1. ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Данное изобретение относится к варианту белка SpoT и способу получения L-аминокислоты с его применением.
2. ОПИСАНИЕ БЛИЖАЙШЕГО УРОВНЯ ТЕХНИКИ
Что касается микроорганизмов, применяющихся для продуцирования L-аминокислот, в первое время в основном использовались отобранные штаммы, демонстрирующие устойчивость к аналогам, полученные в результате мутации, индуцированной химическими или физическими факторами, однако с 1990-х годов возобладало использование рекомбинантных штаммов, полученных при помощи генно-инженерных технологий благодаря быстрому развитию технологии рекомбинации генов и установлению механизмов регуляции на молекулярном уровне.
Строгий ответ представляет собой бактериальный ответ на различные стрессовые ситуации, который был впервые открыт как феномен, при котором синтез белков подавляется аминокислотным голоданием. В настоящее время известно, что строгий ответ позволяет адаптироваться к различным стрессовым окружающим условиям благодаря изменениям в различных физиологических активностях, таких как остановка деления клетки, ингибирование репликации ДНК и так далее. (F.C. Neidhart, ed. (Washington, D.C.: ASM Press), (1996) pp.1458-1496, J. Mol. Microbiol. Biotechnol. (2002) 4:331-340.). Ключевые реакции строгого ответа запускаются гуанозинпентафосфатом (pppGpp) и гуанозинтетрафосфатом (ppGpp). Поскольку не существует больших функциональных различий между этими двумя молекулами, они обычно обозначаются как (p)ppGpp.
У Escherichia coli pppGpp образуется pppGpp-синтетазой под названием Re1A. Белок Re1A контролирует трансляцию белков на рибосоме. Внутриклеточная недостаточность белков повышает концентрацию не заряженной аминокислотой тРНК, которая связывается с рибосомами, приводя к активации Re1A. Re1A продуцирует pppGpp благодаря реакции гуанозинтрифосфата (GTP) с аденозинтрифосфатом (АТР). При повышении внутриклеточной концентрации аминокислот и восстановлении баланса (p)ppGpp подвергается гидролизу белком SpoT. Белок SpoT обладает не только активностью, осуществляющей расщепление (p)ppGpp, но также синтетической активностью (Nat Rev Microbiol. (2012) 10(3):203-12). Однако, взаимоотношение между белком SpoT и образованием аминокислот остается пока неизвестным.
В данном изобретении предложен вариант белка SpoT и способ получения L-аминокислоты с его применением.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Задачей данного изобретения является создание варианта белка SpoT, включающего замену аминокислоты, соответствующей положению 262 в последовательности SEQ ID NO: 1, на другую аминокислоту.
Другой задачей данного изобретения является создание полинуклеотида, кодирующего вариант белка SpoT, и вектора, включающего его.
Еще одной задачей данного изобретения является создание микроорганизма рода Escherichia, включающего в себя один или более вариантов белка SpoT; полинуклеотид, кодирующий вариант белка SpoT, и вектор, включающий полинуклеотид.
Еще одной задачей данного изобретения является разработка способа получения L-аминокислоты, включающего культивирование микроорганизма в среде.
Еще одной задачей данного изобретения является разработка способа повышения способности микроорганизма продуцировать L-аминокислоту, включающего стадию модификации микроорганизма для экспрессии варианта белка SpoT.
СВЕДЕНИЯ, ПОДТВЕРЖДАЮЩИЕ ВОЗМОЖНОСТЬ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Далее данное изобретение будет описано более подробно. При этом, соответствующие описания и воплощения, изложенные в данной заявке, также могут быть применены к другим описаниям и воплощениям. Таким образом, все комбинации различных элементов, изложенных в данной заявке, входят в объем данного изобретения. Более того, объем данного изобретения не ограничивается конкретным описанием, приведенным ниже. Кроме того, в данном описании процитирован ряд публикаций и патентных документов и приведены ссылки на них. Содержание процитированных публикаций и патентных документов включено в данный документ во всей полноте путем ссылки для более точного описания уровня техники, к которой относится данное изобретение, и объекта данного изобретения.
В аспекте данного изобретения предложен вариант белка SpoT, включающий замену аминокислоты, соответствующей положению 262 в последовательности SEQ ID NO: 1, на другую аминокислоту.
Вариант белка SpoT относится к варианту белка, включающего замену аминокислоты, соответствующей положению 262 от N-конца в аминокислотной последовательности SEQ ID NO: 1, на другую аминокислоту, в полипептиде, обладающем активностью белка SpoT, или в белке SpoT.
В одном воплощении другая аминокислота может быть выбрана из глицина, валина, лейцина, изолейцина, метионина, фенилаланина, триптофана, пролина, серина, треонина, цистеина, тирозина, аспарагина, глутамина, аспарагиновой кислоты, глутаминовой кислоты, лизина, аргинина и гистидина.
Применительно к варианту белка по любому из описанных выше воплощений другая аминокислота может быть выбрана из полярных аминокислот.
Применительно к варианту белка по любому из описанных выше воплощений другая аминокислота может быть выбрана из серина, треонина, цистеина, тирозина, аспарагина и глутамина.
Применительно к варианту белка по любому из описанных выше воплощений вариант белка SpoT может иметь или включать любую последовательность из аминокислотных последовательностей, представленных SEQ ID NOS: 19-24, или может по существу состоять из аминокислотной последовательности.
Применительно к варианту белка по любому из описанных выше воплощений вариант белка SpoT может включать замену аминокислоты, соответствующей положениям 262 на основе аминокислотной последовательности SEQ ID NO: 1, на другую аминокислоту и может включать аминокислотную последовательность, по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или более или менее чем на 100% гомологичную или идентичную аминокислотной последовательности, представленной SEQIDNO: 1.
Также, очевидно, что в объем данного изобретения также входит белок, имеющий аминокислотную последовательность с делецией, модификацией, заменой, консервативной заменой или добавлением некоторых последовательностей, при условии, что аминокислотная последовательность обладает такой гомологией или идентичностью и демонстрирует эффективность, которая соответствует таковой варианта белка по данному изобретению.
Его примеры включают имеющие добавление или делецию последовательности на N-конце, С-конце аминокислотной последовательности и/или внутри аминокислотной последовательности, мутацию естественного происхождения, молчащую мутацию или консервативную замену, которая не изменяет функцию варианта белка по данному изобретению.
В данном описании термин «консервативная замена» означает замену одной аминокислоты на другую аминокислоту, имеющую схожие структурные и/или химические свойства. Такая аминокислотная замена, как правило, может возникать на основании сходства в полярности, заряде, растворимости, гидрофобности, гидрофильности и/или амфипатической природы остатков. Как правило, консервативная замена может крайне незначительно влиять или не влияет на активность белков или полипептидов.
Вариант белка по данному изобретению также может быть описан как «вариантный белок».
В данном документе термин «вариант» относится к полипептиду, имеющему аминокислотную последовательность, отличную от таковой варианта до внесения консервативной замены и/или модификации одной или более аминокислот, но сохраняющему функции или свойства. Такой вариант, как правило, может быть обнаружен путем модификации одной или более аминокислот в аминокислотной последовательности полипептида и оценки свойств модифицированного полипептида. Другими словами, способность варианта по сравнению с полипептидом до внесения изменений может быть повышена, оставлена без изменений или снижена. Кроме того, некоторые варианты могут включать варианты, в которых были удалены одна или более частей, такие как N-концевая лидерная последовательность или трансмембранный домен. Другие варианты могут включать варианты, у которых в зрелом белке была удалена N-и/или С-концевая часть. Термин «вариант» может использоваться взаимозаменяемо с такими терминами, как модификация, модифицированный полипептид, модифицированный белок, мутант, мутеин и дивергент, без ограничения, при условии, что этот термин используется в значении вариация. Применительно к задачам данного изобретения, вариант может представлять собой полипептид, включающий аминокислотную последовательность, в которой аминокислота, соответствующая положению 262 в аминокислотной последовательности SEQ ID NO: 1, замещена на другую аминокислоту.
Кроме того, вариант может включать делеции или добавления аминокислот, которые оказывают минимальное влияние на свойства и вторичную структуру полипептида. Например, с N-концом варианта может быть конъюгирована сигнальная (или лидерная) последовательность, которая задействована в транслокации белка одновременно с трансляцией или после завершения трансляции. Вариант может быть конъюгирован с другими последовательностями или линкерами для его обнаружения, очистки или синтеза.
В данном документе термины «гомология» или «идентичность» означают степень сходства между двумя заданными аминокислотными последовательностями или последовательностями оснований и могут выражаться в процентах. Термины «гомология» и «идентичность» зачастую можно использовать взаимозаменяемо.
Гомология или идентичность консервативных полинуклеотидов или полипептидов определяется при помощи стандартных алгоритмов выравнивания, и при этом могут использоваться значения штрафов за открытие гэпа, установленные в используемой программе по умолчанию. По существу гомологичные или идентичные последовательности обычно способны гибридизоваться в условиях умеренной или высокой строгости со всей последовательностью или ее частью. Очевидно, что гибридизация также включает гибридизацию с полинуклеотидом, включающим общие ко доны или ко доны с учетом вырожденности ко до нов.
Установить, обладают ли две любые полинуклеотидные или полипептидные последовательности гомологией, сходством или идентичностью, можно, например, при помощи известных компьютерных алгоритмов, таких как программа FASTA, например, с использованием параметров по умолчанию, как описано Pearson et al. (1988) Proc. Natl. Acad. Sci. USA 85:2444. В альтернативном варианте гомологию, сходство или идентичность можно устанавливать при помощи алгоритма Нидлмана-Вунша (Needleman and Wunsch, 1970, J. Mol. Biol. 48:443-453), как осуществляется в программе Needleman пакета EMBOSS (EMBOSS: The European Molecular Biology Open Software Suite, Rice et al., 2000, Trends Genet. 16:276-277) (версия 5.0.0 или более поздняя) (включая пакет программ GCG (Devereux, J., et al, Nucleic Acids Research 12:387 (1984)), BLASTP, BLASTN, FASTA (Atschul, S. F., et al, J MOLEC BIOL 215:403 (1990); Guide to Huge Computers, Martin J. Bishop, ed., Academic Press, San Diego, 1994, and CARILLO et al. (1988) SIAM J Applied Math 48:1073). Например, для определения гомологии, сходства или идентичности можно использовать BLAST Национального Центра Биотехнологической Информации или ClustalW.
Гомологию, сходство или идентичность полинуклеотидов или полипептидов можно определять путем сравнения информации о последовательности, например, с применением компьютерной программы GAP, такой как у Needleman et al. (1970), J Mol Biol. 48:443, как изложено, например, Smith and Waterman, Adv. Appl. Math (1981) 2:482. В заключение, программу GAP можно охарактеризовать как значение, полученное путем деления общего количества одинаково расположенных символов (то есть нуклеотидов или аминокислот) на общее количество символов в более короткой из двух последовательностей. Параметры по умолчанию для программы GAP могут включать (1) бинарную матрицу сравнения (включающую значение 1 для идентичности и 0 для отсутствия идентичности) и взвешенную матрицу сравнения, предложенную Gribskov et al. (1986) Nucl. Acids Res. 14:6745 (или подстановочную матрицу EDNAFULL (EMBOSS версия NCBI NUC4.4), согласно описанию Schwartz and DayhofF, eds., Atlas Of Protein Sequence And Structure, National Biomedical Research Foundation, pp.353-358 (1979); (2) штраф 3,0 для каждого гэпа и дополнительный штраф 0,10 для каждого символа в каждом гэпе (или штраф за открытие гэпа 10, штраф за продолжение гэпа 0,5); и (3) отсутствие штрафа за концевые гэпы.
В одном примере данного изобретения вариант белка SpoT по данному изобретению может обладать активностью белка SpoT. Кроме того, вариант белка SpoT по данному изобретению может обладать активностью для повышения способности продуцировать L-аминокислоту по сравнению с полипептидом дикого типа, обладающего активностью белка SpoT.
В данном описании термин «белок SpoT» представляет собой полипептид, обладающий расщепляющей (p)ppGpp и/или синтетической активностью. В частности, белок SpoT по данному изобретению может использоваться взаимозаменяемо с бифункциональной (р)ррОрр-синтазой или бифункциональной (р)ррОрр-гидролазой. В данном описании последовательность белка SpoT может быть получена из GenBank NCBI, которая является известной базой данных. В частности, белок SpoT может представлять собой полипептид, кодируемый геном spot, без ограничения.
В одном воплощении белок SpoT по данному изобретению может быть получен из рода Escherichia (Escherichia sp.). В одном воплощении белок SpoT по данному изобретению может представлять собой аминокислотную последовательность SEQ ID NO: 1. В одном воплощении белок SpoT по данному изобретению может включать аминокислотную последовательность, обладающую гомологией или идентичностью 70% или более, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% или более с аминокислотной последовательностью SEQ ID NO: 1, без ограничения.
