ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к антителам, содержащим вариабельный (V) домен и полипептид-вставку в участке каркасной области 3 V-домена. Такие антитела могут обеспечить улучшенную функциональность и/или фармакокинетику полипептида вставки.
УРОВЕНЬ ТЕХНИКИ, К КОТОРОМУ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Высокая специфичность и аффинность антител делает их идеальными диагностическими и терапевтическими средствами. Достижения в области технологии рекомбинантных антител привела к получению фрагментов антител, таких как фрагменты Fv, Fab, Fab' и F(ab')2. Эти молекулы меньшего размера сохраняют антигенсвязывающую активность целых антител и также демонстрируют улучшенное проникновение в ткани и фармакокинетические свойства по сравнению с целыми молекулами иммуноглобулинов. Действительно, доказано, что фрагменты антител являются универсальными терапевтическими средствами. В то время как оказалось, что такие фрагменты демонстрируют ряд преимуществ над целыми иммуноглобулинами, они также обладают увеличенной скоростью выведения из сыворотки, поскольку они лишены Fc-домена, который обеспечивает длительное время полужизни in vivo (Medasan et al., 1997, J. Immunol. 158:2211-2217). Предшествующие подходы для улучшения времени полужизни молекул антител в сыворотке включают конъюгацию с агентами или доменами, которые взаимодействуют с FcRn-рецептором, такими как сывороточный альбумин человека (HSA). Было показано, что ПЭГ также повышает время полужизни в сыворотке.
Ранее были описаны антитела с двойной специфичностью, т.е. которые связываются с двумя различными антигенами (для обзора, см. Segal et al., 1999, Curr. Opin. Immunol. 11:558-562; Pluckthun & Pack, 1997, Immunotechnology, 3:83-105; Fischer and Leger, 2007, Pathobiology, 74: 3-14). В предыдущих подходах получения гетеробиспецифических молекул на основе антител в основном использовались способы химического сшивания или белковой инженерии. Недостатком химического сшивания является низкий выход образования гетеро- и гомодимера и необходимость их последующего хроматографического разделения. Подходы инженерии белков являются либо в высокой степени сложными (например, конструирование выступов-в-углублениях; Ridgway et al., 1996, Proteins Eng. 9(7):617-621) или в них используются молекулы с ненадлежащими характеристиками стабильности (например, диантитела, scFv). В некоторых случаях биспецифические антитела также могут иметь проблемы пространственного препятствования, так что оба антигена не могут одновременно связываться каждым плечом антитела. Антитела с одним вариабельным доменом, также известные как однодоменные антитела или sdAb, соответствуют вариабельным областям либо тяжелой (VH), либо легкой (VL) цепей антитела. Однодоменные антитела мыши описаны Ward et al., 1989, Nature, 341, 544-546. Также описаны однодоменные антитела человека и "камелизованные" однодоменные антитела человека (Holt et al., 2003, Trends in Biotechnology, 21, 484-490). Однодоменные антитела также получают от животных семейства верблюжьих (верблюды и ламы) и хрящевых рыб (ковровая акула и акулы-няньки). У этих организмов в ходе эволюции сформировались высокоаффинные единичные V-подобные домены (называемые VHH у животных семейства верблюжьих и V-NAR у акул), встроенные в каркасную область Fc-эквивалентного константного домена в качестве интегрального и ключевого компонента их иммунной системы (см. Holliger & Hudson, для обзора; 2005, Nature Biotechnology, 23(9):1126-1136).
В связи с этим остается необходимость в предоставлении дополнительных сконструированных антител, таких как слитые белки, содержащие антительную часть, которая демонстрирует одно или несколько желаемых свойств, таких как увеличенная функциональность (например, поливалентное связывание) или улучшенная фармакокинетика (например, увеличенное время полужизни в сыворотке). Задачей настоящего изобретения является предоставление нового формата сконструированного антитела, которой демонстрирует увеличенную функциональность (например, увеличенное связывание с его антигеном, биспецифическое связывание или поливалентное связывание) или улучшенную фармакокинетику (например, увеличенное время полужизни в сыворотке). В частности, настоящее изобретение относится к новому формату биспецифического антитела, в частности, стабильному и способному одновременно связывать два антитела.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В одном аспекте настоящее изобретение относится к антителу, содержащему вариабельный (V) домен и полипептид-вставку, где полипептид-вставка находится в участке каркасной области 3 V-домена.
В некоторых вариантах осуществления V-домен представляет собой VH-домен, и антитело может дополнительно содержать VL-домен.
В некоторых вариантах осуществления V домен представляет собой VL-домен, и антитело может дополнительно содержать VH-домен.
В некоторых вариантах осуществления полипептид-вставка находится между аминокислотными остатками 73 и 76 VH-домена. Например, полипептид-вставка может находиться в VH-домене: (i) между аминокислотными остатками 73 и 74; (ii) между аминокислотными остатками 74 и 75; или (iii) между аминокислотными остатками 75 и 76.
В некоторых вариантах осуществления один или несколько аминокислотных остатков между 73 и 76 VH-домена заменены полипептидом вставки.
В некоторых вариантах осуществления полипептид-вставка находится между аминокислотными остатками 67 и 70 VL-домена. Например, полипептид-вставка может находиться в VL-домене: (i) между аминокислотными остатками 67 и 68; (ii) между аминокислотными остатками 68 и 69; или (iii) между аминокислотными остатками 69 и 70.
В некоторых вариантах осуществления один или несколько аминокислотных остатков между остатками 67 и 70 VL-домена заменены полипептидом вставки.
В некоторых вариантах осуществления антитело по изобретению дополнительно содержит линкерную последовательность, предпочтительно из по меньшей мере одной, двух, трех, четырех, пяти, шести, семи, восьми, девяти или десяти аминокислот, соединяющую N- и/или C-конец полипептида вставки с участком каркасной области 3.
В некоторых вариантах осуществления антитело по изобретению представляет собой полноразмерное антитело или его связывающий фрагмент. В некоторых вариантах осуществления антитело представляет собой полноразмерный IgG или его связывающий фрагмент. В некоторых вариантах осуществления антитело представляет собой Fab, Fab', F(ab')2, VHH или scFv, в частности, dsscFv.
В некоторых вариантах осуществления антитело по изобретению является гуманизированным.
В некоторых вариантах осуществления антитело по изобретению содержит V-домен, который связывается с сывороточным альбумином человека (HSA). В одном варианте осуществления антитело содержит VL-домен, содержащий CDR-L1, имеющую последовательность SEQ ID NO: 3; CDR-L2, имеющую последовательность SEQ ID NO: 4; CDR-L3, имеющую последовательность SEQ ID NO: 5. В одном варианте осуществления антитело содержит VL-домен, содержащий SEQ ID NO: 2. В одном варианте осуществления антитело содержит легкую (L) цепь, содержащую SEQ ID NO: 1. В одном варианте осуществления антитело содержит VH-домен, содержащий CDR-H1, имеющую последовательность SEQ ID NO: 8; CDR-H2, имеющую последовательность SEQ ID NO: 9; CDR-H3, имеющую последовательность SEQ ID NO: 10. В одном варианте осуществления антитело содержит VH-домен, содержащий SEQ ID NO: 7. В одном варианте осуществления антитело содержит тяжелую (H) цепь, содержащую SEQ ID NO: 6.
В некоторых вариантах осуществления полипептид-вставка представляет собой цитокин, предпочтительно где цитокин выбран из IL-10, IL-15, IL-2, G-CSF, GM-CSF и EPO. В других вариантах осуществления полипептид-вставка представляет собой склеростин. В других вариантах осуществления полипептид-вставка представляет собой второе антитело, предпочтительно где второе антитело представляет собой scFv, в частности, dsscFv, или доменное антитело, такое как VH, или VL, или VHH. VHH может содержать последовательность, выбранную из SEQ ID NO: 67, 69 и 89. В таких вариантах осуществления антитело может связывать распознаваемый им антиген посредством указанного V-домена с аффинностью связывания, которая составляет по меньшей мере 50%, 60%, 70%, 80% или 90% по сравнению с антителом без полипептида вставки.
В некоторых вариантах осуществления второе антитело связывается с тем же антигеном, что и V-домен. Предпочтительно, второе антитело связывается с эпитопом, отличным от V-домена. В таких вариантах осуществления антитело может связывать распознаваемый им антиген с более высокой аффинностью связывания по сравнению с антителом без второго антитела. В некоторых вариантах осуществления второе антитело связывается с антигеном, отличным от антигена первого антитела, т.е. антитела, содержащего V-домен. В некоторых вариантах осуществления второе антитело связывается с IL-17 и первое антитело, т.е. антитело, содержащее V-домен, связывается с HSA. В некоторых вариантах осуществления второе антитело связывается с CD3 и первое антитело, т.е. антитело, содержащее V-домен, связывается с CD28. В некоторых вариантах осуществления второе антитело связывается с CD28 и первое антитело, т.е. антитело, содержащее V-домен, связывается с CD3.
Настоящее изобретение также относится к выделенному полинуклеотиду(ам), такому как молекула(ы) ДНК, и клонирующему или экспрессирующему вектору(ам), который кодирует антитело, описанное в настоящем описании. Также предусматривается клетка-хозяин, содержащая указанный полинуклеотид(ы) или вектор(ы). Кроме того, настоящее изобретение относится к способу получения антитела, описанного в настоящем описании, включающему культивирование указанной клетки-хозяина и выделение антитела.
В другом аспекте настоящее изобретение также относится к фармацевтической композиции, содержащей антитело, содержащее V-домен и полипептид-вставку, как описано в настоящем описании, в комбинации с одним или несколькими из фармацевтически приемлемого эксципиента, разбавителя или носителя. Также предусматривается указанное антитело или указанная фармацевтическая композиция для применения в терапии. Также предусматривается способ терапии, включающий введение указанного антитела или указанной фармацевтической композиции индивидууму, нуждающемуся в терапии. Также предусматривается применение указанного антитела для изготовления лекарственного средства.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг. 1: (A) Типичная структура Fab с выделением участка петли каркасной области 3; (B) выделение определенных остатков в участке каркасной области 3 вариабельной области тяжелой цепи, (C) выделение определенных остатков в участке каркасной области 3 вариабельного домена легкой цепи.
Фиг. 2: (A) Сенсограмма SPR, демонстрирующая связывание CA645/IL-15Fwk3 с цепями рецептора IL-15; (B) Смоделированная структура, демонстрирующая вставку IL-15 в каркасную область 3 VH-домена CA645.
Фиг. 3: (A) Сенсограмма SPR, демонстрирующая связывание CA645/IL-2Fwk3 с β-цепью рецептора IL-2 с добавлением IL-15 или без него; (B) смоделированная структура, демонстрирующая вставку IL-2 в каркасную область 3 VH-домена CA645.
Фиг. 4: (A) Сенсограмма SPR, демонстрирующая связывание CA645/G-CSFFwk3 с HSA и/или рецептором G-CSF; (B) Смоделированная структура, демонстрирующая вставку G-CSF в каркасную область 3 VH-домена CA645.
Фиг. 5: (A) Сенсограмма SPR, демонстрирующая связывание CA645/GM-CSFFwk3 (человека) с рецептором GM-CSF по сравнению с конструкцией CA645/GM-CSFCH1 (человека); (B) Смоделированная структура, демонстрирующая вставку GM-CSF в каркасную область 3 VH-домена CA645.
Фиг. 6: Сенсограмма SPR, демонстрирующая связывание CA645/GM-CSFFwk3 (мыши) с рецептором GM-CSF по сравнению с конструкцией CA645/GM-CSFCH1 (мыши).
Фиг. 7: (A) Сенсограмма SPR, демонстрирующая связывание CA645/EPOFwk3 с рецептором EPO с добавлением EPO или без него; (B) Смоделированная структура, демонстрирующая вставку EPO в каркасную область 3 VH-домена CA645.
Фиг. 8: (A) Сенсограмма SPR, демонстрирующая связывание CA645/VHH6Fwk3 с IL-6, HSA, IL-6:gp80 с HSA или без него; (B) Смоделированная структура, демонстрирующая вставку VHH6 в каркасную область 3 VH-домена CA645.
Фиг. 9: (A) Сенсограмма SPR, демонстрирующая связывание CA645/VHH15Fwk3 с HSA и/или IL-6; (B) Смоделированная структура, демонстрирующая вставку VHH15 в каркасную область 3 VH-домена CA645.
Фигура 10: (A) Сенсограмма SPR, демонстрирующая связывание CA645/склеростинаFwk3 (центральная часть или полноразмерный) с внеклеточным доменом LRP6 с различными линкерными конструкциями; (B) Смоделированная структура, демонстрирующая вставку центральной части склеростина в каркасную область 3 VH-домена CA645.
Фиг. 11: Сенсограмма SPR, демонстрирующая связывание CA645/склеростинаFwk3 (центральная часть или полноразмерный) с внеклеточным доменом LRP4 с различными линкерными конструкциями.
Фиг. 12: (A) Сенсограмма SPR, демонстрирующая конструкцию CA645/(IL-10Fwk3)2, имеющую различные линкерные последовательности, связывающиеся с рецептором IL-10; (B) Смоделированная структура, демонстрирующая вставку димера IL-10 в каркасную область 3 VH-домена CA645.
Фиг. 13: Сенсограмма SPR, демонстрирующая связывание CA497/IL-15Fwk3 с β-цепью рецептора IL-2 с добавлением IL-15 или без него.
Фиг. 14: Сенсограмма SPR, демонстрирующая связывание CA240/IL-15Fwk3 с β-цепью рецептора IL-2 с добавлением IL-15 или без него.
Фиг. 15: Сенсограмма SPR, демонстрирующая серии разведений для связывания HyHEL5/VHH D3L11Fwk3 с лизоцимом.
Фиг. 16: Сенсограмма SPR, демонстрирующая серии разведений для конструкции HyHEL5/VHH15Fwk3, связывающейся с лизоцимом.
Фиг. 17: Сенсограмма SPR, демонстрирующая серии разведений для конструкции CA645/VHH D3L11Fwk3, связывающейся c лизоцимом.
Фиг. 18: Представление потенциальной схемы связывания HyHEL5/VHH D3L11Fwk3 с лизоцимом.
Фиг. 19: Сенсограмма SPR, демонстрирующая конструкцию CA645/IL-15Fwk3, имеющую различные линкерные последовательности, связывающуюся с цепью рецептора IL-15.
Фиг. 20: Кривые DSC (сплошная линия: CA645 Fab; точечная линия: CA497 dsscFv; перевернутый треугольник: CA645 Fab- Fwk3 CA497 dsscFv (vLvH); треугольник: CA645 Fab- Fwk3 CA497 dsscFv (vHvL)).
Фиг. 21: Кривые ингибирования IL-17A в отсутствии (закрашенный круг) или в присутствии (незакрашенный круг) HSA; (A) CA497 IgG4; (B) CA645 Fab-Fwk3 CA497 dsscFv (vHvL); (C) CA645 Fab- Fwk3 CA497 dsscFv (vLvH).
Фиг. 22: Анализ SPR, демонстрирующий связывание IL-17A и HSA с иммобилизованной конструкцией CA645 IgG4P, содержащей dsscFv (vHvL) CA497 против IL-17, встроенный в область Fwk3.
Фиг. 23: Кривые DSC. (A) IgG4P против CD28, содержащий dsscFv против CD3 в области Fwk3 (сплошная линия); IgG4P против CD3, содержащий dsscFv против CD3 в области Fwk3 (точечная линия). (B) Fab против CD28, содержащий dsscFv против CD3 в области Fwk3 (сплошная линия); Fab против CD3, содержащий dsscFv против CD3 в области Fwk3 (точечная линия).
Фиг. 24: Анализ активации T-клеток в PBMC. Незакрашенный треугольник: анти-CD28 Fab Fwk3-анти-CD3 dsscFv; закрашенный круг: Fab против CD3; закрашенный перевернутый треугольник: Fab против CD28; закрашенный круг: анти-CD3 Fab Fwk3-анти-CD3 dsscFv; незакрашенный квадрат: анти-CD28 IgG4P Fwk3-анти-CD3 dsscFv; закрашенный треугольник: анти-CD3 IgG4P-Fwk3 анти-CD3 dsscFv; символ *: OKT3; незакрашенный круг: контроль в виде только клеток. (A) активация CD4 T-клеток (пациент 1), (B) активация CD4 T-клеток (пациент 2) (C) активация CD8 T-клеток (пациент 1) (D) активация CD8 T-клеток (пациент 2).
ПОДРОБНОЕ ОПИСАНИЕ
Антитело
Настоящее изобретение относится к антителу, содержащему вариабельный (V) домен и полипептид-вставку, где полипептид-вставка находится в участке каркасной области 3 (Fwk3) V-домена.
Антитело по настоящему изобретению, как правило, конструируют путем включения полипептида вставки в область Fwk3 V-домена антитела. V-домен, например, может представлять собой VH-домен или VL-домен. Антитело, в которое включен полипептид-вставка, может рассматриваться как каркасное антитело и/или, когда полипептид-вставка также представляет собой антитело, антитело, в которое включен полипептид-вставка, альтернативно может называться первым антителом, в то время как полипептид-вставка называется вторым антителом. Термины "каркасное антитело" и "первое антитело" могут использоваться взаимозаменяемо в контексте настоящего изобретения для указания на антитело, в которое включен полипептид-вставка.
В некоторых вариантах осуществления антитело по изобретению представляет собой полноразмерное антитело или его связывающий фрагмент. В некоторых вариантах осуществления антитело представляет собой полноразмерный IgG или его связывающий фрагмент. В некоторых вариантах осуществления антитело представляет собой полноразмерный IgG1. В некоторых вариантах осуществления антитело представляет собой полноразмерный IgG4. В некоторых вариантах осуществления антитело представляет собой полноразмерный IgG4P, где серин в положении 241 (при нумерации согласно системе нумерации Kabat) заменен на пролин. В некоторых вариантах осуществления антитело представляет собой Fab, Fab', F(ab')2, VHH или scFv. В некоторых вариантах осуществления антитело представляет собой стабилизированный дисульфидной связью scFv или "dsscFv". "Стабилизированный дисульфидной связью одноцепоченый вариабельный фрагмент" или "dsscFv", как используют в настоящем описании, относится к одноцепоеченому вариабельному фрагменту, стабилизированному пептидным линкером между вариабельными доминами VH и VL, и также включает междоменную дисульфидную связь между VH и VL. (см., например, Weatherill et al., Protein Engineering, Design & Selection, 25 (321-329), 2012; WO2007109254. В некоторых вариантах осуществления междоменная дисульфидная связь между VH и VL образована между положениями VH44 и VL100 (нумерация согласно системе нумерации Kabat).
Термин "антитело", как используют в рамках изобретения, главным образом, относится к полноразмерным (интактным, целым) антителам, т.е. содержащим элементы из двух тяжелых цепей и двух легких цепей в случае IgG-антитела.
Термины "конструкция антитела", "сконструированное антитела", "слитый белок антитела" могут использоваться взаимозаменяемо в контексте настоящего изобретения для указания на антитело по изобретению.
Связывающие фрагменты антител, как правило, содержат по меньшей мере один вариабельный домен легкой цепи (VL) или вариабельный домен тяжелой цепи (VH) и включают: одноцепочечные антитела (например, полноразмерная тяжелая цепь или легкая цепь), Fab, модифицированный Fab, Fab', модифицированный Fab', F(ab')2, Fv, Fab-Fv, Fab-dsFv, однодоменные антитела (sdAb, например, VH или VL или VHH), scFv, dsscFv, Bis-scFv, диантитела, тритела, триантитела, тетраантитела и эпитоп-связывающие фрагменты любого из описанных выше (см. например Holliger and Hudson, 2005, Nature Biotech. 23(9):1126-1136; Adair and Lawson, 2005, Drug Design Reviews - Online 2(3), 209-217). Способы получения и производства этих антительных связывающих фрагментов хорошо известны в данной области (см., например, Verma et al., 1998, Journal of Immunological Methods, 216, 165-181). Например, антительные связывающие фрагменты можно получать из целого антитела, особенно из целого моноклонального антитела, с использованием любых подходящих способов ферментативного разрезания и/или расщепления, например, обработки пепсином. Альтернативно антительный исходный материал можно получать с использованием способов рекомбинантных ДНК, вовлекающих манипулирование и реэкспрессию ДНК, кодирующей вариабельные и/или константные области антитела. При желании можно использовать стандартные способы молекулярной биологии для модификации, добавления или удаления аминокислот или доменов. Любые изменения вариабельных или константных областей все еще охватываются терминами "вариабельные" и "константные" области, как используют в рамках изобретения. Исходный материал для антительного фрагмента может быть получен из любого вида, включая, например, мышь, крысу, кролика, хомячка, верблюда, ламу, козу или человека. Части фрагмента антитела могут быть получены из более чем одного вида; например, фрагменты антител могут быть химерными. В одном примере, константные области происходят из одного вида, а вариабельные области происходят из другого. Исходный материал для фрагмента антитела также может быть модифицирован. В другом примере вариабельная область фрагмента антитела создана с использованием способов инженерии рекомбинантных ДНК. Такие сконструированные версии включают версии, созданные, например, из природных вариабельных областей антител посредством инсерций, делеций или замен в или из аминокислотных последовательностей природных антител. Конкретные примеры этого типа включают сконструированные домены вариабельной области, содержащие по меньшей мере одну CDR и, необязательно, одну или несколько аминокислот каркасной области из одного антитела, и остальную часть из домена вариабельной области из второго антитела. Формат Fab-Fv впервые был описан в WO2009/040562, и его стабилизированная дисульфидной связью версия Fab-dsFv впервые описана в WO2010/035012. Другие фрагменты антител включают фрагменты Fab и Fab', описанные в международных патентных заявках WO2005/003169, WO2005/003170 и WO2005/003171. Поливалентные антитела могут обладать мультиспецифичностью, как например, биспецифические, или могут быть моноспецифическими (см., например, WO 92/22583 и WO05/113605). Один такой пример последних из них представляет собой Tri-Fab (или TFM), как описано в WO92/22583. Другой мультиспецифический формат описан в WO2015/197772.
Типичная молекула Fab' содержит пару тяжелой цепи и легкой цепи, в которой тяжелая цепь содержит вариабельную область VH, константный домен CH1 и натуральную или модифицированную шарнирную область, и легкая цепь содержит вариабельную область VL и константный домен CL.
Связывающий домен антитела, как правило, содержит 6 CDR, три из тяжелой цепи и три из легкой цепи. В таком антителе CDR находятся в каркасной области и вместе формируют вариабельную область. Таким образом, в одном варианте осуществления антитело содержит связывающий домен, специфический для антигена, содержащего вариабельную область легкой цепи и вариабельную область тяжелой цепи. В других вариантах осуществления, например, когда антитело представляет собой однодоменное антитело, связывающий домен может содержать только 3 CDR из одного V-домена либо тяжелой цепи, либо легкой цепи. Таким образом, в одном варианте осуществления антитело содержит связывающий домен, специфичный к антигену, содержащий одну из вариабельной области легкой цепи (VL-домен) или вариабельной области тяжелой цепи (VH- или VHH-домен). Будет понятно, что V-домен, который связывает антиген, может связывать его самостоятельно или совместно с другим V-доменом, например, в качестве комплементарной пары VH/VL. Таким образом, антитело, содержащее V-домен, может связывать его антиген совместно с комплементарным доменом V-доменом (VH или VL). В одном варианте осуществления V-домен представляет собой VH-домен, и антитело по изобретению дополнительно содержит VL-домен, где VH-домен связывает его антиген совместно с VL-доменом в качестве пары VH/VL, т.е. антитело по изобретению содержит VH-домен и связывает его антиген совместно с комплементарным VL-доменом. В одном варианте осуществления V-домен представляет собой VL-домен, и антитело по изобретению дополнительно содержит VH-домен, где VL связывает его антиген совместно с VH-доменом в качестве пары VH/VL, т.е. антитело по изобретению содержит VL-домен и связывает его антиген совместно с комплементарным VH-доменом.
Связывающий домен антитела по изобретению, главным образом, относится к VH, VL или паре VH/VL, которая связывает антиген.
Антитела, как правило, содержат один или несколько связывающих доменов. Например, немодифицированный Fab-фрагмент содержит один связывающий домен, образованный парой VH/VL, и целый IgG содержит два связывающих домена, каждый из которых образован парой VH/VL, где два связывающих домена являются идентичными (моноспецифическое антитело) или различаются (биспецифическое или бивалентное антитело). В одном примере антитело по изобретению может представлять собой Fab-фрагмент, содержащий VH-домен и полипептид-вставку в участке Fwk3 VH-домена, и дополнительно содержит VL-домен, который образует комплементарную пару с VH-доменом. В одном примере VL-домен может быть немодифицированным (т.е. антитело по изобретению представляет собой Fab-фрагмент, содержащий один полипептид-вставку в участке Fwk3 VH-домена) или может содержать полипептид-вставку в его участке Fwk3 (т.е. антитело по изобретению представляет собой Fab-фрагмент, содержащий два полипептида-вставки, идентичных, сходных или отличающихся, один из которых встроен в участок Fwk3 VH-домена, а другой встроен в участок Fwk3 VL-домена). В одном примере антитело по изобретению может представлять собой Fab-фрагмент, содержащий VL-домен и полипептид-вставку в участке Fwk3 VL-домена, и дополнительно содержащий VH-домен, который образует комплементарную пару с VL-доменом. В одном примере VH-домен может быть немодифицированным (т.е. антитело по изобретению представляет собой Fab-фрагмент, содержащий один полипептид-вставку в участке Fwk3 VL-домена) или может содержать полипептид-вставку в его участке Fwk3 (т.е. антитело по изобретению представляет собой Fab-фрагмент, содержащий два полипептида-вставки, идентичных, сходных или отличающихся, один из которых встроен в участке Fwk3 VL-домена, а другой встроен в участке Fwk3 VH-домена). В другом примере антитело по изобретению представляет собой полный IgG, содержащий VH-домен и полипептид-вставку в участке Fwk3 VH-домена, и дополнительно содержащий VL-домен, который образует комплементарную пару с VH-доменом. В одном примере VL-домен может быть немодифицированным (т.е. антитело по изобретению представляет собой IgG, содержащий два полипептида вставки, каждый из которых встроен в один из двух VH-доменов IgG) или может содержать полипептид-вставку в его области Fwk3 (т.е. антитело по изобретению представляет собой IgG, содержащий четыре полипептида вставки, идентичных, сходных или отличающихся, каждый из которых встроен в область Fwk3 каждого из двух доменов VH и двух доменов VL). В другом примере антитело по изобретению представляет собой полный IgG, содержащий VL-домен и полипептид-вставку в области Fwk3 VL-домена, и дополнительно содержащий VH-домен, который образует комплементарную пару с VL. VH-домен может быть немодифицированным (т.е. антитело по изобретению представляет собой IgG, содержащий два полипептида вставки, каждый из которых встроен в один из двух VL-доменов IgG) или может содержать полипептид-вставку в его участке Fwk3 (т.е. антитело по изобретению представляет собой IgG, содержащий четыре полипептида вставки, идентичных, сходных или отличающихся, каждый из которых встроен в участок Fwk3 каждого из двух доменов VL и двух доменов VH).
Остатки в вариабельных доменах антител обычно нумеруют в соответствии с системой, разработанной Kabat et al. Эта система указана в Kabat et al., 1987, Sequences of Proteins of Immunological Interest, US Department of Health and Human Services, NIH, USA. В настоящем описании используется эта система нумерации за исключением случая, когда указано иное.
Обозначения остатков по Kabat не всегда прямо соответствуют линейной нумерации аминокислотных остатков. Фактическая линейная аминокислотная последовательность может содержать меньше или дополнительные аминокислоты относительно строгой нумерации Kabat, соответствующие укорочению или вставке в структурный компонент - либо каркасную область, либо определяющую комплементарность область (CDR) основной структуры вариабельного домена. Правильная нумерация остатков по Kabat может быть определена для данного антитела посредством выравнивания остатков по гомологии в последовательности антитела со "стандартной" пронумерованной по Kabat последовательностью способами, известными в данной области.
CDR вариабельного домена тяжелой цепи находятся в остатках 31-35 (CDR-H1), остатках 50-65 (CDR-H2) и остатках 95-102 (CDR-H3) в соответствии с системой нумерации Kabat. CDR вариабельного домена легкой цепи находятся в остатках 24-34 (CDR-L1), остатках 50-56 (CDR-L2) и остатках 89-97 (CDR-L3) в соответствии с системой нумерации Kabat. В дополнение к петлям CDR, между CDR-2 и CDR-3 существует четвертая петля, образованная каркасной областью 3 (Fwk3). На фиг. 1A представлено изображение петли каркасной области 3 в контексте Fab, имеющего легкую и тяжелую цепи. На фиг. 1B и 1C также представлены репрезентативные остатки из этих петель. Система нумерации Kabat определяет каркасную область 3 как положения 66-94 в тяжелой цепи и положения 57-88 в легкой цепи.
Таким образом, в одном варианте осуществления антитело по настоящему изобретению содержит вариабельный (V) домен и полипептид-вставку, где полипептид-вставка находится в положениях 66-94, когда V-домен представляет собой VH-домен, или в положениях 57-88, когда V-домен представляет собой VL-домен, в соответствии с системой нумерации Kabat.
В некоторых вариантах осуществления полипептид-вставка находится между аминокислотными остатками 73 и 76 VH-домена в соответствии с системой нумерации Kabat. Предпочтительно, полипептид-вставка может находиться между аминокислотными остатками 73 и 74, 74 и 75, или 75 и 76 VH-домена, в соответствии с системой нумерации Kabat. В некоторых вариантах осуществления полипептид-вставка находится между аминокислотными остатками 76 и 77, или 77 и 78 VH-домена, в соответствии с системой нумерации Kabat. В некоторых вариантах осуществления один или несколько аминокислотных остатков VH-домена заменены полипептидом вставки.
В некоторых вариантах осуществления полипептид-вставка находится между аминокислотными остатками 67 и 70 VL-домена в соответствии с системой нумерации Kabat. Предпочтительно, полипептид-вставка может находиться между аминокислотными остатками 67 и 68, 68 и 69, или 69 и 70 VL-домена в соответствии с системой нумерации Kabat. В некоторых вариантах осуществления один или несколько аминокислотных остатков VL-домена заменены полипептидом вставки.
Антитело по настоящему изобретению может дополнительно содержать домены константной области. Домены константной области молекулы антитела по настоящему изобретению, если присутствуют, могут быть выбраны с учетом предполагаемой функции молекулы антитела и, в частности, эффекторных функций, которые могут требоваться. Например, домены константной области могут представлять собой домены IgA, IgD, IgE, IgG или IgM человека. В частности, можно использовать домены константной области IgG человека, особенно изотипов IgG1 и IgG3, когда требуются эффекторные функции антитела. Альтернативно можно использовать изотипы IgG2 и IgG4, когда эффекторные функции антител не требуются. Будет понятно, что также можно использовать варианты последовательностей этих доменов константных областей. Например, можно использовать IgG4-молекулы, в которых серин в положении 241 заменен на пролин, как описано в Angal et al. Molecular Immunology, 1993, 30 (1), 105-108. Также специалисту в данной области будет понятно, что антитела могут претерпевать различные посттрансляционные модификации. Тип и степень этих модификаций часто зависят от линии клеток-хозяев, используемой для экспрессии антитела, а также от условий культивирования. Такие модификации могут включать варьирование гликозилирования, окисления метионина, образования дикетопиперазина, изомеризации аспартата и дезамидации аспарагина. Частой модификацией является утрата карбокси-концевого основного остатка (такого как лизин или аргинин) вследствие действия карбоксипептидаз (как описано в Harris, RJ. Journal of Chromatography 705:129-134, 1995).
Антитела для применения в рамках настоящего изобретения могут представлять собой полностью человеческие антитела, гуманизированные антитела или химерные антитела.
В одном варианте осуществления антитело по настоящему изобретению не является антителом мыши. Термин "антитело мыши" относится к антителу, имеющему тяжелые и легкие цепи, происходящие исключительно из B-клеток мыши. Таким образом, это антитело состоит из аминокислотных последовательностей мыши, независимо от происхождения клеток, которые обеспечивают их продуцирование.
В одном варианте осуществления антитело по настоящему изобретению представляет собой химерное антитело. Термин "химерное антитело" относится к антителу или его антигенсвязывающему фрагменту, где каждая последовательность легкой цепи и/или тяжелой цепи, из которой оно состоит, содержит или состоит из гибридной последовательности, происходящей из по меньшей мере двух различных животных, особенно млекопитающих, в частности, к антителу, в котором скомбинирована вариабельная область кролика, мыши, крысы или яванского макака с константной областью человека.
В одном варианте осуществления антитело по настоящему изобретению представляет собой гуманизированное антитело. Гуманизированные антитела представляют собой молекулы антител из не являющегося человеком вида, имеющие одну или несколько определяющих комплементарность областей (CDR) из не являющегося человеком вида и каркасную область из молекулы иммуноглобулина человека (см., например, US 5585089). В частности, термин "гуманизированное" относится к антителу или его антигенсвязывающему фрагменту, где тяжелая и/или легкая цепь содержит одну или несколько CDR из не являющегося человеческим антитела (такого как моноклональное антитело кролика, мыши, крысы, яванского макака или ламы), трансплантированные в каркасную область вариабельной области тяжелой и/или легкой цепей акцепторного антитела, такого как антитело человека. Вместо переноса целой CDR, только один или несколько из определяющих специфичность остатков из любой из CDR, описанных в настоящем описании выше, можно переносить в каркасную область антитела человека (см., например, Kashmiri et al., 2005, Methods, 36, 25-34). Отдельные остатки каркасной области гуманизированного антитела могут быть заменены соответствующими остатками (так называемые донорные остатки) из не являющегося человеческим антитела (например, антитело, из которого происходят CDR или определяющие специфичность остатки), например для восстановления или повышения специфичности или аффинности антитела.
Полностью человеческие антитела представляют собой антитела, в которых вариабельная область(и) и константная область(и) (когда присутствуют) тяжелой и/или легкой цепей являются полностью человеческими по происхождению или по существу идентичными последовательностям человеческого происхождения, не обязательно из одного и того же антитела. Примеры полностью человеческих антител могут включать антитела, полученные, например, способами фагового дисплея, и антитела, продуцированные мышами, у которых гены вариабельной и/или константной области иммуноглобулина мыши заменены аналогами из человека, например, как в общем описано в EP0546073 B1, US 5545806, US 5569825, US 5625126, US 5633425, US 5661016, US 5770429, EP 0438474 B1 и EP0463151 B1.
Антитела, содержащие V-домен и полипептид-вставку, как описано в настоящем описании, могут быть далее модифицированы, например, химически конъюгированы с другими соединениями, которые могут влиять, например, на их фармакокинетические свойства или связывание антигена.
В одном примере антитела по настоящему изобретению связаны с частями поли(этиленгликоля) (ПЭГ). Антитела, связанные с ПЭГ, могут демонстрировать, например, дополнительно увеличенное время полужизни в сыворотке. В одном конкретном примере антитело представляет собой фрагмент антитела и молекулы ПЭГ могут быть связаны через любую доступную боковую цепь аминокислоты или функциональную группу концевой аминокислоты, находящейся во фрагменте антитела, например, любую свободную амино, имино, тиольную, гидроксильную или карбоксильную группу. Такие аминокислоты могут быть встречающимися естественным образом во фрагменте антитела или могут быть встроены во фрагмент с использованием способов рекомбинантных ДНК (см., например, US 5219996; US 5667425; WO98/25971, WO2008/038024). В одном примере антитело по настоящему изобретению представляет собой модифицированный Fab-фрагмент, где модификация представляет собой добавление к C-концу его тяжелой цепи одной или нескольких аминокислот для обеспечения присоединения эффекторной молекулы. В подходящем случае дополнительные аминокислоты формируют модифицированную шарнирную область, содержащую один или несколько остатков цистеина, с которым может быть связана эффекторная молекула. Можно использовать множество участков для присоединения двух или более молекул ПЭГ.
В подходящем случае молекулы ПЭГ могут быть ковалентно связаны через тиольную группу по меньшей мере одного остатка цистеина, находящегося во фрагменте антитела. Каждая полимерная молекула, связанная с модифицированным фрагментом антитела, может быть ковалентно связана с атомом серы остатка цистеина, находящегося во фрагменте антитела. Ковалентная связь, как правило, представляет собой дисульфидную связь или, в частности, связь сера-углерод. Когда тиольную группу используют в качестве точки присоединения, можно использовать надлежащим образом активированные эффекторные молекулы, например, тиол-селективные производные, такие как малеинимиды и производные цистеина. В препарате модифицированных полимером фрагментов антител, как описано выше, можно использовать активированный полимер. Активированный полимер может представлять собой любой полимер, содержащий реакционноспособную в отношении тиола группу, такую как α-галогенкарбоновая кислота или сложный эфир, например, йодацетамид, имид, например, малеинимид, винилсульфон или дисульфид. Такие исходные материалы можно получать из коммерческих источников (например, от Nektar, ранее Shearwater Polymers Inc., Huntsville, AL, США) или можно получать из коммерчески доступных исходных материалов с использованием общепринятых химических методик. Конкретные молекулы ПЭГ включают метокси-ПЭГ-амин 20K (получаемый от Nektar, ранее Shearwater; Rapp Polymere; и SunBio) и M-PEG-SPA (получаемый от Nektar, ранее Shearwater).
Предпочтительно, антитело по настоящему изобретению обладает аффинностью связывания (при измерении по его константе диссоциации KD) в отношении распознаваемого им антигена 10-5 M или менее, 10-6 M или менее, 10-7 M или менее, 10-8 M или менее, 10-9 M или менее, 10-10 M или менее, или 10-11 M или менее. Аффинность можно измерять известными способами, такими как способы поверхностного плазмонного резонанса, включая Biacore™. Аффинность можно измерять при комнатной температуре, 25°C или 37°C. Аффинность можно измерять при физиологических значениях pH, т.е. при приблизительно pH 7,4.
Будет понятно, что аффинность антител, предусматриваемых настоящим изобретением, можно изменять с использованием любого подходящего способа, известного в данной области. Настоящее изобретение, таким образом, также относится к вариантам молекул антител по настоящему изобретению, которые обладают увеличенной аффинностью к альбумину. Такие варианты можно получать посредством ряда протоколов созревания аффинности, включающих мутацию CDR (Yang et al., J. Mol. Biol, 254. 392-403, 1995), шаффлинг цепей (Marks et al., Bio/Technology, 10, 779-783, 1992), использование штаммов-мутаторов E. coli (Low et al., J. Mol. Biol., 260. 359-368, 1996), шаффлинг ДНК (Patten et al., Curr. Opin. BiotechnoL, 8, 724-733, 1997), фаговый дисплей (Thompson et al., J. Mol. Biol, 256. 77-88, 1996) и половую ПЦР (Crameri et al.. Nature, 391. 288-291, 1998).
Антитело, связывающее сывороточный альбумин человека
В одном варианте осуществления антитело по изобретению содержит V-домен, который связывается с сывороточным альбумином человека (HSA). В таком варианте осуществления антитело может быть пригодным для продления времени полужизни полипептида вставки в сыворотке. Будет понятно, что V-домен, который связывает альбумин, также может связывать его самостоятельно или совместно с другим V-доменом, например, в качестве комплементарной пары VH/VL. Таким образом, в некоторых вариантах осуществления, антитело, содержащее V-домен, связывает альбумин совместно с комплементарным VH- или VL-доменом. В одном варианте осуществления V-домен представляет собой VH-домен, и антитело по изобретению дополнительно содержит VL-домен, где VH связывает альбумин совместно с VL-доменом в качестве пары VH/VL, т.е. антитело по изобретению содержит VH-домен и связывается с альбумином совместно с комплементарным VL. В одном варианте осуществления V-домен представляет собой VL-домен, и антитело по изобретению дополнительно содержит VH-домен, где VL связывает альбумин совместно с VH-доменом в качестве пары VH/VL, т.е. антитело по изобретению содержит VL-домен и связывается с альбумином совместно с комплементарным VH. Когда антитело по изобретению связывает альбумин, VH/VL, как правило, называют "альбумин-связывающим доменом". В связи с этим, альбуминсвязывающий домен антитела по изобретению относится к VH, VL или паре VH/VL, которая связывается с альбумином.
В одном варианте осуществления антитело по настоящему изобретению связывает сывороточный альбумин яванского макака, сывороточный альбумин мыши и/или сывороточный альбумин крысы.
В одном варианте осуществления антитело по настоящему изобретению, содержащее V-домен, который связывается с сывороточным альбумином человека, является гуманизированным. В одном варианте осуществления антитело по настоящему изобретению, содержащее альбуминсвязывающий домен, представляет собой гуманизированное антитело, где тяжелая и/или легкая цепь содержит одну или несколько CDR из моноклонального антитела кролика, мыши или крысы, трансплантированных в каркасную область вариабельной области тяжелой и/или легкой цепи антитела человека. В одном варианте осуществления антитело по настоящему изобретению, содержащее альбуминсвязывающий домен, представляет собой гуманизированное антитело, где тяжелая и/или легкая цепь содержит одну или несколько CDR из антител кролика, трансплантированных в каркасную область вариабельной области тяжелой и/или легкой цепи антитела человека, и где каркасная область вариабельной области тяжелой и/или легкой цепи содержит по меньшей мере одну замену аминокислоты, такой как аминокислота, которая является донорным остатком.
В одном варианте осуществления антитело по настоящему изобретению, содержащее V-домен, который связывается с сывороточным альбумином человека, содержит последовательность легкой и/или тяжелой цепи, как показано ниже (вариабельный домен подчеркнут; CDR выделены полужирным шрифтом и курсивом). В одном варианте осуществления антитело по настоящему изобретению, содержащее V-домен, который связывается с сывороточным альбумином человека, содержит последовательность легкой и/или тяжелой цепи, выбранную из последовательностей, приведенных ниже. В одном варианте осуществления антитело по настоящему изобретению, содержащее V-домен, который связывается с сывороточным альбумином человека, содержит последовательность вариабельного домена легкой и/или тяжелой цепи, выбранную из приведенных ниже последовательностей. В одном варианте осуществления антитело по настоящему изобретению, содержащее V-домен, который связывается с сывороточным альбумином человека, содержит по меньшей мере одну из последовательностей CDR-L1, CDR-L2 и CDR-L3 и/или по меньшей мере одну из последовательностей CDR-H1, CDR-H2 и CDR-H3, выбранных из приведенных ниже последовательностей. В одном варианте осуществления антитело по настоящему изобретению, содержащее V-домен, который связывается с сывороточным альбумином человека, содержит последовательность легкой и/или тяжелой цепи; и/или последовательность вариабельного домена легкой и/или тяжелой цепи; и/или по меньшей мере одну из последовательностей CDR-L1, CDR-L2 и CDR-L3; и/или по меньшей мере одну из последовательностей CDR-H1, CDR-H2 и CDR-H3, выбранных из приведенных ниже последовательностей.
