Перекрестные ссылки на родственные заявки
Данная международная заявка PCT претендует на преимущество и приоритет от U.S. Provisional Patent Application No. 62/414,785, поданной 30 октября 2016 г. Содержание данной заявки включено сюда путем ссылки.
Область техники, к которой относится изобретение
Изобретение в общем касается антител против PD-L1, их вариантов или мутантов либо их антигенсвязывающих фрагментов, а также способов их применения при лечении раковых заболеваний у человека.
Уровень техники
Лиганд-1 запрограммированной смерти (PD-L1) и лиганд-2 запрограммированной смерти (PD-L2) экспрессируются на антигенпрезентирующих клетках, а также во многих раковых опухолях человека, причем было показано, что они подавляют активацию Т-клеток и секрецию цитокинов при связывании с PD-1 (Freeman et al., 2000; Latchman et al., 2001). В отличие от CTLA-4, PD-1 в основном функционирует в периферических тканях, где активированные T-клетки могут сталкиваться с иммуносупрессивными лигандами PD-L1 (B7-H1) и PD-L2 (B7-DC), экспрессированными в опухолевых и/или стромальных клетках (Flies et al., 2011; Topalian et al., 2012a). Ингибирование взаимодействия PD-1/PD-L1 опосредует сильную противоопухолевую активность на доклинических моделях (U.S. Pat. Nos. 8,008,449 and 7,943,743), а применение антител-ингибиторов взаимодействия PD-1/PD-L1 для лечения рака вошло в клинические испытания (Brahmer et al., 2010; Flies et al., 2011; Topalian et al., 2012b; Brahmer et al., 2012).
По-видимому, активация PD-L1 позволяет раковым опухолям уклоняться от иммунной системы хозяина. Несмотря на то, что многие ингибиторы PD-L1 разрабатываются в качестве иммунотерапии при онкологии и проявляют хорошие результаты при клинических испытаниях, существует потребность в разработке противоопухолевой терапии, направленной против PD-L1. Настоящее изобретение удовлетворяет эту и другие потребности.
Cущность изобретения
Изобретением предусмотрены антитела против PD-L1 и/или их антигенсвязывающие фрагменты. В некоторых воплощениях антитело против PD-L1 по изобретению, т.е. антитело PL1, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 35; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 41; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 44; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 51; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 59.
В других воплощениях антитело против PD-L1 по изобретению, т.е. антитело PL2, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 36; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 42; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 45; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 52; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 56; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 60.
В других воплощениях антитело против PD-L1 по изобретению, т.е. антитело PL3, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 37; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 41; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 46; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 53; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 61.
В других воплощениях антитело против PD-L1 по изобретению, т.е. антитело PL6, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 37; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 41; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 47; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 54; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 57; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 62.
В других воплощениях антитело против PD-L1 по изобретению, т.е. антитело PL8, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 38; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 43; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 48; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 51; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 58; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 63.
В других воплощениях антитело против PD-L1 по изобретению, т.е. антитело PL12, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 39; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 41; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 49; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 51; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 64.
В других воплощениях антитело против PD-L1 по изобретению, т.е. антитело PL15, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 40; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 41; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 50; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 53; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 61.
Изобретением также предусмотрены варианты и/или мутанты антител против PD-L1 либо их антигенсвязывающих фрагментов. В некоторых воплощениях вариант и/или мутант антитела против PD-L1 по изобретению, т.е. антитело PL2#3, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 65; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 42; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 71; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 52; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 56; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 77.
В других воплощениях вариант и/или мутант антитела против PD-L1 по изобретению, т.е. антитело PL3#7, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 37; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 41; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 72; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 75; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 78.
В других воплощениях вариант и/или мутант антитела против PD-L1 по изобретению, т.е. антитело PL3#7-19, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 66; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 68; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 73; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 75; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 79. В некоторых воплощениях мутанты антител против PD-L1 включают в себя CDR-L2, содержащий одну или несколько мутаций по сайтам N-гликозилирования.
В других воплощениях вариант и/или мутант антитела против PD-L1 по изобретению, т.е. антитело PL3#7-43, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 35; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 69; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 74; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 75; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 80. В некоторых воплощениях мутанты антител против PD-L1 включают в себя CDR-L2, содержащий одну или несколько мутаций по сайтам N-гликозилирования.
В других воплощениях вариант и/или мутант антитела против PD-L1 по изобретению, т.е. антитело PL3#7-54, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 67; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 70; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 72; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 75; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 76; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 81.
В других воплощениях вариант и/или мутант антитела против PD-L1 по изобретению, т.е. антитело PL2#4, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 94; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 42; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 95; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 52; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 56; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 96.
В других воплощениях вариант и/или мутант антитела против PD-L1 по изобретению, т.е. антитело PL2#5, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 97; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 42; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 98; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 52; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 56; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 99.
В других воплощениях вариант и/или мутант антитела против PD-L1 по изобретению, т.е. антитело PL2#39, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 100; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 42; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 95; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 52; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 56; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 101.
В других воплощениях вариант и/или мутант антитела против PD-L1 по изобретению, т.е. антитело PL3#1, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 37; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 106; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 107; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 108; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 109.
Аминокислотные последовательности вышеприведенных CDRs (CDR-L1, -L2, -L3; и CDR-H1, -H2, -H3) каждого из антител против PD-L1 и их вариантов/мутантов представлены ниже в табл. 1.
Таблица 1
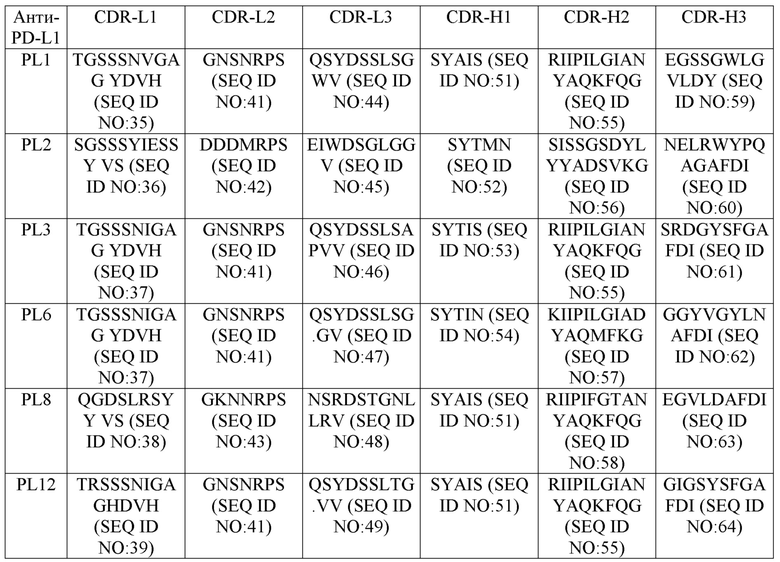
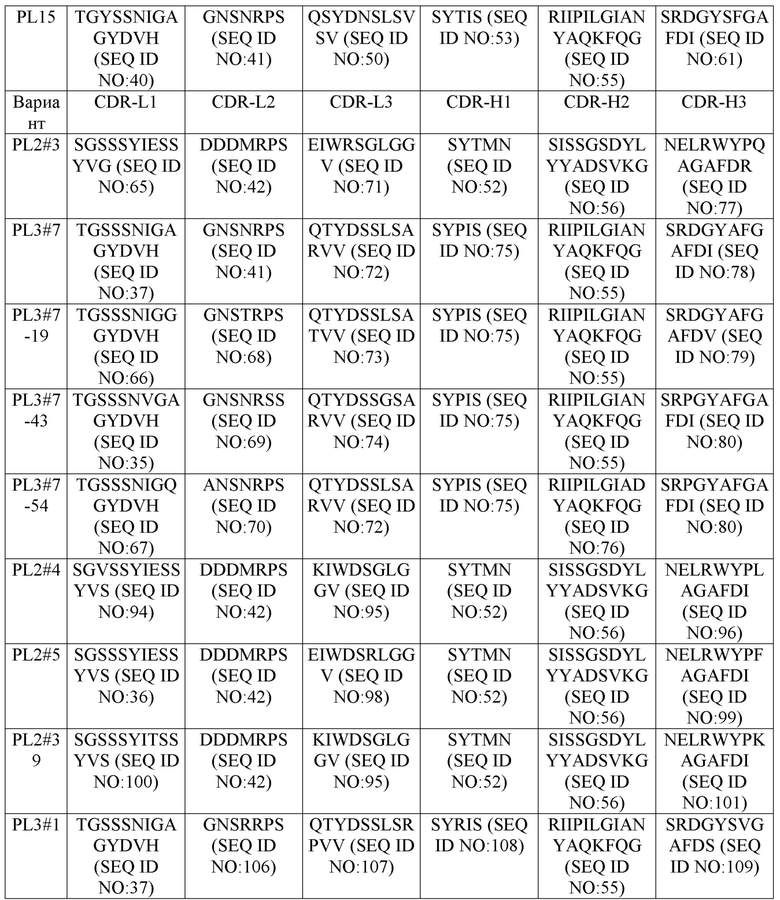
В некоторых воплощениях антитела против PD-L1 и их варианты содержат последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 35-40, 65-67, 94, 97 и 100; (2) CDR-L2, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 41-43, 68-70 и 106; и (3) CDR-L3, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 44-50, 71-74, 95, 98 и 107; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 51-54, 75 и 108; (2) CDR-H2, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 55-58 и 76; и (3) CDR-H3, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 59-64, 77-81, 96, 99, 101 и 109.
Также изобретением предусмотрены антитела против PD-L1, их варианты либо их антигенсвязывающие фрагменты, содержащие последовательность вариабельного домена легкой цепи (LC), включающую аминокислотную последовательность, приведенную в SEQ ID NOs: 1-7, 16, 20, 24, 28, 32, 83, 87, 91 или 103; и последовательность вариабельного домена тяжелой цепи (HC), включающую аминокислотную последовательность, приведенную в SEQ ID NOs: 8-14, 18, 22, 26, 30, 34, 85, 89, 93 или 105. Также предусмотрены последовательности нуклеиновых кислот, кодирующих эти домены LC и HC.
В некоторых воплощениях, согласно (или применительно к) любым из приведенных выше воплощений, антитела по изобретению содержат одну или несколько мутаций по сайтам N-гликозилирования в одном или нескольких CDR вариабельных доменов. Полученные при этом дегликозилированные антитела сохраняют те же функции, что и исходное недегликозилированное антитело.
В некоторых воплощениях, согласно (или применительно к) любым из приведенных выше воплощений, антитела содержат последовательность Fc из IgG человека. В некоторых воплощениях, согласно (или применительно к) любым из приведенных выше воплощений, антигенсвязывающий фрагмент выбирают из группы, состоящей из Fab, Fab′, F(ab′)2, одноцепочечных Fv (scFv), Fv-фрагментов, диател и линейных антител. В некоторых воплощениях, согласно (или применительно к) любым из приведенных выше воплощений, антитела представляют собой полиспецифичные антитела.
В некоторых воплощениях, согласно (или применительно к) любым из приведенных выше воплощений, антитела против PD-L1, их варианты или антигенсвязывающие фрагменты конъюгируют с терапевтическим средством. В некоторых воплощениях, согласно (или применительно к) любым из приведенных выше воплощений, антитела против PD-L1, их варианты или антигенсвязывающие фрагменты конъюгируют с меткой. В некоторых воплощениях, согласно (или применительно к) любым из приведенных выше воплощений, метка выбрана из группы, состоящей из радиоизотопов, флуоресцентных красителей и ферментов.
Изобретением предусмотрены выделенные молекулы нуклеиновой кислоты, кодирующие антитела против PD-L1, их варианты, мутанты или антигенсвязывающие фрагменты согласно (или применительно к) любым из приведенных выше воплощений. Также предусмотрены экспрессирующие векторы, кодирующие молекулы нуклеиновой кислоты согласно (или применительно к) любым из приведенных выше воплощений. Также предусмотрены клетки, содержащие экспрессирующие векторы согласно (или применительно к) любым из приведенных выше воплощений. Изобретением также предусмотрен способ получения антител, их вариантов или их антигенсвязывающих фрагментов, включающий культивирование клеток согласно (или применительно к) любым из приведенных выше воплощений и извлечение антител либо их антигенсвязывающих фрагментов из культуры клеток. В некоторых воплощениях, согласно (или применительно к) любым из приведенных выше воплощений, клетки представлены клетками млекопитающих. В некоторых воплощениях, согласно (или применительно к) любым из приведенных выше воплощений, клетки млекопитающих представлены клетками CHO. В некоторых воплощениях, согласно (или применительно к) любым из приведенных выше воплощений клетки представлены стабильной линией клеток млекопитающих. В некоторых воплощениях, согласно (или применительно к) любым из приведенных выше воплощений стабильная линия клеток млекопитающих представлена клетками линии CHO.
Изобретением предусмотрены композиции, содержащие антитела против PD-L1, их варианты, мутанты или антигенсвязывающие фрагменты согласно (или применительно к) любым из приведенных выше воплощений и фармацевтически приемлемый носитель.
Изобретением предусмотрены способы выявления белка PD-L1 в образцах от пациентов путем контактирования антител против PD-L1, их вариантов, мутантов либо антигенсвязывающих фрагментов согласно (или применительно к) любым из приведенных выше воплощений с образцом и детектирование антител против PD-L1, связавшихся с белком PD-L1. В некоторых воплощениях, согласно (или применительно к) любым из приведенных выше воплощений антитела против PD-L1, их варианты либо антиген-связывающие фрагменты применяются при иммуногистохимическом анализе (IHC) или анализе методом ELISA.
Также предусмотрен способ лечения рака у субъектов, включающий введение им эффективного количества композиции согласно (или применительно к) любым из приведенных выше воплощений. Также предусмотрены композиции, содержащие антитела против PD-L1, их варианты, мутанты или антигенсвязывающие фрагменты согласно (или применительно к) любым из приведенных выше воплощений, для применения при лечении рака. Предусмотрено и применение антител против PD-L1, их вариантов, мутантов или антигенсвязывающих фрагментов согласно (или применительно к) любым из приведенных выше воплощений при изготовлении лекарственных средств для лечения рака. В некоторых воплощениях согласно (или применительно к) любым из приведенных выше воплощений рак выбран из меланомы, рака головы и шеи, рака уротелия, рака молочной железы (напр., тройного отрицательного рака молочной железы, TNBC), рака желудка, классической лимфомы Ходжкина (cHL), неходжкинской лимфомы - первичной B-клеточной лимфомы средостения (NHL PMBCL), мезотелиомы, рака яичников, рака легких (напр., мелкоклеточного рака легких и немелкоклеточного рака легких (NSCLC), рака пищевода, карциномы носоглотки (NPC), рака желчных путей, колоректального рака, рака шейки матки, рака щитовидной железы и рака слюнных желез. В некоторых воплощениях, согласно (или применительно к) любым из приведенных выше воплощений, субъектам дополнительно вводят терапевтическое средство, выбранное из группы, состоящей из противоопухолевых средств, химиотерапевтических средств, средств, ингибирующих рост, и цитотоксических средств. В некоторых воплощениях, согласно (или применительно к) любым из указанных выше воплощений, субъекты дополнительно подвергаются лучевой терапии.
