[0001] Данная заявка испрашивает приоритет по предварительной заявке на патент США с серийным № 62/254,435, поданной 12 ноября 2015 г., которая полностью включена в настоящий документ путем ссылки.
ЗАЯВЛЕНИЕ О ФИНАНСИРОВАНИИ ИССЛЕДОВАНИЙ ИЛИ РАЗРАБОТОК ИЗ ФЕДЕРАЛЬНОГО БЮДЖЕТА
[0002] Данное изобретение было сделано при государственной поддержке в соответствии с грантом № 1RO1EB013584-02, присужденным Национальными институтами здравоохранения США. Государство имеет определенные права на данное изобретение.
ОБЛАСТЬ ТЕХНИКИ
[0003] Варианты реализации данного изобретения относятся по меньшей мере к клеточной биологии, молекулярной биологии, модуляции генов, генной терапии, терапии стволовыми клетками, медицине, молекулярной визуализации, биосенсорам и диагностике.
УРОВЕНЬ ТЕХНИКИ
[0004] Для установления функции продукта конкретного гена или для получения терапевтических белков всегда было необходимо точно контролировать экспрессию трансгена в пределах безопасного диапазона. В настоящее время доступно несколько систем регуляции генов1-3 и они оказались исключительно эффективными экспериментальными инструментами. Тем не менее, несмотря на их полезность, данные системы обладают некоторыми практическими ограничениями из-за их зависимости от гибридных транскрипционных трансактиваторов и специализированных промоторов. Эти ограничения включают: (i) Необходимость наличия двух транскрипционных единиц, одной - для экспрессии активатора транскрипции, другой - для экспрессии подлежащего регуляции трансгена. В результате, использование данных систем требует совместного введения двух экспрессионных конструкций. (ii) Потенциальные токсические эффекты, возникающие вследствие экспрессии гибридного активатора транскрипции (например, индукция иммунных реакций хозяина, направленных против активаторов транскрипции, являющихся чужеродными белками, влияние активаторов транскрипции на эндогенную транскрипцию и т.д.). (iii) Сложности применения таких систем для регуляции тканеспецифическим способом из-за необходимости наличия специализированного промотора. (iv) Ограниченное количество малых индукторных молекул, доступных для экспериментального и терапевтического применения (из-за ограниченного количества доступных систем). Эти ограничения можно преодолевать с помощью системы регуляции генов, содержащей, по существу состоящей из или состоящей только из РНК, в которую не вовлекаются какие-либо белки-активаторы транскрипции и специализированные промоторы. Сообщалось о существовании механизмов контроля экспрессии генов исключительно на основе РНК. В частности, описана система, основанная на модуляции аутокаталитического рибозима4, предоставляющая убедительную концепцию для разработки системы с использованием только РНК, которая обладает намного более широкой сферой применения в генной терапии и биологических исследованиях.
[0005] В противоположность системам регуляции генов, основанным на контроле транскрипции с использованием активаторов транскрипции, система на основе полиА, предложенная в данном документе, не требует экспрессии каких-либо продуктов белков-активаторов транскрипции и не зависит от применения каких-либо специализированных промоторных элементов, и поэтому, теоретически, представляет «портативную» систему регуляции, которая может быть «помещена» в любой эндогенный ген или сконструированную векторную единицу транскрипции. По сути, данная система требует наличия только одной транскрипционной единицы (одной экспрессионной конструкции) и содержит настолько универсальный промотор, что он может использоваться для регуляции трансгенов тканеспецифическим (зависящим от пространства) и зависящим от времени способом. Благодаря тому, что полиаденилирование представляет собой универсальный процесс, происходящий во всех клетках млекопитающих, описанная система может найти широкое применение.
[0006] Другая важная сфера применения регуляции генов, которая еще не полностью разработана из-за ограничений существующей технологии, заключается в способности систем регуляции генов функционировать в качестве биосенсоров для выявления экспрессии конкретных клеточных белков или патологических событий in vivo. Такие биосенсорные платформы могут обеспечивать относящейся ко времени или пространству информацией in vivo, относительно флуктуаций на биомолекулярных уровнях, а вводимую информацию можно использовать для регуляции клеточного поведения или генерирования репортерных сигналов для визуализации или выявления. Например, система регуляции генов связывается с вызывающим рак белком в виде его лиганда и, в ответ на это, включает или выключает конкретный набор генов. Данные гены могут генерировать репортерные сигналы для количественного выявления или визуализации, влиять на ход метаболических путей или экспрессировать терапевтические белки. Молекулярное обнаружение конкретных клеточных белков может обеспечивать отображение биохимических аномалий в живых клетках, лежащих в основе заболевания. Такая технология может использоваться для наблюдения прогрессирования клинического лечения путем отслеживания экспрессии конкретных маркерных белков и, в конкретных вариантах реализации изобретения, формировать важную платформу для обеспечения возможности тестирования и разработки новых терапевтических подходов. Кроме того, такая технология может использоваться для понимания роли конкретных продуктов генов в биологических процессах, а также в развитии заболеваний.
[0007] Современные технологии протеомики предложили несколько подходящих способов визуализации или идентификации эндогенных клеточных белков. Такие способы как использование масс-спектрометрии5 позволяют проводить идентификацию сотен молекул белков, присутствующих в клетках или тканях млекопитающих. Однако такие способы разрушают клетки и ткани в процессе исследования и несовместимы с целями, направленными на выявление конкретных клеточных белков в живых организмах животных или человека. Способы, в которых применяются антитела с конъюгированными генерирующими сигнал мотивами, несмотря на их потенциал для визуализации конкретных белков за пределами мембраны клетки, не подходят для выявления или визуализации внутриклеточных белков, которые составляют большинство экспрессируемых внутри живых клеток белков. Недостаток подходящих способов для выявления конкретных нативных белков in vivo указывает на необходимость новых стратегий для биосенсорного определения, поскольку они будут иметь основное влияние на понимание фундаментальных биологических процессов и болезненных состояний.
[0008 ] Настоящее описание удовлетворяет долго ожидаемую потребность в данной области техники касательно предложения способов, систем и композиций для применения подхода на основе РНК-переключателей для выявления экспрессии нативных белков, а также для регуляции интересующих генов в живых клетках.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
[0009] Варианты реализации данного изобретения относятся к экзогенному контролю регуляции генов с использованием опосредованной аптамерами модуляции полиаденилирования. Варианты реализации данного изобретения относятся к контролю экспрессии генов млекопитающих посредством регуляции расщепления полиA, управляемой контролирующим действием связывания лиганда с аптамером, который содержит сигнал полиA.
[0010] Варианты реализации данного изобретения относятся к молекулярному переключателю, который контролирует экспрессию конкретного полинуклеотида с использованием опосредованной аптамерами модуляции полиаденилирования. В конкретных вариантах реализации изобретения молекулярный переключатель представляет собой основанный на сигнале полиA в 5'-НТО (нетранслируемая область) РНК-переключатель (который в данном документе может называться полиA-переключателем или полиA-сенсором).
[0011] В конкретных вариантах реализации систему данного изобретения используют в качестве биосенсорной системы, которая выявляет одну или более внутриклеточных «сигнатур» в живых клетках, тканях или организмах. «Сигнатура» может отображать экспрессию одного или более обсуждаемых генов; определять наличие или отсутствие одного или более продуцируемых эндогенно соединений, таких как белки или метаболиты; наличие или отсутствие молекулы (молекул), которые являются индикаторами болезненного состояния или нормального состояния, или отображать эффективность терапии, или быть их комбинацией и т.д. Данная система может быть индикатором метаболического или физиологического состояния одной или более клеток или тканей, или организма.
[0012] В конкретных вариантах реализации изобретения система содержит полинуклеотид, который содержит в направлении 5'-3' один или более лиганд-связывающих аптамеров, содержащих в своем составе по меньшей мере один сигнал расщепления полиA; и экспрессируемый полинуклеотид; система подвергается воздействию соответствующих условий таким образом, что когда лиганд отсутствует в системе или ее окружении или лиганд не связывает лиганд-связывающий аптамер, полученная из экспрессируемого полинуклеотида мРНК разрушается. В некоторых вариантах реализации изобретения система подвергается воздействию соответствующих условий таким образом, что когда лиганд связывает аптамер, полученная из экспрессируемого полинуклеотида мРНК не разрушается, а продукт гена экспрессируется на экспрессируемом полинуклеотиде.
[0013] В определенных вариантах реализации система данного изобретения относится к способности модулировать экспрессию одного или более обсуждаемых конкретных генов. Система позволяет манипулировать экспрессией гена(-ов) путем использования конкретного лиганда, который может ингибировать РНК-переключатель на основе сигнала полиA в 5'-НТО, тем самым приводя к экспрессии гена(-ов). Система позволяет осуществлять строго управляемую экспрессию генов с использованием лигандов, таких как малые молекулы, для контроля экспрессии, включая, например, тканеспецифический и/или зависящий от времени способ. В конкретных вариантах реализации изобретения система действует в качестве биосенсорной системы для использования специально подобранной комбинации аптамера/экспрессируемого полинуклеотида для получения информации об эндогенных лигандах в определенном окружении.
[0014] В одном варианте реализации изобретения представлена система для модуляции экспрессии генов, содержащая полинуклеотид, содержащий в направлении 5'-3': a) по меньшей мере один лиганд-связывающий аптамер, который в своем составе, каждый, содержит по меньшей мере один сигнал расщепления полиA (хотя в альтернативных вариантах реализации изобретения не все аптамеры содержат сигнал расщепления полиA); и b) экспрессируемый полинуклеотид. В конкретных вариантах реализации изобретения лиганд-связывающий аптамер, содержащий сигнал расщепления полиA, находится в пределах 5'-нетранслируемой области экспрессируемого полинуклеотида. В конкретных вариантах реализации изобретения система содержит полинуклеотид, который экспрессирует лиганд. В некоторых аспектах изобретения экспрессирующий лиганд полинуклеотид представляет собой тот же полинуклеотид, который содержит аптамер и экспрессируемый полинуклеотид. В других аспектах изобретения экспрессирующий лиганд полинуклеотид представляет собой полинуклеотид, отличающийся от того, который содержит аптамер и экспрессируемый полинуклеотид.
[0015] В некоторых вариантах реализации охватываемые данным изобретением полинуклеотиды содержат два, три или более сигналов полиA в 5'-НТО экспрессируемого полинуклеотида. В конкретном варианте реализации изобретения полинуклеотид содержит: a) по меньшей мере один сигнал полиA; b) лиганд-связывающий аптамер и c) по меньшей мере один богатый U/UG участок, по меньшей мере один богатый G участок или оба варианта из по меньшей мере одного богатого U/UG участка и по меньшей мере одного богатого G участка. В некоторых вариантах реализации изобретения системы лиганд-связывающий аптамер содержит один, два, три или более богатых U/UG участков. В определенных аспектах изобретения в направлении 5'-3' полинуклеотида по меньшей мере один сигнал полиA находится слева (выше) по ходу транскрипции от по меньшей мере одного богатого U/UG участка. В других аспектах изобретения в направлении 5'-3' полинуклеотида лиганд-связывающий аптамер находится слева по ходу транскрипции от одного, двух или более богатых U/UG участков. В конкретных аспектах изобретения в направлении 5'-3' полинуклеотида по меньшей мере один сигнал полиA находится слева по ходу транскрипции от по меньшей мере одного богатого G участка. В некоторых аспектах изобретения в направлении 5'-3' полинуклеотида лиганд-связывающий аптамер находится слева по ходу транскрипции от одного, двух или более богатых G участков. В конкретных аспектах изобретения в направлении 5'-3' полинуклеотида аптамер содержит два сигнала полиA и два богатых U/UG участка. В некоторых аспектах изобретения, в тех случаях, когда полинуклеотид содержит два или более аптамеров, полинуклеотид может содержать только один богатый G участок. В таких случаях богатый G участок может располагаться на наиболее удаленном в 3'-направлении аптамере полинуклеотида, или может располагаться на втором аптамере в направлении 5'-3' полинуклеотида.
[0016] В некоторых вариантах реализации данного изобретения лиганд представляет собой полипептид, пептид, нуклеиновую кислоту, малую молекулу, лекарственное средство, метаболит или их комбинацию. В конкретных аспектах изобретения длина аптамера составляет от 14 до 250 нуклеотидов. В конкретных вариантах реализации изобретения экспрессируемый полинуклеотид представляет собой репортерный ген, терапевтический ген или ген, продукт которого изменяет метаболическое состояние клеток. В некоторых случаях полинуклеотид представляет собой по меньшей мере часть вектора. Полинуклеотид, который экспрессирует лиганд, может представлять собой по меньшей мере часть такого вектора, как плазмида, вирусный вектор или линейная ДНК. В конкретных вариантах реализации изобретения экспрессируемый полинуклеотид кодирует лиганд. В некоторых аспектах изобретения экспрессия экспрессируемого полинуклеотида может регулироваться тканеспецифическим промотором.
[0017] В некоторых вариантах реализации изобретения представлен способ модулирования экспрессии генов, включающий этапы: a) предоставления системы, причем указанная система содержит полинуклеотид, содержащий в направлении 5'-3': 1) по меньшей мере один лиганд-связывающий аптамер, каждый из которых содержит в своем составе по меньшей мере один сигнал расщепления полиA; 2) экспрессируемый полинуклеотид и 3) необязательно лиганд-экспрессирующую конструкцию; и b) воздействия на систему соответствующих условий, при которых, когда не требуется получение мРНК из экспрессируемого полинуклеотида, лиганд не связывает лиганд-связывающий аптамер или отсутствует в системе или ее окружении, а полученная из экспрессируемого полинуклеотида мРНК разрушается; или c) воздействия на систему, содержащую лиганд-экспрессирующую конструкцию, соответствующих условий, при которых, когда требуется экспрессия экспрессионного полинуклеотида, лиганд связывает аптамер и/или присутствует в системе или ее окружении, а полученная из экспрессируемого полинуклеотида мРНК не разрушается.
[0018] В одном варианте реализации изобретения представлен способ модуляции экспрессии генов, включающий этапы: a) предоставления системы, причем указанная система содержит полинуклеотид, содержащий в направлении 5'-3': 1) по меньшей мере один лиганд-связывающий аптамер, который содержит в своем составе по меньшей мере один сигнал расщепления полиA; 2) экспрессируемый полинуклеотид и 3) необязательно лиганд-экспрессирующую конструкцию; и b) воздействия на систему соответствующих условий, при которых в отсутствии лиганда в системе или ее окружении, или в отсутствии связывания ним лиганд-связывающего аптамера, полученная из экспрессируемого полинуклеотида мРНК разрушается; или c) воздействия на систему, содержащую лиганд-экспрессирующую конструкцию, соответствующих условий, при которых, когда лиганд связывает аптамер, полученная из экспрессируемого полинуклеотида мРНК не разрушается, а продукт гена экспрессируется на экспрессируемом полинуклеотиде. В конкретных вариантах реализации изобретения способ осуществляется в такой клетке, как стволовая клетка, раковая клетка или пораженная заболеванием, или дефектная клетка, требующей генной терапии по некоторому гену (такому как ген дистрофина, альбумина или фактора IX). В конкретных аспектах изобретения лиганд по отношению к клетке является эндогенным. В конкретных аспектах изобретения способ осуществляется in vivo, например, в организме млекопитающего, включая человека. В других аспектах изобретения способ осуществляется in vitro. В некоторых вариантах реализации изобретения способ осуществляется в одной или более из клеток индивида, лиганд представляет собой глюкозу, индивид болен диабетом, преддиабетом или имеет осложнения вследствие диабета, и/или экспрессируемый полинуклеотид представляет собой последовательность инсулина. В некоторых аспектах изобретения способ осуществляется в одной или более из клеток индивида, лиганд представляет собой продукт гена ракового биомаркера, а экспрессируемый полинуклеотид представляет собой суицидальный ген. В конкретных аспектах изобретения способ осуществляется в организме индивида, экспрессируемый полинуклеотид представляет собой репортерный ген, а расположение и/или интенсивность экспрессии репортерного гена предоставляет информацию о пространственном распределении, зависящей от времени флуктуации, или обоих этих параметрах для лиганда в одной или более клетках индивида. В некоторых вариантах реализации изобретения способ дополнительно включает этап конструирования аптамера для приемлемого связывания лиганда. В конкретных вариантах реализации изобретения способ осуществляется в организме индивида, ткани или клетке, причем экспрессируемый полинуклеотид кодирует выявляемый продукт гена и при этом визуализируется тело соответствующего индивида, ткань или клетка.
[0019] В некоторых вариантах реализации изобретения представлен способ отслеживания терапии индивида, включающий этап предоставления индивиду: a) вектора, содержащего полинуклеотид, который в направлении 5'-3' содержит: 1) по меньшей мере один лиганд-связывающий аптамер, содержащий в своем составе по меньшей мере один сигнал расщепления полиA; и 2) экспрессируемый полинуклеотид; и/или b) одну или более клеток, содержащих вектор по п. a), причем лиганд представляет собой продукт конкретного гена в виде белка, который является показателем эффективности терапии. В конкретных вариантах реализации изобретения индивиду предоставляют вектор по п. a) и/или клетки по п. b) перед терапией, во время терапии и/или после терапии.
[0020] В одном варианте реализации изобретения представлен способ оценки у индивида наличия, риска возникновения или восприимчивости к патологическому состоянию, включающий этапы предоставления индивиду: a) вектора, содержащего полинуклеотид, который в направлении 5'-3' содержит: 1) по меньшей мере один лиганд-связывающий аптамер, содержащий в своем составе по меньшей мере один сигнал расщепления полиA; и 2) экспрессируемый полинуклеотид; и/или b) одну или более клеток, содержащих вектор по п. a), причем экспрессия экспрессируемого полинуклеотида или отсутствие экспрессии экспрессируемого полинуклеотида идентифицирует связывание присутствующим лигандом аптамера, при этом соответствующее наличие или отсутствие лиганда в организме индивида или его клетках является показателем наличия, восприимчивости или риска возникновения патологического состояния.
[0021] В определенном варианте реализации изобретения представлен полинуклеотид, содержащий в направлении 5'-3': a) по меньшей мере один лиганд-связывающий аптамер, который в своем составе содержит по меньшей мере один сигнал расщепления полиA, и b) экспрессируемый полинуклеотид. В конкретных вариантах реализации изобретения лиганд-связывающий аптамер расположен в 5'-НТО экспрессируемого полинуклеотида. В некоторых аспектах изобретения полинуклеотид дополнительно содержит по меньшей мере один богатый U/UG участок, по меньшей мере один богатый G участок или оба варианта из по меньшей мере одного богатого U/UG участка и по меньшей мере одного богатого G участка. В других аспектах изобретения в направлении 5'-3' полинуклеотида по меньшей мере один сигнал полиA находится слева по ходу транскрипции от по меньшей мере одного богатого U/UG участка. В конкретных аспектах изобретения в направлении 5'-3' полинуклеотида лиганд-связывающий аптамер находится слева по ходу транскрипции от одного, двух или более богатых U/UG участков. В особых случаях в направлении 5'-3' полинуклеотида по меньшей мере один сигнал полиA находится слева по ходу транскрипции от по меньшей мере одного богатого G участка. В особых аспектах изобретения в направлении 5'-3' полинуклеотида лиганд-связывающий аптамер находится слева по ходу транскрипции от одного, двух или более богатых G участков.
[0022] Другие варианты реализации изобретения относятся к клеткам, которые содержат любой полинуклеотид, охватываемый данным описанием. В конкретных вариантах реализации изобретения клетка представляет собой клетку млекопитающего, такую как клетка человека, и данная клетка может располагаться в организме человека. В некоторых вариантах реализации изобретения клетка содержит любую систему, охватываемую данным описанием. В конкретных вариантах реализации изобретения клетка представляет собой клетку млекопитающего, такую как клетка человека, и данная клетка может располагаться в организме человека. В конкретных вариантах реализации изобретения предложен вектор, охватываемый данным описанием и содержащий любой полинуклеотид, охватываемый данным описанием. В конкретных случаях вектор представляет собой вирусный вектор, такой как аденовирусный вектор, аденоассоциированный вирусный вектор, ретровирусный вектор или лентивирусный вектор. В конкретных вариантах реализации изобретения вектор представляет собой плазмиду.
КРАТКОЕ ОПИСАНИЕ ФИГУР
[0023] На ФИГ. 1A изображена примерная стратегия визуализации конкретного внутриклеточного белка посредством модуляции сигнала расщепления полиA. Эффективное расщепление сигнала полиA (pA), сконструированного в пределах 5'-НТО, приводит к разрушению мРНК и потере экспрессии репортерного сигнала. Связывание белка-лиганда с аптамером, связанным с сайтом полиA, блокирует расщепление, приводя к сохранению интактной мРНК, и обеспечивает возможность экспрессии репортерного сигнала. Треугольником обозначен сайт расщепления pA. Для ясности не указан pA, обычно присутствующий в 3'-НТО. Нижнее изображение получено путем котрансфекции ДНК-плазмиды, кодирующей полиA-переключатель с кодируемым плазмидой белком-лигандом (tat) в клетках 293T человека (справа) или без него (слева). Наличие белка-лиганда прямо индуцирует экспрессию биолюминесцентного репортерного сигнала, который легко выявляется путем визуализации. На ФИГ. 1B показано взаимодействие малой молекулы-аптамера, которое регулирует расщепление полиA. Аптамер со встроенным pA, связывающий малую молекулу, вставлен в 5'-НТО гена. Расщепление полиA на 5'-НТО приводит к разрушению 3'-части мРНК и поэтому обеспечивает отсутствие экспрессии гена. Тем не менее связывание введенной малой молекулы с аптамером блокирует полиA 5'-НТО и приводит к образованию интактной мРНК и тем самым к экспрессии белка. Черной линией показана мРНК 5'-НТО, а интересующий ген - оранжевыми прямоугольниками. Низкомолекулярный лиганд показан розовым шестиугольником, а сайт расщепления - черным треугольником. На ФИГ. 1C показан подробный вид компонентов полиA, встроенных внутрь аптамера.
[0024] На ФИГ. 2 проиллюстрировано полиаденилирование, обычно происходящее в 3'-НТО, при котором происходит расщепление мРНК, добавление хвоста полиA к 5'-фрагменту и расщепление 3'-фрагмента.
[0025] На ФИГ. 3 показано, что в случае когда сайт расщепления полиA (синий треугольник) бычьего гормона роста скрыт в стабильном стебле РНК, полиаденилирование в значительной степени снижается, по данным измерения от 100% до 40%. (По материалам Gimmi et al., 1989) [SEQ ID NO: 1; SEQ ID NO: 2].
[0026] На ФИГ. 4 проиллюстрирована полиA-сенсорная система, обеспечивающая возможность использования существующей аптамерной технологии и репортеров визуализации для создания многоцелевой платформы молекулярной визуализации. Сигнал pA: полиA. IFP: белки, флуоресцирующие в инфракрасном спектре. Luc: люцифераза.
[0027] На ФИГ. 5 проиллюстрировано, что сигнал полиA может эффективно расщепляться на 5'-НТО. На ФИГ. 5A проиллюстрирована структура репортерного гена, имеющего вставку сигнала полиA в 5'-НТО репортерного гена зеленого флуоресцентного белка (GFP) совместно с требуемым богатым по U/GU участком. На ФИГ. 5B проиллюстрировано использование одной или двух копий богатого по G-элемента, добавленного после богатого по U/GU участка, и на Фиг. 5C показано включение двух копий сигнала полиA для усиления расщепления полиA. Эффективность расщепления с помощью разных мотивов полиA опосредовано измерена анализом проточной цитометрии. На оси X указаны уровни экспрессии GFP (зеленого флуоресцентного белка). Красная диагональная линия отделяет GFP-позитивные от GFP-негативных клеток. Нижняя конфигурация приводит к меньшему количеству зеленых клеток, вследствие наличия двух копий сигнала полиA, приводящему к значительному снижению уровней GFP [SEQ ID NO: 3].
[0028] На ФИГ. 6 проиллюстрирован подход «фиксации». Связывание лиганда эффективно замыкает аптамер в стабильную двухцепочечную стеблевую структуру и физически блокирует сайт расщепления, указанный треугольником.
