Группа заявляемых изобретений относится к биотехнологии, в частности, к биосинтезу гемицеллюлаз, и представляет собой рекомбинантную секретируемую термостабильную β-глюканазу ThiTeEgh12, рекомбинантный штамм дрожжей Pichia pastoris, способный синтезировать рекомбинантную секретируемую термостабильную β-глюканазу ThiTeEgh12 и полученный путем трансформации штамма Pichia pastoris Х33 плазмидой pPIC-ThiTeEgh12, содержащей ген β-1,3-1,4-глюканазы ThiTeEgh12 Thielavia terrestris VKPM F-144, под контролем промотора АОХ1, а также способ микробирлогического синтеза рекомбинантной секретируемой термостабильной β-глюканазы ThiTeEgh12.
Зерновые культуры составляют основу кормовых смесей для сельскохозяйственных животных. Некрахмалистые полисахариды (НКП), входящие в состав зерна, являются причиной высокой вязкости кормовых смесей, а также кормовой массы в желудочно-кишечном тракте (ЖКТ) животных, значительно снижая усвояемость и перевариваемость кормов. При этом НКП не расщепляются в пищеварительной системе моногастричных животных, таких как свиньи и птицы, а также в пищеварительной системе человека, из-за дефицита соответствующих ферментов. Основными НКП зерна являются β-глюканы - полисахариды, состоящие из единиц целлотриозы и целлотетрозы, связанных β-(1,3)-гликозидными связями. β-Глюканы из различных видов злаков или разных частей растения значительно варьируют по экстрагируемости, молекулярному весу и вязкости (Rykov et al. 2019). Ферментативный гидролиз - эффективный способ снижения вязкости зерновых смесей. Ферменты, способные специфически гидролизовать β-1,4-гликозидные связи β-(1,3)-(1,4)-глюканов, называются β-(1,3)-(1,4)-глюканазами и относятся к группе с ферментативной активностью ЕС 3.2.1.73. Согласно данным базы CAZy ферменты с β-(1,3)-(1,4)-глюканазы найдены в семействах гликозид гидролаз 6, 7, 9, 12, 16, 17, 26, 51 (http://www.cazy.org). Ферменты, специфически гидролизующие β-(1,3)-(1,4)-глюкан, могут проявлять побочную активность на растворимой и нерастворимой целлюлозе, ксилоглюкане, лихенане и ламиранане.
β-Глюканазы входят в состав мультиферментных препаратов для гидролиза НКП в кормовых смесях с целью снижения их вязкости (например, мультиферментные комплексы и композиции Vilzim, Алтавим, Кемзайм, Фидбест VGPro G). Поскольку кормовые смеси на основе комбикормов перед использованием часто запаривают горячей водой, содержащиеся в них кормовые ферменты могут разрушаться. В этом случае кормовые ферменты из термофильных микроорганизмов имеют значительное преимущество перед их аналогами из мезофильных источников.
В источниках информации имеются сведения о грибах - природных продуцентах β-глюканаз: Aspergillus japonicus, Chrysosporium lucknowense, Trichoderma reesei (EP 2041294 A2; Markov et al., 2005; Qi et al., 2013), Aspergillus aculeatus (RU 2303057), Penicillium oxalicum (Zhu et al., 2019), Aspergillus oryzae (Matsuzawa et al., 2020), Malbranchea cinnamomea strain CBS 343.55, Thielavia australiensis strain ATCC 28236, и Paecilomyces byssochlamydoides strain NRRL 3658 (WO 2014138983 A1), Penicillium funiculosum (RU 2323254).
Важнейшим преимуществом рекомбинантных штаммов - продуцентов ферментов перед природными продуцентами является их повышенная продуктивность по целевому белку. В качестве реципиентов для гетерологичной экспрессии грибных β-глюканаз используют штаммы грибов и бактерий. Например, собственные β-глюканазы экспрессированы в Penicillium canescens (RU 2358756) и в Aspergillus niger (Hasper et al., 2002). Описаны также полученные методами генной инженерии штаммы дрожжей Pichia pastoris - продуценты рекомбинантной эндо-бета-1,4-D-глюканазы Aspergillus niger (Shumiao et al., 2010), PoCell2A, PoCell2B и PoCell2C Penicillium oxalicum (Zhu et al., 2019), эндо-бета-1,3(4)-D-глюканазы Paenibacillus sp. F-40 (Yang et al. 2007).
Системы гетерологичной экспрессии на основе бактерий или грибов имеют ряд недостатков. В частности, при внутриклеточном синтезе рекомбинантных ферментов в Е. coli часто образуются нерастворимые тельца включения. Кроме того, в бактериях невозможно воспроизвести все посттрансляционные модификации, характерные для эукариот, например, гликозилирование, фосфорилирование, образование дисульфидных связей между цистеиновыми остатками. Недостатком экспрессионных систем на основе грибов, относящихся к роду Aspergillus или роду Penicillium является отсутствие стандартизированного генетического инструментария, что осложняет проведение генетических манипуляций (Demain and Vaishnav, 2009).
Получение рекомбинантных белков с использованием в качестве штаммов-реципиентов метилотрофных дрожжей, относящихся к роду Pichia, роду Komagataella или роду Hansenula имеет ряд преимуществ по сравнению с биосинтезом в других микроорганизмах. Метилотрофные дрожжи классифицированы как GRAS ("generally recognized as safe") и считаются безопасными (дать ссылку на источник сведений о безопасности). Они непатогенны, нетоксичны, не образуют антибиотиков. В отличие от Е. coli, метилотрофные дрожжи способны осуществлять все посттрансляционные модификации, характерные для эукариотических клеток, включая секрецию рекомбинантных белков в культуральную среду. Низкий уровень секреции собственных белков существенно облегчает выделение и очистку целевого рекомбинантного продукта (Macauley-Patrick and Fazenda, 2005). Культура метилотрофных дрожжей достигает в ферментерах более высокой плотности, чем традиционно используемые дрожжи Saccharomyces cerevisiae, что позволяет получить более высокий уровень продукции целевого белка (Gellissen, 2000).
Thielavia terrestris (анаморф: Acremonium alabamense) - широко распространенный ацидофильный и термофильный гриб, способный расти при повышенных температурах, в том числе в геотермальных источниках, при температуре до 55°С. Т. terrestris является важным источником гликозид гидролаз для промышленной биотехнологии. В заявляемом изобретении ген термостабильной β-глюканазы семейства 12 гликозид гидролаз из штамма Т. terrestris VKPM F-144 интегрирован в хромосому штамма P. pastoris Х33, в результате чего получена секретируемая термостабильная β-глюканаза ThiTeEgh12 (SEQ ID N2), сконструирован рекомбинантный штамм дрожжей P. pastoris ВКПМ Y-4850 - продуцент заявляемой секретируемой термостабильной β-глюканазы и разработан способ микробиологического синтеза термостабильной β-глюканазы на основе этого штамма.
