Область техники
Настоящее изобретение относится к микроорганизму, продуцирующему L-триптофан, где микроорганизм модифицирован так, что экспрессируется белок, обладающий триптофан-экспортирующей активностью, и к способу получения L-триптофана с использованием этого микроорганизма.
Предшествующий уровень техники
L-триптофан, незаменимая аминокислота, широко используется в качестве сырья для кормовых добавок, лекарственных средств (например инфузионных растворов), веществ для здорового питания и т.д. В настоящее время для производства L-триптофана в основном используют метод прямой ферментации с использованием микроорганизма.
Ранее, микроорганизмы, которые в основном использовались в получении L-триптофана, представляли собой отобранные штаммы, проявляющие резистентность к аналогам благодаря химическим или физическим мутациям. Однако быстрое развитие технологии генетической рекомбинации и идентификация регуляторных механизмов на молекулярном уровне в 1990-е годы позволили использовать рекомбинантные штаммы с использованием методов генной инженерии.
Между тем экспрессия гена, способного экспортировать определенную аминокислоту, способствовала увеличению продуктивности соответствующей аминокислоты в микроорганизмах. Усиление экспрессии гена экспортера L-лизина (lysE) в микроорганизме рода Corynebacterium привело к повышению продуктивности лизина (WO 9723597 А2). Кроме того, усиление гена rhtC в Е. coli улучшило резистентность к L-треонину и одновременно повысило продуктивность по L-гомосерину, L-треонину и L-лейцину (ЕР 1013765 А1). В ЕР 1016710 В1 раскрыто, что повышение продуктивности по L-глутаминовой кислоте, L-лизину, L-треонину, L-аланину, L-гистидину, L-пролину, L-аргинину, L-валину и L-изолейцину происходит за счет усиления генов yahN, yeaS, yfiK и yggA, функции которых в Е. coli еще не определены.
Однако до сих пор не сообщалось ни об одном экспортирующем белке, проявляющем специфичность к L-триптофану. Хотя ген yddG Е. coli известен, он проявляет более высокую специфичность к L-фенилаланину, чем к L-триптофану (FEMS Microbiol Lett 275 (2007) 312-318). Кроме того, для микроорганизма рода Corynebacterium, который в основном используется в качестве штамма, обеспечивающего L-аминокислотную ферментацию, не сообщалось ни об одном белке, который может экспортировать L-триптофан или ароматическую аминокислоту (J Ind Microbiol Biotechnol. 2015 May; 42(5): 787-97).
В этих обстоятельствах авторам настоящего изобретения удалось экспрессировать новый триптофан-экспортирующий белок, обладающий специфичностью к L-триптофану в L-триптофан-продуцирующем микроорганизме, и они обнаружили, что количество продуцируемого L-триптофана было значительно увеличено, тем самым осуществив настоящее изобретение.
Область техники
Задачей настоящего изобретения является получение микроорганизма, продуцирующего L-триптофан, где микроорганизм модифицирован таким образом, что экспрессируется белок, имеющий L-триптофан-экспортирующую активность.
Другой задачей настоящего изобретения является предложение способа, который включает культивирование микроорганизма в среде; и выделение L-триптофана из культивируемого микроорганизма или среды.
Еще одной задачей настоящего изобретения является предложение вектора, содержащего полинуклеотид, кодирующий новый белок, имеющий L-триптофан-экспортирующую активность и содержащий аминокислотную последовательность SEQ ID NO: 1.
Еще одной задачей настоящего изобретения является предложение способа увеличения L-триптофан-экспортирующей активности микроорганизма, включающего модификацию микроорганизма, так что белок, имеющий L-триптофан-экспортирующую активность и содержащий аминокислотную последовательность SEQ ID NO: 1, экспрессируется в этом микроорганизме.
Техническое решение
Ниже будут подробно описаны типичные воплощения настоящего изобретения. Между тем, каждое из объяснений и типичных воплощений, раскрытых в настоящем документе, может быть применено к другим объяснениям и типичным воплощениям. То есть все комбинации различных факторов, раскрытых здесь, входят в объем настоящего изобретения. Кроме того, объем настоящего изобретения не следует ограничивать конкретным раскрытием, представленным ниже.
Для достижения вышеуказанных целей один из аспектов настоящего раскрытия относится к микроорганизму, продуцирующему L-триптофан, где микроорганизм модифицирован таким образом, что экспрессируется белок, имеющий L-триптофан-экспортирующую активность и содержащий аминокислотную последовательность SEQ ID NO: 1.
При использовании здесь термин "L-триптофан", α-аминокислота, относится к незаменимой аминокислоте, не синтезируемой in vivo, и относится к ароматической L-аминокислоте, имеющей химическую формулу C11H12N2O2.
В настоящем изобретении "белок, имеющий L-триптофан-экспортирующую активность" относится к мембранному белку, который имеет активность специфического экспорта L-триптофана из клетки.
Белок, имеющий L-триптофан-экспортирующую активность, может быть, например, белком, включающим аминокислотную последовательность SEQ ID NO: 1. Белок, включающий аминокислотную последовательность SEQ ID NO: 1, можно использовать взаимозаменяемо с белком, имеющим аминокислотную последовательность SEQ ID NO: 1, и с белком, состоящим из аминокислотной последовательности SEQ ID NO: 1.
В частности, в настоящем изобретении белок, имеющий L-триптофан-экспортирующую активность, включающий аминокислотную последовательность SEQ ID NO: 1, относится к белку, имеющему L-триптофан-экспортирующую активность, среди белков, происходящих из Herbaspirillum rhizosphaerae. В частности, "Herbaspirillum rhizosphaerae'" представляет собой грамотрицательную бактерию, принадлежащую к роду Herbaspirillum. В Корее его можно выделить из ризосферы в почве, как штамм, выделенный с острова Ulleung и т.д.
Кроме того, хотя белок по настоящему изобретению, имеющий L-триптофан-экспортирующую активность, определен, как белок, включающий аминокислотную последовательность SEQ ID NO: 1, это не исключает добавление последовательности выше или ниже аминокислотной последовательности SEQ ID NO: 1, мутации, которая может произойти естественным путем или ее молчащей мутации, и, когда белок имеет активность, идентичную или соответствующую белку, включающему аминокислотную последовательность SEQ ID NO: 1, специалистам в данной области понятно, что этот белок относится к белку, имеющему L-триптофан-экспортирующую активность, по настоящему изобретению. Например, белок, имеющий L-триптофан-экспортирующую активность по настоящему изобретению, может быть белком, состоящим из аминокислотной последовательности SEQ ID NO: 1 или аминокислотной последовательности, имеющей гомологию с аминокислотной последовательностью SEQ ID NO: 1, составляющую 80%, 90%, 95%, 97% или выше. Кроме того, очевидно, что любой белок, имеющий аминокислотную последовательность с делецией, модификацией, заменой или вставкой в части последовательности, также может входить в объем настоящего изобретения при условии, что белок имеет аминокислотную последовательность с любой из вышеприведенных гомологий и проявляет эффект, соответствующий вышеуказанному белку.
Термин "гомология" относится к проценту идентичности двух полинуклеотидов или полинуклеотидных группировок (то есть к степени совпадения с данной аминокислотной последовательности или нуклеотидной последовательностью) и может быть выражен, как процент. В настоящем описании изобретения гомологичная последовательность, обладающая активностью, идентичной или аналогичной данной аминокислотной или нуклеотидной последовательности, представлена как "% гомологии". Гомология последовательностями одной группировки относительно другой может быть определена методом, известным в данной области техники. Например, гомология может быть определена с помощью стандартного программного обеспечения, в частности BLAST 2.0, для расчета таких параметров, как счет выравнивания, идентичность и сходство, или путем сравнения последовательностей с помощью Саузерн-гибридизации в определенных жестких условиях. Подходящие условия гибридизации могут быть определены в рамках данной области техники и могут быть установлены методом, хорошо известным специалистам в данной области техники (например, J. Sambrook et al., Molecular Cloning, A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory press, Cold Spring Harbor, New York, 1989; F.M. Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., New York).
Любой ген, кодирующий белок, имеющий L-триптофан-экспортирующую активность, происходящий из Herbaspirillum rhizosphaerae, может быть включен без ограничения при условии, что этот ген содержит последовательность, которая кодирует белок. В частности, любая последовательность, которая может кодировать белок, имеющий L-триптофан-экспортирующую активность и содержащий аминокислотную последовательность SEQ ID NO: 1, может быть включена без ограничения. Более конкретно, последовательность может быть полинуклеотидом, содержащим полинуклеотидную последовательность SEQ ID NO: 2. Полинуклеотид, содержащий полинуклеотидную последовательность SEQ ID NO: 2, можно использовать взаимозаменяемо с полинуклеотидом, имеющим полинуклеотидную последовательность SEQ ID NO: 2, и с полинуклеотидом, состоящим из полинуклеотидной последовательности SEQ ID NO: 2.
Кроме того, могут быть выполнены различные модификации в кодирующей области полинуклеотида, кодирующего белок, с учетом вырожденность кодонов и кодоны, предпочитаемые в биоорганизме, где белок должен быть экспрессирован, в объеме, не изменяющем аминокислотную последовательность белка. При этом, полинуклеотид, кодирующий белок, может быть полинуклеотидом, имеющим гомологию с полинуклеотидной последовательностью SEQ ID NO: 2, составляющую 80%, 90%, 95%, 97% или выше. Кроме того, очевидно, что любая полинуклеотидная последовательность с делецией, модификацией, заменой или вставкой в части последовательности также может быть включена в объем настоящего изобретения при условии, что эта полинуклеотидная последовательность имеет вышеописанную гомологию и проявляет эффект, идентичный или соответствующий эффекту белка. Альтернативно, любой зонд, который может быть получен из известных последовательностей генов (например, путем гибридизации в жестких условиях с комплементарными последовательностями со всей или частью вышеуказанной полинуклеотидной последовательности) и любой последовательности, которая кодирует белок, имеющий активность, идентичную или соответствующую активности белка, состоящего из аминокислотной последовательности SEQ ID NO: 1, может быть включены без ограничения. Термин "жесткие условия" относится к условиям, которые обеспечивают специфическую гибридизацию полинуклеотидов. Такие условия конкретно описаны в ссылках (например, J Sambrook et al., выше). Например, эти условия могут включать выполнение гибридизации генов, имеющих высокую гомологию, гомологию 80% или выше, в частности 90% или выше, более конкретно 95% или выше, еще более конкретно 97% или выше, и наиболее конкретно 99% или выше, и при этом не происходит гибридизация генов, имеющих гомологию ниже описанной выше гомологии; или выполнение гибридизации один раз, конкретнее два или три раза, при обычных условиях промывания для Саузерн-гибридизации при концентрации соли и температуре, соответствующих 60°С, 1×SSC (раствор хлорида натрия/цитрата натрия), и 0,1% SDS (додецилсульфат натрия), конкретно 60°С, 0,1×SSC и 0,1% SDS, и наиболее конкретно 68°С, 0,1×SSC и 0,1% SDS. Для гибридизации требуется, чтобы две нуклеиновые кислоты содержали комплементарные последовательности, хотя несоответствия между основаниями возможны в зависимости от жесткости гибридизации. Термин "комплементарный" используется для описания отношения между взаимно гибридизируемыми нуклеотидными основаниями. Например, что касается ДНК, аденозин комплементарен тимину, и цитозин комплементарен гуанину. Соответственно, настоящее изобретение может также включать выделенные фрагменты нуклеиновых кислот, комплементарные всей последовательности, а также по существу аналогичные нуклеотидные последовательности. В частности, полинуклеотиды, имеющие гомологию, могут быть обнаружены при значении Tm 55°C с использованием условий гибридизации, которые включают стадию гибридизации, и с использованием условий, описанных выше. Кроме того, значение Tm может быть 60°С, 63°С или 65°С, но температура не ограничивается ими и может быть соответствующим образом подобрана специалистами в данной области в соответствии с намеченной целью. Жесткость, пригодная для гибридизации полинуклеотидов, зависит от длины и степени комплементарности полинуклеотидов, и соответствующие переменные хорошо известны в данной области техники (см. Sambrook et al., выше, 9.50-9.51 и 11.7-11.8).
При использовании здесь, термин "для экспрессии/экспрессируемый" относится к состоянию, при котором целевой белок введен в микроорганизм, или в случае, когда белок присутствует в микроорганизме, активность белка повышена по сравнению с активностью эндогенного белка или активностью до его модификации.
В частности, термин "введение белка" означает, что микроорганизм демонстрирует активность конкретного белка, которой он первоначально не обладал, или микроорганизм демонстрирует увеличенную активность по сравнению с его эндогенной активностью или с активностью белка до модификации. Например, это может означать, что в хромосому микроорганизма введен полинуклеотид, кодирующий определенный белок, или вектор, содержащий полинуклеотид, кодирующий определенный белок, введен в микроорганизм и при этом он проявляет свою активность. Кроме того, термин "повышение активности" означает, что активность конкретного белка улучшена по сравнению с его эндогенной активностью или с активностью до его модификации. Термин "эндогенный белок" относится к активности определенного белка, первоначально имеющегося у родительского штамма микроорганизма, когда свойство микроорганизма изменяется из-за генетической модификации, вызванной естественным или искусственным фактором.
В частности, в настоящем изобретении повышение активности может быть достигнуто одним или более из следующих способов: способом увеличения внутриклеточного числа копий гена, кодирующего белок, имеющий L-триптофан-экспортирующую активность; способом введения модификации в регуляторную последовательность экспрессии гена на хромосоме, кодирующего белок, имеющий L-триптофан-экспортирующую активность; способом замены последовательности контроля экспрессии гена, кодирующего белок, имеющий L-триптофан-экспортирующую активность на хромосоме, последовательностью, имеющей сильную активность; способом замены гена, кодирующего белок, имеющий L-триптофан-экспортирующую активность на хромосоме, геном, который мутирован таким образом, что активность белка, имеющего L-триптофан-экспортирующую активность, повышается; и способом введения модификации в ген, кодирующий белок, имеющий L-триптофан-экспортирующую активность на хромосоме таким образом, что активность белка, имеющего L-триптофан-экспортирующую активность, повышается.
В указанном выше, способ увеличения числа копий гена может быть выполнен в форме, когда ген функционально связан с вектором или путем вставки гена в хромосому клетки-хозяина, но данный способ не ограничивается этим. В частности, число копий гена может быть увеличено путем введения вектора в клетку-хозяина, где вектор функционально связан с полинуклеотидом, кодирующим белок по настоящему изобретению и способен реплицироваться и функционировать независимо от клетки-хозяина. Альтернативно, способ увеличения числа копий гена может быть осуществлен путем введения вектора, который способен встраивать полинуклеотид в хромосому и функционально связан с полинуклеотидом, в хромосоме клетки-хозяина. Введение полинуклеотид а в хромосому может быть достигнуто известным в данной области методом (например посредством гомологичной рекомбинации).
Затем, чтобы увеличить экспрессию полинуклеотида, последовательность контроля экспрессии может быть модифицирована путем индуцирования модификации в ней посредством делеции, вставки, неконсервативной или консервативной замены или их комбинации, чтобы еще больше усилить активность последовательности контроля экспрессии; или путем замены последовательности контроля экспрессии последовательностью нуклеиновой кислоты с более сильной активностью, но данный метод модификации не ограничивается этим. Последовательность контроля экспрессии может включать промотор, последовательность оператора, последовательность, кодирующую сайт связывания рибосомы, спейсер и последовательности, контролирующие окончанию транскрипции и трансляции, но не ограничивается ими.
Сильный промотор может быть связан с восходящей областью единицы экспрессии полинуклеотид а вместо исходного промотора, без ограничения этим. Примеры сильных промоторов, известных в данной области, могут включать промоторы cj1 - cj7 (KR патент No. 10-0620092), промотор lac, промотор trp, промотор trc, промотор tac, промотор lambda phage PR, промотор PL, промотор tet, промотор gapA, промотор SPL7, промотор SPL13 (sm3) (KR патент No. 10-1783170), промотор 02 (KR патент No. 10-1632642), промотор tkt, промотор уссА, и т.д., но промоторы не ограничены ими.
Кроме того, полинуклеотидная последовательность хромосомы может быть модифицирована посредством индукции модификации последовательности контроля экспрессии посредством делеции, вставки, неконсервативной или консервативной замены или их комбинации, чтобы еще больше усилить активность полинуклеотидной последовательности; или путем замены полинуклеотидной последовательности на полинуклеотидную последовательность, модифицированную таким образом, чтобы иметь более сильную активность, но метод модификации этим не ограничивается.
Вышеописанное введение и повышение активности белка может, как правило, увеличивать активность или концентрацию соответствующего белка по меньшей мере на 1%, по меньшей мере на 10%, по меньшей мере на 25%, по меньшей мере на 50%, по меньшей мере на 75%, по меньшей мере на 100%, по меньшей мере на 150%, по меньшей мере на 200%, по меньшей мере на 300%, по меньшей мере на 400% или по меньшей мере на 500%, и самое большее на 1000% или на 2000% в зависимости от активности или концентрации белка в штамме микроорганизма дикого типа или немодифицированного микроорганизма, но этот диапазон не ограничен указанным.
В другом аспекте настоящего изобретения предложен полинуклеотид, кодирующий новый белок, имеющий L-триптофан-экспортирующую активность, содержащий аминокислотную последовательность SEQ ID NO: 1, или вектор, содержащий полинуклеотид.
При использовании здесь термин "полинуклеотид" относится к нити ДНК или РНК, имеющей длину более определенной, в виде нуклеотидного полимера, который представляет собой длинную цепь нуклеотидных мономеров, соединенных ковалентной связью.