[0029] На ФИГ. 7A-7C показан зависимый от дозы отклик полиA-сенсора, предназначенный для выявления вирусного белка tat. (ФИГ. 7A) Уровень экспрессии репортера, визуализированный прибором для визуализации биолюминесценции IVIS200 при разных дозах конструкций лигандов относительно количества трансфицированной плазмиды. Каждая лунка содержит примерно 10 тысяч клеток. Уровень экспрессии неактивного вектора полиA является эталонной 100% индукцией. (ФИГ. 7B) Индукцию в «кратных» величинах рассчитывали как соотношение репортерного сигнала (люциферазы) в присутствии и отсутствие трансфицированной конструкции лиганда при разных дозах. Репортерный сигнал измеряли с использованием люминометра. Показаны три разных полиA-сенсора (сенсор с активным pA, сенсор с неактивным pA или контрольная плазмида без pA) при разных дозах лиганда относительно количества трансфицированной плазмиды. (ФИГ. 7C) Нозерн-анализом визуализировали экспрессию мРНК люциферазы с помощью зонда к 5'-НТО, проходящую на всех плазмидных векторах. Визуализировали мРНК tat с помощью зонда к данной мРНК. В качестве внутреннего контроля использовали зонд против GAPDH. Сигнал pA: полиA. Данные результаты подтвердили, что присутствие лиганда (tat) приводит к получению интактной полноразмерной мРНК (зона 5, верхняя полоса).
[0030] На ФИГ. 8A-8B показано, что эффективность расщепления полиA зависит от положения сигнала полиA в примере тетрациклин-связывающего аптамера (tc-аптамер cb32). (ФИГ. 8A) (SEQ ID NO: 4). В качестве матрицы использовали tc-аптамер (левое изображение), показаны положения, в которые помещен сигнал полиA (AAUAAA). Мутации, внесенные внутрь tc-аптамера с целью создания сигнала полиA, показаны синими буквами. (ФИГ. 8B) На графиках изображена относительная люциферазная активность, измеренная без тетрациклина (tc), что таким образом отображает эффективность расщепления полиA. Активность исходной плазмиды без полиA-переключателя принята за 100%. В данной серии конфигурация A6 проявляет наибольшую эффективность расщепления полиA. Незакрашенный прямоугольник, соответствующий A6, представляет собой контрольный полиA (CACACA).
[0031] На ФИГ. 9A-9C показано влияние длины стебля P1 и положения богатого GU участка на регуляцию эффективности, на основе конструкции A6. Красные линии показывают два сигнала полиA. Длина P1 и протяженность между богатым GU участком и B1-2 даны в п.н. (парах нуклеотидов). Индукция экспрессии гена показана относительной люциферазной активностью конструкций в отсутствие (черные прямоугольники) и присутствии (белые прямоугольники) 15 мкг/мл tc. Эффективность регуляции определяли как соотношение относительной люциферазной активности, данной цифрами под графиком, с tc и без него. (ФИГ. 9A) Конструкции с разной длиной P1 с протяженностью GU равной 2 п.н. Данные результаты показывают, что длина P1, составляющая 12 п.н., приводит к индуцированной tc экспрессии гена. (ФИГ. 9B) Конструкции с длиной P1, составляющей 12 п.н., но отличающейся протяженностью богатого GU участка. Протяженность богатого GU участка, составляющая 5 п.н. или 8 п.н., приводит к максимальной индуцированной tc экспрессии гена. (ФИГ. 9C) Конструкции с разной длиной P1, но с фиксированной протяженностью GU равной 8 п.н. Длина P1, составляющая 11 п.н. или 12 п.н., приводит к максимальной индуцированной tc экспрессии гена, когда протяженность GU фиксирована 8 п.н.
[0032] На ФИГ. 10A-10B показано, что второй сигнал полиA в петле играл более важную роль в расщеплении полиA. (ФИГ. 10A) (SEQ ID NO: 5; SEQ ID NO: 6; SEQ ID NO: 7) Конструкции с длиной P1, составляющей 12 п. н., с протяженностью богатого GU участка 2 п.н. (ФИГ. 10B) (SEQ ID NO: 5; SEQ ID NO: 6; SEQ ID NO: 7) Конструкции с длиной P1, составляющей 12 п.н., с протяженностью богатого GU участка 8 п.н. Левая сторона: Схематический вид различных конструкций. Активные сигналы полиA показаны красной линией, неактивные сигналы полиA показаны синей линией, а мутации, превращающие активный сигнал полиA в неактивный (CACACA) - синими буквами. Правая сторона: Относительная люциферазная активность в отсутствие (черные прямоугольники) и присутствии (белые прямоугольники) 15 мкг/мл tc. Эффективность регуляции определяли как соотношение относительной люциферазной активности, данной цифрами под графиком, с tc и без него. В обоих случаях второй сигнал полиA в петле играл более важную роль в расщеплении полиA.
[0033] На ФИГ. 11A-11B показан сигнал полиA, частично помещенный в новый стебель P1, это приводит к улучшению эффективности регуляции. (ФИГ. 11A) (SEQ ID NO: 8; SEQ ID NO: 9) Схематический вид исследованных конструкций. Конструкции A8 и A8* отличаются по одной паре A-U, показанной синим прямоугольником. (ФИГ. 11B) Относительная люциферазная активность в отсутствие (закрашенные) и присутствии (незакрашенные прямоугольники) 15 мкг/мл tc. Эффективность регуляции определяли как соотношение относительной люциферазной активности, данной цифрами под графиком, с tc и без него.
[0034] На ФИГ. 12A-12B показаны примеры модификации участка P1 на основе конструкции A8. (ФИГ. 12A) Схематический вид конструкций. В конструкции A8g использовано два аптамера tc, фланкирующих сигнал полиA. В конструкции A8h использован один аптамер tc и один аптамер неомицина, фланкирующие сигнал полиA. (ФИГ. 12B) Относительная люциферазная активность в отсутствие (закрашенные) и присутствии (незакрашенные прямоугольники) 15 мкг/мл tc. Следует отметить, что вставка дополнительных пар оснований G-C непосредственно в 5'-положении сигнала полиA, которая стабилизирует стебель P1, приводит к потере активности полиA (конструкция A8f). Это подтверждает, что сигнал полиA можно инактивировать стабилизацией стебля P1. В дополнение к этому, фланкирование сигнала полиA двумя аптамерами tc дополнительно улучшает регуляцию эффективности (конструкция A8g). Во всех случаях используют 15 мкг/мл tc, за исключением A8h, для которого использовали 15 мкг/мл tc и 15 мкг/мл неомицина.
[0035] На ФИГ. 13A-13B показано, что расщепление полиA происходит возле однонитевой петли B1-2 центра связывания tc. (ФИГ. 13A; [SEQ ID NO: 10; SEQ ID NO: 11; SEQ ID NO: 12]) Секвенирование по Сэнгеру приводит к получению полиаденилированных фрагментов 5'-НТО мРНК. Вертикальная черная линия показывает положение, в котором к фрагменту 5'-НТО добавляли хвост полиA. (ФИГ. 13B; SEQ ID NO:13) Идентифицированные сайты расщепления полиA отмечены черными стрелками возле однонитевой петли B1-2 центра связывания tc.
[0036] На ФИГ. 14A-14B показаны модификации для протяженности полиA-GU, предполагаемые сайты расщепления и их влияние на эффективность регуляции. (ФИГ. 14A; SEQ ID NO:14) Схематический вид производных конструкций, полученных из A8 с разной длиной P1 (A8a, A8b, A8c, A8d). Стрелки указывают на предполагаемые основные сайты расщепления полиA для каждой конструкции, на основе показанных на ФИГ. 13 данных. (ФИГ. 14B) Относительная люциферазная активность конструкций с разной длиной P1 в отсутствие (закрашенные) и в присутствии (незакрашенные прямоугольники) 15 мкг/мл tc. Данный результат показывает, что A8 остается наиболее эффективной конфигурацией.
[0037] На ФИГ. 15A-15B показано, что мутационные исследования в положении 42 выявили конструкцию с лучшей регуляторной эффективностью. Известно, что отмеченные квадратными рамками положения являются неустойчивыми к мутациям, тогда как незакрашенными кругами показаны положения, в которых все нуклеотидные изменения не влияют на связывание tc. Положения A13 и A42 образуют неканоническое спаривание оснований, и их выбрали для мутационных исследований. Данные результаты показывают, что мутация A42G приводит к улучшенной эффективности регуляции (конструкция A8-A42G)[SEQ ID NO:15]. (15B) Относительная люциферазная активность в отсутствие (закрашенные) и присутствии (незакрашенные прямоугольники) 15 мкг/мл tc. Эффективность регуляции определяли как соотношение относительной люциферазной активности, данной цифрами под графиком, с tc и без него.
[0038] На ФИГ. 16 показана визуализация связывания tc с аптамером с использованием геля в неденатурирующих условиях. Транскрибируемый in vitro РНК-аптамер, содержащий сконструированный полиA-переключатель (5 мкM), инкубировали с tc (50 мкM) и без него и наносили на 10% полиакриламидный гель в неденатурирующих условиях. Tc возбуждали УФ-светом при 365 нм и визуализировали непосредственно путем флуоресцентной эмиссии (среднее изображение, синий цвет). Затем гель инкубировали с SYBR Gold для окрашивания РНК (верхнее изображение, красный цвет). Совместную локализацию tc и РНК демонстрируют наложением (нижнее изображение). Интенсивность пикселей из канала tc нормализовали к интенсивности канала РНК, обеспечивая повышение по относительному tc-сигналу. Оказывается, что конструкция A8-A42G имеет более высокую аффинность по сравнению с A8 или с cb32. Конструкция A8-A42G*, которая содержит мутацию в сайте связывания tc внутри аптамера, обеспечивает очень слабое связывание tc.
[0039] На ФИГ. 17 продемонстрирована дозозависимая регуляция A8-A42G. Показана относительная кратность изменения индукции для конструкции A8-A42G (синий круг), неактивного контрольного аптамера A8-A42G* (красный квадрат) и неактивного контроля pA (мутация AAUAAA в CACACA, зеленый треугольник). Каждая точка данных соответствует относительной кратности индукции, определенной как соотношение люциферазной активности с tc или без него.
[0040] На ФИГ. 18A, 18B и 18C проиллюстрированы дополнительные положения полиA-переключателя, а также разные конфигурации аптамеров, которые обеспечивают эффективное расщепление и регуляцию полиA.
[0041] На ФИГ. 19 показаны примеры модификаций типичных аптамеров. Аптамер 103G модифицировали по отмеченным положениям для получения аптамеров 103GP2 и 103GS3 [SEQ ID NO: 16; SEQ ID NO: 17; SEQ ID NO: 18].
[0042] На ФИГ. 20 представлены соответствующие уровни экспрессии и индукции люциферазы типичных аптамеров 103G, 103GP2 и 103GS3. Конструкция 103GP2 показывает наибольший теоретический динамический диапазон, 423-кратный, и наибольший уровень индукции под действием tc, 30-кратный.
[0043] На ФИГ. 21 продемонстрирована вариация в модификации участка P2 из 103GP2 и соответствующее воздействие ее на индукцию экспрессии гена [SEQ ID NO: 19].
[0044] На ФИГ. 22 продемонстрирована вариация в модификации участка L3 из 103GP3 и соответствующее воздействие ее на кратность изменения индукции экспрессии гена [SEQ ID NO: 20].
[0045] На ФИГ. 23 продемонстрирована вариация в модификации участка L1 из 103GP3 и соответствующее воздействие ее на индукцию экспрессии гена [SEQ ID NO: 21; SEQ ID NO: 22; SEQ ID NO: 23; SEQ ID NO: 24; SEQ ID NO: 25].
[0046] На ФИГ. 24 проиллюстрированы примеры модификации в спаривании оснований, которые стабилизируют P1 и таким образом приводят к значительному ингибированию активности полиA. Модифицированные участки находятся в голубой рамке [SEQ ID NO: 26; SEQ ID NO: 27].
[0047] На ФИГ. 25 представлен пример полиA-сенсорной конструкции, GP2, имеющей сигнал полиA, фланкированный 2 аптамерами с разной ориентацией [SEQ ID NO: 28, 29, 30].
[0048] На ФИГ. 26 проиллюстрирован пример полиA-сенсорной конструкции, GP2SLGP2, с конфигурацией с двумя аптамерами GP2, соединенными друг с другом таким путем, который обеспечивает альтернативное сворачивание. Аптамер 5'-GP2 называют «хелперным», аптамер 3'-GP2 называют «центральным». Оба содержат активный сигнал полиA, но только центральный аптамер содержит богатый G участок.
[0049] На ФИГ. 27 продемонстрирована вариация по длине и последовательности участка стебля/петли-II (участок в рамке) хелперного GP2 и центрального GP2 и показано соответствующее влияние на индукцию экспрессии гена. Показана вариация последовательностей между разными конструкциями [SEQ ID NO: 31-58].
[0050] На ФИГ. 28 представлена полная последовательность типичных полиA-сенсоров GP2SLGP2 [SEQ ID NO: 59], C12 [SEQ ID NO: 60] и D11A [SEQ ID NO: 61].
[0051] На ФИГ. 29 продемонстрирована дозозависимая регуляция GP2SLGP2. Каждая точка данных соответствует относительной кратности индукции, определенной как соотношение люциферазной активности с tc или без него. Показана средняя кратность индукции со стандартным отклонением. Индукция достигала 104-кратного увеличения при 15 мкг/мл Tc.
[0052] На ФИГ. 30 продемонстрирована дозозависимая регуляция C12. Каждая точка данных соответствует относительной кратности индукции, определенной как соотношение люциферазной активности с tc или без него. Показана средняя кратность индукции со стандартным отклонением. Индукция достигала 151-кратного увеличения при 15 мкг/мл Tc.
[0053] На ФИГ. 31 продемонстрирована дозозависимая регуляция D11A. Каждая точка данных соответствует относительной кратности индукции, определенной как соотношение люциферазной активности с tc или без него. Показана средняя кратность индукции со стандартным отклонением. Индукция достигала 127-кратного увеличения при 15 мкг/мл Tc.
ПОДРОБНОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
[0054] Аптамеры представляют собой короткие последовательности РНК, которые сворачиваются как рецепторы и связывают конкретные лиганды 19-21. Эффективность методов эволюции in vitro 8, 12 для создания аптамеров с высокой аффинностью с конкретными лигандами хорошо установлена 7, 9, 22. Аффинность связывания аптамеров часто может достигать наномолярного диапазона, сравнимого с антителами. В этом отношении аптамеры можно рассматривать как антитела, созданные из РНК. Что отличает аптамер от антитела, так это его малый размер (часто меньше 50 оснований) и его модульная природа. Эти признаки дают аптамерам возможность интегрироваться со структурами других РНК и контролировать их без потери своей функции связывания. Продемонстрировано, что аптамеры могут трансформироваться в саморасщепляющиеся РНК-рибозимы для функционирования лиганд-зависимым способом и функционировать в исследуемых пробирках подобно молекулярному переключателю23, 24.
[0055] Для регуляции экспрессии генов в природе успешно используется аптамерный механизм в форме «рибопереключателей»25. Множество аптамеров природного происхождения обнаружено в бактериях, причем связывание входящих лигандов с аптамерами модулирует экспрессию генов в дополнение к расщеплению РНК посредством механизмов транскрипционной аттенюации, трансляционного ингибирования и альтернативного сплайсинга25, 26. По оценкам считается, что 2-3% из всех бактериальных генов и неизвестное количество дрожжевых генов должно контролироваться данным способом27. Посредством опосредованного аптамерами механизма, расположение и интенсивность экспрессии репортерного гена с течением времени может отражать пространственное распределение и зависящую от времени флуктуацию конкретного внутриклеточного лиганда in vivo. Продукты репортерного гена, такие как люцифераза, тимидинкиназа, белок (например) ближнего инфракрасного или инфракрасного спектра флуоресценции, можно отслеживать такими современными приборами для визуализации, например, как прибор для визуализации биолюминесценции или позитронно-эмиссионной томографии (ПЭТ). Подходящим для методик визуализации всего тела in vivo является белок ближнего инфракрасного или инфракрасного спектра флуоресценции. При данном подходе не требуется маркирование или манипуляция с нативным белком a priori в процессе исследования, следовательно, он может обеспечивать класс измерений, которые раннее было сложно использовать in vivo.
[0056] РНК-аптамеры распознают конкретные лиганды с высокой аффинностью, но отсутствуют мощные методы, которые усиливают сигнал, созданный в результате связывания аптамеров. Несмотря на то что в нескольких исследованиях сообщалось об использовании аптамера для контроля расщепления рибозимов и возможности его применения для функционирования в качестве молекулярных переключателей, которые отвечают на конкретные лиганды в клетках млекопитающих, малые динамические диапазоны (часто менее 5-кратного индуцируемого диапазона), и высокая фоновая экспрессия 28, 29, обнаруженная для этих сенсоров, в значительной степени ограничила их применение.
[0057] Можно считать, что препятствием для использования аптамеров в качестве воспринимающего устройства в клетках млекопитающих является отсутствие действенного механизма переключения/амплификации, который соединяет связывание лиганда/аптамера с экспрессией репортерного гена. В настоящем описании предложен такой механизм переключения/амплификации, чтобы аптамеры и репортерные гены (или, например, трансгены) можно было эффективно связывать с выявлением конкретных молекулярных сигнатур в живых клетках или контролировать экспрессию трансгена. Как описано в данном документе, в одном варианте реализации изобретения задействована модуляция расщепления сигнала полиA посредством взаимодействия лиганда-аптамера.
[0058] Слова, используемые в настоящем описании в единственном числе согласованно со словом содержащий, включающий в формуле изобретения, обозначают «один или более». В некоторых вариантах реализации изобретения описание может состоять из или состоять по существу из одного или более элементов, этапов способов и/или способов данного описания. Подразумевается, что любой способ или композиция, описанные в данном документе, можно воплощать применительно к любому другому способу или композиции, описанных в данном документе в вариантах реализации изобретения, которые описаны, и при этом может быть получен подобный или аналогичный результат без отступления от сущности и объема данного описания.
[0059] Данное изобретение относится к привлечению способности расщепления полиA для создания эффективного молекулярного переключателя, который будет обеспечивать чувствительное выявление конкретных молекулярных сигнатур или состояний в клетках млекопитающих или который будет контролировать экспрессию трансгена в клетках млекопитающих с использованием малых молекул, таких как лекарственные средства или, например, подобные лекарственным средствам молекулы. Сконструированный молекулярный переключатель может контролировать экспрессию репортерного гена или трансгена (например) путем опосредованной аптамерами модуляции полиаденилирования (ФИГ. 1A и 1B). Полиаденилирование представляет собой важный механизм процессинга мРНК, который присутствует во всех клетках млекопитающих и является универсальным для них. Сигналы полиA млекопитающих обычно расположены на 3'-нетранслируемой области (НТО). Когда на 5'-НТО искусственно создают новый сайт полиA, который никогда не располагался в нормальных транскрипционных единицах, эффективное расщепление этого сигнала полиA приводит к разрушению мРНК и, тем самым, потере экспрессии гена. Однако связывание молекулы-лиганда со сконструированным сигналом полиA эффективно блокирует расщепление, приводя к сохранению интактной мРНК, таким образом обеспечивая возможность индуцированной экспрессии гена.
[0060] Общие варианты реализации переключателей на основе полиA
[0061] Данное описание относится к системам, в которых в целом и намеренно применяют сигнал полиA, имеющийся в экспрессионной конструкции в расположении, отличающемся от 3'-нетранслируемой области (НТО) экспрессируемого полинуклеотида, такого как ген. В конкретных вариантах реализации изобретения эктопическое расположение сигнала полиA позволяет использовать систему настоящего описания для модуляции интересующего экспрессируемого полинуклеотида. В конкретных вариантах реализации изобретения сигнал полиA расположен слева по ходу транскрипции от сайта начала трансляции экспрессируемого полинуклеотида (мРНК) и в конкретных вариантах реализации изобретения сигнал полиA расположен в 5'-НТО мРНК. В конкретных вариантах реализации изобретения экспрессируемый полинуклеотид способен транскрибироваться РНК-полимеразой II. В конкретных вариантах реализации изобретения структура экспрессионной конструкции намеренно располагает сигнал полиA на 5'-НТО экспрессируемого полинуклеотида.
[0062] В некоторых вариантах реализации изобретения наличие сигнала полиA в 5'-НТО направляет мРНК для разрушения и данное свойство используют в системах настоящего описания. В конкретных вариантах реализации изобретения сигнал полиA связан с аптамером, с которым может связываться один или более лигандов, а связывание лиганда с аптамером определяет будет проходить разрушение мРНК или нет. В конкретных вариантах реализации изобретения, в которых требуется экспрессия мРНК, предоставляется лиганд, связывающий аптамер. В некоторых случаях вопрос о присутствии или отсутствии лиганда в конкретном окружении (например, в клетке, ткани или организме) решается тем, экспрессируется ли конкретный экспрессируемый полинуклеотид при регуляции конкретным полиA/аптамером, который может связываться таким лигандом, или нет. В вариантах реализации изобретения, в которых лиганд связывает аптамер и экспрессируется мРНК, система обеспечивает амплификацию сигнала, поскольку на единственной мРНК может продуцироваться множество продуктов генов.
[0063] Как показано на ФИГ. 1A и 1B, связывание конкретного лиганда с аптамером, блокирует расщепление полиA, приводя к сохранению интактной мРНК и тем самым экспрессии репортерного сигнала (такого как в биосенсорных системах данного описания) или конкретного гена, такого как терапевтический ген (такой как в системах регуляции генов данного описания); лиганд может быть любого типа, включая, например, белок или малую молекулу. В конкретных вариантах реализации изобретения лиганд данной системы для биосенсорных вариантов реализации является эндогенным для конкретной клетки, ткани или организма, тогда как лиганд системы для систем регуляции генов не обязательно является эндогенным лигандом.
[0064] В конкретных вариантах реализации изобретения в конструкции, содержащей сигнал полиA, внутри аптамера содержится сигнал полиA. В некоторых вариантах реализации изобретения аптамер содержит один или более сигналов полиA, например один, два, три или более сигналов полиA (см. ФИГ. 11). Замена AAUAAA на CACACA в значительной степени инактивирует сигнал полиA, таким образом возвращая теоретический верхний предел уровня экспрессии гена при блокировании сигнала полиA. Соотношение уровня экспрессии CACACA по сравнению с AAUAAA используют для оценки теоретического динамического диапазона индукции гена в кратном выражении. В некоторых случаях сигнал полиA имеет такую модификацию, что в последовательности содержится одно, два или более изменений по сравнению со стандартной последовательностью AAUAAA. Например, вместо AAUAAA, он может использовать другую последовательность, включая по меньшей мере AUUAAA, AGUAAA, UAUAAA, CAUAAA, GAUAAA, AAUAUA, AAUACA, AAUAGA, AAAAAG или ACUAAA. В вариантах реализации изобретения, в которых в конструкции используют два или более сигналов полиA, данные сигналы полиA могут быть одинаковыми или разными.
[0065] В конкретных вариантах реализации изобретения для усиления активности расщепления полиA и/или усиления связывания лиганда применяют модуляцию структуры РНК возле сайта расщепления полиA. В конкретных вариантах реализации изобретения оптимизировали размещение сигнала полиA в аптамере или возле него для обеспечения улученного расщепления сайта полиA и/или связывания лиганда с аптамером (см. ФИГ. 12, к примеру). Пространственное размещение сайта полиA в пределах аптамера можно оптимизировать и он может располагаться в пределах аптамера обычно по центру или иным образом. Положение сигнала полиA может быть таким, чтобы фланкирующие стороны аптамера могли смежно располагаться рядом друг с другом для возможности связывания лиганда с аптамером (см. ФИГ. 6, просто в качестве примера).
[0066] В вариантах реализации изобретения в отношении конструкции, содержащей сигнал полиA, можно использовать одну или более последовательностей для усиления связывания лиганда и/или для усиления расщепления в сайте расщепления полиA. В конкретных вариантах реализации изобретения конструкция, содержащая сигнал полиA, также содержит один, два, три или более богатых G участков справа по ходу транскрипции от сигнала полиA. В конкретных вариантах реализации изобретения конструкция, содержащая сигнал полиA, также содержит один, два, три или более богатых U/UG участков справа по ходу транскрипции от сигнала полиA. В некоторых вариантах реализации изобретения один или более богатых G участков расположены справа по ходу транскрипции от по меньшей мере одного богатого U/UG участка. В конкретных вариантах реализации изобретения длина одного или более конкретных стеблей или петель в конкретном аптамере и/или положении сигнала полиA, богатого G участка и/или богатого U/UG участка может влиять на эффективность регуляции (ФИГ. 5, 10, 13-25), а для оптимизации подходящей конфигурации специалист в данной области может применять обычные способы.
[0067] Размещение конструкции, содержащей полиA-переключатель, может происходить любыми приемлемыми способами, но в конкретных вариантах реализации изобретения конструкция присутствует в клетке, ткани или организме и может присутствовать экзогенно на векторе (таком как вирусный вектор (аденовирусный, ретровирусный, аденоассоциированный, лентивирусный и подобный) или плазмиде, или она может присутствовать внутри генома клетки. Полинуклеотиды, содержащие полиA-переключатель и экспрессируемый полинуклеотид, можно вводить в целевую клетку, ткань или организм в виде голой нуклеиновой кислоты, они могут находиться внутри приемлемого носителя (такого как липосома или наночастицы), или они могут находиться на векторе, который сам по себе может содержать приемлемый носитель.
[0068] В конкретных вариантах реализации изобретения аптамер сконструирован таким образом, что он связывает конкретный лиганд, или его можно получать в результате отбора из библиотеки аптамеров, или его можно моделировать, например, из существующего аптамера. Для оптимизации связывания лиганда и/или расщепления в сайте полиA можно модифицировать разные участки аптамера. В конкретных вариантах реализации изобретения положение сигнала полиA связано с эффективностью расщепления полиA (см., например, ФИГ. 8). В одном примере модулируют аптамер A8 (см. ФИГ. 13-15) и учитывают получаемое влияние на эффективность. Еще раз, в качестве примера, в таблице 1 показаны результаты мутационного анализа лиганд-связывающего сайта на основе типичного аптамера A8.
[0069]

