ВКЛЮЧЕНИЕ МАТЕРИАЛА, ПОДАННОГО В ЭЛЕКТРОННОЙ ФОРМЕ, ПУТЕМ ССЫЛКИ
Заявитель тем самым включает путем ссылки материал с перечнем последовательностей, поданный с данной заявкой в электронной форме. Данный файл назван UPN-16-7798PCT_ST25.txt.
ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Данная заявка испрашивает приоритет согласно предварительных заявок на патент США: № 62/323,336, поданной 15 апреля 2016 г., № 62/331,807, поданной 4 мая 2016 г., и № 62/428,866, поданной 1 декабря 2016 г. Эти три заявки на патент включены в данный документ в полном объеме путем ссылки.
1. ВВЕДЕНИЕ
Данная заявка относится к вариантам реализации изобретения, используемым для генной терапии для лечения гемофилии A.
2. УРОВЕНЬ ТЕХНИКИ
Гемофилия A (HA или HemA) представляет собой наиболее распространенное наследственное нарушение свертывания крови. В соответствии с Центрами США контроля и профилактики заболеваний, гемофилия A встречается приблизительно в 1 случае на 5000 новорожденных. В США существует около 20 000 людей с гемофилией A. Гемофилия A встречается в четыре раза чаще, чем гемофилия B, и у более чем половины пациентов с гемофилией A наблюдается тяжелая форма гемофилии. HA вызвана дефицитом фактора VIII (FVIII), а также поддается для лечения подходом с замещением генов в первую очередь благодаря тому, что увеличение уровня FVIII (>1 % от нормального) может облегчать фенотип, для которого характерно возникновение тяжелых кровотечений. В настоящее время аденоассоциированные (AAV) вирусные векторы демонстрируют наибольшую перспективу для применения в генной терапии благодаря их превосходному профилю безопасности и способности к прямой долговременной трансгенной экспрессии из постмитотических тканей, таких как печень.
Однако применение векторов AAV для генной терапии HA вызывает новые сложности из-за отличающихся молекулярных и биохимических свойств человеческого FVIII (hFVIII).По сравнению с другими белками сходного размера экспрессия hFVIII является в высшей степени неэффективной. Применение методов биоинженерии к молекуле FVIII привело к улучшению экспрессии FVIII.Например, удаление B-домена, который не нужен для проявления кофакторной активности, и замена его коротким линкером из 14 аминокислот (FVIII SQ) приводила к 17-кратному увеличению уровней мРНК по сравнению с полноразмерным FVIII нормального типа и 30 % увеличению секретированного белка. См., Ward, Natalie J., et al. "Codon optimization of human factor VIII cDNAs leads to high-level expression." Blood 117.3 (2011): 798–807 и патент США № 9,393,323, также опубликованный как WO 2011/005968.Рекомбинантный FVIII-BDD-SQ обеспечивает клиническое применение в качестве заместительного рекомбинантного продукта FVIII (Refacto, Wyeth Pharma; Xyntha, Pfizer).
Другое препятствие для опосредованного AAV переноса генов для генной терапии HA состоит в размере кодирующей FVIII последовательности, которая составляет 7,0 т. н., что намного превышает нормальную емкость упаковки векторов AAV. Сообщалось об упаковке больших экспрессионных кассет в векторы AAV, но это очень нестабильный процесс, приводящий к низким выходам векторных частиц с пониженной инфицирующей способностью и требующий высокой дозы, что может вызывать повреждение печени. См., например, Sarkar, R., W. Xiao, and H. H. Kazazian. "A single adeno‐associated virus (AAV)‐murine factor VIII vector partially corrects the hemophilia A phenotype." Journal of Thrombosis and Haemostasis 1.2 (2003): 220-226; и McIntosh, Jenny, et al. "Therapeutic levels of FVIII following a single peripheral vein administration of rAAV vector encoding a novel human factor VIII variant." Blood 121.17 (2013): 3335–3344.
Таким образом, для лечения HA требуются более эффективные векторы AAV.FVIII.
3. СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Данные варианты реализации изобретения относятся к вектору AAV для генной терапии для доставки нормального человеческого FVIII нуждающемуся в нем субъекту с последующим внутривенным введением такого вектора, приводящим к долговременной, вероятно на 10 лет или более, клинически значимой коррекции нарушения свертывания крови. Популяция субъектов-пациентов представляет собой пациентов, болеющих от умеренной до тяжелой формами гемофилии A. Предполагаемая доза вектора предназначена для увеличения уровней FVIII в крови приблизительно на 3–10 % или 5 %. Цель лечения вектором AAV заключается в переводе тяжелой гемофилии A у пациентов до умеренной или слабой гемофилии A, таким образом облегчая состояние таких пациентов до необходимости нахождения в режиме профилактики.
Продукт генной терапии, описанный в данном документе, обеспечивает множество важных преимуществ по сравнению с доступными в настоящее время профилактическими подходами для контроля тяжелой гемофилии A. Во-первых, результаты доклинических исследований исследуемого продукта согласовывались с его потенциалом достижения уровней циркуляции фактора VIII до 10 % или более от нормы, это уровни, которые могли быть революционными для целевой популяции пациентов. Во-вторых, данный продукт должен приводить к эффективному достижению постоянных уровней фактора VIII в крови, избегая падения уровней, наблюдаемых в настоящее время при введении экзогенного фактора. В-третьих, это достигается только одним введением, причем необходимость частых внутривенных введений может снижаться на продолжительный период времени, вероятно на десятилетие или дольше.
Данная заявка обеспечивает применение репликационно дефектного аденоассоциированного вируса (AAV) для доставки гена человеческого фактора VIII (hFVIII или hF8) в клетки печени пациентов (субъектов-людей) с диагностированной гемофилией A. Рекомбинантный вектор AAV (rAAV), применяемый для доставки гена hFVIII (rAAV.hFVIII), должен обладать тропизмом по отношению к печени (например, rAAV, несущий капсид AAVhu.37 или AAVrh.10), а трансген hFVIII должен управляться специфическими для печени элементами контроля экспрессии. В одном варианте реализации изобретения элементы контроля экспрессии включают один или более из следующих элементов: энхансер транстиретина (enTTR); промотор транстиретина (TTR) и сигнал полиA. В другом варианте реализации изобретения элементы контроля экспрессии включают один или более из следующих элементов: энхансер предшественника укороченного α1-микроглобулина/бикунина (ABPS) и enTTR; промотор транстиретина (TTR) и сигнал полиA. В одном варианте реализации изобретения элементы контроля экспрессии включают один или более из следующих элементов: энхансер транстиретина (enTTR); промотор антитрипсина альфа-1 (A1AT) и сигнал полиA. В другом варианте реализации изобретения элементы контроля экспрессии включают один или более из следующих элементов: энхансер ABPS и enTTR; промотор A1AT и сигнал полиA. Такие элементы описаны далее в данном документе.
В одном варианте реализации изобретения ген hFVIII кодирует форму фактора VIII с удаленным B-доменом (BDD), в котором B-домен замещен коротким аминокислотным линкером (FVIII-BDD-SQ, также называемым в данном документе как hFVIII). В одном варианте реализации изобретения белковая последовательность FVIII-BDD-SQ показана в SEQ ID NO: 3. В одном варианте реализации изобретения кодирующая последовательность FVIII-BDD-SQ показана в SEQ ID NO: 1. В одном варианте реализации изобретения кодирующая последовательность hFVIII является кодон-оптимизированной для экспрессии у людей. Такая последовательность может иметь 80 % идентичности с нативной кодирующей последовательностью hFVIII (SEQ ID NO: 1). В одном варианте реализации изобретения кодирующая последовательность hFVIII показана в SEQ ID NO: 2.
В другом аспекте изобретения в данном документе предложена водная суспензия, подходящая для введения пациенту с гемофилией A, которая содержит rAAV, описанный в данном документе. В некоторых вариантах реализации изобретения данная суспензия содержит водную суспендирующую жидкость и от около 1 x 1012 до около 1 x 1014 геномных копий (ГК) rAAV/мл. В одном варианте реализации изобретения суспензия пригодна для внутривеннной инъекции. В другом варианте реализации изобретения суспензия дополнительно содержит поверхностно-активное вещество, консервант и/или буфер, растворенные в водной суспендирующей жидкости.
В еще одном варианте реализации изобретения в данном документе предложен способ лечения пациента, болеющего гемофилией A, с помощью rAAV, как описано в данном документе. В одном варианте реализации изобретения в водной суспензии в организм пациента доставляют от около 1 x 1011 до около 3 x 1013 геномных копий (ГК) rAAV/кг массы тела пациента.
Цель лечения заключается в функциональном замещении дефектного hFVIII пациента посредством основанной на rAAV направленной на печень генной терапии в качестве целесообразного подхода для лечения данного заболевания и улучшения ответа на существующие виды терапий. Варианты реализации изобретения, описанные в данной заявке, в частности основаны на разработке терапевтических композиций и способов, позволяющих безопасно доставлять эффективные дозы, и усовершенствованных способов промышленного получения для соответствия требованиям по очистке продукции для эффективного введения доз субъектам-людям.
4. КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На ФИГ. 1 показано схематическое представление цис-плазмиды pAAV.E03.P3.hF8co-SQ.PA75.
На ФИГ. 2 показано схематическое представление цис-плазмиды pAAV.E12.P3.hF8co-SQ.PA75.
На ФИГ. 3 показано схематическое представление цис-плазмиды pAAV.E03.P2.hF8co-SQ.PA75.
На ФИГ. 4 показано схематическое представление цис-плазмиды pAAV.E12.P2.hF8co-SQ.PA75.
На ФИГ. 5 показана вариация активности hFVIII перед образованием антител у мышей FVIII KO. Мышам FVIII KO внутривенно (в/в) вводили 1010 ГК векторов AAVrh10, экспрессирующих hFVIIIco-SQ из одной из 42 комбинаций энхансеров/промоторов. Каждый из наборов энхансеров (отмеченных как E01–E14, в таблице 1) комбинировали с промоторами TBG-S1 (левая когорта из каждой группы), A1AT (средняя когорта) и TTR (правая когорта). Анализом COATEST и твердофазным ИФА с IgG против hFVIII определяли, соответственно, активность hFVIII (A) и титры IgG против hFVIII (B). Анализы выполняли на мышиной плазме, выделенной на неделе 2 после введения векторов. Строят график по данным отдельных мышей со средними значениями ± стандартная ошибка среднего (SEM) для показанной активности (n = 10/группу).
На ФИГ. 6 показана активность hFVIII и титр антител против hFVIII на неделю 8 после в/в введения вектора у мышей FVIII KO. Мышам FVIII KO внутривенно (в/в) вводили 1010 ГК векторов AAVrh10, экспрессирующих hFVIIIco-SQ из одной из 42 комбинаций энхансеров/промоторов. Каждый из наборов энхансеров (отмеченных как E01–E14, в таблице 1) комбинировали с промоторами TBG-S1 (левая когорта из каждой группы), A1AT (средняя когорта) и TTR (правая когорта). Анализом COATEST и твердофазным ИФА с IgG против hFVIII определяли, соответственно, активность hFVIII (A) и титры IgG против hFVIII (B). Анализы выполняли на мышиной плазме, выделенной на неделе 8 после введения векторов. Строят график по данным отдельных мышей со средними значениями ± стандартная ошибка среднего (SEM) для показанной активности (n = 10/группу).
На ФИГ. 7 показана в динамике активность hFVIII у мышей FVIII KO с после в/в введение 42 энхансерных/промоторных комбинаций векторов. Мышам FVIII KO внутривенно (в/в) вводили 1010 ГК векторов AAVrh10, экспрессирующих hFVIIIco-SQ из одной из 42 комбинаций энхансеров/промоторов. Каждый из наборов энхансеров (отмеченных как E01–E14, в таблице 1) комбинировали с промоторами TBG-S1 (левая когорта из каждой группы), A1AT (средняя когорта) и TTR (правая когорта). Анализом COATEST определяли активность hFVIII на плазме мышей, выделяемой каждые две недели после введения векторов. Строили график по данным отдельных мышей со средними значениями ± SEM (n = 10/группу).
На ФИГ. 8 показаны в динамике титры антител против hFVIII у мышей FVIII KO с последующим в/в введением 42 энхансерных/промоторных комбинаций векторов. Мышам FVIII KO внутривенно (в/в) вводили 1010 ГК векторов AAVrh10, экспрессирующих hFVIIIco-SQ из одной из 42 комбинаций энхансеров/промоторов. Каждый из наборов энхансеров (отмеченных как E01–E14, в таблице 1) комбинировали с промоторами TBG-S1 (левая когорта), A1AT (средняя когорта) и TTR (правая когорта). Титры IgG против hFVIII определяли твердофазным ИФА с IgG против hFVIII на мышиной плазме, выделяемой каждые две недели. Строили график по данным отдельных мышей (n = 10/группу).
На ФИГ. 9 представлено сравнение активности hFVIII и титра антител против hFVIII с последующим в/в введением генома E06.TTR.hFVIIIco-SQ с помощью различных векторных капсид. Мышам FVIII KO в/в вводили 1010 ГК векторов AAVrh10, AAV8, AAV9, AAVhu37 или AAVrh64R1, экспрессирующих hFVIIIco-SQ из E06.TTR. Собирали плазму каждые две недели и анализом COATEST и твердофазным ИФА с IgG против hFVIII определяли, соответственно, активность hFVIII (A) и титры IgG против hFVIII (B). Строят график по данным отдельных мышей со средними значениями ± стандартная ошибка среднего (SEM) для показанной активности (n = 10/группу).
На ФИГ. 10 представлена экспрессия hFVIII в предварительных исследованиях на приматах, не являющихся людьми (NHP). (A) Двум самцам макака-резус в/в вводили 3x1012 ГК/кг AAVrh10.ABP2.TBG-S1.hFVIIIco-SQ. (B) Двум самцам яванского макака в/в вводили 3x1012 ГК/кг AAVhu37.ABP2.TBG-S1.hFVIIIco-SQ. Для оценки экспрессии hFVIII и наличия антител против трансгена hFVIII у макак еженедельно или раз в две недели отбирали кровь. Твердофазным ИФА измеряли в плазме экспрессию hFVIII (сплошная линия), а значения выражали в виде среднего ± SEM. В плазме твердофазным ИФА также определяли титры IgG против hFVIII (штриховая линия).
На ФИГ. 11 представлена экспрессия hFVIII у яванских макак. Пяти самцам макака-резус в/в вводили 1,2x1013 ГК/кг одного из вариантов AAVrh10.E03.TTR.hFVIIIco-SQ.PA75, AAVrh10.E12.A1AT.hFVIIIco-SQ.PA75, AAVhu37.E03.TTR.hFVIIIco-SQ.PA75, или AAVhu37.E12.A1AT.hFVIIIco-SQ.PA75. У макак каждые две недели отбирали кровь для оценки экспрессии hFVIII в плазме твердофазным ИФА, а значения выражали в виде среднего ± SEM.
На ФИГ. 12 показано образование антител против hFVIII у яванских макак. Пяти самцам макака-резус в/в вводили 1,2x1013 ГК/кг одного из вариантов AAVrh10.E03.TTR.hFVIIIco-SQ.PA75, AAVrh10.E12.A1AT.hFVIIIco-SQ.PA75, AAVhu37.E03.TTR.hFVIIIco-SQ.PA75, или AAVhu37.E12.A1AT.hFVIIIco-SQ.PA75. Для оценки наличия антител против трансгена hFVIII у макак каждые две недели отбирали кровь. В плазме твердофазным ИФА оценивали титры IgG против hFVIII. Статистический анализ показан на ФИГ. 14.
На ФИГ. 13 представлена технологическая схема.
На ФИГ. 14 представлен анализ времени до наступления события образования антител против FVIII, показанного на ФИГ. 12. С использованием лог-рангового критерия (Мантела-Кокса) отмечена статистически значимая разница между AAVrh.10 и AAVhu.37.
На ФИГ. 15 показано сравнение уровней экспрессии rhCG векторами AAVrh10, AAV8, AAV3B и AAV5 (первая инъекция вектора).
На ФИГ. 16A–16D показаны ДНК-копии векторов rhCG в печени в различные моменты времени (AAVrh10, ФИГ. 16A; AAV8, ФИГ. 16B; AAV3B, ФИГ. 16C; AAV5, ФИГ. 16D).
На ФИГ. 17A–17B показаны уровни rhAFP после повторного введения (вторая инъекция вектора) векторов AAV3B (ФИГ. 17A) или AAV5 (ФИГ. 17B), экспрессирующих rhAFP.
На ФИГ. 18A–18B показаны геномные копии вектора rhAFP в печени (ФИГ. 18A, AAV3B.TBG.rhAFP; ФИГ. 18B, AAV5.TBG.rhAFP).
На ФИГ. 19 показаны различные Nab-ответы на AAV у макак.
На ФИГ. 20A–20B представлены уровни векторных ГК (Фиг. 20A) или РНК-транскриптов (Фиг. 20B) в печени мышей, которым инъецировали векторы с энхансером/промотором AAVrh10, экспрессирующие hFVIIIco IV, как описано в разделе 6.3.8.
На ФИГ. 21 представлены уровни РНК-транскриптов hFVIII в мышце (правая икроножная), правом яичке, поджелудочной железе, правой почке, селезенке, правом легком и сердце мышей, которым инъецировали векторы с энхансером/промотором AAVrh10, экспрессирующие hFVIIIco IV, как описано в разделе 6.3.8.
На ФИГ. 22 показан график, демонстрирующий долговременную стабильную экспрессию человеческого FVIII у яванского макака (35 месяцев) после однократной внутривенной инъекции AAVhu37.TBG-S1.hFVIII-SQ.PA75 с дозой 3x1012 ГК/кг.
На ФИГ. 23 показан график, демонстрирующий уровни ферментов в печени (АЛТ, Ед/мл, квадраты; АСТ, Ед/мл, круги) у макак из ФИГ. 22.
На ФИГ. 24 показан график, демонстрирующий ответ в виде нейтрализующих антител (Nab) на капсид AAVhu.37.
На ФИГ. 25 представлено выравнивание последовательности hFVIIIco (SEQ ID NO: 2) по сравнению с нативной последовательностью hFVIII (SEQ ID NO: 1).
5. ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Варианты реализации, описанные в данной заявке, относятся к применению репликационно дефектного аденоассоциированного вируса (AAV) для доставки гена человеческого фактора VIII (hFVIII) в клетки печени пациентов (субъектов-людей) с диагностированной гемофилией A. Рекомбинантный вектор AAV (rAAV), применяемый для доставки гена hFVIII (rAAV.hFVIII), должен обладать тропизмом по отношению к печени (например, rAAV, несущий капсид AAVhu.37 или AAVrh.10), а трансген hFVIII должен управляться специфическими для печени элементами контроля экспрессии. В одном варианте реализации изобретения элементы контроля экспрессии включают один или более из следующих элементов: энхансер транстиретина (TTR); промотор транстиретина (TTR) и сигнал полиA. Такие элементы описаны далее в данном документе.
Как используется в данном документе, «капсид AAVhu.37» относится к hu.37, имеющему аминокислотную последовательность из Genbank, номер регистрации: AAS99285, SEQ ID NO: 17, которая включена в данный документ путем ссылки. Допускается некоторая вариация этой кодирующей последовательности, которая может включать последовательности с около 99 % идентичности к эталонной аминокислотной последовательностью в AAS99285 и US 2015/0315612 (которые включены в данный документ путем ссылки) (т. е. менее чем около 1 % вариации от эталонной последовательности). Поэтому были описаны способы получения капсидных кодирующих последовательностей и способы продукции вирусных векторов rAAV. См., например, Gao, et al, Proc. Natl. Acad. Sci. США 100 (10), 6081-6086 (2003) и US 2015/0315612.
Как используется в данном документе, «капсид AAVrh10» относится к rh.10, имеющему аминокислотную последовательность из Genbank, номер регистрации: AAO88201, SEQ ID NO: 18, которая включена в данный документ путем ссылки. Допускается некоторая вариация этой кодирующей последовательности, которая может включать последовательности с около 99 % идентичности к эталонной аминокислотной последовательностью в AAO88201 и US 2013/0045186A1 (т. е. менее чем около 1 % вариации от эталонной последовательности), при условии, что поддерживается целостность лигандсвязывающего сайта для очистки с аффинным захватом, а изменение в последовательностях существенно не изменяет диапазон pH для капсида для обеспечения очистки на ионообменной смоле (как обсуждается далее в данном документе). Поэтому были описаны способы получения капсидных кодирующих последовательностей и способы продукции вирусных векторов rAAV. См., например, Gao, et al, Proc. Natl. Acad. Sci. США 100 (10), 6081–6086 (2003) и US 2013/0045186A1.
Как используется в данном документе, термином «титр NAb» определяется величина продуцирования нейтрализующего антитела (например, Nab против AAV), которое нейтрализует физиологические действие своего целевого эпитопа (например, AAV). Титры NAb против AAV можно измерять, как описано, например, в Calcedo, R., et al., Worldwide Epidemiology of Neutralizing Antibodies to Adeno-Associated Viruses. Journal of Infectious Diseases, 2009. 199(3): p. 381–390, которая включена в данный документ путем ссылки.
Термины «процент (%) идентичности», «идентичность последовательностей», «процент идентичности последовательностей» или «процентная идентичность», в контексте аминокислотных последовательностей, относится к остаткам в двух последовательностях, которые являются одинаковыми при выравнивании для определения соответствия. Процент идентичности можно легко определять для аминокислотных последовательностей для полной длины белка, полипептида, около 32 аминокислот, около 330 аминокислот или их пептидного фрагмента, или соответствующих кодирующих секвенированных последовательностей нуклеотидных последовательностей. Подходящий аминокислотный фрагмент может в длину содержать по меньшей мере около 8 аминокислот и может содержать до около 700 аминокислот. В целом, при упоминании терминов «идентичность», «гомология» или «сходство» между двумя различными последовательностями, то «идентичность», «гомология» или «сходство» определяют в отношении «выровненных» последовательностей. «Выровненные» последовательности или «выравнивания» относятся к множеству последовательностей нуклеиновых кислот или белков (аминокислот), часто содержащих корректировки по отсутствующим или дополнительным основаниям или аминокислотам, по сравнению с эталонной последовательностью. Выравнивания выполняют с использованием любого из разнообразия общедоступных или коммерчески доступных программ для множественного выравнивания последовательностей. Для аминокислотных последовательностей доступны программы выравнивания последовательностей, например программы Clustal Omega, Clustal X, MAP, PIMA, MSA, BLOCKMAKER, MEME и Match-Box. Как правило, любую из этих программ используют с настройками по умолчанию, хотя специалист в данной области может при необходимости изменять эти настройки. В альтернативном варианте специалист в данной области может использовать другой алгоритм или компьютерную программу, которая обеспечивает по меньшей мере тот же уровень идентичности или выравнивания, которые предоставляются эталонными алгоритмами и программами. См., например, J. D. Thomson et al, Nucl. Acids. Res., “A comprehensive comparison of multiple sequence alignments”, 27(13):2682-2690 (1999).
Как используется в данном документе, термин «функционально связанный» относится как к последовательностям контроля экспрессии, которые примыкают к интересующему гену, так и последовательностям контроля экспрессии, которые действуют в транс-положении или на расстоянии контролируют интересующий ген.
«Дефектный по репликации вирус» или «вирусный вектор» относится к рекомбинантной, синтетической или искусственной вирусной частице, в которой в вирусный капсид или оболочку упакована экспрессионная кассета, содержащая интересующий ген, причем упакованы также в этот вирусный капсид или оболочку любые вирусные геномные последовательности также являются дефектными по репликации, т. е. они не могут создавать вирионы потомства, но сохраняют способность инфицировать клетки-мишени. В одном варианте реализации изобретения геном вирусного вектора не включает гены, кодирующие ферменты, требуемые для репликации (геном может быть сконструирован так, чтобы быть «выпотрошенным» — содержащим только интересующий трансген, фланкированный сигналами, необходимыми для амплификации и упаковки искусственного генома), но эти гены могут предоставляться во время продукции. Поэтому считается, что его безопасно использовать в генной терапии, поскольку не может происходить репликация и инфицирование вирионами потомства, за исключением того случая, когда присутствует требуемый для репликации вирусный фермент.
Следует отметить, что термины в единственном числе относится к одному или более элементам. По существу, термины в единственном числе, термины «один или более» и «по меньшей мере один» применяются в данном документе взаимозаменяемо.
Слова «содержать», «содержит» и «содержащий» должны толковаться как включающие, а не исключающие. Слова «состоит», «состоящий» и их варианты должны толковаться как включающие, а не исключающие. При том что различные варианты реализации изобретения в данном описании представлены с использованием стиля «содержащий», в других обстоятельствах родственный вариант реализации изобретения также подразумевает необходимость интерпретации и описания с использованием стиля «состоящий из» или «состоящий по существу из».
Как используется в настоящем описании термин «около» означает вариабельность на 10 % от данного эталонного значения, если не указано иное.
Если в данном документе не указано иное, технические и научные термины, использованные в данном документе, имеют то же самое значение, которое обычно понимается средним специалистом в данной области, и учитывая ссылки на опубликованные тексты, которые предоставляют специалисту в данной области общим руководством для множества терминов, использованных в настоящей заявке.
5.1 Векторы для генной терапии
В одном аспекте для применения в генной терапии предложен рекомбинантный аденоассоциированный вирусный (rAAV) вектор, несущий ген фактора 8 сворачивания крови человека (hF8 или hFVIII). Вектор rAAV.hFVIII должен обладать тропизмом по отношению к печени (например, rAAV, несущий капсид AAVhu.37 или AAVrh.10), а трансген hFVIII должен управляться специфическими для печени элементами контроля экспрессии. Вектор готовится в виде препарата в буфере/носителе, подходящем для инфузии субъектам-людям. Буфер/носитель должен содержать компонент, который предотвращает прилипание rAAV к трубке для инфузии, но не влияет на активность связывания rAAV in vivo.
5.1.1. Вектор rAAV.hFVIII
5.1.1.1. Последовательность hFVIII
Человеческий фактор коагуляции VIII продуцируется в виде большого гликопротеина массой 330 кДа с доменной структурой A1-A2-B-A3-C1-C2, в которой и A и C домены обладают внутренней гомологией последовательностей и приблизительно 40 % идентичностью последовательностей с доменами A и C фактора V (FV), который несет аналогичную доменную структуру. Для прокоагулянтной активности необязателен B-домен, который составляет 38 % от общей последовательности. FVIII, в котором удален B-домен (BDD) и замещен коротким линкером из 14 аминокислот (FVIII SQ), находится в клиническом применении в качестве заместительного рекомбинантного продукта FVIII, и было показано, что это привело к 17-кратному увеличению уровней мРНК по сравнению с полноразмерным FVIII нормального типа и 30 % увеличению секретированного белка. См., McIntosh et al, Therapeutic levels of FVIII following a single peripheral vein administration of rAAV vector encoding a novel human factor VIII variant, Blood, 121(17):3335-44 (Feb 2013) и Ward et al, Codon optimization of human factor VIII cDNAs leads to high-level expression, Blood, 117(3):798-807 (Jan 2011), которые включены в данный документ путем ссылки.
В одном варианте реализации изобретения ген hFVIII, кодирующий белок hFVIII, показан в SEQ ID NO: 3, являющейся последовательностью FVIII, у которой удален B-домен (BDD) и замещен коротким линкером из 14 аминокислот (FVIII-BDD-SQ). Таким образом, в одном варианте реализации изобретения трансген hFVIII может включать, но не ограничиваться, одной или более последовательностей, представленных SEQ ID NO:1 или SEQ ID NO: 2, которые представлены в приложенном перечне последовательностей, который включен в данный документ путем ссылки. В SEQ ID NO: 1 предложена кДНК для нативного человеческого FVIII-BDD-SQ. В SEQ ID NO: 2 предложена сконструированная кДНК для человеческого FVIII-BDD-SQ, который прошел кодон-оптимизацию для экспрессии у людей (иногда называемый в данном документе как hFVIIIco-SQ или hFVIIIco-BDD-SQ). Необходимо понимать, что ссылка на hFVIII в данном документе, в некоторых вариантах реализации изобретения, относится к нативной или кодон-оптимизированной последовательности hFVIII-BDD-SQ. Альтернативно или дополнительно, могут использоваться доступные в сети или коммерчески доступные компьютерные программы, а также предоставляющие услуги компании для обратной трансляции аминокислотных последовательностей в нуклеотидные кодирующие последовательности, включая как РНК, так и/или кДНК. См., например, обратно транслированные последовательности от компаний EMBOSS, www.ebi.ac.uk/Tools/st/ ; Gene Infinity (www.geneinfinity.org/sms-/sms_backtranslation.html); ExPasy (www.expasy.org/tools/). Предполагается, что охвачены все нуклеиновые кислоты, кодирующие описанные полипептидные последовательности hFVIII, включая нуклеотидные последовательности, которые были оптимизированы для экспрессии у необходимого целевого субъекта (например, методом оптимизации кодонов). В одном варианте реализации изобретения нуклеотидная последовательность, кодирующая hFVIII, имеет по меньшей мере 95 % идентичность с нативной кодирующей hFVIII последовательностью SEQ ID NO: 1. В другом варианте реализации изобретения нуклеотидная последовательность, кодирующая hFVIII, имеет по меньшей мере 90, 85, 80, 75, 70 или 65 % идентичность с нативной кодирующей hFVIII последовательностью SEQ ID NO: 1. В одном варианте реализации изобретения нуклеотидная последовательность, кодирующая hFVIII, имеет около 77 % идентичность с нативной кодирующей hFVIII последовательностью SEQ ID NO: 1. В одном варианте реализации изобретения нуклеотидная последовательность, кодирующая hFVIII представляет собой SEQ ID NO: 2. В другом варианте реализации изобретения нуклеотидная последовательность, кодирующая hFVIII, имеет по меньшей мере 99 %, 97 %, 95 %, 90 %, 85 %, 80 %, 75 %, 70 % или 65 % идентичность с нативной кодирующей hFVIII последовательностью SEQ ID NO: 1 или SEQ ID NO: 2. В другом варианте реализации изобретения нуклеотидная последовательность, кодирующая hFVIII представляет собой SEQ ID NO: 19. В другом варианте реализации изобретения нуклеотидная последовательность, кодирующая hFVIII, имеет по меньшей мере 90, 85, 80, 75, 70 или 65 % идентичность с кодирующей hFVIII последовательностью SEQ ID NO: 19. В еще одном варианте реализации изобретения нуклеотидная последовательность, кодирующая hFVIII, имеет по меньшей мере 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99 % идентичность с кодирующей hFVIII последовательностью SEQ ID NO: 1 или SEQ ID NO: 2. В еще одном варианте реализации изобретения нуклеотидная последовательность, кодирующая hFVIII, имеет по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99 % идентичность с кодирующей hFVIII последовательностью SEQ ID NO: 19. См. Ward et al, Codon optimization of human factor VIII cDNAs leads to high-level expression, Blood, 117(3):798-807 (Jan 2011), которая включена в данный документ путем ссылки для обсуждения различных вариантов FVIII-SQ, включая кодон-оптимизированные варианты.
Кодон-оптимизированные кодирующие области могут конструироваться различными разнообразными методами. Эту оптимизацию можно выполнять, используя методы, которые доступны онлайн (например, GeneArt), опубликованные методы или компании, которые обеспечивают услуги по оптимизации кодонов, например DNA2.0 (г. Менло-Парк, штат Калифорния, США). Один подход для оптимизации кодонов описан, например, в международной публикации патента № WO 2015/012924, которая включена в данный документ путем ссылки. См. также, например, публикацию патента США № 2014/0032186 и публикацию патента США № 2006/0136184. Соответственно модифицируют всю длину открытой рамки считывания (ORF) для данного продукта. Тем не менее, в некоторых вариантах реализации изобретения, можно изменять только фрагмент ORF. Путем использования одного из этих методов, специалист может применять значения частоты встречаемости к любой заданной полипептидной последовательности и получать фрагмент нуклеиновой кислоты кодон-оптимизированного кодирующей области, которая кодирует данный полипептид.
Для выполнения фактических изменений в кодоны или для синтеза сконструированных кодон-оптимизированных кодирующих области доступен ряд вариантов, описанных в данном документе. Такие модификации или синтез можно выполнять с использованием стандартных и обычных молекулярных биологических манипуляций, хорошо известных средним специалистам в данной области. В одном подходе стандартными способами синтезируются наборы комплементарных олигонуклеотидных пар из 80–90 нуклеотидов, каждый по длине и протяженности длины соответствующий требуемой последовательности. Эти олигонуклеотидные пары синтезируют таким образом, чтобы при ренатурации они образовывали двухцепочечные фрагменты из 80–90 пар оснований, содержащие липкие концы, например, каждый олигонуклеотид в паре синтезируют до размера 3, 4, 5, 6, 7, 8, 9, 10 или более оснований больше области, которая является комплементарной к другому олигонуклеотиду в этой паре. Одноцепочечные концы каждой пары олигонуклеотидов разработаны для ренатурации с одноцепочечным концом другой пары олигонуклеотидов. Олигонуклеотидным парам позволяют ренатурировать и затем приблизительно от пяти до шести из этих двухцепочечных фрагментов позволяют ренатурировать вместе посредством липких одноцепочечных концов, а потом их лигируют вместе и клонируют в стандартном бактериальном векторе клонирования, например векторе TOPO®, предлагаемом Thermo Fisher Scientific Inc. Затем полученную конструкцию секвенируют стандартными способами. Получают несколько из этих конструкций, состоящих из от 5 до 6 фрагментов из от 80 до 90 пар лигированных вместе фрагментов, т. е. фрагментов из около 500 пар оснований, таким образом, чтобы в наборе плазмидных конструкций была представлена полная требуемая последовательность. Затем вставки этих плазмид разрезают соответствующими ферментами рестрикции и лигируют вместе с образованием готовой конструкции. Готовую конструкцию потом клонируют в стандартном бактериальном векторе клонирования и секвенируют. Дополнительные способы будут непосредственно очевидны специалисту в данной области. В дополнение к этому, синтез генов легко доступен в коммерческой сфере.
5.1.1.2. Вектор rAAV
Поскольку hFVIII изначально экспрессируется в печени, желательно использовать AAV, который проявляет тропизм к печени. В одном варианте реализации изобретения поставляемый AAV капсид представляет собой AAVrh.37. В другом варианте реализации изобретения поставляемый AAV капсид представляет собой AAVrh.10. Однако можно применять любое количество векторов rAAV с тропизмом к печени.
В конкретном варианте реализации изобретения, описанном в примерах ниже, вектор для генной терапии представляет собой вектор AAVhu.37, экспрессирующий трансген hFVIII под контролем промотора транстиретина, называемый rAAVhu.37.TTR.hFVIII. Внешний компонент вектора AAV представляет собой серотип hu.37, T = 1 икосаэдрального капсида, состоящего из 60 копий трех вирусных белков AAV VP1, VP2 и VP3 в соотношении 1:1:10. Капсид содержит одноцепочечную ДНК векторного генома rAAV.
Геном rAAVhu.37.TTR.hFVIII содержит трансген hFVIII, фланкированный двумя инвертированными концевыми повторами (ITR) AAV. Трансген hFVIII содержит энхансер, промотор, кодирующую hFVIII последовательность и сигнал полиаденилирования (полиA). Эти контрольные последовательности являются «функционально связанными» с генными последовательностями hFVIII. Экспрессионную кассету можно встраивать в плазмиду, которая используется для продукции вирусного вектора.
ITR представляют собой генетические элементы, отвечающие за репликацию и упаковку генома во время продукции вектора, и они являются единственными вирусными цис-элементами, необходимыми для образования rAAV. Минимальные последовательности, требуемые для упаковки экспрессионной кассеты в вирусную частицу AAV, представляют собой 5’ и 3’ ITR AAV, которые могут быть того же происхождения, что и капсид, или которые имеют другое происхождение AAV (для получения псевдотипа AAV). В одном варианте реализации изобретения применяются последовательности ITR из AAV2 или их версия с делецией (∆ITR). Однако могут выбирать ITR из других источников. Если источник ITR происходит из AAV2, а капсид AAV происходит из другого источника AAV, то получаемый вектор может называться псевдотипным. Как правило, экспрессионная кассета для вектора AAV содержит 5’ ITR AAV, кодирующие hFVIII последовательности, любые регуляторные последовательности и 3’ ITR AAV. Тем не менее могут подходить другие конфигурации этих элементов. Была описана укороченная версия 5’ ITR, называемая ∆ITR, у которой удалены D-последовательность и концевой сайт распознавания (trs). В других вариантах реализации изобретения применяют полноразмерные 5’ и 3’ ITR AAV. В одном варианте реализации изобретения последовательность 5' ITR показана в SEQ ID NO: 11. В одном варианте реализации изобретения последовательность 3' ITR показана в SEQ ID NO: 12.
Экспрессия кодирующей hFVIII последовательности управляется специфическим для печени промотором. Из-за размера трансгена hFVIII желательно применение промотора относительно малого размера. Иллюстративная плазмида и вектор, описанные в данном документе, используют промотор транстиретина (TTR) (также называемый в данном документе P3) или его модифицированную версию. В одном варианте реализации изобретения последовательность промотора TTR показана в SEQ ID NO: 7. В альтернативном варианте можно применять специфические для печени промоторы, такие как промотор тироксин-связывающего глобулина (TBG) (также называемый в данном документе как P1) или его укороченную версию, TBG-S1, последовательность которой показана в SEQ ID NO: 8. Другой подходящий промотор представляет собой промотор антитрипсина альфа 1 (A1AT) или его модифицированную версию (также называемый P2), показанную в SEQ ID NO: 9. Другие подходящие промоторы включают промотор альбумина человека (Miyatake et al., J. Virol., 71:5124 32 (1997)), humAlb; и промотор корового белка вируса гепатита B (Sandig et al., Gene Ther., 3:1002 9 (1996). См., например, базу данных промоторов специфических промоторов печени, г. Колд Спринг Харбор, rulai.schl.edu/LSPD, которая включена путем ссылки. Хотя и менее желательно, в описанных в данном документе векторах могут использоваться другие промоторы, такие как вирусные промоторы, конститутивные промоторы, регулируемые промоторы [см., например, WO 2011/126808 и WO 2013/04943] или промотор, восприимчивый к физиологическим сигналам.
