностью покрытой стрептавидином, например, покрытыми стрептавидином магнитными гранулами.
[0135] Если не указано иное, в настоящем изобретении применяют общепринятые способы клеточной биологии, культивирования клеток, молекулярной биологии, трансгенной биологии, микробиологии, рекомбинантной ДНК и иммунология, которые известны специалистам в данной области. Такие способы полностью описаны в литературе. (См., например, Sambrook et al., ed. (1989) Molecular Cloning A Laboratory Manual (2nd ed.; Cold Spring Harbor Laboratory Press); Sambrook et al., ed. (1992) Molecular Cloning: A Laboratory Manual, (Cold Springs Harbor Laboratory, NY); D. N. Glover ed., (1985) DNA Cloning, Volumes I and II; Gait, ed. (1984) Oligonucleotide Synthesis; Mullis et al. U.S. Pat. No. 4,683,195; Hames and Higgins, eds. (1984) Nucleic Acid Hybridization; Hames and Higgins, eds. (1984) Transcription And Translation; Freshney (1987) Culture Of Animal Cells (Alan R. Liss, Inc.); Immobilized Cells And Enzymes (IRL Press) (1986); Perbal (1984) A Practical Guide To Molecular Cloning; the treatise, Methods In Enzymology (Academic Press, Inc., N.Y.); Miller and Calos eds. (1987) Gene Transfer Vectors For Mammalian Cells, (Cold Spring Harbor Laboratory); Wu et al., eds., Methods In Enzymology, Vols. 154 and 155; Mayer and Walker, eds. (1987) Immunochemical Methods In Cell And Molecular Biology (Academic Press, London); Weir and Blackwell, eds., (1986) Handbook Of Experimental Immunology, Volumes I-IV; Manipulating the Mouse Embryo, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., (1986); и у Ausubel et al. (1989) Current Protocols in Molecular Biology (John Wiley and Sons, Baltimore, Md.).
[0136] Общие принципы конструирования антител указаны в Borrebaeck, ed. (1995) Antibody Engineering (2nd ed.; Oxford Univ. Press). General principles of protein engineering are set forth in Rickwood et al., eds. (1995) Protein Engineering, A Practical Approach (IRL Press at Oxford Univ. Press, Oxford, Eng.). Общие принципы связывания антител и антитела-гаптена указаны: в Nisonoff (1984) Molecular Immunology (2nd ed.; Sinauer Associates, Sunderland, Mass.); and Steward (1984) Antibodies, Their Structure and Function (Chapman and Hall, New York, N.Y.). Кроме того, стандартные способы иммунологии известны в данной области и конкретно не описанные, можно выполнять как описано в Current Protocols in Immunology, John Wiley & Sons, New York; Stites et al., eds. (1994) Basic and Clinical Immunology (8th ed; Appleton & Lange, Norwalk, Conn.) и Mishell and Shiigi (eds) (1980) Selected Methods in Cellular Immunology (W.H. Freeman and Co., NY).
[0137] Стандартные справочные издания, в которых описывают основные принципы иммунологии, включают Current Protocols in Immunology, John Wiley & Sons, New York; Klein (1982) J., Immunology: The Science of Self-Nonself Discrimination (John Wiley & Sons, NY); Kennett et al., eds. (1980) Monoclonal Antibodies, Hybridoma: A New Dimension in Biological Analyses (Plenum Press, NY); Campbell (1984) "Monoclonal Antibody Technology" in Laboratory Techniques in Biochemistry and Molecular Biology, ed. Burden et al., (Elsevier, Amsterdam); Goldsby et al., eds. (2000) Kuby Immunology (4th ed.; H. Freeman & Co.); Roitt et al. (2001) Immunology (6th ed.; London: Mosby); Abbas et al. (2005) Cellular and Molecular Immunology (5th ed.; Elsevier Health Sciences Division); Kontermann and Dubel (2001) Antibody Engineering (Springer Verlag); Sambrook and Russell (2001) Molecular Cloning: A Laboratory Manual (Cold Spring Harbor Press); Lewin (2003) Genes VIII (Prentice Hall, 2003); Harlow and Lane (1988) Antibodies: A Laboratory Manual (Cold Spring Harbor Press); Dieffenbach and Dveksler (2003) PCR Primer (Cold Spring Harbor Press).
[0138] Все указанные выше ссылки, а также все цитируемые в настоящем описании ссылки, полностью включены в настоящее описание посредством ссылки. Следующие ниже примеры приведены в качестве иллюстрации, а не в качестве ограничения.
Примеры
Пример 1: Конструирование слитого белка
[0139] IMP вводили в EEV вируса осповакцины с использованием характерных для EEV белков F13L, A56R и B5R следующий ниже способами. Как правило, внеклеточные домены HER2, CD100 (семафорин 4D) и FZD4 вводили в виде слияний с характерными для EEV пронизывающими один раз мембранными белками A56R и B5R как схематически изображено на фиг. 1A. Зрелый слитый белок FZD4-ECD-A56R содержит аминокислоты от 20 до 370 SEQ ID NO:6, зрелый слитый белок HER2-ECD-A56R содержит аминокислоты от 20 до 855 SEQ ID NO:7, и зрелый слитый белок CD100-ECD-A56R содержит аминокислоты от 20 до 935 SEQ ID NO:8. На фиг. 1B и фиг. 1C схематично показано, как пронизывающие несколько раз мембрану белки, такие как GPCRs и CD20, можно вводить в EEV в виде пронизывающих несколько раз мембрану мембранных белков, в виде слияния с EEV мембраносвязанного белка F13L.
Получение слитых белков F13L (FZD4-F13L, CD20-F13L и CXCR4-F13L)
[0140] кДНК, кодирующую IMP, клонировали в рамке считывания с геном F13L вируса осповакцины, кодирующим пальмитоилированный EEV мембранный гликопротеин (SEQ ID NO:1) в плазмиду pJEM1, ранее описанную для целей введения в вирус осповакцины. pJEM1 представляет собой производное p7.5/tk, описанное в публикацию заявки на патент США № 2013/0288927, и при расщеплении NcoI или BssHII и BsiWI содержит фланкирующие области, способные к гомологичной рекомбинации с геном TK вируса осповакцины и промотором H5 вируса осповакцины.
[0141] Открытую рамку считывания гена мембранного белка MS4A1 человека (CD20)(NM_021950.3) клонировали в рамку считывания с F13L вируса осповакцины с использованием SOE (сращивание посредством перекрывающегося удлинения) ЦР в соответствии со стандартными протоколами, в результате чего добавляли участки узнавания рестрикционных эндонуклеаз NcoI и BsiWI к продукту ПЦР путем их кодирования в наиболее фланкирующие 5'- и 3'-праймеры соответственно. Эта стратегия позволяет не вводить лидерный пептид. Конечный продукт ПЦР и pJEM1 расщепляли с помощью NcoI и BsiWI и соединяли две молекулы лигированием согласно стандартные протокола. Затем вводили кольцевую плазмиду в компетентные E. coli с помощью химической трансформации и выбирали колонии на содержащих ампициллин планшетах с агаром.
[0142] Получаемый слитый белок CD20-F13L, кодируемый полинуклеотидом, содержит аминокислотную последовательность SEQ ID NO:4.
[0143] Открытую рамку считывания мембранного белка FZD4 человека (NM_012193.3) клонировали в рамку считывания с F13L вируса осповакцины с использованием SOE (сращивание посредством перекрывающегося удлинения) ПЦР в соответствии со стандартными протоколами, в результате чего добавляли участки узнавания рестрикционных эндонуклеаз NcoI и BsiWI к продукту ПЦР путем их кодирования в наиболее фланкирующие 5'- и 3'-праймеры соответственно. Эта стратегия позволяет использовать лидерный пептид, содержащийся в pJEM1. Конечный продукт ПЦР и pJEM1 расщепляли с помощью BssHII и BsiWI и соединяли две молекулы путем лигирования согласно стандартным протоколам. Затем вводили кольцевую плазмиду в компетентные E. coli путем химической трансформации и отбирали колонии на планшетах с агаром, содержащим ампициллин. Праймеры для ПЦР являлись специфическими для FZD4 и F13L и соответствовали той же общей стратегии, как описано для MS4A1. Получаемый зрелый слитый белок FZD4-F13L, кодируемый полинуклеотидом содержит аминокислоты 20-892 SEQ ID NO:2.
[0144] Открытую рамка считывания мембранного белка CXCR4 человека (NM_001008540.1) клонировали в рамку считывания с F13L вируса осповакцины с использованием SOE (сращивание посредством перекрывающегося удлинения) ПЦР в соответствии со стандартными протоколами в результате чего добавляли участки узнавания рестрикционных эндонуклеаз NcoI и BsiWI к продукту ПЦР путем их кодирования в наиболее фланкирующие 5'- и 3'-праймеры соответственно. Эта стратегия позволяет не вводить лидерный пептид. Конечный продукт ПЦР и pJEM1 расщепляли с помощью NcoI и BsiWI и соединяли две молекулы путем лигирования согласно стандартному протоколу. Затем вводили кольцевую плазмиду в компетентные E. coli путем химической трансформации и отбирали колонии на планшетах с агаром, содержащим ампициллин. Праймеры для ПЦР являлись специфическими к CXCR4 и F13L и соответствовали той же общей стратегии, как описано для MS4A1. Получаемый слитый белок CXCR4-F13L, кодируемый полинуклеотидом, содержит аминокислотную последовательность SEQ ID NO:3.
[0145] Плазмиды, получаемые так, как описано выше, а также аналогичные плазмиды, кодирующие неслитые ("немеченые") варианты CD20, FZD4, CXCR4, CD100 и HER2, линеризовали и вводили в вирус осповакцины путем тримолекулярной рекомбинации.
Пример 2: Экспрессия слитого белка CD20-F13L на EEV
[0146] Клетки BHK инфицировали IMV, кодирующим слитый белок CD20-F13L (SEQ ID NO:4), или штаммом Western Reserve (WR) контрольного вируса при множественности заражения (MOI 1 вирус на клетку в течение двух суток, после чего собирали супернатант, содержащий EEV, и удаляли дебрис низкоскоростным центрифугированием. Белок G DYNABEADS® (110 мкл) осаждали с помощью магнита и добавляли к гранулам 1 мл PBS+20 мкг очищенного антитела к CD20. Раствор инкубировали при комнатной температуре при аккуратном вращении в течение 30-60 минут для обеспечения связывания антитела с белком G на гранулах. К раствору добавляли десять мкг очищенного изотипического контроля mIgG1 для обеспечения полного блокирования и инкубировали раствор при комнатной температуре при аккуратном вращении еще 10-30 минут. Гранулы осаждали с помощью магнита, промывали дин раз 1 мл PBS и ресуспендировали в 110 мкл PBS.
[0147] Добавляли пятьдесят мкл антитела к CD20-Pro G DYNABEADS® к 1 мл супернатанта EEV CD20-F13L или WR и инкубировали при комнатной температуре при аккуратном вращении в течение 1 часа. Гранулы осаждали с использованием магнита и удаляли несвязаный супернатант. Затем гранулы отмывали пять раз 1 мл модифицированной Дульбекко среды Игла (DMEM), дополненной 10% FBS и 1 мМ HEPES (10% DMEM). Все промывания объединяли с несвязанным супернатантом ("несвязанные"). Затем гранулы ("связанные") ресуспендировали в 1 мл 10% DMEM. "несвязанные" и "связанные" титровали на клетках BSC-1 и покрывали средой для выращивания, содержащей метилцеллюлозу. Оставляли до формирования бляшек в течение двух суток, а затем фиксировали клетки и окрашивали 0,1% раствором кристаллического фиолетового. Подсчитывали бляшки для определения количество бляшкообразующих единиц (БОЕ) в "несвязанных" и "связанных", из чего можно рассчитывать % от EEV, связанных с гранулами. Результаты приведены в таблице 2.
Таблица 2: Связывание EEV CD20-F13L
[0148] EEV, связанные с покрытыми антителами к CD20 гранулами, являлся значительно выше для слитого белка CD20-F13L EEV по сравнению с Western Reserve, что указывает на то, что CD20 экспрессируется на поверхности мембраны EEV.
Пример 3: Слияние CD20 с F13L приводит к наиболее эффективной экспрессии на мембране EEV
[0149] Клетки BHK инфицировали вирусом, кодирующим IMV слитый белок CD20-F13L (SEQ ID NO:4), слитый белок CD20-A56R (SEQ ID NO:16), немеченый CD20 (неслитый) или контрольный внеклеточный домен (ECD) HER2-A56R (SEQ ID NO:7), при MOI=1 в течение двух суток, после чего супернатант, содержащий EEV, собирали и удаляли дебрис низкоскоростным центрифугированием. Покрытые стрептавидином DYNABEADS® (200 мкл) осаждали магнитом и добавляли к гранулам 0,2 мл PBS+20 мкг очищенного биотинилированного антитела к CD20 или биотинилированного антитела к HER2. Раствор инкубировали при комнатной температуре при аккуратном вращении в течение 30 минут, чтобы обеспечивать связывание антитела с покрытыми стрептавидином гранулами. Гранулы осаждали магнитом, промывали один раз 1 мл PBS и ресуспендировали в 200 мкл PBS.
CD20-A56R: (SEQ ID NO:16)
MGWSCIILFLVATATGAHT ELIVNTDSESTIDIILSGSTHSPETSSKKPDYIDNSNCSSVFEIATPEPITDNVEDHTDTVTYTSDSINTVSASSGESTTDETPEPITDKEDHTVTDTVSYTTVSTSSGIVTTKSTTDDADLYDTYNDNDTVPPTTVGGSTTSISNYKTKDFVEIFGITALIILSAVAIFCITYYIYNKRSRKYKTENKV MTTPRNSVNGTFPAEPMKGPIAMQSGPKPLFRRMSSLVGPTQSFFMRESKTLGAVQIMNGLFHIALGGLLMIPAGIYAPICVTVWYPLWGGIMYIISGSLLAATEKNSRKCLVKGKMIMNSLSLFAAISGMILSIMDILNIKISHFLKMESLNFIRAHTPYINIYNCEPANPSEKNSPSTQYCYSIQSLFLGILSVMLIFAFFQELVIAGIVENEWKRTCSRPKSNIVLLSAEEKKEQTIEIKEEVVGLTETSSQPKNEEDIEIIPIQEEEEEETETNFPEPPQDQESSPIENDSSP
Одинарное подчеркивание - Сигнальная последовательность (аминокислоты 1-19)
Курсив - Усеченный A56R (аминокислоты 20-190)
Жирный шрифт - Последовательность CD20 (аминокислоты 191-506)
[0150] Пятьдесят мкл подготовленных покрытых стрептавидином гранул добавляли к 1 мл каждого супернатанта с EEV и оставляли вращаться при комнатной температуре в течение 45 минут. Гранулы осаждали с использованием магнита и удаляли несвязавшийся супернатант. Затем гранулы отмывали пять раз 1 мл среды DMEM, дополненной 10% FBS и 1 мМ HEPES (10% DMEM). Все промывания объединяли с несвязанным супернатантом ("несвязанные"). Затем гранулы ("связанные") ресуспендировали в 1 мл 10% DMEM. "Несвязанные" и "связанные" титровали на клетках BSC-1 и покрывали средой для выращивания, содержащей метилцеллюлозу. Оставляли в течение двух суток для формирования бляшек, а затем клетки фиксировали и окрашивали 0,1% раствором кристаллического фиолетового. Подсчитывали бляшки для определения числа бляшкообразующих единиц (БОЕ) в "несвязанных" и "связанных", на основании которого можно было рассчитывать % EEV, связанного с гранулами. Результаты продемонстрированы на фиг. 2.
