Группа заявляемых изобретений относится к биотехнологии, в частности к биосинтезу гемицеллюлаз, и представляет собой рекомбинантный штамм дрожжей Pichia pastoris, способный синтезировать рекомбинантную ксилоглюканазу и полученный путем трансформации штамма Pichia pastoris Х33 плазмидой pPIC-AsCeGH12b, содержащей ген β-1,4-гликозилгидролазы класса GH12 AsCeGH12b из Aspergillus cervinus ВКПМ F-612 под контролем промотора АОХ1.
Растительная биомасса является дешевым и возобновляемым источником сахаров для нужд промышленной биотехнологии. Ферментативный гидролиз - эффективный и экологически безопасный способ конверсии полисахаридов растительной биомассы в сахара. Химическое разнообразие растительных полисахаридов обуславливает необходимость использования широкого спектра ферментов для гидролиза.
Ксилоглюкан - один из основных полисахаридов первичной клеточной стенки двудольных и многих однодольных растений (Hayashi, 1989). Молекулы ксилоглюкана состоят из мономеров 1,4-β-D-глюкотетрозы, в которой три из четырех глюкозных остатка связаны α-1,6 связями с остатками D-ксилозы. Некоторые из остатков глюкозы, в свою очередь, могут быть связаны β-1,2 связями с остатками D-галактозы или L-арабинозы. Остатки D-галактозы могут быть связаны α-1,2 связями с остатками L-фукозы. Модификация остатков ксилозы видоспецифична, что определяет множество типов ксилоглюкана в растениях. В клеточной стенке растений ксилоглюкан выполняет структурную функцию. Ксилоглюкан выступает связующим глюканом, взаимодействуя как с молекулами целлюлозы, так и со своими собственными, и формирует прочную полисахаридную сеть. Также ксилоглюкан может играть роль запасного полисахарида, например в случае с тамариндом индийским Tamarindus indica ксилоглюкан выступает единственным запасным полисахаридом семян (Shankaracharya, 1998).
Ферменты, способные специфически гидролизовать β-1,4-связи глюканового остова молекулы ксилоглюкана, называются ксилоглюканазами. Также известны гликозилгидролазы, которые могут гидролизовать различные связи в молекуле ксилоглюкана: β-галактозидазы, α-ксилозидазы, β-глюкозидазы и α-фукозидазы. Конечными продуктами гидролиза ксилоглюкана ксилоглюканазами является смесь олигосахаридов различного состава и строения (Grishutin et al., 2004). Ксилоглюканазы классифицируют по механизму действия на ксилоглюкановые олигосахариды: экзо- ксилоглюканазы (ЕС 3.2.1.155 и ЕС 3.2.1.150) гидролизуют олигосахариды с редуцирующего конца цепи; эндо-ксилоглюканазы (ксилоглюкан-специфические эндо-β-1,4-глюканазы, ЕС 3.2.1.151) гидролизуют только гликозидную связь у незамещенного остатка глюкозы в молекуле ксилоглюкана и ксилоглюкановых олигосахаридов (Yaoi and Mitsuishi, 2004). Согласно данным базы CAZy (http://www.cazy.org/) ферменты с ксилоглюканазной активностью найдены среди семейств гликозил-гидролаз GH5, GH9, GH12, GH16, GH26, GH44, GH74. Ферменты, специфически гидролизующие ксилоглюкан, также могут проявлять побочную активность на других β-1,4-/1,3-глюканах: растворимой и нерастворимой целлюлозе, β-глюкане, лихенане и ламиранане.
На основе ксилоглюканаз созданы ферментные препараты для ферментативного гидролиза растительных полисахаридов, в особенности биомассы молодых побегов (RU 2358756, RU 2361918). Полный ферментативный гидролиз целлюлозы с помощью только целлюлаз затруднен без предварительного гидролиза связующего ксилоглюкана. Также ксилоглюканазы находят применение как модифицирующие агенты для изменения физических и биохимических свойств растительных волокон (Park et al., 2004), для легирования химически модифицированных олигосахаридов к молекуле ксилоглюкана для изменения функциональности целлюлозных фибрилл (Brumer et al., 2004). Также отмечается возрастающий интерес к применению ксилоглюканаз в пищевой и текстильной индустрии и в фармацевтике (Shankaracharya, 1998; Miyazaki et al., 2003).
В источниках информации имеются сведения о грибах - природных продуцентах ксилоглюканаз: Geotrichum sp.(Yaoi and Mitsuishi, 2004), Aspergillus japonicus, Chrysosporium lucknowense, Trichoderma reesei (Grishutin et al., 2004; Markov et al., 2005; Qi et al., 2013), Phanerochaete chrysosporium (Ishida et al., 2007). Методами селекции и радиационного мутагенеза получены штаммы, синтезирующие повышенные количества ксилоглюканаз: Penicillium verruculosum (RU 2361918), Penicillium funiculosum (RU 2323254), Aspergillus aculeatus (RU 2303057).