Краткое описание фигур
Фиг. 1A-1B. Совмещение аминокислотных последовательностей легкой цепи (A) и тяжелой цепи (B) отобранных лидирующих антител PL1, PL2, PL3, PL6, PL8, PL12, PL15 против PD-L1. Семь отобранных лидирующих антител с активностью связывания и блокирования PD-L1 человека при последующих анализах методами ELISA и проточной цитометрии идентифицировали при скрининге библиотеки фагового дисплея наивных Fab человека с помощью PD-L1_ECD-His человека. Эти отобранные последовательности Fab затем клонировали в Fc-остов IgG1 человека с мутацией N297A, получая полноразмерные антитела. Представлено совмещение последовательностей лидирующих антител, а жирным шрифтом и подчеркиванием отмечены участки, определяющие комплементарность (CDR).
Фиг. 2A-2B. Связывание выбранных антител с рекомбинантными слитыми с Fc белками PD-L1 человека (фиг. 2А) и активированными Т-клетками CD3+ (фиг. 2B). Выбранные антитела тестировали на связывание с рекомбинантными белками PD-L1 человека методом ELISA и с активированными Т-клетками CD3+ методом проточной цитометрии. В качестве положительного и отрицательного контроля использовали контрольное антитело против PD-L1 и контрольное антитело против PD-1, соответственно.
Фиг. 3. Блокирование выбранными антителами связывания PD-1 с PD-L1. Антитела против PD-L1 тестировали на способность блокировать связывание PD-1 с экспрессирующими PD-L1 клетками CHO-S методом проточной цитометрии. В качестве положительного и отрицательного контроля использовали контрольное антитело против PD-L1 и авастин, соответственно. Моноклональные антитела против PD-L1 блокировали связывание PD-1 с трансфецированными PD-L1 клетками CHO-S при измерении по средней интенсивности флуоресценции (MFI) после окрашивания.
Фиг. 4A-4B. Влияние лидирующих антител против PD-L1 на продукцию цитокинов и пролиферацию Т-клеток при реакции смешанных лейкоцитов (MLR). Моноклональные антитела против PD-L1 человека усиливают секрецию IFN-γ и пролиферацию Т-клеток при анализе по реакции смешанных лейкоцитов. В качестве положительного контроля использовали контрольное антитело против PD-L1 и контрольное антитело против PD-1. В качестве отрицательного контроля использовали авастин (антитело против VEGF). На фиг. 4A представлены гистограммы секреции IFN-γ, а на фиг. 4B представлены гистограммы пролиферации Т-клеток CD3+ при указанной концентрации антител.
Фиг. 5A-5B. Активность ингибирования роста опухолей у антител PL2 и PL3 на модели ксенотрансплантатов клеток A375/антиген-специфичных Т-клеток. Мышам (n = 4 на группу) подкожно прививали смесь клеток линии A375 меланомы человека и антиген-специфичных Т-клеток (Т-клетки:раковые клетки = 1:100). Начиная со дня 0, мышам два раза в неделю вводили внутрибрюшинно исследуемые антитела. На фиг. 5A и 5B представлены кривые роста опухолей у мышей, получавших PL2 и PL3, соответственно. Все точки данных представляют средние значения ± SEM.
Фиг. 6A-6B. Влияние вариантов PL2 и PL3 на продукцию цитокинов в реакции смешанных лейкоцитов (MLR). Варианты PL2 и PL3 усиливают секрецию IL-2 при анализе по реакции смешанных лейкоцитов. В качестве антител положительного и отрицательного контроля использовали контрольное антитело против PD-L1 и авастин, соответственно. На фиг. 6A представлены гистограммы секреции IL-2, вызванной исходным PL2 и его вариантами, а на фиг. 6B представлены гистограммы секреции IL-2 в MLR при указанной концентрации исходного антитела PL3 и вариантов.
Фиг. 7A-7B. Активность ингибирования роста опухолей у лучших вариантов PL2 и PL3 на модели ксенотрансплантатов клеток A375/антиген-специфичных Т-клеток. Мышам (n = 4 на группу) прививали подкожно смесь клеток линии A375 меланомы человека и антиген-специфичных Т-клеток (Т-клетки:раковые клетки = 1:100). Начиная со дня 0, мышам два раза в неделю вводили внутрибрюшинно исследуемые антитела. На фиг. 7A представлены кривые роста опухолей у мышей, получавших PL2#3 и PL3#7. На фиг. 7B представлены объемы отдельных опухолей на 35-й день. Все точки данных представляют средние значения ± SEM.
Фиг. 8A-8D. Эффективность различных IgG-форм PL2#3 и PL3#7 при реакции смешанных лейкоцитов (MLR). Исследовали различные IgG-формы PL2#3 и PL3#7 методом MLR. В качестве антитела отрицательного контроля использовали авастин. На фиг. 8A и 8B представлены гистограммы секреции IFN-γ и IL-2, соответственно, индуцированной исследуемыми антителами. На фиг. 8C представлены гистограммы пролиферации Т-клеток CD4+ при различных концентрациях антител. На фиг. 8D представлены гистограммы пролиферации Т-клеток CD8+ при указанных концентрациях антител.
Фиг. 9A-9B. Активность ингибирования роста опухолей у различных IgG-изотипов PL2 и PL3 на модели ксенотрансплантатов клеток A375/антиген-специфичных Т-клеток. На фиг. 9A представлены объемы отдельных опухолей у мышей (n = 4 на группу), получавших различные IgG-формы PL2#3, на 32-й день. На фиг. 9B представлены объемы отдельных опухолей у мышей (n = 4 на группу), получавших PL3#7 различных подклассов IgG, на 32-й день.
Фиг. 10. Активность ингибирования роста опухолей у антител PL2#3-mtIgG1 и PL3#7-mtIgG1 на мышах hPD1 KI. Мышам со вставленным PD-1 человека (hPD1 KI) (n = 6 на группу) подкожно прививали клетки MC38-huPD-L1 (MC38, трансфецированные PD-L1 человека). Обработку антителами начинали при достижении объема опухолей примерно 86 мм3. Исследуемые антитела вводили мышам внутрибрюшинно два раза в неделю в течение 3 недель. Все точки данных представляют средние значения ± SEM.
Фиг. 11A-11B. Совмещение аминокислотных последовательностей легкой цепи (фиг. 11A) (SEQ ID NOs 136-140, соответственно, в порядке появления) и тяжелой цепи (фиг. 11B) (SEQ ID NOs 141-145, соответственно, в порядке появления) у лучших вариантов антител против PD-L1. Лучший вариант PL2 (PL2#3), лучший вариант PL3 (PL3#7) и лучшие варианты PL3#7 (PL3#7-19, -43, -54) с самым высоким сродством и наилучшей функциональной активностью получали в экспериментах in vitro по созреванию сродства на основе фагового дисплея. В общем, проводили три раунда пэннинга, используя биотинилированный hPD-L1-His, конъюгированный с покрытыми стрептавидином магнитными шариками Dynabeads® M-280. Затем Fabs лучших вариантов подвергали скринингу методом ELISA и клонировали в Fc-остов IgG1 человека с мутацией N297A, получая полноразмерные антитела. Представлено совмещение последовательностей лучших вариантов, а жирным шрифтом и подчеркиванием отмечены CDR (определяющие комплементарность участки).
Фиг. 12A-12B. Эффекты вариантов PL3#7 на продукцию цитокинов и пролиферацию Т-клеток при реакции смешанных лейкоцитов (MLR). Модифицированные варианты PL3#7 усиливают секрецию IFN-γ и пролиферацию Т-клеток CD8+ при анализе по реакции смешанных лейкоцитов. В качестве положительного и отрицательного контроля использовали контрольное антитело против PD-L1 и авастин, соответственно. На фиг. 12A представлены гистограммы секреции IFN-γ, а на фиг. 12B представлены гистограммы пролиферации Т-клеток CD8+ при указанной концентрации антител.
Фиг. 13A-13C. Связывание вариантов PL2#3 и PL3#7 с клеточной поверхностью экспрессирующих PD-L1 клеток. Варианты PL3#7 тестировали на связывание с клеточной поверхностью активированных Т-клеток (фиг. 13A), клеток линии A375 меланомы человека (фиг. 13B) и клеток NSCLC линии NCI-H292 человека (фиг. 13C) методом проточной цитометрии. В качестве положительного и отрицательного контроля использовали контрольное антитело против PD-L1 и антитело HLX01 (mAb против CD20), соответственно.
Фиг. 14A-14B. Активность ингибирования роста опухолей у вариантов PL2#3 и PL3#7 на модели ксенотрансплантатов NCI-H292/PBMC. Мышам (n = 4 на группу) подкожно прививали смесь клеток NSCLC линии NCI-H292 человека и свежевыделенных PBMC человека (раковые клетки (T): PBMC (E) = 3:1). Начиная со дня 0, мышам два раза в неделю вводили внутрибрюшинно антитела против PD-L1. На фиг. 14A представлены кривые роста опухолей. На фиг. 14B представлены объемы отдельных опухолей на 28-й день. Все точки данных представляют средние значения ± SEM.
Фиг. 15A-15B. Активность ингибирования роста опухолей у вариантов PL2#3 и PL3#7 на модели ксенотрансплантатов клеток A375/антиген-специфичных Т-клеток. Мышам (n = 4 на группу) подкожно прививали смесь клеток линии A375 меланомы человека и антиген-специфичных Т-клеток (Т-клетки (E):раковые клетки (T) = 1:100). Начиная со дня 0, мышам два раза в неделю вводили внутрибрюшинно исследуемые антитела. На фиг. 15A представлены кривые роста опухолей у получавших mAb мышей. На фиг. 15B представлены объемы отдельных опухолей на 31-й день. Все точки данных представляют средние значения ± SEM.
Фиг. 16. Перекрестное связывание моноклональных антител против PD-L1 человека с клетками меланомы мыши. Варианты PL2#3 и PL3#7 тестировали на связывание с экспрессирующими PD-L1 клетками меланомы мыши (B16-F10) методом проточной цитометрии. В качестве положительного и отрицательного контроля использовали контрольные антитела против PD-L1, соответственно.
Фиг. 17A-17B. Перекрестное связывание PD-L1 макаки-крабоеда (cynomolgus) с вариантами PL2#3 (фиг. 17A) и PL3#7 (фиг. 17B). Варианты PL2#3 и PL3#7 тестировали на связывание с рекомбинантными слитыми с Fc белками PD-L1_ECD макаки-крабоеда методом ELISA. В качестве отрицательного контроля использовали HLX01 (mAb против CD20). Все точки данных представляют средние значения из трех повторов ± SD.
Фиг. 18. Совмещение аминокислотных последовательностей вариабельных областей легкой цепи у дегликозилированных версий варианта PL3#7-19 и варианта PL3#7-43 (SEQ ID NOS 20, 24, 111, 113, 115, 28, 117 и 119, соответственно, в порядке появления). Представлено совмещение последовательностей легких цепей этих дегликозилированных вариантов, а жирным шрифтом и подчеркиванием отмечены CDR (определяющие комплементарность участки). Тяжелые цепи у них были неизменными и идентичны своим исходным вариантам (совмещение последовательностей не приводится).
Фиг. 19A-19B. Связывание дегликозилированных версий варианта PL3#7-19 (фиг. 19A) и варианта PL3#7-43 (фиг. 19B) на целых клетках. Активность связывания на целых клетках у полученных трех дегликозилированных вариантов для PL3#7-19 (т.е. PL3#7-19 деглико1, деглико2, деглико3) и двух дегликозилированных вариантов для PL3#7-43 (т.е. PL3#7-43 деглико1, деглико2) с трансфецированными PD-L1 клетками CHO-S определяли методом проточной цитометрии. Все исследованные варианты дегликозилированных антител были в Fc-остове IgG1 человека с мутацией N297A. В качестве отрицательного контроля использовали наше собственное антитело против PD-1 (т.е. HLX10).
Фиг.20A-20B. Эффективность различных IgG-изотипов PL3#7-19 деглико1 и PL3#7-43 деглико2 в реакции смешанных лейкоцитов (MLR). Тестировали различные IgG-изотипы PL3#7-19 деглико1 и PL3#7-43 деглико2 по методу MLR. В качестве антител отрицательного и положительного контроля использовали HLX04 и HLX20 (PL2#3), соответственно. Представлены гистограммы секреции IFN-γ (фиг. 20A) и IL-2 (фиг. 20B), индуцированной исследуемыми антителами.
Фиг. 21A-21D. Активность ингибирования роста опухолей у дегликозилированных вариантов PL3#7-19 и PL3#7-43 на модели ксенотрансплантатов клеток A375/антиген-специфичных Т-клеток. Мышам (n = 5 на группу) прививали подкожно смесь клеток линии A375 меланомы человека и антиген-специфичных Т-клеток (Т-клетки: раковые клетки = 1:100). Начиная со дня 0, мышам два раза в неделю вводили внутрибрюшинно исследуемые антитела. На фиг. 21A и 21C представлен рост опухолей у мышей, получавших дегликозилированные варианты PL3#7-19 и PL3#7-43, соответственно. На фиг. 21B и 21D, соответственно, представлены объемы отдельных опухолей на 28-й день. Все точки данных представляют средние значения ± SEM.
Фиг. 22A-22B. Активность ингибирования роста опухолей у mAb против PD-L1 плюс mAb против VEGF на модели ксенотрансплантатов NSCLC у мышей. Мышам (n = 5 на группу) подкожно прививали смесь клеток NSCLC линии NCI-H292 человека и свежевыделенных PBMC человека (раковые клетки:PBMC = 3:1). Начиная со дня 1, мышам два раза в неделю вводили внутрибрюшинно антитела против PD-L1 (PL2#3) и против VEGF (HLX04). На фиг. 22A представлены кривые роста опухолей. На фиг. 22B представлены объемы отдельных опухолей на 21-й день. Все точки данных представляют средние значения ± SEM.
Раскрытие сущности изобретения
Настоящим изобретением предусмотрены новые антитела против PD-L1, их варианты, мутанты и/или антигенсвязывающие фрагменты. Авторы изобретения неожиданно обнаружили, что определенные антитела против PD-L1 и их варианты и/или мутанты по сродству усиливают секрецию Т-клетками IL-2 и IFN-γ и пролиферацию Т-клеток CD4+ и CD8+. Описанные здесь антитела против PD-L1 также проявляют повышение эффективности и/или противоопухолевой активности по сравнению с некоторыми эталонными моноклональными антителами против PD-L1, используемыми для лечения рака.
Также предусмотрены иммуноконъюгаты, нуклеиновые кислоты, кодирующие новые антитела против PD-L1, их варианты по сродству либо их антигенсвязывающие фрагменты, как описано здесь, и композиции (типа фармацевтических композиций). Изобретением также предусмотрены способы применения новых антител против PD-L1, их вариантов и/или мутантов по сродству либо их антигенсвязывающих фрагментов для выявления PD-L1 в образцах (типа образцов in vivo или ex vivo), композиции, содержащие такие антитела, их варианты и/или мутанты либо антигенсвязывающие фрагменты, для применения при лечении рака, и применение таких антител, их вариантов или антиген-связывающих фрагментов при изготовлении лекарственных средств для лечения рака.
Определения
В настоящем изобретении термин “лечение” или “терапия” означает способ получения полезных или желательных результатов, включая клинические результаты. Для целей данного изобретения полезные или желательные клинические результаты включают, без ограничения, одно или несколько из следующего: ослабление одного или нескольких симптомов, возникающих при заболевании, уменьшение степени заболевания, стабилизацию заболевания (напр., предотвращение или замедление ухудшения заболевания), предотвращение или замедление распространения (напр., метастазирования) заболевания, предотвращение или замедление рецидива заболевания, задержка или замедление прогрессирования заболевания, улучшение состояния заболевания, обеспечение ремиссии (частичной или полной) заболевания, уменьшение дозы одного или нескольких других лекарственных средств, необходимых для лечения заболевания, замедление прогрессирования заболевания, повышение или улучшение качества жизни, усиление набора веса и/или продление выживаемости. “Лечение” также охватывает уменьшение патологических последствий рака (таких, к примеру, как объем опухоли). Представленные здесь способы предусматривают какой-либо один или несколько из этих аспектов лечения.
