Область техники, к которой относится изобретение
Настоящее изобретение относится к биотехнологии, а именно к способам повышения гомологичной рекомбинации в эукариотических клетках с использованием химерных слитых белков аппарата гомологичной рекомбинации бактериофагов. В частности, изобретение относится к химерной экзонуклеазе слитой на С-конце с SSB белком, связывающим одноцепочечную ДНК. Такой способ будет применяться после направленного внесения двуцепочечного разрыва системой Cas9 в клетке для резекции двуцепочечной ДНК и одновременной защиты одноцепочечных рекомбиногенных концов ДНК.
Уровень техники
Совместное использование экзонуклеаз, ДНК-полимераз и ДНК-лигаз позволяет проводить рекомбинацию последовательностей ДНК in vitro (патент US 20120053087 A1. Daniel G. Gibson, Hamilton О. Smith, Clyde A. Hutchison, Lei Young, J. Craig Venter. Methods for in vitro joning and combinatoral assembly of nuclec acid molecules // 2012). С помощью таких методов можно быстро создавать генетические конструкции различного состава. При этом используются чистые препараты ферментов и ДНК. Все это сложно осуществить при работе на клеточном уровне.
В первых исследованиях с клетками млекопитающих, начатых проводится более 20 лет назад, использовали хоминг-эндонуклеазу I-Scel для индукции двухцепочечного разрыва (DSB) в уникальном положении, что позволило провести точную модификацию последовательности посредством гомологичной рекомбинации (HR) (Rouet Р, Smih F, Jasin М. Expression of a site-specific endonuclease stimulates homologous recombination in mammalian cells. Proc Natl Acad Sci USA. 1994 Jun 21; 91(13): 6064-8. doi: 10.1073/pnas.91.13.6064).
В дальнейшем методы генной терапии стали нацелены на использование генов для лечения генетических дефектов. Такой принцип редактирования заключается в восстановлении функции, утраченной в пораженных клетках больного, например, для лечения первичных иммунодефицитов, вызванных мутациями, инактивирующими ключевые гены в системе кроветворения. Клетки, полученные от пациента, могут быть выделены и трансдуцированы рекомбинантным вектором, модифицированным таким образом, что природные вирусные генетические элементы в геноме удаляются и заменяются кассетой экспрессии, кодирующей ген, который отсутствует в клетках пациента; это приводит к интеграции вирусного генома в геном хозяина, что приводит к экспрессии гена, содержащегося в вирусе. Модифицированные клетки можно использовать в качестве продукта генной терапии и трансплантировать обратно исходному пациенту в качестве терапии. Несмотря на то, что этот подход оказался успешным для лечения множественных иммунодефицитов, он основан на случайном механизме, с помощью которого вирусный геном интегрируется в клетку-хозяин. Это создает серьезные проблемы с безопасностью, поскольку интеграция с онкогенами или онкосупрессорами может привести к неконтролируемой пролиферации модифицированных клеток и, даже если это происходит редко, такие события могут в долгосрочной перспективе привести к возникновению рака.
В последнее десятилетие методы точной модификации собственных клеток пациента без использования случайной интеграции вирусных векторов становятся все более и более привлекательными. Точная модификация генома включает как интеграцию кассеты экспрессии гена в точное место генома-мишени, ранее подтвержденное как безопасное при модификации, так называемые «безопасные гавани», так и точное обращение мутации, вызывающей заболевание, к нормальной последовательности. Оба случая основаны на доставке в клетки фрагмента экзогенной ДНК (так называемой донорной ДНК), гомологичного сайту-мишени и содержащего либо интегрируемую экспрессионную кассету, либо нормальную последовательность для исправления основной мутации.
Восстановление целевых повреждений ДНК, может происходить путем негомологичного соединения концов (Nonhomology End joining, NHEJ). Чтобы обеспечить передачу генетической информации от экзогенной ДНК к точному местоположению изменяемого генома, клетки используют механизм гомологичной рекомбинации (HR), также называемый Homology Directed Repair (HDR), что приводит к удалению, добавлению или модификации генетической информации в клетках практически любого организма.
Обычно HR является очень редким явлением в клетках млекопитающих, но эта частота может быть увеличена, если двухцепочечный разрыв ДНК (DSB) происходит в целевом сайте и в то же время обеспечивается наличие гомологичной донорной ДНК. Таким образом, за последние 20 лет были предприняты огромные усилия для характеристики и улучшения платформ для создания DSB в определенных геномных местах, и это привело к введению в практику нуклеаз цинковых пальцев (ZFN); нуклеаз (TALEN) - эффектора, подобного активатору транскрипции; нуклеаз системы CRISPR-Cas - (РНК-управляемые нуклеазы (RGN)) (Beumer KJ, Carroll D. Targeted genome engineering techniques in Drosophila. Methods. 2014 Jun 15; 68(1): 29-37. doi: 10.1016/j.ymeth.2013.12.002).
Различные системы репарации DSB ДНК могут вступать в игру после расщепления целевой ДНК и определять характер редактирования генома. Классическое негомологичное соединение концов (cNHEJ) и микрогомологичное соединение концов (MMEJ) опосредуют лигирование концов ДНК и приводят к небольшим целевым, но незапрограммированным делециям/вставкам, которые позволяют эффективно инактивировать кодирующие последовательности генов.
При нормальном HDR в клетках гомологичная последовательность ДНК из сестринской хромосомы используется в качестве донорной матрицы для восстановления поврежденной хромосомы, содержащей DSB. В геномной инженерии восстановление DSB с помощью HDR включает добавление матрицы донорной ДНК. Центр экзогенной ДНК содержит желаемый трансген, тогда как фланги содержат сегменты гомологии с ДНК клетки. Донорная ДНК-матрица обычно разрабатывается таким образом, чтобы приводить к модификации конкретных нуклеотидов или вставке последовательности ДНК, такой как ген, в конкретное место генома, например, введение определенных последовательностей-вставок в область двуцепочечного разрыва DSB. Донорная ДНК вводится в клетку стандартными методами, такими как электропорация или трансфекция, опосредованная фосфатом кальция, и рекомбинирует в ДНК клетки, например, с помощью белков, способствующих рекомбинации в клетке.
Для многих приложений важно использовать HDR вместо NHEJ, потому что HDR опосредует модификацию или вставку нужных исследователю последовательностей ДНК в конкретном геномном месте, а не простое нарушение гена, как это обычно происходит с NHEJ. Однако существует конкуренция между использованием NHEJ и HDR во время репарации DSB и гомологичная рекомбинация HDR работает гораздо менее эффективно, чем NHEJ.
NHEJ и HDR представляют собой конкурирующие пути, которые содержат перекрывающиеся, но различные белковые компоненты. NHEJ значительно преобладает и в том случае, когда присутствует донорная ДНК, имеющая гомологичную последовательность для восстановления. Попытки блокировать NHEJ генетически или с помощью белковых и/или химических ингибиторов сдвигают баланс в пользу HDR, но могут привести к риску нежелательной нестабильности генома.
Альтернативный подход - стимулировать путь HDR. Действительно, умеренное улучшение эффективности HDR происходит за счет сверхэкспрессии / активации белков, специфичных для пути HDR, таких как RAD51, или слияния белков репарации ДНК с нуклеазой Cas9. В патенте WO 2020041172 A1 (патент WO 2020041172 A1. Cheng Albert, Jillette, Nathaniel. Methods and compositions for recruiting DNA repair proteins // 2020) авторы изобретения обнаружили, что белки, участвующие в гомологичной рекомбинации можно привязать к нуклеазе, например Cas9, с помощью слияния белка с РНК-связывающим доменом Pumilio-FBF (PUF) (Wang М, Ogé L, Perez-Garcia MD, Hamama L, Sakr S. The PUF Protein Family: Overview on PUF RNA Targets, Biological Functions, and Post Transcriptional Regulation. Int J Mol Sci. 2018 Jan 30; 19(2): 410. doi: 10.3390/ijms19020410). В свою очередь, такой домен имеет сайт связывания на gRNA, и таким образом, данная конструкция оказывается коньюгирована с нуклеазой Cas9. Авторы перечисляют большинство белков пути HR, например FEN-1 эндонуклеазу, EXO1 экзонуклеазу, RPA1-3 белки, связывающие одноцепочечную ДНК. Это позволило авторам увеличить HR, и в некоторых случаях одновременно уменьшить NHEJ.
В работе (Han Yang, Shuling Ren, Siyuan Yu, Haifeng Pan, Tingdong Li, Shengxiang Ge, Jun Zhang and Ningshao Xia. Methods favoring Homology-Directed Repair choice in response to CRISPR/Cas9 induced-double strand breaks. Int. J. Mol. Sci. 2020, 21, 6461; doi: 10.3390/ijms21186461) сообщалось, что имеются другие ортогональные стратегии, которые незначительно улучшают частоту HDR. К ним относятся: контроль клеточного цикла и экспрессии Cas9 или нарушение клеточного цикла, холодовой шок и привязка донорной матрицы ДНК для репарации к нуклеазе. Однако, несмотря на эти достижения, NHEJ остается преобладающим путем репарации. Таким образом, с дополнительной проблемой доставки как нуклеазы, так и репарационной матрицы в клетки in vivo, существует потребность в улучшении эффективности HDR.
Из-за низкой эффективности направленной геномной модификации на основе HDR вероятность события гомологичной репарации часто намного ниже желаемых клинически значимых частот. Таким образом, терапевтический потенциал точного редактирования генома остается неизученной возможностью. Современные стратегии повышения низкой скорости редактирования генов на основе HDR в основном основаны на использовании низкомолекулярных препаратов для блокирования клеток в фазе клеточного цикла S/G2, когда путь HDR наиболее активен, или для ингибирования NHEJ. Гомологичная рекомбинация (HR) активна только во время фаз S/G2 клеточного цикла, когда гомологичная матричная ДНК доступна для репарации.