Рекомбинантные продуценты ксилоглюканаз обладают рядом несомненных преимуществ, основным из которых является повышенная продуктивность рекомбинантных штаммов по целевому белку. В качестве реципиентов для гетерологичной экспрессии грибных ксилоглюканаз используют различные штаммы грибов: в Aspergillus oryzae экспрессированы ксилоглюканазы из Malbranchea cinnamomea (US 6500658), в Fusarium venenatum - ксилоглюканаза из Trichoderma reesei (US 2004067569 A1), в Penicillium canescens - ксилоглюканаза из Penicillium canescens (RU 2358756), в Aspergillus niger - ксилоглюканаза из Thielavia australiensis и собственная ксилоглюканаза (WO 2014138983; Hasper et al., 2002). Описаны также полученные методами генной инженерии штаммы дрожжей Pichia pastoris - продуценты рекомбинантных ксилоглюканаз из Phanerochaete chrysosporium (Ishida et al., 2007).
Используемые для получения рекомбинантного фермента системы гетерологичной экспрессии в бактериях и микроскопических грибах имеют свои технологические и экономические достоинства и недостатки. Способы с применением бактериальных систем экспрессии, например E.coli, имеют такие ограничения, как формирование нерастворимых телец включения и невозможность воспроизвести все посттрансляционные модификации, характерные для эукариот (гликозилирование, фосфорилирование, образование дисульфидных связей между цистеиновыми остатками). К недостаткам штаммов-продуцентов на основе микроскопических грибов родов Aspergillus и Penicillium относят сложности с проведением генетических манипуляций на штаммах вследствие нестандартизированного и недостаточно разработанного генетического инструментария (Demain and Vaishnav, 2009).
Интерес к продукции рекомбинантных белков с использованием метилотрофных дрожжей, в частности, рода Pichia, Komagataella или Hansenula основан на ряде преимуществ этих микроорганизмов как с научной, так и с технологический точки зрения. В отличие от Е.coli, метилотрофные дрожжи обладают способностью ко всем посттрансляционным модификациям, характерным для эукариотических клеток, включая секрецию белков в культуральную среду. Невысокое количество эндогенных секретируемых белков дрожжей существенно облегчает выделение и очистку целевого рекомбинантного продукта (Macauley-Patrick and Fazenda, 2005). Культуру метилотрофных дрожжей можно выращивать в ферментерах до более высоких плотностей, по сравнению с традиционно используемыми дрожжами Saccharomyces cerevisiae, что позволяет получить более высокий уровень продукции целевого белка (Gellissen, 2000). Метилотрофные дрожжи обладают одними из самых мощных в природе промоторов.
В качестве ближайшего аналога заявляемого штамма рассмотрим штамм дрожжей Pichia pastoris, содержащий на плазмиде pPIC9K оптимизированный по кодонному составу ген PoxXEG12A из Penicillium oxalicum под контролем промотора АОХ1 и экспрессирующий гликозил-гидролазу класса GH12, специфически гидролизующую ксилоглюкан (Xian et al., 2016). По результатам сравнительного анализа уровень гомологии используемого в настоящем изобретении гена AsCeGH12b, кодирующего ксилоглюканазу, и гена PoxXEG12A из Penicillium oxalicum, составляет 48,3%. Способ микробиологического синтеза на основе штамма ближайшего аналога приводит к синтезу незначительных количеств фермента и требует стадии выделения белка из клеточного лизата, поскольку генетическая конструкция предназначена для внутриклеточной экспрессии. Так, клон штамма-ближайшего аналога GHP9K13202 после трансформации способен продуцировать только 1,5 единиц активности на мл культуральной жидкости.
Задача заявляемой группы изобретений состоят в расширении арсенала штаммов- продуцентов и способов микробиологического синтеза рекомбинантных ксилоглюканаз.
Задачу решают путем:
- конструирования рекомбинантного штамма дрожжей Pichia pastohs ВКПМ Y-4299 - продуцента секретируемой ксилоглюканазы, кодируемой геном, соответствующим последовательности SEQ ID NO 1;
- разработки способа микробиологического синтеза секретируемой ксилоглюканазы, кодируемой геном, соответствующим последовательности SEQ ID NO 1, путем культивирования рекомбинантного штамма дрожжей Pichia pastoris ВКПМ Y-4299 в аэробных условиях в подходящей питательной среде до максимального накопления целевого продукта.
Процесс конструирования заявляемого штамма состоит из следующих этапов:
- получение полной нуклеотидной последовательности гена AsCeGH12b, кодирующего секретируемую ксилоглюканазу, синтезируемую штаммом Aspergillus cervinus ВКПМ F-612;
- конструирования интегративной плазмидной ДНК (плазмиды) pPIC-AsCeGH12b, содержащей ген AsCeGH12b секретируемой ксилоглюканазы из Aspergillus cervinus ВКПМ F-612;
- конструирования рекомбинантного штамма дрожжей Pichia pastoris ВКПМ Y-4299, способного синтезировать секретируемую ксилоглюканазу, кодируемую геном AsCeGH12b;
- разработки способа микробиологического синтеза секретируемой ксилоглюканазы ASCEGH12B на основе сконструированного рекомбинантного штамма дрожжей Pichia pastoris ВКПМ Y-4299.
