Область техники
Изобретение относится к биотехнологии, в частности, к биосинтезу ксилоглюканазы, и представляет собой рекомбинантный штамм дрожжей Pichia pastoris, несущий на хромосоме мутантный ген ксилоглюканазы из Aspergillus cervinus ВКПМ F-612, содержащий замену (Y37V), которая приводит к повышению термостабильности фермента, и способный синтезировать ксилоглюканазу, а также способ микробиологического синтеза ксилоглюканазы на основе заявленного штамма.
Уровень техники
Лигноцеллюлозная биомасса является дешевым возобновляемым источником ферментируемых Сахаров являющихся субстратами множества биосинтетических процессов. Ферментативный гидролиз - экологически безопасный способ биоконверсии растительной биомассы в сахара. Сложный полисахаридный состав растительных клеточных стенок, содержащих, помимо целлюлозы, множество разнообразных связующих гликанов, обуславливает необходимость использования широкого спектра ферментов для гидролиза.
Основным структурным полисахаридом первичной клеточной стенки всех двудольных и многих однодольных растений является разновидность бета-глюкана - ксилоглюкан (КГ). Наряду с другими связующими гликанами, он формирует аморфный матрикс, в который погружены микрофибриллы целлюлозы. Поэтому полный гидролиз целлюлозы невозможен без предварительного гидролиза полисахаридов матрикса, в том числе КГ.
Основная цепь молекулы КГ состоит из блоков целлотетраозы, соединенных между собой β-(1,4)-связями. В большинстве случаев каждые три из четырех остатков глюкозы в таком блоке связаны α-(1,6)-связями с остатками D-ксилозы. Некоторые из остатков глюкозы, в свою очередь, могут быть связаны β-(1,2)-связями с остатками D-галактозы или, реже, L-арабинозы. В КГ некоторых растений остатки D-галактозы могут быть связаны α-(1,2)-связями с остатками L-фукозы. Видо- и тканеспецифичность модификаций остатков ксилозы определяет разнообразие типов КГ в растениях.
Ферменты, специфически расщепляющие β-(1,4)-связи основной цепи молекулы КГ, называются ксилоглюканазами.
Карбогидразы разделяются на семейства, объединенные общими структурно-функциональными свойствами, на основании сходства аминокислотной последовательностей. В настоящее время в базе данных CAZy находится более 140 семейств гликозил-гидролаз (http://www.Cazy.org). Ферменты, гидролизующие ксилоглюкан, обнаружены в семействах гликозил-гидролаз: GH5, GH7, GH12, GH16, GH26, GH44 и GH12. В отличие от других семейств, содержащих помимо ксилоглюканаз, ферменты с иной субстратной специфичностью, семейство GH12 содержит только ферменты, обладающие активностью на ксилоглюкане.
В источниках информации имеются сведения о микроорганизмах - природных продуцентах ксилоглюканаз: Geotrichum sp.[Yaoi K., Mitsuishi Y. // Purification, characterization, cDNA cloning, and expression of a xyloglucan endoglucanase from Geotrichum sp.M128. // FEBS Lett., 2004], Aspergillus japonicus, Chrysosporium lucknowense, Trichoderma reesei [Grishutin S.G., Gusakov A.V., Markov A.V., Ustinov B.B., Semenova M.V., Sinitsyn A.P. // Specific xyloglucanases as a new class of polysaccharide-degrading enzymes. // Biochim Biophys Acta, 2004.; Markov A.V., Gusakov A.V., Kondratyeva E.G., Okunev O.N., Bekkarevich A.O., Sinitsyn A.P. // New effective method for analysis of the component composition of enzyme complexes from Trichoderma reesei. // Biochemistry (Mosc)., 2005; Qi H., Bai F., Liu A. // Purification and characteristics of xyloglucanase and five other cellulolytic enzymes from Trichoderma reesei QM9414. // Biochemistry (Mosc), 2013], Phanerochaete chrysosporium [Ishida Т., Yaoi K., Hiyoshi A., Igarashi K., Samejima M. // Substrate recognition by glycoside hydrolase family 74 xyloglucanase from the basidiomycete Phanerochaete chrysosporium. // FEBS J., 2007]. Методами селекции и мутагенеза получены штаммы, синтезирующие повышенные количества ксилоглюканаз: Penicillium verruculosum [RU 2361918], Penicillium funiculosum [RU 2323254], Aspergillus aculeatus [RU 2303057]. Подобным образом был получен высокоактивный штамм мицелиального гриба Aspergillus foetidus - продуцента комплекса карбогидраз, содержащего ксилоглюканазу [RU2323973].
