ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к новому варианту белка, обладающего активностью экспортера триптофана, продуцирующему L-триптофан микроорганизму, экспрессирующему указанный вариант белка, и способу получения L-триптофана с применением указанного микроорганизма.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Триптофан, который представляет собой незаменимую аминокислоту, широко применяли в качестве исходного сырья для пищевых добавок, лекарственных средств (например, инфузионных растворов), продуктов лечебного питания и т.д. В настоящее время для получения L-триптофана в основном применяют способ прямой ферментации с использованием микроорганизмов.
Ранее в качестве микроорганизма для получения L-триптофана в основном использовали отобранные штаммы, демонстрирующие устойчивость к аналогам в результате химической или физической мутации. Однако благодаря быстрому развитию технологии генетической рекомбинации и идентификации механизмов регуляции на молекулярном уровне в 1990-е годы в основном используют рекомбинантные штаммы с применением генно-инженерных технологий.
При этом экспрессия гена, кодирующего экспортер конкретной аминокислоты, способствует увеличению продуктивности микроорганизма в отношении соответствующей аминокислоты. Увеличение экспрессии гена, кодирующего экспортер L-лизина (lysE) в микроорганизме рода Corynebacterium улучшило продуктивность в отношении лизина (WO 9723597А2). Кроме того, усиление экспрессии гена rhtC у Е. coli улучшило устойчивость к L-треонину и одновременно улучшило продуктивность в отношении L-гомосерина, L-треонина и L-лейцина (ЕР 1013765 А1). Кроме того, в патенте ЕР 1016710 В1 раскрыто, что продуктивность в отношении L-глутаминовой кислоты, L-лизина, L-треонина, L-аланина, L-гистидина, L-пролина, L-аргинина, L-валина и L-изолейцина улучшалась посредством усиления экспрессии генов yahN, yeaS, yfiK и yggA, функция которых у Е. coli еще не установлена.
Однако до настоящего времени не сообщалось о белках-экспортерах, демонстрирующих специфичность к L-триптофану. Хотя у Е. coli известен ген yddG, он демонстрирует более высокую специфичность в отношении L-фенилаланина, чем в отношении L-триптофана (FEMS Microbiol Lett 275 (2007), 312-318). Кроме того, у микроорганизма рода Corynebacterium, который в основном используют в качестве штамма-продуцента для получения L-аминокислот посредством ферментации, ранее не сообщалось о генах экспортеров L-триптофана или ароматической аминокислоты (J Ind Microbiol Biotechnol. 2015 May; 42(5): 787-797).
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Техническая задача
Авторам настоящего изобретения удалось экспрессировать новый белок-экспортер триптофана, обладающий специфичностью в отношении L-триптофана, в продуцирующем L-триптофан микроорганизме и, в результате, обнаружить, что произошло существенное улучшение продукции L-триптофана в количественном выражении. Кроме того, в результате введения мутаций для дальнейшего улучшения экспортирующей способности соответствующего мембранного белка они подтвердили, что количество продуцируемого L-триптофана существенно увеличилось. Тем самым было выполнено настоящее изобретение.
Техническое решение
Задачей настоящего изобретения является предоставление варианта белка, обладающего активностью экспортера L-триптофана, в котором по меньшей мере одна аминокислота, выбранная из аминокислот, соответствующих аминокислотам в положениях 79-83 аминокислотной последовательности SEQ ID NO: 1, заменена гидрофобной или алифатической аминокислотой.
Другой задачей настоящего изобретения является предоставление полинуклеотида, кодирующего указанный вариант, и вектора, включающего указанный полинуклеотид.
Еще одной задачей настоящего изобретения является предоставление микроорганизма, продуцирующего L-триптофан, экспрессирующего указанный вариант белка.
Еще одной задачей настоящего изобретения является предоставление способа получения L-триптофана, включающего культивирование указанного микроорганизма в среде.
Еще одной задачей настоящего изобретения является предоставление применения указанного варианта белка для увеличения продукции L-триптофана.
Полезные эффекты изобретения
Авторы настоящего изобретения обнаружили ген нового экспортера, обладающего специфичностью в отношении L-триптофана, и предприняли попытку экспрессировать указанный ген в микроорганизме, продуцирующем L-триптофан. В результате они подтвердили, что количество продуцируемого указанным микроорганизмом L-триптофана может существенно увеличиться по сравнению с его родительским штаммом, в котором указанный ген не экспрессируется, а также обнаружили вариант белка, кодируемый указанным геном, который позволяет более значительно увеличить количество продуцируемого микроорганизмом L-триптофана, тем самым подтвердив, что L-триптофан может в нем эффективно продуцироваться.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На Фиг. 1 показана внутриклеточная концентрация триптофана в СА04-8352 и СА04-8405, которые представляют собой модифицированные штаммы Corynebacterium glutamicum, по мере потребления глюкозы.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Далее настоящее изобретение будет описано более подробно. При этом каждое описание и воплощение, раскрытое в данном документе, также может быть применено здесь для описания других описаний и воплощений. Иными словами, все комбинации различных элементов, описанных в данном документе, входят в объем настоящего изобретения. Более того, объем настоящего изобретения не ограничивается конкретным описанием, приведенным ниже.
В одном аспекте настоящего изобретения предложен вариант белка, обладающего способностью экспортировать L-триптофан, где указанный вариант белка включает по меньшей мере одну мутацию в аминокислотной последовательности SEQ ID NO: 1.
Мутация может включать такие, в которых по меньшей мере одна аминокислота, выбранная из аминокислот от 79-й до 83-й от N-конца аминокислотной последовательности SEQ ID NO: 1, заменена на другую аминокислоту.
Вариант белка может представлять собой вариант белка, обладающего активностью экспортера L-триптофана, в котором по меньшей мере одна аминокислота, выбранная из аминокислот в положениях 79-83 аминокислотной последовательности SEQ ID NO: 1, заменена на другую аминокислоту. Конкретно, вариант белка может представлять собой вариант белка, обладающего активностью экспортера L-триптофана, в котором по меньшей мере одна аминокислота, выбранная из аминокислот в положениях 79-83 аминокислотной последовательности SEQ ID NO: 1, заменена гидрофобной аминокислотой или алифатической аминокислотой.
В данном документе термин «L-триптофан», обозначающий одну из α-аминокислот, относится к незаменимой аминокислоте, которая не синтезируется in vivo и представляет собой ароматическую L-аминокислоту, имеющую химическую формулу C11H12N2O2.
В данном документе «белок, обладающий способностью экспортировать L-триптофан» или «белок, обладающий активностью экспортера L-триптофана» относится к мембранному белку, который обладает способностью специфически экспортировать L-триптофан из клетки.
Белок, обладающий активностью экспортера L-триптофана, может представлять собой белок, обладающий активностью экспортера L-триптофана, имеющий происхождение из Herbaspirillum rhizosphaerae. Например, белок, обладающий активностью экспортера L-триптофана, имеющий происхождение из Herbaspirillum rhizosphaerae, может представлять собой белок, включающий аминокислотную последовательность SEQ ID NO: 1. Термин «белок, включающий аминокислотную последовательность SEQ ID NO: 1», может использоваться взаимозаменяемо с терминами «белок, имеющий аминокислотную последовательность SEQ ID NO: 1» и «белок, состоящий из аминокислотной последовательности SEQ ID NO: 1».
В частности, «Herbaspirillum rhizosphaerae» представляет собой грамотрицательную бактерию, относящуюся к роду Herbaspirillum. В Корее штамм Herbaspirillum rhizosphaerae, такой как выделенный на острове Ulleung и т.д., может быть выделен из ризосферы в почве.
Кроме того, несмотря на то, что белок по настоящему изобретению, обладающий активностью экспортера L-триптофана, определен как белок, включающий аминокислотную последовательность SEQ ID NO: 1, он не исключает добавления некодирующей последовательности в 5'- или 3'-направлении относительно аминокислотной последовательности SEQ ID NO: 1, мутации, которая может иметь происхождение естественным образом, или молчащей мутации, и для специалиста в данной области техники очевидно, что любой белок, который обладает активностью, идентичной или соответствующей активности белка, включающего аминокислотную последовательность SEQ ID NO: 1, принадлежит к белкам по настоящему изобретению, обладающим активностью экспортера L-триптофана. В частности, например, белок по настоящему изобретению, обладающий активностью экспортера L-триптофана, может представлять собой белок, состоящий из аминокислотной последовательности SEQ ID NO: 1, или белок, состоящий из аминокислотной последовательности, гомологичной или идентичной аминокислотной последовательности SEQ ID NO: 1 на 80%, 90%, 95%, 97% или более. Кроме того, очевидно, что любой белок, имеющий аминокислотную последовательность с делецией, модификацией, заменой или вставкой в части последовательности, также принадлежит к белкам, входящим в объем настоящего изобретения, подлежащим мутации, при условии, что белок имеет аминокислотную последовательность с любой из указанных выше гомологии или идентичностей, и проявляет эффект, соответствующий эффекту указанного выше белка.
Таким образом, даже если в настоящем изобретении описан «белок или полипептид, имеющий аминокислотную последовательность с конкретным SEQ ID NO» или «белок или полипептид, состоящий из аминокислотной последовательности с конкретным SEQ ID NO», очевидно, что любой белок, имеющий аминокислотную последовательность с делецией, модификацией, заменой или вставкой в части последовательности, также может применяться в настоящем изобретении, при условии, что указанный белок обладает активностью, идентичной или соответствующей активности полипептида, состоящего из аминокислотной последовательности с соответствующим SEQ ID NO. Например, очевидно, что «полипептид, состоящий из аминокислотной последовательности с SEQ ID NO: 1» также может принадлежать к «полипептиду, содержащему аминокислотную последовательность с SEQ ID NO: 1», при условии, что полипептид обладает идентичной или соответствующей активностью.
Вариант белка, обладающего способностью экспортировать L-триптофан, предложенный в настоящем описании, может относиться к варианту описанных выше белков, обладающих способностью экспортировать L-триптофан, в котором осуществлена замена аминокислоты в определенном положении, и итоговая способность экспортировать L-триптофан превосходит 100% по сравнению с активностью белка до мутации.
В данном описании термин «вариант» относится к белку, в котором по меньшей мере одна аминокислота в приведенной последовательности подвергнута консервативной замене и/или модификации, при этом функции или свойства белка сохранены. Вариант отличается от идентифицированной последовательности несколькими аминокислотными заменами, делециями или вставками. Такой вариант может быть идентифицирован путем модификации одной или более аминокислот в аминокислотной последовательности вышеуказанного белка и оценки свойств вышеуказанного модифицированного белка. Таким образом, активность варианта может быть повышена, оставлена без изменений или ослаблена по сравнению с таковой нативного белка. Кроме того, некоторые варианты могут включать варианты, в которых удалена одна или более чем одна часть (например, N-концевая лидерная последовательность или трансмембранный домен). Другие варианты могут включать варианты, в которых была удалена часть N-конца и/или С-конца зрелого белка. Термин «вариант» также может быть использован взаимозаменяемо с «модификацией», «модифицированным белком», «модифицированным полипептидом», «мутантом», «мутеином», «дивергентом», «вариантом» и т.д., однако используемый термин не ограничивается перечисленными и может быть использован любой термин, при условии, что он используется в смысле «мутированный». В настоящем описании вариант может относиться к вариантам, в которых активность мутированного белка повышена по сравнению с таковой белка дикого типа или немодифицированного белка, однако вариант ими не ограничивается.
В данном описании термин «консервативная замена» относится к замене одной аминокислоты на другую аминокислоту, имеющую схожую структуру и/или химические свойства. Например, вариант может иметь одну или более чем одну консервативную замену, при этом сохраняя одну или более чем одну биологическую активность. Такие аминокислотные замены, как правило, могут осуществляться на основании сходства в полярности, заряде, растворимости, гидрофобности, гидрофильности и/или амфипатической природы остатков. Например, среди аминокислот, несущих электрический заряд, положительно заряженные (основные) аминокислоты включают аргинин, лизин и гистидин; а отрицательно заряженные (кислые) аминокислоты включают глутаминовую кислоту и аспарагиновую кислоту. Среди незаряженных аминокислот неполярные аминокислоты включают глицин, аланин, валин, лейцин, изолейцин, метионин, фенилаланин, триптофан и пролин; полярные или гидрофобные аминокислоты включают серин, треонин, цистеин, тирозин, аспарагин и глутамин, а ароматические аминокислоты включают фенилаланин, триптофан и тирозин.
Кроме того, вариант может включать делецию или вставку аминокислот, которые оказывают минимальное влияние на свойства и вторичную структуру полипептида. Например, полипептид может быть конъюгирован с сигнальной (или лидерной) последовательностью на N-конце белка, которая направляет котрансляционный или посттрансляционный транспорт белка. Кроме того, полипептид может также быть конъюгирован с другой последовательностью или линкером для идентификации, очистки или синтеза полипептида.
«Замена на другую аминокислоту» не ограничена, при условии, что заменяющая аминокислота отличается от аминокислоты до замены. Так, «замена на другую аминокислоту» не ограничена, при условии, что 79-я от N-конца аминокислота аминокислотной последовательности SEQ ID NO: 1 (то есть лейцин) заменена на аминокислоту, отличную от лейцина; 80-я от N-конца аминокислота аминокислотной последовательности SEQ ID NO: 1 (то есть серин) заменена на аминокислоту, отличную от серина; 81-я от N-конца аминокислота аминокислотной последовательности SEQ ID NO: 1 (то есть лейцин) заменена на аминокислоту, отличную от лейцина; 82-я от N-конца аминокислота аминокислотной последовательности SEQ ID NO: 1 (то есть серин) заменена на аминокислоту, отличную от серина; 83-я от N-конца аминокислота аминокислотной последовательности SEQ ID NO: 1 (то есть изолейцин) заменена на аминокислоту, отличную от изолейцина. При этом, когда в данном документе используется выражение «конкретная аминокислота заменена», очевидно, что аминокислота заменена на аминокислоту, отличную от аминокислоты до замены, даже если напрямую не указано, что аминокислота была заменена другой аминокислотой.
Альтернативно, вариант белка может представлять собой вариант, в котором по меньшей мере одна аминокислота из аминокислот в положениях 79-83 аминокислотной последовательности SEQ ID NO: 1 заменена на аминокислоту, отличную от аминокислоты до замены, за исключением кислых аминокислот и основных аминокислот. Альтернативно, вариант белка может представлять собой вариант, имеющий незаряженную аминокислоту, в котором заменяющая аминокислота отличается от аминокислоты до замены, но вариант белка не ограничивается этим.
Альтернативно, вариант белка может представлять собой вариант, в котором по меньшей мере одна аминокислота из аминокислот в положениях 79-83 аминокислотной последовательности SEQ ID NO: 1 заменена на аминокислоту, выбранную из гидрофобной аминокислоты и алифатической аминокислоты, отличную от аминокислоты до замены. Конкретно, вариант белка может представлять собой вариант, в котором по меньшей мере одна аминокислота из аминокислот в положениях 79-83 аминокислотной последовательности SEQ ID NO: 1 заменена любой аминокислотой, выбранной из гидрофобной (неполярной) аминокислоты и алифатической аминокислоты. Алифатическая аминокислота может быть, например, аминокислотой, выбранной из группы, состоящей из глицина, аланина, валина, лейцина и изолейцина, но алифатическая аминокислота не ограничена этим. Гидрофобная (неполярная) аминокислота может быть, например, аминокислотой, выбранной из группы, состоящей из глицина, метионина, аланина, валина, лейцина, изолейцина, пролина, фенилаланина, тирозина и триптофана, но гидрофобная (неполярная) аминокислота не ограничена этим.
Альтернативно, вариант белка может представлять собой вариант, в котором по меньшей мере одна из аминокислот в положениях 79-83 аминокислотной последовательности SEQ ID NO: 1 заменена на аминокислоту, отличную от аминокислоты до замены, выбранную из аминокислот малого размера, но вариант белка не ограничен этим.
В данном документе термин «аминокислота малого размера» включает аминокислоты с относительно малым размером, выбранные из 20 аминокислот (то есть глицин, аланин, серин, треонин, цистеин, валин, лейцин, изолейцин, пролин и аспарагин) и, в частности, может относиться к глицину, аланину, серину, треонину, цистеину, валину, лейцину, изолейцину и пролину, но аминокислоты малого размера не ограничены перечисленными; более конкретно, аминокислота малого размера может относиться к глицину, аланину, валину, лейцину, изолейцину, серину и треонину, но аминокислоты малого размера не ограничены перечисленными.
Альтернативно, вариант белка может представлять собой вариант, в котором по меньшей мере одна аминокислота из аминокислот в положениях 79-83 аминокислотной последовательности SEQ ID NO: 1 заменена на аминокислоту, выбранную из группы, состоящей из аланина, валина, лейцина и изолейцина, однако вариант белка не ограничен этим.
В частности, замена на другую аминокислоту в варианте белка может представлять собой одну или более чем одну из замен, выбранных из: замены 79-й аминокислоты в аминокислотной последовательности SEQ ID NO: 1 (то есть лейцина) на аланин, валин или изолейцин; замены 80-й аминокислоты в аминокислотной последовательности SEQ ID NO: 1 (то есть серина) на аланин, валин, лейцин или изолейцин; замены 81-й аминокислоты в аминокислотной последовательности SEQ ID NO: 1 (то есть лейцина) на аланин, валин или изолейцин; замены 82-й аминокислоты в аминокислотной последовательности SEQ ID NO: 1 (то есть серина) на аланин, валин, лейцин или изолейцин; и замены 83-й аминокислоты в аминокислотной последовательности SEQ ID NO: 1 (то есть изолейцина) на аланин, валин или лейцин, но замены не ограничиваются перечисленными.
Варианты белка по настоящему изобретению, как таковые, обладают улучшенной способностью экспортировать L-триптофан по сравнению с белком до мутации.
Очевидно, что варианты белка по настоящему изобретению, в которых по меньшей мере одна аминокислота из аминокислот в положениях с 79 по 83 от N-конца SEQ ID NO: 1 заменена на другую аминокислоту, включают варианты белка, в которых аминокислоты в положениях, соответствующих положениям с 79 по 83, заменены на другую аминокислоту.
Специалист в области техники способен установить, является ли аминокислота в любом положении в любой последовательности аминокислотой, соответствующей аминокислотам в положениях с 79 по 83 SEQ ID NO: 1, путем сравнения любой последовательности с SEQ ID NO: 1 настоящего изобретения, применив способ выравнивания, подтверждения гомологии или идентичности, известный в области техники.
Таким образом, если здесь не указано иное, очевидно, что описание, относящееся к «аминокислотам в положениях с 79 по 83 SEQ ID NO: 1», также можно применять к описанию «аминокислот, соответствующих аминокислотам в положениях с 79 по 83 SEQ ID NO: 1» в любой последовательности, например последовательности, идентичной SEQ ID NO: 1 на 50%, 60%, 70%, 80% или 90% или более.
Например, вариант белка по настоящему изобретению может представлять собой вариант белка, в котором аминокислота, соответствующая аминокислотам в положениях с 79 по 83, заменена на другую аминокислоту, и идентичный SEQ ID NO: 1 на 70%, 80%, 90% или 95%, однако вариант белка по настоящему изобретению не ограничен этим.
Вариант белка, в котором одна или более чем одна аминокислота из аминокислот в положениях 79-83 аминокислотной последовательности SEQ ID NO: 1 заменена на другую аминокислоту, может представлять собой вариант белка, включающий любую аминокислотную последовательность из SEQ ID NO: 131-147, в частности, может представлять собой вариант белка, по существу состоящий из любой аминокислотной последовательности из SEQ ID NO: 131-147, и, более конкретно, может представлять собой вариант белка, состоящий из любой аминокислотной последовательности из SEQ ID NO: 131-147, но вариант белка по настоящему изобретению не ограничен этим.
Кроме того, вариант белка может включать любую аминокислотную последовательность из SEQ ID NO: 131-147 или аминокислотную последовательность, в которой по меньшей мере одна аминокислота, выбранная из аминокислот в положениях 79-83 аминокислотной последовательности SEQ ID NO: 1, фиксирована, и которая гомологична или идентична SEQ ID NO: 1 на 80% или более, однако вариант белка не ограничен этим. В частности, мутантный полипептид по настоящему изобретению может включать полипептид, гомологичный или идентичный любой из аминокислотных последовательностей с SEQ ID NO: 131-147 по меньшей мере на 80%, 90%, 95%, 96%, 97%, 98% или 99%. Кроме того, очевидно, что любой белок, имеющий аминокислотную последовательность с делецией, модификацией, заменой или вставкой в части последовательности, отличной от аминокислотных положений 79-83, также может входить в объем настоящего изобретения, при условии, что белок имеет указанную выше гомологию или идентичность и демонстрирует эффект, соответствующий таковому указанного выше белка.
В данном описании термин «гомология» или «идентичность» относится к степени соответствия двух заданных аминокислотных последовательностей или нуклеотидных последовательностей и может быть выражен в процентах. Указанные термины «гомология» и «идентичность» зачастую могут использоваться взаимозаменяемо.
Гомология или идентичность последовательностей консервативных полинуклеотидов или полипептидов может быть установлена при помощи стандартного алгоритма выравнивания, при этом можно применять значения штрафов за открытие гэпа, установленные в используемой программе по умолчанию. По существу, гомологичные или идентичные последовательности могут гибридизоваться друг с другом в условиях умеренной или высокой жесткости по всей длине их последовательности или по меньшей мере на протяжении приблизительно 50%, 60%, 70%, 80% или 90% от всей длины. При гибридизации также можно учитывать полинуклеотиды, включающие вместо кодона вырожденный кодон.
Установить, обладают ли две любые полинуклеотидные или полипептидные последовательности гомологией, сходством или идентичностью, можно, например, при помощи известных в области техники компьютерных алгоритмов, например программы «FASTA» с использованием параметров по умолчанию, как описано Pearson et al. (1988) Proc. Natl. Acad. Sci. USA 85: 2444]. Альтернативно, для этого можно использовать алгоритм Нидлмана-Вунша (1970, J. Mol. Biol. 48: 443-453), используемый в программе Needleman пакета программ EMBOSS (The European Molecular Biology Open Software Suite, Rice et al., 2000, Trends Genet. 16: 276-277) (версия 5.0.0 или более поздняя) (включая пакет программ GCG (Devereux, J., et al., Nucleic Acids Research 12: 387 (1984)), BLASTP, BLASTN, FASTA (Atschul, [S.] [F.,] [ET AL., J MOLEC BIOL 215]: 403 (1990); Guide to Huge Computers, Martin J. Bishop, [ED.,] Academic Press, San Diego, 1994, and [CARILLO ETA/.] (1988) SIAM J Applied Math 48: 1073). Например, гомологию, сходство или идентичность можно определять при помощи алгоритма BLAST базы данных Национального Центра Биотехнологической Информации (National Center for Biotechnology Information) или ClustalW.
Гомологию, сходство или идентичность полинуклеотидов или полипептидов можно определять, например, путем сравнения имеющейся информации о последовательности с использованием компьютерной программы GAP, такой как программа, разработанная Needleman et al. (J Mol Biol. 48: 443 (1970)), как описано Smith and Waterman (Adv. Appl. Math (1981) 2: 482). Вкратце, программа GAP определяет гомологию, сходство или идентичность как количество символов (то есть нуклеотидов или аминокислот), при выравнивании оказывающихся одинаковыми, поделенное на общее количество символов в более короткой из двух последовательностей. Параметры по умолчанию для программы GAP могут включать: (1) бинарную матрицу сравнения (содержащую значения 1 для идентичных и 0 для неидентичных символов) и взвешенную матрицу сравнения, предложенную Gribskov et al., (Nucl. Acids Res. 14: 6745 (1986)), согласно описанию Schwartz and Dayhoff, eds. (Atlas Of Protein Sequence And Structure, National Biomedical Research Foundation, pp. 353-358 (1979), или подстановочную матрицу EDNAFULL (EMBOSS версия NCBI NUC4.4); (2) штраф 3,0 для каждого гэпа и дополнительный штраф 0,10 для каждого символа в каждом гэпе (или штраф за открытие гэпа 10, штраф за продолжение гэпа 0,5); и (3) отсутствие штрафа за концевые гэпы.
Кроме того, установить, обладают ли любые две полинуклеотидные или полипептидные последовательности гомологией, сходством или идентичностью, можно путем сравнения этих последовательностей посредством экспериментов с гибридизацией по Саузерну, которые выполняют в условиях определенной жесткости, и подходящие условия гибридизации можно установить способом, входящим в объем уровня техники и известным специалисту в области техники (например, J. Sambrook et al., Molecular Cloning, A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory press, Cold Spring Harbor, New York, 1989; F.M. Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., New York).
В данном документе термин «экспрессируемый/экспрессирующийся» применительно к белку означает состояние, в котором целевой белок вводят в микроорганизм или целевой белок модифицируют для экспрессии в микроорганизме. Когда целевой белок представляет собой белок, присутствующий в микроорганизме, термин означает состояние, при котором активность белка усилена по сравнению с активностью эндогенного белка или белка до модификации. В настоящем изобретении «целевой белок» может представлять собой вариант белка, обладающего способностью экспортировать L-триптофан, описанный выше.
В частности, термин «внедрение белка» означает, что микроорганизм демонстрирует активность конкретного белка, которой он изначально не обладал, или микроорганизм демонстрирует активность, которая усилена по сравнению с его эндогенной активностью или активностью белка до модификации. Например, термин «внедрение белка» может означать, что в хромосому микроорганизма внедряют полинуклеотид, кодирующий конкретный белок, или в микроорганизм внедряют вектор, включающий полинуклеотид, кодирующий конкретный белок, и, таким образом, обеспечивают проявление активности конкретного белка. Помимо этого, термин «усиление активности» означает, что активность конкретного белка усиливается по сравнению с его эндогенной активностью или активностью до его модификации. Термин «эндогенная активность» относится к активности конкретного белка, которой исходно обладал родительский штамм до модификации, в случае, когда признак микроорганизма изменен вследствие генетической мутации, вызванной естественным или искусственным фактором.
В частности, в данном описании усиление активности может достигаться одним или более чем одним из способов, выбранных из группы, состоящей из: способа увеличения количества внутриклеточных копий гена, кодирующего вариант белка; способа введения мутации в последовательность, контролирующую экспрессию гена, кодирующего вариант белка; способа замены последовательности, контролирующей экспрессию гена, кодирующего вариант белка, обладающего активностью экспортера L-триптофана, последовательностью, обеспечивающей высокую активность; способа замены в хромосоме гена, кодирующего нативный белок, обладающий активностью экспортера L-триптофана, геном, кодирующим вариант белка; способа внедрения дополнительной мутации в ген, кодирующий белок, обладающий активностью экспортера L-триптофана, так что активность варианта белка усиливается, и способа внедрения варианта белка в микроорганизм, однако способ усиления активности не ограничивается перечисленными.
При этом способ увеличения количества копий гена можно реализовать в форме, где ген функционально связан с вектором, или путем встраивания гена в хромосому клетки-хозяина, однако способ конкретно не ограничивается перечисленными. В частности, количество копий гена может быть увеличено путем введения вектора в клетку-хозяина, когда в клетку-хозяина вводят вектор, с которым функционально связан полинуклеотид, кодирующий белок по настоящему изобретению, и который способен реплицироваться и функционировать независимо от клетки-хозяина. Альтернативно, количество копий гена может быть увеличено путем введения вектора, с которым функционально связан полинуклеотид и который способен встраивать полинуклеотид в хромосому клетки-хозяина. Встраивание полинуклеотида в хромосому можно осуществлять любым известным в области техники способом (например, посредством гомологичной рекомбинации).
Затем, модификацию последовательности, контролирующей экспрессию, для повышения экспрессии полинуклеотида, можно осуществлять путем индуцирования мутации в последовательности нуклеиновой кислоты посредством делеции, вставки, неконсервативной или консервативной замены или их комбинации, так чтобы дополнительно усилить активность последовательности, контролирующей экспрессию, или путем замены последовательности, контролирующей экспрессию, нуклеиновокислотной последовательностью с более высокой активностью, однако способ модификации последовательности, контролирующей экспрессию, не ограничивается указанными. Последовательность, контролирующая экспрессию, может включать промотор, последовательность оператора, последовательность, кодирующую сайт связывания рибосомы, последовательности, контролирующие терминацию транскрипции и трансляции, и т.д., однако последовательность, контролирующая экспрессию, не ограничена этим.
Сильный промотор может быть связан с областью, расположенной в 5'-направлении относительно экспрессионной единицы полинуклеотида, вместо исходного промотора, но не ограничивается этим. Примеры сильных промоторов, известных в области техники, могут включать промоторы cj1-cj7 (патент KR 10-0620092), промотор lac, промотор trp, промотор trc, промотор tac, промотор PR фага лямбда, промотор PL, промотор tet, промотор gapA, промотор SPL7, промотор SPL13 (sm3) (патент KR 10-1783170), промотор O2 (патент KR 10-1632642), промотор tkt, промотор уссА и т.д., однако сильный промотор не ограничен перечисленными.
Кроме того, модификацию полинуклеотидной последовательности в хромосоме можно осуществлять путем индуцирования мутации в последовательности, контролирующей экспрессию, путем делеции, вставки, неконсервативной или консервативной замены или их комбинации, так чтобы дополнительно усилить активность полинуклеотидной последовательности, или путем замены полинуклеотидной последовательности улучшенной полинуклеотидной последовательностью с более высокой активностью, однако способ модификации полинуклеотидной последовательности не ограничивается указанными.
Внедрение и усиление активности белка, как описано выше, обычно позволяет увеличить активность или концентрацию соответствующего белка по меньшей мере на 1%, по меньшей мере на 10%, по меньшей мере на 25%, по меньшей мере на 50%, по меньшей мере на 75%, по меньшей мере на 100%, по меньшей мере на 150%, по меньшей мере на 200%, по меньшей мере на 300%, по меньшей мере на 400% или по меньшей мере на 500% и самое большее на 1000% или 2000%, относительно активности или концентрации белка в штамме микроорганизма дикого типа или немодифицированном штамме микроорганизма, однако диапазон увеличения не ограничен перечисленными.
В другом аспекте настоящего изобретения предложен полинуклеотид, кодирующий вышеупомянутый вариант белка.
В данном описании термин «полинуклеотид» относится к цепи ДНК или РНК, длина которой как нуклеотидного полимера превышает некоторое значение, представляющей собой длинную цепь мономеров нуклеотидов, соединенных ковалентными связями и, в частности, относится к фрагменту полинуклеотида, кодирующему вариант белка.
Полинуклеотид, кодирующий вариант белка по настоящему изобретению, может включать любую полинуклеотидную последовательность, без ограничения, при условии, что полинуклеотидная последовательность кодирует вариант белка, обладающий способностью экспортировать L-триптофан.
В данном документе ген, кодирующий аминокислотную последовательность белка, обладающего способностью экспортировать L-триптофан, может представлять собой ген wex, может иметь происхождение из Herbaspirillum rhizosphaerae, в частности, может представлять собой нуклеотидную последовательность, кодирующую аминокислотную последовательность SEQ ID NO: 1 и, более конкретно, может представлять собой нуклеотидную последовательность, включающую в себя нуклеотидную последовательность SEQ ID NO: 2, однако ген не ограничивается указанным.
С учетом вырожденности кодонов и предпочтения кодонов у биологического организма, в котором будет экспрессироваться полипептид, можно осуществлять различные модификации в кодирующей области полинуклеотида, кодирующего вариант белка по настоящему изобретению, в том объеме, который не меняет аминокислотную последовательность полипептида. В частности, изобретение охватывает любые полинуклеотидные последовательности, кодирующие вариант белка, в которых по меньшей мере одна аминокислота, выбранная из аминокислот в положениях от 79 до 83 в аминокислотной последовательности SEQ ID NO: 1, заменена на другую аминокислоту, без ограничения. Например, полинуклеотид по настоящему изобретению может кодировать вариант белка по настоящему изобретению и, в частности, представлять собой полинуклеотидную последовательность, кодирующую белок, включающий любую аминокислотную последовательность из SEQ ID NO: 131-147, или полипептид, имеющий гомологию или идентичность с указанным белком, однако полинуклеотид по настоящему изобретению не ограничивается указанным, и, более конкретно, может включать любую полинуклеотидную последовательность из полинуклеотидных последовательностей с SEQ ID NO: 80, 81, 82, 89, 90, 91, 92, 101, 102, 103, 110, 111, 112, 113, 122, 123 и 124, однако полинуклеотид по настоящему изобретению не ограничивается этим. Гомология и идентичность являются такими, как описано выше.
Кроме того, в объем изобретения входит любая последовательность, без ограничения, которая кодирует вариант белка, в котором по меньшей мере одна аминокислота, выбранная из аминокислот в положениях с 79 по 83 аминокислотной последовательности SEQ ID NO: 1, заменена на другую аминокислоту, гибридизующаяся в жестких условиях с любым зондом, который может быть получен на основе известных последовательностей гена (например, последовательностей, комплементарных всей вышеупомянутой полинуклеотидной последовательности или ее части).
Термин «жесткие условия» означает условия, при которых обеспечивается специфическая гибридизация между полинуклеотидами. Такие условия подробно описаны в литературе (например, J. Sambrook et al., см. выше). Например, условия могут включать проведение гибридизации между генами, обладающими высокой гомологией, гомологией или идентичностью 40% или более, в частности, 90% или более, более конкретно, 95% или более, еще более конкретно, 97% или более, наиболее конкретно 99% или более, при отсутствии гибридизации между генами, обладающими гомологией меньше указанной, или включать стандартные условия отмывки для гибридизации по Саузерну, то есть, в частности, однократную, двукратную или трехкратную отмывку при концентрации соли и температуре, соответствующих 60°С, IX SSC и 0,1% SDS, в частности, 60°С, 0,1Х SSC и 0,1% SDS, и более конкретно, 68°С, 0,1Х SSC и 0,1% SDS. Однако условия гибридизации не ограничиваются указанными и могут быть соответствующим образом скорректированы специалистом в области техники в зависимости от задачи.
Для гибридизации необходимо, чтобы два полинуклеотида включали комплементарные последовательности, хотя, в зависимости от строгости гибридизации, возможны несовпадения между основаниями. Термин «комплементарный» используется для описания взаимодействия между нуклеотидными основаниями, которые могут гибридизоваться друг с другом. Например, в случае ДНК аденозин комплементарен тимину, а цитозин комплементарен гуанину. Соответственно, настоящее изобретение также может охватывать выделенные полинуклеотидные фрагменты, комплементарные полным последовательностям, а также по существу схожие полинуклеотидные последовательности.
В частности, полинуклеотиды, обладающие гомологией, можно обнаружить при значении Tm 55°С, используя условия гибридизации, включающие стадию гибридизации и условия, описанные выше. Кроме того, значение Tm может составлять 60°С, 63°С или 65°С, однако температура не ограничивается указанными значениями и может быть соответствующим образом скорректирована специалистом в области техники в зависимости от задачи.
Жесткость, подходящая для гибридизации полинуклеотидов, зависит от длины и степени комплементарности полинуклеотидов, и возможные значения хорошо известны в области техники (см. Sambrook et al., выше, 9.50-9.51, 11.7-11.8).
В еще одном аспекте настоящего изобретения предложен вектор, который включает в себя полинуклеотид, кодирующий вариант белка.
В данном описании термин «вектор» означает ДНК-конструкцию, включающую в себя нуклеотидную последовательность полинуклеотида, кодирующего целевой белок, которая функционально связана с подходящей регуляторной последовательностью, так, что целевой белок может экспрессироваться в подходящем хозяине. Регуляторная последовательность включает промотор, способный инициировать транскрипцию, последовательность любого оператора для регуляции транскрипции, последовательность, кодирующую соответствующий сайт связывания рибосомы на мРНК, и последовательность для регуляции терминации транскрипции и трансляции. После трансформации подходящей клетки-хозяина вектор может реплицироваться или функционировать независимо от генома хозяина или может интегрироваться в геном самого хозяина.
Вектор для применения по настоящему изобретению не ограничивается каким-либо конкретным, но можно использовать любой известный в области техники вектор. Примеры обычно используемых векторов могут включать естественные или рекомбинантные плазмиды, космиды, вирусы и бактериофаги. Например, в качестве фагового вектора или космидного вектора можно использовать pWE15, М13, MBL3, MBL4, IXII, ASHII, APII, t10, t11, Charon4A, Charon21A и т.д., а в качестве плазмидного вектора использовать вектор на основе pBR, pUC, pBluescriptII, pGEM, pTZ, pCL, pET и т.д. В частности, можно использовать такие векторы как pDZ, pACYC177, pACYC184, pCL, pECCG117, pUC19, pBR322, pMW118, pCC1BAC и др.
Например, полинуклеотид, кодирующий целевой белок в хромосоме, может быть заменен мутированным полинуклеотидом посредством вектора для внутриклеточного встраивания в хромосому. Встраивание полинуклеотида в хромосому можно осуществлять любым известным в области техники способом, например посредством гомологичной рекомбинации, однако способ этим не ограничивается. Вектор может дополнительно включать в себя селективный маркер для подтверждения его успешного встраивания в хромосому. Селективный маркер используют для отбора клеток, трансформированных вектором, то есть для подтверждения того, встроилась ли целевая молекула нуклеиновой кислоты, и можно использовать маркеры, которые обеспечивают фенотип, позволяющий осуществлять отбор (например, устойчивость к лекарственным средствам, ауксотрофию, устойчивость к цитотоксическим агентам, экспрессию поверхностных белков и т.д.). В условиях с использованием селективных агентов только клетки, способные экспрессировать селективные маркеры, могут выживать или экспрессировать другие фенотипические признаки, что позволяет легко отобрать трансформированные клетки.
Еще одной задачей настоящего изобретения является предоставление продуцирующего L-триптофан микроорганизма, экспрессирующего вариант белка, обладающий активностью экспортера L-триптофана.
В данном описании термин «микроорганизм, продуцирующий L-триптофан» относится к микроорганизму, который способен продуцировать L-триптофан из источников углерода в среде в большем количестве, чем микроорганизм дикого типа или немодифицированный микроорганизм. Кроме того, микроорганизм, продуцирующий L-триптофан, может представлять собой рекомбинантный микроорганизм. В частности, микроорганизм может представлять собой микроорганизм рода Enterobacter, микроорганизм рода Escherichia, микроорганизм рода Erwinia, микроорганизм рода Serratia, микроорганизм рода Providencia, микроорганизм рода Corynebacterium или микроорганизм рода Brevibacterium, однако тип микроорганизма не ограничивается каким-либо конкретным, при условии, что микроорганизм способен продуцировать L-триптофан. Более конкретно, микроорганизм может представлять собой микроорганизм рода Corynebacterium или микроорганизм рода Escherichia.
Еще более конкретно, микроорганизм рода Escherichia может представлять собой Escherichia coli, а микроорганизм рода Corynebacterium может представлять собой Corynebacterium glutamicum, однако изобретение охватывает без ограничения любой микроорганизм рода Escherichia или рода Corynebacterium, в который внедрен белок, обладающий активностью экспортера L-триптофана, или активность которого усилена, благодаря чему может быть увеличено количество продуцируемого L-триптофана.