В данном описании термин «соответствующий» относится к аминокислотным остаткам в положениях, приведенных в составе полипептида, или аминокислотным остаткам, которые схожи, идентичны или гомологичны приведенным в составе полипептида. Обнаружение аминокислоты в соответствующем положении может представлять собой определение специфической аминокислоты в последовательности, относящейся к специфической последовательности. В данном описании «соответствующая область» обычно относится к схожему или соответствующему положению в родственном белке или референсном белке.
Например, произвольную аминокислотную последовательность выравнивают с SEQ ID NO: 1 и на этом основании каждый аминокислотный остаток аминокислотной последовательности можно пронумеровать с указанием выраженного числом положения аминокислотного остатка, соответствующего аминокислотному остатку SEQ ID NO: 1. Например, алгоритм выравнивания последовательностей, описанный в данном изобретении, может устанавливать положение аминокислоты или положение, в котором возникает модификация, такая как замена, вставка или делеция, путем сравнения с таковым в последовательности, вводимой при запросе (также обозначаемой как «референсная последовательность»).
Для таких выравниваний, например, можно использовать алгоритм Нидлмана-Вунша (Needleman and Wunsch, 1970, J. Mol. Biol. 48:443-453), программа Needleman пакета EMBOSS (EMBOSS: The European Molecular Biology Open Software Suite, Rice et al., 2000), Trends Genet. 16:276-277), без ограничения, и можно надлежащим образом использовать программу выравнивания последовательностей, алгоритм попарного сравнения последовательностей и так далее, которые известны в области техники.
В другом аспекте данного изобретения предложен полинуклеотид, кодирующий вариант белка SpoT по данному изобретению.
В данном описании термин «полинуклеотид» представляет собой цепь ДНК или РНК определенной длины или более в виде полимера из нуклеотидов, в котором мономеры нуклеотидов соединены в длинную цепь ковалентными связями и, более конкретно, означает фрагмент полинуклеотида, кодирующий вариант белка SpoT.
Полинуклеотид, кодирующий вариант белка SpoT по данному изобретению может включать нуклеотидную последовательность, кодирующую аминокислотную последовательность, включающую замену аминокислоты, соответствующей положению 262 в последовательности SEQ ID NO: 1, на другую аминокислоту. В одном воплощении полинуклеотид по данному изобретению может включать, состоять из или по существу состоять из нуклеотидных последовательностей, кодирующих аминокислотные последовательности SEQ ID NO: 19 - 24.
Можно осуществлять различные модификации кодирующей области полинуклеотид а по данному изобретению при условии, что аминокислотная последовательность варианта белка по данному изобретению не изменяется с учетом вырожденности кодонов или предпочтения кодонов в организмах, которые намереваются использовать для экспрессии варианта белка по данному изобретению. В частности, полинуклеотид по данному изобретению может иметь или включать в себя последовательность оснований, обладающую гомологией или идентичностью 70% или более, 75% или более, 80% или более, 85% или более, 90% или более, 95% или более, 96% или более, 97% или более, 98% или более и менее чем 100% с последовательностью SEQ ID NO: 2, или может состоять или по существу состоять из последовательности оснований, обладающей гомологией или идентичностью 70% или более, 75% или более, 80% или более, 85% или более, 90% или более, 95% или более, 96% или более, 97% или более или 98% или более и менее чем 100% с последовательностью SEQ ID NO: 2, без ограничения. Так, в данной последовательности, обладающей гомологией или идентичностью, кодон, кодирующий аминокислоту, которая соответствует положению 262 в SEQ ID NO: 1, может быть одним из кодонов, кодирующих аминокислоты, за исключением аланина, в частности, одним из кодонов, кодирующих аминокислоту, выбранную из глицина, валина, лейцина, изолейцина, метионина, фенилаланина, триптофана, пролина, серина, треонина, цистеина, тирозина, аспарагина, глутамина, аспарагиновой кислоты, глутаминовой кислоты, лизина, аргинина и гистидина.
Кроме того, полинуклеотид по данному изобретению может включать зонд, который может быть получен из известной последовательности гена, например, последовательности без ограничения, при условии, что она представляет собой последовательность, которая способна гибрид изо в аться в строгих условиях с комплементарной последовательностью со всей или частью полинуклеотидной последовательности по данному изобретению. «Строгие условия» означают условия, позволяющие полинуклеотидам специфически гибридизоваться между собой. Такие условия подробно описаны в документах (см. Sambrook et al., выше, 9.50-9.51, 11.7-11.8). Их примеры включают условие, когда полинуклеотиды, обладающие более высокой гомологией или идентичностью, то есть, полинуклеотиды, обладающие гомологией или идентичностью 70% или более, 75% или более, 80% или более, 85% или более, 90% или более, 95% или более, 96% или более, 97% или более, 98% или более или 99% или более, гибридизуются друг с другом, тогда как полинуклеотиды, обладающие меньшей гомологией или идентичностью, не гибридизуются друг с другом, или условие, когда осуществляется отмывка один раз, в частности, от двух до трех раз, при концентрации соли и температуре, эквивалентных 60°С, IX SSC, 0,1% SDS, в частности, 60°С, 0,1Х SSC, 0,1% SDS, более конкретно, 68°С, 0,1Х SSC, 0,1% SDS, которые представляют собой условия отмывки для стандартной гибридизации по Саузерну.
Для гибридизации необходимо, чтобы две нуклеиновые кислоты имели комплементарные последовательности, хотя, в зависимости от строгости гибридизации, возможны несовпадения между основаниями. Термин «комплементарный» используется для описания взаимоотношения между нуклеотидными основаниями, способными гибридизоваться друг с другом. Например, в случае ДНК аденин комплементарен тимину, а цитозин комплементарен гуанину. Соответственно, полинуклеотид по данному изобретению может также включать по существу схожие последовательности нуклеиновых кислот, а также выделенные фрагменты нуклеиновых кислот, комплементарные полной последовательности.
В частности, полинуклеотид, обладающий гомологией или идентичностью с полинуклеотидом по данному изобретению, можно обнаружить с применением условий гибридизации, включающих стадию гибридизации при значении Tm 55°С, и описанные выше условия. Кроме того, значение Tm может составлять 60°С, 63°С или 65°С, без ограничения, и может быть соответствующим образом скорректировано специалистом в области техники в зависимости от задачи.
Надлежащая строгость для гибридизации полинуклеотидов зависит от длины и степени комплементарности полинуклеотидов, и эти параметры хорошо известны в области техники (например, J. Sambrook et al., выше).
Еще в одном аспекте данного изобретения предложен вектор, включающий в себя полинуклеотид по данному изобретению. Вектор может представлять собой экспрессирующий вектор для экспрессии полинуклеотида в клетке-хозяине, без ограничения.
В данном описании термин «вектор» может включать ДНК-конструкцию, включающую в себя полинуклеотидную последовательность, кодирующую полипептид, представляющий интерес, которая функционально связана с подходящей областью, регулирующей экспрессию (или последовательностью контроля экспрессии), так чтобы полипептид, представляющий интерес, мог экспрессироваться в подходящем хозяине. Область, регулирующая экспрессию, может включать промотор, способный инициировать транскрипцию, любую последовательность оператора для контролирования транскрипции, последовательность, кодирующую подходящий сайт связывания рибосомы, и последовательность, контролирующую терминацию транскрипции и трансляции. Вектор может быть трансформирован в подходящего хозяина и затем реплицироваться или функционировать независимо от генома хозяина или может интегрироваться собственно в геном хозяина.
Вектор для применения по данному изобретению не ограничивается каким-либо конкретным, но может применяться любой известный в области техники вектор. Примеры обычно используемых векторов включают плазмиды, космиды, вирусы и бактериофаги естественного происхождения или рекомбинантные. Например, в качестве фагового вектора или космидного вектора можно использовать pWE15, М13, MBL3, MBL4, IXII, ASHII, APII, t10, t11, Charon4A, Charon21A и так далее, а в качестве плазмидного вектора можно использовать вектор на основе системы pDZ, системы pBR, системы pUC, системы pBluescript II, системы pGEM, системы pTZ, системы pCL, системы рЕТ и так далее. В частности, можно использовать векторы pDZ, pDC, pDCM2, pACYC177, pACYC184, pCL, pECCG117, pUC19, pBR322, pMW118, pCC1BAC итак далее.
Например, полинуклеотид, кодирующий полипептид, представляющий интерес, может быть встроен в хромосому при помощи вектора для внутриклеточного встраивания в хромосому. Встраивание полинуклеотида в хромосому может достигаться любым известным в области техники способом, например, посредством гомологичной рекомбинации, без ограничения. Вектор может дополнительно включать в себя селективный маркер для подтверждения его встраивания в хромосому. Селективный маркер используют для отбора клеток, трансформированных векторами, то есть для подтверждения встраивания молекулы нуклеиновой кислоты, представляющей интерес, и могут применяться маркеры, придающие фенотипы, позволяющие осуществлять отбор, такие как устойчивость к лекарственным средствам, ауксотрофия, устойчивость к цитотоксическим агентам или экспрессия поверхностных полипептидов. В среде, обработанной агентом для селекции, выживают или демонстрируют другие фенотипические признаки только клетки, экспрессирующие селективный маркер, таким образом, можно осуществлять отбор трансформированных клеток.
В данном описании термин «трансформация» означает, что вектор, включающий в себя полинуклеотид, кодирующий целевой полипептид, внедрен в клетку-хозяина или микроорганизм, так чтобы полипептид, кодируемый полинуклеотидом, мог экспрессироваться в клетке-хозяине. После трансформации полинуклеотид может быть как встроен в хромосому клетки-хозяина, так и располагаться вне хромосомы, при условии, что он может экспрессироваться в клетке-хозяине. Кроме того, полинуклеотид включает ДНК и/или РНК, которые кодируют полипептид, представляющий интерес. Полинуклеотид может быть внедрен в любой форме, при условии, что он может быть внедрен в клетку-хозяина и экспрессироваться. Например, полинуклеотид может быть внедрен в клетку-хозяина в форме экспрессионной кассеты, которая представляет собой генетическую конструкцию, включающую все элементы, необходимые для самостоятельной экспрессии. Обычно экспрессионная кассета может включать в себя промотор, функционально связанный с полинуклеотидом, сигнал терминации транскрипции, сайт связывания рибосомы и сигнал терминации трансляции. Экспрессионная кассета может быть в форме экспрессирующего вектора, способного к самостоятельной репликации. Кроме того, полинуклеотид может быть внедрен в клетку-хозяина как таковой и быть функционально связанным с последовательностью, необходимой для экспрессии в клетке-хозяине, без ограничения.
Кроме того, термин «функционально связанный» означает, что последовательность полинуклеотида функционально связана с последовательностью промотора, который инициирует и опосредует транскрипцию полинуклеотида, кодирующего вариант, представляющий интерес, по данному изобретению.
Еще одной задачей данного изобретения является создание микроорганизма рода Escherichia (Escherichia sp.), включающего в себя один или более вариантов белка SpoT по данному изобретению; полинуклеотид, кодирующий вариант белка SpoT, и вектор, включающий в себя полинуклеотид.
Микроорганизм по данному изобретению может включать вариант белка SpoT по данному изобретению; полинуклеотид, кодирующий вариант белка или вектор, включающий в себя полинуклеотид по данному изобретению.
В одном воплощении микроорганизм по данному изобретению может экспрессировать вариант белка SpoT по данному изобретению, включая в себя вариант белка SpoT по данному изобретению, полинуклеотид, кодирующий вариант белка, или вектор, включающий в себя полинуклеотид по данному изобретению.
В данном описании термин «штамм (или микроорганизм)» включает все микроорганизмы дикого типа или микроорганизмы, генетически модифицированные естественным или искусственным образом, и это может быть микроорганизм, в котором конкретный механизм ослаблен или усилен вследствие вставки чужеродного гена или усиления активности или инактивации эндогенного гена, и это может быть микроорганизм, включающий генетическую модификацию для продуцирования полипептида, белка или продукта, представляющего интерес.
Штамм по данному изобретению может представлять собой штамм, включающий один или более вариантов белка по данному изобретению, полинуклеотид по данному изобретению и вектор, включающий в себя полинуклеотид по данному изобретению; штамм, модифицированный, чтобы экспрессировать вариант белка по данному изобретению или полинуклеотид по данному изобретению; штамм (например, рекомбинантный штамм), экспрессирующий вариант белка по данному изобретению или полинуклеотид по данному изобретению; или штамм (например, рекомбинантный штамм), имеющий активность варианта белка по данному изобретению, без ограничения.
Штамм по данному изобретению может представлять собой штамм, обладающий способностью продуцировать L-аминокислоту.
В одном воплощении L-аминокислота может представлять собой L-триптофан.
