Изобретение относится к экспериментальной биологии, медицине и молекулярной онкологии, и может быть использовано для дистанционной, бесконтактной модуляции с использованием линейного ускорителя частиц транскрипционной активности гена CDKN1B в опухолевых клетках предстательной железы и регуляции их клеточного цикла.
Во всем мире рак предстательной железы (РПЖ) занимает лидирующие позиции в структуре онкологической патологии у мужчин всех возрастных групп. Рост заболеваемости за последние 10 лет составляет 143%, что является самым высоким показателем среди всей онкологической патологии (см. Зинькович М.С., Максимов А.Ю., Розенко Л.Я., Гусарева М.А., Карнаухова Е.А., Фаенсон А.В., Тимошкина Н.Н., Кутилин Д.С.Радиорезистентность как фактор эволюции лучевой терапии рака предстательной железы // Современные проблемы науки и образования. - 2019. - №2; URL: http://www.science-education.ru/ru/article/view?id=28627 (дата обращения: 14.03.2019). Устойчивый рост этого показателя делает как никогда актуальной задачу разработки новых методологических подходов к лечению данной патологии. Основными лечебными подходами при РПЖ являются радикальная простатэктомия и лучевая терапия (см. Chaiswing L., Weiss H.L., Jayswal R.D. Profiles of Radioresistance Mechanisms in Prostate Cancer. Crit Rev Oncog. 2018. Vol. 23(1-2). P. 39-67). Последняя как в самостоятельном варианте, так и в сочетании с другими методиками играет одну из ведущих ролей в лечении рассматриваемой патологии (см. Mottet N., Bellmunt J., Bolla M. EAU-ESTRO-SIOG Guidelines on Prostate Cancer. Part 1: Screening, Diagnosis, and Local Treatment with Curative Intent. Eur. Urol. 2017. V. 71. P. 618-63), однако применение стандартных протоколов облучения приводит к появлению радиорезистентных форм, что диктует необходимость персонифицированного подхода, основанного на данных молекулярно-генетических исследований, которые целесообразно проводить используя в качестве модели клеточную линию человеческих опухолевых клеток предстательной железы, например РС-3 (см. Кутилин Д.С., Зинькович М.С., Гусарева М.А., Фаенсон А.В., Карнаухова Е.А., Розенко Л.Я., Фатькина Н.Б., Удаленкова И.А., Васильева Е.О., Гаппоева М.А. Копийность генов как фактор устойчивости опухолевых клеток предстательной железы к облучению // Современные проблемы науки и образования. - 2020. - №4.; URL: http://science-education.ru/ru/article/view?id=29866 (дата обращения: 28.08.2020).
Облучение используется главным образом у пациентов с локализованными или местно-распространенными формами рака с противопоказаниями для хирургического вмешательства (см. Кутилин Д.С., Сагакянц А.Б., Зинькович М.С., Максимов А.Ю., Гусарева М.А., Бондаренко Е.С., Потемкин Д.С., Васильченко Н.Г. Влияние различных доз лучевой терапии на выживаемость опухолевых клеток предстательной железы линии РС-3 // Современные проблемы науки и образования. - 2019. - №2.;URL: http://science-education.ru/ru/article/view?id=28740 (дата обращения: 11.10.2019).
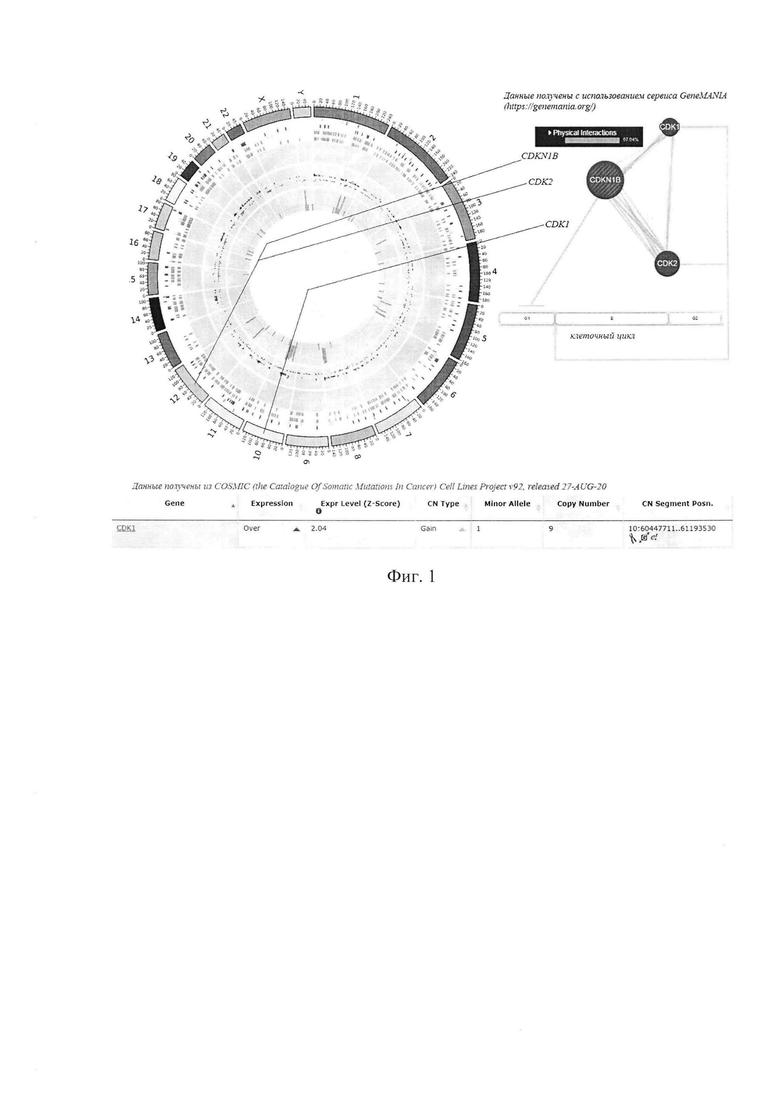