Термины “повторение”, “рецидив” или “возврат” означают возвращение рака или заболевания после клинической оценки его исчезновения. Диагноз удаленного метастазирования или местного повторения можно считать рецидивом.
Термин “рефрактерный” или “резистентный” означает такой рак или заболевание, которое не поддается лечению.
Термин “вспомогательная терапия” означает такое лечение, которое проводится после первичной терапии, обычно хирургии. Вспомогательная терапия рака или заболевания может включать иммунотерапию, химиотерапию, лучевую терапию или гормональную терапию.
Термин “поддерживающая терапия” означает плановое повторное лечение, которое проводится для поддержания эффекта предыдущего лечения. Поддерживающая терапия часто применяется для того, чтобы сохранить рак в стадии ремиссии или продлить ответ на определенную терапию независимо от прогрессирования заболевания.
Термин “инвазивный рак” означает такой рак, который вышел за пределы той ткани, из которой он начал проникать в нормальные окружающие ткани. Инвазивный рак может быть или не быть метастатическим.
Термин “неинвазивный рак” означает очень ранний рак или такой рак, который не вышел за пределы ткани происхождения.
Термин “выживаемость без прогрессирования” в онкологии относится к такому промежутку времени во время и после лечения, когда рак не растет. Выживаемость без прогрессирования включает промежуток времени, при котором пациенты испытывали полный или частичный ответ, а также промежуток времени, при котором заболевание у пациентов было стабильным.
Термин “прогрессирующее заболевание” в онкологии может означать рост опухоли более чем на 20 процентов с момента начала лечения - либо вследствие увеличения массы, либо из-за распространения опухоли.
“Нарушение” означает такое состояние, которое выиграет от лечения с помощью антитела. Например, млекопитающие, которые страдают или нуждаются в профилактике от аномальной активности PD-L1. Это охватывает хронические и острые расстройства или заболевания, включая такие патологические состояния, которые предрасполагают млекопитающих к данному расстройству. Неограничительные примеры заболеваний, подлежащих лечению, включают рак (как-то рак головы и шеи, рак горла, колоректальный рак, рак легких и т.д.).
“Опухоль” в настоящем изобретении относится к росту и пролиферации всех неопластических клеток, будь то злокачественных или доброкачественных, а также ко всем предраковым и раковым клеткам и тканям.
Термин “антитело” применяется в самом широком смысле и конкретно охватывает, к примеру, отдельные моноклональные антитела (включая антитела-агонисты, антагонисты и нейтрализующие антитела), композиции антител с полиэпитопной специфичностью, поликлональные антитела, одноцепочечные антитела и фрагменты антител (см. ниже), если только они специфически связывают нативный полипептид и/или проявляют биологическую активность или иммунологическую активность по изобретению. Согласно одному воплощению, антитело связывается с олигомерной формой целевого белка, напр., тримерной формой. Согласно другому воплощению, антитело специфически связывается с белком, причем связывание может ингибироваться моноклональным антителом по изобретению (напр., депонированным антителом по изобретению и т.д.). Выражение “функциональный фрагмент или аналог” антитела означает соединение, обладающее качественной биологической активностью, общей с тем антителом, с которым оно сопоставляется. Например, функциональный фрагмент или аналог антитела по изобретению может быть таким, который может специфически связываться с PD-L1. В одном воплощении антитело может предотвращать или существенно уменьшать способность PD-L1 индуцировать пролиферацию клеток.
“Выделенное антитело” означает такое антитело, которое было идентифицировано и отделено и/или извлечено из компонента своего природного окружения. Загрязняющие компоненты природного окружения представляют собой материалы, которые могут помешать диагностическому или терапевтическому применению антитела, и могут включать ферменты, гормоны и другие белковые или небелковые вещества. В предпочтительных воплощениях антитело должно быть очищено (1) до содержания антител более 95% масс. при определении методом Лоури, наиболее предпочтительно более 99% масс., (2) до степени, достаточной для получения по меньшей мере 15 аминокислотных остатков N-концевой или внутренней последовательности при использовании секвенатора с вращающейся чашкой, или (3) до гомогенности по SDS-PAGE в восстановительных или невосстановительных условиях с окрашиванием Кумасси синим или, предпочтительно, серебром. Выделенные антитела включают антитела in situ в рекомбинантных клетках, так как не будет присутствовать по меньшей мере один компонент природного окружения антитела. Обычно, однако, выделенные антитела получают по меньшей мере за одну стадию очистки.
Основная 4-цепочечная единица антител представляет собой гетеротетрамерный гликопротеин, состоящий из двух идентичных легких (L) цепей и двух идентичных тяжелых (H) цепей (антитела типа IgM состоят из 5 основных гетеротетрамерных единиц вместе с дополнительным полипептидом, называемым J-цепью, поэтому они содержат 10 сайтов связывания антигена, тогда как секретируемые антитела типа IgA могут полимеризоваться с образованием поливалентных агрегатов, содержащих 2-5 основных 4-цепочечных звеньев вместе с J-цепью). В случае IgG обычно 4-цепочечный комплект составляет около 150 000 дальтон. Каждая L-цепь соединяется с H-цепью одной ковалентной дисульфидной связью, а две H-цепи соединяются друг с другом одной или несколькими дисульфидными связями в зависимости от изотипа H-цепи. Каждая H- и L-цепь также содержит регулярно расположенные внутрицепочечные дисульфидные мостики. Каждая Н-цепь имеет на N-конце вариабельный домен (VH), за которым следуют три константных домена (СН) для каждой из α- и γ-цепей или четыре домена СН для изотипов µ и ε. Каждая L-цепь имеет на N-конце вариабельный домен (VL), за которым следует константный домен (CL) на другом конце. VL совмещается с VH, а CL совмещается с первым константным доменом тяжелой цепи (CH1). Считается, что определенные аминокислотные остатки образуют поверхность раздела между вариабельными доменами легкой цепи и тяжелой цепи. При спаривании VH и VL образуется один антигенсвязывающий сайт. Насчет структуры и свойств различных классов антител см., напр., Basic and Clinical Immunology, 8th edition, Daniel P. Stites, Abba I. Terr and Tristram G. Parslow (eds.), Appleton & Lange, Norwalk, CT, 1994, стр. 71 и глава 6.
L-цепь любого вида позвоночных может относиться к одному из двух четко различимых типов, называемых каппа и лямбда, исходя из аминокислотных последовательностей их константных доменов. В зависимости от аминокислотной последовательности константного домена тяжелых цепей (СН) иммуноглобулины могут относиться к разным классам или изотипам. Существует пять классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, у которых тяжелые цепи обозначаются как α, δ, γ, ε и μ, соответственно. Классы γ и α дополнительно подразделяются на подклассы на основании сравнительно мелких различий в последовательности и функции СН, напр., у человека экспрессируются следующие подклассы: IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2.
Термин “вариабельный” означает то, что определенные сегменты вариабельных доменов сильно различаются по последовательности между антителами. V-домен опосредует связывание антигена и определяет специфичность конкретного антитела к его конкретному антигену. Однако вариабельность распределяется не одинаково по длине вариабельных доменов из 110 аминокислот. Напротив, V-области состоят из относительно инвариантных отрезков, называемых каркасными участками (FR) из 15-30 аминокислот, разделенных более короткими участками чрезвычайной вариабельности, называемыми “гипервариабельными участками” длиной в 9-12 аминокислот. Вариабельные домены нативных тяжелых и легких цепей содержат по четыре FR, в основном принимающие конфигурацию β-листа и соединенные тремя гипервариабельными участками, которые образуют петли, соединяющие и в некоторых случаях входящие в состав структуры β-листа. Гипервариабельные участки в каждой цепи удерживаются в непосредственной близости под действием FR и вместе с гипервариабельными участками из другой цепи участвуют в образовании антигенсвязывающего сайта антител (см. Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD (1991)). Константные домены не принимают прямого участия в связывании антитела с антигеном, но проявляют различные эффекторные функции типа участия антител в антителозависимой клеточной цитотоксичности (ADCC).
В настоящем изобретении термин “CDR” или “ определяющий комплементарность участок” служит для обозначения несмежных сайтов сочетания с антигеном, находящихся в вариабельных областях полипептидов тяжелой и легкой цепи. Эти конкретные участки были описаны в Kabat et al., J. Biol. Chem. 252:6609-6616 (1977); Kabat et al., U.S. Dept. of Health and Human Services, “Sequences of proteins of immunological interest” (1991); Chothia et al., J. Mol. Biol. 196:901-917 (1987); и MacCallum et al., J. Mol. Biol. 262:732-745 (1996), причем их определения включают перекрывающиеся или подмножества аминокислотных остатков при сравнении друг с другом. Тем не менее, применение этих определений в отношении CDR у антител или привитых антител либо их вариантов должно быть в рамках определения и применения этого термина здесь. Аминокислотные остатки, которые составляют участки CDR при определении по каждой из приведенных выше ссылок, представлены ниже в табл. 2 для сравнения.
Таблица 2
Термин “моноклональное антитело” в настоящем изобретении означает антитело, полученное из популяции практически однородных антител, то есть моноклональные антитела, составляющие популяцию, являются идентичными, за исключением возможных естественных мутаций, которые могут присутствовать в незначительном количестве. Моноклональные антитела высокоспецифичны и направлены против одного антигенного сайта. Кроме того, в отличие от препаратов поликлональных антител, которые включают различные антитела, направленные против разных детерминант (эпитопов), каждое моноклональное антитело направлено против единственной детерминанты на антигене. В дополнение к их специфичности, моноклональные антитела имеют преимущество в том, что их можно синтезировать без примеси других антител. Модификатор ”моноклональное” не следует понимать как требующий получения антитела каким-либо конкретным методом. Напр., моноклональные антитела, применимые в настоящем изобретении, могут быть получены гибридомным методом, впервые описанным в Kohler et al., Nature, 256:495 (1975), или же методами рекомбинантной ДНК в бактериальных, эукариотических клетках животных или растений (напр., см. U.S. Patent No. 4,816,567). “Моноклональные антитела” также могут быть выделены из фаговых библиотек антител по методикам, описанным в Clackson et al., Nature, 352: 624-628 (1991); Marks et al., J. Mol. Biol., 222: 581-597 (1991); а также, к примеру, в приведенных ниже примерах.
Моноклональные антитела включают и “химерные” антитела, у которых часть тяжелой и/или легкой цепи идентична или гомологична соответствующим последовательностям у антител, происходящих из определенного вида или принадлежащих к определенному классу или подклассу антител, тогда как остальная часть цепи идентична или гомологична соответствующим последовательностям у антител, происходящих из другого вида или принадлежащих к другому классу или подклассу антител, а также фрагменты таких антител, если только они проявляют биологическую активность по изобретению (см. U.S. Patent No. 4,816,567; и Morrison et al., Proc. Natl. Acad. Sci. USA, 81:6851-6855 (1984)). Представляющие интерес химерные антитела включают “приматизированные” антитела, содержащие антигенсвязывающие последовательности вариабельного домена, происходящие из других приматов (напр., обезьян Старого Света или человекообразных обезьян и т.д.), и последовательности константных областей человека.
“Интактными” являются такие антитела, которые содержат антигенсвязывающий сайт, а также CL и по крайней мере константные домены тяжелой цепи, CH1, CH2 и CH3. Константные домены могут быть представлены константными доменами с нативной последовательностью (напр., константными доменами с нативной последовательностью человека) или вариантами их аминокислотных последовательностей. Предпочтительно интактные антитела обладают одной или несколькими эффекторными функциями.
“Фрагменты антител” содержат часть интактного антитела, предпочтительно антигенсвязывающий участок или вариабельную область интактного антитела. Примеры фрагментов антител включают фрагменты Fab, Fab′, F(ab′)2 и Fv; диатела; линейные антитела (см. U.S. Patent No. 5,641,870, пример 2; Zapata et al., Protein Eng. 8(10): 1057-1062 (1995); одноцепочечные молекулы антител; и мультиспецифичные антитела, состоящие из фрагментов антител. Выражение ”линейные антитела” обычно относится к антителам, описанным в Zapata et al., Protein Eng., 8 (10): 1057-1062 (1995). Вкратце, эти антитела содержат пару тандемных Fd-сегментов (VH-CH1-VH-CH1), которые вместе с комплементарными полипептидами легкой цепи образуют пару антигенсвязывающих участков.
При расщеплении антител папаином образуются два идентичных антигенсвязывающих фрагмента, называемых “Fab”-фрагментами, и остаточный “Fc”-фрагмент, название которого отражает способность к легкой кристаллизации. Fab-фрагмент состоит из всей L-цепи вместе с вариабельной областью H-цепи (VH) и первым константным доменом одной тяжелой цепи (CH1). Каждый Fab-фрагмент является моновалентным в отношении связывания антигена, то есть он содержит один антигенсвязывающий сайт. Обработка антител пепсином дает один большой F(ab′)2-фрагмент, который примерно соответствует двум связанным дисульфидом Fab-фрагментам и является бивалентным по активности связывания антигена, а также способен перекрестно связывать антиген. Fab′-фрагменты отличаются от Fab-фрагментов наличием нескольких дополнительных остатков на карбоксильном конце домена CH1, включая один или несколько цистеинов из шарнирной области антитела. А Fab′-SH означает такой Fab′, у которого остатки цистеина в константных доменах содержат свободные тиоловые группы. F(ab′)2-фрагменты антител первоначально получали в виде пары Fab′-фрагментов, содержащих шарнирные цистеины между ними. Известны и другие химические конъюгаты фрагментов антител.
Fc-фрагмент содержит C-концевые части обеих H-цепей, удерживаемые вместе дисульфидами. Эффекторные функции антител определяются последовательностями Fc-области, которая также распознается Fc-рецепторами (FcR), находящимися на некоторых типах клеток.
“Вариант Fc-области” имеет аминокислотную последовательность, которая отличается от нативной последовательности Fc-области наличием по меньшей мере одной “аминокислотной модификации”, как определено здесь. Предпочтительно вариант Fc-области имеет по меньшей мере одну аминокислотную замену по сравнению с нативной последовательностью Fc-области или с Fc-областью исходного полипептида, напр., от одной до десяти аминокислотных замен, предпочтительно от одной до пяти аминокислотных замен в нативной последовательности Fc-области или в Fc-области исходного полипептида. В одном воплощении вариант Fc-области по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 99% гомологичен нативной последовательности Fc-области. В соответствии с другим воплощением вариант Fc-области по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 99% гомологичен Fc-области исходного полипептида.
Термин “полипептид, содержащий Fc-область” относится к полипептидам типа антител или иммуноадгезинов (см. определения, приведенные в другом месте), которые содержат Fc-область. Из Fc-области может быть удален С-концевой лизин (остаток 447 по системе нумерации EU), к примеру, во время очистки полипептида или путем рекомбинантной инженерии нуклеиновой кислоты, кодирующей полипептид. Соответственно, композиции, содержащие полипептиды, включая антитела, содержащие Fc-область по изобретению, могут содержать популяции полипептидов с удалением всех остатков K447, популяции полипептидов без удаления остатков K447 или же популяции полипептидов, содержащие смесь полипептидов с остатком K447 и без него.
