ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Областью техники, к которой относится изобретение, является оптимизация процесса получения цистеинов, кэпированных 5-тио-2-нитробензоатом (TNB), на антителах посредством манипуляций с условиями роста клеток для обеспечения более эффективного получения конъюгатов антитело-лекарственное вещество (ADC).
УРОВЕНЬ ТЕХНИКИ
Конъюгаты антитело-лекарственное вещество (ADC) относятся к одному из видов направленной терапии, которые обычно состоит из антитела, "вооруженного" сильнодействующими цитотоксическими лекарственными веществами (Chudasama et al. Nature Chemistry, 2016; Junutula and Gerber, ACS medicinal chemistry letters 2016). Таким образом, ADC являются перспективными средствами, позволяющими обеспечивать избирательную доставку токсичной полезной нагрузки в опухоли, избегая распространения токсичности, из-за которой часто ограничивают или исключают химиотерапию в качестве долговременного лечения, за пределы целевой области. В качестве многообещающей терапевтической платформы в настоящее время на рынке представлены как минимум два одобренных для коммерческого применения продукта ADC (брентуксимаб ведотин, и трастузумаб эмтанзин). На фоне экспоненциального роста ADC среди них имеется значительное число терапевтических кандидатов, проходящих клиническую оценку.
Для того чтобы ADC могли достичь своего терапевтического потенциала, требуется выполнение сложных технологических процессов конъюгации для связывания цитотоксических лекарственных веществ с антителом. Большинство нынешних кандидатов на ADC, включая два коммерческих ADC, получают обычными неспецифическими методами конъюгации с участием случайного поверхностного лизина или свободных цистеинов восстановленных четырех межцепочечных дисульфидов. Это приводит к получению высокогетерогенных смесей ADC, что не только создает проблемы с воспроизводимостью их получения, но также значительно снижает терапевтический индекс.
Для решения этой проблемы, исследования в области, относящейся к ADC, движутся в сторону сайт-специфических технологий конъюгации, таких как сайт-специфические ADC на основе цистеина (Cys) (Junutula et al., Nat Biotechnol 2008). Устойчивые нуклеофильные тиоловые боковые цепи, содержащие сконструированные непарные остатки цистеина, позволяют проводить быструю и простую химическую реакцию конъюгации с целью присоединения различных линкеров/полезных нагрузок для обеспечения однородных продуктов ADC. Имеются сообщения о более четко определенных и улучшенных фармакокинетических (PK) профилях полученных таким образом молекул ADC. Сайт-специфическая платформа имеет свои технические проблемы. Если для получения используются клетки млекопитающих, тиоловые группы на введенных остатках цистеина образуют дисульфидные связи со свободными цистеинами или глутатионами (GSH). Эти так называемые модификации Cys-кэпирования должны быть удалены перед осуществлением конъюгации с лекарственным веществом через этап частичного восстановления. Поскольку такая обработка также уменьшает количество межцепочечных дисульфидов в антителе, такие восстановленные межцепочечные дисульфиды в антителе должны быть затем приведены в исходное состояние с помощью процесса повторного окисления, включающего выведение восстанавливающих агентов, цистеина или глутатиона, и обработку окисляющими реагентами (Junutula et al., Nat Biotechnol. 2008). Этот утомительный процесс восстановления и повторного окисления потенциально приводит к перетасовке и перекручиванию дисульфидных связей в антителе, что может отрицательно сказаться укладке и качестве белка, а также вызвать такие проблемы, как ухудшение PK у конечных ADC.
Для решения этой потенциальной проблемы была разработана новая стратегия селективного восстановления Cys-кэппирования с помощью тионитробензоата (TNB) (см. PCT IB2016/054789, включенная в настоящее описание в виде ссылки), не затрагивающего межцепочечные дисульфидные связи антитела. TNB-кэппинг, продукт реакции реагента Эллмана (5,5'-дитиобис(2-нитробензоат), DTNB) со свободной тиольной группой цистеина, представляет собой лабильный кэппинг из-за его слабого окислительно-восстановительного потенциала. Было показано, что восстанавливающий фрагмент трис(3-сульфонатофенил)фосфин (TSPP) может селективно удалять TNB-кэппинг без восстановления эндогенных дисульфидов. Этот процесс TNB/TSPP с последующей прямой конъюгацией устраняет необходимость в жестких условиях традиционных этапов восстановления-повторного окисления, сохраняя в целости исходную укладку антитела. Однако для оптимизации коммерческого производства ADC необходимы более эффективные методы TNB-кэпирования.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение основано на открытии того, что статус кэпирования остатков цистеина в антителах может быть модифицирован путем оптимизации параметров, таких как рост клеток, плотность клеток и/или культивирование клеток. Таким образом, изобретение относится к процессу продуцирования антител в клетках млекопитающих, в которых сконструированные неспаренные остатки цистеина остаются некэпированными тиолом, что обеспечивает эффективную модификацию, как правило, кэпирование тионитробензоатом (TNB) при добавлении различных концентраций дитионитробензоата (DTNB) на разных стадиях культивирования клеток или при разной плотности клеток.
Изобретение также относится к конъюгатам антитело-лекарственное вещество (ADC) или к конъюгатам терапевтический белок-лекарственное вещество, полученным с использованием этих TNB-кэпированных антител, в частности к ADC, образованным путем селективного восстановления остатков цистеина в TNB-кэпированных антителах, что позволяет избежать уменьшения количества межцепочечных дисульфидов и, таким образом, исключает необходимость в этапе (повторного)окисления перед конъюгацией. Кроме того, изобретение также относится к новым антителам, кэпированным тионитробензоатом, полученным способами, описанными в настоящей заявке, которые обеспечивают возможность для селективного восстановления с помощью TSPP или родственных агентов для осуществления прямой конъюгации с улучшенной эффективностью.
Более конкретно, в настоящем изобретении продемонстрировано, что статус кэпирования неспаренных поверхностных цистеинов на антителе можно улучшить путем осторожной манипуляции ростом клеток, плотностью клеток или условиями культивирования клеток. В культуре клеток млекопитающих цистеин является ключевой аминокислотой. Цистеин и его окисленная форма цистин (Ctn) в культуральной среде преимущественно используются для роста клеток, предположительно, с участием транспортера Xc- аминокислот (Sato et al., J Biol Chem 1999; см. фиг. 1). Реакция Cys-кэпирования цистеином/цистином/глутатионом вне клеток является медленным процессом, и, следовательно, непарные поверхностные цистеины остаются некэпированными свободными тиолами во время культивирования клеток с высокой плотностью. При добавлении DTNB к клеточной культуре на определенной стадии, были получены практически гомогенные TNB-кэпированные антитела. Эти результаты обусловили осуществимую стратегию производства TNB-кэпированных антител в больших количествах.
Кроме того, процессы культивирования клеток обычно разрабатываются с гарантией того, что количество аминокислот, таких как цистеин и его эквиваленты, не будет ограничено или истощено, чтобы добиться максимального роста клеток, плотности и титра жизнеспособных клеток и предотвратить аминокислотные замены (т.е. неправильное включение аминокислот) (см. патент США № 8,232,075 В). L-цистеин и его окисленная форма, L-цистин, считаются ключевыми аминокислотами в культуре клеток млекопитающих, поскольку генерация цистеина в результате превращения метионина ограничивает скорость роста клеток, клеточного метаболизма и продуктивности. Однако в настоящем изобретении для получения надлежащего статуса кэпирования остатков цистеина в антителе с помощью TNB использовали несколько способов намеренного ограничения или уменьшения количества цистеина/цистина, доступного для клеточной культуры.
Таким образом, в варианте осуществления изобретения предложен способ получения цистеин-содержащих белков, которые могут быть конъюгированы с химической полезной нагрузкой, причем этот процесс включает этапы: (а) засева ростовой среды клетками, способными экспрессировать цистеин-содержащие белки, причем среда содержит один или более ростовых компонентов, выбранных из цистеина, цистина и глутатиона; (b) инкубации клеток до достижения плотности клеток, достаточной для истощения большинства компонентов, необходимых для роста, присутствующих в ростовой среде; и (с) дальнейшей инкубации клеток для экспрессии цистеин-содержащих белков, имеющих один или более некэпированных остатков цистеина, содержащих свободный тиол. Этот процесс может дополнительно включать этап: (d) введения заданного кэпирующего фрагмента или его предшественника в экспрессированные цистеин-содержащие белки, в результате чего происходит кэпирование одного или более цистеинов в белке(белках) заданным кэпирующим фрагментом. В этом варианте осуществления достигаемая плотность клеток обычно составляет по меньшей мере примерно 1E6 кл./мл и может составлять, например, по меньшей мере примерно 5E6 кл./мл, 10E6 кл./мл, 50E6 кл./мл, 100E6 кл./мл или 500E6 кл./мл, предпочтительно выше 10E6 кл./мл, более предпочтительно выше 50E6 кл./мл.
Следует отметить, что заданный кэпирующий фрагмент может представлять собой алкилирующий агент, в некоторых случаях действующий в качестве химических «ручек», отличных от TNB или аналогичных лабильных фрагментов, полезных для дополнительных типов реакции конъюгации с лекарственным веществом. Эти ручки прикрепляются к антителу путем добавления новых химических алкилирующих спейсеров в культуральную среду. Химические алкилирующие спейсеры содержат химические ручки, такие как альдегиды, кетоны, азиды и алкины. В случае кетонов и альдегидов эти химические ручки могут реагировать с аминоокси-нуклеофилами или гидразидом для дополнительной химии конъюгации, образуя продукты оксима/гидразона. В случае азидов и алкинов эти химические ручки могут обеспечивать конъюгацию через циклоприсоединение. Дополнительные алкилирующие химические спейсеры включают функциональный домен биотина, который обеспечивает специфическое тесное нековалентное взаимодействие между стрепавидином и биотином. См. пример 4 в WO2017/025897, в котором обсуждается химическая ручка - малеимид-триокса-4-формил бензамид (MTFB), дибензоциклооктил-полиэтилен-малеимид (DBCO-ПЭГ4-малеимид) и малеимид-ПЭГ2-биотин (MPB).
В дополнительном варианте осуществления изобретения предложен способ получения цистеин-содержащих белков, которые могут быть конъюгированы с химической полезной нагрузкой, причем этот способ включает этапы: (а) засева ростовой среды клетками, способными экспрессировать цистеин-содержащие белки, причем среда содержит один или более ростовых компонентов, выбранных из цистеина, цистина и глутатиона; (b) инкубации клеток для экспрессии цистеин-содержащих белков, имеющих один или более некэпированных остатков цистеина, содержащих свободный тиол; и (с) на этапе (а), этапе (b) или на обоих этапах (а) и (b) поддержание концентрации одного или более ростовых компонентов на уровне менее 0,4 мМ, менее 0,3 мМ, менее 0,2 мМ, менее 0,1 мМ или менее 0,05 мМ. Этот способ может дополнительно включать этап: (d) введения заданного кэпирующего фрагмента или его предшественника в экспрессированные цистеин-содержащие белки, в результате чего происходит кэпирование одного или более цистеинов в белках заданным кэпирующим фрагментом.
Еще один вариант осуществления относится к способу получения цистеин-содержащих белков, которые могут быть конъюгированы с химической полезной нагрузкой, причем этот способ включает этапы: (а) засева ростовой среды клетками, способными экспрессировать цистеин-содержащие белки, причем среда содержит один или более ростовых компонентов, выбранных из цистеина, цистина и глутатиона, включая, без ограничения, ростовую среду, в которой начальная концентрация ростовых компонентов находится на уровне менее 2 мМ, менее 0,4 мМ, менее 0,3 мМ, менее 0,2 мМ, менее 0,1 мМ или менее 0,05 мМ, затем (b) инкубации клеток для экспрессии цистеин-содержащих белков, имеющих один или более некэпированных остатков цистеина, содержащих свободный тиол. Этот способ может дополнительно включать этап: (c) введения заданного кэпирующего фрагмента или его предшественника в экспрессированные цистеин-содержащие белки, в результате чего происходит кэпирование одного или более цистеинов в белках заданным кэпирующим фрагментом.
Также в рамках настоящего изобретения находятся варианты осуществления, в которых ростовые компоненты в клеточной культуре истощают путем намеренного ограничения в способе цистеина и/или его альтернативных форм. Используя рационально разработанные среды и стехиометрические подходы, изложенные в US 8232075 B, количество цистеина/цистина, необходимое для конкретной пиковой плотности клеток и количества продуцируемого продукта, можно рассчитать по приведенному ниже уравнению (уравнение 1). Это уравнение является общим для культур СНО, и для конкретной клеточной линии можно вывести более частное уравнение путем определения скорости потребления цистеина в этой клеточной линии в заданном способе.
Уравнение 1. Требуемая концентрация цистеина
Требуемая концентрация цистеина=[(x*m)+(x*m*k)+(p*n)]*f
х: концентрация цистеина, необходимая для клеток Е6/мл (0,09 мМ)
m: пиковая плотность клеток (Е6 кл./мл)
k: коэффициент стабилизации (клеток) (10-15%)
р: концентрация цистеина, необходимая для 1 г/л антитела (0,19 мМ)
n: конечная концентрация антитела (1 г/л)
f: коэффициент устойчивости (1,1-1,3)
После определения необходимого количества цистеина для конкретной клеточной линии в данном процессе, для определения количества цистеина/цистина можно использовать уравнение относительного показателя предельного количества цистеина, чтобы обеспечить достижение в процессе определенного показателя предельного количества (уравнение 2), для намеренного ограничения или истощения культуры по цистеину/цистину. В рамках настоящего изобретения представлены варианты, в которых компоненты роста истощают путем ограничения относительного показателя предельного количества цистеина в ростовой среде для клеток до менее чем примерно 1,0x. Эти показатели могут составлять, например, примерно: 0,95x, 0,90x, 0,85x, 0,80x, 0,75x, 0,70x, 0,65x, 0,60x, 0,55x, 0,50x, 0,45x, 0,40x, 0,35x, 0,30. х, 0,25х, 0,20х, 0,15х, 0,10х или 0,05х.
Уравнение 2. Относительный показатель предельного количества цистеина
Относительный показатель предельного количества цистеина =
количество цистеина, обеспечиваемое в процессе/количество цистениа, требуемое по уравнению 1
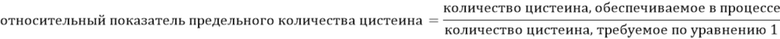
Конечно, цистеин-содержащие белки по изобретению включают широкий спектр фрагментов, включая, без ограничения, антитела и слитые белки. Это могут быть анти-EDB антитела и анти-HER2 антитела, включая трастузумаб.
В способах, описанных в настоящей заявке, заданный кэпирующий фрагмент может быть выбран из группы, состоящей из 5-тио-2-нитробензойной кислоты (TNB), 2-меркаптопиридина, дитиодипиридина (DTDP), 4-тиобензойной кислоты, 2-тиобензойной кислоты, 4-тиобензолсульфокислоты, 2-тиобензолсульфокислоты, метилсульфоната (Ms), п-толуолсульфоната (Ts) и трифторметансульфоната (Tf); и/или заданный кэпирующеий фрагмент выбирают из реакционноспособной группы, состоящей из из малеимид-триокса-4-формилбензамид (MTFB) подобных молекул с альдегидной ручкой или малеимид-азидо-лизин подобных молекул с азидной ручкой или дибензоциклооктил-полиэтилен-малеимид (DBCO-ПЭГ4-малеимид) подобных молекул с алкиновой ручкой. Зачастую заданный кэпирующий фрагмент представляет собой TNB, и часто предшественник представляет собой DTNB.
Также в способах, описанных в настоящей заявке, кэпированные белки подвергают дополнительной обработке, состоящей из одного или более процессов выделения, очистки и концентрации, в одной или более различных точках способа. Таким образом, в некоторых вариантах осуществления изобретения, по меньшей мере, 50% клеток отделяют от экспрессированных цистеин-содержащих белков перед введением заданного кэпирующего фрагмента или его предшественника. Это отделение может быть достигнуто центрифугированием или фильтрацией.
Изобретение также включает варианты осуществления, относящиеся к конъюгации TNB-кэпированного (или кэпированного иным образом) цистеин-содержащего белка, включающей этапы: (a) взаимодействия TNB-кэпированного цистеин-содержащего белка с восстанавливающим агентом, способным отщеплять TNB-кэпирующие фрагменты от белка без существенного уменьшения количества межцепочечных серных связей в антителе; (b) фильтрования реакционной смеси для удаления избытка восстанавливающего агента, отсоединенного TNB или обоих; и (c) без введения окисляющего агента, конъюгации одной или более восстановленных серных связей в антителе с полезной нагрузкой через реакционноспособный линкирующий фрагмент (линкер).
Изобретение также дополнительно включает варианты осуществления конъюгации TNB-кэпированного (или кэпированного иным образом) цистеин-содержащего белка, включающей этапы: (а) взаимодействия TNB-кэпированного цистеин-содержащего белка со стехиометрическим избытком восстанавливающего агента, способного отсоединять TNB-кэпирующие фрагменты от белка без существенного уменьшения количества межцепочечных серных связей в антителе, необязательно в присутствии солей, таких как хлорид натрия или других солей (см. пример 8); (b) фильтрование реакционной смеси для удаления избыточного количества одного или более из восстанавливающего агента, отщепленного TNB; (с) введение окисляющего агента для репарации межцепочечных серных связей, восстановленных за счет избытка восстанавливающего агента; и (d) конъюгации одной или более восстановленных серных связей в антителе с полезной нагрузкой через реакционноспособный линкирующий фрагмент. Стехиометрический избыток восстанавливающего агента к кэпированному остатку цистеина обычно составляет от 4:1 до 6:1 (например, от 16:1 до 24:1 восстанавливающего агента к антителу, содержащему четыре кэпированных цистеина) и предпочтительно примерно 5:1. Этап (с) этого процесса может быть выполнен при температуре окружающей среды, например, примерно 25 градусов Цельсия, чтобы сократить время, требуемое на окисление, и избежать изменения температуры процесса. Производительность при низких температурах, например примерно 4 градусов Цельсия, требует более длительного времени окисления, но менее чувствительна к потере выхода, если превышено целевое время окисления. Вышеописанный процесс может дополнительно включать этап: (е) добавления избытка цистеина после этапа (d) для блокирования реакции линкирующего фрагмента; и (f) отделения блокированного линкера-полезной нагрузки от конъюгата. Блокирование цистеином позволяет улучшить отделение линкера-полезной нагрузки с помощью хроматографии или диафильтрации. Кроме того, в вышеописанном способе отделение может быть выполнено с помощью диафильтрации или колоночной хроматографии, обычно хроматографии гидрофобного взаимодействия (HIC). Использование изопропанол-содержащих буферов для HIC очистки приводит к увеличению извлечения очищенного конъюгата.
ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг. 1. Цистеин и его окисленная форма цистин (Ctn) внутри и вне клетки.
Фиг. 2. Соотношение лекарственное вещество-антитело, равное 4:1 (DAR4).
Фиг. 3. Полностью некэпированное мутантное по цистеину антитело получали в результате стабильной экспрессии в клетках СНО при высокой плотности клеток в обычной среде CD CHO. Клетки CHO-K1, стабильно экспрессирующие мутантный по цистеину трастузумаб K290C-K334C или K392C-K334C, высевали в среду CD CHO с плотностью 3E6 кл./мл или 6E6 кл./мл и культивировали в течение 72 часов при 37 градусах Цельсия. Кондиционированные среды очищали, пропуская через колонку ProA и гель-фильтрационную колонку (SEC). Очищенные белки антител анализировали методом ЖХ/МС, как описано в Примере 1.
Фиг. 4. Условия культивирования клеток с высокой плотностью стабильной клеточной линии СНО в среде СНО с DTNB. Клетки CHO-K1, стабильно экспрессирующие мутантный по цистеину трастузумаб K183C-K290C, высевали в запатентованную минимальную среду с плотностью 0,6E6 кл./мл в биореакторе с контролируемой периодической загрузкой, как описано в Примере 1. На панели A показаны условия культивирования в минимальных средах, питательных средах или DTNB. На панели B показана плотность жизнеспособных клеток (VCD) и жизнеспособность культуры.
Фиг. 5. Полностью TNB-кэпированное антитело мутантное по цистеину в сайте Fc K290C получали в результате стабильной экспрессии в клетках СНО при высокой плотности клеток в среде CHO с DTNB. Кондиционированные среды из условий культивирования, описанных на фиг.4, очищали, пропуская через колонку ProA/SEC, и мутантное по цистеину антитело трастузумаб K183C-K290C расщепляли с помощью IDES (панель A) и анализировали методом ЖХ/МС, как описано в примере 1.
Фиг. 6. Полностью TNB-кэпированное антитело мутантное по цистеину в сайте Fab K183C получали в результате стабильной экспрессии в клетках СНО при высокой плотности клеток в среде CHO с DTNB. Кондиционированные среды из условий культивирования, описанных на фиг.4, очищали, пропуская через колонку ProA/SEC, и мутантное по цистеину антитело трастузумаб K183C-K290C расщепляли IDES и анализировали методом ЖХ/МС, как описано в примере 1.
Фиг. 7. Условия культивирования клеток с высокой плотностью для стабильной экспрессии клеток СНО в HiPDOG и процессах с периодической загрузкой DTNB. Клетки CHO-K1, стабильно экспрессирующие мутантный по цистеину трастузумаб K183C-K290C, высевали в запатентованную минимальную среду с плотностью 2E6 кл./мл или 0,6E6 кл./мл в контролируемый биореактор. На панели A показаны условия культивирования в минимальных средах, питательных средах или DTNB. На панели B показана плотность жизнеспособных клеток (VCD).