Известно, что в опухолевых клетках РС-3 повышена копийность и экспрессия гена CDK1 (см. базу данных COSMIC, Cell Lines Project v. 92, Кутилин Д.С., Зинькович M.C., Гусарева М.А., Фаенсон А.В., Карнаухова Е.А., Розенко Л.Я., Фатькина Н.Б., Удаленкова И.А., Васильева Е.О., Гаппоева М.А. Копийность генов как фактор устойчивости опухолевых клеток предстательной железы к облучению // Современные проблемы науки и образования. - 2020. - №4.; URL: http://science-education.ru/ru/article/view?id=29866 (дата обращения: 28.08.2020), а в резистентных к облучению клетках РС-3 еще и экспрессия гена CDK2 (см. Фиг. 1).
Циклинзависимая киназа 1 (CDK1), является высококонсервативным белком, который является ключевым в регуляции клеточного цикла. При его связывании с циклином происходит прогрессирование клеточного цикла. Связывание с циклином изменяет доступ к активному центру CDK1, обеспечивая его активность; кроме того, циклины могут нацеливать CDK1 на определенные субклеточные локации (см. Morgan D.L. The cell cycle: principles of control. London: New Science Press. 2007. pp. 30-31). Циклинзависимая киназа 2 (CDK2) активна во время фазы G1 и S клеточного цикла и, следовательно, действует как регулятор контрольной точки перехода G1-S. В начале фазы G1 увеличивается синтез циклина Е и активности CDK2. В конце фазы G1 комплекс CDK2/Cyclin Е достигает максимальной активности и играет значительную роль в инициации S фазы (см. Caruso JA, Duong МТ, Carey JP, Hunt KK, Keyomarsi K. Low-Molecular-Weight Cyclin E in Human Cancer: Cellular Consequences and Opportunities for Targeted Therapies. Cancer Research. 2018, 78 (19): 5481-5491).
Хотя CDK2 незаменим в клеточном цикле нормально функционирующих клеток, он также имеет решающее значение для аномальных процессов роста раковых клеток (см. Gopinathan L, Tan SL, Padmakumar VC, Coppola V, Tessarollo L, Kaldis P. Loss of Cdk2 and cyclin A2 impairs cell proliferation and tumorigenesis. Cancer Research. 2014, 74 (14): 3870-9). CDK2 и его партнеры по связыванию циклина представляют собой возможные терапевтические мишени для новых противоопухолевых препаратов (см. Wood DJ, Korolchuk S, Tatum NJ, Wang LZ, Endicott JA, Noble ME, Martin MP. Differences in the Conformational Energy Landscape of CDK1 and CDK2 Suggest a Mechanism for Achieving Selective CDK Inhibition. Cell Chemical Biology. 2018. 26 (1): 121-130).
Доклинические модели показали значительный успех в ограничении роста опухоли при применении ингибиторов CDK2 (см. Xia Р, Liu Y, Chen J, Coates S, Liu D, Cheng Z. Inhibition of cyclin-dependent kinase 2 protects against doxorubicin-induced cardiomyocyte apoptosis and cardiomyopathy. The Journal of Biological Chemistry. 2018. 293 (51): 19672-19685). Однако, идентификация селективных ингибиторов CDK2 затруднена из-за чрезвычайного сходства между активными сайтами CDK2 и CDK1, а ингибирование CDK1 может привести к непредвиденным побочным эффектам (см. Brown NR, Korolchuk S, Martin MP, Stanley WA, Moukhametzianov R, Noble ME, Endicott JA. CDK1 structures reveal conserved and unique features of the essential cell cycle CDK. Nature Communications. 2015.6: 6769).
Известным ингибитором CDK2 является CDKN1B (cyclin-dependent kinase inhibitor 1B, белок p27). p27 регулирует клеточный цикл, отвечая за его остановку в фазе G1. р27 подавляет активность комплексов циклин (А, Е или D) /циклинзависимая киназа (см. Mitrea D.M., Yoon М.K., Ou L., Kriwacki R.W. Disorder-function relationships for the cell cycle regulatory proteins p21 and p27. Biol Chem. 2012. 393(4). P. 259-274).