Штамм по данному изобретению может представлять собой микроорганизм, от природы имеющий белок SpoT или обладающий способностью продуцировать L-аминокислоту, или микроорганизм, в который внедрен вариант белка SpoT по данному изобретению или кодирующий его полинуклеотид (или вектор, включающий в себя полинуклеотид) и/или способность продуцировать L-аминокислоту придана родительскому штамму, который не имеет белка SpoT или не обладает способностью продуцировать L-аминокислоту, без ограничения.
Например, штамм по данному изобретению представляет собой клетку или микроорганизм, который трансформирован полинуклеотидом по данному изобретению или вектором, включающим в себя полинуклеотид, кодирующий вариант белка согласно данному изобретению, чтобы экспрессировать вариант белка по данному изобретению. Штамм по данному изобретению может включать любой микроорганизм, способный продуцировать L-аминокислоты благодаря включению варианта по данному изобретению. Например, штамм по данному изобретению может представлять собой рекомбинантный штамм, в котором полинуклеотид, кодирующий вариант по данному изобретению, внедрен в естественный микроорганизм дикого типа или микроорганизм, продуцирующий L-аминокислоту, чтобы экспрессировать вариант SpoT и обладать повышенной способностью продуцировать L-аминокислоту. Рекомбинантный штамм, обладающий повышенной способностью продуцировать L-аминокислоту, может представлять собой микроорганизм, обладающий повышенной способностью продуцировать L-аминокислоту по сравнению с естественным микроорганизмом дикого типа или микроорганизмом с немодифицированным SpoT (то есть микроорганизмом, экспрессирующим белок SpoT дикого типа), без ограничения. Например, микроорганизмом с немодифицированным SpoT, представляющий собой целевой штамм для сравнения увеличения способности продуцировать L-аминокислоту, может представлять собой штамм СА04-4303 или КССМ11166Р, без ограничения.
Например, рекомбинантный штамм, обладающий повышенной способностью продуцировать L-аминокислоту, может обладать способностью продуцировать L-аминокислоту, повышенной приблизительно на 5% или более, на 6% или более или на 8% или более по сравнению со способностью родительского штамма до изменения или немодифицированного микроорганизма продуцировать L-аминокислоту, но повышенное количество этим не ограничивается, при условии, что продуцирующая способность увеличена на положительную величину по сравнению с продуцирующей способностью родительского штамма до изменения или немодифицированного микроорганизма. Термин «приблизительно» относится к диапазону, включающему все из ±0,5, ±0,4, ±0,3, ±0,2, ±0,1 и так далее, и включает все значения, которые являются эквивалентными или близкими значениям, указанным после термина «приблизительно», однако диапазон ими не ограничивается.
В данном описании термин «немодифицированный микроорганизм» не исключает штаммов, включающих мутацию, которая может возникнуть в микроорганизмах естественным путем, и может представлять собой штамм дикого типа или сам природный штамм или представлять собой штамм до изменения признака вследствие генетической изменчивости, вызванной естественными или искусственными факторами. Например, немодифицированный микроорганизм может представлять собой штамм, в который не внедрен или еще не внедрен вариант белка SpoT, описанный в данном документе. Термин «немодифицированный микроорганизм» может использоваться взаимозаменяемо со «штаммом до модификации», «микроорганизмом до модификации», «неизмененным штаммом», «немодифицированным штаммом», «неизмененным микроорганизмом» или «референсным микроорганизмом».
В другом примере из данного документа микроорганизм по данному изобретению может представлять собой микроорганизм рода Escherichia. В частности, микроорганизм может представлять собой Escherichia coli, без ограничения.
Что касается микроорганизма по данному изобретению, вариант белка SpoT, полинуклеотид, L-аминокислота и так далее являются такими, как описано в других аспектах.
В еще одном аспекте данного изобретения предложен способ получения L-аминокислоты, включающий стадию культивирования в среде микроорганизма рода Escherichia по данному изобретению.
Способ получения L-аминокислоты по данному изобретению может включать стадию культивирования в среде штамма рода Escherichia, включающего в себя вариант белка SpoT по данному изобретению или полинуклеотид по данному изобретению, или вектор по данному изобретению.
В одном воплощении L-аминокислота может представлять собой L-триптофан.
В данном описании термин «культивирование» означает выращивание штамма рода Escherichia по данному изобретению в надлежащим образом контролируемых окружающих условиях. Процесс культивирования по данному изобретению можно осуществлять в соответствии с подходящей средой и условиями культивирования, известными в области техники. Специалист в области техники может легко адаптировать такой процесс культивирования в зависимости от выбранного штамма. В частности, культивирование может представлять собой периодическое культивирование, непрерывное культивирование и/или непрерывное культивирование с подпиткой, без ограничения.
В данном описании термин «среда» означает смешанное вещество, содержащее питательные вещества, необходимые для культивирования штамма рода Escherichia по данному изобретению в качестве основного компонента, и среда снабжает питательными веществами, факторами роста и так далее, включая воду, которые являются незаменимыми для выживания и развития. В частности, в качестве среды и других условий культивирования, применяемых для культивирования штамма рода Escherichia по данному изобретению, можно использовать любые, без конкретного ограничения, при условии, что это среда, используемая для стандартного культивирования микроорганизмов. Штамм рода Escherichia по данному изобретению можно культивировать в аэробных условиях в стандартной среде, содержащей надлежащие источники углерода, источники азота, источники фосфора, неорганические соединения, аминокислоты и/или витамины и так далее, при этом контролируя температуру, рН и так далее.
В данном изобретении источники углерода включают углеводы, такие как глюкоза, сахароза, лактоза, фруктоза, сахароза, мальтоза и так далее; сахароспирты, такие как маннит, сорбит и так далее, органические кислоты, такие как пировиноградная кислота, молочная кислота, лимонная кислота и так далее, аминокислоты, такие как глутаминовая кислота, метионин, лизин и так далее. Можно использовать естественные органические питательные вещества, такие как гидролизат крахмала, меласса, сырая меласса, рисовые отруби, маниок, выжимки сахарного тростника и кукурузный экстракт. В частности, можно использовать такие углеводы как глюкоза и стерилизованная предварительно обработанная меласса (то есть меласса, в которой сахара конвертированы в восстанавливающие) и можно использовать другие источники углерода в надлежащих количествах различным образом, без ограничения. Эти источники углерода можно использовать по-отдельности или в комбинации двух или более из них, без ограничения.
В качестве источников азота можно использовать неорганические источники азота, такие как аммоний, сульфат аммония, хлорид аммония, ацетат аммония, фосфат аммония, карбонат аммония, нитрат аммония и так далее; и органические источники азота, такие как аминокислоты, такие как глутаминовая кислота, метионин, глутамин и так далее, пептон, NZ-амин, мясной экстракт, дрожжевой экстракт, мальтозный экстракт, жидкий кукурузный экстракт, гидролизат казеина, рыба или продукты ее переработки и обезжиренный соевый жмых или продукты его переработки. Эти источники азота можно использовать по-отдельности или в комбинации двух или более из них, без ограничения.
Источники фосфора могут включать дигидрофосфат калия, гидрофосфат калия или соответствующие натрий-содержащие соли. В качестве неорганических соединений можно использовать хлорид натрия, хлорид кальция, хлорид железа, сульфат магния, сульфат железа, сульфат марганца, карбонат кальция и так далее. Кроме того, можно включать аминокислоты, витамины и/или соответствующие предшественники. Эти компоненты или предшественники можно добавлять в среду либо порциями, либо непрерывно, без ограничения.
Кроме того, при культивировании штамма рода Escherichia по данному изобретению рН среды можно корректировать путем добавления в среду должным образом таких соединений, как гидроксид аммония, гидроксид калия, аммиак, фосфорная кислота, серная кислота и так далее. Кроме того, в процессе культивирования можно подавлять образование пены при помощи пеногасителя, такого как сложный полигликолевый эфир жирной кислоты. Кроме того, для поддержания аэробного состояния среды в среду можно вводить кислород или кислород-содержащий газ, либо для поддержания анаэробных или микроаэробных условий газ можно не вводить или можно вводить азот, водород или углекислый газ, без ограничения.
При культивировании по данному изобретению температуру культуры можно поддерживать от 20 до 45°С, в частности, от 25 до 40°С, и штамм можно культивировать приблизительно от 10 до 160 часов, без ограничения.
Продуцируемая в ходе культивирования по данному изобретению L-аминокислота может секретироваться в среду или может оставаться в клетках.
Способ получения L-аминокислоты по данному изобретению может дополнительно включать, например, перед стадией культивирования, стадии получения штамма рода Escherichia по данному изобретению, приготовление среды для культивирования штамма или их комбинацию (в любом порядке).
Способ получения L-аминокислоты по данному изобретению может дополнительно включать стадию выделения L-аминокислоты из среды в соответствии с культивированием (среды, в которой проводилось культивирование) или из штамма рода Escherichia по данному изобретению. После стадии культивирования может быть дополнительно включена стадия выделения.
Выделение может осуществляться для сбора L-аминокислоты, представляющей интерес, с применением подходящего способа, известного в области техники, в зависимости от способа культивирования микроорганизма по данному изобретению, например, способа периодического культивирования, непрерывного культивирования или культивирования с подпиткой. Например, может применяться центрифугирование, фильтрация, обработка агентом для преципитации кристаллизованного белка (высаливание), экстракция, обработка ультразвуком, ультрафильтрация, диализ, различные виды хроматографии, такие как хроматография молекулярных сит (гель-фильтрация), адсорбционная хроматография, ионообменная хроматография и аффинная хроматография, и так далее, ВЭЖХ (высокоэффективная жидкостная хроматография) или их комбинация. L-аминокислоту, представляющую интерес, можно выделять из среды или микроорганизма подходящим способом, известным в области техники.
Способ получения L-аминокислоты по данному изобретению может дополнительно включать стадию очистки. Очистку можно осуществлять подходящим способом, известным в области техники. В одном примере, когда способ получения L-аминокислоты по данному изобретению включает как стадию выделения, так и стадию очистки, стадию выделения и стадию очистки можно осуществлять с перерывами (или непрерывно), вне зависимости от порядка, или можно осуществлять одновременно или путем объединения в одну стадию, без ограничения.
В способе по данному изобретению вариант белка SpoT, полинуклеотид, вектор, штамм и так далее являются такими, как описано в других аспектах.
В еще одном аспекте данного изобретения предложена композиция для получения L-аминокислоты, включающая вариант белка SpoT по данному изобретению.
В одном воплощении композиция может включать вариант белка SpoT по данному изобретению, полинуклеотид, кодирующий вариант белка SpoT, вектор, включающий в себя полинуклеотид, или штамм рода Escherichia, включающий в себя полинуклеотид по данному изобретению; культуральную среду, в которой культивируют штамм, или комбинацию двух или более из перечисленного.
Композиция согласно данному изобретению может дополнительно включать произвольные подходящие эксципиенты, которые обычно применяют в композициях для получения аминокислот. Такие эксципиенты могут представлять собой, например, консервант, увлажняющее вещество, диспергирующее вещество, суспендирующее вещество, буфер, стабилизатор или изотоническое вещество и так далее, без ограничения.
В композиции по данному изобретению вариант белка SpoT, полинуклеотид, вектор, штамм, среда L-аминокислота и так далее являются такими, как описано в других аспектах.
В еще одном аспекте данного изобретения предложен способ повышения способности микроорганизма продуцировать L-аминокислоту, включающий стадию модификации микроорганизма для экспрессии варианта белка SpoT по данному изобретению.
В еще одном аспекте данного изобретения предложено применение варианта белка SpoT в получении L-аминокислот.
Вариант белка SpoT, микроорганизм, L-аминокислоты и так далее являются такими, как описано в других аспектах.
При этом, в данном описании, если из контекста не следует иное, выражение «включать», «включающий», «содержащий» и так далее следует понимать, как включение указанного целого значения или группы целых значений, но не исключение любого другого целого значения или группы целых значений.
Далее данное изобретение будет описано более подробно при помощи приведенных в качестве примера воплощений. Однако, эти приведенные в качестве примера воплощения служат исключительно для иллюстрации данного изобретения, и, таким образом, объем данного изобретения не ограничен этими приведенными в качестве примера воплощениями.
Пример 1. Конструирование рекомбинантного вектора pSKH-spoT
Для получения фрагмента ДНК приблизительно 1,63 тпн (тысяч пар нуклеотидов), содержащего сайт мутации гена spoT (SEQ ID NO: 2), выделяли хромосомную ДНК (гДНК) дикого типа Escherichia coli W3110 при помощи системы колонок для выделения геномной ДНК Genomic-tip от Qiagen и проводили полимеразную цепную реакцию (ПЦР) с использованием гДНК в качестве матрицы и с использованием готовой смеси PCR HL (продукция BIONEER, как и далее). Для амплификации области генного фрагмента, в котором аланин, представляющий собой аминокислоту в положении 262 аминокислотной последовательности SpoT (SEQ ID NO: 1), замещали на другую полярную аминокислоту, такую как серин (S), треонин (Т), цистеин (С), тирозин (Y), аспарагин (N), глутамин (Q), выполняли ПЦР путем повторения 27 циклов, представляющих собой денатурацию при 95°С в течение 30 секунд, отжиг при 56°С в течение 30 секунд и элонгацию при 68°С в течение 1 минуты с использованием набора праймеров, указанных в Таблице 1.