Фиг. 8. Полностью некэпированное мутантное по цистеину антитело получали в стабильно экспрессирующих клетках СНО при высокой плотности клеток в среде CHO с DTNB. Кондиционированные среды из условий культивирования, описанных на фиг.7, очищали, пропуская через колонку ProA/SEC, и мутантное по цистеину антитело трастузумаб K183C-K290C расщепляли IDES и анализировали методом ЖХ/МС (панель А). На панели B показана сводная таблица данных по кэпированию.
Фиг. 9. TNB-кэппинг сгенерированных мутантных по цистеину антител, трастузумаб K183C-K290C. Кэппинг определяли с помощью анализа ЖХ/МС, который позволяет определить виды кэпирования мутированных остатков цистеина. Мутированные по цистеину антитела получали в культуре клеток с высокой плотностью в запатентованных минимальных и питательных сред намеренно истощенных по цистеину/цистину. Во время продуцирования партии в культуру вносили добавку DTNB. Кондиционированные среды из культуры подвергали ProA очистке перед анализом методом ЖХ/МС. Первичные продуцированные виды (> 95%) представляли собой целевые полностью кэпированные mAb с четырьмя TNB; в смеси присутствовали низкие уровни видов, содержащих менее четырех TNB-кэпов.
Фиг. 10. Профили цистеина в условиях питательных сред с высокой и низкой концентрациями цистеина/цистина. Концентрации получали с помощью аминокислотного анализа, выполненного методом СВЭЖХ; концентрации цистина, полученные в результате анализа СВЭЖХ, подвергали стехиометрическому преобразованию в цистеин. В обоих условиях использовали клетки CHO-K1, стабильно экспрессирующие мутантное по цистеину антитело трастузумаб K183C-K290C, которые вводили в биореактор периодического действия объемом 1 л с запатентованной минимальной и питательной средами. В обоих условиях начальные концентрации цистеина/цистина в минимальной среде были одинаковыми, а концентрации цистина в питательной среде различались. Образцы кондиционированных сред из каждого условия анализировали, начиная с 4-го дня, для получения временной динамики истощения цистеина/цистина на протяжении всего периода культивирования партии.
Фиг. 11. Цистеиновые профили условий культивирования с высокой и низкой концентрациями дополнительного цистина. Концентрации получали с помощью аминокислотного анализа, выполненного методом СВЭЖХ; концентрации цистина, полученные в результате анализа СВЭЖХ, подвергали стехиометрическому преобразованию в цистеин. Во всех условиях использовали клетки CHO-K1, стабильно экспрессирующие мутантное по цистеину анти-EDB K183C-K290C антитело, которые вводили в биореактор периодического действия объемом 1 л с запатентованной минимальной и питательной средами. Во всех условиях начальные концентрации цистеина/цистина в минимальной среде были одинаковыми относительно плотности посева. Образцы кондиционированных сред из каждого условия анализировали, начиная с 0-го дня, для получения временной динамики истощения цистеина/цистина на протяжении всего периода культивирования партии.
Фиг. 12. Профили цистеина в условиях с целевыми относительными показателями предельного количества цистеина. Концентрации получали с помощью аминокислотного анализа, выполненного методом СВЭЖХ; концентрации цистина, полученные в результате анализа СВЭЖХ, подвергали стехиометрическому преобразованию в цистеин. Во всех условиях использовали клетки CHO-K1, стабильно экспрессирующие мутантное по цистеину антитело трастузумаб K183C-K290C, которые вводили в биореактор периодического действия объемом 1 л с запатентованной минимальной и питательной средами. Целевые относительные показатели предельного количества цистеина определяли с помощью уравнений 1 и 2, как описано ранее; все условия имели разные уровни цистина в соответствующих средах для достижения целевого относительного показателя предельного количества цистеина. Образцы кондиционированных сред из каждого условия анализировали, начиная с 4-го дня, для получения временной динамики истощения цистеина/цистина на протяжении всего периода культивирования партии.
Фиг. 13. Профили плотности жизнеспособных клеток и концентрации цистеина в условиях высокой и низкой пиковой плотности клеток. Высокие и низкие пиковые плотности клеток получали для клеток CHO-K1, стабильно экспрессирующих мутантное по цистеину антитело трастузумаб K183C-K290C. В обоих условиях использовали биореакторы периодического действия объемом 1 л с запатентованной минимальной и питательной средами; хотя эти два условия различались по некоторым параметрам процесса для достижения и сохранения различных пиковых плотностей, концентрации цистеина/цистина были одинаковыми в минимальной среде и идентичными в питательной среде. На панели A показана плотность жизнеспособных клеток. На панели B показана концентрация цистеина во время культивирования партии. Образцы кондиционированных сред из каждого условия анализировали, начиная с 0-го дня, для получения временной динамики истощения цистеина/цистина на протяжении всего периода культивирования партии. Аминокислотный анализ проводили методом СВЭЖХ; концентрации цистина, полученные из анализа СВЭЖХ, подвергали стехиометрическому преобразованию в цистеин.
Фиг. 14. Результаты анализа исходного DAR4 при добавлении DTNB в кондиционированную среду. Клетки CHO-K1, стабильно экспрессирующие мутантное по цистеину антитело трастузумаб K183C-K290C, культивировали в биореакторе периодического действия объемом 1 л с запатентованной минимальной и питательной средами. После достижения определенной временной точки в процессе культивирования партии в биореакторе, клетки отделяли от кондиционированной среды центрифугированием и фильтровали через 0,2 мкм фильтр. Кондиционированную среду переносили в отдельный сосуд и вводили 2 мМ DTNB, и инкубировали в течение различных промежутков времени. Образцы с разными периодами инкубации подвергали ProA очистке и конъюгировали с помощью базового процесса конъюгации, используя TNB (см. Пример 5), определяли процентное содержание неочищенного DAR4, который использовали в качестве суррогатного маркера полностью TNB-кэпированного антитела.
Фиг. 15. Результаты анализа исходного DAR4 при добавлении DTNB в кондиционированную среду. Клетки CHO-K1, стабильно экспрессирующие мутантное по цистеину антитело анти-EDB K183C-K290C, культивировали в биореакторе периодического действия объемом 1 л с запатентованной минимальной и питательной средами. После достижения определенной временной точки в процессе культивирования партии в биореакторе, клетки отделяли от кондиционированной среды центрифугированием и фильтровали через 0,2 мкм фильтр. Кондиционированную среду переносили в отдельный сосуд и вводили 2 мМ DTNB, и инкубировали в течение различных промежутков времени. Образцы с разными периодами инкубации подвергали ProA очистке и конъюгировали с помощью базового процесса конъюгации, используя TNB (см. Пример 5), определяли процентное содержание неочищенного DAR4, который использовали в качестве суррогатного маркера полностью TNB-кэпированного антитела.
Фиг. 16. Влияние замены буфера после восстановления. TNB-кэпированное антитело трастузумаб K183C-K290C восстанавливали 6 эквивалентами TSPP (37°C в течение 3 часов) и конъюгировали с различными количествами mcvcPABC0101; квадраты: замена буфера после восстановления, ромбы: буфер не меняли. % DAR4 измеряли с помощью аналитической хроматографии гидрофобного взаимодействия.
Фиг. 17. Влияние этапа повторного окисления. Кривые, полученные с помощью аналитической хроматографии гидрофобного взаимодействия (детектирование при 280 нм) после восстановления 20 эквивалентами TSPP, замены буфера и конъюгации с 10 эквивалентами mcvcPABC0101; сплошная линия: отсутствие повторного окисления, пунктирная линия: повторное окисление дегидроаскорбиновой кислотой перед конъюгацией. Сверхконъюгированные виды возникают в результате конъюгации с уменьшенным количеством межцепочечных дисульфидных связей.
Фиг. 18. Сравнение неочищенных конъюгатов мутантного по цистеину анти-EDB K183C-K290C антитела и мутантного по цистеину антитела трастузумаб K183C-K290C. Конъюгаты получали в соответствии с протоколом, описанным ниже в Примере 9, и анализировали с помощью аналитической хроматографии гидрофобного взаимодействия (детектирование при 280 нм).
Фиг. 19. Очистка с помощью хроматографии гидрофобного взаимодействия мутантного по цистеину конъюгата трастузумаба. Неочищенный конъюгат получали согласно процедуре, описанной в Примере 9, и очищали с помощью колонки и условий, описанных в Примере 14.
Фиг. 20. Обработка антитела или конъюгата одним ферментом, который разрезает антитело под шарнирной областью, и вторым ферментом, который разрезает над шарнирной областью.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Общие процедуры
Способы культивирования клеток
Используемые в настоящем описании термины «культура» и «клеточная культура» относятся к популяции клеток, которая суспендирована в среде в условиях, подходящих для выживания и/или роста популяции клеток. Как будет понятно специалистам в данной области техники, в некоторых вариантах осуществления эти термины, используемые в настоящей заявке, относятся к комбинации, содержащей популяцию клеток и среду, в которой суспендирована популяция. В некоторых вариантах осуществления клетки культуры содержат клетки млекопитающих.
В настоящем изобретении могут быть использованы любые способы культивирования клеток, которые обеспечивают нужный процесс (например, продуцирование рекомбинантного белка (например, антитела)). В качестве неограничивающего примера, клетки могут быть выращены в периодической культуре или культуре с подпиткой, при этом культивирование прекращают после получения достаточного количества экспрессированного рекомбинантного белка (например, антитела), после чего экспрессированный белок (например, антитело) собирают. В качестве альтернативы, в качестве другого неограничивающего примера, клетки можно выращивать в условиях с периодической подпиткой, когда культивирование не прекращают, и новые питательные вещества и другие компоненты периодически или непрерывно добавляют в культуру, и во время культивирования экспрессируемый рекомбинантный белок (например, антитело) периодически или непрерывно собирают. В данной области известны и другие подходящие способы (например, культуры в центрифужных пробирках), которые могут быть использованы для практического осуществления настоящего изобретения.
В некоторых вариантах осуществления клеточная культура, подходящая для настоящего изобретения, представляет собой подпитываемую культуру. Используемый в настоящем описании термин «подпитываемая культура» относится к способу культивирования клеток, при котором дополнительные компоненты вводят в культуру во время или после начала процесса культивирования. Такие вводимые компоненты обычно содержат питательные компоненты для клеток, которые истощаются во время процесса культивирования. Культивирование подпитываемой культуры обычно останавливают в определенный момент, и клетки и/или компоненты среды собирают и при необходимости очищают. В некоторых вариантах осуществления подпитываемая культура содержит минимальную среду, дополненную питательными средами. В некоторых вариантах осуществления поддерживают низкие уровни лактата путем доставки глюкозы, регулируемой высоким конечным значением рН (процесс HiPDOG), описанной в Gagnon et al.
В некоторых вариантах осуществления процесс культивирования клеток, подходящий для настоящего изобретения, представляет собой непрерывный процесс. Используемый в настоящем описании термин «непрерывный» относится к способу культивирования клеток, при котором клетки инокулируют в минимальную среду, и в тот момент, когда клетки достигают целевой плотности, переходят на непрерывный процесс культивирования, при котором отработанную среду заменяют свежей средой. Непрерывный процесс позволяет получать культуры с высокой плотностью клеток и, таким образом, позволяет производить большое количество продукта. Однако по меньшей мере некоторые формы непрерывного процесса требуют доставки большого количества среды, в результате чего некоторая часть продукта остается в большом объеме отработанной среды, а не накапливается в едином сборе продукта.
Используемый в настоящем описании термин «биореактор» относится к любому сосуду, используемому для роста культуры прокариотических или эукариотических клеток (например, культуры клеток млекопитающих). Биореактор может быть любого размера при условии, что он подходит для культивирования клеток (например, клеток млекопитающих). Клетки могут быть выращены в любом удобном объеме, выбранном специалистом. Например, клетки могут выращиваться в небольших реакционных сосудах объемом от нескольких миллилитров до нескольких литров. В качестве альтернативы, клетки могут быть выращены в больших коммерческих биореакторах, объем которых составляет примерно от 1 литра до 10, 50, 100, 250, 500, 1000, 2500, 5000, 8000, 10000, 12000, 15000, 20000 или 25000 литров или больше, или любой объем между ними.
В некоторых вариантах осуществления клетки можно выращивать во время начальной фазы роста (или фазы роста) в течение более или менее продолжительного периода времени, в зависимости от потребностей специалиста и потребностей самих клеток. В некоторых вариантах осуществления клетки выращивают в течение периода времени, достаточного для достижения заданной плотности клеток. В некоторых вариантах осуществления клетки выращивают в течение периода времени, достаточного для достижения плотности клеток, которая представляет собой заданный процент от максимальной плотности клеток, которой клетки в конечном итоге достигли бы, если бы росли без помех. Например, клетки можно выращивать в течение периода времени, достаточного для достижения целевой плотности жизнеспособных клеток: 1, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95 или 99 процентов от максимальной плотности клеток. В некоторых вариантах осуществления клетки выращивают до тех пор, пока плотность клеток не станет увеличиваться более чем на 15%, 14%, 13%, 12%, 11%, 10%, 9%, 8%, 7%, 6%, 5%. 4%, 3%, 2% или 1% на каждый день культивирования. В некоторых вариантах осуществления клетки выращивают до тех пор, пока плотность клеток не станет увеличиваться более чем на 5% на каждый день культивирования.
В некоторых вариантах осуществления клетки могут расти в течение определенного периода времени. Например, в зависимости от начальной концентрации культуры клеток, температуры, при которой выращивают клетки, и собственной скорости роста клеток, клетки можно выращивать в течение 0, 1, 2, 3, 4, 5, 6. 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 или более дней, предпочтительно от 4 до 10 дней. В некоторых случаях клетки могут расти в течение месяца или более. Специалист в области к, которой относится настоящее изобретение, сможет выбрать начальную фазу роста в зависимости от требований к продуцированию белка и потребностей самих клеток.
В некоторых вариантах осуществления клетки можно выращивать на следующей фазе продуцирования до тех пор, пока не будет достигнута целевая плотность клеток или титр продукта. В другом варианте осуществления настоящего изобретения клетки выращивают в течение определенного периода времени на следующей фазе продуцирования. Например, в зависимости от концентрации клеток в культуре в начале следующей фазы роста, температуры, при которой выращивают клетки, и собственной скорости роста клеток, клетки можно выращивать в течение 1, 2, 3 г. 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 и более дней. В некоторых случаях клетки могут расти в течение месяца или более. Специалист в области к, которой относится настоящее изобретение, сможет выбрать следующую фазу продуцирования в зависимости от требований к продуцированию полипептида или белка и потребностей самих клеток.
В некоторых вариантах осуществления клетки экспрессируют рекомбинантный белок, и способ культивирования клеток по изобретению включает фазу роста и фазу продуцирования.
Клетки
В соответствии с настоящим изобретением может быть использована любая клетка, чувствительная к клеточной культуре. В некоторых вариантах осуществления клетка представляет собой клетку млекопитающего. Неограничивающие примеры клеток млекопитающих, которые можно использовать в соответствии с настоящим изобретением, включают линию клеток мышиной миеломы BALB/c (NSO/1, ECACC №: 85110503); человеческой ретинобластомы (PER.C6, CruCell, Leiden, Netherlands); линию клеток CV1 почки обезьяны, трансформированная SV40 (COS-7, ATCC CRL 1651); клеточную линию, полученную из эмбриональной почки человека (клетки 293 или 293, субклонированные для роста в суспензионной культуре, Graham et al., J. Gen Virol., 36: 59, 1977); клетки почек детенышей хомяков (BHK, ATCC CCL 10); клетки яичника китайского хомячка +/-DHFR (CHO, Urlaub и Chasin, Proc. Natl. Acad. Sci. USA, 77: 4216, 1980); мышиные клетки Сертоли (TM4, Mather, Biol. Reprod., 23: 243-251, 1980); клетки почки обезьяны (CV1 ATCC CCL 70); клетки почки африканской зеленой мартышки (VERO-76, ATCC CRL-1 587); клетки карциномы шейки матки человека (HeLa, ATCC CCL 2); клетки почки собаки (MDCK, ATCC CCL 34); клетки печени крысы Buffalo (BRL 3A, ATCC CRL 1442); клетки легких человека (W138, ATCC CCL 75); клетки печени человека (Hep G2, HB 8065); клетки мышиной опухоли молочной железы (MMT 060562, ATCC CCL51); клетки TRI (Mather et al., Annals N.Y. Acad. Sci., 383: 44-68, 1982); клетки MRC 5; клетки FS4; и линию клеток гепатомы человека (Hep G2). В некоторых предпочтительных вариантах осуществления клетки представляют собой клетки СНО. В некоторых предпочтительных вариантах осуществления в клетках используется система экспрессии гена глутаминсинтетазы (GS).
Кроме того, в соответствии с настоящим изобретением может быть использовано любое количество коммерчески доступных и некоммерческих линий гибридомных клеток. Термин «гибридома» в контексте настоящего описания относится к клетке или потомству клетки, полученной в результате слияния иммортализованной клетки и клетки, продуцирующей антитело. Полученная таким образом гибридома представляет собой иммортализованную клетку, которая продуцирует антитела. Отдельные клетки, используемые для создания гибридомы, могут быть получены из любого млекопитающего, включая, без ограничения, крысу, свинью, кролика, овцу, свинью, козу и человека. В некоторых вариантах осуществления гибридома представляет собой клеточную линию триомы, которая возникает, когда потомство слитых гетерогибридных миелом, которые являются продуктом слияния человеческой клетки с клеточной линией мышиной миеломы, затем сливают с плазматической клеткой. В некоторых вариантах осуществления гибридома представляет собой любую иммортализованную линию гибридных клеток, которая продуцирует антитела, такую как, например, квадрома (см., например, Milstein et al., Nature, 537: 3053, 1983). Специалист в данной области осведомлен о том, что линии гибридомных клеток могут иметь разные требования к питанию и/или могут требовать разные условия культивирования для оптимального роста, и способен модифицировать условия по мере необходимости.
Рост клеток и продуктивность
Используемая в настоящей заявке высокая плотность клеток относится к плотности клеток выше 1E6 кл./мл, 5E6 кл./мл, 10E6 кл./мл, 50E6 кл./мл, 100E6 кл./мл или 500E6 кл./мл, предпочтительно более 10E6 кл./мл, более предпочтительно более 50E6 кл./мл.
В некоторых вариантах осуществления рост клеток определяется плотностью жизнеспособных клеток (VCD), максимальной плотностью жизнеспособных клеток или общим количеством жизнеспособных клеток (IVCC). В некоторых вариантах осуществления рост клеток определяется максимальной плотностью жизнеспособных клеток.
Используемый в настоящей заявке термин «плотность жизнеспособных клеток» относится к числу клеток, присутствующих в заданном объеме среды. Плотность жизнеспособных клеток может быть измерена любым способом, известным специалисту. Предпочтительно, плотность жизнеспособных клеток измеряется с помощью автоматического счетчика клеток, такого как Bioprofile Flex® (Nova Biomedical, Waltham, MA). Используемый в настоящей заявке термин «максимальная плотность клеток» относится к максимальной плотности клеток, достигнутой во время культивирования клеток. Используемый в настоящей заявке термин «жизнеспособность клеток» относится к способности клеток в культуре выживать при заданном наборе условий культивирования или экспериментальных вариациях. Специалистам в данной области техники должно быть понятно, что в настоящее изобретение включен один из многочисленных способов определения жизнеспособности клеток. Например, для оценки жизнеспособности клетки можно использовать краситель (например, трипановый синий), который не проходит через мембрану живой клетки, но может проходить через разрушенную мембрану мертвой или умирающей клетки.
Используемый в настоящем описании термин «общее количество жизнеспособных клеток (IVCC)» относится к площади под кривой плотности жизнеспособных клеток (VCD). IVCC можно рассчитать по следующей формуле:
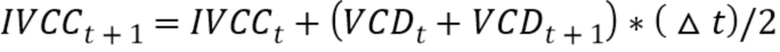
где Δt представляет собой разницу между моментами времени t и t+1. IVCCt=0 можно считать пренебрежимо малой величиной. VCDt и VCDt+1 представляет собой плотность жизнеспособных клеток в моменты времени t и t+1.
В некоторых вариантах осуществления описанных выше способов продуктивность определяется титром и/или объемной продуктивностью.
Используемый в настоящем описании термин «титр» относится, например, к общему количеству рекомбинантно экспрессируемого белка, продуцируемого клеточной культурой, в заданном объеме среды. Титр обычно выражается в единицах граммов белка на литр среды.
В некоторых вариантах осуществления описанных выше способов продуктивность определяется титром. В некоторых вариантах осуществления продуктивность увеличивается по меньшей мере на 5, 10, 15, 20 или 25% по сравнению с продуктивностью контрольной культуры. В некоторых вариантах осуществления продуктивность увеличивается по меньшей мере на 10% по сравнению с таковой контрольной культуры. В некоторых вариантах осуществления продуктивность увеличивается по меньшей мере на 20% по сравнению с таковой контрольной культуры.