Поэтому, для ингибирования CDK2 в клетках рака предстательной железы (в том числе радиорезистентных) допустимо применение воздействия, активирующего транскрипционную активность гена CDKN1B.
Из патентных источников известны следующие изобретения:
1. Лечение связанных с геном-супрессором опухолей заболеваний посредством ингибирования природного транскрипта в антисмысловой ориентации относительно этого гена (см. патент на изобретение RU 2618688, опубл. 10.01.2013, Бюл. №1) Способ основан на повышении экспрессии полинуклеотида PTEN в клетках или тканях пациента in vivo или in vitro, включающий контактирование указанных клеток или тканей с по меньшей мере одним антисмысловым олигонуклеотидом длиной от 10-30 нуклеотида, мишенью которого является природный антисмысловой полинуклеотид PTEN, посредством чего осуществляется повышение экспрессии полинуклеотида PTEN в клетках или тканях пациента in vivo.
2. Средство, индуцирующее гибель клеток, средство, ингибирующее рост клеток, и фармацевтическая композиция для лечения заболевания, вызванного аномальным клеточным ростом (см. патент на изобретение RU 2707746 С2, опубл. 29.11.2019, Бюл. №34). В основе лежит средство, индуцирующее гибель раковых клеток, которое содержит лекарственное вещество, ингибирующее GST-π, и лекарственное вещество, ингибирующее связанный с гомеостазом белок, выбранный из группы, состоящей из белка, регулирующего клеточный цикл, - ATM, CDC25A, р21, PRKDC, RBBP8, SKP2, МСМ10, RNPC1, CCNL1, CENPH, BRSK1, МСМ8, CCNB3, MCMDC1 и MYLK, белка, связанного с подавлением апоптоза, - AATF, ALOX12, ANXA1, ANXA4, API5, ATF5, AVEN, AZU1, BAG1, BCL2L1, BFAR, CFLAR, IL2, MALT1, MCL1, MKL1, МРО, MTL5, MYBL2 и MYO18A, и белка, связанного с сигнальным путем PI3K, MTOR, IRAKI, IRS1, MYD88, NFKB1, PIK3CG, RAC1, AKT3, EIF4B, EIF4E, ILK, МТСР1, PIK3CA и SRF.
3. Средство, вызывающее в экспериментах in vivo замещение опухолевой ткани соединительной и переводящее клетки опухоли из фазы G1 клеточного цикла в состояние покоя G0 (см. патент на изобретение RU 2731105, опубл. 25.10.2019, Бюл. №30). Основано на средстве, вызывающем замещение опухолевой ткани соединительной и переводящее клетки опухоли из фазы G1 клеточного цикла в состояние покоя G0, представляющее собой сухой экстракт листьев аврана лекарственного, полученный путем измельчения их и экстракции.
4. Способ воздействия на пролиферативный статус клеток с помощью специфических нуклеотидных последовательностей G-цепи теломерной ДНК человека (см. патент на изобретение RU 2550267, опубл. 27.06.2013, Бюл. №18). Способ включает внесение в среду, окружающую клетки, и последующее проникновение в них специфических нуклеотидных последовательностей однонитевых оверхенгов G-цепи теломерной ДНК человека в конечной концентрации, не превышающей 30 мкМ. Указанные последовательности различаются вариациями теломерного повтора ДНК и триплетными окончаниями. Предложенное изобретение позволяет контролировать время наступления определенных фаз клеточного цикла и его общую продолжительность.
5. Способ радиосенсибилизации опухолевых клеток (см. патент на изобретение RU 2723393, опубл. 11.06.2020, Бюл. №17). Способ заключается в воздействии на опухолевую клетку веществом, являющимся ингибитором гликолиза и одновременно блокирующим клеточную пролиферацию, в качестве которого используют глюкозамин D гидрохлорид в концентрации 10 мМ. Изобретение обеспечивает повышение радиосенсибилизации и направленной селективности воздействия на опухолевые клетки без влияния на неопухолевые клетки за счет блокирования клеточной пролиферации в радиочувствительной G1 фазе клеточного цикла.
Описанные способы являются более сложными на подготовительных этапах, не нацелены на подавление клеточного цикла именно опухолевых клеток предстательной железы, не нацелены на стимуляцию транскрипционной активности гена ингибитора циклинзависимой киназы 2, т.е. в их основе лежат иные подходы нежели в способе предлагаемом нами.
Техническим результатом заявляемого изобретения является создание нового способа бесконтактной стимуляции транскрипционной активности гена ингибитора циклинзависимой киназы 2 в опухолевых клетках предстательной железы для регулирования клеточного цикла.
Технический результат достигается тем, что проводят облучение опухолевых клеток на линейном ускорителе Novalis ТХ в дискретно-импульсном режиме: 3 импульса по 6,4 Гр, интервал между импульсами 120 секунд и используют высокспецифичные праймеры для CDKN1B, CDK2 и CDK1 для подтверждения эффекта активации ингибитора и ингибирования циклинзависимой киназы 2 при сохраненной транскрипционной активности циклинзависимой киназы 1.