Легкая цепь CA645 (gL5)
DIQMTQSPSSVSASVGDRVTITCQSSPSVWSNFLSWYQQKPGKAPKLLIYEASKLTSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCGGGYSSISDTTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC (SEQ ID NO: 1)
VL-домен (gL5):
DIQMTQSPSSVSASVGDRVTITCQSSPSVWSNFLSWYQQKPGKAPKLLIYEASKLTSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCGGGYSSISDTTFGGGTKVEIK (SEQ ID NO: 2)
CDR-L1: QSSPSVWSNFLS (SEQ ID NO: 3)
CDR-L2: EASKLTS (SEQ ID NO: 4)
CDR-L3: GGGYSSISDTT (SEQ ID NO: 5)
Тяжелая цепь CA645 (gH5)
EVQLLESGGGLVQPGGSLRLSCAVSGIDLSNYAINWVRQAPGKGLEWIGIIWASGTTFYATWAKGRFTISRDNSKNTVYLQMNSLRAEDTAVYYCARTVPGYSTAPYFDLWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC (SEQ ID NO: 6)
VH-домен (gH5): EVQLLESGGGLVQPGGSLRLSCAVSGIDLSNYAINWVRQAPGKGLEWIGIIWASGTTFYATWAKGRFTISRDNSKNTVYLQMNSLRAEDTAVYYCARTVPGYSTAPYFDLWGQGTLVTVSS (SEQ ID NO: 7)
CDR-H1: GIDLSNYAIN (SEQ ID NO: 8)
CDR-H2: IIWASGTTFYATWAKG (SEQ ID NO: 9)
CDR-H3: TVPGYSTAPYFDL (SEQ ID NO: 10)
В некоторых вариантах осуществления антитело по настоящему изобретению содержит варианты VL- и VH-доменов, связывающие сывороточный альбумин человека, как описано выше (SEQ ID NO: 2 и 7, соответственно), которые содержат дополнительный остаток, так что между VL- и VH-доменами может образовываться дисульфидная связь. Дополнительные цистеин-содержащие варианты могут иметь следующие последовательности (где дополнительные остатки цистеина подчеркнуты):
VL-домен CA645-Cys (gL5):
DIQMTQSPSSVSASVGDRVTITCQSSPSVWSNFLSWYQQKPGKAPKLLIYEASKLTSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCGGGYSSISDTTFGCGTKVEIK (SEQ ID NO: 102)
VH-домен CA645-Cys (gH5):
EVQLLESGGGLVQPGGSLRLSCAVSGIDLSNYAINWVRQAPGKCLEWIGIIWASGTTFYATWAKGRFTISRDNSKNTVYLQMNSLRAEDTAVYYCARTVPGYSTAPYFDLWGQGTLVTVSS (SEQ ID NO: 103)
Дополнительные примеры VL- и VH-доменов, которые связывают сывороточный альбумин человека и которые можно использовать в антителе по изобретению, содержат SEQ ID NO: 105, 106, 107, 108 и 109, как описано ниже (CDR выделены полужирным шрифтом и курсивом).
VH-домен CA645 (gH1):
EVQLLESGGGLVQPGGSLRLSCAVSGIDLSNYAINWVRQAPGKGLEWIGIIWASGTTFYATWAKGRFTISRDSTTVYLQMNSLRAEDTAVYYCARTVPGYSTAPYFDLWGQGTLVTVSS (SEQ ID NO: 105)
VH-домен CA645 (gH37):
EVQLLESGGGLVQPGGSLRLSCAVSGIDLSNYAINWVRQAPGKGLEWIGIIWASGTTAYATWAKGRFTI SRDNSKNTVYLQMNSLRAEDTAVYYCARTVPGYSTAPYFDLWGQGTLVTVSS (SEQ ID NO: 106)
VH-домен CA645 (gH47):
EVQLLESGGGLVQPGGSLRLSCAVSGIDLSNYAINWVRQAPGKGLEWIGIIWASGTTFYATWAKGRFTISRDNSKNTVYLQMNSLRAEDAVYYCARTVPGYSAAPYFDLWGQGTLVTVSS (SEQ ID NO: 107)
VL-домен CA645 (gL1):
DIVMTQSPSSVSASVGDRVTITCQSSPSVWSNFLSWYQQKPGKAPKLLIYEASKLTS GVPSRFKGSGSGTDFTLTISSLQPEDFATYYCGGGYSSISDTTFGGGTKVEIK (SEQ ID NO: 108)
VL-домен CA645 (gL4):
DIQMTQSPSSVSASVGDRVTITCQSSPSVWSNFLSWYQQKPGKAPKLLIYEASKLTS GVPSRFSGSGSGTDFTLTISSLQPEDFATYYC GGGYSSISD TTFGGGTKVEIK
(SEQ ID NO: 109)
В некоторых вариантах осуществления антитело по настоящему изобретению содержит варианты VL- и VH-доменов, связывающие сывороточный альбумин человека, как описано выше (SEQ ID NO: 105-SEQ ID NO: 109), которые содержат дополнительный остаток цистеина, так что между VL- и VH-доменами может образовываться дисульфидная связь. Дополнительные цистеинсодержащие варианты могут иметь следующую последовательность (где дополнительный остаток цистеина подчеркнут):
VL CA645-Cys (gL4):
diqmtqspssvsasvgdrvtitcqsspsvwsnflswyqqkpgkapklliyeaskltsgvpsrfsgsgsgtdftltisslqpedfatyycgggyssisdttfgCgtkveikRT(SEQ ID NO: 110)
В некоторых вариантах осуществления каркасная область VH альбуминсвязывающего домена является человеческой (например, VH3, такая как VH3 1-3 3-23), и содержит, например, 1, 2, 3, 4, 5 или 6 аминокислотных замен, таких как аминокислоты, которые являются донорными остатками. В таких вариантах осуществления VH может иметь последовательность, показанную в SEQ ID NO: 7, SEQ ID NO: 103, SEQ ID NO: 105, SEQ ID NO: 106, SEQ ID NO: 107, или вариант любой из них, обладающий по меньшей мере 95, 96, 97, 98 или 99% сходством или идентичностью.
В некоторых вариантах осуществления каркасная область VL альбуминсвязывающего домена является человеческой (например, Vκ1, такая как 2-1- (1) L5), и содержит, например, 1, 2, 3, 4, 5 или 6 замен аминокислот, таких как аминокислоты, которые являются донорными остатками. В таких вариантах осуществления VL может иметь последовательность, показанную в SEQ ID NO: 2, SEQ ID NO: 102, SEQ ID NO: 108, SEQ ID NO: 109, SEQ ID NO: 110, или вариант любой из них, обладающий по меньшей мере 95, 96, 97, 98 или 99% сходством или идентичностью.
В некоторых вариантах осуществления альбуминсвязывающий домен содержит последовательности VH и VL, выбранные из комбинаций SEQ ID NO: 7 и 2, 7 и 102, 7 и 108, 7 и 109, 7 и 110, 103 и 2, 103 и 102, 103 и 108, 103 и 109, 103 и 110, 105 и 2, 105 и 102, 105 и 108, 105 и 109, 105 и 110, 106 и 2, 106 и 102, 106 и 108, 106 и 109, 106 и 110, 107 и 2, 107 и 102, 107 и 108, 107 и 109, 107 и 110, или варианта или вариантов любой из них, обладающих по меньшей мере 95, 96, 97, 98 или 99% сходством или идентичностью.
В некоторых вариантах осуществления последовательности VL и VH альбуминсвязывающего домена представляют собой SEQ ID NO:102 и SEQ ID NO: 103, соответственно. В некоторых вариантах осуществления последовательности VL и VH альбуминсвязывающего домена представляют собой SEQ ID NO:2 и SEQ ID NO: 7, соответственно.
В одном варианте осуществления антитело, содержащее V-домен, который связывает сывороточный альбумин человека, представляет собой Fab или scFv. VL- и VH-домены SEQ ID NO: 2 и 7 являются предпочтительными для Fab-формата. В одном варианте осуществления антитело, содержащее V-домен, который связывает сывороточный альбумин человека, представляет собой scFv, который связывает альбумин совместно с комплементарным V-доменом (VL или VH). В таком варианте осуществления scFv может содержать любой из VH-доменов и любой из VL-доменов, как описано выше, связанных в ориентации VH-VL или VL-VH посредством линкера (G4S)4 (SEQ ID NO:16). VL- и VH-домены SEQ ID NO: 2 и 7 являются предпочтительными для формата scFv. VL-домен SEQ ID NO: 2 может содержать дополнительные две аминокислоты RT на C-конце. В одном варианте осуществления антитело, содержащее V-домен, который связывает сывороточный альбумин человека, представляет собой scFv, который содержит VL- и VH-домены SEQ ID NO: 2 и 7, разделенные линкером (G4S)4. В одном примере scFv имеет ориентацию VH-VL. В другом примере scFv имеет ориентацию VL-VH. В одном варианте осуществления антитело, содержащее V-домен, который связывает сывороточный альбумин человека, представляет собой scFv, который содержит или имеет SEQ ID NO: 143, как показано ниже. В одном конкретном варианте осуществления антитело, содержащее V-домен, который связывает сывороточный альбумин человека, представляет собой dsscFv. В одном варианте осуществления антитело, содержащее V-домен, который связывает сывороточный альбумин человека, представляет собой dsscFv, который связывает альбумин совместно с комплементарным V-доменом (VL или VH). В таком варианте осуществления dsscFv может содержать любые VH-домены и любые VL-домены, как описано выше, содержащие, если он уже не присутствует, дополнительный остаток цистеина, так чтобы между VL- и VH-доменами могла образовываться дисульфидная связь, где VL- и VH-домены связаны в ориентации VH-VL или VL-VH посредством линкера (G4S)4. VL- и VH-домены SEQ ID NO: 102 и 103 являются предпочтительными для формата dsscFv. VL-домен SEQ ID NO: 102 может содержать дополнительные две аминокислоты RT на C-конце. В одном варианте осуществления антитело, содержащее V-домен, который связывает сывороточный альбумин человека, представляет собой dsscFv, который содержит VL- и VH-домены SEQ ID NO: 102 и 103, разделенные линкером (G4S)4 (SEQ ID NO:16). В одном примере dsscFv имеет ориентацию VH-VL. В другом примере dsscFv имеет ориентацию VL-VH. В одном варианте осуществления антитело, содержащее V-домен, который связывает сывороточный альбумин человека, представляет собой dsscFv, который содержит или имеет SEQ ID NO: 144, как показано ниже.
CA645 scFv (VH-VL):
evqllesggglvqpggslrlscavsgidlsnyainwvrqapgkGlewigiiwasgttfyatwakgrftisrdnskntvylqmnslraedtavyycartvpgystapyfdlwgqgtlvtvssggggsggggsggggsggggsdiqmtqspssvsasvgdrvtitcqsspsvwsnflswyqqkpgkapklliyeaskltsgvpsrfsgsgsgtdftltisslqpedfatyycgggyssisdttfgQgtkveik (SEQ ID NO: 143)
dsscFv (VH-VL) CA645, где дополнительные остатки цистеина подчеркнуты:
evqllesggglvqpggslrlscavsgidlsnyainwvrqapgkclewigiiwasgttfyatwakgrftisrdnskntvylqmnslraedtavyycartvpgystapyfdlwgqgtlvtvssggggsggggsggggsggggsdiqmtqspssvsasvgdrvtitcqsspsvwsnflswyqqkpgkapklliyeaskltsgvpsrfsgsgsgtdftltisslqpedfatyycgggyssisdttfgcgtkveik (SEQ ID NO: 144)
В другом варианте осуществления антитело, содержащее V-домен, который связывает сывороточный альбумин человека, представляет собой полноразмерный IgG. VL- и VH-домены SEQ ID NO: 2 и 7 являются предпочтительными для IgG-формата. В одном варианте осуществления антитело, содержащее V-домен, который связывает сывороточный альбумин человека, представляет собой IgG1. В одном варианте осуществления антитело, содержащее V-домен, который связывает сывороточный альбумин человека, представляет собой IgG4. В одном варианте осуществления антитело, содержащее V-домен, который связывает сывороточный альбумин человека, представляет собой IgG4P.
Будет понятно, что одну или несколько (например, 1, 2, 3 или 4) аминокислотных замен, вставок и/или делеций можно вносить в CDR или другие последовательности (например, вариабельные домены), предусматриваемые настоящим изобретением, без значительного изменения способности антитела связываться с распознаваемым им антигеном (например, HSA). Эффект любых аминокислотных замен, вставок и/или делеций может без труда протестировать специалист в данной области. Например, одну или несколько (например, 1, 2, 3 или 4) аминокислотных замен, вставок и/или делеций можно вносить в каркасную область, используемую в антителе или фрагменте, предусматриваемых настоящим изобретением, где аффинность связывания с распознаваемым ими антигеном сохранена или увеличена. В некоторых вариантах осуществления аминокислота в положении 72 каркасной области 3 VH-домена представляет собой аминокислоту дикого типа, естественным образом присутствующую в положении 72 указанного VH-домена.
В связи с этим, настоящее изобретение также относится к антителам, которые содержат последовательности, которые являются по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% сходными или идентичными последовательности, приведенной в настоящем описании. "Идентичность", как используют в рамках изобретения, указывает на любое конкретное положение в выравниваемых последовательностях, где аминокислотный остаток является идентичным между последовательностями. "Сходство", как используют в рамках изобретения, указывает на то, что в любом конкретном положении в выравниваемых последовательностях аминокислотных остаток представляет собой аминокислотный остаток сходного типа между последовательностями. Например, лейцин может быть заменен на изолейцин или валин. Другие аминокислоты, которые часто заменяются на другие аминокислоты, включают, но не ограничиваются ими:
- фенилаланин, тирозин и триптофан (аминокислоты, имеющие ароматические боковые цепи);
- лизин, аргинин и гистидин (аминокислоты, имеющие основные боковые цепи);
- аспартат и глутамат (аминокислоты, имеющие кислотные боковые цепи);
- аспарагин и глутамин (аминокислоты, имеющие амидные боковые цепи); и
- цистеин и метионин (аминокислоты, имеющие содержащие серу боковые цепи).
Степени идентичности и сходства можно без труда вычислять (Computational Molecular Biology, Lesk, A.M., ed., Oxford University Press, New York, 1988; Biocomputing. Informatics and Genome Projects, Smith, D.W., ed., Academic Press, New York, 1993; Computer Analysis of Sequences Data, Part 1, Griffin, A.M., and Griffin, H.G., eds., Humana Press, New Jersey, 1994; Sequence Analysis in Molecular Biology, von Heinje, G., Academic Press, 1987, Sequence Analysis Primer, Gribskov, M. and Devereux, J., eds., M Stockton Press, New York, 1991, the BLAST™ software available from NCBI (Altschul, S.F. et al., 1990, J. Mol. Biol. 215:403-410; Gish, W. & States, D.J. 1993, Nature Genet. 3:266-272. Madden, T.L. et al., 1996, Meth. Enzymol. 266:131-141; Altschul, S.F. et al., 1997, Nucleic Acids Res. 25:3389-3402; Zhang, J. & Madden, T.L. 1997, Genome Res. 7:649-656).
Антитело по настоящему изобретению может связываться с альбумином с аффинностью связывания, достаточной для продления времени полужизни полипептида вставки, такого как терапевтический белок или второе антитело, in vivo. В одном варианте осуществления полипептид-вставка связывается с HSA. В таком варианте осуществления полипептид-вставка может связываться с альбумином с аффинностью связывания, достаточной для продления времени полужизни каркасного антитела. Было описано, что KD для альбумина меньше или ровно 2,5 мкМ будет продлевать время полужизин in vivo (Nguyen, A. et al (2006) Protein Engineering, Design & Selection, 19(7), 291-297). В одном примере антитело по настоящему изобретению может связываться с альбумином с высокой аффинностью связывания, которая является наномолярной или микромолярной, например, с KD, равной или меньшей чем 3 нМ. Аффинность можно измерять с использованием любого подходящего способа, известного в данной области, включая поверхностный плазмонный резонанс с использованием природного или рекомбинантного сывороточного альбумина.
В некоторых вариантах осуществления полипептид-вставка представляет собой альбуминсвязывающий пептид. Примеры альбуминсвязывающих пептидов описаны в WO2007/106120 и включают:
В одном варианте осуществления полипептид-вставка представляет собой второе антитело, которое связывается с HSA. В таком варианте осуществления полипептид-вставка может иметь признаки, описанные выше для антитела, содержащего V-домен, т.е. каркасного антитела. В некоторых вариантах осуществления полипептид-вставка может содержать по меньшей мере одну из CDR, по меньшей мере один из VL- или VH-доменов, или по меньшей мере одну из легкой или тяжелой цепей, которые связывают альбумин, как описано выше. В одном варианте осуществления полипептид-вставка содержит по меньшей мере одну из CDR-L1, CDR-L2 и CDR-L3 с последовательностью SEQ ID NO: 3, SEQ ID NO: 4 и SEQ ID NO: 5, соответственно, и/или одну из CDR-H1, CDR-H2 и CDR-H3 с последовательностью SEQ ID NO: 8, SEQ ID NO: 9 и SEQ ID NO: 10, соответственно.
Полипептид-вставка
В соответствии с настоящим изобретением, настоящее изобретение относится к антителам, содержащим полипептид-вставку в каркасной области 3 V-домена. V-домен может представлять собой VL- или VH-домен. В одном примере антитело по изобретению может содержать полипептид-вставку в участке каркасной области 3 как VL, так и VH, т.е. антитело по изобретению может содержать два полипептида вставки, где один полипептид-вставка встроен в участок каркасной области 3 VL и один полипептид-вставка встроен в участок каркасной области 3 VH, и где полипептиды-вставки являются идентичными, сходными или отличающимися. В одном варианте осуществления полипептид-вставка является гетерологичным для антитела. Как используют в рамках изобретения, "гетерологичный", как правило, означает из происходящий из окружения, генотипически отличного от остальной части окружения, с которой его сравнивают. В этом контексте, это означает, что встроенный полипептид в норме не присутствует в нативной последовательности антитела. В одном варианте осуществления полипептид-вставка представляет собой полноразмерный белок. В одном варианте осуществления полипептид-вставка представляет собой фрагмент полноразмерного белка. В одном варианте осуществления полипептид-вставка представляет собой функциональный фрагмент полноразмерного белка. В одном варианте осуществления полипептид-вставка является функциональным, когда экспрессируется сам по себе. В одном варианте осуществления полипептид-вставка является функциональным и/или сохраняет функциональность, когда он присутствует в участке каркасной области 3 V-домена антитела по изобретению. Таким образом, в одном варианте осуществления настоящее изобретение относится к антителу, содержащему вариабельный (V) домен и полипептид-вставку, где полипептид-вставка находится в участке каркасной области 3 (Fwk3) V-домена и где полипептид-вставка является функциональным и/или сохраняет функциональность, когда он присутствует в участке каркасной области 3 в V-домене антитела по изобретению.
В одном варианте осуществления полипептид-вставка содержит по меньшей мере 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 150 или 200 аминокислот. В одном варианте осуществления полипептид-вставка содержит менее 250, 300 или 350 аминокислот. В одном варианте осуществления полипептид-вставка содержит менее 250 аминокислот. В одном варианте осуществления полипептид-вставка содержит по меньшей мере 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 150 или 200 аминокислот и менее 250 аминокислот. В одном варианте осуществления полипептид-вставка содержит от 50 до 250, предпочтительно от 100 до 250, более предпочтительно от 150 до 250 аминокислот.
В одном варианте осуществления полипептид-вставка может представлять собой терапевтический или диагностический полипептид. Подходящие терапевтические полипептиды могут включать, например, агонисты или антагонисты рецепторов, ингибиторы ферментов, хелаторы металлов, противовирусные средства, противогрибковые средства, сердечно-сосудистые лекарственные средства и химиотерапевтические лекарственные средства. В некоторых вариантах осуществления полипептид-вставка представляет собой цитокин. Цитокин может быть выбран, например, из IL-10, IL-15, IL-2, G-CSF, GM-CSF, EPO. В других вариантах осуществления полипептид-вставка представляет собой склеростин. В некоторых вариантах осуществления полипептид-вставка содержит более одной полипептидной последовательности, например, две копии одного и того же полипептида, необязательно связанных. Преимущественно в таком варианте осуществления полипептид-вставка может образовывать мультимер, в частности, функциональный мультимер. В некоторых вариантах осуществления полипептид-вставка содержит две, три или четыре полипептидных последовательности и способен образовывать димер, тример или тетрамер полипептида. В некоторых вариантах осуществления полипептид-вставка образует гомодимер, гомотример или гомотетрамер. В некоторых вариантах осуществления полипептид-вставка содержит по меньшей мере два цитокина. В одном варианте осуществления полипептид-вставка содержит две последовательности IL-10 человека, необязательно разделенных пептидным линкером, например, (G4S)3 (SEQ ID NO: 15), т.е. две последовательности IL-10 человека встроены в область Fwk3 V-домена антитела по изобретению. В таком варианте осуществления две последовательности IL-10 человека образуют гомодимер IL-10, включенный в антитело по изобретению.
В некоторых вариантах осуществления полипептид-вставка представляет собой хемокин. Хемокин может быть выбран, например, из CCL (лиганды хемокинов с C-C-мотивом), CXCL (лиганды хемокинов с CXC-мотивом). В некоторых вариантах осуществления полипептид-вставка представляет собой гормон. Гормон может быть выбран, например, из инсулина или лептина. В некоторых вариантах осуществления полипептид-вставка представляет собой фактор роста.
Неожиданно авторы настоящего изобретения обнаружили, что вставка полипептида в антитело в участке петли каркасной области 3 V-домена не препятствует в значительной степени функциональности как антитела, так и полипептида вставки. В частности, примеры в настоящем описании показывают, что специфическое и обусловленное распознаванием связывание с соответствующими связывающими партнерами как антитела, так и полипептида вставки, наблюдается для нескольких протестированных конструкций слитых белков.
В некоторых вариантах осуществления антитело по изобретению связывается с распознаваемым им антигеном посредством указанного V-домена с аффинностью связывания, которая составляет по меньшей мере 50%, 60%, 70%, 80% или 90% по сравнению с антителом без полипептида вставки. Будет понятно, что V-домен, который связывает антиген, может связывать его отдельно или совместно с другим V-доменом в зависимости от формата антитела, например, в качестве комплементарной пары VH/VL. Таким образом, антитело, содержащее V-домен, может связывать его антиген совместно с комплементарным V-доменом (VH или VL). В некоторых вариантах осуществления антитело по изобретению связывает распознаваемый им антиген с аффинностью связывания, которая составляет по меньшей мере 50%, 60%, 70%, 80% или 90% по сравнению с антителом без полипептида вставки. В некоторых вариантах осуществления антитело по изобретению содержит VH-домен и связывает его антиген совместно с комплементарным VL с аффинностью связывания, которая составляет по меньшей мере 50%, 60%, 70%, 80% или 90% по сравнению с антителом без полипептида вставки. В некоторых вариантах осуществления антитело по изобретению содержит VL-домен и связывает его антиген совместно с комплементарным VH-доменом с аффинностью связывания, которая составляет по меньшей мере 50%, 60%, 70%, 80% или 90% по сравнению с антителом без полипептида вставки. В некоторых вариантах осуществления часть антитела в виде полипептида вставки связывает распознаваемый ей партнер по связыванию с аффинностью связывания, которая составляет по меньшей мере 50%, 60%, 70%, 80% или 90% по сравнению со свободным полипептидом, когда он не связан с антителом.
Таким образом, в одном варианте осуществления антитело по изобретению содержит вариабельный (V) домен и полипептид-вставку, где полипептид-вставка находится в участке каркасной области 3 (Fwk3) V-домена и где как антитело, содержащее V-домен, так и полипептид-вставка являются функциональными и/или сохраняют функциональность, когда полипептид-вставка присутствует. В таких вариантах осуществления антитело по изобретению обладает функциональностью как каркасного антитела, так и полипептида вставки. Преимущественно, антитело по изобретению может иметь одновременно функциональность как каркасного антитела, так и полипептида вставки.
Антитело, содержащее вариабельный (V) домен, считается функциональным и/или сохраняющим функциональность, когда полипептид-вставка присутствует в участке каркасной области 3 V-домена, где антитело по изобретению демонстрирует по меньшей мере 50%, 60%, 70%, 80% или 90% по меньшей мере одной из функций антитела, содержащего вариабельный (V) домен бе полипептида вставки. Полипептид-вставка считается функциональным и/или сохраняющим функциональность, когда он присутствует в каркасной области 3 V-домена, где полипептид-вставка демонстрирует по меньшей мере 50%, 60%, 70%, 80% или 90% по меньшей мере одной из функций полипептида вставки, когда он экспрессируется самостоятельно, т.е. по сравнению со свободным полипептидом, когда он не связан с антителом.
"Функциональный" или "функциональность" антитела и/или полипептида вставки относится по меньшей мере к одному виду биологической активности указанного антитела и/или полипептида вставки, включая, например, связывание мишени (например, связывание антигена), аффинность связывания, перекрестную реактивность, нейтрализацию, время полужизни в сыворотке, активацию белков клеточной поверхности, например, рецепторов T-клеточной поверхности. Способы определения функциональности, например, аффинности связывания, нейтрализации и активации клеток-мишеней хорошо известны в данной области и включают, например, способы, описанные в примерах, описанных в настоящем описании. Например, стандартные способы анализа, такие как анализ Скэтчарда или способ поверхностного плазмонного резонанса (например, с использованием BIACORE®) можно использовать для определения связывания мишени (например, связывания антигена) и аффинности связывания.
Неожиданно, V-домен (VL- или VH-домен) антитела по изобретению все еще может образовывать функциональный связывающий домен с его соответствующим или комплементарным V-доменом (VH или VL), когда он присутствует.
В некоторых вариантах осуществления полипептид-вставка может представлять собой второе антитело или его связывающий фрагмент. В таких вариантах осуществления полипептид-вставка может иметь любой из признаков антител, как описано выше. В частности, в некоторых вариантах осуществления полипептид-вставка представляет собой scFv, a dsscFv, однодоменное антитело, такое как VH, или VL, или VHH. В некоторых вариантах осуществления полипептид-вставка может представлять собой полностью человеческое, гуманизированное или химерное антитело. Полипептид-вставка может содержать одну или несколько CDR или может состоять из одной или нескольких CDR. Например, полипептид-вставка может содержать или состоять из по меньшей мере одной из CDR-H1, CDR-H2, CDR-H3 и/или CDR-L1, CDR-L2, CDR-L3, необязательно связанных.
Специалистам в данной области будет понятно, что оно может функционировать как двухвалентное антитело или биспецифическое антитело. Таким образом, в одном аспекте изобретение относится к новому бивалентному или биспецифическому формату, способному к одновременному связыванию двух различных эпитопов. В некоторых аспектах эпитопы представляют собой различные эпитопы, присутствующие на одном и том же антигене. В другом аспекте изобретение относится к новому формату биспецифических антител, способных одновременно связывать два различных антигена.
В одном варианте осуществления представляющий интерес антиген, связываемый первым или вторым антителом, или фрагментом антитела, может представлять собой ассоциированный с клеткой белок, например, белок клеточной поверхности на клетках, таких как бактериальные клетки, дрожжевые клетки, T-клетки, эндотелиальные клетки или опухолевые клетки, или он может представлять собой растворимый белок. Представляющие интерес антигены также могут представлять собой любой имеющий медицинское значение белок, такой как белки, уровень которых повышается в ходе заболевания или инфекции, например, рецепторы и/или их соответствующие лиганды. Конкретные примеры белков клеточной поверхности включают молекулы адгезии, например, интегрины, такие как β1-интегрины, например, VLA-4, E-селектин, P-селектин или L-селектин, CD2, CD3, CD4, CD5, CD7, CD8, CDlla, CDllb, GDIS, CD19, CD20, CD23, CD25, CD28, CD33, CD38, CD40, CD45, CDW52, CD69, CD 134 (OX40), ICOS, BCMP7, CD 137, CD27L, CDCP1, DPCR1, DPCR1, дудулин 2, FLJ20584, FLJ40787, HEK2, KIAA0634, KIAA0659, KIAA1246, KIAA1455, FTBP2, FTK, MAF2, MRP2, нектин-подобный 2, NKCC1, PTK7, RAIG1, TCAM1, SC6, BCMP101, BCMP84, BCMP11, DTD, карциноэмбриональный антиген (CEA), человеческий глобулин молочного жира человека (HMFG1 и 2), антигены MHC класса I и MHC класса II, и VEGF, и, когда это целесообразно, их рецепторы. Растворимые антигены включают интерлейкины, такие как IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-8, IL-12, IL-16 или IL-17, вирусные антигены, например, антигены респираторно-синцитиального вируса или цитомегаловируса, иммуноглобулины, такие как IgE, интерфероны, такие как интерферон α, интерферон β или интерферон γ, фактор некроза опухоли α, фактор некроза опухоли β, колониестимулирующие факторы, такие как G-CSF или GM-CSF, и тромбоцитарные факторы роста, такие как PDGF-α и PDGF-β и, когда это целесообразно, их рецепторы. Другие антигены включают антигены поверхности бактериальных клеток, бактериальные токсины, вирусы, такие как вирус гриппа, EBV, Hep A, B и C, агенты биотерроризма, радионуклиды и тяжелые металлы, и яды и токсины змей и пауков.
В некоторых вариантах осуществления ни каркасное антитело, ни полипептид-вставка не связывают гаптен (4-гидрокси-3-нитрофенил)ацетил (NP) или гаптен (4-гидрокси-5-йод-3-нитрофенил)ацетил (NIP). В некоторых вариантах осуществления антитело отличается от антитела мыши BI-8.
В одном варианте осуществления второе антитело связывается с антигеном, отличным от V-домена антитела.
В других вариантах осуществления второе антитело связывается с тем же антигеном, что и V-домен антитела. Предпочтительно, втрое антитело связывается с эпитопом, отличным от эпитопа V-домена антитела. В таких вариантах осуществления антитело может связывать распознаваемый им антиген с аффинностью связывания, которая превышает аффинность связывания антитела без второго антитела.
В одном варианте осуществления полипептид-вставка представляет собой второе антитело, которое связывает IL-17, в частности, IL-17A. В одном варианте осуществления полипептид-вставка представляет собой второе антитело, которое связывает IL-17AA, IL-17AF и IL-17FF. В одном варианте осуществления второе антитело, которое связывает IL-17, представляет собой scFv. В одном варианте осуществления второе антитело, которое связывает IL-17, представляет собой dsscFv. В одном варианте осуществления антитело по настоящему изобретению представляет собой Fab-фрагмент, который связывается как с HSA, так и с IL-17, в частности, антитело, содержащее V-домен, связывается с HSA, и полипептид-вставка представляет собой dsscFv, который связывает IL-17 т.е. каркасное антитело представляет собой Fab-фрагмент, который связывает HSA, и второе антитело, встроенное в участок Fwk3 V-домена каркасного антитела, представляет собой dsscFv, который связывает IL-17. В другом варианте осуществления антитело по изобретению представляет собой целый IgG, который связывается как с HSA, так и с IL-17, в частности, антитело, содержащее V-домен, связывается с HSA, и полипептид-вставка представляет собой dsscFv, который связывает IL-17, т.е. каркасное антитело представляет собой полный IgG, который связывает HSA, и второе антитело, встроенное в участок Fwk3 V-домена каркасного антитела, представляет собой dsscFv, который связывает IL-17. В одном варианте осуществления антитело по изобретению представляет собой Fab-фрагмент, который содержит или имеет тяжелые и легкие цепи SEQ ID NO: 129 (тяжелая цепь Fab CA645 с dsscFv CA497 (vHvL), встроенным в каркасную область 3) и SEQ ID NO: 1, соответственно, или вариант или варианты любого из них, обладающие по меньшей мере 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% сходством или идентичностью, при условии, что они сохраняют их функциональность, например, связывание с HSA и/или IL-17. В одном варианте осуществления антитело по изобретению представляет собой Fab-фрагмент, который содержит или имеет тяжелые и легкие цепи SEQ ID NO: 130 (тяжелая цепь Fab CA645 с dsscFv CA497 (vLvH), встроенным в каркасную область 3) и SEQ ID NO: 1, соответственно, или вариант или варианты любого из них, обладающие по меньшей мере 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% сходством или идентичностью при условии, что они сохраняют их функциональность, например, связывание с HSA и/или IL-17. В одном варианте осуществления антитело по изобретению представляет собой целый IgG, который содержит или имеет тяжелые и легкие цепи SEQ ID NO: 131 (тяжелая цепь CA645 с dsscFv CA497 (vHvL), встроенным в каркасную область 3) и SEQ ID NO: 1, соответственно, или вариант или варианты любого из них, обладающие по меньшей мере 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% сходством или идентичностью при условии, что они сохраняют их функциональность, например, связывание с HSA и/или IL-17. В одном варианте осуществления антитело по изобретению представляет собой целый IgG, который содержит или имеет тяжелые и легкие цепи SEQ ID NO: 132 (тяжелая цепь IgG4P CA645 с dsscFv CA497 (vLvH), встроенным в каркасную область 3) и SEQ ID NO: 1, соответственно, или вариант или варианты любого из них, обладающие по меньшей мере 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% сходством или идентичностью при условии, что они сохраняют их функциональность, например, связывание с HSA и/или IL-17.
В одном варианте осуществления антитело по изобретению связывается как с CD3, так и с CD28. В одном варианте осуществления антитело по изобретению представляет собой Fab-фрагмент, который связывает как CD3, так и CD28, в частности, антитело, содержащее V-домен, связывается с CD3, и полипептид-вставка представляет собой dsscFv, который связывает CD28 т.е. каркасное антитело представляет собой Fab-фрагмент, который связывает CD3, и второе антитело, встроенное в участок Fwk3 V-домена каркасного антитела, представляет собой dsscFv, который связывает CD28. В одном варианте осуществления антитело по изобретению представляет собой Fab-фрагмент, который связывает как CD3, так и CD28, в частности, антитело, содержащее V-домен, связывается с CD28, и полипептид-вставка представляет собой dsscFv, который связывает CD3, т.е. каркасное антитело представляет собой Fab-фрагмент, который связывает CD28, и второе антитело, встроенное в участок Fwk3 V-домена каркасного антитела, представляет собой dsscFv, который связывает CD3. В другом варианте осуществления антитело по изобретению представляет собой целый IgG, который связывает как CD3, так и CD28, в частности, антитело, содержащее V-домен, связывается с CD3, и полипептид-вставка представляет собой dsscFv, который связывает CD28, т.е. каркасное антитело представляет собой полный IgG, который связывает CD3, и второе антитело, встроенное в участок Fwk3 V-домена каркасного антитела, представляет собой dsscFv, который связывает CD28. В другом варианте осуществления антитело по изобретению представляет собой целый IgG, который связывает как CD3, так и CD28, в частности, антитело, содержащее V-домен, связывает CD28, и полипептид-вставка представляет собой dsscFv, который связывает CD3, т.е. каркасное антитело представляет собой полный IgG, который связывает CD28, и второе антитело, встроенное в участок Fwk3 V-домена каркасного антитела, представляет собой dsscFv, который связывает CD3. В одном варианте осуществления антитело по изобретению представляет собой Fab-фрагмент, который содержит или имеет тяжелые и легкие цепи SEQ ID NO: 139 (тяжелая цепь Fab против CD28 с dsscFv против CD3, встроенным в каркасную область 3) и SEQ ID NO: 136, соответственно, или вариант или варианты любого из них, обладающие по меньшей мере 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% сходством или идентичностью при условии, что они сохраняют их функциональность, например, связывание с CD3 и/или CD28. В другом варианте осуществления антитело по изобретению представляет собой целый IgG, который содержит или имеет тяжелые и легкие цепи SEQ ID NO: 141 (тяжелая цепь IgG4P против CD28 с dsscFv против CD3, встроенным в каркасную область 3) и SEQ ID NO: 136, соответственно, или вариант или варианты любого из них, обладающие по меньшей мере 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% сходством или идентичностью при условии, что они сохраняют их функциональность, например, связывание с CD3 и/или CD28.
Иллюстративные последовательности для применения в рамках настоящего изобретения включают SEQ ID NO: 53, SEQ ID NO: 54, SEQ ID NO: 55, SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59, SEQ ID NO: 60, SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, SEQ ID NO: 69, SEQ ID NO: 70, SEQ ID NO: 71, SEQ ID NO: 72, SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 83, SEQ ID NO: 84, SEQ ID NO: 85, SEQ ID NO: 86, SEQ ID NO: 87, SEQ ID NO: 88, SEQ ID NO: 89, SEQ ID NO: 90, SEQ ID NO: 91, SEQ ID NO: 92, SEQ ID NO: 93, SEQ ID NO: 94, SEQ ID NO: 95, SEQ ID NO: 96, SEQ ID NO: 97, SEQ ID NO: 98, SEQ ID NO: 99, SEQ ID NO: 100, SEQ ID NO: 101, SEQ ID NO: 102, SEQ ID NO: 103, SEQ ID NO: 105, SEQ ID NO: 106, SEQ ID NO: 107, SEQ ID NO: 108, SEQ ID NO: 109, SEQ ID NO: 110, SEQ ID NO: 125, SEQ ID NO: 126, SEQ ID NO: 127, SEQ ID NO: 128, SEQ ID NO: 129, SEQ ID NO: 130, SEQ ID NO: 131, SEQ ID NO: 132, SEQ ID NO: 133, SEQ ID NO: 134, SEQ ID NO: 135, SEQ ID NO: 136, SEQ ID NO: 137, SEQ ID NO: 138, SEQ ID NO: 139, SEQ ID NO: 140, SEQ ID NO: 141, SEQ ID NO: 142, SEQ ID NO: 143 и SEQ ID NO: 144. Иллюстративные комбинации тяжелой и легкой цепей для применения в Fab-фрагментах по настоящему изобретению включают SEQ ID NO: 54 и SEQ ID NO: 1, SEQ ID NO: 56 и SEQ ID NO: 1, SEQ ID NO: 58 и SEQ ID NO: 1, SEQ ID NO: 60 и SEQ ID NO: 1, SEQ ID NO: 61 и SEQ ID NO: 1, SEQ ID NO: 63 и SEQ ID NO: 1, SEQ ID NO: 64 и SEQ ID NO: 1, SEQ ID NO: 66 и SEQ ID NO: 1, SEQ ID NO: 68 и SEQ ID NO: 1, SEQ ID NO: 70 и SEQ ID NO: 1, SEQ ID NO: 73 и SEQ ID NO: 1, SEQ ID NO: 74 и SEQ ID NO: 1, SEQ ID NO: 75 и SEQ ID NO: 1, SEQ ID NO: 76 и SEQ ID NO: 1, SEQ ID NO: 93 и SEQ ID NO: 1, SEQ ID NO: 94 и SEQ ID NO: 1, SEQ ID NO: 95 и SEQ ID NO: 1, SEQ ID NO: 101 и SEQ ID NO: 1, SEQ ID NO: 129 и SEQ ID NO: 1, SEQ ID NO: 130 и SEQ ID NO: 1, SEQ ID NO: 139 и SEQ ID NO: 136, SEQ ID NO: 140 и SEQ ID NO: 133. Иллюстративные комбинации тяжелых и легких цепей для применения в целом IgG по настоящему изобретению включают SEQ ID NO: 131 и SEQ ID NO: 1, SEQ ID NO: 132 и SEQ ID NO: 1, SEQ ID NO: 141 и SEQ ID NO: 136, SEQ ID NO: 142 и SEQ ID NO: 133.
Линкерная последовательность
В некоторых вариантах осуществления полипептид-вставка связан прямо с участком каркасной области 3 антитела. Под прямым связыванием, как используют в рамках настоящего изобретения, подразумевают тот факт, что N- и C-концевые аминокислоты гетерологичного полипептида-вставки связаны посредством пептидной связи с участком каркасной области 3 V-домена антитела по настоящему изобретению.
В некоторых вариантах осуществления антитело по изобретению дополнительно содержит линкерную последовательность. В таких вариантах осуществления линкерная последовательность предпочтительно является гетерологичной как для антитела, так и для полипептида вставки, т.е. линкерная последовательность в норме не присутствует в последовательности нативного антитела или полипептиде вставки в этом положении.
В некоторых вариантах осуществления линкерная последовательность содержит по меньшей мере одну, две, три, четыре, пять, шесть, семь, восемь, девять, десять, одиннадцать, двенадцать, тринадцать, четырнадцать, пятнадцать, шестнадцать, семнадцать, восемнадцать, девятнадцать, двадцать, двадцать одну, двадцать две, двадцать три, двадцать четыре, двадцать пять, двадцать шесть, двадцать семь, двадцать восемь, двадцать девять, тридцать или тридцать одну аминокислоту, соединяющие N- и/или C-концы полипептида вставки с участком каркасной области 3. В некоторых вариантах осуществления линкерная последовательность содержит от одной до тридцати одной, или от пяти до тридцати одной, или от шести до двадцати восьми, или от десяти до двадцати шести, или от пятнадцати до двадцати шести, или от десяти до двадцати одной, или от пяти до десяти аминокислот, соединяющих N- и/или C-конце полипептида вставки с участком каркасной области 3.
В некоторых вариантах осуществления линкерная последовательность содержит не более тридцати аминокислот, соединяющих N- и/или C-концы полипептида вставки с участком каркасной области 3.
В некоторых вариантах осуществления линкер на N-конце полипептида вставки отличается по составу и/или длине от линкера на C-конце полипептида вставки. В некоторых вариантах осуществления линкер на N-конце полипептида вставки идентичен или сходен по составу и/или длине с линкером на C-конце полипептида вставки. В некоторых вариантах осуществления линкерная последовательность происходит из последовательности человека. В некоторых вариантах осуществления линкер представляет собой линкер человека. В некоторых вариантах осуществления линкерная последовательность происходит из не являющейся человеческой последовательности. В некоторых вариантах осуществления линкер происходит из последовательности животного подсемейства бычьих. В некоторых вариантах осуществления линкер представляет собой линкер из последовательности животного подсемейства бычьих.
Примеры подходящих линкерных областей для связывания полипептида вставки с участком каркасной области 3 V-домена включают, но не ограничиваются ими, последовательности гибких линкеров и последовательности жестких линкеров. Последовательности гибких линкеров включают последовательности, описанные в Huston et al., 1988, PNAS 85:5879-5883; Wright & Deonarain, Mol. Immunol., 2007, 44(11):2860-2869; Alfthan et al., Prot. Eng., 1995, 8(7):725-731; Luo et al., J. Biochem., 1995, 118(4):825-831; Tang et al., 1996, J. Biol. Chem. 271(26):15682-15686; и Turner et al., 1997, JIMM 205, 42-54 (см. таблицу 1 ниже для репрезентативных примеров).
Таблица 1 - Примеры последовательностей гибких линкеров
(S) является необязательным в последовательностях 13-17. X обозначает любой остаток.
В конкретных вариантах осуществления антитело содержит линкерные последовательности SEQ ID NO: 106 и 107.
