Изобретение относится к биотехнологии, в частности к получению рекомбинантных интерферонов, обладающих антивирусной активностью.
Гамма-интерферон человека представляет собой гликопротеин с молекулярным весом около 15 000 Да. В человеческом организме гамма-интерферон продуцируется индуцированными Т-лимфоцитами. Как природный (гликозилированный), так и рекомбинантный (негликозилированный) гамма-интерфероны обладают значительным противоопухолевым, иммуномодулирующим и противовирусным действием, что существенно повышает защитный ресурс человека в борьбе с онкологическими и вирусными заболеваниями и делает применение рекомбинантного интерферона человека в медицине весьма перспективным.
Однако противовирусное действие гамма-интерферона в десять раз слабее, чем действие лейкоцитарного интеpферона. Такое различие в активности стимулировало работы по реконструкции гена иммунного интерферона таким образом, чтобы кодируемый им белок обладал более высокой антивирусной активностью по сравнению с белком, кодируемым последовательностью природного гена гамма-интерферона.
Известен ген 68, кодирующий аналог зрелого интерферона человека с 19 аминокислотными заменами в районе 120-146 белка и плазмида р652trp, обеспечивающий экспрессию мутантного гена в E. coli под регуляцией триптофанового промотора. Такая конструкция обеспечивает в штамме E. coli JМ101 высокий уровень экспрессии мутантного гена (до 50% от суммарного клеточного белка). В результате одновременных 19 аминокислотных замен в С-области гамма-интерферона антивирусная активность аналога возросла в 2 раза по сравнению с активностью зрелого рекомбинантного гамма-интеpферона.
Известен ген С10L, кодирующий белок с антивирусной активностью в 4 раза выше, чем у зрелого гамма-интерферона. Укороченный ген С10L кодирует белок в 133 аминокислотных заменах остатка (ако), у которого удалены 10 ако с С-конца. Экспрессия гена обеспечивается сильным trc-промотором в векторе рКК233-2. Уровень экспрессии мутантного гена в Е. coli JМ105 составляет 38%
Известна рекомбинантная плазмидная ДНК рIFN- γ-trp2, кодирующая полипептид, обладающий антивирусной активностью до 20 млн. МЕ/мг, аминокислотная последовательность которого совпадает с последова- тельностью рекомбинантного зрелого интерферона (т.е. без CysTyr Cys на N-конце). Высокий уровень синтеза данного полипептида в Е. coli достигается за счет использования синтетического участка связывания рибосом и нуклеотидной последовательности 5'- CCTCТAG между данным участком и инициирующим кодоном ATG гена аналога гамма-интерферона. По принципу конструирования плазмиза pIFN-γ-trp2 является наиболее близким техническим решением к заявляемому.
Однако существенным недостатком плазмиды pIFN- γ -trp2 является то, что она кодирует известный полипептид, обладающий антивирусной активностью, не превышающей 20 млн.МЕ/мг.
Целью изобретения является получение полипептида, обладающего более высокой, чем известные аналоги гамма-интерферона, антивирусной активностью.
Цель достигается получением мутантного гена S54, рекомбинантной плазмиды pS54 и штамма-продуцента Е. coli, обеспечивающего синтез полипептида с высокой антивирусной активностью.
Сущность изобретения состоит в том, что олигонуклеотиднаправленным мутагенезом получают ген S54 и рекомбинацией замещают им ген природного гамма-интерферона в плазмиде pIFN- γ-trp2.
Мутантный ген S54 отличается от природного следующим: два нуклеотида в кодоне, кодирующем аминокислотный остаток Ser в 54 положении (отсчет от N-конца природного белка), заменены таким образом, что новый кодон кодирует Asp.
Плазмида рS54, содержащая мутантный ген S54, состоит из следующих элементов: большого фрагмента ЕсоRI-PstI плазмиды рIFN- γ-trp2 размером 3940 п.н. и фрагмента EcRI PstI, содержащего мутантный ген S54 (481 п.н.).
Плазмида рS54 содержит ген устойчивости к тетрациклину в качестве генетического маркера, имеет молекулярную массу 1,46 Мda (4421 п.н.) и обеспечивает в клетках Е. coli синтез мутантного полипептида, обладающего активностью гамма-интерферона.
Полученная плазмида рS54 обеспечивает синтез мутантного белка в Е.coli. Полученный мутантный белок обладает высокой антивирусной активностью 600 млн. МЕ/мг, т.е. в 30 раз более высокой, чем у зрелого гамма-интерферона, а его температура плавления на 2оС выше.