Например, когда клетки синхронизированы в фазах S/G2, HDR может быть улучшен до 5 раз (Lin S, Staahl ВТ, Alia RK, Doudna JA. Enhanced homology-directed human genome engineering by controlled timing of CRISPR/Cas9 delivery. Elife. 2014 Dec 15; 3: e04766. doi: 10.7554/eLife.04766. PMID: 25497837; PMCID: PMC4383097.) (Yang D, Scavuzzo MA, Chmielowiec J, Sharp R, Bajic A, Borowiak M. Enrichment of G2/M cell cycle phase in human pluripotent stem cells enhances HDR-mediated gene repair with customizable endonucleases. Sci Rep. 2016 Feb 18; 6: 21264. doi: 10.1038/srep21264). Посредством индуцированной перетаскиванием синхронизации клеточного цикла и смогли добиться повышения (в 1,6-3,3 и 3-6 раз соответственно) HDR в клетках млекопитающих. Однако синхронизация клеток может быть сложной для выполнения и, в частности, может часто приводить к нежелательным нарушениям физиологических механизмов клеток. Кроме того, одним из основных недостатков этого метода является то, что синхронизация клеток может быть неприемлемой, когда клетки являются мишенями in vivo. С другой стороны, NHEJ может быть ингибирован посредством инактивации активности Ku70/80 (продукт гена XRCC6), и, следовательно, улучшает HDR (Gandía M, Xu S, Font С, Marcos JF. Disruption of ku70 involved in non-homologous end-joining facilitates homologous recombination but increases temperature sensitivity in the phytopathogenic fungus Penicillium digitatum. Fungal Biol. 2016 Mar; 120(3): 317-23. doi: 10.1016/j.funbio.2015.11.001). Чтобы способствовать HDR по сравнению с NHEJ, эффективность NHEJ была снижена за счет использования мутантных клеток, в которых отсутствует ДНК-лигаза IV, важный фермент в пути NHEJ. Потеря ДНК лигазы IV приводит к существенному увеличению HDR по сравнению с NHEJ у Drosophila (Beumer KJ, Trautman JK, Bozas A, Liu JL, Rutter J, Gall JG, Carroll D. Efficient gene targeting in Drosophila by direct embryo injection with zinc-finger nucleases. Proc Natl Acad Sci USA. 2008 Dec 16; 105(50): 19821-6. doi: 10.1073/pnas.0810475105). Однако для большинства приложений невозможно выбить ген, кодирующий ДНК-лигазу IV.
Обычно такие низкомолекулярные соединения оказывают широкое влияние на клеточную физиологию, и, хотя они могут быть эффективными в некоторых системах (в основном в системах in vitro), их значение для клинически ориентированных исследований может быть подорвано соображениями безопасности и неблагоприятным соотношением риск/польза.
Другой подход был разработан с использованием белка генимин (Geminin) (Gutschner Т, Haemmerle М, Genovese G, Draetta GF, Chin L. Post-translational Regulation of Cas9 during G1 Enhances Homology-Directed Repair. Cell Rep. 2016 Feb 16; 14(6): 1555-1566. doi: 10.1016/j.celrep.2016.01.019). Геминин - естественный субстрат комплекса АРС/Cdhl, который является основным клеточным циклом, контролирующим убиквитинлигазу Е3, и, следовательно, разлагается во время фазы Gl. Уменьшая экспрессию Cas9 в фазе G1 и увеличивая экспрессию Cas9 в фазах S, G2 и М клеточного цикла, исследователи смогли повысить эффективность HDR в 1,87 раза. Часть геминина была слита с каталитически активной нуклеазой Cas9 человека. При использовании комплекса нуклеаза Cas9 - Geminin слитый белок протеолизуется в поздних фазах М и Gl, в то время как во время фаз S/G2/М слитый белок накапливается. Следовательно, частота HDR повышается, а частота незапрограммированных мутаций, индуцированных NHEJ, снижается. Другими словами, этот подход основан на искусственной модуляции присутствия белка Cas9 в определенных фазах клеточного цикла.
Guo et al. (Guo Q, Mintier G, Ma-Edmonds M, Storton D, Wang X, Xiao X, Kienzle B, Zhao D, Feder JN. «Cold shock» increases the frequency of homology directed repair gene editing in induced pluripotent stem cells. Sci Rep. 2018 Feb 1; 8(1): 2080. doi: 10.1038/s41598-018-20358-5) обнаружили, что в условиях холодового шока при +32°С iPS-клетки увеличивают HDR в 1,8-3,6 раза. В том случае, когда базальный уровень HDR был очень низким (<0,2%), авторы наблюдали примерно 10-кратное увеличение эффективности HDR в клетках HEK293 при лечении холодовым шоком.
Альтернативная стратегия - использование небольших молекул (таких как NU7441 и Ku-0060648), которые ингибируют DNA-PK, одного из ключевых ферментов репарации по механизму NHEJ, приводило к 2-4-кратному увеличению HDR в клетках млекопитающих (Robert F, Barbeau М, Éthier S, Dostie J, Pelletier J. Pharmacological inhibition of DNA-PK stimulates Cas9-mediated genome editing. Genome Med. 2015 Aug 27; 7(1): 93. doi: 10.1186/s13073-015-0215-6).
Pinder et al., (Pinder J, Salsman J, Dellaire G. Nuclear domain «knock-in» screen for the evaluation and identification of small molecule enhancers of CRISPR-based genome editing. Nucleic Acids Res. 2015 Oct 30; 43(19): 9379-92. doi: 10.1093/nar/gkv993) показали 3-6-кратное увеличение HDR в клетках млекопитающих за счет сверхэкспрессии или стимуляции Rad51 с помощью небольшой молекулы RSI. В тоже время, Takayama K et al., (Takayama K, Igai K, Hagihara Y, Hashimoto R, Hanawa M, Sakuma T, Tachibana M, Sakurai F, Yamamoto T, Mizuguchi H. Highly efficient biallelic genome editing of human ES/iPS cells using a CRISPR/Cas9 or TALEN system. Nucleic Acids Res. 2017 May 19; 45(9): 5198-5207. doi: 10.1093/nar/gkx130) показали, что комбинация сверхэкспрессии Rad51 и лечения вальпроевой кислотой позволяет достичь более высоких уровней редактирования двуаллельных генов в клетках ES/iPS смогли.
Cas9 дикого типа содержит два нуклеазных домена, обозначенных как RuvC и HNH, каждый из которых разрезает разные цепи ДНК. Домен HNH надрезает цепь ДНК, которая комплементарна crРНК, a RuvC-подобный домен надрезает цепь, которая не комплементарна crRNA. Cas9 расщепляет ДНК на три пары оснований перед соседним мотивом протоспейсера (РАМ), что приводит к расщеплению ДНК по тупым концам. Расщепление ДНК вредно для вторгающейся плазмиды или вируса, приводя к деградации и защите от этих захватчиков. Двухцепочечный разрыв очень эффективен при деградации чужеродной ДНК, поскольку двухцепочечные разрывы, индуцированные Cas9, можно репарировать с помощью негомологичного соединения концов (NHEJ), что может легко привести к вставкам и/или делециям (инделы). Однако для редактирования генома NHEJ невыгоден.
Charpentier et al., (Charpentier M, Khedher AHY, Menoret S, Brion A, Lamribet K, Dardillac E, Boix C, Perrouault L, Tesson L, Geny S, De Cian A, Itier JM, Anegon I, Lopez B, Giovannangeli C, Concordet JP. CtIP fusion to Cas9 enhances transgene integration by homology-dependent repair. Nat Commun. 2018 Mar 19; 9(1): 1133. doi: 10.1038/s41467-018-03475-7) описывают слияние CtlP с Cas9 для усиления интеграции трансгена путем гомологически зависимой репарации (HDR). Они показали двукратное увеличение HDR за счет использования белка Cas9, слитого с фрагментом CtIP, который является ключевым белком на ранних этапах гомологичной рекомбинации. В патенте ЕР 3592852 А1 (Патент ЕР 3592852 А1 Ignacio Anegon, Marine Charpentier, Jean-Paul Concordet, Carine Giovannangeli, Bernard Lopez. Nuclease fusions for enhancing genome editing by homology-directed transgene integration // 2020) авторы изобретения обнаружили, что скорость HDR, опосредованного системой CRISPR/Cas9, может быть существенно улучшена путем предоставления нуклеазы Cas9 в форме слитого белка по крайней мере с N-концевым доменом белка CtIP. Белки CtIP участвуют в ранних стадиях гомологичной рекомбинации. CtIP это эндонуклеаза, которая связывается с комплексом белков MRE11-RAD50-NBN (MRN), участниками гомологичной репарации ДНК, и вовлечена в резекцию концов ДНК - первый шаг в репарации двуцепочечного разрыва через механизм гомологичной рекомбинации (HR). Кроме того, изобретатели идентифицировали субдомены N-концевого домена белка CtIP, которые важны для повышения уровня HDR. Таким образом, изобретение относится к гибридным белкам, содержащим белок Cas9, домен тетрамеризации белка CtIP и домен димеризации белка CtIP. В частности, изобретатели протестировали эти слитые белки клеток HEK293, клеток RG37DR и крыс Sprague-Dawley.
Патент ЕР 3730610 А1 (Патент ЕР 3730610 А1. Claudio Mussolino, Toni Cathomen, Tatjana Cornu, Antonio Carusillo. Modified Cas9 system and its use for improved gene editing // 2019) описывает платформу HDR-CRISPR. Эта платформа относится к модифицированной нуклеазе Cas9, содержащей значительную часть нуклеазы Cas9 и ковалентно связанной с ней по меньшей мере одним из белковых доменов фактора HDR, выбранного из группы, состоящей из RAD51, RAD52, RAD54, MRE11, PALB2, FANCD2 и EXO1. Данный патент охватывает физическое слияние активной нуклеазы Cas9 с одним или несколькими белками или белковыми доменами, участвующими в стимуляции репарации DSB посредством гомологичной рекомбинации. В качестве альтернативы, в патенте также предлагается возможность связать факторы HDR с нуклеазой Cas9 с помощью методов химического связывания. Линкеры, имеющие функциональные концы, которые могут реагировать с факторами HDR, с одной стороны, и с молекулой Cas9, с другой стороны, известны, и такое связывание может быть выгодным при манипуляциях с последовательностями генов в культурах клеток (in vitro).