В качестве реципиентов для гетерологичной экспрессии грибных ксилоглюканаз используют различные штаммы грибов: в Aspergillus oryzae экспрессированы ксилоглюканазы из Malbranchea cinnamomea [US 6500658], в Fusarium venenatum - ксилоглюканаза из Trichoderma reesei [US 2004067569], в Penicillium canescens - ксилоглюканаза из Penicillium canescens [RU 2358756], в Aspergillus niger - ксилоглюканаза из Thielavia australiensis и собственная ксилоглюканаза [WO 2014138983].
Описаны также полученные методами генной инженерии штаммы дрожжей Pichia pastoris - продуценты рекомбинантных ксилоглюканаз из Phanerochaete chrysosporium [Ishida Т., Yaoi K., Hiyoshi A., Igarashi K., Samejima М. // Substrate recognition by glycoside hydrolase family 74 xyloglucanase from the basidiomycete Phanerochaete chrysosporium. // FEBS J., 2007], Aspergillus cervinus ВКПМ F-612 [RU2639248] и эндоксилоглюканазы Penicillium oxalicum [CN104388406A]. Помимо этого, разработан метод микробиологического синтеза в Komagataella phaffii термостабильной рекомбинантной ксилоглюканазы, кодируемой синтетическим геном на основе нуклеотидной последовательности ксилоглюканазуы семейства GH74 из Myceliophthora thermophila ВКПМ F-244 [RU2605629].
Системы гетерологичной экспрессии на основе бактерий и микроскопических грибов имеют свои преимущества и недостатки. К недостаткам штаммов-продуцентов на основе микроскопических грибов родов Aspergillus и Penicillium относят невысокий уровень синтеза рекомбинантных ферментов по сравнению с E.coli и сложности с проведением генетических манипуляций на штаммах вследствие нестандартизированного и недостаточно разработанного генетического инструментария [Demain AL, Vaishnav P. // Production of recombinant proteins by microbes and higher organisms // Biotechnol Adv., 2009]. Метилотрофные дрожжи (Pichia, Komagataella, Hansenula) являются высокоэффективными продуцентами рекомбинантных белков. Преимуществом метилотрофных дрожжей является то, что их культуру можно выращивать в ферментерах до более высокой плотности, по сравнению с традиционно используемыми дрожжами Saccharomyces cerevisiae, что позволяет получить более высокий уровень продукции целевого белка [Gellissen G // Heterologous protein production in methylotrophic yeasts. Appl. Microbiol. Biotechnol. 54(6): 741-750., 2000]. Метилотрофные дрожжи обладают одними из самых мощных в природе промоторов. В отличие от Е. coli, метилотрофные дрожжи обладают способностью ко всем характерным для эукариот посттрансляционным модификациям (гликозилирование, фосфорилирование, образование дисульфидных связей между цистеиновыми остатками, секреция рекомбинантных белков в культивационную среду). Низкий уровень секреции собственных белков существенно облегчает выделение и очистку целевого рекомбинантного продукта [Macauley-Patrick S, Fazenda ML (2005) Heterologous protein production using the Komagataella phaffii expression system. Yeast 22: 249-27]. Рекомбинантные белки в метилотрофных дрожжах, в отличие от Е. coli, не образуют телец включения.
Ближайшим аналогом заявляемого штамма является является штамм дрожжей Pichia pastoris, содержащий на плазмиде pPICZaA ген bgh12A Aspergillus cervinus под контролем промотора АОХ1 и экспрессирующий эндо-(1,4)-β-глюканазу Bgh12A семейства GH12 (Rykov et al., 2019; GenBank: AYM46706.1).
Известным аналогом заявляемого способа является способ микробиологического синтеза секретируемой ксилоглюкан-специфической эндо-(1,4)-β-глюканазы семейства 12 гликозид гидролаз из гриба Aspergillus cervinus рекомбинантным штаммом дрожжей Pichia pastoris ВКПМ Y-4299 (RU 2639248). Уровень синтеза ксилоглюкан-специфической эндо-(1,4)-β-глюканазы AsCeGH12b таким способом, составляет не менее 30 ед/мл культуральной жидкости.
Технической проблемой, на решение которой направлена заявляемая группа изобретений, является расширение арсенала рекомбинантных эндоглюканаз, штаммов-продуцентов и способов микробиологического синтеза рекомбинантных эндоглюканаз.