В описанных выше микроорганизмах количество продуцируемого L-триптофана может быть увеличено путем усиления экспрессии гена tktA или блокирования разветвленных путей биосинтеза L-триптофана для бесперебойного обеспечения предшественниками (например, эритрозо-4-фосфатом, Е4Р) и эффективного использования энергии, или с применением способа использования меньшего количества АТФ (аденозинтрифорсфат) и т.д.
В частности, в настоящем документе родительский штамм микроорганизма, продуцирующего L-триптофан, экспрессирующий белок или вариант белка, обладающий активностью экспортера L-триптофана, или модифицированный таким образом, чтобы экспрессировать белок или вариант белка, обладающий активностью экспортера L-триптофана, не ограничен каким-либо конкретным, при условии, что родительский штамм представляет собой микроорганизм, продуцирующий L-триптофан. Микроорганизм, продуцирующий L-триптофан, может представлять собой микроорганизм, в котором активность гена конкурентного пути, регулятора в направленном пути L-триптофанового оперона, гена импортера L-триптофана или гена импортера и катаболизма L-триптофана ослаблена или инактивирована, чтобы усилить биосинтез L-триптофана, и/или может представлять собой микроорганизм, в котором активность L-триптофанового оперона сверхэкспрессирована. В частности, активность trpR (то есть гена, регулирующего группу ферментов синтеза триптофана, который подавляет экспрессию генов биосинтеза L-триптофана (trpEDCBA)) или активность Mtr (то есть мембранного белка, импортирующего внеклеточный L-триптофан в клетку) может быть ослаблена или устранена по сравнению с их эндогенной активностью.
Для выполнения вышеуказанной задачи в еще одном аспекте настоящего изобретения предложен способ получения триптофана, включающий культивирование в среде микроорганизма, продуцирующего L-триптофан, экспрессирующего вариант белка.
L-триптофан, белок, обладающий активностью экспортера L-триптофана и включающий в себя аминокислотную последовательность SEQ ID NO: 1, экспрессия белка и микроорганизм являются такими, как описано выше.
В настоящем описании термин «культивирование» означает, что микроорганизм выращивают в надлежащим образом контролируемых условиях окружающей среды. Процесс культивирования по настоящему изобретению можно осуществлять в подходящей культуральной среде и условиях культивирования, известных в области техники. Специалист в области техники может легко адаптировать такой процесс культивирования для применения в соответствии с отбираемым штаммом. В частности, процесс культивирования можно осуществлять в виде периодического культивирования, непрерывного культивирования и культивирования с подпиткой, известных в области техники, однако процесс культивирования не ограничивается перечисленными.
В данном описании термин «среда» относится к смеси веществ, содержащих в качестве основного ингредиента питательные вещества, необходимые для культивирования микроорганизма, и наряду с веществом, незаменимым для выживания и роста, она обеспечивает питательными веществами, факторами роста и т.д. В частности, в качестве среды и других условий, применяемых для культивирования микроорганизма по настоящему изобретению, можно использовать любую среду, используемую при стандартном культивировании микроорганизмов, без конкретного ограничения. При этом микроорганизм по настоящему изобретению можно культивировать в аэробных условиях в стандартной среде, содержащей надлежащий источник углерода, источник азота, источник фосфора, неорганические соединения, аминокислоты и/или витамины и т.д., при этом подбирая температуру, рН и т.д.
В данном описании источник углерода может включать углеводы (например, глюкозу, фруктозу, сахарозу, мальтозу и т.д.), сахарные спирты (например, маннит, сорбит и т.д.), органические кислоты (например, пировиноградную кислоту, молочную кислоту, лимонную кислоту и т.д.), аминокислоты (например, глутаминовую кислоту, метионин, лизин и т.д.) и др. Кроме того, источник углерода может включать природные органические питательные вещества (например, гидролизат крахмала, мелассу, сырую мелассу, рисовые отруби, маниок, мелассу тростниково-сахарного производства, кукурузный экстракт и т.д.). В частности, можно использовать такие углеводы как глюкоза и стерилизованная предварительно обработанная меласса (то есть меласса, в которой сахара конвертированы в восстанавливающие) и, кроме того, можно использовать другие источники углерода в соответствующем количестве, без ограничения. Такие источники углерода можно использовать по отдельности или в виде комбинации двух или более видов, но источники углерода не ограничиваются этим.
Примеры источника азота могут включать источники неорганического азота (например, аммоний, сульфат аммония, хлорид аммония, ацетат аммония, фосфат аммония, карбонат аммония, нитрат аммония и т.д.), аминокислоты (например, глутаминовую кислоту, метионин, глутамин и т.д.) и источники органического азота (например, пептон, N-Z амин, мясной экстракт, дрожжевой экстракт, мальтозный экстракт, кукурузный экстракт, гидролизат казеина, рыбу или продукты ее переработки, обезжиренный соевый жмых или продукт его переработки и т.д.). Такие источники азота можно использовать по отдельности или в виде комбинации двух или более видов, но источники азота не ограничиваются этим.
Примеры источника фосфора могут включать однозамещенный фосфат калия, двузамещенный фосфат калия, соответствующую натрий содержащую соль и т.д. Примеры неорганического соединения могут включать хлорид натрия, хлорид кальция, хлорид железа, сульфат магния, сульфат железа, сульфат марганца, карбонат кальция и т.д. Кроме того, можно дополнительно включать аминокислоты, витамины и/или соответствующие предшественники. Такие составляющие ингредиенты или предшественники можно добавлять в среду при периодическом культивировании или непрерывном культивировании, и они не ограничиваются перечисленными.
В настоящем изобретении рН среды можно корректировать в процессе культивирования микроорганизма путем добавления соединения (например, гидроксида аммония, гидроксида калия, аммония, фосфорной кислоты, серной кислоты и т.д.) в среду надлежащим образом. Кроме того, в процессе культивирования для предупреждения образования пены можно добавлять пеногаситель (например, сложный полигликолевый эфир жирной кислоты). Кроме того, для поддержания аэробного состояния среды в среду можно вводить кислород или кислородсодержащий газ, либо для поддержания анаэробного или микроаэробного состояния среды в среду можно вводить азот, водород или диоксид углерода без введения газа, но газ не ограничивается этим.
Температуру среды можно поддерживать в диапазоне от 20°С до 50°С, и в частности, в диапазоне от 30°С до 37°С, однако температура среды не ограничивается указанной. Культивирование можно продолжать до тех пор, пока полезные вещества не будут получены в желаемом количестве и, в частности, на протяжении 10-100 часов, однако период культивирования не ограничивается указанным.
Способ получения может включать выделение L-триптофана из культуральной среды или микроорганизма.
На стадии выделения триптофана необходимый L-триптофан можно выделять из среды, применяя для культивирования микроорганизма способ, изложенный в настоящем описании, например, используя способ, известный в области техники, подходящий для периодического культивирования, непрерывного культивирования или культивирования с подпиткой. Например, такие способы как центрифугирование, фильтрация, обработка белка кристаллизованным осаждающим агентом (метод высаливания), экстрагирование, разрушение ультразвуком, ультрафильтрация, диализ, различные виды хроматографии (например, хроматография на молекулярных ситах (гель-фильтрация), адсорбционная хроматография, ионообменная хроматография, аффинная хроматография и т.д.) и HPLC (высокоэффективная жидкостная хроматография) могут использоваться по отдельности или в комбинации, однако способы не ограничиваются перечисленными.
Способ получения может включать процесс очистки. В процессе очистки выделенный L-триптофан можно очищать с использованием подходящего способа, известного в области техники.
В еще одном аспекте настоящего изобретения предложен способ увеличения способности микроорганизма экспортировать L-триптофан, включающий модификацию микроорганизма таким образом, чтобы вариант белка, обладающий активностью экспортера L-триптофана, в котором по меньшей мере одна аминокислота, выбранная из аминокислот в положениях 79-83 аминокислотной последовательности SEQ ID NO: 1, заменена на другую аминокислоту, мог экспрессироваться в микроорганизме.
В еще одном аспекте настоящего изобретения предложено применение варианта белка для увеличения способности экспортировать L-триптофан.
В еще одном аспекте настоящего изобретения предложено применение варианта белка для увеличения способности продуцировать L-триптофан.
Поскольку вариант белка по настоящему изобретению может увеличивать способность микроорганизма экспортировать L-триптофан, его можно применять для увеличения продукции L-триптофана. Вариант белка и другие аминокислоты являются такими, как описано выше.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Далее настоящее изобретение будет описано более подробно посредством приведенных в качестве примера воплощений. Однако эти приведенные в качестве примера воплощения служат исключительно для иллюстрации и не ограничивают объем настоящего изобретения.
Пример 1. Скрининг и отбор гена экспортера
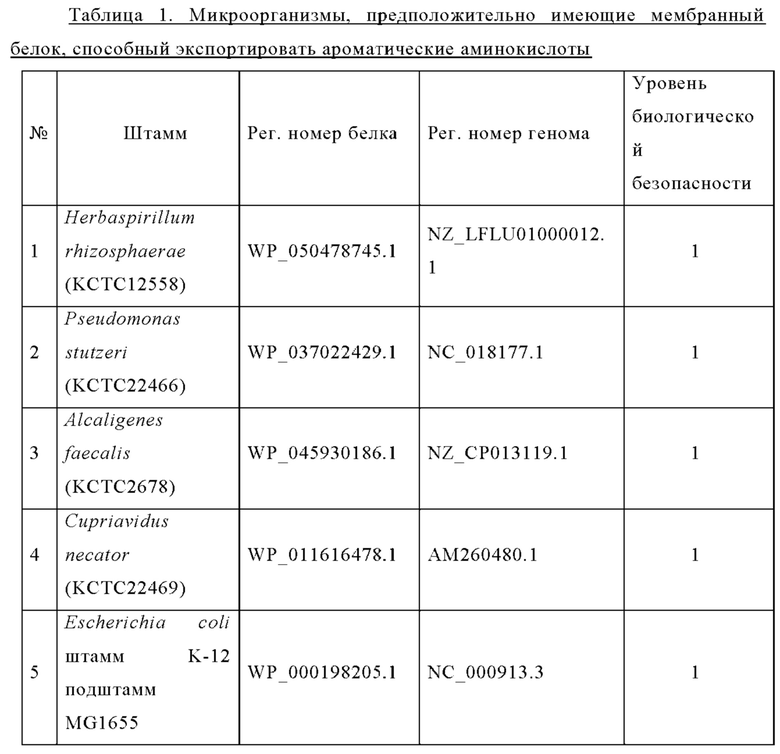
В результате скрининга баз данных NCBI и KEGG с использованием в качестве искомой последовательности аминокислотной последовательности YdeD (имеет происхождение из Е. coli и относится к семейству EamA) при помощи PSI-BLAST отобрали 30 генов-кандидатов, которые считаются мембранными белками, способными экспортировать триптофан, и биологические организмы, обладающие этими генами. Исходя из соображений биологической безопасности, применяемых к штаммам-продуцентам, и доступности этих биологических организмов, из них отобрали пять видов, которые приведены ниже в Таблице 1.
Пример 2. Получение микроорганизма рода Corynebacterium с внедренным геном, имеющим происхождение из Herbaspirillum rhizosphaerae
Мембранный белок, кодируемый геном, имеющим происхождение из Herbaspirillum rhizosphaerae, отобранного в Примере 1, имеет аминокислотную последовательность SEQ ID NO: 1. Информацию о гене, кодирующем мембранный белок, и прилежащих к нему нуклеотидных последовательностях (регистрационный № NZ_LFLU01000012.1) получали из базы данных NIH GenBank.
Праймеры для встраивания гена, имеющего происхождение из Herbaspirillum rhizosphaerae, в геномную ДНК Corynebacterium glutamicum синтезировали на основе полученной информации о нуклеотидных последовательностях. Для амплификации гена, имеющего происхождение из Herbaspirillum rhizosphaerae, выполняли ПЦР (полимеразная цепная реакция) с использованием в качестве матрицы хромосомной ДНК штамма Herbaspirillum rhizosphaerae и праймеров с последовательностями SEQ ID NO: 3 и SEQ ID NO: 4. В качестве полимеразы использовали ДНК-полимеразу Solg™ Pfu-X и проводили ПЦР следующим образом: денатурация при 95°С в течение 2 мин; 27 циклов, включающих денатурацию при 95°С в течение 20 сек, отжиг при 62°С в течение 40 сек и полимеризацию при 72°С в течение 1 мин; и в завершение полимеризация при 72°С в течение 5 мин.
В результате получали генный фрагмент размером 956 п.н., включающий в себя ген размером 924 п.н. (SEQ ID NO: 2).
SEQ ID NO: 3 (wex - 1)
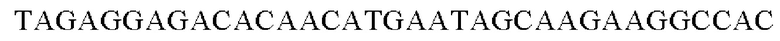
SEQ ID NO: 4 (wex - 2)
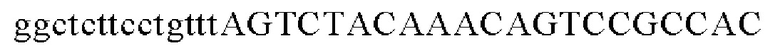
Для получения промотора gapA, имеющего происхождение из Corynebacterium glutamicum, выполняли ПЦР с использованием в качестве матрицы геномной ДНК Corynebacterium glutamicum и праймеров с последовательностями SEQ ID NO: 5 и SEQ ID NO: 6. В качестве полимеразы использовали ДНК-полимеразу Solg™ Pfu-X (SolGent Co., Ltd.) и проводили ПЦР следующим образом: денатурация при 95°С в течение 2 мин; 27 циклов, включающих денатурацию при 95°С в течение 20 сек, отжиг при 62°С в течение 40 сек и полимеризацию при 72°С в течение 30 сек; и в завершение полимеризация при 72°С в течение 5 мин.
SEQ ID NO: 5 (PgapA - 1)
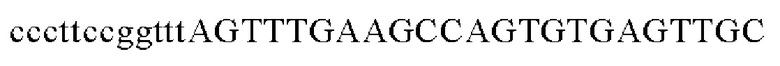
SEQ ID NO: 6 (PgapA(-wex) - 2)
Амплифицированные область промотора gapA и генные фрагменты, имеющие происхождение из Herbaspirillum rhizosphaerae, клонировали в вектор pDZTn (патент KR 10-1126041), расщепленный рестрикционным ферментом ScaI, способом, предложенным Gibson (DG Gibson et al., NATURE METHODS, VOL. 6, №5, MAY 2009, NEBuilder HiFi DNA Assembly Master Mix), и таким образом получали рекомбинантную плазмиду. Рекомбинантную плазмиду обозначали pDZTn-PgapA-Hrh. Клонирование осуществляли путем смешивания реагента Gibson для сборки и каждого генного фрагмента в рассчитанном молярном количестве с дальнейшей инкубацией при 50°С в течение 1 часа.
Полученным вектором pDZTn-PgapA-Hrh трансформировали посредством электропорации штамм Corynebacterium glutamicum АТСС13869 дикого типа (Appl. Microbiol. Biotechnol. (1999) 52: 541-545) и затем осуществляли вторичный кроссинговер для получения штамма, в котором одна копия гена PgapA-Hrh встроена в хромосому между транспозонными генами. Соответствующую генетическую манипуляцию верифицировали при помощи секвенирования генома и метода ПЦР с использованием праймеров SEQ ID NO: 7 и SEQ ID NO: 8, каждый из которых позволяет амплифицировать область, прилежащую к соответствующему встроенному гену в направлении 5' и 3', соответственно, для гомологичной рекомбинации.
SEQ ID NO: 7 (Confirm_PgapA-wex - 1)
SEQ ID NO: 8 (Confirm_PgapA-wex - 2)
Полученный таким образом штамм обозначали Corynebacterium glutamicum ATCC13869::PgapA-Hrh.
Пример 3. Получение микроорганизма рода Corynebacterium с внедренным геном, имеющим происхождение из Pseudomonas stutzeri
Мембранный белок, кодируемый геном, имеющим происхождение из Pseudomonas stutzeri, отобранного в Примере 1, имеет аминокислотную последовательность SEQ ID NO: 9. Информацию о соответствующем гене и прилежащих к нему нуклеотидных последовательностях (регистрационный № NC_018177.1) получали из базы данных NIH GenBank.
Праймеры для встраивания гена, имеющего происхождение из Pseudomonas stutzeri, в геномную ДНК Corynebacterium glutamicum синтезировали на основе полученной информации о нуклеотидных последовательностях. Для амплификации гена, имеющего происхождение из Pseudomonas stutzeri, выполняли ПЦР таким же образом, как описано в Примере 2, с использованием в качестве матрицы хромосомной ДНК штамма Pseudomonas stutzeri и праймеров с последовательностями SEQ ID NO: 11 и SEQ ID NO: 12.
В результате получали генный фрагмент размером 977 п.н., включающий в себя ген экспортера размером 945 п.н. (SEQ ID NO: 10).
SEQ ID NO: 11 (Pst-1)
SEQ ID NO: 12 (Pst-2)
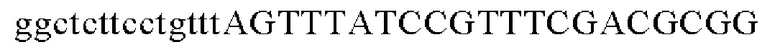
Для использования промотора gapA, имеющего происхождение из Corynebacterium glutamicum, выполняли ПЦР таким же образом, как описано в Примере 2, с использованием в качестве матрицы геномной ДНК Corynebacterium glutamicum АТСС13869 и праймеров с последовательностями SEQ ID NO: 5 и SEQ ID NO: 13.
SEQ ID NO: 13 (PgapA(-Pst)-2)
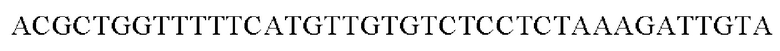
Амплифицированные область промотора gapA и генные фрагменты, имеющие происхождение из Pseudomonas stutzeri, клонировали в вектор pDZTn, расщепленный рестрикционным ферментом ScaI, способом, предложенным Gibson, и, таким образом, получали рекомбинантную плазмиду. Рекомбинантную плазмиду обозначали pDZTn-PgapA-Pst. Клонирование осуществляли путем смешивания реагента Gibson для сборки и каждого генного фрагмента в рассчитанном молярном количестве с дальнейшей инкубацией при 50°С в течение 1 часа.
Полученным вектором pDZTn-PgapA-Pst трансформировали посредством электропорации штамм Corynebacterium glutamicum АТСС13869 дикого типа (Appl. Microbiol. Biotechnol. (1999) 52: 541-545) и затем осуществляли вторичный кроссинговер для получения штамма, в котором одна копия гена PgapA-Pst встроена в хромосому между транспозонными генами. Соответствующую генетическую манипуляцию верифицировали при помощи секвенирования генома и метода ПЦР с использованием праймеров SEQ ID NO: 7 и SEQ ID NO: 8, каждый из которых позволяет амплифицировать область, прилежащую к соответствующему встроенному гену в направлении 5' и 3', соответственно, для гомологичной рекомбинации.
Полученный таким образом штамм обозначали Corynebacterium glutamicum ATCC13869::PgapA-Pst.
Пример 4. Получение микроорганизма рода Corynebacterium с внедренным геном, имеющим происхождение из Alcaligenes faecalis
Мембранный белок, кодируемый геном, имеющим происхождение из Alcaligenes faecalis, отобранного в Примере 1, имеет аминокислотную последовательность SEQ ID NO: 14. Информацию о соответствующем гене и прилежащих к нему нуклеотидных последовательностях (регистрационный № NZ_СР013119.1) получали из базы данных NIH GenBank.
Праймеры для встраивания гена, имеющего происхождение из Alcaligenes faecalis, в геномную ДНК Corynebacterium glutamicum синтезировали на основе полученной информации о нуклеотидных последовательностях. Для амплификации гена, имеющего происхождение из Alcaligenes faecalis, выполняли ПЦР таким же образом, как описано в Примере 2, с использованием в качестве матрицы хромосомной ДНК штамма Alcaligenes faecalis и праймеров с последовательностями SEQ ID NO: 16 и SEQ ID NO: 17.
В результате получали генный фрагмент размером 943 п.н., включающий в себя ген экспортера размером 912 п.н. (SEQ ID NO: 15).
SEQ ID NO: 16 (Afa-1)

SEQ ID NO: 17 (Afa-2)
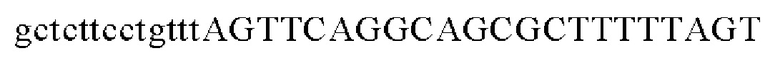
Для получения промотора gapA, имеющего происхождение из Corynebacterium glutamicum, выполняли ПЦР таким же образом, как в Примере 2, с использованием в качестве матрицы геномной ДНК Corynebacterium glutamicum и праймеров с последовательностями SEQ ID NO: 5 и SEQ ID NO: 18.
SEQ ID NO: 18 (PgapA(-Afa)-2)
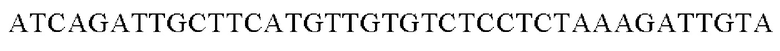
Амплифицированные область промотора gapA и генные фрагменты, имеющие происхождение из Alcaligenes faecalis, клонировали в вектор pDZTn, расщепленный рестрикционным ферментом ScaI, способом, предложенным Gibson, и, таким образом, получали рекомбинантную плазмиду. Рекомбинантную плазмиду обозначали pDZTn-PgapA-Afa. Клонирование осуществляли путем смешивания реагента Gibson для сборки и каждого генного фрагмента в рассчитанном молярном количестве с дальнейшей инкубацией при 50°С в течение 1 часа.
Полученным вектором pDZTn-PgapA-Afa трансформировали штамм Corynebacterium glutamicum АТСС13869 дикого типа посредством электропорации и затем осуществляли вторичный кроссинговер для получения штамма, в котором одна копия гена PgapA-Afa встроена в хромосому между транспозонными генами. Соответствующую генетическую манипуляцию верифицировали при помощи секвенирования генома и метода ПЦР с использованием праймеров SEQ ID NO: 7 и SEQ ID NO: 8, каждый из которых позволяет амплифицировать область, прилежащую к соответствующему встроенному гену в направлении 5' и 3', соответственно, для гомологичной рекомбинации.
Полученный таким образом штамм обозначали Corynebacterium glutamicum ATCC13869::PgapA-Afa.
Пример 5. Получение микроорганизма рода Corynebacterium с внедренным геном, имеющим происхождение из Cupriavidus necator
Мембранный белок, кодируемый геном, имеющим происхождение из Cupriavidus necator, отобранного в Примере 1, имеет аминокислотную последовательность SEQ ID NO: 19. Информацию о соответствующем гене и прилежащих к нему нуклеотидных последовательностях (регистрационный № АМ260480.1) получали из базы данных NIH GenBank.
Праймеры для встраивания гена, имеющего происхождение из Cupriavidus necator, в геномную ДНК Corynebacterium glutamicum синтезировали на основе полученной информации о нуклеотидных последовательностях. Для амплификации гена, имеющего происхождение из Cupriavidus necator, выполняли ПЦР таким же образом, как описано в Примере 2, с использованием в качестве матрицы хромосомной ДНК штамма Cupriavidus necator и праймеров с последовательностями SEQ ID NO: 21 и SEQ ID NO: 22.
В результате получали генный фрагмент размером 977 п.н., включающий в себя ген экспортера, имеющий происхождение из Cupriavidus necator, размером 945 п.н. (SEQ ID NO: 20).
SEQ ID NO: 21 (Cne-1)
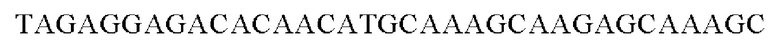
SEQ ID NO: 22 (Cne-2)
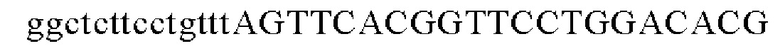
Для получения промотора gapA, имеющего происхождение из Corynebacterium glutamicum, выполняли ПЦР таким же образом, как в Примере 2, с использованием в качестве матрицы геномной ДНК Corynebacterium glutamicum и праймеров с последовательностями SEQ ID NO: 5 и SEQ ID NO: 23.
SEQ ID NO: 23 (PgapA(-Cne)-2)
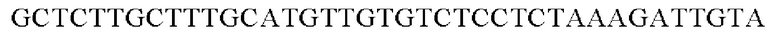
Амплифицированные область промотора gapA и генные фрагменты, имеющие происхождение из Cupriavidus necator, клонировали в вектор pDZTn, расщепленный рестрикционным ферментом ScaI, способом, предложенным Gibson, и, таким образом, получали рекомбинантную плазмиду. Рекомбинантную плазмиду обозначали pDZTn-PgapA-Cne. Клонирование осуществляли путем смешивания реагента Gibson для сборки и каждого генного фрагмента в рассчитанном молярном количестве с дальнейшей инкубацией при 50°С в течение 1 часа.
Полученным вектором pDZTn-PgapA-Cne трансформировали штамм Corynebacterium glutamicum АТСС13869 дикого типа посредством электропорации и затем осуществляли вторичный кроссинговер для получения штамма, в котором одна копия гена PgapA-Cne встроена в хромосому между транспозонными генами. Соответствующую генетическую манипуляцию верифицировали при помощи секвенирования генома и метода ПЦР с использованием праймеров SEQ ID NO: 7 и SEQ ID NO: 8, каждый из которых позволяет амплифицировать область, прилежащую к соответствующему встроенному гену в направлении 5' и 3', соответственно, для гомологичной рекомбинации.
Полученный таким образом штамм обозначали Corynebacterium glutamicum ATCC13869::PgapA-Cne.
Пример 6. Получение микроорганизма рода Corynebacterium с внедренным геном, имеющим происхождение из штамма Escherichia coli K-12 подштамма MG1655
Мембранный белок, кодируемый геном, имеющим происхождение из штамма Escherichia coli K-12 подштамма MG1655, отобранного в Примере 1, имеет аминокислотную последовательность SEQ ID NO: 24. Информацию о соответствующем гене и прилежащих к нему нуклеотидных последовательностях (регистрационный № NC_000913.3) получали из базы данных NIH GenBank.
Праймеры для встраивания гена, имеющего происхождение из Escherichia coli, в геномную ДНК Corynebacterium glutamicum синтезировали на основе полученной информации о нуклеотидных последовательностях. Для амплификации гена, имеющего происхождение из Escherichia coli, выполняли ПЦР таким же образом, как описано в Примере 2, с использованием в качестве матрицы хромосомной ДНК штамма Escherichia coli и праймеров с последовательностями SEQ ID NO: 26 и SEQ ID NO: 27.
В результате получали генный фрагмент размером 913 п.н., включающий в себя ген экспортера размером 882 п.н. (SEQ ID NO: 25).
SEQ ID NO: 26 (Eco-1)
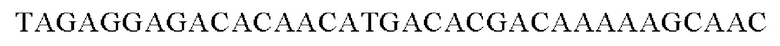
SEQ ID NO: 27 (Eco-2)
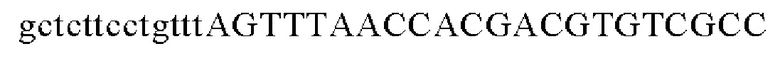
Для получения промотора gapA, имеющего происхождение из Corynebacterium glutamicum, выполняли ПЦР таким же образом, как в Примере 2, с использованием в качестве матрицы геномной ДНК Corynebacterium glutamicum и праймеров с последовательностями SEQ ID NO: 5 и SEQ ID NO: 28.
SEQ ID NO: 28 (PgapA(-Eco)-2)
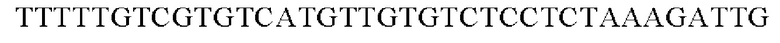
Амплифицированные область промотора gapA и генные фрагменты, имеющие происхождение из Escherichia coli, клонировали в вектор pDZTn, расщепленный рестрикционным ферментом ScaI, способом, предложенным Gibson, и, таким образом, получали рекомбинантную плазмиду. Рекомбинантную плазмиду обозначали pDZTn-PgapA-Eco. Клонирование осуществляли путем смешивания реагента Gibson для сборки и каждого генного фрагмента в рассчитанном молярном количестве с дальнейшей инкубацией при 50°С в течение 1 часа.
Полученным вектором pDZTn-PgapA-Eco трансформировали штамм Corynebacterium glutamicum АТСС13869 дикого типа посредством электропорации и затем осуществляли вторичный кроссинговер для получения штамма, в котором одна копия гена PgapA-Eco встроена в хромосому между транспозонными генами. Соответствующую генетическую манипуляцию верифицировали при помощи секвенирования генома и метода ПЦР с использованием праймеров SEQ ID NO: 7 и SEQ ID NO: 8, каждый из которых позволяет амплифицировать область, прилежащую к соответствующему встроенному гену в направлении 5' и 3', соответственно, для гомологичной рекомбинации.
Полученный таким образом штамм обозначали Corynebacterium glutamicum ATCC13869::PgapA-Eco.
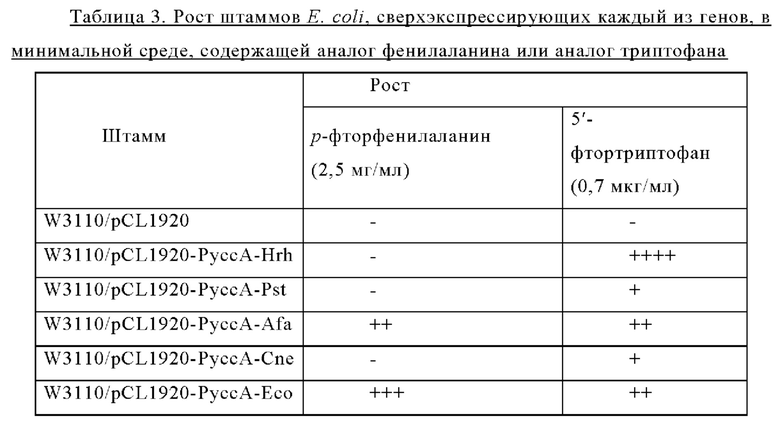
Пример 7. Определение минимальной ингибирующей концентрации (MIC) для штаммов микроорганизмов рода Corynebacterium с внедренными генами, имеющими происхождение из различных микроорганизмов
Для подтверждения наличия триптофан-экспортирующей активности у пяти типов штаммов Corynebacterium glutamicum, полученных в Примерах 2-6 (то есть ATCC13869::PgapA-Hrh, АТСС13869::PgapA-Pst, ATCC13869::PgapA-Afa, АТСС13869::PgapA-Cne и АТСС13869::PgapA-Eco), проводили определение минимальной ингибирующей концентрации (MIC) с использованием аналога триптофана и аналога фенилаланина (то есть другой ароматической аминокислоты). 5 различных штаммов Corynebacterium glutamicum, в каждый из которых был внедрен ген, кодирующий мембранный белок, культивировали в минимальной жидкой среде при 30°С в течение 24 часов, при разведении клеток до концентрации 3×103 и 3×104, соответственно, и затем получали колонии на минимальной твердой среде, в которую добавляли аналог триптофана или аналог фенилаланина.
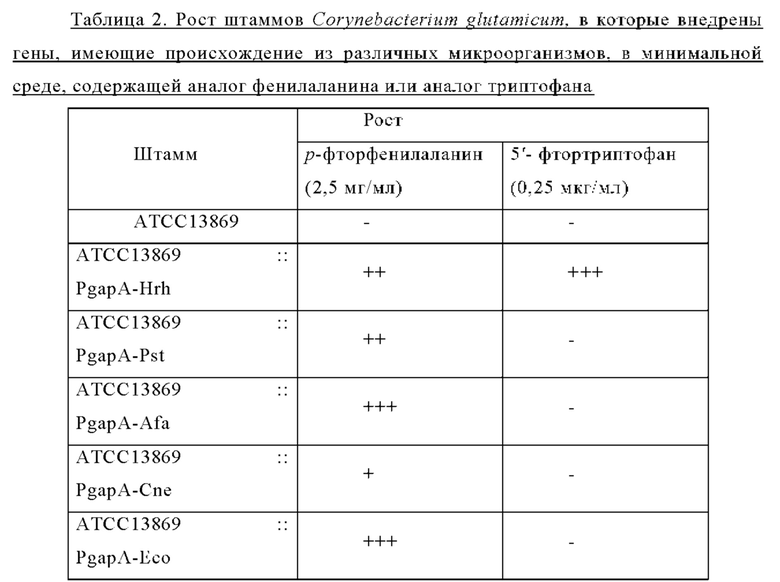
В ходе эксперимента для определения минимальной ингибирующей концентрации (MIC) в минимальную твердую среду добавляли р-фтор-DL-фенилаланин (2,5 мг/мл) или 5-фтор-DL-триптофан (0,25 мкг/мл) и регистрировали рост клеток через 60 часов (Таблица 2).
Внедрение всех пяти типов отобранных генов позволяло клеткам расти в условиях, когда добавляли аналог фенилаланина в концентрации 2,5 мг/мл. Среди них введение генов, имеющих происхождение из Herbaspirillum rhizosphaerae, Alcaligenes faecalis и Escherichia coli, обеспечивало наиболее выраженный рост клеток. Введение гена, имеющего происхождение из Pseudomonas stutzeri, приводило к более слабо выраженному росту клеток по сравнению с вышеупомянутыми тремя типами штаммов, а внедрение гена, имеющего происхождение из Cupriavidus necator, вызывало наиболее слабый рост клеток. В этих же условиях рост штамма АТСС13869 дикого типа отсутствовал. Кроме того, в условиях, когда добавляли аналог триптофана в концентрации 0,25 мкг/мл, только внедрение гена, имеющего происхождение из Herbaspirillum rhizosphaerae, позволяло клеткам расти.
Приведенные выше результаты свидетельствуют, что внедрение всех пяти типов отобранных генов придавало устойчивость к фенилаланину и аналогу фенилаланина, хотя при их внедрении наблюдались различия в активности. Напротив, только внедрение гена, имеющего происхождение из Herbaspirillum rhizosphaera, придавало специфическую и превосходную устойчивость к триптофану и аналогу триптофана. На основании этих результатов можно сделать заключение, что только мембранный белок, имеющий происхождение из Herbaspirillum rhizosphaera, может служить экспортирующим триптофан белком.
Минимальная среда (рН 7,2)
Глюкоза 10 г, KH2PO4 1 г, K2HPO4 2 г, MgSO4×7H2O 0,4 г, мочевина 2 г, (NH4)2SO4 5 г, NaCl 0,5 г, никотинамид 5 мкг, кальция пантотенат 0,1 мкг, биотин 0,2 мкг, тиамин HCl 3 мкг, раствор микроэлементов 1 мл (из расчета на 1 л дистиллированной воды).
Раствор микроэлементов
Na2B4O7×10Н2О 0,09 г, (NH4)6Mo7O27×4H2O 0,04 г, ZnSO4×7H2O 0,01 г, CuSO4×5H2O 0,27 г, MnCl2×4H2O 0,01 г, FeCl3×6H2O 1 г, CaCl2 0,01 г (из расчета на 1 л дистиллированной воды).
Пример 8. Получение экспрессирующего вектора для Escherichia coli, в который внедрены гены, имеющие происхождение из различных микроорганизмов
Для подтверждения устойчивости к триптофану или его аналогу, придаваемой Escherichia coli генами, имеющими происхождение из различных микроорганизмов, отобранными в Примере 1, каждый ген клонировали в pCL1920 (то есть вектор для экспрессии в Е. coli) и экспрессировали под контролем промотора уссА Е. coli W3110.
Для получения генного фрагмента, имеющего происхождение из Herbaspirillum rhizosphaerae, выполняли ПЦР с использованием в качестве матрицы хромосомной ДНК штамма Herbaspirillum rhizosphaerae и праймеров с последовательностями SEQ ID NO: 29 и SEQ ID NO: 30. В качестве полимеразы использовали ДНК-полимеразу Solg™ Pfu-X (SolGent Co., Ltd.) и проводили ПЦР следующим образом: денатурация при 95°С в течение 2 мин; 27 циклов, включающих денатурацию при 95°С в течение 20 сек, отжиг при 62°С в течение 40 сек и полимеризацию при 72°С в течение 1 мин; и в завершение полимеризация при 72°С в течение 5 мин.
SEQ ID NO: 29 (Hrh-3)
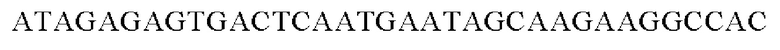
SEQ ID NO: 30 (Hrh-4)

Для получения промотора уссА, имеющего происхождение из Е. coli W3110, выполняли ПЦР с использованием в качестве матрицы геномной ДНК Е. coli W3110 и праймеров с последовательностями SEQ ID NO: 31 и SEQ ID NO: 32. В качестве полимеразы использовали ДНК-полимеразу Solg™ Pfu-X (SolGent Co., Ltd.) и проводили ПЦР следующим образом: денатурация при 95°С в течение 2 мин; 27 циклов, включающих денатурацию при 95°С в течение 20 сек, отжиг при 62°С в течение 40 сек и полимеризацию при 72°С в течение 10 сек; и в завершение полимеризация при 72°С в течение 5 мин.
SEQ ID NO: 31 (РуссА - 1)
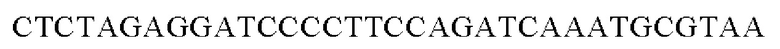
SEQ ID NO: 32 (PyccA(-Hrh)-2)
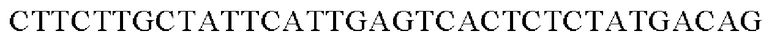
Амплифицированные область промотора уссА и генные фрагменты, имеющие происхождение из Herbaspirillum rhizosphaerae, клонировали в вектор pCL1920 vector (pSC101 ori, Spr), расщепленный рестрикционным ферментом SmaI, способом, предложенным Gibson, и, таким образом, получали рекомбинантную плазмиду. Рекомбинантную плазмиду обозначали pCL1920-PyccA-Hrh. Клонирование осуществляли путем смешивания реагента Gibson для сборки и каждого генного фрагмента в рассчитанном молярном количестве с дальнейшей инкубацией при 50°С в течение 1 часа. Полученную pCL1920-PyccA-Hrh внедряли в Е. coli W3110 дикого типа и, таким образом, получали W3110/pCL1920-PyccA-Hrh (то есть трансформант, экспрессирующий ген).
Для получения генного фрагмента, имеющего происхождение из Pseudomonas stutzeri, выполняли ПЦР с использованием в качестве матрицы хромосомной ДНК штамма Pseudomonas stutzeri и праймеров с последовательностями SEQ ID NO: 33 и SEQ ID NO: 34. Кроме того, выполняли ПЦР таким же образом, как описано выше для получения генного фрагмента из штамма Herbaspirillum rhizosphaerae, за тем исключением, что использовали праймер с последовательностью SEQ ID NO: 35, который использовали для получения промотора уссА, имеющего происхождение из Е. coli W3110.
SEQ ID NO: 33 (Pst-3)
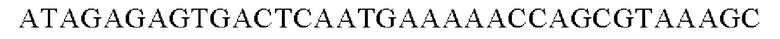
SEQ ID NO: 34 (Pst-4)

SEQ ID NO: 35 (PyccA(-Pst)-2)
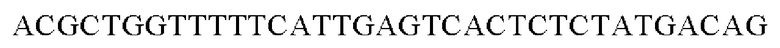
Полученную таким образом рекомбинантную плазмиду обозначали pCL1920-PyccA-Pst. Экспрессирующим вектором pCL1920-PyccA-Pst трансформировали Е. coli W3110 дикого типа и, таким образом, получали W3110/pCL1920-PyccA-Pst (то есть трансформант, экспрессирующий ген).