В этом же патенте авторы рассмотрели возможность слияния Cas9 с белком CtIP. Они уточнили, что CtIP может быть добавлен в конструкцию только в дополнение по крайней мере к одному из факторов HDR, упомянутых выше. CtIP действует как кофактор для эндонуклеазы MRE11 при запуске резекции конца ДНК. В патенте описывается конструкция, содержащая один или несколько предпочтительных факторов HDR, RAD51, RAD52 и Mre11, в дополнение к CtIP. Предполагается, что за счет увеличения концентрации факторов, которые способствуют репарации DSB посредством HR именно в месте двухцепочечного разрыва, достигается повышенная частота HR-опосредованного редактирования генома.
В патенте WO 2020036653 A2 (Патент WO 2020036653 A2. James Kadonaga, Grisel Cruz Becerra. Improved method for homology directed repair in cells // 2019) показано, что эффективность HDR в клетках увеличивается за счет использования экзогенно поставляемой донорной матрицы хроматина, а не голой донорной матрицы ДНК, что не требует использования мутантных клеток, лечения лекарственными препаратами, холодовым шоком, не затрагивает клеточный цикл, регулируемый лекарственными средствами, или экспрессию или добавление экзогенного белка, не являющегося нуклеазой. Успех способа, вероятно, связан с использованием более естественной формы донорной матрицы в качестве хроматина, а не в качестве неестественной голой ДНК. Хроматин - это комплекс ДНК и белков, называемый гистонами ядра. В хроматине ДНК и гистоны организованы в частицы, известные как нуклеосомы. Нуклеосома представляет собой единичный повтор хроматина и состоит из примерно 180-200 п.н. ДНК и двух копий каждого из основных гистонов Н2А, Н2В, Н3 и Н4. При исследовании с помощью электронной микроскопии хроматин имеет вид «бусинок на нитке», где каждая из «бусинок» представляет собой нуклеосому.
Таким образом, предложен способ повышения эффективности гомологически направленной репарации (HDR) в клетках за счет использования донорной матрицы хроматина. Высокие показатели гибели клеток наблюдаются в трансфицированных клетках, подвергающихся редактированию генов с использованием донорной матрицы «голой» ДНК; однако данный способ значительно снижает гибель клеток и увеличивает эффективность HDR в 2-7 раз, что показано положительным эффектом донорных матриц хроматина (по сравнению с матрицами голой ДНК) при редактировании генома (например, HDR-CRISPR-опосредованное редактирование генома, отслеживаемое сайт-направленной вставкой репортерного гена в человеческие клетки MCF10A или HeLa). Другими словами, способ является улучшением производительности существующих методов редактирования генов. Таким образом, использование донорных матриц ДНК, собранных в хроматин, увеличивает скорость HDR для точного редактирования генома (например, при использовании системы CRISPR-Cas9) и увеличивает выживаемость клеток.
Основным недостатком современных инструментов геномной инженерии является отсутствие возможности контролировать переключение между механизмами репарации клеточной ДНК. В результате разрывы ДНК, созданные с помощью этих инструментов, восстанавливаются случайно с помощью NHEJ или HDR. Такой стохастический характер репарации часто приводит к бесполезному результату, который значительно снижает эффективность и точность процесса. Например, обращение вспять патогенного эффекта генетических вариаций, вызывающих заболевание, во многих случаях требует вставки элемента ДНК из внешнего источника в место разрыва. Эта деятельность опосредована HDR. Однако, используя доступные в настоящее время инструменты редактирования генома, большинство разрывов будет подвергнуто репарации через NHEJ, что является доминирующим процессом. В таких случаях NHEJ не только превосходит HDR в восстановлении начальных разрывов ДНК, но также может привести к мутации исходной последовательности.
Раскрытие сущности изобретения
Когда возникает DSB, клетки млекопитающих полагаются в основном на путь репарации ДНК негомологичного соединения концов (NHEJ), чтобы восстановить разрыв и выжить. Этот путь подвержен ошибкам и не приводит к включению генетической информации из экзогенной ДНК в сайт DSB. Даже при использовании высокоэффективных DN, HDR-опосредованное редактирование генов все еще крайне неэффективно, поскольку предпочтительна опосредованная NHEJ коррекция DSB, ведущая к небольшим мутациям в месте разрыва.
Этот дисбаланс между NHEJ и HR сильно затрудняет применимость точного редактирования генома как нового терапевтического варианта лечения заболеваний человека. Действительно, частота событий точного редактирования генома в гемопоэтических стволовых клетках, наиболее подходящем типе клеток в области генной терапии первичного иммунодефицита, сильно варьируется и намного ниже желаемого порога, который был бы значительным для достижения терапевтического эффекта у пациентов. Для дальнейшего увеличения низкой скорости редактирования генов на основе HR было принято несколько стратегий, которые обычно используют химические соединения для изменения физиологического баланса между NHEJ и HDR в пользу последнего. Несмотря на то, что эти стратегии показали себя многообещающими в некоторых клеточных системах, их применимость в клинически значимых условиях вызывает опасения из-за потенциальных побочных эффектов, связанных с их использованием. Следовательно, существует потребность в разработке новой платформы, чтобы сделать HR-опосредованное редактирование генома более эффективным и потенциально применимым в клиниках.
Настоящее изобретение посвящено способам клонирования молекул ДНК с повышенной частотой гомологичной рекомбинации между нуклеиновыми кислотами в клетке.
В настоящем описании представлено создание дополнительного модуля (химерного белка) для стандартных систем редактирования генома, состоящего из 5'-3'-flap-экзонуклеазы (ЕХО) и одноцепочечного ДНК-связывающего полипептида, SSB белка, способного стимулировать гомологичную рекомбинацию. Такие полипептиды, связывающие одноцепочечную ДНК, включают ssbT4 (продукт гена р32), ssbT5, ssbT7, а также функциональные фрагменты и варианты полипептидов, связывающих одноцепочечную ДНК. Такой модуль создается для индукции гомологичной рекомбинации с использованием защищенных одноцепочечных концов ДНК с помощью SSB белка, после направленного двуцепочечного разрыва ДНК, например Cas9, и последующей резекции ДНК с помощью 5'-3' flap-экзонуклеазы (ЕХО).
Один вариант реализации изобретения относится к слитому белку, содержащему по меньшей мере (а) 5'-3'-экзонуклеазу, например, flap-эндонуклеазу FEN1, (b) белок репарации ДНК, например, SSB белок - белок связывающий одноцепочечный конец ДНК, (с) доставку в клетку донорной нуклеиновой кислоты, содержащей интересующую нуклеотидную последовательность, кодирующую этот химерный продукт.
В другом варианте реализации изобретение также относится к нуклеиновой кислоте, кодирующей слитый белок, как определено здесь.
Еще один вариант реализации изобретения относится к созданию вектора нуклеиновой кислоты для экспрессии рекомбинантного белка, содержащему нуклеиновую кислоту, как описано здесь.
Еще один вариант реализации изобретение также относится к слитому белку, нуклеиновой кислоте, вектору нуклеиновой кислоты или частице для доставки, как описано здесь, для использования в качестве лекарственного средства.
В еще одном варианте реализации изобретение также относится к клетке-хозяину, содержащей слитый белок, нуклеиновую кислоту или вектор нуклеиновой кислоты, как описано здесь.
В некоторых вариантах реализации изобретения ДНК-нуклеаза, представляет собой нуклеазу Cas9 или никазу Cas9.
В некоторых вариантах реализации изобретения 5'-3'-flap-экзонуклеаза происходит из Т-четных бактериофагов, например, Т4 или Т-нечетных бактериофагов, например, Т5, Т7.
В некоторых вариантах реализации изобретения SSB белок происходит из Т-четных бактериофагов, например, Т4 или Т-нечетных бактериофагов, например, Т5, Т7.
В некоторых вариантах реализации изобретения SSB белок может происходить из генома млекопитающих RPA1, 2, 3.
Следует понимать, что 5'-3'-flap-экзонуклеаза может быть напрямую (например, слита) или косвенно связана (например, через, по меньшей мере одну, линкерную молекулу) с SSB белком аппарата гомологичной репарации ДНК.
Другие варианты реализации настоящего раскрытия относятся к белкам репарации ДНК, содержащим 5'-3'-flap-экзонуклеазу, связанную с SSB белками репарации ДНК, выбранным из группы, состоящей из: RPA1; RPA2; RPA3; RAD52; XRCC3.
Наконец, еще один вариант реализации изобретения также относится в виде дополнительного модуля к набору для редактирования генома по меньшей мере одной целевой клетки, содержащему:
- слитый белок, нуклеиновую кислоту, кодирующую, указанный слитый белок, вектор, несущий нуклеиновую кислоту, или частицу для доставки, содержащую указанный слитый белок, согласно приведенному здесь описанию; а также:
- одна или несколько сайт-специфичных направляющих РНК (gRNA) или вектор нуклеиновой кислоты для экспрессии одной или нескольких сайт-специфичных направляющих РНК (gRNA).
- ДНК-нуклеаза, которая представляет собой нуклеазу Cas9 или никазу Cas9, или вектор нуклеиновой кислоты для экспрессии ДНК-нуклеазы.
В дополнение к вставке последовательностей этот метод может использоваться для удаления нежелательных последовательностей, таких как повторяющиеся последовательности ДНК, которые встречаются во многих генах, связанных с различными заболеваниями, например ген хантингтина человека (HIT) (болезнь Хантингтона; HD) или человеческий ген фратаксина (FXN) (Атаксия Фридрейха; FRDA).