Способ конструирования мутантного гена S54 включает в себя выделение из плазмиды рIFN- γ-trp2 фрагмента ЕсоRI PstI (481 п.н.) и клонирование его в ДНК фага M13mp8, введение мутации в ген гамма-интерферона с помощью олигонуклеотида S54 по методу Золлера-Смита. Наличие мутации в гене доказано секвенированием ДНК по методу Сэнгера.
Мутантный ген S54 был переклонирован обратно в плазмиду pIFN-γ-trp2, для чего ее фрагмент ЕсоRI PstI (481 п.н.) был замещен на фрагмент ЕсоRI PstI, содержащий мутантный ген S54 и выделенный из репликативной формы ДНК М131FN. Строение полученной плазмиды pS54 подтверждают анализом рестрикционными эндонуклеазами ЕсоRI, PstI и SalI и определением нуклеотидной последовательности гена S54. Сайт SalI появляется в гене S54 в результате мутагенеза.
Для получения штамма-продуцента полипептида с активностью гамма-интерферона плазмидой pS54 трансформируют компетентные клетки Е.соli МН-1. Полученный таким образом штамм Е. сoli МН-1/pS54 характеризуется следующими признаками.
Морфологические признаки. Грамотрицательные, палочковидные клетки, неспороносные.
Культуральные признаки. Клетки хорошо растут на простых питательных средах. На плотной среде образуют круглые, прозрачные, блестящие колонии с ровным краем без пигмента. При росте в жидких средах (на минимальной среде с глюкозой или LB-бульоне) образуют интенсивную ровную муть.
Физико-биологические признаки. Клетки растут при температуре от 4 до 40оС при оптимуме рН от 6,8 до 7. В качестве источника азота используют как минеральные соли в аммонийной форме, так и органические соединения в виде пептона, триптона, дрожжевого экстракта, аминокислот и т.д. Не утилизирует многие углероды, отрицателен по сероводороду, ацетону, образует индол.