При использовании здесь термин "вектор" относится к конструкции ДНК, содержащей нуклеотидную последовательность полинуклеотида, кодирующего целевой белок, в которой целевой белок функционально связан с подходящей регуляторной последовательностью, так что он может быть экспрессирован в соответствующем хозяине. Регуляторная последовательность включает промотор, способный инициировать транскрипцию, любую последовательность оператора для контроля за транскрипцией, последовательность, кодирующую соответствующий домен связывания рибосом на мРНК, и последовательность, контролирующую прекращение транскрипции и трансляции. Вектор, будучи трансформирован в подходящую клетку хозяина, может реплицироваться или может функционировать независимо от генома хозяина, или может быть интегрирован в сам геном хозяина.
Вектор, используемый в настоящем изобретении, конкретно не ограничен, при условии, что он способен реплицироваться в клетке-хозяине, и может быть использован любой вектор, известный в данной области техники. Примеры обычных векторов могут включать природные или рекомбинантные плазмиды, космиды, вирусы и бактериофаги. Например, pWE15, М13, MBL3, MBL4, IXII, ASHII, APII, t10, t11, Charon4A, Charon21A и т.д. могут быть использованы как фаговый вектор или космидный вектор; а те, что основаны на pBR, pUC, pBluescriptII, pGEM, pTZ, pCL, pET, и т.д. могут быть использованы в качестве плазмидного вектора. В частности, могут быть использованы векторы pDZ, pACYC177, pACYC184, pCL, pECCG117, pUC19, pBR322, pMW118, pCC1BAC и т.д.
Например, полинуклеотид, кодирующий целевой белок в хромосоме, может быть заменен вариантом полинуклеотида с помощью вектора для хромосомной вставки в клетку. Введение полинуклеотида в хромосому может быть выполнено любым известным в данной области методом, например посредством гомологичной рекомбинации, но этим способ не ограничивается. Кроме того, может быть дополнительно включен селективный маркер для подтверждения удачной вставки вектора в хромосому. Селективный маркер используют для отбора клеток, трансформированных вектором, то есть для подтверждения, была ли успешно введена молекула целевой нуклеиновой кислоты, также могут быть использованы маркеры, способные обеспечить селектируемые фенотипы (например лекарственную устойчивость, ауксотрофию, резистентность к цитотоксическим агентам и экспрессию поверхностных белков). В условиях применения селективных агентов, только клетки, экспрессирующие селективные маркеры, могут выживать и проявлять другие фенотипические признаки и, таким образом, трансформированные клетки могут быть легко отобраны.
При использовании здесь термин "микроорганизм, продуцирующий L-триптофан" относится к микроорганизму, который может продуцировать L-триптофан из источников углерода, находящегося в среде, в избыточном количестве по сравнению с диким типом или немодифицированным штаммом микроорганизма. Кроме того, микроорганизмом, продуцирующим L-триптофан, может быть рекомбинантный микроорганизм. В частности, микроорганизм может представлять собой микроорганизм рода Enterobacter, микроорганизм рода Escherichia, микроорганизм рода Erwinia, микроорганизм рода Serratia, микроорганизм рода Providencia, микроорганизм рода Corynebacterium, или микроорганизм рода Brevibacterium, но тип микроорганизма не ограничен этим, при условии, что микроорганизм продуцирует L-триптофан. Более конкретно, микроорганизм может быть микроорганизмом рода Corynebacterium или микроорганизмом рода Escherichia, и еще более конкретно микроорганизмом рода Escherichia может быть микроорганизм Escherichia coli и микроорганизмом рода Corynebacterium может быть микроорганизм Corynebacterium glutamicum, но микроорганизм рода Escherichia или рода Corynebacterium, в который введен белок, имеющий L-триптофан-экспортирующую активность, или эта активность усилена и, таким образом, количество продуцируемого L-триптофана может быть увеличено, может быть включен без ограничения.
В вышеописанных микроорганизмах количество продуцируемого L-триптофана может быть повышено путем использования метода увеличения биосинтеза L-триптофана путем усиления экспрессии гена tktA или блокирования разветвленных путей в пути биосинтеза L-триптофана для непрерывной поставки предшественников (например эритроза-4-фосфата (Е4Р)) и эффективного использования энергии; или метода использования меньшего количества АТФ и т.д.
В частности, в настоящем изобретении родительский штамм микроорганизма, продуцирующего L-триптофан, в котором микроорганизм модифицирован таким образом, что экспрессируется белок, имеющий L-триптофан-экспортирующую активность, конкретно не ограничен, при условии, что микроорганизм продуцирует L-триптофан. Микроорганизм, продуцирующий L-триптофан, может быть микроорганизмом, в котором активность гена в конкурентном пути, регулятор в пути регуляции L-триптофанового оперона, или ген для введения и разрушения L-триптофана ослаблен или инактивирован таким образом, чтобы усилить путь биосинтеза L-триптофана; и/или активность оперона L-триптофана сверхэкспрессируется. В частности, активность trpR, гена регуляции ферментной группы синтеза триптофана, который ингибирует экспрессию генов биосинтеза L-триптофана (trpEDCBA), или Mtr (то есть мембранного белка), который импортирует внеклеточный L-триптофан в клетку, может быть ослаблена или удалена по сравнению с их эндогенной активностью.
Для достижения вышеуказанной цели в другом аспекте настоящего изобретения предложен способ получения L-триптофана, который включает культивирование микроорганизма, продуцирующего L-триптофан в среде, в которой микроорганизм модифицируется таким образом, что экспрессируется белок, имеющий L-триптофан-экспортирующую активность, содержащий аминокислотную последовательность SEQ ID NO: 1; и выделение L-триптофана из культивируемого микроорганизма или среды.
L-триптофан, белок, имеющий L-триптофан-экспортирующую активность и содержащий аминокислотную последовательность SEQ ID NO: 1, экспрессия белка и микроорганизм являются такими же, как описано выше.
При использовании здесь термин "культивирование" означает, что микроорганизм выращивают в подходящих контролируемых условиях. Процесс культивирования по настоящему изобретению может быть осуществлен в подходящей питательной среде и условиях культивирования, известных в данной области. Такой процесс культивирования может быть легко приспособлен для использования специалистами в данной области в соответствии с выбранным штаммом. В частности, процесс культивирования может быть выполнен в периодической культуре, в непрерывной культуре и в культуре с подпиткой, известных в данной области, но процесс культивирования не ограничивается ими.
При использовании здесь термин "среда" относится к смеси веществ, которая содержит питательные вещества, необходимые для культивирования микроорганизма в качестве основного ингредиента, и она поставляет питательные вещества, факторы роста и т.д. вместе с водой, которая необходима для выживания и роста. В частности, среда и другие условия культивирования, используемые для роста микроорганизма по настоящему изобретению, могут представлять любую среду, используемую для обычного культивирования микроорганизмов без каких-либо конкретных ограничений. Однако микроорганизм по настоящему изобретению можно культивировать в аэробных условиях в обычной среде, содержащей подходящий источник углерода, источник азота, источник фосфора, неорганическое соединение, аминокислоту и/или витамин и т.д. при регулировке подходящей температуры, рН и т.д.
В настоящем изобретении источник углерода может включать углеводы (например глюкозу, фруктозу, сахарозу, мальтозу и т.д.); сахарные спирты (например маннит, сорбит и т.д.); органическую кислоту (например пировиноградную кислоту, молочную кислоту, лимонную кислоту и т.д.); аминокислоты (например глутаминовую кислоту, метионин, лизин и т.д.) и т.д. Кроме того, источник углерода может включать природные органические питательные вещества, такие как гидролизат крахмала, мелассу, сырую мелассу, рисовые отруби, маниоку, мелассу сахарного тростника, сгущенный кукурузный экстракт и т.д. В частности, можно использовать углеводы, такие как глюкоза и стерилизованная предварительно обработанная меласса (то есть меласса, превращенная в восстанавливающий сахар) и, кроме того, можно использовать без ограничений различные другие источники углерода в подходящем количестве. Эти источники углерода могут использоваться отдельно или в комбинации из двух или более видов, но источники углерода не ограничены ими.
Примеры источника азота могут включать неорганические источники азота (например аммоний, сульфат аммония, хлорид аммония, ацетат аммония, фосфат аммония, карбонат аммония, нитрат аммония и т.д.); аминокислоты (глутаминовую кислоту, метионин, глутамин и т.д.); и органические источники азота (например пептон, N-Z амин, мясной экстракт, дрожжевой экстракт, солодовый экстракт, кукурузный экстракт, гидролизат казеина, рыбу или продукт ее разложения, обезжиренный соевый жмых или продукт его разложения и т.д.). Эти источники азота могут быть использованы отдельно или в комбинации из двух или более видов, но источники азота не ограничены ими.
Примеры источника фосфора могут включать монокалийфосфат, дикалийфосфат, соответствующие натрийсодержащие соли и т.д. Примеры неорганического соединения могут включать хлорид натрия, хлорид кальция, хлорид железа, сульфат магния, сульфат железа, сульфат марганца, карбонат кальция и т.д. Кроме того, могут быть включены аминокислоты, витамины и/или соответствующие предшественники. Эти составляющие ингредиенты или предшественники могут быть добавлены к среде в периодической культуре или в непрерывной культуре, без ограничения ими.
В настоящем изобретении рН среды может быть отрегулирован во время культивирования микроорганизма путем добавления такого соединения, как гидроксид аммония, гидроксид калия, аммиак, фосфорная кислота, серная кислота и т.д., к среде подходящим образом. Кроме того, во время культивирования может быть добавлен пеногаситель (например, полигликолевый эфир жирной кислоты) для предупреждения пенообразования. Кроме того, кислород или кислородсодержащий газ могут быть введены в среду для поддержания аэробного состояния среды; или газообразный азот, водород или двуокись углерода могут быть введены без введения газа для поддержания анаэробного или микроаэробного состояния среды, но газ не ограничен ими.
Температура среды может находиться в диапазоне от 20°С до 50°С, и конкретно, от 30°С до 37°С, но температура не ограничена этими значениями. Культивирование может продолжаться до тех пор, пока полезные вещества не будут получены в нужных количествах, и в частности, в течение 10-100 часов, но культивирование этим не ограничивается.
На стадии выделения триптофана, необходимый триптофан может быть выделен из среды с использованием способа культивирования микроорганизма по настоящему изобретению, например с использованием подходящего метода, известного в данной области, в соответствии с процессом периодической культуры, процессом непрерывной культуры или процессом культуры с подпиткой. Например, такие методы как центрифугирование, фильтрование, обработка осадителем, кристаллизующим белки (метод высаливания), экстракция, ультразвуковое разрушение, ультрафильтрация, диализ, различные виды хроматографий (например молекулярно-ситовая хроматография (гель-фильтрация), адсорбционная хроматография, ионообменная хроматография, аффинная хроматография и т.д.) и ВЭЖХ можно использовать отдельно или в комбинации, но методы не ограничены указанным.
Стадия выделения может дополнительно включать процесс очистки. Выделенный L-триптофан может быть очищен с помощью подходящего способа очистки, известного в данной области техники.
В еще одном аспекте настоящего изобретения предложен способ увеличения L-триптофан-экспортирующей активности микроорганизма, который включает введение белка, имеющего L-триптофан-экспортирующую активность, содержащего аминокислотную последовательность SEQ ID NO: 1, в микроорганизм или повышение его активности.
Еще в одном аспекте настоящего изобретения предлагается применение белка, имеющего L-триптофан-экспортирующую активность и содержащего аминокислотную последовательность SEQ ID NO: 1, для увеличения L-триптофан-экспортирующей активности в микроорганизме.
Выше описаны белок, имеющий L-триптофан-экспортирующую активность и содержащий аминокислотную последовательность SEQ ID NO: 1, введение белка и повышение активности белка.
Полезные эффекты изобретения
Авторы настоящего изобретения открыли новый экспортерный ген, имеющий специфичность к L-триптофану, и попытались экспрессировать ген в микроорганизме, продуцирующем L-триптофан. В результате они подтвердили, что микроорганизм может значительно улучшить количество производимого L-триптофана по сравнению с родительским штаммом, в котором ген не экспрессируется, тем самым подтверждая, что L-триптофан может эффективно продуцироваться в вышеприведенном микроорганизме.
Краткое описание графических материалов
На Фиг. 1 показаны внутриклеточные концентрации триптофана в СА04-8352 и СА04-8405, которые представляют собой модифицированные штаммы Corynebacterium glutamicum, в соответствии с потреблением глюкозы.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Далее настоящее изобретение будет подробно описано посредством типичных воплощений. Однако эти типичные воплощения представлены только для иллюстрации и не предназначены для ограничения объема настоящего изобретения.
Пример 1: Скрининг и отбор экспортирующих генов
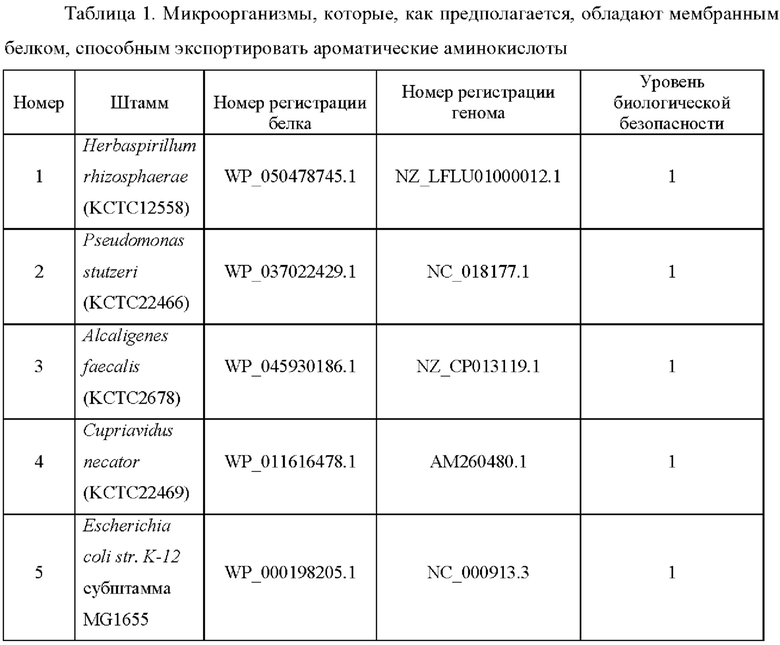
В результате PSI-BLAST скрининга на основе баз данных NCBI и KEGG с аминокислотной последовательностью YdeD (то есть семейства EamA, происходящего из Е. coli) в качестве искомой последовательности, были выбраны 30 генов-кандидатов, которые рассматриваются, как мембранные белки, способные экспортировать триптофан, и были выбраны биоорганизмы, обладающие этими генами. Среди них было выбрано 5 видов биоорганизмов с учетом уровней биобезопасности, которые применимы к продуцирующим штаммам, и доступности, как показано в Таблице 1 ниже.
Пример 2: Получение микроорганизма рода Corynebacterium, в который введен ген, происходящий из Herbaspirillum rhizosphaerae
Ген, кодирующий мембранный белок, происходящий из Herbaspirillum rhizosphaerae, выбранный в Примере 1, имеет аминокислотную последовательность SEQ ID NO: 1. Информация о гене, кодирующем мембранный белок, и прилегающих к нему нуклеотидных последовательностях (номер регистрации NZ_LFLU01000012.1) была получена из NIH GenBank.
На основе полученной информации о нуклеотидных последовательностях были синтезированы праймеры для вставки гена, происходящего из Herbaspirillum rhizosphaerae, в геномную ДНК Corynebacterium glutamicum. Для амплификации гена, происходящего из штамма Herbaspirillum rhizosphaerae, проводили ПЦР с использованием хромосомной ДНК штамма Herbaspirillum rhizosphaerae в качестве матрицы в сочетании с праймерами SEQ ID NO: 3 и SEQ ID NO: 4. ДНК-полимеразу Solg™ Pfu-X DNA использовали в качестве полимеразы и выполняли ПЦР следующим образом: денатурация при 95°С в течение 2 минут; 27 циклов денатурации при 95°С в течение 20 секунд, отжиг при 62°С в течение 40 секунд и полимеризация при 72°С в течение 1 минуты; и полимеризация при 72°С в течение 5 минут.
В результате был получен генный фрагмент из 956 п.о., который содержал ген из 924 п.о. (SEQ ID NO: 2).
SEQ ID NO: 3 (wex - 1)
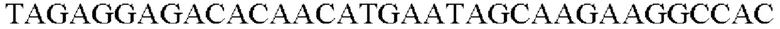
SEQ ID NO: 4 (wex - 2)
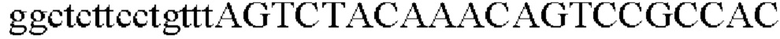
Для получения промотора gapA, происходящего из Corynebacterium glutamicum, выполняли ПЦР с использованием геномной ДНК Corynebacterium glutamicum в качестве матрицы в сочетании с праймерами SEQ ID NO: 5 и SEQ ID NO: 6. ДНК-полимеразу SolgTM Pfu-X (SolGent Co., Ltd.) использовали в качестве полимеразы, и ПЦР выполняли следующим образом: денатурация при 95°С в течение 2 минут; 27 циклов денатурации при 95°С в течение 20 секунд, отжиг при 62°С в течение 40 секунд и полимеризация при 72°С в течение 30 секунд; и полимеризация при 72°С в течение 5 минут.
SEQ ID NO: 5 (PgapA - 1)
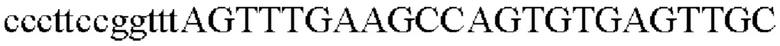
SEQ ID NO: 6 (PgapA(-wex) - 2)
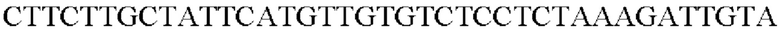
Амплифицированная область промотора gapA, фрагменты гена, происходящего из Herbaspirillum rhizosphaerae и вектор pDZTn (KR патент No. 10-1126041), который был расщеплен с помощью фермента рестрикции Seal, клонировали с помощью метода сборки Гибсона (DG Gibson et al., NATURE METHODS, VOL.6 NO.5, MAY2009, NEBuilder HiFi DNA Assembly Master Mix), и таким образом была получена рекомбинантная плазмида. Рекомбинантная плазмида была названа pDZTn-PgapA-Hrh. Клонирование было выполнено посредством смешивания реагента сборки Гибсона и каждого из генных фрагментов с рассчитанным числом молей с последующей инкубацией при 50°С в течение 1 часа.