Примеры жестких линкеров включают пептидные последовательности GAPAPAAPAPA (SEQ ID NO:51), PPPP (SEQ ID NO:52), PPP, IPFTV (SEQ ID NO: 82) и EYHGLQ (SEQ ID NO: 104). Следующие примеры включают последовательности, происходящие из удлиненного стебля CDRH3 антител животных семейства бычьих (Wang et al., Cell, 2013, 153:1379-1393); предпочтительные последовательности включают длинные линкерные последовательности, такие как TSVHQETKKYQS (SEQ ID NO: 96) и SYTYNYEWHVDV (SEQ ID NO: 97) (содержащие как открытые для растворителя, так и погруженные остатки из стебля CDH3 животного семейства бычьих); и короткие линкерные последовательности, такие как ETKKYQS (SEQ ID NO: 98) и SYTYNYE (SEQ ID NO: 99) (содержащие только открытые для растворителя остатки из стебля CDH3 животного семейства бычьих).
В некоторых вариантах осуществления можно использовать варианты любой из линкерных последовательностей, описанных в настоящем описании, например, варианты, где один, два, три, четыре или более аминокислотных остатков заменены, вставлены или удалены относительно исходной линкерной последовательности. Варианты линкерных последовательностей также могут включать варианты с повторами, где линкерная последовательность удлинена посредством повторения линкерной последовательности. В одном варианте осуществления предпочтительная подлежащая повторению линкерная последовательность представляет собой SEQ ID NO: 13, которая обеспечивает мотив: (GmS)n, где m равен 4 и n может представлять собой любое целое число (например, 1, 2 или 3). В других вариантах осуществления m может представлять собой отличающееся целое число от 1 до 10.
Независимо от того, связан ли полипептид-вставка прямо или через линкерную последовательность, в некоторых вариантах осуществления одна или несколько аминокислот в участке каркасной области 3 могут быть удалены в результате связывания полипептидной вставки. В некоторых вариантах осуществления полипептид-вставка находится между аминокислотными остатками 73 и 75 VH-домена в соответствии с системой нумерации Kabat и аминокислота в положении 74 удалена.
Фармацевтические композиции и способы применения
Антитела по изобретению и как описано в настоящем описании являются пригодными для лечения заболеваний или нарушений, включающих воспалительные заболевания и нарушения, иммунные заболевания и нарушения, фибротические нарушения и злокачественные опухоли.
Термин "воспалительное заболевание" или "нарушение" и "именное заболевание или нарушение" включает ревматоидный артрит, псориатический артрит, болезнь Стилла, болезнь Макла-Уэлса, псориаз, болезнь Крона, язвенный колит, SLE (системная красная волчанка), астму, аллергический ринит, атопический дерматит, рассеянный склероз, васкулит, сахарный диабет типа I, трансплантацию и реакцию "трансплантат" против хозяина.
Термин "фибротическое нарушение" включает идиопатический фиброз легких (IPF), системный склероз (или склеродермию), фиброз почек, диабетическую нефропатию, IgA-нефропатию, гипертензию, терминальную хроническую почечную недостаточность, перитонеальный фиброз (постоянный амбулаторный перитонеальный диализ), цирроз печени, связанную со старением дегенерацию желтого пятна (ARMD), ретинопатию, сердечный реактивный фиброз, рубцы, келоиды, ожоги, кожные язвы, ангиопластику, аорто-коронарное шунтирование, артропластикю и хирургическую операцию по поводу катаракты.
Термин "злокачественная опухоль" включает злокачественный новый рост, который происходит из эпителия, находится на коже или чаще выстилке органов тела, например: молочной железы, яичников, предстательной железы, легкого, почки, поджелудочной железы, желудка, мочевого пузыря или толстого кишечника. Злокачественные опухоли имеют тенденцию к инфильтрации соседних тканей и распространению (метастазированию) в другие органы, например: в кость, печень, легкое или головной мозг.
Таким образом, согласно следующему аспекту изобретения, предусматривается фармацевтическая композиция, которая содержит антитело по изобретению совместно с одним или несколькими фармацевтически приемлемыми носителями, эксципиентами или разбавителями.
Также предусматривается применение антитела по изобретению для изготовления лекарственного средства для лечения заболевания или нарушения. Наиболее предпочтительно, заболевание или нарушение представляет собой воспалительное заболевание или нарушение.
Фармацевтические композиции согласно изобретению могут иметь форму, например, пригодную для парентерального, подкожного или офтальмологического введения. Когда это целесообразно, например, если антитело связывается с альбумином через его V-домен, может быть желательным предварительное составление антитела по изобретению с сывороточным альбумином человека или рекомбинантным сывороточным альбумином с использованием любого подходящего способа, известного в данной области.
Когда фармацевтический состав представляет собой жидкость, например, раствор или суспензию, состав может дополнительно содержать альбумин, например, сывороточный альбумин человека, в частности, рекомбинантный альбумин, такой как рекомбинантный сывороточный альбумин человека. Подходящие количества могут быть в диапазоне менее 2% масс./масс. от всего состава, в частности, менее 1, 0,5 или 0,1% масс./масс. Это может способствовать стабилизации антительного компонента в составе. Фармацевтическая композиция может быть лиофилизированной для восстановления позднее водным растворителем. В одном варианте осуществления предусматривается контейнер для единичной дозы, такой как флакон, содержащий лиофилизированное антитело по изобретению.
Антитела по изобретению можно составлять для парентерального введения посредством инъекции, например, посредством болюсной инъекции или инфузии. Составы для инъекций могут присутствовать в единичной дозированной форме, например, в стеклянных ампулах или многодозовых контейнерах, например, в стеклянных флаконах. Композиции для инъекций могут принимать такие формы в качестве суспензий, растворов или эмульсий в масляных или водных носителях, и могут содержать средства для составления, такие как суспендирующие, стабилизирующие вещества, консерванты и/или диспергирующие средства. Альтернативно активный ингредиент может иметь форму порошка для восстановления подходящим носителем, например, стерильной свободной от пирогенов водой, перед применением.
Если желательно, композиции могут находиться в упаковке или устройстве распределения, которое может содержать одну или несколько единичных дозированных форм, содержащих активный ингредиент. К упаковке или устройству распределения могут прилагаться инструкции по введению.
Форматы антител по изобретению можно доставлять диспергированными в растворителе, например, в форме раствора или суспензии. Их можно суспендировать в подходящем физиологическом растворе, например, солевом растворе или другом фармакологически приемлемом растворителе или буферном растворе. Буферные растворы, известные в данной области, могут содержать от 0,05 мг до 0,15 мг динатрия эдетата, от 8,0 мг до 9,0 мг NaCl, от 0,15 мг до 0,25 мг полисорбата, от 0,25 мг до 0,30 мг безводной лимонной кислоты и от 0,45 мг до 0,55 мг цитрата натрия на 1 мл воды, так чтобы достигнуть pH приблизительно от 4,0 до 5,0. Можно использовать суспензию, например, лиофилизированное антитело.
Терапевтические составы суспензий или растворов также могут содержать один или несколько эксципиентов. Эксципиенты хорошо известны в данной области и включают буферы (например, цитратный буфер, фосфатный буфер, ацетатный буфер и бикарбонатный буфер), аминокислоты, мочевину, спирты, аскорбиновую кислоту, фосфолипиды, белки (например, сывороточный альбумин), EDTA, хлорид натрия, липосомы, маннит, сорбит и глицерин. Растворы или суспензии могут быть инкапсулированы в липосомы или биодеградируемые микросферы. Состав, как правило, предоставляют в по существу стерильной форме с использованием стерильных процессов производства.
Это может включать продуцирование и стерилизацию посредством фильтрации буферного растворителя/раствора, используемого для составления, суспендирование в асептических условиях антитела в стерильном растворе буферного растворителя, и распределение состава в стерильную тару способами, известными специалисту в данной области.
Количество антитела по изобретению, требуемое для профилактики или лечения конкретного состояния, варьируется в зависимости от подвергаемого лечению состояния. Однако, как правило, суточные дозировки могут находиться в диапазоне приблизительно от 10 нг/кг до 1000 мг/кг, от 100 нг/кг до 100 мг/кг, или от 0,1 мг/кг до 10 мг/кг массы тела.
Настоящее изобретение также относится к способам и композициям для доставки антител, как описано в настоящем описании, посредством генной терапии, в частности, посредством аденоассоциированного вирусного (AAV) вектора.
Таким образом, настоящее изобретение относится к фармацевтической композиции, содержащей вирусный вектор, имеющий вирусный капсид и искусственный геном, содержащий экспрессирующую кассету, фланкированную инвертированными концевыми повторами (ITR), где экспрессирующая кассета содержит трансген, содержащий полинуклеотидную последовательность, кодирующую антитело, как описано в настоящем описании. Последовательности ITR можно использовать для упаковывания искусственного генома, содержащего полинуклеотидные последовательности, кодирующие антитело, как описано в настоящем описании, в вирион вирусного вектора.
Трансген в экспрессирующей кассете функционально связан с элементами контроля экспрессии, такими как промоторы, которые контролируют экспрессию трансгена в клетках человека.
Вирусный вектор предпочтительно представляет собой вирусные векторы на основе AAV. Различные капсиды AAV описаны в данной области. Способы получения векторов на основе AAV также широко описаны в литературе (например, WO 2003/042397; WO 2005/033321, WO 2006/110689; патент США № 7588772 B2). Источник капсидов AAV может быть выбран из AAV, который нацелен на желаемую ткань. Например, подходящий AAV может включать, например, AAV9 (патент США № 7906111; US 2011-0236353-A1), rh10 (WO 2003/042397) и/или hu37 (US 7906111B2; US20110236353). Однако также можно выбирать другие AAV, включая, например, AAV1, AAV2, AAV-TT, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8 и AAV.PHP.B (или их варианты) и другие.
Способы получения и выделения вирусных векторов на основе AAV, пригодные для доставки индивидууму, известны в данной области (US7790449B2; US7282199B2; WO 2003/042397; WO 2005/033321, WO 2006/110689; и US7588772 B2).
Фармацевтическую композицию, содержащую вирусный вектор, имеющий вирусный капсид и искусственный геном согласно изобретению, конструируют так, чтоб обеспечить экспрессию как тяжелой, так и легкой цепей. Полинуклеотидные последовательности тяжелой и легкой цепей антител в соответствии с настоящим изобретением можно конструировать в виде одной конструкции, и они могут быть разделены, например, расщепляемым линкером или элементом участка внутренней посадки рибосомы (IRES), так чтобы тяжелая и легкая цепи экспрессировались по отдельности. В некоторых вариантах осуществления расщепляемый линкер представляет собой саморасщепляющийся линкер фурин/F2A (Fang et al., Nature Biotechnology 23: 584-590, 2005 и Fang et al. Mol Ther 15: 1153-9, 2007). Например, линкер фурин/F2A может быть включен в экспрессирующую кассету для разделения последовательностей, кодирующих полинуклеотиды тяжелой и легкой цепей.
В некоторых других вариантах осуществления полинуклеотидные последовательности тяжелой и легкой цепей антител в соответствии с настоящим изобретением соединены через гибкий не расщепляющийся линкер. В некоторых вариантах осуществления такой линкер может представлять собой гибкий пептидный линкер, такой как линкеры, кодирующие scFv (состоящие из Gly и Ser), так чтобы домены тяжелой и легкой цепей свободно двигались относительно друг друга.
Изобретение далее описано с отсылкой на следующие примеры, которые являются только иллюстративными и никоим образом не должны истолковываться как ограничивающие объем настоящего изобретения.
ПРИМЕРЫ
Общая методология
Вставка в каркасную область 3
В дополнение к трем петлям CDR легкие и тяжелые цепи, как общепринятых антител, так и одноцепочечных VHH животных семейства верблюжьих, имеют четвертую петлю, которая образована каркасной областью 3 (фиг. 1A). Система нумерации Kabat определяет каркасную область 3 как положения 66-94 в тяжелой цепи и положения 57-88 в легкой цепи.
Конструирование генов и временная экспрессия клеток
Гены, кодирующие V-области тяжелой и легкой цепей, включающие варианты со вставкой в каркасную область 3, моделировали и конструировали с использованием подхода автоматизированного синтеза (ATUM).
Гены клонировали в экспрессирующие векторы, содержавшие ДНК, кодирующую область Cκ человека (аллотип Km3) и область CH1 тяжелой цепи γ1, соответственно. Затем, экспрессирующими векторами тяжелой и легкой цепей котрансфицировали клетки HEK293 и рекомбинантные молекулы слитых белков антитело/Fab подвергали скринингу с использованием анализа связывания SPR.
V-область легкой цепи или легкая цепь слитых белков Fab CA645, описанных в приведенных ниже примерах, содержала или имела SEQ ID NO: 2 (VL-домен CA645 (gL5)) или SEQ ID NO: 1 (gL5 легкой цепи CA645), соответственно. Можно использовать альтернативные легкие цепи или V-области легкой цепи, например, V-области легкой цепи, содержащие VL-домен SEQ ID NO: 108 или SEQ ID NO: 109.
Определение эффективности связывания слитого белка антитело/Fab посредством поверхностного плазмонного резонанса
Все эксперименты с использованием поверхностного плазмонного резонанса (SPR) проводили при 25°C на системе Biacore 3000 с использованием подвижного буфера при pH 7,4, содержавшего 10 мМ HEPES, 150 мМ NaCl, 2 мМ EDTA и 0,005% (об./об.) P20 (буфер HBS-EP).
Поликлональный F(ab)2-фрагмент козы против F(ab)2 человека (код продукта Jackson Labs #109-006-097) иммобилизовывали на поверхности сенсорных чипов CM5 (GE Healthcare) способом присоединения аминов. В кратком изложении, поверхность карбоксиметилдекстрана активировали свежей смесью 1:1 (об./об.) 50 мМ N-гидроксисукцинимида и 200 мМ 1-этил-3-(3-диметиламинопропил)карбодиимида в течение 5 минут при скорости потока 10 мкл/мин. Противочеловеческий F(ab)2 в 10 мМ ацетатном буфере, pH 5,0, ковалентно связывали с чипами в количестве 50 мкг/мл в ходе обработки в течение 10 минут при той же скорости потока. Наконец, поверхность инактивировали обработкой в течение 10-минут 1 M этаноламином.HCl, pH 8,5. Подготавливали эталонную проточную ячейку на чип путем исключения белка из указанной выше методики.
Супернатанты, содержавшие конструкции вставок Fab-петля, Fab дикого типа или контрольные супернатанты, разбавляли 1 к 5 в буфере HPS-EP перед тестированием в отношении связывания их соответствующего лиганда или рецептора.
Сывороточный альбумин человека (HSA) был предоставлен Jackson ImmunoResearch (каталожный номер #009-000-051), и его разбавляли в буфере HBS-EP до 1000 нМ. Следующие цитокины и рецепторы получали от R&D Systems Ltd.: IL-15, IL-15-Rα-hFc, IL-2, IL-2-sRβ, G-CSF, G-CSF-Rα, GM-CSF, GM-CSF-Rα, EPO, EPO-R, IL-6, LRP4(ecd)-hFc, LRP6(ecd)-hFc, лизоцим, KLK5. Эти белки обычно разбавляли до 100 мкг/мл и хранили при -20°C. Для экспериментальных целей их разбавляли непосредственно перед использованием буфером HBS-EP, как правило, до 50, 100 или 200 нМ.
Типичный цикл SPR включал скорость потока 10 мкл/мин с HBS-EP в качестве подвижного буфера и разбавленный супернатант инжектировали в течение 1 мин для улавливания слитого белка антитело/Fab. После этого следовала инъекция 30 мкл разбавленного распознаваемого лиганда, или рецептора, или контроля в виде буфера. Мониторинг последующей фазы диссоциации проводили в течение по меньшей мере 3 мин. В некоторых экспериментах было включено дополнительное инжектирование 30 мкл соответствующего распознаваемого антителом антигена, HSA в случае конструкций слитого белка с использованием Fab CA645 в качестве полипептидной части антитела. В некоторых экспериментах был включен цикл контроля специфичности, где распознаваемый рецептор/лиганд (часть вставки в слитом белке антитело/Fab) предварительно смешивали с 2-кратным молярным избытком его партнера по связыванию. Каждый цикл завершался стадией регенерации, включавшей два инжектирования 10 мкл 40 мМ HCl, между которыми проводили инжектирование 5 мкл 5 мМ NaOH.
Результаты получали за 10 с до инжектирований супернатанта и лиганда/рецептора для определения улавливания конструкции слитого белка и за 5 с до и через 15 с после окончания инжектирования лиганда/рецептора для оценки связывания распознаваемого лиганда/рецептора. Полученные результаты и данные сенсограмм отражали различие в количестве резонансных единиц (RU) между проточной ячейкой для улавливания с поликлональным антителом и эталонной проточной ячейкой.
Результаты и данные сенсограмм обрабатывали путем вычитания соответствующего контрольного циикла из цикла лиганда/рецептора для учета какой-либо медленной диссоциации конструкции слитого белка от поверхности улавливания. В каждом эксперименте был включен соответствующий супернатант с Fab дикого типа для проверки какого-либо неспецифического связывания лиганда/рецептора. Если оно было значительным, его вычитали из описанных выше величин.
В примерах, в которых оценивали константы диссоциации (KD), описанный выше способ был модифицирован увеличением скорости потока до 30 мкл/мин. После улавливания конструкции Fab, 90 мкл распознаваемого лиганда или контроля в виде буфера инжектировали с последующей фазой диссоциации 180 с. Тестировали связывание лиганда на протяжении диапазона концентраций (0,1, 0,2, 0,5, 1, 2, 5 и 10 нМ). В сенсограммы вносили поправку на дрейф путем вычитания сенсограммы с нулевым содержанием лиганда и на неспецифическое взаимодействие путем вычитания сенсограммы с поправкой на Fab дикого типа. Константы аффинности вычисляли из отдельных констант скорости диссоциации и ассоциации, которые приводили в соответствие с сенсограммами с внесенной поправкой с использованием модели Ленгмюра программного обеспечения BIAevaluation (версия 4.1.1). В примерах, в которых оценивали константы диссоциации (KD), описанный выше способ был модифицирован увеличением скорости потока до 30 мкл/мин. После улавливания конструкции Fab, 90 мкл распознаваемого лиганда или контроля в виде буфера инжектировали с последующей фазой диссоциации 180 с. Тестировали связывание лиганда на протяжении диапазона концентраций (0,1, 0,2, 0,5, 1, 2, 5 и 10 нМ). В сенсограммы вносили поправку на дрейф путем вычитания сенсограммы с нулевым содержанием лиганда и на неспецифическое взаимодействие путем вычитания сенсограммы с поправкой на Fab дикого типа. Константы аффинности вычисляли из отдельных констант скорости диссоциации и ассоциации, которые приводили в соответствие с сенсограммами с внесенной поправкой с использованием модели Ленгмюра программного обеспечения BIAevaluation (версия 4.1.1).
ПРИМЕР 1 - Fab CA645 с полипептидом вставки IL-15
Интерлейкин 15 (IL-15) представляет собой цитокин, который играет роль в иммунном ответе и функционирует, стимулируя пролиферацию T-клеток и натуральных киллеров (NK). Он структурно сходен с IL-2 и его рецептор имеет общие цепи с рецептором IL-2.
Слитый белок получали и экспрессировали в соответствии с описанными выше способами для вставки последовательности IL-15 человека в участок каркасной области 3 конструкции Fab CA645, как показано ниже.
Последовательность IL-15 человека:
NWVNVISDLKKIEDLIQSMHIDATLYTESDVHPSCKVTAMKCFLLELQVISLESGDASIHDTVENLIILANNSLSSNGNVTESGCKECEELEEKNIKEFLQSFVHIVQMFIN (SEQ ID NO: 53)
Тяжелая цепь Fab CA645 с IL-15 человека (полужирным шрифтом и курсивом), встроенным в каркасную область 3, линкеры подчеркнуты:
EVQLLESGGGLVQPGGSLRLSCAVSGIDLSNYAINWVRQAPGKGLEWIGIIWASGTTFYATWAKGRFTISRDNKYQSGGSGMGNWVNVISDLKKIEDLIQSMHIDATLYTESDVHPSCKVTAMKCFLLELQVISLESGDASIHDTVENLIILANNSLSSNGNVTESGCKECEELEEKNIKEFLQSFVHIVQMFINSGGSYTYKNTVYLQMNSLRAEDTAVYYCARTVPGYSTAPYFDLWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC (SEQ ID NO: 54)
На фиг. 2A представлен анализ посредством Biacore эффективности связывания для этого слитого белка антитела CA645/IL-15 при тестировании с100 нМ IL-2-Rβ, IL-15-Rα (оба из которых являются связывающими партнерами для IL-15) и 1000 нМ HSA (лигнад для CA645). Как отчетливо видно из фиг. 2A, связывание наблюдалось для всех трех лигандов, что демонстрирует, что функциональность сохраняется как в части Fab CA645, так и в трансплантате IL-15 в участке каркасной области 3. На фиг. 2B представлено смоделированное представление белка CA645/IL-15Fwk3.
ПРИМЕР 2 - CA645 Fab с полипептидом вставки IL-2
Интерлейкин 2 (IL-2) представляет собой плюрипотентный цитокин, продуцируемый в основном активированными CD4+ T-клетками, который играет ключевую роль в формировании нормального иммунного ответа. IL-2 стимулирует пролиферацию и экспансию активированных T-лимфоцитов, усиливает рост B-клеток и активирует моноциты и натуральные киллеры.
Слитый белок получали и экспрессировали в соответствии с описанными выше способами для вставки последовательности IL-2 человека в участок каркасной области 3 конструкции Fab CA645, как показано ниже.
Последовательность IL-2 человека:
APTSSSTKKTQLQLEHLLLDLQMILNGINNYKNPKLTRMLTFKFYMPKKATELKHLQCLEEELKPLEEVLNLAQSKNFHLRPRDLISNINVIVLELKGSETTFMCEYADETATIVEFLNRWITFCQSIISTLT (SEQ ID NO: 55)
Тяжелая цепь Fab CA645 с IL-2 человека (полужирным шрифтом и курсивом), встроенным в каркасную область 3, линкеры подчеркнуты:
EVQLLESGGGLVQPGGSLRLSCAVSGIDLSNYAINWVRQAPGKGLEWIGIIWASGTTFYATWAKGRFTISRDNSGGGSGGGGAPTSSSTKKTQLQLEHLLLDLQMILNGINNYKNPKLTRMLTFKFYMPKKATELKHLQCLEEELKPLEEVLNLAQSKNFHLRPRDLISNINVIVLELKGSETTFMCEYADETATIVEFLNRWITFCQSIISTLTGGGKNTVYLQMNSLRAEDTAVYYCARTVPGYSTAPYFDLWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC (SEQ ID NO: 56)
На фиг. 3A представлен анализ посредством Biacore эффективности связывания для этого слитого белка антитела CA645/IL-2 при тестировании с100 нМ IL-2-Rβ (партнер по связыванию для IL-2), с предшествующей инкубацией с 200 нМ IL-15 (конкурирующий лиганд) или без нее. Как показано на фиг. 3A, наблюдаемое связывание с IL-2-Rβ устраняется в присутствии конкурирующего лиганда в молярном избытке, демонстрируя, что наблюдаемое связывание с частью IL-2 в каркасной области 3 слитого белка является истинным функциональным взаимодействием, и не является неспецифическим. На фиг. 3B показано смоделированное представление белка CA645/IL-2Fwk3.
ПРИМЕР 3 - Fab CA645 с полипептидом вставки G-CSF
Гранулоцитарный колониестимулирующий фактор (G-CSF) представляет собой цитокин, продуцируемый рядом различных типов клеток (например, эндотелий, макрофаги), который действует, стимулируя продуцирование зрелых гранулоцитов из костного мозга.
Слитый белок получали и экспрессировали в соответствии с описанными выше способами для вставки последовательности G-CSF человека в участок каркасной области 3 конструкции Fab CA645, как показано ниже.
Последовательность G-CSF человека:
ATPLGPASSLPQSFLLKCLEQVRKIQGDGAALQEKLVSECATYKLCHPEELVLLGHSLGIPWAPLSSCPSQALQLAGCLSQLHSGLFLYQGLLQALEGISPELGPTLDTLQLDVADFATTIWQQMEELGMAPALQPTQGAMPAFASAFQRRAGGVLVASHLQSFLEVSYRVLRHLAQP (SEQ ID NO: 57)
Тяжелая цепь Fab CA645 с G-CSF человека (полужирным шрифтом и курсивом), встроенным в каркасную область 3, линкеры Gly-Ser подчеркнуты:
EVQLLESGGGLVQPGGSLRLSCAVSGIDLSNYAINWVRQAPGKGLEWIGIIWASGTTFYATWAKGRFTISRDNSGGGGSATPLGPASSLPQSFLLKCLEQVRKIQGDGAALQEKLVSECATYKLCHPEELVLLGHSLGIPWAPLSSCPSQALQLAGCLSQLHSGLFLYQGLLQALEGISPELGPTLDTLQLDVADFATTIWQQMEELGMAPALQPTQGAMPAFASAFQRRAGGVLVASHLQSFLEVSYRVLRHLAQPSGGGGKNTVYLQMNSLRAEDTAVYYCARTVPGYSTAPYFDLWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC (SEQ ID NO: 58)
На фиг. 4A представлен анализ посредством Biacore эффективности связывания этого слитого белка антитела CA645/G-CSF при тестировании с100 нМ G-CSF-Rα (связывающий партнер для G-CSF), 1000 нМ HSA (лиганд для CA645), и как G-CSF-Rα и HSA. На фиг. 4A показаны наравне с другими примерами, что части как G-CSF, так и Fab слитого белка являются функционально активными и способны связывать их соответствующие партнеры по связыванию. На фиг. 4A, кроме того, показано, что в присутствии обоих партнеров по связыванию сигнал, наблюдаемый на кривой SPR, является суммарным из отдельных кривых связывания, демонстрируя, что связывание обеих функциональных частей может происходить одновременно. На фиг. 4B представлено смоделированное представление белка CA645/G-CSFFwk3.
ПРИМЕР 4 -Fab CA645 с полипептидом вставки GM-CSF
GM-CSF представляет собой цитокин, который стимулирует клетки к продуцированию гранулоцитов (нейтрофилы, эозинофилы и базофилы) и моноцитов.
Слитый белок получали и экспрессировали в соответствии с описанными выше способами для вставки последовательностей GM-CSF человека и мыши в участок каркасной области 3 конструкции Fab CA645, как показано ниже.
Последовательность GM-CSF (hGM-CSF) человека:
APARSPSPSTQPWEHVNAIQEARRLLNLSRDTAAEMNETVEVISEMFDLQEPTCLQTRLELYKQGLRGSLTKLKGPLTMMASHYKQHCPPTPETSCATQIITFESFKENLKDFLLVIPFDCWEPVQE (SEQ ID NO: 59)
Тяжелая цепь Fab CA645 с GM-CSF человека (полужирным шрифтом и курсивом), встроенным в каркасную область 3, линкеры Gly-Ser подчеркнуты:
EVQLLESGGGLVQPGGSLRLSCAVSGIDLSNYAINWVRQAPGKGLEWIGIIWASGTTFYATWAKGRFTISRDNSGGGSGGGGAPARSPSPSTQPWEHVNAIQEARRLLNLSRDTAAEMNETVEVISEMFDLQEPTCLQTRLELYKQGLRGSLTKLKGPLTMMASHYKQHCPPTPETSCATQIITFESFKENLKDFLLVIPFDCWEPVQEGGGKNTVYLQMNSLRAEDTAVYYCARTVPGYSTAPYFDLWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC (SEQ ID NO: 60)
Тяжелая цепь Fab CA645 с GM-CSF человека (полужирным и курсивом) на C-конце CH1, линкер Gly-Ser подчеркнут (для целей сравнения):
EVQLLESGGGLVQPGGSLRLSCAVSGIDLSNYAINWVRQAPGKGLEWIGIIWASGTTFYATWAKGRFTISRDNSKNTVYLQMNSLRAEDTAVYYCARTVPGYSTAPYFDLWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCGGGSGGGGAPARSPSPSTQPWEHVNAIQEARRLLNLSRDTAAEMNETVEVISEMFDLQEPTCLQTRLELYKQGLRGSLTKLKGPLTMMASHYKQHCPPTPETSCATQIITFESFKENLKDFLLVIPFDCWEPVQE (SEQ ID NO: 61)
Последовательность GM-CSF мыши (mGM-CSF):
APTRSPITVTRPWKHVEAIKEALNLLDDMPVTLNEEVEVVSNEFSFKKLTCVQTRLKIFEQGLRGNFTKLKGALNMTASYYQTYCPPTPETDCETQVTTYADFIDSLKTFLTDIPFECKKPGQK (SEQ ID NO: 62)
Тяжелая цепь Fab CA645 с GM-CSF мыши (полужирным шрифтом и курсивом), встроенная в каркасную область 3, линкеры Gly-Ser подчеркнуты:
EVQLLESGGGLVQPGGSLRLSCAVSGIDLSNYAINWVRQAPGKGLEWIGIIWASGTTFYATWAKGRFTISRDNSGGGSGGGGAPTRSPITVTRPWKHVEAIKEALNLLDDMPVTLNEEVEVVSNEFSFKKLTCVQTRLKIFEQGLRGNFTKLKGALNMTASYYQTYCPPTPETDCETQVTTYADFIDSLKTFLTDIPFECKKPGQKGGGKNTVYLQMNSLRAEDTAVYYCARTVPGYSTAPYFDLWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC (SEQ ID NO: 63)
Тяжелая цепь Fab CA645 с GM-CSF мыши (полужирным шрифтом и курсивом) на C-конце CH1, линкеры Gly-Ser подчеркнуты (для целей сравнения):
EVQLLESGGGLVQPGGSLRLSCAVSGIDLSNYAINWVRQAPGKGLEWIGIIWASGTTFYATWAKGRFTISRDNSKNTVYLQMNSLRAEDTAVYYCARTVPGYSTAPYFDLWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCGGGSGGGGAPTRSPITVTRPWKHVEAIKEALNLLDDMPVTLNEEVEVVSNEFSFKKLTCVQTRLKIFEQGLRGNFTKLKGALNMTASYYQTYCPPTPETDCETQVTTYADFIDSLKTFLTDIPFECKKPGQK (SEQ ID NO: 64)
На фиг. 5A и 6 представлен анализ посредством Biacore эффективности связывания для слитых белков антител CA645/hGM-CSF и CA645/mGM-CSF (соответственно) при тестировании с распознаваемым ими рецептором GM-CSF (в концентрации 100 нМ). Результаты демонстрируют, что связывание с GM-CSF является специфическим и зависит от присутствия полипептида вставки. На фиг. также показана отдельная кривая для слитой конструкции константного домена (CH1) пептида GM-CSF, демонстрирующая, что вставка в каркасной области 3 функционирует сравнимым образом. На фиг. 5B показано смоделированное представление белка CA645/GM-CSFFwk3.
ПРИМЕР 5 - Fab CA645 с полипептидом вставки EPO
Эритропоэтин (EPO) представляет собой цитокин, высвобождаемый почкой в ответ на клеточную гипоксию, который стимулирует продуцирование эритроцитов в костном мозге.
Слитый белок получали и экспрессировали в соответствии с описанными выше способами для вставки последовательности EPO человека в участок каркасной области 3 конструкции Fab CA645, как показано ниже.
Последовательность EPO:
APPRLICDSRVLERYLLEAKEAENITTGCAEHCSLNENITVPDTKVNFYAWKRMEVGQQAVEVWQGLALLSEAVLRGQALLVNSSQPWEPLQLHVDKAVSGLRSLTTLLRALGAQKEAISPPDAASAAPLRTITADTFRKLFRVYSNFLRGKLKLYTGEACRTGDR (SEQ ID NO: 65)
Тяжелая цепь Fab CA645 с EPO человека (полужирным шрифтом и курсивом), встроенным в каркасную область 3, линкеры Gly-Ser подчеркнуты:
EVQLLESGGGLVQPGGSLRLSCAVSGIDLSNYAINWVRQAPGKGLEWIGIIWASGTTFYATWAKGRFTISRDNSGGGSGGGGAPPRLICDSRVLERYLLEAKEAENITTGCAEHCSLNENITVPDTKVNFYAWKRMEVGQQAVEVWQGLALLSEAVLRGQALLVNSSQPWEPLQLHVDKAVSGLRSLTTLLRALGAQKEAISPPDAASAAPLRTITADTFRKLFRVYSNFLRGKLKLYTGEACRTGDRGGGKNTVYLQMNSLRAEDTAVYYCARTVPGYSTAPYFDLWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC (SEQ ID NO: 66)
На фиг. 7A представлен анализ посредством Biacore эффективности связывания для этого слитого белка антитела CA645/EPO при тестировании с 50 нМ EPO-R (партнер по связыванию для EPO), с предшествующей инкубацией с 500 нМ растворимым EPO (конкурирующий лиганд) или без нее. На фиг. 7A показано наряду с другими примерами, что EPO-часть слитого является функционально активной и способна связываться с распознаваемым ей рецептором. Более того, снижение сигнала, наблюдаемое для предшествующей инкубации с EPO, демонстрирует специфичность взаимодействия, в то время как немодифицированный Fab CA645 выступает в качестве дальнейшего контроля для демонстрации того, что наблюдаемое связывание происходит с трансплантатом EPO. На фиг. 7B представлено смоделированное представление белка CA645/EPOFwk3.
ПРИМЕР 6 - Слитый белок антитело/Fab с Fab CA645 со вставкой VHH6
VHH6 представляет собой сконструированное однодоменное антитело в виде тяжелой цепи животного семейства верблюжьих, которое связывает объединенный эпитоп, поскольку оно способно связывать комплекс IL-6:gp80, но не индивидуальные компоненты по отдельности.
Слитый белок получали и экспрессировали в соответствии с описанными выше способами для вставки последовательности VHH6 в участок каркасной области 3 конструкции Fab CA645, как показано ниже.
Последовательность VHH6 животного семейства верблюжьих:
DVQFVESGGGSVHAGGSLRLNCATSGYIYSTYCMGWFRQAPGKEREGVAHIYTNSGRTYYADSVKGRFTISQDNAKNTVYLQMNSLKPEDTAIYYCAARPSIRCASFSATEYKDWGQGTQVTVSS (SEQ ID NO: 67)
Тяжелая цепь Fab CA645 с VHH6 (полужирным шрифтом и курсивом), встроенным в каркасную область 3, линкеры Gly-Ser подчеркнуты:
EVQLLESGGGLVQPGGSLRLSCAVSGIDLSNYAINWVRQAPGKGLEWIGIIWASGTTFYATWAKGRFTISRDNSGGGGSGGGGSDVQFVESGGGSVHAGGSLRLNCATSGYIYSTYCMGWFRQAPGKEREGVAHIYTNSGRTYYADSVKGRFTISQDNAKNTVYLQMNSLKPEDTAIYYCAARPSIRCASFSATEYKDWGQGTQVTVSSGGGGSGGGGSKNTVYLQMNSLRAEDTAVYYCARTVPGYSTAPYFDLWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC (SEQ ID NO: 68)
На фиг. 8A представлен анализ посредством Biacore эффективности связывания для этого слитого белка антитела CA645/VHH6 при тестировании с 100 нМ IL-6 (отрицательный контроль для VHH6, поскольку VHH6 связывает только комплекс), 100 нМ слитой конструкцией IL-6/gp80 (партнер по связыванию для VHH6), 1000 нМ HSA (партнер по связыванию для CA645) и как слитой конструкцией IL-6, так и HSA вместе. На фиг. 8A показано, наряду с другими примерами, что обе части слитого белка антитела, VHH6 и CA645, являются функционально активными в качестве связывающих фрагментов и способны связывать их соответствующие партнеры по связыванию. На фиг. 8A, кроме того, показано, что в присутствии обоих партнеров по связыванию сигнал, наблюдаемый на кривой SPR, является суммарным из отдельных кривых связывания, демонстрируя, что связывание с обеими функциональными частями может происходить одновременно. На фиг. 8B показано смоделированное представление белка CA645/VHH6Fwk3.
ПРИМЕР 7 - Слитый белок антитело/Fab с Fab CA645 со вставкой VHH15
VHH15 представляет собой сконструированное однодоменное антитело на основе тяжелой цепи животного семейства верблюжьих, которое распознает и связывает IL-6.
Слитый белок получали и экспрессировали в соответствии с описанными выше способами для вставки последовательности VHH15 в участок каркасной области 3 конструкции Fab CA645, как показано ниже.
Последовательность VHH15 животных семейства верблюжьих:
QVQLVESGGGSVQAGGSLRLSCVAASGYTGCTYDMRWYRQAPGKEREFVSGIDSDGRATYADSVKGRFTISQSNAKIAVYLQMDSLKLEDTAMYYCNLQCLRYPGEYYWGQGTQVTVSS (SEQ ID NO: 69)
Тяжелая цепь Fab CA645 с VHH15, антителом на основе домена животного семейства верблюжьих против IL-6 (полужирным шрифтом и курсивом), встроенным в каркасную область 3, линкеры Gly-Ser подчеркнуты:
EVQLLESGGGLVQPGGSLRLSCAVSGIDLSNYAINWVRQAPGKGLEWIGIIWASGTTFYATWAKGRFTISRDNSGGGGSGGGGSQVQLVESGGGSVQAGGSLRLSCVAASGYTGCTYDMRWYRQAPGKEREFVSGIDSDGRATYADSVKGRFTISQSNAKIAVYLQMDSLKLEDTAMYYCNLQCLRYPGEYYWGQGTQVTVSSGGGGSGGGGSKNTVYLQMNSLRAEDTAVYYCARTVPGYSTAPYFDLWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC (SEQ ID NO: 70)
На фиг. 9A представлен анализ посредством Biacore эффективности связывания для этого слитого белка антитела CA645/VHH15 при тестировании с100 нМ IL-6 (лиганд для VHH15), 1000 нМ HSA (лиганд для CA645) и как IL-6, так и HSA. На фиг. 9A показано, наряду с другими примерами, что обе части слитого белка антитела, VHH15 и Fab, являются функционально активными в качестве связывающих фрагментов и способны связывать их соответствующие партнеры по связыванию. На фиг. 9A, кроме того, показано, что в присутствии обоих партнеров по связыванию сигнал, наблюдаемый на кривой SPR, является суммарным из отдельных кривых связывания, демонстрируя, что связывание с обеими функциональными частями может происходить одновременно. На фиг. 9B показано смоделированное представление белка CA645/VHH15Fwk3.
ПРИМЕР 8 - Fab CA645 с полипептидом-вставкой склеростина
Склеростин является продуктом гена SOST у человека и вовлечен в процесс образования костей, в основном экспрессируясь в остеоцитах. Склеростин функционирует в качестве регуляторного белка в каскаде передачи сигнала Wnt посредством связывания с корецепторами типа LRP.
Слитые белки получали и экспрессировали в соответствии с описанными выше способами для вставки центральной или целой последовательности склеростина в участок каркасной области 3 конструкции Fab CA645, как показано ниже.
Зрелая полноразмерная последовательность склеростина человека:
QGWQAFKNDATEIIPELGEYPEPPPELENNKTMNRAENGGRPPHHPFETKDVSEYSCRELHFTRYVTDGPCRSAKPVTELVCSGQCGPARLLPNAIGRGKWWRPSGPDFRCIPDRYRAQRVQLLCPGGEAPRARKVRLVASCKCKRLTRFHNQSELKDFGTEAARPQKGRKPRPRARSAKANQAELENAY (SEQ ID NO: 71)
Последовательность центрального домена склеростина человека:
CRELHFTRYVTDGPCRSAKPVTELVCSGQCGPARLLPNAIGRGKWWRPSGPDFRCIPDRYRAQRVQLLCPGGEAPRARKVRLVASCKC (SEQ ID NO: 72)
Тяжелая цепь Fab CA645 с центральным доменом склеростина человека (полужирным шрифтом и курсивом), встроенным в каркасную область 3, линкеры из одного S подчеркнуты:
EVQLLESGGGLVQPGGSLRLSCAVSGIDLSNYAINWVRQAPGKGLEWIGIIWASGTTFYATWAKGRFTISRDNSSCRELHFTRYVTDGPCRSAKPVTELVCSGQCGPARLLPNAIGRGKWWRPSGPDFRCIPDRYRAQRVQLLCPGGEAPRARKVRLVASCKCSKNTVYLQMNSLRAEDTAVYYCARTVPGYSTAPYFDLWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC (SEQ ID NO: 73)
Тяжелая цепь Fab CA645 с центральным доменом склеростина человека (полужирным шрифтом и курсивом), встроенным в каркасную область 3, линкеры Gly-Ser (2X G4S) подчеркнуты:
EVQLLESGGGLVQPGGSLRLSCAVSGIDLSNYAINWVRQAPGKGLEWIGIIWASGTTFYATWAKGRFTISRDNSGGGGSGGGGSCRELHFTRYVTDGPCRSAKPVTELVCSGQCGPARLLPNAIGRGKWWRPSGPDFRCIPDRYRAQRVQLLCPGGEAPRARKVRLVASCKCSGGGGSGGGGSKNTVYLQMNSLRAEDTAVYYCARTVPGYSTAPYFDLWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC (SEQ ID NO: 74)
Тяжелая цепь Fab CA645 с центральным доменом склеростина человека (полужирным шрифтом и курсивом), встроенным в каркасную область 3, жесткие линкеры подчеркнуты:
EVQLLESGGGLVQPGGSLRLSCAVSGIDLSNYAINWVRQAPGKGLEWIGIIWASGTTFYATWAKGRFTISRDNSIPFTVCRELHFTRYVTDGPCRSAKPVTELVCSGQCGPARLLPNAIGRGKWWRPSGPDFRCIPDRYRAQRVQLLCPGGEAPRARKVRLVASCKCEYHGLQKNTVYLQMNSLRAEDTAVYYCARTVPGYSTAPYFDLWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC (SEQ ID NO: 75)
Тяжелая цепь Fab CA645 со зрелым полноразмерным склеростина человека (полужирным шрифтом и курсивом), встроенным в каркасную область 3, линкеры Gly-Ser (1X G4S) подчеркнуты:
EVQLLESGGGLVQPGGSLRLSCAVSGIDLSNYAINWVRQAPGKGLEWIGIIWASGTTFYATWAKGRFTISRDNSGGGGSQGWQAFKNDATEIIPELGEYPEPPPELENNKTMNRAENGGRPPHHPFETKDVSEYSCRELHFTRYVTDGPCRSAKPVTELVCSGQCGPARLLPNAIGRGKWWRPSGPDFRCIPDRYRAQRVQLLCPGGEAPRARKVRLVASCKCKRLTRFHNQSELKDFGTEAARPQKGRKPRPRARSAKANQAELENAYSGGGGSKNTVYLQMNSLRAEDTAVYYCARTVPGYSTAPYFDLWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC (SEQ ID NO: 76)
На фиг. 10A и 11 представлен анализ посредством Biacore эффективности этого слитого белка антитела CA645/склеростин при тестировании в качестве супернатанта, разведенного 1 к 5, в отношении связывания с иммобилизованными внеклеточными доменами LRP6 и LRP4, соответственно (партнеры по связыванию для склеростина). Этот эксперимент также исследовал эффекты различных конструкций линкерной области. Было отмечено, что все из протестированных конструкций продемонстрировали связывание с LRP6 и LRP4. На фиг. 10B показано смоделированное представление для белка CA645/Sclerostincore-Fwk3.