В результате получали генные фрагменты 0,82 тпн и 0,88 тпн, соответственно, для мутации в виде замены.
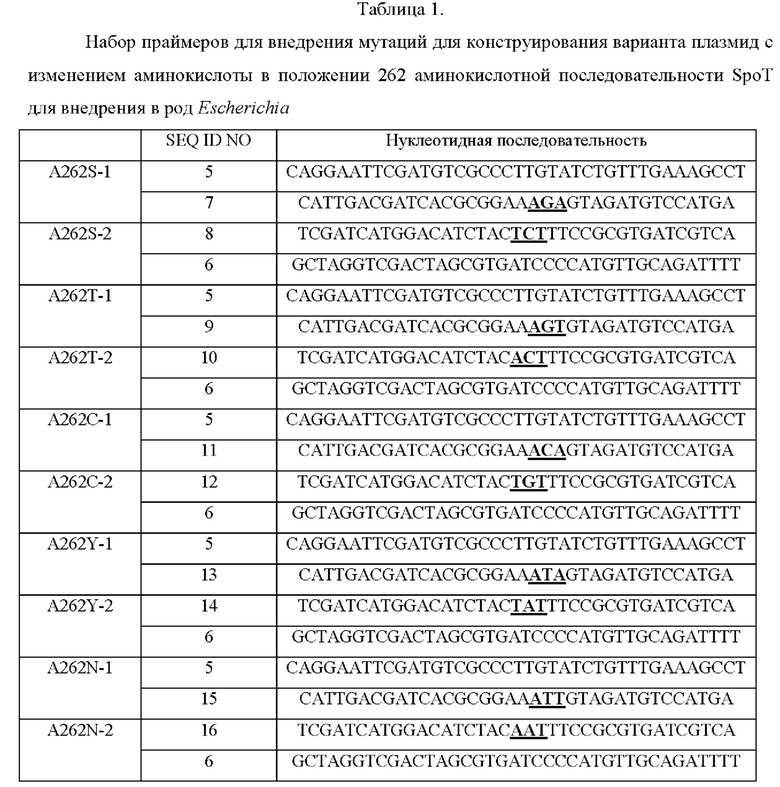

Амплифицированный вариант фрагмента гена spoT и вектор pSKH (US 8569066 В2) для хромосомной трансформации, которые расщепляли рестрикционным ферментом EcoR V, клонировали с применением метода сборки по Гибсону (DG Gibson et al, NATURE METHODS, Vol.6 No. 5, MAY 2009, NEBuilder HiFi DNA Assembly Master Mix) с получением рекомбинантных плазмид, которые обозначали pSKH-spoT(A262S), pSKH-spoT(A262T), pSKH-spoT(A262C), pSKH-spoT(A262Y), pSKH-spoT(A262N) и pSKH-spoT(A262Q) в соответствии с мутациями в виде замены. Клонирование осуществляли путем смешивания реагента для сборки по Гибсону и каждого из генных фрагментов в рассчитанном молярном количестве с последующей инкубацией при 50°С в течение 1 часа.
Пример 2. Создание мутантного штамма, в котором последовательность, кодирующая аминокислоту в положении 262 в SpoT, замещена последовательностью, кодирующей полярную аминокислоту
Каждым из PSKH-spoT(A262S), PSKH-spoT(A262T), PSKH-spoT(A262C), pSKH-spoT(A262Y), pSKH-spoT(A262N) и pSKH-spoT(A262Q), полученными в Примере 1, трансформировали электрокомпетентные клетки продуцирующего L-триптофан штамма КССМ11166Р (US 8835154 В2) посредством электропорации, и затем получали штаммы в результате процесса вторичного кроссинговера, в котором последовательность на хромосоме, кодирующую аланин, являющийся аминокислотой в положении 262 в SpoT, замещали последовательностью, кодирующей полярную аминокислоту, серин, треонин, цистеин, тирозин, аспарагин или глутамин (SEQ ID NO: 19 24). Соответствующую манипуляцию с генами подтверждали при помощи ПЦР и секвенирования генома с использованием праймеров с SEQ ID NO: 3 и SEQ ID NO: 4, которые позволяют амплифицировать области, прилежащие извне в направлении 5' и в направлении 3', соответственно, к области гомологичной рекомбинации, где был встроен соответствующий ген.

Полученные таким образом штаммы обозначали Escherichia coli KCCM11166P::spoT(A262S), KCCM11166P::spoT(A262T), KCCM11166P::spoT(A262C), KCCM11166P::spoT(A262Y), KCCM11166P::spoT(A262N), KCCM11166P::spoT(A262Q), соответственно.
Кроме того, демонстрирует ли внедрение мутации SpoT 262 такой же эффект в других продуцирующих L-триптофан штаммах рода Escherichia, исследовали в другом продуцирующем L-триптофан штамме, СА04-4303 (US 2020-0063219 Al). СА04-4303 представляет собой штамм, полученный путем делеции trpR, mtr и tnaA и замены аминокислоты пролина в положении 21 в trpE на серин в штамме дикого типа W3110. Получали штаммы, в которых последовательность, кодирующую аланин, являющийся аминокислотой в положении 262 в SpoT на хромосоме штамма СА04-4303, замещали последовательностью, кодирующей полярную аминокислоту, серин, треонин, цистеин, тирозин, аспарагин или глутамин. Процедура получения является такой же, как в процедуре внедрения, описанной выше для КССМ11166Р.
Полученные таким образом штаммы обозначали CA04-4303::spoT(A262S), СА04-4303::spoT(A262T), CA04-4303::spoT(A262C), CA04-4303::spoT(A262Y), CA04-4303::spoT(A262N), CA04-4303::spoT(A262Q), соответственно.
Пример 3. Сравнение способности продуцировать L-триптофан у мутантных штаммов, в которых аминокислота в положении 262 в SpoT замещена полярной аминокислотой
Штаммы, полученные в Примере 2 путем замены аминокислоты в положении 262 в SpoT на полярную аминокислоту, на базе продуцирующего L-триптофан штамма КССМ11166Р, культивировали в среде для титрования триптофана, приготовленной в соответствии с составом, показанным в Таблице 2 ниже, и исследовали способность продуцировать L-триптофан.
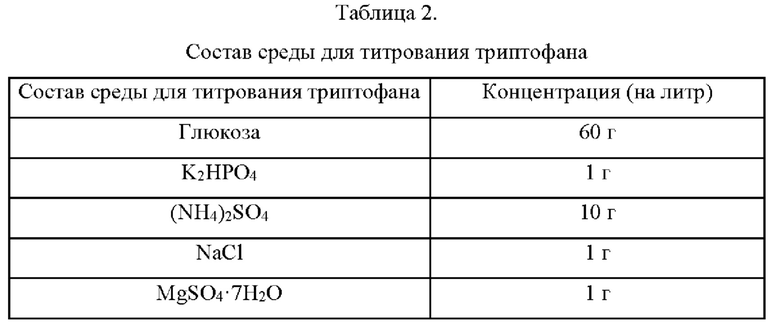
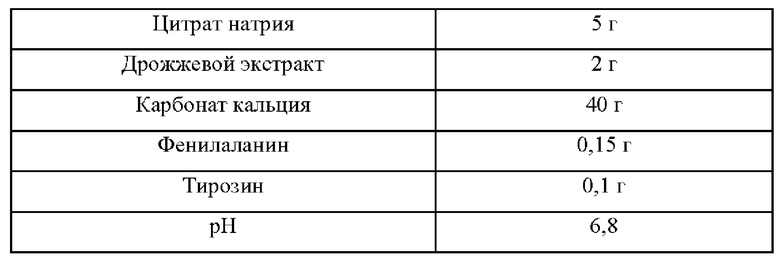
Конкретнее, при помощи платиновой петли осуществляли инокуляцию каждого из Escherichia coli КССМ11166Р и Escherichia coli KCCM11166P::spoT(A262S), KCCM 11166P::spoT(A262T), KCCM 11166P::spoT(A262C), KCCM11166P::spoT(A262Y), KCCM11166P::spoT(A262N) и KCCM11166P::spoT(A262Q), культивировавшимися в течение ночи на твердой среде LB (Лурия-Бертани) в инкубаторе при 37°С, из расчета одна петля на 25 мл среды для титрования, согласно Таблице 2, и затем культивировали в инкубаторе при 37°С и 200 об/мин в течение 48 ч и сравнивали скорости потребления сахара и концентрации L-триптофана.
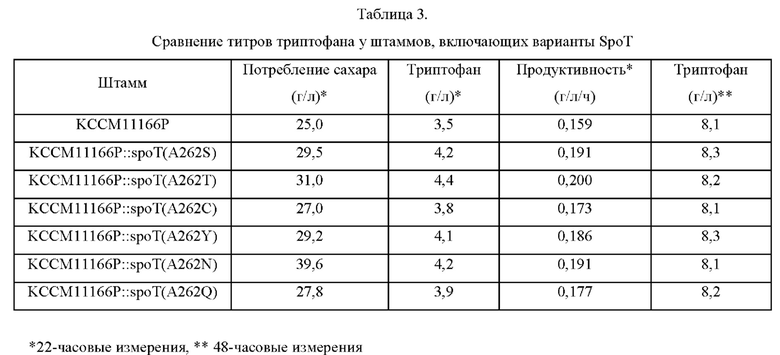
В результате, как показано в Таблице 3, ниже, потребление сахара контрольным штаммом КССМ11166Р, наблюдавшееся через 22 часа культивирования, составляло 25,0 г/л, тогда как мутантные штаммы, у которых аминокислота в положении 262 аминокислотной последовательности белка SpoT замещена полярной аминокислотой, демонстрировал потребление сахара от 27 г/л до 31 г/л в течение такого же количества времени. Это представляет собой величину улучшения продуктивности на 8,8%-25,8%, и, в частности, подтверждали, что KCCM11166P::spoT(A262T) демонстрировал наибольшее улучшение продуктивности.
Эти результаты подтверждают, что аминокислота в положении 262 белка SpoT является основным сайтом для изменения активности соответствующего белка, и замена аминокислоты в положении 262 на полярную аминокислоту может значительно улучшить продуктивность ферментации L-триптофана.
Изменение способности продуцировать L-триптофан вследствие замены аминокислоты в положении 262 в SpoT на полярную аминокислоту вновь подтверждали на штамме СА04-4303, полученном в Примере 2. Способ культивирования и состав среды для сравнения способности продуцировать L-триптофан являются такими же, как и для культуры на основе КССМ11166Р.
Как следует из результатов, приведенных ниже, мутации, выбранные в данном изобретении, значительно улучшали продуктивность L-триптофана даже при замене аминокислоты в положении 262 в SpoT, на основе продуцирующего L-триптофан штамма СА04-4303.
Как показано в Таблице 4, ниже, потребление сахара контрольным штаммом СА04-4303, наблюдавшееся через 22 часа культивирования, составляло 30,0 г/л, тогда как мутантные штаммы, у которых аминокислота в положении 262 аминокислотной последовательности белка SpoT была замещена полярной аминокислотой, демонстрировали потребление сахара от 31,8 г/л до 33,0 г/л в течение такого же интервала времени. Это представляет собой величину улучшения продуктивности на 13,0%-30,4%, и, в частности, подтверждали, что CA04-4303::spoT(A262Y) демонстрировал наибольшее улучшение продуктивности.
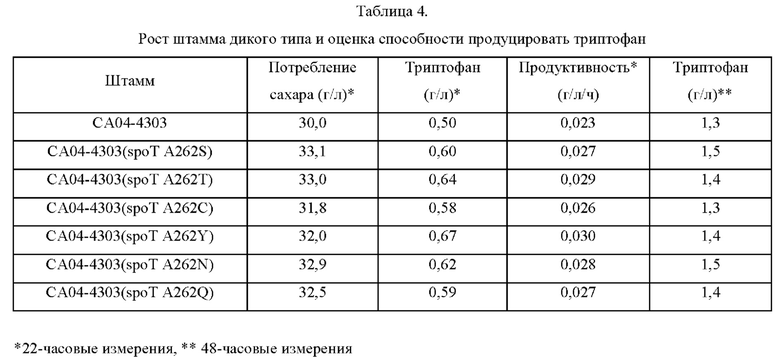
Приведенные выше результаты указывают, что продуктивность L-аминокислоты повышалась у микроорганизмов рода Escherichia, в которые внедрен вариант SpoT, в котором аминокислота в положении 262 аминокислотной последовательности по данному изобретению замещена полярной аминокислотой, что приводит к повышению продуктивности L-аминокислоты по сравнению с немодифицированными штаммами.
