Область техники
Изобретение относится к области биотехнологии, а именно к редактированию генома с помощью CRISPR-Cas9 системы и белка LigD. Данная система позволит редактировать геном бактерии Pseudomonas putida, в том числе для синтеза новых веществ с антибиотической активностью.
Уровень техники
Бактерии являются важным источником новых субстанций, в том числе фармакологического значения. Для этих целей, среди прочих, активно используется бактерия Pseudomonas putida. Различные клоны P. putida, обнаруженные в пробах почвы и пробах воды в акватории Северного Ледовитого океана содержат новые потенциальные антибиотики. Однако, кластеры генов, отвечающих за их продукцию, имеют низкий или вовсе отсутствующий уровень экспрессии. Для наработки этих антибиотиков и их изучения необходимо редактирование генома бактерий, и для этого активно применяется система CRISPR-Cas9.
В последние годы система редактирования генома с короткими палиндромными кластерными повторами (CRISPR) и CRISPR-ассоциированным белком 9 (Cas9) превратилась в универсальный инструмент для точечного внесения мутаций, включая вставки/делеции генов, замены генов и однонуклеотидных мутаций (Zhang & Showalter, 2020). CRISPR-Cas9 был впервые обнаружен как адаптивная система иммунной защиты в бактериальных клетках в качестве механизма защиты от чужеродной ДНК. При использовании для редактирования генома механизм CRISPR-Cas9 в основном состоит из двух частей: гибридной направляющей РНК (sgRNA) и эндонуклеазы Cas9. Длина направляющей части sgRNA составляет 20 нуклеотидов и представляет собой высокоспецифичную последовательность, соответствующую строго определенному участку генома.
Основным преимуществом этой системы является простота её переключения на новые мишени. Для этого достаточно изменения последовательности гибридной направляющей РНК, что позволяет проводить работу одновременно с большим количеством мишеней.
Рекомбинация - это технология, используемая для генной инженерии бактерий in vivo. Она позволяет вносить изменения в масштабе генома, такие как вставки, делеции и точечные мутации (Sharan et al., 2009). Технологии рекомбинации широко используются для генной инженерии in vivo у бактерий, позволяя проводить такие виды геномных модификаций как вставки, делеции или однонуклеотидные замены. Они основаны на гомологичной рекомбинации линейных двуцепочечных или одноцепочечных фрагментов ДНК с помощью ферментов-рекомбиназ, полученных из фагов. Молекулы ДНК, которые должны быть интегрированы в бактериальный геном, сконструированы так, чтобы нести желаемую мутацию и короткие гомологичные последовательности (30-50 п.н.) на флангах. Учитывая низкую эффективность процедур рекомбинации, стандартные методы отбора мутировавших клеток, полученных рекомбинацией двуцепочечной ДНК, основаны на интеграцию в хромосому селективных маркеров, например, генов устойчивости к антибиотикам. Такие методы являются трудоемкими, отнимающими много времени и приводят к образованию “рубца” в мутировавшей области каждый раз, когда маркер удаляется (Ronda et al., 2016). Более того, отбор мутантов, полученных путем рекомбинации одноцепочечной ДНК, ограничен из-за относительно небольшого размера олигонуклеотида, который возможно синтезировать de novo, что сужает выбор возможного маркера. В связи с этим получил развитие метод контрселекции на основе Cas9. Он оказался эффективным для отбора рекомбинантных мутантов E. coli, не приводя к образованию «рубцов» (Jiang et al., 2013b). Методика заключается в том, чтобы направить Cas9 в область, которая была мутирована на стадии рекомбинации. Поскольку распознавание протоспейсера с помощью sgRNA нарушается в мутантных клетках, они будут избегать разрезания ДНК эффектором Cas9. Частота успешных мутантов в конечной популяции клеток увеличится за счет элиминации клеток, не прошедших этап рекомбинации.
Этот метод активно использовался для контрселекции бактерий после геномного редактирования, основанной на гомологичной рекомбинации. Основываясь на этом принципе, несколько исследовательских групп (Aparicio et al., 2018; Bikard et al., 2012; Jiang et al., 2013a; Oh & van Pijkeren, 2014; Penewit et al., 2018) исследовали пригодность методов сопряжения рекомбинации с контрселекцией Cas9 у различных прокариот для проведения эффективного редактирование генома. Этот подход был использован и для редактирования генома P. putida (Aparicio et al., 2018; Ricaurte et al., 2018). Однако такой подход требует этапа подбора последовательности для гомологичной рекомбинации, а также конструирования нескольких необходимых плазмид.
Ранее была разработана стратегия одноэтапной инженерии генома E. coli (Su et al., 2016). Она основана на нацеливании Cas9 с последующим восстановлением области протоспейсера с помощью негомологичного соединения концов (NHEJ). Этот механизм восстановления двунитевых разрывов подвержен ошибкам, что приводит к появлению мутаций. Гены, кодирующие механизм NHEJ и систему CRISPR-Cas9, были доставлены в клетки через плазмиду. Используя этот подход, стало возможным быстро и эффективно инактивировать целевые гены путем удаления больших фрагментов хромосомной ДНК без необходимости создавать гомологичную последовательность и без использования селективных маркеров.
Таким образом, существуют системы для геномного редактирования прокариот на основе гомологичной рекомбинации и CRISPR-Cas9 контрселекции, в том числе и для P. putida. Эта технология была усовершенствована, и на E. coli была показана возможность одностадийного внесения делеций в геном, с использованием только CRISPR-Cas9, без гомологичной рекомбинации. Однако, не было создано аналогичной технологии для редактирования генома P. putida.
Раскрытие изобретения
Технической задачей данного изобретение является разработка и создание нового эффективного способа редактирования генома бактерии Pseudomonas putida, а также упрощение редактирования генома бактерии Pseudomonas putida, для создания возможности одновременного редактирования большого числа геномных мишеней.
Поставленная задача решается в результате применения белка LigD, характеризующегося последовательностью SEQ ID NO:1, или характеризующегося последовательностью, которая, по меньшей мере, на 95% идентична аминокислотной последовательности SEQ ID NO: 1 и имеет отличия по сравнению с SEQ ID NO: 1 только в каталитически неактивных аминокислотных остатках, для редактирования генома бактерии Pseudomonas putida на основе системы CRISPR/Cas9.
Поставленная задача также решается при осуществлении способа редактирования генома бактерии Pseudomonas putida на основе системы CRISPR/Cas9, включающего введение в клетку:
- белка Cas9, матричной РНК (мРНК), кодирующей белок Cas9, или ДНК, кодирующей белок Cas9;
- белка LigD, характеризующегося последовательностью SEQ ID NO:1, или характеризующегося последовательностью, которая, по меньшей мере, на 95% идентична аминокислотной последовательности SEQ ID NO: 1 и имеет отличия по сравнению с SEQ ID NO: 1 только в каталитически неактивных аминокислотных остатках, матричной РНК (мРНК), кодирующей белок LigD, или ДНК, кодирующей белок LigD;
- по меньшей мере, одной направляющей РНК, для нацеливания белка Cas9 на последовательность-мишень нуклеиновой кислоты.