Раскрытие сущности изобретения
Техническим результатом заявляемого изобретения является получение ксилоглюканазы на основе рекомбинантного штамма Pichia pastoris, сочетающего высокий уровень продукции гетерологичного фермента с повышенной термостабильностью, обеспечивающим эффективное снижение вязкости растворов ксилоглюкана.
Для достижения технического результата предложен рекомбинантный штамм дрожжей Pichia pastoris ВКПМ Y-5148 - продуцент секретируемой мутантной ксилоглюканазы AsCeGH12b-mut, кодируемой мутантным геном, соответствующим SEQ ID NO 1.
Предложен способ микробиологического синтеза рекомбинантной мутантной ксилоглюканазы AsCeGH12b-mut, заключающийся в том, что посевной материал, представляющий собой клетки рекомбинантного штамма-продуцента, подготавливают путем инкубации в течение 16-18 часов при температуре 29°С на среде BMGY мас. %: пептон - 2, дрожжевой экстракт - 1, глицерин - 1, фосфатный буфер рН=6.6 - до 100 mM, вода - остальное при постоянной аэрации 200 об/мин на термостатируемой качалке, выросшую культуру переносят в среду BMMY мас. %: пептон - 2, дрожжевой экстракт - 1, метанол - 0,5, фосфатный буфер рН=6.6 - до 100 mM, вода - остальное до ОД600=1, при этом, процесс биосинтеза ведут к колбах Эрленмейера, содержащих 100 мл среды BMMY, в течение 96 часов в ротационном шейкере-термостате со скоростью 200-250 об/мин, при температуре 28°С, при этом, каждые 24 часа проводят индукцию метанолом, путем асептического добавления 50% раствора метанола в пробирки, до конечной концентрации 0,5%, по истечении 96 часов биомассу отделяют центрифугированием, определяют наличие рекомбинантной ксилоглюканазы в культуральной среде при помощи электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия с последующей зимограммой, осуществляют количественную оценку уровня активности рекомбинантной ксилоглюканазы при помощи метода с динитросалициловым реактивом.
Краткое описание чертежей
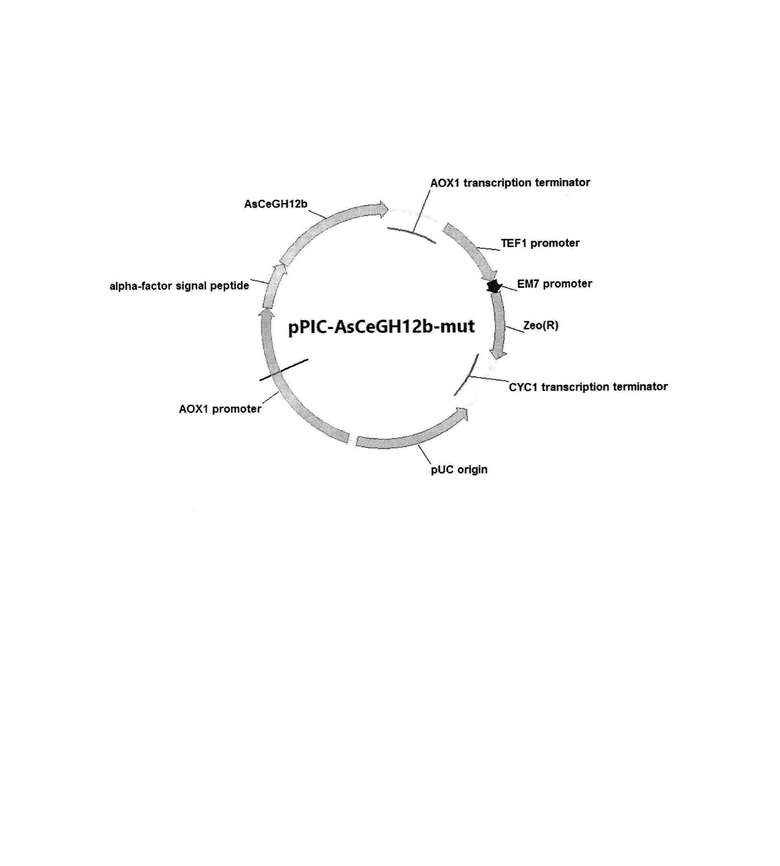
На фигуре показана схема рекомбинантной плазмидной ДНК pPIC-ascegh12b-mut.
