Настоящее изобретение относится к области иммунологии и медицины и может найти применение для лечения онкологических заболеваний с использованием моноклональных человеческих антител, специфичных к интерлейкину-11 человека и ингибирующих его связывание с рецептором IL-11RA и/или с рецептором gp130.
Предшествующий уровень техники
Из патента США US 8518888 В2 известен способ лечения онкологических заболеваний желудочно-кишечного тракта при помощи антител, антагонистичных по отношению к рецептору IL-11R. Однако в данном патенте не раскрывается структура антитела, а в самом способе не описывается каких-либо новых, экспериментально подтвержденных данных.
Из патента РФ RU 2318829 С2 известны химерные анти-IL-6-антитела на основе мышиного антитела CLB-8, каждое из которых содержит константную область из одного или нескольких антител человека. Предпочтительными антителами согласно изобретению являются антитела, которые связываются с эпитопами, распознаваемыми CLB-8 и cCLB-8 и включенными в эпитоп gp130. Описаны варианты нуклеиновых кислот, кодирующих анти-IL-6-антитело. Раскрыто применение анти-IL-6-антитела для лечения иммунологического расстройства или заболевания, опосредуемого IL-6, а также применение анти-IL-6-антитела для модуляции злокачественного расстройства или расстройства иммунной системы, опосредуемого IL-6.
Однако описанные в данном патенте антитела не являются специфичными к интерлейкину-11 и не эффективны при лечении онкологических заболеваний.
В патенте США US 7094402 В2 описано антитело, специфичное к белку рецептора IL-11R человеческого интерлейкина-11, в котором белок рецептора человеческого IL-11R состоит из аминокислотной последовательности, выбранной из группы, состоящей из:
(a) аминокислотной последовательности Seq ID NO: 2;
(b) аминокислотной последовательности Seq ID NO: 2 из аминокислот 24-422;
(c) аминокислотной последовательности Seq ID NO: 2 из аминокислот 24-365;
(d) аминокислотной последовательности Seq ID NO: 2 из аминокислот 391-422;
(e) аминокислотной последовательности Seq ID NO: 2 из аминокислот 112-422;
(f) аминокислотной последовательности Seq ID NO: 2 из аминокислот 112-365;
(g) аминокислотной последовательности Seq ID NO: 2 из аминокислот 24-359;
(h) аминокислотной последовательности Seq ID NO: 2 из аминокислот 24-345; и
(i) аминокислотной последовательности Seq ID NO: 2 из аминокислот 24-324.
В данном патенте не раскрывается способность описанного антитела использоваться для лечения онкологических заболеваний.
Краткое описание существа изобретения
Целью настоящего изобретения является устранение вышеуказанных недостатков аналогов путем создания вариабельных доменов легкой и тяжелой цепей, обладающих в составе полноразмерного антитела или фрагмента антитела, специфичных к интерлейкину-11 человека (IL-11), ингибирующему связывание IL-11 с альфа-субъединицей рецептора интерлейкина-11 (IL-11RA) и/или связывание комплекса IL-11/IL-11RA с бета-субъединицей рецептора интерлейкина-6 (gp130), в котором вариабельные домены тяжелой и легкой цепей иммуноглобулина содержат следующие комбинации аминокислотных последовательностей гипервариабельных участков (CDRs) или гомологичные им комбинации аминокислотных последовательностей гипервариабельных участков легких и\или тяжелых цепей.
В таблице 1. приведены комбинации аминокислотных последовательностей гипервариабельных участков из п. 1 формулы изобретения, а также два примера гомологов.
Отличие клона U#L1 от клона U#4 заключается в трех аминокислотах HCDR1 во 2-м, 3-ми 7-м положениях; отличие клона U#L2 от клона U#4 заключается в одной аминокислоте HCDR3 в 12-м положении. 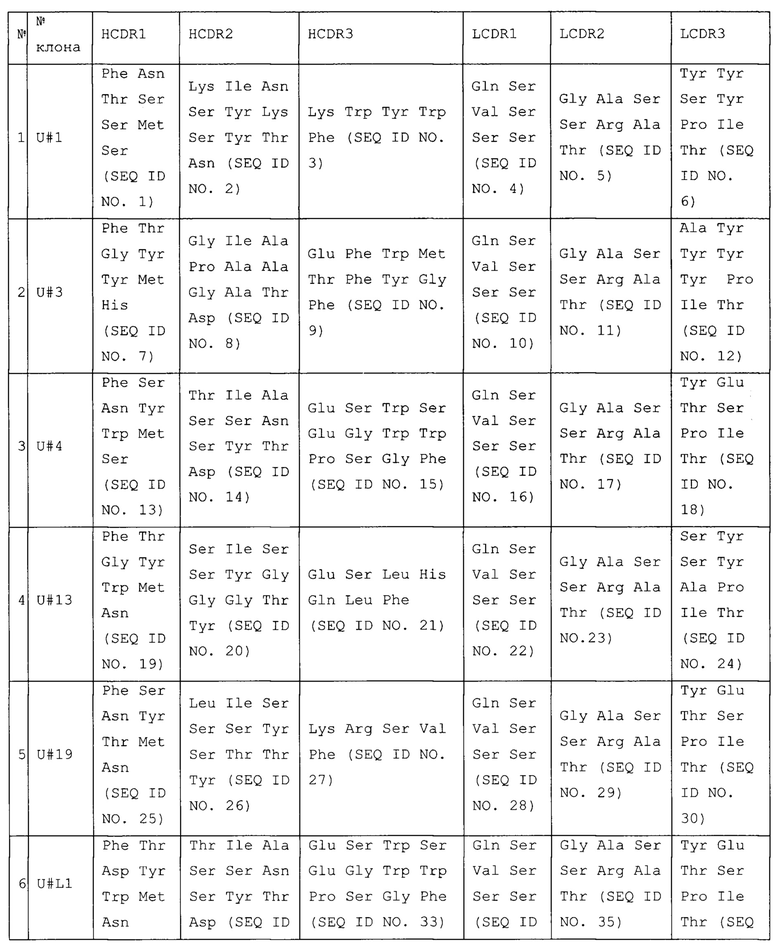
Кроме того, из патента США US 8518888 В2 известен способ использования антитела для лечения пациентов, заключающийся во введении страдающим заболеванием пациентам в составе полноразмерного терапевтического антитела или его фрагмента (Fab-фрагмент, (Fab)2-фрагмент), и обеспечивающие связывание IL-11 и, как следствие, блокирование активации рецептора IL-11RA.
Настоящее изобретение основано на способности IL-11 последовательно связываться с рецепторами IL-11RA и gp130 и тем самым вызывать активацию STAT3 сигналирования. Аномальная активация сигнального белка и фактора транскрипции STAT3 способствует атрофическому гастриту, дисплазии, пролиферации различного рода аденоматозных полипов и пролиферации рака:
Jarnicki, А., Т. Putoczki, and М. Ernst, Stat3: linking inflammation to epithelial cancer - more than a "gut" feeling? Cell Div, 2010. 5: p. 14.;
Guo, X., et al., Induction of innate lymphoid cell-derived interleukin-22 by the transcription factor STAT3 mediates protection against intestinal infection. Immunity, 2014. 40(1): p. 25-39.;
Kortylewski, M., et al., Inhibiting Stat3 signaling in the hematopoietic system elicits multicomponent antitumor immunity. Nat Med, 2005. 11(12): p. 1314-21.;
Schwitalla, S., et al., Intestinal tumorigenesis initiated by dedifferentiation and acquisition of stem-cell-like properties. Cell, 2013. 152(1-2): p. 25-38.;
Bollrath, J., et al., gp130-mediated Stat3 activation in enterocytes regulates cell survival and cell-cycle progression during colitis-associated tumorigenesis. Cancer Cell, 2009. 15(2): p. 91-102.
Putoczki, T.L., et al., Interleukin-11 is the dominant IL-6 family cytokine during gastrointestinal tumorigenesis and can be targeted therapeutically. Cancer Cell, 2013. 24(2): p. 257-71.
Заявленное изобретение представляет собой полноразмерное моноклональное человеческое антитело или фрагменты антитела (Fab-фрагмент, (Tab)2-фрагмент), связывающие IL-11 и стерически или аллостерически блокирующие дальнейшее связывание IL-11 с IL-11RA и/или gp130 и дальнейшее STAT3 сигналирование. Такие антитела-антагонисты могут быть применены в лечении ряда онкологических заболеваний, таких как рак кости, молочной железы, толстого кишечника, рака эндометрия, печени, легкого, яичника, поджелудочной железы, простаты, почки и желудка, а также других видов рака, обусловленных усиленным сигналированием через ось IL-11/IL-11/gp130/STAT3, на основе данных гистохимического анализа опухоли.
В настоящем изобретении описывается способ лечения ряда онкозаболеваний, таких как рак костей, молочной железы, толстого кишечника, рак эндометрия, печени, легкого, яичников, поджелудочной железы, простаты, почек и желудка, в которых наблюдается высокий уровень экспрессии IL-11 и/или IL-11RA, причем способ включает введение субъекту эффективного количества антитела-антагониста IL-11.
В заявленном изобретении используется терапевтически эффективное количество антитела, что означает достаточное количество антитела, обеспечивающее желаемый терапевтический или физиологический эффект, включая ингибирование IL-11/IL-11RA сигналирования. Ингибирующий эффект заключается в ингибировании активации сигнальных белков STAT3 и/или STAT1. Кроме того, терапевтический эффект может быть достигнут в виде ингибирования роста раковой опухоли, или рецессии раковой опухоли, блокирования метастазирования опухоли в другие ткани организма пациента. Настоящее изобретение касается использования описанного антитела-антагониста IL-11 (или фрагмента антитела) как в качестве монотерапии, так и совместно с другими лекарственными средствами, в виде интервенционной терапии или адъювантной терапии, как простой, так и неоадъвантной.
Другим аспектом настоящего изобретения является возможность использования приведенного антитела при лечении больных различными онкологическими заболеваниями. При высоких уровнях экспрессии IL-11 и/или IL-11RA у пациента предполагается использование антитела-антагониста IL-11 в таких концентрациях и условиях, чтобы предотвратить дальнейшее развитие раковой опухоли и метастазирование в другие ткани и органы пациента.
Краткое описание чертежей.
Настоящее изобретение иллюстрируется прилагаемыми чертежами, на которых:
Фигура 1 представляет собой изображение аминокислотных последовательностей IL-11 человека и яванской макаки, взятые из записей Р20809 и Р20808 базы данных UniProtKB, соответственно.
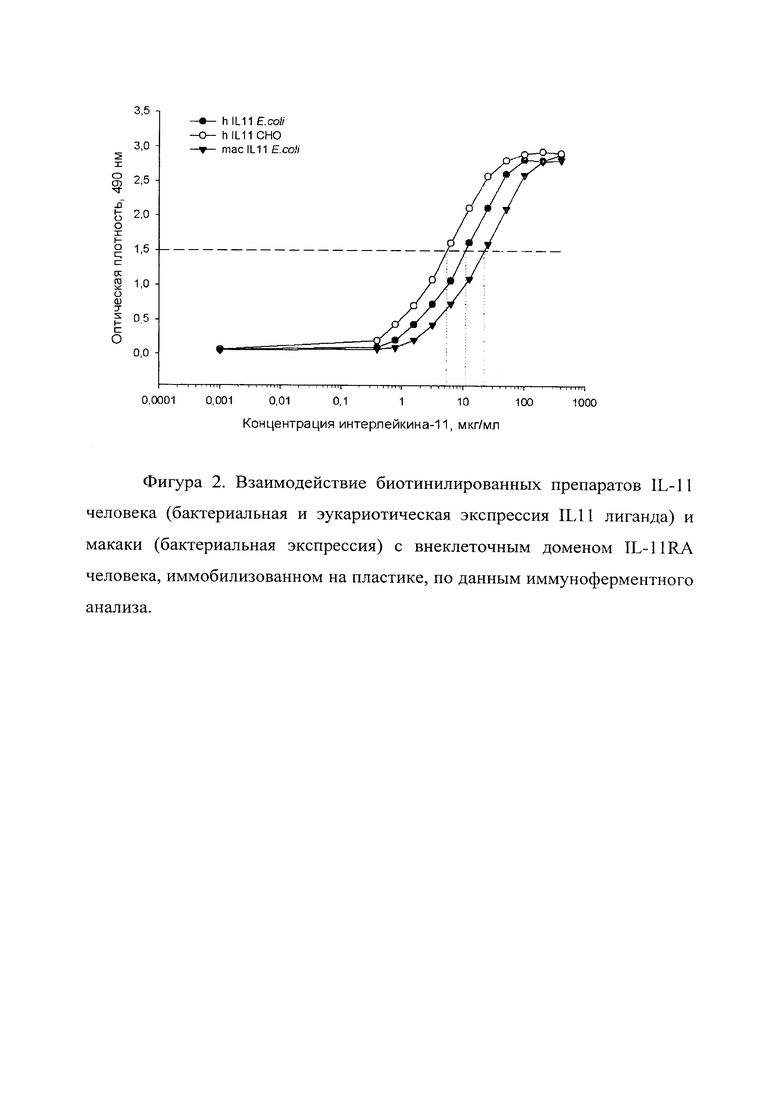
Фигура 2 отражает взаимодействие биотинилированных препаратов IL-11 человека и макаки, экспрессированных в различных экспрессионных системах (бактериальная и эукариотическая экспрессия IL-11 лиганда) с внеклеточным доменом IL-11RA человека, иммобилизованном на пластике, по данным иммуноферментного анализа. Данный пример отражает нативную структуру и функциональную активность лиганда и рецептора.
