Родственная ссылка
По данной заявке испрашивается приоритет и преимущество согласно U.S.S.N 61/381684, поданной 10 сентября 2010 года, которая включена в настоящее описание посредством ссылки во всей своей полноте.
Область техники, к которой относится изобретение
Настоящее изобретение относится к ингибированию форм дикого типа и определенных мутантных форм метилтрансферазы гистонов EZH2 человека - каталитической субъединицы комплекса PRC2, которая катализирует от моно- до триметилирования лизина 27 на гистоне H3 (H3-K27), к способам лечения различных типов рака, включающих фолликулярную лимфому и диффузную крупно-B-клеточную лимфому (DLBCL), и к способам определения способности к ответу на ингибитор EZH2 у индивидуума.
Уровень техники
В эукариотических клетках ДНК упакована с помощью гистонов, образуя хроматин. Приблизительно 150 пар оснований ДНК дважды обернуты вокруг октамера гистонов (по два каждого из гистонов 2A, 2B, 3 и 4), образуя нуклеосому - основной элемент хроматина. Изменения упорядоченной структуры хроматина могут приводить к изменениям транскрипции ассоциированных с ней генов. Этот процесс в высокой степени контролируется, поскольку изменения профилей экспрессии генов могут существенно влиять на фундаментальные клеточные процессы, такие как дифференцировка, пролиферация и апоптоз. Контроль изменений структуры хроматина (и, таким образом, транскрипции) опосредуется ковалентными модификациями гистонов, прежде всего, их N-концов. Эти модификации часто называют эпигенетическими, поскольку они могут приводить к наследственным изменениям экспрессии генов, но они не влияют на саму последовательность ДНК. Ковалентные модификации (например, метилирование, ацетилирование, фосфорилирование и убиквитинилирование) боковых цепей аминокислот опосредуются ферментативно.
Селективное присоединение метильных групп к конкретным аминокислотным участкам на гистонах контролируется действием уникального семейства ферментов, известных как метилтрансферазы гистонов (HMT). На уровень экспрессии конкретного гена влияет присутствие или отсутствие одной или нескольких метильных групп в соответствующем участке гистона. Конкретный эффект метильной группы в конкретном участке сохраняется до тех пор, пока метильная группа не удалится деметилазой гистонов, или до тех пор, пока модифицированный гистон не будет заменен путем обновления нуклеосомы. Подобным образом, другие классы ферментов могут присоединять к ДНК и гистонам другие химические группы, и другие ферменты могут удалять эти группы для обеспечения контроля экспрессии генов.
Организованный набор биохимических систем, обуславливающих регуляцию транскрипции, должен строго контролироваться для оптимального протекания роста и дифференцировки клеток. Когда этот контроль нарушается вследствие нарушения экспрессии и/или активности ферментов, ответственных за модификацию ДНК и гистонов, возникают болезненные состояния. Например, существуют накапливающиеся данные, позволяющие предположить, что при раке человека нарушенная регуляция активности эпигенетических ферментов приводит к неконтролируемой пролиферации клеток, ассоциированной с раком, а также к другим связанным с раком фенотипам, таким как усиленная миграция и инвазия клеток. Помимо рака, существуют накапливающиеся данные о роли эпигенетических ферментов в ряде других заболеваний человека, включая метаболические заболевания (такие как диабет), воспалительные заболевания (такие как болезнь Крона), нейродегенеративные заболевания (такие как болезнь Альцгеймера) и сердечно-сосудистые заболевания. Таким образом, селективное модулирование аберрантного действия эпигенетических ферментов имеет значительную перспективу для лечения ряда заболеваний.
Метилтрансфераза гистонов EZH2
Известно, что белки группы Polycomb (PcG) и группы trithorax (trxG) являются частью системы клеточной памяти. Francis et al. (2001) Nat Rev Mol Cell Biol 2:409-21; Simon et al. (2002) Curr Opin Genet Dev 12:210-8. Обе группы белков вовлечены в поддержание пространственного характера экспрессии генов гомеотического бокса (Hox), который формируются рано в ходе эмбрионального развития с помощью временно экспрессируемых генов сегментации. Как правило, белки PcG представляют собой репрессоры транскрипции, которые поддерживают «выключенное состояние», и белки trxG представляют собой активаторы транскрипции, которые поддерживают «включенное состояние». Поскольку представители белков PcG и trxG обладают собственной активностью метилтрансферазы гистонов (HMTase), белки PcG и trxG могут участвовать в клеточной памяти путем метилирования коровых гистонов. Beisel et al. (2002) Nature 419:857-62; Cao et al. (2002) Science 298: 1039-43; Czermin et al. (2002) Cell 111: 185-96; Kuzmichev et al. (2002) Genes Dev 16:2893-905; Milne et al. (2002) Mol Cell 10: 1107-17; Muller et al. (2002) Cell 111: 197-208; Nakamura et al. (2002) Mol Cell 10: 1119-28.
Биохимические и генетические исследования обеспечили доказательства того, что белки PcG Drosophila функционируют, по меньшей мере, в двух различных белковых комплексах: репрессивный комплекс Polycomb 1 (PRC1) и комплекс ESC-E(Z) (также известный как репрессивный комплекс Polycomb 2 (PRC2)), хотя составы комплексов могут быть динамическими. Otte et al. (2003) Curr Opin Genet Dev 13: 448-54. Studies in Drosophila (Czermin et al. (выше); Muller et al. (выше)) и в клетках млекопитающих (Cao et al. (выше); Kuzmichev et al. (выше)) было показано, что комплексы ESC-E(Z)/EED-EZH2 (т.е. PRC2) обладают собственной активностью метилтрансферазы гистонов. Хотя составы комплексов, выделенных различными группами, немного отличаются, они, главным образом, содержат EED, EZH2, SUZ12 и RbAp48 или их гомологи из Drosophila. Однако воссозданный комплекс, содержащий только EED, EZH2 и SUZ12, сохраняет активность метилтрансферазы гистона в отношении лизина 27 гистона H3. Патент США 7563589 (включен посредством ссылки).
Среди различных белков, составляющих комплексы PRC2, EZH2 (энхансер гомолога 2 Zeste) представляет собой каталитическую субъединицу. Каталитический центр EZH2, в свою очередь, находится в домене SET - мотиве с высоко консервативной последовательностью (назван после Su(var)3-9, энхансер Zeste, Trithorax), которая выявлена в нескольких ассоциированных с хроматином белках, включая представителей как группы Trithorax, так и группы Polycomb. Домен SET характерен для всех известных лизиновых метилтрансфераз гистонов, за исключением метилтрансферазы H3-K79 DOT1.
В дополнение к подавлению гена Hox, было показано, что опосредуемое PRC2 метилирование H3-K27 гистонов участвует в X-инактивации. Plath et al. (2003) Science 300: 131-5; Silva et al. (2003) Dev Cell 4: 481-95. Привлечение комплекса PRC2 в Xi и последующее триметилирование H3-K27 гистонов происходит на стадии инициации X-инактивации и зависит от РНК Xist. Более того, было выявлено, что EZH2 и ассоциированная с ней активность метилтрансферазы H3-K27 гистонов по-разному маркирует плюрипотентные эпибластные клетки и дифференцированную трофэктодерму. Erhardt et al. (2003) Development 130: 4235-48).
В соответствии с ролью EZH2 в поддержании профилей эпигенетической модификации плюрипотентных эпибластных клеток, опосредуемая Cre делеция EZH2 приводит к утрате метилирования H3-K27 гистонов в клетках. Erhardt et al. (выше). Кроме того, исследование в клеточных линиях и тканях рака предстательной железы и рака молочной железы выявило строгую корреляцию между уровнями EZH2 и SUZ12 и инвазивностью этих типов рака (Bracken et al. (2003) EMBO J 22: 5323-35; Kirmizis et al. (2003) Mol Cancer Ther 2: 113-21; Kleer et al. (2003) Proc Natl Acad Sci USA 100: 11606-11; Varambally et al. (2002) Nature 419: 624-9), что указывает на то, что дисфункция комплекса PRC2 может приводить к раку.
Недавно было описано, что соматические мутации тирозина 641 (Y641F, Y641N, Y641S и Y641H) EZH2 ассоциированы с фолликулярной лимфомой (FL) и диффузной крупно-B-клеточной лимфомой (DLBCL) подтипа лимфомы B-клеток герминального центра (GCB). Morin et al. (2010) Nat Genet 42: 181-5. Во всех случаях было выявлено, что встречаемость мутантного гена EZH2 является гетерозиготной, и в мутантных образцах, в которых профиль определяли секвенированием транскриптома, была выявлена экспрессия как аллелей дикого типа, так и мутантных аллелей. Также было продемонстрировано, что все мутантные формы EZH2 могли быть включены в мультибелковый комплекс PRC2, но что полученные комплексы были лишены способности катализировать метилирование эквивалентного H3-K27 остатка пептидного субстрата. Таким образом, было сделано заключение, что ассоциированные с заболеванием изменения Tyr641 в EZH2 приводили к утрате функции в отношении катализируемого EZH2 метилирования H3-K27.
Сущность изобретения
Один аспект настоящего изобретения относится к модулированию активности метилтрансферазы гистонов EZH2 дикого типа и мутантной метилтрансферазы гистонов EZH2, являющейся субъединицей комплекса PRC2, который катализирует от моно- до триметилирования лизина 27 в гистоне H3 (H3-K27). Например, настоящее изобретение относится к ингибированию активности определенных мутантных форм EZH2. Мутантные формы EZH2 включают замену тирозина 641 (Y641, также Tyr641) EZH2 дикого типа другим аминокислотным остатком.
Другой аспект настоящего изобретения относится к определению способности пациента отвечать на ингибитор EZH2, в зависимости от уровня диметилированного H3-K27me2 или предпочтительно в зависимости от уровней диметилированного H3-K27me2 и триметилированного H3-K27me3. Например, клетки с низким или не поддающимся выявлению уровнем диметилированного H3-K27me2 или клетки с низким соотношением H3-K27me2/me3 в значительно большей степени отвечают на антипролиферативный эффект ингибитора EZH2, чем клетки с более типичным более высоким соотношением 3-K27me2/me3.
Один аспект изобретения представляет собой способ ингибирования у индивидуума преобразования H3-K27 в триметилированный H3-K27. Способ включает стадию введения индивидууму, у которого экспрессируется мутант EZH2 по Y641, терапевтически эффективного количества ингибитора EZH2, где ингибитор ингибирует активность метилтрансферазы гистонов EZH2, тем самым ингибируя преобразование H3-K27 в триметилированный H3-K27 у индивидуума.
В этом и других аспектах изобретения в одном варианте осуществления ингибитор ингибирует активность метилтрансферазы гистонов мутанта EZH2 по Y641.
В этом и других аспектах изобретения в одном варианте осуществления ингибитор селективно ингибирует активность метилтрансферазы гистонов у мутанта EZH2 по Y641.
В этом и других аспектах изобретения в одном варианте осуществления мутант EZH2 по Y641 выбран из группы, состоящей из Y641F, Y641H, Y641N и Y641S.
В этом и других аспектах изобретения в одном варианте осуществления ингибитор EZH2 представляет собой S-аденозил-L-гомоцистеин или его фармацевтически приемлемую соль.
В этом и других аспектах изобретения в одном варианте осуществления ингибитор EZH2 представляет собой соединение 75:
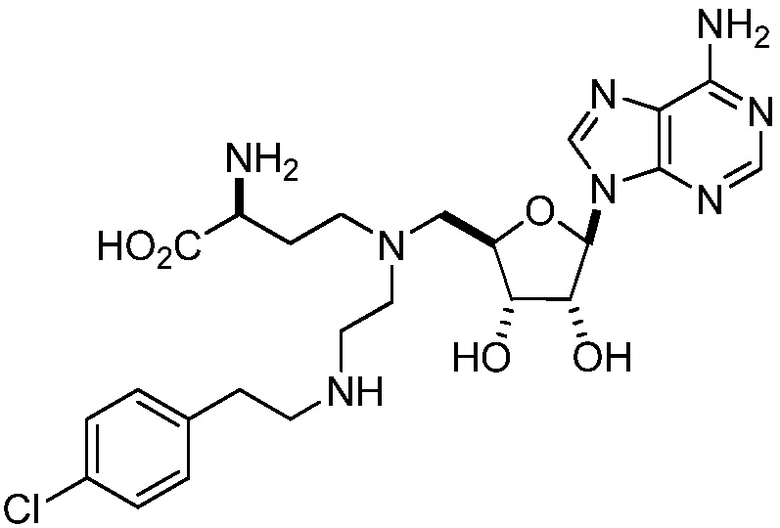 (75)
(75)
или его фармацевтически приемлемую соль.
Один аспект изобретения представляет собой способ ингибирования у индивидуума преобразования H3-K27 в триметилированный H3-K27. Способ включает стадии проведения анализа для выявления мутанта EZH2 по Y641 в образце от индивидуума; и введение индивидууму, у которого экспрессируется мутант EZH2 по Y641, терапевтически эффективного количества ингибитора EZH2, где ингибитор ингибирует активность метилтрансферазы гистонов у EZH2, тем самым ингибируя преобразование H3-K27 в триметилированный H3-K27 у индивидуума.
В этом и других аспектах изобретения в одном варианте осуществления проведение анализа для выявления мутанта EZH2 по Y641 включает полногеномное ресеквенирование или ресеквенирование области-мишени, которое выявляет нуклеиновую кислоту, кодирующую мутант EZH2 по Y641.
В этом и других аспектах изобретения в одном варианте осуществления проведение анализа для выявления мутанта EZH2 по Y641 включает приведение в контакт образца с антителом, которое специфично связывается с полипептидом или его фрагментом, характерным для мутанта EZH2 по Y641.
В этом и других аспектах изобретения в одном варианте осуществления проведение анализа для выявления мутанта EZH2 по Y641 включает приведение в контакт образца в условиях высокой жесткости с зондом нуклеиновой кислоты, который гибридизуется с нуклеиновой кислотой, кодирующей полипептид или его фрагмент, характерный для мутанта EZH2 по Y641.
Один аспект изобретения представляет собой способ ингибирования преобразования H3-K27 в триметилированный H3-K27. Способ включает стадию приведения в контакт мутанта EZH2 по Y641 с гистонным субстратом, включающим H3-K27, и эффективным количеством ингибитора EZH2, где ингибитор ингибирует активность метилтрансферазы гистонов у EZH2, тем самым ингибируя преобразование H3-K27 в триметилированный H3-K27.
Один аспект изобретения представляет собой способ идентификации индивидуума в качестве кандидата для лечения ингибитором EZH2. Способ включает стадии проведения анализа для выявления мутанта EZH2 по Y641 в образце от индивидуума; и идентификации индивидуума, у которого экспрессируется мутант EZH2 по Y641, в качестве кандидата для лечения ингибитором EZH2, где ингибитор ингибирует активность метилтрансферазы гистонов у EZH2.
Один аспект изобретения представляет собой способ идентификации ингибитора мутанта EZH2 по Y641. Способ включает стадии комбинирования выделенного мутанта EZH2 по Y641 с гистонным субстратом, донором метильной группы и тестируемым соединением, где гистонный субстрат включает форму H3-K27, выбранную из группы, состоящей из неметилированного H3-K27, монометилированного H3-K27, диметилированного H3-K27 и любой их комбинации; и проведение анализа для выявления метилирования H3-K27 в гистонном субстрате, тем самым, идентифицируя тестируемое соединение в качестве ингибитора мутанта EZH2 по Y641, когда метилирование H3-K27 в присутствии тестируемого соединения меньше, чем метилирование H3-K27 в отсутствие тестируемого соединения.
В одном варианте осуществления проведение анализа для выявления метилирования H3-K27 в гистонном субстрате включает измерение включения меченных метильных групп.
В одном варианте осуществления меченые метильные группы представляют собой изотопно меченные метильные группы.
В одном варианте осуществления проведение анализа для выявления метилирования H3-K27 в гистонном субстрате включает приведение в контакт гистонного субстрата с антителом, которое специфично связывается с триметилированным H3-K27.
Один аспект изобретения представляет собой способ идентификации ингибитора мутанта EZH2 по Y641. Способ включает стадии комбинирования выделенного мутанта EZH2 по Y641 с гистонным субстратом, донором метильной группы и тестируемым соединением, где гистонный субстрат содержит форму H3-K27, выбранную из группы, состоящей из неметилированного H3-K27, монометилированного H3-K27, диметилированного H3-K27 и любой их комбинации; и проведение анализа для выявления образования триметилированного H3-K27 в гистонном субстрате, тем самым, идентифицируя тестируемое соединение в качестве ингибитора мутанта EZH2 по Y641, когда образование триметилированного H3-K27 в присутствии тестируемого соединения является меньшим, чем образование триметилированного H3-K27 в отсутствие тестируемого соединения.
В одном варианте осуществления проведение анализа для выявления образования триметилированного H3-K27 в гистонном субстрате включает измерение включения меченых метильных групп.
В одном варианте осуществления меченые метильные группы представляют собой изотопно меченные метильные группы.
В одном варианте осуществления проведение анализа для выявления образования триметилированного H3-K27 в гистонном субстрате включает приведение в контакт гистонного субстрата с антителом, которое специфично связывается с триметилированным H3-K27.
Один аспект изобретения представляет собой способ идентификации селективного ингибитора мутанта EZH2 по Y641. Способ включает стадии комбинирования выделенного мутанта EZH2 по Y641 с гистонным субстратом, донором метильной группы и тестируемым соединением, где гистонный субстрат включает форму H3-K27, выбранную из группы, состоящей из монометилированного H3-K27, диметилированного H3-K27 и комбинации монометилированного H3-K27 и диметилированного H3-K27, тем самым формируя тестируемую смесь; комбинирования выделенной EZH2 дикого типа с гистонным субстратом, донором метильной группы и тестируемым соединением, где гистонный субстрат содержит форму H3-K27, выбранную из группы, состоящей из монометилированного H3-K27, диметилированного H3-K27 и комбинации монометилированного H3-K27 и диметилированного H3-K27, тем самым формируя контрольную смесь; проведения анализа для выявления триметилирования гистонного субстрата в каждой из тестируемой смеси и контрольной смеси; вычисления соотношения (a) триметилирования с мутантом EZH2 по Y641 и тестируемым соединением (M+) и (b) триметилирования с мутантом EZH2 по Y641 без тестируемого соединения (M-); вычисления соотношения (c) триметилирования с EZH2 дикого типа и тестируемым соединением (WT+) и (d) триметилирования с EZH2 дикого типа без тестируемого соединения (WT-); сравнения соотношения (a)/(b) с соотношением (c)/(d); и идентификации тестируемого соединения в качестве селективного ингибитора мутанта EZH2 по Y641, когда соотношение (a)/(b) меньше, чем соотношение (c)/(d).
Один аспект изобретения представляет собой способ лечения рака. Способ включает стадию введения индивидууму, имеющему рак, экспрессирующий мутант EZH2 по Y641, терапевтически эффективного количества ингибитора EZH2, где ингибитор ингибирует активность метилтрансферазы гистонов EZH2, тем самым осуществляя лечение рака.
В этом и других аспектах изобретения в одном варианте осуществления рак выбран из группы, состоящей из фолликулярной лимфомы и диффузной крупно-B-клеточной лимфомы (DLBCL) подтипа лимфомы B-клеток герминального центра (GCB).
Один аспект изобретения представляет собой способ лечения рака. Способ включает стадию введения индивидууму, имеющему рак, экспрессирующий мутант EZH2 по Y641, терапевтически эффективного количества ингибитора EZH2, где ингибитор селективно ингибирует активность метилтрансферазы гистонов мутанта EZH2 по Y641, тем самым, осуществляя лечение рака.
Один аспект изобретения представляет собой способ лечения рака. Способ включает стадии проведения анализа для выявления мутанта EZH2 по Y641 в образце, содержащем раковые клетки от индивидуума, имеющего рак; и введения индивидууму, у которого экспрессируется мутант EZH2 по Y641, терапевтически эффективного количества ингибитора EZH2, где ингибитор ингибирует активность метилтрансферазы гистонов EZH2, тем самым, осуществляя лечение рака.
Другой аспект изобретения представляет собой способ определения способности индивидуума отвечать на ингибитор EZH2. В одном варианте осуществления способ включает выделение образца ткани от индивидуума; выявление уровня диметилирования (me2) H3-K27 в образце ткани; сравнение уровня диметилирования (me2) с контрольным уровнем диметилирования (me2); и идентификацию индивидуума как отвечающего на указанный ингибитор EZH2, когда уровень диметилирования (me2) отсутствует или ниже, чем контрольный уровень диметилирования (me2). В одном варианте осуществления способ дополнительно включает выявление уровня триметилирования (me3) H3-K27 в образце ткани; сравнение уровня триметилирования (me3) с контрольным уровнем триметилирования (me3) и уровня диметилирования (me2) с контрольным уровнем диметилирования (me2); и идентификацию указанного индивидуума как отвечающего на ингибитор EZH2, когда уровень триметилирования (me3) является таким же или превышает контрольный уровень триметилирования (me3), и уровень диметилирования (me2) отсутствует или ниже, чем контрольный уровень диметилирования (me2). В другом варианте осуществления способ дополнительно включает получение соотношения уровня диметилирования (me2) и уровня триметилирования (me3) H3-K27 в образце ткани; получение контрольного соотношения контрольного уровня диметилирования (me2) и контрольного уровня триметилирования (me3); сравнение соотношения с контрольным соотношением; и идентификацию индивидуума как отвечающего на указанный ингибитор EZH2, когда указанное соотношение является меньшим, чем указанное контрольное соотношение. В предпочтительном варианте осуществления индивидуум имеет рак. В одном варианте осуществления рак представляет собой фолликулярную лимфому. Альтернативно, рак представляет собой крупно-B-клеточную лимфому (DLBCL). В другом предпочтительном варианте осуществления у индивидуума экспрессируется мутант EZH2 по Y641. В предпочтительном варианте осуществления мутант по Y641 представляет собой Y641F, Y641H, Y641N или Y641S.
Один аспект изобретения представляет собой способ выбора лечения для индивидуума, имеющего рак. Способ включает определение способности индивидуума отвечать на ингибитор EZH2 по уровню диметилированного H3-K27 или предпочтительно по уровням диметилированного H3-K27 и триметилированного H3-K27; и предоставление ингибитора EZH2 индивидууму, когда индивидуум способен отвечать на ингибитор EZH2. В одном варианте осуществления рак представляет собой фолликулярную лимфому. Альтернативно, рак представляет собой диффузную крупно-B-клеточную лимфому (DLBCL). В другом предпочтительном варианте осуществления у индивидуума экспрессируется мутант EZH2 по Y641. В предпочтительном варианте осуществления мутант по Y641 представляет собой Y641F, Y641H, Y641N или Y641S.
Один аспект изобретения представляет собой соединение 75:
(75)
или его фармацевтически приемлемую соль.
Один аспект изобретения представляет собой фармацевтическую композицию, содержащую соединение 75:
(75)
или его фармацевтически приемлемую соль.
Один аспект изобретения представляет собой применение соединения 75:
(75)
или его фармацевтически приемлемой соли при лечении фолликулярной лимфомы.
Один аспект изобретения представляет собой применение соединения 75:
(75)
или его фармацевтически приемлемой соли при лечении диффузной крупно-B-клеточной лимфомы (DLBCL).
Если не указано иное, все технические и научные термины, используемые в настоящем описании, имеют такое же значение, которое обычно подразумевает специалист в области, к которой относится настоящее изобретение. В описании форма единственного числа также включает множественное число, если контекст явно не указывает на иное. Хотя на практике и для исследования настоящего изобретения можно использовать способы и материалы, сходные или эквивалентные способам или материалам, описанным в настоящем описании, подходящие способы и материалы описаны ниже. Все публикации, патентные заявки, патенты и другие ссылки, приведенные в настоящем описании, включены посредством ссылок. Ссылки, цитированные в настоящем описании, не признаются как уровень техники заявленного изобретения. В случае противоречий следует руководствоваться настоящим описанием, включая определения. Кроме того, материалы, способы и примеры являются только иллюстративными, но не ограничивающими.
Другие признаки и преимущества изобретения будут понятны из следующего подробного описания и формулы изобретения.
Краткое описание фигур
На фиг.1 представлены два графика, на которых показано, что ассоциированные с B-клеточной лимфомой мутанты EZH2 являются активными метилтрансферазами гистонов. Активность метилтрансферазы in vitro в комплексах PRC2, содержащих EZH2 дикого типа и различные мутанты EZH2 по Y641, измеряли в виде (A) реакций переноса метильной группы с использованием пептида (H3 21-44) в качестве субстрата, и (B) реакций переноса метильной группы с использованием нуклеосом птиц в качестве субстрата. Обозначения: дикий тип (●), Y641F (o), Y641H (□), Y641N (■) и Y641S (▲). CPM представляет собой количество импульсов в минуту применительно к сцинтилляционному измерению радиационного излучения 3H.
На фиг.2 представлены четыре графика, на которых показано, что комплексы PRC2, содержащие мутант EZH2, предпочтительно катализируют ди- и триметилирование H3-K27 гистонов. (A) Активность метилтрансферазы мутантных комплексов и комплексов дикого типа (WT) в отношении неметилированного пептида (незакрашенные столбики), монометилированного пептида (мозаичные столбики) и диметилированного пептида (закрашенные столбики). (B) Аффинность к пептидным субстратам, исходя из K1/2, является сходной при всех состояниях метилирования пептида у комплексов PRC2, содержащих EZH2 дикого типа (o), Y641F (●), Y641H (□), Y641N (■) и Y641S (▲) EZH2. Следует отметить, что варьирование величин K1/2 среди всех субстратов и всех форм фермента является менее чем 3,5-катным. Для любого конкретного состояния метилирования субстрата варьирование величины K1/2 является менее чем 2-кратным. (C) Число оборотов фермента (kcat) варьирует, в зависимости от состояния метилирования, противоположным образом для WT EZH2 и мутантов EZH2 по Y641. kcat снижается при увеличении состояния метилирования K27 для EZH2 дикого типа (o), но увеличивается для мутантов Y641F (●), Y641H (□), Y641N (■) и Y641S (▲) EZH2. (D) Каталитическая эффективность (kcat/K1/2) снижается при увеличении состояния метилирования K27 для EZH2 дикого типа (o), но возрастает для мутантов Y641F (●), Y641H (□), Y641N (■) и Y641S (▲) EZH2. На панелях B-D линии, соединяющие точки данных, не предполагают какой-либо математической взаимосвязи; вместо этого, они предназначены только в качестве наглядного средства.
На фиг.3A также представлены три графика, на которых показаны предсказанные относительные уровни H3-K27me3 (верхняя панель), H3-K27me2 (средняя панель) и H3-K27me1 (нижняя панель) для клеток, содержащих различные мутанты EZH2. Моделирование проводили с использованием уравнения скорости реакции сопряженных ферментативных реакций в состоянии равновесия и стационарных параметров кинетики, представленных в таблице 1. Все величины представлены относительно гомозиготных содержащих WT EZH2 клеток и предполагают насыщающие концентрации внутриклеточного SAM относительно Km и внутриклеточные концентрации нуклеосом, сходные с Km.
На фиг.3B представлена серия анализов с использованием вестерн-блоттинга относительных профилей состояния метилирования H3-K27 для клеточных линий лимфомы, гомозиготных по WT EZH2 или гетерозиготных по указанной мутации Y641 EZH2. На панелях сверху вниз представлены результаты исследования с помощью антител, специфичных к следующему: общей EZH2; H3-K27me3; H3-K27me2; H3-K27me1; и общему гистону H3 в качестве нагрузочного контроля.