“Эффекторные функции” антител относятся к таким биологическим активностям, которые относятся к Fc-области (Fc-области с нативной последовательностью или Fc-области с вариантом аминокислотной последовательности) антител и зависят от изотипа антител. Примеры эффекторных функций антител: связывание C1q и комплемент-зависимая цитотоксичность; связывание с Fc-рецептором; антителозависимая клеточная цитотоксичность (ADCC); фагоцитоз; понижающая регуляция рецепторов на клеточной поверхности; и активация B-клеток. “Fc-область с нативной последовательностью” содержит аминокислотную последовательность, идентичную аминокислотной последовательности природной Fc-области. Примеры последовательностей Fc описаны, к примеру, но без ограничения, в Kabat et al., Sequences of Immunological Interest. 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991).
“Fv” - это минимальный фрагмент антител, содержащий полный сайт распознавания и связывания антигена. Этот фрагмент состоит из димера одной вариабельной области тяжелой цепи и одной вариабельной области легкой цепи, связанных прочными нековалентными связями. При сворачивании этих двух доменов образуется шесть гипервариабельных петель (по 3 петли из каждой цепи H и L), которые предоставляют аминокислотные остатки для связывания антигена и придают специфичность связыванию антигена с антителом.
Тем не менее, даже один вариабельный домен (то есть половинка Fv, содержащая только три CDRs, специфичные к антигену) обладает способностью к распознаванию и связыванию антигена, хотя и с меньшим сродством, чем полный сайт связывания.
“Одноцепочечный Fv”, также сокращенно “sFv” или “scFv”, представляет собой фрагменты антител, содержащие домены VH и VL антител, соединенные в одну полипептидную цепь. Предпочтительно полипептид sFv также содержит полипептидный линкер между доменами VH и VL, который позволяет sFv образовывать требуемую структуру для связывания антигена. Насчет обзора по sFv см. Pluckthun in The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore, eds., Springer-Verlag, New York, pp. 269-315 (1994); Borrebaeck 1995, infra.
Термин “диатела” относится к небольшим фрагментам антител, полученным при конструировании фрагментов sFv (см. предыдущий абзац) с короткими линкерами (5-10 остатков) между доменами VH и VL, при этом происходит межцепочечное, а не внутрицепочечное спаривание V-доменов, в результате чего образуется бивалентный фрагмент, то есть фрагмент, имеющий два антигенсвязывающих сайта. Биспецифичные диатела представляют собой гетеродимеры двух “перекрестных” фрагментов sFv, у которых домены VH и VL из двух антител находятся на разных полипептидных цепях. Диатела описаны более подробно, к примеру, в ЕР 404097; WO 93/11161; и Hollinger et al., Proc. Natl. Acad. Sci. USA, 90: 6444-6448 (1993).
“Гуманизованные” формы нечеловеческих антител (напр., грызунов) представляют собой химерные антитела, содержащие минимальную последовательность, полученную из нечеловеческого антитела. По большей части гуманизованные антитела представляют собой иммуноглобулины человека (антитела-реципиенты), у которых остатки в гипервариабельной области реципиента заменены на остатки из гипервариабельной области антитела другого вида, чем человек (донорского антитела), типа мыши, крысы, кролика или других приматов, помимо человека, обладающего требуемой специфичностью, сродством и способностью. В некоторых случаях остатки в каркасной области (FR) иммуноглобулинов человека заменяют на соответствующие остатки из другого вида.
Кроме того, гуманизованные антитела могут содержать остатки, которые не встречаются в реципиентном антителе или в донорском антителе. Эти модификации делают для дальнейшего улучшения характеристик антител. В общем, гуманизованное антитело должно содержать практически весь по меньшей мере один, а обычно и два вариабельных домена, у которых все или практически все гипервариабельные петли соответствуют таковым у иммуноглобулина не человека, а все или практически все FRs происходят из последовательности иммуноглобулина человека. Гуманизованное антитело необязательно также должно содержать по меньшей мере часть константной области (Fc) иммуноглобулина, как правило, иммуноглобулина человека. Более подробно см. Jones et al., Nature 321: 522-525 (1986); Riechmann et al., Nature 332: 323-329 (1988); и Presta, Curr. Op. Struct. Biol. 2: 593-596 (1992).
“Степень идентичности (в %)” или “гомологичности аминокислотных последовательностей” в отношении приведенных здесь последовательностей полипептидов и антител определяется как процент таких аминокислотных остатков в рассматриваемой последовательности, которые идентичны аминокислотным остаткам у сравниваемого полипептида после совмещения последовательностей, рассматривая консервативные замены как часть идентичности последовательностей. Совмещение для определения степени идентичности аминокислотных последовательностей может проводиться различными способами, известными специалистам в данной области, к примеру, с помощью общедоступного компьютерного программного обеспечения типа программы BLAST, BLAST-2, ALIGN или Megalign (DNASTAR). Специалисты могут определить соответствующие параметры для измерения степени совмещения, включая любые алгоритмы, необходимые для максимального совмещения по всей длине сравниваемых последовательностей. Однако для целей настоящего изобретения степень идентичности аминокислотных последовательностей в % получают с помощью компьютерной программы для сравнения последовательностей ALIGN-2. Компьютерная программа для сравнения последовательностей ALIGN-2 была разработана в Genentech, Inc., а исходный код был подан вместе с пользовательской документацией в U.S. Copyright Office, Washington D.C., 20559, где он зарегистрирован за номером U.S. Copyright Registration No. TXU510087. Программа ALIGN-2 общедоступна через Genentech, Inc., South San Francisco, Калифорния. Программа ALIGN-2 должна быть скомпилирована для операционной системы UNIX, предпочтительно цифровой системы UNIX V4.0D. Все параметры сравнения последовательностей устанавливаются самой программой ALIGN-2 и не меняются.
Термины “Fc-рецептор” или “FcR” применяются для описания рецептора, который связывается с Fc-областью антител. В одном воплощении FcR по изобретению связывает антитела типа IgG и включает рецепторы подклассов FcγRI, FcγRII и FcγRIII, в том числе аллельные варианты и альтернативные сплайс-формы этих рецепторов. Рецепторы FcγRII включают FcγRIIA (“активирующий рецептор”) и FcγRIIB (“ингибирующий рецептор”), которые имеют близкие аминокислотные последовательности, но отличаются главным образом по цитоплазматическим доменам. Активирующий рецептор FcγRIIA содержит активационный мотив иммунорецептора на основе тирозина (ITAM) в своем цитоплазматическом домене. Ингибирующий рецептор FcγRIIB содержит ингибирующий мотив иммунорецептора на основе тирозина (ITIM) в своем цитоплазматическом домене (см. Rev. Immunol., 15: 203-234 (1997)). Термин охватывает и аллотипы, как-то аллотипы FcγRIIIA: FcγRIIIA-Phe158 и FcγRIIIA-Val158, FcγRIIA-R131 и/или FcγRIIA-H131. Обзоры по FcR см. в Ravetch and Kinet, Annu. Rev. Immunol. 9:457-92 (1991); Capel et al., Immunomethods 4:25-34 (1994); и de Haas et al., J. Lab. Clin. Med. 126:330-41 (1995). Термин “FcR” охватывает и другие FcR, включая те, что будут идентифицированы в будущем. Термин также включает неонатальный рецептор FcRn, который отвечает за перенос материнских IgG к плоду (Guyer et al., J. Immunol. 117: 587 (1976); Kim et al., J. Immunol. 24: 249 (1994).
Термин “FcRn” относится к неонатальному Fc-рецептору (FcRn). FcRn по структуре близок главному комплексу гистосовместимости (MHC) и состоит из α-цепи, нековалентно связанной с β2-микроглобулином. Множественные функции неонатального Fc-рецептора FcRn рассмотрены в обзоре Ghetie and Ward (2000) Annu. Rev. Immunol. 18, 739-766. FcRn играет роль в пассивной доставке иммуноглобулинов типа IgG от матери к плоду и регуляции уровня IgG в сыворотке. FcRn может действовать как спасательный рецептор, связывая и транспортируя захваченные при пиноцитозе IgGs в интактном виде как внутри, так и между клетками, и спасая их от пути деградации по умолчанию.
“Домен CH1” Fc-области IgG человека (также называется доменом “C1” в “H1”) обычно простирается от аминокислоты 118 до аминокислоты 215 (по системе нумерации EU).
“Шарнирный участок” обычно определяется как отрезок от Glu216 до Pro230 в IgG1 человека (Burton, Molec. Immunol. 22: 161-206 (1985)). Шарнирные участки других изотипов IgG можно совмещать с последовательностью IgG1, помещая первый и последний остатки цистеина, образующие связи S-S между тяжелыми цепями, в одних и тех же положениях.
“Нижняя часть шарнирного участка” Fc-области обычно определяется как отрезок из остатков непосредственно на С-конце шарнирного участка, т.е. остатков 233-239 Fc-области. В предыдущих работах связывание FcR обычно связывали с аминокислотными остатками в нижней части шарнирного участка Fc-области IgG.
“Домен CH2” Fc-области IgG человека (также называется доменом “C2” в “H2”) обычно простирается от аминокислоты 231 до аминокислоты 340. Домен CH2 уникален тем, что он не спарен тесно с другим доменом. Скорее, между двумя доменами CH2 интактной нативной молекулы IgG вставлены две цепочки N-связанных разветвленных углеводов. Предполагается, что углеводы могут служить заменой спаривания между доменами и способствовать стабилизации домена CH2: Burton, Molec. Immunol. 22:161-206 (1985).
“Домен СН3” (также называется доменом “С2” или “Н3”) включает отрезок из остатков на С-конце домена СН2 в Fc-области (т.е. от остатка аминокислоты 341 до самого С-конца последовательности антител, обычно до остатка аминокислоты 446 или 447 IgG).
“Функциональная Fc-область” обладает “эффекторной функцией” Fc-области с нативной последовательностью. Типичные “эффекторные функции” включают связывание C1q; комплемент-зависимая цитотоксичность; связывание с Fc-рецептором; антителозависимая клеточная цитотоксичность (ADCC); фагоцитоз; понижающая регуляция рецепторов на клеточной поверхности (напр., B-клеточных рецепторов; BCR) и др. Такие эффекторные функции обычно требуют, чтобы Fc-область сочеталась с доменом связывания (напр., вариабельным доменом антител), и их можно определять различными методами, к примеру, как изложено здесь.
“C1q” представляет собой полипептид, который содержит сайт связывания для Fc-области иммуноглобулина. C1q вместе с двумя сериновыми протеазами, C1r и C1s, образует комплекс C1, первый компонент пути комплемент-зависимой цитотоксичности (CDC). C1q человека коммерчески доступен, напр., от фирмы Quidel, San Diego, CA.
Термин “связывающий домен” относится к той области полипептида, которая связывается с другой молекулой. В случае FcR связывающий домен может содержать ту часть его полипептидной цепи (напр., α-цепи), которая отвечает за связывание Fc-области. Одним из полезных связывающих доменов является внеклеточный домен α-цепи FcR.
Антитело с вариантом Fc IgG с “измененным” сродством связывания с FcR или “измененной” активностью ADCC представляет собой антитело, которое обладает повышенной или пониженной активностью связывания с FcR (напр., FcγR или FcRn) и/или активностью ADCC по сравнению с исходным полипептидом или полипептидом, содержащим Fc-область с нативной последовательностью. Вариант Fc, который “проявляет повышенное связывание” с FcR, связывает по меньшей мере один FcR с более высоким сродством (напр., с меньшим кажущимся значением Kd или IC50), чем у исходного полипептида или нативной последовательности Fc IgG. В некоторых воплощениях улучшение связывания по сравнению с исходным полипептидом составляет примерно в 3 раза, предпочтительно в 5, 10, 25, 50, 60, 100, 150, 200 и вплоть до 500 раз, либо улучшение связывания от 25% до 1000%. Вариант полипептида, который “проявляет пониженное связывание” с FcR, связывается по меньшей мере с одним FcR с более низким сродством (напр., с большим кажущимся Kd или значением IC50), чем исходный полипептид. Снижение связывания по сравнению с исходным полипептидом может означать снижение связывания на 40% или больше.
“Антителозависимая клеточная цитотоксичность” или “ADCC” относится к такой цитотоксичности, при которой секретируемый Ig, связанный с Fc-рецепторами (FcR), находящимися на определенных цитотоксических клетках (напр., клетках натуральных киллеров, нейтрофилах и макрофагах), позволяет этим цитотоксическим эффекторным клеткам специфически связываться с несущими антиген клетками мишени и после этого уничтожать клетки мишени с помощью цитотоксинов. Антитела “вооружают” цитотоксические клетки и абсолютно необходимы для такого уничтожения. Главные клетки, опосредующие ADCC, т.е. NK-клетки, экспрессируют только FcγRIII, тогда как моноциты экспрессируют FcγRI, FcγRII и FcγRIII. Экспрессия FcR на гемопоэтических клетках изложена в табл. 3 на стр. 464 в Ravetch and Kinet, Annu. Rev. Immunol. 9: 457-92 (1991). Для оценки активности ADCC у представляющих интерес молекул можно проводить анализ ADCC in vitro типа описанного в US Patent No. 5,500,362 или 5,821,337 либо в приведенных ниже примерах. Эффекторные клетки, применимые для таких анализов, включают мононуклеарные клетки периферической крови (РВМС) и клетки природных киллеров (NK). С другой стороны, можно оценить активность ADCC у представляющих интерес молекул in vivo, напр., на модели у животных типа изложенной в Clynes et al. PNAS (USA) 95:652-656 (1998).
Полипептид, содержащий вариант Fc-области, который “проявляет повышенную ADCC” или опосредует антителозависимую клеточную цитотоксичность (ADCC) в присутствии эффекторных клеток человека более эффективно, чем полипептид с Fc IgG дикого типа или исходный полипептид, - это такой полипептид, который in vitro или in vivo значительно более эффективно опосредует ADCC при практически одинаковом количестве полипептида с вариантом Fc-области или полипептида с Fc-областью дикого типа (или исходного полипептида) при анализе. Как правило, такие варианты следует идентифицировать любым методом анализа ADCC in vitro, известным в данной области, типа методов определения активности ADCC, напр., на модели у животных и т.д. В одном воплощении предпочтительный вариант от 5 до 100 раз, напр., от 25 до 50 раз более эффективно опосредует ADCC, чем Fc дикого типа (или исходный полипептид).
“Комплемент-зависимая цитотоксичность” или “CDC” означает лизис клеток мишени в присутствии комплемента. Активация классического пути комплемента запускается при связывании первого компонента системы комплемента (C1q) с антителами (соответствующего подкласса), связанными со своим антигеном. Для оценки активации комплемента можно провести анализ CDC, напр., как описано в Gazzano-Santoro et al., J. Immunol. Methods 202:163 (1996). Варианты полипептидов с измененными аминокислотными последовательностями Fc-области и повышенной или пониженной способностью к связыванию C1q описаны в US Patent No. 6,194,551B1 и WO 99/51642. Содержание этих патентных публикаций специально включено сюда путем ссылки. Также см. Idusogie et al., J. Immunol. 164: 4178-4184 (2000).
