Область техники, к которой относится изобретение
Изобретение относится к области биотехнологии и генной инженерии растений, более конкретно к РНК-проводнику (молекуле направляющей РНК со специфичной нуклеотидной последовательностью) для контролирования экспрессии генов растений путем целенаправленного воздействия на нуклеотидную последовательность консервативного участка промоторной области гена VRN-A1 однодольных зерновых культур системами редактирования генома CRISPR/Cas9, в которых молекулы РНК-проводника или сконструированный на его основе экспрессионный ДНК-вектор являются одним из компонентов.
Предшествующий уровень техники
Способность злаков переходить к репродуктивному развитию зависит от функционирования двух основных систем генов развития - VRN (vernalization - ответ на яровизацию) и PPD (photoperiodic - чувствительность к фотопериоду). У представителей Triticeae чувствительность к яровизации контролируется группой скоординировано функционирующих генов VRN1, VRN2 и VRN3. Важнейшая роль в регуляции ответа на яровизацию принадлежит гену VRN1, кодирующему MADS-box транскрипционный фактор (Trevaskis et al., 2007), который необходим для закладки и поддержания флоральной меристемы в точке роста побега пшеницы. У озимых форм потребность в яровизации связана с тем, что VRN1 гены находятся в рецессивном состоянии и их экспрессия блокирована до наступления периода воздействия пониженных температур. Для перехода озимой пшеницы к репродуктивному развитию необходимо, чтобы уровень транскрипта VRN1 достиг определенного порогового значения (Loukoianov et al., 2005).
Отсутствие потребности в яровизации у яровых сортов связано с существенными изменениями нуклеотидной последовательности промотора или первого интрона гена VRN-A1 (Yan et al. 2004; Fu et al. 2005). Показано, что промотор генов VRN1 содержит консервативные цис-элементы (CArG-, VRN- и G-боксы), представляющие собой мишени для транскрипционных факторов, участвующих в ответе на яровизацию. Известен ряд нуклеотидных модификаций промотора VRN-A1 (Konopatskaia et al., 2016; Muterko et al. 2016) (Фиг. 1). Наиболее распространенный доминантный аллель Vrn-A1a отличается от рецессивного аллеля vrn-A1 транспозонными вставками повторяющихся последовательностей в VRN-box участке промотора (Фиг. 1). Другие доминантные аллели содержат делеционные фрагменты разной протяженности в различных участках, таких как VRN-box (аллели Vrn-A1b, Vrn-A1d, Vrn-A1e), CArG-box (аллели Vrn-A1e), G-box (аллели Vrn-A1f), а также делении размером 20 п.н. в участке между -136 и -157 п.н. (аллели Vrn-A1b, Vrn-A1d) или крупную делецию размером 50 п.н. в участке между -62 и -112 п.н. (аллели Vrn-A1f) (Фиг. 1). Помимо делеций, доминантные аллельные варианты Vrn-A1b, Vrn-A1d, Vrn-A1e также могут содержать единичные нуклеотидные замены в VRN-box участке промотора (Фиг. 1). Крупная деления размером 50 п.н. в участке между -62 и -112 п.н. а также 8 п.н. нуклеотидная делеция в G-box была обнаружена только у аллеля Vrn-A1f яровых диких видов пшеницы секции Timopheevii (Фиг. 1), и не присутствует у представителей современных культурных видов мягкой и твердой пшеницы (Muterko et al. 2016). Примечательно, что в настоящий момент не обнаружены какие либо природные яровые и озимые формы пшеницы с измененной последовательностью промотора VRN1 в области между -113 и -122 п.н., а также отсутствуют искусственные мутантные формы зерновых с модификациями промотора VRN1 в областях между -62 и -120 п.н.
Время от посева до колошения имеет важное практическое значение для производителей пшеницы, поэтому значительные усилия селекционеров направлены на сокращение вегетационного периода у культурных сортов пшеницы. Возможность контролировать сроки колошения путем точечного изменения аллелей VRN1 позволит упростить процесс получения новых высокопродуктивных сортов, соответствующих современным требованиям. Получения новых (или аналогичных диким представителям) аллелей генов VRN1, имеющих мутации в регуляторных областях гена на неизменном генетическом фоне, представляет собой техническую проблему, для решения которой необходимо расширение арсенала средств, доступных селекционерам. Настоящее изобретение решает эту проблему путем изменения последовательности промотора VRN1 средствами редактирования геномов растений CRJSPR/Cas9.