Среда для клеточной культуры
Используемые в настоящей заявке термины «среда», «среда для культивирования клеток» и «среда для культивирования» относятся к раствору, содержащему компоненты или питательные вещества, которые необходимы для питания растущих клеткок млекопитающих. Как правило, питательные вещества включают незаменимые и заменимые аминокислоты, витамины, источники энергии, липиды и микроэлементы, необходимые клетке для минимального роста и/или выживания. Такой раствор может также содержать дополнительные питательные вещества или дополнительные компоненты, которые ускоряют рост и/или увеличивают вероятность выживания относительно минимального уровня, включая, без ограничения, гормоны и/или другие факторы роста, в частности ионы (такие как натрий, хлорид, кальций, магний и фосфат), буферы, витамины, нуклеозиды или нуклеотиды, микроэлементы (неорганические соединения, обычно присутствующие в очень низких конечных концентрациях), неорганические соединения, присутствующие в высоких конечных концентрациях (например, железо), аминокислоты, липиды и/или глюкозу или другой источник энергии. В некоторых вариантах осуществления изобретения среду предпочтительно готовят таким образом, чтобы рН и концентрация соли были оптимальными для выживания и пролиферации клеток. В некоторых вариантах осуществления среда представляет собой питательную среду, которая добавляется после начала культивирования клеток.
Методы измерения концентрации аминокислот
Концентрация аминокислоты может быть измерена любым способом, известным специалисту в данной области. Предпочтительные способы измерения концентрации аминокислот в реальном времени или традиционные способы включают, например, жидкостную хроматографию, такую как ВЭЖХ, СВЭЖХ или ЖХ/МС, ЯМР или ГХ/МС.
В некоторых вариантах осуществления концентрацию аминокислоты измеряют традиционным способом, отбирая образец среды для культивирования клеток и измеряя концентрацию указанной по меньшей мере одной аминокислоты в указанном образце. В некоторых вариантах осуществления концентрацию аминокислоты измеряют, как описано в примерах 3.1, 3.2 и 3.3. Предпочтительным способом измерения концентрации аминокислот является традиционный СВЭЖХ способ.
В некоторых вариантах осуществления концентрацию аминокислоты измеряют в режиме реального времени. В некоторых вариантах осуществления концентрацию аминокислоты измеряют в режиме реального времени методом спектроскопии комбинационного рассеяния. В некоторых вариантах осуществления концентрацию аминокислоты измеряют в режиме реального времени методом спектроскопии комбинационного рассеяния. В некоторых вариантах осуществления концентрацию аминокислоты измеряют в режиме реального времени с помощью технологии на основе ВЭЖХ или СВЭЖХ с автоматическим пробоотборником, который отбирает пробу из реактора и передает ее в оборудование программируемым образом.
Дополнительные процедуры, описанные в WO 2015/140708, также могут быть использованы в настоящем изобретении и включены в настоящее описание в виде ссылки.
Методы измерения отношения лекарственное вещество к антителу
Отношение лекарственное вещество-антитело (DAR) может быть измерено любым способом, известным специалисту в данной области. Предпочтительные методы измерения DAR включают, например, жидкостную хроматографию, такую как ВЭЖХ, СВЭЖХ или ЖХ/МС, масс-спектрометрия и ЯМР.
В некоторых вариантах осуществления DAR измеряют путем отбора образца смеси для конъюгации, хроматографической фракции или дополнительного очищенного материала и измерения концентрации указанного в указанном образце. Предпочтительными методами измерения DAR являются хроматография гидрофобного взаимодействия (HIC ВЭЖХ) и ВЭЖХ с обращенной фазой.
Дополнительные определения
В дополнение к приведенным выше определениям приведены следующие дополнительные определения:
СНО описано в настоящей заявке как клетки яичника китайского хомяка (СНО), которые получены из яичника китайского хомяка, используемые для продуцирования терапевтических белков, как описано в приведенных примерах. CHO-K1 описаны в настоящем документе как линия родоначальник клеток-хозяев, субклонированная из родительской клеточной линии CHO, содержащей систему экспрессии глутаминсинтазы (GS), коммерчески доступные от Lonza.
Используемый в настоящей заявке термин «HiPDOG» относится к «доставке глюкозы, контролируемой высоким конечным значением pH» (HiPDOG), представляющей собой метод введения питательных веществ, при котором доставка концентрированного раствора глюкозы инициируется повышением pH с целью подавления накопления лактата в клеточной культуре.
Используемый в настоящем описании термин mAb относится к моноклональному антителу.
Используемый в настоящем описании термин DAR4 относится к соотношению лекарственное вещество-антитело, равному 4:1, при котором четыре молекулы полезной нагрузки-лекарственного вещества связаны с одним белком (например, см. фиг. 2, на которой изображен один такой случай, где «R» представляет собой полезную нагрузку). Термин исходное DAR4 описывает отношение лекарственного вещества к антителу после процесса конъюгации, но до конечной стадии очистки.
Используемый в настоящем описании термин УФ/ДФ относится к ультрафильтрации/диафильтрации.
Примеры
Приведенные ниже примеры иллюстрируют важные признаки изобретения.
Пример 1: Получение полностью некэпированного мутантного по цистеину антитела в результате стабильной экспрессии клеток СНО, культивируемых в обычной CD CHO среде при высокой плотности клеток
Клетки CHO-K1, стабильно экспрессирующие мутантное по цистеину антитело трастузумаб K290C-K334C или K392C-K334C, высевали в обычную среду CD CHO (Thermo Fisher, Waltham, MA) с плотностью 3E6 кл./мл или 6E6 кл./мл и поддерживали в увлажненном инкубаторе с 5% CO2 при 37 градусах Цельсия. В одном из условий к среде CD CHO добавляли 5 мМ цистина. Клетки культивировали в течение 72 часов при 37 градусах Цельсия. Измеряли жизнеспособность клеток, и собирали кондиционированные среды. Клетки были жизнеспособными более чем на 98%.
Белок-антитело очищали, пропуская через колонку ProA и гель-фильтрационную колонку следующим образом. Кондиционированные среды фильтровали через 0,2 мкм фильтры и пропускали через смолу с белком А (GE Healthcare, Piscataway, NJ), предварительно уравновешенную 50 мМ Трис, 150 мМ NaCl, рН 7,5 (TBS). Перед элюированием белка (100% этап) смесью 150 мМ глицина и 40 мМ NaCl (рН 3,5) колонку промывали 2 объемами колонки (CV) TBS, 5CV CaCl2, pH 7,5, 3 CV смеси 10 мМ Трис и 10 мМ NaCl, pH 7,5. Значение рН белка доводили до 7,0 с помощью 2М HEPES, рН 8,0, и белок загружали в колонку Superdex 200, уравновешенную PBS (137 мМ NaCl, 2,7 мМ KCl, 8,1 мМ Na2HPO4, 2,7 мМ KH2PO4, рН 7,2). Пиковые фракции объединяли и концентрировали до 10 мг/мл с помощью 50 кДа MWCO центрифужного устройства.
Образцы белка анализировали vtnjljv масс-спектрометрии для измерения cnfnecf Cys-кэппинга следующим образом. Масс-спектрометрический анализ c помощью жидкостной хроматографии (ЖХ/МС) выполняли на масс-спектрометре Waters Xevo Q-TOF G2 (Waters, Milford, MA) в сочетании с капиллярной ВЭЖХ Agilent (Santa Clara, CA) 1200. Образцы белка обрабатывали протеазой IdeS (Promega, Madison, WI) при комнатной температуре в течение 2 часов. Пробы белка подкисляли, разбавляя в соотношении 1:1 0,05% ТФУ (Sigma-Aldrich, St Louis, MO), с последующим масс-спектрометрическим анализом с помощью жидкостной хроматографии. Образцы разделяли на колонке Waters BEH300 C4, 1,7 мкм (1,0×50 мм), поддерживаемой при 80°C со скоростью потока 65 мкл/мин. Подвижная фаза A представляла собой воду с 0,05% ТФУ, а подвижная фаза B представляла собой ацетонитрил с 0,05% ТФУ. Белки элюировали из колонки, используя градиент: 2%-20% B в течение 0,5 минуты, 20%-40% B в течение 6 минут и 40%-100% B в течение 4 минут. Масс-спектрометр работал в режиме съемки положительных ионов только в пределах 800-3500 m/z, и данные получали с помощью программного обеспечения MassLynx (Waters) 4.1. Сигнал TOF-MS, соответствующий антителу, суммировали и деконволютировали с помощью программы MaxEnt1 (Waters). Виды, кэпированные цистеином и глутатионом, определяли по сдвигу массы (Cys: 119,004 Да, GSH: 305,068 Да).
Как показано на фиг.3, для среды CD CHO мутантное по цистеину антитело было полностью цистеинировано при добавлении в среду 5 мМ цистина. В обычной среде CD CHO клетки CHO продуцировали существенное количество некэпированного мутантного по цистеину белка при плотности клеток 3E6 кл./мл. При увеличении количества клеток CHO до 6E6 кл./мл продуцированные мутантные цистеину антитела были полностью некэпированными, что позволяет предположить, что большая часть цистина в среде используется для роста клеток. Увеличение плотности клеток может истощить среду по цистину и, следовательно, повлиять на состояние кэппинга неспаренных поверхностных цистеинов.
Пример 2: Получение полностью TNB-кэпированных мутантных по цистеину антител при культивировании стабильных СНО клеток в обычной СНО среде при высокой плотности клеток
Как показано в примере 1, во время роста при высокой плотности клетки могли потреблять существенные количества цистеина или цистина, находящегося в среде. В результате можно было получить полностью некэпированное мутантное по цистеину антитело. Результаты примера 1 позволили получить полностью TNB-кэпированные мутантные по цистеину антитела в большом количестве путем добавления различных концентраций DTNB в клеточную культуру в разные моменты времени. DTNB эффективно алкилирует свободные тиолы на некэпированных мутантных антителах, что приводит к образованию TNB-кэпированных антител.
Таким образом, клеточные линии CHO, стабильно экспрессирующие мутантное по цистеину антитело трастузумаб K183C-K290C, высевали в контролируемый биореактор (Applikon, Inc., Schiedam, Netherlands) с начальным рабочим объемом 1 л. Как показано на фиг.4А, была проведена оценка восьми условий с разными концентрациями цистеина и/или цистина в минимальной среде. Культуру с подпиткой засевали с плотностью 0,6E6 кл./мл и контролировали путем подпитки питательными веществами с фиксированным процентным содержанием в зависимости от объема культуры, начиная с дня 3; TNB-кэпированные антитела получали в питательной среде без цистеина и цистина. В день 7 в культуру добавляли 1 мМ DTNB, и культивирование клеток продолжали в течение следующих пяти дней.
Как показано на фиг. 4B, процесс удвоения клеток и их жизнеспособность были нормальными во всех тестируемых условиях.
После очистки белков-антител, их подвергали анализу для определения уровня Cys-кэпирования. Как показано на фиг.5А, антитело расщепляли с помощью протеазы Ides с получением фрагмента Fc и фрагмента Fab2. Смесь анализировали методом ЖХ/МС, как описано в Примере 1 выше. Как показано на фиг. 5В, для участка F2, расположенного в Fc, менее подверженного воздействию растворителя, при всех условиях добавления TNB (опыты 2-8) получали >95% TNB-кэпированных материалов. Как показано на фиг. 6 для сайта K183C, расположенного в Fab легкой цепи, в большей степени подверженного воздействию растворителя, добавление TNB (опыты 5-8) также приводит к образованию преимущественно TNB-кэпированных материалов.
Для дальнейшего повышения эффективности TNB-кэпирования были протестированы условия культивирования с более высокой плотностью клеток. Для улучшенного контроля лактата использовали контролируемый уровнем глюкозы процесс HiPDOG (Gagnon et al., Biotechnol Bioeng 2011). Экспериментальные условия с плотностью посева и концентрациями цистеина/цистина представлены на фиг. 7А. Лактат-управляемый процесс HiPDOG (опыт 3) обеспечивал существенно более высокую пиковую плотность клеток, чем соответствующие условия с подпиткой (опыт 7) (фиг. 7В).
Антитело очищали от кондиционированной среды и подвергали анализу уровня Cys-кэпирования методом ЖХ/МС. Как показано на фиг. 8, сайт K183C Fab был TNB-кэпирован на 95% в опытах 3-7, в то время как сайт Fc K290C был TNB-кэпирован примерно на 98% в опытах 3-7.
Пример 3: Получение полностью TNB-кэпированных мутантных по цистеину антител в процессах с ограниченным количеством цистеина/цистина
В приведенных ниже примерах значения эффективности кэпирования мутантных по цистеину антител представлены в виде процента исходного DAR4 (отношения лекарственное вещество к антителу), полученного после конъюгации линкера-полезной нагрузки с антителом перед окончательной очисткой. Значения исходного DAR4 использовали в этих примерах в качестве суррогатного маркера эффективности кэпирования; однако определение статуса кэппирования каждого мутированного цистеинового остатка на антителе может быть выполнено с помощью более длительного анализа ЖХ/МС (описанного в примере 1) перед процессом конъюгации. Пример анализа ЖХ/МС редставлен фиг. 9, на котором идентифицированы и количественно определены различные виды кэппинга в одном антителе. Образец, использованный для этого анализа представлял собой TNB-кэпированные антитела, полученные в процессе культивирования с низким содержанием цистеина/цистина с DTNB-подпиткой. Требуемый целевой вид представляет собой четыре TNB на антитело, что составляет >95% согласно результатам ЖХ/МС, полученным для этого конкретного примера, с низкими уровнями невосстановленных кэпов и некоторыми видами суб-4 антител.
Пример 3.1: Намеренное ограничение количества цистеина/цистина в культуре путем уменьшения концентрации в питательных средах
Подход 1: В этом примере показано намеренное ограничение концентрации цистеина и/или цистина в культуре путем уменьшения количества любого из этих компонентов, содержащихся в добавляемой питательной среде. Два условия, использованные в этом примере, имели сходные начальные концентрации цистеина и/или цистина в минимальной среде; однако в одном из условий среда для подпитки питательными веществами не содержала цистеинового или цистинового компонента, в то время как во втором условии использовали питательную среду, которая содержала только цистин (таблица 1); все другие параметры процесса двух условий были идентичными. Стандартные процессы культивирования клеток млекопитающих включают цистеин и/или цистин в дополнительных вспомогательных подпитках из-за проблем, связанных с растворимостью этих компонентов в средах для культивирования клеток.
В этом примере клетки CHO-K1, стабильно экспрессирующие мутантное по цистеину антитело трастузумаб K183C-K290C, использовали с запатентованными минимальной и питательной средами в биореакторах Applikon объемом 1 л (Applikon, Inc., Schiedam, Netherlands), работающих с модульными контроллерами BioNet (Broadley-James Corp., Irvine, CA) с модулями перистальтического насоса и регулятора массового расхода газа. Культуру засевали с плотностью примерно 2E6 кл./мл, температуру поддерживали равной примерно 37 градусов Цельсия, и величину рН доводили до примерно 7,0 путем добавления либо раствора карбоната натрия/калия, либо CO2. Уровень растворенного кислорода поддерживали равным >20% путем барботирования чистого кислорода. Для TNB-кэпирования антител подпитку DTNB начинали после фазы роста до достижения в биореакторе целевой концентрации DTNB, равной 4 мМ. Продолжительность культивирования партии для приведенных ниже примеров составляла примерно 12 дней.
Нормализованные исходные значения DAR4 демонстрируют, что удаление цистина из питательной среды оказалось успешным способом истощения культуры по доступному цистеину/цистину, что, в свою очередь, позволило повысить уровень кэпирования антитела с помощью TNB и получить конъюгаты антитело-лекарственное вещество с более высоким процентным значением DAR4 без вредного воздействия на рост или продуктивность клеток.
Таблица 1. Подход 1 - Концентрации цистеина и цистина в среде и нормализованное исходное DAR4 с трастузумабом K183C-K290C
(E6 кл./мл)
(%)
цистина в питательной среде
Acquity UPLC (Waters Corp, Milford, MA) использовали для аминокислотного анализа условий из дополнительного эксперимента, в котором контрольным условием было введение 0,5 мМ цистина в дни 7, 9 и 11, чтобы избежать ограничения по цистину в любой временной точке в партии согласно рациональному дизайну среды, как описано ранее (таблица 2); для TNB-кэпирования используемая целевая концентрация DTNB составляла 5 мМ. Все остальные параметры процесса были идентичны параметрам, использованным в эксперименте, описанном в таблице 1.
Таблица 2. Подход 1 - Концентрации цистеина и цистина в среде и время истощения с трастузумабом K183C-K290C
7, 9, 11
цистина в питательной среде
Результаты ясно демонстрируют, что в этом конкретном примере удаление цистеина/цистина из питательной среды действительно приводит к снижению концентрации цистеина/цистина в культуре в дни 6-7; концентрации цистина, полученные в результате анализа СВЭЖХ, подвергали стехиометрическому преобразованию в цистеин и графически отображали (см. фиг. 10).
Понятно, что если бы длительность культивирования партии в этом конкретном примере составляла только 5 дней, в культуре опыта 4 был бы избыток доступного цистеина/цистина, что привело бы к неоптимальному TNB-кэпированию антитела. В этом случае увеличение продолжительности культивирования партии до 8 дней или дольше было эффективным методом для истощения избытка цистеина/цистина, что привело к получению подходящей среды для оптимального TNB-кэпирования.
Кроме того, следует понимать, что может быть реализована стратегия замены среды, такая как перфузия, для удаления в дальнейшем цистеина/цистина из культуры в опыте 4. Питательная среда с более высокими уровнями цистеина/цистина могла бы быть использована во время фазы роста партии, и после достижения пиковой плотности жизнеспособных клеток можно заменить средой, содержащей низкие уровни цистеина/цистина для имитации очень похожего профиля истощения по цистеину/цистину, сравнимого с фиг. 10.
Подход 2: В этом примере показана такая же стратегия, как в Подходе 1 намеренного ограничения концентраций цистеина и/или цистина в культуре путем уменьшения количества любого из этих компонентов, содержащихся в питательной среде для подпитки, но с использованием второй клеточной линии, продуцирующей антитела. Условия, использованные в этом примере, были сгруппированы на условия с высокой или низкой плотностью посева и имели сходные начальные концентрации цистеина и/или цистина в минимальной среде. В одном из условий с высокой или низкой плотностью посева цистеиновая или цистиновая подпитка отсутствовала, кроме начальных концентраций в минимальной среде, в то время как в остальных условиях использовалась отдельная дополнительная подпитка цистином (таблица 3); во всех условиях подпитка питательными веществами не содержала цистеин или цистин. Все остальные параметры процесса были идентичными для двух условий в соответствии с состоянием посева (например, условия с более высокой плотностью посева требовали более высокой скорости подачи дополнительных питательных веществ).
В этом примере клетки CHO-K1, стабильно экспрессирующие мутантное по цистеину анти-EDB антитело K183C-K290C, использовали с запатентованными минимальной и питательной средами в биореакторах Applikon объемом 1 л (Applikon, Inc., Schiedam, Netherlands), работающих с модульными контроллерами BioNet. (Broadley-James Corp., Irvine, CA) с модулями перистальтического насоса и регулятора массового расхода газа. Культуру засевали с плотностью клеток примерно 0,6E6 кл./мл или 3E6 кл./мл, температуру поддерживали равной примерно 37 градусов по Цельсию, и значение pH доводили до примерно 7,0 путем добавления либо раствора карбоната натрия/калия, либо CO2. Уровень растворенного кислорода поддерживали равным >20% путем барботирования чистого кислорода. Для TNB-кэпирования антител подпитку DTNB начинали после фазы роста до достижения в биореакторе целевой концентрации DTNB, равной 4 мМ. Продолжительность культивирования партии для приведенных ниже примеров составляла примерно 12 дней.
Оценка второй клеточной линии показала, что удаление цистеина и/или цистина из любых дополнительных подпиток является надежной стратегией по истощению цистеина/цистина, доступного в культуре, что впоследствии позволяет осуществлять TNB-кэпирование антитела более эффективно. Эта стратегия в конечном итоге приводит к конъюгатам антитело-лекарственное вещество с более высок процентным значением исходного DAR4 без вредного воздействия на рост или продуктивность клеток (таблица 3).
Таблица 3. Подход 2 - Концентрации цистеина и цистина в средах и нормализованное исходное DAR4 с анти-EDB K183C-K290C
(начиная с дня 4)
(E6 кл./мл)
(%)
Acquity UPLC (Waters Corp, Milford, MA) использовали для аминокислотного анализа условий подхода 2 (таблица 4). Все концентрации цистина, полученные в результате анализа СВЭЖХ подвергали преобразованию в цистеин (см. фиг. 11). Результаты ясно показывают, что удаление подпитки цистеином/цистином приводит к снижению концентрации в культуре уже на 7-й день (фиг. 11), что обеспечивает подходящую среду для оптимального TNB-кэпирования, о чем свидетельствуют значения исходного DAR4.
Таблица 4. Подход 2 - Концентрации цистеина и цистина в среде и время истощения с анти-EDB K183C-K290C
низкая плотность посева
(начиная с дня 4)
цистина
(<0,5 мМ)
1значение в день 7 составило 0,54 мМ; предполагаемое истощение до уровня менее 0,5 мМ в день 8 основано на показателях роста и продуктивности культуры
Пример 3.2: Ограничение уровня цистеина/цистина в культуре с использованием стехиометрического подхода и целевых относительных показателей предельного количества цистеина: Используя рациональный дизайн среды и стехиометрические подходы, описанные выше, можно рассчитать количество цистеина/цистина, необходимое для получения конкретной пиковой плотности клеток, и можно вычислить количество продуцируемого продукта и использовать для разработки оптимальной среды для этого конкретного процесса (уравнение 1). В этом примере требуемое количество цистеина/цистина рассчитывали, исходя из рационального дизайна среды с использованием оцененных пиковой плотности жизнеспособных клеток и титра сбора, получаемых в этом процессе. Выполняли оценку диапазона различных относительных показателей предельного количества цистеина, ниже расчетного требуемого количества цистеина/цистина (например, по сравнению с показателями, описанными в US 8232075 В), с целью определения идеального целевого количество, которое обеспечит ограничение или истощение цистеина/цистина, таким образом способствуя TNB-кэпированию антитела, при этом все еще в условиях приемлемых значений пиковой плотности жизнеспособных клеток и титров сбора (см. Уравнение 2).
