ПЕРЕКРЕСТНЫЕ ССЫЛКИ НА РОДСТВЕННЫЕ ЗАЯВКИ
Настоящая заявка испрашивает приоритет на основании предварительной заявки на патент США №62/387375, поданной 24 декабря 2015 года, содержание которой полностью включено в настоящее описание посредством ссылки.
УРОВЕНЬ ТЕХНИКИ
В настоящее время доступны многие устойчивые высокопродуктивные линии клеток млекопитающих, применяемых при производстве терапевтических лекарственных средств в виде рекомбинантных белков. Были осуществлены улучшение и оптимизация процессов с целью сокращения времени для получения линии клеток с 8-12 месяцев до 4 месяцев с одновременным улучшением продуктивности и стабильности продуцирующих белки линий клеток млекопитающих.
Тем не менее, остается потребность в создании линий клеток млекопитающих с лучшими профилями безопасности.
В частности, существует общая потребность в разработке более эффективных процедур для анализа и описания характеристик линий клеток млекопитающих, продуцирующих рекомбинантные белки, в целях увеличения продуктивности, скорости и эффективности разработки кандидатов на роль лекарственных средств, а также снижения затрат на проведение валидации в регуляторных целях, а также как и рисков непредвиденных и нежелательных воздействий на белковые терапевтические средства. Кроме того, риск закрытия производственного участка по причине необнаруженных загрязняющих агентов линий клеток (например, микоплазмы или адвентивных вирусных агентов) можно было бы уменьшить путем внедрения более эффективного процесса определения клеток в рамках рабочих процедур.
Все охарактеризованные линии клеток и организмы млекопитающих содержат остатки возбудителей ранее перенесенных вирусных инфекций, включая ретровирусную инфекцию, в результате чего вирусные гены были интегрированы в клеточный геном. Такие оставшиеся вирусные последовательности часто повреждаются и мутируют, но некоторые из них, тем не менее, продолжают экспрессироваться и приводят к образованию вирусоподобных частиц, которые можно обнаружить, например, методом электронной микроскопии.
Наиболее широко используются клетки яичников китайского хомячка (СНО), поскольку они могут представлять собой стабильных хозяев для экспрессии гетерологичных генов, их безопасность подтверждена на протяжении длительного периода времени, они продуцируют белки, которые несут человекоподобную посттрансляционную модификацию и их относительно просто адаптировать для независимого от адгезии и быстрого роста в бессывороточной синтетической среде в биореакторах. Тем не менее, процесс разработки и документирования высокоэффективных клональных линий клеток для культивирования в биореакторах зачастую является долгим и трудоемким, и в нем до сих пор имеются «узкие» места и присутствует низкий выход белка.
Клетки СНО считаются безопасными для получения терапевтического белка по сравнению с другими линиями клеток. Тем не менее, они также содержат остатки геномов инфекционных вирусных частиц, для которых необходимо наличие нормативной документации, поскольку такие остатки приводят к высвобождению вирусных частиц клетками СНО (Lie, Y.S. et al., 1994). Кроме того, интегрированные провирусные геномы теоретически могут подвергаться рекомбинации, мутациям и тем самым изменять свойства, хотя и с низкой частотой.
Очень желательны клетки СНО, в геноме которых снижено содержание экспрессируемых остаточных вирусных последовательностей, что таким образом обеспечивает более безопасную клетку-хозяина млекопитающего для получения терапевтического белка.
Недавняя разработка методов на основе высокопроизводительного секвенирования ДНК (секвенирование ДНК следующего поколения или NGS) для характеристики генома клеток, применяемых для получения белков (например, клеток СНО), привела к успешному описанию сайтов интеграции трансгенов (Kostyrko et al, 2016).
Содержание публикаций и других материалы, включая патенты и патентные заявки, упоминаемых в настоящем документе для иллюстрации настоящего изобретения и, в частности, для предоставления дополнительных сведений, относящихся к практической реализации, включено посредством ссылки. Для удобства указанные публикации упоминаются в последующем тексте в виде ссылки на автора и дату и перечислены в алфавитном порядке по автору в прилагаемой библиографии.
С учетом продолжающегося обнаружения адвентивных микроорганизмов в клетках, применяемых на площадках по производству терапевтических средств, растет давление со стороны регулирующих органов, таких как Управление по контролю за пищевыми продуктами и лекарственными средствами США (FDA), в части применения производственных линий клеток, генетические характеристики которых лучшим образом описаны. FDA приводит определение адвентивных микроорганизмов, куда включены ретровирусы, интегрированные в геном клетки (см. Руководство FDA по производственной характеристике и квалификации клеточных субстратов и других биологических материалов, применяемых при производстве вирусных вакцин для инфекционных заболеваний, февраль 2010 г.). Тем не менее, в настоящее время зачастую отсутствуют аналитические и биоинформационные инструменты, необходимые для полной сборки генома различных сублиний клеток СНО, а также для анализа и интерпретации геномных последовательностей.
ОПИСАНИЕ ЧЕРТЕЖЕЙ
На Фигурах 1а и 1b показан процесс образования эндогенного ретровируса (ERV) (а) и его выход после его нахождения в спящем состоянии в геноме хозяина (b). Размножение указанного ретровируса требует интеграции генов, кодирующих обратную транскриптазу и другие компоненты вирусного цикла. ERV-вирусы обнаруживают почти исключительно у представителей царства животных.
На Фигуре 2А показаны компоненты комплекса ESCRT (необходимый для транспортировки эндосомный комплекс сортировки), который содержит комплексы цитозольных белков, известные как ESCRT-0, ESCRT-I, ESCRT-II и ESCRT-III, и ряд вспомогательных белков, включая ALIX, и взаимодействие вирусного белка р6 с таким комплексом приводит к высвобождению вирусных везикул.
На Фигуре 2В показаны различные аминокислотные мотивы ERV, способствующие почкованию («мотивы для почкования»), которые были определены при помощи скрытого моделирования Маркова (НММ) из открытой рамки считывания (ОРС) gag (группового специфичного антигена).
На Фигуре 3 показаны способы определения мотивов для почкования (см. Фигуру 1С) при помощи НММ (скрытая модель Маркова) в рр12, кодируемом участком gag. На основе мотива PPXY, который, как известно, часто характерным для гаммаретровирусов, определяли перекрывающийся мотив PPYP Группы 1 и Группы 2, содержащих ERV СНО, который и служил мишенью для мутагенеза. Также показаны номера первой пары оснований, сравниваемых с перечисленными последовательностями.
На Фигуре 4А схематически показано, как ERV-элементы можно обнаружить в геноме клетки на основе сравнения последовательности клеточного генома полученной из клеток СНО-K1 линии клеток, называемых СНО-М (получена с применением технологии секвенирования и сборки генома PacBio®) с последовательностями обычных ретровирусных элементов (gag, pol, env, LTR (длинные концевые повторы)). Для генома описанных авторами настоящего изобретения клеток СНО-М, полученных с применением технологии PacBio®, было получено 77 совпадений. Совпадения отбирали для дальнейшего исследования и определения консенсусных последовательностей.
На Фигуре 4В показаны различные этапы дальнейшего определения и анализа указанных консенсусных последовательностей и сравнения с опубликованным геномом китайского хомячка.
На Фигуре 4С показано экспериментальное подтверждение присутствия специфических ERV-подобных последовательностей гаммаретровируса, идентифицированных в геноме клеток СНО-М. Подтверждение ERV проводилась с применением праймеров для ПЦР, гибридизующихся с геномной последовательностью СНО с обоих сторон прогнозируемого интегрированного вирусного генома (подтверждение последовательностей гаммаретровируса типа С, обнаруженных с применением биоинформационного анализа). На геле более последовательности большего размера (три верхних круга) представляют собой аллели с интегрированным геномом, тогда как аллели без вирусной интеграции движутся быстрее на геле (три нижних круга).
На Фигуре 5 показано, каким образом можно определить потенциально опасные ERV т.е. ERV, которые могут «проснуться», например, в случае клеточного/геномного стресса. Такой стресс можно применять в разное время, в разных типах культур и/или после нескольких трансфекций. Эпигенетические данные позволяют определить, транскрибируется ли ERV-элемент (эпигенетические данные, включая оценку иРНК и статус метилирования) или нет. В эти периоды стресса инфекционные ERV чаще пробуждаются, и их можно обнаружить.
На Фигуре 6 показаны CRISPR/Cas9-системы, в которых применяется гидовая РНК (также называемая одиночная гидовая РНК или огРНК): (а) системы, предложенные Carrol et al. 2013 и (b) системы, предложенные Guilinger et al. 2014, в которых нацеливание на ген (GT) запускается посредством двухцепочечных разрывов (DSB), демонстрирует, что нуклеазы облегчают GT. Сайт-направленные DSB увеличивают GT вплоть до 10000 раз, но они также могут запускать нежелательные нецелевые мутации. Активность нуклеазы варьируется между 10-80%.
На Фигуре 7 показаны три основных пути репарации, конкурирующих за репарацию DSB, а именно: негомологичное соединение концов (NHEJ), опосредованное микрогомологией соединение концов (MMEJ) и гомологичная рекомбинация (HR). Путь репарации влияет на результат редактирования генома, который может привести к делециям, генной конверсии или к эффективному восстановлению исходной последовательности.
На Фигуре 8 показано восстановление функциональной GFP-кодирующей последовательности из донорской плазмиды, кодирующей часть GFP в геноме клеток СНО DG44, с применением CRISPR/Cas9-системы. Результаты показаны для CRISPR/Cas 9 + донор (а), только для CRISPR/Cas 9 (b) и только для донора (с). Можно заметить, что для восстановления GFP требуется CRISPR/Cas 9, а также донорская плазмида (репарация примерно 0,4% GFP происходит в трансфицированных клетках в (а)). В системе DG44 отсутствует измеримый сигнал GFP, в случае присутствия только CRISPR/Cas9-системы или только донорской плазмиды.
На Фигуре 9 показан процент генной коррекции посредством донорской плазмиды, показанной на Фигуре 8 (клетки СНО DG44), в зависимости от подавления экспрессии (сайленсинга)/нокдауна (посредством миРНК) генов, влияющих на различные пути репарции. Сайленсинг генов в пути NHEJ увеличивает процент коррекции (20% увеличение при сайленсинге/нокдауне лигазы 4 и XRCC4). Сайленсинг/нокдаун генов в HDR (пути репарации, направляемые гомологией, HR и MMEJ) в основном уменьшают процент коррекции. Сайленсинг определенных генов в HDR увеличивает процент коррекции (Nbs4, часть MRN-комплекса). (Ось X показывает различные участвующие в сайленсинге РНК, порядок применения которых зависел от пути репарации, * указывает на значимые различия).
На Фигуре 10 показано восстановление функциональной GFP-кодирующей последовательности из донорской плазмиды, кодирующей часть GFP в геноме клеток СНО-М (СНО-K1), с применением CRISPR/Cas9-системы. Трансфекция компонентов CRISPR/Cas9, а также донорской плазмиды, приводит к репарации 0,7% GFP. Таким образом, клетки СНО-М менее подвержены репарации, чем клетки DG44.
На Фигуре 11 показан процент интеграции донорской плазмиды, показанной на Фиг. 10 (клетки СНО-K1), в зависимости от сайленсинга/нокдауна (посредством миРНК) генов, влияющих на различные пути репарации. Сайленсинг генов HDR (пути репарации, направляемые гомологией, HR и MMEJ) уменьшал процент генной коррекции. На Фигуре также показана высокая степень воспроизводимости результатов, полученных из исследований 3 клонов. Сайленсинг Rad51 (через siRad51) и, таким образом, замедление/остановка HR пути репарации приводит к значимому уменьшению количества GFP-положительных клеток во всех трех клонах, в то время как замедление/остановка пути NHEJ через siXrcc4 практически не влияет на процентили GFP-положительных клеток.
На Фигуре 12А показано увеличение или уменьшение генной коррекции из Фиг. 10 (клетки СНО-K1) в зависимости от сайленсинга (посредством миРНК) генов, влияющих на различные пути репарации. Сайленсинг генов при HR (гомологичная репарация) частично уменьшает процент коррекции (сайленсинг/нокдаун Rad 51) и частично ее увеличивает (сайленсинг/нокдаун Mre11). (Ось X показывает различные участвующие в сайленсинге РНК, порядок применения которых зависел от пути репарации, * указывает на значимые различия).
На Фигуре 12В показано, что увеличение экспрессии ограничивающих скорость HR и/или MMEJ белков стимулирует генную коррекцию зависимым от дозы образом. Как можно видеть, пустой вектор обеспечивает базовый уровень 1, при этом определенные дозы векторов экспрессии, обеспечивающих ограничивающие скорость белки, показывают увеличение (повышающим образом регулируют соответствующий ген) генной коррекции более, чем в 0,05 раз, более, чем в 0,1 раз, в 0,2 раза, 0,3 раза, 0,4 раза или 0,5 раз.
На Фигуре 13 показаны три класса гаммаретровирусов, которые могут быть интегрированы в геном клеток с образованием родственных гаммаретровирусу ERV. На фигуре показаны вирусы в каждом классе (I-III), которые присутствуют и активны, что было обнаружено при анализе генома СНО, полученного методом секвенирования PACBIO. В результате поиска последовательностей GAL, POL, ENV, LTR (80% покрытия, 80% идентичность последовательности): было обнаружено 159 IAP-последовательностей (интрацистернальные частицы А-типа) и 144 ERV-подобных последовательностей мышей типа С (оригинально (boldly) закрученных), а также 6 последовательностей, родственных GALV (вирус лейкоза гиббонов).
На Фигуре 14 показано построение консенсусного дерева методом ближайших соседей, основанного на последовательно связанных 77 последовательностях GAG/POL/ENV подобных гаммаретровирусу ERV, полученных из генома СНО. Группы 1 и 2 различаются, как это описано в другом месте настоящего документа.
На Фигуре 15 показано выравнивание GAG РНК, полученной из выбранных транскриптов подобных гаммаретровирусу ERV, охарактеризованных для клеток СНО-М. Знаки под выровненными последовательностями представляют мутации в открытой рамке считывания GAG. Обе группы хорошо сохранились и представляют собой недавние интеграции в эволюции китайского хомячка.
На Фигуре 16 показан статус метилирования ДНК CpG специфичных ERV-элементов, идентифицированных в клетках СНО-М. 5'-LTR ERV являются гипометилированными в данных конкретных примерах, что свидетельствует о том, что они соответствуют транскрибируемым ERV.
На Фигуре 17 показано построение консенсусного дерева методом ближайших соседей, основанного на 121 последовательности GAG подобного гаммаретровирусу ERV, полученных из генома СНО. Обе группы 1 и 2 содержат транскрипционно активные ERV. Было обнаружено, что окруженная рамкой последовательность в группе 2 является активной, но содержит стоп-кодоны, было обнаружено, что множество помещенных в рамку последовательностей в группе 1 являются активными и не содержат стоп-кодон в кодирующей последовательности. Анализ кДНК Gag и Pol согласовывался с присутствием функциональных ERV. Потенциально экспрессируемые ERV выделены черным прямоугольником. На основе этих последовательностей была определена консенсусная последовательность вирусов группы 1 для процессинга CRISPR/Cas9.
На Фигуре 18 показана стратегия, применяемая для CRISPR/Cas9-опосредованной инактивации ERV последовательностей, представленных на Фигуре 17, либо мотивом PPYP, либо мотивом миристоилирования в качестве мишени (оба препятствуют почкованию ERV). Стрелки показывают, где были сделаны двухцепочечные разрывы. Для модифицирования огРНК см. Материалы и Методы.
На Фигуре 19 показаны схемы двух экспериментов для определения инактивации ERV, один без ингибирования и один с ингибированием путей репарации HDR, в данном случае - посредством миРНК Rad51. Разрушение ERV подтверждают при помощи мРНК (например, кДНК), которую анализируют методом ПЦР с анализом по конечной точке или путем секвенирования для определения рамки считывания.
На Фигуре 20 показано влияние трансфекции трех отдельных, нацеливающихся на РР51 миРНК на уровень мРНК РР51 и активность репарации посредством гомологичной рекомбинации по сравнению с уровнями в клетках, обработанных неспецифическими миРНК, представляющими собой отрицательный контроль. Квадрат: влияние трансфекции смеси из (siRad51 3х) трех нацеливающихся на РР51 миРНК на уровень мРНК Rad51 (ось х) и на активность гомологичной рекомбинационной репарации (ось y) относительно уровней для клеток, обработанных неспецифическими миРНК, представляющими собой отрицательный контроль, которые приняты за значение 1. Закрашенные круги: аналогичные анализы проводили для каждой из трех миРНК, применяемой по отдельности (siRad51_1, siRad51_2, siRad51_3), для демонстрации аналогичного воздействия на ингибирование Rad51 и HR смесью из трех миРНК. Как видно, миРНК Radi являются эффективными и специфичными.
На Фигуре 21 показано влияние CRISPR-Cas9 в случае его применения в сочетании со специфичными огРНК (sgRNA) в ПЦР-анализе на кДНК-матрице, который показывает пониженную экспрессию ERV с применением ERV-специфичных праймеров: первый и второй эксперименты с применением CRISPR показывают воспроизводимость, а третий показывает влияние дополнительного ингибирования, опосредованного миРНК RAD51, перед обработкой CRISPR-Cas9. Наблюдали сильные потери сигнала мРНК ERV в присутствии огРНК PPYP5, а также в случае нокаута RAD51 (верхний гель). На графике ниже: сверху показана консенсусная последовательность ERV группы 1 (SEQ ID NO: 30), контрольная (необработанные клетки) последовательность мотива PPYP группы 1 показана посередине (SEQ ID NO: 31) и внизу показан пример делении вне рамки считывания, полученный из клеток, обработанных огРНК PPYP7 (SEQ ID NO: 32).
На Фигуре 22 показаны данные электрофореза для секвенирования амплифицированных последовательностей GAG ERV, для необработанных СНО-клеток (СНО WT) или из клеток СНО, обработанных Cas9-нуклеазой, кодируемой огРНК PPGP6, полученные по результатам первого эксперимента с применением CRISPR (образец 1) и второго эксперимента с применением CRISPR (образец 2), как это показано на Фиг. 21. Сайт расщепления Cas9 обозначен вертикальной линией, после которой для обработанных клеток наблюдали смесь последовательностей, что говорит о том, что 50% или более последовательностей GAG подверглись мутациям.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение в одном варианте реализации относится к модифицированной клетке, предпочтительно линии клеток млекопитающего, как, например, модифицированной клетке СНО, включая модифицированную СНО-K1, такую как клетку СНО-М, содержащей:
геном клетки, при этом указанный геном содержит по меньшей мере одно изменение, включающее:
- делеции,
- добавления/интеграции и/или
- замены
одной или более нуклеиновых кислот в одном или более, в целом, более, чем в 10, 20, 30, 40, 50, 60, 80, 90 или 100 эндогенных ретровирусных (ERV) элементах, которые являются частью указанного генома.
ERV-элементы могут происходить из гаммаретровирусных ERV, включая ERV вируса эпидемии коал (KoRV), ERV вируса опухоли молочной железы у мышей (MMTV), ERV вируса лейкоза мышей (MLV). По меньшей мере одно изменение может предпочтительно подавлять или исключать высвобождение, или его можно адаптировать для подавления или исключения, по меньшей мере одного или более, предпочтительно более, чем 60%, 70%, 80%, 90%, 95% или 100% указанных ERV.
По меньшей мере один измененный (ERV) элемент может представлять собой или может происходить из гена gag, pol и/или env, предпочтительно из указанного гена gag, кодирующего МА (белок ядерного матрикса), СА (белок капсида), NC (белок нуклеокапсида), белки, кодирующие дополнительный домен, такие как рр12 или р6, и/или он может представлять собой длинные концевые повторы (LTR) ERV.
По меньшей мере один ERV-элемент может, в частности, кодировать белок Gag (групповой антиген), белок Pol (обратная транскриптаза) и/или белок Env (оболочки).
Добавления/вставки, делеции или замены могут содержать более, чем 2, 3, 4, 5, 10, 15, 20, 25, 30, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 150 или 200 нуклеиновых кислот.
Эти изменения могут быть направлены на один или более (ERV) элементов, которые можгут представлять собой или происходить по меньшей мере из одного гена gag, pol и/или env, предпочтительно гена gag, и последовательности указанного гена могут подвергаться направленным интеграциям, например, трансгена, кодирующего маркерный белок, такой как GFP (зеленый флуоресцентный белок), или аминокислотным заменам, приводящим к доминантно-негативному фенотипу.
ERV-элементы могут происходить из гена gag, и по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более или 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 98%, 99% элеиенитов или все указанные элементы могут содержать указанные изменения.
Изменения могут содержаться в консенсусных последовательностях EVR-элементов, и длина указанной консенсусной последовательности может составлять по меньшей мере 5, 10, 15, 20, 25, 30, 35, 40, 45, 50 пар оснований, предпочтительно длина составляет 15-25 или 30-50 пар оснований (см. Фиг. 21В).
Модифицированная клетка может не высвобождать или по существу не высвобождать ERV.
Клетка может также содержать гетерологичные последовательности нуклеиновых кислот, кодирующие, ингибирующие или активирующие по меньшей мере один белок пути рекомбинации, и/или гетерологичные последовательности нуклеиновых кислот, кодирующие по меньшей мере одну последовательность или белок, подавляющий экспрессию по меньшей мере одного белка пути рекомбинации, предпочтительно Nbs1, Mre11, Rad51, лигазы 1 и/или лигазы 3.
Гетерологичные последовательности нуклеиновых кислот могут присутствовать в виде вектора, или могут экспрессироваться вектором, который предпочтительно не интегрирован в геном указанной клетки.
Модифицированная клетка может также дополнительно содержать гетерологичную донорную ДНК, предпочтительно на векторе, кодирующую по меньшей мере один маркерный белок, такой как GFP (зеленый флуоресцентный белок).
Модифицированная клетка может дополнительно содержать трансген. Указанный трансген может представлять собой маркерный ген, кодирующий маркерный белок, такой как GFP, и/или белок, обладающий терапевтической активностью, который может предпочтительно экспрессироваться с вектора, интегрированного в геном.
Указанные одно или более изменений могу представлять собой или содержать замену по меньшей мере одной миристоилированной аминокислоты в мотиве миристоилирования по меньшей мере в одном ERV-элементе, как, например, гене gag или ERV-элементе, который происходит из гена gag, на немиристоилированную аминокислоту.
Для этой цели клетку модифицируют для повышения экспрессии HR/MMEJ (см. Фигуры 12А и 12В) путем обеспечения/экспрессии в указанной клетке, по меньшей мере транзиентно (временным образом), последовательностей, например, в форме транзиентно экспрессируемого вектора, кодирующих белки HR и/или MMEJ, такие как Pad51, Lig3, Ercc1, Pold3; и/или последовательностей, как, например, последовательностей миРНК, таких как миРНК MRE11, или векторов, кодирующих такие последовательности.
ERV-элемент(-ы), такой как ген gag или из гена gag ERV-элемент, может содержать мотив PPYP, и (i) последовательности, кодирующие указанный мотив PPYP, и/или последовательности, содержащие до 20 или 30 нуклеиновых кислот, фланкирующих последовательности в (i), могут содержать изменение.
Модифицированная клетка может представлять собой клетку СНО и может содержать по меньшей мере одну делецию, добавление и/или замену по меньшей мере в одной последовательности, соответствующей SEQ ID NOS: 1, 2, 3, 4, или в последовательностях, которая более, чем на 90% или 95% идентична таким последовательностям, предпочтительно в границах ERV-элементов указанных последовательностей.
Модифицированная клетка может представлять собой клетку СНО и может содержать по меньшей мере одну делецию, добавление и/или замену в консенсусной последовательности ERV группы 1, такой как SEQ ID NO: 30, или в последовательностях, которая более, чем на 90% идентична такой последовательности, и при этом указанные последовательности, содержащие делеции, добавления и/или замены, больше не кодируют функциональный белок Gag.
Настоящее изобретение также относится к способу улучшенного редактирования генома, который включает:
(i) обеспечение или введение не встречающейся в природе/гетерологичной системы для создания одно- или двухцепочечных разрывов в целевой нуклеотидной последовательности клетки, предпочтительно представляющей интерес клетки, такой как консенсусная последовательность ERV-элемента, предпочтительно ERV-элемента, который представляет собой или который происходит из гена gag, и дополнительно
(ii) обеспечение или введение в указанную клетку
(a) гетерологичных нуклеотидных последовательностей, кодирующих или активирующих по меньшей мере один белок по меньшей мере одного пути рекомбинации, и/или
(b) гетерологичных последовательностей, кодирующих по меньшей мере одну последовательность, такую как, например, миРНК, или белок, которые подавляют/снижают экспрессию по меньшей мере одного белка по меньшей мере одного пути рекомбинации, при этом указанная гетерологичная система (i) и/или последовательности (ii) (а) и/или (ii) (b) предпочтительно транзиентно экспрессируются в указанной представляющей интерес клетке.
Гетерологичные последовательности могут кодировать, активировать или инактивировать по меньшей мере один белок пути рекомбинации, в частности по меньшей мере один белок пути гомологичной рекомбинации (HR), в частности Rad51, Nbs1, Mre11, лигазу 1 и/или лигазу 3.
В целевую нуклеотидную последовательность можно вставить маркерный ген, такой как GFP посредством гомологичной рекомбинации. Клетки, содержащие указанный маркерный ген, можно предпочтительно отобрать.
Гетерологичные последовательности (а), приведенные выше, могут кодировать, активировать или инактивировать белки MNR-комплекса, такие как Nbs1 и/или Mre11, и/или указанные гетерологичные последовательности (b) могут кодировать последовательности или белки, подавляющие экспрессию по меньшей мере одного белка пути гомологичной рекомбинации (HR), такого как Rad51, Nbs1, Mre11, лигаза 1 и/или лигаза 3.
Делецию или вставку можно ввести в указанные один или более ERV-элементов посредством негомологичного соединения концов (NHEJ) или опосредованного микрогомологией соединения концов (MMEJ).
Гетерологичные последовательности могут представлять собой часть интегрирующих или предпочтительно неинтегрирующих векторов, или они могут экспрессироваться с них.
Не встречающаяся в природе/гетерологичная система для введения одно- или двухцепочечных разрывов в целевую нуклеотидную последовательность может представлять собой CRISPR/Cas9-систему или может быть основана на ней.
Одну или более миристоилированных аминокислот в мотиве миристоилирования в одном или более ERV-элементов можно заменить на немиристоилированную аминокислоту.
Последовательности, кодирующие мотив PPYP, и/или последовательности, содержащие вплоть до 10, 20 или 30 нуклеиновых кислот, фланкирующих указанный мотив, могут содержать изменение.
Последовательности, подавляющие экспрессию, например, белков HR, таких как Rad51, могут только транзиентно экспрессироваться в клетках.
Настоящее изобретение также относится к модифицированной клетке, предпочтительно клетке млекопитающего, такой как модифицированная клетка СНО, включая клетку СНО-K1, которая предпочтительно не высвобождает вирусные частицы/по существу не высвобождает вирусные частицы, особенно при стандартном и/или стрессовом условии культивирования, и при этом геном указанной клетки содержит одно или более изменений, включающих:
(a) делеции,
(b) добавления/интеграции и/или
(c) замены,
одной или более нуклеиновых кислот в одном или более, в целом в более, чем 10, 20, 30, 40, 50, 60, 80, 90 или 100 эндогенных ретровирусных (ERV) элементах, и которую предпочтительно получают при помощи любого одного из способов, описанных в настоящем документе.
Настоящее изобретение также относится к набору, содержащему:
(i) в одной емкости по меньшей мере один неинтегрирующий вектор, кодирующий по меньшей мере нуклеазу, такую как CRISPR,
(ii) в той же емкости или в другой емкости по меньшей мере одну гидовую РНК или последовательность, кодирующую по меньшей мере одну гидовую РНК, нацеливающую на мотив в ERV-элементе,
(iii) в той же емкости или в дополнительной емкости по меньшей мере одну миРНК или последовательность, кодирующую по меньшей мере одну миРНК, и
в дополнительной емкости инструкцию о том, как обеспечить попадание (i), (ii) и (iii) внутрь клетки.
Последовательности, кодирующие по меньшей мере одну гидовую РНК, нацеливающую на мотив в ERV-элементе, и/или последовательности, кодирующие по меньшей мере одну миРНК, могут представлять собой часть вектора. Любой из описанных в настоящем документе векторов может только транзиентно экспрессироваться (i), и/или может представлять собой неинтегрирующий вектор (i). ERV-элемент может кодировать белок Gag. Мотив может представлять собой мотив миристоилирования или мотив PPXY, в частности, мотив PPYP. миРНК могут быть направлены в противоположном от гена HR-пути направлении.
ОПИСАНИЕ РАЗЛИЧНЫХ И ПРЕДПОЧТИТЕЛЬНЫХ ВАРИАНТОВ РЕАЛИЗАЦИИ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
Клетку, предпочтительно клетку млекопитающего/эукариотическую клетку, которую согласно настоящему изобретению модифицируют для получения модифицированной клетки, можно поддерживать в условиях культивирования клеток. Стандартные условия культивирования клеток составляют от 30 до 40°С, предпочтительно при 37°С или приблизительно при 37°С, например, в полностью синтетической среде для культивирования, применяемой для получения рекомбинантных белков. Неограничивающими примерами такого типа клеток являются эукариотические клетки, полученные не из приматов, такие как клетки яичника китайского хомячка (СНО), включая клетки СНО-K1 (АТСС CCL 61) и клетки SURE СНО-М (производные СНО-K1), и клетки почек детенышей хомяка (BHK, АТСС CCL 10). Полученные от приматов эукариотические клетки-хозяева включают, например, клетки карциномы шейки матки человека (HELA, АТСС CCL 2) и клетки 293 [АТСС CRL 1573], а также клетки 3Т3 [АТСС CCL 163] и клетки линии CV1 почки обезьяны [АТСС CCL 70], также трансформированные при помощи SV40 (COS-7, АТСС CRL-1587). Термин «модифицированный» обозначает клетку, которая была изменена, например, путем трансфекции, например, трансгенной последовательностью, и/или посредством мутации. Поскольку специалист в данной области легко поймет, что эти клетки, даже до описанной в настоящем документе их модификации, представляют собой клетки, не встречающиеся в природе. Вышеуказанные клетки, в частности, различные клетки СНО, обычно применяются в биотехнологических применениях, как, например, для получения терапевтических белков, и в настоящем документе они упоминаются как представляющие интерес клетки. Специалист в данной области техники легко поймет, что другие клетки, помимо тех, которые упомянуты выше, могут представлять собой представляющие интерес клетки, если их применяют в биотехнологических целях, в частности, для экспрессии, например, терапевтических белков.
Три основных белка, кодируемых в ретровирусном геноме, представляют собой белки Gag, Pol и Env. Gag (групповые антигены), кодируемый геном gag, представляет собой полипротеин, который процессируется до белков ядерного матрикса и других белков ядра, включая нуклеопротеиновые ядерные (коровые) частицы, которые определяют ядро ретровируса. Pol представляет собой обратную транскриптазу, кодируемую геном pol, и он выполняет функцию РНКазы Н и интегразы. Его активность приводит к предварительно интегрированной форме двухцепочечной ДНК вируса и, через функцию интегразы для интеграции в геном хозяина, а также функции РНКазы, к обратной транскрипции после интеграции в геном хозяина. Env представляет собой белок оболочки, кодируемый геном env, и он находится в липидном слое вируса, определяющем вирусный тропизм.
Эндогенные ретровирусы (ERV), такие как вирусы из рода гаммаретровирусов, включая вирус эпидемии коал (KoRV), вирус опухоли молочной железы у мышей (MMTV), вирус лейкоза мышей (MLV), являются адвентивными агентами, которые встречаются в геноме любого животного, включая, например, геномы хомячков и геномы родственных им видов. ERV сохраняются в геномах и могут обладать определенными преимуществами в отношении клеток, в геном которых они интегрированы, включая обеспечение источника генетического разнообразия и защиту от других вирусных патогенов. Тем не менее, они могут стать инфекционными. Раковые и/или эпигенетические модификации могут усиливать «пробуждение» ERV. Lewis и др. идентифицировал 403 возможных белков ретровирусов в клетках СНО на основе сравнения гомологий с базой данных из 115 вирусов. Lewis и др. сообщил об экспрессии 40% их мРНК, тем самым предполагая, что многие из них могут все еще синтезироваться ретровирусными компонентами (Lewis et al., 2013, Supplement).
В настоящем контексте в качестве инструмента для сравнения применяли последовательности ретровируса типа С мышей (данные, полученные в публикации 1994 г.). В этой группе четыре вида, как это указано на веб-сайте Viralzone (декабрь 2015 г.), принадлежащем SIB (Швейцарский институт биоинформатики). Они также часто встречаются в геноме мыши и, как известно, вызывают рак у мышей. Тем не менее, специалисту в данной области техники будет понятно, что последовательности многих других ретровирусов можно использовать для поиска ERV в представляющих интерес клетках.
Модифицированная клетка согласно настоящему изобретению может содержать геном, который, по большей части, идентичен геному клетки, из которой она получена такой, как, например, СНО-K1. Тем не менее, по меньшей один, а зачастую множество ERV-элементов, которые являются частью таких геномов, будут содержать описанные в настоящем документе изменения.
ERV-элемент согласно настоящему изобретению описывает в основном участок нуклеотидной последовательности ERV, который обеспечивает в соответствующем неинтегрированном вирусе функциональный объект. Как будет понятно специалисту в данной области техники, зачастую часть таких участков разрушается или удаляется в интегрированной вирусной ДНК. Таким образом, в это определение включены все гены (gag, pol, env) и, во-вторых, их части, приведенные ниже и показанные на Фигуре 2В, но и также их укороченные и/или модифицированные версии, которые присутствуют в геноме клетки. Указывается, что ERV-элемент, например, происходит из gag, если ген gag является их источником, но они не обязательно кодируют полный функциональный белок Gag. ERV-элементы, про которые указано, что они кодируют белок Gag, кодируют функциональный белок Gag.
Ген gag приводит к появлению белка-предшественника Gag, который экспрессируется из несплайсированной вирусной мРНК. Указанный белок-предшественник Gag расщепляется кодируемой вирусом протеазой (продуктом гена pol) в процессе созревания вируса, обычно, до четырех белков меньшего размера, обозначаемые как МА (белок ядерного матрикса), СА (белок капсида), NC (белок нуклеокапсида) и дополнительный белковый домен (например, рр12 в вирусе мышиного лейкоза или р6 в ВИЧ).
Полипептид МА происходит из N-концевого миристоилированного конца указанного белка-предшественника. Большинство молекул МА остаются прикрепленными к внутренней поверхности вирионного липидного бислоя, стабилизирующего частицу.
Белок СА образует коническое ядро вирусных частиц.
NC-область Gag отвечает за специфическое распознавание так называемого сигнала для упаковки ретровируса. Сигнал для упаковки содержит четыре структуры типа «петля-на-стебле», расположенных вблизи 5'-конца вирусной РНК, и его достаточно для опосредования инкорпорации гетерологичной РНК в вирионы. NC связывается с указанным сигналом для упаковки через взаимодействия, опосредствованные двумя цинк-пальцевыми мотивами.
Другой белковый домен опосредует взаимодействия между белком-предшественником Gag и дополнительным белком Vpr и приводит к инкорпорации Vpr в способный к сборке вирионов. Р6-область у вируса ВИЧ также содержит так называемый поздний домен, который необходим для эффективного высвобождения почкующихся вирионов из инфицированной клетки. (Норе & Trono, 2000).
Вирусная протеаза (Pro), интеграза (IN), РНКаза Н и обратная транскриптаза (RT) экспрессируются в контексте гибридного белка Gag-Pol. Предшественник Gag-Pol в целом образуется в результате сдвига рамки считывания рибосом, которое запускается специфичным цис-действующим мотивом РНК (гептануклеотидная последовательность, за которой следует короткая структура типа «петля-на-стебле» в дистальной области РНК Gag). Когда рибосомы сталкиваются с таким мотивом, они сдвигаются приблизительно в 5% случаев по сравнению с рамкой считывания pol без прерывающей трансляции. Частота сдвига рамки считывания рибосом объясняет, почему Gag и предшественник Gag-Pol образуются в соотношении приблизительно 20:1.
Во время созревания вируса кодируемая вирусом протеаза отщепляет полипептид Pol от Gag и дополнительно расщепляет его до белков, по отдельности обладающих протеазной, RTазной, РНКазной Н и интегразной активностями. Все такие расщепления не происходят эффективно, например, примерно 50% белка RT остается связанным с РНКазой Н в виде одного полипептида (р65) (Норе & Trono, 2000).
Ген pol кодирует обратную транскриптазу. В процессе обратной транскрипции полимераза образует двухцепочечную ДНК-копию димера одноцепочечной геномной РНК, присутствующей в вирионе. РНКаза Н удаляет исходный образец РНК из первой цепи ДНК, что позволяет синтезировать комплементарную цепь ДНК. Преобладающими функциональными видами полимеразы являются гетеродимеры. Все продукты гена pol можно обнаружить в капсиде высвобожденных вирионов.
Белок IN опосредует встраивание провирусной ДНК в геномную ДНК инфицированной клетки. Этот процесс опосредуется тремя различными функциями IN.
Белок Env экспрессируется из однократно сплайсированной мРНК. Изначально будучи синтезированным в эндоплазматическом ретикулуме, Env мигрирует через комплекс Гольджи, где он подвергается гликозилированию. Гликозилирование Env в целом требуется для инфекционности. Клеточная протеаза расщепляет указанный белок до трансмембранного домена и поверхностного домена (Норе & Trono, 2000).
Некоторые ERV генома высвобождаются из клеток в виде вирусоподобных частиц, а другие нет. Тем не менее, в целом те ERV, которые высвобождаются, обладают более высоким потенциалом к тому, чтобы стать инфекционными. Таким образом, в целом выгодно иметь модифицированные клетки, описанные в настоящем документе, которые не могут экспрессировать и высвобождать или, по существу, не экспрессировать и не высвобождать ERV, предпочтительно в стандартных или стрессовых условиях культивирования. По существу, ERV не высвобождаются в том случае, если культура клеток, содержащая модифицированную таким образом клетку, высвобождает менее, чем 50%, менее, чем 40%, менее, чем 30%, менее, чем 20%, менее, чем 10%, предпочтительно 5% ERV, по сравнению с ее аналогом, который не был подвергнут описанным в настоящем документе процедурам уменьшения высвобождения ERV. Такой аналог будет, например, представлять собой коммерчески доступную клетку СНО-K1. Отсутствие экспрессии или отсутствие экспрессии по существу означает, что менее, чем 50%, менее, чем 40%, менее, чем 30%, менее, чем 20%, менее, чем 10%, предпочтительно 5% немутированной последовательности мРНК Gag можно обнаружить методом ПЦР и при помощи анализа секвенирования. Отсутствие высвобождения означает, что не происходит или по существу не происходит обнаруживаемого высвобождения вирусной последовательности, которое оценивают путем ПЦР-анализа с применением кДНК, как это показано, например, на Фиг. 21, или которое получают по результатам анализа с применением QuantiTect Rev. Transcription Kit ® от компании QIAGEN. Как будет понятно специалисту в данной области техники, клетки, которые модифицируют таким образом, чтобы они содержали изменения в нуклеотидной последовательности, которые инактивируют образование белка/пептида в одном, двух, трех, четырех, 5, 6, 7, 8, 9, 10 ERV-элементах, также обладают преимуществами и являются частью настоящего изобретения. Фактически, в определенных вариантах реализации целесообразно изменять один или более ERV-элементов, которые являются частью генома представляющей интерес клетки, только путем по меньшей мере одной мутации, чтобы сохранить там, где это необходимо, те положительные функции, которые ERV могли бы выполнять в клетке. Тем не менее, как это обсуждается ниже, определенные варианты реализации настоящего изобретения особенно подходят для большого числа копий ERV (более, чем 30, 40 или 50 ERV/на клетку), и с их помощью можно достигнуть того, что такая модифицированная клетка не будет демонстрировать ни один из уровней экспрессии соответствующего ERV-компонента.