Осуществление изобретения
Процесс получения заявляемого штамма состоит из нескольких этапов:
- конструирования мутированного гена ascegh12b-mut (SEQ ID NO 1) на основе нуклеотидной последовательности гена ascegh12b, кодирующего мутантный вариант ксилоглюканазы семейства GH12 из Aspergillus cervinus ВКПМ F-612;
- конструирования интегративной плазмидной ДНК (плазмиды) pPIC-ascegh12b-mut, содержащей ген ascegh12b-mut, являющийся мутантным вариантом гена ксилоглюканазы ascegh12b;
- конструирования рекомбинантного штамма дрожжей Pichia pastoris/ascegh12b-mut, способного синтезировать ксилоглюканазу семейства GH12 с повышенной термостабильностью;
- разработки способа микробиологического синтеза ксилоглюканазы на основе рекомбинантного штамма дрожжей Pichia pastoris/ascegh12b-mut, сочетающего высокий уровень синтеза гетерологичной ксилоглюканазы семейства GH12 с повышенной термостабильностью.
Этап 1. Конструирование мутированного гена ascegh12b-mut на основе нуклеотидной последовательности гена ascegh12b, интегрированного в геном штамма Pichia pastoris ВКПМ Y-5148 и кодирующего мутантную форму ксилоглюканазы AsCeGH12 с повышенной термостабильностью.
Ксилоглюканаза AsCeGH12b из Aspergillus cervinus ВКПМ F-612 проявляет максимальную активность на ксилоглюкане при 55°С, рН 5.0. AsCeGH12b сохраняет 100% активности после инкубации в течение 2 ч при 50°С, рН 5.0, время полуинактивации при 55°С, рН 5.0, составляет 158 мин, при 60°С - менее 5 мин. Для получения более термостабильного варианта данного фермента конструируют мутантную форму AsCeGH12b-mut, осуществляя в последовательности AsCeGH12b замену остатка тирозина в положении 37 остатком валина путем сайт-направленного мутагенеза. Дизайн нуклеотидной последовательности AsCeGH12b-mut разрабатывают на основании гена ascegh12b (GenBank: MF527262.1), кодирующего ксилоглюканазу AsCeGH12b семейства GH12 из Aspergillus cervinus ВКПМ F-612. Введение мутаций в ген asceghl2b осуществляли методом перекрывающихся ПЦР-продуктов (Overlap PCR) с использованием праймеров и плазмиды pPIC-AsCeGH12b, содержащей ген asceghl2b в качестве матрицы (RU 2639248). Для проведения мутагенеза используют следующие праймеры:
AsCeGH12_EcoRI - GTAGAATTCGCGACTACGTACTGCGGCCA,
AsCeGH12B_XhoI - TAACTCGAGAGCCACACTGGCCGAGTACG,
AsCeXgh_for_Y37V - AGTGCTTCACGGTCAATTCTCTCTC,
AsCeXgh_rev_Y37V - AGAGAATTGACCGTGAAGCACTG.
В результате получают фрагмент ДНК, содержащий мутированный ген ascegh12b-mut. Корректность введенной мутации подтверждают при помощи секвенирования методом Сэнгера.
Этап 2. Конструирование интегративной плазмидной конструкции pPIC-ascegh12b-mut
Плазмиду pPIC-ascegh12b-mut конструируют путем клонирования полученного амплификацией фрагмента ДНК, содержащего структурную часть гена ascegh12b-mut, в вектор pPICZaA разработанный для интеграции в штаммы Pichia pastoris и предназначенный для внеклеточной экспрессии. Вектор pPICZaA размером 3593 пары оснований содержит промотор АОХ1; терминатор АОХ1; кодирующую область гена Zeo, обеспечивающего устойчивость штаммов Е. coli и Pichia pastoris к зеоцину; репликон pUC; последовательность, кодирующую сигнальный пептид α-фактор дрожжей.
Плазмида pPIC-ascegh12b-mut размером 4153 пар оснований, наряду с генами вектора pPICZaA, содержит кодирующую область гена ascegh12b-mut ксилоглюканазы под контролем промотора АОХ1 (Фигура).
Этап 3. Получение рекомбинантного штамма Pichia pastoris/ascegh12b-mut
В качестве штамма-реципиента используют штамм Pichia pastoris Х33 (mut+), не синтезирующий ксилоглюканазу. Компетентные клетки данного штамма трансформируют плазмидой pPIC-ascegh12b-mut. В результате получают рекомбинантный штамм Pichia pastoris, способный синтезировать ксилоглюканазу AsCeGH12b-mut.