На фиг.4 представлены выбранные предложенные механизмы, ведущие к нарушенным высоким уровням триметилирования H3-K27 гистонов при раке. Они включают: a) мутацию Y641 в EZH2, приводящую к изменению предпочтения субстрата из неметилированного на моно- и диметилированный гистон H3-K27; b) сверхэкспрессию EZH2; c) мутации в UTX, которые инактивируют функцию фермента, вызывая снижение деметилирования H3-K27me3; и d) сверхэкспрессию субъединицы PHF19/PCL3 комплекса PRC2, которая приводит к увеличению привлечения комплекса PRC2 к конкретным генам и увеличению триметилирования H3-K27 гистонов. Во всех четырех моделях изменение приводит к нарушенному триметилированию H3-K27 гистонов в проксимальных промоторных областях гена, что приводит к подавлению транскрипции ключевых генов при раке.
На фиг.5 представлена SDS-PAGE на геле, показывающая, что уровни экспрессии каждого из пятикомпонентных комплексов PRC2 сходны в случае мутантной EZH2 и EZH2 дикого типа.
На фиг.6 представлена пара таблиц, на которых показано, что мутантный комплекс PRC2 и комплекс PRC2 дикого типа (WT) обладают строгим предпочтением в качестве субстрата H3-K27-содержащих пептидов. Каждый фермент исследовали против панели перекрывающихся 15-мерных пептидов, охватывающие все H3 и H4. Активность измеряли как скорость (CPM в минуту), и представленная величина представляет собой среднее значение для двух независимых определений в каждой реакции. Для всех комплексов, наиболее предпочтительным пептидом был H3:16-30. Комплекс WT имел более чем в 6 раз более высокую активность против этого пептида, чем любой из мутантных комплексов.
На фиг.7 представлен график, на котором показана эффективность S-аденозил-L-гомоцистеина (SAH) в отношении ингибирования WT EZH2 и мутантов EZH2 по Y641. На оси X представлен логарифм концентрации SAH; на оси Y показано процентное ингибирование.
На фиг.8 представлен график, на котором показана эффективность соединения 75 в отношении ингибирования EZH2 WT и мутантов EZH2 по Y641. На оси X показан логарифм концентрации соединения 75; на оси Y представлено процентное ингибирование.
На фиг.9 представлен анализ вестерн-блотов для относительных уровней H3-K27me1, me2 и me3 на панели клеточных линий, включая множество клеточных линий DLBCL, экспрессирующих WT EZH2 или мутант EZH2 по Y641. a) Гистоны выделяли из представленных клеточных линий, фракционировали с помощью SDS-PAGE на 4-20% геле, переносили на нитроцеллюлозные мембраны и исследовали с помощью антител к гистону H3, H3-K27me1, me2 или me3. Уровни EZH2 определяли путем приготовления лизатов цельных клеток из представленных клеточных линий, обработки, как описано выше, и исследования с помощью антитела к EZH2; b) гистоны выделяли из представленных клеточных линий и обрабатывали, как описано выше, за исключением того, что уровни EZH2 не определяли.
На фиг.10 представлен иммуноцитохимический анализ уровней H3 и H3-K27me3 на панели клеточных линий WT и мутантных по Y641 клеточных линий лимфомы. Клеточные осадки указанных клеточных линий фиксировали и погружали в парафин. Подготавливали препараты и оценивали уровни H3 и H3-K27me3 с помощью иммуноцитохимии с использованием антител к гистону H3 или H3-K27me3.
На фиг.11 представлен иммуноцитохимический анализ уровней H3 и H3-K27me2 на панели клеточных линий WT и мутантных по Y641 клеточных линий лимфомы. Клеточные осадки указанных клеточных линий фиксировали и погружали в парафин. Подготавливали препараты и оценивали уровни H3 и H3-K27me2 с помощью иммуноцитохимии с использованием антител к гистону H3 или H3-K27me2.
На фиг.12 представлен график, на котором показано ингибирование общих уровней H3-K27me3 путем обработки ингибитором EZH2 в мутантных по Y641 клетках WSU-DLCL2. Клетки WSU-DLCL2 обрабатывали в течение 4 суток указанными концентрациями ингибитора EZH2 A или B. После обработки соединением гистоны выделяли, фракционировали с помощью SDS-PAGE на 4-20% геле, переносили на нитроцеллюлозные мембраны и исследовали с помощью антител к гистону H3 или H3-K27me3.
На фиг.13 представлен график, на котором показано, что ингибиторы EZH2 могут блокировать пролиферацию мутантных по Y641 клеток WSU-DLCL2, однако имеют небольшой эффект на не мутантные по Y641 клетки OCI-LY19. Клетки инкубировали в присутствии возрастающих концентраций ингибитора EZH2 A или B в течение одиннадцати суток. В качестве контроля включали клетки, обработанные носителем (ДМСО). Количество и жизнеспособность клеток определяли с использованием анализа Guava Viacount в устройстве Guava EasyCyte Plus. Клетки разделяли, и среду и соединение восполняли каждые 3-4 суток.
На фиг.14 представлен график, на котором показано, что присутствие мутации EZH2 (Y641) и/или высоких уровней H3-K27me3 и низких уровней H3-K27me2 предсказывает чувствительность к ингибиторам EZH2. Клеточные линии поддерживали в присутствии возрастающих концентраций одного ингибитора EZH2 вплоть до 25 мкМ. Для установления величин IC90 использовали количества жизнеспособных клеток после обработки в течение 11 суток. Результаты нанесены на график, где клеточные линии сгруппированы в соответствии с состоянием по мутации EZH2 (A), или сгруппированы в соответствии с уровнями H3-K27me2 и H3-K27me3 (B). На обоих графиках линия указывает на средние величины IC90 для указанной группы клеточных линий.
Подробное описание изобретения
Структура хроматина важна для регуляции генов и эпигенетического наследования. Посттрансляционные модификации гистонов вовлечены в установление и подержание структуры хроматина более высокого порядка; например, концевые части определенных коровых гистонов модифицированы ацетилированием, метилированием, фосфорилированием, риболизированием и/или убиквитинилированием.
EZH2 представляет собой метилтрансферазу гистонов, которая является каталитической субъединицей комплекса PRC2, который катализирует от моно- до триметилирования лизина 27 на гистоне H3 (H3-K27). Триметилирование H3-K27 гистонов представляет собой механизм подавления транскрипции определенных генов, которые расположены близко к участку модификации гистонов. Известно, что триметилирование является маркером рака, экспрессия которого изменяется при раке, таком как рак предстательной железы (см., например, публикацию патентной заявки США № 2003/0175736, включенную в настоящее описание посредством ссылки во всей своей полноте). EZH2 относится к семейству белков группы Polycomb (PcG). Белки группы Polycomb помогают поддерживать характерные признаки клеток путем репрессии транскрипции генов-мишеней. Jacobs et al. (1999) Semin Cell Dev Biol 10(2): 227-35; Jacobs et al. (2002) Biochim Biophys Acta 1602(2): 151-61. На микрочипах ДНК EZH2 был идентифицирован как активированный при гормон-резистентном метастазирующем раке предстательной железы. Dhanasekaran et al. (2001) Nature 412(6849): 822-6; Varambally et al. (2002) Nature 419(6907): 624-9. EZH2 активируется при агрессивных типах рака молочной железы и является медиатором проинвазивного фенотипа. Kleer et al. (2003) Proc Natl Acad Sci USA 100(20): 11606-11. Сверхэкспрессия EZH2 в иммортализованных эпителиальных клеточных линиях молочной железы человека обеспечивает независимый от заякоривания рост и инвазию клеток. Kleer et al. (выше). Для опосредуемой EZH2 инвазии клеток требуется неизмененный домен SET и активность деацетилазы гистонов. Предшествующие испытания обеспечили доказательство функциональной связи между нарушенной регуляцией экспрессии EZH2, подавлением транскрипции и неопластической трансформацией. Varambally et al. (выше); Kleer et al (выше).
Один из аспектов настоящего изобретения относится к ингибированию активности EZH2, включая определенные мутантные формы EZH2. В одном варианте осуществления настоящее изобретение относится к селективному ингибированию активности определенных мутантных форм EZH2.
Было описано, что точечные мутации в гене EZH2 в одном аминокислотном остатке (Tyr641, в настоящем описании обозначаемый, как Y641) EZH2 связаны с подгруппами B-клеточных лимфом человека. Morin et al. (2010) Nat Genet 42(2): 181-5. В частности, Morin et al. описали, что соматические мутации тирозина 641 (Y641F, Y641H, Y641N и Y641S) в EZH2 ассоциированы с фолликулярной лимфомой (FL) и диффузной крупно-B-клеточной лимфомой (DLBCL) подтипа лимфомы B-клеток герминального центра (GCB). Мутантный аллель всегда выявляют в ассоциации с аллелем дикого типа (гетерозиготный) в пораженных заболеванием клетках, и было описано, что мутации устраняют ферментативную активность комплекса PRC2 в отношении метилирования немодифицированного пептидного субстрата.
Недавно неожиданно было открыто, что фермент EZH2 дикого типа (WT) проявляет наиболее высокую каталитическую эффективность (kcat/K) для реакции от нуль- до монометилирования H3-K27 и меньшую эффективность для последующих (от моно- до ди- и от ди- до триметилирования) реакций; в то время как в противоположность этому, ассоциированные с заболеванием мутации Y641 проявляют в значительной степени ограниченную способность к проведению первой реакции метилирования, однако обладают увеличенной каталитической эффективностью в последующих реакциях относительно фермента дикого типа. Эти результаты подразумевают, что злокачественный фенотип заболевания использует комбинированную активность фермента, осуществляющего монометилирование H3-K27 (PRC2, содержащий WT EZH2 или EZH1) вместе с PRC2, содержащим мутантную EZH2 для усиленного преобразования H3-K27 в триметилированную форму (H3-K27me3).
Без связи с какой-либо теорией, полагают, что мутация Y641 на фенилаланин (F), гистидин (H), аспарагин (N) или серин (S) в EZH2 может облегчать множество раундов метилирования H3-K27 путем влияния на характер образования H-связей и/или пространственное скучивание в активном центре фермент-бисубстратного тройного комплекса, влияя на образование надлежащего водного канала для депротонирования реагирующего лизина. Zhang et al. (2008) Proc Natl Acad Sci USA 105: 5728-32. Это препятствование установлено по аналогии с результатами кристаллографического и молекулярного динамического моделирования, выявленного для мутации тирозина в родственных метилтрансферазах лизина белков LSMT, Dim-5 и SET7/9.
Например, когда тирозин 245 рекомбинантной SET7/9 подвергали мутации на аланин, наблюдали изменение субстратной специфичности. Dillon et al. (2005) Genome Biol 6: 227. Способность мутанта Y245A SET7/9 метилировать немодифицированный пептид из 20 остатков, соответствующий последовательности, окружающей H3-K4, была снижена приблизительно до 20% относительно фермента WT. Xiao et al. (2003) Nature 421: 652-6. В то же время способность мутанта Y245A далее метилировать пептиды H3-K4me1 и H3-K4me2 значительно усиливалась (приблизительно в 7 раз и 5 раз, соответственно) в отношении фермента WT. Однако в противоположность настоящему описанию, касающемуся мутаций EZH2 по Y641, мутация Y245 SET7/9 на фенилаланин не усиливала ни от моно- до ди-, ни от ди- до триметилирования пептида; вместо этого мутант Y245F SET7/9 проявлял минимальную каталитическую активность в отношении всех пептидных субстратов. Аналогично, фермент G9a дикого типа может диметилировать H3-K9, но не способен выполнять реакцию от ди- до триметилирования. Тем не менее, когда тирозин 1067 G9a (аналогичный Y641 EZH2) подвергают мутации на фенилаланин, фермент приобретает способность триметилировать H3-K9. Wu, H. et al. (2010) PLoS One 5, e8570, doi:10.1371/journal.pone.0008570).
Нуклеиновые кислоты и полипептиды EZH2 человека были описаны ранее. См., например, Chen et al. (1996) Genomics 38: 30-7 [746 аминокислот]; номер доступа Swiss-Prot Q15910 [746 аминокислот]; номера доступа GenBank NM_004456 и NP_004447 (изоформа a [751 аминокислота]); и номера доступа GenBank NM_152998 и NP_694543 (изоформа b [707 аминокислот]), каждый из которых включен в настоящее описание посредством ссылки во всей своей полноте.








Как указано выше, полагают, что каталитический центр EZH2 расположен в консервативном домене белка, известном как домен SET. Аминокислотная последовательность домена SET EZH2 представлена следующей неполной последовательностью, охватывающей аминокислотные остатки 613-726 последовательности с номером доступа Swiss-Prot Q15910 (SEQ ID NO:1):

SEQ ID NO:6.
Остаток тирозина (Y), показанный подчеркиванием в SEQ ID NO:6, представляет собой Tyr641 (Y641) в последовательности с номером доступа Swiss-Prot Q15910 (SEQ ID NO:1).
Домен SET последовательности с номером доступа GenBank NP 004447 (SEQ ID NO:3) охватывает аминокислотные остатки 618-731 и идентичен SEQ ID NO:6. Остаток тирозина, соответствующий Y641 в последовательности с номером доступа Swiss-Prot Q_15910, показанный подчеркиванием в SEQ ID NO:6, представляет собой Tyr646 (Y646) в последовательности GenBank с номером доступа NP 004447 (SEQ ID NO:3).
Домен SET в последовательности с номером доступа GenBank NP_694543 (SEQ ID NO:5) охватывает аминокислотные остатки 574-687 и идентичен SEQ ID NO:6. Остаток тирозина, соответствующий Y641 в последовательности Swiss-Prot с номером доступа Q_15910, показанный подчеркиванием в SEQ ID NO:6, представляет собой Tyr602 (Y602) в последовательности GenBank с номером доступа NP_694543 (SEQ ID NO:5).
Нуклеотидная последовательность, кодирующая домен SET последовательности с номером доступа GenBank NP_004447 представляет собой
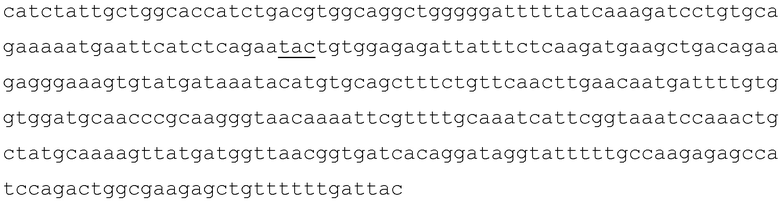
SEQ ID NO:7, где кодон, кодирующий Y641, показан подчеркиванием.
Для целей настоящей заявки следует понимать, что аминокислотный остаток Y641 EZH2 человека относится к остатку тирозина, который представляет собой или соответствует Y641 в последовательности с номером доступа Swiss-Prot Q_15910.

где x может представлять собой любой аминокислотный остаток, отличный от тирозина (Y).
Также для целей настоящей изобретения следует понимать, что мутант EZH2 по Y641 человека и, эквивалентно, мутант EZH2 по Y641 относится к EZH2 человека, в которой аминокислотный остаток, соответствующий Y641 EZH2 человека дикого типа, заменен аминокислотным остатком, отличным от тирозина.
В одном варианте осуществления аминокислотная последовательность мутанта EZH2 по Y641 отличается от аминокислотной последовательности EZH2 человека дикого типа только заменой одного аминокислотного остатка, соответствующего Y641 EZH2 человека дикого типа, аминокислотным остатком, отличным от тирозина.
В одном варианте осуществления аминокислотная последовательность мутанта EZH2 по Y641 отличается от аминокислотной последовательности EZH2 человека дикого типа только заменой фенилаланином (F) одного аминокислотного остатка, соответствующего Y641 EZH2 человека дикого типа. Мутант EZH2 по Y641 в соответствии с данным вариантом осуществления обозначают в настоящем описании как мутант Y641F или, эквивалентно, Y641F.

В одном варианте осуществления аминокислотная последовательность мутанта EZH2 по Y641 отличается от аминокислотной последовательности EZH2 человека дикого типа только заменой гистидином (H) одного аминокислотного остатка, соответствующего Y641 EZH2 человека дикого типа. Мутант EZH2 по Y641 в соответствии с данным вариантом осуществления обозначают в настоящем описании как мутант Y641H или, эквивалентно, Y641H.

В одном варианте осуществления аминокислотная последовательность мутанта EZH2 по Y641 отличается от аминокислотной последовательности EZH2 человека дикого типа только заменой аспарагином (N) одного аминокислотного остатка, соответствующего Y641 EZH2 человека дикого типа. Мутант EZH2 по Y641 в соответствии с данным вариантом осуществления обозначают в настоящем описании как мутант Y641N или, эквивалентно, Y641N.

В одном варианте осуществления аминокислотная последовательность мутанта EZH2 по Y641 отличается от аминокислотной последовательности EZH2 человека дикого типа только заменой серином (S) одного аминокислотного остатка, соответствующего Y641 EZH2 человека дикого типа. Мутант EZH2 по Y641 в соответствии с данным вариантом осуществления обозначают в настоящем описании как мутант Y641S или, эквивалентно, Y641S.

Толерантность к множественным мутациям Y641 в EZH2 позволяет предположить, что устранение пространственного скучивания может обеспечить лучший доступ для надлежащего размещения более крупного диметиллизина в качестве субстрата для реакции от ди- до триметилирования. Кристаллографический анализ метилтрансфераз белков SET7/9 и G9a показывает, что гидроксилы боковых цепей остатков тирозина активного центра прямо вовлечены во взаимодействия для образования H-связей с амином метил-акцептирующего лизина или непрямо через промежуточную молекулу воды. В то время как более крупный активный центр мутантов Y641 способствует ди- и триметилированию, утрата акцептора водородной связи, представляющего собой гидроксил тирозина, может приводить к неблагоприятной ориентации активного центра для первоначального переноса метила на амин лизина.
Значение указанных результатов для заболеваний у человека становятся очевидными с учетом данных, обобщенно представленных в таблице 1 (см. ниже). Можно ожидать, что клетки, гетерозиготные по EZH2, будут проявлять злокачественный фенотип вследствие эффективного образования H3-K27me1 ферментом WT и эффективного последующего преобразования этой молекулы-предшественника в H3-K27me2 и, особенно, в H3-K27me3, с помощью мутантной формы(форм) фермента.
Сообщалось, что образование H3-K27me1 не исключительно зависит от катализа WT-EZH2. Исследования по нокауту EZH2 и другой субъединицы PRC2, EED, продемонстрировали, что образование H3-K27me1 может катализироваться комплексами PRC2, содержащими либо EZH2, либо родственный белок EZH1, в качестве каталитической субъединицы. Shen, X. et al. (2008) Mol Cell 32: 491-502. Таким образом, каталитическое сопряжение между мутантными видами EZH2 и комплексами PRC2, содержащими либо WT-EZH2, либо WT-EZH1, может быть достаточным для усиления образования H3-K27me2/3 и, таким образом, обеспечивает имеющийся злокачественный фенотип. Таким образом, данные указывают на то, что злокачественный фенотип фолликулярной лимфомы (FL) и диффузной крупно-B-клеточной лимфомы (DLBCL) подтипа лимфомы B-клеток герминального центра (GCB), ассоциированный с экспрессией мутантных форм EZH2, является результатом общего приобретения функции образования триметилированной формы H3-K27. Такая интерпретация данных также помогает согласовать существование ассоциированной с раком сверхэкспрессии белков, ассоциированных с EZH2 или PRC2 (например, PHF19/PCL3), а также генотипов с потерей функции деметилазы H3-K27 гистонов UTX. Утрата активности UTX может быть ферментативно эквивалентной приобретению функции EZH2 в любой ситуации, приводящей к более высоким устойчивым уровням триметилированного H3-K27 в раковых клетках (фиг.4).
Состояние моно-, ди- и триметилирования H3-K27 гистонов ассоциировано с различными функциями при контроле транскрипции. Монометилирование H3-K27 гистонов ассоциировано с активной транскрипцией генов, которые готовы к транскрипции. Cui et al. (2009) Cell Stem Cell 4: 80-93; Barski (2007) Cell 129: 823-37. Напротив, триметилирование H3-K27 гистонов ассоциировано либо с генами, которые подверглись транскрипционной репрессии, либо с генами, которые готовы к транскрипции, когда имеется триметилирование H3-K4 гистонов в цис-положении. Cui et al. (выше); Kirmizis et al. (2007) Genes Dev 18: 1592-1605; Bernstein et al. (2006) Cell 125: 315-26. В общем, предсказано, что изменения в активности комплекса PRC2, описанные для рака, включая мутацию Y641 EZH2, приводят к увеличению триметилированного состояния H3-K27 гистонов и, таким образом, приводят к подавлению транскрипции.
Другое открытие согласно настоящему изобретению состоит в том, что клетки, экспрессирующие мутант EZH2 по Y641, являются, как правило, более чувствительными к низкомолекулярным ингибиторам EZH2, чем клетки, экспрессирующие WT EZH2. В частности, клетки, экспрессирующие мутант EZH2 по Y641, проявляют сниженный рост, деление или пролиферацию, или даже претерпевают апоптоз или некроз после обработки ингибиторами EZH2. Напротив, клетки, экспрессирующие WT EZH2, не отвечают на антипролиферативный эффект ингибиторов EZH2 (фиг.13 и 14). Другим неожиданным открытием согласно настоящему изобретению является то, что клетки, экспрессирующие WT EZH2, могут проявлять сходное состояние метилирования H3-K27 гистонов с клетками, экспрессирующими Y641 EZH2, и что такое состояние метилирования также может коррелировать с чувствительностью к ингибитору EZH2 независимо от мутантного состояния EZH2. Как правило, общие уровни H3-K27me3 являются сходными или более высокими в содержащих мутант по Y641 клеточных линиях, чем в клеточных линиях, экспрессирующих WT EZH2; однако уровни H3-K27me2 являются значительно более низкими в мутантных по Y641 клеточных линиях EZH2 и определенных клеточных линиях дикого типа, таких как клеточная линия Pfeiffer, чем в других клеточных линиях дикого типа (фиг.9, 10 и 11). Таким образом, соотношение сигнала H3-K27me2/me3 в мутантных по Y641 линиях и клеточной линии Pfeiffer является значительно более низким, чем соотношение, наблюдаемое в других линиях WT. Настоящие данные далее демонстрируют, что клеточные линии с низким сигналом H3-K27me2 и сходным или более высоким сигналом H3-K27me3 относительно типичных клеточных линий, экспрессирующих WT EZH2, являются более чувствительными к низкомолекулярным ингибиторам EZH2. В частности, клетки с низким сигналом H3-K27me2 и нормальным или высоким сигналом H3K27me3 прекращают делиться или даже погибают после обработки ингибиторами EZH2 (фиг.9, 10, 11, 13 и 14). Напротив, клетки с более высоким соотношением сигнала H3-K27me2/me3 не отвечают на антипролиферативный эффект ингибиторов EZH2 (фиг.9, 10, 11, 13 и 14). Настоящее изобретение обеспечивает ранее неизвестные и неожиданные результаты, состоящие в том, что идентификацию мутаций Y641 EZH2 в опухолях пациента и/или выявление низких уровней H3-K27me2 и нормальных или высоких уровней H3-K27me3 относительно контроля, с использованием таких способов, как вестерн-блоттинг, МС или IHC, у пациента, можно использовать для установления того, какой пациент ответит на лечение ингибитором EZH2.
Было предположено, что EZH2 и другие метилтрансферазы белков являются привлекательными мишенями для разработки лекарственных средств. Copeland et al. (2009) Nat Rev Drug Discov 8: 724-32; Copeland et al. (2010) Curr Opin Chem Biol 14(4): 505-10; Pollock et al. (2010) Drug Discovery Today: Therapeutic Strategies 6(1): 71-9. Настоящие данные также предполагают экспериментальную стратегию для разработки лекарственных средств, специфичных к лимфомам FL и GCB. Поскольку различия в распознавании субстрата между мутантами WT и ассоциированными с заболеванием мутантами являются результатом взаимодействий переходного состояния, низкомолекулярные ингибиторы, которые селективно имитируют переходное состояние мутанта EZH2 относительно переходного состояния фермента WT, должны оказаться эффективными в отношении блокирования метилирования H3-K27 в несущих мутацию клетках. Можно ожидать, что ингибиторы этого типа проявят большой терапевтический индекс, поскольку опосредуемая мишенью токсичность будет минимальной для любых клеток, несущих только фермент WT. Было показано, что мимикрия переходного состояния является эффективной стратегией для конструирования лекарственных средств для многих заболеваний. См., например, Copeland, R. A. Enzymes: A Practical Introduction to Structure, Mechanism и Data Analysis. 2nd ed, (Wiley, 2000).
Настоящие результаты указывают на ранее неизвестную неожиданную зависимость патогенеза фолликулярной лимфомы и диффузной крупно-B-клеточной лимфомы от ферментативного сопряжения между ферментами, которые выполняют монометилирование H3-K27, и определенными мутантными формами EZH2. Без связи с какой-либо теорией, полагают, что эти данные представляют собой первый пример заболевания человека, которое зависит от такого сопряжения каталитической активности между нормальными (WT) и ассоциированными с заболеванием мутантными (Y641) ферментами.
Один аспект изобретения представляет собой способ ингибирования преобразования у индивидуума H3-K27 в триметилированный H3-K27. Ингибирование может вовлекать ингибирование у индивидуума преобразования неметилированного H3-K27 в монометилированный H3-K27, преобразования монометилированного H3-K27 в диметилированный H3-K27, преобразования диметилированного H3-K27 в триметилированный H3-K27 или любую их комбинацию, включая, например, преобразование монометилированного H3-K27 в диметилированный H3-K27 и преобразование диметилированного H3-K27 в триметилированный H3-K27. Как используют в настоящем изобретении, неметилированный H3-K27 относится к гистону H3 без метильной группы, ковалентно связанной с аминогруппой лизина 27. Как используют в настоящем изобретении, монометилированный H3-K27 относится к гистону H3 с одной метильной группой, ковалентно связанной с аминогруппой лизина 27. Монометилированный H3-K27 также обозначают в настоящем описании как H3-K27me1. Как используют в настоящем изобретении, диметилированный H3-K27 относится к гистону H3 с двумя метильными группами, ковалентно связанными с аминогруппой лизина 27. Диметилированный H3-K27 также обозначают в настоящем описании как H3-K27me2. Как используют в настоящем изобретении, триметилированный H3-K27 относится к гистону H3 с тремя метильными группами, ковалентно связанными с аминогруппой лизина 27. Триметилированный H3-K27 также обозначают в настоящем описании как H3-K27me3.
Гистон H3 представляет собой белок длиной 136 аминокислот, последовательность которого известна. См., например, номер доступа GenBank CAB02546, содержание которого включено в настоящее описание посредством ссылки. Как далее описано в настоящем описании, в дополнение к полноразмерному гистону H3, в качестве субстрата для EZH2 (и аналогично для мутантных форм EZH2) можно использовать пептидные фрагменты гистона H3, содержащие остаток лизина, соответствующий K27 полноразмерного гистона H3, в целях оценки преобразования H3-K27m1 в H3-K27m2 и преобразования H3-K27m2 в H3-K27m3. В одном варианте осуществления такой пептидный фрагмент соответствует аминокислотным остаткам 21-44 гистона H3. Такой пептидный фрагмент имеет аминокислотную последовательность LATKAARKSAPATGGVKKPHRYRP (SEQ ID NO:13).
