Данная заявка содержит перечень последовательностей, который был представлен в электронном виде в формате ASCII и в данном документе полностью включен посредством ссылки. Указанная копия ASCII, созданная 11 августа 2017 года, называется TNO-0001-WO_SL. txt и имеет размер 26401 байт.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Данное изобретение относится к трансгенным животным, отличным от человека, продуцирующим модифицированные антитела, содержащие только тяжелые цепи (HCAb - heavy chain-only antibody). В частности, изобретение относится к трансгенным животным, отличным от человека, таким как трансгенные крысы или мыши, продуцирующим модифицированные человеческие или химерные HCAb с пониженной склонностью к агрегации, полученным таким образом антителам и способам их получения и применения.
УРОВЕНЬ ТЕХНИКИ
Антитела, содержащие только тяжелые цепи
Основная четырехцепочечная единица антитела представляет собой гетеротетрамерный гликопротеин, состоящий из двух идентичных легких (L) цепей и двух идентичных тяжелых (H) цепей. В случае IgG 4-цепочечная единица обычно составляет около 150000 дальтон. Каждая L-цепь связана с H-цепью одной ковалентной дисульфидной связью, тогда как две H-цепи связаны друг с другом одной или несколькими дисульфидными связями в зависимости от изотипа H-цепи. Каждая H и L цепь также имеет регулярно расположенные внутрицепочечные дисульфидные мостики. Каждая H-цепь имеет на N-конце вариабельный домен (VH), за которым следуют три константных домена (CH) для каждой из цепей α и γ и четыре домена C H для изотипов μ и ε. Каждая L-цепь имеет на N-конце вариабельный домен (VL), за которым следует константный домен (CL) на ее другом конце. VL выровнен с VH, а CL выровнен с первым константным доменом тяжелой цепи (CH1). Считается, что конкретные аминокислотные остатки образуют поверхность соприкосновения между вариабельными доменами легкой цепи и тяжелой цепи. Спаривание VH и VL вместе образует один антигенсвязывающий сайт. Антитело IgM состоит из 5 основных гетеротетрамерных звеньев вместе с дополнительным полипептидом, называемым J-цепью, и, следовательно, содержит 10 антигенсвязывающих сайтов, в то время как секретируемые IgA-антитела могут полимеризоваться с образованием поливалентных сборок, включающих 2-5 основных 4-цепочечных звеньев вместе с J-цепью. О структуре и свойствах различных классов антител см., например, Basic and Clinical Immunology, 8-е издание, Daniel P. Stites, Abba I. Terr и Tristram G. Parslow (eds.), Appleton & Lange, Norwalk, CT, 1994, стр. 71 и глава 6. В таких антителах взаимодействие доменов VH и VL образует антигенсвязывающую область, хотя связыванию способствуют и домен CH1 и части домена CL.
L-цепь из любого вида позвоночных может быть отнесена к одному из двух различных типов, называемых каппа (κ) и лямбда (λ), что основывается на аминокислотных последовательностях их константных доменов. В зависимости от аминокислотных последовательностей константного домена их тяжелых цепей (CH) иммуноглобулины могут быть отнесены к разным классам или изотипам. Существует пять классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, имеющих тяжелые цепи, обозначенные как α, δ, ε, γ и μ соответственно. Классы γ и α дополнительно подразделяются на подклассы на основе относительно незначительных различий в последовательности и функции CH, например, люди экспрессируют следующие подклассы: IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2.
В обычном IgG-антителе ассоциация тяжелой цепи и легкой цепи частично обусловлена гидрофобным взаимодействием между константной областью легкой цепи и константным доменом CH1 тяжелой цепи. В каркасных областях 2 (FR2) и 4 (FR4) тяжелой цепи имеются дополнительные остатки, которые также способствуют этому гидрофобному взаимодействию между тяжелой и легкой цепями.
Известно, однако, что сыворотка верблюдовых (подотряд Мозоленогие (Tylopoda), который включает верблюдов, дромадеров и лам) содержит в качестве основного тип антител, состоящих исключительно из парных H-цепей (антитела, содержащие только тяжелые цепи или HCAb). HCAb Camelidae ( Camelus dromedarius, Camelus bactrianus, Lama glama, Lama guanaco, Lama alpaca и Lama vicugna) имеют уникальную структуру, состоящую из одного вариабельного домена (VHH), шарнирной области и двух константных доменов (CH2 и CH3), которые высоко гомологичны доменам СН2 и СН3 классических антител. Эти HCAb не имеют первого домена константной области (CH1), который присутствует в геноме, но вырезается при сплайсинге во время процессинга мРНК. Отсутствие домена CH1 объясняет отсутствие легкой цепи в HCAb, поскольку этот домен является местом закрепления константного домена легкой цепи. Такие HCAb естественным образом эволюционировали для придания антигенсвязывающей специфичности и высокой аффинности с помощью трех CDR из обычных антител или их фрагментов (Muyldermans, 2001; J Biotechnol 74: 277-302; Revets et al., 2005; Expert Opin Biol Ther 5: 111-124).
Хрящевые рыбы также выработали особый тип иммуноглобулина, называемого IgNAR, который лишен легких полипептидных цепей и полностью состоит из тяжелых цепей.
Способность антител, содержащих только тяжелые цепи, лишенных легкой цепи, связывать антиген была установлена в 1960-х годах (Jaton et al. (1968) Biochemistry, 7, 4185-4195). Тяжелая цепь иммуноглобулина, физически отделенная от легкой цепи, сохранила 80% антигенсвязывающей активности по сравнению с тетрамерным антителом.
Sitia et al. (1990) Cell, 60, 781-790 продемонстрировали, что удаление домена CH1 из перестроенного гена μ мыши приводит к продуцированию антитела только из тяжелых цепей, лишенного легкой цепи, в культуре клеток млекопитающих. Вырабатываемые антитела сохраняли специфичность связывания VH и эффекторные функции.
Открытие антител, содержащих только тяжелые цепи верблюдовых стимулировало интерес к разработке человеческих однодоменных антител в искусственных системах, таких как фаговый дисплей. Ранние человеческие доменные антитела, идентифицированные таким образом, были склонны к агрегации и имели проблемы с растворимостью, вероятно, из-за экспонированных гидрофобных пятен в каркасных областях, которые обычно скрыты на поверхности соприкосновения с константной областью легкой цепи. Последующие исследования, которые выяснили кристаллическую структуру VH доменных антител человека, выявили поверхностно-экспонированные остатки человеческих доменных антител. Barthelemy et al. (2008) J. Biol. Chem., 283, 3639-3654 сообщают о всестороннем анализе факторов, способствующих стабильности и растворимости автономных доменов VH человека.
Клонированный и выделенный домен VHH представляет собой стабильный полипептид, обладающий полной антигенсвязывающей способностью исходного HCAb. Нанотела представляют собой наименьшие доступные интактные антигенсвязывающие фрагменты (около 12-15 кДа), обладающие полной антигенсвязывающей способностью исходной тяжелой цепи эволюционировавших антител с тяжелыми цепями, которые являются полностью функциональными в отсутствие легких цепей. Эти VHH-домены образуют основу терапевтических антител нового поколения, называемых нанотелами, которые подходят для внутривенного перорального или местного введения, и их можно легко изготовить в моно- или поливалентных формах, проявляющих высокую активность и афинность к одной или большему количеству мишеней.
Однодоменные антитела VHH, включая способы их получения, описаны, например, в WO2004062551.
Мыши, у которых локус легкой (L) цепи λ (лямбда) и/или локусы L цепи λ и κ (каппа) цепи были функционально отключены, и антитела, продуцируемые такими мышами, описаны в патентах США №№ 7541513 и 8367888. Рекомбинантное продуцирование антител, содержащих только тяжелые цепи, у мышей и крыс было описано, например, в WO 2006008548; публикация заявки США № 20100122358; Nguyen et al., 2003, Immunology; 109 (1), 93-101;  и др., Crit. Rev. Immunol.; 2006, 26(5):377-90; and Zou et al., 2007, J Exp Med; 204(13): 3271-3283. Получение нокаутированных крыс с помощью микроинъекций в эмбрионы нуклеаз цинкового пальца описано в Geurts et al., 2009, Science, 325 (5939): 433. Характеризация крыс, нокаутированных по тяжелой цепи иммуноглобулина, описана
и др., Crit. Rev. Immunol.; 2006, 26(5):377-90; and Zou et al., 2007, J Exp Med; 204(13): 3271-3283. Получение нокаутированных крыс с помощью микроинъекций в эмбрионы нуклеаз цинкового пальца описано в Geurts et al., 2009, Science, 325 (5939): 433. Характеризация крыс, нокаутированных по тяжелой цепи иммуноглобулина, описана  et al., 2010, European Journal of Immunology, 40: 2932-2941. Растворимые антитела, содержащие только тяжелые цепи, и трансгенные грызуны, содержащие гетерологичный локус тяжелой цепи, продуцирующий такие антитела, описаны в патентах США №№8883150. Структуры CAR-T, содержащие однодоменные антитела в качестве связывающего (нацеливающего) домена, описаны, например, в Iri-Sofla et al., 2011, Experimental Cell Research 317: 2630-2641 и Jamnani et al., 2014, Biochim Biophys Acta 1840: 378-386.
et al., 2010, European Journal of Immunology, 40: 2932-2941. Растворимые антитела, содержащие только тяжелые цепи, и трансгенные грызуны, содержащие гетерологичный локус тяжелой цепи, продуцирующий такие антитела, описаны в патентах США №№8883150. Структуры CAR-T, содержащие однодоменные антитела в качестве связывающего (нацеливающего) домена, описаны, например, в Iri-Sofla et al., 2011, Experimental Cell Research 317: 2630-2641 и Jamnani et al., 2014, Biochim Biophys Acta 1840: 378-386.
Несмотря на недавние достижения, существует потребность в улучшенных способах получения антител, содержащих только тяжелые цепи, которые имеют меньшую склонность к агрегации и сохраняют высокую афинность к своей мишени.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Данное изобретение основано, по меньшей мере частично, на обнаружении того, что антитела, содержащие только тяжелые цепи (HCAb), с меньшей склонностью к агрегации, можно получить путем замены нативного аминокислотного остатка в первой позиции четвертой каркасной области (FR4) HCAb другим аминокислотным остатком, который способен нарушать целостность экспонированного на поверхности гидрофобного пятна, содержащего нативный аминокислотный остаток или связанного с нативным аминокислотным остатком в этой позиции. Такие гидрофобные пятна обычно скрыты на поверхности соприкосновения с константной областью легкой цепи антитела, но становятся выставленными на поверхности в HCAb и существуют, по меньшей мере частично, для нежелательной агрегации и ассоциации легкой цепи с HCAb.
В одном аспекте данное изобретение относится к изолированному человеческому или химерному антителу, содержащему только тяжелые цепи (HCAb), содержащему вариабельный домен тяжелой цепи (VH), содержащему определяющие комплементарность области (CDR) и каркасные области (FR), имеющему аффинность связывания с антигеном-мишенью в отсутствие легкой цепи антитела, причем в указанном VH домене нативный аминокислотный остаток в первой позиции четвертой каркасной области (FR4) указанного HCAb замещен другим аминокислотным остатком, который способен нарушать целостность экспонированного на поверхности гидрофобного участка, содержащего нативный аминокислотный остаток или связанного с нативным аминокислотным остатком в этой позиции.
В одном варианте осуществления HCAb представляет собой антитело человека.
В другом варианте осуществления в HCAb указанный нативный аминокислотный остаток в первой позиции FR4 замещен полярным аминокислотным остатком.
В еще одном варианте осуществления в HCAb указанный нативный аминокислотный остаток в первой позиции FR4 замещен положительно заряженным аминокислотным остатком, таким как, например, лизин (K), аргинин (R) или гистидин (H), предпочтительно аргинином (R).
В конкретном варианте осуществления HCAb содержит замену триптофана (W) на аргинин (R) в первом аминокислотном остатке в четвертой каркасной области (FR4).
Во всех вариантах осуществления HCAb могут содержать одну или несколько дополнительных мутаций в одной или нескольких каркасных областях.
Во всех вариантах осуществления HCAb могут иметь пониженную склонность к агрегации по сравнению с соответствующим антителом, содержащим нативный аминокислотный остаток в первом аминокислотном остатке в FR4.
Во всех вариантах осуществления HCAb может иметь аффинность связывания от примерно 1 пМ до примерно 1 мкМ со своим антигеном-мишенью.
В другом аспекте данное изобретение относится к изолированному человеческому или химерному антителу, содержащему только тяжелые цепи (HCAb), имеющему аффинность связывания с антигеном-мишенью в отсутствие легкой цепи антитела, содержащему вариабельный домен тяжелой цепи (VH), содержащий определяющие комплементарность области (CDR) и каркасные области (FR), причем указанный HCAb содержит замену триптофана (T) на аргинин (R) в первом аминокислотном остатке в четвертй FR области (FR4) аминокислотной последовательности нативного VH человека.
В одном варианте осуществления HCAb дополнительно содержит константный домен тяжелой цепи (CH), лишенный области CH1, и может представлять собой антитело IgG, такое как антитело IgG1.
В другом варианте осуществления HCAb содержит одну или несколько дополнительных мутаций в одной или большем количестве областей FR.
В еще одном варианте осуществления HCAb обладает пониженной склонностью к агрегации по сравнению с соответствующим антителом, содержащем нативный аминокислотный остаток в первом аминокислотном остатке в FR4.
В другом аспекте данное изобретение относится к химерному антигенному рецептору (CAR), содержащему антитело с исключительно тяжелыми цепями, как описано в данном документе. В одном варианте осуществления указанный CAR содержит один человеческий домен VH.
В дополнительном аспекте данное изобретение относится к изолированному автономному вариабельному домену тяжелой цепи (VH) человеческого антитела, содержащему определяющие комплементарность области (CDR) и каркасные области (FR), обладающие аффинностью связывания с антигеном-мишенью, содержащему замену другого аминокислотного остатка, что способно нарушить целостность экспонированного на поверхности гидрофобного пятна, содержащего нативный аминокислотный остаток или связанного с нативным аминокислотным остатком в этой позиции, для указанного нативного аминокислотного остатка в первом аминокислотном остатке в четвертой каркасной области (FR4).
В одном варианте осуществления в изолированном автономном VH домене человека указанный нативный аминокислотный остаток в первой позиции FR4 замещен полярным аминокислотным остатком.
В другом варианте осуществления указанный нативный аминокислотный остаток в первой позиции FR4 замещен положительно заряженным аминокислотным остатком, таким как остаток лизина (K), аргинина (R) или гистидина (H), предпочтительно остатком аргинина (R).
В дополнительном варианте осуществления выделенный автономный домен VH человека содержит замену триптофана (W) на аргинин (R) в первом аминокислотном остатке в четвертой каркасной области (FR4).
В еще одном варианте осуществления выделенный автономный домен VH человека содержит одну или несколько дополнительных мутаций в одной или нескольких каркасных областях.
В дополнительном аспекте данное изобретение относится к многовалентному связывающему белку, содержащему множество антигенсвязывающих доменов, которые включают в себя по меньшей мере один VH-домен человека, содержащий определяющие комплементарность области (CDR) и каркасные области (FR), имеющие аффинность связывания с антигеном-мишенью, причем в указанном VH домене нативный аминокислотный остаток в первой позиции четвертой каркасной области (FR4) указанного поливалентного связывающего белка замещен другим аминокислотным остатком, который способен нарушать целостность экспонированного на поверхности гидрофобного участка, содержащего или связанного с ним с нативного аминокислотного остатка в этой позиции.
В одном варианте осуществления в многовалентном связывающем белке указанный нативный аминокислотный остаток в первой позиции FR4 замещен полярным аминокислотным остатком.
В другом варианте осуществления в многовалентном связывающем белке указанный нативный аминокислотный остаток в первой позиции FR4 замещен положительно заряженным аминокислотным остатком, таким как остаток лизина (K), аргинина (R) или гистидина (H) предпочтительно остатком аргинина (R).
В дополнительном варианте осуществления поливалентный связывающий белок содержит замену триптофана (W) на аргинин (R) в первом аминокислотном остатке в четвертой каркасной области (FR4).
Во всех вариантах многовалентный связывающий белок может содержать одну или несколько дополнительных мутаций в одной или нескольких каркасных областях.
В дополнительном варианте осуществления данное изобретение относится к рекомбинантному локусу иммуноглобулина (Ig), содержащего только тяжелые цепи, содержащему один или несколько V геных сегментов человека, один или несколько D генных сегментов человека и один или несколько J генных сегментов человека, которые при рекомбинации друг с другом в геноме животного, отличного от человека, и после созревания аффинности, кодируют вариабельную область тяжелой цепи (VH), содержащую определяющие комплементарность области (CDR) и каркасные области (FR), причем по меньшей мере один из указанных J-сегментов человека содержит кодон, кодирующий не нативный аминокислотный остаток в первой позиции четвертой каркасной области (FR4), который способен нарушать целостность экспонированного на поверхности гидрофобного участка, содержащего нативный аминокислотный остаток или связанного с нативным аминокислотным остатком в этой позиции.
В одном варианте осуществления локус рекомбинантного Ig, содержащего только тяжелые цепи, дополнительно содержит генный сегмент константной (C) области, кодирующий константную эффекторную область иммуноглобулина, лишенную функциональности CH1.
В различных вариантах осуществления локус рекомбинантного Ig, содержащего только тяжелые цепи, содержит от двух до 40 D генных сегментов и/или от двух до 20 J генных сегментов.
В другом варианте осуществления более чем один из J-сегментов человека содержит кодон, кодирующий не нативный аминокислотный остаток в первой позиции четвертой каркасной области (FR4), который способен нарушать целостность экспонированного на поверхности гидрофобного участка, содержащего нативный аминокислотный остаток или связанного с нативным аминокислотным остатком в этой позиции.