Этап 1. Получение полной нуклеотидной последовательности гена, кодирующего ксилоглюканазу семейства GH12, синтезируемую штаммом Aspergillus cervinus ВКПМ F- 612
Штамм Aspergillus cervinus ВКПМ F-612 секретирует в культуральную среду ферменты, способные гидролизовать ксилоглюкан. Методом зимографии определено, что культуральная жидкость штамма Aspergillus cervinus ВКПМ F-612 содержит ксилоглюканазу с молекулярной массой 25 кДа. Известно, что ксилоглюканазную активность у видов царства грибов проявляют ферменты семейств GH12 и GH74, которые отчетливо различают по молекулярному весу (около 25-30 кДа и около 75-85 кДа соответственно). Для получения и клонирования нуклеотидной последовательности гена гликозил-гидролазы класса GH12 из Aspergillus cervinus ВКПМ F-612 проводят амплификацию консервативных участков гена с помощью вырожденных праймеров и методами прогулки по хромосоме получают фланкирующие консервативный участок гена последовательности ДНК (Green and Sambrook, 2012). Вырожденные праймеры подобраны на консервативные мотивы в известных аминокислотных последовательностях ксилоглюкан-специфичных гликозил-гидролаз класса GH12 представителей рода Aspergillus. Для определения фланкирующих последовательностей применяют два метода прогулки по хромосоме: FPNI-PCR (Fusion primer and nested integrated PCR) для определения последовательности геномной ДНК и RACE (Rapid Amplification of cDNA Ends) для определения последовательности матричной РНК (Wang et al., 2011). В результате получают последовательность нуклеотидов гена ксилоглюканазы класса GH12 из Aspergillus cervinus ВКПМ F-612, обозначенную как AsCeGH12b.
Этап 2. Конструирование интегративной плазмиды pPIC-AsCeGH12b, содержащей процессированный вариант гена AsCeGH12b, кодирующего ксилоглюканазу
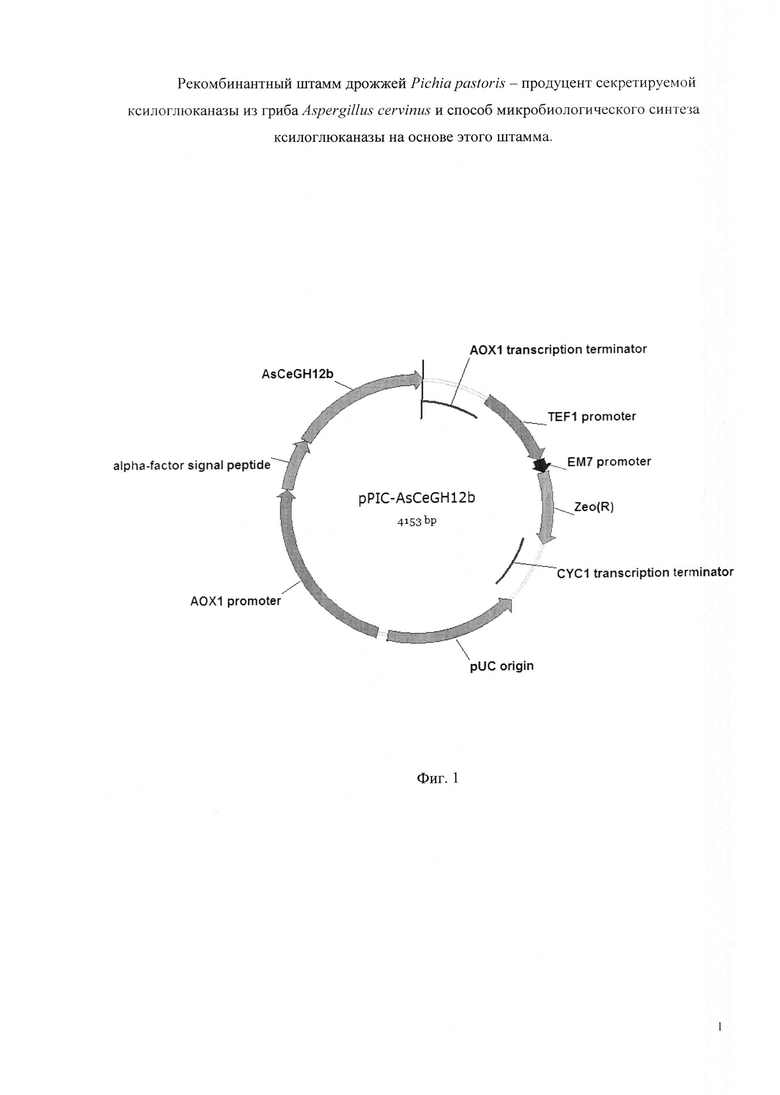
Плазмиду pPIC-AsCeGH12b конструируют путем клонирования полученного амплификацией фрагмента ДНК, содержащего структурную часть гена AsCeGH12b, в вектор pPICZaA, разработанный для интеграции в штаммы Pichia pastoris и предназначенный для внеклеточной экспрессии. Вектор pPICZaA размером 3593 пары оснований содержит промотор АОХ1; терминатор АОХ1; кодирующую область гена Zeo, обеспечивающего устойчивость штаммов Е. coli и Pichia pastoris к зеоцину; репликон pUC; последовательность, кодирующую сигнальный пептид α-фактор дрожжей.