Способ вовлекает введение индивидууму, у которого экспрессируется мутант EZH2 по Y641, терапевтически эффективного количества ингибитора EZH2, где ингибитор ингибирует активность метилтрансферазы гистонов EZH2, тем самым ингибируя преобразование H3-K27 в триметилированный H3-K27 у индивидуума. В одном варианте осуществления индивидуум, у которого экспрессируется мутант EZH2 по Y641, относится к индивидууму, имеющему поддающееся выявлению количество мутанта полипептида EZH2 по Y641. В одном варианте осуществления индивидуум, у которого экспрессируется мутант EZH2 по Y641, относится к индивидууму, имеющему поддающееся выявлению количество нуклеиновой кислоты, кодирующей мутант полипептида EZH2 по Y641.
Мутант полипептида EZH2 по Y641 можно выявлять с использованием любого подходящего способа. Например, мутант полипептида EZH2 по Y641 можно выявлять с использованием антитела, которое специфично связывается с мутантом полипептида EZH2 по Y641 или с пептидным фрагментом, который является характерным для мутанта полипептида EZH2 по Y641. Пептидный фрагмент, который является характерным для мутанта полипептида EZH2 по Y641, может включать, например, домен SET, представленный в SEQ ID NO:6, за исключением замены Y641 аминокислотным остатком, отличным от тирозина. В другом варианте осуществления пептидный фрагмент, который характерен для мутанта полипептида EZH2 по Y641, может включать, например, фрагмент из 10-113 аминокислот домена SET, представленного в SEQ ID NO:6, за исключением замены Y641 аминокислотным остатком, отличным от тирозина, при условии, что фрагмент включает аминокислотный остаток, соответствующий Y641. Ожидается, что эпитоп для такого антитела включает аминокислотный остаток, соответствующий Y641 EZH2 дикого типа. Антитело считается специфично связывающимся с мутантом полипептида EZH2 по Y641 или с пептидным фрагментом, который является характерным для мутанта полипептида EZH2 по Y641, если он связывается с мутантным полипептидом EZH2 или его пептидным фрагментом, но не соответствующим полипептидом EZH2 дикого типа или его пептидным фрагментом. В одном варианте осуществления такое антитело считают специфично связывающимся с мутантом полипептида EZH2 по Y641 или с пептидным фрагментом, который является характерным для мутанта полипептида EZH2 по Y641, если оно связывается с этим мутантным полипептидом EZH2 или его пептидным фрагментом с аффинностью, которая, по меньшей мере, приблизительно в 100 раз превышает аффинность в отношении соответствующего полипептида EZH2 дикого типа или его пептидного фрагмента. В одном варианте осуществления такое антитело считают специфично связывающимся с мутантом полипептида EZH2 по Y641 или с пептидным фрагментом, который является характерным для мутанта полипептида EZH2 по Y641, если оно связывается с мутантным полипептидом EZH2 или его пептидным фрагментом с аффинностью, которая, по меньшей мере, приблизительно в 1000 раз превышает аффинность в отношении соответствующего полипептида EZH2 дикого типа или его пептидного фрагмента. Антитело можно использовать, например, в твердофазном иммуноферментном анализе (ELISA) или анализе с использованием вестерн-блоттинга.
В одном варианте осуществления антитело представляет собой моноклональное антитело. Моноклональное антитело можно получать общепринятыми способами, хорошо известными в данной области. См., например, Kohler and Milstein (1975) Nature 256 (5517): 495-7.
В качестве другого примера мутант полипептида EZH2 по Y641 можно выявлять с использованием масс-спектрометрии (МС), например, времяпролетной электрораспылительной ионизации (ESI-TOF) или времяпролетной ионизации лазерной десорбцией с использованием матрицы (MALDI-TOF). Такие способы хорошо известны в данной области. Анализ вовлекает идентификацию одного или нескольких пептидных фрагментов, содержащих представляющую интерес мутацию, например, пептида длиной 12-24 аминокислот, содержащего последовательность, охватывающую аминокислоту, соответствующую Y641 в EZH2 дикого типа.
Нуклеиновая кислота, кодирующая мутант полипептида EZH2 по Y641 или пептидный фрагмент, который является характерным для мутанта полипептида EZH2 по Y641, можно выявлять с использованием любого подходящего способа. Например, нуклеиновую кислоту, кодирующую мутант полипептида EZH2 по Y641, можно выявлять с использованием полногеномного ресеквенирования или ресеквенирования области-мишени (последнее также известно, как нацеленное ресеквенирование) с использованием подходящим образом выбранных источников ДНК и праймеров для полимеразной цепной реакции (ПЦР) в соответствии со способами, хорошо известными в данной области. См., например, Bentley (2006) Curr Opin Genet Dev. 16: 545-52, и Li et al. (2009) Genome Res 19: 1124-32. Способ, обычно и, как правило, включает стадии очистки геномной ДНК, амплификации способом ПЦР представляющей интерес области, циклического секвенирования, очистки после реакции секвенирования, капиллярного электрофореза и анализа данных. Высококачественные праймеры ПЦР для охвата представляющей интерес области конструируют с использованием инструментов для конструирования праймеров in silico. Циклическое секвенирование представляет собой простой способ, в котором последовательные раунды денатурации, отжига и удлинения в термоблоке приводят к линейной амплификации продуктов удлинения. Продукты, как правило, оканчиваются флуоресцентной меткой, которая идентифицирует концевое нуклеотидное основание как G, A, T или C. Не встроившиеся красящие концевые метки и соли, которые могут конкурировать при капиллярной электрофоретической инжекции, удаляют промыванием. В ходе капиллярного электрофореза продукты реакции циклического секвенирования мигрируют через капилляры, заполненные полимером. Отрицательно заряженные фрагменты ДНК разделяются по размеру по мере того, как они продвигаются через капилляры в направлении положительного электрода. После электрофореза программное обеспечение для сбора данных создает файл для образца с необработанными данными. С использованием последующих приложений программного обеспечения проводят дальнейший анализ данных для преобразования собранных данных в виде цветовых изображений в соответствующие нуклеотидные основания. Альтернативно или дополнительно, способ может включать использование улавливания области-мишени из геномной ДНК на основе микрочипов и/или секвенирование. Наборы, реагенты и способы выбора соответствующих праймеров для ПЦР и проведения ресеквенирования являются коммерчески доступными, например, от Applied Biosystems, Agilent, и NimbleGen (Roche Diagnostics GmbH). Способы, такие как указанные, используют для выявления мутаций JAK2 и гена миелопролиферативного лейкоза (MPL) и диагностики истинной полицитемии, эссенциальной тромбоцитемии и идиопатического миелофиброза. Для применения в настоящем изобретении праймеры для ПЦР можно выбрать таким образом, чтобы амплифицировать, например, по меньшей мере, соответствующую часть SEQ ID NO:7 (выше).
Альтернативно или дополнительно, нуклеиновую кислоту, кодирующую мутант полипептида EZH2 по Y641, можно выявлять с использованием саузерн-блоттинга в соответствии со способами, хорошо известными в данной области. В одном варианте осуществления последовательность ДНК, кодирующую мутант полипептида EZH2 по Y641, выявляют с использованием гибридизации нуклеиновых кислот, проводимой в условиях высокой жесткости. Зонд нуклеиновой кислоты выбирают таким образом, чтобы его последовательность была комплементарна являющейся мишенью последовательности нуклеиновой кислоты, которая включает кодон для мутантной аминокислоты, соответствующий Y641 EZH2 дикого типа.
Специфичный к последовательности зонд комбинируют с образцом, подлежащим исследованию, в условиях высокой жесткости. Термин «условия высокой жесткости», как используют в настоящем изобретении, относится к параметрам, известным в данной области. Параметры гибридизации нуклеиновых кислот могут быть найдены в ссылках, в которых собраны такие способы, например, J. Sambrook, et al, eds., Molecular Cloning: A Laboratory Manual, Second Edition, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1989, или F. M. Ausubel, et al, eds., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., New York. Более конкретно, условия высокой жесткости, как используют в настоящем изобретении, относится, например, к гибридизации при 65°C в буфере для гибридизации (3,5×SSC, 0,02% Ficoll, 0,02% поливинилпирролидон, 0,02% бычий сывороточный альбумин (BSA), 2,5 мМ NaH2PO4 (pH 7), 0,5% SDS, 2 мМ EDTA). SSC представляет собой 0,15M хлорид натрия/0,015M цитрат натрия, pH 7; SDS представляет собой додецилсульфат натрия; и EDTA представляет собой этилендиаминтетрауксусную кислоту. После гибридизации мембрану, на которую перенесена ДНК, промывают, например, в 2×SSC при комнатной температуре, и затем при 0,1-0,5×SSC/0,1×SDS при температурах вплоть до 68°C.
Существуют другие условия, реагенты и т.д., которые можно использовать, обеспечивающие сходную степень жесткости. Специалисту в данной области известны такие условия, и, таким образом, они не приведены в настоящем описании. Однако следует понимать, что специалист в данной области способен манипулировать условиями таким образом, чтобы обеспечить четкую идентификацию ассоциированных с EZH2 нуклеиновых кислот по изобретению, включая, в частности, нуклеиновые кислоты, кодирующие мутанты EZH2 по Y641 (например, с использованием условий более низкой жесткости). Специалисту в данной области также известны технологии скрининга клеток и библиотек для экспрессии таких молекул, которые затем выделяют общепринятыми способами, с последующим выделением подходящей молекулы нуклеиновой кислоты и секвенированием.
Индивидууму вводят терапевтически эффективное количество ингибитора EZH2. Как используют в настоящем изобретении, ингибитор EZH2 относится, главным образом, к низкомолекулярному соединению, т.е. к молекуле с молекулярной массой менее чем приблизительно 1,5 килодальтон (кДа), которое способно препятствовать ферментативной активности метилтрансферазы гистонов EZH2.
В одном варианте осуществления ингибитор EZH2 ингибирует активность метилтрансферазы гистонов EZH2 дикого типа. В одном варианте осуществления ингибитор EZH2 ингибирует активность метилтрансферазы гистонов мутанта EZH2 по Y641. В одном варианте осуществления ингибитор EZH2 ингибирует активность метилтрансферазы гистонов EZH2 дикого типа и активность метилтрансферазы гистонов мутанта EZH2 по Y641. В одном варианте осуществления ингибитор EZH2 селективно ингибирует активность метилтрансферазы гистонов мутанта EZH2 по Y641.
Как описано в настоящем описании, определенные мутанты EZH2 по Y641 являются относительно слабыми катализаторами преобразования неметилированного H3-K27 в H3-K27me1 и, тем не менее, являются неожиданно эффективными катализаторами для преобразования H3-K27me2 в H3-K27me3. Напротив, EZH2 дикого типа является относительно эффективным катализатором для преобразования неметилированного H3-K27 в H3-K27me1 и, тем не менее, является неожиданно неэффективным катализатором для преобразования H3-K27me2 в H3-K27me3. Это является важным, поскольку моно-, ди- и триметилированные состояния H3-K27 проявляют различные функции в контроле транскрипции. Например, H3-K27me1 ассоциирован с активной транскрипцией генов, которые готовы к транскрипции, в то время как H3-K27me3 ассоциирован с генами, подвергнутыми транскрипционной репрессии, или генами, которые готовы к транскрипции, когда имеется триметилирование H3-K4 в цис-положении. Таким образом, селективное ингибирование активности метилтрансферазы гистонов мутанта EZH2 по Y641 обеспечивает селективное ингибирование продуцирования триметилированной формы H3-K27, тем самым способствуя транскрипции, ассоциированной с H3-K27me1, и препятствуя подавлению транскрипции, ассоциированной с H3-K27me3.
Ингибитор EZH2 «селективно ингибирует» активность метилтрансферазы гистонов мутанта EZH2 по Y641, когда он ингибирует активность метилтрансферазы гистонов мутанта EZH2 по Y641 более эффективно, чем он ингибирует активность метилтрансферазы гистонов EZH2 дикого типа. Например, в одном варианте осуществления селективный ингибитор имеет IC50 для мутанта EZH2 по Y641, которая, по меньшей мере, на 40 процентов ниже, чем IC50 для EZH2 дикого типа. В одном варианте осуществления селективный ингибитор имеет IC50 для мутанта EZH2 по Y641, которая, по меньшей мере, на 50 процентов ниже, чем IC50 для EZH2 дикого типа. В одном варианте осуществления селективный ингибитор имеет IC50 для мутанта EZH2 по Y641, которая, по меньшей мере, на 60 процентов ниже, чем IC50 для EZH2 дикого типа. В одном варианте осуществления селективный ингибитор имеет IC50 для мутанта EZH2 по Y641, которая, по меньшей мере, на 70 процентов ниже, чем IC50 для EZH2 дикого типа. В одном варианте осуществления селективный ингибитор имеет IC50 для мутанта EZH2 по Y641, которая, по меньшей мере, на 80 процентов ниже, чем IC50 для EZH2 дикого типа. В одном варианте осуществления селективный ингибитор имеет IC50 для мутанта EZH2 по Y641, которая, по меньшей мере, на 90 процентов ниже, чем IC50 для EZH2 дикого типа.
В одном варианте осуществления селективный ингибитор мутанта EZH2 по Y641 по существу не обладает ингибиторным эффектом в отношении EZH2 дикого типа.
Ингибитор ингибирует преобразование H3-K27me2 в H3-K27me3. В одном варианте осуществления ингибитор считают ингибирующим триметилирование H3-K27. Поскольку преобразование H3-K27me1 в H3-K27me2 предшествует преобразованию H3-K27me2 в H3-K27me3, ингибитор преобразования H3-K27me1 в H3-K27me2 естественным образом также ингибирует преобразование H3-K27me2 в H3-K27me3, т.е. он ингибирует триметилирование H3-K27. Также возможно ингибировать преобразование H3-K27me2 в H3-K27me3 без ингибирования преобразования H3-K27me1 в H3-K27me2. Ингибирование этого типа также может приводить к ингибированию триметилирования H3-K27, хотя и без ингибирования диметилирования H3-K27.
В одном варианте осуществления ингибитор ингибирует преобразование H3-K27me1 в H3-K27me2 и преобразование H3-K27me2 в H3-K27me3. Такой ингибитор может прямо ингибировать только преобразование H3-K27me1 в H3-K27me2. Альтернативно, такой ингибитор может прямо ингибировать как преобразование H3-K27me1 в H3-K27me2, так и преобразование H3-K27me2 в H3-K27me3.
Ингибитор ингибирует активность метилазы гистонов. Ингибирование активности метилазы гистонов можно выявлять с использованием любого подходящего способа. Ингибирование можно определить, например, либо в значениях уровня активности метилазы гистонов, либо в качестве активности продукта метилазы гистонов. Способы, подходящие для любых из полученных данных, включены в примеры ниже.
Ингибирование представляет собой поддающееся измерению ингибирование по сравнению с подходящим отрицательным контролем. В одном варианте осуществления ингибирование представляет собой ингибирование, по меньшей мере, на 10 процентов по сравнению с подходящим отрицательным контролем. Иными словами, уровень ферментативной активности или количество продукта с ингибитором являются меньшими или равны 90 процентам соответствующего уровня или количества без ингибитора. В различных других вариантах осуществления ингибирование представляет собой ингибирование, по меньшей мере, на 20, 25, 30, 40, 50, 60, 70, 75, 80, 90 или 95 процентов по сравнению с подходящим отрицательным контролем. В одном варианте осуществления ингибирование представляет собой ингибирование, по меньшей мере, на 99 процентов по сравнению с подходящим отрицательным контролем. Иными словами, уровень ферментативной активности или количество продукта с ингибитором являются меньшими или равны 1 проценту от соответствующего уровня или количества без ингибитора.
В одном варианте осуществления ингибитор представляет собой S-аденозил-L-гомоцистеин (SAH). SAH имеет структурную формулу:
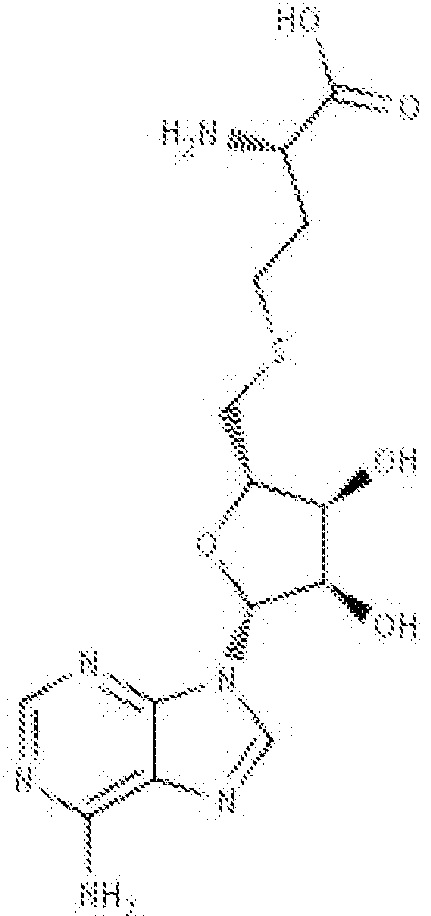 (SAH)
(SAH)
и коммерчески доступен от ряда поставщиков, включая, например, Sigma-Aldrich, St. Louis, MO. SAH описан в качестве ингибитора трансметилирования S-аденозилметионин-зависимыми метилтрансферазами.
В одном варианте осуществления ингибитор представляет собой соединение 75:
(75)
или его фармацевтически приемлемую соль.
В определенных вариантах осуществления изобретение включает стадию проведения анализа для выявления мутанта EZH2 по Y641 в образце от индивидуума. Анализы данного типа описаны выше. Как используют в настоящем изобретении, «образец от индивидуума» относится к любому подходящему образцу, содержащему клетки или компоненты клеток, полученные или происходящие из индивидуума. В одном варианте осуществления образец включает клетки, предположительно экспрессирующие мутант EZH2 по Y641, например, раковые клетки. В одном варианте осуществления образец представляет собой образец крови. В одном варианте осуществления образец представляет собой биоптат, полученный, например, из лимфатической ткани (например, лимфатический узел) или костного мозга. В одном варианте осуществления образец представляет собой биоптат, полученный из ткани, отличной от или в дополнение к лимфатической ткани (например, лимфатический узел), или костного мозга. Например, в одном варианте осуществления образец представляет собой биоптат рака, например, опухоли, состоящей из раковых клеток. Клетки в образце можно выделять из других компонентов образца. Например, мононуклеарные клетки периферической крови (PBMC) можно выделять в виде лейкоцитарной пленки из образца крови, центрифугированного способами, известными специалистам в данной области.
Когда результат анализа образца индивидуума указывает на то, что мутант EZH2 по Y641 присутствует в образце, то утверждают, что у индивидуума экспрессируется мутант EZH2 по Y641. Действительно, в одном варианте осуществления, когда результат анализа образца от индивидуума указывает на то, что мутант EZH2 по Y641 присутствует в образце, индивидуума идентифицируют в качестве кандидата для лечения ингибитором EZH2, где ингибитор селективно ингибирует активность метилтрансферазы гистонов мутанта EZH2 по Y641.
Когда результат анализа образца рака указывает на то, что в данном типе рака присутствует мутант EZH2 по Y641, то утверждают, что рак экспрессирует мутант EZH2 по Y641.
Аналогично, когда результат анализа образца, содержащего раковые клетки индивидуума, имеющего рак, указывают на то, что в образце присутствует мутант EZH2 по Y641, то утверждают, что у индивидуума экспрессируется мутант EZH2 по Y641.
Настоящее изобретение также относится к ранее неизвестной, неожиданной корреляции способности пациента отвечать на ингибитор EZH2 с уровнем H3-K27me2 или предпочтительно с уровнями H3-K27me и H3-K27me3. Например, клетки с низкими уровнями H3-K27me2 и нормальными или высокими уровнями me3 относительно контроля в значительно большей степени отвечают на антипролиферативный эффект ингибитора EZH2, чем клетки с нормальными уровнями H3-K27me2 и me3.
Один из аспектов изобретения представляет собой способ определения способности индивидуума отвечать на ингибитор EZH2. В одном варианте осуществления способ включает выделение образца ткани из индивидуума; выявление уровня диметилирования (me2) H3-K27 в образце ткани; сравнение уровня диметилирования (me2) с контрольным уровнем диметилирования (me2); и идентификацию того, что индивидуум отвечает на указанный ингибитор EZH2, когда уровень диметилирования (me2) отсутствует или ниже контрольного уровня диметилирования (me2). В одном варианте осуществления способ дополнительно включает выявление уровня триметилирования (me3) H3-K27 в образце ткани; сравнение уровня триметилирования (me3) с контрольным уровнем триметилирования (me3) и уровня диметилирования (me2) с контрольным уровнем диметилирования (me2); и идентификацию того, что указанный индивидуум отвечает на ингибитор EZH2, когда уровень триметилирования (me3) является таким же или превышает контрольный уровень триметилирования (me3), и уровень диметилирования (me2) отсутствует или является более низким, чем контрольный уровень диметилирования (me2). В другом варианте осуществления способ дополнительно включает получение соотношения уровня диметилирования (me2) и уровня триметилирования (me3) H3-K27 в образце ткани; получение контрольного соотношения контрольного уровня диметилирования (me2) и контрольного уровня триметилирования (me3); сравнение соотношения с контрольным соотношением; и идентификацию того, что индивидуум отвечает на указанный ингибитор EZH2, когда указанное соотношение является более низким, чем указанное контрольное соотношение. В предпочтительном варианте осуществления индивидуум имеет рак. В одном варианте осуществления рак представляет собой фолликулярную лимфому. Альтернативно, рак представляет собой диффузную крупно-B-клеточную лимфому (DLBCL). В другом предпочтительном варианте осуществления у индивидуума экспрессируется мутант EZH2 по Y641. В предпочтительном варианте осуществления мутант по Y641 представляет собой Y641F, Y641H, Y641N или Y641S.
Выявление диметилированного H3-K27 или триметилированного H3-K27 можно проводить с использованием любого подходящего способа в данной области. В одном варианте осуществления уровень метилирования выявляют с использованием антител, специфичных к диметилированному H3-K27 или триметилированному H3-K27. Например, выделенную ткань фиксируют в формалине и погружают в парафиновые блоки для длительного хранения. Блоки можно использовать для получения препаратов для иммуногистохимического окрашивания или флуоресцентного окрашивания антителами против метилированного H3-K27. Альтернативно, из выделенного образца ткани можно получать лизаты цельных клеток и экстракты гистонов и затем использовать их для иммуногистохимического окрашивания, анализа с использованием вестерн-блоттинга или флуоресцентного окрашивания. В другом варианте осуществления уровень метилирования выявляют с использованием полипептида или аптамера, специфичного к диметилированному H3-K27 или триметилированному H3-K27. В другом варианте осуществления уровень метилирования определяют с использованием масс-спектрометрии (МС).
Контрольный диметилированный H3-K27 или контрольный триметилированный H3-K27 можно получать из контрольного образца, например, соседней неопухолевой ткани, выделенной из индивидуума, или здоровой ткани от здорового индивидуума. Альтернативно, контрольный уровень метилирования H3-K27me2 или H3-K27me3 может быть установлен патологом способами, известными в данной области.
Способы скрининга
Один аспект изобретения представляет собой способ идентификации тестируемого соединения в качестве ингибитора мутанта EZH2 по Y641. В одном варианте осуществления способ включает комбинирование выделенного мутанта EZH2 по Y641 с гистонным субстратом, донором метильных групп (таким как S-аденозилметионин (SAM)) и тестируемым соединением, где гистонный субстрат содержит форму H3-K27, выбранную из группы, состоящей из неметилированного H3-K27, монометилированного H3-K27, диметилированного H3-K27 и любой их комбинации; и проведение анализа для выявления метилирования H3-K27 в гистонном субстрате, тем самым идентифицируя тестируемое соединение в качестве ингибитора мутанта EZH2 по Y641, когда метилирование H3-K27 в присутствии тестируемого соединения является меньшим, чем метилирование H3-K27 в отсутствие тестируемого соединения. Анализ для выявления метилирования H3-K27 может быть выбран таким образом, чтобы измерять скорость метилирования, степень метилирования или как скорость, так и степень метилирования.
Мутант EZH2 по Y641 выделяют в виде комплекса PRC2 или его функционального эквивалента. Как используют в настоящем изобретении, термин «выделенный» означает по существу отделенный от других компонентов, с которыми комплекс может быть выявлен, как он встречается в природе. Соединение можно выделять, необязательно очищая его. В одном варианте осуществления мутант EZH2 выделяют в виде комплекса мутанта EZH2 по Y641 вместе с EED и SUZ12. В другом варианте осуществления мутант EZH2 выделяют в виде комплекса мутанта EZH2 по Y641 вместе с EED, SUZ12 и RbAp48. В соответствующих условиях комплекс PRC2 или его функциональный эквивалент проявляет активность метилтрансферазы гистонов в отношении H3-K27. В одном варианте осуществления комплекс состоит из рекомбинантно экспрессируемых составляющих полипептидов, например, EZH2, EED, SUZ12, с RbAp48 или без него.
Выделенный мутант EZH2 по Y641 комбинируют с гистонным субстратом. Гистонный субстрат включает любой подходящий источник полипептидов гистонов или их фрагментов, которые могут служить в качестве субстрата для EZH2. В одном варианте осуществления гистонный субстрат включает выделенные гистоны от индивидуума. Гистоны можно выделять из клеток индивидуума с использованием любого подходящего способа; такие способы хорошо известны специалистам в данной области и нет необходимости в их дальнейшем уточнении в настоящем описании. См., например, Fang et al. (2004) Methods Enzymol 377: 213-26. В соответствии с примерами ниже, в одном варианте осуществления гистонный субстрат предоставляют в виде нуклеосом. В соответствии с примерами ниже, в одном варианте осуществления гистонный субстрат предоставляют в виде нуклеосом эритроцитов птиц (курицы).
Гистонный субстрат, предоставленный таким образом, может включать смесь состояний модификаций гистонов, включая различные состояния метилирования H3-K27, исходя из вестерн-блоттинга с антителами, специфичными к состоянию метилирования H3-K27. В одном варианте осуществления гистонный субстрат может быть предоставлен в виде очищенного полноразмерного гистона H3. Такой очищенный полноразмерный гистон H3 может быть предоставлен в виде гомогенного препарата в отношении состояний метилирования H3-K27 или в виде смеси различных состояний метилирования H3-K27. Гомогенные препараты выделенного гистона H3 в отношении состояний метилирования H3-K27 можно получать, частично путем пропускания через иммуноаффинную колонку, нагруженную подходящими антителами, специфичными к состоянию метилирования H3-K27, или путем иммунопреципитации с использованием магнитных гранул, покрытых подходящими антителами, специфичными к состоянию метилирования H3-K27. Альтернативно или дополнительно, состояние метилирования H3-K27 может быть охарактеризовано как часть проведения анализа. Например, исходный материал гистонного субстрата может быть охарактеризован, как содержащий 50 процентов неметилированного H3-K27, 40 процентов монометилированного H3-K27, 10 процентов диметилированного H3-K27 и 0 процентов триметилированного H3-K27.