Все необходимые концентрации цистеина/цистина рассчитывали на основе целевого относительного показателя предельного количества цистеина, которые добавляли только в минимальную среду.
Клетки CHO-K1, стабильно экспрессирующие мутантное по цистеину антитело трастузумаб K183C-K290C, использовали с запатентованными минимальной и питательной средами в биореакторах Applikon объемом 1 л (Applikon, Inc., Schiedam, Netherlands), работающих с модульными контроллерами BioNet (Broadley-James Corp., Irvine, CA) с модулями перистальтического насоса и регулятора массового расхода газа. Культуру засевали с плотностью клеток примерно 2E6 кл./мл, температуру поддерживали, равной примерно 37 градусов по Цельсию, и значение pH доводили до примерно 7,0 путем добавления либо раствора карбоната натрия/калия, либо CO2. Уровень растворенного кислорода поддерживали равным >20% путем барботирования чистого кислорода. Для TNB-кэппирования антитела подачу DTNB начинали после фазы роста до достижения в биореакторе концентрации DTNB, равной 4 мМ. Продолжительность культивирования партии для приведенных ниже примеров составляла примерно 12 дней. Помимо концентраций цистеина/цистина в минимальной среде, все параметры процесса были идентичными для всех оцениваемых условий.
Из таблицы 5 ясно видно, что с увеличением относительного показателя предельного количества цистеина исходное DAR4 (и, следовательно, кэпирование антитела) уменьшается; это указывает на то, что более низкий относительный показатель предельного количества цистеина является наиболее оптимальным для кэпирования антитела, и в этом примере отношения 0,63x и 0,76x обеспечивали приемлемые значения пиковой плотности жизнеспособных клеток, титры сбора и аналогичные результаты кэпирования (таблица 5). Титр сбора и значения исходного DAR4 были нормализованы.
Таблица 5. Сравнение относительный показатель предельного количества цистеина
(E6 кл./мл)
(%)
Acquity UPLC (Waters Corp, Milford, MA) использовали для аминокислотного анализа условий, сходных с описанными в таблице 5. Все концентрации цистина, полученные в результате анализа СВЭЖХ подвергали стехиометрическому преобразованию в цистеин. Результаты ясно показывают, что более низкие относительные показатели предельного количества цистеина раньше приводят к истощению культуры по цистеину/цистину по сравнению с условием 1,15x, что свидетельствует о том, что использование целевого относительного показателя предельного количества цистеина может привести к согласованному и предсказуемому низкому уровню Cys-кэпирования и высокой эффективности TNB-кэпирования (фиг. 12).
Пример 3.3: Ограничение концентрации цистеина/цистина в культуре путем увеличения пиковой плотности клеток: Как было продемонстрировано ранее на двух клеточных линиях в Примере 3.1, если цистеин/цистин ограничен низкими концентрациями или полностью отсутствует во время культивирования партии, эффективность TNB-кэпирования увеличивается. Согласно второму примеру, намеренное снижение концентрации цистеина/цистина в культуре или истощение культуры по цистеину/цистину может быть получено путем использования пиковой плотности жизнеспособных клеток. По мере увеличения плотности жизнеспособных клеток в данной культуре, как правило, увеличивается потребление и потребность в аминокислотах (US 8232075 B). В этом примере проведена оценка двух отдельных условий, в которых были достигнуты различные пиковые плотности жизнеспособных клеток.
Клетки CHO-K1, стабильно экспрессирующие мутантное по цистеину антитело трастузумаб K183C-K290C, использовали с запатентованной минимальной и питательной средами в биореакторах Applikon объемом 1 л (Applikon, Inc., Schiedam, Netherlands), работающих с модульными контроллерами BioNet (Broadley-James Corp., Irvine CA, Калифорния) с модулями перистальтического насоса и регулятора массового расхода газа. Культуру засевали с плотностью клеток примерно 0,6E6 кл./мл или 2E6 кл./мл, температуру поддерживали, равной примерно 37 градусов по Цельсию, и значение pH доводили до примерно 7,0 путем добавления либо раствора карбоната натрия/калия, либо CO2. Уровень растворенного кислорода поддерживали равным >20% путем барботирования чистого кислорода. Продолжительность культивирования партии для приведенных ниже примеров составляла примерно 12 дней; сбор в одном из условий был выполнен рано из-за низкой жизнеспособности клеток. Некоторые параметры процесса, такие как скорость подачи питательных веществ, для двух условий различались для достижения и поддержания различных пиковых плотностей клеток. Однако концентрации цистеина и цистина в минимальной среде были одинаковыми, и концентрации цистина в питательной среде были одинаковыми (Таблица 6).
Таблица 6: Концентрации цистеина и цистина в среде
(E6 кл./мл)
Acquity UPLC (Waters Corp, Milford, MA) использовали для аминокислотного анализа условий, описанных в таблице 6. Все концентрации цистина, полученные в результате анализа СВЭЖХ, подвергали стехиометрическому преобразованию в цистеин. Аминокислотный анализ этих условий показал, что в условии с более высокой пиковой плотностью жизнеспособных клеток произошло успешное истощение цистеина/цистина в партии раньше по сравнению с условием с более низкой пиковой плотностью жизнеспособных клеток (фиг. 13B), в котором остаточная концентрация цистеина/цистина сохранилась до момента сбора. Исходное DAR4 не было проанализировано для этих образцов, но получено на основании данных аминокислотного анализа; в условии с посева с высокой плотностью истощение цистеина/цистина произошло раньше. Из этих данных можно сделать вывод, что более высокая пиковая плотность жизнеспособных клеток обеспечила оптимальную среду с низким содержанием цистеина/цистина для эффективного TNB-кэпирования. Это свидетельствует о том, что поддержание условий, которые могут стимулировать рост культуры с достижением более высоких плотностей клеток может быть эффективным альтернативным способом поддержания низких или истощенных уровней цистеина/цистина в клеточной культуре для создания требуемой среды для TNB-кэпирования антитела.
Пример 4. Получение TNB-кэпированного мутантного по Cys антитела в отсутствие клеток. Антитело или слитый белок в форме, позволяющей впоследствии использовать его на этапе конъюгации для получения целевого конъюгата лекарственное вещество-антитело, обычно получают как часть разработки получения клеточной линии и/или как часть процесса выращивания/культивирования клеток. В случае получения TNB-кэпированного антитела, описанный выше подход заключался во включении подпитки DTNB как части процесса культивирования клеток наряду с различными дополнительными модификациями процесса культивирования клеток (например, без ограничения, корректировки концентраций цистеина/цистина в питательной среде, модификации длительности культивирования партии), как описано ранее. Хотя это позволяет получать целевое кэпированное антитело, эти подходы зависят от той части производственного процесса, которая связана с культивированием клеток и может снизить общую эффективность производственного оборудования. Таким образом, выход TNB-кэпированного антитела ограничен неоптимальным использованием установки, в частности, производственного биореактора или ферментера, используемого для клеточной культуры.
В этом примере мутантный по цистеину рекомбинантный белок, такой как мутантное по цистеину антитело, мутантный по цистеину белок или т.п., получают с помощью методов культивирования/ферментации клеток. Белок-содержащая кондиционированная среда затем может быть отделена от клеток центрифугированием, микрофильтрацией или другим подходящим способом отделения клеток. Отделение клеток может быть полным или частичным. Следующим этапом является экспозиция/инкубация мутантного по cys рекомбинантного белка с DTNB для получения TNB-кэпированного белка. Путем разделения этапов: а) генерации мутантного по цистеину рекомбинантного белка и b) кэпирование мутантного по цистеину рекомбинантного белка, производственный процесс и использование оборудования могут быть оптимизированы для достижения максимальной продуктивности путем исключения использования биореактора/ферментера для культивирования клеток в относящейся к кэпированию части процесса и, следовательно, минимизации времени цикла, в котором необходимо использование биореактора/ферментера. Минимизация времени цикла, в котором необходимо использование биореактора/ферментера, таким образом, обеспечивает более высокую производительность (благодаря большему количеству циклов, выполненных за один период времени).
Подход 1: В этом примере клетки CHO-K1, стабильно экспрессирующие мутантное по цистеину антитело трастузумаб K183C-K290C, использовали с запатентованными питательной средами в биореакторах Applikon объемом 1 л (Applikon, Inc., Schiedam, Netherlands), работающих с модульными контроллерами BioNet (Broadley-James Corp., Irvine, CA) с модулями перистальтического насоса и регулятора массового расхода газа. Температуру культуры клеток поддерживали, равной примерно 37 градусов по Цельсию, и величину pH доводили до 7,0 путем добавления либо раствора карбоната натрия/калия, либо CO2. Уровни растворенного кислорода поддерживали равными >20% путем барботирования чистого кислорода. После культивирования клеточных культур белок-содержащую кондиционированную среду отделяли центрифугированием и фильтрованием через 0,2 мкм фильтр. Затем кондиционированную среду переносили в отдельный сосуд. Конкретную концентрацию DTNB добавляли непосредственно в белок-содержащую кондиционированную среду, находящуюся в этом сосуде. Реакцию проводили в течение разных периодов времени, чтобы продемонстрировать надежность процедуры получения целевого материала. Как видно из фиг. 14, целевой продукт (трастузумаб K183C-K290C) получали, используя диапазон периодов инкубации, причем более длительное время приводило к улучшению производительности.
Подход 2: В этом примере использовали вторую клеточную линию CHO-K1, стабильно экспрессирующую мутантное по цистеину анти-EDB антитело K183C-K290C, с запатентованными минимальной и питательной средами в биореакторах Applikon объемом 1 л (Applikon, Inc., Schiedam, Netherlands), работающих с модульными контроллерами BioNet. (Broadley-James Corp., Irvine, CA) с модулями перистальтического насоса и регулятора массового расхода газа. Температуру культуры клеток поддерживали равной примерно 37 градусов по Цельсию, и величину pH доводили до 7,0 путем добавления либо раствора карбоната натрия/калия, либо CO2. Уровень растворенного кислорода поддерживали равным >20% путем барботирования чистого кислорода. После культивирования клеточных культур белок-содержащую кондиционированную среду отделяли центрифугированием и фильтрованием через 0,2 мкм фильтр. Затем кондиционированную среду переносили в отдельный сосуд. Конкретную концентрацию DTNB добавляли непосредственно в белок-содержащую кондиционированную среду, находящуюся в этом сосуде. Реакцию проводили в течение разных периодов времени, чтобы продемонстрировать надежность процедуры получения целевого материала. Как видно на фиг. 15 целевой продукт (анти-EDB K183C-K290C) получали, используя диапазон периодов инкубации, причем более длительные периоды времени приводили к улучшению производительности, демонстрируя одинаковую тенденцию, наблюдаемую в двух разных клеточных линиях, экспрессирующих антитела.
Пример 5: Процесс конъюгации с помощью TNB. TNB-кэпированное мутантное по цистеину антитело избирательно восстанавливали с помощью TSPP. Образующиеся свободные тиоловые группы обеспечивали прямую конъюгацию с лекарственным веществом без этапа ультрафильтрации/диафильтрации (UF/DF) и повторного окисления, что упрощает процесс. Мутантное по цистеину антитело трастузумаб K183C-K290C, полностью кэпированное TNB в форме 4 кэппингов на антитело (DAR4), генерировали и подвергали процессу прямой конъюгации после обработки TSPP с эффективностью 70%.
В качестве дополнительного примера в настоящем описании обсуждается TNB-кэпирование и конъюгация (на участке K183C-K290C). Протокол конъюгации TNB-кэпированного мутантного по цистеину антитела состоит из двух этапов, ведущих к получению неочищенного конъюгата: селективное восстановление и конъюгация. На первом этапе осуществляют селективное восстановление мутантных цистеинов для удаления защитной группы(групп) из мутантных остатков цистеина с минимальным или без восстановления межцепочечных дисульфидов. Обычно этот процесс осуществляется с использованием избытка (~7 эквивалентов) восстанавливающего агента, такого как трис(3-сульфонатофенил)фосфин (TSPP), при температуре окружающей среды в течение 2 часов. На втором этапе некэпированные мутантные цистеины конъюгируют с линкером-полезной нагрузкой. Обычно в реакционную смесь добавляют избыток (~12 эквивалентов) линкера-полезной нагрузки, и реакцию проводят при температуре окружающей среды в течение 1 часа с получением неочищенного конъюгата. В конечном конъюгате сохраняются нативные межцепочечные дисульфидные связи, поскольку они не были разорваны на этапе восстановления.
В конкретном случае: к 1,0 г (6,9 мкмоль; 25 мг/мл в 60 мМ гистидина, pH 7; 38,9 мл) антитела трастузумаб K183C-K290C добавляли 27,5 мг TSPP (7 эквивалентов; 48,3 мкмоль; 10 мМ в воде; 4,83 мл). Реакционную смесь инкубировали при температуре окружающей среды в течение 2 часов. К этой реакционной смеси добавляли 111 мг полезной нагрузки-линкера mcvcPABC0101 (12 эквивалентов, 82,7 мкмоль; 25 мМ в диметилсульфоксиде; 3,31 мл). Реакционную смесь инкубировали при температуре окружающей среды в течение 1 часа, получая неочищенный конъюгат.
Пример 6: Процесс конъюгации с помощью TNB с диафильтрацией после восстановления. Избыточное количество полезной нагрузки-линкера, используемой в процессе, описанном в примере 5, может быть уменьшено путем добавления этапа диафильтрации (замены буфера) после восстановления TSPP для удаления видов, которые реагируют с полезной нагрузкой-линкером. Как показано на фиг. 16, 70% DAR4 достигается с 6 эквивалентами полезной нагрузки-линкера после замены буфера, в то время как в случаях, когда замена буфера отсутствует, необходимы значительно более высокие эквиваленты. Кроме того, количество TSPP может быть немного уменьшено путем увеличения температуры и/или времени восстановления.
В конкретном случае: к 4,0 г (27,6 мкмоль; 25 мг/мл в 60 мМ гистидина, pH 7; 156 мл) антитела трастузумаб K183C-K290C добавляли 93,8 мг TSPP (6 эквивалентов; 165 мкмоль; 10 мМ в воде; 16,5 мл). Реакционную смесь инкубировали при 37°C в течение 3 часов, а затем выполняли замену буфера путем диафильтрации (мембрана TangenX ProStream 50 кД, 110-210 г/м2, 10 диаобъемов 60 мМ гистидина, рН 7). После диафильтрации добавляли 222 мг полезной нагрузки-линкера mcvcPABC0101 (6 эквивалентов, 165 мкмоль; 25 мМ в диметилсульфоксиде; 6,62 мл). Реакционную смесь инкубировали при 25°C в течение 1 часа, получая неочищенный конъюгат.
Пример 7: Процесс конъюгации с помощью TNB с повышенным стехиометрическим количеством TSPP, диафильтрацией после восстановления и повторным окислением. Количество агрегата, полученного способом, описанным в Примере 6, может быть уменьшено путем увеличения стехиометрического количества TSPP. Повышенное стехиометрическое количество TSPP позволяет ускорить удаление TNB из мутированных цистеинов и предотвращает образование дисульфидных связей между антителами. Однако увеличение стехиометрического количества TSPP также приводит к уменьшению небольшой фракции межцепочечных дисульфидных связей. Присутствие хлорида натрия и других солей (см. Пример 8) снижает степень восстановления дисульфидов между цепями при более высоком стехиометрическом количестве TSPP. Добавление этапа повторного окисления после диафильтрации регенерирует некоторые из восстановленных межцепочечных дисульфидных связей. Как показано на фиг. 17, конъюгация после восстановления 20 эквивалентами TSPP и замены буфера без этапа повторного окисления приводит к низким уровням сверхконъюгированных соединений из-за уменьшения количества межцепочечных дисульфидных связей. Когда в процесс добавляют этап повторного окисления, сверхконъюгированные вещества не образуются.
В конкретном случае: к 24,7 г (0,170 ммоль; 26 мг/мл в 60 мМ гистидина, 150 мМ NaCl, pH 7; 961 мл) антитела трастузумаб K183C-K290C добавляли 1,94 г TSPP (20 эквивалентов; 3,41 ммоль; 100 мМ в воде; 34,1 мл). Реакционную смесь перемешивали при 37°C в течение 3 часов, а затем заменяли буфер путем диафильтрации (мембрана TangenX ProStream 50 кД, 110-210 г/м2, 10 диамолей 60 мМ гистидина, 150 мМ NaCl, pH 7). После диафильтрации смесь охлаждали до 4°C, добавляли 0,45 г дегидроаскорбиновой кислоты (15 эквивалентов; 2,56 ммоль; 50 мМ в 1:1 ДМСО/вода; 51,2 мл), и смесь перемешивали при 4°C в течение 16 часов. Смесь нагревали до 25°C, добавляли 1,37 г полезной нагрузки-линкера mcvcPABC0101 (6 эквивалентов, 1,02 ммоль; 25 мМ в диметилсульфоксиде; 40,9 мл) и перемешивали при 25°C в течение 1,5 часов с получением неочищенного конъюгата.
Пример 8: Процесс конъюгации с помощью TNB с повышенным стехиометрическим количеством TSPP, диафильтрацией после восстановления, повторным окислением и блокированием линкера-полезной нагрузки. Присутствие солей в восстановительной смеси, такой как хлорид натрия в примере 7, снижает степень восстановления межцепочечных дисульфидов при более высоком стехиометрическом количестве TSPP.
В качестве еще одного примера, используя приведенный на фиг. 17 показатель для сверхконъюгированных соединений, образовавшихся без повторного окисления, показало уменьшение уровня восстановления межцепочечных дисульфидов с ~33%, для случая в отсутствие соли, до 10-20% в присутствии солей 150-500 мМ. Соли, такие как хлорид натрия, ацетат натрия, нитрат калия, гидрофосфат калия, нитрат магния, сульфат магния, гуанидиния хлорид и сульфат аммония, снижают уровень восстановления межцепочечных дисульфидов.
В конкретном случае уровень восстановления межцепочечных дисульфидов снизился с 33%, в случае без добавления солей, до 15, 14 и 12% в присутствии хлорида натрия 150, 250 и 500 мМ соответственно.
Пример 9: Процесс конъюгации с помощью TNB с повышенным стехиометрическим количеством TSPP, диафильтрацией после восстановления, повторным окислением и блокированием линкера-полезной нагрузки. Этот пример аналогичен примеру 7, но с признаками (1) повторного окисления при 25°C, что значительно сокращает время повторного окисления и устраняет трудоемкое охлаждение после диафильтрации и нагревания перед процессом конъюгации, и (2) блокирования линкера-полезной нагрузки путем реакции с цистеином, что улучшает последующую очистку при определенных условиях (см. пример 14 ниже).
В конкретном случае: к 80,0 г (0,541 ммоль; 26 мг/мл в 60 мМ гистидина, 150 мМ NaCl, pH 7; 3053 мл) антитела трастузумаб K183C-K290C добавляли 6,14 г TSPP (20 эквивалентов; 10,8 ммоль; 100 мМ в воде; 108 мл). Реакционную смесь перемешивали при 37°C в течение 3 часов, а затем заменяли буфер путем диафильтрации (мембрана TangenX ProStream 50 кД, 110-210 г/м2, 10 диамолей 60 мМ гистидина, 150 мМ NaCl, pH 7). После диафильтрации добавляли 0,19 г дегидроаскорбиновой кислоты (2 эквивалента; 1,08 ммоль; 50 мМ в 1:1 ДМСО/вода; 21,6 мл), и смесь перемешивали при 25°C в течение 0,5 часа. Затем добавляли 4,35 г полезной нагрузки-линкера mcvcPABC0101 (6 эквивалентов, 3,24 ммоль; 25 мМ в диметилсульфоксиде; 130 мл), и смесь перемешивали при 25°C в течение 1 часа. Наконец, добавляли 0,79 г цистеина (12 эквивалентов, 6,48 ммоль; 100 мМ в воде; 64,9 мл), и смесь перемешивали при 25°C в течение 1 часа с получением неочищенного конъюгата.
Пример 10: Процесс конъюгации с помощью TNB - Регенерация фрагмента антитела путем повторного окисления. Добавление этапа повторного окисления, как в примерах 7 и 9, приводило к регенерации фрагментов, присутствующих во вводимом антителе, таким образом улучшая качество и выход конечного конъюгата. Фрагменты возникали из антител, в которых одна или более межцепочечных дисульфидных связей не были целыми. Таким образом, конъюгация партий антител, содержащих 15-35% фрагментов, с помощью процесса, описанного в примере 9, дает конъюгат с низкими и сопоставимыми уровнями фрагментов.
В конкретном случае: антитело с 35% фрагментов приводило к образованию неочищенного конъюгата с примерно 10% фрагментов; большинство фрагментов было регенерировано путем повторного окисления. Конечный очищенный конъюгат содержал менее 3% фрагментов.