Процесс получения трансформанта, в котором экспрессировался ген, имеющий происхождение из штамма Alcaligenes faecalis, был таким же, как описано выше, за тем исключением, что ПЦР выполняли с использованием в качестве матрицы хромосомной ДНК штамма Alcaligenes faecalis и праймеров с последовательностями 36 и SEQ ID NO: 37, а также праймера с последовательностью SEQ ID NO: 38 для получения промотора уссА.
SEQ ID NO: 36 (Afa-3)
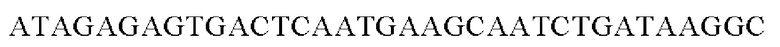
SEQ ID NO: 37 (Afa-4)
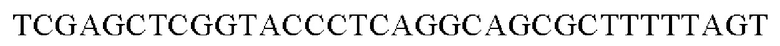
SEQ ID NO: 38 (PyccA(-Afa)-2)

Таким образом, получали рекомбинантную плазмиду, в которую клонировали ген, имеющий происхождение из Alcaligenes faecalis, и обозначали pCL1920-PyccA-Afa. Экспрессирующим вектором pCL1920-PyccA-Afa трансформировали Е. coli W3110 дикого типа и, таким образом, получали W3110/pCL1920-PyccA-Afa (то есть трансформант, экспрессирующий ген).
Для получения генного фрагмента, имеющего происхождение из Cupriavidus necator, выполняли ПЦР с использованием в качестве матрицы хромосомной ДНК штамма Cupriavidus necator и праймеров с последовательностями SEQ ID NO: 39 и SEQ ID NO: 40. Кроме того, выполняли ПЦР таким же образом, как описано выше для получения генного фрагмента из штамма Herbaspirillum rhizosphaerae, за тем исключением, что использовали праймер с последовательностью SEQ ID NO: 41, который использовали для получения промотора уссА, имеющего происхождение из Е. coli W3110.
SEQ ID NO: 39 (Cne-3)
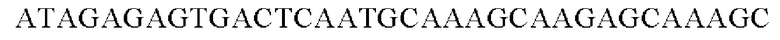
SEQ ID NO: 40 (Cne-4)
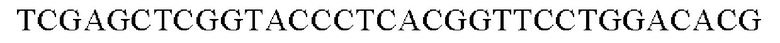
SEQ ID NO: 41 (PyccA(-Cne)-2)
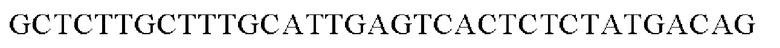
Полученную таким образом рекомбинантную плазмиду обозначали pCL1920-PyccA-Cne. Экспрессирующим вектором pCL1920-PyccA-Cne трансформировали Е. coli W3110 дикого типа и, таким образом, получали W3110/pCL1920-PyccA-Cne (то есть трансформант, экспрессирующий ген).
Для получения генного фрагмента, имеющего происхождение из штамма Escherichia coli, выполняли ПЦР с использованием в качестве матрицы хромосомной ДНК штамма Escherichia coli K-12 подштамма MG1655 и праймеров с последовательностями SEQ ID NO: 42 и SEQ ID NO: 43. Кроме того, выполняли ПЦР таким же образом, как описано выше для получения генного фрагмента из штамма Herbaspirillum rhizosphaerae, за тем исключением, что использовали праймер с последовательностью SEQ ID NO: 44, который использовали для получения промотора уссА, имеющего происхождение из Е. coli W3110.
SEQ ID NO: 42 (Eco-3)
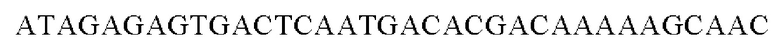
SEQ ID NO: 43 (Eco-4)
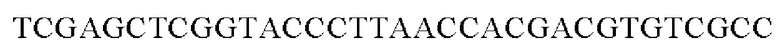
SEQ ID NO: 44 (PyccA(-Eco)-2)
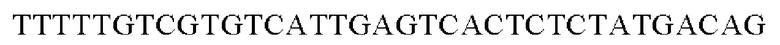
Полученную таким образом рекомбинантную плазмиду обозначали pCL1920-РуссА-Есо. Экспрессионный вектор pCL1920-РуссА-Есо внедряли в Е. coli W3110 дикого типа и, таким образом, получали W3110/pCL1920-PyccA-Cne (то есть трансформант, экспрессирующий ген).
Пример 9. Определение MIC для Е. coli со сверхэкспрессией генов мембранных белков, имеющих происхождение из различных микроорганизмов
Для подтверждения устойчивости штаммов Е. coli, сверхэкспрессирующих пять типов генов, полученных в Примере 8 (то есть W3110/pCL1920-PyccA-Hrh, W3110/pCL1920-PyccA-Pst, W3110/pCL1920-PyccA-Afa, W3110/pCL1920-PyccA-Cne и W3110/pCL1920-PyccA-Eco), проводили определение минимальной ингибирующей концентрации (MIC) с использованием аналога триптофана и аналога фенилаланина. Штаммы Е. coli, в которых сверхэкспрессировали пять типов генов, культивировали в минимальной жидкой среде М9, содержащей спектиномицин (50 мкг/мл) при 37°С в течение 15 часов, при разведении клеток до концентрации 104 и 105, соответственно, и затем получали колонии на минимальной твердой среде М9 с глюкозой, содержащей спектиномицин (50 мкг/мл), в которую добавляли аналог триптофана или аналог фенилаланина. В ходе эксперимента для определения минимальной ингибирующей концентрации (MIC) в минимальную твердую среду М9 добавляли р-фтор-DL-фенилаланин (2 мг/мл) или 5-фтор-DL-триптофан (0,7 мкг/мл) и регистрировали рост клеток через 48 часов (Таблица 3).
Как и штаммы Corynebacterium glutamicum, штаммы Е. coli, сверхэкспрессирующие гены, имеющие происхождение из Е. coli, демонстрировали превосходный рост в условиях добавления аналога фенилаланина, а сверхэкспрессия генов, имеющих происхождение из Alcaligenes faecalis, также обеспечивала выраженный рост. Однако, сверхэкспрессия генов, имеющих происхождение из Herbaspirillum rhizosphaerae, Pseudomonas stutzeri и Cupriavidus necator, не приводила к сопоставимому росту, как и в случае W3110/pCL1920 (то есть в контрольной группе). Напротив, сверхэкспрессия всех пяти типов отобранных генов позволяла клеткам расти в условиях, когда добавляли аналог триптофана. Среди них сверхэкспрессия гена, имеющего происхождение из Herbaspirillum rhizosphaerae, обеспечивала наиболее сильный рост, а на втором месте по степени выраженности был рост, обеспечиваемый при сверхэкспрессии генов экспортеров, имеющих происхождение из Alcaligenes faecalis и Е. coli. Сверхэкспрессия генов экспортеров, имеющих происхождение из Pseudomonas stutzeri и Cupriavidus necator, не приводила к заметному росту.
Результаты эксперимента по определению MIC с пятью типами генов в штамме Е. coli были аналогичны результатам, наблюдавшимся с Corynebacterium glutamicum. Ген, имеющий происхождение из Herbaspirillum rhizosphaerae, обеспечивал специфическую и превосходную устойчивость к триптофану и его аналогу как у штаммов Corynebacterium glutamicum, так и у штаммов Е. coli, а ген экспортера, имеющий происхождение из Е. coli, обеспечивал более устойчивый экспорт фенилаланина и его аналога по сравнению с триптофаном. На основании этих результатов установили, что ген, имеющий происхождение из Herbaspirillum rhizosphaerae, придает специфическую и превосходную способность экспортировать триптофан как штаммам Corynebacterium glutamicum, так и штаммам Е. coli.
Справочный Пример 1. Получение микроорганизма рода Corynebacterium, продуцирующего L-триптофан
Штаммы, продуцирующие L-триптофан, были производными Corynebacterium glutamicum ATCC13869 дикого типа. Поскольку Corynebacterium glutamicum дикого типа не способен продуцировать L-триптофан или если и способен, то продуцирует его лишь в небольшом количестве, предпринимали попытку в качестве родительского штамма использовать штамм, в котором усилен путь биосинтеза, необходимый для продукции L-триптофана. В частности, повышали экспрессию генов L-триптофанового оперона за счет усиления промотора. Кроме того, для устранения ингибирования по принципу обратной связи белка TrpE, 38-ю аминокислоту в TrpE (то есть серин) заменяли на аргинин (Journal of Bacteriology, Nov. 1987, p. 5330-5332).
Для осуществления указанной генетической манипуляции вначале для встраивания в хромосому посредством гомологичной рекомбинации получали область, расположенную в направлении 5' от промотора trpE, и 3'-концевую область, кодирующую часть TrpE дальше мутированной 38-й аминокислоты. В частности, генный фрагмент, расположенный в направлении 5' от промотора trpE, получали при помощи ПЦР с использованием в качестве матрицы хромосомной ДНК Corynebacterium glutamicum и праймеров с последовательностями SEQ ID NO: 45 и SEQ ID NO: 46, тогда как 3'-концевой генный фрагмент, кодирующий часть TrpE дальше мутированной 38-й аминокислоты, получали при помощи ПЦР с использованием в качестве матрицы хромосомной ДНК Corynebacterium glutamicum и праймеров с последовательностями SEQ ID NO: 47 и SEQ ID NO: 48.
SEQ ID NO: 45 (Pspl7-trpE(S38R)_L-1)
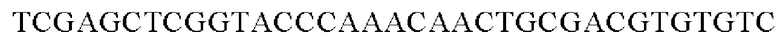
SEQ ID NO: 46 (Pspl7-trpE(S38R)_L-2)
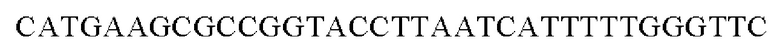
SEQ ID NO: 47 (Pspl7-trpE(S38R)_R-1)
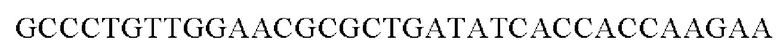
SEQ ID NO: 48 (Pspl7-trpE(S38R)_R-2)
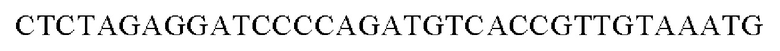
В качестве полимеразы использовали ДНК-полимеразу Solg™ Pfu-X (SolGent Co., Ltd.) и проводили ПЦР следующим образом: денатурация при 95°С в течение 5 мин; 30 циклов, включающих денатурацию при 95°С в течение 30 сек, отжиг при 60°С в течение 30 сек и полимеризацию при 72°С в течение 60 сек; и в завершение полимеризация при 72°С в течение 5 мин.
Выполняли ПЦР, используя в качестве матрицы синтетический промотор SPL7 (SEQ ID NO: 49) и праймеры с последовательностями SEQ ID NO: 50 и SEQ ID NO: 51.
SEQ ID NO: 50 (Pspl7 - 1)
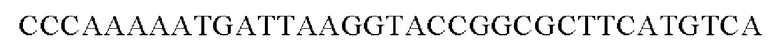
SEQ ID NO: 51 (Pspl7 - 2)
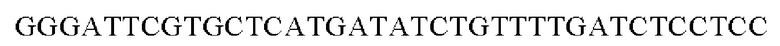
В качестве полимеразы использовали ДНК-полимеразу Solg™ Pfu-X (SolGent Co., Ltd.) и проводили ПЦР следующим образом: денатурация при 95°С в течение 5 мин; 30 циклов, включающих денатурацию при 95°С в течение 30 сек, отжиг при 60°С в течение 30 сек и полимеризацию при 72°С в течение 30 сек; и в завершение полимеризация при 72°С в течение 5 мин.
Для получения начального фрагмента TrpE, имеющего происхождение из Corynebacterium glutamicum, включавшего аминокислотную последовательность от 1-й аминокислоты до 38-й мутированной аминокислоты, выполняли ПЦР с использованием в качестве матрицы геномной ДНК Corynebacterium glutamicum и праймеров с последовательностями SEQ ID NO: 52 и SEQ ID NO: 53.
SEQ ID NO: 52 (trpE (S38R) - 1)
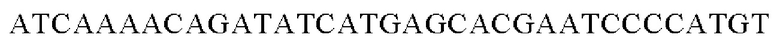
SEQ ID NO: 53 (trpE (S38R) - 2)
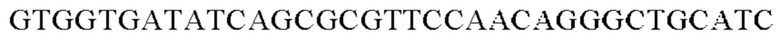
В качестве полимеразы использовали ДНК-полимеразу Solg™ Pfu-X (SolGent Co., Ltd.) и проводили ПЦР следующим образом: денатурация при 95°С в течение 5 мин; 30 циклов, включающих денатурацию при 95°С в течение 30 сек, отжиг при 60°С в течение 30 сек и полимеризацию при 72°С в течение 30 сек; и в завершение полимеризация при 72°С в течение 5 мин.
Рекомбинантную плазмиду получали способом, предложенным Gibson, путем клонирования амплифицированных: области, расположенной в направлении 5' от промотора trpE, 3'-концевой области, кодирующей часть TrpE дальше мутированной 38-й аминокислоты, промотора SPL7 и области, кодирующей начальный фрагмент TrpE, в вектор pDZ, расщепленный рестрикционным ферментом SmaI. Рекомбинантную плазмиду обозначали pDZ-PSPL7-trpE (S38R). Клонирование осуществляли путем смешивания реагента Gibson для сборки и каждого генного фрагмента в рассчитанном молярном количестве с дальнейшей инкубацией при 50°С в течение 1 часа.
Полученным вектором pDZ-PSPL7-trpE (S38R) трансформировали штамм Corynebacterium glutamicum АТСС13869 посредством электропорации и затем осуществляли вторичный кроссинговер. Таким образом, получали штамм, в хромосоме которого промотор trpE заменен промотором SPL7 (то есть более сильным промотором) и 38-я аминокислота TrpE (то есть серин) заменена на аргинин. Соответствующую генетическую манипуляцию верифицировали при помощи секвенирования генома и метода ПЦР с использованием праймеров SEQ ID NO: 54 и SEQ ID NO: 55, которые позволяют амплифицировать область, расположенную в направлении 5', и область, расположенную в направлении 3' от встраиваемого гена для гомологичной рекомбинации, и полученный штамм обозначали СА04-8325.
SEQ ID NO: 54 (Confirm_Pspl7-trpE(S38R) - 1)
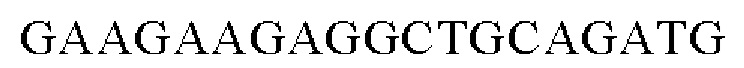
SEQ ID NO: 55 (Confirm_Pspl7-trpE(S38R) - 2)

Продуцирование триптофана осуществляется в метаболическом пути ароматических аминокислот, и данный метаболический путь начинается реакцией конденсации между фосфоенолпируватом и эритрозо-4-фосфатом. Соответственно, бесперебойное обеспечение этими двумя предшественниками является ключевым для продуцирования триптофана, и для бесперебойного обеспечения эритрозо-4-фосфатом, об относительной недостаточности которого известно, осуществляли сверхэкспрессию гена tkt.
Для вышеуказанной генетической манипуляции выполняли ПЦР с использованием в качестве матрицы хромосомной ДНК Corynebacterium glutamicum и праймеров с последовательностями SEQ ID NO: 56 и SEQ ID NO: 57 для получения расположенной в направлении 5' области для вставки дополнительного гена tkt, а также с праймерами SEQ ID NO: 58 и SEQ ID NO: 59 для получения расположенной в направлении 3' области для вставки дополнительного гена tkt.
SEQ ID NO: 56 (Pn-tkt_L - 1)
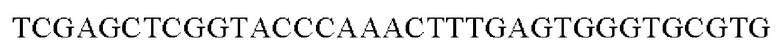
SEQ ID NO: 57 (Pn-tkt_L - 2)
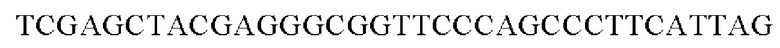
SEQ ID NO: 58 (Pn-tkt_R - 1)

SEQ ID NO: 59 (Pn-tkt_R - 2)
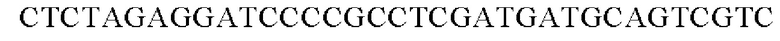
В качестве полимеразы использовали ДНК-полимеразу Solg™ Pfu-X и проводили ПЦР следующим образом: денатурация при 95°С в течение 2 мин; 27 циклов, включающих денатурацию при 95°С в течение 20 сек, отжиг при 62°С в течение 40 сек и полимеризацию при 72°С в течение 30 сек; и в завершение полимеризация при 72°С в течение 5 мин.
Для получения гена tkt и его промотора выполняли ПЦР с использованием в качестве матрицы хромосомной ДНК Corynebacterium glutamicum дикого типа АТСС13869 и праймеров с последовательностями SEQ ID NO: 60 и SEQ ID NO: 61 и, таким образом, получали ген tkt, включая его промотор.
SEQ ID NO: 60 (Pn-tkt - 1)
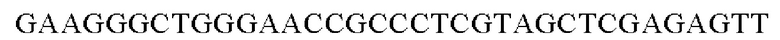
SEQ ID NO: 61 (Pn-tkt - 2)
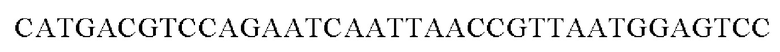
В качестве полимеразы использовали ДНК-полимеразу Solg™ Pfu-X и проводили ПЦР следующим образом: денатурация при 95°С в течение 2 мин; 27 циклов, включающих денатурацию при 95°С в течение 20 сек, отжиг при 62°С в течение 40 сек и полимеризацию при 72°С в течение 1 мин 20 сек; и в завершение полимеризация при 72°С в течение 5 мин.
Рекомбинантную плазмиду для трансформации хромосомы получали способом, предложенным Gibson, путем клонирования амплифицированных: расположенной в направлении 5' области для вставки дополнительного гена tkt и расположенной в направлении 3' области для вставки дополнительного гена tkt, а также гена tkt, включая промотор tkt, в вектор pDZ, расщепленный рестрикционным ферментом SmaI, и полученную рекомбинантную плазмиду обозначали pDZ-Pn-tkt. Клонирование осуществляли путем смешивания реагента Gibson для сборки и каждого генного фрагмента в рассчитанном молярном количестве с дальнейшей инкубацией при 50°С в течение 1 часа.
Полученным вектором pDZ-Pn-tkt трансформировали штамм СА04-8325 посредством электропорации и затем осуществляли вторичный кроссинговер для получения штамма, у которого ген tkt, включая промотор tkt, встроен в хромосому. Соответствующую генетическую манипуляцию верифицировали при помощи секвенирования генома и метода ПЦР с использованием праймеров SEQ ID NO: 62 и SEQ ID NO: 63, которые позволяют амплифицировать область, прилежащую к встроенному гену в направлении 5' и в направлении 3', соответственно, для гомологичной рекомбинации. Полученный штамм обозначали СА04-8352. Международное депонирование штамма СА04-8352 осуществляли 2 февраля 2018 в Корейском центре культур микроорганизмов (KCCM), являющемся международным депозитарием, в соответствии с Будапештским договором, с присвоением ему регистрационного номера KCCM12218.
SEQ ID NO: 62 (Confirm_Pn-tkt - 1)
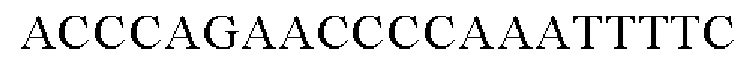
SEQ ID NO: 63 (Confirm_Pn-tkt - 2)
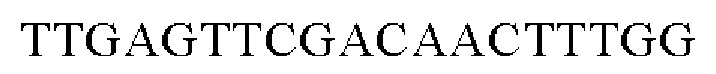
Пример 10. Продукция триптофана микроорганизмом рода Corynebacterium в который внедрен ген, имеющий происхождение из Herbaspirillum rhizosphaerae и Е. coli
Ген, имеющий происхождение из Herbaspirillum rhizosphaerae, демонстрирующий превосходную активность при минимальной ингибирующей концентрации аналога триптофана в Примере 7, внедряли в СА04-8352, который представляет собой триптофан-продуцирующий штамм, полученный в справочном Примере 1. С этой целью, вектором для внедрения гена, имеющего происхождение из Herbaspirillum rhizosphaerae, pDZTn-PgapA-Hrh, полученным в Примере 2, трансформировали СА04-8352 (то есть триптофан-продуцирующий штамм) посредством электропорации и осуществляли процесс, как описано в Примере 2, и, таким образом, получали штамм, в котором одна копия гена, имеющего происхождение из Herbaspirillum rhizosphaerae, встроена между транспозонными генами. Полученный штамм обозначали СА04-8405.
Кроме того, ген, имеющий происхождение из Е. coli, внедряли в СА04-8352 (то есть триптофан-продуцирующий штамм) и применяли в качестве контрольной группы. Вектором для внедрения гена, имеющего происхождение из Е. coli, pDZTn-PgapA-Eco, полученным в Примере 6, трансформировали СА04-8352 (то есть триптофан-продуцирующий штамм) посредством электропорации и осуществляли процесс, как описано в Примере 6, и, таким образом, получали штамм, в котором одна копия гена, имеющего происхождение из Е. coli, встроена между транспозонными генами. Полученный штамм обозначали СА04-8406.
Штаммы СА04-8405 и СА04-8406, полученные описанным выше способом, культивировали следующим образом, чтобы проверить количество продуцируемого триптофана в сравнении со штаммом СА04-8352, полученным в справочном Примере 1, служившим в качестве контрольного штамма. Каждым штаммом инокулировали 25 мл среды для посева, находящейся в конической колбе с дефлекторами объемом 250 мл, и культивировали при встряхивании со скоростью 200 об/мин при температуре 30°С в течение 20 часов. Затем 1 мл каждой среды для посева инокулировали 25 мл среды для продуцирования, находящейся в конической колбе с дефлекторами объемом 250 мл, и культивировали при встряхивании со скоростью 200 об/мин при температуре 30°С в течение 24 часов. После окончания культивирования измеряли количество продуцируемого L-триптофана при помощи HPLC.
Среда для посева (рН 7,0)
Глюкоза 20 г, пептон 10 г, дрожжевой экстракт 5 г, мочевина 1,5 г, KH2PO4 4 г, K2HPO4 8 г, MgSO4×7H2O 0,5 г, биотин 100 мкг, тиамин HCl 1000 мкг, кальция пантотенат 2000 мкг, никотинамид 2000 мкг (из расчета на 1 л дистиллированной воды).
Среда для продуцирования (рН 7,0)
Глюкоза 30 г, (NH4)2SO4 15 г, MgSO4×7H2O 1,2 г, KH2PO4 1 г, дрожжевой экстракт 5 г, биотин 900 мкг, тиамин HCl 4500 мкг, кальция пантотенат 4500 мкг, СаСО3 30 г (из расчета на 1 л дистиллированной воды).
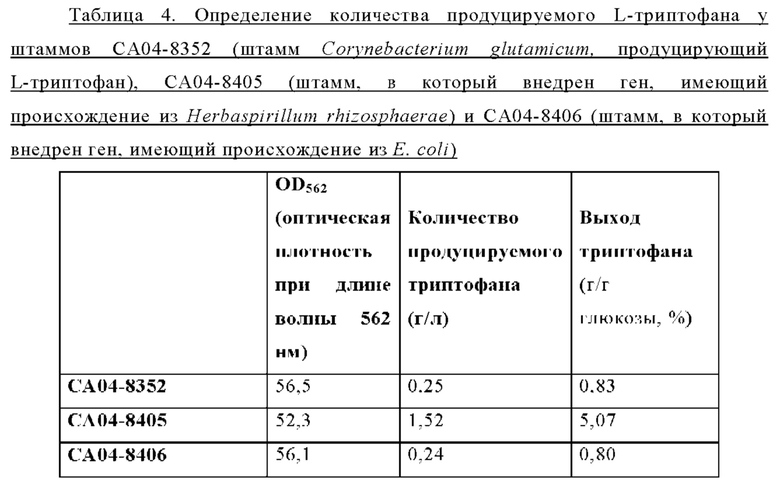
Результаты, демонстрирующие продукцию L-триптофана штаммами СА04-8352, СА04-8405 и СА04-8406 в среде, приведены в Таблице 4 выше. Штамм, в который внедрен ген, имеющий происхождение из Herbaspirillum rhizosphaerae, продуцировал L-триптофан в конечной концентрации 1,52 г/л при культивировании во флаконе, что соответствует приблизительно 5-кратному увеличению по сравнению со штаммом СА04-8352 (то есть контрольной группой). Это указывает, что ген, имеющий происхождение из Herbaspirillum rhizosphaerae, может существенно улучшать продукцию L-триптофана штаммом Corynebacterium glutamicum. Напротив, штамм СА04-8406, в который внедрен ген, имеющий происхождение из Е. coli, продуцировал L-триптофан в концентрации 0,23 г/л, что почти совпадает с количеством L-триптофана, продуцируемым штаммом СА04-8352 (то есть родительским штаммом для штамма СА04-8406). Как подтвердил эксперимент, в котором определяли минимальную ингибирующую концентрацию (MIC) аналога триптофана и аналога фенилаланина в Примерах 7 и 9, ген, имеющий происхождение из Е. coli, считается геном экспортера, демонстрирующего более высокую специфичность к фенилаланину, чем к триптофану. Международное депонирование штамма СА04-8405 осуществляли 21 августа 2017 в Корейском центре культур микроорганизмов (KCCM), являющемся международным депозитарием, в соответствии с Будапештским договором, с присвоением ему регистрационного номера KCCM12099P (СА04-8405).
Пример 11. Анализ внутриклеточных метаболитов триптофана в Corynebacterium slutamicum, в который внедряли ген, имеющий происхождение из Herbaspirillum rhizosphaerae
Для однозначного подтверждения, что внутриклеточная концентрация триптофана у штамма СА04-8405 (то есть штамма, продуцирующего триптофан) снижается по мере увеличения способности экспортировать триптофан, определяли внутриклеточную концентрации триптофана у штамма СА04-8405 и штамма СА04-8352 (то есть родительского штамма для штамма СА04-8405) методом экстракции с использованием органического растворителя.
Анализ внутриклеточных метаболитов выполняли способом, описанным Nakamura J et al., (Appl. Environ. Microbiol. 73(14): 4491-4498, 2007).
Что касается мутированных штаммов Corynebacterium glutamicum СА04-8352 и СА04-8405, каждым из них вначале инокулировали 25 мл среды для посева, находящейся в конической колбе с дефлекторами объемом 250 мл, и культивировали при встряхивании со скоростью 200 об/мин при температуре 30°С в течение 20 часов. Затем 1 мл каждой среды для посева инокулировали 25 мл среды для продуцирования, находящейся в конической колбе с дефлекторами объемом 250 мл, и культивировали при встряхивании со скоростью 200 об/мин при температуре 30°С. Внутриклеточную концентрацию триптофана определяли три раза, по мере потребления глюкозы. На каждой стадии культивируемые клетки отделяли от культурального раствора посредством быстрой вакуум-фильтрации (Durapore HV, 0.45 m; Millipore, Billerica, MA). Фильтр с адсорбированными на нем клетками промывали двукратно дистиллированной водой (10 мл) и вымачивали в метаноле, содержащем 5 мкМ морфолинэтансульфоновую кислоту и 5 мкМ метионина сульфона в течение 10 минут. К полученному выше клеточному экстракту (1,6 мл) добавляли хлороформ (1,6 мл) и дистиллированную воду (0,64 мкл), тщательно перемешивали и на микроцентрифужную колонку наносили только водную фазу для удаления белковых примесей. Профильтрованный экстракт анализировали при помощи капиллярного электрофореза и масс-спектрометрии, результаты показаны на Фиг. 1.
Как показано на Фиг. 1, подтвердили, что штамм СА04-8405 демонстрировал снижение внутриклеточной концентрации триптофана по сравнению со своим родительским штаммом СА04-8352 до уровня 33% - 41%. Эти результаты свидетельствуют, что поскольку триптофан, продуцируемый внутри клеток штамма Corynebacterium glutamicum, беспрепятственно экспортировался из клеток благодаря экспрессии гена; имеющего происхождение из Herbaspirillum rhizosphaerae, внутриклеточная концентрация триптофана у штамма СА04-8405 снижалась. Эти результаты подтвердили, что ген, имеющий происхождение из Herbaspirillum rhizosphaerae, представляет собой ген, кодирующий мембранный белок, который обладает экспортирующей способностью, специфической в отношении триптофана.
Справочный Пример 2. Получение микроорганизма рода Escherichia, продуцирующего L-триптофан
Микроорганизм рода Escherichia, продуцирующий L-триптофан, получали на основе Е. coli W3110 дикого типа. Чтобы проверить, может ли количество продуцируемого триптофана существенно повышаться при экспрессии модифицированного белка, обладающего активностью экспортера L-триптофана, в качестве родительского штамма использовали штамм, приготовленный для продуцирования L-триптофана. В частности, экспрессия генов биосинтеза L-триптофана (trpEDCBA), задействованных в продукции L-триптофана из хоризмата, ингибируется TrpR. Поэтому ген trpR, кодирующий TrpR, удаляли. Кроме того, для устранения ингибирования по принципу обратной связи полипептида TrpE с целью улучшения продукции L-триптофана 21-ю аминокислоту от N-конца TrpE (то есть пролин) заменяли на серин (J. Biochem. Mol. Biol. 32, 20 - 24 (1999)).
Мембранный белок Mtr играет роль при внедрении внутриклеточного L-триптофана в клетку, а белок TnaA играет роль при распаде внутриклеточного L-триптофана при участии молекул воды на индол, пируват и аммоний (NH3). Следовательно, гены mtr и tnaA, ингибирующие продуцирование L-триптофана и его распад, удаляли.
Для удаления этих генов использовали способ с рекомбиназой фага λ-red (One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products, Datsenko KA, Wanner BL., Proc Nati Acad Sci USA. 2000 Jun 6; 97(12): 6640-6645). Для удаления гена mtr выполняли ПЦР с использованием в качестве матрицы вектора pKD4 и праймеров с последовательностями SEQ ID NO: 64 и SEQ ID NO: 65. В результате получали генный фрагмент размером 1580 п.н., в котором кассета FRT-ген устойчивости к канамицину-FRT связана с двумя нуклеотидными последовательностями размером 50 п.н., гомологичными участкам, фланкирующими ген mtr в хромосоме, между которыми происходит гомологичная рекомбинация. Маркер резистентности к антибиотику канамицину вектора pKD4 служил для подтверждения удаления целевого гена и встраивания гена антибиотикорезистентности, а области FRT служили для удаления маркера антибиотикорезистентности после удаления целевого гена.
В качестве полимеразы использовали ДНК-полимеразу Solg™ Pfu-X и проводили ПЦР следующим образом: денатурация при 95°С в течение 2 мин; 27 циклов, включающих денатурацию при 95°С в течение 20 сек, отжиг при 62°С в течение 40 сек и полимеризацию при 72°С в течение 1 мин; и в завершение полимеризация при 72°С в течение 5 мин.
SEQ ID NO: 64 (Δmtr cassette - 1)
TGCAATGCATAACAACGCAGTCGCACTATTTTTCACTGGAGAGAAGCCCTGTGTAGGCTGGAGCTGCTTC
SEQ ID NO: 65 (Δmtr cassette - 2)
TGCAATGCATAACAACGCAGTCGCACTATTTTTCACTGGAGAGAAGCCCTGTCCATATGAATATCCTCCT
Вектором pKD46, экспрессирующим рекомбиназу λ red (гены gam, bet и ехо) трансформировали штамм Е. coli W3110 посредством электропорации и осуществляли посев штамма на твердой среде LB, содержащей канамицин (50 мг/л). Экспрессию рекомбинантного фермента в штамме Е. coli W3110, у которого была подтверждена трансформация вектором pKD46, индуцировали добавлением 10 мМ L-арабинозы при достижении OD600 (оптическая плотность при длине волны 600 нм) приблизительно 0,1. При достижении OD600 приблизительно 0,6 приготавливали компетентные клетки штамма и трансформировали их линеаризованным генным фрагментом, полученным в ходе вышеуказанного процесса, в котором кассета FRT-ген устойчивости к канамицину-FRT была связана с двумя нуклеотидными последовательностями размером 50 п.н., гомологичными участкам, фланкирующим ген mtr. Для колоний, выросших на твердой среде LB, содержащей канамицин (25 мг/л), выполняли ПЦР с использованием праймеров с последовательностями SEQ ID NO: 66 и SEQ ID NO: 67, и отбирали колонии, содержащие генный фрагмент размером 782 п.н.
SEQ ID NO: 66 (Confirm_Cassette - 1)
GGGCAGGATCTCCTGTCATC
SEQ ID NO: 67 (Confirm_Δmtr - 2)
AAATGTCGGATAAGGCACCG
Для удаления маркера устойчивости к антибиотику канамицину из штамма, в котором ген mtr был удален в результате гомологичной рекомбинации, готовили компетентные клетки, а затем трансформировали вектором рСР20 посредством электропорации. Вектор рСР20 экспрессирует белок FLP, за счет чего происходит распознавание сайтов FRT, фланкирующих ген маркера устойчивости к антибиотику канамицину в хромосоме, связывание с ними и удаление маркера устойчивости к антибиотику между сайтами FRT. Штамм, трансформированный вектором рСР20, который выращивали на твердой среде LB, содержащей ампициллин (100 мг/л) и хлорамфеникол (25 мг/л), культивировали в жидкой среде LB при 30°С в течение одного часа, затем культивировали при 42°С в течение 15 часов и выполняли рассев шпателем на твердой среде LB. Полученные колонии культивировали на твердой среде LB, содержащей ампициллин (100 мг/л) и хлорамфеникол (25 мг/л), твердой среде LB, содержащей канамицин (12,5 мг/л), и твердой среде LB, не содержащей антибиотик. Отбирали только колонии, выросшие на твердой среде LB, не содержащей антибиотик. Удаление гена mtr окончательно подтверждали путем секвенирования генома и обозначали штамм СА04-9300.
Для удаления гена tnaA генетические манипуляции проводили способом, описанным выше. Выполняли ПЦР с использованием в качестве матрицы вектора pKD4 и праймеров с последовательностями SEQ ID NO: 68 и SEQ ID NO: 69 и, таким образом, получали генный фрагмент размером 1580 п.н., в котором кассета FRT-ген устойчивости к канамицину-FRT связана с двумя нуклеотидными последовательностями размером 50 п.н., гомологичными участкам, фланкирующим ген tnaA в хромосоме, между которыми происходила гомологичная рекомбинация. В качестве полимеразы использовали ДНК-полимеразу Solg™ Pfu-X и проводили ПЦР следующим образом: денатурация при 95°С в течение 2 мин; 27 циклов, включающих денатурацию при 95°С в течение 20 сек, отжиг при 62°С в течение 40 сек и полимеризацию при 72°С в течение 1 мин; и в завершение полимеризация при 72°С в течение 5 мин.
SEQ ID NO: 68 (ΔtnaA cassette - 1)
TGTAATATTCACAGGGATCACTGTAATTAAAATAAATGAAGGATTATGTAGTGTAGGCTGGAGCTGCTTC
SEQ ID NO: 69 (ΔtnaA cassette - 2)
TGTAGGGTAAGAGAGTGGCTAACATCCTTATAGCCACTCTGTAGTATTAAGTCCATATGAATATCCTCCT
Подтверждали трансформацию вектором pKD46. CA04-9300, в котором индуцировали экспрессию рекомбиназ путем добавления 10 мМ L-арабинозы, трансформировали посредством электропорации линеаризованным генным фрагментом, полученным в ходе вышеуказанного процесса, в котором кассета FRT-ген устойчивости к канамицину-FRT была связана с двумя нуклеотидными последовательностями размером 50 п.н., гомологичными участкам, фланкирующим ген tnaA. Для колоний, выросших на твердой среде LB, содержащей канамицин (25 мг/л), выполняли ПЦР с использованием праймеров с последовательностями SEQ ID NO: 66 и SEQ ID NO: 70, и отбирали колонии, в которых был получен генный фрагмент размером 787 п.н.
SEQ ID NO: 70 (Confirm_ΔtnaA - 2)
ACATCCTTATAGCCACTCTG
Для удаления маркера устойчивости к антибиотику канамицину из штамма, в котором ген tnaA был удален в результате гомологичной рекомбинации, готовили компетентные клетки, а затем трансформировали вектором рСР20 посредством электропорации и получали штамм, в котором маркер устойчивости к антибиотику канамицину был удален благодаря экспрессии белка FLP. Удаление гена tnaA окончательно подтверждали путем секвенирования генома и обозначали штамм СА04-9301.
Для удаления гена trpR выполняли ПЦР с использованием в качестве матрицы вектора pKD4 и праймеров с последовательностями SEQ ID NO: 71 и SEQ ID NO: 72 и, таким образом, получали генный фрагмент (1580 п.н.), в котором кассета FRT-ген устойчивости к канамицину-FRT связана с двумя нуклеотидными последовательностями размером 50 п.н., гомологичными участкам, фланкирующим ген trpR в хромосоме, между которыми происходила гомологичная рекомбинация. В качестве полимеразы использовали ДНК-полимеразу Solg™ Pfu-X и проводили ПЦР следующим образом: денатурация при 95°С в течение 2 мин; 27 циклов, включающих денатурацию при 95°С в течение 20 сек, отжиг при 62°С в течение 40 сек и полимеризацию при 72°С в течение 1 мин; и в завершение полимеризация при 72°С в течение 5 мин.
SEQ ID NO: 71 (ΔtrpR cassette - 1)
TACAACCGGGGGAGGCATTTTGCTTCCCCCGCTAACAATGGCGACATATTGTGTAGGCTGGAGCTGCTTC
SEQ ID NO: 72 (ΔtrpR cassette - 2)
GCATTCGGTGCACGATGCCTGATGCGCCACGTCTTATCAGGCCTACAAAAGTCCATATGAATATCCTCCT
Подтверждали трансформацию вектором pKD46. СА04-9301, в котором индуцировали экспрессию рекомбиназ путем добавления 10 мМ L-арабинозы, трансформировали посредством электропорации линеаризованным генным фрагментом, полученным в ходе вышеуказанного процесса, в котором кассета FRT-ген устойчивости к канамицину-FRT была связана с двумя нуклеотидными последовательностями размером 50 п.н., гомологичными участкам, фланкирующим ген trpR. Для колоний, выросших на твердой среде LB, содержащей канамицин (25 мг/л), выполняли ПЦР с использованием праймеров с последовательностями SEQ ID NO: 66 и SEQ ID NO: 73, и отбирали колонии, в которых был получен генный фрагмент размером 838 п.н.