Устойчивость к антибиотикам. Клетки проявляют устойчивость к тетрациклину (до 10 мкг/мл), обусловленную наличием плазмиды.
Штамм Е. соli МН-1/pS54 обеспечивает индуцибельный синтез полипептида с антивирусной активностью и уровнем экспрессии до 40% от суммарного клеточного белка бактерии.
Штамм Е. соli MH-1/pS54 депонирован во Всесоюзной коллекции промышленных микроорганизмов ВНИИгенетика под коллекционным номером ВКПМ В-5943.
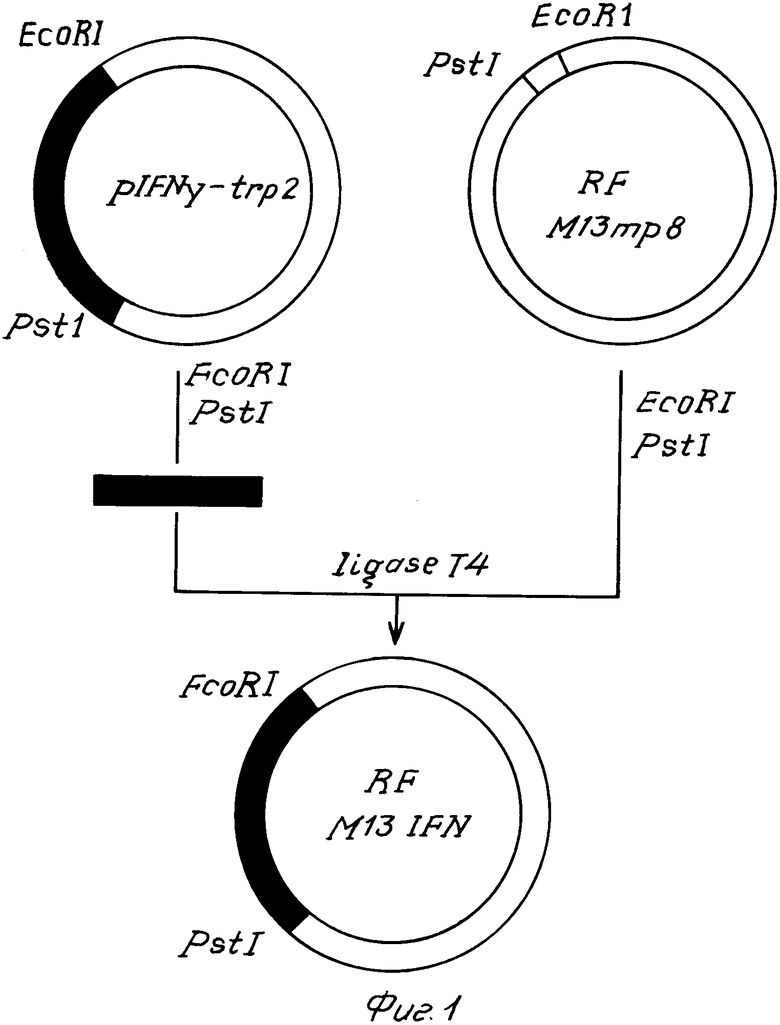
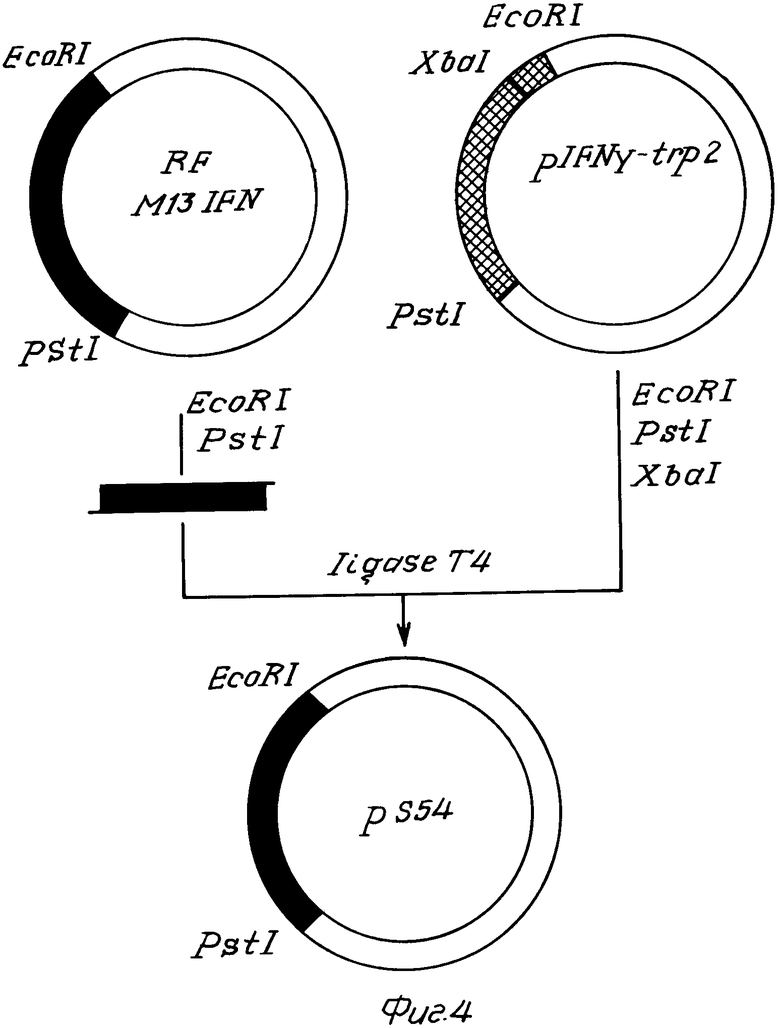
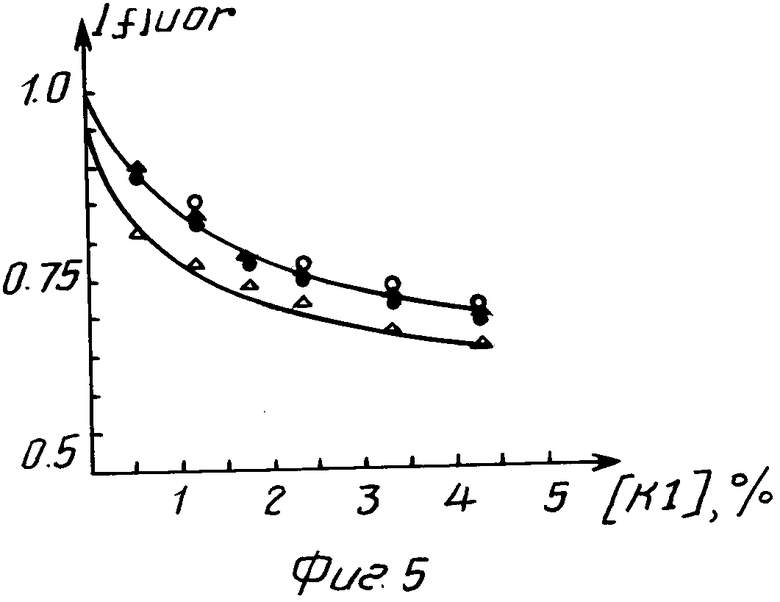
На фиг. 1 показана схема получения ДНК фага М13 IFN; на фиг. 2 схема введения мутации в ген гамма-интерферона, где А мутируемый участок гена гамма-интерферона; сверху показана аминокислотная последовательность кодируемого белка, цифрами отмечена нумерация аминокислотных остатков и оснований в белке и гене, Б олигонуклеотид-мутаген, звездами отмечены неиспаренные основания; на фиг. 3 нуклеотидная последовательность гена S54 и кодируемого им белка, подчеркнуты нуклеотидные замены в гене S54, отличающие его от прототипа; на фиг. 4 схема получения плазмиды рS54; на фиг. 5 зависимость интенсивности флуоресценции раствора интерферона от концентрации К1, где при возбуждении светом с длиной волны 268 нм и измерении флуоресценции на 320 нм ·- зрелый рекомбинантный гамма-интерферон человека, ° мутантный интерферон; при возбуждении светом с длиной волны 296 нм и измерении флуоресценции на 365 нм  - зрелый интерферон,
- зрелый интерферон,  мутантный интерферон.