Пример 11: Процесс конъюгации с помощью TNB с анти-EDB антителом K183C-K290C. Процесс конъюгации с помощью TNB подходит для TNB-кэпированных антител, отличных от трастузумаба.
В конкретном случае: обработка TNB-кэпированного анти-EDB антитела K183C-K290C с помощью процедуры, описанной в примере 9, давала неочищенный конъюгат, сравнимый с конъюгатом, полученным с антителом трастузумаб K183C-K290C. Как показано на фиг. 18, неочищенные конъюгаты этих антител очень похожи с величиной DAR4 ~75%.
Пример 12: Очистка конъюгатов, полученных способом конъюгации с помощью TNB. Неочищенный конъюгат, полученный этим способом, может быть очищен с помощью хроматографии гидрофобного взаимодействия (HIC). Очистка HIC удаляет или значительно уменьшает количество свободного линкера-полезной нагрузки, агрегатов, фрагментов и конъюгатов с более низким значением DAR.
В качестве еще одного примера, очистка HIC может быть выполнена на колонке CaptoТМ Butyl ImpRes (GE) и колонке Toyopearl® PPG-600M (Tosoh), соединенных последовательно. CaptoТМ Butyl ImpRes обеспечивает подходящее удаление агрегатов, фрагментов и конъюгатов с более низким значением DAR, а Toyopearl® PPG-600M задерживает свободную полезную нагрузку mcvcPABC0101. Очищенный конъюгат может быть сконцентрирован, а буфер может быть заменен ультрафильтрацией/диафильтрацией (УФ/ДФ). УФ/ДФ также позволяет удалять остаточный свободный линкер-полезную нагрузку, если он присутствует после очистки HIC.
В конкретном случае: неочишенный конъюгат (4,0 г, 26 мг/мл), полученный способом, описанным в примере 6, разбавляли одним объемом 10 мМ фосфата натрия, pH 7. Разбавленную неочищенную смесь дополнительно разбавляли до соотношения 1:1 смесью 20 мМ фосфата натрия и 1 М сульфатом аммония, рН 7, для получения загрузочного раствора для HIC. Очистку HIC выполняли на колонке CaptoТМ Butyl ImpRes (GE) (24 см (h) x 2,6 см (d)) и колонке Toyopearl® PPG-600M (Tosoh) ((4 см (h) x 2,6 см (d)), соединенных последовательно. После введения загрузочного раствора в колонку CaptoТМ Butyl ImpRes очищенный конъюгат элюировали из двух последовательно соединенных колонок 50-100% градиентом буфера B в буфере A на 25 объемов колонки; буфер A: 20 мМ фосфат натрия, 1 М сульфат аммония, pH 7; буфер B: 10 мМ фосфат натрия, pH 7. Фракции, содержащие целевой конъюгат, объединяли и подвергали УФ/ДФ для осуществления концентрации и замены буфера (мембрана TangenX ProStream 50 кД, 110-210 г/м2, 10 диаобъемов 20 мМ гистидина, рН 5,8).
Пример 13: Очистка конъюгатов, полученных в результате процесса конъюгации с помощью TNB с увеличенным выходом. При очистке методом HIC восстановление ADC DAR4 может быть улучшено путем добавления в буфер B подвижной фазы изопропанола, таким образом увеличивая выход процесса.
В конкретном случае: неочищенный конъюгат (12,4 г, 26 мг/мл), полученный способом, описанным в примере 7, разбавляли одним объемом 10 мМ фосфата натрия, pH 7, и 5% (об./об.) изопропанола. Разбавленную неочищенную смесь дополнительно разбавляли до соотношения 1:1 смесью 20 мМ фосфата натрия и 1 М сульфатом аммония, рН 7, с получением загрузочного раствора для HIC. Очистку HIC выполняли на колонке CaptoТМ Butyl ImpRes (GE) (22 см (h) x 5 см (d)) и колонке Toyopearl® PPG-600M (Tosoh) ((4 см (h) x 5 см (d)), соединенных последовательно. После введения загрузочного раствора в колонку CaptoТМ Butyl ImpRes очищенный конъюгат элюировали из двух последовательно соединенных колонок 50-100% градиентом буфера B в буфере A на 25 объемов колонки, буфер A: 20 мМ фосфат натрия, 1 М сульфат аммония, pH 7; буфер B: 10 мМ фосфат натрия, pH 7,5% (об./об.) изопропанол. Фракции, содержащие целевой конъюгат, объединяли и подвергали УФ/ДФ для осуществления концентрации и замены буфера (TangenX ProStream 50kD мембрана, 110-210 г/м2, 10 диаобъемов 20 мМ гистидина, рН 5,8). Выход ADC для всего процесса, в котором в процессе очистки HIC в буфере В присутствовал изопропанол, улучшился примерно на 50% по сравнению с процессом очистки в Примере 12.