В одном варианте реализации изобретения последовательности контроля экспрессии включают один или более энхансеров. В одном варианте реализации изобретения включена последовательность энхансера транстиретина (enTTR) (100 п. н. последовательности энхансера транстиретина), данная последовательность показана в SEQ ID NO: 5. См. Wu et al, Molecular Therapy, 16(2):280–289, Feb. 2008, которая включена в данный документ путем ссылки. В другом варианте реализации изобретения включен энхансер En34 (коровый энхансер размером 34 п. н. аполипопротеина человека из контрольной области печени), который показан в SEQ ID NO: 4. В еще одном варианте реализации изобретения включен энхансер ABPS (укороченная до 42 п. н. версия дистального энхансера размером 100 п. н. предшественника α1-микроглобулина/букинина [ABP]). Такая последовательность показана в SEQ ID NO: 6. В другом варианте реализации изобретения представлено более одного энхансера. Такая комбинация может включать более одной копии любого из описанных в данном документе энхансеров и/или более одного типа энхансеров. В различных вариантах реализации изобретения энхансеры представлены в одной из следующих комбинаций.
Таблица 1. Комбинации энхансеров
В одном варианте реализации изобретения энхансеры комбинируют в следующей последовательности: 5’-EnTTR-ABPS-En34-промотор-3’. В другом варианте реализации изобретения энхансеры комбинируют в следующей последовательности: 5’-промотор-EnTTR-ABPS-En34-3’. В одном варианте реализации изобретения последовательности контроля экспрессии включают enTTR. В другом варианте реализации изобретения последовательности контроля экспрессии включают две копии ABPS и 1 копию enTTR.
В дополнение к этому промотор, экспрессионная кассета и/или вектор могут содержать соответствующие сигналы инициации транскрипции, терминации, энхансерные последовательности и сигналы эффективного процессинга РНК. Такие последовательности включают сигналы сплайсинга и полиаденилирования (полиA); стабилизирующие цитоплазматическую мРНК последовательности; усиливающие эффективность трансляции последовательности (т. е. консенсусная последовательность Козак); усиливающие белковою стабильность последовательности и, при необходимости, последовательности, которые усиливают секрецию кодированного продукта. В одном варианте реализации изобретения включен сигнал полиаденилирования (полиA) для опосредования терминации мРНК-транскриптов hFVIII. Сигнал полиA, используемый в данном документе, представляет собой искусственную полиA, которая представляет собой по размеру около 75 п. н. (PA75), показанную в SEQ ID NO: 10. Примеры других подходящих последовательностей полиA включают в себя среди прочих, например, полиA бычьего гормона роста, SV40, бета-глобина кролика и TK.
В одном варианте реализации изобретения выбирают регуляторные последовательности таким образом, чтобы полный геном вектора rAAV составлял по размеру от около 5 до 5,5 тысяч пар нуклеотидов. В другом варианте реализации изобретения выбирают регуляторные последовательности таким образом, чтобы полный геном вектора rAAV составлял по размеру около 5,1 т. п. н. В другом варианте реализации изобретения выбирают регуляторные последовательности таким образом, чтобы полный геном вектора rAAV составлял по размеру около 5,2 т. п. н. В другом варианте реализации изобретения полный геном вектора rAAV составляет по размеру менее 5 т. п. н.
В одном варианте реализации изобретения геном вектора содержится от нуклеотида 1 до нуклеотида 5110 SEQ ID NO: 13. В одном варианте реализации изобретения геном вектора содержится от нуклеотида 1 до нуклеотида 5194 SEQ ID NO: 14. В одном варианте реализации изобретения геном вектора содержится от нуклеотида 1 до нуклеотида 5138 SEQ ID NO: 15. В другом варианте реализации изобретения геном вектора содержится от нуклеотида 1 до нуклеотида 5222 SEQ ID NO: 16.
5.1.2. Лекарственная форма rAAV.hFVIII
В одном варианте реализации изобретения предложен вектор rAAV.hFVIII в фармацевтической композиции, которая содержит водный носитель, наполнитель, разбавитель или буфер. В одном варианте реализации буфер представляет собой фосфатно-солевой буфер (ФСБ). В конкретном варианте реализации изобретения лекарственная форма rAAV.hFVIII представляет собой суспензию, содержащую эффективное количество вектора rAAV.hFVIII, суспензированного в водном растворе, содержащем 0,001 % плюроник F-68 в TMN200 (200 мM хлорида натрия, 1 мM хлорида магния, 20 мM трис, pH 8,0).Однако известны различные подходящие растворы, включая те, которые содержат один или более из следующих компонентов:забуференный солевой раствор, поверхностно-активное вещество и физиологически совместимую соль или смесь солей, подобранных по ионной силе, эквивалентной раствору от около 100 мМ хлорида натрия (NaCl) до около 250 мМ хлорида натрия, или физиологически совместимую соль, подобранную до эквивалентной ионной концентрации.
Например, предложенная в данном документе суспензия может содержать как NaCl, так и KCl. Значение pH может находиться в диапазоне от 6,5 до 8,5 или от 7 до 8,5, или от 7,5 до 8. Подходящее поверхностно-активное вещество или комбинацию поверхностно-активных веществ можно выбирать из полоксамеров, т. е. неионогенных триблоксополимеров, состоящих из центральной гидрофобной цепи полиоксипропилена (поли(пропиленоксид)), фланкированного двумя гидрофильными цепями полиоксиэтилена (поли(этиленоксид)), СОЛЮТОЛА HS 15 (макрогол-15 гидроксистеарат), ЛАБРАСОЛА (глицерид полиоксикаприловой кислоты), полиокси-10-олеилового эфира, ТВИНА (сложные эфиры жирных кислот и полиоксиэтиленсорбитана), этанола и полиэтиленгликоля. В одном варианте реализации изобретения лекарственная форма содержит полоксамер. Эти сополимеры обычно обозначаются буквой «P» (полоксамер) с последующими тремя цифрами: первые две цифры x 100 дают приблизительную молекулярную массу полиоксипропиленового ядра, а последняя цифра x 10 дает процентное содержание полиоксиэтилена. В одном варианте реализации изобретения выбран полоксамер 188. Поверхностно-активное вещество может присутствовать в количестве от около 0,0005 % до около 0,001 % суспензии. В другом варианте реализации изобретения вектор суспендируется в водном растворе, содержащем 180 мМ хлорид натрия, 10 мМ фосфат натрия, 0,001 % полоксамер 188, pH 7,3.
В одном варианте реализации изобретения лекарственная форма пригодна для применения для субъектов-людей и вводится внутривенно. В одном варианте реализации изобретения лекарственную форму доставляют через периферическую вену посредством болюсной инъекции. В одном варианте реализации изобретения лекарственную форму доставляют через периферическую вену посредством инфузии в течение около 10 минут (±5 минут). В одном варианте реализации изобретения лекарственную форму доставляют через периферическую вену посредством инфузии в течение около 90 минут (±10 минут). В другом варианте реализации изобретения лекарственную форму доставляют через периферическую вену посредством инфузии в течение около 20 минут (±5 минут). В другом варианте реализации изобретения лекарственную форму доставляют через периферическую вену посредством инфузии в течение около 30 минут (±5 минут). В другом варианте реализации изобретения лекарственную форму доставляют через периферическую вену посредством инфузии в течение около 40 минут (±5 минут). В другом варианте реализации изобретения лекарственную форму доставляют через периферическую вену посредством инфузии в течение около 50 минут (±5 минут). В другом варианте реализации изобретения лекарственную форму доставляют через периферическую вену посредством инфузии в течение около 60 минут (±5 минут). В другом варианте реализации изобретения лекарственную форму доставляют через периферическую вену посредством инфузии в течение около 70 минут (±5 минут). В другом варианте реализации изобретения лекарственную форму доставляют через периферическую вену посредством инфузии в течение около 80 минут (±5 минут). Однако данный период времени может, при необходимости или желании, корректироваться. Для введения содержащей AAV композиции, описанной в данном документе, и необязательно для совместного введения других активных лекарственных средств или применения терапий в сочетании с опосредованной AAV доставки hFVIII, описанной в данном документе, может применяться любой подходящий способ или путь. Пути введения включают, например, системный, пероральный, ингаляционный, интраназальный, внутритрахеальный, внутриартериальный, интраокулярный, внутривенный, внутримышечный, подкожный, внутрикожный и другие парентеральные пути введения.
В одном варианте реализации изобретения состав может содержать, например, от около 1,0 x 1011 геномных копий на килограмм массы тела пациента (ГК/кг) до около 1 x 1014 ГК/кг, от около 5 x 1011 геномных копий на килограмм массы тела пациента (ГК/кг) до около 3 x 1013 ГК/кг или от около 1 x 1012 до около 1 x 1014 ГК/кг, при измерении методом окПЦР или цифровой капельной ПЦР (цкПЦР), как описано, например, в M. Lock et al, Hum Gene Ther Methods. 2014 Apr;25(2):115–25. doi: 10.1089/hgtb.2013.131. Epub 2014 Feb 14, которая включена в данный документ путем ссылки. В одном варианте реализации изобретения лекарственная форма rAAV.hFVIII представляет собой суспензию, содержащую по меньшей мере 1x1013 геномных копий (ГК)/мл или больше, при измерении окПКЦ или цифровой капельной ПЦР (цкПЦР), как описано в, например, в M. Lock et al, выше.
С целью обеспечения удаления пустых капсидов из дозы AAV.hFVIII, которую вводили пациентам, пустые капсиды отделяют от векторных частиц во время процесса очистки вектора, используя, например, обсуждаемый в данном документе метод. В одном варианте реализации изобретения векторные частицы, содержащие упакованные геномы, очищают от пустых капсидов с использованием процесса, описанного в международной заявке на патент № PCT/US2016/066013, поданной 9 декабря 2016 г. и заявке на патент США № 62/322,055, поданной 13 апреля 2016 г. и озаглавленной «Scalable Purification Method for AAVrh.10 (масштабируемый метод очистки AAVrh.10)», которые включены в данный документ путем ссылки. Вкратце, описана двухстадийная схема очистки, которая избирательно захватывает и выделяет содержащие геном векторные частицы rAAV из осветленного концентрированного супернатанта клеточной культуры, продуцирующей rAAV. В процессе используется метод аффинного захвата, проводимый при высокой концентрации солей, с последующим методом с использованием анионообменной смолы, проводимым при высоком значении pH, что обеспечивает получение векторных частиц rAAV практически не содержащих промежуточных продуктов rAAV. Для векторов на основе AAVhu.37 могут применяться аналогичные методы очистки. Другие методы очистки описаны, например, в заявках на патент США № 62/266,347, 62/266,357, 62/322,071, 62/266,351, 62/322,083, 62/266,341 и 62/322,098, каждая из которых включена в данный документ путем ссылки.
Поскольку может применяться любой традиционный технологический процесс, то описанный в данном документе процесс (и в международной заявке на патент № PCT/US2016/066013) обеспечивает выходы векторных препаратов, в которых от 50 до 70 % частиц содержат векторный геном, т. е. от 50 до 70 % заполненных частиц. Таким образом примерная доза составляет 1,6x1012 ГК/кг, а общая доза частиц будет составлять от 2,3x1012 до 3x1012 частиц. В другом варианте реализации изобретения предложенная доза оказывается на половину логарифма выше или равна 5x1012 ГК/кг, а общая доза частиц будет составлять от 7,6x1012 до 1,1x1013 частиц. В одном варианте реализации изобретения лекарственная форма характеризуется содержанием концентрата rAAV, имеющего соотношение «пустых» к «полным» равным 1 или меньше, предпочтительно менее 0,75, предпочтительнее 0,5, предпочтительно менее 0,3.
Вкратце, в одном варианте реализации изобретения предложен способ отделения вирусных частиц AAV от капсидных промежуточных продуктов AAV, который включает: проведение для смеси, содержащей рекомбинантные вирусные частицы AAV и капсидные промежуточные продукты AAV, жидкостной хроматографии быстрого разрешения, в ходе которой вирусные частицы AAV и промежуточные продукты AAV связываются с анионообменной смолой, уравновешенной при pH около 10,0, и проведение солевого градиента при контроле элюата на поглощение в ультрафиолете при длине волны от около 260 до около 280, причем полные капсиды AAV собирают из фракции, которая элюируется, когда соотношение A260/A280 достигает точки перегиба.
В одном варианте реализации изобретения данный метод дополнительно включает (a) смешивание суспензии, содержащей рекомбинантные вирусные частицы AAV и капсидные промежуточные продукты AAV, и буфера A, содержащего от 20 мМ до 50 мМ бис-триспропана (BTP) и pH около 10,0; (b) нанесение суспензии из (a) на колонку с сильной анионообменной смолой; (c) промывку нагруженной анионообменной смолы 1 % буфером B, который содержит соль с ионной силой от 10 мМ до 40 мМ NaCl и BTP с pH около 10,0; (d) приложение градиента с возрастающей концентрацией соли к нагруженной и промытой анионообменной смоле, причем солевой градиент эквивалентен от около 10 мМ до около 40 мМ NaCl, и (e) сбор частиц rAAV из элюата, полученного при концентрации солей, эквивалентной по меньшей мере 70 мМ NaCl, при этом частицы rAAV являются по меньшей мере на около 90 % очищенными от промежуточных продуктов AAV. В одном варианте реализации изобретения это значение определяется по числу геномных копий.
В одном варианте реализации изобретения промежуточные продукты элюируют из анионообменной смолы, когда концентрация солей эквивалентна более чем около 50 мМ NaCl. В еще одном варианте реализации изобретения буфер A дополнительно смешан с NaCl до конечной концентрации 1M с целью образования или получения буфера B. В еще одном варианте реализации изобретения солевой градиент имеет ионную силу эквивалентную от 10 мМ до около 190 мМ NaCl. Градиент элюирования может составлять от 1 % буфера B до около 19 % буфера B. Необязательно емкость, содержащая анионообменную смолу представляет собой монолитную колонку, причем буфер A, буфер B и солевой градиент составляют около 60 объемов колонки.
Концентрат или препарат частиц rAAV (с упакованным геномом) является «практически свободным» от пустых капсидов AAV (и других промежуточных продуктов), когда частицы rAAV в концентрате составляют по меньшей мере от около 75 % до около 100 %, по меньшей мере около 80 %, по меньшей мере около 85 %, по меньшей мере около 90 %, по меньшей мере около 95 % или по меньшей мере 99 % rAAV в концентрате, а «пустые капсиды» составляют в концентрате или препарате менее около 1 %, менее около 5 %, менее около 10 %, менее около 15 % rAAV.
В дополнительном варианте реализации изобретения средний выход частиц rAAV составляет по меньшей мере около 70 %. Это значение можно рассчитывать путем определения титра (геномных копий) в данной смеси, нагруженной на колонку, и количества, присутствующего в конечных элюатах. Дополнительно это можно определять на основе к-ПЦР анализа и/или методик ДСН-ПААГ-электрофореза, таких как описанные в данном документе, или таких, которые описаны в данной области.
Например, для расчёта содержания пустых и полных частиц, объемы в полосе VP3 выбранного образца (например, препарата очищенного на градиенте йодиксанола, в котором число ГК = числу частиц) отмечали на графике по сравнению с внесенным количеством ГК-частиц. Полученное линейное уравнение (y = mx+c) используют для расчета количества частиц в объемах полос пиков исследуемого препарата. Затем количество частиц (част.) во внесенных 20 мкл умножают на 50 для получения количества частиц (част.) /мл. Значение частиц/мл, деленное на ГК/мл, дает соотношение частиц к геномным копиям (част./ГК). Вычисление част./мл–ГК/мл дает величину пустых част./мл. Значение пустых част./мл, деленное на част./мл и x 100, дает процентное содержание пустых частиц.
В целом, методы для оценки количества пустых капсидов и векторных частиц AAV с упакованными геномами, известны в данной области.См., например, Grimm et al., Gene Therapy (1999) 6:1322-1330; Sommer et al., Molec. Ther. (2003) 7:122–128. Для испытания на денатурированные капсиды, методы включают проведение для обработанного концентрата AAV электрофореза на ДСН-полиакриламидном геле, состоящем из любого геля, способного разделять три капсидных белка, например гель с градиентом, содержащим в буфере 3–8 % трис-ацетата, затем обработка геля до разделения исследуемого материала и блоттинг геля на нейлоновые или нитроцеллюлозные мембраны, предпочтительно нейлоновые.Затем антитела против капсидов AAV применяют как первичные антитела, которые связываются с денатурированными капсидными белками, предпочтительно моноклональное антитело против капсида AAV, наиболее предпочтительно моноклональное антитело B1 против AAV-2 (Wobus et al., J. Virol. (2000) 74:9281–9293). Затем используют вторичное антитело, которое связывается с первичным антителом и содержит средство для определения связывания с первичным антителом, предпочтительнее антитело против IgG, содержащее ковалентно связанную с ним молекулу детекции, наиболее предпочтительно антитело овцы против мышиного IgG, ковалентно связанное с пероксидазой хрена. Метод для выявления связывания используется для полуколичественного определения связывания между первичными и вторичными антителами, предпочтительно метод выявления способен определять эмиссию радиоактивных изотопов, электромагнитное излучение или колориметрические изменения, наиболее предпочтительно — это набор для выявления хемилюминесценции. Например, можно брать образцы из фракций колонки для ДСН-ПААГ-электрофореза и нагревать в буфере для внесения, содержащем восстанавливающий агент (например, ДТТ) и разделять капсидные белки на готовых градиентных полиакриламидных гелях (например, Novex).Можно выполнять окрашивание серебром с использованием набора SilverXpress (Invitrogen, штат Калифорния) в соответствии с инструкциями производителя. В одном варианте реализации изобретения концентрацию векторных геномов (вг) в фракциях колонки можно измерять количественным ПЦР в реальном времени (К-ПЦР).Образцы разбавляют и обрабатывают ДНКазой I (или другой подходящей нуклеазой) для удаления экзогенной ДНК.После инактивации нуклеазы, образцы дополнительно разбавляют и амплифицируют с использованием праймеров и флуорогенного зонда TaqMan™, специфического к последовательности ДНК между праймерами. Количество циклов, требуемое для достижения определенного уровня флуоресценции (пороговый цикл, Ct), измеряется для каждого образца на системе выявления последовательностей Prism 7700, Applied Biosystems.Для получения стандартной кривой в реакции К-ПЦР применяют плазмидную ДНК, содержащую последовательности, идентичные к последовательностям, содержащимся в векторе AAV.Значения порогового цикла (Ct), полученные из образцов, используют для определения титра векторного генома путем нормализации его со значением Ct стандартной кривой для плазмиды.Можно также использовать анализы по конечной точке, основанные на цифровой ПЦР.
В одном аспекте в данном документе предложен оптимизированный метод к-ПЦР, в котором используется широкий спектр сериновых протеаз, например протеиназа K (такая как коммерчески доступная у Qiagen). Более конкретно, анализ титра генома оптимизированной кПЦР аналогичен стандартному анализу, за исключением того, что после расщепления ДНКазой I образцы разбавляют буфером для протеиназы K и обрабатывают протеиназой K, с последующей ее инактивацией нагреванием. Подходящие образцы разбавляют буфером для протеиназы K в количестве равном объему образца. Буфер для протеиназы K можно концентрировать в 2 раза или выше. Как правило, обработка протеиназой K составляет около 0,2 мг/мл, но может варьировать от 0,1 мг/мл до около 1 мг/мл. Этап обработки обычно проводят при температуре около 55 °C в течение около 15 минут, но могут выполнять при более низкой температуре (например, от около 37 °C до около 50 °C) в течении более длительного периода времени (например, от около 20 минут до около 30 минут) или при более высокой температуре (например, до около 60 °C) в течение более короткого периода времени (например, от около 5 до 10 минут). Аналогичным образом инактивацию нагреванием обычно проводят при температуре около 95 °C в течение около 15 минут, но могут выполнять при более низкой температуре (например, от около 70 до около 90 °C) в течении продолжительного периода времени (например, от около 20 минут до около 30 минут). Затем образцы разбавляют (например, 1000-кратно) и подвергают анализу TaqMan, как описано в стандартном анализе.
Дополнительно или альтернативно можно использовать капельный цифровой ПЦР (кцПЦР). Например, были описаны методы для определения титров одноцепочечного и самокомплементарного генома вектора AAV с помощью кцПЦР. См., например, M. Lock et al, Hu Gene Therapy Methods, Hum Gene Ther Methods. 2014 Apr;25(2):115–25. doi: 10.1089/hgtb.2013.131. Epub 2014 Feb 14.
5.1.3 Промышленное получение
Вектор rAAV.hFVIII можно промышленно получать как показано на технологической схеме, показанной на ФИГ. 13. Вкратце, клетки (например, клетки HEK 293) размножают в подходящей системе для культивирования и трансфицируют для получения векторов. Затем собирают вектор rAAV.hFVIII, концентрируют и очищают для получения субстанции вектора, которую потом разливают и готовят готовый препарат в финишном технологическом процессе.
Способы промышленного получения векторов для генной терапии, описанные в данном документе, включают способы, хорошо известные в данной области, такие как получение плазмидной ДНК, используемой для продукции векторов для генной терапии, получения и очистки этих векторов. В некоторых вариантах реализации изобретения вектор для генной терапии представляет собой вектор AAV, а полученные плазмиды представляют собой цис-плазмиду AAV, кодирующую геном AAV и интересующий ген, транс-плазмиду AAV, содержащую гены rep и cap AAV, и аденовирусную хелперную плазмиду. Процесс получения вектора может включать такие стадии способа, как размножение клеточной культуры, пересев клеток, высевание клеток, трансфекция клеток плазмидной ДНК, замена среды после трансфекции на бессывороточную среду и сбор содержащих вектор клеток и культуральной среды. Собранные содержащие вектор клетки и культуральную среду называют в данном документе неочищенным урожаем клеток.
После этого неочищенный урожай клеток подвергают таким стадиям способа, как концентрирование урожая векторов, диафильтрация урожая векторов, микрофлюидизация урожая векторов, расщепление нуклеазами урожая векторов, фильтрация микрофлюидизированного промежуточного продукта, очистка хроматографией, очистка ультрацентрифугированием, замена буфера фильтрацией с тангенциальным потоком и приготовление лекарственной формы и фильтрация для получения субстанции вектора.
В одном варианте реализации изобретения продуцирующая плазмида, является такой как показана в SEQ ID NO: 13. В одном варианте реализации изобретения продуцирующая плазмида, является такой как показана в SEQ ID NO: 14. В одном варианте реализации изобретения продуцирующая плазмида, является такой как показана в SEQ ID NO: 15. В другом варианте реализации изобретения продуцирующая плазмида является такой, как показана в SEQ ID NO: 16.
В конкретном варианте реализации изобретения способы, используемые для промышленного получения векторов для генной терапии описаны в разделе 8, ниже.
5.2 Популяция пациентов
По нескольким причинам выбранной исследуемой популяцией являются пациенты с тяжелой или умеренной гемофилией A (HemA). Пациенты с тяжелой гемофилией A определены как имеющие менее 1 % от нормальной активности фактора VIII (FVIII), таким образом, требующие частых инфузий FVIII для контроля их предрасположенности к кровотечениям. Это представляет значительное затруднение в отношении ведения нормального образа жизни и, в дополнение к этому, уровни FVIII в крови проходят через хорошо известные пики и имеют снижающийся профиль, что неоптимально. Тот факт, что уровни FVIII в крови у тяжелых пациентов оказываются меньше 1 %, дает возможность легко измерять даже от низкого до умеренного увеличения уровней FVIII в крови после введения rAAV.hVIII. Недавние клинические исследования подтвердили действенность этого подхода. Пациенты с умеренной HemA определены, как имеющие уровни от 1 % вплоть до 5 % FVIII в крови.
Пациенты, которые являются кандидатами для проведения лечения предпочтительно представляют собой взрослых мужчин возрастом ≥18 с диагностированной умеренной/тяжелой или тяжелой гемофилией A. В одном варианте реализации изобретения пациент имеет базовую активность FVIII ≤2 % от нормальной или задокументированной истории активности FVIII ≤2 %. В некоторых вариантах реализации изобретения можно лечить пациента возрастом <18 лет. Кандидаты для лечения включают субъектов, у которых было по меньшей мере 3 эпизода с кровотечением за год, которые требуют лечения FVIII по необходимости. Другие кандидаты для лечения, включают субъектов, которых лечат FVIII в профилактическом режиме. Другие критерии демонстрируют, что для субъекта подходит лечение, включающее по меньшей мере 100 дней истории лечения FVIII; в истории болезни отсутствует образование ингибиторов (нейтрализирующих антител) к экзогенному FVIII; отсутствуют известные аллергические реакции к экзогенному FVIII или любому компоненту векторной композиции rAAV.FVIII.
Перед лечением пациента с гемофилией A необходимо оценивать на наличие NAb к серотипу AAV, используемому для доставки гена hFVIII, (например, AAVhu.37 или AAVrh.10). Такие NAb могут влиять на эффективность трансдукции и снижать терапевтическую эффективность. Пациенты с гемофилией A, которые имеют базовый титр NAb ≤ 1:5 являются хорошими кандидатами для лечения с помощью протокола генной терапии rAAV.hFVIII.
Субъектам могут позволять продолжать их стандартные виды лечения (-ий) (например, терапия рекомбинантным FVIII) перед и во время лечения генной терапией по усмотрению лечащего врача. В альтернативном варианте врач может предпочесть прекратить стандартные виды лечения перед проведением лечения генной терапией и, необязательно, возобновить стандартные виды лечения в качестве котерапии после проведения генной терапии.
Желательные конечные критерии оценки схемы генной терапии определяются по увеличению активности FVIII до 3 % от нормы от базового уровня в течение до 52 недель после проведения лечения генной терапией. В одном варианте реализации изобретения у пациентов достигаются уровни циркулирующего FVIII (например, 5 % или выше) после лечения с помощью rAAV.hFVIII, самостоятельно и/или в комбинации с использованием дополнительных видов лечения. В другом варианте реализации изобретения у пациентов достигаются уровни циркулирующего FVIII 10 %, 15 %, 20 % или выше после лечения с помощью rAAV.hFVIII самостоятельно и/или в комбинации с использованием дополнительных видов лечения. В другом варианте реализации изобретения у пациентов достигаются уровни циркулирующего FVIII 25 %, 30 %, 35 %, 40 %, 45 %, 50 %, 55 %, 60 %, 65 %, 70 %, 75 %, 80 %, 85 %, 95 %, 95 % или выше после лечения с помощью rAAV.hFVIII самостоятельно и/или в комбинации с использованием дополнительных видов лечения.
Несмотря на это, пациенты имеющие одну или более из следующих особенностей могут исключаться из лечения по усмотрению их лечащего врача:
1. В истории болезни серьезное заболевание печени (т. е. портальная гипертензия).
2. Серьезное воспаление или цирроз печени.
3. Подтверждение инфекции с активным вирусом гепатита B (ВГB) или вирусом гепатита C (ВГC).
4. В истории болезни инфекция вирусом иммунодефицита человека (ВИЧ) И любой анализ из следующего: число клеток CD4+ <350 клеток/мм3, изменение режима антиретровирусной терапии в пределах 6 месяцев до дня 0 или вирусная нагрузка в плазме >200 копий/мл в 2 отдельных случаях при измерении ПЦР.
5. Титр нейтрализирующих антител против AAVhu.37 (или против AAVrh10, соответственно) >1:5 или ≥1:10.
6. Участие (текущее или предыдущее) в другом исследовании генной терапии.
7. Участие в исследовании другого исследуемого лекарственного средства в течение 3 месяцев до скрининга.
В других вариантах реализации изобретения лечащий врач может определить, что наличие одной или более из этих физических особенностей (история болезни), не должно исключать лечение, предложенное в данном документе.
5.3. Дозирование и путь введения
В одном варианте реализации изобретения вектор rAAV.hFVIII доставляется в виде однократной дозы для одного пациента. В другом варианте реализации изобретения вектор rAAV.hFVIII доставляется в виде множества доз для одного пациента. В дополнительном варианте реализации изобретения вектор rAAV.hFVIII доставляется в виде двух доз для одного пациента. В одном варианте реализации изобретения субъекту доставляется минимальная эффективная доза (МЭД) (определяемая доклиническим исследованием, описанным в данном документе в примерах). Как используется в данном документе, МЭД относится к дозе rAAV.hFVIII, требуемой для достижения 5 % от нормальной активности фактора VIII.
Как принято, титр векторов определяют на основе содержания ДНК векторного препарата. В одном варианте реализации изобретения для определения содержания ДНК препаратов векторов rAAV.hFVIII используют количественную ПЦР или оптимизированную ПЦР, как описано в примерах. В одном варианте реализации изобретения для определения содержания ДНК препаратов векторов rAAV.hFVIII используют цифровую капельную ПЦР, как описано в примерах. В одном варианте реализации изобретения дозировка составляет от около 1x1011 геномных копий (ГК)/кг массы тела до около 1x1013 ГК/кг, включая конечные критерии оценки. В одном варианте реализации изобретения дозировка составляет 5x1011 ГК/кг. В другом варианте реализации изобретения дозировка составляет 5x1012 ГК/кг. В конкретных вариантах реализации изобретения дозировка rAAV.hFVIII, введенного пациенту составляет по меньшей мере 5 x 1011 ГК/кг, 1 x 1012 ГК/кг, 1,5 x 1012 ГК/кг, 2,0 x 1012 ГК/кг, 2,5 x 1012 ГК/кг, 3,0 x 1012 ГК/кг, 3,5 x 1012 ГК/кг, 4,0 x 1012 ГК/кг, 4,5 x 1012 ГК/кг, 5,0 x 1012 ГК/кг, 5,5 x 1012 ГК/кг, 6,0 x 1012 ГК/кг, 6,5 x 1012 ГК/кг, 7,0 x 1012 ГК/кг или 7,5 x 1012 ГК/кг. Композиции с дефектными по репликации вирусами также можно готовить в виде дозированных единиц, содержащих количество дефектного по репликации вируса, которое находится в диапазоне от около 1,0 x 109 ГК до около 1,0 x 1015 ГК. Как используется в данном документе, термин «дозировка» может относится к общей дозировке, доставляемой субъекту в ходе лечения, или количеству, доставляемому в ходе однократного (или многократных) введения (-ий).
В другом варианте реализации изобретения композицию повторно вводят в более поздний срок. Необязательно допускается более одного повторного введения. Такое повторное введение может происходить с тем же типом вектора или с другим вирусным вектором, как описано в данном документе. В одном варианте реализации изобретения вектор повторно вводят через около 6 месяцев после первого введения. В другом варианте реализации изобретения вектор повторно вводят через около 1 год после первого введения. В другом варианте реализации изобретения вектор повторно вводят через около 2 года после первого введения. В другом варианте реализации изобретения вектор повторно вводят через около 3 года после первого введения. В другом варианте реализации изобретения вектор повторно вводят через около 4 года после первого введения. В другом варианте реализации изобретения вектор повторно вводят через около 5 лет после первого введения. В другом варианте реализации изобретения вектор повторно вводят через около 6 лет после первого введения. В другом варианте реализации изобретения вектор повторно вводят через около 7 лет после первого введения. В другом варианте реализации изобретения вектор повторно вводят через около 8 лет после первого введения. В другом варианте реализации изобретения вектор повторно вводят через около 9 лет после первого введения. В другом варианте реализации изобретения вектор повторно вводят через около 10 лет после первого введения.
В одном варианте реализации изобретения дозировка достаточна для увеличения уровней фактора VIII у пациента до 1 % от нормального. В одном варианте реализации изобретения дозировка достаточна для увеличения уровней фактора VIII у пациента до 2% от нормального. В одном варианте реализации изобретения дозировка достаточна для увеличения уровней фактора VIII у пациента до 3% от нормального. В другом варианте реализации изобретения дозировка достаточна для увеличения уровней фактора VIII у пациента до 4 % от нормального. В другом варианте реализации изобретения дозировка достаточна для увеличения уровней фактора VIII у пациента до 5% от нормального. В другом варианте реализации изобретения дозировка достаточна для увеличения уровней фактора VIII у пациента до 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95 % или больше от нормального.
В некоторых вариантах реализации изобретения rAAV.hFVIII вводят в комбинации с одним или более видами терапии для лечения гемофилии A, например с введением рекомбинантного FVIII.
5.4. Измерение клинических критериев
Определение эффективности лечения можно измерять экспрессией и активностью трансгена, что определяется по уровням фактора VIII в плазме и активности фактора VIII. Дополнительную оценку эффективности можно определять клинической оценкой потребности в заместительной терапии фактора VIII и частотой эпизодов спонтанных кровотечений. Такие оценивания можно проводить дважды в неделю в течение 4 недель после введения продукта, еженедельно с недели 6 до недели 12, ежемесячно в течение оставшегося периода времени в первый год и с интервалами по 6 месяцев в течение общего периода времени в 5 лет.
Безопасность вектора для генной терапии можно оценивать по количеству нежелательных явлений, отмеченных при медицинском осмотре изменений и/или параметров, определяемых в клинической лаборатории, оцениваемых в множество моментов времени вплоть до около 52 недели после введения вектора. Хотя физиологическое действие можно наблюдать раньше, например, через около одну неделю, в одном варианте реализации изобретения постоянные устоявшиеся уровни экспрессии достигаются до около 12 недель. Последующие оценивания можно проводить дважды в неделю в течение 4 недель после введения продукта, еженедельно с недели 6 до недели 12, ежемесячно в течение оставшегося периода времени в первый год и с интервалами по 6 месяцев в течение общего периода времени в 5 лет. Такие оценивания включают:
a. Медицинский осмотр
b. Электрокардиограмму (ЭКГ)
c. Биохимические анализы: Уровни электролитов в сыворотке, АМК, креатинина, кальция, фосфата, общего белка, альбумина, ЛДГ, КФК, АСТ, АЛТ, щелочной фосфатазы, билирубина
d. Гематологический анализ: ОАК и лейкограмма, профиль коагуляции
e. Общий анализ мочи
f. Иммунологическую оценку:
g. Серологическая реакция на капсид hu.37 (или капсид rh.10, соответственно) и на фактор VIII
h. Реакция T-клеток на капсид hu.37 (или капсид rh.10, соответственно) и антигены фактора VIII
i. Оценка векторной ДНК; измерения кПЦР в плазме, моче и слюне.
Увеличение hFVIII, достигаемое вместе с введением rAAV.hFVIII, можно оценивать в виде определенного процентного изменения уровня hFVIII в течение около 12 недель или в другие требуемые моменты времени, по сравнению с уровнями hFVIII у пациента, не болеющего гемофилией A, т. е. так называемыми нормальными уровнями hFVIII равными около 100 %. В другом варианте реализации изобретения изменение сравнивают с базовыми уровнями hFVIII у пациента. В одном варианте реализации изобретения желаемая эффективность заключается в увеличении у пациента уровней фактора VIII до 3 % от нормального. В одном варианте реализации изобретения желаемая эффективность заключается в увеличении у пациента уровней фактора VIII до 4% от нормального. В одном варианте реализации изобретения желаемая эффективность заключается в увеличении у пациента уровней фактора VIII до 5% от нормального. В одном варианте реализации изобретения желаемая эффективность заключается в увеличении у пациента уровней фактора VIII до 6% от нормального. В одном варианте реализации изобретения желаемая эффективность заключается в увеличении у пациента уровней фактора VIII до 7% от нормального. В одном варианте реализации изобретения желаемая эффективность заключается в увеличении у пациента уровней фактора VIII до 8% от нормального. В одном варианте реализации изобретения желаемая эффективность заключается в увеличении у пациента уровней фактора VIII до 9% от нормального. В другом варианте реализации изобретения дозировка достаточна для увеличения уровней фактора VIII у пациента до 10% от нормального. В другом варианте реализации изобретения дозировка достаточна для увеличения уровней фактора VIII у пациента до 15% от нормального. В другом варианте реализации изобретения дозировка достаточна для увеличения уровней фактора VIII у пациента до 20 % или больше от нормального. В одном варианте реализации изобретения проводили исследования на панелях коагуляции как часть стандартного испытания по установлению активности FVIII.
Как используется в данном документе, вектор rAAV.hFVIII «функционально замещает» или «функционально дополняет» организм пациентов с недостаточностью по FVIII активным FVIII, когда в организме пациентов экспрессируется достаточный уровень FVIII для достижения по меньшей мере одного из этих клинических конечных критериев. Функциональное замещение могут обеспечивать уровни экспрессии hFVIII, которые достигаются всего лишь от около 1 % до менее 100 % от нормальных уровней клинических конечных критериев нормального типа у пациента без гемофилии.
В одном варианте реализации изобретения экспрессию можно наблюдать как ранний ответ через от около 8 часов до около 24 часов после введения дозы. Один или более из желаемых клинических эффектов, описанных выше, могут наблюдаться в течение от нескольких дней до нескольких недель после введения дозы.
Долговременную (до 260 недель) безопасность и эффективность можно оценивать после введения rAAV.hFVIII.