В частных вариантах воплощения изобретения способ осуществляется в условиях in vitro или in vivo.
В частных вариантах воплощения изобретения в способе осуществляют введение двух или более последовательностей направляющей РНК, комплементарных разным последовательностям-мишеням нуклеиновой кислоты, причем две или более последовательности направляющей РНК связываются с разными последовательностями-мишенями нуклеиновой кислоты.
В частных вариантах воплощения изобретения в клетке белок Cas9 взаимодействует с двумя или более последовательностями направляющей РНК.
В частных вариантах воплощения изобретения белок Cas9 представляет собой модифицированный белок Cas9 или гомолог белка Cas9.
В частных вариантах воплощения изобретения белок Cas9, матричную РНК (мРНК), кодирующую белок Cas9, или ДНК, кодирующую белок Cas9; белок LigD, характеризующийся последовательностью SEQ ID NO:1, матричную РНК (мРНК), кодирующую белок LigD, или ДНК, кодирующую белок LigD, а также, по меньшей мере, одну направляющую РНК для нацеливания белка Cas9 на последовательность-мишень нуклеиновой кислоты или ДНК ее кодирующей вводят одновременно или последовательно в клетку.
В частных вариантах воплощения изобретения бактерия Pseudomonas putida представляет собой штамм Pseudomonas putida KT2440.
В частных вариантах воплощения изобретения направляющая РНК состоит из крРНК (crisprRNA), способной гибридизоваться с целевой последовательностью, и трейсерной РНК (tracrRNA), совместно способных образовать комплекс с участком целевой ДНК и белком Cas9.
В частных вариантах воплощения изобретения в качестве направляющей РНК может быть использована гибридная направляющая РНК, сконструированная на основе крРНК и трейсерной РНК.
В частных вариантах воплощения изобретения ДНК, кодирующая белок Cas9, характеризуется последовательностью SEQ ID NO: 14.
В частных вариантах воплощения изобретения ДНК, кодирующая направляющую РНК, для нацеливания белка Cas9 на последовательность-мишень нуклеиновой кислоты, характеризуется последовательностью SEQ ID NO: 15.
В частных вариантах воплощения изобретения белок Cas9 представляет собой белок Cas9 Streptococcus pyogenes.
В результате осуществления изобретения достигаются следующие технические результаты:
- разработка нового и эффективного способа редактирования генома на основе системы CRISPR-Cas9 и белка LigD;
- способ по изобретению позволяет редактировать геном бактерии Pseudomonas putida, в частности штамма P. putida KT2440, делать это с меньшим количеством подготовительных этапов и проще подбирать новые геномные мишени;
- снижение количества необходимых этапов для осуществления способа по изобретению позволяет ускорить и упростить редактирование генома указанной бактерии, что позволит осуществлять высокопроизводительное редактирование различных штаммов P. Putida, в частности штамма P. putida KT2440;
- способ по изобретению позволяет вносить изменения сразу в несколько генов-мишеней;
- группа изобретений расширяет арсенал доступных средств для редактирования генома бактерии Pseudomonas putida.
Краткое описание чертежей
Фиг. 1А - Процент мутаций для каждого нуклеотида геномного сайта mcb1 после работы Cas9 без сверхэкспрессии белка LigD. Фиг. 1Б - Процент мутаций каждого нуклеотида геномного сайта mcb1 после работы Cas9 совместно со сверхэкспрессией белка LigD.
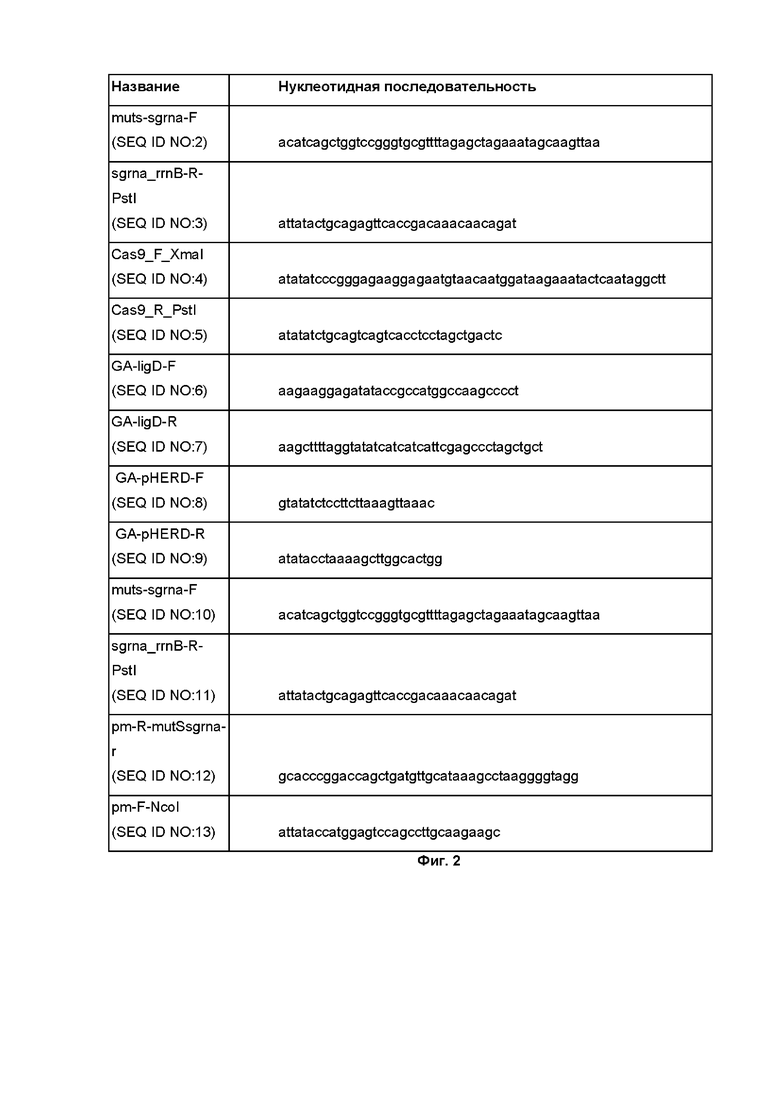
Фиг. 2 - Список праймеров, использованных для получения обозначенных в данной заявке плазмид.