В еще одном варианте осуществления в рекомбинантном локусе Ig, содержащего только тяжелые цепи, все J-сегменты человека содержат кодон, кодирующий не нативный аминокислотный остаток в первой позиции четвертой каркасной области (FR4), который способен нарушать целостность экспонированного на поверхности гидрофобного пятна, содержащего нативный аминокислотный остаток или связанного с нативным аминокислотным остатком в этой позиции.
В дополнительном варианте осуществления в кодируемой области тяжелой цепи VH нативный аминокислотный остаток в первой позиции FR4 замещен полярным аминокислотным остатком, таким как лизин (K), аргинин (R) или гистидин (H), предпочтительно остатком аргинина (R).
В еще одном следующем варианте осуществления кодированная VH область содержит замену триптофана (W) на аргинин (R) в первом аминокислотном остатке в четвертой каркасной области (FR4).
В еще одном варианте осуществления локус рекомбинантного Ig, содержащего только тяжелые цепи, содержит J4 генный сегмент, в котором кодон для W заменен на R.
Во всех вариантах осуществления локус рекомбинантного Ig, содержащего только тяжелые цепи, кодирует VH область, которая содержит одну или несколько дополнительных мутаций в одной или нескольких каркасных областях.
В дополнительном варианте осуществления локус рекомбинантного Ig только с тяжелой цепью кодирует человеческое или гуманизированное антитело только с тяжелой цепью, содержащее VH область, как описано выше.
В другом аспекте данное изобретение относится к трансгенному животному, отличному от человека, содержащему локус рекомбинантного Ig, содержащего только тяжелые цепи, как описано выше.
В различных вариантах осуществления трансгенное животное, отличное от человека, представляет собой млекопитающее, отличное от человека, такое как позвоночное, отличное от человека, грызуна, мышь или крысу, например UniRat™.
В дополнительном аспекте данное изобретение относится к трансгенному животному, отличному от человека, которое не экспрессирует никаких функциональных генов легкой цепи иммуноглобулина и содержит гетерологичный локус Ig исключительно с тяжелыми цепями, содержащий один или несколько V генных сегментов, один или несколько D генных сегментов, и один или несколько J генных сегментов, которые при рекомбинации друг с другом и после созревания аффинности кодируют домен VH, содержащий определяющие комплементарность области (CDR) и каркасные области (FR), в котором нативный аминокислотный остаток находящийся в первой позиции четвертой каркасной области (FR4) указанного домена VH замещен другим аминокислотным остатком, который способен нарушать целостность экспонированного на поверхности гидрофобного участка, содержащего нативный аминокислотный остаток или связанного с нативным аминокислотным остатком в этой позиции, и один или несколько константных сегментов гена эффекторной области, каждый из которых кодирует константную эффекторную область антитела, включающую в себя функциональность CH1, причем генные сегменты расположены так, что V, D и J генные сегменты и генный сегмент константной области рекомбинируют для получения перестроенного аффинно-зрелого локуса гена, содержащего только тяжелые цепи, кодирующего антитело исключительно с тяжелыми цепями (HCAb).
В одном варианте осуществления в VH домене указанного трансгенного животного, отличного от человека, нативный аминокислотный остаток в первой позиции FR4 замещен полярным аминокислотным остатком или положительно заряженными аминокислотными остатками, такими как лизин (K), остаток аргинина (R) или гистидина (H), предпочтительно остатком аргинина (R).
В другом варианте осуществления у трансгенного животного, отличного от человека, домен VH включает замену триптофана (W) на аргинин (R) в первом аминокислотном остатке в четвертой каркасной области (FR4).
В еще одном варианте осуществления у трансгенного животного, отличного от человека, гетерологичный локус, содержащий только тяжелые цепи, содержит J4 сегмент, в котором кодон для W заменен кодоном для R.
Во всех вариантах реализации закодированное антитело, содержащее только тяжелые цепи, содержит одну или несколько дополнительных мутаций в одной или нескольких каркасных областях.
В дополнительном варианте осуществления трансгенное животное, отличное от человека, представляет собой млекопитающее, позвоночное, грызуна, мышь или крысу, такую как UniRat™.
Во всех аспектах в определенных вариантах осуществления антитела, содержащие только тяжелые цепи в данном документе не содержат мутаций в других каркасных областях, включая области FR1, FR2 и FR3.
Во всех аспектах в определенных вариантах осуществления антитела, содержащие только тяжелые цепи в данном документе не содержат дополнительных каркасных мутаций, обычно присутствующих у верблюдовых, таких как верблюд, лама, дромедар, альпака или гуанако.
Во всех аспектах в некоторых вариантах осуществления антитела, содержащие только тяжелые цепи в данном документе могут содержать одну или несколько дополнительных мутаций в одной или нескольких каркасных областях, включая области FR1, FR2, FR3 и/или FR4, такие как, например, в FR2 область или в областях FR2 и FR4.
Во всех аспектах и вариантах осуществления целевые антигены, к которым HCAb по данному изобретению обладает аффинностью связывания, включают, без ограничения, рецепторы клеточной поверхности и опухолевые антигены, такие как, например, EGFR, ErbB2 (HER2), ErbB3 (HER3), ErbB4 (HER4), CTLA-4/CD152, RANKL, TNF-α, CD20, IL-12/IL-23, IL1-β, IL-17A, IL-17F, CD38, NGF, IGF-1, IL-12, CD20, CD30, CD39, CD73, CD40, PD-1, PDL-1, PD-L2, BCMA, BTLA, тимусный стромальный лимфопоэтин (TSP), рецептор фолликулостимулирующего гормона (FSHR), простат-специфический мембранный антиген (PSMA), антиген стволовых клеток простаты (PSCA), CD137, OX-40 и IL-33.
Во всех аспектах и вариантах осуществления связывающие домены HCAb могут быть частью мультиспецифического связывающего белка, который связывается с несколькими различными эпитопами одного и того же антигена-мишени или несколькими различными эпитопами на более чем одном антигене-мишени. В частности, включены такие мультиспецифические, как биспецифические HCAb, включая, например, биспецифические структуры HCAb, имеющие следующие аффинности связывания: молекула адгезии эпителиальных клеток (EpCam) × CD3; CD19 × CD3; EpCam × CD3; TNF-α x IL-17; IL-1α × IL-1β; CD30 × CD16A; рецептор 2 эпидермального фактора роста человека (HER2) × HER3; IL-4 × IL-13; ангиопоэтин 2 (Ang-2) × фактор a роста эндотелия сосудов (VEGF-A); Фактор IXa × Фактор X; рецептор эпидермального фактора роста (EGFR) × HER3; IL-17A × IL-17F; HER2 × HER3; карциноэмбриональный антиген × CD3; CD20 × CD3; CD123 × CD3; BCMA × CD3, PSMA × PSCA × CD3, PSMA × CD3, PSCA × CD3, CD19 × CD22 × CD3, CD22 × CD3, CD38 × PD1, CD38 × PD-L1, CD38 × CD73, CD38 × CD39, PD1 × CD39 × CD73 и PD1 × CD73.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг. 1 представляет собой диаграмму человеческого трансгена с J4 генным сегментом (SEQ ID NO: 40), экспрессирующего R в позиции 101 в UniRat™ (SEQ ID NO: 41), который экспрессирует антитела, содержащие только тяжелые цепи, как описано в примерах.
Фиг. 2 представляет собой диаграмму человеческого трансгена со всеми J-сегментами (SEQ ID NOS 42-47, соответственно, в порядке появления), экспрессирующими R в позиции 101 в UniRat™, которые экспрессируют антитела, содержащие только тяжелые цепи, как описано в примерах.
Фиг. 3 демонстрирует использование гена J в UniRat™ и OmniFlic™, причем последняя является трансгенной крысой с тем же V генным кластером человека, что и UniRat™, но экспрессирует фиксированные легкие цепи каппа.
Фиг. 4 иллюстрирует то, что мутация W → R в первом аминокислотном остатке четвертой каркасной области (FR4) в антителах человека, содержащих исключительно тяжелые цепи, ингибирует ассоциацию с легкой цепью лямбда.
Фиг. 5 Свободная связь белка лямбда с антителами тяжелой цепи в том же семействе CDR3. На фигуре показано множественное выравнивание последовательностей 11 последовательностей VH антител с тяжелыми цепями в одном и том же семействе CDR3 (SEQ ID NOS 48-58, соответственно, в порядке появления). Все эти последовательности содержат W в позиции 101.
Фиг. 6 иллюстрирует две структуры химерного антигенного рецептора с использованием внеклеточного связывающего домена VH человека, сравнивая структуру scFv CAR-T (панель A) и структуру CAR-T (панель B), используя антитело человека, содержащее исключительно тяжелые цепи по данному изобретению.
На Фиг. 7 изображены различные мультиспецифические конструкции HCAb, содержащие связывающие домены VH человека.
ПОДРОБНОЕ ОПИСАНИЕ ПРЕДПОЧТИТЕЛЬНЫХ ВЫРИАНТОВ ОСУЩЕСТВЛЕНИЯ
Практика данного изобретения будет использовать, если не указано иное, обычные методы молекулярной биологии (включая рекомбинантные методы), микробиологии, клеточной биологии, биохимии и иммунологии, которые известны специалистам в данной области. Такие методы полностью описаны в литературе, такой как “Molecular Cloning: A Laboratory Manual”, второе издание (Sambrook et al., 1989); “Oligonucleotide Synthesis” (MJ Gait, ed., 1984); “Animal Cell Culture” (R. I. Freshney, ed., 1987); “Methods in Enzymology” (Academic Press, Inc.); “Current Protocols in Molecular Biology” (F. M. Ausubel et al., eds., 1987, и периодические обновления); “PCR: The Polymerase Chain Reaction”, (Mullis et al., ed., 1994); “A Practical Guide to Molecular Cloning” (Perbal Bernard V., 1988); “Phage Display: A Laboratory Manual” (Barbas et al., 2001).
Все источники, цитируемые в данном документе, включая заявки на патент и публикации, включены посредством ссылки во всей их полноте.
I. Определения
Используемый в данном документе термин «трансгенное животное, отличное от человека», как определено в данном документе, представляет собой животное, отличное от человека, способное продуцировать человеческое или гуманизированное антитело, содержащее только тяжелые цепи, в котором аминокислотный остаток находящийся в первой позиции четвертой каркасной области (FR4) заменяется другим остатком, который способен нарушить целостность экспонированного на поверхности гидрофобного пятна, содержащего или связанного с таким остатком. В одном варианте осуществления указанный нативный аминокислотный остаток заменен заряженным аминокислотным остатком, таким как положительно заряженный аминокислотный остаток. Трансгенное животное, отличное от человека, предпочтительно представляет собой млекопитающее, включая, без ограничения, крыс, мышей, крупный рогатый скот, обезьян, свиней, овец, коз, кроликов, собак, кошек, морских свинок, хомяков и тому подобное. Предпочтительно трансгенное животное, отличное от человека, представляет собой грызуна, предпочтительно крысу или мышь, наиболее предпочтительно UniRat™. Выбор трансгенного животного ограничен только способностью продуцировать человеческое или химерное антитело человека или химерное антитело, содержащее только тяжелые цепи, с мутацией в FR4, описанной в данном документе.
Используемый в данном документе термин «генетическая модификация» представляет собой одно или несколько изменений в последовательностях генов животного, отличного от человека. Неограничивающим примером является вставка трансгена в геном трансгенного животного.
Используемый в данном документе термин «трансген» относится к экзогенной ДНК, содержащей промотор, репортерный ген, сигнал полиаденилирования и другие элементы для усиления экспрессии (инсуляторы, интроны). Эта экзогенная ДНК интегрируется в геном одноклеточного эмбриона, из которого развивается трансгенное животное, и трансген остается в геноме зрелого животного. Интегрированная трансгенная ДНК может встречаться в одном или нескольких местах в геноме яйцеклетки или мыши, а также от одной до нескольких (нескольких сотен) тандемных копий трансгена может интегрироваться в каждом геномном местоположении.
«Обычные антитела» обычно представляют собой гетеротетрамерные гликопротеины составляющие около 150000 дальтон, состоящие из двух идентичных легких (L) цепей и двух идентичных тяжелых (H) цепей. Каждая легкая цепь связана с тяжелой цепью одной ковалентной дисульфидной связью, в то время как число дисульфидных связей варьируется среди тяжелых цепей разных изотипов иммуноглобулина. Каждая тяжелая и легкая цепь также имеет регулярно расположенные внутрицепочечные дисульфидные мостики. Каждая тяжелая цепь имеет на одном конце вариабельный домен (VH), за которым следует ряд константных доменов. Каждая легкая цепь имеет вариабельный домен на одном конце (VL) и константный домен на другом конце; константный домен легкой цепи выровнен с первым константным доменом тяжелой цепи, а вариабельный домен легкой цепи выровнен с вариабельным доменом тяжелой цепи. Считается, что конкретные аминокислотные остатки образуют поверхность соприкосновения между вариабельными доменами легкой и тяжелой цепи.
Остатки антител в данном документе пронумерованы в соответствии с системой нумерации Кабата (например, Kabat et al., Sequences of Immunological Interest. 5th Ed. Public Health Service, National Institutes of Health, Бэтэсда, Мэриленд. (1991)). Согласно этой нумерации первый аминокислотный остаток области FR4 находится в аминокислотной позиции 101.
Термин «вариабельный» относится к тому факту, что определенные части вариабельных доменов сильно различаются по последовательности между антителами и используются в связывании и специфичности каждого конкретного антитела к его конкретному антигена. Однако эта вариабельность неравномерно распределена по вариабельным доменам антител. Она сконцентрирована в трех сегментах, называемых областями, определяющими комплементарность (CDR) или гипервариабельными областями как в вариабельных доменах легкой цепи, так и в вариабельных доменах тяжелой цепи. Более высоко консервативные части вариабельных доменов называются каркасом (FR). Каждый из вариабельных доменов нативных тяжелых и легких цепей содержит четыре области FR, в значительной степени принимающих конфигурацию β-листа, соединенных тремя CDR, которые образуют петли, соединяющие и в некоторых случаях являющиеся частью структуры β-листа. CDR в каждой цепи удерживаются вместе в непосредственной близости областями FR и вместе с CDR из другой цепи способствуют образованию антигенсвязывающего сайта антител (см. Kabat et al., NIH Publ. No. 91-3242, Vol. 1, pages 647-669 (1991)). Константные домены не участвуют непосредственно в связывании антитела с антигеном, но проявляют различные эффекторные функции, такие как участие антитела в антителозависимой клеточной токсичности.
Используемый в данном документе термин «моноклональное антитело» относится к антителу, полученному из популяции по существу гомогенных антител, то есть отдельные антитела, составляющие популяцию, являются идентичными, за исключением возможных встречающихся в природе мутаций, которые могут присутствовать в незначительных количествах. Моноклональные антитела являются высокоспецифичными и направлены против одного антигенного сайта. Кроме того, в отличие от обычных (поликлональных) препаратов антител, которые обычно включают разные антитела, направленные против разных детерминант (эпитопов), каждое моноклональное антитело направлено против одной детерминанты на антигене.
Термины «антитело, содержащее только тяжелые цепи», «антитело с тяжелой цепью» и «HCAb» используются взаимозаменяемо и относятся в самом широком смысле к антителам, лишенным легкой цепи обычного антитела. Так как гомодимерные HCAb не имеют легкой цепи и, следовательно, домена VL, антиген распознается одним единственным доменом, то есть вариабельным доменом тяжелой цепи антитела тяжелой цепи (VH или VHH, при упоминании вариабельного домена тяжелой цепи верблюдовых). Этот термин, в частности, включает, без ограничения, гомодимерные антитела, содержащие антигенсвязывающий домен VHH и константные домены CH2 и CH3 или антитело верблюдовых, в отсутствие домена CH1; функциональные (антигенсвязывающие) варианты таких антител, растворимые варианты VH, Ig-NAR, содержащие гомодимер одного вариабельного домена (V-NAR) и пять C-подобных константных доменов (C-NAR) и их функциональные фрагменты; и растворимые однодоменные антитела (sdAb) или нанотела. Антитела, содержащие только тяжелые цепи, по данному изобретению содержат по меньшей мере один вариабельный домен (VH) тяжелой цепи, в котором аминокислотный остаток в первой позиции FR4 (аминокислотный остаток 101 согласно нумерации Кабата) заменен другим остатком, который способен нарушать целостность экспонированного на поверхности гидрофобного пятна, содержащего или связанного с таким остатком. В одном варианте осуществления указанный нативный аминокислотный остаток заменен заряженным аминокислотным остатком, таким как положительно заряженный аминокислотный остаток. Антитела, содержащие только тяжелые цепи, по данному изобретению предпочтительно представляют собой антитела человека или химеры, предпочтительно содержащие мутацию Trp (W) - Arg (R) в аминокислотной позиции 101 (мутация W101R). В одном варианте осуществления антитела, содержащие только тяжелые цепи, в данном документе используются в качестве связывающего (нацеливающего) домена химерного антигенного рецептора (CAR).
Термин «растворимое однодоменное антитело (sdAb)» используется для обозначения в самом широком смысле полипептидов, содержащих вариабельный домен тяжелой цепи антитела с тяжелыми цепями или обычного IgG, в отсутствие константных доменов. Базовая структура sdAb обычно состоит из четырех каркасных областей (FR1-FR4), прерываемых тремя областями, определяющими комплементарность (CDR1-CDR3). Таким образом, sdAb может быть представлен следующей структурой: FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4. Для дальнейшего ознакомления, см., например, Holt et al, "Domain antibodies:proteins for therapy" Trends in Biotechnology (2003):Vol. 21, No. 11:484-490.
Антитела по данному изобретению включают мультиспецифичные антитела. Мультиспецифичные антитела имеют более чем одну специфичность связывания. Термин «мультиспецифический», в частности, включает «биспецифический» и «триспецифический», а также независимые аффинности специфического связывания более высокого порядка, такие как полиэпитопная специфичность высшего порядка, а также четырехвалентные антитела и фрагменты антител. Термины «мультиспецифическое антитело», «мультиспецифическое только одноцепочечное антитело» и «мультиспецифическое HCAb» используются в данном документе в самом широком смысле и охватывают все антитела с более чем одной специфичностью связывания.