В одном варианте осуществления гистонный субстрат включает пептидную библиотеку или подходящий пептид, содержащий одну или более аминокислотных последовательностей, родственных гистону H3, включая, в частности, последовательность, которая охватывает H3-K27. Например, в одном варианте осуществления гистонный субстрат представляет собой пептидный фрагмент, который соответствует аминокислотным остаткам 21-44 гистона H3. Такой пептидный фрагмент имеет аминокислотную последовательность LATKAARKSAPATGGVKKPHRYRP (SEQ ID NO:13). Библиотеку пептидов или пептид можно получать путем пептидного синтеза способами, хорошо известными в данной области, и необязательно модифицировать так, чтобы они включали любую желаемую степень метилирования лизина, соответствующего H3-K27. Как описано в примерах ниже, такие пептиды также могут быть модифицированы, чтобы они включали метку, такую как биотин, пригодную для проведения последующих анализов. В одном варианте осуществления метка присоединена к амино(N)-концу пептида(ов). В одном варианте осуществления метка присоединена к карбокси(C)-концу пептида(ов).
Антитела, специфичные к метилированию H3-K27, доступны из различных коммерческих источников, включая, например, Cell Signaling Technology (Danvers, MA) и Active Motif (Carlsbad, CA).
Выделенный мутант EZH2 по Y641 комбинируют с тестируемым соединением. Как используют в настоящем изобретении, «тестируемое соединение» относится к низкомолекулярному органическому соединению, имеющему молекулярную массу менее чем приблизительно 1,5 кДа. В одном варианте осуществления тестируемое соединение представляет собой известное соединение. В одном варианте осуществления тестируемое соединение представляет собой новое соединение. В одном варианте осуществления тестируемое соединение может быть предоставлено в виде части библиотеки таких соединений, где библиотека включает, например, десятки, сотни, тысячи или даже более соединений. Библиотека соединений может быть подвергнута преимущественно скринингу в высокопроизводительном скрининговом анализе, например, с использованием матриц тестируемых соединений и роботизированного манипулирования в соответствии с общими способами, хорошо известными в данной области.
В определенных вариантах осуществления тестируемое соединение представляет собой соединение, которое является производным SAH или производным соединения 75.
Выявление метилирования H3-K27 можно проводить с использованием любого подходящего способа. В одном варианте осуществления источник донорных метильных групп включает метильные группы, которые мечены поддающейся детекции меткой. Поддающаяся детекции метка в одном варианте осуществления представляет собой изотопную метку, например, тритий. Другие типы меток могут включать, например, флуоресцентные метки.
Выявление образования триметилированного H3-K27 можно проводить с использованием любого подходящего способа. Например, выявление образования триметилированного H3-K27 можно проводить с использованием анализа для выявления включения меченых метильных групп, такого как описано выше, необязательно в комбинации с хроматографическим или другим способом разделения меченых продуктов по размеру, например, полиакриламидным гельэлектрофорезом (PAGE), капиллярным электрофорезом (CE) или высокоэффективной жидкостной хроматографией (ВЭЖХ). Альтернативно или дополнительно, выявление образования триметилированного H3-K27 можно проводить с использованием антител, которые являются специфичными к триметилированному H3-K27.
Выявление преобразования монометилированного H3-K27 в диметилированный H3-K27 можно проводить с использованием любого подходящего способа. В одном варианте осуществления преобразование измеряют с использованием антител, специфичных к монометилированному H3-K27 и диметилированному H3-K27. Например, исходные количества или концентрации монометилированного H3-K27 и диметилированного H3-K27 можно определять с использованием соответствующих антител, специфичных к монометилированному H3-K27 и диметилированному H3-K27. После комбинирования фермента, субстрата, донора метильных групп и тестируемого соединения, полученные количества или концентрации монометилированного H3-K27 и диметилированного H3-K27 можно определять с использованием соответствующих антител, специфичных к монометилированному H3-K27 и диметилированному H3-K27. Затем можно сравнить начальные и конечные количества или концентрации монометилированного H3-K27 и диметилированного H3-K27. Альтернативно или дополнительно, начальные и конечные количества или концентрации монометилированного H3-K27 и диметилированного H3-K27 затем можно сравнить с соответствующими количествами или концентрациями отрицательного контроля. Реакцию отрицательного контроля, в которой в анализ не включено тестируемое вещество, можно проводить параллельно или в качестве исторического контроля. Результаты такой контрольной реакции необязательно можно вычесть из соответствующих результатов экспериментальной реакции до или одновременно с проведением сравнения, указанного выше.
Поскольку диметилированная форма H3-K27 может далее метилироваться в указанном анализе, может оказаться, что уменьшение количества или концентрации монометилированного H3-K27 не соответствует прямо увеличению количества диметилированного H3-K27. В этом случае, однако, можно предположить, что снижение количества или концентрации монометилированного H3-K27 само по себе отражает преобразование монометилированного H3-K27 в диметилированный H3-K27.
Выявление преобразования диметилированного H3-K27 в триметилированный H3-K27 можно проводить с использованием любого подходящего способа. В одном варианте осуществления преобразование измеряют с использованием антител, специфичных к диметилированному H3-K27 и триметилированному H3-K27. Например, исходные количества или концентрации диметилированного H3-K27 и триметилированного H3-K27 можно определить с использованием соответствующих антител, специфичных к диметилированному H3-K27 и триметилированному H3-K27. После комбинирования фермента, субстрата и тестируемого соединения, полученные количества или концентрации диметилированного H3-K27 и триметилированного H3-K27 можно определить с использованием соответствующих антител, специфичных к диметилированному H3-K27 и триметилированному H3-K27. Затем можно сравнить начальные и конечные количества или концентрации диметилированного H3-K27 и триметилированного H3-K27. Альтернативно или дополнительно, начальные и конечные количества или концентрации диметилированного H3-K27 и триметилированного H3-K27 затем можно сравнить с соответствующими количествами или концентрациями отрицательного контроля. Реакцию отрицательного контроля, в которой в анализ не включено тестируемое вещество, можно проводить параллельно или в качестве исторического контроля. Результаты такой контрольной реакции необязательно можно вычесть из соответствующих результатов экспериментальной реакции до или одновременно с проведением сравнения, указанного выше.
Тестируемое вещество идентифицируют в качестве ингибитора мутанта EZH2 по Y641, когда метилирование H3-K27 с тестируемым соединением является меньшим, чем метилирование H3-K27 без тестируемого соединения. В одном варианте осуществления тестируемое вещество идентифицируют в качестве ингибитора мутанта EZH2 по Y641, когда образование триметилированного H3-K27 в присутствии тестируемого соединения является меньшим, чем образование триметилированного H3-K27 в отсутствие тестируемого соединения.
Один аспект изобретения представляет собой способ идентификации селективного ингибитора мутанта EZH2 по Y641. В одном варианте осуществления способ включает комбинирование выделенного мутанта EZH2 по Y641 с гистонным субстратом, донором метильной группы (например, SAM) и тестируемым соединением, где гистонный субстрат включает форму H3-K27, выбранную из группы, состоящей из монометилированного H3-K27, диметилированного H3-K27 и комбинации монометилированного H3-K27 и диметилированного H3-K27, тем самым формируя тестируемую смесь; комбинирование выделенного EZH2 дикого типа с гистонным субстратом, донором метильной группы (например, SAM) и тестируемым соединением, где гистонный субстрат включает форму H3-K27, выбранную из группы, состоящей из монометилированного H3-K27, диметилированного H3-K27 и комбинации монометилированного H3-K27 и диметилированного H3-K27, тем самым образуя контрольную смесь; проведение анализа для выявления триметилирования гистонного субстрата в каждой из тестируемой смеси и контрольной смеси; вычисление соотношения (a) триметилирования с мутантом EZH2 по Y641 и тестируемым соединением (M+) и (b) триметилирования с мутантом EZH2 по Y641 без тестируемого соединения (M-); вычисление соотношения (c) триметилирования с EZH2 дикого типа и тестируемым соединением (WT+) и (d) триметилирования с EZH2 дикого типа без тестируемого соединения (WT-); сравнение соотношения (a)/(b) с соотношением (c)/(d); и идентификацию тестируемого соединения в качестве селективного ингибитора мутанта EZH2 по Y641, когда соотношение (a)/(b) является меньшим, чем соотношение (c)/(d). В одном варианте осуществления способ дополнительно включает учет отрицательного контроля без тестируемого соединения для любой или обеих из тестируемой смеси и контрольной смеси.
Фармацевтические композиции
Один или более антагонистов EZH2 можно вводить отдельно пациенту-человеку или в фармацевтических композициях, где они смешаны с подходящими носителями или эксципиентом(ами) в дозах, осуществляющих лечение или смягчающих заболевание или состояние, как описано в настоящем описании. Смеси антагонистов EZH2 также можно вводить пациенту в виде простой смеси или в подходящих составленных фармацевтических композициях. Например, один аспект изобретения относится к фармацевтической композиции, содержащей терапевтически эффективную дозу антагониста EZH2 или его фармацевтически приемлемой соли, гидрата, энантиомера или стереоизомера; и фармацевтически приемлемый разбавитель или носитель.
Способы составления и введения антагонистов EZH2 могут быть найдены в источниках литературы, хорошо известных специалисту в данной области, таких как Remington's «The Science and Practice of Pharmacy», 21st ed., Lippincott Williams & Wilkins 2005.
Подходящие пути введения могут, например, включать пероральное, ректальное или внутрикишечное введение; парентеральную доставку, включая внутривенную, внутримышечную, внутрибрюшинную, подкожную или внутрикостномозговую инъекции, а также интратекальную, прямую внутрижелудочковую или внутриглазную инъекции; местную доставку, включая глазные капли и чрескожное введение; и интраназальный и другие способы доставки через слизистую оболочку.
Альтернативно, можно вводить антагонист EZH2 локальным в отличие от системного пути, например, инъекцией антагониста EZH2 прямо в отечную область, часто в депо-составе или составе для замедленного высвобождения.
В одном варианте осуществления антагонист EZH2 вводят путем прямой инъекции в опухоль или лимфатический узел.
Более того, антагонист EZH2 можно вводить в системе направленной доставки лекарственных средств, например, в липосоме, покрытой антителом, специфичным к раковой клетке.
Фармацевтические композиции по настоящему изобретению можно изготавливать, например, путем общепринятых способов смешения, растворения, грануляции, получения драже, растирания в порошок, эмульгации, инкапсулирования, захвата или лиофилизации.
Фармацевтические композиции для применения в соответствии с настоящим изобретением, таким образом, могут быть изготовлены общепринятым способом с использованием одного или более физиологически приемлемых носителей, содержащих эксципиенты и вспомогательные вещества, которые способствуют переработке активных антагонистов EZH2 в препараты, которые можно использовать фармацевтически. Надлежащий состав зависит от выбранного пути введения.
Для инъекции средства по изобретению можно составлять в водных растворах, предпочтительно в физиологически совместимых буферах, таких как раствор Хенкса, раствор Рингера или забуференный физиологический солевой раствор. Для чрескожного введения в составе используют обеспечивающие проникновение вещества, подходящие для барьера, предназначенного для прохождения. Такие обеспечивающие проникновение вещества, главным образом, известны в данной области.
Для перорального введения антагонисты EZH2 можно без труда составлять путем комбинирования активных антагонистов EZH2 с фармацевтически приемлемыми носителями, хорошо известными в данной области. Такие носители обеспечивают составление антагонистов EZH2 по изобретению в виде таблеток, пилюль, драже, капсул, жидкостей, гелей, сиропов, взвесей, суспензий и т.п., для перорального приема пациентом, подлежащим лечению. Фармацевтические препараты для перорального введения можно получать путем комбинирования активного антагониста EZH2 с твердым эксципиентом, необязательно растирания полученной смеси и переработки смеси гранул, после добавления подходящих вспомогательных веществ, если желательно, с получением таблеток или сердцевин драже. Подходящие эксципиенты включают наполнители, такие как сахара, включая лактозу, сахарозу, маннит или сорбит; препараты целлюлозы, например, такие как кукурузный крахмал, пшеничный крахмал, рисовый крахмал, картофельный крахмал, желатин, трагакантовая камедь, метилцеллюлоза, гидроксипропилметилцеллюлоза, натрийкарбоксиметилцеллюлоза и/или поливинилпирролидон (PVP). Если желательно, можно добавлять дезинтегрирующие вещества, такие как сшитый поливинилпирролидон, агар или альгиновая кислота, или их соль, такая как альгинат натрия.
Сердцевины драже предоставляют с подходящими покрытиями. Для этой цели можно использовать концентрированные растворы сахаров, которые необязательно могут содержать гуммиарабик, тальк, поливинилпирролидон, карбопол-гель, полиэтиленгликоль и/или диоксид титана, глазурирующие растворы и подходящие органические растворители или смеси растворителей. В таблетки или покрытия драже можно добавлять красители или пигменты для обозначения или характеризации различных комбинаций доз активного антагониста EZH2.
Фармацевтические препараты, которые можно использовать перорально, включают твердые капсулы из желатина, а также мягкие запаянные капсулы, изготовленные из желатина и пластификатора, такого как глицерин или сорбит. Твердые капсулы могут содержать активные ингредиенты в смеси с наполнителем, таким как лактоза, связующие вещества, такие как крахмалы, и/или лубриканты, такие как тальк или стеарат магния и, необязательно, стабилизаторы. В мягких капсулах активные антагонисты EZH2 можно растворять или суспендировать в подходящих жидкостях, таких как жирные масла, вазелиновое масло или жидкие полиэтиленгликоли. Кроме того, можно добавлять стабилизаторы.
Для букального введения композиции могут иметь форму таблеток или пастилок, составленных общепринятым способом.
Для введения путем ингаляции антагонисты EZH2 для применения в соответствии с настоящим изобретением удобно доставлять в форме распыляемого аэрозоля из находящихся под давлением упаковок или небулайзера, с использованием подходящего пропеллента, например, дихлордифторметана, трихлорфторметана, дихлортетрафторэтана, диоксида углерода или другого подходящего газа. В случае находящегося под давлением аэрозоля дозированная единица может определяться путем предоставления клапана для доставки отмеренного количества. Можно составлять капсулы и кассеты, например, из желатина, содержащие порошковую смесь антагониста EZH2 и подходящую порошковую основу, такую как лактоза или крахмал, для применения в ингаляторе или инсуфляторе.
Антагонисты можно составлять для парентерального введения путем инъекции, например, болюсной инъекции или непрерывной инфузии. Составы для инъекции могут быть предоставлены в единичной дозированной форме, например, в ампулах или многодозовых контейнерах, с добавлением консерванта. Композиции могут иметь такие формы, как суспензии, растворы или эмульсии, в масляных или водных носителях, и они могут содержать компоненты состава, такие как суспендирующие, стабилизирующие и/или диспергирующие вещества.
Фармацевтические составы для парентерального введения включают водные растворы активных антагонистов EZH2 в растворимой в воде форме. Кроме того, суспензии активных антагонистов EZH2 можно получить в виде подходящих масляных инъекционных суспензий. Подходящие липофильные растворители или носители включают жирные масла, такие как кунжутное масло или синтетические сложные эфиры жирных кислот, такие как этилолеат или триглицериды, или липосомы. Водные инъекционные суспензии могут содержать вещества, которые повышают вязкость суспензии, такие как натрийкарбоксиметилцеллюлоза, сорбит или декстран. Необязательно суспензия также может содержать подходящие стабилизаторы или вещества, которые повышают растворимость антагонистов EZH2, чтобы обеспечить получение высококонцентрированных растворов.
Альтернативно, активный ингредиент может быть в форме порошка для восстановления перед введением подходящим носителем, например, стерильной не содержащей пирогенов водой.
Антагонисты EZH2 также можно составлять в виде ректальных композиций, таких как суппозитории или удерживающие клизмы, например, содержащие общепринятые основы суппозиториев, такие как масло какао или другие глицериды.
В дополнение к составам, описанным выше, антагонисты EZH2 также можно составлять в виде депо-препаратов. Такие составы длительного действия можно вводить путем имплантации (например, подкожно или внутримышечно, или путем внутримышечной инъекции). Таким образом, например, антагонисты EZH2 можно составлять с подходящими полимерными или гидрофобными материалами (например, в виде эмульсий в приемлемом масле) или ионообменными смолами, или в виде умеренно растворимых производных (например, в виде умеренно растворимой соли).
Альтернативно, можно использовать другие системы для доставки гидрофобных фармацевтических антагонистов EZH2. Липосомы и эмульсии являются примерами переносчиков или носителей для гидрофобных лекарственных средств. Также можно использовать определенные органические растворители, такие как диметилсульфоксид. Кроме того, антагонисты EZH2 можно доставлять с использованием системы с замедленным высвобождением, такой как полупроницаемые матрицы твердых гидрофобных полимеров, содержащих лекарственное средство. Различные материалы для замедленного высвобождения разработаны и хорошо известны специалистам в данной области. Капсулы с замедленным высвобождением могут, в зависимости от их химической природы, высвобождать антагонисты EZH2 в течение от нескольких недель вплоть до свыше 100 суток. В зависимости от химической природы и биологической стабильности терапевтического реагента, можно использовать дополнительные стратегии для стабилизации белка.
Фармацевтические композиции также могут содержать подходящие носители или эксципиенты в твердой или гелевой фазе. Примеры таких носителей или эксципиентов включают, но не ограничиваются ими, карбонат кальция, фосфат кальция, различные сахара, крахмалы, производные целлюлозы, желатин, и полимеры, такие как полиэтиленгликоли.
Способы лечения
В настоящем описании предусмотрены способы лечения или профилактики состояний и заболеваний, на течение которых может влиять модулирование состояния метилирования гистонов или других белков, где указанное состояние метилирования опосредуется, по меньшей мере, частично активностью EZH2. Модулирование состояния метилирования гистонов может, в свою очередь, влиять на уровень экспрессии генов-мишеней, активируемых метилированием, и/или генов-мишеней, подавляемых метилированием.
Например, один аспект изобретения относится к способу лечения рака. Способ включает стадию введения индивидууму, имеющему рак, экспрессирующий мутант EZH2 по Y641, терапевтически эффективного количества ингибитора EZH2, где ингибитор ингибирует активность метилтрансферазы гистонов EZH2, тем самым осуществляя лечение рака. В одном варианте осуществления ингибитор ингибирует активность метилтрансферазы гистонов у мутанта EZH2 по Y641. В одном варианте осуществления ингибитор селективно ингибирует активность метилтрансферазы гистонов у мутанта EZH2 по Y641. В одном варианте осуществления рак представляет собой фолликулярную лимфому. В одном варианте осуществления рак представляет собой диффузную крупно-B-клеточную лимфому (DLBCL).
Один аспект изобретения относится к способу лечения рака. Способ включает стадии проведения анализа для выявления мутанта EZH2 по Y641 в образце, содержащем раковые клетки от индивидуума, имеющего рак; и введения индивидууму, у которого экспрессируется мутант EZH2 по Y641, терапевтически эффективного количества ингибитора EZH2, где ингибитор ингибирует активность метилтрансферазы гистонов EZH2, тем самым осуществляя лечение рака. В одном варианте осуществления ингибитор ингибирует активность метилтрансферазы гистонов мутанта EZH2 по Y641. В одном варианте осуществления ингибитор селективно ингибирует активность метилтрансферазы гистонов мутанта EZH2 по Y641. В одном варианте осуществления рак представляет собой фолликулярную лимфому. В одном варианте осуществления рак представляет собой диффузную крупно-B-клеточную лимфому (DLBCL).
Заболевания, такие как рак и неврологическое заболевание, можно лечить путем ведения модуляторов метилирования белков (например, гистонов), например, модуляторов метилтрансферазы гистонов или активности фермента деметилазы гистонов. Описано, что метилирование гистонов вовлечено в нарушенную экспрессию определенных генов при различных типах рака и в подавление нейрональных генов в не являющихся нейрональными клетках. Модуляторы, описанные в настоящем описании, можно использовать для лечения таких заболеваний, т.е. для ингибирования метилирования гистонов в пораженных клетках.
Исходя, по меньшей мере, из того факта, что было обнаружено, что увеличенное метилирование гистонов ассоциировано с определенными типами рака, способ лечения рака у индивидуума включает введение индивидууму, нуждающемуся в этом, терапевтически эффективного количества соединения, которое ингибирует метилирование или восстанавливает метилирование приблизительно до его уровня в аналогичных нормальных клетках. В одном варианте осуществления способ лечения рака у индивидуума включает введение индивидууму, нуждающемуся в этом, терапевтически эффективного количества соединения, которое ингибирует преобразование неметилированного H3-K27 в монометилированный H3-K27 (H3-K27me1). В одном варианте осуществления способ лечения рака у индивидуума включает введение индивидууму, нуждающемуся в этом, терапевтически эффективного количества соединения, которое ингибирует преобразование монометилированного H3-K27 (H3-K27me1) в диметилированный H3-K27 (H3-K27me2). В одном варианте осуществления способ лечения рака у индивидуума включает введение индивидууму, нуждающемуся в этом, терапевтически эффективного количества соединения, которое ингибирует преобразование H3-K27me2 в триметилированный H3-K27 (H3-K27me3). В одном варианте осуществления способ лечения рака у индивидуума включает введение индивидууму, нуждающемуся в этом, терапевтически эффективного количества соединения, которое ингибирует как преобразование H3-K27me1 в H3-K27me2, так и преобразование H3-K27me2 в H3-K27me3. Важно отметить, что специфическое для заболевания увеличение метилирования может происходить в ключевых геномных локусах хроматина в отсутствие общего увеличения клеточных уровней метилирования гистонов или белков. Например, возможно, чтобы неправильное гиперметилирование в ключевых связанных с заболеванием генах происходило на фоне общего гипометилирования гистонов или белков.
Модуляторы метилирования можно использовать для модулирования, главным образом, пролиферации клеток. Например, в некоторых случаях, избыточная пролиферация может быть снижена с помощью средств, которые снижают метилирование, в то время как недостаточную пролиферацию можно стимулировать с помощью средств, которые повышают метилирование. Таким образом, заболевания, которые можно лечить, включают гиперпролиферативные заболевания, такие как доброкачественный рост клеток и злокачественный рост клеток (рак).
Иллюстративные типы рака, которые можно лечить, включают лимфомы, включающие фолликулярную лимфому (FL) и диффузную крупно-B-клеточную лимфому (DLBCL).
Другие типы рака включают острый лимфобластный лейкоз; острый миелоидный лейкоз; карциному надпочечников; обусловленные СПИДом типы рака; обусловленную СПИДом лимфому; рак анального канала; детскую мозжечковую астроцитому; детскую церебральную астроцитому; базально-клеточную карцииному, см. рак кожи (не меланома); внепеченочный рак желчных протоков; рак мочевого пузыря; рак кости, остеосаркому/злокачественную фиброзную гистиоцитому; глиому ствола головного мозга; опухоль головного мозга; опухоль головного мозга, астроцитому мозжечка; опухоль головного мозга, церебральную астроцитому/злокачественную глиому; опухоль головного мозга, эпендимому; опухоль головного мозга, медуллобластому; опухоль головного мозга, супратенториальную примитивную нейроэктодермальную опухоль; опухоль головного мозга, глиому зрительного пути и гипоталамуса; рак молочной железы; аденомы/карциноиды бронхов; лимфому беркитта; карциноидную опухоль; желудочно-кишечную карциноидную опухоль; первичную карциному неясного происхождения; лимфому центральной нервной системы, первичную; астроцитому мозжечка; рак шейки матки; типы рака детского возраста; хронический лимфоцитарный лейкоз; хронический миелогенный лейкоз; хронический миелогенный волосатоклеточный лейкоз; хронические миелопролиферативные нарушения; рак толстого кишечника; рак ободочной и прямой кишки; т-клеточную лимфому кожи, см. фунгоидный микоз и синдром Сезари; рак эндометрия; рак пищевода; семейство опухолей Юинга; внепеченочный рак желчных протоков; рак глаза, внутриглазную меланому; рак глаза, ретинобластому; рак желчного пузыря; гастральный рак (рак желудка); желудочно-кишечную карциноидную опухоль; внечерепную герминому; герминому, внегонадную; яичниковую герминому; гестационную трофобластическую опухоль; глиому; детскую глиому ствола головного мозга; детскую глиому, церебральную астроцитому; детскую глиому, зрительного пути и гипоталамуса; волосатоклеточный лейкоз; рак головы и шеи; печеночно-клеточный рак (печени) взрослых (первичный); печеночно-клеточный рак (печени) детского возраста (первичный); лимфому Ходжкина; лимфому Ходжкина в ходе беременности; гипофарингеальный рак; глиому гипоталамуса и зрительного пути; внутриглазную меланому; карциному островковых клеток (эндокринная часть поджелудочной железы); саркому Капоши; рак почки (почечно-клеточный рак); рак почки; рак гортани; лейкоз; рак губы и полости рта; рак печени взрослых (первичный); рак печени детского возраста (первичный); немелкоклеточный рак легкого; мелкоклеточный рак легкого; первичную лимфому центральной нервной системы; макроглобулинемию Вальденстрема; злокачественную фиброзную гистиоцитому костей/остеосаркому; медуллобластому; меланому; карциному клеток Меркеля; мезотелиому; злокачественную мезотелиому взрослых; метастатический плоскоклеточный рак шеи неясного происхождения; синдром множественной эндокринной неоплазии; множественную миелому; множественную миелому/фунгоидный микоз с новообразованием из плазматических клеток; миелодиспластические синдромы; миелодиспластические/миелопролиферативные заболевания; острый миелоидный лейкоз взрослых; острый миелоидный лейкоз детского возраста; хронические миелопролиферативные нарушения; рак носовой полости и околоносовой пазухи; рак носоглотки; нейробластому; неходжскинскую лимфому; неходжскинскую лимфому в ходе беременности; рак полости рта; рак полости рта, рак губы и ротоглотки; остеосаркому/злокачественную фиброзную гистиоцитому кости; рак яичника; эпителиальный рак яичника; опухоль яичника с низким раковым потенциалом; рак поджелудочной железы; рак островковых клеток поджелудочной железы; рак околоносовой пазухи и носовой полости; рак паращитовидной железы; рак полового члена; феохромацитому; пинеобластому и супратенториальные примитивные нейроэктодермальные опухоли; опухоль гипофиза; новообразование из плазматических клеток/множественную миелому; плевролегочную бластому; рак молочной железы при беременности; рак предстательной железы; рак прямой кишки; ретинобластому; рабдомиосаркому; рак слюнных желез; саркому, семейство опухолей Юинга; саркому мягких тканей; саркому матки; синдром Сезари; рак кожи; рак кожи (не меланома); рак тонкого кишечника; саркому мягких тканей; плоскоклеточную карциному, см. рак кожи (не меланома); плоскоклеточный рак шеи неясного происхождения, метастатический; рак желудка (гастральный рак); рак яичка; тимому; тимому и тимическую карциному; рак щитовидной железы; рак переходных клеток почечной лоханки и мочеточника; гестационную трофобластическую опухоль; рак с неизвестным первичным очагом; необычные типы рака детского возраста; уретральный рак; рак тела матки, эндометриальный; саркому матки; рак влагалища; глиому зрительного пути и гипоталамуса; рак женских наружных половых органов; макроглобулинемию Вальденстрема; опухоль Вильмса и женские типы рака.
С использованием соединений и способов, описанных в настоящем описании, можно лечить или предупреждать любое другое заболевание, при котором играет роль эпигенетическое метилирование, опосредуемое EZH2.
Например, неврологические заболевания, которые можно лечить, включают эпилепсию, шизофрению, биполярное нарушение или другие психологические и/или психиатрические нарушения, невропатии, атрофию скелетных мышц и нейродегенеративные заболевания, например, нейродегенеративное заболевание.