мутантный интерферон.
П р и м е р 1. Способ получения мутантного гена S54.
Для проведения олигонуклеотиднаправленного мутагенеза необходимо было переклонировать ген γ -IFN из плазмиды pIFN- γ -trp2 в ДНК фага М13mp8. Схема клонирования представлена на фиг. 1.
5 мкг ДНК плазмиды pIFN- γ-trp2 расщепляли одновременно двумя эндонуклеазами рестрикции EcoRI и РstI (по десять единиц активности каждого фермента) в буфере, содержащем 50 мМ NaCl, 10 мМ MgCl, 10 мМ трис-НСl рН 8,1 мМ меркаптоэтанола. Гидролиз проводили при 37оС 3 ч Полноту гидролиза анализировали с помощью электрофореза.
Реакцию останавливали добавлением 0,5 М ЭДТА рН 7,5 до конечной концентрации 10 мМ. ДНК осаждали добавлением 1/10 объема ацетона натрия рН 7 и 2 объемов спирта. После высушивания осадок растворяли в необходимом объеме ТЕ-буфера, содержащем 10 мМ трис-HCl рН 8 и 1 мМ ЭДТА.
Фрагмент ДНК с геном IFN (481 п.н.) выделяли путем электрофоретического разделения реакционной смеси в 6%-ном полиакриламидном геле (ПААГ) с последующей электроэлюцией.
Лигирование смеси гидролизованной по ЕсоRI- и РstI-сайтам репликативной формы ДНК фага 13mp8 (80 нг) и фрагмента проводили с помощью ДНК-лигазы фага Т4 (4 ед.акт.) в объеме 30 мкл буферного раствора состава: 70 мМ трис-НСl рН 7,6. 5 мМ MgCl, 1 мМ дитиотрейтола, 2 мМ АТФ при 4-10оС в течение 12 ч Реакцию останавливали добавлением раствора ЭДТА до концентрации 20 мМ. ДНК осаждали этанолом из раствора 0,3 М ацетата натрия рН 7. Осадок растворяли после высушивания в 20 мкл стерильной воды. Контроль сшивки проводили электрофорезом в 1% -ном агарозном геле. Полученной ДНК трансформировали компетентные клетки Е. соli JM103, отбирали клоны с белой краской на индикаторной агаризованной среде с IPТG и Ygal, выделяли из них одноцепочечную ДНК и определяли нуклеотидную последовательность клонированного фрагмента по методу Сэнгера. Полученную одноцепочечную ДНК М13 IFN использовали для мутагенеза.
Для введения одновременно двух нуклеотидных замен в ген гамма-интерферона (фиг. 2) был синтезирован олигонуклеотид S54:5' -gaagtaaaagtcgacaatttgg.
Мутагенез проводили следующим способом.