[0070] Специалист в данной области осведомлен о стандартных способах для создания аптамеров для нацеливания соединений с высокой аффинностью 7-12.
[0071] В конкретных вариантах реализации изобретения применяют систему, в которой использовано два или более отдельных полиA-переключателей. Одна конструкция полиA может быть чувствительной к первому аптамеру, который связывает первый лиганд, а другая конструкция полиA может быть чувствительной ко второму аптамеру, который связывает второй лиганд. Причем в конкретных вариантах реализации изобретения первый и второй аптамеры не являются идентичными и при этом первый и второй лиганды не являются идентичными. В других случаях конструкция полиA содержит множество аптамеров, которые могут быть чувствительными к одним и тем же лигандам.
[0072] Варианты реализации биосенсорных систем
[0073] В данном описании предложены РНК-переключатели на основе полиA в 5'-НТО, применяемые в данном документе для биосенсорного подхода, которые предоставляют несколько ключевых преимуществ по отношению к существующим технологиям выявления внутриклеточных сигнатур в живых клетках. Во-первых, в данном документе показано, что репортерный сигнал от сенсора проявляет предельно низкую фоновую экспрессию в живых клетках человека и при выявлении конкретного белка-лиганда, данный сигнал эффективно индуцировали со стократным повышением. Соотношение сигнал-шум составляет по меньшей мере величину на порядок выше, чем было достигнуто в живых клетках человека раннее, давая динамический диапазон, который может обеспечивать новые применения в разнообразных экспериментальных условиях. Во-вторых, полиA-сенсор дает возможность неинвазивного выявления/визуализации эндогенных белков в живых клетках без необходимости в метках, маркерах или красителях, обеспечивая класс измерений, которые было раннее трудно выполнить. Наконец, путем эффективного соединения существующей аптамерной технологии с существующими репортерными системами визуализации, данный способ обеспечивает платформу для выявления широкого диапазона белковых лигандов путем молекулярной визуализации. Таким образом полиA-сенсор предлагает уникальную возможность для выявления молекулярных сигнатур in vivo для значительно более широкого применения в молекулярном обнаружении, чем возможно в настоящее время, и может даже быть перенесен в клинические условия, используя соответствующий репортерный зонд для визуализации (т. е. ПЭТ, МРТ, NIRF (флуоресценция в ближнем ИК-спектре)).
[0074] Такой биосенсер обеспечивает получение пространственной, а также относящейся ко времени информации, например, касательно уровней конкретных лигандов при заболевании, а входящую информацию можно использовать для регуляции клеточного поведения для достижения терапевтических целей. Например, примером полиA-биосенсора является тот, который сконструирован для распознавания в качестве своего лиганда глюкозы, и в ответ на нее, регулирует экспрессию сконструированного белка инсулина для модуляции уровней глюкозы у пациентов с диабетом. Сходные биосенсоры также могут функционировать как безопасные переключатели. Например, можно конструировать биосенсор для выявления наличия ракового биомаркера в стволовых клетках. Когда нормальная стволовая клетка ошибочно превращается в раковую клетку, биосенсор может включать суицидальный ген для ее саморазрушения.
[0075] В конкретных вариантах реализации изобретения примеры молекул-лигандов для переключателя на основе полиA, при применении в биосенсорной системе, включают по меньшей мере клеточные метаболиты; нуклеиновые кислоты (включая регуляторные нуклеиновые кислоты, такие как миРНК или интерферирующие молекулы РНК (мшРНК или киРНК)); малые молекулы; клеточные белки (например, связанные с болезненным состоянием белки, включая, например, раковые белки) или вирусные белки, продуцируемые вирусной инфекцией.
[0076] В некоторых вариантах реализации изобретения экспрессируемый полинуклеотид кодирует продукт репортерного гена или продукт терапевтического гена. В некоторых вариантах реализации изобретения продукт репортерного гена может быть гибридизирован с продуктом терапевтического гена в виде гибридного белка. В других вариантах реализации изобретения экспрессия продукта репортерного гена и продукта терапевтического гена транслируется отдельно от единственной мРНК с использованием IRES (участка внутренней посадки рибосомы). Примеры репортерных генов включают ген люциферазы, зеленого флуоресцентного белка, белка красной флуоресценции, β-галактозидазы и т. п. Примеры терапевтических генов включают гены инсулина, гормонов роста, дистрофина, альбумина, фактора IX и т.п. В других случаях в системе используют экспрессируемый полинуклеотид, кодирующий продукт репортерного гена, и отдельный экспрессируемый полинуклеотид, кодирующий продукт терапевтического гена, а их экспрессия может управляться одним и тем же или разными лиганд-связывающими аптамерами. Конструкция(-и) для продукта репортерного гена и продукта терапевтического гена может (могут) располагаться на одном векторе или разных векторах.
[0077] В некоторых вариантах реализации изобретения полинуклеотид содержит 2, 3, 4, 5 или более аптамеров, функционально связанных линейным образом в направлении 5'-3'. Данные аптамеры могут обладать практически одинаковой последовательностью или структурой, или нет, и каждый из них может содержать сигнал полиA или не содержать его. Когда в пределах одного аптамера или в пределах множества аптамеров присутствует множество сигналов полиA, то данные сигналы полиA могут быть идентичными или нет. В конкретных вариантах реализации изобретения единственный полинуклеотид содержит множество аптамеров, но только один богатый G участок и, в некоторых случаях, данный богатый G участок может присутствовать на любом аптамере на полинуклеотиде, но в конкретных случаях он присутствует на втором аптамере в направлении 5'-3' полинуклеотида или находится на наиболее удаленном в 3'-направлении аптамере молекулы. Сворачивание полинуклеотида, содержащего два или более аптамеров, может изменяться в зависимости от различных факторов, включающих длину полинуклеотида и его последовательность, но в конкретных случаях посредством более одной конфигурации сворачивания способен действовать единственный полинуклеотид. В конкретных вариантах реализации изобретения полинуклеотид сконфигурирован таким образом, что аптамеры не содержат какого-либо спаривания оснований между аптамерами (например, левое изображение на ФИГ. 26), хотя в других случаях существует по меньшей мере несколько спариваний оснований между аптамерами (включая расположение вдоль основной части последовательности аптамеров), например, правое изображение на ФИГ. 26. В некоторых конфигурациях сворачивания стеблевая петля одного аптамера имеет конфигурацию, противоположную стеблевой петле другого аптамера в той же молекуле (см. правое изображение на ФИГ. 26).
[0078] В некоторых вариантах реализации изобретения полинуклеотид, содержащий два или более аптамеров разделен линейно в направлении 5'-3' определенной последовательностью. Определенная последовательность может быть случайной или может быть определена как богатый G участок. В конкретных случаях длина между двумя петлями линейно в направлении 5'-3' составляет конкретное значение длины. Например, количество нуклеотидов между двумя петлями может составлять 10-25, 10-24, 10-23, 10-22, 10-21, 10-20, 10-19, 10-18, 10-17, 10-16, 10-15, 10-14, 10-13, 10-12, 10-11, 11-25, 11-24, 11-23, 11-22, 11-21, 11-20, 11-19, 11-18, 11-17, 11-16, 11-15, 11-14, 11-13, 11-12, 12-25, 12-24, 12-23, 12-22, 12-21, 12-20, 12-19, 12-18, 12-17, 12-16, 12-15, 12-14, 12-13, 13-25, 13-24, 13-23, 13-22, 13-21, 13-20, 13-19, 13-18, 13-17, 13-16, 13-15, 13-14, 14-25, 14-24, 14-23, 14-22, 14-21, 14-20, 14-19, 14-18, 14-17, 14-16, 14-15, 15-25, 15-24, 15-23, 15-22, 15-21, 15-20, 15-19, 15-18, 15-17, 15-16, 16-25, 16-24, 16-23, 16-22, 16-21, 16-20, 16-19, 16-18, 16-17, 17-25, 17-24, 17-23, 17-22, 17-21, 17-20, 17-19, 17-18, 18-25, 18-24, 18-23, 18-22, 18-21, 18-20, 18-19, 19-25, 19-24, 19-23, 19-22, 19-21, 19-20, 20-25, 20-24, 20-23, 20-22, 20-21, 21-25, 21-24, 21-23, 21-22, 22-25, 22-24, 22-23, 23-25, 23-24 или 24-25 нуклеотидов. Количество нуклеотидов между двумя петлями в аптамере или в полинуклеотиде может составлять 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25. В конкретных случаях в конфигурации множества аптамеров, в которых существует спаривание оснований между по меньшей мере частями двух разных аптамеров, расстояние между одной и другой петлями, включая последовательность стволовой петли 2 аптамера (см. ФИГ. 26), составляет между 10-25 нуклеотидами, включая, например, 18-20 нуклеотидов.
[0079] Способы применения биосенсорных систем
[0080] В вариантах реализации изобретения существуют способы использования систем данного описания для обнаружения или выявления конкретного требуемого биологического состояния, включая, например, выявление наличия или отсутствия одной или более композиций в конкретном расположении или окружении, включающем определенные клетки, ткани и/или организм. Биосенсорная система может предоставлять информацию зависящим от пространства и/или времени способом о конкретном окружении или расположении. Биосенсорная система может предоставлять информацию о конкретном болезненном состоянии, восприимчивости к нему или риску его возникновения. Биосенсорную систему можно применять в способах определения наличия у индивида конкретного заболевания или существования риска заболеть конкретным заболеванием, или возможности ответа индивида на терапию заболевания. Биосенсорная система может предоставлять информацию о том, будет ли определенная терапия эффективной для индивида.
[0081] В конкретных вариантах реализации изобретения лиганд для биосенсорной системы является эндогенным по отношению к клетке или индивиду. Лиганд может экспрессироваться эндогенно все время и во всех тканях, или не экспрессироваться таким образом. Эндогенный лиганд может экспрессироваться тканеспецифическим или зависящим от времени образом. В конкретных вариантах реализации изобретения вектор, содержащий полинуклеотиды данной системы, нацелен на конкретную ткань или область в организме индивида, причем предполагается, что данная ткань или область содержат или не содержат конкретный эндогенный лиганд.
[0082] В вариантах реализации изобретения, в которых используется данная биосенсорная система, экспрессия экспрессируемого полинуклеотида может регулироваться одним или более элементами, отличающимися от полиA-переключателя. А именно, в некоторых случаях может существовать один или более транскрипционных элементов, которые обеспечивают или ингибируют экспрессию в определенных окружениях или в определенных временными рамками условиях (таких как определенные состояния биологического развития стадий заболеваний). В случаях, в которых требуется чтобы экспрессия системы с полиA-переключателем проходила в определенном окружении, экспрессию полинуклеотида можно регулировать тканеспецифическим промотором. Выбор тканеспецифического промотора может быть продиктован обсуждаемым окружением, а примеры тканеспецифических промоторов известны в данной области и могут быть получены, например, из таких баз данных, как TiProD.
[0083] В случаях, в которых требуется определять является ли один или более видов терапии эффективным для индивида, можно применять по отношению к индивиду систему перед проведением терапии, чтобы таким образом выявлять присутствие или отсутствие конкретного индикаторного соединения, важного для терапии, а затем после проведения терапии, один или более раз можно применять по отношению к индивиду систему для выявления присутствия или отсутствия конкретного индикаторного соединения. В других вариантах реализации изобретения систему не применяют для отслеживания терапии пока по отношению к индивиду ее не проведут один или более раз для того чтобы идентифицировать присутствие или отсутствие конкретного индикаторного соединения, являющегося показателем эффективности терапии.
[0084] В некоторых случаях биосенсорная система используется для получения информации, но в других вариантах реализации изобретения биосенсорную систему можно применять в терапевтических целях (и необязательно она также может предоставлять информацию). Например, система может давать возможность распознавать метаболит в виде лиганда и при связывании лиганда-метаболита с аптамером обеспечивать экспрессию продукта гена, который предоставляет терапию патологическому состоянию, для которого метаболит является маркером (или его присутствие или уровни являются показателем патологического состояния или восприимчивости к нему). В других вариантах реализации изобретения экспрессируемый полинуклеотид не является продуктом терапевтического гена сам по себе, но является продуктом суицидального гена, который экспрессируется, когда существует лиганд, являющимся показателем пагубного патологического состояния, восприимчивости к нему или риска его возникновения. В конкретных вариантах реализации изобретения лиганд начинает образовываться в клетках, которые являются, например, злокачественными или предзлокачественными.
[0085] Примеры лигандов, которые могут управлять аптамерами в биосенсорных системах данного описания, включают по меньшей мере клеточные метаболиты, малые РНК (такие как миРНК), нормальные и аномальные клеточные белки, чужеродные белки, экспрессируемые вирусом и другими патогенами, суицидальные белки, индуцирующие гибель клеток, и т.п.
[0086] Примеры лигандов, которые могут управлять аптамерами в биосенсорных системах данного описания включают по меньшей мере клеточные метаболиты, малые РНК (такие как миРНК), клеточные белки и т.п.
[0087] Варианты реализации систем регуляции генов
[0088] Для определения функции продукта конкретного гена или манипулирования уровнями конкретного белка для достижения терапевтических воздействий всегда требовалась возможность контролировать экспрессию гена. В вариантах реализации данного изобретения, в которых полиA-переключатель помещен в 5'-НТО мРНК, расщепление сигнала полиA ведет к разрушению мРНК и, тем самым, к потере экспрессии трансгена. Молекулы, (например) подобные низкомолекулярным лекарственным средствам, способные ингибировать полиA-переключатель, приводят к сохранению интактной мРНК и тем самым индуцируют экспрессию гена. Как описано в другом месте данного документа, экспрессия трансгена, контролируемая таким переключателем, проявляет предельно низкую фоновую экспрессию в живых клетках человека, а при введении малой молекулы (в данном случае тетрациклина) эффективно индуцировалась экспрессия трансгена с 30-кратным повышением. Оптимизацию и генерализацию данной системы регуляции генов можно выполнять в направлении создания множества строго управляемых систем регуляции генов, каждая из которых контролировалась таким лигандом, например, как одобренное FDA (Управление по контролю за продуктами питания и лекарственными средствами США) низкомолекулярное лекарственное средство. В противоположность существующим системам регуляции генов, основанным на контроле транскрипции с использованием активаторов транскрипции, система на основе полиА, описанная в данном документе, не требует экспрессии каких-либо продуктов белков-активаторов транскрипции (которые могут вызывать тяжелую иммунную реакцию хозяина), не зависит от применения каких-либо специализированных промоторных элементов и поэтому представляет «портативную» систему регуляции, которая может быть «помещена» в любой эндогенный ген или сконструированную векторную единицу транскрипции. По сути, данная система требует наличия только одной транскрипционной единицы (одной экспрессионной конструкции) и содержит настолько универсальный промотор, что он может использоваться для регуляции трансгенов тканеспецифическим (зависящим от пространства) и зависящим от времени способом. Такие системы регуляции генов, комбинирующие безопасные малые молекулы с неиммуногенными полиA-переключателями на основе РНК, будут в значительной степени более безопасными для применения в клинической практике, а также в биологических исследованиях.
[0089] В конкретных вариантах реализации изобретения в системах регуляции генов для переключателей на основе полиA в качестве молекул-лигандов используют низкомолекулярные соединения. Примеры низкомолекулярных соединений включают тетрациклин или аналоги тетрациклина (например, такие как доксициклин, демеклоциклин, миноциклин, хлортетрациклин, санциклин, метациклин или тигециклин) или его функциональные производные; аминогликозиды или их функциональные производные; рапамицин или его функциональные производные (например, эверолимус, темсиролимус, дефоролимус, ридафоролимус) и одобренные FDA лекарственные средства или их функциональные производные. В других вариантах реализации систем регуляции генов применяют отличающийся от малой молекулы лиганд, такой как белок, пептид, нуклеиновая кислота и т.п. И в конкретных вариантах реализации изобретения лиганды данной системы предоставлены в приемлемом носителе.
[0090] В некоторых вариантах реализации изобретения в системе регуляции генов используют два или более аптамеров для независимого ответа на разные лиганды, например, разные аналоги тетрациклина. А именно, если одну систему с полиA-переключателем можно сделать отвечающей на такой лиганд, как тетрациклин, затем можно создать множество строго управляемых систем регуляции генов с помощью мутировавшей последовательности аптамерной РНК для ответа на разные аналоги тетрациклина (доксициклина и т.д.).
[0091] В некоторых вариантах реализации изобретения экспрессируемый полинуклеотид кодирует продукт репортерного гена или продукт терапевтического гена. В некоторых вариантах реализации изобретения продукт репортерного гена может быть гибридизирован с продуктом терапевтического гена в виде гибридного белка. В других вариантах реализации изобретения экспрессия продукта репортерного гена и продукта терапевтического гена транслируется отдельно из единственной мРНК с использованием IRES (участка внутренней посадки рибосомы). Примеры репортерных генов включают ген люциферазы, зеленого флуоресцентного белка, белка красной флуоресценции, β-галактозидазы и т.п. Примеры терапевтических генов включают гены инсулина, гормонов роста, дистрофина, альбумина, фактора IX и т.п. В других случаях в системе используют экспрессируемый полинуклеотид, кодирующий продукт репортерного гена, и отдельный экспрессируемый полинуклеотид, кодирующий продукт терапевтического гена, а их экспрессия может управляться одним и тем же или разными лиганд-связывающими аптамерами. Конструкция(-и) для продукта репортерного гена и продукта терапевтического гена может (могут) располагаться на одном векторе или разных векторах.
[0092] Способы применения систем регуляции генов
[0093] В конкретных вариантах реализации изобретения систему данного описания можно использовать для задач регуляции генов, в которых требуется, чтобы конкретный экспрессируемый полинуклеотид экспрессировался в конкретном расположении и/или при конкретном событии или в конкретный момент времени. В некоторых вариантах реализации изобретения контроль за экспрессией гена позволяет проводить определение функции продукта конкретного гена, тогда как в других вариантах реализации изобретения контроль за экспрессией гена обеспечивает терапевтическую пользу. В некоторых случаях не требуется, чтобы экспрессировался экспрессируемый полинуклеотид трансгена (например, в определенные моменты времени и/или в определенных расположениях), а лиганд либо может не предоставляться вместе с трансгеном, либо его экспрессия может игибироваться, пока не потребуется экспрессия лиганда для обеспечения связывания с аптамером трансгена для экспрессии последнего (например, с определенными регуляторными элементами).
[0094] В некоторых вариантах реализации изобретения лиганд для вариантов реализаций регуляторной системы генов не является эндогенным по отношению к конкретной клетке, ткани или организму. В конкретных вариантах реализации изобретения лиганд в системах для регуляции генов представляет собой лекарственное средство или подобную лекарственному средству молекулу (в по меньшей мере некоторых вариантах реализации изобретения подобную лекарственному средству молекулу определяют как низкомолекулярные соединения обычно с молекулярными массами ниже 500 Дальтон). В конкретных вариантах реализации изобретения лиганд в системах регуляции генов предоставляют индивиду одним из любых приемлемых способов, например пероральным, внутримышечно, путем ингаляции и т.п.
[0095] В некоторых вариантах реализации изобретения системы регуляции генов на основе полиA-переключателей можно конфигурировать в виде систем ауторегуляции с петлями негативной и позитивной обратной связи. Регуляция гена своим собственным продуктом известна как ауторегуляция, которая может создавать уникальные свойства для практического применения. В конкретных вариантах реализации изобретения полиA-переключатель можно конфигурировать для функционирования в качестве системы ауторегуляции. А именно, продукт гена, находящийся под контролем полиA-переключателя, может позитивно или негативно регулировать свою собственную экспрессию. Негативная ауторегуляция происходит, когда продукт гена репрессирует свою собственную экспрессию. Известно, что она увеличивает устойчивость установившейся экспрессии и снижает флуктуации уровней экспрессии генов в клетках. (1) В одном примере негативная ауторегуляция, которую можно конфигурировать с полиA-переключателем, представляет собой биосенсорную систему, предназначенную для выявления глюкозы. В данном случае полиA-переключатель сконструирован с аптамером, распознающим в качестве своего лиганда глюкозу, и в ответ на связывание глюкозы включает экспрессию трансгена, кодирующего белок инсулин. Индуцированная экспрессия инсулина снижает уровни глюкозы, что в свою очередь снижает экспрессию инсулина. В противоположность этому, позитивная ауторегуляция происходит, когда продукт гена стимулирует свою собственную экспрессию. Известно, что это создает состояние с двойной устойчивостью. А именно, сразу после активации трансгена его лигандом, он может блокироваться в состоянии с высокой экспрессией и поддерживать себя во «включенном» состоянии даже после удаления исходного входящего лиганда (2-6). Один пример системы с позитивной обратной связью, которую можно конфигурировать с полиA-переключателем, представляет собой биосенсорную систему, предназначенную для выявления вирусного белка tat. В данном случае полиA-переключатель сконструирован с аптамером, распознающим в качестве своего лиганда tat, и в ответ на связывание tat включает экспрессию трансгена, кодирующего белок tat. Начальное введение белка tat индуцирует экспрессию большего количества белков tat. Это обеспечивает петлю позитивной обратной связи, блокирует систему в состоянии с высокой экспрессией и поддерживает ее во «включенном» состоянии.
[0096] Наборы данного описания
[0097] Любая из композиций, описанных в данном документе, может входить в состав набора. В неограничивающем примере в наборе могут содержаться любые полинуклеотиды, лиганды или векторы, охватываемые данным описанием.
[0098] Набор может содержать подходящим образом разделенную(-ые) на порции композицию(-ии) настоящего изобретения. Компонент(-ы) наборов можно фасовать либо в водной среде, либо в лиофилизированной форме. В отношении наборов контейнер означает, что они обычно будут содержать по меньшей мере один флакон, пробирку для исследования, колбу, бутылку, шприц или другие средства для хранения, в которые может быть помещен компонент и, предпочтительно, подходящим образом разделен на порции. Когда в наборе имеется более одного компонента, набор обычно также будет содержать второй, третий или другой дополнительный контейнер, в который раздельно можно помещать дополнительные компоненты. Однако во флаконе могут содержаться различные комбинации компонентов. Наборы настоящего изобретения, как правило, также будут содержать средства для хранения контейнеров в закрытой оболочке для коммерческой продажи. Такие контейнеры могут включать контейнеры для инъекций или пластмассовые контейнеры, изготовленные выдувным формованием, в которых, например, содержатся требуемые флаконы.
[0099] Когда компоненты набора представлены в одном и/или более жидких растворах, жидкий раствор представляет собой водный раствор, особенно предпочтительный стерильный водный раствор. Композиции также можно готовить в виде композиции в шприце. В таком случае средство для хранения может само представлять собой шприц, пипетку и/или другое подобное устройство, из которого состав можно вводить в инфицированную область тела, инъецировать в организм животного и/или даже применять и/или смешивать с другими компонентами набора. Однако компоненты набора также можно предоставлять в виде высушенного(-ых) порошка (-ов). Когда реактивы и/или компоненты предоставлены в виде сухого порошка, его можно растворять добавлением подходящего растворителя. Предусмотрено, что растворитель также можно предоставлять в другом средстве для хранения.
Примеры