Гетерологичная последовательность нуклеиновой кислоты представляет собой последовательность нуклеиновой кислоты, которая не встречается в клетках до их модификации согласно настоящему изобретению, в то время как родственные типы последовательностей нуклеиновых кислот могут очень хорошо существовать в клетке. В контексте настоящего изобретения трансген представляет собой такую гетерологичную последовательность нуклеиновой кислоты, в частности дезоксирибонуклеотидную последовательность (ДНК), кодирующую: некоторый зрелый белок (также упоминаемая в настоящем документе как кодирующая белок ДНК), белок-предшественник или для функциональную РНК, которая не кодирует белок (некодирующая РНК). Трансген выделяют и вводят в клетку для получения продукта такого трансгена. Некоторые предпочтительные трансгены согласно настоящему изобретению кодируют маркерные белки, такие как GFP (зеленый флуоресцентный белок). Такие белки можно применять для обнаружения успешной интеграции в ERV-элементы, а следовательно, и для обнаружения их изменения/инактивации. Другими трансгенами являются те, которые кодируют, например, белки, которые в конечном итоге будут продуцироваться рассматриваемой клеткой, такие как иммуноглобулины (Igs) и Fc-гибридные белки и другие белки, в частности обладающие терапевтической активностью белки («биотерапевтические средства»). Согласно настоящему изобретению дополнения включают интеграции, как, например, направленные интеграции. Тем не менее, специалисту в данной области техники будет понятно, что во время процесса интеграции определенные нуклеотиды могут «потеряться» в геноме реципиента. Такие интеграции являются частью настоящего изобретения и считаются дополнениями.
В контексте настоящего описания термин «трансген» в контексте кодирующей белок ДНК не включает нетранскрибируемые фланкирующие области, такие как сигналы инициации транскрипции РНК, промоторы или энхансеры. Другие предпочтительные трансгены включают последовательности ДНК, кодирующие функциональные РНК. Таким образом, указанный термин трансген применяется в настоящем контексте в случае, когда речь идет о последовательности ДНК, которую вводят в клетку, такую как эукариотическую клетку-хозяин, посредством трансфекции (которая в контексте настоящего изобретения также включает трансдукцию, т.е. введение через вирусные векторы), и которая кодирует представляющий интерес продукт («продукт экспрессии трансгена», например, «гетерологичный белок»). Указанный трансген можно функционально присоединить к кодирующей сигнальный пептид последовательности, которая кодирует сигнальный пептид, который в свою очередь опосредует и/или способствует транслокации и/или секреции через эндоплазматический ретикулум и/или цитоплазматическую мембрану и удаляется до или во время секреции.
Последовательность промотора или просто промотор представляет собой нуклеотидную последовательность, которая распознается клеткой-хозяином для экспрессии по меньшей мере одной последовательности нуклеиновой кислоты. Указанная последовательность промотора содержит последовательности контроля транскрипции, которые регулируют экспрессию указанного полинуклеотида. Промотор может представлять собой любую последовательность нуклеиновой кислоты, которая проявляет транскрипционную активность в выбранной клетке-хозяине, включая мутантные, укороченные и гибридные промоторы, и ее можно получить из генов, кодирующих внеклеточные или внутриклеточные полипептиды, которые либо гомологичны, либо гетерологичны указанной клетке-хозяину. Промоторы согласно настоящему изобретению включают индуцируемые и неиндуцируемые промоторы. Последовательность нуклеиновой кислоты находится под контролем промотора, когда указанный промотор выполняет свою функцию в указанной нуклеиновой кислоте. Клетки/векторы согласно настоящих изобретений зачастую содержат такие промоторы.
Изменения последовательности нуклеиновой кислоты представляют собой изменения, такие как добавления/вставки, делеции и/или замены, которые не встречаются в таких клетках до их модификации согласно настоящему изобретению.
В контексте настоящего описания «редактирование генома» относится к модификации («редактированию») геномных последовательностей и может включать делецию по меньшей мере одного нуклеотида, добавление/вставку по меньшей мере одного нуклеотида или замещение по меньшей мере одного нуклеотида. Редактированная геномная последовательность упоминается в настоящем документе как последовательность целевой нуклеиновой кислоты. Направленные вставки представляют собой вставки, которые происходят в специфичном, заранее определенном сайте-мишени. Инструменты для редактирования генома вводят двух- или одноцепочечные разрывы в геном, например, через нуклеазы или никазы, и по меньшей мере частично зависят от механизмов клеточной рекомбинации (см. обсуждение ниже) для восстановления таких разрывов. Такие средства также содержат в целом сиквенс-специфичные ДНК-связывающие модули.
ZFN (цинк-пальцевые нуклеазы) и TALEN (подобные активатору транскрипции эффекторные нуклеазы) позволяют проводить широкий спектр генетических модификаций путем индуцирования двухцепочечных разрывов ДНК, которые способствуют подверженному ошибкам негомологичному соединению концов (NHEJ) или репарации, направляемой гомологией (HDR), в определенных местах локализации гена в геноме.
Сиквенс-специфичность CRISPR-систем (короткие палиндромные повторы, регулярно расположенные группами) определяется малыми РНК. Локусы CRISPR состоят из серии повторов, разделенных спейсерными последовательностями, которые соответствуют геномам бактериофагов и других мобильных генетических элементов. Матрица повторов-спейсеров транскрибируется в виде длинного предшественника и процессируется в пределах повторяющихся последовательностях с образованием малой crРНК, которая определяет целевые последовательности (также известные как протоспейсеры), расщепляемые CRISPR-системами. Для расщепления зачастую требуется наличие мотива последовательности, располагающегося непосредственно за областью-мишенью, который известен как мотив, смежный с протоспейсером (РАМ). CRISPR-ассоциированные (cas) гены обычно фланкируют матрицу повторов-спейсеров и кодируют ферментативный механизм, ответственный за биогенез и нацеливание crРНК (CRISPR РНК). Cas9 представляет собой эндонуклеазу дцДНК, которая использует гидовую crРНК для определения сайта расщепления. Загрузка гидовой crРНК на Cas9 происходит во время процессинга предшественника crРНК и требует присутствия малой антисмысловой РНК для указанного предшественника, tracrРНК, и РНКазы III. В отличие от редактирования генома с применением ZFN или TALEN, изменение специфичности мишени для Cas9 не требует модифицирования белка, а только разработки гидовой crРНК, также называемой огРНК.
На сегодняшний день в протоколах редактирования генома применяют три различных варианта Cas9-нуклеазы. Первый - это Cas9 дикого типа, которая может сайт-специфичным образом расщеплять двухцепочечную ДНК, что приводит к активации механизма репарации двухцепочечных разрывов (DSB). Репарация DSB может осуществляться посредством клеточного пути негомологичного соединения концов (NHEJ), что приводит к вставкам и/или делециям (вставки-делеции (indels)), которые нарушают целевой локус. В альтернативном варианте, в случае, если предоставляется донорская матрица (см. Фиг. 7), гомологичная указанному целевому локусу, репарация DSB может осуществляться посредством пути репарации, направляемой гомологией (HDR), который позволяет производить точные замещающие мутации.
Cas9-система дополнительно модифицировали для повышения точности за счет разработки мутантной формы, известной как Cas9D10A, обладающей только никазной активностью. Это означает, что она расщепляет только одну цепь ДНК и не активирует NHEJ. Вместо этого, в том случае, когда имеется матрица для гомологичной репарации, репарации ДНК осуществляются только через высокоточный HDR-путь, что приводит к уменьшению числа мутаций вида вставки-делеции. Поэтому Cas9D10A во многих практических применениях является более привлекательной с точки зрения специфичности мишени, когда локусы нацеливаются на парные комплексы Cas9, предназначенные для образования смежных одноцепочечных разрывов ДНК.
В контексте настоящего изобретения специфичная последовательность или консенсусная последовательность ERV-элементов обозначает определенный сайт расщепления посредством, например, одной из вышеуказанных систем. Длина такой специфичной или консенсусной последовательности предпочтительно составляет от 5 до 50 пар оснований, предпочтительно 10-50 или 15-25, или 25-50, или 30-50. Указанные консенсусные последовательности могут содержать, например, 1, 2, 3, 4 или 5 несоответствий (относительно друга они комплементарны на более, чем 60%, 70%, 80%, 90% или 95%) до тех пор, пока еще можно проводить расщепление. См., например, Фиг. 21. Вышеуказанные системы называются не встречающимися в природе системами или гетерологичными системами, что означает, что они введены в клетку, а не являются частью указанной клетки, до ее модификации согласно настоящему изобретению.
Вектор согласно настоящему изобретению представляет собой молекулу нуклеиновой кислоты, способную транспортировать другую нуклеиновую кислоту такую, как, например, трансген, которая будет экспрессироваться данным вектором, с которым она была связана, в целом в который она была интегрирована. Например, плазмида представляет собой разновидность вектора, ретровирус или лентивирус представляют собой другой тип вектора. В предпочтительном варианте реализации настоящего изобретения вектор линеаризуют до начала трансфекции. Вектор экспрессии содержит гетерологичные регуляторные элементы или находится под контролем таких регуляторных элементов, которые предназначены для дальнейшей транскрипции и/или экспрессии нуклеотидной последовательности, как, например, трансгена, переносимой указанным вектором экспрессии. Регуляторные элементы включают энхансеров и/или промоторы, а также множество других описанных в настоящем документе элементов.
Среди числа невирусных векторов транспозоны являются особенно привлекательными из-за их способности интегрировать одиночные копии последовательностей ДНК с высокой частотой в несколько локусов генома хозяина (интегрирующий вектор). В отличие от вирусных векторов, сообщалось, что некоторые транспозоны не интегрируются в предпочтительной близости к клеточным генам, и, следовательно, они с меньшей долей вероятности вводят вредные мутации. Кроме того, транспозоны легко получить и с ними легко работать, и они состоят в целом из вектора-донора транспозона, содержащего ДНК-переносчик (cargo DNA), фланкированную последовательностями инвертированных повторов, и экспрессирующих транспозазу вспомогательной (хелперной) плазмиды или мРНК. Было разработано несколько транспозонных систем для мобилизации ДНК в различных линиях клеток без вмешательства в копии эндогенных транспозонов. Например, транспозон PiggyBac (РВ), первоначально выделенный из металловидки серой (cabbage looper moth), эффективно переносит ДНК-переносчик в различные клетки млекопитающих. В контексте настоящего изобретения векторы, в частности неинтегрирующие векторы, также можно применять для транзиентной (ограниченной по времени) экспрессии гена или функциональной РНК. Транзиентная экспрессия представляет собой экспрессию в течение ограниченного периода времени, и период времени экспрессии зависит от конструкции вектора и условий культивирования. Тем не менее, транзиентная экспрессия означает экспрессию в течение по меньшей мере 24 часового периода, но в целом не более, чем в течение 7 дней.
Эпигенетические регуляторные элементы можно использовать для защиты переносимую ДНК от нежелательных эпигенетических воздействий в случае ее размещения вблизи трансгена на плазмидных векторах. Например, были предложены элементы, называемые участком прикрепления к ядерному матриксу (MAR), для увеличения геномной интеграции и транскрипции ДНК-переносчика с одновременным предотвращением сайленсинга гетерохроматина, примером которого является эффективный белок MAR 1-68 человека. Они также могут выступать в качестве инсуляторов и тем самым предотвращать активацию соседних клеточных генов. Таким образом, MAR-элементы применяли для опосредования высокой и устойчивой экспрессии в контексте плазмид или вирусных векторов. Для транзиентной экспрессии генов можно применять неинтегрирующие векторы (иногда упоминаемые как эписомальные векторы), такие как плазмиды, или неинтегрирующие лентивирусные (NIL) векторы. Они могут стабильно или транзиентно сохраняться и реплицироваться внутри клетки-хозяина.
Векторная последовательность вектора представляет собой последовательность ДНК или РНК указанного вектора за исключением любых «других» нуклеиновых кислот, таких как трансгены, а также генетических элементов, таких как MAR-элементы.
Термин идентичность последовательности относится к измерению идентичности нуклеотидных последовательностей или аминокислотных последовательностей. В целом последовательности выравнивают так, чтобы получить соответствие наивысшего порядка. «Идентичность» сама по себе имеет признанное определение в данной области техники, и ее можно рассчитать с применением опубликованных методов. (См., например: Computational Molecular Biology, Lesk, A.M., ed., Oxford University Press, New York, 1988; Biocomputing: Informatics and Genome Projects, Smith, D.W., ed., Academic Press, New York, 1993; Computer Analysis of Sequence Data, Part I, Griffin, A.M., and Griffin, H.G., eds., Humana Press, New Jersey, 1994; Sequence Analysis in Molecular Biology, von Heinje, G., Academic Press, 1987; и Sequence Analysis Primer, Gribskov, M. and Devereux, J., eds., M. Stockton Press, New York, 1991). Хотя и существует целый ряд способов измерения идентичности между двумя полинуклеотидными или полипептидными последовательностями, термин «идентичность» хорошо известен специалистам в данной области техники как определение идентичных нуклеотидов или аминокислот в рассматриваемом положении в последовательности (Carillo, Н. & Lipton, D., SIAM J. Applied Math. 48: 1073 (1988)).
Является ли какая-либо конкретная молекула нуклеиновой кислоты по меньшей мере на 50%, 60%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичной, например, к последовательности, подобной последовательности гаммаретровируса, SEQ ID NOS: 1, 2, 3 или 4 или их части (см., например, последовательности, описанные на Фиг. 18), можно традиционно определить с применением известных компьютерных программ, таких как программное обеспечение DNAsis (Hitachi Software, Сан Бруно, штат Калифорния) для начального выравнивания последовательности с последующим применением программного обеспечения ESEE версии 3.0 для последовательностей ДНК/белков (cabot@trog.mbb.sfu.ca) для множественного выравнивания последовательностей.
Является ли аминокислотная последовательность по меньшей мере на 50%, 60%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичной к, например, последовательности белка, экспрессируемого последовательностью SEQ ID NOS: 1, 2, 3 или 4 или ее частью, можно традиционно определить с применением известных компьютерных программ, таких как программа BESTFIT (пакет анализа последовательности Висконсина, версия 8 для Unix, Genetics Computer Group, University Research Park, 575 Science Drive, Мадисон, штат Висконсин, 53711). BESTFIT использует алгоритм локальной гомологии Смита-Уотермана, Advances in Applied Mathematics 2: 482-489 (1981), для того, чтобы найти наилучший сегмент гомологии между двумя последовательностями.
В случае применения DNAsis, ESEE, BESTFIT или любой другой программы выравнивания последовательностей для определения того, является ли конкретная последовательность, например, на 95% идентичной к эталонной (референсной) последовательности согласно настоящему изобретению, параметры устанавливаются таким образом, чтобы процент идентичности вычислялся по полной длине эталонной нуклеотидной или аминокислотной последовательности, и чтобы допускались гэпы (пропуски) в гомологии вплоть до 5% от общего количества нуклеотидов в эталонной последовательности.
Другой предпочтительный способ определения наилучшего общего соответствия (совпадения) между запрашиваемой (искомой, от англ. query) последовательностью (последовательностью согласно настоящему изобретению) и исследуемой (от англ. subject) последовательностью, также упоминаемый как глобальное выравнивание последовательностей, можно определить с применением компьютерной программы FASTDB на основе алгоритма Brutlag et al. (Comp. Арр. Biosci. (1990) 6: 237-245). При выравнивании последовательностей обе из указанных запрашиваемой и исследуемой последовательностей представляют собой последовательности ДНК. Последовательность РНК можно сравнить путем преобразования U в Т. Результат указанного глобального выравнивания последовательностей указывается в процентах идентичности. Предпочтительные параметры, применяемые в выравнивании последовательностей ДНК с использованием FASTDB для расчета процента идентичности, составляют: Матрица = Унитарная, Участок максимального совпадения = 4, Штраф за несовпадение = 1, Штраф за присоединение = 30, Длина группы рандомизации = 0, Пограничный вес выравнивания = 1, Штраф за гэп (пропуск) = 5, Штраф за размер гэпа (пропуска) = 0,05, Размер окна = 500 или длина исследуемой нуклеотидной последовательности, в зависимости от того, что короче.
Например, полинуклеотид, обладающий 95% «идентичностью» к эталонной нуклеотидной последовательности согласно настоящему изобретению, идентичен к эталонной последовательности, за исключением того, что указанная полинуклеотидная последовательность может содержать в среднем вплоть до пяти точечных мутаций на каждые 100 нуклеотидов эталонной нуклеотидной последовательности, кодирующей полипептид. Другими словами, чтобы получить полинуклеотид, у которого нуклеотидная последовательность по меньшей мере на 95% идентична к эталонной нуклеотидной последовательности, вплоть до 5% нуклеотидов в указанной эталонной последовательности можно удалить или заменить на другой нуклеотид, или ряд нуклеотидов вплоть до 5% от общего числа нуклеотидов в указанной эталонной последовательности можно вставить в эталонную последовательность. Запрашиваемая последовательность может представлять собой полную последовательность, ОРС (открытую рамку считывания) или любой определенный фрагмент, описанный в настоящем документе.
Средство поиска основного локального выравнивания NCBI (BLAST) (Altschul et al., J. Mol. Biol., 215: 403-410, 1990) доступно в нескольких источниках, включая Национальный центр биотехнологической информации (NCBI, Бетесда, штат Мэриленд) и Интернет, для применения в сочетании с программами для анализа последовательностей blastp, blastn, blastx, tblastn и tblastx. Доступ к ней можно получить на веб-сайте NCBI совместно с описанием того, каким образом определить идентичность последовательности и сходство последовательностей при помощи данной программы.
Настоящее изобретение включает молекулы нуклеиновой кислоты и/или аминокислоты, обладающие 80%, 85%, 90% или более, чем 95% или более, чем 98% идентичностью последовательности или полной идентичностью последовательности к раскрытым в настоящем документе последовательностями и любыми их фрагментами, в частности фрагментами, расположенными на расстоянии вплоть до 40 п.о., 30 п.о. или 20 п.о. по обеим сторонам от мотива PPYP.
ПУТИ РЕКОМБИНАЦИ
Пути рекомбинации, также известные как пути рекомбинации ДНК, представляют собой клеточные пути, которые приводят к репарации повреждений ДНК, такие как соединение концов молекул ДНК после хромосомных двухцепочечных разрывов (DSB), а также к обмену или гибридизации последовательностей ДНК между молекулами хромосомной и нехромосомной ДНК, такой как, например, перекрывание хромосом при мейозе или перегруппировка генов иммуноглобулина в лимфоцитарных клетках. Основными путями рекомбинации являются путь гомологичной рекомбинации (HR), путь негомологичного соединения концов (NHEJ) и опосредованное микрогомологией соединение концов (MMEJ) и пути альтернативного соединения концов (Alt-EJ).
В контексте настоящего изобретения нокауты определенных членов по меньшей мере одного пути зачастую происходят посредством комплементарных нуклеотидных последовательностей. Нуклеотидная последовательность, такая как ДНК или РНК, комплементарна последовательности другой ДНК или РНК в случае, если нуклеотиды, например, две цепи одноцепочечных ДНК или две цепи одноцепочечных РНК, могут образовывать стабильные водородные связи, такие как водородная связь между гуанином (G) и цитозином (С). В клетке спаривание комплементарных оснований позволяет, например, клеткам копировать информацию из одного поколения в другое. При РНК-интерференции (РНКи) спаривание комплементарных оснований способствует сайленсингу или полному нокауту определенных генов-мишеней. По существу, последовательности малых интерфирирующих РНК (миРНК), коротких шпилечных РНК (кшРНК) или микроРНК специфически понижают или нокаутируют экспрессию гена-мишени при помощи содержащейся в них одиночной цепи РНК (например, антисмысловая цепь в миРНК), соответствующей РНК, в частности мРНК, клетки-хозяина. Степень комплементарности между двумя цепями нуклеиновых кислот может варьироваться от полной комплементарности (каждый нуклеотид комплементарен нуклеотиду напротив) до частичной комплементарности (50%, 60%, 70%, 80%, 90% или 95%). Степень комплементарности определяет стабильность комплекса и, следовательно, то, насколько успешно ген можно, например, нокаутировать. Таким образом, предпочтительна полная или по меньшей мере 95% комплементарность.
МЕХАНИЗМЫ ГОМОЛОГИЧНОЙ РЕКОМБИНАЦИИ (HR), NHEJ И MMEJ
Трансгены используют механизмы рекомбинации для интеграции в двухцепочечные разрывы в геноме хозяина
Двухцепочечные разрывы (DSB) представляют собой с биологической точки зрения самый вредный тип повреждения генома, потенциально приводящий к гибели клеток или широкому спектру генетических перестроек. Точная репарация необходима для успешного сохранения и распространения генетической информации.
Существует два основных механизма репарации DSB: NHEJ и HR. Третий механизм, называемый MMEJ, зачастую запускается в случае, когда два основных механизма репарации DSB оказываются неэффективными. Гомологичная рекомбинация представляет собой процесс генетического обмена между гомологичными последовательностями ДНК, и он действует преимущественно во время S/G2-фазы клеточного цикла, тогда как NHEJ просто соединяет вместе два «разорванных» (нарушенных, от англ. broken) конца ДНК, в которых обычно гомология последовательностей отсутствует, и он функционирует во всех фазах клеточного цикла, но имеет особое значение во время фазы G0-G1 и ранней S-фазы митотических клеток. У позвоночных HR, NHEJ и MMEJ вносят различный вклад в репарацию DSB, в зависимости от природы DSB и фазы клеточного цикла.
NHEJ: основные механизмы
По существу, молекулярный механизм процесса NHEJ представляется простым: 1) набор ферментов захватывает «разорванную» молекулу ДНК, 2) образуется молекулярный мост, который объединяет два конца ДНК и 3) «разорванные» молекулы повторно лигируют. Для проведения таких реакций механизм NHEJ в клетках млекопитающих включает в себя два белковых комплекса: гетеродимера Ku80/Ku70, ассоциированного с ДНК-PKcs (каталитической субъединицей ДНК-зависимой протеинкиназы), и ДНК-лигазы IV с ее кофактором XRCC4 (комплементирующий ген репарации радиационных повреждений в клетках китайского хомячка 4) и многими белковыми факторами, такими как Artemis и XLF (XRCC4-подобный фактор или Cernunnos). NHEJ часто рассматривают как подверженную ошибкам репарацию DSB, поскольку она просто соединяет два «разорванных» конца ДНК, в которых обычно гомология последовательностей отсутствует, и образует небольшие вставки и/или удаления. NHEJ обеспечивает механизм для репарации DSB в течение клеточного цикла, однако это путь имеет особое значение во время фазы G0-G1 и ранней S-фазы митотических клеток. Репарацию DSB посредством NHEJ наблюдают в организмах от бактерий до млекопитающих, что свидетельствует о том, что он сохраняется на протяжении эволюции.
После образования DSB ключевой стадией в пути репарации NHEJ является физическое соединение «разорванных» концов ДНК. NHEJ инициируется ассоциацией белкового комплекса гетеродимера Ku70/80 с обоими концами «разорванной» молекулы ДНК для захвата, соединения концов и создания каркаса для сборки других ключевых факторов NHEJ. ДНК-связанный комплекс Ku-гетеродимер осуществляет рекрутинг ДНК-PKcs к DSB, белка с массой 460 кДа, принадлежащему к семейству PIKK (фосфоинозитид-3-киназоподобному семейству протеинкиназ), и активирует его серин/треонинкиназную функцию. Две молекулы ДНК-PKcs взаимодействуют друг с другом через DSB, тем самым формируя молекулярный мост между обоими «разорванными» концами ДНК и ингибирует их разрушение. Затем концы ДНК можно непосредственно лигировать, хотя для большей части концевых участков, полученных из DSB, следует осуществить надлежащим образом процессинг до начала лигирования. В зависимости от природы разрыва может потребоваться действие различных комбинаций ферментов процессинга для образования совместимых выступов (overhand) путем заполнения гэпов (пробелов), удаления поврежденных ДНК или вторичных структур, окружающих указанный разрыв. Считается, что данная стадия в процессе NHEJ ответственна за случайную потерю нуклеотидов, ассоциированную с путем репарации NHEJ. Одним из ключевых ферментов конечного процессинга в пути NHEJ у млекопитающих является фермент Artemis, член суперсемейства металло-β-лактамазных ферментов, который был обнаружен как мутированный ген у большинства пациентов с радиочувствительным тяжелым комбинированным иммунодефицитом (SCID). Artemis обладает как от 5' до 3' экзонуклеазной активностью, так и ДНК-PKcs-зависимой эндонуклеазной активностью в отношении ДНК-содержащих ds-ss-транзиций и ДНК-шпилек (Ma et al., 2002). Его активность также регулируется ATM. Таким образом, Artemis, вероятнее всего, участвует во множественных ответах на повреждение ДНК. Тем не менее, Artemis осуществляет, по-видимому, репарацию только подмножества повреждений ДНК, поскольку в клетках, не содержащих Artemis, никаких серьезных дефектов в репарации DSB не наблюдали.
Гэпы (пробелы) в ДНК должны быть заполнены для того, чтобы обеспечить репарацию. Добавление нуклеотидов в DSB ограничено полимеразами μ и λ. Путем взаимодействия с XRCC4 также происходит рекрутинг полинуклеотидкиназы (PNK) к концам ДНК, что позволяет как полимеризовать, так и лигировать ДНК. Наконец, NHEJ завершается лигированием концов ДНК, стадией, осуществляемой комплексом, содержащим XRCC4, ДНК-лигазу IV и XLF. Другие лигазы могут частично замещать ДНК-лигазу IV, поскольку NHEJ может происходить в отсутствие XRCC4 и лигазы IV. Кроме того, исследования показали, что XRCC4 и лигаза IV не выполняют вне NHEJ никаких иных функций, тогда как, напротив, KU функционирует в других процессах, таких как транскрипция, апоптоз и ответы на микроокружение.
Путь NHEJ можно ослабить или «отключить» при помощи разных способов, многие из которых непосредственно влияют на вышеуказанные белки (например, на гетеродимер Ku80/Ku70, DNA-PKcs, но, в частности, на ДНК-лигазу IV, XRCC4, Artemis и XLF (XRCC4-подобный фактор или Cernunnos), PIKK (фосфоинозитид-3-киназоподобное семейство протеинкиназ). Тем не менее, хотя NHEJ в большинстве случаев является нежелательным, неточная репарация двухцепочечного разрыва в целом нарушает функциональность гена, в котором происходит двухцепочечный разрыв, и зачастую такой разрыв полностью представляет собой тот разрыв, который требуется в контексте настоящего изобретения. Соответственно, настоящее изобретение также включает варианты реализации, в которых желательно усиление NHEJ.
HR: основные механизмы
Гомологичная рекомбинация (HR) представляет собой очень точный механизм репарации. Гомологичная хроматида служит в качестве матрицы для репарации «разорванной» цепи. HR происходит во время S-фазы и G2-фазы клеточного цикла, когда доступны сестринские хроматиды. Классический механизм HR в основном характеризуется тремя стадиями: 1) отрезанием 5'-конца от «разорванных» концов, 2) внедрением (инвазией) цепи и обменом с гомологичным ДНК-дуплексом и 3) разрешением промежуточных продуктов рекомбинации. Различные пути могут завершать репарацию DSB, в зависимости от способности осуществлять внедрение цепи, и включают синтез-зависимый отжиг цепи (SDSA-путь), классическую репарацию двухцепочечного разрыва (DSBR), репликацию, индуцированную разрывом (BIR) и, в альтернативном варианте, отжиг одиночной цепи (SSA-путь). Все HR-механизмы взаимосвязаны и включают множество опосредуемых ферментами стадий.
Первая стадия всех HR-реакций соответствует отрезанию 5'-концевой «разорванной» цепи ДНК-нуклеазами при помощи MRN-комплекса (MRE11, RAD50, NBN (ранее NBS1 для синдрома неймегеновского повреждения 1)) и CtIP (взаимодействующий с CtBP белок). Получающееся в результате поколение 3'-одноцепочечных DSB может искать гомологичную последовательность. Внедрение указанного гомологичного дуплекса осуществляется посредством нуклеофиламента, состоящего из 3'-фрагмента одноцепочечной ДНК (оцДНК), покрытого белком рекомбиназы RAD51. Присутствие репликативного белка A (RPA), гетеротримерного оцДНК-связывающего белка, участвующего в метаболических процессах ДНК, связанного с оцДНК у эукариот, необходимо для сборки RAD51-филамента. Затем RAD51 взаимодействует с RAD52, который имеет кольцеобразную структуру, с вытеснением молекул RPA и содействием нагрузки RAD51. Rad52 важен для процессов рекомбинации у дрожжей. Тем не менее, предполагается, что у позвоночных BRCA2 (белок восприимчивости к раку молочной железы типа 2), а не RAD52, играет важную роль в процессах внедрения цепи и обмена. Взаимодействие RAD51/RAD52 стабилизируется связыванием RAD54. RAD54 также играет роль в созревании промежуточных продуктов рекомбинации после образования D-петли. С другой стороны, BRCA1 (ген рака молочной железы 1) взаимодействует с BARD1 (BRCA1-ассоциированный домен RING 1) и ВАСН1 (ген гомолога 1 ВТВ и CNC) для осуществления лигазной и геликазной активности репарации DSB соответственно. BRCA1 также взаимодействует с CtIP CDK-зависимом образом и подвергается убиквитинированию в ответ на повреждение ДНК. Как следствие, BRCA1, CtIP и MRN-комплекс играют роль в активации HR-опосредованной репарации ДНК в S-фазе и G2-фазе клеточного цикла.
Внедрение нуклеофиламента приводит к образованию гетеродуплекса, называемого петлей смещения (D-петля), и включает смещение одной цепи дуплекса посредством внедренной цепи и спаривания с другой. Затем несколько HR-путей могут завершить репарацию, применяя гомологичную последовательность в качестве матрицы для замены последовательности, окружающей DSB. В зависимости от применяемого механизма взаимные обмены (кроссинговеры) между указанной гомологичной матрицей и «разорванной» молекулой ДНК могут быть ассоциированы или могут быть не ассоциированы с HR-репарацией. Кроссинговеры могут иметь важные генетические последствия, такие как перегруппировка генома или потеря гетерозиготности.
Пять Rad51-паралогов также участвуют в гомологичной рекомбинации: Xrcc2, Xrcc3, Rad51B, Rad51C, Rad51D. Rad51-паралоги образуют два типа комплексов: один, называемый BCDX2, содержит Rad51B, Rad51C, Rad51D и Xrcc2; другой содержит Rad51C и Xrcc3 (СХ3). Было высказано предположение, что указанный первый комплекс участвует в образовании и/или стабилизации комплекса Rad51-ДНК. Роль второго комплекса, по всей видимости, заключается в миграции ветви и разрешения узла Холлидея.
Как сообщалось ранее, усиление HR по отношению к NHEJ (см. публикацию патента США №20030231449, содержание которого полностью включено в настоящее описание посредством ссылки) можно применять для улучшения и/или содействия экспрессии трансгена.
Были также описаны преимущества ослабления или «отключения» HR (WO 2014/118619, публикация патента США №20150361451, содержание которого полностью включено в настоящее описание посредством ссылки). HR можно ослабить или «отключить» при помощи разных способов, многие из которых непосредственно влияют на вышеуказанные белки (см. также Таблицу С, тем не менее, отмечается, что не существует четкого различия между различными путями HDR). РНК, такие как миРНК или кшРНК, в целом применяют для осуществления ослабления или «отключения».
Опосредованное микрогомологией соединение концов (MMEJ)
В том случае, когда другие пути рекомбинации являются неэффективными или неактивными, репарацию DSB можно осуществить другим, подверженному ошибкам механизму репарации, а именно MMEJ. Полная характеристика данного пути до сих пор отсутствует, и иногда он также упоминается как альтернативное соединение концов (alt-EJ), хотя и остается непонятным, основаны ли эти два процесса на одном и том же механизме или нет. Наиболее характерной особенностью указанного пути, которая отличает его от NHEJ, является использование микрогомологий размером в 5-25 п.о. во время выравнивания «разорванных» цепей ДНК, что приводит, в отличие от пути NHEJ, к делеции более крупных участков нуклеиновых кислот в целевом геноме, например, с размером более, чем 20, 40, 60, 80, 100, 150, 200 п.о., что является преимуществом во многих вариантах реализации настоящего изобретения.
MMEJ может происходить в любое время клеточного цикла и не зависит от ядерных NHEJ- и HR-факторов, т.е. генов Ku70, лигазы IV и Rad52. Вместо этого инициация MMEJ зависит от собственного набора белков, наиболее важными из которых являются компоненты MRN-комплекса (MRX у дрожжей), включающие Mre11, Rad50 и Nbs1 (Xrs2 у дрожжей), также участвующие в первых стадиях HR (Ma et al., 2003). Было высказано предположение, что помимо MRN-комплекса множество других факторов принимает участие в MMEJ и в связанном механизме SD-MMEJ, который зависит от синтеза ДНК, например, СТВР-взаимодействующий белок, поли(ADP-рибоза)полимераза 1 (PARP1), комплекс лигаза III/Xrcc1, лигаза I, ДНК-полимераза θ (Yu и McVey, 2010) и комплекс ERCC1/XPF. Тем не менее, в этом процессе также может участвовать множество других белков.
Было высказано предположение, что при отсутствии других связывающих концы ДНК белков (таких как Ku или Rad51) DSB распознаются белком PARP1, который затем инициирует их репарацию посредством MMEJ. Процесс репарации, аналогично HR, начинается с 5'-3' отрезания, которое воздействует на короткие области гомологии с каждой стороны разрыва. Эта стадия процессинга осуществляется MRN-комплексом и регулируется CtIP. Комплементарные области (присутствуют в 3'-фрагментах оцДНК) спариваются, а некомплементарные сегменты (флэпы) удаляются, возможно, при помощи комплекса ERCC1/XPF. Затем гэпы (промежутки) (в том случае, если они присутствуют) заполняются полимеразой (например, ДНК-полимеразой θ или δ и разрывами, соединенными комплексом лигазы I или лигазы III с Xrcc1.
При отсутствии расположенных рядом участков микрогомологии на концах ДНК, что бывает наиболее часто, более отдаленный фрагмент подвергнувшейся репарации молекулы можно копировать с применением точной ДНК-полимеразы (например, полимеразы θ). Затем этот дублированный участок принимает участие в выравнивании ДНК-концов, что приводит к вставке в созданный узел. Этот более сложный вариант опосредованного микрогомологией репарации был назван синтез-зависимым MMEJ (SD-MMEJ).
Хотя было высказано предположение, что MMEJ действует в качестве альтернативного пути рекомбинационной репарации, было показано, что он является очень эффективным в процессе рекомбинации переключателей класса IgH в В-лимфоцитах, что свидетельствует о том, что он может представлять собой больше, чем резервный механизм. Также возможно, что репарацию некоторых DSB, например, несовместимых выступов или тупых концов (которые представляют собой плохие мишени для NHEJ и/или HR), можно более эффективно осуществить посредством MMEJ.
На Фиг. 12В показано, что повышение экспрессии ограничивающих скорость HR и/или MMEJ белков стимулирует коррекцию генов зависимым от дозы образом. Как можно видеть, пустой вектор обеспечивает базовый уровень 1, при этом определенные дозы векторов экспрессии, обеспечивающих ограничивающие скорость белки, показывают увеличение, следовательно, повышающим образом регулируют генную коррекцию более, чем в 0,05 раз, более, чем в 0,1 раз, в 0,2 раза, 0,3 раза, 0,4 раза или 0,5 раз. Тем не менее, как это будет понятно специалисту в данной области техники, можно повышающим образом регулировать любой из представленных белков, например, в Таблице С.
ОБНАРУЖЕНИЕ ВОЗМОЖНЫХ НЕОЖИДАННЫХ АДВЕНТИВНЫХ АГЕНТОВ, ТАКИХ КАК ВИРУСЫ
Характеристика всех вирусоподобных элементов в геноме представляющей интерес клетки, таком как геноме СНО, позволяет охарактеризовать потенциально новые адвентивные агенты, для линий, полученных из линий маточного банка клеток. Такой подход может заменить многие из многочисленных способов анализа, которые в настоящее время применяют для характеристики клона клетки-продуцента по мере необходимости для получения одобрения контролирующих органов на любой белок, обладающий терапевтической активностью, продуцируемый указанными линиями клеток. Характеристика также может применяться в контексте анализа производственных партий на регулярной основе. Эффективное редактирование генома СНО путем направленного редактирования гена посредством, например, гомологичной рекомбинации, можно применять для удаления экспрессируемых ретровирусных элементов, которые присутствуют в геномах культивируемых клеток, представляющих интерес, включая клетки СНО, такие как клетки СНО-К1, что приводит к пониженной вирусной нагрузке на геном.
В настоящее время существует несколько доступных коммерческих услуг для редактирования генов или направляемой трансгеном интеграции, которые основаны на механизмах гомологичной рекомбинации. К ним относятся цинк-пальцевые нуклеазы, подобные активаторам транскрипции эффекторные нуклеазы Talen и CRISPR/Cas9-нуклеазы, и аденовирусные векторы, например, SAFC-SANGAMO в США, CELLECTIS во Франции и HORIZON в Великобритании. Тем не менее, двое из таких средств, цинк-пальцевые нуклеазы и нуклеазы Talen, по-прежнему труднодоступны и сложны в применении, поскольку конкретная нуклеаза и/или вектор необходимо модифицировать для каждой редактируемой геномной последовательности, тогда как доступность третьего средства (CRISPR) остается весьма неопределенной. Кроме того, гомологичная рекомбинация остается довольно неэффективным процессом в клетке, для которого необходим скрининг многих линий клеток-кандидатов. Таким образом, удаление двух аллелей рассматриваемого гена зачастую требует множественных циклов мутагенеза и выделения и характеристика клона клеток. Таким образом, эти современные технологии зачастую являются слишком медленными и слишком дорогостоящими, чтобы обеспечить модификацию клеточных геномов на системной основе, как это требуется, например, для удаления множественных экспрессируемых остатков последовательностей вирусного генома, рассредоточенных по геному СНО. Модификация метаболических путей и механизмов рекомбинации в клетке СНО обеспечивает возможность для более эффективных подходов редактирования генома посредством одного из путей рекомбинации, причем в большинстве случаев гомологичная рекомбинация является предпочтительной.
Для идентификации экспрессируемых ERV-элементов, геномную последовательность линии клеток СНО (например, клеток СНО-М, полученных из SELEXIS'S СНО-K1) определяли с приблизительно 120-кратным покрытием с применением технологии PACBIO (Pacific Bioscience Inc.). Секвенирование генома может быть, например, выполнено в секвенаторе для прочтения лонг-ридов PacBio RS II™.
При сборке генома СНО было получено примерно 7200 контигов с размером N50, равным 6,1 Мб.
ИДЕНТИФИКАЦИЯ И ХАРАКТЕРИСТИКА ЭКСПРЕССИРУЕМЫ ERV-ЭЛЕМЕНТОВ В ГЕНОМЕ КЛЕТОК СНО
Геномные последовательности клеток СНО, отображающие сходство последовательностей с известными ретровирусными последовательностями мышей, идентифицировали с применением алгоритма BLAST для поиска кодирующих последовательностей вирусных GAG-, POL- и ENV-элементов с сохранением последовательности по меньшей мере на 80%, а также присутствием вирусных длинных концевых повторов (LTR).
Как показано на Фиг. 13, было идентифицировано три класса гаммаретровирусов, которые могут способствовать образованию ERV или ERV-элементов, обнаруживаемых в представляющих интерес клетках. ERV-элементы, которые экспрессируются с образованием функционального белка, представляют особый интерес в контексте настоящего изобретения. Отправной точкой является база данных вирусных, в частности ретровирусных и/или микробоподобных, последовательностей в представляющих интерес клетках. Соответствующая база данных соответствующих последовательностей РНК таких клеток обеспечивает информацию о том, какой из элементов ДНК фактически экспрессируется и может кодировать функциональный белок.
Построение базы данных соответствующих ДНК-элементов
Были идентифицированы рассредоточенные по геному представляющих интерес клеток повторяющиеся последовательности, образованные в результате прошлых вирусной и ретровирусной интеграции и ретротранспозиции, например, клеток, применяемых при производстве обладающих терапевтической активностью белков, в частности, тех элементов, которые остаются экспрессируемыми.
В качестве первого этапа идентификации экспрессируемых ERV-элементов определяли геномную последовательность линии клеток СНО (клетки СНО-М, полученные из СНО-K1 с применением платформы SELEXIS) с приблизительно 120-кратным покрытием. Для этого применяли технологию PACBIO (Pacific Bioscience Inc.). Такое обширное секвенирование можно, например, осуществить в оригинальном секвенаторе для прочтения лонг-ридов PacBio RS II™. Тем не менее, специалисту в данной области техники будет понятно, что одинаково успешно можно применять и другие технологии и устройства для одномолекулярного секвенирования ДНК. Также осуществляли специфичные амплификации методом ПЦР и направленное ДНК-секвенирование соответствующих локусов для получения, например, геномной последовательности СНО-М версии 3.0.