ПРИМЕР 9 - Fab CA645 с полипептидом-вставкой IL-10 (димер IL-10)
Интерлейкин 10 (IL-10), также известный как инкубирующий синтез цитокинов фактор (CSIF) человека, представляет собой противовоспалительный цитокин. Он в основном функционирует как гомодимер.
Слитый белок получали и экспрессировали в соответствии с описанными выше способами для вставки двух последовательностей IL-10 человека в участок каркасной области 3 конструкции Fab CA645, как показано ниже.
Последовательность мономера IL-10 человека:
NCWLRQAKNGRCQVLYKTELSKEECCSTGRLSTSWTEEDVNDNTLFKWMIFNGGAPNCIPCKEPRCVCAPDCSNITWKGPVCGLDGKTYRNECALLKARCKEQPELEVQYQGRCKKTCRDVFCPGSSTCVVDQTNNAYCVTCNRICPEPASSEQYLCGNDGVTYSSACHLRKATCLLGRSIGLAGEYK (SEQ ID NO: 100)
Тяжелая цепь Fab CA645 с двумя мономерами IL-10 человека (полужирным шрифтом и курсивом), встроенными в каркасную область 3, линкеры подчеркнуты:
EVQLLESGGGLVQPGGSLRLSCAVSGIDLSNYAINWVRQAPGKGLEWIGIIWASGTTFYATWAKGRFTISRDNSGGGGSGSNCWLRQAKNGRCQVLYKTELSKEECCSTGRLSTSWTEEDVNDNTLFKWMIFNGGAPNCIPCKEPRCVCAPDCSNITWKGPVCGLDGKTYRNECALLKARCKEQPELEVQYQGRCKKTCRDVFCPGSSTCVVDQTNNAYCVTCNRICPEPASSEQYLCGNDGVTYSSACHLRKATCLLGRSIGLAGEYKGGGGSGGGGSGGGGSNCWLRQAKNGRCQVLYKTELSKEECCSTGRLSTSWTEEDVNDNTLFKWMIFNGGAPNCIPCKEPRCVCAPDCSNITWKGPVCGLDGKTYRNECALLKARCKEQPELEVQYQGRCKKTCRDVFCPGSSTCVVDQTNNAYCVTCNRICPEPASSEQYLCGNDGVTYSSACHLRKATCLLGRSIGLAYEGGGGSGSKNTVYLQMNSLRAEDTAVYYCARTVPGYSTAPYFDLWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC (SEQ ID NO: 101)
На фиг. 12A представлен анализ посредством Biacore эффективности связывания в отношении этого слитого белка антитела CA645/IL-10 при тестировании с 100 нМ рецептором IL-10 (партнер по связыванию для IL-10), с предшествующей инкубацией с 200 нМ IL-10 (конкурирующий лиганд) или без нее. Как показано на фиг. 12A, связывание, наблюдаемое для рецептора IL-10, устранялось в присутствии конкурирующего лиганда в молярном избытке, демонстрируя, что связывание, наблюдаемое с частью IL-10 в каркасной области 3 слитого белка, является истинным функциональным взаимодействием и не является неспецифическим. Этот результат также указывает на то, что встроенные мономеры IL-10 также способны образовывать функциональный димер в конструкции каркасной области 3. На фиг. 12B показано смоделированное представление белка CA645/IL-10Fwk3.
ПРИМЕР 10 - конструкции вставок в каркасную область 3 антитела с другими Fab
Слитые белки получали и экспрессировали в соответствии с описанными выше способами для вставки последовательности IL-15 человека в участок каркасной области 3 конструкций Fab CA497 и CA240, как показано ниже.
Последовательность тяжелой цепи Fab CA497 против IL-17A (Fab, CH1 из IgG1; вариабельный домен подчеркнут; CDR полужирным шрифтом и курсивом)
EVQLVESGGGLVKPGGSLRLSCAASGVIFSDYYMAWVRQAPGKGLEWVASINFNADISYYRESVKGRFTISRDDSKNTLYLQMNSLKTEDTAVYYCTTDANRQNYDWFAYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC (SEQ ID NO: 125)
Тяжелая цепь Fab CA497 против IL-17A с IL-15 человека (полужирным шрифтом и курсивом), встроенным в каркасную область 3, линкеры Gly-Ser подчеркнуты:
EVQLVESGGGLVKPGGSLRLSCAASGVIFSDYYMAWVRQAPGKGLEWVASINFNADISYYRESVKGRFTISRDDSGGGSGGGGNWVNVISDLKKIEDLIQSMHIDATLYTESDVHPSCKVTAMKCFLLELQVISLESGDASIHDTVENLIILANNSLSSNGNVTESGCKECEELEEKNIKEFLQSFVHIVQMFINGGGKNTLYLQMNSLKTEDTAVYYCTTDANRQNYDWFAYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC (SEQ ID NO: 83)
Последовательность легкой цепи Fab CA497 против IL-17A:
AIQLTQSPSSLSASVGDRVTITCKASESVSSSMYSYMHWYQQKPGKAPKLLIYRASNLESGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQSWTAPRTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC (SEQ ID NO: 84)
Последовательность тяжелой цепи Fab CA240 против IL-6 (Fab, CH1 из IgG1; вариабельный домен подчеркнут; CDR выделены полужирным шрифтом и курсивом):
EVQLVESGGGLVQPGGSLRLSCAASGFNFNDYFMNWVRQAPGKGLEWVAQMRNKNYQYGTYYAESLEGRFTISRDDSKNSLYLQMNSLKTEDTAVYYCARESYYGFTSYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC (SEQ ID NO: 126)
Тяжелая цепь Fab CA240 против IL-6 с IL-15 человека (полужирным шрифтом и курсивом), встроенным в каркасную область 3, линкеры Gly-Ser подчеркнуты:
EVQLVESGGGLVQPGGSLRLSCAASGFNFNDYFMNWVRQAPGKGLEWVAQMRNKNYQYGTYYAESLEGRFTISRDDSGGGSGGGGNWVNVISDLKKIEDLIQSMHIDATLYTESDVHPSCKVTAMKCFLLELQVISLESGDASIHDTVENLIILANNSLSSNGNVTESGCKECEELEEKNIKEFLQSFVHIVQMFINGGGKNSLYLQMNSLKTEDTAVYYCARESYYGFTSYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC (SEQ ID NO: 85)
Последовательность легкой цепи Fab CA240 против IL-6:
DIQMTQSPSSLSASVGDRVTITCQASQDIGISLSWYQQKPGKAPKLLIYNANNLADGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCLQHNSAPYTFGQGTKLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC (SEQ ID NO: 86)
На фиг. 13 и 14 представлен анализ посредством Biacore эффективности связывания для слитых белков антител CA497/IL-15 и CA240/IL-15 (соответственно) при тестировании с 100 нМ IL-2-Rβ (партнер по связыванию для IL-15), с предшествующей инкубацией с IL-15 (конкурирующий лиганд) или без нее. Как показано в обоих экспериментах, наблюдаемое связывание с IL-2-Rβ устранялось в присутствии конкурирующего лиганда в молярном избытке, демонстрируя, что наблюдаемое связывание с частью IL-15 в каркасной области 3 слитых белков является истинным функциональным взаимодействием и не является неспецифическим.
ПРИМЕР 11 - Конструкции Fab с VHH, встроенным в каркасную область 3
HyHEL5 представляет собой Fab против лизоцима со следующими последовательностями:
Тяжелая цепь HyHEL5:
EVQLQQSGAELMKPGASVKISCKASGYTFSDYWIEWVKQRPGHGLEWIGEILPGSGSTNYHERFKGKATFTADTSSSTAYMQLNSLTSEDSGVYYCLHGNYDFDGWGQGTTLTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC (SEQ ID NO: 87)
Легкая цепь HyHEL5:
DIVLTQSPAIMSASPGEKVTMTCSASSSVNYMYWYQQKSGTSPKRWIYDTSKLASGVPVRFSGSGSGTSYSLTISSMETEDAATYYCQQWGRNPTFGGGTKLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC (SEQ ID NO: 88)
D3L11 представляет собой VHH против лизоцима со следующей последовательностью:
D3L11:
DVQLVESGGGSVQAGGSLRLSCAASGSTDSIEYMTWFRQAPGKAREGVAALYTHTGNTYYTDSVKGRFTISQDKAKNMAYLRMDSVKSEDTAIYTCGATRKYVPVRFALDQSSYDYWGQGTQVTVSS (SEQ ID NO: 89)
Следующие скрытые конструкции получали для тестирования того, приведет ли вставка второй молекулы антитела в участок каркасной области 3 первого антитела, где антитела, связывающиеся с одним и тем же антигеном-мишенью, иметь более высокую аффинность к антигену, чем каждое из индивидуальных антител.
Легкая цепь HyHEL5 против лизоцима с VHH D3L11 против лизоцима (полужирным шрифтом), встроенным в каркасную область 3, линкеры Gly-Ser подчеркнуты:
DIVLTQSPAIMSASPGEKVTMTCSASSSVNYMYWYQQKSGTSPKRWIYDTSKLASGVPVRFSGSGSGSGGGGSDVQLVESGGGSVQAGGSLRLSCAASGSTDSIEYMTWFRQAPGKAREGVAALYTHTGNTYYTDSVKGRFTISQDKAKNMAYLRMDSVKSEDTAIYTCGATRKYVPVRFALDQSSYDYWGQGTQVTVSSGGGGSGGGGSGGGGSTSYSLTISSMETEDAATYYCQQWGRNPTFGGGTKLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC (SEQ ID NO: 90)
Легкая цепь HyHEL5 против лизоцима с VHH15 против IL-6 (полужирным шрифтом), встроенным в каркасную область 3, линкеры Gly-Ser подчеркнуты:
DIVLTQSPAIMSASPGEKVTMTCSASSSVNYMYWYQQKSGTSPKRWIYDTSKLASGVPVRFSGSGSGSGGGGSQVQLVESGGGSVQAGGSLRLSCVAASGYTGCTYDMRWYRQAPGKEREFVSGIDSDGRATYADSVKGRFTISQSNAKIAVYLQMDSLKLEDTAMYYCNLQCLRYPGEYYWGQGTQVTVSSGGGGSGGGGSGGGGSTSYSLTISSMETEDAATYYCQQWGRNPTFGGGTKLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC (SEQ ID NO: 91)
Легкая цепь CA645 с VHH D3L11 против лизоцима (полужирным шрифтом), встроенным в каркасную область 3, линкеры Gly-Ser подчеркнуты:
DIQMTQSPSSVSASVGDRVTITCQSSPSVWSNFLSWYQQKPGKAPKLLIYEASKLTSGVPSRFSGSGSGSGGGGSDVQLVESGGGSVQAGGSLRLSCAASGSTDSIEYMTWFRQAPGKAREGVAALYTHTGNTYYTDSVKGRFTISQDKAKNMAYLRMDSVKSEDTAIYTCGATRKYVPVRFALDQSSYDYWGQGTQVTVSSGGGGSGGGGSGGGGSTDFTLTISSLQPEDFATYYCGGGYSSISDTTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC (SEQ ID NO: 92)
Кривые SPR для серии разведений каждой из конструкций при тестировании в отношении связывания с лизоцимом представлены на фиг. 15-17. Из последующего анализа с использованием способов, описанных выше, в следующей таблице представлена аффинность связывания в отношении лизоцима, вычисленная для каждой конструкции:
Таблица 2 - Аффинность связывания для конструкций антител
Вставка в каркасную область 3: VHH D3L11
Вставка в каркасную область 3: VHH15
Вставка в каркасную область 3: VHH D3L11
Как отчетливо видно из приведенных выше результатов, аффинность связывания в отношении лизоцима возрастает (т.е. наблюдается снижение KD) для конструкции, где как Fab, так и вставка VHH связываются с лизоцимом. На фиг. 18 проиллюстрировано, как это может происходить на молекулярном уровне. Это также позволяет предположить, что длина линкера может быть важной для обеспечения поливалентных взаимодействий на одной и той же молекуле-мишени в таких антителах.
ПРИМЕР 12 - Fab CA645 с полипептидом-вставкой IL-15 с использованием линкеров из животных подсемейства бычьих
Получали варианты белка CA645/IL-15Fwk3, описанного в примере 1, где линкеры заменяли на другие линкеры, включая линкеры из животных семейства бычьих, и тестировали их эффекты на связывание. Помимо исходной конструкции тяжелой цепи (SEQ ID NO: 54), получали дополнительные конструкции, как показано ниже:
Тяжелая цепь Fab CA645 с IL-15 человека (полужирным шрифтом и курсивом), встроенным в каркасную область 3, с длинным линкером из животного семейства бычьих (подчеркнут)
EVQLLESGGGLVQPGGSLRLSCAVSGIDLSNYAINWVRQAPGKGLEWIGIIWASGTTFYATWAKGRFTISRDNSTSVHQETKKYQSNWVNVISDLKKIEDLIQSMHIDATLYTESDVHPSCKVTAMKCFLLELQVISLESGDASIHDTVENLIILANNSLSSNGNVTESGCKECEELEEKNIKEFLQSFVHIVQMFINSYTYNYEWHVDVKNTVYLQMNSLRAEDTAVYYCARTVPGYSTAPYFDLWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC (SEQ ID NO: 93)
Тяжелая цепь Fab CA645 с IL-15 человека (полужирным и курсивом), встроенным в каркасную область 3, с коротким линкером животного семейства бычьих (подчеркнут)
EVQLLESGGGLVQPGGSLRLSCAVSGIDLSNYAINWVRQAPGKGLEWIGIIWASGTTFYATWAKGRFTISRDNSETKKYQSNWVNVISDLKKIEDLIQSMHIDATLYTESDVHPSCKVTAMKCFLLELQVISLESGDASIHDTVENLIILANNSLSSNGNVTESGCKECEELEEKNIKEFLQSFVHIVQMFINSYTYNYEKNTVYLQMNSLRAEDTAVYYCARTVPGYSTAPYFDLWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC (SEQ ID NO: 94)
Тяжелая цепь Fab CA645 с IL-15 человека (полужирным шрифтом и курсивом), встроенным в каркасную область 3 с линкером 2xG4S (подчеркнут)
EVQLLESGGGLVQPGGSLRLSCAVSGIDLSNYAINWVRQAPGKGLEWIGIIWASGTTFYATWAKGRFTISRDNSGGGGSGGGGSNWVNVISDLKKIEDLIQSMHIDATLYTESDVHPSCKVTAMKCFLLELQVISLESGDASIHDTVENLIILANNSLSSNGNVTESGCKECEELEEKNIKEFLQSFVHIVQMFINGGGGSGGGGSKNTVYLQMNSLRAEDTAVYYCARTVPGYSTAPYFDLWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC (SEQ ID NO: 95)
На фиг. 19 представлен анализ связывания Biacore для описанных выше конструкций CA645/IL-15Fwk3 при тестировании с 100 нМ IL-2-Rβ, и при сравнении против конструкции согласно примеру 1 (исходный линкер) и Fab CA645 дикого типа, который не имеет вставки IL-15. Следует отметить, что все из протестированных конструкций продемонстрировали успешное связывание.
ПРИМЕР 13 - фрагмент Fab CA645 с a dsscFv, встроенным в каркасную область 3
Конструирование генов
Два антитела по изобретению получали и экспрессировали в соответствии с описанными выше способами для вставки dsscFv CA497 против IL17A (vHvL или vLvH) в участок каркасной области 3 Fab-фрагмента CA645 (каркасное антитело), как показано ниже. Полученные антитела могут быть названы CA645 Fab-Fwk3 CA497 dsscFv (vHvL) или CA645 Fab-Fwk3 CA497 dsscFv (vLvH) соответственно. В качестве контролей получали dsscFv CA497 и Fab CA645.
Гены, кодирующие V-области легкой и тяжелой цепей, включая варианты со вставкой в каркасную область 3, моделировали и конструировали посредством подхода автоматизированного синтеза (ATUM). Гены V-области легкой цепей (например, кодирующие VL-домен CA645 SEQ ID NO: 2), клонировали в экспрессирующие векторы, содержавшие ДНК, кодирующую область Cκ человека (аллотип Km3). Гены V-области тяжелой цепи клонировали в экспрессирующие векторы, содержавшие ДНК, кодирующую область CH1 IgG1.
dsscFv CA497 (vHvL)
EVQLVESGGGLVKPGGSLRLSCAASGVIFSDYYMAWVRQAPGKCLEWVASINFNADISYYRESVKGRFTISRDDSKNTLYLQMNSLKTEDTAVYYCTTDANRQNYDWFAYWGQGTLVTVSSGGGGSGGGGSGGGGSAIQLTQSPSSLSASVGDRVTITCKASESVSSSMYSYMHWYQQKPGKAPKLLIYRASNLESGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQSWTAPRTFGCGTKVEIK (SEQ ID NO: 127)
dsscFv CA497 (vLvH)
AIQLTQSPSSLSASVGDRVTITCKASESVSSSMYSYMHWYQQKPGKAPKLLIYRASNLESGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQSWTAPRTFGCGTKVEIKGGGGSGGGGSGGGGSEVQLVESGGGLVKPGGSLRLSCAASGVIFSDYYMAWVRQAPGKCLEWVASINFNADISYYRESVKGRFTISRDDSKNTLYLQMNSLKTEDTAVYYCTTDANRQNYDWFAYWGQGTLVTVSS (SEQ ID NO: 128)
Тяжелая цепь Fab CA645 с dsscFv CA497 (vHvL) (полужирным шрифтом и курсивом), встроенным в каркасную область 3, линкеры подчеркнуты
EVQLLESGGGLVQPGGSLRLSCAVSGIDLSNYAINWVRQAPGKGLEWIGIIWASGTTFYATWAKGRFTISRDNSGGGGSGGGGSEVQLVESGGGLVKPGGSLRLSCAASGVIFSDYYMAWVRQAPGKCLEWVASINFNADISYYRESVKGRFTISRDDSKNTLYLQMNSLKTEDTAVYYCTTDANRQNYDWFAYWGQGTLVTVSSGGGGSGGGGSGGGGSAIQLTQSPSSLSASVGDRVTITCKASESVSSSMYSYMHWYQQKPGKAPKLLIYRASNLESGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQSWTAPRTFGCGTKVEIKGGGGSGGGGSKNTVYLQMNSLRAEDTAVYYCARTVPGYSTAPYFDLWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC (SEQ ID NO: 129)
Тяжелая цепь Fab CA645 с dsscFv CA497 (vLvH) (полужирным шрифтом и курсивом), встроенным в каркасную область 3, линкеры подчеркнуты:
EVQLLESGGGLVQPGGSLRLSCAVSGIDLSNYAINWVRQAPGKGLEWIGIIWASGTTFYATWAKGRFTISRDNSGGGGSGGGGSAIQLTQSPSSLSASVGDRVTITCKASESVSSSMYSYMHWYQQKPGKAPKLLIYRASNLESGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQSWTAPRTFGCGTKVEIKGGGGSGGGGSGGGGSEVQLVESGGGLVKPGGSLRLSCAASGVIFSDYYMAWVRQAPGKCLEWVASINFNADISYYRESVKGRFTISRDDSKNTLYLQMNSLKTEDTAVYYCTTDANRQNYDWFAYWGQGTLVTVSSGGGGSGGGGSKNTVYLQMNSLRAEDTAVYYCARTVPGYSTAPYFDLWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC (SEQ ID NO: 130)
Временная экспрессия в клетках
Сконструированные Fab-фрагменты временно экспрессировали либо в клетках HEK-293 с использованием набора для трансфекции ExpiFectamine 293 (Life Technologies, в соответствии с инструкциями изготовителя) либо в клетках CHO-S XE, происходящей из CHO-K1 клеточной линии (Cain K et al, A CHO cell line engineered to express XBP1 and ERO1-Lα has increased levels of transient protein expression. Biotechnology Progress 2013; 29: 697-706), с использованием набора для трансфекции ExpiFectamine CHO (Life Technologies, в соответствии с инструкциями изготовителя). Клетки HEK-293 использовали для мелкомасштабной экспрессии (2 мл) для получения конструкций антител для прямого анализа с использованием SPR. Клетки CHO-S XE использовали для крупномасштабной экспрессии (200 мл) для получения конструкций антител для очистки перед дальнейшей охарактеризацией.
Очистка
Аффинную хроматографию использовали для очистки Fab-фрагментов из культуральных супернатантов. Супернатанты, содержавшие антитела, пропускали через колонку HiTrap с белком G (GE Healthcare). После стадии промывания фосфатно-солевым буфером (PBS), pH 7,4, связавшийся материал элюировали с использованием градиента PBS, pH 7,4, против возрастающего процента по объему 0,1 M фосфатно-цитратного буфера, pH 2,6. Элюат нейтрализовывали 2 M Tris-HCl (pH 8). Фракции объединяли, количественно определяли по поглощению при 280 нм, и концентрировали с использованием центрифужных фильтров Amicon Ultra (Merck Millipore). Для выделения мономерных фракций конструкций использовали эксклюзионную хроматографию на колонке HiLoad 16/60, Superdex 200 (GE Healthcare), уравновешенной PBS, pH 7,4. Фракции, содержавшие мономерный Fab, объединяли, количественно определяли, концентрировали и хранили при 4°C.
UPLC (Сверхвысокоэффективная жидкостная хроматография):
2 мкг образца инжектировали на колонку SEC BEH200, предварительно уравновешенную PBS, pH7,4, и прогоняли в течение 10 минут. Интенсивность флуоресценции регистрировали против времени элюирования и площади пиков отбирали вручную в программном обеспечении EMpower.
Профили элюирования UPLC получали и анализировали. Пики выглядели на >98% мономерными (99,42% для CA645 Fab-Fwk3 CA497 dsscFv (vLvH), 99,51% для CA645 Fab-Fwk3 CA497 dsscFv (vHvL)). Контроли dsscFv CA497 и Fab CA645 продемонстрировали, соответственно, 99,99% и 98,43% мультимерных форм (данные не представлены). Таким образом, очищенные белки были однородными и в них отсутствовали мультимерные формы или агрегаты.
Молекулярная стабильность
Молекулярную стабильность определяли посредством анализа температуры плавления (Tm) (показатель разворачивания) способом дифференциальной сканирующей калориметрии (DSC). Термограммы получали на автоматизированном MicroCal VP DSC (Malvern Panalytical) при температуре 0-100°C, с использованием шага 1°C/мин, с временем сканирования перед инкубацией 15 мин, периодом фильтрации 5 с и пассивной обратной связью. Из данных вычитали данные для буфера перед вычитанием исходного уровня вручную и внесения поправки на концентрацию. Термограммы аппроксимировали к модели не 2 состояний в Origin7.0.
Результаты представлены на фиг. 20 (кривые DSC; вертикальная ось: (Cp) удельная теплота; горизонтальная ось: температура в градусах цельсия C (°C)) и в таблице 3 ниже. dsscFv CA497 и Fab CA645 продемонстрировали единичные переходы при 58,40°C и 83,57°C, соответственно. При комбинировании в формате Fab-Fwk3 с вставкой dsscFv, Tm dsscFv CA497 является практически идентичной (разность 2-3°C) и Fab CA645 имеет варьирование Tm приблизительно 4-5°C. Таким образом, образование пар dsscFv CA497 и Fab CA645 в этом формате антитела не оказывает значительного влияния на конформационную стабильность каждого из доменов.
Таблица 3 - Температуры перехода для разворачивания
Анализ нейтрализации
Эффективность антитела IgG4 CA497, CA645 Fab-Fwk3 CA497 dsscFv (vHvL) и CA645 Fab-Fwk3 CA497 dsscFv (vLvH) против рекомбинантного IL-17A человека в клетках Hela тестировали в присутствии или в отсутствии HSA. Клетки Hela получали из банка клеток ATCC (ATCC CCL-2). Клетки выращивали в модифицированной способом Дульбекко среде Игла (DMEM), дополненной 10% эмбриональной телячьей сывороткой, пенициллином, гентамицином и глутамином. 1×104 клеток высевали в 96-луночные планшеты для культивирования тканей с плоским дном. Клетки инкубировали в течение ночи и промывали один раз в буфере для анализа. IL-17A человека (25 нг мл-1) инкубировали в присутствии фиксированной концентрации TNF-α человека, и эту смесь предварительно инкубировали с тестируемым антителом. Затем Цитокины и антитело добавляли к клеткам Hela для стимуляции клеток, которые инкубировали в течение ночи. В клеточной линии Hela IL-17A действует синергично с TNF-альфа, индуцируя продуцирование IL-6, который может быть количественно определен, например, посредством ELISA. Продуцирование IL-6 в супернатанте клеточной культуры пропорционально количеству IL-17A, добавляемого к клеткам. Уровни IL-6 человека измеряли посредство ELISA и количественно определяли посредством сравнения с концентрациями известного стандарта IL-6 человека. В этом анализе ингибирование секреции IL-6 коррелирует с ингибированием IL-17A.
Кривые ингибирования представлены на фиг. 21 (горизонтальная ось: концентрация антитела (ингибитора) в нг/мл, вертикальная ось: ингибирование секреции IL-6 (%)) и указывает на то, что родительское антитело IgG4 CA497 и обе слитых конструкции антитела эффективно нейтрализовывали рекомбинантный IL-17A человека. Кроме того, результаты указывают на то, что dsscFv против IL-17 сохранял функциональность при встраивании в каркасную область Fwk3 Fab против HSA. Величины EC50, вычисленные из этих экспериментов (таблица 4 ниже), показали, что антитело CA645 Fab-Fwk3 CA497 dsscFv (vLvH) и CA645 Fab-Fwk3 CA497 dsscFv (vHvL) обеспечили EC50 10,5 нг/мл и 10,6 нг/мл, соответственно, против рекомбинантного IL-17A человека (25 нг.мл-1) и что присутствие HSA не влияло на эффективность нейтрализации (EC50 10,8 и 8,7 нг/мл, соответственно, в присутствии HSA). Также конструкции CA645 Fab-Fwk3 CA497 dsscFv имели эффективность нейтрализации, сравнимую с одним из исходных IgG4-антител CA497.
Таблица 4. Величины EC50 для нейтрализации IL-17A в присутствии или в отсутствии HSA
ПРИМЕР 14 - Полный IgG CA645 с dsscFv, встроенным в каркасную область 3
Слитый белок получали и экспрессировали в соответствии с описанными выше способами для вставки dsscFv (vH-vL) CA497 или dsscFv (vL-vH) CA497 в участок каркасной области 3 конструкции IgG4P CA645, как показано ниже. Полученные слитые белки антител могут быть названы CA645 IgG4P Fwk3-CA497 dsscFv (vHvL) и CA645 IgG4P Fwk3-CA497 dsscFv (vLvH).
Конструирование гена
гены, кодирующие V-области тяжелой и легкой цепи, включая варианты со вставкой в каркасную область 3, моделировали и конструировали посредством подхода автоматизированного синтеза (ATUM).
Гены V-области легкой цепи (например, кодирующие VL-домен CA645 SEQ ID NO: 2) клонировали в экспрессирующие векторы, содержавшие ДНК, кодирующую область Cκ человека (аллотип Km3). Гены V-области тяжелой цепи клонировали в экспрессирующие векторы, содержавшие ДНК, кодирующую константные области IgG4P для полноразмерного антитела.
Тяжелая цепь IgG4P CA645 с dsscFv (vHvL) CA497 (полужирным шрифтом и курсивом), встроенным в каркасную область 3, линкеры подчеркнуты:
EVQLLESGGGLVQPGGSLRLSCAVSGIDLSNYAINWVRQAPGKGLEWIGIIWASGTTFYATWAKGRFTISRDNSGGGGSGGGGSEVQLVESGGGLVKPGGSLRLSCAASGVIFSDYYMAWVRQAPGKCLEWVASINFNADISYYRESVKGRFTISRDDSKNTLYLQMNSLKTEDTAVYYCTTDANRQNYDWFAYWGQGTLVTVSSGGGGSGGGGSGGGGSAIQLTQSPSSLSASVGDRVTITCKASESVSSSMYSYMHWYQQKPGKAPKLLIYRASNLESGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQSWTAPRTFGCGTKVEIKGGGGSGGGGSKNTVYLQMNSLRAEDTAVYYCARTVPGYSTAPYFDLWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 131)
Тяжелая цепь IgG4P CA645 с dsscFv (vLvH) CA497 (полужирным шрифтом и курсивом), встроенным в каркасную область 3, линкеры подчеркнуты:
EVQLLESGGGLVQPGGSLRLSCAVSGIDLSNYAINWVRQAPGKGLEWIGIIWASGTTFYATWAKGRFTISRDNSGGGGSGGGGSAIQLTQSPSSLSASVGDRVTITCKASESVSSSMYSYMHWYQQKPGKAPKLLIYRASNLESGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQSWTAPRTFGCGTKVEIKGGGGSGGGGSGGGGSEVQLVESGGGLVKPGGSLRLSCAASGVIFSDYYMAWVRQAPGKCLEWVASINFNADISYYRESVKGRFTISRDDSKNTLYLQMNSLKTEDTAVYYCTTDANRQNYDWFAYWGQGTLVTVSSGGGGSGGGGSKNTVYLQMNSLRAEDTAVYYCARTVPGYSTAPYFDLWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 132)
Временная экспрессия в клетках
Антитела временно экспрессировали либо в клетках HEK-293 с использованием набора для трансфекции ExpiFectamine 293 (Life Technologies, в соответствии с инструкциями изготовителя) либо в клетках CHO-S XE, происходящей из CHO-K1 клеточной линии (Cain K et al, A CHO cell line engineered to express XBP1 and ERO1-Lα has increased levels of transient protein expression. Biotechnology Progress 2013; 29: 697-706), с использованием набора для трансфекции ExpiFectamine CHO (Life Technologies, в соответствии с инструкциями изготовителя). Клетки HEK-293 использовали для мелкомасштабной экспрессии (2 мл) для получения конструкций антител для прямого анализа с использованием SPR. Клетки CHO-S XE использовали для крупномасштабной экспрессии (200 мл) для получения конструкций антител для очистки перед дальнейшей охарактеризацией.
Очистка
Аффинную хроматографию использовали для очистки IgG из культуральных супернатантов. Супернатанты, содержавшие IgG, пропускали через колонку MabSelect SuRe (GE Healthcare). После стадии промывания фосфатно-солевым буфером (PBS), pH 7,4, связавшийся материал элюировали с использованием градиента PBS, pH 7,4, против возрастающего процента по объему 0,1 M фосфатно-цитратного буфера, pH 2,6. Элюат нейтрализовывали 2 M Tris-HCl (pH 8). Фракции объединяли, количественно определяли по поглощению при 280 нм, и концентрировали с использованием центрифужных фильтров Amicon Ultra (Merck Millipore). Для выделения мономерных фракций конструкций на основе IgG использовали эксклюзионную хроматографию на колонке HiLoad 16/60, Superdex 200 (GE Healthcare), уравновешенной PBS, pH 7,4. Фракции, содержавшие мономерный Fab, объединяли, количественно определяли, концентрировали и хранили при 4°C.
Одновременное связывание с IL-17 и HSA
Для тестирования одновременного связывания IL-17A и HSA с CA645 IgG4P Fwk3-CA497 dsscFv (vHvL), последний иммобилизовывали на чипе Biacore вместо поликлонального F(ab)2 козы по той же методике, которая описана выше. Три концентрации как IL-17A, так и HSA выбирали для обеспечения низких, средних и высоких уровней связывания с иммобилизованным антителом, в частности, они соответствовали стехиометрическим соотношениям лиганда и антитела в диапазонах от 0,15 до 0,3, от 0,4 до 0,6 и от 0,65 до 0,75, соответственно. В случае IL-17A, концентрации составляли 4, 20 и 100 нМ, и в случае HSA, концентрации составляли 40, 200 и 1000 нМ, соответственно. Указанные выше низкие средние и высокие концентрации тестировали посредством SPR, как описано ранее, в индивидуальных циклах либо в качестве отдельных лигнадов, либо в качестве смесей IL17A/HSС, и в результаты вносили поправку путем вычитания циклов для контроля в виде буфера. Результаты представлены на фиг. 22 и показывают, что CA645 IgG4P Fwk3-CA497 dsscFv (vHvL) связывает как IL-17A, так и HSA, зависимым от концентрации образом. На фиг. 22, кроме того, показано, что в присутствии обоих партнеров по связыванию сигнал является аддитивным, демонстрируя, что связывание как с IL-17A, так и с HSA может происходить одновременно.
ПРИМЕР 15 - Антитела по изобретению, которые связываются как с CD3, так и с CD28
Конструирование гена
Антитела получали и экспрессировали в соответствии со способами, описанными выше, для вставки dsscFv (vH-vL) против CD3 в участок каркасной области 3 каркасного антитела против CD3 или против CD28 (IgG4P или Fab-фрагмент), как показано ниже в таблице 5. Полученные конструкции антител согласно изобретению могут быть названы анти-CD28 Fab Fwk3-анти-CD3 dsscFv (LC SEQ ID NO: 136, HC SEQ ID NO: 139), анти-CD3 Fab Fwk3-анти-CD3 dsscFv (LC SEQ ID NO: 133, HC SEQ ID NO: 140), анти-CD28 IgG4P Fwk3-анти-CD3 dsscFv (LC SEQ ID NO: 136, HC SEQ ID NO: 141), анти-CD3 IgG4P Fwk3-анти-CD3 dsscFv (LC SEQ ID NO: 133, HC SEQ ID NO: 142).
Таблица 5 - Конструкции антител
Тяжелая цепь SEQ ID NO: 134
Тяжелая цепь SEQ ID NO: 137
Тяжелая цепь SEQ ID NO: 139
Тяжелая цепь SEQ ID NO: 135
Тяжелая цепь SEQ ID NO: 138
Тяжелая цепь SEQ ID NO: 141
Тяжелая цепь SEQ ID NO: 142
Легкая цепь антитела против CD3, муромонаб (гуманизированный OKT3) (вариабельный домен подчеркнут; CDR полужирным шрифтом и курсивом):
DIQMTQSPSSLSASVGDRVTITCSASSSVSYMNWYQQTPGKAPKLLIYDTSKLASGVPSRFSGSGSGTDYTFTISSLQPEDIATYYCQQWSSNPFTFGQGTKLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC (SEQ ID NO: 133)
Тяжелая цепь Fab против CD3, CH1 из IgG1, муромонаб (гуманизированный OKT3) (вариабельный домен подчеркнут; CDR полужирным шрифтом и курсивом):
QVQLVQSGGGVVQPGRSLRLSCKASGYTFTRYTMHWVRQAPGKGLEWIGYINPSRGYTNYNQKFKDRFTISRDNSKNTAFLQMDSLRPEDTGVYFCARYYDDHYCLDYWGQGTPVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC (SEQ ID NO: 134)
Тяжелая цепь IgG4P против CD3, муромонаб (гуманизированный OKT3) (вариабельный домен подчеркнут; CDR полужирным шрифтом и курсивом):
QVQLVQSGGGVVQPGRSLRLSCKASGYTFTRYTMHWVRQAPGKGLEWIGYINPSRGYTNYNQKFKDRFTISRDNSKNTAFLQMDSLRPEDTGVYFCARYYDDHYCLDYWGQGTPVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTKTYTCNVDHKPSNTKVDKRVESKYGPPCPPCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK (SEQ ID NO: 135)
Легкая цепь антитела против CD28, терализумаб (вариабельный домен подчеркнут; CDR полужирным шрифтом и курсивом):
DIQMTQSPSSLSASVGDRVTITCHASQNIYVWLNWYQQKPGKAPKLLIYKASNLHTGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQGQTYPYTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC (SEQ ID NO: 136)
Тяжелая цепь Fab против CD28, CH1 из IgG1, терализумаб (вариабельный домен подчеркнут; CDR полужирным шрифтом и курсивом):
QVQLVQSGAEVKKPGASVKVSCKASGYTFTSYYIHWVRQAPGQGLEWIGCIYPGNVNTNYNEKFKDRATLTVDTSISTAYMELSRLRSDDTAVYFCTRSHYGLDWNFDVWGQGTTVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC (SEQ ID NO: 137)
Тяжелая цепь IgG4P против CD28, терализумаб (вариабельный домен подчеркнут; CDR полужирным шрифтом и курсивом):
QVQLVQSGAEVKKPGASVKVSCKASGYTFTSYYIHWVRQAPGQGLEWIGCIYPGNVNTNYNEKFKDRATLTVDTSISTAYMELSRLRSDDTAVYFCTRSHYGLDWNFDVWGQGTTVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTKTYTCNVDHKPSNTKVDKRVESKYGPPCPPCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK (SEQ ID NO: 138)
Тяжелая цепь Fab против CD28 с dsscFv (vHvL) против CD3 (полужирным шрифтом и курсивом), встроенным в каркасную область 3, линкеры подчеркнуты:
QVQLVQSGAEVKKPGASVKVSCKASGYTFTSYYIHWVRQAPGQGLEWIGCIYPGNVNTNYNEKFKDRATLTVDTSGGGGSGGGGSQVQLVQSGGGVVQPGRSLRLSCKASGYTFTRYTMHWVRQAPGKCLEWIGYINPSRGYTNYNQKFKDRFTISRDNSKNTAFLQMDSLRPEDTGVYFCARYYDDHYCLDYWGQGTPVTVSSGGGGSGGGGSGGGGSGGGGSDIQMTQSPSSLSASVGDRVTITCSASSSVSYMNWYQQTPGKAPKLLIYDTSKLASGVPSRFSGSGSGTDYTFTISSLQPEDIATYYCQQWSSNPFTFGCGTKLEIKGGGGSGGGGSISTAYMELSRLRSDDTAVYFCTRSHYGLDWNFDVWGQGTTVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC (SEQ ID NO: 139)
Тяжелая цепь Fab против CD3 с dsscFv (vHvL) против CD3 (полужирным шрифтом и курсивом), встроенным в каркасную область 3, линкеры подчеркнуты:
QVQLVQSGGGVVQPGRSLRLSCKASGYTFTRYTMHWVRQAPGKGLEWIGYINPSRGYTNYNQKFKDRFTISRDNSGGGGSGGGGSQVQLVQSGGGVVQPGRSLRLSCKASGYTFTRYTMHWVRQAPGKCLEWIGYINPSRGYTNYNQKFKDRFTISRDNSKNTAFLQMDSLRPEDTGVYFCARYYDDHYCLDYWGQGTPVTVSSGGGGSGGGGSGGGGSGGGGSDIQMTQSPSSLSASVGDRVTITCSASSSVSYMNWYQQTPGKAPKLLIYDTSKLASGVPSRFSGSGSGTDYTFTISSLQPEDIATYYCQQWSSNPFTFGCGTKLEIKGGGGSGGGGSKNTAFLQMDSLRPEDTGVYFCARYYDDHYCLDYWGQGTPVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC (SEQ ID NO: 140)
Тяжелая цепь IgG4P против CD28 с dsscFv (vHvL) против CD3 (полужирным шрифтом и курсивом), встроенным в каркасную область 3, линкеры подчеркнуты:
QVQLVQSGAEVKKPGASVKVSCKASGYTFTSYYIHWVRQAPGQGLEWIGCIYPGNVNTNYNEKFKDRATLTVDTSGGGGSGGGGSQVQLVQSGGGVVQPGRSLRLSCKASGYTFTRYTMHWVRQAPGKCLEWIGYINPSRGYTNYNQKFKDRFTISRDNSKNTAFLQMDSLRPEDTGVYFCARYYDDHYCLDYWGQGTPVTVSSGGGGSGGGGSGGGGSGGGGSDIQMTQSPSSLSASVGDRVTITCSASSSVSYMNWYQQTPGKAPKLLIYDTSKLASGVPSRFSGSGSGTDYTFTISSLQPEDIATYYCQQWSSNPFTFGCGTKLEIKGGGGSGGGGSISTAYMELSRLRSDDTAVYFCTRSHYGLDWNFDVWGQGTTVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTKTYTCNVDHKPSNTKVDKRVESKYGPPCPPCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK (SEQ ID NO: 141)
Тяжелая цепь IgG4P против CD3 с dsscFv (vHvL) против CD3 (полужирным шрифтом и курсивом), встроенным в каркасную область 3, линкеры подчеркнуты:
QVQLVQSGGGVVQPGRSLRLSCKASGYTFTRYTMHWVRQAPGKGLEWIGYINPSRGYTNYNQKFKDRFTISRDNSGGGGSGGGGSQVQLVQSGGGVVQPGRSLRLSCKASGYTFTRYTMHWVRQAPGKCLEWIGYINPSRGYTNYNQKFKDRFTISRDNSKNTAFLQMDSLRPEDTGVYFCARYYDDHYCLDYWGQGTPVTVSSGGGGSGGGGSGGGGSGGGGSDIQMTQSPSSLSASVGDRVTITCSASSSVSYMNWYQQTPGKAPKLLIYDTSKLASGVPSRFSGSGSGTDYTFTISSLQPEDIATYYCQQWSSNPFTFGCGTKLEIKGGGGSGGGGSKNTAFLQMDSLRPEDTGVYFCARYYDDHYCLDYWGQGTPVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTKTYTCNVDHKPSNTKVDKRVESKYGPPCPPCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK (SEQ ID NO: 142)
UPLC (сверхвысокоэффективная жидкостная хроматография):
2 мкг образца инжектировали на колонку SEC BEH200, предварительно уравновешенную PBS, pH7,4, и прогоняли в течение 10 мин. Регистрировали интенсивность флуоресценции против времени элюирования и площади пиков отбирали вручную в программном обеспечении EMpower.
Профили элюирования UPLC получали и анализировали. Пики выглядели на >98% мономерными (анти-CD3 IgG4P Fwk3-анти-CD3 dsscFv: 99,02%, анти-CD3 Fab Fwk3-анти-CD3 dsscFv: 99,79%, анти-CD28 Fab Fwk3-анти-CD3 dsscFv: 99,48%) за исключением более низкомолекулярного плеча 12,12% в анти-CD28 IgG4P Fwk3-анти-CD3 dsscFv. Не было признаков мультимеризации ни в одном из форматов.
Молекулярная стабильность
Молекулярную стабильность определяли посредством анализа температуры плавления (Tm) (показатель разворачивания) способом дифференциальной сканирующей калориметрии (DSC), как описано выше. Кривые DSC представлены на фиг. 23 (A) и (B), и величины Tm представлены в таблице 6 и 7 ниже.
Таблица 6 - Fab-Fwk3-dsscFv
Кривые DSC для анти-CD3 Fab Fwk3-анти-CD3 dsscFv продемонстрировали единичный переход разворачивания при 70,04°C. Напротив, разворачивание конструкции, содержавшей конструкцию против CD28, не может быть точно описано посредством одного перехода, а лучше соответствует модели не 2 состояний с двумя переходами; наиболее низкий из которых может быть отнесен к компоненту dsscFv. Примечательно, что результаты демонстрируют, что встраивание Fab в положение каркасной области 3 приводило к стабильным конструкциям, поскольку величины 65°C-75°C сопоставимы с величинами для типичного IgG.