Полученный вектор pDZTn-PgapA-Hrh трансформировали в штамм дикого типа Corynebacterium glutamicum АТСС13869 с помощью электропорации (Appl. Microbiol. Biotechnol. (1999) 52:541-545) и подвергали вторичному кроссинговеру с получением штамма, в котором одна копия гена PgapA-Hrh встроена в транспозон на хромосоме. Соответствующие генетические манипуляции были подтверждены с помощью секвенирования генома и метода ПЦР с использованием праймеров SEQ ID NO: 7 и SEQ ID NO: 8, каждый из которых может соответственно амплифицировать внешнюю область восходящей области и нисходящей области участка гомологичной рекомбинации, куда встроен соответствующий ген.
SEQ ID NO: 7 (Confirm_PgapA-wex - 1)
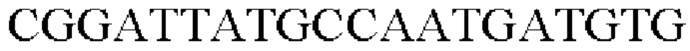
SEQ ID NO: 8 (Confirm_PgapA-wex - 2)
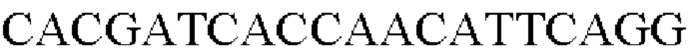
Полученный таким образом штамм был назван Corynebacterium glutamicum АТСС13869::PgapA-Hrh.
Пример 3: Получение микроорганизма рода Corynebacterium, в который введен ген, происходящий из Pseudomonas stutzeri
Ген, кодирующий мембранный белок, происходящий из Pseudomonas stutzeri, выбранный в Примере 1, имеет аминокислотную последовательность SEQ ID NO: 9. Информация о соответствующем гене и смежных с ним нуклеотидных последовательностях (номер регистрации NC_018177.1) была получена из NIH GenBank.
На основе полученной информации о нуклеотидных последовательностях были синтезированы праймеры для встраивания гена, происходящего из Pseudomonas stutzeri, в геномную ДНК Corynebacterium glutamicum. Для амплификации гена, происходящего из Pseudomonas stutzeri, проводили ПЦР таким же образом, как и в Примере 2, используя хромосомную ДНК штамма Pseudomonas stutzeri в качестве матрицы в сочетании с праймерами SEQ ID NO: 11 и SEQ ID NO: 12.
В результате был получен генный фрагмент из 977 п.о., который содержит экспортерный ген из 945 п.о. (SEQ ID NO: 10).
SEQ ID NO: 11 (Pst-1)
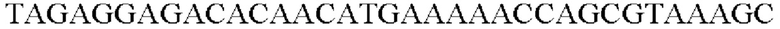
SEQ ID NO: 12 (Pst-2)
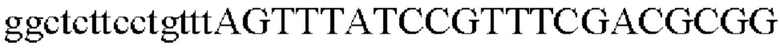
Для использования промотора gapA, происходящего из Corynebacterium glutamicum, выполняли ПЦР таким же образом, как в Примере 2, с использованием геномной ДНК Corynebacterium glutamicum АТСС13869 в качестве матрицы в сочетании с праймерами SEQ ID NO: 5 и SEQ ID NO: 13.
SEQ ID NO: 13 (PgapA(-Pst)-2)
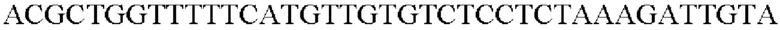
Амплифицированная область промотора gapA, генные фрагменты, происходящие из Pseudomonas stutzeri, и вектор pDZTn, который был расщеплен рестрикционным ферментом Seal, клонировали с помощью метода сборки Гибсона, и таким образом была получена рекомбинантная плазмида. Рекомбинантная плазмида была названа pDZTn-PgapA-Pst. Клонирование было выполнено посредством смешивания реагента сборки Гибсона и каждого из генных фрагментов с рассчитанным числом молей и с последующей инкубацией при 50°С в течение 1 часа.
Полученный вектор pDZTn-PgapA-Pst трансформировали в штамм дикого типа Corynebacterium glutamicum АТСС13869 посредством электропорации (Appl. Microbiol. Biotechnol. (1999) 52:541-545) и подвергали вторичному кроссинговеру с получением штамма, в котором одна копия гена PgapA-Pst встроена в транспозон в хромосоме. Соответствующие генетические манипуляции были подтверждены с помощью секвенирования генома и метода ПЦР с использованием праймеров SEQ ID NO: 7 и SEQ ID NO: 8, каждый из которых может соответственно амплифицировать внешнюю область восходящей области и нисходящей области участка гомологичной рекомбинации, куда встроен соответствующий ген
Полученный таким образом штамм был назван Corynebacterium glutamicum ATCC13869::PgapA-Pst.
Пример 4: Получение микроорганизма рода Corynebacterium, в который введен ген, происходящий из Alcaligenes faecalis
Ген, кодирующий мембранный белок, происходящий из Alcaligenes faecalis, выбранный в Примере 1, имеет аминокислотную последовательность SEQ ID NO: 14. Информация о соответствующем гене и смежных с ним нуклеотидных последовательностях (номер регистрации NZ_CP013119.1) была получена из NIH GenBank.
Праймеры для вставки гена, происходящего из Alcaligeness faecalis, в геномную ДНК Corynebacterium glutamicum синтезировали на основе полученной информации о нуклеотидных последовательностях. Для амплификации гена, происходящего из Alcaligeness faecalis, ПЦР выполняли так же, как в Примере 2, с использованием хромосомной ДНК штамма Alcaligenes faecalis в качестве матрицы в сочетании с праймерами SEQ ID NO: 16 и SEQ ID NO: 17.
В результате был получен генный фрагмент из 943 п.о., который содержит экспортерный ген из 912 п.о. (SEQ ID NO: 15).
SEQ ID NO: 16 (Afa-1)
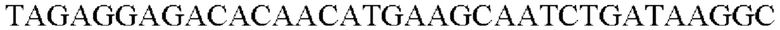
SEQ ID NO: 17 (Afa-2)
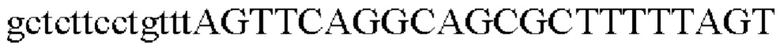
Для получения промотора gapA, происходящего из Corynebacterium glutamicum, выполняли ПЦР так же, как в Примере 2, с использованием геномной ДНК Corynebacterium glutamicum в качестве матрицы в сочетании с праймерами SEQ ID NO: 5 и SEQ ID NO: 18.
SEQ ID NO: 18 (PgapA(-Afa)-2)
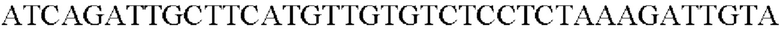
Амплифицированную область промотора gap А, генные фрагменты, происходящие из Alcaligenes faecalis, и вектор pDZTn, который был расщеплен рестрикционным ферментом Seal, клонировали с помощью метода сборки Гибсона, и таким образом была получена рекомбинантная плазмида. Рекомбинантная плазмида была названа pDZTn-PgapA-Afa. Клонирование выполняли посредством смешивания реагента сборки Гибсона и каждого из генных фрагментов с рассчитанным числом молей и с последующей инкубации при 50°С в течение 1 часа.
Полученный вектор pDZTn-PgapA-Afa трансформировали в штамм дикого типа Corynebacterium glutamicum АТСС13869 посредством электропорации и подвергали вторичному кроссинговеру с получением штамма, в котором одна копия гена PgapA-Afa встроена в транспозон на хромосоме. Соответствующие генетические манипуляции были подтверждены с помощью секвенирования генома и метода ПЦР с использованием праймеров SEQ ID NO: 7 и SEQ ID NO: 8, каждый из которых может соответственно амплифицировать внешнюю область восходящей области и нисходящей области участка гомологичной рекомбинации, куда был встроен соответствующий ген.
Полученный таким образом штамм был назван Corynebacterium glutamicum ATCC13869::PgapA-Afa.
Пример 5: Получение микроорганизма рода Corynebacterium, в который встроен ген, происходящий из Cupriavidus necator
Ген, кодирующий мембранный белок, происходящий из Cupriavidus necator, выбранный в Примере 1, имеет аминокислотную последовательность SEQ ID NO: 19. Информация о соответствующем гене и смежных с ним нуклеотидных последовательностях (номер регистрации АМ260480.1) была получена из NIH GenBank.
Праймеры для вставки гена, происходящего из Cupriavidus necator, в геномную ДНК Corynebacterium glutamicum синтезировали на основе полученной информации о нуклеотидных последовательностях. Для амплификации гена, происходящего из Cupriavidus necator, выполняли ПЦР так же, как в Примере 2, с использованием хромосомной ДНК штамма Cupriavidus necator в качестве матрицы в сочетании с праймерами SEQ ID NO: 21 и SEQ ID NO: 22.
В результате получали генный фрагмент из 977 п.о., который содержит экспортерный ген из 945 п.о., происходящий из Cupriavidus necator (SEQ ID NO: 20).
SEQ ID NO: 21 (Cne-1)
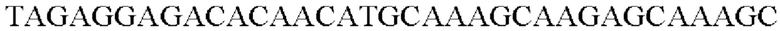
SEQ ID NO: 22 (Cne-2)
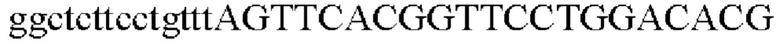
Для получения промотора gapA, происходящего из Corynebacterium glutamicum, выполняли ПЦР так же, как в Примере 2, с использованием геномной ДНК Corynebacterium glutamicum в качестве матрицы в сочетании с праймерами SEQ ID NO: 5 и SEQ ID NO: 23.
SEQ ID NO: 23 (PgapA(-Cne)-2)
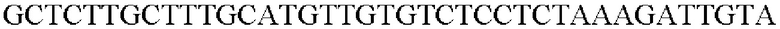
Амплифицированную область промотора gap А, генные фрагменты, происходящие из Cupriavidus necator, и вектор pDZTn, который был расщеплен рестрикционным ферментом SeaI, клонировали с помощью метода сборки Гибсона, и таким образом была получена рекомбинантная плазмида. Рекомбинантная плазмида была названа pDZTn-PgapA-Cne. Клонирование было выполнено посредством смешивания реагента сборки Гибсона и каждого из генных фрагментов с рассчитанным числом молей и с последующей инкубацией при 50°С в течение 1 часа.
Полученный вектор pDZTn-PgapA-Cne трансформировали в штамм дикого типа Corynebacterium glutamicum АТСС13869 посредством электропорации и подвергали вторичному кроссинговеру с получением штамма, в котором одна копия гена PgapA-Cne встроена между генами транспозона на хромосоме. Соответствующие генетические манипуляции были подтверждены с помощью секвенирования генома и метода ПЦР с использованием праймеров SEQ ID NO: 7 и SEQ ID NO: 8, каждый из которых может соответственно амплифицировать внешнюю область восходящей области и нисходящей области участка гомологичной рекомбинации, куда был встроен соответствующий ген.
Полученный таким образом штамм был назван Corynebacterium glutamicum ATCC13869::PgapA-Cne.
Пример 6: Получение микроорганизма рода Corynebacterium, в который встроен ген, происходящий из субштамма MG1655 штамма Escherichia coli K-12.
Ген, кодирующий мембранный белок, полученный из субштамма MG1655 штамма Escherichia coli K-12, выбранный в Примере 1, имеет аминокислотную последовательность SEQ ID NO: 24. Информация о соответствующем гене и смежных с ним нуклеотидных последовательностях (номер регистрации NC_000913.3) была получена из NIH GenBank.
Праймеры для вставки гена, происходящего из Escherichia coli, в геномную ДНК Corynebacterium glutamicum синтезировали на основе полученной информации о нуклеотидных последовательностях. Для амплификации гена, происходящего из Escherichia coli, выполняли ПЦР так же, как в Примере 2, с использованием хромосомной ДНК штамма Escherichia coli в качестве матрицы в сочетании с праймерами SEQ ID NO: 26 и SEQ ID NO: 27.
В результате был получен генный фрагмент из 913 п.о., который содержит экспортерный ген из 882 п.о. (SEQ ID NO: 25).
SEQ ID NO: 26 (Eco-1)
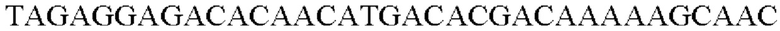
SEQ ID NO: 27 (Eco-2)
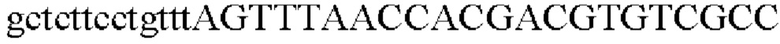
Для получения промотора gapA, происходящего из Corynebacterium glutamicum, выполняли ПЦР так же, как в Примере 2, с использованием геномной ДНК Corynebacterium glutamicum в качестве матрицы в сочетании с праймерами SEQ ID NO: 5 и SEQ ID NO: 28.
SEQ ID NO: 28 (PgapA(-Eco)-2)
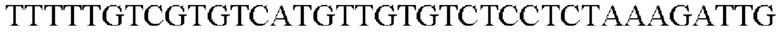
Амплифицированную область промотора gapA, фрагменты гена, происходящие из Escherichia coli, и вектор pDZTn, который был расщеплен рестрикционным ферментом Seal, клонировали с помощью метода сборки Гибсона, и таким образом была получена рекомбинантная плазмида. Рекомбинантная плазмида была названа pDZTn-PgapA-Eco. Клонирование было выполнено посредством смешивания реагента сборки Гибсона и каждого из фрагментов с рассчитанным числом молей с последующей инкубацией при 50°С в течение 1 часа.
Полученный вектор pDZTn-PgapA-Eco трансформировали в штамм дикого типа Corynebacterium glutamicum АТСС13869 посредством электропорации и подвергали вторичному кроссинговеру с получением штамма, в котором одна копия гена PgapA-Eco встроена между генами транспозона на хромосоме. Соответствующие генетические манипуляции были подтверждены с помощью секвенирования генома и способа ПЦР с использованием праймеров SEQ ID NO: 7 и SEQ ID NO: 8, каждый из которых может соответственно амплифицировать внешнюю область восходящей области и нисходящей области участка гомологичной рекомбинации, куда встроен соответствующий ген.
Полученный таким образом штамм был назван Corynebacterium glutamicum ATCC13869::PgapA-Eco.
Пример 7: Определение MIC в штаммах микроорганизма рода Corynebacterium, в который введены гены, происходящие из различных микроорганизмов
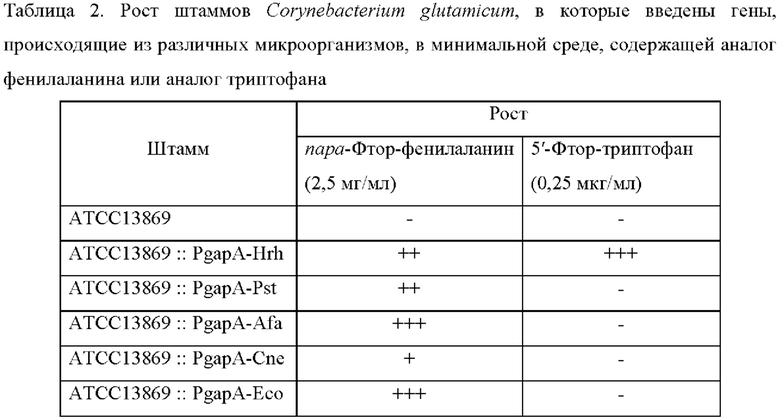
Для подтверждения присутствия триптофан-экспортирующей активности у 5 типов штаммов Corynebacterium glutamicum, полученных в Примерах 2-6, (то есть ATCC13869::PgapA-Hrh, ATCC13869::PgapA-Pst, ATCC13869::PgapA-Afa, ATCC13869::PgapA-Cne и ATCC13869::PgapA-Eco), проводили эксперимент с минимальной ингибирующей концентрацией (MIC) с использованием аналога триптофана и аналога фенилаланина (то есть другой ароматической аминокислоты). 5 разных штаммов Corynebacterium glutamicum, каждый с введенным геном, кодирующим мембранный белок, культивировали в минимальной жидкой среде при 30°С в течение 24 часов, разбавляли до концентрации 3×103 клеток и 3×104 клеток, соответственно, и затем культивировали пятнами в минимальной твердой среде, куда добавляли аналог триптофана и аналог фенилаланина.
Для эксперимента с минимальной ингибирующей концентрацией (MIC) в минимальную твердую среду добавляли пара-фтор-DL-фенилаланин (2,5 мг/мл) или 5-фтор-DL-триптофан (0,25 мкг/мл), и рост клеток наблюдали через 60 часов (Таблица 2).
Все введения выбранных 5 типов генов обеспечивали рост клеток в условиях, когда аналог фенилаланина добавляли в концентрации 2,5 мг/мл. Среди них, введение генов, происходящих из Herbaspirillum rhizosphaerae, Alcaligenes faecalis и Escherichia coli, показало самый высокий рост клеток. Введение гена, происходящего из Pseudomonas stutzeri, показало немного сниженный рост клеток, а введение гена, происходящего из Cupriavidus necator, показало самый низкий рост клеток. В таких же условиях штамм АТСС13869 дикого типа не рос. Кроме того, в случае добавления аналога триптофана в концентрации 0,25 мкг/мл, только введение гена, происходящего из Herbaspirillum rhizosphaerae, обеспечивало рост клеток.
Из приведенных выше результатов было замечено, что все из введений выбранных 5 типов генов, показали резистентность к фенилаланину и к аналогу фенилаланина даже несмотря на то, что были различия в активностях между введениями. В отличие от этого, в отношении триптофана и аналога триптофана, только введение гена, происходящего из Herbaspirillum rhizosphaera, показало специфическую и отличную резистентность к ним. Основываясь на этих результатах, можно интерпретировать, что только мембранный белок, кодируемый геном, происходящим из Herbaspirillum rhizosphaera, может выступать в качестве белка-экспортера для триптофана.