Основными компонентами системы редактирования CRISPR/Cas9 являются белок Cas9, обладающий нуклеазной активностью, и РНК-проводник (sgRNA, направляющая РНК специфичной композиции), которая направляет белок Cas9 к специфичному участку-мишени генома растений. РНК-проводник функционирует в качестве матрицы при редактировании ДНК и представляет собой химерную последовательность с элементами распознавания белка Cas9 и участка, комплементарного ДНК-мишени генома растения. Вследствие этого РНК-проводник состоит из двух областей - константной, длиной около 70 нуклеотидов, и вариабельной, длиной 17-25 нуклеотидов. Первая отвечает за образование функционального комплекса Cas9-sgRNA, а вариабельная - за специфичное связывания этого комплекса с целевой последовательностью ДНК в геноме (Jinek et al., 2012; Nishimasu et al., 2014). Наиболее распространенная система использует РАМ, состоящий из трех оснований NGG, он распознается нуклеазой Cas9 из Streptococcus pyogenes (Hsu et al., 2014; Mao et al., 2013). Разрезание целевой мишени возможно только при условии совпадения мишени и наличия проксимального участка 5' РАМ-мотива, прилегающего к протоспейсеру.
Сущность изобретения
Задачей изобретения является разработка РНК-проводников (направляющих РНК со специфичной нуклеотидной последовательностью), которые могут быть использованы в системах CRISPR/Cas9 для внесения изменений в консервативный участок протяженностью 50 п.н. между -62 и -112 п.н. промоторной области гена VRN-A1 (Фиг. 1) без изменения других участков генома зерновых.
Задача решается, а технический результат достигается созданием молекулы РНК-проводника, где указанный РНК-проводник содержит первую нуклеотидную последовательность, выбранную из SEQ ID NO: 1 или SEQ ID NO: 2 или SEQ ID NO: 3 и вторую нуклеотидную последовательность SEQ ID NO: 4, фланкирующую первую последовательность с 3'-конца и представляющую собой РНК-шпильку, при этом указанная первая последовательность обеспечивает связывание с целевым высококонсервативным участком промотора гена VRN-A1, а указанная вторая последовательность способна распознаваться РНК-направляемой ДНК-эндонуклеазой Cas9 с обеспечением внесения мутаций в целевой участок.
Принцип функционирования предлагаемых РНК-проводников базируется на особенностях работы системы CRISPR/Cas9. Нуклеаза Cas9 взаимодействует с представленными в настоящей заявке направляющими молекулами РНК-проводников, в результате чего образуется функциональный комплекс, который нацеливает белок Cas9 в специфическое местоположение внутри целевой последовательности промотора гена VRN-A1, комплементарной участку РНК-проводника. В результате такого взаимодействия функциональные домены комплексы CRISPR/Cas9 вносят разрывы в обе цепи ДНК, которые в дальнейшем восстанавливаются репарационными системами клетки. В отсутствии матрицы репарация проводится за счет негомологичного соединения концов (NHEJ, non-homologous end joining). При этом в месте разрыва могут происходить вставки или делеции нуклеотидов, что вероятно приведет к образованию мутантных вариантов последовательности промотора гена VRN-A1.
В одном аспекте настоящее изобретение предусматривает возможность применения разрабатываемого РНК-проводника непосредственно в виде молекулы РНК и использование его в работах по геномному редактированию в составе рибонуклеопротеиновых комплексов (РНП), т.е. в системах CPJSPR/Cas9 где комплекс Cas9-sgRNA может быть представлен в виде очищенного функционального белка Cas9 в смеси с молекулами РНК-проводника.
В другом аспекте настоящее изобретение предусматривает создание ДНК-векторов для интродукции в геном однодольных зерновых культур последовательности разрабатываемого РНК-проводника с целью его стабильной экспрессии в отдельных клетках, тканях и растениях в результате генетической трансформации.
Краткое описание чертежей
Фигура 1. Сравнительная характеристика нуклеотидной последовательности доминантных (Vrn) и рецессивных (vrn) аллелей гена VRN-A1, несущих мутации в области промотора, идентифицированных у видов и сортов полиплоидной пшеницы (согласно Konopatskaia et al. 2016; с модификациями).
Фигура 2. Нуклеотидная последовательность амплифицированного с геномной ДНК пшеницы 'Chinese Spring' фрагмента промоторной последовательности гена VRN-A1 (699 п.н.) (рецессивный вариант) использованная для разработки РНК-проводников.
Фигура 3. Положение таргетных последовательностей на гене VRN-A1, являющихся целевыми мишенями для разработанных РНК-проводников в консервативной области (выделено зеленым) и в VRN-box участке (выделено желтым).
Фигура 4. Нуклеотидные последовательности мишеней для РНК-проводников указанных на Фиг. 3.