[0100] Следующие примеры включены для демонстрации предпочтительных вариантов реализации данного изобретения. Специалисты в данной области должны понимать, что описанные в примерах методики, которое соответствуют раскрытым автором изобретения представленным методикам, должны надлежащим образом функционировать при практическом осуществлении данного изобретения, и таким образом могут считаться основными предпочтительными способами его практического осуществления. Тем не менее в свете настоящего описания специалисты в данной области должны понимать, что в описанные конкретные варианты реализации изобретения можно внести множество изменений и при этом может быть получен подобный или аналогичный результат без отступления от сущности и объема данного описания.
Пример 1
Новшество данного ИЗОБРЕТЕНИЯ
[0101] Привлечение способности расщепления полиA для создания внутриклеточной сенсорной системы
[0102] Полиаденилирование представляет собой важный механизм процессинга мРНК, который присутствует во всех клетках млекопитающих и является универсальным для них. За исключением генов гистонов, все кодирующие белки мРНК млекопитающих содержат 3′-конец, состоящий из около 200-300 аденозиновых остатков 30, 31. Образование хвоста полиA включает два последовательных этапа: расщепление пре-мРНК и последующее добавление хвоста полиA к вновь расщепленному 3'-концу. Процесс полиаденилирования направляется элементами последовательности, имеющимися на пре-мРНК, под действием комплекса полиаденилирования, состоящего из множества мультимерных белковых факторов. Перед добавлением хвоста полиA необходимо расщепление пре-мРНК. Сайт расщепления находится между высококонсервативным сигналом AAUAAA и правым по ходу транскрипции богатым U или GU мотивом (ФИГ. 2). Предпочтительно расщепление происходит после нуклеотида A 32. Важно отметить, что расщепленные фрагменты 3'-РНК быстро разрушаются из-за отсутствия кэп-структуры.
[0103] В нескольких сообщениях указывали, что вторичные структуры РНК внутри сайта полиA могут сильно влиять на эффективность полиаденилирования в клетках млекопитающих 6, 33, 34. В одном исследовании сообщали, что простым маскированием сайта расщепления внутри стабилизированного стебля РНК в значительной степени снижается степень расщепления/полиаденилирования 6 (ФИГ. 3). Это дает основание предполагать, что расщепление можно эффективно контролировать локальными структурами РНК. В одном варианте реализации изобретения выявление связывания белков и контроль экспрессии репортерного гена основано на этой простой, но при этом эффективной модуляции структуры РНК возле сайта расщепления полиA. Как проиллюстрировано на ФИГ. 1, когда на 5'-НТО искусственно создают новый сайт полиA, который никогда не располагался в нормальных транскрипционных единицах, эффективное расщепление сигнала полиA приводит к разрушению мРНК и потере экспрессии репортерного гена из-за быстрой деградации расщепленного и некэпированного 3'-фрагмента мРНК. В отличие от нормального сигнала полиA, расположенного после гена на 3'-НТО, искусственно вставленный в 5'-НТО сигнал полиA, функционирует в мРНК как «суицидальная единица». Можно спасти мРНК от «суицида» вставкой в данный сайт расщепления аптамера. Связывание конкретного лиганда с аптамером блокирует расщепление, приводя к сохранению интактной мРНК и, тем самым, к экспрессии репортерного сигнала или продукта трансгена. Как описано в другом месте данного документа, такой опосредованный расщеплением полиA сенсор обеспечивает как специфичность посредством динамического обнаружения лиганда посредством связывания аптамера, так и чувствительность благодаря лиганд-зависимой экспоненциальной амплификации репортерного гена или трансгена.
[0104] Преимущество полиA-сенсора в качестве средства визуализации с возможностью генетического кодирования
[0105] Сенсорную последовательность полиA можно кодировать как ДНК в формате плазмидного или вирусного вектора (например) для доставки или генетически встраивать в геном клеток (таких как стволовые клетки или раковые клетки) или в организм мышей для трансплантации и трансгенного применения. Одно ключевое преимущество сенсора с генетическим кодированием заключается в возможности ограничивать его пространственные распределения и контролировать временные интервалы in vivo. В отличие от зондов с химическими красителями, которые непреднамеренно могут окрашивать большую область нецелевых тканей, экспрессию РНК-сенсоров можно эффективно ограничивать целевыми клетками или тканями путем использования тканеспецифических промоторов, включенных в ДНК-векторы. В конкретных вариантах реализации изобретения эти дополнительные признаки безопасности могут в значительной степени минимизировать потенциальные побочные эффекты, возникающие в результате связывания сенсора со своим целевым лигандом, а также снижать фоновый шум in vivo. Кроме того, в отличие от способов выявления, в которых отсутствует амплификация сигнала, единственная кодированная копия ДНК в клетках может генерировать сотни идентичных РНК-сенсоров, которые в свою очередь генерируют в тысячи раз больше репортерных белков, и посредством последующих ферментативных реакций с помощью репортерных белков, это может приводить к экспоненциальной амплификации сигнала.
[0106] ПолиA-сенсор может преодолевать критический барьер для отслеживания молекулярных сигнатур in vivo
[0107] ПолиA-сенсор обеспечивает в высшей степени востребованный механизм для отслеживания нативного белка сочетанием обнаружения, амплификации с выявлением/визуализацией сигнала in vivo. Как упоминается выше, поскольку белки содержат избыточные функциональные группы для химических взаимодействий, способы создания аптамеров для нацеливания на белки с высокой аффинностью хорошо установлены 7-12. Например, аптамеры создавали для специфического связывания с белком tat ВИЧ 35, 36, тромбином 37, фактором роста эндотелия сосудов 38, белка Ku восстановления ДНК39 и опухолевого регулятора остеопонтина40, называя некоторых из них. Специфичность аптамеров позволяет распознавать даже разные изоформы одного и того же белка, например изоформы P50 и P65 транскрипционного фактора NF-κB41, 42 или нефосфорилированную и фосфорилированную форму протеинкиназы ERK243. Данные примеры демонстрируют эффективность аптамеров в распознавании связанных с заболеваниями белков и их метаболических состояний.
[0108] Быстро развивается также сфера репортера для визуализации. Например, GFP произвел революцию во многих областях биологии в качестве репортера экспрессии генов, хотя такие применения для визуализации in vivo были ограничены прозрачными тканями из-за ограничений проницаемости тканей для оптической визуализации. Разработаны сконструированные флуоресцентные в инфракрасном спектре белки (IFP) с длинами волн возбуждения/эмиссии, способными проникать глубоко в ткани и пригодными для визуализации всего тела 44. Более того, продемонстрировано, что тимидинкиназу, связанную с радиоактивным субстратом [18F]FHBG, можно применять для визуализации ПЭТ in vivo в клинических условиях45-47. Относительно недавно для отслеживания экспрессии GFP in vivo разработали новый метод магнито-резонансной томографии (МРТ), предлагающий новое использование GFP для неинвазивной визуализации методом магнитного резонанса48.
[0109] Таким образом обнаружение на основе аптамеров и выявление/визуализация на основе репортеров являются по своим предполагаемым функциям эффективными, но в комбинации они еще более эффективны, например для выявления молекулярных сигнатур in vivo. Недостает только эффективного биологического усилителя, способного соединить эти две функциональные возможности и перевести «обнаружение аптамером» в «экспрессию репортера», которого можно выявлять существующим в настоящее время способом визуализации или другими способами выявления. Обобщенная схема вариантов реализации изобретения, в которых выполняется данное описание, проиллюстрирована на ФИГ. 4. Для отслеживания конкретного белкового лиганда сначала посредством эволюции in vitro создают аптамер с высокой аффинностью. Для образования воспринимающего устройства полиA-переключатель, функционирующий в качестве биологического усилителя, связывается с аптамером для визуализации репортера. Такое воспринимающее устройство может кодироваться в ДНК-векторе и доставляться в целевую клетку/ткань с использованием соответствующих вирусных и невирусных способов переноса. В присутствии клеточного лиганда связывание аптамера генерирует сильный сигнал посредством лиганд-зависимой экспоненциальной амплификации репортерного гена. Важно отметить, что универсальность аптамеров обеспечивает возможность выявления широкого диапазона нативных белков, а универсальность выбора репортеров обеспечивает визуализацию посредством разных способов выявления. Таким образом, предложенный полиA-сенсор дает возможность использовать существующие технологии для получения многоцелевой неинвазивной стратегии молекулярного обнаружения in vivo, которая зависит от наличия и концентрации нативной молекулярной сигнатуры.
[0110] В последнее десятилетие всплеск данных по экспрессии генов, полученных с помощью высокопроизводительного секвенирования и ДНК-микрочипов, предоставил сотни белков и их мутантных изоформ, которые можно применять в будущем в качестве биомаркеров заболеваний. Зондирование молекулярных сигнатур in vivo не только помогает в диагностике, но также обеспечивает измерения клеточной пролиферации, изменений в метаболизме и терапевтического ответа, (например) на основе увеличения уровней конкретных белковых биомаркеров. В конкретных вариантах реализации изобретения полиA-сенсор обеспечивает данный класс измерений, которые раннее было трудно проводить, и предоставляет значительно более широкое применение при молекулярном обнаружении, чем возможно в настоящее время.
[0111] В конкретных вариантах реализации способы и композиции настоящего изобретения обеспечивают возможность для точного контроля экспрессии трансгена, избегая применения используемых в настоящее время реактивных элементов экспрессии генов, таких как гибридные транскрипционные трансактиваторы и специализированные промоторы. В конкретных вариантах реализации изобретения в настоящих способах обходится необходимость применения более одной экспрессионной конструкции, обходятся потенциальные токсические эффекты благодаря экспрессии гибридного активатора транскрипции (включая индукцию иммунных реакций хозяина, направленных против активаторов транскрипции, представляющих собой чужеродные белки), обходятся сложности, связанные с требованиями наличия специализированного промотора и обходится необходимость использования ограниченного количества малых индукторных молекул, доступных для экспериментального и терапевтического применения (из-за ограниченного количества доступных систем).
[0112] Вместо этого, настоящая система на основе полиA данного описания не требует экспрессии каких-либо белковых продуктов-активаторов транскрипции и не зависит от использования каких-либо специализированных промоторных элементов. Таким образом, предложена «портативная» система регуляции, которая может быть «помещена» в любой эндогенный ген или сконструированную векторную единицу транскрипции. По сути, данная система требует наличия только одной транскрипционной единицы (одной экспрессионной конструкции) и ее промотор настолько универсален, что он может использоваться для регуляции трансгенов тканеспецифическим (зависящим от пространства) и зависящим от времени способом. Универсальная природа существующего во всех клетках млекопитающих полиаденилирования обеспечивает широкое применение данной системы.
Пример 2
Типичные начальные исследования
[0113] Начальные исследования, описанные в данном примере, демонстрируют развитие систем с полиA-сенсорами. Конкретно, посредством анализа ряда сигналов полиA разработан сенсор, который опосредуется расщеплением полиA, эффективно расщепляемый в клетках человека при размещении в 5'-НТО стандартного экспрессионного вектора. Важно отметить, что описанная в данном документе аптамерная последовательность предназначена для связывания конкретного лиганда, как части сенсора, без нарушения эффективности расщепления полиA. Наконец, путем совместной экспрессии белка-лиганда в живых клетках человека сенсор легко выявляет свой лиганд, приводя к более чем стократному увеличению репортерного сигнала. Данные результаты демонстрируют, что в живых клетках человека легко достигается выявление конкретных немеченых внутриклеточных белков, а степень индукции, измеряемая соотношением сигнал-шум, представлена диапазоном, который может находить использование во множестве применений.
[0114] Идентификация мотивов полиA, эффективно функционирующих в 5'-НТО в клетках человека
[0115] Общая стратегия выявления и визуализации белков посредством модуляции расщепления полиA в критической степени зависима от высокой эффективности расщепления сигнала полиA в 5'-НТО (ФИГ. 1). Первым этапом было исследование потенциальных сигналов полиA и связанных с ними мотивов слева и справа по ходу транскрипции, которые обеспечивают эффективное расщепление полиA в клетках. Для исследования эффективности расщепления полиA использовали анализ транзиентной трансфекции, с вовлечением экспрессионного вектора49 млекопитающих с GFP или люциферазным репортером (просто в качестве примеров). Потенциальные последовательности сигнала полиA клонировали в 5'-НТО репортерного гена или трансгена внутри экспрессионного вектора. Для обеспечения верхнего предела уровней экспрессии также получали контрольные векторы с неактивным сигналом полиA (мутации AAUAAA в CACACA)50, с которым после трансфекции клеток HEK293T человека сравнивали эффективность расщепления активными сигналами полиA.
[0116] Для начального анализа использовали ряд разных сигналов полиA и связанных с ними слева и справа по ходу транскрипции мотивов. Поскольку оказалось, что большинство из этих исследованных сигналов полиA в некоторой степени функционируют в клетках человека, как отражено их способностью в различной степени подавлять экспрессию репортерных генов в 5'-НТО, в условиях исследования в конкретных вариантах реализации изобретения особенно подходящей оказалась одна синтетическая последовательность полиA, опубликованная Proudfoot lab 51. Данную полиA использовали для дальнейших целей исследования, основываясь на ее очевидно более высоком уровне активности расщепления. Для улучшения эффективности активности расщепления выполняли и оценивали ряд модификаций последовательности полиA. Прежде всего положение богатой G последовательности 52 справа по ходу транскрипции сайта полиA значительно усиливает расщепление, вероятно посредством транзиентной приостановки транскрипции, вызванной богатой G последовательностью 53, 54. Увеличение количества копий богатого G участка от одной до двух дополнительно усиливало расщепление. Размещение двух копий AAUAAA в тандеме также заметно увеличивало расщепление (см. ФИГ. 5). В качестве общего шаблона в последующих исследованиях использовали результирующую конфигурацию полиA, содержащую комбинацию из 2 AAUAAA+спейсер+богатый G/U участок+2 богатых G участка.
[0117] В конкретных вариантах реализации изобретения изменение длины спейсера между AAUAAA и богатым G/U участком от 14 до 25 оснований или замена спейсера аптамером оказывали слабый эффект на эффективность расщепления полиA, указывая на то, что длина спейсера универсальна в пределах исследованного диапазона и может вмещать аптамерную последовательность разной длины. Таким образом, сигнал полиA может эффективно расщепляться в клетках человека в 5'-НТО и легко воспринимаются некоторые изменения последовательности в пределах спейсера.
[0118] Разработки по конструированию аптамера в сенсор на основе полиA для выявления внутриклеточного лиганда в клетках человека
[0119] Продемонстрировав способность сигнала полиA расщепляться в 5'-НТО в клетках, в конкретных вариантах реализации изобретения конструировали лиганд-связывающий аптамер как часть полиA-сенсора. Это дает сенсору возможность связывать конкретный белковый лиганд, приводя к ингибированию расщепления и, тем самым, к индуцированной экспрессии репортера. В качестве конкретного варианта реализации изобретения использовали аптамер, связывающий белок tat ВИЧ. Данное связывание представляет собой простое взаимодействие один-к-одному, в которое не вовлечен никакой другой кофактор, а константа диссоциации Kd составляет 0,1 нM35, 36, что сравнимо с достигаемой антителами аффинностью. Для создания эффективного сенсора использовали подход «рациональной структуры», используя принцип конструирования, называемый «фиксацией» (ФИГ. 6). В данном документе аптамер рационально располагается сбоку от сигнала AAUAAA и заменяет большую часть спейсерного участка. Результирующую РНК конструировали для обеспечения переключения между двумя первичными конформациями: в одной лиганд связывает сенсор и, тем самым, «фиксирует» участок спейсера; в другой сенсор не связан, а спейсер, главным образом, существует в виде одной цепи. В первой конформации связывание лиганда эффективно замыкает спейсер в стабильной двухцепочечной стеблевой структуре, которая в значительной степени снижает опосредованное полиA расщепление6, 33, 34. Кроме того, лиганд физически присоединяется к сайту расщепления, по существу блокируя доступ для связанных с полиA клеточных белков, требующихся для инициирования расщепления.
[0120] На основании данного принципа «фиксации» разработали более 40 разных конструкций, в которых стабильность и длина спейсера, а также положение аптамера в пределах спейсера, подбирали для получения максимального соотношения активности репортера между «связанным» (в присутствии лиганда, без расщепления) и «несвязанным» (в отсутствие лиганда, расщепление) состояниями. Исследование данной серии структуры в клетках человека выявило, что данная стратегия «фиксации» привела к получению высокочувствительных сенсоров с превосходным поведением при переключении. Одна структура в клетках показала очень низкую фоновую экспрессию в несвязанном состоянии и сильную индукцию репортерного сигнала в присутствии лиганда (см. ниже). В сконструированных выше вариантах реализации изобретения создана эффективная сенсорная система на основе полиA, как установлено примером, в котором сконструированные РНК можно эффективно приспособить для построения функционального сенсора/усилителя с целью выявления внутриклеточного лиганда в живых клетках.
[0121] Демонстрация функционирования полиA-сенсора в клетках в ответ на клеточный лиганд
[0122] В отношении сконструированного полиA-сенсора показано, что трансфекция экспрессирующего лиганд вектора вместе с вектором сенсора в клетках 293T приводит к резкому увеличению люциферазного репортерного сигнала (ФИГ. 7). В отсутствие лиганда, репортерный сигнал из такого сенсора проявлял очень низкую фоновую экспрессию (ФИГ. 7A, ниже 1% от уровня экспрессии неактивного контроля полиA). Данная фоновая экспрессия обеспечивает основной уровень для расчета индукции в «кратном» выражении в виде соотношения репортерного сигнала в присутствии и отсутствие лиганда. Важно отметить, что отклик сенсора был «настраиваемым» и отображал количество лиганда в клетках дозозависимым образом. Кроме того, индукция достигала вплоть до 120-кратных увеличений (ФИГ. 7B) или приблизительно 76% от теоретического индуцируемого диапазона, используя в качестве эталона уровень экспрессии неактивного полиA-сенсора (ФИГ. 7A). Данная степень индукции оказывается по величине на около два порядка выше, чем было достигнуто для живых клеток человека раннее, и является достаточной для формирования основы для сенсорной платформы. В противоположность этому не наблюдали значительной индукции в клетках, трансфицированных неактивным полиA или исходным вектором без помещенного в него полиA-сенсора (ФИГ. 7B), что указывает на то, что данный отвечающий за индукцию механизм действительно опосредован сигналом полиA.
[0123] Для определения того, что при некотором уровне мРНК отклик также согласуется с предложенным механизмом, выполняли нозерн-анализ мРНК люциферазы в ответ на связывание лиганда. Как показано на ФИГ. 7C, в отсутствие лиганда (дорожка 4) не наблюдали мРНК люциферазы, что дает основание предполагать быстрое разрушение расщепленных мРНК. В противоположность этому, наличие лиганда значительно повышало количество мРНК люциферазы (дорожка 5). Кроме того, полосу, которая находилась слегка выше чем для эталонного расщепленного 5'-фрагмента, наблюдали в клетках, несущих активный сенсор (дорожки 4, 5 по сравнению с дорожкой 1), что дает основание предполагать, что расщепленный 5'-фрагмент был полиаденилирован. Для подтверждения того, что в предполагаемый сайт расщепления действительно добавлен хвост полиA в конце расщепленного 5'-фрагмента, применяли ОТ-ПЦР (полимеразная цепная реакция с обратной транскрипцией), а затем секвенирование кДНК. Вместе данные результаты близко совпадают с механизмом, в котором связывание лиганда блокирует расщепление полиA и сохраняет интактную мРНК для индукции.
[0124] Предварительная демонстрация применимости сенсора на основе РНК in vivo
[0125] Выполняли исследования in vivo другого РНК-сенсора на основе саморасщепления рибозима типа «головка молотка», который эффективно функционировал in vivo4, 55. Данный рибозим использовали для конструирования РНК-сенсора, в котором спонтанное саморасщепление рибозима, помещенного в 5'-НТО, приводит к разрушению мРНК и, тем самым, потере экспрессии репортерного гена. Присутствие тойокамицина, низкомолекулярного ингибитора рибозима, блокирует саморасщепление рибозима и приводит к сохранению интактной мРНК и, тем самым, индуцирует экспрессию репортера. Для демонстрации функционирования сенсора на основе рибозима in vivo создавали рекомбинантный аденоассоциированный вирус (AAV), кодирующий РНК-сенсор на основе рибозима, и доставляли его в ткань хозяина в виде глаз бестимусных мышей. В качестве контроля доставляли вирус AAV, кодирующий неактивный рибозим, для инфицирования задней мышцы бедра. Затем вводили молекулу тойокамицина посредством гранулы лекарственного средства, помещаемой мышам под кожу дорсальной области. Проводили визуализацию для мышей с использованием прибора для визуализации IVIS200 для обеспечения количественного измерения экспрессии люциферазы на основе детекции фотонов 56, 57. Репрезентативные изображения мышей, полученные перед и после введения тойокамицина, показали, что сенсор способен выявлять присутствие тойокамицина в сетчатке и такое введение приводило к индуцированным экспрессиям люциферазного репортера вплоть до их 180-кратного увеличения4. В противоположность этому, в несущей неактивный рибозим мышце не наблюдали увеличения экспрессии люциферазы. Данные результаты продемонстрировали, что РНК-сенсоры, кодируемые в вирусных векторах и доставляемые в ткани-мишени, были способные эффективно функционировать in vivo, а интенсивность индуцированного репортерного сигнала легко выявлялась путем визуализации всего тела. Поскольку в данных исследованиях в качестве механизма не использовалось расщепление аптамера или полиA, общую экспериментальную настройку для визуализации in vivo можно модифицировать для демонстрации применимости полиA-сенсора у живых мышей.
[0126] Оптимизированный сенсор, разработанный для tat ВИЧ, представляет собой пример, который можно было использовать для выявления и визуализации инфицированных ВИЧ клеток крови или тканей in vivo. Альтернативно, данный или другой сенсор можно встраивать в линию клеток для создания стабильной «сенсорной клетки» для титрования количества инфекционных частиц ВИЧ в крови пациента. Недавний всплеск исследования биомаркеров предоставил сотни дополнительных белков, которых можно использовать аналогичным образом в качестве молекулярных сигнатур заболеваний. Зондирование данных молекулярных сигнатур in vivo помогает не только при диагностике, но также отслеживает терапевтический ответ, основанный на увеличении уровней конкретных белков. Данная технология пригодна для выявления/визуализации этих или других конкретных клеточных белков и обеспечивает класс измерений, проведение которых раннее было трудно достижимо. Спектр молекулярных сигнатур, которые можно выявлять полиA-сенсорами, может включать не только белки, но и другие биомолекулы, а также одобренные FDA малые молекулы. Вместе с будущими поколениями применимых в клинике репортеров, которые можно визуализировать в инфракрасной области спектра44, ПЭТ45-47 или МРТ48, полиA-сенсор имеет широкое применение в молекулярной визуализации.
[0127] Демонстрация применимости полиA-переключателя для регуляции генов с использованием одобренного FDA лекарственного средства
[0128] Как показано на ФИГ. 19, сигнал полиA можно эффективно встраивать внутрь аптамера, который распознает в качестве своего лиганда тетрациклин. Тетрациклин представляет собой одобренное FDA лекарственное средство с длительной историей клинического применения и данными по безопасности. В отсутствие тетрациклина трансген или репортерный ген проявлял очень низкую фоновую экспрессию (ФИГ. 20), ниже 0,2% от уровня экспрессии неактивного контроля полиA. При введении тетрациклина включалась экспрессия трансгена или репортерного гена и возрастала до ~30 раз. Индукция тетрациклином является дозозависимой (ФИГ. 17), позволяя точно контролировать экспрессию трансгена с помощью одобренного FDA лекарственного средства.
Пример 3
Аптамер полиA, содержащий множество аптамеров