Минимальная среда (рН 7.2)
10 г глюкозы, 1 г KH2PO4, 2 г K2HPO4, 0,4 г MgSO4⋅7H2O, 2 г мочевины, 5 г (NH4)2SO4, 0,5 г NaCl, 5 мкг никотинамида, 0,1 мкг пантотената кальция, 0,2 мкг биотина, 3 мкг тиамина HCl, 1 мл раствора микроэлементов (в расчете на 1 л дистиллированной воды).
Раствор микроэлементов
0,09 г Na2B4O7⋅10H2O, 0,04 г (NH4)6Mo7O27 4H2O, 0,01 г ZnSO4⋅7H2O, 0,27 г CuSO4⋅5H2O, 0,01 г MnCl2⋅4H2O, 1 г FeCl3⋅6H2O, 0,01 г CaCl2 (в расчете на 1 л дистиллированной воды)
Пример 8: Получение экспрессионного вектора для Escherichia coli, в который введены гены, происходящие из различных микроорганизмов
Чтобы подтвердить резистентность генов, происходящих из различных микроорганизмов, выбранных в Примере 1, к триптофану или его аналогу в Escherichia coli, каждый из генов клонировали в pCL1920 (то есть экспрессионный вектор для Е. coli) и экспрессирован с промотором yccA E. coli W3110.
Для получения фрагмента гена, происходящего из Herbaspirillum rhizosphaerae, выполняли ПЦР с использованием хромосомной ДНК штамма Herbaspirillum rhizosphaerae в качестве матрицы в сочетании с праймерами SEQ ID NO: 29 и SEQ ID NO: 30. ДНК-полимеразу SolgTM Pfu-X (SolGent Co., Ltd.) использовали в качестве полимеразы, и ПЦР выполняли следующим образом: денатурация при 95°С в течение 2 минут; 27 циклов денатурации при 95°С в течение 20 секунд, отжиг при 62°С в течение 40 секунд и полимеризация при 72°С в течение 1 минуты; и полимеризация при 72°С в течение 5 минут.
SEQ ID NO: 29 (Hrh-3)
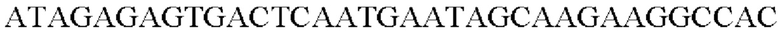
SEQ ID NO: 30 (Hrh-4)
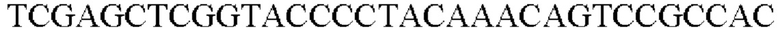
Для получения промотора уссА, происходящего из Е. coli W3110, ПЦР выполняли с использованием геномной ДНК Е. coli W3110 в качестве матрицы в сочетании с праймерами SEQ ID NO: 31 и SEQ ID NO: 32. ДНК-полимеразу SolgTM Pfu-X (SolGent Co., Ltd.) использовали в качестве полимеразы, и ПЦР выполняли следующим образом: денатурация при 95°С в течение 2 минут; 27 циклов денатурации при 95°С в течение 20 секунд, отжиг при 62°С в течение 40 секунд и полимеризация при 72°С в течение 10 секунд; и полимеризация при 72°С в течение 5 минут.
SEQ ID NO: 31 (РуссА - 1)
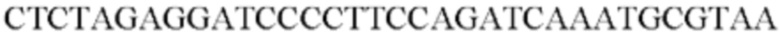
SEQ ID NO: 32 (PyccA(-Hrh)-2)
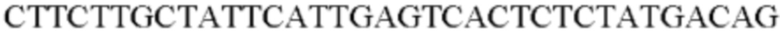
Амплифицированная область промотора уссА, фрагменты гена, происходящего из Herbaspirillum rhizosphaerae, и вектор pCL1920 (pSC101 ori, Spr), который расщепляли рестрикционным ферментом SmaI, клонировали с помощью метода сборки Гибсона, и таким образом была получена рекомбинантная плазмида. Рекомбинантная плазмида была названа pCL1920-PyccA-Hrh. Клонирование было выполнено посредством смешивания реагента сборки Гибсона и каждого из генных фрагментов с рассчитанным числом молей с последующей инкубацией при 50°С в течение 1 часа. Полученный pCL1920-PyccA-Hrh вводили в штамм дикого типа Е. coli W3110, и таким образом был получен W3110/pCL1920-PyccA-Hrh (то есть трансформант, в котором экспрессируется ген).
Для получения фрагмента гена, происходящего из Pseudomonas stutzeri, ПЦР выполняли с использованием хромосомной ДНК штамма Pseudomonas stutzeri в качестве матрицы в сочетании с праймерами SEQ ID NO: 33 и SEQ ID NO: 34. Кроме того, ПЦР выполняли таким же образом, как и при получении фрагмента гена из штамма Herbaspirillum rhizosphaerae, описанного выше, за исключением использования праймера SEQ ID NO: 35, который использовали для получения промотора усе А, происходящего из Е. coli W3110.
SEQ ID NO: 33 (Pst-3)
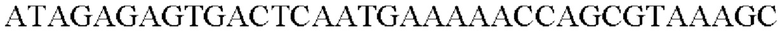
SEQ ID NO: 34 (Pst-4)
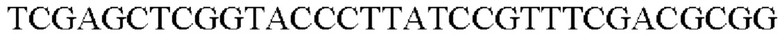
SEQ ID NO: 35 (PyccA(-Pst)-2)
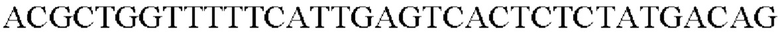
Таким образом, была получена рекомбинантная плазмида, получившая название pCL1920-PyccA-Pst. Экспрессионный вектор pCL1920-PyccA-Pst трансформировали в штамм дикого типа Е. coli W3110, и таким образом был получен W3110/pCL1920-PyccA-Pst (то есть трансформант, в котором экспрессируется ген).
Процесс получения трансформанта, где экспрессируется ген, происходящий из штамма Alcaligenes faecalis, был таким же, как описано выше, за исключением того, что ПЦР выполняли с использованием хромосомной ДНК Alcaligenes faecalis в качестве матрицы в сочетании с праймерами SEQ ID NO: 36 и SEQ ID NO: 37, и праймером SEQ ID NO: 38 для получения промотора уссА.
SEQ ID NO: 36 (Afa-3)
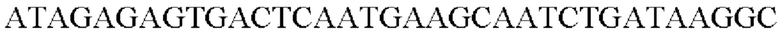
SEQ ID NO: 37 (Afa-4)
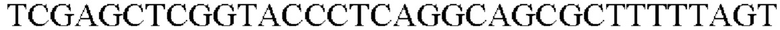
SEQ ID NO: 38 (PyccA(-Afa)-2)
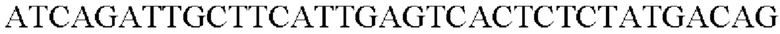
Таким образом, была получена рекомбинантная плазмида, в которую клонирован ген, происходящий из Alcaligenes faecalis, и названа pCL1920-PyccA-Afa. Экспрессионный вектор pCL1920-PyccA-Afa трансформировали в Е. coli W3110 дикого типа и, таким образом, был получен W3110/pCL1920-PyccA-Afa (то есть трансформант).
Для получения фрагмента гена, происходящего из штамма Cupriavidus necator, ПЦР выполняли с использованием хромосомной ДНК штамма Cupriavidus necator в качестве матрицы в сочетании с праймерами SEQ ID NO: 39 и SEQ ID NO: 40. Кроме того, ПЦР выполняли таким же образом, как и при получении фрагмента гена из штамма Herbaspirillum rhizosphaerae, описанного выше, за исключением использования праймера SEQ ID NO: 41, который использовали для получения промотора уссА, происходящего из Е. coli W3110.
SEQ ID NO: 39 (Cne-3)
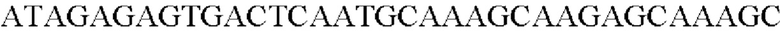
SEQ ID NO: 40 (Cne-4)
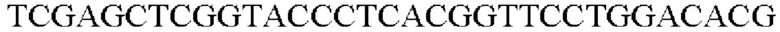
SEQ ID NO: 41 (PyccA(-Cne)-2)
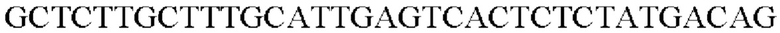
Таким образом, получали рекомбинантную плазмиду, которую назвали pCL1920-PyccA-Cne. Экспрессионный вектор pCL1920-PyccA-Cne трансформировали в Е. coli W3110 дикого типа и, таким образом, получали W3110/pCL1920-PyccA-Cne (то есть трансформант, в котором экспрессируется ген).
Для получения фрагмента гена, происходящего из штамма Escherichia coli, ПЦР выполняли с использованием хромосомной ДНК субштамма MG1655 штамма. Escherichia coli K-12 в качестве матрицы в сочетании с праймерами SEQ ID NO: 42 и SEQ ID NO: 43. Кроме того, ПЦР выполняли таким же образом, как и при получении фрагмента гена из штамма Herbaspirillum rhizosphaerae, описанного выше, за исключением того, что был использован праймер SEQ ID NO: 44, который использовали для получения промотора уссА, полученного из E. coli W3110.
SEQ ID NO: 42 (Eco-3)
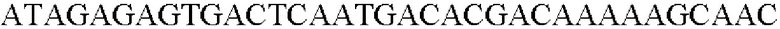
SEQ ID NO: 43 (Eco-4)
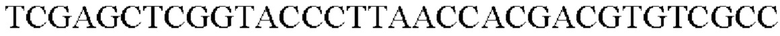
SEQ ID NO: 44 (PyccA(-Eco)-2)
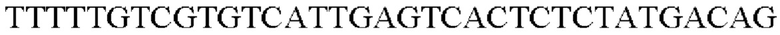
Таким образом, была получена рекомбинантная плазмида и названа pCL1920-PyccA-Eco. Экспрессионный вектор pCL1920-PyccA-Eco вводили в Е. coli W3110 дикого типа, и, таким образом, получали W3110/pCL1920-PyccA-Cne (то есть трансформант, в котором экспрессируется ген).
Пример 9: Измерение MIC Е. coli, в котором сверхэкспрессируются гены мембранных белков из различных микроорганизмов.
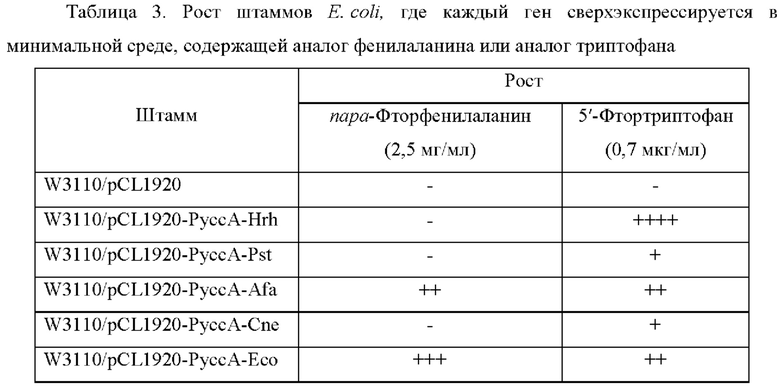
Для подтверждения резистентности штаммов Е. coli, у которых сверхэкспрессируются 5 типов генов, полученных в Примере 8, (то есть W3110/pCL1920-PyccA-Hrh, W3110/pCL1920-PyccA-Pst, W3110/pCL1920-PyccA-Afa, W3110/pCL1920-PyccA-Cne, и W3110/pCL1920-PyccA-Eco), выполняли эксперимент с минимальной ингибирующей концентрацией (MIC) с использованием аналога триптофана и аналога фенилаланина. Штаммы Е. coli, у которых сверхэкспрессировались 5 типов генов, культивировали в минимальной жидкой среде М9, содержащей спектиномицин (50 мкг/мл) при 37°С в течение 15 часов, разбавляли до концентрации 104 и 105 клеток, соответственно, и затем культивировали пятнами в М9 минимальной твердой среде с глюкозой, содержащей спектиномицин (50 мкг/мл), куда были добавлены аналог триптофана или аналог фенилаланина. Для эксперимента с минимальной ингибирующей концентрацией (MIC) добавляли пара-фтор-DL-фенилаланин (2 мг/мл) или 5-фтор-DL-триптофан (0,7 мкг/мл) к М9 минимальной твердой среде и рост клеток определяли через 48 часов (Таблица 3).
Штаммы Е. coli показали прекрасный рост в условиях добавления аналога фенилаланина при сверхэкспрессии генов, происходящих из Е. coli, и сверхэкспрессия гена, происходящего из Alcaligeness faecalis, также показала существенный рост. Однако при сверхэкспрессии генов, происходящих из Herbaspirillum rhizosphaerae, Pseudomonas stutzeri и Cupriavidus necator, не удалось показать рост, сопоставимый с W3110/pCL1920 (то контрольной группой). Наоборот, сверхэкспрессия всех 5 типов выбранных генов позволила выращивать все клетки в условиях, когда был добавлен аналог триптофана. Среди них наибольший рост показала сверхэкспрессия гена, происходящего из Herbaspirillum rhizosphaerae, а сверхэкспрессия экспортерных генов, происходящих из Alcaligenes faecalis и Е. coli, показала второй по величине рост. Сверхэкспрессия экспортерных генов, происходящих из Pseudomonas stutzeri и Cupriavidus necator, показала незначительный рост.
Результаты эксперимента с MIC для 5 типов генов в штамме E. coli были аналогичны результатам для С. glutamicum. Ген, происходящий из Herbaspirillum rhizosphaerae, показал специфическую и превосходную резистентность к триптофану и его аналогу в обоих штаммах Corynebacterium glutamicum и Е. coli, и экспортерный ген, происходящий из Е. coli, показал более высокую резистентность экспорта к фенилаланину и его аналогу, чем к триптофану. Исходя из этих результатов, было высказано предположение, что ген, происходящий из Herbaspirillum rhizosphaerae, показывает специфическую и существенную экспортирующую способность к триптофану в обоих штаммах Corynebacterium glutamicum и Е. coli.
Сравнительный пример 1: Получение микроорганизма рода Corynebacterium glutamicum, продуцирующего L-триптофан
L-триптофан-продуцирующие штаммы были получены из штамма дикого типа Corynebacterium glutamicum АТСС13869. Поскольку штамм дикого типа Corynebacterium glutamicum не может производить L-триптофан, или может производить только очень небольшое его количество, была сделана попытка использовать штамм, у которого усилен путь биосинтеза, необходимый для производства L-триптофана, в качестве родительского штамма. В частности, экспрессия оперона генов биосинтеза L-триптофана была увеличена за счет усиления промотора. Кроме того, для высвобождения ингибирования по типу обратной связи белка TrpE, 38-ая аминокислота trpE (то есть серии) была заменена аргинином (Journal of Bacteriology, Nov. 1987, p. 5330-5332).
Для вышеуказанных генетических манипуляций сначала были получены восходящая область промотора trpE и нисходящая область мутации 38-ой аминокислоты trpE для гомологичной рекомбинации на хромосоме. В частности, генетический фрагмент восходящей области промотора trpE получали, выполняя ПЦР с использованием хромосомной ДНК Corynebacterium glutamicum в качестве матрицы в сочетании с праймерами SEQ ID NO: 45 и SEQ ID NO: 46, в то время как генетический фрагмент нисходящей области мутации 38-ой аминокислоты trpE получали посредством выполнения ПЦР с использованием хромосомной ДНК Corynebacterium glutamicum в качестве матрицы в сочетании с праймерами SEQ ID NO: 47 и SEQ ID NO: 48.
SEQ ID NO: 45 (Pspl7-trpE(S38R)_L-1)
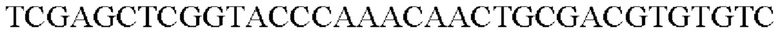
SEQ ID NO: 46 (Pspl7-trpE(S38R) L-2)
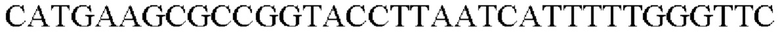
SEQ ID NO: 47 (Pspl7-trpE(S38R)_R-l)
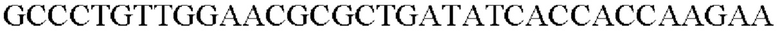
SEQ ID NO: 48 (Pspl7-trpE(S38R)_R-2)
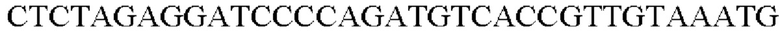
ДНК-полимеразу SolgTM Pfu-X (SolGent Co., Ltd.) использовали в качестве полимеразы, и ПЦР выполняли следующим образом: денатурация при 95°С в течение 5 минут; 30 циклов денатурации при 95°С в течение 30 секунд, отжиг при 60°С в течение 30 секунд и полимеризация при 72°С в течение 60 секунд; и полимеризация при 72°С в течение 5 минут.
ПЦР выполняли с использованием синтезированного промотора SPL7 (SEQ ID NO: 49) в качестве матрицы в сочетании с праймерами SEQ ID NO: 50 и SEQ ID NO: 51.
SEQ ID NO: 50 (Pspl7 - 1)
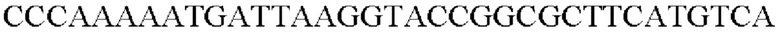
SEQ ID NO: 51 (Pspl7 - 2)
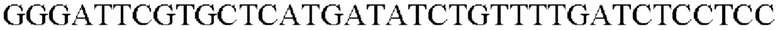
ДНК-полимеразу SolgTM Pfu-X (SolGent Co., Ltd.) использовали в качестве полимеразы, и ПЦР выполняли следующим образом: денатурация при 95°С в течение 5 минут; 30 циклов денатурации при 95°С в течение 30 секунд, отжиг при 60°С в течение 30 секунд, и полимеризация при 72°С в течение 30 секунд; и полимеризация при 72°С в течение 5 минут.