При сборке генома СНО было получено примерно 7200 контигов с размером N50, равным 6,1 Мб.
Для создания базы данных индикаторных последовательностей ДНК или кДНК для потенциальных адвентивных загрязняющих микробных агентов, основанной на списке адвентивных агентов, результаты скрининга которых в настоящее время запрашиваются такими агентствами, как FDA, были идентифицированы геномные последовательности, отображающие сходство последовательностей с известными ретровирусными последовательностями мышей.
Для картирования их позиции внутри геномных каркасов СНО-М, были идентифицированы геномные последовательности СНО-М, отображающие сходство последовательностей с известными ретровирусными последовательностями мышей (например, последовательностями ML2G, Lie et al, 2014), с применением алгоритма BLAST, направленного на поиск кодирующих вирусных последовательностей GAG, POL и ENV с по меньшей мере 80% сохранением последовательности. Выравнивание с применением алгоритма Blast выполняли отдельно для каждой из указанных кодирующих последовательностей. Кроме того, также через алгоритм Blast с 80% пороговым значением исследовали присутствие вирусных длинных концевых повторов (LTR). Средний процент идентичности последовательностей между последовательностями ML2G и вирусным последовательностями GAG, POL и ENV был определен на уровне 92%. Средний процент идентичности последовательностей между вирусными последовательностями GAG, POL и ENV и последовательностью вируса мышиного лейкоза составляет 66%. В некоторых случаях ERV не были полными, и были обнаружены только определенные ERV-элементы.
Идентифицированные ERV-последовательности включали 159 членов семейства ретровирусов класса II, формирующих интрацистернальные частицы А-типа (IAP), 144 гаммаретровирусов типа С (ERV класса 1), а также 8 других гаммаретровирусов, в частности GALV (см. Фигуру 13 для основных классов и филогенетической связи этих вирусных последовательностей). Обычный поиск последовательности мышиного ретровируса типа С в клетке СНО показан на Фиг. 4А.
Дальнейшее исследование гаммаретровирусных ERV-последовательностей типа С и число обнаруженных ERV-элементов показано в таблице А:
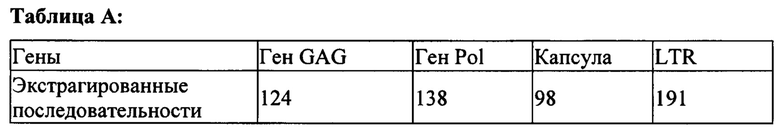
После того, как последовательности обнаружили путем выравнивания, были разработаны праймеры, и с применением таких праймеров был осуществлен поиск соответствующих последовательностей для экспериментального их подтверждения с применением метода ПЦР и секвенирования ДНК. Был осуществлен поиск граничных последовательностей генов, специфических меток интеграции, метилирования и экспрессии ДНК и т.д.
Только 77 анализируемых ERV-последовательностей содержали все элементы (гены gag, pol, env) для создания ERV. Эти результаты были дополнительно проанализированы и результаты показаны на Фигуре 14. Эти 77 ERV-последовательностей также проанализировали с филогенетической точки зрения, и их можно было разделить на две группы, а именно на ERV группы 1 (52) и группы 2 (14). В пределах этих групп идентичность последовательностей составляла 97-99%, а между группами идентичность последовательностей составляла 84-97%. Указанные две группы отличались по своим LTR. GAG-кодирующая последовательность была хорошо сохранена. Сравнение последовательностей выявило, что две вспышки ретровирусных инфекций могли произойти сравнительно недавно в течение эволюции китайского хомячка, что привело к возникновению указанных хорошо сохранившихся и, возможно, функциональных ERV (Фиг. 15).
Кроме того, было обнаружено, что также присутствует последовательность ДНК, кодирующая связанный с PPXY мотив PPYP, важный для функционирования GAG. ERV группы 1 демонстрировали от одного до двенадцати различий или интеграции с размером в 30 п.о. в геноме, ERV группы 2 не демонстрировали вообще или демонстрировали до 6 различий в гене gag по сравнению с родственными функциональными генами gag.
Исследование потенциальной экспрессии идентифицированных ДНК-элементов
Для оценки того, какой ERV-элемент(-ы) может транскрибироваться, статус их ДНК-метилирования CpG анализировали с применением базы данных последовательностей PACBIO (Suzuki et al., (2016)). Это позволило идентифицировать несколько ERV, чьи LTR-последовательности, как и ожидалось, были гипометилированы для экспрессии разрешающей структуры хроматина и транскрипционно активных LTR-промоторных последовательностей, как это показано на Фиг. 16.
Создание базы данных РНК
Дальнейшее секвенирование и анализ нескольких независимых мРНК-препаратов из находящихся в банке клеток, таких как клеток-хозяева СНО-М, можно осуществить для создания базы данных последовательностей РНК (т.е. кДНК), которые в виде таблицы отображают базы данных геномных и/или индикаторных адвентивных последовательностей. Продукт представляет собой коллекцию экспрессируемых ретровирусных/вирусных, ретротранспозонных и бактериальных маркерных генов и семейств, представленных в виде таблицы относительных уровней экспрессии для каждого типа элемента.
Последовательности многокопийных РНК, которые невозможно отнести к геномной или митохондриальной геномной ДНК, экспериментально подтверждают методом количественной ПЦР в режиме реального времени (RT-qPCR). Экспериментально подтвержденные последовательности ищут возможные варианты для транс-сплайсинга. Оставшиеся неидентифицированные (unexplained) последовательности подвергаются скринингу на наличие гомологий с доступными геномами вирусных, прокариотических и эукариотических последовательностей для того, чтобы их можно было отнести к идентифицированным видам микроорганизмов. Соответствующие совпадения добавляются в базу данных экспрессируемых последовательностей.
Примеры специфичного создания базы данных РНК, экспрессируемой идентифицированными элементами ДНК
В настоящем документе транскрипция специфичных ERV-элементов дополнительно оценивали посредством их прямого сравнения с последовательностью кДНК GAG, полученной с применением обратной транскрипции всей РНК клетки СНО, в настоящем документе кДНК СНО-М, РНК клетки, и амплификации методом ПЦР. Это свидетельствовало о том, что ERV-элементы из обеих групп 1 и 2 являлись транскрипционно активными. Сравнение геномных и кДНК-последовательностей также выявило, что транскрибировались ERV только одной группы 2, которые, тем не менее, не могли экспрессировать функциональный белок GAG вследствие наличия стоп-кодонов (Фиг. 17). Тем не менее, экспрессировались некоторые идентифицированные ERV-последовательности группы 1, последовательность их РНК не содержала преждевременных стоп-кодонов, и, таким образом, они, по всей видимости, опосредуют экспрессию функционального белка GAG. Анализ смежных POL-кодирующих последовательностей показал, что они также были функциональными, что привело к идентификации нескольких экспрессируемых и консервативных ERV в качестве очень эффективных кандидатов для экспрессии и высвобождения ретровирусных частиц из клеток СНО. Полная консенсусная последовательность ДНК таких функциональных ERV Группы 1 представлена SEQ ID NO: 1, тогда как консенсусные последовательности ERV Группы 2, IAP-элементы и другие гаммаретровирусные последовательности, идентифицированные в настоящем документе, представлены SEQ ID NOS: 2, 3 и 4.
ОПРЕДЕЛЕНИЕ ХАРАКТЕРИСТИК АДВЕНТИВНЫХ ЭЛЕМЕНТОВ, ПОЛУЧЕННЫХ В РЕЗУЛЬТАТЕ ПОЛНОГЕНОМНОГО/ЭПИСОМНОГО СЕКВЕНИРОВАНИЯ
Биоинформационный пакет предназначен для быстрой идентификации (i) новых мобильных генетических и адвентивных элементов, полученных в результате полногеномного и эписомного секвенирования, и (ii) значимых изменений в уровнях экспрессии известных микробоподобных генов, поскольку они могут указывать на загрязнение культуры клеток адвентивными агентами. Загрязнение следует отличать от хромосомных перегруппировок.
В настоящем документе программы сравнения генома применяют для разработки автоматизированных процессов, чтобы отличать небольшие изменения в геноме (такие как транспозиция или интеграция вирусного генома) от крупных хромосомных перегруппировок (таких как крупные хромосомные транслокации или явления дупликации) или от загрязнения другими культивированными клетками.
Параметры вышеуказанного процесса распознавания определяют с применением геномных и РНК-последовательностей, полученных из банка клеток, таких как клетки СНО-М, и из клеток клонов, полученных из таких клеток, применяя клоны, в которых ранее подтверждалось отсутствие адвентивных агентов при помощи обычных средств.
Вышеуказанный процесс применяют в сочетании с полученными выше базами данных, для идентификации потенциальных изменений в геноме, вызванных в ходе выделения клонов клеток СНО. Соответствующие совпадения подтверждают экспериментально методом ПЦР и мелкомасштабным секвенированием ДНК.
Аналогичный анализ осуществляют с применением транскриптома, например, клонов клеток СНО-М. Новые последовательности РНК или последовательности РНК, уровень экспрессии которых изменяется значимым образом, идентифицируют с применением биоинформационного анализа. Совпадения подтверждают экспериментально методом RT-qPCR.
Процесс обнаружения сначала подтверждают с точки зрения чувствительности при помощи биоинформационного моделирования с последующим экспериментальных введением препаратов нуклеиновой кислоты СНО-М с известным количеством бактериальных (например, микоплазменных) последовательностей или последовательностей, подобных последовательностям ретровирусов, и эталонных материалов. Это обеспечивает порог чувствительности с точки зрения количества вирусного генома на эквивалент клеточного генома, который можно обнаружить после заданной глубины (или геномного покрытия) «секвенирования следующего поколения».
Способ более эффективного редактирования генома клеток СНО и/или транскриптомы
Предложен количественный способ анализа для определения направляемой HR и HDR интеграции и удаления трансгена из генома клеток СНО посредством нокдауна или гиперэкспрессии белков, участвующих в рекомбинантных действиях.
В настоящем документе сконструировано несколько независимых индикаторных линий клеток СНО, которые содержат один интегрированный в геном трансген GFP (например, через мобильные векторы), и интеграцию трансгена подтверждают методом количественной ПЦР и картированием сайта интеграции. Индикаторную плазмиду конструируют методом брэкетинга кассеты экспрессии dsRed при помощи последовательностей, соответствующих концам GFP-кодирующей последовательности. Частоты спонтанной мутации последовательности GFP и нецелевой интеграции кассеты экспрессии dsRed подтверждают документально относительно надлежащей направленной интеграции с применением флуорометрического и qPCR-анализов. В альтернативном варианте одна или несколько удаленных GFP-кодирующих последовательностей интегрируют в геном клетки, и их восстанавливают с применением механизмов, связанных с HR или HDR, для восстановления функциональной GFP-кодирующей последовательности и флуоресцентных клеток.
Вышеуказанный анализ применяют после транзиентного миРНК-опосредованного нокдауна белков MMEJ и NHEJ с тем, чтобы идентифицировать предельные активности, которые могут препятствовать эффективным HR-механизмам. Аналогичный анализ осуществляют в присутствии вектора экспрессии для Cas9 и нацеленной на GFP гидовой РНК CRISPR для оценки HDR. Параллельно с этим, кДНК, кодирующие белки HR и HDR в СНО, клонируют и интегрируют в векторы экспрессии. Эти кДНК совместно трансфицируют индикаторной плазмидой dsRed для идентификации активности, которая ограничивает эффективность HR или HDR в клетках СНО-М.
Комбинации вышеуказанного нокдауна или гиперэкспрессии осуществляют для улучшения частоты HR-подобных явлений (см. Фиг. 9-12). Наиболее эффективные комбинации оценивают и выбирают на основе i) результатов вышеуказанных анализов отдельных активностей, ii) уровней мРНК конкретных генов рекомбинации в клетках СНО-М и iii) относительных положений в пути рекомбинации и мультимерных структур, образованных белками. Такие комбинации анализируют для удаления последовательности dsRed, интегрированной в GFP, для восстановления функционального GFP, для идентификации процесса, который эффективно удаляет экспрессируемые последовательности из генома СНО.
Генетическое или эпигенетическое редактирование СНО-М для удаления инфекционных агентов, представляющих собой остатки ДНК или РНК
В настоящем документе экспрессируемые остатки вирусного генома из линий клеток СНО удаляют или осуществляют их сайленсинг. Вирусные и подобные ретровирусам элементы, экспрессируемые клетками СНО-М, определенные выше, нацеливаются на HR или HDR с применением оптимизированного вышеуказанного подхода, чтобы нарушить функциональные вирусные последовательности, предпочтительно последовательности gag, которые опосредуют высвобождение вирусных частиц. В альтернативном варианте или одновременно механизмы, опосредующие HR или HDR, ингибируют для того, чтобы способствовать восстановлению DSB в вирусных последовательностях посредством механизмов репарации, которые способствуют делеции на участке расщепления ДНК, таких как механизмы NHEJ и MMEJ, чтобы удалить последовательности gag, которые необходимы для высвобождения вирусных частиц.
Проводят несколько циклов редактирования генома, а после уменьшения количества геномных вирусных последовательностей и последовательностей РНК следует количественная ПЦР. После каждого цикла будет определено время деления клеток, чтобы гарантировать, что дополнительные изменения в геноме не ухудшают метаболические свойства указанных клеток.
Подходящим образом модифицированные клетки, такие как популяции клеток СНО-М, исследуют на их стабильность и способность экспрессировать маркер (например, GFP) или терапевтические (например, иммуноглобулины) белки. Получают субклоны и аналогичным образом их анализируют. Документально подтверждают, что в подходящих субклонах отсутствуют возможные дополнительные адвентивные агенты, применяя описанные в настоящем документе способы обнаружения.
Геномную последовательность наиболее эффективных клеток, таких как субклоны СНО-М, получают и осуществляют ее сборку для документального подтверждения изменений в геноме, а также уменьшения количества адвентивных вирусоподобных последовательностей.
Для того, чтобы специфическим образом расщепить и произвести мутацию в экспрессируемых ERV Группы 1 с применением подхода для редактирования генома CREMPR/Cas9, били разработаны гидовые РНК для распознавания предпочтительных последовательностей вокруг мотива PPYP или последовательности миристоилирования последовательности GAG (Фиг. 18 и SEQ ID NOS: 5-12). Мотив PPYP был выбран для применения подхода с нокаутом, при котором не являющиеся HR механизмы репарации ДНК могут приводить к делециям в GAG-кодирующей последовательности, что может ухудшать экспрессию функциональной версии указанного белка, который опосредует почкование вирусных частиц. На мотив миристоилирования нацеливаются, чтобы заменить миристоилированную аминокислоту на немодифицируемый аналог путем направляемой гомологией HR-репарации ДНК для образования доминантно-негативной версии GAG, которая может быть как нефункциональной, так и может ингибировать функцию вероятно оставшихся немутированных белков GAG, что основывается на известном требовании к миристолированию белка GAG ВИЧ1 для почкования ВИЧ (Abdusetir Cerfoglio et al. (2014)). Одиночные гидовые РНК (огРНК) были разработаны так, чтобы соответствовать мономерной версии CRIPR/Cas9, как это показано на Фиг. 6 (Carrol 2013).
Векторы экспрессии Cas9 и огРНК были совместно трансфицированы вектором экспрессии dsRed в клетки СНО, и клетки, экспрессирующие dsRed, сортировали для приумножения числа (обогащения) эффективно трансфицированных и трансген-экспрессирующих клеток (Фиг. 19). В альтернативном варианте, клетки трансфицировали смесью из трех нацеливающихся на РР51 миРНК, чтобы понизить экспрессию Rad51 и уменьшить репарацию ДНК посредством механизмов гомологичной рекомбинации. Эффективность нокдауна Rad51 оценивали как по уровням мРНК Rad51, так и по результатам ингибирования восстановления функции мутированного трансгена GFP посредством гомологичной рекомбинационной репарации, что свидетельствует о том, что все разработанные миРНК специфическим образом ингибировали экспрессию Rad51 и HR (Фиг. 20).
Чтобы непосредственно оценить, была ли надлежащим образом обработка CRISPR/Cas9 нацелена на экспрессируемые ERV, РНК GAG подвергали обратной транскрипции и амплифицировали методом ПЦР. В то время как РНК GAG можно было легко обнаружить в контрольных клетках, как и ожидалось клетки, обработанные огРНК Cas9 PPGP5, стабильно приводили к получению низких или неопределяемых уровней в экспериментах по независимой трансфекции (1-й и 2-й эксперименты с применением CRISPR на Фиг. 21). Поскольку обработка РР9Р6 и PPYP13-кодируемой Cas9 значимым образом не изменяла уровни РНК GAG в случае только ее применения, оценивали, может ли миРНК-опосредованный нокдаун активности Rad51 увеличить эффект такой обработки CRISPR/Cas9, поскольку можно было ожидать, что это предотвратит эффективную репарацию расщепленных генов gag посредством HR-механизмов и, таким образом, может привести к альтернативному пути репарации ДНК, подверженному делециям. Такой случай уже наблюдали, поскольку предыдущий нокдаун Rad51 привел к исчезновению сигнала мРНК GAG при обработке PPYP6 или PPYP13 и Cas9 (3 эксперимента с применением CRISPR на Фиг. 21).
Предыдущие попытки мутировать гены с применением мутагенных систем на основе CRISPR/Cas или других нуклеаз не привели к исчезновению кодированной мРНК даже для низкокопийных генов. Ожидалось, что такая ситуация усложнится в случае большого количества копий ERV, которые включают ERV, присутствующие в клетке в количестве копий более, чем 30, 40 или даже 50, как это обнаруживают в геномах млекопитающих. Таким образом, совершенно неожиданно наблюдали, что ретровирусные РНК невозможно было обнаружить в клетках, обработанных огРНК Cas9-нуклеазы PPGP5 (1-й эксперимент с применением CRISPR на Фиг. 21). В предыдущей попытке удалить или мутировать ген POL из 62 связанных эндогенных ретровирусных элементов не удалось достигнуть их удаления из клеток свиней с применением транзиентных трансфекций, что привело к стабильной экспрессии элементов CRISPR/CAS9 с применением мобильных или вирусных векторов (Yang L. et al., 2015). Тем не менее, стабильная активность CRISPR/Cas на протяжении только 17 дней приводила к частоте максимального нацеливания в 37% от числа клеток, вероятно потому, что пролонгированная активность CRISPR/CAS приводила к токсическим нецелевым эффектам расщепления.
Проведенные эксперименты показывают, что нокдаун белков HR, таких как Rad51, можно применять для увеличения частоты инактивации гена GAG при помощи вредных делеций с частотой, которую не наблюдают при отсутствии трансфекции миРНК.
В то время как нокдаун Rad51 представлен в качестве примера, специалисту в данной области техники будет понятно, что при нокдауне других белков HR будут получены сопоставимые результаты.
Соответственно, клетки СНО, содержащие другие белки HR, подвергнувшиеся нокдауну, входят в объем настоящего изобретения. Однако также клетки, содержащие 53ВР1, CtIP, Mre11, Rad50, лигазу III, Pold3 (дельта-субъединица 3 ДНК-полимеразы), Xpf и Blm (RecQ-подобная геликаза синдрома Блума), подвергнувшиеся нокдауну (см. Фиг. 9), особым образом включены в настоящее изобретение. В Таблице С приводятся примеры белков HR, для которых можно произвести нокдаун, а также белки других путей рекомбинации, для которых можно произвести нокдаун или осуществить их гиперэкспрессию. Тем не менее, специалисту в данной области техники будет понятно, что Таблица С упрощена: некоторые из белков, упоминаемые в Таблице С, обладают двойной функцией (например, лигаза III), и их можно расположить в разных категориях.
Не ограничиваясь какой-либо конкретной теорией, для расщепляемых Cas9 ERV можно осуществить репарацию посредством гомологичной рекомбинации, взяв один ERV из множества других ERV в качестве гомологичной матрицы, которую можно ингибировать путем инактивации белков путей, направляемых гомологией, таких как Rad51. Напротив, некоторые предыдущие попытки осуществить нокдаун экспрессии Rad51 до экспрессии компонентов CRISPR/Cas9 привели к усилению путей репарации ДНК, направляемых гомологией, чего следует избегать в случае, когда предпочтительны делеции, приводящие к инактивации гена (Davis L. и Maizels, N. 2014 и 2016). В других исследованиях было обнаружено, что гиперэкспрессия Rad51 может усиливать путь репарации ДНК, направляемый гомологией, при исследованиях направленной интеграции генов (т.е. нокину) после опосредованного CRISPR/Cas9 или TALEN расщепления ДНК (Song J. et al., (2016)). В настоящем документе показано, что RAD51 способствует генной коррекции по путям HDR в различных типах клеток СНО (Фиг. 9 и 12), и что ингибирование активности гомологичной рекомбинации может значительно усилить мутацию ERV при применении в сочетании с ERV-специфичной ДНК-нуклеазой. Таким образом в том случае, когда следует инактивировать множество аллелей, а значит, в случае ERV с большим количеством копий, зачастую предпочтителен вариант реализации, который объединяет нокдаун миРНК, в частности, HR-пути, в сочетании, в частности, с подходом применения CRISPR/Cas9, нежели чем применение только CRISPR/Cas9, что является еще одним вариантом реализации настоящего изобретения.
В целом, очень высокая мутагенная активность компонентов CRISPR/Cas9 неожиданно оказалась повышенной по сравнению с уровнем техники. Не ограничиваясь какой-либо конкретной теорией, причиной может быть то, что Cas9-нуклеаза специфически нацеливается на экспрессируемые ERV, а не на неактивные ERV. Тем не менее, это вряд ли объясняет уменьшение вирусных РНК до неопределяемых уровней при самых эффективных настройках. Таким образом, также может быть, что специфичные расщепления ДНК могут привести к транскрипционному сайленсингу экспрессируемых ERV.
В конечном итоге, некодирующие мутированные РНК, содержащие преждевременные стоп-кодоны, зачастую нарушаются внутри клетки посредством механизма нонсенс-опосредованного распада РНК (NMD) (Baker and Parker, 2004). Таким образом, затем оценивали, может ли огРНК PPYP опосредовать мутации вне рамки считывания, что может привести к NMD-опосредованному разложению РНК GAG. Такая мутация сдвига рамки считывания, образуемая в результате делеции 11 нуклеотидов из мотива PPYP ERV Группы 1, показана на Фиг. 21 в качестве примера.
На Фиг. 22 показан результат амплификации методом ПЦР и секвенирования генов GAG, содержащихся в поликлональной популяции клеток СНО, обработанных CRISPR/Cas. Результаты четко свидетельствуют о том, что большинство GAG-кодирующих последовательностей мутировали после транзиентной трансфекции векторов экспрессии для компонентов CRISPR/Cas9-нуклеазы, даже при использовании умеренно активной огРНК PPGP6 и отсутствии обработки siRAD51.
В целом, был сделан вывод о том, что предлагаемый подход с применением CRISPR/Cas9, нацеливающейся на клетки СНО с эндогенными ERV, можно применять для исключения или понижения экспрессии белка GAG, который опосредует высвобождение вирусных частиц, и в этом случае отсутствует необходимость в пролонгированной экспрессии CRISPR/Cas или стабильной интеграции векторов их экспрессии в клеточный геном и потенциальное негативное воздействие с их стороны на культуру клеток. Таким образом, данный подход можно применять для создания более безопасных линий клеток СНО, которые не обладают способностью экспрессировать такие ретровирусные белки.
Материалы и Способы
Линия клеток
Редактированной линией клеток была линия клеток SURE СНО-М™ (SELEXIS SA, Швейцария) (см. патенты США №№7129062 и 8252917 и опубликованные заявки №№20110061117, 20120231449 и 20130143264, содержание которых включено в настоящее описание посредством ссылки).
CRISPR/Cas9-система
Для редактирования генома авторы настоящего изобретения применяли кодон-оптимизированную для млекопитающих Cas9-нуклеазу Streptococcus pyogenes. Последовательности одиночных гидовых РНК (огРНК) клонировали в вектор экспрессии гРНК млекопитающих MLM3636, описанный ранее (Fu et al, 2013). Для достижения высокой эффективности нокаута исследовали несколько сайтов-мишеней для огРНК на сайт-мишень (PPYP: n=5, Myr: n=3). Пакет CRISPRseek R (Zhu et al., 2014) применяли для идентификации ERV-специфичных огРНК с минимальным числом нецелевых сайтов, применяя находящийся в собственности авторов настоящего изобретения геном СНО-М в качестве эталонной последовательности. Последовательности огРНК разработали для гибридизации с прилежащими к сайтам-мишеням участками, а точнее с мотивами Myr и PPYP, для максимизации потенциала мутагенеза CRISPR/Cas9 для данных локусов. Все возможные последовательности огРНК получили с применением пакета CRISPRseek R (Zhu et al., 2014), который содержит структуру последовательности вида 5'-20 нуклеотидов NGG-3' и который опосредует DSB размером не более, чем 25 п.о. вне зависимости от сайта мутагенеза. Среди числа всех потенциальных последовательностей огРНК, эффективности огРНК были предсказаны с применением различных инструментов оценки, включая CRISPRseek (Zhu et al., 2014), Sequence Scan для CRISPR (SSC, веб-сайт Dana Faber Institute crispr) и sgRNA scorer 1.0 (веб-сайт Dana Faber Institute crispr). Кроме того, пакет CRISPRseek R (Zhu et al., 2014) применяли для идентификации ERV-специфичных огРНК с минимальными числом нецелевых сайтов, применяя находящийся в собственности авторов настоящего изобретения геном СНО-М в качестве эталонного генома. В конечном итоге, было отобрано и исследовано несколько последовательностей огРНК на сайт-мишень (Myr: n=3, PPYP: n=5) для идентификации огРНК с максимальным потенциалом мутации. Cas9 (№43861) и плазмиды экспрессии гРНК (№43860) доступны от ADDGENE (Кембридж, Массачусетс, США).

РНК-интерференция
Малую интерферирующую РНК (миРНК) против гомологичной Rad51 клеток СНО разработали и синтезировали при помощи MICROSYNTH AG (Бальгах, Швейцария). Сайленсинг уровней экспрессии мРНК Rad51 осуществили с применением смеси из трех Rad51-специфичных миРНК для минимизации нецелевых эффектов. Анализ BLAST исключал гомологию для других мишеней. В качестве контроля применяли смесь из трех ненацеливающихся миРНК (siNeg).
Модификация генома СНО-М
Клетки СНО-М трансфицировали с применением системы Neon Transfection System® (INVITROGEN) согласно протоколу производителя. Вкратце, 400000 клеток трансфицировали 100 нМ смесью миРНК, и через 48 часов 700000 клеток повторно трансфицировали CRISPR/Cas9-системой с применением dsRed-кодирующей плазмиды в качестве контроля трансфекции. Для приумножения числа (обогащения) трансфицированных клеток, примерно 100000 dsRed-положительных клеток отсортировали через 48-72 часа после трансфекции методом проточной цитометрии.
Специалисту в данной области техники будет понятно, что приведенное выше описание не является ограничивающим, однако обеспечивает примеры определенных вариантов реализации настоящего изобретения. В соответствии с приведенным выше руководством, специалист в данной области техники может разработать множество альтернативных вариантов, конкретно не указанных в настоящем документе.
В Таблице С перечислены некоторые из ключевых генов в каждом из трех путей (см. также публикацию патента США №20120231449, содержание которого полностью включено в настоящее описание посредством ссылки). Также в таблицу включены репарационные белки ДНК, такие как MDC1 и MHS2. MDC1 необходим для активации контрольных точек интра-S фазы и G2/М-фазы клеточного цикла в ответ на повреждение ДНК. Тем не менее, MDC1 также функционирует в Rad51-опосредованной гомологичной рекомбинации, удерживая Rad51 в хроматине. (Для последовательностей см. публикацию WO 2014/118619, публикацию патента США №20150361451, содержание которого полностью включено в настоящее описание посредством ссылки).
ВЫБРАННЫЕ ГЕНЫ ОПРЕДЕЛЕННЫХ ПУТЕЙ РЕКОМБИНАЦИИ

БИБЛИОГАРАФИЯ
Abdusetir Cerfoglio et al., Structural elements in the Gag polyprotein of feline immunodeficiency virus involved in Gag self-association and assembly, Journal of General Virology 95, 2050-2059 (2014).
Baker, K.E.; Parker, R., Nonsense-mediated mRNA decay: Terminating erroneous gene expression". Current Opinion in Cell Biology 16: 293-299 (2004).
Carrol et al., Staying on target with CRISPR-Cas, Nature Biotechnology 31, 807-809 (2013).
Davis, L., and Maizels N., Homology-directed repair of DNA nicks via pathways distinct from canonical double-strand break repair. Proc. Natl. Acad. Sci. USA 111, E924-E932 (2014).
Davis, L., and Maizels, N., Two Distinct Pathways Support Gene Correction by Single-Stranded Donors at DNA Nicks. Cell Reports 17, 1872-1881 (2016).
Food and Drug Administration (FDA), FDA's Guidance for Industry Characterization and Qualification of Cell Substrates and Other Biological Materials Used in the Production of Viral Vaccines for Infectious Disease Indications, February 2010, available at the Fda.gov website.
Fu et al., High-frequency off-target mutagenesis induced by CRISPR-Cas nucleases in human cells. Nat. Biotechnol. 31, 822-6 (2013).
Guilinger et al., Fusion of catalytically inactive Cas9 to FokI nuclease improves the specificity of genome modification, Nature Biotechnology 32, 577-582 (2014)
Hope & Trono, HIV, Structure, Expression, and Regulation of the HIV Genome, Site Knowledge Base Chapter November 2000.
Публикация международного патента WO 2014/118619 (2014).
Kostyrko et al., MAR-Mediated transgene integration into permissive chromatin and increased expression by recombination pathway engineering, Biotechnology & Bioengineering, 3 October 2016 online edition.
Lewis et al., Genomic landscapes of Chinese hamster ovary cell lines as revealed by the Cricetulus griseus draft genome, Nature Biotechnology 31, 759-765 (2013), Supplement)
Lie, Y.S. et al., Chinese hamster ovary cells contain transcriptionally active full-length type С proviruses. J. Virol. 68:7840-7849(1994).
Ma et al., Hairpin opening and overhang processing by an Artemis:DNA-PKcs complex in V(D)J recombination and in nonhomologous end joining. Cell 108, 781-794 (2002).
Song J et al., RS-1 enhances CRISPR/Cas9- and TALEN-mediated knock-in efficiency, Nature Communications 7:10548 (2016).
Suzuki et al., AgIn: measuring the landscape of CpG methylation of individual repetitive elements. Bioinformatics 52:2911-2919 (2016).
Публикация патента США №20120231449 (2012).
Публикация патента США 20150361451 (2015).
Yang L. et al., Genome-wide inactivation of porcine endogenous retroviruses (PERVs), Science 350:1101-1104 (2015).
Zhu et al., CRISPRseek: A Bioconductor package to identify target-specific guide RNAs for CRISPR-Cas9 genome-editing systems. PLoS One. 9 (2014).