[0151] % EEV, связанного с покрытыми антителом к CD20 гранулами, для CD20-F13L являлся больше, чем % EEV, связанного с немеченым (неслитым) CD20 или слитым с A56 CD20, что указывает на более высокую экспрессию CD20-F13L на мембране EEV. Отсутствие связывания с покрытыми антителом к HER2 гранулами, подтверждало специфичность анализа.
[0152] Указанный выше эксперимент повторяли с использованием слитого белка CD20-F13L (SEQ ID NO:4), немеченого CD20 (неслитого), слитого белка FZD-F13L (SEQ ID NO:2) и немеченого FZD (неслитого). Вирус осаждали с использованием гранул, покрытых антителами к CD20 или антителами к FZD, как описано выше. Данные на фиг. 3A (покрытые антителом к CD20 гранулы) и на фиг. 3B (покрытые антителом к FZD гранулы) демонстрирует, что слитые белки F13L специфически осаждались их соответствующими антителами, и они более эффективно встраивались в вирус осповакцины чем немеченые (неслитые) белки.
Пример 4: Вирус осповакцина можно конструировать для экспрессии различных конструкций антиген-EEV
[0153] Клетки BHK инфицировали при MOI=1, вирусом, экспрессирующим следующие ниже антигенные конструкции: CD20-F13L (SEQ ID NO:4), CXCR4-F13L (SEQ ID NO:3), HER2-ECD-A56R (SEQ ID NO:7) и CD100-ECD-A56R (SEQ ID NO:8). Через двое суток супернатант, содержащий EEV, собирали и удаляли дебрис низкоскоростным центрифугированием. Покрытые стрептавидином DYNABEADS® осаждали с помощью магнита и для каждого образца 50 мкл гранул ресуспендировали в 0,1 мл PBS+5 мкг очищенного биотинилированного антитела к CD20, биотинилированного антитела к CXCR4 (12G5), биотинилированного антитела к CD100 (2503) или биотинилированного антитела к HER2. Растворы инкубировали при комнатной температуре при аккуратном вращении в течение 30 минут для обеспечения связывания антитела с покрытыми стрептавидином гранулами. Гранулы осаждали с помощью магнита, промывали один раз 1 мл PBS и ресуспендировали в 100 мкл PBS на образец.
[0154] Тысячу мкл подготовленных покрытых стрептавидином гранул добавляли к 1 мл каждого супернатанта с EEV и оставляли вращаться при комнатной температуре в течение 45 минут. Гранулы осаждали с использованием магнита и удаляли несвязавшийся супернатант. Затем гранулы отмывали пять раз 1 мл среды DMEM, дополненной 10% FBS и 1 мМ HEPES (10% DMEM). Все промывания объединяли с несвязанным супернатантом ("несвязанные"). Затем гранулы ("связанные") ресуспендировали в 1 мл 10% DMEM. "Несвязанные" и "связанные" титровали на клетках BSC-1 и покрывали средой для выращивания, содержащей метилцеллюлозу. Оставляли в течение двух суток для формирования бляшек, а затем клетки фиксировали и окрашивали 0,1% раствором кристаллического фиолетового. Подсчитывали бляшки для определения числа бляшкообразующих единиц (БОЕ) в "несвязанных" и "связанных", на основании которого можно было рассчитывать % EEV, связанного с гранулами. Результаты продемонстрированы на фиг. 4.
[0155] Все из антигенов-EEV специфически связывались с гранулами, покрытыми соответствующим им антителом, указывая на эффективную экспрессию антигена на мембране EEV.
Пример 5: Антиген-EEV можно непосредственно связывать с магнитными гранулами для селекции антител
[0156] Клетки BHK (2×108 клеток) инфицировали при MOI=1 вирусом, экспрессирующим HER2-ECD-A56R (SEQ ID NO:7), FZD-F13L (SEQ ID NO:2), CXCR4-F13L (SEQ ID NO:3) или CD100 (семафорин 4D)-ECD-A56R (SEQ ID NO:8), каждый в камере для культивирования клеток cellSTACK (Corning). Через двое суток супернатант, содержащий EEV, собирали и удаляли дебрис низкоскоростным центрифугированием. Затем осветленный супернатант центрифугировали при 13000 об./мин. (28000×g) в течение 1 часа для осаждения антигена-EEV. Супернатант аспирировали и ресуспендировали осадок в 1,5 мл 1× PBS. Различные вирусы переносили в свежие пробирки и добавляли просален (триоксален, 4'-аминометилгидрохлорид; Sigma) до конечной концентрации 20 мкг/мл. EEV и просален инкубировали при комнатной температуре в течение 10 минут перед облучением в УФ-сшивателе STRATALINKER® (Stratagene) 99999 микроджоулей. Процедура обработки просаленом/УФ обеспечивает то, что антиген-EEV является инактивированным и, таким образом, неспособен образовывать бляшки или размножаться в любом последующем тестировании.
[0157] Активированные тозилом MyOne DYNABEADS® (100 мкл) осаждали с помощью магнита и промывали 1 мл PBS. Гранулы осаждали с помощью магнита, удаляли PBS и добавляли 1 мл каждого инактивированного просаленом/УФ антигена-EEV к отдельной аликвоте гранул. Гранулы и антиген-EEV оставляли вращаться при 37°C в течение 16-20 часов. Гранулы осаждали и удаляли супернатант. Гранулы блокировали 1 мл 1× PBS, 10% FBS и 0,5% BSA при 37°C в течение 1 часа. Перед тем как ресуспендировать в 200 мкл 1× PBS гранулы осаждали и промывали 1 мл 1× PBS.
[0158] Тысячу микролитров связанных с антигеном-EEV гранул добавляли к 1 мл каждого соответствующего супернатанта антитело-EEV, экспрессирующего антитело к FZD4, антитело к CXCR4, антитело к CD100 или антитело к HER2, а также к контрольному антителу-EEV. Антитело к EEV получали инфицированием клеток BHK при MOI=1 каждый в течение 2 суток вирусом осповакцины, кодирующим тяжелые и легкие цепи соответствующих антител, и собирали супернатанты с последующим центрифугированием для удаления любых клеток. Вирус, связанный с гранулами и антителом к EEV,оставляли вращаться при комнатной температуре в течение 2 часов. Гранулы осаждали с использованием магнита и удаляли несвязанные супернатанты. Затем гранулы отмывали пять раз 1 мл среды DMEM, дополненной 10% FBS и 1 мМ HEPES (10% DMEM). Все промывания объединяли с несвязанным супернатантом ("несвязанные"). Затем гранулы ("связанные") ресуспендировали в 1 мл 10% DMEM. "Несвязанные" и "связанные" титровали на клетках BSC-1 и покрывали средой для выращивания, содержащей метилцеллюлозу. Оставляли в течение двух суток для формирования бляшек, а затем клетки фиксировали и окрашивали 0,1% раствором кристаллического фиолетового. Подсчитывали бляшки для определения числа бляшкообразующих единиц (БОЕ) в "несвязанных" и "связанных", на основании которого можно было рассчитывать % EEV, связанного с гранулами. Схематическое изображение способа приведено на фиг. 5, и результаты приведены на фиг. 6A (HER2), фиг. 6B (FZD4), фиг. 6C (CXCR4) и фиг. 6D (CD100 ("Sema")).
[0159] Антитело-EEV, экспрессирующие антитело к HER2 специфически осаждали гранулами, связанными с антигеном EEV HER2-ECD-A56R, антитело-EEV, экспрессирующее антитело к FZD, специфически осаждали гранулами, связанными с антигеном EEV FZD-F13L, антитело-EEV, экспрессирующее антитело к CXCR4, специфически осаждали гранулами, связанными с антигеном EEV CXCR4-F13L, и антитело-EEV, экспрессирующее антитело к SEMA, специфически осаждали гранулами, связанными с антигеном EEV Sema-ECD-A56R.
Пример 6: Скрининг библиотеки антител
[0160] Клетки BHK инфицировали при MOI=1 каждым из библиотеки антител (H-IgG-A56R) и L48 (производным VK1-39 зародышевой линии) в четырех камерах для культивирования клеток cellSTACK (Corning) (2×108 клеток на штатив). Библиотека антител содержала различную популяцию вариабельных доменов тяжелых цепей в формате полноразмерного IgG, слитых в рамке считывания с A56R (см. публикацию заявки на патент США № 2013-0288927, полностью включенную в настоящее описание посредством ссылки). Разнообразие этой библиотеки составляло приблизительно 400 миллионов независимых клонов. Через двое суток, супернатант, содержащий EEV, собирали и удаляли дебрис низкоскоростным центрифугированием. Затем осветленный супернатант центрифугировали при 13000 об./мин. (28000×g) в течение 1 часа для осаждения антитела-EEV. Антитело-EEV ресуспендировали в 1 мл EMEM с 10% FBS и хранили при 4 градусах до момента использования. Для подготовки антигена вирус для пэннинга клетки BHK (2×108 клеток) инфицировали при MOI=1,5 вирусом, экспресисрующим FZD4-ECD-A56R (SEQ ID NO:6) в двух камерах для культивирования клеток cellSTACK (Corning). Через двое суток супернатант, содержащий EEV, собирали и удаляли дебрис низкоскоростным центрифугированием. Затем осветленный супернатант центрифугировали при 13000 об./мин. (28000×g) в течение 1 часа для осаждения антигена-EEV. Супернатант аспирировали и ресуспендировали осадок в 1,0 мл 1× PBS. Один мл EEV FZD4-ECD-A56R переносили в свежую пробирку и добавляли просален (триоксален, 4'-аминометилгидрохлорид; Sigma) до конечной концентрации 40 мкг/мл. EEV и просален инкубировали при комнатной температуре в течение 10 минут перед облучением в УФ-сшивателе STRATALINKER® (Stratagene) при 99999 микроджоулей. Процедура обработки просаленом/УФ обеспечивает то, что антиген-EEV является инактивированным и, таким образом, неспособен образовывать бляшки или размножаться в любом последующем тестировании.
[0161] Активированные тозилом MyOne DYNABEADS® (150 мкл) осаждали с помощью магнита и промывали 1 мл PBS два раза. Гранулы осаждали с помощью магнита, удаляли PBS и 1 мл инактивированного просаленом/УФ FZD4-ECD-A56R добавляли к гранулам. Гранулы и антиген-EEV оставляли вращаться при 37°C в течение 18-20 часов. Гранулы осаждали и удаляли супернатант. Гранулы блокировали 1 мл 1× PBS, 10% FBS и 0,5% BSA при 37°C в течение 2 часов. Перед ресуспендированием в 150 мкл 1× PBS гранулы осаждали и промывали 1 мл 1× PBS.
[0162] Пятьдесят микролитров связанных с FZD4-ECD-A56R гранул добавляли к 1 мл библиотеке антитело-EEV. Связанные с FZD4-ECD-A56R гранулы и антитело к EEV оставляли вращаться при комнатной температуре в течение 2 часов. Гранулы осаждали с использованием магнита и удаляли несвязавшийся супернатант. Затем гранулы отмывали пять раз 1 мл среды DMEM, дополненной 10% FBS и 1 мМ HEPES (10% DMEM). Все промывания объединяли с несвязанным супернатантом ("несвязанные"). Затем гранулы ("связанные") ресуспендировали в 1 мл 10% DMEM. "Несвязанные" и "связанные" титровали на клетках BSC-1 и покрывали средой для выращивания, содержащей метилцеллюлозу. Оставляли на двое суток для формирования бляшек, а затем клетки фиксировали и окрашивали 0,1% раствором кристаллического фиолетового. Оставшийся связанный вирус (990 мкл) распределяли по 5 колбам T175, содержащим конфлюэнтные клетки BSC1 и оставляли размножаться в DMEM 2,5%, содержащей 1 мг/мл G418, в течение 3 суток. Затем собирали клетки, и высвобождали вирус с помощью трех циклов заморозки/оттаивания и титровали вирус.
[0163] Для второго цикла селекции (Rd2) амплифицированные тяжелые цепи из цикла 1 совместно инфицировали со свежей L48 в одну камеру cellSTACK BHK. Антитело к EEV собирали, как описано выше. Для каждого цикла пэннинга получали, концентрировали, инактивировали и связывали с гранулами свежий вирус с антигеном FZD-ECD-A56R, как описано выше. Пятьдесят микролитров связанных с FZD4-ECD-A56R гранул добавляли к 1 мл антитела-EEV Rd2. Связанные с FZD4-ECD-A56R гранулы и антитело к EEV оставляли вращаться при комнатной температуре в течение 2 часов. Гранулы осаждали с использованием магнита и удаляли несвязавшийся супернатант. Затем гранулы отмывали пять раз 1 мл среды DMEM, дополненной 10% FBS и 1 мМ HEPES (10% DMEM). Все промывания объединяли с несвязанным супернатантом ("несвязанные"). Затем гранулы ("связанные") ресуспендировали в 1 мл 10% DMEM. "Несвязанные" и "связанные" титровали на клетках BSC-1 и покрывали средой для выращивания, содержащей метилцеллюлозу. Оставляли на двое суток для формирования бляшек, а затем клетки фиксировали и окрашивали 0,1% раствором кристаллического фиолетового. Оставшийся связанный вирус (990 мкл) распределяли по 5 колбам T175, содержащим конфлюэнтные клетки BSC1 и оставляли размножаться в DMEM 2,5%, содержащей 1 мг/мл G418, в течение 3 суток. Затем собирали клетки, и высвобождали вирус с помощью трех циклов заморозки/оттаивания, и титровали вирус.
[0164] Три дополнительных цикла пэннинга (Rd3, Rd4 и Rd5) проводили так, как описано выше.