Известным аналогом заявляемой β-глюканазы яввляется эндо-(1,4)-β-глюканаза Bgh12A семейства GH12 (Rykov et al., 2019; GenBank: AYM46706.1). Температурный оптимум рекомбинантной β-глюканазы Bgh12A, составляет 55°С, рН 5,0; время полуинактивации фермента при 50°С составляет 31 мин (Rykov et al., 2019).
Известным аналогом заявляемого штамма является штамм дрожжей Pichia pastoris, содержащий на плазмиде pPICZaA ген bgh12A Aspergillus cervinus под контролем промотора АОХ1 и экспрессирующий эндо-(1,4)-β-глюканазу Bgh12A семейства GH12 (Rykov et al., 2019; GenBank: AYM46706.1).
Известным аналогом заявляемого способа является способ микробиологического синтеза секретируемой ксилоглюкан-специфической эндо-(1,4)-β-глюканазы семейства 12 гликозид гидролаз из гриба Aspergillus cervinus рекомбинантным штаммом дрожжей Pichia pastoris ВКПМ Y-4299 (RU 2639248). Уровень синтеза ксилоглюкан-специфической эндо-(1,4)-β-глюканазы AsCeGH12b таким способом, составляет не менее 30 ед/ мл культуральной жидкости.
Задачи заявляемой группы изобретений состоят в расширении арсенала рекомбинантных β-глюканаз, штаммов-продуцентов и способов микробиологического синтеза рекомбинантных β-глюканаз.
Задачи решают путем:
- получения рекомбинантной секретируемой термостабильной β-глюканазы ThiTeEgh12, имеющей последовательность SEQ ID NO 2;
- конструирования рекомбинантного штамма дрожжей Pichia pastoris ВКПМ Y-4850 - продуцента заявляемой β-глюканазы.
- разработки способа микробиологического синтеза заявляемой β-глюканазы путем культивирования заявляемого штамма дрожжей в аэробных условиях в подходящей питательной среде с периодическим добавлением в качестве индуктора- метанола до максимального накопления целевого продукта.
Процесс конструирования заявляемого штамма и разработки заявляемого способа микробиологического синтеза заявляемой β-глюканазы на основе этого штамма состоит из следующих этапов:
- конструирования интегративной плазмидной ДНК (плазмиды) pPIC-ThiTeEgh12, содержащей последовательность SEQ ID NO 1, включающую кодирующую часть гена (исключая интроны и сигнальную последовательность) β-глюканазы семейства 12, из Thielavia terrestris VKPM F-144);
- конструирования рекомбинантного штамма дрожжей Pichia pastoris ВКПМ Y-4850, способного синтезировать β-глюканазу ThiTeEgh12, имеющую последовательность SEQ ID NO 2;
- разработки способа микробиологического синтеза β-глюканазы ThiTeEgh12 на основе сконструированного рекомбинантного штамма дрожжей Pichia pastoris ВКПМ Y-4850.
Этап 1. Конструирование интегративной плазмидной ДНК (плазмиды) pPIC-ThiTeEgh12, содержащей последовательность SEQ ID NO 1
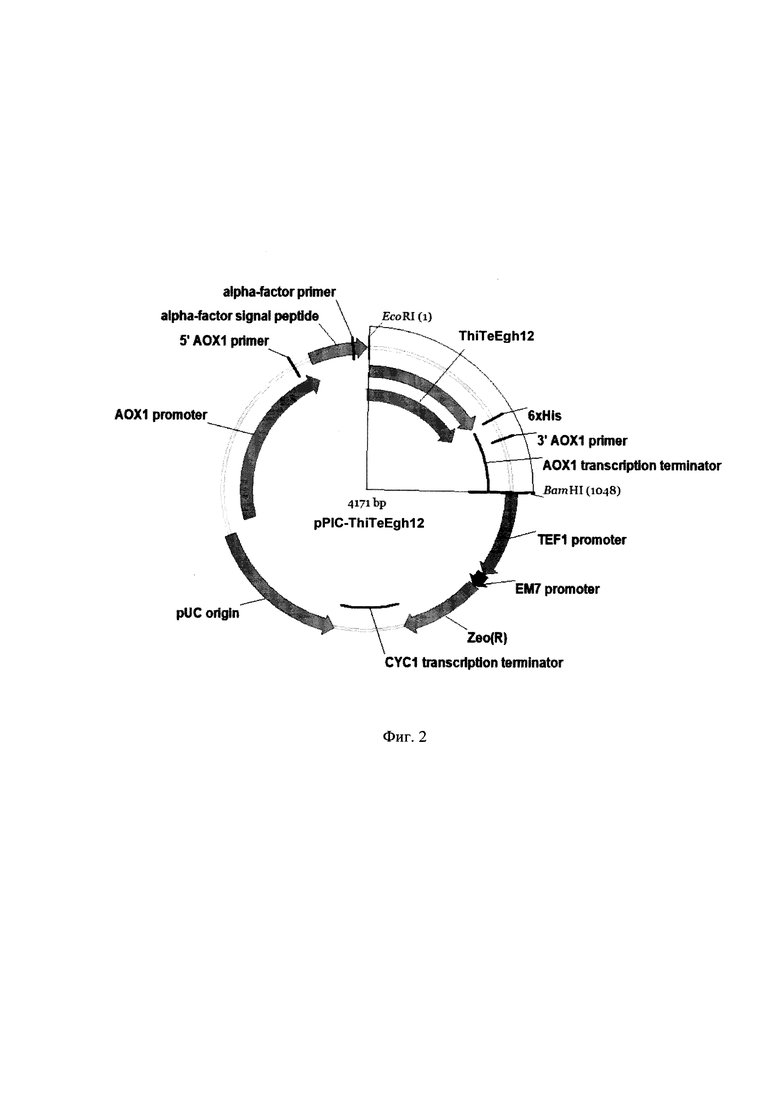
Анализ транскриптома Т. terrestris (синоним Thermothielavioides terrestris, штамм NRRL 8126) выявил наличие последовательности THITE_2117762 (NCBI Ref. Seq. ХМ_003654584), гомологичной генам гликозид гидролаз из семейства 12. На основе последовательности гена THITE_2117762 конструируют праймеры для амплификации кодирующей (не содержащей интроны и сигнальную последовательность) части гена β-глюканазы семейства 12 из Т. terrestris VKPM F-144. В состав прямого и обратного праймеров вводят сайты рестриктаз EcoRI и XhoI, соответственно, для клонирования в вектор pPICZaA (Thermo Fisher Scientific, США). Интегративный вектор pPICZaA размером 3593 пары оснований предназначен для индуцированной экспрессии рекомбинантных генов в Pichia pastoris и секреции рекомбинантных белков во внеклеточную среду. Этот вектор содержит репликон pUC, промотор АОХ1, последовательность, кодирующую альфа-фактор дрожжей, сайт множественного клонирования, последовательность, кодирующую с-myc эпитоп, последовательность, кодирующую 6xHis-tag, терминатор транскрипции АОХ1, ген Zeo, обеспечивающий устойчивость штаммов Е. coli и P. pastoris к зеоцину под контролем промоторов TEF1 и ЕМ7, терминатор транскрипции CYC1 (Фиг. 1).