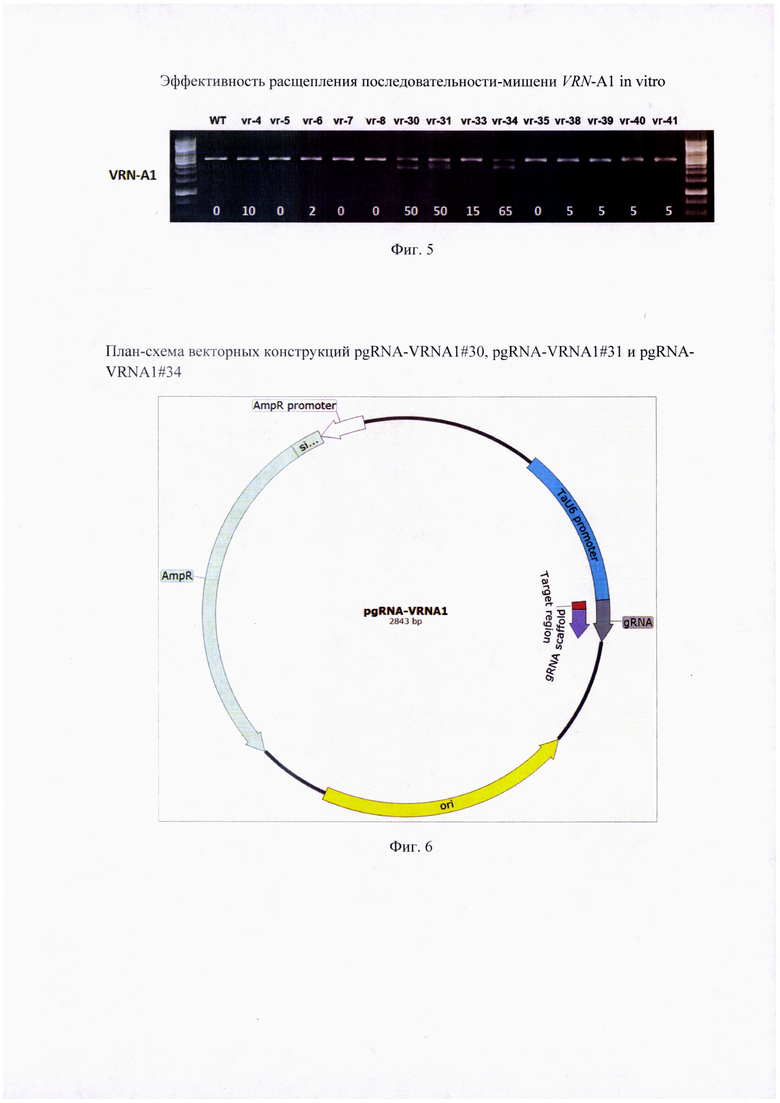
Фигура 5. Эффективность расщепления последовательностей-мишеней промоторной области гена VRN-A1. Представлена активность in vitro комплекса Cas9-РНК-проводник при разрезании целевого ДНК-субстрата. Визуализация реакции разрезания с помощью фрагментного анализа в 1,2% агарозном геле. В качестве отрицательного контроля (WT) использовали линеаризованный ДНК-субстрат мягкой пшеницы 'Chinese Spring' в соответствующей концентрации, к которому не был добавлен комплекс эндонуклеазы и РНК-проводника. Интенсивность полосы нерасщепленного контроля принимали за 100% и сравнивали со значения экспериментальных полос денситометрически; эффективность расщепления (указана в нижней части геля) определяли как разность в % между WT и полученным значением для каждого отдельного РНК-проводника.
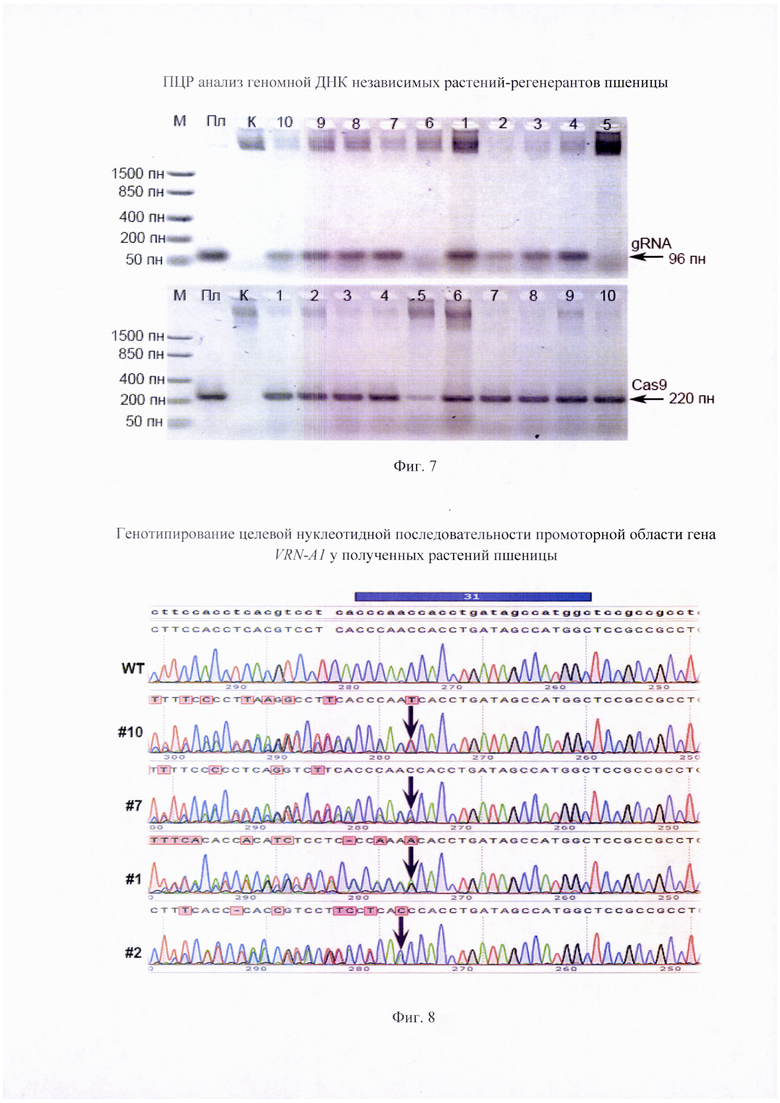
Фигура 6. План-схема векторных конструкций pgRNA-VRNA1#30, pgRNA-VRNA1#31 и pgRNA-VRNA1#34
Фигура 7. ПЦР анализ геномной ДНК независимых растений-регенерантов пшеницы, полученных в результате биобаллистического переноса разработанного РНК-проводника vr-31 в составе экспрессионного вектора pgRNA-VRNA1#31 в смеси с вектором несущим последовательность Cas9 для внесения мутаций в промоторную область гена пшеницы VRN- A1. Наличие искомых фрагментов последовательности РНК-проводника размером 96 п.н. подтверждена у восьми образцов содержащих последовательность гена Cas9 (202 п.н.). На Фигуре 7. представлены агарозные гели, содержащие ПЦР-продукты Cas9 и РНК-проводника vr-31, где М - маркер молекулярного веса; Пл - ДНК плазмиды pGCB (положительный контроль для Cas9) или pgRNA-VRNA1#31 (положительный контроль для РНК-проводника vr-31); К - нетрансгенное растение 'Chinese Spring' (отрицательный контроль), 1-10 - отобранные независимые растения.