[0165] Циклы 3, 4 и 5 тестировали на обогащение инфицированными клетками A431 в 6-луночном планшете при moi=1 с каждым циклом амплификации VH и L48. После инфицирования в течение ночи клетки собирали и разделяли напополам. Одну половину окрашивали 10 мкг/мл FZD-His с последующим окрашиванием антителом к His-Dyelight650 и антителом к Fab-FITC. Другую половину окрашивали 10 мкг/мл CD100-His (отрицательный контроль) с последующим окрашиванием антителом к His-Dyelight650 и антителом к Fab-FITC. Данные, представленные на фиг. 7, демонстрируют постоянное обогащение для каждого цикла селекции. Антитела из 5 цикла субклонировали в экспрессирующий вектор млекопитающих для экспрессии в виде полноразмерного растворимого IgG и трансфицировали (с L48 в экспрессирующий вектор млекопитающего). Получаемые антитела, присутствующие в супернатанте, тестировали проточной цитометрией на связывание с трансфицированными FZD4 клетками CHO и отсутствие связывания с трансфицированными CXCR4 клетками CHO. Идентифицировали ряд антител, которые специфически связывались с FZD.
Пример 7: Двойной EEV метка/антиген можно связывать с магнитными гранулами
[0166] Клетки BHK инфицировали по два вириона на клетку, где один вирион представлял собой гемагглютининовую метку (HA)-A56R (SEQ ID NO:17), а второй представлял собой FZD4-F13L (SEQ ID NO:2), для сбора EEV, экспрессирующего так метку HA, так и антиген FZD4 на своей поверхности, или инфицировали по одному вириону на клетку каждым отдельным вирусом. Через двое суток супернатант, содержащий EEV, собирали и удаляли дебрис низкоскоростным центрифугированием. Магнитные гранулы с белком G (150 мкл) осаждали с помощью магнита и добавляли к гранулам 1 мл PBS+30 мкг очищенного антитела к FZD4 (C6073). Раствор инкубировали при комнатной температуре при аккуратном вращении в течение 25 минут для обеспечения связывания антитела с белком G на гранулах. Гранулы осаждали с помощью магнита, промывали один раз 1 мл PBS и ресуспендировали в 300 мкл DMEM+10% FBS. Перед ресуспендированием в 150 мкл PBS покрытые антителом к HA-метка магнитные гранулы (ThermoFisher, 150 мкл) осаждали с помощью магнита и промывали один раз 1 мл PBS.
HA-A56R (SEQ ID NO:14)
MGWSCIILFLVATATGAHS YPYDVPDYA TSTTNDTDKVDYEEYSTELIVNTDSESTIDIILSGSTHSPETSSKKPDYIDNSNCSSVFEIATPEPITDNVEDHTDTVTYTSDSINTVSASSGESTTDETPEPITDKEDHTVTDTVSYTTVSTSSGIVTTKSTTDDADLYDTYNDNDTVPPTTVGGSTTSISNYKTKDFVEIFGITALIILSAVAIFCITYYIYNKRSRKYKTENKV
Одинарное подчеркивание - Сигнальная последовательность (аминокислоты 1-19)
Жирный шрифт - Метка HA (аминокислоты 20-29)
Курсив - Усеченный A56R (аминокислоты 30-235)
[0167] Пятьдесят мкл подготовленных покрытых антителом к HA-метка грагул или 100 мкл подготовленного белка G с антителом к FZD4 добавляли к 1 мл каждого супернатанта с EEV и оставляли вращаться при комнатной температуре в течение 60 минут. Гранулы осаждали с использованием магнита и удаляли несвязавшийся супернатант. Затем гранулы отмывали пять раз 1 мл среды DMEM, дополненной 10% FBS и 1 мМ HEPES (10% DMEM). Все промывания объединяли с несвязанным супернатантом ("несвязанные"). Затем гранулы ("связанные") ресуспендировали в 1 мл 10% DMEM. "Несвязанные" и "связанные" титровали на клетках BSC-1 и покрывали средой для выращивания, содержащей метилцеллюлозу. Оставляли на двое суток для формирования бляшек, а затем клетки фиксировали и окрашивали 0,1% раствором кристаллического фиолетового. Подсчитывали бляшки для определения числа бляшкообразующих единиц (БОЕ) в "несвязанных" и "связанных", на основании которого можно было рассчитывать % EEV, связанного с гранулами. Результаты приведены на фиг. 8. EEV, экспрессирующий оба слитых белка, осаждали любым антителом.
Пример 8: Двойной EEV метка/антиген можно связывать с магнитными гранулами и использовать здля захвата mAb EEV
[0168] Клетки BHK (2×108 клеток) инфицировали по два вириона на клетку, где один вирион представлял собой HA-A56R (SEQ ID NO:17), а второй представлял собой CXCR4-F13L (SEQ ID NO:3), для сбора EEV, экспрессирующего так метку HA, так и антиген CXCR4 на своей поверхностьи. Через двое суток супернатант, содержащий EEV, собирали и удаляли дебрис низкоскоростным центрифугированием. Затем осветленный супернатант центрифугировали при 13000 об./мин. (28000×g) в течение 1 часа для осаждения метка/антиген-EEV. Супернатант аспирировали и осадок ресуспендировали в 1 мл 1× PBS. Добавляли просален (триоксален, 4'-аминометилгидрохлорид; Sigma) до конечной концентрации 40 мкг/мл. EEV и просален инкубировали при комнатной температуре в течение 10 минут перед облучением в УФ-сшивателе Stratalinker (Stratagene) при 99999 микродхоулей дважды. Процедура обработки просаленом/УФ обеспечивает то, что антиген-EEV является инактивированным и, таким образом, неспособен образовывать бляшки или размножаться в любом последующем тестировании.
[0169] Антитела к CXCR4 EEV и mAb EEV к HER2 получали посредством инфицирования клеток BHK по два вириона на клетку, где один вирион представлял собой конкретную тяжелую цепь, и второй представлял собой конкретную легкую цепь. Через двое суток супернатанты, содержащие антитело к CXCR4 EEV и антитело к HER2 EEV, собирали и удаляли дебрис низкоскоростным центрифугированием.
[0170] Три тысячи микролитров магнитных гранул, покрытых антителом к HA, отмывали 1 мл PBS, а затем ресуспендировали в одном миллилитре инактивированных просаленом/УФ HA/CXCR4 EEV. Гранулы и EEV инкубировали при комнатной температуре при аккуратном вращении в течение 90 минут для обеспечения связвания EEV с гранулами, покрытыми антителом к HA. Гранулы осаждали с помощью магнита, промывали один раз 1 мл PBS и ресуспендировали в 300 мкл PBS.
[0171] Тысячу мкл HA/CXCR4 EEV, связанных с покрытыми антителом к HA гранулами, добавляли к 1 мл каждого супернатанта с mAb EEV и инкубировали при комнатной температуре при аккуратном вращении в течение 1-1,5 часов. Гранулы осаждали с использованием магнита и удаляли несвязавшийся супернатант. Затем гранулы отмывали пять раз 1 мл среды DMEM, дополненной 10% FBS и 1 мМ HEPES (10% DMEM). Все промывания объединяли с несвязанным супернатантом ("есвязанные"). Затем гранулы ("связанные") ресуспендировали в 1 мл 10% DMEM. "Несвязанные" и "связанные" титровали на клетках BSC-1 и покрывали средой для выращивания, содержащей метилцеллюлозу. Оставляли на двое суток для формирования бляшек, а затем клетки фиксировали и окрашивали 0,1% раствором кристаллического фиолетового. Подсчитывали бляшки для определения числа бляшкообразующих единиц (БОЕ) в "несвязанных" и "связанных", на основании которого можно было рассчитывать % EEV, связанного с гранулами. Результаты приведены на фиг. 9. Гранулы, покрытые EEV, коэкспрессирующим HA-A45R и CXCR4-F13L, специфически захватывали EEV с антителом к CXCR4.
Пример 9: Антиген-EEV можно биотинилировать для связывания с магнитными гранулами
[0172] Клетки BHK инфицировали при MOI=1, вирусом, экспрессирующим FZD4-F13L, FZD4-ECD-A56R или CD20-F13L. Через двое суток супернатант, содержащий EEV, собирали и удаляли дебрис низкоскоростным центрифугированием. Затем осветленный супернатант центрифугировали при 13000 об./мин. в течение 1 часа для осаждения антигена-EEV. Супернатант аспирировали и ресуспендировали осадок в 1-2 мл 1× PBS. Для биотинилирования EEV 2,5 мкл исходного раствора биотин-XX SSE в 1× PBS (FLUOREPORTERВ® набор Cell Surface Biotinylation, Molecular Probes) добавляли к 1 мл каждого EEV в PBS и инкубировали на льду в течение 30 минут. Для гашения реакции добавляли пятьдесят мкл 1M Tris pH 8.
[0173] Для связывания биотин-EEV с гранулами 150 мкл покрытыех стрептавидином DYNABEADS® осаждали и промывали один раз 1 мл 1× PBS. Гранулы ресуспендировали в 150 мкл и добавляли 50 мкл к 1 мл минимальной поддерживающей среды Игла (EMEM), содержащей 10% FBS и 1 мМ HEPES (10% EMEM). Пятьдесят мкл биотин-EEV добавляли к гранулам и среде и оставляли вращаться при комнатной температуре в течение 1 часа. Гранулы осаждали с использованием магнита и удаляли несвязавшийся супернатант. Затем гранулы отмывали пять раз 1 мл среды DMEM, дополненной 10% FBS и 1 мМ HEPES (10% DMEM). Все промывания объединяли с несвязанным супернатантом ("несвязанные"). Затем гранулы ("связанные") ресуспендировали в 1 мл 10% DMEM. "Несвязанные" и "связанные" титровали на клетках BSC-1 и покрывали средой для выращивания, содержащей метилцеллюлозу. Оставляли на двое суток для формирования бляшек, а затем клетки фиксировали и окрашивали 0,1% раствором кристаллического фиолетового. Подсчитывали бляшки для определения числа бляшкообразующих единиц (БОЕ) в "несвязанных" и "связанных", на основании которого можно было рассчитывать % EEV, связанного с гранулами. Результаты продемонстрированы на фиг. 10.
[0174] Антиген-EEV являлся пригодным для биотинилирования и связывания с магнитными гранулами, покрытыми стрептавидином.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Smith, Ernest S.
Paris, Mark
Scrivens, Maria G.M.
Kirk, Renee A.
Moksa-Cornelison, Angelica A.