В таблице 2 приведены рассчитанные по данным ИФА из примера 2 величины ЕС50, свидетельствующие об их специфическом взаимодействии и функциональной активности:

Эти данные показывают, что белки IL-11 человека и макаки, полученные в бактериальной и эукариотической системах экспрессии, функциональны и способны взаимодействовать с рецептором IL-11, т.е. антитела получены к нативному лиганду.
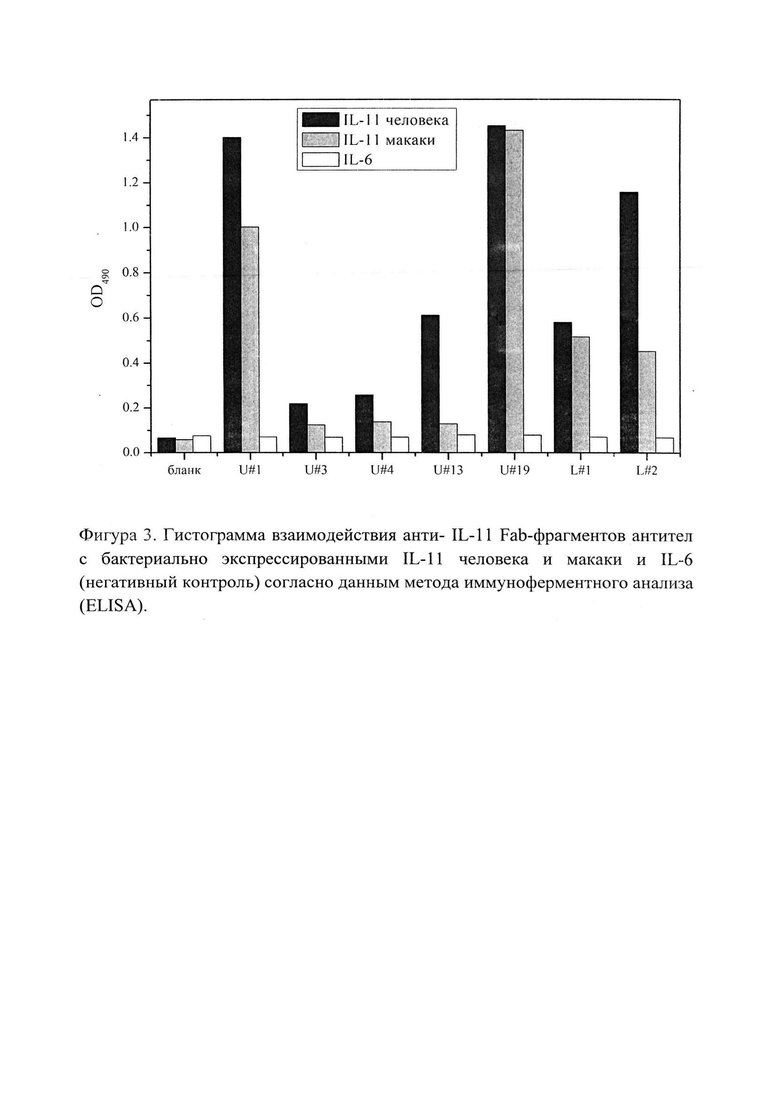
Фигура 3 отражает гистограмму взаимодействия анти-IL-11 Fab-фрагментов антител с бактериально экспрессированными IL-11 человека и макаки и IL-6 в качестве негативного контроля, согласно данным метода иммуноферментного анализа (ELISA). Лиганды иммобилизованы на поверхности пластика (1 мкг/мл), антитело выступает в качестве аналита.
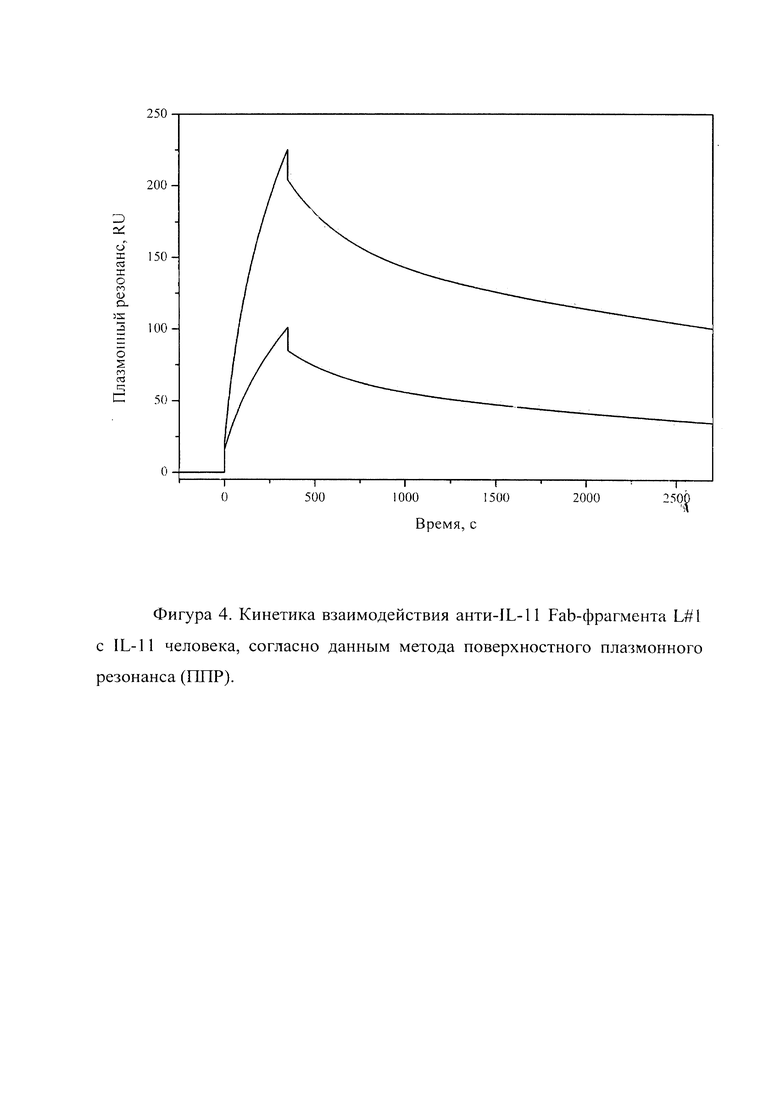
Фигура 4 отражает взаимодействие анти-IL-11 Fab-фрагмента L#1 с IL-11 человека, ковалентно пришит на поверхность GLH сенсор-чипа за аминогруппы, согласно данным метода поверхностного плазмонного резонанса (ППР). Концентрации нанесения Fab-фрагментов в рабочем буфере: 20, 10 нМ. Отображены полученные сенсограммы и теоретические кривые, построенные в программе ProteOn Manager 3.0 в рамках схемы гетерогенного лиганда.
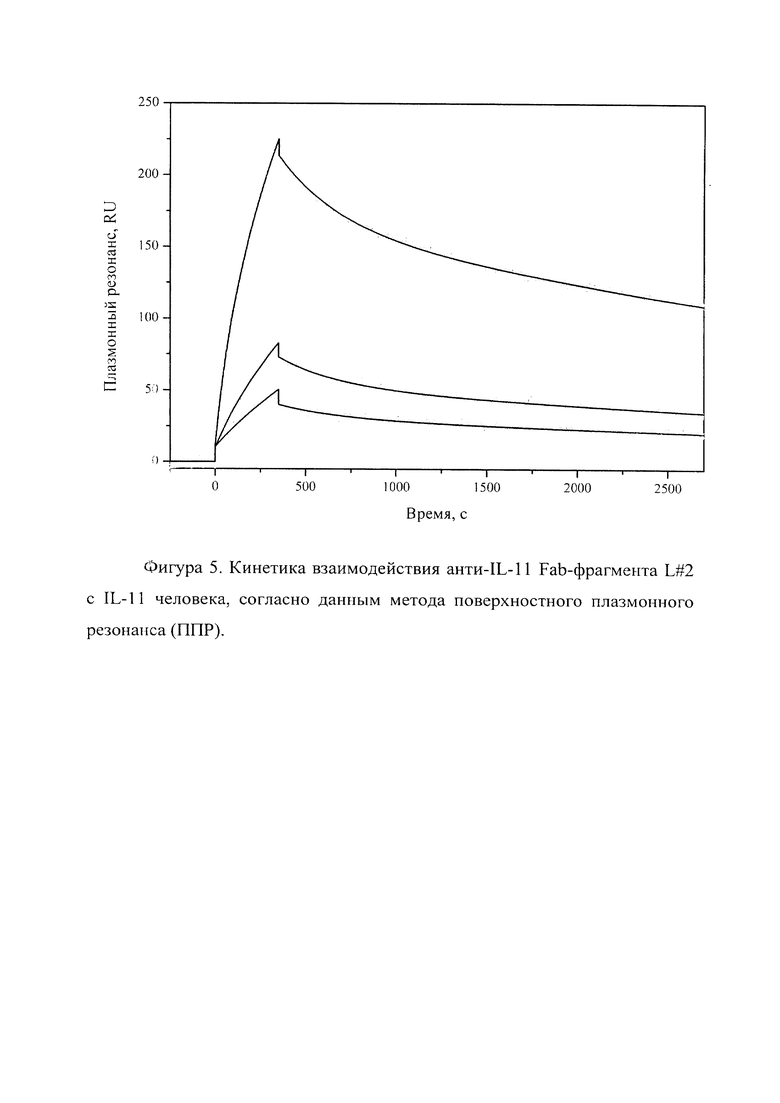
Фигура 5 отражает взаимодействие анти-IL-11 Fab-фрагмента L#2 с IL-11 человека, ковалентно пришитого на поверхность GLH сенсор-чипа за аминогруппы, согласно данным метода поверхностного плазмонного резонанса (ППР). Концентрации нанесения Fab-фрагментов в рабочем буфере: 20, 5, 2.5 нМ. Отображены полученные сенсограммы и теоретические кривые, построенные в программе ProteOn Manager 3.0 в рамках схемы гетерогенного лиганда.
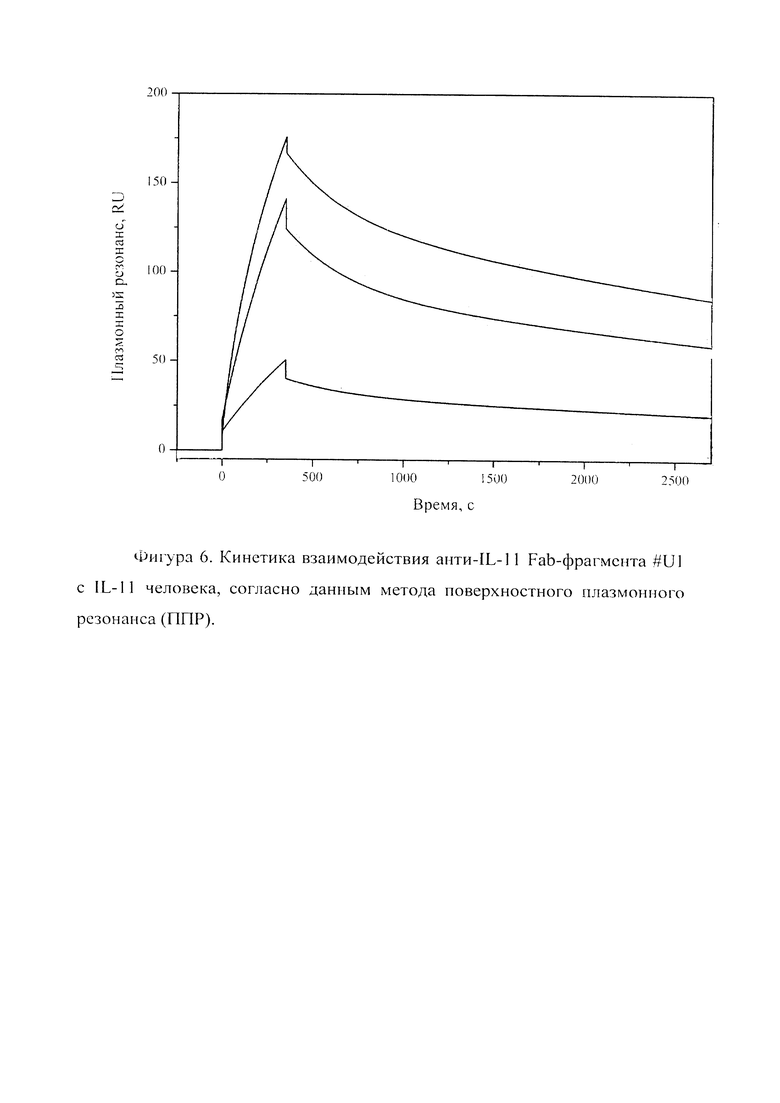
Фигура 6 отражает взаимодействие анти-IL-11 Fab-фрагмента U#1 с IL-11 человека, ковалентно пришитого на поверхность GLH сенсор-чипа за аминогруппы, согласно данным метода поверхностного плазмонного резонанса (ППР). Концентрации нанесения Fab-фрагментов в рабочем буфере: 20, 5, 2.5 нМ. Отображены полученные сенсограммы и теоретические кривые, построенные в программе ProteOn Manager 3.0 в рамках схемы гетерогенного лиганда.