Используемый в данном документе термин «валентность» относится к указанному количеству сайтов связывания в молекуле антитела.
«Поливалентное» антитело имеет два или более сайтов связывания. Таким образом, термины «двухвалентный», «трехвалентный» и «четырехвалентный» относятся к наличию двух сайтов связывания, трех сайтов связывания и четырех сайтов связывания, соответственно. Биспецифическое антитело по данному изобретению является по меньшей мере двухвалентным и может быть трехвалентным, четырехвалентным или иным образом поливалентным. Мультиспецифичные одноцепочечные антитела по данному изобретению, например биспецифические антитела, включают поливалентные одноцепочечные антитела.
Термин «химерный антигенный рецептор» или «CAR» (Chimeric Antigen Receptor) используется в данном описании в самом широком смысле для обозначения сконструированного рецептора, который прививает желаемую специфичность связывания (например, антигенсвязывающую область моноклонального антитела или другого лиганда) к трансмембранным и внутриклеточным сигнальным доменам. Как правило, рецептор используется для прививания специфичности моноклонального антитела на Т-клетку для создания J Natl Cancer Inst, 2015; 108 (7): dvj439; и Jackson et al., Nature Reviews Clinical Oncology, 2016; 13: 370-383. Типичная конструкция CAR-T, содержащая внеклеточный связывающий домен VH человека, продемонстрирована на Фиг. 6.
Под «рекомбинантным локусом иммуноглобулина (Ig)» подразумевается локус Ig, в котором отсутствует часть эндогенного локуса Ig и/или который содержит по меньшей мере один фрагмент, который не является эндогенным по отношению к локусу Ig у используемого млекопитающего. Такой фрагмент может быть человеческим или нечеловеческим и может включать в себя любой генный сегмент Ig или его часть или может составлять весь локус Ig. Локус рекомбинантного Ig предпочтительно представляет собой функциональный локус, способный подвергаться перестройке генов и продуцировать репертуар иммуноглобулинов у трансгенного животного. Рекомбинантные локусы Ig включают рекомбинантные локусы легкой цепи Ig и рекомбинантные локусы тяжелой цепи Ig. После включения в геном хозяина искусственный Ig-локус можно назвать рекомбинантным Ig-локусом.
Под «трансгенным антителом» подразумевается антитело, кодируемое рекомбинантным локусом Ig и полученное из трансгенного млекопитающего, отличного от человека, или полученное иным образом, содержащее локус рекомбинантного Ig в соответствии с данным изобретением. Трансгенное антитело, полученное от используемого трансгенного млекопитающего, включает трансгенное антитело, полученное с использованием изолированной клетки или нуклеиновой кислоты, полученной от используемого трансгенного животного, или с использованием клетки или нуклеиновой кислоты, полученной из изолированной клетки или нуклеиновой кислоты, полученной от используемого трансгенного животного. В предпочтительном варианте осуществления трансгенное антитело содержит аминокислотную последовательность, кодируемую интегрированным донорным полинуклеотидом или его частью.
Используемый в данном документе термин «генный сегмент Ig» относится к сегментам ДНК, кодирующим различные части молекулы Ig, которые присутствуют в зародышевой линии животных и людей, не являющихся людьми, и которые объединяются в В-клетках с образованием перестроенных генов Ig. Таким образом, «генные сегменты Ig», как используется в данном документе, может относиться к V генным сегментам, D генным сегментам J генным сегментам и генам С-области, а также их частям.
Используемый в данном документе термин «генный сегмент Ig человека» включает как встречающиеся в природе последовательности генного сегмента Ig человека, вырожденные формы встречающихся в природе последовательностей генного сегмента Ig человека, так и синтетические последовательности, которые кодируют полипептидную последовательность, практически идентичную полипептиду, который кодируется природной последовательностью генного сегмента Ig человека. «По существу» означает, что степень идентичности аминокислотной последовательности составляет, по меньшей мере, около 85%-95%. Предпочтительно, степень идентичности аминокислотной последовательности составляет более 90%, более предпочтительно, более 95%.
Термин «локус, содержащий только тяжелые цепи», как он определено в данном документе, относится к локусу, кодирующему домен VH, в котором первый аминокислотный остаток области FR4 антитела является положительно заряженным, включающим один или несколько V генных сегментов, один или несколько D генных сегментов и один или несколько J генных сегментов, необязательно связанных с одним или несколькими генными сегментами эффекторной области тяжелой цепи, каждый из которых кодирует константную эффекторную область антитела, лишенную функциональности домена CH1. Предпочтительно, локус, содержащий только тяжелые цепи, содержит от около пяти до около двадцати V генных фрагментов, от около двух до около 40 D генных фрагментов и от около двух до около двадцати J генных фрагментов, причем V/D/J фрагменты предпочтительно происходят от человека. Термины «D генный сегмент» и «J генный сегмент» также охватывают производные, гомологи и их фрагменты до тех пор, пока результирующий сегмент может рекомбинировать с остальными компонентами локуса антитела с тяжелыми цепями, как описано в данном документе, с образованием антитела, содержащего только тяжелые цепи. D и J генные сегменты могут быть получены из природных источников или они могут быть синтезированы с использованием методов, знакомых специалистам в данной области техники и описанных в данном документе. В одном варианте осуществления в J4 генном сегменте кодон для W (TGG) заменен кодоном для R (CGG) для кодирования R вместо W в первом аминокислотном остатке FR4 (позиция 101 антитела, содержащего только тяжелые цепи, согласно нумерации по Кабату). D и J генные сегменты могут включать кодоны для определенных дополнительных аминокислотных остатков или определенных аминокислотных замен или делеций для увеличения разнообразия CDR3. Термин «V генный сегмент» охватывает встречающиеся в природе V генные сегменты, происходящие от животного, отличного от человека, такого как млекопитающее, отличное от человека, например грызун, сконструированный с введением положительно заряженного аминокислотного остатка в первой позиции области FR4. «V генный сегмент» должен быть способен рекомбинировать с D генным сегментом, J генным сегментом и константной областью тяжелой цепи, которая исключает экзон CH1, чтобы генерировать антитело только с тяжелой цепью в данном случае, когда экспрессируется нуклеиновая кислота.
Под «человеческим идиотипом» подразумевается эпитоп полипептидной последовательности, присутствующий на антителе человека в вариабельной области тяжелой и/или легкой цепи иммуноглобулина. Используемый в данном документе термин «человеческий идиотип» включает как встречающиеся в природе последовательности человеческого антитела, так и синтетические последовательности, практически идентичные полипептиду, обнаруженному в встречающихся в природе человеческих антителах. «По существу» означает, что степень идентичности аминокислотной последовательности составляет, по меньшей мере, около 85%-95%. Предпочтительно, степень идентичности аминокислотной последовательности составляет более 90%, более предпочтительно, более 95%.
Под «химерным антителом» или «химерным иммуноглобулином» подразумевается молекула иммуноглобулина, содержащая аминокислотные последовательности по меньшей мере из двух разных локусов Ig, например, трансгенное антитело, содержащее часть, кодируемую локусом Ig человека, и часть, кодируемую локусом Ig крысы. Химерные антитела включают трансгенные антитела с не-человеческими Fc-областями или искусственными Fc-областями и человеческие идиотипы. Такие иммуноглобулины могут быть выделены из животных по данному изобретению, которые были сконструированы для получения таких химерных антител.
«Аффинность связывания» относится к силе общей суммы нековалентных взаимодействий между одним сайтом связывания молекулы (например, антителом) и его партнером по связыванию (например, антигеном). Если не указано иное, как используется в данном документе, «аффинность связывания» относится к внутренней аффинности связывания, которая отражает взаимодействие 1:1 между членами пары связывания (например, антителом и антигеном). Сродство молекулы X к ее партнеру Y обычно может быть представлено константой диссоциации (Kd). Kd HCAb по данному изобретению обычно составляет от около 1 пм до около 1 мкм. Например, Kd может составлять около 200 нМ, 150 нМ, 100 нМ, 60 нМ, 50 нМ, 40 нМ, 30 нМ, 20 нМ. 10 нМ, 8 нМ, 6 нМ, 4 нМ, 2 нМ, 1 нМ или быть сильнее. Сродство может быть измерено общепринятыми способами, известными в данной области. Антитела с низким афинностьм обычно связывают антиген медленно и имеют тенденцию легко диссоциировать, тогда как антитела с высоким афинностьм обычно связывают антиген быстрее и имеют тенденцию оставаться.
Используемый в данном документе термин «Kd» или «значение Kd» относится к константе диссоциации, измеренной с использованием анализов поверхностного плазмонного резонанса, например, с использованием BIAcore™ -2000 или BIAcore™ -3000 (BIAcore, Inc., Piscataway, NJ.) при 25°С с иммобилизованными антигенными чипами СМ5 при ~ 10 единицах ответа (RU - response unit). Для получения дополнительной информации см., например, Chen et al., J. Mol. Biol. 293:865-881 (1999).
«Эпитоп» представляет собой место на поверхности молекулы антигена, с которым связывается одна молекула антитела. Обычно антиген имеет несколько или много разных эпитопов и реагирует со многими различными антителами. Этот термин конкретно включает линейные эпитопы и конформационные эпитопы.
«Полиэпитопная специфичность» относится к способности специфически связываться с двумя или более разными эпитопами на одной или разных мишенях.
Антитело связывает «по существу тот же самый эпитоп», что и эталонное антитело, когда два антитела распознают идентичные или стерически перекрывающиеся эпитопы. Наиболее широко используемыми и быстрыми методами для определения того, связываются ли два эпитопа с идентичными или стерически перекрывающимися эпитопами, являются конкурентные анализы, которые могут быть сконфигурированы во всем количестве различных форматов с использованием либо меченого антигена, либо меченого антитела. Обычно антиген иммобилизуют на 96-луночном планшете, и способность немеченых антител блокировать связывание меченых антител измеряют с использованием радиоактивных или ферментных меток.
«Эпитопное картирование» представляет собой процесс идентификации сайтов связывания или эпитопов антител с их антигенами-мишенями. Эпитопы антител могут быть линейными или конформационными эпитопами. Линейные эпитопы образованы непрерывной последовательностью аминокислот в белке. Конформационные эпитопы образуются из аминокислот, которые расположены на удалении друг от друга в последовательности белка, но которые объединяются при сворачивании белка в его трехмерную структуру.
«Опухоль» в контексте данного описания относится ко всему неопластическому росту и пролиферации, как злокачественному, так и доброкачественному, а также ко всем предраковым и раковым клеткам и тканям. Термин «опухоль» включает как солидные (твердые) опухоли, так и гематологический рак.
Термины «рак» и «раковый» относятся или описывают физиологическое состояние у млекопитающих, которое обычно характеризуется нерегулируемым ростом клеток. Примеры рака включают, но не ограничиваются ими, карциному, лимфому, бластому, саркому и лейкоз. Более конкретные примеры рака включают рак молочной железы, рак желудка, плоскоклеточный рак, глиобластому, рак шейки матки, рак яичников, рак печени, рак мочевого пузыря, гепатому, рак толстой кишки, колоректальный рак, рак эндометрия, рак слюнной железы, рак почки, рак эпителия почечных канальцев, рак простаты, рак влагалища, рак щитовидной железы, рак печени, рак головы и шеи, рак прямой кишки, рак толстой кишки, рак легких, включая мелкоклеточный рак легких, немелкоклеточный рак легких, аденокарциному легкого и плоскоклеточный рак легких, плоскоклеточный рак (например, эпителиально-плоскоклеточный рак), рак простаты, перитонеальный рак, гепатоцеллюлярный рак, рак желудка, включая рак желудочно-кишечного тракта, рак поджелудочной железы, глиобластому, ретинобластому, астроцитому, текомы, арренобластомы, гепатомы, гематологические злокачественные новообразования, в том числе не-Ходжкинскую лимфому (НХЛ), множественную миелому и острые гематологические злокачественные новообразования, рак эндометрия или матки, эндометроз, фибросаркомы, хориокарциному, рак слюнных желез, рак влагалища, рак щитовидной железы, рак пищевода, рак печени, рак полового члена, назофарингеальную карциному, карциному гортани, саркому Капоши, меланому, карциномы кожи, Шванному, олигодендроглиому, нейробластомы, рабдомиосаркому, остеогенную саркому, лейомиосаркомы, рак уретры, рак щитовидной железы, опухоль Вильмса, а также В-клеточную лимфому (включая малоразвитую/фолликулярную неходжкинскую лимфому (НХЛ), малую лимфоцитарную (МЛ) НХЛ, среднеразвитую (промежуточную)/фолликулярную НХЛ; среднеразвитую (промежуточную) НХЛ; сильноразвитую иммунобластную НХЛ; сильноразвитую лимфобластную НХЛ; сильноразвитую НХЛ с нерассеченными клетками; крупноразмерную НХЛ; лимфому мантийных клеток; связанную со СПИДом лимфому; и макроглобулинемию Вальденстрема; хронический лимфолейкоз (ХЛЛ); острый лимфобластный лейкоз (ALL); волосатоклеточный лейкоз; хронический миелобластный лейкоз; и посттрансплантационное лимфопролиферативное расстройство (PTLD), а также аномальную сосудистую пролиферацию, связанную с факоматозами, и синдром Мейгса.
II. ПОДРОБНОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
HCAb по данному изобретению являются человеческими или химерными, в которых нативный аминокислотный остаток в первой позиции области FR4 (аминокислотная позиция 101 согласно системе нумерации Кабата) замещен другим аминокислотным остатком, который способен нарушать целостность экспонированного на поверхности гидрофобного пятна, содержащего нативный аминокислотный остаток или связанного с нативным аминокислотным остатком в этой позиции. Такие гидрофобные пятна обычно скрыты в поверхности соприкосновения с константной областью легкой цепи антитела, но экспонируются на поверхности в HCAb и, по меньшей мере, частично, являются причиной нежелательной агрегации и ассоциации с легкими цепями HCAb. Замещенный аминокислотный остаток предпочтительно является заряженным и, более предпочтительно, является положительно заряженным. Полученные HCAb предпочтительно имеют высокую антигенсвязывающую аффинность и растворимость в физиологических условиях в отсутствие агрегации.
В частности, включены антитела, содержащие только тяжелые цепи, в которых отсутствует VHH каркас и мутации верблюдовых, и их функциональные VH-области. Такие антитела, содержащие только тяжелые цепи, можно, например, продуцировать в трансгенных крысах или мышах, которые содержат полностью человеческие локусы генов антител, содержащих только тяжелые цепи, как описано, например, в WO 2006/008548, но и другие трансгенные млекопитающие, такие как кролик, морская свинка, крыса также могут быть использованы, при этом предпочтение отдается крысам и мышам. Антитела, содержащие только тяжелые цепи, включая их функциональные фрагменты VHH или VH, также могут быть получены с помощью технологии рекомбинантных ДНК путем экспрессии кодирующей нуклеиновой кислоты в подходящем эукариотическом или прокариотическом хозяине, включая E. coli или дрожжи.
Домены антител, содержащих только тяжелые цепи, объединяют преимущества антител и низкомолекулярных лекарств: могут быть одно- или многовалентными; имеют низкую токсичность; и рентабельны в производстве. Из-за своего небольшого размера эти домены легко вводить, включая пероральное или местное введение, характеризуются высокой стабильностью, включая желудочно-кишечную стабильность; и их период полураспада может быть адаптирован к желаемому использованию или показаниям. Кроме того, домены VH и VHH HCAb могут быть изготовлены экономически эффективным способом.
В одном варианте осуществления домены HCAb представляют собой нанотела, как определено в данном документе выше.
В одном варианте осуществления HCAb-связывающие домены являются частью мультиспецифического связывающего белка, который связывается с несколькими различными эпитопами одного и того же антигена-мишени или несколькими различными эпитопами на более чем одном антигене-мишени. В одном варианте осуществления антитело представляет собой биспецифическое антитело. Различные мультиспецифические структуры, содержащие VH-связывающие домены человека, изображены на Фиг. 7. Мультиспецифические или биспецифические HCAb по данному изобретению могут, например, связываться с двумя или более сайтами на одной и той же растворимой мишени или двумя или более сайтами на одной и той же клеточной поверхности (рецепторе)-мишени, такой как опухолевый антиген или одна или несколько растворимых мишеней и одна или несколько мишеней-рецепторов на клеточной поверхности. В определенных вариантах осуществления биспецифические структуры HCAb в данном документе имеют следующие аффинности связывания: молекула адгезии эпителиальных клеток (EpCam) × CD3; CD19 × CD3; EpCam × CD3; TNF-α x IL-17; IL-1α × IL-1β; CD30 × CD16A; рецептор 2 эпидермального фактора роста человека (HER2) × HER3; IL-4 × IL-13; ангиопоэтин 2 (Ang-2) × фактор роста эндотелия сосудов a (VEGF-A); Фактор IXa × Фактор X; рецептор эпидермального фактора роста (EGFR) × HER3; IL-17A × IL-17F; HER2 × HER3; карциноэмбриональный антиген × CD3; CD20 × CD3; CD123 × CD3; BCMA × CD3, PSMA × PSCA × CD3, PSMA × CD3, PSCA × CD3, CD19 × CD22 × CD3, CD22 × CD3, CD38 × PD1, CD38 × PD-L1, CD38 × CD73, CD38 × CD39, PD1 × CD39 × CD73 и PD1 × CD73.