Иллюстративные нейродегенеративные заболевания включают: болезнь Альцгеймера, боковой амиотрофический склероз (ALS) и болезнь Паркинсона. Другой класс нейродегенеративных заболеваний включает заболевания, вызываемые, по меньшей мере, частично, агрегацией полиглутамина. Заболевания этого класса включают: болезнь Гентингтона, мышечную атрофию спинного и продолговатого мозга (SBMA или болезнь Кеннеди), дентато-рубро-паллидо-льюисову атрофию (DRPLA), спинально-церебеллярную атаксию 1 (SCA1), спинально-церебеллярную атаксию 2 (SCA2), болезнь Мачадо—Джозефа (MJD; SCA3), спинально-церебеллярную атаксию 6 (SCA6), спинально-церебеллярную атаксию 7 (SCA7) и спинально-церебеллярную атаксию 12 (SCA12).
Также в настоящем описании предусмотрены способы выбора лечения для индивидуума, имеющего рак. Способ включает определение способности индивидуума отвечать на ингибитор EZH2 по уровню диметилированного H3-K27, предпочтительно по уровням диметилированного H3-K27 и триметилированного H3-K27; и предоставление ингибитора EZH2 индивидууму, когда индивидуум способен отвечать на ингибитор EZH2. В одном варианте осуществления рак представляет собой фолликулярную лимфому. Альтернативно, рак представляет собой диффузную крупно-B-клеточную лимфому (DLBCL). В другом предпочтительном варианте осуществления у индивидуума экспрессируется мутант EZH2 по Y641. В предпочтительном варианте осуществления мутант по Y641 представляет собой Y641F, Y641H, Y641N или Y641S.
Комбинированная терапия
В одном аспекте изобретения антагонист EZH2 или его фармацевтически приемлемую соль можно использовать в комбинации с другим лекарственным средством для лечения заболеваний, таких как рак и/или неврологические нарушения. Например, дополнительное средство может представлять собой лекарственное средство, которое признано в данной области в качестве полезного для лечения заболевания или состояния, подвергаемого лечению соединением по настоящему изобретению. Дополнительное средство также может представлять собой средство, которое придает полезное свойство терапевтической композиции (например, средство, которое влияет на вязкость композиции).
Комбинированная терапия, предусматриваемая согласно изобретению, включает, например, введение соединения по изобретению или его фармацевтически приемлемой соли и дополнительного средства (средств) в одном фармацевтическом составе, а также введение соединения по изобретению или его фармацевтически приемлемой соли и дополнительного средства(средств) в отдельных фармацевтических составах. Иными словами, совместное введение означает введение, по меньшей мере, двух средств индивидууму, чтобы обеспечить полезные эффекты комбинации обоих средств. Например, средства можно вводить одновременно или последовательно на протяжении периода времени.
Средства, указанные ниже, представлены для иллюстративных целей и не предназначены для ограничения. Комбинации, которые являются частью настоящего изобретения, могут представлять собой соединения по настоящему изобретению и, по меньшей мере, одно дополнительное средство, выбранное из перечня ниже. Комбинация также может включать более одного дополнительного средства, например, два или три дополнительных средства, если комбинация является такой, что полученная композиция может выполнять ее предполагаемую функцию.
Например, один аспект изобретения относится к применению антагониста EZH2 в комбинации с другим средством для лечения рака и/или неврологического нарушения. В одном варианте осуществления дополнительное средство представляет собой средство против рака, т.е. соединение, которое влияет на модификации гистонов, такое как ингибитор HDAC. В определенных вариантах осуществления дополнительное средство против рака выбрано из группы, состоящей из химиотерапевтических средств (таких как 2CdA, 5-FU, 6-меркаптопурин, 6-TG, Abraxane™, Accutane®, актиномицин-D, Adriamycin®, Alimta®, полностью ретиноевая кислота, аметоптерин, Ara-C, азацитадин, BCNU, Blenoxane®, Camptosar®, CeeNU®, клофарабин, Clolar™, Cytoxan®, даунорубицина гидрохлорид, DaunoXome®, Dacogen®, DIC, Doxil®, Ellence®, Eloxatin®, Emcyt®, этопозида фосфат, Fludara®, FUDR®, Gemzar®, Gleevec®, гексаметилмеламин, Hycamtin®, Hydrea®, Idamycin®, Ifex®, иксабепилон, Ixempra®, L-аспарагиназа, Leukeran®, липосомальный Ara-C, L-PAM, лизодрен, Matulane®, митрацин, митомицин-C, Myleran®, Navelbine®, Neutrexin®, нилотиниб, Nipent®, азотистый иприт, Novantrone®, Oncaspar®, Panretin®, Paraplatin®, Platinol®, пролифероспан 20 с иплантатом кармустина, Sandodtatin®, Targretin®, Tasigna®, Taxotere®, Temodar®, TESPA, Trisenox®, Valstar®, Velban®, Vidaza™, винкристина сульфат, VM 26, Xeloda® и Zanosar®); биологические препараты (такие как альфа-интерферон, бацилла Кальметта-Герена, Bexxar®, Capath®, Ergamisol®, эрлотиниб, Herceptin®, интерлейкин-2, Iressa®, леналидомид, Mylotarg®, Ontak®, Pegasys®, Revlimid®, Rituxan®, Tarceva™, Thalomid®, Tykerb®, Velcade® и Zevalin™); кортикостероиды (такие как дексаметазон фосфат натрия, DeltaSone® и Delta-Cortef®); средства гормональной терапии (такие как Arimidex®, Aromasin®, Casodex®, Cytadren®, Eligard®, Eulexin®, Evista®, Faslodex®, Femara®, Halotestin®, Megace®, Nilandron®, Nolvadex®, Plenaxis™ и Zoladex®); и радиофармацевтические препараты (такие как Iodotope®, Metastron®, Phosphocol® и самарий SM-153).
Дозировка
Как используют в настоящем изобретении, «терапевтически эффективное количество» или «терапевтически эффективная доза» представляет собой количество антагониста EZH2 или комбинации двух или более таких соединений, которое ингибирует полностью или частично прогрессирование состояния или смягчает, по меньшей мере, частично, один или более симптомов состояния. Терапевтически эффективное количество также может представлять собой количество, которое является профилактически эффективным. Количество, которое является терапевтически эффективным, зависит от размера и пола пациента, состояния, подвергаемого лечению, тяжести состояния и искомого результата. В одном варианте осуществления терапевтически эффективная доза относится к количеству антагонистов EZH2, которое приводит к смягчению симптомов у пациента. Для данного пациента терапевтически эффективное количество можно определить способами, известными специалистам в данной области.
Токсичность и терапевтическую эффективность антагонистов EZH2 можно определить с помощью стандартных фармацевтических методик в клеточных культурах или на экспериментальных животных, например, путем определения максимальной переносимой дозы (MTD) и ED50 (эффективная доза для 50% максимального ответа). Соотношение дозы между токсическим и терапевтическим эффектами представляет собой терапевтический индекс, и он может быть выражен в виде соотношения между MTD и ED50. Данные, полученные в данных анализах клеточных культур и в исследованиях на животных, можно использовать при составлении ряда дозировок для введения человеку. Дозировку также можно определить путем мониторинга эффекта антагониста EZH2 на фармакодинамические маркеры ингибирования фермента (например, метилирование гистонов или экспрессия гена-мишени) в пораженной ткани или заместительной ткани. Эксперименты на культуре клеток или на животных можно использовать для определения взаимосвязи между дозами, требуемыми для изменения фармакодинамических маркеров, и дозы, требуемые для терапевтической эффективности, можно определить в клеточной культуре или в экспериментах на животных или в ранних клинических испытаниях. Дозировка таких антагонистов EZH2 предпочтительно находится в диапазоне концентраций в крови, которые включают ED50 с небольшой токсичностью или без токсичности. Дозировка может варьировать в этом диапазоне, в зависимости от используемой дозированной формы и используемого пути введения. Точный состав, путь введения и дозировку могут быть выбраны конкретным врачом с учетом состояния пациента. При лечении криза может потребоваться введение неотложного болюса или инфузии, достигающих MTD, для получения быстрого ответа.
Уровень дозировки и интервал между введениями можно корректировать индивидуально для обеспечения уровней активного соединения в плазме, которые являются достаточными для поддержания эффектов модулирования метилтрансферазы или минимальной эффективной концентрация (MEC) в течение требуемого периода времени для достижения терапевтической эффективности. MEC варьирует для каждого антагониста EZH2, однако она может быть оценена из данных in vitro и экспериментов на животных. Дозировки, необходимые для достижения MEC, зависят от индивидуальных характеристик и пути введения. Однако для определения концентраций в плазме можно использовать анализы с использованием высокоэффективной жидкостной хроматографии (ВЭЖХ) или биоанализы.
Интервалы между введениями также могут быть определены с использованием величины MEC. В определенных вариантах осуществления антагонисты EZH2 следует вводить с использованием режима, который поддерживает уровни в плазме выше MEC в течение 10-90% времени, предпочтительно 30-90% и наиболее предпочтительно 50-90%, до тех пор, пока не будет достигнуто желаемое смягчение симптомов. В других вариантах осуществления различные уровни MEC в плазме можно поддерживать в течение различных периодов времени. В случаях местного введения или селективного захвата, эффективная локальная концентрация лекарственного средства может быть не связана с концентрацией в плазме.
Специалист в данной области сможет выбрать различные режимы введения, и вводимое количество антагониста EZH2 будет зависеть, безусловно, от индивидуума, подвергаемого лечению, массы тела индивидуума, тяжести заболевания, способа введения и мнения назначающего врача.
Соединения и фармацевтические композиции
Аспекты изобретения относятся к соединениям, которые являются пригодными согласно способам по изобретению. Эти соединения приведены в настоящем описании как «ингибиторы EZH2» и, эквивалентно, «антагонисты EZH2». Соединения могут быть представлены в виде самих соединений, фармацевтически приемлемых солей соединений или в виде фармацевтических композиций.
Такие соединения, в частности, включают соединение 75:
(75)
и его фармацевтически приемлемые соли.
Кроме того, изобретение включает фармацевтическую композицию, содержащую соединение 75:
(75)
или его фармацевтически приемлемую соль.
Антагонист EZH2 и необязательно другие лекарственные средства можно вводить сами по себе (в чистом виде) или в форме фармацевтически приемлемой соли. При использовании в медицине соли должны быть фармацевтически приемлемыми, однако фармацевтически неприемлемые соли можно удобным образом использовать для получения из них фармацевтически приемлемых солей.
Соединения, пригодные согласно изобретению, могут быть предоставлены в виде солей с фармацевтически совместимыми противоионами (т.е. фармацевтически приемлемые соли). «Фармацевтически приемлемая соль» означает любую нетоксичную соль, которая при введении реципиенту способна обеспечивать, прямо или косвенно, соединение или пролекарство соединения, пригодное согласно настоящему изобретению. «Фармацевтически приемлемый противоион» представляет собой ионную часть соли, которая является нетоксичной при высвобождении из соли при введении индивидууму. Фармацевтически совместимые соли могут быть образованы со многими кислотами, включая, но, не ограничиваясь ими, хлористоводородную, серную, уксусную, молочную, винную, яблочную и янтарную кислоты. Соли имеют тенденцию быть более растворимыми в воде или других протонных растворителях, чем их соответствующие формы свободного основания. Настоящее изобретение относится к применению таких солей.
Фармацевтически приемлемые кислотно-аддитивные соли включают соли, образованные с неорганическими кислотами, такими как хлористоводородная кислота и бромистоводородная кислота, а также соли, образованные с органическими кислотами, такими как малеиновая кислота. Например, кислоты, обычно используемые для получения фармацевтически приемлемых солей, включают неорганические кислоты, такие как персульфид водорода, хлористоводородная, бромистоводородная, серная и фосфорная кислоты, а также органические кислоты, такие как паратолуолсульфоновая, салициловая, винная, бивинная, аскорбиновая, малеиновая, бесиловая, фумаровая, глюконовая, глюкуроновая, муравьиная, глутаминовая, метансульфоновая, этансульфоновая, бензолсульфоновая, молочная, щавелевая, пара-бромфенилсульфоновая, угольная, янтарная, лимонная, бензойная и уксусная кислоты, и родственные неорганические и органические кислоты. Такие фармацевтически приемлемые соли, таким образом, включают сульфат, пиросульфат, бисульфат, сульфит, бисульфит, фосфат, моногидрофосфат, дигидрофосфат, метафосфат, пирофосфат, хлорид, бромид, йодид, ацетат, пропионат, деканоат, каприлат, акрилат, формиат, изобутират, капрат, гептаноат, пропиолят, оксалат, малонат, сукцинат, суберат, себакат, фумарат, малеат, бутин-1,4-диоат, гексин-1,6-диоат, бензоат, хлорбензоат, метилбензоат, динитробензоат, гидроксибензоат, метоксибензоат, фталат, терефталат, сульфонат, ксилолсульфонат, фенилацетат, фенилпропионат, фенилбутират, цитрат, лактат, β-гидроксибутират, гликолят, малеат, тартрат, метансульфонат, пропансульфонат, нафталин-1-сульфонат, нафталин-2-сульфонат, манделат и т.п.
Пригодные основания для образования фармацевтически приемлемых солей с кислотными функциональными группами включают, но не ограничиваются ими, гидроксиды щелочных металлов, таких как натрий, калий и литий; гидроксиды щелочноземельных металлов, таких как кальций и магний; гидроксиды других металлов, таких как алюминий и цинк; аммиак и органические амины, такие как незамещенные или гидрокси-замещенные моно-, ди- или триалкиламины; дициклогексиламин; трибутиламин; пиридин; N-метил-N-этиламин; диэтиламин; триэтиламин; моно-, бис- или трис-(2-гидрокси-низшие алкиламины), такие как моно-, бис- или трис-(2-гидроксиэтил)амин, 2-гидрокси-трет-бутиламин или трис-(гидроксиметил)метиламин, N,N-диалкил-N-(гидроксиалкил)амины, такие как N,N-диметил-N-(2-гидроксиэтил)амин или три(2-гидроксиэтил)амин; N-метил-D-глюкамин; и аминокислоты, такие как аргинин, лизин и т.п.
Определенные соединения, пригодные согласно изобретению, и их соли могут существовать более чем в одной кристаллической форме (т.е. полиморф); настоящее изобретение включает использование каждой из кристаллических форм и их смесей.
Определенные соединения, пригодные согласно изобретению, могут содержать один или более хиральных центров и существовать в различных оптически активных формах. Когда соединения, пригодные согласно изобретению, содержат один хиральный центр, соединения существуют в двух энантиомерных формах, и настоящее изобретение включает использование обоих энантиомеров и смесей энантиомеров, таких как их рацемические смеси. Энантиомеры можно разделить способами, известными специалистам в данной области; например, энантиомеры могут быть разделены путем образования диастереоизомерных солей, которые можно разделять, например, кристаллизацией; путем образования диастереоизомерных производных или комплексов, которые можно разделять, например, кристаллизацией, газожидкостной или жидкостной хроматографией; селективной реакцией одного энантиомера с энантиомер-специфическим реагентом, например, путем ферментативной этерификации; или газожидкостной или жидкостной хроматографией в хиральной среде, например, на хиральной подложке (например, диоксиде кремния со связанным с ним хиральным лигандом) или в присутствии хирального растворителя. Когда желаемый энантиомер преобразовывают в другую химическую структуру способами разделения, описанными выше, можно использовать дополнительную стадию для высвобождения желаемого очищенного энантиомера. Альтернативно, определенные энантиомеры можно синтезировать путем асимметричного синтеза с использованием оптически активных реагентов, субстратов, катализаторов или растворителей, или путем преобразования одного энантиомера в другой путем асимметричной трансформации.
Когда соединение, пригодное согласно изобретению, содержит более одного хирального центра, оно может существовать в диастереоизомерных формах. Диастереоизомерные соединения могут быть разделены способами, известными специалистам в данной области (например, хроматографией или кристаллизацией) и индивидуальные энантиомеры могут быть разделены, как описано выше. Настоящее изобретение включает использование различных диастереоизомеров соединений, пригодных согласно изобретению, и их смесей. Соединения, пригодные согласно изобретению, могут существовать в различных таутомерных формах или в виде различных геометрических изомеров, и настоящее изобретение включает использование каждого таутомера и/или геометрического изомера соединений, пригодных согласно изобретению, и их смесей. Соединения, пригодные согласно изобретению, могут существовать в цвиттерионной форме. Настоящее изобретение охватывает использование каждой цвиттерионной формы соединений, пригодной согласно изобретению, и их смесей.
Наборы
Антагонист EZH2 может, если желательно, присутствовать в виде набора (например, упаковки или распределяющего устройства), который может содержать одну или более единичных дозированных форм, содержащих антагонист EZH2. Упаковка, например, может содержать металлическую или пластмассовую фольгу, как например, блистерная упаковка. К упаковке или распределяющему устройству могут прилагаться инструкции по введению. Композиции, содержащие антагонист EZH2 по изобретению, составленный в совместимом фармацевтическом носителе, также может быть получен, помещен в соответствующий контейнер и маркирован для лечения указанного состояния. Также могут быть предоставлены инструкции по применению.
Также в настоящем изобретении предусмотрены наборы, содержащие несколько реагентов для выявления метилирования, которые выявляют метилированный H3-K27. Например, набор включает реагенты для выявления монометилированного H3-K27, диметилированного H3-K27 и триметилированного H3-K27. Реагентом для выявления являются, например, антитела или их фрагменты, полипептид или аптамеры. Набор может содержать в отдельных контейнерах аптамер или антитело, контрольные составы (положительный и/или отрицательный) и/или поддающуюся детекции метку, такую как флуоресцеин, зеленый флуоресцентный белок, родамин, цианиновые красители, красители Alexa, люцифераза, радиоактивные метки, среди прочего. В набор могут быть включены инструкции (например, написанные, напечатанные, VCR, CD-ROM и т.д.) для проведения анализа. Анализ может быть в форме, например, вестерн-блоттинга, иммуногистохимии (IHC), иммунофлуоресценции (IF) и масс-спектрометрии (МС), как известно в данной области.
Определения
Для удобства, в этом разделе собраны определенные термины, используемые в описании, примерах и прилагаемой формуле изобретения. Все определения, определенные и используемые в настоящем описании, заменяют определения в словарях, определения в документах, включенных посредством ссылок, и/или обычные значения определяемых терминов.
Форму единственного числа используют в настоящем описании для обозначения одного или более чем одного (т.е., по меньшей мере, одного) грамматического объекта. В качестве примера, «элемент» означает один или более элементов.
Выражение «и/или», как используют в настоящем изобретении в описании и в формуле изобретения, следует понимать как «любой или оба» из элементов, объединенных таким образом, т.е. элементы, которые совместно присутствуют в некоторых случаях и по отдельности присутствуют в других случаях. Множество элементов, перечисленных с помощью «и/или», следует истолковывать аналогичным образом, т.е. как «один или более» элементов, объединенных таким образом. Необязательно могут присутствовать другие элементы, отличные от элементов, конкретно идентифицированных условием «и/или», как родственные, так и не родственные конкретно указанным элементам. Таким образом, в качестве неограничивающего примера, указание на «A и/или B», когда его используют вместе с неограничивающей формулировкой, такой как «содержащий», может относиться в одном варианте осуществления только к A (необязательно включая элементы, отличные от B); в другом варианте осуществления только к B (необязательно включая элементы, отличные от A); в другом варианте осуществления, как к A, так и к B (необязательно включая другие элементы); и т.д.
Как используют в настоящем изобретении в описании и в формуле изобретения, «или» следует понимать как имеющий такое же значение, как «и/или», как определено выше. Например, при разделении объектов в перечне, «или» или «и/или» следует интерпретировать как включительные, т.е. включающие, по меньшей мере, один, но также включающие более одного из ряда или перечня элементов и необязательно дополнительных не перечисленных объектов. Только термины, явно указывающие на противоположное, такие как «только один из» или «абсолютно один из», или при использовании в формуле изобретения, «состоящий из», относится к включению абсолютно одного элемента из ряда или перечня элементов. Как правило, термин «или», как используют в настоящем изобретении, следует интерпретировать только как указывающий на исключающие альтернативы (т.е. «один или другой, но не оба»), когда ему предшествуют термины исключительности, такие как «любой из», «один из», «только один из» или «абсолютно один из». «По существу состоящий», когда его используют в формуле изобретения, имеет обычное значение, используемое в области патентного закона.
Как используют в настоящем изобретении в описании и формуле изобретения, выражение, «по меньшей мере, один» в отношении перечня из одного или более элементов, следует понимать как означающее, по меньшей мере, один элемент, выбранный из одного или нескольких элементов в перечне элементов, но необязательно включающий, по меньшей мере, один каждый элемент, конкретно включенный в перечень элементов, но не исключающее какие-либо комбинации элементов в перечне элементов. Это определение также допускает, что необязательно могут присутствовать элементы, отличные от элементов, конкретно указанных в перечне элементов, к которому относится выражение «по меньшей мере, один», либо родственные, либо не родственные конкретно указанным элементам. Таким образом, в качестве неограничивающего примера, «по меньшей мере, один из A и B» (или, эквивалентно, «по меньшей мере, один из A или B», или, эквивалентно, «по меньшей мере, один A и/или B») может относиться, в одном варианте осуществления, по меньшей мере, к одному, необязательно включая более одного A без присутствия B (и необязательно включая элементы, отличные от B); в другом варианте осуществления, по меньшей мере, к одному, необязательно включая более одного B без присутствия A (и необязательно включая элементы, отличные от A); в другом варианте осуществления, по меньшей мере, к одному, необязательно включая более одного A и, по меньшей мере, к одному, необязательно включая более одного B (и необязательно включая другие элементы); и т.д.
Также следует понимать, что, если явно не указано иное, в любых способах, представленных в настоящем описании, которые включают более одной стадии или действия, порядок стадий или действий способа необязательно ограничивается порядком, в котором стадии или действия указаны.
В формуле изобретения, а также в представленном выше описании, все переходные выражения, такие как «содержащий», «включающий», «несущий», «имеющий», «охватывающий», «вовлекающий», «удерживающий», «составленный из» и т.п. следует понимать как открытые, т.е. означающие «включая, но, не ограничиваясь ими». Только переходные выражения «состоящий из» и «по существу состоящий из» являются закрытыми или полузакрытыми переходными выражениями, соответственно, как указано в United States Patent Office Manual of Patent Examining Procedures, Section 2111.03.
Термины «совместное введение» и «совместное проведение введения» относятся как к одновременному введению (введение двух или более лекарственных средств в одно и то же время), так и к введению в различное время (введение одного или более лекарственных средств во время, отличное от введения дополнительного лекарственного средства или средств), при условии, что лекарственные средства до некоторой степени присутствуют у пациента в одно и то же время.
Термин «проведение лечения», как используют в настоящем изобретении, относится к смягчению, по меньшей мере, одного симптома заболевания, нарушения или состояния. Этот термин охватывает введение и/или применение одного или более соединений, описанных выше, индивидууму, для целей обеспечения управления течением или излечением состояния. «Лечение» для целей настоящего описания может, но необязательно, обеспечивать излечение; вместо этого, «лечение» может быть в форме управления течением состояния. Когда соединения, описанные в настоящем описании, используют для лечения нежелательных пролиферирующих клеток, включая рак, «лечение» включает частичное или полное уничтожение нежелательных пролиферирующих клеток с минимальными воздействиями на нормальные клетки. Желаемым механизмом лечения нежелательных быстро пролиферирующих клеток, включая рак, на клеточном уровне является апоптоз.
Термин «профилактика», как используют в настоящем изобретении, включает либо предупреждение, либо замедление появления прогрессирования клинически очевидного заболевания в целом, либо предупреждение или замедление возникновения доклинически очевидной стадии заболевания у имеющего риск индивидуума. Это включает профилактическое лечение индивидуумов, имеющих риск развития заболевания.
Термин «индивидуум», как используют в настоящем изобретении для целей лечения, включает любого человека, у которого было диагностировано, который имеет симптомы или который имеет риск развития нарушения. Для способов профилактики индивидуумом является любой человек. Для иллюстрации, для целей изобретения индивидуумом может быть человек, который имеет риск или генетически предрасположен к возникновению нарушения, характеризующегося нежелательной быстрой пролиферацией клеток, такой как рак. Индивидуумом может иметь риск вследствие воздействия канцерогенных веществ, наличия генетической предрасположенности к нарушениям, характеризующимся нежелательной быстрой пролиферацией клеток, и т.д.
Если не указано иное, для получения рекомбинантных и синтетических полипептидов, слитых белков, антител или их антигенсвязывающих фрагментов, манипулирования с последовательностями нуклеиновых кислот, получения трансформированных клеток и т.п. можно использовать стандартные способы. Такие способы известны специалистам в данной области. См., например, Sambrook et al, Molecular Cloning: A Laboratory Manual, 3rd Ed. (Cold Spring Harbor, N.Y., 2001); F.M. Ausubel et al. Current Protocols in Molecular Biology (Green Publishing Associates, Inc. and John Wiley & Sons, Inc., New York).
Термин «полипептид EZH2» охватывает функциональные фрагменты полноразмерных полипептидов и функциональные эквиваленты любого из указанных выше, которые имеют по существу сходные или по существу идентичные аминокислотные последовательности (сходство, идентичность аминокислотных последовательностей, по меньшей мере, приблизительно на 75%, 80%, 85%, 90%, 95% 98% или более), где функциональный фрагмент или функциональный эквивалент сохраняет одно или более функциональных свойств нативного полипептида.
Под «функциональным» подразумевают, что полипептид (или нуклеиновая кислота) имеет такую же или по существу сходную активность в отношении одного или более биологических свойств нативного полипептида (или нуклеиновой кислоты), например, по меньшей мере, приблизительно 50%, 75%, 85%, 90%, 95% или 98% или более активности нативного полипептида (или нуклеиновой кислоты).
Термин «модулировать» (и грамматические эквиваленты) относится к увеличению или снижению активности. В конкретных вариантах осуществления термин «увеличивать» или «усиливать» (и грамматические эквиваленты) означает увеличение, по меньшей мере, приблизительно на 25%, 50%, 75%, в 2 раза, в 3 раза, в 5 раз, в 10 раз, в 15 раз, 20 раз или более. В конкретных вариантах осуществления термины «уменьшать» или «снижать» (и грамматические эквиваленты) означают снижение, по меньшей мере, приблизительно на 25%, 40%, 50%, 60%, 75%, 80%, 85%, 90%, 95%, 98% или более. В некоторых вариантах осуществления указанная активность, вещество или другой параметр не поддаются выявлению. Конкретно предусмотрены антагонисты EZH2.
Термин «фармакодинамический маркер» относится к молекулярному маркеру ответа на лекарственное средство, который может быть измерен у пациентов, которым вводят лекарственное средство. Маркер должен представлять собой прямую меру модулирования мишени лекарственного средства и быть способен продемонстрировать количественные изменения в ответе на дозу. Потенциальным фармакодинамическим маркером для антагонистов EZH2 могут быть уровни метилирования H3-K27 гистонов в пораженной или заместительной ткани.
Как используют в настоящем изобретении, термин «способность отвечать» является взаимозаменяемым с терминами «отвечающий», «чувствительный» и «чувствительность», и он означает, что индивидуум демонстрирует терапевтический ответ при введении ингибитора EZH, например, опухолевые клетки или опухолевые ткани индивидуума претерпевают апоптоз и/или некроз, и/или проявляют сниженный рост, деление или пролиферацию.
Термин «контроль» или «эталон» относится к уровням метилирования (например, уровень монометилирования, уровень диметилирования или уровень триметилирования), выявляемым в соседней неопухолевой ткани, выделенной от индивидуума, выявляемым в здоровой ткани от здорового индивидуума или установленным патологом с помощью способов, стандартных в данной области.