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Selexis SA
<120> УЛУЧШЕННЫЕ ЭУКАРИОТИЧЕСКИЕ КЛЕТКИ ДЛЯ ПОЛУЧЕНИЯ БЕЛКА И
СПОСОБЫ ИХ ПОЛУЧЕНИЯ
<130> 3024-232/ SEL WO 070
<140> PCT/EP2016/082567
<141> 2016-12-23
<150> US62/387375
<151> 2015-12-24
<160> 32
<170> PatentIn version 3.5
<210> 1
<211> 8366
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<222> (1)..(8366)
<223> Тип ERV группы 1 клеток CHO-K1
<400> 1
tgtaccagac accagacctt gagaatatgc tgatctggaa tggctctgtg tctcatttga 60
accatccaat ggaaatgatt ctgtatttcg cctcatttga aagactctgt gtttcacctc 120
atttgaataa ctctgtactt tacctcattt gaataaccct gtatagcgcc tcatatacat 180
tgaccaatgg gaatagctct gtataatgcc tcattagaat tatccaatag aatccttgct 240
cctagcttgc gccttttttc ctatataagg accccttttc ccttggctcg gggcgcttag 300
ccacacagaa gctaagtcgc cccaggtacc tgcgtctcca ataaagcctc ttgtttttac 360
atccagttcg tggcctcgct gattcctggg tgtgtgggtc tccctctacg aaagtgcctc 420
ttcggggtct ttcatttggg ggctcgtccg ggattgagac ccgcccaggg accaccgacc 480
cacgtctggg aggtaagtgt tgtgcggatc cgctgttttg tcttgtctgg tctgagtctg 540
tcttgtgaat tgcgcttgcg tttgtagtat acagctgtgt acatttgtag gcggatccga 600
ggagggactg acgggtccga actcccgacc gcggctccag gagacgtcct ggtagcgttt 660
gaagccctca ggaagaggga tttgtatttt gaacttggga agccctcagg gtgagagatt 720
tgtactttga acttagatct atgactggac attttcccag tctctttgga gaaggccctc 780
ggcttgaggg atttgcaatc tttactgggg acgaggaagg agggccccct tcctcgactc 840
tctctcaatt ccttctgtcg actctctgtc gaaaccgcgc tgcgaaagtc tgttctgtgt 900
tattcggtct ttgtcttgta gctgtcattt gtgccctcct aagcctagaa actatgggca 960
aactgtcacc actcctttgt ccctaacact ctcccactgg aaagatgtac aggaatatgc 1020
tcataaccaa tctgttaatg tgcgtaaacg caaatggatt actctttgtt cttcagaatg 1080
gccgaccttt gatgtaggct ggccgcgaga tggtaccttt aacccccaga ctatattcca 1140
gataaaagag aagattatgg atcctggacc acacgggcat cccgatcaag tggcttatat 1200
cgtcacttgg gaggctttgg ttcaggaccc ccctccctgg gtacgtcctt tcttacatcc 1260
caagggcccc tctctccttc cccctctaac cgctccaacc gacccattcc ttcggcccta 1320
cacctcccac tcctttgatt cctcccaacc cccttcccat tccaaccttt accctaccgt 1380
gatgaaagac actaaggcta aagaaaagaa gacacctaag gtactccctc cgggagaaga 1440
ccagttggtt gatctattaa cggaggagcc cccgccatat ccgccactgc cgcccccacc 1500
agaggcagaa gcggactccg ccgctgcctt ggcggaagcg gccctgaccc ttcaccaatg 1560
gcttatcgac taagaggtcg tagggagcag cccgttccag attcaaccac tctgcccctc 1620
cgaactgggc tgaacggcca acctcagtat tggccattct cagcatcgga cctctataac 1680
tggaaaaata ataatccttc tttttctgca gaccccgtga ggctgacatc tctcatagag 1740
tcggtactca cgactcacca acccacctgg gatgattgtc agcagctttt gcaggtcctt 1800
ttaacctcgg aagagaaaca gcgcgtgcta ctagaagcac gaaaaaatgt cccaggagta 1860
aacgggcagc ccacccagct acccaatgaa attgatgcgg cttgccctct tgaaagacct 1920
gaatgggatt ttaccaccga agcaggtagg acccacctgc gtctctatcg ccagttgctg 1980
gtagcgggac tccgggggca ggacgcagac ccactaattt ggcccaggtg aagcagataa 2040
tacagggtgc ggaggaatca cctgccgctt ttctagagag attgaaagaa gcgtacagga 2100
tgtatactcc ctataatccg gaagatccag gtcaggccac caacgtttct atgtccttta 2160
tttggcaatc agccccggac ataagaaaca agcttcaaag gctagaaaat ctacagggat 2220
atacactcca agatttgttg aaggaagcag aacgtatttt taataagagg gaaacacaga 2280
cagaaagaga agaacgttgg agaaggaaac tcaggagaga gaggaaagac taagacagga 2340
agctgaggaa aaagaggttg cgagagaccg taagcggaat aaagaaatga gcaggttatt 2400
ggccacagta gtgacaggcc agagacagaa tagacagagg gatgacagaa gggggcccca 2460
cctggacagg gaccaatgtg cttactgtaa agaaaaagga cattgggcaa gagaatgccc 2520
taagaacccc cggccaagct tccaccgcca agggtttctg acctcctgaa cctagaagat 2580
tagaggagtc ggggccagga gcccccccct gagccaggat aacactgcaa gtcggggggc 2640
atccggtcac cttcctagtc gatacagggg cacaacattc cgttctgaat cggtcacccg 2700
gacccctgag tcacaggact gcatgggtac agggagctac aggcggaaag cagtaccatt 2760
ggactacaaa tcggcagctc cagctcgcga ccggtaaggt tatgcattct ttcctccatg 2820
tgccagactg cccctacccc ttactagacg ggacctattg accaaattaa aagctcaaat 2880
acactttgag aggtcagaag tcaaagtcac agggccagag ggaattcccc ttaccatctt 2940
gacaatgtcc atagaagatg aatatagact ccatgaaaag aggactaatt cgaacaatca 3000
ggaaaccctt gatcactggc ttgcggaatt tccccaagcc tgggctgaaa caggaggaat 3060
gggccttgcc attaaccagg ccccaattat agtaacctta aaagctgcca tccttcctgc 3120
atccgtcaga cagtatccaa tgcctaaaga agcccgcgaa ggaattcggc cacatattaa 3180
aaggttactt gaacaaggga ttctggtgcc ctgtaaatct ccttggaata cacccttgct 3240
acccgttagg aagccgggaa ctaatgacta ccggccagta caggacctga gagaagtcaa 3300
taaaaggata gaggacatac accctactgt ccccaaccct tacaatttgc tgagtggatt 3360
gccacctaac tatacctggt acacagtctt agatcttaaa gacgctttct tctgcctccg 3420
cctgcatccc accagccagc ctatatttgc ctttgaatgg caggacgcgg cccttggaat 3480
ctctgggcag ctgacttgga ctaggctacc tcaagggttt aagaacagcc ctaccctttt 3540
tgatgaagct ttacatcagg acctggcaga attccggtta ggtaccccgc tctaatcctc 3600
ttacaatatg tagatgacat tctcctggca gccaaaacca aagggaaatg caaggaaggc 3660
actcaagccc tcctccagac tcttgggagc ctaggtaccg ggcatccgcc aagaaggccc 3720
agatatgtca gaaacaggtg acctatttag gatacaagat aaaggatgga cgtcgatggc 3780
taacggaagc ccgtatgcga gccatcttag acattcccac cccacaaaat ccccgccaac 3840
tgagagaatt cttgggaacg gcaggcttct gccgcctatg gatccctggg tttgccgaaa 3900
tggcggctcc cctctacccc ctcactcggc caggggttgc ttttaaatgg gaagagcccc 3960
aaaagaaagc cttcaccgac atcaaaaagg ctctccttga atcaccagcc ctgggtctac 4020
cggacttagc taagccattt gaacttttta tagatgagaa ggagggctat gctaagggag 4080
tcctcaccca aaatctgggg ccttggagaa ggcccactgc atacctctcc aagaaattgg 4140
atcctgtggc atcgggatgg ccaccctgcc ttcgaatgat tgctgctata gccctgctgg 4200
taaaagattc tcacaagcta accttggggc agcctttgac catacatgcc cctcatgcag 4260
tagaggcagt catcagacag cctccagata gatggcttac taatgcccga atgactcatt 4320
accagactat gctgttagac aaagaccggg tccacttcgg gcctttggtg actctgaacc 4380
cagccaccct gctccccctc cctggggagc ccgaggctca tgattgctta caggtattgg 4440
ccgaggccca tggagcgaga tccgacctga ctgaccagcc tctacctagc ccggaccaca 4500
tctggttcac ggatggaagc agctttttgc atcaaggaga acgaaaggcg ggcgcggcag 4560
tcaccacaga gaatcaggtc gtctgggccc aggcactccc cctggaactt ccgcacagag 4620
ggcagaactc atagcactca cgcaggctct aaaattggca gaaggtaaga ggctcaccgt 4680
gtatacagac agtcgttatg cctttgccac tgcccatata catggagaaa tttacagacg 4740
gagggggctg cttacctccg aagggaaaga cattaaaaat aaggaggaaa tcctcgctct 4800
cttaagggct cttcatctgc ccgctgcctt aagtatcata cattgccctg gacaccaaaa 4860
agggattctt tcgaagcaag ggcaatcgaa gggcagactt ggctgcccga gaggcggccc 4920
tgaccacaga caccactaac ctcctggctc tagagcccac caacgaccat cccttcccct 4980
catgggacta tgaacaaaga gacatccaaa ccctagagaa attgggagcc gcaaaggaac 5040
caaacgggga ttggacttat gaaggaaaga ctgtcatccc ctaccgggta accaagtacc 5100
tagtgacatt tttacataag atgacacatc tgagctccaa gaagatgcgg gagctcctcg 5160
aacgagaaga ggaattcaat ttccttttgg gaaagaacga tattctaaaa caggtaactg 5220
agcaatgtga tgcgtgcgcc cgagtcaacg catccagact gaagcttcct cccgggaacc 5280
gggtcagagg ctaccggccc ggaacacatt gggagataga tttcactgag attaaaccag 5340
gaaaatatgg atacaagtat ctattaattt ttgtagacac cttttcagga tgggttaaag 5400
ccttccctac taaacatgaa acagccaaga tcgttactaa gaaattgctt gaagaaatct 5460
ttccccgtta tgggatgcct caggtattgg gaacagacaa tgggcccgcc ttcgtctccc 5520
aggtaagtca gtcagtggcc accttattgg ggattgattg gaaattacat tgtgcttata 5580
gaccccaaag ttcaggacag gtagaaagga tgaatagaac aatcaaggag actttaacaa 5640
aattgtcgct tgcaactggc actagagact gggtcctcct actcccccta gcactctacc 5700
gcgctcgtaa tacccctgga ccacatgggc tcacaccctt tgagatcctg tatggagtac 5760
ctactcctat cattaacttt cttgatcaag atgtctcaga ttttgctaac tccccttctc 5820
tccaagctca tttacaggcc tccaactagt acaacgggag gtctggaaac cccttgctca 5880
agcttataaa gaccagaggg accatcccac catcccccat tcctaccaga tcgggacact 5940
gtttgggtcc ggcgtcacca ggccaagaac cttgaacccc gctggaaggg accctacatc 6000
gttttgctta ccactcccac cgcactcaag gtagacggca ttgcagcttg gatacatgct 6060
tcacatgtaa agccagccca acccaccgat tcagccactg catcagaatg gaccgcacac 6120
cgcactcaaa atcctttaaa gataagactc tctcgtacac cctcctgttg attggttgtc 6180
tgtttacccc ccatgtagca actaaccccc acagggttta taatatcacc tggaaaatag 6240
ccaatctagg gaccggggaa atagccaacc tcagcactta tatagggact ctacatgatg 6300
ggttccctcc tctctatgtc gacctatatg acttagtagg gtctgattgg gatccctctg 6360
accaggaacc attcccaggg tacggatgcc accaccctgg ggaaggatag gaacaagaag 6420
caaggatttt tatgtttgcc ccggccataa accaactcat ggctgcgggg gccgcaggaa 6480
gggtactgtg caagatgggg atgtgaaacc acaggggagg cttactggaa accctcttcc 6540
tcttgggatt tcatcactct caaacggagg gagatcccag ggtacgcagg gaaaggacca 6600
tggagatgtg ggcaaagagc ctgcggacct tgttatgata gtgccggagg gggaggtttt 6660
caaggcgcca cccccggagg aaaatgcaac cctctcatcc taaggttcac agatgctgga 6720
aaaagaacta cttgggatag tcctaaggtc tggggactca ggctgcaccg agcagggaaa 6780
gatccggtga ctttattctc cctgtacaga caaattactc ccctaagcca acaatcagtt 6840
gggccaaaca tagtaatagc ggaccagaga tcccaaccca ttttcaagtc cctaaacccc 6900
ctaccgttcc taaagctatc actcctacac caggtgctgt caccttctcc cccaccccag 6960
atgccctaaa catcgagata accagagacc ctccaggtac cagagataga ttattacaat 7020
taatccaagg agtttaccaa gccttaaatt tttcagaccc caacaagact caggaatgct 7080
ggttatgcct agtttcccgg cccccatatt atgaaggcgt ggcaatactg ggcaactact 7140
ccaaccagac ctcagcacct accagttgcg gagctgctat gcagcacaag ctcacaatat 7200
ctgaggtctc aggaaagggg ctatgcatag gcaggattcc ttcctcacat caagaattat 7260
gtaaccaagt agagccatta tctcaggaca gccgatacct tgttgcccct tatggaactt 7320
attgggcttg cagtactggg ttgactccct gtgtctctac cactgttctc aacaccacca 7380
ttgacttttg tatattgata gaactttggc ccaaagtcac ataccaccaa cctgaatatg 7440
tttacagcgt actagagaaa tcaacccgat ataagaggga gccactaaag caaatagatc 7500
cagagtatct agagacctct gaatgaaaga ttccattcag ttacaagaga aatgggggaa 7560
tgaaagaccc ctctccctta gccctttctt tctcaagttt gtctcctctt cctcctgtcg 7620
gcggcttccc cgatccccac ccccggtggc ctttccccgc ccggcccgag aacaagcacc 7680
gggtggggcc ggcccgagaa caagcaccgg gtggggccgg cccgagaatg agcaccaggt 7740
gggccagccc gagaacgagc accgggtggg ctggcacgag ggcgagcacc aggtgggtca 7800
gcacgaggac aaacaccgag tgagccggcc tggagctctg cccctgagcc cccgccccgc 7860
ccgaagagaa acactccgtc ccaaggtctc cgccccaagg tcagccatca ggaaaagggg 7920
ggaattgagt ctgctgtacc agacaccaga ccttgagaat atgctgatct ggaatggctc 7980
tgtgtctcat ttgaaccatc caatggaaat gattctgtat ttcgcctcat ttgaaagact 8040
ctgtgtttca cctcatttga ataactctgt actttgcctc atttgaataa ccctgtatag 8100
cgcctcatat acattgacca atgggaatag ctctgtataa tgcctcatta gaattatcca 8160
atagaatcct tgctcctagc ttgcgccttt tttcctatat aaggacccct tttcccttgg 8220
ctcgggcgct tagccacaca gaagctaagt cgcccaggta cctgcgtctc caataaagcc 8280
tcttgttttt acatccagtt cgtggcctcg ctgattcctg ggtgtgtggg tctccctcta 8340
cgaaagtgcc tcttcggggt ctttca 8366
<210> 2
<211> 8667
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<222> (1)..(8667)
<223> Тип ERV группы 2 клеток CHO-K1
<400> 2
ctctctctct ctctcttggg ctagctaaat tacagaccaa gagaccaaga ccacccagaa 60
ccatttctga gaatagaagg tcagggcgtt atgactactt gtttgccaca caggatatct 120
gtggtcagcc atctggacgc ggagggagga actgccgggt ctgttgtaac cactgtgtac 180
ccaagataga gccaagtcag cttcgtctga ttaccagacc ttgagaactt gctaacctga 240
aataactctg tgcctcattt gaattgtcca atagagtccc tgtaactatg cttttcgctt 300
ctgtactatg ctttttgcta tataaggacc cctctccctt ggctcggcgt gcttggcctc 360
acagaagcta agtcgcccca ggtacccgtg tatccaataa agcctcttgc tgtttgcatc 420
caagttcatg gtctcgctga ttcctgggta cgggtctcct tctgtgaaag tacctcttct 480
gggggtcttt catttggggg ctcgtccggg atcgaaaccc cccgcccagg gatcaccgac 540
ccacgtctgg gaggtaagcg ttgtgcggat ccgctgtctt gtctgtcttg tcttgcctgt 600
ctgtctgaat ctgtcttatg aactgcgctt gcgtctgtag tacacagctg cgtacatctg 660
tatctggcgg ctccgaggaa aaactgaaga gttcagactc ccgaccacgg ctccaggaga 720
cgtcctggta gcgtctagag cctcagtggg ctcactcttt tttctgacta gggtagatgc 780
ctaggcatct tcccaatctg tttggggaag cccctcagat tgagggattt gctatcctca 840
tcggggacga ggaagggagg cccccttctt cggccccatc tgaagtttta ctttcgacct 900
tgtgtcgaaa ccgcgccgca aaagcctgtt ctgtgttgct cggtctttgt cttgtaacca 960
tcatttgtgt cctcctaggt ctagaaacta tggggcaaac tgtcaccact cctttgtccc 1020
taacactctc gcactggaga gatgtgcagg attatgctca taaccagtct gttgatgtgc 1080
ataaacgcaa atggattact ctttgttcct cagaatggcc aacctttgac gtgggctggc 1140
cgcgagatgg tacctttaac ccccaaacta tatttcaggt gaaagagaag actatggatc 1200
ctggaccaca ggggcatccc gaccaggtgg attacatagt cacttgggag gccttggttc 1260
aggaccctcc ccctgggtac gtcctttctt acaccccaag ggcccctcct tcctttcccc 1320
ctctaactgc tccagccgac ccactccctc ggcccctacc cttcccaatc cctcgacacc 1380
cactcctccc taccctcctc cccattccaa cctttaccct acagcggtaa aagacgctaa 1440
agctgaagaa aagaagacac ctgaggtact cccccgggag aagaactgtt ggttgactta 1500
ctgactgagg aacccccgcc atatccagca ccgccaccgc ccccaccaga ggcagaagcg 1560
gcccccgccg cctcggtgga ggtggcccca gctcccccac ctgacccttc cccaatggct 1620
caccgactga gaggtcgcag ggagcagccc gctccagatt caaccactct gcccctcaga 1680
actggaccga acggccattc tcagcctcgg acctctataa ctggaaaaat aacaatcctc 1740
ctttctctgc agaccctgtg aggctgacat ctctcataga atcggtcctc acaactcacc 1800
aacctacctg ggatgattgt gaacagattt tgcaggttct cttaacctcg gaggagaaac 1860
agcgcgttct actagaggca cgaaagaatg tcccaggagc caacgggcag cccacccagc 1920
tgcccaatga aatcgatgcg gcttgccctc ttgaaagacc tgaatgggat tttaccactg 1980
aagcaggtag gacccatcta cgtctctatc gccagttgct cgtagcgggt ctccgggggg 2040
caggacaccc acccaccaat ttggcccagg taaagcaggt aatacagggg gcggaggaat 2100
cgcccgccac ttttctagag agactaaaag aagcgtatag gatgtatact ccctatgatc 2160
cggaagatcc agggcaggcc accagcgttt ctatgtcctt tatttggcag tcagccccgg 2220
acataagaaa caagcttcaa aggctagaaa atttgcaggg atatacactc caagatttgt 2280
tgaaggaagc agaacgtatt tttaataaga gagagacaca gacagaaaga gaagaacgct 2340
ggagaaagga aacccaggag agagaggaaa gactgagaaa agaagctgag gaaaaagagg 2400
ctgcaagaga ccgtaagcgg aatagagaga tgagcaggct attggccaca gtagtgacag 2460
gtcagagaca gaataggcag agggatggca gaagggggcc ccacctggac agggatcaat 2520
gtgcttactg caaggaaaaa ggacattggg caagagattg ccctaagaac cccgggccaa 2580
gcctccacgg ccaagaacct ctgaccttct aaacctagaa gattagagaa gtcagggcca 2640
ggagcccccc cccctgagcc caggataaca ctgcaagtcg gggggcatcc ggtcaccttc 2700
ctagtcgata caggggcaca acattccgtt ctgaatcagt cacccggacc cctgagtcac 2760
cggactgcat gggtacaggg ggctacaggc ggaaagcagt accgttggac cacagatcgg 2820
cagctccagc tcgcgaccgg taaggtcatg cactctttcc tccatgtgcc tgattgcccc 2880
taccctttac taggacggga cctattgacc aaactaaaag ctcaaataca ctttgagaaa 2940
tcaaaagtca aagtcacagg gccagaagga attcccctta ccatcttgac aatgtccata 3000
gaagatgaat atagactcca tgaaaaagag actaactcgg gcaaccagga aacccttgac 3060
cactggctcg cggaatttcc ccaaacctgg gctgagacag gcggaatggg ccttgccagc 3120
aaccaggccc caattatagt aaccttaaaa gctgccgccc ttcctgcatc cgtcaagcag 3180
tacccgatgc ctaaagaggc ccgcgaagga aacatattaa aaggttactt gaacaagaga 3240
ttctgacgcc ctgtaaatct ccttggaata cacccttgtt gcccgttagg aagccaggaa 3300
ccaatgacta tagaccagtg caggatctga gagaagtcaa taaaaggata gaggacatac 3360
accctactgt ccctaaccct tacaatttgc tgagcggatt gccacctgac tatacctggt 3420
atacagtctt agatcttaaa gatgccttct tctgcctcca cctgcatccc accagccagc 3480
ctatatttgc ctttgaatgg caggacgctg accttggaat ctctgggcag ctaacttgga 3540
ctaggttacc tcaagggttt aagaacagcc ctaccctttt tgatgaagct ttacatcagg 3600
acctggcagg attccaggtt cggtacccgg ctctaatcct cttacaatat gtagatgaca 3660
tcctcctggc agccaaaacc caggggaatg taaggagggc actcaagccc tcctccagac 3720
tcttgggagc ctagggtacc gagcatccgc caagaagtcc cagatatgtc agaagcaggt 3780
gacctattta ggatacaaga taaaggacgg actgcgatgg ctaacggaag cccgtatgtg 3840
agccatctta gaccttccca ccccgcaaaa tccccaccaa ctaagagagt tcttgggaac 3900
ggcaggtttc tgccgcctat ggatccccgg gtttgctgaa atggcggctc ccctctaccc 3960
cctcactcgt ccagggattg cttttaaatg ggaagagccc caaaagaaag ccttcaccaa 4020
catcaaaaag gctctccttg aatcaccagc cctgggacta ccggacttgg ctaagccatt 4080
tgagttcttt gtagatgaga aagagggcta tgccaaggga gtcctcaccc aaaaactggg 4140
gccttggaga aggcccactg catacctctc caagaaattg gatcctgtgg catcgggatg 4200
gccaccctgc ctccaaatga ttgccgccat agccttgtta gtaaaagact ctcacaaact 4260
aaccttgggg cagcctttga ccatacatgc ccctcatgcg gttgaggcag tcatcagaca 4320
gcctccagac agatggctca ctaatgcccg aatgactcat taccagacta tgctgttgga 4380
caaagaccgg gtccactttg gccctttggt gactctgaac ccagccacct tgcccccctc 4440
cctggggagc ccgaggctca tgattgcttg caggtactgg ctgaggccca tgggacaaga 4500
cccgacctaa ctgaccagcc tctacccagc ccggaccaca tctggttcac ggatggaagc 4560
agctttttgc atcaaggaga acggagggct ggtgcagcgg tcacctcaga ggatcaggtc 4620
gtctgggccc aggcactccc tcccggaacc tccgcgcaga gggcggagct cattgcactc 4680
acgcaagctc taaagttggc agaaggtaag aggctcaccg tgtacacaga cagtcgttat 4740
gccttcgcca ctgcccacat acatggagaa atttacagac ggagggggta gcttacctca 4800
gaagggaagg acattaagaa taaggaggaa atcctcgctc tcctaagagc tcttcatctg 4860
ccctccgcct taagtatcat acattgcccc ggacatcaaa aaggggattc tcttgaagcg 4920
aggggcaatc gaagggcaga tttggccgcc caagaggcgg ccctggccac agaaaccact 4980
aacctcctgg ctctagaacc caccaacgac cgtccctccc cctcatggga ctatgaacaa 5040
agagacatcc agaccctaga gaaattggga gccaccaagg aatcaaacgg ggattggact 5100
tatgaaggaa agactatcat cccctaacgg gtaaccaagt acctagtgac atttttacat 5160
aagatgacac atctgagctc caagaagatg cgagaactcc tcgagcgaga agacgaattc 5220
aatttccttt tggggaagaa taacattcta aaacaggtaa ctgaacaatg tgatgcgtgc 5280
gcccgagtca acgcatccag actgaagctt cctcccggga accgggtcag aggctaccgg 5340
cccggaacac attgggagat agatttcact gagattaaac caggtaaata tggatacaag 5400
tatctattag tttttataga caccttttca ggatgggttg aagccttccc tactaaacat 5460
gaaacagcca agatcgttac taagaaattg cttgaagaaa tctttccccg ttatgggatg 5520
cctcaggtac tggggacaga caatgggccc gccttcgtct cccaggtaag tcagtcagtg 5580
gccaccttgt tggggattga ttggaaatta cattgtgctt atagacccca aagttcagga 5640
caggtagaaa ggatgaatag aacaattaag gagactttaa caaaattgtc gcttgcaact 5700
ggcactagag actgggtcct cctactcccc ctagcactct accgcgctcg caatacccct 5760
gggccacatg ggctcacacc ttttgagatc ctgcatggag tatctacccc tatcattaac 5820
ttccttgatc aagatgtctc agattttgct aactcccctt ctctccaagc tcatttacag 5880
gccctccaac ttgtacaacg ggaggtctgg aaaccccttg ctcaagctta taaagaccag 5940
agggaccatc ctaccattcc ccattcctac cagatcgggg acaccgtctg ggtccggcgt 6000
caccagtcca agaacctcga accccgctgg aagggaccct acatcgtttt gcttaccacc 6060
cccaccgcac tcaaagtaga cagcattgca gcttggatac atgcttctca tgtaaagcca 6120
gcccgaccca ccgattcagc cactgcatcg gaatggactg cgcactgcac tcaaaatcct 6180
ttaaagataa gactctctcg tacaccctcc tattgattgt ttgtctgttt atcccccatg 6240
tagcaaccag cccctatacc attcatgcgt taacctggca ggtactttcc cagtcagggg 6300
aggtcattta tgagaccacc aaaaaccatg cccttggtgt atggtggcct acattgacac 6360
ccgatttctg tgacttggtc gctggtctag acacctggga catctcacac tgttacccta 6420
aggcttgtag caacgaaaac gatttgccac gatttcgcag gtcactccaa agttcgagag 6480
acaccactcc tggatgtagc agtcaacagg ccaaagacca actgtcgggt agtgactttt 6540
atgtctgtcc cagagacgga cgggcctgga agaagactca gacttgtggg ggttatgaac 6600
agtttttctg tgccacttgg ggatgtgaaa cgacagggga tgcttactgg aagccttcat 6660
cctcatggga caagattcaa gtaaccagag gatggcaaaa gggctccacc ggcaccacct 6720
ctatgcctct ccttatttcc tttacggatg agggtaagaa agcaacagaa tggcagaacg 6780
gatatgcttg gggccttcgc tggtacctgt cgggaccaga ccggggggcc atctttaaaa 6840
ttagactcaa agtaaaaact atcaccaaaa ccatgggccc aaacccagtt ctggcagacc 6900
agaaaccccc accaagcctc atgggtctcg gccccatagg cctctatccc atctcccacc 6960
ctcatcaaca ccctcatcca atactgccac ccttactctt acgactccag ttcacacccc 7020
cacctcccca tatccaggga caagggacag attacttgaa ctaatacaag gggcttactt 7080
ggccctcaat tcctcagacc ccaacaagac tcaggaatgc tggctatgcc tagtttcccg 7140
gcccccatat tatgaaggca tagcagtact gggcaactac tctaaccaga cctcagcgcc 7200
taccagttgt ggagctgcca tgcagcacaa gctcacaatg tctgaggtct caggaaaggg 7260
gctatgcata ggcagggttc ctccctcaca tcaagagtta tgtaaccaaa cagagccatt 7320
atctcaagac agccgatacc ttgttgcccc ttatggaact tattgggctt gcagtactgg 7380
tttgacaccc tgcgtctcta ccactgtcct caatgccacc attgactttt gtatattgac 7440
agaactttgg cccaaagtca cataccacca acctgagtat gtttacagcg tactagagaa 7500
atcaacccga catcagaggg agccaatgtc cttaaccgtg gccctattat taggaggaat 7560
aacaatgggg ggtatagcag ctggaagagg gaccagtacg gttgccttgc aggaaactag 7620
ttatttcaaa cttctacaac aagccatgca tacagatatc caggccctag aagagtcagt 7680
cagcgcactc gagaaatcct taacatcact ctctgaggta gtcctgcaga acagacgaga 7740
gttagattta ttatttctac aggaacgggg actatgtgct gccctcaagg aagaatgctg 7800
cttttatgca gaccacacag gaatagttag ggacagcatg gccaaactta gagaaagact 7860
taagcagagg caacaactat ttgagtctca acagggatgg ttcgaaggat ggttcgctaa 7920
gtccccctgg ttcactaccc ttatatccat tctcatggga cctctgatta ttctattttt 7980
aatccttata tttggtacct gcattctaaa taagatgact caattcatca gagaacgact 8040
atctgtcata caggccttag tcttaactca acaataccat cagctaaagc aaatagatcc 8100
agaatatcca gagacctctg aatgaaagat tccattcagt tacaagagaa atgggggaat 8160
gaaagaccac tgccccttaa gctctctctc tctctctctt gggctagcta aattacagac 8220
caagagacca agaccaccca gaaccgtttc tgagaataga aggtcagggc gttatgacta 8280
cttgtttgcc acacaggata tctgtggtca gccatctgga cgcggaggga ggaactaccg 8340
ggtctgttgt aaccactgtg tacccaagat agagccaagt cagcttcgtc tgattaccag 8400
accttgagaa cttgctaacc tgaaataact ctgtgcctca tttgaattgt ccaatagaat 8460
ccctgtaact atgcttttcg cttctgtact atgctttttg ctatataagg acccctctcc 8520
cttggctcgg tgcgcttggc ctcacagaag ctaagtcgcc ccgggtaccc gtgtatccaa 8580
taaagcctct tgctgtttgc atccaagttc gtggtctcgc tgattcctgg gtacgggtct 8640
ccttctgtga aagtacctct tctgggg 8667
<210> 3
<211> 8181
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<222> (1)..(8181)
<223> Ретровирус-подобные IAP-последовательности клетки CH0-K1
<400> 3
ctgtagagag ctgcagaaaa ccgcacccaa aagatggcgc tggtttccgc cttccgccag 60
cccgacggcg agcgctctct gtggtaaaca actccaaata tggtagaggt catgtatcct 120
cttaattctg cttggagaca acctatcctg gcgcgccacg tagggttagg tgattggtag 180
atgtagacta tatcaggccc cgtctccctc ggcctggggc cgccgcctcc attatacatt 240
gtacaaaagc aaagaggttc ccgattaaac tgtgtttgaa gaagattcct cggtgtggcg 300
tctttcttgc tggtcaaggg tggacgccgc aagtggtggc ccgtacgggg aattcagaac 360
ttctcacagt cggggcggcg ccggtaagtt cccaaggtaa gtggaactgt aaagtccgcg 420
gtgaatgcgg agataggtct ccggtaaagg agcaccaagg tcctcaggaa agaggagata 480
ggtctcagta aaagagtacc aaagccctcg gggaagaggc agtaaagtct ccggaaagga 540
gcaacattgt tcgcgacaaa agcgaaccga aagtaaaaaa ccctcgcttc tgctttaatt 600
tgcagagtga aagtaagtta atggaccccg ctattatagt ggtgatttgg ttgcttttta 660
atatcgcagt gtgggctttt gttttctggt ggtgttatcc gacttgcaca ccttcggagc 720
gcgctaatta agacaaaagc acaatgggat cgtcacaatc acatccaatt tttctggctc 780
ttcaagagct acttcattct aacaatttaa ggttaactaa agccacatta cagaggttcc 840
tgagtgaatg cgacaccatt gcaccctggt ttgcagcctc gggcaatctc actgtggata 900
gttgggataa attagggaaa gacctggatt ttgcctggga gcaaggagcg ttgaagccga 960
gtgttcgccc ggtttggcgc ctggtgcgca gctgtctaga ggacaaaggg ggaaataaga 1020
aagctatagc aaatggacag gctgcccttg aaatgctcca agaagaaagg tcagaacagt 1080
cagaaagtga gagaacaaag gagaaacaga aaagggataa gaagcgagtc tatccttctc 1140
taggaaggtt gaggaaggaa ttaaaagaac caggagagga ttcagaatca gaggaggata 1200
tatgtgagga tgaggacatc atggaggaag aggaggaaga gattattgag ctcatggaaa 1260
aacattcatt aaaagtatca gaaaaacagc gtcctaaaat ggcgactgta aagcaagggc 1320
accgccctat cgctcctccc cctactgctc gaaagaggaa gtaggaggtc ctggttgttc 1380
caccttctgt ccggaagtgt ggcgtacggt tcgcactgag tttcgggtta ctcgaccact 1440
ggcttatcca gtttttacag acggtaacca acagagatat catgaaccaa ttgactttaa 1500
aatagtcaaa gctctggcag aatcggtacg cacctatggg gtgacggcag cttttacaat 1560