В предпочтительном варианте осуществления данного изобретения HCAb продуцируются трансгенными животными, включая трансгенных мышей и крыс, предпочтительно крыс, у которых эндогенные гены иммуноглобулина нокаутированы или инактивированы. В предпочтительном варианте осуществления данного изобретения HCAb получают в UniRat™. У UniRat™ эндогенные гены иммуноглобулинов инактивированы и используют транслокус тяжелой цепи человеческого иммуноглобулина для экспрессии разнообразного, естественно оптимизированного репертуара полностью человеческих HCAb. В то время как эндогенные локусы иммуноглобулина у крыс могут быть нокаутированы или инактивированы с использованием различных технологий, в UniRat™ технология (эндо)нуклеаз цинкового пальца (ZNF) была использована для инактивации эндогенного J-локуса тяжелой цепи крысы, локуса Cκ легкой цепи и локуса Cλ легкой цепи. ZNF конструкции для микроинъекции в яйцеклетку могут продуцировать IgH и IgL-нокаутные (KO) линии. Подробности см., например, в Geurts et al., 2009, Science 325: 433 Characterization of Ig heavy chain knockout rats has been reported by Menoret et al., 2010, Eur. J. Immunol. 40:2932-2941. Преимущества технологии ZNF заключаются в том, что негомологичное соединение концов для инактивации гена или локуса посредством делеций размером до нескольких т.п.н. также может обеспечить целевой сайт для гомологичной интеграции (Cui et al., 2011, Nat Biotechnol 29: 64-67. UniRat™ HCAb связывают эпитопы, которые нельзя атаковать обычными антителами. Их высокая специфичность, афинность и небольшой размер делают их идеальными для моно- и поли-специфических применений.
В антителах, содержащих только тяжелые цепи, по данному изобретению, нативный аминокислотный остаток в первой позиции четвертой каркасной области (FR4) заменен другим аминокислотным остатком, который способен нарушать целостность экспонированного на поверхности гидрофобного участка, содержащего нативный аминокислотный остаток или связанного с нативным аминокислотным остатком в этой позиции. В одном варианте осуществления замещенный аминокислотный остаток является заряженным. В другом варианте осуществления замещенный аминокислотный остаток является положительно заряженным, таким как лизин (Lys, K), аргинин (Arg, R) или гистидин (His, H), предпочтительно аргинином (R). Как показано на Фиг. 5, все последовательности VH из антител с тяжелыми цепями в одном и том же семействе CDR3 содержат Trp (W) в позиции 101, таким образом, в предпочтительном варианте осуществления антитела, содержащие только тяжелые цепи, полученные из трансгенных животных по данному изобретению, содержат мутацию Trp-Arg по позиции 101.
Антитела человека или только химерные тяжелые цепи по данному изобретению могут быть получены против любого желаемого антигена-мишени и имеют большой потенциал для различных клинических применений. Целевые антигены для терапевтического применения включают, без ограничения, EGFR, ErbB2 (HER2), ErbB3 (HER3), ErbB4 (HER4), CTLA-4/CD152, RANKL, TNF-α, CD20, IL-12/IL-23, IL1 -β, IL-17A, IL-17F, CD38, NGF, IGF-1, IL-12, CD20, CD30, CD39, CD73, CD40, PD-1, PDL-1, PD-L2, BCMA, BTLA, стромальный лимфопоэтин тимуса (TSP), рецептор фолликулостимулирующего гормона (FSHR), простат-специфический мембранный антиген (PSMA), антиген стволовых клеток простаты (PSCA), CD137, OX-40 и IL-33. Терапевтические показания включают, без ограничения, лечение солидных опухолей, гематологических опухолей, воспалительных заболеваний, таких как ревматоидный артрит, псориаз, болезнь Крона, язвенный колит, нарушения обмена веществ, сердечно-сосудистых заболеваний, респираторных, дерматологических заболеваний, заболеваний центральной нервной системы, гематологических заболеваний, заболеваний глаз/уха, печени.
Опухоли-мишени включают, например, рак молочной железы, рак желудка, плоскоклеточный рак, глиобластому, рак шейки матки, рак яичников, рак печени, рак мочевого пузыря, гепатому, рак толстой кишки, колоректальный рак, рак эндометрия, рак слюнной железы, рак почки, рак эпителия почечных канальцев, рак простаты, рак влагалища, рак щитовидной железы, рак печени, рак головы и шеи, рак прямой кишки, рак толстой кишки, рак легких, включая мелкоклеточный рак легких, немелкоклеточный рак легких аденокарциному легкого и плоскоклеточный рак легких, плоскоклеточный рак (например, эпителиально-плоскоклеточный рак), рак простаты, перитонеальный рак, гепатоцеллюлярный рак, рак желудка, включая рак желудочно-кишечного тракта, рак поджелудочной железы, глиобластому, ретинобластому, астроцитому, текомы, арренобластомы, гепатомы, гематологические злокачественные новообразования, в том числе не-Ходжкинскую лимфому (НХЛ), множественную миелому (ММ) и острые гематологические злокачественные новообразования, рак эндометрия или матки, эндометроз, фибросаркомы, хориокарциному, рак слюнных желез, рак влагалища, рак щитовидной железы, рак пищевода, рак печени, рак полового члена, назофарингеальную карциному, карциному гортани, саркому Капоши, меланому, карциномы кожи, Шванному, олигодендроглиому, нейробластомы, рабдомиосаркому, остеогенную саркому, лейомиосаркомы, рак уретры, рак щитовидной железы, опухоль Вильмса, а также В-клеточную лимфому (включая малоразвитую/фолликулярную неходжкинскую лимфому (НХЛ), малую лимфоцитарную (МЛ) НХЛ, среднеразвитую (промежуточную)/фолликулярную НХЛ; среднеразвитую (промежуточную) НХЛ; сильноразвитую иммунобластную НХЛ; сильноразвитую лимфобластную НХЛ; сильноразвитую НХЛ с нерассеченными клетками; крупноразмерную НХЛ; лимфому мантийных клеток; связанную со СПИДом лимфому; и макроглобулинемию Вальденстрема; хронический лимфолейкоз (ХЛЛ); острый лимфобластный лейкоз (ALL); волосатоклеточный лейкоз; хронический миелобластный лейкоз; и посттрансплантационное лимфопролиферативное расстройство (PTLD), а также аномальную сосудистую пролиферацию, связанную с факоматозами, и синдром Мейгса.
Дальнейшие подробности изобретения иллюстрируются следующими неограничивающими примерами, в которых используются следующие сокращения:
BAC Бактериальная искусственная хромосома
YAC дрожжевая искусственная хромосома
ZFN нуклеаза цинкового пальца
H тяжелая цепь
С константная область
V вариабельная область
D сегмент разнообразия
J соединяющий сегмент
ПРИМЕРЫ
Пример 1: Получение генетически модифицированных крыс, экспрессирующих антитела, содержащие только тяжелые цепи (HCAb)
Ранее идентифицированные, охарактеризованные и, частично, модифицированные BAC и YAC вмещают гены вариабельной области тяжелой цепи человека и гены константной области крысы (Osborn et al., 2013, J. Immunol. 190:1481-1490; Ma et al., 2013, J. Immunol. Methods 400-401:78-86). Чтобы обеспечить экспрессию антител с тяжелыми цепями, константную область BAC крысы модифицировали путем удаления Cμ и делеции экзонов CH1 во всех Cγ. Экспрессия только тяжелых цепей была затем усилена путем подавления эндогенных локусов тяжелых и легких цепей (каппа и лямбда).
Конструирование модифицированных человеческих локусов IgH на YAC и BAC.
«Человеческий-крысиный» локус IgH был сконструирован и собран в нескольких частях. Это включало изменение и присоединение генов области C крысы ниже человеческих JH и, последующим добавлением выше области человеческого VH 6-D-сегмента. Два BAC с отдельными кластерами человеческих VH- генов [BAC6 и BAC3] были затем совместно инъецированы с BAC, называемой Georg, кодирующей собранную и модифицированную область, содержащую человеческий VH 6 - все D - все JH - Cγ2a/1/2b (ΔCH 1) крысы.
Для введения модификаций в точных местоположениях в последовательности ДНК и для одновременного соединения множества больших участков ДНК были разработаны технологии для сборки последовательностей с перекрывающимися концами в S. cerevisiae в виде кольцевого YAC (cYAC) и, впоследствии, для преобразования таких cYAC в BAC. Преимущества YAC включают их способность удерживать большие вставки ДНК, легкость гомологичных изменений в дрожжевом хозяине и поддержание стабильности последовательности, особенно в сильно повторяющихся областях (например, областях переключения, энхансерах). С другой стороны, BAC, размножающиеся в E. coli, обладают преимуществами простоты приготовления и большой продуктивности. Кроме того, подробное рестрикционное картирование и анализ последовательности могут быть лучше достигнуты с BAC, чем с YAC. Были сконструированы два самореплицирующихся челночных вектора S. cerevisiae/E. Coli, pBelo-CEN-URA и pCAU. Вкратце, S. cerevisiae CEN4 был вырезан как фрагмент AvrII из pYAC-RC (Marchuk и Collins, 1988; Nucleic Acids Res. 16 (15): 7743) и лигирован с SpeI-линеаризованным pAP599 (Ma et al. Mol Microbiol. 2007; 66(1):14-25). Полученная плазмида содержит CEN4, клонированный ниже URA3. Из этого был вырезан фрагмент ApaLI-BamHI URA3-CEN4 и лигирован в расщепленный ApaLI и BamHI pBACBelo11 (New England Biolabs) с получением pBelo-CEN-URA. Автономно реплицирующаяся последовательность ARS209 S. cerevisiae была синтезирована и клонирована в уникальный сайт SexAI в pBelo-CEN-URA с получением pCAU.
Чтобы облегчить модификации областей человеческого JH4, Cμ и Cγ1 крысы, SacII-фрагмент размером ~ 37 т.п.н., простирающийся от ~ 2. 2 т.п.н. выше от человеческих JH до ~ 5. 5 т.п.н. ниже от кодирующей области Cγ1 крысы, был вырезан из конструкции ВАС Annabel (Osborn et al., 2013; J. Immunol. 190: 1481-1490) и клонирован в уникальный сайт SacII в pBelo-CEN-URA [pBelo + SacII, 37 т.п.н.]. Кроме того, для модификации области Cγ2b крысы фрагмент SacII - SwaI размером ~ 19 т.п.н. из Annabel, охватывающий ~ 6.9 т.п.н. выше от области переключения γ2b до ~ 2.0 т.п.н. ниже от кодирующей области Cγ2b, был клонирован в расщепленный двумя рестриктазами SacII и HpaI pBelo-CEN-URA [pBelo + SacII - SwaI, 19 т.п.н.]. Обе плазмиды были использованы в качестве матриц для амплификации различных областей генома человека и крысы и для создания необходимых рестрикционных фрагментов.
Область ДНК, простирающаяся от ~ 3,1 т.п.н. выше от человеческих JH и включающая крысиный Cμ с участком 3' области, была модифицирована и собрана в pCAU в виде фрагмента SnaBI-FspI размером 16,7 т.п.н. Модифицированная область включает все подлинные человеческие JH, кроме точечной мутации T → C, вводимой в JH4 (приводящей к аминокислотной замене W → R), за которой следует межгенная область крысы от J H до μCH1, которая была удалена вместе с остальной областью кодирования Cμ и точно заменена крысиной последовательностью Cγ2a, в которой отсутствует CH1 (начиная с интрона непосредственно перед шарниром до 3'-конца мембранных экзонов). Эта конструкция была получена путем сборки следующих 5 перекрывающихся фрагментов в дрожжах в виде cYAC, а затем преобразована в BAC: амплифицированный фрагмент размером ~ 4,3 т.п.н. с использованием праймеров HC27 - 1 и - 2, покрывающих область выше от человеческого JH до мутированного JH4 (точечная мутация, введенная через последний праймер, обозначенный  ), амплифицированный фрагмент размером ~ 3,4 т.п.н. с использованием праймеров HC27 -3 и -4, охватывающий мутированный JH4 (обозначенный
), амплифицированный фрагмент размером ~ 3,4 т.п.н. с использованием праймеров HC27 -3 и -4, охватывающий мутированный JH4 (обозначенный  ) до расположения выше области переключателя μ, ~ 5. 2 т.п.н. AflII - фрагмент, охватывающий область переключения μ и фланкирующие последовательности, вырезанные из pBelo+SacII 37 т.п.н., амплифицированный Cγ2a крысы без CH1, слитый с последовательностями, фланкирующими Cμ крысы с использованием длинных праймеров HC27 - 5 и - 6, и амплификации вектора pCAU с использованием праймеров HC27 - 7 и - 8. Это привело к pCAU + HuJ - Cγ2a (-CH1) крысы. Все измененные области были проверены сиквенированием для подтверждения точности.
) до расположения выше области переключателя μ, ~ 5. 2 т.п.н. AflII - фрагмент, охватывающий область переключения μ и фланкирующие последовательности, вырезанные из pBelo+SacII 37 т.п.н., амплифицированный Cγ2a крысы без CH1, слитый с последовательностями, фланкирующими Cμ крысы с использованием длинных праймеров HC27 - 5 и - 6, и амплификации вектора pCAU с использованием праймеров HC27 - 7 и - 8. Это привело к pCAU + HuJ - Cγ2a (-CH1) крысы. Все измененные области были проверены сиквенированием для подтверждения точности.
Крысиный Cγ1 без CH1 и Cγ2b без CH1 индивидуально генерировали с помощью ПЦР. Фрагмент размером ~1,7 т.п.н., расположенный непосредственно перед кодирующей областью Cγ1 с 3'-хвостом, совпадающим с 5'-концом интрона между CH1 и шарниром, амплифицировали с использованием праймеров HC27 - 9 и - 10. Cγ1 амплифицировали как фрагмент размером ~ 3,9 т.п.н. из интрона между CH1 и шарниром до 3 'конца кодирующей области используя праймеры HC27 - 11 и - 12. Впоследствии фрагменты ~ 1,7 т.п.н. и ~ 3,9 т.п.н. оба были очищены в геле и объединены посредством перекрывающейся ПЦР с использованием праймеров HC27-9 и -12 с получением фрагмента ~ 5,6 т.п.н. Аналогично, для Cγ2b без CH1 фрагмент ~ 0,3 т.п.н. выше от кодирующей области Cγ2b амплифицировали с использованием праймеров HC27 - 13 и - 14 и фрагмент ~ 5,4 т.п.н., охватывающийо область от интрона между CH1 и шарниром до 3'-конца кодирующей области амплифицировали с использованием праймеров HC27 - 15 и - 16, и впоследствии эти два фрагмента объединяли посредством перекрывающейся ПЦР с использованием праймеров HC27 - 13 и - 16 с получением фрагмента ~ 5,7 т.п.н. pCAU + Cγ1, 2b (-CH1) крысы был сконструирован так, чтобы содержать следующее: область гомологии 100 п. н., совпадающую с 3'-концом Cμ крысы, за которой следуют Cγ1 и Cγ2b в геномной конфигурации, за исключением того, что CH1 обоих были удалены. Шесть перекрывающихся фрагментов были использованы для конструирования pCAU +Cγ1, 2b (-CH1)крысы: фрагмент SpeI - NarI ~ 10,2 т.п.н., охватывающий область гомологии 3 'Cμ, за которым следует область переключения γ1, вырезанная из pBelo + SacII, 37 т.п.н., фрагмент ПЦР ~ 5,6 т.п.н., содержащий Cγ1 без C H 1, как описано выше, амплифицированный фрагмент ~ 7,4 т.п.н., охватывающий межгенную область между Cγ1 и Cγ2b с использованием праймеров HC27 - 17 и - 18, фрагмент XhoI ~ 11,3 т.п.н., включающий область переключателя Cγ2b крысы вырезанный из pBelo + SacII - SwaI 19 т.п.н., фрагмента ПЦР ~ 5,7 т.п.н., содержащий Cγ2b без CH1, и амплифицированный вектор pCAU с использованием праймеров HC27 - 19 и - 20. Геномная область крысы в pCAU + Cγ1, 2b (-CH1) крысы может быть вырезана как один FspI фрагмент размером ~ 40 т.п.н.
Наконец, BAC (Georg), кодирующий C-области человеческий VH6-D-JH-крысы со всеми модификациями, был собран с использованием следующих четырех перекрывающихся фрагментов: очищенный фрагмент FspAI - MluI размером ~ 78,2 т.п.н., охватывающий область человеческих VH6-D, вырезанную из BAC10 (CTD-3216M13, Invitrogen), фрагмент SnaBI - FspI размером 16,7 т.п.н., вырезанный из pCAU + HuJ - Cγ2a (-CH1) крысы, как описано выше, фрагмент FspI размером ~ 40 т.п.н., вырезанный из pCAU + Cγ1, 2b (-CH1) крысы и очищенный фрагмент ~ 77,2 т.п.н. SwaI - SacII, вырезанный из конструкции Annabel, который включает межгенную область между Cγ2b и Cε, за которой следуют Cε, Cα, 3'-энхансерная область, вектор pBelo-CEN-URA и 5'-область выше человеческого VH 6. Эта последняя конструкция была тщательно проверена с помощью рестрикционного картирования и частичного секвенирования. Область (человеческий VH6-D - JH-крыса C) может быть вырезана и очищена как фрагмент NotI размером ~ 201 т.п.н.
BAC6 содержит область генома человека от VH4-39 до VH3-23, тогда как BAC3 содержит нижерасположенную область от VH3-11 до VH6-1 (наиболее D-проксимальный ген VH). Чтобы обеспечить перекрытие между BAC6 и BAC3, фрагмент размером 10,6 т.п.н., расположенный на 5'-конце VH- локусов человека в BAC3, был интегрирован ниже VH3-23 в BAC6, как описано ранее (Osborn et al. 2013, выше). Гены VH человека в BAC6 были вырезаны в виде фрагмента AsiSI -AscI размером ~ 182 т.п.н. BAC3 не был модифицирован, и гены VH человека в этом BAC были вырезаны как NotI-фрагмент размером ~ 173 т.п.н.
Олигонуклеотиды:



Конструкция Georg II
«Человеческий - крысиный» локус IgH был сконструирован и собран в нескольких частях. Это включало модификацию и объединение генов С-области крысы ниже мутированных человеческих JH1 - JH6, каждый из которых содержит точечную мутацию, кодирующую W → R, и, впоследствии, добавление в расположении выше области VH6-1-D-сегмента человека. Этот BAC назван Georg II.
Во-первых, мутированный человеческий JH1 - JH6 синтезируется в ScaI-фрагменте размером 2,3 т.п.н. (Thermo Fisher Scientific, см. ниже).