В одном аспекте предложена схема доставки генного продукта hFVIII в организм пациента-человека. Данная схема включает (a) доставку вектора rAAV.hFVIII, содержащего экспрессионную кассету, как описано в данном документе, и (b) доставку второго вектора rAAV.hFVIII, содержащего экспрессионную кассету, как описано в данном документе, причем первый рекомбинантный вектор AAV или второй рекомбинантный вектор AAV содержит капсид AAV3B. Последовательность AAV3B показана в SEQ ID NO: 20, имеющая № регистрации AAB95452.1. В одном варианте реализации изобретения один из первого или второго вектора AAV содержит капсид rh.10. В одном варианте реализации изобретения один из первого или второго вектора AAV содержит капсид AAVhu.37. Такие схемы описаны в международной заявке на патент № PCT/US16/42472, которая включена в данный документ путем ссылки.
Вирусные векторы, описанные в данном документе, можно использовать для получения лекарственного средства для доставки hFVIII нуждающемуся в этом субъекту (например, пациенту-человеку), как поставляющие функциональный hFVIII субъекту и/или для лечения заболевания гемофилией A.
В другом аспекте для лечения гемофилии A предложен вектор rAAV.hFIII, описанный в данном документе. В одном варианте реализации изобретения для лечения гемофилии A предложено введение множества доз. В другом аспекте предложен вектор rAAV.hFIII, описанный в данном документе, для производства лекарственного средства для лечения гемофилии A.
В одном варианте реализации изобретения обеспечивается второе введение вектора rAAV.hFVIII. В одном варианте реализации изобретения вектор rAAV.hFVIII для второго введения имеет тот же капсид AAV, который дается с первой дозировкой. В одном варианте реализации изобретения вектор rAAV.hFVIII для второго введения имеет капсид AAVrh.10. В другом варианте реализации изобретения вектор rAAV.hFVIII для второго введения имеет отличающийся от вектора первой дозы капсид AAV. В одном варианте реализации изобретения вектор rAAV.hFVIII для второго введения обладает тропизмом в отношении печени. В одном варианте реализации изобретения вектор rAAV.hFVIII для второго введения имеет капсид AAV3B.
В дополнительном аспекте в изобретении обеспечивается нацеливание на гепатоциты пациента.
В одном аспекте доставка первого rAAV и второго rAAV временно разделяется периодом в по меньшей мере один месяц, по меньшей мере около трех месяцев или от около 1 года до около 10 лет.
Следующие примеры предназначены для иллюстрации и не предназначены для ограничения настоящего изобретения.
ПРИМЕРЫ
6. Пример 1. Доклиническое исследование
6.1 Вектор hFVIII
В отличие от человеческого фактора FIX (hFIX) кДНК hFVIII намного больше и необходимо вносить изменения для подгонки этого трансгена к стандартному геному AAV. Так как трансген hFVIII с удаленным B-доменом (BDD) и с исключением других необходимых элементов транскрипции содержит 1457 аминокислот, то вектор AAV все еще имеет ограниченную емкость упаковки. Следовательно, были предприняты шаги для снижения размера других элементов, включая элементы контроля экспрессии трансгена.
Для ограничения экспрессии hFVIII в печени при сохранении размера элементов насколько возможно меньшими с целью получения 42 комбинаций энхансеров/промоторов было укорочено и скомбинировано несколько сильных специфических для печени промоторов с сочетаниями до трех специфических для печени энхансерных последовательностей. После введения векторов AAV на мышах FVIII KO оценивали активность и иммуногенность hFVIII данного трансгена.
6.1.1 Продукция векторов AAV для доклинического исследования
Получали 42 плазмиды, содержащие одну из комбинаций энхансера/промотора. Получали 14 энхансерных комбинаций, используя три энхансерные последовательности; En34 (коровый энхансер размером 34 п. н. аполипопротеина человека из контрольной области печени), ABPS (укороченная до 42 п. н. версия дистального энхансера размером 100 п. н. предшественника α1-микроглобулина/букинина [ABP]) и EnTTR (энхансерная последовательность размером 100 п. н. из транстиретина). Ряд энхансерных комбинаций ограничен из-за общего размера ITR-ITR и комбинации энхансеров в следующей последовательности: 5’-EnTTR-ABPS-En34-промотор-3’. Таблица 1. Каждую из 14 энхансерных комбинаций вставляли выше по ходу транскрипции от одного из трех промоторов: TBG-S1 (P1, укороченная версия специфического для печени промотора тироксин-связывающего глобулина или TBG), A1AT (P2, модифицированный промотор SERINA1 [α1-антитрипсин]) и TTR (P3, промотор транстиретина). Полученные конструкции были сконструированы для экспрессии кодон-оптимизированной версии человеческого белка-фактора FVIII, у которой удален B-домен (BDD) и замещен коротким линкером из 14 аминокислот hFVIIIco-SQ (SEQ ID NO: 2).
Все векторы AAV получали, как описано в Gao G, Lu Y, Calcedo R, et al. Biology of AAV serotype vectors in liver-directed gene transfer to nonhuman primates. Mol Ther. 2006;13(1):77-87, которая включена в данный документ путем ссылки. Вкратце, экспрессирующие hFVIII плазмиды из одной из 42 комбинаций энхансеров/промоторов упаковывали с вирусным капсидом AAVrh10. Экспрессирующие hFVIII плазмиды из E06.TTR также упаковывали в вирусные капсиды AAV8, AAV9, AAVhu37 и AAVrh64R1.
6.1.2 Анализ векторов методом ДСН-ПААГ-электрофореза
Серии векторов AAVrh10 с комбинациями энхансеров/промоторов, использованные в исследовании, подвергали оценке на чистоту методом электрофореза в полиакриламидном геле с додецилсульфатом натрия (ДСН-ПААГ), как описано в Lock M, Alvira M, Vandenberghe LH, et al. Rapid, simple, and versatile manufacturing of recombinant adeno-associated viral vectors at scale. Hum Gene Ther. 2010;21(10):1259–1271, которая включена в данный документ путем ссылки. Вкратце, на ДСН-ПААГ наносили денатурированные и восстановленные векторные образцы, содержащие 5x109 ГК. Белки окрашивали красным красителем SYPRO (Invitrogen, г. Карслбад, штат Калифорния, США) с последующей фиксацией, визуализацией, а затем количественным определением с использованием системы для анализа изображений Syngene и программного обеспечения GeneTool (Syngene, г. Фредерик, штат Мэриленд, США). Рассчитывали процент чистоты капсида (белки (VP1, VP2 и VP3, определяемые к общему белку). Процентное значение чистоты 42 векторов находилось в диапазоне от 29 % (AAVrh10.E12.P3) до 100 %, со средним значением чистоты равным 90 % (данные не показаны).
6.1.3 Мыши
Размножающиеся пары мышей FVIII KO получали у The Jackson Laboratory (г. Бар Харбор, штат Мэн, США) и колонию поддерживали в Лабораториях исследования трансляции университета штата Пенсильвания, размещая в специальных условиях без проникновения патогенов). Все процедуры и протоколы обхождения с животными утверждены Институциональным комитетом по содержанию и использованию животных (IACUC) университета штата Пенсильвания. Самцам FVIII KO в возрасте 6–12 недель в/в в хвостовую вену инъецировали 1010 ГК вектора на мышь. Разбавляли вектор в фосфатно-солевом растворе (ФСБ) и инъецировали 100 мкл этого разведения вектора. Каждые две недели собирали плазму в пробирки для сбора с цитратом натрия путем отборов крови из ретроорбитального синуса.
6.1.4 Определение активности hFVIII
В плазме измеряли активность hFVIII с помощью набора COATEST SP4 в соответствии с протоколом производителя (DiaPharma, штат Огайо, США). На неделю 2 после инъекции у мышей обнаруживали широкий диапазон активности hFVIII 0,12–2,12 ИЕ/мл (ФИГ. 5).
Среди прочих пять конструкций демонстрировали значительное увеличение уровней активности: E03.TTR, E05.A1AT, E05.TTR, E06.TTR и E12.A1AT (ФИГ. 5A). Вариация уровней активности hFVIII, отмеченная на неделе 2, возникла перед образованием антител к трансгену (ФИГ. 5B). Следовательно существовали зависимые от конструкции значительные отличия в уровнях активности.
6.1.5 Выявление IgG против hFVIII в плазме мышей
Антитела-IgG против hFVIII в плазме мышей измеряли методом твердофазного ИФА, причем все реактивы были получены, если не указано иное, у Sigma-Aldrich (г. Сент-Луис, штат Миссури, США). Планшеты для твердофазного ИФА покрывали 1 мкг/мл BDD-hFVIII-SQ (Xyntha, Wyeth Pharmaceuticals Inc., г. Даллас, штат Техас, США) в 0,1 M карбонатном буфере (pH 9,6) и инкубировали в течение ночи при 4 °C. Промывали лунки пять раз с помощью 0,05 % Твина 20 в ФСБ и блокировали их с помощью 5 % обезжиренного молока (Bio Rad, г. Геркулес, штат Калифорния, США) в ФСБ в течение одного часа при комнатной температуре. После удаления блокирующего буфера образцы плазмы, разбавленные в 5 % обезжиренном молоке, добавляли в планшеты и инкубировали в течение одного часа при комнатной температуре. В качестве контроля использовали образцы плазмы, полученные от наивных мышей. Затем планшеты промывали пять раз и добавляли конъюгированный с HRP IgG против антител мыши при разведении 1:1000 в обезжиренном молоке. После инкубации при комнатной температуре в течение 90 минут планшеты промывали восемь раз и для выявления добавляли 3,3',5,5'-тетраметилбензидин (ТМБ). После выдержки в течение 5 минут при комнатной температуре останавливали реакцию с использованием 2N серной кислоты, и планшеты считывали при 450 нм с использованием устройства для считывания планшетов BioTek µQuant (г. Уинуски, штат Вермонт, США).
У мышей FVIII KO отмечено образование антител к hFVIII на неделю 4 после введения вектора, а на неделю 8 у мышей в большинстве из 42 групп векторов обнаружили выявляемые уровни IgG против hFVIII с соответствующим снижением уровней активности hFVIII (ФИГ. 6). Временная динамика активности hFVIII и образования антител, количественно определенная по титру, представлена на ФИГ. 7 и 8, соответственно. Более 50 % мышей, которым инъецировали конструкции E05.A1AT, E10.A1AT и все промоторы с комбинацией энхансеров E06, синтезировали антитела к трансгену на неделю 8. Однако антитела к трансгену были обнаружены не во всех группах. Для 6 из 42 векторных групп (E11.TTR, E13.TBG-S1, E13.A1AT и все конструкции с использованием E01) на неделе 8 не было выявлено антител к hFVIII (ФИГ. 6), а у двух групп не было обнаружено выявляемых антител на протяжении 12 недель исследования (E01.A1AT и E11.TTR). Выполняли также анализ времени до наступления события для образования антител к hFVIII. У мышей, которым инъецировали вектор с использованием для экспрессии E01.A1AT, E11.TTR, E01.TBG-S1, E11.A1AT и E13.TBG-S1, оказалось самое длительное время до экспрессии антител, тогда как конструкции, содержащие E06.TTR, E06.A1AT, E05.A1AT, E09.TBG-S1 и E14.TTR показали самое короткое время до экспрессии антител.
6.1.6 Статистический анализ
Для выявления сходных групп лечения, в соответствии с активностью hFVIII в плазме на неделю 2, анализировали данные с использованием модели дисперсионного анализа (ANOVA) с одним фиксированным критерием с ретроспективными анализами с критерием Тьюки для идентификации средних уровней активности, которые отличаются друг от друга. Выполняли анализ времени до наступления события — образования антител против hFVIII.
6.1.9 Сравнение активности и иммуногенности для различных капсидов AAV
Далее определяли разницу в уровнях активности и устанавливали потенциальное распределение по иммуногенности использованных капсидов AAV. Для этого исследования был отобран наиболее иммуногенный геном из предыдущих исследований — E06.TTR. Интересно, что эта конструкция продуцировала значительно более высокую экспрессию, чем большинство других конструкций на неделю 2, но в последующие недели образовывались антитела против трансгена hFVIII у 80 % мышей с инъекциями конструкций.
Тот же самый векторный геном с использованием энхансерной комбинации E06 со специфическим для печени промотором TTR упаковали с одним из пяти капсидов AAV; AAVrh10, AAV8, AAV9, AAVhu37 и AAVrh64R1. Снова мышам FVIII KO в/в инъецировали дозу 1010 ГК на мышь и в течение всего 12-недельного исследования отслеживали уровни активности hFVIII в плазме и титры IgG против hFVIII. Четкие различия в экспрессии и иммуногенности трансгена видны на основе вектора AAV, используемого для переноса гена (ФИГ. 9). На неделю 2 после введения вектора активность hFVIII в плазме варьировала от 0,51 ИЕ/мл после введения AAVrh64R1 до 1,26 ИЕ/мл после AAVrh10 (ФИГ. 9A). В течение проведения исследования у нескольких мышей синтезировались антитела против hFVIII в диапазоне от 20 % мышей, которым ввели векторы AAV8 или AAV9, до 63 % мышей, которые получили AAVrh10 (ФИГ. 9B). Следовательно даже с высокоиммуногенной комбинацией энхансера и промотора иммунный ответ к трансгену может варьировать в зависимости от капсида AAV, использованного для переноса гена.
6.1.10 Обсуждение
Предыдущие исследования мышей FVIII KO, у которых для экспрессии использовался промотор HLP, не выявили антитела к трансгену на протяжении всего исследования. Данная промоторная последовательность HLP сходна с такой у комбинации энхансера/промотора E01.A1AT, причем у мышей, которым вводили этот вектор AAVrh10 не обнаружили выявляемых антител на протяжении 12 недель исследования. К сожалению, активность этого вектора у мышей FVIII KO была относительно низкой только с выявляемыми 0,189 ИЕ/мл в плазме на неделе 2 после введения вектора и пиковым уровнем 0,303 ИЕ/мл на неделе 6.
Интересно, что капсид AAV, используемый для переноса гена из той же трансгенной кассеты значимо влиял как на иммуногенность к hFVIIII, так и на пиковую активность hFVIII. Для того чтобы исследовать влияние капсида на образование антител против hFVIII была использована наиболее иммуногенная трансгенная кассета (E06.TTR). Пять исследованных векторных капсидов можно разделить на две группы: в одной у ≤ 20 % мышей синтезировались антитела против hFVIII (AAV8 и AAV9), а в другой у > 20 % мышей вырабатывалась гуморальная иммунная реакция (AAVrh10, AAVhu37 и AAVrh64R1). Переносимость по отношению к трансгену hFVIII, связанная с капсидом AAV8, вероятно оказывается ожидаемой благодаря предыдущим исследованиям, которые демонстрируют, что в/в доставка этого капсида может активировать специфические к трансгену регуляторные T-клетки в контексте толерогенной природы печени. В дополнение к этому авторы раннее показали, что не существует выявляемых ингибиторов FIX после в/м инъекции AAV8 у мышей с гемофилией B. Следовательно даже с высокоиммуногенной комбинацией энхансера и промотора иммунный ответ к трансгену может варьировать в зависимости от капсида AAV, использованного для переноса гена.
Уровни активности hFVIII в плазме после введения пяти различных капсидов AAV также значительно варьировали от 0,51 ИЕ/мл с AAVrh64R1 до 1,26 ИЕ/мл с AAVrh10 на неделю 2 после введения векторов. Экспрессия вектора AAVrh10 была значительно увеличенной по сравнению с AAV8 и совпадала с образованием антител против hFVIII у 63 % мышей. Эта сравнительное исследование капсидов выполняли с использованием высокоиммуногенной трансгенной кассеты, и вероятно оно не моделирует то, что наблюдается у людей, для которых у ~30 % пациентов образуются антитела к рекомбинантному белку. Следовательно высокие уровни экспрессии, продуцируемые после введения вектора AAVrh10, могут быть более выгодными для клинической ситуации, при которой может требоваться низкая доза вектора для значимых улучшений в отношении возникновения событий кровотечения и дополнительного введения рекомбинантного белка hFVIII.
По результатам этого исследования для дополнительного изучения были выбраны конструкции E03.TTR (ФИГ. 1; SEQ ID NO: 13), E03.A1AT (ФИГ. 3; SEQ ID NO: 15), E12.TTR (ФИГ. 2; SEQ ID NO: 14) и E12.A1AT (ФИГ. 4; SEQ ID NO: 16).
6.2 Исследования по определению дозировок
6.2.1 Исследования на мышах FVIII KO для определения приблизительной МЭД
Мыши FVIII KO линий C57BL/6 и 129 исходно получают в хвостовую вену инъекцию одной из четырех доз AAVhu37.E03.TTR.hFVIIIco-SQ.PA75. Дозы таких векторов составляют 5x1010 ГК/кг, 5x1011 ГК/кг, 5x1012 ГК/кг и 5x1013 ГК/кг. В качестве контроля носителя включена когорта животных, получающая только контрольный препарат (буфер-носитель). После введения вектора ежедневно исследуют животных путем общего осмотра. У животных в соответствующие моменты времени отбирают кровь для определения уровней активности hFVIII. Животных из подгруппы A умерщвляют на день 60 после введения доз, животных из подгруппы B умерщвляют на день 28 после введения доз, а животных из подгруппы C умерщвляют на день 3 после введения доз. Кровь также собирают при аутопсии для химического анализа сыворотки с помощью панелей и гематологического анализа. Умерщвленных животных вскрывают и отбирают органы для изучения биораспределения и гистопатологии, такие как правый паховый лимфоидный узел, правое яичко, поджелудочная железа, двенадцатиперстная кишка, толстая кишка, головной мозг, правая икроножная мышца, желудок, правая почка, правое легкое, селезенка, сердце, печень и обширные поражения, если они есть. Из органов мышей, которые получали самую высокую дозу вектора и которые получали контрольный препарат, экстрагируют общую клеточную ДНК и РНК. Выполняют анализы кПЦР и ОТ-кПЦР на экстрагированной ДНК/РНК для измерения количества копий векторного генома и уровни транскриптов в этих органах, соответственно. Эффективность исследуемого препарата определяют по уровням активности в плазме белка hFVIII анализом COATEST. Антитела против hFVIII также повторно определяют анализом твердофазного ИФА с IgG против hFVIII, а внешний путь коагуляции оценивают в анализе определения протромбинового времени (PT).
6.3 Исследования на нечеловеческих приматах (NHP)
6.3.1 Исследования экспрессии на NHP
Первичной целью этого не предусматривающего требований GLP исследования является оценивание потенциального вектора относительно токсичности и биораспределения у NHP.
Для этого исследования использовали самцов макака-резус и яванского макака. В этом исследовании использовали только животных самцов поскольку гемофилия A является сцепленным с X-хромосомой генетическим нарушением. В начале исследования у всех макак наблюдали титры NAb <1:5, определенные как описано раннее(CALCEDO et al. (2009) Worldwide epidemiology of neutralizing antibodies to adeno-associated viruses. J Infect Dis, 199, 381-90).Перед введением вектора макаки получали анестезию смесью кетамина (10–15 мг/кг) и дексмедетомидина (0,05–0,10 мг/кг), инъецируемую в/м. Макакам в/в вводили векторы через подкожную вену бедра.Перед началом исследования и дважды в неделю в течение исследования путем пункции бедренной вены отбирали образцы крови. Все клинические анализы по определению патологии в образцах крови проводили в Antech Diagnostics (г. Ирвайн, штат Калифорния, США), включая полный анализ крови и лейкограмму, полные клинические химические показатели и панели коагуляции.
На NHP выполняли пилотные исследования экспрессии hFVIII. Двум макакам-резус и двум яванским макакам в/в вводили 3x1012 ГК/кг AAVrh10 (ФИГ. 10A) и AAVhu37 (ФИГ. 10B) векторы, экспрессирующие hFVIII из комбинации энхансера/промотора ABP2.TGB-S1, соответственно. У всех животных отмечена высокая пиковая экспрессия, но до недель 6–8. У макак, которым инъецировали AAVrh10, наблюдали гуморальный иммунный ответ к hFVIII. У одного животного, которое получило AAVhu37, была задержка в образовании антител против hFVIII, которая происходила на 12 неделю после введения вектора, и они не образовывались у других животных в течение всего хода исследования. За макаком, который получал AAVhu37.TBG-S1.hFVIII-SQ.PA7, наблюдали в течение 35 месяцев после введения (ФИГ. 22–24).
На основании исследований на мышах FVIII KO и этого пилотного исследования на макаках-резус, отобрали две из исходных 42 комбинаций энхансеров/промоторов для дополнительной оценки на яванский макаках, используя для экспрессии разные капсиды клада E.
6.3.2 Дополнительные исследования на NHP
Далее 20 самцам яванского макака вводили дозы одного из четырех векторов: AAVrh10.E03.TTR.hFVIIIco-SQ.PA75, AAVrh10.E12.A1AT.hFVIIIco-SQ.PA75, AAVhu37.E03.TTR.hFVIIIco-SQ.PA75 и AAVhu37.E12.A1AT.hFVIIIco-SQ.PA75 (n = 5 макак на вектор). Вектор в/в вводили в дозе 1,2x1013 ГК/кг (на основе среднего титра по окПЦР). На неделю 2 после введения векторов наблюдали пиковую экспрессию равную 37 % от нормальных уровней FVIII для одного капсида плюс комбинации энхансера/промотора, которая потом выходила на плато 20 % от нормы (ФИГ. 11). В то время когда антитела к hFVIII выявляли у большинства макак на неделю 8, антитела не выявлялись у двух животных на неделю 30 после введения вектора (ФИГ. 12). Эти методы обсуждаются ниже.Применяя анализ времени до наступления события для образования антител, определили, что существует значимая разница между AAVrh10 и AAVhu37 (ФИГ.14).
6.3.3 Определение экспрессии hFVIII в плазме NHP
Экспрессию hFVIII измеряли методом твердофазного ИФА, причем все реактивы были получены, если не указано иное, у Sigma-Aldrich (г. Сент-Луис, штат Миссури, США). Планшеты для твердофазного ИФА покрывали IgG против hFVIII (Green Mountain Antibodies, штат Вермонт, США) при разведении 1:500 в 0,1 M карбонатном буфере (pH 9,6) и инкубировали в течение ночи при 4 °C. Промывали лунки четыре раза с помощью 0,1 % твина 20 в ФСБ и блокировали их с помощью 5 % обезжиренного молока (Bio Rad, г. Геркулес, штат Калифорния, США) в ФСБ в течение одного часа при комнатной температуре. После удаления блокирующего буфера образцы плазмы, разбавленные в 5 % обезжиренном молоке, добавляли в планшеты и инкубировали в течение одного часа при комнатной температуре. Затем планшеты промывали четыре раза и добавляли IgG против hFVIII (ThermoFisher Scientific, штат Массачусетс, США) при разведении 1:1000 в обезжиренном молоке. После инкубации в течение одного часа при комнатной температуре планшеты промывали четыре раза и добавляли конъюгированный с HRP IgG против антител овцы при разведении 1:1000 в обезжиренном молоке. После инкубации при комнатной температуре в течение 90 минут планшеты промывали пять раз и для выявления добавляли 3,3',5,5'-тетраметилбензидин (ТМБ). После выдержки в течение 5 минут при комнатной температуре останавливали реакцию с использованием 2N серной кислоты, и планшеты считывали при 450 нм с использованием устройства для считывания планшетов BioTek µQuant (г. Уинуски, штат Вермонт, США). На ФИГ. 11.
Результаты долговременной стабильной экспрессии человеческого FVIII у яванского макака (35 месяцев) после однократной внутривенной инъекции AAVhu37.TBG-S1.hFVIII-SQ.PA75 показаны на ФИГ. 22. Результаты показывают стабильную экспрессию до аутопсии через 35 месяцев. Результаты исследования активности ферментов печени показаны на ФИГ. 23. Данные результаты показывают, что уровни активности ферментов печени находятся в пределах диапазона нормы, за исключением временного повышения после биопсии печени. Реакция образования нейтрализующих антител (Nab) на капсид AAVhu.37 показана на ФИГ. 24.
Применяли технологию с использованием одиночных клеток для выявления ДНК и РНК вектора AAV в гепатоцитах NHP M11269, упомянутого выше, который получил внутривенное введение AAVhu37.TBG-S1.hFVIII-SQ.PA75 в дозе 3E12 ГК/кг. Для оценки наличия геномов и экспрессии AAV выделяли одиночные клетки гепатоцитов. Через 150 недель после введения вектора выделяли гепатоциты из клиновидного фрагмента печени путем перфузии со смесью коллагеназы/протеазы. Одиночные клетки распределяли в отдельные лунки 96-луночного планшета. Для клеток из одного 96-луночного планшета провели амплификацию целого генома (WGA), а геномы AAV в клетках количественно определяли методом цифровой ПЦР, используя зонд к FVIII человека. Второй 96-луночный планшет был использован для целых амплифицированных транскриптом (WTA) для оценки экспрессии FVIII.
Результаты показаны в таблице 3. Геномы AAV можно было выявить у ~25 % выделенных одиночных клеток. Экспрессию РНК обнаружили только у ~4 % или 20 % клеток, которые приняли ДНК. Экспрессирующие РНК клетки можно разделить на два типа с низкой и высокой экспрессией. Остается неясно, почему значительное количество клеток приняло вектор, но не способно экспрессировать трансген. Эти результаты были подтверждены исследованиями с гибридизацией in situ, выполненными на подложке CMC (данные не показаны).
6.3.4 Выявление IgG против hFVIII в плазме NHP
Антитела-IgG против hFVIII в плазме NHP измеряли методом твердофазного ИФА, как описано раннее, за исключением добавления для выявления конъюгированного с HRP IgG против антител NHP в разведении 1:2000 в обезжиренном молоке (ФИГ. 12).
6.3.5 Выявление титров анализом Бетезда в плазме NHP
Ингибирующее антитело против FVIII человека измеряли модифицированным Ниджмегеном (Nijmegen) анализом Бетезда (Giles et al., 1998). Одна единица Бетезда представляет ингибирование коагуляционной активности нормальной человеческой плазмы на 50 %.
6.3.6 Проведение биопсии печени
Двум NHP из каждой группы проводили биопсию печени, выполняемую минилапаротомией на неделю 5 после введения вектора. Выбор животных основывался на экспрессии hFVIII в плазме на неделю 4. Первое отобранное животное представляло собой животное с медианным уровнем hFVIII, второе отобранное животное представляло собой животное с ближайшим уровнем hFVIII или вторым ближайшим (если ближайший был медианным) к среднему значению. Отбирали образцы ткани печени для гистопатологического исследования, биораспределения вектора и анализа трансгенной мРНК. На день 3 после биопсии печени отбирали кровь для общего анализа крови и лейкограммы, полного анализа клинических химических показателей и панелей коагуляции.
6.3.7 Протокол иммуносупрессии
Начинали схему иммуносупрессии, в объеме необходимом животным, когда после введения вектора возможность выявлять экспрессию hFVIII была утрачена в присутствии выявляемых антител к hFVIII (единица Бетезда >1). Схему иммуносупрессии выполняли с ритуксимабом (250 мг/м2, в/в с интервалами в 4 недели, всего 4 инфузии) и циклофосфамидом (300 мг/м2, медленная внутривенная инфузия каждые 15 дней, всего 8 доз в течение 4 месяцев), как описано раннее(Mcintosh et al. (2013) Therapeutic levels of FVIII following a single peripheral vein administration of rAAV vector encoding a novel human factor VIII variant. Blood, 121, 3335-44.).
6.3.8 Биораспределение вектора
Во время проведения аутопсии быстро замораживали образцы тканей (паховые лимфоузлы, поясничные лимфоузлы, мышца [правая икроножная], правое яичко, поджелудочная железа, правая почка, селезенка, правое легкое, сердце и печень) мышей C57BL/6J и экстрагировали ДНК с использованием мининабора для выделения ДНК QIAamp (Qiagen, г. Валенсия, штат Калифорния, США). Выполняли выявление и количественное определение геномных копий (ГК) векторов в экстрагированной ДНК и относительную экспрессию транскриптов hFVIII в экстрагированной РНК методом ПЦР в реальном времени, как описано раннее. Вкратце, векторные ГК и РНК количественно определяли с использованием праймеров/зонда, предназначенных для трансгенной последовательности hFVIII вектора. Количественное определение ГК из печени выполняли на одном образце печени из каждой мыши (n = 3/группу). Относительную экспрессию транскриптов РНК определяли с использованием ΔΔCT каждого образца, нормализованного по экспрессии 18S.
Биораспределение векторов оценивали для субпопуляции из 18 векторов AAVrh10 с вариантами энхансеров/промоторов. Векторы AAVrh10, экспрессировавшие hFVIIIco IV, вводили в дозе 1011 ГК на мышь в организм от 6 до 8 недельных мышей C57BL/6J дикого типа. Мышам проводили аутопсию на день 14 после введения векторов и отбирали мышцу (правую икроножную), правое яичко, поджелудочную железу, правую почку, селезенку, правое легкое, сердце и печень. Экстрагировали ДНК и РНК и количественно определяли уровни векторных ГК и транскриптов РНК, соответственно, с использованием праймеров/зонда, предназначенных для трансгенной последовательности hFVIII вектора. Не обнаружено значимых отличий в печени для уровней векторных ГК (Фиг. 20A) или РНК-транскриптов (ФИГ. 20B) среди получавших векторы групп, и отсутствовали выявляемые количества ГК и РНК в получавших контрольный препарат (ФСБ) групп. Однако отмечена тенденция к повышенным уровням транскриптов РНК в печени для векторов, в которых для экспрессии использован промотор A1AT, независимо от энхансерных последовательностей (ФИГ. 20B). Для других отобранных тканей уровни РНК-транскриптов hFVIII были в среднем в 1000 раз ниже, чем в печени, но высокая экстрагепатическая экспрессия отмечалась в мышцах и сердцах мышей C57BL/6J после введения E01.TBG-S1, E02.A1AT, E09.A1AT и E09.TTR (ФИГ. 21).
6.4. Исследование вектора
Выполняли анализы по определению характеристик, включающие идентификацию серотипа, содержание пустых частиц и идентификацию продукта трансгена. Описания всех этих анализов приведены ниже.
6.4.1 Титр геномных копий (ГК)
Для определения титра геномных копий использовали оптимизированную количественную ПЦР (окПЦР) путем сравнения распознаваемого плазмидного стандарта. В анализе окПЦР применяли последовательное расщепление с помощью ДНКазы I и протеиназы K с последующим проведением анализа кПЦР для измерения инкапсидированных векторных геномных копий. Выявления ДНК выполняли с использованием специфических к последовательности праймеров, таргетирующих полиA-область PA75, в комбинации с флуоресцентно меченным зондом, гибридизирующимся с той же областью. Сравнение стандартной кривой плазмидной ДНК позволяет проводить определение титров без необходимости какой-либо манипуляции с образцом после ПЦР. В данном анализе были применены ряд стандартов, образцов для валидации и контролей (для фонового уровня и загрязнения ДНК). Для этого анализа провели квалификацию путем принятия и определения аналитических характеристик, включающих чувствительность, предел обнаружения, диапазон количественного определения и межлабораторную и внутрилабораторную прецизионность анализа. Была квалифицирована внутренняя эталонная серия AAVrh.10 и использована для выполнения количественных исследований.
6.4.2 Идентификация векторных капсидов Масс-спектрометрия капсида AAV из VP3
Подтверждение серотипа AAV2/hu.37 или AAV2/rh.10 вектора достигали исследованием на основе анализа пептидов белка капсида VP3 методом масс-спектрометрии (МС). В данном методе использовали мультиферментное расщепление (трипсин, химотрипсин и эндопротеаза Glu-C) полосы белка VP3, вырезанной из гелей ДСН-ПААГ с последующим определением характеристик СВЭЖХ-МС/МС на масс-спектрометре Q-Exactive Orbitrap для секвенирования капсидного белка. Разработали метод тандемной масс-спектрометрии (МС), позволяющий проводить идентификацию определенных загрязняющих белков и выводить пептидную последовательность из масс-спектров.
6.4.3 Соотношение пустых-полных частиц
Профили векторных частиц получали с использованием скорости седиментации при аналитическом ультрацентрифугировании (AUC), измеренной на аналитической центрифуге, что является превосходным методом для получения информации о гетерогенности макромолекулярной структуры, отличия при подтверждении и состояния ассоциации или агрегации. Образец вносили в ячейки и проводили седиментацию при 12 000 об/мин на аналитической центрифуге Beckman Coulter Proteomelab XL-I. Каждые две минуты в течение 3,3 часов записывали сканы показателей преломления. Анализировали данные с помощью модели c(s) (программа Sedfit) и рассчитывали коэффициенты седиментации, построенные по отношению к нормализованным значениям c(s). Должен наблюдаться основной пик, представляющий мономерный вектор. Появление пиков, мигрирующих медленнее основного мономерного пика, показывает пустые/неправильно собранные частицы. Коэффициент седиментации пика пустых частиц определяют с использованием препаратов с пустыми частицами AAV8. Прямое количественное определение основного мономерного пика и предшествующих пиков обеспечивает определение соотношения пустых-полных частиц.
6.4.4 Инфекционный титр
Для определения продуктивного поглощения и репликации вектора в клетках RC32 (клетки HeLa, экспрессирующие rep2) использовали анализ инфекционных единиц (ИЕ). Вкратце, клетки RC32 в 96-луночных планшетах совместно инфицировали серийными разведениями вектора и одинаковым разведением Ad5 с 12 репликатами для каждого разведения rAAV. Через 72 часа после инфицирования клетки лизировали и выполняли кПЦР для выявления амплификации векторов rAAV превышающей исходное введение. Для определения репликативного титра, выраженного в ИЕ/мл, выполняли расчет конечного разведения TCID50 (Спирмена-Кербера). Поскольку значения «инфекционности» зависят от вида частиц, входящих в контакт с клетками, связывания рецепторов, интернализации, транспорта в ядро и репликации генома, они находятся под влиянием конфигурации анализа и наличия соответствующих рецепторов и механизмов ответа после связывания в используемой линии клеток. Рецепторы и механизмы ответа после связывания, критически важные для импорта векторов AAV, обычно поддерживаются в иммортализированных линиях клеток, и поэтому анализы титров инфекционности не являются абсолютным показателем количества присутствующих «инфекционных» частиц. Однако соотношение инкапсидированных ГК к «инфекционным единицам» (описанным как соотношение ГК/ИЕ) можно применять как показатель однородности продукта от серии к серии.
7. Пример 2. Протокол лечения людей
Этот пример относится к лечению генной терапией пациентов с генетически подтвержденной сцепленной с Х-хромосомой гемофилией A из-за мутаций в гене фактора свертывания 8 (FVIII). В этом примере вектор для генной терапии, AAVhu.37.hFVIII, дефектный по репликации аденоассоциированный вирусный вектор hu.37 (AAVhu.37), экспрессирующий hFVIII, вводили пациентам с гемофилией A. Эффективность лечения можно оценивать с использованием уровней FVIII, как заменителе показателя трансгенной экспрессии. Первичные оценки эффективности включают уровни FVIII через около 12 недель после лечения с сохранением эффекта после этого в течение по меньшей мере 1 года. Долгосрочную безопасность и сохранение трансгенной экспрессии можно определять в образцах биопсии печени после лечения.
7.1. Вектор для генной терапии AAV.hFVIII
7.1.1. AAVhu.37.hFVIII
Вектор AAVhu.37.hFVIII состоит из активного ингредиента вектора AAV и буфера лекарственной формы. Внешний компонент вектора AAV представляет собой серотип hu.37, T = 1 икосаэдрального капсида, состоящего из 60 копий трех вирусных белков AAV VP1, VP2 и VP3 в ожидаемом соотношении 1:1:10. Капсид содержит рекомбинантный геном вектора AAV (rAAV) из одноцепочечной ДНК (ФИГ.1–ФИГ.4).
Геном содержит трансген человеческого фактора VIII (FVIII), фланкированный двумя инвертированными концевыми повторами (ITR) AAV. Кодон-оптимизированный трансген FVIII человека с удаленным B-доменом содержит энхансер, промотор, кодирующая человеческий фактор VIII (hFVIII) последовательность и сигнал полиаденилирования (полиA). ITR представляют собой генетические элементы, отвечающие за репликацию и упаковку генома во время продукции вектора, и они являются единственными вирусными цис-элементами, необходимыми для образования rAAV. В одном варианте реализации изобретения экспрессия кодирующей человеческий FVIII последовательности управляется промотором транстиретина (SEQ ID NO: 7). В другом варианте реализации изобретения экспрессия кодирующей человеческий FVIII последовательности управляется модифицированным промотором A1AT (SEQ ID NO: 9). Данная конструкции включает по меньшей мере один энхансерный элемент для стимуляции промоторной активности. В одном варианте реализации изобретения включен энхансер EnTTR (SEQ ID NO: 5). В другом варианте реализации изобретения две копии энхансера ABP-S (SEQ ID NO: 6) идут за одной копией энхансера EnTTR (SEQ ID NO: 5). Включен синтетический сигнал полиA из около 75 нуклеотидов (SEQ ID NO:10) для опосредования терминации транскриптов мРНК человеческого FVIII.
Вектор представлен в виде суспензии вектора AAVhu.37.hFVIII в буфере лекарственной формы. В одном варианте реализации изобретения буфер лекарственной формы представляет собой 0,001 % плюроник F-68 в TMN200 (200 мM хлорида натрия, 1 мM хлорида магния, 20 мM трис, pH 8,0).
Подробная информация о промышленном получении и определении характеристик векторов описана ниже в разделах.
7.1.2. AAVrh.10.hFVIII
Вектор AAVrh.10.hFVIII состоит из активного ингредиента вектора AAV и буфера лекарственной формы. Внешний компонент вектора AAV представляет собой серотип rh.10, T = 1 икосаэдрального капсида, состоящего из 60 копий трех вирусных белков AAV VP1, VP2 и VP3 в ожидаемом соотношении 1:1:10. Капсид содержит рекомбинантный геном вектора AAV (rAAV) из одноцепочечной ДНК (ФИГ.1–ФИГ.4).