Амплификацию кодирующей части гена β-глюканазы семейства 12 из Т. terrestris VKPM F-144 (без интронов и сигнальной последовательности) проводят методом ПЦР на матрице кДНК штамма Т. terrestris VKPM F-144. Препарат кДНК получают синтезом одно- и двухцепочечных ДНК на матрице тотальной РНК штамма Т. terrestris VKPM F-144 методом обратной транскрипции. Полученный амплификацией фрагмент ДНК размером 678 пар нклеотидов содержит кодирующую часть гена β-глюканазы семейства 12 из штамма Т. terrestris VKPM F-144.
Плазмиду pPIC-ThiTeEgh12 конструируют путем клонирования полученного амплификацией фрагмента ДНК, содержащего кодирующую часть гена β-глюканазы семейства 12 из T. terrestris VKPM F-144, в вектор pPICZaA. Плазмида pPIC-ThiTeEgh12 размером 4171 пар нуклеотидов (SEQ ID NO 3), наряду с генами вектора pPICZaA, содержит кодирующую область гена β-глюканазы семейства 12 из T. terrestris VKPM F-144. С 5'-конца гена β-глюканазы в той же рамке считывания расположена последовательность, кодирующая α-фактор дрожжей под контролем промотора АОХ1, а с 3'-конца гена β-глюканазы в той же рамке считывания расположена последовательность, кодирующая гистидиновый хвост (Фиг. 2). Таким образом, плазмида pPIC-ThiTeEgh12 содержит открытую рамку считывания, кодирующую заявляемую рекомбинантную β-глюканазу, обозначенную ThiTeEgh12, которая содержит на N-конце сигнальный пептид, обеспечивающий секрецию в Pichia pastoris, а на С-конце - гистидиновый хвост, предназначенный для очистки заявляемого фермента β-глюканазы (SEQ ID NO 2).
Этап 2. Конструирование рекомбинантного штамма дрожжей Pichia pastoris, способного синтезировать заявляемую β-глюканазу
В качестве штамма-реципиента используют штамм Pichia pastoris Х33 (mut+), компетентные клетки которого трансформируют линеаризованной плазмидой pPIC-ThiTeEgh12. В результате получают рекомбинантный штамм Pichia pastoris, содержащий поседовательность SEQ ID NO 1 и способный синтезировать заявляемую β-глюканазу ThiTeEgh12. Сконструированный рекомбинантный штамм Pichia pastoris депонирован во Всероссийской коллекции промышленных микроорганизмов как Pichia pastoris ВКПМ Y-4850.
Заявляемый штамм Pichia pastoris ВКПМ Y-4850 имеет следующие морфологические и физиолого-биохимические характеристики:
Морфологические признаки: при культивировании при температуре 28°С в течение 48 часов на агаризованной среде YPD следующего состава (мас. %: пептон - 2, дрожжевой экстракт - 1, глюкоза - 2, агар - 2, вода - остальное) формируются колонии светло-бежевого цвета с ровным краем, матовой поверхностью, линзовидным профилем и пастообразной консистенцией. При росте в жидкой среде YPD при 28°С в течение 24 ч культивирования - жидкость мутная, осадок белый, коагуляции не наблюдается, пристеночных пленок культура не образует. Клетки имеют овальную форму, 3-4 мкм в диаметре; почкуются; почкование истинное, многостороннее; истинного мицелия не образуют.
Физиолого-биохимические признаки: штамм является факультативным анаэробом с температурой роста - 20-33°С (оптимум - 28°С) и рН среды культивирования - 4,8-7,4 (оптимум - 6,0). В качестве источников углерода штамм может использовать глюкозу, глицерин, метанол, олеат, сорбитол, рамнозу. Не утилизирует галактозу, ксилозу, арабинозу. В качестве источников азота штамм может использовать аминокислоты, сернокислый аммоний, азотнокислый аммоний. Штамм Pichia pastoris ВКПМ Y-4850 синтезирует рекомбинантную секретируемую термостабильную β-глюканазу ThiTeEgh12.
Способ микробиологического синтеза заявляемой β-глюканазы в общем виде
Посевной материал, представляющий собой клетки заявляемого рекомбинантного штамма Pichia pastoris ВКПМ Y-4850, подготавливают путем инкубации в течение 16-18 часов при температуре 29°С на среде BMGY (мас. %: пептон - 2, дрожжевой экстракт - 1, глицерин - 1, фосфатный буфер рН 6.6 - до 100 мМ, вода - остальное) при постоянной аэрации на термостатируемой качалке (200 об/мин). Затем выросшую культуру переносят в среду BMMY (мас. %: пептон - 2, дрожжевой экстракт - 1, метанол - 0,5, фосфатный буфер рН 6.6 - до 100 мМ, вода - остальное) и продолжают культивировать до достижения уровня оптической плотности ОД600=1.
Процесс биосинтеза ведут к колбах Эрленмейера, содержащих 100 мл среды BMMY, в течение 96 часов в ротационном шейкере-термостате (200-250 об/мин) при температуре 28°С. Каждые 24 часа проводят индукцию метанолом путем асептического добавления 50% раствора метанола в пробирки до конечной концентрации 0,5%. По истечении 96 часов биомассу отделяют центрифугированием. Наличие рекомбинантной β-глюканазы в культуральной среде определяют при помощи электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия. Количественную оценку уровня активности рекомбинантной термостабильной β-глюканазы осуществляют при помощи ДНС-метода (Miller, 1959).