tgctcaggta gaggcgctcg gtcgctactg tatgacaccc agtgattgga caaatttggt 1620
aagagcttgc ttatcaccag gacaatattt ggattggaga gcttttcaaa ttgagtttgc 1680
caatgatcaa ggtgcgatta atcgtgcagc tggtgatcaa tattgggatg tggacatgct 1740
tcttggtcaa ggacgttttg cacagcaaca aacaggatat cctcctcaag tgtatgatca 1800
gattaatcaa atagccatta gggcttggag gtctctgcct aacaaaggtg aggtcagtgg 1860
aaatcttaca aagatattac agggtcctat ggaacccttt tcagactttg tggcaagaat 1920
ggtagaagct gcaggaagag tgtttgggga tccagatgct gctatgcctt tgatcaaaca 1980
gctgatttat gaacagtgta caaaagagtg tagagctgca ataactccat ataagggtaa 2040
aggccttgaa gtctggatga aagtctgtag ggagttaggg ggtccgctga ctaatgctgg 2100
actagcagct gctgtgatgc aattaactaa gaaaggtgga ggttcaggag cttgctttaa 2160
atgcggcaag caagggcatt tgaaaaagca atgccccgag ggaggaaaca ctaaagtcaa 2220
taactttgct ccgcgcccta agcaacctgg cttatgtcct agatgtaaaa aaggaaatca 2280
ttgggctaag gattgtagat cagtaaaaga catcagtgga cagcctcttg ttcaggggta 2340
tggaggagcc cgttcaaaaa acggacgacg gggcccacga ccccagggcc cacaaatata 2400
tggggccatg gaggatcaga accaggagca gagtcccgaa acctggccct ctcttcgtca 2460
tccgagggac cgaggagagc cactacaggc tccgcggggc tggacttacg ctccaccacc 2520
agactcgtat taactcccag aatgggggtc cagcttgttg acaccgattt taagggaccc 2580
cttgagcctg gcacagtagg tttgcttata ggaagatcat ctgcagcatt gaaaggttta 2640
cgagtacatc ctggagttat agatcctgat tacatgggtg tagtaaagat catggtagaa 2700
tctcctagag ggattacggc catttctcct ggagacagga tagcacagtt actgcttttg 2760
ccaagcttgc atgacaagtt tccagcacaa gccagagaga gaggagaggg aaactttggc 2820
tccactggat caaacttaac tttcctagct ttagaccttg atcaacgtcc aacccttgag 2880
ttaatagtga atggtaagaa aatcttaggc ttactagatt ctggagctga taagagcatc 2940
atagccacta aagattggcc ctctggctgg cctatacagg tttcttctca aagtttacaa 3000
ggtttaggct atgctaaggc tcctgatatg agtgctagac aattgccttg gaaagatcag 3060
gaagggcatt cagggaccat gcaaccttat gtgttagact taccaatttc attatgggga 3120
agagatttgt taaaggatat gggttttaaa ctcacaaatg aatactcaga aacatctcaa 3180
ggtatcatga aacgaatggg atacgtccca ggccgaggcc tcgggaaaca tctgcagggt 3240
cgtaccagtc ctattattcc acaaccgaga ccaaagaatc taggtctggg tttttcctag 3300
gggccactga gggaggtatt cctattacct ggaaaacaga ggagccggta tgggttcctc 3360
agtggccact ttcctctgaa aaactggaag ctgctaagac tctagtgcgg gagcagctgg 3420
atctggggca tataaaatcc tctgtatctc catggaatac tcctattttt gtcattaaga 3480
aaaaatctgg taaatggaga ctgcttcacg atcttagagc tattaatcaa cagatgcaaa 3540
ttatgggccc tgtacaacgt ggtcttccac ttttaacttc tttacctgca tcatggccta 3600
tcatctctat agatattaaa gattgcttct tttccatacc tttgtgtgcc aaggattcag 3660
ggcgttttgc gtttacgctg ccctcttgta atcatgaaca acctgattta aggtatgaat 3720
ggatagtgtt gccacagggg atggccaata gtcctactat gtgtcagttg tttgtagcag 3780
aagcaattgc tcctttgaga gtggactttc caaagattag atgtgttcat tatatggatg 3840
atattttatt ggctgccaaa gatgataaaa cgcttaataa ggcatataca aaattggtaa 3900
aattgcttga gatgcataat ttagtcatag cctcagaaaa ggtacaaaag gacactgttg 3960
ttaactatct aggggctaag attctccctc atacaattat tccacaaaag atagagatta 4020
gaaaagataa tttaaaaact cttaatgatt ttcaaaagtt gttgggagat ataaattgga 4080
taagatgtta tttaaaatta ccaaattatg agttgaagcc attgtataat attctcaatg 4140
gtgattcagc attagattca cctaggcagt taactgctga agccagagaa gctttaaaga 4200
aagtggagac agaattacaa aatgccagcc ttaaaaggct aatagaaaat atggatattc 4260
ttttgtgtgt gcttcctaca ttctcccagc ctacagcttt gctgtggcaa gatggtccgt 4320
tactatggat ttattctaaa tcctccccag gaaaaactat tgaatactat cctgcagcgg 4380
ttgctgactt ggctcaaaat gggatacagc aatgtattca atattttggg aaattaccaa 4440
ttaagatcat aatcccttat actgctcaac aagttaagat actttgtgga actgtggatg 4500
attgggcaat tcttcaatgt ggattttcag gagaaataga taatcattat ccaaaacacc 4560
ctctaatgtc attttttaaa gaacatccag tcatttttcc taaaatgact gcagcagctc 4620
ctattgctgg agccgcaaat attttcactg atggatccaa gacgggctgt ggagcctaca 4680
tgatagagca tcaggatcca gtgcaatttc aatatcagcc tggctccccc caaattattg 4740
aatgtaaaat tgttcttgaa gtgtttagaa attgcccgtt ctcctttaat ctgatttctg 4800
attcggcata tgttgtgaat gctgttaagg ccctggaggt tgcaggacct attaaaccta 4860
atagtacagt ttgcaaactc ttacaggagt tgcaaaagtt gatctggcat cgcgatcaga 4920
aattttttat acaacatatt agagctcata ccaacttgcc tggacctcta tcaaaaggca 4980
atgagatcgt agatctctgt actagagggg aatatgtatt tttcgcttcc tctatggaac 5040
gagcacagca ctttcacaaa caatttcatg tatctgcaaa aactttacag caaaggtttc 5100
agctatctcg agcagaagct agacagatag tattgaattg tcaacagtgt ataaccttct 5160
tgcaccccct agcctggggg tcaatcccag agggttattg cccttgaaga tctggcaaat 5220
ggacgtaact catttttcag gatttggaac tcttaaatat atccatgtgt ctgtggatac 5280
ctgttcaggc atcatccatg ccacccctat gagtggggaa aaggctcgca atgtcattgg 5340
acattgcctt gaggcgtggg ctgcctgggg aaaacctcag caattaaaga ctgacaatgg 5400
tcctgcttat acagctcaat cttttacctc cttctgtaaa cagatggaag tccaattaaa 5460
ccatggcctg ccttataatc cccaaggaca aggaatagtt gaaagagccc atcgcacatt 5520
gaaagaatgt ttactaaaac aaaaagggga ataggccatg gtagaacacc aaaggaacaa 5580
ctatctttgg ctctctttac tctgaatttt ttaaatttgg attctcaagg cctttctgct 5640
gccgataggc accagactcg ggcacctgct cagaagggct atgtaaaatg gaaagatatc 5700
cttacgggtc tatggcatgg accggacccg gtattagcat gggcgagagg ttctgtttgt 5760
gttttccctc aggatcagca agacccggtg tggattccag agagactaac gaggaggtgc 5820
aacaaacaag atgaagcctc cgatattgat cctgctgctg atcttgacaa caccgtggct 5880
gatccaggcc agagaacaaa tcctgtgggc catagcgaaa gcgtggccaa ttcctttgcc 5940
tgttcatagc acatctaaag ttttgcctgt tttcttttct actagttgtg aattgggttt 6000
gccttgtgtc aatttagatc ctgacactgc ccagtattca gccactaata tttctctcaa 6060
tggagcatta tgcttctccc ttattaataa ttctaaccca tgcatttggt tgaagaatgg 6120
ttctattggc aattggcttg atcctcttac taataaccag atctcgagtg ctatgctcac 6180
tgaagccctg tctcagttta gtacaggcgc tagttctggc tctggcactg gtacaaatgg 6240
aaccaaagat gtaaatatca ctacatttct gatgctgatg gaggggctcc gaccctcaaa 6300
ttcacgcttt agtcaagaga actccactgc tcgacaagtc ttgccttatt gtgcacccaa 6360
tcgaggctat ccacctcatt ggacgccttg tcagagtccg gatcacaaac ttaagcaaat 6420
agctccaggt tttgaattta ctccccctct tggcaagtac cagtataacc taactcaggg 6480
aggttcccaa agaacaggca gtaaaaacac ctggccatgg tttcaatggg tactctccaa 6540
cgagcgtgaa acatttactt ctcttgaacc ttttgcagtt ctccagaatc tctgcataag 6600
acttcttaat gtttctggta caagggatga gaagggacct tctcttaaga atctatcaat 6660
acataaagct ttgaataatc tcactgttcc agctagccca gtatgcttaa aagctccttt 6720
tttctttctg ctctctaatg ctattgatgc taccaaagaa gtgatctcgt gcaattcgac 6780
tgatgtctcc tgcattgcta gccaatgttg gaatggttct gctaatacag cagtagtgat 6840
gaggattcct atgtatgttc ctattccagt caaagtggat acaggaacgt ttcccattac 6900
ggaaatcatc agaaccaaac gggattttgg cataactgca gcattggtta ctgccattac 6960
cctttctgca gcagcggcta cgactgccac cgttgccatg gctgctcagg tccaatcagc 7020
tgagacagtc aatgatattg tggaaaaaac agctacaaca ttaactacct tgagatctat 7080
tgatggccac ttaaaagctg gcattctaac tgtgaaccaa agagtggacc tgttgcagga 7140
acaggtggat gatttagtaa ccttaacttc cataggatgc attcattcct tttcttcttt 7200
gtgtatcacc tccaggatgg ccaataattt cacggagaac agcaacctga gttggcagct 7260
gagtgcctac cttcagggaa attggagtca acagttcgag aacctgacag acaccctgac 7320
acaacagatt attgccgtga atgctacacg cctgagcctg cccactgtta ataccctctt 7380
aactaccctt aagcgagcct ttagtttagt taaggaatgg gctggaatag gggttatgtt 7440
ttccatgatg ctgttagcca tttttgtttg cttgtggtgc ctatgcagga tcaggaagtc 7500
gcaaaaatcc caagctgcta tgttggtaca ggcctttgct gctgtggaag caggacagtc 7560
ccctcaagca tggttatcca tgctgacaga caaatagatg tctgtctcgc ccttgctgag 7620
tgagtaatgc gtatctgaat cagtcatgct caatgacggg cacctaccct catgctgagc 7680
gaataagcat acgtgtgttg ccttcaacct aagacatgag cctgtgaggg cgacacagtg 7740
actctttgac gggtaagata gggacactgg ggttgaacct aagacagaga tgactgaaat 7800
ctgacaacag atagatactg ctactcctat aatattaaaa caaaaagggg gaactgtagg 7860
gagctgcaga aaaccgcacc caaaagatgg cgccggtttc cgccttccgc cagcccgacg 7920
gcgagcgctc tctgtggtaa acaactccaa atatggtaga ggtcatgtat cctcttaatt 7980
ctgcttggag acaacctatc ctggcgcgcc acgtagggtt aggtgattgg tagatgtaga 8040
ctatatcagg ccccgtctcc ctcggcccgg ggccgccgcc tccattatac attgtacaaa 8100
gcaaagaggt tcccgattaa actgtgtttg aagaagattc ctcggtgtgg cgtctttctt 8160
gctggtcaag ggtggacgcc g 8181
<210> 4
<211> 8369
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<222> (1)..(8369)
<223> Подобный вирусу лейкоза гиббонов ERV клетки CHO-K1
<400> 4
aaaaaaaaaa aaagaaaaga aaagaaaaga aaaaacagtt cccagaataa ctacccgagg 60
ttctgatgac ctctatggaa tgtttcaacg ccctaagtgt gcctcaggtg tatgattcat 120
gtgacacatg gtttatggtt taactgaaga atactgtatg actctgatat ttccccattc 180
ttttgtgggt tttttgcctt tataagcctg ttacaaattc agaccggggt cgaactcctc 240
tacccctgtg tggtgtatgg gtttcgaccc cagcatgctg gtctgagatc tcgttccgtt 300
tcactctcaa taaacttcct cgtgtgattg cagcaagtct ggtcttggtg cctactgggt 360
gcgcgccttt cccgaggctt gagtgagggg tctcccttcg ggggtctttc atttgggggc 420
tcgtccggga tttgcgcgac cacccagggg tcctagaccc acttggaggt aaggattttc 480
attcctcgtt gttctgttgt gattctgtgt ttgtggattg ttcagagatc tggtgcgatc 540
gcagttctgg tttttgcaaa cgctctaatc gagtctgctc ctccagaggg gggagcagag 600
tggacgggga tagacgtatc cgggtgtcca ccgtttgttc gccctgggag acgtcccagg 660
aggacagggg aaggaccagg gacgcctggt ggaccccacg aggaaagtcc ggggattagt 720
ttcctccttg acaatctgta gaccgtctga ctcttttcag gatttctggt cgactcaagg 780
tcggcgattt tgtctctctg ttttgttgtc tttgtgattg ttctttttgt gaagtgtgat 840
agaaccagga atgggacaga ccgtgacgac cccgctaaca ctgaccctgg accattggac 900
ggacgttaag acgagggctc ataatctttc tgttgaagtc agaaaagggc catggcagac 960
tttctgctcc tctgagtggc cgagttttgg cgtaggatgg cctccggagg gaacttttaa 1020
cctgtctctc atttccgctg tcaagcgaat tatctttcag gcctcagggg gacaccctga 1080
tcaggttcca tacattatcg tatgggagga cctggtccaa aatccgacac cctgggtgag 1140
accctggacg acaggggcag aaacggtgac ggtggccgtg gccgccaaac cgaaagtacc 1200
ccctccagat aaaccggtac ctgctccatc ggcccctacg aagatttatc ctgaaattga 1260
tgatggttcc cttctcctcg actaccctcc acccccgtat cctcaaccag cttcccacgc 1320
acgtccccca caggtgtcgc cccccgccga ctcccggacg gcctctgaaa caggaccggt 1380
ggccggaacc cgaagccgcc ggggtcgtag cccaggggag aaggcgcggg gtttgactct 1440
gctactgccc tacctttacg agcttatgga cccgccccgg ctccgggaga actggtcccg 1500
ctacagtact ggccgttctc atcagccgat atgtataact ggaaaactaa ccactcctct 1560
ttttcagaga atccctcagg cctgacaggg cttcttgagt ccctaatgtt ttctcatcag 1620
cccacctggg atgattgcca gcagcttttg caagtccttt ttaccacgga agagagggag 1680
agaatcctcc tagaagctag aaaaaatgtt ccgggaccag atggagcccc caccaacctc 1740
cctaacctta tagatgcagc ttttcccttg atccgccctg actgggatta taactctgca 1800
gaaggtaggg agcgtctcac ggtctaccgc cgggctctag tggcaggtct caagggagct 1860
gcaaggcgac ccacgaattt ggctaaggta agggaggtcc tacaaggtcc ggtggagcca 1920
ccctctgtct ttttagagcg cttaatggag gcttacagga gatatactcc atttgatccc 1980
tcctcagagg gacagtgggc ggctgtagct atggctttta taggacaatc agcccctgat 2040
atcaagaaga agttacaaag attagagggg ctccaagatt atactttaca ggatttagtg 2100
aaggaagcag agaaagtgta tcacaagagg gagactgagg aagagaagca ggagagagaa 2160
aaaagagaga cggaagaaag ggagaaacgg cgtgaccgcc gccaagagaa gaatctgact 2220
aggattttgg ccgcagtagt aggtgaagaa agggtagaga aaagacagtt agggcacctg 2280
ggcaacaaag caaatagacc cgtgggcgga cagagaccac gacttgaaaa ggaccaatgt 2340
gcatactgca aagagaaagg acattgggcc agagaatgcc ccaaaaagaa acagaagggc 2400
cctaaggtac tggcccttga agaagattag gggagtcggg gctcggtccc cctccccgag 2460
cccagggtaa ctctctctgt ggaggggaac cccgttgact tcttagtgga cactggggct 2520
gaacactcgg tcttgactga accattagga caattaggat caagaaggac tctggtcaca 2580
ggagccactg gtagtaaact ttatccctgg acaacaaaaa gaagtttaaa aataggaaaa 2640
agccaagtga ctcactcttt ccttgtaatc cctgagtgcc ctgcaccgct tctgggcagg 2700
gacttgctga ctaaactaaa ggctcaggtc cagttcacct cgacagggcc ggaagtcacc 2760
tggggggaga cacctatagc atgtttagtc ctaaatctag aagaagaata tcgcctccat 2820
gaacccaaga ttgagaattt gccctccaag gaatggttgg tagcttttcc aggagtttgg 2880
gcagagcaag cgggaatggg actggctaag cgggtaccgc cagtggtggt cgaattaaaa 2940
gcagctgcta cccccatctc agtcaggcag taccccatga ctcaggaggc taaagaaggc 3000
attcgaccac acatccagag gctattacaa caaggtatac tgattcattg ccagtcccct 3060
ggaatactcc cctcttgccg gtacgtaagc ctggaactaa tgactatcga cctgtccagg 3120
acttgagaga ggtcaacaag cgggtactgg acatccaccc gacagtgccc aatccctaca 3180
atttgctaag ttccctgcca ccggagcggt cctggtacac tgttctggat ttaaaagatg 3240
cttttttctg cctcagacta catccaagta gccaactcct gtttgccttc gaatggcggg 3300
accccgacgg aggacacacc ggtcagctga cctggaccag gcttccacag ggattcaaga 3360
actcgccaac tctcttcgat gaagcgctcc actgtgacct tgcgcccttc agggcacgaa 3420
acccccaaat ctctctctta cagtatgtag atgatttgct gcttgcagcc tcgacctggg 3480
aactgtgcct taatgggacc aaagagcttc taaaagaatt gggtgagttg gggtaccggg 3540
tgtccgcaaa aaaggcccaa ttatgctgtc cagaagttac ctacctggga tacaccctcc 3600
gagaagggaa gcggtggctc actgaagccc gaaagaaaac tgtgatgcag atcccgaccc 3660
ccaccactcc acgacaagta cgtgagtttc tgggaacggc gggcttttgt agactctgga 3720
taccaggatt cgccactttg gcggcccctt tgtaccctct gactaaagac aaagtcccat 3780
ttacatggaa agaagaacat cagaaagcct ttgaaaaaat aaagactgcc ctgctcacag 3840
cccctgcttt gactctgcca gacttgacca aaactttcac cttatacgtt gatgaacggg 3900
ctggaatagc tcggggagtc ctgactcagg ccctgggtcc ttggaagcgg ccggtggctt 3960
acttgtcaaa gaaactagac ccggtagcca gcggttggcc ctcttgtctg aaagccattg 4020
ccgcagtagc cttgcttgtc aaagatgctg acaagctgat cctgggtcaa catgtgacta 4080
taatcgcccc tcatgcctta gagagtattg tacggcagcc ccccgaccgc tggatgacaa 4140
actctcgaat gactcattac cagagcttgt tactaactga tcgaataact tttgctaccc 4200
ctgctatcct caaccctgcc accctactcc ctgaagcaga tgactctact cctgtgcatc 4260
gatgcgctga tatcctggcg gaggagactg gagtcagaac ggacctgact gatcaacctt 4320
ggccaggtgt gcctagctgg tatacggatg gcagcagctt cgtggtggaa ggaaaaagaa 4380
aagcaggagc agcggtggta gatgggaaac aggtgctgtg ggccagcagc ctcccagagg 4440
ggacgtctgc tcaaaaagcc gaacttttgg cactaattca agctctgcgc ctggcagaag 4500
gtaaggctgt caacatctat actgatagcc gctacgcctt tgctaccgcc cacattcatg 4560
gagctatcta taggcagagg gcctactcac atcggcaggg aaagacatta aaaacaaaga 4620
agaaatcctg gctcttctag aagccataca cctacccaag aaattggcca ttgtccattg 4680
tccaggacat caaaagggga ctgacccagt tacacgggga aaccagatgg cagatcagac 4740
agccaagcaa gctgcccatg gaactacggt gctaatagaa gaaatcagaa atcacccacc 4800
agagcccccg gagcacacac ataaaggctg tgccttgact gaagatgggc aaattgtctt 4860
accagctaaa gaagggcagg actatgtaaa aaggttacat caactaactc acctcggaac 4920
tgaaaaactt aaacaactaa ttaaaagttc caaatactgt gttcaagacc tacacaccac 4980
aacaaaacag gtagtggagg catgccaagc ctgtactatg accaatgcgg cccgaccgtt 5040
caaggagcct ggaaaaagaa tgaggggaga ccgaccagga gtctattggg aagtagactt 5100
tactgaagtt aagccaggaa aatatggtaa taaatatctt ctagttttta tagacacttt 5160
ttcaggatgg gtggaagcct ttcccacaaa atctgaaact gctcagatgg tgaccaagaa 5220
gatactagaa gaaattctgc ccagattcgg gattcccaag gtaatcgggt ctgataacgg 5280
cccagctttt gttgcccagg taagtcaaga attggccact caactgggga ttaattggaa 5340
attacattgt gcttacagac cccagagctc aggacaggta gaaaggatga atagaactct 5400
aaaagagacc cttactaaat tagctattga gaccggcggc aaggactggg tgaccctcct 5460
cccctttgca cttcttagag tccgaaactc gcccgggcgt ttcggcctca ccccttatga 5520
aatcctacat gggggaccgc cccccttaac ggagtcgtgc gggatattgg atcctagcac 5580
ttactccccc tcctcttctg ctttatttac tcacttaaag gccttagaag ttattaaaac 5640
acagatctgg agtcgaatta aagaagccta cgctccagga accatcacgg tgccccacgg 5700
gttccaggtc ggagactcag tcctggtcag acgacatcgc gctggcacgc ttgagccccg 5760
gtggaaggga ccctacctgg tgctgttgac taccccctca gcagtcaagg tcgacgggat 5820
tgctgcctgg gtccatgctt cccatgtcaa gaagacaccc agtcagggca agaataacca 5880
tgaagaaaat tggacagtgg caaccagtga caatcctctt aagctgcgcc ttcgccgtag 5940
ccccaacctt gggtaataac ccccatgccc ccaaaaacta acctgggaag tgcttaacga 6000
agagggcgac gtcgtatggt caacaaccgc agtgcatccc ctttggactt ggtggcctga 6060
tcttgcaccc gatgtctgta agttagcagc cggctctatg gcttgggatc tttccagtta 6120
cactgactta gaaaagccac ccccagaaga acagtgtgtc cctcatggca tagggagcac 6180
attggggtgt tcaggacagt ttttccgagc caacctacag gcagcagact tctatgtctg 6240
ccccgggcag ggacaatcca gagaaatgaa gagacggtgc ggaggagctg cagactattt 6300
ttgtaataaa tggggatgtg agactacagg agaggcctac tggaaccctt cctcctcttg 6360
ggatctaatt atggttaaga agggtcgtaa gccagaagag ccagaaaaag gagaaagaga 6420
tccctccaaa tacccagctt ataggactgg gtgtgcctct ggaagctcgg atggccctca 6480
ggggccttgc aaaaacaatt attgtaaccc cttaaatatc agctttaccc cacagggaaa 6540
acaggatcgt caatgggtaa gaggaaatcg gtggggatgg agagtctata ctggcagcgg 6600
ggaccctgga ttcgtttttg tcattaagat aaaaatagaa agtccaccca cacgacccgt 6660
aggccccaat tcggtgcttg ataaccccat acccaggcct gagaggcctc ctcctcccac 6720
cctgtcacca aaggaaggct catattctct ctggactgca cccccacgtg tccctatggg 6780
gactgaggac cttctgttca atctgcttga tagtgcattt gttgttttaa atcgcactaa 6840
ccctgctgct actcagtcct gctggctgtg ttttacttcc aatccccctt actatgaagg 6900
aatagctcag ctacgtagct ataatgttac ttctgatcac tcacattgcc cctggggaag 6960
ccaacgaaag ttaaccctgt ctgcagtgtc aggaactgga ctgtgtatag gaaaagtacc 7020
taccaggcat cagcccctct gtaatgtaac tatctccaac ctgacaactg gcccagataa 7080
atacttagta ccccctctca atacctggtg ggcttgtggt accagcttga cccctgcctc 7140
catacttcac tttttgacaa tactaaagac ttttgtattt tagtccaatt ggtacctagg 7200
gtcatttatc atgatgatgc ctcgtttata gatgaatttg accaccgtgc tcgctacaag 7260
agagaaccta taaccttgac cttagctgtt ttgttaggac tcggagtggc ggcaggaatt 7320
ggaacaggag cagctgccct catccaacag ccctattact ataatgaact cagagcagct 7380
atggatgcgg acttgggagc cttagaacag tccataacca aactggaaga gtcattgact 7440
tcattatcag aggtagtatt acagaacagg aggggattag acctgctttt tctaaaagaa 7500
ggagggctct gtgctgccct aaaagaagaa tgttgctttt acgtagatca ttctggtgtc 7560
ataagagact ctatggcaaa gctcagagat agactagaaa ccagaaaaaa agaaagagaa 7620
acacaccagg gatggtatga aaattggttt aaccggtcac cctggttctc tacgctagtc 7680
tccactttgg tcgggccttt actcatattg ttgctaatcc tgacctttgg gccgtgcatc 7740
ctcaataaac tagtagcctt tgtaaaagaa agagtgagcg ctgtgcaggt aatggtccta 7800
agacaacaat atcaggtatt acaagaagct gagggatcgc tctaagatta gagcttttac 7860
aaaaagaaaa aggagggaat gaagagtaaa aacaaaactt tcaaagaaca ctagcccctg 7920
ccctagggcc ctcttctctg taggtaatca cacccggaac tttcttaaaa aaaaaaaaaa 7980
aaaagaaaag aaaagaaaaa acagttccca gaataactac ccgaggttct gatgacctct 8040
atggaatgtt tcaacgccct aagtgtgcct caggtgtatg attcatgtga cacatggttt 8100
atggtttaac tgaagaatac tgtatgactc tgatatttcc ccattctttt gtgggttttt 8160
tgcctttata agcctgttac aaattcagac cggggtcgaa ctcctctacc cctgtgtggt 8220
gtatgggttt cgaccccagc atgctggtct gagatctcgt tccgtttcac tctcaagaaa 8280
cttcctcgtg tgattgcagc aagtctggtc ttggtgccta ctgggtgcgc gcctttcccg 8340
aggcttgagt gaggggtctc ccttcgggg 8369
<210> 5
<211> 24
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<222> (2)..(24)
<223> Целевая последовательность миристоилирования 2 (Myr2), прямая цепь
<220>
<221> прочий_признак
<222> (22)..(24)
<223> Мотив, смежный с протоспейсером
<400> 5
gtcctaagcc tagaaactat gggg 24
<210> 6
<211> 24
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<222> (2)..(24)
<223> Целевая последовательность миристоилирования 4 (Myr4), обратная цепь
<220>
<221> прочий_признак
<222> (22)..(24)
<223> Мотив, смежный с протоспейсером
<400> 6
gcatagtttc taggcttagg aggg 24
<210> 7
<211> 23
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<222> (1)..(23)
<223> Целевая последовательность миристоилирования 8 (Myr8), обратная цепь
<220>
<221> прочий_признак
<222> (21)..(23)
<223> Мотив, смежный с протоспейсером
<400> 7
gagtgttagg gacaaaggag tgg 23
<210> 8
<211> 23
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<222> (1)..(23)
<223> PPYP5-мишень, прямая цепь
<220>
<221> прочий_признак
<222> (21)..(23)
<223> Мотив, смежный с протоспейсером
<400> 8
gttggttgat ctattaacgg agg 23
<210> 9
<211> 23
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<222> (1)..(23)
<223> PPYP6-мишень, прямая цепь
<220>
<221> прочий_признак
<222> (21)..(23)
<223> Мотив, смежный с протоспейсером
<400> 9
gccactgccg cccccaccag agg 23
<210> 10
<211> 23
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<222> (1)..(23)
<223> PPYP7-мишень, прямая цепь
<220>
<221> прочий_признак
<222> (21)..(23)
<223> Мотив, смежный с протоспейсером
<400> 10
gcccccacca gaggcagaag cgg 23
<210> 11
<211> 23
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<222> (1)..(23)
<223> PPYP13-мишень, обратная цепь
<220>
<221> прочий_признак
<222> (21)..(23)
<223> Мотив, смежный с протоспейсером
<400> 11
ggcagtggcg gatatggcgg ggg 23
<210> 12
<211> 23
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<222> (1)..(23)
<223> PPYP20-мишень, обратная цепь
<220>
<221> прочий_признак
<222> (21)..(23)
<223> Мотив, смежный с протоспейсером
<400> 12
gcttctgcct ctggtggggg cgg 23
<210> 13
<211> 1020
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<223> Rad51
<400> 13
atggctatgc agatgcagct tgaagcaaat gcagatactt cagtggaaga agaaagcttt 60
ggtccacaac ctatttcacg gttagagcaa tgtggcataa gtgccaatga tgtgaagaaa 120
ttagaagaag ctggtttcca tacggtggag gctgttgctt atgcaccaaa gaaggaactc 180
ataaatatta agggaattag tgaagccaaa gcagacaaaa ttctggctga ggccgctaaa 240
ttagttccaa tgggtttcac cactgcaact gaatttcacc aaaggcgttc agaaatcata 300
cagattacta ctggctccaa agagcttgac aaactgcttc aaggtggaat tgagactgga 360
tctatcacag agatgtttgg agaattccga actgggaaga cacagatctg tcatacattg 420
gctgtaacat gccagcttcc cattgatcgt ggtggaggtg aaggaaaggc catgtacatt 480
gacaccgagg gtacgtttag gccagaacgg ctgctagcag tggctgagag gtatggtctg 540
tctggcagcg atgtcctaga taatgtagca tatgctcgag ggttcaacac agaccaccaa 600
acccagctcc tttatcaagc atcagccatg atggtagaat ccagatatgc actgcttatt 660
gtagacagtg ctactgccct ctacagaaca gactactcag gtcgaggaga gctttcagcc 720
aggcaaatgc atttggccag atttctgagg atgctgctgc gacttgctga tgagtttggt 780
gtagcagtgg taatcaccaa ccaggtagta gcccaagtgg atggagcagc catgttcact 840
gcagatccca aaaaacccat tggaggaaac atcattgccc atgcatcaac aaccaggctg 900
tacctgagga aaggaagagg ggagaccaga atctgcaaag tctatgactc tccctgtctc 960
cctgaagctg aagccatgtt tgccattaat gcagatggag taggagatgc caaggactga 1020
<210> 14
<211> 1053
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<223> Rad51b
<400> 14
atgagcaaca agaaactaag acgagtaggt ttatcacaag agctgtgtga ccgtttgagc 60
agacatcaga ttgttaattg tcaggacttt ttaggtcttt ccccactgga acttatgaaa 120
gtgactggcc tgagttatgg aggtgtccag gagcttctgt atatggtcag cagggcctgt 180
gccccacaga tgcaaacagc ctatgatata aagatgcgga ggtctgctga actctcccca 240
gcgttcctgt ctactaccct ttctgctttg gacaaagccc tgcatggtgg tgtggcttgt 300
ggatcgctca cagagattac aggtccgcca ggttgtggga aaactcagtt ttgtataatg 360
atgagtgttt tagctacatt gcccaccaac atgggaggat tagaagggac tgtcgtgtat 420
attgacaccg agtctgcgtt tactgctgaa agactggtcg agattgcaga atcccgtttt 480
ccactctatt ttaacacaga agaaaaactg cttttgatga gcagtaaagt tcatcttcac 540
cgggaactca gctgtgaggc agttctgcaa aggcttgaat ctttggagga agagattatt 600
tccaaaggag ttaagcttgt gattgttgac tctattgctt ctgtggtcag aaaggagttc 660
gatcctcagc ttcagggcaa catcaaagaa aggaacaagt tcttggccaa acaagcatcc 720
ttgctgaagt acctggctga ggaattttca ctcccagtta tcttgacgaa tcaaattacg 780
acccatctga gtggagccct cccttctcaa gcagacctgg tgtctccggc tgatgatttg 840
tccctgtctg aaggcacttc tggatccagc tgtgtggtag ccgcactggg aaactcatgg 900
agtcactgtg tgaacacccg gctgattctc cagtaccttg actcagagag aaggcagatt 960
ctcattgcca agtcacctct ggctgccttc acctccttca tctataccat caagggggaa 1020
ggcctagttc ttcaaggcca agaaaggcca tag 1053
<210> 15
<211> 1101
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<223> Rad51c
<400> 15