SEQ ID NO: 73 (Confirm_ΔtrpR - 2)
AGGACGGATAAGGCGTTCAC
Для удаления маркера устойчивости к антибиотику канамицину из штамма, в котором ген trpR был удален в результате гомологичной рекомбинации, готовили компетентные клетки, а затем трансформировали вектором рСР20 и получали штамм, в котором маркер устойчивости к антибиотику канамицину был удален благодаря экспрессии белка FLP. Удаление гена trpR окончательно подтверждали путем секвенирования генома и обозначали штамм СА04-9307.
Чтобы получить штамм СА04-9307, у которого trpE обладает устойчивостью к ингибированию по принципу обратной связи, выполняли ПЦР с использованием в качестве матрицы геномной ДНК Е. coli W3110 и праймеров с последовательностями SEQ ID NO: 74 и SEQ ID NO: 75 (оба из которых включали рестрикционный сайт EcoRI), и, таким образом, получали генный фрагмент trpE размером 1575 п.п., содержащий рестрикционные сайты EcoRI. В качестве полимеразы использовали ДНК-полимеразу Solg™ Pfu-X и проводили ПЦР следующим образом: денатурация при 95°С в течение 2 мин; 27 циклов, включавших денатурацию при 95°С в течение 20 сек, отжиг при 62°С в течение 1 мин и полимеризацию при 72°С в течение 1 мин; и в завершение полимеризация при 72°С в течение 5 мин.
SEQ ID NO: 74 (trpE - 1)
GAATTCATGCAAACACAAAAACCGAC
SEQ ID NO: 75 (trpE - 2)
GAATTCTCAGAAAGTCTCCTGTGCA
Клонирование выполняли после обработки генного фрагмента trpE, полученного вышеуказанным способом, и плазмиды pSG76-C (Journal of Bacteriology, July 1997, p.4426-4428), соответственно, рестрикционным ферментом EcoRI. Клонированной плазмидой трансформировали Е. coli DH5a посредством электропорации и отбирали трансформированные штаммы Е. coli DH5a на чашках со средой LB, содержащей хлорамфеникол (25 мкг/мл) и, таким образом, получали плазмиду pSG76-C-trpE.
PSG76-C-trpE(P21S) получали посредством сайт-специфического мутагенеза с использованием плазмиды pSG76-C-trpE, полученной выше, и праймеров с последовательностями SEQ ID NO: 76 и SEQ ID NO: 77 (Stratagene, USA).
SEQ ID NO: 76 (trpE(P21S) - 1)
CGCTTATCGCGACAATTCCACCGCGCTTTTTCACCAG
SEQ ID NO: 77 (trpE(P21S) - 2)
CTGGTGAAAAAGCGCGGTGGAATTGTCGCGATAAGCG
Штамм СА04-9307 трансформировали плазмидой pSG76-C-trpE(P21S), культивировали в среде LB-Cm (дрожжевой экстракт 10 г/л, NaCl 5 г/л, триптон 10 г/л и хлорамфеникол 25 мкг/л) и отбирали колонии, обладающие устойчивостью к хлорамфениколу. Отобранные трансформанты представляют собой штаммы, в геном которых в результате первой вставки в области trpE встроена плазмида pSG76-C-trpE(P21S). Штамм, в который встроен полученный ген trpE(P21S), трансформировали плазмидой pAScep (Journal of Bacteriology, July 1997, p.4426 4428), экспрессирующей рестрикционный фермент 1-SceI, который расщепляет области I-SceI, присутствующие в плазмиде pSG76-C, и отбирали штамм, растущий на среде LB-Ap (дрожжевой экстракт 10 г/л, NaCl 5 г/л, триптон 10 г/л и ампициллин 100 мкг/л). Амплифицировали ген trpE отобранного штамма с использованием праймеров с последовательностями SEQ ID NO: 74 и SEQ ID NO: 75 и при помощи секвенирования подтверждали, что в амплифицированном гене trpE была произведена замена trpE(P21S). Полученный таким образом штамм обозначали СА04-4303.
Пример 12. Продуцирование L-триптофана микроорганизмом рода Escherichia, в который внедрен ген, имеющий происхождение из Herbaspirillum rhizosphaerae
PCL1920-PyccA-Hrh, полученный в Примере 8, внедряли в штамм СА04-4303, полученный в справочном Примере 2, и, таким образом, получали штамм СА04-4306, сверхэкспрессирующий ген, имеющий происхождение из Herbaspirillum rhizosphaerae. Кроме того, в качестве контрольной группы, штамм СА04-4303 трансформировали вектором pCL1920. Для определения количества продуцируемого L-триптофана у двух штаммов (то есть CA04-4303/pCL1920 и СА04-4306) указанные два штамма культивировали в жидкой среде LB, содержащей спектиномицин (50 мг/л) в течение 12 часов. Затем каждым штаммом инокулировали 25 мл среды для продуцирования, находящейся в конической колбе с дефлекторами объемом 250 мл, при этом начальное значение OD600 достигало 0,01, и затем культивировали при встряхивании со скоростью 200 об/мин при температуре 37°С в течение 48 часов. По окончании культивирования измеряли количество продуцированного L-триптофана при помощи HPLC.
Результаты, демонстрирующие продукцию L-триптофана штаммами CA04-4303/pCL1920 и СА04-4306 в среде, приведены в Таблице 5 ниже. При культивировании в колбе штамма СА04-4306, в который был внедрен и сверхэкспрессировался ген, имеющий происхождение из Herbaspirillum rhizosphaerae, итоговая концентрация L-триптофана составляла 2,1 г/л, что было приблизительно на 50% выше по сравнению с контрольной группой. Это свидетельствует, что ген, имеющий происхождение из Herbaspirillum rhizosphaerae, также осуществляет экспорт L-триптофана у Е. coli, в результате чего продукция L-триптофана существенно улучшается.
Среда для продуцирования (рН 7.0)
Глюкоза 70 г, (NH4)2SO4 20 г, MgSO4×7Н2О 1 г, KH2PO4 2 г, дрожжевой экстракт 2,5 г, цитрат натрия 5 г, NaCl 1 г, СаСО3 40 г (из расчета на 1 л дистиллированной воды).
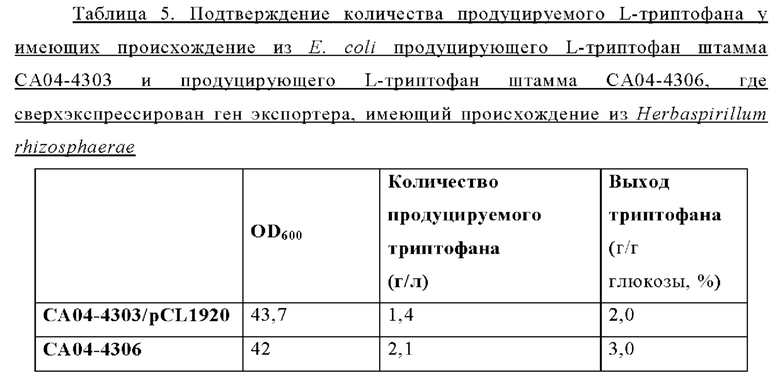
Следовательно, как показывают результаты в Примерах 7 и 9, ген, имеющий происхождение из Herbaspirillum rhizosphaerae, демонстрировал высокую специфичность и превосходную устойчивость к L-триптофану и его аналогу. Как показывают результаты в Примерах 10 и 12, ген, имеющий происхождение из Herbaspirillum rhizosphaerae, улучшал продукцию L-триптофана как у штамма Corynebacterium glutamicum, так и у штамма Е. coli. Кроме того, как было показано в Примере 11, ген, имеющий происхождение из Herbaspirillum rhizosphaerae, обеспечивал выраженный экспорт триптофана из клеток. В связи с этим, ген, имеющий происхождение из Herbaspirillum rhizosphaerae, обозначили wex (экспортер триптофана (W)).
Пример 13. Улучшение экспортирующей способности Wex с использованием случайного мутагенеза
В данном примере для случайного мутагенеза гена wex дикого типа использовали ПЦР пониженной точности, и при выполнении ПЦР пониженной точности применяли набор Diversify PCR Random Mutagenesis Kit (Clontech, USA).
Для получения варианта гена wex с внедренными случайными мутациями проводили ПЦР пониженной точности с использованием в качестве матрицы pCL1920-PyccA-wex, полученный в Примере 8, и праймеров с последовательностями SEQ ID NO: 78 и SEQ ID NO: 79.
Для подбора условий ПЦР пониженной точности, обеспечивающих желаемую частоту мутаций, использовали две композиции со следующими концентрациями MnSO4.
SEQ ID NO: 78 (wex - 1)
ACTCTAGAGGATCCCCTTCCAGATCAAATGCGTAA
SEQ ID NO: 79 (wex - 2)
ATTCGAGCTCGGTACCCCTACAAACAGTCCGCCAC
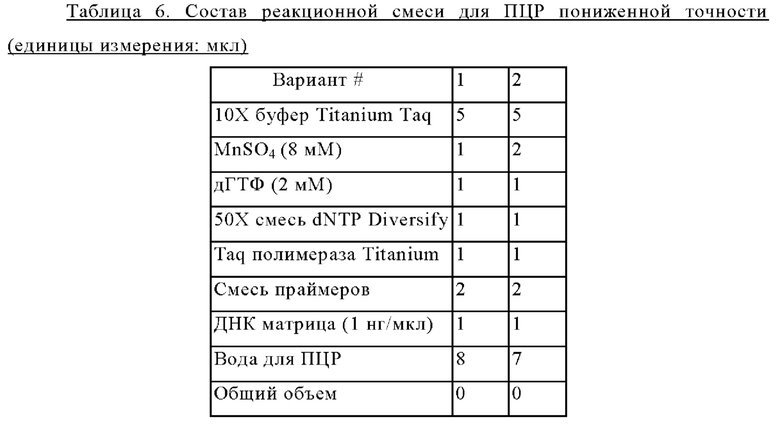
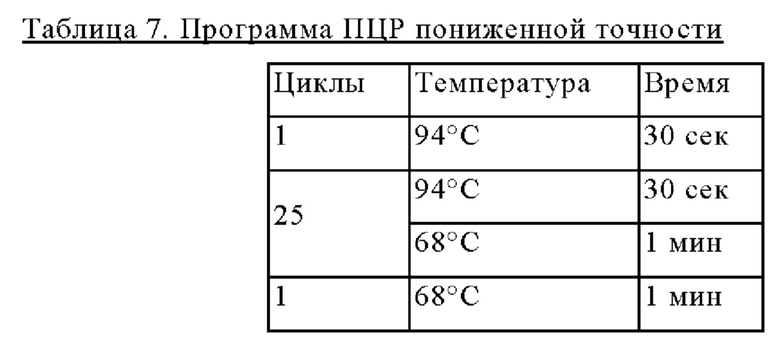
Библиотеку рекомбинантных мутантных плазмид получали методом сборки по Gibson (DG Gibson et al., Nature Methods, VOL. 6, №5, MAY 2009, NEBuilder HiFi DNA Assembly Master Mix) с использованием продуктов ПЦР пониженной точности (которую проводили в условиях, приведенных в Таблице 6 и Таблице 7) и pCL1920 (которую расщепляли рестрикционным ферментом SmaI). Клетки Escherichia coli W3110 трансформировали мутантной библиотекой, полученной вышеуказанным способом, pCL1920-PyccA-Hrh и pCL1920, культивировали на чашках со средой LB, содержащей спектиномицин (50 мкг/л) и отбирали из мутантной библиотеки 50 колоний. Выполняли секвенирование для определения в них частоты мутаций и наличие/отсутствие мутаций в различных положениях. В результате секвенирования частота мутаций в условиях #1 составляла 1,3 т.п.н.-1, а в условиях #2 составляла 2,5 т.п.н.-1. И в условиях #1, и в условиях #2 частота мутаций оказалась подходящей для получения мутантной библиотеки, и процесс скрининга эффективных мутаций проводили с использованием библиотеки, полученной в вышеупомянутых условиях.
300 мкл минимальной среды М9, содержащей 50 мкг/мл 5'-фтортриптофана (то есть аналога L-триптофана) разливали по лункам 96-луночного глубокодонного планшета, после чего инокулировали каждой из колоний ранее трансформированных мутантных библиотек (то есть мутантных библиотек W3110/pCL1920-PyccA-Hrh, W3110/pCL1920 и W3110/pCL1920-PyccA-wex). Культивирование осуществляли при 1200 об/мин при 37°С, и через 16 часов культивирования измеряли оптическую плотность (OD) при длине волны 600 нм. У большинства мутантов библиотеки рост в глубокодонных планшетах отсутствовал, как и в случае контрольных штаммов (W3110/pCL1920 и W3110/pCL1920-PyccA-Hrh). Из них у 51 колонии наблюдался улучшенный рост, их отбирали и из колоний выделяли мутантные плазмиды pCL1920-PyccA-wex и вновь трансформировали ими штаммы Е. coli W3110 для оценки воспроизводимости. Регистрировали значения OD у 10 типов штаммов, демонстрирующих специфическое улучшение роста в минимальной среде М9, в которой 5'-фтортриптофан содержался в обычной концентрации 50 мкг/мл.
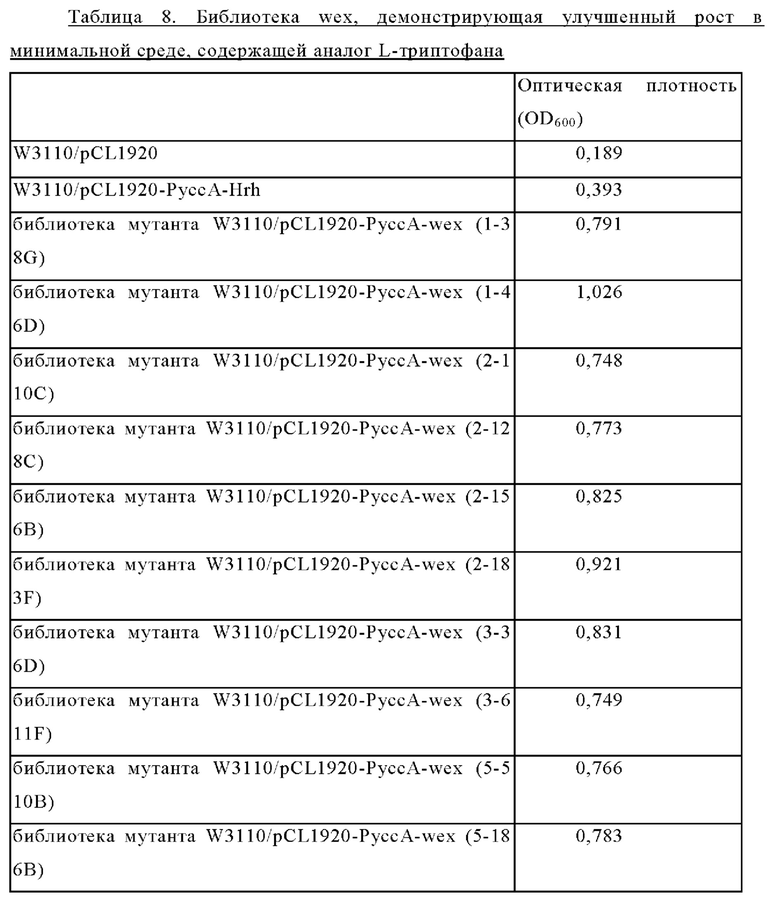
После выделения мутантной плазмиды pCL1920-PyccA-wex из 10 отобранных мутантных штаммов проводили секвенирование для подтверждения мутаций и в результате подтвердили, что мутации возникали в кодирующих последовательностях (CDS), которые не затрагивали области промотора. Кроме того, показали, что у 8 из 10 мутантных штаммов сайты мутаций были сгруппированы в области между 79-й аминокислотой (то есть лейцином) и 83-й аминокислотой (то есть изолейцином) аминокислотной последовательности мембранного белка Wex, и большинство мутаций представляли собой замену гидрофобной аминокислотой, алифатической аминокислотой или L-аминокислотой относительно малого размера. Таким образом, установили, что положения от 79 до 83 в аминокислотной последовательности мембранного белка Wex могут представлять собой ключевую область для внедрения мутаций, улучшающих способность экспортировать L-триптофан. В этой связи, предприняли попытку улучшить его активность путем замены аминокислот в соответствующей последовательности различными гидрофобными аминокислотами, алифатическими аминокислотами или L-аминокислотами относительно малого размера.
Пример 14. Замена 79-й аминокислоты в последовательности Wex (лейцина) у микроорганизма рода Corynebacterium на гидрофобную аминокислоту
Проверяли повышение эффективности экспорта L-триптофана при замене 79-й аминокислоты лейцина (далее будет обозначаться как лейцин в положении 79 или в 79-м положении) в аминокислотной последовательности мембранного белка Wex на другую гидрофобную аминокислоту. Выполняли сайт-специфический мутагенез с использованием в качестве матрицы pDZTn-PgapA-Hrh, полученного в Примере 2, для создания мутаций с заменой тремя различными аминокислотами, отличными от лейцина. Сайт-специфический мутагенез осуществляли следующим образом.
Для замены аминокислоты в положении 79 (то есть лейцина) аминокислотной последовательности Wex каждой из других гидрофобных аминокислот (то есть аланином (A) (SEQ ID NO: 80, SEQ ID NO: 131), валином (V) (SEQ ID NO: 81, SEQ ID NO: 132) и изолейцином (I) (SEQ ID NO: 82, SEQ ID NO: 133)); готовили реакционную смесь для ПЦР согласно Таблице 9 с использованием каждого набора праймеров для внедрения мутаций, приведенных в Таблице 11, и выполняли ПЦР способом, изложенным в Таблице 10. После завершения ПЦР добавляли 1 мкл рестрикционного фермента DpnI и инкубировали смесь в течение 1 часа при 37°С. 3 мкл ДНК, обработанной DpnI, трансформировали компетентные клетки DH5a для получения мутантных плазмид pDZTn-PgapA-wex и каждую из мутаций, приведенных в Таблице 11, подтверждали посредством секвенирования.
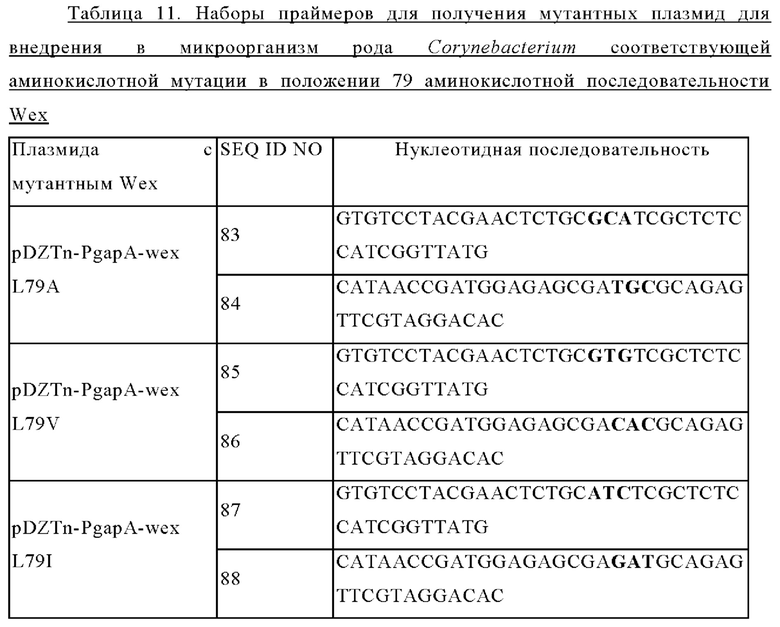
Каждым из векторов pDZTn-PgapA-wex L79A, pDZTn-PgapA-wex L79V и pDZTn-PgapA-wex L79I, полученных, как показано в Таблице 11, трансформировали посредством электропорации штамм СА04-8352, который получали в справочном Примере 1, и затем осуществляли вторичный кроссинговер для получения трех типов штаммов; соответственно; у которых мутированный ген wex внедрен в хромосому. Соответствующую генетическую манипуляцию верифицировали при помощи секвенирования генома и метода ПЦР с использованием праймеров SEQ ID NO: 7 и SEQ ID NO: 8, каждый из которых позволяет амплифицировать область, прилежащую к соответствующему встроенному гену в направлении 5' и 3', соответственно, для гомологичной рекомбинации.
Полученные мутантные штаммы обозначали CA04-8352(wex L79A), CA04-8352(wex L79V) и CA04-8352(wex L79I), соответственно.
Для определения количества триптофана, продуцируемого штаммами СА04-8352, СА04-8405, CA04-8352(wex L79A), CA04-8352(wex L79V) и CA04-8352(wex L79I), указанные штаммы культивировали тем же способом, как и в Примере 10. После окончания культивирования количество L-триптофана, продуцируемого каждым штаммом, измеряли при помощи HPLC.
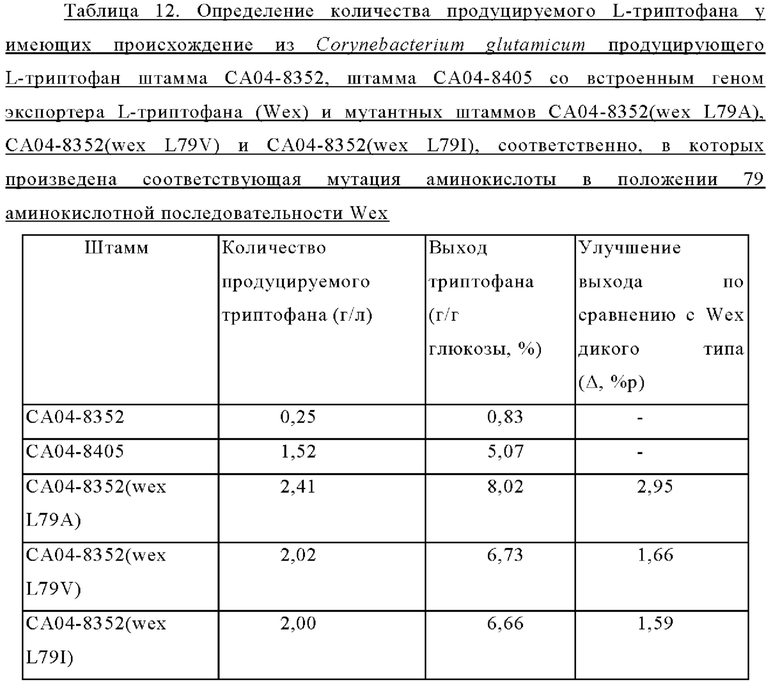
Как показывают результаты, приведенные в Таблице 12, все три типа мутантов, в которых лейцин в положении 79 аминокислотной последовательности Wex заменен на другую гидрофобную аминокислоту, демонстрировали при ферментации увеличение выхода на 1,59%р - 2,95%р по сравнению со штаммом СА04-8405, в который внедрен Wex дикого типа. В частности; мутантный штамм wex L79A демонстрировал наибольшее увеличение выхода L-триптофана при ферментации, и эти результаты указывают, что замена лейцина в положении 79 аминокислотной последовательности Wex на другую гидрофобную аминокислоту может существенно повысить способность Wex экспортировать L-триптофан.
Пример 15. Замена 80-й аминокислоты в последовательности Wex (серина) у микроорганизма рода Corynebacterium на гидрофобную аминокислоту
Проверяли повышение эффективности экспорта L-триптофана при замене 80-й аминокислоты серина (далее будет обозначаться как серии в положении 80) в аминокислотной последовательности мембранного белка Wex на гидрофобную аминокислоту. Выполняли сайт-специфический мутагенез с использованием в качестве матрицы pDZTn-PgapA-wex, полученного таким же способом, как и в Примере 14, для создания мутаций с заменой четырьмя различными гидрофобными аминокислотами.
Кроме того, серии в положении 80 аминокислотной последовательности Wex заменяли каждой из других гидрофобных аминокислот (то есть аланином (A) (SEQ ID NO: 89, SEQ ID NO: 134), валином (V) (SEQ ID NO: 90, SEQ ID NO: 135), лейцином (L) (SEQ ID NO: 91, SEQ ID NO: 136) и изолейцином (I) (SEQ ID NO: 92, SEQ ID NO: 137)), таким же способом, как и в Примере 14. Наборы праймеров для внедрения мутаций, а также формы мутаций, использованные для получения мутантных плазмид pDZTn-PgapA-wex, приведены в Таблице 13 ниже.

Каждым из векторов pDZTn-PgapA-wex S80A, pDZTn-PgapA-wex S80V, pDZTn-PgapA-wex S80L и pDZTn-PgapA-wex S80I, полученных, как показано в Таблице 13, трансформировали штамм СА04-8352 посредством электропорации, и затем осуществляли вторичный кроссинговер для получения четырех типов штаммов, соответственно, у которых мутированный ген wex внедрен в хромосому. Соответствующую генетическую манипуляцию верифицировали при помощи секвенирования генома и метода ПЦР с использованием праймеров SEQ ID NO: 7 и SEQ ID NO: 8, каждый из которых позволяет амплифицировать область, прилежащую к соответствующему встроенному гену в направлении 5' и 3', соответственно, для гомологичной рекомбинации.
Полученные мутантные штаммы обозначали CA04-8352(wex S80A), CA04-8352(wex S80V), CA04-8352(wex S80L) и CA04-8352(wex S80I), соответственно.
Для определения количества триптофана, продуцируемого штаммами СА04-8352, СА04-8405, CA04-8352(wex S80A), CA04-8352(wex S80V), CA04-8352(wex S80L) и CA04-8352(wex S80I), указанные штаммы культивировали тем же способом, как и в Примере 10. После окончания культивирования количество L-триптофана, продуцируемого каждым штаммом, измеряли при помощи HPLC.
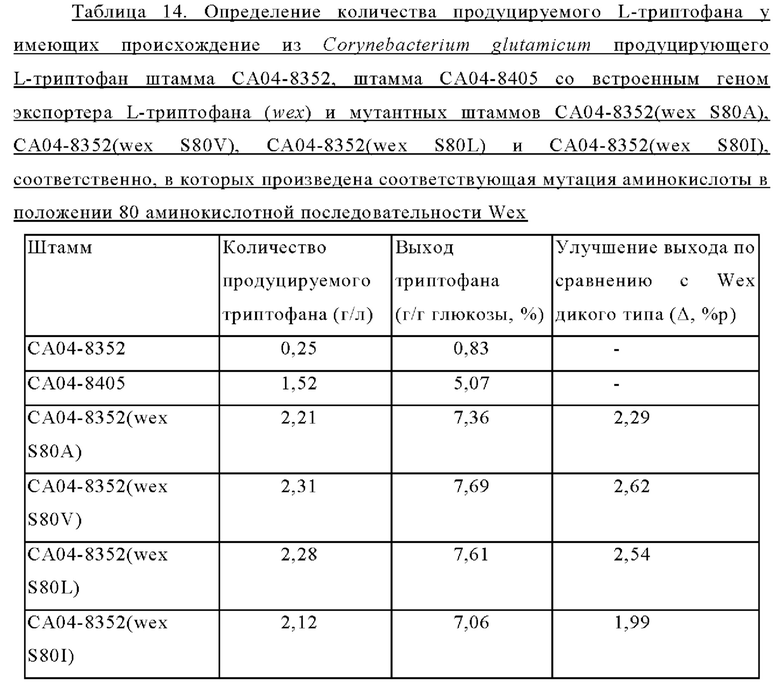
Как показывают результаты, приведенные в Таблице 14, все четыре типа мутантов, в которых аминокислота в положении 80 аминокислотной последовательности Wex заменена на другую гидрофобную аминокислоту, демонстрировали при ферментации увеличение выхода на 1,99%р - 2,62%р по сравнению со штаммом СА04-8405, в который внедрен Wex дикого типа. В частности, мутантный штамм wex S80V демонстрировал наибольшее увеличение выхода L-триптофана при ферментации, и эти результаты указывают, что замена серина в положении 80 аминокислотной последовательности Wex на другую гидрофобную аминокислоту может существенно повысить способность Wex экспортировать L-триптофан.
Пример 16. Замена 81-й аминокислоты в последовательности Wex (лейцина) у микроорганизма рода Corynebacterium на гидрофобную аминокислоту
Проверяли повышение эффективности экспорта L-триптофана при замене 81-й аминокислоты лейцина (далее будет обозначаться как лейцин в положении 81) в аминокислотной последовательности мембранного белка Wex на другую гидрофобную аминокислоту. Выполняли сайт-специфический мутагенез с использованием в качестве матрицы pDZTn-PgapA-wex, полученного таким же способом, как и в Примере 14, для создания мутаций с заменой тремя различными гидрофобными аминокислотами, отличными от лейцина.
Кроме того, лейцин в положении 81 аминокислотной последовательности Wex заменяли каждой из других гидрофобных аминокислот (то есть аланином (А) (SEQ ID NO: 101, SEQ ID NO: 138), валином (V) (SEQ ID NO: 102, SEQ ID NO: 139) и изолейцином (I) (SEQ ID NO: 103, SEQ ID NO: 140)), таким же способом, как и в Примере 14. Наборы праймеров для внедрения мутаций, а также формы мутаций, использованные для получения мутантных плазмид pDZTn-PgapA-wex, приведены в Таблице 15 ниже.
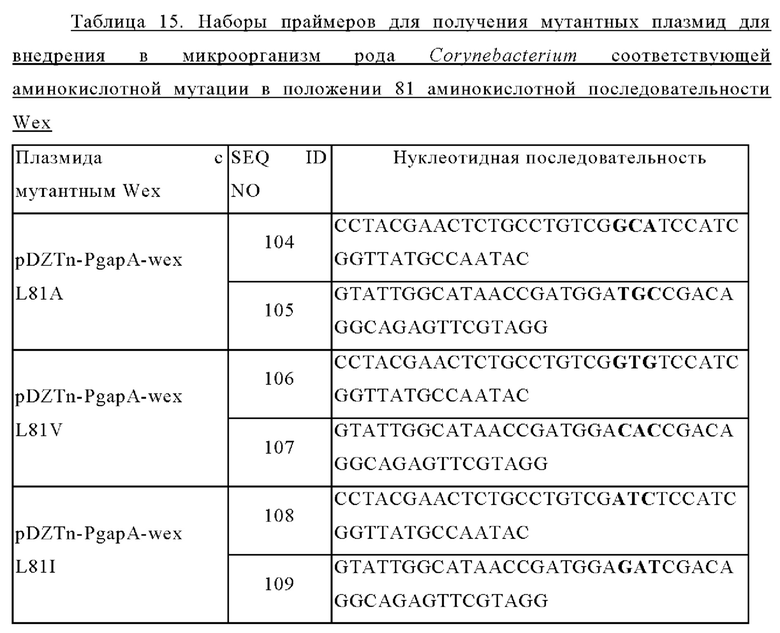
Каждым из векторов pDZTn-PgapA-wex L81A, pDZTn-PgapA-wex L81V и pDZTn-PgapA-wex L81I, полученных, как показано в Таблице 15, трансформировали посредством электропорации штамм СА04-8352 и затем осуществляли вторичный кроссинговер для получения трех типов штаммов, соответственно, у которых мутированный ген wex встроен в хромосому. Соответствующую генетическую манипуляцию верифицировали при помощи секвенирования генома и метода ПЦР с использованием праймеров SEQ ID NO: 7 и SEQ ID NO: 8, каждый из которых позволяет амплифицировать область, прилежащую к соответствующему встроенному гену в направлении 5' и 3', соответственно, для гомологичной рекомбинации.
Полученные мутантные штаммы обозначали CA04-8352(wex L81A), CA04-8352(wex L81V) и CA04-8352(wex L81I), соответственно.
Для определения количества триптофана, продуцируемого штаммами СА04-8352, СА04-8405, CA04-8352(wex L81A), CA04-8352(wex L81V) и CA04-8352(wex L81I), указанные штаммы культивировали тем же способом, как и в Примере 10. После окончания культивирования количество L-триптофана, продуцируемого каждым штаммом, измеряли при помощи HPLC.
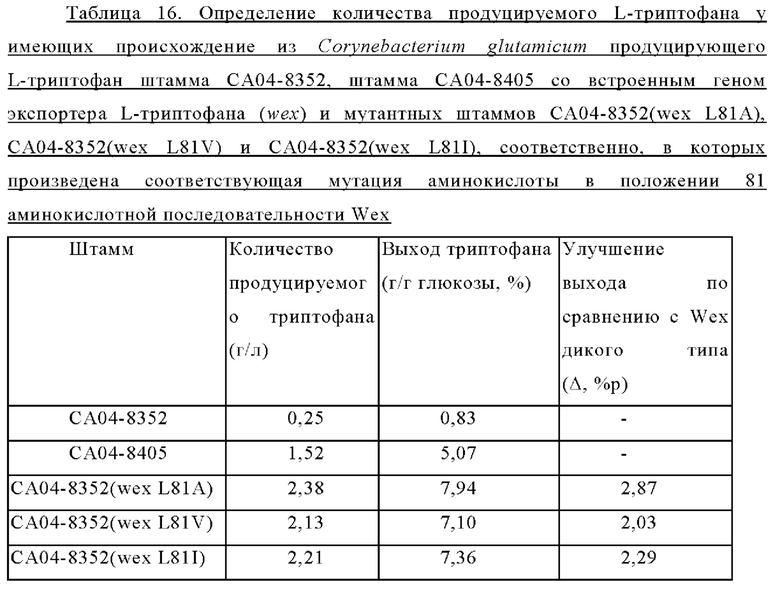
Как показывают результаты, приведенные в Таблице 16, все три типа мутантов, в которых аминокислота в положении 81 аминокислотной последовательности Wex заменена на другую гидрофобную аминокислоту, демонстрировали при ферментации увеличение выхода на 2,03%р - 2,87%р по сравнению со штаммом СА04-8405, в который внедрен Wex дикого типа. В частности, мутантный штамм wex L81A демонстрировал наибольшее увеличение выхода L-триптофана при ферментации, и эти результаты указывают, что замена лейцина в положении 81 аминокислотной последовательности Wex на другую гидрофобную аминокислоту может существенно повысить способность Wex экспортировать L-триптофан.
Пример 17. Замена 82-й аминокислоты в последовательности Wex (серина) у микроорганизма рода Corynebacterium на гидрофобную аминокислоту
Проверяли повышение эффективности экспорта L-триптофана при замене 82-й аминокислоты серина (далее будет обозначаться как серии в положении 82) в аминокислотной последовательности мембранного белка Wex на гидрофобную аминокислоту. Выполняли сайт-специфический мутагенез с использованием в качестве матрицы pDZTn-PgapA-wex, полученного таким же способом, как и в Примере 14, для создания мутаций с заменой четырьмя различными гидрофобными аминокислотами.
Кроме того, лейцин в положении 82 аминокислотной последовательности Wex заменяли каждой из других гидрофобных аминокислот (то есть аланином (А) (SEQ ID NO: 110, SEQ ID NO: 141), валином (V) (SEQ ID NO: NO: 111, SEQ ID NO: 142), лейцином (SEQ ID N0:112, SEQ ID NO: 143) и изолейцином (I) (SEQ ID NO: 113, SEQ ID NO: 144)), таким же способом, как и в Примере 14. Наборы праймеров для внедрения мутаций, а также формы мутаций, использованные для получения мутантных плазмид pDZTn-PgapA-wex, приведены в Таблице 17 ниже.
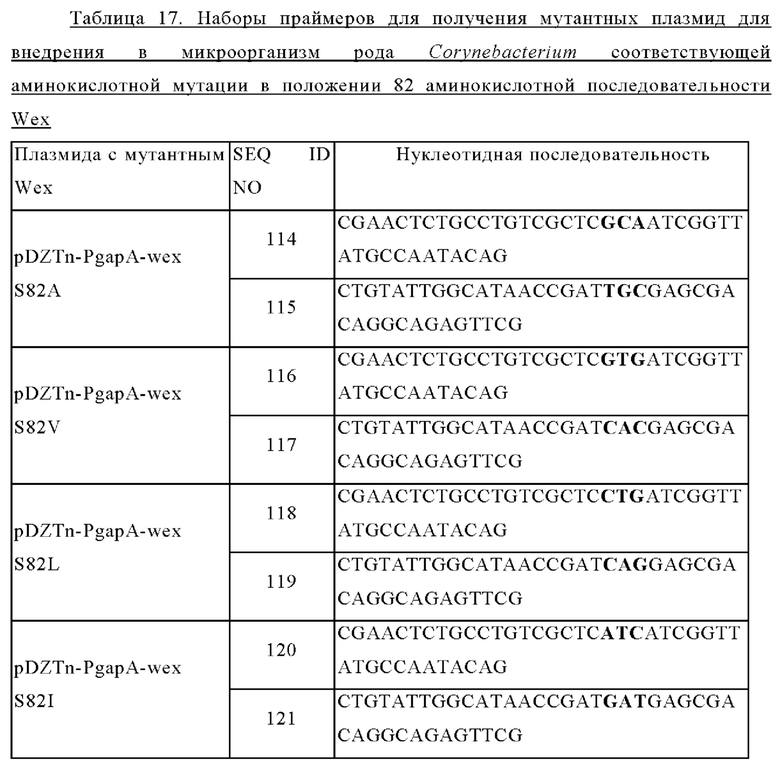
Каждым из векторов pDZTn-PgapA-wex S82A, pDZTn-PgapA-wex S82V, pDZTn-PgapA-wex S82L и pDZTn-PgapA-wex S82I, полученных, как показано в Таблице 17, трансформировали штамм СА04-8352 посредством электропорации, и затем осуществляли вторичный кроссинговер для получения четырех типов штаммов, соответственно, у которых мутированный ген wex встроен в хромосому. Соответствующую генетическую манипуляцию верифицировали при помощи секвенирования генома и метода ПЦР с использованием праймеров SEQ ID NO: 7 и SEQ ID NO: 8, каждый из которых позволяет амплифицировать область, прилежащую к соответствующему встроенному гену в направлении 5' и 3', соответственно, для гомологичной рекомбинации.
Полученные мутантные штаммы обозначали CA04-8352(wex S82A), CA04-8352(wex S82V), CA04-8352(wex S82L) и CA04-8352(wex S82I), соответственно.
Для определения количества триптофана, продуцируемого штаммами СА04-8352, СА04-8405, CA04-8352(wex S82A), CA04-8352(wex S82V), CA04-8352(wex S82L) и CA04-8352(wex S82I), указанные штаммы культивировали тем же способом, как и в Примере 10. После окончания культивирования количество L-триптофана, продуцируемого каждым штаммом, измеряли при помощи HPLC.