Под «образцом» подразумевают любой биологический образец, происходящий из индивидуума, включая, но, не ограничиваясь ими, образцы клеток, тканей и жидкостей организма (включая, но, не ограничиваясь ими, слизь, кровь, плазму, сыворотку, мочу, слюну и семенную жидкость).
ПРИМЕРЫ
После общего описания изобретения оно будет более понятно с помощью следующих примеров, которые включены только для целей иллюстрации определенных аспектов и вариантов осуществления настоящего изобретения, и не предназначены для ограничения изобретения.
Пример 1 - Рекомбинантный пятикомпонентный комплекс PRC2
EZH2 дикого типа (номер доступа GenBank NM 004456) или мутанты по Tyr641 коэкспрессировали с AEBP2 дикого типа (номер доступа GenBank NM_153207), EED (номер доступа GenBank NM_003797), SUZ12 (номер доступа GenBank NM_015355) и RbAp48 (номер доступа GenBank NM_005610) в клетках Spodoptera frugiperda (Sf9) с использованием бакуловирусной экспрессирующей системы. Для очистки активного комплекса PRC2 из клеточных лизатов использовали N-концевую FLAG-метку на EED (BPS Bioscience, каталожный номер 51004). Чистоту конечных препаратов PRC2 оценивали с помощью SDS-PAGE окрашиванием кумасси синим.
Пример 2 - Панель пептидов H3, H4
Библиотеку, состоящую из 44 пептидов по 15 аминокислот в каждом, синтезировали в 21st Century Biochemicals (Marlboro, MA). Данная панель пептидов охватывала все аминокислоты гистонов H3 и H4 человека с перекрыванием 5 остатков между последовательными пептидными последовательностями. К N-концу каждого пептида присоединяли биотин, и C-концы присутствовали в виде амида. Чистоту (>95%) и идентичность подтверждали жидкостной хроматографией/масс-спектральным анализом.
Для исследования зависимости состояния метилирования H3-K27 от активности фермента синтезировали пептиды, соответствующие аминокислотной последовательности H3 человека в остатках 21-44 (H3:21-44), где лизин 27 представлен в виде немодифицированного, монометилированного, диметилированного или триметилированного амина боковой цепи. Данные пептиды приобретали от New England Peptide (Gardner, MA) с биотином, присоединенным к C-концу каждого пептида.
Пример 3 - оценка состояния метилирования H3-K27 в клетках
Клеточные линии OCI-LY19 (ACC 528), KARPAS-422 (ACC 32) и WSU-DLCL2 (ACC 575) приобретали в DSMZ. Клеточные линии DB (CRL-2289) и SU-DHL2 (CRL-2959) приобретали в ATCC. Клеточные линии OCI-LY19, WSU-DLCL2 и DB выращивали в RPMI-1640 с 10% FBS, и клеточные линии KARPAS-422 и SU-DHL2 выращивали в RPMI-1640 с 20% FBS. Клетки выращивали до плотности 1,5-2×106 клеток/мл и 1×107 клеток собирали центрифугированием при 264×g, промывали ледяным PBS и лизировали ресуспендированием в 10X объеме осадка лизирующим буфером RIPA, содержащим 50 мМ Tris-HCl, 150 мМ NaCl, 0,25% DOC, 1% NP-40, и 1 мМ EDTA (Millipore #20-188), вместе с 0,1% SDS и таблетками ингибиторов протеаз (Roche # 1836153). Лизаты обрабатывали ультразвуком в течение 2 раундов с 10 импульсами по 1 секунде в положении 3 устройства Misonix XL-2000 для обеспечения эффективной экстракции гистонов, и очищали центрифугированием при 4°C с использованием настольной центрифуги при 14000 об./мин. в течение 10 минут. Концентрацию белка определяли анализом BCA (Pierce). Четыре микрограмма каждого лизата фракционировали на 4-20% Tris-глициновом геле (Invitrogen), переносили на PVDF и исследовали с помощью следующих антител в блокирующем буфере Odyssey: антитела мыши против EZH2 (CST 3147; разведение 1:2000), антитела кролика против H3-K27me3 (CST 9733; разведение 1:10000), антитела кролика против 3-K27me2 (CST 9755; разведение 1:5000), антитела кролика против H3-K27me1 (Active Motif 39377; разведение 1:5000) и антитела мыши против тотального H3 (CST 3638; разведение 1:20000). После первичной инкубации Ab, мембраны исследовали с помощью вторичных Ab: IRDye 800CW-антитело осла против IgG мыши (LiCOR #926-32212) или Alexa Fluor 680-антитело козы против IgG кролика (Invitrogen #A-21076) и визуализировали с использованием системы LiCOR Odyssey.
Пример 4 - Ферментология
Как указано выше, ранее было сделано заключение, что ассоциированные с заболеванием изменения Tyr641 приводили к утрате функции катализируемого EZH2 метилирования H3-K27. Однако предполагаемое снижение уровня метилирования H3-K27 вследствие гетерозиготности по ферменту было трудно обосновать как основу злокачественного фенотипа, особенно ввиду предшествующих данных о том, что сверхэкспрессия EZH2, мутации с утратой функции в соответствующей деметилазе H3-K27 UTX или сверхэкспрессия компонентов PRC2, таких как PHF19/PCL3, вовлеченные в повышение триметилирования H3-K27, приводят к злокачественным фенотипам в конкретных типах рака человека. Morin et al. (2010) Nat Genet 42: 181-5; Martinez-Garcia et al. (2010) Nat Genet 42: 100-1; Bracken et al. (2003) EMBO J 22: 5323-35; Kleer et al. (2003) Proc Natl Acad Sci USA 100: 11606-11; Varambally et al. (2002) Nature 419: 624-9; Simon et al. (2008) Mutat Res 647: 21-9; van Haaften et al. (2009) Nat Genet 41: 521-3; Wang et al. (2004) Gene 343: 69-78; Cao et al. (2008) Mol Cell Biol 28: 1862-72; и Sarma et al. (2008) Mol Cell Biol 28: 2718-31). Таким образом, ферментологию этих мутаций исследовали более детально.
Рекомбинантные комплексы PRC2 получали с WT и мутантными по Tyr641 вариантами EZH2 человека (см. пример 1 выше; Cao et al. (2004) Mol Cell 15: 57-67). Равные концентрации (номинально 8 нМ, на основе определения белков) каждого комплекса первоначально исследовали на способность катализировать перенос H-метила с меченного S-аденозилметионина (SAM) на немодифицированный пептид, соответствующий аминокислотной последовательности, окружающей H3-K27 (H3:21-44) или на нативные олигонуклеосомы эритроцитов птиц. Как описано ранее (Morin et al. (2010) Nat Genet 42: 181-5), было обнаружено, что фермент WT проявлял устойчивую активность переноса метильной группы в отношении этого неметилированного пептидного субстрата, но ни один из мутантных ферментов не проявлял существенной активности метилтрансферазы (фиг.1A). В противоположность ранее описанным данным и данным на фиг.1A, было обнаружено, что все мутантные конструкции EZH2 представляли собой активные метилтрансферазы в отношении субстрата в виде нуклеосом птиц (фиг.1B). Нуклеосомы, выделенные из природного источника птиц, представляют собой смесь состояний модификаций гистонов, включая различные состояния метилирования H3-K27, исходя из вестерн-блоттинга с антителами, специфичными к метилированию H3-K27.
Существует несколько возможных объяснений противоречивой активности мутантных комплексов PRC2 в отношении пептидных и нуклеосомных субстратов. Одной из возможностей является то, что участки распознавания субстрата дистальные активного центра (т.е. экзоучастки) являются важными детерминантами связывания субстрата и обновления; эти участки могут связываться с комплементарными распознаваемыми элементами на нуклеосоме, которые являются недоступными на небольших пептидных субстратах. Однако, когда экспрессированный в E. coli рекомбинантный гистон H3 человека исследовали в качестве субстрата для WT и мутантных комплексов PRC2, полученный профиль активности был идентичен профилю, наблюдаемому для пептидного субстрата; т.е. фермент WT продемонстрировал устойчивую активность метилтрансферазы в отношении субстрата H3, мутант Y641F продемонстрировал 7% активности комплекса WT, и все другие мутанты проявляли ≤1% активности комплекса WT. Таким образом, связывание экзоучастка оказалось маловероятным объяснением текущих результатов. Нуклеосома предоставляет множество остатков лизина, помимо H3-K27, в качестве потенциальных участков метилирования, которые могут не присутствовать в небольшом пептидном субстрате. Таким образом, другая возможность состоит в том, что мутация Y641 изменяет субстратную специфичность EZH2, что приводит к метилированию остатков лизина, отличных от H3-K27. Эта возможность является маловероятной, учитывая превосходную согласованность между активностью мутанта в отношении субстратов в виде малых пептидов и рекомбинантного белка H3.
Очевидное несоответствие между представленными результатами и результатами, описанными ранее, было разрешено, когда ферментативную активность WT и мутантных комплексов PRC2 исследовали в отношении панели пептидных субстратов, которые представляют все возможные остатки лизина (K) на гистоне H3 и гистоне H4 (см. пример 2 выше). Все формы фермента продемонстрировали значительную активность только против пептидов, содержащих эквивалент остатка H3-K27. Удельная активность мутантов, однако, была значительно снижена в отношении WT в порядке WT>>Y641F>Y641S~Y641H>Y641N, что вновь согласуется с предшествующими описанными данными.
Пример 5 - Ферментология
Для дальнейшего понимания ферментативной активности этих мутантов и для устранения очевидного противоречия между активностью в отношении пептидных и нуклеосомных субстратов, исследовали способность форм фермента катализировать дальнейшее метилирование различных состояний метилирования H3-K27 в контексте пептида H3:21-44. Как указано выше, было обнаружено, что все мутантные ферменты были дефектными катализаторами метилирования немодифицированного пептида H3-K27 в отношении фермента WT. Однако примечательно, что было обнаружено, что все мутантные ферменты превосходили фермент WT в отношении катализа дальнейшего метилирования моно- и особенно диметилированных пептидов H3-K27 (фиг.2). Таким образом, данные указывают на то, что фермент WT является наиболее эффективным в отношении катализа реакции от нуль- до монометилирования. Мутантные ферменты являются дефектными в отношении катализа этой первоначальной стадии, однако они являются более эффективными, чем фермент WT, в отношении катализа последующих стадий, ведущих от монометилированного к ди- и триметилированному H3-K27.
Источники дифференциальной субстратной специфичности WT и мутантной EZH2 исследовали с помощью ферментативной кинетики устойчивого состояния. Как обобщенно представлено в таблице 1, мутации имеют минимальные эффекты на распознавание субстрата в основном состоянии, как продемонстрировано с помощью сходных величин Km для нуклеосомных субстратов и K1/2 для пептидных субстратов. Во всех случаях пептидные субстраты проявляли сигмоидальное поведение связывания; таким образом, концентрация пептида, приводящая к полумаксимальной скорости, описана в настоящем описании как K1/2 вместо более распространенной константы Михаэлеса, Km. Copeland (2005) Evaluation of Enzyme Inhibitors in Drug Discovery: A Guide to Medicinal Chemists and Pharmacologists, Wiley. Km для SAM аналогично проявляла минимальное варьирование среди форм фермента, находясь в диапазоне от 208±50 до 304±64 нМ. Вместо этого, различия в утилизации субстрата, по-видимому, являлись следствием распознавания переходного состояния, как продемонстрировано по различиям в величинах kcat среди ферментов для различных субстратов (таблица 1). В результате, каталитическая эффективность, количественно определенная как соотношение kcat/K (где K представляет собой либо Km, либо K1/2, в зависимости от типа субстрата; см. выше), варьирует между WT и мутантными ферментами для различных состояний метилирования H3-K27 (таблица 1).
Кинетические параметры устойчивого состояния для реакций метилирования, катализируемых PRC2, содержащим EZH2 дикого типа или мутанты EZH2 по Y641
H3-K27
(нМ)
(ч-1×10-2)
(ч-1.нМ-1×10-4)
Пример 6 - Ферментология
Параметры кинетики устойчивого состояния, приведенные в таблице 1, дают возможность вычислить ожидаемые уровни различных состояний метилирования H3-K27 для клеток, гетерозиготных по различным мутантным формам EZH2, относительно клеток, гомозиготных по ферменту WT. Для проведения такого моделирования был сделан ряд упрощающих допущений: (1) что стационарная кинетика фермента соответствует катализируемому PRC2 метилированию H3-K27 в клеточном контексте, и что все измерения проводят в один и тот же момент времени при росте клеток; (2) что мутантный и WT ферменты экспрессировались на равных уровнях в гетерозиготных клетках, и что общий уровень EZH2 является равным во всех клетках; (3) что клеточная концентрация SAM относительно его Km является насыщающей и не изменяется среди клеток; (4) что клеточная концентрация нуклеосом сходна с ее Km и аналогично не изменяется среди клеток; (5) что катализируемое EZH1 метилирование H3-K27 было незначительным и постоянным среди клеток; и (6) что любая активность деметилазы H3-K27 также была постоянной среди клеток.
Учитывая эти допущения, были сделаны предсказания, проиллюстрированные на фиг.3A, для относительных уровней H3-K27me3 (верхняя панель), H3-K27me2 (средняя панель) и H3-K27me1 (нижняя панель). Из такого моделирования выявляется четкий профиль. Уровень H3-K27me3 возрастает относительно клеток WT для всех содержащих мутант клеток, находясь в диапазоне от 30% увеличения для мутанта Y641H до >400% для мутанта Y641N. В то же время, уровни H3-K27me2 снижены до <50% относительно WT для всех мутантов, и уровни H3-K27me1 снижены приблизительно наполовину для всех мутантов относительно WT.
Затем измеряли относительные уровни состояний метилирования H3-K27 в клеточных линиях B-клеточной лимфомы, которые, как известно, являются гомозиготными по WT EZH2 (OCI-LY19) или гетерозиготными по EZH2 Y641N (DB, KARPAS 422, и SU-DHL-6) или EZH2 Y641F (WSU-DLCL2) с помощью вестерн-блоттинга (фиг.3B). Профиль относительных состояний метилирования H3-K27, наблюдаемый на фиг.3b, превосходно согласуется с результатами моделирования на основе стационарных кинетических параметров in vitro, несмотря на допущения, использованные в моделировании, и использование нефизиологического пептидного заместителя в качестве субстрата.
Таким образом, увеличение H3-K27me3 наблюдали для всех клеток, содержащих мутант по Y641, относительно WT, снижение H3-K27me2 наблюдали для всех клеток, содержащих мутант по Y641, относительно WT, и снижение H3-K27me1 наблюдали, по меньшей мере, для двух из четырех мутантных клеточных линий. Практически сравнимые уровни H3-K27me1 в клетках WT и KARPAS 422 и SU-DHL-6 могли отражать различные уровни экспрессии WT и мутантного EZH2, различный вклад EZH1 или другие факторы, не учитываемые в моделировании. Тем не менее соответствие между предсказанными и экспериментальными профилями состояния метилирования H3-K27 является существенным и подтверждает точку зрения, что ферментативное сопряжение между WT и мутантной EZH2 приводит к увеличению H3-K27me3, что, таким образом, приводит к злокачественному фенотипу клеток, которые являются гетерозиготными по этим мутантам.
Пример 7 - Анализы активности метилтрансферазы PRC2 in vitro
Анализ на Flashplate с пептидным субстратом. Для первоначального сравнения WT и мутантов EZH2 по Y641, биотинилированный пептид гистона H3:21-44, содержащий неметилированный K27 (New England Peptides), монометилированный K27 (Millipore) или диметилированный K27 (Millipore) в концентрации 800 нМ, комбинировали со смесью S-аденозилметионин-Cl (SAM) в концентрации 1700 нМ и тритированного SAM в концентрации 300 нМ (Perkin Elmer). Затем данную комбинацию субстратов добавляли в буфер для анализа PRC2 (20 мМ BICINE, 1 мМ DTT, 0,002% Tween 20, 0,005% бычий желатин кожи (BSG), pH 7,6). Реакциям давали возможность протекать в течение указанного интервала и затем гасили добавлением избытка холодного SAM (конечная концентрация 600 мкМ). Реакционные смеси после гашения переносили в покрытый стрептавидином Flashplate (Perkin Elmer, каталожный номер SMP410), давали возможность связываться в течение одного часа и затем подвергали детекции на сцинтилляционном и люминесцентном счетчике TopCount NXT HTS (Perkin Elmer). Каждый момент времени соответствовал в среднем шести отдельным реакциям. Стационарные параметры кинетики определяли в идентичных условиях реакции, за исключением того, что концентрацию пептида или SAM варьировали в насыщающих условиях другого субстрата. Скорость наносили на график в виде функции варьирующей концентрации субстрата, и данные приводили в соответствие с непреобразованной версией уравнения Михаэлеса-Ментен или непреобразованной версией уравнения сигмоидальной кинетики для вычисления величин K и kcat. Стандартные ошибки приведенных в соответствие параметров приведены в таблице 1 и их использовали для построения планок погрешностей, проиллюстрированных на фиг.2 панели B и C. Ошибки, ассоциированные с kcat/K (таблица 1) вычисляли в соответствии со стандартными способами размножения ошибок; относительную погрешность kcat/K определяли как:
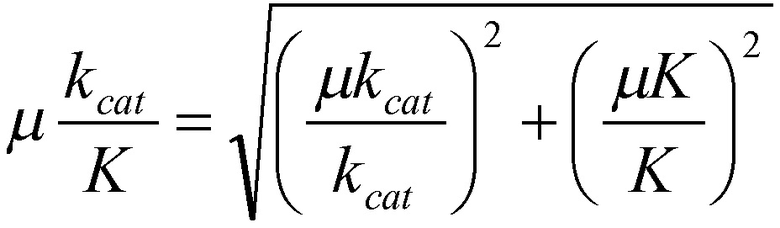 (1),
(1),
где μkcat представляет собой стандартную ошибку kcat, и μΚ представляет собой стандартную ошибку K.
Анализ с олигонуклеосомами на фильтровальных планшетах. Олигонуклеосомы эритроцитов курицы очищали, как описано ранее. Fang et al. (2004) Methods Enzymol 377:213-26. Нуклеосомы комбинировали со смесью SAM и тритированного SAM и добавляли к PRC2 в буфере для анализа (20 мМ BICINE, 100 мМ KC1, 1 мМ DTT, 0,002% Tween 20, 0,005% BSG, pH 7,6). Реакции проводили и гасили, как описано выше. Реакционную смесь переносили в фильтровальный планшет из стекловолокна (Millipore, каталожный номер MSFBN6B), промывали три раза 10% трихлоруксусной кислотой и давали возможность высохнуть. Добавляли Microscint Zero (30 мкл) и включение трития выявляли на сцинтилляционном и люминесцентном счетчике TopCount. Стационарные параметры определяли в идентичных условиях реакции, за исключением того, что концентрацию нуклеосом или SAM варьировали в насыщающих условиях другого субстрата. Скорость наносили на график как функцию варьирующей концентрации субстрата в соответствие с непреобразованной версией уравнения Михаэлеса-Ментен для установления величин Km и kcat, как описано выше.
Пример 8 - Получение соединения 75
A. Получение соединения 37
К раствору 9-((3aR,4R,6R,6aR)-6-(аминометил)-2,2-диметилтетрагидрофтор[3,4-d][1,3]диоксол-4-ил)-9H-пурин-6-амина (Townsend, A. P. et al. (2009) Org. Let. 11: 2976-2979) (3,05 г, 9,96 ммоль) в DCE (250 мл) добавляли (9H-флуорен-9-ил)метил-(2-оксоэтил)карбамат (2,8 г, 9,96 ммоль) и NaB(OAc)3H (2,96 г, 13,95 ммоль), смесь перемешивали в течение 4 ч при комнатной температуре. Добавляли раствор K2CO3 до pH 8-9. Добавляли DCM, органический слой сушили над Na2SO4, концентрировали и очищали SGC (DCM MeOH=30:1), с получением соединения 37 (2,9 г, выход: 50,9%).
B. Получение соединения 65
К раствору соединения 37 (2,9 г, 5,08 ммоль) в DCE (250 мл) добавляли (S)-бензил 2-((трет-бутоксикарбонил)амино)-4-оксобутаноат (1,56 г, 5,08 ммоль) и NaB(OAc)3H (1,51 г, 7,11 ммоль), смесь перемешивали в течение 4 ч при комнатной температуре. Добавляли раствор K2CO3 до pH 8-9. Добавляли DCM, органический слой сушили над Na2SO4, концентрировали и очищали SGC (DCM: MeOH=100:1), с получением соединения 65 (2,8 г, выход: 63,9%).
C. Получение соединения 75
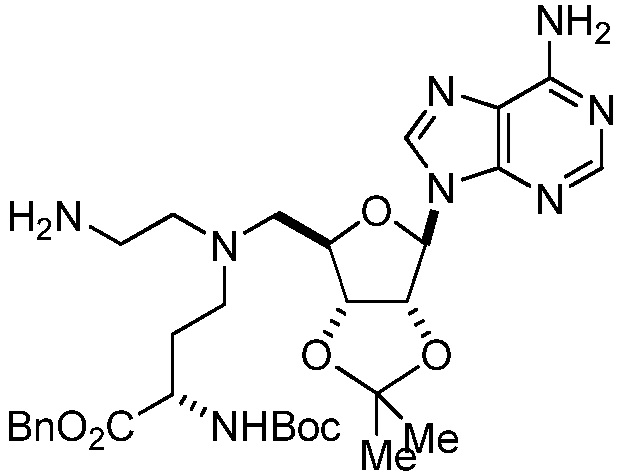
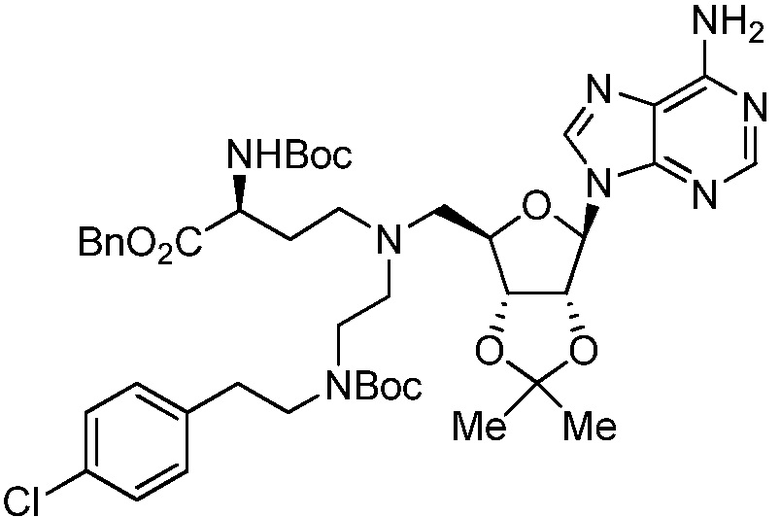
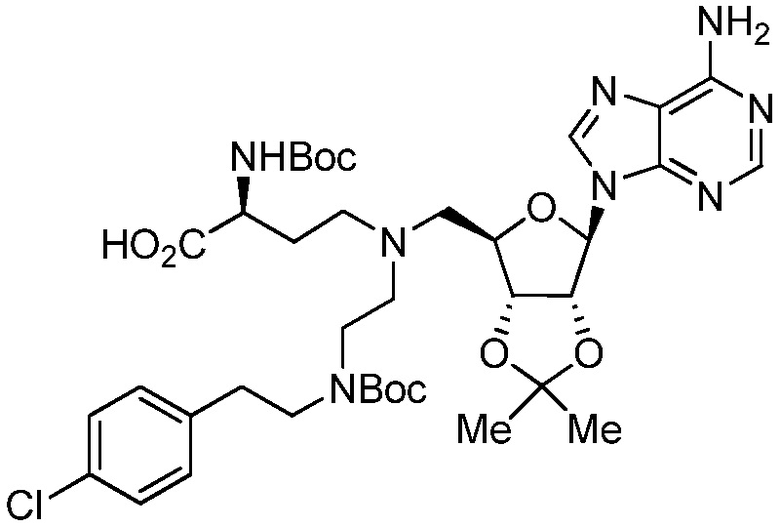
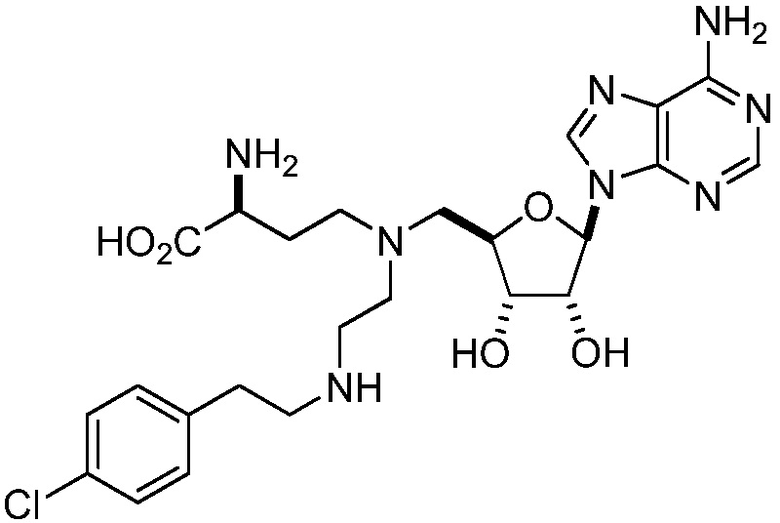
Стадия 1. К раствору соединения 65B (2,2 г, 2,55 ммоль) в DCM (10 мл) добавляли Et2NH (1,1 г, 15,3 ммоль), смесь перемешивали в течение 4 ч при комнатной температуре. Смесь концентрировали, с получением неочищенного соединения 72 (2,2 г).
Стадия 2. К перемешиваемому раствору соединения 72 (167 мг, 0,26 ммоль) в MeOH (4 мл) добавляли 2-(4-хлорфенил)ацетальдегид (40 мг, 0,26 ммоль) и перемешивали при комнатной температуре в течение 20 мин. Затем добавляли Na(OAc)3BH (83 мг, 0,39 ммоль) и HOAc (0,4 мл) и перемешивали в течение ночи. Затем добавляли NaHCO3 (водн.) и экстрагировали DCM (25 мл×3), промывали насыщенным раствором соли, сушили над Na2SO4 и концентрировали. Неочищенный продукт очищали препаративной ТСХ (DCM/MeOH=10:1), с получением соединения 73 (30 мг, выход: 14%) в виде белого порошка. ЖХ/МС (m/z): 779,7 [M+1]+.
Стадия 3. Смесь соединения 73 (30 мг, 0,038 ммоль) и 10% Pd/C (15 мг) в MeOH (2 мл) перемешивали при комнатной температуре в атмосфере H2 в течение ночи. Смесь фильтровали, и фильтрат концентрировали, с получением неочищенного продукта. Неочищенный продукт очищали препаративной ТСХ (DCM/MeOH=8:1), с получением соединения 74 (20 мг, выход: 69%) в виде белого порошка. ЖХ/МС (m/z): 689,7 [M+1]+.
Стадия 4. Раствор соединения 74 (20 мг, 0,028 ммоль) в 90% TFA (1 мл) перемешивали при комнатной температуре в течение 1 ч и концентрировали до состояния твердого вещества, удаляя TFA, с получением соединения 75 (соль TFA) в виде бесцветного масла без очистки. ЖХ/МС (m/z): 549,7 [M+1]+.