Краткое описание фигур и таблиц
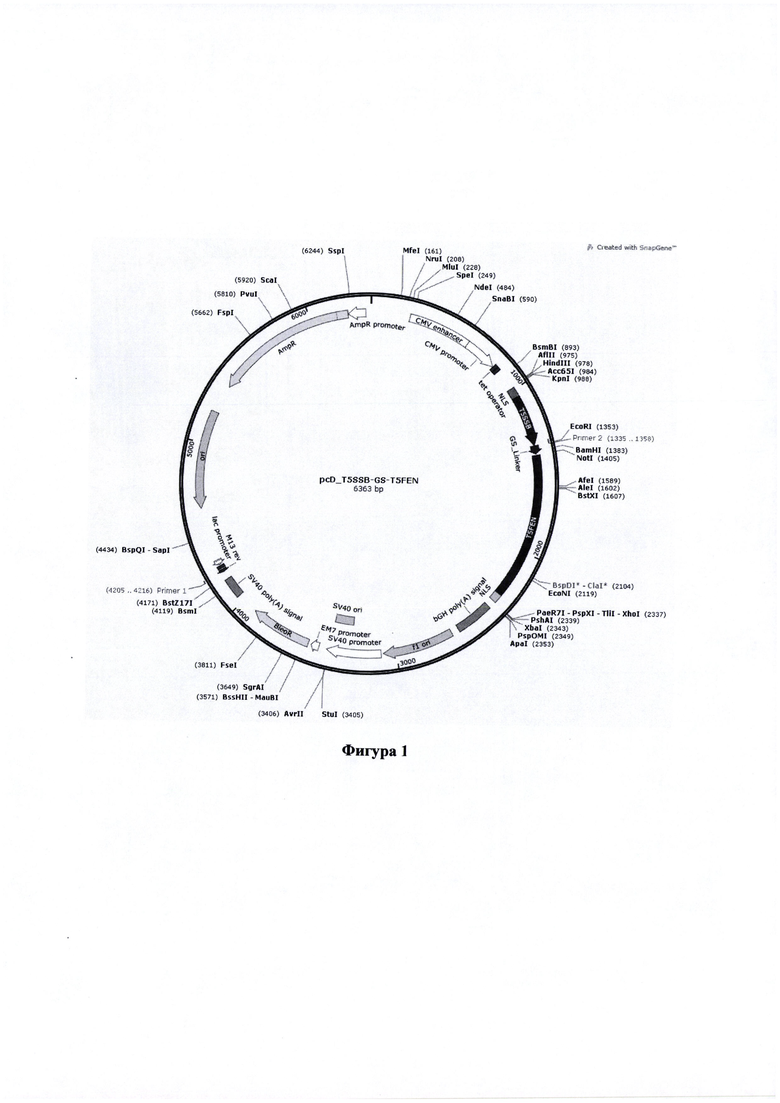
Фигура 1. Карта вектора pcD_T5SSB-GS-T5FEN, содержащего последовательность, кодирующую химерный белок - SSB белок и экзонуклеазу (ЕХО) аппарата гомологичной рекомбинации бактериофага Т5
Основные элементы конструкций:
NLS - сигнал ядерной локализации - nt 989-1046
T5SSB - nt 1047-1353
GS_Linker - nt 1356-1403
T5FEN - nt 1413-2285
NLS - nt 2286-2336
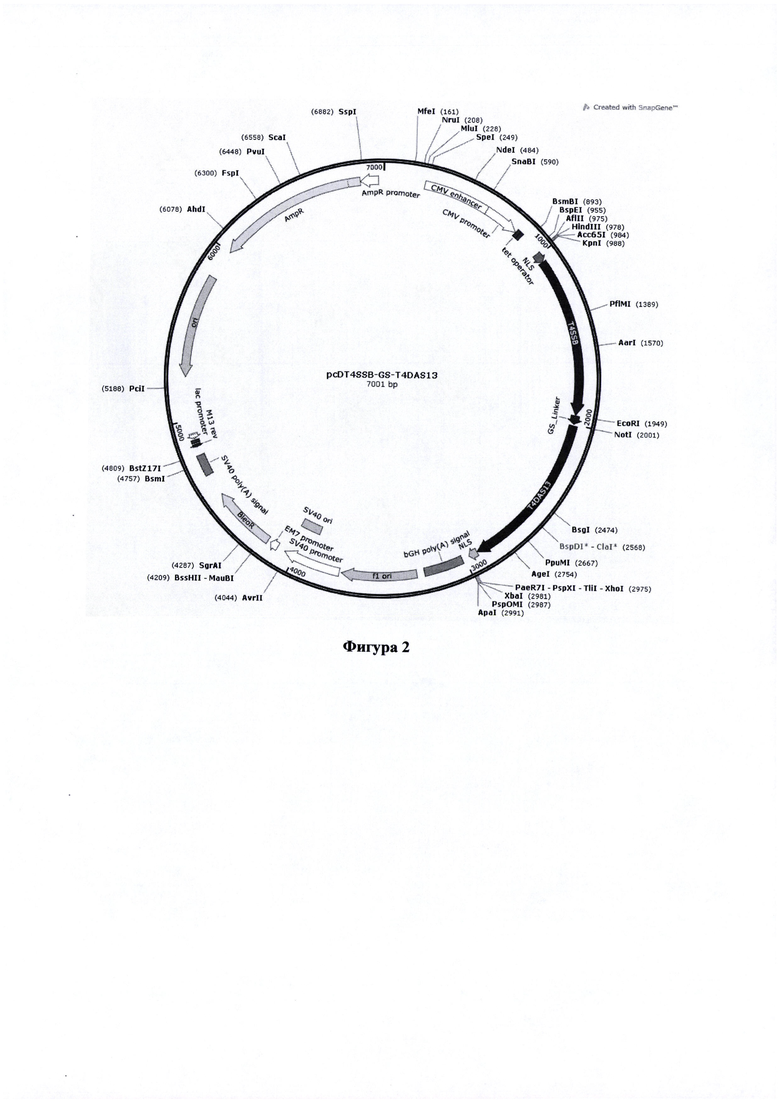
Фигура 2. Карта вектора pcD_T4SSB-GS-T4DAS13, содержащего последовательность, кодирующую химерный белок - SSB белок и экзонуклеазу (ЕХО) аппарата гомологичной рекомбинации бактериофага Т4.
Основные элементы конструкций:
T4SSB - nt 1046-1949
GS_Linker - nt 1952-1999
T4DAS13 - nt 2009-2923
NLS - nt 2924-2974
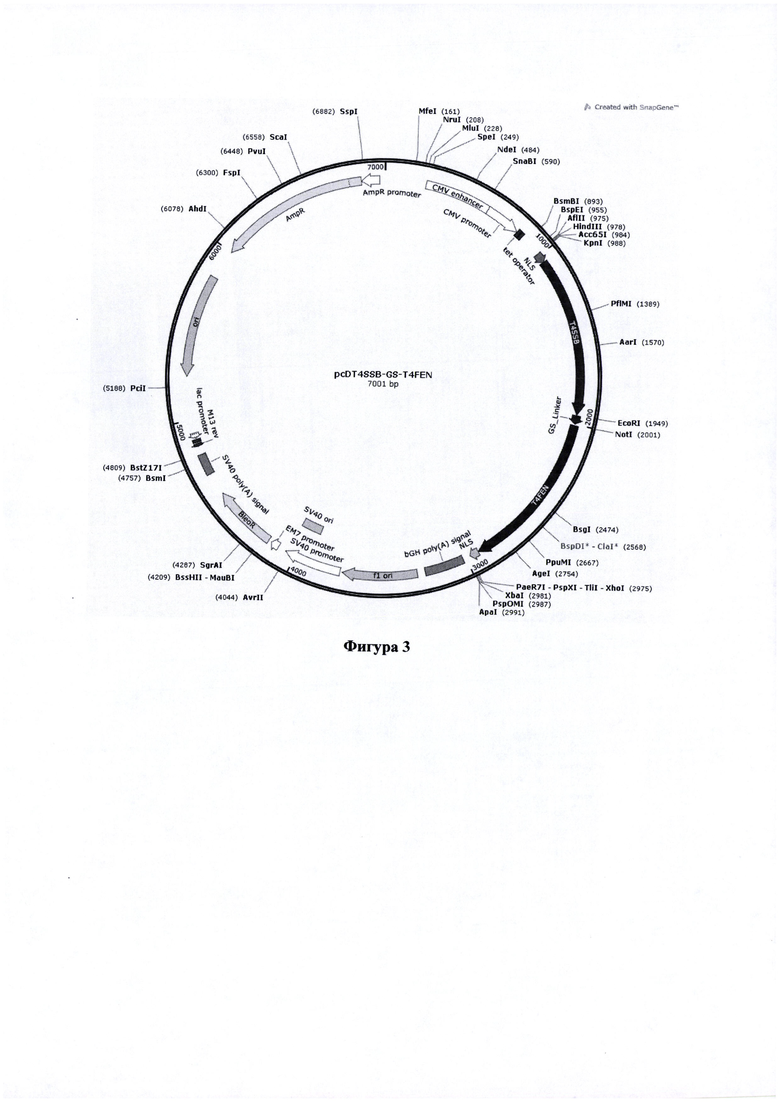
Фигура 3. Карта вектора pcD_T4SSB-GS-T4FEN, содержащего последовательность, кодирующую химерный белок - SSB белок и экзонуклеазу (ЕХО) аппарата гомологичной рекомбинации бактериофага Т4.
Основные элементы конструкций:
NLS - сигнал ядерной локализации - nt 989-1045
T4SSB - nt 1046-1949
GS_Linker - nt 1952-1999
T4FEN - nt 2009-2923
NLS - nt 2924-2974
Осуществление изобретения
Целью изобретения являлось создание способа одновременной резекции и защиты рекомбиногенных концов ДНК после направленного внесения двуцепочечного разрыва в последовательность ДНК-мишени для повышения уровня гомологичной рекомбинации. Данная технология может быть применена в качестве дополнительного модуля при редактировании генома эукариотической клетки.