После отжига с матричной ДНК М13mp8 IFN мутантого олигонуклеотида, фосфорилированного по 5'-концу, синтезировали с помощью фрагмента Кленова ДНК-полимеразы 1 и лигазы фага Т4 гетеродуплексную двухцепочечную ДНК. После трансформации такой ДНК компетентных клеток JМ 103 клоны, несущие мутантный ген γ -IFN, выявляли методом гибридизации с радиоактивно меченным олигонуклеотидом. Мутантный олигонуклеотид, меченный [32P] АТФ по 5'-концу, гибридизовали с репликами примерно ста клонов на нитроцеллюлозном (НЦ) фильтре при комнатной температуре. Далее НЦ-фильтр отмывали в 6*SSC (0,9 М NaCl, 0,09 М цитрата натрия) при различных температурах, получая после каждой отмывки его радиоавтограф. Поскольку олигонуклеотид-зонд полностью комплементарен лишь мутантной ДНК, то температура плавления дуплекса мутантной одноцепочечной ДНК с зондом выше, чем у немутантной ДНК с олигонуклеотидом-мутагеном. ДНК клонов, которые имели более сильный гибридизационный сигнал на радиографической пленке после отмывки фильтра при различных температурах, подвергали рестрикционному анализу и секвенированию по методу Сэнгера. ДНК фага M13mp8 IFN, содержащую мутантный ген гамма-интерферона (фиг. 3), назвали MIFN54.
П р и м е р 2. Получение плазмиды рS54.
Для этого репликативную форму ДНК MIFN54 гидролизовали по сайтам EcoRI-PstI (фиг. 4), рестрикционную смесь фракци- онировали в 6%-ной ПААГ. Полученный фрагмент длиной 481 п.н. клонировали в плазмиду pIFN-γ-trp2, обработанную рестриктазами ЕсоRI, PstI, XbaI для исключения возможности встраивания немутантного гена γ -IFN обратно в плазмиду. Компетентные клетки Е. coli JM 103 трансформировали реакционной смесью и высевали на среду с тетрациклином. Бактериальные колонии, содержащие плазмиду с мутантным геном интерферона, выявляли гибридизацией с радиоактивным олигонуклеотидом-зондом S54 и окончательно подтверждали структуру мутантного гена секвенированием ДНК плазмиды по методу Максама-Гилберта. Полученную плазмиду назвали pS54.
П р и м е р 3. Получение штамма-продуцента полипептида с активностью гамма-интерферона.
Плазмидой рS54 трансформируют компетентные клетки Е. coli МН-1 и получают штамм-продуцент полипептида с антивирусной активностью 600 млн. МЕ/мг.
П р и м е р 4. Выделение мутантного белка S54.
10 г биомассы ресуспендируется в 50 мл раствора А (7М гуанидина гидрохлорида, 25 мМ трис-HCl рН 7,3), суспензия перемешивается 30 мин при 0оС, обрабатывается ультразвуком 3 мин во льду и центрифугируется 30 мин со скоростью 17 000 об./мин при 4оС (центрифуга J-21 "Beckman"). Супернатант отбирается и разбавляется в 10 раз раствором В (25 мМ трис-HCl рН 7,3, 2 мM CuCl, 2 мМ ZnCl), перемешивается 2 ч на магнитной мешалке при 4оС, затем центрифугируется 60 мин со скоростью 12000 об./мин. Супернатант удаляется, а осадок растворяется в 50 мл буфера С (6М гуанидина гидрохлорида, 0,05 М ацетат аммония рН 8,5. 0,05% твин 80). С раствором проводится гель-фильтрация на Сефадексе G75, уравновешенном буфером (7М мочевины, 20 мМ трис-HCl рН 7,3). рН выделенной фракции доводится до 5 уксусной кислотой и проводится хроматография на сорбенте "Ольвагель". Элюция ведется последовательно растворами:
7М мочевина, 01 М ацетат натрия рН 5
7М мочевина, 0,1 М ацетат натрия рН 7
7М мочевина, 0,1 М ацетат аммония рН 7, 0,1 М NaCl
7М мочевина, 0,1 М ацетат аммония рН 7, 0,2 М NaCl.
Затем фракции, в которых находится мутантный интерферон, объединяются и диализуются против раствора 7М мочевины с 0,1 М ацетатом натрия рН 5 в течение 12 ч. Раствор рехроматографируется на сорбенте "Ольвагель", а затем выделенная фракция диализуется против 0,1 М ацетате аммония рН 7. Полученный раствор высокоочищенного мутантного интерферона стерильно фильтруется и распределяется по 100 мкл в пробирки "Eppendorf" для хранения при -20оС.
П р и м е р 5. Определение антивирусной активности мутантного белка S54.