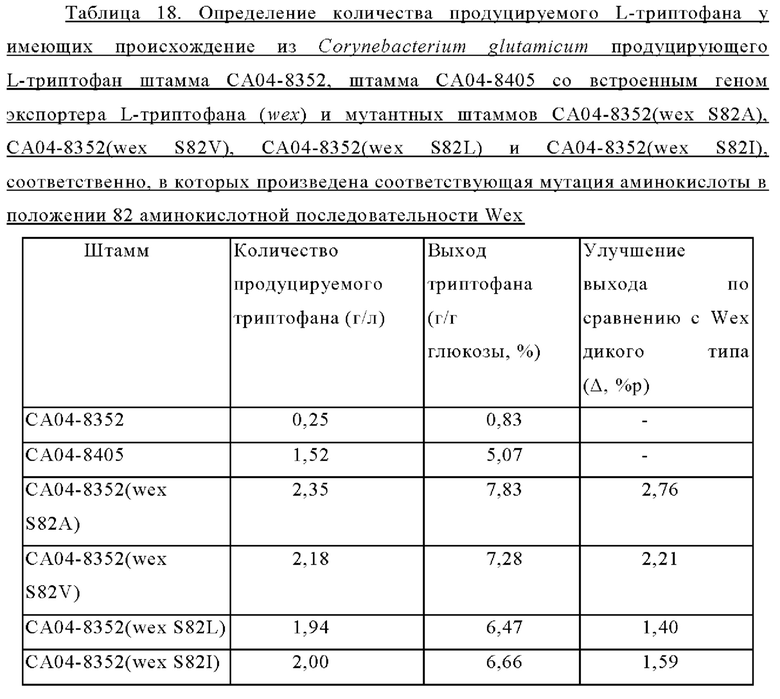
Как показывают результаты, приведенные в Таблице 18, все четыре типа мутантов, в которых аминокислота в положении 82 аминокислотной последовательности Wex заменена на другую гидрофобную аминокислоту, демонстрировали при ферментации увеличение выхода на 1,40%р - 2,76%р по сравнению со штаммом СА04-8405, в который внедрен Wex дикого типа. В частности, мутантный штамм wex S82A демонстрировал наибольшее увеличение выхода L-триптофана при ферментации, и эти результаты указывают, что замена серина в положении 82 аминокислотной последовательности Wex на другую гидрофобную аминокислоту может существенно повысить способность Wex экспортировать L-триптофан.
Пример 18. Замена 83-й аминокислоты в последовательности Wex (изолейцина) у микроорганизма рода Corynebacterium на гидрофобную аминокислоту
Проверяли повышение эффективности экспорта L-триптофана при замене 83-й аминокислоты изолейцина (далее будет обозначаться как изолейцин в положении 83) в аминокислотной последовательности мембранного белка Wex на другую гидрофобную аминокислоту. Выполняли сайт-специфический мутагенез с использованием в качестве матрицы pDZTn-PgapA-wex, полученного таким же способом, как и в Примере 14, для создания мутаций с заменой тремя различными гидрофобными аминокислотами, отличными от изолейцина.
Кроме того, изолейцин в положении 83 аминокислотной последовательности Wex заменяли каждой из других гидрофобных аминокислот (то есть аланином (А) (SEQ ID NO: 122, SEQ ID NO: 145), валином (V) (SEQ ID NO: 123, SEQ ID NO: 146) и лейцином (L) (SEQ ID NO: 124, SEQ ID NO: 147)), таким же способом, как и в Примере 14. Наборы праймеров для внедрения мутаций, а также формы мутаций, использованные для получения мутантных плазмид pDZTn-PgapA-wex, приведены в Таблице 19 ниже.
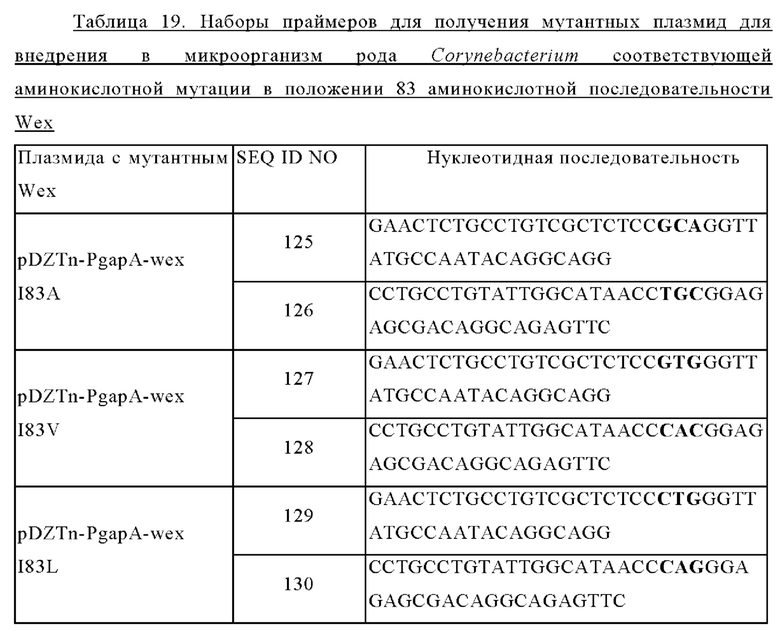
Каждым из векторов pDZTn-PgapA-wex I83A, pDZTn-PgapA-wex I83V и pDZTn-PgapA-wex I83L, полученных, как показано в Таблице 19, трансформировали посредством электропорации штамм СА04-8352 и затем осуществляли вторичный кроссинговер для получения трех типов штаммов, соответственно, у которых мутированный ген wex встроен в хромосому. Соответствующую генетическую манипуляцию верифицировали при помощи секвенирования генома и метода ПЦР с использованием праймеров SEQ ID NO: 7 и SEQ ID NO: 8, каждый из которых позволяет амплифицировать область, прилежащую к соответствующему встроенному гену в направлении 5' и 3', соответственно, для гомологичной рекомбинации.
Полученные мутантные штаммы обозначали CA04-8352(wex I83A), CA04-8352(wex I83V) и CA04-8352(wex I83L), соответственно.
Для определения количества триптофана, продуцируемого штаммами СА04-8352, СА04-8405, CA04-8352(wex I83A), CA04-8352(wex I83V) и CA04-8352(wex I83L), указанные штаммы культивировали тем же способом, как и в Примере 10. После окончания культивирования количество L-триптофана, продуцируемого каждым штаммом, измеряли при помощи HPLC.
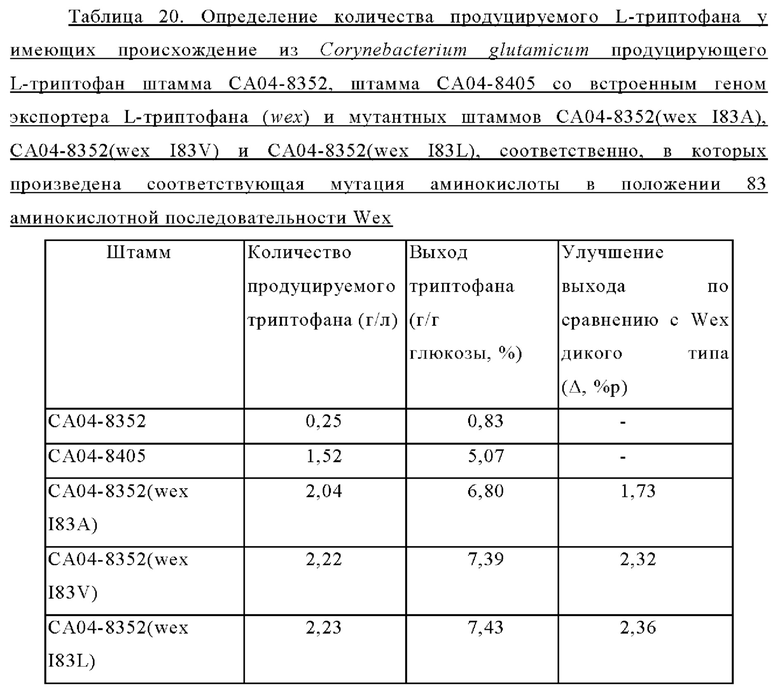
Как показывают результаты, приведенные в Таблице 20, все три типа мутантов, в которых аминокислота в положении 83 аминокислотной последовательности Wex заменена на другую гидрофобную аминокислоту, демонстрировали при ферментации увеличение выхода на 1,73%р - 2,36%р по сравнению со штаммом СА04-8405, в который внедрен Wex дикого типа. В частности, мутантный штамм wex I83L демонстрировал наибольшее увеличение выхода L-триптофана при ферментации, и эти результаты указывают, что замена изолейцина в положении 83 аминокислотной последовательности Wex на другую гидрофобную аминокислоту может существенно повысить способность Wex экспортировать L-триптофан. Результаты, приведенные выше в Примерах 2-6, подтверждают, что замена аминокислот в положениях 79 - 83 аминокислотной последовательности мембранного белка Wex на другую гидрофобную аминокислоту может существенно повысить способность штамма Corynebacterium glutamicum экспортировать L-триптофан.
Штаммы CA04-8352(wex L79I), CA04-8352(wex S80V), CA04-8352(wex L81V), CA04-8352(wex S82V) и CA04-8352(wex I83L), в которые был внедрен мутантный wex по настоящему изобретению, обозначали СМ05-9022, СМ05-9023, СМ05-9024, СМ05-9025 и СМ05-9026, соответственно. Международное депонирование указанных штаммов осуществляли 29 марта 2019 в Корейском центре культур микроорганизмов (КССМ), являющемся международным депозитарием, в соответствии с Будапештским договором, с присвоением им регистрационных номеров KCCM12475P, KCCM12476P, KCCM12477P, KCCM12478P и KCCM12479P, соответственно.
Пример 19. Замена аминокислоты в положении 79 последовательности Wex (лейцина) на гидрофобную аминокислоту у микроорганизма рода Escherichia
Эффект от введения мутации Wex, проявляющийся в улучшении способности транспортировать L-триптофан у микроорганизма рода Corynebacterium, продемонстрированный в Примерах 14 - 18 выше, еще раз подтвердили на микроорганизме рода Escherichia.
Как и в случае микроорганизма рода Corynebacterium, у микроорганизма рода Escherichia проверяли увеличение эффективности экспорта L-триптофана при замене аминокислоты в положении 79 (то есть лейцина) в аминокислотной последовательности мембранного белка Wex на другую гидрофобную аминокислоту. Выполняли сайт-специфический мутагенез с использованием в качестве матрицы pCL1920-PyccA-Hrh, полученного в Примере 8, для создания мутаций с заменой тремя различными гидрофобными аминокислотами, отличными от лейцина. Сайт-специфический мутагенез осуществляли следующим образом.
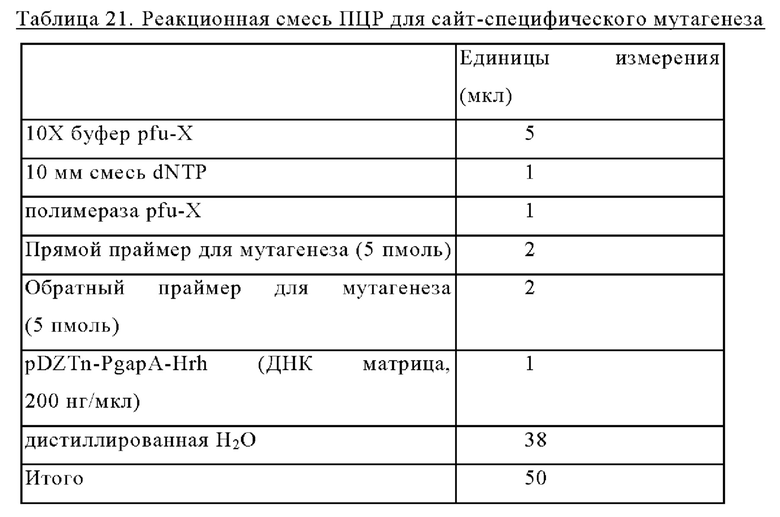
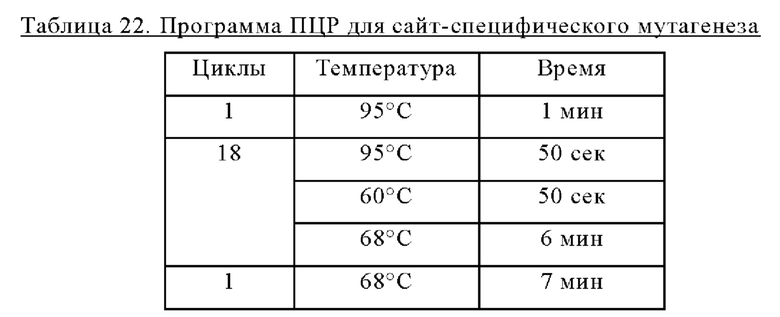
Для замены аминокислоты в положении 79 аминокислотной последовательности Wex (то есть лейцина) каждой из других гидрофобных аминокислот (то есть аланином (A) (SEQ ID NO: 80, SEQ ID NO: 131), валином (V) (SEQ ID NO: 81, SEQ ID NO: 132) и изолейцином (I) (SEQ ID NO: 82, SEQ ID NO: 133)); готовили смесь для ПЦР, согласно Таблице 21 с использованием каждого набора праймеров для внедрения мутаций, приведенных в Таблице 23, и выполняли ПЦР способом, изложенным в Таблице 22. После завершения ПЦР добавляли 1 мкл рестрикционного фермента DpnI и инкубировали смесь в течение 1 часа при 37°С. 3 мкл ДНК, обработанной DpnI, трансформировали компетентные клетки DH5a для получения мутантных плазмид pCL1920-PgapA-wex и каждую из мутаций, приведенных в Таблице 23, подтверждали посредством секвенирования.
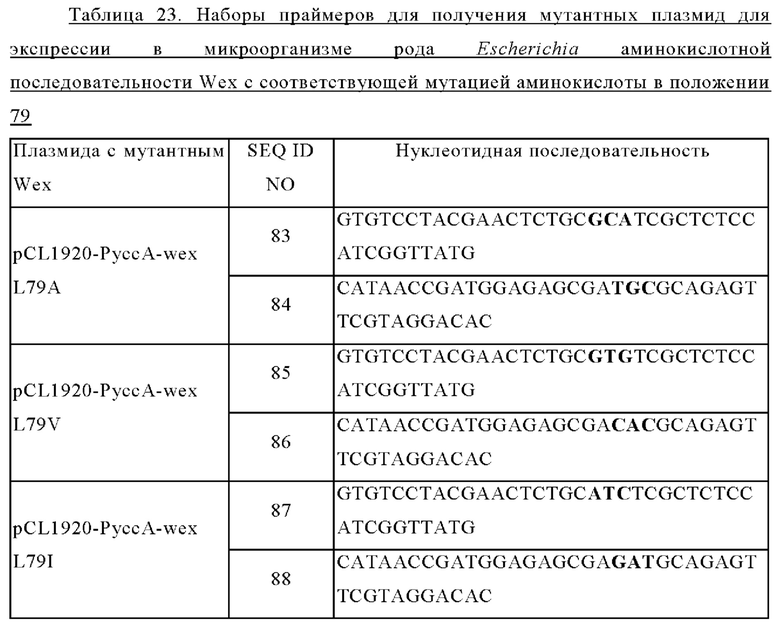
Каждым из векторов pCL1920-PyccA-wex L79A, pCL1920-PyccA-wex L79V и pCL1920-PyccA-wex L79I, полученных, как показано в Таблице 23, трансформировали посредством электропорации штамм СА04-4303 и, таким образом, получали три типа штаммов, в которые был внедрен каждый из мутированных генов wex, отличавшихся друг от друга кодируемой аминокислотой в положении 79. Полученные мутантные штаммы обозначали CA04-4303(wex L79A), CA04-4303(wex L79V) и CA04-4303(wex L79I), соответственно.
Для определения количества триптофана, продуцируемого штаммами CA04-4303(wex L79A), CA04-4303(wex L79V) и CA04-4303(wex L79I), с использованием в качестве контроля штаммов CA04-4303(pCL1920) и СА04-4306, полученных в Примере 12, указанные штаммы культивировали тем же способом, как и в Примере 12. После окончания культивирования количество L-триптофана, продуцируемого каждым штаммом, измеряли при помощи HPLC.

Как показывают результаты, приведенные в Таблице 24, все три типа мутантов, в которых аминокислота в положении 79 аминокислотной последовательности Wex заменена на другую гидрофобную аминокислоту, демонстрировали при ферментации увеличение выхода на 0,6%р - 1,5%р по сравнению со штаммом СА04-4306, в который внедрен Wex дикого типа. В частности, как и в случае продуцирующего L-триптофан штамма Corynebacterium glutamicum, штамм Е. coli с мутацией wex L79A также демонстрировал наибольшее увеличение выхода L-триптофана при ферментации, и эти результаты указывают, что замена лейцина в положении 79 аминокислотной последовательности Wex на другую гидрофобную аминокислоту может существенно повысить способность Wex экспортировать L-триптофан у микроорганизма рода Escherichia, как и у микроорганизма рода Corynebacterium.
Пример 20. Замена аминокислоты в положении 80 последовательности Wex (серина) на гидрофобную аминокислоту у микроорганизма рода Escherichia
Проверяли повышение эффективности экспорта L-триптофана у микроорганизма рода Escherichia при замене серина в положении 80 в аминокислотной последовательности мембранного белка Wex на другую гидрофобную аминокислоту. Выполняли сайт-специфический мутагенез с использованием в качестве матрицы pCL1920-PyccA-wex, полученного таким же способом, как и в Примере 19, для создания мутаций с заменой четырьмя различными гидрофобными аминокислотами.
Кроме того, серии в положении 80 аминокислотной последовательности Wex заменяли каждой из других гидрофобных аминокислот (то есть аланином (A) (SEQ ID NO: 89, SEQ ID NO: 134), валином (V) (SEQ ID NO: 90, SEQ ID NO: 135), лейцином (L) (SEQ ID NO: 91, SEQ ID NO: 136) и изолейцином (I) (SEQ ID NO: 92, SEQ ID NO: 137)), таким же способом, как и в Примере 19. Наборы праймеров для внедрения мутаций, а также формы мутаций, использованные для получения мутантных плазмид pCL1920-PyccA-wex, приведены в Таблице 25 ниже.
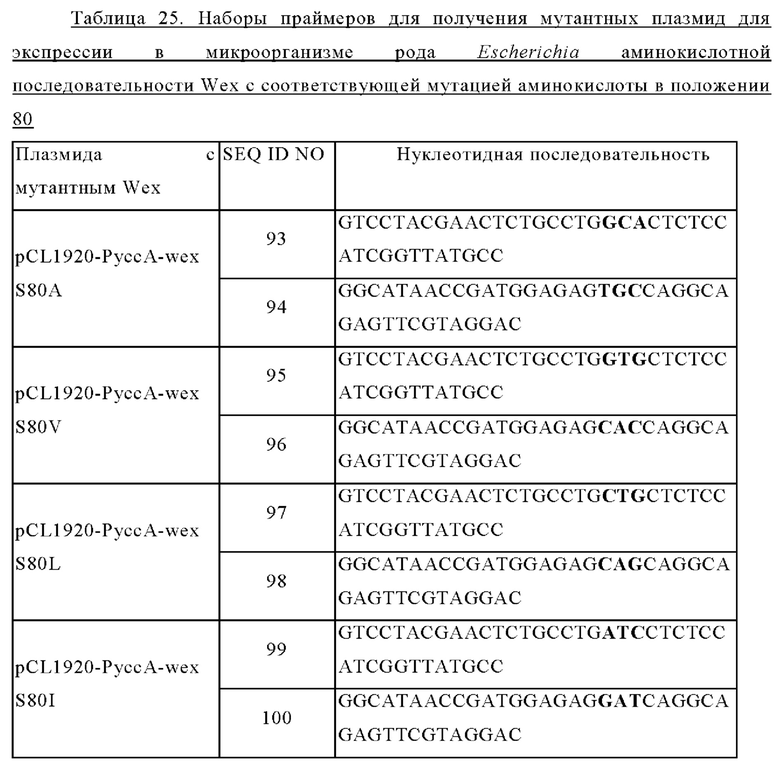
Каждым из векторов pCL1920-PyccA-wex S80A, pCL1920-PyccA-wex S80V, pCL1920-PyccA-wex S80L и pCL1920-PyccA-wex S80I, полученных, как показано в Таблице 15, трансформировали посредством электропорации штамм СА04-4303 и, таким образом, получали четыре типа штаммов, в которые был внедрен каждый из мутированных генов wex, отличавшихся друг от друга кодируемой аминокислотой в положении 80. Полученные мутантные штаммы обозначали CA04-4303(wex S80A), CA04-4303(wex S80V), CA04-4303(wex S80L) и CA04-4303(wex S80I), соответственно.
Для определения количества триптофана, продуцируемого штаммами CA04-4303(pCL1920), СА04-4306, CA04-4303(wex S80A), CA04-4303(wex S80V), CA04-4303(wex S80L) и CA04-4303(wex S80I), указанные штаммы культивировали тем же способом, как и в Примере 12. После окончания культивирования количество L-триптофана, продуцируемого каждым штаммом, измеряли при помощи HPLC.
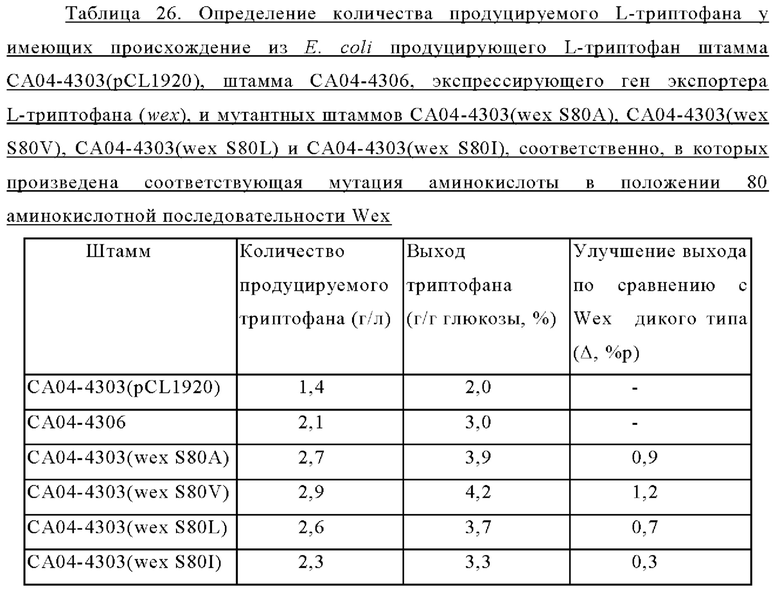
Как показывают результаты, приведенные в Таблице 16, все четыре типа мутантов, в которых аминокислота в положении 80 аминокислотной последовательности Wex заменена на другую гидрофобную аминокислоту, демонстрировали при ферментации увеличение выхода на 0,3%р - 1,2%р по сравнению со штаммом СА04-4306, в который внедрен Wex дикого типа. В частности, как и в случае продуцирующего L-триптофан штамма Corynebacterium glutamicum, штамм Е. coli с мутацией wex S80V также демонстрировал наибольшее увеличение выхода L-триптофана при ферментации, и эти результаты указывают, что замена серина в положении 80 аминокислотной последовательности Wex на другую гидрофобную аминокислоту может существенно повысить способность Wex экспортировать L-триптофан у микроорганизма рода Escherichia, как и у микроорганизма рода Corynebacterium.
Пример 21. Замена аминокислоты в положении 81 последовательности Wex (лейцина) на гидрофобную аминокислоту у микроорганизма рода Escherichia
Проверяли повышение эффективности экспорта L-триптофана у микроорганизма рода Escherichia при замене лейцина в положении 81 в аминокислотной последовательности мембранного белка Wex на другую гидрофобную аминокислоту. Выполняли сайт-специфический мутагенез с использованием в качестве матрицы pCL1920-PyccA-wex, полученного таким же способом, как и в Примере 19, для создания мутаций с заменой тремя различными гидрофобными аминокислотами, отличными от лейцина.
Кроме того, лейцин в положении 81 аминокислотной последовательности Wex заменяли каждой из других гидрофобных аминокислот (то есть аланином (А) (SEQ ID NO: 101, SEQ ID NO: 138), валином (V) (SEQ ID NO: 102, SEQ ID NO: 139) и изолейцином (I) (SEQ ID NO: 103, SEQ ID NO: 140)), таким же способом, как и в Примере 19. Наборы праймеров для внедрения мутаций, а также формы мутаций, использованные для получения мутантных плазмид pCL1920-PyccA-wex, приведены в Таблице 27 ниже.

Каждым из векторов pCL1920-PyccA-wex L81A, pCL1920-PyccA-wex L81V и pCL1920-PyccA-wex L81I, полученных, как показано в Таблице 27, трансформировали посредством электропорации штамм СА04-4303 и, таким образом, получали три типа штаммов, в которые был внедрен каждый из мутированных генов wex, отличавшихся друг от друга кодируемой аминокислотой в положении 81. Полученные мутантные штаммы обозначали CA04-4303(wex L81A), CA04-4303(wex L81V) и CA04-4303(wex L81I), соответственно.
Для определения количества триптофана, продуцируемого штаммами CA04-4303(pCL1920), СА04-4306, CA04-4303(wex L81A), CA04-4303(wex L81V) и CA04-4303(wex L81I), указанные штаммы культивировали тем же способом, как и в Примере 12. После окончания культивирования количество L-триптофана, продуцируемого каждым штаммом, измеряли при помощи HPLC.
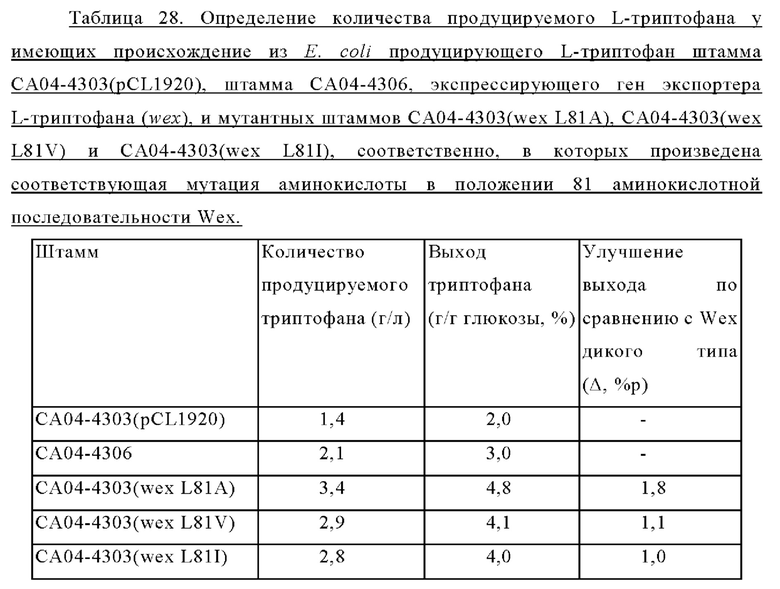
Как показывают результаты, приведенные в Таблице 28, все три типа мутантов, в которых аминокислота в положении 81 аминокислотной последовательности Wex заменена на другую гидрофобную аминокислоту, демонстрировали при ферментации увеличение выхода на 1,0%р - 1,8%р по сравнению со штаммом СА04-4306, в который внедрен Wex дикого типа. В частности, как и в случае продуцирующего L-триптофан штамма Corynebacterium glutamicum, штамм Е. coli с мутацией wex L81A также демонстрировал наибольшее увеличение выхода L-триптофана при ферментации, и эти результаты указывают, что замена лейцина в положении 81 аминокислотной последовательности Wex на другую гидрофобную аминокислоту может существенно повысить способность Wex экспортировать L-триптофан у микроорганизма рода Escherichia, как и у микроорганизма рода Corynebacterium.
Пример 22. Замена аминокислоты в положении 82 последовательности Wex (серина) на гидрофобную аминокислоту у микроорганизма рода Escherichia
Проверяли повышение эффективности экспорта L-триптофана у микроорганизма рода Escherichia при замене аминокислоты в положении 82 в аминокислотной последовательности мембранного белка Wex (то есть серина) на другую гидрофобную аминокислоту. Выполняли сайт-специфический мутагенез с использованием в качестве матрицы pCL1920-PyccA-wex, полученного таким же способом, как и в Примере 19, для создания мутаций с заменой четырьмя различными гидрофобными аминокислотами.
Кроме того, серии в положении 82 аминокислотной последовательности Wex заменяли каждой из других гидрофобных аминокислот (то есть аланином (A) (SEQ ID NO: 110, SEQ ID NO: 141), валином (V) (SEQ ID NO: NO: 111, SEQ ID NO: 142), лейцином (L) (SEQ ID N0:112, SEQ ID NO: 143) и изолейцином (I) (SEQ ID NO:
113, SEQ ID NO: 144)), таким же способом, как и в Примере 19. Наборы праймеров для внедрения мутаций, а также формы мутаций, использованные для получения мутантных плазмид pCL1920-PyccA-wex, приведены в Таблице 29 ниже.
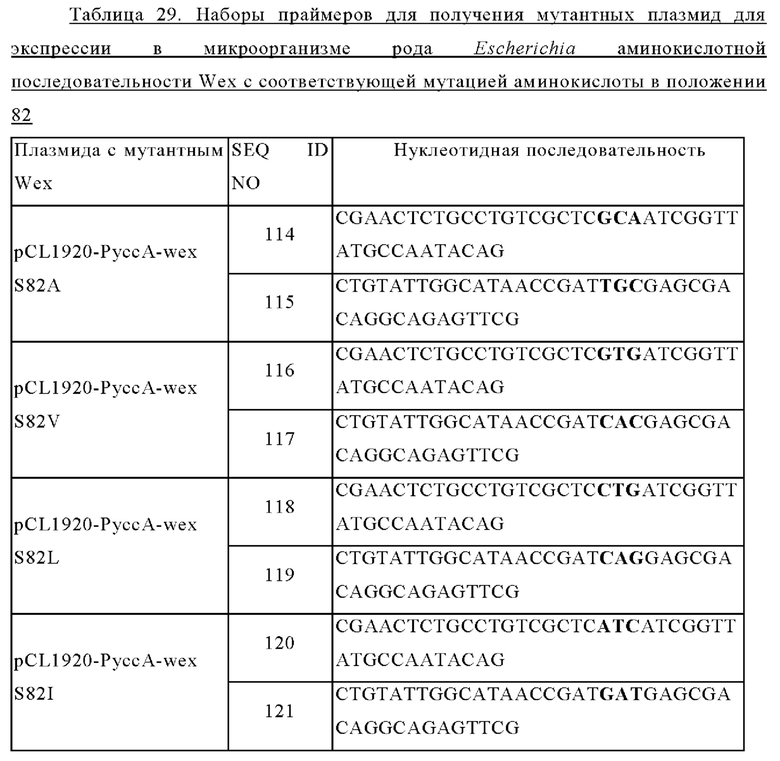
Каждым из векторов pCL1920-PyccA-wex S82A, pCL1920-PyccA-wex S82V, pCL1920-PyccA-wex S82L и pCL1920-PyccA-wex S82I, полученных, как показано в Таблице 29, трансформировали посредством электропорации штамм СА04-4303 и, таким образом, получали четыре типа штаммов, в которые был внедрен каждый из мутированных генов wex, отличавшихся друг от друга кодируемой аминокислотой в положении 82. Полученные мутантные штаммы обозначали CA04-4303(wex S82A), CA04-4303(wex S82V), CA04-4303(wex S82L) и CA04-4303(wex S82I), соответственно.
Для определения количества триптофана, продуцируемого штаммами CA04-4303(pCL1920), СА04-4306, CA04-4303(wex S82A), CA04-4303(wex S82V), CA04-4303(wex S82L) и CA04-4303(wex S82I), указанные штаммы культивировали тем же способом, как и в Примере 12. После окончания культивирования количество L-триптофана, продуцируемого каждым штаммом, измеряли при помощи HPLC.

Как показывают результаты, приведенные в Таблице 30, все четыре типа мутантов, в которых аминокислота в положении 82 аминокислотной последовательности Wex заменена на другую гидрофобную аминокислоту, демонстрировали при ферментации увеличение выхода на 0,2%р - 1,4%р по сравнению со штаммом СА04-4306, в который внедрен Wex дикого типа. В отличие от продуцирующего L-триптофан штамма Corynebacterium glutamicum, наибольшее увеличение выхода L-триптофана при ферментации демонстрировал штамм Е. coli с мутацией wex S82V, и эти результаты указывают, что замена серина в положении 82 аминокислотной последовательности Wex на другую гидрофобную аминокислоту может существенно повысить способность Wex экспортировать L-триптофан у микроорганизма рода Escherichia, как и у микроорганизма рода Corynebacterium.
Пример 23. Замена аминокислоты в положении 83 последовательности Wex (изолейцина) на гидрофобную аминокислоту у микроорганизма рода Escherichia
Проверяли повышение эффективности экспорта L-триптофана у микроорганизма рода Escherichia при замене аминокислоты в положении 83 в аминокислотной последовательности мембранного белка Wex (то есть изолейцина) на другую гидрофобную аминокислоту. Выполняли сайт-специфический мутагенез с использованием в качестве матрицы pCL1920-PyccA-wex, полученного таким же способом, как и в Примере 19, для создания мутаций с заменой тремя различными гидрофобными аминокислотами, отличными от изолейцина.
Кроме того, аминокислоту в положении 83 аминокислотной последовательности Wex (то есть изолейцин) заменяли каждой из других гидрофобных аминокислот (то есть аланином (A) (SEQ ID NO: 122, SEQ ID NO: 145), валином (V) (SEQ ID NO: 123, SEQ ID NO: 146) и лейцином (L) (SEQ ID NO: 124, SEQ ID NO: 147)), таким же способом, как и в Примере 19. Наборы праймеров для внедрения мутаций, а также формы мутаций, использованные для получения мутантных плазмид pCL1920-PyccA-wex, приведены в Таблице 31 ниже.

Каждым из векторов pCL1920-PyccA-wex I83A, pCL1920-PyccA-wex I83V и pCL1920-PyccA-wex I83L, полученных, как показано в Таблице 31, трансформировали посредством электропорации штамм СА04-4303 и, таким образом, получали три типа штаммов, в которые был внедрен каждый из мутированных генов wex, отличавшихся друг от друга кодируемой аминокислотой в положении 83. Полученные мутантные штаммы обозначали CA04-4303(wex I83A), CA04-4303(wex I83V) и CA04-4303(wex I83L), соответственно.
Для определения количества триптофана, продуцируемого штаммами CA04-4303(pCL1920), СА04-4306, CA04-4303(wex I83A), CA04-4303(wex I83V) и CA04-4303(wex I83L), указанные штаммы культивировали тем же способом, как и в Примере 12. После окончания культивирования количество L-триптофана, продуцируемого каждым штаммом, измеряли при помощи HPLC.
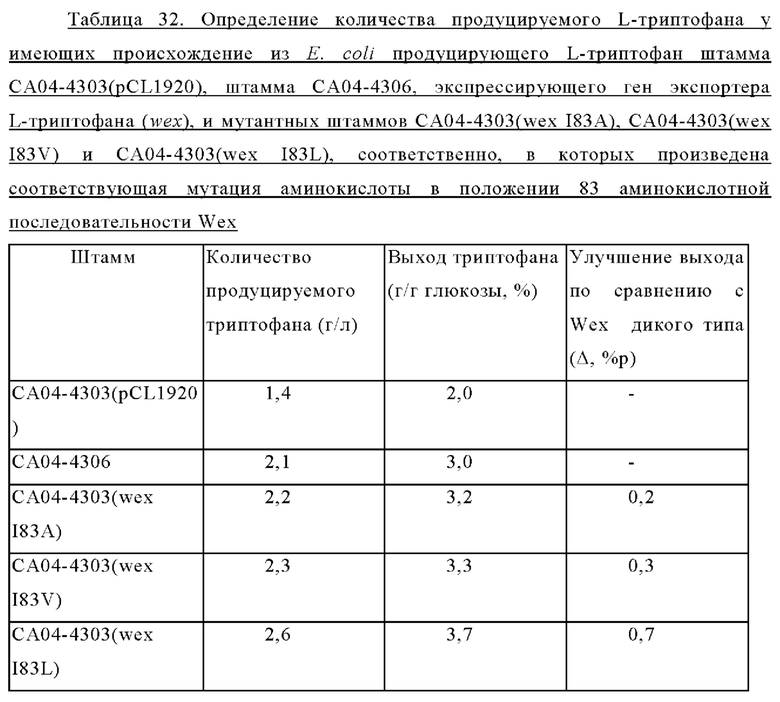
Как показывают результаты, приведенные в Таблице 32, все три типа мутантов, в которых аминокислота в положении 83 аминокислотной последовательности Wex заменена на другую гидрофобную аминокислоту, демонстрировали при ферментации увеличение выхода на 0,2%р - 0,7%р по сравнению со штаммом СА04-4306, в который внедрен Wex дикого типа. В частности, как и в случае продуцирующего L-триптофан штамма Corynebacterium glutamicum, штамм Е. coli с мутацией wex I83L также демонстрировал наибольшее увеличение выхода L-триптофана при ферментации, и эти результаты указывают, что замена изолейцина в положении 83 аминокислотной последовательности Wex на другую гидрофобную аминокислоту может существенно повысить способность Wex экспортировать L-триптофан у микроорганизма рода Escherichia, как и у микроорганизма рода Corynebacterium.
На основании вышеизложенного специалисты в области техники, к которой относится настоящее изобретение, поймут, что изобретение может быть реализовано и в других специфических формах, без изменения технической концепции или существенных признаков настоящего изобретения. В этой связи приведенные в качестве примера воплощения служат исключительно для иллюстративных целей и не должны рассматриваться как ограничивающие объем изобретения. Следует понимать, что объем настоящего изобретения включает значение и объем прилагаемой формулы изобретения и все измененные и модифицированные формы, следующие из эквивалентных концепций, а не подробное описание изобретения.