Для подтверждения эффекта активации CDKN1B и ингибирования CDK2 при сохраненной транскрипционной активности CDK1 проводится анализ экспрессии соответствующих генов в клетках подвергнутых облучению и интактных клетках РС-3. Также проводится подсчет общего количества клеток и анализ соотношения живых и мертвых клеток в камере Горяева с использованием 0,4%-ного раствора красителя трипанового синего и оценка количества клеток, находящихся на различных стадиях апоптоза, на проточном цитофлюориметре FacsCantoII с использованием Annexin V-FITC Apoptosis Detection Kit, а также ДНК-цитометрия для оценки клеточного цикла и подтверждения его остановки в стадии G1.
Заявленный способ, основан на использовании линейного ускорителя частиц Novalis ТХ для генерации облучения направленного на клеточную линию РС-3, что приводит к активации транскрипционной активности гена CDKN1B и ингибированию транскрипционной активности гена CDK2 и остановке клеточного цикла в фазе G1.
Представляем рисунки для понимания сущности способа.
На фигуре 1 изображена схема, отражающая геномную локализацию, транскрипционную активность и копийность гена CDK1, а также его взаимосвязи с генами CDK2 и CDKN1B у клеток РС-3.
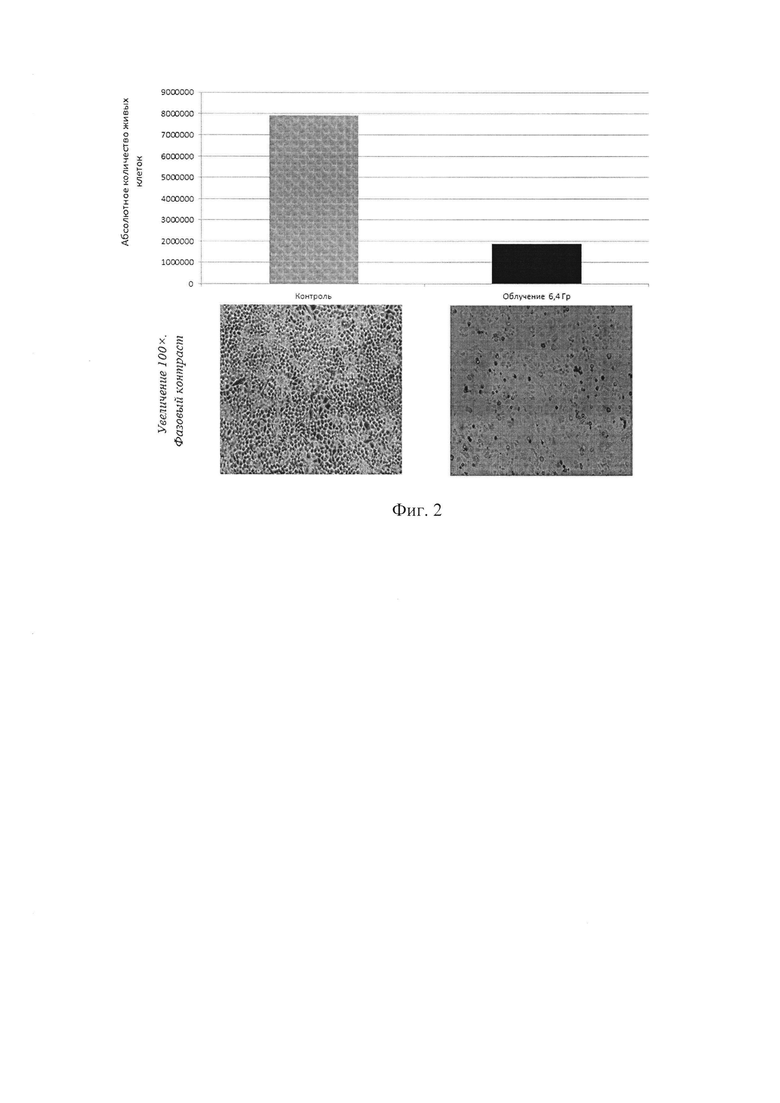
На фигуре 2 изображен результат микроскопирования и подсчета абсолютного количества клеток (* - статистически значимые отличия относительно контроля (интактных клеток), р<0.05).
На фигуре 3 изображены результаты ДНК-цитометрии интактных и облученных клеток.