Геном содержит трансген человеческого фактора VIII (FVIII), фланкированный двумя инвертированными концевыми повторами (ITR) AAV. Кодон-оптимизированный трансген FVIII человека с удаленным B-доменом содержит энхансер, промотор, кодирующая человеческий фактор VIII (hFVIII) последовательность и сигнал полиаденилирования (полиA). ITR представляют собой генетические элементы, отвечающие за репликацию и упаковку генома во время продукции вектора, и они являются единственными вирусными цис-элементами, необходимыми для образования rAAV. В одном варианте реализации изобретения экспрессия кодирующей человеческий FVIII последовательности управляется промотором транстиретина (SEQ ID NO: 7). В другом варианте реализации изобретения экспрессия кодирующей человеческий FVIII последовательности управляется модифицированным промотором A1AT (SEQ ID NO: 9). Данная конструкции включает по меньшей мере один энхансерный элемент для стимуляции промоторной активности. В одном варианте реализации изобретения включен энхансер EnTTR (SEQ ID NO: 5). В другом варианте реализации изобретения две копии энхансера ABP-S (SEQ ID NO: 6) идут за одной копией энхансера EnTTR (SEQ ID NO: 5). Включен синтетический сигнал полиA из около 75 нуклеотидов (SEQ ID NO:10) для опосредования терминации транскриптов мРНК человеческого FVIII.
Вектор представлен в виде суспензии вектора AAVrh.10.hFVIII в буфере лекарственной формы. В одном варианте реализации изобретения буфер лекарственной формы представляет собой 0,001 % плюроник F-68 в TMN200 (200 мM хлорида натрия, 1 мM хлорида магния, 20 мM трис, pH 8,0).
7.2. Популяция пациентов
По нескольким причинам выбранной исследуемой популяцией являются пациенты с тяжелой гемофилией A. Пациенты с тяжелой гемофилией A определены как имеющие менее 1 % от нормальной активности фактора VIII (FVIII), таким образом, требующие частых инфузий FVIII для контроля их предрасположенности к кровотечениям. Это представляет значительное затруднение в отношении ведения нормального образа жизни и, в дополнение к этому, уровни FVIII в крови проходят через хорошо известные пики и имеют снижающийся профиль, что неоптимально. Тот факт, что уровни FVIII в крови у тяжелых пациентов оказываются меньше 1 %, дает возможность легко измерять даже от низкого до умеренного увеличения уровней FVIII в крови после введения AAV.hFVIII. Недавние клинические исследования подтвердили действенность этого подхода.
Пациенты, которые являются кандидатами для проведения лечения предпочтительно представляют собой взрослых мужчин возрастом ≥18 с диагностированной умеренной/тяжелой или тяжелой гемофилией A. В одном варианте реализации изобретения пациент имеет базовую активность FVIII ≤2 % от нормальной или задокументированной истории активности FVIII ≤2 %. В некоторых вариантах реализации изобретения можно лечить пациента возрастом <18 лет. Кандидаты для лечения включают субъектов, у которых было по меньшей мере 3 эпизода с кровотечением за год, которые требуют лечения FVIII по необходимости. Другие кандидаты для лечения, включают субъектов, которых лечат FVIII в профилактическом режиме. Другие критерии демонстрируют, что для субъекта подходит лечение, включающее по меньшей мере 100 дней истории воздействия к FVIII; в истории болезни отсутствует образование ингибиторов (нейтрализирующих антител) к экзогенному FVIII; отсутствуют известные аллергические реакции к экзогенному FIX или любому компоненту AAV.hFVIII.
Получающие лечение пациенты могут иметь в сыворотке базовый уровень нейтрализующего антитела к AAVhu.37 или AAVrh.10 (соответствуют выбранному вектору) ≤ 1:5.
Субъектам могут позволять продолжать их стандартные виды лечения (-ий) (например, введение заместительного FVIII) перед и во время лечения генной терапией по усмотрению лечащего врача. В альтернативном варианте врач может предпочесть прекратить стандартные виды лечения перед проведением лечения генной терапией и, необязательно, возобновить стандартные виды лечения в качестве котерапии после проведения генной терапии.
7.3. Дозирование и путь введения
Пациенты получают однократную дозу AAVrh.10.hFVIII или AAVhu.37.hFVIII, вводимые посредством инфузии через периферическую вену. Вводимая пациенту доза AAVrh.10.hFVIII или AAVhu.37.hFVIII составляет около 5x1011 ГК/кг или 1,6x1012 ГК/кг, или 5x1012 ГК/кг, или 1x1013 ГК/кг. С целью обеспечения удаления пустых капсидов из дозы AAVrh.10.hFVIII или AAVhu.37.hFVIII, которую вводили пациентам, пустые капсиды отделяют от векторных частиц путем ультрацентрифугирования в градиенте хлорида цезия или ионообменной хроматографии во время процесса очистки вектора, как обсуждается выше.
7.4. Измерение клинических критериев
Первичное оценивание безопасности вводимого продукта. Последующие оценивания проводят дважды в неделю в течение 4 недель после введения продукта, еженедельно с недели 6 до недели 12, ежемесячно в течение оставшегося времени в первый год и с интервалами по 6 месяцев в течение общего периода времени в 5 лет.
a. Медицинский осмотр
b. Электрокардиограмма (ЭКГ)
c. Биохимические анализы: Уровни электролитов в сыворотке, АМК, креатинина, кальция, фосфата, общего белка, альбумина, ЛДГ, КФК, АСТ, АЛТ, щелочной фосфатазы, билирубина
d. Гематологический анализ: ОАК и лейкограмма, профиль коагуляции
e. Общий анализ мочи
f. Иммунологическая оценка:
g. Серологическая реакция на капсид hu.37 или rh.10 и на фактор VIII
h. Реакция T-клеток на капсид hu.37 или rh.10 и антигены фактора VIII
i. Оценка векторной ДНК; измерения кПЦР в плазме, моче и слюне.
Вторичное оценивание основано на измерениях экспрессии и активности трансгена, определяемых по
a. Уровням фактора VIII в плазме и активности фактора VIII;
b. Клинической оценке потребности в заместительной терапии фактора VIII и частотой эпизодов спонтанных кровотечений.
8. Пример 3. Промышленное получение AAV.hFVIII
8.1. Используемые для продукции AAV.hFVIII плазмиды
AAVrh.10.hFVIII продуцируют трансфекцией 3 плазмидных ДНК в человеческие клетки HEK 293 MCB с помощью:
(i) векторной плазмиды, описанной в разделах 8.2.1.1–8.2.1.4
(ii) хелперной плазмиды AAV, называемой pAAV2.rh10.KanR и содержащей гены rep2 и cap rh10 AAV дикого типа, описанной в разделе 8.2.2.1, и
(iii) хелперной аденовирусной плазмиды, называемой pAdDeltaF6(Kan), описанной в разделе 8.2.3.
AAVhu.37.hFVIII продуцируют трансфекцией 3 плазмидных ДНК в человеческие клетки HEK 293 MCB с помощью:
(i) векторной плазмиды, описанной в разделах 8.2.1.1–8.2.1.4
(ii) хелперной плазмиды AAV, называемой pAAV2.hu.37.KanR и содержащей гены rep2 и cap hu.37 AAV дикого типа, описанной в разделе 8.2.2.2, и
(iii) хелперной аденовирусной плазмиды, называемой pAdDeltaF6(Kan), описанной в разделе 8.2.3.
8.2.1 Цис-плазмиды (экспрессионная конструкция векторного генома)
8.2.1.1 pAAV.E03.p3.hF8co-SQ.PA75, содержащая экспрессионную кассету человеческого FVIII (ФИГ. 1). Эта цис-плазмида кодирует геном вектора rAAV. Экспрессия кДНК человеческого FVIII-SQco управляется промотором TTR с энхансером enTTR. Сигнал полиA экспрессионной кассеты является искусственной последовательностью полиA из около 75 нуклеотидов.
Описание элементов последовательностей
1. Инвертированные концевые повторы (ITR): ITR AAV представляют собой последовательности, идентичные на обоих концах, но расположенные в противоположной ориентации. Функции последовательностей ITR (GenBank № NC001401) AAV2 как во отношении точки репликации векторной ДНК, так и сигнала упаковки для векторного генома, при которой функционируют AAV и аденовирусный (ad) хелпер, представлены в положении транс. По сути последовательности ITR представляют только цис-действующие последовательности, требуемые для репликации и упаковки векторного генома. Последовательность 5' ITR используется в примере вектора, показанном в SEQ ID NO: 11. Последовательность 3' ITR используется в примере вектора, показанном в SEQ ID NO: 12.
2. Промотор TTR: Транстиретиновый промотор (SEQ ID NO:7) используется для управления высокоэффективной специфической для печени экспрессии гена hFVIII.
3. Энхансер TTR (enTTR): Энхансерная последовательность размером 100 п. н. (SEQ ID NO:5) из транстиретина присутствует в экспрессионной кассете вектора для увеличения экспрессии FVIII.
4. кДНК человеческого фактора коагуляции VIII (FVIII) (SEQ ID NO: 1 демонстрирует нативную последовательность; SEQ ID NO: 2 демонстрирует кодон-оптимизированную последовательность). кДНК человеческого фактора коагуляции 8 (FVIII) кодирует фактор коагуляции, необходимый для образования сгустков крови. hFVIII представляет собой последовательность с удаленным B-доменом, в которой домен B был замещен короткой последовательностью SQ, как описано в данном документе. кДНК hFVIII является кодон-оптимизированной для экспрессии у людей.
5. Искусственный сигнал полиаденилирования: (SEQ ID NO: 10) В искусственном сигнале полиаденилирования размером 75 п. н. представлены цис-последовательности для эффективного полиаденилирования мРНК hFVIII. Этот элемент функционирует как сигнал для транскрипционной терминации, специфического события отщепления на 3’-конце образующегося транскрипта после добавления полиаденильного хвоста.
8.2.1.2 pAAV.E12.p3.hF8co-SQ.PA75, содержащая экспрессионную кассету человеческого FVIII (ФИГ. 2). Эта цис-плазмида кодирует геном вектора rAAV. Экспрессия кДНК человеческого FVIII-SQco управляется промотором TTR с энхансером ABPS и enTTR. Сигнал полиA экспрессионной кассеты является искусственной последовательностью полиA из около 75 нуклеотидов.
Описание элементов последовательностей
1. Инвертированные концевые повторы (ITR): Такие как у 8.2.1.1
2. Промотор TTR:Такой как у 8.2.1.1
3. Энхансер: Укороченная до 42 п. н. версия дистального энхансера размером 100 п. н. предшественника α1-микроглобулина/букинина [ABP] (SEQ ID NO: 6) с двумя копиями энхансерной последовательности размером 100 п. н. из энхансера транстиретина (enTTR) (SEQ ID NO:5) присутствует в экспрессионной кассете вектора для увеличения экспрессии FVIII.
4. кДНК человеческого фактора коагуляции VIII (FVIII): Такая как у 8.2.1.1
5. Искусственный сигнал полиаденилирования: Такой как у 8.2.1.1
8.2.1.3 pAAV.E03.p2.hF8co-SQ.PA75, содержащая экспрессионную кассету человеческого FVIII (ФИГ. 3). Эта цис-плазмида кодирует геном вектора rAAV. Экспрессия кДНК человеческого FVIII-SQco управляется модифицированным промотором A1AT с энхансером enTTR. Сигнал полиA экспрессионной кассеты является искусственной последовательностью полиA из около 75 нуклеотидов.
Описание элементов последовательностей
1. Инвертированные концевые повторы (ITR): Такие как у 8.2.1.1
2. Промотор A1AT: Модифицированный промотор SERINA1 [α1-антитрипсин] (SEQ ID NO:9) используется для управления высокоэффективной специфической для печени экспрессии гена hFVIII.
3. Энхансер TTR (enTTR): Энхансерная последовательность размером 100 п. н. из транстиретина присутствует в экспрессионной кассете вектора для увеличения экспрессии FVIII.
4. кДНК человеческого фактора коагуляции VIII (FVIII): Такая как у 8.2.1.1
5. Искусственный сигнал полиаденилирования: Такой как у 8.2.1.1
8.2.1.4 pAAV.E12.p2.hF8co-SQ.PA75, содержащая экспрессионную кассету человеческого FVIII (ФИГ. 4). Эта цис-плазмида кодирует геном вектора rAAV. Экспрессия кДНК человеческого FVIII-SQco управляется промотором TTR с энхансером ABPS и enTTR. Сигнал полиA экспрессионной кассеты является искусственной последовательностью полиA из около 75 нуклеотидов.
Описание элементов последовательностей
1. Инвертированные концевые повторы (ITR): Такие как у 8.2.1.1
2. Промотор A1AT: Такой же как у 8.2.1.3
3. Энхансер: Такой как у 8.2.1.1
4. кДНК человеческого фактора коагуляции VIII (FVIII): Такая как у 8.2.1.1
5. Искусственный сигнал полиаденилирования: Такой как у 8.2.1.1
8.2.2 Хелперная плазмида
8.2.2.1 AAVrh10-хелперная плазмида pAAV2.rh10.KanR
Эта хелперная плазмида AAVrh10 (8036 п. н.) кодирует 4 белка rep AAV2 дикого типа и 3 капсидных белка VP AAV дикого типа серотипа rh10. Новую последовательность AAV получали из ДНК ткани печени макака-резус и установили AAV-серотип rh10. Для образования химерной упаковывающей конструкции ген cap AAV2 удаляли из плазмиды p5E18 и замещали ПЦР-фрагментом гена cap AAVrh10, амплифицированного из ДНК печени примата для получения плазмиды p5E18VD2/rh10. Следует отметить, что промотор p5 AAV, который в норме управляет экспрессией rep, перемещен в этой конструкции из 5’-конца rep к 3’-концу гена cap rh10. Это расположение служит для введения спейсера между промотором и геном rep (т. е. в остов плазмиды) для снижения регуляции экспрессии rep и увеличения способности поддерживать продукцию вектора с высокими титрами. Остов плазмиды в p5E18 взят у pBluescript KS. Все части компонентов плазмиды были проверены прямым секвенированием. Наконец, для получения pAAV2/rh10 (Kan) ген устойчивости к ампициллину был замещен геном устойчивости к канамицину.
8.2.2.2 AAVhu.37-хелперная плазмида pAAV2.hu.37.KanR
Эта хелперная плазмида AAVhu.37 (8036 п. н.) кодирует 4 белка rep AAV2 дикого типа и 3 капсидных белка VP AAV дикого типа серотипа hu.37. Схематический вид плазмиды pAAV2.rh10.KanR показан ниже. Для образования химерной упаковывающей конструкции ген cap AAV2 удаляли из плазмиды p5E18 и замещали ПЦР-фрагментом гена cap AAVhu.37, амплифицированного из ДНК печени примата для получения плазмиды p5E18VD2/hu.37. Остов плазмиды в p5E18 взят у pBluescript KS. Все части компонентов плазмиды были проверены прямым секвенированием. Наконец, для получения pAAV2/hu.37 (Kan) ген устойчивости к ампициллину был замещен геном устойчивости к канамицину.
8.2.3 Аденовирусная хелперная плазмида pAdDeltaF6(Kan)
Плазмида pAdDeltaF6(Kan) имеет размер 15 774 п. н. Данная плазмида содержит участки аденовирусного генома, который является важным для репликации AAV, а именно РНК E2A, E4 и VA (функции аденовируса E1 обеспечиваются клетками 293), но не содержит другие гены для репликации или структурные гены аденовируса. Плазмида не содержит цис-элементы, критически важные для репликации, такие как аденовирусные инвертированные концевые повторы и, поэтому, не предполагается возможный синтез инфекционного аденовируса. Она получена из молекулярного клона Ad5 с удаленными E1, E3 (pBHG10, на основе плазмиды pBR322). В ДНК Ad5 проводили делеции для удаления экспрессии ненужных аденовирусных генов и снижения количества аденовирусной ДНК от 32 т. н. до ~12 т. н. Наконец, для получения pAdΔF6(kan) ген устойчивости к ампициллину был замещен геном устойчивости к канамицину. Идентичность этих 3 аденовирусных генов подтверждали секвенированием плазмидной ДНК, выполненным Qiagen Genomic Services на исходном плазмидном материале, который отправили в Aldevron Inc. для промышленной наработки плазмидной ДНК. Анализ ДНК выявил 100 % гомологию с 3 генными областями аденовируса типа 5 (GenBank, номер регистрации AF369965).
8.2.4 Главный банк клеток (ГБК) бактерий
ГБК бактерии для продукции этих трех продуцирующих ДНК плазмид, которые используются для поддержания производства DTX101, получали с помощью Aldevron Inc. Банки клеток получали в результате размножения выбранных культур и проводили дорогостоящие исследования для квалификации каждого ГБК бактерий, следуя СОП Aldevron в соответствии с рекомендациями CBER. Выполняли тестирования для каждой из трех плазмид и фиксировали информацию, касающуюся особенностей получения ГБК бактерий.
8.2.5 Промышленное получение плазмидной ДНК
Все плазмиды, используемые в технологическом процессе, получали с помощью Aldevron Inc. под контролем системы контроля качества GMP-S™ и инфраструктуры, использующей актуальные особенности производства cGMP; прослеживаемость, контроль за документацией и разделение материалов. Выполняли тестирования для каждой плазмиды и фиксировали информацию, касающуюся особенностей получения плазмидной ДНК.
8.2.6 Главный банк клеток (ГБК) почки эмбриона человека (HEK) 293
Клетки HEK 293 исходно созданы путем трансформации клеток HEK расщепленной ДНК аденовируса типа 5 Фрэнком Грэхемом (Frank Graham) с коллегами. Для продуцирования rAAV с высокими титрами требуются клетки, экспрессирующие генные продукты E1a и E1b. Клетки HEK293 являются адгезивными и в высшей степени трансфицируемыми с образованием высоких титров rAAV при трансфекции плазмидной ДНК.
8.3 Промышленное получение вектора AAV
8.3.1 Описание производственного процесса
1. Высаживание клеток. Для технологического процесса используют квалифицированную линию клеток 293 почки эмбриона человека. Клетки культивируют в среде, состоящей из среды Игла в модификации Дюльбекко (DMEM) с добавкой 10 % фетальной бычьей сыворотки (FBS), облученной гамма-радиацией. Клетки являются субстратзависимыми и диссоциацию клеток выполняют с использованием TrypLE Select, реактива для диссоциации неживотных клеток. Клетки поддерживают при 37 °C (+/- 1 °C) в атмосфере 5 % (+/- 0,5 %) CO2.
2. Временная трансфекция. Через 3 дня выращивания (среда DMEM + 10 % FBS), клеточную культуральную среду в камере Hyperstack замещают свежей бессывороточной средой DMEM и трансфецируют 3 продуцирующими плазмидами с использованием оптимизированного метода преципитации с PEI.
Полный комплекс для трансфекции ДНК-плазмид готовят в BSC для трансфекции двадцати 36-слойных камер HyperStack Corning (на серию BDS). Вначале готовят смесь ДНК/PEI, содержащую 3,0 мг векторной плазмиды pDTX.hFIX.101, 60 мг pAdDeltaF6(Kan), 30 мг хелперной плазмиды pAAV2.rh10.KanR AAV и PEI, уровня качества для GMP (PEIPro, PolyPlus Transfection SA). После интенсивного перемешивания раствору дают отстояться при комнатной температуре в течение 25 мин, а затем добавляют к бессывороточной среде для гашения реакции и потом добавляют его в 36-слойные камеры Hyperstack Corning. Среду для трансфекции распределяют между всеми 36 слоями камеры Hyperstack и клетки поддерживают при 37 °C (+/- 2 °C) в атмосфере 5 % (+/- 0,5 %) CO2 5 дней.
3. Сбор среды с клетками. Трансфицированные клетки и среду собирают из каждой камеры Hypertack, используя одноразовые пакеты для биопроцессов путем асептического слива среды из модулей. После сбора среды в объем ~ 80 литров добавляют MgCl2 до конечной концентрации 2 мМ (кофактор бензоназы) и добавляют нуклеазу бензоназу (№ 1.016797.0001 по кат. Merck Group) до конечной концентрации 25 единиц/мл. Продукт (в одноразовом пакете для биопроцессов) инкубируют при 37 °C в течение 2–3 ч в инкубаторе для предоставления достаточного времени для ферментативного расщепления остаточной клеточной и плазмидной ДНК в урожае, полученном в результате процедуры трансфекции.Эту стадию выполняют для минимизации количества остаточной ДНК в готовом векторе DP.После инкубационного периода добавляют NaCl до конечной концентрации 500 мМ, чтобы способствовать выделению продукта во время фильтрации и последующей фильтрации в тангенциальном потоке.
4. Осветление. Удаляют клетки и клеточный дебрис из продукта с использованием погружной фильтровальной капсулы (1,2 мкм/0,22 мкм), присоединенной последовательно в виде набора из стерильной закрытой трубки и пакета, используемого с перистальтическим насосом. Среда проходит через капсульный фильтр Sartorius Sartoguard из PES (1,2 мкм/0,22 мкм) (Sartorius Stedim Biotech Inc.).
5. Крупномасштабная фильтрация в тангенциальном потоке. Снижение объема (в 10–20 раз) осветленного продукта достигают с использованием фильтрации в тангенциальном потоке (TFF), используя специальную набор из стерильной закрытой трубки для биопроцессов, пакет и мембрану, изготовленный Spectrum Labs.
8.4 Повторное введение второго вектора
8.4.1 Повторное введение AAV3B или AAV5
Оценивали эффективность повторного введения вектора с использованием AAV3B или AAV5 у макак-резус, которых ранее лечили векторами AAVrh10 или AAV8. Векторы, показанные в таблице 4, получали, как описано ранее, когда вектор выделяли из супернатанта после тройной трансфекции в клетках HEK293 и очищали на градиенте йодиксанола. Определяли титр векторов методом цифровой ПЦР.
В исследовании на 8 группах (n=3/группу; таблица 1) было задействовано 24 макака-резус (возрастом 3–5 лет) на основании их статуса по предварительно имеющимся NAb. На день 0 макакам инъецировали 1,0x1013 ГК/кг AAV.TBG.hCG.WPRE с вектором AAV, как показано в таблице 4. На неделю 12 макаки получали вторую инъекцию 1,0x1013 ГК/кг AAV.TBG.hCG.WPRE с вектором AAV, как показано в таблице 4. Выполняли биопсию печени на неделю 2 и неделю 14, а аутопсию выполняли на неделю 26.