Фигура 8. Генотипирование целевой нуклеотидной последовательности промоторной области VRN-A1 у полученных растений пшеницы методом секвенирования по Сэнгеру. Представлены нуклеотидные последовательности аллелей в целевом фрагменте контрольного растения (WT) и четырех независимых растений пшеницы #1, #2, #7, #10 с изменениями (отклонениями в сравнении с WT), которые отмечены стрелкой.
Фигура 9. Генотипирование мутантных аллелей пшеницы относительно целевой последовательности в промоторной области рецессивного гена VRN-A1, являющейся мишенью для РНК-проводника vr-31. Представлена последовательность целевой мишени, которой комплементарна последовательность РНК-проводника (заглавные буквы) в которой указана РАМ последовательность и стрелкой указан предпочтительный сайт разрезания ДНК. Красным цветом выделены нуклеотидные изменения (замены и вставки) у пшеницы #1, #2, #7, #10; при этом отсутствующие в редактированных растениях последовательности (делетированные нуклеотиды) изображены пунктирной линией.
Фигура 10. Анализ возможных изменений нуклеотидной последовательности промоторной области генов VRN-1B и VRN-1D у растений пшеницы с отредактированной последовательностью VRN-A1. Представлен порядок нуклеотидных последовательностей растений пшеницы #1, #2. #7. #10 согласно результатам секвенирования по Сэнгеру в сравнении с референсными последовательностями (WT) промоторной области генов VRN-1 В и VRN-1D, включая фрагмент имеющий сходство с последовательностью РНК-проводника vr-31.
Примеры
Далее приводятся примеры, которые иллюстрируют возможные варианты осуществления изобретения.