atgcagcggg agctagtgag tttcccgctg tctcccacgg tgcgagtgaa gctggtggct 60
gcgggtttcc agacggccga ggacgtcctg ggggtgaagc cctctgagct cagcaaagaa 120
gttgggatat ctaaagagga agctttagaa actctacaaa ttgtaagaag agagagtctc 180
acagacaaac ccagatgtgc tggtgcatct gtggcaggca agaagtacac cgcactggaa 240
cttcttgagc aggagcacac ccagggcttc ataatcacct tctgctcagc gctagataac 300
attcttgggg gtggaatacc cctaatgaaa acaacggaag tttgtggtgt accaggtgtt 360
ggaaaaacac agttatgtat gcagttggca gtggatgtgc agattccaga gtgttttgga 420
ggagtggcag gtgaagcagt gtttattgat acggagggaa gttttatggt tgatagagtg 480
gtcacccttg caaatgcctg cattcagcac cttcacctta tagcaggaac acacaaggat 540
gaagaacacc agaaagcctt ggagggcttt actcttgaaa atattctttc tcatatttat 600
tatttccgtt gtcatgatta tactgagttg ctggcacaag tctatctcct tccagatttc 660
ctttcaaatc attcaaaggt gcagttagtg ataatagatg gcattgctct tccttttcga 720
catgaccttg atgatctatc ccttcgtact cgattactaa atggccttgc ccaacaaatg 780
atcagcctgg caaataatca cagattagct gttattttaa ctaatcagat gacaacaaag 840
attgataaaa atcaagcatt gcttgttcct gcattagggg aaagctgggg acatgctgct 900
acaataagac ttatttttca ctgggagcaa aagcaaaggt ttgcaacatt gtacaagtca 960
ccaagccaga aggagtccac aataccattt cagatcacac cacagggatt tagagatgct 1020
gctgtcactg cttcttcatc acagacagaa ggttcttcaa atctccggaa acggtcacga 1080
gaaccagagg aaggatgctg a 1101
<210> 16
<211> 1002
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<223> Rad51d
<400> 16
aaacatgggc gtgctcaggg tggggctgtg cccgggcctc accgaagaga tcgtccagct 60
tctgaagggc caaaggatca agacagtggc ggacctggca gctgctaacc tggaggaggt 120
agcccagaag tgtggcttgt cctacaaggc cctggttgcc ctgaggaggg tattgctggc 180
gcagttctcg gctttcccat taaacggagc agacctatat gaggaactga agacttccac 240
tgccatcctg tccacaggca ttggaagcct ggacaaacta cttgatgctg gcctctatac 300
aggggaggtg actgaaattg taggaggtcc aggtagcgga aaaacccagg tgtgcctgtg 360
tgtggctgca aatgtggccc atagcctgca gcagaatgta ctgtatgttg attccagtgg 420
aggaatgaca gcatcccgcc tcctgcagct actacaggct agaacccaag atgaggagaa 480
acaggcaggt gctctccaga ggatacaggt ggtgcatgta tttgacatct tccagatgct 540
ggatatgcta caggaccttc ggggctccat ggcccagcag tcgacatctt cttcaggcac 600
tgtgaaggtt gtgattgtgg attctgtcac tgccgtgatt gccccacttc tgggaggtca 660
gcagagggaa ggcctggcct tgatgatgca gctggcccga gagctcaaga tcctggcccg 720
ggacctagct gtggcagtgg tggtgaccaa ccacttgacc cgagacaggg atggtaggag 780
gttcaaacct gctctgggac gctcctggag ctttgtgccc agtacccgga ttctcctgga 840
tgtcaccaaa ggggctggaa cattaggcag aggccaacgc acagtgtgtc tgaccaagtc 900
tccccgccag ccaacaggtc tacaggaggt gatagacatt gggacattgg ggactgagga 960
gcagagccca gaattgcctg ccaaacagac atgatgctgt tt 1002
<210> 17
<211> 1257
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<223> Rad52
<400> 17
atggctggag ctgaggaaac agtccgtgga ggctgtgaca cccatcctcc ctttgctggt 60
gggaaatctg tgctgtgctt tgggcagagc cagtacacag cagaggaata ccaggctatc 120
cagagggctc tgaggcagcg gctgggcccg gagtacatca gcagccgcat ggctggagga 180
ggccagaagg tgtgttatat tgaaggtcat cgagtaatta acctggccaa tgagatgttt 240
ggttacaatg gctgggcaca ctccatcacc cagcagaatg tggattttgt tgacttcaac 300
aatggcaaat tctacgtggg agtctgtgca tttgtaaggg tgcagttaaa ggatggttcc 360
tatcatgagg acgtgggtta tggagttagt gagggcctaa ggtcaaaggc cttgtcactg 420
gagaaggcca ggaaggaggc tgtgactgac gggctgaagc gggcactcag gagttttgga 480
aatgcacttg gaaactgcat tctggacaaa gactatctga ggtcactaaa taagcttcca 540
cgacagctcc ctcttgaagt ggatttaact aaagcaaaga gagaagattt tgaaccatct 600
gtggaacagg caagatataa tagctgccta cagaatgaag caccgggacc cccaaaacca 660
caagaagcgg cttccccttg cagaccaagc cacccacatg attcgaacat taggctgcag 720
ggggctaagg actgcagcag ctcctgcagt ctggccgccc ccatggagag tgatgccatt 780
caccagcgca agctccggaa gctccggcag aaacagctgc agcagcagtt ccggcagcag 840
atggaggccc acctacaggg ccacacacct gccgtaaaag tgaaagccga gcgtgaggca 900
gtgcttccag accttcctcc aaaacacagt acccctgtaa ctgctgcctc agaactcctc 960
agggagaaag ccatttttcc agataaccct gaagacaacc ttgaaatgtg ggacctgact 1020
ccggatttag aggacatcat taagcccttg tctagaccag aaccacctca aacctctgcc 1080
accagagtcc aggtgatcca ggatggtgtc ctacacggcc tttgccacca gatgccacca 1140
gaaaaacatg aagctggtca cctgcaggcc cacagcactc accagcatgt attaggaaac 1200
tctgactctc ataggaagag ccaggacctg aagaaaagga aactggatcc atcctga 1257
<210> 18
<211> 2235
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<223> Rad54
<400> 18
atgaggagga gcttagctcc cagccagttg gccaggagga aaccagaagg cagatcatct 60
gatgatgaag actggcagcc tgggacagta actcctaaga aacaaaaatc cagtaatgag 120
acccagtgct tcctgtctcc ttttcggaaa cctttgactc agttaatcaa ccgaccgcct 180
tgtctggata gcagtcaaca tgaagcgttt attcgaagca ttttgtcaaa gcctttcaag 240
gtccccattc caaattatca aggtcctctg ggctgtcgag cattgggctt gaaaaaggct 300
ggtattcgtc gtgccctcca tgaccctctg gaagaaggtg ccttggttct gtatgagcct 360
cccccactaa gcgtccatga ccaactgaag ctggacaagg agaaactccc tgtccacgtg 420
gttgttgatc ctattctcag taaggtgttg cggcctcatc agagagaggg agtgaagttc 480
ctatgggagt gtgtcaccag tcgtcgaatc cctggaagcc atggctgtat catggctgat 540
gagatgggcc tgggaaagac actacagtgc atcacattga tgtggacact tttacgccag 600
agcccagagt gcaagccaga aattgagaaa gcagtggtag tgtcaccttc cagcttggtg 660
aagaactggt acaatgaggt tggaaagtgg cttggaggca ggatccaacc tctggccatc 720
gacggaggct ctaaggacga gatagaccga aaactggaag gattcatgaa ccagcgtgga 780
gcgagagtgc cttctcctat tctcatcatt tcctatgaga cttttcgcct tcatgtcgga 840
gtccttaaaa aaggaaatgt tggactggtc atatgtgatg agggacacag gcttaaaaac 900
tctgagaatc agacttacca ggccctggac agcttgaata ccagtcgtcg ggtgctcatc 960
tctgggaccc ccatccaaaa tgacttgctt gaatatttca gcttggtgca cttcgttaat 1020
tcaggcattt tgggaactgc ccaggagttc aagaagcatt ttgagctgcc aattttgagg 1080
agtcgagatg cagctgccag cgaggcagac aggcagctag gggaggaacg tcttcgagag 1140
ctcatcagta tagtgaatag gtgcctgata cggagaacat cagatatcct ctctaaatat 1200
ctgccagtga agattgagca ggtggtttgt tgtaggctga caccccttca aactgagtta 1260
tataagagat ttctgagaca ggctaagcct gaagaagaat tgcgtgaagg caagatgagc 1320
gtgtcttccc tgtcttctat cacctctcta aagaagctat gtaaccatcc agccctaatc 1380
tatgacaagt gtgtgtcagg ggaagatggc tttgaggata ctttggatat cttcccacct 1440
ggttatactt ctaaagctgt agaaccacag ctttcaggta aaatgctggt ccttgattac 1500
attctggcca tgactcgaag ccgcagcagt gataaagttg tgctggtgtc taattatact 1560
caaacgttgg atctctttga aaagctgtgc cgagctcgaa ggtacttgta tgttcgcctg 1620
gatggtacaa tgtccattaa gaagcgagcc aaggttgtgg agcgcttcaa tagtccatcg 1680
agtcccgatt ttgtcttcat gctgagcagc aaagctgggg gctgtggact taatctcatt 1740
ggtgctaacc ggctggtcat gtttgatcct gactggaacc cagccaatga tgaacaagct 1800
atggcccgag tctggcgtga tggtcaaaag aagacctgct atatttaccg actgctatct 1860
gctggaacca tagaagagaa gatctttcag cggcagagcc ataagaaggc actgagcagc 1920
tgtgtggttg atgaggagca agatgtagag agacatttct ctcttggcga gctcaaagag 1980
ctgtttaccc tggatgaagc tagcctcagc gacacacatg acaggctgca ttgccgccgt 2040
tgtgtaaaca gacgccaggt ctggccaccc cctgacggtt ctgactgtac ttcagatctg 2100
gctcagtgga accatagcac agataaacgg ggactccagg atgaggtact ccaggctgcc 2160
tgggatgctt cacctacagc catcaccttc gtcttccatc atcgttctca tgaggagcag 2220
cggggtctcc actga 2235
<210> 19
<211> 837
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<223> Xrcc2
<400> 19
atgtgcagcg actttcgtaa ggccgagtcc gggacggagc tccttgcccg gcttgaaggc 60
agaagctccc tgaaagaact agaacccaac ctgtttgctg atgaagattc accagtacat 120
ggtgatgttc ttgaatttca tggtccagaa ggaacaggaa aaacagaaat gctttatcat 180
ttaacagccc gatgtatact tccaaaatca gagggtggac tgcaaataga agtcttattt 240
attgacacag attaccactt tgacatgctc cggcttgtga cagtgctcga gcacagactg 300
tctcgaagct ccgaggagac catcaagctc tgcctgggaa gattgttcct ggcctactgc 360
agcagcagct tgcagctact gctcacgctg cactcactgg aagccctgtt ctgtagtcac 420
ccctctctct gccttctcat tgtggatagc ctgtcggctt tttactggac agaccgcgcc 480
agtggaggag agagtgtggc cctgcaggaa tccactctga agaagtgttc tcagctccta 540
gagaggcttg tcactgagta ccgcttggtg cttttcacaa caacacaaag tctaatgcag 600
aaagcctctg actcagcgga gcagcctgct tcctccaagc tcccaggtga cggagacaca 660
gactacagag cctatctctg caaggcctgg cagaaggtgg tgaagcacag agtcatcttc 720
tccagagagg acgaggctaa gagcagccgc ttctcattag tttcacgtca tttaaaaagt 780
aacagtttaa aaaaacatgc ttttatgatc agagaaagtg gggtggaatt ctgttga 837
<210> 20
<211> 1050
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<223> Xrcc3
<400> 20
atggacttgg atcaactgga cctaaatccc agaattactg ctgcaattaa gaaggggaga 60
ctgaggtcag tgaaggaggt tctgtgctac tcgggaccag acctgcagag gctcaccagc 120
ctgtccaccc acgatgtgca gcacctactg agagtggccg ctctgcacct ccagggcagc 180
cgggtcctca cagcactgca gctgttccag cagagggaga gcttccccga gcagcatcaa 240
cgcctgagcc tgggctgccc ggtcttggat cagttcctgg gtggcggcct gcccctggag 300
ggcatcactg acctggctgg tcgaagctct gcagggaaga cccagctggg gctacagctc 360
tgcttgactg tgcagttccc acgacaatat ggaggcctgg aggctggggc tgtctacatc 420
tgcacagagg atgccttccc cagcaagcgg ctgtggcagc tcattgaaca gcaacagcag 480
ctgcggacgg atgttcctgg ggaggtgatc cagaagatca gattcagcaa ccacatcttc 540
atcgagcatg cggccgacgt ggacgccttg ctggagtgtg tgagcaagag ggttcccatt 600
ctgctgtcaa gggggatggc ccgcctggta gtggttgact ctgttgctgc cccattccgt 660
tgtgagtatg atgctcaggc cttggccacc agggccaagc acctgcagtc tctgggagcc 720
gcgctccgca gactgagcag taccttccgg agccctgtgc tgtgcatcaa ccaggtgatg 780
gaaacggtgg aggagcaaga gtctatgccc aggccactgg gggcctggga tgagcacctc 840
tctccagccc ttggcatcac ctgggccaac cagatcctga tgagactgat ggttgaccgg 900
gcacatgagg acgatgcctc catgggctta cccagaagcc cggcacggac catacgggtg 960
ctctctgccc cgcacctgcc cctctcctcc tgctgctaca cggtcagtgc ggaaggcatc 1020
agagggatac cgggaaccga gtcctgctaa 1050
<210> 21
<211> 5373
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<223> Brca1
<400> 21
atggatttat ctgctgttca aatcgaagaa gtacaaaatg tccttcatgc tatgcagaaa 60
atcctggagt gtccaatctg tttggaattg atcaaagaac ccatttccac aaagtgtgac 120
cacatatttt gcaaattttg tatgctgaaa cttcttaacc agaagaaagg gccttcacaa 180
tgtcctttgt gtaagaatga gataaccaaa aggagcctgc aaggaagcac gaggtttagt 240
cagcttgttg aagagctgtt gaaaatcact gatgcttttg agcttgacac aggaatacag 300
tatgcaaacg gttacagtat ttcaaaattg aaaaattctt ctgaaccttt gaatgaggaa 360
gcttccatca tccagagtgt gggctaccga aaccgcggca aaagacttag acagattgaa 420
tctggaaatg ccaccttgaa ggacagtctc agtgtccagc tgtctaacct tggaattgtg 480
agatcaacaa agaaaaatca tcagacacag cctcgaaata aatctgtgta cattgaatta 540
gagtctgatt cttctgaaga gacagttagt aaaccagatg actgcagtgt gagagaccag 600
gaactgttac aaaccacctc tctgggagct ggagatgaag tcagtttgga ttcttcaacc 660
agagctgctt gtgagttttc tgaggacgta acaaatattg atctgcacca gtgcagtaat 720
aaagatttgg accccattga gaatcatgca actggaagat atccagaaaa atgtcaggat 780
atttctgttt caaacttaca tgtggagcca tgtggcacag atattcatgc cagctcgtta 840
cagcatgaga acagcagctt attactcact gaagacagaa tgaatgtaga aaaggctgag 900
ttctataata aaagcaaaca gtctggctta gccaggagcc aacagaacag atgggctgaa 960
agtaaagaaa catgtaatga taggcagatt cccagcactg agaaaaaggt agatccaaat 1020
actgattcct tctgtgggag aaaaaaatgg aaaaatcaga aaagtctgtg tcctgagaat 1080
tctagagcta cccaggatgt tccttggatg acactgaata gcagcattca gaaagtcaac 1140
gagtggtttt ccaaaactgg tgaaatgcta acttgtgatg gcacatctga caggaggcat 1200
gagtcaaatg ctggagcagc tgttgtgtta gaagtttcag atgaagtaga tggatgttcc 1260
agttcttcaa aagaaatgga cttagtggcc tctgatcccc ataatgcttt aaagtgtaaa 1320
agtgaaagcg acctctccaa accagtagag aacaatatcc gagataaaat atttgggaaa 1380
acgtatcaga gaaagggaag tctccctcac ttgaaccacg taactgaaat tataggcaca 1440
tttattgcag aaccacagat aacacaagaa caccccttca caaataaatt aaaacgtaaa 1500
aggagaacta catgccttca tcctgaggat tttatcaaga aagcagattt aacagttact 1560
caaaagactt ctaaaaatgt aaatcaagga actgaccaaa tggagccaga tggcctagtg 1620
atgggtatta ccagtaatgg tcaagagaat gaaacacaag gtaataatct tcagaaagag 1680
aaaaatgcta atccagtgaa gtccttggaa aaggggtctg tttccacaac taaagccaaa 1740
actataagca acagtataag tgatttggag ctagaattaa atgtccacaa ttcacaggca 1800
cctaagaaaa gtcggctgag gaggaagtct tcaaccagat gtgttcttgc actggaacca 1860
gtcagtagga atccaagccc acctgcttgt actgaatttc aaattgatag ttgtagcagc 1920
agtgaagaaa cagagaaaaa caattccaac ccaacaccag tcaagaacat tagaaagcct 1980
caactcacgg aggacacaga acctgcagca gattccaaga agaataaaga gccaaatgaa 2040
caaagaagga agagaagggc cagtgatcct ttcccagaag agaaatttat gcctggctta 2100
ttaactaact gtccaagttc tagtaaacct caaggacctg ccaatcctag ccctcagaga 2160
aaggaagtag agaaacttga aacaagccaa atgtctgaca gtaccaaaga cctgagggat 2220
ctgttgctgg atggagaaca gggtttgccc actgagaggt cggaggagag taccagtgtt 2280
tcattggtgc ctgacactga ttatgacact cagaacagtg tgtcattgct ggaagctaat 2340
gctgtcagat atgcaaaaac agtatcaagc cagtgtatga cccagtttgt agcaagtgat 2400
aaccctaacg aactggtcca tggttctaaa gatgctggaa gtggcacaga gtgcttcaag 2460
catccattga gacacgaact tagccacatt caggagacca tagaaatgga agagagtgaa 2520
cttgatactc agtatttaca gaatacattt caagcttcaa agcgtcagtc atttacttta 2580
ttttcaaaac caagagatcc ccaagaggag tgtgtaacag cttgtgcttc ctctgtgtcc 2640
ttaagggaca tgagtccaaa agtgacttct gaaggtgaac aacaagaaga aagtcgggga 2700
cacgaagagg ctgaaatcag tcacatacag gcagggcctg cgacagtggg cgtgcctctg 2760
cctcgtcagg aaggtgatcc aggcgctgat acaatgcatg ctgcagtgtg cagcctttgt 2820
ccatcatctc agtacagaag caatgaaaac tcacatccta aacaatcagc ttctcccatc 2880
atgtcttcta taaaaactgg ccatgggaaa accctgtcag aggaacaatc tgagaaacat 2940
acattgtcaa atgaaaaggc aatgggaaat gagacctttg ttcaaagcac aatgcacaca 3000
attagccaaa accacagaaa aaatggttgt caagaagcca actcaggcag tattaatgaa 3060
gtgccttcca gtggtgaaaa cttccaagga cagctaggcc gaaacagaag gactaagtta 3120
aacactgtgc ttccactagg tcttatgcaa cctggagtct gtaagcatag ttttcctgtc 3180
agtgattata aatatcttga aataaaaaag caggaaggtg agaatgttgg tgcacacttc 3240
tcttcgtgtc tgttttcaga taagctcgaa caacctgtag ggagtggtaa tgtttttcag 3300
atttgttctg agacacctga tgacctgttg gatgatgttg aaatacagga aaatactagc 3360
tttggtgaag ttgacataat ggagaagtct gctgttttta acgggagtgt ccagagacga 3420
gagctcagta ggagccctag ccctttaacc catgcatcgc tggctcggaa tctccagaga 3480
cggtctagga agttagagtc ctcagaagac agcggatctt gtgaggatga agaccttccc 3540
tgcttccaac acttacttgg ccaagtaagc aaaacacctg aacctagcag tgttgtgaca 3600
cagcctctgt cagagaaagc agaggggacc caagtgccat ggaagagtag tgttggtgac 3660
tctgataacg aggtgatctt gatagaggca tctcaggaac atcaccctag tgaggatgca 3720
aaatactctg gcagcatgtt ctcttctcag cccagtgctg tacaaggttc aactgcaaat 3780
acaagctccc aggacccctt gtttaacctt tccaaacaaa agagtcacca gtctgaggat 3840
gaggaagatt ttctaagtga caaggaattg atttcagatg atgaggaaat gggaacttgc 3900
ctagaagaag ataatgatca agaagaggat attataatcc cagattcagc tgaagcggca 3960
tctggatatg agagtgaaac aaacctttct gaagattgct cacagagtga tattttaacc 4020
actcagcaga gggccaccat gaaggataac ctgataaagc tccagcagga gatggcccac 4080
ctggaggctg tgctagaaca gcaaggggac cagccttctg tccactcctc gtctctcaca 4140
gctgaacctt gtgcccctga agacctacca aacgcagaac aaaacttatc aggaacagca 4200
attttaactt caaagaatat taatgaaaat cctgtaagcc ggaatccagc atgcgtttct 4260
gctgacaagt tccaaccaca acctccagac agttccagca gtgaaaataa agagtcaagg 4320
gaaggaaggc cttccctttt taaatctccg ttggcaggca gtaggtgctc tgcacacagc 4380
cgctctggga gtcttcaaaa cagaaactgc ccatctcaag aggagctcct ttggactgtt 4440
gaagcaggga agtcagaagg aacgccatac ctggaatctg gaatcagcct tttctctaat 4500
agagaccctg aatctgagtc ccctaaagag ccagcttatg tttgcaccac accagcttca 4560
acctctgcac ggacaatatc ccagtatcag gtttctgact ctttcgagag tccggctgct 4620
actcatgctc atactgcagt gatagaaact gtgagcaaga aaaagccaga attgacatct 4680
tcaaaaggaa gagccaataa aggaatatcc atggtggtgt caggcttgac ccccaaagaa 4740
gtaatgattg tgcaaaagtt tgctgaaaaa taccgcctca ctttaactga tgcaattact 4800
gaggagacta cccatgtcat tatgaaaaca gatgctgagc ttgtgtgtga acggacactg 4860
aaatattttc tgggaattgc aggaggaaaa tggatagtta gctattcatg ggtgattcag 4920
tctattcaag aaagaaaact tctgagtgtg catgaatttg aagtcagagg agatgttgtg 4980
actggaagaa accaccaagg tccaaagcga tccagagaat cccaggaaaa gctcttcaaa 5040
ggcctaacaa tctgttgttg tgagcccttc accaacatgc ccaaagatga gctggagaag 5100
atgctgcagc tgtgtggggc ttccgtggtg aaggagcttt cgtcactcac ccctgacacc 5160
ggtgttcatc caattgtgat tgtacagcca agtgcctgga cagaagagaa tggctgccct 5220
gagattgacc agctgtgtgg ggcgcacctg gtgatgtggg actgggtgtt ggacagtata 5280
tccgtctacc ggtgtcggga tctggacgcc tacctggtac agaatatcac ccatggccac 5340
gacagcagcg agccacagga ccccaatgat tag 5373
<210> 22
<211> 10098
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<223> Brca2
<400> 22
atgtccaatg aatacaaaag gagaccaact ttttttgaaa tttttaaggc acgatgcagc 60
acagcagact taggacctat aagcctgaat tggtttgaag aactttcttc agaagcccca 120
ccatataatt ttgagcctcc agaagaatat gaatataaga ccaacaatta tgagccacag 180
ctgtttaaaa cgccacagag gaagctctct taccatctgt tggcttcaac tccaataata 240
ttcaatgaac aaggtcaaac tctaccagta gaccagtcgc ctttcaaaga actagggaag 300
actcttggaa atagtagaca taaaaatcac cacagaacca aggccaaaat ggactccatg 360
gttgatgttg ccagtccacc tctgaagtct tgtctcagtg aaagccctct tactctgcga 420
tgcacacagg ttgtaccaca aagagaaaag ccagtgatgt gtggaagttt attttctaca 480
ccaaaaatcg aggagggtca gacacctaaa cctatttctg aaagtctggg agttgaggtg 540
gatcctgata tgtcttggtc cagttcatta gctacaccac caacccttag ttccactgtg 600
ctcatagccc gagatgggga agcacgagga attgtgtttc ctgaagactc ccccgctgtt 660
ttgaaaagct acttttccag ccacaatgaa agtctgaaaa agagtggtat atcagttccc 720
tctgtgactg acagcgagaa caaaaaccag agggaagctt ttagtcatgg attggggaaa 780
acgttagagg attcatctgg caaaacaaac agcttcaaag attgccttag gcagtcaata 840
ccaaatgttc tagaagatgg ggagacagct gcagatactt ctgaagaaga tagtttctca 900
ttatgttttc ctaagcataa aaccagaaaa atgcaaaaaa tgagaatggg caagaccagg 960
aagaaaattt tcagtgaaat aagaactgat gaattaagtg aagaaactag aagtggagct 1020
gatggaaaac attcatttgc acttgaaatt gacccaagag atagtgatcc cttagatcca 1080
gatgtgacaa accagaaatg ctttgacaat gggaatgagg aaatctgcga ggaagttgta 1140
cagtcttcag acactcgatg gtctcagcta accctttctc gtctaactgg tacccagaag 1200
ggaaaaatac ctctacctcc tatttcttct tgtaaccaaa ataattcaga aaaagacttc 1260
atagatacga aggaagaagg tattgactct gttactttag aaaattcttt gcctcatatt 1320
tctagttttc cagaaccaga aaagatgttc tgtgagaaaa ctctggtaga taaggaacat 1380
gagggacagc atcttgaatc acatgagaac tccattgcag ggaagcaagt ggtatctgga 1440
acttgtcaag cagcttgcct attgagagag ccacttgaag agtctctggg taatctcttc 1500
tcagagagtg tgactagctc agcctttaca gaagaaccca gtgcctctgc aagtggattg 1560
ggaatatgtg ctgtgtcctc acagagagag gattctttgt gtcctagttc aggtgacact 1620
ggaaactggc catcaactct cactcacact tctgcaactg tgaacaatac aggtttaata 1680
tccagtctaa aaactaaaag aagaaagttt atttactctg tcggtgacag tgcatctcat 1740
caaggaaaaa tactacaggc agacagaaag tcagagctca ctaacacttc tgctcagttc 1800
gaaacaagtg cttttgaagc gccattcacg tttacaaatg ggaattcagg tttatcagat 1860
tcttctctcc aaagaagctg tttacagaat gatgctgaag agccatcttt gtccttatcc 1920
acctcatttg tgactgcttc caagaaagaa agtagttata gtaatgcatt gatatctcag 1980
gatctccatg acaaagaagc aatagtcagt gaagaaaaac tgcagccaca tacagccctg 2040
gaaactgatt gtctgtcgtg cttgccagaa agacaatgtg aatatgatcc aaagggtccc 2100
aaagtttcag atggaaaaga agaagtctta gtctcagcat gtcatcctgc aggacagcac 2160
acagcagcag cacagcccag cagcattagc tttgaattac aggaagaccc tgtcaatggc 2220
cacaatagta caagtcctaa agaaactcct agcttgaagg tgcttctgtc aaagccagtt 2280
gtgctttcta gaggaaaagt gtcatgtaaa atgccagaga aactgcagtg tgagagttat 2340
aaagataata ctgaattaag caaaagcatt cccttgggag gaaataaaat acacatccta 2400
agtgaaaatt ctaaacctcc tgagcttctg ccacctggaa aatatgtaac agaagcatcc 2460
cccacagtga agtctcagtt caatcagaat acaaatctag cagtcaaaaa aaatgaccaa 2520
gaagaaacgc cttttatttc agaagtaaca gtcaatgtga gttctggaga acttttccca 2580
gacaatgaga ataatttcgc ctttcaagta actcatgaaa ataataagac tgccttagga 2640
agtactgtgg aactgcagga agaagacctc cgccatgcta aagggcctaa tctcaacaac 2700
tctcccacag cagtagatgg agacataggt gatgagcaag cagctcatgc actgattatg 2760
gaagactccg attcctcagc tttagtccat gagtgtgcaa agaagagcag aaatactata 2820
gagcagcatc tgaaaggaac cacagacaaa gatttcaatt cctccttgga tgtgaaatca 2880
gatgggaaca atgactatac agacaaatgg ccaggatttt tggatccagt ttttaagcat 2940
aaatttggag gtagcttcag aacagcttcc aataaagaaa taaaactttc agaacataat 3000
gttaagaaaa gcaaaatgtt cttcaaagat attgaagagg agtatcctac tagtttaacc 3060
tgtattgaca tagttaatgc ctcaccatta gcaaaccagg agatactaag tggaccttat 3120
acatttgatt tgcagtcagt cactaccatg tctgcccatc cacagagtca ggcatctgtt 3180
tcttgtgaag atactcacac atcacttcag gtgttacctt caaagcaaga ttttcattcg 3240
aatcacaatt taacacccag ccaaaaggca gaaattacag aactttctac tatcttggaa 3300
gaatcaggaa gtcagtttga attcacacag ttcaaaaagc caagccatgt agcacagaat 3360
aatatacctg aagtgcctgg aaaacagacg gttgccataa atactacttc tgaggggtgg 3420
aaaaggattg gtcttcatct cacagtggat cctgcctctg tagctcagac agatgacagc 3480
aagaaatttg aaggttctgc tggatttaga caaagctttc cttgcctgtt gaaaagcagt 3540
tgtaacaaaa atacatctag ttttttagca aatgtaaatg aaatggagtt tcgaggattt 3600
cgttctgctc ttggcacaaa gcttagtgtg tctagtgagg ctctgcaaaa agctgtgaaa 3660
ctcttcagtg acattgaaag tggtagtgag gagacttcca caaaagtcga cccaagagct 3720
ttgtcttcag gtgctcgtca tgattctggt gcttctgtgt ttaagataag gaaacaaaac 3780
agtggtaaaa gttttgatga gaaaactagt aagtgccaag taacattaca gaataatacc 3840
gaagtgacta ctggtatttt tgttgacaga aatcctgaaa attatgcaag aaatacaaaa 3900
tgtgaagata acaactctac tggttttcaa agaagtcctt ataaattaaa aaactcggag 3960
gatagtgaat caagtacaag cggcacagtt tctgttcatc aagatgatgg tgacttacca 4020
tgtgctgctg atcactgcag caagtaccct gagtcgtgtt cccaatatgt aagggaggaa 4080
aacacacaaa ttaaggaatg tgtatcagat ttaacatgtt tggaagtcat gaaagctgag 4140
gaaacatgct atattcaacc ttcagataaa gaacaattac cttcaggtaa gatggaacaa 4200
aatagaaaag attttaatat atcctttcag actgcaagtg ggaaaaatgt cagagtctct 4260
gaggagtcat taagtaaaag tatgaatatt ttaaatcaag taacagatga atcgatcatc 4320
tcttcagatt ctttgaattc taaatttcat tgtggcacaa ataacaacaa gatgggtatt 4380
tcacatcaca aggaaactac cagtactaaa aaggtatttg aagaacgttt cccagttggg 4440
actgtcagtc aattaccaac tctccagcag catcctagat gtgaaataga aagtatcaaa 4500
gaacctgctc tgttgggttt tcataccgct agtggaaaaa aagtcaaaat tatgcagaaa 4560
tctttagaca aagtgaaaaa cctttttgat gagacacagt atgttagtca tcaagggtca 4620
aaacccttga aggacagaga gaactgcaaa gaaggacttg cattaggatg tgagacaatt 4680
gaaatacctg cctccaagtg tgaagaaatg cagaagagct ttctctctaa ggagtctgaa 4740
gtgctatcta agcaaagtga tcgtttgtat aggtcgactg aaaatctcag aacatcaaat 4800
ggtacctctt ctaaagctaa tgtacatgga aacatagaaa gtgaaataga aaaaagtcct 4860
acaacttgct gcattagtca tttatcttat tcagtcactg aagattctgc tttgacatgt 4920
gacacaggac acggtagaaa aacttgtgtc agtgagtctt ctctatccaa agacagaaaa 4980
tggcttagag accgactggg tgataagctt caaaaaagag atgctgccga aattgaatgt 5040
gtaaaagaac atactgaggg ttatgctgga gatgcctcat gtgagcatag tttagacagt 5100
atcagaaatg aagttgatat aaattgtgtc tctgaaaatc aaacttcagc cctctttagt 5160
gaccctagca tgtgtcatag ctgtccatcc cattttggtt gtcattgtga taacaagcat 5220
aatgactcag gatatttctc aaaaaataaa atttattctg atactcagcc agacacgaag 5280
aatgaagaca ctgccaattt ttccagtgta tatgctacaa aagaagtaaa tatatacccg 5340
ccaactgtaa atgaagatat ttgtgttcag aaacttgaga ctaactcttc accacataca 5400
aataaaaatg tagccattga cttggctata gcagattcaa ggaattgtaa ggtatgccca 5460
tccaagttca ttacagatca ctcacaagaa actgtgaaaa cagtaaaagc aatatttaca 5520
cataacagtg ataaaacaat taagcaaaac acaaagagta aaccagatac ttgccggaca 5580
agctgtcaga aagcattgga taattcagag gattttatat gtcctagctc tttagaagat 5640
gactatatga actcacataa gacttctgtt tatacccacg atgaacaaat attacagcat 5700
aacctaagtg tgtctggact ggagaaagct caaataccac ctgttcactt ggaaacttgg 5760
gataagtgta aatctacaag ggaacttgca caggcagcct gttcttcaca catgccgggg 5820