Плазмида pPIC-AsCeGH12b размером 4153 пар оснований, наряду с генами вектора pPICZaA, содержит кодирующую область гена AsCeGH12b ксилоглюканазы под контролем промотора АОХ1 (Фиг. 1).
Этап 3. Конструирование рекомбинантного штамма дрожжей Pichia pastoris, способного синтезировать секретируемую ксилоглюканазу, кодируемую геном AsCeGH12b
В качестве штамма-реципиента используют штамм Pichia pastoris Х33 (mut+), не синтезирующий ксилоглюканазу. Компетентные клетки данного штамма трансформируют плазмидой pPIC-AsCeGH12b. В результате получают рекомбинантный штамм Pichia pastoris, способный синтезировать ксилоглюканазу семейства GH12 из Aspergillus cervinus ВКПМ F-612.
Рекомбинантный штамм Pichia pastoris депонирован во Всероссийской коллекции промышленных микроорганизмов как Pichia pastoris ВКПМ Y-4299.
Заявляемый штамм Pichia pastoris ВКПМ Y-4299 имеет следующие морфологические и физиолого-биохимические характеристики:
Морфологические признаки: при культивировании при температуре 28°С в течение 48 часов на агаризованной среде YPD следующего состава (мас.%: пептон - 2, дрожжевой экстракт - 1, глюкоза - 2, агар - 2, вода - остальное) формируются колонии светло-бежевого цвета с ровным краем, матовой поверхностью, линзовидным профилем и пастообразной консистенцией. При росте в жидкой среде YPD при 28°С в течение 24 ч культивирования - жидкость мутная, осадок белый, коагуляции не наблюдается, пристеночных пленок культура не образует. Клетки имеют овальную форму, 3-4 мкм в диаметре; почкуются; почкование истинное, многостороннее; истинного мицелия не образуют.
Физиолого-биохимические признаки: штамм является факультативным анаэробом с температурой роста - 20-33°C (оптимум - 28°C) и рН среды культивирования - 4,8-7,4 (оптимум - 6,0). В качестве источников углерода штамм может использовать глюкозу, глицерин, метанол, олеат, сорбитол, рамнозу. Не утилизирует галактозу, ксилозу, арабинозу. В качестве источников азота штамм может использовать аминокислоты, сернокислый аммоний, азотнокислый аммоний. Штамм Pichia pastoris ВКПМ Y-4299 синтезирует рекомбинантную секретируемую ксилоглюканазу, кодируемую геном AsCeGH12b.
Способ микробиологического синтеза ксилоглюканазы ASCEGH12B в общем виде
Посевной материал, представляющий собой клетки рекомбинантного штамма - продуцента, подготавливают путем инкубации в течение 16-18 часов при температуре 29°C на среде BMGY (мас.%: пептон - 2, дрожжевой экстракт - 1, глицерин - 1, фосфатный буфер рН=6.6 - до 100 mM, вода - остальное) при постоянной аэрации на термостатируемой качалке (200 об/мин). Затем выросшую культуру переносят в среду BMMY (мас.%: пептон - 2, дрожжевой экстракт - 1, метанол - 0,5, фосфатный буфер рН=6.6 - до 100 mM, вода - остальное) до ОД600=1.
Процесс биосинтеза ведут в колбах Эрленмейера, содержащих 100 мл среды BMMY, в течение 96 часов в ротационном шейкере-термостате (200-250 об/мин), при температуре 28°C. Каждые 24 часа проводят индукцию метанолом, путем асептического добавления 50% раствора метанола в пробирки, до конечной концентрации 0,5%. По истечении 96 часов биомассу отделяют центрифугированием. Наличие рекомбинантной ксилоглюканазы в культуральной среде определяют при помощи электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия с последующей зимограммой. Количественную оценку уровня активности рекомбинантной ксилоглюканазы осуществляют при помощи ДНС-метода (Miller, 1959).
Уровень синтеза ксилоглюканазы заявляемым способом составляет не менее 30 ед/мл культуральной жидкости, что значительно превосходит ближайший аналог. Оптимум активности рекомбинантной ксилоглюканазы, продуцируемой штаммом Pichia pastoris ВКПМ Y-4299, - 55°C, рН 5.0. Рекомбинантная ксилоглюканаза, продуцируемая штаммом Pichia pastoris ВКПМ Y-4299, является термостабильной: при инкубации препарата фермента в течение часа при 50°C, рН 5.0 сохраняется 100% ферментативной активности.
Изобретение проиллюстрировано следующими фигурами графических изображений:
Фигура 1. Рекомбинантная плазмида pPIC-AsCeGH12b.