Во-вторых, мутированный человеческий JН1-JН6 соединяется с областью ДНК ~3,1 т.п.н. выше от человеческих JH и областью ~ 11,4 т.п.н., охватывающей область от крысиной последовательности непосредственно ниже JH до Cμ крысы, чей кодирующий участок был заменен на кодирующую область Cγ2a крысы, но не содержащую CH1 в BAC, называемой pCAU + HuJ (WR) -Cγ2a (-CH1) крысы. Вся модифицированная область в этом BAC может быть вырезана как фрагмент SnaBI - FspI размером 16,7 т.п.н. Этот BAC был получен путем сборки следующих 4 перекрывающихся фрагментов в дрожжах в качестве cYAC и затем преобразованных в BAC: амплифицированный ~ фрагмент 2,7 т.п.н. с использованием праймеров HC32 - 1 и - 2, покрывающий область выше человеческого JH, 2,3 т.п.н. мутированный человеческий фрагмент JH1 - JH6 - ScaI, амплифицированный фрагмент ~ 2,1 т.п.н. с использованием праймеров HC32 - 3 и - 4, покрывающих область крысы непосредственно ниже JH, и фрагмент MluI - HpaI ~ 18,4 т.п.н., вырезанный из pCAU + HuJ - Cγ2a (-CH1) крысы (содержащий крысиный локус Cμ, в которой кодирующая область заменена кодирующей областью Cγ2a крысы, в которой отсутствует CH 1 и pCAU вектор, описанный в «методе конструирования HC27»). Все измененные области были проверены сиквенированием для подтверждения точности.
Наконец, Georg II, кодирующий человеческие VH6-1-D-мутированные JH-модифицированные C-области крысы, был собран с использованием следующих 4 перекрывающихся фрагментов: очищенного фрагмента FspAI - MluI размером ~ 78,2 т.п.н., охватывающего человеческую VH6-1-D область вырезанную из BAC10 (CTD-3216M13, Invitrogen), фрагмента SnaBI-FspI размером 16,7 т.п.н., вырезанного из pCAU + HuJ (WR) -Cγ2a (-CH1) крысы, как описано выше, фрагмента FspI размером ~ 40 т.п.н. вырезанного из pCAU + Cγ1, 2b (-CH1) крысы (описано в «способе конструирования HC27») и очищенного фрагмента SwaI-SacII ~ 77,2 т.п.н., вырезанного из конструкции Annabel, которая включает межгенную область между Cγ2b и Cε, за которой следует Cε, Cα, 3'-энхансерная область, вектор pBelo-CEN-URA и 5'-область выше человеческого VH 6-1. Эта последняя конструкция была тщательно проверена с помощью рестрикционного картирования и частичного сиквенирования. Область (человеческий VH 6-1-D - мутированный J H -модифицированный C крысы) может быть вырезана и очищена как NotI-фрагмент размером ~ 201 т.п.н.
Микроинъекция для получения трансгенных крыс HC32 и HC33.
BAC6 содержит область генома человека от VH4-39 до VH3-23, тогда как BAC3 содержит нижерасположенную область от VH3-11 до VH6-1 (наиболее D-проксимальный ген VH). Чтобы обеспечить перекрытие между BAC6 и BAC3, фрагмент размером 10,6 т.п.н., расположенный на 5'-конце локуса VH человека в BAC3, был интегрирован ниже VH3-23 в BAC6, как описано ранее (Osborn et al. 2013, J. Immunol. 190:1481-1490). Гены VH человека в BAC6 были вырезаны в виде фрагмента AsiSI -AscI размером ~ 182 т.п.н. BAC3 не был модифицирован, и человеческие гены VH в этом BAC были вырезаны как NotI-фрагмент размером ~173 т.п.н. Оба фрагмента были очищены и совместно инъецированы с фрагментом NotI ~ 201 т.п.н. из Georg II в эмбрионы крысы для конструирования HC32.
BAC9 содержит геномную область человека от VH3-74 до VH3-53. BAC (14 + 5) содержит нижерасположенную область от VH3-53 до VH3-13, и область 6,1 т.п.н., расположенную непосредственно перед VH6-1, добавили к ее 3'-области, чтобы обеспечить перекрытие для Georg II. Область VH человека в BAC9 была вырезана как фрагмент NotI размером ~ 185 т.п.н., а область BAC (14 + 5) была вырезана как фрагмент BsiwI размером ~ 209 т.п.н. Оба фрагмента были очищены и совместно введены с фрагментом NotI ~ 201 т.п.н. из Georg II в эмбрионы крысы для конструирования HC33.
Очистка ДНК
Линейные YAC, кольцевые YAC и фрагменты BAC после расщепления очищали электроэлюцией с использованием Elutrap™ (Schleicher and Schuell) (Gu et al., J. Biochem. Biophys Methods.,1992, 24:45-50) из полосок, вырезанных из 0,8% агарозных гелей, прогнанных обычным способом, или из электрофореза в импульсном геле (PFGE - pulsed-field-gel electrophoresis). Концентрация ДНК обычно составляла несколько нг/мкл в объеме ~ 100 мкл. Для фрагментов до ~200 т.п.н. ДНК осаждали и повторно растворяли в буфере для микроинъекции (10 мМ Трис-HCl рН 7,5, 100 мМ ЭДТА рН 8 и 100 мМ NaCl, но без спермина/спермидина) до желаемой концентрации.
Очистку циркулярных YAC из дрожжей осуществляли с использованием анионообменной смолы на основе диоксида кремния Nucleobond AX (Macherey-Nagel, Германия). Вкратце, сферопласты получали с использованием зимолазы или литиказы и осаждали (Davies et al., 1996, Human antibody repertoires in transgenic mice: manipulation of transfer of YACs. В Antibody Engineering: A Practical Approach. J. McCafferty, HR Hoogenboom and DJ Chiswell eds. IRL, Oxford, U. K., p. 59-76). Затем клетки подвергали щелочному лизису, связыванию с колонкой AX100 и элюированию, как описано в методе Nucleobond для плазмиды с низкой копийностью. Загрязняющую хромосомную ДНК дрожжей гидролизовали с использованием АТФ-зависимой ДНКазы Plasmid-Safe™ (Epicenter Biotechnologies) с последующим этапом очистки с использованием SureClean (Bioline). Аликвоту электрокомпетентных клеток DH10 (Invitrogen) затем трансформировали кольцевым YAC для получения колоний BAC. Для отделения ДНК-вставки для микроинъекции, 150-200 т.п.н., от ДНК-вектора ВАС, ~ 10 т.п.н., использовали стадию фильтрации с сефарозой 4B-CL (Yang et al., 1997, Nat. Biotechnol. 1997, 15;859-865).
Анализ на геле
Очищенную ДНК YAC и BAC анализировали путем рестрикционного расщепления и разделения на обычных 0,7% агарозных гелях (Sambrook and Russell, 2001). Более крупные фрагменты, 50-200 т.п.н., были отделены друг от друга с помощью PFGE (Biorad Chef Mapper™) при 8°C, с использованием 0,8% PFC агарозы в 0,5% TBE, при 2-20 сек времени переключения в течение 16 ч, 6V/см, 10 мА. Очистка позволила провести прямое сравнение полученных фрагментов с предсказанным размером, полученным из анализа последовательности. Изменения были проанализированы с помощью ПЦР и сиквенирования.
Микроинъекция
Беспородных животных линии SD/Hd содержали в стандартных клетках для микроизоляторов в соответствии с утвержденными протоколами по уходу за животными в помещении для животных, которое аккредитовано Ассоциацией по оценке и аккредитации для ухода за лабораторными животными (AAALAC). Крыс содержали при циклах 14-10 ч свет/темнота с неограниченной свободой доступа к еде и воде. Самкам крыс SD/Hd в возрасте от четырех до пяти недель вводили 20-25 МЕ PMSG (Sigma-Aldrich), а спустя 48 часов - 20-25 МЕ ХГЧ (Sigma-Aldrich) перед скрещиванием с беспородными самцами SD/Hd. Оплодотворенные эмбрионы 1-клеточной стадии собирали для последующей микроинъекции. Измененные эмбрионы переносили в псевдобеременных самок крыс SD/Hd для родов.
Очищенную ДНК, кодирующую рекомбинантные локусы иммуноглобулина, ресуспендировали в буфере для микроинъекции с 10 мМ спермина и 10 мМ спемидина (Spemidine). ДНК вводили в оплодотворенные ооциты в различных концентрациях от 0,5 до 3 нг/мкл.
Плазмидную ДНК или мРНК, кодирующую ZFN, специфичные для генов иммуноглобулина крысы, инъецировали в оплодотворенные ооциты в различных концентрациях от 0,5 до 10 нг/мкл.
Нуклеазы цинкового пальца (ZFN)
Были получены ZFN, специфичные для генов иммуноглобулинов крысы.
ZFN, специфичный для Ckappa крысы, имел следующий сайт связывания: ATGAGCAGCACCCTCtcgttgACCAAGGCTGACTATGAA (SEQ ID NO: 22)
ZFN, специфичные для последовательностей J-локуса крысы, имели следующие сайты связывания:
CAGGTGTGCCCATCCagctgaGTTAAGGTGGAG (SEQ ID NO: 23)
и
CAGGACCAGGACACCTGCAgcagcTGGCAGGAAGCAGGT (SEQ ID NO: 24)
ZFN, специфичные для последовательностей крысиного Cγ-локуса, имели следующие сайты связывания:
AACAGCCATTTGcagaccAAAGGGAAGGAAAGA (SEQ ID NO: 25)
и
TTCTACCCTGGTGTTATGacagtgGTCTGGAAGGCAGATGGT (SEQ ID NO: 26)
Крысы с транслокусами.
Были получены трансгенные крысы, несущие искусственные локусы иммуноглобулинов тяжелой цепи в неупорядоченной конфигурации. ОТ-ПЦР и анализ сыворотки (ИФА) трансгенных крыс выявили продуктивную перестройку локусов трансгенного иммуноглобулина и экспрессию в сыворотке антител, содержащих только тяжелые цепи, различных изотипов. Иммунизация трансгенных крыс приводила к получению высокоаффинных антиген-специфических антител, содержащих только тяжелые цепи.
Новая технология нокаутирования с помощью нуклеаз цинковых пальцев.
Для дальнейшей оптимизации генерации антител, содержащих только тяжелые цепи, у трансгенных крыс были получены нокаутные крысы с инактивированными эндогенными локусами иммуноглобулинов крыс.
Для инактивации экспрессии тяжелых цепей тяжелых иммуноглобулинов крыс и экспрессии легких цепей λ крыс ZFN микроинъецировали в одноклеточные эмбрионы крыс. Впоследствии эмбрионы были перенесены в псевдобеременных самок крыс и выношены до родов. Животных с мутированными локусами тяжелой цепи и легкой цепи идентифицировали с помощью ПЦР. Анализ таких животных продемонстрировал инактивацию экспрессии тяжелой и легкой цепи иммуноглобулина у крыс у мутантных животных.
Пример 2: Получение антигенспецифических антител, содержащих только тяжелые цепи, (HCAb) у крыс
Для получения антиген-специфических антител, содержащих только тяжелые цепи, у генетически сконструированных крыс, экспрессирующих антитела, содержащие только тяжелые цепи, таких как те, которые описаны в Примере 1, крыс иммунизируют различными способами.
Иммунизация инактивированным вирусом
Вирусы гриппа с различными генами гемагглютинина и нейраминидазы предоставлены Отделением иммунологии и патогенеза Отдела гриппа, CDC (Centers for Disease Control and Prevention), Атланта, Джорджия. Исходный материал вируса размножают в аллантоисных полостях 10-дневных куриных яиц с эмбрионами и очищают через градиент сахарозы 10-50% посредством ультрацентрифугирования. Вирусы ресуспендируют в забуференном фосфато-солевом буфере и инактивируют обработкой 0,05% формалином при 4°C в течение 2 недель. Инактивированный вирус и раствор для разведения (Pierce) смешивают в соотношении 3:1 и инкубируют при комнатной температуре в течение 1 часа перед иммунизацией. Генетически модифицированные крыс, экспрессирующих антитела, содержащие только тяжелые цепи, иммунизируют цельными неактивными (вирусами).
Иммунизация белками или пептидами
Обычно иммуногены (белки или пептиды, такие как легкая цепь каппа человеческого иммуноглобулина, тяжелая цепь человеческого IgM или IgG, человеческий сывороточный альбумин, белково-пептидные конъюгаты) разводят до 0,05-0,15 мл стерильным физиологическим раствором и объединяют с адъювантом до конечного объема 0,1-0,3 мл. Доступны многие подходящие адъюванты (т. е. инактивированная нагреванием Bordetella pertussis, гель гидроксида алюминия, Quil A или сапонин, бактериальный липополисахарид или анти-CD40), но ни один из них не обладает активностью полного адъюванта Фрейнда (CFA - Complete Freund's Adjuvant) и неполного адъюванта Фрейнда (IFA - Incomplete Freund's Adjuvant). Концентрация растворимых иммуногенов, таких как белки и пептиды, может варьироваться от 5 мкг до 5 мг в конечном препарате. Первую иммунизацию (прайминг) иммуногеном в CFA вводят внутрибрюшинно и/или подкожно и/или внутримышечно. Если в качестве иммуногенов используются интактные клетки, их лучше всего вводить внутрибрюшинно и/или внутривенно. Клетки разводят физиологическим раствором и вводят от 1 до 20 миллионов клеток за одну инъекцию. Клетки, которые выживают в организме крысы, дадут лучшие результаты иммунизации. После первой иммунизации иммуногенами в CFA вторая иммунизация в IFA (бустерная) обычно проводится через 4 недели. Эта последовательность приводит к развитию В-клеток, продуцирующих антитела, имеющие высокую афинность. Если иммуноген является слабым, бустер-иммунизация проводится каждые 2 недели, пока не будет достигнут сильный гуморальный ответ. Концентрации иммуногена могут быть ниже при бустерной иммунизации и могут быть использованы внутривенные пути. Сыворотку собирают у крыс каждые 2 недели для определения гуморального ответа.
Иммунизацию пентапептидами Липида II, белками, связывающими пенициллин, бета-лактамазами, сортазами и другими мембранными белками прокариот проводят, как описано выше.
Иммунизация липидами, гликолипидами и углеводными полимерами
Иммунизация поли-N-ацетил-β-(1-6) глюкозамином (PNAG-poly-N-acetyl-β-(1-6)-glucosamine) и другими углеводными полимерами
Очищенные dPNAG, PNAG или другие углеводные полимеры конъюгируют с дифтерийным токсином, альбумином или другим белком-носителем путем восстановительного аминирования. Альдегидные группы сначала вводятся на поверхность белка-носителя обработкой глутаральдегидом. Активированные белки-носители впоследствии реагируют с dPNAG (или другими углеводными полимерами) через его свободные аминогруппы в присутствии восстановительного агента NaCNBH3. Животных иммунизируют PNAG и dPNAG-DT конъюгатами подкожно или в ступни в день 0 и бустируются в последующие недели с помощью 0,15-100- мкг дозами конъюгированного PNAG или dPNAG в полном адъюванте Фрейнда или других адъювантах. Бустерные иммунизации желательно проводить в неполных Фрейндах. Кровь отбирается еженедельно, титры специфических антител определяются методом ИФА. В-клетки извлекаются из дренирующих лимфатических узлов или селезенки и гибридомы получаются стандартными методами. Альтернативно, дрожжевые клетки используются для отбора антигенсвязывающих антител или фрагментов.
Иммунизация Липидом II
Очищенный Липид II с С55 или более коротким липидным хвостом смешивают с Titermax, липосомами, адъювантом Риби или другими адъювантами. Кроме того, в смесь добавляют белки, такие как KLH или овальбумин, и иммуностимулирующие соединения, такие как липид А или полный адъювант Фрейнда. Первичную иммунизацию проводят 0,1-1 мг Липида II в подушечку стопы или подкожно в день 0 и бустируют в последующие недели дозами 0,15-100 мкг конъюгированного PNAG или dPNAG в полном адъюванте Фрейнда или других адъювантах. Бустерные иммунизации желательно проводить в неполных Фрейндах. Кровь отбирается еженедельно, титры специфических антител определяются методом ИФА. В-клетки извлекаются из дренирующих лимфатических узлов или селезенки и гибридомы получаются стандартными методами. Альтернативно, дрожжевые клетки используются для отбора антигенсвязывающих антител или фрагментов.
Иммунизация Липидом А
Базовый-липид А получают по методу Bogard et al., 1987, Infection and Immunity, 55: 4, 899-908. Acinetobacter baumannii ATCC 19606 или другие грамотрицательные бактерии культивируют в среде Лурия (Luria broth), собирают центрифугированием, суспендируют в лиофилизированной среде до плотности клеток 109 КОЕ/мл и помещают в предварительно взвешенную ампулу или стеклянный флакон, заполняя не более 1/3 их объема. Стерильную вату или стекловату вставляют в горловину ампулы или в пробирку вставляют пробку. Образец медленно замораживают в ультранизкотемпературном морозильнике, а затем вакуум применяют в течение ночи для первичной сушки. Вторичная сушка осуществляется путем повышения температуры до 20°С в течение нескольких часов. Лиофилизированные образцы хранят при 4°С. 10 мг лиофилизированных клеток суспендируют в 400 мкл изомасляной кислоты и 1 М гидроксида аммония (5: 3, об. /Об.) и инкубируют в пробирке с завинчивающейся крышкой с мешалкой при 100°С в течение 2 часов. Образцы охлаждают в ледяной воде и центрифугируют при 2000х g в течение 15 минут. Надосадочную жидкость переносят в новую пробирку, разбавляют равной частью воды и лиофилизируют. Образец дважды промывают 400 мкл метанола и центрифугируют при 2000g в течение 15 минут, а нижние органические слои и их поверхности раздела сохраняют и сушат в токе азота. Затем пять миллиграммов липида А суспендируют в 5 мл 0,5% (вес/объем) триэтиламида (Sigma, St. Louis, МО), после чего добавляют 5 мг обработанных кислотой бактерий. Смесь медленно перемешивают в течение 30 минут при комнатной температуре и сушат в вакууме с помощью центрифуги Speed Vac.