[0129] В некоторых вариантах реализации данного описания аптамерная молекула полиA содержит 2, 3, 4, 5 или более аптамеров, а в конкретных случаях аптамеры соединены линейно вдоль направления 5'-3' единственной молекулы. Между разными аптамерами могут находится неаптамерные последовательности или нет. В конкретных вариантах реализации изобретения, когда имеется множество аптамеров, один аптамер, связанный с другим аптамером, может иметь противоположную ориентацию аптамера (а именно, направление 5'-3' одного аптамера обращено в направление 3'-5' для другого аптамера) (ФИГ. 18A (среднее изображение) и 25), хотя в альтернативных случаях второй аптамер находится в той же ориентации (ФИГ. 18A (крайнее правое изображение). В конкретных случаях конструкции, содержащей один или более аптамеров, богатый G участок находится на наиболее удаленном в 3'-направлении конце молекулы или возле него, а примером богатого G участка является участок MAZ (ФИГ. 25). Таким образом на ФИГ. 25 проиллюстрирована новая конструкция GP2, в которой сигнал полиA фланкирован 2 аптамерами в разной ориентации. Данная структура основана на одной из конфигураций, описанных на Фиг. 18. При обработке 5 мкг/мл Tc GP2 показала улучшенную индукцию с 12,6-кратным увеличением по сравнению с 10-кратным увеличением индукции 103GP2, описанной на Фиг. 21. Показана также вариация в модификации участка P2b из GP2 и соответствующее воздействие ее на индукцию экспрессии гена.
[0130] В некоторых вариантах реализации изобретения молекула аптамера полиA содержит два аптамера, соединенные линейно, хотя данная молекула может иметь трехмерную форму с разнообразными формами. На ФИГ. 26 иллюстрируется новая конструкция GP2SLGP2, в которой два аптамера GP2, соединены вплотную друг с другом. Аптамер 5'-GP2 называют «хелперным», аптамер 3'-GP2 называют «центральным». Оба содержат активный сигнал полиA, но только центральный аптамер содержит богатый G участок. Данная конфигурации предназначена для обеспечения альтернативного сворачивания между двумя разными структурами: структурой по умолчанию и сверхструктурой C. При обработке 5 мкг/мл Tc GP2SLGP2 показала кардинально улучшенную индукцию с 70-кратным увеличением по сравнению с 12,6-кратным увеличением индукции GP2. В случаях, в которых имеется множество аптамеров, образующих трехмерную структуру, могут выравниваться последовательности двух петель (см. правое изображение ФИГ. 26). В случаях, в которых на молекуле имеется множество аптамеров, на данной молекуле присутствует только один богатый G участок. В конкретных вариантах реализации изобретения богатый G участок может присутствовать на втором аптамере (в направлении 5'-3') или, в определенных случаях, присутствовать на наиболее удаленном в 3'-направлении аптамере молекулы.
[0131] На ФИГ. 27 показана конкретная молекула в виде сверхструктуры, такая же как на правом изображении ФИГ. 26 и содержащая последовательность GP2 петли P2b, указанной на ФИГ. 25. Показаны вариация по длине и последовательности участка стебля/петли-II (участок в рамке) хелперного GP2 и центрального GP2 и соответствующее влияние на индукцию экспрессии гена. Красным цветом показана вариация последовательностей между разными конструкциями. При сворачивании как структура сверхструктуры C центральный стебель GP2SLGP2 имеет эквивалентную длину из 18 п. н., тогда как C12 имеет эквивалентную длину из 20 п. н. Конструкция C12 при 5 мкг/мл Tc достигала индукции с 94-кратным повышением, тогда как конструкция D11A при 5 мкг/мл Tc достигала индукции с 84-кратным повышением. Обе значительно превышали 70-кратную индукцию GP2SLGP2 при 5 мкг/мл Tc. В конкретных вариантах реализации изобретения последовательность петли 2 (участок в рамке на ФИГ. 27) вызывает переход участка в двухцепочечный участок вместо петли. В молекуле со сверхструктурой, показанной на ФИГ. 26 (правое изображение) и ФИГ. 27, длина участка между «гантелями» полиA (pA), включая участок L2 в рамке, составляет между 10-25 нуклеотидами, например, 18-20 нуклеотидами.
[0132] Просто в качестве примера на ФИГ. 28 показана полная последовательность полиA-сенсорной конструкции GP2SLGP2, содержащей последовательность стеблевой петли GP2 (см. ФИГ. 25), последовательности стволовой петли C12 (см. ФИГ. 25) и последовательности стволовой петли D11A (см. ФИГ. 25). Для определения их способности индуцировать экспрессию исследовали три молекулы при различных концентрациях тетрациклина (Tc, в качестве примера лиганда для кармана связывания). На ФИГ. 29 продемонстрирована дозозависимая регуляция GP2SLGP2. Каждая точка данных соответствует относительной кратности индукции, определенной как соотношение люциферазной активности с tc или без него. Показана средняя кратность индукции со стандартным отклонением. Индукция достигала 104-кратного увеличения при 15 мкг/мл Tc. На ФИГ. 30 продемонстрирована дозозависимая регуляция конструкции, содержащей последовательность C12. Каждая точка данных соответствует относительной кратности индукции, определенной как соотношение люциферазной активности с tc или без него. Показана средняя кратность индукции со стандартным отклонением. Индукция достигала 151-кратного увеличения при 15 мкг/мл Tc. На ФИГ. 31 продемонстрирована дозозависимая регуляция конструкции, содержащей последовательность D11A. Каждая точка данных соответствует относительной кратности индукции, определенной как соотношение люциферазной активности с tc или без него. Показана средняя кратность индукции со стандартным отклонением. Индукция достигала 127-кратного увеличения при 15 мкг/мл Tc.
[0133] Не предполагается ограничения объема настоящей заявки конкретными вариантами реализации процесса, механизма, способа изготовления, композиции объекта изобретения, средств, способов и этапов, описанных в данном описании.
ССЫЛКИ НА ЛИТЕРАТУРУ
[0134] Все упомянутые в данном описании публикации являются показателем уровня специалистов в данной области, к которой принадлежит данное изобретение. Все публикации в данном документе включены в него путем ссылки в той же степени, как если бы каждая отдельная публикация была конкретно и отдельно указана, как полностью включенная путем ссылки.
[0135] 1. Gossen, M. & Bujard, H. Tight control of gene expression in mammalian cells by tetracycline-responsive promoters. Proc Natl Acad Sci U S A 89, 5547-5551 (1992).
[0136] 2. Rivera, V.M. et al. A humanized system for pharmacologic control of gene expression. Nat Med 2, 1028-1032 (1996).
[0137] 3. Suhr, S.T., Gil, E.B., Senut, M.C. & Gage, F.H. High level transactivation by a modified Bombyx ecdysone receptor in mammalian cells without exogenous retinoid X receptor. Proc Natl Acad Sci U S A 95, 7999-8004 (1998).
[0138] 4. Yen, L. et al. Exogenous control of mammalian gene expression through modulation of RNA self-cleavage. Nature 431, 471-476 (2004).
[0139] 5. Warnock, D.E., Fahy, E. & Taylor, S.W. Identification of protein associations in organelles, using mass spectrometry-based proteomics. Mass Spectrom Rev 23, 259-280 (2004).
[0140] 6. Gimmi, E.R., Reff, M.E. & Deckman, I.C. Alterations in the pre-mRNA topology of the bovine growth hormone polyadenylation region decrease poly(A) site efficiency. Nucleic acids research 17, 6983-6998 (1989).
[0141] 7. Ciesiolka, J. et al. Affinity selection-amplification from randomized ribooligonucleotide pools. Methods Enzymol 267, 315-335 (1996).
[0142] 8. Ellington, A.D. & Szostak, J.W. In vitro selection of RNA molecules that bind specific ligands. Nature 346, 818-822 (1990).
[0143] 9. Fitzwater, T. & Polisky, B. A SELEX primer. Methods Enzymol 267, 275-301 (1996).
[0144] 10. Lee, J.F., Hesselberth, J.R., Meyers, L.A. & Ellington, A.D. Aptamer database. Nucleic acids research 32, D95-100 (2004).
[0145] 11. Nimjee, S.M., Rusconi, C.P. & Sullenger, B.A. Aptamers: an emerging class of therapeutics. Annu Rev Med 56, 555-583 (2005).
[0146] 12. Tuerk, C. & Gold, L. Systematic evolution of ligands by exponential enrichment: RNA ligands to bacteriophage T4 DNA polymerase. Science 249, 505-510 (1990).
[0147] 13. Becskei, A. & Serrano, L. Engineering stability in gene networks by autoregulation. Nature 405, 590-593 (2000).
[0148] 14. Becskei, A., Seraphin, B. & Serrano, L. Positive feedback in eukaryotic gene networks: cell differentiation by graded to binary response conversion. The EMBO journal 20, 2528-2535 (2001).
[0149] 15. Carrier, T.A. & Keasling, J.D. Investigating autocatalytic gene expression systems through mechanistic modeling. Journal of theoretical biology 201, 25-36 (1999).
[0150] 16. Demongeot, J., Kaufman, M. & Thomas, R. Positive feedback circuits and memory. Comptes rendus de l'Academie des sciences. Serie III, Sciences de la vie 323, 69-79 (2000).
[0151] 17. Ferrell, J.E., Jr. Self-perpetuating states in signal transduction: positive feedback, double-negative feedback and bistability. Current opinion in cell biology 14, 140-148 (2002).
[0152] 18. Isaacs, F.J., Hasty, J., Cantor, C.R. & Collins, J.J. Prediction and measurement of an autoregulatory genetic module. Proc Natl Acad Sci U S A 100, 7714-7719 (2003).
[0153] 19. Famulok, M., Hartig, J.S. & Mayer, G. Functional aptamers and aptazymes in biotechnology, diagnostics, and therapy. Chem Rev 107, 3715-3743 (2007).
[0154] 20. Jenison, R.D., Gill, S.C., Pardi, A. & Polisky, B. High-resolution molecular discrimination by RNA. Science 263, 1425-1429 (1994).
[0155] 21. Navani, N.K. & Li, Y. Nucleic acid aptamers and enzymes as sensors. Curr Opin Chem Biol 10, 272-281 (2006).
[0156] 22. Hesselberth, J.R., Robertson, M.P., Knudsen, S.M. & Ellington, A.D. Simultaneous detection of diverse analytes with an aptazyme ligase array. Anal Biochem 312, 106-112 (2003).
[0157] 23. Breaker, R.R. Engineered allosteric ribozymes as biosensor components. Current opinion in biotechnology 13, 31-39 (2002).
[0158] 24. Tang, J. & Breaker, R.R. Rational design of allosteric ribozymes. Chem Biol 4, 453-459 (1997).
[0159] 25. Mandal, M. & Breaker, R.R. Gene regulation by riboswitches. Nature reviews 5, 451-463 (2004).
[0160] 26. Winkler, W.C., Nahvi, A., Roth, A., Collins, J.A. & Breaker, R.R. Control of gene expression by a natural metabolite-responsive ribozyme. Nature 428, 281-286 (2004).
[0161] 27. Winkler, W.C. & Breaker, R.R. Regulation of bacterial gene expression by riboswitches. Annual review of microbiology 59, 487-517 (2005).
[0162] 28. Win, M.N. & Smolke, C.D. A modular and extensible RNA-based gene-regulatory platform for engineering cellular function. Proc Natl Acad Sci U S A 104, 14283-14288 (2007).
[0163] 29. Win, M.N. & Smolke, C.D. Higher-order cellular information processing with synthetic RNA devices. Science 322, 456-460 (2008).
[0164] 30. Proudfoot, N. New perspectives on connecting messenger RNA 3' end formation to transcription. Current opinion in cell biology 16, 272-278 (2004).
[0165] 31. Proudfoot, N.J., Furger, A. & Dye, M.J. Integrating mRNA processing with transcription. Cell 108, 501-512 (2002).
[0166] 32. Chen, F., MacDonald, C.C. & Wilusz, J. Cleavage site determinants in the mammalian polyadenylation signal. Nucleic acids research 23, 2614-2620 (1995).
[0167] 33. Klasens, B.I., Thiesen, M., Virtanen, A. & Berkhout, B. The ability of the HIV-1 AAUAAA signal to bind polyadenylation factors is controlled by local RNA structure. Nucleic acids research 27, 446-454 (1999).
[0168] 34. Zarudnaya, M.I., Kolomiets, I.M., Potyahaylo, A.L. & Hovorun, D.M. Downstream elements of mammalian pre-mRNA polyadenylation signals: primary, secondary and higher-order structures. Nucleic acids research 31, 1375-1386 (2003).
[0169] 35. Yamamoto, R., Baba, T. & Kumar, P.K. Molecular beacon aptamer fluoresces in the presence of Tat protein of HIV-1. Genes Cells 5, 389-396 (2000).
[0170] 36. Yamamoto, R. et al. A novel RNA motif that binds efficiently and specifically to the Ttat protein of HIV and inhibits the trans-activation by Tat of transcription in vitro and in vivo. Genes Cells 5, 371-388 (2000).
[0171] 37. Long, S.B., Long, M.B., White, R.R. & Sullenger, B.A. Crystal structure of an RNA aptamer bound to thrombin. RNA (New York, N.Y 14, 2504-2512 (2008).
[0172] 38. Ng, E.W. et al. Pegaptanib, a targeted anti-VEGF aptamer for ocular vascular disease. Nat Rev Drug Discov 5, 123-132 (2006).
[0173] 39. Zhang, L., Yoo, S., Dritschilo, A., Belyaev, I. & Soldatenkov, V. Targeting Ku protein for sensitizing of breast cancer cells to DNA-damage. Int J Mol Med 14, 153-159 (2004).
[0174] 40. Mi, Z. et al. RNA aptamer blockade of osteopontin inhibits growth and metastasis of MDA-MB231 breast cancer cells. Mol Ther 17, 153-161 (2009).
[0175] 41. Cassiday, L.A. & Maher, L.J., 3rd Yeast genetic selections to optimize RNA decoys for transcription factor NF-kappa B. Proc Natl Acad Sci U S A 100, 3930-3935 (2003).
[0176] 42. Reiter, N.J., Maher, L.J., 3rd & Butcher, S.E. DNA mimicry by a high-affinity anti-NF-kappaB RNA aptamer. Nucleic acids research 36, 1227-1236 (2008).
[0177] 43. Vaish, N.K. et al. Monitoring post-translational modification of proteins with allosteric ribozymes. Nature biotechnology 20, 810-815 (2002).
[0178] 44. Shu, X. et al. Mammalian expression of infrared fluorescent proteins engineered from a bacterial phytochrome. Science 324, 804-807 (2009).
[0179] 45. Hung, S.C. et al. Mesenchymal stem cell targeting of microscopic tumors and tumor stroma development monitored by noninvasive in vivo positron emission tomography imaging. Clin Cancer Res 11, 7749-7756 (2005).
[0180] 46. Yaghoubi, S.S. & Gambhir, S.S. PET imaging of herpes simplex virus type 1 thymidine kinase (HSV1-tk) or mutant HSV1-sr39tk reporter gene expression in mice and humans using [18F]FHBG. Nat Protoc 1, 3069-3075 (2006).
[0181] 47. Yaghoubi, S.S. et al. Noninvasive detection of therapeutic cytolytic T cells with 18F-FHBG PET in a patient with glioma. Nat Clin Pract Oncol 6, 53-58 (2009).
[0182] 48. Perez-Torres, C.J., Massaad, C.A., Hilsenbeck, S.G., Serrano, F. & Pautler, R.G. In vitro and in vivo magnetic resonance imaging (MRI) detection of GFP through magnetization transfer contrast (MTC). Neuroimage 50, 375-382 (2010).
[0183] 49. Ory, D.S., Neugeboren, B.A. & Mulligan, R.C. A stable human-derived packaging cell line for production of high titer retrovirus/vesicular stomatitis virus G pseudotypes. Proc Natl Acad Sci U S A 93, 11400-11406 (1996).
[0184] 50. Sheets, M.D., Ogg, S.C. & Wickens, M.P. Point mutations in AAUAAA and the poly (A) addition site: effects on the accuracy and efficiency of cleavage and polyadenylation in vitro. Nucleic acids research 18, 5799-5805 (1990).
[0185] 51. Levitt, N., Briggs, D., Gil, A. & Proudfoot, N.J. Definition of an efficient synthetic poly(A) site. Genes & development 3, 1019-1025 (1989).
[0186] 52. Ashfield, R. et al. MAZ-dependent termination between closely spaced human complement genes. The EMBO journal 13, 5656-5667 (1994).
[0187] 53. Gromak, N., West, S. & Proudfoot, N.J. Pause sites promote transcriptional termination of mammalian RNA polymerase II. Molecular and cellular biology 26, 3986-3996 (2006).
[0188] 54. Yonaha, M. & Proudfoot, N.J. Specific transcriptional pausing activates polyadenylation in a coupled in vitro system. Molecular cell 3, 593-600 (1999).
[0189] 55. Yen, L., Magnier, M., Weissleder, R., Stockwell, B.R. & Mulligan, R.C. Identification of inhibitors of ribozyme self-cleavage in mammalian cells via high-throughput screening of chemical libraries. RNA (New York, N.Y 12, 797-806 (2006).
[0190] 56. Contag, C.H. et al. Visualizing gene expression in living mammals using a bioluminescent reporter. Photochem Photobiol 66, 523-531 (1997).
[0191] 57. Contag, P.R., Olomu, I.N., Stevenson, D.K. & Contag, C.H. Bioluminescent indicators in living mammals. Nat Med 4, 245-247 (1998).
[0192] Хотя настоящее изобретение и его преимущества подробно описаны, следует понимать, что для данного изобретения можно выполнять различные изменения, замены и модификации без отклонения от сущности и объема данного описания, определенных прилагаемой формулой изобретения. Более того, не предполагается ограничения объема настоящей заявки конкретными вариантами реализации процесса, механизма, способа изготовления, композиции объекта изобретения, средств, способов и этапов, описанных в данном описании. Специалисту в данной области будет понятно, что по описанию настоящего изобретения можно разработать процессы, механизмы, способ изготовления, композиции объекта изобретения, средства, способы или этапы, существующие в настоящее время или доступные позже, которые выполняют практически такую же функцию или достигают практически такого же результата, для чего в соответствии с настоящим описанием можно использовать соответствующие варианты реализации, описанные в данном документе. Соответственно подразумевается, что прилагаемая формула изобретения включает в пределы своего объема такие процессы, механизмы, способ изготовления, композиции объекта изобретения, средства, способы или этапы.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Yen, Laising
Luo, Liming
Chao, Pei-Wen
<120> Экзогенный контроль экспрессии генов млекопитающих с
помощью опосредованной аптамерами модуляции полиаденилирования
<130> BAYM.P0165WO
<140> НЕ УСТАНОВЛЕНО
<141> 10-11-2016
<150> 62/254,435
<151> 12-11-2015
<160> 61
<170> Версия PatentIn 3.5
<210> 1
<211> 50
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 1
ucccacuguc cuuuccuaau aaaaugagga aauugcaucg cauugucuga 50
<210> 2
<211> 93
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 2
ucagacaaug cgaugcaauu ucgagcucuc gcgaggugcc acuccccacu guccuuuccu 60
aauaaaauga ggaaauugca ucgcauuguc uga 93
<210> 3
<211> 12
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 3
aauaaaaaua aa 12
<210> 4
<211> 71
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 4
gggccuaaaa cauaccagau cgccacccgc gcuuuaaucu ggagagguga agaauacgac 60
caccuaggcu c 71
<210> 5
<211> 12
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 5
aauaaaaaua aa 12
<210> 6
<211> 12
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 6
cacacaaaua aa 12
<210> 7
<211> 12
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 7
aaauaaacac ac 12
<210> 8
<211> 92
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 8
cacagaucug gagaggugaa gaauacgacc accugcguuu uauacuuugu aacacaaaua 60
aaguauaaag ugcaaaacau accagaucug ug 92
<210> 9
<211> 92
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 9
cacagaucug gagaggugaa gaauacgacc accugcguua uauacuuugu aacacaaaua 60
aaguauauag ugcaaaacau accagaucug ug 92
<210> 10
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 10
gtgcaaaaca aaaaaaaaaa aaaaaaa 27
<210> 11
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 11
gtgcaaaaca taaaaaaaaa aaaaaaa 27
<210> 12
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 12
gtgcaaaaca taccaaaaaa aaaaaaaa 28
<210> 13
<211> 92
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 13
cacagaucug gagaggugaa gaauacgacc accugcguuu uauacuuugu aacacaaaua 60
aaguauaaag ugcaaaacau accagaucug ug 92
<210> 14
<211> 11
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 14
aaaacauacc a 11
<210> 15
<211> 92
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 15
cacagaucug gagaggugaa gaauacgacc accugcguuu uauacuuugu aacacaaaua 60
aaguauaaag ugcaaaacau accagaucug ug 92
<210> 16
<211> 92
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 16
cacagaucug gggaggugaa gaauacgacc accugcguuu uauacuuugu aacacaaaua 60
aaguauaaag ugcaaaacau accagaucug ug 92
<210> 17
<211> 92
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 17
cacaaaucug gggaggugaa gaauacgacc accugcguuu uauacuuugu aacacaaaua 60
aaguauaaag ugcaaaacau accagaucug ug 92
<210> 18
<211> 92
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 18
cacaaaucug gggaggugaa gaauacgacc accugcuagu uauacuuugu aacacaaaua 60
aaguauagct agcaaaacau accagaucug ug 92
<210> 19
<211> 92
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 19
cacaaaucug gggaggugaa gaauacgacc accugcguuu uauacuuugu aacacaaaua 60
aaguauaaag ugcaaaacau accagaucug ug 92
<210> 20
<211> 92
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 20
cacaaaucug gggaggugaa gaauacgacc accugcuagu uauacuuugu aacacaaaua 60
aaguauagct agcaaaacau accagaucug ug 92
<210> 21
<211> 92
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 21
cacaaaucug gggaggugaa gaauacgacc accugcuagu uauacuuugu aacacaaaua 60
aaguauagct agcaaaacau accagaucug ug 92
<210> 22
<211> 12
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 22
ccaacccccg ca 12
<210> 23
<211> 11
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 23
uuugcuuaga a 11
<210> 24
<211> 12
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 24
ccgucucccc ca 12
<210> 25
<211> 12
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 25
ggggccucga ga 12
<210> 26
<211> 97
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 26
cacagaucug gggaggugaa gaauacgacc accugcguuu uauacuuugu uggaacacac 60
caauaaagua uaaagugcaa aacauaccag aucugug 97
<210> 27
<211> 92
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 27
cacaaaucug gggaggugaa gaauacgacc accugcuagu uacgcuuugu aacacaaaua 60
aagcguagct agcaaaacau accagaucug ug 92
<210> 28
<211> 136
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 28
cacaaaucug gggaggugaa gaauacgacc accugcguuu uauacuuugu uaaaacauac 60
cagaucgaaa gaucugggga ggugaagaau acgaccaccu aauaaaguau aaagugcaaa 120
acauaccaga ucugug 136
<210> 29
<211> 10
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 29
aucgaaagau 10
<210> 30
<211> 10
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 30
auugaaagau 10
<210> 31
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 31
accagatcga aagatctggg g 21
<210> 32
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 32
accagatcga tcgatctggg g 21
<210> 33
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 33
accagatcgc gatctgggg 19
<210> 34
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 34
accagatcga tctgggg 17
<210> 35
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 35
accagatcga ttcgatctgg gg 22
<210> 36
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 36
accagatatc tgggg 15
<210> 37
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 37
accagatcga attcgatctg ggg 23
<210> 38
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 38
accagatcga atttcgatct gggg 24
<210> 39
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 39
accagatcga atattcgatc tgggg 25
<210> 40
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 40
accagatcga atcattcgat ctgggg 26
<210> 41
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 41
accagatcga atcgattcga tctgggg 27
<210> 42
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 42
accagattac ccttgggtga tctgggg 27
<210> 43
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 43
accagatcga atctgattcg atctgggg 28
<210> 44
<211> 41
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 44
accatcctaa gcctaaggca aacgctatgg tatggatggg g 41
<210> 45
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 45
accagatcga aagatctggg g 21
<210> 46
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 46
accagatcga tcgatctggg g 21
<210> 47
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 47
accagatcgc gatctgggg 19
<210> 48
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 48
accagatcga tctgggg 17
<210> 49
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 49
accagatcga atcgatctgg gg 22
<210> 50
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 50
accagatatc tgggg 15
<210> 51
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 51
accagatcga attcgatctg ggg 23
<210> 52
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 52
accagatcga aattcgatct gggg 24
<210> 53
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 53
accagatcga atattcgatc tgggg 25
<210> 54
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 54
accagatcga atgattcgat ctgggg 26
<210> 55
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 55
accagatcga atcgattcga tctgggg 27
<210> 56
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 56
accagattac ccgagggtga tctgggg 27
<210> 57
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 57
accagatcga atcagattcg atctgggg 28
<210> 58
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 58
accagatcga aagatctggg g 21
<210> 59
<211> 370
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 59
gctagccaca cacacaaatc tggggaggtg aagaatacga ccacctgcgt tttatacttt 60
gttaaaacat accagatcga aagatctggg gaggtgaaga atacgaccac ctaataaagt 120
ataaagtgca aaacatacca gatctgtgtg ttggtttttt gtgtggaatt ccacacacac 180
aaatctgggg aggtgaagaa tacgaccacc tgcgttttat actttgttaa aacataccag 240
atcgaaagat ctggggaggt gaagaatacg accacctaat aaagtataaa gtgcaaaaca 300
taccagatct gtgtgttggt tttttgtgtg aacgggggag ggggaggaaa gggggagggg 360
gagcggccgc 370
<210> 60
<211> 374
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 60
ctgcagcaca cacacaaatc tggggaggtg aagaatacga ccacctgcgt tttatacttt 60
gttaaaacat accagatcga attcgatctg gggaggtgaa gaatacgacc acctaataaa 120
gtataaagtg caaaacatac cagatctgtg tgttggtttt ttgtgtggct agccacacac 180
acaaatctgg ggaggtgaag aatacgacca cctgcgtttt atactttgtt aaaacatacc 240
agatcgaatt cgatctgggg aggtgaagaa tacgaccacc taataaagta taaagtgcaa 300
aacataccag atctgtgtgt tggttttttg tgtgaacggg ggagggggag gaaaggggga 360
gggggagcgg ccgc 374
<210> 61
<211> 396
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический праймер
<400> 61
gctagcgaat tgcacacaca caaatctggg gaggtgaaga atacgaccac ctgcgtttta 60
tactttgtta aaacatacca tcctaagcct aaggcaaacg ctatggtatg gatggggagg 120
tgaagaatac gaccacctaa taaagtataa agtgcaaaac ataccagatc tgtgtgttgg 180
ttttttgtgt ggaattccac acacacaaat ctggggaggt gaagaatacg accacctgcg 240
ttttatactt tgttaaaaca taccagatcg aaagatctgg ggaggtgaag aatacgacca 300
cctaataaag tataaagtgc aaaacatacc agatctgtgt gttggttttt tgtgtgaacg 360
ggggaggggg aggaaagggg gagggggagc ggccgc 396
<---