Таблица 7- IgG4P-Fwk3-dsscFv
Кривые DSC для анти-CD3 IgG4P Fwk3-анти-CD3 dsscFv продемонстрировали единичный переход разворачивания при 68,55°C. Напротив, разворачивание конструкций, содержащих конструкцию против CD28, не может быть точно описано посредством одного перехода, а лучше соответствует модели не 2 состояний с двумя переходами; наиболее низкий из которых может быть отнесен к компоненту dsscFv. Примечательно, что результаты демонстрируют, что встраивание IgG в положение каркасной области 3 приводило к стабильным конструкциям, поскольку величины 65°C-75°C являются типичными для IgG-подобных молекул.
Оценка конструкций Fab и IgG4P со вставками в каркасную область 3 в функциональном анализе с PBMC человека
Конструкции тестировали в отношении их способности стимулировать активацию T-клеток в культуре мононуклеарных клеток периферической крови (PBMC) в соответствии со способом, описанным ниже.
Реагенты
Антитела, использованные для детекции CD4 и CD8 T-клеток, различают живые и погибшие клетки и определяют маркер активации CD69, и их получали от BD Biosciences (антитело против CD4, антитело против CD8 и антитело против CD69, различающие живые/погибшие клетки, были связаны, соответственно, с флуороформами FITC, PE, BV421 и Near Infrared. Все культивирование клеток проводили в RPMI1640 (Thermo) плюс 5% инактивированная нагреванием сыворотка AB человека (Sigma) и 2 мМ глутамин (Thermo). Все окрашивание посредством проточной цитометрии проводили с использованием стандартного буфера для проточной цитометрии, содержавшего PBS Дульбекко (Thermo) плюс 1% бычий сывороточный альбумин (Sigma), 2 мМ EDTA (Sigma) и 0,1% азид натрия (Sigma), OKT3 человека (Ebioscience) и T cell Transact (Miltenyi), коллоидную суспензию антител против CD3 и CD28. Все данные проточной цитометрии получали с использованием iQUEplus (Sartorius). T-клетки идентифицировали с использованием программного обеспечения Forecyt (Sartorius) и данные импортировали в GraphPad Prism 7 (GraphPad) для преобразования и графического представления данных.
Способ
Конструкции антител в таблице 5 разбавляли, начиная с 25 мкг/мл с восемью шагами разведения 1 к 5. На последующей стадии анализа все эти величины конвертировали в концентрации в нМ для обеспечения равных сравнений всех конструкций. В 96-луночных планшетах для культивирования тканей с "U"-образным дном 1,5×105 PBMC (от двух различных доноров, анонимно называемых донорами 1 и 2) смешивали с разбавленными антителами. OKT3 (5 мкг/мл), Transact (CD3/CD28, разбавленное 1 к 100) и среда отдельно были эталонными контролями для эксперимента. После смешения планшеты инкубировали при 37 градусах Цельсия в течение 24 часов. После этого планшеты охлаждали на льду и центрифугировали (все последующее центрифугирование проводили при 500g в течение 5 мин при 4 градусах Цельсия) и супернатант удаляли из каждой лунки. Планшеты вновь промывали, супернатант аспирировали и 20 мкл коктейля антител добавляли в каждую лунку (все антитела разбавляли 1:20 в буфере для проточной цитометрии и 1 мкл красителя для определения живых/погибших добавляли на 1 мл коктейля). Планшеты оставляли на льду в темноте в течение 1 часа. После этого планшеты промывали дополнительно два раза и супернатант извлекали из всех лунок. В конце планшеты ресуспендировали в 20 мкл буфера для проточной цитометрии и получали по меньшей мере 10000 клеток на лунку.
Результаты
Результаты представлены на фиг. 24 (A-D; по вертикальной оси представлен уровень экспрессии CD69 (MFI) и по горизонтальной оси представлена концентрация антительной конструкции в нМ). Эксперимент считался успешным, поскольку положительный контроль (OKT3) индуцировал ранний маркер активации CD69 как на CD4, так и на CD8 T-клетках. Минимальный (только клетки) уровень активации определялся контролем, содержавшим только клетки. Как можно видеть из фиг. 24(A) и (B), индукция CD69 на CD4 T-клеток для протестированных конструкций была сопоставимой между донорами.
Антитела по изобретению, содержавшие CD3 и CD28 (либо в формате Fab, либо в формате IgG4P) были наиболее мощными активаторами CD4 T-клеток. Как и ожидалось в этом анализе, конструкции, содержащие Fc, также могут обеспечивать дополнительную активность через Fc-рецепторы на других (не) T-клетках, в частности, моноцитах, что дает усиленные костимулирующие сигналы. В этом отношении анти-CD3 IgG4P Fwk3-анти-CD3dsscFv также продемонстрировал активность в этом анализе (как и контроль OKT3, который представляет собой целое IgG-антитело против CD3). Эти данные демонстрируют, что антитела по изобретению, которые связываются как с CD3, так и с CD28, являются функциональными активаторами CD4 T-клеток человека в анализе со смешанными PBMC, и по существу способны выполнять функцию биспецифического антитела.
Кроме того, также измеряли активацию CD8 T-клеток в том же анализе. Как описано ранее, все положительные и отрицательные контроли функционировали, как и ожидалось. Как можно видеть из фиг. 24 (C) и (D), индукция CD69 на CD8 T-клетках для протестированных конструкций была сопоставимой между донорами. Антитела по изобретению, связывающие CD3 и CD28 (либо в формате Fab, либо в формате IgG4P) были наиболее мощными активаторами CD8 T-клеток. Анти-CD3 IgG4P Fwk3 анти-CD3dsscFv также продемонстрировал активность в этом анализе (как и контроль в виде OKT3, который представляет собой целое IgG-антитело против CD3). Эти данные демонстрируют, что антитела по изобретению, которые связывают как CD3, так и CD28, являются функциональными активаторами CD8 T-клеток человека в анализе смешанных PBMC и по существу способны выполнять функцию биспецифического антитела.
Результаты показывают, что антитела по изобретению обладают функциональностью как каркасного антитела (например, Fab или IgG4P против CD28), так и полипептида вставки, т.е. второго антитела (например, dsscFv против CD3), когда полипептид-вставка присутствует в области Fwk3 каркасного антитела.
"Содержащий" в контексте настоящего описания означает "включающий".
Когда это технически возможно, варианты осуществления изобретения можно комбинировать. Варианты осуществления описаны в настоящем описании в качестве включающих определенные признаки/элементы. Изобретение также распространяется на отдельные варианты осуществления, состоящие или по существу состоящие из указанных признаков/элементов.
Безусловно, будет понятно, что настоящее изобретение описано в качестве примера, подразумевается, что примеры никоим образом не являются ограничивающими, и в объем формулы изобретения, прилагаемой далее, могут быть внесены модификации. Предпочтительные признаки каждого варианта осуществления изобретения являются такими, как и для других вариантов осуществления с соответствующими поправками. Все публикации, включая, но не ограничиваясь ими, патенты и патентные заявки, цитированные в настоящем описании, включены в настоящее описание в качестве ссылок, как если было конкретно и индивидуально указано, что каждая индивидуальная публикация включена в качестве ссылки.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ЮСБ Биофарма СРЛ
<120> АНТИТЕЛА, СОДЕРЖАЩИЕ ПОЛИПЕПТИД, ВСТРОЕННЫЙ В УЧАСТОК КАРКАСНОЙ ОБЛАСТИ 3
<130> PF0155-WO
<150> GB1811368.8
<151> 2018-07-11
<160> 144
<170> PatentIn version 3.5
<210> 1
<211> 217
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь gL5 CA645
<400> 1
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ser Ser Pro Ser Val Trp Ser Asn
20 25 30
Phe Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Glu Ala Ser Lys Leu Thr Ser Gly Val Pro Ser Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln
65 70 75 80
Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gly Gly Gly Tyr Ser Ser Ile
85 90 95
Ser Asp Thr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr
100 105 110
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
115 120 125
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
130 135 140
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
145 150 155 160
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
165 170 175
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
180 185 190
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
195 200 205
Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 2
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-домен (gL5) CA645
<400> 2
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ser Ser Pro Ser Val Trp Ser Asn
20 25 30
Phe Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Glu Ala Ser Lys Leu Thr Ser Gly Val Pro Ser Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln
65 70 75 80
Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gly Gly Gly Tyr Ser Ser Ile
85 90 95
Ser Asp Thr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 3
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR L1 CA645
<400> 3
Gln Ser Ser Pro Ser Val Trp Ser Asn Phe Leu Ser
1 5 10
<210> 4
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR L2 CA645
<400> 4
Glu Ala Ser Lys Leu Thr Ser
1 5
<210> 5
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR L3 CA645
<400> 5
Gly Gly Gly Tyr Ser Ser Ile Ser Asp Thr Thr
1 5 10
<210> 6
<211> 224
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> gH5 тяжелой цепи CA645
<400> 6
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
<210> 7
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-домен (gH5) CA645
<400> 7
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 8
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR H1 CA645
<400> 8
Gly Ile Asp Leu Ser Asn Tyr Ala Ile Asn
1 5 10
<210> 9
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR H2 CA645
<400> 9
Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys Gly
1 5 10 15
<210> 10
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR H3 CA645
<400> 10
Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu
1 5 10
<210> 11
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 11
Ser Gly Gly Gly Gly Ser Glu
1 5
<210> 12
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 12
Asp Lys Thr His Thr Ser
1 5
<210> 13
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 13
Ser Gly Gly Gly Gly Ser
1 5
<210> 14
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 14
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210> 15
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 15
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 16
<211> 21
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 16
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
Gly Gly Gly Gly Ser
20
<210> 17
<211> 26
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 17
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
20 25
<210> 18
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 18
Ala Ala Ala Gly Ser Gly Gly Ala Ser Ala Ser
1 5 10
<210> 19
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<220>
<221> дополнительный признак
<222> (7)..(7)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<400> 19
Ala Ala Ala Gly Ser Gly Xaa Gly Gly Gly Ser Gly Ala Ser Ala Ser
1 5 10 15
<210> 20
<211> 21
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<220>
<221> дополнительный признак
<222> (7)..(7)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<220>
<221> дополнительный признак
<222> (12)..(12)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<400> 20
Ala Ala Ala Gly Ser Gly Xaa Gly Gly Gly Ser Xaa Gly Gly Gly Ser
1 5 10 15
Gly Ala Ser Ala Ser
20
<210> 21
<211> 26
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<220>
<221> дополнительный признак
<222> (7)..(7)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<220>
<221> дополнительный признак
<222> (12)..(12)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<220>
<221> дополнительный признак
<222> (17)..(17)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<400> 21
Ala Ala Ala Gly Ser Gly Xaa Gly Gly Gly Ser Xaa Gly Gly Gly Ser
1 5 10 15
Xaa Gly Gly Gly Ser Gly Ala Ser Ala Ser
20 25
<210> 22
<211> 31
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<220>
<221> дополнительный признак
<222> (7)..(7)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<220>
<221> дополнительный признак
<222> (12)..(12)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<220>
<221> дополнительный признак
<222> (17)..(17)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<220>
<221> дополнительный признак
<222> (22)..(22)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<400> 22
Ala Ala Ala Gly Ser Gly Xaa Gly Gly Gly Ser Xaa Gly Gly Gly Ser
1 5 10 15
Xaa Gly Gly Gly Ser Xaa Gly Gly Gly Ser Gly Ala Ser Ala Ser
20 25 30
<210> 23
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<220>
<221> дополнительный признак
<222> (7)..(7)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<400> 23
Ala Ala Ala Gly Ser Gly Xaa Ser Gly Ala Ser Ala Ser
1 5 10
<210> 24
<211> 28
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 24
Pro Gly Gly Asn Arg Gly Thr Thr Thr Thr Arg Arg Pro Ala Thr Thr
1 5 10 15
Thr Gly Ser Ser Pro Gly Pro Thr Gln Ser His Tyr
20 25
<210> 25
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 25
Ala Thr Thr Thr Gly Ser Ser Pro Gly Pro Thr
1 5 10
<210> 26
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 26
Ala Thr Thr Thr Gly Ser
1 5
<210> 27
<211> 21
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 27
Glu Pro Ser Gly Pro Ile Ser Thr Ile Asn Ser Pro Pro Ser Lys Glu
1 5 10 15
Ser His Lys Ser Pro
20
<210> 28
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 28
Gly Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
1 5 10 15
<210> 29
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 29
Gly Gly Gly Gly Ile Ala Pro Ser Met Val Gly Gly Gly Gly Ser
1 5 10 15
<210> 30
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 30
Gly Gly Gly Gly Lys Val Glu Gly Ala Gly Gly Gly Gly Gly Ser
1 5 10 15
<210> 31
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 31
Gly Gly Gly Gly Ser Met Lys Ser His Asp Gly Gly Gly Gly Ser
1 5 10 15
<210> 32
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 32
Gly Gly Gly Gly Asn Leu Ile Thr Ile Val Gly Gly Gly Gly Ser
1 5 10 15
<210> 33
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 33
Gly Gly Gly Gly Val Val Pro Ser Leu Pro Gly Gly Gly Gly Ser
1 5 10 15
<210> 34
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 34
Gly Gly Glu Lys Ser Ile Pro Gly Gly Gly Gly Ser
1 5 10
<210> 35
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 35
Arg Pro Leu Ser Tyr Arg Pro Pro Phe Pro Phe Gly Phe Pro Ser Val
1 5 10 15
Arg Pro
<210> 36
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 36
Tyr Pro Arg Ser Ile Tyr Ile Arg Arg Arg His Pro Ser Pro Ser Leu
1 5 10 15
Thr Thr
<210> 37
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 37
Thr Pro Ser His Leu Ser His Ile Leu Pro Ser Phe Gly Leu Pro Thr
1 5 10 15
Glu Asn
<210> 38
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 38
Arg Pro Val Ser Pro Phe Thr Phe Pro Arg Leu Ser Asn Ser Trp Leu
1 5 10 15
Pro Ala
<210> 39
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 39
Ser Pro Ala Ala His Phe Pro Arg Ser Ile Pro Arg Pro Gly Pro Ile
1 5 10 15
Arg Thr
<210> 40
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 40
Ala Pro Gly Pro Ser Ala Pro Ser His Arg Ser Leu Pro Ser Arg Ala
1 5 10 15
Phe Gly
<210> 41
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 41
Pro Arg Asn Ser Ile His Phe Leu His Pro Leu Leu Val Ala Pro Leu
1 5 10 15
Gly Ala
<210> 42
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 42
Met Pro Ser Leu Ser Gly Val Leu Gln Val Arg Tyr Leu Ser Pro Pro
1 5 10 15
Asp Leu
<210> 43
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 43
Ser Pro Gln Tyr Pro Ser Pro Leu Thr Leu Thr Leu Pro Pro His Pro
1 5 10 15
Ser Leu
<210> 44
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 44
Asn Pro Ser Leu Asn Pro Pro Ser Tyr Leu His Arg Ala Pro Ser Arg
1 5 10 15
Ile Ser
<210> 45
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 45
Leu Pro Trp Arg Thr Ser Leu Leu Pro Ser Leu Pro Leu Arg Arg Arg
1 5 10 15
Pro
<210> 46
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 46
Pro Pro Leu Phe Ala Lys Gly Pro Val Gly Leu Leu Ser Arg Ser Phe
1 5 10 15
Pro Pro
<210> 47
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 47
Val Pro Pro Ala Pro Val Val Ser Leu Arg Ser Ala His Ala Arg Pro
1 5 10 15
Pro Tyr
<210> 48
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 48
Leu Arg Pro Thr Pro Pro Arg Val Arg Ser Tyr Thr Cys Cys Pro Thr
1 5 10 15
Pro
<210> 49
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 49
Pro Asn Val Ala His Val Leu Pro Leu Leu Thr Val Pro Trp Asp Asn
1 5 10 15
Leu Arg
<210> 50
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 50
Cys Asn Pro Leu Leu Pro Leu Cys Ala Arg Ser Pro Ala Val Arg Thr
1 5 10 15
Phe Pro
<210> 51
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Жесткий линкер
<400> 51
Gly Ala Pro Ala Pro Ala Ala Pro Ala Pro Ala
1 5 10
<210> 52
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Жесткий линкер
<400> 52
Pro Pro Pro Pro
1
<210> 53
<211> 112
<212> БЕЛОК
<213> Homo sapiens
<400> 53
Asn Trp Val Asn Val Ile Ser Asp Leu Lys Lys Ile Glu Asp Leu Ile
1 5 10 15
Gln Ser Met His Ile Asp Ala Thr Leu Tyr Thr Glu Ser Asp Val His
20 25 30
Pro Ser Cys Lys Val Thr Ala Met Lys Cys Phe Leu Leu Glu Leu Gln
35 40 45
Val Ile Ser Leu Glu Ser Gly Asp Ala Ser Ile His Asp Thr Val Glu
50 55 60
Asn Leu Ile Ile Leu Ala Asn Asn Ser Leu Ser Ser Asn Gly Asn Val
65 70 75 80
Thr Glu Ser Gly Cys Lys Glu Cys Glu Glu Leu Glu Glu Lys Asn Ile
85 90 95
Lys Glu Phe Leu Gln Ser Phe Val His Ile Val Gln Met Phe Ile Asn
100 105 110
<210> 54
<211> 352
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CA645 Fab HC Fwk3 IL-15
<400> 54
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Lys Tyr Gln Ser Gly Gly Ser
65 70 75 80
Gly Met Gly Asn Trp Val Asn Val Ile Ser Asp Leu Lys Lys Ile Glu
85 90 95
Asp Leu Ile Gln Ser Met His Ile Asp Ala Thr Leu Tyr Thr Glu Ser
100 105 110
Asp Val His Pro Ser Cys Lys Val Thr Ala Met Lys Cys Phe Leu Leu
115 120 125
Glu Leu Gln Val Ile Ser Leu Glu Ser Gly Asp Ala Ser Ile His Asp
130 135 140
Thr Val Glu Asn Leu Ile Ile Leu Ala Asn Asn Ser Leu Ser Ser Asn
145 150 155 160
Gly Asn Val Thr Glu Ser Gly Cys Lys Glu Cys Glu Glu Leu Glu Glu
165 170 175
Lys Asn Ile Lys Glu Phe Leu Gln Ser Phe Val His Ile Val Gln Met
180 185 190
Phe Ile Asn Ser Gly Gly Ser Tyr Thr Tyr Lys Asn Thr Val Tyr Leu
195 200 205
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
210 215 220
Arg Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly
225 230 235 240
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
245 250 255
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
260 265 270
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
275 280 285
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
290 295 300
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
305 310 315 320
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
325 330 335
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
340 345 350
<210> 55
<211> 133
<212> БЕЛОК
<213> Homo sapiens
<400> 55
Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His
1 5 10 15
Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys
20 25 30
Asn Pro Lys Leu Thr Arg Met Leu Thr Phe Lys Phe Tyr Met Pro Lys
35 40 45
Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu Glu Leu Lys
50 55 60
Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His Leu
65 70 75 80
Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu
85 90 95
Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala
100 105 110
Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile
115 120 125
Ile Ser Thr Leu Thr
130
<210> 56
<211> 368
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CA645 Fab HC Fwk3 IL-2
<400> 56
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Gly Gly Gly Ser Gly Gly
65 70 75 80
Gly Gly Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu
85 90 95
Glu His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn
100 105 110
Tyr Lys Asn Pro Lys Leu Thr Arg Met Leu Thr Phe Lys Phe Tyr Met
115 120 125
Pro Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu Glu
130 135 140
Leu Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe
145 150 155 160
His Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu
165 170 175
Glu Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu
180 185 190
Thr Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln
195 200 205
Ser Ile Ile Ser Thr Leu Thr Gly Gly Gly Lys Asn Thr Val Tyr Leu
210 215 220
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
225 230 235 240
Arg Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly
245 250 255
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
260 265 270
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
275 280 285
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
290 295 300
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
305 310 315 320
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
325 330 335
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
340 345 350
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
355 360 365
<210> 57
<211> 178
<212> БЕЛОК
<213> Homo sapiens
<400> 57
Ala Thr Pro Leu Gly Pro Ala Ser Ser Leu Pro Gln Ser Phe Leu Leu
1 5 10 15
Lys Cys Leu Glu Gln Val Arg Lys Ile Gln Gly Asp Gly Ala Ala Leu
20 25 30
Gln Glu Lys Leu Val Ser Glu Cys Ala Thr Tyr Lys Leu Cys His Pro
35 40 45
Glu Glu Leu Val Leu Leu Gly His Ser Leu Gly Ile Pro Trp Ala Pro
50 55 60
Leu Ser Ser Cys Pro Ser Gln Ala Leu Gln Leu Ala Gly Cys Leu Ser
65 70 75 80
Gln Leu His Ser Gly Leu Phe Leu Tyr Gln Gly Leu Leu Gln Ala Leu
85 90 95
Glu Gly Ile Ser Pro Glu Leu Gly Pro Thr Leu Asp Thr Leu Gln Leu
100 105 110
Asp Val Ala Asp Phe Ala Thr Thr Ile Trp Gln Gln Met Glu Glu Leu
115 120 125
Gly Met Ala Pro Ala Leu Gln Pro Thr Gln Gly Ala Met Pro Ala Phe
130 135 140
Ala Ser Ala Phe Gln Arg Arg Ala Gly Gly Val Leu Val Ala Ser His
145 150 155 160
Leu Gln Ser Phe Leu Glu Val Ser Tyr Arg Val Leu Arg His Leu Ala
165 170 175
Gln Pro
<210> 58
<211> 412
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CA645 Fab HC Fwk3 G-CSF
<400> 58
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Gly Gly Gly Gly Ser Ala
65 70 75 80
Thr Pro Leu Gly Pro Ala Ser Ser Leu Pro Gln Ser Phe Leu Leu Lys
85 90 95
Cys Leu Glu Gln Val Arg Lys Ile Gln Gly Asp Gly Ala Ala Leu Gln
100 105 110
Glu Lys Leu Val Ser Glu Cys Ala Thr Tyr Lys Leu Cys His Pro Glu
115 120 125
Glu Leu Val Leu Leu Gly His Ser Leu Gly Ile Pro Trp Ala Pro Leu
130 135 140
Ser Ser Cys Pro Ser Gln Ala Leu Gln Leu Ala Gly Cys Leu Ser Gln
145 150 155 160
Leu His Ser Gly Leu Phe Leu Tyr Gln Gly Leu Leu Gln Ala Leu Glu
165 170 175
Gly Ile Ser Pro Glu Leu Gly Pro Thr Leu Asp Thr Leu Gln Leu Asp
180 185 190
Val Ala Asp Phe Ala Thr Thr Ile Trp Gln Gln Met Glu Glu Leu Gly
195 200 205
Met Ala Pro Ala Leu Gln Pro Thr Gln Gly Ala Met Pro Ala Phe Ala
210 215 220
Ser Ala Phe Gln Arg Arg Ala Gly Gly Val Leu Val Ala Ser His Leu
225 230 235 240
Gln Ser Phe Leu Glu Val Ser Tyr Arg Val Leu Arg His Leu Ala Gln
245 250 255
Pro Ser Gly Gly Gly Gly Lys Asn Thr Val Tyr Leu Gln Met Asn Ser
260 265 270
Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Thr Val Pro
275 280 285
Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly Gln Gly Thr Leu
290 295 300
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
305 310 315 320
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
325 330 335
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
340 345 350
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
355 360 365
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
370 375 380
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
385 390 395 400
Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
405 410
<210> 59
<211> 127
<212> БЕЛОК
<213> Homo sapiens
<400> 59
Ala Pro Ala Arg Ser Pro Ser Pro Ser Thr Gln Pro Trp Glu His Val
1 5 10 15
Asn Ala Ile Gln Glu Ala Arg Arg Leu Leu Asn Leu Ser Arg Asp Thr
20 25 30
Ala Ala Glu Met Asn Glu Thr Val Glu Val Ile Ser Glu Met Phe Asp
35 40 45
Leu Gln Glu Pro Thr Cys Leu Gln Thr Arg Leu Glu Leu Tyr Lys Gln
50 55 60
Gly Leu Arg Gly Ser Leu Thr Lys Leu Lys Gly Pro Leu Thr Met Met
65 70 75 80
Ala Ser His Tyr Lys Gln His Cys Pro Pro Thr Pro Glu Thr Ser Cys
85 90 95
Ala Thr Gln Ile Ile Thr Phe Glu Ser Phe Lys Glu Asn Leu Lys Asp
100 105 110
Phe Leu Leu Val Ile Pro Phe Asp Cys Trp Glu Pro Val Gln Glu
115 120 125
<210> 60
<211> 362
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CA645 Fab HC Fwk3 GM-CSF
<400> 60
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Gly Gly Gly Ser Gly Gly
65 70 75 80
Gly Gly Ala Pro Ala Arg Ser Pro Ser Pro Ser Thr Gln Pro Trp Glu
85 90 95
His Val Asn Ala Ile Gln Glu Ala Arg Arg Leu Leu Asn Leu Ser Arg
100 105 110
Asp Thr Ala Ala Glu Met Asn Glu Thr Val Glu Val Ile Ser Glu Met
115 120 125
Phe Asp Leu Gln Glu Pro Thr Cys Leu Gln Thr Arg Leu Glu Leu Tyr
130 135 140
Lys Gln Gly Leu Arg Gly Ser Leu Thr Lys Leu Lys Gly Pro Leu Thr
145 150 155 160
Met Met Ala Ser His Tyr Lys Gln His Cys Pro Pro Thr Pro Glu Thr
165 170 175
Ser Cys Ala Thr Gln Ile Ile Thr Phe Glu Ser Phe Lys Glu Asn Leu
180 185 190
Lys Asp Phe Leu Leu Val Ile Pro Phe Asp Cys Trp Glu Pro Val Gln
195 200 205
Glu Gly Gly Gly Lys Asn Thr Val Tyr Leu Gln Met Asn Ser Leu Arg
210 215 220
Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Thr Val Pro Gly Tyr
225 230 235 240
Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly Gln Gly Thr Leu Val Thr
245 250 255
Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro
260 265 270
Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val
275 280 285
Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala
290 295 300
Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly
305 310 315 320
Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly
325 330 335
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
340 345 350
Val Asp Lys Lys Val Glu Pro Lys Ser Cys
355 360
<210> 61
<211> 359
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CA645 Fab HC с GM-CSF на C-конце CH1
<400> 61
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Gly Gly Gly Ser Gly Gly Gly Gly Ala Pro Ala Arg Ser Pro Ser Pro
225 230 235 240
Ser Thr Gln Pro Trp Glu His Val Asn Ala Ile Gln Glu Ala Arg Arg
245 250 255
Leu Leu Asn Leu Ser Arg Asp Thr Ala Ala Glu Met Asn Glu Thr Val
260 265 270
Glu Val Ile Ser Glu Met Phe Asp Leu Gln Glu Pro Thr Cys Leu Gln
275 280 285
Thr Arg Leu Glu Leu Tyr Lys Gln Gly Leu Arg Gly Ser Leu Thr Lys
290 295 300
Leu Lys Gly Pro Leu Thr Met Met Ala Ser His Tyr Lys Gln His Cys
305 310 315 320
Pro Pro Thr Pro Glu Thr Ser Cys Ala Thr Gln Ile Ile Thr Phe Glu
325 330 335
Ser Phe Lys Glu Asn Leu Lys Asp Phe Leu Leu Val Ile Pro Phe Asp
340 345 350
Cys Trp Glu Pro Val Gln Glu
355
<210> 62
<211> 124
<212> БЕЛОК