“Эффективное количество” антитела против PD-L1 (либо его фрагмента) или композиции, как описано здесь, означает количество, достаточное для достижения конкретно заявленной цели. “Эффективное количество” может быть определено эмпирически и известными способами, относящимися к заявленной цели. Термин “терапевтически эффективное количество” означает такое количество антитела против PD-L1 (либо его варианта или антигенсвязывающего фрагмента) или композиции, как описано здесь, которое эффективно для “лечения” заболеваний или расстройств у млекопитающих (то есть пациентов). В случае рака терапевтически эффективное количество антитела против PD-L1 (либо его варианта или антигенсвязывающего фрагмента) или композиции, как описано здесь, может уменьшать количество раковых клеток; уменьшать размер или вес опухолей; ингибировать (то есть замедлять в некоторой степени и предпочтительно останавливать) инфильтрацию раковых клеток в периферические органы; ингибировать (то есть замедлять в некоторой степени и предпочтительно останавливать) метастазирование опухолей; ингибировать в некоторой степени рост опухолей; и/или ослаблять в некоторой степени один или несколько симптомов, связанных с раком. В той степени, в которой антитело против PD-L1 (либо его вариант или антигенсвязывающий фрагмент) или композиция, как описано здесь, может предотвращать рост и/или уничтожать существующие раковые клетки, оно может быть цитостатическим и/или цитотоксическим. В одном воплощении терапевтически эффективное количество означает количество, ингибирующее рост. В другом воплощении терапевтически эффективное количество означает количество, которое продлевает выживаемость пациентов. В другом воплощении терапевтически эффективное количество означает количество, которое улучшает выживаемость пациентов без прогрессирования заболевания.
“Ингибирующее рост количество” антитела против PD-L1 (либо его варианта или антигенсвязывающего фрагмента) или композиции по изобретению, как описано здесь, означает количество, способное ингибировать рост клеток, особенно опухолей, напр., раковых клеток, in vitro либо in vivo. “Ингибирующее рост количество” полипептида, антитела, антагониста или композиции по изобретению для целей ингибирования роста неопластических клеток можно определить эмпирически и известными способами либо по приведенным здесь примерам.
“Цитотоксическое количество” антитела против PD-L1 (либо его варианта или антигенсвязывающего фрагмента) или композиции по изобретению означает количество, способное вызвать разрушение клеток, особенно опухолей, напр., раковых клеток, in vitro или in vivo. “Цитотоксическое количество” антитела против PD-L1 (либо его варианта или антигенсвязывающего фрагмента) или композиции по изобретению для целей ингибирования роста опухолевых клеток можно определить эмпирически и известными в данной области способами.
“Ингибирующее рост количество” антитела против PD-L1 (либо его варианта или антигенсвязывающего фрагмента) или композиции по изобретению означает количество, способное ингибировать рост клеток, особенно опухолей, напр., раковых клеток, in vitro либо in vivo. “Ингибирующее рост количество” антитела против PD-L1 (либо его варианта или антигенсвязывающего фрагмента) или композиции по изобретению для целей ингибирования роста неопластических клеток можно определить эмпирически и известными способами либо по приведенным здесь примерам.
В настоящем изобретении “фармацевтически приемлемый” или “фармакологически совместимый” означает, что материал не является биологически или иным образом нежелательным, напр., материал может быть включен в фармацевтическую композицию, вводимую пациентам, не вызывая значительных нежелательных биологических эффектов или не взаимодействуя зловредным образом с какими-либо другими компонентами той композиции, в которой он содержится. Фармацевтически приемлемые носители или эксципиенты предпочтительно соответствуют требуемым стандартам токсикологических и производственных испытаний и/или включены в Руководство по неактивным ингредиентам, подготовленное Управлением по контролю за продуктами и лекарствами США.
Термин “обнаружение” служит для обозначения определения наличия или отсутствия вещества или определения количества вещества (типа PD-L1). Таким образом, термин относится к применению материалов, композиций и способов настоящего изобретения для качественных и количественных определений. В общем, для практического применения изобретения не имеет значения, какая конкретно методика применяется для обнаружения.
Например, “обнаружение” согласно изобретению может включать в себя: выявление наличия или отсутствия продукта гена PD-L1, молекул мРНК или полипептида PD-L1; изменение уровня полипептида PD-L1 или его количества, связанного с мишенью; изменение биологической функции/активности полипептида PD-L1. В некоторых воплощениях “обнаружение” может включать в себя выявление уровня PD-L1 дикого типа (напр., уровня мРНК или полипептида). Обнаружение может включать количественную оценку изменения (увеличения или уменьшения) любого значения от 10% до 90% или любого значения от 30% до 60% или более 100% по сравнению с контролем. Обнаружение может включать количественную оценку изменения любого значения от 2 до 10 раз включительно или больше, напр., в 100 раз.
Слово “метка” в настоящем изобретении означает детектируемое соединение или композицию, которая прямо или косвенно конъюгирована с антителом. Сама метка может быть детектируемой сама по себе (напр., радиоизотопные метки или флуоресцентные метки) или же, в случае ферментативной метки, она может катализировать химическое изменение соединения или композиции субстрата, которое детектируется.
Ссылка “примерно” на приведенное здесь значение или параметр относится к обычному диапазону ошибок для соответствующего значения, хорошо известному специалистам в данной области. Ссылка “примерно” на значение или параметр включает в себя (и описывает) аспекты, которые касаются этого значения или параметра per se. Например, описание со ссылкой на “примерно X” включает и значение “X”.
Предусматривается, что описанные здесь аспекты и воплощения изобретения включают в себя “содержащие”, “состоящие” и “состоящие в основном из” аспекты и воплощения.
Все ссылки, приведенные здесь, включая патентные заявки и публикации, тем самым включены сюда посредством ссылки во всей полноте.
Антитела против PD-L1 и их варианты/мутанты по сродству
Настоящее изобретение основывается на идентификации новых антител, которые связываются с рецептором PD-L1 (PD-L1). Антитела против PD-L1 и их варианты и/или мутанты по сродству либо их антигенсвязывающие фрагменты могут применяться в различных способах терапии и диагностики. Например, антитела против PD-L1 и их варианты и/или мутанты по сродству либо их или антигенсвязывающие фрагменты могут применяться по отдельности или в комбинации с другими средствами при лечении заболеваний, характеризующихся аномальной экспрессией PD-L1 или аномальной активностью PD-L1, в том числе, напр., меланомы, NSCLC, рака головы и шеи, рака уротелия, рака молочной железы (напр., тройного отрицательного рака молочной железы, TNBC), рака желудка, классической лимфомы Ходжкина (cHL), неходжкинской лимфомы - первичной B-клеточной лимфомы средостения (NHL PMBCL), мезотелиомы, рака яичников, рака легких (напр., мелкоклеточного рака легких), рака пищевода, карциномы носоглотки (NPC), рака желчных путей, колоректального рака, рака шейки матки, рака щитовидной железы. Представленные здесь антитела также могут применяться для обнаружения белка PD-L1 у пациентов или в образцах пациентов путем введения пациентам антител против PD-L1 и/или их вариантов/мутантов по сродству либо их антигенсвязывающих фрагментов и детектирования антител и/или вариантов/мутантов против PD-L1 либо их антиген-связывающих фрагментов, связавшихся с белком PD-L1 в образцах от пациентов (напр., in vivo или ex vivo), или путем контактирования антител и/или вариантов/мутантов против PD-L1 либо их антигенсвязывающих фрагментов с образцами от пациентов и качественного или количественного определения антител против PD-L1 и/или вариантов/мутантов по сродству либо их антигенсвязывающих фрагментов, связавшихся с белком PD-L1.
Лиганд-1 запрограммированной смерти (PD-L1), также известный как кластер дифференцировки 274 (CD274) или гомолог-1 B7 (B7-H1), представляет собой белок, который у человека кодируется геном CD274 (PD-L1). PD-L1 является трансмембранным белком 1-го типа и играет главную роль в подавлении иммунной системы во время определенных событий типа беременности, аллотрансплантации тканей, аутоиммунных заболеваний и других заболеваний типа гепатита. При связывании PD-L1 с PD-1 или B7.1 передается ингибирующий сигнал, который снижает пролиферацию T-клеток CD8+ в лимфатических узлах, а вместе с этим PD-1 также способен контролировать накопление специфичных к чужеродным антигенам T-клеток в лимфатических узлах посредством апоптоза, который в дальнейшем опосредуется понижающей регуляцией гена Bcl-2.
Антитела против PD-L1 - это антитела, которые связываются с PD-L1 с достаточным сродством и специфичностью. Предпочтительно представленные здесь антитела против PD-L1 (или их варианты или мутанты либо их антигенсвязывающие фрагменты) могут применяться в качестве терапевтического средства при воздействии и вмешательстве в такие заболевания, в которых вовлечена активность PD-L1. Антитела против PD-L1 (или их варианты/мутанты либо антигенсвязывающие фрагменты) обычно не связываются с другими представителями суперсемейства иммуноглобулинов. Предпочтительно антитела против PD-L1 (либо их варианты/мутанты или антигенсвязывающие фрагменты) представляют собой человеческие или рекомбинантные гуманизованные моноклональные антитела против PD-L1.
Согласно определенным воплощениям, антитела против PD-L1 содержат участки CDR, вариабельную область тяжелой цепи и/или вариабельную область легкой цепи любого из приведенных здесь антител.
Изобретением предусмотрены антитела против PD-L1, их варианты/мутанты по сродству и/или их антигенсвязывающие фрагменты. В некоторых воплощениях антитело против PD-L1 по изобретению, т.е. антитело PL1, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 35; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 41; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 44; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 51; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 59.
В других воплощениях антитело против PD-L1 по изобретению, т.е. антитело PL2, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 36; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 42; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 45; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 52; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 56; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 60.
В других воплощениях антитело против PD-L1 по изобретению, т.е. антитело PL3, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 37; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 41; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 46; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 53; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 61.
В других воплощениях антитело против PD-L1 по изобретению, т.е. антитело PL6, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 37; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 41; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 47; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 54; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 57; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 62.
В других воплощениях антитело против PD-L1 по изобретению, т.е. антитело PL8, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 38; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 43; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 48; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 51; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 58; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 63.
В других воплощениях антитело против PD-L1 по изобретению, т.е. антитело PL12, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 39; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 41; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 49; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 51; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 64.
В других воплощениях антитело против PD-L1 по изобретению, т.е. антитело PL15, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 40; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 41; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 50; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 53; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 61.
Изобретением предусмотрено, что отмеченные здесь вариабельные домены и участки CDR тяжелой и легкой цепи комбинируются во всевозможных парных комбинациях, образуя целый ряд антител против PD-L1.
В некоторых воплощениях аминокислотные замены представляют собой консервативные замены аминокислот. В некоторых воплощениях аминокислотные замены не снижают существенно способность антител к связыванию с антигеном. Например, можно проводить консервативные изменения (напр., консервативные замены, как предусмотрено здесь), которые не снижают существенно сродство связывания с PD-L1. Сродство связывания вариантов антител против PD-L1 можно оценить способами, описанными ниже в Примерах.
Консервативные замены представлены в табл. 3 под заголовком «Консервативные замены». Более существенные изменения представлены в табл. 3 под заголовком «Типичные замены», которые более подробно описаны ниже со ссылкой на классы боковых цепей аминокислот. В представляющие интерес антитела можно вводить аминокислотные замены, а продукты подвергать скринингу на требуемую активность, напр., сохранение/улучшение связывания PD-L1, уменьшение иммуногенности либо улучшение ADCC или CDC.
Таблица 3. Консервативные замены
Неконсервативные замены означают замену представителя одного из этих классов на представителя другого класса. Типичными вариантами с заменой являются антитела, прошедшие созревание сродства, которые можно удобно получить, напр., методами созревания сродства на основе фагового дисплея типа описанных здесь. Вкратце, подвергают мутации один или несколько остатков CDR, варианты антител выставляются на фагах и подвергаются скринингу на определенную биологическую активность (напр., сродство связывания). Изменения (напр., замены) могут проводиться в HVR, напр., для улучшения сродства антитела. Такие изменения могут проводиться в “горячих точках” HVR, то есть в остатках, кодируемых теми кодонами, которые подвергаются мутации с высокой частотой в процессе соматического созревания (напр., см. Chowdhury, Methods Mol. Biol. 207: 179-196 (2008)), и/или в SDR (a-CDR), причем полученные варианты VH или VL тестируют на сродство связывания. Созревание сродства путем конструирования и повторного отбора из вторичных библиотек описано, напр., в Hoogenboom et al., Methods in Molecular Biology 178: 1-37 (O’Brien et al., ed., Human Press, Totowa, NJ (2001).
В некоторых воплощениях созревания сродства вводится разнообразие в вариабельные гены, выбранные для созревания, различными способами (напр., подверженной ошибкам ПЦР, перетасовки цепей или направляемого олигонуклеотидами мутагенеза). Затем создается вторичная библиотека. Затем проводится скрининг библиотеки для выявления вариантов антител с требуемым сродством. Другой способ введения разнообразия включает HVR-ориентированные подходы, в которых несколько остатков HVR (напр., 4-6 остатков за 1 раз) подвергаются случайному мутагенезу. Конкретно можно идентифицировать остатки HVR, участвующие в связывании антигена, напр., методом сканирующего мутагенеза по аланинам или моделирования.
Изобретением также предусмотрены варианты и/или мутанты антител против PD-L1 либо их антигенсвязывающих фрагментов. В некоторых воплощениях вариант и/или мутант антитела против PD-L1 по изобретению, т.е. антитело PL2#3, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 65; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 42; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 71; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 52; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 56; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 77.
В других воплощениях вариант и/или мутант антитела против PD-L1 по изобретению, т.е. антитело PL3#7, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 37; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 41; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 72; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 75; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 78.
В других воплощениях вариант и/или мутант антитела против PD-L1 по изобретению, т.е. антитело PL3#7-19, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 66; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 68; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 73; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 75; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 79. В некоторых воплощениях мутантное антитело PL3#7-19 против PD-L1 включают в себя CDR-L2, содержащий одну или несколько мутаций по сайтам N-гликозилирования. В некоторых воплощениях сайты гликозилирования находятся в районе CDR-L2, напр., в последовательности N-X-S/T. У антител против PD-L1 с одной или несколькими мутациями в последовательности N-X-S/T устраняется сайт N-гликозилирования, но функция остается та же.
В других воплощениях вариант и/или мутант антитела против PD-L1 по изобретению, т.е. антитело PL3#7-43, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 35; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 69; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 74; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 75; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 80. В некоторых воплощениях мутантное антитело PL3#7-19 против PD-L1 включают в себя CDR-L2, содержащий одну или несколько мутаций по сайтам N-гликозилирования. В некоторых воплощениях сайты гликозилирования находятся в районе CDR-L2, напр., в последовательности N-X-S/T. У антител против PD-L1 с одной или несколькими мутациями в последовательности N-X-S/T устраняется сайт N-гликозилирования, но функция остается та же.
В других воплощениях вариант и/или мутант антитела против PD-L1 по изобретению, т.е. антитело PL3#7-54, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 67; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 70; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 72; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 75; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 76; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 81.
В других воплощениях вариант и/или мутант антитела против PD-L1 по изобретению, т.е. антитело PL2#4, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 94; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 42; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 95; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 52; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 56; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 96.
В других воплощениях вариант и/или мутант антитела против PD-L1 по изобретению, т.е. антитело PL2#5, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 97; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 42; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 98; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 52; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 56; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 99.
В других воплощениях вариант и/или мутант антитела против PD-L1 по изобретению, т.е. антитело PL2#39, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 100; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 42; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 95; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 52; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 56; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 101.
В других воплощениях вариант и/или мутант антитела против PD-L1 по изобретению, т.е. антитело PL3#1, содержит последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 37; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 106; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 107; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 108; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 109.
Изобретением предусмотрено, что отмеченные здесь вариабельные домены и участки CDR тяжелой и легкой цепи комбинируются во всевозможных парных комбинациях, образуя целый ряд вариантов антител против PD-L1.