Уровень синтеза заявляемой β-глюканазы ThiTeEgh12 при получении заявляемым способом составляет не менее 230 ед/мл.
Изобретение проиллюстрировано следующими фигурами графических изображений.
Фигура 1. Интегративный плазмидный вектор pPICZaA
Фигура 2. Рекомбинантная плазмида pPIC- ThiTeEgh12
Пример 1. Конструирование интегративной плазмидной ДНК (плазмиды) pPIC-ThiTeEgh12, содержащей последовательность SEQ ID NO 1
Все стандартные генно-инженерные и микробиологические манипуляции проводят по известным методикам (Green and Sambrook, 2012).
Праймеры для амплификации кодирующей части гена β-глюканазы семейства 12 из Т. terrestris VKPM F-144 конструируют на основе последовательности м-РНК THITE_2117762 (NCBI Ref. Seq. ХМ_003654584) из транскриптома Thermothielavioides terrestris NRRL 8126, гомологичной генам гликозид гидролаз семейства 12. Наличие сигнального пептида определяют с помощью сервиса SignalP 4.1 (http://www.cbs.dtu.dk/services/SignalP/). Прямой праймер ThiTeEgh12_F 5'-GTAGAATTCGAACCCAGGCAGCAGGCAAC-3' и обратный праймер - ThiTeEgh12_R ATACTCGAGCGAAACGTTGGCCGAGAACT содержат, соответственно, сайты рестриктаз EcoIII и XhoI и предназначены для клонирования в интегративный вектор pPICZaA. Праймеры для амплификации конструируют таким образом, чтобы кодирующая часть гена β-глюканазы семейства 12 из Т. terrestris VKPM F-144 после клонирования в вектор pPICZaA находилась в одной рамке считывания с последовательностями, кодирующими α-фактор дрожжей под контролем промотора АОХ1 и гистидиновый хвост и расположенными со стороны 5'-конца и 3'-конца гена β-глюканазы, соответственно. С использованием этих праймеров, кодирующую часть гена β-глюканазы семейства 12 из Т. terrestris VKPM F-144 амплифицируют методом ПЦР на матрице кДНК штамма Thielavia terrestris ВКПМ F-144 Амплификацию проводят с помощью полимеразы Phusion (Thermo Fisher Scientific, США) по протоколам производителя. кДНК получают путем синтеза одно- и двухцепочечной ДНК методом обратной транскрипции с матрицы тотальной РНК. Выделение тотальной РНК производят с помощью реагента ExtractRNA (Евроген, Россия). Очистку полученного амплификацией фрагмента ДНК, содержащего кодирующую часть гена β-глюканазы семейства 12 из Т. terrestris VKPM F-144, проводят с помощью гель-электрофореза в 1,8% агаровом геле. Последовательность амплифицированного фрагмента определяют путем секвенирования по методу Сэнгера на генетическом анализаторе ABI 3500 (Thermo Fisher Scientific, США).
Кодирующая часть гена β-глюканазы семейства 12 из Т. terrestris VKPM F-144 имеет длину 678 пар нуклеотидов (SEQ ID NO1).
Плазмиду pPIC-ThiTeEgh12 конструируют путем клонирования амплифицированного фрагмента, содержащего кодирующую часть гена β-глюканазы семейства 12 из T. terrestris VKPM F-144, в вектор pPICZaA. Гидролиз плазмиды pPICZaA и амплифицированного фрагмента ДНК проводят рестриктазами EcoRI/SalI и EcoRI/XhoI соответственно (Thermo Fisher Scientific, США). Очистку полученных фрагментов ДНК проводят с помощью гель-электрофореза в 1,8% агарозном геле с последующим выделением нужной полосы с помощью набора для выделения ДНК из агарозных гелей (Биосилика, Россия). Лигирование проводят в реакционной смеси объемом 20 мкл, содержащей 20 нг ДНК вектора, 20 нг ДНК фрагмента и 5 ед. Т4 ДНК лигазы (СибЭнзим, Россия), согласно методике производителя. Полученной лигазной смесью в количестве 5 мкл трансформируют компетентные клетки штамма Escherichia coli XL1 blue MRF' (Agilent Technologies, США) (генотип - Δ(mcrA)183 Δ(mcrCB-hsdSMR-mrr)173 endA1 supE44 thi-1 recA1 gyrA96 relA1 lac [F' proAB lacIqZAM15 Tn10 (Tetr)]) и высевают на чашки с агаризованной низкосолевой средой LB (мас. %: триптон - 1, дрожжевой экстракт - 0,5, хлорид натрия - 0,5, агар - 2, вода - остальное), содержащей антибиотик зеоцин в концентрации 25 мкг/см. Плазмидную ДНК полученных трансформантов анализируют путем гидролиза рестриктазами BamHI и HindIII. В результате отбирают клоны, содержащие BamHI/HindIII фрагменты расчетного размера. Правильность клонирования подтверждают секвенированием вставки методом Сэнгера на генетическом анализаторе ABI 3500 (Life technology, США). В результате получают рекомбинантную плазмиду pPIC-ThiTeEgh12 (SEQ ID NO 3) размером 4171 пару оснований, включающую ген, кодирующий термостабильную β-глюканазу ThiTeEgh12 (Фиг. 1).
Последовательность рекомбинантного белка ThiTeEgh12 (SEQ ID NO 2), состоит из 236 аминокислотных остатков, и отличается от природного белка двумя дополнительными аминокислотами с N-конца и 6 аминокислотами, включающими полигистидиновый хвост с С-конца. Расчетная масса рекомбинантного белка ThiTeEgh12 - 25,7 кДа.
Пример 2. Конструирование штамма Pichia pastoris ВКПМ Y-4850 - продуцента заявляемой β-глюканазы
С целью конструирования заявляемого рекомбинантного штамма Pichia pastoris ВКПМ Y-4850 - продуцента β-глюканазы ThiTeEgh12, клетки штамма Pichia pastoris Х33 (mut+) (Invitrogen) трансформируют плазмидой pPIC-ThiTeEgh12. Трансформацию проводят 5 мг линеаризованной плазмидной ДНК, для получения которой исходную плазмиду обрабатывают эндонуклеазой рестрикции MssI.