Рекомбинантный штамм Pichia pastoris депонирован во Всероссийской коллекции промышленных микроорганизмов как Pichia pastoris ВКПМ Y-5148.
Заявляемый штамм Pichia pastoris ВКПМ Y-5148 имеет следующие морфологические и физиолого-биохимические характеристики:
Морфологические признаки
При культивировании при температуре 28°С в течение 48 часов на агаризованной среде YPD следующего состава (мас. %):
пептон – 2;
дрожжевой экстракт – 1;
глюкоза – 2;
агар – 2;
вода - остальное.
Клетки имеют овальную форму, 3-4 мкм в диаметре; почкуются; почкование истинное, многостороннее; истинного мицелия не образуют. На агаризованной среде YPD формируются колонии светло-бежевого цвета с ровным краем, матовой поверхностью, линзовидным профилем и пастообразной консистенцией. При росте в жидкой среде YPD при 28°С в течение 24 ч культивирования - жидкость мутная, осадок белый, коагуляции не наблюдается, пристеночных пленок культура не образует.
Физиолого-биохимические признаки
Штамм является факультативным анаэробом. Температура роста - 20-33°С (оптимум - 28°С). рН среды культивирования - 4,8-7,4 (оптимум - 6,0). В качестве источников углерода штамм может использовать глюкозу, глицерин, метанол, олеат, сорбитол, рамнозу. Не утилизирует галактозу, ксилозу, арабинозу. В качестве источников азота штамм может использовать аминокислоты, сернокислый аммоний, азотнокислый аммоний. Существенными признаками штамма является отсутствие потребности в гистидине. Штамм Pichia pastoris ВКПМ Y-5148 синтезирует рекомбинантную секретируемую ксилоглюканазу с повышенной термостабильностью.
Способ микробиологического синтеза ксилоглюканазы в общем виде.
Посевной материал, представляющий собой клетки рекомбинантного штамма-продуцента, подготавливают путем инкубации в течение 16-18 часов при температуре 29°С на среде BMGY (мас. %: пептон - 2, дрожжевой экстракт - 1, глицерин - 1, фосфатный буфер рН=6.6 - до 100 mM, вода -остальное) при постоянной аэрации на термостатируемой качалке (200 об/мин). Затем выросшую культуру переносят в среду BMMY (мас. %: пептон - 2, дрожжевой экстракт - 1, метанол - 0,5, фосфатный буфер рН=6.6 - до 100 mM, вода - остальное) до ОД600=1.
Процесс биосинтеза ведут к колбах Эрленмейера, содержащих 100 мл среды BMMY, в течение 96 часов в ротационном шейкере-термостате (200-250 об/мин), при температуре 28°С. Каждые 24 часа проводят индукцию метанолом, путем асептического добавления 50% раствора метанола в пробирки, до конечной концентрации 0,5%. По истечении 96 часов биомассу отделяют центрифугированием. Наличие рекомбинантной ксилоглюканазы в культуральной среде определяют при помощи электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия с последующей зимограммой. Количественную оценку уровня активности рекомбинантной ксилоглюканазы осуществляют при помощи метода с динитросалициловым реактивом [ГОСТ 31662-2012. Препараты ферментные. Методы определения ферментативной активности целлюлазы. Москва, 2012. 9 с.]. Уровень синтеза ксилоглюканазы заявляемым способом составляет не менее 43 ед/мл культуральной жидкости, что превосходит ближайший аналог. Количество целевого белка составляет не менее 40 единиц активности физиологически активного белка на 1 мл культуральной жидкости, измеренной при 50°С на ксилоглюкане.
Пример 1. Конструирование рекомбинантной плазмидной ДНК pPIC-ascegh12b-mut, содержащей ген ascesh12b-mut, кодирующий ксилоглюканазу семейства GH12 с повышенной термостабильностью
Мутантный ген ascegh12b-mut (SEQ ID NO 1) получают путем введения мутаций в ген ascegh12b (GenBank: MF527262.1) методом перекрывающихся ПЦР-продуктов (Overlap PCR) с использованием плазмиды pPIC-AsCeGH12b, содержащей ген ascegh12b в качестве матрицы (RU 2639248).
Для этого используют следующие праймеры:
прямой - AsCeXgh_for_BamHI - ATAGGATCCGCGACTACGTACTGC и обратный - AsCeXgh_rev_Y37V - AGAGAATTGACCGTGAAGCACTG для получения первого фрагмента размером 576 п.о.;
прямой - AsCeXgh_rev_Sal - ATAGTCGACTTAAGCCACACTGGCCGAG и обратный AsCeXgb_for_Y37V - AGTGCTTCACGGTCAATTCTCTCTC для получения второго фрагмента размером 129 п.о.