<120> ДИСПЛЕЙ ИНТЕРАЛЬНОГО МЕМБРАННОГО БЕЛКА НА ВНЕКЛЕТОЧНЫХ
ОБОЛОЧЕЧНЫХ ВИРИОНАХ ПОКСВИРУСА
<130> 58008-165443
<140> PCT/US2017/028787
<141> 2017-04-21
<160> 17
<170> PatentIn version 3.5
<210> 1
<211> 372
<212> Белок
<213> Искусственная последовательность
<220>
<223> F13L
<400> 1
Met Trp Pro Phe Ala Ser Val Pro Ala Gly Ala Lys Cys Arg Leu Val
1 5 10 15
Glu Thr Leu Pro Glu Asn Met Asp Phe Arg Ser Asp His Leu Thr Thr
20 25 30
Phe Glu Cys Phe Asn Glu Ile Ile Thr Leu Ala Lys Lys Tyr Ile Tyr
35 40 45
Ile Ala Ser Phe Cys Cys Asn Pro Leu Ser Thr Thr Arg Gly Ala Leu
50 55 60
Ile Phe Asp Lys Leu Lys Glu Ala Ser Glu Lys Gly Ile Lys Ile Ile
65 70 75 80
Val Leu Leu Asp Glu Arg Gly Lys Arg Asn Leu Gly Glu Leu Gln Ser
85 90 95
His Cys Pro Asp Ile Asn Phe Ile Thr Val Asn Ile Asp Lys Lys Asn
100 105 110
Asn Val Gly Leu Leu Leu Gly Cys Phe Trp Val Ser Asp Asp Glu Arg
115 120 125
Cys Tyr Val Gly Asn Ala Ser Phe Thr Gly Gly Ser Ile His Thr Ile
130 135 140
Lys Thr Leu Gly Val Tyr Ser Asp Tyr Pro Pro Leu Ala Thr Asp Leu
145 150 155 160
Arg Arg Arg Phe Asp Thr Phe Lys Ala Phe Asn Ser Ala Lys Asn Ser
165 170 175
Trp Leu Asn Leu Cys Ser Ala Ala Cys Cys Leu Pro Val Ser Thr Ala
180 185 190
Tyr His Ile Lys Asn Pro Ile Gly Gly Val Phe Phe Thr Asp Ser Pro
195 200 205
Glu His Leu Leu Gly Tyr Ser Arg Asp Leu Asp Thr Asp Val Val Ile
210 215 220
Asp Lys Leu Lys Ser Ala Lys Thr Ser Ile Asp Ile Glu His Leu Ala
225 230 235 240
Ile Val Pro Thr Thr Arg Val Asp Gly Asn Ser Tyr Tyr Trp Pro Asp
245 250 255
Ile Tyr Asn Ser Ile Ile Glu Ala Ala Ile Asn Arg Gly Val Lys Ile
260 265 270
Arg Leu Leu Val Gly Asn Trp Asp Lys Asn Asp Val Tyr Ser Met Ala
275 280 285
Thr Ala Arg Ser Leu Asp Ala Leu Cys Val Gln Asn Asp Leu Ser Val
290 295 300
Lys Val Phe Thr Ile Gln Asn Asn Thr Lys Leu Leu Ile Val Asp Asp
305 310 315 320
Glu Tyr Val His Ile Thr Ser Ala Asn Phe Asp Gly Thr His Tyr Gln
325 330 335
Asn His Gly Phe Val Ser Phe Asn Ser Ile Asp Lys Gln Leu Val Ser
340 345 350
Glu Ala Lys Lys Ile Phe Glu Arg Asp Trp Val Ser Ser His Ser Lys
355 360 365
Ser Leu Lys Ile
370
<210> 2
<211> 892
<212> Белок
<213> Искусственная последовательность
<220>
<223> Слитый белок FZD (FL) - F13L
<400> 2
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Ala His Ser Phe Gly Asp Glu Glu Glu Arg Arg Cys Asp Pro Ile Arg
20 25 30
Ile Ser Met Cys Gln Asn Leu Gly Tyr Asn Val Thr Lys Met Pro Asn
35 40 45
Leu Val Gly His Glu Leu Gln Thr Asp Ala Glu Leu Gln Leu Thr Thr
50 55 60
Phe Thr Pro Leu Ile Gln Tyr Gly Cys Ser Ser Gln Leu Gln Phe Phe
65 70 75 80
Leu Cys Ser Val Tyr Val Pro Met Cys Thr Glu Lys Ile Asn Ile Pro
85 90 95
Ile Gly Pro Cys Gly Gly Met Cys Leu Ser Val Lys Arg Arg Cys Glu
100 105 110
Pro Val Leu Lys Glu Phe Gly Phe Ala Trp Pro Glu Ser Leu Asn Cys
115 120 125
Ser Lys Phe Pro Pro Gln Asn Asp His Asn His Met Cys Met Glu Gly
130 135 140
Pro Gly Asp Glu Glu Val Pro Leu Pro His Lys Thr Pro Ile Gln Pro
145 150 155 160
Gly Glu Glu Cys His Ser Val Gly Thr Asn Ser Asp Gln Tyr Ile Trp
165 170 175
Val Lys Arg Ser Leu Asn Cys Val Leu Lys Cys Gly Tyr Asp Ala Gly
180 185 190
Leu Tyr Ser Arg Ser Ala Lys Glu Phe Thr Asp Ile Trp Met Ala Val
195 200 205
Trp Ala Ser Leu Cys Phe Ile Ser Thr Ala Phe Thr Val Leu Thr Phe
210 215 220
Leu Ile Asp Ser Ser Arg Phe Ser Tyr Pro Glu Arg Pro Ile Ile Phe
225 230 235 240
Leu Ser Met Cys Tyr Asn Ile Tyr Ser Ile Ala Tyr Ile Val Arg Leu
245 250 255
Thr Val Gly Arg Glu Arg Ile Ser Cys Asp Phe Glu Glu Ala Ala Glu
260 265 270
Pro Val Leu Ile Gln Glu Gly Leu Lys Asn Thr Gly Cys Ala Ile Ile
275 280 285
Phe Leu Leu Met Tyr Phe Phe Gly Met Ala Ser Ser Ile Trp Trp Val
290 295 300
Ile Leu Thr Leu Thr Trp Phe Leu Ala Ala Gly Leu Lys Trp Gly His
305 310 315 320
Glu Ala Ile Glu Met His Ser Ser Tyr Phe His Ile Ala Ala Trp Ala
325 330 335
Ile Pro Ala Val Lys Thr Ile Val Ile Leu Ile Met Arg Leu Val Asp
340 345 350
Ala Asp Glu Leu Thr Gly Leu Cys Tyr Val Gly Asn Gln Asn Leu Asp
355 360 365
Ala Leu Thr Gly Phe Val Val Ala Pro Leu Phe Thr Tyr Leu Val Ile
370 375 380
Gly Thr Leu Phe Ile Ala Ala Gly Leu Val Ala Leu Phe Lys Ile Arg
385 390 395 400
Ser Asn Leu Gln Lys Asp Gly Thr Lys Thr Asp Lys Leu Glu Arg Leu
405 410 415
Met Val Lys Ile Gly Val Phe Ser Val Leu Tyr Thr Val Pro Ala Thr
420 425 430
Cys Val Ile Ala Cys Tyr Phe Tyr Glu Ile Ser Asn Trp Ala Leu Phe
435 440 445
Arg Tyr Ser Ala Asp Asp Ser Asn Met Ala Val Glu Met Leu Lys Ile
450 455 460
Phe Met Ser Leu Leu Val Gly Ile Thr Ser Gly Met Trp Ile Trp Ser
465 470 475 480
Ala Lys Thr Leu His Thr Trp Gln Lys Cys Ser Asn Arg Leu Val Asn
485 490 495
Ser Gly Lys Val Lys Arg Glu Lys Arg Gly Asn Gly Trp Val Lys Pro
500 505 510
Gly Lys Gly Ser Glu Thr Val Val Met Trp Pro Phe Ala Ser Val Pro
515 520 525
Ala Gly Ala Lys Cys Arg Leu Val Glu Thr Leu Pro Glu Asn Met Asp
530 535 540
Phe Arg Ser Asp His Leu Thr Thr Phe Glu Cys Phe Asn Glu Ile Ile
545 550 555 560
Thr Leu Ala Lys Lys Tyr Ile Tyr Ile Ala Ser Phe Cys Cys Asn Pro
565 570 575
Leu Ser Thr Thr Arg Gly Ala Leu Ile Phe Asp Lys Leu Lys Glu Ala
580 585 590
Ser Glu Lys Gly Ile Lys Ile Ile Val Leu Leu Asp Glu Arg Gly Lys
595 600 605
Arg Asn Leu Gly Glu Leu Gln Ser His Cys Pro Asp Ile Asn Phe Ile
610 615 620
Thr Val Asn Ile Asp Lys Lys Asn Asn Val Gly Leu Leu Leu Gly Cys
625 630 635 640
Phe Trp Val Ser Asp Asp Glu Arg Cys Tyr Val Gly Asn Ala Ser Phe
645 650 655
Thr Gly Gly Ser Ile His Thr Ile Lys Thr Leu Gly Val Tyr Ser Asp
660 665 670
Tyr Pro Pro Leu Ala Thr Asp Leu Arg Arg Arg Phe Asp Thr Phe Lys
675 680 685
Ala Phe Asn Ser Ala Lys Asn Ser Trp Leu Asn Leu Cys Ser Ala Ala
690 695 700
Cys Cys Leu Pro Val Ser Thr Ala Tyr His Ile Lys Asn Pro Ile Gly
705 710 715 720
Gly Val Phe Phe Thr Asp Ser Pro Glu His Leu Leu Gly Tyr Ser Arg
725 730 735
Asp Leu Asp Thr Asp Val Val Ile Asp Lys Leu Lys Ser Ala Lys Thr
740 745 750
Ser Ile Asp Ile Glu His Leu Ala Ile Val Pro Thr Thr Arg Val Asp
755 760 765
Gly Asn Ser Tyr Tyr Trp Pro Asp Ile Tyr Asn Ser Ile Ile Glu Ala
770 775 780
Ala Ile Asn Arg Gly Val Lys Ile Arg Leu Leu Val Gly Asn Trp Asp
785 790 795 800
Lys Asn Asp Val Tyr Ser Met Ala Thr Ala Arg Ser Leu Asp Ala Leu
805 810 815
Cys Val Gln Asn Asp Leu Ser Val Lys Val Phe Thr Ile Gln Asn Asn
820 825 830
Thr Lys Leu Leu Ile Val Asp Asp Glu Tyr Val His Ile Thr Ser Ala
835 840 845
Asn Phe Asp Gly Thr His Tyr Gln Asn His Gly Phe Val Ser Phe Asn
850 855 860
Ser Ile Asp Lys Gln Leu Val Ser Glu Ala Lys Lys Ile Phe Glu Arg
865 870 875 880
Asp Trp Val Ser Ser His Ser Lys Ser Leu Lys Ile
885 890
<210> 3
<211> 728
<212> Белок
<213> Искусственная последовательность
<220>
<223> Слитый белок CXCR4-F13L
<400> 3
Met Ala Ile Pro Leu Pro Leu Leu Gln Ile Tyr Thr Ser Asp Asn Tyr
1 5 10 15
Thr Glu Glu Met Gly Ser Gly Asp Tyr Asp Ser Met Lys Glu Pro Cys
20 25 30
Phe Arg Glu Glu Asn Ala Asn Phe Asn Lys Ile Phe Leu Pro Thr Ile
35 40 45
Tyr Ser Ile Ile Phe Leu Thr Gly Ile Val Gly Asn Gly Leu Val Ile
50 55 60
Leu Val Met Gly Tyr Gln Lys Lys Leu Arg Ser Met Thr Asp Lys Tyr
65 70 75 80
Arg Leu His Leu Ser Val Ala Asp Leu Leu Phe Val Ile Thr Leu Pro
85 90 95
Phe Trp Ala Val Asp Ala Val Ala Asn Trp Tyr Phe Gly Asn Phe Leu
100 105 110
Cys Lys Ala Val His Val Ile Tyr Thr Val Asn Leu Tyr Ser Ser Val
115 120 125
Leu Ile Leu Ala Phe Ile Ser Leu Asp Arg Tyr Leu Ala Ile Val His
130 135 140
Ala Thr Asn Ser Gln Arg Pro Arg Lys Leu Leu Ala Glu Lys Val Val
145 150 155 160
Tyr Val Gly Val Trp Ile Pro Ala Leu Leu Leu Thr Ile Pro Asp Phe
165 170 175
Ile Phe Ala Asn Val Ser Glu Ala Asp Asp Arg Tyr Ile Cys Asp Arg
180 185 190
Phe Tyr Pro Asn Asp Leu Trp Val Val Val Phe Gln Phe Gln His Ile
195 200 205
Met Val Gly Leu Ile Leu Pro Gly Ile Val Ile Leu Ser Cys Tyr Cys
210 215 220
Ile Ile Ile Ser Lys Leu Ser His Ser Lys Gly His Gln Lys Arg Lys
225 230 235 240
Ala Leu Lys Thr Thr Val Ile Leu Ile Leu Ala Phe Phe Ala Cys Trp
245 250 255
Leu Pro Tyr Tyr Ile Gly Ile Ser Ile Asp Ser Phe Ile Leu Leu Glu
260 265 270
Ile Ile Lys Gln Gly Cys Glu Phe Glu Asn Thr Val His Lys Trp Ile
275 280 285
Ser Ile Thr Glu Ala Leu Ala Phe Phe His Cys Cys Leu Asn Pro Ile
290 295 300
Leu Tyr Ala Phe Leu Gly Ala Lys Phe Lys Thr Ser Ala Gln His Ala
305 310 315 320
Leu Thr Ser Val Ser Arg Gly Ser Ser Leu Lys Ile Leu Ser Lys Gly
325 330 335
Lys Arg Gly Gly His Ser Ser Val Ser Thr Glu Ser Glu Ser Ser Ser
340 345 350
Phe His Ser Ser Met Trp Pro Phe Ala Ser Val Pro Ala Gly Ala Lys
355 360 365
Cys Arg Leu Val Glu Thr Leu Pro Glu Asn Met Asp Phe Arg Ser Asp
370 375 380
His Leu Thr Thr Phe Glu Cys Phe Asn Glu Ile Ile Thr Leu Ala Lys