На основании приведенного выше описания специалист в области техники поймет, что данное изобретение может быть воплощено в различных частных формах без изменения его технического замысла или существенных признаков. В этой связи, следует понимать, что приведенное выше воплощение не ограничивает объем изобретения, а является иллюстративным во всех аспектах. Объем данного изобретения определяется прилагаемой формулой изобретения, а не предшествующим описанием, и таким образом, все изменения и модификации, подпадающие под рамки формулы изобретения, или их эквиваленты, считаются входящими в объем формулы изобретения.
ТЕХНИЧЕСКИЙ ЭФФЕКТ ИЗОБРЕТЕНИЯ
L-аминокислоты могут эффективно получены с применением варианта белка по данному изобретению.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> CJ CheilJedang Corporation
<120> Мутированный белок SpoT и способ получения L-аминокислот
с его применением
<130> OPA22080
<150> KR 10-2021-0055169
<151> 2021-04-28
<160> 24
<170> KoPatentIn 3.0
<210> 1
<211> 702
<212> PRT
<213> Artificial Sequence
<220>
<223> SpoT_WT
<400> 1
Met Tyr Leu Phe Glu Ser Leu Asn Gln Leu Ile Gln Thr Tyr Leu Pro
1 5 10 15
Glu Asp Gln Ile Lys Arg Leu Arg Gln Ala Tyr Leu Val Ala Arg Asp
20 25 30
Ala His Glu Gly Gln Thr Arg Ser Ser Gly Glu Pro Tyr Ile Thr His
35 40 45
Pro Val Ala Val Ala Cys Ile Leu Ala Glu Met Lys Leu Asp Tyr Glu
50 55 60
Thr Leu Met Ala Ala Leu Leu His Asp Val Ile Glu Asp Thr Pro Ala
65 70 75 80
Thr Tyr Gln Asp Met Glu Gln Leu Phe Gly Lys Ser Val Ala Glu Leu
85 90 95
Val Glu Gly Val Ser Lys Leu Asp Lys Leu Lys Phe Arg Asp Lys Lys
100 105 110
Glu Ala Gln Ala Glu Asn Phe Arg Lys Met Ile Met Ala Met Val Gln
115 120 125
Asp Ile Arg Val Ile Leu Ile Lys Leu Ala Asp Arg Thr His Asn Met
130 135 140
Arg Thr Leu Gly Ser Leu Arg Pro Asp Lys Arg Arg Arg Ile Ala Arg
145 150 155 160
Glu Thr Leu Glu Ile Tyr Ser Pro Leu Ala His Arg Leu Gly Ile His
165 170 175
His Ile Lys Thr Glu Leu Glu Glu Leu Gly Phe Glu Ala Leu Tyr Pro
180 185 190
Asn Arg Tyr Arg Val Ile Lys Glu Val Val Lys Ala Ala Arg Gly Asn
195 200 205
Arg Lys Glu Met Ile Gln Lys Ile Leu Ser Glu Ile Glu Gly Arg Leu
210 215 220
Gln Glu Ala Gly Ile Pro Cys Arg Val Ser Gly Arg Glu Lys His Leu
225 230 235 240
Tyr Ser Ile Tyr Cys Lys Met Val Leu Lys Glu Gln Arg Phe His Ser
245 250 255
Ile Met Asp Ile Tyr Ala Phe Arg Val Ile Val Asn Asp Ser Asp Thr
260 265 270
Cys Tyr Arg Val Leu Gly Gln Met His Ser Leu Tyr Lys Pro Arg Pro
275 280 285
Gly Arg Val Lys Asp Tyr Ile Ala Ile Pro Lys Ala Asn Gly Tyr Gln
290 295 300
Ser Leu His Thr Ser Met Ile Gly Pro His Gly Val Pro Val Glu Val
305 310 315 320
Gln Ile Arg Thr Glu Asp Met Asp Gln Met Ala Glu Met Gly Val Ala
325 330 335
Ala His Trp Ala Tyr Lys Glu His Gly Glu Thr Ser Thr Thr Ala Gln
340 345 350
Ile Arg Ala Gln Arg Trp Met Gln Ser Leu Leu Glu Leu Gln Gln Ser
355 360 365
Ala Gly Ser Ser Phe Glu Phe Ile Glu Ser Val Lys Ser Asp Leu Phe
370 375 380
Pro Asp Glu Ile Tyr Val Phe Thr Pro Glu Gly Arg Ile Val Glu Leu
385 390 395 400
Pro Ala Gly Ala Thr Pro Val Asp Phe Ala Tyr Ala Val His Thr Asp
405 410 415
Ile Gly His Ala Cys Val Gly Ala Arg Val Asp Arg Gln Pro Tyr Pro
420 425 430
Leu Ser Gln Pro Leu Thr Ser Gly Gln Thr Val Glu Ile Ile Thr Ala
435 440 445
Pro Gly Ala Arg Pro Asn Ala Ala Trp Leu Asn Phe Val Val Ser Ser
450 455 460
Lys Ala Arg Ala Lys Ile Arg Gln Leu Leu Lys Asn Leu Lys Arg Asp
465 470 475 480
Asp Ser Val Ser Leu Gly Arg Arg Leu Leu Asn His Ala Leu Gly Gly
485 490 495
Ser Arg Lys Leu Asn Glu Ile Pro Gln Glu Asn Ile Gln Arg Glu Leu
500 505 510
Asp Arg Met Lys Leu Ala Thr Leu Asp Asp Leu Leu Ala Glu Ile Gly
515 520 525
Leu Gly Asn Ala Met Ser Val Val Val Ala Lys Asn Leu Gln His Gly
530 535 540
Asp Ala Ser Ile Pro Pro Ala Thr Gln Ser His Gly His Leu Pro Ile
545 550 555 560
Lys Gly Ala Asp Gly Val Leu Ile Thr Phe Ala Lys Cys Cys Arg Pro
565 570 575
Ile Pro Gly Asp Pro Ile Ile Ala His Val Ser Pro Gly Lys Gly Leu
580 585 590
Val Ile His His Glu Ser Cys Arg Asn Ile Arg Gly Tyr Gln Lys Glu
595 600 605
Pro Glu Lys Phe Met Ala Val Glu Trp Asp Lys Glu Thr Ala Gln Glu
610 615 620
Phe Ile Thr Glu Ile Lys Val Glu Met Phe Asn His Gln Gly Ala Leu
625 630 635 640
Ala Asn Leu Thr Ala Ala Ile Asn Thr Thr Thr Ser Asn Ile Gln Ser
645 650 655
Leu Asn Thr Glu Glu Lys Asp Gly Arg Val Tyr Ser Ala Phe Ile Arg
660 665 670
Leu Thr Ala Arg Asp Arg Val His Leu Ala Asn Ile Met Arg Lys Ile
675 680 685
Arg Val Met Pro Asp Val Ile Lys Val Thr Arg Asn Arg Asn
690 695 700
<210> 2
<211> 2109
<212> DNA
<213> Artificial Sequence
<220>
<223> spoT_WT
<400> 2
ttgtatctgt ttgaaagcct gaatcaactg attcaaacct acctgccgga agaccaaatc 60
aagcgtctgc ggcaggcgta tctcgttgca cgtgatgctc acgaggggca aacacgttca 120
agcggtgaac cctatatcac gcacccggta gcggttgcct gcattctggc cgagatgaaa 180
ctcgactatg aaacgctgat ggcggcgctg ctgcatgacg tgattgaaga tactcccgcc 240
acctaccagg atatggaaca gctttttggt aaaagcgtcg ccgagctggt agagggggtg 300
tcgaaacttg ataaactcaa gttccgcgat aagaaagagg cgcaggccga aaactttcgc 360
aagatgatta tggcgatggt gcaggatatc cgcgtcatcc tcatcaaact tgccgaccgt 420
acccacaaca tgcgcacgct gggctcactt cgcccggaca aacgtcgccg catcgcccgt 480
gaaactctcg aaatttatag cccgctggcg caccgtttag gtatccacca cattaaaacc 540
gaactcgaag agctgggttt tgaggcgctg tatcccaacc gttatcgcgt aatcaaagaa 600
gtggtgaaag ccgcgcgcgg caaccgtaaa gagatgatcc agaagattct ttctgaaatc 660
gaagggcgtt tgcaggaagc gggaataccg tgccgcgtca gtggtcgcga gaagcatctt 720
tattcgattt actgcaaaat ggtgctcaaa gagcagcgtt ttcactcgat catggacatc 780
tacgctttcc gcgtgatcgt caatgattct gacacctgtt atcgcgtgct gggccagatg 840
cacagcctgt acaagccgcg tccgggccgc gtgaaagact atatcgccat tccaaaagcg 900
aacggctatc agtctttgca cacctcgatg atcggcccgc acggtgtgcc ggttgaggtc 960
cagatccgta ccgaagatat ggaccagatg gcggagatgg gtgttgccgc gcactgggct 1020
tataaagagc acggcgaaac cagtactacc gcacaaatcc gcgcccagcg ctggatgcaa 1080
agcctgctgg agctgcaaca gagcgccggt agttcgtttg aatttatcga gagcgttaaa 1140
tccgatctct tcccggatga gatttacgtt ttcacaccgg aagggcgcat tgtcgagctg 1200
cctgccggtg caacgcccgt cgacttcgct tatgcagtgc ataccgatat cggtcatgcc 1260
tgcgtgggcg cacgcgttga ccgccagcct tacccgctgt cgcagccgct taccagcggt 1320
caaaccgttg aaatcattac cgctccgggc gctcgcccga atgccgcttg gctgaacttt 1380
gtcgttagct cgaaagcgcg cgccaaaatt cgtcagttgc tgaaaaacct caagcgtgat 1440
gattctgtaa gcctgggccg tcgtctgctc aaccatgctt tgggtggtag ccgtaagctg 1500
aatgaaatcc cgcaggaaaa tattcagcgc gagctggatc gcatgaagct ggcaacgctt 1560
gacgatctgc tggcagaaat cggacttggt aacgcaatga gcgtggtggt cgcgaaaaat 1620
ctgcaacatg gggacgcctc cattccaccg gcaacccaaa gccacggaca tctgcccatt 1680
aaaggtgccg atggcgtgct gatcaccttt gcgaaatgct gccgccctat tcctggcgac 1740
ccgattatcg cccacgtcag ccccggtaaa ggtctggtga tccaccatga atcctgccgt 1800
aatatccgtg gctaccagaa agagccagag aagtttatgg ctgtggaatg ggataaagag 1860
acggcgcagg agttcatcac cgaaatcaag gtggagatgt tcaatcatca gggtgcgctg 1920
gcaaacctga cggcggcaat taacaccacg acttcgaata ttcaaagttt gaatacggaa 1980
gagaaagatg gtcgcgtcta cagcgccttt attcgtctga ccgctcgtga ccgtgtgcat 2040
ctggcgaata tcatgcgcaa aatccgcgtg atgccagacg tgattaaagt cacccgaaac 2100
cgaaattaa 2109
<210> 3
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 3
ccaggaacag caagagca 18
<210> 4
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 4
acggatatta cggcagga 18
<210> 5
<211> 39
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 5
caggaattcg atgtcgccct tgtatctgtt tgaaagcct 39
<210> 6
<211> 38
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 6
gctaggtcga ctagcgtgat ccccatgttg cagatttt 38
<210> 7
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 7
cattgacgat cacgcggaaa gagtagatgt ccatga 36
<210> 8
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 8
tcgatcatgg acatctactc tttccgcgtg atcgtca 37
<210> 9
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 9
cattgacgat cacgcggaaa gtgtagatgt ccatga 36
<210> 10
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 10
tcgatcatgg acatctacac tttccgcgtg atcgtca 37
<210> 11
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 11
cattgacgat cacgcggaaa cagtagatgt ccatga 36
<210> 12
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 12
tcgatcatgg acatctactg tttccgcgtg atcgtca 37
<210> 13
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 13
cattgacgat cacgcggaaa tagtagatgt ccatga 36
<210> 14
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 14
tcgatcatgg acatctacta tttccgcgtg atcgtca 37
<210> 15