--->
<110> CJ CheilJedang Corporation
<120> NOVEL L-TRYPTOPHAN-EXPORTING PROTEIN VARIANT AND METHOD FOR
PRODUCING L-TRYPTOPHAN USING THE SAME
<130> OPA19299
<150> KR 10-2019-0040397
<151> 2019-04-05
<160> 147
<170> KoPatentIn 3.0
<210> 1
<211> 307
<212> PRT
<213> Artificial Sequence
<220>
<223> аминокислотная последовательность wex
<400> 1
Met Asn Ser Lys Lys Ala Thr Leu Ile Gly Leu Thr Ala Val Val Leu
1 5 10 15
Trp Ser Ser Ile Val Gly Leu Ile Arg Gly Val Ser Glu His Leu Gly
20 25 30
Ala Thr Gly Gly Ala Ala Met Met Tyr Ser Val Ala Ser Leu Phe Leu
35 40 45
Leu Leu Ser Val Gly Phe Pro Lys Leu Gly Ser Phe Pro Lys Lys Tyr
50 55 60
Leu Leu Trp Gly Ser Leu Leu Phe Val Ser Tyr Glu Leu Cys Leu Ser
65 70 75 80
Leu Ser Ile Gly Tyr Ala Asn Thr Gly Arg Gln Ala Ile Glu Val Ser
85 90 95
Met Val Asn Tyr Leu Trp Pro Ala Phe Thr Leu Ile Ala Ala Ile Ala
100 105 110
Phe Asn Arg Gln Arg Ala Asn Trp Met Val Val Pro Gly Phe Ile Leu
115 120 125
Ser Ile Ile Gly Ile Cys Trp Val Leu Gly Gly Asp Gln Gly Leu Asp
130 135 140
Leu Ala Gly Met Leu Gly Asn Val Gln Asp Asn Pro Leu Ser Tyr Gly
145 150 155 160
Leu Ala Phe Leu Gly Ala Val Ile Trp Ala Ala Tyr Cys Thr Val Thr
165 170 175
Ala Arg Leu Ala Lys Gly Lys Asn Gly Val Thr Leu Phe Phe Ile Leu
180 185 190
Val Ala Leu Thr Leu Trp Val Lys Phe Phe Phe Gly Asp His Arg Pro
195 200 205
Met Ser Phe Ser Leu Pro Ala Ile Val Tyr Leu Leu Leu Ala Ala Ala
210 215 220
Ala Met Gly Phe Gly Tyr Ala Ala Trp Asn Val Gly Ile Leu His Gly
225 230 235 240
Asn Val Thr Val Leu Ala Gly Val Ser Tyr Phe Ile Pro Val Phe Ser
245 250 255
Ala Ala Leu Ser Ala Met Val Leu His Ala Pro Leu Pro Arg Ser Phe
260 265 270
Trp Val Gly Ala Ser Leu Val Cys Ala Gly Ser Ile Leu Cys Trp Leu
275 280 285
Ala Thr Arg Ala Arg Arg Ala Ser Ala Ala Gln Glu Asp Ala Val Ala
290 295 300
Asp Cys Leu
305
<210> 2
<211> 924
<212> DNA
<213> Artificial Sequence
<220>
<223> нуклеотидная последовательность wex
<400> 2
atgaatagca agaaggccac gctgatcgga cttactgcag tggtcctctg gagttccatt 60
gtcggattga ttcgcggcgt cagcgaacat ctcggcgcta ccgggggcgc ggcgatgatg 120
tatagcgttg cctcgctgtt tctgctgttg tcggtcggtt tcccgaaact gggttccttt 180
ccgaaaaaat acctcttgtg gggcagcctg ctgtttgtgt cctacgaact ctgcctgtcg 240
ctctccatcg gttatgccaa tacaggcagg caagcaatcg aagtcagcat ggtcaactat 300
ctgtggccgg cattcacgct catcgccgcc attgcattca accggcagag agcgaactgg 360
atggtggtgc ccggattcat cctctcgatt atcggtatct gctgggtgct gggcggtgac 420
caggggctgg acctggcggg catgcttggc aacgtgcagg acaatccgct cagttatggg 480
ctggcttttt tgggcgccgt gatctgggcc gcctattgca ctgtgacggc ccgcctcgcg 540
aaggggaaga acggagtgac gctgttcttc attcttgtgg cattgacgct ctgggtgaag 600
tttttcttcg gcgatcaccg cccgatgtct ttcagcctgc cggcaattgt ctacctgctc 660
ctggcggcgg ccgcgatggg cttcggctat gcggcatgga atgtcgggat cttgcacggt 720
aacgtgaccg tgctggcggg cgtgtcgtac tttatcccgg ttttttcggc ggcgttgtcc 780
gcgatggtgt tgcatgcgcc gttgccgcga tcgttttggg tgggggcgtc gctggtatgc 840
gccggttcga tactgtgctg gctggcaacc agggccaggc gcgcttcggc cgcgcaagaa 900
gatgcggtgg cggactgttt gtag 924
<210> 3
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> wex - 1
<400> 3
tagaggagac acaacatgaa tagcaagaag gccac 35
<210> 4
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> wex - 2
<400> 4
ggctcttcct gtttagtcta caaacagtcc gccac 35
<210> 5
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> PgapA(-wex) - 1
<400> 5
cccttccggt ttagtttgaa gccagtgtga gttgc 35
<210> 6
<211> 39
<212> DNA
<213> Artificial Sequence
<220>
<223> PgapA(-wex) - 2
<400> 6
cttcttgcta ttcatgttgt gtctcctcta aagattgta 39
<210> 7
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Confirm_PgapA-wex - 1
<400> 7
cggattatgc caatgatgtg 20
<210> 8
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Confirm_PgapA-wex - 2
<400> 8
cacgatcacc aacattcagg 20
<210> 9
<211> 314
<212> PRT
<213> Artificial Sequence
<220>
<223> аминокислотная последовательность Pst
<400> 9
Met Lys Asn Gln Arg Lys Ala Thr Leu Ile Gly Leu Val Ala Ile Val
1 5 10 15
Leu Trp Ser Ser Ile Val Gly Leu Ile Arg Gly Val Ser Glu Ser Leu
20 25 30
Gly Ala Thr Gly Gly Ala Val Met Met Tyr Ser Val Ala Ser Val Met
35 40 45
Leu Leu Phe Thr Val Gly Phe Pro Arg Ile Arg Glu Phe Pro Arg Arg
50 55 60
Tyr Leu Val Trp Gly Ser Leu Leu Phe Val Ser Tyr Glu Leu Cys Leu
65 70 75 80
Ala Leu Ser Ile Gly Tyr Ala Asn Ser Ser Arg Gln Ala Ile Glu Val
85 90 95
Gly Met Val Asn Tyr Leu Trp Pro Ala Phe Thr Ile Leu Ala Ala Ile
100 105 110
Leu Phe Asn Arg Gln Gln Ala Asn Leu Leu Ile Val Pro Gly Phe Leu
115 120 125
Ile Ala Ile Leu Gly Ile Cys Trp Val Leu Gly Gly Glu Gln Gly Leu
130 135 140
Asp Leu Ser Gly Met Thr Ala Asn Ile Arg Asp Asn Pro Leu Ser Tyr
145 150 155 160
Gly Leu Ala Phe Ala Gly Ala Val Ile Trp Ala Ala Tyr Cys Thr Val
165 170 175
Thr Thr Arg Ile Ala Gly Gly Lys Asn Gly Val Thr Leu Phe Phe Met
180 185 190
Leu Thr Ala Leu Ala Leu Trp Ala Lys Tyr Leu Ala Ile Gly Gly Glu
195 200 205
Thr Met Glu Phe Ser Tyr His Ala Leu Ile Tyr Leu Val Leu Ala Ala
210 215 220
Ser Ala Met Gly Phe Gly Tyr Ala Ala Trp Asn Val Gly Ile Leu His
225 230 235 240
Gly Asn Val Thr Val Leu Ala Gly Ala Ser Tyr Phe Ile Pro Val Leu
245 250 255
Ser Ala Ala Leu Ala Ala Val Leu Leu Arg Thr Pro Leu Ser Leu Ser
260 265 270
Phe Trp Gln Gly Ala Ala Met Val Cys Ile Gly Ser Ile Leu Cys Trp
275 280 285
Phe Ala Thr Arg Ala Lys Pro Pro Glu Ser Ala Gln Ser Gly Asp Gln
290 295 300
Ala Ser Ala Thr Thr Pro Arg Arg Asn Gly
305 310
<210> 10
<211> 945
<212> DNA
<213> Artificial Sequence
<220>
<223> нуклеотидная последовательность Pst
<400> 10
atgaaaaacc agcgtaaagc gaccctcatc gggctcgttg caattgtctt gtggagctcg 60
atcgtcggcc tgatccgggg cgtcagcgag agcctcggcg cgaccggtgg tgccgtcatg 120
atgtacagcg tcgcatcggt aatgctgttg ttcacggtcg gctttccgcg gatacgggag 180
ttcccccgac gctatcttgt ctggggcagc ctgctgttcg tctcgtacga gctgtgcctt 240
gccctgtcca tcggctacgc caacagcagc cgacaggcca tcgaggtcgg catggtcaat 300
tacctgtggc cggccttcac gatcctggcg gcgatcctgt tcaacaggca gcaggccaac 360
ctgctcatcg ttcccggctt cctcatcgcg atcctcggga tctgctgggt gctcggcggg 420
gaacaggggc tggacctgtc cgggatgacg gcgaacatcc gcgacaatcc cctcagctac 480
gggctggcct tcgccggcgc ggtgatctgg gcggcatact gcacggtgac cacgcggatc 540
gccggcggca agaacggtgt cacgctgttc ttcatgctga cggcattggc gctatgggcc 600
aagtacctgg ccatcggcgg ggaaacgatg gaattcagct accacgcgct gatctacctg 660
gtgctggccg cctccgcgat gggcttcggc tatgcggcgt ggaacgtcgg catcctgcac 720
ggcaatgtca ccgtcctcgc tggtgcttcg tatttcatcc cggtgctgtc cgccgccctg 780
gcggccgtac tgttgcgtac gccgctgtcg ctgtcgttct ggcagggtgc cgccatggtc 840
tgcatcgggt cgatcctctg ctggttcgcc acccgtgcga aaccgccaga atcggcgcag 900
tcgggtgacc aggccagcgc aaccacgccg cgtcgaaacg gataa 945
<210> 11
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Pst-1
<400> 11
tagaggagac acaacatgaa aaaccagcgt aaagc 35
<210> 12
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Pst-2
<400> 12
ggctcttcct gtttagttta tccgtttcga cgcgg 35
<210> 13
<211> 39
<212> DNA
<213> Artificial Sequence
<220>
<223> PgapA(-Pst)-2
<400> 13
acgctggttt ttcatgttgt gtctcctcta aagattgta 39
<210> 14
<211> 303
<212> PRT
<213> Artificial Sequence
<220>
<223> аминокислотная последовательность Afa
<400> 14
Met Lys Gln Ser Asp Lys Ala Thr Leu Ile Gly Leu Ile Ala Ile Val
1 5 10 15
Leu Trp Ser Thr Ile Val Gly Leu Ile Arg Ser Val Ser Asp Ser Leu
20 25 30
Gly Val Thr Gly Gly Ala Ala Leu Ile Tyr Thr Leu Ala Ser Val Phe
35 40 45
Leu Leu Leu Ser Val Gly Trp Val Arg Leu Arg Asp Phe Pro Arg Arg
50 55 60
Tyr Leu Ile Trp Gly Ser Val Leu Phe Val Cys Tyr Glu Leu Cys Leu
65 70 75 80
Ala Leu Ser Ile Gly Tyr Ala His Asn Ser Gln Gln Ala Ile Glu Val
85 90 95
Gly Met Val Asn Tyr Leu Trp Pro Thr Phe Thr Ile Val Ala Ala Ile
100 105 110
Leu Phe Asn Lys Gln Lys Ala Asn Gly Leu Leu Ala Pro Gly Leu Leu
115 120 125
Leu Ser Met Met Gly Ile Ser Trp Ile Leu Gly Gly Glu Gln Gly Leu
130 135 140
Ser Leu His Asn Ile Trp Leu Asn Val Gln Asp Asn Pro Leu Ser Tyr
145 150 155 160
Gly Leu Ala Phe Ser Gly Ala Leu Ile Trp Ala Gly Tyr Ser Thr Met
165 170 175
Thr Ala Arg Ile Ala Gln Gly Lys Asn Gly Ile Thr Leu Phe Phe Met
180 185 190
Leu Thr Ala Ala Ala Leu Trp Val Lys Tyr Leu Val Gln Gly Ala Pro
195 200 205
Ala Met Thr Phe Thr Val Pro Ala Leu Val Tyr Leu Leu Leu Ala Ala
210 215 220
Met Ala Met Gly Phe Gly Tyr Ala Ala Trp Asn Val Gly Ile Leu His
225 230 235 240
Gly Asn Val Thr Ile Leu Ala Gly Ala Ser Tyr Phe Ile Pro Val Phe
245 250 255
Ser Ala Ala Leu Ser Thr Val Leu Leu Gln Ala Pro Leu Thr Leu Thr
260 265 270
Phe Trp Gln Gly Ser Ser Met Val Cys Leu Gly Ala Leu Leu Cys Trp
275 280 285
Leu Ala Ile Arg Val Arg Lys Pro Arg Ser Leu Lys Ser Ala Ala
290 295 300
<210> 15
<211> 912
<212> DNA
<213> Artificial Sequence
<220>
<223> нуклеотидная последовательность Afa
<400> 15
atgaagcaat ctgataaggc aaccctgatc gggctgatcg ccattgtcct ttggagcacg 60
attgtcggcc tgatacgcag cgtcagcgac tctctgggcg taaccggcgg cgctgccctg 120
atttacaccc tggcctcggt ctttcttctt ttatcagtgg gctgggtacg cttgcgcgac 180
ttcccgcgtc gctacctgat ctggggcagt gtgctgtttg tctgctatga actctgcctg 240
gccctgtcca tcggctatgc ccacaacagc cagcaggcaa ttgaagtggg gatggtcaac 300
tatctgtggc cgacctttac cattgtggcc gccatcttgt tcaataagca aaaagccaat 360
gggctgcttg cacccggcct gctcttgtcc atgatgggaa tcagctggat tctgggcggc 420
gagcaaggct tgagcctgca caacatctgg ctgaatgtgc aggacaatcc cttgagctac 480
ggcctggcct ttagcggcgc gctgatctgg gccggctaca gcaccatgac cgcccgcatc 540
gcccagggca aaaatggcat caccctgttt ttcatgctga cggcagcggc cttgtgggtg 600
aagtacctgg tccaaggtgc tcctgccatg acgtttacgg ttcccgcctt ggtgtatttg 660
ctgctggcgg ccatggcgat gggctttggc tatgccgcct ggaatgtcgg tattttgcat 720
ggcaatgtca ccatcctggc cggcgcttcc tactttattc cggtattttc agccgccctg 780
tccaccgttt tgctgcaagc tccgttgacg ctgaccttct ggcaaggctc gtccatggtg 840
tgtttgggtg ccctgctatg ctggctggcc atccgggttc gcaaaccccg gtcactaaaa 900
agcgctgcct ga 912
<210> 16
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Afa-1
<400> 16
tagaggagac acaacatgaa gcaatctgat aaggc 35
<210> 17
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Afa-2
<400> 17
gctcttcctg tttagttcag gcagcgcttt ttagt 35
<210> 18
<211> 39
<212> DNA
<213> Artificial Sequence
<220>
<223> PgapA(-Afa)-2
<400> 18
atcagattgc ttcatgttgt gtctcctcta aagattgta 39
<210> 19
<211> 314
<212> PRT
<213> Artificial Sequence
<220>
<223> аминокислотная последовательность Cne
<400> 19
Met Gln Ser Lys Ser Lys Ala Thr Leu Ile Gly Leu Ile Ala Ile Leu
1 5 10 15
Leu Trp Ser Ser Ile Val Gly Leu Ile Arg Gly Val Ser Glu Asn Leu
20 25 30
Gly Ala Thr Gly Gly Ala Ala Met Ile Tyr Thr Val Ala Ser Ala Leu
35 40 45
Leu Leu Leu Thr Val Gly Phe Val Arg Met Gln Asp Phe Pro Arg Arg
50 55 60
Tyr Leu Val Trp Gly Ser Ile Leu Phe Val Ser Tyr Glu Leu Cys Leu
65 70 75 80
Ser Leu Ser Ile Gly Tyr Ala Asn Ser Ser Arg Gln Ala Ile Glu Val
85 90 95
Gly Met Val Asn Tyr Leu Trp Pro Ser Phe Thr Met Leu Cys Ala Ile
100 105 110
Ala Phe Asn Lys Gln Lys Ala Asn Leu Leu Ile Ile Pro Gly Phe Leu
115 120 125
Ile Ala Ile Leu Gly Ile Cys Trp Val Leu Gly Gly Asp Gln Gly Leu
130 135 140
Asp Phe Ala Gly Met Ala Glu Asn Ile Gln Asp Asn Pro Leu Ser Tyr
145 150 155 160
Gly Leu Ala Phe Leu Gly Ala Leu Ile Trp Ala Ala Tyr Cys Thr Val
165 170 175
Thr Asn Arg Ile Ala Glu Gly Arg Asn Gly Ile Thr Leu Phe Phe Met
180 185 190
Leu Thr Ala Leu Ala Leu Trp Ile Lys Tyr Phe Ala Thr Glu Ser Gly
195 200 205
Ser Met Glu Phe Ser Tyr Gln Ala Val Ile Tyr Leu Ala Leu Ala Ala
210 215 220
Ser Ala Met Gly Phe Gly Tyr Ala Ala Trp Asn Val Gly Ile Leu His
225 230 235 240
Gly Asn Val Thr Val Leu Ala Gly Ala Ser Tyr Phe Ile Pro Val Leu
245 250 255
Ser Ala Ala Leu Ala Ala Met Leu Leu Arg Thr Pro Leu Ser Ile Ala
260 265 270
Phe Trp Lys Gly Ala Ser Met Val Cys Ala Gly Ser Ile Leu Cys Trp
275 280 285
Leu Ala Thr Arg Gly Gln Arg Ser Lys Ala Pro Pro Leu Pro Glu Leu
290 295 300
Pro Gln Ser Arg Glu Arg Val Gln Glu Pro
305 310
<210> 20
<211> 945
<212> DNA
<213> Artificial Sequence
<220>
<223> нуклеотидная последовательность Cne
<400> 20
atgcaaagca agagcaaagc aactctcatc gggctcatcg cgattctgtt atggagctcg 60
attgtcggcc tgattcgcgg tgtcagcgaa aaccttgggg caaccggtgg ggcggcaatg 120
atctataccg tcgcctcggc cctgctcttg ctgacagtcg gtttcgtcag aatgcaggat 180
tttccccggc gctatctggt ttggggaagc attctgttcg tttcgtacga gctgtgtctt 240
tccttgtcca ttggctacgc caacagcagc aggcaagcca ttgaggtggg gatggtcaac 300
tacttgtggc cgagcttcac catgctgtgt gccatcgcat tcaacaagca gaaggccaac 360
ttgctgatca ttcccggctt cctgatcgcc attctcggga tctgctgggt gcttggcggg 420
gatcagggcc tggacttcgc cgggatggcg gagaacatcc aggacaatcc gctcagctat 480
gggctggcct tccttggtgc cctgatctgg gcggcgtatt gcactgtgac caaccggatt 540
gccgaaggca ggaatggcat cacgctgttc ttcatgctga cagcgctggc gttgtggatc 600
aagtatttcg ccacagagag cgggtcgatg gaatttagct atcaggcagt gatttatctt 660
gcgttggccg cctctgcgat gggattcggc tatgcggcct ggaatgttgg catcctgcat 720
ggcaatgtca ccgtccttgc cggcgcttcc tacttcattc cggtactttc cgccgccctg 780
gcggccatgc tcttgcgtac acccctgtcg atcgccttct ggaagggcgc atccatggta 840
tgtgcggggt cgatcctctg ttggctggca acacgtgggc aacgttccaa ggcacctccg 900
ttgccggaat taccgcagtc gcgcgaacgt gtccaggaac cgtga 945
<210> 21
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Cne-1
<400> 21
tagaggagac acaacatgca aagcaagagc aaagc 35
<210> 22
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Cne-2
<400> 22
ggctcttcct gtttagttca cggttcctgg acacg 35
<210> 23
<211> 39
<212> DNA
<213> Artificial Sequence
<220>
<223> PgapA(-Cne)-2
<400> 23
gctcttgctt tgcatgttgt gtctcctcta aagattgta 39
<210> 24
<211> 293
<212> PRT
<213> Artificial Sequence
<220>
<223> аминокислотная последовательность Eco
<400> 24
Met Thr Arg Gln Lys Ala Thr Leu Ile Gly Leu Ile Ala Ile Val Leu
1 5 10 15
Trp Ser Thr Met Val Gly Leu Ile Arg Gly Val Ser Glu Gly Leu Gly
20 25 30
Pro Val Gly Gly Ala Ala Ala Ile Tyr Ser Leu Ser Gly Leu Leu Leu
35 40 45
Ile Phe Thr Val Gly Phe Pro Arg Ile Arg Gln Ile Pro Lys Gly Tyr
50 55 60
Leu Leu Ala Gly Ser Leu Leu Phe Val Ser Tyr Glu Ile Cys Leu Ala
65 70 75 80
Leu Ser Leu Gly Tyr Ala Ala Thr His His Gln Ala Ile Glu Val Gly
85 90 95
Met Val Asn Tyr Leu Trp Pro Ser Leu Thr Ile Leu Phe Ala Ile Leu
100 105 110
Phe Asn Gly Gln Lys Thr Asn Trp Leu Ile Val Pro Gly Leu Leu Leu
115 120 125
Ala Leu Val Gly Val Cys Trp Val Leu Gly Gly Asp Asn Gly Leu His
130 135 140
Tyr Asp Glu Ile Ile Asn Asn Ile Thr Thr Ser Pro Leu Ser Tyr Phe
145 150 155 160
Leu Ala Phe Ile Gly Ala Phe Ile Trp Ala Ala Tyr Cys Thr Val Thr
165 170 175
Asn Lys Tyr Ala Arg Gly Phe Asn Gly Ile Thr Val Phe Val Leu Leu
180 185 190
Thr Gly Ala Ser Leu Trp Val Tyr Tyr Phe Leu Thr Pro Gln Pro Glu
195 200 205
Met Ile Phe Ser Thr Pro Val Met Ile Lys Leu Ile Ser Ala Ala Phe
210 215 220
Thr Leu Gly Phe Ala Tyr Ala Ala Trp Asn Val Gly Ile Leu His Gly
225 230 235 240
Asn Val Thr Ile Met Ala Val Gly Ser Tyr Phe Thr Pro Val Leu Ser
245 250 255
Ser Ala Leu Ala Ala Val Leu Leu Ser Ala Pro Leu Ser Phe Ser Phe
260 265 270
Trp Gln Gly Ala Leu Met Val Cys Gly Gly Ser Leu Leu Cys Trp Leu
275 280 285
Ala Thr Arg Arg Gly
290
<210> 25
<211> 882
<212> DNA
<213> Artificial Sequence
<220>
<223> нуклеотидная последовательность Eco
<400> 25
atgacacgac aaaaagcaac gctcataggg ctgatagcga tcgtcctgtg gagcacgatg 60
gtaggattga ttcgcggtgt cagtgagggg ctcggcccgg tcggcggcgc agctgctatc 120
tattcattaa gcgggctgct gttaatcttc acggttggat ttccgcgtat tcggcaaatc 180
ccgaaaggct atttactcgc cgggagtctg ttattcgtca gctatgaaat ctgtctggcg 240
ctttccttag ggtatgcggc gacccatcat caggcgattg aagtgggtat ggtgaactat 300
ctgtggccca gcctgacaat tctctttgcc attctgttta atggtcagaa aaccaactgg 360
ttgattgtac ctggattatt attagccctc gtcggcgtct gttgggtgtt aggcggtgac 420
aatgggttac attatgatga aatcatcaat aatatcacca ccagcccatt gagttatttc 480
ctggcgttca ttggtgcgtt tatctgggca gcctattgca cagtaacgaa taaatacgca 540
cgcggattta atggaattac cgtttttgtc ctgctaacgg gagcaagtct gtgggtttac 600
tattttctta cgccacaacc agaaatgata tttagcacgc ccgtcatgat taaactcatc 660
tctgcggcat ttaccttagg atttgcttat gctgcatgga atgtcggtat attgcatggc 720
aatgtcacca ttatggcggt aggttcgtat tttacgcctg tactttcctc agcgcttgca 780
gccgtgctgc tcagcgcccc gctgtcgttc tcgttctggc aaggcgcgct gatggtctgc 840
ggcggttccc tgctctgctg gctggcgaca cgtcgtggtt aa 882
<210> 26
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Eco-1
<400> 26
tagaggagac acaacatgac acgacaaaaa gcaac 35
<210> 27
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Eco-2
<400> 27
gctcttcctg tttagtttaa ccacgacgtg tcgcc 35
<210> 28
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> PgapA(-Eco)-2
<400> 28
tttttgtcgt gtcatgttgt gtctcctcta aagattg 37
<210> 29
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Hrh-3
<400> 29
atagagagtg actcaatgaa tagcaagaag gccac 35
<210> 30
<211> 33
<212> DNA
<213> Artificial Sequence
<220>
<223> Hrh-4
<400> 30
tcgagctcgg tacccctaca aacagtccgc cac 33
<210> 31
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<223> PyccA - 1
<400> 31
ctctagagga tccccttcca gatcaaatgc gtaa 34
<210> 32
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> PyccA(-Hrh)-2
<400> 32
cttcttgcta ttcattgagt cactctctat gacag 35
<210> 33
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Pst-3
<400> 33
atagagagtg actcaatgaa aaaccagcgt aaagc 35
<210> 34
<211> 33
<212> DNA
<213> Artificial Sequence
<220>
<223> Pst-4
<400> 34
tcgagctcgg tacccttatc cgtttcgacg cgg 33
<210> 35
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> PyccA(-Pst)-2
<400> 35
acgctggttt ttcattgagt cactctctat gacag 35
<210> 36
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Afa-3
<400> 36
atagagagtg actcaatgaa gcaatctgat aaggc 35
<210> 37
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<223> Afa-4
<400> 37
tcgagctcgg taccctcagg cagcgctttt tagt 34
<210> 38
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> PyccA(-Afa)-2
<400> 38
atcagattgc ttcattgagt cactctctat gacag 35
<210> 39
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Cne-3
<400> 39
atagagagtg actcaatgca aagcaagagc aaagc 35
<210> 40
<211> 33
<212> DNA
<213> Artificial Sequence
<220>
<223> Cne-4
<400> 40
tcgagctcgg taccctcacg gttcctggac acg 33
<210> 41
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> PyccA(-Cne)-2
<400> 41
gctcttgctt tgcattgagt cactctctat gacag 35
<210> 42
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Eco-3
<400> 42
atagagagtg actcaatgac acgacaaaaa gcaac 35
<210> 43
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<223> Eco-4
<400> 43
tcgagctcgg tacccttaac cacgacgtgt cgcc 34
<210> 44
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> PyccA(-Eco)-2
<400> 44
tttttgtcgt gtcattgagt cactctctat gacag 35
<210> 45
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Pspl7-trpE(S38R)_L-1
<400> 45
tcgagctcgg tacccaaaca actgcgacgt gtgtc 35
<210> 46
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Pspl7-trpE(S38R)_L-2
<400> 46
catgaagcgc cggtacctta atcatttttg ggttc 35
<210> 47
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Pspl7-trpE(S38R)_R-1
<400> 47
gccctgttgg aacgcgctga tatcaccacc aagaa 35
<210> 48
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Pspl7-trpE(S38R)_R-2
<400> 48
ctctagagga tccccagatg tcaccgttgt aaatg 35
<210> 49
<211> 294
<212> DNA
<213> Artificial Sequence
<220>
<223> последовательность промотора spl7
<400> 49
ggcgcttcat gtcaacaatc tttaacgttt tcaagttcac aagtcgtgtt caaatggtga 60
caagattgga cactgtgctg aattggcacc aagccctcat aaatgataga tctaaatcga 120
atatcaatat atggtctgtt tattggaacg cgtcccagtg gctgagacgc atccgctaaa 180
gccccaggaa ccctgtgcag aaagaacaaa taatcgtgaa ttttggcagc aacagcaatt 240
cctgctacaa ttgaaaacgt gcaaaagcat agattattgg aggagatcaa aaca 294
<210> 50
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Pspl7 - 1
<400> 50
cccaaaaatg attaaggtac cggcgcttca tgtca 35
<210> 51
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> Pspl7 - 2
<400> 51
gggattcgtg ctcatgatat ctgttttgat ctcctcc 37
<210> 52
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> trpE (S38R) - 1
<400> 52
atcaaaacag atatcatgag cacgaatccc catgt 35
<210> 53
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> trpE (S38R) - 2
<400> 53
gtggtgatat cagcgcgttc caacagggct gcatc 35
<210> 54
<211> 19
<212> DNA
<213> Artificial Sequence
<220>
<223> Confirm_Pspl7-trpE(S38R) - 1
<400> 54
gaagaagagg ctgcagatg 19
<210> 55
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> Confirm_Pspl7-trpE(S38R) - 2
<400> 55
gatcagcgcc atcatgtt 18
<210> 56
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Pn-tkt_L - 1
<400> 56
tcgagctcgg tacccaaact ttgagtgggt gcgtg 35
<210> 57
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Pn-tkt_L - 2
<400> 57
tcgagctacg agggcggttc ccagcccttc attag 35
<210> 58
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Pn-tkt_R - 1
<400> 58
attaacggtt aattgattct ggacgtcatg actac 35
<210> 59
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Pn-tkt_R - 2
<400> 59
ctctagagga tccccgcctc gatgatgcag tcgtc 35
<210> 60
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Pn-tkt - 1
<400> 60
gaagggctgg gaaccgccct cgtagctcga gagtt 35
<210> 61
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> Pn-tkt - 2
<400> 61
catgacgtcc agaatcaatt aaccgttaat ggagtcc 37
<210> 62
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Confirm_Pn-tkt - 1
<400> 62
acccagaacc ccaaattttc 20
<210> 63
<211> 19
<212> DNA
<213> Artificial Sequence
<220>
<223> Confirm_Pn-tkt - 2
<400> 63
ttgagttcga caactttgg 19
<210> 64
<211> 70
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 64
tgcaatgcat aacaacgcag tcgcactatt tttcactgga gagaagccct gtgtaggctg 60
gagctgcttc 70
<210> 65
<211> 70
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 65
tgcaatgcat aacaacgcag tcgcactatt tttcactgga gagaagccct gtccatatga 60
atatcctcct 70
<210> 66
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 66
gggcaggatc tcctgtcatc 20
<210> 67
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 67
aaatgtcgga taaggcaccg 20
<210> 68
<211> 70
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 68
tgtaatattc acagggatca ctgtaattaa aataaatgaa ggattatgta gtgtaggctg 60
gagctgcttc 70
<210> 69
<211> 70
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 69
tgtagggtaa gagagtggct aacatcctta tagccactct gtagtattaa gtccatatga 60
atatcctcct 70
<210> 70
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 70
acatccttat agccactctg 20
<210> 71
<211> 70
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 71
tacaaccggg ggaggcattt tgcttccccc gctaacaatg gcgacatatt gtgtaggctg 60
gagctgcttc 70
<210> 72
<211> 70
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 72
gcattcggtg cacgatgcct gatgcgccac gtcttatcag gcctacaaaa gtccatatga 60
atatcctcct 70
<210> 73
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 73
aggacggata aggcgttcac 20
<210> 74
<211> 26
<212> DNA
<213> Artificial Sequence
<220>
<223> trpE - 1
<400> 74
gaattcatgc aaacacaaaa accgac 26
<210> 75
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> trpE - 2
<400> 75
gaattctcag aaagtctcct gtgca 25
<210> 76
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> trpE(P21S) - 1
<400> 76
cgcttatcgc gacaattcca ccgcgctttt tcaccag 37
<210> 77
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> trpE(P21S) - 2
<400> 77
ctggtgaaaa agcgcggtgg aattgtcgcg ataagcg 37
<210> 78
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex-1)
<400> 78
actctagagg atccccttcc agatcaaatg cgtaa 35
<210> 79
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex-2)
<400> 79
attcgagctc ggtaccccta caaacagtcc gccac 35
<210> 80
<211> 924
<212> DNA
<213> Artificial Sequence
<220>
<223> wex(L79A)
<400> 80
atgaatagca agaaggccac gctgatcgga cttactgcag tggtcctctg gagttccatt 60
gtcggattga ttcgcggcgt cagcgaacat ctcggcgcta ccgggggcgc ggcgatgatg 120
tatagcgttg cctcgctgtt tctgctgttg tcggtcggtt tcccgaaact gggttccttt 180
ccgaaaaaat acctcttgtg gggcagcctg ctgtttgtgt cctacgaact ctgcgcatcg 240
ctctccatcg gttatgccaa tacaggcagg caagcaatcg aagtcagcat ggtcaactat 300
ctgtggccgg cattcacgct catcgccgcc attgcattca accggcagag agcgaactgg 360
atggtggtgc ccggattcat cctctcgatt atcggtatct gctgggtgct gggcggtgac 420
caggggctgg acctggcggg catgcttggc aacgtgcagg acaatccgct cagttatggg 480
ctggcttttt tgggcgccgt gatctgggcc gcctattgca ctgtgacggc ccgcctcgcg 540
aaggggaaga acggagtgac gctgttcttc attcttgtgg cattgacgct ctgggtgaag 600
tttttcttcg gcgatcaccg cccgatgtct ttcagcctgc cggcaattgt ctacctgctc 660
ctggcggcgg ccgcgatggg cttcggctat gcggcatgga atgtcgggat cttgcacggt 720
aacgtgaccg tgctggcggg cgtgtcgtac tttatcccgg ttttttcggc ggcgttgtcc 780
gcgatggtgt tgcatgcgcc gttgccgcga tcgttttggg tgggggcgtc gctggtatgc 840
gccggttcga tactgtgctg gctggcaacc agggccaggc gcgcttcggc cgcgcaagaa 900
gatgcggtgg cggactgttt gtag 924
<210> 81
<211> 924
<212> DNA
<213> Artificial Sequence
<220>
<223> wex(L79V)
<400> 81
atgaatagca agaaggccac gctgatcgga cttactgcag tggtcctctg gagttccatt 60
gtcggattga ttcgcggcgt cagcgaacat ctcggcgcta ccgggggcgc ggcgatgatg 120
tatagcgttg cctcgctgtt tctgctgttg tcggtcggtt tcccgaaact gggttccttt 180
ccgaaaaaat acctcttgtg gggcagcctg ctgtttgtgt cctacgaact ctgcgtgtcg 240
ctctccatcg gttatgccaa tacaggcagg caagcaatcg aagtcagcat ggtcaactat 300
ctgtggccgg cattcacgct catcgccgcc attgcattca accggcagag agcgaactgg 360
atggtggtgc ccggattcat cctctcgatt atcggtatct gctgggtgct gggcggtgac 420
caggggctgg acctggcggg catgcttggc aacgtgcagg acaatccgct cagttatggg 480
ctggcttttt tgggcgccgt gatctgggcc gcctattgca ctgtgacggc ccgcctcgcg 540
aaggggaaga acggagtgac gctgttcttc attcttgtgg cattgacgct ctgggtgaag 600
tttttcttcg gcgatcaccg cccgatgtct ttcagcctgc cggcaattgt ctacctgctc 660
ctggcggcgg ccgcgatggg cttcggctat gcggcatgga atgtcgggat cttgcacggt 720
aacgtgaccg tgctggcggg cgtgtcgtac tttatcccgg ttttttcggc ggcgttgtcc 780
gcgatggtgt tgcatgcgcc gttgccgcga tcgttttggg tgggggcgtc gctggtatgc 840
gccggttcga tactgtgctg gctggcaacc agggccaggc gcgcttcggc cgcgcaagaa 900
gatgcggtgg cggactgttt gtag 924
<210> 82
<211> 924
<212> DNA
<213> Artificial Sequence
<220>
<223> wex(L79I)
<400> 82
atgaatagca agaaggccac gctgatcgga cttactgcag tggtcctctg gagttccatt 60