385 390 395 400
Lys Tyr Ile Tyr Ile Ala Ser Phe Cys Cys Asn Pro Leu Ser Thr Thr
405 410 415
Arg Gly Ala Leu Ile Phe Asp Lys Leu Lys Glu Ala Ser Glu Lys Gly
420 425 430
Ile Lys Ile Ile Val Leu Leu Asp Glu Arg Gly Lys Arg Asn Leu Gly
435 440 445
Glu Leu Gln Ser His Cys Pro Asp Ile Asn Phe Ile Thr Val Asn Ile
450 455 460
Asp Lys Lys Asn Asn Val Gly Leu Leu Leu Gly Cys Phe Trp Val Ser
465 470 475 480
Asp Asp Glu Arg Cys Tyr Val Gly Asn Ala Ser Phe Thr Gly Gly Ser
485 490 495
Ile His Thr Ile Lys Thr Leu Gly Val Tyr Ser Asp Tyr Pro Pro Leu
500 505 510
Ala Thr Asp Leu Arg Arg Arg Phe Asp Thr Phe Lys Ala Phe Asn Ser
515 520 525
Ala Lys Asn Ser Trp Leu Asn Leu Cys Ser Ala Ala Cys Cys Leu Pro
530 535 540
Val Ser Thr Ala Tyr His Ile Lys Asn Pro Ile Gly Gly Val Phe Phe
545 550 555 560
Thr Asp Ser Pro Glu His Leu Leu Gly Tyr Ser Arg Asp Leu Asp Thr
565 570 575
Asp Val Val Ile Asp Lys Leu Lys Ser Ala Lys Thr Ser Ile Asp Ile
580 585 590
Glu His Leu Ala Ile Val Pro Thr Thr Arg Val Asp Gly Asn Ser Tyr
595 600 605
Tyr Trp Pro Asp Ile Tyr Asn Ser Ile Ile Glu Ala Ala Ile Asn Arg
610 615 620
Gly Val Lys Ile Arg Leu Leu Val Gly Asn Trp Asp Lys Asn Asp Val
625 630 635 640
Tyr Ser Met Ala Thr Ala Arg Ser Leu Asp Ala Leu Cys Val Gln Asn
645 650 655
Asp Leu Ser Val Lys Val Phe Thr Ile Gln Asn Asn Thr Lys Leu Leu
660 665 670
Ile Val Asp Asp Glu Tyr Val His Ile Thr Ser Ala Asn Phe Asp Gly
675 680 685
Thr His Tyr Gln Asn His Gly Phe Val Ser Phe Asn Ser Ile Asp Lys
690 695 700
Gln Leu Val Ser Glu Ala Lys Lys Ile Phe Glu Arg Asp Trp Val Ser
705 710 715 720
Ser His Ser Lys Ser Leu Lys Ile
725
<210> 4
<211> 669
<212> Белок
<213> Искусственная последовательность
<220>
<223> Слитый белок CD20-F13L
<400> 4
Met Ala Thr Pro Arg Asn Ser Val Asn Gly Thr Phe Pro Ala Glu Pro
1 5 10 15
Met Lys Gly Pro Ile Ala Met Gln Ser Gly Pro Lys Pro Leu Phe Arg
20 25 30
Arg Met Ser Ser Leu Val Gly Pro Thr Gln Ser Phe Phe Met Arg Glu
35 40 45
Ser Lys Thr Leu Gly Ala Val Gln Ile Met Asn Gly Leu Phe His Ile
50 55 60
Ala Leu Gly Gly Leu Leu Met Ile Pro Ala Gly Ile Tyr Ala Pro Ile
65 70 75 80
Cys Val Thr Val Trp Tyr Pro Leu Trp Gly Gly Ile Met Tyr Ile Ile
85 90 95
Ser Gly Ser Leu Leu Ala Ala Thr Glu Lys Asn Ser Arg Lys Cys Leu
100 105 110
Val Lys Gly Lys Met Ile Met Asn Ser Leu Ser Leu Phe Ala Ala Ile
115 120 125
Ser Gly Met Ile Leu Ser Ile Met Asp Ile Leu Asn Ile Lys Ile Ser
130 135 140
His Phe Leu Lys Met Glu Ser Leu Asn Phe Ile Arg Ala His Thr Pro
145 150 155 160
Tyr Ile Asn Ile Tyr Asn Cys Glu Pro Ala Asn Pro Ser Glu Lys Asn
165 170 175
Ser Pro Ser Thr Gln Tyr Cys Tyr Ser Ile Gln Ser Leu Phe Leu Gly
180 185 190
Ile Leu Ser Val Met Leu Ile Phe Ala Phe Phe Gln Glu Leu Val Ile
195 200 205
Ala Gly Ile Val Glu Asn Glu Trp Lys Arg Thr Cys Ser Arg Pro Lys
210 215 220
Ser Asn Ile Val Leu Leu Ser Ala Glu Glu Lys Lys Glu Gln Thr Ile
225 230 235 240
Glu Ile Lys Glu Glu Val Val Gly Leu Thr Glu Thr Ser Ser Gln Pro
245 250 255
Lys Asn Glu Glu Asp Ile Glu Ile Ile Pro Ile Gln Glu Glu Glu Glu
260 265 270
Glu Glu Thr Glu Thr Asn Phe Pro Glu Pro Pro Gln Asp Gln Glu Ser
275 280 285
Ser Pro Ile Glu Asn Asp Ser Ser Pro Met Trp Pro Phe Ala Ser Val
290 295 300
Pro Ala Gly Ala Lys Cys Arg Leu Val Glu Thr Leu Pro Glu Asn Met
305 310 315 320
Asp Phe Arg Ser Asp His Leu Thr Thr Phe Glu Cys Phe Asn Glu Ile
325 330 335
Ile Thr Leu Ala Lys Lys Tyr Ile Tyr Ile Ala Ser Phe Cys Cys Asn
340 345 350
Pro Leu Ser Thr Thr Arg Gly Ala Leu Ile Phe Asp Lys Leu Lys Glu
355 360 365
Ala Ser Glu Lys Gly Ile Lys Ile Ile Val Leu Leu Asp Glu Arg Gly
370 375 380
Lys Arg Asn Leu Gly Glu Leu Gln Ser His Cys Pro Asp Ile Asn Phe
385 390 395 400
Ile Thr Val Asn Ile Asp Lys Lys Asn Asn Val Gly Leu Leu Leu Gly
405 410 415
Cys Phe Trp Val Ser Asp Asp Glu Arg Cys Tyr Val Gly Asn Ala Ser
420 425 430
Phe Thr Gly Gly Ser Ile His Thr Ile Lys Thr Leu Gly Val Tyr Ser
435 440 445
Asp Tyr Pro Pro Leu Ala Thr Asp Leu Arg Arg Arg Phe Asp Thr Phe
450 455 460
Lys Ala Phe Asn Ser Ala Lys Asn Ser Trp Leu Asn Leu Cys Ser Ala
465 470 475 480
Ala Cys Cys Leu Pro Val Ser Thr Ala Tyr His Ile Lys Asn Pro Ile
485 490 495
Gly Gly Val Phe Phe Thr Asp Ser Pro Glu His Leu Leu Gly Tyr Ser
500 505 510
Arg Asp Leu Asp Thr Asp Val Val Ile Asp Lys Leu Lys Ser Ala Lys
515 520 525
Thr Ser Ile Asp Ile Glu His Leu Ala Ile Val Pro Thr Thr Arg Val
530 535 540
Asp Gly Asn Ser Tyr Tyr Trp Pro Asp Ile Tyr Asn Ser Ile Ile Glu
545 550 555 560
Ala Ala Ile Asn Arg Gly Val Lys Ile Arg Leu Leu Val Gly Asn Trp
565 570 575
Asp Lys Asn Asp Val Tyr Ser Met Ala Thr Ala Arg Ser Leu Asp Ala
580 585 590
Leu Cys Val Gln Asn Asp Leu Ser Val Lys Val Phe Thr Ile Gln Asn
595 600 605
Asn Thr Lys Leu Leu Ile Val Asp Asp Glu Tyr Val His Ile Thr Ser
610 615 620
Ala Asn Phe Asp Gly Thr His Tyr Gln Asn His Gly Phe Val Ser Phe
625 630 635 640
Asn Ser Ile Asp Lys Gln Leu Val Ser Glu Ala Lys Lys Ile Phe Glu
645 650 655
Arg Asp Trp Val Ser Ser His Ser Lys Ser Leu Lys Ile
660 665
<210> 5
<211> 314
<212> Белок
<213> Искусственная последовательность
<220>
<223> A56R
<400> 5
Met Thr Arg Leu Pro Ile Leu Leu Leu Leu Ile Ser Leu Val Tyr Ala
1 5 10 15
Thr Pro Phe Pro Gln Thr Ser Lys Lys Ile Gly Asp Asp Ala Thr Leu
20 25 30
Ser Cys Asn Arg Asn Asn Thr Asn Asp Tyr Val Val Met Ser Ala Trp
35 40 45
Tyr Lys Glu Pro Asn Ser Ile Ile Leu Leu Ala Ala Lys Ser Asp Val
50 55 60
Leu Tyr Phe Asp Asn Tyr Thr Lys Asp Lys Ile Ser Tyr Asp Ser Pro
65 70 75 80
Tyr Asp Asp Leu Val Thr Thr Ile Thr Ile Lys Ser Leu Thr Ala Arg
85 90 95
Asp Ala Gly Thr Tyr Val Cys Ala Phe Phe Met Thr Ser Thr Thr Asn
100 105 110
Asp Thr Asp Lys Val Asp Tyr Glu Glu Tyr Ser Thr Glu Leu Ile Val
115 120 125
Asn Thr Asp Ser Glu Ser Thr Ile Asp Ile Ile Leu Ser Gly Ser Thr
130 135 140
His Ser Pro Glu Thr Ser Ser Lys Lys Pro Asp Tyr Ile Asp Asn Ser
145 150 155 160
Asn Cys Ser Ser Val Phe Glu Ile Ala Thr Pro Glu Pro Ile Thr Asp
165 170 175
Asn Val Glu Asp His Thr Asp Thr Val Thr Tyr Thr Ser Asp Ser Ile
180 185 190
Asn Thr Val Ser Ala Ser Ser Gly Glu Ser Thr Thr Asp Glu Thr Pro
195 200 205
Glu Pro Ile Thr Asp Lys Glu Asp His Thr Val Thr Asp Thr Val Ser
210 215 220
Tyr Thr Thr Val Ser Thr Ser Ser Gly Ile Val Thr Thr Lys Ser Thr
225 230 235 240
Thr Asp Asp Ala Asp Leu Tyr Asp Thr Tyr Asn Asp Asn Asp Thr Val
245 250 255
Pro Pro Thr Thr Val Gly Gly Ser Thr Thr Ser Ile Ser Asn Tyr Lys
260 265 270
Thr Lys Asp Phe Val Glu Ile Phe Gly Ile Thr Ala Leu Ile Ile Leu
275 280 285
Ser Ala Val Ala Ile Phe Cys Ile Thr Tyr Tyr Ile Tyr Asn Lys Arg
290 295 300
Ser Arg Lys Tyr Lys Thr Glu Asn Lys Val
305 310
<210> 6
<211> 370
<212> Белок
<213> Искусственная последовательность
<220>
<223> Слитый белок FZD-ECD-A56R
<400> 6
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Ala His Ser Phe Gly Asp Glu Glu Glu Arg Arg Cys Asp Pro Ile Arg
20 25 30
Ile Ser Met Cys Gln Asn Leu Gly Tyr Asn Val Thr Lys Met Pro Asn
35 40 45
Leu Val Gly His Glu Leu Gln Thr Asp Ala Glu Leu Gln Leu Thr Thr
50 55 60
Phe Thr Pro Leu Ile Gln Tyr Gly Cys Ser Ser Gln Leu Gln Phe Phe
65 70 75 80
Leu Cys Ser Val Tyr Val Pro Met Cys Thr Glu Lys Ile Asn Ile Pro
85 90 95
Ile Gly Pro Cys Gly Gly Met Cys Leu Ser Val Lys Arg Arg Cys Glu
100 105 110
Pro Val Leu Lys Glu Phe Gly Phe Ala Trp Pro Glu Ser Leu Asn Cys
115 120 125
Ser Lys Phe Pro Pro Gln Asn Asp His Asn His Met Cys Met Glu Gly
130 135 140
Pro Gly Asp Glu Glu Val Pro Leu Pro His Lys Thr Pro Ile Gln Pro
145 150 155 160
Gly Glu Glu Thr Ser Thr Thr Asn Asp Thr Asp Lys Val Asp Tyr Glu
165 170 175
Glu Tyr Ser Thr Glu Leu Ile Val Asn Thr Asp Ser Glu Ser Thr Ile
180 185 190
Asp Ile Ile Leu Ser Gly Ser Thr His Ser Pro Glu Thr Ser Ser Lys
195 200 205
Lys Pro Asp Tyr Ile Asp Asn Ser Asn Cys Ser Ser Val Phe Glu Ile
210 215 220
Ala Thr Pro Glu Pro Ile Thr Asp Asn Val Glu Asp His Thr Asp Thr
225 230 235 240
Val Thr Tyr Thr Ser Asp Ser Ile Asn Thr Val Ser Ala Ser Ser Gly
245 250 255
Glu Ser Thr Thr Asp Glu Thr Pro Glu Pro Ile Thr Asp Lys Glu Asp
260 265 270
His Thr Val Thr Asp Thr Val Ser Tyr Thr Thr Val Ser Thr Ser Ser
275 280 285
Gly Ile Val Thr Thr Lys Ser Thr Thr Asp Asp Ala Asp Leu Tyr Asp
290 295 300
Thr Tyr Asn Asp Asn Asp Thr Val Pro Pro Thr Thr Val Gly Gly Ser
305 310 315 320
Thr Thr Ser Ile Ser Asn Tyr Lys Thr Lys Asp Phe Val Glu Ile Phe
325 330 335
Gly Ile Thr Ala Leu Ile Ile Leu Ser Ala Val Ala Ile Phe Cys Ile
340 345 350
Thr Tyr Tyr Ile Tyr Asn Lys Arg Ser Arg Lys Tyr Lys Thr Glu Asn
355 360 365
Lys Val
370
<210> 7
<211> 855
<212> Белок