Для получения фрагмента фронтальных последовательностей trpE, включая мутацию от 1-ой до 38-ой аминокислоты, происходящую из Corynebacterium glutamicum, ПЦР выполняли с использованием геномной ДНК Corynebacterium glutamicum в качестве матрицы в сочетании с праймерами SEQ ID NO: 52 и SEQ ID NO: 53.
SEQ ID NO: 52 (trpE (S38R) - 1)
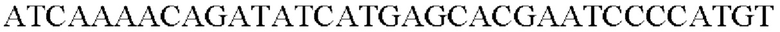
SEQ ID NO: 53 (trpE (S38R) - 2)
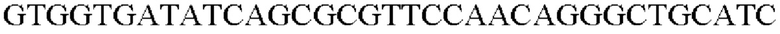
ДНК-полимеразу SolgTM Pfu-X (SolGent Co., Ltd.) использовали в качестве полимеразы, и ПЦР выполняли следующим образом: денатурация при 95°С в течение 5 минут; 30 циклов денатурации при 95°С в течение 30 секунд, отжиг при 60°С в течение 30 секунд и полимеризация при 72°С в течение 30 секунд; и полимеризация при 72°С в течение 5 минут.
Рекомбинантную плазмиду получили путем клонирования амплифицированной восходящей области промотора trpE и нисходящей области мутации 38-й аминокислоты trpE, промотора SPL7 и фрагмента фронтальной последовательности trpE и вектора pDZ, расщепленного с помощью рестрикционного фермента SmaI, с использованием метода сборки Гибсона. Рекомбинантная плазмида была названа DZ-PSPL7-trpE (S38R). Клонирование было выполнено посредством смешивания реагента сборки Гибсона и каждого из генных фрагментов с рассчитанным числом молей с последующей инкубацией при 50°С в течение 1 часа.
Полученный вектор pDZ-PSPL7-trpE (S38R) трансформировали в штамм Corynebacterium glutamicum АТСС13869 посредством электропорации и подвергали второму кроссинговеру. Затем был получен штамм, в котором промотор trpE заменен промотором SPL7 (то есть более сильным промотором) и 38-ая аминокислота trpE (то есть серии) замещена аргинином на хромосоме. Соответствующая генетическая манипуляция была подтверждена с помощью секвенирования генома и метода ПЦР с использованием праймеров SEQ ID NO: 54 и SEQ ID NO: 55, которые могут амплифицировать восходящую область и нисходящую область гомологичной рекомбинации, куда встроен ген, и полученный штамм назвали СА04-8325.
SEQ ID NO: 54 (Confirm_Pspl7-trpE(S38R) - 1)
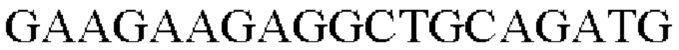
SEQ ID NO: 55 (Confirm_Pspl7-trpE(S38R) - 2)

Продуцирование триптофана происходит через метаболический путь ароматических аминокислот, и этот метаболический путь начинается с реакции конденсации между фосфоенолпируватом и эритрозо-4-фосфатом. Соответственно, бесперебойная поставка этих двух предшественников является существенной для продуцирования триптофана, и сверхэкспрессию гена tkt выполняли для бесперебойной поставки эритрозо-4-фосфата, который, как известно, является относительно дефицитным.
Для вышеуказанной генетической манипуляции выполняли ПЦР с использованием хромосомной ДНК Corynebacterium glutamicum в качестве матрицы в сочетании с праймерами SEQ ID NO: 56 и SEQ ID NO: 57 с получением восходящей области для дополнительной вставки гена tkt, и в сочетании с праймерами SEQ ID NO: 58 и SEQ ID NO: 59 для получения нисходящей области для дополнительной вставки гена tkt.
SEQ ID NO: 56 (Pn-tkt_L - 1)
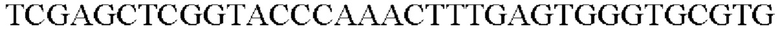
SEQ ID NO: 57 (Pn-tkt_L - 2)
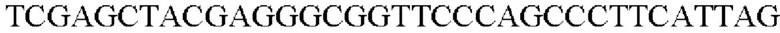
SEQ ID NO: 58 (Pn-tkt_R - 1)
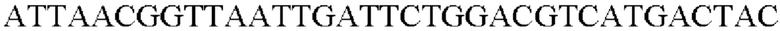
SEQ ID NO: 59 (Pn-tkt_R - 2)
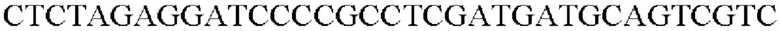
ДНК-полимеразу SolgTM Pfu-X использовали в качестве полимеразы, и ПЦР выполняли следующим образом: денатурация при 95°С в течение 2 минут; 27 циклов денатурации при 95°С в течение 20 секунд, отжиг при 62°С в течение 40 секунд и полимеризация при 72°С в течение 30 секунд; и полимеризация при 72°С в течение 5 минут.
Для получения гена tkt и его промотора ПЦР выполняли с использованием хромосомной ДНК штамма дикого типа Corynebacterium glutamicum АТСС13869 в качестве матрицы, в сочетании с праймерами SEQ ID NO: 60 и SEQ ID NO: 61, и, таким образом, был получен ген tkt, содержащий его промотор.
SEQ ID NO: 60 (Pn-tkt - 1)
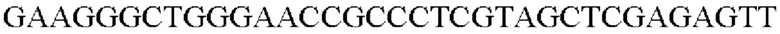
SEQ ID NO: 61 (Pn-tkt - 2)
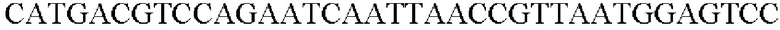
ДНК-полимеразу SolgTM Pfu-X использовали в качестве полимеразы и ПЦР выполняли следующим образом: денатурация при 95°С в течение 2 минут; 27 циклов денатурации при 95°С в течение 20 секунд, отжиг при 62°С в течение 40 секунд, и полимеризация при 72°С в течение 1 минуты и 20 секунд; и полимеризация при 72°С в течение 5 минут.
Рекомбинантную плазмиду получали путем клонирования амплифицированной восходящей области для дополнительной вставки гена tkt и нисходящей области для дополнительной вставки гена tkt, содержащего промотор tkt, и вектора pDZ для хромосомной трансформации, который расщеплен рестрикционным ферментом SmaI, с использованием метода сборки Гибсона, и полученная рекомбинантная плазмида была названа pDZ-Pn-tkt. Клонирование выполняли путем смешивания реагента сборки Гибсона и каждого из генных фрагментов с рассчитанным числом молей с последующей инкубацией при 50°С в течение 1 часа.
Полученный вектор pDZ-Pn-tkt трансформировали в штамм CJ04-8325 посредством электропорации и подвергали вторичному кроссинговеру с получением штамма, в котором ген tkt, содержащий промотор tkt, вставлен в хромосому. Соответствующая генетическая манипуляция была подтверждена с помощью секвенирования генома и метода ПЦР с использованием праймеров SEQ ID NO: 62 и SEQ ID NO: 63, которые могут, соответственно, амплифицировать внешнюю область восходящей области и нисходящей области гомологичной рекомбинации, куда встроен соответствующий ген. Полученный штамм назвали СА04-8352.
SEQ ID NO: 62 (Confirm_Pn-tkt - 1)

SEQ ID NO: 63 (Confirm Pn-tkt - 2)
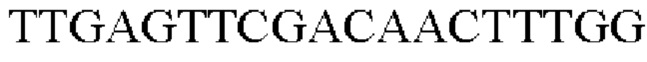
Пример 10: Продуцирование триптофана микроорганизмом рода Corynebacterium, в который введены гены, происходящие из Herbaspirillum rhizosphaerae и Е. coli
Ген, происходящий из Herbaspirillum rhizosphaerae, который показал существенную активность в эксперименте определения минимальной ингибирующей концентрации аналога триптофана в Примере 7, был введен в СА04-8352, который представляет собой триптофан-продуцирующий штамм, полученный в Сравнительном примере 1. Для этой цели вектор pDZTn-PgapA-Hrh для введения гена, происходящего из Herbaspirillum rhizosphaerae, трансформировали в СА04-8352 (то есть триптофан-продуцирующий штамм) посредством электропорации и подвергали такому же процессу, как в Примере 2, и, таким образом, был получен штамм, в котором одна копия гена, происходящего из Herbaspirillum rhizosphaerae, вставлена между генами транспозона. Полученный штамм назвали СА04-8405.
Кроме того, ген, происходящий из Е. coli, вводили в СА04-8352 (то есть триптофан-продуцирующий штамм), в качестве контрольной группы. Вектор pDZTn-PgapA-Eco для введения гена, происходящего из Е. coli, полученный в Примере 6, трансформировали в СА04-8352 (то есть триптофан-продуцирующий штамм) посредством электропорации и подвергали такому же процессу, что и в Примере 6, и, таким образом, был получен штамм, в котором одна копия гена, происходящего из Е. coli, вставлена между генами транспозона. Полученный штамм назвали СА04-8406.
Штаммы СА04-8405 и СА04-8406, полученные посредством вышеописанного процесса, культивировали следующим методом для подтверждения количества произведенного триптофана по сравнению со штаммом СА04-8352, полученным в Сравнительном примере 1, в качестве контрольной группы. Каждый штамм инокулировали в 250 мл колбу с угловыми перегородками, содержащую среду для посева (25 мл) и культивировали при встряхивании при 30°С, при 200 об/мин в течение 20 часов. Затем каждый раствор посевной культуры (1 мл) инокулировали в 250 мл колбу с угловыми перегородками, содержащую среду для продуцирования (25 мл) и культивировали при встряхивании при 30°С, при 200 об/мин в течение 24 часов. По завершении культивирования количество продуцированного L-триптофана измеряли с помощью ВЭЖХ.
Среда для посева (рН 7.0)
20 г глюкозы, 10 г пептона, 5 г дрожжевого экстракте, 1,5 г мочевины, 4 г KH2PO4, 8 г K2HPO4, 0,5 г MgSO4⋅7H2O, 100 мкг биотина, 1000 мкг тиамина HCl, 2000 мкг пантотената кальция, 2000 мкг никотинамида (на 1 л дистиллированной воды). Среда для продуцирования (рН 7.0)
30 г глюкозы, 15 г (NH4)2SO4, 1,2 г MgSO4⋅7H2O, 1 г KH2PO4, 5 г дрожжевого экстракта, 900 мкг биотина, 4500 мкг тиамина HCl, 4500 мкг пантотената кальция, 30 г СаСО3 (на 1 л дистиллированной воды)
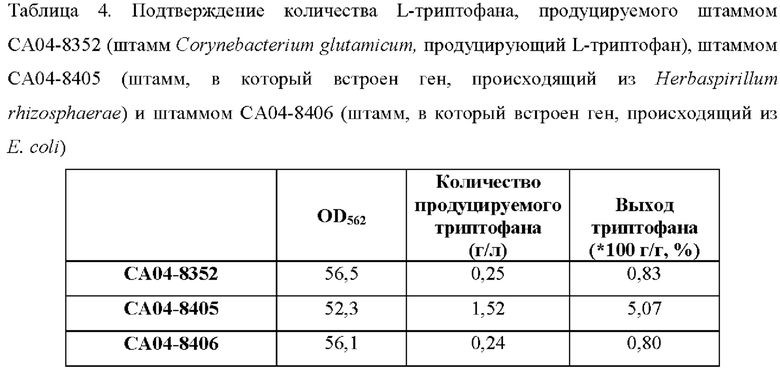
Результаты продуцирования L-триптофана штаммами СА04-8352, СА04-8405 и СА04-8406 в среде показаны в Таблице 4 выше.
Штамм СА04-8405, в который встроен ген, происходящий из Herbaspirillum rhizosphaerae, продуцирует L-триптофан в конечной концентрации 1,52 г/л при культивировании в колбе и это улучшение составляет примерно 5 раз по сравнению со штаммом СА04-8352, контрольной группой. Это указывает на то, что ген, происходящий из Herbaspirillum rhizosphaerae, значительно улучшает продуцирование L-триптофана в штамме Corynebacterium glutamicum. Напротив, штамм СА04-8406, в который введен ген, происходящий из Е. coli, продуцирует L-триптофан в концентрации 0,23 г/л, что почти совпадает с количеством L-триптофана, продуцируемого штаммом СА04-8352 (то есть родительским штаммом для СА04-8406). Как подтверждено в эксперименте с минимальной ингибирующей концентрацией (MIC) для аналога триптофана и для аналога фенилаланина, подтвержденного в Примерах 7 и 9, ген, происходящий из Е. coli, считается экспортерным геном, который проявляет более высокую специфичность к фенилаланину, чем к триптофану.
Штамм СА04-8405 был депонирован на международном уровне в Корейском Центре Культур Микроорганизмов (KCCM), международном органе депонирования, 21 августа 2017 года, согласно Будапештскому договору, с присвоенным регистрационным номером KCCM12099P (СА04-8405).
Пример 11: Анализ внутриклеточных метаболитов триптофана в Corynebacterium glutamicum, в которую введен ген, происходящий из Herbaspirillum rhizosphaerae
Чтобы явно подтвердить, уменьшается ли внутриклеточная концентрация триптофана по мере улучшения триптофан-экспортирующей способности штамма СА04-8405 (т.е. триптофан-продуцирующего штамма), измеряли внутриклеточную концентрацию триптофана для штамма СА04-8405 и его родительского штамма (т.е. СА04-8352) методом экстракции с использованием органического растворителя.
Способ анализа внутриклеточных метаболитов выполняли в соответствии с методом, описанным в ссылке (Nakamura J et al., Appl. Environ. Microbiol. 73(14): 4491-4498, 2007).
Во-первых, что касается модифицированных штаммов Corynebacterium glutamicum СА04-8352 и СА04-8405, то каждый штамм инокулировали в 250 мл колбу с угловыми перегородками, содержащую среду для посева (25 мл) и культивировали при встряхивании при 30°С, 200 об/мин в течение 20 часов. Затем каждый раствор посевной культуры (1 мл) инокулировали в 250 мл колбу с угловыми перегородками, содержащую среду для продуцирования (25 мл), и культивировали при встряхивании при 30°С, 200 об/мин. Внутриклеточную концентрацию триптофана анализировали три раза в соответствии с потреблением глюкозы. Культивируемые клетки на каждой стадии отделяли от культуральной жидкости посредством быстрой вакуумной фильтрации (Durapore HV, 0.45 m; Millipore, Billerica, MA, USA). Фильтр, на который адсорбировались клетки, дважды промывали дистиллированной водой (10 мл) и погружали в метанол, содержащий 5 мкм морфолин-этансульфоновой кислоты и 5 мкм метионинсульфона, на 10 минут. К полученному выше клеточному экстракту (1,6 мл) добавляли хлороформ (1,6 мл) и дистиллированную воду (0,64 мкл) и тщательно перемешивали, и только водную фазу наносили на спин-колонку для удаления белковых примесей. Отфильтрованный экстракт анализировали с помощью капиллярного электрофореза и масс-спектрометрии, и результаты представлены на Фиг. 1.
Как показано на Фиг. 1, было подтверждено, что штамм СА04-8405 демонстрировал уменьшение внутриклеточной концентрации триптофана до уровня 33%-41%, по сравнению с родительским штаммом СА04-8352. Исходя из этого, можно заключить, что, по мере того, как триптофан, продуцируемый в клетках штамма Corynebacterium glutamicum благодаря экспрессии гена, происходящего из Herbaspirillum rhizosphaerae, беспрепятственно экспортировался внеклеточно, внутриклеточная концентрация триптофана для штамма СА04-8405 уменьшалась. Из приведенных выше результатов, было подтверждено, что ген, происходящий из Herbaspirillum rhizosphaerae, представляет собой ген, кодирующий мембранный белок, имеющий экспортирующую способность, специфичную к триптофану.
Сравнительный пример 2: Получение микроорганизма рода Escherichia, продуцирующего L-триптофан
Микроорганизм рода Escherichia, продуцирующий L-триптофан, был получен из штамма дикого типа Е. coli W3110. Для подтверждения того, что количество продукции триптофана значительно увеличивается, когда белок, обладающий L-триптофан-экспортирующей активностью, модифицирован для экспрессии, в качестве родительского штамма использовали штамм, подготовленный для продуцирования L-триптофана. В частности, экспрессия генов биосинтеза L-триптофана (trpEDCBA), которые вовлечены в производство L-триптофана из хоризмата, ингибируется TrpR. Соответственно, ген trpR, кодирующий TrpR, был удален. Кроме того, для высвобождения ингибирования по типу обратной связи полипептидом TrpE в соответствии с улучшением продуцирования L-триптофана, 21-ую аминокислоту с N-конца TrpE (то есть пролин) заменяли серином (J. Biochem. Mol. Biol. 32, 20-24 (1999)).
Мембранный белок Mtr играет важную роль во введении внеклеточного L-триптофана в клетку и белок TnaA играет важную роль разделения внутриклеточного L-триптофана и молекул воды в индол, пируват и аммиак (NH3). Соответственно, были удалены гены mtr и tnaA, которые ингибируют продуцирование L-триптофана и разлагают его.
Для удаления этих генов использовали метод рекомбинации λ-red (One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products, Datsenko KA, Wanner BL., Proc Natl Acad Sci USA. 2000 Jun 6; 97(12): 6640-6645). Для удаления гена mtr, ПЦР выполняли с использованием вектора pKD4 в качестве матрицы в сочетании с праймерами SEQ ID NO: 64 и SEQ ID NO: 65, и, таким образом, был получен генный фрагмент (1580 п.о.), в котором соединены кассета FRT-канамицин-FRT и пара гомологичных нуклеотидов из 50 п.о., фланкирующих ген mtr, где происходит хромосомная гомологичная рекомбинация между ними. Маркерный антибиотик канамицин вектора pKD4 использовали для подтверждения удаления целевого гена и вставки гена антибиотика, а область FRT участвовала в удалении антибиотического маркера после удаления целевого гена. ДНК-полимеразу SolgTM Pfu-X использовали в качестве полимеразы, и ПЦР выполняли следующим образом: денатурация при 95°С в течение 2 минут; 27 циклов денатурации при 95°С в течение 20 секунд, отжиг при 62°С в течение 40 секунд, и полимеризация при 72°С в течение 1 минуты; и полимеризация при 72°С в течение 5 минут.