Конструкции pcDT4SSB-GS-T4DAS13, pcDT4SSB-GS-T4FEN, pcDT5SSB-GS-T5FEN создавали на основе вектора pcDNA™4/TO Mammalian Expression Vector (ThermoFischer, USA) с применение стандартных молекулярно-биологических методов. На первом этапе была создана промежуточная конструкция, содержащая последовательность глицин-серинового полилинкера (Перечень последовательностей 1), клонированная в указанный вектор по сайтам рестрикции EcorI и NotI, расположенным в мультиклонинг сайте плазмидного вектора pcDNA™4/TO Mammalian Expression Vector. Для этого проводили обработку ферментами рестрикции EcorI (ThermoFischer, USA) и NotI (ThermoFischer, USA) указанного вектора и отжиг друг на друга олигонуклеотидных праймеров GS4_F и GS4_R (SEQ ID NO: 5 и SEQ ID NO: 6, соответственно, Таблица 1), затем проводили реакцию лигирования двух реакционных смесей с использованием Т4 DNA ligase (ThermoFischer, USA) при +4°С в течение ночи.
Кодон-оптимизированные ДНК последовательности, кодирующие SSB-белок бактериофага Т4, РС4 домен-содержащий SSB-белок бактериофага Т5 и белки экзонуклеаз системы гомологичной рекомбинации бактериофагов Т4 и Т5 (Ribonuclease Н, мутантная форма Ribonuclease Н, Flap endonuclease), были синтезированы и клонированы в плазмидный вектор pUC57 компанией Евроген (Россия).
Клонированные таким образом последовательности кодировали белки, соответствующие последовательностям, представленным в базе данных UniProt:
Идентификационный номер SSB-белка бактериофага Т4 (T4SSB): Р03695 (VHED_BPT4)
Идентификационный номер Рибонуклеазы Н бактериофага Т4 (T4FEN): Р13319 (RNH_BPT4)
Идентификационный номер мутантной формы рибонуклеазы Н бактериофага Т4 (T4DAS13): M1XLP1 (M1XLP1_BPT4)
Идентификационный номер РС4 домен-содержащего белка (РС4 domain-containing protein) бактериофага Т5 (T5SSB): Q6QGH2 (Q6QGH2_BPT5)
Идентификационный номер Flap endonuclease бактериофага Т5 (T5FEN): Р06229 (FEN_BPT5)
К последовательностям, кодирующим SSB-белок бактериофага Т4 и РС4 домен-содержащий белок бактериофага Т5 добавляли последовательность ядерной локализации. Для этого проводили ПЦР реакцию с использованием олигонуклеотидных праймеров: для гена, кодирующего SSB-белок фага Т4 (T4SSB) T4SSBCnlsRev (SEQ ID NO: 10) и CnlsinvF (SEQ ID NO: 8); для гена, кодирующего SSB-белок фага Т5 (T5SSB) T5SCnlsinR (SEQ ID NO: 11) и CnlsinvF (SEQ ID NO: 8) (Таблица 1). После проведения ПЦР продукты анализировали в 1% агарозном геле в 1х ТАЕ буфере с последующим выделением целевого продукта с помощью набора для очистки ДНК из агарозного геля и реакционных смесей (Евроген, Россия) согласно протоколу производителя. Далее полученные ПЦР-продукты и плазмидный вектор, содержащий глицин-сериновый полилинкер обрабатывали ферментами рестрикции NotI (NEB, USA) и XhoI (NEB, USA) при +37°C в течение 2 часов, и проводили независимое лигирование обработанных ферментами рестрикции ПЦР-продуктов T4SSB и T5SSB с плазмидным вектором pcDNA™4/TO, содержащим глицин-сериновый полилинкер. Реакцию проводили с использованием Т4 DNA лигазы (ThermoFischer, USA) согласно протоколу производителя. Продукты лигирования использовали для проведения трансформации компетентных клеток ТОР10 (ThermoFischer, USA). Отбор целевых клонов осуществляли методом рестриктного анализа с использованием ферментов рестрикции NotI (NEB, USA) и XhoI (NEB, USA). Таким образом были получены две конструкции:
1) pcDNA™4/TO, содержащие сигнал ядерной локализации, T4SSB и серин-глициновый полилинкер
2) pcDNA™4/TO, содержащие сигнал ядерной локализации, T5SSB и серин-глициновый полилинкер.
К последовательностям, кодирующим экзонуклеазы системы гомологичной рекомбинации бактериофага Т4 (5'-3'-flap эндонуклеазы, также называемой рибонуклеаза Н бактериофага Т4 (T4FEN)), ее мутантной формы Т4 (T4DAS13) и 5'-3'-flap эндонуклеазы бактериофага Т5 (T5FEN) добавляли последовательность ядерной локализации. Для этого проводили ПЦР реакцию с использованием олигонуклеотидных праймеров: для T4FEN и T4DAS13 были использованы олигонуклеотидные праймеры T4FnnlsinF и NnlsinvR (SEQ ID NO: 7 и SEQ ID NO: 18, соответственно, Таблица 1). Для 5'-3'-flap эндонуклеазы бактериофага Т5 (T5FEN) были использованы олигонуклеотидные праймеры T5FNnlsinF и NnlsinvR (SEQ ID NO: 9 и SEQ ID NO: 18, соответственно, Таблица 1). После проведения ПЦР продукты анализировали в 1% агарозном геле в 1х ТАЕ буфере с последующим выделением целевого продукта с помощью набора для очистки ДНК из агарозного геля и реакционных смесей (Евроген, Россия) согласно протоколу производителя. Далее полученные ПЦР-продукты и плазмидный вектор, содержащий глицин-сериновый полилинкер обрабатывали ферментами рестрикции EcorIR (NEB, USA) и KpnIF (NEB, USA) при +37°C в течение 2 часов. Проводили независимое лигирование обработанных ферментами рестрикции ПЦР-продуктов T4FEN и T4DAS13 сплазмидным вектором pcDNA™4/TO, содержащим T4SSB и глицин-сериновый полилинкер, а также лигирование обработанного ферментами рестрикции ПЦР-продукта T5FEN с плазмидным вектором pcDNA™4/TO, содержащим T5SSB и глицин-сериновый полилинкер, Т4 DNA лигазой (ThermoFischer, USA) согласно протоколу производителя. Продукты лигирования затем использовали для проведения трансформации компетентных клеток ТОР10 (ThermoFischer, USA). Отбор целевых клонов осуществляли методом рестриктного анализа с использованием ферментов рестрикции EcorIR (NEB, USA) и KpnIF (NEB, USA). Таким образом были получены три конструкции:
1) pcDNA™4/TO, содержащие сигнал ядерной локализации, T4SSB, серин-глициновый полилинкер, T4FEN и сигнал ядерной локализации (Фигура 3, Перечень последовательностей 3);
2) pcDNA™4/TO, содержащие сигнал ядерной локализации, T4SSB, серин-глициновый полилинкер, T4DAS13 и сигнал ядерной локализации (Фигура 2, Перечень последовательностей 2);
3) pcDNA™4/TO, содержащие сигнал ядерной локализации, T5SSB, серин-глициновый полилинкер, T5FEN и сигнал ядерной локализации (Фигура 1, Перечень последовательностей 4).
Далее изобретение будет проиллюстрировано примерами, которые предназначены для обеспечения лучшего понимания сущности заявленного изобретения, но не должны рассматриваться как ограничивающие данное изобретение.
ПРИМЕР 1
Кодон-оптимизированные ДНК последовательности, кодирующие SSB-белок бактериофага Т4, РС4 домен-содержащего S SB-белка бактериофага Т5 и экзонуклеазы системы гомологичной рекомбинации бактериофагов Т4 и Т5 (5'-3'flap эндонуклеазы и мутантная форма 5'-3'flap эндонуклеазы бактериофага Т4), были синтезированы и клонированы в плазмидный вектор pUC57 компанией Евроген (Россия).
Клонированные таким образом последовательности кодировали белки, соответствующие последовательностям, представленным в базе данных UniProt:
Идентификационный номер SSB-белка бактериофага Т4 (T4SSB): Р03695 (VHED_BPT4);
Идентификационный номер Рибонуклеазы Н бактериофага Т4 (T4FEN): Р13319 (RNH_BPT4);
Идентификационный номер мутантной формы рибонуклеазы Н бактериофага Т4 (T4DAS13): M1XLP1 (M1XLP1_BPT4);
Идентификационный номер РС4 домен-содержащего белка (РС4 domain-containing protein) бактериофага Т5 (T5SSB): Q6QGH2 (Q6QGH2_BPT5);
Идентификационный номер Flap endonuclease бактериофага Т5 (T5FEN): Р06229 (FEN_BPT5).
Конструкции создавали на основе вектора pcDNA™4/TO Mammalian Expression Vector (ThermoFischer, USA). Для этого была создана промежуточная конструкция pcDNA4/TO-4GS, включающая в себя последовательность глицин-серинового полилинкера (Gly-Gly-Ser-Gly)x3. Для этого были синтезированы два олигонуклеотидных праймера:
GS4_F (SEQ ID NO: 5) и GS4_R (SEQ ID NO: 6) (Таблица 1)
Указанные олигонуклеотидные праймеры отжигали друг на друга. Параметры отжига: 94°С - 2 минуты, затем ступенчатое снижение температуры до 25°С с шагом 5°С/минута. Использованное количество праймеров на реакцию - 100 мкмоль каждого.