<213> Искусственная последовательность
<220>
<223> Слитый белок Her2-A56R
<400> 7
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Ala His Ser Ser Thr Gln Val Cys Thr Gly Thr Asp Met Lys Leu Arg
20 25 30
Leu Pro Ala Ser Pro Glu Thr His Leu Asp Met Leu Arg His Leu Tyr
35 40 45
Gln Gly Cys Gln Val Val Gln Gly Asn Leu Glu Leu Thr Tyr Leu Pro
50 55 60
Thr Asn Ala Ser Leu Ser Phe Leu Gln Asp Ile Gln Glu Val Gln Gly
65 70 75 80
Tyr Val Leu Ile Ala His Asn Gln Val Arg Gln Val Pro Leu Gln Arg
85 90 95
Leu Arg Ile Val Arg Gly Thr Gln Leu Phe Glu Asp Asn Tyr Ala Leu
100 105 110
Ala Val Leu Asp Asn Gly Asp Pro Leu Asn Asn Thr Thr Pro Val Thr
115 120 125
Gly Ala Ser Pro Gly Gly Leu Arg Glu Leu Gln Leu Arg Ser Leu Thr
130 135 140
Glu Ile Leu Lys Gly Gly Val Leu Ile Gln Arg Asn Pro Gln Leu Cys
145 150 155 160
Tyr Gln Asp Thr Ile Leu Trp Lys Asp Ile Phe His Lys Asn Asn Gln
165 170 175
Leu Ala Leu Thr Leu Ile Asp Thr Asn Arg Ser Arg Ala Cys His Pro
180 185 190
Cys Ser Pro Met Cys Lys Gly Ser Arg Cys Trp Gly Glu Ser Ser Glu
195 200 205
Asp Cys Gln Ser Leu Thr Arg Thr Val Cys Ala Gly Gly Cys Ala Arg
210 215 220
Cys Lys Gly Pro Leu Pro Thr Asp Cys Cys His Glu Gln Cys Ala Ala
225 230 235 240
Gly Cys Thr Gly Pro Lys His Ser Asp Cys Leu Ala Cys Leu His Phe
245 250 255
Asn His Ser Gly Ile Cys Glu Leu His Cys Pro Ala Leu Val Thr Tyr
260 265 270
Asn Thr Asp Thr Phe Glu Ser Met Pro Asn Pro Glu Gly Arg Tyr Thr
275 280 285
Phe Gly Ala Ser Cys Val Thr Ala Cys Pro Tyr Asn Tyr Leu Ser Thr
290 295 300
Asp Val Gly Ser Cys Thr Leu Val Cys Pro Leu His Asn Gln Glu Val
305 310 315 320
Thr Ala Glu Asp Gly Thr Gln Arg Cys Glu Lys Cys Ser Lys Pro Cys
325 330 335
Ala Arg Val Cys Tyr Gly Leu Gly Met Glu His Leu Arg Glu Val Arg
340 345 350
Ala Val Thr Ser Ala Asn Ile Gln Glu Phe Ala Gly Cys Lys Lys Ile
355 360 365
Phe Gly Ser Leu Ala Phe Leu Pro Glu Ser Phe Asp Gly Asp Pro Ala
370 375 380
Ser Asn Thr Ala Pro Leu Gln Pro Glu Gln Leu Gln Val Phe Glu Thr
385 390 395 400
Leu Glu Glu Ile Thr Gly Tyr Leu Tyr Ile Ser Ala Trp Pro Asp Ser
405 410 415
Leu Pro Asp Leu Ser Val Phe Gln Asn Leu Gln Val Ile Arg Gly Arg
420 425 430
Ile Leu His Asn Gly Ala Tyr Ser Leu Thr Leu Gln Gly Leu Gly Ile
435 440 445
Ser Trp Leu Gly Leu Arg Ser Leu Arg Glu Leu Gly Ser Gly Leu Ala
450 455 460
Leu Ile His His Asn Thr His Leu Cys Phe Val His Thr Val Pro Trp
465 470 475 480
Asp Gln Leu Phe Arg Asn Pro His Gln Ala Leu Leu His Thr Ala Asn
485 490 495
Arg Pro Glu Asp Glu Cys Val Gly Glu Gly Leu Ala Cys His Gln Leu
500 505 510
Cys Ala Arg Gly His Cys Trp Gly Pro Gly Pro Thr Gln Cys Val Asn
515 520 525
Cys Ser Gln Phe Leu Arg Gly Gln Glu Cys Val Glu Glu Cys Arg Val
530 535 540
Leu Gln Gly Leu Pro Arg Glu Tyr Val Asn Ala Arg His Cys Leu Pro
545 550 555 560
Cys His Pro Glu Cys Gln Pro Gln Asn Gly Ser Val Thr Cys Phe Gly
565 570 575
Pro Glu Ala Asp Gln Cys Val Ala Cys Ala His Tyr Lys Asp Pro Pro
580 585 590
Phe Cys Val Ala Arg Cys Pro Ser Gly Val Lys Pro Asp Leu Ser Tyr
595 600 605
Met Pro Ile Trp Lys Phe Pro Asp Glu Glu Gly Ala Cys Gln Pro Cys
610 615 620
Pro Ile Asn Cys Thr His Ser Cys Val Asp Leu Asp Asp Lys Gly Cys
625 630 635 640
Pro Ala Glu Gln Arg Ala Ser Pro Thr Ser Thr Thr Asn Asp Thr Asp
645 650 655
Lys Val Asp Tyr Glu Glu Tyr Ser Thr Glu Leu Ile Val Asn Thr Asp
660 665 670
Ser Glu Ser Thr Ile Asp Ile Ile Leu Ser Gly Ser Thr His Ser Pro
675 680 685
Glu Thr Ser Ser Lys Lys Pro Asp Tyr Ile Asp Asn Ser Asn Cys Ser
690 695 700
Ser Val Phe Glu Ile Ala Thr Pro Glu Pro Ile Thr Asp Asn Val Glu
705 710 715 720
Asp His Thr Asp Thr Val Thr Tyr Thr Ser Asp Ser Ile Asn Thr Val
725 730 735
Ser Ala Ser Ser Gly Glu Ser Thr Thr Asp Glu Thr Pro Glu Pro Ile
740 745 750
Thr Asp Lys Glu Asp His Thr Val Thr Asp Thr Val Ser Tyr Thr Thr
755 760 765
Val Ser Thr Ser Ser Gly Ile Val Thr Thr Lys Ser Thr Thr Asp Asp
770 775 780
Ala Asp Leu Tyr Asp Thr Tyr Asn Asp Asn Asp Thr Val Pro Pro Thr
785 790 795 800
Thr Val Gly Gly Ser Thr Thr Ser Ile Ser Asn Tyr Lys Thr Lys Asp
805 810 815
Phe Val Glu Ile Phe Gly Ile Thr Ala Leu Ile Ile Leu Ser Ala Val
820 825 830
Ala Ile Phe Cys Ile Thr Tyr Tyr Ile Tyr Asn Lys Arg Ser Arg Lys
835 840 845
Tyr Lys Thr Glu Asn Lys Val
850 855
<210> 8
<211> 935
<212> Белок
<213> Искусственная последовательность
<220>
<223> Слитый белок CD100-A56R
<400> 8
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Ala His Ser Phe Ala Pro Ile Pro Arg Ile Thr Trp Glu His Arg Glu
20 25 30
Val His Leu Val Gln Phe His Glu Pro Asp Ile Tyr Asn Tyr Ser Ala
35 40 45
Leu Leu Leu Ser Glu Asp Lys Asp Thr Leu Tyr Ile Gly Ala Arg Glu
50 55 60
Ala Val Phe Ala Val Asn Ala Leu Asn Ile Ser Glu Lys Gln His Glu
65 70 75 80
Val Tyr Trp Lys Val Ser Glu Asp Lys Lys Ala Lys Cys Ala Glu Lys
85 90 95
Gly Lys Ser Lys Gln Thr Glu Cys Leu Asn Tyr Ile Arg Val Leu Gln
100 105 110
Pro Leu Ser Ala Thr Ser Leu Tyr Val Cys Gly Thr Asn Ala Phe Gln
115 120 125
Pro Ala Cys Asp His Leu Asn Leu Thr Ser Phe Lys Phe Leu Gly Lys
130 135 140
Asn Glu Asp Gly Lys Gly Arg Cys Pro Phe Asp Pro Ala His Ser Tyr
145 150 155 160
Thr Ser Val Met Val Asp Gly Glu Leu Tyr Ser Gly Thr Ser Tyr Asn
165 170 175
Phe Leu Gly Ser Glu Pro Ile Ile Ser Arg Asn Ser Ser His Ser Pro
180 185 190
Leu Arg Thr Glu Tyr Ala Ile Pro Trp Leu Asn Glu Pro Ser Phe Val
195 200 205
Phe Ala Asp Val Ile Arg Lys Ser Pro Asp Ser Pro Asp Gly Glu Asp
210 215 220
Asp Arg Val Tyr Phe Phe Phe Thr Glu Val Ser Val Glu Tyr Glu Phe
225 230 235 240
Val Phe Arg Val Leu Ile Pro Arg Ile Ala Arg Val Cys Lys Gly Asp
245 250 255
Gln Gly Gly Leu Arg Thr Leu Gln Lys Lys Trp Thr Ser Phe Leu Lys
260 265 270
Ala Arg Leu Ile Cys Ser Arg Pro Asp Ser Gly Leu Val Phe Asn Val
275 280 285
Leu Arg Asp Val Phe Val Leu Arg Ser Pro Gly Leu Lys Val Pro Val
290 295 300
Phe Tyr Ala Leu Phe Thr Pro Gln Leu Asn Asn Val Gly Leu Ser Ala
305 310 315 320
Val Cys Ala Tyr Asn Leu Ser Thr Ala Glu Glu Val Phe Ser His Gly
325 330 335
Lys Tyr Met Gln Ser Thr Thr Val Glu Gln Ser His Thr Lys Trp Val
340 345 350
Arg Tyr Asn Gly Pro Val Pro Lys Pro Arg Pro Gly Ala Cys Ile Asp
355 360 365
Ser Glu Ala Arg Ala Ala Asn Tyr Thr Ser Ser Leu Asn Leu Pro Asp
370 375 380
Lys Thr Leu Gln Phe Val Lys Asp His Pro Leu Met Asp Asp Ser Val
385 390 395 400
Thr Pro Ile Asp Asn Arg Pro Arg Leu Ile Lys Lys Asp Val Asn Tyr
405 410 415
Thr Gln Ile Val Val Asp Arg Thr Gln Ala Leu Asp Gly Thr Val Tyr
420 425 430
Asp Val Met Phe Val Ser Thr Asp Arg Gly Ala Leu His Lys Ala Ile
435 440 445
Ser Leu Glu His Ala Val His Ile Ile Glu Glu Thr Gln Leu Phe Gln
450 455 460
Asp Phe Glu Pro Val Gln Thr Leu Leu Leu Ser Ser Lys Lys Gly Asn
465 470 475 480
Arg Phe Val Tyr Ala Gly Ser Asn Ser Gly Val Val Gln Ala Pro Leu
485 490 495
Ala Phe Cys Gly Lys His Gly Thr Cys Glu Asp Cys Val Leu Ala Arg
500 505 510
Asp Pro Tyr Cys Ala Trp Ser Pro Pro Thr Ala Thr Cys Val Ala Leu
515 520 525
His Gln Thr Glu Ser Pro Ser Arg Gly Leu Ile Gln Glu Met Ser Gly
530 535 540
Asp Ala Ser Val Cys Pro Asp Lys Ser Lys Gly Ser Tyr Arg Gln His
545 550 555 560
Phe Phe Lys His Gly Gly Thr Ala Glu Leu Lys Cys Ser Gln Lys Ser
565 570 575
Asn Leu Ala Arg Val Phe Trp Lys Phe Gln Asn Gly Val Leu Lys Ala
580 585 590
Glu Ser Pro Lys Tyr Gly Leu Met Gly Arg Lys Asn Leu Leu Ile Phe
595 600 605
Asn Leu Ser Glu Gly Asp Ser Gly Val Tyr Gln Cys Leu Ser Glu Glu
610 615 620
Arg Val Lys Asn Lys Thr Val Phe Gln Val Val Ala Lys His Val Leu
625 630 635 640
Glu Val Lys Val Val Pro Lys Pro Val Val Ala Pro Thr Leu Ser Val
645 650 655
Val Gln Thr Glu Gly Ser Arg Ile Ala Thr Lys Val Leu Val Ala Ser
660 665 670
Thr Gln Gly Ser Ser Pro Pro Thr Pro Ala Val Gln Ala Thr Ser Ser
675 680 685
Gly Ala Ile Thr Leu Pro Pro Lys Pro Ala Pro Thr Gly Thr Ser Cys
690 695 700
Glu Pro Lys Ile Val Ile Asn Thr Val Pro Gln Leu His Ser Glu Lys
705 710 715 720
Thr Met Tyr Leu Lys Ser Ser Asp Thr Ser Thr Thr Asn Asp Thr Asp
725 730 735
Lys Val Asp Tyr Glu Glu Tyr Ser Thr Glu Leu Ile Val Asn Thr Asp
740 745 750
Ser Glu Ser Thr Ile Asp Ile Ile Leu Ser Gly Ser Thr His Ser Pro
755 760 765
Glu Thr Ser Ser Lys Lys Pro Asp Tyr Ile Asp Asn Ser Asn Cys Ser
770 775 780
Ser Val Phe Glu Ile Ala Thr Pro Glu Pro Ile Thr Asp Asn Val Glu
785 790 795 800
Asp His Thr Asp Thr Val Thr Tyr Thr Ser Asp Ser Ile Asn Thr Val
805 810 815
Ser Ala Ser Ser Gly Glu Ser Thr Thr Asp Glu Thr Pro Glu Pro Ile
820 825 830
Thr Asp Lys Glu Asp His Thr Val Thr Asp Thr Val Ser Tyr Thr Thr
835 840 845
Val Ser Thr Ser Ser Gly Ile Val Thr Thr Lys Ser Thr Thr Asp Asp
850 855 860