Пример 1. Определение полной нуклеотидной последовательности гена ксилоглюканазы Aspergillus cervinus ВКПМ F-612
Для определения полной нуклеотидной последовательности гена гликозил-гидролазы класса GH12 из Aspergillus cervinus ВКПМ F-612 проводят амплификацию консервативных участков последовательности гена с помощью вырожденных праймеров. Методами прогулки по хромосоме получают фланкирующие консервативный участок гена последовательности ДНК. Вырожденные праймеры подбирают на консервативные мотивы в аминокислотных последовательностях белков класса GH12, охарактеризованных как ксилоглюкан-специфичные гликозил-гидролазы, описанные у представителей рода Aspergillus: A. clavatus, A. fumigatus, A. aculeatus, A. niger, A. flavus, А. lerreus, A. nidulans. Амплификацию проводят методом ПЦР с помощью высокоточной полимеразы Phusion (Thermo Scientific) по протоколам производителя. Геномную ДНК выделяют по стандартной методике (Green and Sambrook, 2012). В качестве матрицы используют геномную ДНК штамма Aspergillus cervinus ВКПМ F-612. Амплификация с применением полученных вырожденных праймеров: прямого - F_140_gr.2 - GCYYTGGGCGGYGCKGGHCC, и обратного - R_230_gr.2 GAAKGGCTCRGTWCCGGCCTG, приводит к получению продукта ПЦР с длиной 280 пар оснований. Полученный фрагмент очищают с помощью гель-электрофореза в 1,8% агаровом геле с последующим выделением нужной полосы с помощью набора для выделения ДНК из агарозных гелей (Биосилика, Россия). Полученный амплификат секвенируют методом Сэнгера на генетическом анализаторе ABI 3500 (Life technology, США) и проверяют последовательность на соответствие ожидаемому классу ферментов поиском по гомологии с помощью BLAST.
На основе полученной последовательности синтезируют серию праймеров, элонгация которых приводит к амплификации ДНК фланговых последовательностей, лежащих за пределами полученного фрагмента. Для определения фланкирующих последовательностей применяют два метода прогулки по хромосоме: FPN1-PCR для определения последовательности геномной ДНК и RACE для определения последовательности матричной РНК (Wang et al., 2011; Lukyanov et al., 1999; Matz et al., 1999). Синтезируемые праймеры подбирают с таким расчетом, чтобы соответствовать требованиям обоих методов: длина 20-25 пар оснований, температура плавления 58-62°С, GC состав не более 50%. Получают праймеры: 3'for_1 - AGCATTGCAGGGCAGACGTGG, 3'for_2 - CACGACGGTGTACAGTTTCGTGG, 3'for_3 - AGCTTCTCGGCTGATCTGATGG, 5'rev_1 - ACGTGTAGAAGTCCATCAGATCAGCC, 5'rev_2, CACGAAACTGTACACCGTCGTGG, 5'rev_3 - CGAGTACAGTTTCCACGTCTGCC. Метод FPN1-PCR проводят на матрице геномной ДНК штамма Aspergillus cervinus ВКПМ F-612 согласно методике (Wang et al., 2011). В результате получают продукты амплификации, приходящиеся на фланкирующие последовательности геномной ДНК в направлении 3' начала гена и 5' конца гена. Метод RACE проводят на матрице кДНК, полученной с генетического материала культуры штамма Aspergillus cervinus ВКПМ F-612, индуцированной добавлением ксилоглюкана в культуральную среду. Метод RACE осуществляют с использованием набора RACE Primer Set (Евроген, Россия) по протоколу производителя. кДНК получают синтезом одно- и двухцепочечных ДНК с матрицы тотальной РНК методом обратной транскрипции с использованием набора реагентов Mint (Евроген, Россия) по протоколам производителя. Выделение тотальной РНК осуществляют с помощью реагента ExtractRNA (Евроген, Россия) по протоколам производителя. Амплификацию проводят методом ПЦР с помощью высокоточной полимеразы Phusion (Thermo Scientific) по протоколам производителя. В результате получают продукты амплификации, приходящиеся на фланкирующие последовательности матричной РНК в направлении 3' начала гена и 5' конца гена. Полученные методами FPNI-PCR и RACE продукты амплификации секвенируют методом Сэнгера на генетическом анализаторе ABI 3500 (Life technology, США) и собирают единую последовательность ДНК. В результате получена последовательность ДНК длиной 908 пар оснований, которая содержит ген ксилоглюканазы класса GH12 длиной 768 пар оснований, который назван AsCeGH12b и содержит один интрон длиной 53 пары оснований и два экзона длиной 314 и 400 пар оснований. Процессированный транскрипт гена AsCeGH12b длиной 714 пар нуклеотидов (SEQ ID NO 1) кодирует белок ASCEGH12B (SEQ ID NO 2). Белок ASCEGH12B состоит из 238 аминокислотных остатков и содержит на N-конце предсказанный программой SignalP 4.1 (http://www.cbs.dtu.dk/services/SignalP/) сигнальный пептид длиной 17 аминокислотных остатков. Процессированный секретируемый зрелый белок имеет массу 24,5 кДа. Поиск по гомологии аминокислотной последовательности с помощью программы BLAST выявляет ближайшего гомолога белка ASCEGH12B - вероятную ксилоглюкан - специфичную эндо-бета-1,4-глюканазу из Aspergillus lentulus с идентичностью 72%.
Пример 2. Конструирование рекомбинантной плазмидной ДНК, содержащей процессированный вариант гена AsCeGH12b
Все стандартные генно-инженерные и микробиологические манипуляции проводят по известным методикам (Green and Sambrook, 2012).