<213> Животные семейства мышиных
<400> 62
Ala Pro Thr Arg Ser Pro Ile Thr Val Thr Arg Pro Trp Lys His Val
1 5 10 15
Glu Ala Ile Lys Glu Ala Leu Asn Leu Leu Asp Asp Met Pro Val Thr
20 25 30
Leu Asn Glu Glu Val Glu Val Val Ser Asn Glu Phe Ser Phe Lys Lys
35 40 45
Leu Thr Cys Val Gln Thr Arg Leu Lys Ile Phe Glu Gln Gly Leu Arg
50 55 60
Gly Asn Phe Thr Lys Leu Lys Gly Ala Leu Asn Met Thr Ala Ser Tyr
65 70 75 80
Tyr Gln Thr Tyr Cys Pro Pro Thr Pro Glu Thr Asp Cys Glu Thr Gln
85 90 95
Val Thr Thr Tyr Ala Asp Phe Ile Asp Ser Leu Lys Thr Phe Leu Thr
100 105 110
Asp Ile Pro Phe Glu Cys Lys Lys Pro Gly Gln Lys
115 120
<210> 63
<211> 359
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CA645 Fab HC Fwk3 mGM-CSF
<400> 63
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Gly Gly Gly Ser Gly Gly
65 70 75 80
Gly Gly Ala Pro Thr Arg Ser Pro Ile Thr Val Thr Arg Pro Trp Lys
85 90 95
His Val Glu Ala Ile Lys Glu Ala Leu Asn Leu Leu Asp Asp Met Pro
100 105 110
Val Thr Leu Asn Glu Glu Val Glu Val Val Ser Asn Glu Phe Ser Phe
115 120 125
Lys Lys Leu Thr Cys Val Gln Thr Arg Leu Lys Ile Phe Glu Gln Gly
130 135 140
Leu Arg Gly Asn Phe Thr Lys Leu Lys Gly Ala Leu Asn Met Thr Ala
145 150 155 160
Ser Tyr Tyr Gln Thr Tyr Cys Pro Pro Thr Pro Glu Thr Asp Cys Glu
165 170 175
Thr Gln Val Thr Thr Tyr Ala Asp Phe Ile Asp Ser Leu Lys Thr Phe
180 185 190
Leu Thr Asp Ile Pro Phe Glu Cys Lys Lys Pro Gly Gln Lys Gly Gly
195 200 205
Gly Lys Asn Thr Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp
210 215 220
Thr Ala Val Tyr Tyr Cys Ala Arg Thr Val Pro Gly Tyr Ser Thr Ala
225 230 235 240
Pro Tyr Phe Asp Leu Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
245 250 255
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
260 265 270
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
275 280 285
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
290 295 300
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
305 310 315 320
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
325 330 335
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
340 345 350
Lys Val Glu Pro Lys Ser Cys
355
<210> 64
<211> 356
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CA645 Fab HC с mGM-CSF на C-конце CH1
<400> 64
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Gly Gly Gly Ser Gly Gly Gly Gly Ala Pro Thr Arg Ser Pro Ile Thr
225 230 235 240
Val Thr Arg Pro Trp Lys His Val Glu Ala Ile Lys Glu Ala Leu Asn
245 250 255
Leu Leu Asp Asp Met Pro Val Thr Leu Asn Glu Glu Val Glu Val Val
260 265 270
Ser Asn Glu Phe Ser Phe Lys Lys Leu Thr Cys Val Gln Thr Arg Leu
275 280 285
Lys Ile Phe Glu Gln Gly Leu Arg Gly Asn Phe Thr Lys Leu Lys Gly
290 295 300
Ala Leu Asn Met Thr Ala Ser Tyr Tyr Gln Thr Tyr Cys Pro Pro Thr
305 310 315 320
Pro Glu Thr Asp Cys Glu Thr Gln Val Thr Thr Tyr Ala Asp Phe Ile
325 330 335
Asp Ser Leu Lys Thr Phe Leu Thr Asp Ile Pro Phe Glu Cys Lys Lys
340 345 350
Pro Gly Gln Lys
355
<210> 65
<211> 166
<212> БЕЛОК
<213> Homo sapiens
<400> 65
Ala Pro Pro Arg Leu Ile Cys Asp Ser Arg Val Leu Glu Arg Tyr Leu
1 5 10 15
Leu Glu Ala Lys Glu Ala Glu Asn Ile Thr Thr Gly Cys Ala Glu His
20 25 30
Cys Ser Leu Asn Glu Asn Ile Thr Val Pro Asp Thr Lys Val Asn Phe
35 40 45
Tyr Ala Trp Lys Arg Met Glu Val Gly Gln Gln Ala Val Glu Val Trp
50 55 60
Gln Gly Leu Ala Leu Leu Ser Glu Ala Val Leu Arg Gly Gln Ala Leu
65 70 75 80
Leu Val Asn Ser Ser Gln Pro Trp Glu Pro Leu Gln Leu His Val Asp
85 90 95
Lys Ala Val Ser Gly Leu Arg Ser Leu Thr Thr Leu Leu Arg Ala Leu
100 105 110
Gly Ala Gln Lys Glu Ala Ile Ser Pro Pro Asp Ala Ala Ser Ala Ala
115 120 125
Pro Leu Arg Thr Ile Thr Ala Asp Thr Phe Arg Lys Leu Phe Arg Val
130 135 140
Tyr Ser Asn Phe Leu Arg Gly Lys Leu Lys Leu Tyr Thr Gly Glu Ala
145 150 155 160
Cys Arg Thr Gly Asp Arg
165
<210> 66
<211> 401
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CA645 Fab HC Fwk3 EPO
<400> 66
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Gly Gly Gly Ser Gly Gly
65 70 75 80
Gly Gly Ala Pro Pro Arg Leu Ile Cys Asp Ser Arg Val Leu Glu Arg
85 90 95
Tyr Leu Leu Glu Ala Lys Glu Ala Glu Asn Ile Thr Thr Gly Cys Ala
100 105 110
Glu His Cys Ser Leu Asn Glu Asn Ile Thr Val Pro Asp Thr Lys Val
115 120 125
Asn Phe Tyr Ala Trp Lys Arg Met Glu Val Gly Gln Gln Ala Val Glu
130 135 140
Val Trp Gln Gly Leu Ala Leu Leu Ser Glu Ala Val Leu Arg Gly Gln
145 150 155 160
Ala Leu Leu Val Asn Ser Ser Gln Pro Trp Glu Pro Leu Gln Leu His
165 170 175
Val Asp Lys Ala Val Ser Gly Leu Arg Ser Leu Thr Thr Leu Leu Arg
180 185 190
Ala Leu Gly Ala Gln Lys Glu Ala Ile Ser Pro Pro Asp Ala Ala Ser
195 200 205
Ala Ala Pro Leu Arg Thr Ile Thr Ala Asp Thr Phe Arg Lys Leu Phe
210 215 220
Arg Val Tyr Ser Asn Phe Leu Arg Gly Lys Leu Lys Leu Tyr Thr Gly
225 230 235 240
Glu Ala Cys Arg Thr Gly Asp Arg Gly Gly Gly Lys Asn Thr Val Tyr
245 250 255
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
260 265 270
Ala Arg Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp
275 280 285
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
290 295 300
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
305 310 315 320
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
325 330 335
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
340 345 350
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
355 360 365
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
370 375 380
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
385 390 395 400
Cys
<210> 67
<211> 125
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VHH6
<400> 67
Asp Val Gln Phe Val Glu Ser Gly Gly Gly Ser Val His Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Asn Cys Ala Thr Ser Gly Tyr Ile Tyr Ser Thr Tyr
20 25 30
Cys Met Gly Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Gly Val
35 40 45
Ala His Ile Tyr Thr Asn Ser Gly Arg Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Gln Asp Asn Ala Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Ile Tyr Tyr Cys
85 90 95
Ala Ala Arg Pro Ser Ile Arg Cys Ala Ser Phe Ser Ala Thr Glu Tyr
100 105 110
Lys Asp Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120 125
<210> 68
<211> 369
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CA645 Fab HC Fwk3 VHH6
<400> 68
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Gly Gly Gly Gly Ser Gly
65 70 75 80
Gly Gly Gly Ser Asp Val Gln Phe Val Glu Ser Gly Gly Gly Ser Val
85 90 95
His Ala Gly Gly Ser Leu Arg Leu Asn Cys Ala Thr Ser Gly Tyr Ile
100 105 110
Tyr Ser Thr Tyr Cys Met Gly Trp Phe Arg Gln Ala Pro Gly Lys Glu
115 120 125
Arg Glu Gly Val Ala His Ile Tyr Thr Asn Ser Gly Arg Thr Tyr Tyr
130 135 140
Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Gln Asp Asn Ala Lys
145 150 155 160
Asn Thr Val Tyr Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala
165 170 175
Ile Tyr Tyr Cys Ala Ala Arg Pro Ser Ile Arg Cys Ala Ser Phe Ser
180 185 190
Ala Thr Glu Tyr Lys Asp Trp Gly Gln Gly Thr Gln Val Thr Val Ser
195 200 205
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Lys Asn Thr Val Tyr
210 215 220
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
225 230 235 240
Ala Arg Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp
245 250 255
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
260 265 270
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
275 280 285
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
290 295 300
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
305 310 315 320
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
325 330 335
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
340 345 350
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
355 360 365
Cys
<210> 69
<211> 119
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VHH15
<400> 69
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ala Ser Gly Tyr Thr Gly Cys Thr
20 25 30
Tyr Asp Met Arg Trp Tyr Arg Gln Ala Pro Gly Lys Glu Arg Glu Phe
35 40 45
Val Ser Gly Ile Asp Ser Asp Gly Arg Ala Thr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Gln Ser Asn Ala Lys Ile Ala Val Tyr
65 70 75 80
Leu Gln Met Asp Ser Leu Lys Leu Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Asn Leu Gln Cys Leu Arg Tyr Pro Gly Glu Tyr Tyr Trp Gly Gln Gly
100 105 110
Thr Gln Val Thr Val Ser Ser
115
<210> 70
<211> 363
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CA645 Fab HC Fwk3 VHH15
<400> 70
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Gly Gly Gly Gly Ser Gly
65 70 75 80
Gly Gly Gly Ser Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val
85 90 95
Gln Ala Gly Gly Ser Leu Arg Leu Ser Cys Val Ala Ala Ser Gly Tyr
100 105 110
Thr Gly Cys Thr Tyr Asp Met Arg Trp Tyr Arg Gln Ala Pro Gly Lys
115 120 125
Glu Arg Glu Phe Val Ser Gly Ile Asp Ser Asp Gly Arg Ala Thr Tyr
130 135 140
Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Gln Ser Asn Ala Lys
145 150 155 160
Ile Ala Val Tyr Leu Gln Met Asp Ser Leu Lys Leu Glu Asp Thr Ala
165 170 175
Met Tyr Tyr Cys Asn Leu Gln Cys Leu Arg Tyr Pro Gly Glu Tyr Tyr
180 185 190
Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser Gly Gly Gly Gly Ser
195 200 205
Gly Gly Gly Gly Ser Lys Asn Thr Val Tyr Leu Gln Met Asn Ser Leu
210 215 220
Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Thr Val Pro Gly
225 230 235 240
Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly Gln Gly Thr Leu Val
245 250 255
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
260 265 270
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
275 280 285
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
290 295 300
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
305 310 315 320
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
325 330 335
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
340 345 350
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
355 360
<210> 71
<211> 190
<212> БЕЛОК
<213> Homo sapiens
<400> 71
Gln Gly Trp Gln Ala Phe Lys Asn Asp Ala Thr Glu Ile Ile Pro Glu
1 5 10 15
Leu Gly Glu Tyr Pro Glu Pro Pro Pro Glu Leu Glu Asn Asn Lys Thr
20 25 30
Met Asn Arg Ala Glu Asn Gly Gly Arg Pro Pro His His Pro Phe Glu
35 40 45
Thr Lys Asp Val Ser Glu Tyr Ser Cys Arg Glu Leu His Phe Thr Arg
50 55 60
Tyr Val Thr Asp Gly Pro Cys Arg Ser Ala Lys Pro Val Thr Glu Leu
65 70 75 80
Val Cys Ser Gly Gln Cys Gly Pro Ala Arg Leu Leu Pro Asn Ala Ile
85 90 95
Gly Arg Gly Lys Trp Trp Arg Pro Ser Gly Pro Asp Phe Arg Cys Ile
100 105 110
Pro Asp Arg Tyr Arg Ala Gln Arg Val Gln Leu Leu Cys Pro Gly Gly
115 120 125
Glu Ala Pro Arg Ala Arg Lys Val Arg Leu Val Ala Ser Cys Lys Cys
130 135 140
Lys Arg Leu Thr Arg Phe His Asn Gln Ser Glu Leu Lys Asp Phe Gly
145 150 155 160
Thr Glu Ala Ala Arg Pro Gln Lys Gly Arg Lys Pro Arg Pro Arg Ala
165 170 175
Arg Ser Ala Lys Ala Asn Gln Ala Glu Leu Glu Asn Ala Tyr
180 185 190
<210> 72
<211> 88
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Центральный домен склеростина человека
<400> 72
Cys Arg Glu Leu His Phe Thr Arg Tyr Val Thr Asp Gly Pro Cys Arg
1 5 10 15
Ser Ala Lys Pro Val Thr Glu Leu Val Cys Ser Gly Gln Cys Gly Pro
20 25 30
Ala Arg Leu Leu Pro Asn Ala Ile Gly Arg Gly Lys Trp Trp Arg Pro
35 40 45
Ser Gly Pro Asp Phe Arg Cys Ile Pro Asp Arg Tyr Arg Ala Gln Arg
50 55 60
Val Gln Leu Leu Cys Pro Gly Gly Glu Ala Pro Arg Ala Arg Lys Val
65 70 75 80
Arg Leu Val Ala Ser Cys Lys Cys
85
<210> 73
<211> 314
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь Fab CA645 с центральным доменом склеростина человека, встроенным в каркасную область 3, линкеры из единичного S
<400> 73
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Ser Cys Arg Glu Leu His
65 70 75 80
Phe Thr Arg Tyr Val Thr Asp Gly Pro Cys Arg Ser Ala Lys Pro Val
85 90 95
Thr Glu Leu Val Cys Ser Gly Gln Cys Gly Pro Ala Arg Leu Leu Pro
100 105 110
Asn Ala Ile Gly Arg Gly Lys Trp Trp Arg Pro Ser Gly Pro Asp Phe
115 120 125
Arg Cys Ile Pro Asp Arg Tyr Arg Ala Gln Arg Val Gln Leu Leu Cys
130 135 140
Pro Gly Gly Glu Ala Pro Arg Ala Arg Lys Val Arg Leu Val Ala Ser
145 150 155 160
Cys Lys Cys Ser Lys Asn Thr Val Tyr Leu Gln Met Asn Ser Leu Arg
165 170 175
Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Thr Val Pro Gly Tyr
180 185 190
Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly Gln Gly Thr Leu Val Thr
195 200 205
Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro
210 215 220
Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val
225 230 235 240
Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala
245 250 255
Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly
260 265 270
Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly
275 280 285
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
290 295 300
Val Asp Lys Lys Val Glu Pro Lys Ser Cys
305 310
<210> 74
<211> 333
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь Fab CA645 с центральным доменом склеростина человека, встроенным в каркасную область 3, линкеры Gly-Ser (2X G4S)
<400> 74
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Gly Gly Gly Gly Ser Gly
65 70 75 80
Gly Gly Gly Ser Cys Arg Glu Leu His Phe Thr Arg Tyr Val Thr Asp
85 90 95
Gly Pro Cys Arg Ser Ala Lys Pro Val Thr Glu Leu Val Cys Ser Gly
100 105 110
Gln Cys Gly Pro Ala Arg Leu Leu Pro Asn Ala Ile Gly Arg Gly Lys
115 120 125
Trp Trp Arg Pro Ser Gly Pro Asp Phe Arg Cys Ile Pro Asp Arg Tyr
130 135 140
Arg Ala Gln Arg Val Gln Leu Leu Cys Pro Gly Gly Glu Ala Pro Arg
145 150 155 160
Ala Arg Lys Val Arg Leu Val Ala Ser Cys Lys Cys Ser Gly Gly Gly
165 170 175
Gly Ser Gly Gly Gly Gly Ser Lys Asn Thr Val Tyr Leu Gln Met Asn
180 185 190
Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Thr Val
195 200 205
Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly Gln Gly Thr
210 215 220
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
225 230 235 240
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
245 250 255
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
260 265 270
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
275 280 285
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
290 295 300
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
305 310 315 320
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
325 330
<210> 75
<211> 323
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь Fab CA645 с центральным доменом склеростина человека, встроенным в каркасную область 3, жесткие линкеры
<400> 75
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Ile Pro Phe Thr Val Cys
65 70 75 80
Arg Glu Leu His Phe Thr Arg Tyr Val Thr Asp Gly Pro Cys Arg Ser
85 90 95
Ala Lys Pro Val Thr Glu Leu Val Cys Ser Gly Gln Cys Gly Pro Ala
100 105 110
Arg Leu Leu Pro Asn Ala Ile Gly Arg Gly Lys Trp Trp Arg Pro Ser
115 120 125
Gly Pro Asp Phe Arg Cys Ile Pro Asp Arg Tyr Arg Ala Gln Arg Val
130 135 140
Gln Leu Leu Cys Pro Gly Gly Glu Ala Pro Arg Ala Arg Lys Val Arg
145 150 155 160
Leu Val Ala Ser Cys Lys Cys Glu Tyr His Gly Leu Gln Lys Asn Thr
165 170 175
Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
180 185 190
Tyr Cys Ala Arg Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp
195 200 205
Leu Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
210 215 220
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
225 230 235 240
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
245 250 255
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
260 265 270
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
275 280 285
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
290 295 300
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
305 310 315 320
Lys Ser Cys
<210> 76
<211> 425
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь Fab CA645 со зрелым полноразмерным склеростином человека, встроенным в каркасную область 3, линкеры Gly-Ser (1X G4S)
<400> 76
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Gly Gly Gly Gly Ser Gln
65 70 75 80
Gly Trp Gln Ala Phe Lys Asn Asp Ala Thr Glu Ile Ile Pro Glu Leu
85 90 95
Gly Glu Tyr Pro Glu Pro Pro Pro Glu Leu Glu Asn Asn Lys Thr Met
100 105 110
Asn Arg Ala Glu Asn Gly Gly Arg Pro Pro His His Pro Phe Glu Thr
115 120 125
Lys Asp Val Ser Glu Tyr Ser Cys Arg Glu Leu His Phe Thr Arg Tyr
130 135 140
Val Thr Asp Gly Pro Cys Arg Ser Ala Lys Pro Val Thr Glu Leu Val
145 150 155 160
Cys Ser Gly Gln Cys Gly Pro Ala Arg Leu Leu Pro Asn Ala Ile Gly
165 170 175
Arg Gly Lys Trp Trp Arg Pro Ser Gly Pro Asp Phe Arg Cys Ile Pro
180 185 190
Asp Arg Tyr Arg Ala Gln Arg Val Gln Leu Leu Cys Pro Gly Gly Glu
195 200 205
Ala Pro Arg Ala Arg Lys Val Arg Leu Val Ala Ser Cys Lys Cys Lys
210 215 220
Arg Leu Thr Arg Phe His Asn Gln Ser Glu Leu Lys Asp Phe Gly Thr
225 230 235 240
Glu Ala Ala Arg Pro Gln Lys Gly Arg Lys Pro Arg Pro Arg Ala Arg
245 250 255
Ser Ala Lys Ala Asn Gln Ala Glu Leu Glu Asn Ala Tyr Ser Gly Gly
260 265 270
Gly Gly Ser Lys Asn Thr Val Tyr Leu Gln Met Asn Ser Leu Arg Ala
275 280 285
Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Thr Val Pro Gly Tyr Ser
290 295 300
Thr Ala Pro Tyr Phe Asp Leu Trp Gly Gln Gly Thr Leu Val Thr Val
305 310 315 320
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
325 330 335
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
340 345 350
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
355 360 365
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
370 375 380
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
385 390 395 400
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
405 410 415
Asp Lys Lys Val Glu Pro Lys Ser Cys
420 425
<210> 77
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 77
Lys Tyr Gln Ser Gly Gly Ser Gly Met Gly
1 5 10
<210> 78
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 78
Ser Gly Gly Ser Tyr Thr Tyr
1 5
<210> 79
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 79
Gly Gly Gly Ser Gly Gly Gly Gly
1 5
<210> 80
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 80
Gly Ser Ser Ser Ser Gly Ser
1 5
<210> 81
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 81
Ser Gly Gly Gly Gly
1 5
<210> 82
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Жесткий линкер
<400> 82
Ile Pro Phe Thr Val
1 5
<210> 83
<211> 347
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь Fab CA497 с IL-15 человека, встроенным в каркасную область 3, линкеры Gly-Ser
<400> 83
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Val Ile Phe Ser Asp Tyr
20 25 30
Tyr Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Ser Ile Asn Phe Asn Ala Asp Ile Ser Tyr Tyr Arg Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Gly Gly Gly Ser Gly
65 70 75 80
Gly Gly Gly Asn Trp Val Asn Val Ile Ser Asp Leu Lys Lys Ile Glu
85 90 95
Asp Leu Ile Gln Ser Met His Ile Asp Ala Thr Leu Tyr Thr Glu Ser
100 105 110
Asp Val His Pro Ser Cys Lys Val Thr Ala Met Lys Cys Phe Leu Leu
115 120 125
Glu Leu Gln Val Ile Ser Leu Glu Ser Gly Asp Ala Ser Ile His Asp
130 135 140
Thr Val Glu Asn Leu Ile Ile Leu Ala Asn Asn Ser Leu Ser Ser Asn
145 150 155 160
Gly Asn Val Thr Glu Ser Gly Cys Lys Glu Cys Glu Glu Leu Glu Glu
165 170 175
Lys Asn Ile Lys Glu Phe Leu Gln Ser Phe Val His Ile Val Gln Met
180 185 190
Phe Ile Asn Gly Gly Gly Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser
195 200 205
Leu Lys Thr Glu Asp Thr Ala Val Tyr Tyr Cys Thr Thr Asp Ala Asn
210 215 220
Arg Gln Asn Tyr Asp Trp Phe Ala Tyr Trp Gly Gln Gly Thr Leu Val
225 230 235 240
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
245 250 255
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
260 265 270
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
275 280 285
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
290 295 300
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
305 310 315 320
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
325 330 335
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
340 345
<210> 84
<211> 218
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь Fab CA497
<400> 84
Ala Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Glu Ser Val Ser Ser Ser
20 25 30
Met Tyr Ser Tyr Met His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Arg Ala Ser Asn Leu Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Trp
85 90 95
Thr Ala Pro Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 85
<211> 346
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь Fab CA240 с IL-15 человека, встроенным в каркасную область 3, линкеры Gly-Ser
<400> 85
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Phe Asn Asp Tyr
20 25 30
Phe Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Met Arg Asn Lys Asn Tyr Gln Tyr Gly Thr Tyr Tyr Ala Glu
50 55 60
Ser Leu Glu Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Gly Gly Gly
65 70 75 80
Ser Gly Gly Gly Gly Asn Trp Val Asn Val Ile Ser Asp Leu Lys Lys
85 90 95
Ile Glu Asp Leu Ile Gln Ser Met His Ile Asp Ala Thr Leu Tyr Thr
100 105 110
Glu Ser Asp Val His Pro Ser Cys Lys Val Thr Ala Met Lys Cys Phe
115 120 125
Leu Leu Glu Leu Gln Val Ile Ser Leu Glu Ser Gly Asp Ala Ser Ile
130 135 140
His Asp Thr Val Glu Asn Leu Ile Ile Leu Ala Asn Asn Ser Leu Ser
145 150 155 160
Ser Asn Gly Asn Val Thr Glu Ser Gly Cys Lys Glu Cys Glu Glu Leu
165 170 175
Glu Glu Lys Asn Ile Lys Glu Phe Leu Gln Ser Phe Val His Ile Val
180 185 190
Gln Met Phe Ile Asn Gly Gly Gly Lys Asn Ser Leu Tyr Leu Gln Met
195 200 205
Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Glu
210 215 220
Ser Tyr Tyr Gly Phe Thr Ser Tyr Trp Gly Gln Gly Thr Leu Val Thr
225 230 235 240
Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro
245 250 255
Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val
260 265 270
Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala
275 280 285
Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly
290 295 300
Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly
305 310 315 320
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
325 330 335
Val Asp Lys Lys Val Glu Pro Lys Ser Cys
340 345
<210> 86
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь Fab CA240
<400> 86
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Asp Ile Gly Ile Ser
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asn Ala Asn Asn Leu Ala Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln His Asn Ser Ala Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 87
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь HyHEL5
<400> 87
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Met Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Ser Asp Tyr
20 25 30
Trp Ile Glu Trp Val Lys Gln Arg Pro Gly His Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Leu Pro Gly Ser Gly Ser Thr Asn Tyr His Glu Arg Phe
50 55 60
Lys Gly Lys Ala Thr Phe Thr Ala Asp Thr Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Thr Ser Glu Asp Ser Gly Val Tyr Tyr Cys
85 90 95
Leu His Gly Asn Tyr Asp Phe Asp Gly Trp Gly Gln Gly Thr Thr Leu
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215
<210> 88
<211> 212
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь HyHEL5
<400> 88
Asp Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Asn Tyr Met
20 25 30
Tyr Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Val Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Thr Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Gly Arg Asn Pro Thr Phe
85 90 95
Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro Ser
100 105 110
Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala
115 120 125
Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val
130 135 140
Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser
145 150 155 160
Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr
165 170 175
Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys
180 185 190
Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn
195 200 205
Arg Gly Glu Cys
210
<210> 89
<211> 127
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> D3L11
<400> 89
Asp Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ser Thr Asp Ser Ile Glu
20 25 30
Tyr Met Thr Trp Phe Arg Gln Ala Pro Gly Lys Ala Arg Glu Gly Val
35 40 45
Ala Ala Leu Tyr Thr His Thr Gly Asn Thr Tyr Tyr Thr Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Gln Asp Lys Ala Lys Asn Met Ala Tyr
65 70 75 80
Leu Arg Met Asp Ser Val Lys Ser Glu Asp Thr Ala Ile Tyr Thr Cys
85 90 95
Gly Ala Thr Arg Lys Tyr Val Pro Val Arg Phe Ala Leu Asp Gln Ser
100 105 110
Ser Tyr Asp Tyr Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120 125
<210> 90
<211> 360
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь HyHEL5 против лизоцима с VHH D3L11 против лизоцима, встроенным в каркасную область 3, линкеры Gly-Ser
<400> 90
Asp Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Asn Tyr Met
20 25 30
Tyr Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Val Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Ser Gly Gly Gly Gly Ser Asp Val Gln Leu Val Glu Ser
65 70 75 80
Gly Gly Gly Ser Val Gln Ala Gly Gly Ser Leu Arg Leu Ser Cys Ala
85 90 95
Ala Ser Gly Ser Thr Asp Ser Ile Glu Tyr Met Thr Trp Phe Arg Gln
100 105 110
Ala Pro Gly Lys Ala Arg Glu Gly Val Ala Ala Leu Tyr Thr His Thr
115 120 125
Gly Asn Thr Tyr Tyr Thr Asp Ser Val Lys Gly Arg Phe Thr Ile Ser
130 135 140
Gln Asp Lys Ala Lys Asn Met Ala Tyr Leu Arg Met Asp Ser Val Lys
145 150 155 160
Ser Glu Asp Thr Ala Ile Tyr Thr Cys Gly Ala Thr Arg Lys Tyr Val
165 170 175
Pro Val Arg Phe Ala Leu Asp Gln Ser Ser Tyr Asp Tyr Trp Gly Gln
180 185 190
Gly Thr Gln Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly
195 200 205
Gly Ser Gly Gly Gly Gly Ser Thr Ser Tyr Ser Leu Thr Ile Ser Ser
210 215 220
Met Glu Thr Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Gly Arg
225 230 235 240
Asn Pro Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Thr Val
245 250 255
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
260 265 270
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
275 280 285
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
290 295 300
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
305 310 315 320
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
325 330 335
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr
340 345 350
Lys Ser Phe Asn Arg Gly Glu Cys
355 360
<210> 91
<211> 352
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь HyHEL5 против лизоцима с VHH15 против IL-6, встроенным в каркасную область 3, линкеры Gly-Ser
<400> 91
Asp Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Asn Tyr Met
20 25 30
Tyr Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Val Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Val Glu Ser
65 70 75 80
Gly Gly Gly Ser Val Gln Ala Gly Gly Ser Leu Arg Leu Ser Cys Val
85 90 95
Ala Ala Ser Gly Tyr Thr Gly Cys Thr Tyr Asp Met Arg Trp Tyr Arg
100 105 110
Gln Ala Pro Gly Lys Glu Arg Glu Phe Val Ser Gly Ile Asp Ser Asp
115 120 125
Gly Arg Ala Thr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser
130 135 140
Gln Ser Asn Ala Lys Ile Ala Val Tyr Leu Gln Met Asp Ser Leu Lys