В некоторых воплощениях изобретения предусмотрено, что антитела против PD-L1, их варианты или антигенсвязывающие фрагменты содержат последовательность вариабельного домена легкой цепи (VL), включающую (1) CDR-L1, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 35-40, 65-67, 94, 97 и 100; (2) CDR-L2, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 41-43, 68-70 и 106; и (3) CDR-L3, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 44-50, 71-74, 95, 98 и 107; и последовательность вариабельного домена тяжелой цепи (VH), включающую (1) CDR-H1, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 51-54, 75 и 108; (2) CDR-H2, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 55-58 и 76; и (3) CDR-H3, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 59-64, 77-81, 96, 99, 101 и 109. Вариабельные домены тяжелой и легкой цепи комбинируются во всевозможных парных комбинациях, образуя целый ряд антител против PD-L1.
Также изобретением предусмотрены антитела против PD-L1, их варианты и/или мутанты либо их антигенсвязывающие фрагменты, содержащие последовательность вариабельного домена легкой цепи (LC), включающую аминокислотную последовательность, приведенную в SEQ ID NOs: 1-7, 16, 20, 24, 28, 32, 83, 87, 91 или 103; и последовательность вариабельного домена тяжелой цепи (HC), включающую аминокислотную последовательность, приведенную в SEQ ID NOs: 8-14, 18, 22, 26, 30, 34, 85, 89, 93 или 105. Также предусмотрены последовательности нуклеиновых кислот, кодирующих эти домены LC и HC. Вариабельные домены тяжелой и легкой цепи комбинируются во всевозможных парных комбинациях, образуя целый ряд антител против PD-L1 и/или их вариантов.
В некоторых воплощениях у антител против PD-L1, их вариантов или мутантов либо антигенсвязывающих фрагментов может отсутствовать мотив N-гликозилирования в вариабельной области тяжелой цепи или легкой цепи, что может вызвать отличия в пределах одной партии антител, вызывающие изменения функции, иммуногенности или стабильности. Методы анализа гликозилирования антител включают, без ограничения, напр., хроматографию (типа катионообменной хроматографии (CEX) или жидкостной хроматографии), масс-спектрометрию (типа масс-спектрометрии с ионизацией электрораспылением) и капиллярный электрофорез с додецилсульфатом натрия. Такие методы описаны, напр., в Jung et al. (2011) Curr Op Biotechnol. 22(6):858-67; Cummings RD, Etzler ME. Antibodies and Lectins in Glycan Analysis. In: Varki A, Cummings RD, Esko JD, et al., editors. Essentials of Glycobiology, 2nd edition. Cold Spring Harbor (NY): Cold Spring Harbor Laboratory Press, 2009, Chapter 45; Mulloy B, Hart GW, Stanley P. Structural Analysis of Glycans. In: Varki A, Cummings RD, Esko JD, et al., editors. Essentials of Glycobiology, 2nd edition. Cold Spring Harbor (NY): Cold Spring Harbor Laboratory Press, 2009, Chapter 47; Leymarie et al. (2012) Anal Chem. 84(7): 3040-3048; Fernandez (2005) European Biopharmaceutical Review, pp 106-110; and Raju T. (2013) Methods Mol. Biol. 988: 169-180.
В некоторых воплощениях антитела против PD-L1, их варианты и/или мутанты либо антигенсвязывающие фрагменты имеют большее сродство связывания с лигандом PD-L1, чем с гомологом этого PD-L1. Обычно антитела против PD-L1 и/или их варианты либо антигенсвязывающие фрагменты “специфически связываются” с PD-L1 (то есть у них значение сродства связывания (Kd) составляет не более 1×10−7 М, предпочтительно не более 1×10−8 и наиболее предпочтительно не более 1×10−9 М), но сродство связывания с другими представителями семейства PD-L1 по меньшей мере в 50 раз или по меньшей мере в 500 раз или по меньшей мере в 1000 раз меньше, чем сродство связывания с PD-L1. Антитела против PD-L1, которые специфически связываются с PD-L1, могут представлять собой любые из различных типов антител, как определено выше, но предпочтительно они представляют собой гуманизованные или человеческие антитела.
В некоторых воплощениях степень связывания антител против PD-L1 с нецелевым белком составляет менее 10% от связывания антител с PD-L1 при определении такими известными в данной области методами, как ELISA, активируемая флуоресценцией сортировка клеток (FACS) или радиоиммунопреципитация (RIA). Специфическое связывание можно измерить, к примеру, путем определения связывания с молекулой по сравнению со связыванием с контрольной молекулой, которая обычно представляет собой молекулу с близкой структурой, но не обладает активностью связывания. Например, специфическое связывание можно определять по конкуренции с контрольной молекулой, похожей на мишень, к примеру, с избытком немеченой мишени. При этом специфическое связывание проявляется, если связывание меченой мишени с зондом конкурентно ингибируется избытком немеченой мишени. Термин “специфическое связывание” или “специфически связывается” или “специфично к” определенному полипептиду или эпитопу на определенной полипептидной мишени в настоящем изобретении может проявляться, к примеру, тем, что молекула имеет значение Kd для мишени по меньшей мере 10−4 М или же по меньшей мере 10−5 М или же по меньшей мере 10−6 М или же по меньшей мере 10−7 М или же по меньшей мере 10−8 М или же по меньшей мере 10−9 М или же по меньшей мере 10−10 М или же по меньшей мере 10−11 М или же по меньшей мере 10−12 М или больше. В одном воплощении термин “специфическое связывание” означает такое связывание, когда молекула связывается с определенным полипептидом или эпитопом на определенном полипептиде без существенного связывания с каким-либо другим полипептидом или полипептидным эпитопом.
Механизм действия терапевтических антител против опухолевых клеток составляет антителозависимая клеточная цитотоксичность (ADCC). ADCC - опосредованная клетками иммунная защита, посредством которой эффекторные клетки иммунной системы активно лизируют такие клетки мишени (напр., раковые клетки), у которых антигены мембранной поверхности связались со специфичными антителами (напр., типа описанных здесь антител против PD-L1 и/или их вариантов по сродству). В некоторых воплощениях антитела против PD-L1 и/или их варианты по сродству проявляют аналогичную эффекторную функцию типа антителозависимой клеточной цитотоксичности (ADCC) в качестве контрольных моноклональных антител против PD-L1, как показано, напр., при анализах, описанных в Примерах.
Например, в некоторых воплощениях активность эффекторной функции ADCC у описанных здесь антител против PD-L1 и/или их вариантов или мутантов по сродству составляет по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99%, по меньшей мере 100% или более 100% (напр., примерно 105%, 106%, 107%, 108%, 109%, 110%, 111%, 112%, 113%, 114%, 115%, 116%, 117%, 118%, 119%, 120%, 121%, 122%, 123%, 124%, 125% или 130%) от активности эффекторной функции ADCC у контрольных антител против PD-L1, включая любые интервалы между этими значениями.
В некоторых воплощениях антитела против PD-L1 и/или их варианты или мутанты по сродству проявляют сродство связывания с PD-L1, близкое контрольным антителам против PD-L1. В некоторых воплощениях связывание с PD-L1 определяется методом ELISA, как описано в примерах. Например, сродство связывания антител против PD-L1 и/или их вариантов или мутантов по сродству к PD-L1 примерно на 1%, 5%, 10%, 15%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98%, 99%, 100% или более чем на 100% (напр., примерно на 105%, 106%, 107%, 108%, 109%, 110%, 111%, 112%, 113%, 114%, 115%, 116%, 117%, 118%, 119%, 120%, 121%, 122%, 123%, 124%, 124%, 125% или более чем на 125%) выше, чем сродство связывания с PD-L1 у контрольных антител против PD-L1.
В некоторых воплощениях антитела против PD-L1 и/или их варианты или мутанты по сродству связывают PD-L1 человека со значением Kd от 0,1 до 200 пМ (0,2 нМ), напр., примерно 0,1 пМ, 0,25 пМ, 0,5 пМ, 0,75 пМ, 1 пМ, 5 пМ, 10 пМ, 20 пМ, 30 пМ, 40 пМ, 50 пМ, 60 пМ, 70 пМ, 80 пМ, 90 пМ, 100 пМ, 110 пМ, 120 пМ, 130 пМ, 140 пМ, 150 пМ, 160 пМ, 170 пМ, 180 пМ, 190 пМ или более 190 пМ, включая любые интервалы между этими значениями. В некоторых воплощениях сродство связывания с PD-L1 у антител против PD-L1 и/или их вариантов или мутантов по сродству примерно на 1%, 5%, 10%, 15%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98%, 99%, 100% или более чем на 100% (напр., примерно на 105%, 110%, 120% или 130%) выше, чем сродство связывания с PD-L1 у контрольных антител против PD-L1. В некоторых воплощениях сродство связывания с PD-L1 у антител против PD-L1 и/или их вариантов или мутантов по сродству примерно в 1,1, 1,2, 1,3, 1,4, 1,5, 1,6, 1,7, 1,8, 1,9, 2, 2,25, 2,5, 2,75, 3, 3,25, 3,5, 3,75, 4, 4,25, 4,5, 4,75 раза или более чем в 4,75 раза выше, чем сродство связывания с PD-L1 у контрольных антител против PD-L1, включая любые интервалы между этими значениями.
В некоторых воплощениях представленные здесь антитела против PD-L1 и их варианты или мутанты имеют больший период полужизни in vivo по сравнению с контрольными антителами против PD-L1. В некоторых воплощениях период полужизни in vivo у описанных здесь антител против PD-L1 и/или их вариантов или мутантов не меньше, чем период полужизни in vivo у контрольных антител против PD-L1.
В некоторых воплощениях представленные здесь антитела против PD-L1 и их варианты или мутанты проявляют фармакокинетические свойства, близкие свойствам контрольных антител против PD-L1. В некоторых воплощениях представленные здесь антитела против PD-L1 и их варианты или мутанты проявляют значения AUC (площади под кривой), которые составляют примерно 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или более 95% (напр., примерно 96%, 97%, 98%, 99% или более 99%) от профиля концентрации в сыворотке от времени у контрольных антител против PD-L1, включая любые интервалы между этими значениями.
В некоторых воплощениях антитела содержат последовательность Fc человеческого IgG, напр., IgG1 человека или IgG4 человека. В некоторых воплощениях последовательность Fc была изменена или иным образом заменена таким образом, что она лишена эффекторной функции антителозависимой клеточной цитотоксичности (ADCC), которая часто связана со связыванием их с Fc-рецепторами (FcR). Существует много примеров изменений или мутаций в последовательностях Fc, которые могут изменять эффекторную функцию. Напр., в WO 00/42072 и в Shields et al., J Biol. Chem. 9(2): 6591-6604 (2001) описаны варианты антител с улучшенным или уменьшенным связыванием с FcR. Содержание этих публикаций специально включено сюда путем ссылки. Антитела могут иметь вид Fab, Fab′, F(ab′)2, одноцепочечных Fv (scFv), фрагментов Fv; диател и линейных антител. Кроме того, антитела и их варианты или мутанты могут представлять собой мультиспецифичные антитела либо такие варианты или мутанты, которые связываются с PD-L1, но также связываются с одной или несколькими другими мишенями и ингибируют их функции. Антитела и/или их варианты или мутанты могут быть конъюгированы с терапевтическими средствами (напр., цитотоксическими средствами, радиоизотопами и химиотерапевтическими средствами) или метками для обнаружения PD-L1 в образцах пациентов или in vivo методами визуализации (напр., радиоизотопами, флуоресцентными красителями и ферментами). Другие модификации включают конъюгирование токсинов с представленными здесь антителами против PD-L1 и/или их вариантами или мутантами.
Также предусмотрены молекулы нуклеиновой кислоты, кодирующие антитела против PD-L1, их варианты и/или мутанты, экспрессирующие векторы, содержащие молекулы нуклеиновой кислоты, кодирующие описанные здесь участки CDR и/или вариабельные домены тяжелой цепи и/или вариабельные домены легкой цепи, и клетки, содержащие молекулы нуклеиновой кислоты. Эти антитела и их варианты или мутанты могут применяться в описанных здесь способах лечения и для выявления белка PD-L1 в образцах пациентов (напр., методом FACS, иммуногистохимии (IHC), методами ELISA) или у пациентов.
Моноклональные антитела
Моноклональные антитела могут быть получены, напр., гибридомными методами типа описанных в Kohler and Milstein, Nature, 256: 495 (1975) или могут быть получены методами рекомбинантной ДНК (US Patent No. 4,816,567) или же способами, описанными здесь в примерах ниже. В гибридомном методе обычно иммунизируют хомяков, мышей или других подходящих животных-хозяев иммунизирующим средством для индукции лимфоцитов, вырабатывающих или способных вырабатывать антитела, которые будут специфически связываться с иммунизирующим средством. С другой стороны, можно иммунизировать лимфоциты in vitro.
Иммунизирующие средства обычно включают в себя полипептид или белок, слитый с представляющим интерес белком, или композиции, содержащие этот белок. Как правило, используют лимфоциты периферической крови (“PBLs”), если нужны клетки человеческого происхождения, либо клетки селезенки или лимфатических узлов, если нужны клетки из других млекопитающих, а не человека. Затем лимфоциты сливают с клетками иммортализованной линии с помощью подходящего реагента для слияния типа полиэтиленгликоля, получая клетки гибридомы (Goding, Monoclonal Antibodies: Principles and Practice, New York: Academic Press, 1986, pp. 59-103). Бессмертные клеточные линии обычно представляют собой трансформированные клетки млекопитающих, в частности, клетки миеломы от грызунов, бычьи и человеческие. Обычно используют линии клеток миеломы крыс или мышей. Клетки гибридомы можно культивировать в подходящей культуральной среде, которая предпочтительно содержит одно или несколько веществ, ингибирующих рост или выживаемость не слившихся иммортализованных клеток. Например, если исходные клетки лишены фермента гипоксантин-гуанин-фосфорибозилтрансферазы (HGPRT или HPRT), то культуральная среда для гибридом обычно включает в себя гипоксантин, аминоптерин и тимидин (“среда HAT”), то есть вещества, которые предотвращают рост дефектных по HGPRT клеток.
Предпочтительными являются такие иммортализованные линии клеток, которые эффективно сливаются, поддерживают стабильную экспрессию антител на высоком уровне выбранными продуцирующими антитела клетками и чувствительны к средам типа среды HAT. Более предпочтительно иммортализованные линии клеток представлены линиями клеток миеломы мыши, которые можно получить, к примеру, из Центра по распределению клеток Salk Institute, San Diego, Калифорния, и Американской коллекции типовых культур, Manassas, Вирджиния. Также для получения человеческих моноклональных антител были описаны линии клеток миеломы человека и гетеромиеломы мыши-человека (Kozbor, J. Immunol., 133:3001 (1984); Brodeur et al. Monoclonal Antibody Production Techniques and Applications, Marcel Dekker, Inc.: New York, 1987, pp. 51-63).
Культуральную среду, в которой культивируются клетки гибридомы, можно затем проанализировать на наличие моноклональных антител, направленных против полипептида. Специфичность связывания моноклональных антител, вырабатываемых клетками гибридомы, можно определить методом иммунопреципитации или по связыванию in vitro типа радиоиммуноанализа (RIA) или ферментного иммуносорбционного анализа (ELISA). Такие методы и анализы известны в данной области. Сродство связывания моноклональных антител можно определить, к примеру, методом анализа по Скэтчарду типа Munson and Pollard, Anal. Biochem., 107: 220 (1980).