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 15
cattgacgat cacgcggaaa ttgtagatgt ccatga 36
<210> 16
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 16
tcgatcatgg acatctacaa tttccgcgtg atcgtca 37
<210> 17
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 17
cattgacgat cacgcggaat tggtagatgt ccatga 36
<210> 18
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 18
tcgatcatgg acatctacca attccgcgtg atcgtca 37
<210> 19
<211> 702
<212> PRT
<213> Artificial Sequence
<220>
<223> SpoT_A262S
<400> 19
Met Tyr Leu Phe Glu Ser Leu Asn Gln Leu Ile Gln Thr Tyr Leu Pro
1 5 10 15
Glu Asp Gln Ile Lys Arg Leu Arg Gln Ala Tyr Leu Val Ala Arg Asp
20 25 30
Ala His Glu Gly Gln Thr Arg Ser Ser Gly Glu Pro Tyr Ile Thr His
35 40 45
Pro Val Ala Val Ala Cys Ile Leu Ala Glu Met Lys Leu Asp Tyr Glu
50 55 60
Thr Leu Met Ala Ala Leu Leu His Asp Val Ile Glu Asp Thr Pro Ala
65 70 75 80
Thr Tyr Gln Asp Met Glu Gln Leu Phe Gly Lys Ser Val Ala Glu Leu
85 90 95
Val Glu Gly Val Ser Lys Leu Asp Lys Leu Lys Phe Arg Asp Lys Lys
100 105 110
Glu Ala Gln Ala Glu Asn Phe Arg Lys Met Ile Met Ala Met Val Gln
115 120 125
Asp Ile Arg Val Ile Leu Ile Lys Leu Ala Asp Arg Thr His Asn Met
130 135 140
Arg Thr Leu Gly Ser Leu Arg Pro Asp Lys Arg Arg Arg Ile Ala Arg
145 150 155 160
Glu Thr Leu Glu Ile Tyr Ser Pro Leu Ala His Arg Leu Gly Ile His
165 170 175
His Ile Lys Thr Glu Leu Glu Glu Leu Gly Phe Glu Ala Leu Tyr Pro
180 185 190
Asn Arg Tyr Arg Val Ile Lys Glu Val Val Lys Ala Ala Arg Gly Asn
195 200 205
Arg Lys Glu Met Ile Gln Lys Ile Leu Ser Glu Ile Glu Gly Arg Leu
210 215 220
Gln Glu Ala Gly Ile Pro Cys Arg Val Ser Gly Arg Glu Lys His Leu
225 230 235 240
Tyr Ser Ile Tyr Cys Lys Met Val Leu Lys Glu Gln Arg Phe His Ser
245 250 255
Ile Met Asp Ile Tyr Ser Phe Arg Val Ile Val Asn Asp Ser Asp Thr
260 265 270
Cys Tyr Arg Val Leu Gly Gln Met His Ser Leu Tyr Lys Pro Arg Pro
275 280 285
Gly Arg Val Lys Asp Tyr Ile Ala Ile Pro Lys Ala Asn Gly Tyr Gln
290 295 300
Ser Leu His Thr Ser Met Ile Gly Pro His Gly Val Pro Val Glu Val
305 310 315 320
Gln Ile Arg Thr Glu Asp Met Asp Gln Met Ala Glu Met Gly Val Ala
325 330 335
Ala His Trp Ala Tyr Lys Glu His Gly Glu Thr Ser Thr Thr Ala Gln
340 345 350
Ile Arg Ala Gln Arg Trp Met Gln Ser Leu Leu Glu Leu Gln Gln Ser
355 360 365
Ala Gly Ser Ser Phe Glu Phe Ile Glu Ser Val Lys Ser Asp Leu Phe
370 375 380
Pro Asp Glu Ile Tyr Val Phe Thr Pro Glu Gly Arg Ile Val Glu Leu
385 390 395 400
Pro Ala Gly Ala Thr Pro Val Asp Phe Ala Tyr Ala Val His Thr Asp
405 410 415
Ile Gly His Ala Cys Val Gly Ala Arg Val Asp Arg Gln Pro Tyr Pro
420 425 430
Leu Ser Gln Pro Leu Thr Ser Gly Gln Thr Val Glu Ile Ile Thr Ala
435 440 445
Pro Gly Ala Arg Pro Asn Ala Ala Trp Leu Asn Phe Val Val Ser Ser
450 455 460
Lys Ala Arg Ala Lys Ile Arg Gln Leu Leu Lys Asn Leu Lys Arg Asp
465 470 475 480
Asp Ser Val Ser Leu Gly Arg Arg Leu Leu Asn His Ala Leu Gly Gly
485 490 495
Ser Arg Lys Leu Asn Glu Ile Pro Gln Glu Asn Ile Gln Arg Glu Leu
500 505 510
Asp Arg Met Lys Leu Ala Thr Leu Asp Asp Leu Leu Ala Glu Ile Gly
515 520 525
Leu Gly Asn Ala Met Ser Val Val Val Ala Lys Asn Leu Gln His Gly
530 535 540
Asp Ala Ser Ile Pro Pro Ala Thr Gln Ser His Gly His Leu Pro Ile
545 550 555 560
Lys Gly Ala Asp Gly Val Leu Ile Thr Phe Ala Lys Cys Cys Arg Pro
565 570 575
Ile Pro Gly Asp Pro Ile Ile Ala His Val Ser Pro Gly Lys Gly Leu
580 585 590
Val Ile His His Glu Ser Cys Arg Asn Ile Arg Gly Tyr Gln Lys Glu
595 600 605
Pro Glu Lys Phe Met Ala Val Glu Trp Asp Lys Glu Thr Ala Gln Glu
610 615 620
Phe Ile Thr Glu Ile Lys Val Glu Met Phe Asn His Gln Gly Ala Leu
625 630 635 640
Ala Asn Leu Thr Ala Ala Ile Asn Thr Thr Thr Ser Asn Ile Gln Ser
645 650 655
Leu Asn Thr Glu Glu Lys Asp Gly Arg Val Tyr Ser Ala Phe Ile Arg
660 665 670
Leu Thr Ala Arg Asp Arg Val His Leu Ala Asn Ile Met Arg Lys Ile
675 680 685
Arg Val Met Pro Asp Val Ile Lys Val Thr Arg Asn Arg Asn
690 695 700
<210> 20
<211> 702
<212> PRT
<213> Artificial Sequence
<220>
<223> SpoT_A262T
<400> 20
Met Tyr Leu Phe Glu Ser Leu Asn Gln Leu Ile Gln Thr Tyr Leu Pro
1 5 10 15
Glu Asp Gln Ile Lys Arg Leu Arg Gln Ala Tyr Leu Val Ala Arg Asp
20 25 30
Ala His Glu Gly Gln Thr Arg Ser Ser Gly Glu Pro Tyr Ile Thr His
35 40 45
Pro Val Ala Val Ala Cys Ile Leu Ala Glu Met Lys Leu Asp Tyr Glu
50 55 60
Thr Leu Met Ala Ala Leu Leu His Asp Val Ile Glu Asp Thr Pro Ala
65 70 75 80
Thr Tyr Gln Asp Met Glu Gln Leu Phe Gly Lys Ser Val Ala Glu Leu
85 90 95
Val Glu Gly Val Ser Lys Leu Asp Lys Leu Lys Phe Arg Asp Lys Lys
100 105 110
Glu Ala Gln Ala Glu Asn Phe Arg Lys Met Ile Met Ala Met Val Gln
115 120 125
Asp Ile Arg Val Ile Leu Ile Lys Leu Ala Asp Arg Thr His Asn Met
130 135 140
Arg Thr Leu Gly Ser Leu Arg Pro Asp Lys Arg Arg Arg Ile Ala Arg
145 150 155 160
Glu Thr Leu Glu Ile Tyr Ser Pro Leu Ala His Arg Leu Gly Ile His
165 170 175
His Ile Lys Thr Glu Leu Glu Glu Leu Gly Phe Glu Ala Leu Tyr Pro
180 185 190
Asn Arg Tyr Arg Val Ile Lys Glu Val Val Lys Ala Ala Arg Gly Asn
195 200 205
Arg Lys Glu Met Ile Gln Lys Ile Leu Ser Glu Ile Glu Gly Arg Leu
210 215 220
Gln Glu Ala Gly Ile Pro Cys Arg Val Ser Gly Arg Glu Lys His Leu
225 230 235 240
Tyr Ser Ile Tyr Cys Lys Met Val Leu Lys Glu Gln Arg Phe His Ser
245 250 255
Ile Met Asp Ile Tyr Thr Phe Arg Val Ile Val Asn Asp Ser Asp Thr
260 265 270
Cys Tyr Arg Val Leu Gly Gln Met His Ser Leu Tyr Lys Pro Arg Pro
275 280 285
Gly Arg Val Lys Asp Tyr Ile Ala Ile Pro Lys Ala Asn Gly Tyr Gln
290 295 300
Ser Leu His Thr Ser Met Ile Gly Pro His Gly Val Pro Val Glu Val
305 310 315 320
Gln Ile Arg Thr Glu Asp Met Asp Gln Met Ala Glu Met Gly Val Ala
325 330 335
Ala His Trp Ala Tyr Lys Glu His Gly Glu Thr Ser Thr Thr Ala Gln
340 345 350
Ile Arg Ala Gln Arg Trp Met Gln Ser Leu Leu Glu Leu Gln Gln Ser
355 360 365
Ala Gly Ser Ser Phe Glu Phe Ile Glu Ser Val Lys Ser Asp Leu Phe
370 375 380
Pro Asp Glu Ile Tyr Val Phe Thr Pro Glu Gly Arg Ile Val Glu Leu
385 390 395 400
Pro Ala Gly Ala Thr Pro Val Asp Phe Ala Tyr Ala Val His Thr Asp
405 410 415
Ile Gly His Ala Cys Val Gly Ala Arg Val Asp Arg Gln Pro Tyr Pro
420 425 430
Leu Ser Gln Pro Leu Thr Ser Gly Gln Thr Val Glu Ile Ile Thr Ala
435 440 445
Pro Gly Ala Arg Pro Asn Ala Ala Trp Leu Asn Phe Val Val Ser Ser
450 455 460
Lys Ala Arg Ala Lys Ile Arg Gln Leu Leu Lys Asn Leu Lys Arg Asp
465 470 475 480
Asp Ser Val Ser Leu Gly Arg Arg Leu Leu Asn His Ala Leu Gly Gly
485 490 495
Ser Arg Lys Leu Asn Glu Ile Pro Gln Glu Asn Ile Gln Arg Glu Leu
500 505 510
Asp Arg Met Lys Leu Ala Thr Leu Asp Asp Leu Leu Ala Glu Ile Gly
515 520 525
Leu Gly Asn Ala Met Ser Val Val Val Ala Lys Asn Leu Gln His Gly
530 535 540
Asp Ala Ser Ile Pro Pro Ala Thr Gln Ser His Gly His Leu Pro Ile
545 550 555 560
Lys Gly Ala Asp Gly Val Leu Ile Thr Phe Ala Lys Cys Cys Arg Pro
565 570 575
Ile Pro Gly Asp Pro Ile Ile Ala His Val Ser Pro Gly Lys Gly Leu
580 585 590
Val Ile His His Glu Ser Cys Arg Asn Ile Arg Gly Tyr Gln Lys Glu
595 600 605
Pro Glu Lys Phe Met Ala Val Glu Trp Asp Lys Glu Thr Ala Gln Glu
610 615 620
Phe Ile Thr Glu Ile Lys Val Glu Met Phe Asn His Gln Gly Ala Leu
625 630 635 640
Ala Asn Leu Thr Ala Ala Ile Asn Thr Thr Thr Ser Asn Ile Gln Ser
645 650 655
Leu Asn Thr Glu Glu Lys Asp Gly Arg Val Tyr Ser Ala Phe Ile Arg
660 665 670
Leu Thr Ala Arg Asp Arg Val His Leu Ala Asn Ile Met Arg Lys Ile
675 680 685
Arg Val Met Pro Asp Val Ile Lys Val Thr Arg Asn Arg Asn
690 695 700
<210> 21
<211> 702
<212> PRT
<213> Artificial Sequence
<220>
<223> SpoT_A262C
<400> 21
Met Tyr Leu Phe Glu Ser Leu Asn Gln Leu Ile Gln Thr Tyr Leu Pro
1 5 10 15
Glu Asp Gln Ile Lys Arg Leu Arg Gln Ala Tyr Leu Val Ala Arg Asp
20 25 30
Ala His Glu Gly Gln Thr Arg Ser Ser Gly Glu Pro Tyr Ile Thr His
35 40 45
Pro Val Ala Val Ala Cys Ile Leu Ala Glu Met Lys Leu Asp Tyr Glu
50 55 60
Thr Leu Met Ala Ala Leu Leu His Asp Val Ile Glu Asp Thr Pro Ala
65 70 75 80
Thr Tyr Gln Asp Met Glu Gln Leu Phe Gly Lys Ser Val Ala Glu Leu
85 90 95
Val Glu Gly Val Ser Lys Leu Asp Lys Leu Lys Phe Arg Asp Lys Lys
100 105 110
Glu Ala Gln Ala Glu Asn Phe Arg Lys Met Ile Met Ala Met Val Gln
115 120 125
Asp Ile Arg Val Ile Leu Ile Lys Leu Ala Asp Arg Thr His Asn Met
130 135 140
Arg Thr Leu Gly Ser Leu Arg Pro Asp Lys Arg Arg Arg Ile Ala Arg
145 150 155 160
Glu Thr Leu Glu Ile Tyr Ser Pro Leu Ala His Arg Leu Gly Ile His
165 170 175
His Ile Lys Thr Glu Leu Glu Glu Leu Gly Phe Glu Ala Leu Tyr Pro