gtcggattga ttcgcggcgt cagcgaacat ctcggcgcta ccgggggcgc ggcgatgatg 120
tatagcgttg cctcgctgtt tctgctgttg tcggtcggtt tcccgaaact gggttccttt 180
ccgaaaaaat acctcttgtg gggcagcctg ctgtttgtgt cctacgaact ctgcatctcg 240
ctctccatcg gttatgccaa tacaggcagg caagcaatcg aagtcagcat ggtcaactat 300
ctgtggccgg cattcacgct catcgccgcc attgcattca accggcagag agcgaactgg 360
atggtggtgc ccggattcat cctctcgatt atcggtatct gctgggtgct gggcggtgac 420
caggggctgg acctggcggg catgcttggc aacgtgcagg acaatccgct cagttatggg 480
ctggcttttt tgggcgccgt gatctgggcc gcctattgca ctgtgacggc ccgcctcgcg 540
aaggggaaga acggagtgac gctgttcttc attcttgtgg cattgacgct ctgggtgaag 600
tttttcttcg gcgatcaccg cccgatgtct ttcagcctgc cggcaattgt ctacctgctc 660
ctggcggcgg ccgcgatggg cttcggctat gcggcatgga atgtcgggat cttgcacggt 720
aacgtgaccg tgctggcggg cgtgtcgtac tttatcccgg ttttttcggc ggcgttgtcc 780
gcgatggtgt tgcatgcgcc gttgccgcga tcgttttggg tgggggcgtc gctggtatgc 840
gccggttcga tactgtgctg gctggcaacc agggccaggc gcgcttcggc cgcgcaagaa 900
gatgcggtgg cggactgttt gtag 924
<210> 83
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex L79A-1)
<400> 83
gtgtcctacg aactctgcgc atcgctctcc atcggttatg 40
<210> 84
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex L79A-2)
<400> 84
cataaccgat ggagagcgat gcgcagagtt cgtaggacac 40
<210> 85
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex L79V-1)
<400> 85
gtgtcctacg aactctgcgt gtcgctctcc atcggttatg 40
<210> 86
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex L79V-2)
<400> 86
cataaccgat ggagagcgac acgcagagtt cgtaggacac 40
<210> 87
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex L79I-1)
<400> 87
gtgtcctacg aactctgcat ctcgctctcc atcggttatg 40
<210> 88
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex L79I-2)
<400> 88
cataaccgat ggagagcgag atgcagagtt cgtaggacac 40
<210> 89
<211> 924
<212> DNA
<213> Artificial Sequence
<220>
<223> wex(S80A)
<400> 89
atgaatagca agaaggccac gctgatcgga cttactgcag tggtcctctg gagttccatt 60
gtcggattga ttcgcggcgt cagcgaacat ctcggcgcta ccgggggcgc ggcgatgatg 120
tatagcgttg cctcgctgtt tctgctgttg tcggtcggtt tcccgaaact gggttccttt 180
ccgaaaaaat acctcttgtg gggcagcctg ctgtttgtgt cctacgaact ctgcctggca 240
ctctccatcg gttatgccaa tacaggcagg caagcaatcg aagtcagcat ggtcaactat 300
ctgtggccgg cattcacgct catcgccgcc attgcattca accggcagag agcgaactgg 360
atggtggtgc ccggattcat cctctcgatt atcggtatct gctgggtgct gggcggtgac 420
caggggctgg acctggcggg catgcttggc aacgtgcagg acaatccgct cagttatggg 480
ctggcttttt tgggcgccgt gatctgggcc gcctattgca ctgtgacggc ccgcctcgcg 540
aaggggaaga acggagtgac gctgttcttc attcttgtgg cattgacgct ctgggtgaag 600
tttttcttcg gcgatcaccg cccgatgtct ttcagcctgc cggcaattgt ctacctgctc 660
ctggcggcgg ccgcgatggg cttcggctat gcggcatgga atgtcgggat cttgcacggt 720
aacgtgaccg tgctggcggg cgtgtcgtac tttatcccgg ttttttcggc ggcgttgtcc 780
gcgatggtgt tgcatgcgcc gttgccgcga tcgttttggg tgggggcgtc gctggtatgc 840
gccggttcga tactgtgctg gctggcaacc agggccaggc gcgcttcggc cgcgcaagaa 900
gatgcggtgg cggactgttt gtag 924
<210> 90
<211> 924
<212> DNA
<213> Artificial Sequence
<220>
<223> wex(S80V)
<400> 90
atgaatagca agaaggccac gctgatcgga cttactgcag tggtcctctg gagttccatt 60
gtcggattga ttcgcggcgt cagcgaacat ctcggcgcta ccgggggcgc ggcgatgatg 120
tatagcgttg cctcgctgtt tctgctgttg tcggtcggtt tcccgaaact gggttccttt 180
ccgaaaaaat acctcttgtg gggcagcctg ctgtttgtgt cctacgaact ctgcctggtg 240
ctctccatcg gttatgccaa tacaggcagg caagcaatcg aagtcagcat ggtcaactat 300
ctgtggccgg cattcacgct catcgccgcc attgcattca accggcagag agcgaactgg 360
atggtggtgc ccggattcat cctctcgatt atcggtatct gctgggtgct gggcggtgac 420
caggggctgg acctggcggg catgcttggc aacgtgcagg acaatccgct cagttatggg 480
ctggcttttt tgggcgccgt gatctgggcc gcctattgca ctgtgacggc ccgcctcgcg 540
aaggggaaga acggagtgac gctgttcttc attcttgtgg cattgacgct ctgggtgaag 600
tttttcttcg gcgatcaccg cccgatgtct ttcagcctgc cggcaattgt ctacctgctc 660
ctggcggcgg ccgcgatggg cttcggctat gcggcatgga atgtcgggat cttgcacggt 720
aacgtgaccg tgctggcggg cgtgtcgtac tttatcccgg ttttttcggc ggcgttgtcc 780
gcgatggtgt tgcatgcgcc gttgccgcga tcgttttggg tgggggcgtc gctggtatgc 840
gccggttcga tactgtgctg gctggcaacc agggccaggc gcgcttcggc cgcgcaagaa 900
gatgcggtgg cggactgttt gtag 924
<210> 91
<211> 924
<212> DNA
<213> Artificial Sequence
<220>
<223> wex(S80L)
<400> 91
atgaatagca agaaggccac gctgatcgga cttactgcag tggtcctctg gagttccatt 60
gtcggattga ttcgcggcgt cagcgaacat ctcggcgcta ccgggggcgc ggcgatgatg 120
tatagcgttg cctcgctgtt tctgctgttg tcggtcggtt tcccgaaact gggttccttt 180
ccgaaaaaat acctcttgtg gggcagcctg ctgtttgtgt cctacgaact ctgcctgctg 240
ctctccatcg gttatgccaa tacaggcagg caagcaatcg aagtcagcat ggtcaactat 300
ctgtggccgg cattcacgct catcgccgcc attgcattca accggcagag agcgaactgg 360
atggtggtgc ccggattcat cctctcgatt atcggtatct gctgggtgct gggcggtgac 420
caggggctgg acctggcggg catgcttggc aacgtgcagg acaatccgct cagttatggg 480
ctggcttttt tgggcgccgt gatctgggcc gcctattgca ctgtgacggc ccgcctcgcg 540
aaggggaaga acggagtgac gctgttcttc attcttgtgg cattgacgct ctgggtgaag 600
tttttcttcg gcgatcaccg cccgatgtct ttcagcctgc cggcaattgt ctacctgctc 660
ctggcggcgg ccgcgatggg cttcggctat gcggcatgga atgtcgggat cttgcacggt 720
aacgtgaccg tgctggcggg cgtgtcgtac tttatcccgg ttttttcggc ggcgttgtcc 780
gcgatggtgt tgcatgcgcc gttgccgcga tcgttttggg tgggggcgtc gctggtatgc 840
gccggttcga tactgtgctg gctggcaacc agggccaggc gcgcttcggc cgcgcaagaa 900
gatgcggtgg cggactgttt gtag 924
<210> 92
<211> 924
<212> DNA
<213> Artificial Sequence
<220>
<223> wex(S80I)
<400> 92
atgaatagca agaaggccac gctgatcgga cttactgcag tggtcctctg gagttccatt 60
gtcggattga ttcgcggcgt cagcgaacat ctcggcgcta ccgggggcgc ggcgatgatg 120
tatagcgttg cctcgctgtt tctgctgttg tcggtcggtt tcccgaaact gggttccttt 180
ccgaaaaaat acctcttgtg gggcagcctg ctgtttgtgt cctacgaact ctgcctgatc 240
ctctccatcg gttatgccaa tacaggcagg caagcaatcg aagtcagcat ggtcaactat 300
ctgtggccgg cattcacgct catcgccgcc attgcattca accggcagag agcgaactgg 360
atggtggtgc ccggattcat cctctcgatt atcggtatct gctgggtgct gggcggtgac 420
caggggctgg acctggcggg catgcttggc aacgtgcagg acaatccgct cagttatggg 480
ctggcttttt tgggcgccgt gatctgggcc gcctattgca ctgtgacggc ccgcctcgcg 540
aaggggaaga acggagtgac gctgttcttc attcttgtgg cattgacgct ctgggtgaag 600
tttttcttcg gcgatcaccg cccgatgtct ttcagcctgc cggcaattgt ctacctgctc 660
ctggcggcgg ccgcgatggg cttcggctat gcggcatgga atgtcgggat cttgcacggt 720
aacgtgaccg tgctggcggg cgtgtcgtac tttatcccgg ttttttcggc ggcgttgtcc 780
gcgatggtgt tgcatgcgcc gttgccgcga tcgttttggg tgggggcgtc gctggtatgc 840
gccggttcga tactgtgctg gctggcaacc agggccaggc gcgcttcggc cgcgcaagaa 900
gatgcggtgg cggactgttt gtag 924
<210> 93
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex S80A-1)
<400> 93
gtcctacgaa ctctgcctgg cactctccat cggttatgcc 40
<210> 94
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex S80A-2)
<400> 94
ggcataaccg atggagagtg ccaggcagag ttcgtaggac 40
<210> 95
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex S80V-1)
<400> 95
gtcctacgaa ctctgcctgg tgctctccat cggttatgcc 40
<210> 96
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex S80V-2)
<400> 96
ggcataaccg atggagagca ccaggcagag ttcgtaggac 40
<210> 97
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex S80L-1)
<400> 97
gtcctacgaa ctctgcctgc tgctctccat cggttatgcc 40
<210> 98
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex S80L-2)
<400> 98
ggcataaccg atggagagca gcaggcagag ttcgtaggac 40
<210> 99
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex S80I-1)
<400> 99
gtcctacgaa ctctgcctga tcctctccat cggttatgcc 40
<210> 100
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex S80I-2)
<400> 100
ggcataaccg atggagagga tcaggcagag ttcgtaggac 40
<210> 101
<211> 924
<212> DNA
<213> Artificial Sequence
<220>
<223> wex(L81A)
<400> 101
atgaatagca agaaggccac gctgatcgga cttactgcag tggtcctctg gagttccatt 60
gtcggattga ttcgcggcgt cagcgaacat ctcggcgcta ccgggggcgc ggcgatgatg 120
tatagcgttg cctcgctgtt tctgctgttg tcggtcggtt tcccgaaact gggttccttt 180
ccgaaaaaat acctcttgtg gggcagcctg ctgtttgtgt cctacgaact ctgcctgtcg 240
gcatccatcg gttatgccaa tacaggcagg caagcaatcg aagtcagcat ggtcaactat 300
ctgtggccgg cattcacgct catcgccgcc attgcattca accggcagag agcgaactgg 360
atggtggtgc ccggattcat cctctcgatt atcggtatct gctgggtgct gggcggtgac 420
caggggctgg acctggcggg catgcttggc aacgtgcagg acaatccgct cagttatggg 480
ctggcttttt tgggcgccgt gatctgggcc gcctattgca ctgtgacggc ccgcctcgcg 540
aaggggaaga acggagtgac gctgttcttc attcttgtgg cattgacgct ctgggtgaag 600
tttttcttcg gcgatcaccg cccgatgtct ttcagcctgc cggcaattgt ctacctgctc 660
ctggcggcgg ccgcgatggg cttcggctat gcggcatgga atgtcgggat cttgcacggt 720
aacgtgaccg tgctggcggg cgtgtcgtac tttatcccgg ttttttcggc ggcgttgtcc 780
gcgatggtgt tgcatgcgcc gttgccgcga tcgttttggg tgggggcgtc gctggtatgc 840
gccggttcga tactgtgctg gctggcaacc agggccaggc gcgcttcggc cgcgcaagaa 900
gatgcggtgg cggactgttt gtag 924
<210> 102
<211> 924
<212> DNA
<213> Artificial Sequence
<220>
<223> wex(L81V)
<400> 102
atgaatagca agaaggccac gctgatcgga cttactgcag tggtcctctg gagttccatt 60
gtcggattga ttcgcggcgt cagcgaacat ctcggcgcta ccgggggcgc ggcgatgatg 120
tatagcgttg cctcgctgtt tctgctgttg tcggtcggtt tcccgaaact gggttccttt 180
ccgaaaaaat acctcttgtg gggcagcctg ctgtttgtgt cctacgaact ctgcctgtcg 240
gtgtccatcg gttatgccaa tacaggcagg caagcaatcg aagtcagcat ggtcaactat 300
ctgtggccgg cattcacgct catcgccgcc attgcattca accggcagag agcgaactgg 360
atggtggtgc ccggattcat cctctcgatt atcggtatct gctgggtgct gggcggtgac 420
caggggctgg acctggcggg catgcttggc aacgtgcagg acaatccgct cagttatggg 480
ctggcttttt tgggcgccgt gatctgggcc gcctattgca ctgtgacggc ccgcctcgcg 540
aaggggaaga acggagtgac gctgttcttc attcttgtgg cattgacgct ctgggtgaag 600
tttttcttcg gcgatcaccg cccgatgtct ttcagcctgc cggcaattgt ctacctgctc 660
ctggcggcgg ccgcgatggg cttcggctat gcggcatgga atgtcgggat cttgcacggt 720
aacgtgaccg tgctggcggg cgtgtcgtac tttatcccgg ttttttcggc ggcgttgtcc 780
gcgatggtgt tgcatgcgcc gttgccgcga tcgttttggg tgggggcgtc gctggtatgc 840
gccggttcga tactgtgctg gctggcaacc agggccaggc gcgcttcggc cgcgcaagaa 900
gatgcggtgg cggactgttt gtag 924
<210> 103
<211> 924
<212> DNA
<213> Artificial Sequence
<220>
<223> wex(L81I)
<400> 103
atgaatagca agaaggccac gctgatcgga cttactgcag tggtcctctg gagttccatt 60
gtcggattga ttcgcggcgt cagcgaacat ctcggcgcta ccgggggcgc ggcgatgatg 120
tatagcgttg cctcgctgtt tctgctgttg tcggtcggtt tcccgaaact gggttccttt 180
ccgaaaaaat acctcttgtg gggcagcctg ctgtttgtgt cctacgaact ctgcctgtcg 240
atctccatcg gttatgccaa tacaggcagg caagcaatcg aagtcagcat ggtcaactat 300
ctgtggccgg cattcacgct catcgccgcc attgcattca accggcagag agcgaactgg 360
atggtggtgc ccggattcat cctctcgatt atcggtatct gctgggtgct gggcggtgac 420
caggggctgg acctggcggg catgcttggc aacgtgcagg acaatccgct cagttatggg 480
ctggcttttt tgggcgccgt gatctgggcc gcctattgca ctgtgacggc ccgcctcgcg 540
aaggggaaga acggagtgac gctgttcttc attcttgtgg cattgacgct ctgggtgaag 600
tttttcttcg gcgatcaccg cccgatgtct ttcagcctgc cggcaattgt ctacctgctc 660
ctggcggcgg ccgcgatggg cttcggctat gcggcatgga atgtcgggat cttgcacggt 720
aacgtgaccg tgctggcggg cgtgtcgtac tttatcccgg ttttttcggc ggcgttgtcc 780
gcgatggtgt tgcatgcgcc gttgccgcga tcgttttggg tgggggcgtc gctggtatgc 840
gccggttcga tactgtgctg gctggcaacc agggccaggc gcgcttcggc cgcgcaagaa 900
gatgcggtgg cggactgttt gtag 924
<210> 104
<211> 43
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex L81A-1)
<400> 104
cctacgaact ctgcctgtcg gcatccatcg gttatgccaa tac 43
<210> 105
<211> 43
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex L81A-2)
<400> 105
gtattggcat aaccgatgga tgccgacagg cagagttcgt agg 43
<210> 106
<211> 43
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex L81V-1)
<400> 106
cctacgaact ctgcctgtcg gtgtccatcg gttatgccaa tac 43
<210> 107
<211> 43
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex L81V-2)
<400> 107
gtattggcat aaccgatgga caccgacagg cagagttcgt agg 43
<210> 108
<211> 43
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex L81I-1)
<400> 108
cctacgaact ctgcctgtcg atctccatcg gttatgccaa tac 43
<210> 109
<211> 43
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex L81I-2)
<400> 109
gtattggcat aaccgatgga gatcgacagg cagagttcgt agg 43
<210> 110
<211> 924
<212> DNA
<213> Artificial Sequence
<220>
<223> wex(S82A)
<400> 110
atgaatagca agaaggccac gctgatcgga cttactgcag tggtcctctg gagttccatt 60
gtcggattga ttcgcggcgt cagcgaacat ctcggcgcta ccgggggcgc ggcgatgatg 120
tatagcgttg cctcgctgtt tctgctgttg tcggtcggtt tcccgaaact gggttccttt 180
ccgaaaaaat acctcttgtg gggcagcctg ctgtttgtgt cctacgaact ctgcctgtcg 240
ctcgcaatcg gttatgccaa tacaggcagg caagcaatcg aagtcagcat ggtcaactat 300
ctgtggccgg cattcacgct catcgccgcc attgcattca accggcagag agcgaactgg 360
atggtggtgc ccggattcat cctctcgatt atcggtatct gctgggtgct gggcggtgac 420
caggggctgg acctggcggg catgcttggc aacgtgcagg acaatccgct cagttatggg 480
ctggcttttt tgggcgccgt gatctgggcc gcctattgca ctgtgacggc ccgcctcgcg 540
aaggggaaga acggagtgac gctgttcttc attcttgtgg cattgacgct ctgggtgaag 600
tttttcttcg gcgatcaccg cccgatgtct ttcagcctgc cggcaattgt ctacctgctc 660
ctggcggcgg ccgcgatggg cttcggctat gcggcatgga atgtcgggat cttgcacggt 720
aacgtgaccg tgctggcggg cgtgtcgtac tttatcccgg ttttttcggc ggcgttgtcc 780
gcgatggtgt tgcatgcgcc gttgccgcga tcgttttggg tgggggcgtc gctggtatgc 840
gccggttcga tactgtgctg gctggcaacc agggccaggc gcgcttcggc cgcgcaagaa 900
gatgcggtgg cggactgttt gtag 924
<210> 111
<211> 924
<212> DNA
<213> Artificial Sequence
<220>
<223> wex(S82V)
<400> 111
atgaatagca agaaggccac gctgatcgga cttactgcag tggtcctctg gagttccatt 60
gtcggattga ttcgcggcgt cagcgaacat ctcggcgcta ccgggggcgc ggcgatgatg 120
tatagcgttg cctcgctgtt tctgctgttg tcggtcggtt tcccgaaact gggttccttt 180
ccgaaaaaat acctcttgtg gggcagcctg ctgtttgtgt cctacgaact ctgcctgtcg 240
ctcgtgatcg gttatgccaa tacaggcagg caagcaatcg aagtcagcat ggtcaactat 300
ctgtggccgg cattcacgct catcgccgcc attgcattca accggcagag agcgaactgg 360
atggtggtgc ccggattcat cctctcgatt atcggtatct gctgggtgct gggcggtgac 420
caggggctgg acctggcggg catgcttggc aacgtgcagg acaatccgct cagttatggg 480
ctggcttttt tgggcgccgt gatctgggcc gcctattgca ctgtgacggc ccgcctcgcg 540
aaggggaaga acggagtgac gctgttcttc attcttgtgg cattgacgct ctgggtgaag 600
tttttcttcg gcgatcaccg cccgatgtct ttcagcctgc cggcaattgt ctacctgctc 660
ctggcggcgg ccgcgatggg cttcggctat gcggcatgga atgtcgggat cttgcacggt 720
aacgtgaccg tgctggcggg cgtgtcgtac tttatcccgg ttttttcggc ggcgttgtcc 780
gcgatggtgt tgcatgcgcc gttgccgcga tcgttttggg tgggggcgtc gctggtatgc 840
gccggttcga tactgtgctg gctggcaacc agggccaggc gcgcttcggc cgcgcaagaa 900
gatgcggtgg cggactgttt gtag 924
<210> 112
<211> 924
<212> DNA
<213> Artificial Sequence
<220>
<223> wex(S82L)
<400> 112
atgaatagca agaaggccac gctgatcgga cttactgcag tggtcctctg gagttccatt 60
gtcggattga ttcgcggcgt cagcgaacat ctcggcgcta ccgggggcgc ggcgatgatg 120
tatagcgttg cctcgctgtt tctgctgttg tcggtcggtt tcccgaaact gggttccttt 180
ccgaaaaaat acctcttgtg gggcagcctg ctgtttgtgt cctacgaact ctgcctgtcg 240
ctcctgatcg gttatgccaa tacaggcagg caagcaatcg aagtcagcat ggtcaactat 300
ctgtggccgg cattcacgct catcgccgcc attgcattca accggcagag agcgaactgg 360
atggtggtgc ccggattcat cctctcgatt atcggtatct gctgggtgct gggcggtgac 420
caggggctgg acctggcggg catgcttggc aacgtgcagg acaatccgct cagttatggg 480
ctggcttttt tgggcgccgt gatctgggcc gcctattgca ctgtgacggc ccgcctcgcg 540
aaggggaaga acggagtgac gctgttcttc attcttgtgg cattgacgct ctgggtgaag 600
tttttcttcg gcgatcaccg cccgatgtct ttcagcctgc cggcaattgt ctacctgctc 660
ctggcggcgg ccgcgatggg cttcggctat gcggcatgga atgtcgggat cttgcacggt 720
aacgtgaccg tgctggcggg cgtgtcgtac tttatcccgg ttttttcggc ggcgttgtcc 780
gcgatggtgt tgcatgcgcc gttgccgcga tcgttttggg tgggggcgtc gctggtatgc 840
gccggttcga tactgtgctg gctggcaacc agggccaggc gcgcttcggc cgcgcaagaa 900
gatgcggtgg cggactgttt gtag 924
<210> 113
<211> 924
<212> DNA
<213> Artificial Sequence
<220>
<223> wex(S82I)
<400> 113
atgaatagca agaaggccac gctgatcgga cttactgcag tggtcctctg gagttccatt 60
gtcggattga ttcgcggcgt cagcgaacat ctcggcgcta ccgggggcgc ggcgatgatg 120
tatagcgttg cctcgctgtt tctgctgttg tcggtcggtt tcccgaaact gggttccttt 180
ccgaaaaaat acctcttgtg gggcagcctg ctgtttgtgt cctacgaact ctgcctgtcg 240
ctcatcatcg gttatgccaa tacaggcagg caagcaatcg aagtcagcat ggtcaactat 300
ctgtggccgg cattcacgct catcgccgcc attgcattca accggcagag agcgaactgg 360
atggtggtgc ccggattcat cctctcgatt atcggtatct gctgggtgct gggcggtgac 420
caggggctgg acctggcggg catgcttggc aacgtgcagg acaatccgct cagttatggg 480
ctggcttttt tgggcgccgt gatctgggcc gcctattgca ctgtgacggc ccgcctcgcg 540
aaggggaaga acggagtgac gctgttcttc attcttgtgg cattgacgct ctgggtgaag 600
tttttcttcg gcgatcaccg cccgatgtct ttcagcctgc cggcaattgt ctacctgctc 660
ctggcggcgg ccgcgatggg cttcggctat gcggcatgga atgtcgggat cttgcacggt 720
aacgtgaccg tgctggcggg cgtgtcgtac tttatcccgg ttttttcggc ggcgttgtcc 780
gcgatggtgt tgcatgcgcc gttgccgcga tcgttttggg tgggggcgtc gctggtatgc 840
gccggttcga tactgtgctg gctggcaacc agggccaggc gcgcttcggc cgcgcaagaa 900
gatgcggtgg cggactgttt gtag 924
<210> 114
<211> 41
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex S82A-1)
<400> 114
cgaactctgc ctgtcgctcg caatcggtta tgccaataca g 41
<210> 115
<211> 41
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex S82A-2)
<400> 115
ctgtattggc ataaccgatt gcgagcgaca ggcagagttc g 41
<210> 116
<211> 41
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex S82V-1)
<400> 116
cgaactctgc ctgtcgctcg tgatcggtta tgccaataca g 41
<210> 117
<211> 41
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex S82V-2)
<400> 117
ctgtattggc ataaccgatc acgagcgaca ggcagagttc g 41
<210> 118
<211> 41
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex S82L-1)
<400> 118
cgaactctgc ctgtcgctcc tgatcggtta tgccaataca g 41
<210> 119
<211> 41
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex S82L-2)
<400> 119
ctgtattggc ataaccgatc aggagcgaca ggcagagttc g 41
<210> 120
<211> 41
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex S82I-1)
<400> 120
cgaactctgc ctgtcgctca tcatcggtta tgccaataca g 41
<210> 121
<211> 41
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex S82I-2)
<400> 121
ctgtattggc ataaccgatg atgagcgaca ggcagagttc g 41
<210> 122
<211> 924
<212> DNA
<213> Artificial Sequence
<220>
<223> wex(I83A)
<400> 122
atgaatagca agaaggccac gctgatcgga cttactgcag tggtcctctg gagttccatt 60
gtcggattga ttcgcggcgt cagcgaacat ctcggcgcta ccgggggcgc ggcgatgatg 120
tatagcgttg cctcgctgtt tctgctgttg tcggtcggtt tcccgaaact gggttccttt 180
ccgaaaaaat acctcttgtg gggcagcctg ctgtttgtgt cctacgaact ctgcctgtcg 240
ctctccgcag gttatgccaa tacaggcagg caagcaatcg aagtcagcat ggtcaactat 300
ctgtggccgg cattcacgct catcgccgcc attgcattca accggcagag agcgaactgg 360
atggtggtgc ccggattcat cctctcgatt atcggtatct gctgggtgct gggcggtgac 420
caggggctgg acctggcggg catgcttggc aacgtgcagg acaatccgct cagttatggg 480
ctggcttttt tgggcgccgt gatctgggcc gcctattgca ctgtgacggc ccgcctcgcg 540
aaggggaaga acggagtgac gctgttcttc attcttgtgg cattgacgct ctgggtgaag 600
tttttcttcg gcgatcaccg cccgatgtct ttcagcctgc cggcaattgt ctacctgctc 660
ctggcggcgg ccgcgatggg cttcggctat gcggcatgga atgtcgggat cttgcacggt 720
aacgtgaccg tgctggcggg cgtgtcgtac tttatcccgg ttttttcggc ggcgttgtcc 780
gcgatggtgt tgcatgcgcc gttgccgcga tcgttttggg tgggggcgtc gctggtatgc 840
gccggttcga tactgtgctg gctggcaacc agggccaggc gcgcttcggc cgcgcaagaa 900
gatgcggtgg cggactgttt gtag 924
<210> 123
<211> 924
<212> DNA
<213> Artificial Sequence
<220>
<223> wex(I83V)
<400> 123
atgaatagca agaaggccac gctgatcgga cttactgcag tggtcctctg gagttccatt 60
gtcggattga ttcgcggcgt cagcgaacat ctcggcgcta ccgggggcgc ggcgatgatg 120
tatagcgttg cctcgctgtt tctgctgttg tcggtcggtt tcccgaaact gggttccttt 180
ccgaaaaaat acctcttgtg gggcagcctg ctgtttgtgt cctacgaact ctgcctgtcg 240
ctctccgtgg gttatgccaa tacaggcagg caagcaatcg aagtcagcat ggtcaactat 300
ctgtggccgg cattcacgct catcgccgcc attgcattca accggcagag agcgaactgg 360
atggtggtgc ccggattcat cctctcgatt atcggtatct gctgggtgct gggcggtgac 420
caggggctgg acctggcggg catgcttggc aacgtgcagg acaatccgct cagttatggg 480
ctggcttttt tgggcgccgt gatctgggcc gcctattgca ctgtgacggc ccgcctcgcg 540
aaggggaaga acggagtgac gctgttcttc attcttgtgg cattgacgct ctgggtgaag 600
tttttcttcg gcgatcaccg cccgatgtct ttcagcctgc cggcaattgt ctacctgctc 660
ctggcggcgg ccgcgatggg cttcggctat gcggcatgga atgtcgggat cttgcacggt 720
aacgtgaccg tgctggcggg cgtgtcgtac tttatcccgg ttttttcggc ggcgttgtcc 780
gcgatggtgt tgcatgcgcc gttgccgcga tcgttttggg tgggggcgtc gctggtatgc 840
gccggttcga tactgtgctg gctggcaacc agggccaggc gcgcttcggc cgcgcaagaa 900
gatgcggtgg cggactgttt gtag 924
<210> 124
<211> 924
<212> DNA
<213> Artificial Sequence
<220>
<223> wex(I83L)
<400> 124
atgaatagca agaaggccac gctgatcgga cttactgcag tggtcctctg gagttccatt 60
gtcggattga ttcgcggcgt cagcgaacat ctcggcgcta ccgggggcgc ggcgatgatg 120
tatagcgttg cctcgctgtt tctgctgttg tcggtcggtt tcccgaaact gggttccttt 180
ccgaaaaaat acctcttgtg gggcagcctg ctgtttgtgt cctacgaact ctgcctgtcg 240
ctctccctgg gttatgccaa tacaggcagg caagcaatcg aagtcagcat ggtcaactat 300
ctgtggccgg cattcacgct catcgccgcc attgcattca accggcagag agcgaactgg 360
atggtggtgc ccggattcat cctctcgatt atcggtatct gctgggtgct gggcggtgac 420
caggggctgg acctggcggg catgcttggc aacgtgcagg acaatccgct cagttatggg 480
ctggcttttt tgggcgccgt gatctgggcc gcctattgca ctgtgacggc ccgcctcgcg 540
aaggggaaga acggagtgac gctgttcttc attcttgtgg cattgacgct ctgggtgaag 600
tttttcttcg gcgatcaccg cccgatgtct ttcagcctgc cggcaattgt ctacctgctc 660
ctggcggcgg ccgcgatggg cttcggctat gcggcatgga atgtcgggat cttgcacggt 720
aacgtgaccg tgctggcggg cgtgtcgtac tttatcccgg ttttttcggc ggcgttgtcc 780
gcgatggtgt tgcatgcgcc gttgccgcga tcgttttggg tgggggcgtc gctggtatgc 840
gccggttcga tactgtgctg gctggcaacc agggccaggc gcgcttcggc cgcgcaagaa 900
gatgcggtgg cggactgttt gtag 924
<210> 125
<211> 45
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex I83A-1)
<400> 125
gaactctgcc tgtcgctctc cgcaggttat gccaatacag gcagg 45
<210> 126
<211> 45
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex I83A-2)
<400> 126
cctgcctgta ttggcataac ctgcggagag cgacaggcag agttc 45
<210> 127
<211> 45
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex I83V-1)
<400> 127
gaactctgcc tgtcgctctc cgtgggttat gccaatacag gcagg 45
<210> 128
<211> 45
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex I83V-2)
<400> 128
cctgcctgta ttggcataac ccacggagag cgacaggcag agttc 45
<210> 129
<211> 45
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex I83L-1)
<400> 129
gaactctgcc tgtcgctctc cctgggttat gccaatacag gcagg 45
<210> 130
<211> 45
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер(wex I83L-2)
<400> 130
cctgcctgta ttggcataac ccagggagag cgacaggcag agttc 45
<210> 131
<211> 307
<212> PRT
<213> Artificial Sequence
<220>
<223> wex L79A
<400> 131
Met Asn Ser Lys Lys Ala Thr Leu Ile Gly Leu Thr Ala Val Val Leu
1 5 10 15
Trp Ser Ser Ile Val Gly Leu Ile Arg Gly Val Ser Glu His Leu Gly
20 25 30
Ala Thr Gly Gly Ala Ala Met Met Tyr Ser Val Ala Ser Leu Phe Leu
35 40 45
Leu Leu Ser Val Gly Phe Pro Lys Leu Gly Ser Phe Pro Lys Lys Tyr
50 55 60
Leu Leu Trp Gly Ser Leu Leu Phe Val Ser Tyr Glu Leu Cys Ala Ser
65 70 75 80
Leu Ser Ile Gly Tyr Ala Asn Thr Gly Arg Gln Ala Ile Glu Val Ser
85 90 95
Met Val Asn Tyr Leu Trp Pro Ala Phe Thr Leu Ile Ala Ala Ile Ala
100 105 110
Phe Asn Arg Gln Arg Ala Asn Trp Met Val Val Pro Gly Phe Ile Leu
115 120 125
Ser Ile Ile Gly Ile Cys Trp Val Leu Gly Gly Asp Gln Gly Leu Asp
130 135 140
Leu Ala Gly Met Leu Gly Asn Val Gln Asp Asn Pro Leu Ser Tyr Gly
145 150 155 160
Leu Ala Phe Leu Gly Ala Val Ile Trp Ala Ala Tyr Cys Thr Val Thr
165 170 175
Ala Arg Leu Ala Lys Gly Lys Asn Gly Val Thr Leu Phe Phe Ile Leu
180 185 190
Val Ala Leu Thr Leu Trp Val Lys Phe Phe Phe Gly Asp His Arg Pro
195 200 205
Met Ser Phe Ser Leu Pro Ala Ile Val Tyr Leu Leu Leu Ala Ala Ala
210 215 220
Ala Met Gly Phe Gly Tyr Ala Ala Trp Asn Val Gly Ile Leu His Gly
225 230 235 240
Asn Val Thr Val Leu Ala Gly Val Ser Tyr Phe Ile Pro Val Phe Ser
245 250 255
Ala Ala Leu Ser Ala Met Val Leu His Ala Pro Leu Pro Arg Ser Phe
260 265 270
Trp Val Gly Ala Ser Leu Val Cys Ala Gly Ser Ile Leu Cys Trp Leu
275 280 285
Ala Thr Arg Ala Arg Arg Ala Ser Ala Ala Gln Glu Asp Ala Val Ala
290 295 300
Asp Cys Leu
305
<210> 132
<211> 307
<212> PRT
<213> Artificial Sequence
<220>
<223> wex L79V
<400> 132
Met Asn Ser Lys Lys Ala Thr Leu Ile Gly Leu Thr Ala Val Val Leu
1 5 10 15
Trp Ser Ser Ile Val Gly Leu Ile Arg Gly Val Ser Glu His Leu Gly
20 25 30
Ala Thr Gly Gly Ala Ala Met Met Tyr Ser Val Ala Ser Leu Phe Leu
35 40 45
Leu Leu Ser Val Gly Phe Pro Lys Leu Gly Ser Phe Pro Lys Lys Tyr
50 55 60
Leu Leu Trp Gly Ser Leu Leu Phe Val Ser Tyr Glu Leu Cys Val Ser
65 70 75 80
Leu Ser Ile Gly Tyr Ala Asn Thr Gly Arg Gln Ala Ile Glu Val Ser
85 90 95
Met Val Asn Tyr Leu Trp Pro Ala Phe Thr Leu Ile Ala Ala Ile Ala
100 105 110
Phe Asn Arg Gln Arg Ala Asn Trp Met Val Val Pro Gly Phe Ile Leu
115 120 125
Ser Ile Ile Gly Ile Cys Trp Val Leu Gly Gly Asp Gln Gly Leu Asp
130 135 140
Leu Ala Gly Met Leu Gly Asn Val Gln Asp Asn Pro Leu Ser Tyr Gly
145 150 155 160
Leu Ala Phe Leu Gly Ala Val Ile Trp Ala Ala Tyr Cys Thr Val Thr
165 170 175
Ala Arg Leu Ala Lys Gly Lys Asn Gly Val Thr Leu Phe Phe Ile Leu
180 185 190
Val Ala Leu Thr Leu Trp Val Lys Phe Phe Phe Gly Asp His Arg Pro
195 200 205
Met Ser Phe Ser Leu Pro Ala Ile Val Tyr Leu Leu Leu Ala Ala Ala
210 215 220
Ala Met Gly Phe Gly Tyr Ala Ala Trp Asn Val Gly Ile Leu His Gly
225 230 235 240
Asn Val Thr Val Leu Ala Gly Val Ser Tyr Phe Ile Pro Val Phe Ser
245 250 255
Ala Ala Leu Ser Ala Met Val Leu His Ala Pro Leu Pro Arg Ser Phe
260 265 270
Trp Val Gly Ala Ser Leu Val Cys Ala Gly Ser Ile Leu Cys Trp Leu
275 280 285
Ala Thr Arg Ala Arg Arg Ala Ser Ala Ala Gln Glu Asp Ala Val Ala
290 295 300
Asp Cys Leu
305
<210> 133
<211> 307
<212> PRT
<213> Artificial Sequence
<220>
<223> wex L79I
<400> 133
Met Asn Ser Lys Lys Ala Thr Leu Ile Gly Leu Thr Ala Val Val Leu
1 5 10 15
Trp Ser Ser Ile Val Gly Leu Ile Arg Gly Val Ser Glu His Leu Gly
20 25 30
Ala Thr Gly Gly Ala Ala Met Met Tyr Ser Val Ala Ser Leu Phe Leu
35 40 45
Leu Leu Ser Val Gly Phe Pro Lys Leu Gly Ser Phe Pro Lys Lys Tyr