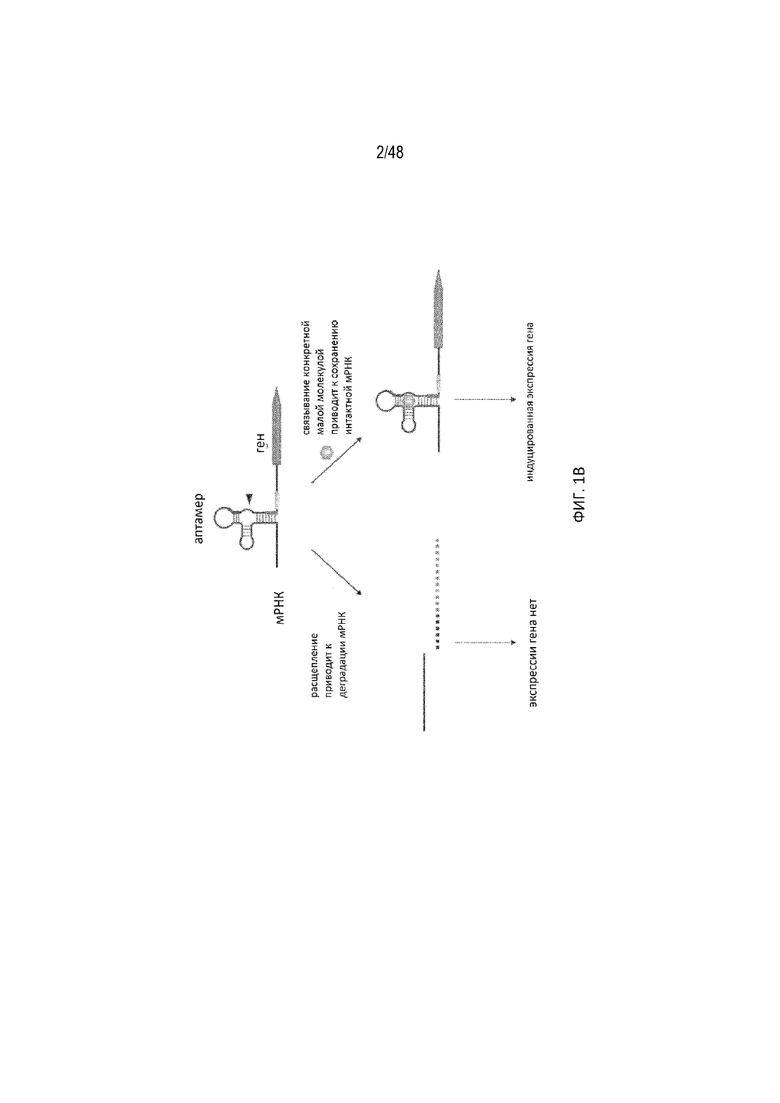


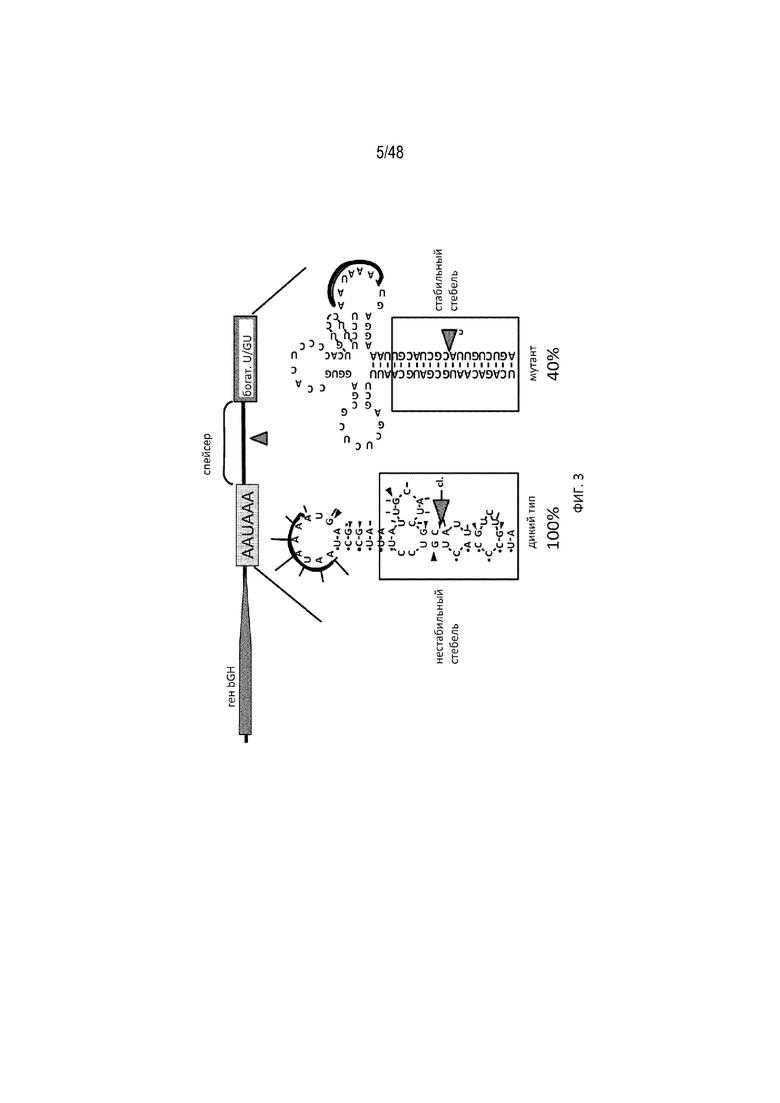

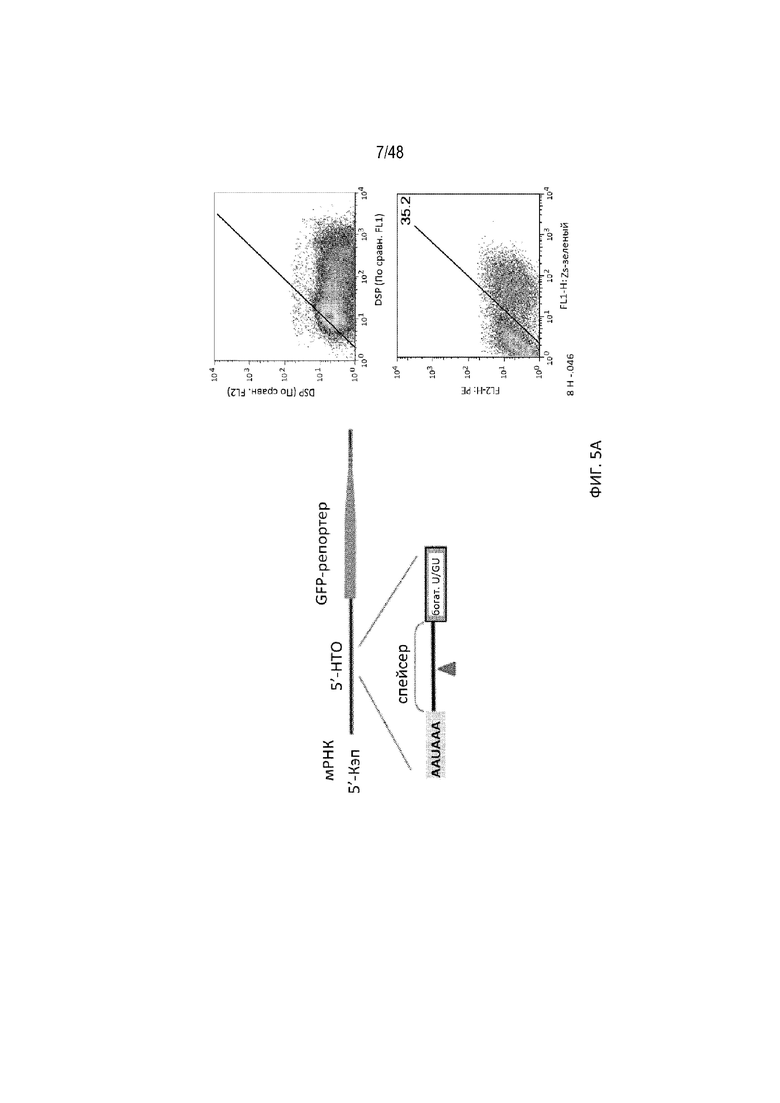

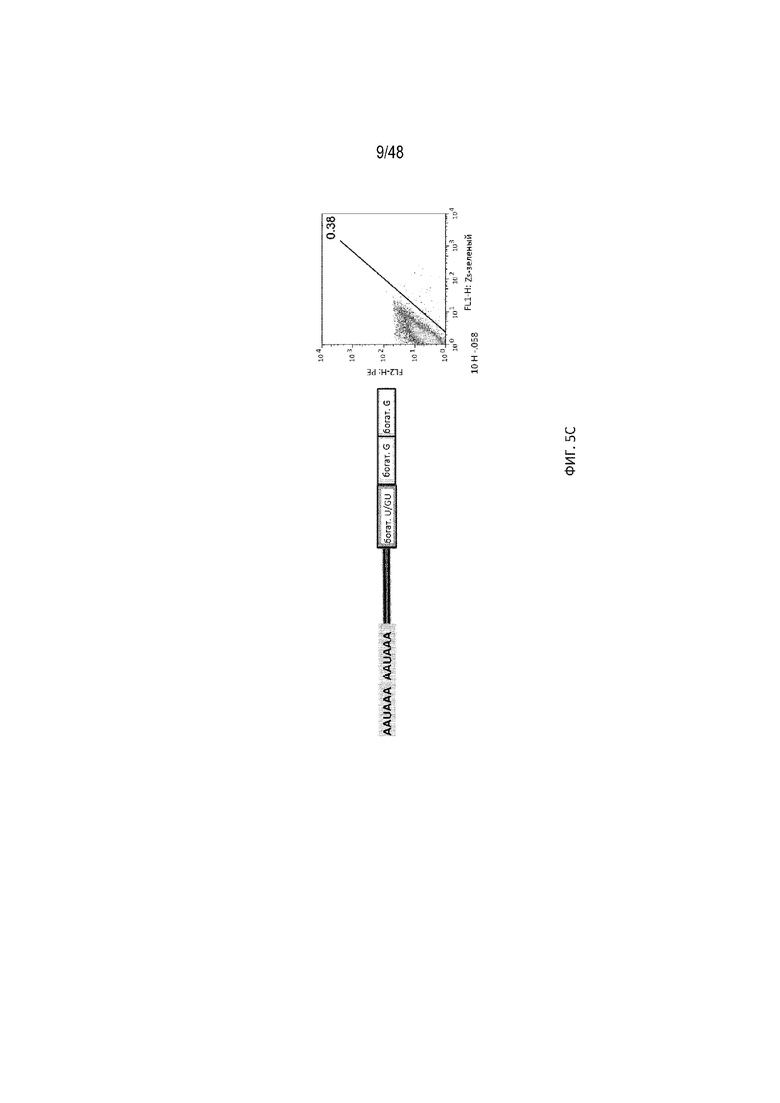
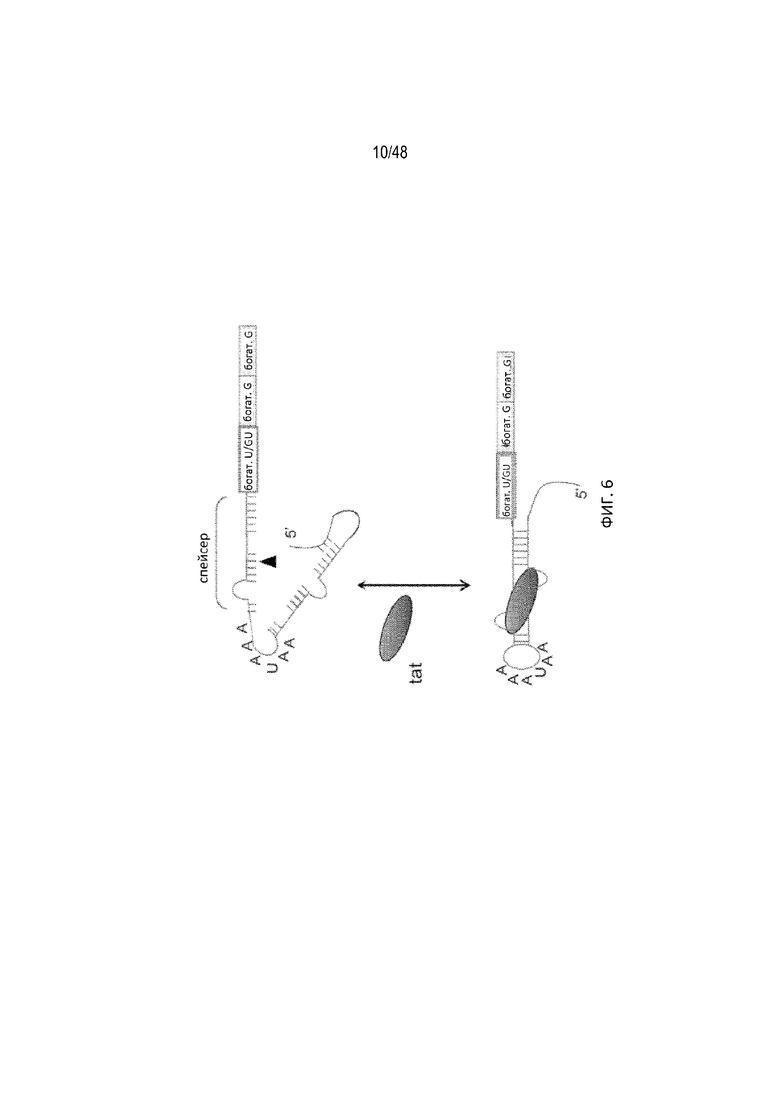
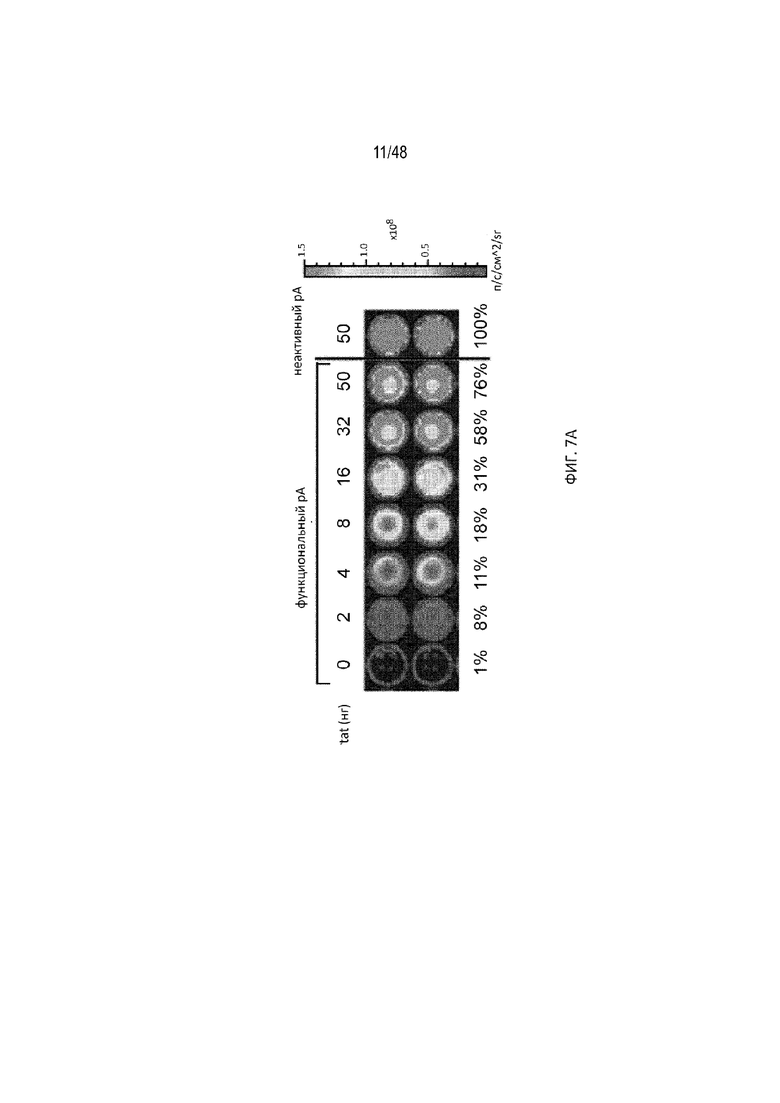
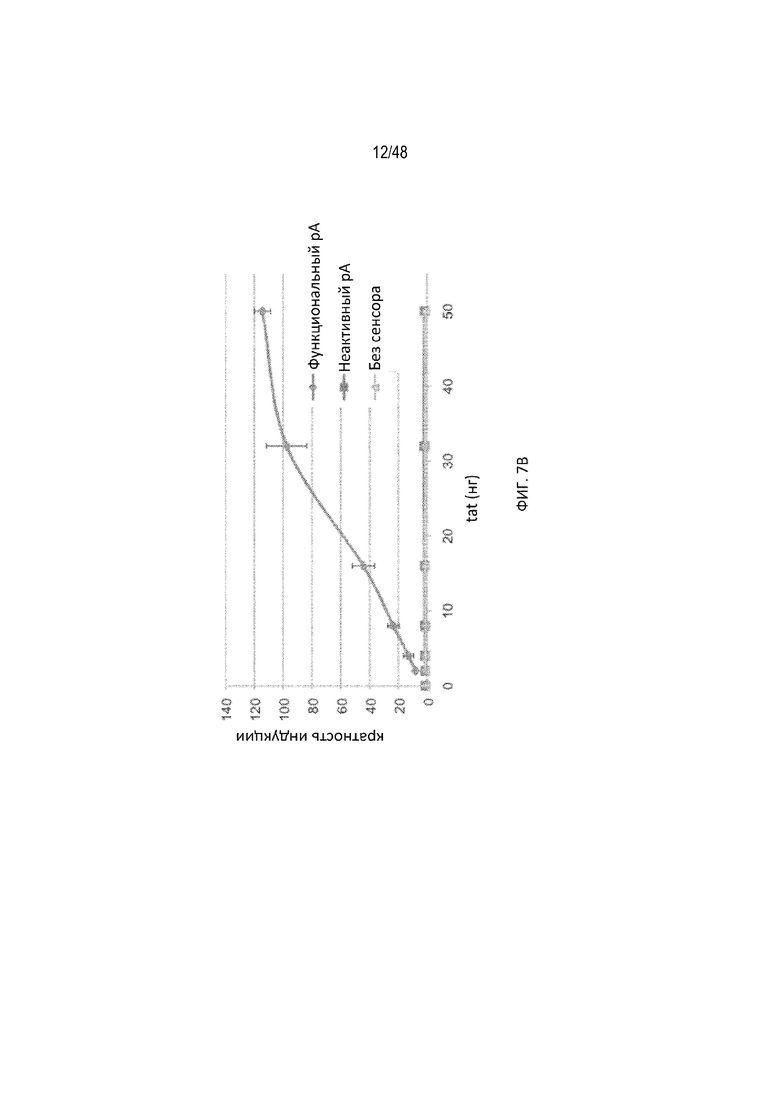
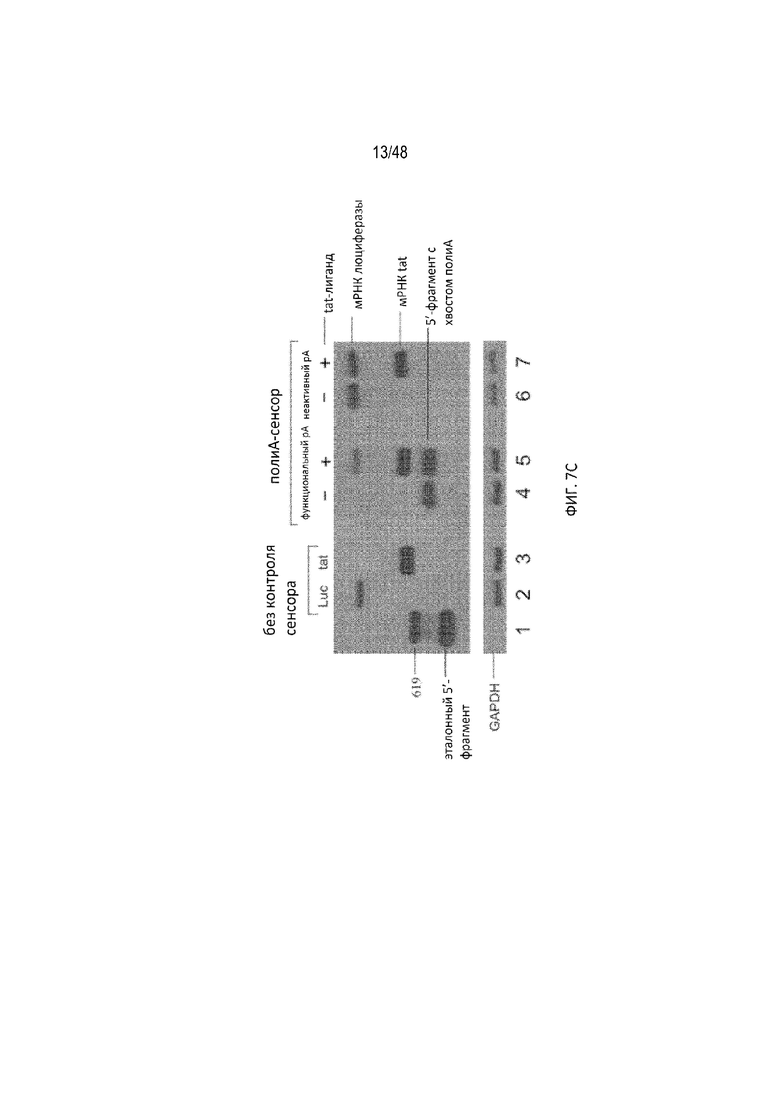
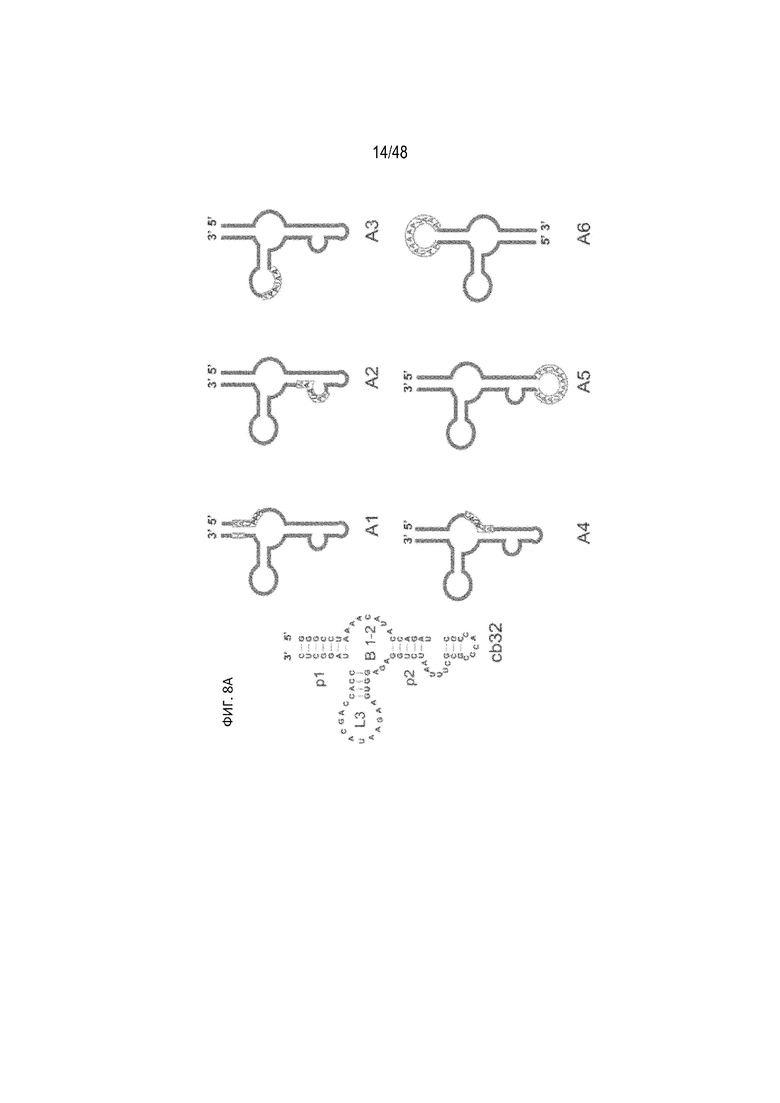
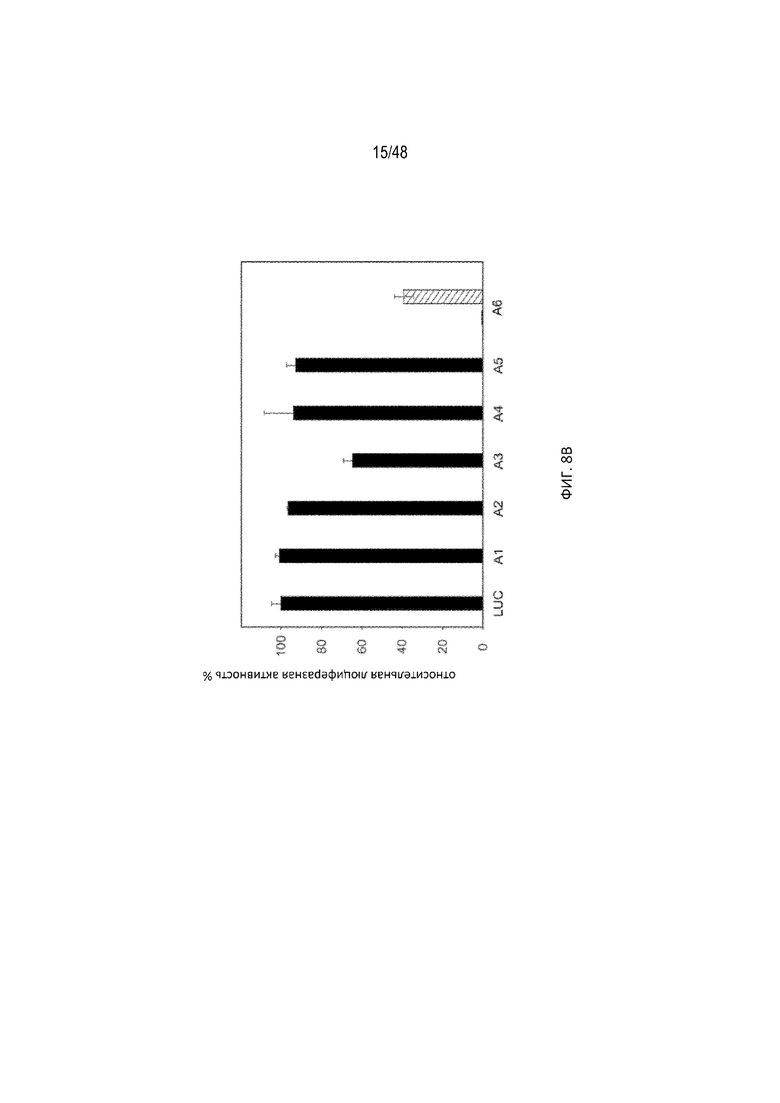



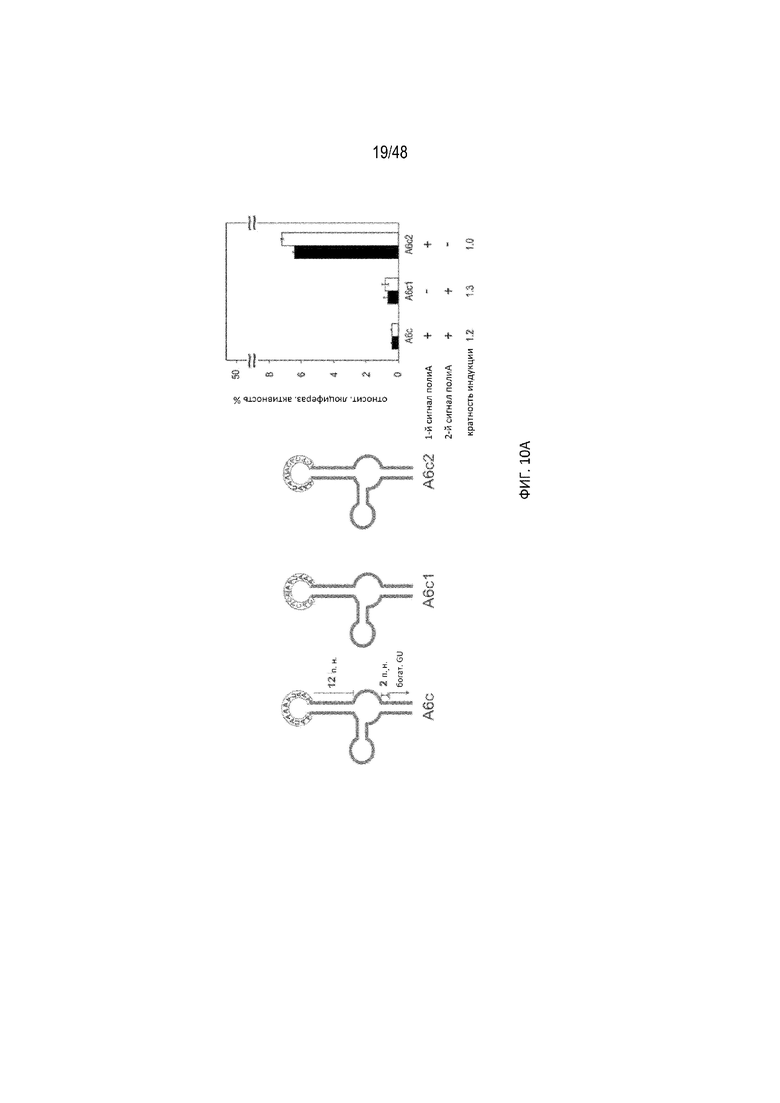

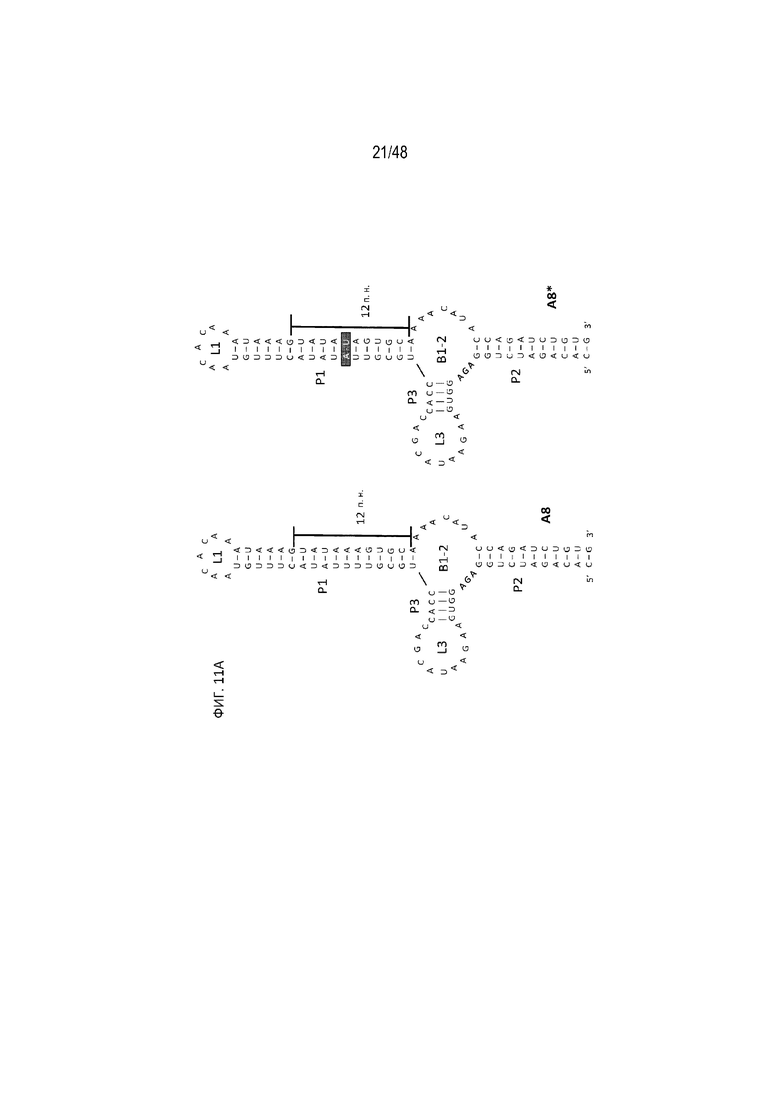
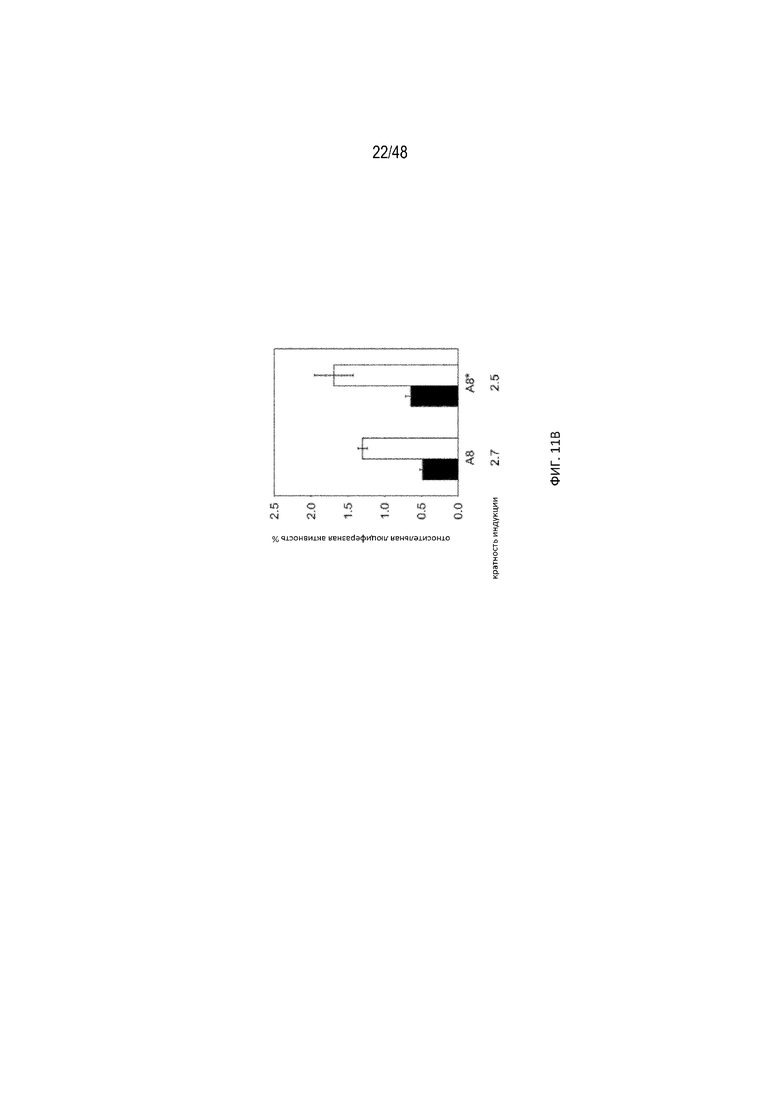
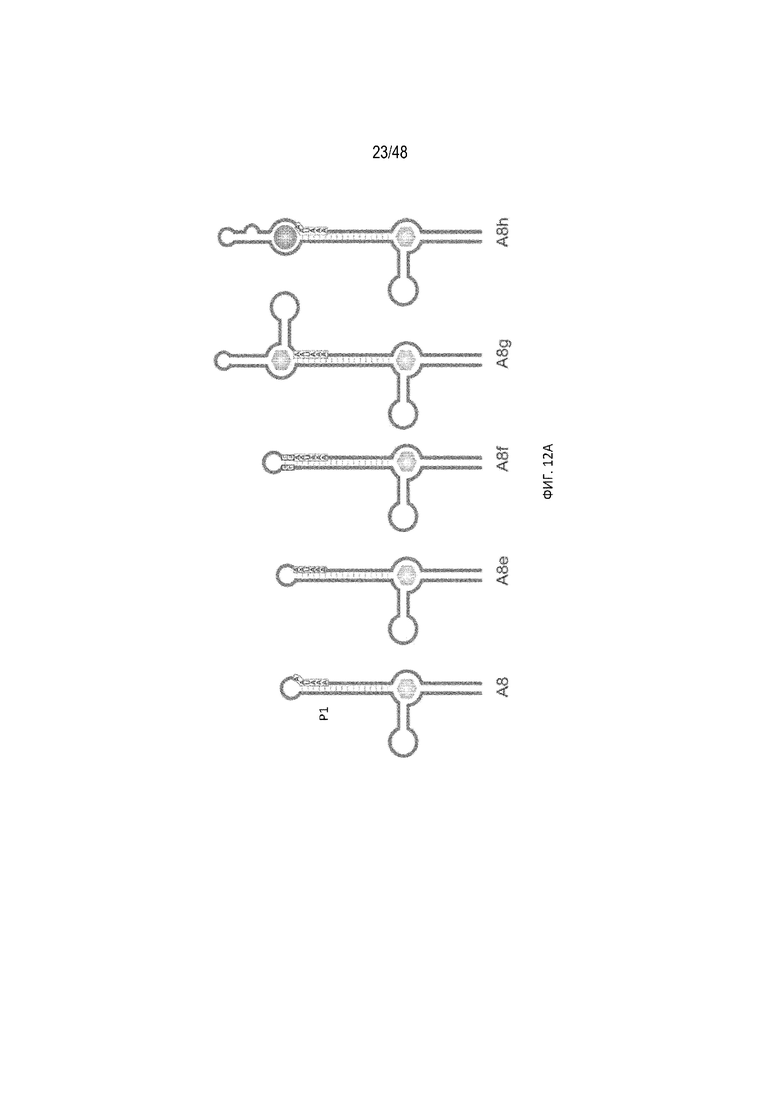