Таблица 4. Краткое описание когорт и векторов 
Уровни экспрессии трансгенов (rhCG — хорионический гонадотропин макака-резус, b-субъединица; rhAFP — альфа-фетопротеин макака-резус) в сыворотке измеряли твердофазным иммуноферментным анализом (твердофазный ИФА). Для измерения ДНК-копий векторов в печени выполняли анализы кПЦР на общей клеточной ДНК, экстрагированной из образцов печени, собранных во время биопсии и аутопсии печени. Выполняли анализ на NAb к AAV как описано раннее. Для визуализации окрашивали срезы печени антителом против CG.
На ФИГ. 15 показано сравнение уровней экспрессии rhCG векторами AAVrh10, AAV8, AAV3B и AAV5 (первая инъекция вектора). На ФИГ. 16A–16D показаны ДНК-копии векторов rhCG в печени в различные моменты времени. На ФИГ. 17A–17B показаны уровни rhAFP после повторного введения (вторая инъекция вектора) векторов AAV3B (ФИГ. 17A) или AAV5 (ФИГ. 17B), экспрессирующих rhAFP. На ФИГ. 18A и ФИГ. 18B показаны геномные копии вектора rhAFP в печени. На ФИГ. 19 показаны различные Nab-ответы на AAV у макак.
У наивных животных векторы клада E (AAVrh10 и AAV8) продемонстрировали наибольшие уровни перипортального переноса генов по сравнению с векторами AAV5 с наименьшими уровнями; перипортальная зона располагается ближе всего к входу кровотока, получает наиболее оксигенированную кровь и является самой важной областью печени для обеспечения метаболических процессов. Векторы AAVrh10 и AAV5 проявили более высокие уровни нейтрализирующих антител (NAb), чем AAV8 и AAV3B. У серонегативных животных отмечена значимая вариация экспрессии трансгенов, включая AAV3B, среди разных животных. В течение исследованного короткого временного промежутка оказывается, что образующееся к AAVrh10 NAb, впоследствии ингибировало трансдукцию in vivo серологически отличающегося серотипа AAV3B; а предыдущее воздействие AAV8 не влияло на трансдукцию AAV3B.
Все публикации, цитируемые в этом описании, а также предварительных заявках на патенты США с номерами 62/323,336, 62/331,807 и 62/428,866, включены в данный документ путем ссылки. Аналогичным образом, номера SEQ ID NO, которые упомянутые в данном документе и оказываются в прилагаемом перечне последовательностей и самом перечне последовательностей, включены путем ссылки. Несмотря на то, что изобретение было описано со ссылкой на конкретные варианты реализации изобретения, следует понимать, что можно выполнять различные модификации, не отступая от сущности данного изобретения. Подразумевается, что такие модификации входят в объем прилагаемой формулы изобретения.
(Перечень последовательностей в произвольном формате)
Следующая информация представлена для последовательностей в произвольном формате под числовым идентификатором <223>.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110>ЗЕ ТРАСТИС ОФ ЗЕ ЮНИВЕРСИТИ ОФ ПЕНСИЛЬВАНИЯ
<120> ГЕННАЯ ТЕРАПИЯ ДЛЯ ЛЕЧЕНИЯ ГЕМОФИЛИИ A
<130> UPN-16-7798PCT
<150> US 62/323,336
<151> 2016-04-15
<150> US 62/331,807
<151> 2016-05-04
<150> US 62/428,866
<151> 2016-12-01
<160> 20
<170> PatentIn, версия 3.5
<210> 1
<211> 4371
<212>ДНК
<213> Homo sapiens
<400> 1
atgcaaatag agctctccac ctgcttcttt ctgtgccttt tgcgattctg ctttagtgcc 60
accagaagat actacctggg tgcagtggaa ctgtcatggg actatatgca aagtgatctc 120
ggtgagctgc ctgtggacgc aagatttcct cctagagtgc caaaatcttt tccattcaac 180
acctcagtcg tgtacaaaaa gactctgttt gtagaattca cggatcacct tttcaacatc 240
gctaagccaa ggccaccctg gatgggtctg ctaggtccta ccatccaggc tgaggtttat 300
gatacagtgg tcattacact taagaacatg gcttcccatc ctgtcagtct tcatgctgtt 360
ggtgtatcct actggaaagc ttctgaggga gctgaatatg atgatcagac cagtcaaagg 420
gagaaagaag atgataaagt cttccctggt ggaagccata catatgtctg gcaggtcctg 480
aaagagaatg gtccaatggc ctctgaccca ctgtgcctta cctactcata tctttctcat 540
gtggacctgg taaaagactt gaattcaggc ctcattggag ccctactagt atgtagagaa 600
gggagtctgg ccaaggaaaa gacacagacc ttgcacaaat ttatactact ttttgctgta 660
tttgatgaag ggaaaagttg gcactcagaa acaaagaact ccttgatgca ggatagggat 720
gctgcatctg ctcgggcctg gcctaaaatg cacacagtca atggttatgt aaacaggtct 780
ctgccaggtc tgattggatg ccacaggaaa tcagtctatt ggcatgtgat tggaatgggc 840
accactcctg aagtgcactc aatattcctc gaaggtcaca catttcttgt gaggaaccat 900
cgccaggcgt ccttggaaat ctcgccaata actttcctta ctgctcaaac actcttgatg 960
gaccttggac agtttctact gttttgtcat atctcttccc accaacatga tggcatggaa 1020
gcttatgtca aagtagacag ctgtccagag gaaccccaac tacgaatgaa aaataatgaa 1080
gaagcggaag actatgatga tgatcttact gattctgaaa tggatgtggt caggtttgat 1140
gatgacaact ctccttcctt tatccaaatt cgctcagttg ccaagaagca tcctaaaact 1200
tgggtacatt acattgctgc tgaagaggag gactgggact atgctccctt agtcctcgcc 1260
cccgatgaca gaagttataa aagtcaatat ttgaacaatg gccctcagcg gattggtagg 1320
aagtacaaaa aagtccgatt tatggcatac acagatgaaa cctttaagac tcgtgaagct 1380
attcagcatg aatcaggaat cttgggacct ttactttatg gggaagttgg agacacactg 1440
ttgattatat ttaagaatca agcaagcaga ccatataaca tctaccctca cggaatcact 1500
gatgtccgtc ctttgtattc aaggagatta ccaaaaggtg taaaacattt gaaggatttt 1560
ccaattctgc caggagaaat attcaaatat aaatggacag tgactgtaga agatgggcca 1620
actaaatcag atcctcggtg cctgacccgc tattactcta gtttcgttaa tatggagaga 1680
gatctagctt caggactcat tggccctctc ctcatctgct acaaagaatc tgtagatcaa 1740
agaggaaacc agataatgtc agacaagagg aatgtcatcc tgttttctgt atttgatgag 1800
aaccgaagct ggtacctcac agagaatata caacgctttc tccccaatcc agctggagtg 1860
cagcttgagg atccagagtt ccaagcctcc aacatcatgc acagcatcaa tggctatgtt 1920
tttgatagtt tgcagttgtc agtttgtttg catgaggtgg catactggta cattctaagc 1980
attggagcac agactgactt cctttctgtc ttcttctctg gatatacctt caaacacaaa 2040
atggtctatg aagacacact caccctattc ccattctcag gagaaactgt cttcatgtcg 2100
atggaaaacc caggtctatg gattctgggg tgccacaact cagactttcg gaacagaggc 2160
atgaccgcct tactgaaggt ttctagttgt gacaagaaca ctggtgatta ttacgaggac 2220
agttatgaag atatttcagc atacttgctg agtaaaaaca atgccattga accaagaagc 2280
ttctcccaga atccaccagt cttgaaacgc catcaacggg aaataactcg tactactctt 2340
cagtcagatc aagaggaaat tgactatgat gataccatat cagttgaaat gaagaaggaa 2400
gattttgaca tttatgatga ggatgaaaat cagagccccc gcagctttca aaagaaaaca 2460
cgacactatt ttattgctgc agtggagagg ctctgggatt atgggatgag tagctcccca 2520
catgttctaa gaaacagggc tcagagtggc agtgtccctc agttcaagaa agttgttttc 2580
caggaattta ctgatggctc ctttactcag cccttatacc gtggagaact aaatgaacat 2640
ttgggactcc tggggccata tataagagca gaagttgaag ataatatcat ggtaactttc 2700
agaaatcagg cctctcgtcc ctattccttc tattctagcc ttatttctta tgaggaagat 2760
cagaggcaag gagcagaacc tagaaaaaac tttgtcaagc ctaatgaaac caaaacttac 2820
ttttggaaag tgcaacatca tatggcaccc actaaagatg agtttgactg caaagcctgg 2880
gcttatttct ctgatgttga cctggaaaaa gatgtgcact caggcctgat tggacccctt 2940
ctggtctgcc acactaacac actgaaccct gctcatggga gacaagtgac agtacaggaa 3000
tttgctctgt ttttcaccat ctttgatgag accaaaagct ggtacttcac tgaaaatatg 3060
gaaagaaact gcagggctcc ctgcaatatc cagatggaag atcccacttt taaagagaat 3120
tatcgcttcc atgcaatcaa tggctacata atggatacac tacctggctt agtaatggct 3180
caggatcaaa ggattcgatg gtatctgctc agcatgggca gcaatgaaaa catccattct 3240
attcatttca gtggacatgt gttcactgta cgaaaaaaag aggagtataa aatggcactg 3300
tacaatctct atccaggtgt ttttgagaca gtggaaatgt taccatccaa agctggaatt 3360
tggcgggtgg aatgccttat tggcgagcat ctacatgctg ggatgagcac actttttctg 3420
gtgtacagca ataagtgtca gactcccctg ggaatggctt ctggacacat tagagatttt 3480
cagattacag cttcaggaca atatggacag tgggccccaa agctggccag acttcattat 3540
tccggatcaa tcaatgcctg gagcaccaag gagccctttt cttggatcaa ggtggatctg 3600
ttggcaccaa tgattattca cggcatcaag acccagggtg cccgtcagaa gttctccagc 3660
ctctacatct ctcagtttat catcatgtat agtcttgatg ggaagaagtg gcagacttat 3720
cgaggaaatt ccactggaac cttaatggtc ttctttggca atgtggattc atctgggata 3780
aaacacaata tttttaaccc tccaattatt gctcgataca tccgtttgca cccaactcat 3840
tatagcattc gcagcactct tcgcatggag ttgatgggct gtgatttaaa tagttgcagc 3900
atgccattgg gaatggagag taaagcaata tcagatgcac agattactgc ttcatcctac 3960
tttaccaata tgtttgccac ctggtctcct tcaaaagctc gacttcacct ccaagggagg 4020
agtaatgcct ggagacctca ggtgaataat ccaaaagagt ggctgcaagt ggacttccag 4080
aagacaatga aagtcacagg agtaactact cagggagtaa aatctctgct taccagcatg 4140
tatgtgaagg agttcctcat ctccagcagt caagatggcc atcagtggac tctctttttt 4200
cagaatggca aagtaaaggt ttttcaggga aatcaagact ccttcacacc tgtggtgaac 4260
tctctagacc caccgttact gactcgctac cttcgaattc acccccagag ttgggtgcac 4320
cagattgccc tgaggatgga ggttctgggc tgcgaggcac aggacctcta c 4371
<210> 2
<211> 4374
<212> ДНК
<213> Искусственная последовательность
<220>
<223> сконструированная последовательность
<400> 2
atgcagatcgagctgagcacctgcttcttcctgtgcctgctgcggttctgcttctccgcc 60
acccggcggtactacctgggagccgtggagctgagctgggattacatgcagagcgatctg 120
ggagagctgccagtggatgcccggttcccaccacgggtgccaaagagcttcccattcaac 180
accagcgtggtgtacaagaagaccctgttcgtggagttcaccgatcacctgttcaacatc 240
gccaagccacggccaccctggatgggactgctgggaccaaccatccaggccgaggtgtac 300
gataccgtggtgatcaccctgaagaacatggcctctcatcctgtgtccctgcacgccgtg 360
ggagtgagctactggaaggccagcgagggagccgagtacgatgatcagaccagccagcgg 420
gagaaggaggatgataaggtgttcccaggaggaagccacacctacgtgtggcaggtgctg 480
aaggagaacggaccaatggccagcgatccactgtgcctgacctacagctacctgagccac 540
gtggatctggtgaaggatctgaacagcggactgatcggagccctgctggtgtgccgggag 600
ggaagcctggccaaggagaagacccagaccctgcacaagttcatcctgctgttcgccgtg 660
ttcgatgagggaaagagctggcacagcgagaccaagaacagcctgatgcaggatcgggat 720
gccgccagcgcccgggcctggccaaagatgcacaccgtgaacggatacgtgaaccggagc 780
ctgccaggactgatcggatgccaccggaagagcgtgtactggcacgtgatcggaatggga 840
accaccccagaggtgcactctatcttcctggagggacacacctttctggtgcggaaccac 900
cggcaggccagcctggagatcagcccaatcaccttcctgaccgcccagaccctgctgatg 960
gatctgggacagttcctgctgttctgccatatcagcagccaccagcacgatggaatggag 1020
gcctacgtgaaggtggatagctgcccagaggagccacagctgcggatgaagaacaacgag 1080
gaggccgaggattacgatgatgatctgaccgatagcgagatggatgtggtgcggttcgat 1140
gatgataacagcccaagcttcatccagatccggagcgtggccaagaagcacccaaagacc 1200
tgggtgcactacatcgccgccgaggaggaggattgggattacgccccactggtgctggcc 1260
cctgatgatcggagctacaagagccagtacctgaacaacggaccacagcggatcggacgg 1320
aagtacaaaaaagtgcggttcatggcctacaccgatgagaccttcaagacccgggaggcc 1380
atccagcacgagagcggaatcctgggaccactgctgtacggagaggtgggagataccctg 1440
ctgatcatcttcaagaaccaggccagccggccatacaacatctacccacacggaatcacc 1500
gatgtgcggccactgtacagccggcggctgccaaagggagtgaagcacctgaaggatttc 1560
ccaatcctgccaggagagatcttcaagtacaagtggacagtgacagtggaggatggacca 1620
accaagtctgatccaagatgcctgaccagatactacagcagctttgtgaacatggagaga 1680
gacctggcctctggactgattggaccactgctgatctgctacaaggagtctgtggatcag 1740
agaggaaaccagatcatgtctgataagagaaatgtgatcctgttctctgtgtttgatgag 1800
aacagaagctggtacctgacagagaacatccagagattcctgccaaacccagccggagtg 1860
cagctggaggatccagagttccaggccagcaacatcatgcacagcatcaacggatacgtg 1920
ttcgatagcctgcagctgagcgtgtgcctgcacgaggtggcctattggtatatcctgagc 1980
atcggagcccagaccgatttcctgagcgtgttcttcagcggatacaccttcaagcacaag 2040
atggtgtacgaggataccctgaccctgttcccattctccggagagaccgtgttcatgagc 2100
atggagaacccaggactgtggatcctgggatgccacaactctgatttcagaaacagagga 2160
atgactgccctgctgaaagtgtccagctgtgataagaacactggagattactatgaggat 2220
agctatgaggatatctctgcctacctgctgagcaagaacaatgccattgagccaagaagc 2280
ttcagccagaacccaccagtgctgaagagacaccagagagagatcaccagaaccaccctg 2340
cagtctgatcaggaggagattgattatgatgataccatctctgtggagatgaagaaggag 2400
gattttgatatctatgatgaggatgagaaccagagcccaagaagcttccagaagaagacc 2460
agacactacttcatcgctgcagtggagagactgtgggattatggaatgagcagcagccca 2520
cacgtgctgagaaacagagcccagagcggatctgtgccacagttcaagaaggtggtgttc 2580
caggagttcaccgatggaagcttcacccagccactgtaccggggagagctgaacgagcac 2640
ctgggactgctgggaccatacatccgggccgaggtggaggataacatcatggtgaccttc 2700
cggaaccaggccagccggccatacagcttctacagcagcctgatcagctacgaggaggat 2760
cagcggcagggagccgagccacggaagaacttcgtgaagccaaacgagaccaagacctac 2820
ttctggaaggtgcagcaccacatggccccaaccaaggatgagttcgattgcaaggcctgg 2880
gcctacttcagcgatgtggatctggagaaggatgtgcacagcggactgatcggaccactg 2940
ctggtgtgccacaccaacaccctgaacccagcccacggacggcaggtgaccgtgcaggag 3000
ttcgccctgttcttcaccatcttcgatgagaccaagagctggtacttcaccgagaacatg 3060
gagcggaactgccgggccccttgcaacatccagatggaggatccaaccttcaaggagaac 3120
taccggttccacgccatcaacggatacatcatggataccctgccaggactggtgatggcc 3180
caggatcagcggatccggtggtacctgctgagcatgggaagcaacgagaacatccacagc 3240
atccacttcagcggacacgtgttcaccgtgcggaagaaggaggagtacaagatggccctg 3300
tacaacctgtacccaggagtgttcgagaccgtggagatgctgccaagcaaggccggaatc 3360
tggcgggtggagtgcctgatcggagagcacctgcacgccggaatgagcaccctgttcctg 3420
gtgtacagcaacaagtgccagaccccactgggaatggccagcggacacatccgggatttc 3480
cagatcaccgccagcggacagtacggacagtgggccccaaagctggcccggctgcactac 3540
agcggaagcatcaacgcctggagcaccaaggagccattcagctggatcaaagtggatctg 3600
ctggccccaatgatcatccacggaatcaagacccagggagcccggcagaagttcagcagc 3660
ctgtacatcagccagttcatcatcatgtacagcctggatggaaagaagtggcagacctac 3720
cggggaaacagcaccggaaccctgatggtgttcttcggaaacgtggatagcagcggaatc 3780
aagcacaacatcttcaacccaccaatcatcgcccgatacatccggctgcacccaacccac 3840
tacagcatcagaagcaccctgcggatggagctgatgggatgtgatctgaacagctgctcc 3900
atgccactgggaatggagagcaaggccatcagcgatgcccagatcaccgccagcagctac 3960
ttcaccaacatgttcgccacctggagcccaagcaaggcccggctgcacctgcagggacgg 4020
agcaacgcctggcggccacaggtgaataacccaaaggagtggctgcaggtggatttccag 4080
aagaccatgaaggtgaccggagtgaccacccagggagtgaagagcctgctgactagcatg 4140
tatgtgaaggagttcctgatcagcagcagccaggatggacaccagtggaccctgttcttc 4200
cagaacggaaaggtgaaggtgttccagggaaaccaggatagcttcaccccagtggtgaac 4260
agcctggatccaccactgctgacccgatacctgcggatccacccacagagctgggtgcac 4320
cagatcgccctgagaatggaggtgctgggatgcgaggcccaggatctgtactga 4374
<210> 3
<211> 1457
<212>ПРТ
<213> Homo sapiens
<400> 3
Met Gln Ile Glu Leu Ser Thr Cys Phe Phe Leu Cys Leu Leu Arg Phe
1 5 10 15
Cys Phe Ser Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser
20 25 30
Trp Asp Tyr Met Gln Ser Asp Leu Gly Glu Leu Pro Val Asp Ala Arg
35 40 45
Phe Pro Pro Arg Val Pro Lys Ser Phe Pro Phe Asn Thr Ser Val Val
50 55 60
Tyr Lys Lys Thr Leu Phe Val Glu Phe Thr Asp His Leu Phe Asn Ile
65 70 75 80
Ala Lys Pro Arg Pro Pro Trp Met Gly Leu Leu Gly Pro Thr Ile Gln
85 90 95
Ala Glu Val Tyr Asp Thr Val Val Ile Thr Leu Lys Asn Met Ala Ser
100 105 110
His Pro Val Ser Leu His Ala Val Gly Val Ser Tyr Trp Lys Ala Ser
115 120 125
Glu Gly Ala Glu Tyr Asp Asp Gln Thr Ser Gln Arg Glu Lys Glu Asp
130 135 140
Asp Lys Val Phe Pro Gly Gly Ser His Thr Tyr Val Trp Gln Val Leu
145 150 155 160
Lys Glu Asn Gly Pro Met Ala Ser Asp Pro Leu Cys Leu Thr Tyr Ser
165 170 175
Tyr Leu Ser His Val Asp Leu Val Lys Asp Leu Asn Ser Gly Leu Ile
180 185 190
Gly Ala Leu Leu Val Cys Arg Glu Gly Ser Leu Ala Lys Glu Lys Thr
195 200 205
Gln Thr Leu His Lys Phe Ile Leu Leu Phe Ala Val Phe Asp Glu Gly
210 215 220
Lys Ser Trp His Ser Glu Thr Lys Asn Ser Leu Met Gln Asp Arg Asp
225 230 235 240
Ala Ala Ser Ala Arg Ala Trp Pro Lys Met His Thr Val Asn Gly Tyr
245 250 255
Val Asn Arg Ser Leu Pro Gly Leu Ile Gly Cys His Arg Lys Ser Val
260 265 270
Tyr Trp His Val Ile Gly Met Gly Thr Thr Pro Glu Val His Ser Ile
275 280 285
Phe Leu Glu Gly His Thr Phe Leu Val Arg Asn His Arg Gln Ala Ser
290 295 300
Leu Glu Ile Ser Pro Ile Thr Phe Leu Thr Ala Gln Thr Leu Leu Met
305 310 315 320
Asp Leu Gly Gln Phe Leu Leu Phe Cys His Ile Ser Ser His Gln His
325 330 335
Asp Gly Met Glu Ala Tyr Val Lys Val Asp Ser Cys Pro Glu Glu Pro
340 345 350
Gln Leu Arg Met Lys Asn Asn Glu Glu Ala Glu Asp Tyr Asp Asp Asp
355 360 365
Leu Thr Asp Ser Glu Met Asp Val Val Arg Phe Asp Asp Asp Asn Ser
370 375 380
Pro Ser Phe Ile Gln Ile Arg Ser Val Ala Lys Lys His Pro Lys Thr
385 390 395 400
Trp Val His Tyr Ile Ala Ala Glu Glu Glu Asp Trp Asp Tyr Ala Pro
405 410 415
Leu Val Leu Ala Pro Asp Asp Arg Ser Tyr Lys Ser Gln Tyr Leu Asn
420 425 430
Asn Gly Pro Gln Arg Ile Gly Arg Lys Tyr Lys Lys Val Arg Phe Met
435 440 445
Ala Tyr Thr Asp Glu Thr Phe Lys Thr Arg Glu Ala Ile Gln His Glu
450 455 460
Ser Gly Ile Leu Gly Pro Leu Leu Tyr Gly Glu Val Gly Asp Thr Leu
465 470 475 480
Leu Ile Ile Phe Lys Asn Gln Ala Ser Arg Pro Tyr Asn Ile Tyr Pro
485 490 495
His Gly Ile Thr Asp Val Arg Pro Leu Tyr Ser Arg Arg Leu Pro Lys
500 505 510
Gly Val Lys His Leu Lys Asp Phe Pro Ile Leu Pro Gly Glu Ile Phe
515 520 525
Lys Tyr Lys Trp Thr Val Thr Val Glu Asp Gly Pro Thr Lys Ser Asp
530 535 540
Pro Arg Cys Leu Thr Arg Tyr Tyr Ser Ser Phe Val Asn Met Glu Arg
545 550 555 560
Asp Leu Ala Ser Gly Leu Ile Gly Pro Leu Leu Ile Cys Tyr Lys Glu
565 570 575
Ser Val Asp Gln Arg Gly Asn Gln Ile Met Ser Asp Lys Arg Asn Val
580 585 590
Ile Leu Phe Ser Val Phe Asp Glu Asn Arg Ser Trp Tyr Leu Thr Glu
595 600 605
Asn Ile Gln Arg Phe Leu Pro Asn Pro Ala Gly Val Gln Leu Glu Asp
610 615 620
Pro Glu Phe Gln Ala Ser Asn Ile Met His Ser Ile Asn Gly Tyr Val
625 630 635 640
Phe Asp Ser Leu Gln Leu Ser Val Cys Leu His Glu Val Ala Tyr Trp
645 650 655
Tyr Ile Leu Ser Ile Gly Ala Gln Thr Asp Phe Leu Ser Val Phe Phe
660 665 670
Ser Gly Tyr Thr Phe Lys His Lys Met Val Tyr Glu Asp Thr Leu Thr
675 680 685
Leu Phe Pro Phe Ser Gly Glu Thr Val Phe Met Ser Met Glu Asn Pro
690 695 700
Gly Leu Trp Ile Leu Gly Cys His Asn Ser Asp Phe Arg Asn Arg Gly
705 710 715 720
Met Thr Ala Leu Leu Lys Val Ser Ser Cys Asp Lys Asn Thr Gly Asp
725 730 735
Tyr Tyr Glu Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys
740 745 750
Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser Gln Asn Pro Pro Val Leu
755 760 765
Lys Arg His Gln Arg Glu Ile Thr Arg Thr Thr Leu Gln Ser Asp Gln
770 775 780
Glu Glu Ile Asp Tyr Asp Asp Thr Ile Ser Val Glu Met Lys Lys Glu
785 790 795 800
Asp Phe Asp Ile Tyr Asp Glu Asp Glu Asn Gln Ser Pro Arg Ser Phe
805 810 815
Gln Lys Lys Thr Arg His Tyr Phe Ile Ala Ala Val Glu Arg Leu Trp
820 825 830
Asp Tyr Gly Met Ser Ser Ser Pro His Val Leu Arg Asn Arg Ala Gln
835 840 845
Ser Gly Ser Val Pro Gln Phe Lys Lys Val Val Phe Gln Glu Phe Thr
850 855 860
Asp Gly Ser Phe Thr Gln Pro Leu Tyr Arg Gly Glu Leu Asn Glu His
865 870 875 880
Leu Gly Leu Leu Gly Pro Tyr Ile Arg Ala Glu Val Glu Asp Asn Ile
885 890 895
Met Val Thr Phe Arg Asn Gln Ala Ser Arg Pro Tyr Ser Phe Tyr Ser
900 905 910
Ser Leu Ile Ser Tyr Glu Glu Asp Gln Arg Gln Gly Ala Glu Pro Arg
915 920 925
Lys Asn Phe Val Lys Pro Asn Glu Thr Lys Thr Tyr Phe Trp Lys Val
930 935 940
Gln His His Met Ala Pro Thr Lys Asp Glu Phe Asp Cys Lys Ala Trp
945 950 955 960
Ala Tyr Phe Ser Asp Val Asp Leu Glu Lys Asp Val His Ser Gly Leu
965 970 975
Ile Gly Pro Leu Leu Val Cys His Thr Asn Thr Leu Asn Pro Ala His
980 985 990
Gly Arg Gln Val Thr Val Gln Glu Phe Ala Leu Phe Phe Thr Ile Phe
995 1000 1005
Asp Glu Thr Lys Ser Trp Tyr Phe Thr Glu Asn Met Glu Arg Asn
1010 1015 1020
Cys Arg Ala Pro Cys Asn Ile Gln Met Glu Asp Pro Thr Phe Lys
1025 1030 1035
Glu Asn Tyr Arg Phe His Ala Ile Asn Gly Tyr Ile Met Asp Thr
1040 1045 1050
Leu Pro Gly Leu Val Met Ala Gln Asp Gln Arg Ile Arg Trp Tyr
1055 1060 1065
Leu Leu Ser Met Gly Ser Asn Glu Asn Ile His Ser Ile His Phe
1070 1075 1080
Ser Gly His Val Phe Thr Val Arg Lys Lys Glu Glu Tyr Lys Met
1085 1090 1095
Ala Leu Tyr Asn Leu Tyr Pro Gly Val Phe Glu Thr Val Glu Met
1100 1105 1110
Leu Pro Ser Lys Ala Gly Ile Trp Arg Val Glu Cys Leu Ile Gly
1115 1120 1125
Glu His Leu His Ala Gly Met Ser Thr Leu Phe Leu Val Tyr Ser
1130 1135 1140
Asn Lys Cys Gln Thr Pro Leu Gly Met Ala Ser Gly His Ile Arg
1145 1150 1155
Asp Phe Gln Ile Thr Ala Ser Gly Gln Tyr Gly Gln Trp Ala Pro
1160 1165 1170
Lys Leu Ala Arg Leu His Tyr Ser Gly Ser Ile Asn Ala Trp Ser
1175 1180 1185
Thr Lys Glu Pro Phe Ser Trp Ile Lys Val Asp Leu Leu Ala Pro
1190 1195 1200
Met Ile Ile His Gly Ile Lys Thr Gln Gly Ala Arg Gln Lys Phe
1205 1210 1215
Ser Ser Leu Tyr Ile Ser Gln Phe Ile Ile Met Tyr Ser Leu Asp
1220 1225 1230
Gly Lys Lys Trp Gln Thr Tyr Arg Gly Asn Ser Thr Gly Thr Leu
1235 1240 1245
Met Val Phe Phe Gly Asn Val Asp Ser Ser Gly Ile Lys His Asn
1250 1255 1260
Ile Phe Asn Pro Pro Ile Ile Ala Arg Tyr Ile Arg Leu His Pro
1265 1270 1275
Thr His Tyr Ser Ile Arg Ser Thr Leu Arg Met Glu Leu Met Gly
1280 1285 1290
Cys Asp Leu Asn Ser Cys Ser Met Pro Leu Gly Met Glu Ser Lys
1295 1300 1305
Ala Ile Ser Asp Ala Gln Ile Thr Ala Ser Ser Tyr Phe Thr Asn
1310 1315 1320
Met Phe Ala Thr Trp Ser Pro Ser Lys Ala Arg Leu His Leu Gln
1325 1330 1335
Gly Arg Ser Asn Ala Trp Arg Pro Gln Val Asn Asn Pro Lys Glu
1340 1345 1350
Trp Leu Gln Val Asp Phe Gln Lys Thr Met Lys Val Thr Gly Val
1355 1360 1365
Thr Thr Gln Gly Val Lys Ser Leu Leu Thr Ser Met Tyr Val Lys
1370 1375 1380
Glu Phe Leu Ile Ser Ser Ser Gln Asp Gly His Gln Trp Thr Leu
1385 1390 1395
Phe Phe Gln Asn Gly Lys Val Lys Val Phe Gln Gly Asn Gln Asp
1400 1405 1410
Ser Phe Thr Pro Val Val Asn Ser Leu Asp Pro Pro Leu Leu Thr
1415 1420 1425