В праймер AsCeXgh_for_BamHI был введен сайт BamHI, а в праймер AsCeXgh_rev_Sal - сайт SalI. Фрагмент конструируют с таким расчетом, чтобы рамка считывания начиналась с полигистидиновой последовательности.
Плазмиду pPIC-ascegh12b-mut конструируют путем клонирования фрагмента, содержащего ген ascegh12b-mut в вектор pPICZaA.
Гидролиз плазмиды pPICZaA и фрагмента ДНК, содержащего ген AsCeGH12b-mut, проводят рестриктазами BamHI и SalI (Thermo Fisher Scientific, США). Очистку полученных фрагментов ДНК проводят с помощью гель-электрофореза в 1,8% агарозном геле с последующим выделением нужной полосы с помощью набора для выделения ДНК из агарозных гелей (Евроген, Россия) согласно инструкции производителя. Лигирование проводят в реакционной смеси объемом 20 мкл, содержащей 20 нг ДНК вектора, 1,5 нг ДНК фрагмента и 5 ед. Т4 ДНК лигазы (Thermo Fisher Scientific, США), согласно методике производителя. Полученной лигазной смесью в количестве 2,5 мкл трансформируют компетентные клетки штамма Е. coli ТОР10 (Invitrogen, США) (генотип - F- mcrA Δ(mrr-hsdRMS-mcrBC) ϕ801acZΔM15 ΔlacX74 nupG recA1 araD139 Δ(ara-leu)7697 galE15 galK16 rpsL(StrR) endA1 λ-). Плазмидную ДНК полученных трансформантов анализируют путем гидролиза рестриктазами BamHI и SalI. В результате отбирают клоны, содержащие BamHI/SalI фрагменты размером, равным исходному фрагменту вставки. Правильность клонирования подтверждают секвенированием каждой из рекомбинантных вставок методом Сэнгера на генетическом анализаторе ABI 3500 (Life technology, США).
В результате получают рекомбинантную плазмиду pPIC-ascegh12b-mut размером 4153 п. о., содержащую наряду с генами вектора pPICZaA, также мутированный ген ascegh12b-mut, кодирующий рекомбинантную ксилоглюканазу AsCeGH12b-mut (Фигура).
Пример 2. Получение заявляемого штамма Pichia pastoris ВКПМ Y-5148 - продуцента ксилоглюканазы семейства GH12 с повышенной термостабильностью.
С целью получения рекомбинантного штамма Pichia pastoris ВКПМ Y-5148 - продуцента ксилоглюканазы, клетки штамма Pichia pastoris Х33 (mut+) (Invitrogen) трансформируют плазмидой pPIC-ascegh12b-mut. Трансформацию проводят 5 мг линеаризованной плазмидной ДНК, для получения которой исходную плазмиду обрабатывают эндонуклеазой рестрикции SacI.
Культуру дрожжей штамма Pichia pastoris Х33 (mut+) выращивают на среде YPD в аэробных условиях (200-250 об/мин) при 29 °С в ночь. Инокулируют свежую среду YPD объемом 500 мл ночной культурой в соотношении 1:1000 и растят в ночь до ОД600=1,3-1,5. Центрифугируют культуру при 1500 об/мин 5 минут при 4 °С. Промывают осадок клеток 500 мл стерильной дистиллированной воды. Центрифугируют культуру при 1500 об/мин 5 минут при 4 °С. Промывают осадок клеток 250 мл стерильной дистиллированной воды. Центрифугируют культуру при 1500 об/мин 5 минут при 4 °С. Промывают осадок клеток 25 мл 1 М сорбитола. Центрифугируют культуру при 1500 об/мин 5 минут при 4 °С и осадок клеток ресуспендируют в 1 мл 1 М сорбитола. Суспензию разделяют на аликвоты по 80 мкл. Электропорацию проводят на приборе GenePulser Xcell (Biorad) в кюветах с зазором 2 мм при 2000 В, 25 мкФ, 200 Ом. После электропорации быстро добавляют 1 мл холодного раствора 1 М сорбитола, инкубируют 1 час при температуре 30°С и высевают на селективную среду для отбора трансформантов. Отбор трансформантов проводят на среде YPDS (мас. %: пептон - 2, дрожжевой экстракт - 1, глюкоза - 2, сорбитол - до 1 М, агар - 2, вода - остальное), содержащей зеоцин в концентрации 100 мкг/см3.