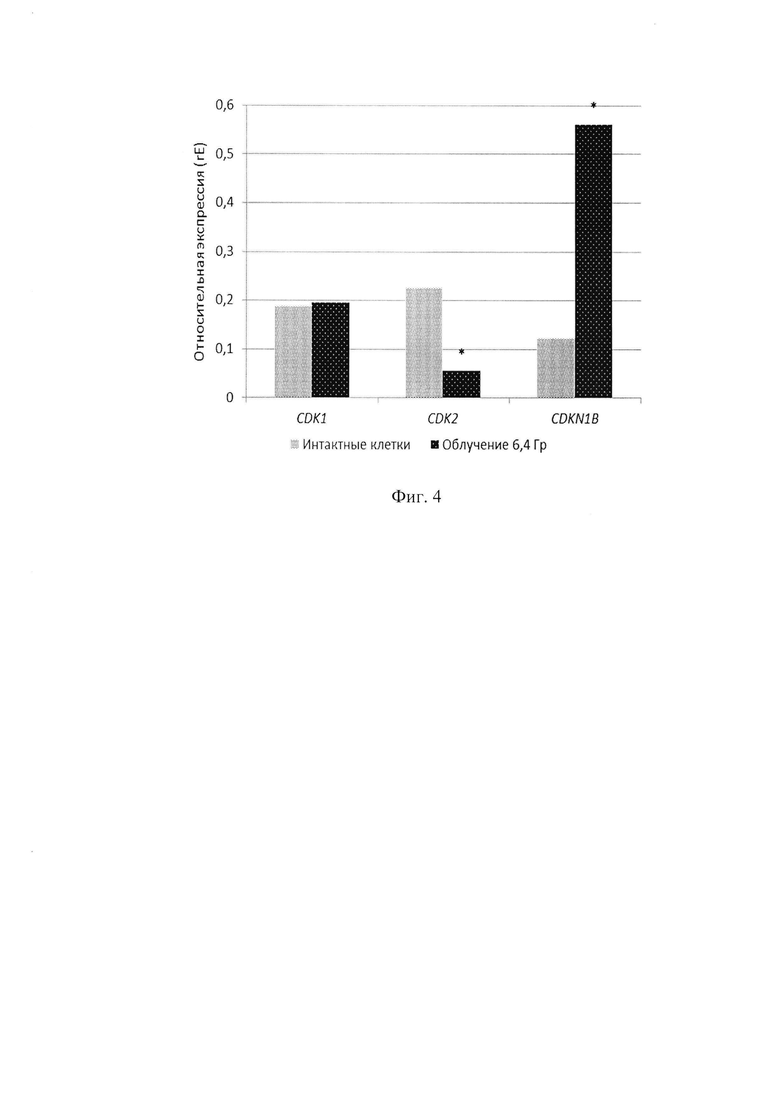
На фигуре 4 изображены данные по относительной экспрессии генов ингибитора циклинзависимой киназы 2, циклинзависимой киназы 2 и 1 в интактных и облученных клетках.
Заявленный способ включает следующие приемы:
• облучение в дозе 6,4 Гр на линейном ускорителе Novalis ТХ (Varian, США) в дискретно-импульсном режиме (3 импульса по 6,4 Гр, интервал между импульсами 120 секунд),
• топометрию на компьютерном томографе Somatom Definition AS Siemens (Германия),
• сегментирование с использованием ПО Eclips (Varian, США),
• выделение препаратов тотальной РНК из суспензии клеток РС-3 с помощью метода фенол-хлороформной экстракции,
• подтверждение эффекта активации CDKN1B и ингибирования CDK2 при сохраненной транскрипционной активности CDKI с использованием метода ПЦР в режиме реального времени (RT-qPCR),
• оценку стадии апоптоза на проточном цитофлюориметре FacsCantoII с использованием Annexin V-FITC Apoptosis Detection Kit,
• ДНК-цитометрию для оценки клеточного цикла.
Для осуществления способа были разработаны специфичные олигонуклеотидные прямые и обратные праймеры для генов CDKN1B, CDK2, CDK1 и GAPDH. Дизайн специфичных олигонуклеотидных праймеров (см. Таблица 1) осуществлялся с использованием референсных последовательностей NCBI GenBank.

Заявленный способ осуществляется следующим образом: культивирование клеток РС-3 проводится в стерильных плоскодонных флаконах в среде RPMI-1640 с 10% фетальной телячьей сывороткой и концентрацией гентамицина 50 мкг/мл в условиях 5% СО2 и 95% влажности при 37°С (см. Кутилин Д.С., Сагакянц А.Б., Зинькович М.С., Максимов А.Ю., Гусарева М.А., Бондаренко Е.С., Потемкин Д.С., Васильченко Н.Г. Влияние различных доз лучевой терапии на выживаемость опухолевых клеток предстательной железы линии РС-3. Современные проблемы науки и образования. 2019. №2. URL: http://www.science-education.ru/article/view?id=28740).
РС-3 облучают в дозе 6,4 Гр на линейном ускорителе Novalis ТХ (Varian, США) в дискретно-импульсном режиме (3 импульса по 6,4 Гр, интервал между импульсами 120 секунд), для топометрии используют компьютерный томограф Somatom Definition AS Siemens (Германия), для сегментирования и планирования используют программное обеспечения Eclips (Varian, США).
После облучения проводят микроскопирование клеточной линии и снимают ее с подложки флакона путем трипсинизации раствором Трипсин/Версена.
Далее проводят:
- оценку количества клеток, находящихся на различных стадиях апоптоза, на проточном цитофлюориметре с использованием Annexin V-FITC Apoptosis Detection Kit;
- оценку количества клеток, находящихся на различных стадиях/ фазах клеточного цикла (ДНК-цитометрия).
Для подтверждения эффекта активации CDKN1B и ингибирования CDK2 при сохраненной транскрипционной активности CDK1 проводится анализ экспрессии соответствующих генов в клетках подвергнутых облучению и интактных клетках РС-3 (через 24 часа после облучения).