Пример 1
Разработка РНК-проводника
Первый этап - подбор последовательностей-мишеней для внесения изменений в консервативный участок протяженностью 50 п.н. между -62 и -112 п.н. промоторной области рецессивного гена VRN-A1 (Фиг. 1) с помощью разрабатываемых РНК-проводников. Для этого амплифицировали фрагменты промоторной области гена VRN-A1 с геномной ДНК пшеницы 'Chinese Spring' с использованием праймеров CTGAATTCTGAAAGGAAAAATTCTGCTCG и ACTGGTACCGAAGGCGTATTGGGGAAC. ПЦР-продукты обрабатывали эндонуклеазами рестрикции EcoRI и KpnI (Thermo Fisher Scientific, США), клонировали по соответствующим сайтам в векторе pUC18 и секвенировали. Последовательность участка VRN-A1 приведена на Фиг. 2. Далее подбирали потенциальные мишени для РНК-проводников в целевой последовательности промотора гена VRN-A1. С этой целью использовали современные алгоритмы и программы, находящиеся в открытом доступе. (Doenchet al., 2016). Основные критерии для выбора эффективных sgRNA были следующими: содержание GC от 30% до 80%, сохранение интактной вторичной структуры, отсутствие определенных мотивов в последовательности, отсутствие оффтергетных участков в геноме пшеницы. Поскольку выбор нуклеотидной последовательности РНК-проводника ограничен наличием РАМ последовательности (NGG) в целевой нуклеотидной последовательности промотора гена VRN-A1, авторы настоящего изобретения составили перечень из 9 потенциальных мишеней в участке между -62 и -112 п.н. промоторной области рецессивного гена VRN-A1 (Фиг. 3, Фиг. 4). Дополнительно подбирали 5 потенциальных мишеней в участке VRN-box (Фиг. 3, Фиг. 4) для проверки эффективности разрезания in vitro в значимых участках промотора. Матрицы для синтеза РНК-проводников в пробирке изготавливали с помощью олигонуклеотидов. Для этого к последовательности являющейся мишенью РНК-проводников без РАМ сайта добавляли к 5'-концу последовательность нуклеотидов TTCTAATACGACTCACTATAG, а к 3'-концу - GTTTTAGAGCTAGAAATAGC. Получившиеся олигонуклеотиды смешивали с общим праймером AAAAGCACCGACTCGGTGCCACTTTTTCAAGTTGATAACGGACTAGCCTTATTTTAACTTGCTATTTCTAGCTCTAAAAC и достраивали на амплификаторе. Затем на созданных ДНК матрицах синтезировали РНК с помощью набора HiScribe Т7 Quick High Yield RNA Synthesis Kit (New England Biolabs, США) по инструкции производителя.
Далее проводили оценку эффективности РНК-проводников путем расщепления последовательности-мишени in vitro. На реакцию объемом 30 мкл брали молекулы РНК-проводника до концентрации 30 нМ, белок Cas9 (NEB) до концентрации 30 нМ и после совместной 10 минутной инкубации при комнатной температуре добавляли линеаризованный по SspI (Thermo) вектор с геном мишенью до 3 нМ. В качестве контроля служили смеси вектора в той же концентрации без добавления Cas9 и РНК-проводника, или в присутствии Cas9, но без РНК-проводника. Инкубировали при 37°С в течение 120 мин. реакцию останавливали добавлением 1 мкл протеиназы К (30 мг/мл). Продукты реакции разделяли в 1,2% агарозном геле и фотографировали. Затем проводили денситометрический анализ для сравнительной оценки активности РНК-проводников. За 100% принимали интенсивность полосы нерасщепленного контроля; с этим значением сравнивали значения экспериментальных полос и определяли эффективность вычитая полученное значение из 100 (Фиг. 5). Анализ показал, что для внесения мутаций в целевые мишени в консервативном участке протяженностью 50 п.н. между -62 и -112 п.н. промоторной области рецессивного гена VRN-A1 среди девяти изучаемых РНК-проводников эффективными являются РНК-проводники vr-30 (SEQ ID NO: 1), vr-31 (SEQ ID NO: 2) и vr-34 (SEQ ID NO: 3). При их использовании, эффективность расщепления соответствующей целевой последовательности in vitro составила 50%, 50% и 65%, соответственно. Тогда как результативность комплексов Cas9 и РНК-проводников №33, №35, №38, №39, №40 и №41 варьировала лишь в диапазоне 0-15%. Эффективность расщепления альтернативных последовательностей в участке VRN-box рецессивного гена VRN-A1 также была низкой, и не превысила 10% при использовании РНК-проводников №4, №5, №6, №7 и №8. Сравнительная оценка расщепляющей активности индивидуальных РНК-проводников показала, что для реализации технического результата изобретения предпочтительным вариантом является использование молекул РНК-проводников vr-30 (SEQ ID NO: 1), vr-31 (SEQ ID NO: 2) и vr-34 (SEQ ID NO: 3).
Пример 2
Разработка векторов для экспрессии в клетках однодольных зерновых культур
Одним из вариантов осуществления изобретения валяется применение РНК-проводников в составе экспрессирующих векторов. В данном примере последовательности трех наиболее эффективных РНК-проводников SEQ ID NO: 1, SEQ ID NO: 2 и SEQ ID NO: 3 использовали для разработки генетических конструкций, способных экспрессироваться в клетках однодольных зерновых культур. Указанные последовательности клонировали в вектор pU6-gRNA (Addgene plasmid #53062; http://n2t.net/addgene:53062; Shan et al., 2013), расщепленном по сайту рестрикции BpiI (Thermo). Исходя их этого, РНК-проводник для внесения мутаций в геном однодольных зерновых культур содержит первую нуклеотидную последовательность, выбранную из SEQ ID NO: 1 или SEQ ID NO: 2 или SEQ ID NO: 3, и вторую нуклеотидную последовательность SEQ ID NO: 4, фланкирующую первую последовательность с 3'-конца и при этом указанная первая последовательность обеспечивает связывание с целевым высококонсервативным участком промотора гена VRN-A1, а указанная вторая последовательность способна распознаваться РНК-направляемой ДНК-эндонуклеазой Cas9 с обеспечением внесения мутаций в целевой участок. Каждый из плазмидных векторов, получивших название pgRNA-VRNA1#30, pgRNA-VRNAl#31 и pgRNA-VRNA1#34 (Фиг. 6), обеспечивает возможность трансформации компетентных клеток E. coli с последующей наработкой большого количества копий. Очищенный препарат плазмидной ДНК указанных векторов может быть использован для биолистической трансформации или трансфекции однодольных зерновых культур при условии ко-трансформации с вектором, кодирующим последовательность нуклеазы Cas9, либо при условии ко-переноса в смеси с белковым препаратом нуклеазы Cas9, либо при условии, что Cas9 уже присутствует (нарабатывается) в переносимых клетках в результате предварительной стабильной трансформации векторами, кодирующими последовательность Cas9 нуклеазы.