Клетки инкубируют при 29°С в течение 3 суток. Трансформанты выявляют методом ПЦР-скрининга. В результате получают заявляемый штамм Pichia pastoris ВКПМ Y-5148, продуцирующий секретируюмую ксилоглюканазу AsCeGH12b-mut.
Пример 3. Микробиологический синтез термостабильной секретируемой ксилоглюканазы заявляемым штаммом Pichia pastoris ВКПМ Y-5148
Посевной материал, представляющий собой клетки рекомбинантного штамма-продуцента Pichia pastoris ВКПМ Y-5148, подготавливают путем инкубации в течение 16-18 часов при температуре 29°С на среде BMGY при постоянной аэрации на термостатируемой качалке (200-250 об/мин). Затем выросшую культуру переносят в среду BMMY до ОД600=1.
Процесс биосинтеза ведут к колбах Эрленмейера, содержащих 100 мл среды BMMY, в течение 96 часов в ротационном шейкере-термостате (200-250 об/мин), при температуре 28°С. Каждые 24 часа проводят индукцию метанолом, путем асептического добавления 50% раствора метанола в пробирки, до конечной концентрации 0,5%. По истечении 96 часов биомассу отделяют центрифугированием. Наличие рекомбинантной ксилоглюканазы в культуральной среде определяют при помощи электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия с последующей зимограммой (Green and Sambrook, 2012). Количественную оценку уровня активности рекомбинантной ксилоглюканазы осуществляют при помощи ДНС-метода.
Уровень синтеза ксилоглюканазы заявляемым способом составляет не менее 43 ед/мл культуральной жидкости, что превосходит ближайший аналог. Рекомбинантная ксилоглюканаза AsCeGH12b-mut, продуцируемая штаммом Pichia pastoris ВКПМ Y-5148, проявляет максимум активности при 60°С, рН 5.0. Дикий вариант фермента AsCeGH12b, сконструированный и очищенный аналогичным образом, а также ближайший аналог -рекомбинантная ксилоглюканаза AsCeGH12b [Rykov S.V., Kornberger P., Herlet J., Tsurin N.V., Zorov I.N., Zverlov V.V., Liebl W., Schwarz W.H., Yarotsky S.V., Berezina O.V. Novel endo-(1,4)-β-glucanase Bgh12A and xyloglucanase Xgh12B from Aspergillus cervinus belong to GH12 subgroup I and II, respectively. Appl. Microbiol. Biotechnol., 2019, 103(18), 7553-7566], имеют более низкий максимум активности - 55°С, рН 5.0. При инкубации препарата фермента AsCeGH12b-mut в течение 10 минут при 65 и 70°С рН 5.0 сохраняется 80% и 28% ферментативной активности соответственно. В таких же условиях у дикого варианта фермента AsCeGH12b, сконструированного и очищенного аналогичным образом, сохраняется 30% и 12% активности, соответственно.
Таким образом, получен рекомбинантный штамм Pichia pastoris ВКПМ Y-5148, способный к гетерологичному биосинтезу ксилоглюканазы семейства GH12 из A. cervinus, кодируемой мутантным геном. Получена рекомбинантная ксилоглюканаза AsCeGH12b-mut, которая отличается от фермента дикого типа более высокими температурным максимумом активности и более высокой термостабильностью в интервале температур 60-70°С. При культивировании заявляемым способом уровень синтеза целевого продукта у заявляемого штамма превышает таковой у ближайшего аналога. Также, в отличие от ближайшего аналога, продуцируемый фермент имеет более высокий температурный максимум активности и более эффективно гидролизует ксилоглюкан при температурах 55-70°С. Разработан способ микробиологического синтеза ксилоглюканазы AsCeGH12b-mut на основе штамма Pichia pastoris ВКПМ Y-5148, отличающийся тем, что в качестве продуцента используют заявляемый рекомбинантный штамм, а целевым продуктом является мутантная ксилоглюканаза AsCeGH12b-mut, обладающая повышенной по сравнению с ферментом дикого типа термостабильностью.