Параллельно вектор обрабатывали ферментами рестрикции EcoRI и NotI (ThermoFischer, USA) согласно протоколу производителя. Продукты рестрикции далее подвергали электрофорезу в 1% агарозном геле в 1х ТАЕ буфере (ThermoFischer, USA) и выделяли из геля с использованием набора для очистки ДНК из агарозного геля и реакционных смесей (Евроген, Россия) согласно протоколу производителя. Далее производили лигирование двуцепочеченого фрагмента глицин-серинового полилинкера с обработанной ферментами ресктрикции EcorI и NotI плазмидой pcDNA4/TO (по 10 мкмоль каждого с использованием Т4 DNA лигазы (ThermoFischer, USA) при +4°С в течение ночи. Продукт лигирования далее трансформировали в компетентные клетки ТОР10 (ThermoFischer, USA). Для этого 10 мкл лигазной смеси на льду добавляли к 100 мкл объема компетентных клеток, проводили инкубацию в течение 20 минут при 0°С и проводили тепловой шок при +42°С в течение 2 минут. Далее к клеткам добавляли 1 мл среды LB и инкубировали при +37°С в течение одного часа при постоянном перемешивании, после чего клетки рассеивали на твердую среду LB-агар, содержащую антибиотик в концентрации 100 μг/мл, и инкубировали в течение ночи при +37°С. Колонии клеток пересеивали в 3 мл жидкой среды LB, содержащую антибиотик в концентрации 100 мкг/мл, и инкубировали с потряхиванием в течение ночи при +37°С. Далее производили выделение плазмидной ДНК с использованием набора для выделения плазмидной ДНК (Евроген, Россия) согласно протоколу производителя. Анализ наличия целевого фрагмента, кодирующего глицин-сериновый полилинкер производили с помощью рестриктного анализа. Для этого плазмидные ДНК обрабатывали ферментами рестрикции EcoRI и NotI (ThermoFischer, USA) согласно протоколу производителя и анализировали результаты на 1% агарозном геле в 1х буфере ТАЕ (ThermoFischer, USA). Отбор целевых плазмид осуществляли по наличию фрагмента ДНК ожидаемой длины после реакции рестрикции.
На 5'-концы используемых последовательностей экзонуклеаз и на 3'-концы использованных SSB-белков добавляли последовательность ядерной локализации. Для этого производили ПЦР с использованием следующих олигонуклеотидных праймеров: для добавления последовательности ядерной локализации к экзонуклеазам фага Т4 (T4FEN и T4DAS13) - T4FnnlsinF (SEQ ID NO: 7) и NnlsinvR (SEQ ID NO: 18) (Таблица 1)
Для экзонуклеазы фага Т5 - T5FNnlsinF (SEQ ID NO: 9) и NnlsinvR (SEQ ID NO: 18) (Таблица 1)
Для SSB фага T4 (T4SSB) - T4SSBCnlsRev (SEQ ID NO: 10) и CnlsinvF (SEQ ID NO: 8) (Таблица 1)
Для SSB фага T5 (T5SSB) - T5SCnlsinR (SEQ ID NO: 11) и CnlsinvF (SEQ ID NO: 8) (Таблица 1)
ПЦР-реакцию проводили с использованием полимеразы Q5 (NEB, USA) согласно протоколу производителя. Параметры амплификации: +98°С - 2 минуты, +98°С - 30 секунд, +60°С - 15 секунд, +72°С - 4 минуты, количество циклов - 25. Продукты амплификации анализировали с использованием 1% агарозного геля. Целевой продукт амплификации вырезали из геля и выделяли с помощью набора для очистки ДНК из агарозного геля и реакционных смесей (Евроген, Россия). Выделенный фрагмент ДНК далее обрабатывали Т4 полинуклеотидой киназой (NEB, USA) согласно протоколу производителя и производили очистку набором для очистки ДНК из агарозного геля и реакционных смесей (Евроген, Россия) и проводили лигирование с использованием Т4 DNA лигазой (ThermoFischer, USA) при +24°С в течение ночи. Продукт лигирования трансформировали в компетентные клетки ТОР10 (ThermoFischer, USA)). Для этого 10 мкл лигазной смеси на льду добавляли к 100 мкл объема компетентных клеток, проводили инкубацию в течение 20 минут при 0°С и проводили тепловой шок при +42°С в течение 2 минут. Далее к клеткам добавляли 1 мл среды LB и инкубировали при +37°С в течение одного часа при постоянном перемешивании, после чего клетки рассеивали на твердую среду LB-агар, содержащую антибиотик ампицилин в концентрации 100 мкг/мл, и инкубировали в течение ночи при +37°С. Колонии клеток пересеивали в 3 мл жидкой среды LB, содержащую антибиотик в концентрации 100 мкг/мл, и инкубировали с потряхиванием в течение ночи при +37°С. Далее производили выделение плазмидной ДНК с использованием набора для выделения плазмидной ДНК (Евроген, Россия) согласно протоколу производителя. Далее, полученные таким образом плазмидные ДНК использовали в качестве матрицы для ПЦР. Для этого были использованы следующие олигонуклеотидные праймеры: для экзонуклеаз фага T4FEN, T4DAS13 - T4FEcorIR (SEQ ID NO: 12) и NnlsKpnIF (SEQ ID NO: 13) (Таблица 1);
Для экзонуклеазы фага Т5 (T5FEN) - T5FEcoRIR (SEQ ID NO: 14) и NnlsKpnIF (SEQ ID NO: 13) (Таблица 1);
Для SSB фага T4 (T4SSB) - T4SNotIF (SEQ ID NO: 15) и CnlsXhoIR (SEQ ID NO: 16) (Таблица 1);
Для SSB фага T5 (T5SSB) - T5SNotIF (SEQ ID NO: 17) и CnlsXhoIR (SEQ ID NO: 16) (Таблица 1).
ПЦР-реакцию проводили с использованием полимеразы Q5 (NEB, USA). Параметры амплификации: +98°С - 2 минуты, +98°С - 30 секунд, +60°С - 15 секунд, +72°С - 4 минуты, количество циклов - 27. Продукты амлификации анализировали с использованием 1% агарозного геля. Целевой продукт амплификации вырезали из геля и выделяли с использованием набора для очистки ДНК из агарозного геля и реакционных смесей (Евроген, Россия). Выделенный фрагменты ДНК далее обрабатывали ферментами рестрикции:
Для экзонуклеаз фага T4FEN, T4DAS13:
EcorIR (NEB, USA) и KpnIF (NEB, USA) при +37°C в течение 2 часов.
Для экзонуклеазы фага Т5 (T5FEN):
EcorIR (NEB, USA) и KpnIF (NEB, USA) при +37°C в течение 2 часов.
Для SSB фага Т4 (T4SSB):
NotI (NEB, USA) и XhoI (NEB, USA) при +37°C в течение 2 часов.
Для SSB фага Т5 (T5SSB):
NotI (NEB, USA) и XhoI (NEB, USA) при +37°C в течение 2 часов.
ПРИМЕР 2.
Обработку вектора pcDNA4/TO-4GS производили ферментами рестрикции EcorIR (NEB, USA) и KpnIF (NEB, USA) при +37°C в течение 2 часов. Продукты рестрикции подвергали электрофорезу в 1% агарозном геле в 1х ТАЕ буфере (ThermoFischer, USA) и выделяли из геля с использованием набора для очистки ДНК из агарозного геля и реакционных смесей (Евроген, Россия) согласно протоколу производителя и, после чего проводили независимое лигирование обработанных ферментами рестрикции EcoRI и KpnI ПЦР продуктов T4FEN и T4DAS и обработанного ферментами рестрикции EcoRI и KpnI вектора pcDNA4\TO-4GS с использованием Т4 DNA лигазы (ThermoFischer, USA) при +24°С в течение ночи. Продукт лигирования далее трансформировали в компетентные клетки ТОР (ThermoFischer, USA). Для этого 10 мкл лигазной смеси на льду добавляли к 100 мкл объема компетентных клеток, проводили инкубацию в течение 20 минут и проводили тепловой шок при +42°С в течение 2 минут, затем к клеткам добавляли 1 мл среды LB и инкубировали при +37°С в течение одного часа с перемешиванием. Далее клетки рассеивали на твердую среду LB-агар, содержащую антибиотик в концентрации 100 мкг/мл и инкубировали в течение ночи при +37°С. Колонии клеток затем пересеивали в 3 мл жидкой среды LB, содержащую антибиотик ампицилин в концентрации 100 мкг/мл, и инкубировали с потряхиванием в течение ночи при +37°С. Далее производили выделение плазмидной ДНК с использованием набора для выделения плазмидной ДНК (Евроген, Россия) согласно протоколу производителя. Наличие целевого фрагмента проводили с помощью рестриктного анализа с использованием ферментов рестрикции EcoRI и KpnI и идентификацией полученных продуктов в 1%-агарозном геле.
Далее полученные вектора pcDNA4\TO-GS, содержащие последовательность T4FEN (pcDNA4/TO-T4FEN-GS), T4DAS13 (pcDNA4\TO-T4DAS13-GS), T5FEN (pcDNA4/TO-T5FEN-GS) обрабатывали ферментами рестрикции NotI и XhoI (NEB, USA) при +37°С в течение 2 часов. Продукты рестрикции подвергали электрофорезу в 1% агарозном геле в 1х ТАЕ буфере (ThermoFischer, USA) и выделяли из геля с использованием набора для очистки ДНК из агарозного геля и реакционных смесей (Евроген, Россия) согласно протоколу производителя и проводили лигирование:
Вектор pcDNA4\TO-T4FEN-GS лигировали с обработанным ферментами рестрикции продуктом T4SSB.
Вектор pcDNA4\TO-T4DAS13-GS лигировали с обработанным ферментами рестрикции продуктом T4SSB.
Вектор pcDNA4\TO-T5FEN-GS лигировали с обработанным ферментами рестрикции продуктом T5SSB.
Лигирование проводили при +24°С в течение ночи с использованием Т4 DNA лигазы (ThermoFischer, USA).
Продукт лигирования далее трансформировали в компетентные клетки ТОР10 (ThermoFischer, USA). Для этого 10 мкл лигазной смеси на льду добавляли к 100 мкл объема компетентных клеток, проводили инкубацию в течение 20 минут и проводили тепловой шок при +42°С в течение 2 минут. К клеткам далее добавляли 1 мл среды LB и инкубировали при +37°С в течение одного часа с перемешиванием. Далее клетки рассеивали на твердую среду LB-агар и инкубировали в течение ночи при 37°С. Колонии клеток затем пересеивали в 3 мл жидкой среды LB и инкубировали с потряхиванием в течение ночи при +37°С, затем производили выделение плазмидной ДНК с использованием набора для выделения плазмидной ДНК (Евроген, Россия) согласно протоколу производителя. Анализ наличия целевого фрагмента проводили с помощью рестриктного анализа с использованием ферментов рестрикции EcoRI и KpnI и анализом в 1%-агарозном геле.