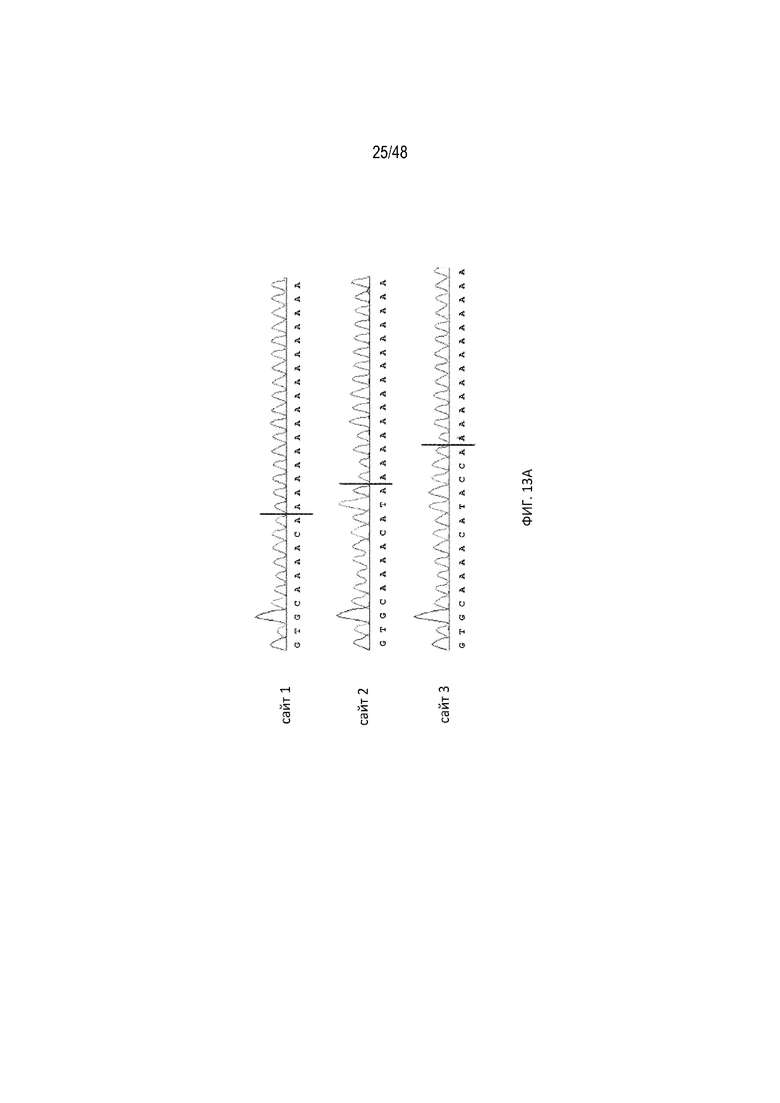
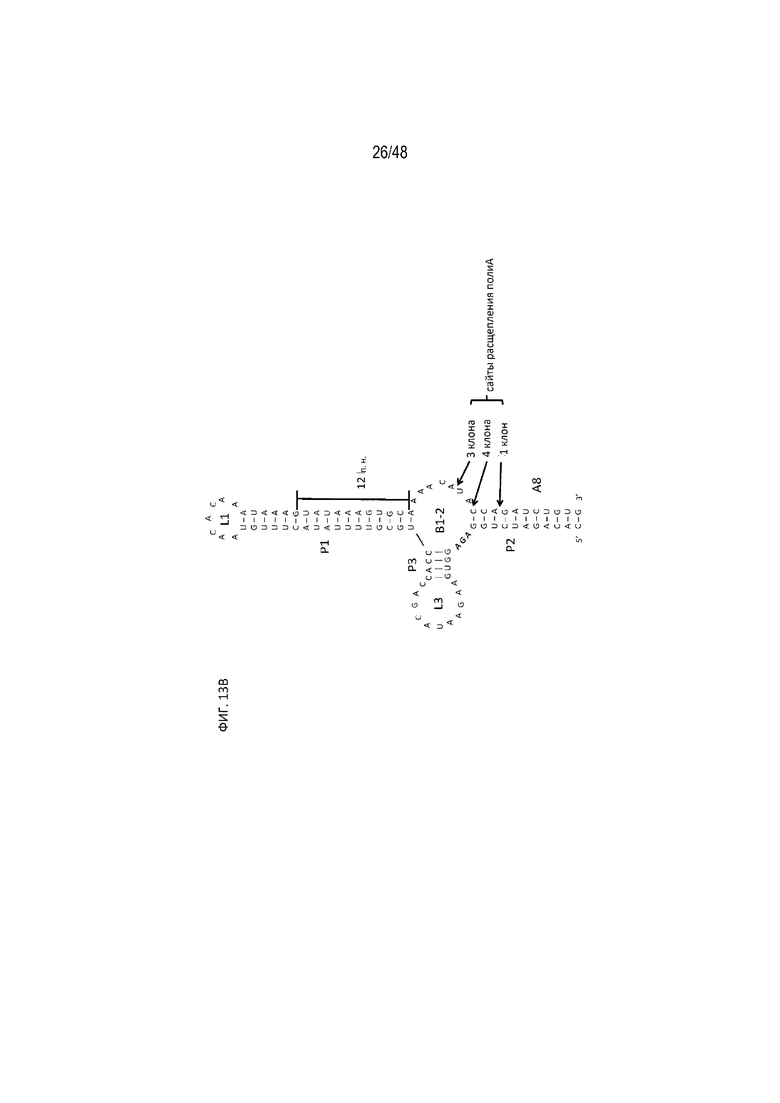
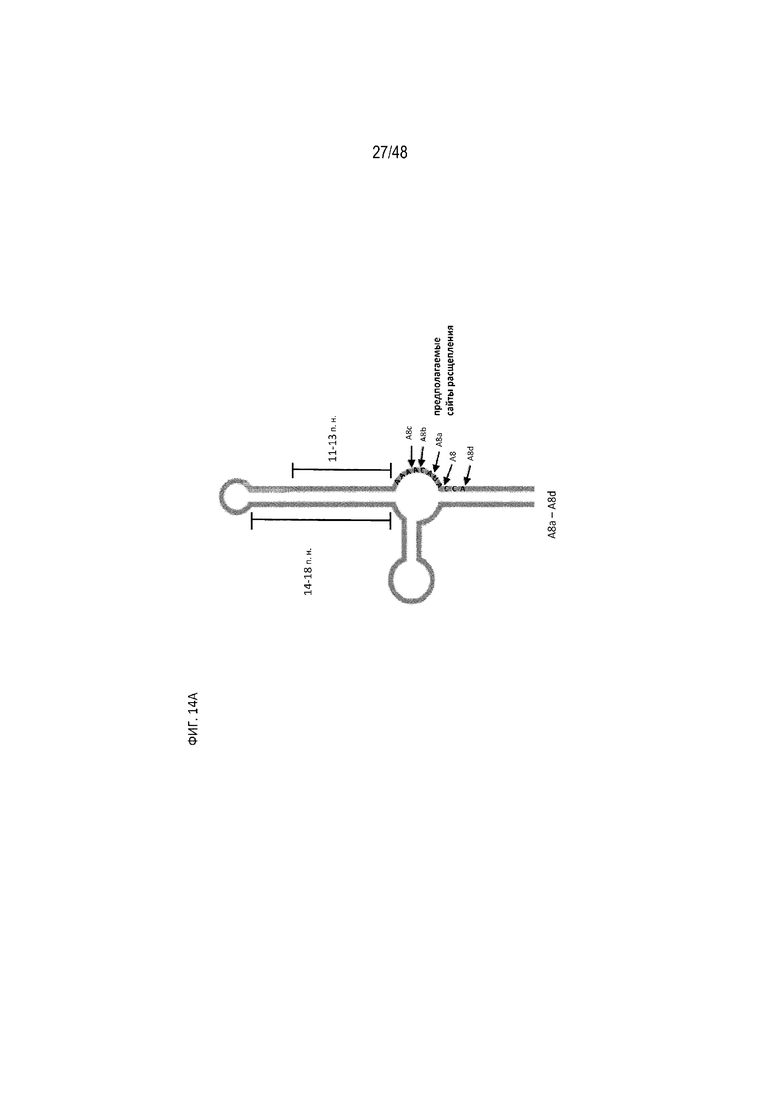
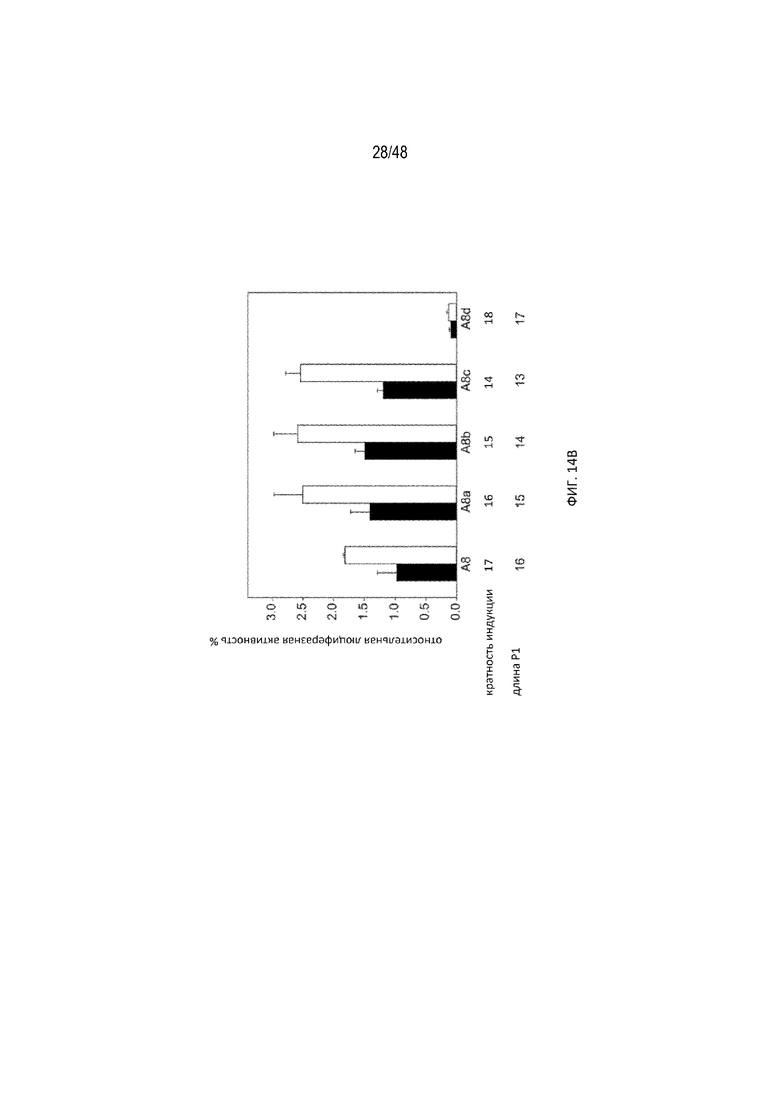
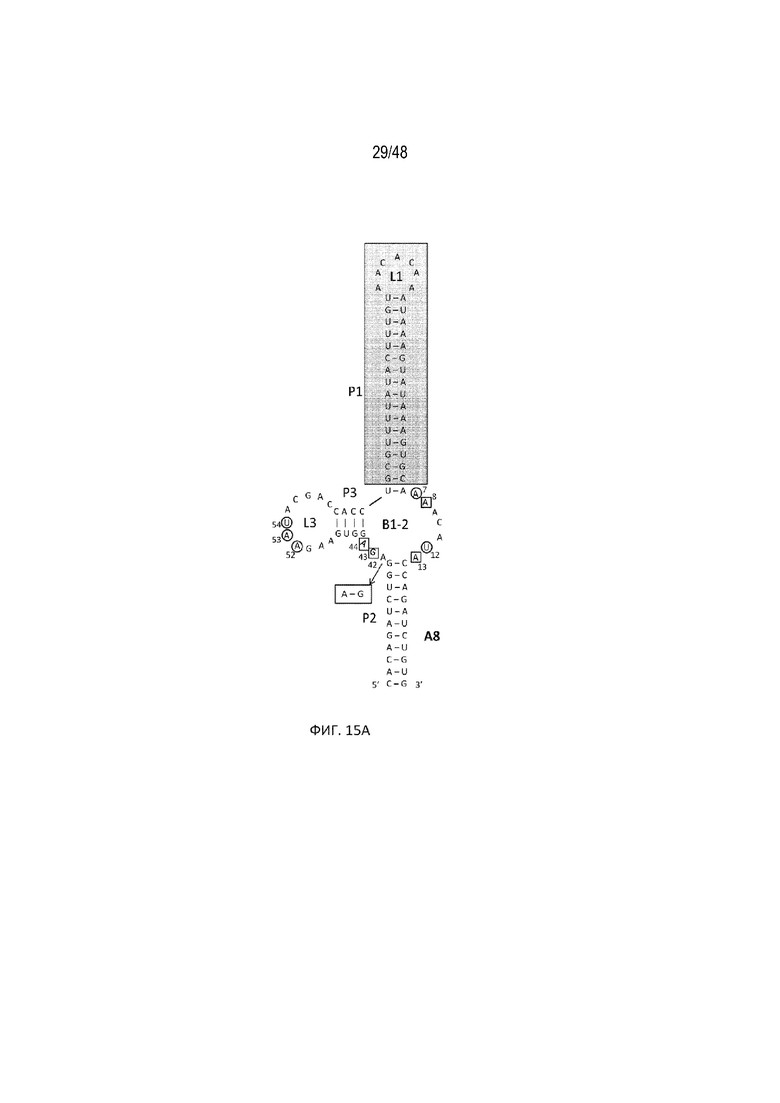
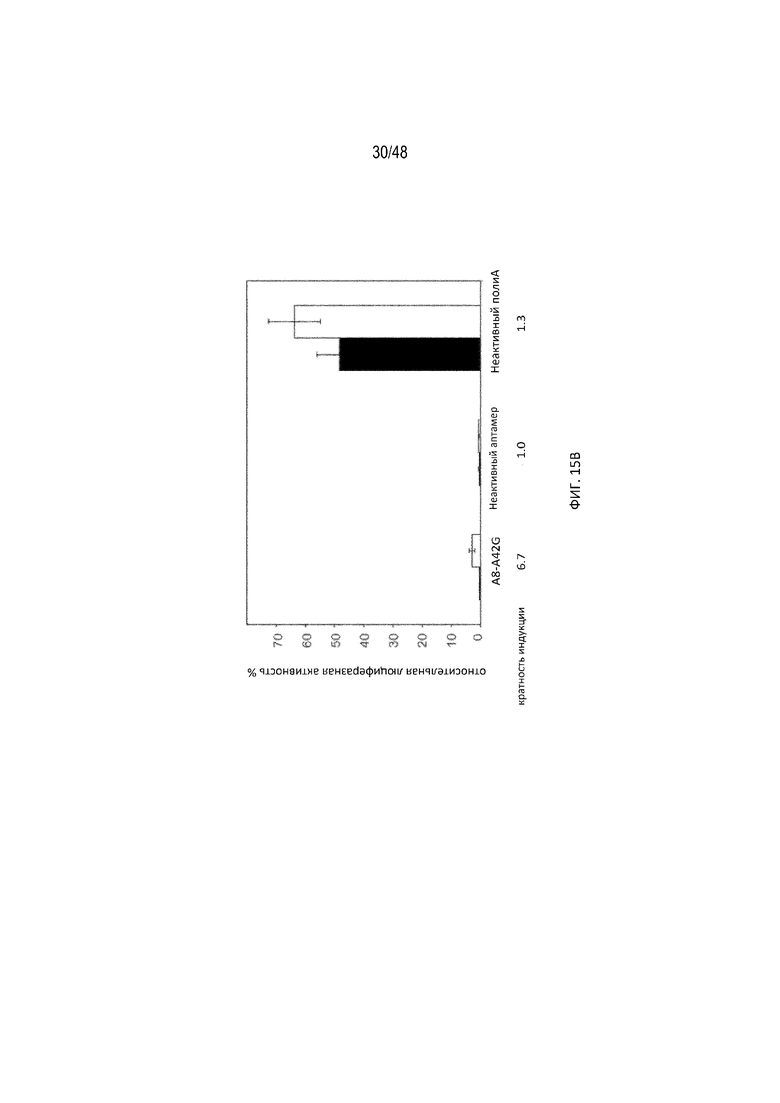


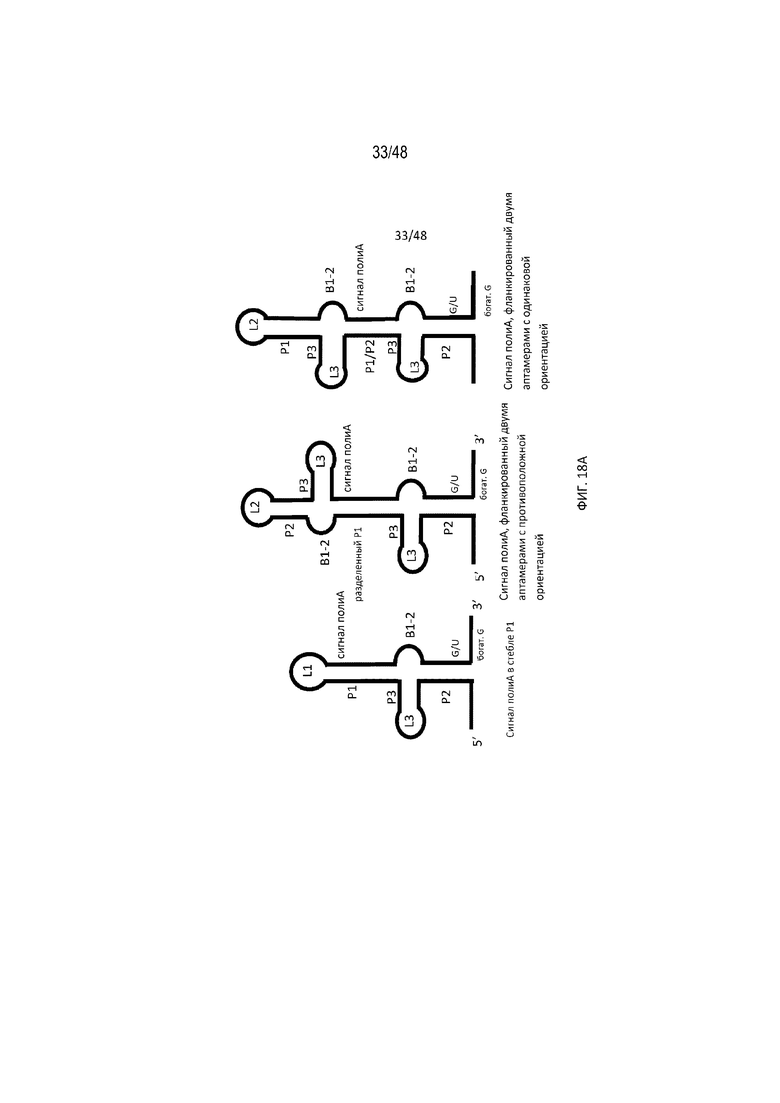
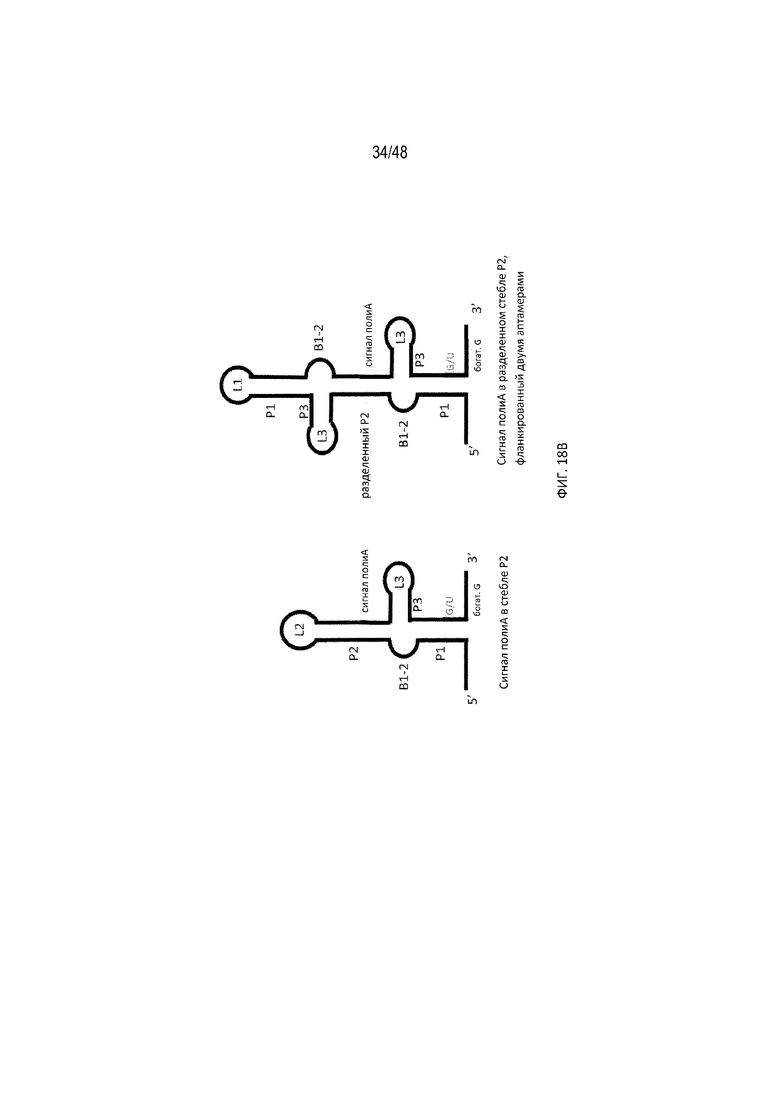