SEQ ID NO: 64 (Δmtr cassette - 1)
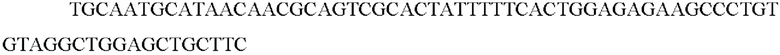
SEQ ID NO: 65 (Δmtr cassette - 2)
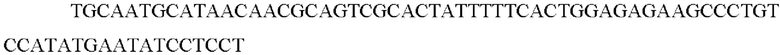
Вектор pKD46, который экспрессирует рекомбиназу λ-red (гены gam, bet и exo), трансформировали в штамм E. coli W3110 посредством электропорации, и каждый штамм наносили на твердую среду LB, содержащую канамицин (50 мг/л). В штамме Е. coli W3110, в котором была подтверждена трансформация вектором pKD46, индуцировали экспрессию рекомбинантного фермента путем добавления 10 мМ L-арабинозы, когда значение OD600 достигало примерно 0,1. Когда значение OD600 достигало примерно 0,6, штаммы получали в виде компетентных клеток и линейный генный фрагмент, полученный в вышеприведенном процессе, в котором соединены кассета FRT-канамицин-FRT и пара гомологичных нуклеотидов из 50 п.о., фланкирующих ген mtr, трансформировали посредством электропорации. Для колоний, выращенных на твердой среде LB, содержащей канамицин (25 мг/л), выполняли ПЦР с использованием праймеров SEQ ID NO: 66 и SEQ ID NO: 67, и были отобраны колонии, в которых был получен генный фрагмент из 782 п.о..
SEQ ID NO: 66 (Confirm_Cassette - 1)
SEQ ID NO: 67 (Confirm_Δmtr - 2)
Штаммы, у которых ген mtr был удален в результате гомологичной рекомбинации, были получены в виде компетентных клеток таким образом, чтобы удалить маркер антибиотика канамицина, и затем трансформированы с помощью вектора рСР20 посредством электропорации. Вектор рСР20 экспрессирует белок FLP и таким образом распознает сайты FRT, фланкирующие антибиотик канамицин, и связывается с ними на хромосоме, тем самым удаляя маркер антибиотика между сайтами FRT. Штамм, трансформированный вектором рСР20, выросший на твердой среде LB, содержащей ампициллин (100 мг/л) и хлорамфеникол (25 мг/л), культивировали в жидкой среде LB при 30°С в течение 1 часа, затем культивировали при 42°С в течение 15 часов и наносили на твердую среду LB. Выросшие колонии культивировали на твердой среде LB, содержащей ампициллин (100 мг/л) и хлорамфеникол (25 мг/л); твердой среде LB, содержащей канамицин (12,5 мг/л); и твердой среде, не содержащей антибиотик. Отбирались только те колонии, которые росли на твердой среде LB, не содержащей антибиотик. Удаление гена mtr было окончательно подтверждено секвенированием генома, и штамм был назван СА04-9300.
Генетическую манипуляцию выполняли вышеописанным способом для того, чтобы удалить ген tnaA. ПЦР выполняли с использованием вектора pKD4 в качестве матрицы в сочетании с праймерами SEQ ID NO: 68 и SEQ ID NO: 69, и таким образом получали генный фрагмент (1580 п.о.), в котором соединены кассета FRT-канамицин-FRT и пара гомологичных нуклеотидов из 50 п.о., фланкирующих ген tnaA, где происходит гомологичная рекомбинация. ДНК-полимеразу SolgTM Pfu-X использовали в качестве полимеразы и ПЦР выполняли следующим образом: денатурация при 95°С в течение 2 минут; 27 циклов денатурации при 95°С в течение 20 секунд, отжиг при 62°С в течение 40 секунд и полимеризация при 72°С в течение 1 минуты; и полимеризация при 72°С в течение 5 минут.
SEQ ID NO: 68 (ΔtnaA cassette - 1)
SEQ ID NO: 69 (ΔtnaA cassette - 2)
Плазмиду pKD46 трансформировали в СА04-9300. Штамм СА04-9300, в котором экспрессировались рекомбиназы посредством добавления 10 мМ L-арабинозы, трансформировали путем электропорации линейным генным фрагментом, полученным в вышеуказанном процессе, в котором соединены кассета FRT-канамицин-FRT и пара гомологичных нуклеотидов из 50 п.о., фланкирующих ген tnaA. Для роста колоний на твердой среде LB, содержащей канамицин (25 мг/л), выполняли ПЦР колоний с использованием праймеров SEQ ID NO: 66 и SEQ ID NO: 70 и отбирали колонии, в которых получен генный фрагмент из 787 п.о.
SEQ ID NO: 70 (Confirm_ΔtnaA - 2)
Штаммы, у которых ген tnaA был удален в результате гомологичной рекомбинации, были получены в виде компетентных клеток таким образом, чтобы удалить маркер антибиотика канамицина, и затем трансформированы вектором рСР20, и был получен штамм, у которого маркер антибиотика канамицина удален, посредством экспрессии белка FLP. Удаление гена tnaA было окончательно подтверждено секвенированием генома и штамм был назван СА04-9301.
Для удаления гена trpR ПЦР выполняли с использованием вектора pKD4 в качестве матрицы в сочетании с праймерами SEQ ID NO: 71 и SEQ ID NO: 72, и таким образом получали генный фрагмент (1580 п.о.), в котором соединены кассета FRT-канамицин-FRT и пара гомологичных нуклеотидов из 50 п.о., фланкирующих ген trpR, где происходит гомологичная рекомбинация между ними. ДНК-полимеразу SolgTM Pfu-X использовали в качестве полимеразы, и ПЦР выполняли следующим образом: денатурация при 95°С в течение 2 минут; 27 циклов денатурации при 95°С в течение 20 секунд, отжиг при 62°С в течение 40 секунд, и полимеризация при 72°С в течение 1 минуты; и полимеризация при 72°С в течение 5 минут.
SEQ ID NO: 71 (ΔtrpR cassette - 1)
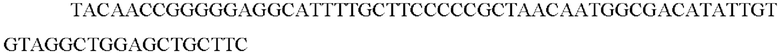
SEQ ID NO: 72 (ΔtrpR cassette - 2)
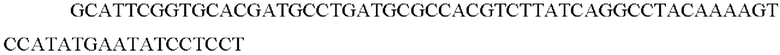
Плазмиду pKD46 трансформировали в CA04-9301. Штамм CA04-9301, в котором рекомбиназы экспрессировались посредством добавления 10 мМ L-арабинозы, трансформировали посредством электропорации с линейным фрагментом гена, полученным в вышеприведенным процессе, в котором соединены кассета FRT-канамицин-FRT и пара гомологичных нуклеотидов из 50 п.о., фланкирующих ген trpR. Колоний, выращенных на твердой среде LB, содержащей канамицин (25 мг/л), выполняли ПЦР колоний, используя праймеры SEQ ID NO: 66 и SEQ ID NO: 73, и были отобраны колонии, где получен генный фрагмент из 838 п.о.
SEQ ID NO: 73 (Confirm_ΔtrpR - 2)
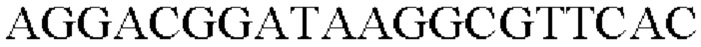
Штаммы, в которых ген trpR был удален в результате гомологичной рекомбинации, получили в виде компетентных клеток, таким образом, чтобы удалить маркер антибиотика канамицина, и затем трансформировали вектором рСР20, и был получен штамм, где маркер антибиотика канамицина был удален посредством экспрессии белка FLP. Удаление гена trpR было окончательно подтверждено секвенированием генома, и штамм был назван СА04-9307.
Чтобы обеспечить штамм СА04-9307 свойством резистентности trpE к обратной связи, ПЦР выполняли с использованием Е. coli W3110 gDNA в качестве матрицы в сочетании с праймерами SEQ ID NO: 74 и SEQ ID NO: 75, которые содержали сайт рестрикции EcoRI и, таким образом, был получен генный фрагмент trpE, содержащий сайт рестрикции EcoRI (1575 п.о.). ДНК-полимеразу SolgTM Pfu-X использовали в качестве полимеразы, и ПЦР выполняли следующим образом: денатурация при 95°С в течение 2 минут; 27 циклов денатурации при 95°С в течение 20 секунд, отжиг при 62°С в течение 1 минуты и полимеризация при 72°С в течение 1 минуты; и полимеризация при 72°С в течение 5 минут.
SEQ ID NO: 74 (trpE - 1)
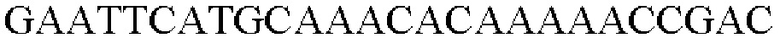
SEQ ID NO: 75 (trpE - 2)
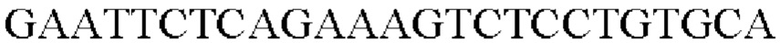
Клонирование выполняли после обработки генного фрагмента trpE, полученного вышеприведенным способом, и плазмиды pSG76-C (Journal of Bacteriology, July 1997, p. 4426-4428) рестриктазой EcoRI соответственно. Клонированную плазмиду трансформировали в Е. coli DH5α посредством электропорации и трансформированные штаммы Е. coli DH5α отбирали на LB чашках, содержащих хлорамфеникол (25 мкг/мл); таким образом была получена плазмида pSG76-C-trpE.
pSG76-C-trpE(P21S) получали с использованием полученной плазмиды pSG76-C-trpE в сочетании с праймерами SEQ ID NO: 76 и SEQ ID NO: 77 посредством сайт-направленного мутагенеза (Stratagene, USA).
SEQ ID NO: 76 (trpE(P21S) - 1)
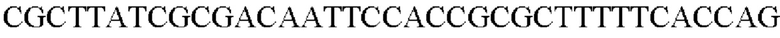
SEQ ID NO: 77 (trpE(P21S) - 2)
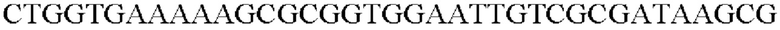
Плазмиду pSG76-C-trpE(P21S) трансформировали в штамм CA04-9307, культивировали в среде LB-Cm (10 г/л дрожжевого экстракта, 5 г/л NaCl, 10 г/л триптона и 25 мкг/л хлорамфеникол а), и отбирали колонии, имеющие резистентность к хлорамфениколу. Отобранные трансформанты представляют собой штаммы, в которых плазмида pSG76-C-trpE(P21S) встроена в область trpE генома посредством первой вставки. Штамм, в который введен полученный ген trpE(P21S), трансформировали плазмидой pAScep (Journal of Bacteriology, July 1997, p. 4426-4428), которая экспрессирует рестриктазу I-SceI, расщепляющую область I-SceI, присутствующую в плазмиде pSG76-C, и отбирали штамм, который рос в LB-Ap (10 г/л дрожжевого экстракта, 5 г/л NaCl, 10 г/л триптона и 100 мкг/л ампициллина). Ген trpE в отобранном штамме амплифицировали с использованием праймеров SEQ ID NO: 74 и SEQ ID NO: 75, и посредством секвенирования было подтверждено, что амплифицированный ген trpE был заменен геном trpE(P21S). Полученный таким образом штамм был назван СА04-4303.
Пример 12: Продуцирование L-триптофана микроорганизмом рода Escherichia, в который введен ген, происходящий из Herbaspirillum rhizosphaerae
pCL1920-PyccA-Hrh, полученный в Примере 8, вводили в штамм СА04-4303, полученный в Сравнительном примере 2, и таким образом получали штамм СА04-4306, в котором сверхэкспрессируется ген, происходящий из Herbaspirillum rhizosphaerae. Кроме того, вектор pCL1920, в качестве контрольной группы, трансформировали в штамм СА04-4303. Для анализа количества продуцированного L-триптофана в двух штаммах (то есть CA04-4303/pCL1920 и СА04-4306), эти штаммы культивировали в жидкой среде LB, содержащей спектиномицин (50 мг/л) в течение 12 часов. Затем каждый из этих штаммов инокулировали в 250 мл колбу с угловыми перегородками, содержащую 25 мл среды для продуцирования, так что исходное значение OD600 составило 0,01 и затем культивировали при встряхивании при 37°С при 200 об/мин в течение 48 часов. По завершении культивирования количество продуцированного L-триптофана измеряли с помощью ВЭЖХ.
Результаты в отношении продуцирования L-триптофана в штаммах са04-4303/pcl1920 и са04-4306 в среде приведены в Таблице 5 ниже. Штамм СА04-4306, в который был введен и сверхэкспрессировался ген, происходящий из Herbaspirillum rhizosphaerae, продемонстрировал конечное количество L-триптофана 2,1 г/л при культивировании в колбе, что примерно на 50% выше, чем в контрольной группе. Это указывает на то, что ген, происходящий из Herbaspirillum rhizosphaerae, экспортирует L-триптофан даже в Е. coli, тем самым значительно улучшая продуцирование L-триптофана.
Среда для продуцирования (рН 7.0)
70 г глюкозы, 20 г (NH4)2SO4, 1 г MgSO4⋅7H2O, 2 г KH2PO4, 2,5 г дрожжевого экстракта, 5 г цитрата Na, 1 г NaCl, 40 г СаСО3 (на 1 л дистиллированной воды)
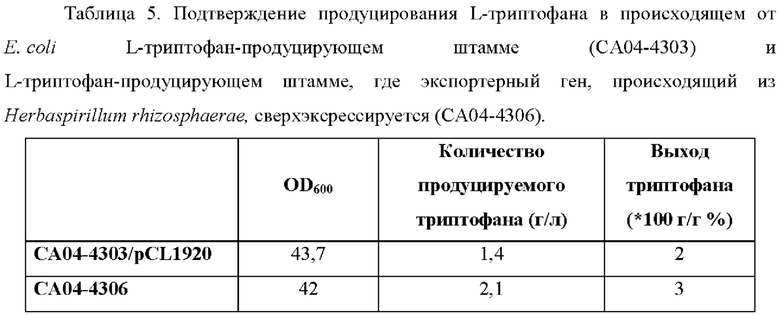
Соответственно, как видно из результатов Примеров 7 и 9, ген, происходящий из Herbaspirillum rhizosphaerae, продемонстрировал высокую специфичность и превосходную резистентность к L-триптофану и его аналогу, в то время как, это видно из результатов Примеров 10 и 12, ген, происходящий из Herbaspirillum rhizosphaerae улучшал продуцирование L-триптофана в обоих штаммах Corynebacterium glutamicum и Е. coli. Кроме того, в Примере 11 наблюдалось, что ген, происходящий из Herbaspirillum rhizosphaerae, по существу экспортировал триптофана внеклеточно. В результате, ген, происходящий из Herbaspirillum rhizosphaerae, был назван wex (триптофан (W) экспортер).
Исходя из вышеизложенного, специалист в области, к которой относится настоящее изобретение, должен понимать, что настоящее изобретение может быть воплощено в других конкретных формах без изменения технических концепций или существенных характеристик настоящего изобретения. В связи с этим, типичные воплощения, раскрытые здесь, предназначены только для иллюстративных целей и их не следует истолковывать как ограничивающие объем настоящего изобретения. Напротив, предполагается, что настоящее изобретение охватывает не только типичные воплощения но также различные альтернативы, модификации, эквиваленты и другие воплощения, которые могут быть включены в сущность и объем настоящего изобретения, как это определено в прилагаемой формуле изобретения.
--->
<110> CJ CheilJedang Corporation
<120> Novel L-tryptophan exporter and the method of producing
L-tryptophan using the same
<130> OPA18282
<150> KR 10-2018-0022054
<151> 2018-02-23
<160> 77
<170> KoPatentIn 3.0
<210> 1
<211> 307
<212> PRT
<213> Artificial Sequence
<220>
<223> wex AA последовательность.
<400> 1
Met Asn Ser Lys Lys Ala Thr Leu Ile Gly Leu Thr Ala Val Val Leu
1 5 10 15
Trp Ser Ser Ile Val Gly Leu Ile Arg Gly Val Ser Glu His Leu Gly
20 25 30
Ala Thr Gly Gly Ala Ala Met Met Tyr Ser Val Ala Ser Leu Phe Leu
35 40 45
Leu Leu Ser Val Gly Phe Pro Lys Leu Gly Ser Phe Pro Lys Lys Tyr
50 55 60
Leu Leu Trp Gly Ser Leu Leu Phe Val Ser Tyr Glu Leu Cys Leu Ser
65 70 75 80
Leu Ser Ile Gly Tyr Ala Asn Thr Gly Arg Gln Ala Ile Glu Val Ser
85 90 95
Met Val Asn Tyr Leu Trp Pro Ala Phe Thr Leu Ile Ala Ala Ile Ala
100 105 110
Phe Asn Arg Gln Arg Ala Asn Trp Met Val Val Pro Gly Phe Ile Leu
115 120 125
Ser Ile Ile Gly Ile Cys Trp Val Leu Gly Gly Asp Gln Gly Leu Asp
130 135 140
Leu Ala Gly Met Leu Gly Asn Val Gln Asp Asn Pro Leu Ser Tyr Gly
145 150 155 160
Leu Ala Phe Leu Gly Ala Val Ile Trp Ala Ala Tyr Cys Thr Val Thr
165 170 175
Ala Arg Leu Ala Lys Gly Lys Asn Gly Val Thr Leu Phe Phe Ile Leu
180 185 190
Val Ala Leu Thr Leu Trp Val Lys Phe Phe Phe Gly Asp His Arg Pro
195 200 205
Met Ser Phe Ser Leu Pro Ala Ile Val Tyr Leu Leu Leu Ala Ala Ala
210 215 220
Ala Met Gly Phe Gly Tyr Ala Ala Trp Asn Val Gly Ile Leu His Gly
225 230 235 240
Asn Val Thr Val Leu Ala Gly Val Ser Tyr Phe Ile Pro Val Phe Ser
245 250 255
Ala Ala Leu Ser Ala Met Val Leu His Ala Pro Leu Pro Arg Ser Phe
260 265 270
Trp Val Gly Ala Ser Leu Val Cys Ala Gly Ser Ile Leu Cys Trp Leu
275 280 285
Ala Thr Arg Ala Arg Arg Ala Ser Ala Ala Gln Glu Asp Ala Val Ala
290 295 300
Asp Cys Leu
305
<210> 2
<211> 924
<212> DNA
<213> Artificial Sequence
<220>
<223> последовательность wex NT
<400> 2
atgaatagca agaaggccac gctgatcgga cttactgcag tggtcctctg gagttccatt 60
gtcggattga ttcgcggcgt cagcgaacat ctcggcgcta ccgggggcgc ggcgatgatg 120
tatagcgttg cctcgctgtt tctgctgttg tcggtcggtt tcccgaaact gggttccttt 180
ccgaaaaaat acctcttgtg gggcagcctg ctgtttgtgt cctacgaact ctgcctgtcg 240
ctctccatcg gttatgccaa tacaggcagg caagcaatcg aagtcagcat ggtcaactat 300
ctgtggccgg cattcacgct catcgccgcc attgcattca accggcagag agcgaactgg 360
atggtggtgc ccggattcat cctctcgatt atcggtatct gctgggtgct gggcggtgac 420
caggggctgg acctggcggg catgcttggc aacgtgcagg acaatccgct cagttatggg 480
ctggcttttt tgggcgccgt gatctgggcc gcctattgca ctgtgacggc ccgcctcgcg 540
aaggggaaga acggagtgac gctgttcttc attcttgtgg cattgacgct ctgggtgaag 600
tttttcttcg gcgatcaccg cccgatgtct ttcagcctgc cggcaattgt ctacctgctc 660
ctggcggcgg ccgcgatggg cttcggctat gcggcatgga atgtcgggat cttgcacggt 720
aacgtgaccg tgctggcggg cgtgtcgtac tttatcccgg ttttttcggc ggcgttgtcc 780
gcgatggtgt tgcatgcgcc gttgccgcga tcgttttggg tgggggcgtc gctggtatgc 840
gccggttcga tactgtgctg gctggcaacc agggccaggc gcgcttcggc cgcgcaagaa 900
gatgcggtgg cggactgttt gtag 924
<210> 3
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> wex - 1
<400> 3
tagaggagac acaacatgaa tagcaagaag gccac 35
<210> 4
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> wex - 2
<400> 4
ggctcttcct gtttagtcta caaacagtcc gccac 35
<210> 5
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> PgapA(-wex) - 1
<400> 5
cccttccggt ttagtttgaa gccagtgtga gttgc 35
<210> 6
<211> 39
<212> DNA
<213> Artificial Sequence
<220>
<223> PgapA(-wex) - 2
<400> 6
cttcttgcta ttcatgttgt gtctcctcta aagattgta 39
<210> 7
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Confirm_PgapA-wex - 1
<400> 7
cggattatgc caatgatgtg 20
<210> 8
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Confirm_PgapA-wex - 2
<400> 8
cacgatcacc aacattcagg 20
<210> 9
<211> 314
<212> PRT
<213> Artificial Sequence
<220>
<223> последовательность Pst AA
<400> 9
Met Lys Asn Gln Arg Lys Ala Thr Leu Ile Gly Leu Val Ala Ile Val
1 5 10 15
Leu Trp Ser Ser Ile Val Gly Leu Ile Arg Gly Val Ser Glu Ser Leu
20 25 30
Gly Ala Thr Gly Gly Ala Val Met Met Tyr Ser Val Ala Ser Val Met
35 40 45
Leu Leu Phe Thr Val Gly Phe Pro Arg Ile Arg Glu Phe Pro Arg Arg
50 55 60
Tyr Leu Val Trp Gly Ser Leu Leu Phe Val Ser Tyr Glu Leu Cys Leu
65 70 75 80
Ala Leu Ser Ile Gly Tyr Ala Asn Ser Ser Arg Gln Ala Ile Glu Val
85 90 95
Gly Met Val Asn Tyr Leu Trp Pro Ala Phe Thr Ile Leu Ala Ala Ile
100 105 110
Leu Phe Asn Arg Gln Gln Ala Asn Leu Leu Ile Val Pro Gly Phe Leu
115 120 125
Ile Ala Ile Leu Gly Ile Cys Trp Val Leu Gly Gly Glu Gln Gly Leu
130 135 140
Asp Leu Ser Gly Met Thr Ala Asn Ile Arg Asp Asn Pro Leu Ser Tyr
145 150 155 160
Gly Leu Ala Phe Ala Gly Ala Val Ile Trp Ala Ala Tyr Cys Thr Val
165 170 175
Thr Thr Arg Ile Ala Gly Gly Lys Asn Gly Val Thr Leu Phe Phe Met
180 185 190
Leu Thr Ala Leu Ala Leu Trp Ala Lys Tyr Leu Ala Ile Gly Gly Glu
195 200 205
Thr Met Glu Phe Ser Tyr His Ala Leu Ile Tyr Leu Val Leu Ala Ala
210 215 220
Ser Ala Met Gly Phe Gly Tyr Ala Ala Trp Asn Val Gly Ile Leu His
225 230 235 240
Gly Asn Val Thr Val Leu Ala Gly Ala Ser Tyr Phe Ile Pro Val Leu
245 250 255
Ser Ala Ala Leu Ala Ala Val Leu Leu Arg Thr Pro Leu Ser Leu Ser
260 265 270
Phe Trp Gln Gly Ala Ala Met Val Cys Ile Gly Ser Ile Leu Cys Trp
275 280 285
Phe Ala Thr Arg Ala Lys Pro Pro Glu Ser Ala Gln Ser Gly Asp Gln
290 295 300
Ala Ser Ala Thr Thr Pro Arg Arg Asn Gly
305 310
<210> 10
<211> 945
<212> DNA
<213> Artificial Sequence
<220>
<223> последовательность Pst NT
<400> 10
atgaaaaacc agcgtaaagc gaccctcatc gggctcgttg caattgtctt gtggagctcg 60
atcgtcggcc tgatccgggg cgtcagcgag agcctcggcg cgaccggtgg tgccgtcatg 120
atgtacagcg tcgcatcggt aatgctgttg ttcacggtcg gctttccgcg gatacgggag 180
ttcccccgac gctatcttgt ctggggcagc ctgctgttcg tctcgtacga gctgtgcctt 240
gccctgtcca tcggctacgc caacagcagc cgacaggcca tcgaggtcgg catggtcaat 300
tacctgtggc cggccttcac gatcctggcg gcgatcctgt tcaacaggca gcaggccaac 360
ctgctcatcg ttcccggctt cctcatcgcg atcctcggga tctgctgggt gctcggcggg 420
gaacaggggc tggacctgtc cgggatgacg gcgaacatcc gcgacaatcc cctcagctac 480
gggctggcct tcgccggcgc ggtgatctgg gcggcatact gcacggtgac cacgcggatc 540
gccggcggca agaacggtgt cacgctgttc ttcatgctga cggcattggc gctatgggcc 600
aagtacctgg ccatcggcgg ggaaacgatg gaattcagct accacgcgct gatctacctg 660
gtgctggccg cctccgcgat gggcttcggc tatgcggcgt ggaacgtcgg catcctgcac 720
ggcaatgtca ccgtcctcgc tggtgcttcg tatttcatcc cggtgctgtc cgccgccctg 780
gcggccgtac tgttgcgtac gccgctgtcg ctgtcgttct ggcagggtgc cgccatggtc 840
tgcatcgggt cgatcctctg ctggttcgcc acccgtgcga aaccgccaga atcggcgcag 900
tcgggtgacc aggccagcgc aaccacgccg cgtcgaaacg gataa 945
<210> 11
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Pst-1
<400> 11
tagaggagac acaacatgaa aaaccagcgt aaagc 35
<210> 12
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Pst-2
<400> 12
ggctcttcct gtttagttta tccgtttcga cgcgg 35
<210> 13
<211> 39
<212> DNA
<213> Artificial Sequence
<220>
<223> PgapA(-Pst)-2
<400> 13
acgctggttt ttcatgttgt gtctcctcta aagattgta 39
<210> 14
<211> 303
<212> PRT
<213> Artificial Sequence
<220>
<223> последовательность Afa AA
<400> 14
Met Lys Gln Ser Asp Lys Ala Thr Leu Ile Gly Leu Ile Ala Ile Val
1 5 10 15
Leu Trp Ser Thr Ile Val Gly Leu Ile Arg Ser Val Ser Asp Ser Leu
20 25 30
Gly Val Thr Gly Gly Ala Ala Leu Ile Tyr Thr Leu Ala Ser Val Phe
35 40 45
Leu Leu Leu Ser Val Gly Trp Val Arg Leu Arg Asp Phe Pro Arg Arg
50 55 60
Tyr Leu Ile Trp Gly Ser Val Leu Phe Val Cys Tyr Glu Leu Cys Leu
65 70 75 80
Ala Leu Ser Ile Gly Tyr Ala His Asn Ser Gln Gln Ala Ile Glu Val
85 90 95
Gly Met Val Asn Tyr Leu Trp Pro Thr Phe Thr Ile Val Ala Ala Ile
100 105 110
Leu Phe Asn Lys Gln Lys Ala Asn Gly Leu Leu Ala Pro Gly Leu Leu
115 120 125
Leu Ser Met Met Gly Ile Ser Trp Ile Leu Gly Gly Glu Gln Gly Leu
130 135 140
Ser Leu His Asn Ile Trp Leu Asn Val Gln Asp Asn Pro Leu Ser Tyr
145 150 155 160
Gly Leu Ala Phe Ser Gly Ala Leu Ile Trp Ala Gly Tyr Ser Thr Met
165 170 175
Thr Ala Arg Ile Ala Gln Gly Lys Asn Gly Ile Thr Leu Phe Phe Met
180 185 190
Leu Thr Ala Ala Ala Leu Trp Val Lys Tyr Leu Val Gln Gly Ala Pro
195 200 205
Ala Met Thr Phe Thr Val Pro Ala Leu Val Tyr Leu Leu Leu Ala Ala
210 215 220
Met Ala Met Gly Phe Gly Tyr Ala Ala Trp Asn Val Gly Ile Leu His
225 230 235 240
Gly Asn Val Thr Ile Leu Ala Gly Ala Ser Tyr Phe Ile Pro Val Phe
245 250 255
Ser Ala Ala Leu Ser Thr Val Leu Leu Gln Ala Pro Leu Thr Leu Thr
260 265 270
Phe Trp Gln Gly Ser Ser Met Val Cys Leu Gly Ala Leu Leu Cys Trp
275 280 285
Leu Ala Ile Arg Val Arg Lys Pro Arg Ser Leu Lys Ser Ala Ala
290 295 300
<210> 15
<211> 912
<212> DNA
<213> Artificial Sequence
<220>
<223> последовательность Afa NT
<400> 15
atgaagcaat ctgataaggc aaccctgatc gggctgatcg ccattgtcct ttggagcacg 60
attgtcggcc tgatacgcag cgtcagcgac tctctgggcg taaccggcgg cgctgccctg 120
atttacaccc tggcctcggt ctttcttctt ttatcagtgg gctgggtacg cttgcgcgac 180
ttcccgcgtc gctacctgat ctggggcagt gtgctgtttg tctgctatga actctgcctg 240
gccctgtcca tcggctatgc ccacaacagc cagcaggcaa ttgaagtggg gatggtcaac 300
tatctgtggc cgacctttac cattgtggcc gccatcttgt tcaataagca aaaagccaat 360
gggctgcttg cacccggcct gctcttgtcc atgatgggaa tcagctggat tctgggcggc 420
gagcaaggct tgagcctgca caacatctgg ctgaatgtgc aggacaatcc cttgagctac 480
ggcctggcct ttagcggcgc gctgatctgg gccggctaca gcaccatgac cgcccgcatc 540
gcccagggca aaaatggcat caccctgttt ttcatgctga cggcagcggc cttgtgggtg 600
aagtacctgg tccaaggtgc tcctgccatg acgtttacgg ttcccgcctt ggtgtatttg 660
ctgctggcgg ccatggcgat gggctttggc tatgccgcct ggaatgtcgg tattttgcat 720
ggcaatgtca ccatcctggc cggcgcttcc tactttattc cggtattttc agccgccctg 780
tccaccgttt tgctgcaagc tccgttgacg ctgaccttct ggcaaggctc gtccatggtg 840
tgtttgggtg ccctgctatg ctggctggcc atccgggttc gcaaaccccg gtcactaaaa 900
agcgctgcct ga 912
<210> 16
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Afa-1
<400> 16
tagaggagac acaacatgaa gcaatctgat aaggc 35
<210> 17
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Afa-2
<400> 17
gctcttcctg tttagttcag gcagcgcttt ttagt 35
<210> 18
<211> 39
<212> DNA
<213> Artificial Sequence
<220>
<223> PgapA(-Afa)-2
<400> 18
atcagattgc ttcatgttgt gtctcctcta aagattgta 39
<210> 19
<211> 314
<212> PRT
<213> Artificial Sequence
<220>
<223> последовательность Cne AA
<400> 19
Met Gln Ser Lys Ser Lys Ala Thr Leu Ile Gly Leu Ile Ala Ile Leu
1 5 10 15
Leu Trp Ser Ser Ile Val Gly Leu Ile Arg Gly Val Ser Glu Asn Leu
20 25 30
Gly Ala Thr Gly Gly Ala Ala Met Ile Tyr Thr Val Ala Ser Ala Leu
35 40 45
Leu Leu Leu Thr Val Gly Phe Val Arg Met Gln Asp Phe Pro Arg Arg
50 55 60
Tyr Leu Val Trp Gly Ser Ile Leu Phe Val Ser Tyr Glu Leu Cys Leu
65 70 75 80
Ser Leu Ser Ile Gly Tyr Ala Asn Ser Ser Arg Gln Ala Ile Glu Val
85 90 95
Gly Met Val Asn Tyr Leu Trp Pro Ser Phe Thr Met Leu Cys Ala Ile
100 105 110
Ala Phe Asn Lys Gln Lys Ala Asn Leu Leu Ile Ile Pro Gly Phe Leu
115 120 125
Ile Ala Ile Leu Gly Ile Cys Trp Val Leu Gly Gly Asp Gln Gly Leu
130 135 140
Asp Phe Ala Gly Met Ala Glu Asn Ile Gln Asp Asn Pro Leu Ser Tyr
145 150 155 160
Gly Leu Ala Phe Leu Gly Ala Leu Ile Trp Ala Ala Tyr Cys Thr Val
165 170 175
Thr Asn Arg Ile Ala Glu Gly Arg Asn Gly Ile Thr Leu Phe Phe Met
180 185 190
Leu Thr Ala Leu Ala Leu Trp Ile Lys Tyr Phe Ala Thr Glu Ser Gly
195 200 205
Ser Met Glu Phe Ser Tyr Gln Ala Val Ile Tyr Leu Ala Leu Ala Ala
210 215 220
Ser Ala Met Gly Phe Gly Tyr Ala Ala Trp Asn Val Gly Ile Leu His
225 230 235 240
Gly Asn Val Thr Val Leu Ala Gly Ala Ser Tyr Phe Ile Pro Val Leu
245 250 255
Ser Ala Ala Leu Ala Ala Met Leu Leu Arg Thr Pro Leu Ser Ile Ala
260 265 270
Phe Trp Lys Gly Ala Ser Met Val Cys Ala Gly Ser Ile Leu Cys Trp
275 280 285
Leu Ala Thr Arg Gly Gln Arg Ser Lys Ala Pro Pro Leu Pro Glu Leu
290 295 300
Pro Gln Ser Arg Glu Arg Val Gln Glu Pro
305 310
<210> 20
<211> 945
<212> DNA
<213> Artificial Sequence
<220>
<223> последовательность Cne NT
<400> 20
atgcaaagca agagcaaagc aactctcatc gggctcatcg cgattctgtt atggagctcg 60
attgtcggcc tgattcgcgg tgtcagcgaa aaccttgggg caaccggtgg ggcggcaatg 120
atctataccg tcgcctcggc cctgctcttg ctgacagtcg gtttcgtcag aatgcaggat 180
tttccccggc gctatctggt ttggggaagc attctgttcg tttcgtacga gctgtgtctt 240
tccttgtcca ttggctacgc caacagcagc aggcaagcca ttgaggtggg gatggtcaac 300
tacttgtggc cgagcttcac catgctgtgt gccatcgcat tcaacaagca gaaggccaac 360
ttgctgatca ttcccggctt cctgatcgcc attctcggga tctgctgggt gcttggcggg 420
gatcagggcc tggacttcgc cgggatggcg gagaacatcc aggacaatcc gctcagctat 480
gggctggcct tccttggtgc cctgatctgg gcggcgtatt gcactgtgac caaccggatt 540
gccgaaggca ggaatggcat cacgctgttc ttcatgctga cagcgctggc gttgtggatc 600
aagtatttcg ccacagagag cgggtcgatg gaatttagct atcaggcagt gatttatctt 660
gcgttggccg cctctgcgat gggattcggc tatgcggcct ggaatgttgg catcctgcat 720
ggcaatgtca ccgtccttgc cggcgcttcc tacttcattc cggtactttc cgccgccctg 780
gcggccatgc tcttgcgtac acccctgtcg atcgccttct ggaagggcgc atccatggta 840
tgtgcggggt cgatcctctg ttggctggca acacgtgggc aacgttccaa ggcacctccg 900
ttgccggaat taccgcagtc gcgcgaacgt gtccaggaac cgtga 945
<210> 21
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Cne-1
<400> 21
tagaggagac acaacatgca aagcaagagc aaagc 35
<210> 22
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Cne-2
<400> 22
ggctcttcct gtttagttca cggttcctgg acacg 35
<210> 23
<211> 39