Для получения полноразмерной, не содержащей интронов и сигнального пептида, кодирующей части гена AsCeGH12b проводят амплификацию на матрице кДНК, полученной с генетического материала культуры штамма Aspergillus cervinus ВКПМ F-612, индуцированной добавлением ксилоглюкана в культуральную среду. кДНК получают синтезом одно- и двухцепочечных ДНК с матрицы тотальной РНК методом обратной транскрипции с использованием набора реагентов Mint (Евроген, Россия) по протоколам производителя. Выделение тотальной РНК производят с помощью реагента ExtractRNA (Евроген, Россия) по протоколам производителя. Амплификацию фрагмента, содержащего ген AsCeGH12b, проводят методом ПЦР с помощью высокоточной полимеразы Phusion (Thermo Scientific) по протоколам производителя с использованием следующих праймеров: прямого - F_AsCeGH12_B_wo_sp_EcoRI_rev.2 - GTAGAATTCGCGACTACGTACTGCGGCCA, и обратного - ASP_AsCeGH12B_XhoI_his_tag - TAACTCGAGAGCCACACTGGCCGAGTACG.
В прямой праймер вводят сайт EcoRI, а в обратный - сайт XhoI. Фрагмент конструируют с таким расчетом, чтобы рамка считывания продолжилась полигистидиновой последовательностью на С-конце рекомбинантного белка, кодируемого последовательностью на векторе pPICZaA. Очистку полученного амплификацией фрагмента ДНК проводят с помощью гель-электрофореза в 1,8% агаровом геле с последующим выделением нужной полосы с помощью набора для выделения ДНК из агарозных гелей (Биосилика, Россия).
Плазмиду pPIC-AsCeGH12b конструируют путем клонирования фрагмента, содержащего ген AsCeGH12b в вектор pPICZaA.
Гидролиз плазмиды pPICZaA и фрагмента ДНК, содержащего ген AsCeGH12b, проводят рестриктазами EcoRI/Sall и EcoRI/XhoI соответственно (Thermo scientific, EU). Очистку полученных фрагментов ДНК проводят с помощью гель-электрофореза в 1,8% агаровом геле с последующим выделением нужной полосы с помощью набора для выделения ДНК из агарозных гелей (Биосилика, Россия). Легирование проводят в реакционной смеси объемом 20 мкл, содержащей 20 нг ДНК вектора, 20 нг ДНК фрагмента и 5 ед. Т4 ДНК лигазы (СибЭнзим, Россия), согласно методике производителя. Полученной лигазной смесью в количестве 5 мкл трансформируют компетентные клетки штамма Escherichia coli XL1 blue MRF' (Stratagene, США) (генотип - Δ(mcrA)183 Δ(mcrCB-hsdSMR-mrr)173 endA1 supE44 thi-1 recA1 gyrA96 relA1 lac [F' proAB laclqZΔM15 Tn10 (Tetr)]) и высевают на чашки с агаризованной низкосолевой LB (мас.%: гриптон - 1, дрожжевой экстракт - 0,5, хлорид натрия - 0,5, агар - 2, вода - остальное), содержащей зеоцин в концентрации 25 мкг/см3. Плазмидную ДНК полученных трансформантов анализируют путем гидролиза рестриктазами BamHI и HindIII. В результате отбирают клоны, содержащие BamHI/HindIII фрагменты расчетного размера. Правильность клонирования подтверждают секвенированием вставки методом Сэнгера на генетическом анализаторе ABI 3500 (Life technology, США).
В результате получают рекомбинантную плазмиду pPIC-AsCeGH12b (SEQ ID NO 3) размером 4153 пар оснований, включающую ген AsCeGH12b, кодирующий ксилоглюканазу ASCEGH12B (Фиг. 1).
Пример 3. Получение заявляемого штамма - продуцента ксилоглюканазы ASCEGH12B
С целью получения рекомбинантного штамма Pichia pastoris ВКПМ Y-4299 - продуцента ксилоглюканазы, клетки штамма Pichia pastoris Х33 (mut+) (Invitrogen) трансформируют плазмидой pPIC-AsCeGH12b. Трансформацию проводят 5 мг линеаризованной плазмидной ДНК, для получения которой исходную плазмиду обрабатывают эндонуклеазой рестрикции MssI.
Культуру дрожжей штамма Pichia pastoris Х33 (mut+) выращивают на среде YPD в аэробных условиях (200-250 об/мин) при 29°С в ночь. Инокулируют свежую среду YPD объемом 500 мл ночной культурой в соотношении 1:1000 и растят в ночь до ОД600=1,3-1,5. Центрифугируют культуру при 1500 об/мин 5 минут при 4 С. Промывают осадок клеток 500 мл стерильной дистиллированной воды. Центрифугируют культуру при 1500 об/мин 5 минут при 4 С. Промывают осадок клеток 250 мл стерильной дистиллированной воды. Центрифугируют культуру при 1500 об/мин 5 минут при 4°С. Промывают осадок клеток 25 мл 1 М сорбитола. Центрифугируют культуру при 1500 об/мин 5 минут при 4°С и осадок клеток ресуспендируют в 1 мл 1 М сорбитола. Суспензию разделяют на аликвоты по 80 мкл. Электропорацию проводят на приборе GenePulser Xcell (Biorad) в кюветах с зазором 2 мм при 2000 В, 25 мкФ, 200 Ом. После электропорации быстро добавляют 1 мл холодного раствора 1 М сорбитола, инкубируют 1 час при температуре 30°C и высевают на селективную среду для отбора трансформантов. Отбор трансформантов проводят на среде YPDS (мас.%: пептон - 2, дрожжевой экстракт - 1, глюкоза - 2, сорбитол - до 1 М, агар - 2, вода - остальное), содержащей зеоцин в концентрации 100 мкг/см3.