Полученные таким образом конструкции pcDT5FEN-GS-T5SSB (Фигура 1), pcD_T4SSB-GS-T4DAS13 (Фигура 2), pcDT4FEN-GS-T4SSB (Фигура 3) далее секвенировали на автоматическом секвенаторе. Секвенирование ДНК проводили с помощью набора реактивов ABI PRISM BigDye Terminator v. 3.1 с анализом на автоматическом секвенаторе ДНК ABI PRISM 3100-AVANT в ЦКП «ГЕНОМ» ИМБ РАН.

--->
ГЕНЕТИЧЕСКИЕ КОНСТРУКЦИИ pcDEXO-GS-SSB, СОДЕРЖАЩИЕ ПОСЛЕДОВАТЕЛЬНОСТИ, КОДИРУЮЩИЕ
ХИМЕРНЫЕ БЕЛКИ SSB И ЭКЗОНУКЛЕАЗЫ АППАРАТА ГОМОЛОГИЧНОЙ РЕКОМБИНАЦИИ БАКТЕРИОФАГОВ,
ОБЪЕДИНЕННЫЕ ЧЕРЕЗ ГЛИЦИН-СЕРИНОВЫЙ ПОЛИЛИНКЕР
<210> 1
<211> 48
<212> Глицин-сериновый спейсер (Gly-Gly-Ser-Gly) x3
<213> Искусственная последовательность
<220>
<223>
<400> 1
ttcggtggag gcggttcagg cggaggtgga tccggcggtg gcggatcg (SEQ ID NO: 1)
<210> 2
<211> 2176
<212> NLS - T4SSB - GS-linker - T4DAS13 – NLS
<213> Искусственная последовательность
<220>
<223> Увеличение вероятности HDR при трансфекции
<400> 2
atggccccaa agaagaagcg gaaggtcggt atccacggag tcccagcagc atgttcaagc ggaagagcac
cgcagagctg gcagcccaga tggccaaact gaacggcaac aagggcttca gcagcgagga caaaggcgag
tggaagctga agctggacaa cgcaggcaac ggacaggccg tgatcagatt cctgccctct aagaacgacg
agcaggcccc tttcgccatc ctggtgaacc acggcttcaa gaagaacggc aagtggtaca tcgagacttg
cagcagcacc cacggcgatt acgactcttg tcccgtctgc cagtacatca gcaagaacga cctgtataac
accgacaaca aggagtacag cctggtgaag cggaagacca gctattgggc caacatcctc gtggtgaagg
atccagccgc cccagaaaac gagggcaagg tgttcaagta ccgcttcggc aagaagattt gggacaagat
caacgccatg atcgccgtgg acgtggaaat gggagagacc ccagtggacg tgacttgtcc ttgggaggga
gccaacttcg tgctgaaggt caagcaggtg tccggcttca gcaactacga cgagagcaag ttcctgaacc
agagcgccat ccccaacatc gacgacgaga gcttccagaa ggagctgttc gagcagatgg tggacctgag
cgagatgacc agcaaggaca agttcaagag cttcgaggag ctgaacacca agttcgggca ggtcatgggc
acagcagtga tgggaggagc agcagccaca gccgctaaga aggccgataa agtggccgac gacctggacg
ccttcaacgt ggacgacttc aacaccaaga ccgaggacga cttcatgagc agcagcagcg gctcttcttc
tagcgccgat gacaccgacc tggacgatct gctgaacgac ctggaattcg gtggaggcgg ttcaggcgga
ggtggatccg gcggtggcgg atcggcggcc gctatggacc tggagatgat gctggacgag gactacaagg
agggcatttg cctgatcgac ttcagccaga tcgctctgtc tacagccctg gtgaacttcc ccgacaagga
gaagatcaac ctgagcatga tccggcacct gatcctgaac agcatcaagt tcaacgtgaa gaaggccaag
accctgggct acaccaagat cgtcctctgc atcgacaacg ccaagagcgg atattggagg cgggacttcg
cctactacta caagaagaac cggggcaagg ccagagagga gagcacttgg gattgggagg gctacttcga
gagcagccac aaggtcatcg acgagctgaa ggcctacatg ccctacatcg tgatggacat cgacaagtac
gaggccgacg atcacattgc cgtgctggtg aagaagttca gcctggaggg ccacaagatc ctgatcatca
gcagcgacgg cgacttcacc cagctgcaca agtaccccaa cgtcaagcag tggagcccca tgcacaagaa
gtgggtgaag atcaagagcg gcagcgccga gatcgattgc atgaccaaga tcctgaaggg cgacaagaag
gacaacgtgg ccagcgtgaa agtgcgctca gacttctggt tcaccagggt ggagggcgaa aggaccccta
gcatgaagac cagcatcgtg gaggccatcg ccaacgatag ggagcaggcc aaagtgatcc tgaccgagag
cgagtacaac cggtacaagg agaacctggt gctgatcgac ttcgactaca tccccgacaa catcgccagc
aacatcgtga actactacaa cagctacaag ctgcccccca ggggcaagat ctacagctac ttcgtgaagg
ccggcctgag caagctgaca aacagcatca acgagttcaa aaggccggcg gccacgaaaa agccggccag
gcaaaaaaga aaaagttga (SEQ ID NO: 2)
<210> 3
<211> 2176
<212> NLS - T4SSB - GS-linker - T4FEN - NLS
<213> Искусственная последовательность
<220>
<223> Увеличение вероятности HDR при трансфекции
<400> 3
atggccccaa agaagaagcg gaaggtcggt atccacggag tcccagcagc atgttcaagc ggaagagcac
cgcagagctg gcagcccaga tggccaaact gaacggcaac aagggcttca gcagcgagga caaaggcgag
tggaagctga agctggacaa cgcaggcaac ggacaggccg tgatcagatt cctgccctct aagaacgacg
agcaggcccc tttcgccatc ctggtgaacc acggcttcaa gaagaacggc aagtggtaca tcgagacttg
cagcagcacc cacggcgatt acgactcttg tcccgtctgc cagtacatca gcaagaacga cctgtataac
accgacaaca aggagtacag cctggtgaag cggaagacca gctattgggc caacatcctc gtggtgaagg
atccagccgc cccagaaaac gagggcaagg tgttcaagta ccgcttcggc aagaagattt gggacaagat
caacgccatg atcgccgtgg acgtggaaat gggagagacc ccagtggacg tgacttgtcc ttgggaggga
gccaacttcg tgctgaaggt caagcaggtg tccggcttca gcaactacga cgagagcaag ttcctgaacc
agagcgccat ccccaacatc gacgacgaga gcttccagaa ggagctgttc gagcagatgg tggacctgag
cgagatgacc agcaaggaca agttcaagag cttcgaggag ctgaacacca agttcgggca ggtcatgggc
acagcagtga tgggaggagc agcagccaca gccgctaaga aggccgataa agtggccgac gacctggacg
ccttcaacgt ggacgacttc aacaccaaga ccgaggacga cttcatgagc agcagcagcg gctcttcttc
tagcgccgat gacaccgacc tggacgatct gctgaacgac ctggaattcg gtggaggcgg ttcaggcgga
ggtggatccg gcggtggcgg atcggcggcc gctatggacc tggagatgat gctggacgag gactacaagg
agggcatttg cctgatcgac ttcagccaga tcgctctgtc tacagccctg gtgaacttcc ccgacaagga
gaagatcaac ctgagcatgg tccggcacct gatcctgaac agcatcaagt tcaacgtgaa gaaggccaag
accctgggct acaccaagat cgtcctctgc atcgacaacg ccaagagcgg atattggagg cgggacttcg
cctactacta caagaagaac cggggcaagg ccagagagga gagcacttgg gattgggagg gctacttcga
gagcagccac aaggtcatcg acgagctgaa ggcctacatg ccctacatcg tgatggacat cgacaagtac
gaggccgacg atcacattgc cgtgctggtg aagaagttca gcctggaggg ccacaagatc ctgatcatca
gcagcgacgg cgacttcacc cagctgcaca agtaccccaa cgtcaagcag tggagcccca tgcacaagaa
gtgggtgaag atcaagagcg gcagcgccga gatcgattgc atgaccaaga tcctgaaggg cgacaagaag
gacaacgtgg ccagcgtgaa agtgcgctca gacttctggt tcaccagggt ggagggcgaa aggaccccta
gcatgaagac cagcatcgtg gaggccatcg ccaacgatag ggaacaggcc aaggtgctgc tgaccgaaag
cgagtacaac cggtacaag gagaacctgg tgctgatcga cttcgactac atccccgaca acatcgccag
caacatcgtg aactactaca acagctacaa gctgcccccc aggggcaaga tctacagcta cttcgtgaag
gccggcctga gcaagctgac aaacagcatc aacgagttca aaaggccggc ggccacgaaa aagccggcca
ggcaaaaaag aaaaagttga (SEQ ID NO: 3)
<210> 4
<211> 1473
<212> NLS - T5SSB - GS-linker - T5FEN - NLS
<213> Искусственная последовательность
<220>
<223> Увеличение вероятности HDR при трансфекции
<400> 4
atggccccaa agaagaagcg gaaggtcggt atccacggag tcccagcagc atgagcgagc aggtcaacca
gaactacgag ggccacgtgg acgatcagag catcatcctc tgggagaagg agggagaaca ggtccgactg
accgtgtcag agttccgggg caacctgtat atgggcatcc gctattggct gctggacatc aacgacgagt
ggttccccac caagtccgga ttcagcttcc cctacaccct ggagaccacc agccagctgt tctacgcctt
cacccagatc ctgagcgaga gcgaagtgct gcacgaagtg cagaagaggg ccgaggaact gaaggccaag
aacgccgaat tcggtggagg cggttcaggc ggaggtggat ccggcggtgg cggatcggcg gccgctatga
gcaagtcttg gggcaagttc atcgaggagg aggaagccga gatggccagc agaagaaacc tgatgatcgt