50 55 60
Leu Leu Trp Gly Ser Leu Leu Phe Val Ser Tyr Glu Leu Cys Ile Ser
65 70 75 80
Leu Ser Ile Gly Tyr Ala Asn Thr Gly Arg Gln Ala Ile Glu Val Ser
85 90 95
Met Val Asn Tyr Leu Trp Pro Ala Phe Thr Leu Ile Ala Ala Ile Ala
100 105 110
Phe Asn Arg Gln Arg Ala Asn Trp Met Val Val Pro Gly Phe Ile Leu
115 120 125
Ser Ile Ile Gly Ile Cys Trp Val Leu Gly Gly Asp Gln Gly Leu Asp
130 135 140
Leu Ala Gly Met Leu Gly Asn Val Gln Asp Asn Pro Leu Ser Tyr Gly
145 150 155 160
Leu Ala Phe Leu Gly Ala Val Ile Trp Ala Ala Tyr Cys Thr Val Thr
165 170 175
Ala Arg Leu Ala Lys Gly Lys Asn Gly Val Thr Leu Phe Phe Ile Leu
180 185 190
Val Ala Leu Thr Leu Trp Val Lys Phe Phe Phe Gly Asp His Arg Pro
195 200 205
Met Ser Phe Ser Leu Pro Ala Ile Val Tyr Leu Leu Leu Ala Ala Ala
210 215 220
Ala Met Gly Phe Gly Tyr Ala Ala Trp Asn Val Gly Ile Leu His Gly
225 230 235 240
Asn Val Thr Val Leu Ala Gly Val Ser Tyr Phe Ile Pro Val Phe Ser
245 250 255
Ala Ala Leu Ser Ala Met Val Leu His Ala Pro Leu Pro Arg Ser Phe
260 265 270
Trp Val Gly Ala Ser Leu Val Cys Ala Gly Ser Ile Leu Cys Trp Leu
275 280 285
Ala Thr Arg Ala Arg Arg Ala Ser Ala Ala Gln Glu Asp Ala Val Ala
290 295 300
Asp Cys Leu
305
<210> 134
<211> 307
<212> PRT
<213> Artificial Sequence
<220>
<223> wex S80A
<400> 134
Met Asn Ser Lys Lys Ala Thr Leu Ile Gly Leu Thr Ala Val Val Leu
1 5 10 15
Trp Ser Ser Ile Val Gly Leu Ile Arg Gly Val Ser Glu His Leu Gly
20 25 30
Ala Thr Gly Gly Ala Ala Met Met Tyr Ser Val Ala Ser Leu Phe Leu
35 40 45
Leu Leu Ser Val Gly Phe Pro Lys Leu Gly Ser Phe Pro Lys Lys Tyr
50 55 60
Leu Leu Trp Gly Ser Leu Leu Phe Val Ser Tyr Glu Leu Cys Leu Ala
65 70 75 80
Leu Ser Ile Gly Tyr Ala Asn Thr Gly Arg Gln Ala Ile Glu Val Ser
85 90 95
Met Val Asn Tyr Leu Trp Pro Ala Phe Thr Leu Ile Ala Ala Ile Ala
100 105 110
Phe Asn Arg Gln Arg Ala Asn Trp Met Val Val Pro Gly Phe Ile Leu
115 120 125
Ser Ile Ile Gly Ile Cys Trp Val Leu Gly Gly Asp Gln Gly Leu Asp
130 135 140
Leu Ala Gly Met Leu Gly Asn Val Gln Asp Asn Pro Leu Ser Tyr Gly
145 150 155 160
Leu Ala Phe Leu Gly Ala Val Ile Trp Ala Ala Tyr Cys Thr Val Thr
165 170 175
Ala Arg Leu Ala Lys Gly Lys Asn Gly Val Thr Leu Phe Phe Ile Leu
180 185 190
Val Ala Leu Thr Leu Trp Val Lys Phe Phe Phe Gly Asp His Arg Pro
195 200 205
Met Ser Phe Ser Leu Pro Ala Ile Val Tyr Leu Leu Leu Ala Ala Ala
210 215 220
Ala Met Gly Phe Gly Tyr Ala Ala Trp Asn Val Gly Ile Leu His Gly
225 230 235 240
Asn Val Thr Val Leu Ala Gly Val Ser Tyr Phe Ile Pro Val Phe Ser
245 250 255
Ala Ala Leu Ser Ala Met Val Leu His Ala Pro Leu Pro Arg Ser Phe
260 265 270
Trp Val Gly Ala Ser Leu Val Cys Ala Gly Ser Ile Leu Cys Trp Leu
275 280 285
Ala Thr Arg Ala Arg Arg Ala Ser Ala Ala Gln Glu Asp Ala Val Ala
290 295 300
Asp Cys Leu
305
<210> 135
<211> 307
<212> PRT
<213> Artificial Sequence
<220>
<223> wex S80V
<400> 135
Met Asn Ser Lys Lys Ala Thr Leu Ile Gly Leu Thr Ala Val Val Leu
1 5 10 15
Trp Ser Ser Ile Val Gly Leu Ile Arg Gly Val Ser Glu His Leu Gly
20 25 30
Ala Thr Gly Gly Ala Ala Met Met Tyr Ser Val Ala Ser Leu Phe Leu
35 40 45
Leu Leu Ser Val Gly Phe Pro Lys Leu Gly Ser Phe Pro Lys Lys Tyr
50 55 60
Leu Leu Trp Gly Ser Leu Leu Phe Val Ser Tyr Glu Leu Cys Leu Val
65 70 75 80
Leu Ser Ile Gly Tyr Ala Asn Thr Gly Arg Gln Ala Ile Glu Val Ser
85 90 95
Met Val Asn Tyr Leu Trp Pro Ala Phe Thr Leu Ile Ala Ala Ile Ala
100 105 110
Phe Asn Arg Gln Arg Ala Asn Trp Met Val Val Pro Gly Phe Ile Leu
115 120 125
Ser Ile Ile Gly Ile Cys Trp Val Leu Gly Gly Asp Gln Gly Leu Asp
130 135 140
Leu Ala Gly Met Leu Gly Asn Val Gln Asp Asn Pro Leu Ser Tyr Gly
145 150 155 160
Leu Ala Phe Leu Gly Ala Val Ile Trp Ala Ala Tyr Cys Thr Val Thr
165 170 175
Ala Arg Leu Ala Lys Gly Lys Asn Gly Val Thr Leu Phe Phe Ile Leu
180 185 190
Val Ala Leu Thr Leu Trp Val Lys Phe Phe Phe Gly Asp His Arg Pro
195 200 205
Met Ser Phe Ser Leu Pro Ala Ile Val Tyr Leu Leu Leu Ala Ala Ala
210 215 220
Ala Met Gly Phe Gly Tyr Ala Ala Trp Asn Val Gly Ile Leu His Gly
225 230 235 240
Asn Val Thr Val Leu Ala Gly Val Ser Tyr Phe Ile Pro Val Phe Ser
245 250 255
Ala Ala Leu Ser Ala Met Val Leu His Ala Pro Leu Pro Arg Ser Phe
260 265 270
Trp Val Gly Ala Ser Leu Val Cys Ala Gly Ser Ile Leu Cys Trp Leu
275 280 285
Ala Thr Arg Ala Arg Arg Ala Ser Ala Ala Gln Glu Asp Ala Val Ala
290 295 300
Asp Cys Leu
305
<210> 136
<211> 307
<212> PRT
<213> Artificial Sequence
<220>
<223> wex S80L
<400> 136
Met Asn Ser Lys Lys Ala Thr Leu Ile Gly Leu Thr Ala Val Val Leu
1 5 10 15
Trp Ser Ser Ile Val Gly Leu Ile Arg Gly Val Ser Glu His Leu Gly
20 25 30
Ala Thr Gly Gly Ala Ala Met Met Tyr Ser Val Ala Ser Leu Phe Leu
35 40 45
Leu Leu Ser Val Gly Phe Pro Lys Leu Gly Ser Phe Pro Lys Lys Tyr
50 55 60
Leu Leu Trp Gly Ser Leu Leu Phe Val Ser Tyr Glu Leu Cys Leu Leu
65 70 75 80
Leu Ser Ile Gly Tyr Ala Asn Thr Gly Arg Gln Ala Ile Glu Val Ser
85 90 95
Met Val Asn Tyr Leu Trp Pro Ala Phe Thr Leu Ile Ala Ala Ile Ala
100 105 110
Phe Asn Arg Gln Arg Ala Asn Trp Met Val Val Pro Gly Phe Ile Leu
115 120 125
Ser Ile Ile Gly Ile Cys Trp Val Leu Gly Gly Asp Gln Gly Leu Asp
130 135 140
Leu Ala Gly Met Leu Gly Asn Val Gln Asp Asn Pro Leu Ser Tyr Gly
145 150 155 160
Leu Ala Phe Leu Gly Ala Val Ile Trp Ala Ala Tyr Cys Thr Val Thr
165 170 175
Ala Arg Leu Ala Lys Gly Lys Asn Gly Val Thr Leu Phe Phe Ile Leu
180 185 190
Val Ala Leu Thr Leu Trp Val Lys Phe Phe Phe Gly Asp His Arg Pro
195 200 205
Met Ser Phe Ser Leu Pro Ala Ile Val Tyr Leu Leu Leu Ala Ala Ala
210 215 220
Ala Met Gly Phe Gly Tyr Ala Ala Trp Asn Val Gly Ile Leu His Gly
225 230 235 240
Asn Val Thr Val Leu Ala Gly Val Ser Tyr Phe Ile Pro Val Phe Ser
245 250 255
Ala Ala Leu Ser Ala Met Val Leu His Ala Pro Leu Pro Arg Ser Phe
260 265 270
Trp Val Gly Ala Ser Leu Val Cys Ala Gly Ser Ile Leu Cys Trp Leu
275 280 285
Ala Thr Arg Ala Arg Arg Ala Ser Ala Ala Gln Glu Asp Ala Val Ala
290 295 300
Asp Cys Leu
305
<210> 137
<211> 307
<212> PRT
<213> Artificial Sequence
<220>
<223> wex S80I
<400> 137
Met Asn Ser Lys Lys Ala Thr Leu Ile Gly Leu Thr Ala Val Val Leu
1 5 10 15
Trp Ser Ser Ile Val Gly Leu Ile Arg Gly Val Ser Glu His Leu Gly
20 25 30
Ala Thr Gly Gly Ala Ala Met Met Tyr Ser Val Ala Ser Leu Phe Leu
35 40 45
Leu Leu Ser Val Gly Phe Pro Lys Leu Gly Ser Phe Pro Lys Lys Tyr
50 55 60
Leu Leu Trp Gly Ser Leu Leu Phe Val Ser Tyr Glu Leu Cys Leu Ile
65 70 75 80
Leu Ser Ile Gly Tyr Ala Asn Thr Gly Arg Gln Ala Ile Glu Val Ser
85 90 95
Met Val Asn Tyr Leu Trp Pro Ala Phe Thr Leu Ile Ala Ala Ile Ala
100 105 110
Phe Asn Arg Gln Arg Ala Asn Trp Met Val Val Pro Gly Phe Ile Leu
115 120 125
Ser Ile Ile Gly Ile Cys Trp Val Leu Gly Gly Asp Gln Gly Leu Asp
130 135 140
Leu Ala Gly Met Leu Gly Asn Val Gln Asp Asn Pro Leu Ser Tyr Gly
145 150 155 160
Leu Ala Phe Leu Gly Ala Val Ile Trp Ala Ala Tyr Cys Thr Val Thr
165 170 175
Ala Arg Leu Ala Lys Gly Lys Asn Gly Val Thr Leu Phe Phe Ile Leu
180 185 190
Val Ala Leu Thr Leu Trp Val Lys Phe Phe Phe Gly Asp His Arg Pro
195 200 205
Met Ser Phe Ser Leu Pro Ala Ile Val Tyr Leu Leu Leu Ala Ala Ala
210 215 220
Ala Met Gly Phe Gly Tyr Ala Ala Trp Asn Val Gly Ile Leu His Gly
225 230 235 240
Asn Val Thr Val Leu Ala Gly Val Ser Tyr Phe Ile Pro Val Phe Ser
245 250 255
Ala Ala Leu Ser Ala Met Val Leu His Ala Pro Leu Pro Arg Ser Phe
260 265 270
Trp Val Gly Ala Ser Leu Val Cys Ala Gly Ser Ile Leu Cys Trp Leu
275 280 285
Ala Thr Arg Ala Arg Arg Ala Ser Ala Ala Gln Glu Asp Ala Val Ala
290 295 300
Asp Cys Leu
305
<210> 138
<211> 307
<212> PRT
<213> Artificial Sequence
<220>
<223> wex L81A
<400> 138
Met Asn Ser Lys Lys Ala Thr Leu Ile Gly Leu Thr Ala Val Val Leu
1 5 10 15
Trp Ser Ser Ile Val Gly Leu Ile Arg Gly Val Ser Glu His Leu Gly
20 25 30
Ala Thr Gly Gly Ala Ala Met Met Tyr Ser Val Ala Ser Leu Phe Leu
35 40 45
Leu Leu Ser Val Gly Phe Pro Lys Leu Gly Ser Phe Pro Lys Lys Tyr
50 55 60
Leu Leu Trp Gly Ser Leu Leu Phe Val Ser Tyr Glu Leu Cys Leu Ser
65 70 75 80
Ala Ser Ile Gly Tyr Ala Asn Thr Gly Arg Gln Ala Ile Glu Val Ser
85 90 95
Met Val Asn Tyr Leu Trp Pro Ala Phe Thr Leu Ile Ala Ala Ile Ala
100 105 110
Phe Asn Arg Gln Arg Ala Asn Trp Met Val Val Pro Gly Phe Ile Leu
115 120 125
Ser Ile Ile Gly Ile Cys Trp Val Leu Gly Gly Asp Gln Gly Leu Asp
130 135 140
Leu Ala Gly Met Leu Gly Asn Val Gln Asp Asn Pro Leu Ser Tyr Gly
145 150 155 160
Leu Ala Phe Leu Gly Ala Val Ile Trp Ala Ala Tyr Cys Thr Val Thr
165 170 175
Ala Arg Leu Ala Lys Gly Lys Asn Gly Val Thr Leu Phe Phe Ile Leu
180 185 190
Val Ala Leu Thr Leu Trp Val Lys Phe Phe Phe Gly Asp His Arg Pro
195 200 205
Met Ser Phe Ser Leu Pro Ala Ile Val Tyr Leu Leu Leu Ala Ala Ala
210 215 220
Ala Met Gly Phe Gly Tyr Ala Ala Trp Asn Val Gly Ile Leu His Gly
225 230 235 240
Asn Val Thr Val Leu Ala Gly Val Ser Tyr Phe Ile Pro Val Phe Ser
245 250 255
Ala Ala Leu Ser Ala Met Val Leu His Ala Pro Leu Pro Arg Ser Phe
260 265 270
Trp Val Gly Ala Ser Leu Val Cys Ala Gly Ser Ile Leu Cys Trp Leu
275 280 285
Ala Thr Arg Ala Arg Arg Ala Ser Ala Ala Gln Glu Asp Ala Val Ala
290 295 300
Asp Cys Leu
305
<210> 139
<211> 307
<212> PRT
<213> Artificial Sequence
<220>
<223> wex L81V
<400> 139
Met Asn Ser Lys Lys Ala Thr Leu Ile Gly Leu Thr Ala Val Val Leu
1 5 10 15
Trp Ser Ser Ile Val Gly Leu Ile Arg Gly Val Ser Glu His Leu Gly
20 25 30
Ala Thr Gly Gly Ala Ala Met Met Tyr Ser Val Ala Ser Leu Phe Leu
35 40 45
Leu Leu Ser Val Gly Phe Pro Lys Leu Gly Ser Phe Pro Lys Lys Tyr
50 55 60
Leu Leu Trp Gly Ser Leu Leu Phe Val Ser Tyr Glu Leu Cys Leu Ser
65 70 75 80
Val Ser Ile Gly Tyr Ala Asn Thr Gly Arg Gln Ala Ile Glu Val Ser
85 90 95
Met Val Asn Tyr Leu Trp Pro Ala Phe Thr Leu Ile Ala Ala Ile Ala
100 105 110
Phe Asn Arg Gln Arg Ala Asn Trp Met Val Val Pro Gly Phe Ile Leu
115 120 125
Ser Ile Ile Gly Ile Cys Trp Val Leu Gly Gly Asp Gln Gly Leu Asp
130 135 140
Leu Ala Gly Met Leu Gly Asn Val Gln Asp Asn Pro Leu Ser Tyr Gly
145 150 155 160
Leu Ala Phe Leu Gly Ala Val Ile Trp Ala Ala Tyr Cys Thr Val Thr
165 170 175
Ala Arg Leu Ala Lys Gly Lys Asn Gly Val Thr Leu Phe Phe Ile Leu
180 185 190
Val Ala Leu Thr Leu Trp Val Lys Phe Phe Phe Gly Asp His Arg Pro
195 200 205
Met Ser Phe Ser Leu Pro Ala Ile Val Tyr Leu Leu Leu Ala Ala Ala
210 215 220
Ala Met Gly Phe Gly Tyr Ala Ala Trp Asn Val Gly Ile Leu His Gly
225 230 235 240
Asn Val Thr Val Leu Ala Gly Val Ser Tyr Phe Ile Pro Val Phe Ser
245 250 255
Ala Ala Leu Ser Ala Met Val Leu His Ala Pro Leu Pro Arg Ser Phe
260 265 270
Trp Val Gly Ala Ser Leu Val Cys Ala Gly Ser Ile Leu Cys Trp Leu
275 280 285
Ala Thr Arg Ala Arg Arg Ala Ser Ala Ala Gln Glu Asp Ala Val Ala
290 295 300
Asp Cys Leu
305
<210> 140
<211> 307
<212> PRT
<213> Artificial Sequence
<220>
<223> wex L81I
<400> 140
Met Asn Ser Lys Lys Ala Thr Leu Ile Gly Leu Thr Ala Val Val Leu
1 5 10 15
Trp Ser Ser Ile Val Gly Leu Ile Arg Gly Val Ser Glu His Leu Gly
20 25 30
Ala Thr Gly Gly Ala Ala Met Met Tyr Ser Val Ala Ser Leu Phe Leu
35 40 45
Leu Leu Ser Val Gly Phe Pro Lys Leu Gly Ser Phe Pro Lys Lys Tyr
50 55 60
Leu Leu Trp Gly Ser Leu Leu Phe Val Ser Tyr Glu Leu Cys Leu Ser
65 70 75 80
Ile Ser Ile Gly Tyr Ala Asn Thr Gly Arg Gln Ala Ile Glu Val Ser
85 90 95
Met Val Asn Tyr Leu Trp Pro Ala Phe Thr Leu Ile Ala Ala Ile Ala
100 105 110
Phe Asn Arg Gln Arg Ala Asn Trp Met Val Val Pro Gly Phe Ile Leu
115 120 125
Ser Ile Ile Gly Ile Cys Trp Val Leu Gly Gly Asp Gln Gly Leu Asp
130 135 140
Leu Ala Gly Met Leu Gly Asn Val Gln Asp Asn Pro Leu Ser Tyr Gly
145 150 155 160
Leu Ala Phe Leu Gly Ala Val Ile Trp Ala Ala Tyr Cys Thr Val Thr
165 170 175
Ala Arg Leu Ala Lys Gly Lys Asn Gly Val Thr Leu Phe Phe Ile Leu
180 185 190
Val Ala Leu Thr Leu Trp Val Lys Phe Phe Phe Gly Asp His Arg Pro
195 200 205
Met Ser Phe Ser Leu Pro Ala Ile Val Tyr Leu Leu Leu Ala Ala Ala
210 215 220
Ala Met Gly Phe Gly Tyr Ala Ala Trp Asn Val Gly Ile Leu His Gly
225 230 235 240
Asn Val Thr Val Leu Ala Gly Val Ser Tyr Phe Ile Pro Val Phe Ser
245 250 255
Ala Ala Leu Ser Ala Met Val Leu His Ala Pro Leu Pro Arg Ser Phe
260 265 270
Trp Val Gly Ala Ser Leu Val Cys Ala Gly Ser Ile Leu Cys Trp Leu
275 280 285
Ala Thr Arg Ala Arg Arg Ala Ser Ala Ala Gln Glu Asp Ala Val Ala
290 295 300
Asp Cys Leu
305
<210> 141
<211> 307
<212> PRT
<213> Artificial Sequence
<220>
<223> wex S82A
<400> 141
Met Asn Ser Lys Lys Ala Thr Leu Ile Gly Leu Thr Ala Val Val Leu
1 5 10 15
Trp Ser Ser Ile Val Gly Leu Ile Arg Gly Val Ser Glu His Leu Gly
20 25 30
Ala Thr Gly Gly Ala Ala Met Met Tyr Ser Val Ala Ser Leu Phe Leu
35 40 45
Leu Leu Ser Val Gly Phe Pro Lys Leu Gly Ser Phe Pro Lys Lys Tyr
50 55 60
Leu Leu Trp Gly Ser Leu Leu Phe Val Ser Tyr Glu Leu Cys Leu Ser
65 70 75 80
Leu Ala Ile Gly Tyr Ala Asn Thr Gly Arg Gln Ala Ile Glu Val Ser
85 90 95
Met Val Asn Tyr Leu Trp Pro Ala Phe Thr Leu Ile Ala Ala Ile Ala
100 105 110
Phe Asn Arg Gln Arg Ala Asn Trp Met Val Val Pro Gly Phe Ile Leu
115 120 125
Ser Ile Ile Gly Ile Cys Trp Val Leu Gly Gly Asp Gln Gly Leu Asp
130 135 140
Leu Ala Gly Met Leu Gly Asn Val Gln Asp Asn Pro Leu Ser Tyr Gly
145 150 155 160
Leu Ala Phe Leu Gly Ala Val Ile Trp Ala Ala Tyr Cys Thr Val Thr
165 170 175
Ala Arg Leu Ala Lys Gly Lys Asn Gly Val Thr Leu Phe Phe Ile Leu
180 185 190
Val Ala Leu Thr Leu Trp Val Lys Phe Phe Phe Gly Asp His Arg Pro
195 200 205
Met Ser Phe Ser Leu Pro Ala Ile Val Tyr Leu Leu Leu Ala Ala Ala
210 215 220
Ala Met Gly Phe Gly Tyr Ala Ala Trp Asn Val Gly Ile Leu His Gly
225 230 235 240
Asn Val Thr Val Leu Ala Gly Val Ser Tyr Phe Ile Pro Val Phe Ser
245 250 255
Ala Ala Leu Ser Ala Met Val Leu His Ala Pro Leu Pro Arg Ser Phe
260 265 270
Trp Val Gly Ala Ser Leu Val Cys Ala Gly Ser Ile Leu Cys Trp Leu
275 280 285
Ala Thr Arg Ala Arg Arg Ala Ser Ala Ala Gln Glu Asp Ala Val Ala
290 295 300
Asp Cys Leu
305
<210> 142
<211> 307
<212> PRT
<213> Artificial Sequence
<220>
<223> wex S82V
<400> 142
Met Asn Ser Lys Lys Ala Thr Leu Ile Gly Leu Thr Ala Val Val Leu
1 5 10 15
Trp Ser Ser Ile Val Gly Leu Ile Arg Gly Val Ser Glu His Leu Gly
20 25 30
Ala Thr Gly Gly Ala Ala Met Met Tyr Ser Val Ala Ser Leu Phe Leu
35 40 45
Leu Leu Ser Val Gly Phe Pro Lys Leu Gly Ser Phe Pro Lys Lys Tyr
50 55 60
Leu Leu Trp Gly Ser Leu Leu Phe Val Ser Tyr Glu Leu Cys Leu Ser
65 70 75 80
Leu Val Ile Gly Tyr Ala Asn Thr Gly Arg Gln Ala Ile Glu Val Ser
85 90 95
Met Val Asn Tyr Leu Trp Pro Ala Phe Thr Leu Ile Ala Ala Ile Ala
100 105 110
Phe Asn Arg Gln Arg Ala Asn Trp Met Val Val Pro Gly Phe Ile Leu
115 120 125
Ser Ile Ile Gly Ile Cys Trp Val Leu Gly Gly Asp Gln Gly Leu Asp
130 135 140
Leu Ala Gly Met Leu Gly Asn Val Gln Asp Asn Pro Leu Ser Tyr Gly
145 150 155 160
Leu Ala Phe Leu Gly Ala Val Ile Trp Ala Ala Tyr Cys Thr Val Thr
165 170 175
Ala Arg Leu Ala Lys Gly Lys Asn Gly Val Thr Leu Phe Phe Ile Leu
180 185 190
Val Ala Leu Thr Leu Trp Val Lys Phe Phe Phe Gly Asp His Arg Pro
195 200 205
Met Ser Phe Ser Leu Pro Ala Ile Val Tyr Leu Leu Leu Ala Ala Ala
210 215 220
Ala Met Gly Phe Gly Tyr Ala Ala Trp Asn Val Gly Ile Leu His Gly
225 230 235 240
Asn Val Thr Val Leu Ala Gly Val Ser Tyr Phe Ile Pro Val Phe Ser
245 250 255
Ala Ala Leu Ser Ala Met Val Leu His Ala Pro Leu Pro Arg Ser Phe
260 265 270
Trp Val Gly Ala Ser Leu Val Cys Ala Gly Ser Ile Leu Cys Trp Leu
275 280 285
Ala Thr Arg Ala Arg Arg Ala Ser Ala Ala Gln Glu Asp Ala Val Ala
290 295 300
Asp Cys Leu
305
<210> 143
<211> 307
<212> PRT
<213> Artificial Sequence
<220>
<223> wex S82L
<400> 143
Met Asn Ser Lys Lys Ala Thr Leu Ile Gly Leu Thr Ala Val Val Leu
1 5 10 15
Trp Ser Ser Ile Val Gly Leu Ile Arg Gly Val Ser Glu His Leu Gly
20 25 30
Ala Thr Gly Gly Ala Ala Met Met Tyr Ser Val Ala Ser Leu Phe Leu
35 40 45
Leu Leu Ser Val Gly Phe Pro Lys Leu Gly Ser Phe Pro Lys Lys Tyr
50 55 60
Leu Leu Trp Gly Ser Leu Leu Phe Val Ser Tyr Glu Leu Cys Leu Ser
65 70 75 80
Leu Leu Ile Gly Tyr Ala Asn Thr Gly Arg Gln Ala Ile Glu Val Ser
85 90 95
Met Val Asn Tyr Leu Trp Pro Ala Phe Thr Leu Ile Ala Ala Ile Ala
100 105 110
Phe Asn Arg Gln Arg Ala Asn Trp Met Val Val Pro Gly Phe Ile Leu
115 120 125
Ser Ile Ile Gly Ile Cys Trp Val Leu Gly Gly Asp Gln Gly Leu Asp
130 135 140
Leu Ala Gly Met Leu Gly Asn Val Gln Asp Asn Pro Leu Ser Tyr Gly
145 150 155 160
Leu Ala Phe Leu Gly Ala Val Ile Trp Ala Ala Tyr Cys Thr Val Thr
165 170 175
Ala Arg Leu Ala Lys Gly Lys Asn Gly Val Thr Leu Phe Phe Ile Leu
180 185 190
Val Ala Leu Thr Leu Trp Val Lys Phe Phe Phe Gly Asp His Arg Pro
195 200 205
Met Ser Phe Ser Leu Pro Ala Ile Val Tyr Leu Leu Leu Ala Ala Ala
210 215 220
Ala Met Gly Phe Gly Tyr Ala Ala Trp Asn Val Gly Ile Leu His Gly
225 230 235 240
Asn Val Thr Val Leu Ala Gly Val Ser Tyr Phe Ile Pro Val Phe Ser
245 250 255
Ala Ala Leu Ser Ala Met Val Leu His Ala Pro Leu Pro Arg Ser Phe
260 265 270
Trp Val Gly Ala Ser Leu Val Cys Ala Gly Ser Ile Leu Cys Trp Leu
275 280 285
Ala Thr Arg Ala Arg Arg Ala Ser Ala Ala Gln Glu Asp Ala Val Ala
290 295 300
Asp Cys Leu
305
<210> 144
<211> 307
<212> PRT
<213> Artificial Sequence
<220>
<223> wex S82I
<400> 144
Met Asn Ser Lys Lys Ala Thr Leu Ile Gly Leu Thr Ala Val Val Leu
1 5 10 15
Trp Ser Ser Ile Val Gly Leu Ile Arg Gly Val Ser Glu His Leu Gly
20 25 30
Ala Thr Gly Gly Ala Ala Met Met Tyr Ser Val Ala Ser Leu Phe Leu
35 40 45
Leu Leu Ser Val Gly Phe Pro Lys Leu Gly Ser Phe Pro Lys Lys Tyr
50 55 60
Leu Leu Trp Gly Ser Leu Leu Phe Val Ser Tyr Glu Leu Cys Leu Ser
65 70 75 80
Leu Ile Ile Gly Tyr Ala Asn Thr Gly Arg Gln Ala Ile Glu Val Ser
85 90 95
Met Val Asn Tyr Leu Trp Pro Ala Phe Thr Leu Ile Ala Ala Ile Ala
100 105 110
Phe Asn Arg Gln Arg Ala Asn Trp Met Val Val Pro Gly Phe Ile Leu
115 120 125
Ser Ile Ile Gly Ile Cys Trp Val Leu Gly Gly Asp Gln Gly Leu Asp
130 135 140
Leu Ala Gly Met Leu Gly Asn Val Gln Asp Asn Pro Leu Ser Tyr Gly
145 150 155 160
Leu Ala Phe Leu Gly Ala Val Ile Trp Ala Ala Tyr Cys Thr Val Thr
165 170 175
Ala Arg Leu Ala Lys Gly Lys Asn Gly Val Thr Leu Phe Phe Ile Leu
180 185 190
Val Ala Leu Thr Leu Trp Val Lys Phe Phe Phe Gly Asp His Arg Pro
195 200 205
Met Ser Phe Ser Leu Pro Ala Ile Val Tyr Leu Leu Leu Ala Ala Ala
210 215 220
Ala Met Gly Phe Gly Tyr Ala Ala Trp Asn Val Gly Ile Leu His Gly
225 230 235 240
Asn Val Thr Val Leu Ala Gly Val Ser Tyr Phe Ile Pro Val Phe Ser
245 250 255
Ala Ala Leu Ser Ala Met Val Leu His Ala Pro Leu Pro Arg Ser Phe
260 265 270
Trp Val Gly Ala Ser Leu Val Cys Ala Gly Ser Ile Leu Cys Trp Leu
275 280 285
Ala Thr Arg Ala Arg Arg Ala Ser Ala Ala Gln Glu Asp Ala Val Ala
290 295 300
Asp Cys Leu
305
<210> 145
<211> 307
<212> PRT
<213> Artificial Sequence
<220>
<223> wex I83A
<400> 145
Met Asn Ser Lys Lys Ala Thr Leu Ile Gly Leu Thr Ala Val Val Leu
1 5 10 15
Trp Ser Ser Ile Val Gly Leu Ile Arg Gly Val Ser Glu His Leu Gly
20 25 30
Ala Thr Gly Gly Ala Ala Met Met Tyr Ser Val Ala Ser Leu Phe Leu
35 40 45
Leu Leu Ser Val Gly Phe Pro Lys Leu Gly Ser Phe Pro Lys Lys Tyr
50 55 60
Leu Leu Trp Gly Ser Leu Leu Phe Val Ser Tyr Glu Leu Cys Leu Ser
65 70 75 80
Leu Ser Ala Gly Tyr Ala Asn Thr Gly Arg Gln Ala Ile Glu Val Ser
85 90 95
Met Val Asn Tyr Leu Trp Pro Ala Phe Thr Leu Ile Ala Ala Ile Ala
100 105 110
Phe Asn Arg Gln Arg Ala Asn Trp Met Val Val Pro Gly Phe Ile Leu
115 120 125
Ser Ile Ile Gly Ile Cys Trp Val Leu Gly Gly Asp Gln Gly Leu Asp
130 135 140
Leu Ala Gly Met Leu Gly Asn Val Gln Asp Asn Pro Leu Ser Tyr Gly
145 150 155 160
Leu Ala Phe Leu Gly Ala Val Ile Trp Ala Ala Tyr Cys Thr Val Thr
165 170 175
Ala Arg Leu Ala Lys Gly Lys Asn Gly Val Thr Leu Phe Phe Ile Leu
180 185 190
Val Ala Leu Thr Leu Trp Val Lys Phe Phe Phe Gly Asp His Arg Pro
195 200 205
Met Ser Phe Ser Leu Pro Ala Ile Val Tyr Leu Leu Leu Ala Ala Ala
210 215 220
Ala Met Gly Phe Gly Tyr Ala Ala Trp Asn Val Gly Ile Leu His Gly
225 230 235 240
Asn Val Thr Val Leu Ala Gly Val Ser Tyr Phe Ile Pro Val Phe Ser
245 250 255
Ala Ala Leu Ser Ala Met Val Leu His Ala Pro Leu Pro Arg Ser Phe
260 265 270
Trp Val Gly Ala Ser Leu Val Cys Ala Gly Ser Ile Leu Cys Trp Leu
275 280 285
Ala Thr Arg Ala Arg Arg Ala Ser Ala Ala Gln Glu Asp Ala Val Ala
290 295 300
Asp Cys Leu
305
<210> 146
<211> 307
<212> PRT
<213> Artificial Sequence
<220>
<223> wex I83V
<400> 146
Met Asn Ser Lys Lys Ala Thr Leu Ile Gly Leu Thr Ala Val Val Leu
1 5 10 15
Trp Ser Ser Ile Val Gly Leu Ile Arg Gly Val Ser Glu His Leu Gly
20 25 30
Ala Thr Gly Gly Ala Ala Met Met Tyr Ser Val Ala Ser Leu Phe Leu
35 40 45
Leu Leu Ser Val Gly Phe Pro Lys Leu Gly Ser Phe Pro Lys Lys Tyr
50 55 60
Leu Leu Trp Gly Ser Leu Leu Phe Val Ser Tyr Glu Leu Cys Leu Ser
65 70 75 80
Leu Ser Val Gly Tyr Ala Asn Thr Gly Arg Gln Ala Ile Glu Val Ser
85 90 95
Met Val Asn Tyr Leu Trp Pro Ala Phe Thr Leu Ile Ala Ala Ile Ala
100 105 110
Phe Asn Arg Gln Arg Ala Asn Trp Met Val Val Pro Gly Phe Ile Leu
115 120 125
Ser Ile Ile Gly Ile Cys Trp Val Leu Gly Gly Asp Gln Gly Leu Asp
130 135 140
Leu Ala Gly Met Leu Gly Asn Val Gln Asp Asn Pro Leu Ser Tyr Gly
145 150 155 160
Leu Ala Phe Leu Gly Ala Val Ile Trp Ala Ala Tyr Cys Thr Val Thr
165 170 175
Ala Arg Leu Ala Lys Gly Lys Asn Gly Val Thr Leu Phe Phe Ile Leu
180 185 190
Val Ala Leu Thr Leu Trp Val Lys Phe Phe Phe Gly Asp His Arg Pro
195 200 205
Met Ser Phe Ser Leu Pro Ala Ile Val Tyr Leu Leu Leu Ala Ala Ala
210 215 220
Ala Met Gly Phe Gly Tyr Ala Ala Trp Asn Val Gly Ile Leu His Gly
225 230 235 240
Asn Val Thr Val Leu Ala Gly Val Ser Tyr Phe Ile Pro Val Phe Ser
245 250 255
Ala Ala Leu Ser Ala Met Val Leu His Ala Pro Leu Pro Arg Ser Phe
260 265 270
Trp Val Gly Ala Ser Leu Val Cys Ala Gly Ser Ile Leu Cys Trp Leu
275 280 285
Ala Thr Arg Ala Arg Arg Ala Ser Ala Ala Gln Glu Asp Ala Val Ala
290 295 300
Asp Cys Leu
305
<210> 147
<211> 307
<212> PRT
<213> Artificial Sequence
<220>
<223> wex I83L
<400> 147
Met Asn Ser Lys Lys Ala Thr Leu Ile Gly Leu Thr Ala Val Val Leu
1 5 10 15
Trp Ser Ser Ile Val Gly Leu Ile Arg Gly Val Ser Glu His Leu Gly
20 25 30
Ala Thr Gly Gly Ala Ala Met Met Tyr Ser Val Ala Ser Leu Phe Leu
35 40 45
Leu Leu Ser Val Gly Phe Pro Lys Leu Gly Ser Phe Pro Lys Lys Tyr
50 55 60
Leu Leu Trp Gly Ser Leu Leu Phe Val Ser Tyr Glu Leu Cys Leu Ser
65 70 75 80
Leu Ser Leu Gly Tyr Ala Asn Thr Gly Arg Gln Ala Ile Glu Val Ser
85 90 95
Met Val Asn Tyr Leu Trp Pro Ala Phe Thr Leu Ile Ala Ala Ile Ala
100 105 110
Phe Asn Arg Gln Arg Ala Asn Trp Met Val Val Pro Gly Phe Ile Leu
115 120 125
Ser Ile Ile Gly Ile Cys Trp Val Leu Gly Gly Asp Gln Gly Leu Asp
130 135 140
Leu Ala Gly Met Leu Gly Asn Val Gln Asp Asn Pro Leu Ser Tyr Gly
145 150 155 160
Leu Ala Phe Leu Gly Ala Val Ile Trp Ala Ala Tyr Cys Thr Val Thr
165 170 175
Ala Arg Leu Ala Lys Gly Lys Asn Gly Val Thr Leu Phe Phe Ile Leu
180 185 190
Val Ala Leu Thr Leu Trp Val Lys Phe Phe Phe Gly Asp His Arg Pro
195 200 205
Met Ser Phe Ser Leu Pro Ala Ile Val Tyr Leu Leu Leu Ala Ala Ala
210 215 220
Ala Met Gly Phe Gly Tyr Ala Ala Trp Asn Val Gly Ile Leu His Gly
225 230 235 240
Asn Val Thr Val Leu Ala Gly Val Ser Tyr Phe Ile Pro Val Phe Ser
245 250 255
Ala Ala Leu Ser Ala Met Val Leu His Ala Pro Leu Pro Arg Ser Phe
260 265 270
Trp Val Gly Ala Ser Leu Val Cys Ala Gly Ser Ile Leu Cys Trp Leu
275 280 285
Ala Thr Arg Ala Arg Arg Ala Ser Ala Ala Gln Glu Asp Ala Val Ala
290 295 300
Asp Cys Leu
305
<---
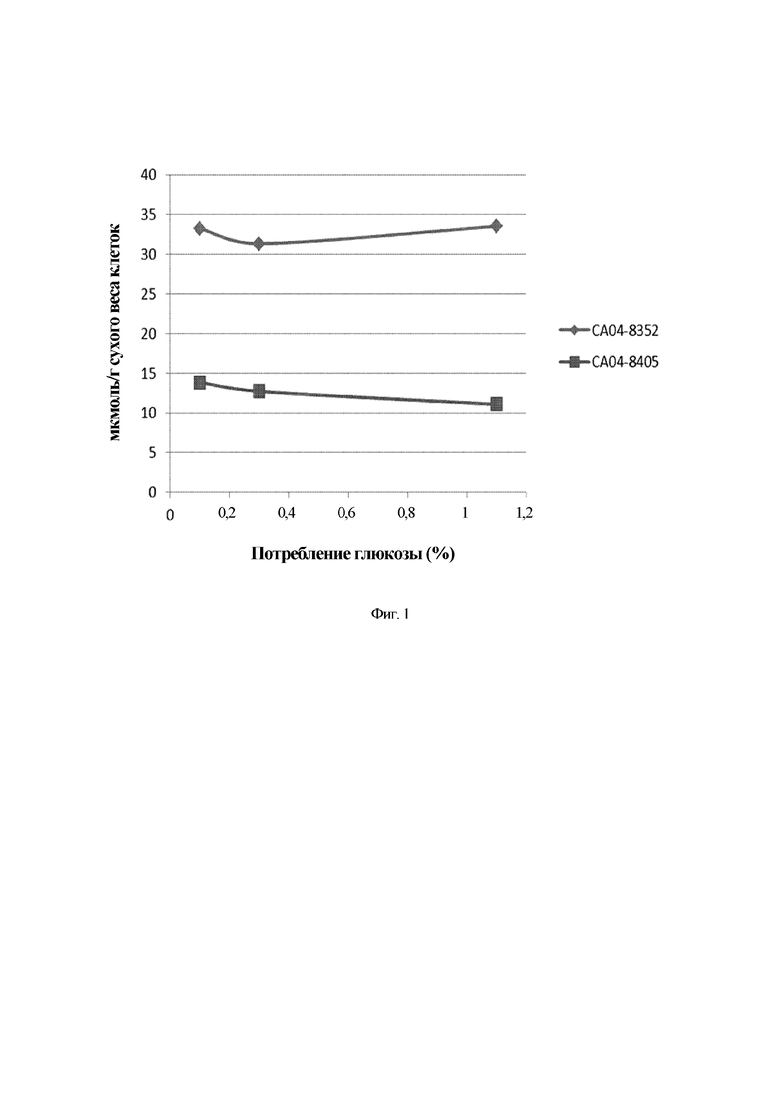
Изобретение относится к биотехнологии, в частности к варианту белка, обладающему активностью экспортера триптофана, микроорганизму, продуцирующему L-триптофан, экспрессирующему указанный вариант белка, и способу получения L-триптофана с применением указанного микроорганизма. Изобретение позволяет получать L-триптофан с высокой степенью эффективности. 6 н. и 5 з.п. ф-лы, 1 ил., 32 табл., 23 пр.
1. Вариант белка, обладающий активностью экспортера L-триптофана, где по меньшей мере одна аминокислота, выбранная из аминокислот в положениях 79 – 83 аминокислотной последовательности SEQ ID NO: 1, заменена гидрофобной или алифатической аминокислотой.
2. Вариант белка по п. 1, где гидрофобная аминокислота представляет собой аминокислоту, выбранную из группы, состоящей из глицина, метионина, аланина, валина, лейцина, изолейцина, пролина, фенилаланина, тирозина и триптофана.
3. Вариант белка по п. 1, где алифатическая аминокислота представляет собой аминокислоту, выбранную из группы, состоящей из глицина, аланина, валина, лейцина и изолейцина.
4. Вариант белка по п. 1, где гидрофобная или алифатическая аминокислота представляет собой аминокислоту, выбранную из группы, состоящей из аланина, валина, лейцина и изолейцина.
5. Вариант белка по п. 1, содержащий любую аминокислотную последовательность, выбранную из SEQ ID NO: 131 – 147.
6. Полинуклеотид, кодирующий вариант белка по любому из пп. 1 – 5.
7. Вектор экспрессии, содержащий полинуклеотид по п. 6.
8. Микроорганизм, продуцирующий L-триптофан, содержащий вариант белка по любому из пп. 1 – 5, полинуклеотид по п.6 или вектор экспрессии по п.7.
9. Микроорганизм по п. 8, относящийся к роду Corynebacterium или роду Escherichia.
10. Способ получения L-триптофана, включающий
культивирование микроорганизма по п. 8 в среде и выделение L-триптофана из культуральной среды или культивируемого микроорганизма.
11. Применение варианта белка, обладающего активностью экспортера L-триптофана, где по меньшей мере одна аминокислота, выбранная из аминокислот в положениях 79 – 83 аминокислотной последовательности SEQ ID NO: 1, заменена на другую аминокислоту, для увеличения продукции L-триптофана.
| МИКРООРГАНИЗМ ИЗ РОДА Escherichia, ОБЛАДАЮЩИЙ УСИЛЕННОЙ СПОСОБНОСТЬЮ К ПРОДУКЦИИ L-ТРИПТОФАНА, И СПОСОБ ПОЛУЧЕНИЯ L-ТРИПТОФАНА С ЕГО ИСПОЛЬЗОВАНИЕМ | 2013 |
|
RU2593957C2 |
| Установка для градуировки и проверки расходомеров (счетчиков) жидкости | 1981 |
|
SU1013765A2 |
| JIAN WANG et.al | |||
| Genetic engineering of Escherichia coli to enhance production of L-tryptophan, Appl Microbiol Biotechnol 2013 Sep;97(17):7587-96, doi: 10.1007/s00253-013-5026-3 | |||
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |