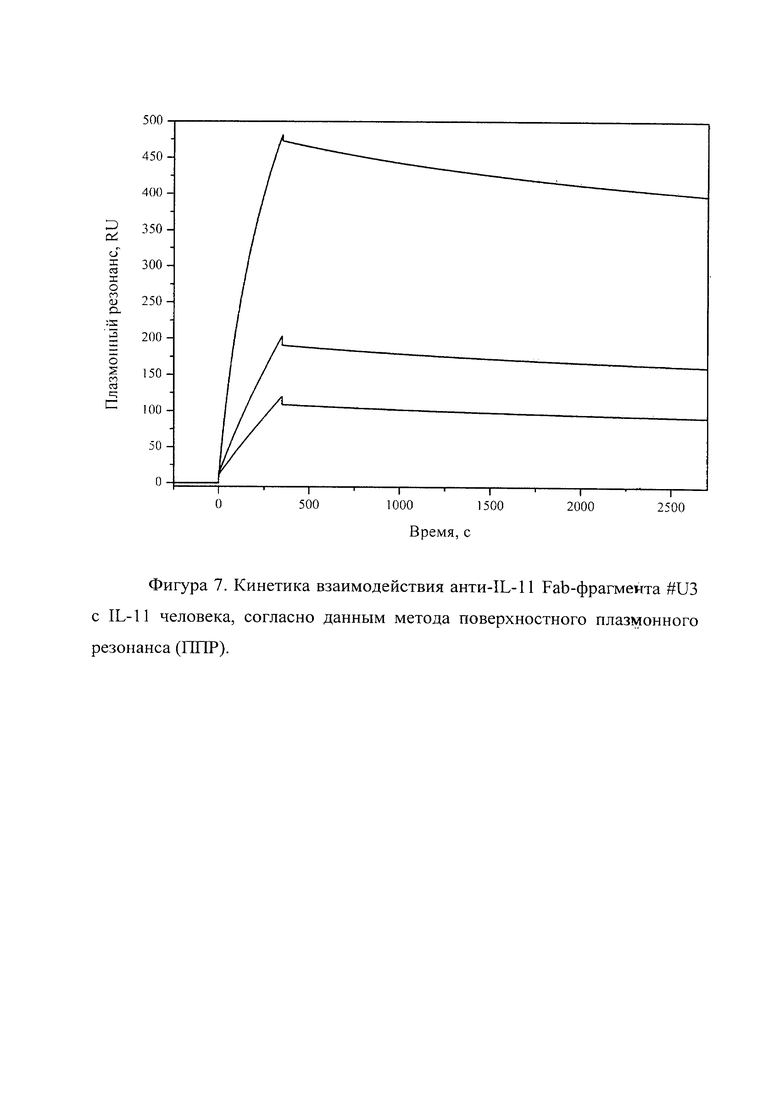
Фигура 7 отражает взаимодействие анти-IL-11 Fab-фрагмента U#3 с IL-11 человека, ковалентно пришитого на поверхность GLH сенсор-чипа за аминогруппы, согласно данным метода поверхностного плазмонного резонанса (ППР). Концентрации нанесения Fab-фрагментов в рабочем буфере: 20, 5, 2.5 нМ. Отображены полученные сенсограммы и теоретические кривые, построенные в программе ProteOn Manager 3.0 в рамках схемы гетерогенного лиганда.
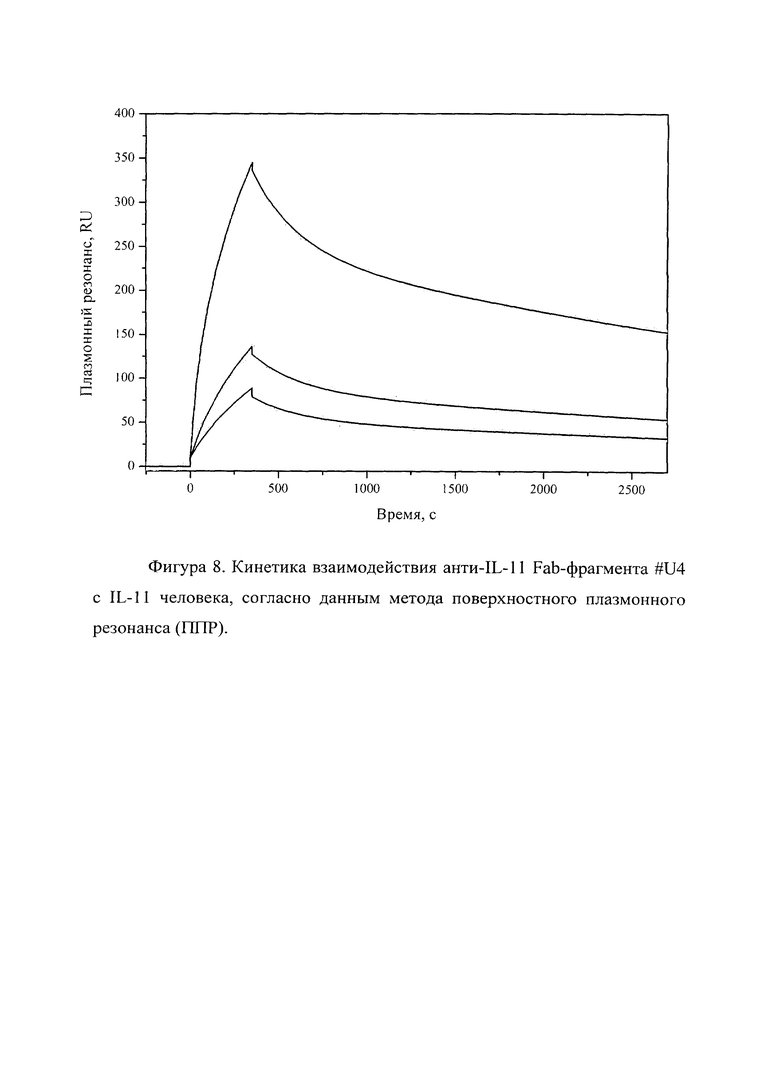
Фигура 8 отражает взаимодействие анти-IL-11 Fab-фрагмента U#4 с IL-11 человека, ковалентно пришитого на поверхность GLH сенсор-чипа за аминогруппы, согласно данным метода поверхностного плазмонного резонанса (ППР). Концентрации нанесения Fab-фрагментов в рабочем буфере: 20, 5, 2.5 нМ. Отображены полученные сенсограммы и теоретические кривые, построенные в программе ProteOn Manager 3.0 в рамках схемы гетерогенного лиганда.
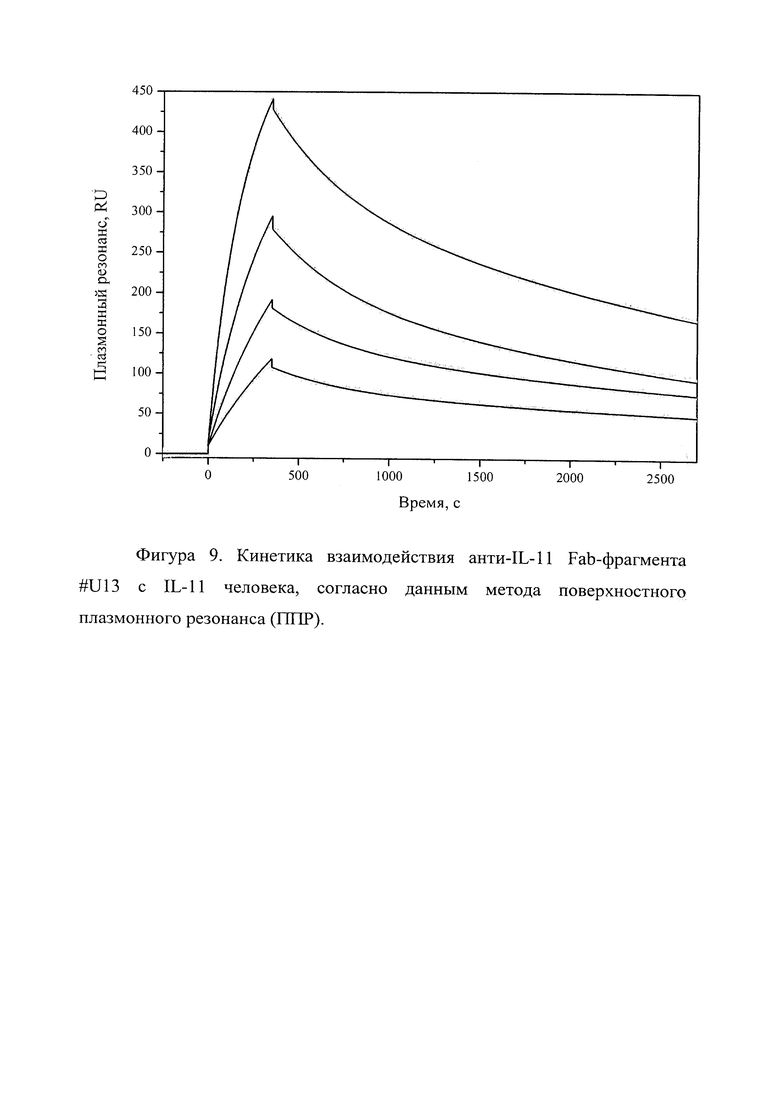
Фигура 9 отражает взаимодействие анти-IL-11 Fab-фрагмента U#13 с IL-11 человека, ковалентно пришитого на поверхность GLH сенсор-чипа за аминогруппы, согласно данным метода поверхностного плазмонного резонанса (ППР). Концентрации нанесения Fab-фрагментов в рабочем буфере: 20, 10, 5, 2.5 нМ. Отображены полученные сенсограммы и теоретические кривые, построенные в программе ProteOn Manager 3.0 в рамках схемы гетерогенного лиганда.
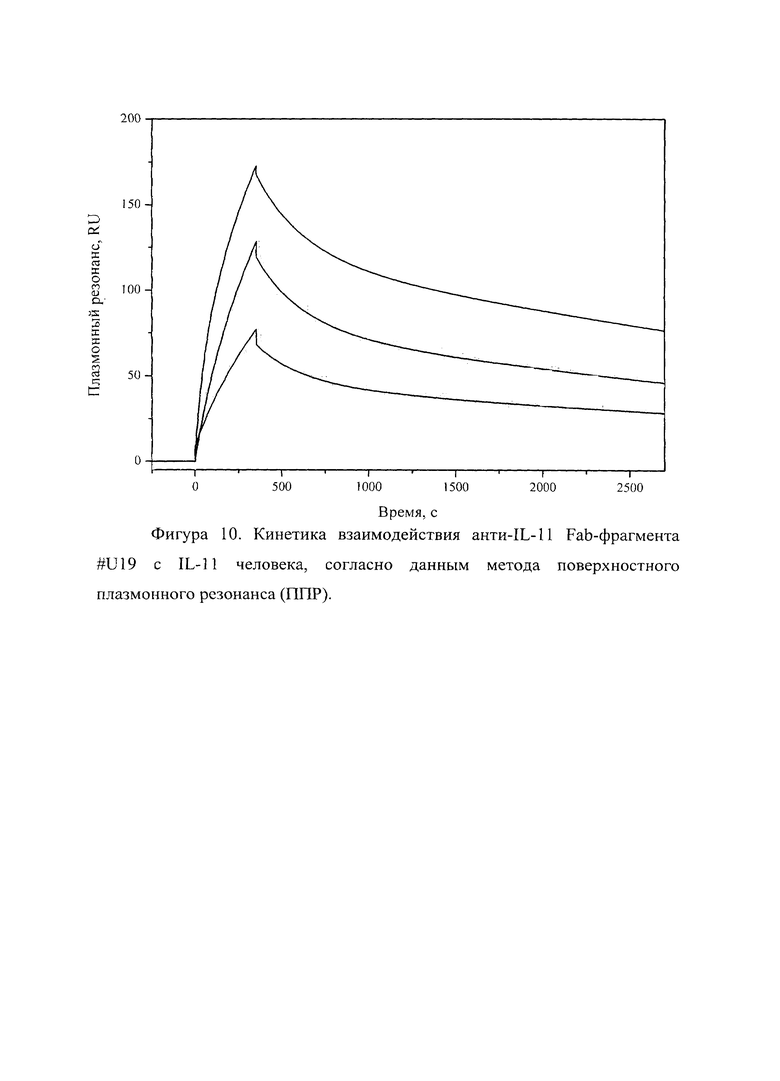
Фигура 10 отражает взаимодействие анти-IL-11 Fab-фрагмента U#19 с IL-11 человека, ковалентно пришитого на поверхность GLH сенсор-чипа за аминогруппы, согласно данным метода поверхностного плазмонного резонанса (ППР). Концентрации нанесения Fab-фрагментов в рабочем буфере: 20, 5, 2.5 нМ. Отображены полученные сенсограммы и теоретические кривые, построенные в программе ProteOn Manager 3.0 в рамках схемы гетерогенного лиганда.
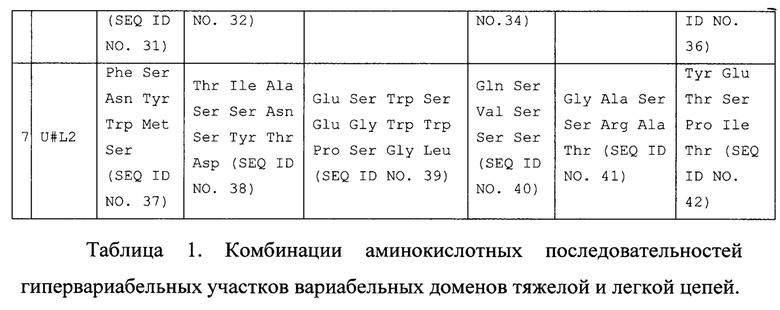
В таблице 3 приведены константы взаимодействия анти-IL-11 Fab-фрагментов с IL-11 человека. Расчет проводился в программе ProteOn Manager 3.0 в рамках схемы гетерогенного лиганда.
kon - кинетическая константа ассоциации. koff - кинетическая константа диссоциации. KD - равновесная константа диссоциации.