Пример 9 - Ингибирование EZH2 дикого типа и мутантов по Y641 с помощью SAH
S-Аденозил-L-гомоцистеин (SAH) серийно разводили в 3 раза в ДМСО для получения 10 точек, и 1 мкл наносили в 384-луночный микропланшет для титрования. Положительный контроль (стандарт со 100% ингибированием) представлял собой SAH в конечной концентрации 100 мкМ, и отрицательный контроль (стандарт с 0% ингибированием) содержал 1 мкл ДМСО. Затем SAH инкубировали в течение 30 минут с 40 мкл на лунку EZH2 дикого типа и мутантами в концентрации 8 нМ в буфере для анализа с pH 7,6 (20 мМ BICINE, 100 мМ KCl, 1 мМ DTT, 0,002% Tween 20, 0,005% BSG). Добавляли 10 мкл на лунку смеси субстратов, которая содержала S-аденозилметионин-Cl (SAM) в концентрации 150 нМ и тритированный SAM в концентрации 100 нМ, и биотинилированную олигонуклеосому в концентрации 150 нМ в буфере для анализа с pH 7,6. Ферментативную реакцию после гашения переносили в покрытый стрептавидином Flashplate (Perkin Elmer, каталожный номер SMP410), давали возможность связываться в течение одного часа и подвергали детекции на TopCount NXT HTS (Perkin Elmer).
Результаты представлены на фиг.7. Величины IC50 представлены в таблице 2.
Ингибирование WT EZH2 и мутантов EZH2 по Y641 с помощью SAH
Пример 10 - Ингибирование EZH2 дикого типа и мутантов по Y641 с помощью соединения 75
Соединение 75 серийно разводили в 3 раза в ДМСО для получения 10 точек, и 1 мкл наносили в 384-луночный микропланшет для титрования. Положительный контроль (стандарт со 100% ингибированием) представлял собой SAH в конечной концентрации 100 мкМ, и отрицательный контроль (стандарт с 0% ингибированием) содержал 1 мкл ДМСО. Затем соединение 75 инкубировали в течение 30 минут с 40 мкл на лунку EZH2 дикого типа и мутантами в концентрации 8 нМ в буфере для анализа с pH 7,6 (20 мМ BICINE, 100 мМ KCl, 1 мМ DTT, 0,002% Tween 20, 0,005% BSG). Добавляли 10 мкл на лунку смеси субстратов, которая содержала S-аденозилметионин-Cl (SAM) в концентрации 150 нМ и тритированный SAM в концентрации 100 нМ, и биотинилированную олигонуклеосому в концентрации 150 нМ в буфере для анализа с pH 7,6. Ферментативную реакцию после гашения переносили в покрытый стрептавидином Flashplate (Perkin Elmer, каталожный номер SMP410), давали возможность связываться в течение одного часа и подвергали детекции на TopCount NXT HTS (Perkin Elmer).
Результаты представлены на фиг.8. Величины IC50 представлены в таблице 3.
Ингибирование WT EZH2 и мутантов EZH2 по Y641 с помощью соединения 75
Пример 11 - Соотношения H3-K27me2/me3 предсказывают чувствительность к ингибитору EZH2
Опухолевые клеточные линии, гетерозиготные по мутации (Y641) EZH2 проявляют увеличенные уровни H3-K27me3 - состояние метилирования H3-K27, как полагают, являющееся важным при образовании опухоли. Оценивали уровни моно- (H3-K27me1), ди- (H3-K27me2) или триметилированных (H3-K27me3) форм H3-K27 в панели клеточных линий, которые были WT по EZH2 или гетерозиготными по мутациям (Y641) EZH2. Использованные клеточные линии приведены в таблице 4. Большинство линий представляют собой линии B-клеточной лимфомы, однако также были включены две линии меланомы. IGR1 представляет собой линию меланомы, которая, как недавно было обнаружено, содержит мутацию Y641N в EZH2, и клетки A375 были включены в качестве контрольной линии меланомы с WT EZH2. На фиг.9A и B представлены результаты анализа с использованием вестерн-блоттинга гистонов, выделенных из этой панели клеточных линий, исследованной с помощью антител, распознающих H3-K27me1, H3-K27me2 или H3-K27me3. Как правило, общие уровни H3-K27me3 являются более высокими в клеточных линиях, содержащих мутант по Y641, чем в клеточных линиях, экспрессирующих исключительно WT EZH2. Двумя исключениями являются клетки Farage и Pfeiffer, где уровни H3-K27me3 являются сходными с уровнями в линиях WT. Более поразительными являются значительно более низкие уровни H3-K27me2 в мутантных по Y641 клеточных линиях EZH2 относительно клеточных линий дикого типа. В вестерн-блотах гистонов, экстрагированных из мутантных по Y641 клеточных линий, наблюдали небольшой или отсутствующий сигнал H3-K27me2, в то время как сигнал, наблюдаемый с тем же антителом в клеточных линиях WT, был более интенсивным, чем сигнал, наблюдаемый с антителом, специфичным к H3-K27me3. В целом, в клеточных линиях WT сигнал вестерн-блота с антителом HK27me2 был более высоким, чем сигнал, наблюдаемый с антителом H3-K27me3, в то время как обратное было справедливым в мутантных по Y641 клеточных линиях. Таким образом, соотношение сигнала H3-K27me3/me2 в мутантных по Y641 клеточных линиях является более высоким, чем соотношение, наблюдаемое в линиях WT. Одним исключением из этого является клеточная линия Pfeiffer, которая не содержит мутацию Y641 EZH2, но имеет высокий сигнал H3-K27me3 и небольшой или отсутствующий сигнал H3-K27me2. Таким образом, клетки Pfeiffer имеют соотношение H3-K27me3/me2, сходное с мутантными по Y641 клеточными линиями.
Состояние метилирования H3-K27 также можно исследовать с помощью масс-спектрометрии (МС) - независимого способа, который не основан на антительных реагентах. Анализ МС продемонстрировал, что уровни H3-K27me3 являются более высокими в мутантных по Y641 линиях и линии Pfeiffer, чем в других линиях WT, в то время как для уровней H3-K27me2 является справедливым обратное. В мутантных по Y641 линиях и линии Pfeiffer уровни H3-K27me3 были более высокими, чем уровни H3-K27me2, в то время как в других клеточных линиях WT было справедливым обратное. Эти результаты согласуются с результатами, наблюдаемыми в анализе с использованием вестерн-блоттинга, представленными на фиг.9A и B.
Различия в состоянии метилирования H3-K27 также можно выявлять с помощью иммуноцитохимии с использованием антител к H3-K27me2 или H3-K27me3. Иммуногистохимический анализ используют для выявления аберрантных соотношений H3-K27me2/3, ассоциированных с мутантом по Y641 EZH2, в фиксированных формалином погруженных в парафин образцах опухолевой ткани пациента. Панель из пяти WT и пяти мутантных по Y641 осадков клеточных линий лимфомы фиксировали и погружали в парафиновые блоки и окрашивали антителами против H3-K27me2 или против H3-K27me3. Антитело к гистону H3 включали в качестве положительного контроля, поскольку все клетки должны содержать ядерный гистон H3. На фиг.10 показано, что все клеточные линии были положительными в 100% клеток по окрашиванию как на H3-K27me3, так и на H3. В этих условиях не было выявлено явных различий в интенсивности окрашивания на H3-K27me3 между клеточными линями WT и мутантными по Y641 клеточными линиями. Это может отражать динамически ограниченный диапазон хромогенного иммуноцитохимического окрашивания по сравнению с другими способами выявления. Однако, как показано на фиг.11, клеточные линии можно было четко разделить на клеточные линии, положительно или отрицательно окрашивающиеся по H3-K27me2. Все клеточные линии WT, за исключением клеток Pfeiffer, окрашивались положительно на H3-K27me2, в то время как все мутантные по Y641 клеточные линии и клетки Pfeiffer не продемонстрировали окрашивания антителом против H3-K27me2. Эти результаты согласуются с результатами, полученными с помощью вестерн-блоттинга и МС-анализа.
Без связи с какой-либо теорией, увеличенные уровни H3-K27me3, ассоциированные с мутациями приобретения функции EZH2 (Y641), могут делать клетки, несущие мутации EZH2, более чувствительными к низкомолекулярным ингибиторам EZH2. Для оценки того, коррелируют ли также увеличенные уровни H3-K27me3 и/или сниженные уровни H3-K27me2, наблюдаемые в клетках Pfeiffer в отсутствие мутации Y641 EZH2 с чувствительностью к ингибиторам EZH2, исследовали два соединения, которые демонстрируют мощное ингибирование EZH2 в биохимических анализах с IC50 85 и 16 нМ, соответственно. Обработка клеток WSU-DLCL2 любым из соединений приводила к ингибированию общих уровней H3-K27me3, подтверждая их способность входить в клетки и ингибировать активность клеточной метилтрансферазы EZH2 (фиг.12).
Чувствительность панели клеточных линий WT и мутантных по Y641 клеточных линий к каждому соединению оценивали в анализах пролиферации. Поскольку проявление антипролиферативной активности ингибиторов EZH2 занимает несколько суток, соединения оценивали в анализах пролиферации в течение 11 суток. На фиг.13 представлены конкретные кривые роста для клеточных линий WT (OCI-LY19) или мутантных по Y641 клеточных линий (WSU-DLCL2), обработанных тестируемыми соединениями. Оба соединения продемонстрировали антипролиферативную активность против клеток WSU-DLCL2, однако, небольшую активность против клеток OCI-LY19. Ингибитор A был более мощным ингибитором пролиферации WSU-DLCL2, чем ингибитор B, и это согласуется с тем, что ингибитор A является более мощным ингибитором EZH2 в биохимических анализах. Анализы пролиферации проводили на панели клеточных линий лимфомы WT и мутантных по Y641 клеточных линий лимфомы, с ингибитором B, и получали величины IC90 на 11 сутки. На фиг.14A представлены величины IC90 для клеточных линий лимфомы, сгруппированных по состоянию Y641 EZH2. В целом, мутантные по Y641 клеточные линии продемонстрировали чувствительность к ингибиторам EZH2 относительно клеточных линий WT, хотя клетки RL и SUDHL4 были значительно менее чувствительными, чем другие клеточные линии. Клетки Pfeiffer являются исключением, поскольку они являются WT, но они высокочувствительны к антипролиферативным эффектам обоих соединений с IC90 в низком или субнаномолярном диапазоне. Клетки Pfeiffer демонстрируют высокие уровни H3-K27me3 и низкие уровни H3-K27me2, и, таким образом, группирование клеточных линий по высоким уровням H3-K27me3 и низким уровням H3-K27me2 обеспечивает лучшую дифференциацию чувствительности к ингибитору EZH2, как показано для ингибитора B на фиг.14B. Таким образом, высокие уровни H3-K27me3 и низкие уровни H3-K27me2 можно использовать для предсказания чувствительности к ингибиторам EZH2, независимо от знания о мутантном состоянии. Неправильное соотношение метилирования, выявленное в клетках Pfeiffer, возникает вследствие отдельного механизма, который обеспечивает зависимость от активности EZH2.