Пример 14: Очистка конъюгатов, полученных в результате конъюгации с помощью TNB с блокированием линкера-полезной нагрузки. Когда на этапе блокируют полезную нагрузку-линкер mcvcPABC0101 кэпировали цистеином, как в примере 9, очистку HIC можно выполнять только на колонке CaptoТМ Butyl ImpRes (GE). Цистеин-кэпированный mcvcPABC0101 элюируется раньше, чем конъюгат DAR4, и хорошо отделяется на колонке CaptoТМ Butyl ImpRes. Как показано на фиг. 19, конъюгат DAR4 отделяют от цистеин-кэпированного mcvcPABC0101, конъюгатов с более низким значением DAR, агрегатов и фрагментов, используя условия, описанные в этом примере. Цистеин-кэпированный McvcPABC0101 также легко очищается путем диафильтрации.
В конкретном случае: неочищенный конъюгат (40,0 г, 26 мг/мл), полученный способом, описанным в примере 9, разбавляли одним объемом 10 мМ фосфата натрия, pH 7, и 5% (об./об.) изопропанола. Разбавленную неочищенную смесь дополнительно разбавляли до соотношения 1:1 смесью 20 мМ фосфата натрия и 1 М сульфата аммония, рН 7, с получением загрузочного раствора для HIC. Очистку HIC выполняли на колонке CaptoТМ Butyl ImpRes (GE) (24,5 см (h) х 10 см (d)). После введения загрузочного раствора в колонку очищенный конъюгат элюировали 50-100% градиентом буфера В в буфере А на 10 объемов колонки; буфер А: 20 мМ фосфат натрия, 1 М сульфат аммония, рН 7; буфер В: 10 мМ фосфат натрия, рН 7, 5% (об./об.) изопропанол. Фракции, содержащие целевой конъюгат, объединяли и подвергали УФ/ДФ для осуществления концентрации и замены буфера (TangenX ProStream 50kD мембрана, 110-210 г/м2, 10 диаобъемов 20 мМ гистидина, рН 5,8).
В отдельном конкретном случае: неочищенный конъюгат (0,83 г, 23 мг/мл), полученный способом, описанным в примере 9, разбавляли одним объемом 10 мМ фосфата натрия, pH 7, и 5% (об./об.) изопропанола. Разбавленную неочищенную смесь дополнительно разбавляли до соотношения 1:1 смесью 20 мМ фосфата натрия, 1 М сульфата аммония, рН 7, с получением загрузочного раствора для HIC. Очистку HIC выполняли на колонке с бутиловой смолой (GE) (24 см (h) х 1,6 см (d)) и колонке Toyopearl® PPG-600M (Tosoh) ((5,3 см (h) х 1,6 см (d)), соединенных последовательно. После введения загрузочного раствора в колонку очищенный конъюгат элюировали 50-100% градиентом буфера В в буфере А на 10 объемов колонки, буфер А: 20 мМ фосфат натрия, 1 М сульфат аммония, рН 7 буфер В: 10 мМ фосфат натрия, рН 7, 5% (об./об.) изопропанола. Фракции, содержащие целевой конъюгат, объединяли. При отделении полезной нагрузки-линкера mcvcPABC0101 от конъюгата DAR4 достигалось высокое извлечение.
Пример 15: Анализ дисульфидного скремблирования в шарнирной области антитела. Преимущество процесса конъюгации с TNB заключается в существенном уменьшении или полном устранении восстановления дисульфидных связей в шарнирной области антитела и, следовательно, в получении конъюгата, имеющего нативные межцепочечные дисульфидные связи. Был разработан анализ для измерения уровня скремблирования в шарнирной области как в антителе, так и в конъюгате. В этом анализе антитело или конъюгат обрабатывают одним ферментом, который разрезает антитело под шарнирной областью, и вторым ферментом, который разрезает над шарнирной областью, как показано на фиг. 20.
Затем фрагмент шарнирной области анализируют с помощью ВЭЖХ или ЖХ/МС для определения количества нативных и скремблированных связей в шарнире, присутствующих в антителе или конъюгате.
Образец ADC в количестве 1 мг/мл обрабатывали IdeS (фермент 1, 1 ед./мкг ADC) при 37°C в течение 30 минут с последующей обработкой Lys-C (фермент 2, 1 мкг/150 мкг ADC) при 37°C в течение 5 минут. Затем реакцию блокируют трифторуксусной кислотой. Образец анализировали с помощью ВЭЖХ: колонка Waters XBridge BEH C18, температура колонки: 60°C, подвижная фаза A: 0,1% ТФУ в воде, подвижная фаза B: 0,1% ТФУ в ацетонитриле, градиент: 20-30% подвижной фазы B в течение 30 минут, скорость потока: 0,2 мл/мин, УФ-детекция при 214 нм. Пики подтверждали МС.