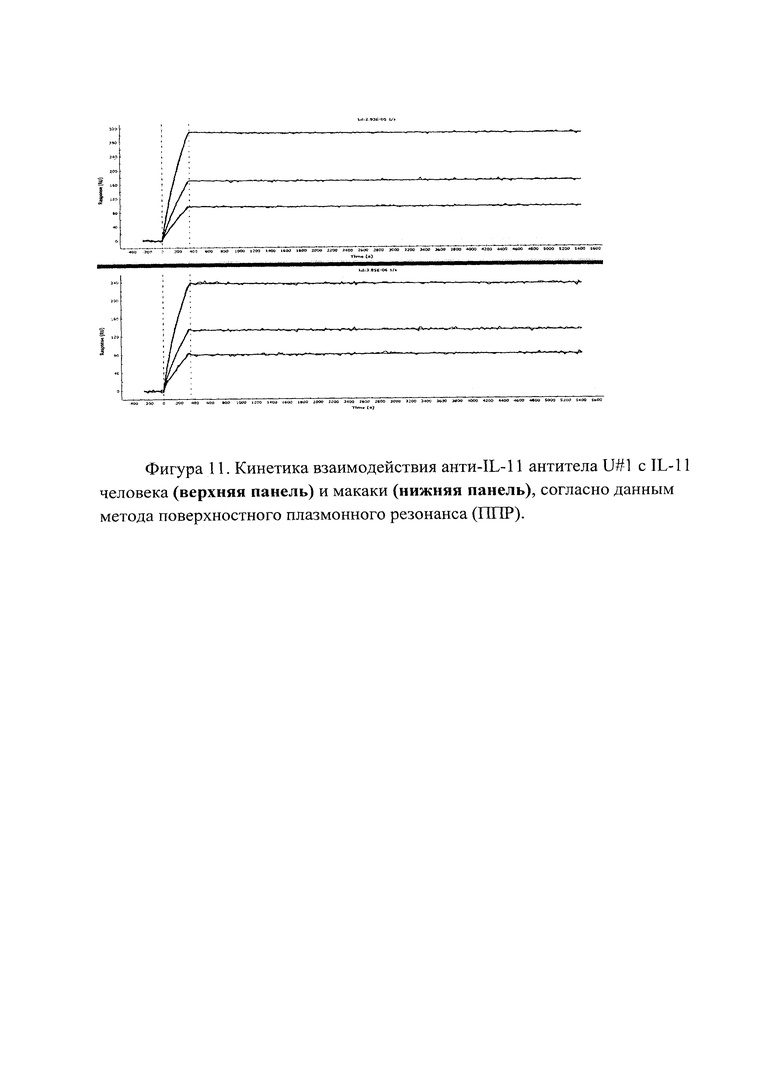
Фигура 11 отражает взаимодействие анти-IL-11 полноразмерного антитела U#1 с IL-11 человека (верхняя панель) и макаки (нижняя панель). Лиганды ковалентно пришиты на поверхности сенсорного чипа GLH за аминогруппы, согласно методике проведения поверхностного плазмонного резонанса (ПГТР). Концентрации нанесения антител в рабочем буфере: 5.71, 2.86, 1.43 нМ. Отображены полученные сенсограммы и теоретические кривые, построенные в программе ProteOn Manager 3.0 в рамках схемы бивалентного аналита.
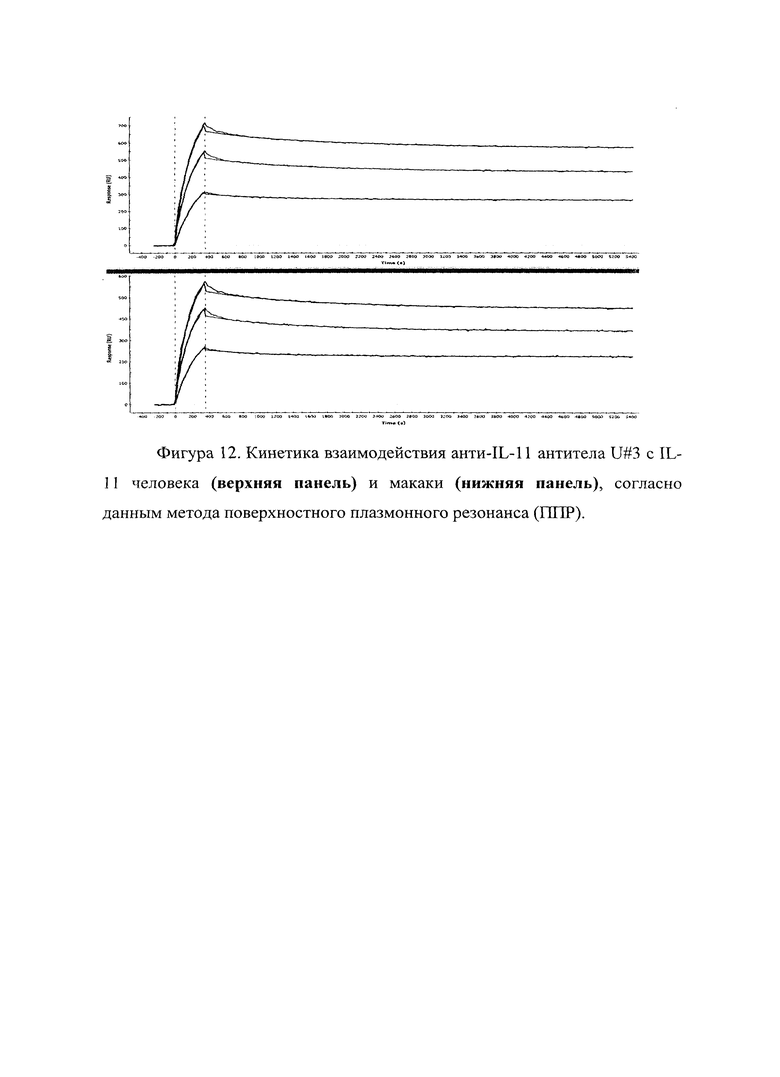
Фигура 12 отражает взаимодействие анти-IL-11 полноразмерного антитела U#3 с IL-11 человека (верхняя панель) и макаки (нижняя панель). Лиганды ковалентно пришиты на поверхности сенсорного чипа GLH за аминогруппы, согласно методике проведения поверхностного плазмонного резонанса (ПГТР). Концентрации нанесения антител в рабочем буфере: 3, 2, 1 мкМ. Отображены полученные сенсограммы и теоретические кривые, построенные в программе ProteOn Manager 3.0 в рамках схемы бивалентного аналита.
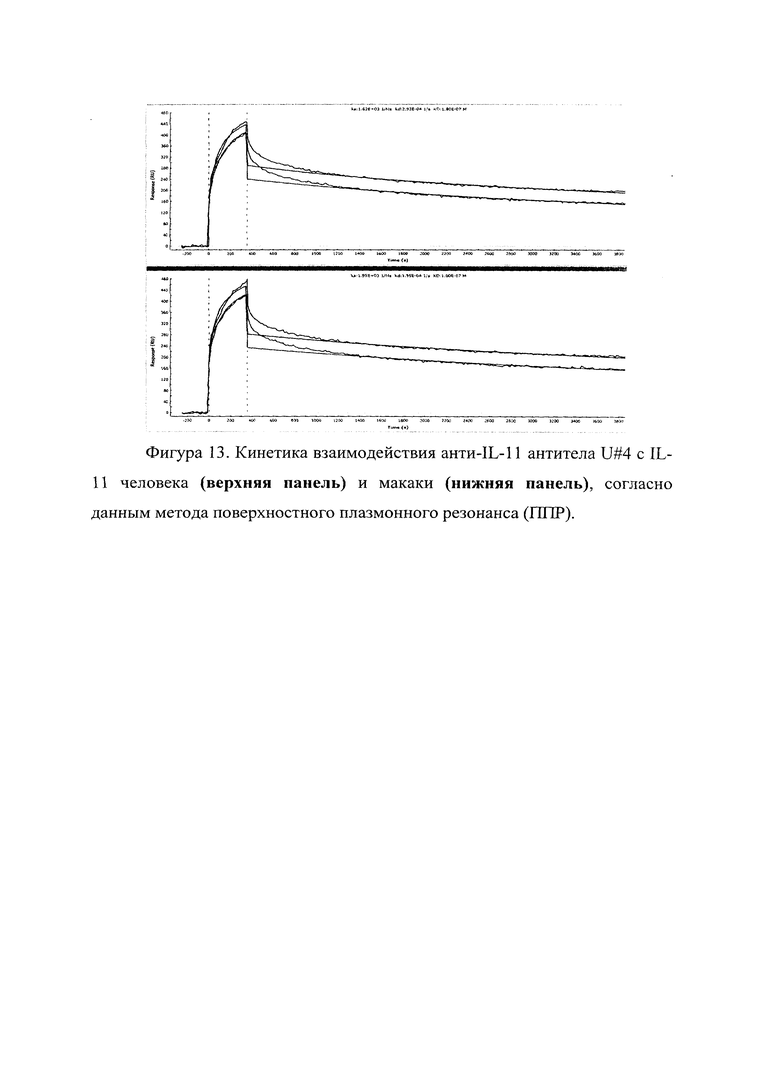
Фигура 13 отражает взаимодействие анти-IL-11 полноразмерного антитела U#4 с IL-11 человека (верхняя панель) и макаки (нижняя панель). Лиганды ковалентно пришиты на поверхности сенсорного чипа GLH за аминогруппы, согласно методике проведения поверхностного плазмонного резонанса (ППР). Концентрации нанесения антител в рабочем буфере: 3,2 мкМ. Отображены полученные сенсограммы и теоретические кривые, построенные в программе ProteOn Manager 3.0 в рамках схемы бивалентного аналита.
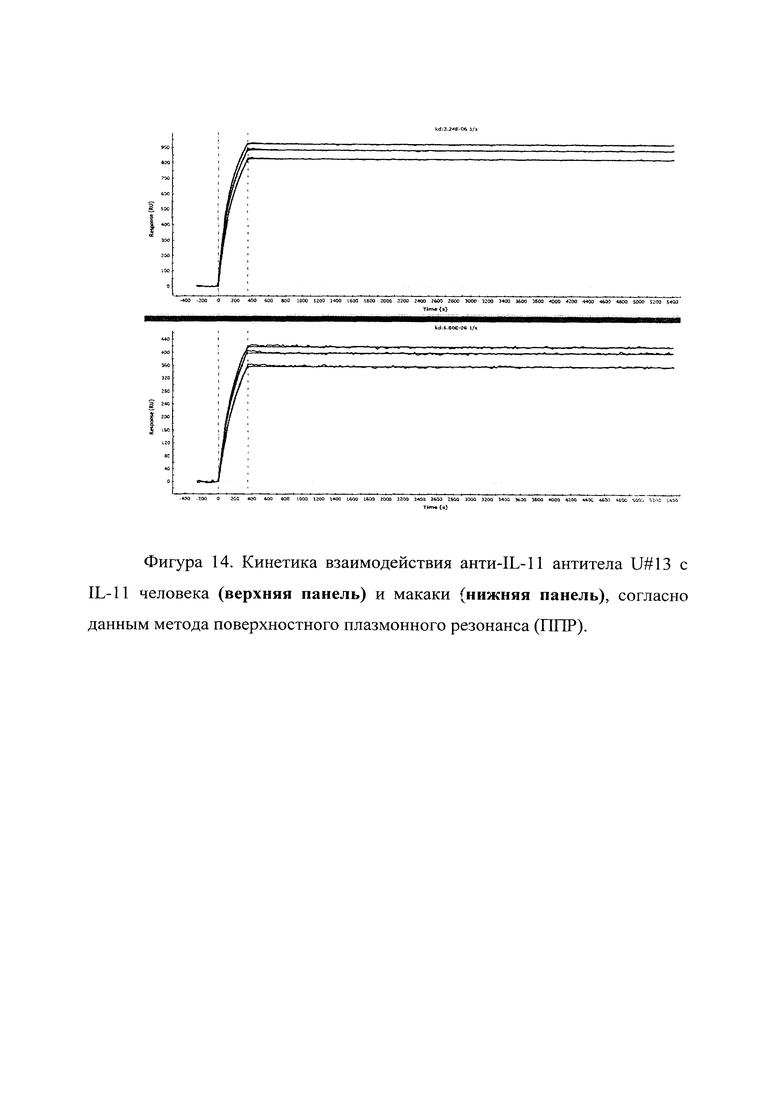
Фигура 14 отражает взаимодействие анти-IL-11 полноразмерного антитела U#13 с IL-11 человека (верхняя панель) и макаки (нижняя панель). Лиганды ковалентно пришиты на поверхности сенсорного чипа GLH за аминогруппы, согласно методике проведения поверхностного плазмонного резонанса (ППР). Концентрации нанесения антител в рабочем буфере: 15, 12.9, 8.57 нМ. Отображены полученные сенсограммы и теоретические кривые, построенные в программе ProteOn Manager 3.0 в рамках схемы бивалентного аналита.