Клетки инкубируют при 29°C в течение 3 суток. Трансформанты выявляют методом ПЦР-скрининга. В результате получают заявляемый штамм Pichia pastoris ВКПМ Y-4299, продуцирующий секретируюмую ксилоглюканазу ASCEGH12B.
Пример 4. Микробиологический синтез ксилоглюканазы ASCEGH12B
Посевной материал, представляющий собой клетки рекомбинантного штамма- продуцента Pichia pastoris ВКПМ Y-4299, подготавливают путем инкубации в течение 16-18 часов при температуре 29°С на среде BMGY при постоянной аэрации на термостатируемой качалке (200-250 об/мин). Затем выросшую культуру переносят в среду ВММУ до ОД600=1.
Процесс биосинтеза ведут к колбах Эрленмейера, содержащих 100 мл среды BMMY, в течение 96 часов в ротационном шейкере-термостате (200-250 об/мин), при температуре 28°C. Каждые 24 часа проводят индукцию метанолом, путем асептического добавления 50% раствора метанола в пробирки, до конечной концентрации 0,5%. По истечении 96 часов биомассу отделяют центрифугированием. Наличие рекомбинантной ксилоглюканазы в культуральной среде определяют при помощи электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия с последующей зимограммой (Green and Sambrook, 2012). Количественную оценку уровня активности рекомбинантной ксилоглюканазы осуществляют при помощи ДНС-метода (Miller, 1959).
Уровень синтеза ксилоглюканазы заявляемым способом составляет не менее 30 ед/мл культуральной жидкости, что превышает уровень ближайшего аналога и природного штамма-продуцента Aspergillus cervinus ВКПМ F-612. Оптимум активности рекомбинантной ксилоглюканазы, продуцируемой штаммом Pichia pastoris ВКПМ Y-4299, - 55°C, рН 5.0. Рекомбинантная ксилоглюканаза, продуцируемая штаммом Pichia pastoris ВКПМ Y-4299 является термостабильной: при инкубации препарата фермента в течении часа при 50°C, рН 5.0 сохраняется 100% ферментативной активности.
Таким образом, получен рекомбинантный штамм дрожжей Pichia pastoris ВКПМ Y-4299, способный к биосинтезу секретируемой ксилоглюканазы ASCEGH12B. Целевой продукт заявляемого штамма - ксилоглюканаза ASCEGH12B отличается от целевого продукта штамма-ближайшего аналога такими свойствами как термостабильность и способность секретироваться в культуральную среду. Заявляемый штамм при культивировании заявляемым способом значительно превосходит ближайший аналог по уровню синтеза целевого фермента.
Источники информации
1. Brumer Н., Zhou Q., Baumann М.J., Carlsson K., Teeri Т.Т. // Activation of crystalline cellulose surfaces through the chemoenzymatic modification of xyloglucan // J. Am. Chem. Soc, 2004.
2. Demain AL, Vaishnav P. // Production of recombinant proteins by microbes and higher organisms // Biotechnol Adv., 2009.
3. Gellissen G (2000) Heterologous protein production in methylotrophic yeasts. Appl. Microbiol. Biotechnol. 54(6): 741-750.
4. Green M.R., Sambrook J. (2012) Molecular Cloning: a Laboratory Manual. (Fourth Edition) Cold Spring Harbor Laboratory Press.
5. Grishutin S.G., Gusakov A.V., Markov A.V., Ustinov B.B., Semenova M.V., Sinitsyn A.P. // Specific xyloglucanases as a new class of polysaccharide-degrading enzymes. // Biochim Biophys Acta, 2004.
6. Hasper A.A., Dekkers E., van Mil M., van de Vondervoort P.J., de Graaff L.H. // EglC, a new eadoglucanase from Aspergillus niger with major activity towards xyloglucan. // Appl Environ Microbiol., 2002.
7. Hayashi T. // Xyloglucans in the primary-cell wall // Annu. Rev. Plant Physiol. Plant Mol.: Biol., 1989.
8. Ishida Т., Yaoi K., Hiyoshi A., Igarashi K., Samejima M. // Substrate recognition by glycoside hydrolase family 74 xyloglucanase from the basidiomycete Phanerochaete chrysosporium. II FEBS J., 2007.
9. Lukyanov K., Gurskaya N., Bogdanova E., Lukyanov S. // Selective suppression of polymerase chain reaction // Bioorganicheskaya Khimiya (Russ), 1999.
10. Macauley-Patrick S, Fazenda ML (2005) Heterologous protein production using the Pichia pastoris expression system. Yeast 22: 249-270.
11. Markov A.V., Gusakov A.V., Kondratyeva E.G., Okunev O.N., Bekkarevich A.O., Sinitsyn A.P.// New effective method for analysis of the component composition of enzyme complexes from Trichoderma reesei. // Biochemistry (Mosc)., 2005.