Пример 3
Геномное редактирование промоторной области гена VRN-A1 однодольных зерновых с помощью РНК-проводника на примере мягкой пшеницы 'Chinese Spring'
Для получения растений с отредактированным геномом требуется взаимодействие комплексов Cas9-РНК-проводник с геномной ДНК. Полученную по примеру 2 конструкцию pgRNA-VRNA1#31 смешивали с вектором, кодирующим последовательность Cas9 (вектор также содержит дополнительные селективные гены), и переносили в клетки пшеницы с помощью генной пушки для внесения мутаций в промоторную область гена VRN-A1. В качестве эксплантов использовали эмбриогенные каллусы, инициированные из тканей незрелых зиготических зародышей пшеницы 'Chinese Spring' (Triticum aestivum L.). В результате генетической трансформации отобрали 10 побегов-регенерантов. Наличие вставки последовательности РНК-проводника подтверждали ПЦР анализом тотальной ДНК, выделенной из растений-регенерантов. На Фиг. 7 представлен образец агарозных гелей после проведения ПЦР анализа. Подтвердили, что геном восьми первичных предполагаемых трансформантов содержит помимо последовательности гена Cas9 (наблюдали амплификацию характерного фрагмента размером 220 п.н.) также содержит последовательность РНК-проводника из вектора pgRNA-VRNA1#31 (наблюдали амплификацию характерного фрагмента размером 96 п.н.) (Фиг. 7).
Оценку эффективности РНК-проводника в способности обеспечить внесение мутаций в консервативную последовательность промотора VRN-A1 гена осуществляли через анализ трансгенных растений пшеницы #1, #2, #3, #4, #7, #8, #9, #10, содержащих одновременно в геноме последовательности нуклеазы Cas9 и РНК-проводника. Для этого фрагмент гена VRN-A1 амплифицировали с помощью праймеров CTGAATTCTGAAAGGAAAAATTCTGCTCG/ACTGGTACCGAAGGCGTATTGGGGAAC. ПЦР-продукты очищали набором GeneJET PCR Purification Kit (Thermo) и секвенировали с применением секвенирующего праймера TTACCATGACTCGGTGGAG. Дополнительно к этому, для подтверждения избирательности и точности разработанного РНК-проводника проводили анализ аналогичного участка промотора гомеологичных VRN-1 генов присутствующих в элементарных геномах В и D пшеницы. Для этого предварительно секвенировали фрагмент промоторной области генов Vrn-B1 и Vrn-D1, полученный путем его амплификации с геномной ДНК пшеницы 'Chinese Spring' с использованием праймеров CTGAATTCATAGTAGTATAAAAAGGACAATTG/CTGGTACCACCGAATCAACCAAACAGTG и CTGAATTCGTATAAAAGGAAAATTGTGCTCT/ACTGGTACCATCAACCAAACAGCCCCG, соответственно. Полученные фрагменты очищали набором GeneJET PCR Purification Kit (Thermo) и секвенировали с применением секвенирующего праймера TTACCATGACTCGGTGGAG. Сиквенсы фрагментов Vrn-A1, Vrn-B1 и Vrn-D1 трансгенных растений расшифровывали; последовательности раскладывали на аллели. В качестве контроля (референсная последовательность) использовали геномную ДНК пшеницы 'Chinese Spring'.
Анализ нуклеотидной последовательности показал, что четыре из восьми независимых трансгенных растений, а именно растения #1, #2, #7, #10, содержали изменения нуклеотидной последовательности в сравнении с немодифицированной последовательностью рецессивного гена VRN-A1, присутствующего в геноме 'Chinese Spring' (Фиг. 8). Эти изменения возникли в результате функциональной активности РНК-проводника vr-31, поскольку вставки/делеции преимущественно обнаружены в ожидаемом сайте расщепления молекулы ДНК комплексом Cas9-РНК-проводник (Фиг. 9). Анализ аналогичной нуклеотидной последовательности гомеологичных генов Vrn-B1 и Vrn-D1 подтвердил отсутствие каких-либо изменений в растениях #1, #2, #7, #10 (Фиг. 10). Эффективность получения растений с мутантным вариантом консервативного участка промоторной области гена VRN-A1 составила 50% (4 из 8 растений-регенерантов). Полученные растения содержали различные изменения, включающие единичную нуклеотидную делению, единичную нуклеотидную вставку, делецию из четырех нуклеотидов; у одной из линий одновременно достигнута и вставка, и делеция нуклеотидов (Фиг. 9).