ggacggcacc aacctgggct tccggttcaa gcacaacaac agcaagaagc ccttcgccag cagctacgtg
tctaccatcc agagcctggc caagagctac agcgctagaa ccaccatcgt gctgggcgat aagggcaaga
gcgtgttcag gctggagcac ctgccagagt acaagggcaa ccgggacgag aagtacgctc agagaaccga
agaggagaag gccctggacg agcagttctt cgagtacctc aaggacgcct tcgagctctg caagaccacc
ttccccacct tcaccatcag gggagtggaa gccgacgaca tggcagccta catcgtgaag ctgatcggcc
acctgtacga ccacgtctgg ctgatcagca ccgacggaga ttgggacaca ctgctgaccg acaaggtgtc
ccggttcagc ttcaccacca ggagagagta ccacctgcgg gacatgtacg agcaccacaa cgtggacgac
gtggagcagt tcatcagcct gaaggccatc atgggcgacc tgggagacaa catcagaggc gtggagggca
tcggagccaa gaggggctac aacatcatcc gggagttcgg gaacgtgctg gacatcatcg atcagctgcc
tctgccaggc aagcagaagt acatccagaa cctgaacgcc agcgaggagc tgctgttccg gaacctgatc
ctggtggacc tgcccaccta ttgcgtggac gctatcgcag cagtgggaca ggacgtgctg gataagttca
ccaaggacat cctggagatc gccgagcaga aaaggccggc ggccacgaaa aagccggcca ggcaaaaaag
aaaaagttga (SEQ ID NO: 4)
<210> 5
<211> 52
<212> GS4_F
<213> Искусственная последовательность
<220>
<223> праймер прямой
<400> 5
aattcggtgg aggcggttca ggcggaggtg gatccggcgg tggcggatcg gc (SEQ ID NO: 5)
<210> 6
<211> 52
<212> GS4_R
<213> Искусственная последовательность
<220>
<223> праймер обратный
<400> 6
ggccgccgat ccgccaccgc cggatccacc tccgcctgaa ccgcctccac cg (SEQ ID NO: 6)
<210> 7
<211> 49
<212> T4FnnlsinF
<213> Искусственная последовательность
<220>
<223> праймер прямой
<400> 7
ggtcggtatc cacggagtcc cagcagccat ggacctggag atgatgctg (SEQ ID NO: 7)
<210> 8
<211> 47
<212> CnlsinvF
<213> Искусственная последовательность
<220>
<223> праймер прямой
<400> 8
gccggccagg caaaaaagaa aaagtgaggc tcgagcatgg tcatagc (SEQ ID NO: 8)
<210> 9
<211> 46
<212> T5FNnlsinF
<213> Искусственная последовательность
<220>
<223> праймер прямой
<400> 9
ggtcggtatc cacggagtcc cagcagccat gagcaagtct tggggc (SEQ ID NO: 9)
<210> 10
<211> 46
<212> T4SSBCnlsRev
<213> Искусственная последовательность
<220>
<223> праймер обратный
<400> 10
cttttttcgt ggccgccggc cttttcaggt cgttcagcag atcgtc (SEQ ID NO: 10)
<210> 11
<211> 44
<212> T5SCnlsinR
<213> Искусственная последовательность
<220>
<223> праймер обратный
<400> 11
ctttttcgtg gccgccggcc ttttggcgtt cttggccttc agtt (SEQ ID NO: 11)
<210> 12
<211> 43
<212> T4FEcorIR
<213> Искусственная последовательность
<220>
<223> праймер обратный
<400> 12
tttgaattcg aactcgttga tgctgtttgt cag (SEQ ID NO: 12)
<210> 13
<211> 31
<212> NnlsKpnIF
<213> Искусственная последовательность
<220>
<223> праймер прямой
<400> 13
TTTGGTACCG CCACCATGGC CCCAAAGAAG A (SEQ ID NO: 13)
<210> 14
<211> 29
<212> T5FEcorIR
<213> Искусственная последовательность
<220>
<223> праймер обратный
<400> 14
tttgaattcc tgctcggcga tctccagga (SEQ ID NO: 14)
<210> 15
<211> 29
<212> T4SNotIF
<213> Искусственная последовательность
<220>
<223> праймер обратный
<400> 15
tttgaattcc tgctcggcga tctccagga (SEQ ID NO: 15)
<210> 16
<211> 34
<212> CnlsXhoIR
<213> Искусственная последовательность
<220>
<223> праймер обратный
<400> 16
tttctcgagt cactttttct tttttgcctg gccg (SEQ ID NO: 16)
<210> 17
<211> 32
<212> T5SNotIF
<213> Искусственная последовательность
<220>
<223> праймер прямой
<400> 17
tttgcggccg ctatgagcga gcaggtcaac ca (SEQ ID NO: 17)
<210> 18
<211> 46
<212> NnlsinvR
<213> Искусственная последовательность
<220>
<223> праймер обратный
<400> 18
ttccgcttct tctttggggc catggtggca ctggccgtcg ttttac (SEQ ID NO: 18)
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ЛОКУСЫ ФУНКЦИОНАЛЬНОСТИ FAD2 И СООТВЕТСТВУЮЩИЕ СПЕЦИФИЧНЫЕ К УЧАСТКУ-МИШЕНИ СВЯЗЫВАЮЩИЕ БЕЛКИ, СПОСОБНЫЕ ИНДУЦИРОВАТЬ НАПРАВЛЕННЫЕ РАЗРЫВЫ | 2013 |
|
RU2656159C2 |
| Средство редактирования генома на основе белка LigD из бактерии Pseudomonas putida и Cas9 комплекса | 2022 |
|
RU2797049C1 |
| ЛОКУСЫ FAD3 ДЛЯ ВЫПОЛНЕНИЯ ОПЕРАЦИЙ И СООТВЕТСТВУЮЩИЕ СВЯЗЫВАЮЩИЕСЯ СО СПЕЦИФИЧЕСКИМИ САЙТАМИ-МИШЕНЯМИ БЕЛКИ, СПОСОБНЫЕ К ВЫЗОВУ НАПРАВЛЕННЫХ РАЗРЫВОВ | 2013 |
|
RU2665811C2 |
| РЕДАКТИРОВАНИЕ ГЕНОВ С ИСПОЛЬЗОВАНИЕМ МОДИФИЦИРОВАННОЙ ДНК С ЗАМКНУТЫМИ КОНЦАМИ (зкДНК) | 2018 |
|
RU2811724C2 |
| СКОНСТРУИРОВАННАЯ СПОСОБАМИ ИНЖЕНЕРИИ ПЛАТФОРМА ДЛЯ ВСТРАИВАНИЯ ТРАНСГЕНА (ETIP) ДЛЯ НАЦЕЛИВАНИЯ ГЕНОВ И СТЭКИНГА ПРИЗНАКОВ | 2013 |
|
RU2666916C2 |
| ГЕНОМНАЯ ИНЖЕНЕРИЯ | 2014 |
|
RU2688462C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ОПОСРЕДОВАННОЙ НУКЛЕАЗОЙ НАПРАВЛЕННОЙ ИНТЕГРАЦИИ ТРАНСГЕНОВ | 2013 |
|
RU2650819C2 |
| Рекомбинантные плазмиднные ДНК PX458D10A_Myc, обеспечивающие нокаут гена MYC в клетках человека, способ получения линии клеток человека с нокаутом гена MYC и линия клеток рака молочной железы человека MDA-MB231_Myc с подавленной экспрессией гена MYC | 2022 |
|
RU2818348C1 |
| ФУНКЦИОНАЛЬНЫЕ ЛОКУСЫ FAD2 И СООТВЕТСТВУЮЩИЕ СПЕЦИФИЧНЫЕ ДЛЯ САЙТА-МИШЕНИ СВЯЗЫВАЮЩИЕСЯ БЕЛКИ, СПОСОБНЫЕ ИНДУЦИРОВАТЬ НАПРАВЛЕННЫЕ РАЗРЫВЫ | 2013 |
|
RU2656158C2 |
| УЛУЧШЕННЫЕ ЭУКАРИОТИЧЕСКИЕ КЛЕТКИ ДЛЯ ПОЛУЧЕНИЯ БЕЛКА И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2016 |
|
RU2752529C2 |
Изобретение относится к биотехнологии, а именно к получению рекомбинантной белковой конструкции SSB-GS-EXO. Предложен способ получения рекомбинантной белковой конструкции SSB-GS-EXO, предусматривающий трансформацию компетентных клеток и экспрессию в них генетических конструкций на основе плазмидной ДНК, в которых генно-инженерным путем объединены 5’-3'Flap-экзонуклеаза (ЕХО) и белок SSB через глицин-сериновый спейсер, представленный последовательностью SEQ ID NO: 1. Изобретение обеспечивает клонирование молекул ДНК с повышенной частотой гомологичной рекомбинации между нуклеиновыми кислотами в клетке. 3 ил., 1 табл., 2 пр.
Способ получения рекомбинантной белковой конструкции SSB-GS-EXO, предусматривающий трансформацию компетентных клеток и экспрессию в них генетических конструкций на основе плазмидной ДНК, в которых генно-инженерным путем объединены 5'-3'Flap-экзонуклеаза (ЕХО) и белок SSB через глицин-сериновый спейсер, представленный последовательностью SEQ ID NO: 1.
| EP 3728589 A1, 28.10.2020 | |||
| WO 2020036653 A2, 20.02.2020 | |||
| EP 3730610 A1, 28.10.2020 | |||
| ЛИТВИНОВ С.В | |||
| Основные пути репарации двойных разрывов геномной ДНК и взаимодействия между ними | |||
| Цитология и генетика, 2014, Т | |||
| Приспособление для автоматической односторонней разгрузки железнодорожных платформ | 1921 |
|
SU48A1 |
| Нефтяной конвертер | 1922 |
|
SU64A1 |