atttttagca cagcaagtgg aaaagctgta caggtatcag acgcttcatt agaaaaggca 5880
aggcaagtgt tttctgagat ggatggtggt gctaaacagt tactttccac attgtctctg 5940
gaaagtcatg aacaatcaga ccactctggg agaagagaaa actctgtgac acataaccct 6000
gaggatgtat tgtcactccc aaaaaccttt gcaagcaatg ccaattcatc tgtattctct 6060
ggatttagta cagcaggtgg taaacgggtc acagtttcag aaagtgcctt acacaaagtt 6120
aagggaatgt tagaggaatt tgatttgatc ggaactgaac atactctcca gtgtccacct 6180
acatctgaag gtgtatcaaa aatacttcct caatattgtg ttgaaaagag aaccccagaa 6240
taccctataa actctaaatt gcagaaaacc tatgatgata aattcagttt accaaacaac 6300
tataaagaaa gtgcttcttt ggggaatact cattctcttg aagcttctcc ccaactctct 6360
cagtttaagc aagacacacg gttggtatta ggaaccaaag tatcccttct ggaaaaagaa 6420
caaaccttcc ctcaaaatat aaaaacagaa agtggtgtaa tggaaacttg tcctgatgtc 6480
cctgtgagaa caaatgtagg agactgttct gctttcgaca aaaagccaga gaactgtttt 6540
gaaacagaag cggtggagat tgccaaagct tttatggaag atgatgagct gacggattct 6600
gaaccaagtc acgacaaata ctcattgttt acctgccccc aaaatgaggc tttgttaaat 6660
ttgagaacta gaaagagaag aggaatggct gttgatgcag ttggacaacc cccaatcaaa 6720
agaagcttat taaatgagtt tgacaggata atagaaaata aaggaaaatc cttaaagcct 6780
tcaaaaagtg ctccagatgg tacaataaaa gacagaagat tgttcacgca ccacatttct 6840
ctagagccag ttacctgtgg acccttctgc tctaccagag aaaggcaaga aatgcagagt 6900
ccacatttca ctgcacctgc tcaaggactt ctgtctaaag ggcatccttc tgtgcgttcg 6960
gctttggaaa aatcttcaag caattctaca gtttcagtcc agccgactca taaagtttct 7020
gctacaagga atgaaaggac aggatgctca gtcacaggca aatcccacaa agtcttcgtc 7080
ccacctttca aaatgaaatc acagtttcac agagataaac atttgaataa caagaatatt 7140
aattcagagg aaaagaacca aaagagcaga gatggagaca gtgaagatgt gaacgacggt 7200
gacattcgtc aatttaaaaa aggcagcttc catcaagagg ccgctagaat tgtcacagag 7260
tgtgaagaag agcctttaga tttaatgaca agccttcaga atgccagaga cctacagaat 7320
atacgaatta aaaagaaaga aaggcatcag ctctgtccac agccaggcag tctgtatctt 7380
accaagtcat ccactctgcc tcggatctct ctgcaggccg cagtaggagg ccggcttccc 7440
tctgcgtgtt ctcataagca gctctatatg tatggcgttt ctaaagagtg cataagcatt 7500
aacagcaaaa atgcggaata ttttcagttt gacattcagg attattttgg taaagaggac 7560
ctgtgtgctg gaaagggcat tcagctggct gatggtggat ggctgatccc ctccaatgaa 7620
gggaaggctg gaaaagaaga attctatagg gctctgtgtg acactccagg tgtggatcca 7680
aagcttattt ctagcctctg ggtctctaat cactaccggt ggattgtgtg gaaactggca 7740
gctatggaat ttgcttttcc taaggaattt gctaacagat gcctaacccc agaaagagtg 7800
ctgcttcaac taaaatacag atacgatgtg gaaattgaca acagcagcag atcagcccta 7860
aagaagatcc tggaaaggga tgatactgct gcaaaaacac tcgttctctg tgtttctgac 7920
gtcatgtcac taagcacaac tggatcagaa acttcaggcg gcaaaactag tggtgcagat 7980
cccaagagca tagacacgat cgaactcaca gacggatggt atgctgtcaa ggcccagcta 8040
gaccctccac ttgtggctct tgtaaagagc aggagactga ctgtgggtca gaaaatcatt 8100
acccaagggg cagaactggt gggatctcct gatgcctgcg cacctctgga agcaccagac 8160
tctcttagac tgaagatttc tgccaacagc acacggcctg ctcgctggca cagcaaactg 8220
gggttctttc atgaccccag gcctttccct ctgcccttgt cctcactgtt cggtgatgga 8280
ggaagcgttg gctgtgtgga catcattgtt cagagagtgt accctttaca gtgggtggag 8340
aagacggcat caggattgta catattccgt aatgagagag aagaggagaa ggaagcagtg 8400
agatttgcag cggcccaaca gaagaaactg gaggccttgt tcaccaaaat ccaggcagag 8460
tttaaagacc atgaagaaga ctcagcgcag cggcgcgtgc tgtcccatgc actcacaagg 8520
cagcaggtct atgctctgca agatggtgca gagctctatg cagcggtgca gagtgcattg 8580
gatccagatc accttgaggg ttgtttcagt gaagagcagc tgagagcctt gaacaaacac 8640
agacaaatgt tgaatgataa gaagcaagca cagatccaat cagaattccg gaaggccgtg 8700
gagtccgctg agcaagagga gggcttatca agggatgttt caactgtgtg gaagcttcga 8760
gttacaagct acgagagaaa agaaaagtca gctctgctga gtatctggcg tccatcatca 8820
gacttacact ccctgttaac agaaggaaag agatacagaa tctatcatct ggcagtatca 8880
aaatctagga gtacatttga acgacctagc atccagttaa cagctacgaa aagaactcag 8940
tatcagcagc taccggctgc cagtgaaacc gtattccagg tttaccagcc gagggagccg 9000
cttgacttca gcagactgtt agagacagcc tttcagcctc cttgttctga agtggacctc 9060
gtaggagctg tagtttctgt tgtaagaaca acaggtcttg ctcctttggt ctacctatca 9120
gacgaatgcc ttaatttatt agtggtaaag tttgggatag accttaatga agacataaag 9180
ccacatgtgc taattgctgc aagcaacctc cagtggcacc cagaatccag atcaggagtg 9240
ccaacgttat ttgccgggaa tctttccata gtttctgcca gtccaaagga ggtctacttc 9300
cagaagcgag tcaacagaat gaggcaaact gttgagaata tcgatacatt ttacaaggag 9360
gcagaaaaga agcttttaca tctgcttaat ggaaacagtc ccaagtggtc caccccaaat 9420
aaagactcta ctcagccggc ctacacttgc cctgcttcag agctccttgc tacaggaggt 9480
cagctaatga ggttctcacc taatagtgag caaagttatc caagtccttt atcacattgc 9540
acaccaaaag aaaagtctac acccctggct cggtcagccc agatggcatc aaagtcttgt 9600
aacagggaga gagagactga tgaccgaaaa acctgcagaa aaagaagagc cttggatttt 9660
ctgagtcggc tgcccttacc tcccccggtc agtcccattt gtacctttgt ctctccggct 9720
gcacagaagg cttttcagcc gccacggagt tgtggcacca aatatgcaac acctatgaag 9780
aaaaaagaac ccagttcccc tcagaggagg acgccatttc agaaggacag tggcatttct 9840
cttctggaac aagattcggt agctgatgaa gaacttgcct tcctcaatac ccaagctctt 9900
gtacctggct caccagaaga aaatcaacaa gtacttcctg gtgactccac aagaacctct 9960
gtgccccaca acaaaagaac ccccagccag cacagaggcc agaccagcat gtaggacccc 10020
gatccaggaa ggggagtcta tctcagggat tacagaggtg gcaagggcag tgatggaaaa 10080
ttagctgtcg agtcttag 10098
<210> 23
<211> 888
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<223> Циклин D1
<400> 23
atggaacacc agctcctgtg ctgcgaagtg gagaccatcc gccgcgcgta ccctgacacc 60
aacctcctga acgaccgagt gctgcgagcc atgctcaaga ccgaggagac ctgcgcgccc 120
tccgtgtctt acttcaagtg cgtgcagagg gagattgtgc cgtccatgcg gaaaatcgtg 180
gccacctgga tgctggaggt ctgcgaggag cagaagtgcg aagaggaggt cttccctctg 240
gccatgaact acctggaccg tttcctgtct ctggagcccc taaaaaagag ccgcctgcag 300
ctgctagggg ccacctgcat gttcgtggcc tccaagatga aggagaccat tcctttgacc 360
gccgagaagt tgtgcatcta cactgacaac tctatccggc ccgaggagct gctgcaaatg 420
gaactccttc tggtgaacaa acttaagtgg aacctggccg ccatgactcc ccacgatttc 480
atcgaacact tcctctccaa aatgccagag gcggatgaga acaagcagat catccgcaag 540
catgcgcaga cctttgtggc cctctgtgcc acagacgtga agttcatttc caacccgccc 600
tccatggtgg ctgccgggag cgtggtggct gcaatgcaag gcctgaacct gggcagcccc 660
aacaactacc tgtcctgcta ccgcacaacg cactttcttt ccagagtaat caagtgtgac 720
ccggactgcc tccgcgcctg ccaggaacag attgaggccc ttctggagtc cagcctgcgc 780
caggcccagc agaacatcaa ccccaaggcc accgaggaag agggagaagc agagggagag 840
actgacctgg cctgcacacc caccgatgta cgagatgtgg acatctga 888
<210> 24
<211> 882
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<223> Ercc1
<400> 24
atggaccttg ggaaagacga gggaagcctg ccgcagccca ccaggaagaa gtttgtgatc 60
ccactggaag acgaggcccc tcctgcaggg gccaagccct tattcagatc ctcacggaac 120
cccagcacca cggccccctc ggtcccagcg gcccctcaga cgtacgccga gtatgccatt 180
gcccagcctc caggaggggc tgggcccaca gggcccacag gctctgaacc tgtgaaggga 240
gagaaccccg gccagacggt gaaaacggga gcgaaatcca atagcatcct tgtgagcccc 300
cggcagaggg gcaaccctgt gttgaagttc gtgcgcaacg tgccctggga attcggcgag 360
gtgacccctg actatgtgct gggacagagc acttgcgccc ttttcctcag cctccgctac 420
cacaatctcc atccagacta catccacgaa cggctgcaga gcctggggaa gagctttgcc 480
ctgcgtgtgc tgttggtcca agtggatgtg aaagatcctc agaaggccct gaaggacctg 540
gctaaaatgt gtatcttagc ggactgcacc ctggtcctgg cctggagtgc cgaggaagca 600
ggacggtacc tggagaccta caaggcatat gagcagaagc ccgctgacct cctcatggag 660
aagctggagc agaacttcct gtcccgggcc accgagtgtc tgaccaccgt gaagtcagtc 720
aacaaaaccg acagccagac cctcctggct acatttggat cccttgaaca gctcttgacg 780
gcatcacggg aggacctagc cttgtgcccc ggcctgggcc cccagaaggc ccgcaggctc 840
tttgacgtcc tccatgaacc cttcctcaaa gtgccccgat ga 882
<210> 25
<211> 4473
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<223> MDC1
<400> 25
atggaaaaca cccaggttat tgactgggat gctgaagagg aggaggagac agaaataccc 60
agtgggtcct tggggtatag cttggagcct atagggcagc tgcgtctctt cagtagtact 120
catggaccag aaagagattt cccactctac cttggcaaga atgtaattgg ccgaagcccc 180
gactgctctg tggccctgcc ttttccatcc atctctaaac agcatgcagt aattgaaatc 240
tcagcttgca acaaagcccc tatcctccag gattgtggga gcctcaatgg cactcaactg 300
gtaaagcccc caaaggtcct aacccctgga gtgagccatc gtctgaagga ccaggaatta 360
attctgtttg cagactttcc ctgccagtac cgtcgcttgg atgtccctcc acctttggtc 420
tctcggggac ttctaactat agagaagacc cccagaatac agggagggtc ccaaacctcc 480
agagttttgt tggctgagga ttcggaggag gaagtagatt ttccttctgg aaggtatgtg 540
gcaaatggat caaggaatcc aacgtctcca tcagcaacag tagtaccaga aagtgatgaa 600
gaggggtctt ctccagccca aagtgtccgt gggccgtctt tgccctttga cttgggcagt 660
gacacagatg aagagctatg tccacagcca gcagttgggg agtcctcttc agttgccagg 720
gatggtgctg ctgcagaggc caagcagctg gaagctaatg gagtgacatc tggtatccag 780
ctggtactgg ctcagcctgc tgagcaaaag ctcagagatg caaaagtcaa gagtgaggct 840
ggcaatggag cggctgtggt tgggaggggc tccactgtgg gtaaggacag caacacggaa 900
atgggtgaag aacaccagcc ttccggcttt gtggacagcg atacagacgt ggaagaagac 960
aggatccctg tgaccccccc tgtagttccg gggaagaagc aagtcctgct tggagttggt 1020
aaaaaggacc ctggagcacc tgtcgtgacc catctgcagg acagcccagc tggtcgtggc 1080
acaggtgtgg aagaaggcaa gactccagtg gctggccctc cagagagaaa ccacacggcc 1140
atggtgatca gcagcgacac agatgaggaa gatggagtct cagcagcagt caccttggcc 1200
catctgaaag acagggggat ggctctgtgg agcagggagc caggcatgga agaggtcaag 1260
tcccaaccac aggtcctcgt agaacgaagc cagagtgcct ctgggaaaga cagtgacaca 1320
gacgtggagg aggaaaagag agaagtggtc cctgacagcc ccatggacac agatgaggct 1380
cttactgtcc cacattcaga gagccaagct ccccatagag ccagtgatga tgtagaccag 1440
ggtgtggata tgggctcccc tggtggtcag ctagagggaa accaggcctc ctctgccaca 1500
gtagaagaca gtagtgcaca agcagggaag gaagtcctgc ctcctgagga ggcctgggaa 1560
acagctgtgg aggaaggctc atcttcagca gcggcagatg taagacaaag caagcagcca 1620
ggagtagaag atgctgggac agagtgggct gcaactgttt gtgagcaggc gagcatgctt 1680
gaggtggggg tccaaggcag gtcacctgct acactggtag agccagtggt ggtgcctaca 1740
gctactctgg gggatcccac tcagccacag agagagggag cccagacccc cacaggaaag 1800
gagagaggag tacaaatggg caggaccaag aatgccaaag actgccgtga tgcagagtct 1860
gaagatgtgt gccttccggc tacccagtgc tttgtagaca gggagggcca gagctcagaa 1920
gctgtccaga gtttggaaga tgagcctacg caagtctttc catgcactct tccccaagag 1980
cctgggcctt cccacctcag cctgcagacg ccaggtccag gtgcccagga tgtgccttgg 2040
gaagtcttag ctacacagcc gttttgtctg agagaagctg aggcctctga actgcagctc 2100
atggacaccc accctgcagc tcatgaatcc cacccatctg tgtctagtgc atcagcagga 2160
cagcagcatc tggttcacac agagccgctg ggaattgaag gcggagagat gcaaactgtg 2220
gagaaagcca tgggccaatt gagttgcaag atggcatctg ctggagagga ctcaaggggt 2280
gatccagaac cctcggccca tcgcctgctt tctccagttc ctgaagcttc ttctccacct 2340
cagagtctgc tcacctctca gagccaaaag ccgtctacac cccagtctct gttacttacc 2400
tctccccctt ctgagctaca ccttcctgaa actcctcaca ccaagcctaa tgtcaggcct 2460
cggcggtcct ccaggatgac ctcctcccca cactcctctg ctgcccttaa gccctatact 2520
acctgcccca caaacctgcc tgctgcccct agaccaacat ctcgggctac tcggggcagg 2580
ggcagggcaa ataggtcctc taccaggacc ccggaaccag ttgtccctac agactctgag 2640
cttcaacctc ccacctccac agaacagtct gtcatcccca aacccacatc tccagtcact 2700
cagggcagca taaatggttc ctttgttaaa acgcctgaac cagttgttct cacaggtccc 2760
aaaatccagc ctcccacctc cacagaacag cctgttaccc caaaccccac acctcaggcc 2820
actcggggca ggccacatag gtcttccatc cagaccccag aaccacttac tcccactggt 2880
cctgacctcc agcctcccac ctccacagaa cagcctgtca tacctaaacc cacatctcgg 2940
gctgctcggg gcaggtcacg taagtcttct accaggaccc cagaaccagc tgtccccact 3000
ggtcctgacc tccagcctcc cacctccaca gaacagcctg tcacacctaa acccacatct 3060
cgggccactc ggggcaggtc acgtaagtct gtcagaaccc cagaaccagc tgtccccact 3120
ggtcctgacc tccagcttcc cacctccaca gctctgggca ctttgggaag ggcatgtaag 3180
tcctccattg aggattctga atcagttgga ccagtagcct ctgattttga acctcccatc 3240
tccacagacc ttgttgcccc tgaggtgaca ggtcagagca taacactaaa gtcttcacca 3300
ctaagtgctt ctccagtttc tgccacctct gaactccagc cacctgtccc cacagcccag 3360
cctgttctcc tggagcccat tcctcaagcc aaccaccgaa ggcggcggaa ggctgctggg 3420
aaacggggct cccacacagt tcccattggc caaaagcctt actctgcacc ctctgaacct 3480
ggttcccaat cttcaatcaa tcaaggcttg gcattggaag ccgctgagtc tattactgtt 3540
gctcctgagc ctgctgtttc ccaggctcca gagacctcca ctcagaatcc cgtggtgcaa 3600
aatgaagcag ctgggagatc agggctcatt cccaagcccc agcctgaggt ttctcgatcc 3660
cgcaagaagc cttctactac cacaacctca ccaattcaaa aacgtccccg aagacaaata 3720
ccccagaaga caatagtccc caaggaagaa gatcctgagg aaatgccagt gaaggaagag 3780
cctcaggaga tagcaattcc aacacccggc aaaagaaaga gggaccgtgt agaggatgag 3840
acccagggaa acccaagtcg gagccggcgg gctaaaccta accaagaagc agcagccccc 3900
aaggtactgt tcacaggagt tgtggattct cggggagagc gtgcagtcct ggctctgggt 3960
ggcagtctag ccagctcagt aaatgaggcc tcccacttgg tcactgatcg catccgaaga 4020
acggtcaagt tcttgtgtgc cctggggaag ggcatcccta tcctctccct gaactggctg 4080
tatcagtcca gaaaggctgg tcacttcctg ccacctgatg actacttggt gactgaccct 4140
gaacaagaga agaactttag cttcagcctt cgggactccc tgagccgggc tcggcaacaa 4200
aaactgctgg agggctatga gatttacgtg acccctggag tgcagccacc cccacctcag 4260
atgggcgaga tcatcagctg ctgtgggggc accaacctgc ccagcatgcc ccagtcctat 4320
aagccttacc gagttgtcat aacttgcact gaagacctac ctcgctgcgc tatcccatct 4380
cgactggggc tgcccctcct ctctcctgag ttcctcctga ctggagtgct aaagcaggaa 4440
gtcacaccag aggcctttgt cctttcaaat tag 4473
<210> 26
<211> 2412
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<223> Bard1
<400> 26
atgcaacttt ctgagtctgt ggttggacta ctaggtaggt acacaagccc tgctcgggaa 60
cgaaacccac gaggagcccc gccccttggc gttgagggcg tggccttggg ccgcgctcct 120
ctcagaaatt cgagcgcgct ctcattgcct ctcccctccc tcttacttgc tataaaggag 180
caacccagga tggtctgctg tctgcttcgg ctcatcatca tccagagtgt cgagttcttt 240
gaaatcatca ctaatattct gaaggagcca gtatgcttag gagggtgtga gcacatcttc 300
tgtagtgtgt gtataagtga ttgcgttgga tcaggatgtc ccgtgtgtta cacaccagcc 360
tggatcctag acctcaagat aaacagacaa ttggacaaca tgattcagct ttataggaag 420
cttcaaaatt tgctacatga caataaactt tcagatccaa aagacaacac atctaggaca 480
ggttcatttg atgatgcaga aagcaagaag aattcaataa aaatgtggtt tagtcctcga 540
agtaagaagg ttagatgtgt tgtgaataaa gtatcagtgc caacccagcc tcaaaagaca 600
aaagatgaca aagcccagga agcctccata tacgaatttg tttccacaag tccccctgaa 660
gttgtttcta agagggctaa aacagcttct agaacatctg caaaaaagca gcagaagaaa 720
tctttagctg agatcaacaa gaaggggaat tcaaggccag aaacagaaga tagcaggttt 780
gattctaaag aggaactgaa agaggagagg gttgtctcct gtagccaaat accagttatg 840
gaaagtccac aggtaaatgg tgaaatagac ttgttagcta gtggttctgt tgcagaatct 900
gaatgttctg gaagattgac cgaagtttct ttaccattgg ctgagcatat agtatctcca 960
ggcactgtga gcaagagtga agagactcct gagaagaaag tctgtgtaaa agatcttctt 1020
tcagtagggc gtaatgaaaa tcacaaatac tgtagcaggc ctcctgatcc tacttctaag 1080
aattgtgaga gaagcattcc gagcaccagc agagatgtca ttaaaccaac agtgcttgca 1140
gaaaatatac tgttggttga ctgttcttca ctgccttcag accagcttca agttgatgtc 1200
acactcagga gaaagagtaa catatcagat aactccctta gcctttcacc aggcacaccc 1260
ccaccactgc tgaataattc aactcccaga caaatgatgt caaaaccctc catagtgaag 1320
ctgtcaccca gcattaccgc caggaaaaga aaccacagag gagagactct gcttcatatt 1380
gcttctatta agggcgatat accttctgtt gaatacctct tgcaaaacgg aaacgacccc 1440
aatgttaaag accatgctgg atggacacca ttgcatgaag cctgcagtca tgggcacctg 1500
aaggtagtgg aattgctgct ccagcataac gctttggtga ataccacagg ctatcagaat 1560
gacacaccac tgcacgatgc agtcaagaac ggccatgtgg atatagtcag ggtgttactg 1620
tccaatggcg cctccaggaa tgctgttaac atatttggtg tacggcctgt ggattataca 1680
gacaatgaaa atataagatc attattgctg ctaccagaga agagtgaatc atcctcaact 1740
agccagtgtg cagttgtgac tgctggacag cgaagaaatg ggcctctggt gcttataggc 1800
agtgggcttt cttcccaaca gcagaaaatg ctcagcaaac ttgagacagt tctcaaggct 1860
aaaagatgtg ctgagtttga cagtacagta actcatgtca ttgttcctga tgatgaagct 1920
caaagtactc tgaagtgtat gcttgggatt ctcaatgggt gttggatcct gaagtttgac 1980
tgggtgaaag cttgtttgca cagcaaagta cgtgaacagg aagaaaagta cgaagttcct 2040
ggaggtccgc agaagagcag gctcaacaga gagcagctgt tgccaaagct atttgatgga 2100
tgctacttct ttttgggggg aaacttcaaa catcatccaa agaatgatct ccttaagctc 2160
attactgcag caggaggcaa ggtgctcagt agaaagccca agccagacag tgacgtgact 2220
cagaccatca acactgttgc ataccatgcc aaccctgact ctgaccagcg cttctgtaca 2280
cagtacatcg tctatgaaga tctgtttaat tgtcgcccag aaagggttcg gcagggcaaa 2340
gtctggatgg ctccttccac ttggcttatc agctgtgtga tggcctttga attgcttcct 2400
cttgacagct ga 2412
<210> 27
<211> 2796
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<223> Lig1 (Лигаза 1)
<400> 27
atgagaaaaa aagaaccaga gaggaaaggg gagaactctg ctgccaccat gcagaaaagt 60
atcaggtcat ttttccaacc catgaaagag ggtaaagcac agaagccgga gaaggagaca 120
gctaacagca ccaaagagaa ggagccacct ccaaaggtgg cactgaagga gaggaatcga 180
gcagtgcctg agagtgattc tccagtgaag aggcctggaa ggaaggcagc ccaggtccta 240
agcagcgaag gggaggagga agatgaagcc cccagcaccc ctaaagtcca gaagtctgtg 300
tcagactcca aacaaagctc tcctcccagc cctgacgcat gtcctgagaa cagtcctttc 360
cacagtagcc cctccatgga gatctcccca tcaggattcc cgaagcgtcg cactgctcgg 420
aagcagctcc cgaaacggac aattgaggac actgtggagg agcagaatga ggacaaaggc 480
agagcagcca agaaaaggaa gaaggaagaa gaagcacaga ctccaatgga aagcctcaca 540
gagagtgaag atgtaaaacc caaggaagaa aaggaggagg gcaagcatgc tgaggcttcc 600
aagtcccctg agtcgggaac cttgacaaag acagagacca tcccagtgtg taaggccggc 660
gtgaaacaga agcctcagga agaggagcag agcaagcctc ctgccagagg cgccaagaca 720
ctcagcagct tcttcactcc ccggaagcca gcagagaaag ccatagtgaa acaagaagag 780
ccaggtactc cagggaagga agagaccaag ggagccctgg atccaacaaa ttacaatcct 840
tccaagagaa actaccaccc cattgaagat gcctgctgga aacatggcca gaaagtccct 900
tttctcgctg tggcccggac ctttgagaag attgaggagg tttctgctcg gctcaagatg 960
gtggagacac tgagcaactt gctgcgctcg gtggtggccc tgtcacctcc agacctgctt 1020
cctgttcttt acctcagcct caaccgcctt gggccacctc agcagggact agagctgggt 1080
gttggtgatg gtgtcctcct taaggcagtt gcccaggcca caggccgtca gctggagtcc 1140
atccgggctg aggtagctga gaagggtgac gtgggactgg tggccgagaa cagccgcagc 1200
actcagagac tcatgctgcc ccctcctccg ctcaccacct ccggggtctt taccaaattc 1260
tgtgacattg cccggctcac tggcagtgct tccatggcca agaagttgga tgtcatcaag 1320
ggcctgtttg ttgcctgccg tcactcggaa gcccggttca ttgccaggtc cctaagtgga 1380
cgcctgcgcc tcgggctggc tgagcagtcc gtcttggctg cccttgccct ggctgtgagc 1440
ctcacacccc ctggccaaga atttccccca gctgttgtgg atgctgggaa gggcaagacc 1500
acagaggcca gaaagacatg gttggaagaa caaggcatga tcttgaagca gaccttctgt 1560
gaggtacctg acctggaccg aatcatcccg gtgctgctgg aacatggcct ggaacgcctc 1620
ccagagcact gcaggctgag cccaggggtc cctcttaaac caatgctggc tcatcccact 1680
cggggtgtca gcgaggtact gaaacgcttt gaggaggtgg actttacctg cgagtacaaa 1740
tatgacgggc agcgggccca gattcatgtt ctggaaggtg gagaggtgaa gatcttcagt 1800
aggaaccagg aagacaacac aggaaagtac ccggacatca tcagccgcat ccccaagatt 1860
aaactcccct cggtcacctc ctttatcctg gacactgagg ctgtggcctg ggaccgggaa 1920
aagaagcaga tccagccatt ccaagtgctc accacacgca agcgcaagga ggttgacgcc 1980
tcggagatac aggtgcaggt gtgtctgtat gcctttgatc tcatctacct caacggagag 2040
tccctgattc gccagcccct gtctcgacgt cggcagctgc tccgggagaa ctttgtggag 2100
acagagggtg agtttgtctt cgccacctcc ctggacacca aggacatcga gcagatcgct 2160
gagttcttgg agcagtccgt gaaggactcc tgtgagggac tgatggtgaa gaccctggat 2220
gttgatgcca cctatgagat tgccaagagg tctcacaact ggctcaagct aaagaaggac 2280
taccttgacg gtgtgggcga cactctggac cttgtggtga ttggcgccta cctgggccgg 2340
gggaagcgtg ccggccggta tgggggcttc ctcttggctg cctatgatga ggagagtgaa 2400
gagctgcagg ccatatgcaa gctgggaact ggattcagtg atgaagagct ggaggagcat 2460
caccagagcc taaaggccct agtgttgccg accccacgcc cctatgtgag gatcgatggg 2520
gcagttgccc cagaccactg gctggaccca aaggtcgtat gggaggtgaa gtgtgcggat 2580
ctctccctgt cccctatcta ccctgctgcg cggggcctgg tggacaaaga gaaagggatc 2640
tcccttcgtt tccctcggtt cattcgtgtc cgtgaagaca agcagccaga gcaggccacc 2700
accagtgacc aggtggcctg tttgtaccgg aagcagagtc agatacagaa ccagcacaac 2760
tcagacttgg actccgactt tgaggactgc tattaa 2796
<210> 28
<211> 2121
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<223> Mre11
<400> 28
atgagccctg cagatccact tgatgatgaa gacactttta aaatcctggt tgccactgat 60
attcaccttg gatttatgga gaaagatgca gttcgaggaa atgatacatt tgtgacactt 120
aatgaaattt taaaacttgc cctggaaaat gaagtggatt ttattttgtt aggtggtgat 180
cttttccatg aaaacaagcc ctcaaggaaa actcttcata gttgcttgga gttgcttagg 240
aagtactgta tgggtgatcg ccctgtgcag tttgagatca tcagtgacca gtcagtcaac 300
tttggtttta gtaggtttcc atgggtgaac tatcaggatg gcaatctcaa catttccatt 360
ccagtgttta gtatccatgg caaccatgac gatcccacag gggcagatgc cctctgtgcc 420
ctggatattt taagctgtgc tgggtttgtg aatcactttg gacggtcaat gtctgtggag 480
aagattgaca ttagtccagt tctgctgcag aaaggaagca caaaacttgc tctgtacggc 540
ctagggtcca ttcctgatga aaggctctat cggatgtttg tcaataaaaa agtaacaatg 600
ctgagaccaa aggaagatga gaactcatgg tttaacttat ttgtgattca tcagaacagg 660
agtaaacatg gaagtaccaa cttcatccca gaacagtttt tggatgactt cattgacctc 720
gttatctggg gccatgaaca cgagtgtaaa attggcccaa ccaaaaatga gcagcagctc 780
ttctatgtgt ctcagcctgg aagctcagtg gtgacttctc tttcccctgg agaagctgtt 840
aagaaacacg tgggcttgct gcgcgttaaa gggaggaaga tgaacctgca gaagctgcct 900
ctccgcacag tgcggcagtt cttcatggag gatgtggttc tcgctaacca cccaaacctg 960
ttcaaccctg acaatcctaa ggtgacccag gccatccaga gcttctgctt ggagaagatt 1020
gaagaaatgc ttgaaaatgc cgagcgcgaa cggctaggga attctcttca accagagaag 1080
cctcttatcc gactacgggt ggactacagt ggaggctttg aaccttttag tgttcttcgc 1140
tttagccaga aatttgtgga tcgggtcgct aaccccaaag atatcatcca ctttttcagg 1200
catagggagc aaaagggaaa aacaggtgaa gagatcaact ttgggaagct tgtttcaaaa 1260
tctccttcgg aaggaacgac actcagagta gaagacctgg tgaagcagta tttccagact 1320
gcggagaaga atgttcagct ctcactgctg acagaaagag ggatgggtga agccgttcaa 1380
gaatttgtgg acaaggaaga aaaagatgcc atcgaggaat tagtgaagta ccagctggag 1440
aaaacacagc ggtttcttaa ggagcgccat atcgatgctc tggaagacaa gattgacgaa 1500
gaggtccggc gtttcagaga aagcagacag agaaatacca acgaagaaga cgatgaagtg 1560
cgagaggcca tgagcagggc ccgggcgctc agatcgcagt cagagaactc tgcctcagcc 1620
ttcagtgctg acgacctgag tttcgatata gcagaacaga cagcaaatga ctctgatgac 1680
agcctgtcag cagtgcccag cagaggccgg ggccgaggcc gaggtcgaag aggaggcaga 1740
gggcagagca ccgcatcaag aggaggatct cagagaggcc gagacactgg gcaggagacg 1800
gctactcgag gcagatgctc aaaggccact acatcgacct ctagaaatat gtccattcta 1860
gacgctttca gatctactcg acagcagcct tccagaaaca ctgccactaa aaattactca 1920
gagactattg aagtggatga atctgatgaa gatgacattt ttcctaccag ttccaaggct 1980
gatcaaaggt ggccaggcac aacatctagc aaacggatgt cccagagcca gatagccaag 2040
ggggttgact ttgaatcaga tgaggacgat gacgatgacc ctttcatgag cagtagttct 2100
ctaagaagcc gaagataata a 2121
<210> 29
<211> 5973
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<223> 53BP1
<400> 29
atgccagggg agcagatgga ccctactgga agtcagttgg attcagattt ctctcagcaa 60
gatactcctt gcctgataat tgaagattct cagcctgaaa gccaggttct ggaagaagat 120
gcgggctctc acttcagtgt gctgtctcga caccttccca atctgcagat acacaaagag 180
aaccctgtgt tggatattgt gtccaatccg gaacaagcag ctgcagaaga gcaaggagac 240
aataatagct ccttcaatga acatctgaaa gagaacaaag ctgctgcaga tcctgtggat 300
tccactcatt tgggcacatg tgattccatc agtcaggtca tagaacagtt gcctcagcca 360
aacaggacaa gcagtgttct gggagtgaca gtggaagctg cttctcttcc agaagaggag 420
aaggaggaag agctggaaga ggagaatgaa gaggtgggag cagatgctgc tgatgccgct 480
ccgtgctccc ttggtgctga agattctgct tcatcacagt tgggctttgg ggttctggaa 540
ctgtcccaga gccaggatgt tgaagaacat acagtgccat atgatgtcaa ccaagaacat 600
atgcagttgg ttaccaccaa ctcgggttct tcccagctgt ctgacatgga tgcaaataat 660
gttaaatgtg aagaacagtc cactgaagat acctccatgg cagaacaacc taacaaagac 720
atccctgtta cagtcgagca cagtaaaggt atccctgtgg tagatgagca aaatctacca 780
ccggcaaggc cagaggatct gccttccagt cctcaagcct ctgctgcagc tgtggaaaca 840