Для этого клеточную массу отмывают от среды фосфатно-солевым буферным раствором Дюльбекко (DPBS) («Биолот», Россия) и осаждают центрифугированием. К осадку пробирки добавляют 900 мкл реагента QIAzol (QIAGEN). Дальнейшее выделение и очистку фракции тотальной РНК производят с помощью набора RNeasy Plus Universal Kits (QIAGEN) согласно протоколу производителя. При необходимости препараты готовой РНК обрабатывали ДНКазой I для устранения следов геномной ДНК согласно протоколу производителя фермента.
В конечный препарат тотальной РНК добавляют РНКазин до конечной концентрации в растворе 1 ед.акт/мкл. Для наработки кДНК готовят реакционную смесь, содержащую 5 мкМ рандомных гексонуклеотидных праймеров, 1x RT буфер, 0,5 мМ dNTP микс, 0,5 ед.акт./мкл ингибитора РНКаз и 5 U/мкл ReverseTranscriptase MMLV.
Полученную смесь инкубируют в термостате при 44°С в течение 1 часа, далее обратную транскриптазу инактивируют 10 минут при 92°С.
Методом количественной ПЦР в режиме реального времени (RT-qPCR) определяют величины относительной экспрессии 3 генетических локусов (CDK2, CDK1 и CDKN1B). В качестве референсного гена используют GAPDH.
Постановку каждого локуса проводят в трех технических повторах. Для амплификации используют реакционную смесь, содержащую 1х PCR буфер, смесь 0,2 мМ dNTP, 1,5 мМ MgCl2, 550 нМ праймеров, 0,1 U/мкл Taq-полимераза и 10 нг кДНК. Полученные смеси инкубируют в RT-амплификаторе CFX 96 (Bio-Rad, USA) по программе: первичная денатурация: t=95°C в течение 240 с; 40 циклов: t=95°C в течение 10 с, t=58°C в течение 30 с, t=72°C в течение 30 с.
Относительную экспрессию (rE) рассчитывают по формуле rE=2-ΔCt, где AC(t)=C(t)target-C(t)reference (см. Kutilin D.S., Mogushkova К.А. Effect of anthracycline antitumor antibiotics upon transcription activity of cancer-testis antigens in model experiments with HeLa cells. Medical Immunology (Russia). 2019;21(3):539-546).
Сравнивают значения rE для интактных клеток и клеток подвергнутых облучению на линейном ускорителе частиц. Наблюдают увеличение экспрессии гена CDKN1B в 4 раза, снижение экспрессии гена CDK2 в 4 раза и отсутствие отличий в экспрессии гена CDK1 в облученных клетках относительно интактных.
Для доказательства возможности осуществления предлагаемого способа приводим данные модельного эксперимента.
Было подготовлено 6 культуральных флаконов (3 для контроля и 3 для облучения) площадью 25 см2 с адгезионной поверхностью и вентилируемыми крышками (Sarstedt, Германия). Инкубирование осуществляли в условиях контролируемого 5% CO2 и влажности 95% при температуре 37 градусов по Цельсию в мультигазовом инкубаторе СВ 150 (Binder, Германия) в среде RPMI-1640 (Биолот, Россия) с 10%-ной фетальной телячьей сывороткой (Thermo Scientific hyclone, США) в присутствие 50 мкг/мл гентамицина (Биолот, Россия). По достижению количества клеток во всех флаконах уровня в 7000000, культуру РС-3 в трех флаконах облучают в дозе 6,4 Гр на линейном ускорителе Novalis ТХ в дискретно-импульсном режиме (3 импульса по 6,4 Гр, интервал между импульсами 120 секунд).
Через 24 часа после облучения проводят микроскопирование клеточной линии (как интактной, так и облученной) (см. Фиг. 2) и снимают ее с подложки флаконов путем трипсинизации раствором Трипсин/Версена.
Далее проводят:
- оценку количества живых клеток на проточном цитофлюориметре с использованием Annexin V-FITC Apoptosis Detection Kit,
- оценку количества клеток, находящихся на различных стадиях/ фазах клеточного цикла (ДНК-цитометрия),
- анализ экспрессии генов в клетках подвергнутых облучению и интактных клетках РС-3 (CDKN1B, CDK2, CDK1) (см. Фиг. 3).
Данные модельного эксперимента представлены на рисунках 2-4. Показано, что облучение клеток в дозе 6,4 Гр на линейном-ускорителе Novalis ТХ в дискретно-импульсном режиме (3 импульса по 6,4 Гр, интервал между импульсами 120 секунд) приводит увеличению транскрипционной активности гена CDKN1B (rE увеличивается 4,6 раза с 0,121 до 0,561) и снижению транскрипционной активности CDK2 (rE снижается 4,2 раза с 0,224 до 0,054), при этом траскрипционная активность CDK1 не изменяется относительно интактных клеток (см. Фиг. 4).