Ala Asp Leu Tyr Asp Thr Tyr Asn Asp Asn Asp Thr Val Pro Pro Thr
865 870 875 880
Thr Val Gly Gly Ser Thr Thr Ser Ile Ser Asn Tyr Lys Thr Lys Asp
885 890 895
Phe Val Glu Ile Phe Gly Ile Thr Ala Leu Ile Ile Leu Ser Ala Val
900 905 910
Ala Ile Phe Cys Ile Thr Tyr Tyr Ile Tyr Asn Lys Arg Ser Arg Lys
915 920 925
Tyr Lys Thr Glu Asn Lys Val
930 935
<210> 9
<211> 317
<212> Белок
<213> Искусственная последовательность
<220>
<223> Белок WR B5R
<400> 9
Met Lys Thr Ile Ser Val Val Thr Leu Leu Cys Val Leu Pro Ala Val
1 5 10 15
Val Tyr Ser Thr Cys Thr Val Pro Thr Met Asn Asn Ala Lys Leu Thr
20 25 30
Ser Thr Glu Thr Ser Phe Asn Asp Lys Gln Lys Val Thr Phe Thr Cys
35 40 45
Asp Gln Gly Tyr His Ser Ser Asp Pro Asn Ala Val Cys Glu Thr Asp
50 55 60
Lys Trp Lys Tyr Glu Asn Pro Cys Lys Lys Met Cys Thr Val Ser Asp
65 70 75 80
Tyr Ile Ser Glu Leu Tyr Asn Lys Pro Leu Tyr Glu Val Asn Ser Thr
85 90 95
Met Thr Leu Ser Cys Asn Gly Glu Thr Lys Tyr Phe Arg Cys Glu Glu
100 105 110
Lys Asn Gly Asn Thr Ser Trp Asn Asp Thr Val Thr Cys Pro Asn Ala
115 120 125
Glu Cys Gln Pro Leu Gln Leu Glu His Gly Ser Cys Gln Pro Val Lys
130 135 140
Glu Lys Tyr Ser Phe Gly Glu Tyr Met Thr Ile Asn Cys Asp Val Gly
145 150 155 160
Tyr Glu Val Ile Gly Ala Ser Tyr Ile Ser Cys Thr Ala Asn Ser Trp
165 170 175
Asn Val Ile Pro Ser Cys Gln Gln Lys Cys Asp Met Pro Ser Leu Ser
180 185 190
Asn Gly Leu Ile Ser Gly Ser Thr Phe Ser Ile Gly Gly Val Ile His
195 200 205
Leu Ser Cys Lys Ser Gly Phe Thr Leu Thr Gly Ser Pro Ser Ser Thr
210 215 220
Cys Ile Asp Gly Lys Trp Asn Pro Val Leu Pro Ile Cys Val Arg Thr
225 230 235 240
Asn Glu Glu Phe Asp Pro Val Asp Asp Gly Pro Asp Asp Glu Thr Asp
245 250 255
Leu Ser Lys Leu Ser Lys Asp Val Val Gln Tyr Glu Gln Glu Ile Glu
260 265 270
Ser Leu Glu Ala Thr Tyr His Ile Ile Ile Val Ala Leu Thr Ile Met
275 280 285
Gly Val Ile Phe Leu Ile Ser Val Ile Val Leu Val Cys Ser Cys Asp
290 295 300
Lys Asn Asn Asp Gln Tyr Lys Phe His Lys Leu Leu Pro
305 310 315
<210> 10
<211> 243
<212> Белок
<213> Искусственная последовательность
<220>
<223> FZD-B5R (короткий)
<400> 10
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Ala Tyr Ala Phe Gly Asp Glu Glu Glu Arg Arg Cys Asp Pro Ile Arg
20 25 30
Ile Ser Met Cys Gln Asn Leu Gly Tyr Asn Val Thr Lys Met Pro Asn
35 40 45
Leu Val Gly His Glu Leu Gln Thr Asp Ala Glu Leu Gln Leu Thr Thr
50 55 60
Phe Thr Pro Leu Ile Gln Tyr Gly Cys Ser Ser Gln Leu Gln Phe Phe
65 70 75 80
Leu Cys Ser Val Tyr Val Pro Met Cys Thr Glu Lys Ile Asn Ile Pro
85 90 95
Ile Gly Pro Cys Gly Gly Met Cys Leu Ser Val Lys Arg Arg Cys Glu
100 105 110
Pro Val Leu Lys Glu Phe Gly Phe Ala Trp Pro Glu Ser Leu Asn Cys
115 120 125
Ser Lys Phe Pro Pro Gln Asn Asp His Asn His Met Cys Met Glu Gly
130 135 140
Pro Gly Asp Glu Glu Val Pro Leu Pro His Lys Thr Pro Ile Gln Pro
145 150 155 160
Gly Glu Glu Cys His Ser Val Gly Thr Asn Ser Asp Gln Tyr Ile Trp
165 170 175
Val Lys Arg Ser Leu Asn Cys Val Leu Lys Cys Gly Tyr Asp Ala Gly
180 185 190
Leu Tyr Ser Arg Ser Ala Lys Glu Cys Ala Thr Tyr His Ile Ile Ile
195 200 205
Val Ala Leu Thr Ile Met Gly Val Ile Phe Leu Ile Ser Val Ile Val
210 215 220
Leu Val Cys Ser Cys Asp Lys Asn Asn Asp Gln Tyr Lys Phe His Lys
225 230 235 240
Leu Leu Pro
<210> 11
<211> 281
<212> Белок
<213> Искусственная последовательность
<220>
<223> FZD-B5R (длинный)
<400> 11
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Ala Tyr Ala Phe Gly Asp Glu Glu Glu Arg Arg Cys Asp Pro Ile Arg
20 25 30
Ile Ser Met Cys Gln Asn Leu Gly Tyr Asn Val Thr Lys Met Pro Asn
35 40 45
Leu Val Gly His Glu Leu Gln Thr Asp Ala Glu Leu Gln Leu Thr Thr
50 55 60
Phe Thr Pro Leu Ile Gln Tyr Gly Cys Ser Ser Gln Leu Gln Phe Phe
65 70 75 80
Leu Cys Ser Val Tyr Val Pro Met Cys Thr Glu Lys Ile Asn Ile Pro
85 90 95
Ile Gly Pro Cys Gly Gly Met Cys Leu Ser Val Lys Arg Arg Cys Glu
100 105 110
Pro Val Leu Lys Glu Phe Gly Phe Ala Trp Pro Glu Ser Leu Asn Cys
115 120 125
Ser Lys Phe Pro Pro Gln Asn Asp His Asn His Met Cys Met Glu Gly
130 135 140
Pro Gly Asp Glu Glu Val Pro Leu Pro His Lys Thr Pro Ile Gln Pro
145 150 155 160
Gly Glu Glu Cys His Ser Val Gly Thr Asn Ser Asp Gln Tyr Ile Trp
165 170 175
Val Lys Arg Ser Leu Asn Cys Val Leu Lys Cys Gly Tyr Asp Ala Gly
180 185 190
Leu Tyr Ser Arg Ser Ala Lys Glu Tyr Val Arg Thr Asn Glu Glu Phe
195 200 205
Asp Pro Val Asp Asp Gly Pro Asp Asp Glu Thr Asp Leu Ser Lys Leu
210 215 220
Ser Lys Asp Val Val Gln Tyr Glu Gln Glu Ile Glu Ser Leu Glu Ala
225 230 235 240
Thr Tyr His Ile Ile Ile Val Ala Leu Thr Ile Met Gly Val Ile Phe
245 250 255
Leu Ile Ser Val Ile Val Leu Val Cys Ser Cys Asp Lys Asn Asn Asp
260 265 270
Gln Tyr Lys Phe His Lys Leu Leu Pro
275 280
<210> 12
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> MS4A1S
<400> 12
tataccatgg caacacccag aaattcagta aatg 34
<210> 13
<211> 47
<212> ДНК
<213> Искусственная последовательность
<220>
<223> MS4A1AS
<400> 13
ggtaccgatg caaatggcca cataggagag ctgtcatttt ctattgg 47
<210> 14
<211> 47
<212> ДНК
<213> Искусственная последовательность
<220>
<223> F13S
<400> 14
ccaatagaaa atgacagctc tcctatgtgg ccatttgcat cggtacc 47
<210> 15
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> F13AS
<400> 15
tatacgtacg ttaatggtga tggtgatgat gaatttttaa cgatttactg tg 52
<210> 16
<211> 506
<212> Белок
<213> Искусственная последовательность
<220>
<223> Слитый белок CD20-A56R
<400> 16
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Ala His Thr Glu Leu Ile Val Asn Thr Asp Ser Glu Ser Thr Ile Asp
20 25 30
Ile Ile Leu Ser Gly Ser Thr His Ser Pro Glu Thr Ser Ser Lys Lys
35 40 45
Pro Asp Tyr Ile Asp Asn Ser Asn Cys Ser Ser Val Phe Glu Ile Ala
50 55 60
Thr Pro Glu Pro Ile Thr Asp Asn Val Glu Asp His Thr Asp Thr Val
65 70 75 80
Thr Tyr Thr Ser Asp Ser Ile Asn Thr Val Ser Ala Ser Ser Gly Glu
85 90 95
Ser Thr Thr Asp Glu Thr Pro Glu Pro Ile Thr Asp Lys Glu Asp His
100 105 110
Thr Val Thr Asp Thr Val Ser Tyr Thr Thr Val Ser Thr Ser Ser Gly
115 120 125
Ile Val Thr Thr Lys Ser Thr Thr Asp Asp Ala Asp Leu Tyr Asp Thr
130 135 140
Tyr Asn Asp Asn Asp Thr Val Pro Pro Thr Thr Val Gly Gly Ser Thr
145 150 155 160
Thr Ser Ile Ser Asn Tyr Lys Thr Lys Asp Phe Val Glu Ile Phe Gly
165 170 175
Ile Thr Ala Leu Ile Ile Leu Ser Ala Val Ala Ile Phe Cys Ile Thr
180 185 190
Tyr Tyr Ile Tyr Asn Lys Arg Ser Arg Lys Tyr Lys Thr Glu Asn Lys
195 200 205
Val Met Thr Thr Pro Arg Asn Ser Val Asn Gly Thr Phe Pro Ala Glu
210 215 220
Pro Met Lys Gly Pro Ile Ala Met Gln Ser Gly Pro Lys Pro Leu Phe
225 230 235 240
Arg Arg Met Ser Ser Leu Val Gly Pro Thr Gln Ser Phe Phe Met Arg
245 250 255
Glu Ser Lys Thr Leu Gly Ala Val Gln Ile Met Asn Gly Leu Phe His
260 265 270
Ile Ala Leu Gly Gly Leu Leu Met Ile Pro Ala Gly Ile Tyr Ala Pro
275 280 285
Ile Cys Val Thr Val Trp Tyr Pro Leu Trp Gly Gly Ile Met Tyr Ile
290 295 300
Ile Ser Gly Ser Leu Leu Ala Ala Thr Glu Lys Asn Ser Arg Lys Cys
305 310 315 320
Leu Val Lys Gly Lys Met Ile Met Asn Ser Leu Ser Leu Phe Ala Ala
325 330 335
Ile Ser Gly Met Ile Leu Ser Ile Met Asp Ile Leu Asn Ile Lys Ile
340 345 350
Ser His Phe Leu Lys Met Glu Ser Leu Asn Phe Ile Arg Ala His Thr
355 360 365
Pro Tyr Ile Asn Ile Tyr Asn Cys Glu Pro Ala Asn Pro Ser Glu Lys
370 375 380
Asn Ser Pro Ser Thr Gln Tyr Cys Tyr Ser Ile Gln Ser Leu Phe Leu
385 390 395 400
Gly Ile Leu Ser Val Met Leu Ile Phe Ala Phe Phe Gln Glu Leu Val
405 410 415
Ile Ala Gly Ile Val Glu Asn Glu Trp Lys Arg Thr Cys Ser Arg Pro
420 425 430
Lys Ser Asn Ile Val Leu Leu Ser Ala Glu Glu Lys Lys Glu Gln Thr
435 440 445
Ile Glu Ile Lys Glu Glu Val Val Gly Leu Thr Glu Thr Ser Ser Gln
450 455 460
Pro Lys Asn Glu Glu Asp Ile Glu Ile Ile Pro Ile Gln Glu Glu Glu
465 470 475 480
Glu Glu Glu Thr Glu Thr Asn Phe Pro Glu Pro Pro Gln Asp Gln Glu
485 490 495
Ser Ser Pro Ile Glu Asn Asp Ser Ser Pro
500 505
<210> 17
<211> 235
<212> Белок
<213> Искусственная последовательность
<220>
<223> Слитый белок HA-A56R
<400> 17
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Ala His Ser Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Thr Ser Thr Thr
20 25 30
Asn Asp Thr Asp Lys Val Asp Tyr Glu Glu Tyr Ser Thr Glu Leu Ile
35 40 45
Val Asn Thr Asp Ser Glu Ser Thr Ile Asp Ile Ile Leu Ser Gly Ser
50 55 60
Thr His Ser Pro Glu Thr Ser Ser Lys Lys Pro Asp Tyr Ile Asp Asn
65 70 75 80
Ser Asn Cys Ser Ser Val Phe Glu Ile Ala Thr Pro Glu Pro Ile Thr
85 90 95
Asp Asn Val Glu Asp His Thr Asp Thr Val Thr Tyr Thr Ser Asp Ser
100 105 110
Ile Asn Thr Val Ser Ala Ser Ser Gly Glu Ser Thr Thr Asp Glu Thr
115 120 125