Arg Tyr Leu Arg Ile His Pro Gln Ser Trp Val His Gln Ile Ala
1430 1435 1440
Leu Arg Met Glu Val Leu Gly Cys Glu Ala Gln Asp Leu Tyr
1445 1450 1455
<210> 4
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> сконструированная последовательность
<400> 4
tgtttgctgcttgcaatgtttgcccattttaggg 34
<210> 5
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> сконструированная последовательность
<400> 5
ctacctcgtgatcgcccggcccctgttcaaacatgtcctaatactctgtctctgcaaggg 60
tcatcagtagttttccatcttactcaacatcctcccagtg 100
<210> 6
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> сконструированная последовательность
<400> 6
aggttaatttttaaactgtttgctctggttaataatctcagg 42
<210> 7
<211> 190
<212> ДНК
<213> Искусственная последовательность
<220>
<223> сконструированная последовательность
<400> 7
atttcatagaacgaatgttccgatgctctaatctctctagacaaggttcatatttgtatg 60
ggttacttattctctctttgttgactaagtcaataatcagaatcagcaggtttgcagtca 120
gattggcagggataagcagcctagctcaggagaagtgagtataaaagccccaggctggga 180
gcagccatca 190
<210> 8
<211> 176
<212> ДНК
<213> Искусственная последовательность
<220>
<223> сконструированная последовательность
<400> 8
actcaaagttcaaaccttatcattttttgctttgttcctcttggccttggttttgtacat 60
cagctttgaaaataccatcccagggttaatgctggggttaatttataactaagagtgctc 120
tagttttgcaatacaggacatgctataaaaatggaaagatgttgctttctgagaga 176
<210> 9
<211> 218
<212> ДНК
<213> Искусственная последовательность
<220>
<223> сконструированная последовательность
<400> 9
tggacacaggacgctgtggtttctgagccagggggcgactcagatcccagccagtggact 60
tagcccctgtttgctcctccgataactggggtgaccttggttaatattcaccagcagcct 120
cccccgttgcccctctggatccactgcttaaatacggacgaggacagggccctgtctcct 180
cagcttcaggcaccaccactgacctgggacagtgaata 218
<210> 10
<211> 75
<212> ДНК
<213> Искусственная последовательность
<220>
<223> сконструированная последовательность
<400> 10
aataaagtctgagtgggcggcagcctgtgtgtgcctgggttctctctgtcccggaatgtg 60
caaacaatggaggtg 75
<210> 11
<211> 168
<212> ДНК
<213> Искусственная последовательность
<220>
<223> сконструированная последовательность
<400> 11
ctgcgcgctcgctcgctcactgaggccgcccgggcaaagcccgggcgtcgggcgaccttt 60
ggtcgcccggcctcagtgagcgagcgagcgcgcagagagggagtggccaactccatcact 120
aggggttccttgtagttaatgattaacccgccatgctacttatctact 168
<210> 12
<211> 164
<212> ДНК
<213> Искусственная последовательность
<220>
<223> сконструированная последовательность
<400> 12
gataagtagcatggcgggttaatcattaactacaaggaacccctagtgatggagttggcc 60
actccctctctgcgcgctcgctcgctcactgaggccgggcgaccaaaggtcgcccgacgc 120
ccgggctttgcccgggcggcctcagtgagcgagcgagcgcgcag 164
<210> 13
<211> 7920
<212> ДНК
<213> Искусственная последовательность
<220>
<223> сконструированная последовательность
<400> 13
ctgcgcgctcgctcgctcactgaggccgcccgggcaaagcccgggcgtcgggcgaccttt 60
ggtcgcccggcctcagtgagcgagcgagcgcgcagagagggagtggccaactccatcact 120
aggggttccttgtagttaatgattaacccgccatgctacttatctacttaagctacctcg 180
tgatcgcccggcccctgttcaaacatgtcctaatactctgtctctgcaagggtcatcagt 240
agttttccatcttactcaacatcctcccagtggaattcatttcatagaacgaatgttccg 300
atgctctaatctctctagacaaggttcatatttgtatgggttacttattctctctttgtt 360
gactaagtcaataatcagaatcagcaggtttgcagtcagattggcagggataagcagcct 420
agctcaggagaagtgagtataaaagccccaggctgggagcagccatcagcggccgccacc 480
atgcagatcgagctgagcacctgcttcttcctgtgcctgctgcggttctgcttctccgcc 540
acccggcggtactacctgggagccgtggagctgagctgggattacatgcagagcgatctg 600
ggagagctgccagtggatgcccggttcccaccacgggtgccaaagagcttcccattcaac 660
accagcgtggtgtacaagaagaccctgttcgtggagttcaccgatcacctgttcaacatc 720
gccaagccacggccaccctggatgggactgctgggaccaaccatccaggccgaggtgtac 780
gataccgtggtgatcaccctgaagaacatggcctctcatcctgtgtccctgcacgccgtg 840
ggagtgagctactggaaggccagcgagggagccgagtacgatgatcagaccagccagcgg 900
gagaaggaggatgataaggtgttcccaggaggaagccacacctacgtgtggcaggtgctg 960
aaggagaacggaccaatggccagcgatccactgtgcctgacctacagctacctgagccac 1020
gtggatctggtgaaggatctgaacagcggactgatcggagccctgctggtgtgccgggag 1080
ggaagcctggccaaggagaagacccagaccctgcacaagttcatcctgctgttcgccgtg 1140
ttcgatgagggaaagagctggcacagcgagaccaagaacagcctgatgcaggatcgggat 1200
gccgccagcgcccgggcctggccaaagatgcacaccgtgaacggatacgtgaaccggagc 1260
ctgccaggactgatcggatgccaccggaagagcgtgtactggcacgtgatcggaatggga 1320
accaccccagaggtgcactctatcttcctggagggacacacctttctggtgcggaaccac 1380
cggcaggccagcctggagatcagcccaatcaccttcctgaccgcccagaccctgctgatg 1440
gatctgggacagttcctgctgttctgccatatcagcagccaccagcacgatggaatggag 1500
gcctacgtgaaggtggatagctgcccagaggagccacagctgcggatgaagaacaacgag 1560
gaggccgaggattacgatgatgatctgaccgatagcgagatggatgtggtgcggttcgat 1620
gatgataacagcccaagcttcatccagatccggagcgtggccaagaagcacccaaagacc 1680
tgggtgcactacatcgccgccgaggaggaggattgggattacgccccactggtgctggcc 1740
cctgatgatcggagctacaagagccagtacctgaacaacggaccacagcggatcggacgg 1800
aagtacaaaaaagtgcggttcatggcctacaccgatgagaccttcaagacccgggaggcc 1860
atccagcacgagagcggaatcctgggaccactgctgtacggagaggtgggagataccctg 1920
ctgatcatcttcaagaaccaggccagccggccatacaacatctacccacacggaatcacc 1980
gatgtgcggccactgtacagccggcggctgccaaagggagtgaagcacctgaaggatttc 2040
ccaatcctgccaggagagatcttcaagtacaagtggacagtgacagtggaggatggacca 2100
accaagtctgatccaagatgcctgaccagatactacagcagctttgtgaacatggagaga 2160
gacctggcctctggactgattggaccactgctgatctgctacaaggagtctgtggatcag 2220
agaggaaaccagatcatgtctgataagagaaatgtgatcctgttctctgtgtttgatgag 2280
aacagaagctggtacctgacagagaacatccagagattcctgccaaacccagccggagtg 2340
cagctggaggatccagagttccaggccagcaacatcatgcacagcatcaacggatacgtg 2400
ttcgatagcctgcagctgagcgtgtgcctgcacgaggtggcctattggtatatcctgagc 2460
atcggagcccagaccgatttcctgagcgtgttcttcagcggatacaccttcaagcacaag 2520
atggtgtacgaggataccctgaccctgttcccattctccggagagaccgtgttcatgagc 2580
atggagaacccaggactgtggatcctgggatgccacaactctgatttcagaaacagagga 2640
atgactgccctgctgaaagtgtccagctgtgataagaacactggagattactatgaggat 2700
agctatgaggatatctctgcctacctgctgagcaagaacaatgccattgagccaagaagc 2760
ttcagccagaacccaccagtgctgaagagacaccagagagagatcaccagaaccaccctg 2820
cagtctgatcaggaggagattgattatgatgataccatctctgtggagatgaagaaggag 2880
gattttgatatctatgatgaggatgagaaccagagcccaagaagcttccagaagaagacc 2940
agacactacttcatcgctgcagtggagagactgtgggattatggaatgagcagcagccca 3000
cacgtgctgagaaacagagcccagagcggatctgtgccacagttcaagaaggtggtgttc 3060
caggagttcaccgatggaagcttcacccagccactgtaccggggagagctgaacgagcac 3120
ctgggactgctgggaccatacatccgggccgaggtggaggataacatcatggtgaccttc 3180
cggaaccaggccagccggccatacagcttctacagcagcctgatcagctacgaggaggat 3240
cagcggcagggagccgagccacggaagaacttcgtgaagccaaacgagaccaagacctac 3300
ttctggaaggtgcagcaccacatggccccaaccaaggatgagttcgattgcaaggcctgg 3360
gcctacttcagcgatgtggatctggagaaggatgtgcacagcggactgatcggaccactg 3420
ctggtgtgccacaccaacaccctgaacccagcccacggacggcaggtgaccgtgcaggag 3480
ttcgccctgttcttcaccatcttcgatgagaccaagagctggtacttcaccgagaacatg 3540
gagcggaactgccgggccccttgcaacatccagatggaggatccaaccttcaaggagaac 3600
taccggttccacgccatcaacggatacatcatggataccctgccaggactggtgatggcc 3660
caggatcagcggatccggtggtacctgctgagcatgggaagcaacgagaacatccacagc 3720
atccacttcagcggacacgtgttcaccgtgcggaagaaggaggagtacaagatggccctg 3780
tacaacctgtacccaggagtgttcgagaccgtggagatgctgccaagcaaggccggaatc 3840
tggcgggtggagtgcctgatcggagagcacctgcacgccggaatgagcaccctgttcctg 3900
gtgtacagcaacaagtgccagaccccactgggaatggccagcggacacatccgggatttc 3960
cagatcaccgccagcggacagtacggacagtgggccccaaagctggcccggctgcactac 4020
agcggaagcatcaacgcctggagcaccaaggagccattcagctggatcaaagtggatctg 4080
ctggccccaatgatcatccacggaatcaagacccagggagcccggcagaagttcagcagc 4140
ctgtacatcagccagttcatcatcatgtacagcctggatggaaagaagtggcagacctac 4200
cggggaaacagcaccggaaccctgatggtgttcttcggaaacgtggatagcagcggaatc 4260
aagcacaacatcttcaacccaccaatcatcgcccgatacatccggctgcacccaacccac 4320
tacagcatcagaagcaccctgcggatggagctgatgggatgtgatctgaacagctgctcc 4380
atgccactgggaatggagagcaaggccatcagcgatgcccagatcaccgccagcagctac 4440
ttcaccaacatgttcgccacctggagcccaagcaaggcccggctgcacctgcagggacgg 4500
agcaacgcctggcggccacaggtgaataacccaaaggagtggctgcaggtggatttccag 4560
aagaccatgaaggtgaccggagtgaccacccagggagtgaagagcctgctgactagcatg 4620
tatgtgaaggagttcctgatcagcagcagccaggatggacaccagtggaccctgttcttc 4680
cagaacggaaaggtgaaggtgttccagggaaaccaggatagcttcaccccagtggtgaac 4740
agcctggatccaccactgctgacccgatacctgcggatccacccacagagctgggtgcac 4800
cagatcgccctgagaatggaggtgctgggatgcgaggcccaggatctgtactgatgagca 4860
tgcaataaagtctgagtgggcggcagcctgtgtgtgcctgggttctctctgtcccggaat 4920
gtgcaaacaatggaggtgctcgagtagataagtagcatggcgggttaatcattaactaca 4980
aggaacccctagtgatggagttggccactccctctctgcgcgctcgctcgctcactgagg 5040
ccgggcgaccaaaggtcgcccgacgcccgggctttgcccgggcggcctcagtgagcgagc 5100
gagcgcgcagccttaattaacctaattcactggccgtcgttttacaacgtcgtgactggg 5160
aaaaccctggcgttacccaacttaatcgccttgcagcacatccccctttcgccagctggc 5220
gtaatagcgaagaggcccgcaccgatcgcccttcccaacagttgcgcagcctgaatggcg 5280
aatgggacgcgccctgtagcggcgcattaagcgcggcgggtgtggtggttacgcgcagcg 5340
tgaccgctacacttgccagcgccctagcgcccgctcctttcgctttcttcccttcctttc 5400
tcgccacgttcgccggctttccccgtcaagctctaaatcgggggctccctttagggttcc 5460
gatttagtgctttacggcacctcgaccccaaaaaacttgattagggtgatggttcacgta 5520
gtgggccatcgccctgatagacggtttttcgccctttgacgttggagtccacgttcttta 5580
atagtggactcttgttccaaactggaacaacactcaaccctatctcggtctattcttttg 5640
atttataagggattttgccgatttcggcctattggttaaaaaatgagctgatttaacaaa 5700
aatttaacgcgaattttaacaaaatattaacgcttacaatttaggtggcacttttcgggg 5760
aaatgtgcgcggaacccctatttgtttatttttctaaatacattcaaatatgtatccgct 5820
catgagacaataaccctgataaatgcttcaataatattgaaaaaggaagagtatgagtat 5880
tcaacatttccgtgtcgcccttattcccttttttgcggcattttgccttcctgtttttgc 5940
tcacccagaaacgctggtgaaagtaaaagatgctgaagatcagttgggtgcacgagtggg 6000
ttacatcgaactggatctcaacagcggtaagatccttgagagttttcgccccgaagaacg 6060
ttttccaatgatgagcacttttaaagttctgctatgtggcgcggtattatcccgtattga 6120
cgccgggcaagagcaactcggtcgccgcatacactattctcagaatgacttggttgagta 6180
ctcaccagtcacagaaaagcatcttacggatggcatgacagtaagagaattatgcagtgc 6240
tgccataaccatgagtgataacactgcggccaacttacttctgacaacgatcggaggacc 6300
gaaggagctaaccgcttttttgcacaacatgggggatcatgtaactcgccttgatcgttg 6360
ggaaccggagctgaatgaagccataccaaacgacgagcgtgacaccacgatgcctgtagc 6420
aatggcaacaacgttgcgcaaactattaactggcgaactacttactctagcttcccggca 6480
acaattaatagactggatggaggcggataaagttgcaggaccacttctgcgctcggccct 6540
tccggctggctggtttattgctgataaatctggagccggtgagcgtgggtctcgcggtat 6600
cattgcagcactggggccagatggtaagccctcccgtatcgtagttatctacacgacggg 6660
gagtcaggcaactatggatgaacgaaatagacagatcgctgagataggtgcctcactgat 6720
taagcattggtaactgtcagaccaagtttactcatatatactttagattgatttaaaact 6780
tcatttttaatttaaaaggatctaggtgaagatcctttttgataatctcatgaccaaaat 6840
cccttaacgtgagttttcgttccactgagcgtcagaccccgtagaaaagatcaaaggatc 6900
ttcttgagatcctttttttctgcgcgtaatctgctgcttgcaaacaaaaaaaccaccgct 6960
accagcggtggtttgtttgccggatcaagagctaccaactctttttccgaaggtaactgg 7020
cttcagcagagcgcagataccaaatactgttcttctagtgtagccgtagttaggccacca 7080
cttcaagaactctgtagcaccgcctacatacctcgctctgctaatcctgttaccagtggc 7140
tgctgccagtggcgataagtcgtgtcttaccgggttggactcaagacgatagttaccgga 7200
taaggcgcagcggtcgggctgaacggggggttcgtgcacacagcccagcttggagcgaac 7260
gacctacaccgaactgagatacctacagcgtgagctatgagaaagcgccacgcttcccga 7320
agggagaaaggcggacaggtatccggtaagcggcagggtcggaacaggagagcgcacgag 7380
ggagcttccagggggaaacgcctggtatctttatagtcctgtcgggtttcgccacctctg 7440
acttgagcgtcgatttttgtgatgctcgtcaggggggcggagcctatggaaaaacgccag 7500
caacgcggcctttttacggttcctggccttttgctggccttttgctcacatgttctttcc 7560
tgcgttatcccctgattctgtggataaccgtattaccgcctttgagtgagctgataccgc 7620
tcgccgcagccgaacgaccgagcgcagcgagtcagtgagcgaggaagcggaagagcgccc 7680
aatacgcaaaccgcctctccccgcgcgttggccgattcattaatgcagctggcacgacag 7740
gtttcccgactggaaagcgggcagtgagcgcaacgcaattaatgtgagttagctcactca 7800
ttaggcaccccaggctttacactttatgcttccggctcgtatgttgtgtggaattgtgag 7860
cggataacaatttcacacaggaaacagctatgaccatgattacgccagatttaattaagg 7920
<210> 14
<211> 8004
<212> ДНК
<213> Искусственная последовательность
<220>
<223> сконструированная последовательность
<400> 14
ctgcgcgctcgctcgctcactgaggccgcccgggcaaagcccgggcgtcgggcgaccttt 60
ggtcgcccggcctcagtgagcgagcgagcgcgcagagagggagtggccaactccatcact 120
aggggttccttgtagttaatgattaacccgccatgctacttatctacttaagctacctcg 180
tgatcgcccggcccctgttcaaacatgtcctaatactctgtctctgcaagggtcatcagt 240
agttttccatcttactcaacatcctcccagtgaggttaatttttaaactgtttgctctgg 300
ttaataatctcaggaggttaatttttaaactgtttgctctggttaataatctcagggaat 360
tcatttcatagaacgaatgttccgatgctctaatctctctagacaaggttcatatttgta 420
tgggttacttattctctctttgttgactaagtcaataatcagaatcagcaggtttgcagt 480
cagattggcagggataagcagcctagctcaggagaagtgagtataaaagccccaggctgg 540
gagcagccatcagcggccgccaccatgcagatcgagctgagcacctgcttcttcctgtgc 600
ctgctgcggttctgcttctccgccacccggcggtactacctgggagccgtggagctgagc 660
tgggattacatgcagagcgatctgggagagctgccagtggatgcccggttcccaccacgg 720
gtgccaaagagcttcccattcaacaccagcgtggtgtacaagaagaccctgttcgtggag 780
ttcaccgatcacctgttcaacatcgccaagccacggccaccctggatgggactgctggga 840
ccaaccatccaggccgaggtgtacgataccgtggtgatcaccctgaagaacatggcctct 900
catcctgtgtccctgcacgccgtgggagtgagctactggaaggccagcgagggagccgag 960
tacgatgatcagaccagccagcgggagaaggaggatgataaggtgttcccaggaggaagc 1020
cacacctacgtgtggcaggtgctgaaggagaacggaccaatggccagcgatccactgtgc 1080
ctgacctacagctacctgagccacgtggatctggtgaaggatctgaacagcggactgatc 1140
ggagccctgctggtgtgccgggagggaagcctggccaaggagaagacccagaccctgcac 1200
aagttcatcctgctgttcgccgtgttcgatgagggaaagagctggcacagcgagaccaag 1260
aacagcctgatgcaggatcgggatgccgccagcgcccgggcctggccaaagatgcacacc 1320
gtgaacggatacgtgaaccggagcctgccaggactgatcggatgccaccggaagagcgtg 1380
tactggcacgtgatcggaatgggaaccaccccagaggtgcactctatcttcctggaggga 1440
cacacctttctggtgcggaaccaccggcaggccagcctggagatcagcccaatcaccttc 1500
ctgaccgcccagaccctgctgatggatctgggacagttcctgctgttctgccatatcagc 1560
agccaccagcacgatggaatggaggcctacgtgaaggtggatagctgcccagaggagcca 1620
cagctgcggatgaagaacaacgaggaggccgaggattacgatgatgatctgaccgatagc 1680
gagatggatgtggtgcggttcgatgatgataacagcccaagcttcatccagatccggagc 1740
gtggccaagaagcacccaaagacctgggtgcactacatcgccgccgaggaggaggattgg 1800
gattacgccccactggtgctggcccctgatgatcggagctacaagagccagtacctgaac 1860
aacggaccacagcggatcggacggaagtacaaaaaagtgcggttcatggcctacaccgat 1920
gagaccttcaagacccgggaggccatccagcacgagagcggaatcctgggaccactgctg 1980
tacggagaggtgggagataccctgctgatcatcttcaagaaccaggccagccggccatac 2040
aacatctacccacacggaatcaccgatgtgcggccactgtacagccggcggctgccaaag 2100
ggagtgaagcacctgaaggatttcccaatcctgccaggagagatcttcaagtacaagtgg 2160
acagtgacagtggaggatggaccaaccaagtctgatccaagatgcctgaccagatactac 2220
agcagctttgtgaacatggagagagacctggcctctggactgattggaccactgctgatc 2280
tgctacaaggagtctgtggatcagagaggaaaccagatcatgtctgataagagaaatgtg 2340
atcctgttctctgtgtttgatgagaacagaagctggtacctgacagagaacatccagaga 2400
ttcctgccaaacccagccggagtgcagctggaggatccagagttccaggccagcaacatc 2460
atgcacagcatcaacggatacgtgttcgatagcctgcagctgagcgtgtgcctgcacgag 2520
gtggcctattggtatatcctgagcatcggagcccagaccgatttcctgagcgtgttcttc 2580
agcggatacaccttcaagcacaagatggtgtacgaggataccctgaccctgttcccattc 2640
tccggagagaccgtgttcatgagcatggagaacccaggactgtggatcctgggatgccac 2700
aactctgatttcagaaacagaggaatgactgccctgctgaaagtgtccagctgtgataag 2760
aacactggagattactatgaggatagctatgaggatatctctgcctacctgctgagcaag 2820
aacaatgccattgagccaagaagcttcagccagaacccaccagtgctgaagagacaccag 2880
agagagatcaccagaaccaccctgcagtctgatcaggaggagattgattatgatgatacc 2940
atctctgtggagatgaagaaggaggattttgatatctatgatgaggatgagaaccagagc 3000
ccaagaagcttccagaagaagaccagacactacttcatcgctgcagtggagagactgtgg 3060
gattatggaatgagcagcagcccacacgtgctgagaaacagagcccagagcggatctgtg 3120
ccacagttcaagaaggtggtgttccaggagttcaccgatggaagcttcacccagccactg 3180
taccggggagagctgaacgagcacctgggactgctgggaccatacatccgggccgaggtg 3240
gaggataacatcatggtgaccttccggaaccaggccagccggccatacagcttctacagc 3300
agcctgatcagctacgaggaggatcagcggcagggagccgagccacggaagaacttcgtg 3360
aagccaaacgagaccaagacctacttctggaaggtgcagcaccacatggccccaaccaag 3420
gatgagttcgattgcaaggcctgggcctacttcagcgatgtggatctggagaaggatgtg 3480
cacagcggactgatcggaccactgctggtgtgccacaccaacaccctgaacccagcccac 3540
ggacggcaggtgaccgtgcaggagttcgccctgttcttcaccatcttcgatgagaccaag 3600
agctggtacttcaccgagaacatggagcggaactgccgggccccttgcaacatccagatg 3660
gaggatccaaccttcaaggagaactaccggttccacgccatcaacggatacatcatggat 3720
accctgccaggactggtgatggcccaggatcagcggatccggtggtacctgctgagcatg 3780
ggaagcaacgagaacatccacagcatccacttcagcggacacgtgttcaccgtgcggaag 3840
aaggaggagtacaagatggccctgtacaacctgtacccaggagtgttcgagaccgtggag 3900
atgctgccaagcaaggccggaatctggcgggtggagtgcctgatcggagagcacctgcac 3960
gccggaatgagcaccctgttcctggtgtacagcaacaagtgccagaccccactgggaatg 4020
gccagcggacacatccgggatttccagatcaccgccagcggacagtacggacagtgggcc 4080
ccaaagctggcccggctgcactacagcggaagcatcaacgcctggagcaccaaggagcca 4140
ttcagctggatcaaagtggatctgctggccccaatgatcatccacggaatcaagacccag 4200
ggagcccggcagaagttcagcagcctgtacatcagccagttcatcatcatgtacagcctg 4260
gatggaaagaagtggcagacctaccggggaaacagcaccggaaccctgatggtgttcttc 4320
ggaaacgtggatagcagcggaatcaagcacaacatcttcaacccaccaatcatcgcccga 4380
tacatccggctgcacccaacccactacagcatcagaagcaccctgcggatggagctgatg 4440
ggatgtgatctgaacagctgctccatgccactgggaatggagagcaaggccatcagcgat 4500
gcccagatcaccgccagcagctacttcaccaacatgttcgccacctggagcccaagcaag 4560
gcccggctgcacctgcagggacggagcaacgcctggcggccacaggtgaataacccaaag 4620
gagtggctgcaggtggatttccagaagaccatgaaggtgaccggagtgaccacccaggga 4680
gtgaagagcctgctgactagcatgtatgtgaaggagttcctgatcagcagcagccaggat 4740
ggacaccagtggaccctgttcttccagaacggaaaggtgaaggtgttccagggaaaccag 4800
gatagcttcaccccagtggtgaacagcctggatccaccactgctgacccgatacctgcgg 4860
atccacccacagagctgggtgcaccagatcgccctgagaatggaggtgctgggatgcgag 4920
gcccaggatctgtactgatgagcatgcaataaagtctgagtgggcggcagcctgtgtgtg 4980
cctgggttctctctgtcccggaatgtgcaaacaatggaggtgctcgagtagataagtagc 5040
atggcgggttaatcattaactacaaggaacccctagtgatggagttggccactccctctc 5100
tgcgcgctcgctcgctcactgaggccgggcgaccaaaggtcgcccgacgcccgggctttg 5160
cccgggcggcctcagtgagcgagcgagcgcgcagccttaattaacctaattcactggccg 5220
tcgttttacaacgtcgtgactgggaaaaccctggcgttacccaacttaatcgccttgcag 5280
cacatccccctttcgccagctggcgtaatagcgaagaggcccgcaccgatcgcccttccc 5340
aacagttgcgcagcctgaatggcgaatgggacgcgccctgtagcggcgcattaagcgcgg 5400
cgggtgtggtggttacgcgcagcgtgaccgctacacttgccagcgccctagcgcccgctc 5460
ctttcgctttcttcccttcctttctcgccacgttcgccggctttccccgtcaagctctaa 5520
atcgggggctccctttagggttccgatttagtgctttacggcacctcgaccccaaaaaac 5580
ttgattagggtgatggttcacgtagtgggccatcgccctgatagacggtttttcgccctt 5640
tgacgttggagtccacgttctttaatagtggactcttgttccaaactggaacaacactca 5700
accctatctcggtctattcttttgatttataagggattttgccgatttcggcctattggt 5760
taaaaaatgagctgatttaacaaaaatttaacgcgaattttaacaaaatattaacgctta 5820
caatttaggtggcacttttcggggaaatgtgcgcggaacccctatttgtttatttttcta 5880
aatacattcaaatatgtatccgctcatgagacaataaccctgataaatgcttcaataata 5940
ttgaaaaaggaagagtatgagtattcaacatttccgtgtcgcccttattcccttttttgc 6000
ggcattttgccttcctgtttttgctcacccagaaacgctggtgaaagtaaaagatgctga 6060
agatcagttgggtgcacgagtgggttacatcgaactggatctcaacagcggtaagatcct 6120
tgagagttttcgccccgaagaacgttttccaatgatgagcacttttaaagttctgctatg 6180
tggcgcggtattatcccgtattgacgccgggcaagagcaactcggtcgccgcatacacta 6240
ttctcagaatgacttggttgagtactcaccagtcacagaaaagcatcttacggatggcat 6300
gacagtaagagaattatgcagtgctgccataaccatgagtgataacactgcggccaactt 6360
acttctgacaacgatcggaggaccgaaggagctaaccgcttttttgcacaacatggggga 6420
tcatgtaactcgccttgatcgttgggaaccggagctgaatgaagccataccaaacgacga 6480
gcgtgacaccacgatgcctgtagcaatggcaacaacgttgcgcaaactattaactggcga 6540
actacttactctagcttcccggcaacaattaatagactggatggaggcggataaagttgc 6600
aggaccacttctgcgctcggcccttccggctggctggtttattgctgataaatctggagc 6660
cggtgagcgtgggtctcgcggtatcattgcagcactggggccagatggtaagccctcccg 6720
tatcgtagttatctacacgacggggagtcaggcaactatggatgaacgaaatagacagat 6780
cgctgagataggtgcctcactgattaagcattggtaactgtcagaccaagtttactcata 6840
tatactttagattgatttaaaacttcatttttaatttaaaaggatctaggtgaagatcct 6900
ttttgataatctcatgaccaaaatcccttaacgtgagttttcgttccactgagcgtcaga 6960
ccccgtagaaaagatcaaaggatcttcttgagatcctttttttctgcgcgtaatctgctg 7020
cttgcaaacaaaaaaaccaccgctaccagcggtggtttgtttgccggatcaagagctacc 7080
aactctttttccgaaggtaactggcttcagcagagcgcagataccaaatactgttcttct 7140
agtgtagccgtagttaggccaccacttcaagaactctgtagcaccgcctacatacctcgc 7200
tctgctaatcctgttaccagtggctgctgccagtggcgataagtcgtgtcttaccgggtt 7260
ggactcaagacgatagttaccggataaggcgcagcggtcgggctgaacggggggttcgtg 7320
cacacagcccagcttggagcgaacgacctacaccgaactgagatacctacagcgtgagct 7380
atgagaaagcgccacgcttcccgaagggagaaaggcggacaggtatccggtaagcggcag 7440
ggtcggaacaggagagcgcacgagggagcttccagggggaaacgcctggtatctttatag 7500
tcctgtcgggtttcgccacctctgacttgagcgtcgatttttgtgatgctcgtcaggggg 7560
gcggagcctatggaaaaacgccagcaacgcggcctttttacggttcctggccttttgctg 7620
gccttttgctcacatgttctttcctgcgttatcccctgattctgtggataaccgtattac 7680
cgcctttgagtgagctgataccgctcgccgcagccgaacgaccgagcgcagcgagtcagt 7740
gagcgaggaagcggaagagcgcccaatacgcaaaccgcctctccccgcgcgttggccgat 7800
tcattaatgcagctggcacgacaggtttcccgactggaaagcgggcagtgagcgcaacgc 7860
aattaatgtgagttagctcactcattaggcaccccaggctttacactttatgcttccggc 7920
tcgtatgttgtgtggaattgtgagcggataacaatttcacacaggaaacagctatgacca 7980
tgattacgccagatttaattaagg 8004