Пример 16: Сравнение с конъюгации с цистеин-кэпированным антителом. ADC, полученный из цистеин-кэпированного антитела, содержит продукт со скремблированным шарниром. Для конъюгации цистеин-кэпированного антитела, антитело должно быть полностью восстановлено избытком трис(2-карбоксиэтил)фосфин гидрохлорида (ТСЕР) и восстановленные межцепочечные дисульфидные связи повторно окисляют дегидроаскорбиновой кислотой, оставляя только мутированные остатки цистеина, доступные для конъюгации в виде свободных тиолов. Затем мутированные остатки цистеина конъюгируют с линкером-полезной нагрузкой с получением неочищенного ADC. Этап повторного окисления дает как нативные межцепочечные дисульфидные связи, так и скремблированные дисульфидные связи в шарнирной области антитела. Для ADC, полученного в результате этого процесса полного восстановления/повторного окисления, уровень скремблирования в шарнирной области, измеренный с помощью анализа, описанного в примере 15, обычно составляет 5-20%. Следует отметить, что все случаи скремблирования в шарнирной области происходят в результате процесса конъюгации ADC, поскольку в исходном антителе скремблирование в шарнирной области не обнаружено.
К 4,0 г (27,6 мкмоль; 27 мг/мл в 60 мМ гистидина, pH 7; 144 мл) цистеин-кэпированного антитела трастузумаб K183C-K290C добавляли 1,65 мл 0,5 М TCEP (30 эквивалентов; 0,827 ммоль; 0,5 М раствор в воды). Реакционную смесь инкубировали при 37°C в течение 5 часов, а затем выполняли замену буфера путем диафильтрации (мембрана TangenX ProStream 50 кД, 110-210 г/м2, 10 диаобъемов 60 мМ гистидина, рН 7). После диафильтрации смесь охлаждали до 4°C, добавляли 0,144 г дегидроаскорбиновой кислоты (30 эквивалентов; 0,827 ммоль; 50 мМ в 1:1 ДМСО/вода; 16,5 мл), и смесь инкубировали при 4°C в течение примерно 16 часов. Смесь нагревали до 25°C, добавляли 0,185 г полезной нагрузки-линкера mcvcPABC0101 (5 эквивалентов, 0,138 ммоль; 25 мМ в диметилсульфоксиде; 5,51 мл), и инкубировали при 25°C в течение 1 часа с получением неочищенного конъюгата. После очистки HIC уровень скремблирования в шарнирной области ADC, измеренный анализом, описанным в Примере 15, составлял 18%.
Для сравнения, ADC, полученные из TNB-кэпированного антитела, как описано в Примерах 6, 7 и 9 выше, не содержат детектируемого скремблирования в шарнирной области после очистки HIC, согласно результатам анализа, описанным в Примере 15.
| название | год | авторы | номер документа |
|---|---|---|---|
| КЭПИРОВАННЫЕ И НЕКЭПИРОВАННЫЕ ЦИСТЕИНЫ АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ В КОНЪЮГАЦИИ АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО | 2016 |
|
RU2742260C2 |
| АНТИТЕЛА И ФРАГМЕНТЫ АНТИТЕЛ ДЛЯ САЙТ-СПЕЦИФИЧЕСКОГО КОНЪЮГИРОВАНИЯ | 2016 |
|
RU2757815C2 |
| СПОСОБЫ КУЛЬТИВИРОВАНИЯ КЛЕТОК | 2018 |
|
RU2758674C2 |
| САЙТ-СПЕЦИФИЧЕСКИЕ КОНЪЮГАТЫ АНТИТЕЛА К HER2 И ЛЕКАРСТВЕННОГО СРЕДСТВА | 2016 |
|
RU2745565C2 |
| КОНЪЮГАТЫ МЕЧЕННЫХ РАДИОАКТИВНЫМ ЦИРКОНИЕМ СКОНСТРУИРОВАННЫХ АНТИТЕЛ С ЦИСТЕИНОВЫМИ ЗАМЕНАМИ | 2010 |
|
RU2562862C2 |
| СПОСОБ КУЛЬТИВИРОВАНИЯ КЛЕТОК МЛЕКОПИТАЮЩИХ И ПОЛУЧЕНИЕ ПОЛИПЕПТИДОВ С ИСПОЛЬЗОВАНИЕМ КОМПОЗИЦИИ КЛЕТОЧНОЙ КУЛЬТУРЫ | 2013 |
|
RU2678142C2 |
| СРЕДА ДЛЯ КУЛЬТИВИРОВАНИЯ КЛЕТОК БЕЗ БЕЛКОВ И БЕЗ СЫВОРОТКИ | 2009 |
|
RU2536244C2 |
| СПОСОБ ДВОЙНОЙ КОНЪЮГАЦИИ ДЛЯ ПОЛУЧЕНИЯ КОНЪЮГАТОВ АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО | 2018 |
|
RU2771310C2 |
| КОНЪЮГАТ АНТИТЕЛА И ЛЕКАРСТВЕННОГО СРЕДСТВА, КОМПОЗИЦИИ НА ЕГО ОСНОВЕ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2015 |
|
RU2728235C2 |
| АНТИТЕЛА ПРОТИВ EDB И КОНЪЮГАТЫ АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО | 2017 |
|
RU2758632C2 |
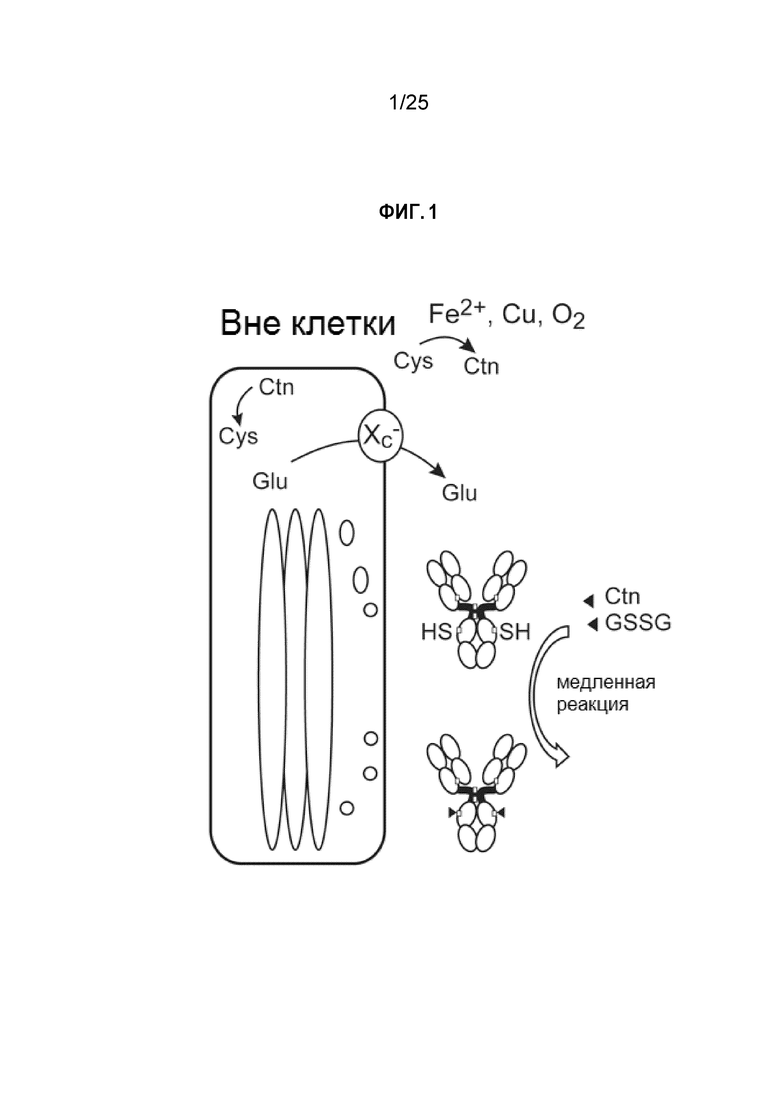
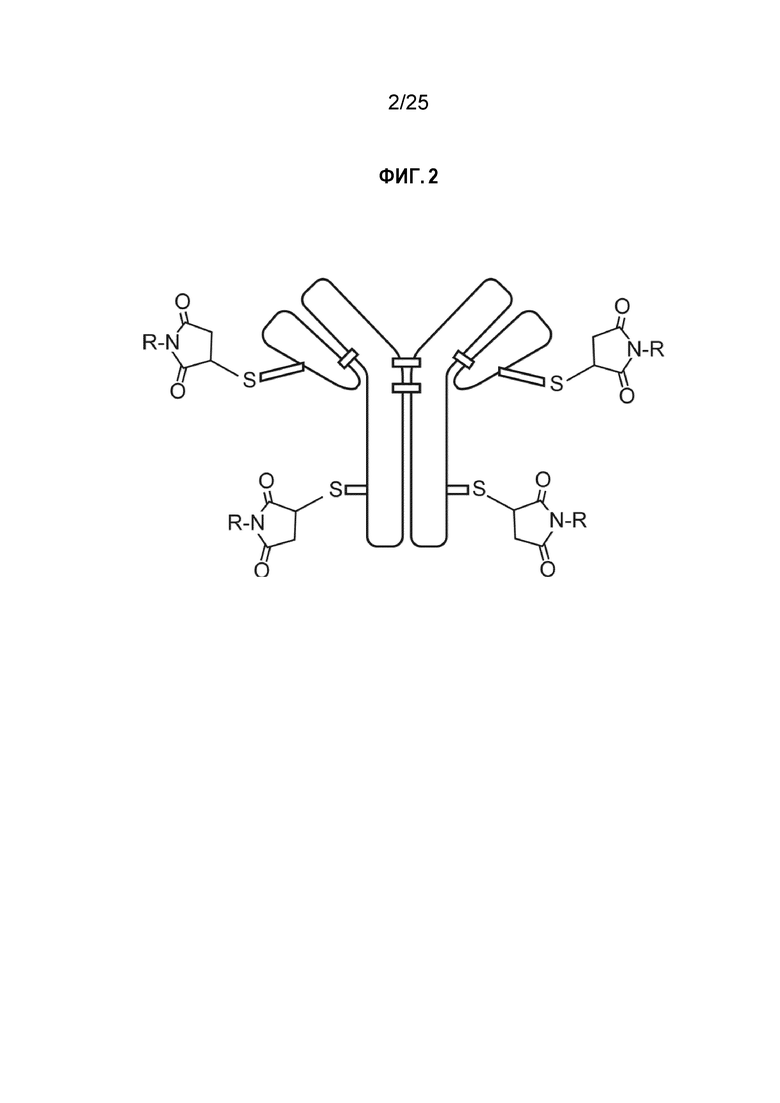
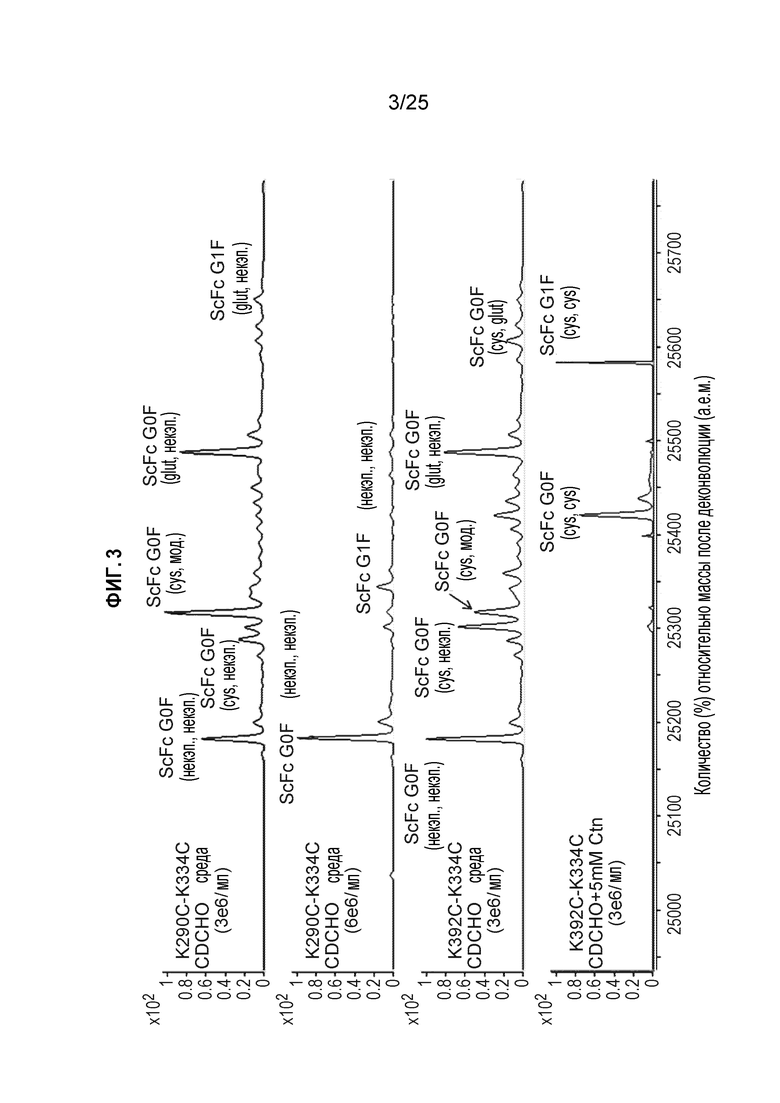