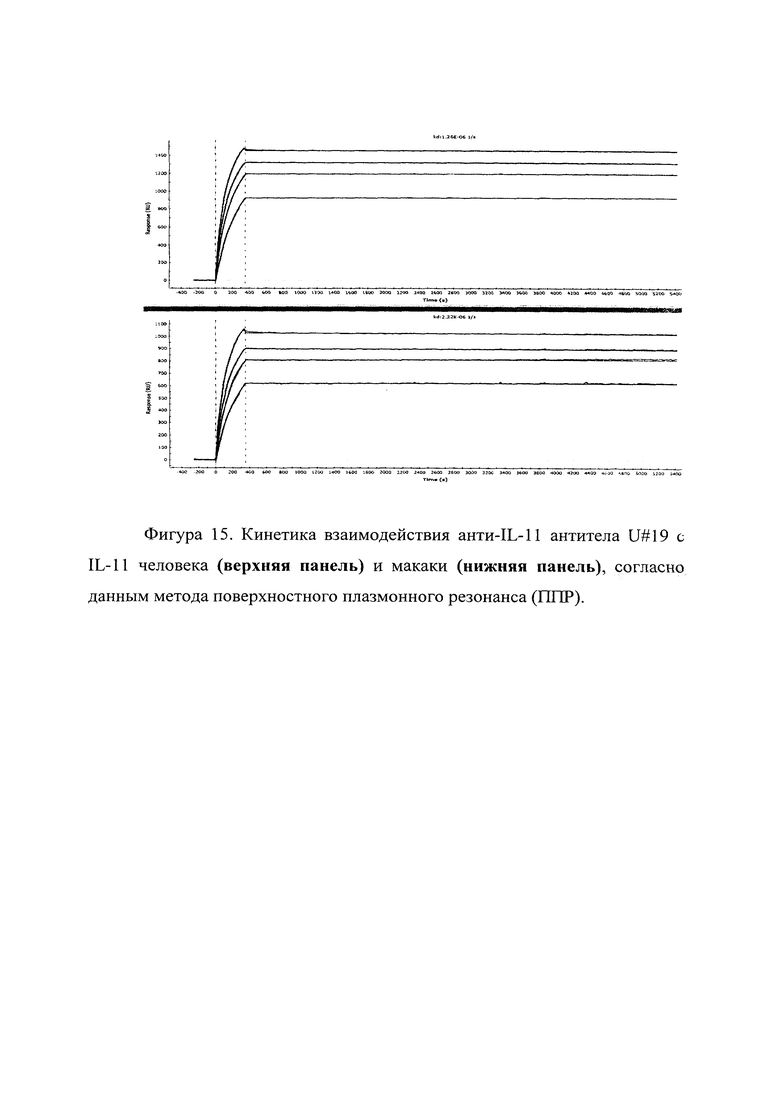
Фигура 15 отражает взаимодействие анти-IL-11 полноразмерного антитела U#19 с IL-11 человека (верхняя панель) и макаки (нижняя панель). Лиганды ковалентно пришиты на поверхности сенсорного чипа GLH за аминогруппы, согласно методике проведения поверхностного плазмонного резонанса (ППР). Концентрации нанесения антител в рабочем буфере: 30, 22.9, 14.4, 5.71 нМ. Отображены полученные сенсограммы и теоретические кривые, построенные в программе ProteOn Manager 3.0 в рамках схемы бивалентного аналита.
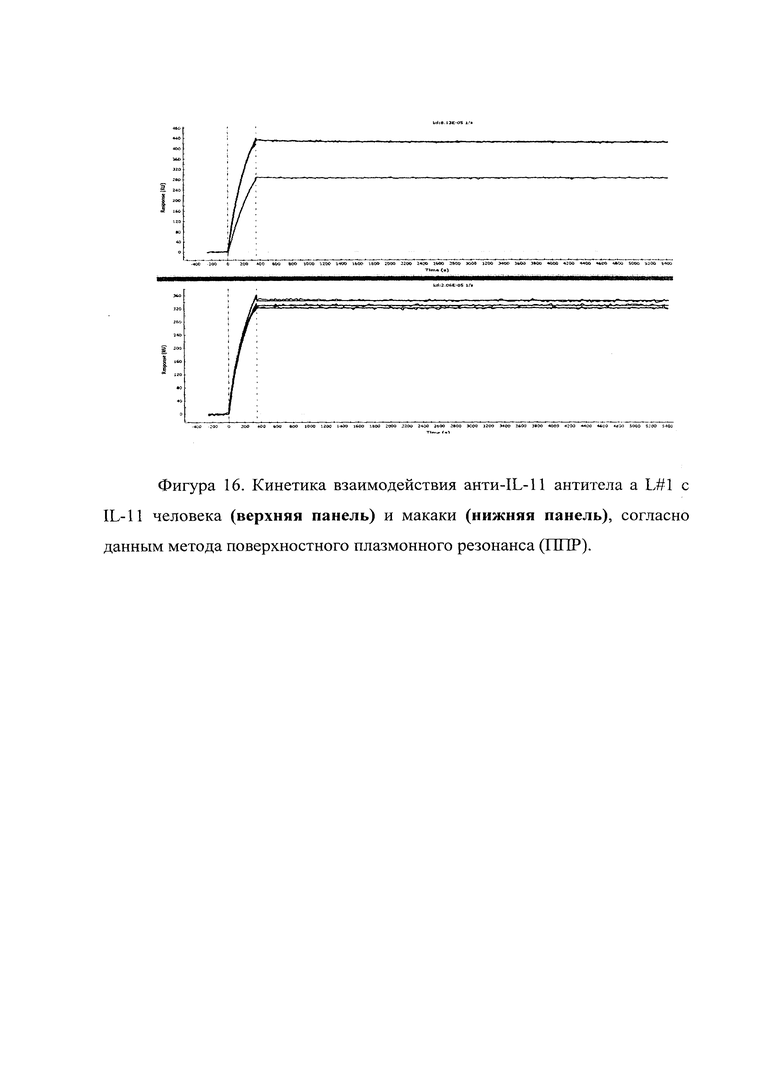
Фигура 16 отражает взаимодействие анти-IL-11 полноразмерного антитела L#1 с IL-11 человека (верхняя панель) и макаки (нижняя панель). Лиганды ковалентно пришиты на поверхности сенсорного чипа GLH за аминогруппы, согласно методике проведения поверхностного плазмонного резонанса (ППР). Концентрации нанесения антител в рабочем буфере: 10, 8.57, 2.58 нМ. Отображены полученные сенсограммы и теоретические кривые, построенные в программе ProteOn Manager 3.0 в рамках схемы бивалентного аналита.
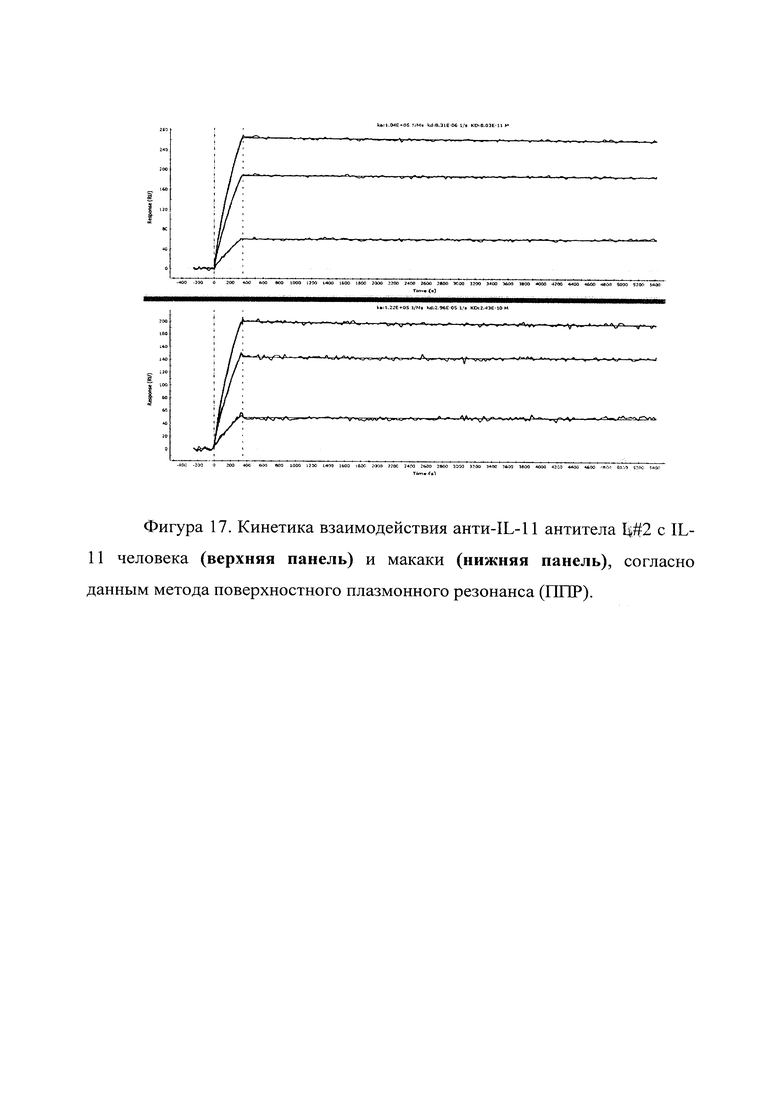
Фигура 17 отражает взаимодействие анти-IL-11 полноразмерного антитела L#2 с IL-11 человека (верхняя панель) и макаки (нижняя панель). Лиганды ковалентно пришиты на поверхности сенсорного чипа GLH за аминогруппы, согласно методике проведения поверхностного плазмонного резонанса (ПНР). Концентрации нанесения антител в рабочем буфере: 8.57, 5.71, 2.58 нМ. Отображены полученные сенсограммы и теоретические кривые, построенные в программе ProteOn Manager 3.0 в рамках схемы бивалентного аналита.
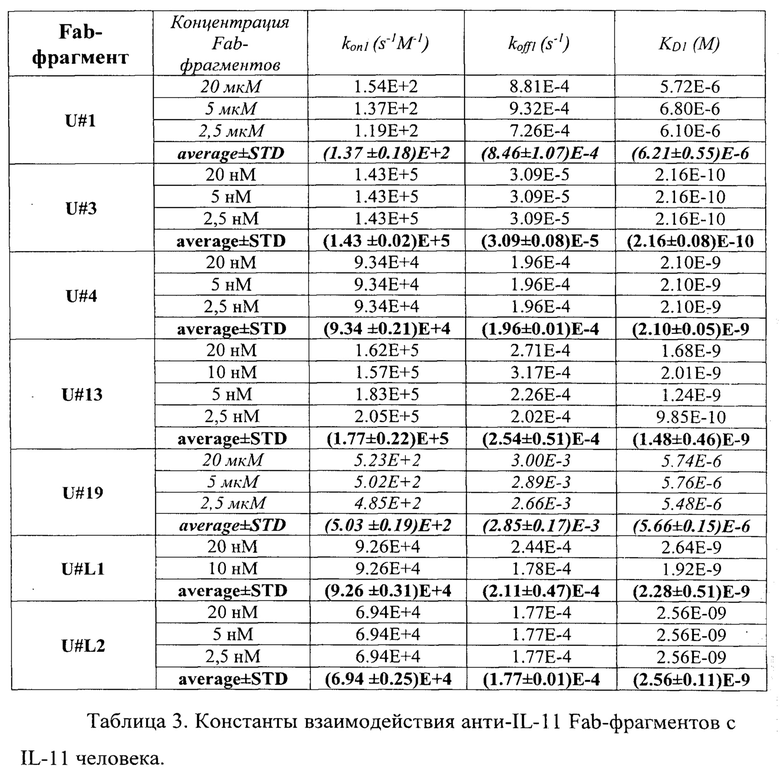
В таблице 4 приведены константы взаимодействия анти-IL-11 полноразмерных антител с IL-11 человека. Расчет проводился в программе ProteOn Manager 3.0 в рамках схемы бивалентного аналита. kon - кинетическая константа ассоциации. koff - кинетическая константа диссоциации. KD - равновесная константа диссоциации.