Культуру дрожжей штамма Pichia pastoris Х33 (mut+) выращивают на среде YPD в аэробных условиях (200-250 об/мин) при 29°С в течение ночи. В 500 мл свежей среды YPD вносят ночную культуру в соотношении 1:1000 и выращивают в течение ночи до ОД600=1,3-1,5, после чего клетки отделяют центрифугированием при 1500 об/мин 5 минут при 4°С. Затем клеточный осадок дважды промывают стерильной дистиллированной водой (в объеме 500 мл и 250 мл, соответственно) и один раз 1 М сорбитолом (в объеме 25 мл), после каждой промывки осаждая суспензию клеток при 1500 об/мин 5 минут при 4°С. Далее осадок клеток ресуспендируют в 1 мл 1 М сорбитола, а полученную суспензию разделяют на аликвоты по 80 мкл. Электропорацию проводят на приборе GenePulser Xcell (Biorad) в кюветах с зазором 2 мм при 2000 В, 25 мкФ, 200 Ом. После электропорации быстро добавляют 1 мл холодного раствора 1 М сорбитола, инкубируют 1 час при температуре 30°С и высевают на твердую селективную среду для отбора трансформантов. Отбор трансформантов проводят на среде YPDS (мас. %: пептон - 2, дрожжевой экстракт -1, глюкоза - 2, сорбитол - до 1 М, агар - 2, вода - остальное), содержащей зеоцин в концентрации 100 мкг/см3.
Клетки инкубируют при 29°С в течение 3 суток. Полученные трансформанты выявляют методом ПЦР-скрининга с помощью следующих праймеров: a-Factor sequencing primer - TACTATTGCCAGCATTGCTGC; 3 AOX1 sequencing primer - GCAAATGGCATTCTGACATCC. В результате получают заявляемый штамм Pichia pastoris ВКПМ Y-4850, несущий ген, кодирующий заявляемую β-глюканазу ThiTeEgh12.
Пример 3. Получение заявляемой β-глюканазы ThiTeEgh12
Посевной материал, представляющий собой клетки заявляемого штамма Pichia pastoris ВКПМ Y-4850 - продуцента β-глюканазы ThiTeEgh12 подготавливают путем инкубации в течение 18 часов при температуре 29°С на среде BMGY при постоянной аэрации на термостатируемой качалке (250 об/мин). Затем выросшую культуру переносят в среду BMMY до ОД600=1.
Процесс биосинтеза ведут к колбах Эрленмейера, содержащих 100 мл среды BMMY, в течение 96 часов в ротационном шейкере-термостате (250 об/мин), при температуре 28°С. Каждые 24 часа проводят индукцию метанолом, путем асептического добавления 50% раствора метанола в пробирки, до конечной концентрации 0,5%. По истечении 96 часов биомассу отделяют центрифугированием. Наличие β-глюканазы в культуральной среде определяют при помощи электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия с последующей зимограммой (Green and Sambrook, 2012). Количественную оценку уровня активности β-глюканазы осуществляют при помощи ДНС-метода (Miller, 1959).
Рекомбинантная секретируемая β-глюканаза ThiTeEgh12, продуцируемая штаммом Pichia pastoris ВКПМ Y-4850, является термостабильной: при инкубации препарата фермента в течении 2 часов при 55, 60 и 65°С, рН 4.6, фермент при тестировании на β-глюкане сохраняет 100%, 80% и 60% ферментативной активности, соответственно,.
Уровень идентичности белковых последовательностей заявляемой β-глюканазы ThiTeEgh12 и аналога β-глюканазы Bghl2A из A. cervinus составляет 52%.
Рекомбинантная секретируемая термостабильная β-глюканаза ThiTeEgh12, продуцируемая заявляемым штаммом Pichia pastoris ВКПМ Y-4850, проявляет максимальную активность при 70°С, рН 4.6, в то время как аналог внеклеточная рекомбинантная β-глюканаза Bgh12A, имеет оптимум активности при гораздо более низкой температуре (55°С, рН 5,0). По сравнению с β-глюканазой ThiTeEgh12, β-глюканаза Bgh12A является менее термостабильной - время полуинактивации фермента при 50°С составляет 31 мин (Rykov et al., 2019). Помимо активности на β-глюкане, заявляемая β-глюканаза проявляет активность на лихенане, ксилоглюкане и карбоксиметилцеллюлозе, то есть обладает более широким спектром субстратной специфичности, чем аналог β-глюканаза Bgh12A.
Уровень синтеза β-глюканазы ThiTeEgh12 заявляемым способом составляет не менее 230 ед/мл, что значительно превышает уровень синтеза рекомбинантной ксилоглюкан-специфической эндо-(1,4)-β-глюканазы AsCeGH12b аналогичным способом (30 ед/мл).
Таким образом,
- получена рекомбинантная секретируемая термостабильная β-глюканаза ThiTeEgh12, которая отличается от аналога такими свойствами как более высокие температурный максимум активности и термостабильность;
- сконструирован рекомбинантный штамм дрожжей Pichia pastoris ВКПМ Y-4850 - продуцент заявляемой β-глюканазы ThiTeEgh12;
- разработан способ микробиологического синтеза заявляемой β-глюканазы ThiTeEgh12, позволяющий получать β-глюканазу в количестве не менее 230 ед/мл, что в 7.7 раза превосходит уровень биосинтеза β-глюканазы AsCeGH12b, полученной аналогичным способом.
Источники информации
1. Demain AL, Vaishnav P. // Production of recombinant proteins by microbes and higher organisms // Biotechnol Adv., 2009.
2. Gellissen G (2000) Heterologous protein production in methylotrophic yeasts. Appl. Microbiol. Biotechnol. 54(6): 741-750.
3. Green M.R., Sambrook J. (2012) Molecular Cloning: a Laboratory Manual. (Fourth Edition) Cold Spring Harbor Laboratory Press.
4. Hasper A.A., Dekkers E., van Mil M., van de Vondervoort P.J., de Graaff L.H. // EglC, a new endoglucanase from Aspergillus niger with major activity towards xyloglucan. // Appl Environ Microbiol., 2002
5. Macauley-Patrick S, Fazenda ML (2005) Heterologous protein production using the Pichia pastoris expression system. Yeast 22: 249-270.
6. Markov A.V., Gusakov A.V., Kondratyeva E.G., Okunev O.N., Bekkarevich A.O., Sinitsyn A.P. // New effective method for analysis of the component composition of enzyme complexes from Trichoderma reesei. // Biochemistry (Mosc)., 2005
7. Miller, G.L. // Use of dinitrosalicylic acid reagent for determination of reducing sugar // Analytical Chemistry. 1959.