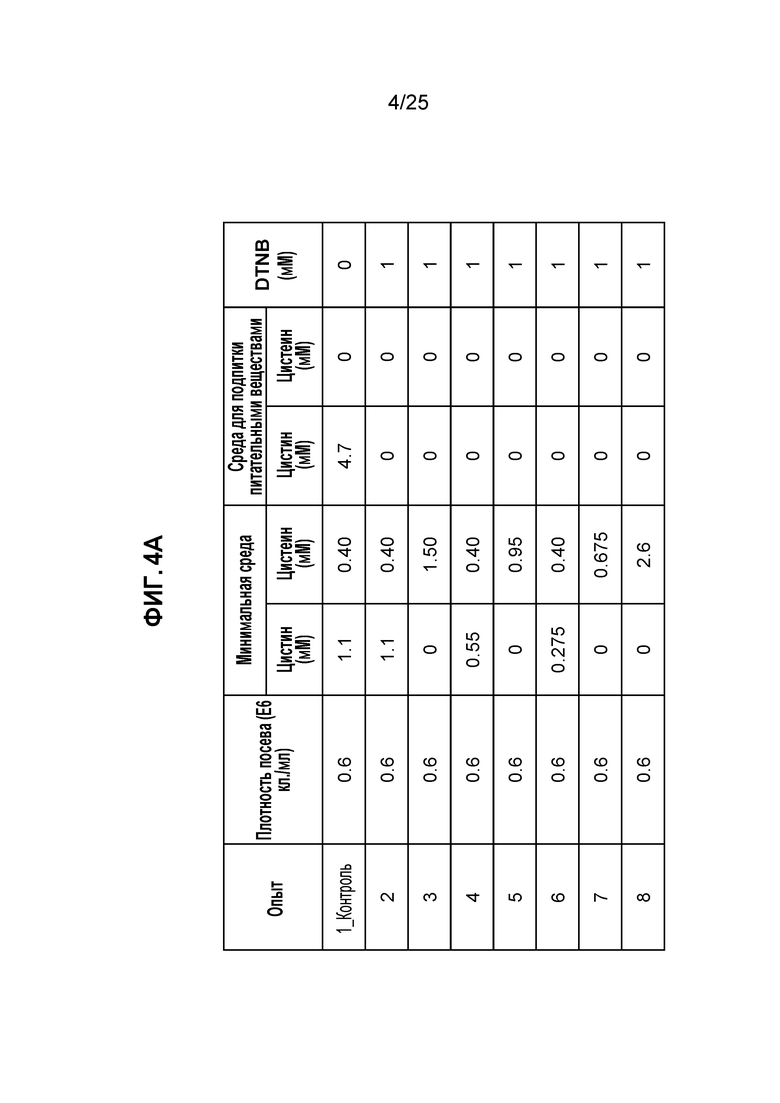
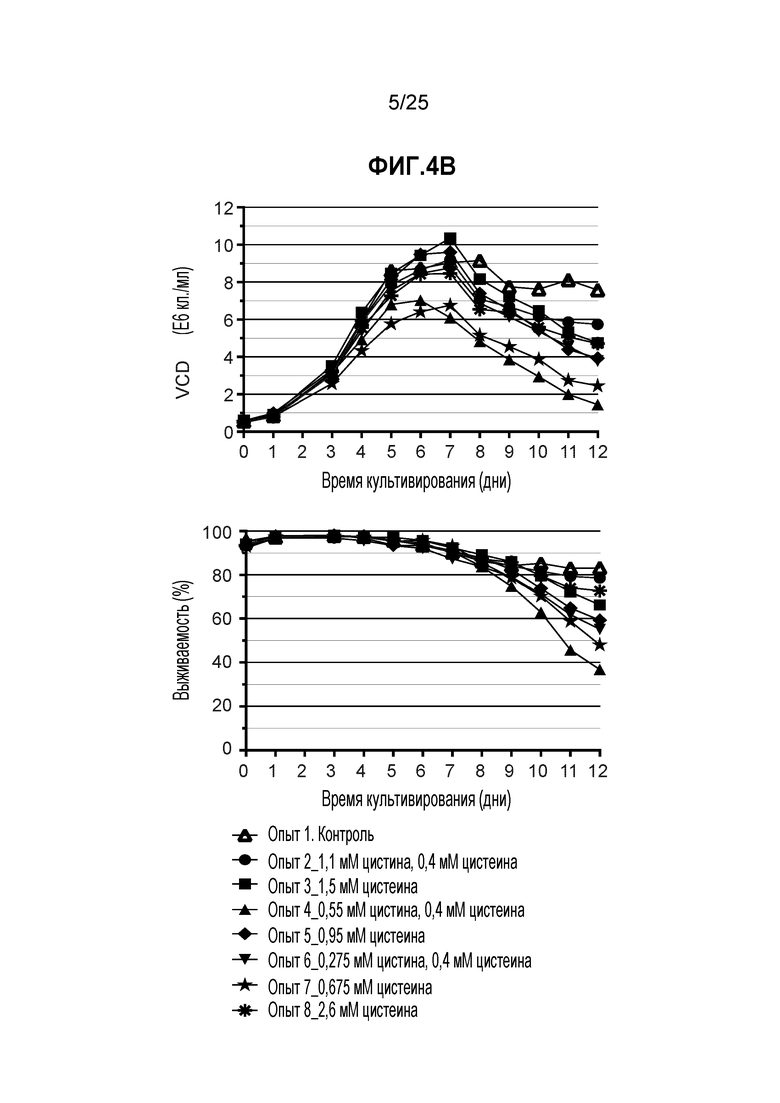
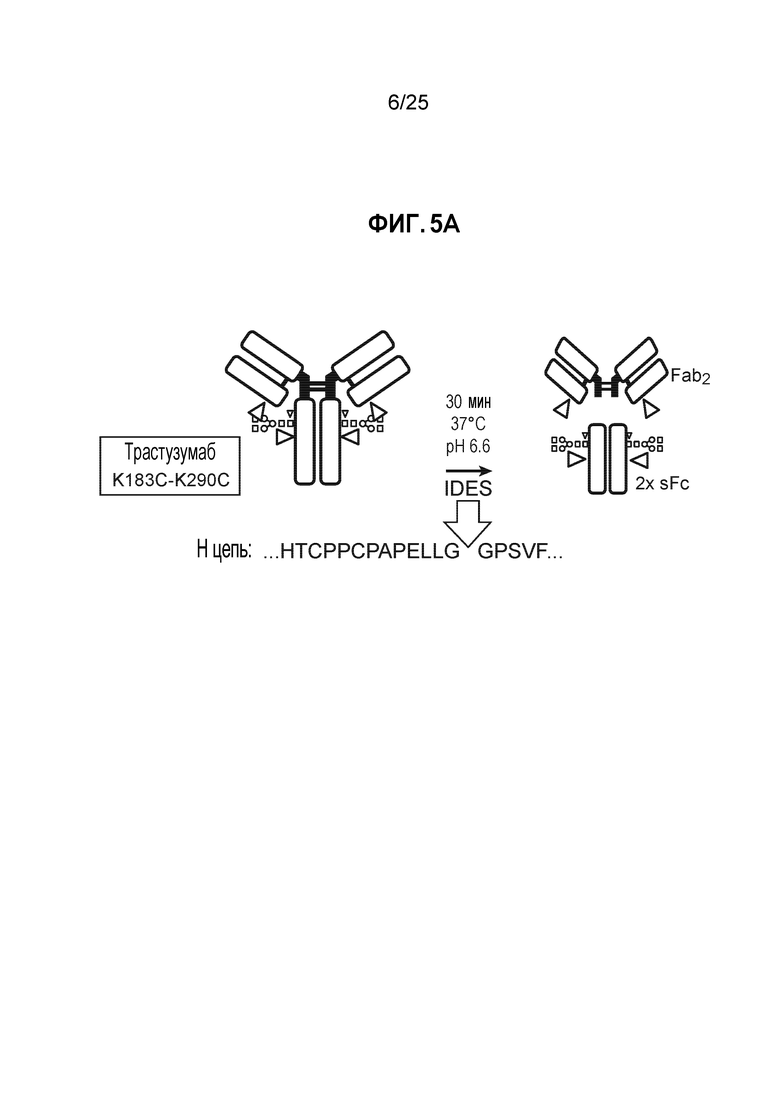
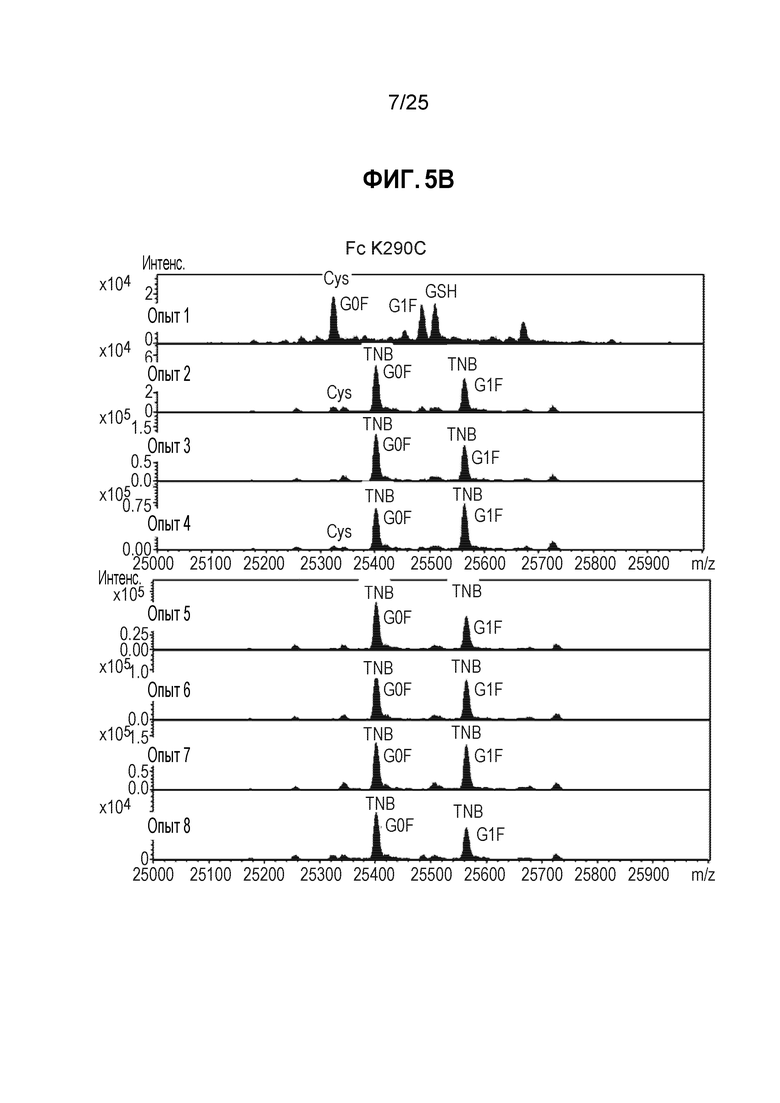
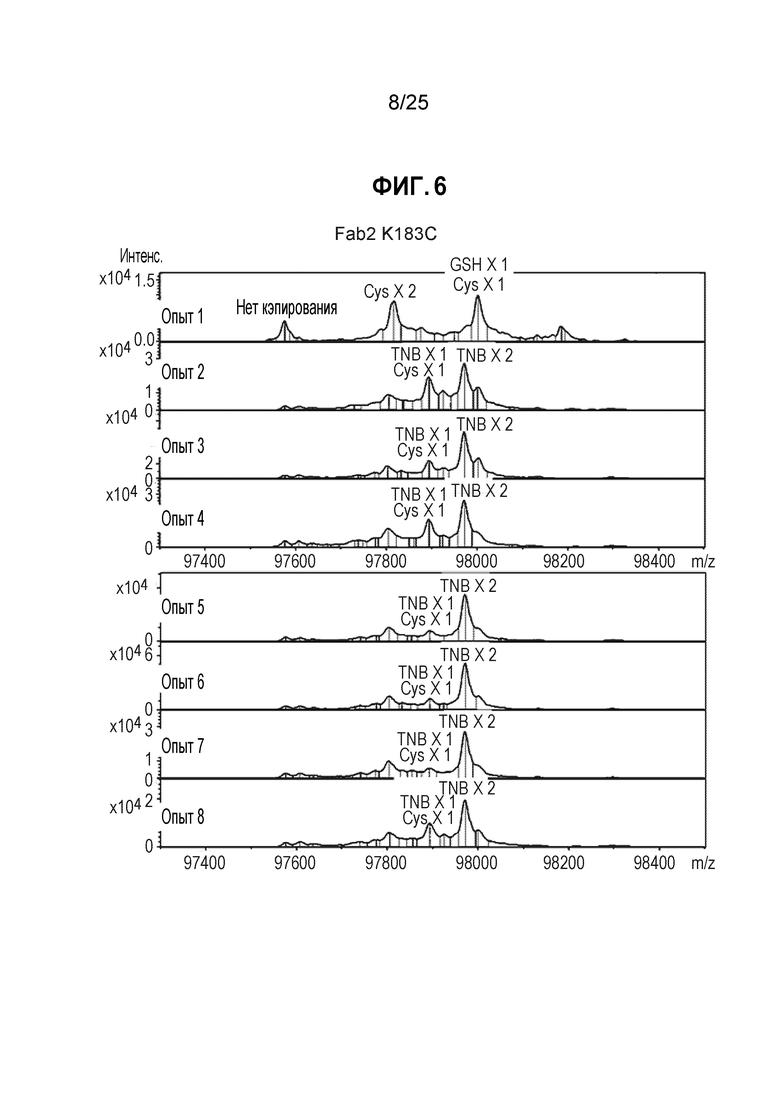
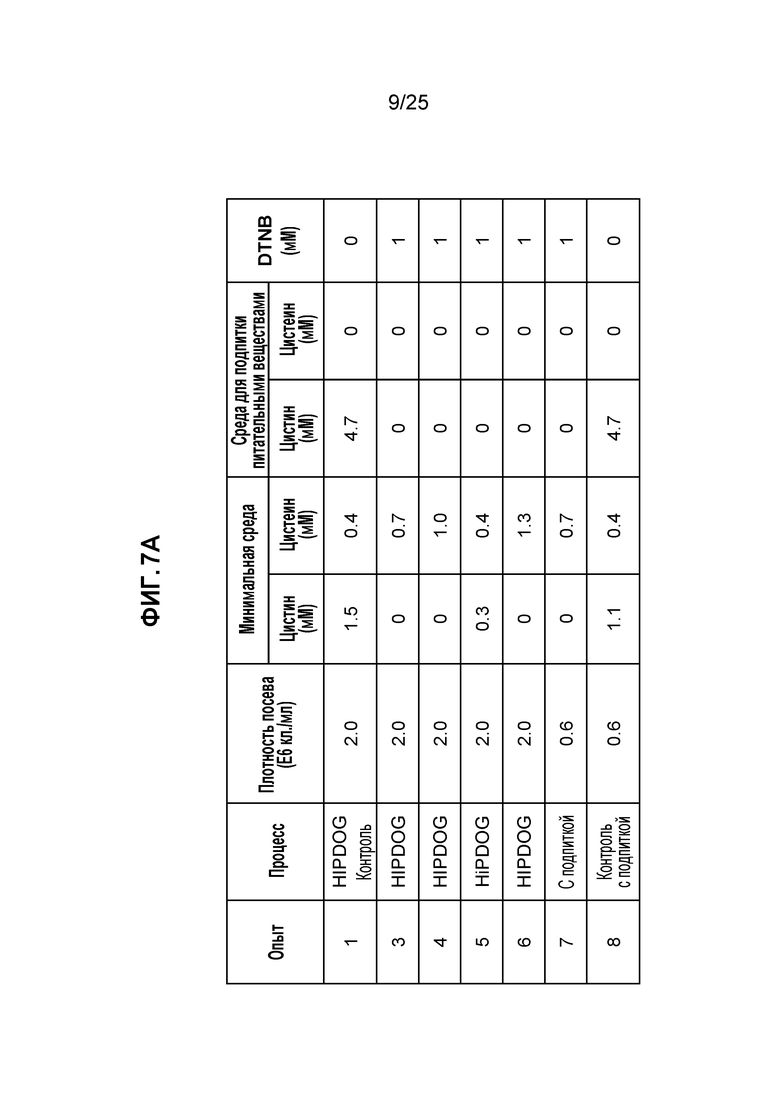
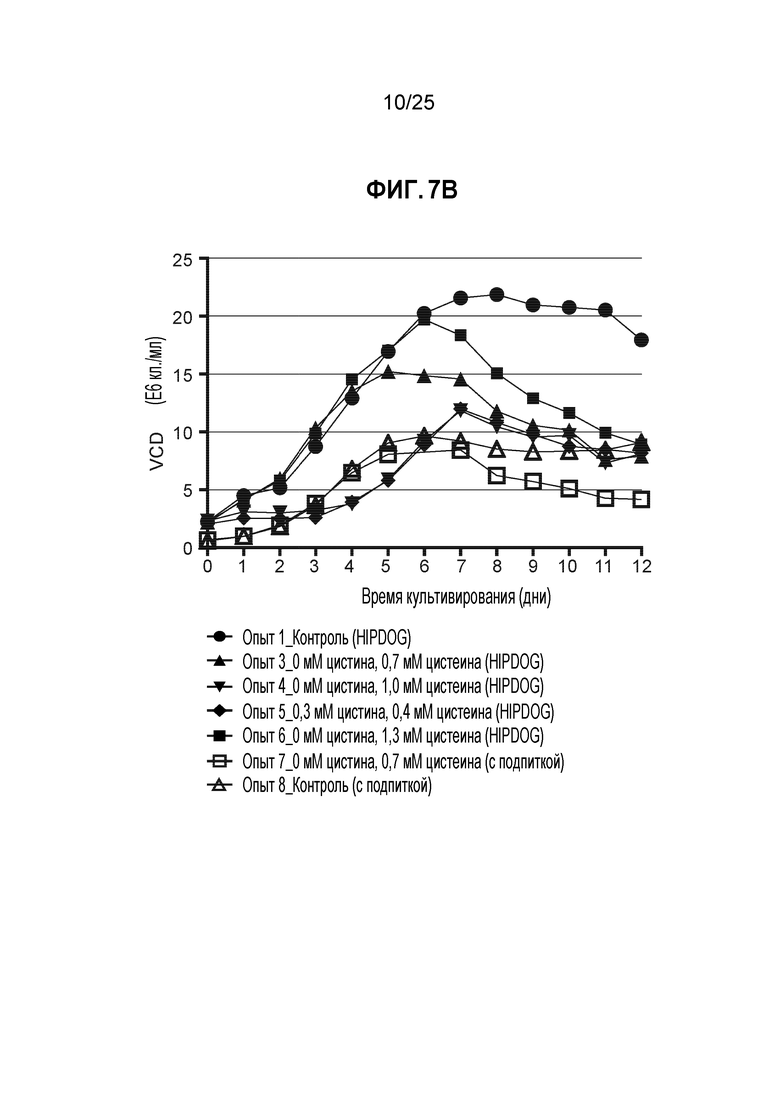
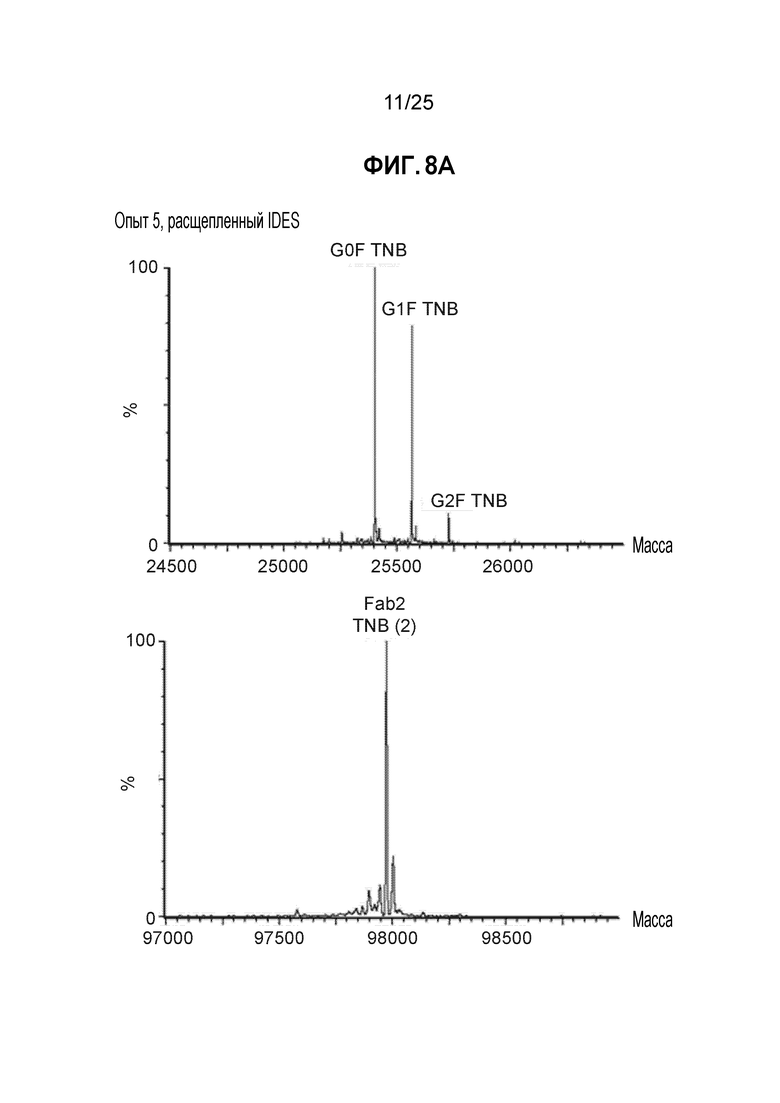
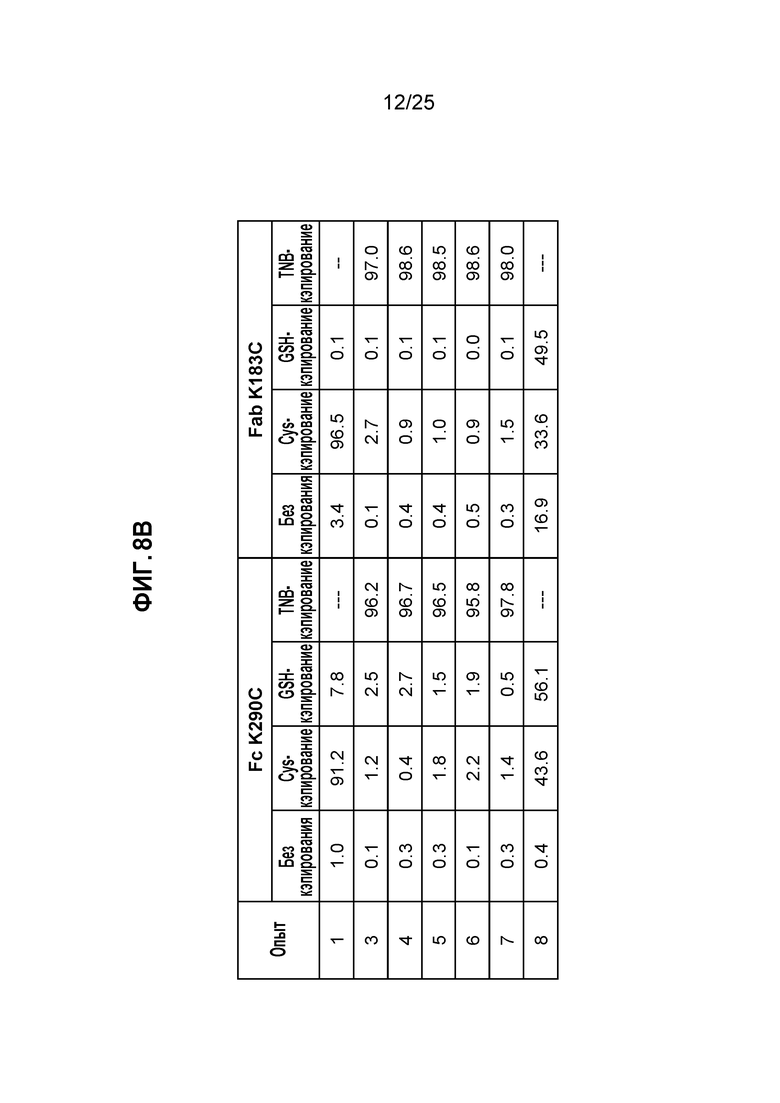
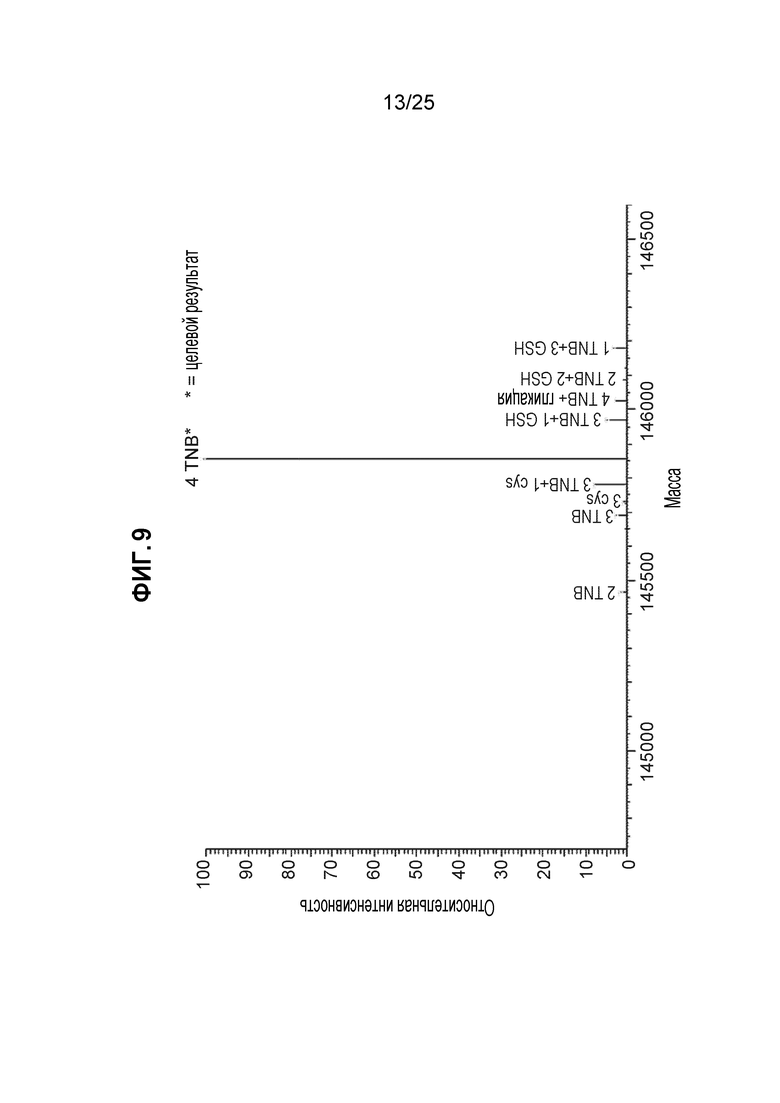
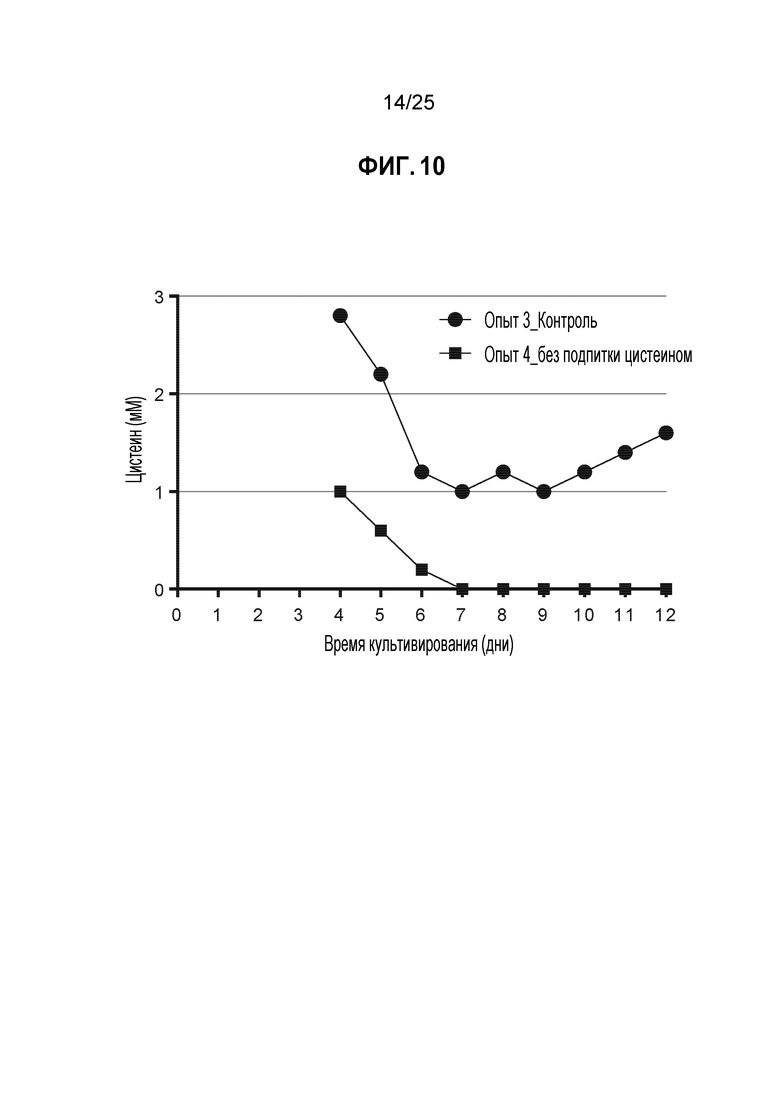
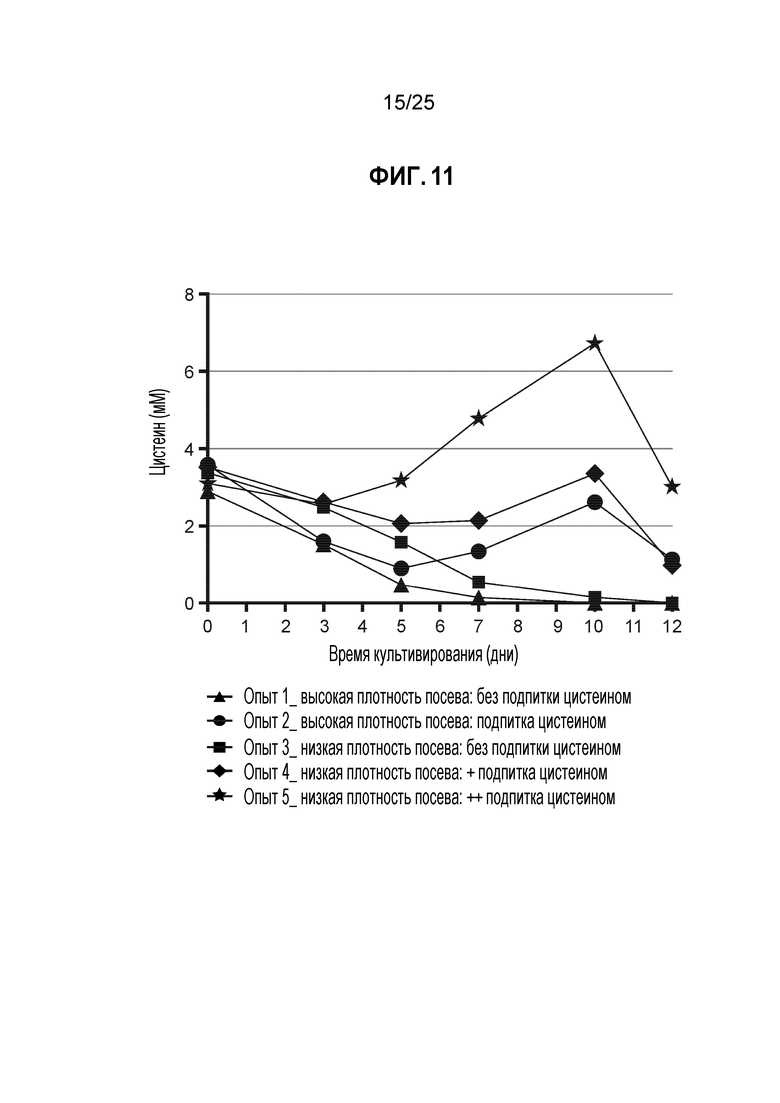
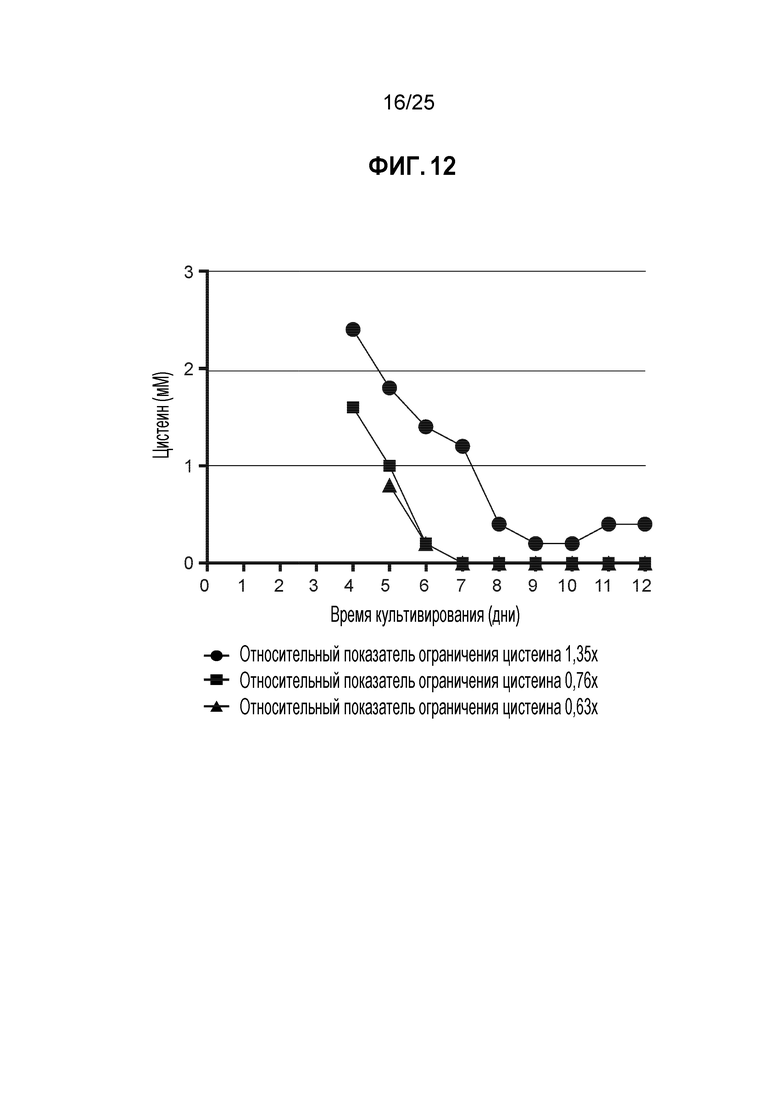
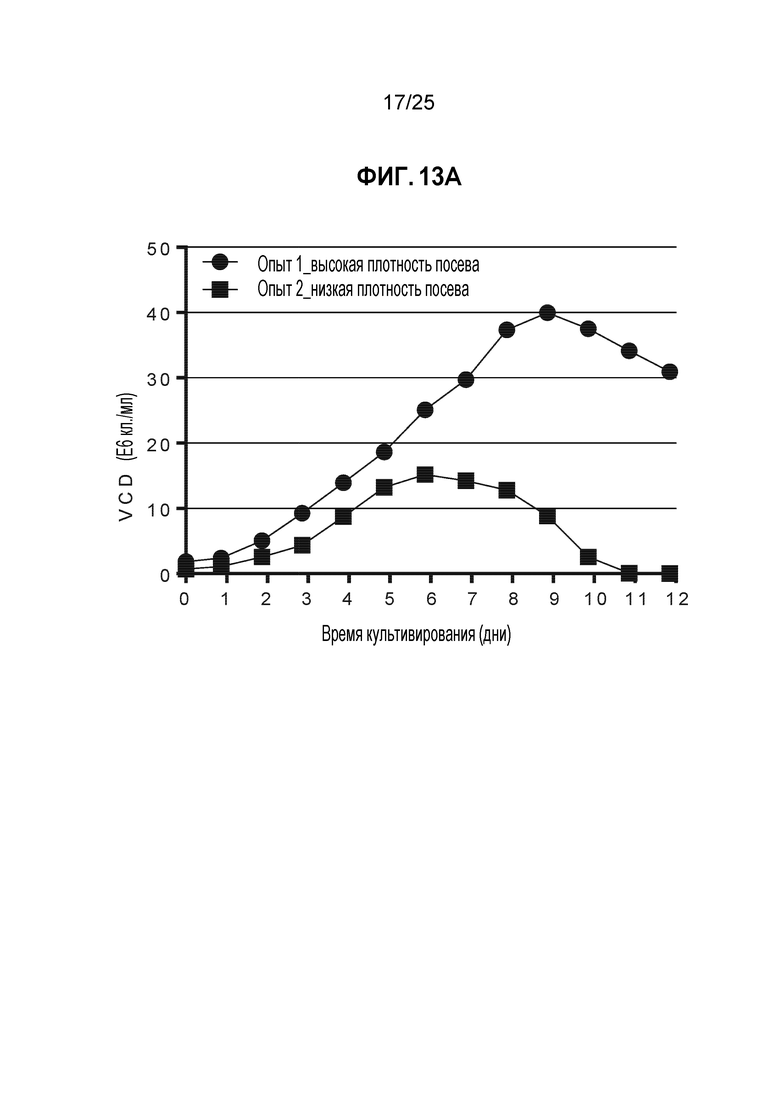
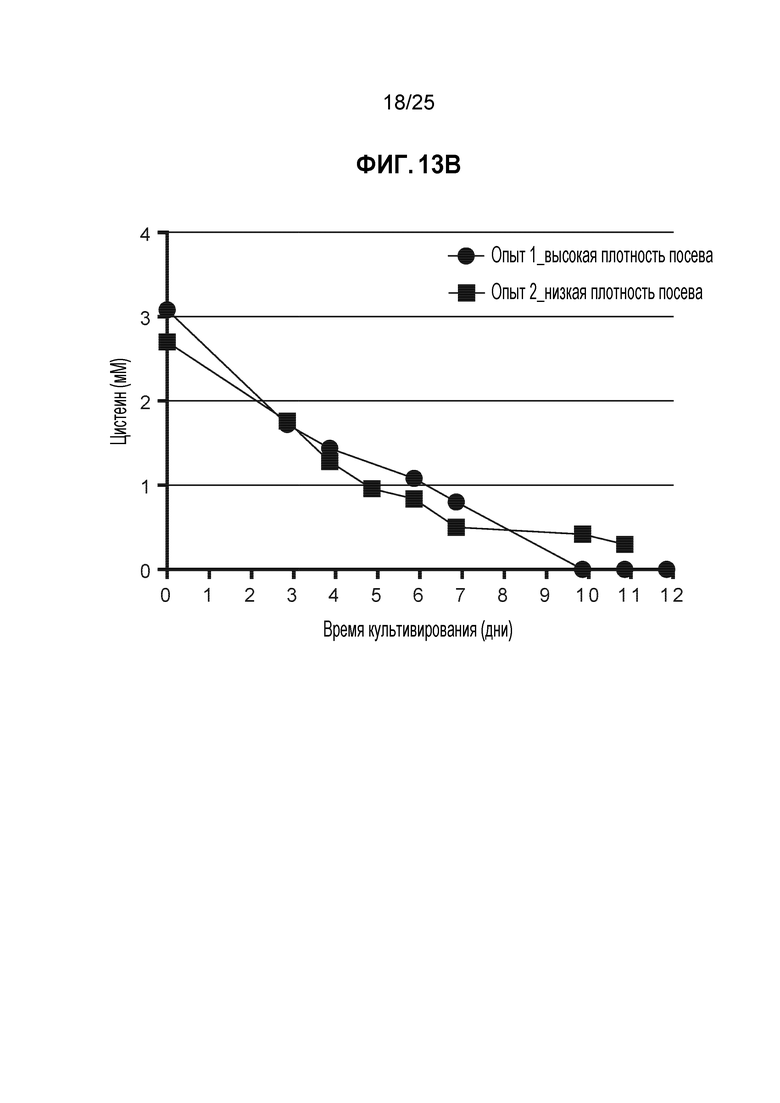
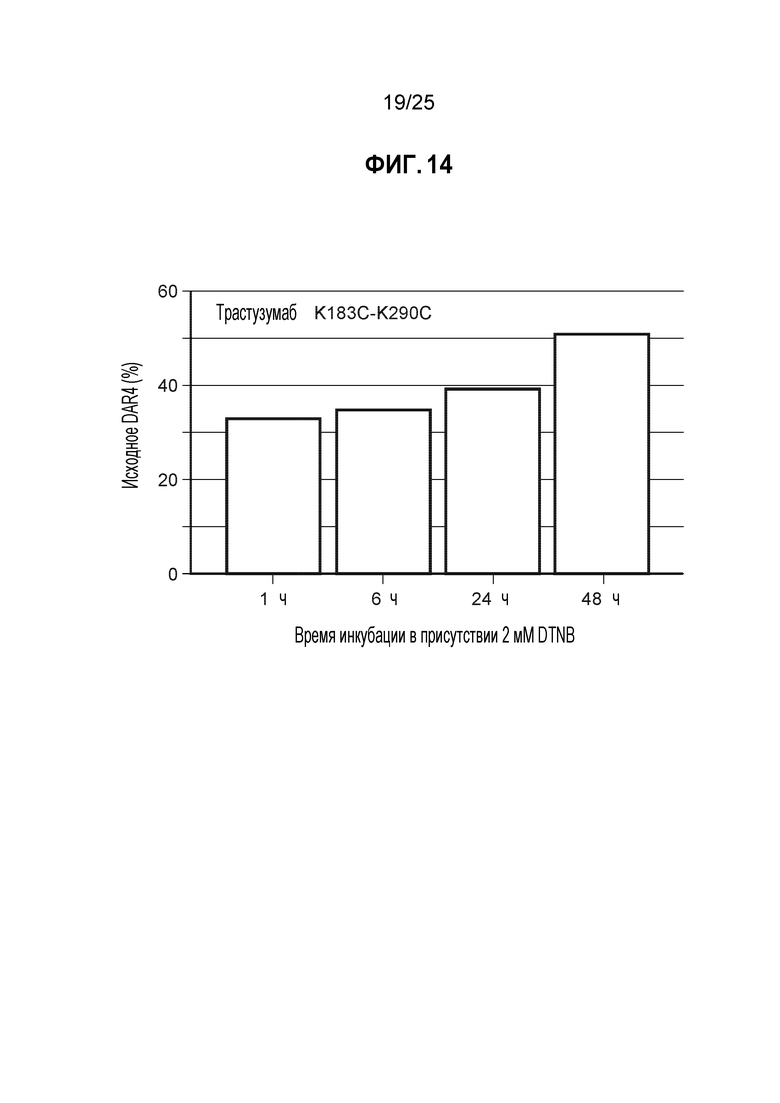
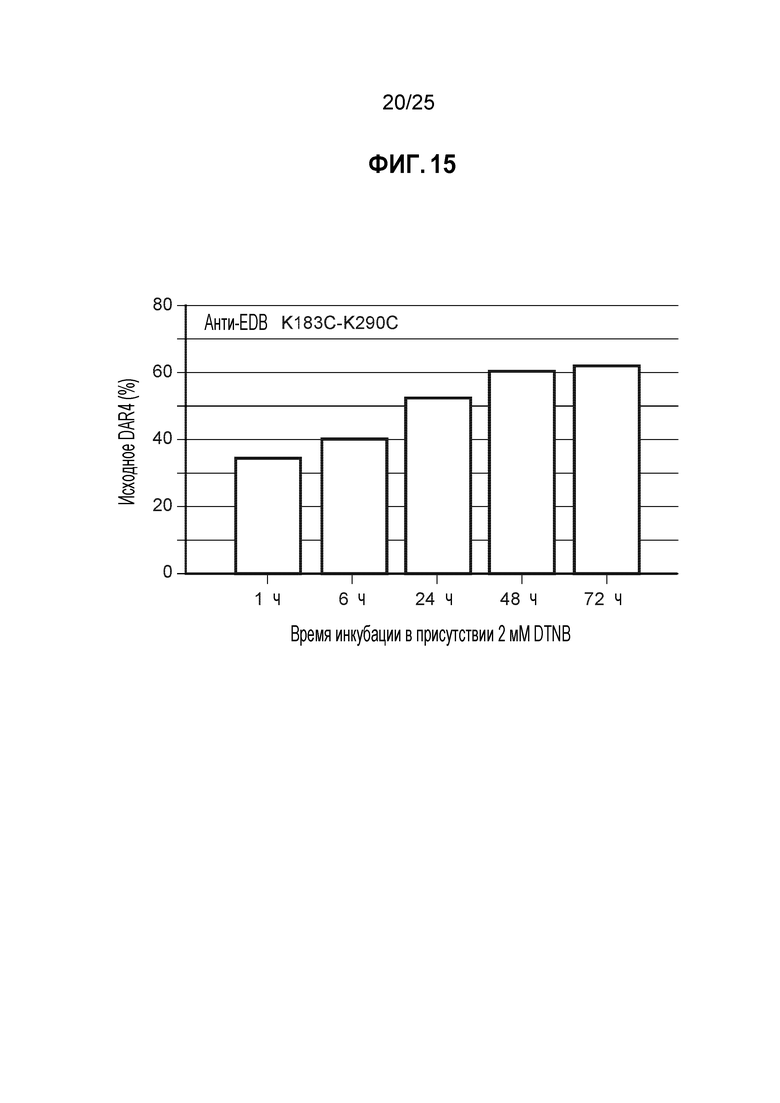
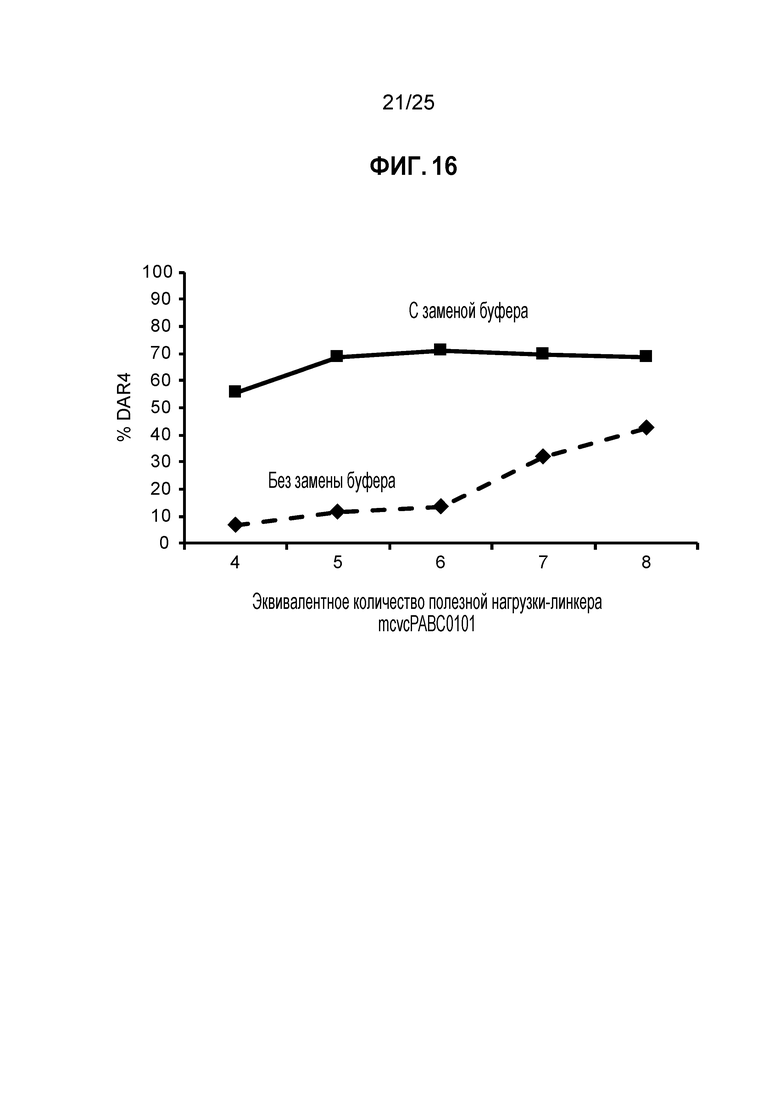
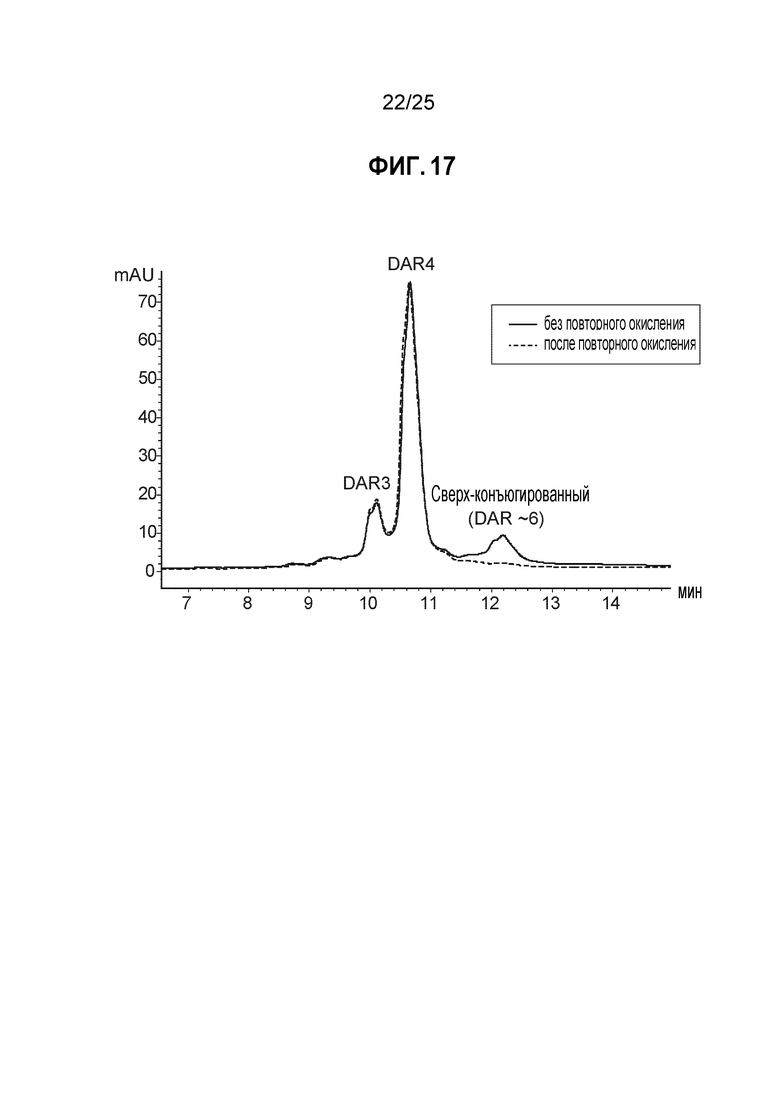
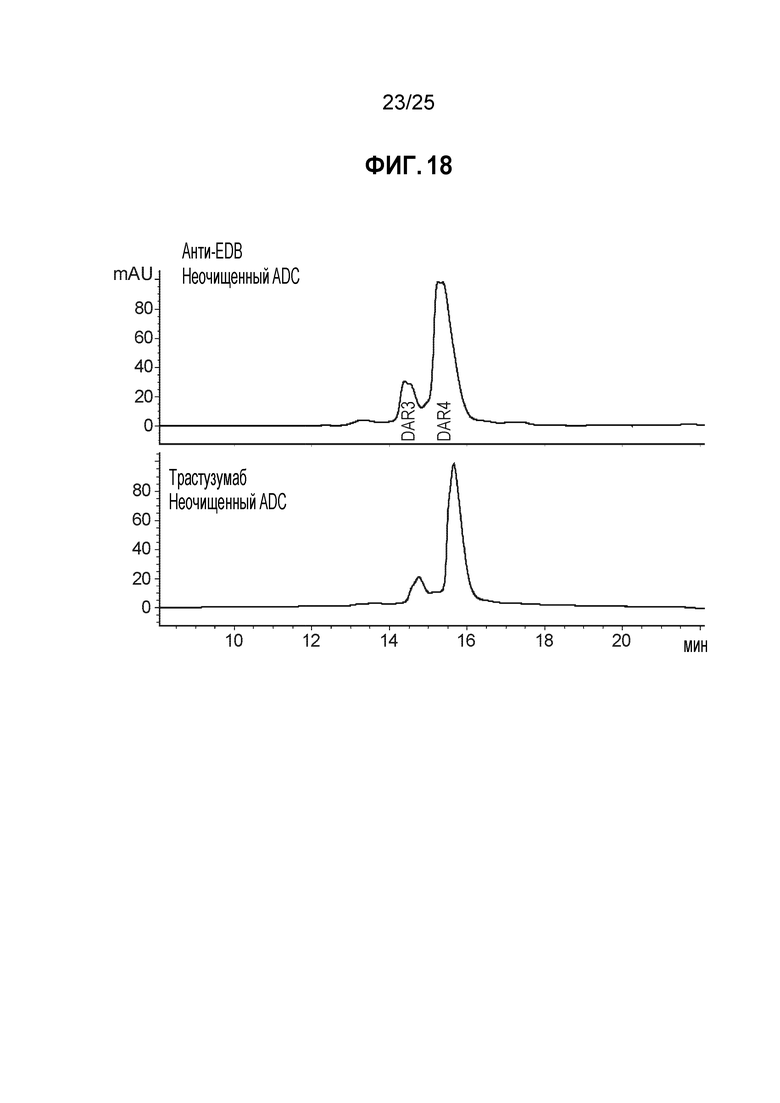
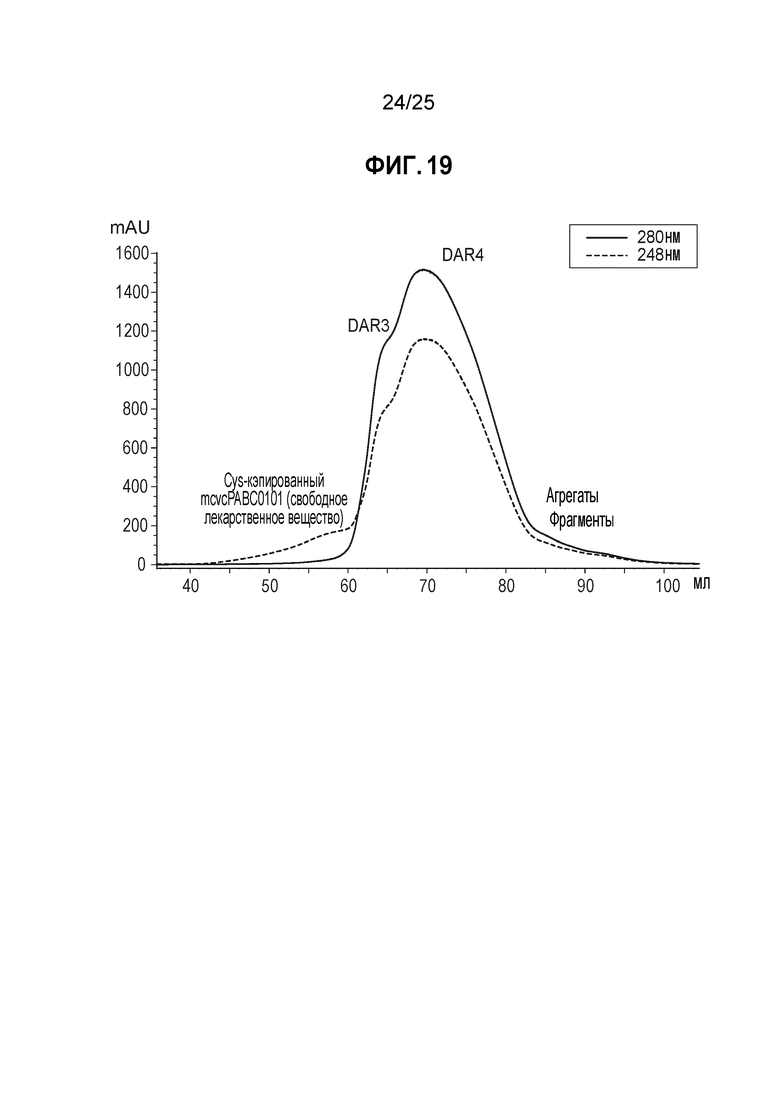
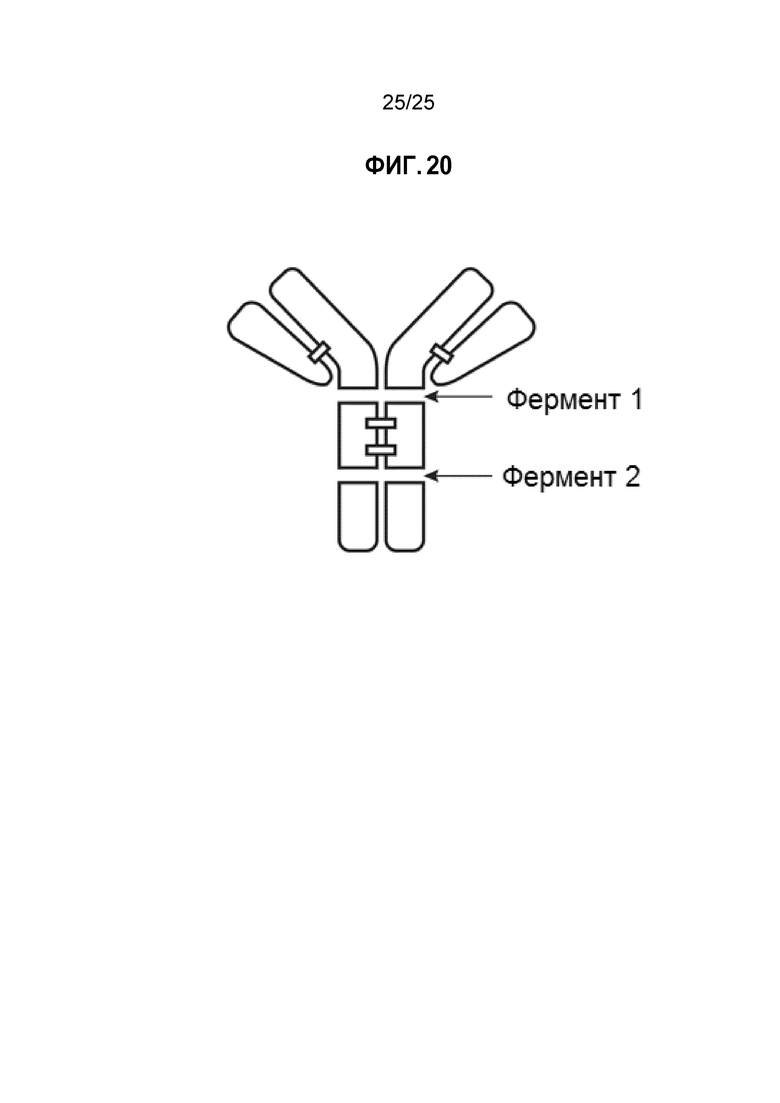
Настоящее изобретение относится к области биотехнологии, в частности к способам получения антител. Изобретение направлено на оптимизацию получения селективно кэпированных и некэпированных цистеинов в антителах путем манипуляции условиями роста клеток, включая намеренное истощение цистеина и/или цистина в процессе культивирования клеток с помощью компонентов среды, продолжительности культивирования партии или плотности клеток, для достижения эффективного продуцирования белка. Изобретение позволяет получить мутантное по цистеину антитело трастузумаб K290C-K334C или K392C-K334C, которое в дальнейшем может быть конъюгировано с химической полезной нагрузкой. 2 з.п. ф-лы, 20 ил., 6 табл., 16 пр.
1. Способ получения цистеин-содержащих белков, которые могут быть конъюгированы с химической полезной нагрузкой, включающий этапы:
(а) засева ростовой среды для клеток, содержащей один или более ростовых компонентов, выбранных из цистеина, цистина и глутатиона, клетками, способными экспрессировать цистеин-содержащие белки;
(b) инкубации указанных клеток до достижения плотности клеток, достаточной для истощения большинства указанных ростовых компонентов, присутствующих в указанной ростовой среде; и
(с) дальнейшей инкубации указанных клеток для экспрессии цистеин-содержащих белков, имеющих один или более некэпированных остатков цистеина, содержащих свободный тиол; и
где указанный цистеин-содержащий белок представляет собой мутантное по цистеину антитело трастузумаб K290C-K334C или K392C-K334C;
где указанные клетки, способные экспрессировать цистеин-содержащие белки, являются клетками млекопитающих CHO-K1, стабильно экспрессирующими мутантное по цистеину антитело трастузумаб K290C-K334C или K392C-K334C;
где указанная ростовая среда для клеток представляет собой среду CD CHO;
где указанную ростовую среду для клеток засевают с плотностью 3E6 кл./мл или 6E6 кл./мл;
где указанные клетки культивировали в увлажненном инкубаторе с 5% CO2 при 37 градусах Цельсия в течение 72 часов.
2. Способ по п.1, дополнительно включающий этап:
(d) введения заданного кэпирующего фрагмента или его предшественника в указанные экспрессированные цистеин-содержащие белки;
в результате чего происходит кэпирование одного или более цистеинов в указанных белках указанным заданным кэпирующим фрагментом, и где указанный заданный кэпирующий фрагмент представляет собой 5-тио-2-нитробензойную кислоту (TNB) или ее предшественник 5,5'-дитиобис(2-нитробензоат) (DTNB).
3. Способ по п.2, в котором указанные кэпированные белки подвергают дальнейшей обработке, состоящей из одного или более из: выделения, очистки и концентрации.
| DUPPATLA V | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| BIOCONJUGATE CHEMISTRY, 2012, v.23, p | |||
| Санный велосипед | 1924 |
|
SU1396A1 |
| SEZONOV G | |||
| et al.: Escherichia Coli Physiology in Luria-Bertani Broth | |||
| JOURNAL OF BACTERIOLOGY, 2007, v.189, n.23, p.8746-8749 | |||
| US 2008108553 A1, 08.05.2008 | |||
| WO 2012064733 A2, 18.05.2012 | |||