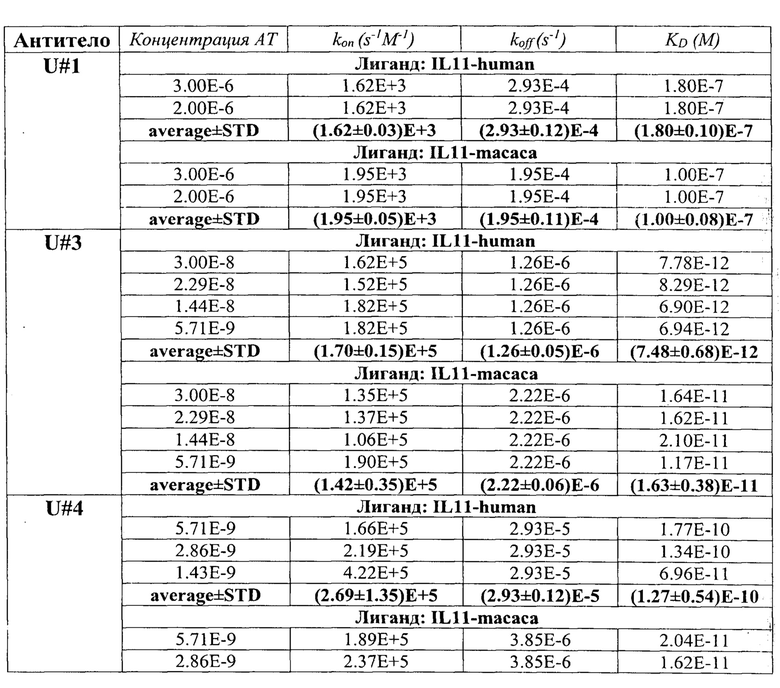
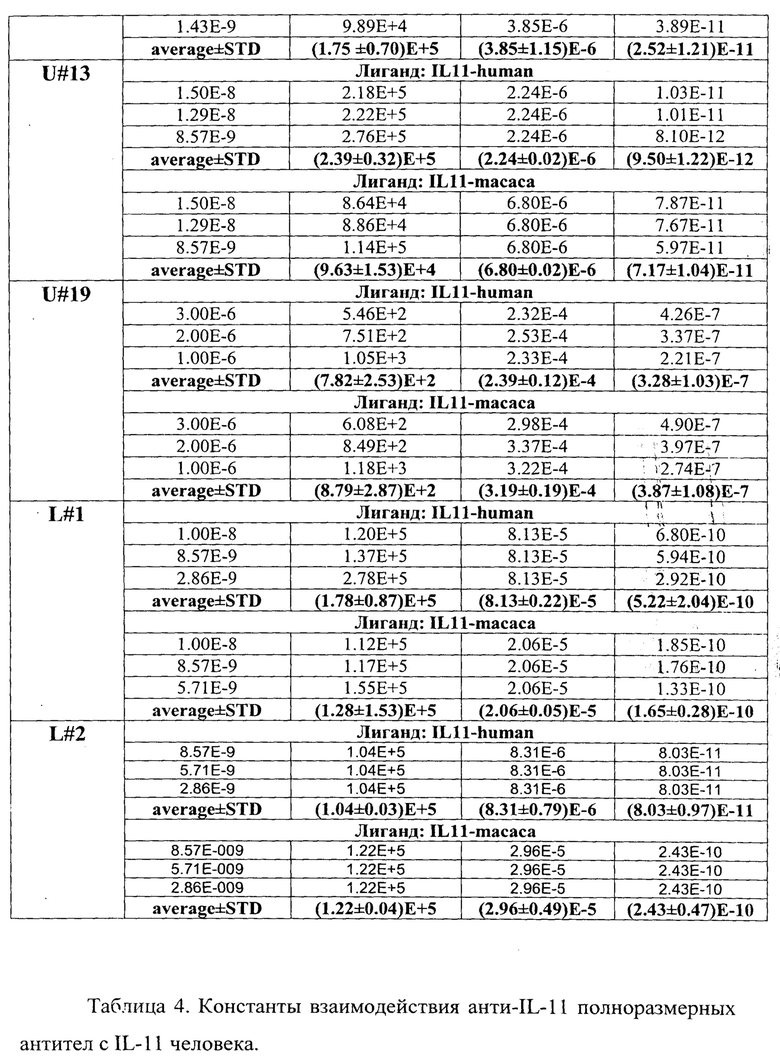
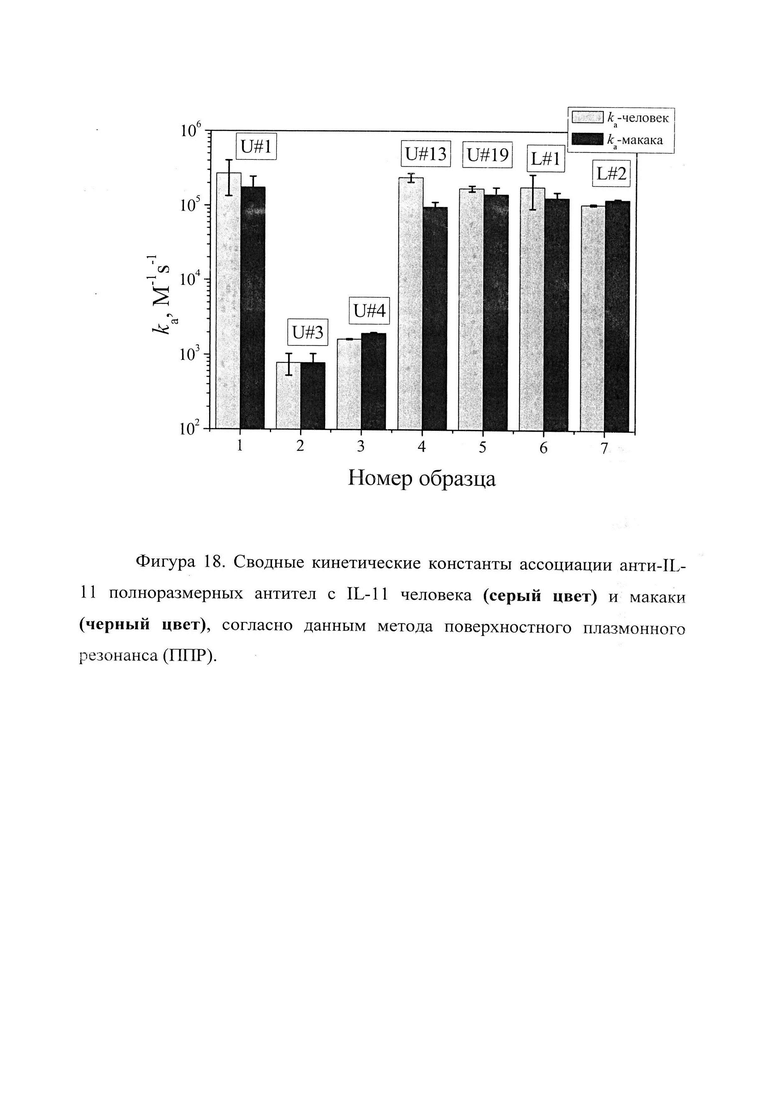
Кинетические константы характеризуют временные характеристики взаимодействия. Чем выше кинетическая константа ассоциации (kon), тем быстрее образуется комплекс антитела с IL-11. Чем ниже кинетическая константа диссоциации (koff), тем дольше происходит взаимодействие антитела с IL-11. Равновесная константа диссоциации (KD) характеризует концентрационные параметры взаимодействия. Чем ниже равновесная константа диссоциации, тем более низкий концентрационный диапазон, при котором происходит взаимодействие антитела с IL-11. Таким образом, наиболее быстро комплекс с IL-11 образуется у Fab-фрагментов U#3 и U#13 (таблица 3), а также у антитела U#3 (таблица 4). Наиболее долгоживущие комплексы между IL-11 и Fab-фрагментами образуются с U#3 и U#L2 (таблица 3), а между IL-11 и антителами с U#13 и U#3 (таблица 4). Исходя из равновесных констант диссоциации, наиболее сильное взаимодействие IL-11 происходит с Fab-фрагментом U#3 (таблица 3) и антителами U#13 и U#3 (таблица 4).
Фигура 18. Отображены сводные кинетические константы ассоциации анти-IL-11 полноразмерных антител с IL-11 человека (серый цвет) и макаки (черный цвет), согласно данным метода поверхностного плазмонного резонанса (ПГТР) (см. таблицу 4).
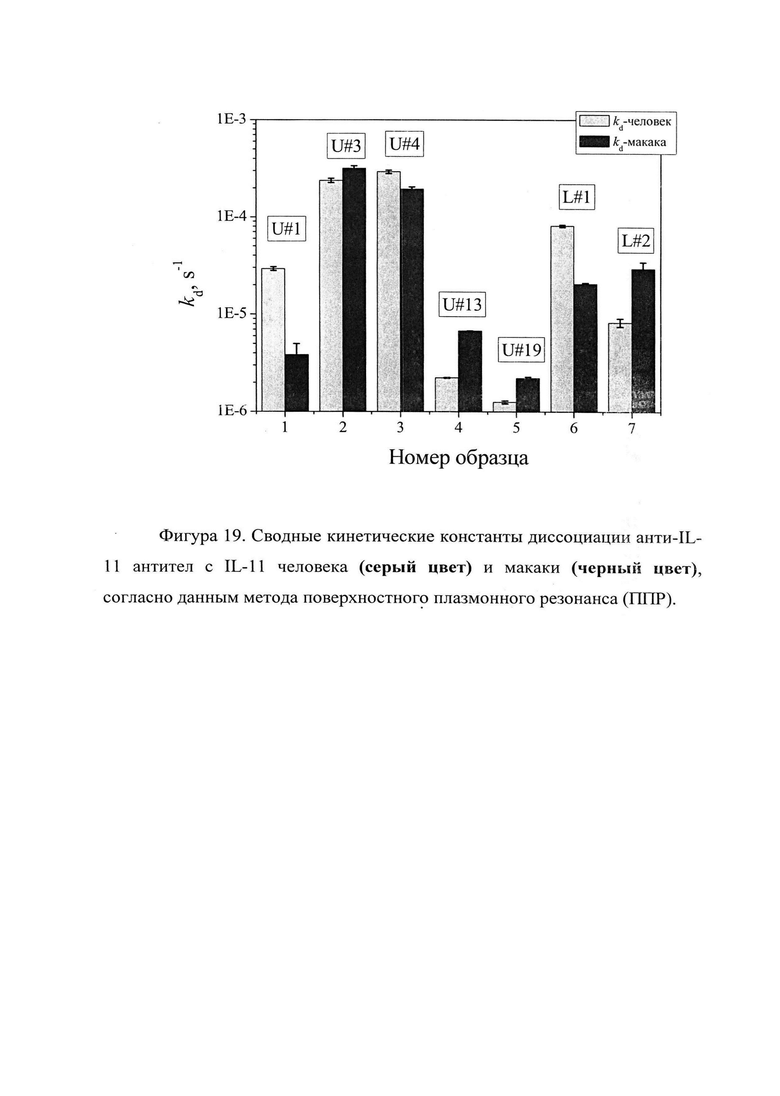
Фигура 19. Отображены сводные кинетические константы диссоциации анти-IL-11 полноразмерных антител с IL-11 человека (серый цвет) и макаки (черный цвет), согласно данным метода поверхностного плазмонного резонанса (ППР) (см. таблицу 4).
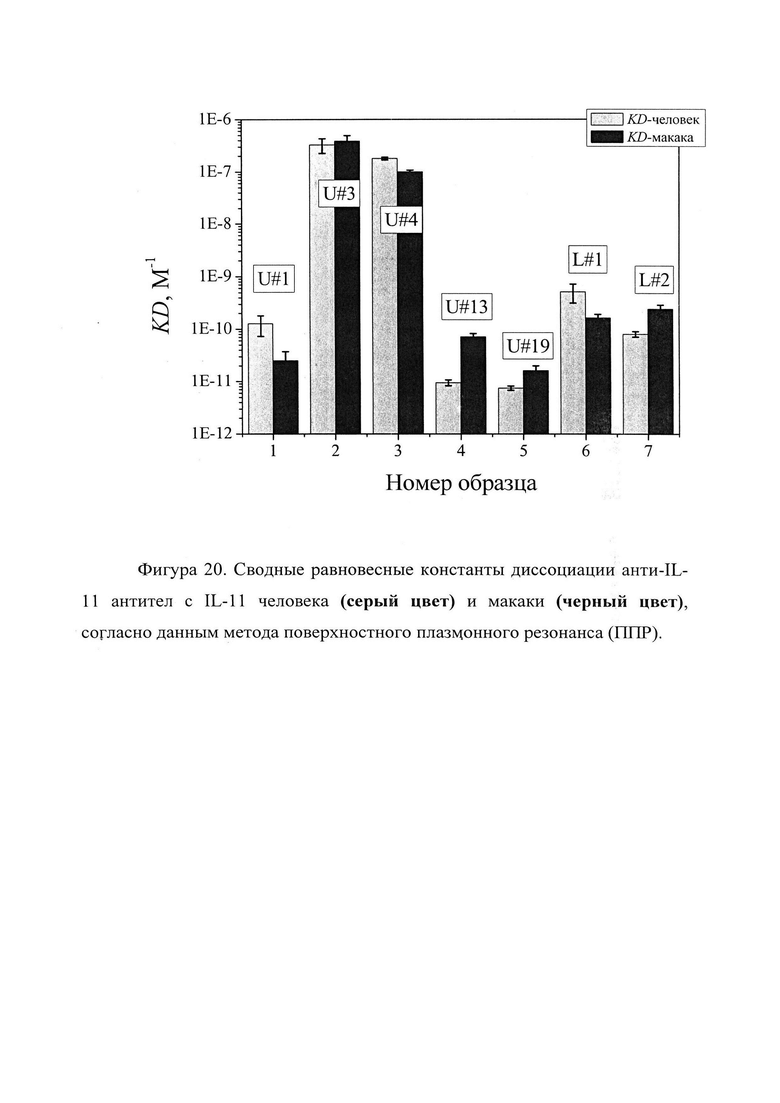
Фигура 20. Отображены сводные равновесные константы диссоциации анти-IL-11 антител с IL-11 человека (серый цвет) и макаки (черный цвет), согласно данным метода поверхностного плазмонного резонанса (ППР) (см. таблицу 4).