180 185 190
Asn Arg Tyr Arg Val Ile Lys Glu Val Val Lys Ala Ala Arg Gly Asn
195 200 205
Arg Lys Glu Met Ile Gln Lys Ile Leu Ser Glu Ile Glu Gly Arg Leu
210 215 220
Gln Glu Ala Gly Ile Pro Cys Arg Val Ser Gly Arg Glu Lys His Leu
225 230 235 240
Tyr Ser Ile Tyr Cys Lys Met Val Leu Lys Glu Gln Arg Phe His Ser
245 250 255
Ile Met Asp Ile Tyr Cys Phe Arg Val Ile Val Asn Asp Ser Asp Thr
260 265 270
Cys Tyr Arg Val Leu Gly Gln Met His Ser Leu Tyr Lys Pro Arg Pro
275 280 285
Gly Arg Val Lys Asp Tyr Ile Ala Ile Pro Lys Ala Asn Gly Tyr Gln
290 295 300
Ser Leu His Thr Ser Met Ile Gly Pro His Gly Val Pro Val Glu Val
305 310 315 320
Gln Ile Arg Thr Glu Asp Met Asp Gln Met Ala Glu Met Gly Val Ala
325 330 335
Ala His Trp Ala Tyr Lys Glu His Gly Glu Thr Ser Thr Thr Ala Gln
340 345 350
Ile Arg Ala Gln Arg Trp Met Gln Ser Leu Leu Glu Leu Gln Gln Ser
355 360 365
Ala Gly Ser Ser Phe Glu Phe Ile Glu Ser Val Lys Ser Asp Leu Phe
370 375 380
Pro Asp Glu Ile Tyr Val Phe Thr Pro Glu Gly Arg Ile Val Glu Leu
385 390 395 400
Pro Ala Gly Ala Thr Pro Val Asp Phe Ala Tyr Ala Val His Thr Asp
405 410 415
Ile Gly His Ala Cys Val Gly Ala Arg Val Asp Arg Gln Pro Tyr Pro
420 425 430
Leu Ser Gln Pro Leu Thr Ser Gly Gln Thr Val Glu Ile Ile Thr Ala
435 440 445
Pro Gly Ala Arg Pro Asn Ala Ala Trp Leu Asn Phe Val Val Ser Ser
450 455 460
Lys Ala Arg Ala Lys Ile Arg Gln Leu Leu Lys Asn Leu Lys Arg Asp
465 470 475 480
Asp Ser Val Ser Leu Gly Arg Arg Leu Leu Asn His Ala Leu Gly Gly
485 490 495
Ser Arg Lys Leu Asn Glu Ile Pro Gln Glu Asn Ile Gln Arg Glu Leu
500 505 510
Asp Arg Met Lys Leu Ala Thr Leu Asp Asp Leu Leu Ala Glu Ile Gly
515 520 525
Leu Gly Asn Ala Met Ser Val Val Val Ala Lys Asn Leu Gln His Gly
530 535 540
Asp Ala Ser Ile Pro Pro Ala Thr Gln Ser His Gly His Leu Pro Ile
545 550 555 560
Lys Gly Ala Asp Gly Val Leu Ile Thr Phe Ala Lys Cys Cys Arg Pro
565 570 575
Ile Pro Gly Asp Pro Ile Ile Ala His Val Ser Pro Gly Lys Gly Leu
580 585 590
Val Ile His His Glu Ser Cys Arg Asn Ile Arg Gly Tyr Gln Lys Glu
595 600 605
Pro Glu Lys Phe Met Ala Val Glu Trp Asp Lys Glu Thr Ala Gln Glu
610 615 620
Phe Ile Thr Glu Ile Lys Val Glu Met Phe Asn His Gln Gly Ala Leu
625 630 635 640
Ala Asn Leu Thr Ala Ala Ile Asn Thr Thr Thr Ser Asn Ile Gln Ser
645 650 655
Leu Asn Thr Glu Glu Lys Asp Gly Arg Val Tyr Ser Ala Phe Ile Arg
660 665 670
Leu Thr Ala Arg Asp Arg Val His Leu Ala Asn Ile Met Arg Lys Ile
675 680 685
Arg Val Met Pro Asp Val Ile Lys Val Thr Arg Asn Arg Asn
690 695 700
<210> 22
<211> 702
<212> PRT
<213> Artificial Sequence
<220>
<223> SpoT_A262Y
<400> 22
Met Tyr Leu Phe Glu Ser Leu Asn Gln Leu Ile Gln Thr Tyr Leu Pro
1 5 10 15
Glu Asp Gln Ile Lys Arg Leu Arg Gln Ala Tyr Leu Val Ala Arg Asp
20 25 30
Ala His Glu Gly Gln Thr Arg Ser Ser Gly Glu Pro Tyr Ile Thr His
35 40 45
Pro Val Ala Val Ala Cys Ile Leu Ala Glu Met Lys Leu Asp Tyr Glu
50 55 60
Thr Leu Met Ala Ala Leu Leu His Asp Val Ile Glu Asp Thr Pro Ala
65 70 75 80
Thr Tyr Gln Asp Met Glu Gln Leu Phe Gly Lys Ser Val Ala Glu Leu
85 90 95
Val Glu Gly Val Ser Lys Leu Asp Lys Leu Lys Phe Arg Asp Lys Lys
100 105 110
Glu Ala Gln Ala Glu Asn Phe Arg Lys Met Ile Met Ala Met Val Gln
115 120 125
Asp Ile Arg Val Ile Leu Ile Lys Leu Ala Asp Arg Thr His Asn Met
130 135 140
Arg Thr Leu Gly Ser Leu Arg Pro Asp Lys Arg Arg Arg Ile Ala Arg
145 150 155 160
Glu Thr Leu Glu Ile Tyr Ser Pro Leu Ala His Arg Leu Gly Ile His
165 170 175
His Ile Lys Thr Glu Leu Glu Glu Leu Gly Phe Glu Ala Leu Tyr Pro
180 185 190
Asn Arg Tyr Arg Val Ile Lys Glu Val Val Lys Ala Ala Arg Gly Asn
195 200 205
Arg Lys Glu Met Ile Gln Lys Ile Leu Ser Glu Ile Glu Gly Arg Leu
210 215 220
Gln Glu Ala Gly Ile Pro Cys Arg Val Ser Gly Arg Glu Lys His Leu
225 230 235 240
Tyr Ser Ile Tyr Cys Lys Met Val Leu Lys Glu Gln Arg Phe His Ser
245 250 255
Ile Met Asp Ile Tyr Tyr Phe Arg Val Ile Val Asn Asp Ser Asp Thr
260 265 270
Cys Tyr Arg Val Leu Gly Gln Met His Ser Leu Tyr Lys Pro Arg Pro
275 280 285
Gly Arg Val Lys Asp Tyr Ile Ala Ile Pro Lys Ala Asn Gly Tyr Gln
290 295 300
Ser Leu His Thr Ser Met Ile Gly Pro His Gly Val Pro Val Glu Val
305 310 315 320
Gln Ile Arg Thr Glu Asp Met Asp Gln Met Ala Glu Met Gly Val Ala
325 330 335
Ala His Trp Ala Tyr Lys Glu His Gly Glu Thr Ser Thr Thr Ala Gln
340 345 350
Ile Arg Ala Gln Arg Trp Met Gln Ser Leu Leu Glu Leu Gln Gln Ser
355 360 365
Ala Gly Ser Ser Phe Glu Phe Ile Glu Ser Val Lys Ser Asp Leu Phe
370 375 380
Pro Asp Glu Ile Tyr Val Phe Thr Pro Glu Gly Arg Ile Val Glu Leu
385 390 395 400
Pro Ala Gly Ala Thr Pro Val Asp Phe Ala Tyr Ala Val His Thr Asp
405 410 415
Ile Gly His Ala Cys Val Gly Ala Arg Val Asp Arg Gln Pro Tyr Pro
420 425 430
Leu Ser Gln Pro Leu Thr Ser Gly Gln Thr Val Glu Ile Ile Thr Ala
435 440 445
Pro Gly Ala Arg Pro Asn Ala Ala Trp Leu Asn Phe Val Val Ser Ser
450 455 460
Lys Ala Arg Ala Lys Ile Arg Gln Leu Leu Lys Asn Leu Lys Arg Asp
465 470 475 480
Asp Ser Val Ser Leu Gly Arg Arg Leu Leu Asn His Ala Leu Gly Gly
485 490 495
Ser Arg Lys Leu Asn Glu Ile Pro Gln Glu Asn Ile Gln Arg Glu Leu
500 505 510
Asp Arg Met Lys Leu Ala Thr Leu Asp Asp Leu Leu Ala Glu Ile Gly
515 520 525
Leu Gly Asn Ala Met Ser Val Val Val Ala Lys Asn Leu Gln His Gly
530 535 540
Asp Ala Ser Ile Pro Pro Ala Thr Gln Ser His Gly His Leu Pro Ile
545 550 555 560
Lys Gly Ala Asp Gly Val Leu Ile Thr Phe Ala Lys Cys Cys Arg Pro
565 570 575
Ile Pro Gly Asp Pro Ile Ile Ala His Val Ser Pro Gly Lys Gly Leu
580 585 590
Val Ile His His Glu Ser Cys Arg Asn Ile Arg Gly Tyr Gln Lys Glu
595 600 605
Pro Glu Lys Phe Met Ala Val Glu Trp Asp Lys Glu Thr Ala Gln Glu
610 615 620
Phe Ile Thr Glu Ile Lys Val Glu Met Phe Asn His Gln Gly Ala Leu
625 630 635 640
Ala Asn Leu Thr Ala Ala Ile Asn Thr Thr Thr Ser Asn Ile Gln Ser
645 650 655
Leu Asn Thr Glu Glu Lys Asp Gly Arg Val Tyr Ser Ala Phe Ile Arg
660 665 670
Leu Thr Ala Arg Asp Arg Val His Leu Ala Asn Ile Met Arg Lys Ile
675 680 685
Arg Val Met Pro Asp Val Ile Lys Val Thr Arg Asn Arg Asn
690 695 700
<210> 23
<211> 702
<212> PRT
<213> Artificial Sequence
<220>
<223> SpoT_A262N
<400> 23
Met Tyr Leu Phe Glu Ser Leu Asn Gln Leu Ile Gln Thr Tyr Leu Pro
1 5 10 15
Glu Asp Gln Ile Lys Arg Leu Arg Gln Ala Tyr Leu Val Ala Arg Asp
20 25 30
Ala His Glu Gly Gln Thr Arg Ser Ser Gly Glu Pro Tyr Ile Thr His
35 40 45
Pro Val Ala Val Ala Cys Ile Leu Ala Glu Met Lys Leu Asp Tyr Glu
50 55 60
Thr Leu Met Ala Ala Leu Leu His Asp Val Ile Glu Asp Thr Pro Ala
65 70 75 80
Thr Tyr Gln Asp Met Glu Gln Leu Phe Gly Lys Ser Val Ala Glu Leu
85 90 95
Val Glu Gly Val Ser Lys Leu Asp Lys Leu Lys Phe Arg Asp Lys Lys
100 105 110
Glu Ala Gln Ala Glu Asn Phe Arg Lys Met Ile Met Ala Met Val Gln
115 120 125
Asp Ile Arg Val Ile Leu Ile Lys Leu Ala Asp Arg Thr His Asn Met
130 135 140
Arg Thr Leu Gly Ser Leu Arg Pro Asp Lys Arg Arg Arg Ile Ala Arg
145 150 155 160
Glu Thr Leu Glu Ile Tyr Ser Pro Leu Ala His Arg Leu Gly Ile His
165 170 175
His Ile Lys Thr Glu Leu Glu Glu Leu Gly Phe Glu Ala Leu Tyr Pro
180 185 190
Asn Arg Tyr Arg Val Ile Lys Glu Val Val Lys Ala Ala Arg Gly Asn
195 200 205
Arg Lys Glu Met Ile Gln Lys Ile Leu Ser Glu Ile Glu Gly Arg Leu
210 215 220
Gln Glu Ala Gly Ile Pro Cys Arg Val Ser Gly Arg Glu Lys His Leu
225 230 235 240
Tyr Ser Ile Tyr Cys Lys Met Val Leu Lys Glu Gln Arg Phe His Ser
245 250 255
Ile Met Asp Ile Tyr Asn Phe Arg Val Ile Val Asn Asp Ser Asp Thr
260 265 270
Cys Tyr Arg Val Leu Gly Gln Met His Ser Leu Tyr Lys Pro Arg Pro
275 280 285
Gly Arg Val Lys Asp Tyr Ile Ala Ile Pro Lys Ala Asn Gly Tyr Gln
290 295 300
Ser Leu His Thr Ser Met Ile Gly Pro His Gly Val Pro Val Glu Val
305 310 315 320
Gln Ile Arg Thr Glu Asp Met Asp Gln Met Ala Glu Met Gly Val Ala
325 330 335
Ala His Trp Ala Tyr Lys Glu His Gly Glu Thr Ser Thr Thr Ala Gln
340 345 350
Ile Arg Ala Gln Arg Trp Met Gln Ser Leu Leu Glu Leu Gln Gln Ser
355 360 365
Ala Gly Ser Ser Phe Glu Phe Ile Glu Ser Val Lys Ser Asp Leu Phe
370 375 380
Pro Asp Glu Ile Tyr Val Phe Thr Pro Glu Gly Arg Ile Val Glu Leu
385 390 395 400
Pro Ala Gly Ala Thr Pro Val Asp Phe Ala Tyr Ala Val His Thr Asp
405 410 415
Ile Gly His Ala Cys Val Gly Ala Arg Val Asp Arg Gln Pro Tyr Pro
420 425 430
Leu Ser Gln Pro Leu Thr Ser Gly Gln Thr Val Glu Ile Ile Thr Ala
435 440 445
Pro Gly Ala Arg Pro Asn Ala Ala Trp Leu Asn Phe Val Val Ser Ser
450 455 460
Lys Ala Arg Ala Lys Ile Arg Gln Leu Leu Lys Asn Leu Lys Arg Asp
465 470 475 480