8. Rykov SV, Kornberger P, Herlet J, Tsurin NV, Zorov IN, Zverlov VV, Liebl W, Schwarz WH, Yarotsky SV, Berezina OV (2019) Novel endo-(1,4)-β-glucanase Bgh12A and xyloglucanase Xgh12B from Aspergillus cervinus belong to GH12 subgroup I and II, respectively. Appl Microbiol Biotechnol 103, 7553-7566. https://doi.org/10.1007/s00253-019-10006-x
9. Qi H., Bai F., Liu A.// Purification and characteristics of xyloglucanase and five other cellulolytic enzymes from Trichoderma reesei QM9414.// Biochemistry (Mosc), 2013
10. Shumiao Z., Huang J., Zhang C, Deng L, Hu N, Liang Y. // High-level expression of an Aspergillus niger endo-beta-1,4-glucanase in Pichia pastoris through gene codon optimization and synthesis // J. Microbiol. Biotechnol.
11. Yang P., Shi P., Wang Y., Bai Y., Meng K., Luo H., Yuan Т., Yao В // Cloning and overexpression of a Paenibacillus beta-glucanase in Pichia pastoris: purification and characterization of the recombinant enzyme // J Microbiol Biotechnol, 2007
12. Zhu, Z., Qu, J., Yu, L. et al. // Three glycoside hydrolase family 12 enzymes display diversity in substrate specificities and synergistic action between each other. // Mol Biol Rep, 2019.
13. EP 2041294 A2 // Construction of highly efficient cellulase compositions for enzymatic hydrolysis of cellulose // Priority to US11/487,547 2006-07-13
14. RU 2303057 // Штамм мицелиального гриба Aspergillus aculeatus - продуцент комплекса карбогидраз, содержащего ксиланазы, бета-глюканазы, пектиназы и ксилоглюканазы // Дата приоритета: 14.11.2005
15. RU 2323254 // Штамм мицелиального гриба Penicillium funiculosum - продуцент комплекса карбогидраз, содержащего целлюлазы, глюканазы, глюкозидазы, ксиланазы и ксилоглюканазы, и способ получения ферментного препарата комплекса карбогидраз для осахаривания лигноцеллюлозных материалов // Дата приоритета: 04.04.2006.
16. RU 2358756 // Способ получения ферментного препарата для расщепления гемицеллюлозных гетерополисахаридов клеточной стенки растений и ферментный препарат (варианты) // Дата приоритета: 26.11.2007
17. RU 2639248 С1 // Рекомбинантный штамм дрожжей Pichia pastoris - продуцент секретируемой ксилоглюканазы из гриба Aspergillus cervinus и способ микробиологического синтеза ксилоглюканазы на основе этого штамма // Дата приоритета: 15.12.2016
18. WO 2014138983 А1 // Novel cell wall deconstruction enzymes of Malbranchea cinnamomea, Thielavia australiensis, and Paecilomyces byssochlamydoides, and uses thereof // Priority date: US201361783222P 20130314; US201361783313P 20130314; US201361783485P 20130314.
--->
Перечень последовательностей
<110> НИЦ "Курчатовский институт"- ГосНИИгенетика
<120> Рекомбинантная секретируемая термостабильная β-глюканаза, рекомбинантный
штамм дрожжей Pichia pastoris – продуцент этой β-глюканазы,
способ микробиологического синтеза β-глюканазы на основе этого штамма
<160> 3
<210> 1
<211> 678
<212> DNA
<213> Thielavia terrestris
<400> 1
gaacccaggc agcaggcaac actctgcgac cagtatggct actggtccgg caatggttac 60
gaggttaaca acaacctttg gggcgagagc gcagccacct caggctcgca gtgcacgtac 120
gtggacggca gctcttctgg cggcgtccaa tggcacacga cgtggacgtg gaacggcgga 180
gacaacaacg tgaagagctt cgcctactcg ggcaggcaga tcaccaaggg ccagaagatc 240
tcctccatca gcagcataca gacgtcggtc tcgtggtcgt acagcaacac caacatccgc 300
gctgacgtcg cgtacgacat cttcaccgcc gcggacccca accactccac cagcagcggc 360
gactacgagc tcatgatctg gctcgccaag tacggcagca tctccccgat cggctcgtcg......420
gtcggcacgg tcaacgtggg cggccgcagc tgggacctgt gggtgggcta caacggcgcc......480
atgaaggtgt tcagcttcgt cgcgccgagc cccgtcacca gcttcagcgc caacgtcaag......540
gacttcttca actacctgca gaacaatcag ggcttccccg ccagcagcca gaatctgctt......600
actttccaaa tcggcacgga acccttcacg ggcggaccgg ccaccttcac ggtgtcgcag......660
ttctcggcca acgtttcg 678
<210> 2
<211> 236
<212> PRT
<213> Thielavia terrestris
<400> 2
Glu Phe Glu Pro Arg Gln Gln Ala Thr Leu Cys Asp Gln Tyr Gly Tyr
1 5 10 15
Trp Ser Gly Asn Gly Tyr Glu Val Asn Asn Asn Leu Trp Gly Glu Ser
20 25 30
Ala Ala Thr Ser Gly Ser Gln Cys Thr Tyr Val Asp Gly Ser Ser Ser
35 40 45
Gly Gly Val Gln Trp His Thr Thr Trp Thr Trp Asn Gly Gly Asp Asn
50 55 60
Asn Val Lys Ser Phe Ala Tyr Ser Gly Arg Gln Ile Thr Lys Gly Gln
65 70 75 80
Lys Ile Ser Ser Ile Ser Ser Ile Gln Thr Ser Val Ser Trp Ser Tyr
85 90 95
Ser Asn Thr Asn Ile Arg Ala Asp Val Ala Tyr Asp Ile Phe Thr Ala
100 105 110
Ala Asp Pro Asn His Ser Thr Ser Ser Gly Asp Tyr Glu Leu Met Ile
115 120 125
Trp Leu Ala Lys Tyr Gly Ser Ile Ser Pro Ile Gly Ser Ser Val Gly
130 135 140
Thr Val Asn Val Gly Gly Arg Ser Trp Asp Leu Trp Val Gly Tyr Asn
145 150 155 160
Gly Ala Met Lys Val Phe Ser Phe Val Ala Pro Ser Pro Val Thr Ser
165 170 175
Phe Ser Ala Asn Val Lys Asp Phe Phe Asn Tyr Leu Gln Asn Asn Gln
180 185 190
Gly Phe Pro Ala Ser Ser Gln Asn Leu Leu Thr Phe Gln Ile Gly Thr
195 200 205
Glu Pro Phe Thr Gly Gly Pro Ala Thr Phe Thr Val Ser Gln Phe Ser
210 215 220
Ala Asn Val Ser Leu Asp His His His His His His
225 230 235
<210> 3
<211> 4171
<212> DNA
<213> Artificial sequence
<400> 3
aattcgaacc caggcagcag gcaacactct gcgaccagta tggctactgg tccggcaatg 60
gttacgaggt taacaacaac ctttggggcg agagcgcagc cacctcaggc tcgcagtgca 120
cgtacgtgga cggcagctct tctggcggcg tccaatggca cacgacgtgg acgtggaacg 180
gcggagacaa caacgtgaag agcttcgcct actcgggcag gcagatcacc aagggccaga 240
agatctcctc catcagcagc atacagacgt cggtctcgtg gtcgtacagc aacaccaaca 300
tccgcgctga cgtcgcgtac gacatcttca ccgccgcgga ccccaaccac tccaccagca 360
gcggcgacta cgagctcatg atctggctcg ccaagtacgg cagcatctcc ccgatcggct 420
cgtcggtcgg cacggtcaac gtgggcggcc gcagctggga cctgtgggtg ggctacaacg 480
gcgccatgaa ggtgttcagc ttcgtcgcgc cgagccccgt caccagcttc agcgccaacg 540
tcaaggactt cttcaactac ctgcagaaca atcagggctt ccccgccagc agccagaatc 600
tgcttacttt ccaaatcggc acggaaccct tcacgggcgg accggccacc ttcacggtgt 660
cgcagttctc ggccaacgtt tcgctcgacc atcatcatca tcatcattga gtttgtagcc 720
ttagacatga ctgttcctca gttcaagttg ggcacttacg agaagaccgg tcttgctaga 780
ttctaatcaa gaggatgtca gaatgccatt tgcctgagag atgcaggctt catttttgat 840
acttttttat ttgtaaccta tatagtatag gatttttttt gtcattttgt ttcttctcgt 900
acgagcttgc tcctgatcag cctatctcgc agctgatgaa tatcttgtgg taggggtttg 960
ggaaaatcat tcgagtttga tgtttttctt ggtatttccc actcctcttc agagtacaga 1020
agattaagtg agaccttcgt ttgtgcggat cccccacaca ccatagcttc aaaatgtttc 1080
tactcctttt ttactcttcc agattttctc ggactccgcg catcgccgta ccacttcaaa 1140
acacccaagc acagcatact aaattttccc tctttcttcc tctagggtgt cgttaattac 1200
ccgtactaaa ggtttggaaa agaaaaaaga gaccgcctcg tttctttttc ttcgtcgaaa 1260
aaggcaataa aaatttttat cacgtttctt tttcttgaaa tttttttttt tagttttttt 1320
ctctttcagt gacctccatt gatatttaag ttaataaacg gtcttcaatt tctcaagttt 1380
cagtttcatt tttcttgttc tattacaact ttttttactt cttgttcatt agaaagaaag 1440
catagcaatc taatctaagg ggcggtgttg acaattaatc atcggcatag tatatcggca 1500
tagtataata cgacaaggtg aggaactaaa ccatggccaa gttgaccagt gccgttccgg 1560
tgctcaccgc gcgcgacgtc gccggagcgg tcgagttctg gaccgaccgg ctcgggttct 1620
cccgggactt cgtggaggac gacttcgccg gtgtggtccg ggacgacgtg accctgttca 1680
tcagcgcggt ccaggaccag gtggtgccgg acaacaccct ggcctgggtg tgggtgcgcg 1740
gcctggacga gctgtacgcc gagtggtcgg aggtcgtgtc cacgaacttc cgggacgcct 1800
ccgggccggc catgaccgag atcggcgagc agccgtgggg gcgggagttc gccctgcgcg 1860
acccggccgg caactgcgtg cacttcgtgg ccgaggagca ggactgacac gtccgacggc 1920
ggcccacggg tcccaggcct cggagatccg tccccctttt cctttgtcga tatcatgtaa 1980
ttagttatgt cacgcttaca ttcacgccct ccccccacat ccgctctaac cgaaaaggaa 2040
ggagttagac aacctgaagt ctaggtccct atttattttt ttatagttat gttagtatta 2100
agaacgttat ttatatttca aatttttctt ttttttctgt acagacgcgt gtacgcatgt 2160
aacattatac tgaaaacctt gcttgagaag gttttgggac gctcgaaggc tttaatttgc 2220
aagctggaga ccaacatgtg agcaaaaggc cagcaaaagg ccaggaaccg taaaaaggcc 2280
gcgttgctgg cgtttttcca taggctccgc ccccctgacg agcatcacaa aaatcgacgc 2340
tcaagtcaga ggtggcgaaa cccgacagga ctataaagat accaggcgtt tccccctgga 2400
agctccctcg tgcgctctcc tgttccgacc ctgccgctta ccggatacct gtccgccttt 2460
ctcccttcgg gaagcgtggc gctttctcaa tgctcacgct gtaggtatct cagttcggtg 2520
taggtcgttc gctccaagct gggctgtgtg cacgaacccc ccgttcagcc cgaccgctgc 2580
gccttatccg gtaactatcg tcttgagtcc aacccggtaa gacacgactt atcgccactg 2640
gcagcagcca ctggtaacag gattagcaga gcgaggtatg taggcggtgc tacagagttc 2700
ttgaagtggt ggcctaacta cggctacact agaaggacag tatttggtat ctgcgctctg 2760
ctgaagccag ttaccttcgg aaaaagagtt ggtagctctt gatccggcaa acaaaccacc 2820
gctggtagcg gtggtttttt tgtttgcaag cagcagatta cgcgcagaaa aaaaggatct 2880
caagaagatc ctttgatctt ttctacgggg tctgacgctc agtggaacga aaactcacgt 2940
taagggattt tggtcatgag atcagatcta acatccaaag acgaaaggtt gaatgaaacc 3000
tttttgccat ccgacatcca caggtccatt ctcacacata agtgccaaac gcaacaggag 3060
gggatacact agcagcagac cgttgcaaac gcaggacctc cactcctctt ctcctcaaca 3120
cccacttttg ccatcgaaaa accagcccag ttattgggct tgattggagc tcgctcattc 3180
caattccttc tattaggcta ctaacaccat gactttatta gcctgtctat cctggccccc 3240
ctggcgaggt tcatgtttgt ttatttccga atgcaacaag ctccgcatta cacccgaaca 3300
tcactccaga tgagggcttt ctgagtgtgg ggtcaaatag tttcatgttc cccaaatggc 3360
ccaaaactga cagtttaaac gctgtcttgg aacctaatat gacaaaagcg tgatctcatc 3420
caagatgaac taagtttggt tcgttgaaat gctaacggcc agttggtcaa aaagaaactt 3480
ccaaaagtcg gcataccgtt tgtcttgttt ggtattgatt gacgaatgct caaaaataat 3540
ctcattaatg cttagcgcag tctctctatc gcttctgaac cccggtgcac ctgtgccgaa 3600
acgcaaatgg ggaaacaccc gctttttgga tgattatgca ttgtctccac attgtatgct 3660
tccaagattc tggtgggaat actgctgata gcctaacgtt catgatcaaa atttaactgt 3720
tctaacccct acttgacagc aatatataaa cagaaggaag ctgccctgtc ttaaaccttt 3780
ttttttatca tcattattag cttactttca taattgcgac tggttccaat tgacaagctt 3840
ttgattttaa cgacttttaa cgacaacttg agaagatcaa aaaacaacta attattcgaa 3900
acgatgagat ttccttcaat ttttactgct gttttattcg cagcatcctc cgcattagct 3960
gctccagtca acactacaac agaagatgaa acggcacaaa ttccggctga agctgtcatc 4020
ggttactcag atttagaagg ggatttcgat gttgctgttt tgccattttc caacagcaca 4080
aataacgggt tattgtttat aaatactact attgccagca ttgctgctaa agaagaaggg 4140
gtatctctcg agaaaagaga ggctgaagct g 4171
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ PICHIA PASTORIS - ПРОДУЦЕНТ СЕКРЕТИРУЕМОЙ КСИЛОГЛЮКАНАЗЫ СЕМЕЙСТВА GH12, КОДИРУЕМОЙ МУТИРОВАННЫМ ГЕНОМ, И СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА СЕКРЕТИРУЕМОЙ КСИЛОГЛЮКАНАЗЫ НА ОСНОВЕ ЭТОГО ШТАММА | 2023 |
|
RU2833908C1 |
| Рекомбинантный штамм дрожжей Pichia pastoris - продуцент секретируемой ксилоглюканазы из гриба Aspergillus cervinus и способ микробиологического синтеза ксилоглюканазы на основе этого штамма | 2016 |
|
RU2639248C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ Pichia pastoris - ПРОДУЦЕНТ СЕКРЕТИРУЕМОЙ ТЕРМОСТАБИЛЬНОЙ КСИЛОГЛЮКАНАЗЫ, КОДИРУЕМОЙ СИНТЕТИЧЕСКИМ ГЕНОМ, И СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА СЕКРЕТИРУЕМОЙ ТЕРМОСТАБИЛЬНОЙ КСИЛОГЛЮКАНАЗЫ НА ОСНОВЕ ЭТОГО ШТАММА | 2015 |
|
RU2605629C1 |
| Рекомбинантный штамм дрожжей Pichia pastoris - продуцент бета-глюканазы | 2018 |
|
RU2701494C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ESCHERICHIA COLI - ПРОДУЦЕНТ ЭНДОГЛЮКАНАЗЫ ИЗ ГРИБА THIELAVIA TERRESTRIS И СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА ЭНДОГЛЮКАНАЗЫ НА ОСНОВЕ ЭТОГО ШТАММА | 2022 |
|
RU2811434C1 |
| Рекомбинантный штамм дрожжей Komagataella kurtzmanii - продуцент бета-глюканазы из Paenibacillus jamilae | 2019 |
|
RU2730577C1 |
| Рекомбинантный штамм дрожжей Komagataella kurtzmanii - продуцент бета-глюканазы | 2018 |
|
RU2701640C1 |
| Трансформант дрожжей Pichia pastoris, продуцирующий бета-глюканазу | 2019 |
|
RU2720914C1 |
| Штамм дрожжей Komagataella kurtzmanii, продуцирующий бета-глюканазу из Bacillus pumilus и бета-глюканазу из Paenibacillus jamilae | 2019 |
|
RU2736441C1 |
| Трансформант дрожжей Komagataella kurtzmanii, продуцирующий бета-глюканазу | 2019 |
|
RU2722563C1 |

Группа изобретений относится к биотехнологии. Предложена рекомбинантная секретируемая термостабильная β-глюканаза, включающая последовательность кодирующей части гена β-глюканазы семейства 12 из штамма Thielavia terrestris ВКПМ F-144 и имеющая последовательность SEQ ID NO 2. Также предложен рекомбинантный штамм дрожжей Pichia pastoris ВКПМ Y-4850, продуцирующий вышеуказанную β-глюканазу и полученный путем введения кодирующей части гена β-глюканазы из штамма Thielavia terrestris ВКПМ F-144, соответствующей последовательности SEQ ID NO 1 в штамм-реципиент Pichia pastoris Х33 в составе плазмиды pPIC-ThiTeEgh12. Также предложен способ микробиологического синтеза вышеуказанной β-глюканазы, предусматривающий культивирование штамма Pichia pastoris ВКПМ Y-4850 в аэробных условиях в подходящей питательной среде с периодическим добавлением в качестве индуктора метанола до максимального накопления целевого продукта. Группа изобретений обеспечивает расширение арсенала рекомбинантных β-глюканаз, штаммов-продуцентов и способов микробиологического синтеза рекомбинантных β-глюканаз. 3 н.п. ф-лы, 2 ил., 3 пр.
1. Рекомбинантная секретируемая термостабильная β-глюканаза, включающая последовательность кодирующей части гена β-глюканазы семейства 12 из штамма Thielavia terrestris ВКПМ F-144 и имеющая последовательность SEQ ID NO 2.
2. Рекомбинантный штамм дрожжей Pichia pastoris ВКПМ Y-4850 - продуцент β-глюканазы по п. 1, полученный путем введения кодирующей части гена β-глюканазы из штамма Thielavia terrestris ВКПМ F-144, соответствующей последовательности SEQ ID NO 1 в штамм-реципиент Pichia pastoris Х33 в составе плазмиды pPIC-ThiTeEgh12.
3. Способ микробиологического синтеза β-глюканазы по п. 1 путем культивирования штамма дрожжей Pichia pastoris по п. 2 в аэробных условиях в подходящей питательной среде с периодическим добавлением в качестве индуктора метанола до максимального накопления целевого продукта.
| RYKOV S.V | |||
| ET AL | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Appl Microbiol Biotechnol | |||
| Станок для придания концам круглых радиаторных трубок шестигранного сечения | 1924 |
|
SU2019A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| база данных NCBI Reference Sequence: XP_003654632.1, 29.11.2019 | |||
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |