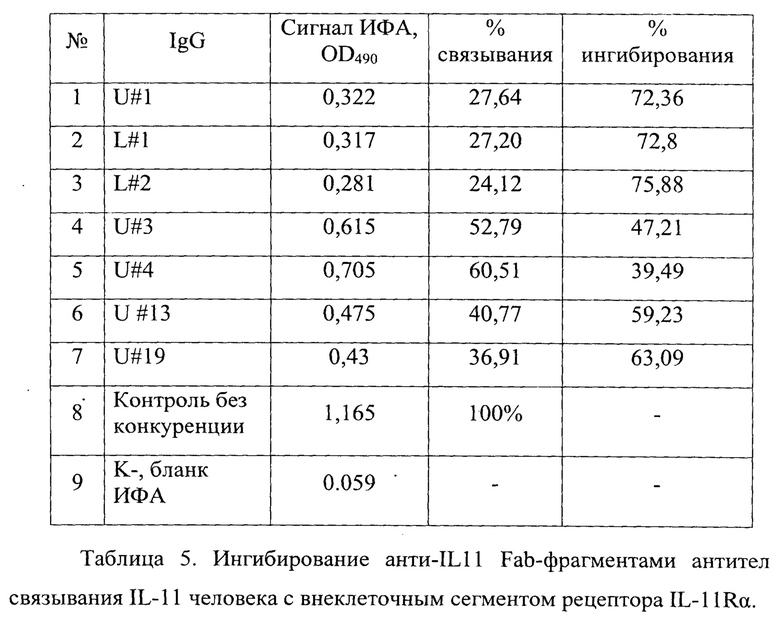
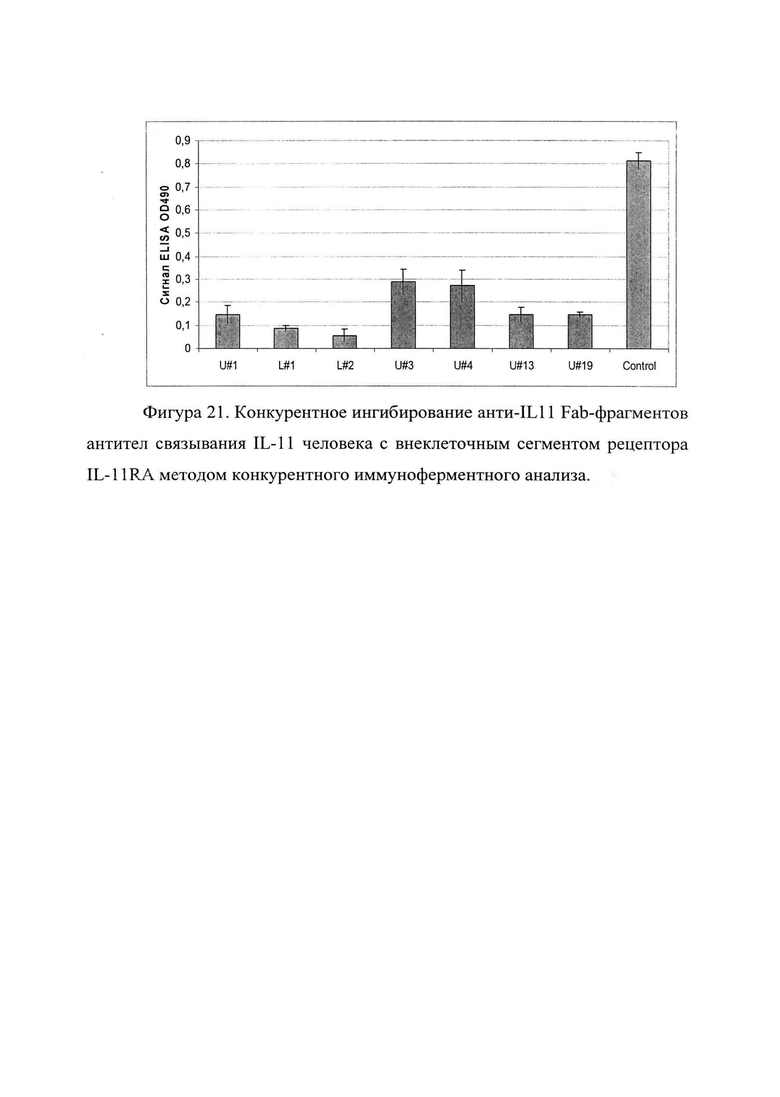
Фигура 21. Отображено конкурентное ингибирование анти-IL11 Fab-фрагментов антител связывания бактериально экспрессированного лиганда IL-11 человека с внеклеточным сегментом рецептора IL-11RA (Sino Biological Inc. cat #10252-H08H). методом конкурентного иммуноферментного анализа.
В таблице 5 приведены рассчитанные значения ингибирования от сигнала взаимодействия лиганда IL-11 с IL-11RA рецептором, принятых за 100%.
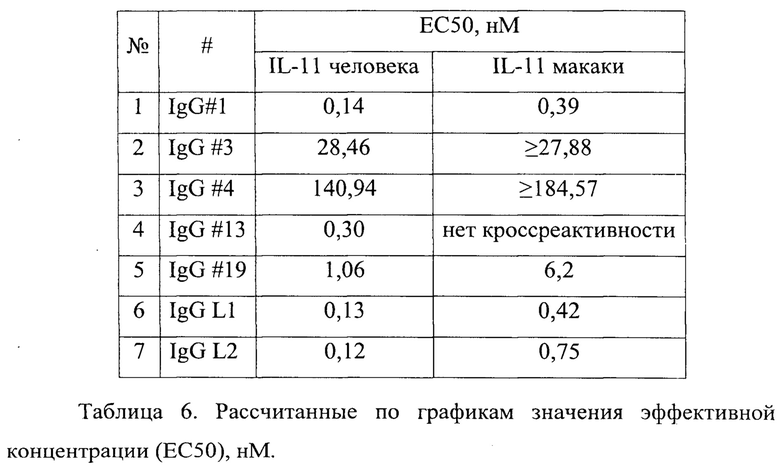
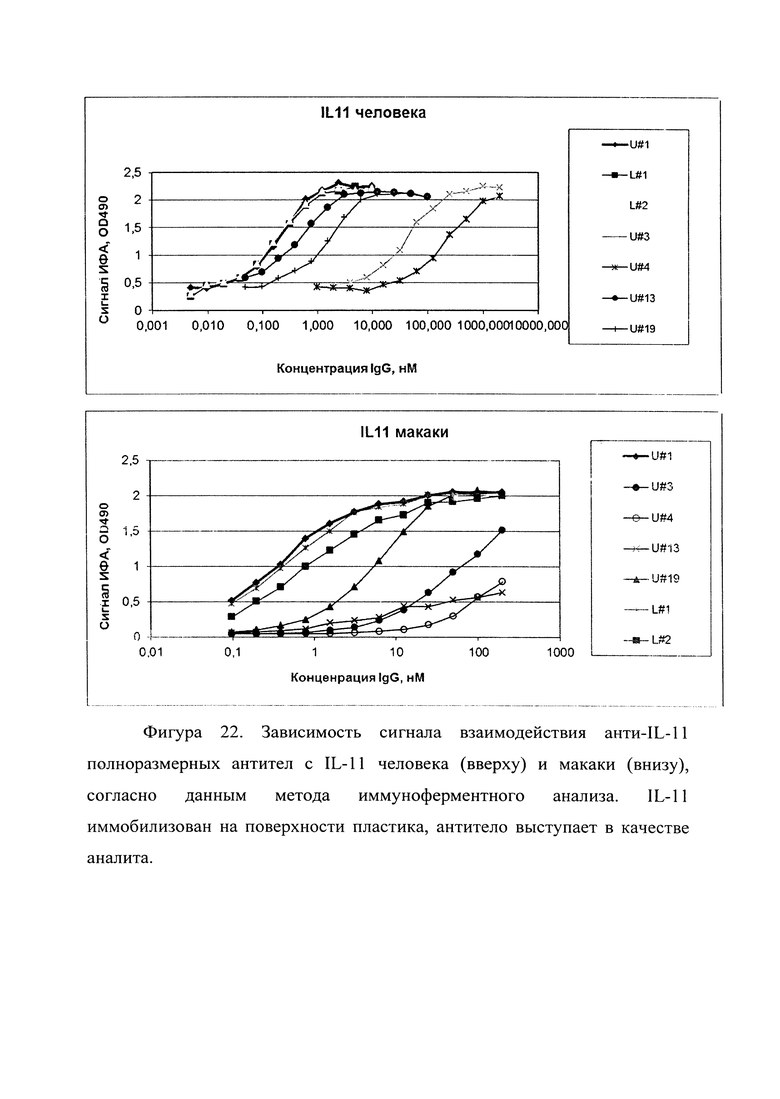
Фигура 22. Отображен график взаимодействия анти-IL-11 полноразмерных IgG1 антител с бактериально экспрессированным IL-11 человека (вверху) и макаки (внизу), согласно данным метода иммуноферментного анализа (ELISA). IL-11 иммобилизован на поверхности пластика (1 мкг/мл), антитело выступает в качестве аналита.
В таблице 6 приведены рассчитанные по графикам фигуры 22 значения эффективной концентрации (ЕС50).
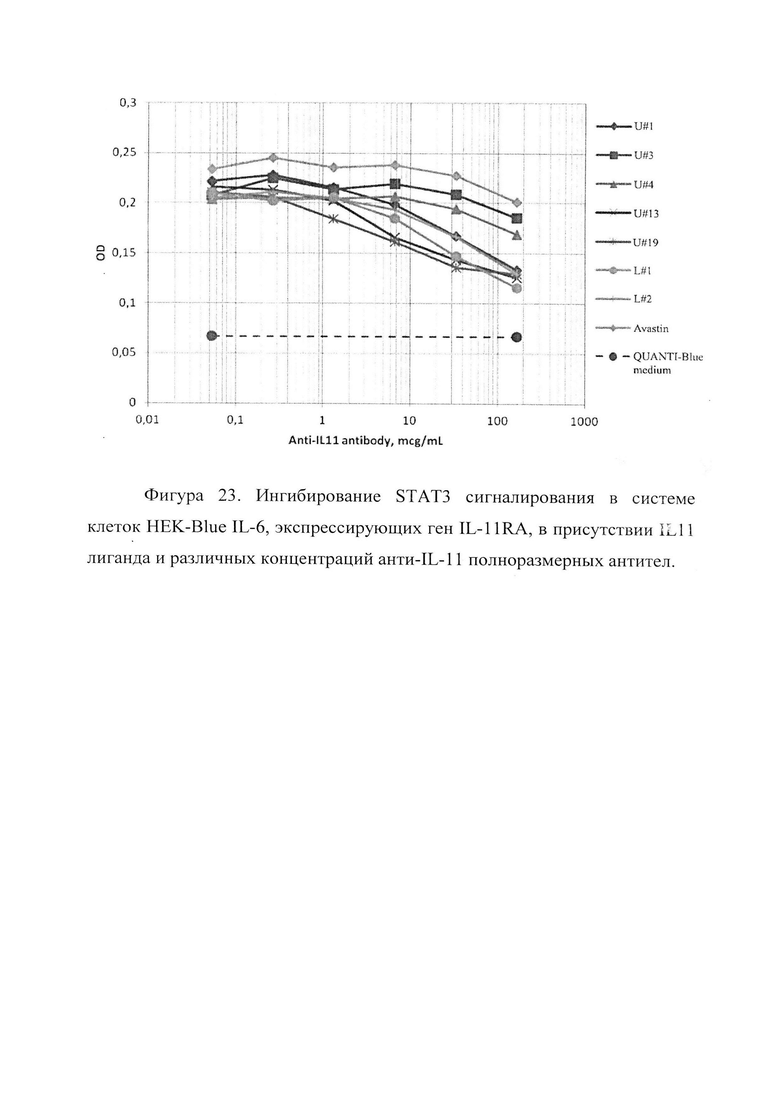
На Фигуре 23 отображено ингибирование STAT3 сигналирования в линии клеток HEK-Blue IL-11, созданной на основе реагентного набора HEK-Blue™ IL-6 Cells компании InvivoGen (http://www.invivogen.com/hekblue-il6), и экспрессирующей полноразмерный ген IL-11RA, в присутствии IL-11 лиганда и различных концентраций анти-IL-11 полноразмерных IgG1 антител. В качестве контрольного антитела использовалось коммерческое терапевтическое полноразмерное антитело препарата Авастин®, не связывающееся с IL-11. Концентрация IL-11 человека 1 нг/мл (52 пМ). Каждая точка соответствует величине, усредненной по двум измерениям. Негативный контроль реакции приведен по уровню поглощения среды без добавления IL-11 лиганда.
Настоящее изобретение далее описано с помощью следующих, не ограничивающих его примеров. В примерах использованы следующие методы:
Пример 1.
Получение рекомбинантного IL-11 человека в бактериальной системе экспрессии описано в источнике [Kazakov, A.S., et al., High-affinity interaction between interleukin-11 and S100P protein. Biochem Bibphys Res Commum, 2015]. Рекомбинантный IL-11 человека в эукариотической системе экспрессии нарабатывался в виде гибридного белка с Fc доменом иммуноглобулина IgG1 человека. Белок был продуцирован в суспензионной культуре клеток линии СНО Ebna при помощи временной (транзиентной) системы экспрессии под управлением CMV промотора. Выделение белка из культуральной жидкости проводилось методом хроматографии на аффинном сорбенте ProteinA-sepharose (метод представляет собой стандартную рутинную операцию).
Пример 2.
Препарат рекомбинантного внеклеточного домена IL-11RA человека приобретен у компании Sino Biological Inc. (#10252-Н08Н).
Пример 3. Получение линии клеток млекопитающих (HEK-Blue IL-11), стабильно экспрессирующих IL-11RA, описано в источнике [Kazakov, A.S., et al., High-affinity interaction between interleukin-11 and S100P protein. Biochem Biophys Res Commum, 2015].