При этом наблюдается снижение абсолютного количества живых клеток в 4,2 раза после облучения (см. Фиг. 2), а также остановка клеточного цикла в фазе G1 (см. Фиг. 3), по данным ДНК-цитометрии после облучения в фазе G1 остается 95% клеток, в фазе G2 0% клеток и в фазе S 5% клеток).
Заявляемый способ, включает разработанный нами алгоритм облучения на линейном ускорителе частиц, разработанные нами синтетические нуклеотиды (праймеры) и является экономически оправданным для регуляции клеточного цикла опухолевых клеток предстательной железы в условиях модельного эксперимента, осуществляется в условиях стандартной лаборатории молекулярной биологии и отделения радиологии, оснащенного линейным ускорителем частиц.
--->
Перечень последовательностей
<110>Kutilin, Denis; Natsional'nyy meditsinskiy
issledovatel'skiy tsentr onkologii
<120>Method of non-contact stimulation of inhibitor
cyclin-dependent kinase 2 gene transcriptional
activity in prostatic tumor cells.
<160>1
<210>1
<211>20
<212>DNA
<213>Homo sapiens
<400>1
AAGCCGGGAT CTACCATACC 20
<110>Kutilin, Denis; Natsional'nyy meditsinskiy
issledovatel'skiy tsentr onkologii
<120>Method of non-contact stimulation of inhibitor
cyclin-dependent kinase 2 gene transcriptional
activity in prostatic tumor cells.
<160>2
<210>1
<211>21
<212>DNA
<213>Homo sapiens
<400>1
CATGGCTACC ACTTGACCTG T 21
<110>Kutilin, Denis; Natsional'nyy meditsinskiy
issledovatel'skiy tsentr onkologii
<120>Method of non-contact stimulation of inhibitor
cyclin-dependent kinase 2 gene transcriptional
activity in prostatic tumor cells.
<160>3
<210>1
<211>20
<212>DNA
<213>Homo sapiens
<400>1
TAATTGGGGC TCCGGCTAAC 20
<110>Kutilin, Denis; Natsional'nyy meditsinskiy
issledovatel'skiy tsentr onkologii
<120>Method of non-contact stimulation of inhibitor
cyclin-dependent kinase 2 gene transcriptional
activity in prostatic tumor cells.
<160>4
<210>1
<211>21
<212>DNA
<213>Homo sapiens
<400>1
GAAGAATCGT CGGTTGCAGG T 21
<110>Kutilin, Denis; Natsional'nyy meditsinskiy
issledovatel'skiy tsentr onkologii
<120>Method of non-contact stimulation of inhibitor
cyclin-dependent kinase 2 gene transcriptional
activity in prostatic tumor cells.
<160>5
<210>1
<211>22
<212>DNA
<213>Homo sapiens
<400>1
ATCTTTGCTG AGATGGTGAC ТС 22
<110>Kutilin, Denis; Natsional'nyy meditsinskiy
issledovatel'skiy tsentr onkologii
<120>Method of non-contact stimulation of inhibitor
cyclin-dependent kinase 2 gene transcriptional
activity in prostatic tumor cells.
<160>6
<210>1
<211>21
<212>DNA
<213>Homo sapiens
<400>1
CACTTGGGGA AACTTGGCTT G 21
<110>Kutilin, Denis; Natsional'nyy meditsinskiy
issledovatel'skiy tsentr onkologii
<120>Method of non-contact stimulation of inhibitor
cyclin-dependent kinase 2 gene transcriptional
activity in prostatic tumor cells.
<160>7
<210>1
<211>20
<212>DNA
<213>Homo sapiens
<400>1
GTCAAGGCTG AGAACGGGAA 20
<110>Kutilin, Denis; Natsional'nyy meditsinskiy
issledovatel'skiy tsentr onkologii
<120>Method of non-contact stimulation of inhibitor
cyclin-dependent kinase 2 gene transcriptional
activity in prostatic tumor cells.