Клетки, покрытые Липидом А, используются для иммунизации животных внутрибрюшинно или в подушечки лап. Дозу 50 мкл суспензии клеток, покрытых базовым LPS, на одну инъекцию используют в смеси 1: 1 с неполным адъювантом Фрейнда (Difco) (De Kievit&Lam, Journal of Bacteriology, Dec. 1994, p. 7129-7139). Животных иммунизируют в дни 0, 4, 9, 14 и 28. Линии гибридомных клеток подвергают скринингу на продуцирование антител против LPS посредством иммуноферментного анализа (ИФА) и LPS, очищенных из разных источников (Avanti Polar Lipids, Алабастер, Алабама). Супернатанты также тестируют на убитых теплом или живых грамотрицательных бактериях, таких как Klebsiella, E. coli, Pseudomonas aeruginosa, Acitenobacter Baumani i и Salmonella. Широко реактивные антитела отбирают и проводят конкурентные анализы с полимиксином В.
Минимальная ингибирующая концентрация (MIC) HCAb
Различные лабораторные методы могут быть использованы для измерения восприимчивости бактерий к противомикробным агентам in vitro. Для количественного измерения in vitro активности антимикробного агента против данного бактериального изолята можно использовать методы разведения среды (бульона) или агара. Для проведения испытаний готовят серию пробирок или чашек с бульоном или агаровой средой, в которую добавляют различные концентрации антимикробных агентов. Пробирки или чашки затем инокулируют стандартизированной суспензией тестового организма. После инкубации в течение ночи при 35 ±2°C тесты проверяются и определяется минимальная ингибирующая концентрация (MIC - minimal inhibitory concentration). На конечный результат существенное влияние оказывает методология, которая должна тщательно контролироваться для достижения воспроизводимых результатов (внутрилабораторных и межлабораторных). MIC, полученная с использованием теста на разбавление, указывает на концентрацию антимикробного агента, необходимую для подавления заражающего организма. MIC, однако, не представляет собой абсолютное значение. «Истинная» MIC находится где-то между самой низкой тестовой концентрацией, которая подавляет рост организма (то есть показание MIC), и следующей более низкой тестовой концентрацией. Если, например, были использованы двукратные разведения, а MIC составляет 16 мкг/мл, то «истинная» MIC была бы между 16 и 8 мкг/мл. Даже при самых лучших контролируемых условиях тест на разбавление может не давать одну и ту же конечную точку каждый раз, когда он проводится. Как правило, приемлемая воспроизводимость теста находится в пределах двухкратного разбавления фактической конечной точки.
In vitro активность антител или их фрагментов и сравнительных антибиотиков (ванкомицина, полимиксинов, цефалоспоринов или других бета-лактамов) будет проверяться с помощью анализа MIC с микроразведениями среды (Broth Microdilution MIC) в соответствии со стандартной методологией CLSI (Институт клинических и лабораторных стандартов. 2003. Методы для разбавительных тестов на чувствительность к противомикробным препаратам для бактерий, которые растут аэробно. Утвержденный стандарт М7-А7).
Активность антител или их фрагментов тестируют против Staphylococcus aureus ATCC 29213, грамположительных и грамотрицательных бактерий. MIC определяется следующим образом:
1. Приготовление бактериального инокулята. Выберите отдельную колонию каждого штамма из свежей чашки (возраст менее 1 недели) и перенесите в 5 мл скорректированной по катионам среды Мюллера-Хинтона (CAMHB). Инкубируйте в шейкере при 37°C в течение ночи.
2. На следующее утро сделайте разведение ночных культур 1:100 (50 мкл в 5 мл). Выдержите 2-4 часа в шейкере при 37°C.
3. Через 2-4 часа (при оптической плотности около 0,3-0,5 при 600 нм) отцентрифугируйте бактерии (5000 об/мин, 5 мин), ресуспендирйте в PBS, доведите культуры до МакФарланд 0,5 и перенесите 50 мкл доведенной культуры в 9950 мкл CAMHB, перемешивают встряхиванием.
4. Через 2-4 часа (при оптической плотности около 0,3-0,5 при 600 нм) отцентрифугируйте бактерии (5000 об/мин, 5 мин), ресуспендируйте в PBS, доведите культуры до МакФарланд 0,5 и перенесите 50 мкл доведенной культуры в 9950 мкл 1,1x CAMHB (доведенная по катионам среда Мюллера - Хинтона), перемешайте на вортексе.
5. Подготовка материнского планшета [один материнский планшет на 10 дочерних планшетов, один дочерний планшет на штамм]: (приготовьте, пока бактерии растут, этап 5): Добавьте 200 мкл стоков соединений в первые лунки планшета с глубокими лунками и 100 мкл ДМСО во все другие лунки. Сделайте 2-кратные серийные разведения соединений в лунках 2-11 (перенесите 100 мкл из 1-й во 2-ю, перемешайте 4 раза пипетированием, перенесите 100 мкл из 2-й в 3-ю, перемешайте и т. д., удалите 100 мкл из лунки 11). Лунка 12 - это контроль только с ДМСО.
Добавте 900 мкл стерилизованной дистиллированной воды в каждую лунку, перемешайте пипетированием.
6. Подготовка планшета для MIC: Перенесите 10 мкл из каждой лунки материнского планшета в соответствующие лунки дочерних планшетов. Добавте 90 мкл бактериального инокулята в каждую лунку, перемешайте пипетированием.
7. Инкубируйте 24 часа при 37°C и оцените рост. Наличие роста определяется либо по мутной лунке, либо по очевидному осадку, либо по > 3 скоплениям бактерий, присутствующих в лунке. Самая низкая концентрация соединения, при которой нет роста, оценивается как MIC.
MIC ванкомицина для Staphylococcus aureus варьируют от 0,1 мкг/мл до 10 мкг/мл с использованием стандартного анализа MIC с микроразведениями среды. MIC интактных HCAb для Staph. aureus варьируют от 0,025 мкг/мл до 25 мкг/мл. Если используются фрагменты вариабельных областей или нанотела, MIC для Staph. aureus варьирует от 0,008 мкг/мл до 8 мкг/мл.
(MIC ванкомицина Staph. Aureus: 0,25-4 мкг/мл, MIC ванкомицина MRSA: 1-138 мкг/мл, мол. масса ванкомицина составляет 1450 дальтон, мол. масса HCAb составляет 80000 дальтон, HCAb примерно в 50 раз тяжелее ванкомицина, афинность ванкомицина к Липиду II составляет 50 нМ, афинность HCAb будет от 0,5 до 10 нМ. HCAb будут иметь MIC от 0,025 до 25 мкг/мл).
(J. Clin. Microbiol. 2006, 44(11):3883. DOI: A. Bruckner Guiqing Wang, Janet F. Hindler, Kevin W. Ward and David. Increased Vancomycin MICs for Staphylococcus aureus Clinical Isolates from a University Hospital during a 5-Year Period)
MIC HCAb в присутствии иммуноглобулинов, комплемента и фагоцитарных клеток
Фагоцитарные анализы опсонизации (OPA - opsonization phagocytic assays) проводят, как описано в патенте США № 8410249. OPA измеряют уничтожение бактерий в присутствии антител, комплемента и фагоцитирующих клеток. Комплемент и клетки человека широко используются. HCAb или фрагменты тестируют в концентрациях от 0,01 мкг/мл до 10 мкг/мл. Уничтожение бактерий измеряется путем определения колониеобразующих единиц до и после инкубации со всеми компонентами. Опсонизация также визуализируется под микроскопом. В качестве эффекторных клеток используют либо полиморфно-ядерные лейкоциты человека (PMN - polymorphonuclear leukocyte), либо дифференцированные клетки промиелоцитарной лейкемии HL60 (HL60). Комплемент в человеческой сыворотке или сыворотке крови кролика используется в качестве источника комплемента. Вкратце, HCAb или другие образцы серийно разводят в два этапа в 96-луночном планшете для микротитрования сбалансированным солевым раствором Хэнкса, а затем инкубируют с бактериями (~ 2000 КОЕ на лунку) и комплементом в течение 30 минут при 37°C на орбитальном шейкере. Оптимальная скорость встряхивания определяется для каждого бактериального штамма, чтобы минимизировать неспецифическое уничтожение или чрезмерный рост. Свеже выделенные человеческие PMN или дифференцированные клетки HL60 (эффекторные клетки) добавляют в соотношении 400:1 к смеси бактерий (мишень) -комплемент-сыворотка и смесь инкубируют при 37°С в течение 45 мин. Титр ОРА - это разведение сыворотки, которое вызывает снижение КОЕ (уничтожение) на 50% по сравнению с КОЕ из контрольных лунок, содержащих все реагенты, кроме HCAb или их фрагментов. HCAb убивает бактерии от 10 нг/мл до 10 мкг/мл.
Иммунизация клетками
Способ иммунизации клетками хорошо известен в данной области. Например, для экспрессии анти-CD3e-антител клетки человека Jurkat выращивают в культуре ткани. Экспрессию желаемого человеческого антитела анализируют путем инкубации клеток Jurkat с моноклональным антителом OKT3. Затем несвязанный OKT3 удаляют путем отмывки клеток, и связанный OKT3 обнаруживают с помощью антимышиного IgG, конъюгированного с флуоресцеином и анализом проточной цитометрией.
Клетки гибридомы T-клеток крысы трансфицируют электропорацией с помощью эукариотической экспрессионной плазмиды, кодирующей CD3 человека, как описано (Transy et al., 1989, Eur J. Immunol. 19 (5): 947-50). Трансфектанты, экспрессирующие CD3 человека, обогащаются FACS и размножаются в культуре ткани.
Генетически сконструированные крысы, экспрессирующие антитела, содержащие только тяжелые цепи (HCO - heavy chain only), иммунизируют путем инъекции 30×10 (6) клеток Jurkat внутрибрюшинно. Через четыре и восемь недель после первичной иммунизации крыс иммунизируют Т-клетками крысы, экспрессирующими CD3 человека. Животных, экспрессирующих антитела против CD3e человека, содержащие только тяжелые цепи, используют для выделения моноклональных антител против CD3e, содержащих только тяжелые цепи.
Протоколы иммунизации на основе ДНК
Генные вакцины или использование антиген-кодирующих ДНК для иммунизации представляют собой альтернативный подход к развитию сильных ответов антител у крыс.
Путем инокуляции ДНК, как правило, является кожа, мышцы и любой другой путь, который поддерживает трансфекцию и экспрессию антигена. Используют очищенные плазмидные ДНК, которые были сконструированы для экспрессии антигенов, таких как гликопротеин гемагглютинина вируса гриппа или другие человеческие или вирусные антигены. Пути инокуляции ДНК включают следующие: внутривенный (хвостовая вена), внутрибрюшинный; внутримышечный (оба квадрицепса), интраназальный, внутрикожный (например, подушечка стопы) и подкожный (например, загривок шеи). Как правило, 10-100 мкг ДНК вводят в 100 мкл физиологического раствора на сайт инокуляции, или ДНК вводят с помощью соответствующих носителей, таких как частицы золота или определенные составы (http://www.incellart.com/index.php?page=genetic-immunization&menu=3.3), которые облегчают поглощение и трансфекцию клеток. Схема иммунизации аналогична протоколу, описанному выше; первичная иммунизация с последующей бустерной иммунизацией.
Очистка антител, содержащих только тяжелые цепи
Для очистки антител кровь собирают у иммунизированных крыс, а сыворотку или плазму получают центрифугированием, которое отделяет коагулированный клеточный осадок от жидкой верхней фазы, содержащей антитела сыворотки. Антитела из сыворотки плазмы очищают стандартными процедурами. Такие процедуры включают осаждение, ионообменную хроматографию и/или аффинную хроматографию. Для очистки IgG можно использовать белок А или белок G ( et al., JI, 142, 3145, 1989).
et al., JI, 142, 3145, 1989).
Пример 3: Выделение В клеток, экспрессирующих антитела из крыс.
Выделение В-клеток из селезенки, лимфатических узлов или периферической крови
Одноклеточную суспензию получают из селезенки или лимфатических узлов иммунизированной крысы. Клетки можно использовать без дополнительного обогащения, после удаления эритроцитов или после выделения В-клеток, В-клеток памяти, антиген-специфических В-клеток или плазматических клеток. Обогащение может привести к лучшим результатам и, как минимум, рекомендуется удаление эритроцитов. В-клетки памяти выделяют деплетированием нежелательных клеток и последующим положительным отбором. Нежелательные клетки, например, Т-клетки, NK-клетки, моноциты, дендритные клетки, гранулоциты, тромбоциты и эритроидные клетки деплетируются, используя набор антител против CD2, CD14, CD16, CD23, CD36, CD43 и CD235a (гликофорина A). Положительный отбор с помощью антител, специфичных к IgG или CD19, приводит к высокообогащенным В-клеткам (от 50% до 95%). Антиген-специфические В-клетки получают путем воздействия на клетки антигеном(ами), меченным флуоресцентными маркерами и/или магнитными гранулами. Затем клетки, меченные флуорохромными и/или магнитными гранулами, разделяют с использованием (проточной цитометрии или проточного цитометра [FACS - fluorescence activated cell sorter]), сортировщика FACS и/или магнитов. Поскольку плазматические клетки могут экспрессировать небольшое количество поверхностного Ig, может применяться внутриклеточное окрашивание.
Выделение В-клеток методом проточной цитометрии с сортировкой клкток
Методы FACS используются для разделения клеток по их индивидуальным свойствам. Важно, чтобы клетки находились в одноклеточной суспензии. Суспензии отдельных клеток, полученные из периферической крови, селезенки или других иммунных органов иммунизированных крыс, смешивают с меченными флуорохромом антителами, специфичными для маркеров В-клеток, таких как CD19, CD138, CD27 или IgG. Альтернативно, клетки инкубируют с меченными флуорохромами антигенами. Концентрация клеток составляет от 1 до 20 миллионов клеток/мл в подходящем буфере, таком как PBS. Например, В-клетки памяти могут быть выделены путем отбора клеток, положительных по CD27 и отрицательных по CD45R. Плазматические клетки могут быть выделены путем отбора клеток, положительных по CD138 и отрицательных по CD45R. Клетки загружают в прибор FACS, и гейтированные клетки распределяются в 96-луночные планшеты или пробирки, содержащие среду. При необходимости в эксперименте используются положительные контроли для каждого флуорохрома, что позволяет вычитать фон для расчета компенсации.
Выделение В-клеток из костного мозга
Плазматические клетки костного мозга (BMPC - bone marrow plasma cells) выделяют из иммунизированных животных, как описано (Reddy et al., 2010, Nature Biotechnology 28, 965-969). Мышечные и жировые ткани удаляются из собранных голеней и бедер. Концы большеберцовой кости и бедренной кости обрезают хирургическими ножницами, а костный мозг вымывают с помощью инсулинового шприца 26 калибра (0,45 мм) (Becton Dickinson, BD). Костный мозг собирают в стерильно отфильтрованный буфер №. 1 (PBS, 0,1% BSA, 2 мМ ЭДТА). Клетки костного мозга собирают фильтрованием через клеточный фильтр (BD) с механическим воздействием, промывают 20 мл PBS и собирают в пробирку на 50 мл (Falcon, BD). Клетки костного мозга центрифугируют при 335g в течение 10 минут при 4°C. Надосадочную жидкость удаляют и осадок клеток ресуспендируют в 3 мл буфера для лизиса эритроцитов (eBioscience) и осторожно встряхивают при 25°C в течение 5 минут. Суспензию клеток разбавляют 20 мл PBS и центрифугируют при 335g в течение 10 мин при 4°C. Надосадочную жидкость удаляют и клеточный осадок ресуспендируют в 1 мл буфера №1.
Суспензии клеток костного мозга инкубируют с биотинилированными анти-CD45R и анти-CD49b антителами. Суспензию клеток затем вращают при 4°C в течение 20 мин. Затем следует центрифугирование при 930g в течение 6 мин при 4°C, удаление супернатанта и повторное суспендирование осадка клеток в 1,5 мл буфера №.1. Конъюгированные со стрептавидином магнитные гранулы М28 (Invitrogen) промывают и ресуспендируют в соответствии с протоколом производителя. Магнитные гранулы (50 мкл) добавляют к каждой клеточной суспензии и смесь вращают при 4°C в течение 20 мин. Затем клеточные суспензии помещают на магниты Dynabead (Invitrogen) и собирают супернатант (отрицательная фракция, клетки, не конъюгированные с гранулами), и клетки, связанные с гранулами, отбрасывают.
Предварительно промытые магнитные гранулы со стрептавидином M280 инкубируют в течение 30 мин при 4°C с биотинилированным анти-CD138 с 0,75 мкг антитела на 25 мкл смеси магнитных гранул. Затем гранулы промывают в соответствии с протоколом производителя и ресуспендируют в буфере №1. Отрицательную клеточную фракцию (обедненную клетками CD45R + и CD49b +), собранную как указано выше, инкубируют с 50 мкл CD138-конъюгированных магнитных гранул и суспензию вращают при 4°C в течение 30 минут. Гранулы с CD138 + связанными клетками выделяют магнитом, промывают 3 раза буфером №1, и отрицательные (CD138-) клетки, не связанные с гранулами, отбрасывают. Положительные CD138 + связанные с гранулами клетки собирают и хранят при 4°C до дальнейшей обработки.
Альтернативно, может быть использован метод, описанный в Ouisse et al., BMC Biotechnology, 2017, 17: 3, 1-17.
Пример 4: Генерация гибридом
Выделенные В-клетки иммортализуют путем слияния с клетками миеломы, такими как клетки X63 или YB2/0, как описано ( and Milstein, Nature, 256, 495, 1975). Клетки гибридомы культивируют в селективных средах, а клетки гибридомы, продуцирующие антитела, получают путем лимитированного разведения или сортировки отдельных клеток.
and Milstein, Nature, 256, 495, 1975). Клетки гибридомы культивируют в селективных средах, а клетки гибридомы, продуцирующие антитела, получают путем лимитированного разведения или сортировки отдельных клеток.