12. Matz M., Shagin D., Bogdanova E., Britanova O., Lukyanov S., Diatchenko L., Chenchik A. // Amplification of cDNA ends based on template-switching effect and step-out PCR // Nucleic Acids Research, 1999.
13. Miller, G.L. // Use of dinitrosalicylic acid reagent for determination of reducing sugar // Analytical Chemistry. 1959.
14. Miyazaki S., Endo K., Kawasaki N., Kubo W., Watanabe H., Attwood D. // Oral sustained delivery of paracetamol from in situ gelling xyloglucan formulations // Drug Dev. Ind. Pharm., 2003.
15. Park Y.W., Baba K., Furuta Y., Iida I., Sameshima K., Arai M., Hayashi T. //. Enhancement of growth and cellulose accumulation by overexpression of xyloglucanase in poplar // FEBS Lett., 2004.
16. Qi H., Bai F., Liu A.// Purification and characteristics of xyloglucanase and five ether cellulolytic enzymes from Trichoderma reesei QM9414. // Biochemistry (Mosc), 2013.
17. Shankaracharya N.B. // Tamarind - chemistry, technology and uses - a critical approach // J. Food. Sci. Technol., 1998.
18. Wang Z., Ye S., Li J., Zheng В., Bao M., Ning G. // Fusion primer and nested integrated PGR (FPNI-PCR): a new high-efficiency strategy for rapid chromosome walking or flanking sequence cloning // BMC Biotechnol., 2011.
19. Xian L., Wang F., Yin X., Feng J.X. // Identification and characterization of an acidic and acid-stable endoxyloglucanase from Penicillium oxalicum. // Int. J Biol. Macromol., 2016.
20. Yaoi K.., Mitsuishi Y. // Purification, characterization, cDNA cloning, and expression of a xyloglucan endoglucanase from Geotrichum sp. M128. // FEBS Lett., 2004.
21. RU 2303057 // Штамм мицелиального гриба Aspergillus aculeatus - продуцент комплекса карбогидраз, содержащего ксиланазы, бета-глюканазы, пектиназы и ксилоглюканазы // Дата приоритета: 14.11.2005.
22. RU 2323254 // Штамм мицелиального гриба Penicillium funiculosum - продуцент комплекса карбогидраз, содержащего целлюлазы, глюканазы, глюкозидазы, ксиланазы и ксилоглюканазы, и способ получения ферментного препарата комплекса карбогидраз для осахаривания лигноцеллюлозных материалов // Дата приоритета: 04.04.2006.
23. RU 2358756 // Способ получения ферментного препарата для расщепления гемицеллюлозных гетерополисахаридов клеточной стенки растений и ферментный препарат (варианты) // Дата приоритета: 26.11.2007.
24. RU 2361918 // Штамм мицелиального гриба Penicillium verruculosum - продуцент комплекса целлюлаз, ксиланазы и ксилоглюканазы и способ получения ферментного препарата комплекса целлюлаз, ксиланазы и ксилоглюканазы для гидролиза целлюлозы и гемицеллюлозы // Дата приоритета: 26.02.2008.
25. US 2004067569 А1 // Polypeptides having xyloglucanase activity and nucleic acids encoding same // Priority date: US 20030420191 20030418; US 20020373987P 20020419.
26. US 6500658 // Xyloglucanase from Malbranchea II Priority date: US 20000653778 20000901; WO 2000DK00450 20000811; US 19990149397P 19990817.
27. WO 2014138983 A1 // Novel cell wall deconstruction enzymes of Malbranchea cinnamomea, Thielavia australiensis, and Paecilomyces byssochlamydoides, and uses thereof // Priority date: US 201361783222P 20130314; US 201361783313P 20130314; US 201361783485P 20130314.




Группа изобретений относится к области биотехнологии. Предложен рекомбинантный штамм дрожжей Pichia pastoris ВКПМ Y-4299 – продуцент секретируемой ксилоглюканазы, кодируемой геном AsCeGH12b (SEQ ID NO: 1), клонированным из Aspergillus cervinus ВКПМ F-612. Предложен способ микробиологического синтеза ксилоглюканазы, кодируемой указанным геном, путем культивирования указанного рекомбинантного штамма в аэробных условиях в подходящей питательной среде до максимального накопления целевого продукта. Группа изобретений позволяет получать термостабильную ксилоглюканазу семейства GH12, с оптимумом активности при 55°C, рН 5.0, в количестве не менее 30 единиц на 1 мл культуральной жидкости. 2 н.п. ф-лы, 1 ил., 4 пр.
1. Рекомбинантный штамм дрожжей Pichia pastoris ВКПМ Y-4299 - продуцент секретируемой ксилоглюканазы, кодируемой геном, соответствующим последовательности SEQ ID NO 1.
2. Способ микробиологического синтеза ксилоглюканазы, путем культивирования рекомбинантного штамма дрожжей Pichia pastoris, содержащего ген ксилоглюканазы, в аэробных условиях в подходящей питательной среде до максимального накопления целевого продукта, отличающийся тем, что в качестве продуцента используют рекомбинантный штамм по п.1, а целевым продуктом является ксилоглюканаза, соответствующая последовательности SEQ ID NO 2.