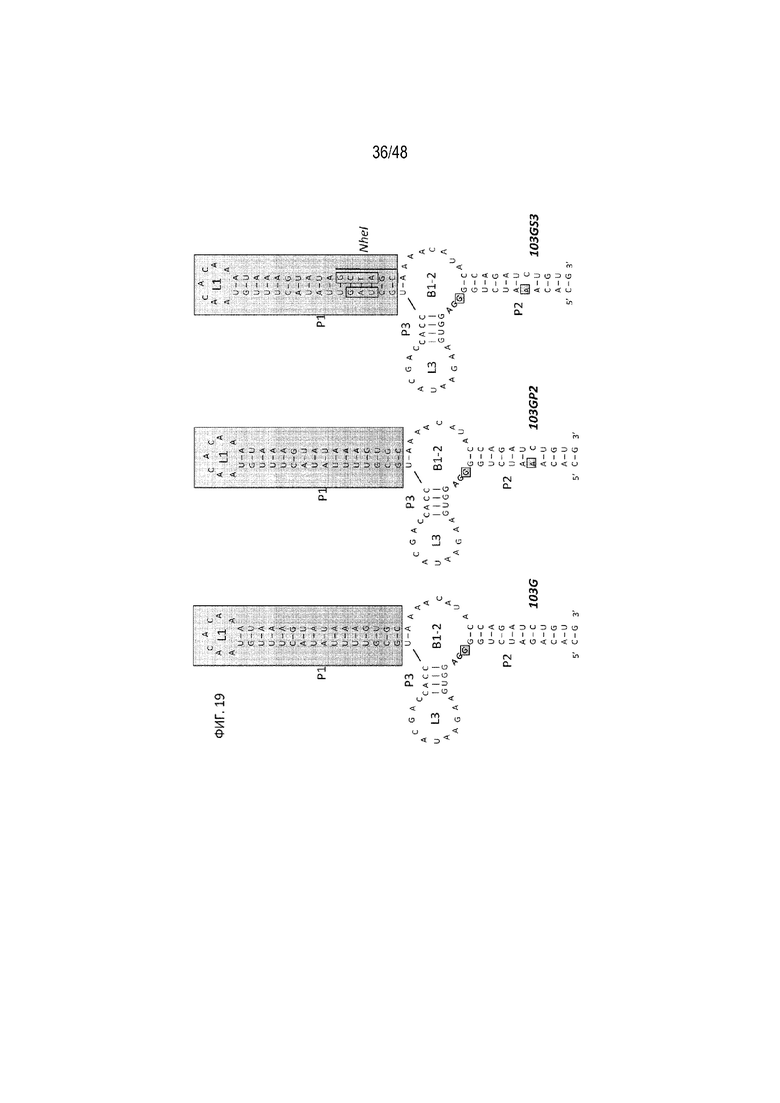


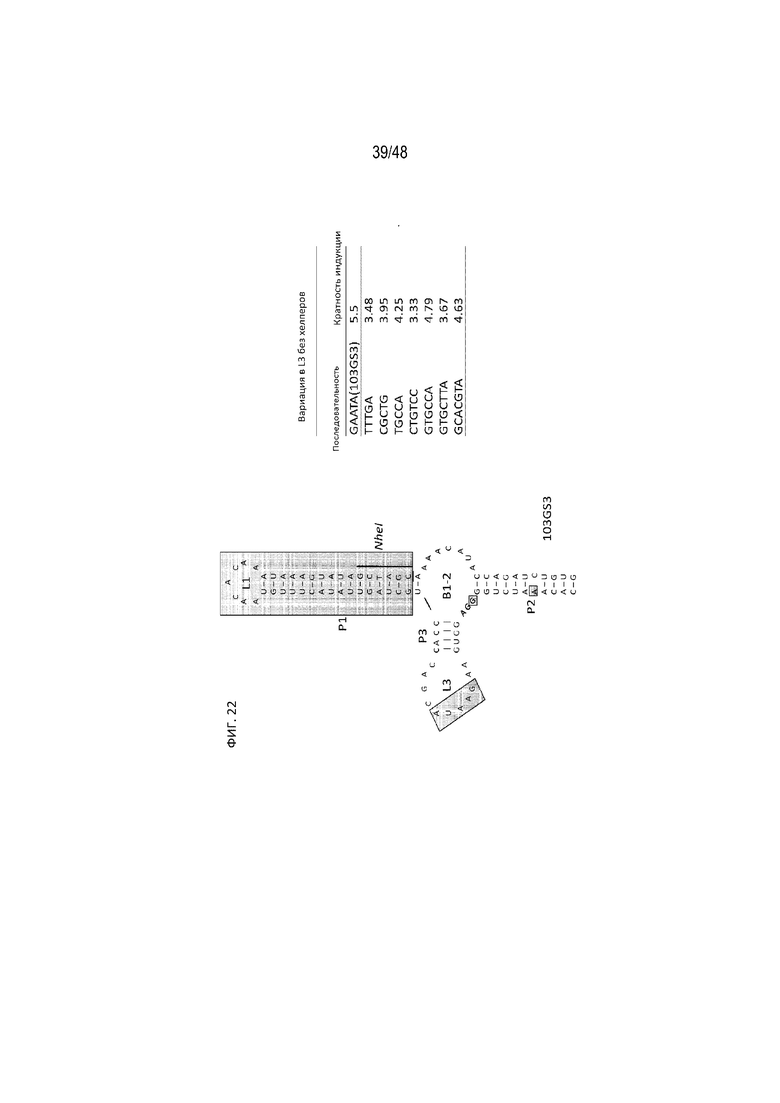


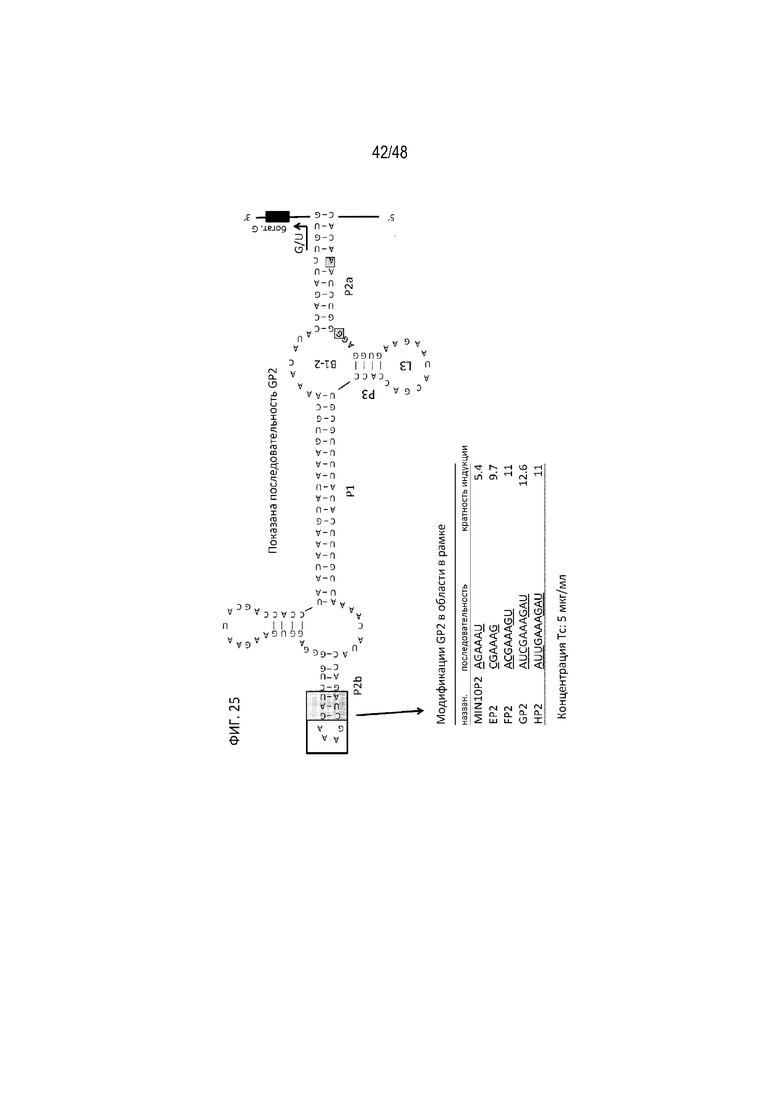

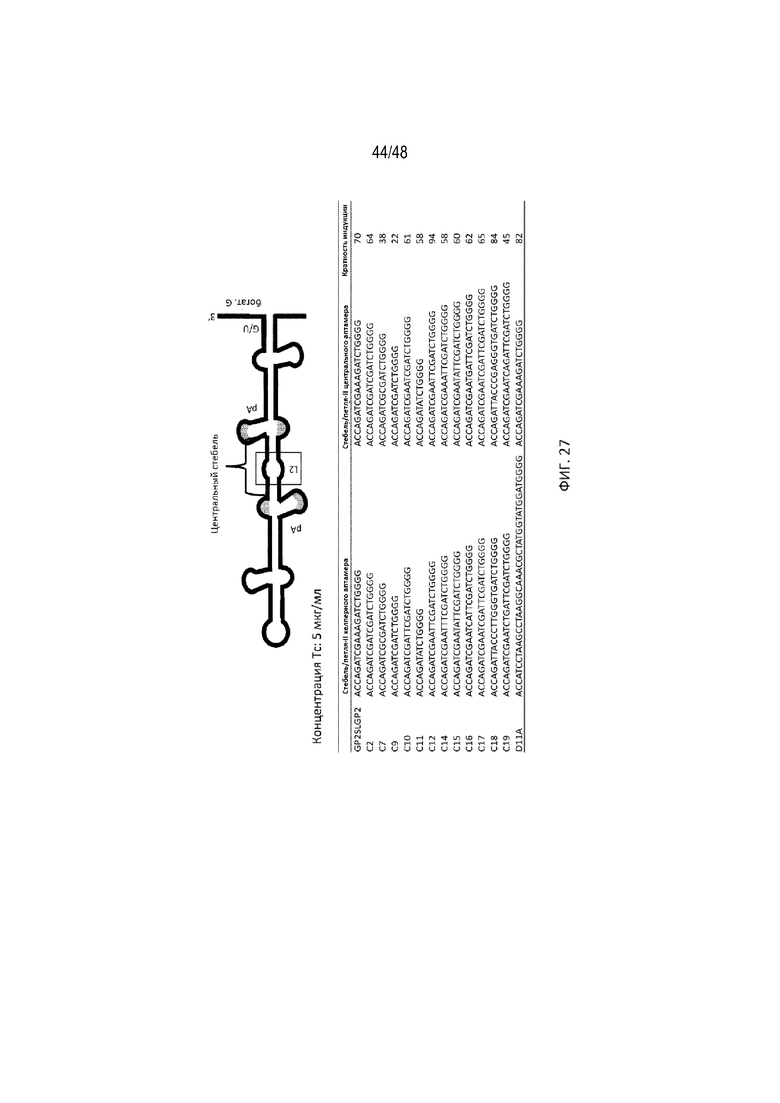

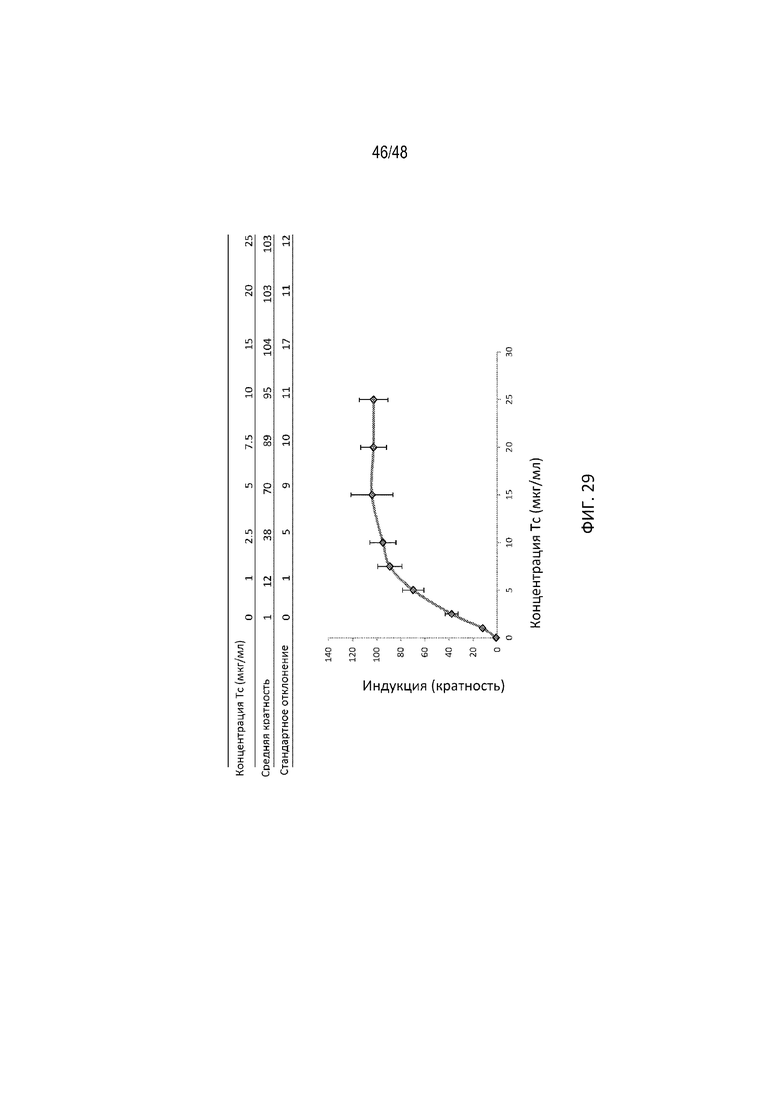
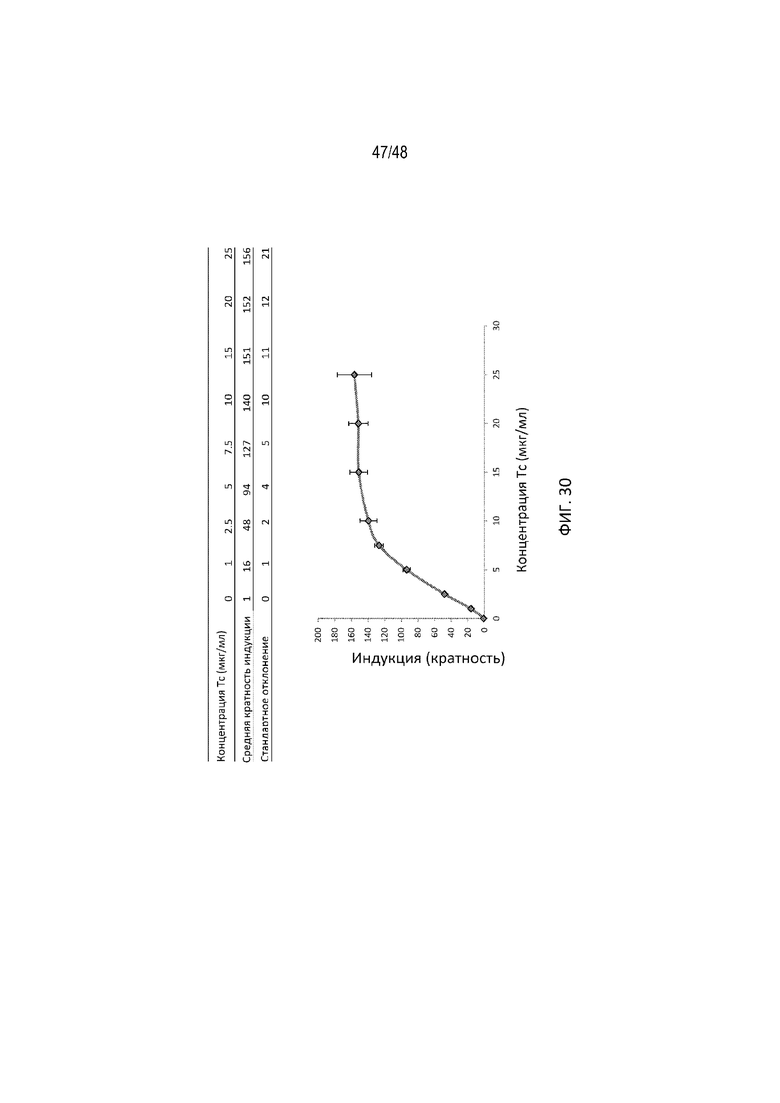

Изобретение относится к биотехнологии, в частности к применению экспрессионных конструкций, в которых по меньшей мере один сигнал полиA помещен слева по ходу транскрипции экспрессируемого транскрипта как, например, в пределах 5'-НТО транскрипта. В некоторых вариантах реализации изобретения сигнал полиA содержится в пределах лиганд-связывающего аптамера, а связывание лиганда с аптамером или отсутствие связывания определяет выход экспрессируемого транскрипта. В конкретных вариантах реализации изобретения отсутствие лиганда вызывает экспрессию экспрессируемого транскрипта, содержащего полиA в своей 5'-НТО, но затем происходит его разрушение, тогда как присутствие лиганда вызывает ингибирование разрушения при экспрессии экспрессируемого транскрипта. На одной и той же экспрессионной конструкции может присутствовать более одного лиганд-связывающего аптамера. 9 н. и 55 з.п. ф-лы, 31 ил., 1 табл., 3 пр.
1. Система для модуляции экспрессии генов, содержащая полинуклеотид аптамера полиA, который в направлении 5'-3' содержит:
a) по меньшей мере один лиганд-связывающий аптамер, содержащий в себе по меньшей мере один сигнал расщепления полиA, и
b) экспрессируемый полинуклеотид,
где:
(i) по меньшей мере один лиганд-связывающий аптамер находится в пределах 5'-нетранслируемой области (UTR) экспрессируемого полинуклеотида; и
(ii) в направлении 5'-3' полинуклеотида аптамера полиA по меньшей мере один лиганд-связывающий аптамер находится слева по ходу транскрипции от одного, двух или более богатых G участков.
2. Система по п. 1, содержащая полинуклеотид, который экспрессирует лиганд, который связывает лиганд-связывающий аптамер.
3. Система по п. 2, отличающаяся тем, что экспрессирующий лиганд полинуклеотид представляет собой тот же полинуклеотид аптамера полиA, который содержит лиганд-связывающий аптамер и экспрессируемый полинуклеотид.
4. Система по п. 2, отличающаяся тем, что экспрессирующий лиганд полинуклеотид представляет собой полинуклеотид, отличающийся от полинуклеотида аптамера полиA, который содержит аптамер и экспрессируемый полинуклеотид.
5. Система по любому из пп. 2-4, отличающаяся тем, что полинуклеотид аптамера полиA содержит два, три или более сигналов полиA в 5'-НТО экспрессируемого полинуклеотида.
6. Система по любому из пп. 1-5, отличающаяся тем, что полинуклеотид аптамера полиA содержит:
a) по меньшей мере один сигнал полиA;
b) по меньшей мере один лиганд-связывающий аптамер, содержащий по меньшей мере один сигнал полиA; и
c) по меньшей мере один богатый U/UG участок, по меньшей мере один богатый G участок или оба варианта из по меньшей мере одного богатого U/UG участка и по меньшей мере одного богатого G участка.
7. Система по п. 6, отличающаяся тем, что лиганд-связывающий аптамер содержит один, два, три или более богатых U/UG участков.
8. Система по п. 6, отличающаяся тем, что в направлении 5'-3' полинуклеотида аптамера полиA по меньшей мере один сигнал полиA находится слева (выше) по ходу транскрипции от по меньшей мере одного богатого U/UG участка.
9. Система по п. 6, отличающаяся тем, что в направлении 5'-3' полинуклеотида аптамера полиA лиганд-связывающий аптамер находится слева по ходу транскрипции от одного, двух или более богатых U/UG участков.
10. Система по п. 6, отличающаяся тем, что в направлении 5'-3' полинуклеотида аптамера полиA по меньшей мере один сигнал полиA находится слева по ходу транскрипции от по меньшей мере одного богатого G участка.
11. Система по любому из пп. 1-10, отличающаяся тем, что в направлении 5'-3' полинуклеотида аптамера полиA аптамер содержит два сигнала полиA и два богатых U/UG участка.
12. Система по любому из пп. 1-11, отличающаяся тем, что полинуклеотид аптамера полиA содержит 2, 3, 4, 5 или более аптамеров.
13. Система по п. 12, отличающаяся тем, что первый аптамер и второй аптамер находятся в одной и той же ориентации в линейном направлении 5'-3'.
14. Система по п. 12, отличающаяся тем, что первый аптамер и второй аптамер находятся в разной ориентации в линейном направлении 5'-3'.
15. Система по любому из пп. 12-14, отличающаяся тем, что полинуклеотид аптамера полиA содержит один богатый G участок.
16. Система по п. 15, отличающаяся тем, что богатый G участок находится на наиболее удаленном в 3'-направлении аптамере в направлении 5'-3' полинуклеотида аптамера полиA.
17. Система по п. 15, отличающаяся тем, что богатый G участок находится на втором аптамере в направлении 5'-3' полинуклеотида аптамера полиA.
18. Система по любому из пп. 12-17, отличающаяся тем, что полинуклеотид аптамера полиA не содержит спаривания оснований между двумя или более аптамерами.
19. Система по любому из пп. 12-17, отличающаяся тем, что полинуклеотид аптамера полиA содержит спаривание оснований между двумя или более аптамерами.
20. Система по любому из пп. 12-19, отличающаяся тем, что в полинуклеотиде аптамера полиA существует спаривание оснований между по меньшей мере частью петли из двух разных аптамеров.
21. Система по любому из пп. 1-20, отличающаяся тем, что количество нуклеотидов между двумя петлями в пределах аптамера составляет 10-25 нуклеотидов.
22. Система по любому из пп. 1-20, отличающаяся тем, что количество нуклеотидов между двумя петлями в пределах аптамера составляет 18-20 нуклеотидов.
23. Система по любому из пп. 1-22, отличающаяся тем, что лиганд представляет собой полипептид, пептид, нуклеиновую кислоту, малую молекулу, лекарственное средство, метаболит или их комбинацию.
24. Система по любому из пп. 1-23, отличающаяся тем, что длина аптамера составляет от 14 до 250 нуклеотидов.
25. Система по любому из пп. 1-24, отличающаяся тем, что экспрессируемый полинуклеотид представляет собой репортерный ген, терапевтический ген или ген, продукт которого изменяет метаболическое состояние клеток.
26. Система по любому из пп. 1-25, отличающаяся тем, что полинуклеотид аптамера полиA является по меньшей мере частью вектора.
27. Система по п. 2, отличающаяся тем, что полинуклеотид, экспрессирующий лиганд, является по меньшей мере частью вектора.
28. Система по п. 26 или 27, отличающаяся тем, что вектор представляет собой плазмиду, вирусный вектор или линейную ДНК.
29. Система по любому из пп. 1-28, отличающаяся тем, что экспрессируемый полинуклеотид кодирует лиганд.
30. Система по любому из пп. 1-29, отличающаяся тем, что экспрессия экспрессируемого полинуклеотида регулируется тканеспецифическим промотором.
31. Способ модуляции экспрессии генов, включающий этапы:
a) обеспечения системы, содержащей полинуклеотид аптамера полиA, который в направлении 5'-3' содержит:
1) по меньшей мере один лиганд-связывающий аптамер, содержащий в себе по меньшей мере один сигнал расщепления полиA,
2) экспрессируемый полинуклеотид и
3) необязательно лиганд-экспрессирующую конструкцию; и
b) когда не требуется получение мРНК из экспрессируемого полинуклеотида, воздействия на систему соответствующих условий, при которых лиганд не связывает лиганд-связывающий аптамер или отсутствует в системе или ее окружении, что приводит к разрушению полученной из экспрессируемого полинуклеотида мРНК; или
c) когда требуется экспрессия экспрессируемого полинуклеотида, воздействия на систему условий, при которых лиганд связывает лиганд-связывающий аптамер и/или присутствует в системе или ее окружении, в результате чего полученная из экспрессируемого полинуклеотида мРНК не разрушается.
32. Способ модуляции экспрессии генов, включающий этапы:
a) обеспечения системы, содержащей полинуклеотид аптамера полиA, который в направлении 5'-3' содержит:
1) по меньшей мере один лиганд-связывающий аптамер, содержащий в себе по меньшей мере один сигнал расщепления полиA,
2) экспрессируемый полинуклеотид и
3) необязательно лиганд-экспрессирующую конструкцию; и
b) воздействия на систему условий, при которых лиганд отсутствует в системе или ее окружении или не связывает лиганд-связывающий аптамер, приводит к разрушению полученной из экспрессируемого полинуклеотида мРНК; или
c) воздействия на систему условий, при которых лиганд связывает лиганд-связывающий аптамер, в результате чего полученная из экспрессируемого полинуклеотида мРНК не разрушается, а продукт гена экспрессируется на экспрессируемом полинуклеотиде.
33. Способ по п. 31 или 32, осуществляемый в клетке.
34. Способ по п. 33, отличающийся тем, что лиганд по отношению к клетке является эндогенным.
35. Способ по п. 33 или 34, отличающийся тем, что клетка представляет собой стволовую клетку, раковую клетку, или пораженную заболеванием или дефектную клетку, требующую генной терапии по некоторому гену.
36. Способ по п. 35, отличающийся тем, что ген представляет собой ген дистрофина, альбумина или фактора IX.
37. Способ по любому из пп. 31-36, осуществляемый in vivo.
38. Способ по любому из пп. 31-37, осуществляемый в организме млекопитающего.
39. Способ по п. 38, отличающийся тем, что млекопитающее является человеком.
40. Способ по п. 31 или 32, осуществляемый in vitro.
41. Способ по любому из пп. 31-39, осуществляемый в одной или более клеток индивида, лиганд представляет собой глюкозу, индивид болен диабетом, преддиабетом или имеет осложнения вследствие диабета, и/или экспрессируемый полинуклеотид представляет собой последовательность инсулина.
42. Способ по любому из пп. 31-39, осуществляемый в одной или более клеток индивида, лиганд представляет собой продукт гена ракового биомаркера, а экспрессируемый полинуклеотид представляет собой суицидальный ген.
43. Способ по любому из пп. 31-39, осуществляемый в организме индивида, экспрессируемый полинуклеотид представляет собой репортерный ген, а расположение и/или интенсивность экспрессии репортерного гена предоставляет информацию о пространственном распределении зависящей от времени флуктуации или обоих этих параметрах для определенного лиганда в одной или более клетках индивида.
44. Способ по любому из пп. 31-43, дополнительно включающий этап конструирования аптамера для приемлемого связывания лиганда.
45. Способ по любому из пп. 31-44, осуществляемый в организме индивида, ткани или клетке, причем экспрессируемый полинуклеотид кодирует выявляемый продукт гена и при этом визуализируют тело соответствующего индивида, ткань или клетку.
46. Способ отслеживания терапии индивида, включающий этапы предоставления индивиду:
a) вектора, содержащего полинуклеотид аптамера полиA, который в направлении 5'-3' содержит:
1) по меньшей мере один лиганд-связывающий аптамер, содержащий в себе по меньшей мере один сигнал расщепления полиA, и
2) экспрессируемый полинуклеотид; и/или
b) одной или более клеток, содержащих вектор по этапу a),
причем лиганд, который связывает лиганд-связывающий аптамер, представляет собой продукт конкретного гена в виде белка, который является показателем эффективности терапии.
47. Способ по п. 46, отличающийся тем, что индивиду предоставляют вектор по этапу a) и/или клетки по этапу b) перед терапией, во время терапии и/или после терапии.
48. Способ анализа у индивида наличия, существования риска или восприимчивости к патологическому состоянию, включающий этапы предоставления индивиду:
a) вектора, содержащего полинуклеотид аптамера полиA, который в направлении 5'-3' содержит:
1) по меньшей мере один лиганд-связывающий аптамер, содержащий в себе по меньшей мере один сигнал расщепления полиA, и
2) экспрессируемый полинуклеотид; и/или
b) одну или более клеток, содержащих вектор по этапу a),
причем экспрессия экспрессируемого полинуклеотида или отсутствие экспрессии экспрессируемого полинуклеотида идентифицирует то, присутствует ли лиганд, который связывает лиганд-связывающий аптамер, чтобы связывать аптамер, при этом соответствующее наличие или отсутствие лиганда в организме индивида или его клетках является показателем наличия, восприимчивости или риска возникновения патологического состояния.
49. Полинуклеотид аптамера полиA для модуляции или отслеживания экспрессии генов, содержащий в направлении 5'-3':
a) по меньшей мере один лиганд-связывающий аптамер, содержащий в себе по меньшей мере один сигнал расщепления полиA, и
b) экспрессируемый полинуклеотид,
где:
(i) по меньшей мере один лиганд-связывающий аптамер находится в пределах 5'-нетранслируемой области (UTR) экспрессируемого полинуклеотида; и
(ii) в направлении 5'-3' полинуклеотида аптамера полиA по меньшей мере один лиганд-связывающий аптамер находится слева по ходу транскрипции от одного, двух или более богатых G участков.
50. Полинуклеотид по п. 49, отличающийся тем, что полинуклеотид аптамера полиA дополнительно содержит по меньшей мере один богатый U/UG участок, по меньшей мере один богатый G участок или оба варианта из по меньшей мере одного богатого U/UG участка и по меньшей мере одного богатого G участка.
51. Полинуклеотид по п. 50, отличающийся тем, что в направлении 5'-3' полинуклеотида аптамера полиA по меньшей мере один сигнал полиA находится слева по ходу транскрипции от по меньшей мере одного богатого U/UG участка.
52. Полинуклеотид по п. 50, отличающийся тем, что в направлении 5'-3' полинуклеотида аптамера полиA лиганд-связывающий аптамер находится слева по ходу транскрипции от одного, двух или более богатых U/UG участков.
53. Полинуклеотид по п. 50, отличающийся тем, что в направлении 5'-3' полинуклеотида аптамера полиA по меньшей мере один сигнал полиA находится слева по ходу транскрипции от по меньшей мере одного богатого G участка.
54. Клетка, содержащая полинуклеотид по любому из пп. 49-53, для модуляции или отслеживания экспрессии генов.
55. Клетка по п. 54, представляющая собой клетку млекопитающего.
56. Клетка по п. 55, причем клетка млекопитающего является клеткой человека.
57. Клетка по любому из пп. 54-56, находящаяся в организме человека.
58. Клетка, содержащая систему по любому из пп. 1-30, для модуляции экспрессии генов.
59. Клетка по п. 58, представляющая собой клетку млекопитающего.
60. Клетка по п. 59, причем клетка млекопитающего является клеткой человека.
61. Вектор, содержащий полинуклеотид по любому из пп. 49-53, для модуляции или отслеживания экспрессии генов.
62. Вектор по п. 61, представляющий собой вирусный вектор.
63. Вектор по п. 62, причем вирусный вектор представляет собой аденовирусный вектор, аденоассоциированный вирусный вектор, ретровирусный вектор или лентивирусный вектор.
64. Вектор по п. 61, представляющий собой плазмиду.
| Способ защиты переносных электрических установок от опасностей, связанных с заземлением одной из фаз | 1924 |
|
SU2014A1 |
| US 2011190386 A1, 09.08.2011 | |||
| Зарудная М | |||
| И., Полиаденилирование про-мРНК | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Образование З-концов мРНК позвоночных, Биополимеры и клетка, 2001, 17(2), стр.93-108. | |||