Антивирусную активность аналога S54 определяли на культуре клеток фибробластов человека L68 по ингибированию цитопатического действия вируса везикулярного стоматита (ВВС) стандартным методом. К монослою клеток добавляли различные разведения препарата аналога S54, инкубировали 24 ч при 37оС в атмосфере углекислого газа, затем добавляли к каждому монослою 100 ЛД50 ВВС и инкубировали в тех же условиях еще 24 ч. после чего проводили учет. Состояние монослоя изучали с помощью инвертированного микроскопа. За единицу активности принимали то разведение аналога, которое обеспечивало сохранение 50% клеток монослоя. Для определения активности аналога в международных единицах (МЕ) пользовались референс-препаратом гамма-интерферона, изготовленным в НПО "Фермент" (г. Вильнюс).
Для расчета удельной антивирусной активности аналога S54 определяли также содержание белка в высокоочищенном препарате (не менее 99%) по методу Лоури или Бредфорда. Удельная антивирусная активность аналога S54 составляла 600 млн.МЕ/мг, тогда как у гамма-интерферона всего лишь 20 млн.МЕ/мг.
П р и м е р 6. Исследование пространственной структуры зрелого и мутантного (S54) гамма-интерферонов.
На спектрофлуориметре Hitachi 650 60 измерялась интенсивность флуоресценции растворов интерферонов с концентрацией 0,1 мг/мл в 0,1 М ацетата аммония (рН 7) в зависимости от концентрации К1 в интервале от 0,5 до 4%
Для изучения экспонированности тирозинов измерялась интенсивность флуоресценции на длине волны 320 нм при возбуждении светом с длиной волны 268 нм. Для изучения экспонированности триптофанов измерялась интенсивность флуоресценции на длине волны 365 нм при возбуждении светом с длиной волны 296 нм.
На фиг. 5 представлены зависимости интенсивностей флуоресценции растворов исходного и мутантного интерферонов от концентрации К1. Видно, что при совпадении всех остальных кривых кривая флуоресценции на длине волны 320 нм при возбуждении светом с длиной волны 268 нм у мутантного интерферона лежит ниже, чем у исходного интерферона. Это свидетельствует о том, что по крайней мере один остаток тирозина у молекулы мутантного интерферона в большей степени экспонирован, чем у молекулы исходного интерферона.
П р и м е р 7. Определение температуры плавления мутантного белка S54.
Калориметрические измерения проводились на сканирующем микрокалориметре ДАСМ-1М при нагревании 2оС/мин. Использовались растворы интерферонов с концентрацией 2 мг/мл в натрий-фосфатном буфере (20 мМ, рН 7). Температуру плавления определяли по максимуму приращения энтальпии.
Температура плавления исходного интерферона оказалась равной 62,3оС, а мутантного интерферона S54 63,6оС.
В настоящее время проводится работа по получению образцов полипептида, кодируемого геном S54, для доклинических испытаний. В перспективе планируется организация производства лекарственных препаратов на основе данного аналога гамма-интерферона человека.


Изобретение относится к биотехнологии, в частности к получению рекомбинантных интерферонов, обладающих активируемой активностью. Сущность изобретения: полипептид, обладающий более высокой, чем известные аналоги гамма-интерферона активностью, получают в результате замены двух нуклеотидов в кодоне, кодирующем Ser54 отсчет от Cys на N-конце непроцессированного гамма-интерферона. В результате мутантный ген кодирует полипептид, отличающийся от природного гамма-интерферона заменой Ser54 на Asp. Для экспрессии мутантного гена в E.coli была сконструирована плазмида pS54 с молекулярной массой 2,92 Md (4421 п. о.), содержащая большой фрагмент EcoR I Pst I плазмиды pBR 322 размером 3940 п. о. фрагмент EcoR I Pst I с геном (481 п. о.) в качестве генетического маркера геном устойчивости к тетрациклину, а также сайты узнавания рестрикционными эндонуклеазами, расположенными на следующих расстояниях от EcoR I-сайта: Hind III 29 п. о. BamH I 375 п. о. Pvu II 2068 н. о, Pst I 3609 п. о. EcoR I 4079 п. о. и участки узнавания Sal I расположены от EcoR I на 651 и 3941 п. о. Штамм-продуцент данного полипептида E.coli ВКПМ 5943 получается в результате трансформации E.coli МН-1 плазмидой pS54. Уровень экспрессии мутантного интерферона составляет 38% от суммарного клеточного белка. 3 с. п. ф-лы, 5 ил.
| Рекомбинантная плазмидная ДНК pIFN- @ тRр, кодирующая синтез человеческого иммунного интерферона | 1986 |
|
SU1433019A1 |