Пример 5: Выделение кДНК, кодирующих антитела, содержащие только тяжелые цепи
Генерация последовательностей кДНК из изолированных клеток
Выделенные клетки центрифугируют при 930g при 4°C в течение 5 мин. Клетки лизируют реагентом TRI, и общую РНК выделяют в соответствии с протоколом производителя в наборе для выделения РНК Ribopure (Ambion). мРНК выделяют из общей РНК с помощью олиго dT матрикса и Poly (A) набора для очистки (Ambion) в соответствии с протоколом производителя. Концентрация мРНК измеряется с помощью спектрофотометра ND-1000 (Nanodrop).
Выделенную мРНК используют для синтеза первой цепи кДНК путем обратной транскрипции с помощью обратной транскриптазы вируса мышиного лейкоза Малони (MMLV-RT, Ambion). Синтез кДНК осуществляют путем ОТ-ПЦР-праймирования с использованием 50 нг мРНК-матрицы и олиго-dT праймеров в соответствии с протоколом производителя Retroscript (Ambion). После конструирования кДНК проводят амплификацию ПЦР для амплификации антител, содержащих только тяжелые цепи. Список праймеров приведен в Таблице 1:
Таблица 1. Демонстрирует SEQ ID NOS 27-39, соответственно, в порядке появления.

50 мкл ПЦР-реакция состоит из 0,2 мМ смеси прямого и обратного праймера, 5 мкл буфера Thermopol (NEB), 2 мкл неочищенной кДНК, 1 мкл Taq ДНК-полимеразы (NEB) и 39 мкл дважды дистиллированной H2O. Программа термо циклеров ПЦР следующая 92°C в течение 3 мин; 4 цикла (92°С в течение 1 мин, 50°С в течение 1 мин, 72oC в течение 1 мин) ; 4 цикла (92°C в течение 1 минуты, 55°C в течение 1 минуты, 72°C в течение 1 минуты), 20 циклов (92°C в течение 1 минуты, 63°C в течение 1 минуты, 72°C в течение 1 минуты); 72°C в течение 7 мин, хранение при 4°C. Генные продукты ПЦР очищают в геле и сиквенируют ДНК.
Пример 6: Клонирование и экспрессия рекомбинантных антител, содержащих только тяжелые цепи
Продукты ПЦР субклонируют в плазмидный вектор. Для экспрессии в эукариотических клетках. кДНК, кодирующую антитело, содержащее только тяжелые цепи, клонируют в вектор экспрессии, как описано ( Tiller et al., 2008; 329 (1-2): 112-124).
Альтернативно, гены, кодирующие антитела, содержащие только тяжелые цепи, клонируют в продуцирующую минициркулярную плазмиду, как описано (Kay et al., 2010; Nature Biotechnology 28. 12 (2010): 1287-1289).
Альтернативно, гены, кодирующие антитела, содержащие только тяжелые цепи, синтезируют из перекрывающихся олигонуклеотидов с использованием модифицированной термодинамически сбалансированной ПЦР с нуклеацией изнутри (thermodynamically balanced inside-out nucleation PCR) (Gao at al., 2003; Nucleic Acids Research.; 31 (22): e143) и клонируют в вектор эукариотической экспрессии.
Альтернативно, гены, кодирующие антитела, содержащие только тяжелые цепи, синтезируют и клонируют в плазмиду.
Для сборки множественных экспрессионных кассет, кодирующих различные антитела, содержащие только тяжелые цепи, в искусственной хромосоме, множественные экспрессионные кассеты лигируют друг с другом и затем клонируют в вектор BAC, который размножается в бактериях. Для трансфекции используются клетки ElectroMAX™ DH10B™ от Invitrogen (http://tools.invitrogen.com/content/sfs/manuals/18290015.pdf). Альтернативно, лигированные экспрессионные кассеты дополнительно лигируют с плечами искусственных хромосом дрожжей, которые размножаются в дрожжевых клетках (Davies et al., 1996, выше).
Очистка плазмиды
GenElute™ наборы для мини-приготовлений плазмид от компании Sigma-Aldrich используются для выделения плазмид из ~5 мл (или больше) ночной бактериальной культуры (http://www.sigmaaldrich.com/life-science/molecular-biology/dna-and-rna-purification/plasmid-miniprep-kit.html). Это включает сбор бактериальных клеток центрифугированием с последующим щелочным лизисом. Затем ДНК связывается на колонке, промывается и элюируется и становится готовой к расщеплению или секвенированию.
Очистка ВАС
NucleoBondR BAC100 от Clontech - это набор, разработанный для очистки BAC (http://www.clontech.com/products/detail.asp?tabno=2&product_id=186802). Для этого бактерии собирают из 200 мл культуры и лизируют с использованием модифицированной щелочной/SDS процедуры. Бактериальный лизат очищают фильтрацией и загружают в уравновешенную колонку, причем плазмидная ДНК связывается с анионообменным матриксом. После последующих стадий промывки очищенную плазмидную ДНК элюируют в высокосолевом буфере и осаждают изопропанолом. Плазмидную ДНК восстанавливают в буфере ТЕ для дальнейшего использования.
Очистка YAC
Линейные YAC, кольцевые YAC и фрагменты BAC после расщепления очищают электроэлюцией с использованием Elutrap™ (Schleicher и Schuell) (Gu et al., 1992, выше) из полосок, нарезанных из 0,8% агарозных гелей, которые обычно используются или из гель-электрофореза в импульсном поле (PFGE - pulsed-field-gel electrophoresis). Очищенную ДНК преципитируют и повторно растворяют в буфере до желаемой концентрации.
Очистку кольцевых YAC из дрожжей проводят с использованием анионообменного матрикса на основе диоксида кремния Nucleobond AX (Macherey-Nagel, Германия). Вкратце, сферопласты получают с использованием зимолазы или литиказы и осаждают ( Davies et al., 1996, выше). Затем клетки подвергаются щелочному лизису, связыванию с колонкой AX100 и элюированию, как описано в методе Nucleobond для плазмиды с низкой копийностью. Загрязняющая дрожжевая хромосомная ДНК гидролизируется с использованием Plasmid -Safe™ АТФ-зависимой ДНКазы (Epicenter Biotechnologies), а затем следует конечная стадия очистки с использованием SureClean (Bioline). Аликвоту электрокомпетентных клеток DH10 (Invitrogen) затем трансформируют кольцевой YAC для получения колоний BAC (см. выше). Для отделения ДНК-вставки размером 150-200 т.п.н. от ДНК-вектора ВАС ~ 10 т.п.н. используют стадию фильтрации с сефарозой 4B-CL (Yang et al., выше).
Трансфекция клеток плазмидной или ВАС ДНК
Для экспрессии рекомбинантных антител, содержащих только тяжелые цепи, эукариотические клетки трансфицируют, как описано (Andreason and Evans, 1989, Anal. Biochem. 180(2):269-75; Baker and Cotten, 1997, Nucleic Acid Res., 25(10):1950-6; http://www.millipore.com/cellbiology/cb3/mammaliancell). Клетки, экспрессирующие антитела, содержащие только тяжелые цепи, выделяют с использованием различных методов отбора. Лимитирующие разведения или сортировка клеток используются для выделения отдельных клеток. Клоны анализируют на экспрессию антител, содержащих только тяжелые цепи.
Пример 7: использование гена J в UniRat™ и OmniFlic™
На Фиг. 1 продемонстрирован человеческий трансген, используемый в UniRat™ для экспрессии антител, содержащих только тяжелые цепи, как описано в предыдущих примерах. OmniFlic™ использует тот же кластер V-генов человека, что и UniRat, но экспрессирует фиксированную легкую цепь κ.
На Фиг. 2 продемонстрирован человеческий трансген, используемый в UniRat™ для экспрессии антител, содержащих только тяжелые цепи со всеми J-генами, экспрессирующими аргинин в позиции 101, как описано в предыдущих примерах.
Репертуар экспрессированных антител UniRat™ и OmniFlic™ определяли сиквенированием следующего поколения (next-generation sequencing) полной области VH из мРНК, выделенной из В-клеток, полученных из лимфатических узлов, от иммунизированных животных. Полные последовательности VH из экспрессированных антител были выровнены с последовательностями зародышевой линии IGHV и IGHJ человека. Частота использования гена J была рассчитана по крайней мере для 6 независимых животных UniRat™ и OmniFlic™. Фиг. 3 демонстрирует, что UniRat™ используют IGHJ4, содержащий мутацию W101R с более высокой частотой, чем OmniFlic™, которые имеют последовательность IGHJ4 дикого типа. OmniFlic™ также использует IGHJ6 с гораздо более высокой частотой, чем UniRat™
Пример 8: Мутация W101 ингибирует ассоциацию с λ
Ассоциацию лямбда с большой коллекцией из 1058 антител с тяжелыми цепями измеряли стандартным ИФА. Из общего количества 1058 антител, содержащих только тяжелые цепи, 859 содержали R в позиции 101, а 199 содержали W в позиции 101. Фиг. 4 демонстрирует, что только 2,7% антител с тяжелой цепью R101 показали значительную ассоциацию со свободным лямбда-белком, определяемую как сигнал ИФА в 10 раз или более сильный по сравнению с фоновым сигналом. Напротив, 31,2% антител с тяжелой цепью W101 показали значительную связь со свободным лямбда-белком, используя те же критерии. Эти результаты показывают, что мутация W101R дает высокую степень защиты от ассоциации с лямбда и что антитела с тяжелой цепью W101 имеют гораздо более высокую вероятность ассоциирования с лямбда, чем R101.
Пример 9: Ассоциация свободного белка λ с антителами, содержащими только тяжелые цепи в одном и том же семействе CDR3
Фиг. 5 демонстрирует множественное выравнивание 11 последовательностей VH антител с тяжелыми цепями одного семейства CDR3. Все эти последовательности содержат W в позиции 101. Все верхние 7 последовательностей в выравнивании были положительными для ассоциации с лямбда, измеренной с помощью ИФА. Все нижние 4 последовательности в выравнивании были отрицательными для ассоциации с лямбда, также измеренной с помощью ИФА. Это семейство последовательностей VH демонстрирует дополнительные мутации в позициях a и b, которые различают лямбда-положительные и отрицательные последовательности. Ser или Glu в любой позиции a или b устраняет ассоциацию с лямбда. Однако Ser или Glu в этих позициях в других семействах CDR3 не имели такой же связи с лямбда-связыванием. Результаты из этого семейства позволяют предположить, что существуют компенсаторные мутации в последовательностях VH W101, которые предотвращают ассоциации с лямбда, но эти компенсаторные мутации специфичны для семейства CDR3.
Пример 10: Разнообразие D-J соединений отличается между UniRat™ и OmniFlic™, когда используется IGHJ6.
Как продемонстрировано в Примере 6, IGHJ6 используется в 3 раза чаще в OmniFlic™ по сравнению с UniRat™. Кроме того, как продемонстрировано на Фиг. 3, когда IGHJ6 используется в UniRat™, участок из 5 остатков Tyr, наблюдаемый в последовательности зародышевой линии IGHJ6, наиболее часто укорачивается до 1 Tyr. Напротив, участок из 4 Tyr является наиболее распространенной длиной в OmniFlic™ при использовании IGHJ6. Это говорит о том, что существует селективное давление, чтобы укоротить участок из 5 остатков Tyr, присутствующих в последовательности зародышевой линии IGHJ6 в антителах тяжелой цепи, содержащих W101, когда используется IGHJ6.
Пример 11: Химерный антигенный рецептор с использованием внеклеточного связывающего домена VH человека
Экспрессия химерного антигенного рецептора на первичной Т-клетке требует экспрессии одного химерного белка, который содержит внеклеточный антигенсвязывающий домен и внутриклеточный сигнальный домен. Одноцепочечные фрагменты Fv обычно используются в качестве антигенсвязывающего домена. Пример химерного антигенного рецептора scFv продемонстрирован на панели А на Фиг. 6. Альтернативно, один связывающий домен VH человека используется в качестве внеклеточного связывающего домена (панель B или Фиг. 6). Использование одного VH человека имеет то преимущество, что он является меньшим и менее сложным белком для экспрессии и является менее иммуногенным.
Хотя предпочтительные варианты осуществления данного изобретения были продемонстрированы и описаны в данном документе, специалистам в данной области техники будет очевидно, что такие варианты осуществления представлены только в качестве примера. Специалистам в данной области техники будут известны многочисленные вариации, изменения и замены без отступления от данного изобретения. Следует понимать, что различные альтернативы вариантам осуществления данного изобретения, описанным в данном документе, могут использоваться при практическом применении данного изобретения. Предполагается, что нижеследующая формула изобретения определяет объем данного изобретения и что таким образом охватываются способы и структуры в пределах объема этой формулы изобретения и их эквивалентов.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> TENEOBIO, INC.
<120> ТРАНСГЕННЫЕ ЖИВОТНЫЕ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА, ПРОДУЦИРУЮЩИЕ
МОДИФИЦИРОВАННЫЕ АНТИТЕЛА, СОДЕРЖАЩИЕ ТОЛЬКО ТЯЖЕЛЫЕ ЦЕПИ
<130> TNO-0001-WO
<140>
<141>
<150> 62/379,075
<151> 2016-08-24
<160> 58
<170> PatentIn версия 3.5
<210> 1
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
олигонуклеотид"
<400> 1
gtattacaca caaaatggga aaagctg 27
<210> 2
<211> 32
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
олигонуклеотид"
<400> 2
ccggtagtca aagtagtcac attgtgggag gc 32
<210> 3
<211> 64
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
олигонуклеотид"
<400> 3
ccttaatggg gcctcccaca atgtgactac tttgactacc ggggccaggg aaccctggtc 60
accg 64
<210> 4
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
олигонуклеотид"
<400> 4
gaatcctagg attgccttct tagcctg 27
<210> 5
<211> 176
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
полинуклеотид"
<400> 5
ccatagacca aacttaccta ctatctagtc ctgccaacct taagagcagc aacatggaga 60
cagcagagtg tagagagatc tcctgactgg caggaggcaa gaagatggat tcttactcgt 120
ccatttctct tttatccctc tctggtccta gagaacaacc aggggatgag gggctc 176
<210> 6
<211> 208
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
полинуклеотид"
<400> 6
gcacaagtgg acaaagtctt tggccagtct agaaagaagc ccgtctcaga gatcaaagct 60
ggagggcaac acaggaaaga tgtgggaata agtttactag tcatacaggc aggaacccca 120
ggcccagagg tagtgtccct gtgggagggt ctcttgctct ctgatgtcct tccatgctga 180
gagttagggc ccttgtccaa tcatgttc 208
<210> 7
<211> 71
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
олигонуклеотид"
<400> 7
gaattttgcc caagtttttt cagcttttcc cattttgtgt gtaatacgta cacaccgcag 60
ggtaataact g 71
<210> 8
<211> 74
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
олигонуклеотид"
<400> 8
gacgggcttc tttctagact ggccaaagac tttgtccact tgtgcgcagt tatctatgct 60
gtctcaccat agag 74
<210> 9
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
олигонуклеотид"
<400> 9
ggaggtctag gctggagctg atccag 26
<210> 10
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
олигонуклеотид"
<400> 10
cctcgtcccc tggttgttct ctcaagaaaa agtatgcgtg atcattttgt c 51
<210> 11
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
олигонуклеотид"
<400> 11
agagaacaac caggggacga gg 22
<210> 12
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
олигонуклеотид"
<400> 12
gtccacatag tcctccagag agagaag 27
<210> 13
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
олигонуклеотид"
<400> 13
gacccaagtc cagttcccaa caaccac 27
<210> 14
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
олигонуклеотид"
<400> 14
cctcgtcccc tggttgtcct ctcaagagag gagggagtgt gagcttttcc 50
<210> 15
<400> 15
000
<210> 16
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
олигонуклеотид"
<400> 16
agaggacaac caggggacga ggggctc 27
<210> 17
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
олигонуклеотид"
<400> 17
gcatggggaa ggggcattgt atgtagg 27
<210> 18
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
олигонуклеотид"
<400> 18
cagatcacac tgtctgctca cttcac 26
<210> 19
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
олигонуклеотид"
<400> 19
aaggcagcag gatggaagct gatgtcg 27
<210> 20
<211> 124
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
полинуклеотид"
<400> 20
gctggagggc aacacaggaa agatgtggga ataagtttac tagtcataca ggcaggaacc 60
ccaggcccag aggtagtgtc cctgtgggag ggtctcttgc gcacacaccg cagggtaata 120
actg 124
<210> 21
<211> 91
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
олигонуклеотид"
<400> 21
gatttaaatg tcaattggtg agtcttctgg ggcttcctac atacaatgcc ccttccccat 60
gcgcagttat ctatgctgtc tcaccataga g 91
<210> 22
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
олигонуклеотид"
<400> 22
atgagcagca ccctctcgtt gaccaaggct gactatgaa 39
<210> 23
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
олигонуклеотид"
<400> 23
caggtgtgcc catccagctg agttaaggtg gag 33
<210> 24
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
олигонуклеотид"
<400> 24
caggaccagg acacctgcag cagctggcag gaagcaggt 39
<210> 25
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
олигонуклеотид"
<400> 25
aacagccatt tgcagaccaa agggaaggaa aga 33
<210> 26
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
олигонуклеотид"
<400> 26
ttctaccctg gtgttatgac agtggtctgg aaggcagatg gt 42
<210> 27
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
праймер"
<400> 27
atggactgga cctggaggat cc 22
<210> 28
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
праймер"
<400> 28
atggactgca cctggaggat cc 22
<210> 29
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
праймер"
<400> 29
tccacgctcc tgctgctgac 20
<210> 30
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
праймер"
<400> 30
gctacacact cctgctgctg acc 23
<210> 31
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
праймер"
<400> 31
atggagtttg ggctgagctg g 21
<210> 32
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
праймер"
<400> 32
atggaattgg ggctgagctg 20
<210> 33
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
праймер"
<400> 33
atggagttgg gactgagctg ga 22
<210> 34
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
праймер"
<400> 34
atggaatttg gcctgagctg g 21
<210> 35
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
праймер"
<400> 35
atgcagtttg tgctgagctg g 21
<210> 36
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
праймер"
<400> 36
tgaaacacct gtggttcttc c 21
<210> 37
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
праймер"
<400> 37
tgaagcacct gtggttcttc c 21
<210> 38
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
праймер"
<400> 38
tcatcttcct gcccgtgctg g 21
<210> 39
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
праймер"
<400> 39
gctttcagtg atggtcagtg tgcttatgac 30
<210> 40
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
олигонуклеотид"
<400> 40
actactttga ctacyggggc cagggaaccc tggtcaccgt ctcctcag 48
<210> 41
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
пептид"
<220>
<221> ВАРИАНТ
<222> (5)..(5)
<223> /replace="Arg"
<220>
<221> MISC_FEATURE
<222> (5)..(5)
<223> /note="Остаток приведенный в этой последовательности не является
предпочтительным по отношению к остаткам в аннотациях для указанной
позиции"
<400> 41
Tyr Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
1 5 10 15
<210> 42
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
олигонуклеотид"
<400> 42
gctgaatact tccagcaccg gggccagggc accctggtca ccgtctcctc ag 52
<210> 43
<211> 53
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
олигонуклеотид"
<400> 43
ctactggtac ttcgatctcc ggggccgtgg caccctggtc actgtctcct cag 53
<210> 44
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
олигонуклеотид"
<400> 44
tgatgctttt gatatccggg gccaagggac aatggtcacc gtctcttcag 50
<210> 45
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
олигонуклеотид"
<400> 45
actactttga ctaccggggc cagggaaccc tggtcaccgt ctcctcag 48
<210> 46
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
олигонуклеотид"
<400> 46
acaactggtt cgacccccgg ggccagggaa ccctggtcac cgtctcctca g 51
<210> 47
<211> 63
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
олигонуклеотид"
<400> 47
attactacta ctactactac atggacgtcc ggggcaaagg gaccacggtc accgtctcct 60
cag 63
<210> 48
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
полипептид"
<400> 48
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Thr Val Ser Gly Thr Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Arg Phe Gly Thr Gly Thr Asn Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 49
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
полипептид"
<400> 49
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Arg Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Arg Phe Gly Ser Gly Thr Asn Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 50
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
полипептид"
<400> 50
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Thr Val Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Lys Trp Gly Thr Gly Thr Asn Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 51
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
полипептид"
<400> 51
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Thr Val Ser Gly Thr Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Gln Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Arg Phe Gly Thr Gly Thr Asn Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 52
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
полипептид"
<400> 52
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Ser
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Lys Asp Arg Phe Gly Thr Gly Thr Asn Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 53
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
полипептид"
<400> 53
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Val Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Ser
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Lys Asp Arg Phe Gly Thr Gly Thr Asn Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 54
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
полипептид"
<400> 54
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Thr Val Ser Gly Thr Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Gln Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Ile Tyr Tyr Cys
85 90 95
Ala Lys Asp Arg Phe Gly Thr Gly Thr Asn Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 55
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
полипептид"
<400> 55
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Ser Cys
85 90 95
Ala Lys Asp Arg Phe Gly Thr Gly Thr Asn Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 56
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
полипептид"
<400> 56
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Glu
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Trp Ala Pro Gly Thr Gly Leu Glu Trp Val
35 40 45
Ser Thr Val Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Glu Tyr Tyr Cys
85 90 95
Ala Lys Asp Arg Phe Gly Thr Gly Thr Asn Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 57
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
полипептид"
<400> 57
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Arg Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Ser
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Ser Cys
85 90 95
Ala Lys Asp Arg Phe Gly Thr Gly Thr Asn Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 58
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<221> источник
<223> /note="Описание искусственной последовательности: Синтетический
полипептид"
<400> 58
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Leu Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Thr Val Ser Gly Thr Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Ser Cys
85 90 95
Ala Lys Asp Arg Phe Gly Ser Gly Thr Asn Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА ПРОТИВ ANG-2, СОДЕРЖАЩИЕ ТОЛЬКО ТЯЖЕЛУЮ ЦЕПЬ | 2016 |
|
RU2749674C2 |
| АНТИТЕЛА К BCMA, СОДЕРЖАЩИЕ ТОЛЬКО ТЯЖЕЛУЮ ЦЕПЬ | 2018 |
|
RU2816994C2 |
| АНТИТЕЛА, СОДЕРЖАЩИЕ ТОЛЬКО ТЯЖЕЛЫЕ ЦЕПИ, КОТОРЫЕ СВЯЗЫВАЮТСЯ С CD19 | 2019 |
|
RU2824170C2 |
| АНТИ-BCMA АНТИТЕЛА, СОДЕРЖАЩИЕ ТОЛЬКО ТЯЖЁЛУЮ ЦЕПЬ | 2017 |
|
RU2781301C2 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ НА ОСНОВЕ АКТИВИРУЮЩИХ Т-КЛЕТКИ БИСПЕЦИФИЧЕСКИХ АНТИГЕНСВЯЗЫВАЮЩИХ МОЛЕКУЛ ПРОТИВ CD3 И ФОЛАТНОГО РЕЦЕПТОРА 1 (FOLR1) И АНТАГОНИСТОВ, СВЯЗЫВАЮЩИХСЯ С ОСЬЮ PD-1 | 2015 |
|
RU2753902C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕ SIGLEC15, И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ | 2021 |
|
RU2839071C1 |
| KIR3DL3 - ИНГИБИРУЮЩИЙ РЕЦЕПТОР ИММУННОЙ СИСТЕМЫ И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2827900C1 |
| АНТИТЕЛА ДЛЯ ЛЕЧЕНИЯ ИНФЕКЦИИ ГЕПАТИТОМ В И СВЯЗАННЫХ С НЕЙ ЗАБОЛЕВАНИЙ | 2018 |
|
RU2765878C2 |
| АНТИТЕЛА, СОДЕРЖАЩИЕ ТОЛЬКО ТЯЖЕЛЫЕ ЦЕПИ, КОТОРЫЕ СВЯЗЫВАЮТСЯ С CD22 | 2018 |
|
RU2797348C2 |
| АНТИТЕЛА ПРОТИВ AXL И КОМПОЗИЦИИ | 2021 |
|
RU2837590C1 |



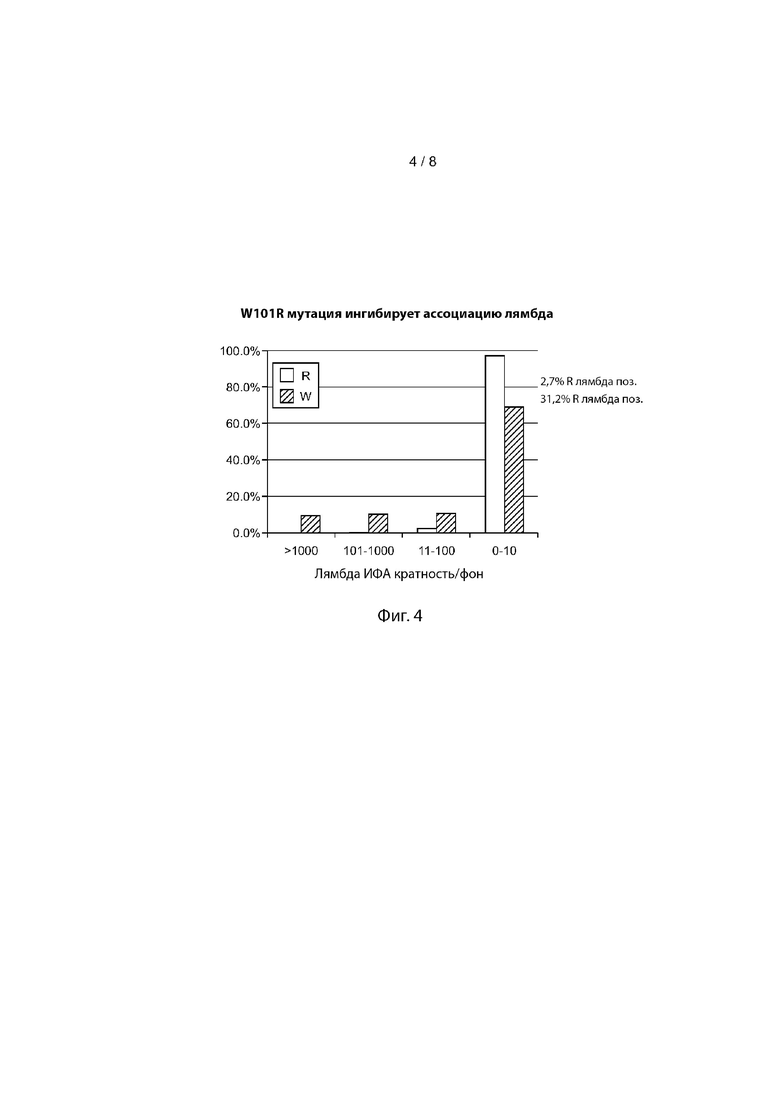




Изобретение относится к области биотехнологии, в частности к нуклеиновой кислоте, кодирующей рекомбинантный иммуноглобулин (Ig), содержащий только тяжелую цепь, которая содержит один или несколько V генных сегментов человека, один или несколько D генных сегментов человека и один или несколько J генных сегментов человека, при этом по меньшей мере один из указанных J-сегментов человека содержит кодон, кодирующий другой аминокислотный остаток по сравнению с нативным аминокислотным остатком в первой позиции четвертой каркасной области (FR4), который способен нарушить целостность экспонированного на поверхности гидрофобного пятна. Также раскрыто трансгенное животное, содержащее вышеуказанную нуклеиновую кислоту. Изобретение эффективно для получения антитела, содержащего только тяжелую цепь (HCAb). 3 н. и 29 з.п. ф-лы, 7 ил., 1 табл., 11 пр.
1. Нуклеиновая кислота, кодирующая рекомбинантный иммуноглобулин (Ig), содержащий только тяжелую цепь, содержащая:
(i) один или несколько V генных сегментов человека, один или несколько D генных сегментов человека и один или несколько J генных сегментов человека, которые при рекомбинации друг с другом в геноме животного, отличного от человека, и после созревания аффинности кодируют вариабельную область тяжелой цепи (VH), содержащую определяющие комплементарность области (CDR) и каркасные области (FR), при этом по меньшей мере один из указанных J-сегментов человека содержит кодон, кодирующий другой аминокислотный остаток по сравнению с нативным аминокислотным остатком в первой позиции четвертой каркасной области (FR4), который способен нарушить целостность экспонированного на поверхности гидрофобного пятна, содержащего нативный аминокислотный остаток или связанного с нативным аминокислотным остатком в этой позиции, и
(ii) сегмент константной (С) области гена, кодирующий константную эффекторную область иммуноглобулина, лишенную функциональности CH1.
2. Нуклеиновая кислота по п. 1, содержащая от 2 до 40 D генных сегментов и от 2 до 20 J генных сегментов.
3. Нуклеиновая кислота по п. 2, отличающаяся тем, что более чем один из указанных J-сегментов человека содержит кодон, кодирующий другой аминокислотный остаток в первой позиции четвертой каркасной области (FR4), который способен нарушить целостность экспонированного на поверхности гидрофобного пятна, содержащего нативный аминокислотный остаток или связанного с нативным аминокислотным остатком в этой позиции.
4. Нуклеиновая кислота по п. 3, отличающаяся тем, что все указанные J-сегменты человека содержат кодон, кодирующий другой аминокислотный остаток в первой позиции четвертой каркасной области (FR4), который способен нарушить целостность экспонированного на поверхности гидрофобного пятна, содержащего нативный аминокислотный остаток или связанного с нативным аминокислотным остатком в этой позиции.
5. Нуклеиновая кислота по п. 4, отличающаяся тем, что в кодируемой области VH указанный нативный аминокислотный остаток в первой позиции FR4 замещен полярным аминокислотным остатком.
6. Нуклеиновая кислота по п. 5, отличающаяся тем, что в кодируемой области VH нативный аминокислотный остаток в первом положении FR4 замещен положительно заряженным аминокислотным остатком.
7. Нуклеиновая кислота по п. 6, отличающаяся тем, что в кодируемой области VH положительно заряженный аминокислотный остаток выбран из группы, состоящей из лизина (K), аргинина (R) и гистидина (H).
8. Нуклеиновая кислота по п. 7, отличающаяся тем, что в кодируемой области VH положительно заряженный аминокислотный остаток представляет собой аргинин (R).
9. Нуклеиновая кислота по п. 8, отличающаяся тем, что кодируемая VH область содержит замену триптофана (W) на аргинин (R) в первом аминокислотном остатке в четвертой каркасной области (FR4).
10. Нуклеиновая кислота по п. 9, содержащая J4 генный сегмент, в котором кодон для W заменен кодоном для R.
11. Нуклеиновая кислота по любому из пп. 1-10, отличающаяся тем, что кодируемая область VH содержит одну или несколько дополнительных мутаций в одной или нескольких каркасных областях.
12. Нуклеиновая кислота по любому из пп. 1-11, кодирующая человеческое или гуманизированное антитело, содержащее только тяжелую цепь, содержащее указанную область VH.
13. Трансгенное животное, отличное от человека, содержащее нуклеиновую кислоту, кодирующую рекомбинантный Ig, содержащий только тяжелую цепь, по любому из пп. 1-12 для получения антитела, содержащего только тяжелую цепь (HCAb).
14. Трансгенное животное, отличное от человека, по п. 13, отличающееся тем, что является млекопитающим, отличным от человека.
15. Трансгенное животное, отличное от человека, по п. 14, отличающееся тем, что представляет собой грызуна.
16. Трансгенное животное, отличное от человека, по п. 15, отличающееся тем, что представляет собой крысу или мышь.
17. Трансгенное животное, отличное от человека, по п. 16, отличающееся тем, что представляет собой крысу.
18. Трансгенное животное, отличное от человека, по п. 17, отличающееся тем, что представляет собой UniRat™.
19. Трансгенное животное, отличное от человека, которое не экспрессирует никаких функциональных генов легкой цепи иммуноглобулина и содержит нуклеиновую кислоту, кодирующую гетерологичный Ig, содержащий только тяжелую цепь, содержащую один или несколько V генных сегментов, один или несколько D генных сегментов и один или несколько J генных сегментов, которые при рекомбинации друг с другом и после созревания аффинности кодируют область VH, содержащую определяющие комплементарность области (CDR) и каркасные области (FR), в которой указанный нативный аминокислотный остаток в первой позиции четвертой каркасной области (FR4) указанной области VH замещен другим аминокислотным остатком, который способен нарушить целостность экспонированного на поверхности гидрофобного пятна, содержащего нативный аминокислотный остаток или связанного с нативным аминокислотным остатком в этой позиции, и один или несколько генных сегментов константной эффекторной области, кодирующие константную эффекторную область антитела, лишенную функциональности CH1, при этом генные сегменты расположены так, что V, D и J генные сегменты и генные сегменты константной области рекомбинируют с образованием перестроенного аффинно-зрелого гена только тяжелой цепи, кодирующего антитело, содержащее только тяжелую цепь (HCAb), для получения антитела, содержащего только тяжелую цепь (HCAb).
20. Трансгенное животное, отличное от человека, по п. 19, отличающееся тем, что в указанной области VH нативный аминокислотный остаток в первой позиции FR4 замещен полярным аминокислотным остатком.
21. Трансгенное животное, отличное от человека, по п. 19, отличающееся тем, что в указанной области VH нативный аминокислотный остаток в первой позиции FR4 замещен положительно заряженным аминокислотным остатком.
22. Трансгенное животное, отличное от человека, по п. 21, отличающееся тем, что в указанной области VH положительно заряженный аминокислотный остаток выбран из группы, состоящей из лизина (K), аргинина (R) и гистидина (H).
23. Трансгенное животное, отличное от человека, по п. 22, отличающееся тем, что в указанной области VH положительно заряженный аминокислотный остаток представляет собой аргинин (R).
24. Трансгенное животное, отличное от человека, по п. 23, отличающееся тем, что указанная область VH содержит замену триптофана (W) на аргинин (R) в первом аминокислотном остатке в четвертой каркасной области (FR4).
25. Трансгенное животное, отличное от человека, по п. 24, отличающееся тем, что указанная нуклеиновая кислота, кодирующая гетерологичный Ig, содержащий только тяжелую цепь, содержит J4 сегмент, в котором кодон для W заменен кодоном для R.
26. Трансгенное животное, отличное от человека, по любому из пп. 19-25, отличающееся тем, что кодируемое антитело, содержащее только тяжелую цепь, содержит одну или несколько дополнительных мутаций в одной или нескольких каркасных областях.
27. Трансгенное животное, отличное от человека, по любому из пп. 19-26, отличающееся тем, что является млекопитающим.
28. Трансгенное животное, отличное от человека, по п. 27, отличающееся тем, что является грызуном.
29. Трансгенное животное, отличное от человека, по п. 28, отличающееся тем, что является крысой или мышью.
30. Трансгенное животное, отличное от человека, по п. 29, отличающееся тем, что является крысой.
31. Трансгенное животное, отличное от человека, по п. 30, отличающееся тем, что является UniRat™.
| WO03035694 A2, 01.05.2003 | |||
| ALINE DESMYTER et al., Antigen Specificity and High Affinity Binding Provided by One Single Loop of a Camel Single-domain Antibody, THE JOURNAL OF BIOLOGICAL CHEMISTRY, 2001, Vol | |||
| СПОСОБ СОСТАВЛЕНИЯ ЗВУКОВОЙ ЗАПИСИ | 1921 |
|
SU276A1 |
| Видоизменение прибора с двумя приемами для рассматривания проекционные увеличенных и удаленных от зрителя стереограмм | 1919 |
|
SU28A1 |
| РАЗРЫВНАЯ ЗАСТЕЖКА | 1930 |
|
SU26285A1 |
| RICK JANSSENS et al., Generation of heavy-chain-only antibodies in mice, PNAS, | |||