Определения и термины
Для лучшего понимания настоящего изобретения ниже приведены некоторые термины, использованные в настоящем описании изобретения. Следующие определения применяются в данном документе, если иное не указано явно.
В описании данного изобретения термины «включает» и «включающий» интерпретируются как означающие «включает, помимо всего прочего». Указанные термины не предназначены для того, чтобы их истолковывали как «состоит только из».
Также здесь перечисление числовых диапазонов по конечным точкам включает все числа, входящие в этот диапазон.
Использованные научные и технические термины, если это не оговорено отдельно, имеют значения, общепринятые в научной и технической литературе.
Идентичность последовательностей определяется на основании референсной последовательности. Алгоритмы для анализа последовательности известны в данной области, такие как BLAST, описанный в Altschul et al., J. Mol. Biol., 215, pp. 403-10 (1990). Для целей настоящего изобретения для определения уровня идентичности и сходства между нуклеотидными последовательностями и аминокислотными последовательностями может быть использовано сравнение нуклеотидных и аминокислотных последовательностей, производимое с помощью пакета программ Blast, предоставляемого National Center for Biotechnology Information (http://www.ncbi.nlm.nih.gov/blast) с использованием содержащего разрывы выравнивания со стандартными параметрами. Процент идентичности двух последовательностей определяется числом положений идентичных аминокислот в этих двух последовательностях с учетом числа пробелов и длины каждого пробела, которые необходимо ввести для оптимального сопоставления двух последовательностей путем выравнивания. Процент идентичности равен числу идентичных аминокислот в данных положениях с учетом выравнивания последовательностей, разделенному на общее число положений и умноженному на 100.
В некоторых воплощениях LigD настоящего изобретения характеризуются аминокислотной последовательностью, которая имеет не менее 95% идентичности, например, не менее 96% идентичности, или не менее 97% идентичности, или не менее 98% идентичности, или не менее 99% идентичности, (например, по крайней мере 96%, 97%, 98%, 99% идентичности) с аминокислотной последовательностью SEQ ID NOs: 1.
Термин "специфически гибридизуется" относится к ассоциации между двумя одноцепочечными молекулами нуклеиновых кислот или в достаточной степени комплементарными последовательностями, что разрешает такую гибридизацию в предопределенных условиях, обычно использующихся в данной области.
Фраза "двунитевой разрыв" означает, что двунитевой разрыв в таргетной последовательности ДНК будет произведен без образования одноцепочечных участков ДНК, характерное место внесения подобного разрыва для Cas9 белка - между 3 и 4 нуклеотидами перед нуклеотидной последовательностью РАМ.
Под эффективным количеством вводимых в клетку белка и/или нуклеиновых кислот следует понимать такое количество белка и/или нуклеиновых кислот, которое при попадании в указанную клетку будет способно образовать функциональный комплекс, то есть комплекс, который будет специфически связываться с целевой ДНК и производить в ней двунитевой разрыв в месте, определяемом направляющей РНК и РАМ (A protospacer adjacent motif) последовательностью на ДНК. Эффективность этого процесса может быть оценена при помощи анализа целевой ДНК, выделенной из указанной клетки с помощью стандартных методов, известных специалистам.
Подробное раскрытие изобретения
Задачей настоящего изобретения является создание нового инструмента для изменения геномной последовательности на основе системы CRISPR-Cas9 и белка LigD. Текущие системы модификации генома прокариот используют метод гомологичной рекомбинации, включающий в себя этап конструирования гомологичной последовательности и её ведения в клетку. Это ограничивает возможности по одновременному редактирования большого количества сайтов генома. Отказ от этого этапа открывает возможность к высокопроизводительному редактированию генома P. putida.
Эта задача была решена авторами настоящего изобретения за счет использования белка P. putida LigD, характеризующегося последовательностью SEQ ID NO: 1, или характеризующегося аминокислотной последовательностью, которая по меньшей мере на 95% идентична аминокислотной последовательности SEQ ID NO: 1 и имеет отличия по сравнению с SEQ ID NO: 1 только в каталитически неактивных аминокислотных остатках. Данный белок ранее был охарактеризован как участник системы репарации двунитевых разрывов с помощью негомологичного соединения концов. В отличие от многих прокариот, P. putida обладает системой негомологичного соединений концов, что позволяет ей восстанавливать двунитевые разрывы ДНК без использования гомологичной ДНК-последовательности (Paris et al., 2015).
Таким образом, указанная задача решается:
1) за счет введения в клетку аминокислотной последовательности, идентичной SEQ ID NO:1, или последовательности, которая по меньшей мере на 95% идентична аминокислотной последовательности SEQ ID NO: 1 и имеет отличия по сравнению с SEQ ID NO: 1 только в каталитически неактивных аминокислотных остатках, либо доставки в клетку нуклеиновой кислоты, кодирующей белок с последовательностью SEQ ID NO:1 или белок с последовательностью, которая по меньшей мере на 95% идентична аминокислотной последовательности SEQ ID NO: 1 и имеет отличия по сравнению с SEQ ID NO: 1 только в каталитически неактивных аминокислотных остатках;
2) за счет введения в клетку Cas9 белка или нуклеиновой кислоты, кодирующей его аминокислотную последовательность, и введения гибридных направляющих РНК, или ДНК их кодирующих, обеспечивающих связывание Cas9 эффектора с необходимой последовательностью генома или иной ДНК, находящейся в клетке, и непосредственно примыкающей к нуклеотидной последовательности 5’-NGG-3’. Возможно использование одновременно нескольких направляющих РНК, для внесения множественных одновременных разрывов в геном, которые будут восстановлены LigD белком с внесением мутаций.
Изобретение может быть использовано как для редактирования ДНК внутри живой клетки (in vivo), так и in vitro. Модификация генома будет осуществляться за счет работы белка LigD, соединяющего двухнитевые разрывы ДНК и, одновременно, вносящего мутации в места восстановления. В качестве мишеней для редактирования генома могут быть выбраны гены-супрессоры, подавляющие работу известных биосинтетических генных кластеров (BGC). Известны примеры (Aigle & Corre, 2012), когда биосинтетические генные кластеры бактерий контролируются одним белком или одной молекулой РНК, и прекращение их экспрессии приводило к активации кластера. Внесение мутаций белком LigD в места восстановления разрывов, созданных Cas9, может привести к необратимому отключению подобных репрессеров, что позволит эффективно нарабатывать продукты подобных кластеров.
Также возможно использование изобретения для внесения мутаций в кодирующую часть интересующих белков, для получения библиотеки мутантных белков. Подобную библиотеку возможно использовать для последующего скрининга и отбора мутаций, повышающих функциональность белка, либо для изучения функций отдельных доменов белка.