Asp Ser Val Ser Leu Gly Arg Arg Leu Leu Asn His Ala Leu Gly Gly
485 490 495
Ser Arg Lys Leu Asn Glu Ile Pro Gln Glu Asn Ile Gln Arg Glu Leu
500 505 510
Asp Arg Met Lys Leu Ala Thr Leu Asp Asp Leu Leu Ala Glu Ile Gly
515 520 525
Leu Gly Asn Ala Met Ser Val Val Val Ala Lys Asn Leu Gln His Gly
530 535 540
Asp Ala Ser Ile Pro Pro Ala Thr Gln Ser His Gly His Leu Pro Ile
545 550 555 560
Lys Gly Ala Asp Gly Val Leu Ile Thr Phe Ala Lys Cys Cys Arg Pro
565 570 575
Ile Pro Gly Asp Pro Ile Ile Ala His Val Ser Pro Gly Lys Gly Leu
580 585 590
Val Ile His His Glu Ser Cys Arg Asn Ile Arg Gly Tyr Gln Lys Glu
595 600 605
Pro Glu Lys Phe Met Ala Val Glu Trp Asp Lys Glu Thr Ala Gln Glu
610 615 620
Phe Ile Thr Glu Ile Lys Val Glu Met Phe Asn His Gln Gly Ala Leu
625 630 635 640
Ala Asn Leu Thr Ala Ala Ile Asn Thr Thr Thr Ser Asn Ile Gln Ser
645 650 655
Leu Asn Thr Glu Glu Lys Asp Gly Arg Val Tyr Ser Ala Phe Ile Arg
660 665 670
Leu Thr Ala Arg Asp Arg Val His Leu Ala Asn Ile Met Arg Lys Ile
675 680 685
Arg Val Met Pro Asp Val Ile Lys Val Thr Arg Asn Arg Asn
690 695 700
<210> 24
<211> 702
<212> PRT
<213> Artificial Sequence
<220>
<223> SpoT_A262Q
<400> 24
Met Tyr Leu Phe Glu Ser Leu Asn Gln Leu Ile Gln Thr Tyr Leu Pro
1 5 10 15
Glu Asp Gln Ile Lys Arg Leu Arg Gln Ala Tyr Leu Val Ala Arg Asp
20 25 30
Ala His Glu Gly Gln Thr Arg Ser Ser Gly Glu Pro Tyr Ile Thr His
35 40 45
Pro Val Ala Val Ala Cys Ile Leu Ala Glu Met Lys Leu Asp Tyr Glu
50 55 60
Thr Leu Met Ala Ala Leu Leu His Asp Val Ile Glu Asp Thr Pro Ala
65 70 75 80
Thr Tyr Gln Asp Met Glu Gln Leu Phe Gly Lys Ser Val Ala Glu Leu
85 90 95
Val Glu Gly Val Ser Lys Leu Asp Lys Leu Lys Phe Arg Asp Lys Lys
100 105 110
Glu Ala Gln Ala Glu Asn Phe Arg Lys Met Ile Met Ala Met Val Gln
115 120 125
Asp Ile Arg Val Ile Leu Ile Lys Leu Ala Asp Arg Thr His Asn Met
130 135 140
Arg Thr Leu Gly Ser Leu Arg Pro Asp Lys Arg Arg Arg Ile Ala Arg
145 150 155 160
Glu Thr Leu Glu Ile Tyr Ser Pro Leu Ala His Arg Leu Gly Ile His
165 170 175
His Ile Lys Thr Glu Leu Glu Glu Leu Gly Phe Glu Ala Leu Tyr Pro
180 185 190
Asn Arg Tyr Arg Val Ile Lys Glu Val Val Lys Ala Ala Arg Gly Asn
195 200 205
Arg Lys Glu Met Ile Gln Lys Ile Leu Ser Glu Ile Glu Gly Arg Leu
210 215 220
Gln Glu Ala Gly Ile Pro Cys Arg Val Ser Gly Arg Glu Lys His Leu
225 230 235 240
Tyr Ser Ile Tyr Cys Lys Met Val Leu Lys Glu Gln Arg Phe His Ser
245 250 255
Ile Met Asp Ile Tyr Gln Phe Arg Val Ile Val Asn Asp Ser Asp Thr
260 265 270
Cys Tyr Arg Val Leu Gly Gln Met His Ser Leu Tyr Lys Pro Arg Pro
275 280 285
Gly Arg Val Lys Asp Tyr Ile Ala Ile Pro Lys Ala Asn Gly Tyr Gln
290 295 300
Ser Leu His Thr Ser Met Ile Gly Pro His Gly Val Pro Val Glu Val
305 310 315 320
Gln Ile Arg Thr Glu Asp Met Asp Gln Met Ala Glu Met Gly Val Ala
325 330 335
Ala His Trp Ala Tyr Lys Glu His Gly Glu Thr Ser Thr Thr Ala Gln
340 345 350
Ile Arg Ala Gln Arg Trp Met Gln Ser Leu Leu Glu Leu Gln Gln Ser
355 360 365
Ala Gly Ser Ser Phe Glu Phe Ile Glu Ser Val Lys Ser Asp Leu Phe
370 375 380
Pro Asp Glu Ile Tyr Val Phe Thr Pro Glu Gly Arg Ile Val Glu Leu
385 390 395 400
Pro Ala Gly Ala Thr Pro Val Asp Phe Ala Tyr Ala Val His Thr Asp
405 410 415
Ile Gly His Ala Cys Val Gly Ala Arg Val Asp Arg Gln Pro Tyr Pro
420 425 430
Leu Ser Gln Pro Leu Thr Ser Gly Gln Thr Val Glu Ile Ile Thr Ala
435 440 445
Pro Gly Ala Arg Pro Asn Ala Ala Trp Leu Asn Phe Val Val Ser Ser
450 455 460
Lys Ala Arg Ala Lys Ile Arg Gln Leu Leu Lys Asn Leu Lys Arg Asp
465 470 475 480
Asp Ser Val Ser Leu Gly Arg Arg Leu Leu Asn His Ala Leu Gly Gly
485 490 495
Ser Arg Lys Leu Asn Glu Ile Pro Gln Glu Asn Ile Gln Arg Glu Leu
500 505 510
Asp Arg Met Lys Leu Ala Thr Leu Asp Asp Leu Leu Ala Glu Ile Gly
515 520 525
Leu Gly Asn Ala Met Ser Val Val Val Ala Lys Asn Leu Gln His Gly
530 535 540
Asp Ala Ser Ile Pro Pro Ala Thr Gln Ser His Gly His Leu Pro Ile
545 550 555 560
Lys Gly Ala Asp Gly Val Leu Ile Thr Phe Ala Lys Cys Cys Arg Pro
565 570 575
Ile Pro Gly Asp Pro Ile Ile Ala His Val Ser Pro Gly Lys Gly Leu
580 585 590
Val Ile His His Glu Ser Cys Arg Asn Ile Arg Gly Tyr Gln Lys Glu
595 600 605
Pro Glu Lys Phe Met Ala Val Glu Trp Asp Lys Glu Thr Ala Gln Glu
610 615 620
Phe Ile Thr Glu Ile Lys Val Glu Met Phe Asn His Gln Gly Ala Leu
625 630 635 640
Ala Asn Leu Thr Ala Ala Ile Asn Thr Thr Thr Ser Asn Ile Gln Ser
645 650 655
Leu Asn Thr Glu Glu Lys Asp Gly Arg Val Tyr Ser Ala Phe Ile Arg
660 665 670
Leu Thr Ala Arg Asp Arg Val His Leu Ala Asn Ile Met Arg Lys Ile
675 680 685
Arg Val Met Pro Asp Val Ile Lys Val Thr Arg Asn Arg Asn
690 695 700
<---
Изобретение относится к области биотехнологии, конкретно - к мутантным белкам, вовлеченным в продуцирование L-триптофана, и может быть использовано для получения L-триптофана в Escherichia coli. Предложен мутантный белок SpoT с аминокислотной последовательностью, по меньшей мере на 90% идентичной SEQ ID NO: 1, в которой аминокислота, соответствующая положению 262, заменена на серин, цистеин, тирозин, аспарагин или глутамин. Изобретение обеспечивает эффективное получение L-триптофана в Escherichia coli с применением предложенного мутантного SpoT. 7 н. и 4 з.п. ф-лы, 4 табл., 3 пр.
1. Полипептид, вовлеченный в продуцирование L-триптофана, обладающий активностью белка SpoT,
где указанный полипептид имеет 90% или более идентичности последовательности с аминокислотной последовательностью SEQ ID NO: 1 и аминокислоту, выбранную из серина, цистеина, тирозина, аспарагина и глутамина в положении, соответствующем положению 262 в последовательности SEQ ID NO: 1,
где указанный полипептид обладает активностью для повышения способности продуцировать L-триптофан по сравнению с полипептидом дикого типа.
2. Полипептид по п. 1, где указанный полипептид состоит из любой последовательности из аминокислотных последовательностей, представленных SEQ ID NO: 19, 21-24.
3. Полинуклеотид, кодирующий полипептид по п. 1 или п. 2.
4. Микроорганизм Escherichia coli для получения L-триптофана, содержащий один или более полипептидов по п. 1 или п. 2;
полинуклеотид, кодирующий указанный полипептид;
и экспрессирующий вектор, включающий в себя указанный полинуклеотид.
5. Микроорганизм по п. 4, где указанный полипептид состоит из любой последовательности из аминокислотных последовательностей, представленных SEQ ID NO: 19, 21-24.
6. Способ получения L-триптофана, включающий стадию культивирования микроорганизма Escherichia coli по п. 4 в среде, и
стадию выделения L-триптофана из микроорганизма или из среды.
7. Композиция для получения L-триптофана, содержащая полипептид по п. 1 или п. 2, микроорганизм по п. 4 или культуру микроорганизма по п. 4 и эксципиенты.
8. Композиция по п. 7, где указанный полипептид состоит из любой последовательности из аминокислотных последовательностей, представленных SEQ ID NO: 19, 21-24.
9. Способ повышения способности микроорганизма Escherichia coli продуцировать L-триптофан, включающий стадию трансформации микроорганизма вектором экспрессии, содержащим полинуклеотид, кодирующий полипептид по п. 1 или п. 2.
10. Способ по п. 9, где указанный полипептид состоит из любой последовательности из аминокислотных последовательностей, представленных SEQ ID NO: 19, 21-24.
11. Применение композиции для получения L-триптофана, где указанная композиция содержит полипептид,
где указанный полипептид вовлечен в продуцирование L-триптофана, обладает активностью белка SpoT, где указанный полипептид имеет 90% или более идентичности последовательности с аминокислотной последовательностью SEQ ID NO: 1 и аминокислоту, выбранную из серина, цистеина, тирозина, аспарагина и глутамина в положении, соответствующем положению 262 в последовательности SEQ ID NO: 1, и где указанный полипептид обладает активностью для повышения способности продуцировать L-триптофан по сравнению с полипептидом дикого типа,
микроорганизм Escherichia coli, содержащий указанный полипептид или полинуклеотид, кодирующий указанный полипептид, или культуру указанного микроорганизма и эксципиенты.
| JP 2010187552 A, 02.09.2010, RU 2549689 C2, 27.04.2015, CN 107267568 А, 20.10.2017, CN 110592109 А, 20.12.2019, YAMPOLSKY L.Y | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| et al., Genetic analysis of protein stability and function | |||
| Anna | |||
| Rev | |||
| Genet | |||
| Механизм для сообщения поршню рабочего цилиндра возвратно-поступательного движения | 1918 |
|
SU1989A1 |

