РОДСТВЕННЫЕ ЗАЯВКИ И ВКЛЮЧЕНИЕ ПОСРЕДСТВОМ ССЫЛКИ
[0001] Данная заявка испрашивает приоритет согласно предварительной заявке на патент США под регистрационным №63/000566, поданной 27 марта 2020 г.
[0002] Вышеуказанные заявки и все документы, цитируемые в них или во время их рассмотрения («цитируемые в заявках документы») и все документы, цитируемые или упоминаемые в данном документе (включая без ограничения все литературные документы, патенты, опубликованные патентные заявки, цитируемые в данном документе) («документы, цитируемые в данном документе»), а также все документы, цитируемые или упоминаемые в цитируемых в данном документе документах, вместе с любыми инструкциями производителя, описаниями, спецификациями продуктов и описаниями продуктов, имеющимися для любых продуктов, упомянутых в данном документе, или приведенными в любом документе, включенном в данный документ в качестве ссылки, настоящим включены в данный документ посредством ссылки и могут быть использованы при практическом осуществлении настоящего изобретения. Более конкретно, все ссылочные документы включены посредством ссылки в той же степени, как если бы каждый отдельный документ был конкретно и индивидуально указан как включенный посредством ссылки. Любые последовательности из Genbank, упомянутые в данном документе, включены посредством ссылки на последовательность из Genbank по состоянию на самую раннюю эффективную дату подачи данного документа.
ОБЛАСТЬ ТЕХНИКИ
[0003] Настоящее изобретение в целом относится к выделенному моноклональному антителу, такому как полностью человеческое, мышиное, химерное или гуманизированное моноклональное антитело, или его антигенсвязывающей части, которые специфично связываются с Siglec15 с высокой аффинностью и функциональностью. Также представлены молекула нуклеиновой кислоты, кодирующая антитело или его антигенсвязывающую часть, вектор экспрессии, клетка-хозяин и способ обеспечения экспрессии антитела или его антигенсвязывающей части.
В настоящем изобретении дополнительно представлены иммуноконъюгат, биспецифическая молекула, химерный антигенный рецептор, онколитический вирус и фармацевтическая композиция, содержащие антитело или его антигенсвязывающую часть, а также способ диагностики и лечения с применением антитела к Siglec15 согласно настоящему изобретению.
УРОВЕНЬ ТЕХНИКИ
[0004] Иммунотерапия является революционным терапевтическим подходом, обеспечивающим повышение эффективности иммунной системы для борьбы с такими заболеваниями, как рак. Она применима ко многим показаниям и обуславливает менее высокую степень токсичности по сравнению с другими стандартными видами терапии. Сигнальный путь PD-1/PD-L1 является важной мишенью в иммунотерапии опухолей, и было клинически одобрено несколько ингибиторов для каждого из PD-1 или PD-L1, таких как антитела к PD-1 Opdivo® и Keytruda®, а также антитело к PD-L1 Tecentriq®. Однако, у части пациентов не наблюдается наличия ответа на такие средства лечения. Как показало недавнее исследование, нацеливание на Siglec15 может являться дополнительным подходом для пациентов, страдающих от рака, у которых не наблюдается ответа на средства терапии, нацеливающиеся на PD-1/PD-L1 (Jun Wang et al., (2019) Nature Medicine 25:656-666).
[0005] Siglec15 является членом семейства белков Siglec, характеризующихся структурой лектинов иммуноглобулинового типа, связывающих сиаловую кислоту. Он содержит два внеклеточных иммуноглобулиноподобных домена, трансмембранный домен с остатком лизина, который необходим для взаимодействия с адапторным белком DAP 12, и цитоплазматический хвостовой участок (Takashi Angata et al., (2007) Glycobiology 17(8): 83 8-846).
[0006] Siglec15 экспрессируется на остеокластах и играет роль в дифференцировке остеокластов и ремоделировании кости (Hiruma Y et al., (2011) Biochemical and Biophysical Research Communications 409(3):424-429; Takashi Angata (2020) Journal of Biomedical Science 27:10). Введение антител к Siglec15 подавляло резорбцию кости остеокластами и увеличивало костную массу в моделях на грызунах (Stuible M et al., (2014) Journal of Biological Chemistry 289(10):6498-6512; Sato D et al., (2018) Вопе 116:172-180).
[0007] Siglec15 также экспрессируется на опухоль-ассоциированных макрофагах и предпочтительно распознает антиген сиалил-Tn, представляющий собой опухоль-ассоциированную гликановую структуру. Совместное культивирование сиалил-Tn/- линии раковых клеток и M-CSF-индуцированных макрофагов человека или Siglec15+ линии миелоидных клеток индуцировало продуцирование трансформирующего фактора роста-β, который способствовал эпителиально-мезенхимальному переходу и метастазированию раковых клеток (Takamiya R et al., (2013) Glycobiology 23(2): 178-187). Lieping Cheng et al. недавно обнаружили, что Siglec15 дополнительно экспрессируется на опухолевых клетках и/или опухоль-ассоциированных стромальных клетках в клинических образцах немелкоклеточного рака легкого. Они также обнаружили, что белки Siglec15 подавляли пролиферацию и активацию Т-клеток, а антитела к Siglec15 обеспечивали реверсию подавления активности Т-клеток и ослабление роста раковых клеток in vivo. Поскольку Siglec15 и PD-L1 являются взаимоисключающими в раковых тканях, Siglec15 может служить в качестве дополнительной терапевтической мишени и обеспечивать альтернативные средства лечения для пациентов, у которых наблюдается устойчивость к блокаде PD-1/PD-L1, как упоминалось выше (Jun Wang et al., (2019) выше). NC318, гуманизированное антитело к Siglec15, было клинически протестировано у пациентов с солидными опухолями на поздней стадии, включая немелкоклеточный рак легкого, рак яичника, меланому, колоректальный рак и рак молочной железы, и при этом длительную стабилизацию заболевания наблюдали у 54% пациентов, а объективный ответ у 5,4% (Sun J et al., (2021) Clin Cancer Res. 27(3):680-688).
[0008] Ввиду участия Siglec15 в ремоделировании кости и развитии опухоли Siglect15 определенно является многообещающей терапевтической мишенью. Имеется необходимость в антителах к Siglec15 с улучшенными фармацевтическими характеристиками.
[0009] Цитирование или указание любого документа в данной заявке не является признанием того, что такой документ доступен в качестве предшествующего уровня техники для настоящего изобретения.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0010] В настоящем изобретении представлено выделенное мышиное, химерное, человеческое или гуманизированное моноклональное антитело или его антигенсвязывающая часть, которые связываются с Siglec15 (например, Siglec15 человека и Siglec15 обезьяны) и характеризуется сравнимой, если не более высокой, аффинностью/способностью связывания с Siglec15 и блокирующей активностью в отношении связывания Siglec15 с лигандом, таким как LRRC4C, по сравнению с антителами к Siglec15 из предшествующего уровня техники, такими как Siglec15-ch5G9 (Nextcure). Антитело или его антигенсвязывающая часть способны обеспечивать реверсию Т-клеточной супрессии, опосредованной Siglec15.
[0011] Антитело или его антигенсвязывающая часть по настоящему изобретению могут быть использованы для ряда различных вариантов применения, включая обнаружение белка Siglec15, а также лечение и предупреждение связанных с Siglec15 заболеваний, таких как опухоли и остеопороз.
[0012] Соответственно, в одном аспекте изобретение относится к выделенному моноклональному антителу (например, человеческому, мышиному, химерному или гуманизированному антителу) или его антигенсвязывающей части, которые связывают Siglec15, характеризующимся I) вариабельной областью тяжелой цепи, которая может содержать участок CDR1 VH, участок CDR2 VH и участок CDR3 VH, где участок CDR1 VH, участок CDR2 VH и участок CDR3 VH могут содержать аминокислотные последовательности, характеризующиеся по меньшей мере 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичностью с (l) SEQ ID NO: 1, 2 (X1 представляет собой D, Х2 представляет собой Q) и 3, соответственно; (2) SEQ ID NO: 1, 2 (X1 представляет собой Е, Х2 представляет собой Q) и 3, соответственно; (3) SEQ ID NO: 1, 2 (X1 представляет собой D, Х2 представляет собой K) и 3, соответственно; (4) SEQ ID NO: 9, 10 и 11, соответственно, или (5) SEQ ID NO: 33, 34 и 35, соответственно; и/или II) вариабельной областью легкой цепи, которая может содержать участок CDR1 VL, участок CDR2 VL и участок CDR3 VL, где участок CDR1 VL, участок CDR2 VL и участок CDR3 VL могут содержать аминокислотные последовательности, характеризующиеся по меньшей мере 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичностью с (1) SEQ ID NO: 4, 5 и 6, соответственно; (2) SEQ ID NO: 12, 13 и 14, соответственно, или (3) SEQ ID NO: 36, 37 и 38, соответственно.
[0013] Антитело или его антигенсвязывающая часть по настоящему изобретению могут содержать вариабельную область тяжелой цепи, которая может содержать участок CDR1 VH, участок CDR2 VH и участок CDR3 VH, и вариабельную участок легкой цепи, которая может содержать участок CDR1 VL, участок CDR2 VL и участок CDR3 VL, где участок CDR1 VH, участок CDR2 VH, участок CDR3 VH, участок CDR1 VL, участок CDR2 VL и участок CDR3 VL могут содержать аминокислотные последовательности, характеризующиеся по меньшей мере 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичностью с (1) SEQ ID NO: 1, 2 (X1 представляет собой D, Х2 представляет собой Q), 3, 4, 5 и 6, соответственно; (2) SEQ ID NO: 1, 2 (X1 представляет собой Е, Х2 представляет собой Q), 3, 4, 5 и 6, соответственно; (3) SEQ ID NO: 1, 2 (X1 представляет собой D, Х2 представляет собой K), 3, 4, 5 и 6, соответственно; (4) SEQ ID NO: 9, 10, 11, 12, 13 и 14, соответственно или (5) SEQ ID NO: 33, 34, 35, 36, 37 и 38, соответственно, при этом антитело или его антигенсвязывающий фрагмент связываются с Siglec15.
[0014] Вариабельная область тяжелой цепи антитела или его антигенсвязывающая часть по настоящему изобретению может содержать аминокислотную последовательность, характеризующуюся по меньшей мере 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичностью с SEQ ID NO: 7 (X1 представляет собой D, Х2 представляет собой Q; X1 представляет собой Е, Х2 представляет собой Q; или X1 представляет собой D, Х2 представляет собой K), 15, 39, 40 или 41, где антитело или его антигенсвязывающий фрагмент связываются с Siglec15. Аминокислотная последовательность под SEQ ID NO: 7 (X1 представляет собой D, Х2 представляет собой Q) может кодироваться нуклеотидной последовательностью под SEQ ID NO: 27, а аминокислотная последовательность под SEQ ID NO: 15 может кодироваться нуклеотидной последовательностью под SEQ ID NO: 29.
[0015] Вариабельная область легкой цепи антитела или его антигенсвязывающая часть по настоящему изобретению могут содержать аминокислотную последовательность, характеризующуюся по меньшей мере 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичностью с SEQ ID NO: 8, 16, 42 или 43, где антитело или его антигенсвязывающий фрагмент связываются с Siglec15. Аминокислотные последовательности под SEQ ID NO: 8 и 16 могут кодироваться нуклеотидными последовательностями под SEQ ID NO: 28 и 30.
[0016] Антитело или его антигенсвязывающая часть по настоящему изобретению могут содержать вариабельную область тяжелой цепи и вариабельную область легкой цепи, характеризующиеся аминокислотными последовательностями, характеризующимися по меньшей мере 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичностью с (1) SEQ ID NO: 7 (X1 представляет собой D, Х2 представляет собой Q) и 8, соответственно; (2) SEQ ID NO: 7 (X1 представляет собой Е, Х2 представляет собой Q) и 8, соответственно; (3) SEQ ID NO: 7 (X1 представляет собой D, Х2 представляет собой K) и 8, соответственно; (4) SEQ ID NO: 15 и 16, соответственно; (5) SEQ ID NO: 39 и 42, соответственно; (6) SEQ ID NO: 40 и 42, соответственно или (7) SEQ ID NO: 41 и 43, соответственно, где антитело или его антигенсвязывающий фрагмент связываются с Siglec15.
[0017] Выделенное моноклональное антитело или его антигенсвязывающая часть по настоящему изобретению могут содержать тяжелую цепь и легкую цепь, связанные дисульфидными связями, причем тяжелая цепь может содержать вариабельную область тяжелой цепи и константную область тяжелой цепи, легкая цепь может содержать вариабельную область легкой цепи и константную область легкой цепи, где С-конец вариабельной области тяжелой цепи соединен с N-концом константной области тяжелой цепи, а С-конец вариабельной области легкой цепи соединен с N-концом константной области легкой цепи, при этом вариабельная область тяжелой цепи и вариабельная область легкой цепи могут содержать аминокислотные последовательности, описанные выше, и антитело или его антигенсвязывающая часть связываются с Siglec15. Константная область тяжелой цепи может представлять собой константную область тяжелой цепи IgG1, IgG2 или IgG4, например, константную область тяжелой цепи IgG1, IgG2 или IgG4 человека, характеризующуюся аминокислотной последовательностью, указанной, например, под SEQ ID NO: 17. Константная область тяжелой цепи, такая как Fc-фрагмент, может быть сконструирована с приданием ей сниженной или повышенной аффинности связывания FcR. Константная область легкой цепи может представлять собой константную область каппа-цепи, например, константную область каппа-цепи человека, характеризующуюся аминокислотными последовательностями, указанными, например, под SEQ ID NO.: 18. Аминокислотные последовательности под SEQ ID NO: 17 и 18 могут кодироваться нуклеотидными последовательностями под SEQ ID NO: 31 и 32, соответственно.
[0018] Антитело по настоящему изобретению в некоторых вариантах осуществления может содержать две тяжелые цепи и две легкие цепи или состоять из них, где каждая тяжелая цепь может содержать последовательности константной области тяжелой цепи, вариабельной области тяжелой цепи или CDR, упомянутые выше, и каждая легкая цепь может содержать последовательности константной области легкой цепи, вариабельной области легкой цепи или CDR, упомянутые выше, при этом антитело связывается с Siglec15. Антитело или его антигенсвязывающая часть по настоящему изобретению могут представлять собой полноразмерное антитело, например, изотипа IgG1, IgG2 или IgG4. Антитело или его антигенсвязывающая часть по настоящему изобретению в других вариантах осуществления может представлять собой антитело, представленное одноцепочечным вариабельным фрагментом (scFv), или фрагмент антитела, такой как фрагмент Fab или F(ab')2.
[0019] В настоящем изобретении также представлена биспецифическая молекула, которая может предусматривать антитело или его антигенсвязывающую часть по настоящему изобретению, связанная со вторым функциональным фрагментом (например, вторым антителом), характеризующимся специфичность связывания, отличной от таковой у указанного антитела или его антиген-связывающего фрагмента. В настоящем изобретении также представлен иммуноконъюгат, такой как конъюгат антитело-лекарственное средство, который может содержать антитело или его антигенсвязывающую часть по настоящему изобретению, связанные с терапевтическим средством, таким как цитотоксин. В другом аспекте антитело или его антигенсвязывающая часть по настоящему изобретению могут быть представлены в виде части химерного антигенного рецептора (CAR). Также представлена иммунная клетка, которая может содержать химерный антигенный рецептор, такая как Т-клетка и NK-клетка. Антитело или его антигенсвязывающая часть по настоящему изобретению также могут кодироваться онколитическим вирусом или применяться в сочетании с ним.
[0020] Молекулы нуклеиновой кислоты, кодирующие антитело или его антигенсвязывающую часть по настоящему изобретению, также охватываются настоящим изобретением, а также векторы экспрессии, которые могут содержать такие нуклеиновые кислоты, и клетки-хозяева, которые могут содержать такие векторы экспрессии. Также представлен способ получения антитела к Siglec15 или его антигенсвязывающей части по настоящему изобретению с использованием клетки-хозяина, который может включать стадии (I) экспрессии антитела в клетке-хозяине и (II) выделения антитела из клетки-хозяина или ее клеточной культуры.
[0021] Также представлены композиции, которые могут содержать антитело или его антигенсвязывающую часть, иммуноконъюгат, биспецифическую молекулу, онколитический вирус, CAR, CAR-T-клетку, молекулу нуклеиновой кислоты, вектор экспрессии или клетки-хозяева по настоящему изобретению и фармацевтически приемлемый носитель. В некоторых вариантах осуществления фармацевтическая композиция может дополнительно содержать терапевтическое средство, такое как противораковое средство.
[0022] В еще одном аспекте настоящего изобретения представлен способ модулирования иммунного ответа у субъекта, включающий введение субъекту терапевтически эффективного количества антитела или его антигенсвязывающей части по настоящему изобретению или, в качестве альтернативы, молекулы нуклеиновой кислоты, способной обеспечивать экспрессию вышеуказанного у субъекта, вследствие чего происходит модуляция иммунного ответа у субъекта. Предпочтительно антитело или его антигенсвязывающая часть по настоящему изобретению усиливают, стимулируют или повышают иммунный ответ у субъекта, например, путем обеспечения реверсии Т-клеточной супрессии, опосредованной Siglec15. В некоторых вариантах осуществления способ включает введение биспецифической молекулы, иммуноконъюгата, CAR-T-клетки или онколитического вируса, кодирующего антитело или несущего антитело, по настоящему изобретению.
[0023] В еще одном аспекте настоящего изобретения представлен способ подавления потери костной ткани или увеличения костной массы у субъекта, включающий введение субъекту терапевтически эффективного количества антитела или его антигенсвязывающей части по настоящему изобретению или, в качестве альтернативы, молекулы нуклеиновой кислоты, способной обеспечивать экспрессию вышеуказанного.
[0024] В дополнительном аспекте в настоящем изобретении представлен способ подавления роста опухоли у субъекта, нуждающегося в этом, включающий введение субъекту терапевтически эффективного количества антитела или его антигенсвязывающей части по настоящему изобретению или, в качестве альтернативы, молекулы нуклеиновой кислоты, способной обеспечивать экспрессию вышеуказанного. В некоторых вариантах осуществления способ включает введение биспецифической молекулы, иммуноконъюгата, CAR-T-клетки или онколитического вируса, кодирующего антитело или несущего антитело, по настоящему изобретению. Опухоль может быть солидной или несолидной опухолью. В некоторых вариантах осуществления опухоль представляет собой солидную опухоль, включая без ограничения немелкоклеточный рак легкого, рак яичника, меланому, колоректальный рак, рак молочной железы (включая трижды негативный рак молочной железы), плоскоклеточную карциному головы и шеи, рак эндометрия и плоскоклеточную карциному. В некоторых вариантах осуществления вместе с антителом или его антигенсвязывающей частью по настоящему изобретению можно вводить по меньшей мере одно дополнительное противораковое антитело, такое как антитело к VISTA, антитело к PD-1, антитело к PD-L1, антитело к LAG-3, антитело к CTLA-4, антитело к TIM-3, антитело к STAT3 и/или антитело к ROR1. В еще одном варианте осуществления антитело или его антигенсвязывающую часть по настоящему изобретению вводят с цитокином (например, IL-2, IL-21, GM-CSF и/или IL-4) или костимулирующим антителом, (например, антителом к CD137 и/или к GITR). В другом варианте осуществления антитело или его антигенсвязывающая часть по настоящему изобретению вводят с химиотерапевтическим средством, которое может представлять собой цитотоксическое средство, такое как эпирубицин, оксалиплатин и/или 5-фторурацил (5-FU). Антитело или его антигенсвязывающая часть по настоящему изобретению могут являться, например, мышиными, человеческими, химерными или гуманизированными.
[0025] Другие признаки и преимущества настоящего изобретения будут очевидны из следующего подробного описания и примеров, которые не следует рассматривать как ограничивающие. Содержание всех ссылок, записей в Genbank, патентов и опубликованных патентных заявок, цитируемых по всему тексту данной заявки, непосредственно включено в данный документ посредством ссылки.
[0026] Соответственно, цель изобретения состоит в том, чтобы изобретением не охватывался какой-либо ранее известный продукт, процедура изготовления продукта или способ применения продукта, вследствие чего заявители сохраняют за собой право и настоящим заявляют об отказе от ответственности в отношении любого из ранее известных продукта, процедуры или способа. Кроме того, следует отметить, что изобретение не предусматривает включение в объем изобретения какого-либо продукта, процедуры или способа изготовления продукта или способа применения продукта, которые не соответствуют требованиям к письменному описанию и условиям практической реализации изобретения согласно USPTO (35 U.S.С. §112, первый абзац) или ЕРО (статья 83 ЕРС), вследствие чего заявители сохраняют за собой право и настоящим заявляют об отказе от ответственности в отношении любого из ранее описанных продукта, процедуры изготовления продукта или способа применения продукта. При практическом осуществлении настоящего изобретения преимущественным может являться соответствие условиям, изложенным в ст. 53(c) ЕРС и Правиле 28 (b) и (с) ЕРС. Все права явного отказа от любых вариантов осуществления, которые являются предметом любого(-ых) выданного(-ых) патента(-ов) заявителя в серии заявок, родственных данной заявке, или в любой другой серии родственных заявок, или в любой ранее поданной заявке любой третьей стороны, явным образом сохранены. Ничто в данном документе не должно быть истолковано как обещание.
[0027] Следует отметить, что в настоящем изобретении и, в частности, в формуле изобретения и/или абзацах термины, такие как «содержит», «содержащий», «содержание» и т.п., могут иметь значение, приписываемое им в патентном законодательстве США; например, они могут означать «включает», «включающий», «включение» и т.п.; и что термины, такие как «состоящий по существу из» и «состоит по существу из» имеют значение, приписываемое им в патентном законодательстве США, например, они допускают элементы, не указанные явным образом, но исключают элементы, которые встречаются в предыдущем уровне техники или которые влияют на основные или новые характеристики изобретения.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0028] Наилучшее понимание следующего подробного описания, приводимого в качестве примера, но не предназначенного для ограничения изобретения исключительно описанными конкретными вариантами осуществления, может быть обеспечено в сочетании с прилагаемыми графическими материалами.
[0029] На фиг. 1А-1С показаны уровни связывающей способности антител А1С8С6Н1, A1D1B7H9, A1D5E2H1 и A1D11A7H10 (A), A1E7G5D1, A1E10G7H9, A2A1D2F1 и А2А5С7Е8 (В), А2А6 В1С2, A2G4C8G7 и A2H5F1A1 (С) по отношению к Siglec15 человека в ELISA с захватом.
[0030] На фиг. 2А-2С показаны уровни связывающей способности антител А1С8С6Н1, A1D1B7H9, A1D5E2H1 и A1D11A7H10 (A), A1E7G5D1, A1E10G7H9, A2A1D2F1 и А2А5С7Е8 (В), А2А6 В1С2, A2G4C8G7 и A2H5F1A1 (С) по отношению к клеткам человеческий-siglec15-2D3-1E1, экспрессирующим Siglec15, в клеточном FACS-анализе связывания.
[0031] На фиг. 3А-3С показаны уровни связывающей способности антител А1С8С6Н1, A1D1B7H9, A1D5E2H1 и A1D11A7H10 (A), A1E7G5D1, A1E10G7H9, A2A1D2F1 и А2А5С7Е8 (В), А2А6 В1С2, A2G4C8G7 и A2H5F1A1 (С) по отношению к Siglec15 яванского макака в непрямом ELISA.
[0032] На фиг. 4А-4С показаны уровни связывающей способности антител А1С8С6Н1, A1D1B7H9, A1D5E2H1 и A1D11A7H10 (A), A1E7G5D1, A1E10G7H9, A2A1D2F1 и А2А5С7Е8 (В), А2А6 В1С2, A2G4C8G7 и A2H5F1A1 (С) по отношению к Siglec15 мыши в непрямом ELISA.
[0033] На фиг. 5А-5С показаны значения способности антител А1С8С6Н1, A1D1B7H9, A1D5E2H1 и A1D11A7H10 (A), A1E7G5D1, A1E10G7H9, A2A1D2F1 и А2А5С7Е8 (В), А2А6 В1С2, A2G4C8G7 и A2H5F1A1 (С) блокировать связывание Siglec15 человека с LRRC4C в конкурентном ELISA.
[0034] На фиг. 6А-6С показаны значения способности антител А1С8С6Н1, A1D1B7H9, A1D5E2H1 и A1D11A7H10 (A), A1E7G5D1, A1E10G7H9, A2A1D2F1 и А2А5С7Е8 (В), А2А6 В1С2, A2G4C8G7 и A2H5F1A1 (С) блокировать связывание эталонного антитела с Siglec15 человека в конкурентном ELISA.
[0035] На фиг. 7А-7С показаны значения способности антител А1С8С6Н1, A1D1B7H9, A1D5E2H1 и A1D11A7H10 (A), A1E7G5D1, A1E10G7H9, A2A1D2F1 и А2А5С7Е8 (В), А2А6 В1С2, A2G4C8G7 и A2H5F1A1 (С) блокировать Siglec15 человека при его взаимодействии с LRRC4C человека на клеточной поверхности в клеточном FACS-анализе блокирования.
[0036] На фиг. 8А и 8В показано, что антитела А2А5С7Е8 и A1E10G7H9 обеспечивали реверсию индуцированной Siglec15 супрессии CD8+ клеток (А) и CD4+ клеток (В) в клеточном функциональном анализе.
[0037] На фиг. 9 показаны уровни связывающей способности антител А2А5С7Е8-1, А2А5С7Е8-2 и А2А5С7Е8-3 по отношению к Siglec15 человека в ELISA с захватом.
[0038] На фиг. 10 показаны уровни связывающей способности антител А2А5С7Е8-1, А2А5С7Е8-2 и А2А5С7Е8-3 по отношению к клеткам человеческий-siglec15-2D3-1E1, экспрессирующим Siglec15 человека, в клеточном FACS-анализе связывания.
[0039] На фиг. 11 показаны уровни связывающей способности антител А2А5С7Е8-1, А2А5С7Е8-2 и А2А5С7Е8-3 по отношению к Siglec15 яванского макака в непрямом ELISA.
[0040] На фиг. 12 показаны уровни связывающей способности антител А2А5С7Е8-1, А2А5С7Е8-2 и А2А5С7Е8-3 по отношению к Siglec15 мыши в непрямом ELISA.
[0041] На фиг. 13 показаны значения способности антител А2А5С7Е8-1, А2А5С7Е8-2 и А2А5С7Е8-3 блокировать связывание Siglec15 человека с LRRC4C в конкурентном ELISA.
[0042] На фиг. 14 показаны значения способности антител А2А5С7Е8-1, А2А5С7Е8-2 и А2А5С7Е8-3 блокировать связывание эталонного антитела с Siglec15 человека в конкурентном ELISA.
[0043] На фиг. 15А-15D показано, что антитела А2А5С7Е8-1, А2А5С7Е8-2 и А2А5С7Е8-3 обеспечивали реверсию подавления CD8+ клеток (А) и CD4+ клеток (В), индуцированного 93,5 нМ Siglec15, и подавление CD8+ клеток (С) и CD4+ клеток (D), индуцированного 186,9 нМ Siglec15, в клеточном функциональном анализе.
[0044] На фиг. 16 показаны уровни связывающей способности мышиных антител B2D7H7A3C1, B2G12H3E8 и В2Н2Н1Н7 по отношению к Siglec15 человека в ELISA с захватом.
[0045] На фиг. 17 показаны уровни связывающей способности мышиных антител B2D7H7A3C1, B2G12H3E8 и В2Н2Н1Н7 по отношению к клеткам человеческий-siglec15-2D3-1E1, экспрессирующим Siglec15 человека, в клеточном FACS-анализе связывания.
[0046] На фиг. 18 показаны уровни связывающей способности мышиных антител B2D7H7A3C1, B2G12H3E8 и В2Н2Н1Н7 по отношению к Siglec15 яванского макака в непрямом ELISA.
[0047] На фиг. 19 показаны уровни связывающей способности мышиных антител B2D7H7A3C1, B2G12H3E8 и В2Н2Н1Н7 по отношению к Siglec15 мыши в непрямом ELISA.
[0048] На фиг. 20 показаны значения способности мышиных антител B2D7H7A3C1, B2G12H3E8 и В2Н2Н1Н7 блокировать связывание Siglec15 человека с LRRC4C в конкурентном ELISA.
[0049] На фиг. 21 показаны значения способности мышиных антител B2D7H7A3C1, B2G12H3E8 и В2Н2Н1Н7 блокировать связывание эталонного антитела с Siglec15 человека в конкурентном ELISA.
[0050] На фиг. 22А и 22В показано, что мышиные антитела B2D7H7A3C1, B2G12H3E8 и В2Н2Н1Н7 обеспечивали реверсию опосредованной Siglec15 супрессии CD8+ клеток (А) и CD4+ клеток (В) в клеточном функциональном анализе.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0051] Чтобы обеспечить более легкое понимание настоящего изобретения, сначала даны определения некоторых терминов. Дополнительные определения приведены на протяжении всего подробного описания.
[0052] Термин « Siglec15» включает варианты, изоформы, гомологи, ортологи и паралоги. Например, антитело, специфичное по отношению к белку Siglec15 человека, может в некоторых случаях перекрестно реагировать с белком Siglec15 представителя другого вида, отличного от человека, такого как обезьяна. В других вариантах осуществления антитело, специфичное по отношению к белку Siglec15 человека, может быть полностью специфичным по отношению к белку Siglec15 человека и не проявлять перекрестной реактивности по отношению к другим видам или другим типам или может перекрестно реагировать с Siglec15 представителей некоторых других видов, но не представителей всех других видов.
[0053] Термин «Siglec15 человека» относится к белку Siglec15, характеризующемуся аминокислотной последовательностью человека, такой как аминокислотная последовательность Siglec15 человека, имеющая номер доступа в Genbank Q6ZMC9. Термины «Siglec15 обезьяны или макака-резуса» и «Siglec15 мыши» относятся к последовательностям Siglec15 обезьяны и мыши, соответственно, например к тем, аминокислотные последовательности которых имеют номера доступа в Genbank ХР_028694069.1 и NP_001094508.1, соответственно.
[0054] Термин «иммунный ответ» относится к действию, например, лимфоцитов, антигенпрезентирующих клеток, фагоцитарных клеток, гранулоцитов и растворимых форм макромолекул, продуцируемых указанными выше клетками или печенью (включая антитела, цитокины и компоненты комплемента), которое приводит к селективному повреждению, разрушению или элиминации из организма человека вторгшихся патогенов, клеток или тканей, инфицированных патогенами, раковых клеток или, в случаях аутоиммунной реакции или патологического воспаления, нормальных клеток или тканей человека.
[0055] Термин «антитело», используемый в данном документе, относится к молекуле иммуноглобулина, которая распознает и специфично связывает мишень, такую как Siglec15, посредством по меньшей мере одного антигенсвязывающего участка, где антигенсвязывающий участок обычно находится в пределах вариабельной области молекулы иммуноглобулина. Как используется в данном документе, термин охватывает интактные поликлональные антитела, интактные моноклональные антитела, антитела, представленные одноцепочечным Fv (scFv), антитела с тяжелыми цепями (HCAb), антитела с легкими цепями (LCAb), мультиспецифические антитела, биспецифические антитела, моноспецифические антитела, моновалентные антитела, слитые белки, содержащие антигенсвязывающий участок антитела, и любую другую модифицированную молекулу иммуноглобулина, содержащую антигенсвязывающий участок {например, молекулу иммуноглобулина с двойным вариабельным доменом), при условии, что антитела проявляют необходимую биологическую активность. Антитела также включают без ограничения мышиные антитела, химерные антитела, гуманизированные антитела и человеческие антитела. Антитело может относиться к любому из пяти основных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM или их подклассам (изотипам) (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2), исходя из идентичности константных доменов их тяжелых цепей, обозначаемых как альфа, дельта, эпсилон, гамма и мю, соответственно. Различные классы иммуноглобулинов имеют различные и хорошо известные структуры и трехмерные конфигурации субъединиц. Антитела могут являться «голыми» или конъюгированными с другими молекулами, включая без ограничения токсины и радиоизотопы. Если явно не указано иное, термин «антитело», используемый в данном документе, включает «антигенсвязывающую часть» интактных антител. IgG представляет собой гликопротеин, который может содержать две тяжелые (Н) цепи и две легкие (L) цепи, соединенные между собой дисульфидными связями. Каждая тяжелая цепь может состоять из вариабельной области тяжелой цепи (сокращенно обозначаемой в данном документе как VH) и константной области тяжелой цепи. Константная область тяжелой цепи может состоять из трех доменов, представляющих собой CH1, CH2 и CH3. Каждая легкая цепь может состоять из вариабельной области легкой цепи (сокращенно обозначаемой в данном документе как VL) и константной области легкой цепи. Константная область легкой цепи может состоять из одного домена, представляющего собой CL. Области VH и VL могут быть дополнительно подразделены на области гипервариабельности, называемые определяющими комплементарность участками (CDR), чередующиеся с более консервативными областями, называемыми каркасными областями (FR). Каждая VH и VL состоит из трех CDR и четырех FR, расположенных от амино-конца к карбокси-концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Вариабельные области тяжелой и легкой цепей содержат связывающий домен, взаимодействующий с антигеном. Константные области антител могут опосредовать связывание иммуноглобулина с тканями или факторами хозяина, включая различные клетки иммунной системы (например, эффекторные клетки) и первый компонент (C1q) классической системы комплемента.
[0056] Термин «антигенсвязывающая часть» антитела (или просто «часть антитела»), как используется в данном документе, относится к одному или более фрагментам антитела, которые сохраняют способность специфически связываться с антигеном (например, белком Siglec15). Было показано, что антигенсвязывающую функцию антитела могут выполнять фрагменты полноразмерного антитела. Примеры связывающих фрагментов, охватываемых термином «антигенсвязывающая часть» антитела, включают (i) Fab-фрагмент, представляющий собой моновалентный фрагмент, состоящий из доменов VL, VH, CL и CH1; (ii) фрагмент F(ab')2, представляющий собой бивалентный фрагмент, который может содержать два фрагмента Fab, связанных дисульфидным мостиком в шарнирной области; (iii) фрагмент Fd, состоящий из доменов VH и CH1; (iv) фрагмент Fv, состоящий из доменов VL и VH одного плеча антитела, (v) фрагмент dAb (Ward et al., (1989) Nature 341:544-546), который состоит из домена VH; (vi) выделенную определяющую комплементарность область (CDR); а также (viii) нанотело, представляющее собой вариабельную область тяжелой цепи, содержащую один вариабельный домен и два константных домена. Кроме того, хотя два домена Fv-фрагмента, VL и VH, кодируются отдельными генами, их можно соединить посредством рекомбинантных способов с помощью синтетического линкера, что позволяет получить их в виде единой белковой цепи, в которой области VL и VH образуют пары с образованием моновалентных молекул (известных как одноцепочечные Fv (scFv); см., например, Bird et al., (1988) Science 242:423-426; and Huston et al., (1988) Proc. Natl Acad. Sci. USA 85:5879-5883). Предусматривается, что такие одноцепочечные антитела также охватываются термином «антигенсвязывающая часть» антитела. Эти фрагменты антител получают с использованием традиционных методик, известных специалистам в данной области, и фрагменты подвергают скринингу на применимость таким же образом, как и интактные антитела.
[0057] Термин «выделенное антитело», как используется в данном документе, предназначен для обозначения антитела, которое по сути не содержит других антител, обладающих другими видами антигенной специфичности (например, выделенное антитело, которое специфично связывает белок Siglec15, по сути не содержит антител, которые специфично связывают отличные от белка Siglec15 антигены). Выделенное антитело, которое специфично связывается с белком Siglec15 человека, может, однако, характеризоваться перекрестной реактивностью с другими антигенами, такими как белки Siglec15 представителей других видов. Кроме того, выделенное антитело может по сути не содержать другого клеточного материала и/или химических веществ.
[0058] Термины «моноклональное антитело» или «композиция на основе моноклонального антитела», используемые в настоящем документе, относятся к препарату молекул антитела на основе мономолекулярной композиции. Композиция на основе моноклонального антитела демонстрирует один вид специфичности связывания и аффинности к конкретному эпитопу.
[0059] Термин «человеческое антитело», используемый в данном документе, предназначен для включения антител, имеющих вариабельные области, в которых как каркасные области, так и участки CDR получены из человеческих последовательностей иммуноглобулинов зародышевого типа. Кроме того, если антитело содержит константную область, константная область также происходит из человеческих последовательностей иммуноглобулинов зародышевого типа. Человеческие антитела по настоящему изобретению могут содержать аминокислотные остатки, не кодируемые человеческими последовательностями иммуноглобулинов зародышевого типа (например, мутации, введенные путем случайного или сайт-специфического мутагенеза in vitro или при соматической мутации in vivo). Однако термин «человеческое антитело», используемый в данном документе, не предназначен для включения антител, в которых последовательности CDR, происходящие из последовательностей зародышевого типа другого вида млекопитающих, были привиты на человеческие каркасные последовательности.
[0060] Термин «мышиное антитело», используемый в данном документе, предназначен для включения антител, имеющих вариабельные области, в которых как каркасные области, так и участки CDR получены из мышиных последовательностей иммуноглобулинов зародышевого типа. Кроме того, если антитело содержит константную область, константная область также происходит из мышиных последовательностей иммуноглобулинов зародышевого типа. Мышиные антитела по настоящему изобретению могут содержать аминокислотные остатки, не кодируемые мышиными последовательностями иммуноглобулинов зародышевого типа (например, мутации, введенные путем случайного или сайт-специфического мутагенеза in vitro или при соматической мутации in vivo). Однако термин «мышиное антитело», используемый в данном документе, не предназначен для включения антител, в которых последовательности CDR, происходящие из последовательностей зародышевого типа другого вида млекопитающих, были привиты на мышиные каркасные последовательности.
[0061] Термин «химерное антитело» относится к антителу, полученному путем комбинирования генетического материала из источника, не относящегося к организму человека, с генетическим материалом из организма человека. Или, в более общем смысле, химерное антитело представляет собой антитело, характеризующееся генетическим материалом из организма представителя определенного вида с генетическим материалом из организма представителя другого вида.
[0062] Термин «гуманизированное антитело», используемый в данном документе, относится к антителу представителя вида, отличного от человека, чьи белковые последовательности были модифицированы для повышения сходства с вариантами антител, продуцируемыми у людей естественным образом.
[0063] Термин «изотип» относится к классу антител (например, IgM или IgG1), который кодируется генами константной области тяжелой цепи.
[0064] Фразы «антитело, распознающее антиген» и «антитело, специфичное в отношении антигена» используются в данном документе взаимозаменяемо с термином «антитело, которое специфично связывается с антигеном».
[0065] Используемый в настоящем документе термин «антитело, которое специфично связывается с Siglec15 человека» предназначен для обозначения антитела, которое связывается с белком Siglec 5 человека (и, возможно, белком Siglec15 представителя одного или более видов, отличного от человека), но по сути не связывается с белками не относящимися к Siglec15. Предпочтительно антитело связывается с белком Siglec15 человека с «высокой аффинностью», а именно с KD 5,0×10-9 М или меньше, более предпочтительно 1,0×10-9 М или меньше и более предпочтительно 1,0×10-10 М или меньше.
[0066] Термин «по сути не связывается» с белком или клетками, используемый в данном документе, означает, что молекула не связывается или не связывается с высокой аффинностью с белком или клетками, т.е. связывается с белком или клетками с KD 1,0×10-6 М или больше, более предпочтительно 1,0×10-5 М или больше, более предпочтительно 1,0×10-4 М или больше, более предпочтительно 1,0×10-3 М или больше, еще более предпочтительно 1,0×10-2 М или больше.
[0067] Термин «высокая аффинность» для антитела класса IgG относится к антителу, характеризующемуся KD 5,0×10-9 М или меньше, более предпочтительно 1,0×10-9 М или меньше, еще более предпочтительно 5,0×10-10 М или меньше, еще более предпочтительно 1,0×10-10 М или меньше и еще более предпочтительно 5,0×10-11 М или меньше в отношении антигена-мишени. Однако связывание с «высокой аффинностью» может варьироваться для других изотипов антител. Например, связывание с «высокой аффинностью» в случае изотипа IgM относится к антителу, характеризующемуся KD 10-6 М или меньше, более предпочтительно 10-7 М или меньше, еще более предпочтительно 10-8 М или меньше.
[0068] Термин «Kassoc» или «Ka», используемый в данном документе, предназначен для обозначения скорости ассоциации при конкретном взаимодействии антитело-антиген, тогда как термин «Kdis» или «Kd», используемый в данном документе, предназначен для обозначения скорости диссоциации при конкретном взаимодействии антитело-антиген. Термин «KD», используемый в данном документе, предназначен для обозначения константы диссоциации, которую получают из отношения Kd к Ka (т.е., Kd/Ka) и выражают в виде молярной концентрации (М). Значения KD для антител можно определить с использованием способов, хорошо известных в данной области техники. Предпочтительным способом определения KD антитела является использование поверхностного плазмонного резонанса, предпочтительно с использованием биосенсорной системы, такой как система Biacore™.
[0069] Термин «ЕС50», также известный как полумаксимальная эффективная концентрация, относится к концентрации антитела, которая индуцирует ответ при величине, находящейся посередине между исходным уровнем и максимумом, после определенного времени воздействия.
[0070] Термин «IC50», также известный как полумаксимальная ингибирующая концентрация, относится к концентрации антитела, которая подавляет конкретную биологическую или биохимическую функцию на 50% по сравнению с отсутствием антитела.
[0071] Термин «субъект» включает любого человека или животное, отличное от человека. Термин «животное, отличное от человека» включает всех позвоночных, например, млекопитающих и не относящихся к млекопитающим, таких как приматы, овцы, собаки, кошки, коровы, лошади, куры, земноводные и рептилии, хотя предпочтительными являются млекопитающие, такие как приматы, отличные от человека, овцы, собаки, кошки, коровы и лошади.
[0072] Термин «терапевтически эффективное количество» означает количество антитела или его антигенсвязывающей части по настоящему изобретению, достаточное для предупреждения или облегчения симптомов, связанных с заболеванием или состоянием (таким как рак), и/или уменьшения тяжести заболевания или состояния. Терапевтически эффективное количество понимается в контексте состояния, которое подвергается лечению, где фактическое эффективное количество легко определяется специалистами в данной области техники.
[0073] Различные аспекты настоящего изобретения описаны более подробно в следующих подразделах.
[0074] Антитело или его антигенсвязывающая часть по настоящему изобретению специфично связываются с Siglec15 человека или обезьяны со сравнимой, если не лучшей, аффинностью связывания по сравнению с ранее описанными антителами к Siglec15, такими как Siglec15-ch5G9.
[0075] Дополнительные функциональные свойства включают способность блокировать связывание Siglec15 с его лигандом и обеспечивать реверсию Т-клеточной супрессии, опосредованной Siglec15.
[0076] Иллюстративные антитело или его антигенсвязывающая часть по настоящему изобретению структурно и химически охарактеризованы, как описано ниже и в примерах. Идентификационные номера аминокислотных последовательностей вариабельных областей тяжелой/легкой цепи антител сведены в таблице 1 ниже. Константная область тяжелой цепи для антител может представлять собой константную область тяжелой цепи IgG1 человека, характеризующуюся аминокислотной последовательностью, указанной, например, под SEQ ID NO: 17, а константная область легкой цепи для антител может представлять собой константную область каппа-цепи человека, характеризующуюся аминокислотной последовательностью, указанной, например, под SEQ ID NO: 18. Антитело по настоящему изобретению может быть человеческим, мышиным, химерным или гуманизированным антителом.
[0077] CDR вариабельной области тяжелой цепи и CDR вариабельной области легкой цепи в таблице 1 были определены с помощью системы нумерации согласно Kabat. Однако, как хорошо известно в данной области техники, участки CDR можно также определять с помощью других систем, как например согласно Chothia и IMGT, AbM или контактной системе/способу нумерации, основанным на последовательностях вариабельных областей тяжелой/легкой цепи.

[0078] Последовательности VH и VL (или последовательности CDR) других антител к Siglec15, которые связываются с Siglec15 человека, можно «смешивать и совмещать» с последовательностями VH и VL (или последовательностями CDR) антитела к Siglec15 по настоящему изобретению. Предпочтительно, когда цепи VH и VL (или CDR в таких цепях) смешивают и совмещают, последовательность VH из конкретной пары VH/VL заменяется структурно сходной последовательностью VH. Аналогичным образом, последовательность VL из конкретной пары VH/VL предпочтительно заменяется структурно сходной последовательностью VL.
[0079] Соответственно, в одном варианте осуществления антитело по настоящему изобретению или его антигенсвязывающая часть содержат
(а) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, указанную выше в таблице 1; и
(b) вариабельную область легкой цепи, содержащую аминокислотную последовательность, указанную выше в таблице 1, или VL другого антитела к Siglec15, где антитело специфично связывает Siglec15 человека.
[0080] В другом варианте осуществления антитело по настоящему изобретению или его антигенсвязывающая часть содержат
(a) участки CDR1, CDR2 и CDR3 вариабельной области тяжелой цепи, перечисленные выше в таблице 1; и
(b) участки CDR1, CDR2 и CDR3 вариабельной области легкой цепи, перечисленные выше в таблице 1, или CDR другого антитела к Siglec15, где антитело специфично связывает Siglec15 человека.
[0081] В еще одном варианте осуществления антитело или его антигенсвязывающая часть содержат CDR2 вариабельной области тяжелой цепи антитела к Siglec15 в сочетании с CDR других антител, которые связывают Siglec15 человека, например, CDR1 и/или CDR3 из вариабельной области тяжелой цепи и/или CDR1, CDR2 и/или CDR3 из вариабельной области легкой цепи другого антитела к Siglec15.
[0082] Кроме того, в данной области техники хорошо известно, что домен CDR3, независимо от домена(-ов) CDR1 и/или CDR2, сам по себе может определять специфичность связывания антитела с когнатным антигеном, а также что можно предсказуемым образом генерировать несколько антител, имеющих такую же специфичность связывания, основанную на общей последовательности CDR3. См., например, Klimka et a.l, British J. of Cancer 83(2):252-260 (2000); Beiboer et al., J. Mol Biol 296:833-849 (2000); Rader et al., Proc. Natl Acad. Sci. U.S.A. 95:8910-8915 (1998); Barbas et al., J. Am. Chem. Soc. 116:2161-2162 (1994); Barbas et al., Proc. Natl Acad. Sci. U.S.A. 92:2529-2533 (1995); Ditzel et al., J. Immunol 157:739-749 (1996); Berezov et al., BIAjournal 8: Scientific Review 8 (2001); Igarashi et al., J. Biochem (Tokyo) 117:452-7 (1995); Bourgeois et al., J. Virol 72:807-10 (1998); Levi et al., Proc. Natl Acad. Sci. U.S.A. 90:4374-8 (1993); Polymenis and Stoller, J. Immunol 152:5218-5329 (1994) and Xu and Davis, Immunity 13:37-45 (2000). See also, U.S. Pat. Nos. 6,951,646; 6,914,128; 6,090,382; 6,818,216; 6,156,313; 6,827,925; 5,833,943; 5,762,905 и 5760185. Каждая из этих ссылок настоящим включена в данный документ посредством ссылки во всей своей полноте.
[0083] Соответственно, в другом варианте осуществления антитела по настоящему изобретению содержат CDR2 вариабельной области тяжелой цепи антитела к Siglec15 и по меньшей мере CDR3 вариабельной области тяжелой и/или легкой цепи антитела к Siglec15 или CDR3 вариабельной области тяжелой и/или легкой цепи другого антитела к Siglec15, при этом антитело способно специфически связываться с Siglec15 человека. Эти антитела предпочтительно (а) конкурируют за связывание с Siglec15; (b) сохраняют функциональные характеристики; (с) связываются с одним и тем же эпитопом и/или (d) обладают такой же аффинностью связывания, что и антитело к Siglec15 по настоящему изобретению. В еще одном варианте осуществления антитела дополнительно могут содержать CDR2 вариабельной области легкой цепи антитела к Siglec15 или CDR2 вариабельной области легкой цепи другого антитела к Siglec15, где антитело способно специфически связываться с Siglec15 человека. В другом варианте осуществления антитела по настоящему изобретению могут содержать CDR1 вариабельной области тяжелой и/или легкой цепи антитела к Siglec15 или CDR1 вариабельной области тяжелой и/или легкой цепи другого антитела к Siglec15, где антитело способно специфически связываться с Siglec15 человека.
[0084] В другом варианте осуществления антитело по настоящему изобретению содержит последовательности CDR1, CDR2 и CDR3 из последовательностей вариабельной области тяжелой и/или легкой цепи, которые отличаются от таковых из антител к Siglec15 по настоящему изобретению одной или более консервативными модификациями. В данной области техники известно, что может быть осуществлена определенная консервативная модификация последовательности, которая не устраняет способность к связыванию антигена. См., например, Brummell et al., (1993) Biochem 32:1180-8; de Wildt et al., (1997) Prot. Eng. 10:835-41; Komissarov et al., (1997) J. Biol Chem. 272:26864-26870; Hall et al., (1992) J. Immunol 149:1605-12; Kelley and O'Connell (1993).Biochem.32:6862-35; Adib-Conquy et al., (1998) Int. Immunol 10:341-6 и Beers et al., (2000) Clin. Can. Res. 6:2835-43.
[0085] Соответственно, в одном варианте осуществления антитело содержит вариабельную область тяжелой цепи, содержащую последовательности CDR1, CDR2 и CDR3, и/или вариабельную область легкой цепи, содержащую последовательности CDR1, CDR2 и CDR3, где
(a) последовательность CDR1 вариабельной области тяжелой цепи предусматривает последовательность, указанную в таблице 1 выше, и/или ее консервативные модификации; и/или
(b) последовательность CDR2 вариабельной области тяжелой цепи предусматривает последовательность, указанную в таблице 1 выше, и/или ее консервативные модификации; и/или
(c) последовательность CDR3 вариабельной области тяжелой цепи предусматривает последовательность, указанную в таблице 1 выше, и/или ее консервативные модификации; и/или
(d) последовательности CDR1, и/или CDR2, и/или CDR3 вариабельной области легкой цепи предусматривают последовательность(-и), указанную(-ые) в таблице 1 выше; и/или ее(их) консервативные модификации; и
(e) антитело специфично связывает Siglec15 человека.
[0086] Антитело или его антигенсвязывающая часть по настоящему изобретению обладают одним или более из следующих функциональных свойств, описанных выше, таких как связывание с высокой аффинностью с Siglec15 человека и способность обеспечивать реверсию опосредованной Siglec15 Т-клеточной супрессии.
[0087] В различных вариантах осуществления антитело или его антигенсвязывающая часть могут являться, например, человеческими, мышиными, гуманизированными или химерными.
[0088] Используемый в данном документе термин «консервативные модификации последовательности» предназначен для обозначения аминокислотных модификаций, которые не влияют на характеристики связывания антитела, содержащего аминокислотную последовательность, или не изменяют их в значительной степени. Такие консервативные модификации включают аминокислотные замены, добавления и делеции. Модификации могут быть введены в антитело по настоящему изобретению посредством стандартных методик, известных в данной области техники, таких как сайт-направленный мутагенез и мутагенез, опосредованный ПЦР. Консервативные аминокислотные замены являются таковыми, в которых аминокислотный остаток заменен на аминокислотный остаток, имеющий аналогичную боковую цепь. В уровне техники определены семейства аминокислотных остатков, имеющих сходные боковые цепи. Эти семейства включают аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислотными боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота), незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серии, треонин, тирозин, цистеин, триптофан), неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин), бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Таким образом, один или более аминокислотных остатков в пределах участков CDR антитела по настоящему изобретению могут быть заменены другими аминокислотными остатками из того же семейства боковых цепей, и измененное антитело может быть протестировано в отношении сохранения функции (т.е. функций, указанных выше) с использованием описанных в данном документе функциональных анализов.
[0089] Антитела по настоящему изобретению могут быть получены с применением антитела, имеющего одну или более последовательностей VH/VL антитела к Siglec15 по настоящему изобретению, в качестве исходного материала для конструирования модифицированного антитела. Антитело можно конструировать путем модификации одного или более остатков в пределах одной или обеих вариабельных областей (т.е. VH и/или VL), например, в пределах одной или более участков CDR и/или в пределах одной или более каркасных областей. Дополнительно или в качестве альтернативы, антитело можно конструировать путем модификации остатков в пределах константной(-ых) области(-ей), например, для изменения эффекторной(-ых) функции(-й) антитела.
[0090] В некоторых вариантах осуществления трансплантацию CDR можно использовать для конструирования вариабельных областей антител. Антитела взаимодействуют с антигенами-мишенями главным образом посредством аминокислотных остатков, расположенных в шести определяющих комплементарность областях (CDR) тяжелой и легкой цепей. По этой причине аминокислотные последовательности в пределах CDR характеризуются более высоким разнообразием среди отдельных антител, чем последовательности за пределами CDR. Поскольку последовательности CDR ответственны за большинство взаимодействий антитело-антиген, имеется возможность экспрессировать рекомбинантные антитела, которые имитируют свойства специфических встречающихся в природе антител, путем конструирования векторов экспрессии, которые содержат последовательности CDR из специфического встречающегося в природе антитела, привитые на каркасные последовательности из другого антитела с отличающимися свойствами (см., например, Riechmann et al., (1998) Nature 332:323-327; Jones et al., (1986) Nature 321:522-525; Queen et al., (1989) Proc. Natl Acad. See also U.S.A. 86:10029-10033; U.S. Pat. Nos. 5,225,539; 5,530,101; 5,585,089; 5,693,762 и 6180370).
[0091] Соответственно, другой вариант осуществления настоящего изобретения относится к выделенному моноклональному антителу или его антигенсвязывающей части, содержащим вариабельную область тяжелой цепи, содержащую последовательности CDR1, CDR2 и CDR3, предусматривающие последовательности по настоящему изобретению, как описано выше, и/или вариабельную область легкой цепи, содержащую последовательности CDR1, CDR2 и CDR3, предусматривающие последовательности по настоящему изобретению, как описано выше. Хотя эти антитела содержат последовательности CDR VH и VL моноклонального антитела по настоящему изобретению, они могут содержать отличающиеся каркасные последовательности.
[0092] Такие каркасные последовательности можно получать из общедоступных баз данных ДНК или опубликованных ссылочных документов, в которых содержатся последовательности генов антител зародышевого типа. Например, последовательности ДНК зародышевого типа для человеческих генов вариабельных областей тяжелой и легкой цепи можно найти в базе данных человеческих последовательностей зародышевого типа «VBase» (доступной в сети Интернет по адресу www.mrc-cpe.cam.ac.uk/vbase), а также в Kabat et al., (1991), приведенном выше; Tomlinson et al., (1992) J. Mol. Biol. 227:776-798; и Cox et al., (1994) Eur. J. Immunol 24:827-836; содержание каждого из которых явным образом включено в данный документ посредством ссылки. В качестве другого примера, последовательности ДНК зародышевого типа для человеческих генов вариабельных областей тяжелых и легких цепей можно найти в базе данных Genbank. Например, следующие последовательности тяжелой цепи зародышевого типа, обнаруженные у мыши НСо7 HuMAb, доступны под прилагаемыми номерами доступа в Genbank: 1-69 (NG--0010109, NT--024637 и ВС070333), 3-33 (NG--0010109 и NT--024637) и 3-7 (NG-0010109 и NT--024637). В качестве другого примера, следующие последовательности тяжелых цепей зародышевого типа, обнаруженные у мыши НСо12 HuMAb, доступны под прилагаемыми номерами доступа в Genbank: 1-69 (NG--0010109, NT--024637 & ВС070333), 5-51 (NG-0010109 & NT-024637), 4-34 (NG-0010109 & NT-024637), 3-30.3 (CAJ556644) & 3-23 (AJ406678).
[0093] Последовательности белков антител сравнивают со скомпилированной базой данных последовательностей белков с использованием одного из способов поиска подобия последовательностей, называемого Gapped BLAST (Altschul et al., (1997), см. выше), который хорошо известен специалистам в данной области техники.
[0094] Предпочтительными каркасными последовательностями для использования в антителах по настоящему изобретению являются те, которые структурно аналогичны каркасным последовательностям, используемым в антителах по настоящему изобретению. Последовательности CDR1, CDR2 и CDR3 VH могут быть привиты на каркасные области, которые имеют последовательность, идентичную последовательности, обнаруженной в гене иммуноглобулина зародышевого типа, из которого происходит каркасная последовательность, или последовательности CDR могут быть привиты на каркасные области, которые содержат одну или более мутаций по сравнению с последовательностями зародышевого типа. Например, было обнаружено, что в некоторых случаях полезным является мутирование остатков в каркасных областях для поддержания или усиления антигенсвязывающей способности антитела (см., например, патенты США №№5530101; 5585089; 5693762 и 6180370).
[0095] Другой тип модификации вариабельной области заключается в мутировании аминокислотных остатков в пределах участков CDR1, CDR2 и/или CDR3 VH и/или VL для улучшения, таким образом, одного или более свойств связывания (например, аффинности) представляющего интерес антитела. Сайт-направленный мутагенез или мутагенез, опосредованный ПЦР, можно проводить для введения мутации(-ий), а эффект в отношении связывания антител или другое представляющее интерес функциональное свойство можно оценить в анализах in vitro или in vivo, как известно в данной области техники. Предпочтительно вводят консервативные модификации (как известно в данной области техники). Мутации могут являться аминокислотными заменами, добавлениями или делециями, но предпочтительно являются заменами. Кроме того, обычно изменяют не более одного, двух, трех, четырех или пяти остатков в пределах участка CDR.
[0096] Соответственно, в другом варианте осуществления в настоящем изобретении представлены выделенные моноклональные антитела к Siglec15 или их антигенсвязывающие части, содержащие вариабельную область тяжелой цепи, содержащую (а) участок CDR1 VH, содержащую последовательность по настоящему изобретению или аминокислотную последовательность, характеризующуюся одной, двумя, тремя, четырьмя или пятью аминокислотными заменами, делециями или добавлениями; (b) участок CDR2 VH, содержащую последовательность по настоящему изобретению или аминокислотную последовательность, характеризующуюся одной, двумя, тремя, четырьмя или пятью аминокислотными заменами, делециями или добавлениями; (с) участок CDR3 VH, содержащую последовательность по настоящему изобретению или аминокислотную последовательность, характеризующуюся одной, двумя, тремя, четырьмя или пятью аминокислотными заменами, делециями или добавлениями; (d) участок CDR1 VL, содержащую последовательность по настоящему изобретению или аминокислотную последовательность, характеризующуюся одной, двумя, тремя, четырьмя или пятью аминокислотными заменами, делециями или добавлениями; (е) участок CDR2 VL, содержащую последовательность по настоящему изобретению или аминокислотную последовательность, характеризующуюся одной, двумя, тремя, четырьмя или пятью аминокислотными заменами, делециями или добавлениями; и (f) участок CDR3 VL, содержащую последовательность по настоящему изобретению или аминокислотную последовательность, характеризующуюся одной, двумя, тремя, четырьмя или пятью аминокислотными заменами, делециями или добавлениями.
[0097] Сконструированные антитела по настоящему изобретению включают таковые, в которые были внесены модификации каркасных остатков в пределах VH и/или VL, например, для улучшения свойств антитела. Обычно такие модификации каркасной области делают для снижения иммуногенности антитела. Например, один из подходов заключается в подвергании «обратной мутации» одного или более каркасных остатков с получением соответствующей последовательности зародышевого типа. Более конкретно, антитело, подвергшееся соматической мутации, может содержать каркасные остатки, которые отличаются от последовательности зародышевого типа, из которой получено антитело. Такие остатки могут быть идентифицированы путем сравнения каркасных последовательностей антитела с последовательностями зародышевого типа, из которых получено антитело.
[0098] Другой тип модификации каркасной области предусматривает мутирование одного или более остатков в пределах каркасной области или даже в пределах одной или более участков CDR для удаления Т-клеточных эпитопов со снижением таким образом потенциальной иммуногенности антитела. Этот подход также называется «деиммунизацией» и более подробно описан в публикации патента США №20030153043.
[0099] Кроме того, или в качестве альтернативы модификациям, выполненным в пределах каркасных областей или участков CDR, антитела по настоящему изобретению могут быть сконструированы таким образом, чтобы включать модификации в пределах Fc-области, как правило, с целью изменения одного или более функциональных свойств антитела, таких как время полужизни в сыворотке крови, фиксация компонентов комплемента, связывание рецептора Fc и/или антиген-зависимая клеточная цитотоксичность. Кроме того, антитело по настоящему изобретению может быть химически модифицировано (например, к антителу может быть присоединен один или более химических фрагментов) или модифицировано с изменением его гликозилирования, опять же, с изменением одного или более функциональных свойств антитела.
[00100] В одном варианте осуществления шарнирная область CH1 модифицирована таким образом, что количество цистеиновых остатков в шарнирной области изменено, например, увеличено или уменьшено. Этот подход дополнительно описан в патенте США №№5677425. Количество остатков цистеина в шарнирной области CH1 изменяют, например, для способствования сборке легкой и тяжелой цепей или для увеличения или уменьшения стабильности антитела.
[00101] В другом варианте осуществления шарнирная область Fc антитела мутирована с уменьшением биологического периода полужизни антитела. Более конкретно, одна или более аминокислотных мутаций вводятся в граничную область доменов CH2-CH3 фрагмента, представленного Fc-шарнирной областью, вследствие чего антитело характеризуется нарушенным связыванием стафилококкового белка А (SpA) по сравнению со связыванием SpA нативным фрагментом, представленным Fc-шарнирной областью. Этот подход более подробно описан в патенте США №№6165745.
[00102] В еще одном варианте осуществления модифицирование осуществлено в отношении гликозилирования антитела. Например, может быть получено гликозилированное антитело (т.е. антитело не предусматривающее гликозилирования). Гликозилирование может быть изменено, например, для увеличения аффинности антитела по отношению к антигену. Такие углеводные модификации могут быть выполнены, например, путем изменения одного или более сайтов гликозилирования в последовательности антитела. Например, можно выполнить одну или более аминокислотных замен, которые приводят к устранению одного или более сайтов гликозилирования каркасной области вариабельной области, тем самым устраняя гликозилирование в этом сайте. Такое гликозилирование может увеличить аффинность антитела по отношению к антигену. См., например, патенты США №№5714350 и 6350861.
[00103] Дополнительно или в качестве альтернативы, может быть получено антитело с измененным типом гликозилирования, такое как гипофукозилированное антитело, имеющее уменьшенное количество фукозильных остатков, или антитело, имеющее повышенное количество структур GlcNac в точках ветвления. Было продемонстрировано, что такие измененные паттерны гликозилирования повышают способность антител к индуцированию ADCC. Такие углеводные модификации могут быть осуществлены, например, путем экспрессии антитела в клетке-хозяине с измененным механизмом гликозилирования. Клетки с измененным механизмом гликозилирования были описаны в данной области техники, и их можно использовать в качестве клеток-хозяев, в которых можно экспрессировать рекомбинантные антитела по настоящему изобретению, чтобы таким образом получить антитело с измененным гликозилированием. Например, в клеточных линиях Ms704, Ms705 и Ms709 отсутствует ген фукозилтрансферазы, FUT8 (α-(1,6)-фукозилтрансфераза), так что антитела, экспрессируемые в клеточных линиях Ms704, Ms705 и Ms709, не содержат фукозы на своих углеводных компонентах. FUT8-/- клеточные линии Ms704, Ms705 и Ms709 получали путем направленного разрушения гена FUT8 в клетках CHO/DG44 с использованием двух векторов замещения (см. публикацию патента США №20040110704 и Yamane-Ohnuki et al., (2004) Biotechnol Bioeng 87:614-22). В качестве другого примера, в ЕР 1176195 описана клеточная линия с функционально нарушенным геном FUT8, который кодирует фукозилтрансферазу, вследствие чего антитела, экспрессируемые в такой клеточной линии, демонстрируют гипофукозилирование за счет снижения или устранения фермента, активность которого связана с α-1,6-связью. В ЕР 1176195 также описаны клеточные линии, которые характеризуются низкой ферментативной активностью в отношении добавления фукозы к N-ацетилглюкозамину, который связывается с Fc-областью антитела, или не обладают ферментативной активностью, например, линия миеломных клеток крысы YB2/0 (АТСС CRL 1662). В РСТ-публикации WO 03/035835 описан вариант линии клеток СНО, представляющий собой клетки Lec13, со сниженной способностью присоединять фукозу к углеводам, связанным с Asn (297), что также приводит к гипофукозилированию антител, экспрессируемых в этой клетке-хозяине (см. также Shields et al., (2002) J. Biol. Chem. 277:26733-26740). Антитела с модифицированным профилем гликозилирования также могут быть получены в куриных яйцах, как описано в РСТ-публикации WO 06/089231. В качестве альтернативы, антитела с модифицированным профилем гликозилирования могут быть получены в клетках растений, как например Lemna. Способы получения антител в растительной системе раскрыты в заявке на патент США, соответствующей делу патентного поверенного Alston & Bird LLP №040989/314911, поданной 11 августа 2006 г. В РСТ-публикации WO 99/54342 описаны клеточные линии, сконструированные для экспрессии гликопротеин-модифицирующих гликозилтрансфераз (например, β(l,4)-N-ацетилглюкозаминилтрансферазы III (GnTIII)), вследствие чего антитела, экспрессируемые в сконструированных клеточных линиях, проявляют повышенное количество структур GlcNac в точках ветвления, что приводит к повышенной ADCC-активности антител (см. также Umana et al., (1999) Nat. Biotech. 17:176-180). В качестве альтернативы, фукозные остатки на антителе могут быть отщеплены с использованием фермента фукозидазы; например, фукозидаза, представленная α-L-фукозидазой, удаляет фукозильные остатки из антител (Tarentino et al., (1975) Biochem. 14:5516-23).
[00104] Другой модификацией антител, рассматриваемой в данном документе, является пэгилирование. Антитело может быть пэгилировано, например, для увеличения биологического (например, в сыворотке крови) периода полужизни антитела. Для пэгилирования антитела антитело или его фрагмент обычно вводят в реакцию с полиэтиленгликолем (PEG), таким как реакционноспособное сложноэфирное или альдегидное производное PEG, в условиях, в которых одна или более групп PEG присоединяются к антителу или фрагменту антитела. Предпочтительно пэгилирование осуществляют с помощью реакции ацилирования или реакции алкилирования с реакционноспособной молекулой PEG (или аналогичным реакционноспособным водорастворимым полимером). Используемый в данном документе термин «полиэтиленгликоль» предназначен для охвата любой из форм PEG, которые использовались для получения производных других белков, таких как моно-(C1-С10) алкокси- или арилоксиполиэтиленгликоль или полиэтиленгликольмалеимид. В некоторых вариантах осуществления антитело, подлежащее пэгилированию, представляет собой агликозилированное антитело. Способы пэгилирования белков известны в данной области и могут быть применены к антителам по настоящему изобретению. См., например, ЕРО 154316 и ЕР 0401384.
[00105] Антитела по настоящему изобретению можно охарактеризовать по их различным физическим свойствам для обнаружения и/или дифференциации их различных классов.
[00106] Например, антитела могут содержать один или более сайтов гликозилирования в вариабельной области либо легкой, либо тяжелой цепи. Такие сайты гликозилирования могут приводить к повышенной иммуногенности антитела или изменению pK антитела ввиду измененного связывания антигена (Marshall et al (1972) Annu Rev Biochem 41:673-702; Gala and Morrison (2004) J Immunol 172:5489-94; Wallick et al (1988) J Exp Med 168:1099-109; Spiro (2002) Glycobiology 12:43R-56R; Parekh et al (1985) Nature 316:452-7; Mimura et al., (2000) Mol Immunol 37:697-706). Известно, что гликозилирование происходит в мотивах, содержащих последовательность N-X-S/T. В некоторых случаях предпочтительно иметь антитело к Siglec15, которое не предусматривает гликозилирования вариабельной области. Этого можно достичь либо путем выбора антител, не содержащих мотив гликозилирования в вариабельной области, либо путем мутирования остатков в пределах области гликозилирования.
[00107] В предпочтительном варианте антитела не содержат аспарагиновых сайтов изомерии. Деамидирование аспарагина может происходить в последовательностях N-G или D-G и приводить к образованию остатка изоаспарагиновой кислоты, который вводит связь в полипептидную цепь и снижает ее стабильность (эффект изоаспарагиновой кислоты).
[00108] Каждое антитело будет иметь уникальную изоэлектрическую точку (pI), которая обычно находится в диапазоне рН от 6 до 9,5. pI для антитела изотипа IgG1 обычно находится в диапазоне рН 7-9,5, a pI для антитела изотипа IgG4 обычно находится в диапазоне рН 6-8. Есть предположение, что антитела с pI за пределами нормального диапазона могут в некоторой степени подвергаться разворачиванию и характеризоваться нестабильностью в условиях in vivo. Таким образом, предпочтительно иметь антитело к Siglec15, которое характеризуется значением pI, находящимся в нормальном диапазоне. Это может быть достигнуто либо путем выбора антител с pI в пределах нормального диапазона значений, либо путем мутирования заряженных остатков, находящихся на поверхности.
[00109] В другом аспекте в настоящем изобретении представлены молекулы нуклеиновой кислоты, которые кодируют вариабельные области тяжелой и/или легкой цепей или CDR антител по настоящему изобретению. Нуклеиновые кислоты могут быть представлены в целых клетках, в лизате клеток или в частично очищенной или по сути чистой форме. Нуклеиновая кислота является «выделенной» или «представленной в по сути чистом виде», когда она является очищенной от других клеточных компонентов или других загрязняющих веществ, например, других клеточных нуклеиновых кислот или белков, с помощью стандартных методик. Нуклеиновая кислота по настоящему изобретению может представлять собой, например, ДНК или РНК и может содержать или не содержать интронные последовательности. В предпочтительном варианте нуклеиновая кислота представляет собой молекулу кДНК.
[00110] Нуклеиновые кислоты по настоящему изобретению могут быть получены с использованием стандартных методик молекулярной биологии. В случае антител, экспрессируемых гибридомами (например, гибридомами, полученными из трансгенных мышей, несущими человеческие гены иммуноглобулинов, как описано далее ниже), кДНК, кодирующие легкую и тяжелую цепи антитела, продуцируемого гибридомой, могут быть получены посредством стандартных методик ПЦР-амплификации или клонирования кДНК. В случае антител, получаемых из библиотеки генов иммуноглобулинов (например, с использованием методик фагового дисплея), нуклеиновая кислота, кодирующая такие антитела, может быть извлечена из генной библиотеки.
[00111] Предпочтительные молекулы нуклеиновых кислот по настоящему изобретению включают таковые, кодирующие последовательности VH и VL моноклонального антитела к Siglec15 или CDR. После получения фрагментов ДНК, кодирующих сегменты VH и VL, эти фрагменты ДНК можно подвергать дополнительным манипуляциям с использованием стандартных методик рекомбинантной ДНК, например, для преобразования генов вариабельной области в гены цепей полноразмерного антитела, в гены Fab-фрагментов или в ген scFv. В ходе этих манипуляций фрагмент ДНК, кодирующий VL или VH, функционально связывают с другим фрагментом ДНК, кодирующим другой белок, такой как константная область антитела или гибкий линкер. Термин «функционально связанный», используемый в данном контексте, предназначен для обозначения того, что два фрагмента ДНК соединены таким образом, что аминокислотные последовательности, кодируемые двумя фрагментами ДНК, остаются внутри рамки считывания.
[00112] Выделенная ДНК, кодирующая область VH, может быть преобразована в ген полноразмерной тяжелой цепи путем функционального связывания ДНК, кодирующей VH, с другой молекулой ДНК, кодирующей константные области тяжелой цепи (CH1, CH2 и CH3). Последовательности генов константной области тяжелой цепи человека известны в данной области, а фрагменты ДНК, охватывающие эти области, могут быть получены с помощью стандартной ПЦР-амплификации. Константная область тяжелой цепи может представлять собой константную область IgG1, IgG2, IgG3, IgG4, IgA, IgE, IgM или IgD, но наиболее предпочтительно представляет собой константную область IgG1 или IgG4. В случае гена тяжелой цепи Fab-фрагмента ДНК, кодирующая Vh, может быть функционально связана с другой молекулой ДНК, кодирующей только константную область тяжелой цепи CH1.
[00113] Выделенная ДНК, кодирующая область VL, может быть преобразована в ген полноразмерной легкой цепи (а также ген легкой цепи Fab) путем функционального связывания ДНК, кодирующей VL, с другой молекулой ДНК, кодирующей константную область легкой цепи CL. Последовательности генов константной области легкой цепи человека известны в данной области, а фрагменты ДНК, охватывающие эти области, могут быть получены с помощью стандартной ПЦР-амплификации. В предпочтительных вариантах осуществления константная область легкой цепи может представлять собой константную область каппа-цепи или лямбда-цепи.
[00114] Для получения гена scFv фрагменты ДНК, кодирующие VH и VL, функционально связывают с другим фрагментом, кодирующим гибкий линкер, например, кодирующий аминокислотную последовательность (Gly4-Ser)3, так что последовательности VH и VL могут экспрессироваться в виде непрерывного одноцепочечного белка с областями VL и VH, соединенными гибким линкером (см., например, Bird et al., (1988) Science 242:423-426; Huston et al., (1988) Proc. Natl Acad Sci. USA 85:5879-5883; McCafferty et al., (1990) Nature 348:552-554).
[00115] Моноклональные антитела (mAb) по настоящему изобретению могут быть получены с использованием платформы на основе трансгенных мышей (например, CAMouseHG, B000.60.01T(G15), HG5042, Chongqing Camab Biotech Ltd.) путем иммунизации трансгенных мышей, генетически сконструированных с обеспечением продуцирования полностью человеческих антител, с помощью антигена-мишени, т.е. Siglec15, особенно Siglec15 человека. Клетки селезенки от иммунизированных трансгенных мышей могут быть слиты с миеломными клетками в соответствии с способом, описанным в Kohler G, and Milstein С, «Continuous cultures of fused cells secreting antibody of predefined specificity,» Nature, 256: 495-497 (1975). Слитые «гибридные клетки» затем распределяют по чашкам, и выжившие колонии гибридом исследуют под микроскопом через семь-десять дней после слияния. Через, например, две недели надосадочную жидкость из каждой лунки можно подвергать тестированию на связывание антигена, и положительную гибридому, секретирующую необходимые антитела, субклонируют с использованием предельного разведения для обеспечения клональности клеточной линии, а затем моноклональные антитела очищают.
[00116] Антитела по настоящему изобретению также могут быть получены посредством других методик, хорошо известных в данной области техники, таких как вирусная или онкогенная трансформация В-лимфоцитов и методики фагового дисплея.
[00117] Антитела по настоящему изобретению также могут быть получены в клетке-хозяине, являющейся трансфектомой, с использованием, например, комбинации методик рекомбинантной ДНК и методик генной трансфекции, как хорошо известно в данной области техники (например, Morrison, S. (1985) Science 229:1202). В одном варианте осуществления ДНК, кодирующая частичные или полноразмерные легкие и тяжелые цепи, полученные посредством стандартных методик молекулярной биологии, встраивают в один или более векторов экспрессии таким образом, чтобы гены были функционально связаны с последовательностями, регулирующими транскрипцию и трансляцию. В данном контексте термин «функционально связанный» предназначен для обозначения того, что ген антитела лигирован в вектор таким образом, что последовательности, регулирующие транскрипцию и трансляцию в векторе, выполняют свою предусматриваемую функцию регуляции транскрипции и трансляции гена антитела.
[00118] Предполагается, что термин «регуляторная последовательность» включает промоторы, энхансеры и другие элементы контроля экспрессии (например, сигналы полиаденилирования), которые контролируют транскрипцию или трансляцию генов антител. Такие регуляторные последовательности описаны, например, у Goeddel (Gene Expression Technology. Methods in Enzymology 185, Academic Press, San Diego, Calif. (1990)). Предпочтительные регуляторные последовательности для обеспечения экспрессии в клетке-хозяине млекопитающего включают вирусные элементы, которые способствуют высоким уровням экспрессии белка в клетках млекопитающих, такие как промоторы и/или энхансеры, полученные из цитомегаловируса (CMV), вируса обезьян 40 (SV40), аденовируса, например, главного позднего промотора аденовируса (AdMLP) и полиомы. В качестве альтернативы, можно использовать невирусные регуляторные последовательности, такие как промотор убиквитина или промотор β-глобина. Более того, регуляторные элементы, состоящие из последовательностей из разных источников, таких как промоторная система SRα, которая содержит последовательности из раннего промотора SV40 и длинный концевой повтор вируса Т-клеточного лейкоза человека 1 типа (Takebe et al., (1988) Mol. Cell. Biol. 8:466-472). Вектор экспрессии и последовательности контроля экспрессии выбирают таким образом, чтобы они были совместимы с используемой для экспрессии клеткой-хозяином.
[00119] Ген легкой цепи антитела и ген тяжелой цепи антитела могут быть вставлены в один и тот же или в отдельные векторы экспрессии. В предпочтительных вариантах осуществления вариабельные области используют для создания генов полноразмерных антител, относящихся к антителу любого изотипа, путем встраивания их в векторы экспрессии, уже кодирующие константные области тяжелой цепи и константные области легкой цепи необходимого изотипа, вследствие чего сегмент VH является функционально связанным с сегментом(-ами) CH в пределах вектора, а сегмент VL является функционально связанным с сегментом CL в пределах вектора. Дополнительно или в качестве альтернативы, рекомбинантный вектор экспрессии может кодировать сигнальный пептид, который способствует секреции цепи антитела из клетки-хозяина. Ген цепи антитела может быть встроен в вектор таким образом, чтобы сигнальный пептид был связан в пределах рамки считывания с амино-концом гена цепи антитела. Сигнальный пептид может представлять собой сигнальный пептид иммуноглобулина или гетерологичный сигнальный пептид (т.е. сигнальный пептид из неиммуноглобулинового белка).
[00120] В дополнение к генам цепи антитела и регуляторным последовательностям рекомбинантные векторы экспрессии по настоящему изобретению могут нести дополнительные последовательности, такие как последовательности, регулирующие репликацию вектора в клетках-хозяевах (например, точки начала репликации), и селектируемые маркерные гены. Селектируемый маркерный ген облегчает отбор клеток-хозяев, в которые был введен вектор (см., например, патенты США №№4399216; 4634665 и 5179017). Например, обычно селективный маркерный ген придает устойчивость к лекарственным средствам, таким как G418, гигромицин или метотрексат, клетке-хозяину, в которую был введен вектор. Предпочтительные селектируемые маркерные гены включают ген дигидрофолатредуктазы (DHFR) (для использования в dhfr- клетках-хозяевах при селекции/амплификации, опосредованной метотрексатом) и ген пео (для селекции, опосредованной G418).
[00121] Для экспрессии легкой и тяжелой цепей вектором(-ами) экспрессии, кодирующим(-и) тяжелую и легкую цепи, трансфицируют клетку-хозяина посредством стандартных методик. Различные формы термина «трансфекция» предназначены для охвата широкого ряда методик, обычно используемых для введения экзогенной ДНК в прокариотическую или эукариотическую клетку-хозяина, например, электропорацию, осаждение фосфатом кальция, трансфекцию, опосредованную DEAE-декстраном, и т.п.Хотя теоретически возможно экспрессировать антитела по настоящему изобретению как в прокариотических, так и в эукариотических клетках-хозяевах, экспрессия антител в эукариотических клетках и наиболее предпочтительно в клетках-хозяевах млекопитающих является наиболее предпочтительной, поскольку такие эукариотические клетки а в частности клетки млекопитающих, с наибольшей вероятностью будут обеспечивать сборку и секретирование правильно свернутого и иммунологически активного антитела в отличие от прокариотических клеток.
[00122] Предпочтительные клетки-хозяева млекопитающих для экспрессии рекомбинантных антител по настоящему изобретению включают клетки яичника китайского хомячка (клетки СНО) (включая dhfr- клетки СНО, описанные в Urlaub and Chasin, (1980) Proc. Natl. Acad. Sci. USA 77:4216-4220, используемые с селектируемым маркером DHFR, например, как описано в R. J. Kaufman and P.A. Sharp (1982) J. Mol. Biol. 159:601-621), миеломные клетки NSO, клетки COS и клетки SP2. В частности, для использования с миеломными клетками NSO другой предпочтительной системой экспрессии является система экспрессии на основе гена GS, раскрытая в WO 87/04462, WO 89/01036 и ЕР 338841. Когда рекомбинантные векторы экспрессии, кодирующие гены антител, вводят в клетки-хозяева млекопитающих, антитела продуцируют путем культивирования клеток-хозяев в течение периода времени, достаточного для обеспечения экспрессии антитела в клетках-хозяевах или, что более предпочтительно, секреции антитела в культуральную среду, в которой выращивают клетки-хозяева. Антитела могут быть выделены из культуральной среды с использованием стандартных способов очистки белков.
[00123] Антитела по настоящему изобретению могут быть конъюгированы с терапевтическим средством с образованием иммуноконъюгата, такого как конъюгат антитело-лекарственное средство (ADC). Подходящие терапевтические средства включают цитотоксины, алкилирующие средства, вещества, связывающие малую бороздку ДНК, интеркаляторы ДНК, ДНК-сшивающие средства, ингибиторы гистондеацетилазы, ингибиторы ядерного экспорта, ингибиторы протеасом, ингибиторы топоизомеразы I или II, ингибиторы белков теплового шока, ингибиторы тирозинкиназы, антибиотики и антимитотические средства. В ADC антитело и терапевтическое средство предпочтительно конъюгированы посредством расщепляемого линкера, такого как пептидильный, дисульфидный или гидразоновый линкер. Более предпочтительно линкер представляет собой пептидильный линкер, такой как Val-Cit, Ala-Val, Val-Ala-Val, Lys-Lys, Ala-Asn-Val, Val-Leu-Lys, Ala-Ala-Asn, Cit-Cit, Val-Lys, Lys, Cit, Ser или Glu. ADC можно получать, как описано в патентах США №№7087600; 6989452; и 7129261; РСТ-публикациях WO 02/096910; WO 07/038658; WO 07/051081; WO 07/059404; WO 08/083312 и WO 08/103693; публикациях патентов США 20060024317; 20060004081; и 20060247295; раскрытия которых включены в данный документ посредством ссылки.
[00124] В другом аспекте настоящее изобретение относится к биспецифическим молекулам, содержащим одно или более антител по настоящему изобретению, связанных по меньшей мере с одной другой функциональной молекулой, например, с другим пептидом или белком (например, с другим антителом или лигандом для рецептора), с образованием биспецифической молекулы, которая связывается по меньшей мере с двумя разными сайтами связывания или молекулами-мишенями. Таким образом, используемый в данном документе термин «биспецифическая молекула» включает молекулы, обладающие тремя или более видами специфичности.
[00125] Биспецифические молекулы могут иметь множество различных форматов и размеров. На одном конце спектра размеров биспецифическая молекула сохраняет традиционный формат антитела, за исключением того, что вместо двух связывающих плеч с одинаковой специфичностью она имеет два связывающих плеча, каждое из которых характеризуется различной специфичностью. Другим крайним случаем являются биспецифические молекулы, состоящие из двух одноцепочечных фрагментов антитела (scFv), связанных пептидной цепью, так называемая конструкция Bs(scFv) 2. Биспецифические молекулы промежуточного размера включают два разных фрагмента F(ab), связанных пептидильным линкером. Биспецифические молекулы этих и других форматов могут быть получены способами генной инженерии, соматической гибридизации или химическими способами. См., например, Kufer et al., приведенный выше; Cao and Suresh, Bioconjugate Chemistry, 9 (6), 635-644 (1998); и van Spriel et al., Immunology Today, 21 (8), 391-397 (2000), а также приведенные в нем ссылки.
[00126] Также в настоящем документе представлен онколитический вирус, который предпочтительно инфицирует и уничтожает раковые клетки. Антитела по настоящему изобретению можно использовать в сочетании с онколитическими вирусами. В качестве альтернативы, в организм человека можно вводить онколитические вирусы, кодирующие антитела по настоящему изобретению.
[00127] В данном документе также представлен химерный антигенный рецептор (CAR), содержащий scFv к Siglec15, scFv к Siglec15, содержащий CDR и вариабельные области тяжелой/легкой цепи, описанные в настоящем документе.
[00128] CAR к Siglec15 может содержать (а) внеклеточный антигенсвязывающий домен, содержащий scFv к Siglec15; (b) трансмембранный домен и (с) внутриклеточный сигнальный домен.
[00129] CAR может содержать сигнальный пептид на N-конце внеклеточного антигенсвязывающего домена, направляющий формирующийся рецептор в эндоплазматический ретикулум, и шарнирный пептид на N-конце внеклеточного антигенсвязывающего домена, который делает рецептор более доступным для связывания. CAR предпочтительно содержит во внутриклеточном сигнальном домене первичный внутриклеточный сигнальный домен и один или более костимулирующих сигнальных доменов. Используемый в большинстве случаев и наиболее эффективный первичный внутриклеточный сигнальный домен представляет собой цитоплазматический домен CD3-зета, который содержит ITAM, фосфорилирование которых приводит к активации Т-клеток. Костимулирующий сигнальный домен может быть получен из костимулирующих белков, таких как CD28, CD137 и ОХ40.
[00130] CAR могут дополнительно добавлять факторы, которые усиливают экспансию, персистенцию и противоопухолевую активность Т-клеток, такие как цитокины и костимулирующие лиганды.
[00131] Также представлены сконструированные иммунные эффекторные клетки, содержащие CAR, представленный в данном документе. В некоторых вариантах осуществления иммунная эффекторная клетка представляет собой Т-клетку, NK-клетку, мононуклеарную клетку периферической крови (РВМС), гемопоэтическую стволовую клетку, плюрипотентную стволовую клетку или эмбриональную стволовую клетку. В некоторых вариантах осуществления иммунная эффекторная клетка представляет собой Т-клетку.
[00132] В другом аспекте настоящее изобретение относится к фармацевтической композиции, которая может содержать одно или более антител или их антигенсвязывающих частей, биспецифические антитела, CAR-T-клетки, онколитические вирусы, иммуноконъюгаты, молекулы нуклеиновой кислоты, векторы экспрессии или клетки-хозяева по настоящему изобретению, составленные совместно с фармацевтически приемлемым носителем. Антитела или их антигенсвязывающую часть, биспецифические антитела, CAR-T-клетки, онколитические вирусы, иммуноконъюгаты, молекулы нуклеиновой кислоты, векторы экспрессии или клетки-хозяева можно дозированно вводить раздельно, если композиция содержит более чем одно антитело (или его антигенсвязывающую часть, биспецифические антитела, CAR-T-клетки, онколитические вирусы, иммуноконъюгаты, молекулы нуклеиновой кислоты, векторы экспрессии или клетки-хозяева). Композиция может необязательно содержать один или более дополнительных фармацевтически активных ингредиентов, таких как другое антитело или лекарственное средство, такое как противоопухолевое лекарственное средство.
[00133] Фармацевтическая композиция может содержать любое количество вспомогательных веществ. Вспомогательные вещества, которые можно использовать, включают носители, поверхностно-активные вещества, загущающие или эмульгирующие средства, твердые связующие вещества, вещества, способствующие диспергированию или суспендированию, солюбилизаторы, красящие вещества, ароматизирующие средства, покрытия, разрыхлители, смазывающие вещества, подсластители, консерванты, средства для обеспечения изотоничности и их комбинации. Выбор и применение подходящих вспомогательных веществ описаны в Gennaro, ed., Remington: The Science and Practice of Pharmacy, 20th Ed. (Lippincott Williams & Wilkins 2003), содержание которого включено в данный документ посредством ссылки.
[00134] Предпочтительно фармацевтическая композиция является подходящей для внутривенного, внутримышечного, подкожного, парентерального, спинального или эпидермального введения (например, путем инъекции или инфузии). В зависимости от пути введения активный ингредиент может быть покрыт материалом для его защиты от действия кислот и других естественных условий, которые могут обуславливать его инактивацию. Фраза «парентеральное введение», используемая в настоящем документе, означает способы введения, отличные от энтерального и местного введения, обычно осуществляемые путем инъекции, и включает без ограничения внутривенные, внутримышечные, внутриартериальные, интратекальные, внутрикапсульные, интраорбитальные, внутри сердечные, внутрикожные, внутрибрюшинные, транстрахеальные, подкожные, подкутикулярные, внутрисуставные, подкапсульные, субарахноидальные, интраспинальные, эпидуральные и интрастернальные инъекцию и инфузию. В качестве альтернативы, антитело по настоящему изобретению можно вводить непарентеральным путем, таким как местный, эпидермальный или слизистый путь введения, например, интраназально, перорально, вагинально, ректально, сублингвально или местно.
[00135] Фармацевтические композиции могут быть представлены в виде стерильных водных растворов или дисперсий. Они также могут быть составлены в виде микроэмульсии, липосомы или другой упорядоченной структуры, подходящей для высокой концентрации лекарственного средства.
[00136] Количество активного ингредиента, которое можно комбинировать с материалом-носителем для получения единичной лекарственной формы, будет варьироваться в зависимости от субъекта, подвергаемого лечению, и конкретного способа введения, и обычно будет являться таким количеством композиции, которое обеспечивает терапевтический эффект. Как правило, в расчете на сто процентов это количество будет варьироваться от приблизительно 0,01% до приблизительно девяноста девяти процентов активного ингредиента, предпочтительно от приблизительно 0,1% до приблизительно 70%, наиболее предпочтительно от приблизительно 1% до приблизительно 30% активного ингредиента в комбинации с фармацевтически приемлемым носителем.
[00137] Схемы введения дозы корректируются для обеспечения оптимального необходимого ответа (например, терапевтического ответа). Например, можно вводить одну болюсную дозу, можно вводить несколько разделенных доз с течением времени или дозу можно пропорционально уменьшать или увеличивать в соответствии с требованиями терапевтической ситуации. Особенно преимущественным является составление парентеральных композиций в виде единичной дозированной формы для простоты введения и однородности дозировки. Единичная дозированная форма, как используется в данном документе, относится к физически дискретным единицам, подходящим в качестве единичных доз для субъектов, подлежащих лечению; каждая единица содержит заранее определенное количество активного ингредиента, рассчитанное для получения необходимого терапевтического эффекта в сочетании с требуемым фармацевтическим носителем. В качестве альтернативы, антитело можно вводить в виде состава с замедленным высвобождением, в случае чего требуется менее частое введение.
[00138] Для введения композиции доза может составлять от приблизительно 0,0001 до 100 мг/кг.
[00139] «Терапевтически эффективная доза» антитела к Siglec15 или его антигенсвязывающей части или биспецифических антител, CAR-T-клеток, онколитических вирусов, иммуноконъюгатов по настоящему изобретению предпочтительно приводит к уменьшению тяжести симптомов заболевания, увеличению частоты и продолжительности бессимптомных периодов заболевания или предупреждению нарушений или инвалидности ввиду поражения заболеванием. Например, для лечения субъектов с опухолями «терапевтически эффективная доза» предпочтительно подавляет рост опухоли на по меньшей мере приблизительно 20%, более предпочтительно на по меньшей мере приблизительно 40%), еще более предпочтительно на по меньшей мере приблизительно 60% и даже еще более предпочтительно на по меньшей мере приблизительно 80%) по сравнению с субъектами, которых не подвергали лечению. Терапевтически эффективное количество терапевтического антитела может обеспечивать уменьшение размера опухоли или иным образом облегчать симптомы у субъекта, которым обычно является человек или который может являться другим млекопитающим.
[00140] Фармацевтическая композиция может представлять собой состав с контролируемым высвобождением, включая имплантаты, трансдермальные пластыри и микроинкапсулированные системы доставки. Можно использовать биоразлагаемые биосовместимые полимеры, такие как этиленвинилацетат, полиангидриды, полигликолевая кислота, коллаген, сложные полиортоэфиры и полимолочная кислота. См., например, Sustained and Controlled Release Drug Delivery Systems, J. R. Robinson, ed., Marcel Dekker, Inc., New York, 1978.
[00141] Терапевтические композиции можно вводить с помощью устройств медицинского назначения, таких как (1) безыгольные устройства для подкожных инъекций (например, патенты США №№5399163, 5383851, 5312335, 5064413,4941880, 4790824 и 4596556); (2) микроинфузионные помпы (патент США №4487603); (3) устройства для трансдермальной доставки (патент США №4486194); (4) инфузионные приборы (патенты США №№4447233 и 4447224) и (5) осмотические устройства (патенты США №№4439196 и 4475196); раскрытия которых включены в данный документ посредством ссылки.
[00142] В некоторых вариантах осуществления моноклональные антитела по настоящему изобретению могут быть составлены таким образом, чтобы обеспечить надлежащее распределение in vivo. Например, чтобы гарантировать, что терапевтическое антитело по настоящему изобретению будет преодолевать гематоэнцефалический барьер, его можно составлять в липосомы, которые могут дополнительно содержать нацеливающие фрагменты для усиления избирательного транспорта к специфическим клеткам или органам. См., например, патенты США №№4522811; 5374548; 5416016 и 5,399,331; V.V. Ranade (1989) J. Clin. Pharmacol. 29:685; Umezawa et al., (1988) Biochem. Biophys. Res. Commun. 153:1038; Bloeman et al., (1995) FEBS Lett.357:140; M. Owais et al., (1995) Antimicrob. Agents Chemother. 39:180; Briscoe et al., (1995) Am. J. Physiol 1233:134; Schreier et al., (1994) J. Biol Chem. 269:9090; Keinanen and Laukkanen (1994) FEBS Lett. 346:123; and Killion and Fidler (1994) Immunomethods 4:273.
[00143] Композиция по настоящему изобретению характеризуется множеством видов применимости in vitro и in vivo, предусматривающих, например, лечение видов рака и остеопороза. Антитела можно вводить субъектам-людям, например in vivo, с целью подавления роста опухоли или подавления потери костной ткани.
[00144] Учитывая способность антител к Siglec15 или антигенсвязывающих частей по настоящему изобретению обеспечивать реверсию Т-клеточной супрессии, опосредованной Siglec15, и подавлять пролиферацию и выживание раковых клеток, в настоящем изобретении представлены способы подавления роста опухолевых клеток у субъекта, включающие введение субъекту композиции по настоящему изобретению, вследствие чего происходит подавление роста опухоли у субъекта. Неограничивающие примеры опухолей, которые можно лечить с помощью композиции по настоящему изобретению, включают без ограничения немелкоклеточный рак легкого, рак яичника, меланому, колоректальный рак, рак молочной железы (включая трижды негативный рак молочной железы), плоскоклеточную карциному головы и шеи, рак эндометрия и плоскоклеточную карциному. Кроме того, рефрактерные или рецидивирующие злокачественные новообразования, рост которых можно подавлять с применением антител по настоящему изобретению.
[00145] В другом аспекте в настоящем изобретении представлен способ подавления потери костной ткани или увеличения массы костной ткани, включающий введение субъекту терапевтически эффективного количества антитела или его антигенсвязывающей части по настоящему изобретению.
[00146] В другом аспекте в настоящем изобретении представлены способы комбинированной терапии, в которых антитела к Siglec15 или их антигенсвязывающая часть или биспецифические антитела, CAR-T-клетки, онколитические вирусы, иммуноконъюгаты по настоящему изобретению вводят совместно с одним или более дополнительными антителами, которые эффективно подавляют рост опухоли у субъекта. В одном варианте осуществления в настоящем изобретении представлен способ подавления роста опухоли у субъекта, включающий введение субъекту антитела к Siglec15 (или его антигенсвязывающей части или CAR-T-клетки, онколитического вируса, иммуноконъюгата) и одного или более дополнительных антител, таких как антитело к VISTA, антитело к LAG-3, антитело к PD-L1 и антитело к PD-1 и/или антитело к CTLA-4. В некоторых вариантах осуществления субъектом является человек.
[00147] Активация передачи сигналов Siglec15 также может быть дополнительно объединена со стандартными средствами лечения рака. Например, блокада передачи сигналов Siglec15 может сочетаться с блокадой CTLA-4, и/или LAG-3, и/или PD-1, а также химиотерапевтическими режимами. Например, с антителами к Siglec15 можно вводить химиотерапевтическое средство, которое может являться цитотоксическим средством. Например, эпирубицин, оксалиплатин и 5-FU вводят пациентам, получающим средство терапии, направленное против Siglec15.
[00148] Необязательно, комбинация антитела к Siglec15 и одного или более дополнительных антител (например, антитела к CTLA-4, и/или антитела к LAG-3, и/или антитела к PD-1) может быть дополнительно объединена с иммуногенным средством, таким как раковые клетки, очищенные опухолевые антигены (включая рекомбинантные белки, пептиды и молекулы углеводов) и клетки, трансфицированные генами, кодирующими иммуностимулирующие цитокины (Не et al., (2004) J. Immunol 173:4919-28). Неограничивающие примеры противоопухолевых вакцин, которые можно использовать, включают пептиды меланомных антигенов, такие как пептиды gp100, антигенов MAGE, Trp-2, MART1 и/или тирозиназы, или опухолевые клетки, трансфицированные для экспрессии цитокина GM-CSF.
[00149] Другие виды терапии, которые можно комбинировать со средством терапии, направленным против Siglec15, включают, помимо прочего, введение интерлейкина-2 (IL-2), облучение, хирургическое вмешательство или гормональную депривацию.
[00150] В другом аспекте в настоящем изобретении представлены способы комбинированной терапии, в которых антитела к Siglec15 или их антигенсвязывающая часть или биспецифические антитела, CAR-T-клетки, онколитические вирусы, иммуноконъюгаты по настоящему изобретению вводят совместно с одним или более дополнительными средствами, эффективными в отношении подавления потери костной ткани. В одном варианте осуществления в настоящем изобретении представлен способ подавления потери массы костной ткани у субъекта, включающий введение субъекту антитела к Siglec15 (или его антигенсвязывающей части или CAR-T-клетки, онколитического вируса, иммуноконъюгатов) и одного или более дополнительных антител для лечения остеопороза, таких как антитело к RANKL и антитело к IL-11. В некоторых вариантах осуществления субъектом является человек.
[00151] Комбинацию терапевтических средств, обсуждаемых в данном документе, можно вводить параллельно в виде одной композиции в фармацевтически приемлемом носителе или параллельно в виде отдельных композиций с каждым средством в фармацевтически приемлемом носителе. В другом варианте осуществления комбинацию терапевтических средств можно вводить последовательно.
[00152] Кроме того, если более одной дозы средства комбинированной терапии вводят последовательно, порядок последовательного введения может быть изменен на обратный или сохранен в том же порядке для каждого момента времени введения, последовательные введения могут быть объединены с параллельными введениями, или предусматривается любая их комбинацией.
[00153] Настоящее раскрытие дополнительно проиллюстрировано следующими примерами, которые не следует рассматривать как налагающие дополнительные ограничения. Содержание всех графических материалов и всех ссылок, последовательностей Genbank, патентов и опубликованных патентных заявок, приводимых по всему тексту данной заявки, явным образом включено в данный документ посредством ссылки.
Примеры
Пример 1. Получение человеческих моноклональных антител к Siglec15 с использованием гибридомной технологии
Иммунизация
[00154] Платформу на основе трансгенных мышей CAMouseHG (HG5042, Chongqing САМАВ Biotech Ltd.) использовали для получения полностью человеческих антител. Трансгенных мышей иммунизировали в соответствии со способом, описанным в Е Harlow, D. Lane, Antibody: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1998. В качестве иммуногена использовали самостоятельно полученный рекомбинантный человеческий белок Siglec15 с человеческим Fc IgG1 на С-конце (аминокислотная последовательность, указанная под SEQ ID NO: 19), а самостоятельно полученный белок Siglec15 яванского макака (аминокислотная последовательность, указанная под SEQ ID NO: 21) использовали для определения титра антисывороток и для скрининга гибридом, секретирующих антигенспецифические антитела. Иммунизирующие дозы содержали 50 мкг человеческого белка Siglec15-Fc/мышь/инъекция как для первичной, так и для бустерной процедур иммунизации. Для повышения иммунного ответа использовали полный адъювант Фрейда и неполный адъювант Фрейда (Сигма, Сент-Луис, Миссури, США), соответственно, для первичной и бустерной процедур иммунизации. Вкратце, смесь адъювант-антиген получали путем начального осторожного перемешивания адъюванта во флаконе с использованием вортекса. Необходимое количество адъюванта переносили в автоклавированную микроцентрифужную пробирку объемом 1,5 мл. Антиген получали в PBS или физиологическом растворе с концентрацией в диапазоне 0,5-0,67 мг/мл. Рассчитанное количество антигена затем добавляли в микроцентрифужную пробирку с адъювантом, и полученную смесь перемешивали путем осторожного центрифугирования на вортексе в течение 2 минут с получением эмульсий по типу «вода в масле». Эмульсию на основе адъюванта-антигена затем набирали в соответствующий шприц для инъекции животным. В общей сложности вводили 50 мкг антигена в объеме 150-200 мкл. Каждое животное иммунизировали, а затем подвергали бустерной иммунизации, выполняемой от 3 до 4 раз в зависимости от титра антисыворотки. Животных с хорошими уровнями титров перед выполнением процедуры слияния подвергали окончательной стимуляции путем внутрибрюшинной инъекции.
Слияние и скрининг гибридом
[00155] Клетки линии мышиных миеломных клеток (SP2/0-Ag14, ATCC#CRL-1581) культивировали до достижения стадии логарифмической фазы непосредственно перед слиянием. Клетки селезенки от иммунизированных мышей получали в стерильных условиях и осуществляли их слияние с миеломными клетками в соответствии с способом, описанным в Kohler G, and Milstein С, «Continuous cultures of fused cells secreting antibody of predefined specificity», Nature, 256: 495-497 (1975). Слитые «гибридные клетки» затем распределяли в 96-луночные планшеты в среде DMEM/20% FCS/HAT. Выжившие колонии гибридом исследовали под микроскопом через семь-десять дней после слияния. Через две недели надосадочную жидкость из каждой лунки подвергали непрямому ELISA с использованием рекомбинантного белка Siglec15-his яванского макака. Затем отбирали положительные гибридомы, секретирующие антитела, которые связывались с белком Siglec15-his яванского макака, и осуществляли их перенос в 24-луночные планшеты. Эти гибридомные клоны дополнительно тестировали на их активность по блокированию связывания человеческого Siglec15 с LRRC4C. Гибридомные клоны, продуцирующие антитела, которые демонстрировали высокоспецифичное связывание Siglec15 яванского макака и активность по блокированию взаимодействия Siglec15-LRRC4C, субклонировали путем предельного разведения для обеспечения клональности клеточной линии, а затем моноклональные антитела очищали. Вкратце, колонки с белок А-сефарозой (от Bestchrom (Шанхай) Biosciences, № по каталогу АА0273) промывали с использованием буфера PBS в объеме, составляющем от 5 до 10 объемов колонки. Надосадочные жидкости гибридомных моноклонов пропускали через колонки, а затем колонки промывали с использованием буфера PBS до тех пор, пока показатель поглощения для белка не достигал исходного уровня. Колонки элюировали элюирующим буфером (0,1 М глицин-HCl, рН 2,7) и полученное незамедлительно собирали в пробирки объемом 1,5 мл с нейтрализующим буфером (1 М трис-HCl, рН 9,0). Фракции, содержащие иммуноглобулины, объединяли и подвергали диализу в PBS в течение ночи при 4°С. Впоследствии характеристики форм функциональной активности очищенных моноклональных антител in vitro определяли следующим образом.
Пример 2. Определение аффинности связывания моноклональных антител к Siglec15 с использованием поверхностного плазмонного резонанса BIACORE.
[00156] Очищенные моноклональные антитела (mAb) к Siglec15, полученные в примере 1, характеризовали по аффинности связывания и кинетике связывания с помощью системы Biacore Т200 (GE Healthcare, Питтсбург, Пенсильвания, США).
[00157] Вкратце, козье антитело к человеческому иммуноглобулину класса IgG (GE Healthcare, № по каталогу BR 100839, набор для захвата человеческих антител) ковалентно связывали с чипом СМ5 (чип, покрытый карбоксиметилдекстраном, GE Healthcare, № по каталогу BR-1005-30) посредством первичных аминов с использованием стандартного набора для связывания через аминогруппу, предоставленного Biacore (GE Healthcare, Питтсбург, Пенсильвания, США). Непрореагировавшие фрагменты на поверхности биосенсора блокировали этаноламином. Затем очищенные антитела к Siglec15 по настоящему изобретению в концентрации 13,3 нМ и референсное антитело к Siglec15 (ch5G12, также обозначаемое как ВМ, см. публикацию патента США №20190202912 А1, аминокислотные последовательности тяжелой цепи и легкой цепи указаны под SEQ ID NO: 24 и 25, соответственно) при 13,3 Нм, соответственно, наносили проточным способом на чип со скоростью потока 10 мкл/мин. Затем серийно разведенный рекомбинантный человеческий белок Siglec15-his (получен самостоятельно, аминокислотная последовательность указана под SEQ ID NO: 20) или белок яванского макака Siglec15-his (получен самостоятельно, аминокислотная последовательность указана под SEQ ID NO: 21) при 2-кратном серийном разведении в буфере HBS-EP+, начиная с 80 нМ, соответственно, наносили проточным способом на чип со скоростью потока 30 мкл/мин. За кинетикой ассоциации антиген-антитело следили в течение 2 минут, а за кинетикой диссоциации следили в течение 10 минут.Кривые ассоциации и диссоциации аппроксимировали согласно модели связывания Ленгмюра 1:1 с использованием программного обеспечения для оценки BIAcore. Определяли значения KD, Ka и Kd, и они в обобщенном виде указаны в таблице 2 ниже.
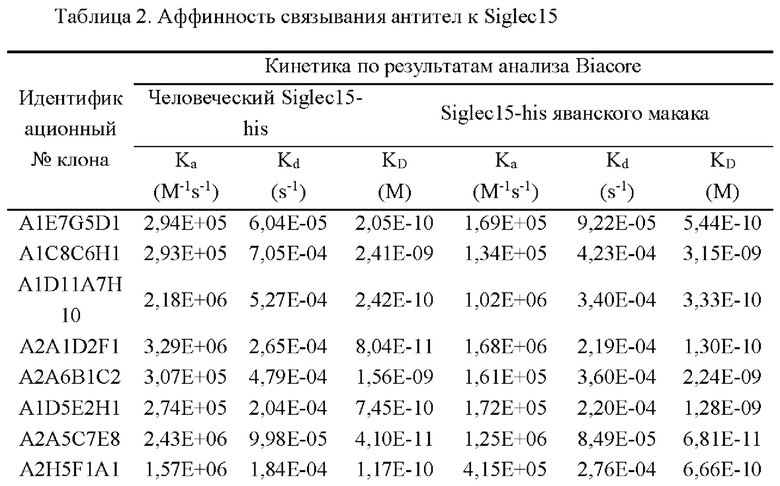
[00158] Все антитела к Siglec15 по настоящему изобретению специфически связывались с человеческим Siglec15 и Siglec15 яванского макака со сравнимой или более высокой аффинностью связывания по сравнению с эталонным антителом. Антитела A2A1D2F1, А2А5С7Е8, A1E10G7H9 и A1D1B7H9 показали самую высокую аффинность связывания.
Пример 3. Активность по связыванию Siglec15 антителами к Siglec15
[00159] Антитела по настоящему изобретению дополнительно тестировали на активность по связыванию с Siglec15 с помощью ELISA с захватом, проточной цитометрии (FACS) и непрямого ELISA.
3.1. ELISA с захватом
[00160] Вкратце, 96-луночные планшеты покрывали 100 мкл находящегося в концентрации 2 мкг/мл козьего антитела к человеческому иммуноглобулину класса IgG AffiniPure, специфичного к фрагменту F(ab')2 (Jackson Immuno Research, № по каталогу 109-005-097), в PBS в течение 2 часов при 37°С. Планшеты однократно промывали промывочным буфером (PBS+0,05% об./об. Tween-20, PBST) и затем блокировали 200 мкл блокирующего буфера (5% вес/об. обезжиренного молока в PBST) в течение ночи при 4°С. Планшеты промывали 4 раза и, соответственно, инкубировали со 100 мкл серийно разбавленных антител к Siglec15 по настоящему изобретению, эталонного антитела и hIgG (человеческий иммуноглобулин (рН4) для внутривенной инъекции, Hualan Biological Engineering Inc.), предусматривающих 5-кратное разведение в 2,5% вес/об. обезжиренного молока в PBST, начиная с 66,7 нМ, в течение 40 минут при 37°С, а затем снова промывали 4 раза. Планшеты, содержащие захваченные антитела, инкубировали со 100 мкл меченного биотином человеческого белка Siglec15-his (получен самостоятельно с SEQ ID NO: 20, 145 нг/мл 2,5% вес/об. обезжиренного молока в PBST) в течение 40 минут при 37°С, промывали 4 раза и инкубировали со стрептавидином, конъюгированным с HRP (разведение 1:10000 в PBST, Jackson Immuno Research, № по каталогу 016-030-084, 100 мкл/лунка), в течение 40 минут при 37°С. После окончательной промывки планшеты инкубировали со 100 мкл/лунка ТМВ-субстрата для ELISA (Innoregents, № по каталогу TMB-S-002). Реакцию останавливали через 4 минуты при комнатной температуре с помощью 50 мкл/лунка 1 М H2SO4, и показатель поглощения для каждой лунки считывали на микропланшетном ридере с использованием двухволнового режима с 450 нм для ТМВ и 630 нм в качестве референсной длины волны, затем значения OD (450-630) наносили на график в зависимости от концентрации антител. Данные анализировали с использованием программного обеспечения Graphpad Prism и устанавливали значения ЕС50.
3.2. Клеточный FACS-анализ связывания
[00161] Показатели активности антител к Siglec15 по связыванию с Siglec15 на клеточной поверхности тестировали с помощью проточной цитометрии (FACS) с использованием клеток человеческий-siglec15-2D3-1E1, полученных самостоятельно, экспрессирующих человеческий Siglec15 (аминокислотные остатки Metl-Pro328 в uniprot №Q6ZMC9) на клеточных мембранах. Клетки человеческий-siglec15-2D3-1E1 получали путем трансфекции клеток HEK-293 (№ в АТСС CRL-1573) плазмидами pCMV-T-P со вставленной в них последовательностью, кодирующей человеческий Siglec15, между сайтами EcoRI и XbaI, следуя инструкции по применению реагента для трансфекции липофектамин 3000 (Thermo Fisher). Клетки человеческий-siglec15-2D3-1Е1 собирали из колб для культивирования клеток, дважды промывали и повторно суспендировали в забуференном фосфатом солевом растворе (PBS), содержащем 2% об./об. фетальной бычьей сыворотки (буфер для FACS). 2×105 клеток на лунку в 96-луночных планшетах инкубировали в 100 мкл серийно разведенных антител к Siglec15 или контролей (начиная с 66,67 нМ, 5-кратное серийное разведение) в буфере для FACS в течение 40 минут на льду. Клетки дважды промывали буфером для FACS и добавляли 100 мкл конъюгированного с R-фикоэритрином козьего антитела к человеческому иммуноглобулину класса IgG AffiniPure, специфичного к фрагменту Fcγ (разведение 1:1000 в буфере для FACS, Jackson Immunoresearch, № по каталогу 109-115-098). После 40-минутной инкубации при 4°С в темноте клетки трижды промывали и повторно суспендировали в буфере для FACS. Флуоресценцию измеряли с использованием оборудования Becton Dickinson FACS Canto II-HTS. Данные анализировали с использованием программного обеспечения Graphpad Prism и устанавливали значения ЕС50.
3.3. Непрямой ELISA
[00162] Измеряли перекрестные реакции антител к Siglec15 с белками Siglec15 яванского макака или мыши. Вкратце, 96-луночные микропланшеты покрывали 100 мкл белка Siglec15-his яванского макака в концентрации 2 мкг/мл (получен самостоятельно с SEQ ID NO: 21) или белка Siglec15-his мыши в концентрации 2 мкг/мл (получен самостоятельно с SEQ ID NO: 22) в карбонатном/бикарбонатном буфере (рН 9,6) на 2 часа при 37°С.Планшеты для ELISA однократно промывали промывочным буфером (PBS +0,05% об./об. Tween-20, PBST) и затем блокировали 200 мкл/лунка блокирующего буфера (5% вес/об. обезжиренного молока в PBST) в течение ночи при 4°С. Планшеты промывали 4 раза и инкубировали со 100 мкл серийно разбавленных антител к Siglec15 по настоящему изобретению или контролей (начиная с 66,7 нМ, 5-кратное серийное разведение в PBST с 2,5% вес/об. обезжиренного молока) в течение 40 минут при 37°С. Планшеты для ELISA снова промывали 4 раза и инкубировали с конъюгированным с пероксидазой козьим антителом к человеческому иммуноглобулину класса IgG AffiniPure, специфичному к фрагменту F(ab')2 (разведение 1:5000 в буфере PBST, Jackson Immunoresearch, № по каталогу 109-035-097, 100 мкл/лунка), в течение 40 минут при 37°С. После окончательной промывки планшеты инкубировали со 100 мкл/лунка ТМВ (Innoregents, № по каталогу TMB-S-002). Реакцию останавливали через 4 минуты при комнатной температуре с помощью 50 мкл 1 М H2SO4, и показатель поглощения для каждой лунки считывали на микропланшетном ридере с использованием двухволнового режима с 450 нм для ТМВ и 630 нм в качестве референсной длины волны, затем значения OD (450-630) наносили на график в зависимости от концентрации антител. Данные анализировали с использованием программного обеспечения Graphpad Prism и устанавливали значения ЕС50.
[00163] Результаты для трех анализов показаны на фиг.1А-1С-4А-4С.
[00164] На фиг. 1А-1С видно, что все антитела по настоящему изобретению, за исключением А1С8С6Н1, специфически связывались с человеческим Siglec15 с более низким значением ЕС50 и более высоким значением Bmax (максимальное связывание), чем референсное антитело.
[00165] Как показано на фиг. 2А-2С, антитела к Siglec15 A1D1B7H9, A1D11A7H10, A1E10G7H9, A2A1D2F1, A2G4C8G7 и A2H5F1A1 более эффективно связывались с человеческим Siglec15 на клеточной поверхности (с более низкой ЕС50) с более высоким Bmax по сравнению с эталонным антителом.
[00166] На фиг. 3А-3С показано, что большинство антител по настоящему изобретению связывается с белком Siglec15 яванского макака с аналогичной активностью по связыванию по сравнению с эталонным антителом. В то время как только несколько антител по настоящему изобретению показали аналогичную или лучшую активность по связыванию с Siglec15 мыши, как показано на фиг. 4А-4С. Например, антитела A1E10G7H9 и A2A1D2F1 показали более высокие значения Bmax, чем референсное антитело.
Пример 4. Блокирующая активность антител к Siglec15 в отношении связывания Siglec15-LRRC4C или Siglec15-референсное антитело
4.1. ELISA с блокированием лиганда
[00167] Активность антител к Siglec15 по настоящему изобретению по блокированию связывания Siglec15-LRRC4C измеряли в конкурентном анализе ELISA. LRRC4C является лигандом Siglec15 и может экспрессироваться раковыми клетками (WO 2018057735). Вкратце, 100 мкл человеческого белка Siglec15-Fc (полученного самостоятельно с аминокислотной последовательностью SEQ ID NO: 19) наносили на 96-луночные микропланшеты в концентрации 2 мкг/мл в карбонатном/бикарбонатном буфере на 2 часа при 37°С. Планшеты однократно промывали промывочным буфером (PBS +0,05% об./об. Tween-20, PBST) и блокировали 5% вес/об. обезжиренного молока в PBST в течение ночи при 4°С. Затем планшеты промывали 4 раза с использованием промывочного буфера.
[00168] В планшеты со связанным Siglec15-Fc добавляли в количестве по 100 мкл на лунку серийно разведенные антитела к Siglec15 или контроли (начиная с 66,67 нМ при 5-кратном серийном разведении) в PBST с 2,5% вес/об. обезжиренного молока и их инкубировали с человеческим белком Siglec15-Fc при 37°С в течение 40 минут. Планшеты снова промывали 4 раза с использованием промывочного буфера, затем добавляли 100 мкл/лунка меченного биотином человеческого белка LRRC4C-Fc (полученного самостоятельно с SEQ ID NO: 23) в количестве 290 нг/мл и осуществляли инкубирование в течение 40 минут при 37°С. Планшеты снова промывали с использованием промывочного буфера. После этого в планшеты добавляли 100 мкл/лунка HRP, конъюгированной со стрептавидином (разведение 1:5000 в буфере PBST, Jackson Immunoresearch, № по каталогу 016-030-084), и инкубировали в течение 40 минут при 37°С. Планшеты снова промывали с использованием промывочного буфера. Наконец, добавляли ТМВ и реакцию останавливали с использованием 1 М H2SO4, и показатель поглощения для каждой лунки считывали на микропланшетном ридере с использованием двухволнового режима с 450 нм для ТМВ и 630 нм в качестве референсной длины волны, затем значения OD (450-630) наносили на график в зависимости от концентрации антител. Данные анализировали с использованием программного обеспечения Graphpad Prism и устанавливали значения IC50.
4.2. Блокирующий ELISA для эталонного антитела
[00169] Способность антител к Siglec15 по настоящему изобретению блокировать связывание эталонного антитела с человеческим Siglec15 измеряли в конкурентном анализе ELISA. Вкратце, эталонным антителом при концентрации 2 мкг/мл в PBS в количестве 100 мкл на лунку покрывали 96-луночные микропланшеты и их инкубировали в течение 2 часов при 37°С. Затем планшеты один раз промывали промывочным буфером и блокировали 5% вес/об. обезжиренным молоком в PBST в течение ночи при 4°С. На следующий день антитела к Siglec15 по настоящему изобретению или контроли разводили меченным биотином человеческим белком Siglec15-Fc (полученным самостоятельно с SEQ ID NO: 19, 37 нг/мл в PBST с 2,5% вес/об. обезжиренного молока), начиная с 80 нМ с 4-кратным серийным разведением, и инкубировали при комнатной температуре в течение 40 минут. После 4-кратного промывания планшета смеси атитело/Siglec15-Fc добавляли в количестве 100 мкл на лунку в покрытые эталонным антителом планшеты. После инкубации при 37°С в течение 40 минут планшеты снова промывали 4 раза с использованием промывочного буфера. Затем в планшеты добавляли 100 мкл/лунка HRP, конъюгированной со стрептавидином, и инкубировали в течение 40 минут при 37°С. Планшеты снова промывали с использованием промывочного буфера. Наконец, добавляли ТМВ и реакцию останавливали с использованием 1 М H2SO4, и показатель поглощения для каждой лунки считывали на микропланшетном ридере с использованием двухволнового режима с 450 нм для ТМВ и 630 нм в качестве референсной длины волны, затем значения OD (450-630) наносили на график в зависимости от концентрации антител. Данные анализировали с использованием программного обеспечения Graphpad Prism и устанавливали значения IC50.
4.3. Клеточный FACS-анализ блокирования лиганда
[00170] Активность антител к Siglec15 по блокированию связывания белка Siglec15 с LRRC4C на клеточной поверхностью оценивали с помощью проточной цитометрии (FACS) с использованием полученных самостоятельно клеток LRRC4C-3F12-1B9. Вкратце, клетки HEK-293 (№ в АТСС CRL-1573) трансфицировали плазмидными конструкциями pCMV-T-P с нуклеотидной последовательностью, кодирующей человеческий LRRC4C (аминокислотные остатки Met1-lle640 в uniprot под № Q9HCJ2), вставленной между EcoRI и XbaI, следуя инструкции по применению реагента для трансфекции липофектамин 3000 (Thermo Fisher). Стабильную клетку, названную LRRC4C-3F12-1B9, выбирали для последующего клеточного анализа блокирования лиганда.
[00171] Вкратце, антитела к Siglec15 по настоящему изобретению или контроли разводили человеческим белком Siglec15 с мышиным Fc (полученным самостоятельно с SEQ ID NO: 26, 8 мкг/мл, нМ, в буфере для FACS), начиная с 66,67 нМ с 5-кратным серийным разведением, и инкубировали при комнатной температуре в течение 40 минут.Затем клетки LRRC4C-3F12-1B9 собирали из колб для культивирования клеток, дважды промывали и повторно суспендировали в PBS, содержащем 2% об/об. фетальной бычьей сыворотки (буфер для FACS). Затем, 1×105 клеток на лунку в 96-луночных планшетах инкубировали в 100 мкл смесей антитело/Siglec15-мышиный Fc в течение 40 минут при 4°С. Планшеты дважды промывали буфером для FACS, а затем добавляли в количестве 100 мкл/лунка конъюгированное с R-фикоэритрином козье антитело к человеческому иммуноглобулину класса IgG AffiniPure, специфичное к F(ab')2 (H+L) (разведение 1:1000 в буфере для FACS, Jackson Immunoresearch, № по каталогу 115-116-146), и инкубировали в течение 40 минут при 4°С в темноте. Клетки дважды промывали и повторно суспендировали в буфере для FACS. Флуоресценцию измеряли с использованием оборудования Becton Dickinson FACS Canto II-HTS. Данные анализировали с использованием программного обеспечения Graphpad Prism и устанавливали значения IC50.
[00172] Результаты трех анализов представлены на фиг.5А-5С-7А-7С.
[00173] Из фиг. 5А-5С видно, что все антитела к Siglec15 по настоящему изобретению были способны блокировать связывание человеческого Siglec15 с человеческим LRRC4C, а блокирующая активность являлась аналогичной таковой для эталонного антитела или более низкой, чем у него.
[00174] На фиг. 6А-6С показано, что антитела A1E7G5D1, А2А5С7Е8, A2G4C8G7 и A2H5F1A1 были способны блокировать связывание человеческого Siglec15 с эталонным антителом, что указывает на то, что они связывались с тем же эпитопом или эпитопом, являющемся аналогичным тому, с которым связывалось референсное антитело. Антитела А1С8С6Н1, A1D1B7H9, A1D5E2H1, A1D11A7H10, A1E10G7H9, A2A1D2F1 и А2А6 В1С2 не характеризовались способностью блокировать связывание человеческого Siglec15 с эталонным антителом, что указывает на то, что А1С8С6Н1, A1D1B7H9, A1D5E2H1, A1D11A7H10, A1E10G7H9, A2A1D2F1 и А2А6 В1С2 могут связываться с другими эпитопами по сравнению с эталонным антителом.
[00175] Кроме того, как показано на фиг. 7А-7С, все антитела к Siglec15 по настоящему изобретению были способны блокировать связывание человеческого Siglec15 с человеческим LRRC4C с мышиным Fc на клеточной поверхности, характеризуясь блокирующей активностью, аналогичной таковой для эталонного антитела или меньшей, чем у него.
Пример 5. Термостабильность антител к Siglec15
[00176] Антитела к Siglec15 также тестировали в отношении их показателей термостабильности. Вкратце, для определения Tm (температуры плавления) использовали анализ белкового теплового сдвига с использованием набора для определения стабильности белка при тепловом сдвиге GloMelt™ (Biotium, № по каталогу 33022-Т). Затем обеспечивали оттаивание красителя GloMelt™ и достижение им комнатной температуры. Флакон, содержащий краситель, встряхивали на вортексе и центрифугировали. Затем получали 10-кратный краситель путем добавления 5 мкл 200-кратного красителя к 95 мкл PBS. Добавляли 2 мкл 10-кратного красителя и 10 мкг антител и добавляли PBS до достижения общего реакционного объема 20 мкл. Пробирки, содержащие краситель и антитела, кратковременно центрифугировали и помещали в термоциклер для ПЦР в реальном времени (Roche, LightCycler 480 II) с установленной программой кривой плавления, характеризующейся параметрами, указанными в таблице 3.
[00177] Температуры плавления антител по настоящему изобретению представлены в обобщенном виде в таблице 4, при этом они указывают на то, что антитела по настоящему изобретению, вероятно, стабильны в организме человека.
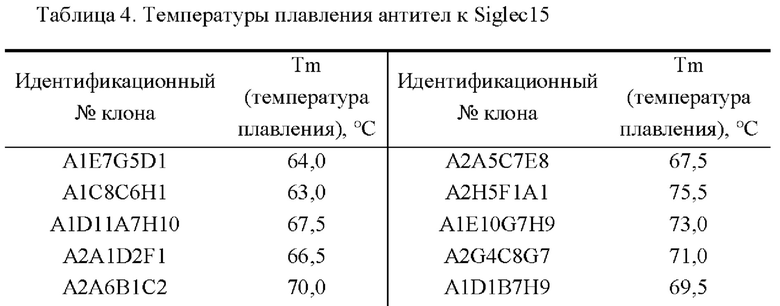

Пример 6. Антитела к Siglec15 обеспечивают реверсию опосредованной Siglec15 Т-клеточной супрессии
[00178] Моноклональными антителами к CD3 (ОКТЗ) (eBioscience Inc., № по каталогу 16-0037-85), представленными в концентрации 50 нг/мл в DPBS, в течение ночи при 4°С покрывали 96-луночные микропланшеты в количестве 100 мкл на лунку. На следующий день несвязанные моноклональные антитела к CD3 аспирировали непосредственно перед добавлением РВМС.
[00179] Общие РВМС от здорового донора-человека промывали средой RPMI-1640 (Gibco, № по каталогу А10491-01), дополненной 10% FBS (Gibco, № по каталогу 10099-141), и центрифугировали при 200 g в течение 15 минут с удалением надосадочную жидкости. Затем клетки РВМС метили 0,5 мкМ CFSE (Invitrogen, № по каталогу С1157) в течение 20 минут на льду, и конечная плотность клеток составляла 1×106 клеток/мл. К клеткам добавляли четырехкратный объем среды RPMI-1640 с добавлением 10% FBS и инкубировали в течение 5 минут при комнатной температуре. Планшеты центрифугировали при 300 g в течение 10 минут и клетки ресуспендировали в среде RPMI-1640 с добавлением 10% FBS при плотности 6×106 клеток/мл. В то же время, серийно разведенные антитела к Siglec15 по настоящему изобретению или контроли (4-кратное разведение в среде RPMI-1640 с добавлением 10% FBS, начиная с 120 мкг/мл) смешивали с человеческим белком Siglec15-Fc (полученным самостоятельно с SEQ ID NO: 19, 40 мкг/мл в среде RPMI-1640 с добавлением 10% FBS) при объемном соотношении 1:1 и инкубировали при комнатной температуре в течение 30 минут. Затем, 50 мкл среды, содержащей клетки РМВС, и 50 мкл смесей антитела/Siglec15-Fc добавляли в планшеты со связанными антителами к CD3 и инкубировали в инкубаторе с СО2 при 37°С в течение 3 дней. Затем в каждую лунку добавляли по 100 мкл буфера для FACS, клетки пипетировали несколько раз и переносили в U-образные планшеты. U-образные планшеты центрифугировали с удалением надосадочной жидкости. И затем, клетки инкубировали в течение 15 минут при 4°С в темноте с 50 мкл/лунка HFCR (разведение 1:10 в буфере для FACS, Biolegend Inc, № по каталогу 422302). Клетки окрашивали флуоресцентными mAb к CD4 (Biolegend Inc., № по каталогу 357410) и к CD8 (Biolegend Inc., № по каталогу 301066) (разведение 1:10 в буфере для FACS) в течение 30 минут при 4°С. Клетки дважды промывали в буфере для FACS (200 мкл/лунка) и повторно суспендировали в буфере для FACS (200 мкл/лунка). Флуоресценцию измеряли с использованием оборудования Becton Dickinson FACS Canto II-HTS. Данные анализировали с использованием программного обеспечения Graphpad Prism и устанавливали значения EC50.
[00180] Результаты показаны на фиг. 8А и 8В.
[00181] Можно увидеть, что антитела А2А5С7Е8 и A1E10G7H9 были способны обеспечивать реверсию опосредованной Siglec15 супрессии CD8+ Т-клеток и CD4+ Т-клеток при более высоких значениях ЕС50 по сравнению с эталонным антителом. Примечательно, что антитело А2А5С7Е8 в высоких дозах было более эффективным, чем референсное антитело в отношении обеспечения реверсии подавления Т-клеток, что приводило к более высоким процентным значениям уровня пролиферации CD4+ и CD8+ Т-клеток.
Пример 7. Секвенирование антител к Siglec15
[00182] Два антитела А2А5С7Е8 и A1E10G7H9 секвенировали и получали полные последовательности вариабельных областей тяжелой цепи и легкой цепи и последовательности константных областей. Идентификационные номера последовательностей вариабельных областей тяжелой цепи и легкой цепи перечислены в таблице 1, а изотип тяжелой/легкой цепи определяли путем выравнивания последовательностей в базе данных.
Пример 8. Генно-инженерное конструирование антитела к Siglec15 А2А5С7Е8
[00183] Чтобы избежать или уменьшить посттрансляционные модификации, такие как дезамидирование и изомеризация определенных аминокислотных остатков в, например, областях CDR, которые могут неблагоприятно влиять на выработку, стабильность, безопасность и/или эффективность антитела, антитело А2А5С7Е8 (также далее обозначаемое в данном документе как А2А5С7Е8-1) дополнительно модифицировали в участке CDR2 тяжелой цепи. Всего получали 2 модифицированных варианта, а именно А2А5С7Е8-2 и А2А5С7Е8-3, идентификационные номера последовательностей CDR и вариабельной области тяжелой/легкой цепи которых перечислены в таблице 1.
[00184] Векторами, каждый из которых содержал нуклеотид, кодирующий вариабельную область тяжелой цепи А2А5С7Е8-2 или А2А5С7Е8-3, связанную с константной областью тяжелой цепи человеческого иммуноглобулина изотипа IgGl (SEQ ID NO: 17), и векторами, каждый из которых содержал нуклеотид, кодирующий вариабельную область легкой цепи, связанную с константной областью легкой каппа-цепи человеческого иммуноглобулина (SEQ ID NO: 18), временно трансфицировали 50 мл суспензионных культур клеток 293F при отношении конструкции с легкой цепью к конструкции с тяжелой цепью 1,1:1 с использованием PEI в количестве 1 мг/мл.
Пример 9. Определение характеристик модифицированных вариантов А2А5С7Е8
[00185] Модифицированные варианты А2А5С7Е8-2 и А2А5С7Е8-3 очищали, как описано выше, и тестировали в ходе анализа Biacore, ELISA с захватом, непрямого ELISA, клеточного FACS-анализа связывания, конкурентного ELISA и клеточного функционального анализа, следуя протоколам согласно предыдущим примерам с модификациями или без них, а также протоколам, описанным ниже.
[00186] Для BIAcore-теста, измеряющего значения аффинности связывания модифицированных вариантов А2А5С7Е8 с мышиным Siglec15, использовали мышиный белок Siglec15-his (полученный самостоятельно с SEQ ID NO: 22).
[00187] В тесте на реверсию супрессии Т-клеток варианты А2А5С7Е8 по настоящему изобретению или контроли в концентрации 48 мкг/мл смешивали с человеческим белком Siglec15-Fc (SEQ ID NO: 19, 20 мкг/мл в среде RPMI-1640 с добавлением 10% FBS) при объемном соотношении 1:1; а варианты А2А5С7Е8 или контроли в концентрации 120 мкг/мл смешивали с человеческим белком Siglec15-Fc (40 мкг/мл в среде RPMI-1640 с добавлением 10% FBS) в объемном соотношении 1:1.
[00188] Результаты тестирования с помощью BIAcore в обобщенном виде представлены в таблицах 5-1 и 5-2. Результаты других анализов показаны на фиг. 9-14 и 15А-15D.
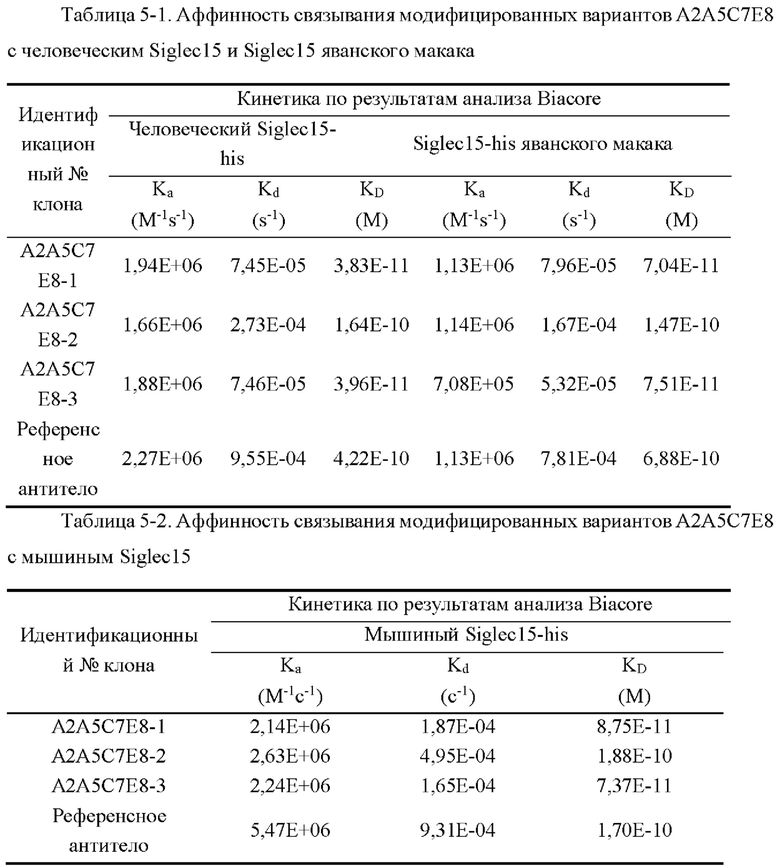
[00189] Как показано в таблицах 5-1 и 5-2, модифицированные варианты А2А5С7Е8-2 и А2А5С7Е8-3 специфически связывались с человеческим Siglec15 и Siglec15 яванского макака с более высокой аффинностью связывания, чем референсное антитело. Антитело А2А5С7Е8-3 также продемонстрировало более высокую аффинность связывания с мышиным Siglec15, чем референсное антитело. Антитело А2А5С7Е8-2 показало сравнимую аффинность связывания с мышиным Siglec15 по сравнению с эталонным антителом.
[00190] Модифицированные варианты А2А5С7Е8-2 и А2А5С7Е8-3 связывались с человеческим Siglec15 с большей эффективностью или с более высоким Втах, чем референсное антитело, как показано на фиг. 9 и 10, и более эффективно подавляли связывание Siglec15-LRRC4C, чем референсное антитело, как показано на фиг. 13. Кроме того, в соответствии с фиг. 11 и 12 модифицированные варианты А2А5С7Е8-2 и А2А5С7Е8-3 более эффективно связывались с Siglec15 яванского макака и мышиным Siglec15, чем референсное антитело.
[00191] Как показано на фиг. 14, модифицированные варианты А2А5С7Е8-2 и А2А5С7Е8-3 были способны блокировать связывание эталонного антитела с Siglec15, что указывает на то, что модифицированные варианты А2А5С7Е8-2 и А2А5С7Е8-3 связывались с эпитопом, являющимся аналогичным таковому для эталонного антитела.
[00192] Как показано на фиг. 15А-15D, в клеточном функциональном анализе модифицированные варианты А2А5С7Е8-2 и А2А5С7Е8-3 обеспечивали реверсию опосредованной Siglec15 супрессии CD8+ Т-клеток и CD4+ Т-клеток, что приводило к аналогичным или более высоким процентным значениям уровня пролиферации клеток по сравнению с эталонным антителом. Примечательно, что когда человеческий белок Siglec15-Fc использовали в относительно низкой дозе, антитела А2А5С7Е8-2 и А2А5С7Е8-3 являлись более эффективными, чем референсное антитело, в отношении подавления CD4+ и CD8+ Т-клеток.
Пример 10. Получение мышиных моноклональных антител к Siglec15 с использованием гибридомной технологии
Иммунизация, слияние и скрининг гибридом
[00193] Иммунизацию мышей, слияние и скрининг гибридом проводили в соответствии с протоколом, приведенным в примере 1, со следующими модификациями. В частности, использовали нормальных мышей, иммунизирующие дозы содержали 50 мкг рекомбинантного человеческого белка Siglec15-Fc на мышь на инъекцию для первичной иммунизации и 25 мкг человеческого белка Siglec15-Fc на мышь на инъекцию для бустерных процедур иммунизации. Антиген получали в PBS или физиологическом растворе с концентрацией в диапазоне 0,25-0,67 мг/мл. В общей сложности вводили 50 или 25 мкг антигена в объеме 150-200 мкл. Каждое животное иммунизировали, а затем подвергали бустерной иммунизации, выполняемой от 3 до 4 раз в зависимости от титра антисыворотки. Надосадочную жидкость культуры слитых клеток подвергали непрямому ELISA с использованием полученного самостоятельно человеческого белка Siglec15-his. Положительные гибридомы, секретирующие антитела, которые связываются с человеческим белком Siglec15-his, отбирали и переносили в 24-луночные планшеты. Эти гибридомы также подвергали клеточному FACS-анализу связывания, ELISA с захватом, непрямому ELISA и ELISA с блокированием лиганда. Гибридомные клоны, продуцирующие антитела, которые демонстрировали высокоспецифичное связывание человеческого Siglec15, связывание Siglec15 яванского макака, связывания мышиного Siglec15 и активность по блокированию взаимодействия человеческого Siglec15 с LRRC4C, субклонировали путем предельного разведения для обеспечения клональности клеточной линии, а затем моноклональные антитела очищали.
Пример 11. Определение аффинности связывания мышиного моноклонального антитела к Siglec15 с использованием поверхностного плазмонного резонанса BIACORE
[00194] Очищенные мышиные моноклональные антитела к Siglec15, полученные в примере 10, характеризовали по их аффинности связывания и кинетике связывания с помощью системы Biacore Т200 (GE Healthcare, Питтсбург, Пенсильвания, США).
[00195] Вкратце, козье антитело к мышиному иммуноглобулину класса IgG (GE Healthcare, № по каталогу BR100838, набор для захвата мышиных антител) ковалентно связывали с чипом СМ5 (чип, покрытый карбоксиметилдекстраном, от GE Healthcare, № по каталогу BR100530) посредством первичных аминов с использованием стандартного набора для связывания через аминогруппу (GE Healthcare, Питтсбург, Пенсильвания, США), предоставленного Biacore. Непрореагировавшие фрагменты на поверхности биосенсора блокировали этаноламином. Очищенные мышиные антитела к Siglec15 по настоящему изобретению в концентрации 13,3 нМ, соответственно, наносили проточным способом на чип со скоростью потока 10 мкл/мин. Затем серийно разводили человеческий белок Siglec15 (получен самостоятельно, аминокислотная последовательность указана под SEQ ID NO: 20), белок Siglec15 яванского макака (получен самостоятельно, аминокислотная последовательность указана под SEQ ID NO: 21) или мышиный белок Siglec15-his (получен самостоятельно, аминокислотная последовательность указана под SEQ ID NO: 22) при 2-кратном серийном разведении в буфере HBS-EP+ (предоставлен Biacore), начиная с 80 нМ, полученное наносили проточным способом на чип со скоростью потока 30 мкл/мин. За кинетикой ассоциации антиген-антитело следили в течение 2 минут, а за кинетикой диссоциации следили в течение 10 минут.Кривые ассоциации и диссоциации аппроксимировали согласно модели связывания Ленгмюра 1:1 с использованием программного обеспечения для оценки BIAcore. Определяли значения KD, Ka и Kd, и они в обобщенном виде указаны в таблицах 6-1 и 6-2 ниже.
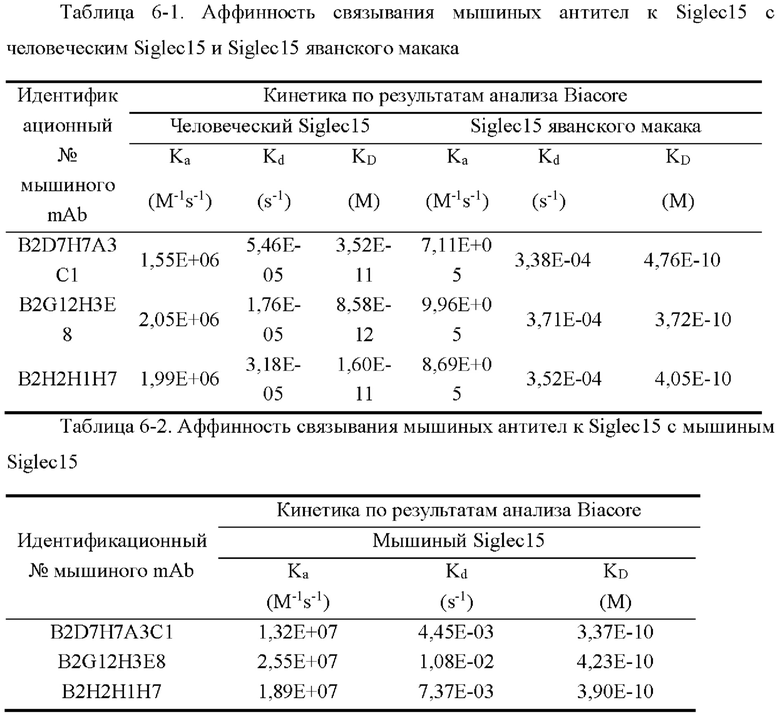
[00196] Все мышиные антитела к Siglec15 по настоящему изобретению специфически связывались с человеческим Siglec15, Siglec15 яванского макака и мышиным Sigle 15 с высокой аффинностью связывания.
Пример 12. Активность по связыванию мышиных моноклональных антител к Siglec15
[00197] Активность по связыванию мышиных антител к Siglec15 с человеческим Siglec15, Siglec15 яванского макака и мышиным Siglec15 дополнительно определяли с помощью ELISA с захватом, непрямого ELISA и проточной цитометрии (FACS).
[00198] ELISA с захватом выполняли в соответствии с протоколом, приведенном в примере 3, за исключением того, что вместо козьего антитела к человеческому иммуноглобулину класса IgG AffiniPure, специфичного к фрагменту F(ab')2, использовали козье антитело к мышиному иммуноглобулину класса IgG, специфичное к фрагменту F(ab')2 (Jackson ImmunoResearch, № по каталогу 115-005-072), в количестве 100 мкл/лунка. Результаты показаны на фиг. 16.
[00199] Клеточный FACS-анализ проводили в соответствии с протоколом, приведенным в примере 3, за исключением того, что 1,5×105 клеток на лунку в 96-луночных планшетах инкубировали в 100 мкл серийно разведенных антител к Siglec15 или контролей (начиная с 66,67 нМ, 5-кратное серийное разведение) в буфере для FACS в течение 40 минут на льду, и к клеткам добавляли 100 мкл конъюгированного с R-фикоэритрином козьего антитела к мышиному иммуноглобулину класса IgG AffiniPure, специфичного к фрагменту F(ab')2 (H+L) (разведение 1:1000 в буфере для FACS, Jackson Immunoresearch, № по каталогу 115-116-146). Результаты показаны на фиг. 17.
[00200] Непрямой ELISA проводили в соответствии с протоколом, приведенным в примере 3, за исключением того, что вместо конъюгированного с R-фикоэритрином козьего антитела к человеческому иммуноглобулину класса IgG AffiniPure, специфичного к фрагменту Fcγ, использовали конъюгированное с пероксидазой козье антитело к мышиному иммуноглобулину класса IgG AffiniPure, специфичное к фрагменту Fcγ (Jackson Immunoresearch, № по каталогу 115-035-071), в количестве 100 мкл/лунка. Результаты показаны на фиг. 18 и 19.
[00201] Из фиг. 16 видно, что мышиные антитела к Siglec15 по настоящему изобретению специфично связываются с человеческим Siglec15 с более высоким значением Bmax (максимальное связывание) и более низким значением ЕС50 по сравнению с эталонным антителом (ch5G12, также обозначаемое как ВМ, см. публикацию патента США №20190202912 А1, аминокислотные последовательности тяжелой цепи и легкой цепи представлены под SEQ ID NO: 24 и 25, соответственно), что указывает на то, что они более эффективно связываются с большим количеством человеческого белка Siglec15. Согласно фиг. 17, мышиные антитела к Siglec15 по настоящему изобретению специфически связывались с человеческим Siglec15 на клеточной поверхности с немного меньшей активностью в FACS-тесте.
[00202] На фиг. 18 показано, что мышиные антитела к Siglec15 по настоящему изобретению специфично связываются с белком Siglec15 яванского макака с более высокой активностью по связыванию, чем у эталонного антитела, в то время как на фиг. 19 показано, что мышиные антитела к Siglec15 по настоящему изобретению специфично связываются с мышиным белком Siglec15 с немного более низкой активностью по связыванию по сравнению с эталонным антителом.
Пример 13. Блокирующая активность мышиных антител к Siglec15 в отношении связывания Siglec15-LRRC4C или Siglec15-референсное антитело
[00203] Мышиные антитела к Siglec15 по настоящему изобретению также тестировали в ELISA с блокированием лиганда и в ELISA с блокированием эталонного антитела в соответствии с протоколами, описанными выше. Результаты показаны на фиг. 20 и 21.
[00204] Из фиг. 20 можно увидеть, что мышиные антитела к Siglec15 по настоящему изобретению были способны блокировать связывание человеческого Siglec15 с человеческим LRRC4C, а блокирующая активность была немного выше, чем таковая у эталонного антитела.
[00205] На фиг. 21 показано, что мышиные антитела к Siglec15 по настоящему изобретению были способны блокировать связывание человеческого Siglec15 с эталонным антителом, что указывает на то, что мышиные антитела к Siglec15 по настоящему изобретению могут связываться с эпитопом, являющемся аналогичным тому, с которым связывалось референсное антитело.
Пример 14. Мышиные антитела к Siglec15 обеспечивали реверсию опосредованной Siglec15 Т-клеточной супрессии
[00206] Мышиные антитела к Siglec15 по настоящему изобретению дополнительно тестировали на формы биологической активности в отношении реверсии опосредованной Siglec15 супрессии Т-клеток в соответствии с протоколом, указанным в предыдущем примере, с модификацией, а также с протоколом, описанным ниже.
[00207] В тесте на реверсию супрессии Т-клеток мышиные антитела к Siglec15 по настоящему изобретению или контроли в концентрации 400 мкг/мл смешивали с человеческим белком Siglec15-Fc (SEQ ID NO: 19, 40 мкг/мл в среде RPMI-1640 с добавлением 10% FBS) при объемном соотношении 1:1. Результаты показаны на фиг. 22А и 22В.
[00208] Можно увидеть, что мышиные антитела к Siglec15 по настоящему изобретению были способны обеспечивать реверсию опосредованной Siglec15 супрессии CD8+ Т-клеток и CD4+ Т-клеток при более низкой или аналогичной ЕС50 по сравнению с эталонным антителом, и обработка антителами по настоящему изобретению в высоких дозах приводила к гораздо более высоким процентным значениям уровня пролиферации клеток.
[00209] Затем мышиные антитела по настоящему изобретению секвенировали, и идентификационные номера последовательностей вариабельных областей тяжелой/легкой цепи обобщены в таблице 1. Интересно отметить, что три антитела B2D7H7A3C1, B2G12H3E8 и В2Н2Н1А7 имели одинаковые последовательности CDR тяжелой/легкой цепи.
[00210] Хотя настоящее изобретение было описано выше в связи с одним или более вариантами осуществления, следует понимать, что настоящее изобретение не ограничивается этими вариантами осуществления, и предполагается, что описание охватывает все альтернативы, модификации и эквиваленты, которые могут быть включены в сущность и объем прилагаемой формулы изобретения. Все ссылочные материалы, приведенные в данном документе, дополнительно включены в качестве ссылки во всей своей полноте.
[00211] Последовательности, представленные в настоящей заявке, обобщены ниже.






[00212] Таким образом, с учетом приведенного подробного описания предпочтительных вариантов осуществления настоящего изобретения следует понимать, что настоящее изобретение, определенное в приведенных выше абзацах, не должно ограничиваться конкретными деталями, изложенными в приведенном выше описании, поскольку возможны многие его очевидные вариации без отступления от сущности или объема настоящего изобретения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Биосион, Инк.
<120> АНТИТЕЛА, СВЯЗЫВАЮЩИЕ SIGLEC15, И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ
<130> 55532 00027
<150> 63/000566
<151> 2020-03-27
<160> 43
<170> PatentIn версия 3.5
<210> 1
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR1 VH для A2A5C7E8, A2A5C7E8-2 и A2A5C7E8-3
<400> 1
Thr Tyr Trp Ile Ser
1 5
<210> 2
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR2 VH для A2A5C7E8, A2A5C7E8-2 и A2A5C7E8-3
<220>
<221> прочий_признак
<222> (6)..(6)
<223> Xaa может представлять собой Asp или Glu
<220>
<221> прочий_признак
<222> (16)..(16)
<223> Xaa может представлять собой Gln или Lys
<400> 2
Leu Ile Asp Pro Ser Xaa Ser Tyr Thr Asn Tyr Ser Pro Ser Phe Xaa
1 5 10 15
Gly
<210> 3
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR3 VH для A2A5C7E8, A2A5C7E8-2 и A2A5C7E8-3
<400> 3
Gly Gly Tyr Tyr Gly Ser Glu Glu Asp Tyr
1 5 10
<210> 4
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR1 VL для A2A5C7E8, A2A5C7E8-2 и A2A5C7E8-3
<400> 4
Arg Ala Ser Gln Ser Val Ser Ser Ser Arg Leu Ala
1 5 10
<210> 5
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR2 VL для A2A5C7E8, A2A5C7E8-2 и A2A5C7E8-3
<400> 5
Asp Ala Ser Ser Arg Ala Thr
1 5
<210> 6
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR3 VL для A2A5C7E8, A2A5C7E8-2 и A2A5C7E8-3
<400> 6
Gln Gln Tyr Gly Ser Ser Pro Arg Thr
1 5
<210> 7
<211> 119
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH для A2A5C7E8, A2A5C7E8-2 и A2A5C7E8-3
<220>
<221> прочий_признак
<222> (55)..(55)
<223> Xaa может представлять собой Asp или Glu
<220>
<221> прочий_признак
<222> (65)..(65)
<223> Xaa может представлять собой Gln или Lys
<400> 7
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Arg Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Thr Thr Tyr
20 25 30
Trp Ile Ser Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Leu Ile Asp Pro Ser Xaa Ser Tyr Thr Asn Tyr Ser Pro Ser Phe
50 55 60
Xaa Gly His Val Thr Ile Ser Thr Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Tyr Tyr Gly Ser Glu Glu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 8
<211> 108
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL для A2A5C7E8, A2A5C7E8-2 и A2A5C7E8-3
<400> 8
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Arg Leu Ala Trp Phe Gln Gln Lys Ser Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Phe Asp Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Phe Lys
100 105
<210> 9
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR1 VH для A1E10G7H9
<400> 9
Ser Ser Asn Trp Trp His
1 5
<210> 10
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR2 VH для A1E10G7H9
<400> 10
Glu Ile Tyr His Ser Gly Asn Thr Asn Tyr Lys Pro Ser Leu Lys Ser
1 5 10 15
<210> 11
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR3 VH для A1E10G7H9
<400> 11
Asp Glu Gly Asn Gly Trp Ser Asn Ala Phe Asp Ile
1 5 10
<210> 12
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR1 VL для A1E10G7H9
<400> 12
Arg Ala Ser Gln Ser Val Ser Ser Ser Tyr Leu Ala
1 5 10
<210> 13
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR2 VL для A1E10G7H9
<400> 13
Gly Ala Ser Gly Gly Ala Thr
1 5
<210> 14
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR3 VL для A1E10G7H9
<400> 14
Gln Gln Tyr Gly Ser Ser Pro Ile Thr
1 5
<210> 15
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH для A1E10G7H9
<400> 15
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gly
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly Gly Ser Ile Ser Ser Ser
20 25 30
Asn Trp Trp His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp
35 40 45
Ile Gly Glu Ile Tyr His Ser Gly Asn Thr Asn Tyr Lys Pro Ser Leu
50 55 60
Lys Ser Arg Val Thr Ile Ser Val Asp Lys Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Glu Gly Asn Gly Trp Ser Asn Ala Phe Asp Ile Trp Gly
100 105 110
Gln Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 16
<211> 108
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL для A1E10G7H9
<400> 16
Glu Asn Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Val Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Phe Leu
35 40 45
Ile Tyr Gly Ala Ser Gly Gly Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
Ile Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105
<210> 17
<211> 330
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Константная область тяжелой цепи
<400> 17
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 18
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Константная область легкой цепи
<400> 18
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 19
<211> 502
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Человеческий Siglec15-Fc
<400> 19
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Phe Val Arg Thr Lys Ile Asp Thr Thr Glu Asn Leu Leu
20 25 30
Asn Thr Glu Val His Ser Ser Pro Ala Gln Arg Trp Ser Met Gln Val
35 40 45
Pro Pro Glu Val Ser Ala Glu Ala Gly Asp Ala Ala Val Leu Pro Cys
50 55 60
Thr Phe Thr His Pro His Arg His Tyr Asp Gly Pro Leu Thr Ala Ile
65 70 75 80
Trp Arg Ala Gly Glu Pro Tyr Ala Gly Pro Gln Val Phe Arg Cys Ala
85 90 95
Ala Ala Arg Gly Ser Glu Leu Cys Gln Thr Ala Leu Ser Leu His Gly
100 105 110
Arg Phe Arg Leu Leu Gly Asn Pro Arg Arg Asn Asp Leu Ser Leu Arg
115 120 125
Val Glu Arg Leu Ala Leu Ala Asp Asp Arg Arg Tyr Phe Cys Arg Val
130 135 140
Glu Phe Ala Gly Asp Val His Asp Arg Tyr Glu Ser Arg His Gly Val
145 150 155 160
Arg Leu His Val Thr Ala Ala Pro Arg Ile Val Asn Ile Ser Val Leu
165 170 175
Pro Ser Pro Ala His Ala Phe Arg Ala Leu Cys Thr Ala Glu Gly Glu
180 185 190
Pro Pro Pro Ala Leu Ala Trp Ser Gly Pro Ala Leu Gly Asn Ser Leu
195 200 205
Ala Ala Val Arg Ser Pro Arg Glu Gly His Gly His Leu Val Thr Ala
210 215 220
Glu Leu Pro Ala Leu Thr His Asp Gly Arg Tyr Thr Cys Thr Ala Ala
225 230 235 240
Asn Ser Leu Gly Arg Ser Glu Ala Ser Val Tyr Leu Phe Arg Phe His
245 250 255
Gly Ala Ser Gly Ala Ser Thr Glu Asn Leu Tyr Phe Gln Gly Glu Pro
260 265 270
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
275 280 285
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
290 295 300
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
305 310 315 320
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
325 330 335
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
340 345 350
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
355 360 365
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
370 375 380
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
385 390 395 400
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
405 410 415
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
420 425 430
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
435 440 445
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
450 455 460
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
465 470 475 480
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
485 490 495
Ser Leu Ser Pro Gly Lys
500
<210> 20
<211> 271
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Человеческий Siglec15-his
<400> 20
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Phe Val Arg Thr Lys Ile Asp Thr Thr Glu Asn Leu Leu
20 25 30
Asn Thr Glu Val His Ser Ser Pro Ala Gln Arg Trp Ser Met Gln Val
35 40 45
Pro Pro Glu Val Ser Ala Glu Ala Gly Asp Ala Ala Val Leu Pro Cys
50 55 60
Thr Phe Thr His Pro His Arg His Tyr Asp Gly Pro Leu Thr Ala Ile
65 70 75 80
Trp Arg Ala Gly Glu Pro Tyr Ala Gly Pro Gln Val Phe Arg Cys Ala
85 90 95
Ala Ala Arg Gly Ser Glu Leu Cys Gln Thr Ala Leu Ser Leu His Gly
100 105 110
Arg Phe Arg Leu Leu Gly Asn Pro Arg Arg Asn Asp Leu Ser Leu Arg
115 120 125
Val Glu Arg Leu Ala Leu Ala Asp Asp Arg Arg Tyr Phe Cys Arg Val
130 135 140
Glu Phe Ala Gly Asp Val His Asp Arg Tyr Glu Ser Arg His Gly Val
145 150 155 160
Arg Leu His Val Thr Ala Ala Pro Arg Ile Val Asn Ile Ser Val Leu
165 170 175
Pro Ser Pro Ala His Ala Phe Arg Ala Leu Cys Thr Ala Glu Gly Glu
180 185 190
Pro Pro Pro Ala Leu Ala Trp Ser Gly Pro Ala Leu Gly Asn Ser Leu
195 200 205
Ala Ala Val Arg Ser Pro Arg Glu Gly His Gly His Leu Val Thr Ala
210 215 220
Glu Leu Pro Ala Leu Thr His Asp Gly Arg Tyr Thr Cys Thr Ala Ala
225 230 235 240
Asn Ser Leu Gly Arg Ser Glu Ala Ser Val Tyr Leu Phe Arg Phe His
245 250 255
Gly Ala Ser Gly Ala Ser Thr His His His His His His His His
260 265 270
<210> 21
<211> 271
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Siglec15-his яванского макака
<400> 21
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Phe Val Arg Thr Lys Ile Asp Thr Thr Glu Asn Leu Leu
20 25 30
Asn Thr Glu Val His Ser Ser Pro Ala Gln Arg Trp Ser Met Gln Val
35 40 45
Pro Ala Glu Val Ser Ala Ala Ala Gly Asp Ala Ala Val Leu Pro Cys
50 55 60
Thr Phe Thr His Pro His Arg His Tyr Asp Gly Pro Leu Thr Ala Ile
65 70 75 80
Trp Arg Ala Gly Glu Pro Tyr Ala Gly Pro Gln Val Phe Arg Cys Ala
85 90 95
Ala Ala Arg Gly Ser Glu Leu Cys Gln Thr Ala Leu Ser Leu His Gly
100 105 110
Arg Phe Arg Leu Leu Gly Asn Pro Arg Arg Asn Asp Leu Ser Leu Arg
115 120 125
Val Glu Arg Leu Ala Leu Ala Asp Asp Arg Arg Tyr Phe Cys Arg Val
130 135 140
Glu Phe Ala Gly Asp Val His Asp Arg Tyr Glu Ser Arg His Gly Val
145 150 155 160
Arg Leu His Val Thr Ala Ala Pro Arg Ile Ile Asn Ile Ser Val Leu
165 170 175
Pro Gly Pro Ala His Ala Phe Arg Ala Leu Cys Thr Ala Glu Gly Glu
180 185 190
Pro Pro Pro Ala Leu Ala Trp Ser Gly Pro Ala Leu Gly Asn Gly Ser
195 200 205
Ala Ala Val Pro Ser Ser Gly Gln Gly His Gly His Leu Val Thr Ala
210 215 220
Glu Leu Pro Ala Leu Asn His Asp Gly Arg Tyr Thr Cys Thr Ala Ala
225 230 235 240
Asn Ser Leu Gly Arg Ser Glu Ala Ser Val Tyr Leu Phe Arg Phe His
245 250 255
Gly Ala Ser Gly Ala Ser Thr His His His His His His His His
260 265 270
<210> 22
<211> 266
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Мышиный Siglec15-his
<400> 22
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Arg Arg Asp Ala Ser Gly Asp Leu Leu Asn Thr Glu Ala
20 25 30
His Ser Ala Pro Ala Gln Arg Trp Ser Met Gln Val Pro Ala Glu Val
35 40 45
Asn Ala Glu Ala Gly Asp Ala Ala Val Leu Pro Cys Thr Phe Thr His
50 55 60
Pro His Arg His Tyr Asp Gly Pro Leu Thr Ala Ile Trp Arg Ser Gly
65 70 75 80
Glu Pro Tyr Ala Gly Pro Gln Val Phe Arg Cys Thr Ala Ala Pro Gly
85 90 95
Ser Glu Leu Cys Gln Thr Ala Leu Ser Leu His Gly Arg Phe Arg Leu
100 105 110
Leu Gly Asn Pro Arg Arg Asn Asp Leu Ser Leu Arg Val Glu Arg Leu
115 120 125
Ala Leu Ala Asp Ser Gly Arg Tyr Phe Cys Arg Val Glu Phe Thr Gly
130 135 140
Asp Ala His Asp Arg Tyr Glu Ser Arg His Gly Val Arg Leu Arg Val
145 150 155 160
Thr Ala Ala Ala Pro Arg Ile Val Asn Ile Ser Val Leu Pro Gly Pro
165 170 175
Ala His Ala Phe Arg Ala Leu Cys Thr Ala Glu Gly Glu Pro Pro Pro
180 185 190
Ala Leu Ala Trp Ser Gly Pro Ala Pro Gly Asn Ser Ser Ala Ala Leu
195 200 205
Gln Gly Gln Gly His Gly Tyr Gln Val Thr Ala Glu Leu Pro Ala Leu
210 215 220
Thr Arg Asp Gly Arg Tyr Thr Cys Thr Ala Ala Asn Ser Leu Gly Arg
225 230 235 240
Ala Glu Ala Ser Val Tyr Leu Phe Arg Phe His Gly Ala Pro Gly Thr
245 250 255
Ser Thr His His His His His His His His
260 265
<210> 23
<211> 759
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Человеческий LRRC4C-Fc
<400> 23
Met Leu Asn Lys Met Thr Leu His Pro Gln Gln Ile Met Ile Gly Pro
1 5 10 15
Arg Phe Asn Arg Ala Leu Phe Asp Pro Leu Leu Val Val Leu Leu Ala
20 25 30
Leu Gln Leu Leu Val Val Ala Gly Leu Val Arg Ala Gln Thr Cys Pro
35 40 45
Ser Val Cys Ser Cys Ser Asn Gln Phe Ser Lys Val Ile Cys Val Arg
50 55 60
Lys Asn Leu Arg Glu Val Pro Asp Gly Ile Ser Thr Asn Thr Arg Leu
65 70 75 80
Leu Asn Leu His Glu Asn Gln Ile Gln Ile Ile Lys Val Asn Ser Phe
85 90 95
Lys His Leu Arg His Leu Glu Ile Leu Gln Leu Ser Arg Asn His Ile
100 105 110
Arg Thr Ile Glu Ile Gly Ala Phe Asn Gly Leu Ala Asn Leu Asn Thr
115 120 125
Leu Glu Leu Phe Asp Asn Arg Leu Thr Thr Ile Pro Asn Gly Ala Phe
130 135 140
Val Tyr Leu Ser Lys Leu Lys Glu Leu Trp Leu Arg Asn Asn Pro Ile
145 150 155 160
Glu Ser Ile Pro Ser Tyr Ala Phe Asn Arg Ile Pro Ser Leu Arg Arg
165 170 175
Leu Asp Leu Gly Glu Leu Lys Arg Leu Ser Tyr Ile Ser Glu Gly Ala
180 185 190
Phe Glu Gly Leu Ser Asn Leu Arg Tyr Leu Asn Leu Ala Met Cys Asn
195 200 205
Leu Arg Glu Ile Pro Asn Leu Thr Pro Leu Ile Lys Leu Asp Glu Leu
210 215 220
Asp Leu Ser Gly Asn His Leu Ser Ala Ile Arg Pro Gly Ser Phe Gln
225 230 235 240
Gly Leu Met His Leu Gln Lys Leu Trp Met Ile Gln Ser Gln Ile Gln
245 250 255
Val Ile Glu Arg Asn Ala Phe Asp Asn Leu Gln Ser Leu Val Glu Ile
260 265 270
Asn Leu Ala His Asn Asn Leu Thr Leu Leu Pro His Asp Leu Phe Thr
275 280 285
Pro Leu His His Leu Glu Arg Ile His Leu His His Asn Pro Trp Asn
290 295 300
Cys Asn Cys Asp Ile Leu Trp Leu Ser Trp Trp Ile Lys Asp Met Ala
305 310 315 320
Pro Ser Asn Thr Ala Cys Cys Ala Arg Cys Asn Thr Pro Pro Asn Leu
325 330 335
Lys Gly Arg Tyr Ile Gly Glu Leu Asp Gln Asn Tyr Phe Thr Cys Tyr
340 345 350
Ala Pro Val Ile Val Glu Pro Pro Ala Asp Leu Asn Val Thr Glu Gly
355 360 365
Met Ala Ala Glu Leu Lys Cys Arg Ala Ser Thr Ser Leu Thr Ser Val
370 375 380
Ser Trp Ile Thr Pro Asn Gly Thr Val Met Thr His Gly Ala Tyr Lys
385 390 395 400
Val Arg Ile Ala Val Leu Ser Asp Gly Thr Leu Asn Phe Thr Asn Val
405 410 415
Thr Val Gln Asp Thr Gly Met Tyr Thr Cys Met Val Ser Asn Ser Val
420 425 430
Gly Asn Thr Thr Ala Ser Ala Thr Leu Asn Val Thr Ala Ala Thr Thr
435 440 445
Thr Pro Phe Ser Tyr Phe Ser Thr Val Thr Val Glu Thr Met Glu Pro
450 455 460
Ser Gln Asp Glu Ala Arg Thr Thr Asp Asn Asn Val Gly Pro Thr Pro
465 470 475 480
Val Val Asp Trp Glu Thr Thr Asn Val Thr Thr Ser Leu Thr Pro Gln
485 490 495
Ser Thr Arg Ser Thr Glu Lys Thr Phe Thr Ile Pro Val Thr Asp Ile
500 505 510
Asn Ser Gly Ile Pro Gly Ile Asp Glu Val Met Lys Thr Thr Lys Glu
515 520 525
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
530 535 540
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
545 550 555 560
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
565 570 575
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
580 585 590
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
595 600 605
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
610 615 620
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
625 630 635 640
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
645 650 655
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys
660 665 670
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
675 680 685
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
690 695 700
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
705 710 715 720
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
725 730 735
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
740 745 750
Leu Ser Leu Ser Pro Gly Lys
755
<210> 24
<211> 452
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь Siglec15-ch5G9
<400> 24
Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Ile Thr Trp Val Ile Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Asp Ile Tyr Cys Gly Ser Asp Thr Met His Tyr Asn Glu Lys Phe
50 55 60
Lys Asn Lys Ala Thr Leu Thr Val Asp Thr Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Trp Asp Tyr Gly Ser Ser Tyr Asp Tyr Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 25
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь Siglec15-ch5G9
<400> 25
Asp Ile Lys Met Thr Gln Ser Pro Ser Ser Met Tyr Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Ile Asn Ser Tyr
20 25 30
Leu Ser Trp Phe Gln Gln Lys Pro Gly Lys Ser Pro Lys Thr Leu Ile
35 40 45
Tyr Arg Ala Asn Arg Leu Val Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Gln Asp Tyr Ser Leu Thr Ile Ser Ser Leu Glu Tyr
65 70 75 80
Glu Asp Met Gly Ile Tyr Tyr Cys Leu Gln Tyr Asp Glu Phe Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 26
<211> 503
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Человеческий Siglec15 с мышиным Fc
<400> 26
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Phe Val Arg Thr Lys Ile Asp Thr Thr Glu Asn Leu Leu
20 25 30
Asn Thr Glu Val His Ser Ser Pro Ala Gln Arg Trp Ser Met Gln Val
35 40 45
Pro Pro Glu Val Ser Ala Glu Ala Gly Asp Ala Ala Val Leu Pro Cys
50 55 60
Thr Phe Thr His Pro His Arg His Tyr Asp Gly Pro Leu Thr Ala Ile
65 70 75 80
Trp Arg Ala Gly Glu Pro Tyr Ala Gly Pro Gln Val Phe Arg Cys Ala
85 90 95
Ala Ala Arg Gly Ser Glu Leu Cys Gln Thr Ala Leu Ser Leu His Gly
100 105 110
Arg Phe Arg Leu Leu Gly Asn Pro Arg Arg Asn Asp Leu Ser Leu Arg
115 120 125
Val Glu Arg Leu Ala Leu Ala Asp Asp Arg Arg Tyr Phe Cys Arg Val
130 135 140
Glu Phe Ala Gly Asp Val His Asp Arg Tyr Glu Ser Arg His Gly Val
145 150 155 160
Arg Leu His Val Thr Ala Ala Pro Arg Ile Val Asn Ile Ser Val Leu
165 170 175
Pro Ser Pro Ala His Ala Phe Arg Ala Leu Cys Thr Ala Glu Gly Glu
180 185 190
Pro Pro Pro Ala Leu Ala Trp Ser Gly Pro Ala Leu Gly Asn Ser Leu
195 200 205
Ala Ala Val Arg Ser Pro Arg Glu Gly His Gly His Leu Val Thr Ala
210 215 220
Glu Leu Pro Ala Leu Thr His Asp Gly Arg Tyr Thr Cys Thr Ala Ala
225 230 235 240
Asn Ser Leu Gly Arg Ser Glu Ala Ser Val Tyr Leu Phe Arg Phe His
245 250 255
Gly Ala Ser Gly Ala Ser Thr Glu Asn Leu Tyr Phe Gln Gly Glu Pro
260 265 270
Arg Gly Pro Thr Ile Lys Pro Cys Pro Pro Cys Lys Cys Pro Ala Pro
275 280 285
Asn Leu Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Ile Lys
290 295 300
Asp Val Leu Met Ile Ser Leu Ser Pro Ile Val Thr Cys Val Val Val
305 310 315 320
Asp Val Ser Glu Asp Asp Pro Asp Val Gln Ile Ser Trp Phe Val Asn
325 330 335
Asn Val Glu Val His Thr Ala Gln Thr Gln Thr His Arg Glu Asp Tyr
340 345 350
Asn Ser Thr Leu Arg Val Val Ser Ala Leu Pro Ile Gln His Gln Asp
355 360 365
Trp Met Ser Gly Lys Glu Phe Lys Cys Lys Val Asn Asn Lys Asp Leu
370 375 380
Pro Ala Pro Ile Glu Arg Thr Ile Ser Lys Pro Lys Gly Ser Val Arg
385 390 395 400
Ala Pro Gln Val Tyr Val Leu Pro Pro Pro Glu Glu Glu Met Thr Lys
405 410 415
Lys Gln Val Thr Leu Thr Cys Met Val Thr Asp Phe Met Pro Glu Asp
420 425 430
Ile Tyr Val Glu Trp Thr Asn Asn Gly Lys Thr Glu Leu Asn Tyr Lys
435 440 445
Asn Thr Glu Pro Val Leu Asp Ser Asp Gly Ser Tyr Phe Met Tyr Ser
450 455 460
Lys Leu Arg Val Glu Lys Lys Asn Trp Val Glu Arg Asn Ser Tyr Ser
465 470 475 480
Cys Ser Val Val His Glu Gly Leu His Asn His His Thr Thr Lys Ser
485 490 495
Phe Ser Arg Thr Pro Gly Lys
500
<210> 27
<211> 357
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH для A2A5C7E8
<400> 27
gaagtgcagc tggtgcagtc cggagcagag gtgaaaaagc ccggggagtc tctgaggatc 60
tcctgtaagg gttctggata cagctttacc acctactgga tcagctgggt gcgccagatg 120
cccgggaaag gcctggagtg gatgggtttg attgatccta gtgactctta taccaactac 180
agtccgtcct tccaaggcca cgtcaccatc tcaactgaca agtccatcag cactgcctac 240
ctgcagtgga gcagcctgaa ggcctcggac accgccatgt attactgtgc gagagggggt 300
tactatggtt cggaagagga ctactggggc cagggaaccc tggtcaccgt ctcctca 357
<210> 28
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VL для A2A5C7E8, A2A5C7E8-2 и A2A5C7E8-3
<400> 28
gaaattgtgt tgacgcagtc tccaggcacc ctgtctttgt ctccagggga aagagccacc 60
ctctcctgca gggccagtca gagtgttagc agcagccggt tagcctggtt ccagcagaaa 120
tctggccagg ctcccagact cctcatcttt gatgcatcca gcagggccac tggcatccca 180
gacaggttca gtggcagtgg gtctgggaca gacttcactc tcaccatcag cagactggag 240
cctgaagatt ttgcagtgta ttactgtcag cagtatggta gctcacctcg gacgttcggc 300
caagggacca aggtggaatt caaa 324
<210> 29
<211> 363
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH для A1E10G7H9
<400> 29
caggtgcagc tgcaggagtc gggcccagga ctggtgaagc cttcggggac cctgtccctc 60
acctgcgctg tctctggtgg ctccatcagc agtagtaact ggtggcattg ggtccgccag 120
cccccaggga aggggctgga gtggattggg gaaatctatc atagtgggaa caccaactac 180
aaaccgtccc tcaagagtcg agtcaccata tcagtggaca agtccaagaa ccagttctcc 240
ctgaagctga gctctgtgac cgccgcggac acggccgtct attattgtgc gagagacgag 300
ggcaatggct ggtccaatgc ttttgatatc tggggccaag ggacaatggt caccgtctct 360
tca 363
<210> 30
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VL для A1E10G7H9
<400> 30
gaaaatgtgt tgacgcagtc tccaggcacc ctgtctctgt ctccagggga aagagtcacc 60
ctctcctgca gggccagtca gagtgttagc agcagctact tagcctggta tcagcagaaa 120
cctggccagg ctcccaggtt cctcatctat ggtgcatccg gcggggccac tggcatccca 180
gacaggttca gtggcagtgg gtctgggaca gacttcactc tcaccatcag cagactggag 240
cctgaagatt ttgcagtgta ttactgtcag cagtatggta gctcaccgat caccttcggc 300
caagggacac gactggagat taaa 324
<210> 31
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константная область тяжелой цепи
<400> 31
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gggtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 32
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Константная область легкой цепи
<400> 32
cgtacggtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagctcgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttag 324
<210> 33
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR1 VH для мышиных B2D7H7A3C1, B2G12H3E8 и B2H2H1A7
<400> 33
Asn Tyr Trp Ile Gly
1 5
<210> 34
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR2 VH для мышиных B2D7H7A3C1, B2G12H3E8 и B2H2H1H7
<400> 34
Asp Ile His Pro Gly Ile Asn Tyr Thr Asn Asn Asn Glu Lys Phe Arg
1 5 10 15
Gly
<210> 35
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR3 VH для мышиных B2D7H7A3C1, B2G12H3E8 и B2H2H1H7
<400> 35
Val Asp Tyr Asp Tyr Asp Gly Ser Tyr Ile Met Asp Tyr
1 5 10
<210> 36
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR1 VL для мышиных B2D7H7A3C1, B2G12H3E8 и B2H2H1H7
<400> 36
Arg Ala Ser Ser Ser Val Ser Tyr Met Tyr
1 5 10
<210> 37
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR2 VL для мышиных B2D7H7A3C1, B2G12H3E8 и B2H2H1H7
<400> 37
Ala Thr Ser Asn Leu Thr Ser
1 5
<210> 38
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR3 VL для мышиных B2D7H7A3C1, B2G12H3E8 и B2H2H1H7
<400> 38
Gln Gln Trp Asn Ser Lys Pro Trp Thr
1 5
<210> 39
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH для мышиного B2D7H7A3C1
<400> 39
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Thr
1 5 10 15
Ser Val Gln Met Ser Cys Lys Ala Ala Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Trp Ile Gly Trp Val Lys Gln Arg Pro Gly His Gly Leu Glu Trp Val
35 40 45
Gly Asp Ile His Pro Gly Ile Asn Tyr Thr Asn Asn Asn Glu Lys Phe
50 55 60
Arg Gly Lys Ala Thr Leu Thr Ala Asp Thr Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Arg Gly Leu Thr Ser Asp Asp Ser Ala Ile Tyr Tyr Cys
85 90 95
Ala Arg Val Asp Tyr Asp Tyr Asp Gly Ser Tyr Ile Met Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 40
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH для мышиного B2G12H3E8
<400> 40
Gln Val Gln Leu Lys Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Thr
1 5 10 15
Ser Val Gln Met Ser Cys Lys Ala Val Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Trp Ile Gly Trp Val Lys Gln Arg Pro Gly His Gly Leu Glu Trp Val
35 40 45
Gly Asp Ile His Pro Gly Ile Asn Tyr Thr Asn Asn Asn Glu Lys Phe
50 55 60
Arg Gly Lys Ala Thr Leu Thr Ala Asp Thr Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Arg Gly Leu Thr Ser Asp Asp Ser Ala Ile Tyr Tyr Cys
85 90 95
Ala Arg Val Asp Tyr Asp Tyr Asp Gly Ser Tyr Ile Met Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 41
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH для мышиного B2H2H1H7
<400> 41
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Thr
1 5 10 15
Ser Val Gln Met Ser Cys Lys Ala Ala Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Trp Ile Gly Trp Val Lys Gln Arg Pro Gly His Gly Leu Glu Trp Val
35 40 45
Gly Asp Ile His Pro Gly Ile Asn Tyr Thr Asn Asn Asn Glu Lys Phe
50 55 60
Arg Gly Lys Ala Thr Leu Thr Ala Asp Thr Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Arg Gly Leu Thr Ser Asp Asp Ser Ala Ile Tyr Tyr Cys
85 90 95
Ala Arg Val Asp Tyr Asp Tyr Asp Gly Ser Tyr Ile Met Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 42
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL для мышиных B2D7H7A3C1 и B2G12H3E8
<400> 42
Gln Ile Val Leu Ser Gln Ser Pro Ala Leu Leu Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Ala Thr Ser Asn Leu Thr Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Phe Cys Gln Gln Trp Asn Ser Lys Pro Trp Thr
85 90 95
Phe Gly Gly Gly Ser Lys Leu Glu Ile Arg
100 105
<210> 43
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL для мышиного B2H2H1H7
<400> 43
Gln Ile Ile Leu Ser Gln Ser Pro Ala Leu Leu Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Ala Thr Ser Asn Leu Thr Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Phe Cys Gln Gln Trp Asn Ser Lys Pro Trp Thr
85 90 95
Phe Gly Gly Gly Ser Arg Leu Glu Ile Arg
100 105
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ИНДУЦИРУЮЩИЙ ЦИТОТОКСИЧНОСТЬ ТЕРАПЕВТИЧЕСКИЙ АГЕНТ | 2015 |
|
RU2743464C2 |
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2837821C1 |
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2808030C2 |
| АНТИ-PCSK9 АНТИТЕЛО, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ИХ МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2016 |
|
RU2739208C2 |
| БИСПЕЦИФИЧЕСКИЙ БЕЛОК | 2019 |
|
RU2784486C1 |
| ПРОКОАГУЛЯНТНЫЕ АНТИТЕЛА | 2018 |
|
RU2810094C2 |
| ТЕРАПЕВТИЧЕСКИЕ АНТИТЕЛА ПРОТИВ ЛИГАНДА CD40 | 2018 |
|
RU2770209C2 |
| АМАТОКСИНОВЫЕ КОНЪЮГАТЫ АНТИТЕЛА С ЛЕКАРСТВЕННЫМ СРЕДСТВОМ И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2826004C2 |
| АНТИТЕЛО ПРОТИВ LILRB1 И ЕГО ПРИМЕНЕНИЯ | 2021 |
|
RU2813373C1 |
| МУЛЬТИВАЛЕНТНЫЕ FV-АНТИТЕЛА | 2016 |
|
RU2785766C2 |
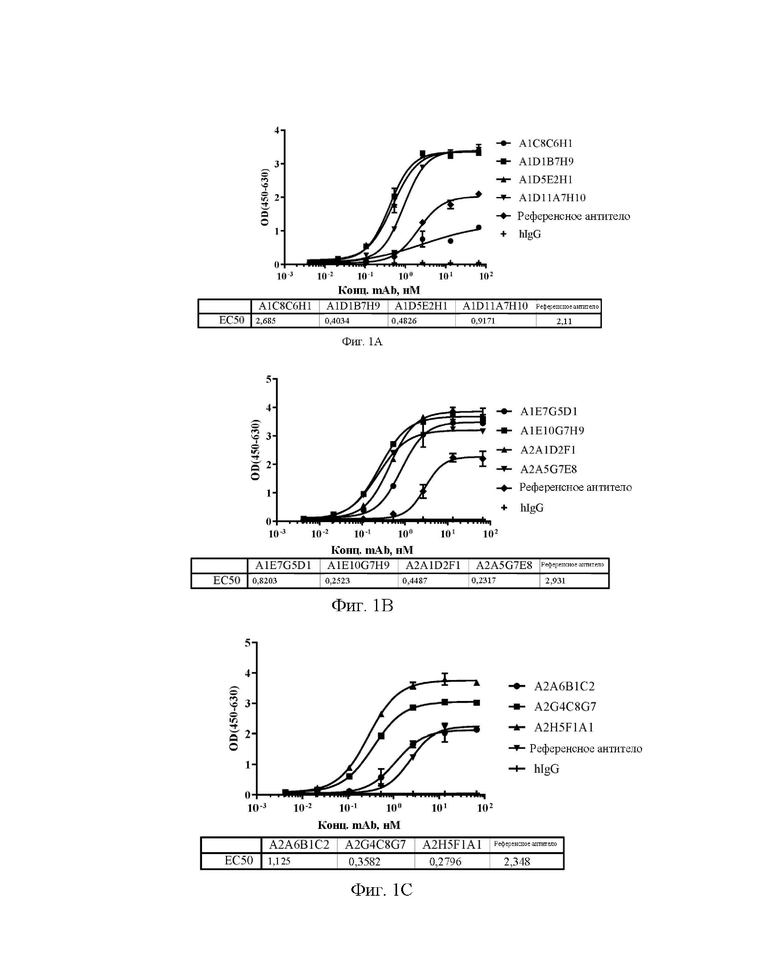
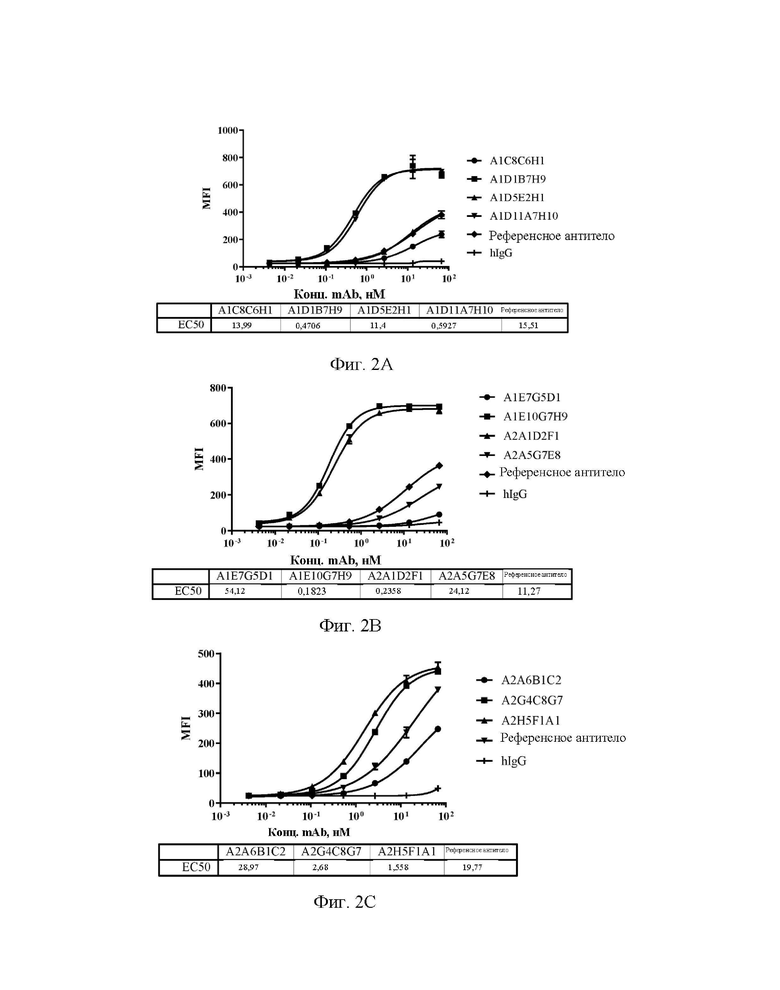
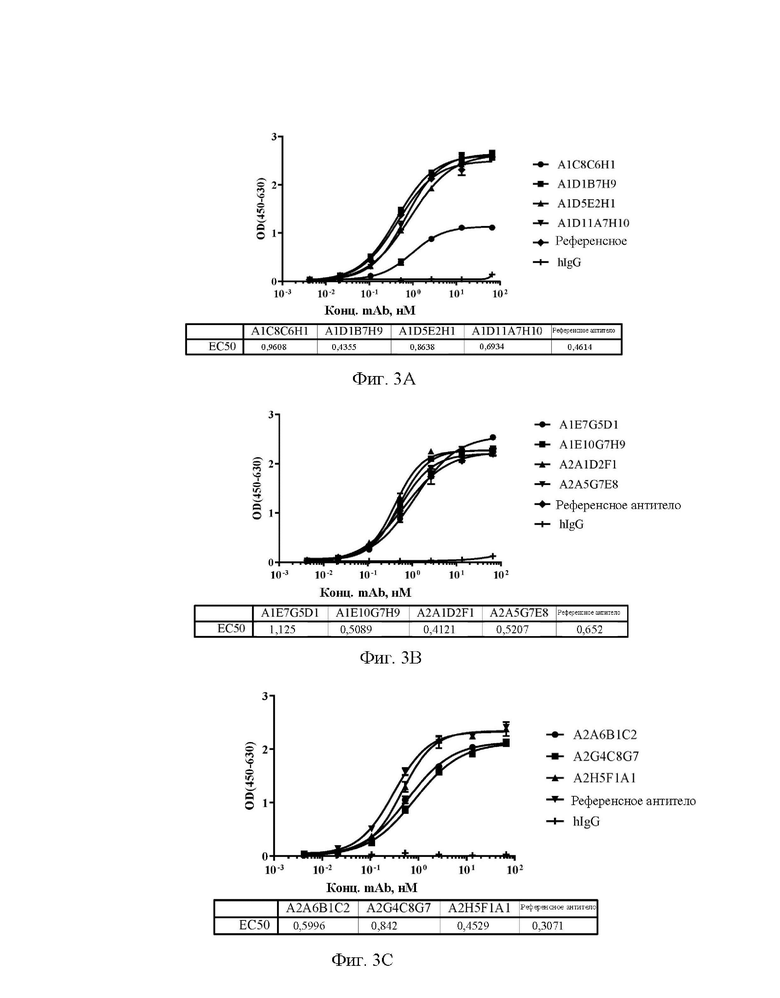
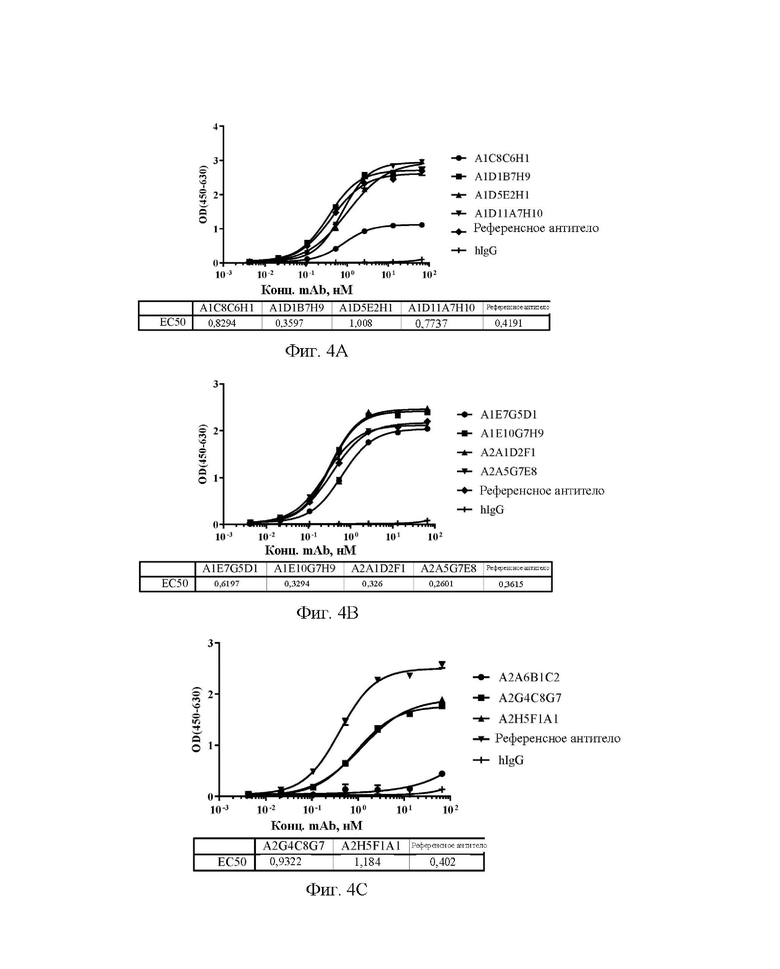
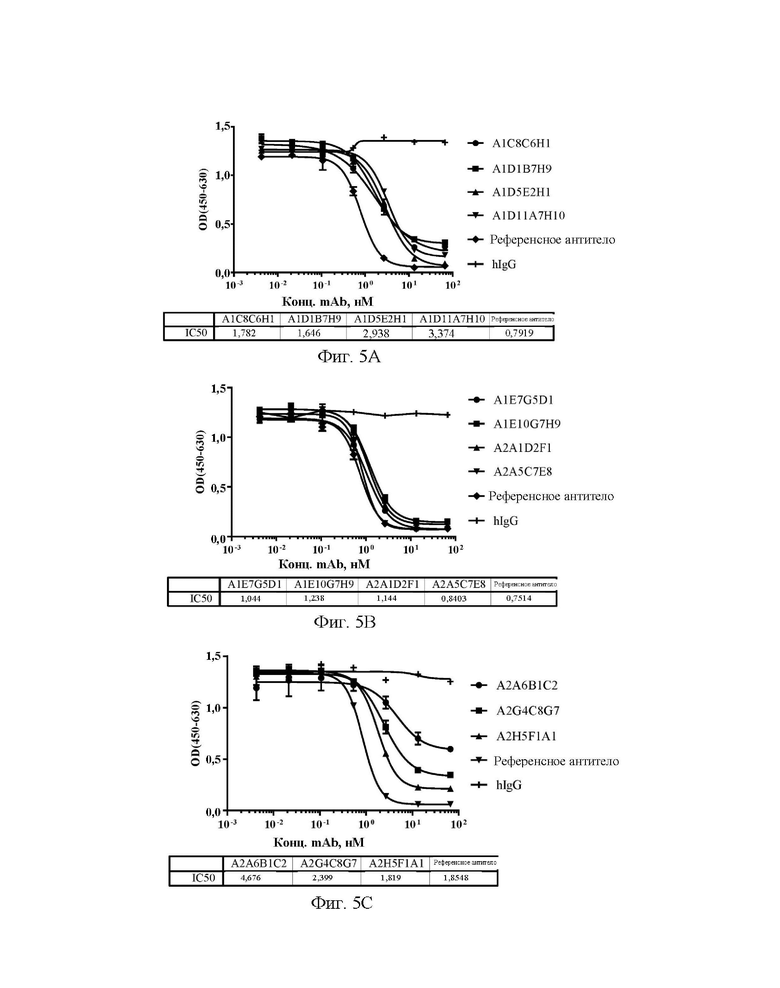
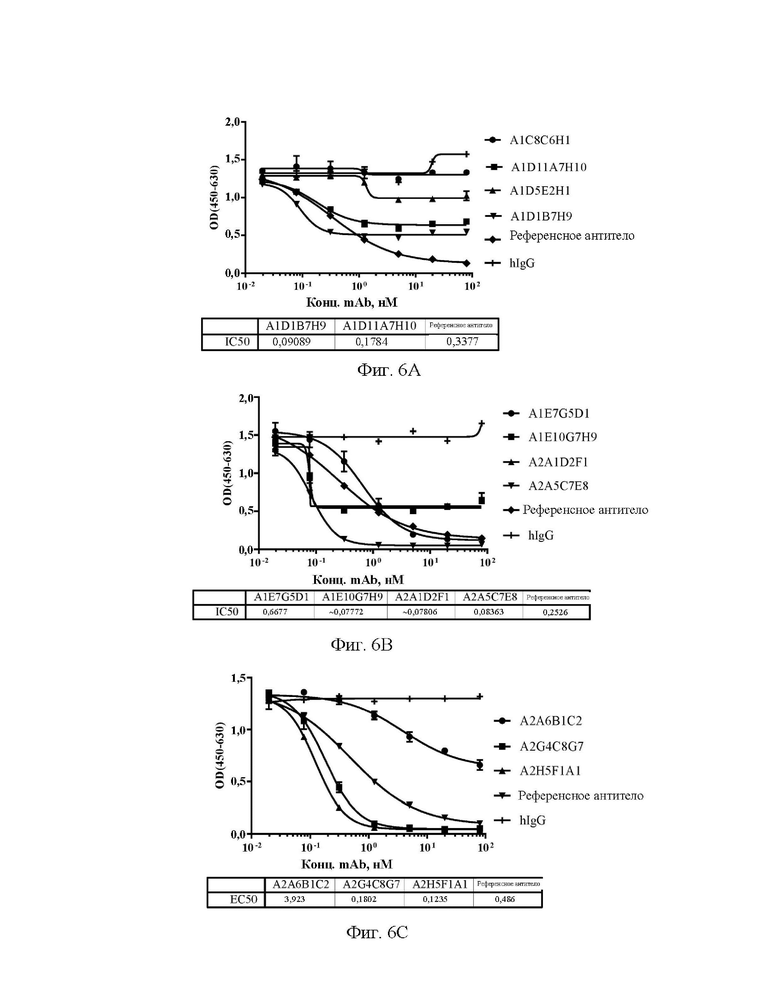
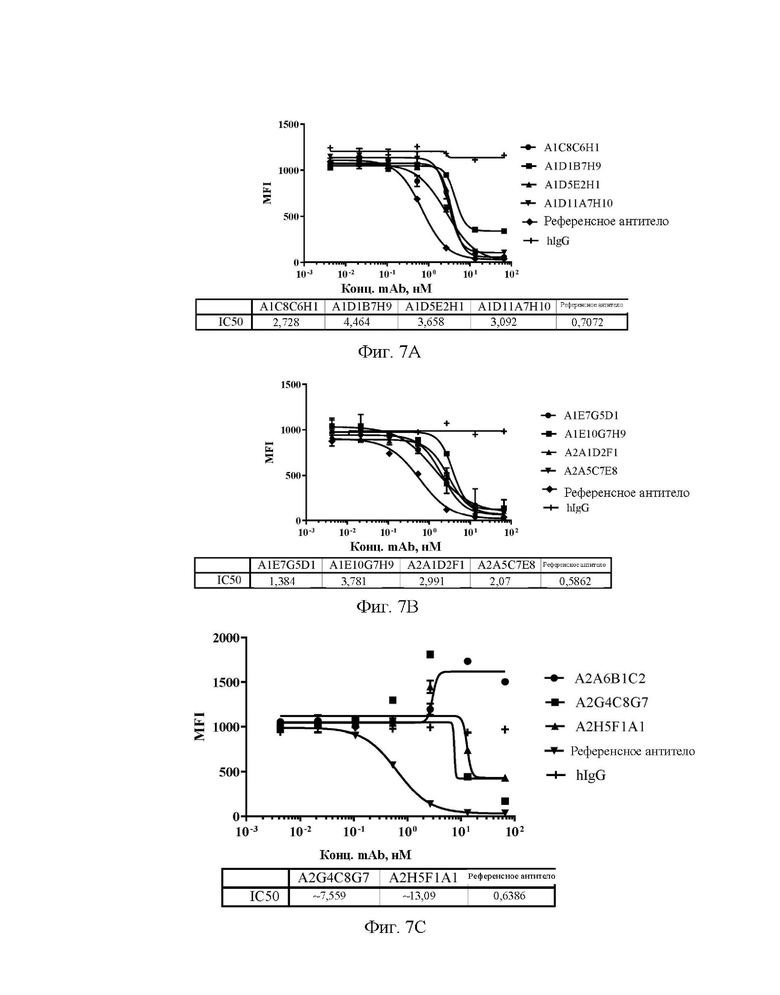
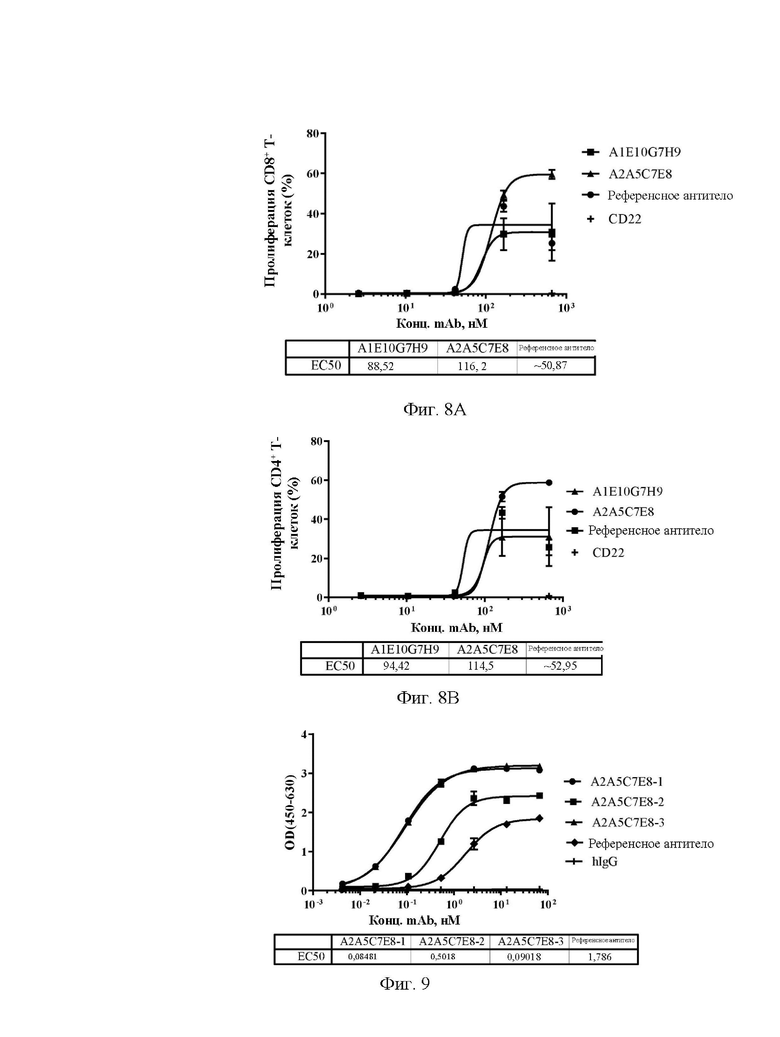
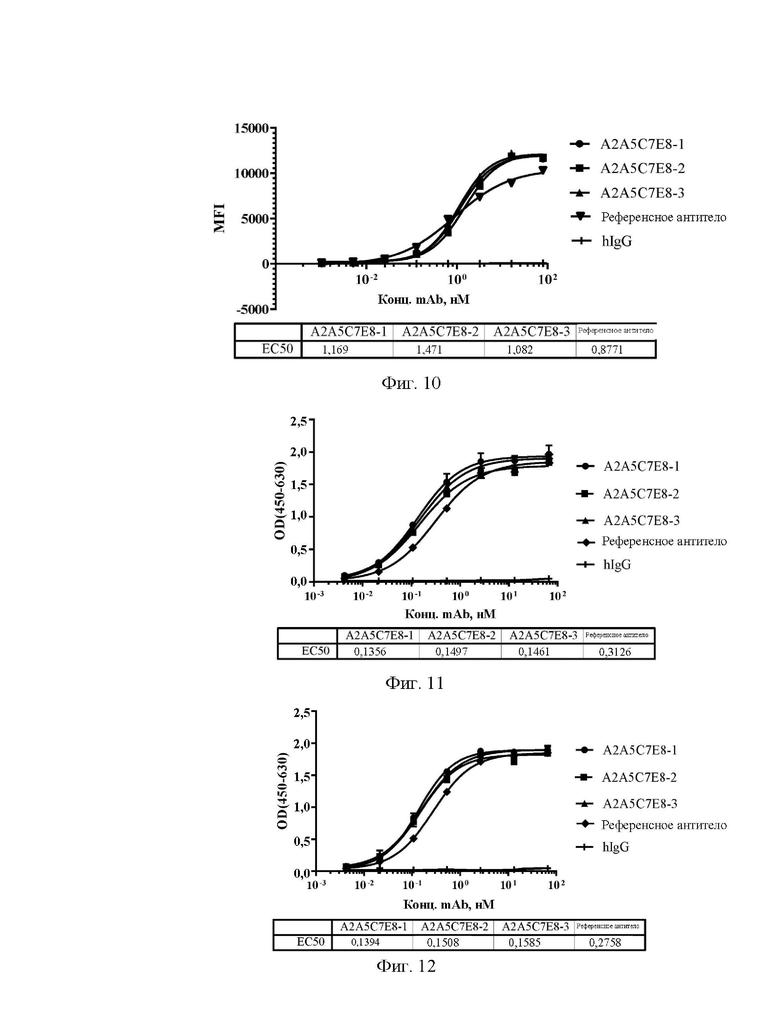
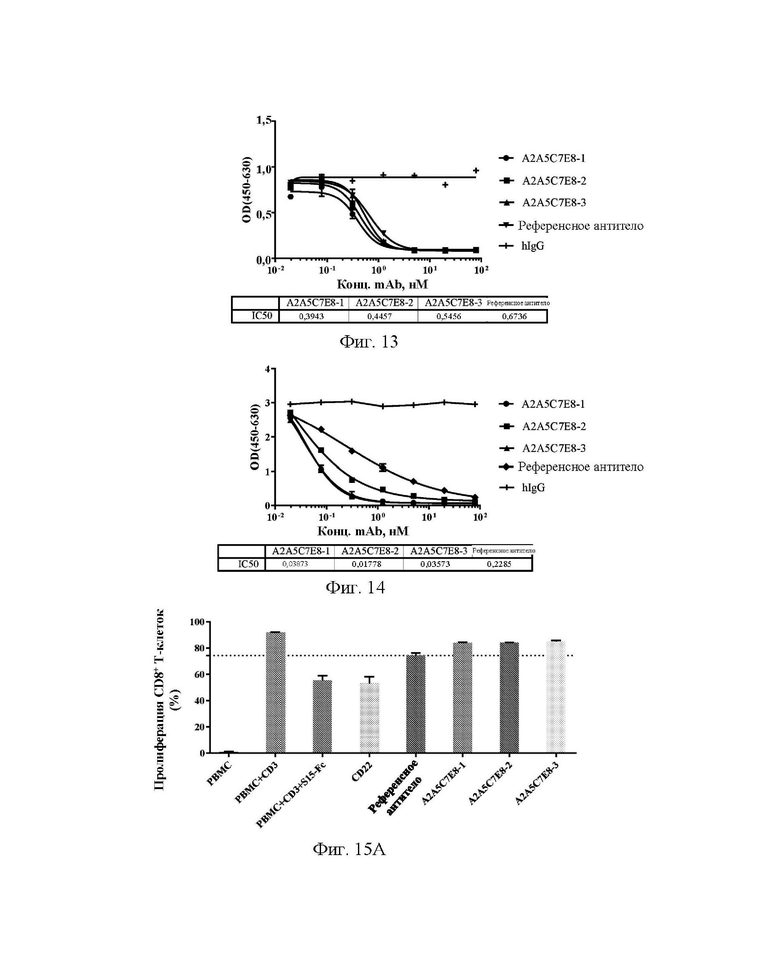
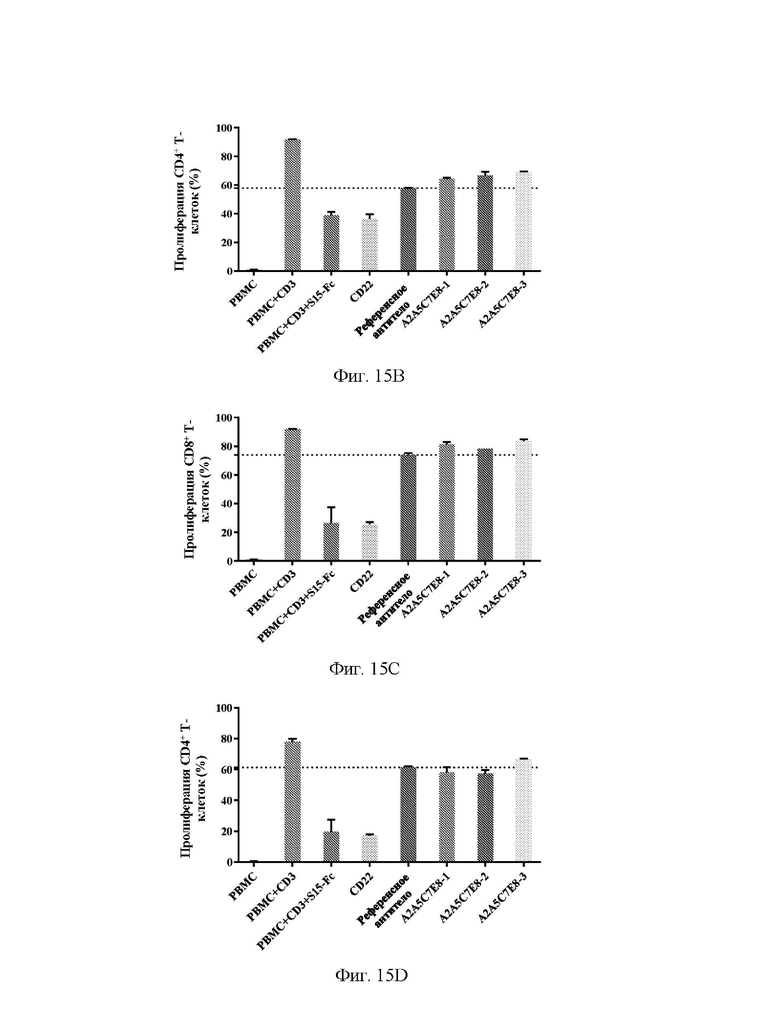
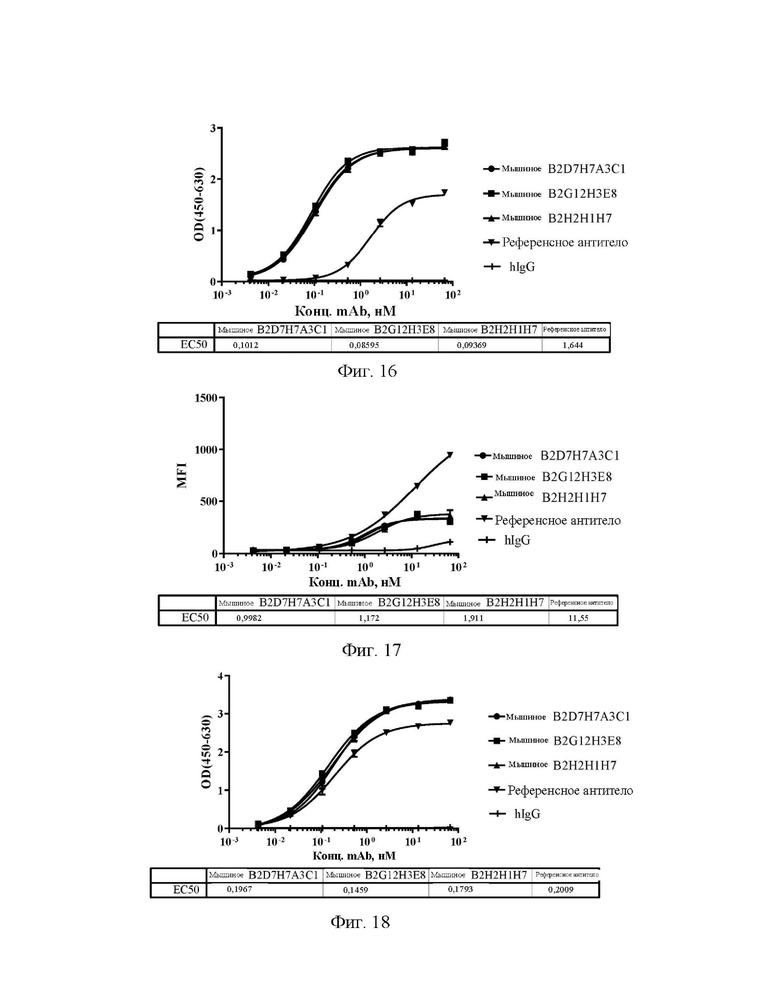
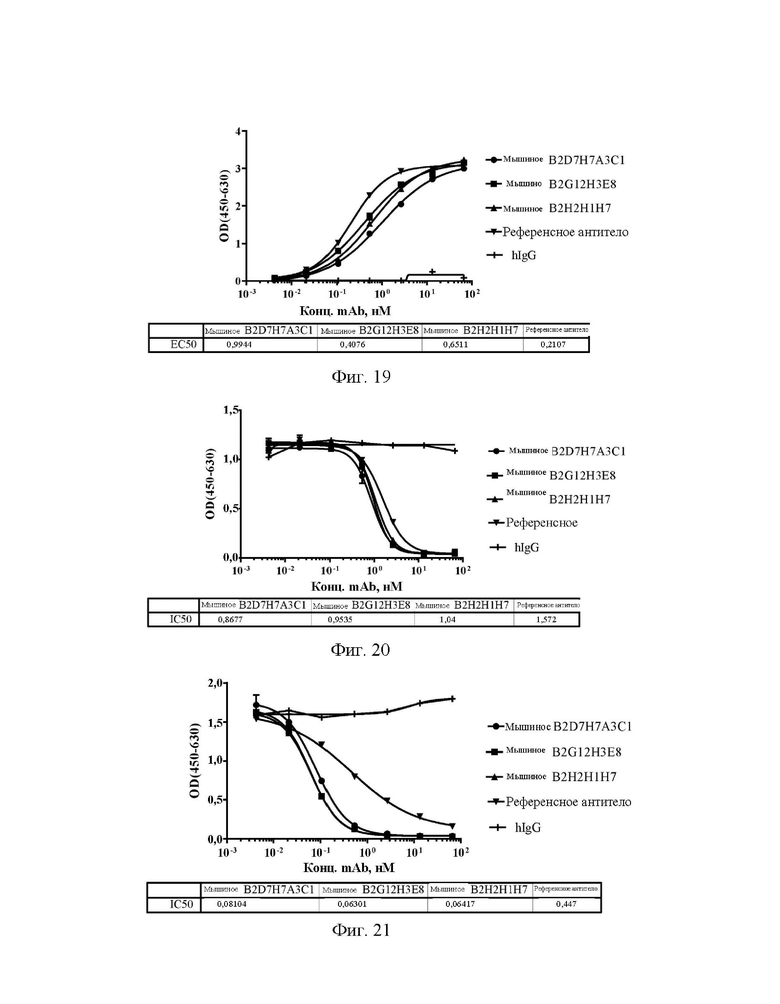

Группа изобретений относится к биотехнологии. Раскрыты выделенные моноклональные антитела, которые специфично связывают Siglec15 человека, или их антигенсвязывающие части и их применение. Также предложены полинуклеотид, кодирующий вариабельную область тяжелой цепи и вариабельную область легкой цепи антитела, фармацевтическая композиция, содержащая антитело или его антигенсвязывающую часть, а также способ лечения рака и способ подавления потери костной ткани у субъекта. Изобретение может использоваться для лечения рака у субъекта. 9 н. и 12 з.п. ф-лы, 22 ил., 6 табл., 14 пр.
1. Выделенное антитело, которое специфично связывается с Siglec15, содержащее вариабельную область тяжелой цепи, которая содержит участок CDR1 VH, участок CDR2 VH и участок CDR3 VH аминокислотной последовательности VH SEQ ID NO: 7, где X1=D и X2=K, X1=D и X2=Q или X1=E и X2=Q, и вариабельную область легкой цепи, которая содержит участок CDR1 VL, участок CDR2 VL и участок CDR3 VL аминокислотной последовательности VL SEQ ID NO: 8.
2. Выделенное антитело по п. 1, где участок CDR1 VH, участок CDR2 VH, участок CDR3 VH, участок CDR1 VL, участок CDR2 VL и участок CDR3 VL содержат аминокислотные последовательности, представленные в SEQ ID NO: 1, 2, 3, 4, 5 и 6, соответственно.
3. Выделенное антитело по п. 2, где участок CDR2 VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 2, где X1 представляет собой D и X2 представляет собой K.
4. Выделенное антитело по любому из пп. 1-3, где вариабельная область тяжелой цепи содержит аминокислотную последовательность, характеризующуюся по меньшей мере 85% идентичностью с SEQ ID NO: 7, и/или вариабельная область легкой цепи содержит аминокислотную последовательность, характеризующуюся по меньшей мере 85% идентичностью с SEQ ID NO: 8.
5. Выделенное антитело по любому из пп. 1-4, где вариабельная область тяжелой цепи содержит аминокислотную последовательность, представленную в SEQ ID NO: 7; и/или вариабельная область легкой цепи содержит аминокислотную последовательность, представленную в SEQ ID NO: 8.
6. Выделенное антитело по п. 5, где вариабельная область тяжелой цепи содержит аминокислотную последовательность, представленную в SEQ ID NO: 7, где X1 представляет собой D и X2 представляет собой K.
7. Выделенное антитело по любому из пп. 1-6, которое представляет собой изотип IgG1, IgG2 или IgG4.
8. Выделенное антитело по любому из пп. 1-7, содержащее константную область тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 17, связанную с вариабельной областью тяжелой цепи.
9. Выделенное антитело по любому из пп. 1-8, где антитело содержит константную область легкой цепи каппа человека или лямбда человека.
10. Выделенное антитело по любому из пп. 1-9, содержащее константную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 18, связанную с вариабельной областью легкой цепи.
11. Выделенное антитело, которое специфично связывается с Siglec15, содержащее вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 7, где X1=D и X2=K, X1=D и X2=Q или X1=E и X2=Q, и вариабельную область легкой цепи, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 8.
12. Выделенное антитело по п. 11, где вариабельная область тяжелой цепи содержит аминокислотную последовательность, представленную в SEQ ID NO: 7, где X1 представляет собой D и X2 представляет собой K.
13. Выделенное антитело по п. 11 или 12, содержащее константную область тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 17, связанную с вариабельной областью тяжелой цепи, и/или константную область легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 18, связанную с вариабельной областью легкой цепи.
14. Выделенное антитело, которое специфично связывается с Siglec15, содержащее вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 7, где X1 представляет собой D и X2 представляет собой K, и константную область тяжелой цепи, имеющую аминокислотную последовательность, представленную в SEQ ID NO: 17, и вариабельную область легкой цепи, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 8, и константную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 18.
15. Выделенное антитело по любому из пп. 1-14, где указанное антитело представляет собой мышиное, человеческое, химерное или гуманизированное антитело.
16. Полинуклеотид, кодирующий:
вариабельную область тяжелой цепи и вариабельную область легкой цепи антитела по любому из пп. 1-15.
17. Фармацевтическая композиция для лечения рака у субъекта, содержащая антитело по любому из пп. 1-15 или полинуклеотид по п. 16, и фармацевтически приемлемый носитель или вспомогательное вещество, где указанный рак представляет собой немелкоклеточный рак легкого, рак яичника, меланому, колоректальный рак, рак молочной железы, рак эндометрия или плоскоклеточную карциному.
18. Способ лечения рака у субъекта, включающий введение субъекту терапевтически эффективного количества антитела по любому из пп. 1-15, полинуклеотида по п. 16 или фармацевтической композиции по п. 17, где рак представляет собой немелкоклеточный рак легкого, рак яичника, меланому, колоректальный рак, рак молочной железы, рак эндометрия или плоскоклеточную карциному.
19. Способ подавления потери костной ткани у субъекта, включающий введение субъекту терапевтически эффективного количества антитела по любому из пп. 1-15, полинуклеотида по п. 16 или фармацевтической композиции по п. 17.
20. Применение антитела по любому из пп. 1-15, полинуклеотида по п. 16 или фармацевтической композиции по п. 17 для изготовления лекарственного средства для лечения рака у субъекта, нуждающегося в этом, где рак представляет собой немелкоклеточный рак легкого, рак яичника, меланому, колоректальный рак, рак молочной железы, рак эндометрия или плоскоклеточную карциному.
21. Применение антитела по любому из пп. 1-15, полинуклеотида по п. 16 или фармацевтической композиции по п. 17 для лечения рака у субъекта, нуждающегося в этом, где рак представляет собой немелкоклеточный рак легкого, рак яичника, меланому, колоректальный рак, рак молочной железы, рак эндометрия или плоскоклеточную карциному.
| CN 102803293 A, 28.11.2012 | |||
| CN 104321430 A, 28.01.2015 | |||
| CN 108430498 A, 21.08.2018 | |||
| АНТИТЕЛА ПРОТИВ СИГЛЕКА-15 ДЛЯ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЯ, СВЯЗАННОГО С ПОТЕРЕЙ КОСТНОЙ МАССЫ | 2010 |
|
RU2596392C2 |