В отличие от ранее описанных подходов, представленный способ редактирования использует белок LigD для восстановления двунитевых разрывов ДНК, возникающих после работы Cas9 белка. Прочие подходы для восстановления подобных разрывов используют гомологичные последовательности ДНК, содержащие требуемые мутации, и эти гомологичные последовательности обеспечивают восстановление двунитевых разрывов, возникающих в результате работы Cas9 белка. Подобные подходы требуют заблаговременного подбора гомологичной ДНК. Предложенный авторами настоящего изобретения подход позволяет отказаться от этого этапа, так как белок LigD при восстановлении разрывов вносит мутации без использования гомологичной ДНК.
Аминокислотная последовательность SEQ ID NO: 1 белка LigD Pseudomonas putida представляет собой:
MAKPLQEYARKRDFNATPEPSGKRKRGKRAHALQFCIQKHDASHLHYDFRLELDGTLKSWAIPKGPSLDPKVRRLAVHVEDHPLDYADFEGHIPEGHYGAGDVIVWDRGIWEPEGDAHEAYAKGKLRFRLLGEKLGGVWNLFRTHLAGKKEQWMLVKSHDAQACSETEYSIVEALPDSVLSERTLVPCKSAAKKATAPRSSRKRADKVQAAPLPAQLQPQLATLVDSPPSGDWRYEVKFDGYRILARIEGQDIRLFTRNGHDWSSKMPRQVAALRALGIDSAWLDGEMVVADENGMADFQALQNAFDTEHDERITYYLFDLPFLGGQDLRQLPLQNRRNTLRQLLEHNHCDVIKYSADFDEPVQSLLDSACRLKLEGLIGKRVDSVYSGRRSPDWIKLKCKQRQEFVIVGYTDPKGSRSGFGALLLALHDNDSGQLRYAGKVGTGFNATTLASIHARLKPLEIARPALPSPPAGAEARGVHWLRPQLLAEVAFAQMTRDGIVRHSVFHGLRDDKAATAIDLERAMPAKTVSNRNKQAKASENLGELRLTHPDRVIDPTTGVTKREVAEYYAGISQWILPQLKHRPVALVRAPDGLGGELFFQKNAGQLYIPNVLSYEKAEAGQAAMVINRADTLLGAVQMNMLELHTWNATDKDFDKPDRFVLDLDPDPALPWKAMVEATQLTLTLLDELGLKVFLKTSGGKGIHVVVPLTRRAGWDEVKDFSHAIVNYLAKLFPERLSAVSGPKNRVGRIFIDYLRNGKGATTACAYSLRARDGLPVSVPIWREELAQIKAANQWNITNLHEHLAEVDDPWADMGKVRQSITARMRKQLGLE.
Осуществление изобретения
Настоящее изобретение охватывает применение белка LigD совместно с работой CRISPR-Cas9 комплекса в клетках бактерии Pseudomonas putida. Во-первых, использование Cas9 эффектора и гибридных направляющих РНК позволяет, используя один тип нуклеазы, вносить разрывы в различные генетические последовательности-мишени. Во-вторых, использование одновременно нескольких направляющих РНК позволяет вносить изменения сразу в несколько генов-мишеней. В-третьих, сверхэкпрессия белка LigD в клетках P. putida приводит к делециям в местах восстановления двунитевых разрывов, что позволяет отказаться от гомологичной рекомбинации и сократить затраты времени на внесение делеций в геном клетки.
Введение белка и РНК в клетку возможно осуществить различными способами. Белок может быть доставлен в клетку в виде нуклеотидной последовательности ДНК, кодирующей мРНК, для её последующей трансляции в аминокислотную последовательность искомого белка. Доставка нуклеиновой кислоты может быть осуществлена трансформацией, трансфекцией или любым иным известным специалистам способом. Доставка белка и/или комплекса белок-РНК в клетку напрямую также возможна любым известным специалистам способом.
ДНК, введенная в клетку, для последующего синтеза белка или гибридной направляющей РНК, может быть включена в состав клеточной хромосомы или находиться в виде внехромосомно реплицирующейся ДНК.
Двуцепочечные разрывы в геномной ДНК смертельны для клетки, если их не восстановить. Чтобы устранить этот тип опасного повреждения, клетки полагаются на два основных механизма репарации: гомологичную рекомбинацию (HR) и негомологичное соединение концов (NHEJ). В то время как гомологичная рекомбинация требует генетической последовательности с высокой степенью сходства в качестве шаблона для репарации, негомологичное соединение концов не требует шаблона: поврежденные концы ДНК соединяются вместе и лигируются напрямую, обычно с внесением ошибок в восстанавливаемую последовательность. В прокариотических клетках гомологичная рекомбинация преобладает во время экспоненциальной фазы роста, когда клетки быстро делятся и доступны дополнительные копии геномной ДНК. Негомологичное соединение концов можно использовать независимо от того, имеется ли гомологичная ДНК-матрица или нет.
Механистически NHEJ состоит из следующих этапов: обнаруживаются сломанные концы ДНК, при необходимости обрабатываются, и лигируются, восстанавливая целостность ДНК. В P. putida в этом пути репарации задействованы только два белка: ДНК-связывающий белок Ku, который связывает и объединяет оба конца разорванной ДНК, и многофункциональная лигаза LigD, которая отвечает за обработку концов ДНК (если концы не комплементарны и/или химически модифицированы) и лигирование. Хотя Ku непосредственно не участвует в обработке разрывов, он необходим для рекрутирования LigD к поврежденным концам и остается физически связанным с ДНК в течение всего процесса репарации (McGovern et al., 2016).
Минимальные компоненты NHEJ были идентифицированы у нескольких видов Pseudomonas (Paris et al., 2015; Zhu & Shuman, 2005). Ферменты Ku и LigD были тщательно изучены в человеческом патогене P. aeruginosa. Штамм P. putida KT2440 (далее именуемый KT2440), еще один представитель рода, привлек внимание научного сообщества благодаря своему высокому потенциалу для метаболической инженерии и биоремедиации (Loeschcke & Thies, 2015). Как следствие, существует высокий спрос на методы манипулирования геномом этой бактерии. Геном KT2440 содержит гены ku и ligD, которые обладают высокой гомологией с соответствующими генами Pseudomonas aeruginosa. Учитывая, что некоторые из наиболее популярных стратегий генной инженерии, доступных в настоящее время, включают внесение двунитевых разрывов в геном, и учитывая мутагенный потенциал NHEJ, использование компонентов этого пути репарации увеличит продуктивность существующих подходов к редактированию генома P. putida. В результате проведенных исследований авторами настоящего изобретения показано, что клетки KT2440 могут преодолевать токсичность, опосредованную действием Cas9 на геном, при этом сверхэкспрессия белка LigD во время стационарной фазы роста клеток приводит к делециям в месте действия Cas9 нуклеазы.