Полученные результаты демонстрируют, что идентификация мутаций Y641 EZH2 в опухолях пациента и/или выявление низких уровней H3-K27me2 относительно H3-K27me3 с использованием способов, таких как вестерн-блоттинг, МС или IHC, у пациента можно использовать для идентификации того, какой пациент будет отвечать на лечение ингибитором EZH2.
Клеточные линии, использованные в данном исследовании
DLBCL (диффузная крупно-В-клеточная лимфома) и другая B-клетка
Эквиваленты
Хотя несколько вариантов осуществления настоящего изобретения были описаны и проиллюстрированы в настоящем описании, специалистам в данной области без труда будут очевидны различные другие средства и/или структуры для выполнения функций и/или получения результатов, и/или одного или более преимуществ, описанных в настоящем описании, и каждый из таких вариантов и/или модификаций считается входящим в объем настоящего изобретения. В более общем смысле, специалистам в данной области без труда будет понятно, что все параметры, размеры, материалы и конфигурации, описанные в настоящем описании, являются иллюстративными, и что истинные параметры, размеры, материалы и/или конфигурации будут зависеть от конкретного применения или применений, для которых используют идеи настоящего изобретения. Специалистам в данной области будут понятны или они смогут установить с использованием не более чем общепринятого экспериментирования многие эквиваленты конкретных вариантов осуществления изобретения, описанных в настоящем описании. Таким образом, следует понимать, что указанные выше варианты осуществления представлены только в качестве примера и что в объеме прилагаемой формулы изобретения и ее эквивалентов изобретение можно осуществлять на практике иначе, чем конкретно описано и заявлено. Настоящее изобретение относится к каждому конкретному признаку, системе, изделию, материалу, набору и/или способу, описанным в настоящем описании. Кроме того, любая комбинация двух или более таких признаков, систем, изделий, материалов, наборов и/или способов, если такие признаки, системы, изделия, материалы, наборы и/или способы не являются взаимно несовместимыми, включена в объем настоящего изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ИНГИБИТОРЫ EZH2 ЧЕЛОВЕКА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2011 |
|
RU2618475C2 |
| ИНГИБИТОРЫ EZH2 ЧЕЛОВЕКА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2704445C2 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ ДЛЯ ЛЕЧЕНИЯ РАКА | 2013 |
|
RU2699546C2 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ ДЛЯ ЛЕЧЕНИЯ РАКА | 2013 |
|
RU2827936C2 |
| РЕПРОГРАММИРОВАНИЕ ЭФФЕКТОРНЫХ БЕЛКОВЫХ ВЗАИМОДЕЙСТВИЙ ДЛЯ ИСПРАВЛЕНИЯ ЭПИГЕНЕТИЧЕСКИХ ДЕФЕКТОВ В ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ | 2013 |
|
RU2637441C2 |
| АРИЛ-ИЛИ ГЕТЕРОАРИЛЗАМЕЩЕННЫЕ БЕНЗОЛЬНЫЕ СОЕДИНЕНИЯ | 2012 |
|
RU2632193C2 |
| СПОСОБЫ ЛЕЧЕНИЯ РАКА | 2013 |
|
RU2677276C2 |
| ЗАМЕЩЕННЫЕ БЕНЗОЛЬНЫЕ СОЕДИНЕНИЯ | 2012 |
|
RU2629118C2 |
| СОЛЕВАЯ ФОРМА ИНГИБИТОРА ГИСТОН-МЕТИЛТРАНСФЕРАЗЫ EZH2 ЧЕЛОВЕКА | 2013 |
|
RU2658911C2 |
| СОДЕРЖАЩИЕ ЗАМЕСТИТЕЛИ БЕНЗОЛЬНЫЕ СОЕДИНЕНИЯ | 2013 |
|
RU2662436C2 |
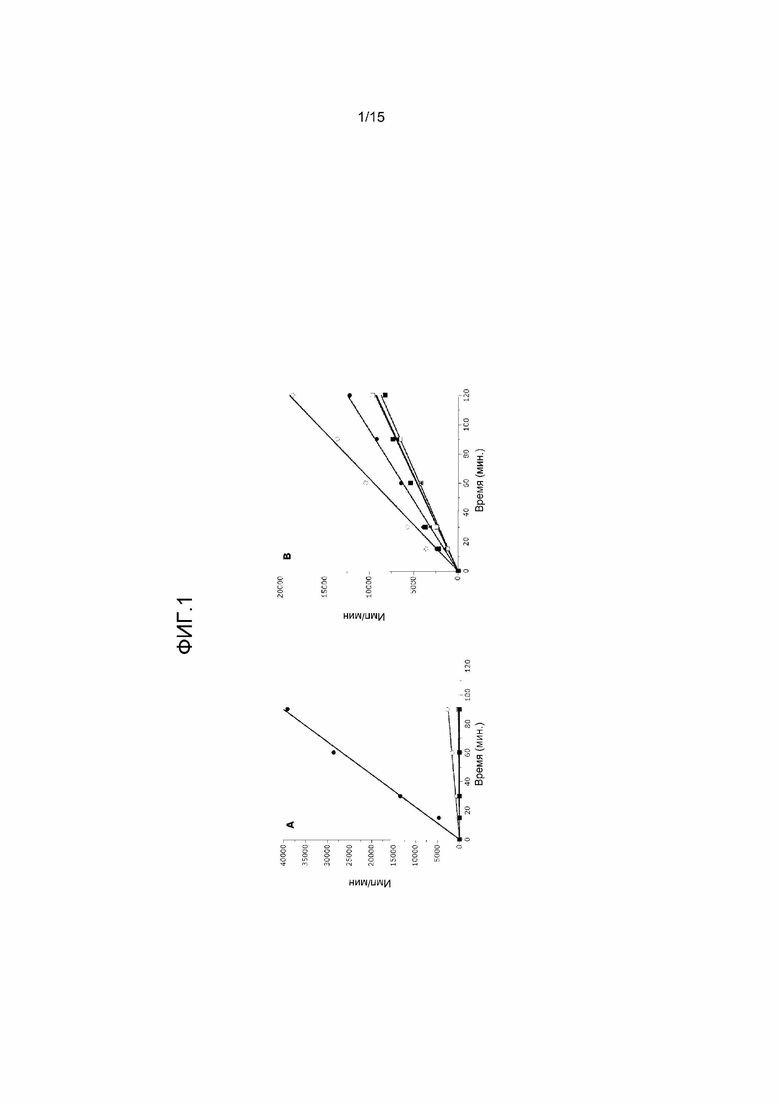
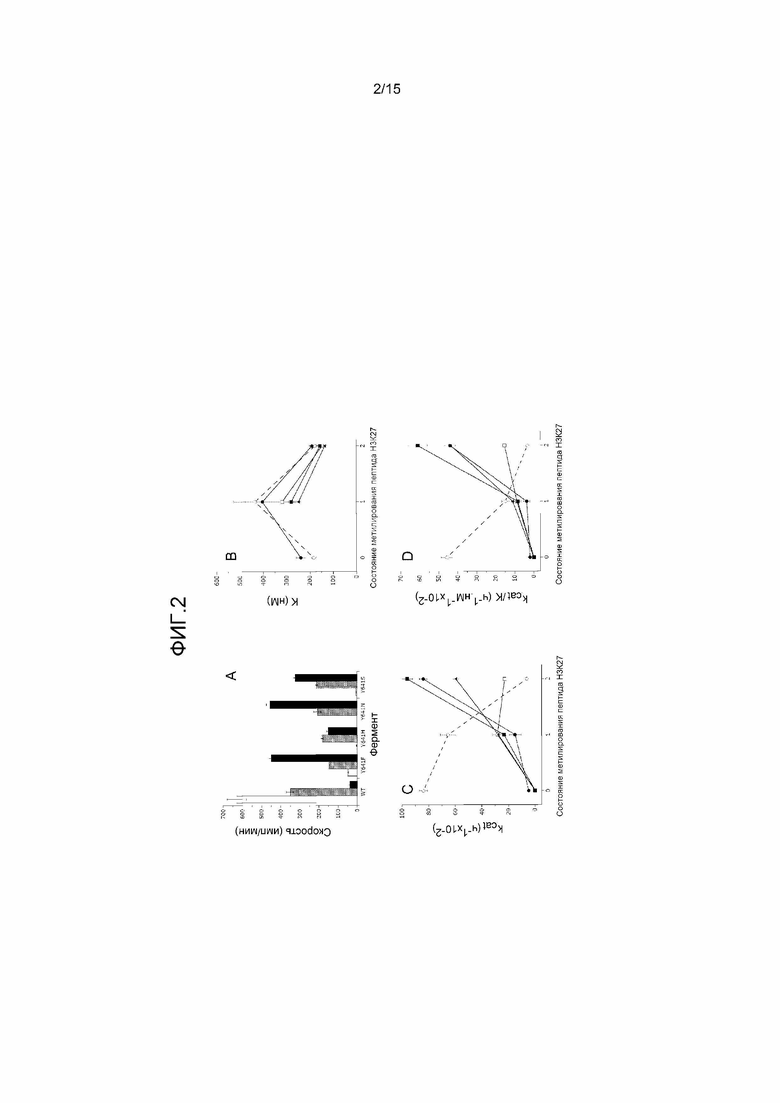
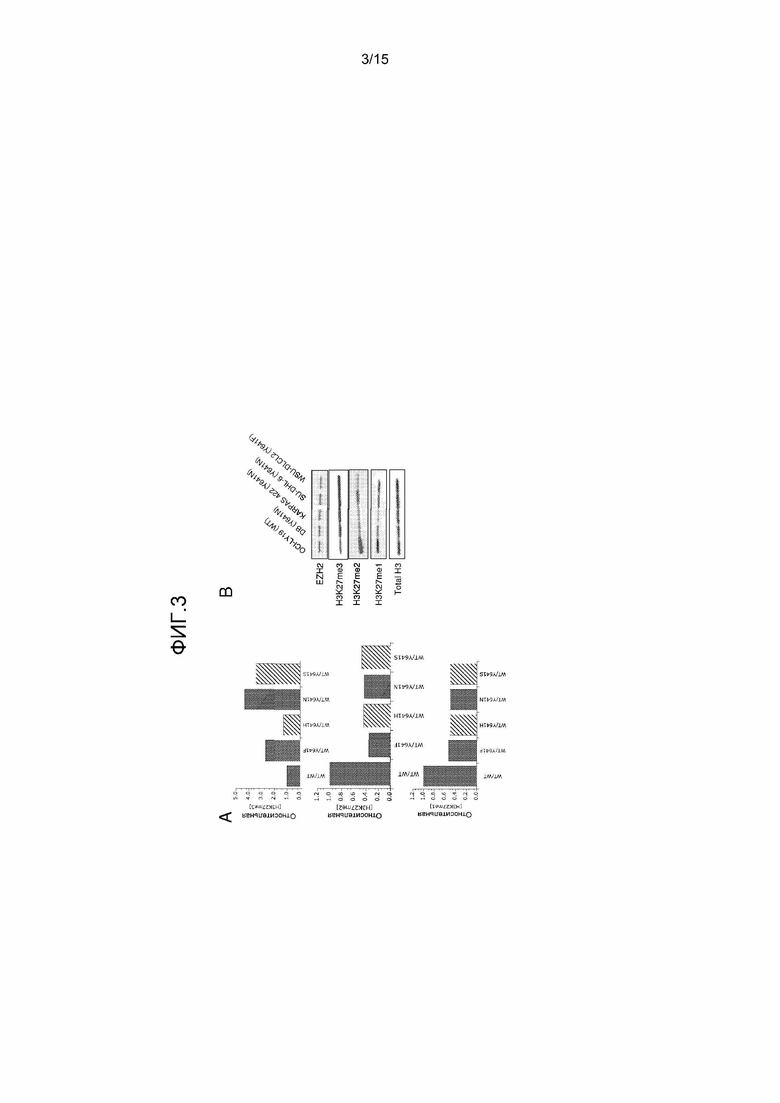
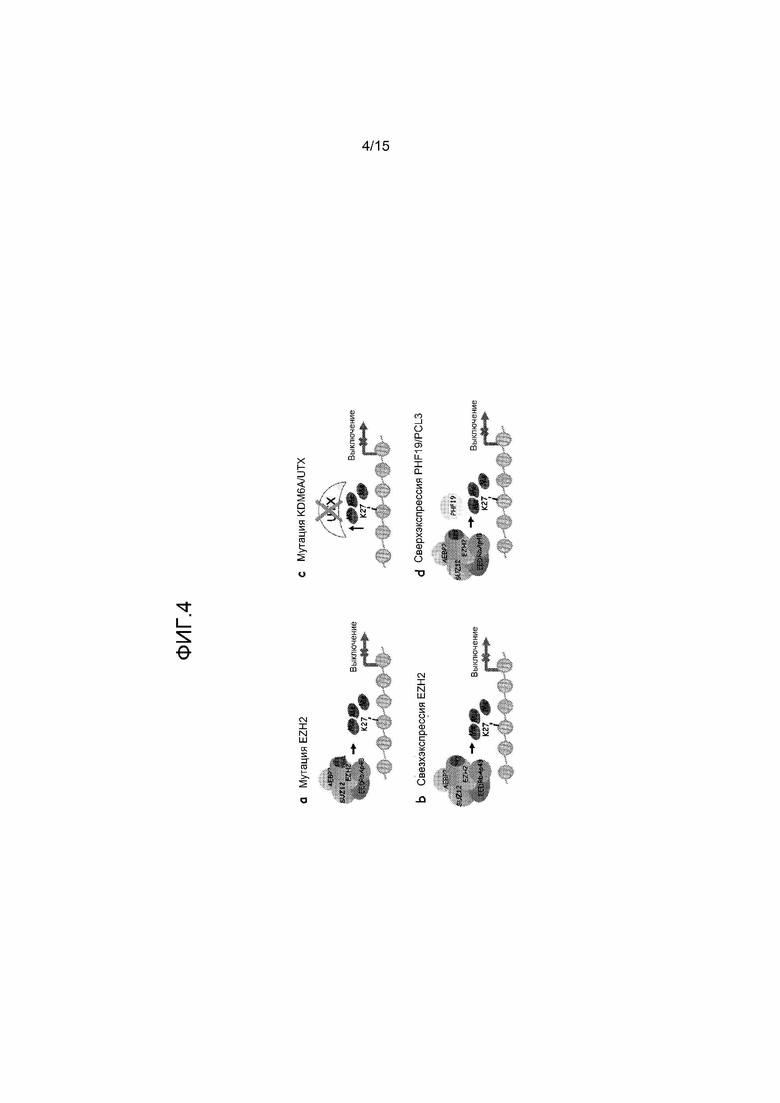
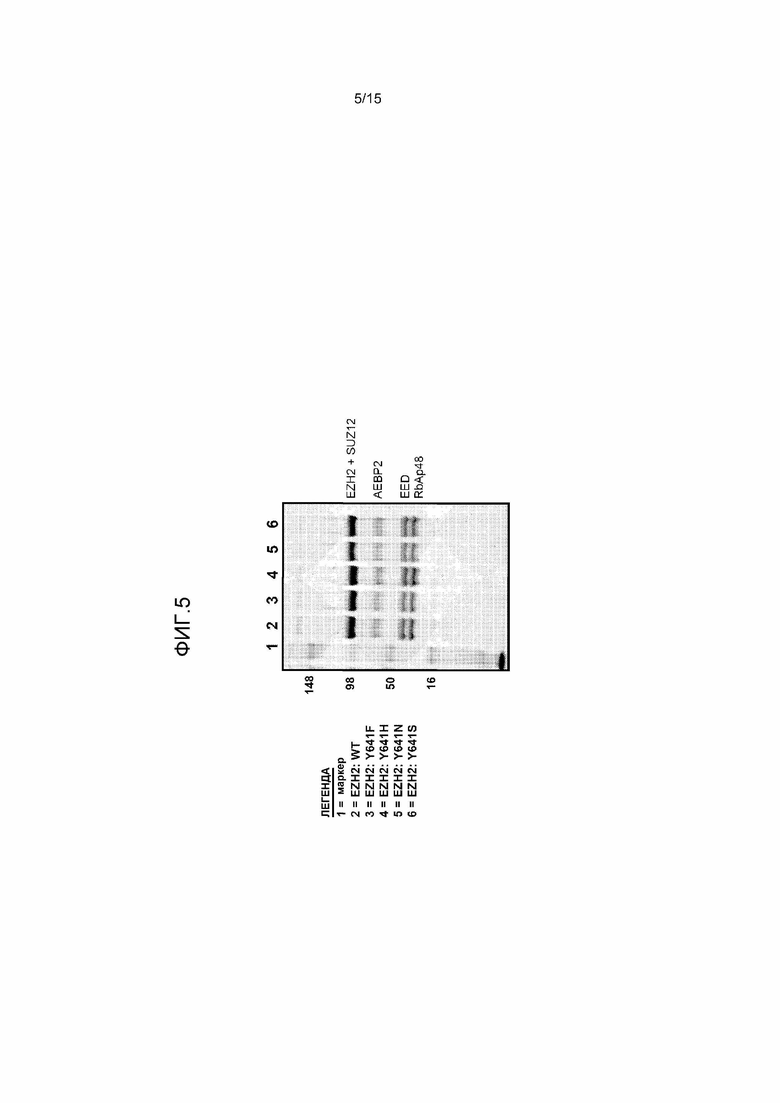
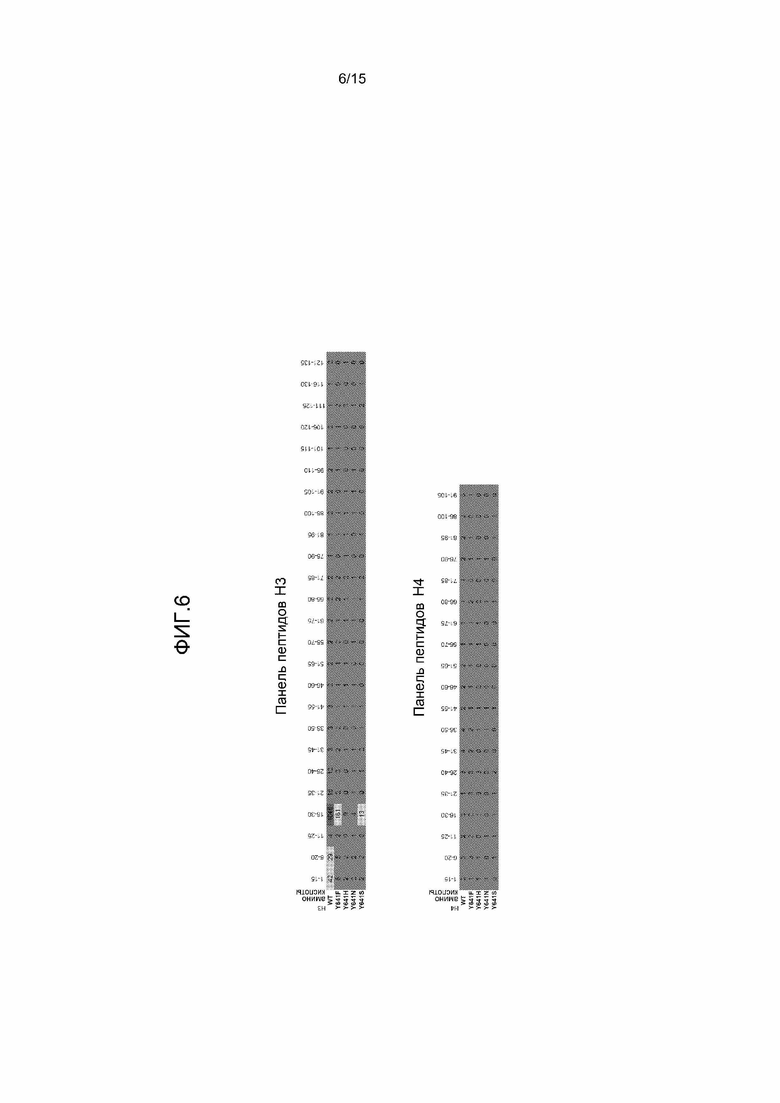

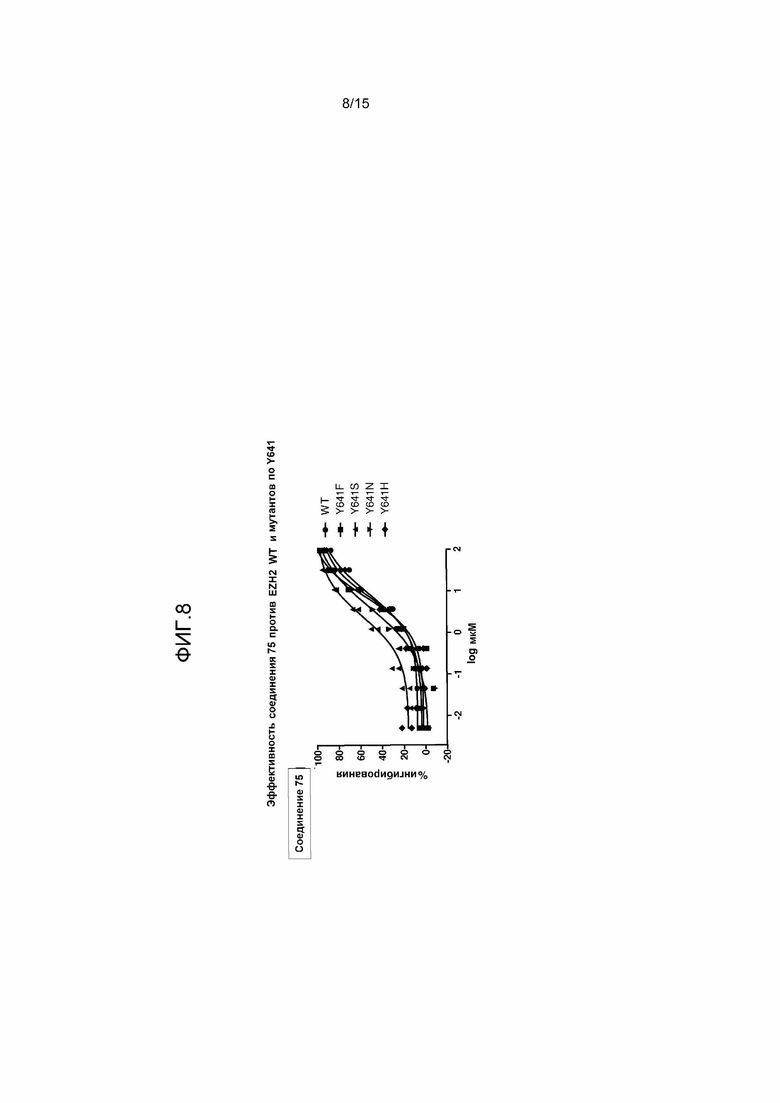
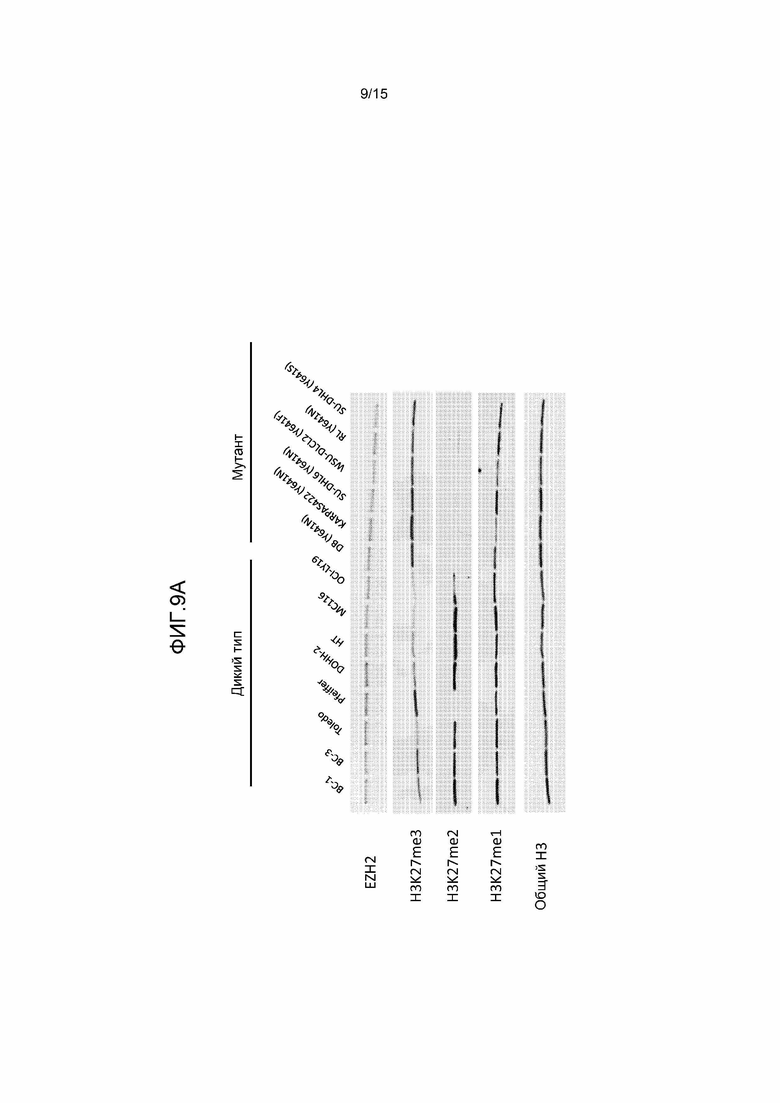
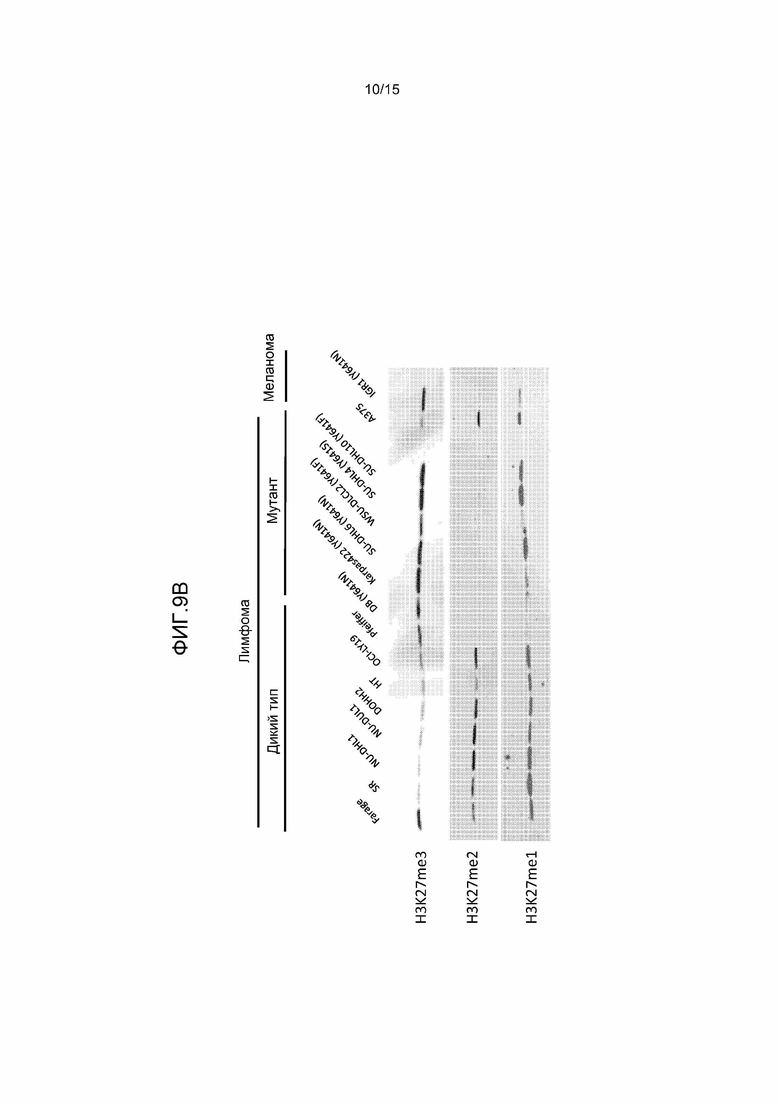
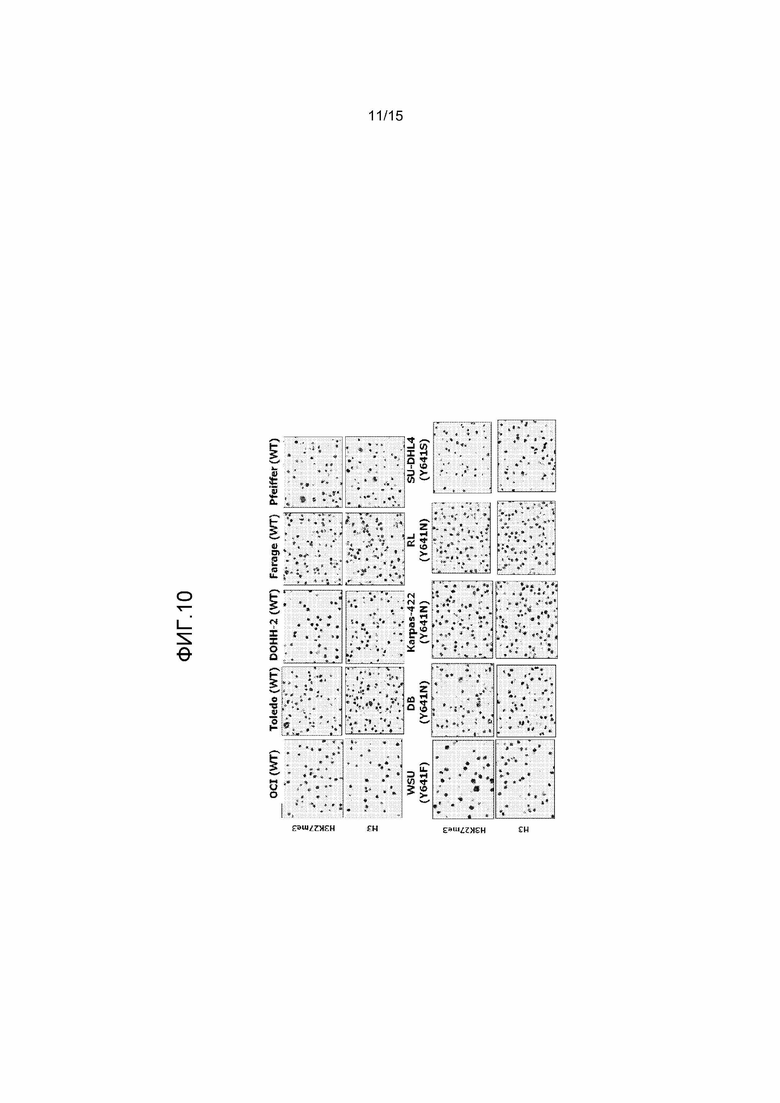
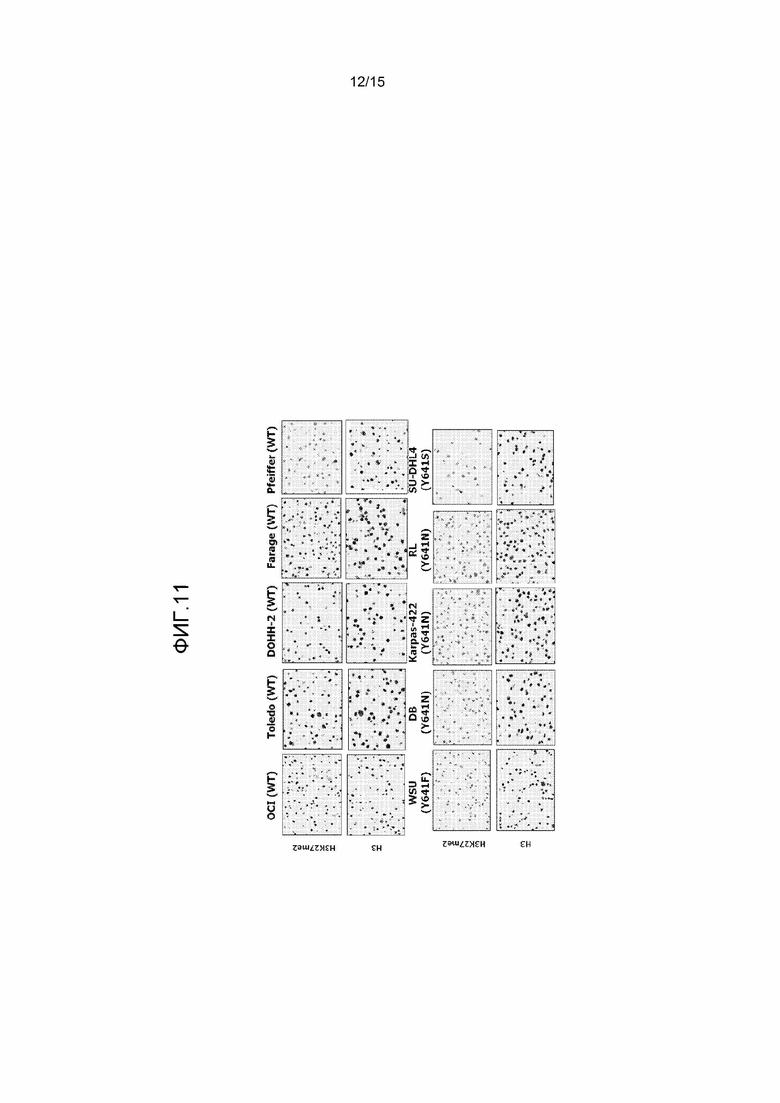
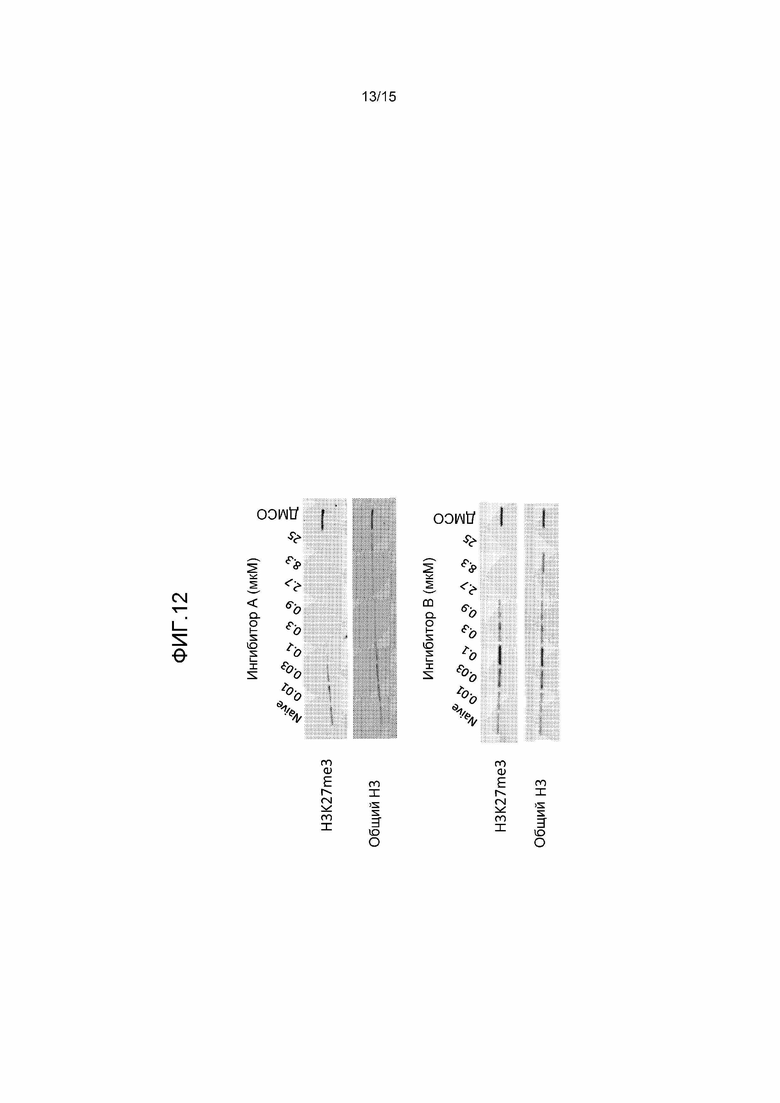
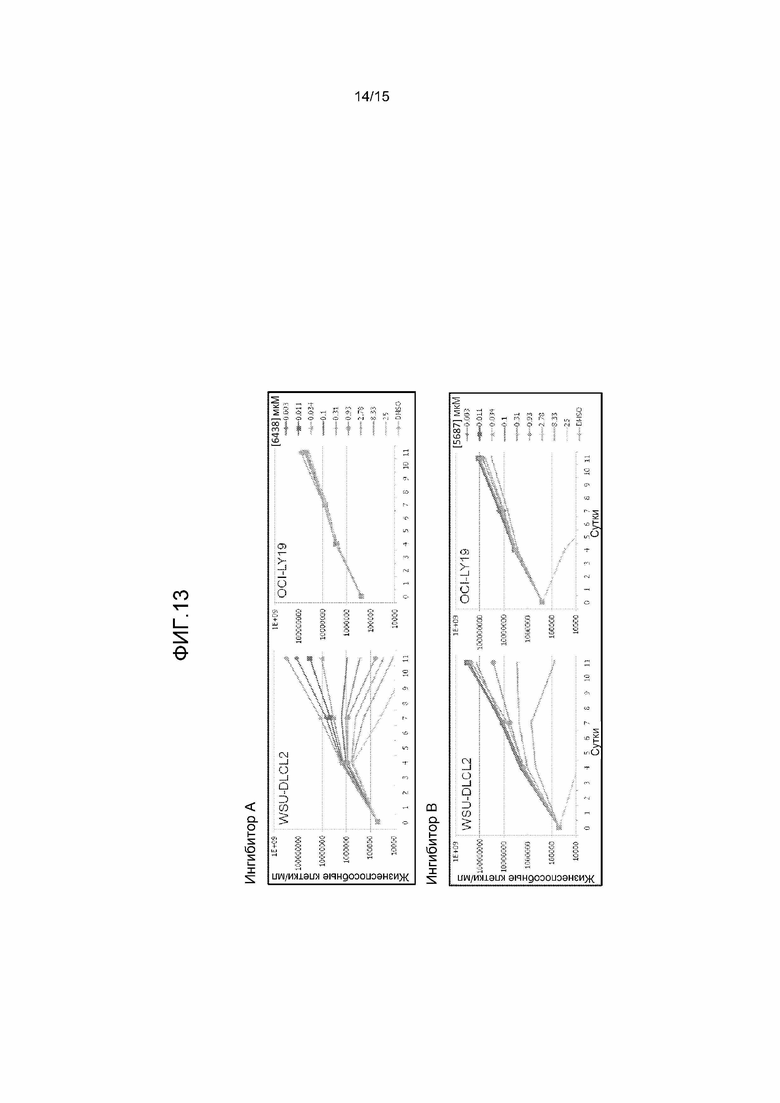

Настоящее изобретение относится к ингибированию форм дикого типа и определенных мутантных форм метилтрансферазы гистонов EZH2 человека - каталитической субъединицы комплекса PRC2, которая катализирует от моно- до триметилирования лизина 27 на гистоне H3 (H3-K27), к способам лечения различных типов рака, включающих фолликулярную лимфому и диффузную крупно-B-клеточную лимфому (DLBCL), и к способам определения способности к ответу на ингибитор EZH2 у индивидуума. Способ идентификации индивидуума как отвечающего на селективный ингибитор мутанта EZH2 по Y641 включает a) получение образца ткани от индивидуума; b) выявление уровня диметилирования (me2) H3-K27, если оно присутствует в указанном образце ткани, и сравнение указанного уровня диметилирования (me2) с контрольным уровнем диметилирования (me2); где указанный индивидуум является отвечающим на селективный ингибитор мутанта EZH2 по Y641, когда указанный уровень диметилирования (me2) отсутствует или ниже указанного контрольного уровня диметилирования (me2), и где контрольный уровень представляет собой уровень диметилирования (me2) H3-K27 в образце ткани от индивидуума, у которого не экспрессируется мутант EZH2 по Y641. 9 з.п. ф-лы, 4 табл., 14 ил., 11 пр.
1. Способ идентификации индивидуума как отвечающего на селективный ингибитор мутанта EZH2 по Y641, включающий:
a) получение образца ткани от индивидуума;
b) выявление уровня диметилирования (me2) H3-K27, если оно присутствует в указанном образце ткани, и сравнение указанного уровня диметилирования (me2) с контрольным уровнем диметилирования (me2);
где указанный индивидуум является отвечающим на селективный ингибитор мутанта EZH2 по Y641, когда указанный уровень диметилирования (me2) отсутствует или ниже указанного контрольного уровня диметилирования (me2), и где контрольный уровень представляет собой уровень диметилирования (me2) H3-K27 в образце ткани от индивидуума, у которого не экспрессируется мутант EZH2 по Y641.
2. Способ по п. 1, дополнительно включающий:
c) выявление уровня триметилирования (me3) H3-K27, если оно присутствует в указанном образце ткани, и сравнение указанного уровня триметилирования (me3) с контрольным уровнем триметилирования (me3),
где указанный индивидуум является отвечающим на селективный ингибитор мутанта EZH2 по Y641, когда указанный уровень триметилирования (me3) является таким же или более высоким, чем указанный контрольный уровень триметилирования (me3), и указанный уровень диметилирования (me2) отсутствует или ниже указанного контрольного уровня диметилирования (me2); и где контрольный уровень триметилирования (me3) представляет собой уровень триметилирования (me3) H3-K27 в образце ткани от индивидуума, у которого не экспрессируется мутант EZH2 по Y641.
3. Способ по п.1, дополнительно включающий:
d) получение соотношения уровня диметилирования (me2) и уровня триметилирования (me3) H3-K27 в указанном образце ткани, для получения тестируемого соотношения, и сравнение тестируемого соотношения с контрольным соотношением;
где указанный индивидуум является отвечающим на ингибитор EZH2, когда указанное тестируемое соотношение является более низким, чем указанное контрольное соотношение, и где контрольное соотношение представляет собой соотношение уровня диметилирования (me2) и уровня триметилирования (me3) H3-K27 в образце ткани от индивидуума, у которого не экспрессируется мутант EZH2 по Y641.
4. Способ по п.1, где указанный индивидуум имеет рак.
5. Способ по п.1, где указанное выявление проводят с помощью анализа, выбранного из анализа с использованием вестерн-блоттинга, иммуногистохимии (IHC), иммунофлуоресценции (IF) и масс-спектрометрии (МС).
6. Способ по п.1, где указанное выявление включает приведение в контакт указанного образца ткани со средством, которое специфично связывается с диметилированным H3-K27.
7. Способ по п.2, где указанное выявление включает приведение в контакт указанного образца ткани со средством, которое специфично связывается с триметилированным H3-K27.
8. Способ по п.6, где указанное средство представляет собой антитело, полипептид, аптамер или их фрагмент.
9. Способ по п.4, где указанный рак выбран из фолликулярной лимфомы и диффузной крупно-B-клеточной лимфомы (DLBCL).
10. Способ по любому из пп.1-9, где указанный мутант по Y641 выбран из Y641F, Y641H, Y641N и Y641S.
| D.B | |||
| YAP et al., Somatic mutations at EZH2 Y641 act dominantly through a mechanism of selectively altered PRC2 catalytic activity, to increase H3K27 trimethylation, BLOOD, 2011, V | |||
| Аппарат для испытания прессованных хлебопекарных дрожжей | 1921 |
|
SU117A1 |
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| Способ получения мочевин ароматического ряда | 1914 |
|
SU2451A1 |
| Гидравлический динамометр | 1914 |
|
SU2452A1 |
| Способ образования части, служащей для скрепления косы с косовищем у кос, изготовляемых штамповкой из листовой или тонкой полосовой стали | 1919 |
|
SU2457A1 |
| Приспособление для убивания крыс и мышей электрическим током | 1921 |
|
SU2461A1 |