Для получение линии клеток млекопитающих HEK-Blue IL-11, стабильно экспрессирующих IL-11RA, ген рецептора IL-11Rα человека (Sino Biological Inc., #HG10252-М) был внесен в вектор pCMV-Script (Agilent Technologies) между сайтами рестрикции NotI и HindIII. Трансфекцию проводили используя TurboFect Transfection Reagent (Thermo Scientific, # R0532) с последующим отбором позитивных клонов с использованием Генетицина.
Пример 4.
Получение Fab-фрагментов антител проводили с использованием технологии фагового дисплея [Bazan, J., I. Calkosinski, and A. Gamian, Phage display-a powerful technique for immunotherapy: 1. Introduction and potential of therapeutic applications. Hum Vaccin Immunother, 2012. 8(12): p. 1817-28] на основе фаговых библиотек ООО «Антерикс», г. Пущино. Из исходного разнообразия 3-х фаговых библиотек, в ходе трех раундов селекции были отобраны фаговые частицы, содержащие на своей поверхности Fab-фрагменты антител, специфически связывающихся с IL-11 человека и макаки. Были использованы синтетические фаговые библиотеки Fab2h-library, Fab5h-library, Fab7h-library с разнообразием 7,2*Е9, 1,4*Е11 и 2,3*10Е11, соответственно) компании ООО «Антерикс», г. Пущино. Использованы две схемы селекции, отличающиеся порядком презентации антигенов (IL-11 человека - IL-11 макаки - IL-11 человека и IL-11 макаки - IL-11 человека - IL-11 человека). Полученный поликлональный фаг был переклонирован в вектор pLL4 для экспрессии в прокариотах Fab-фрагментов, несущих с-myc и 6XHis метки на С-конце. Проведены ИФА-скрининги отобранных Fab-фрагментов, секретированных в клетках Е. coli штамма BL-21(DE3), на предмет их специфичности к IL-11 человека и макаки. Выделение и очистку белка проводили с помощью металл-хелатной аффинной хроматографии на аффинном сорбенте Profinity IMAC (Bio-Rad Laboratories, Inc.) согласно рекомендациям производителя.
Использованные в работе рутинные методы широко описаны в литературе (Barbas CF, Wagner J (October 1995). "Synthetic Human Antibodies: Selecting and Evolving Functional Proteins". Methods. 8(2): 94-103. doi:10.1006/meth.1995.9997; Barbas CF (August 1995). "Synthetic human antibodies". Nat. Med. 1(8): 837-9. doi: 10.1038/nm0895-837. PMID 7585190)
Пример 5.
Выбор функционально активных антител, блокирующих связывание IL-11 лиганда с внеклеточным доменом рецептора IL-11RA, проводили методом конкурентного ИФА [Wild, D., ed. The Immunbassay Handbook (Fourth edition). Theory and applications of ligand binding, ELISA and related techniques. 4 ed. 2013, Elsevier.].
Специфичность антитела определяется Fab-фрагментами, их функциональной активностью и специфичностью.
Белок IL11Ra рецептор (2 мкг/мл, 50 мкл/лунку) сорбировался на пластик (96-луночный планшет для иммуноферментного анализа). Меченый биотином лиганд IL-11 (50 нМ=1 мкг/мл) прединкубировался с соответствующим Fab-фрагментом антитела (300 нМ) в течение 30 минут и раствор переносился на Elisa-планшет. В контрольном образце раствор Fab-фрагмента заменен на буферный раствор. Сигнал Elisa взаимодействия лиганда IL-11 с рецептором в контрольном образце (без конкуренции) принимался за 100%. Падение сигнала Elisa в образцах в присутствии Fab-фрагментов антител от контрольного образца было выражено в процентах и отражало ингибирующую активность фрагментов антител (конкурентное связывание IL-11 лиганда фрагментом антитела, блокирующее связывание лиганда с рецептором). После подтверждения функциональной активности (способности блокировать взаимодействие лиганда с рецептором) Fab-фрагменты антител были конвертированы в формат полноразмерного антитела, как описано в примере 6.
Пример 6. Получение полноразмерных антител
Клонирование нуклеотидных последовательностей, кодирующих Fab-фрагменты антител, рассмотренных в Примере 4, проводили отдельно для легкой цепи (Lc) в вектор рТТ по сайтам рестрикции SalI, BsiWI (Pfl23II) и отдельно для тяжелой цепи (Hc) в вектор рТТ IgG1 по сайтам рестрикции NheI, SalI. Наработку антител проводили в системе транзиентной экспрессии в эукариотических клетках линии СНО-3Е7, адаптированных для продукции белков млекопитающих. Клетки СНО-3Е7 трансфицировали соответствующими плазмидами рТТ Lc и рТТ IgG1 Hc с использованием агента LPEI. Выделение и очистку антител из культуральной среды проводили с помощью аффинной хроматографии с использованием сорбента с иммобилизованным белком A (HiTrap Protein А FF, GE Healthcare Life Sciences), имеющим высокое сродство к иммуноглобулину G. Полученные антитела были диализованы и стерилизованы фильтрацией через 0,22 мкм фильтр. Чистота препаратов была оценена с помощью электрофореза в полиакриламидном геле (ПААГ) в денатурирующих условиях. Сродство антител к IL-11 было проверено с помощью ИФА и спектроскопии поверхностного плазмонного резонанса.
Использованные в работе рутинные методы широко описаны в литературе, в том числе [Wild, D., ed. The Immunoassay Handbook (Fourth edition). Theory and applications of ligand binding, ELISA and related techniques. 4 ed. 2013, Elsevier. Greenfield, E.A., Antibodies. A laboratory manual. Second edition, in A laboratory manual 2014.
Пример 7.
Исследование способности полноразмерных IgG1 антител подавлять IL-11-индуцированную активацию STAT3 в клетках HEK-Blue IL-11Ra человека, проводили аналогично методу, описанному в инструкции к набору HEK-Blue™ IL-6 Cells компании InvivoGen (http://www.invivogen.com/hekblue-il6).
Была приобретена коммерческая линия клеток HEK-Blue™ IL-6 (InvivoGen), в которых экспресс ируется SEAP репортерный ген, секретирующийся при активации STAT3 системы внутриклеточного сигналирования при активации IL6 рецептора в присутствии IL6 лиганда. Уровень экспрессии SEAP детектируется с помощью QUANTI-Blue™, т.е. при стимуляции клеток рекомбинантным человеческим IL-6 клетки вырабатывают и секретируют белок SEAP, катализирующий образование цветного продукта реакции.
На основе этой линии клеток мы разработали собственную линию клеток, экспрессирующих IL11Ra рецептор. Т.к. механизм внутриклеточного сигналирования у IL-6 и IL-11 рецепторов общий (STAT3 сигналирование), то клетки приобрели способность проявлять активацию обоих рецепторов (вместе или раздельно). В нашем эксперименте была показана активация IL-11 рецептора в присутствии лиганда IL-11 и ингибирование его активации в присутствии полноразмерных антител с аналогичной контрольной концентрации лиганда. Данные результаты демонстрируют способность разработанных антител ингибировать взаимодействие лиганда IL-11 с IL11Ra рецептором и таким образом понижать STAT3 внутриклеточное сигналирование.
| название | год | авторы | номер документа |
|---|---|---|---|
| Антитела для лабораторной диагностики концентрации интерлейкина-11 | 2020 |
|
RU2763178C2 |
| АНТИ-IL-22R-АНТИТЕЛА | 2017 |
|
RU2758721C2 |
| АНТИТЕЛА, СОДЕРЖАЩИЕ ПОЛИПЕПТИД, ВСТРОЕННЫЙ В УЧАСТОК КАРКАСНОЙ ОБЛАСТИ 3 | 2019 |
|
RU2796254C2 |
| АНТИТЕЛА К ЧЕЛОВЕЧЕСКОМУ NKG2D И ИХ ПРИМЕНЕНИЯ | 2008 |
|
RU2563343C2 |
| Моноклональное антитело, которое специфически связывается с GITR | 2019 |
|
RU2734432C1 |
| PD-L1 специфические антитела | 2017 |
|
RU2756236C2 |
| Анти-PD-L1 и IL-2 цитокины | 2017 |
|
RU2769282C2 |
| АНТИ-LAG-3 АНТИТЕЛА И ИХ КОМПОЗИЦИИ | 2017 |
|
RU2755503C2 |
| АНТИТЕЛО, КОТОРОЕ СВЯЗЫВАЕТСЯ С VEGF И IL-1БЕТА, И СПОСОБЫ ПРИМЕНЕНИЯ | 2019 |
|
RU2816476C2 |
| АНТИ-ОХ40 АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2017 |
|
RU2753493C2 |

Настоящее изобретение относится к области иммунологии и медицины и представляет собой терапевтическое антитело, ингибирующее связывание IL-11 с альфа-субъединицей рецептора IL-11RA и сигналирование через его рецептор и/или связывание комплекса IL-11/IL-11RA с бета-субъединицей рецептора интерлейкина-6, а также его применение для лечения пациентов с онкологическими заболеваниями, при которых наблюдается высокий уровень экспрессии IL-11 и/или IL-11RA. 2 н.п. ф-лы, 7 пр., 23 ил., 6 табл.
1. Терапевтическое антитело, ингибирующее связывание IL-11 с альфа-субъединицей рецептора интерлейкина-11 (IL-11RA) и сигналирование через его рецептор и связывание комплекса IL-11/IL-11RA с бета-субъединицей рецептора интерлейкина-6 (gpl30), в котором вариабельные домены тяжелой и легкой цепей содержат одну из следующих комбинаций аминокислотных последовательностей гипервариабельных участков:
Клон U#l: HCDR1: Phe Asn Thr Ser Ser Met Ser (SEQ ID NO: 1); HCDR2: Lys Ile Asn Ser Tyr Lys Ser Tyr Thr Asn (SEQ ID NO: 2); HCDR3: Lys Trp Tyr Trp Phe (SEQ ID NO: 3); LCDR1: Gln Ser Val Ser Ser Ser (SEQ ID NO: 4); LCDR2: Gly Ala Ser Ser Arg Ala Thr (SEQ ID NO: 5); LCDR3: Tyr Tyr Ser Tyr Pro Ile Thr (SEQ ID NO: 6), или
Клон U#3: HCDR1: Phe Thr Gly Tyr Tyr Met His (SEQ ID NO: 7); HCDR2: Gly Ile Ala Pro Ala Ala Gly Ala Thr Asp (SEQ ID NO: 8); HCDR3: Glu Phe Trp Met Thr Phe Tyr Gly Phe (SEQ ID NO: 9); LCDR1: Gin Ser Val Ser Ser Ser (SEQ ID NO: 10); LCDR2: Gly Ala Ser Ser Arg Ala Thr (SEQ ID NO: 11); LCDR3: Ala Tyr Tyr Tyr Tyr Pro Ile Thr (SEQ ID NO: 12), или
Клон U#4: HCDR1: Phe Ser Asn Tyr Trp Met Ser (SEQ ID NO: 13); HCDR2: Thr Ile Ala Ser Ser Asn Ser Tyr Thr Asp (SEQ ID NO: 14); HCDR3: Glu Ser Trp Ser Glu Gly Trp Trp Pro Ser Gly Phe (SEQ ID NO: 15); LCDR1: Gln Ser Val Ser Ser Ser (SEQ ID NO: 16); LCDR2: Gly Ala Ser Ser Arg Ala Thr (SEQ ID NO: 17); LCDR3: Tyr Glu Thr Ser Pro Ile Thr (SEQ ID NO: 18), или
Клон U#13: HCDR1: Phe Thr Gly Tyr Trp Met Asn (SEQ ID NO: 19); HCDR2: Ser Ile Ser Ser Tyr Gly Gly Gly Thr Tyr (SEQ ID NO: 20); HCDR3: Glu Ser Leu His Gln Leu Phe (SEQ ID NO: 21); LCDR1: Gln Ser Val Ser Ser Ser (SEQ ID NO: 22); LCDR2: Gly Ala Ser Ser Arg Ala Thr (SEQ ID NO: 23); LCDR3: Ser Tyr Ser Tyr Ala Pro Ile Thr (SEQ ID NO: 24), или
Клон U#19: HCDR1: Phe Ser Asn Tyr Thr Met Asn (SEQ ID NO: 25); HCDR2: Leu Ile Ser Ser Ser Tyr Ser Thr Thr Tyr (SEQ ID NO: 26); HCDR3: Lys Arg Ser Val Phe (SEQ ID NO: 27); LCDR1: Gln Ser Val Ser Ser Ser (SEQ ID NO. 28); LCDR2: Gly Ala Ser Ser Arg Ala Thr (SEQ ID NO: 29); LCDR3: Tyr Glu Thr Ser Pro Ile Thr (SEQ ID NO: 30).
2. Применение терапевтического антитела по пп.1, 2 для лечения пациентов с онкологическими заболеваниями, при которых наблюдается высокий уровень экспрессии IL-11 и/или IL-11RA, заключающееся во введении страдающим заболеванием пациентам терапевтического антитела, обеспечивающего связывание IL11 лиганда и стерическую или аллостерическую блокировку дальнейшего связывания IL-11 с IL-11RA и/или gp130 и дальнейшего STAT3 сигналирования.
| CAMERON N.J | |||
| et al | |||
| Походная разборная печь для варки пищи и печения хлеба | 1920 |
|
SU11A1 |
| PUTOCZKI T | |||
| L | |||
| et al | |||
| Походная разборная печь для варки пищи и печения хлеба | 1920 |
|
SU11A1 |
| Кинематографический аппарат | 1918 |
|
SU441A1 |
| PUTOCZKI T.L | |||
| et al | |||
| Походная разборная печь для варки пищи и печения хлеба | 1920 |
|
SU11A1 |