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3" fileName="Project 4.xml"
softwareName="WIPO Sequence" softwareVersion="2.3.0"
productionDate="2023-09-06">
<ApplicantFileReference>1234567890</ApplicantFileReference>
<ApplicantName languageCode="ru">Национальный исследовательский
центр «Курчатовский институт»</ApplicantName>
<ApplicantNameLatin>National Research Center Kurchatov
Institute</ApplicantNameLatin>
<InventionTitle languageCode="ru">РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ
PICHIA PASTORIS – ПРОДУЦЕНТ СЕКРЕТИРУЕМОЙ КСИЛОГЛЮКАНАЗЫ СЕМЕЙСТВА
GH12, КОДИРУЕМОЙ МУТИРОВАННЫМ ГЕНОМ, И СПОСОБ МИКРОБИОЛОГИЧЕСКОГО
СИНТЕЗА СЕКРЕТИРУЕМОЙ КСИЛОГЛЮКАНАЗЫ НА ОСНОВЕ ЭТОГО
ШТАММА.</InventionTitle>
<SequenceTotalQuantity>1</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>693</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..693</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>gaattcgcgactacgtactgcggccaatgggactcagtgactgagggca
actacatcctctataatgacctctggggcgaatccgctgctaccagcggctcccagtgcttcacgtacaa
ttctctctccggaaacacactctcctggagtacctcctggacctgggccggaggctcaagcagcgtgaag
agctttgccaatgccgctcttcagtttaccccgaaacagttgagcagtattagtagtattgatactacgt
ggacgtggagctacaccggcacctccattgtcgccgacgtcgcctatgacatgttcctagccgctagcgc
aagcggatctagcgagtacgaaatcatggtctggcttgctgcgttgggcggcgcaggcccgatctcgtct
accggctcgaccatcgcgactcccagcattgcagggcagacgtggaaactgtactcgggcccgaacggcg
ccacgacggtgtacagtttcgtggcagagtctgagacgacgagcttctcggctgatctgatggacttcta
cacgtatttgattgataatcaggggctgtcgtcggcgttgtatttgactaatgtgcaggcgggtacggag
ccatttaccggaagcaatgcggtgctgacggtgtcatcgtactcggccagtgtggctctcgaccatcatc
atcatcatcattga</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
Изобретение относится к биотехнологии. Предложен рекомбинантный штамм дрожжей Pichia pastoris ВКПМ Y-5148, продуцирующий секретируемую мутантную ксилоглюканазу AsCeGH12b-mut, кодируемую мутантным геном, соответствующим SEQ ID NO 1. Также предложен способ микробиологического синтеза рекомбинантной мутантной ксилоглюканазы AsCeGH12b-mut, кодируемой мутантным геном, соответствующим SEQ ID NO 1, с использованием указанного штамма. Изобретение позволяет получить термостабильную мутантную ксилоглюканазу AsCeGH12b-mut с высоким уровнем продукции, использование которой обеспечивает эффективное снижение вязкости растворов ксилоглюкана. 2 н.п. ф-лы, 1 ил., 3 пр.
1. Рекомбинантный штамм дрожжей Pichia pastoris ВКПМ Y-5148 - продуцент секретируемой мутантной ксилоглюканазы AsCeGH12b-mut, кодируемой мутантным геном, соответствующим SEQ ID NO 1.
2. Способ микробиологического синтеза рекомбинантной мутантной ксилоглюканазы AsCeGH12b-mut, кодируемой мутантным геном, соответствующим SEQ ID NO 1, заключающийся в том, что посевной материал, представляющий собой клетки рекомбинантного штамма-продуцента по п. 1, подготавливают путем инкубации в течение 16-18 ч при температуре 29°С на среде BMGY, мас. %: пептон - 2, дрожжевой экстракт - 1, глицерин - 1, фосфатный буфер рН=6.6 - до 100 mM, вода - остальное при постоянной аэрации 200 об/мин на термостатируемой качалке, выросшую культуру переносят в среду BMMY, мас. %: пептон - 2, дрожжевой экстракт - 1, метанол - 0,5, фосфатный буфер рН=6.6 - до 100 mM, вода - остальное до ОД600=1, при этом процесс биосинтеза ведут к колбах Эрленмейера, содержащих 100 мл среды BMMY, в течение 96 ч в ротационном шейкере-термостате со скоростью 200-250 об/мин, при температуре 28°С, при этом каждые 24 ч проводят индукцию метанолом, путем асептического добавления 50% раствора метанола в пробирки, до конечной концентрации 0,5%, по истечении 96 ч биомассу отделяют центрифугированием, определяют наличие рекомбинантной ксилоглюканазы в культуральной среде при помощи электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия с последующей зимограммой, осуществляют количественную оценку уровня активности рекомбинантной ксилоглюканазы при помощи метода с динитросалициловым реактивом.