Для осуществления редактирования генома способом по изобретению были сконструированы следующие плазмиды. Для работы Cas9 нуклеазы была сконструирована плазмида pSEVA258-Cas9. Кодирующий Cas9 ген (SEQ ID NO: 14) амплифицировали методом ПЦР с использованием плазмиды pCas9 (Jiang et al., 2013b) в качестве матрицы и праймеров Cas9_F_XmaI и Cas9_R_PstI (Фиг. 2). Амплифицированный фрагмент ПЦР сначала клонировали в вектор pJET1.2/blunt (“ThermoFisher Scientific”) с использованием протокола производителя. Полученную плазмиду pJET-Cas9 и плазмиду pSEVA258 разрезали с помощью XmaI и PstI эндонуклеаз. Фрагмент, кодирующий Cas9, и разрезанную плазмиду pSEVA258 лигировали.
ДНК-последовательность гена белка Cas9 Streptococcus pyogenes, использованная для конструирования плазмиды pSEVA258-Cas9 (SEQ ID NO: 14):
ATGGATAAGAAATACTCAATAGGCTTAGATATCGGCACAAATAGCGTCGGATGGGCGGTGATCACTGATGAATATAAGGTTCCGTCTAAAAAGTTCAAGGTTCTGGGAAATACAGACCGCCACAGTATCAAAAAAAATCTTATAGGGGCTCTTTTATTTGACAGTGGAGAGACAGCGGAAGCGACTCGTCTCAAACGGACAGCTCGTAGAAGGTATACACGTCGGAAGAATCGTATTTGTTATCTACAGGAGATTTTTTCAAATGAGATGGCGAAAGTAGATGATAGTTTCTTTCATCGACTTGAAGAGTCTTTTTTGGTGGAAGAAGACAAGAAGCATGAACGTCATCCTATTTTTGGAAATATAGTAGATGAAGTTGCTTATCATGAGAAATATCCAACTATCTATCATCTGCGAAAAAAATTGGTAGATTCTACTGATAAAGCGGATTTGCGCTTAATCTATTTGGCCTTAGCGCATATGATTAAGTTTCGTGGTCATTTTTTGATTGAGGGAGATTTAAATCCTGATAATAGTGATGTGGACAAACTATTTATCCAGTTGGTACAAACCTACAATCAATTATTTGAAGAAAACCCTATTAACGCAAGTGGAGTAGATGCTAAAGCGATTCTTTCTGCACGATTGAGTAAATCAAGACGATTAGAAAATCTCATTGCTCAGCTCCCCGGTGAGAAGAAAAATGGCTTATTTGGGAATCTCATTGCTTTGTCATTGGGTTTGACCCCTAATTTTAAATCAAATTTTGATTTGGCAGAAGATGCTAAATTACAGCTTTCAAAAGATACTTACGATGATGATTTAGATAATTTATTGGCGCAAATTGGAGATCAATATGCTGATTTGTTTTTGGCAGCTAAGAATTTATCAGATGCTATTTTACTTTCAGATATCCTAAGAGTAAATACTGAAATAACTAAGGCTCCCCTATCAGCTTCAATGATTAAACGCTACGATGAACATCATCAAGACTTGACTCTTTTAAAAGCTTTAGTTCGACAACAACTTCCAGAAAAGTATAAAGAAATCTTTTTTGATCAATCAAAAAACGGATATGCAGGTTATATTGATGGGGGAGCTAGCCAAGAAGAATTTTATAAATTTATCAAACCAATTTTAGAAAAAATGGATGGTACTGAGGAATTATTGGTGAAACTAAATCGTGAAGATTTGCTGCGCAAGCAACGGACCTTTGACAACGGCTCTATTCCCCATCAAATTCACTTGGGTGAGCTGCATGCTATTTTGAGAAGACAAGAAGACTTTTATCCATTTTTAAAAGACAATCGTGAGAAGATTGAAAAAATCTTGACTTTTCGAATTCCTTATTATGTTGGTCCATTGGCGCGTGGCAATAGTCGTTTTGCATGGATGACTCGGAAGTCTGAAGAAACAATTACCCCATGGAATTTTGAAGAAGTTGTCGATAAAGGTGCTTCAGCTCAATCATTTATTGAACGCATGACAAACTTTGATAAAAATCTTCCAAATGAAAAAGTACTACCAAAACATAGTTTGCTTTATGAGTATTTTACGGTTTATAACGAATTGACAAAGGTCAAATATGTTACTGAAGGAATGCGAAAACCAGCATTTCTTTCAGGTGAACAGAAGAAAGCCATTGTTGATTTACTCTTCAAAACAAATCGAAAAGTAACCGTTAAGCAATTAAAAGAAGATTATTTCAAAAAAATAGAATGTTTTGATAGTGTTGAAATTTCAGGAGTTGAAGATAGATTTAATGCTTCATTAGGTACCTACCATGATTTGCTAAAAATTATTAAAGATAAAGATTTTTTGGATAATGAAGAAAATGAAGATATCTTAGAGGATATTGTTTTAACATTGACCTTATTTGAAGATAGGGAGATGATTGAGGAAAGACTTAAAACATATGCTCACCTCTTTGATGATAAGGTGATGAAACAGCTTAAACGTCGCCGTTATACTGGTTGGGGACGTTTGTCTCGAAAATTGATTAATGGTATTAGGGATAAGCAATCTGGCAAAACAATATTAGATTTTTTGAAATCAGATGGTTTTGCCAATCGCAATTTTATGCAGCTGATCCATGATGATAGTTTGACATTTAAAGAAGACATTCAAAAAGCACAAGTGTCTGGACAAGGCGATAGTTTACATGAACATATTGCAAATTTAGCTGGTAGCCCTGCTATTAAAAAAGGTATTTTACAGACTGTAAAAGTTGTTGATGAATTGGTCAAAGTAATGGGGCGGCATAAGCCAGAAAATATCGTTATTGAAATGGCACGTGAAAATCAGACAACTCAAAAGGGCCAGAAAAATTCGCGAGAGCGTATGAAACGAATCGAAGAAGGTATCAAAGAATTAGGAAGTCAGATTCTTAAAGAGCATCCTGTTGAAAATACTCAATTGCAAAATGAAAAGCTCTATCTCTATTATCTCCAAAATGGAAGAGACATGTATGTGGACCAAGAATTAGATATTAATCGTTTAAGTGATTATGATGTCGATCACATTGTTCCACAAAGTTTCCTTAAAGACGATTCAATAGACAATAAGGTCTTAACGCGTTCTGATAAAAATCGTGGTAAATCGGATAACGTTCCAAGTGAAGAAGTAGTCAAAAAGATGAAAAACTATTGGAGACAACTTCTAAACGCCAAGTTAATCACTCAACGTAAGTTTGATAATTTAACGAAAGCTGAACGTGGAGGTTTGAGTGAACTTGATAAAGCTGGTTTTATCAAACGCCAATTGGTTGAAACTCGCCAAATCACTAAGCATGTGGCACAAATTTTGGATAGTCGCATGAATACTAAATACGATGAAAATGATAAACTTATTCGAGAGGTTAAAGTGATTACCTTAAAATCTAAATTAGTTTCTGACTTCCGAAAAGATTTCCAATTCTATAAAGTACGTGAGATTAACAATTACCATCATGCCCATGATGCGTATCTAAATGCCGTCGTTGGAACTGCTTTGATTAAGAAATATCCAAAACTTGAATCGGAGTTTGTCTATGGTGATTATAAAGTTTATGATGTTCGTAAAATGATTGCTAAGTCTGAGCAAGAAATAGGCAAAGCAACCGCAAAATATTTCTTTTACTCTAATATCATGAACTTCTTCAAAACAGAAATTACACTTGCAAATGGAGAGATTCGCAAACGCCCTCTAATCGAAACTAATGGGGAAACTGGAGAAATTGTCTGGGATAAAGGGCGAGATTTTGCCACAGTGCGCAAAGTATTGTCCATGCCCCAAGTCAATATTGTCAAGAAAACAGAAGTACAGACAGGCGGATTCTCCAAGGAGTCAATTTTACCAAAAAGAAATTCGGACAAGCTTATTGCTCGTAAAAAAGACTGGGATCCAAAAAAATATGGTGGTTTTGATAGTCCAACGGTAGCTTATTCAGTCCTAGTGGTTGCTAAGGTGGAAAAAGGGAAATCGAAGAAGTTAAAATCCGTTAAAGAGTTACTAGGGATCACAATTATGGAAAGAAGTTCCTTTGAAAAAAATCCGATTGACTTTTTAGAAGCTAAAGGATATAAGGAAGTTAAAAAAGACTTAATCATTAAACTACCTAAATATAGTCTTTTTGAGTTAGAAAACGGTCGTAAACGGATGCTGGCTAGTGCCGGAGAATTACAAAAAGGAAATGAGCTGGCTCTGCCAAGCAAATATGTGAATTTTTTATATTTAGCTAGTCATTATGAAAAGTTGAAGGGTAGTCCAGAAGATAACGAACAAAAACAATTGTTTGTGGAGCAGCATAAGCATTATTTAGATGAGATTATTGAGCAAATCAGTGAATTTTCTAAGCGTGTTATTTTAGCAGATGCCAATTTAGATAAAGTTCTTAGTGCATATAACAAACATAGAGACAAACCAATACGTGAACAAGCAGAAAATATTATTCATTTATTTACGTTGACGAATCTTGGAGCTCCCGCTGCTTTTAAATATTTTGATACAACAATTGATCGTAAACGATATACGTCTACAAAAGAAGTTTTAGATGCCACTCTTATCCATCAATCCATCACTGGTCTTTATGAAACACGCATTGATTTGAGTCAGCTAGGAGGTGACTGA.
Плазмида pHERD26T-LigD, несущая копию белка LigD для сверхэкспрессии, была сконструирована путем клонирования гена P. putida KT2440 ligD, амплифицированных GA-ligD-F и GA-ligD-R праймеров (Фиг. 2). Праймеры GA-pHERD-F и GA-pHERD-R (Фиг. 2) использовали для амплификации коммерческого вектора pHERD26T, а продукт реакции обрабатывали эндонуклеазой DpnI для разрушения метилированных молекул матрицы. Затем продукты ПЦР инкубировали вместе с HiFi DNA Assembly Master-mix (“New England Biolabs”).
Плазмида pBBR5-sgRNA-mcb1, несущая гибридную направляющую РНК для нацеливания Cas9 комплекса на геномный сайт mcb1, была получена на основе плазмиды pBBR5-sgRNA-mutS. Для создания pBBR5-sgRNA-mutS был амплифицирован с помощью ПЦР фрагмент гибридной направляющей РНК плазмиды pKDsgRNA-ack, отвечающий за связывание с Cas9 эффектором и праймеров muts-sgrna-F и sgrna_rrnB-R-PstI (Фиг. 2), которые вводят спейсер, нацеленный на ген P. putida mutS. Область, кодирующую Pm промотор, амплифицировали с использованием pSEVA258 в качестве матрицы и праймеров pm-R-mutssgrna-r и pm-F-NcoI (Фиг. 2). Полученные фрагменты амплифицировали в одной реакции с использованием праймеров pm-F-NcoI и sgrna_rrnB-R-PstI. Полученные Pm-sgRNA и плазмиду pBBR5 (Kovach et al., 1995) разрезали ферментами NcoI и PstI, а затем лигировали. Плазмида pBBR5-sgRNA-mcb1 была сконструирована с использованием полученной плазмиды pBBR5-sgRNA-mutS в качестве матрицы.
ДНК-последовательность, кодирующая направляющую часть гибридной направляющей РНК mcb1 (SEQ ID NO: 15):
CTTTAAACCAAAAACTCCAATACCATTCCAAGCCACTTACCCTCCTATAATAGATTGACGTTGAAGAATCTAACGCCTAAACCTCCTGATGCCCGC.
Плазмида pHERD26T-LigD совместима с плазмидами pSEVA258-Cas9 и pBBR-sgRNA. Клетки, несущие три разные плазмиды, получали и выращивали в присутствии 0,2% L-арабинозы, чтобы индуцировать экспрессию генов ku или ligD, что приводило к ~65-кратному и ~100-кратному увеличению уровней транскриптов ku или ligD в стационарной фазе роста клеток соответственно, что было оценено с помощью RT-qPCR.
Для внесения мутаций в геном, клетки штамма KT2440 трансформировали плазмидами pSEVA258-Cas9, pBBr5-sgRNA и pHERD26T-ligD одновременно. Трансформированные клетки инкубировались в течение двух часов при 30°C в среде SOC. Для использования клеток на стационарной фазе роста их культивировали в течение ночи в жидкой среде LB, дополненной канамицином (50 мкг/мл), гентамицином (50 мкг/мл) и тетрациклином (12 мкг/мл). На следующий день аликвоты от 0,5 до 2 мл из ночных культур высевали на LB-агар, добавляли канамицин, гентамицин, тетрациклин и бензоат натрия (2 мМ) и инкубировали при 30°C в течение ночи. Конечное количество клеток варьировалось в зависимости от эффективности работы Cas9 нуклеазы, однако объем клеток для посева подбирался таким образом, чтобы обеспечить КОЕ>105.