Использование заявляемого РНК-проводника, в составе экспрессионного вектора pgRNA-VRNA1#31 в описываемом примере, позволило эффективно внести мутации в целевую нуклеотидную последовательность консервативного участка промоторной области гена VRN-A1 мягкой пшеницы, не затрагивая гомеологичные варианты гена VRN-1 других элементарных геномов.
Финансирование работ по созданию настоящего изобретения проводилось из средств Соглашения №075-15-2019-1667 от «31» октября 2019 г. о предоставлении из федерального бюджета грантов в форме субсидий в соответствии с пунктом 4 статьи 78.1 Бюджетного кодекса Российской Федерации на осуществление государственной поддержки создания и развития центра геномных исследований мирового уровня «Курчатовский геномный центр» в рамках реализации федерального проекта «Развитие научной и научно-производственной кооперации» национального проекта «Наука».
Список литературы
Doench JG, Fusi N, Sullender M, et al. (2016). Optimized sgRNA design to maximize activity and minimize off-target effects of CRISPR-Cas9. Nature biotechnology. 34(2), 184-191.
Fu D, Szucs P, Yan L, et al. (2005). Large deletions within the first intron in VRN-1 are associated with spring growth habit in barley and wheat. Mol. Genet. Genomics. 273, 54-65.
Hsu PD, Lander ES, Zhang F. (2014). Development and applications of CRISPR-Cas9 for genome engineering. Cell. 157, 1262-1278.
Jinek M, Chylinski K, Fonfara I, et al. (2012). A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science, 337, 816-821.
Konopatskaia I, Vavilova V, Kondratenko EY, et al. (2016). VRN1 genes variability in tetraploid wheat species with a spring growth habit. BMC Plant Biol. 16(Suppl 3):244.
Loukoianov A, Yan L, Blechl A, et al. (2005). Regulation of VRN-1 vernalization genes in normal and transgenic polyploid wheat. Plant Physiol. 138(4), 2364-2373.
Mao Y, Zhang H, Xu N, et al. (2013). Application of the CRISPR-Cas system for efficient genome engineering in plants. Mol. Plant. 6, 2008-2011.
Muterko A, Kalendar R, Salina E (2016). Novel alleles of the VERNALIZATION1 genes in wheat are associated with modulation of DNA curvature and flexibility in the promoter region. BMC Plant Biol, 16(Suppl 1), 9.
Nishimasu HFA, Ran PD, Hsu S et al. (2014). Crystal structure of Cas9 in complex with guide RNA and target DNA. Cell, 156, 935-949.
Shan Q, Wang Y, Li J et al. (2013). Targeted genome modification of crop plants using a CRISPR-Cas system. Nat. Biotechnol. 2013, 31(8), 686-688.
Trevaskis B, Hemming MN, Dennis ES, et al. (2007). The molecular basis of vernalization-induced flowering in cereals. Trends Plant Sci. 12, 352-357.
Yan L, Helguera M, Kato K. et al. (2004). Allelic variation at the VRN-1 promoter region in polyploid wheat. Theor. Appl. Genet. 109:1677-1186.
--->
Перечень последовательностей
<110> ФГБНУ ВНИИСБ
<120> МОЛЕКУЛА РНК-ПРОВОДНИКА ДЛЯ ГЕНОМНОГО РЕДАКТИРОВАНИЯ ПРОМОТОРНОЙ ОБЛАСТИ
ГЕНА VRN-A1 ОДНОДОЛЬНЫХ ЗЕРНОВЫХ С ПРИМЕНЕНИЕМ СИСТЕМЫ CRISPR/CAS9
<160> 4
<210> 1
<211> 20
<212> РНК
<213> Искусственная последовательность
<400> 20
ACCCAACCACCUGAUAGCCA
<210> 2
<211> 20
<212> РНК
<213> Искусственная последовательность
<400> 20
GCCAUGGCUAUCAGGUGGUU
<210> 3
<211> 20
<212> РНК
<213> Искусственная последовательность
<400> 20
GCGGCGGAGCCAUGGCUAUC
<210> 4
<211> 76
<212> РНК
<213> Искусственная последовательность
<400> 76
GUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGC
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЯ ПШЕНИЦЫ С НУКЛЕОТИДНОЙ ДЕЛЕЦИЕЙ В ПРОМОТОРНОЙ ОБЛАСТИ ГЕНА VRN-A1 ПРИ ПОМОЩИ ТЕХНОЛОГИИ РЕДАКТИРОВАНИЯ ГЕНОМА РАСТЕНИЙ CRISPR/CAS9 | 2020 |
|
RU2772577C2 |
| СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЯ ПШЕНИЦЫ С НУКЛЕОТИДНОЙ ВСТАВКОЙ В ПРОМОТОРНОЙ ОБЛАСТИ ГЕНА VRN-A1 ПРИ ПОМОЩИ ТЕХНОЛОГИИ РЕДАКТИРОВАНИЯ ГЕНОМА РАСТЕНИЙ CRISPR/CAS9 | 2020 |
|
RU2772578C2 |
| СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЯ ПШЕНИЦЫ С БИАЛЛЕЛЬНЫМИ МУТАЦИЯМИ В ПРОТОМОТОРНОЙ ОБЛАСТИ ГЕНА VRN-A1 ПРИ ПОМОЩИ ТЕХНОЛОГИИ РЕДАКТИРОВАНИЯ ГЕНОМА РАСТЕНИЙ CRISPR/CAS9 | 2020 |
|
RU2772575C2 |
| Молекула РНК-проводника sgRNA для внесения мутаций в консервативный участок промоторной области гена PPD-D1 мягкой пшеницы с применением системы редактирования генома CRISPR/Cas9 | 2024 |
|
RU2822358C1 |
| ГЕНЕТИЧЕСКАЯ КОНСТРУКЦИЯ НА ОСНОВЕ СИСТЕМЫ РЕДАКТИРОВАНИЯ ГЕНОМА CRISPR/CAS9, КОДИРУЮЩАЯ НУКЛЕАЗУ CAS9, ДЛЯ ГЕНОМНОГО РЕДАКТИРОВАНИЯ ОДНОДОЛЬНЫХ ЗЕРНОВЫХ КУЛЬТУР НА ОСНОВЕ ДВОЙНОГО ОТБОРА РАСТЕНИЙ | 2020 |
|
RU2762830C1 |
| КОМПОЗИЦИИ ДЛЯ ИНДУКЦИИ ГАПЛОИДИИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2771141C2 |
| РНК-проводники для подавления репликации вируса гепатита B и для элиминации вируса гепатита B из клетки-хозяина | 2017 |
|
RU2652899C1 |
| ГЕТЕРОЗИГОТНЫЕ ПО CENH3 ОДНОДОЛЬНЫЕ РАСТЕНИЯ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ ДЛЯ ИНДУКЦИИ ГАПЛОИДОВ И ОДНОВРЕМЕННОГО РЕДАКТИРОВАНИЯ ГЕНОМА | 2021 |
|
RU2832578C1 |
| СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЯ КАРТОФЕЛЯ С ТЕТРААЛЛЕЛЬНЫМИ МУТАЦИЯМИ В КОДИРУЮЩЕЙ ОБЛАСТИ ГЕНА EDR1 ПРИ ПОМОЩИ ТЕХНОЛОГИИ РЕДАКТИРОВАНИЯ ГЕНОМА РАСТЕНИЙ CRISPR/CAS9 | 2022 |
|
RU2817384C2 |
| СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЯ КАРТОФЕЛЯ С ТРИАЛЛЕЛЬНЫМИ МУТАЦИЯМИ В КОДИРУЮЩЕЙ ОБЛАСТИ ГЕНА EDR1 ПРИ ПОМОЩИ ТЕХНОЛОГИИ РЕДАКТИРОВАНИЯ ГЕНОМА РАСТЕНИЙ CRISPR/CAS9 | 2022 |
|
RU2817374C2 |




Изобретение относится к области биотехнологии, в частности к молекуле РНК-проводника для целенаправленного воздействия на нуклеотидную последовательность консервативного участка промоторной области гена VRN-A1 однодольных зерновых культур системами редактирования генома CRISPR/Cas9. Изобретение позволяет эффективно получать новые аллели генов VRN1, имеющих мутации в регуляторных областях гена на неизменном генетическом фоне. 10 ил., 3 пр.
Молекула РНК-проводника (sgRNA) для внесения мутаций в консервативный участок промоторной области гена VRN-A1 однодольных зерновых с применением системы редактирования генома CRISPR/Cas9, где указанная молекула РНК-проводника содержит первую нуклеотидную последовательность, выбранную из SEQ ID NO: 1, SEQ ID NO: 2 или SEQ ID NO: 3, и вторую нуклеотидную последовательность SEQ ID NO: 4, фланкирующую первую последовательность с 3'-конца, при этом указанная первая последовательность обеспечивает связывание с целевым консервативным участком промотора гена VRN-A1, а указанная вторая последовательность способна распознаваться РНК-направляемой ДНК-эндонуклеазой Cas9 с обеспечением внесения мутаций в целевой участок.
| KONOPATSKAIA I | |||
| et al., VRN1 genes variability in tetraploid wheat species with a spring growth habit, The Author(s) BMC Plant Biology, 2016, 16(Suppl 3):244 | |||
| MUTERKO A | |||
| et al., Novel alleles of the VERNALIZATION1 genes in wheat are associated with modulation of DNA curvature and flexibility in the promoter region, BMC Plant Biology, 2016, |