aaggaagagg tacctgccca agagttgcca gaaggggcgc tggaggttca gatgtcctcg 900
gagcctgagg tctcgtccac tcaggaggac ttgtttgagc agagtagtaa aacagcttct 960
gatggttgtt ctactccttc aagggaggaa ggtgggtgtt ctccggtttc cacacccgcc 1020
accaccctgc aactcctgca gctgtctggt cagaggcccc ttgtccagga cagtctttcc 1080
acgaattcct cagatcttgt tgctccttcc cctgatgctt tccgacctac cccttttatc 1140
gttcctagca gtcccacaga gcaaggaggg agaaaagttg agcccttgga tatgtcagtg 1200
atgcctgaag aaggagggga gactttgcag aagcttcagg atgacgaagc agtggagata 1260
gaaaagcccc atctcccatc tcagccggct gtttccccac aagtgtcaac accagtgtct 1320
cagagcacac ctgtcttcac tccaggctct cttcccatcc cgtcccagcc tgaattttct 1380
catgacattt tcattccatc accaagtttg gaagaaccat caagtgatgg gaagaaaggt 1440
ggcgatttgc acagctcatc tttgactgtg gagtgttcta agacttcaga gagtgaacca 1500
aagaatttca ctgctgatct tgggctctcc ttgacagggg agtcttgcaa actgatgctt 1560
tcttcaagtg agtatagtca atcctcaaag atggagagct tggcttctcc caggagtgag 1620
ggagatggag agaataccca gattgaggac actgaaccgt tgtctccagt caccaattct 1680
aaacttcctg ctgaaagtga tgatgtcctg atgaatccag caccagatga ccaagtagaa 1740
atgaatcaga atgatgacaa agtaaaagag ggtgacacag agaacacagg tgaccatggt 1800
gttttagctg ctggtagtaa aggcagagaa gaaaccgttg ctgaagatgt ttgcattgat 1860
ctcacttgtg attctgggag tcaggcagtt ccgtctcccg ctactcggtc cgaggcactt 1920
tctagtgtct tagatcagga ggaagctatg gaaattaaag aacagcatcc agaggagggg 1980
ttttcgggat ctgaggtaga agaagtcccc gagactccct gtgaaagtca cagagaggag 2040
cccaaggaag aaacgatgga gagtatccca ctgcaccttt ctcttactga aacacagtcc 2100
caggcattgt gtcttcagaa ggaaatgccc aaagaagaat gcccagaggc catggaagtg 2160
gaaccctctg tgattagtgt tgactctccc cagaagctgc cggtactcga ccaggagtca 2220
gagcataagg agccagaggc ctgggaagaa gccgcgtcgg aggactcaag tgtggaggac 2280
tcaagtgtgg ttatcgttga tgtgaaggag ccctcgccaa gagttgtccc ctgcgaacct 2340
ttggagggag cagagaaatg ctcagattcc cactcctggg aggatgtggt gcccgaggtg 2400
gaaccgtgtg ctgcaaatag agtagacact ccggaggaaa agattgtaga atgtgacgga 2460
gatgtgaaag cagagaccac aagaaaggac tctgtagagg aagactcccc acagcctcct 2520
ttgccttcag tgaaagacga gcctcccaga gacgagcctc ccagacccga ccaggagatg 2580
cagcagtccc agcttcaaga gaaagagagc ccagtgacca tagatgcaaa agtggctgat 2640
gccaagcagc tggagtcaga gggagcatcc cagcagcttt cgaaagcccc tgcccgcgac 2700
tcacaaagtt tctgtgaaag ttctagtgaa accccatttc atttcacttt gcctaaagaa 2760
ggtgatatta tcccaccatt gactggtgca accccacctc acattgggca cctaaaattg 2820
gatcgcaaca gacatagtac tccaattggg attggcaact atccagaaag caccatagca 2880
accagtgatg tcatgtctga aagcatggtg gagatcaatg atcctctact tgggagtgaa 2940
aaaggagatt ctgagtctgc cccagaaatg gacggaaaac tgtatctgaa aatgaaactg 3000
gttagtcctg agacagaggc cagtgaagaa tctttgcagt ttagcctgga aaagcctgct 3060
actgctgaga gaaaaaatgg atcaactgct gttgctgagc ctgttgcaaa aaatggatca 3120
actgctgttg ctgagcctgt tgccagtccc cagaagaccg tgcctgtgtt tagctgcagg 3180
caagaggagg tttggagtga ggaccctccc tctgtaccca tcagggcaaa cttgctccat 3240
tttccaagtg ttgaagaaga ggacaaagaa aaactggatg gtaccccaaa gcttaggcag 3300
agtgaacagc ctgtgaggcc cgttgggctg gtcaaggatg ctgctacttc tgaggactct 3360
gcttcttctg ttccccagca gagagcaaca caggggtcat tcagccctca aggagaagtg 3420
atggaaacag acctgctaga aggactgagt gctaaccagg aaaaaccgtg taaggtcttg 3480
atggaaaggc ccacccagag taacatagga atccagacca tggaccattc cctgtgtgct 3540
ccagaaactg tttcagcagc aacccagact gtgaagagtg tatgtgaaca agggaccagt 3600
acagtggacc agaactctgg gaaacaagat gccactgtgc agaccgagag ggggggtgtc 3660
gagaaacagg cccctgtgga cgatacagaa tccctccaca gccagggaga agaagaattt 3720
gaaatgcccc agcctccaca tggccatgtc ttgcatcgtc acatgagaac catccgtgaa 3780
gtccggacac ttgtcacccg tgtcatcaca gatgtttact atgtggatgg gacagaagtg 3840
gaaaggaaag taactgagga gactgaagaa ccaattgtag aatgtcagga atgtgaaaca 3900
gaggtttccc cttcccagac tggaggctct tctggagacc tgggagacat cagttccttc 3960
tcctccaaag catccagctc acaccataca tcaagtggga caagtctctc agccatgcat 4020
agcagtggca gctcaggacg aggagccggg ccactcaaag ggaaaaccag cgggacagaa 4080
cctgcagatt ttgctttacc cagttcccga ggaggcccag gaaaactgag tcctagaaaa 4140
gggatcaatc agacaggggc accagtgtgt gaggaagatg gtgatgcagg ccttggcatc 4200
agacagggag ggaaggctcc tgtcacacct cgtggtcgtg gtcgaagggg ccgcccacct 4260
tctcggacca ctggaacaag agatgcagtt gtgtctggtc cgttgggcat agaagacatt 4320
tcacctagca tgtcaccaga tgacaagtcc ttcacccgaa ttgtgccccg tgtaccagac 4380
tctaccaaac gaattgatac cagttctact gttttgaggc ggagtgattc cccagagatt 4440
ccttttcagg ctgctactgg gtcttctgat ggcttggatg cctcatctcc aggaaatagc 4500
tttgtcgggc tccgtgttgt agccaagtgg tcatccaatg gctattttta ctctgggaaa 4560
atcacacggg atgttggagc tgggaagtat aagctgctct tcgatgatgg gtatgaatgt 4620
gatgtgctgg gcaaagacat tctcctgtgt gaccctatac ccctggacac tgaagtgaca 4680
gccctctcag aagatgagta tttcagtgca ggagtggtca aaggacacag aaaggagtct 4740
ggggagctgt actacagcat tgaaaaagaa ggccaaagga agtggtataa gcggatggca 4800
gtcatcctgt ccttggaaca aggaaacagg ttaagagagc aatatgggct tggcccatat 4860
gaagctgtta cacccctcac aaaggcagca gacatcagcc tagataattt ggtggaagga 4920
aagcggaaac gtcgcagtaa catcagctcc ccagccaccc ccactgcctc cagcagcagc 4980
agcagcagca gcacaacacc cacccgtaaa accacagaga gtccccgtgc ttccacggga 5040
gttccatcag gcaaaaggaa actcatcact tctgaagagg aacggtcccc agctaagcga 5100
ggccgcaagt ctgtcaccgt gaaacctggt acagtggggg caggagaatt tgtgagcccc 5160
tgcgagagtg gagacaacac aggtgaacct tctgtcctgg aagagccaag agggcctttg 5220
cccctcaaca agaccttgtt tctgggctat gcctttctcc ttaccatggc cacaaccaat 5280
gacaagctgg ccagccgctc taaactgcta gatggtccta caggaagcag tgaagaagag 5340
gaggaatttt tagaaattcc tcctttcaac aagcagtata cagaatgcca gcttcgagca 5400
ggagctgggt atatccttga agacttcaat gaagcccagt gtaacacagc ctaccagtgt 5460
ctcctaattg ctgatcagca ttgtcgaacc cggaagtact tcctgtgcct tgccagtgga 5520
attccttgtg tgtctcatgt ctgggtccat gacagctgcc atgccaacca gctccaaaac 5580
taccgtaatt atctgctgcc tgctgggtat agccttgagg agcagcgaat tctggattgg 5640
caacctcgtg aaaacccttt ccagaatctg aaggtactct tggtgtcaga tcaacaacag 5700
aacttcctgg agctctggtc tgagatcctc atgactggag gggcagcctc tgtgaagcag 5760
caccattcaa gtgcccataa caaagacatt gctttagggg tatttgatgt ggtggtgaca 5820
gacccctcat gcccagcctc ggtgctcaag tgtgctgaag ccttgcagct gcctgtggtg 5880
tcacaagagt gggtgatcca gtgcctcatt gttggggaga gaattggatt caagcagcat 5940
ccaaaatata aacatgatta tgtttctcac taa 5973
<210> 30
<211> 47
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<222> (1)..(47)
<223> Консенсусная последовательность ERV группы 1
<400> 30
gcccccgcca tatccgccac tgccgccccc accagaggca gaagcgg 47
<210> 31
<211> 47
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<222> (1)..(47)
<223> Последовательность мотива PPYP группы 1
<400> 31
gcccccgcca tatccgccac tgccgccccc accagaggca gaagcgg 47
<210> 32
<211> 37
<212> ДНК
<213> Cricetulus griseus
<220>
<221> прочий_признак
<222> (1)..(36)
<223> Последовательность мотива PPYP с делециями вне рамки считывания
посредством огРНК PPYP7
<400> 32
gcccccgcca tatccgcccc accagaggca gaagcgg 37
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ГЕНЕТИЧЕСКИЕ КОНСТРУКЦИИ pcDEXO-GS-SSB, СОДЕРЖАЩИЕ ПОСЛЕДОВАТЕЛЬНОСТИ, КОДИРУЮЩИЕ ХИМЕРНЫЕ БЕЛКИ SSB И ЭКЗОНУКЛЕАЗЫ АППАРАТА ГОМОЛОГИЧНОЙ РЕКОМБИНАЦИИ БАКТЕРИОФАГОВ, ОБЪЕДИНЕННЫЕ ЧЕРЕЗ ГЛИЦИН-СЕРИНОВЫЙ ПОЛИЛИНКЕР | 2020 |
|
RU2763747C1 |
| РНК-НАПРАВЛЯЕМАЯ ИНЖЕНЕРИЯ ГЕНОМА ЧЕЛОВЕКА | 2013 |
|
RU2699523C2 |
| Средство редактирования генома на основе белка LigD из бактерии Pseudomonas putida и Cas9 комплекса | 2022 |
|
RU2797049C1 |
| АДРЕСНОЕ ВСТРАИВАНИЕ В ПОСЛЕДОВАТЕЛЬНОСТИ МЛЕКОПИТАЮЩИХ, УСИЛИВАЮЩЕЕ ЭКСПРЕССИЮ ГЕНОВ | 2020 |
|
RU2829170C1 |
| РНК-НАПРАВЛЯЕМАЯ ИНЖЕНЕРИЯ ГЕНОМА ЧЕЛОВЕКА | 2013 |
|
RU2766685C2 |
| СИСТЕМЫ CRISPR-CAS И СПОСОБЫ ИЗМЕНЕНИЯ ЭКСПРЕССИИ ПРОДУКТОВ ГЕНОВ | 2013 |
|
RU2687451C1 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ НАЦЕЛЕННОЙ ГЕНЕТИЧЕСКОЙ МОДИФИКАЦИИ С ИСПОЛЬЗОВАНИЕМ ПАРНЫХ ГИДОВЫХ РНК | 2015 |
|
RU2734770C2 |
| НАПРАВЛЯЕМАЯ РНК РЕГУЛЯЦИЯ ТРАНСКРИПЦИИ | 2014 |
|
RU2690935C2 |
| КОМПОНЕНТЫ СИСТЕМЫ CRISPR-CAS, СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ МАНИПУЛЯЦИИ С ПОСЛЕДОВАТЕЛЬНОСТЯМИ | 2013 |
|
RU2701662C2 |
| ДОСТАВКА, КОНСТРУИРОВАНИЕ И ОПТИМИЗАЦИЯ СИСТЕМ, СПОСОБОВ И КОМПОЗИЦИЙ ДЛЯ МАНИПУЛЯЦИИ С ПОСЛЕДОВАТЕЛЬНОСТЯМИ И ПРИМЕНЕНИЯ В ТЕРАПИИ | 2013 |
|
RU2721275C2 |
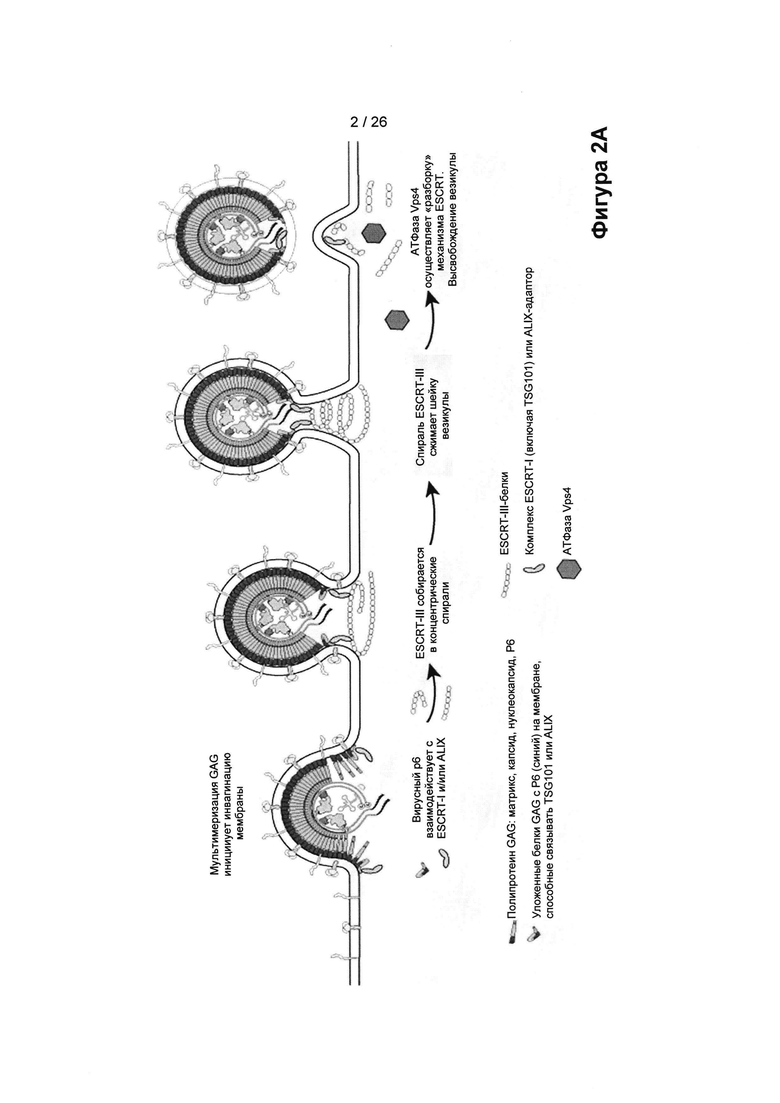
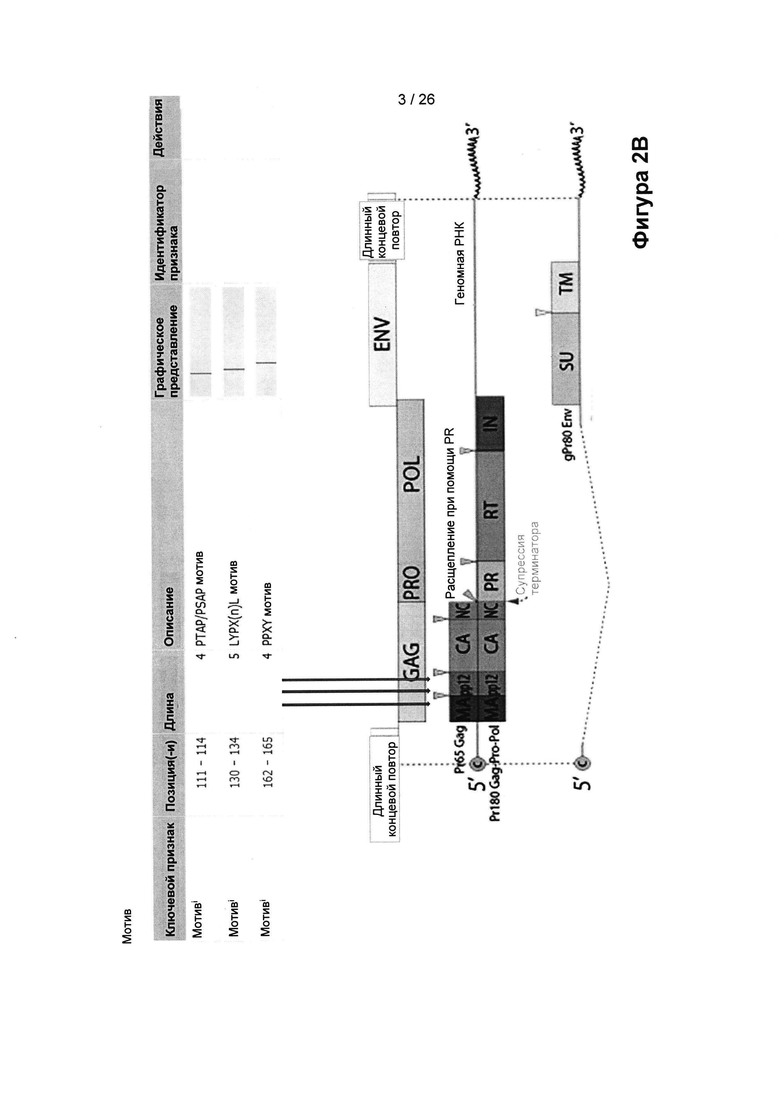

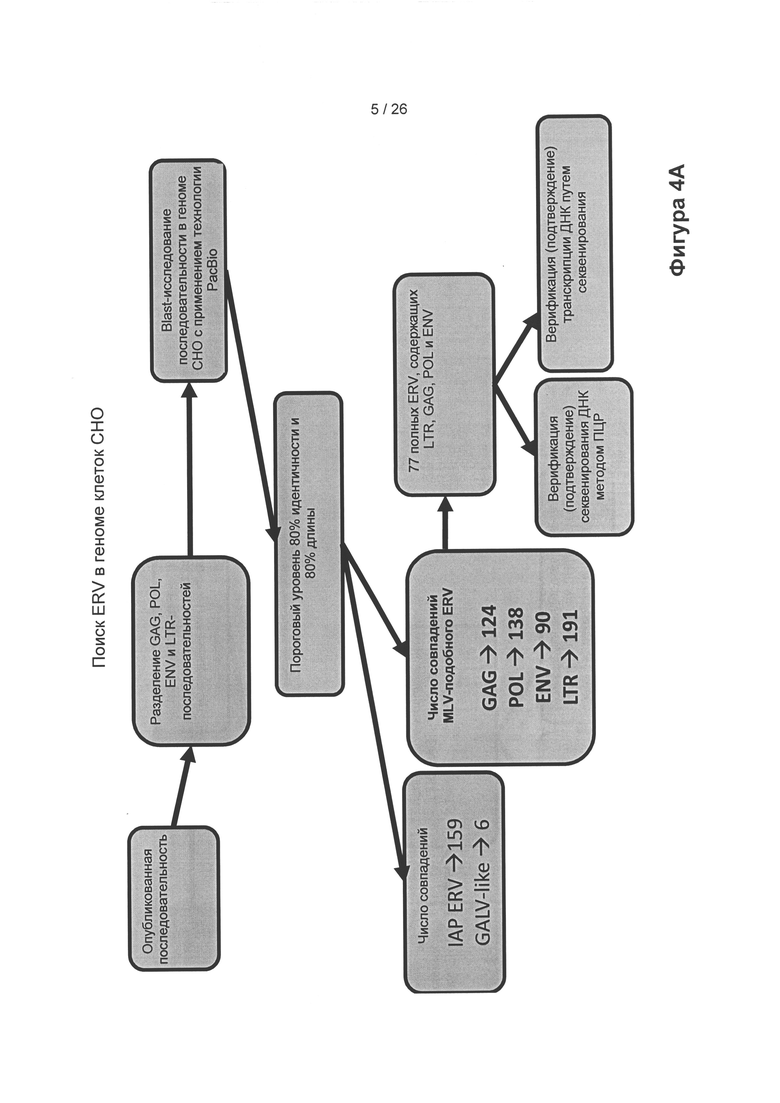


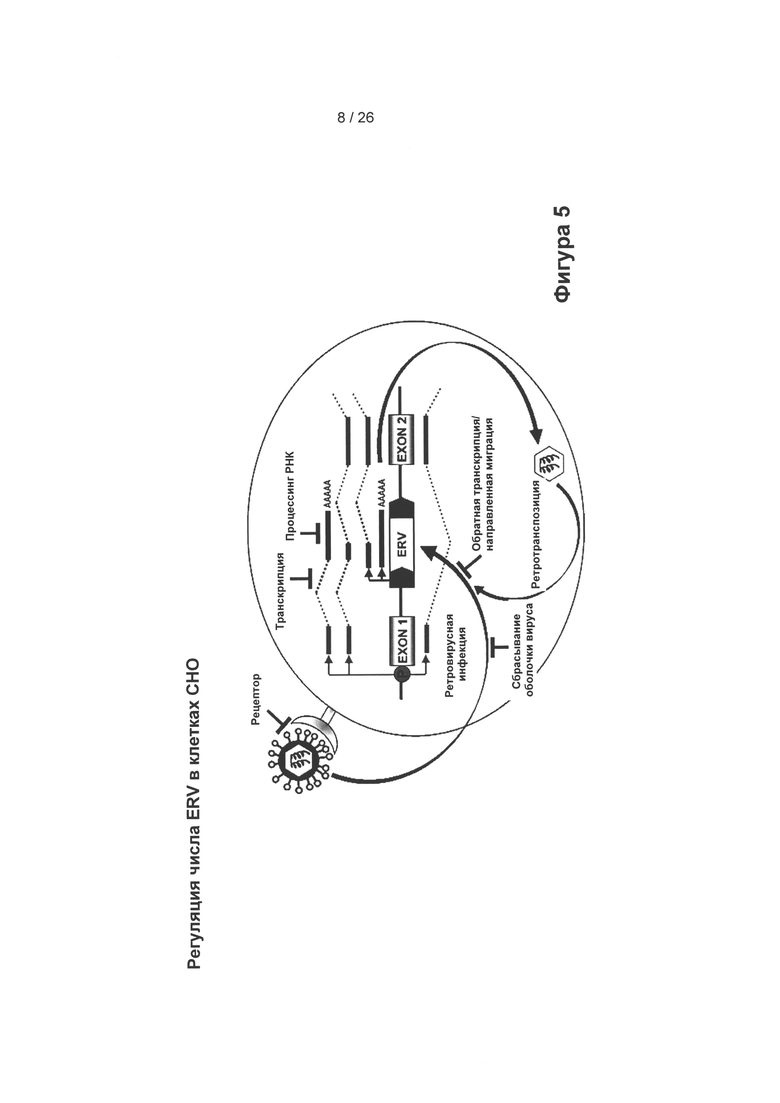
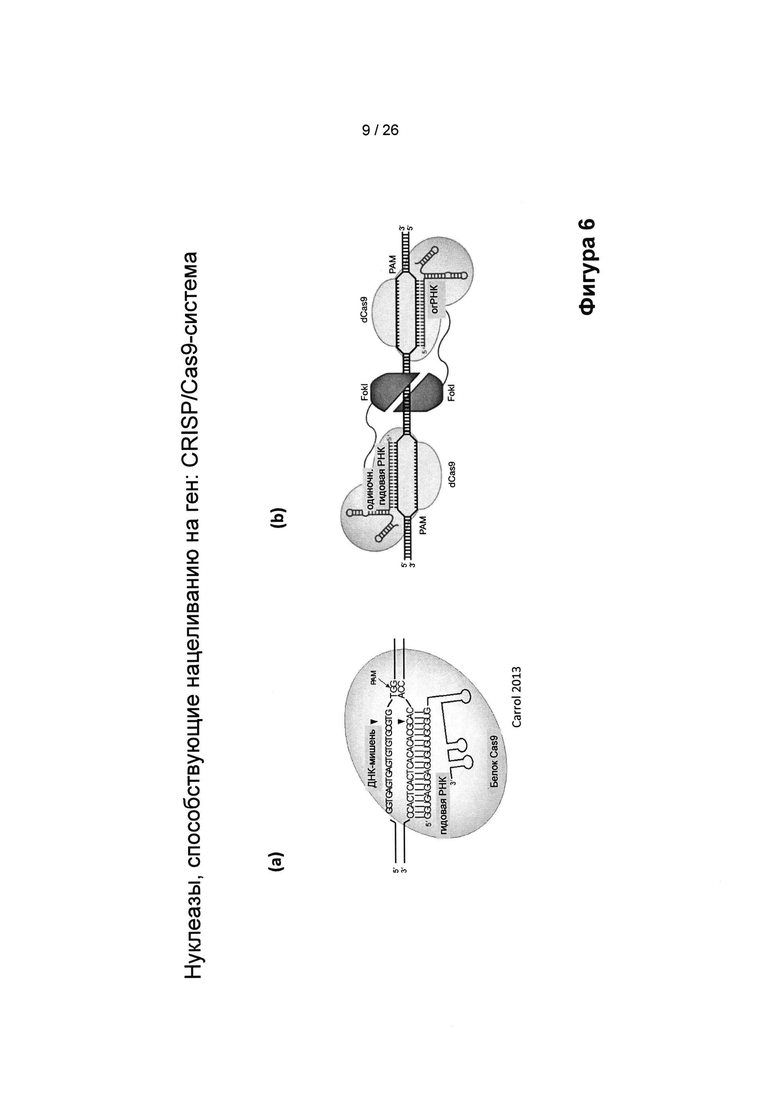
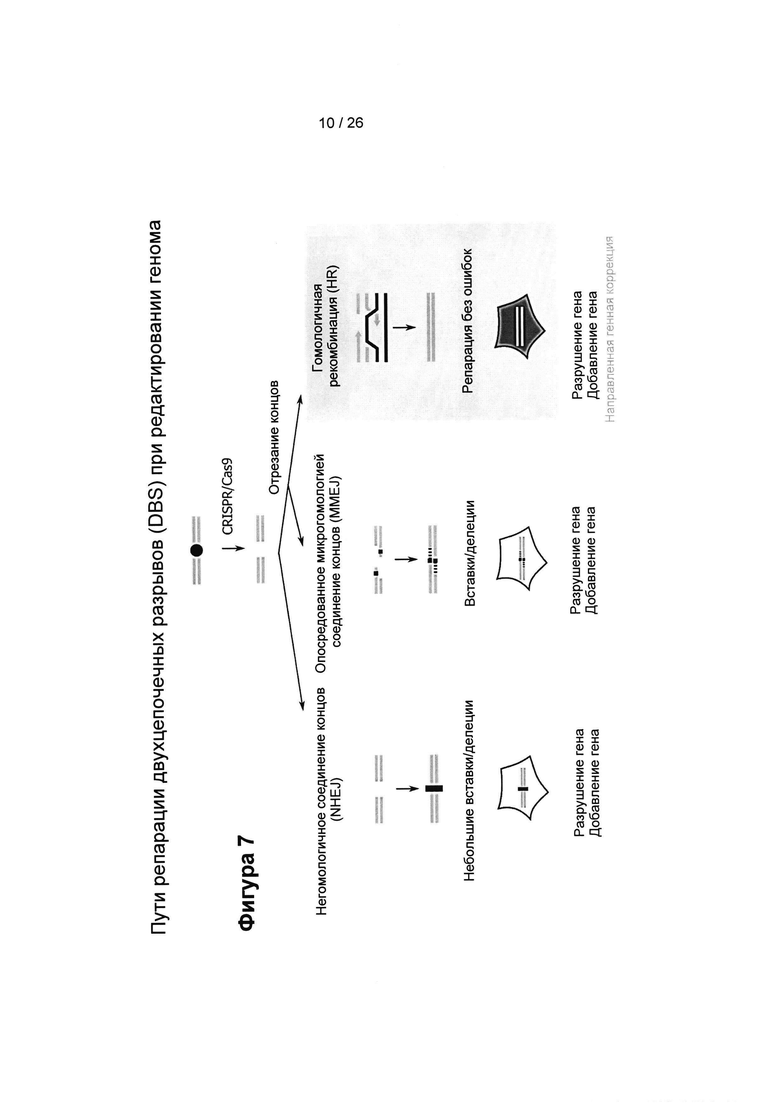
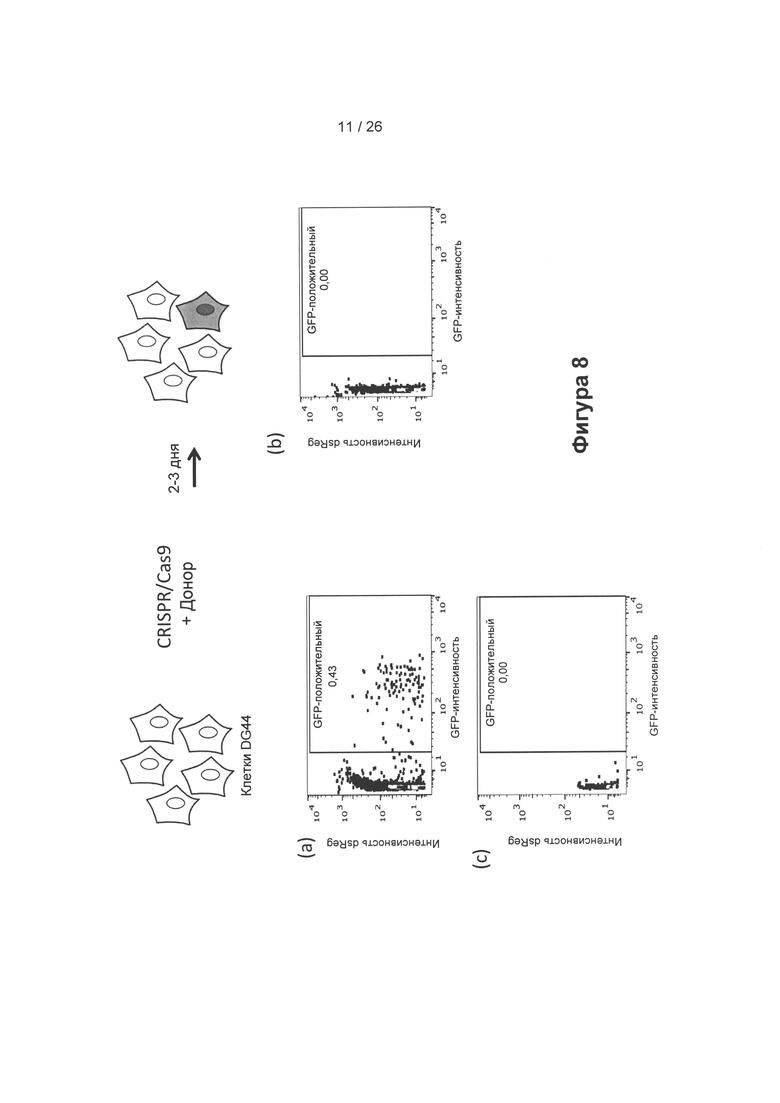
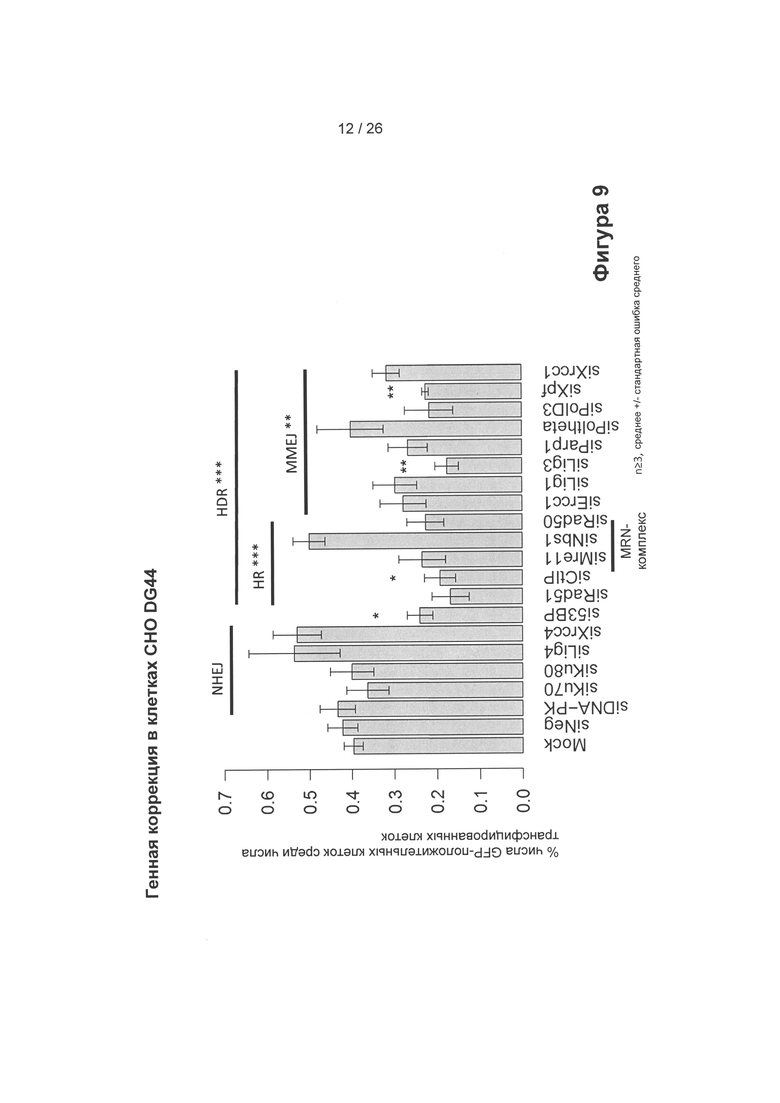
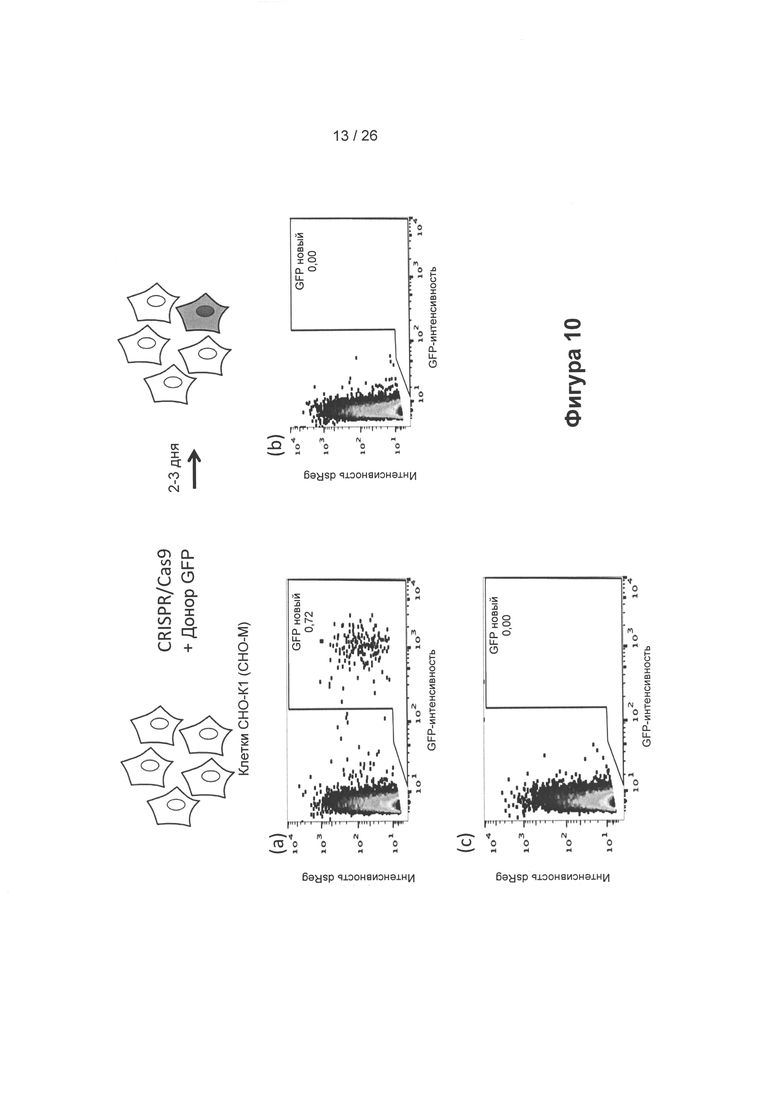
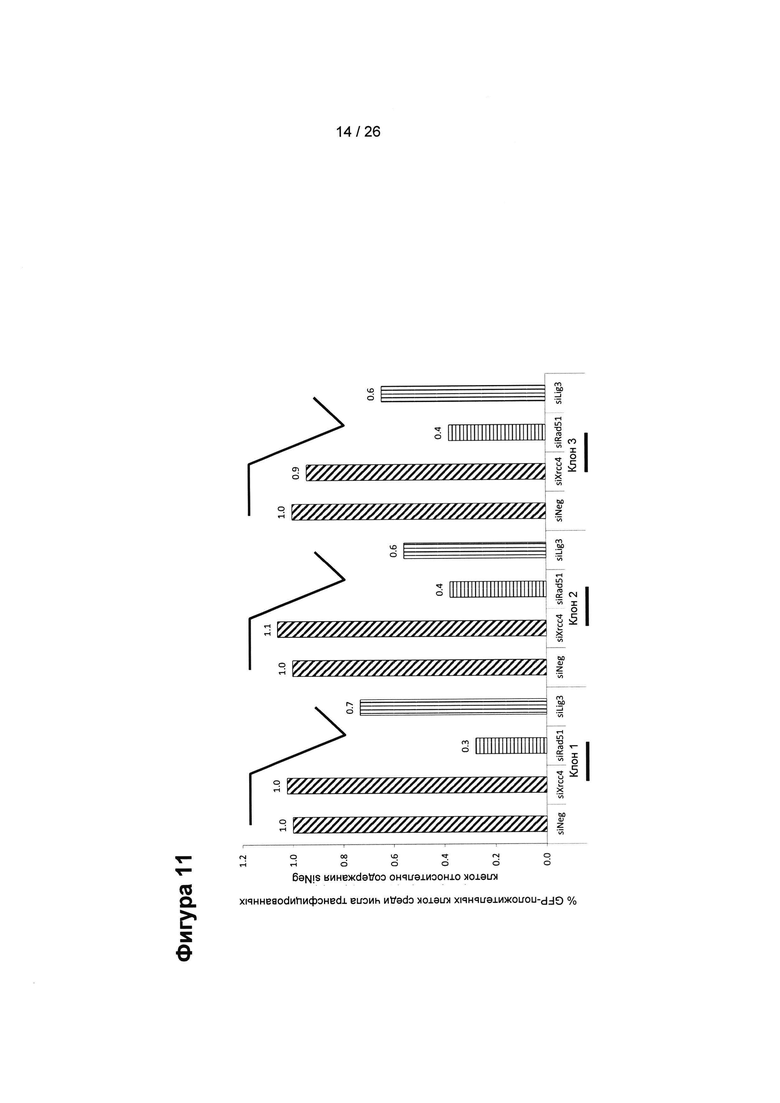
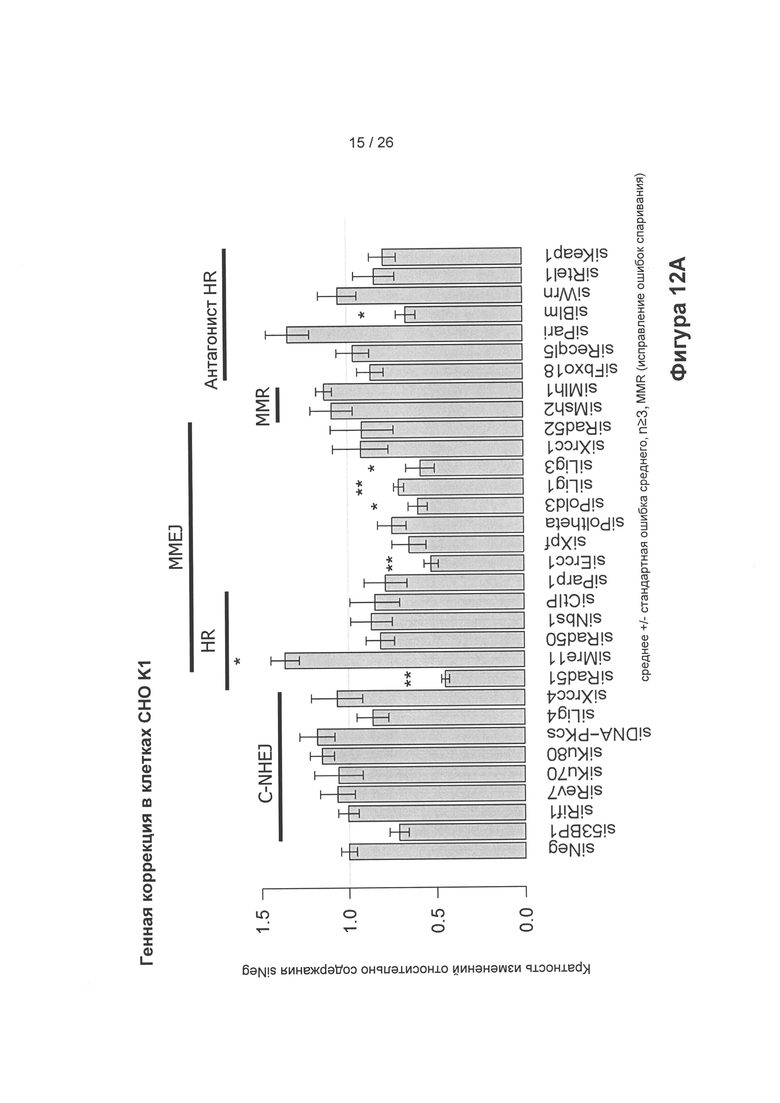
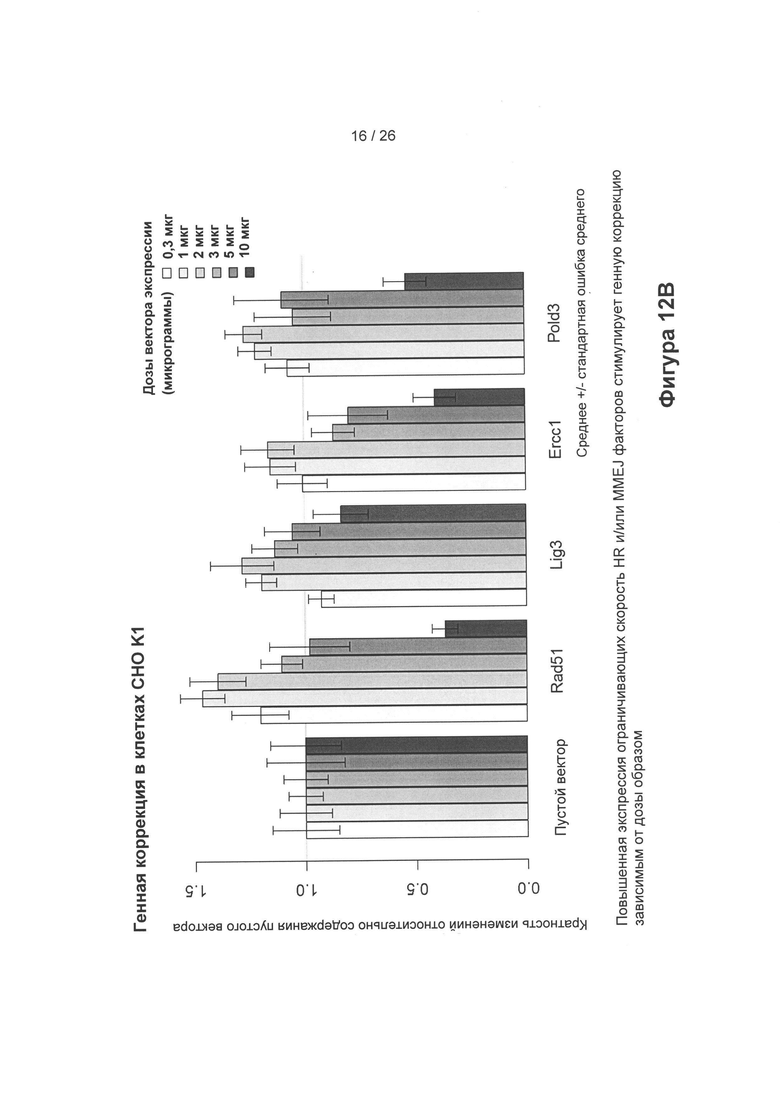
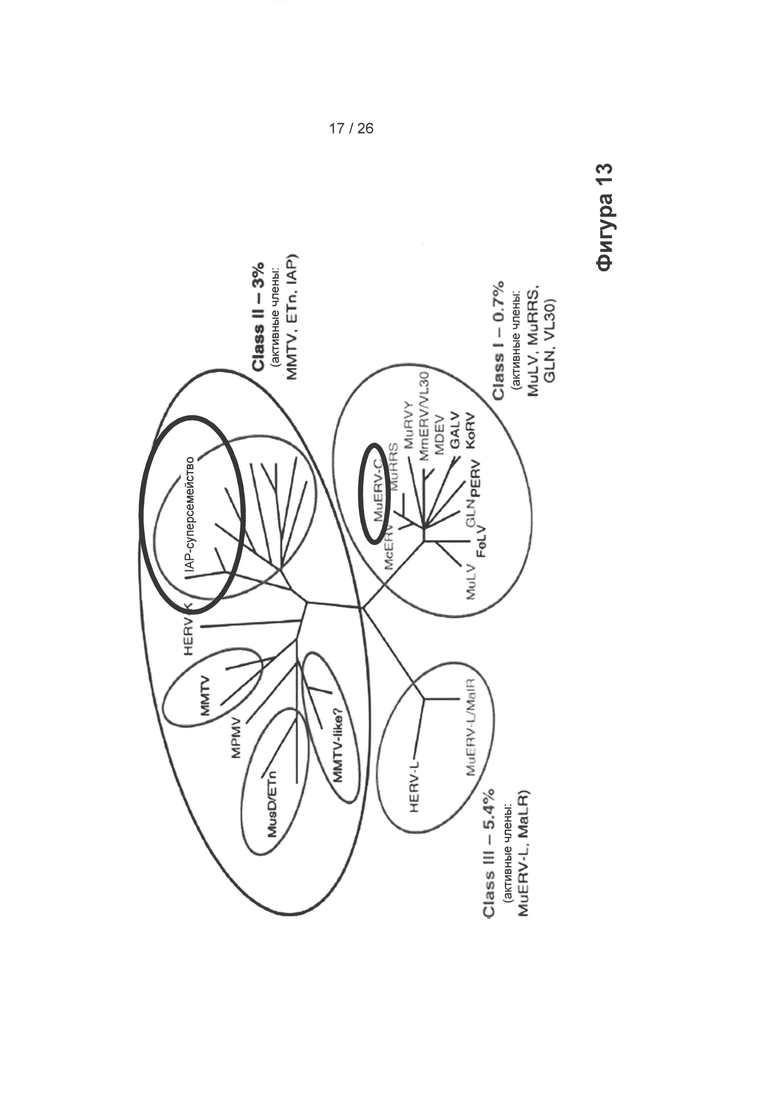
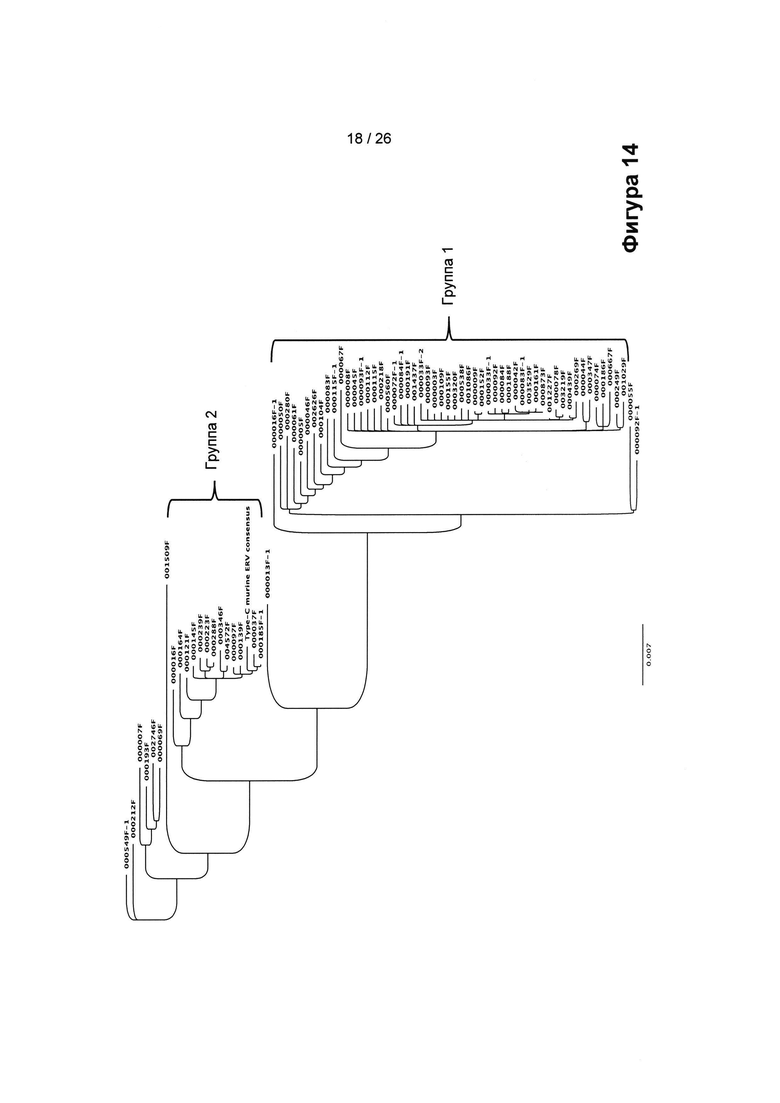


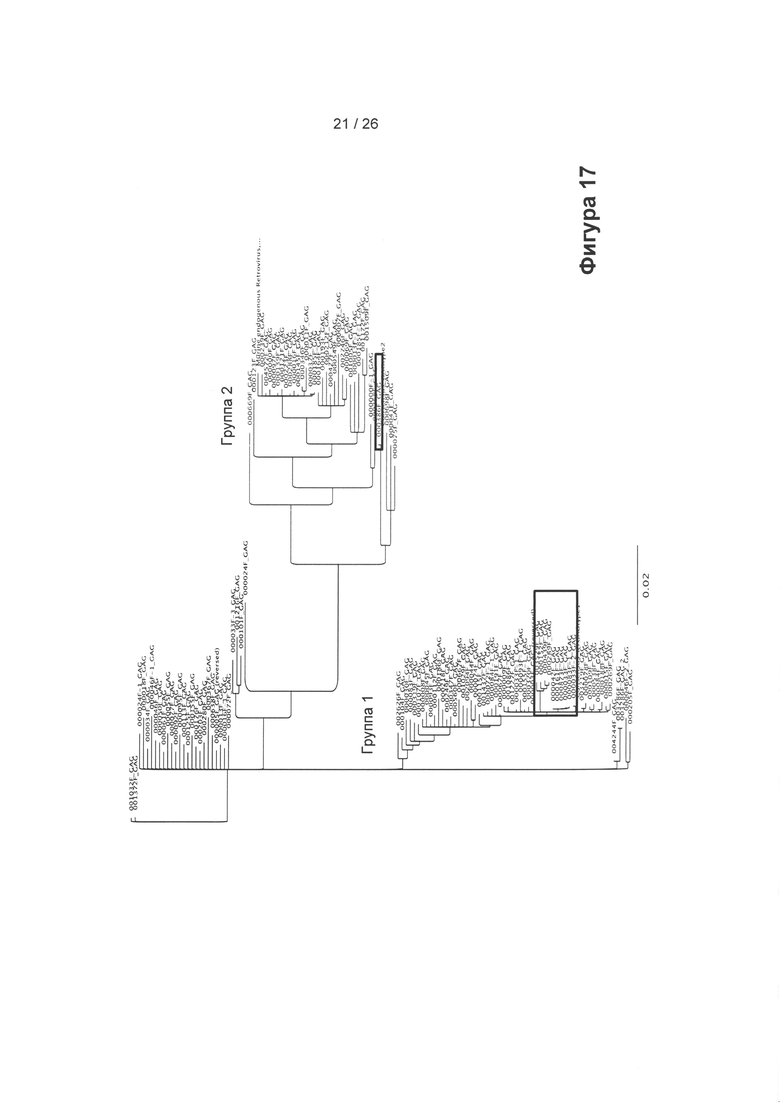
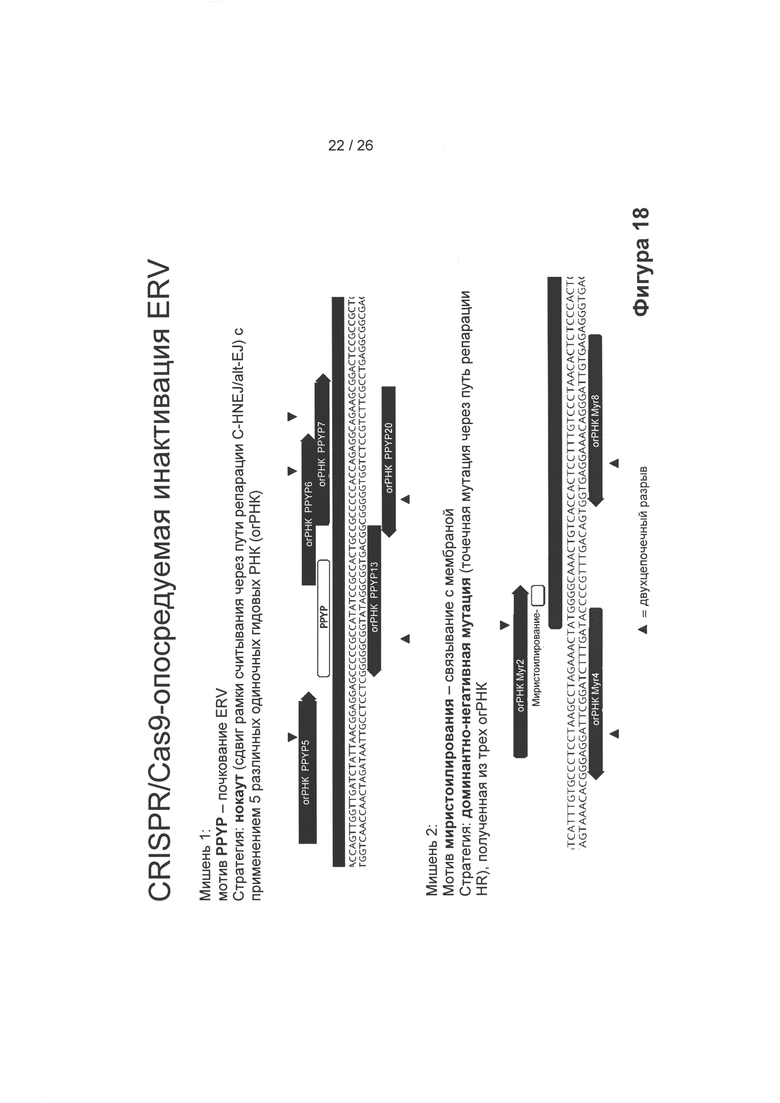

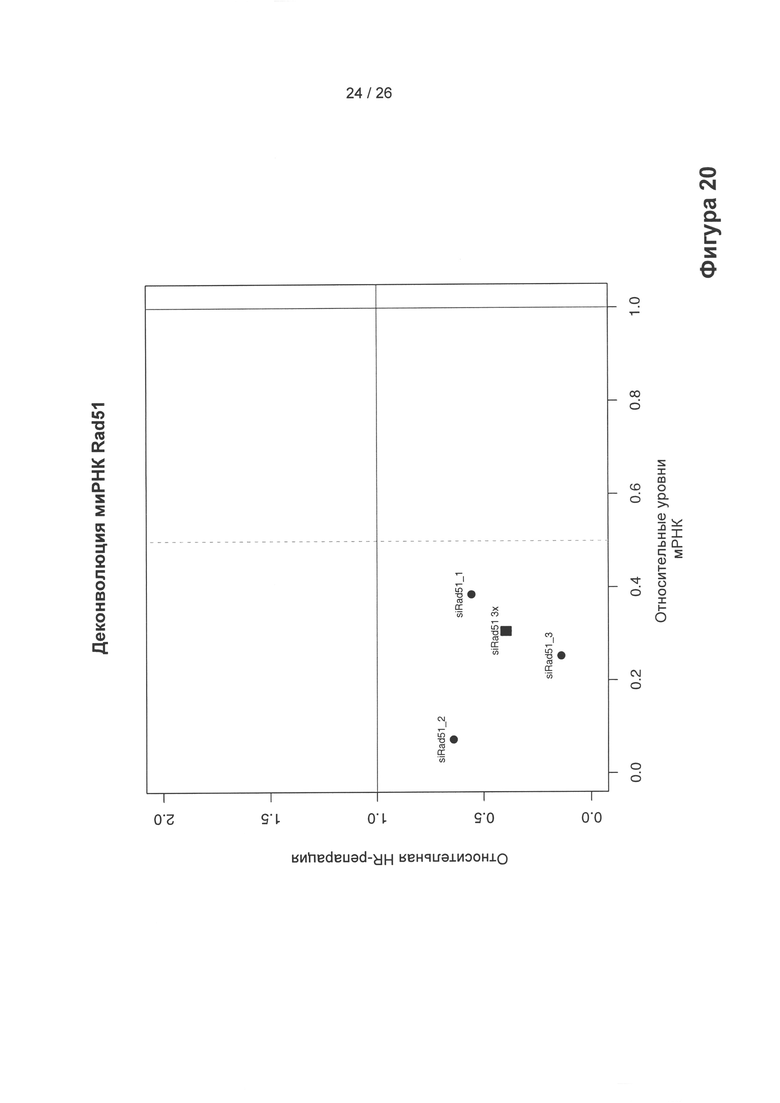
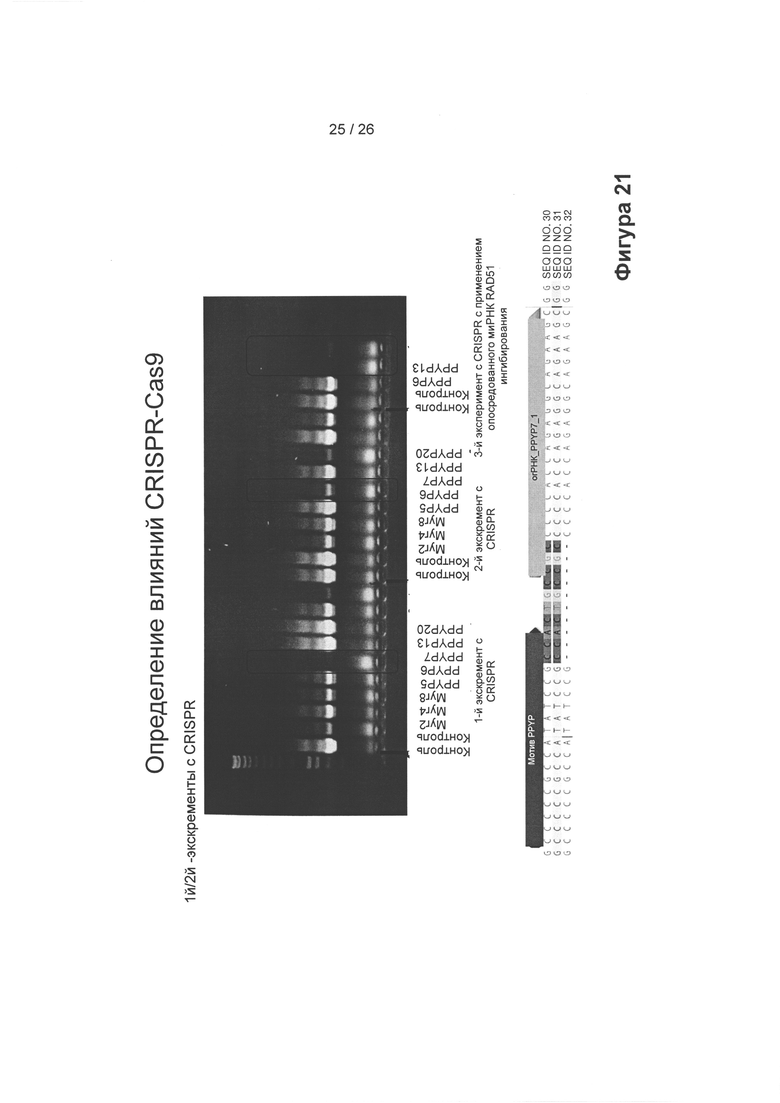
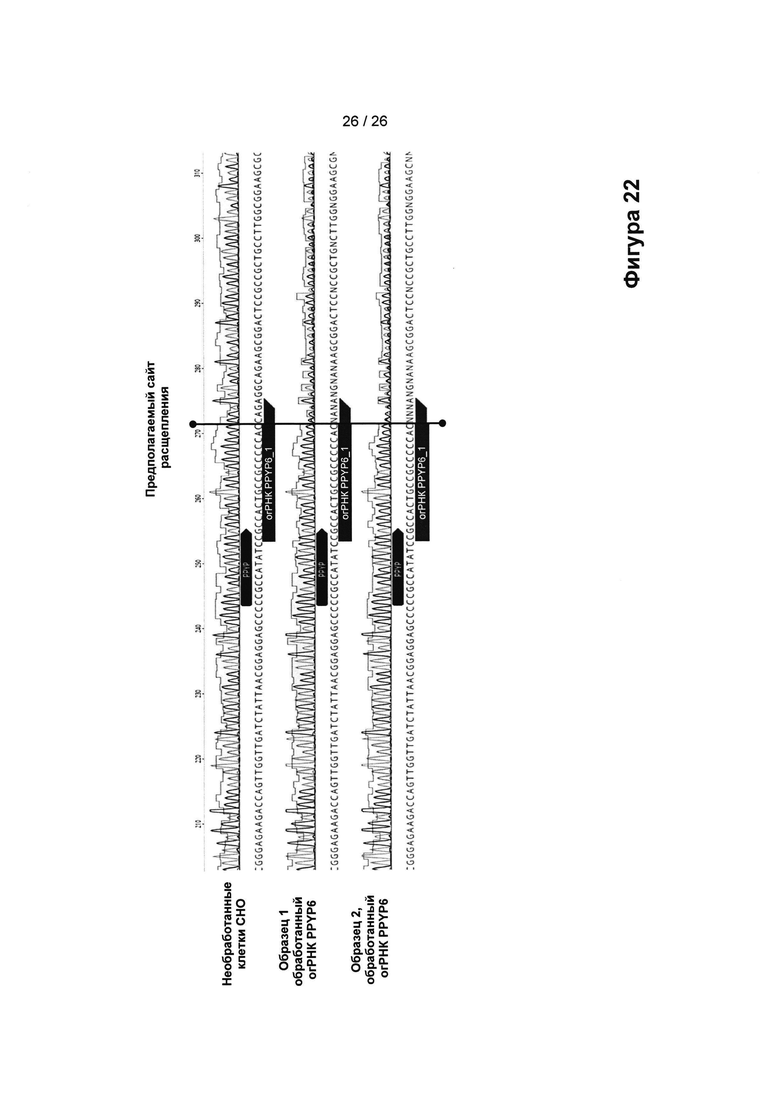
Изобретение относится к биотехнологии, в частности описаны клетки млекопитающих и линии клеток млекопитающих с уменьшенной нагрузкой остатков возбудителей ранее перенесенных вирусных/ретровирусных инфекций и способы получения и применения таких клеток. 4 н. и 32 з.п. ф-лы, 22 ил., 3 табл., 1 пр.
1. Модифицированная клетка млекопитающего для продукции терапевтического белка, содержащая:
геном указанной клетки, при этом указанный геном содержит одно или более изменений, включающих:
- делеции,
- добавления и/или
- замены,
по меньшей мере одной нуклеиновой кислоты в одном или более, в целом более, чем в 10, 20, 30, 40, 50, 60, 80, 90 или 100 эндогенных ретровирусных (ERV) элементах, которые являются частью указанного генома.
2. Модифицированная клетка млекопитающего по п. 1, отличающаяся тем, что указанная клетка представляет собой клетку линии клеток млекопитающего, такую как модифицированная клетка CHO, включая модифицированную CHO-K1.
3. Модифицированная клетка млекопитающего по п. 1 или 2, отличающаяся тем, что ERV-элементы происходят из гамма-ретровирусных ERV, включая ERV вируса эпидемии коал (KoRV), ERV вируса опухоли молочной железы у мышей (MMTV), ERV вируса лейкоза мышей (MLV), и при этом указанные одно или более изменений адаптированы для подавления или исключения высвобождения одного или более, предпочтительно более чем 60%, 70%, 80%, 90%, 95% или 100% указанных ERV.
4. Модифицированная клетка млекопитающего по любому из предшествующих пунктов, отличающаяся тем, что указанные один или более (ERV) элементов представляют собой или происходят из гена gag, pol и/или env, предпочтительно из указанного гена gag, кодирующего белок MA (ядерного матрикса), CA (белок капсида), NC (белок нуклеокапсида), белки, кодирующие дополнительный домен, такие как pp12 или p6, и/или представляют собой длинные концевые повторы (LTR) ERV.
5. Модифицированная клетка млекопитающего по п. 4, отличающаяся тем, что указанный один или более ERV-элементов кодирует белок Gag (групповой антиген), белок Pol (обратная транскриптаза) и/или белок Env (оболочки).
6. Модифицированная клетка млекопитающего по любому из предшествующих пунктов, отличающаяся тем, что указанные делеции, добавления или замены содержат более чем 2, 3, 4, 5, 10, 15, 20, 25, 30, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 150 или 200 нуклеиновых кислот.
7. Модифицированная клетка млекопитающего по любому из предшествующих пунктов, отличающаяся тем, что указанные изменения представляют собой добавления в форме направленных интеграций и указанные один или более (ERV) элементов представляют собой или происходят по меньшей мере из одного гена gag, pol и/или env, предпочтительно генов gag, и последовательности указанного гена подвергнуты указанным направленным интеграциям трансгена, предпочтительно кодирующего маркерный белок, такой как GFP (зеленый флуоресцентный белок), и/или указанное изменение(-я) приводит к доминантно-негативному фенотипу.
8. Модифицированная клетка млекопитающего по любому из предшествующих пунктов, отличающаяся тем, что указанные ERV-элементы получены из гена gag и по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 98%, 99% или все указанные элементы содержат указанные изменения.
9. Модифицированная клетка млекопитающего по любому из предшествующих пунктов, отличающаяся тем, что указанные изменения содержатся в консенсусных последовательностях указанных EVR-элементов, при этом длина такой консенсусной последовательности составляет по меньшей мере 5, 10, 15 или 20 пар оснований, предпочтительно длина составляет 15-25 или 30-50 пар оснований.
10. Модифицированная клетка млекопитающего по любому из предшествующих пунктов, отличающаяся тем, что указанная модифицированная клетка не высвобождает или по существу не высвобождает ERV.
11. Модифицированная клетка млекопитающего по любому из предшествующих пунктов, отличающаяся тем, что указанная модифицированная клетка содержит гетерологичные последовательности нуклеиновых кислот, которые кодируют, ингибируют или активируют по меньшей мере один белок пути рекомбинации, и/или гетерологичные последовательности нуклеиновых кислот, которые кодируют по меньшей мере одну последовательность или белок, подавляющие экспрессию по меньшей мере одного белка пути рекомбинации, предпочтительно HR, еще более предпочтительно Nbs1, Mre11, Rad51, лигазы 1 и/или лигазы 3.
12. Модифицированная клетка млекопитающего по п. 11, отличающаяся тем, что указанные гетерологичные последовательности нуклеиновых кислот присутствуют в виде вектора, который не интегрирован в геном указанной клетки.
13. Модифицированная клетка млекопитающего по любому из предшествующих пунктов, дополнительно содержащая гетерологичную донорную ДНК, предпочтительно в векторе, кодирующую по меньшей мере один маркерный белок, такой как GFP (зеленый флуоресцентный белок).
14. Модифицированная клетка млекопитающего по любому из предшествующих пунктов, отличающаяся тем, что указанная клетка дополнительно содержит трансген.
15. Модифицированная клетка млекопитающего по любому из предшествующих пунктов, отличающаяся тем, что указанный трансген представляет собой маркерный ген, кодирующий маркерный белок, такой как GFP (зеленый флуоресцентный белок), и/или обладающий терапевтической активностью белок, который предпочтительно интегрирован в геном.
16. Модифицированная клетка млекопитающего по любому из предшествующих пунктов, отличающаяся тем, что указанная клетка представляет собой клетку СНО, и при этом по меньшей мере одно изменение представляет собой замену по меньшей мере одной миристоилированной аминокислоты в мотиве миристоилирования в одном или более ERV-элементе, таком как, например, ген gag или ERV-элементе, полученном из гена gag, на немиристоилированную аминокислоту, при этом указанная модифицированная клетка содержит по меньшей мере транзиентно:
- последовательности, как, например, в форме транзиентно экспрессированного вектора, кодирующего белки HR и/или MMEJ, такие как Pad51, Lig3, Ercc1, Pold3; и/или
- последовательности, как, например, последовательности миРНК, такие как siMRel1, или векторы, кодирующие указанные последовательности.
17. Модифицированная клетка млекопитающего по любому из предшествующих пунктов, отличающаяся тем, что указанная клетка представляет собой клетку СНО, содержащую один или более указанных ЕRV-элементов, таких как ген gag или ERV-элемент, полученный из гена gag, который содержит мотив PPYP, и при этом (i) последовательности, кодирующие указанный мотив PPYP, и/или последовательности, содержащие вплоть до 20 нуклеиновых кислот, фланкирующие последовательности в (i), содержат указанное изменение.
18. Модифицированная клетка млекопитающего по любому из предшествующих пунктов, отличающаяся тем, что указанная модифицированная клетка представляет собой клетку СНО и содержит делеции, добавления и/или замены по меньшей мере в одной последовательности SEQ ID NOS: 1, 2, 3, 4, или в последовательностях с более чем 90% или 95% идентичностью последовательности к указанным последовательностям, предпочтительно в границах ERV-элементов указанных последовательностей.
19. Модифицированная клетка млекопитающего по любому из предшествующих пунктов, отличающаяся тем, что указанная модифицированная клетка представляет собой клетку СНО и содержит делеции, добавления и/или замену в консенсусной последовательности ERV группы 1, такой как SEQ ID NO: 30, или в последовательностях с более чем 90% идентичностью последовательности к указанной последовательности, и при этом указанная последовательность, содержащая такие делеции, добавления и/или замены, не кодирует функциональный белок Gag.
20. Способ улучшенного редактирования генома для получения клетки по любому из пп. 1-19, включающий:
(i) введение гетерологичной системы для введения одно- или двухцепочечных разрывов в последовательность целевой нуклеиновой кислоты, такой как консенсусная последовательность ERV-элемента, предпочтительно ERV-элемента, который представляет собой или который получен из гена gag, в клетку, предпочтительно в представляющую интерес клетку, и дополнительно
(ii) введение в указанную клетку
(а) гетерологичных последовательностей нуклеиновых кислот, кодирующих или активирующих по меньшей мере один белок по меньшей мере одного указанного пути рекомбинации, и/или
(b) гетерологичных последовательностей, кодирующих одну или более последовательностей, таких как, например, миРНК, или белки, подавляющие/снижающие экспрессию по меньшей мере одного белка по меньшей мере одного указанного пути рекомбинации, при этом указанная гетерологичная система (i) и/или последовательности (ii) (а) и/или (ii) (b) предпочтительно по меньшей мере транзиентно экспрессируются в представляющей интерес клетке.
21. Способ по п.20, согласно которому указанные гетерологичные последовательности кодируют или активируют по меньшей мере один белок пути рекомбинации, в частности по меньшей мере один белок гомологичной рекомбинации (HR), в частности Rad51.
22. Способ по п. 20, согласно которому маркерный ген, такой как GFP, вставлен в указанную целевую последовательность нуклеиновой кислоты посредством гомологичной рекомбинации, и при этом клетки, содержащие указанный маркерный ген, предпочтительно являются отобранными.
23. Способ по любому из пп. 20-22, согласно которому указанные гетерологичные последовательности нуклеиновых кислот (а) кодируют или активируют белки MNR-комплекса, такие как Nbs1 и/или Mre 11, и/или при этом указанные гетерологичные последовательности (b) кодируют последовательности или белки, подавляющие экспрессию по меньшей мере одного белка пути HR, такого как Rad51.
24. Способ по любому из пп. 20-23, согласно которому делеция или вставка введены в указанные один или более ERV-элементов посредством негомологичного соединения концов (NHEJ) или опосредованного микрогомологией соединения концов (MMEJ).
25. Способ по любому из пп. 20-24, согласно которому указанные гетерологичные последовательности представляют собой часть интегрирующих или неинтегрирующих векторов.
26. Способ по любому из пп. 20-25, согласно которому указанная не встречающаяся в природе система для введения одно- или двухцепочечных разрывов в целевую последовательность нуклеиновой кислоты представляет собой CRISPR/Cas9-систему или основанную на ней систему.
27. Способ по любому из пп. 20-26, согласно которому указанная клетка представляет собой клетку СНО, и при этом по меньшей мере одна миристоилированная аминокислота в мотиве миристоилирования в одном или более ERV-элементах заменена на немиристоилированную аминокислоту, причем указанный способ предпочтительно осуществляют во время транзиентной экспрессии последовательностей, кодирующих белки HR и/или MMEJ, такие как Pad51, Lig3, Ercc1, Pold3, и/или последовательностей, как, например, последовательности миРНК, такие как siMRel1, или кодирующих их векторов.
28. Способ по любому из пп. 20-27, согласно которому указанная клетка представляет собой клетку СНО, содержащую в одном или более указанных ERV-элементах мотив PPYP, и при этом (i) последовательности, кодирующие указанный мотив PPYP, и/или последовательности, содержащие вплоть до 20 нуклеиновых кислот, фланкирующие указанные последовательности (i), содержат изменение, причем указанный способ предпочтительно осуществляют во время транзиентной экспрессии последовательностей РНК, подавляющих экспрессию белков HR, таких как Rad51.
29. Модифицированная клетка млекопитающего для продукции терапевтического белка, при этом геном указанной клетки содержит одно или более изменений, включающих:
(а) делеции,
(b) добавления и/или
(с) замены,
по меньшей мере одной нуклеиновой кислоты в одном или более, в целом в более чем 10, 20, 30, 40, 50, 60, 80, 90 или 100 эндогенных ретровирусных (ERV) элементах, и которая получена при помощи способа по любому из пп. 20-28.
30. Модифицированная клетка млекопитающего по п. 29, отличающаяся тем, что указанная клетка представляет собой клетку линии клеток млекопитающего, такую как модифицированная клетка CHO, включая модифицированную CHO-K1.
31. Набор для получения клетки по любому из пп. 1-19 или 29-30, содержащий:
(i) в одной емкости по меньшей мере один неинтегрирующий вектор, кодирующий по меньшей мере CRISPR нуклеазу,
(ii) в той же или в дополнительной емкости по меньшей мере одну гидовую РНК или последовательность, кодирующую по меньшей мере одну гидовую РНК, нацеливающую на мотив в ERV-элементе,
(iii) в той же или в дополнительной емкости по меньшей мере одну миРНК или последовательность, кодирующую по меньшей мере одну миРНК, и
в дополнительной емкости инструкцию о том, как обеспечить попадание (i), (ii) и (iii) внутрь клетки.
32. Набор по п. 31, отличающийся тем, что указанные последовательности, кодирующие по меньшей мере одну гидовую РНК, нацеливающую на указанный мотив в ERV-элементе, и/или указанные последовательности, кодирующие по меньшей мере одну миРНК, являются частью вектора.
33. Набор по п. 32, отличающийся тем, что указанный вектор представляет собой неинтегрирующий вектор (i).
34. Набор по любому из пп. 31-33, отличающийся тем, что указанный ERV-элемент кодирует белок Gag.
35. Набор по п. 32, отличающийся тем, что указанный мотив представляет собой мотив миристоилирования или мотив PPXY, в частности мотив PPYP.
36. Набор по любому из пп. 31-35, отличающийся тем, что указанные миРНК направлены против гена пути HR.
| Hsu P | |||
| D., Lander E | |||
| S., Zhang F | |||
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
| Соломорезка | 1918 |
|
SU157A1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| Пишущая машина с ножной педалью для передвижения каретки и бумажного валика | 1923 |
|
SU1262A1 |
| Lee J | |||
| S | |||
| et al | |||
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| Устройство для избирательного управления двумя реле | 1918 |
|
SU979A1 |
| Рымарь С | |||
| Е | |||
| и др | |||
| Инкапсулированные | |||