145 150 155 160
Leu Glu Asp Thr Ala Met Tyr Tyr Cys Asn Leu Gln Cys Leu Arg Tyr
165 170 175
Pro Gly Glu Tyr Tyr Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser
180 185 190
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr
195 200 205
Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Thr Glu Asp Ala Ala Thr
210 215 220
Tyr Tyr Cys Gln Gln Trp Gly Arg Asn Pro Thr Phe Gly Gly Gly Thr
225 230 235 240
Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe
245 250 255
Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys
260 265 270
Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val
275 280 285
Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln
290 295 300
Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser
305 310 315 320
Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His
325 330 335
Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
340 345 350
<210> 92
<211> 365
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь CA645 с VHH D3L11 против лизоцима, встроенным в каркасную область 3, линкеры Gly-Ser
<400> 92
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ser Ser Pro Ser Val Trp Ser Asn
20 25 30
Phe Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Glu Ala Ser Lys Leu Thr Ser Gly Val Pro Ser Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Ser Gly Gly Gly Gly Ser Asp Val Gln Leu Val
65 70 75 80
Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly Ser Leu Arg Leu Ser
85 90 95
Cys Ala Ala Ser Gly Ser Thr Asp Ser Ile Glu Tyr Met Thr Trp Phe
100 105 110
Arg Gln Ala Pro Gly Lys Ala Arg Glu Gly Val Ala Ala Leu Tyr Thr
115 120 125
His Thr Gly Asn Thr Tyr Tyr Thr Asp Ser Val Lys Gly Arg Phe Thr
130 135 140
Ile Ser Gln Asp Lys Ala Lys Asn Met Ala Tyr Leu Arg Met Asp Ser
145 150 155 160
Val Lys Ser Glu Asp Thr Ala Ile Tyr Thr Cys Gly Ala Thr Arg Lys
165 170 175
Tyr Val Pro Val Arg Phe Ala Leu Asp Gln Ser Ser Tyr Asp Tyr Trp
180 185 190
Gly Gln Gly Thr Gln Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly
195 200 205
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Asp Phe Thr Leu Thr Ile
210 215 220
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gly Gly Gly
225 230 235 240
Tyr Ser Ser Ile Ser Asp Thr Thr Phe Gly Gly Gly Thr Lys Val Glu
245 250 255
Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser
260 265 270
Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn
275 280 285
Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala
290 295 300
Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys
305 310 315 320
Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp
325 330 335
Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu
340 345 350
Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
355 360 365
<210> 93
<211> 360
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь Fab CA645 с IL-15 человека, встроенным в каркасную область 3, с длинным линкером животного семейства бычьих
<400> 93
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Thr Ser Val His Gln Glu
65 70 75 80
Thr Lys Lys Tyr Gln Ser Asn Trp Val Asn Val Ile Ser Asp Leu Lys
85 90 95
Lys Ile Glu Asp Leu Ile Gln Ser Met His Ile Asp Ala Thr Leu Tyr
100 105 110
Thr Glu Ser Asp Val His Pro Ser Cys Lys Val Thr Ala Met Lys Cys
115 120 125
Phe Leu Leu Glu Leu Gln Val Ile Ser Leu Glu Ser Gly Asp Ala Ser
130 135 140
Ile His Asp Thr Val Glu Asn Leu Ile Ile Leu Ala Asn Asn Ser Leu
145 150 155 160
Ser Ser Asn Gly Asn Val Thr Glu Ser Gly Cys Lys Glu Cys Glu Glu
165 170 175
Leu Glu Glu Lys Asn Ile Lys Glu Phe Leu Gln Ser Phe Val His Ile
180 185 190
Val Gln Met Phe Ile Asn Ser Tyr Thr Tyr Asn Tyr Glu Trp His Val
195 200 205
Asp Val Lys Asn Thr Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu
210 215 220
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Thr Val Pro Gly Tyr Ser Thr
225 230 235 240
Ala Pro Tyr Phe Asp Leu Trp Gly Gln Gly Thr Leu Val Thr Val Ser
245 250 255
Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser
260 265 270
Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp
275 280 285
Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr
290 295 300
Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr
305 310 315 320
Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln
325 330 335
Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp
340 345 350
Lys Lys Val Glu Pro Lys Ser Cys
355 360
<210> 94
<211> 350
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь Fab CA645 с IL-15 человека, встроенным в каркасную область 3, с бычьим коротким линкером
<400> 94
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Glu Thr Lys Lys Tyr Gln
65 70 75 80
Ser Asn Trp Val Asn Val Ile Ser Asp Leu Lys Lys Ile Glu Asp Leu
85 90 95
Ile Gln Ser Met His Ile Asp Ala Thr Leu Tyr Thr Glu Ser Asp Val
100 105 110
His Pro Ser Cys Lys Val Thr Ala Met Lys Cys Phe Leu Leu Glu Leu
115 120 125
Gln Val Ile Ser Leu Glu Ser Gly Asp Ala Ser Ile His Asp Thr Val
130 135 140
Glu Asn Leu Ile Ile Leu Ala Asn Asn Ser Leu Ser Ser Asn Gly Asn
145 150 155 160
Val Thr Glu Ser Gly Cys Lys Glu Cys Glu Glu Leu Glu Glu Lys Asn
165 170 175
Ile Lys Glu Phe Leu Gln Ser Phe Val His Ile Val Gln Met Phe Ile
180 185 190
Asn Ser Tyr Thr Tyr Asn Tyr Glu Lys Asn Thr Val Tyr Leu Gln Met
195 200 205
Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Thr
210 215 220
Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly Gln Gly
225 230 235 240
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
245 250 255
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
260 265 270
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
275 280 285
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
290 295 300
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
305 310 315 320
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
325 330 335
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
340 345 350
<210> 95
<211> 356
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь Fab CA645 с IL-15, встроенным в каркасную область 3, с линкером 2xG4S
<400> 95
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Gly Gly Gly Gly Ser Gly
65 70 75 80
Gly Gly Gly Ser Asn Trp Val Asn Val Ile Ser Asp Leu Lys Lys Ile
85 90 95
Glu Asp Leu Ile Gln Ser Met His Ile Asp Ala Thr Leu Tyr Thr Glu
100 105 110
Ser Asp Val His Pro Ser Cys Lys Val Thr Ala Met Lys Cys Phe Leu
115 120 125
Leu Glu Leu Gln Val Ile Ser Leu Glu Ser Gly Asp Ala Ser Ile His
130 135 140
Asp Thr Val Glu Asn Leu Ile Ile Leu Ala Asn Asn Ser Leu Ser Ser
145 150 155 160
Asn Gly Asn Val Thr Glu Ser Gly Cys Lys Glu Cys Glu Glu Leu Glu
165 170 175
Glu Lys Asn Ile Lys Glu Phe Leu Gln Ser Phe Val His Ile Val Gln
180 185 190
Met Phe Ile Asn Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Lys Asn
195 200 205
Thr Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
210 215 220
Tyr Tyr Cys Ala Arg Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe
225 230 235 240
Asp Leu Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
245 250 255
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
260 265 270
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
275 280 285
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
290 295 300
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
305 310 315 320
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
325 330 335
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
340 345 350
Pro Lys Ser Cys
355
<210> 96
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 96
Thr Ser Val His Gln Glu Thr Lys Lys Tyr Gln Ser
1 5 10
<210> 97
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 97
Ser Tyr Thr Tyr Asn Tyr Glu Trp His Val Asp Val
1 5 10
<210> 98
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 98
Glu Thr Lys Lys Tyr Gln Ser
1 5
<210> 99
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 99
Ser Tyr Thr Tyr Asn Tyr Glu
1 5
<210> 100
<211> 188
<212> БЕЛОК
<213> Homo sapiens
<400> 100
Asn Cys Trp Leu Arg Gln Ala Lys Asn Gly Arg Cys Gln Val Leu Tyr
1 5 10 15
Lys Thr Glu Leu Ser Lys Glu Glu Cys Cys Ser Thr Gly Arg Leu Ser
20 25 30
Thr Ser Trp Thr Glu Glu Asp Val Asn Asp Asn Thr Leu Phe Lys Trp
35 40 45
Met Ile Phe Asn Gly Gly Ala Pro Asn Cys Ile Pro Cys Lys Glu Pro
50 55 60
Arg Cys Val Cys Ala Pro Asp Cys Ser Asn Ile Thr Trp Lys Gly Pro
65 70 75 80
Val Cys Gly Leu Asp Gly Lys Thr Tyr Arg Asn Glu Cys Ala Leu Leu
85 90 95
Lys Ala Arg Cys Lys Glu Gln Pro Glu Leu Glu Val Gln Tyr Gln Gly
100 105 110
Arg Cys Lys Lys Thr Cys Arg Asp Val Phe Cys Pro Gly Ser Ser Thr
115 120 125
Cys Val Val Asp Gln Thr Asn Asn Ala Tyr Cys Val Thr Cys Asn Arg
130 135 140
Ile Cys Pro Glu Pro Ala Ser Ser Glu Gln Tyr Leu Cys Gly Asn Asp
145 150 155 160
Gly Val Thr Tyr Ser Ser Ala Cys His Leu Arg Lys Ala Thr Cys Leu
165 170 175
Leu Gly Arg Ser Ile Gly Leu Ala Gly Glu Tyr Lys
180 185
<210> 101
<211> 627
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь Fab CA645 с двумя мономерами IL-10 человека, встроенными в каркасную область 3
<400> 101
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Gly Gly Gly Gly Ser Gly
65 70 75 80
Ser Asn Cys Trp Leu Arg Gln Ala Lys Asn Gly Arg Cys Gln Val Leu
85 90 95
Tyr Lys Thr Glu Leu Ser Lys Glu Glu Cys Cys Ser Thr Gly Arg Leu
100 105 110
Ser Thr Ser Trp Thr Glu Glu Asp Val Asn Asp Asn Thr Leu Phe Lys
115 120 125
Trp Met Ile Phe Asn Gly Gly Ala Pro Asn Cys Ile Pro Cys Lys Glu
130 135 140
Pro Arg Cys Val Cys Ala Pro Asp Cys Ser Asn Ile Thr Trp Lys Gly
145 150 155 160
Pro Val Cys Gly Leu Asp Gly Lys Thr Tyr Arg Asn Glu Cys Ala Leu
165 170 175
Leu Lys Ala Arg Cys Lys Glu Gln Pro Glu Leu Glu Val Gln Tyr Gln
180 185 190
Gly Arg Cys Lys Lys Thr Cys Arg Asp Val Phe Cys Pro Gly Ser Ser
195 200 205
Thr Cys Val Val Asp Gln Thr Asn Asn Ala Tyr Cys Val Thr Cys Asn
210 215 220
Arg Ile Cys Pro Glu Pro Ala Ser Ser Glu Gln Tyr Leu Cys Gly Asn
225 230 235 240
Asp Gly Val Thr Tyr Ser Ser Ala Cys His Leu Arg Lys Ala Thr Cys
245 250 255
Leu Leu Gly Arg Ser Ile Gly Leu Ala Gly Glu Tyr Lys Gly Gly Gly
260 265 270
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asn Cys Trp Leu
275 280 285
Arg Gln Ala Lys Asn Gly Arg Cys Gln Val Leu Tyr Lys Thr Glu Leu
290 295 300
Ser Lys Glu Glu Cys Cys Ser Thr Gly Arg Leu Ser Thr Ser Trp Thr
305 310 315 320
Glu Glu Asp Val Asn Asp Asn Thr Leu Phe Lys Trp Met Ile Phe Asn
325 330 335
Gly Gly Ala Pro Asn Cys Ile Pro Cys Lys Glu Pro Arg Cys Val Cys
340 345 350
Ala Pro Asp Cys Ser Asn Ile Thr Trp Lys Gly Pro Val Cys Gly Leu
355 360 365
Asp Gly Lys Thr Tyr Arg Asn Glu Cys Ala Leu Leu Lys Ala Arg Cys
370 375 380
Lys Glu Gln Pro Glu Leu Glu Val Gln Tyr Gln Gly Arg Cys Lys Lys
385 390 395 400
Thr Cys Arg Asp Val Phe Cys Pro Gly Ser Ser Thr Cys Val Val Asp
405 410 415
Gln Thr Asn Asn Ala Tyr Cys Val Thr Cys Asn Arg Ile Cys Pro Glu
420 425 430
Pro Ala Ser Ser Glu Gln Tyr Leu Cys Gly Asn Asp Gly Val Thr Tyr
435 440 445
Ser Ser Ala Cys His Leu Arg Lys Ala Thr Cys Leu Leu Gly Arg Ser
450 455 460
Ile Gly Leu Ala Tyr Glu Gly Gly Gly Gly Ser Gly Ser Lys Asn Thr
465 470 475 480
Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
485 490 495
Tyr Cys Ala Arg Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp
500 505 510
Leu Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
515 520 525
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
530 535 540
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
545 550 555 560
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
565 570 575
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
580 585 590
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
595 600 605
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
610 615 620
Lys Ser Cys
625
<210> 102
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-домен (gL5) CA645-Cys
<400> 102
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ser Ser Pro Ser Val Trp Ser Asn
20 25 30
Phe Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Glu Ala Ser Lys Leu Thr Ser Gly Val Pro Ser Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln
65 70 75 80
Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gly Gly Gly Tyr Ser Ser Ile
85 90 95
Ser Asp Thr Thr Phe Gly Cys Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 103
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-домен (gH5) CA645-Cys
<400> 103
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 104
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Жесткий линкер
<400> 104
Glu Tyr His Gly Leu Gln
1 5
<210> 105
<211> 119
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-домен (gH1) CA645
<400> 105
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Ser Thr Thr Val Tyr Leu Gln Met
65 70 75 80
Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Thr
85 90 95
Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 106
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-домен (gH37) CA645
<400> 106
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Ala Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 107
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-домен (gH47) CA645
<400> 107
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Ala Val Tyr Tyr Cys Ala Arg
85 90 95
Thr Val Pro Gly Tyr Ser Ala Ala Pro Tyr Phe Asp Leu Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 108
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-домен (gL1) CA645
<400> 108
Asp Ile Val Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ser Ser Pro Ser Val Trp Ser Asn
20 25 30
Phe Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Glu Ala Ser Lys Leu Thr Ser Gly Val Pro Ser Arg Phe Lys
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln
65 70 75 80
Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gly Gly Gly Tyr Ser Ser Ile
85 90 95
Ser Asp Thr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 109
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-домен (gL4) CA645
<400> 109
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ser Ser Pro Ser Val Trp Ser Asn
20 25 30
Phe Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Glu Ala Ser Lys Leu Thr Ser Gly Val Pro Ser Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln
65 70 75 80
Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gly Gly Gly Tyr Ser Ser Ile
85 90 95
Ser Asp Thr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 110
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-домен (gL4) CA645-Cys
<400> 110
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ser Ser Pro Ser Val Trp Ser Asn
20 25 30
Phe Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Glu Ala Ser Lys Leu Thr Ser Gly Val Pro Ser Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln
65 70 75 80
Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gly Gly Gly Tyr Ser Ser Ile
85 90 95
Ser Asp Thr Thr Phe Gly Cys Gly Thr Lys Val Glu Ile Lys Arg Thr
100 105 110
<210> 111
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Альбуминсвязывающий пептид
<400> 111
Asp Leu Cys Leu Arg Asp Trp Gly Cys Leu Trp
1 5 10
<210> 112
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Альбуминсвязывающий пептид
<400> 112
Asp Ile Cys Leu Pro Arg Trp Gly Cys Leu Trp
1 5 10
<210> 113
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Альбуминсвязывающий пептид
<400> 113
Met Glu Asp Ile Cys Leu Pro Arg Trp Gly Cys Leu Trp Gly Asp
1 5 10 15
<210> 114
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Альбуминсвязывающий пептид
<400> 114
Gln Arg Leu Met Glu Asp Ile Cys Leu Pro Arg Trp Gly Cys Leu Trp
1 5 10 15
Glu Asp Asp Glu
20
<210> 115
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Альбуминсвязывающий пептид
<400> 115
Gln Gly Leu Ile Gly Asp Ile Cys Leu Pro Arg Trp Gly Cys Leu Trp
1 5 10 15
Gly Arg Ser Val
20
<210> 116
<211> 21
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Альбуминсвязывающий пептид
<400> 116
Gln Gly Leu Ile Gly Asp Ile Cys Leu Pro Arg Trp Gly Cys Leu Trp
1 5 10 15
Gly Arg Ser Val Lys
20
<210> 117
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Альбуминсвязывающий пептид
<400> 117
Glu Asp Ile Cys Leu Pro Arg Trp Gly Cys Leu Trp Glu Asp Asp
1 5 10 15
<210> 118
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Альбуминсвязывающий пептид
<400> 118
Arg Leu Met Glu Asp Ile Cys Leu Pro Arg Trp Gly Cys Leu Trp Glu
1 5 10 15
Asp Asp
<210> 119
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Альбуминсвязывающий пептид
<400> 119
Met Glu Asp Ile Cys Leu Pro Arg Trp Gly Cys Leu Trp Glu Asp Asp
1 5 10 15
<210> 120
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Альбуминсвязывающий пептид
<400> 120
Met Glu Asp Ile Cys Leu Pro Arg Trp Gly Cys Leu Trp Glu Asp
1 5 10 15
<210> 121
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Альбуминсвязывающий пептид
<400> 121
Arg Leu Met Glu Asp Ile Cys Leu Ala Arg Trp Gly Cys Leu Trp Glu
1 5 10 15
Asp Asp
<210> 122
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Альбуминсвязывающий пептид
<400> 122
Glu Val Arg Ser Phe Cys Thr Arg Trp Pro Ala Glu Lys Ser Cys Lys
1 5 10 15
Pro Leu Arg Gly
20
<210> 123
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Альбуминсвязывающий пептид
<400> 123
Arg Ala Pro Glu Ser Phe Val Cys Tyr Trp Glu Thr Ile Cys Phe Glu
1 5 10 15
Arg Ser Glu Gln
20
<210> 124
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Альбуминсвязывающий пептид
<400> 124
Glu Met Cys Tyr Phe Pro Gly Ile Cys Trp Met
1 5 10
<210> 125
<211> 224
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь Fab CA497
<400> 125
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Val Ile Phe Ser Asp Tyr
20 25 30
Tyr Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Ser Ile Asn Phe Asn Ala Asp Ile Ser Tyr Tyr Arg Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Thr Asp Ala Asn Arg Gln Asn Tyr Asp Trp Phe Ala Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
<210> 126
<211> 223
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь Fab CA240
<400> 126
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Phe Asn Asp Tyr
20 25 30
Phe Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Met Arg Asn Lys Asn Tyr Gln Tyr Gly Thr Tyr Tyr Ala Glu
50 55 60
Ser Leu Glu Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Glu Ser Tyr Tyr Gly Phe Thr Ser Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
<210> 127
<211> 247
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> dsscFv (vHvL) CA497
<400> 127
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Val Ile Phe Ser Asp Tyr
20 25 30
Tyr Met Ala Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val
35 40 45
Ala Ser Ile Asn Phe Asn Ala Asp Ile Ser Tyr Tyr Arg Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Thr Asp Ala Asn Arg Gln Asn Tyr Asp Trp Phe Ala Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gly Gly Gly Gly Ser Ala Ile Gln Leu Thr Gln Ser Pro
130 135 140
Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Lys
145 150 155 160
Ala Ser Glu Ser Val Ser Ser Ser Met Tyr Ser Tyr Met His Trp Tyr
165 170 175
Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Arg Ala Ser
180 185 190
Asn Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly
195 200 205
Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala
210 215 220
Thr Tyr Tyr Cys Gln Gln Ser Trp Thr Ala Pro Arg Thr Phe Gly Cys
225 230 235 240
Gly Thr Lys Val Glu Ile Lys
245
<210> 128
<211> 247
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> dsscFv (vLvH) CA497
<400> 128
Ala Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Glu Ser Val Ser Ser Ser
20 25 30
Met Tyr Ser Tyr Met His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Arg Ala Ser Asn Leu Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Trp
85 90 95
Thr Ala Pro Arg Thr Phe Gly Cys Gly Thr Lys Val Glu Ile Lys Gly
100 105 110
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val
115 120 125
Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly Ser Leu
130 135 140
Arg Leu Ser Cys Ala Ala Ser Gly Val Ile Phe Ser Asp Tyr Tyr Met
145 150 155 160
Ala Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val Ala Ser
165 170 175
Ile Asn Phe Asn Ala Asp Ile Ser Tyr Tyr Arg Glu Ser Val Lys Gly
180 185 190
Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Leu Tyr Leu Gln
195 200 205
Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr Tyr Cys Thr Thr
210 215 220
Asp Ala Asn Arg Gln Asn Tyr Asp Trp Phe Ala Tyr Trp Gly Gln Gly
225 230 235 240
Thr Leu Val Thr Val Ser Ser
245
<210> 129
<211> 491
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CA645 Fab HC Fwk3 CA497 dsscFv (vHvL)
<400> 129
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Gly Gly Gly Gly Ser Gly
65 70 75 80
Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val
85 90 95
Lys Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Val Ile
100 105 110
Phe Ser Asp Tyr Tyr Met Ala Trp Val Arg Gln Ala Pro Gly Lys Cys
115 120 125
Leu Glu Trp Val Ala Ser Ile Asn Phe Asn Ala Asp Ile Ser Tyr Tyr
130 135 140
Arg Glu Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys
145 150 155 160
Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala
165 170 175
Val Tyr Tyr Cys Thr Thr Asp Ala Asn Arg Gln Asn Tyr Asp Trp Phe
180 185 190
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly
195 200 205
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Ala Ile Gln Leu
210 215 220
Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr
225 230 235 240
Ile Thr Cys Lys Ala Ser Glu Ser Val Ser Ser Ser Met Tyr Ser Tyr
245 250 255
Met His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
260 265 270
Tyr Arg Ala Ser Asn Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
275 280 285
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
290 295 300
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Trp Thr Ala Pro Arg
305 310 315 320
Thr Phe Gly Cys Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser
325 330 335
Gly Gly Gly Gly Ser Lys Asn Thr Val Tyr Leu Gln Met Asn Ser Leu
340 345 350
Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Thr Val Pro Gly
355 360 365
Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly Gln Gly Thr Leu Val
370 375 380
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
385 390 395 400
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
405 410 415
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
420 425 430
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
435 440 445
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
450 455 460
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
465 470 475 480
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
485 490
<210> 130
<211> 491
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CA645 Fab HC Fwk3 CA497 dsscFv (vLvH)
<400> 130
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Gly Gly Gly Gly Ser Gly
65 70 75 80
Gly Gly Gly Ser Ala Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser
85 90 95
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Glu Ser
100 105 110
Val Ser Ser Ser Met Tyr Ser Tyr Met His Trp Tyr Gln Gln Lys Pro
115 120 125
Gly Lys Ala Pro Lys Leu Leu Ile Tyr Arg Ala Ser Asn Leu Glu Ser
130 135 140
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
145 150 155 160
Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
165 170 175
Gln Gln Ser Trp Thr Ala Pro Arg Thr Phe Gly Cys Gly Thr Lys Val
180 185 190
Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
195 200 205
Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro
210 215 220
Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Val Ile Phe Ser
225 230 235 240
Asp Tyr Tyr Met Ala Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu
245 250 255
Trp Val Ala Ser Ile Asn Phe Asn Ala Asp Ile Ser Tyr Tyr Arg Glu
260 265 270
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
275 280 285
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
290 295 300
Tyr Cys Thr Thr Asp Ala Asn Arg Gln Asn Tyr Asp Trp Phe Ala Tyr
305 310 315 320
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser
325 330 335
Gly Gly Gly Gly Ser Lys Asn Thr Val Tyr Leu Gln Met Asn Ser Leu
340 345 350
Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Thr Val Pro Gly
355 360 365
Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly Gln Gly Thr Leu Val
370 375 380
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
385 390 395 400
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
405 410 415
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
420 425 430
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
435 440 445
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
450 455 460
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
465 470 475 480
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
485 490
<210> 131
<211> 718
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CA645 IgG4P HC Fwk3 CA497 dsscFv (vHvL)
<400> 131
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Gly Gly Gly Gly Ser Gly
65 70 75 80
Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val
85 90 95
Lys Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Val Ile
100 105 110
Phe Ser Asp Tyr Tyr Met Ala Trp Val Arg Gln Ala Pro Gly Lys Cys
115 120 125
Leu Glu Trp Val Ala Ser Ile Asn Phe Asn Ala Asp Ile Ser Tyr Tyr
130 135 140
Arg Glu Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys
145 150 155 160
Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala
165 170 175
Val Tyr Tyr Cys Thr Thr Asp Ala Asn Arg Gln Asn Tyr Asp Trp Phe
180 185 190
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly
195 200 205
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Ala Ile Gln Leu
210 215 220
Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr
225 230 235 240
Ile Thr Cys Lys Ala Ser Glu Ser Val Ser Ser Ser Met Tyr Ser Tyr
245 250 255
Met His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
260 265 270
Tyr Arg Ala Ser Asn Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
275 280 285
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
290 295 300
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Trp Thr Ala Pro Arg
305 310 315 320
Thr Phe Gly Cys Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser
325 330 335
Gly Gly Gly Gly Ser Lys Asn Thr Val Tyr Leu Gln Met Asn Ser Leu
340 345 350
Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Thr Val Pro Gly
355 360 365
Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly Gln Gly Thr Leu Val
370 375 380
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
385 390 395 400
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
405 410 415
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
420 425 430
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
435 440 445
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
450 455 460
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
465 470 475 480
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
485 490 495
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
500 505 510
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