<210> 15
<211> 7948
<212> ДНК
<213> Искусственная последовательность
<220>
<223> сконструированная последовательность
<400> 15
ctgcgcgctcgctcgctcactgaggccgcccgggcaaagcccgggcgtcgggcgaccttt 60
ggtcgcccggcctcagtgagcgagcgagcgcgcagagagggagtggccaactccatcact 120
aggggttccttgtagttaatgattaacccgccatgctacttatctacttaagctacctcg 180
tgatcgcccggcccctgttcaaacatgtcctaatactctgtctctgcaagggtcatcagt 240
agttttccatcttactcaacatcctcccagtggaattctggacacaggacgctgtggttt 300
ctgagccagggggcgactcagatcccagccagtggacttagcccctgtttgctcctccga 360
taactggggtgaccttggttaatattcaccagcagcctcccccgttgcccctctggatcc 420
actgcttaaatacggacgaggacagggccctgtctcctcagcttcaggcaccaccactga 480
cctgggacagtgaatagcggccgccaccatgcagatcgagctgagcacctgcttcttcct 540
gtgcctgctgcggttctgcttctccgccacccggcggtactacctgggagccgtggagct 600
gagctgggattacatgcagagcgatctgggagagctgccagtggatgcccggttcccacc 660
acgggtgccaaagagcttcccattcaacaccagcgtggtgtacaagaagaccctgttcgt 720
ggagttcaccgatcacctgttcaacatcgccaagccacggccaccctggatgggactgct 780
gggaccaaccatccaggccgaggtgtacgataccgtggtgatcaccctgaagaacatggc 840
ctctcatcctgtgtccctgcacgccgtgggagtgagctactggaaggccagcgagggagc 900
cgagtacgatgatcagaccagccagcgggagaaggaggatgataaggtgttcccaggagg 960
aagccacacctacgtgtggcaggtgctgaaggagaacggaccaatggccagcgatccact 1020
gtgcctgacctacagctacctgagccacgtggatctggtgaaggatctgaacagcggact 1080
gatcggagccctgctggtgtgccgggagggaagcctggccaaggagaagacccagaccct 1140
gcacaagttcatcctgctgttcgccgtgttcgatgagggaaagagctggcacagcgagac 1200
caagaacagcctgatgcaggatcgggatgccgccagcgcccgggcctggccaaagatgca 1260
caccgtgaacggatacgtgaaccggagcctgccaggactgatcggatgccaccggaagag 1320
cgtgtactggcacgtgatcggaatgggaaccaccccagaggtgcactctatcttcctgga 1380
gggacacacctttctggtgcggaaccaccggcaggccagcctggagatcagcccaatcac 1440
cttcctgaccgcccagaccctgctgatggatctgggacagttcctgctgttctgccatat 1500
cagcagccaccagcacgatggaatggaggcctacgtgaaggtggatagctgcccagagga 1560
gccacagctgcggatgaagaacaacgaggaggccgaggattacgatgatgatctgaccga 1620
tagcgagatggatgtggtgcggttcgatgatgataacagcccaagcttcatccagatccg 1680
gagcgtggccaagaagcacccaaagacctgggtgcactacatcgccgccgaggaggagga 1740
ttgggattacgccccactggtgctggcccctgatgatcggagctacaagagccagtacct 1800
gaacaacggaccacagcggatcggacggaagtacaaaaaagtgcggttcatggcctacac 1860
cgatgagaccttcaagacccgggaggccatccagcacgagagcggaatcctgggaccact 1920
gctgtacggagaggtgggagataccctgctgatcatcttcaagaaccaggccagccggcc 1980
atacaacatctacccacacggaatcaccgatgtgcggccactgtacagccggcggctgcc 2040
aaagggagtgaagcacctgaaggatttcccaatcctgccaggagagatcttcaagtacaa 2100
gtggacagtgacagtggaggatggaccaaccaagtctgatccaagatgcctgaccagata 2160
ctacagcagctttgtgaacatggagagagacctggcctctggactgattggaccactgct 2220
gatctgctacaaggagtctgtggatcagagaggaaaccagatcatgtctgataagagaaa 2280
tgtgatcctgttctctgtgtttgatgagaacagaagctggtacctgacagagaacatcca 2340
gagattcctgccaaacccagccggagtgcagctggaggatccagagttccaggccagcaa 2400
catcatgcacagcatcaacggatacgtgttcgatagcctgcagctgagcgtgtgcctgca 2460
cgaggtggcctattggtatatcctgagcatcggagcccagaccgatttcctgagcgtgtt 2520
cttcagcggatacaccttcaagcacaagatggtgtacgaggataccctgaccctgttccc 2580
attctccggagagaccgtgttcatgagcatggagaacccaggactgtggatcctgggatg 2640
ccacaactctgatttcagaaacagaggaatgactgccctgctgaaagtgtccagctgtga 2700
taagaacact ggagattact atgaggatag ctatgaggat atctctgcct acctgctgag 2760
caagaacaat gccattgagc caagaagctt cagccagaac ccaccagtgc tgaagagaca 2820
ccagagagag atcaccagaa ccaccctgca gtctgatcag gaggagattg attatgatga 2880
taccatctct gtggagatga agaaggagga ttttgatatc tatgatgagg atgagaacca 2940
gagcccaaga agcttccaga agaagaccag acactacttc atcgctgcag tggagagact 3000
gtgggattat ggaatgagca gcagcccaca cgtgctgaga aacagagccc agagcggatc 3060
tgtgccacag ttcaagaagg tggtgttcca ggagttcacc gatggaagct tcacccagcc 3120
actgtaccgg ggagagctga acgagcacct gggactgctg ggaccataca tccgggccga 3180
ggtggaggat aacatcatgg tgaccttccg gaaccaggcc agccggccat acagcttcta 3240
cagcagcctg atcagctacg aggaggatca gcggcaggga gccgagccac ggaagaactt 3300
cgtgaagcca aacgagacca agacctactt ctggaaggtg cagcaccaca tggccccaac 3360
caaggatgag ttcgattgca aggcctgggc ctacttcagc gatgtggatc tggagaagga 3420
tgtgcacagc ggactgatcg gaccactgct ggtgtgccac accaacaccc tgaacccagc 3480
ccacggacgg caggtgaccg tgcaggagtt cgccctgttc ttcaccatct tcgatgagac 3540
caagagctgg tacttcaccg agaacatgga gcggaactgc cgggcccctt gcaacatcca 3600
gatggaggat ccaaccttca aggagaacta ccggttccac gccatcaacg gatacatcat 3660
ggataccctg ccaggactgg tgatggccca ggatcagcgg atccggtggt acctgctgag 3720
catgggaagc aacgagaaca tccacagcat ccacttcagc ggacacgtgt tcaccgtgcg 3780
gaagaaggag gagtacaaga tggccctgta caacctgtac ccaggagtgt tcgagaccgt 3840
ggagatgctg ccaagcaagg ccggaatctg gcgggtggag tgcctgatcg gagagcacct 3900
gcacgccgga atgagcaccc tgttcctggt gtacagcaac aagtgccaga ccccactggg 3960
aatggccagc ggacacatcc gggatttcca gatcaccgcc agcggacagt acggacagtg 4020
ggccccaaag ctggcccggc tgcactacag cggaagcatc aacgcctgga gcaccaagga 4080
gccattcagc tggatcaaag tggatctgct ggccccaatg atcatccacg gaatcaagac 4140
ccagggagcc cggcagaagt tcagcagcct gtacatcagc cagttcatca tcatgtacag 4200
cctggatgga aagaagtggc agacctaccg gggaaacagc accggaaccc tgatggtgtt 4260
cttcggaaac gtggatagca gcggaatcaa gcacaacatc ttcaacccac caatcatcgc 4320
ccgatacatc cggctgcacc caacccacta cagcatcaga agcaccctgc ggatggagct 4380
gatgggatgt gatctgaaca gctgctccat gccactggga atggagagca aggccatcag 4440
cgatgcccag atcaccgcca gcagctactt caccaacatg ttcgccacct ggagcccaag 4500
caaggcccgg ctgcacctgc agggacggag caacgcctgg cggccacagg tgaataaccc 4560
aaaggagtgg ctgcaggtgg atttccagaa gaccatgaag gtgaccggag tgaccaccca 4620
gggagtgaag agcctgctga ctagcatgta tgtgaaggag ttcctgatca gcagcagcca 4680
ggatggacac cagtggaccc tgttcttcca gaacggaaag gtgaaggtgt tccagggaaa 4740
ccaggatagc ttcaccccag tggtgaacag cctggatcca ccactgctga cccgatacct 4800
gcggatccac ccacagagct gggtgcacca gatcgccctg agaatggagg tgctgggatg 4860
cgaggcccag gatctgtact gatgagcatg caataaagtc tgagtgggcg gcagcctgtg 4920
tgtgcctggg ttctctctgt cccggaatgt gcaaacaatg gaggtgctcg agtagataag 4980
tagcatggcg ggttaatcat taactacaag gaacccctag tgatggagtt ggccactccc 5040
tctctgcgcg ctcgctcgct cactgaggcc gggcgaccaa aggtcgcccg acgcccgggc 5100
tttgcccggg cggcctcagt gagcgagcga gcgcgcagcc ttaattaacc taattcactg 5160
gccgtcgttt tacaacgtcg tgactgggaa aaccctggcg ttacccaact taatcgcctt 5220
gcagcacatc cccctttcgc cagctggcgt aatagcgaag aggcccgcac cgatcgccct 5280
tcccaacagt tgcgcagcct gaatggcgaa tgggacgcgc cctgtagcgg cgcattaagc 5340
gcggcgggtg tggtggttac gcgcagcgtg accgctacac ttgccagcgc cctagcgccc 5400
gctcctttcg ctttcttccc ttcctttctc gccacgttcg ccggctttcc ccgtcaagct 5460
ctaaatcggg ggctcccttt agggttccga tttagtgctt tacggcacct cgaccccaaa 5520
aaacttgatt agggtgatgg ttcacgtagt gggccatcgc cctgatagac ggtttttcgc 5580
cctttgacgt tggagtccac gttctttaat agtggactct tgttccaaac tggaacaaca 5640
ctcaacccta tctcggtcta ttcttttgat ttataaggga ttttgccgat ttcggcctat 5700
tggttaaaaa atgagctgat ttaacaaaaa tttaacgcga attttaacaa aatattaacg 5760
cttacaattt aggtggcact tttcggggaa atgtgcgcgg aacccctatt tgtttatttt 5820
tctaaataca ttcaaatatg tatccgctca tgagacaata accctgataa atgcttcaat 5880
aatattgaaa aaggaagagt atgagtattc aacatttccg tgtcgccctt attccctttt 5940
ttgcggcatt ttgccttcct gtttttgctc acccagaaac gctggtgaaa gtaaaagatg 6000
ctgaagatca gttgggtgca cgagtgggtt acatcgaact ggatctcaac agcggtaaga 6060
tccttgagag ttttcgcccc gaagaacgtt ttccaatgat gagcactttt aaagttctgc 6120
tatgtggcgc ggtattatcc cgtattgacg ccgggcaaga gcaactcggt cgccgcatac 6180
actattctca gaatgacttg gttgagtact caccagtcac agaaaagcat cttacggatg 6240
gcatgacagt aagagaatta tgcagtgctg ccataaccat gagtgataac actgcggcca 6300
acttacttct gacaacgatc ggaggaccga aggagctaac cgcttttttg cacaacatgg 6360
gggatcatgt aactcgcctt gatcgttggg aaccggagct gaatgaagcc ataccaaacg 6420
acgagcgtga caccacgatg cctgtagcaa tggcaacaac gttgcgcaaa ctattaactg 6480
gcgaactact tactctagct tcccggcaac aattaataga ctggatggag gcggataaag 6540
ttgcaggacc acttctgcgc tcggcccttc cggctggctg gtttattgct gataaatctg 6600
gagccggtga gcgtgggtct cgcggtatca ttgcagcact ggggccagat ggtaagccct 6660
cccgtatcgt agttatctac acgacgggga gtcaggcaac tatggatgaa cgaaatagac 6720
agatcgctga gataggtgcc tcactgatta agcattggta actgtcagac caagtttact 6780
catatatact ttagattgat ttaaaacttc atttttaatt taaaaggatc taggtgaaga 6840
tcctttttga taatctcatg accaaaatcc cttaacgtga gttttcgttc cactgagcgt 6900
cagaccccgt agaaaagatc aaaggatctt cttgagatcc tttttttctg cgcgtaatct 6960
gctgcttgca aacaaaaaaa ccaccgctac cagcggtggt ttgtttgccg gatcaagagc 7020
taccaactct ttttccgaag gtaactggct tcagcagagc gcagatacca aatactgttc 7080
ttctagtgta gccgtagtta ggccaccact tcaagaactc tgtagcaccg cctacatacc 7140
tcgctctgct aatcctgtta ccagtggctg ctgccagtgg cgataagtcg tgtcttaccg 7200
ggttggactc aagacgatag ttaccggata aggcgcagcg gtcgggctga acggggggtt 7260
cgtgcacaca gcccagcttg gagcgaacga cctacaccga actgagatac ctacagcgtg 7320
agctatgaga aagcgccacg cttcccgaag ggagaaaggc ggacaggtat ccggtaagcg 7380
gcagggtcgg aacaggagag cgcacgaggg agcttccagg gggaaacgcc tggtatcttt 7440
atagtcctgt cgggtttcgc cacctctgac ttgagcgtcg atttttgtga tgctcgtcag 7500
gggggcggag cctatggaaa aacgccagca acgcggcctt tttacggttc ctggcctttt 7560
gctggccttt tgctcacatg ttctttcctg cgttatcccc tgattctgtg gataaccgta 7620
ttaccgcctt tgagtgagct gataccgctc gccgcagccg aacgaccgag cgcagcgagt 7680
cagtgagcga ggaagcggaa gagcgcccaa tacgcaaacc gcctctcccc gcgcgttggc 7740
cgattcatta atgcagctgg cacgacaggt ttcccgactg gaaagcgggc agtgagcgca 7800
acgcaattaa tgtgagttag ctcactcatt aggcacccca ggctttacac tttatgcttc 7860
cggctcgtat gttgtgtgga attgtgagcg gataacaatt tcacacagga aacagctatg 7920
accatgattacgccagatttaattaagg 7948
<210> 16
<211> 8032
<212> ДНК
<213> Искусственная последовательность
<220>
<223> сконструированная последовательность
<400> 16
ctgcgcgctcgctcgctcactgaggccgcccgggcaaagcccgggcgtcgggcgaccttt 60
ggtcgcccggcctcagtgagcgagcgagcgcgcagagagggagtggccaactccatcact 120
aggggttccttgtagttaatgattaacccgccatgctacttatctacttaagctacctcg 180
tgatcgcccggcccctgttcaaacatgtcctaatactctgtctctgcaagggtcatcagt 240
agttttccatcttactcaacatcctcccagtgaggttaatttttaaactgtttgctctgg 300
ttaataatctcaggaggttaatttttaaactgtttgctctggttaataatctcagggaat 360
tctggacacaggacgctgtggtttctgagccagggggcgactcagatcccagccagtgga 420
cttagcccctgtttgctcctccgataactggggtgaccttggttaatattcaccagcagc 480
ctcccccgttgcccctctggatccactgcttaaatacggacgaggacagggccctgtctc 540
ctcagcttcaggcaccaccactgacctgggacagtgaatagcggccgccaccatgcagat 600
cgagctgagcacctgcttcttcctgtgcctgctgcggttctgcttctccgccacccggcg 660
gtactacctgggagccgtggagctgagctgggattacatgcagagcgatctgggagagct 720
gccagtggatgcccggttcccaccacgggtgccaaagagcttcccattcaacaccagcgt 780
ggtgtacaagaagaccctgttcgtggagttcaccgatcacctgttcaacatcgccaagcc 840
acggccaccctggatgggactgctgggaccaaccatccaggccgaggtgtacgataccgt 900
ggtgatcaccctgaagaacatggcctctcatcctgtgtccctgcacgccgtgggagtgag 960
ctactggaaggccagcgagggagccgagtacgatgatcagaccagccagcgggagaagga 1020
ggatgataaggtgttcccaggaggaagccacacctacgtgtggcaggtgctgaaggagaa 1080
cggaccaatggccagcgatccactgtgcctgacctacagctacctgagccacgtggatct 1140
ggtgaaggatctgaacagcggactgatcggagccctgctggtgtgccgggagggaagcct 1200
ggccaaggagaagacccagaccctgcacaagttcatcctgctgttcgccgtgttcgatga 1260
gggaaagagctggcacagcgagaccaagaacagcctgatgcaggatcgggatgccgccag 1320
cgcccgggcctggccaaagatgcacaccgtgaacggatacgtgaaccggagcctgccagg 1380
actgatcggatgccaccggaagagcgtgtactggcacgtgatcggaatgggaaccacccc 1440
agaggtgcactctatcttcctggagggacacacctttctggtgcggaaccaccggcaggc 1500
cagcctggagatcagcccaatcaccttcctgaccgcccagaccctgctgatggatctggg 1560
acagttcctgctgttctgccatatcagcagccaccagcacgatggaatggaggcctacgt 1620
gaaggtggatagctgcccagaggagccacagctgcggatgaagaacaacgaggaggccga 1680
ggattacgatgatgatctgaccgatagcgagatggatgtggtgcggttcgatgatgataa 1740
cagcccaagcttcatccagatccggagcgtggccaagaagcacccaaagacctgggtgca 1800
ctacatcgccgccgaggaggaggattgggattacgccccactggtgctggcccctgatga 1860
tcggagctacaagagccagtacctgaacaacggaccacagcggatcggacggaagtacaa 1920
aaaagtgcggttcatggcctacaccgatgagaccttcaagacccgggaggccatccagca 1980
cgagagcggaatcctgggaccactgctgtacggagaggtgggagataccctgctgatcat 2040
cttcaagaaccaggccagccggccatacaacatctacccacacggaatcaccgatgtgcg 2100
gccactgtacagccggcggctgccaaagggagtgaagcacctgaaggatttcccaatcct 2160
gccaggagagatcttcaagtacaagtggacagtgacagtggaggatggaccaaccaagtc 2220
tgatccaagatgcctgaccagatactacagcagctttgtgaacatggagagagacctggc 2280
ctctggactgattggaccactgctgatctgctacaaggagtctgtggatcagagaggaaa 2340
ccagatcatgtctgataagagaaatgtgatcctgttctctgtgtttgatgagaacagaag 2400
ctggtacctgacagagaacatccagagattcctgccaaacccagccggagtgcagctgga 2460
ggatccagagttccaggccagcaacatcatgcacagcatcaacggatacgtgttcgatag 2520
cctgcagctgagcgtgtgcctgcacgaggtggcctattggtatatcctgagcatcggagc 2580
ccagaccgatttcctgagcgtgttcttcagcggatacaccttcaagcacaagatggtgta 2640
cgaggataccctgaccctgttcccattctccggagagaccgtgttcatgagcatggagaa 2700
cccaggactgtggatcctgggatgccacaactctgatttcagaaacagaggaatgactgc 2760
cctgctgaaagtgtccagctgtgataagaacactggagattactatgaggatagctatga 2820
ggatatctctgcctacctgctgagcaagaacaatgccattgagccaagaagcttcagcca 2880
gaacccaccagtgctgaagagacaccagagagagatcaccagaaccaccctgcagtctga 2940
tcaggaggagattgattatgatgataccatctctgtggagatgaagaaggaggattttga 3000
tatctatgatgaggatgagaaccagagcccaagaagcttccagaagaagaccagacacta 3060
cttcatcgctgcagtggagagactgtgggattatggaatgagcagcagcccacacgtgct 3120
gagaaacagagcccagagcggatctgtgccacagttcaagaaggtggtgttccaggagtt 3180
caccgatggaagcttcacccagccactgtaccggggagagctgaacgagcacctgggact 3240
gctgggaccatacatccgggccgaggtggaggataacatcatggtgaccttccggaacca 3300
ggccagccggccatacagcttctacagcagcctgatcagctacgaggaggatcagcggca 3360
gggagccgagccacggaagaacttcgtgaagccaaacgagaccaagacctacttctggaa 3420
ggtgcagcaccacatggccccaaccaaggatgagttcgattgcaaggcctgggcctactt 3480
cagcgatgtggatctggagaaggatgtgcacagcggactgatcggaccactgctggtgtg 3540
ccacaccaacaccctgaacccagcccacggacggcaggtgaccgtgcaggagttcgccct 3600
gttcttcaccatcttcgatgagaccaagagctggtacttcaccgagaacatggagcggaa 3660
ctgccgggccccttgcaacatccagatggaggatccaaccttcaaggagaactaccggtt 3720
ccacgccatcaacggatacatcatggataccctgccaggactggtgatggcccaggatca 3780
gcggatccggtggtacctgctgagcatgggaagcaacgagaacatccacagcatccactt 3840
cagcggacacgtgttcaccgtgcggaagaaggaggagtacaagatggccctgtacaacct 3900
gtacccaggagtgttcgagaccgtggagatgctgccaagcaaggccggaatctggcgggt 3960
ggagtgcctgatcggagagcacctgcacgccggaatgagcaccctgttcctggtgtacag 4020
caacaagtgccagaccccactgggaatggccagcggacacatccgggatttccagatcac 4080
cgccagcggacagtacggacagtgggccccaaagctggcccggctgcactacagcggaag 4140
catcaacgcctggagcaccaaggagccattcagctggatcaaagtggatctgctggcccc 4200
aatgatcatccacggaatcaagacccagggagcccggcagaagttcagcagcctgtacat 4260
cagccagttcatcatcatgtacagcctggatggaaagaagtggcagacctaccggggaaa 4320
cagcaccggaaccctgatggtgttcttcggaaacgtggatagcagcggaatcaagcacaa 4380
catcttcaacccaccaatcatcgcccgatacatccggctgcacccaacccactacagcat 4440
cagaagcaccctgcggatggagctgatgggatgtgatctgaacagctgctccatgccact 4500
gggaatggagagcaaggccatcagcgatgcccagatcaccgccagcagctacttcaccaa 4560
catgttcgccacctggagcccaagcaaggcccggctgcacctgcagggacggagcaacgc 4620
ctggcggccacaggtgaataacccaaaggagtggctgcaggtggatttccagaagaccat 4680
gaaggtgaccggagtgaccacccagggagtgaagagcctgctgactagcatgtatgtgaa 4740
ggagttcctgatcagcagcagccaggatggacaccagtggaccctgttcttccagaacgg 4800
aaaggtgaaggtgttccagggaaaccaggatagcttcaccccagtggtgaacagcctgga 4860
tccaccactgctgacccgatacctgcggatccacccacagagctgggtgcaccagatcgc 4920
cctgagaatggaggtgctgggatgcgaggcccaggatctgtactgatgagcatgcaataa 4980
agtctgagtgggcggcagcctgtgtgtgcctgggttctctctgtcccggaatgtgcaaac 5040
aatggaggtgctcgagtagataagtagcatggcgggttaatcattaactacaaggaaccc 5100
ctagtgatggagttggccactccctctctgcgcgctcgctcgctcactgaggccgggcga 5160
ccaaaggtcgcccgacgcccgggctttgcccgggcggcctcagtgagcgagcgagcgcgc 5220
agccttaattaacctaattcactggccgtcgttttacaacgtcgtgactgggaaaaccct 5280
ggcgttacccaacttaatcgccttgcagcacatccccctttcgccagctggcgtaatagc 5340
gaagaggcccgcaccgatcgcccttcccaacagttgcgcagcctgaatggcgaatgggac 5400
gcgccctgtagcggcgcattaagcgcggcgggtgtggtggttacgcgcagcgtgaccgct 5460
acacttgccagcgccctagcgcccgctcctttcgctttcttcccttcctttctcgccacg 5520
ttcgccggctttccccgtcaagctctaaatcgggggctccctttagggttccgatttagt 5580
gctttacggcacctcgaccccaaaaaacttgattagggtgatggttcacgtagtgggcca 5640
tcgccctgatagacggtttttcgccctttgacgttggagtccacgttctttaatagtgga 5700
ctcttgttccaaactggaacaacactcaaccctatctcggtctattcttttgatttataa 5760
gggattttgccgatttcggcctattggttaaaaaatgagctgatttaacaaaaatttaac 5820
gcgaattttaacaaaatattaacgcttacaatttaggtggcacttttcggggaaatgtgc 5880
gcggaacccctatttgtttatttttctaaatacattcaaatatgtatccgctcatgagac 5940
aataaccctgataaatgcttcaataatattgaaaaaggaagagtatgagtattcaacatt 6000
tccgtgtcgcccttattcccttttttgcggcattttgccttcctgtttttgctcacccag 6060
aaacgctggtgaaagtaaaagatgctgaagatcagttgggtgcacgagtgggttacatcg 6120
aactggatctcaacagcggtaagatccttgagagttttcgccccgaagaacgttttccaa 6180
tgatgagcacttttaaagttctgctatgtggcgcggtattatcccgtattgacgccgggc 6240
aagagcaactcggtcgccgcatacactattctcagaatgacttggttgagtactcaccag 6300
tcacagaaaagcatcttacggatggcatgacagtaagagaattatgcagtgctgccataa 6360
ccatgagtgataacactgcggccaacttacttctgacaacgatcggaggaccgaaggagc 6420
taaccgcttttttgcacaacatgggggatcatgtaactcgccttgatcgttgggaaccgg 6480
agctgaatgaagccataccaaacgacgagcgtgacaccacgatgcctgtagcaatggcaa 6540
caacgttgcgcaaactattaactggcgaactacttactctagcttcccggcaacaattaa 6600
tagactggatggaggcggataaagttgcaggaccacttctgcgctcggcccttccggctg 6660
gctggtttattgctgataaatctggagccggtgagcgtgggtctcgcggtatcattgcag 6720
cactggggccagatggtaagccctcccgtatcgtagttatctacacgacggggagtcagg 6780
caactatggatgaacgaaatagacagatcgctgagataggtgcctcactgattaagcatt 6840
ggtaactgtcagaccaagtttactcatatatactttagattgatttaaaacttcattttt 6900
aatttaaaaggatctaggtgaagatcctttttgataatctcatgaccaaaatcccttaac 6960
gtgagttttcgttccactgagcgtcagaccccgtagaaaagatcaaaggatcttcttgag 7020
atcctttttttctgcgcgtaatctgctgcttgcaaacaaaaaaaccaccgctaccagcgg 7080
tggtttgtttgccggatcaagagctaccaactctttttccgaaggtaactggcttcagca 7140
gagcgcagataccaaatactgttcttctagtgtagccgtagttaggccaccacttcaaga 7200
actctgtagcaccgcctacatacctcgctctgctaatcctgttaccagtggctgctgcca 7260
gtggcgataagtcgtgtcttaccgggttggactcaagacgatagttaccggataaggcgc 7320
agcggtcgggctgaacggggggttcgtgcacacagcccagcttggagcgaacgacctaca 7380
ccgaactgagatacctacagcgtgagctatgagaaagcgccacgcttcccgaagggagaa 7440
aggcggacaggtatccggtaagcggcagggtcggaacaggagagcgcacgagggagcttc 7500
cagggggaaacgcctggtatctttatagtcctgtcgggtttcgccacctctgacttgagc 7560
gtcgatttttgtgatgctcgtcaggggggcggagcctatggaaaaacgccagcaacgcgg 7620
cctttttacggttcctggccttttgctggccttttgctcacatgttctttcctgcgttat 7680
cccctgattctgtggataaccgtattaccgcctttgagtgagctgataccgctcgccgca 7740
gccgaacgaccgagcgcagcgagtcagtgagcgaggaagcggaagagcgcccaatacgca 7800
aaccgcctctccccgcgcgttggccgattcattaatgcagctggcacgacaggtttcccg 7860
actggaaagcgggcagtgagcgcaacgcaattaatgtgagttagctcactcattaggcac 7920
cccaggctttacactttatgcttccggctcgtatgttgtgtggaattgtgagcggataac 7980
aatttcacacaggaaacagctatgaccatgattacgccagatttaattaagg 8032
<210> 17
<211> 738
<212> ПРТ
<213> Неизвестный
<220>
<223> Капсид AAVhu.37
<400> 17
MetAlaAlaAspGlyTyrLeuProAspTrpLeuGluAspAsnLeuSer
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Asp Leu Lys Pro Gly Ala Pro Lys Pro
20 25 30
Lys Ala Asn Gln Gln Lys Gln Asp Asp Gly Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Arg Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Gln Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Ala Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Pro Ser Pro Gln Arg Ser Pro Asp Ser Ser Thr Gly Ile
145 150 155 160
Gly Lys Lys Gly Gln Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln
165 170 175
Thr Gly Asp Ser Glu Ser Val Pro Asp Pro Gln Pro Ile Gly Glu Pro
180 185 190
Pro Ala Gly Pro Ser Gly Leu Gly Ser Gly Thr Met Ala Ala Gly Gly
195 200 205
Gly Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Ser
210 215 220
Ser Ser Gly Asn Trp His Cys Asp Ser Thr Trp Leu Gly Asp Arg Val
225 230 235 240
Ile Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His
245 250 255
Leu Tyr Lys Gln Ile Ser Asn Gly Thr Ser Gly Gly Ser Thr Asn Asp
260 265 270
Asn Thr Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn
275 280 285
Arg Phe His Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn
290 295 300
Asn Asn Trp Gly Phe Arg Pro Lys Arg Leu Ser Phe Lys Leu Phe Asn
305 310 315 320
Ile Gln Val Lys Glu Val Thr Gln Asn Glu Gly Thr Lys Thr Ile Ala
325 330 335
Asn Asn Leu Thr Ser Thr Ile Gln Val Phe Thr Asp Ser Glu Tyr Gln
340 345 350
Leu Pro Tyr Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe
355 360 365
Pro Ala Asp Val Phe Met Ile Pro Gln Tyr Gly Tyr Leu Thr Leu Asn
370 375 380
Asn Gly Ser Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr
385 390 395 400
Phe Pro Ser Gln Met Leu Arg Thr Gly Asn Asn Phe Glu Phe Ser Tyr
405 410 415
Thr Phe Glu Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser
420 425 430
Leu Asp Arg Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu
435 440 445
Ser Arg Thr Gln Ser Thr Gly Gly Thr Gln Gly Thr Gln Gln Leu Leu