Pro Glu Pro Ile Thr Asp Lys Glu Asp His Thr Val Thr Asp Thr Val
130 135 140
Ser Tyr Thr Thr Val Ser Thr Ser Ser Gly Ile Val Thr Thr Lys Ser
145 150 155 160
Thr Thr Asp Asp Ala Asp Leu Tyr Asp Thr Tyr Asn Asp Asn Asp Thr
165 170 175
Val Pro Pro Thr Thr Val Gly Gly Ser Thr Thr Ser Ile Ser Asn Tyr
180 185 190
Lys Thr Lys Asp Phe Val Glu Ile Phe Gly Ile Thr Ala Leu Ile Ile
195 200 205
Leu Ser Ala Val Ala Ile Phe Cys Ile Thr Tyr Tyr Ile Tyr Asn Lys
210 215 220
Arg Ser Arg Lys Tyr Lys Thr Glu Asn Lys Val
225 230 235
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ПРОМОТОРЫ ДЛЯ УСИЛЕНИЯ ЭКСПРЕССИИ В ПОКСВИРУСАХ | 2016 |
|
RU2753884C2 |
| МУЛЬТИВАЛЕНТНЫЕ PD-L1-СВЯЗЫВАЮЩИЕ СОЕДИНЕНИЯ ДЛЯ ЛЕЧЕНИЯ ЗЛОКАЧЕСТВЕННЫХ НОВООБРАЗОВАНИЙ | 2020 |
|
RU2816646C2 |
| МОДИФИЦИРОВАННЫЕ НЕПРИРОДНЫЕ ЛИГАНДЫ NKG2D, КОТОРЫЕ ИЗБИРАТЕЛЬНО ДОСТАВЛЯЮТ ПРИСОЕДИНЕННЫЕ ГЕТЕРОЛОГИЧНЫЕ МОЛЕКУЛЫ К НЕПРИРОДНЫМ РЕЦЕПТОРАМ NKG2D НА CAR-КЛЕТКАХ | 2020 |
|
RU2823728C2 |
| Вакцина против герпеса | 2019 |
|
RU2731073C1 |
| Конструкции для направленного редактирования генов и способы с их применением | 2020 |
|
RU2832109C2 |
| ЛЕЧЕНИЕ И ПРОФИЛАКТИКА АЛЛЕРГИИ НА КЛЕЩЕЙ ДОМАШНЕЙ ПЫЛИ | 2019 |
|
RU2815386C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО ПРОТИВ ВИРУСА БЕШЕНСТВА И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2764740C1 |
| СЛИТЫЙ ПОЛИПЕПТИД, ВКЛЮЧАЮЩИЙ GDF15 И ПОЛИПЕПТИДНУЮ ОБЛАСТЬ, СПОСОБНУЮ К О-ГЛИКОЗИЛИРОВАНИЮ | 2020 |
|
RU2812049C1 |
| ВАКЦИННАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ РАКА | 2018 |
|
RU2782261C2 |
| АДЕНОВИРУСЫ И СПОСОБЫ ПРИМЕНЕНИЯ АДЕНОВИРУСОВ | 2019 |
|
RU2782528C1 |
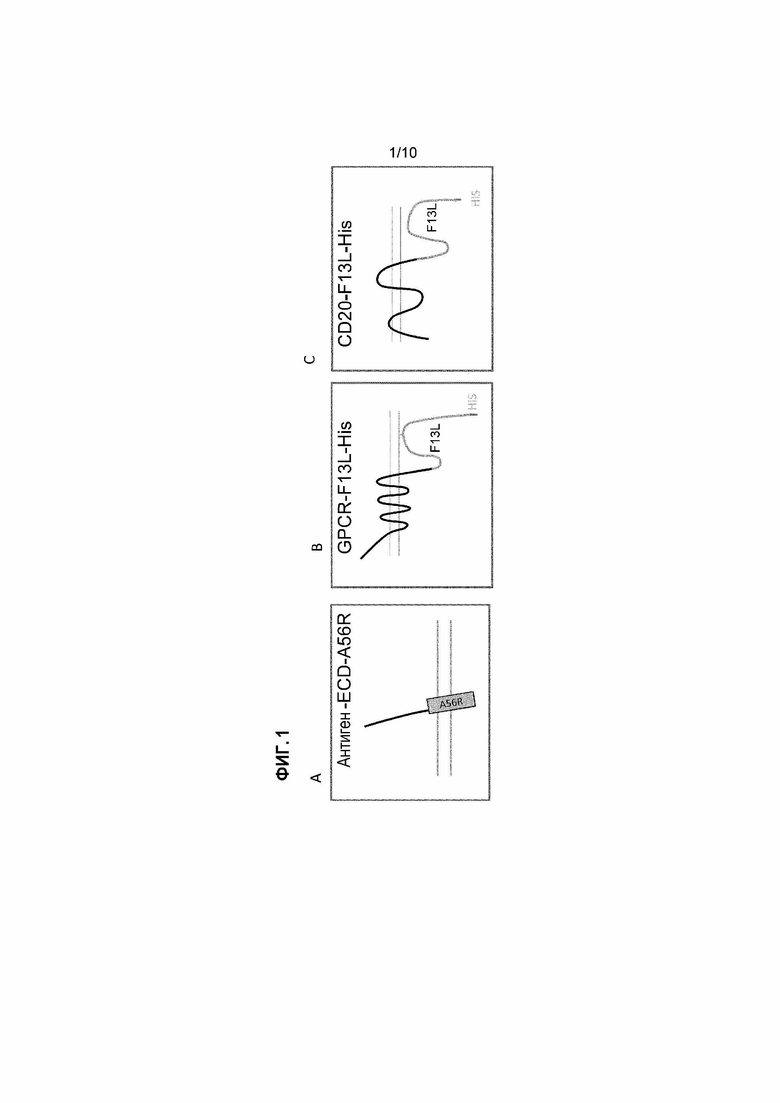


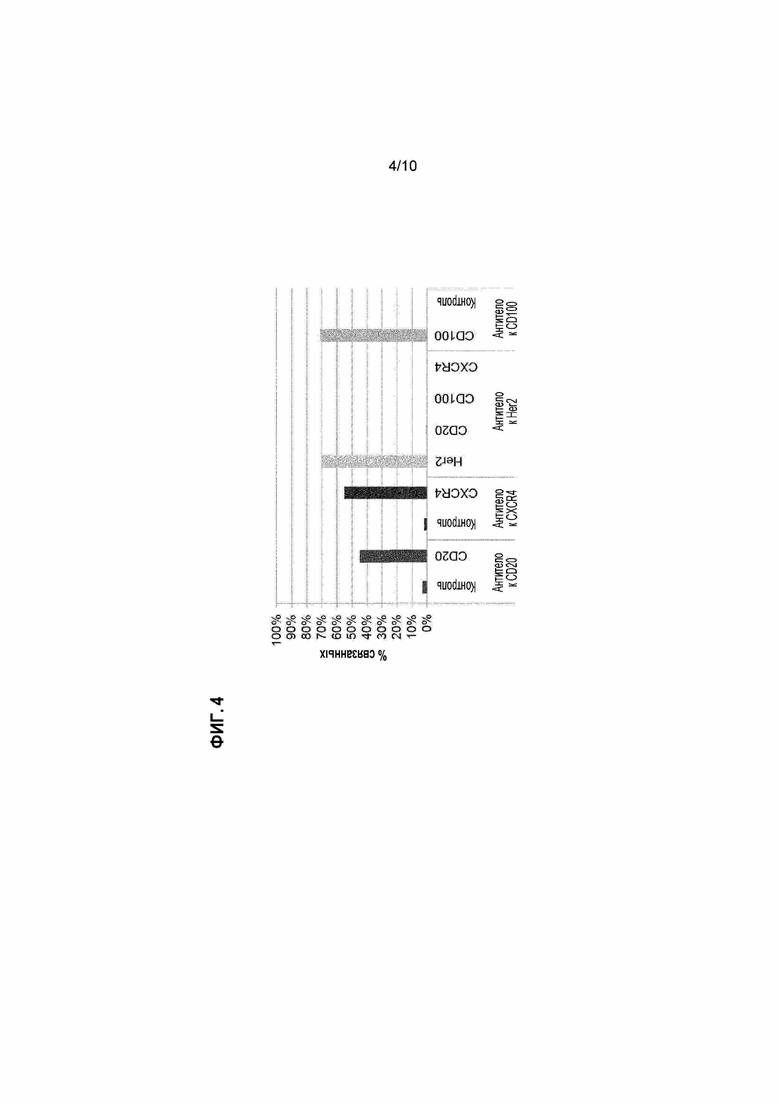
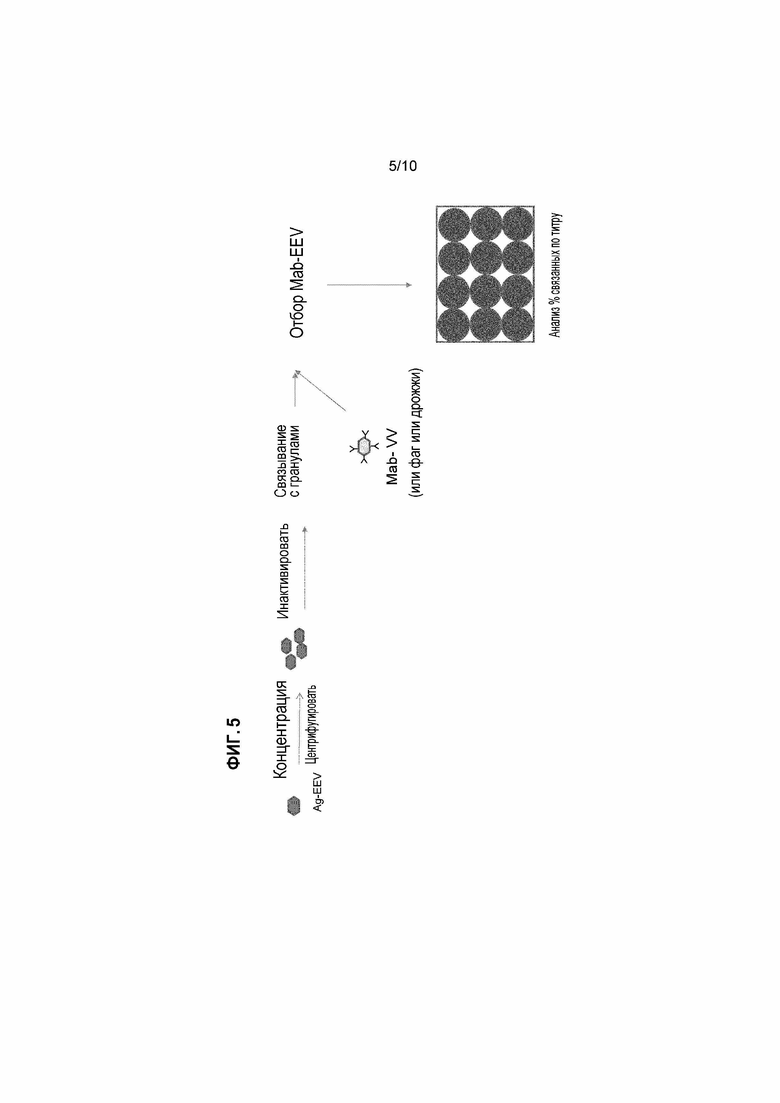

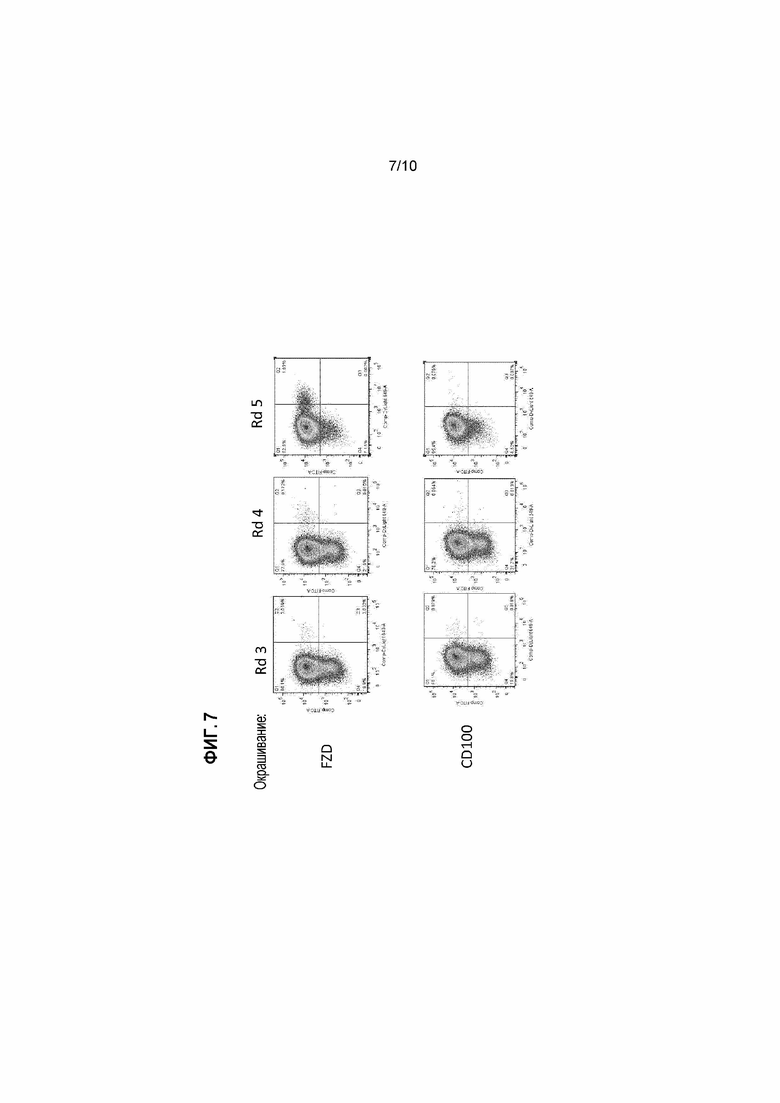



Настоящее изобретение относится к биотехнологии, в частности к композициям и способам экспрессии и получения выделенных интегральных мембранных белков (IMP) или их фрагментов в нативной конформации для применения в скрининге, селекции и идентификации антител или антителоподобных молекул, которые связываются с представляющей интерес IMP-мишенью. 6 н. и 28 з.п. ф-лы, 10 ил., 1 табл., 9 пр.
1. Выделенный полинуклеотид для экспрессии выделенного интегрального мембранного белка-мишени (IMP) или его фрагмента, содержащий:
(a) фрагмент первой нуклеиновой кислоты, кодирующей интегральный мембранный белок-мишень (IMP) или его фрагмент, где IMP-мишень или его фрагмент содержит по меньшей мере одну внемембранную область, по меньшей мере один трансмембранный домен и по меньшей мере одну внутримембранную область и где участок фрагмента первой нуклеиновой кислоты, кодирующей по меньшей мере одну внутримембранную область, располагается на 5'- или 3'-конце фрагмента первой нуклеиновой кислоты; и
(b) фрагмент второй нуклеиновой кислоты, кодирующий белок F13L вируса осповакцины или его функциональный фрагмент, где фрагмент второй нуклеиновой кислоты является слитым в рамке считывания с участком фрагмента первой нуклеиновой кислоты, кодирующей внутримембранную область IMP-мишени;
где инфицированная поксвирусом клетка, содержащая полинуклеотид, может экспрессировать слитый белок IMP-F13L в виде части внешней мембраны оболочки внеклеточного оболочечного вириона (EEV).
2. Полинуклеотид по п.1, где белок F13L или его функциональный фрагмент содержит аминокислотную последовательность SEQ ID NO:1 или его функциональный фрагмент.
3. Полинуклеотид по п.1 или 2, где IMP представляет собой пронизывающий несколько раз мембрану белок, содержащий по меньшей мере два, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять, по меньшей мере шесть или по меньшей мере семь трансмембранных доменов.
4. Полинуклеотид по п.3, где IMP содержит нечетное число трансмембранных доменов, где 5'-конец фрагмента первой нуклеиновой кислоты кодирует внемембранную область, где 3'-конец фрагмента первой нуклеиновой кислоты кодирует внутримембранную область и где 5'-конец второго полинуклеотида является слитым с 3'-концом фрагмента первой нуклеиновой кислоты.
5. Полинуклеотид по п.4, где IMP содержит сопряженный с G-белком рецептор (GPCR).
6. Полинуклеотид по п.5, где IMP представляет собой белок Фрайзлед 4 человека (FZD4) или его фрагмент.
7. Полинуклеотид по п.6, который кодирует полипептид, содержащий аминокислоты от 20 до 892 SEQ ID NO:2.
8. Полинуклеотид по п.7, дополнительно содержащий сигнальный пептид.
9. Полинуклеотид по п.8, где сигнальный пептид содержит аминокислоты от 1 до 19 SEQ ID NO:2.
10. Полинуклеотид по п.5, где IMP представляет собой рецептор хемокинов CXC.
11. Полинуклеотид по п.10, где рецептор хемокинов CXC представляет собой CXCR4 или его фрагмент.
12. Полинуклеотид по п.11, который кодирует полипептид, содержащий аминокислотную последовательность SEQ ID NO:3.
13. Полинуклеотид по п.3, где IMP содержит четное число трансмембранных доменов и где 5'- и 3'-конец фрагмента первой нуклеиновой кислоты кодируют внутримембранные области.
14. Полинуклеотид по п.13, где фрагмент второй нуклеиновой кислоты является слитым с 3'-концом фрагмента первой нуклеиновой кислоты.
15. Полинуклеотид по п.14, где IMP представляет собой белок CD20 человека или его фрагмент.
16. Полинуклеотид по п.15, который кодирует белок, содержащий аминокислотную последовательность SEQ ID NO:4.
17. Полинуклеотид по любому из пп.1-16, где первый и второй фрагменты нуклеиновой кислоты являются непосредственно слитыми.
18. Полинуклеотид по любому из пп.1-17, дополнительно содержащий третий фрагмент нуклеиновой кислоты, кодирующий гетерологичный пептид.
19. Полинуклеотид по п.18, где гетерологичный пептид содержит линкерную последовательность, аминокислотную метку или пептидную или полипептидную последовательность, которая облегчает очистку.
20. Полинуклеотид по п.19, где гетерологичный пептид содержит гистидиновую метку.
21. Полинуклеотид по любому из пп.1-20, который является функционально связанным с промотором поксвируса.
22. Полинуклеотид по п.21, где промотор поксвируса представляет собой p7.5, или H5, или T7.
23. Слитый белок F13L, кодируемый полинуклеотидом по любому из пп. 1-22, для скрининга, селекции и идентификации антител, которые связываются с IMP-мишенью.
24. Геном поксвируса, содержащий полинуклеотид по любому из пп.1-22, для получения EEV, которые представляют слитый белок IMP-F13L в виде части мембраны внешней оболочки EEV.
25. Геном поксвируса по п.24, который представляет собой геном вируса осповакцины.
26. Рекомбинантный EEV вируса осповакцины для представления слитого белка IMP-F13L в виде части мембраны внешней оболочки EEV, содержащий геном поксвируса по п.25.
27. Способ получения рекомбинантного EEV вируса осповакцины по п.26, включающий:
(a) инфицирование клетки-хозяина, чувствительной к инфекции вирусом осповакцины, вирусом осповакцины, содержащим геном поксвируса по п.24, и
(b) выделение EEV, высвобождаемого клеткой-хозяином.
28. Способ селекции антител, которые связываются с пронизывающим несколько раз мембрану белком, включающий:
(a) связывание рекомбинантного EEV по п.26 с твердой подложкой;
(b) получение библиотеки дисплея антител, где библиотека содержит пакеты дисплея, предоставляющие совокупность антигенсвязывающих доменов;
(c) приведение библиотеки дисплея в контакт с EEV таким образом, что пакеты дисплея, предоставляющие антигенсвязывающие домены, которые специфически связываются с IMP, экспрессируемым на EEV, могут его связывать;
(d) удаление несвязанных пакетов дисплея и
(e) выделение пакетов дисплея, которые предоставляют антигенсвязывающий домен, специфический к IMP, экспрессируемому на EEV.
29. Способ по п.28, где рекомбинантный EEV инактивируют перед связыванием с твердой подложкой.
30. Способ по п.29, где EEV инактивируют инкубацией с просаленом (триоксаленом, 4'-аминометилгидрохлоридом) в присутствии УФ-облучения.
31. Способ по любому из пп.28-30, где EEV связывают с твердой поверхностью с помощью реакции с тозильными группами, связанными с поверхностью.
32. Способ по п.31, где твердая поверхность представляет собой активированные тозилом магнитные гранулы.
33. Способ по любому из пп.28-30, где EEV подвергают битионилированию и связывают с покрытой стрептавидином твердой поверхностью.
34. Способ по п.33, где твердая поверхность представляет собой покрытые стрептавидином магнитные гранулы.
| WO 2015193143 A1, 23.12.2015 | |||
| US 2004014033 A1, 22.01.2004 | |||
| WO 2013163602 A1, 31.10.2013 | |||
| RU 2441071 C2, 27.01.2012. |