Резкое изменение паттерна мутации наблюдалось в участке mcb1 (Фиг. 1), что говорит о том, что двухнитевые разрывы, производимые Cas9 белком, восстанавливались системой репарации, подверженной ошибкам, в состав которой входит LigD. В условиях сверхэкспрессии белка LigD преобладающими становились длинные (до 60 п.н.) делеции, охватывающие область мишени.
Примером использования технологии является получение мутантов с делециями ku и ligD в геноме P. putida. Использование гибридных направляющих РНК, гомологичных началу кодирующей части этих генов, с одновременной сверхэкспрессией белка LigD, позволило получить делецию от нуклеотида #59 до нуклеотида #210 кодирующей области гена ku, и делецию от нуклеотида #112 до нуклеотида #1335 кодирующей области гена ligD, что в обоих генах привело к сдвигу рамки считывания и нарушению синтеза продуктов этих генов. Данный пример демонстрирует возможности изобретения для внесения делеций в геном P. putida. Специалистам в области генетической инженерии очевидны применения изобретения для редактирования прочих участков генома, в том числе с целью реактивировать молчащие гены P. putida, находящиеся под контролем различных репрессоров.
Несмотря на приведенную ссылку на вариант применения изобретения, для специалистов в данной области очевидно, что конкретный пример не может ограничивать возможности применения изобретения. Возможно внесение различных модификаций без отступления от сути изобретения.
Список литературы:
1. Aigle, B., & Corre, C. (2012). Waking up Streptomyces secondary metabolism by constitutive expression of activators or genetic disruption of repressors. Methods in Enzymology, 517, 343-366. https://doi.org/10.1016/B978-0-12-404634-4.00017-6;
2. Aparicio, T., de Lorenzo, V., & Martínez-García, E. (2018). CRISPR/Cas9-Based Counterselection Boosts Recombineering Efficiency in Pseudomonas putida. Biotechnology Journal, 13(5), e1700161. https://doi.org/10.1002/biot.201700161;
3. Bikard, D., Hatoum-Aslan, A., Mucida, D., & Marraffini, L. A. (2012). CRISPR interference can prevent natural transformation and virulence acquisition during in vivo bacterial infection. Cell Host & Microbe, 12(2), 177-186. https://doi.org/10.1016/j.chom.2012.06.003;
4. Jiang, W., Bikard, D., Cox, D., Zhang, F., & Marraffini, L. A. (2013a). RNA-guided editing of bacterial genomes using CRISPR-Cas systems. Nature Biotechnology, 31(3), 233-239. https://doi.org/10.1038/nbt.2508;
5. Jiang, W., Bikard, D., Cox, D., Zhang, F., & Marraffini, L. A. (2013b). RNA-guided editing of bacterial genomes using CRISPR-Cas systems. Nature Biotechnology 2013 31:3, 31(3), 233-239. https://doi.org/10.1038/nbt.2508;
6. Kovach, M. E., Elzer, P. H., Steven Hill, D., Robertson, G. T., Farris, M. A., Roop, R. M., & Peterson, K. M. (1995). Four new derivatives of the broad-host-range cloning vector pBBR1MCS, carrying different antibiotic-resistance cassettes. Gene, 166(1), 175-176. https://doi.org/10.1016/0378-1119(95)00584-1;
7. Loeschcke, A., & Thies, S. (2015). Pseudomonas putida-a versatile host for the production of natural products. Applied Microbiology and Biotechnology, 99(15), 6197-6214. https://doi.org/10.1007/s00253-015-6745-4;
8. McGovern, S., Baconnais, S., Roblin, P., Nicolas, P., Drevet, P., Simonson, H., Pietrement, O., Charbonnier, J. B., le Cam, E., Noirot, P., & Lecointe, F. (2016). C-terminal region of bacterial Ku controls DNA bridging, DNA threading and recruitment of DNA ligase D for double strand breaks repair. Nucleic Acids Research, 44(10), 4785-4806. https://doi.org/10.1093/nar/gkw149;
9. Oh, J. H., & van Pijkeren, J. P. (2014). CRISPR-Cas9-assisted recombineering in Lactobacillus reuteri. Nucleic Acids Research, 42(17). https://doi.org/10.1093/NAR/GKU623;
10. Paris, Ü., Mikkel, K., Tavita, K., Saumaa, S., Teras, R., & Kivisaar, M. (2015). NHEJ enzymes LigD and Ku participate in stationary-phase mutagenesis in Pseudomonas putida. DNA Repair, 31, 11-18. https://doi.org/10.1016/j.dnarep.2015.04.005;Penewit, K., Holmes, E. A., McLean, K., Ren, M., Waalkes, A., & Salipante, S. J. (2018). Efficient and Scalable Precision Genome Editing in Staphylococcus aureus through Conditional Recombineering and CRISPR/Cas9-Mediated Counterselection. MBio, 9(1). https://doi.org/10.1128/MBIO.00067-18;
11. Ricaurte, D. E., Martínez-García, E., Nyerges, Á., Pál, C., de Lorenzo, V., & Aparicio, T. (2018). A standardized workflow for surveying recombinases expands bacterial genome-editing capabilities. Microbial Biotechnology, 11(1), 176-188. https://doi.org/10.1111/1751-7915.12846;
12. Ronda, C., Pedersen, L. E., Sommer, M. O. A., & Nielsen, A. T. (2016). CRMAGE: CRISPR Optimized MAGE Recombineering. Scientific Reports, 6. https://doi.org/10.1038/SREP19452;
13. Sharan, S. K., Thomason, L. C., Kuznetsov, S. G., & Court, D. L. (2009). Recombineering: a homologous recombination-based method of genetic engineering. Nature Protocols, 4(2), 206-223. https://doi.org/10.1038/NPROT.2008.227;Su, T., Liu, F., Gu, P., Jin, H., Chang, Y., Wang, Q., Liang, Q., & Qi, Q. (2016). A CRISPR-Cas9 Assisted Non-Homologous End-Joining Strategy for One-step Engineering of Bacterial Genome. Scientific Reports 2016 6:1, 6(1), 1-11. https://doi.org/10.1038/srep37895;
14. Zhang, Y., & Showalter, A. M. (2020). CRISPR/Cas9 Genome Editing Technology: A Valuable Tool for Understanding Plant Cell Wall Biosynthesis and Function. Frontiers in Plant Science, 11, 1779. https://doi.org/10.3389/FPLS.2020.589517/BIBTEX;
15. Zhu, H., & Shuman, S. (2005). A primer-dependent polymerase function of pseudomonas aeruginosa ATP-dependent DNA ligase (LigD). The Journal of Biological Chemistry, 280(1), 418-427. https://doi.org/10.1074/JBC.M410110200.