515 520 525
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
530 535 540
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
545 550 555 560
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
565 570 575
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
580 585 590
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
595 600 605
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
610 615 620
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
625 630 635 640
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
645 650 655
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
660 665 670
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
675 680 685
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
690 695 700
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
705 710 715
<210> 132
<211> 718
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CA645 IgG4P HC Fwk3 CA497 dsscFv (vLvH)
<400> 132
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Gly Gly Gly Gly Ser Gly
65 70 75 80
Gly Gly Gly Ser Ala Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser
85 90 95
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Glu Ser
100 105 110
Val Ser Ser Ser Met Tyr Ser Tyr Met His Trp Tyr Gln Gln Lys Pro
115 120 125
Gly Lys Ala Pro Lys Leu Leu Ile Tyr Arg Ala Ser Asn Leu Glu Ser
130 135 140
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
145 150 155 160
Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
165 170 175
Gln Gln Ser Trp Thr Ala Pro Arg Thr Phe Gly Cys Gly Thr Lys Val
180 185 190
Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
195 200 205
Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro
210 215 220
Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Val Ile Phe Ser
225 230 235 240
Asp Tyr Tyr Met Ala Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu
245 250 255
Trp Val Ala Ser Ile Asn Phe Asn Ala Asp Ile Ser Tyr Tyr Arg Glu
260 265 270
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
275 280 285
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
290 295 300
Tyr Cys Thr Thr Asp Ala Asn Arg Gln Asn Tyr Asp Trp Phe Ala Tyr
305 310 315 320
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser
325 330 335
Gly Gly Gly Gly Ser Lys Asn Thr Val Tyr Leu Gln Met Asn Ser Leu
340 345 350
Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Thr Val Pro Gly
355 360 365
Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly Gln Gly Thr Leu Val
370 375 380
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
385 390 395 400
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
405 410 415
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
420 425 430
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
435 440 445
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
450 455 460
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
465 470 475 480
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
485 490 495
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
500 505 510
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
515 520 525
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
530 535 540
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
545 550 555 560
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
565 570 575
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
580 585 590
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
595 600 605
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
610 615 620
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
625 630 635 640
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
645 650 655
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
660 665 670
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
675 680 685
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
690 695 700
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
705 710 715
<210> 133
<211> 213
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь против CD3
<400> 133
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
Asn Trp Tyr Gln Gln Thr Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Tyr Thr Phe Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Phe Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 134
<211> 222
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HC Fab против CD3
<400> 134
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Arg Tyr
20 25 30
Thr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Ala Phe
65 70 75 80
Leu Gln Met Asp Ser Leu Arg Pro Glu Asp Thr Gly Val Tyr Phe Cys
85 90 95
Ala Arg Tyr Tyr Asp Asp His Tyr Cys Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Pro Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
<210> 135
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HC IgG4P против CD3
<400> 135
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Arg Tyr
20 25 30
Thr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Ala Phe
65 70 75 80
Leu Gln Met Asp Ser Leu Arg Pro Glu Asp Thr Gly Val Tyr Phe Cys
85 90 95
Ala Arg Tyr Tyr Asp Asp His Tyr Cys Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Pro Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val
260 265 270
Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
<210> 136
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь против CD28
<400> 136
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys His Ala Ser Gln Asn Ile Tyr Val Trp
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Asn Leu His Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Gln Thr Tyr Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 137
<211> 223
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HC Fab против CD28
<400> 137
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Cys Ile Tyr Pro Gly Asn Val Asn Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Asp Arg Ala Thr Leu Thr Val Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Phe Cys
85 90 95
Thr Arg Ser His Tyr Gly Leu Asp Trp Asn Phe Asp Val Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
<210> 138
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HC IgG4P против CD28
<400> 138
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Cys Ile Tyr Pro Gly Asn Val Asn Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Asp Arg Ala Thr Leu Thr Val Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Phe Cys
85 90 95
Thr Arg Ser His Tyr Gly Leu Asp Trp Asn Phe Asp Val Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
<210> 139
<211> 488
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Анти-CD28 Fab HC Fwk3 анти-CD3 dsscFv (vHvL)
<400> 139
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Cys Ile Tyr Pro Gly Asn Val Asn Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Asp Arg Ala Thr Leu Thr Val Asp Thr Ser Gly Gly Gly Gly Ser
65 70 75 80
Gly Gly Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly Gly Gly Val
85 90 95
Val Gln Pro Gly Arg Ser Leu Arg Leu Ser Cys Lys Ala Ser Gly Tyr
100 105 110
Thr Phe Thr Arg Tyr Thr Met His Trp Val Arg Gln Ala Pro Gly Lys
115 120 125
Cys Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn
130 135 140
Tyr Asn Gln Lys Phe Lys Asp Arg Phe Thr Ile Ser Arg Asp Asn Ser
145 150 155 160
Lys Asn Thr Ala Phe Leu Gln Met Asp Ser Leu Arg Pro Glu Asp Thr
165 170 175
Gly Val Tyr Phe Cys Ala Arg Tyr Tyr Asp Asp His Tyr Cys Leu Asp
180 185 190
Tyr Trp Gly Gln Gly Thr Pro Val Thr Val Ser Ser Gly Gly Gly Gly
195 200 205
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
210 215 220
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
225 230 235 240
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
245 250 255
Asn Trp Tyr Gln Gln Thr Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
260 265 270
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
275 280 285
Gly Ser Gly Thr Asp Tyr Thr Phe Thr Ile Ser Ser Leu Gln Pro Glu
290 295 300
Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Phe Thr
305 310 315 320
Phe Gly Cys Gly Thr Lys Leu Glu Ile Lys Gly Gly Gly Gly Ser Gly
325 330 335
Gly Gly Gly Ser Ile Ser Thr Ala Tyr Met Glu Leu Ser Arg Leu Arg
340 345 350
Ser Asp Asp Thr Ala Val Tyr Phe Cys Thr Arg Ser His Tyr Gly Leu
355 360 365
Asp Trp Asn Phe Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser
370 375 380
Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser
385 390 395 400
Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp
405 410 415
Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr
420 425 430
Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr
435 440 445
Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln
450 455 460
Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp
465 470 475 480
Lys Lys Val Glu Pro Lys Ser Cys
485
<210> 140
<211> 487
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Анти-CD3 Fab HC Fwk3 анти-CD3 dsscFv (vHvL)
<400> 140
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Arg Tyr
20 25 30
Thr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Arg Phe Thr Ile Ser Arg Asp Asn Ser Gly Gly Gly Gly Ser
65 70 75 80
Gly Gly Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly Gly Gly Val
85 90 95
Val Gln Pro Gly Arg Ser Leu Arg Leu Ser Cys Lys Ala Ser Gly Tyr
100 105 110
Thr Phe Thr Arg Tyr Thr Met His Trp Val Arg Gln Ala Pro Gly Lys
115 120 125
Cys Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn
130 135 140
Tyr Asn Gln Lys Phe Lys Asp Arg Phe Thr Ile Ser Arg Asp Asn Ser
145 150 155 160
Lys Asn Thr Ala Phe Leu Gln Met Asp Ser Leu Arg Pro Glu Asp Thr
165 170 175
Gly Val Tyr Phe Cys Ala Arg Tyr Tyr Asp Asp His Tyr Cys Leu Asp
180 185 190
Tyr Trp Gly Gln Gly Thr Pro Val Thr Val Ser Ser Gly Gly Gly Gly
195 200 205
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
210 215 220
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
225 230 235 240
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
245 250 255
Asn Trp Tyr Gln Gln Thr Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
260 265 270
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
275 280 285
Gly Ser Gly Thr Asp Tyr Thr Phe Thr Ile Ser Ser Leu Gln Pro Glu
290 295 300
Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Phe Thr
305 310 315 320
Phe Gly Cys Gly Thr Lys Leu Glu Ile Lys Gly Gly Gly Gly Ser Gly
325 330 335
Gly Gly Gly Ser Lys Asn Thr Ala Phe Leu Gln Met Asp Ser Leu Arg
340 345 350
Pro Glu Asp Thr Gly Val Tyr Phe Cys Ala Arg Tyr Tyr Asp Asp His
355 360 365
Tyr Cys Leu Asp Tyr Trp Gly Gln Gly Thr Pro Val Thr Val Ser Ser
370 375 380
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
385 390 395 400
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
405 410 415
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
420 425 430
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
435 440 445
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
450 455 460
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
465 470 475 480
Lys Val Glu Pro Lys Ser Cys
485
<210> 141
<211> 712
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Анти-CD28 IgG4P HC Fwk3 анти-CD3 dsscFv (vHvL)
<400> 141
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Cys Ile Tyr Pro Gly Asn Val Asn Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Asp Arg Ala Thr Leu Thr Val Asp Thr Ser Gly Gly Gly Gly Ser
65 70 75 80
Gly Gly Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly Gly Gly Val
85 90 95
Val Gln Pro Gly Arg Ser Leu Arg Leu Ser Cys Lys Ala Ser Gly Tyr
100 105 110
Thr Phe Thr Arg Tyr Thr Met His Trp Val Arg Gln Ala Pro Gly Lys
115 120 125
Cys Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn
130 135 140
Tyr Asn Gln Lys Phe Lys Asp Arg Phe Thr Ile Ser Arg Asp Asn Ser
145 150 155 160
Lys Asn Thr Ala Phe Leu Gln Met Asp Ser Leu Arg Pro Glu Asp Thr
165 170 175
Gly Val Tyr Phe Cys Ala Arg Tyr Tyr Asp Asp His Tyr Cys Leu Asp
180 185 190
Tyr Trp Gly Gln Gly Thr Pro Val Thr Val Ser Ser Gly Gly Gly Gly
195 200 205
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
210 215 220
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
225 230 235 240
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
245 250 255
Asn Trp Tyr Gln Gln Thr Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
260 265 270
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
275 280 285
Gly Ser Gly Thr Asp Tyr Thr Phe Thr Ile Ser Ser Leu Gln Pro Glu
290 295 300
Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Phe Thr
305 310 315 320
Phe Gly Cys Gly Thr Lys Leu Glu Ile Lys Gly Gly Gly Gly Ser Gly
325 330 335
Gly Gly Gly Ser Ile Ser Thr Ala Tyr Met Glu Leu Ser Arg Leu Arg
340 345 350
Ser Asp Asp Thr Ala Val Tyr Phe Cys Thr Arg Ser His Tyr Gly Leu
355 360 365
Asp Trp Asn Phe Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser
370 375 380
Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser
385 390 395 400
Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp
405 410 415
Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr
420 425 430
Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr
435 440 445
Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys
450 455 460
Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp
465 470 475 480
Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala
485 490 495
Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
500 505 510
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
515 520 525
Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val
530 535 540
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
545 550 555 560
Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
565 570 575
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly
580 585 590
Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
595 600 605
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr
610 615 620
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
625 630 635 640
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
645 650 655
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
660 665 670
Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe
675 680 685
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
690 695 700
Ser Leu Ser Leu Ser Leu Gly Lys
705 710
<210> 142
<211> 711
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Анти-CD3 IgG4P HC Fwk3 анти-CD3 dsscFv (vHvL)
<400> 142
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Arg Tyr
20 25 30
Thr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Arg Phe Thr Ile Ser Arg Asp Asn Ser Gly Gly Gly Gly Ser
65 70 75 80
Gly Gly Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly Gly Gly Val
85 90 95
Val Gln Pro Gly Arg Ser Leu Arg Leu Ser Cys Lys Ala Ser Gly Tyr
100 105 110
Thr Phe Thr Arg Tyr Thr Met His Trp Val Arg Gln Ala Pro Gly Lys
115 120 125
Cys Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn
130 135 140
Tyr Asn Gln Lys Phe Lys Asp Arg Phe Thr Ile Ser Arg Asp Asn Ser
145 150 155 160
Lys Asn Thr Ala Phe Leu Gln Met Asp Ser Leu Arg Pro Glu Asp Thr
165 170 175
Gly Val Tyr Phe Cys Ala Arg Tyr Tyr Asp Asp His Tyr Cys Leu Asp
180 185 190
Tyr Trp Gly Gln Gly Thr Pro Val Thr Val Ser Ser Gly Gly Gly Gly
195 200 205
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
210 215 220
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
225 230 235 240
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
245 250 255
Asn Trp Tyr Gln Gln Thr Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
260 265 270
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
275 280 285
Gly Ser Gly Thr Asp Tyr Thr Phe Thr Ile Ser Ser Leu Gln Pro Glu
290 295 300
Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Phe Thr
305 310 315 320
Phe Gly Cys Gly Thr Lys Leu Glu Ile Lys Gly Gly Gly Gly Ser Gly
325 330 335
Gly Gly Gly Ser Lys Asn Thr Ala Phe Leu Gln Met Asp Ser Leu Arg
340 345 350
Pro Glu Asp Thr Gly Val Tyr Phe Cys Ala Arg Tyr Tyr Asp Asp His
355 360 365
Tyr Cys Leu Asp Tyr Trp Gly Gln Gly Thr Pro Val Thr Val Ser Ser
370 375 380
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
385 390 395 400
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
405 410 415
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
420 425 430
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
435 440 445
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
450 455 460
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
465 470 475 480
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
485 490 495
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
500 505 510
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
515 520 525
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
530 535 540
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
545 550 555 560
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
565 570 575
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
580 585 590
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
595 600 605
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
610 615 620
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
625 630 635 640
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
645 650 655
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
660 665 670
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
675 680 685
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
690 695 700
Leu Ser Leu Ser Leu Gly Lys
705 710
<210> 143
<211> 251
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> scFv (VH-VL) CA645
<400> 143
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln
130 135 140
Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly Asp Arg Val
145 150 155 160
Thr Ile Thr Cys Gln Ser Ser Pro Ser Val Trp Ser Asn Phe Leu Ser
165 170 175
Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Glu
180 185 190
Ala Ser Lys Leu Thr Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly
195 200 205
Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp
210 215 220
Phe Ala Thr Tyr Tyr Cys Gly Gly Gly Tyr Ser Ser Ile Ser Asp Thr
225 230 235 240
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
245 250
<210> 144
<211> 251
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> dsscFv (VH-VL) CA645
<400> 144
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln
130 135 140
Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly Asp Arg Val
145 150 155 160
Thr Ile Thr Cys Gln Ser Ser Pro Ser Val Trp Ser Asn Phe Leu Ser
165 170 175
Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Glu
180 185 190
Ala Ser Lys Leu Thr Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly
195 200 205
Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp
210 215 220
Phe Ala Thr Tyr Tyr Cys Gly Gly Gly Tyr Ser Ser Ile Ser Asp Thr
225 230 235 240
Thr Phe Gly Cys Gly Thr Lys Val Glu Ile Lys
245 250
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| C3-СВЯЗЫВАЮЩИЕ АГЕНТЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2802307C2 |
| АНТИТЕЛА К РЕЦЕПТОРУ IL4 ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2019 |
|
RU2837141C2 |
| МУЛЬТИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО СО СПЕЦИФИЧНОСТЬЮ СВЯЗЫВАНИЯ IL-13 И IL-17 ЧЕЛОВЕКА | 2020 |
|
RU2827946C1 |
| АНТИ-IL31 АНТИТЕЛА ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2020 |
|
RU2835543C1 |
| СЛИТЫЙ БЕЛОК, ВКЛЮЧАЮЩИЙ IL-12 И АНТИТЕЛО ПРОТИВ FAP, И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2831612C1 |
| КОНСТРУКЦИИ СЛИТОГО БЕЛКА ДЛЯ ЗАБОЛЕВАНИЯ, СВЯЗАННОГО С КОМПЛЕМЕНТОМ | 2019 |
|
RU2824402C2 |
| АНТИТЕЛО ПРОТИВ ФАКТОРА РОСТА СОЕДИНИТЕЛЬНОЙ ТКАНИ И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2819228C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО ПРОТИВ ВИРУСА БЕШЕНСТВА И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2764740C1 |
| ИММУНОЦИТОКИНЫ И ИХ ПРИМЕНЕНИЕ | 2021 |
|
RU2818371C1 |
| ИНДУЦИРУЮЩИЙ ЦИТОТОКСИЧНОСТЬ ТЕРАПЕВТИЧЕСКИЙ АГЕНТ | 2015 |
|
RU2743464C2 |
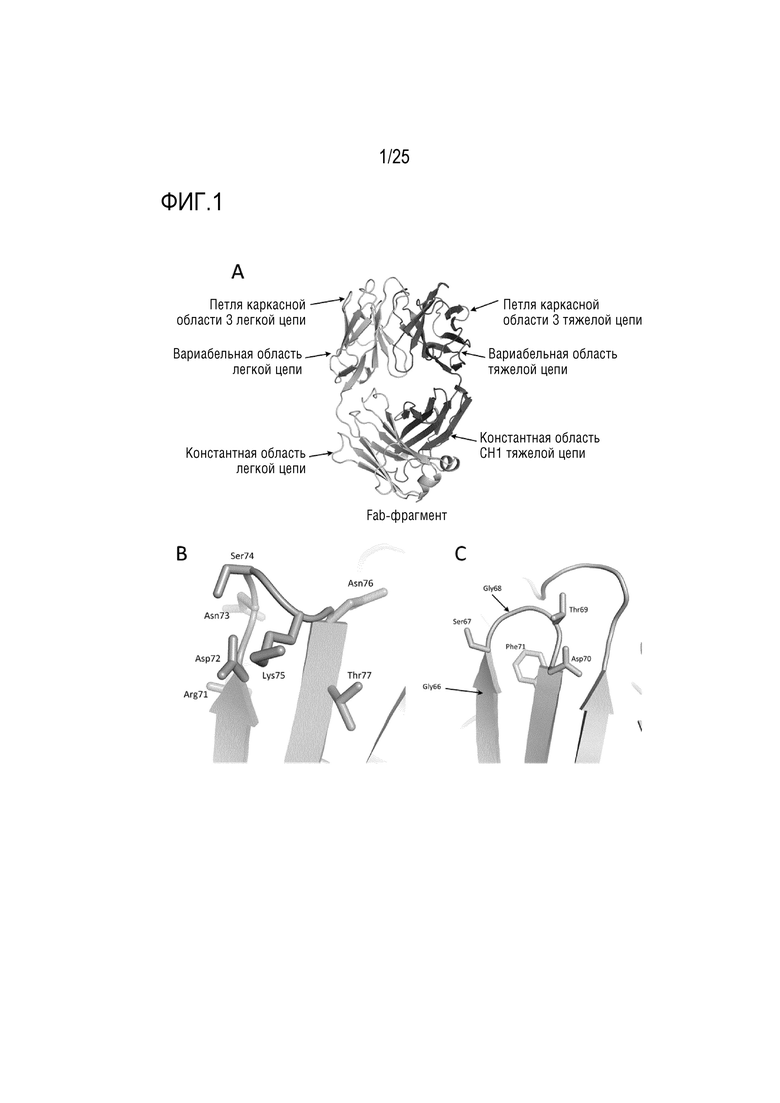
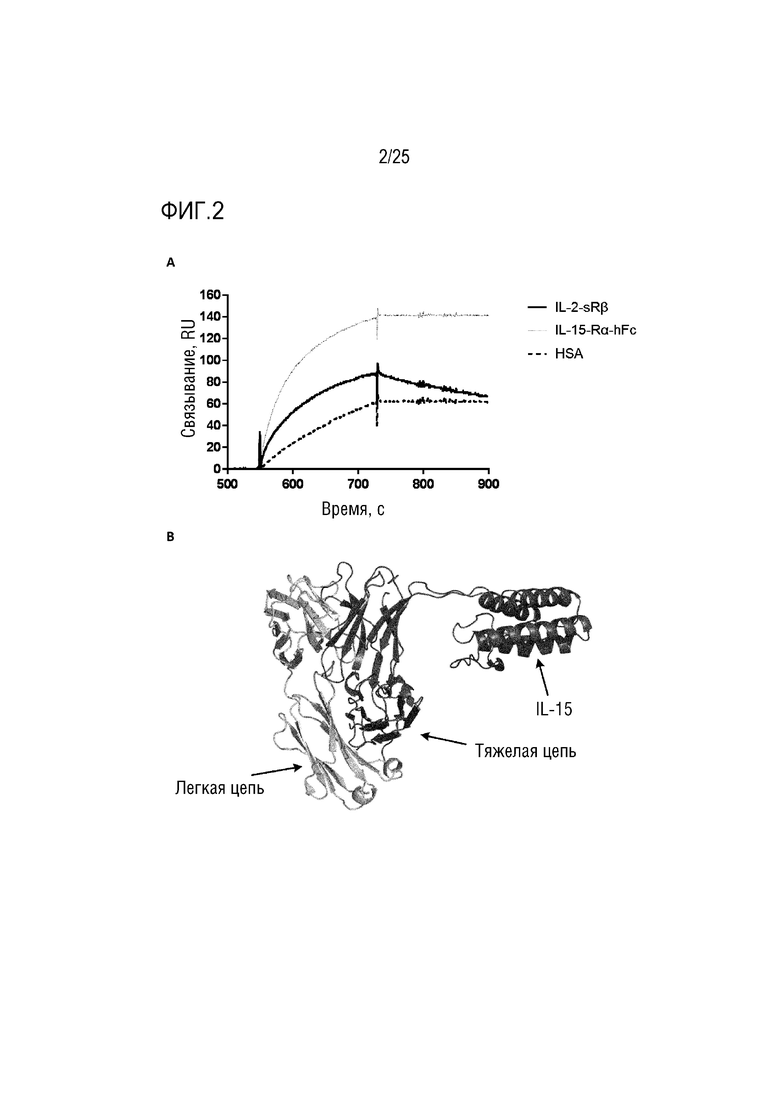
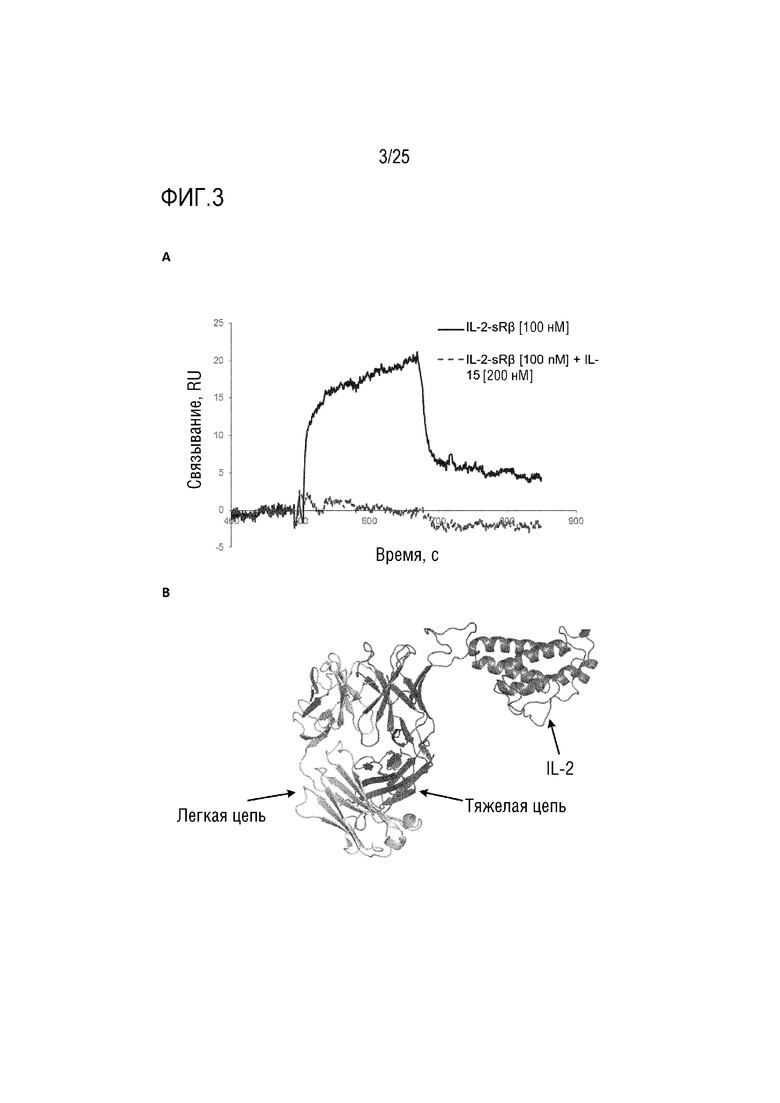
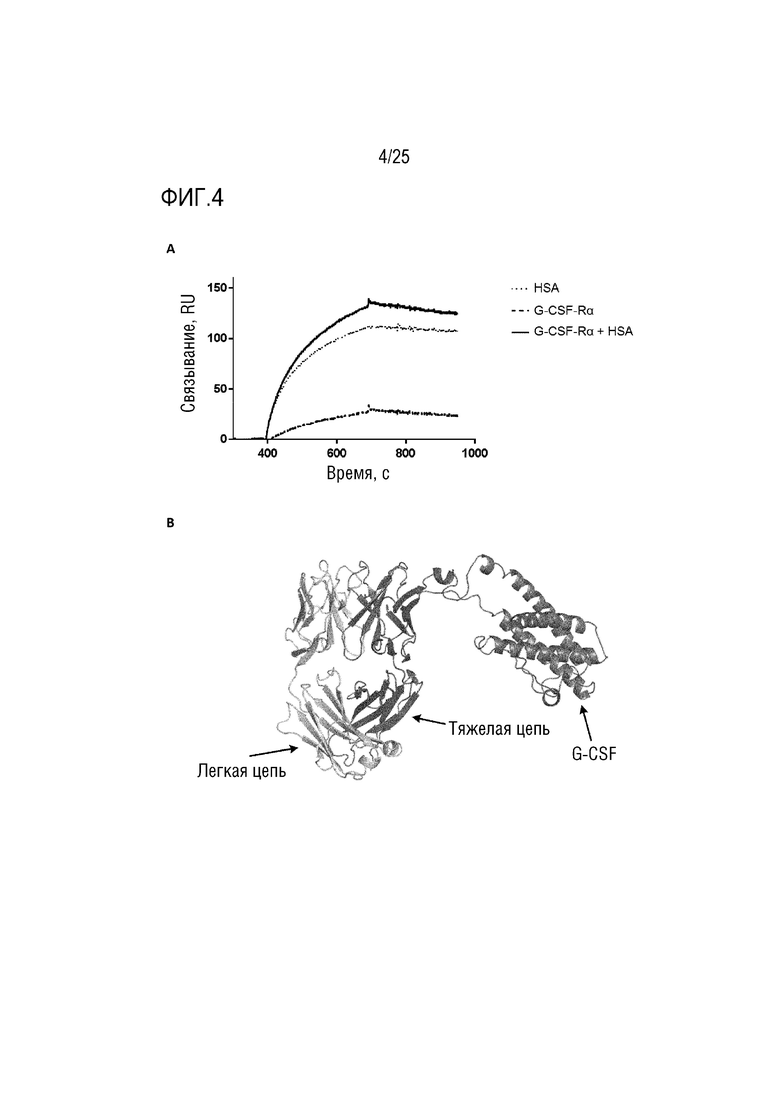
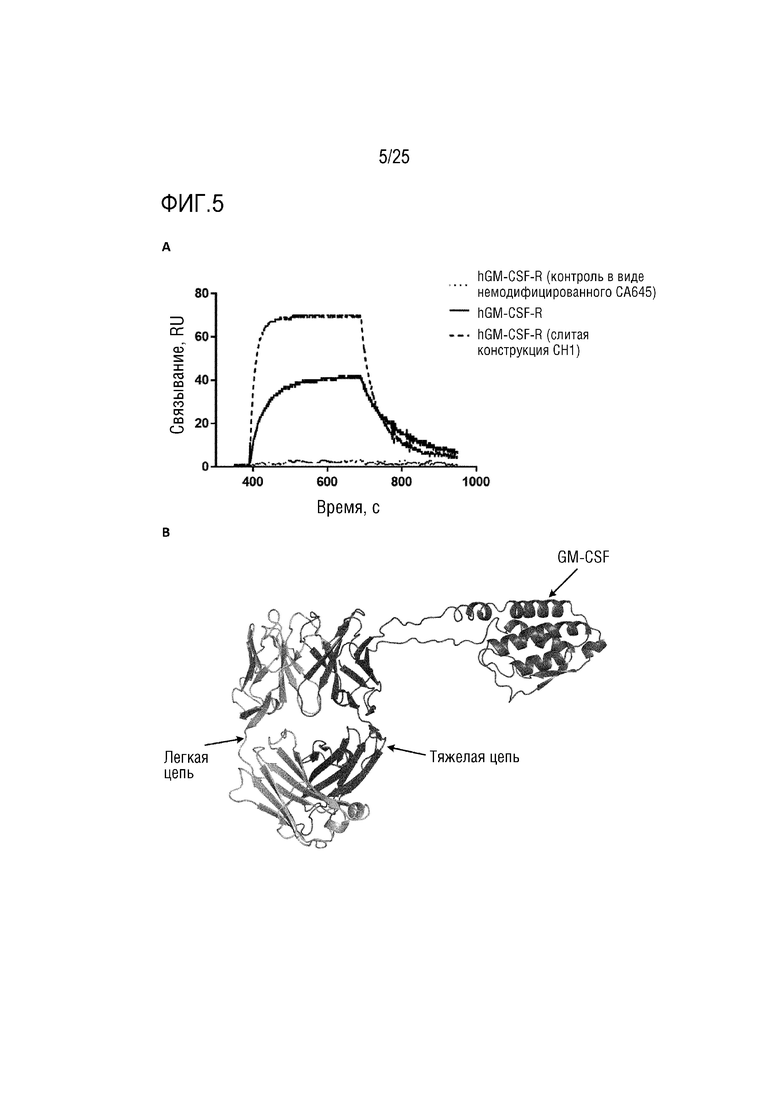
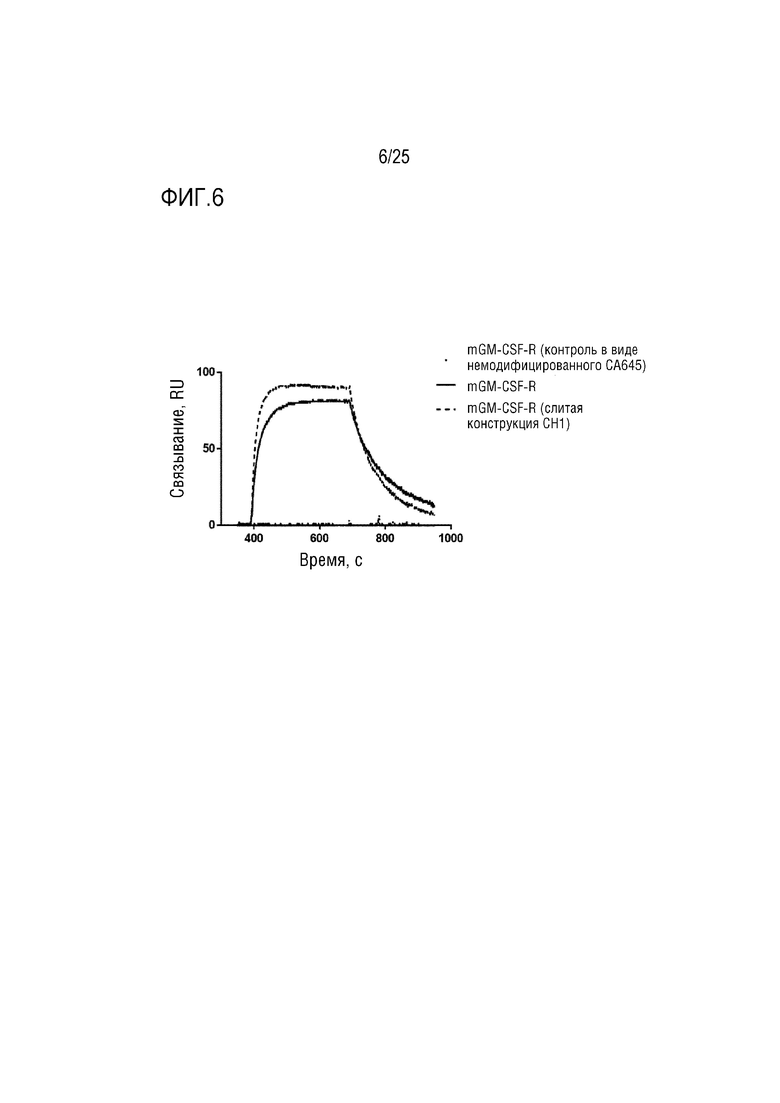
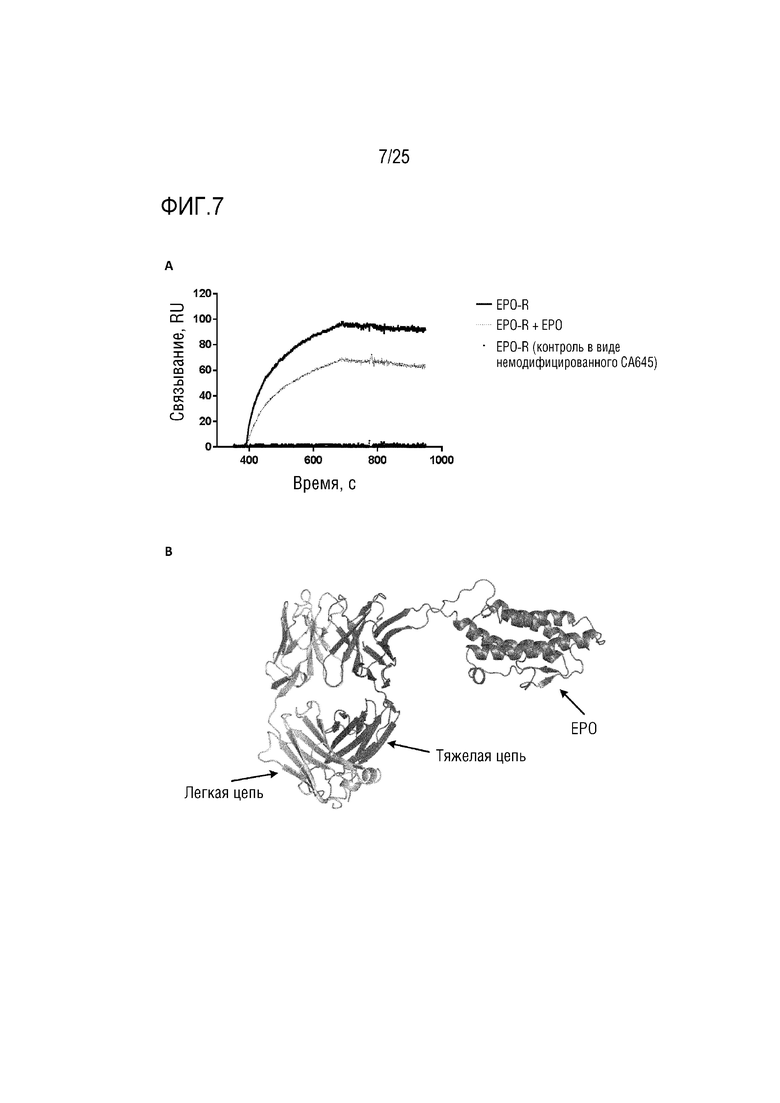
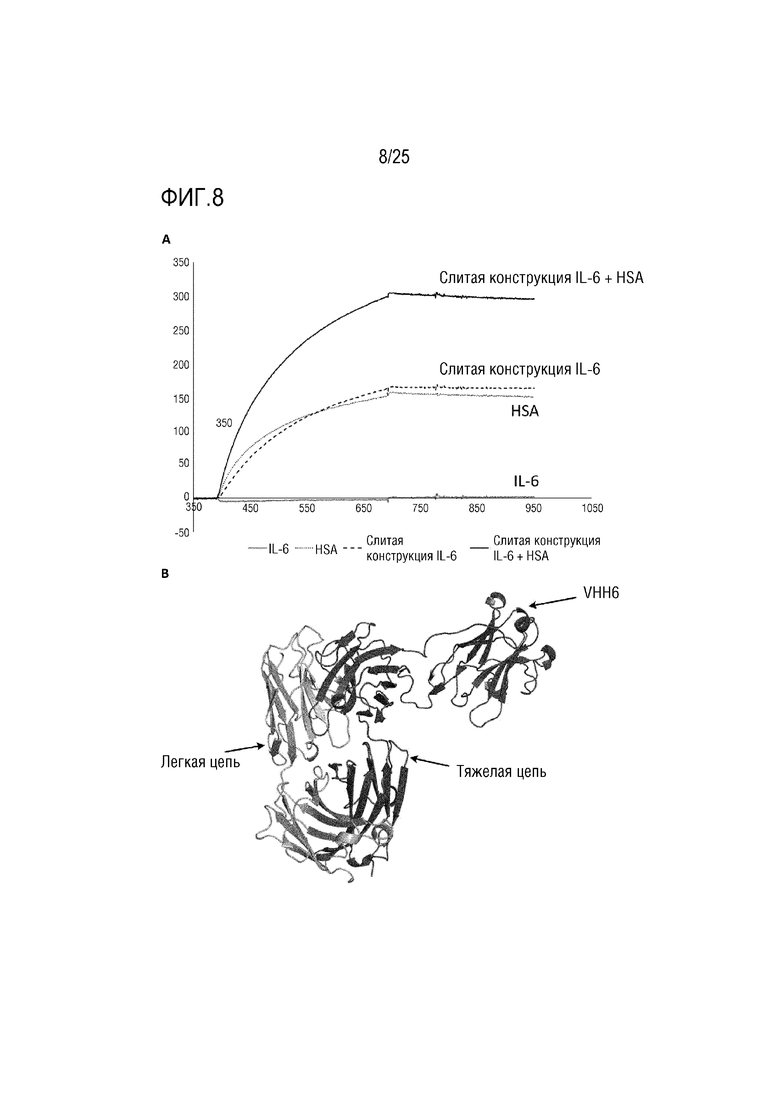
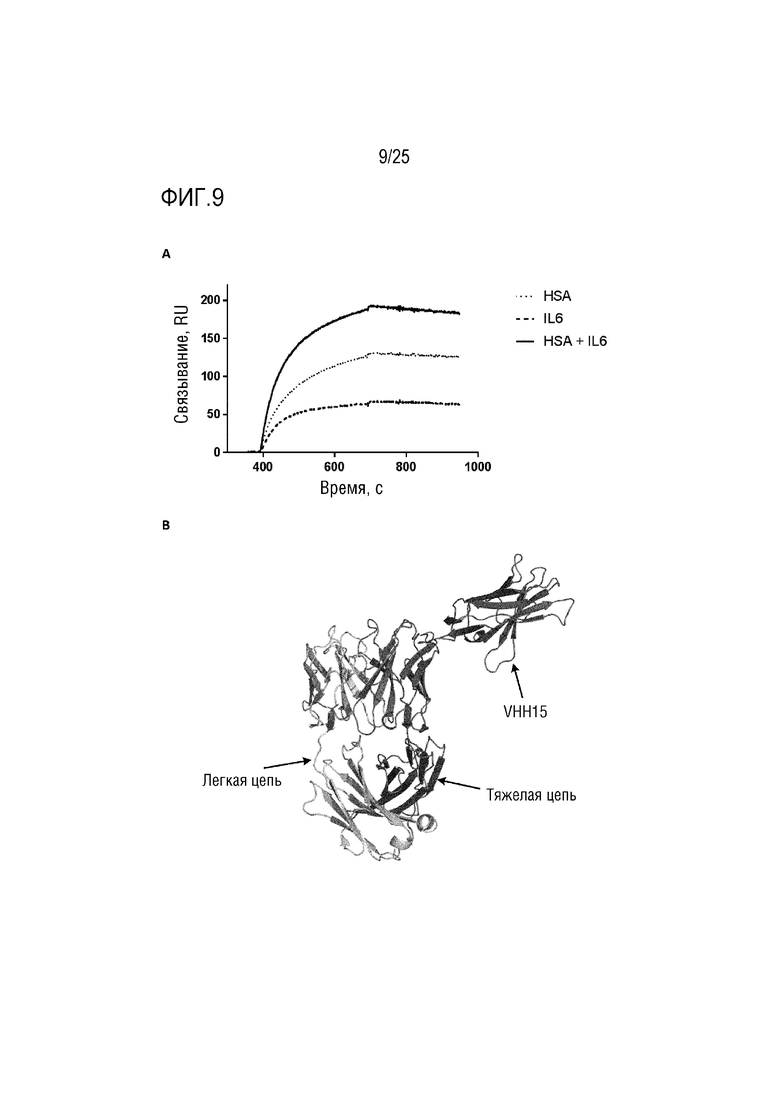
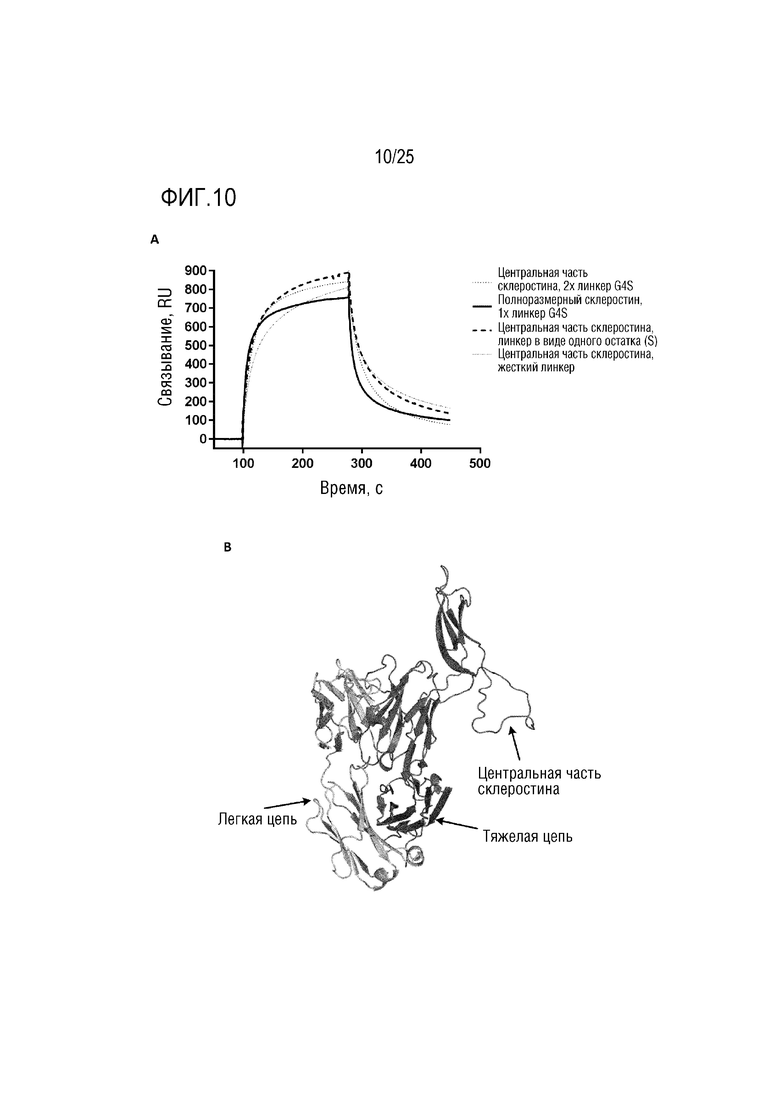
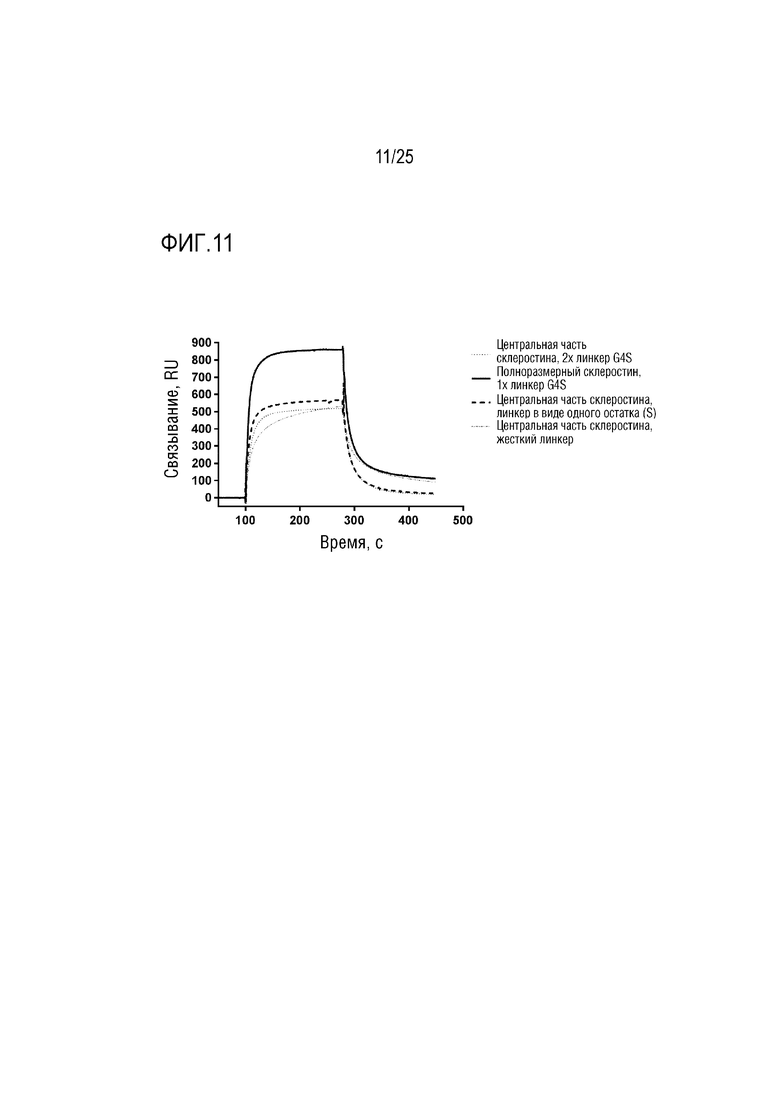
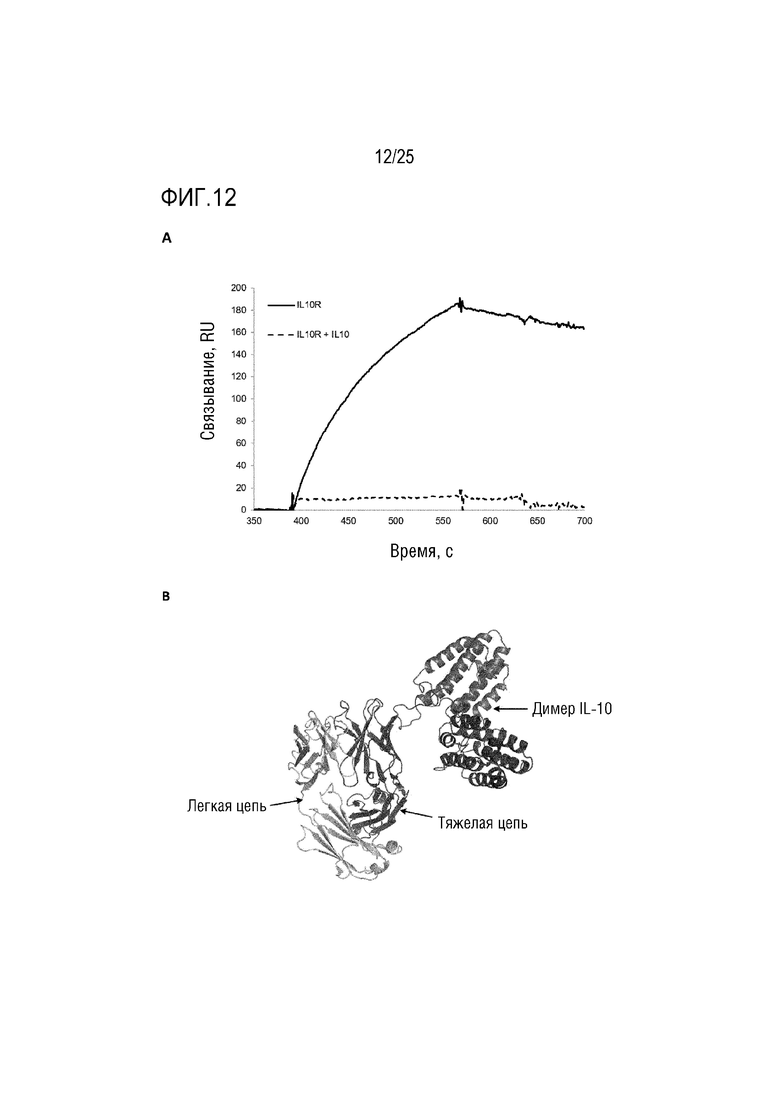
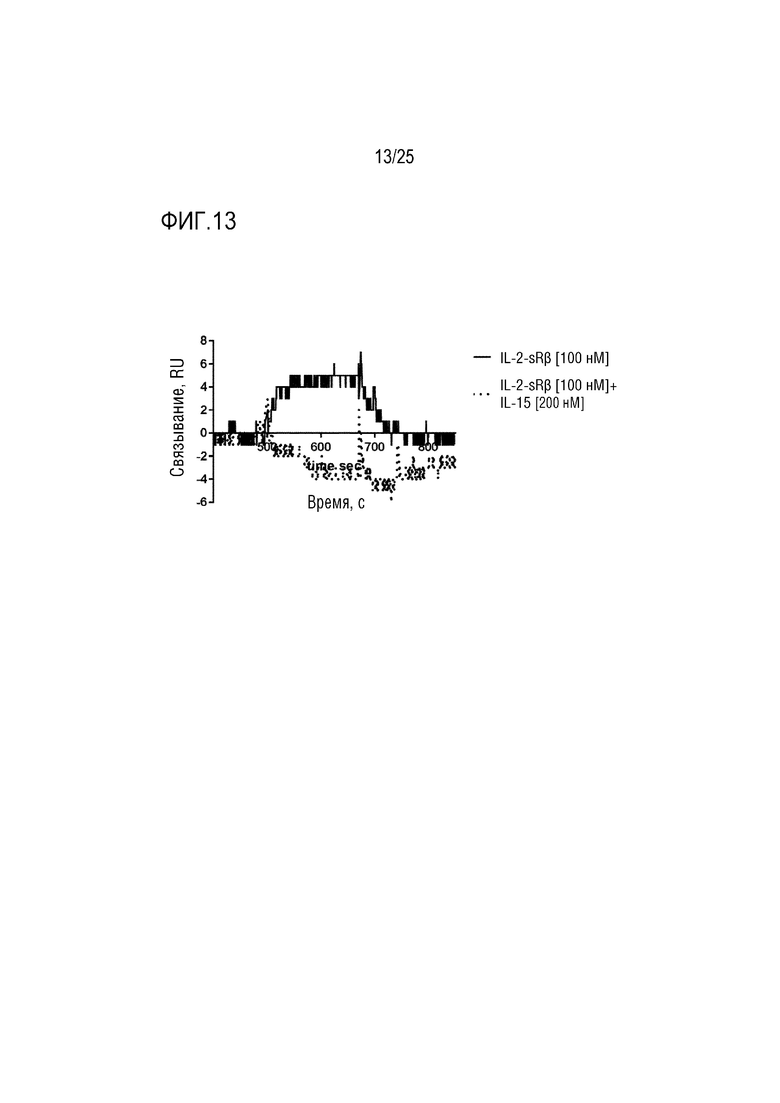
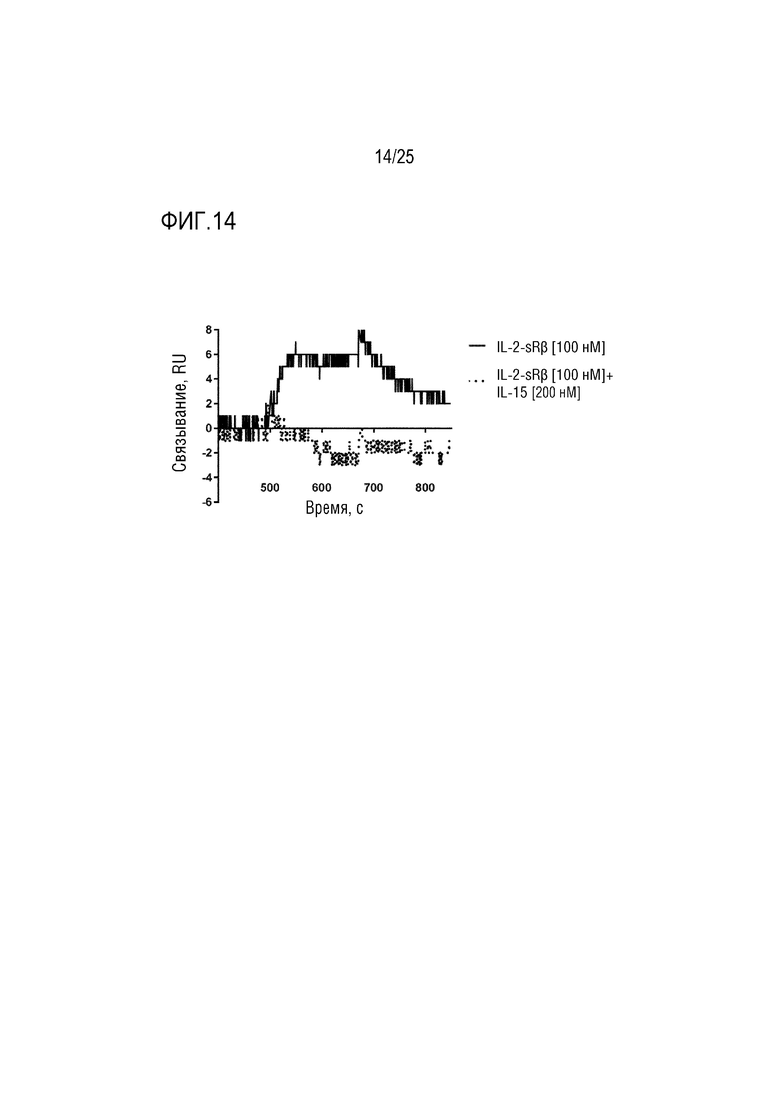
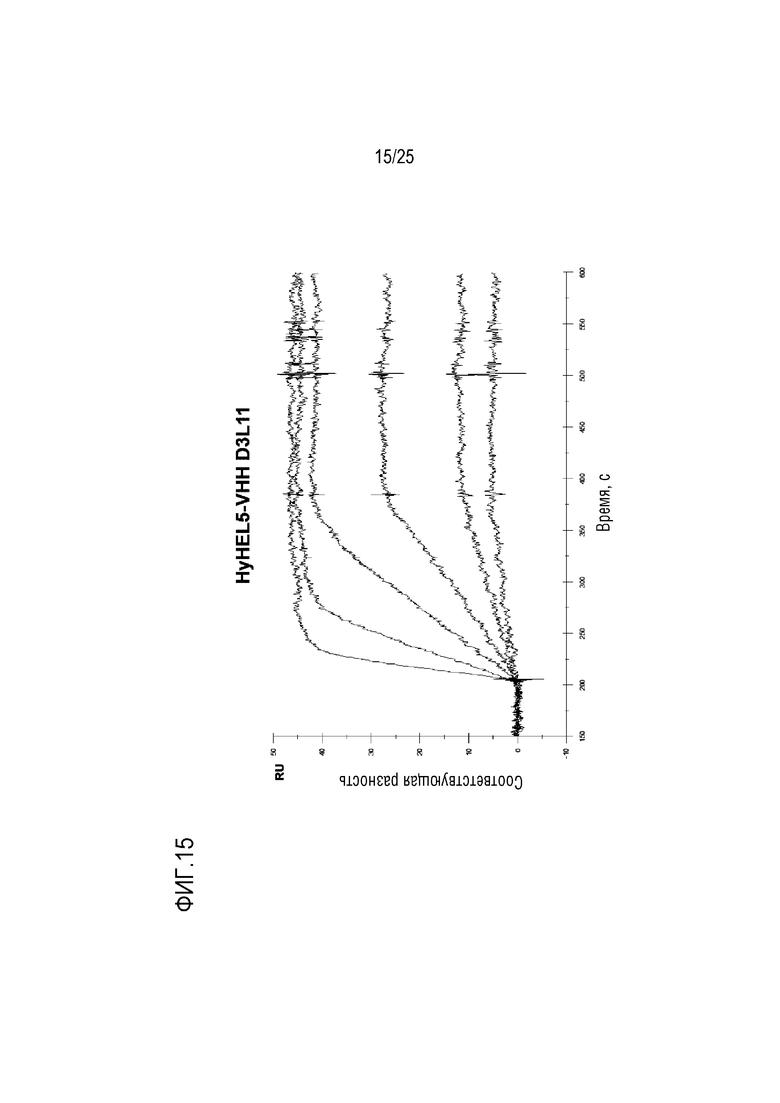
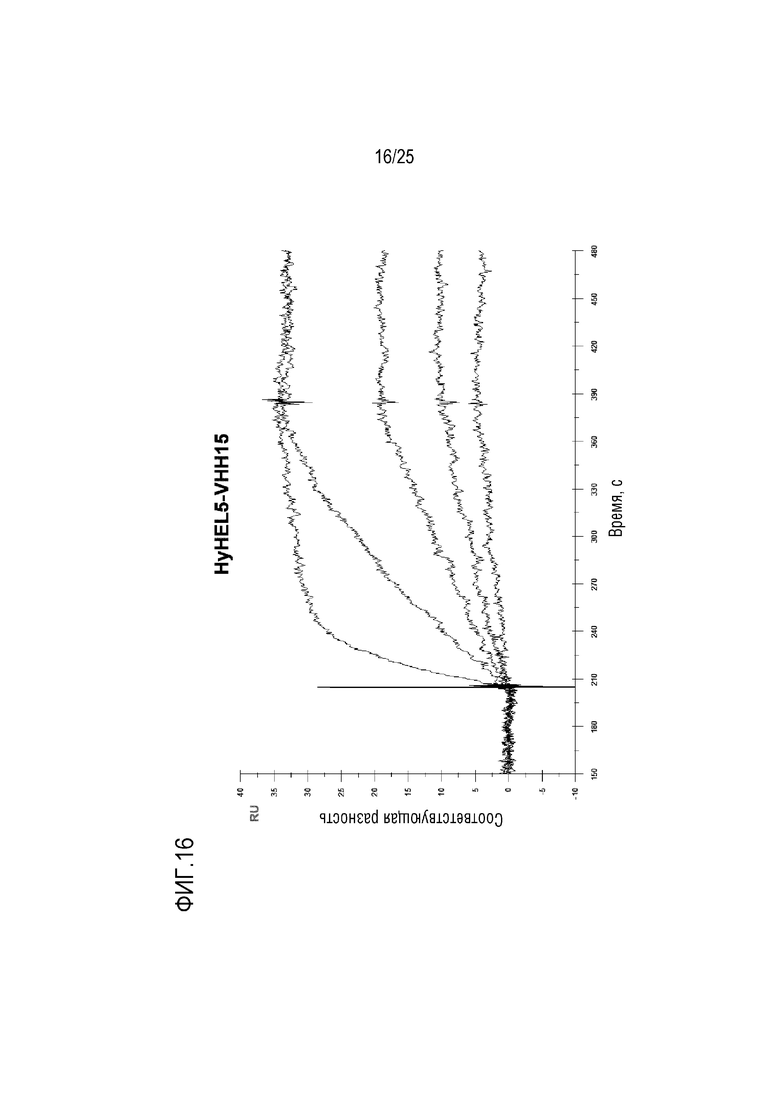
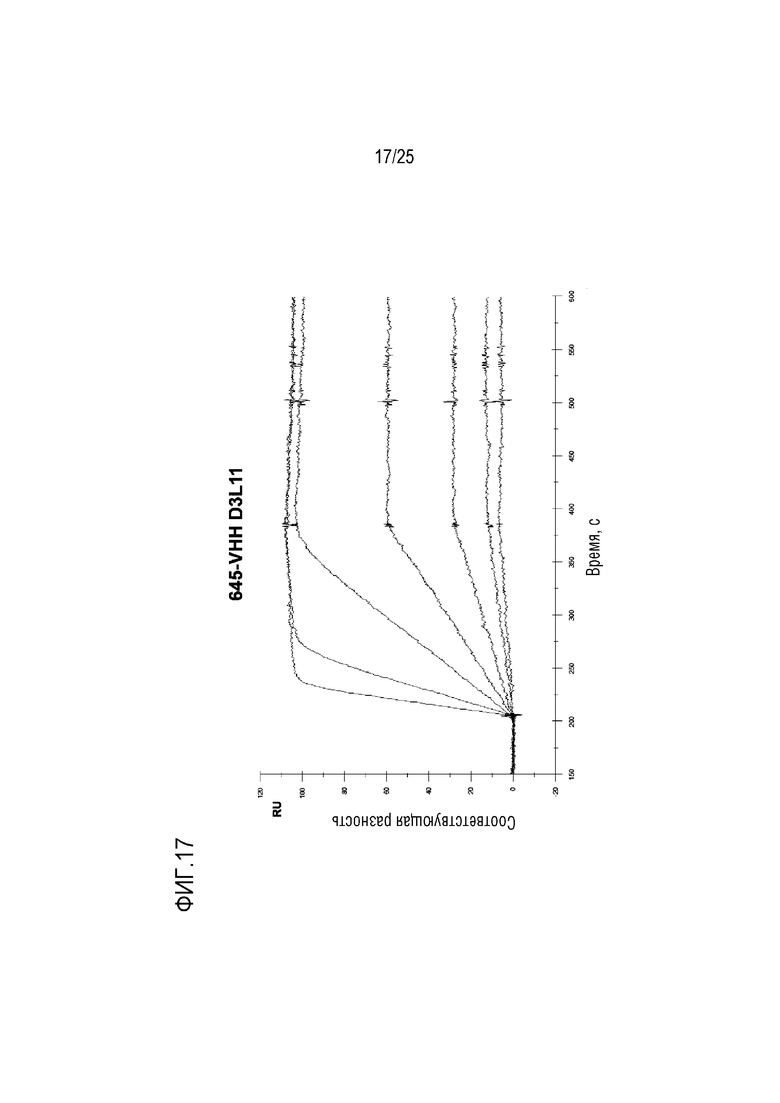
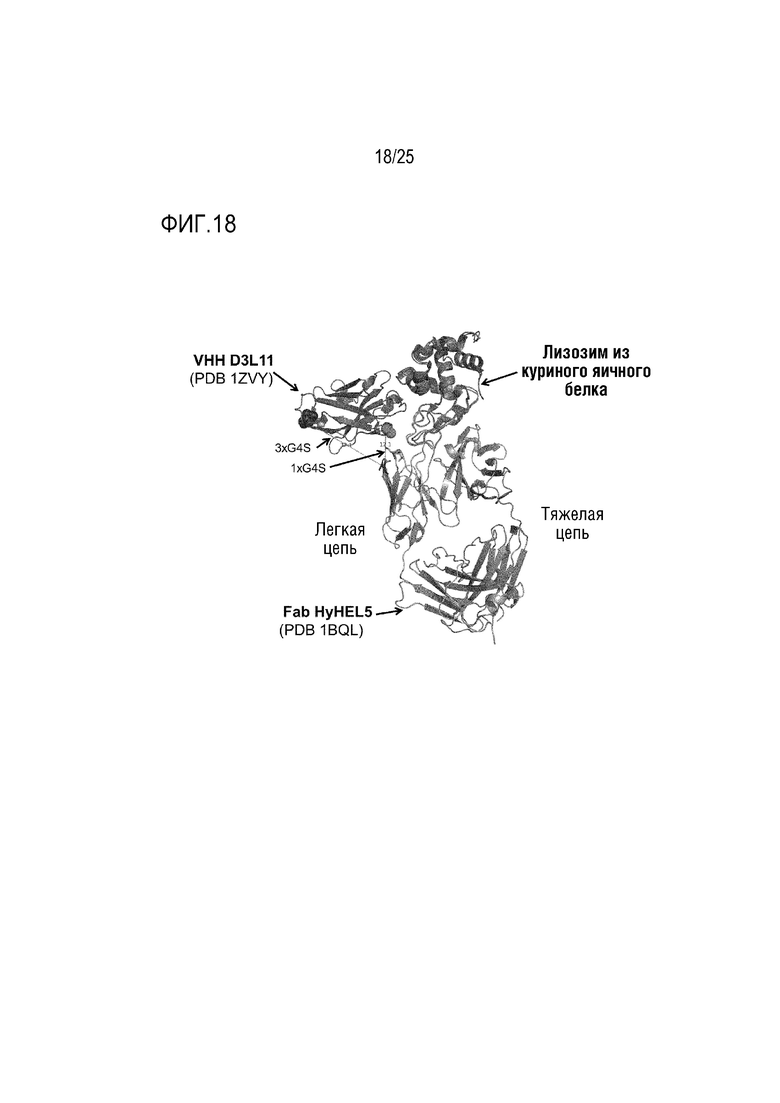
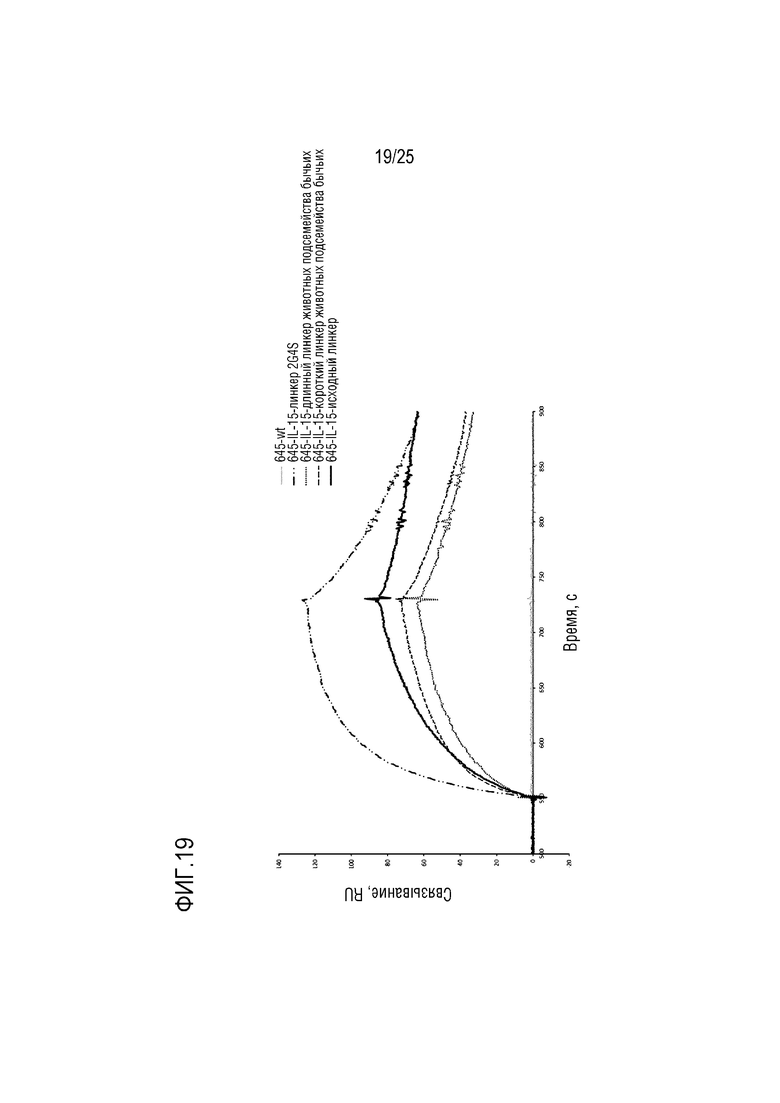
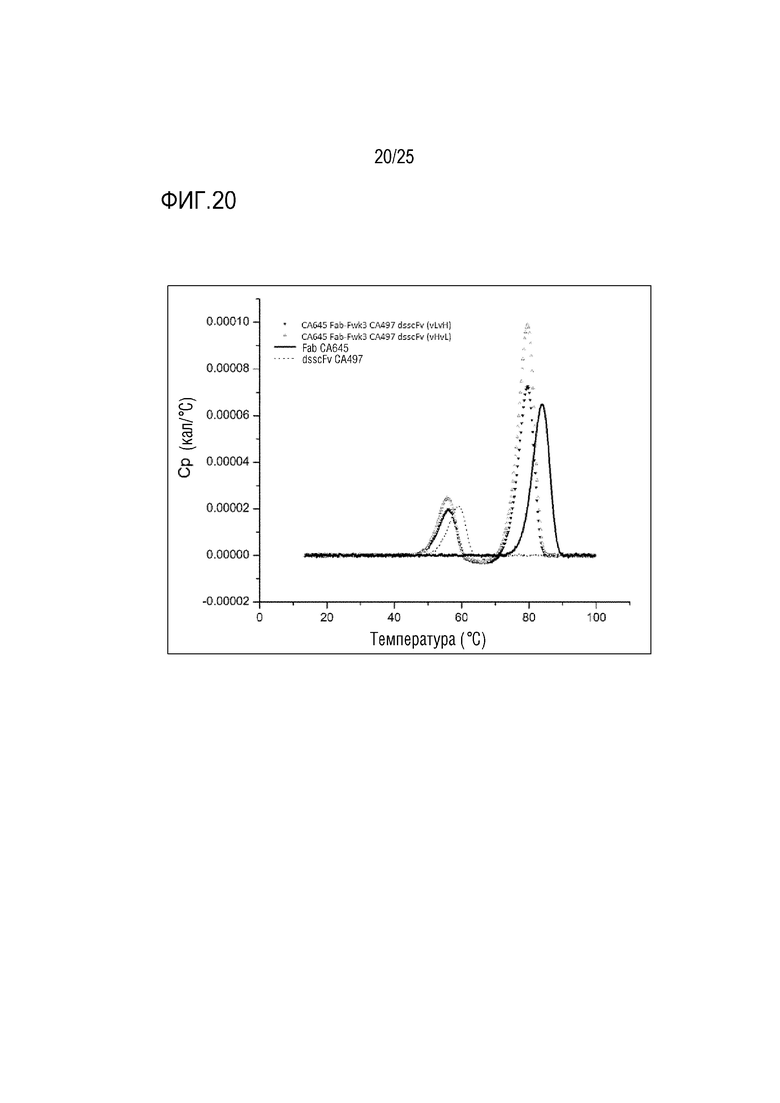
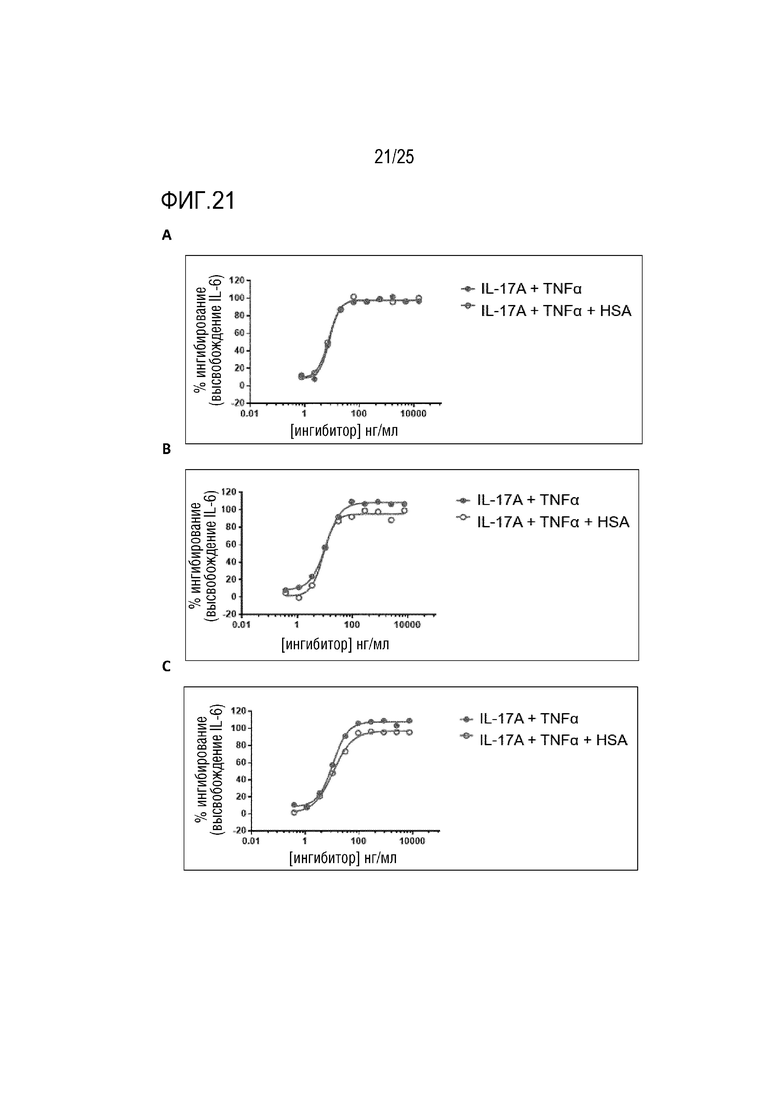
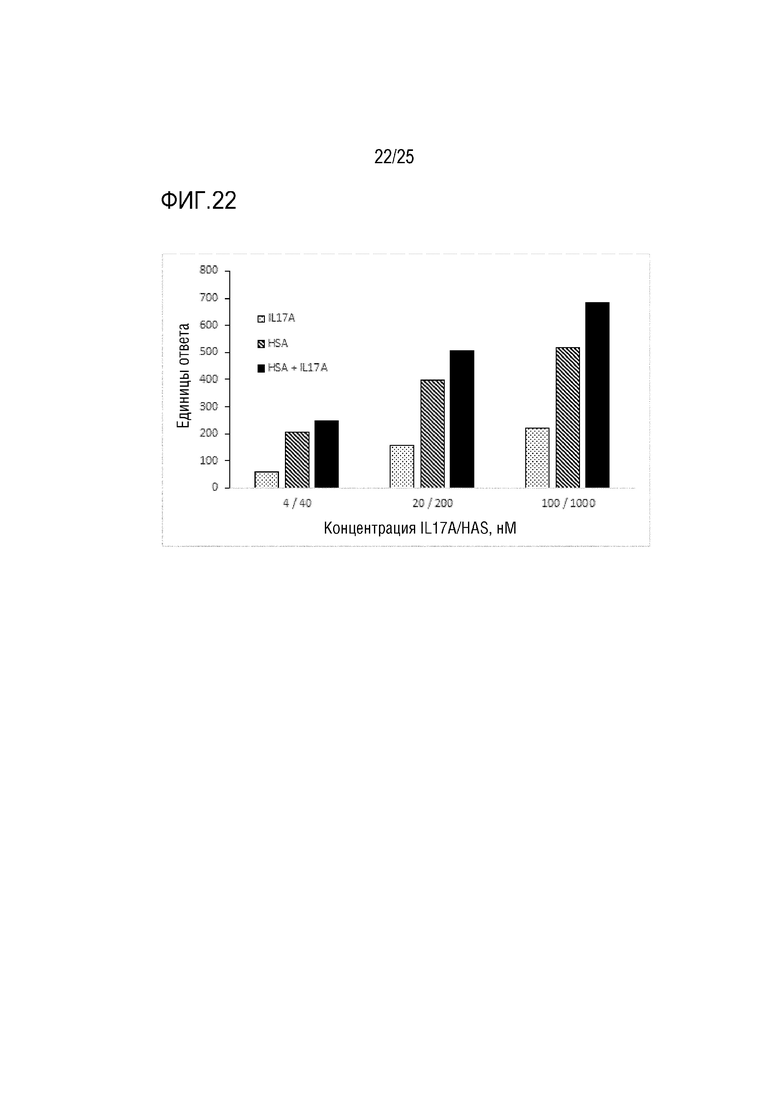
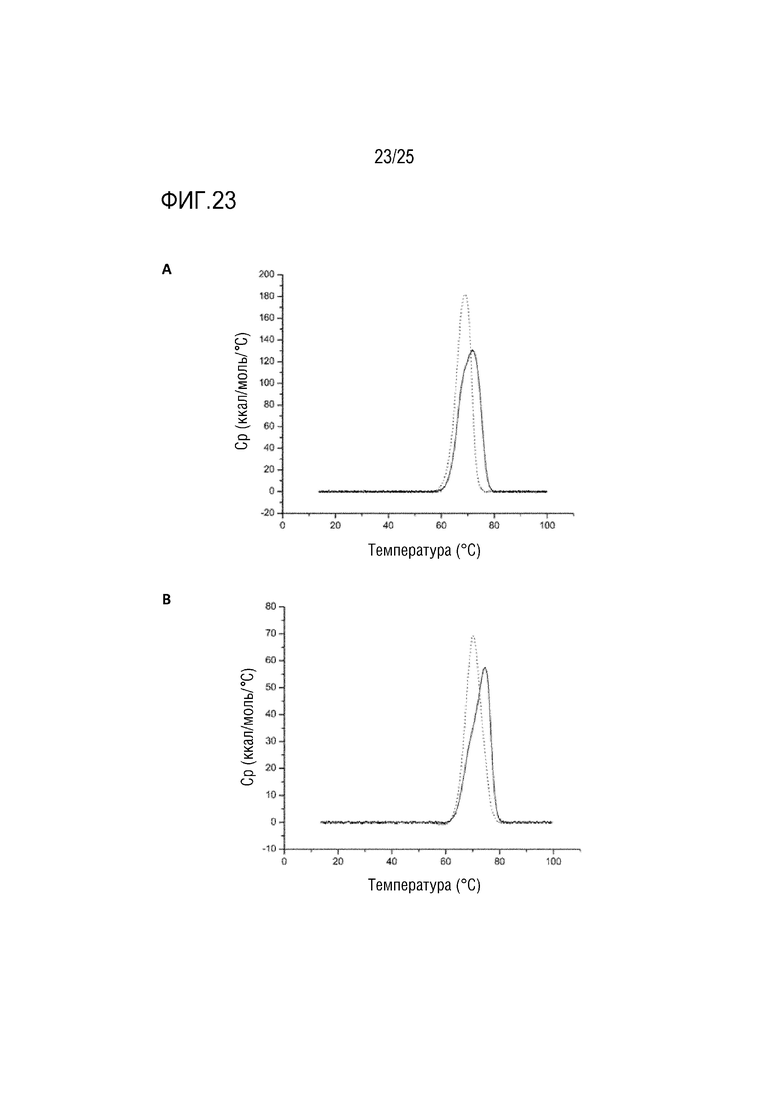
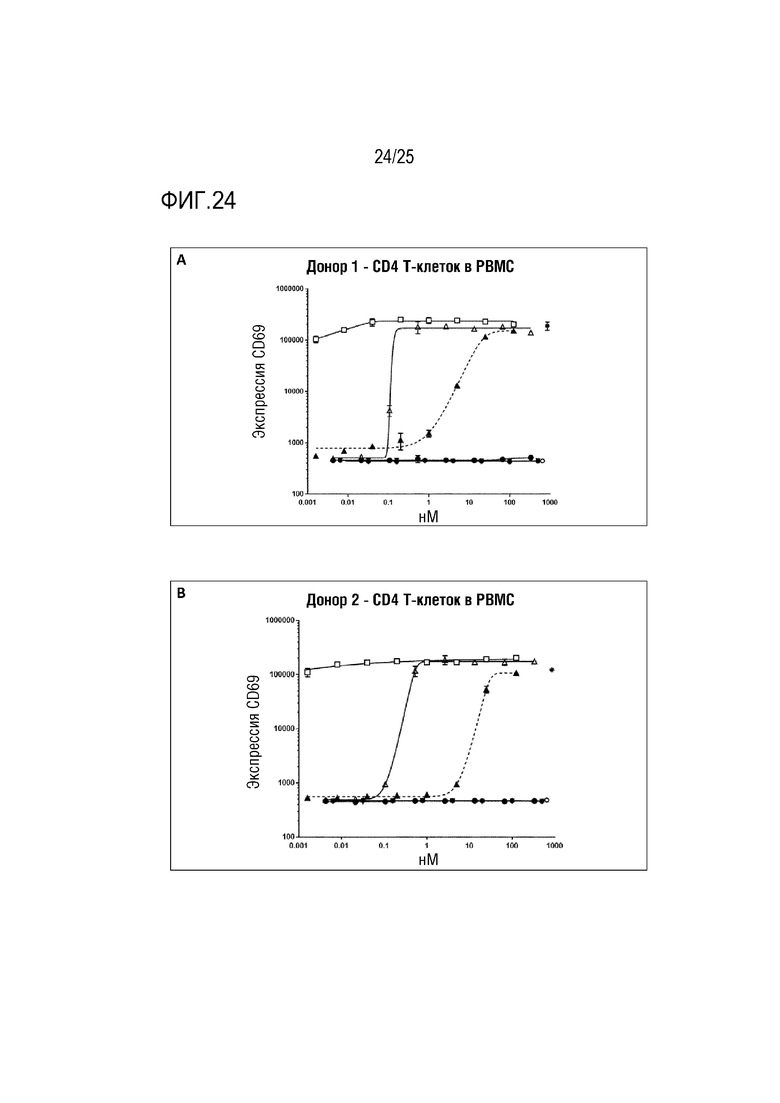
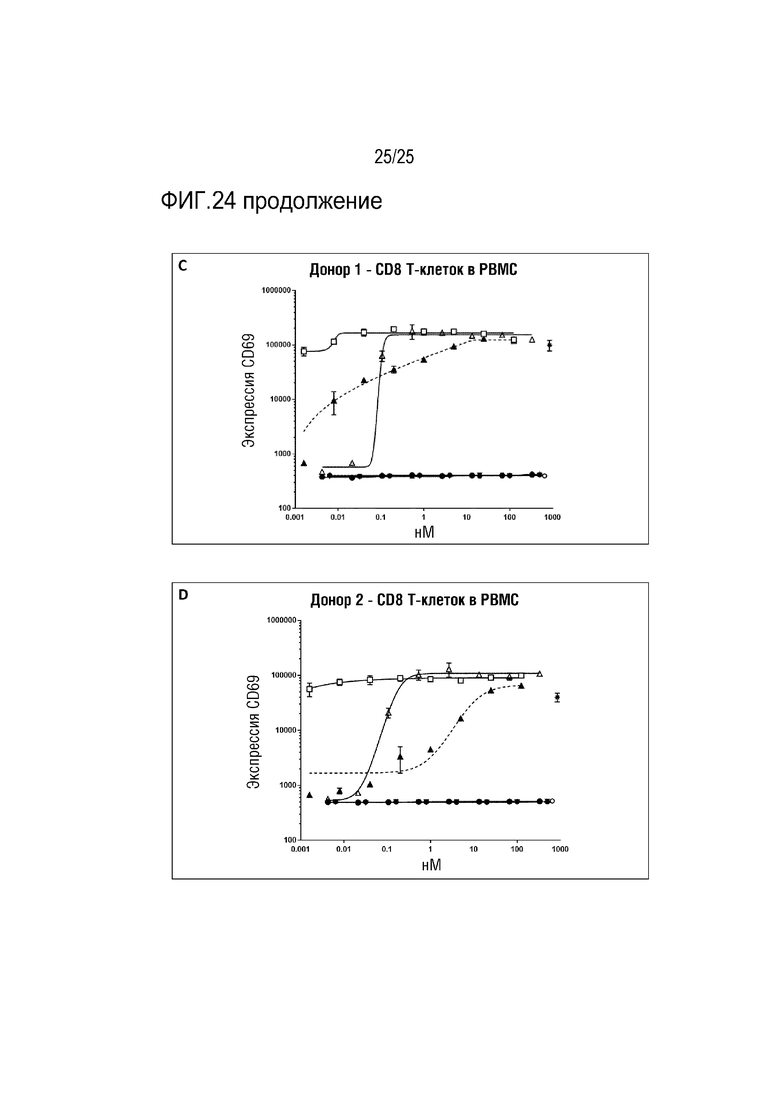
Изобретение относится к области биотехнологии. Предложена конструкция антитела для улучшения фармакокинетики полипептида-вставки, содержащая каркасное антитело, которое содержит вариабельный (V) домен и полипептид-вставку. Полипептид-вставка находится в участке каркасной области 3 V-домена. Также предложены нуклеиновые кислоты, векторы экспрессии, векторы клонирования и клетки-хозяева для получения указанной конструкции антитела. Изобретение обеспечивает получение нового формата сконструированного антитела, которой демонстрирует улучшенную фармакокинетику, например, увеличенное время полужизни в сыворотке. 12 н. и 20 з.п. ф-лы, 24 ил., 7 табл., 15 пр.
1. Конструкция антитела для улучшения фармакокинетики полипептида-вставки,
где конструкция антитела содержит каркасное антитело, которое содержит вариабельный (V) домен и полипептид-вставку, где полипептид-вставка находится в участке каркасной области 3 V-домена,
где V-домен представляет собой VH-домен и каркасное антитело также содержит VL-домен,
где каркасное антитело представляет собой полноразмерное антитело, Fab, Fab’, F(ab’)2, VHH или scFv, которое связывается с альбумином сыворотки и содержит альбумин-связывающий домен, и
где полипептид-вставка представляет собой полипептид или второе антитело, которое связывается с другим антигеном по сравнению с каркасным антителом, и где каркасное антитело обеспечивает улучшение время полураспада полипептида-вставки при наличии полипептида-вставки,
где конструкция антитела содержит VL-домен, содержащий CDR-L1 с последовательностью SEQ ID NO:3; CDR-L2 с последовательностью SEQ ID NO:4; CDR-L3 с последовательностью SEQ ID NO:5; и VH-домен, содержащий CDR-H1 с последовательностью SEQ ID NO:8; CDR-H2 с последовательностью SEQ ID NO:9; CDR-H3 с последовательностью SEQ ID NO:10.
2. Конструкция антитела по п.1, где полипептид-вставка находится между аминокислотными остатками 73 и 76 VH-домена.
3. Конструкция антитела по п.2, где полипептид-вставка находится в VH-домене:
(i) между аминокислотными остатками 73 и 74;
(ii) между аминокислотными остатками 74 и 75; или
(iii) между аминокислотными остатками 75 и 76.
4. Конструкция антитела по п.2 или 3, где один или несколько аминокислотных остатков между остатками 73 и 76 VH-домена заменены полипептидом-вставкой.
5. Конструкция антитела по любому из пп.1-4, дополнительно содержащая линкерную последовательность из по меньшей мере одной аминокислоты, соединяющую N- и/или C-концы полипептида-вставки с участком каркасной области 3.
6. Конструкция антитела по п.5, где линкерная последовательность представляет собой по меньшей мере две, три, четыре, пять, шесть, семь или восемь или более аминокислот на N-конце части, представляющей собой полипептид-вставку.
7. Конструкция антитела по п.5 или 6, где линкерная последовательность представляет собой по меньшей мере две, три, четыре, пять, шесть, семь или восемь, или более аминокислот на C-конце части, представляющей собой полипептид-вставку.
8. Конструкция антитела по любому из пп.1-7, которая является полноразмерным IgG.
9. Конструкция антитела по любому из пп.1-7, которая является dsscFv.
10. Конструкция антитела по любому из пп.1-7, которая является Fab.
11. Конструкция антитела по любому из предшествующих пунктов, которое является гуманизированным.
12. Конструкция антитела по любому из предшествующих пунктов, где альбумин сыворотки представляет собой альбумин сыворотки человека.
13. Конструкция антитела по любому из предшествующих пунктов, где каркасное антитело содержит VL-домен, содержащий SEQ ID NO:2 или SEQ ID NO:102.
14. Конструкция антитела по п.13, где каркасное антитело содержит легкую цепь, содержащую SEQ ID NO:1.
15. Конструкция антитела по любому из предыдущих пунктов, где каркасное антитело содержит VH-домен, содержащий SEQ ID NO:7 или SEQ ID NO:103.
16. Конструкция антитела по п.15, где каркасное антитело содержит тяжелую цепь, содержащую SEQ ID NO:6.
17. Конструкция антитела по любому из предшествующих пунктов, где полипептид-вставка представляет собой цитокин.
18. Конструкция антитела по п.17, где цитокин выбран из IL-10, IL-15, IL-2, G-CSF, GM-CSF и EPO.
19. Конструкция антитела по любому из пп.1-16, где полипептид-вставка представляет собой второе антитело.
20. Конструкция антитела по п.19, где второе антитело представляет собой scFv, в частности, dsscFv, или доменное антитело, такое как VH, или VL, или VHH.
21. Конструкция антитела по п.19 или 20, где второе антитело связывается с IL-17.
22. Выделенная молекула ДНК, кодирующая конструкцию антитела по любому из пп.1-21.
23. Пара молекул ДНК для получения конструкции антитела по любому из пп.1-21.
24. Клонирующий вектор для получения конструкции антитела по любому из пп.1-21.
25. Пара клонирующих векторов для получения конструкции антитела по любому из пп.1-21.
26. Вектор экспрессии, кодирующий конструкцию антитела по любому из пп.1-21.
27. Пара векторов экспрессии, кодирующая конструкцию антитела по любому из пп.1-21.
28. Клетка-хозяин для получения конструкции антитела по любому из пп.1-21, содержащая клонирующий вектор по п.24.
29. Клетка-хозяин для получения конструкции антитела по любому из пп.1-21, содержащая пару клонирующих векторов по п.25.
30. Клетка-хозяин для получения конструкции антитела по любому из пп.1-21, содержащая вектор экспрессии по п.26.
31. Клетка-хозяин для получения конструкции антитела по любому из пп.1-21, содержащая пару векторов экспрессии по п.27.
32. Способ получения конструкции антитела по любому из пп.1-21, включающий культивирование клетки-хозяина по любому из пп.28-31 и выделение антитела.
| DIMITRIS NIKOLOUDIS et al., A complete, multi-level conformational clustering of antibody complementarity-determining regions, PEERJ, 2014, vol.2, page e456 | |||
| SIMON T., RAJEWSKY K., A functional antibody mutant with an insertion in the framework region 3 loop of the VH domain: implications for antibody engineering, PROTEIN ENGINEERING, 1992, vol |