450 455 460
Phe Ser Gln Ala Gly Pro Ala Asn Met Ser Ala Gln Ala Lys Asn Trp
465 470 475 480
Leu Pro Gly Pro Cys Tyr Arg Gln Gln Arg Val Ser Thr Thr Leu Ser
485 490 495
Gln Asn Asn Asn Ser Asn Phe Ala Trp Thr Gly Ala Thr Lys Tyr His
500 505 510
Leu Asn Gly Arg Asp Ser Leu Val Asn Pro Gly Val Ala Met Ala Thr
515 520 525
His Lys Asp Asp Glu Glu Arg Phe Phe Pro Ser Ser Gly Val Leu Met
530 535 540
Phe Gly Lys Gln Gly Ala Gly Arg Asp Asn Val Asp Tyr Ser Ser Val
545 550 555 560
Met Leu Thr Ser Glu Glu Glu Ile Lys Thr Thr Asn Pro Val Ala Thr
565 570 575
Glu Gln Tyr Gly Val Val Ala Asp Asn Leu Gln Gln Thr Asn Thr Gly
580 585 590
Pro Ile Val Gly Asn Val Asn Ser Gln Gly Ala Leu Pro Gly Met Val
595 600 605
Trp Gln Asn Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile
610 615 620
Pro His Thr Asp Gly Asn Phe His Pro Ser Pro Leu Met Gly Gly Phe
625 630 635 640
Gly Leu Lys His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val
645 650 655
Pro Ala Asp Pro Pro Thr Thr Phe Ser Gln Ala Lys Leu Ala Ser Phe
660 665 670
Ile Thr Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu
675 680 685
Leu Gln Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr
690 695 700
Ser Asn Tyr Tyr Lys Ser Thr Asn Val Asp Phe Ala Val Asn Thr Glu
705 710 715 720
Gly Thr Tyr Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg
725 730 735
AsnLeu
<210> 18
<211> 738
<212> ПРТ
<213> Неизвестный
<220>
<223> Капсид AAVrh.10
<400> 18
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Asp Leu Lys Pro Gly Ala Pro Lys Pro
20 25 30
Lys Ala Asn Gln Gln Lys Gln Asp Asp Gly Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Arg Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Gln Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Gly Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Pro Ser Pro Gln Arg Ser Pro Asp Ser Ser Thr Gly Ile
145 150 155 160
Gly Lys Lys Gly Gln Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln
165 170 175
Thr Gly Asp Ser Glu Ser Val Pro Asp Pro Gln Pro Ile Gly Glu Pro
180 185 190
Pro Ala Gly Pro Ser Gly Leu Gly Ser Gly Thr Met Ala Ala Gly Gly
195 200 205
Gly Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Ser
210 215 220
Ser Ser Gly Asn Trp His Cys Asp Ser Thr Trp Leu Gly Asp Arg Val
225 230 235 240
Ile Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His
245 250 255
Leu Tyr Lys Gln Ile Ser Asn Gly Thr Ser Gly Gly Ser Thr Asn Asp
260 265 270
Asn Thr Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn
275 280 285
Arg Phe His Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn
290 295 300
Asn Asn Trp Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn
305 310 315 320
Ile Gln Val Lys Glu Val Thr Gln Asn Glu Gly Thr Lys Thr Ile Ala
325 330 335
Asn Asn Leu Thr Ser Thr Ile Gln Val Phe Thr Asp Ser Glu Tyr Gln
340 345 350
Leu Pro Tyr Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe
355 360 365
Pro Ala Asp Val Phe Met Ile Pro Gln Tyr Gly Tyr Leu Thr Leu Asn
370 375 380
Asn Gly Ser Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr
385 390 395 400
Phe Pro Ser Gln Met Leu Arg Thr Gly Asn Asn Phe Glu Phe Ser Tyr
405 410 415
Gln Phe Glu Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser
420 425 430
Leu Asp Arg Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu
435 440 445
Ser Arg Thr Gln Ser Thr Gly Gly Thr Ala Gly Thr Gln Gln Leu Leu
450 455 460
Phe Ser Gln Ala Gly Pro Asn Asn Met Ser Ala Gln Ala Lys Asn Trp
465 470 475 480
Leu Pro Gly Pro Cys Tyr Arg Gln Gln Arg Val Ser Thr Thr Leu Ser
485 490 495
Gln Asn Asn Asn Ser Asn Phe Ala Trp Thr Gly Ala Thr Lys Tyr His
500 505 510
Leu Asn Gly Arg Asp Ser Leu Val Asn Pro Gly Val Ala Met Ala Thr
515 520 525
His Lys Asp Asp Glu Glu Arg Phe Phe Pro Ser Ser Gly Val Leu Met
530 535 540
Phe Gly Lys Gln Gly Ala Gly Lys Asp Asn Val Asp Tyr Ser Ser Val
545 550 555 560
Met Leu Thr Ser Glu Glu Glu Ile Lys Thr Thr Asn Pro Val Ala Thr
565 570 575
Glu Gln Tyr Gly Val Val Ala Asp Asn Leu Gln Gln Gln Asn Ala Ala
580 585 590
Pro Ile Val Gly Ala Val Asn Ser Gln Gly Ala Leu Pro Gly Met Val
595 600 605
Trp Gln Asn Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile
610 615 620
Pro His Thr Asp Gly Asn Phe His Pro Ser Pro Leu Met Gly Gly Phe
625 630 635 640
Gly Leu Lys His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val
645 650 655
Pro Ala Asp Pro Pro Thr Thr Phe Ser Gln Ala Lys Leu Ala Ser Phe
660 665 670
Ile Thr Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu
675 680 685
Leu Gln Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr
690 695 700
Ser Asn Tyr Tyr Lys Ser Thr Asn Val Asp Phe Ala Val Asn Thr Asp
705 710 715 720
Gly Thr Tyr Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg
725 730 735
AsnLeu
<210> 19
<211> 4371
<212> ДНК
<213> Искусственная последовательность
<220>
<223> сконструированная последовательность
<400> 19
atgcagatcgagctgtctacctgcttcttcctgtgcctgctgcggttctgcttcagcgcc 60
accagacggtactatctgggcgccgtggaactgagctgggactacatgcagagcgacctg 120
ggcgagctgcccgtggacgccagattccctccaagagtgcccaagagcttccccttcaac 180
acctccgtggtgtacaagaaaaccctgttcgtggaattcaccgaccacctgttcaatatc 240
gccaagcccagacccccctggatgggcctgctgggacctacaattcaggccgaggtgtac 300
gacaccgtcgtgatcaccctgaagaacatggccagccaccccgtgtctctgcacgccgtg 360
ggagtgtcctactggaaggcctctgagggcgccgagtacgacgatcagaccagccagcgc 420
gagaaagaggacgacaaggtgttccctggcggcagccacacctacgtgtggcaggtgctg 480
aaagaaaacggccccatggcctccgaccctctgtgcctgacatacagctacctgagccac 540
gtggacctcgtgaaggacctgaacagcggcctgatcggagccctgctcgtgtgtagagag 600
ggcagcctggccaaagagaaaacccagaccctgcacaagttcatcctgctgttcgccgtg 660
ttcgacgagggcaagagctggcacagcgagacaaagaacagcctgatgcaggaccgggac 720
gccgcctctgctagagcctggcccaaaatgcacaccgtgaacggctacgtgaacagaagc 780
ctgcccggactgatcggctgccaccggaagtctgtgtactggcacgtgatcggcatgggc 840
accacccctgaggtgcacagcatctttctggaaggacacacctttctcgtgcggaaccac 900
cggcaggccagcctggaaatcagccctatcaccttcctgaccgcccagacactgctgatg 960
gacctgggccagtttctgctgttctgccacatcagctcccaccagcacgacggcatggaa 1020
gcctacgtgaaggtggacagctgccccgaggaaccccagctgcggatgaagaacaacgag 1080
gaagccgaggactacgacgacgacctgaccgacagcgagatggacgtggtgcgcttcgac 1140
gacgataacagccccagcttcatccagatcagaagcgtggccaagaagcaccccaagacc 1200
tgggtgcactatatcgccgccgaggaagaggactgggattacgcccctctggtgctggcc 1260
cccgacgacagaagctacaagagccagtacctgaacaacggcccccagcggatcggccgg 1320
aagtataagaaagtgcggttcatggcctacaccgacgagacattcaagaccagagaggcc 1380
atccagcacgagagcggcatcctgggccctctgctgtatggcgaagtgggcgacaccctg 1440
ctgatcatcttcaagaaccaggccagcagaccctacaacatctaccctcacggcatcacc 1500
gacgtgcggcccctgtactctagaaggctgcccaagggcgtgaaacacctgaaggacttc 1560
cccatcctgcccggcgagatcttcaagtacaagtggaccgtgaccgtggaagatggcccc 1620
accaagagcgaccccagatgcctgacacggtactatagcagcttcgtgaacatggaacgg 1680
gacctggcctccggcctgattggcccactgctgatctgctacaaagaaagcgtggaccag 1740
cggggcaaccagatcatgagcgacaagcggaacgtgatcctgtttagcgtgttcgatgag 1800
aaccggtcctggtatctgaccgagaatatccagcggttcctgcccaaccctgccggcgtg 1860
cagctggaagatcctgagttccaggcctccaacatcatgcactccatcaatggctatgtg 1920
ttcgacagcctgcagctgagcgtgtgcctgcacgaggtggcctactggtacatcctgagc 1980
atcggggcccagaccgacttcctgtccgtgttcttctccggctacaccttcaagcacaag 2040
atggtgtacgaggataccctgaccctgttcccctttagcggcgaaaccgtgttcatgagc 2100
atggaaaaccccggcctgtggatcctgggctgccacaacagcgacttccggaacagaggc 2160
atgaccgccctgctgaaggtgtccagctgcgacaagaacaccggcgactactacgaggac 2220
agctatgaggacatcagcgcctacctgctgagcaagaacaacgccatcgagcccagaagc 2280
ttcagccagaacccccccgtgctgaagcggcaccagagagagatcacccggaccaccctg 2340
cagtccgaccaggaagagatcgattacgacgacaccatcagcgtggaaatgaagaaagaa 2400
gatttcgacatctacgacgaggacgagaaccagagcccccggtcctttcagaaaaagacc 2460
cggcactacttcattgccgctgtggaacggctgtgggactacggcatgagcagcagccct 2520
cacgtgctgagaaacagggcccagagcggcagcgtgccccagttcaagaaagtggtgttc 2580
caggaattcacagacggcagcttcacccagcctctgtaccgcggcgagctgaacgagcac 2640
ctgggactgctgggcccctatatcagagccgaagtggaagataacatcatggtcaccttc 2700
cggaatcaggcctcccggccctacagcttctacagctccctgatcagctacgaagaggac 2760
cagagacagggcgctgagccccggaagaacttcgtgaagcccaacgagactaagacctac 2820
ttttggaaggtgcagcaccacatggcccctacaaaggacgagttcgactgcaaggcctgg 2880
gcctacttctccgacgtggacctggaaaaggacgtgcactctgggctgatcggccccctg 2940
ctcgtgtgccacaccaacaccctgaatcccgcccacggcagacaggtgacagtgcaggaa 3000
ttcgccctgttcttcaccatcttcgacgaaacaaagagctggtacttcaccgaaaacatg 3060
gaaagaaactgccgggctccctgcaacatccagatggaagatcccaccttcaaagagaac 3120
taccggttccacgccatcaacggctacatcatggacacactgcccggcctcgtgatggct 3180
caggatcagcggatccggtggtatctgctgtccatgggctccaacgagaacatccacagc 3240
atccacttcagcggccacgtgttcaccgtgcggaaaaaagaagagtacaaaatggccctg 3300
tacaacctgtaccctggggtgttcgagacagtggaaatgctgcccagcaaggccggcatc 3360
tggcgggtggagtgtctgatcggcgagcacctgcacgctgggatgagcacactgtttctg 3420
gtgtacagcaacaagtgccagacacctctgggcatggcctctggccacatccgggacttt 3480
cagatcacagccagcggccagtacggccagtgggccccaaaactggccagactgcactac 3540
agcggcagcatcaacgcctggtccaccaaagagcccttcagctggatcaaggtggacctg 3600
ctggctcccatgatcatccacggaatcaagacccagggcgccagacagaagttcagcagc 3660
ctgtacatcagccagttcatcatcatgtacagcctggacggcaagaagtggcagacctac 3720
cggggcaatagcaccggcaccctgatggtgttcttcggcaacgtggactccagcggcatt 3780
aagcacaacatcttcaacccccccatcattgcccggtacatccggctgcaccccacccac 3840
tacagcatccggtccaccctgagaatggaactgatgggctgcgacctgaactcctgctcc 3900
atgcccctggggatggaaagcaaggccatctccgacgcccagatcaccgcctccagctac 3960
ttcaccaacatgttcgccacctggtccccatccaaggcccggctgcacctgcagggcaga 4020
agcaatgcttggaggcctcaggtgaacaaccccaaagagtggctgcaggtggacttccag 4080
aaaaccatgaaagtgaccggcgtgaccacccagggcgtgaagtctctgctgacctctatg 4140
tacgtgaaagagttcctgatctccagcagccaggacggccaccagtggaccctgtttttc 4200
cagaacggcaaagtgaaagtgtttcaggggaaccaggactccttcacccccgtcgtgaat 4260
agcctggaccctccactgctgaccagatacctgcggatccaccctcagagttgggtgcac 4320
cagattgctctgcggatggaagtgctgggatgcgaggcccaggacctgtac 4371
<210> 20
<211> 736
<212> ПРТ
<213> Неизвестный
<220>
<223> Капсид AAV3B
<400> 20
MetAlaAlaAspGlyTyrLeuProAspTrpLeuGluAspAsnLeuSer
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Ala Leu Lys Pro Gly Val Pro Gln Pro
20 25 30
Lys Ala Asn Gln Gln His Gln Asp Asn Arg Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Gly Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Glu Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Gln Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Ile Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Ala Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Asp Gln Ser Pro Gln Glu Pro Asp Ser Ser Ser Gly Val Gly
145 150 155 160
Lys Ser Gly Lys Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ser Glu Ser Val Pro Asp Pro Gln Pro Leu Gly Glu Pro Pro
180 185 190
Ala Ala Pro Thr Ser Leu Gly Ser Asn Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Gln Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Gln Ser Gly Ala Ser Asn Asp Asn His Tyr
260 265 270
Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe His
275 280 285
Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn Trp
290 295 300
Gly Phe Arg Pro Lys Lys Leu Ser Phe Lys Leu Phe Asn Ile Gln Val
305 310 315 320
Lys Glu Val Thr Gln Asn Asp Gly Thr Thr Thr Ile Ala Asn Asn Leu
325 330 335
Thr Ser Thr Val Gln Val Phe Thr Asp Ser Glu Tyr Gln Leu Pro Tyr
340 345 350
Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala Asp
355 360 365
Val Phe Met Val Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly Ser
370 375 380
Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro Ser
385 390 395 400
Gln Met Leu Arg Thr Gly Asn Asn Phe Gln Phe Ser Tyr Thr Phe Glu
405 410 415
Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp Arg
420 425 430
Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Asn Arg Thr
435 440 445
Gln Gly Thr Thr Ser Gly Thr Thr Asn Gln Ser Arg Leu Leu Phe Ser
450 455 460
Gln Ala Gly Pro Gln Ser Met Ser Leu Gln Ala Arg Asn Trp Leu Pro
465 470 475 480
Gly Pro Cys Tyr Arg Gln Gln Arg Leu Ser Lys Thr Ala Asn Asp Asn
485 490 495
Asn Asn Ser Asn Phe Pro Trp Thr Ala Ala Ser Lys Tyr His Leu Asn
500 505 510
Gly Arg Asp Ser Leu Val Asn Pro Gly Pro Ala Met Ala Ser His Lys
515 520 525
Asp Asp Glu Glu Lys Phe Phe Pro Met His Gly Asn Leu Ile Phe Gly
530 535 540
Lys Glu Gly Thr Thr Ala Ser Asn Ala Glu Leu Asp Asn Val Met Ile
545 550 555 560
Thr Asp Glu Glu Glu Ile Arg Thr Thr Asn Pro Val Ala Thr Glu Gln
565 570 575
Tyr Gly Thr Val Ala Asn Asn Leu Gln Ser Ser Asn Thr Ala Pro Thr
580 585 590
Thr Arg Thr Val Asn Asp Gln Gly Ala Leu Pro Gly Met Val Trp Gln
595 600 605
Asp Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His
610 615 620
Thr Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu
625 630 635 640
Lys His Pro Pro Pro Gln Ile Met Ile Lys Asn Thr Pro Val Pro Ala
645 650 655
Asn Pro Pro Thr Thr Phe Ser Pro Ala Lys Phe Ala Ser Phe Ile Thr
660 665 670
Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln
675 680 685
Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr Ser Asn
690 695 700
Tyr Asn Lys Ser Val Asn Val Asp Phe Thr Val Asp Thr Asn Gly Val
705 710 715 720
Tyr Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn Leu
725 730 735
<---
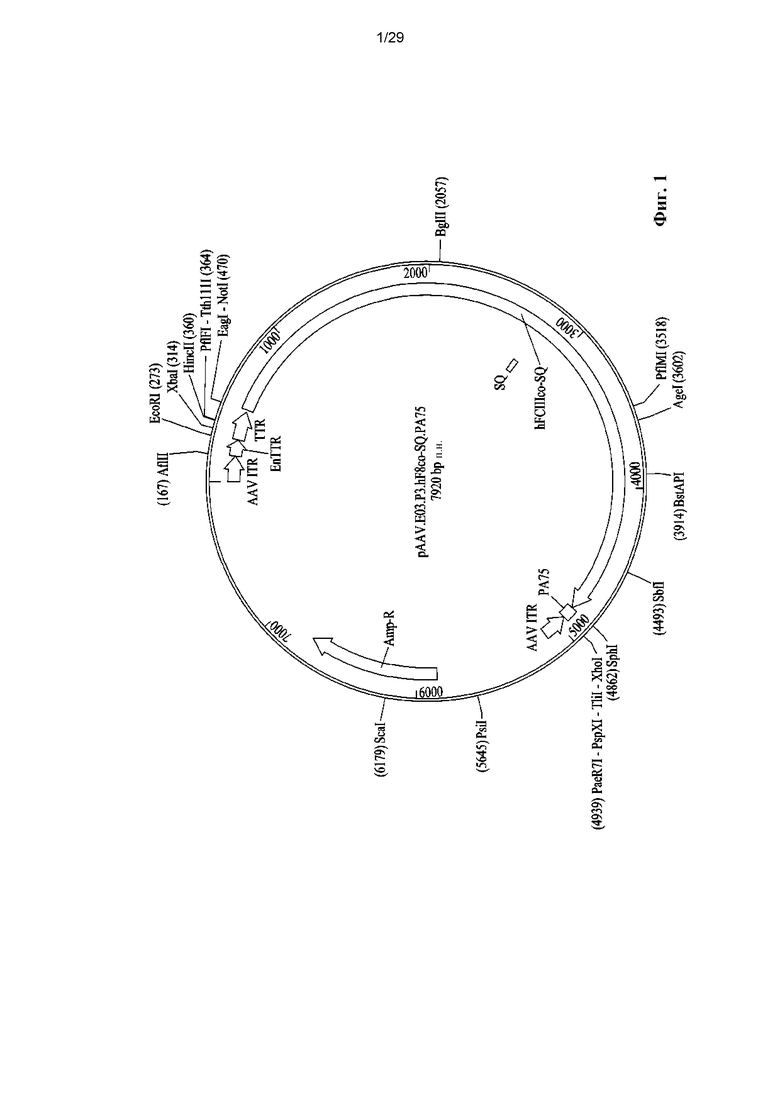
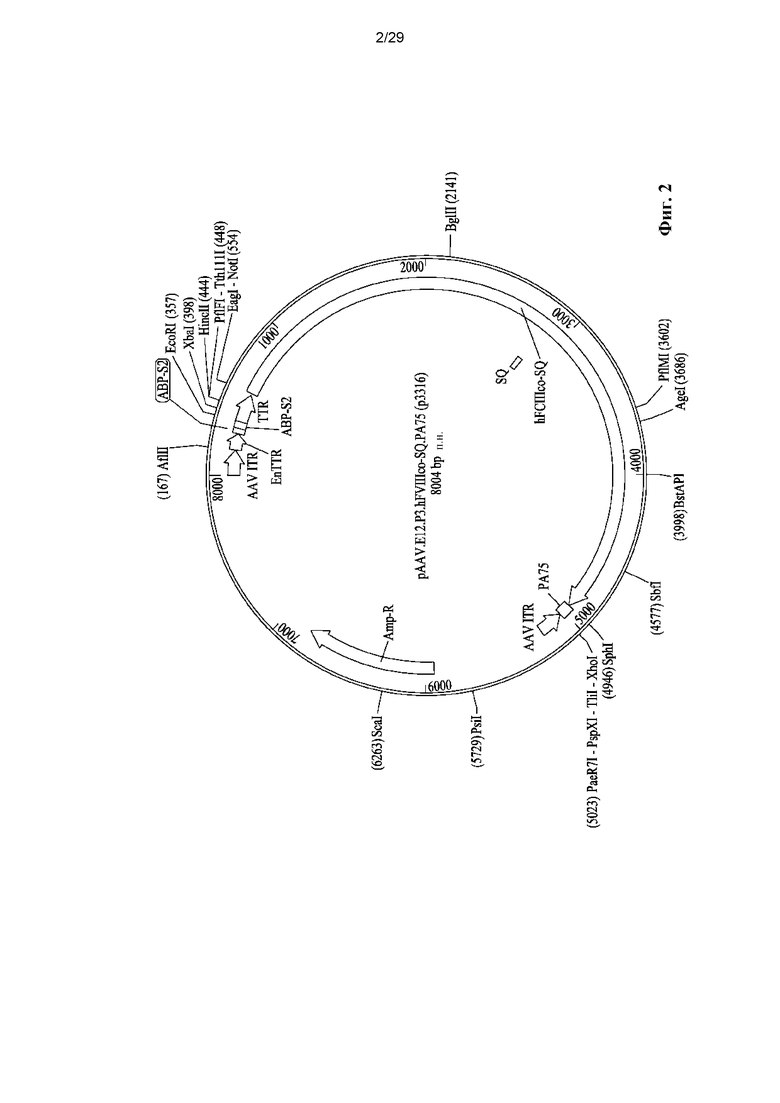
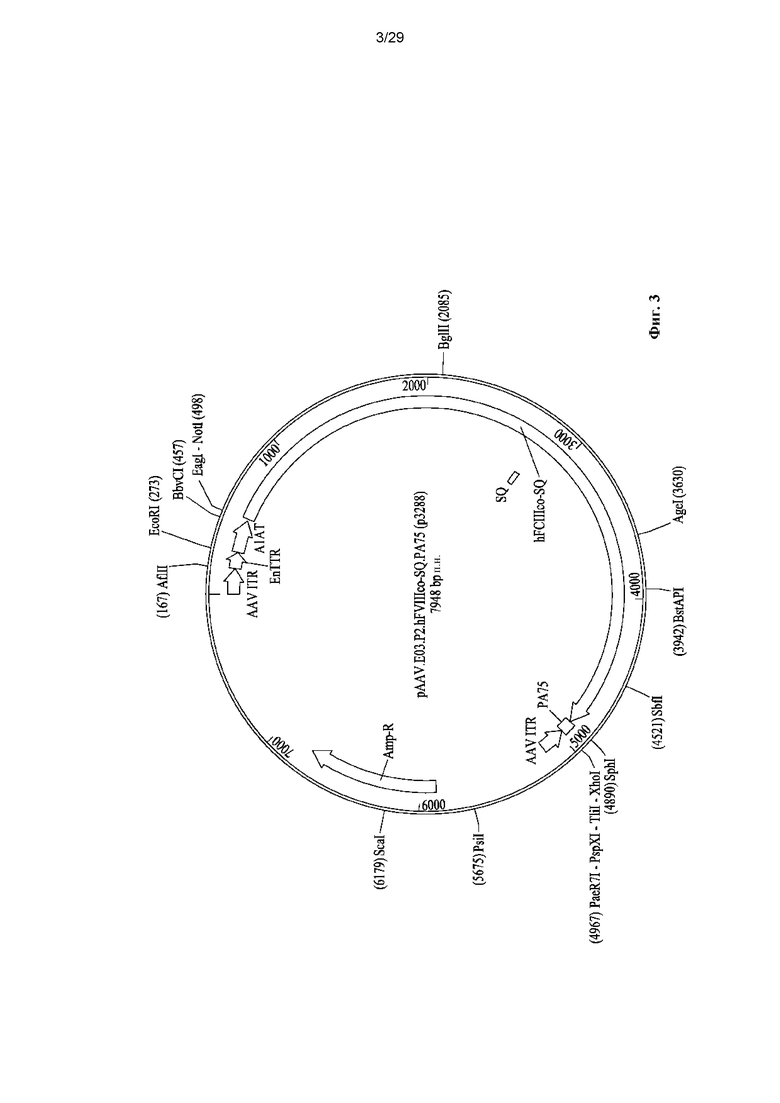
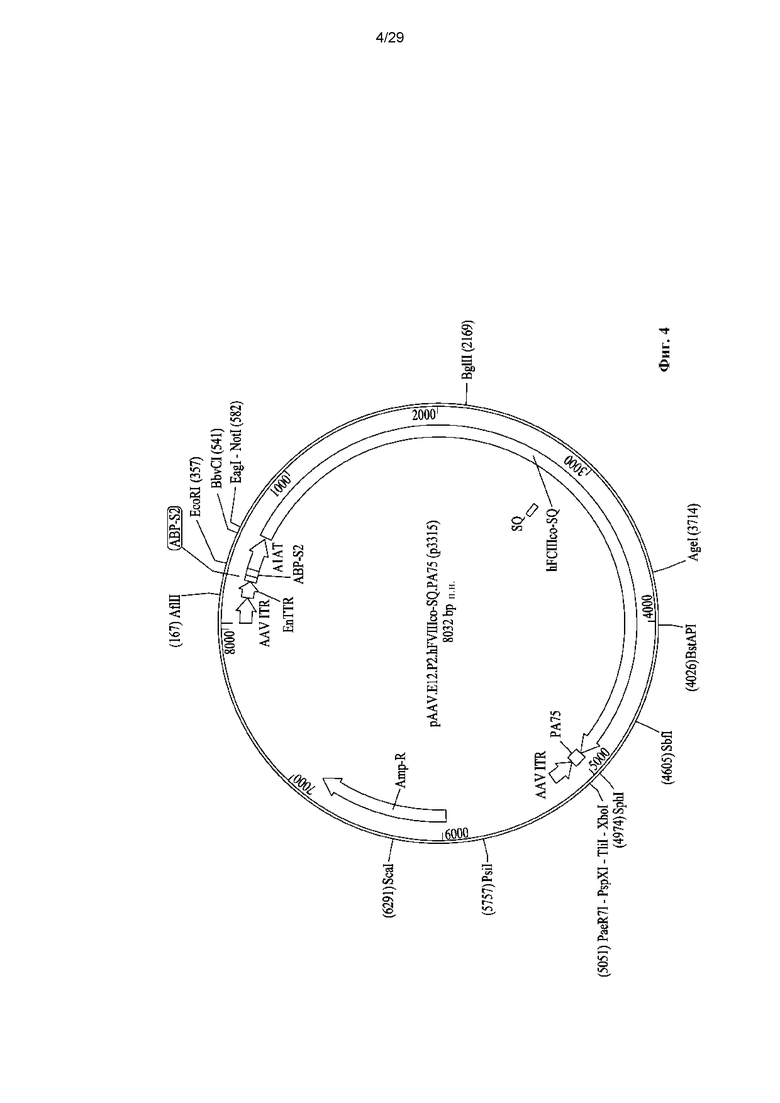





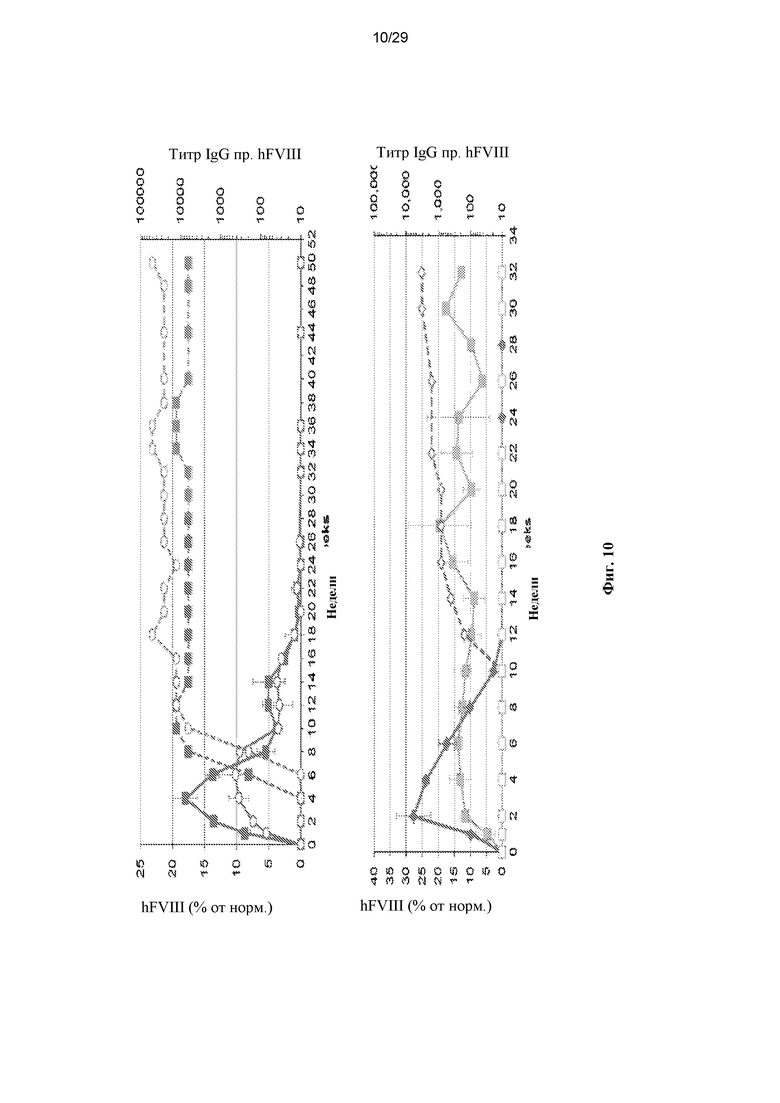
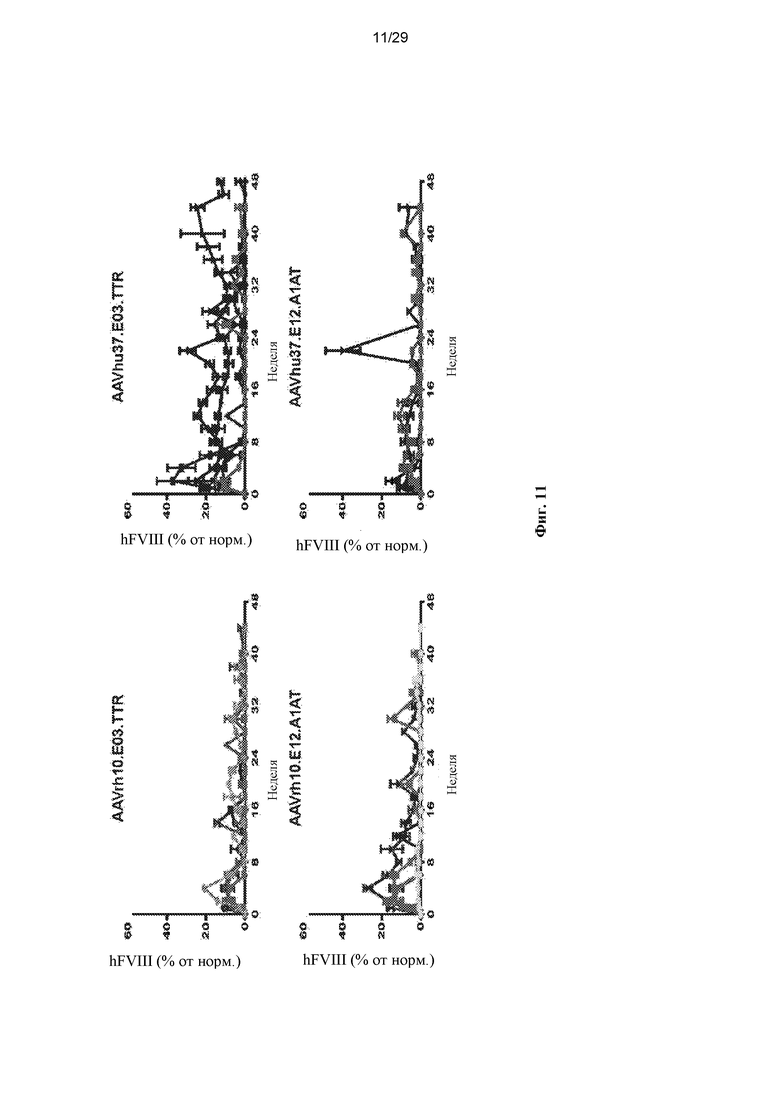
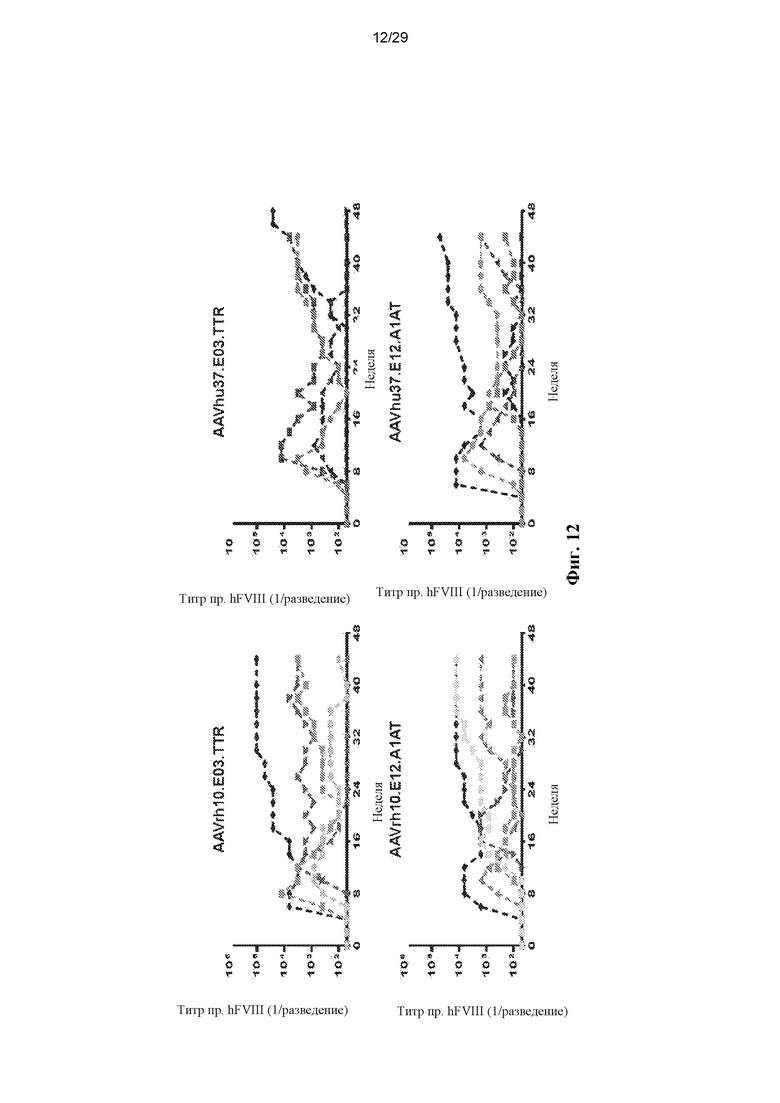
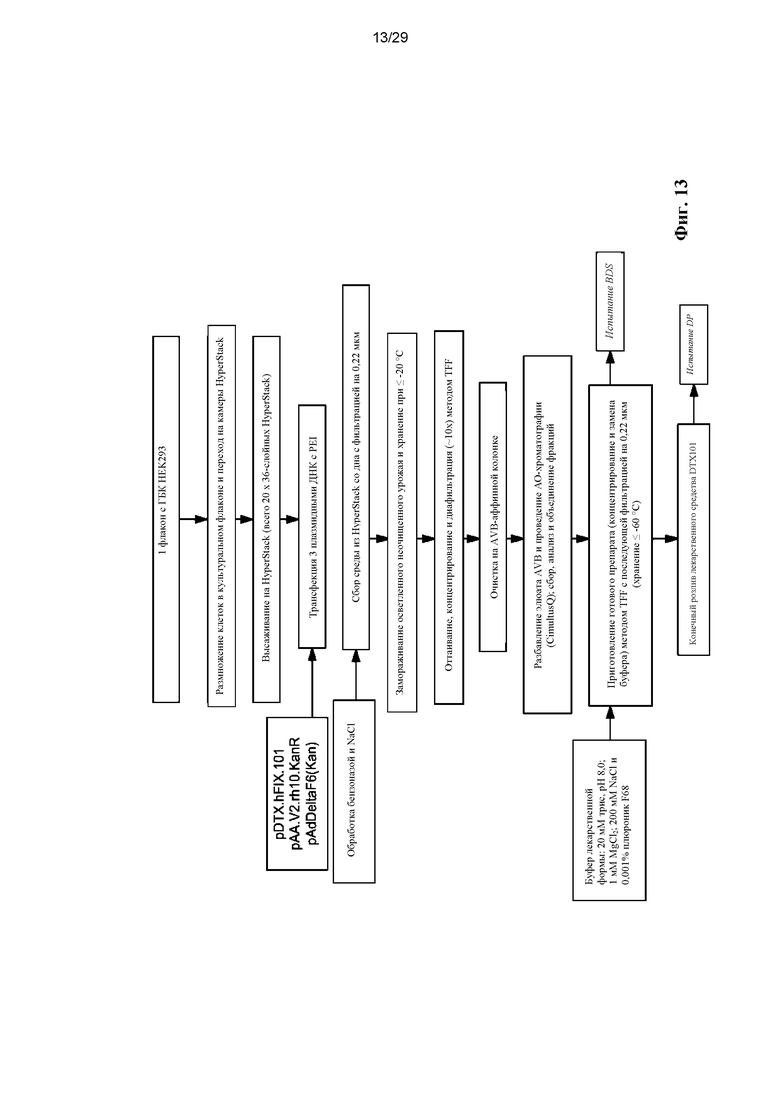
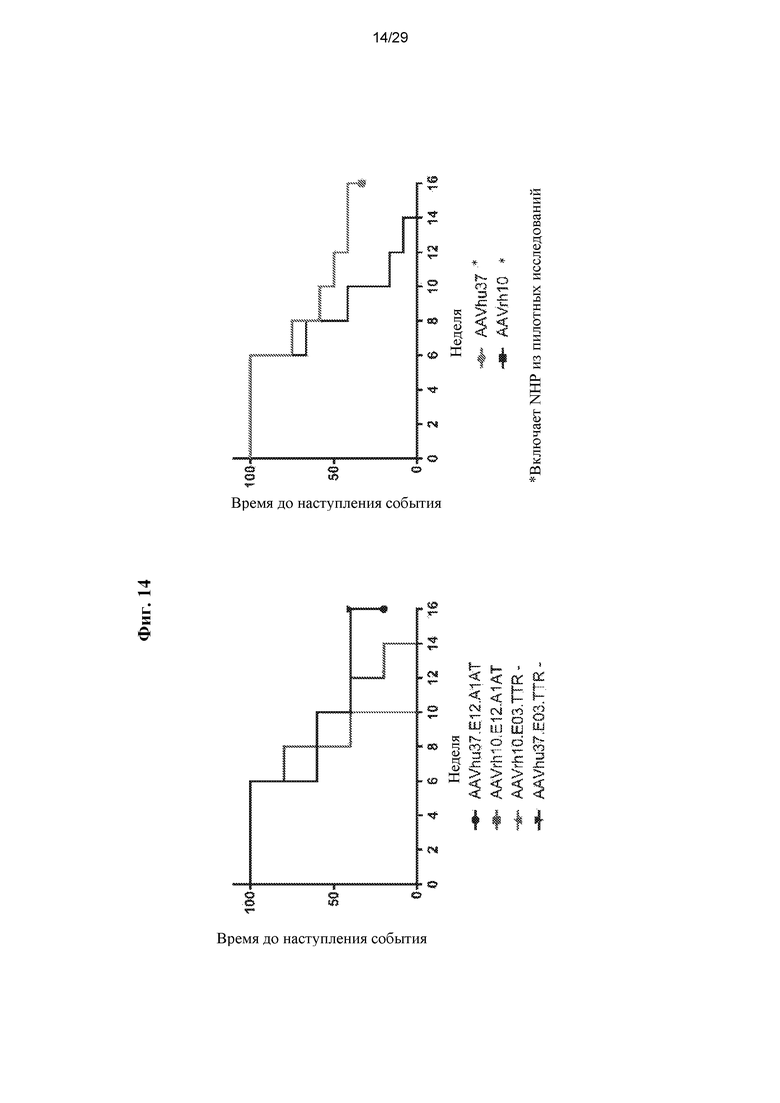
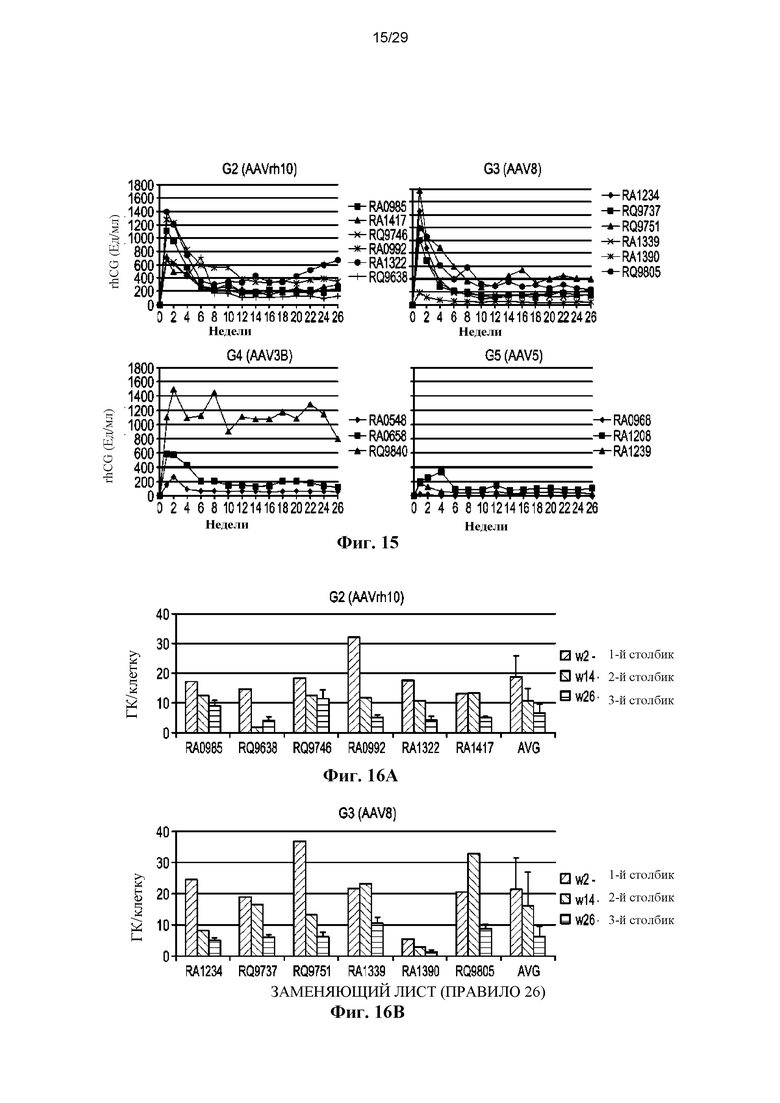
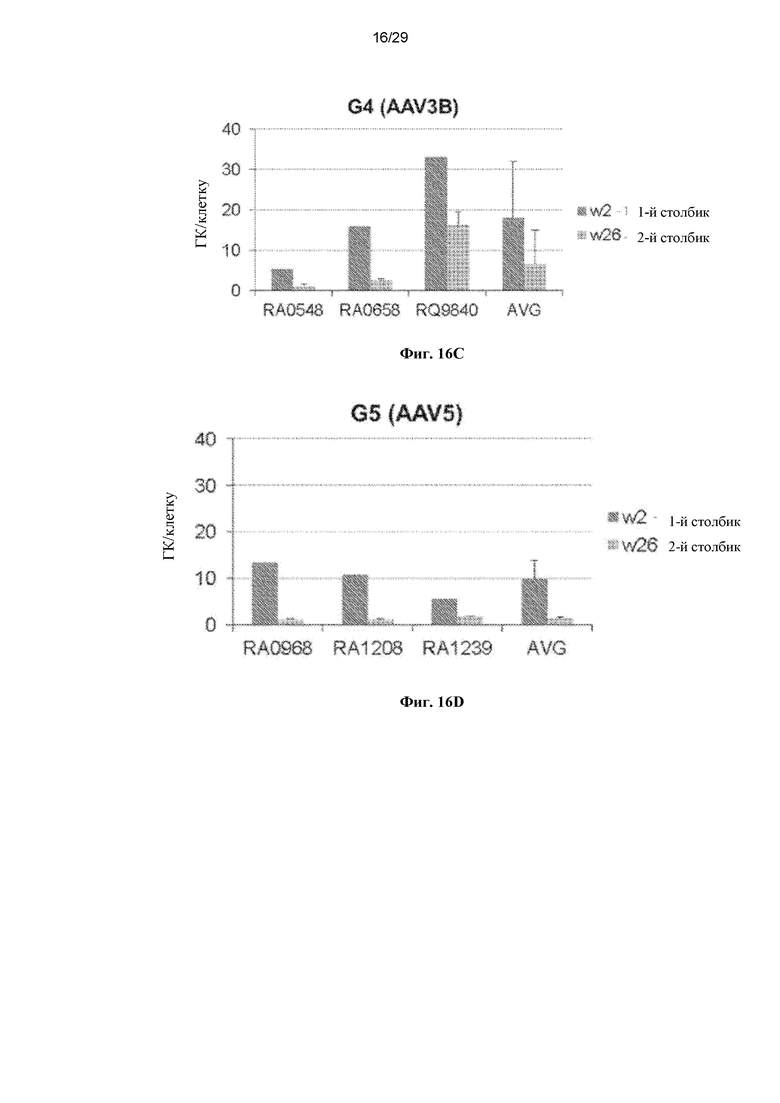
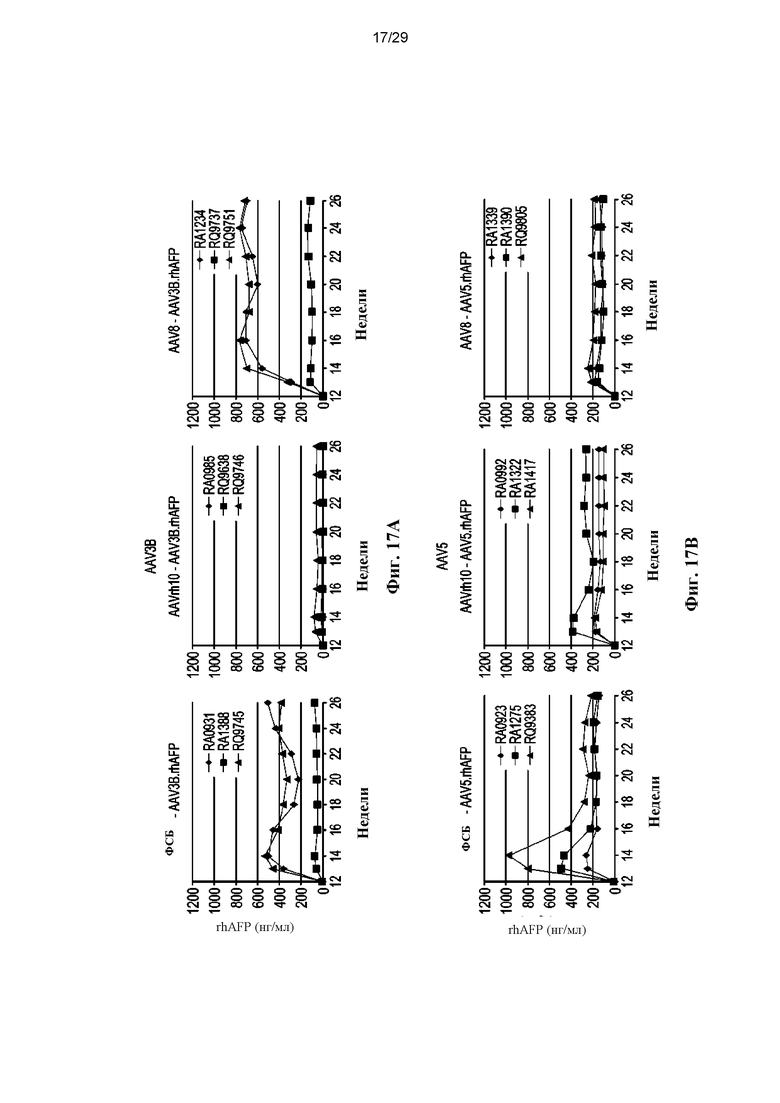
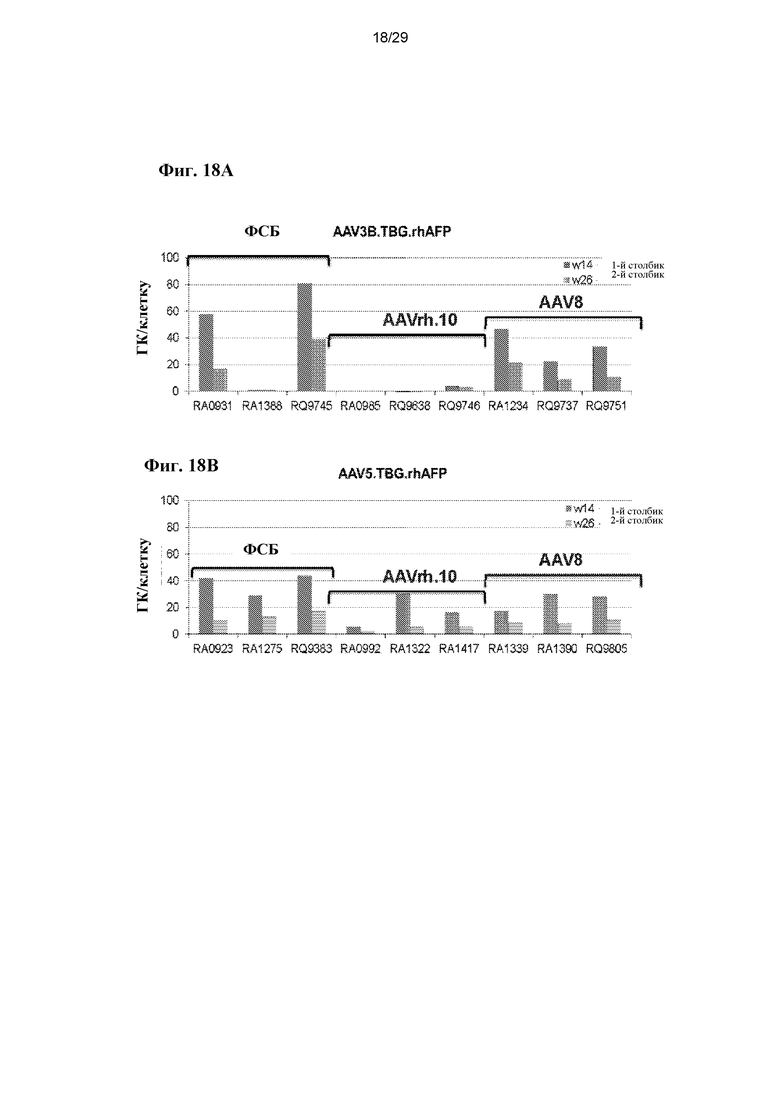
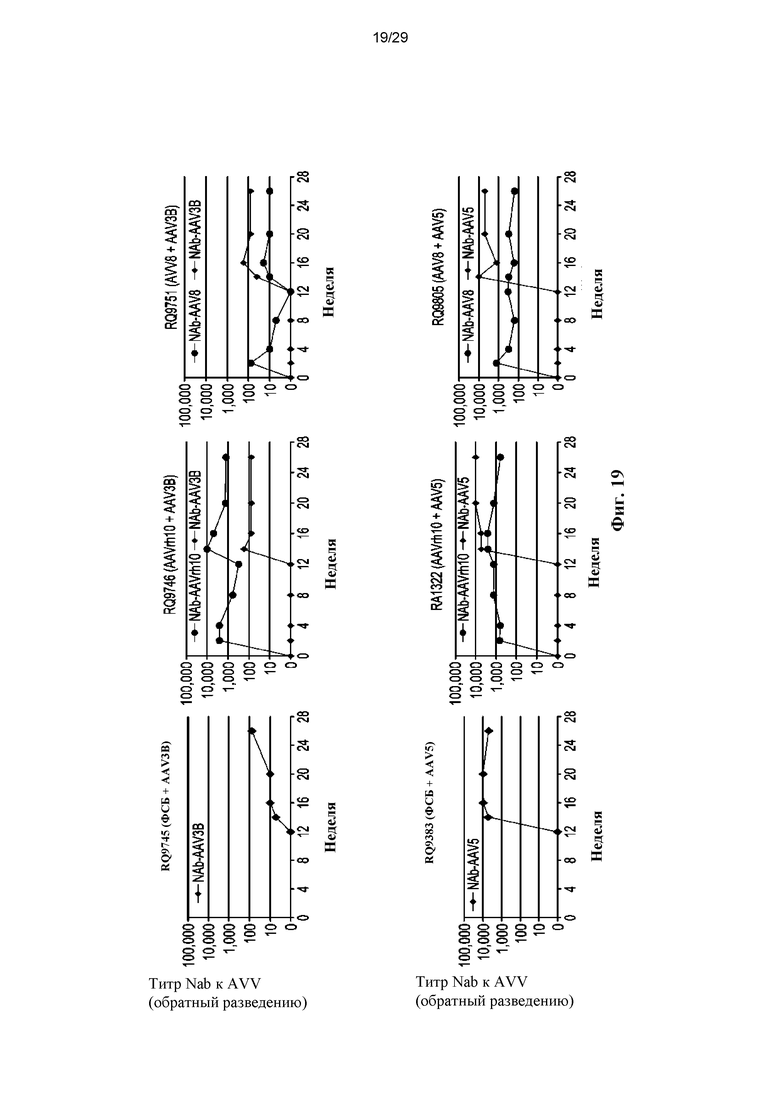
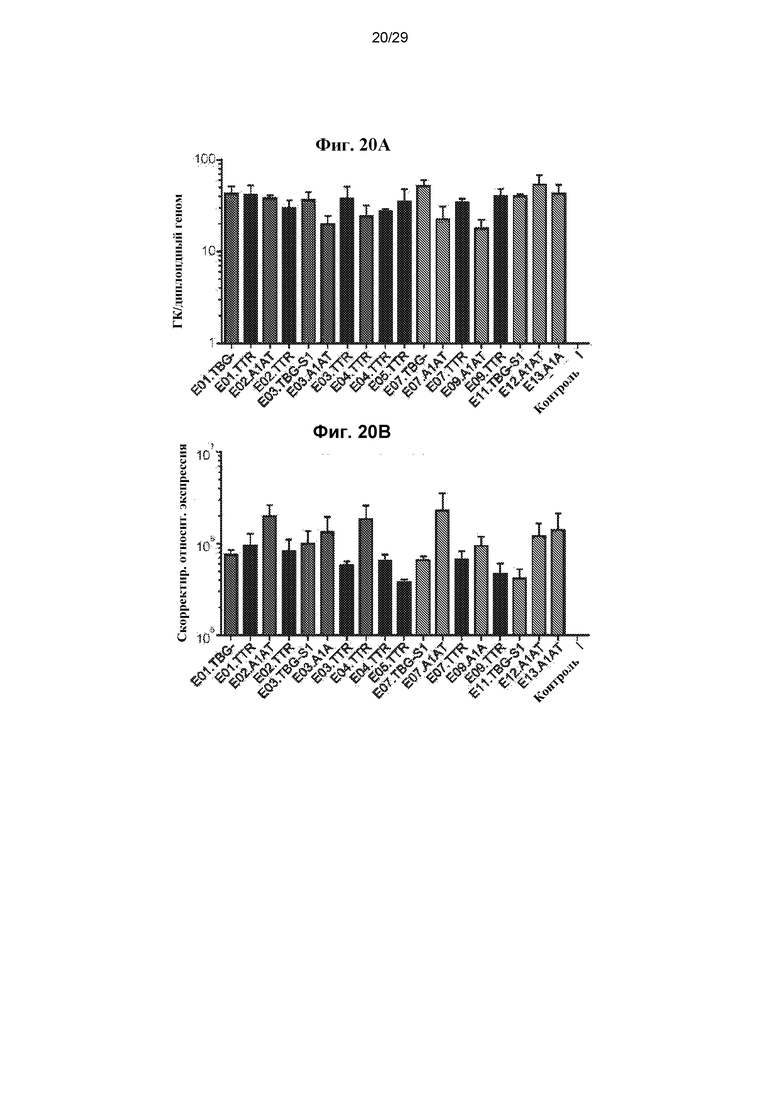
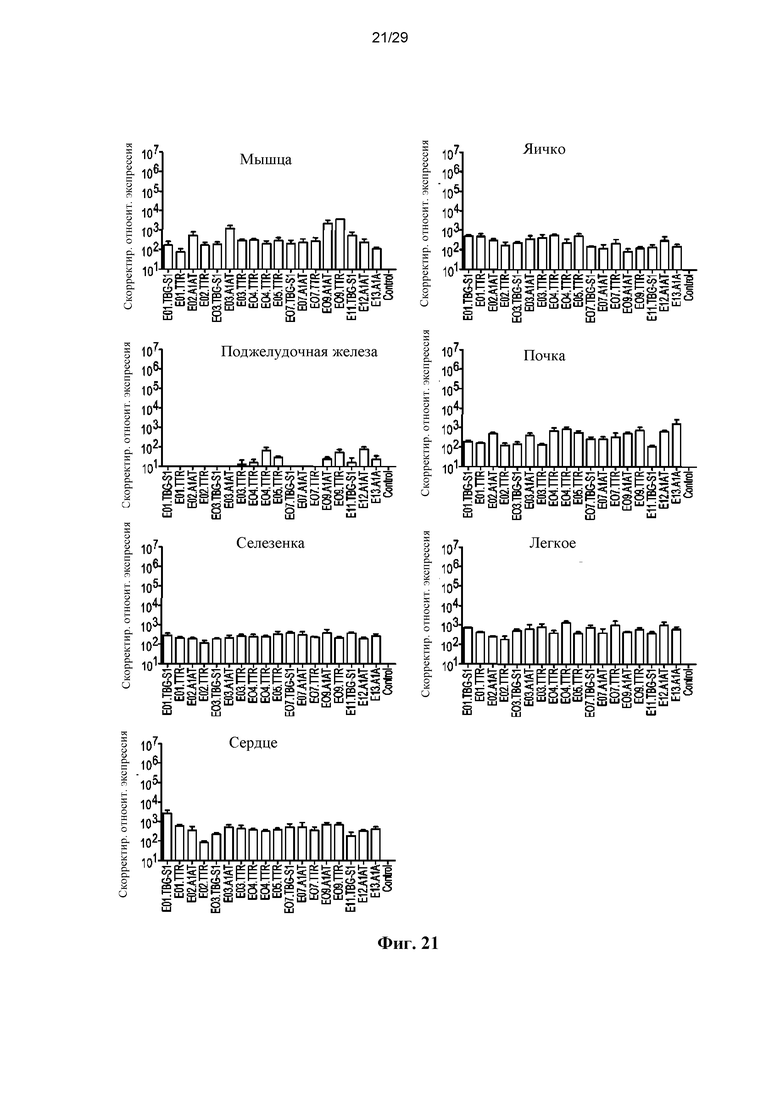
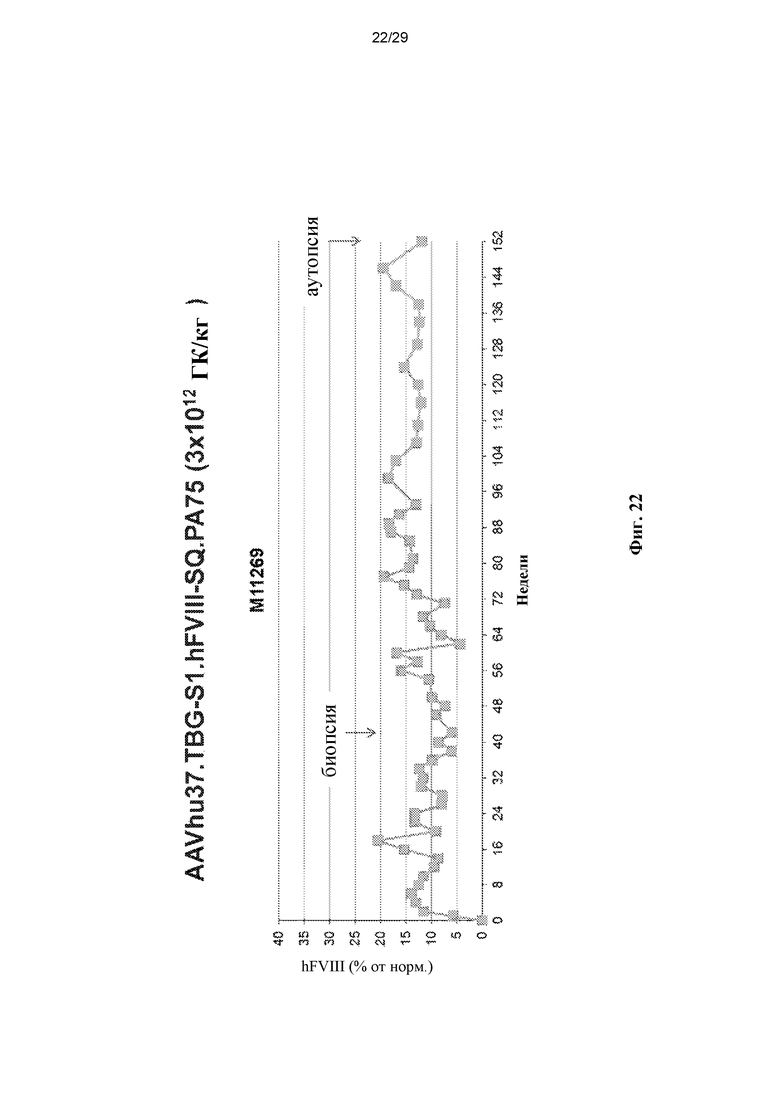
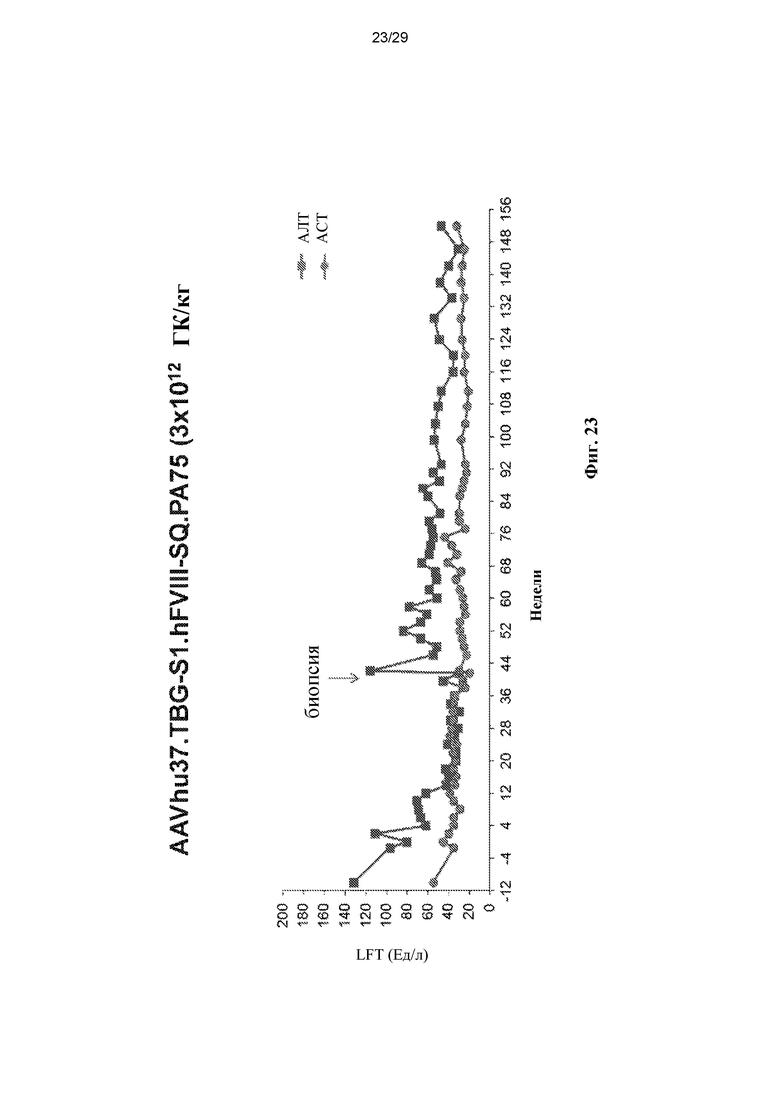
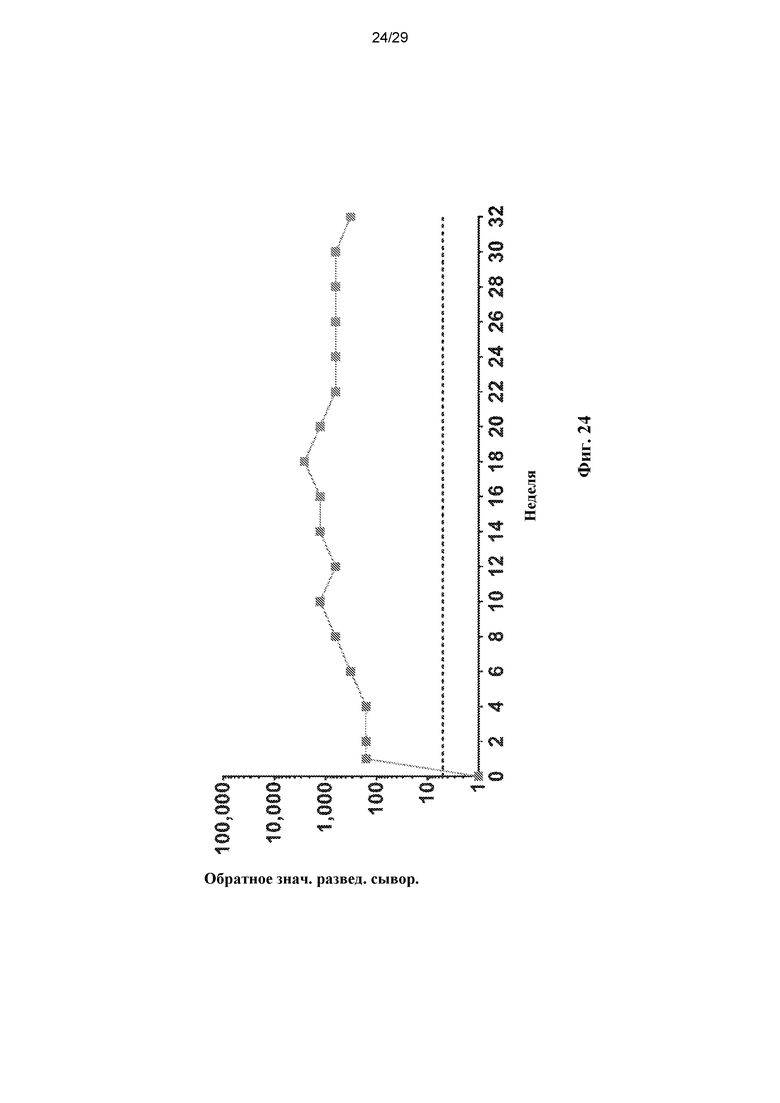





Изобретение относится к биотехнологии. Описаны рекомбинатный аденоассоциированный вирус (rAAV) для доставки гена человеческого фактора VIII пациенту, содержащий капсид AAV и упакованный в него векторный геном, при этом указанный векторный геном содержит: (a) AAV с 5' инвертированным концевым повтором (ITR); (b) специфический для печени промотор; (c) кодирующую последовательность, кодирующую человеческий фактор VIII, обеспечивающий функцию обеспечения коагуляции; и (d) 3' ITR AAV, причем указанная кодирующая последовательность содержит нуклеотидную последовательность, представленную в SEQ ID NO: 2, или последовательность, идентичную ей по меньшей мере на 95%. Представлен вирус rAAV для доставки гена человеческого фактора VIII пациенту, содержащий капсид AAVhu37 и упакованный в него векторный геном, при этом указанный векторный геном содержит: (a) последовательность AAV с 5' инвертированным концевым повтором (ITR); (b) промотор транстиретина (TTR); (с) энхансер транстиретина (enTTR); (d) кодирующую последовательность, кодирующую человеческий фактор VIII, обеспечивающий функцию обеспечения коагуляции; и (e) последовательность 3' ITR AAV, причем указанная кодирующая последовательность содержит нуклеотидную последовательность, представленную в SEQ ID NO: 2. Также описана водная суспензия, пригодная для введения пациенту с гемофилией A, причем указанная суспензия содержит водную суспендирующую жидкость и от около 1 x 1012 геномных копий (ГК)/мл до около 1 x 1014 ГК/мл вируса rAAV. Также представлена нуклеиновая кислота, кодирующая человеческий фактор VIII, обеспечивающий функцию обеспечения коагуляции, причем указанная нуклеиновая кислота содержит нуклеотидную последовательность, представленную в SEQ ID NO: 2, или последовательность, идентичную ей по меньшей мере на 95%. Изобретение расширяет арсенал средств для лечения гемофилии A. 5 н. и 17 з.п. ф-лы, 25 ил., 4 табл., 3 пр.
1. Рекомбинатный аденоассоциированный вирус (rAAV) для доставки гена человеческого фактора VIII пациенту, содержащий капсид AAV и упакованный в него векторный геном, при этом указанный векторный геном содержит:
(a) AAV с 5' инвертированным концевым повтором (ITR);
(b) специфический для печени промотор;
(c) кодирующую последовательность, кодирующую человеческий фактор VIII, обеспечивающий функцию обеспечения коагуляции; и
(d) 3' ITR AAV,
причем указанная кодирующая последовательность содержит нуклеотидную последовательность, представленную в SEQ ID NO: 2, или последовательность, идентичную ей по меньшей мере на 95%.
2. Вирус rAAV по п. 1, отличающийся тем, что человеческий фактор VIII представляет собой фактор VIII SQ с удаленным B-доменом, содержащий в длину около 1457 аминокислот.
3. Вирус rAAV по п. 1 или 2, отличающийся тем, что кодирующая последовательность из (с) представляет собой нуклеотидную последовательность, представленную в SEQ ID NO: 2.
4. Вирус rAAV по любому из пп. 1–3, отличающийся тем, что капсид AAV представляет собой капсид hu37.
5. Вирус rAAV по любому из пп. 1–4, отличающийся тем, что 5' ITR AAV и/или 3' ITR AAV взяты из AAV2.
6. Вирус rAAV по п. 5, отличающийся тем, что 5' ITR AAV содержит нуклеотидную последовательность, представленную в SEQ ID NO: 11, и 3' ITR AAV содержит нуклеотидную последовательность, представленную в SEQ ID NO: 12.
7. Вирус rAAV по любому из пп. 1–6, отличающийся тем, что специфический для печени промотор представляет собой промотор транстиретина (TTR).
8. Вирус rAAV по п. 7, отличающийся тем, что промотор TTR содержит нуклеотидную последовательность, представленную в SEQ ID NO: 7.
9. Вирус rAAV по любому из пп. 1–8, отличающийся тем, что векторный геном дополнительно содержит энхансер.
10. Вирус rAAV по п. 9, отличающийся тем, что энхансер представляет собой специфический для печени энхансер.
11. Вирус rAAV по п. 10, отличающийся тем, что специфический для печени энхансер представляет собой энхансер транстиретина (enTTR).
12. Вирус rAAV по п. 11, отличающийся тем, что энхансер enTTR содержит нуклеотидную последовательность, представленную в SEQ ID NO: 5.
13. Вирус rAAV по любому из пп. 1–12, отличающийся тем, что векторный геном дополнительно содержит последовательность полиA.
14. Вирус rAAV по п. 13, отличающийся тем, что последовательность полиA содержит нуклеотидную последовательность, представленную в SEQ ID NO: 10.
15. Вирус rAAV по любому из пп. 1–14, отличающийся тем, что векторный геном имеет размер от около 5 т. п. н. до около 5,5 т. п. н.
16. Вирус rAAV для доставки гена человеческого фактора VIII пациенту, содержащий капсид AAVhu37 и упакованный в него векторный геном, при этом указанный векторный геном содержит:
(a) последовательность AAV с 5' инвертированным концевым повтором (ITR);
(b) промотор транстиретина (TTR);
(с) энхансер транстиретина (enTTR);
(d) кодирующую последовательность, кодирующую человеческий фактор VIII, обеспечивающий функцию обеспечения коагуляции; и
(e) последовательность 3' ITR AAV, причем указанная кодирующая последовательность содержит нуклеотидную последовательность, представленную в SEQ ID NO: 2.
17. Вирус rAAV по п. 16, отличающийся тем, что векторный геном содержит нуклеотиды 1-5110 нуклеотидной последовательности, представленной в SEQ ID NO: 13.
18. Водная суспензия, пригодная для введения пациенту с гемофилией A, причем указанная суспензия содержит водную суспендирующую жидкость и от около 1 x 1012 геномных копий (ГК)/мл до около 1 x 1014 ГК/мл вируса rAAV по любому из пп. 1–17.
19. Водная суспензия по п. 18, отличающаяся тем, что суспензия дополнительно содержит поверхностно-активное вещество, консервант и/или буфер, в водной суспендирующей жидкости.
20. Применение rAAV по любому из пп. 1-17 в лечении пациента с гемофилией A, в котором пациенту доставляют водную суспензию от около 1 x 1012 до около 1 x 1014 геномных копий (ГК)/кг указанного rAAV.
21. Применение по п. 20, отличающийся тем, что rAAV вводят повторно в более поздний момент времени.
22. Нуклеиновая кислота, кодирующая человеческий фактор VIII, обеспечивающий функцию обеспечения коагуляции, причем указанная нуклеиновая кислота содержит нуклеотидную последовательность, представленную в SEQ ID NO: 2, или последовательность, идентичную ей по меньшей мере на 95%.
| WO 2014064277 A1, 01.05.2014 | |||
| US 20070243168 A1, 18.10.2007 | |||
| ZHIJIAN WU ET AL, "Optimization of Self-complementary AAV Vectors for Liver-directed Expression Results in Sustained Correction of Hemophilia B at Low Vector Dose", MOLECULAR THERAPY,Vol | |||
| Устройство для электрической сигнализации | 1918 |
|
SU16A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| СТАБИЛЬНОЕ ПОЛУЧЕНИЕ ЛЕНТИВИРУСНЫХ ВЕКТОРОВ | 2011 |
|
RU2577979C2 |