| название | год | авторы | номер документа |
|---|---|---|---|
| СРЕДСТВО РАЗРЕЗАНИЯ ДНК НА ОСНОВЕ CAS9 БЕЛКА ИЗ DEFLUVIIMONAS SP. | 2018 |
|
RU2712492C1 |
| Средство разрезания ДНК на основе Cas9 белка из биотехнологически значимой бактерии Clostridium cellulolyticum | 2018 |
|
RU2712497C1 |
| ПРИМЕНЕНИЕ CAS9 БЕЛКА ИЗ БАКТЕРИИ PASTEURELLA PNEUMOTROPICA ДЛЯ МОДИФИКАЦИИ ГЕНОМНОЙ ДНК В КЛЕТКАХ | 2019 |
|
RU2724470C1 |
| СРЕДСТВО РАЗРЕЗАНИЯ ДНК НА ОСНОВЕ CAS9 БЕЛКА ИЗ БАКТЕРИИ PASTEURELLA PNEUMOTROPICA | 2019 |
|
RU2722934C1 |
| Способ получения препарата рибонуклеопротеинового комплекса CRISPR/Cas и препарат для выявления гена антибиотикоустойчивости bla VIM-2 (металло-бета-лактамаза класс B VIM-2) Pseudomonas aeruginosa в ультранизких концентрациях | 2020 |
|
RU2745637C1 |
| Новая компактная нуклеаза CAS9 II типа из Anoxybacillus flavithermus | 2020 |
|
RU2749307C1 |
| Средство разрезания двунитевой ДНК с помощью Cas12d белка из Katanobacteria и гибридной РНК, полученной путем слияния направляющей CRISPR РНК и scout РНК | 2020 |
|
RU2771626C1 |
| Способ обнаружения гена антибиотикоустойчивости blaVIM-2 (металло-бета-лактамаза класс B VIM-2) Pseudomonas aeruginosa в ультранизких концентрациях и специфические олигонуклеотиды для использования в способе | 2020 |
|
RU2734520C1 |
| Система для увеличения эффективности редактирования генома млекопитающего за счет гомологичной репарации | 2022 |
|
RU2808045C1 |
| Система привлечения белка к месту разрыва ДНК для увеличения эффективности редактирования генома млекопитающего | 2022 |
|
RU2808601C1 |

Изобретение относится к области биотехнологии, а именно к редактированию генома с помощью CRISPR-Cas9 системы и белка LigD. Данная система позволит редактировать геном бактерии Pseudomonas putida, в том числе для синтеза новых веществ с антибиотической активностью. Изобретение раскрывает новый способ упрощенного редактирования генома бактерии Pseudomonas putida, который, в частности, позволяет одновременного редактировать большое число геномных мишеней. 2 н. и 11 з.п. ф-лы, 2 ил., 1 пр.
1. Применение белка LigD, характеризующегося последовательностью SEQ ID NO:1, или характеризующегося последовательностью, которая, по меньшей мере, на 95% идентична аминокислотной последовательности SEQ ID NO: 1 и имеет отличия по сравнению с SEQ ID NO: 1 только в каталитически неактивных аминокислотных остатках, для редактирования генома бактерии Pseudomonas putida на основе системы CRISPR/Cas9.
2. Способ редактирования генома бактерии Pseudomonas putida на основе системы CRISPR/Cas9, включающий введение в клетку:
- белка Cas9, матричной РНК (мРНК), кодирующей белок Cas9, или ДНК, кодирующей белок Cas9;
- белка LigD, характеризующегося последовательностью SEQ ID NO:1, или характеризующегося последовательностью, которая, по меньшей мере, на 95% идентична аминокислотной последовательности SEQ ID NO: 1 и имеет отличия по сравнению с SEQ ID NO: 1 только в каталитически неактивных аминокислотных остатках, матричной РНК (мРНК), кодирующей белок LigD, или ДНК, кодирующей белок LigD;
- по меньшей мере, одной направляющей РНК, для нацеливания белка Cas9 на последовательность-мишень нуклеиновой кислоты.
3. Способ по п. 2, в котором способ осуществляется в условиях in vitro или in vivo.
4. Способ по п. 2, в котором осуществляют введение двух или более последовательностей направляющей РНК, комплементарных разным последовательностям-мишеням нуклеиновой кислоты, причем две или более последовательности направляющей РНК связываются с разными последовательностями-мишенями нуклеиновой кислоты.
5. Способ по п. 2, в котором в клетке белок Cas9 взаимодействует с двумя или более последовательностями направляющей РНК.
6. Способ по п. 2, в котором белок Cas9 представляет собой модифицированный белок Cas9 или гомолог белка Cas9.
7. Способ по п. 2, в котором белок Cas9, матричную РНК (мРНК), кодирующую белок Cas9, или ДНК, кодирующую белок Cas9; белок LigD, характеризующийся последовательностью SEQ ID NO:1, матричную РНК (мРНК), кодирующую белок LigD, или ДНК, кодирующую белок LigD, а также, по меньшей мере, одну направляющую РНК для нацеливания белка Cas9 на последовательность-мишень нуклеиновой кислоты или ДНК ее кодирующей вводят одновременно или последовательно в клетку.
8. Способ по п. 2, в котором бактерия Pseudomonas putida представляет собой штамм Pseudomonas putida KT2440.
9. Способ по п. 2, в котором направляющая РНК состоит из крРНК (crisprRNA), способной гибридизоваться с целевой последовательностью, и трейсерной РНК (tracrRNA), совместно способных образовать комплекс с участком целевой ДНК и белком Cas9.
10. Способ по п. 9, в котором в качестве направляющей РНК может быть использована гибридная РНК, сконструированная на основе крРНК и трейсерной РНК.
11. Способ по п. 2, в котором ДНК, кодирующая белок Cas9, характеризуется последовательностью SEQ ID NO: 14.
12. Способ по п. 2, в котором ДНК, кодирующая направляющую РНК, для нацеливания белка Cas9 на последовательность-мишень нуклеиновой кислоты, характеризуется последовательностью SEQ ID NO: 15.
13. Способ по п. 2, в котором белок Cas9 представляет собой белок Cas9 Streptococcus pyogenes.
| US20210207134 A1, 08.07.2021 | |||
| WU Z | |||
| et al., Combination of ssDNA recombineering and CRISPR-Cas9 for Pseudomonas putida KT2440 genome editing, Appl Microbiol Biotechnol, 2019, vol | |||
| Клапанный регулятор для паровозов | 1919 |
|
SU103A1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| Электрическая лампа накаливания с несколькими нитями, как дополнительный патент к патенту № 1073 | 1925 |
|
SU2783A1 |
| БЛАТОВ И | |||
| А | |||
| и др., CRISPR/Cas-системы: характеристика и возможности использования для редактирования геномов бактерий, Бактериология, 2020, | |||