<160>8
<210>1
<211>20
<212>DNA
<213>Homo sapiens
<400>1
TCGCCCCACT TGATTTTGGA 20
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ определения радиочувствительности злокачественных опухолей прямой кишки | 2020 |
|
RU2754154C1 |
| Малоинвазивный способ диагностики серозной аденокарциномы яичников высокой степени злокачественности на основании показателя копийности генов SULT1E1, CYP1B1 и ESR1 | 2021 |
|
RU2750472C1 |
| Тест-система "ESSC-tipe-1" для молекулярно-генетического типирования плоскоклеточного рака пищевода | 2020 |
|
RU2742468C1 |
| СПОСОБ ПРОГНОЗИРОВАНИЯ РЕЦИДИВИРОВАНИЯ СЕРОЗНОЙ КАРЦИНОМЫ ЯИЧНИКОВ | 2020 |
|
RU2749361C1 |
| МАЛОИНВАЗИВНЫЙ СПОСОБ ОПРЕДЕЛЕНИЯ ЧУВСТВИТЕЛЬНОСТИ ОПУХОЛЕЙ ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ К ЛУЧЕВОЙ ТЕРАПИИ НА ОСНОВАНИИ ИЗМЕНЕНИЯ КОПИЙНОСТИ ГЕНОВ BRCA2 И RAD50 | 2019 |
|
RU2728428C1 |
| Малоинвазивный способ дифференциальной ранней диагностики гепатоцеллюлярной карциномы и жирового гепатоза | 2023 |
|
RU2822224C1 |
| Тест-система для определения статуса генов IDH ½ в тканях глиальных опухолей | 2023 |
|
RU2823028C1 |
| КОМПОЗИЦИЯ И СПОСОБЫ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ РАКА ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ ЧЕЛОВЕКА | 2002 |
|
RU2346685C2 |
| АГЕНТЫ, КОТОРЫЕ РЕГУЛИРУЮТ, ИНГИБИРУЮТ ИЛИ МОДУЛИРУЮТ АКТИВНОСТЬ И/ИЛИ ЭКСПРЕССИЮ ФАКТОРОВ РОСТА СОЕДИНИТЕЛЬНОЙ ТКАНИ (ФРСТ), В КАЧЕСТВЕ УНИКАЛЬНЫХ СРЕДСТВ ДЛЯ СНИЖЕНИЯ ВНУТРИГЛАЗНОГО ДАВЛЕНИЯ И ЛЕЧЕНИЯ ГЛАУКОМАТОЗНЫХ РЕТИНОПАТИЙ/ОПТИЧЕСКИХ НЕВРОПАТИЙ | 2003 |
|
RU2332213C2 |
| Способ дифференциальной диагностики плоскоклеточного рака языка и доброкачественного новообразования языка | 2024 |
|
RU2841122C1 |

Настоящее изобретение относится к медицине и представляет собой способ бесконтактной стимуляции транскрипционной активности гена CDKN1B. Для осуществления настоящего изобретения проводят облучение опухолевых клеток предстательной железы на линейном ускорителе Novalis ТХ в дискретно-импульсном режиме, 3 импульса по 6,4 Гр, интервал между импульсами 120 секунд, и оценивают уровень экспрессии генов CDKN1B, CDK2 и CDK1. Способ позволяет осуществить процедуру регуляции клеточного цикла опухолевых клеток предстательной железы путем активации ингибитора циклинзависимой киназы 2 при сохраненной транскрипционной активности циклинзависимой киназы 1. 4 з.п. ф-лы, 4 ил., 1 табл., 1 пр.
1. Способ бесконтактной стимуляции транскрипционной активности гена ингибитора циклинзависимой киназы 2 в опухолевых клетках предстательной железы, заключающийся в том, что проводят облучение опухолевых клеток предстательной железы на линейном ускорителе Novalis ТХ в дискретно-импульсном режиме: 3 импульса по 6,4 Гр, интервал между импульсами 120 секунд.
2. Способ по п. 1, отличающийся тем, что проводят оценку уровня экспрессии генов CDKN1B, CDK2, CDK1.
3. Способ по п. 2, отличающийся тем, что для оценки уровня экспрессии гена CDKN1B используют праймеры: SEQ ID 3 и SEQ ID 4.
4. Способ по п. 2, отличающийся тем, что для оценки уровня экспрессии гена CDK2 используют праймеры: SEQ ID 5 и SEQ ID 6.
5. Способ по п. 2, отличающийся тем, что для оценки уровня экспрессии гена CDK1 используют праймеры: SEQ ID 1 и SEQ ID 2.
| ЛЕЧЕНИЕ СВЯЗАННЫХ С ГЕНОМ-СУПРЕССОРОМ ОПУХОЛЕЙ ЗАБОЛЕВАНИЙ ПОСРЕДСТВОМ ИНГИБИРОВАНИЯ ПРИРОДНОГО ТРАНСКРИПТА В АНТИСМЫСЛОВОЙ ОРИЕНТАЦИИ ОТНОСИТЕЛЬНО ЭТОГО ГЕНА | 2009 |
|
RU2618688C2 |
| СПОСОБ ВОЗДЕЙСТВИЯ НА ПРОЛИФЕРАТИВНЫЙ СТАТУС КЛЕТОК С ПОМОЩЬЮ СПЕЦИФИЧЕСКИХ НУКЛЕОТИДНЫХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ G-ЦЕПИ ТЕЛОМЕРНОЙ ДНК ЧЕЛОВЕКА | 2012 |
|
RU2550267C2 |
| МАЛОИНВАЗИВНЫЙ СПОСОБ ОПРЕДЕЛЕНИЯ ЧУВСТВИТЕЛЬНОСТИ ОПУХОЛЕЙ ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ К ЛУЧЕВОЙ ТЕРАПИИ НА ОСНОВАНИИ ИЗМЕНЕНИЯ КОПИЙНОСТИ ГЕНОВ BRCA2 И RAD50 | 2019 |
|
RU2728428C1 |
| LANG J-W., et al., Identification of novel CDK2 inhibitors by a multistage virtual screening method based on SVM, pharmacophore and docking model, Journal of Enzyme Inhibition and Medicinal Chemistry, 35:1,pp | |||
| Упругая металлическая шина для велосипедных колес | 1921 |
|
SU235A1 |
| De MARCO C., et al., The T197A Knock-in | |||