<212> DNA
<213> Artificial Sequence
<220>
<223> PgapA(-Cne)-2
<400> 23
gctcttgctt tgcatgttgt gtctcctcta aagattgta 39
<210> 24
<211> 293
<212> PRT
<213> Artificial Sequence
<220>
<223> последовательность Eco AA
<400> 24
Met Thr Arg Gln Lys Ala Thr Leu Ile Gly Leu Ile Ala Ile Val Leu
1 5 10 15
Trp Ser Thr Met Val Gly Leu Ile Arg Gly Val Ser Glu Gly Leu Gly
20 25 30
Pro Val Gly Gly Ala Ala Ala Ile Tyr Ser Leu Ser Gly Leu Leu Leu
35 40 45
Ile Phe Thr Val Gly Phe Pro Arg Ile Arg Gln Ile Pro Lys Gly Tyr
50 55 60
Leu Leu Ala Gly Ser Leu Leu Phe Val Ser Tyr Glu Ile Cys Leu Ala
65 70 75 80
Leu Ser Leu Gly Tyr Ala Ala Thr His His Gln Ala Ile Glu Val Gly
85 90 95
Met Val Asn Tyr Leu Trp Pro Ser Leu Thr Ile Leu Phe Ala Ile Leu
100 105 110
Phe Asn Gly Gln Lys Thr Asn Trp Leu Ile Val Pro Gly Leu Leu Leu
115 120 125
Ala Leu Val Gly Val Cys Trp Val Leu Gly Gly Asp Asn Gly Leu His
130 135 140
Tyr Asp Glu Ile Ile Asn Asn Ile Thr Thr Ser Pro Leu Ser Tyr Phe
145 150 155 160
Leu Ala Phe Ile Gly Ala Phe Ile Trp Ala Ala Tyr Cys Thr Val Thr
165 170 175
Asn Lys Tyr Ala Arg Gly Phe Asn Gly Ile Thr Val Phe Val Leu Leu
180 185 190
Thr Gly Ala Ser Leu Trp Val Tyr Tyr Phe Leu Thr Pro Gln Pro Glu
195 200 205
Met Ile Phe Ser Thr Pro Val Met Ile Lys Leu Ile Ser Ala Ala Phe
210 215 220
Thr Leu Gly Phe Ala Tyr Ala Ala Trp Asn Val Gly Ile Leu His Gly
225 230 235 240
Asn Val Thr Ile Met Ala Val Gly Ser Tyr Phe Thr Pro Val Leu Ser
245 250 255
Ser Ala Leu Ala Ala Val Leu Leu Ser Ala Pro Leu Ser Phe Ser Phe
260 265 270
Trp Gln Gly Ala Leu Met Val Cys Gly Gly Ser Leu Leu Cys Trp Leu
275 280 285
Ala Thr Arg Arg Gly
290
<210> 25
<211> 882
<212> DNA
<213> Artificial Sequence
<220>
<223> последовательность Eco NT
<400> 25
atgacacgac aaaaagcaac gctcataggg ctgatagcga tcgtcctgtg gagcacgatg 60
gtaggattga ttcgcggtgt cagtgagggg ctcggcccgg tcggcggcgc agctgctatc 120
tattcattaa gcgggctgct gttaatcttc acggttggat ttccgcgtat tcggcaaatc 180
ccgaaaggct atttactcgc cgggagtctg ttattcgtca gctatgaaat ctgtctggcg 240
ctttccttag ggtatgcggc gacccatcat caggcgattg aagtgggtat ggtgaactat 300
ctgtggccca gcctgacaat tctctttgcc attctgttta atggtcagaa aaccaactgg 360
ttgattgtac ctggattatt attagccctc gtcggcgtct gttgggtgtt aggcggtgac 420
aatgggttac attatgatga aatcatcaat aatatcacca ccagcccatt gagttatttc 480
ctggcgttca ttggtgcgtt tatctgggca gcctattgca cagtaacgaa taaatacgca 540
cgcggattta atggaattac cgtttttgtc ctgctaacgg gagcaagtct gtgggtttac 600
tattttctta cgccacaacc agaaatgata tttagcacgc ccgtcatgat taaactcatc 660
tctgcggcat ttaccttagg atttgcttat gctgcatgga atgtcggtat attgcatggc 720
aatgtcacca ttatggcggt aggttcgtat tttacgcctg tactttcctc agcgcttgca 780
gccgtgctgc tcagcgcccc gctgtcgttc tcgttctggc aaggcgcgct gatggtctgc 840
ggcggttccc tgctctgctg gctggcgaca cgtcgtggtt aa 882
<210> 26
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Eco-1
<400> 26
tagaggagac acaacatgac acgacaaaaa gcaac 35
<210> 27
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Eco-2
<400> 27
gctcttcctg tttagtttaa ccacgacgtg tcgcc 35
<210> 28
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> PgapA(-Eco)-2
<400> 28
tttttgtcgt gtcatgttgt gtctcctcta aagattg 37
<210> 29
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Hrh-3
<400> 29
atagagagtg actcaatgaa tagcaagaag gccac 35
<210> 30
<211> 33
<212> DNA
<213> Artificial Sequence
<220>
<223> Hrh-4
<400> 30
tcgagctcgg tacccctaca aacagtccgc cac 33
<210> 31
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<223> PyccA - 1
<400> 31
ctctagagga tccccttcca gatcaaatgc gtaa 34
<210> 32
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> PyccA(-Hrh)-2
<400> 32
cttcttgcta ttcattgagt cactctctat gacag 35
<210> 33
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Pst-3
<400> 33
atagagagtg actcaatgaa aaaccagcgt aaagc 35
<210> 34
<211> 33
<212> DNA
<213> Artificial Sequence
<220>
<223> Pst-4
<400> 34
tcgagctcgg tacccttatc cgtttcgacg cgg 33
<210> 35
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> PyccA(-Pst)-2
<400> 35
acgctggttt ttcattgagt cactctctat gacag 35
<210> 36
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Afa-3
<400> 36
atagagagtg actcaatgaa gcaatctgat aaggc 35
<210> 37
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<223> Afa-4
<400> 37
tcgagctcgg taccctcagg cagcgctttt tagt 34
<210> 38
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> PyccA(-Afa)-2
<400> 38
atcagattgc ttcattgagt cactctctat gacag 35
<210> 39
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Cne-3
<400> 39
atagagagtg actcaatgca aagcaagagc aaagc 35
<210> 40
<211> 33
<212> DNA
<213> Artificial Sequence
<220>
<223> Cne-4
<400> 40
tcgagctcgg taccctcacg gttcctggac acg 33
<210> 41
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> PyccA(-Cne)-2
<400> 41
gctcttgctt tgcattgagt cactctctat gacag 35
<210> 42
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Eco-3
<400> 42
atagagagtg actcaatgac acgacaaaaa gcaac 35
<210> 43
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<223> Eco-4
<400> 43
tcgagctcgg tacccttaac cacgacgtgt cgcc 34
<210> 44
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> PyccA(-Eco)-2
<400> 44
tttttgtcgt gtcattgagt cactctctat gacag 35
<210> 45
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Pspl7-trpE(S38R)_L-1
<400> 45
tcgagctcgg tacccaaaca actgcgacgt gtgtc 35
<210> 46
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Pspl7-trpE(S38R)_L-2
<400> 46
catgaagcgc cggtacctta atcatttttg ggttc 35
<210> 47
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Pspl7-trpE(S38R)_R-1
<400> 47
gccctgttgg aacgcgctga tatcaccacc aagaa 35
<210> 48
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Pspl7-trpE(S38R)_R-2
<400> 48
ctctagagga tccccagatg tcaccgttgt aaatg 35
<210> 49
<211> 294
<212> DNA
<213> Artificial Sequence
<220>
<223> последовательность промотора spl7
<400> 49
ggcgcttcat gtcaacaatc tttaacgttt tcaagttcac aagtcgtgtt caaatggtga 60
caagattgga cactgtgctg aattggcacc aagccctcat aaatgataga tctaaatcga 120
atatcaatat atggtctgtt tattggaacg cgtcccagtg gctgagacgc atccgctaaa 180
gccccaggaa ccctgtgcag aaagaacaaa taatcgtgaa ttttggcagc aacagcaatt 240
cctgctacaa ttgaaaacgt gcaaaagcat agattattgg aggagatcaa aaca 294
<210> 50
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Pspl7 - 1
<400> 50
cccaaaaatg attaaggtac cggcgcttca tgtca 35
<210> 51
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> Pspl7 - 2
<400> 51
gggattcgtg ctcatgatat ctgttttgat ctcctcc 37
<210> 52
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> trpE (S38R) - 1
<400> 52
atcaaaacag atatcatgag cacgaatccc catgt 35
<210> 53
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> trpE (S38R) - 2
<400> 53
gtggtgatat cagcgcgttc caacagggct gcatc 35
<210> 54
<211> 19
<212> DNA
<213> Artificial Sequence
<220>
<223> Confirm_Pspl7-trpE(S38R) - 1
<400> 54
gaagaagagg ctgcagatg 19
<210> 55
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> Confirm_Pspl7-trpE(S38R) - 2
<400> 55
gatcagcgcc atcatgtt 18
<210> 56
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Pn-tkt_L - 1
<400> 56
tcgagctcgg tacccaaact ttgagtgggt gcgtg 35
<210> 57
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Pn-tkt_L - 2
<400> 57
tcgagctacg agggcggttc ccagcccttc attag 35
<210> 58
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Pn-tkt_R - 1
<400> 58
attaacggtt aattgattct ggacgtcatg actac 35
<210> 59
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Pn-tkt_R - 2
<400> 59
ctctagagga tccccgcctc gatgatgcag tcgtc 35
<210> 60
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Pn-tkt - 1
<400> 60
gaagggctgg gaaccgccct cgtagctcga gagtt 35
<210> 61
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> Pn-tkt - 2
<400> 61
catgacgtcc agaatcaatt aaccgttaat ggagtcc 37
<210> 62
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Confirm_Pn-tkt - 1
<400> 62
acccagaacc ccaaattttc 20
<210> 63
<211> 19
<212> DNA
<213> Artificial Sequence
<220>
<223> Confirm_Pn-tkt - 2
<400> 63
ttgagttcga caactttgg 19
<210> 64
<211> 70
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 64
tgcaatgcat aacaacgcag tcgcactatt tttcactgga gagaagccct gtgtaggctg 60
gagctgcttc 70
<210> 65
<211> 70
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 65
tgcaatgcat aacaacgcag tcgcactatt tttcactgga gagaagccct gtccatatga 60
atatcctcct 70
<210> 66
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 66
gggcaggatc tcctgtcatc 20
<210> 67
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 67
aaatgtcgga taaggcaccg 20
<210> 68
<211> 70
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 68
tgtaatattc acagggatca ctgtaattaa aataaatgaa ggattatgta gtgtaggctg 60
gagctgcttc 70
<210> 69
<211> 70
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 69
tgtagggtaa gagagtggct aacatcctta tagccactct gtagtattaa gtccatatga 60
atatcctcct 70
<210> 70
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 70
acatccttat agccactctg 20
<210> 71
<211> 70
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 71
tacaaccggg ggaggcattt tgcttccccc gctaacaatg gcgacatatt gtgtaggctg 60
gagctgcttc 70
<210> 72
<211> 70
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 72
gcattcggtg cacgatgcct gatgcgccac gtcttatcag gcctacaaaa gtccatatga 60
atatcctcct 70
<210> 73
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 73
aggacggata aggcgttcac 20
<210> 74
<211> 26
<212> DNA
<213> Artificial Sequence
<220>
<223> trpE - 1
<400> 74
gaattcatgc aaacacaaaa accgac 26
<210> 75
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> trpE - 2
<400> 75
gaattctcag aaagtctcct gtgca 25
<210> 76
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> trpE(P21S) - 1
<400> 76
cgcttatcgc gacaattcca ccgcgctttt tcaccag 37
<210> 77
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> trpE(P21S) - 2
<400> 77
ctggtgaaaa agcgcggtgg aattgtcgcg ataagcg 37
<---
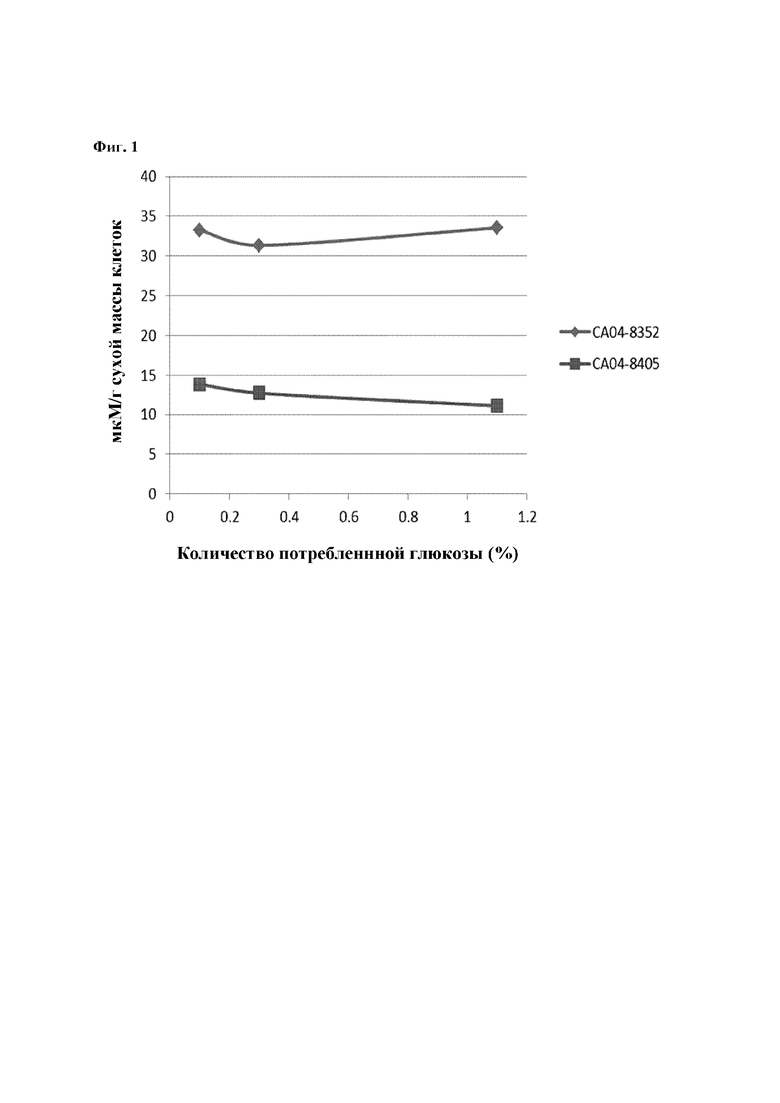
Изобретения относятся к продуцированию L-триптофана. Предложен микроорганизм для продуцирования L-триптофана, модифицированный так, что экспрессируется белок, содержащий аминокислотную последовательность SEQ ID NO: 1. При этом модификации представляют собой введение и/или усиление активности белка, имеющего L-триптофан-экспортирующую активность. Также предложен способ получения L-триптофана, в котором используется указанный микроорганизм. Также предложен экспрессионный вектор, содержащий полинуклеотид, кодирующий белок, имеющий L-триптофан-экспортирующую активность и содержащий аминокислотную последовательность SEQ ID NO: 1. Также предложен способ увеличения L-триптофан-экспортирующей активности микроорганизма. Изобретения обеспечивают увеличение количества продуцируемого L-триптофана. 4 н. и 4 з.п. ф-лы, 1 ил., 5 табл., 12 пр.
1. Микроорганизм для продуцирования L-триптофана, где микроорганизм модифицирован так, что экспрессируется белок, содержащий аминокислотную последовательность SEQ ID NO: 1, где модификации представляют собой введение и/или усиление активности белка, имеющего L-триптофан-экспортирующую активность.
2. Микроорганизм по п. 1, который принадлежит к роду Corynebacterium или к роду Escherichia.
3. Микроорганизм по п. 2, который представляет собой Corynebacterium glutamicum или Escherichia coli.
4. Способ получения L-триптофана, включающий:
культивирование в среде микроорганизма, продуцирующего L-триптофан, где микроорганизм модифицирован так, что экспрессируется белок, имеющий L-триптофан-экспортирующую активность и содержащий аминокислотную последовательность SEQ ID NO: 1, где модификации представляют собой введение и/или усиление активности белка, имеющего L-триптофан-экспортирующую активность; и
выделение L-триптофана из культивируемого микроорганизма или культуральной среды.
5. Способ по п. 4, где микроорганизм принадлежит к роду Corynebacterium или к роду Escherichia.
6. Способ по п. 5, где микроорганизм представляет собой Corynebacterium glutamicum или Escherichia coli.
7. Экспрессионный вектор, содержащий полинуклеотид, кодирующий белок, имеющий L-триптофан-экспортирующую активность и содержащий аминокислотную последовательность SEQ ID NO: 1.
8. Способ увеличения L-триптофан-экспортирующей активности микроорганизма, включающий модификацию микроорганизма таким образом, что белок, имеющий L-триптофан-экспортирующую активность и содержащий аминокислотную последовательность SEQ ID NO: 1, экспрессируется в этом микроорганизме, и где модификации представляют собой введение и/или усиление активности белка, имеющего L-триптофан-экспортирующую активность.
| WO 2003044192 A1, 30.05.2003 | |||
| База данных NCBI Reference Sequence: WP_050478745.1, 16.10.2017, "drug/metabolite DMT transporter permease [Herbaspirillum rhizosphaerae]" Найдено онлайн: https://www.ncbi.nlm.nih.gov/protein/913733518?sat=48&satkey=55716412; | |||
| DOROSHENKO V | |||
| еt al., YddG from Escherichia coli promotes export of aromatic amino acids |