После идентификации требуемых клеток гибридомы можно субклонировать кло-ны методом предельных разведений и культивировать их стандартными способами, см. Goding, supra. Подходящие культуральные среды для этой цели включают, к примеру, модифицированную Дюльбекко среду Игла и среду RPMI-1640. С другой стороны, клетки гибридомы можно культивировать in vivo в виде асцитов у млекопитающих.
Моноклональные антитела, секретируемые субклонами, можно выделить или очистить из культуральной среды или асцитной жидкости стандартными методами очистки иммуноглобулинов, такими, к примеру, как хроматография на сефарозе с белком A или на гидроксиапатите, гель-электрофорез, диализ или аффинная хроматография.
Моноклональные антитела также можно получить методами рекомбинантной ДНК типа описанных в U.S. Patent No. 4,816,567. ДНК, кодирующую представленные здесь моноклональные антитела, можно легко выделить и секвенировать по стандартным методикам (напр., с помощью олигонуклеотидных зондов, способных специфически связываться с генами, кодирующими тяжелые и легкие цепи мышиных антител). Предпочтительным источником такой ДНК служат представленные здесь клетки гибридомы. После выделения ДНК можно вставить в экспрессирующие векторы, а затем трансфецировать в клетки хозяина типа клеток COS обезьяны, клеток яичников китайского хомячка (CHO) или клеток миеломы, которые иначе не вырабатывают белок иммуноглобулина, получая синтез моноклональных антител в рекомбинантных клетках хозяина. ДНК также можно модифицировать, к примеру, путем замены кодирующей последовательности для константных доменов тяжелой и легкой цепи человека вместо гомологичных последовательностей мыши (U.S. Patent No. 4,816,567; Morrison et al., supra) или путем ковалентного присоединения к кодирующей последовательности иммуноглобулина всей или части кодирующей последовательности другого полипептида, не иммуноглобулина. Такой неиммуноглобулиновый полипептид можно вставить вместо константных доменов представленных здесь антител или вариабельных доменов одного антигенсвязывающего сайта представленных здесь антител, получая химерные бивалентные антитела.
В некоторых воплощениях представленные здесь антитела против PD-L1 и/или их варианты или мутанты экспрессируются в стабильных линиях клеток млекопитающих. В некоторых воплощениях представленные здесь антитела против PD-L1 и/или их варианты или мутанты экспрессируются в стабильных линиях клеток млекопитающих при титре в 2,0 г/л, 2,5 г/л, 3,0 г/литр, 3,5 г/л, 4,0 г/л, 4,5 г/л, 5,0 г/л, 5,5 г/л, 6 г/л, 6,5 г/л, 7,0 г/л или более 7,0 г/л, включая любые интервалы между этими значениями. В некоторых воплощениях стабильная линия клеток млекопитающих, в которой экспрессируется антитело против PD-L1 и/или его вариант или мутант, предусмотренный изобретением, представлена линией клеток СНО.
В некоторых воплощениях антитела представляют собой моновалентные антитела. Способы получения моновалентных антител известны в данной области. Например, один способ включает рекомбинантную экспрессию легкой цепи иммуноглобулина и модифицированной тяжелой цепи. Тяжелая цепь обычно укорачивается в любой точке Fc-области с тем, чтобы предотвратить образование сшивок тяжелой цепи. С другой стороны, чтобы предотвратить сшивание, соответствующие остатки цистеина заменяют на другие аминокислотные остатки или удаляют.
Для получения моновалентных антител также подходят способы in vitro. Расщепление антител для получения их фрагментов, в частности, Fab-фрагментов, может осуществляться, без ограничения, известными в данной области методами.
Человеческие и гуманизованные антитела
Антитела (и/или их варианты) могут представлять собой гуманизованные антитела или человеческие антитела. Гуманизированные формы нечеловеческих (напр., мышиных) антител представляют собой химерные иммуноглобулины, цепи иммуноглобулинов или их фрагменты (типа Fv, Fab, Fab′, F(ab′)2 или других антигенсвязывающих частей антител), которые обычно содержат минимальную последовательность, происходящую из иммуноглобулина не человека. Гуманизованные антитела включают иммуноглобулины человека (из антитела-реципиента), в которых остатки участка CDR реципиента заменены на остатки из CDR другого вида (донорского антитела) типа мыши, крысы или кролика, обладающего требуемой специфичностью, сродством и емкостью. В некоторых случаях каркасные остатки Fv иммуноглобулина человека заменяют на соответствующие остатки не человека. Гуманизованные антитела также могут содержать остатки, которые не встречаются ни в реципиентном антителе, ни в импортированных последовательностях CDR или каркасных участков. В общем, гуманизированные антитела могут содержать практически весь по меньшей мере один, а обычно два вариабельных домена, в которых все или практически все участки CDR соответствуют участкам иммуноглобулина не человека, а все или практически все участки FR имеют консенсусные последовательности иммуноглобулина человека. Гуманизованные антитела предпочтительно также должны содержать по меньшей мере часть константной области (Fc) иммуноглобулина, как правило, иммуноглобулина человека: Jones et al., Nature, 321: 522-525 (1986); Riechmann et al., Nature, 332: 323-329 (1988); Presta, Curr. Op. Struct. Biol., 2:593-596 (1992).
Как правило, гуманизованные антитела содержат один или несколько аминокислотных остатков, введенных в них из других источников, чем человек. Эти аминокислотные остатки не человека часто называют “импортными” остатками, так как обычно они происходят из “импортного” вариабельного домена. Согласно одному воплощению. гуманизация может проводиться в основном по методу Winter and co-workers (Jones et al., Nature, 321: 522-525 (1986); Riechmann et al., Nature, 332: 323-327 (1988); Verhoeyen et al., Science, 239: 1534-1536 (1988)), путем замены участков или последовательностей CDR грызунов вместо соответствующих последовательностей антител человека. Соответственно, “гуманизованными” являются такие антитела (U.S. Patent No. 4,816,567), у которых значительно меньшая часть интактного вариабельного домена человека заменена на соответствующую последовательность из другого вида. На практике гуманизованные антитела обычно представляют собой антитела человека, у которых некоторые остатки CDR, а возможно и некоторые остатки FR заменены на остатки из аналогичных сайтов в антителах грызунов.
В качестве альтернативы гуманизации можно получать человеческие антитела. Например, теперь уже можно получать трансгенных животных (напр., мышей), которые способны после иммунизации вырабатывать полный репертуар человеческих антител в отсутствие продукции эндогенных иммуноглобулинов. Например, было описано, что гомозиготная делеция гена соединительного участка тяжелой цепи антител (JH) у химерных и гаметных мутантных мышей приводит к полному ингибированию продукции эндогенных антител. Перенос комплекта гаметных генов иммуноглобулина человека таким гаметных мутантным мышам приводит к продукции человеческих антител при стимуляции антигеном. Напр., см., Jakobovits et al., PNAS USA, 90: 2551 (1993); Jakobovits et al., Nature, 362: 255-258 (1993); Bruggemann et al., Year in Immunol., 7:33 (1993); U.S. Patent Nos. 5,545,806, 5,569,825, 5,591,669; 5,545,807; и WO 97/17852.
С другой стороны, человеческие антитела можно получать путем введения локусов иммуноглобулина человека трансгенным животным, напр., мышам, у которых эндогенные гены иммуноглобулина были частично или полностью инактивированы. При стимуляции антигеном наблюдается вырабатывание человеческих антител, которое очень похоже на то, что наблюдается у людей во всех отношениях, включая перегруппировку генов, сбор-ку и репертуар антител. Этот подход описан, к примеру, в U.S. Patent Nos. 5,545,807; 5,545,806; 5,569,825; 5,625,126; 5,633,425; и 5,661,016; Marks et al., Bio/Technology, 10: 779-783 (1992); Lonberg et al., Nature, 368: 856-859 (1994); Morrison, Nature, 368: 812-813 (1994); Fishwild et al., Nature Biotechnology, 14: 845-851 (1996); Neuberger, Nature Biotechnology, 14: 826 (1996); Lonberg and Huszar, Intern. Rev. Immunol., 13: 65-93 (1995).
А с другой стороны, для получения человеческих антител и фрагментов антител in vitro из репертуара генов вариабельного (V) домена иммуноглобулина от неиммунизированных доноров можно использовать технологию фагового дисплея (McCafferty et al., Nature 348: 552-553, 1990). Согласно одному варианту этого метода, последовательности V-домена антител клонируют в одной рамке в ген главного или второстепенного белка оболочки филаментного бактериофага типа M13 или fd и экспрессируют в виде функциональных фрагментов антител на поверхности фаговых частиц. Фаговый дисплей может осуществляться в различных форматах, напр., как описано ниже в разделе «Примеры» или как изложено, напр., в Johnson, Kevin S. and Chiswell, David J., Current Opinion in Structural Biology 3: 564-571 (1993). Для фагового дисплея можно использовать несколько источников сегментов V-генов. Clackson et al., Nature, 352: 624-628 (1991) выделили многообразный набор антител против оксазолона из небольшой случайной комбинаторной библиотеки V-генов, полученных из селезенки иммунизированных мышей. Можно создать репертуар V-генов от неиммунизированных доноров и выделить антитела к разнообразным антигенам (включая аутоантигены), в основном по методам, описанным в Marks et al., J. Mol. Biol. 222: 581-597 (1991); или Griffith et al., EMBO J. 12: 725-734 (1993). Также см. U.S. Patent Nos. 5,565,332 и 5,573,905.
Как обсуждалось выше, человеческие антитела также могут вырабатываться активированными in vitro B-клетками (см. U.S. Patents 5,567,610 и 5,229,275).
Человеческие антитела также могут быть получены различными методами, известными в данной области, включая библиотеки фагового дисплея: Hoogenboom and Winter, J. Mol. Biol., 227: 381 (1991); Marks et al., J. Mol. Biol., 222: 581 (1991). Для получения моноклональных человеческих антител также доступны методы Cole et al. и Boerner et al., см. Cole et al., Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, p. 77 (1985); Boerner et al., J. Immunol., 147(1): 86-95 (1991).
Мультиспецифичные антитела
Мультиспецифичные антитела - моноклональные, предпочтительно человеческие или гуманизованные антитела, которые обладают специфичностью связывания для двух или нескольких разных антигенов (напр., биспецифичные антитела обладают специфичностью связывания для по меньшей мере двух антигенов). Например, одна специфичность связывания может быть для белка a5~1, а другая - для какого-то другого антигена. В одном предпочтительном воплощении другой антиген представляет собой белок клеточной поверхности или рецептор или субъединица рецептора. Например, белок клеточной поверхности может представлять собой рецептор природных клеток-киллеров (NK). Так, в одном воплощении биспецифичные антитела по изобретению могут связывать и PD-L1, и, напр., второй рецептор клеточной поверхности.
Подходящие способы получения биспецифичных антител хорошо известны в данной области. Например, рекомбинантная продукция биспецифичных антител основана на совместной экспрессии двух пар тяжелых цепей/легких цепей иммуноглобулина, причем две тяжелые цепи имеют разные специфичности: Milstein and Cuello, Nature, 305: 537-539 (1983). Вследствие случайного ассортимента тяжелых и легких цепей иммуноглобулина эти гибридомы (квадромы) могут вырабатывать смесь из десяти различных молекул антител, из которых только одна имеет правильную биспецифичную структуру. Очистка правильных молекул обычно проводится методом аффинной хроматографии. Подобные процедуры изложены в WO 93/08829 и в Traunecker et al., EMBO, 10: 3655-3659 (1991).
Вариабельные домены антител с требуемой специфичностью связывания (сайты связывания антитело-антиген) можно сливать с последовательностями константных доменов иммуноглобулина. Слияние предпочтительно происходит с константными доменами тяжелой цепи иммуноглобулина, содержащими по меньшей мере часть шарнира и участки CH2 и CH3. Предпочтительно по меньшей мере в одном из слияний должен находиться первый константный участок тяжелой цепи (СН1), содержащий сайт, необходимый для связывания легкой цепи. ДНК, кодирующие слияния тяжелой цепи иммуноглобулина и, если нужно, легкой цепи иммуноглобулина, вставляют в отдельные экспрессирующие векторы и совместно трансфецируют в подходящий организм хозяина. Более подробно о получении биспецифичных антител см., к примеру, Suresh et al., Methods in Enzymology, 121: 210 (1986).
Также были описаны различные способы получения и выделения биспецифичных фрагментов антител прямо из культуры рекомбинантных клеток. Например, биспецифичные антитела получали с помощью лейциновых застежек: Kostelny et al., J. Immunol., 148 (5): 1547-1553 (1992). Пептиды лейциновых молний из белков Fos и Jun соединяли с Fab′-фрагментами двух разных антител путем слияния генов. Гомодимеры антител подвергали восстановлению в шарнирной области с получением мономеров, а затем снова окисляли с получением гетеродимеров антител. Этот метод также может применяться для получения гомодимеров антител. Технология “диател”, описанная в Hollinger et al., PNAS USA, 90: 6444-6448 (1993), обеспечила альтернативный механизм получения биспецифичных фрагментов антител. Фрагменты содержат VH, соединенный с VL при помощи линкера, слишком короткого для того, чтобы происходило спаривание между двумя доменами на одной и той же цепи. Соответственно, домены VH и VL одного фрагмента вынуждены спариваться с комплементарными доменами VL и VH из другого фрагмента, образуя при этом два антигенсвязывающих сайта. Также сообщалось о другой стратегии получения биспецифичных фрагментов антител с помощью димеров одноцепочечных Fv (sFv). См. Gruber et al., J. Immunol., 152: 5368 (1994).
Предусмотрены антитела с более чем двумя валентностями. К примеру, могут быть получены триспецифичные антитела: Tutt et al. J. Immunol. 147: 60 (1991).
Гетероконъюгаты антител
Гетероконъюгаты антител состоят из двух ковалентно связанных антител. Такие антитела, к примеру, были предложены для наведения клеток иммунной системы на нежелательные клетки (U.S. Patent No. 4,676,980) и для лечения ВИЧ-инфекции: WO 91/00360; WO 92/200373; EP 03089. Предусматривается, что антитела могут быть получены in vitro известными в химии белков методами синтеза, в том числе с использованием сшивающих агентов. Например, можно создавать иммунотоксины по реакции дисульфидного обмена или путем образования тиоэфирной связи. Примеры подходящих реагентов для этой цели включают иминотиолат и метил-4-меркаптобутиримидат и те реагенты, которые приведены, к примеру, в U.S. Patent No. 4,676,980.
Инженерия эффекторных функций
Может потребоваться модифицировать представленные здесь антитела в отношении эффекторных функций, например, чтобы повысить эффективность антител при лечении рака. Например, можно ввести остатки цистеина в Fc-область, что позволит образовывать межцепочечные дисульфидные связи в этом районе. Полученные при этом гомодимерные антитела могут обладать лучшей способностью к интернализации и/или повышенным опосредованным комплементом уничтожением клеток и антителозависимой клеточной цитотоксичностью (ADCC). См. Caron et al., J. Exp. Med., 176: 1191-1195 (1992); и Shapes, J. Immunol., 148: 2918-2922 (1992). Гомодимерные антитела с повышенной противоопухолевой активностью также можно получить с помощью гетеробифункциональных сшивающих реагентов, как описано в Wolff et al., Cancer Research, 53: 2560-2565 (1993). С другой стороны, можно создать антитела, содержащие сдвоенные Fc-области, при этом улучшатся способности к лизису при помощи комплемента и ADCC. См. Stevenson et al., Anti-Cancer Drug Design 3: 219-230 (1989).
Можно проводить мутации или изменения в последовательности Fc-области для улучшения связывания FcR (напр., FcγR, FcRn). В одном воплощении антитела по изобретению имеют по меньшей мере одну измененную эффекторную функцию, выбранную из группы, состоящей из ADCC, CDC и улучшения связывания с FcRn по сравнению с нативным IgG или исходным антителом. Примеры нескольких полезных специфических мутаций описаны, напр., в Shields RL et al. (2001) JBC 276 (6) 6591-6604; Presta L.G. (2002) Biochemical Society Transactions 30(4): 487-490; и WO 00/42072.
В соответствии с одним воплощением мутация Fc-рецептора представляет собой замену по меньшей мере в одном положении, выбранном из группы, состоящей из: 238, 239, 246, 248, 249, 252, 254, 255, 256, 258, 265, 267, 268, 269, 270, 272, 276, 278, 280, 283, 285, 286, 289, 290, 292, 293, 294, 295, 296, 298, 301, 303, 305, 307, 309, 312, 315, 320, 322, 324, 326, 327, 329, 330, 331, 332, 333, 334, 335, 337, 338, 340, 360, 373, 376, 378, 382, 388, 389, 398, 414, 416, 419, 430, 434, 435, 437, 438 или 439 в Fc-области, причем нумерация остатков в Fc-области соответствует системе нумерации EU. В некоторых воплощениях мутация Fc-рецептора представляет собой замену D265A. В некоторых воплощениях мутация Fc-рецептора представляет собой замену N297A. Другие подходящие мутации изложены в U.S. Patent No. 7,332,581.
Иммуноконъюгаты
Изобретение также касается иммуноконъюгатов, содержащих антитела либо их варианты или мутанты, конъюгированные с цитотоксическими средствами типа химиотерапевтических средств, токсинов (напр., ферментативно активных токсинов бактериального, грибкового, растительного или животного происхождения либо их фрагментов) или радиоактивных изотопов (т.е. радиоконъюгатов).
Ферментативно активные токсины и их фрагменты, которые можно использовать, включают цепь A дифтерийного токсина, несвязывающие активные фрагменты дифтерийного токсина, цепь A экзотоксина (из Pseudomonas aeruginosa), цепь A рицина, цепь A абрина, цепь A модецина, альфа-сарцин, белки Aleurites fordii, белки диантина, белки Phytolaca americana (PAPI, PAPII и PAP-S), ингибитор из Momordica charantia, курцин, кротин, ингибитор из Sapaonaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трихотецены. Для получения радиоконъюгированных антител доступны различные радионуклиды. Их примеры включают 212Bi, 131I, 131In, 90Y и 186Re. Типичные химиотерапевтические средства, применимые для получения таких иммуноконъюгатов, включают описанные здесь в другом месте.
В некоторых воплощениях представленные здесь антитела против PD-L1 и/или их варианты или мутанты конъюгированы с майтанзином, майтансиноидом или калихеамицином. В некоторых воплощениях представленные здесь антитела против PD-L1 и/или их варианты или мутанты конъюгированы с мейтансиноидом DM1.
Конъюгаты антител и цитотоксических средств получают с помощью различных бифункциональных реагентов для конъюгирования белков типа N-сукцинимидил-3-(2-пиридилдитиол)пропионата (SPDP), иминотиолана (IT), бифункциональных производных имидоэфиров (типа диметиладипимидата⋅HCl), активных сложных эфиров (типа дисукцинимидилсуберата), альдегидов (типа глутаральдегида), бис-азидосоединений (типа бис-(п-азидобензоил)гександиамина), производных бисдиазония (типа бис-(п-диазонийбензоил)этилендиамина), диизоцианатов (типа толуол-2,6-диизоцианата) и бис-активных соединений фтора (типа 1,5-дифтор-2,4-динитробензола). Например, иммунотоксин рицина можно получить, как описано в Vitetta et al., Science, 238: 1098 (1987). Типичным хелатирующим реагентом для конъюгирования радионуклеотидов с антителами является меченная 14C 1-изотиоцианатобензил-3-метилдиэтилентриаминпентауксусная кислота (MX-DTPA). См. WO 94/11026.
В другом воплощении антитела либо их варианты или мутанты могут быть конъюгированы с “рецептором” (типа стрептавидина) для применения при предварительном наведении на опухоль, при этом пациенту вводят конъюгат антитело-рецептор с последующим удалением несвязавшегося конъюгата из кровотока с помощью связывающего реагента, а затем вводят “лиганд” (напр., авидин), конъюгированный с цитотоксическим средством (напр., радионуклеотидом).
Ковалентные модификации
В рамки изобретения входят ковалентные модификации антител против PD-L1, их вариантов, мутантов и фрагментов. Один тип ковалентных модификаций включает реакции целевых аминокислотных остатков полипептида с органическим дериватизирующим реагентом, способным реагировать с выбранными боковыми цепями либо N- или С-концевыми остатками полипептида. Дериватизация с помощью бифункциональных реагентов применима, к примеру, для сшивания полипептида с нерастворимой в воде подложкой или поверхностью для применения в способе очистки антител и наоборот. К распространенным сшивающим реагентам относятся, напр., 1,1-бис-(диазоацетил)-2-фенилэтан, глутаральдегид, N-гидроксисукцинимидные эфиры, к примеру, сложные эфиры с 4-азидосалициловой кислотой, гомобифункциональные имидоэфиры, включая дисукцинимидиловые эфиры типа 3,3′-дитиобис(сукцинимидил-пропионата), бифункциональные малеимиды типа бис-N-малеимидо-1,8-октана, и такие реагенты, как метил-3-[(п-азидофенил)-дитио]пропиоимидат.
Другие модификации включают дезамидирование остатков глутамина и аспарагина до соответствующих остатков глутаминовой и аспарагиновой кислоты, соответственно, гидроксилирование пролина и лизина, фосфорилирование гидроксильных групп остатков серина или треонина, метилирование α-аминогрупп боковых цепей лизина, аргинина и гистидина (T.E. Creighton. Proteins: Structure and Molecular Properties. W.H. Freeman & Co., San Francisco, pp. 79-86 (1983)), ацетилирование N-концевых аминогрупп и амидирование C-концевых карбоксигрупп.
Другой тип ковалентных модификаций полипептидов составляет присоединение к полипептидам одного из различных небелковых полимеров, например, полиэтиленгликоля (ПЭГ), полипропиленгликоля или полиоксиалкиленов, способом, изложенным в U.S. Patent Nos. 4,640,835; 4,496,689; 4,301,144; 4,670,417; 4,791,192 или 4,179,337.
Химерные молекулы
Антитела против PD-L1, их варианты, мутанты и/или фрагменты по настоящему изобретению также могут быть модифицированы, если это выгодно, путем образования химерных молекул, содержащих полипептид, слитый с другим, гетерологичным полипептидом или аминокислотной последовательностью (например, иммуноадгезинов или пептидоантител).
В одном воплощении такие химерные молекулы включают слияние полипептида с доменом трансдукции белка, который наводит полипептид для доставки в различные ткани, в особенности через гематоэнцефалический барьер, используя, к примеру, домен трансдукции белка из белка TAT вируса иммунодефицита человека (Schwarze et al., 1999, Science 285: 1569-72).
В другом воплощении такие химерные молекулы включают слияние полипептида с полипептидом-меткой, который обеспечивает эпитоп, с которым антитело против метки может избирательно связываться. Эпитопная метка обычно размещается на N- или C-конце полипептида. Присутствие таких форм полипептида с эпитопной меткой можно обнаружить с помощью антител против полипептида-метки. Кроме того, наличие эпитопной метки облегчает очистку полипептида аффинными методами с помощью антител против метки или аффинного матрикса другого типа, связывающегося с эпитопной меткой. В данной области известны различные полипептиды-метки и соответствующие им антитела. Примеры включают метки типа полигистидина (поли-His) или полигистидин-глицина (поли-His-Gly); метки на основе полипептида HA вируса гриппа и его антитело 12CA5 (Field et al., Mol. Cell. Biol., 8: 2159-2165 (1988)]; метки c-myc и антитела 8F9, 3C7, 6E10, G4, B7 и 9E10 к нему (Evan et al., Molecular and Cellular Biology, 5: 3610-3616 (1985)] и метки на основе гликопротеина D (gD) вируса Herpes simplex и антитела к нему (Paborsky et al., Protein Engineering, 3(6): 547-553 (1990)]. Другие полипептидные метки включают пептид Flag (Hopp et al., BioTechnology, 6: 1204-1210 (1988)]; эпитопный пептид KT3 (Martin et al., Science, 255: 192-194 (1992)], эпитопный пептид α-тубулина (Skinner et al., J. Biol. Chem., 266: 15163-15166 (1991)] и пептидная метка на основе белка гена 10 T7 (Lutz-Freyermuth et al., Proc. Natl. Acad. Sci. USA, 87: 6393-6397 (1990)).
В альтернативном воплощении химерные молекулы могут включать слияние полипептида с иммуноглобулином или определенной областью иммуноглобулина. Для бивалентных форм химерных молекул (напр., “иммуноадгезинов”) такое слияние может происходить с Fc-областью молекулы IgG. Слияния с Ig по изобретению включают полипептиды, содержащие примерно или только остатки 94-243, остатки 33-53 или остатки 33-52 Ig человека вместо по меньшей мере одной вариабельной области в молекуле Ig. В особенно предпочтительном воплощении слияние с иммуноглобулином включает шарнир, СН2 и СН3 либо шарнир и участки СН1, СН2 и СН3 молекулы IgG1. Насчет получения слияний с иммуноглобулином также см. U.S. Patent No. 5,428,130 от 27 июня 1995 г.
Иммунолипосомы
Представленные здесь антитела также могут быть составлены в виде иммунолипосом. Липосомы, содержащие антитела, получают известными в данной области способами типа описанных в Epstein et al., PNAS USA, 82: 3688 (1985); Hwang et al., PNAS USA, 77: 4030 (1980); и U.S. Pat. Nos. 4,485,045 и 4,544,545. Липосомы с повышенным временем циркуляции раскрыты в U.S. Patent No. 5,013,556.
Особенно полезные липосомы можно получить методом обратнофазового испарения с липидной композицией, содержащей фосфатидилхолин, холестерин и дериватизированный PEG фосфатидилэтаноламин (PEG-PE). Липосомы пропускают через фильтры с заданным размером пор, получая липосомы требуемого диаметра. Fab-фрагменты антител настоящего изобретения можно конъюгировать с липосомами, как описано в Martinet al., J. Biol. Chem., 257: 286-288 (1982), по реакции дисульфидного обмена. Липосомы также могут содержать противоопухолевые средства, средства, ингибирующие рост, или химиотерапевтические средства (типа доксорубицина). См. Gabizon et al., J. National Cancer Inst., 81(19): 1484 (1989).
Лечение с помощью антител против PD-L1 и их вариантов
Представленные здесь антитела против PD-L1, их варианты, мутанты и/или фрагменты и/или композиции можно вводить субъектам (напр., млекопитающим типа человека) для лечения заболеваний и расстройств, связанных с аномальной активностью PD-L1, включая, к примеру, рак (как-то рак головы и шеи, рак горла, колоректальный рак, рак легких и т.п.). В некоторых воплощениях изобретения предусмотрены описанные здесь антитела против PD-L1 и/или их варианты (либо фрагменты) для применения при изготовлении лекарственных средств для лечения рака (как-то меланомы, NSCLC, рака головы и шеи, рака уротелия, рака молочной железы (напр., тройного отрицательного рака молочной железы, TNBC), рака желудка, классической лимфомы Ходжкина (cHL), неходжкинской лимфомы - первичной B-клеточной лимфомы средостения (NHL PMBCL), мезотелиомы, рака яичников, рака легких (напр., мелкоклеточного рака легких), рака пищевода, карциномы носоглотки (NPC), рака желчных путей, колоректального рака, рака шейки матки, рака щитовидной железы) у субъектов. В некоторых воплощениях изобретения предусмотрены описанные здесь антитела против PD-L1 и/или их варианты (либо фрагменты) для применения при лечении рака (типа меланомы, NSCLC, рака головы и шеи, рака уротелия, рака молочной железы (напр., тройного отрицательного рака молочной железы, TNBC), рака желудка, классической лимфомы Ходжкина (cHL), неходжкинской лимфомы - первичной B-клеточной лимфомы средостения (NHL PMBCL), мезотелиомы, рака яичников, рака легких (напр., мелкоклеточного рака легких), рака пищевода, карциномы носоглотки (NPC), рака желчных путей, колоректального рака, рака шейки матки, рака щитовидной железы) у субъектов.
В некоторых воплощениях изобретения предусмотрены фармацевтические композиции, содержащие представленные здесь антитела против PD-L1 и/или их варианты или мутанты (либо фрагменты), для применения при лечении рака (меланомы, NSCLC, рака головы и шеи, рака уротелия, рака молочной железы (напр., тройного отрицательного рака молочной железы, TNBC), рака желудка, классической лимфомы Ходжкина (cHL), неходжкинской лимфомы - первичной B-клеточной лимфомы средостения (NHL PMBCL), мезотелиомы, рака яичников, рака легких (напр., мелкоклеточного рака легких), рака пищевода, карциномы носоглотки (NPC), рака желчных путей, колоректального рака, рака шейки матки, рака щитовидной железы) у субъектов. В некоторых воплощениях подлежащими лечению субъектами являются млекопитающие (напр., человек, другие приматы, крысы, мыши, коровы, лошади, свиньи, овцы, козы, собаки, кошки и т.д.). В некоторых воплощениях субъектом является человек. В некоторых воплощениях субъектами являются клинические пациенты, добровольцы клинических испытаний, экспериментальные животные и т.п. В некоторых воплощениях есть подозрение на наличие у субъекта или риск заболевания раком (типа меланомы, NSCLC, рака головы и шеи, рака уротелия, рака молочной железы (напр., тройного отрицательного рака молочной железы, TNBC), рака желудка, классической лимфомы Ходжкина (cHL), неходжкинской лимфомы - первичной B-клеточной лимфомы средостения (NHL PMBCL), мезотелиомы, рака яичников, рака легких (напр., мелкоклеточного рака легких), рака пищевода, карциномы носоглотки (NPC), рака желчных путей, колоректального рака, рака шейки матки, рака щитовидной железы) или поставлен диагноз рака или какого-либо другого заболевания с аномальной экспрессией или активностью PD-L1.
Многие методы диагностики рака (как-то меланомы, NSCLC, рака головы и шеи, рака уротелия, рака молочной железы (напр., тройного отрицательного рака молочной железы, TNBC), рака желудка, классической лимфомы Ходжкина (cHL), неходжкинской лимфомы - первичной B-клеточной лимфомы средостения (NHL PMBCL), мезотелиомы, рака яичников, рака легких (напр., мелкоклеточного рака легких), рака пищевода, карциномы носоглотки (NPC), рака желчных путей, колоректального рака, рака шейки матки, рака щитовидной железы) или каких-либо других заболеваний с аномальной активностью PD-L1 и клинические описания этих заболеваний известны в данной области. Такие методы включают, без ограничения, напр., иммуногистохимию, ПЦР, флуоресцентную гибридизацию in situ (FISH). Более подробно методы диагностики аномальной активности или экспрессии PD-L1 описаны, напр., в Gupta et al. (2009) Mod Pathol. 22(1): 128-133; Lopez-Rios et al. (2013) J Clin Pathol. 66(5): 381-385; Ellison et al. (2013) J Clin Pathol 66(2): 79-89; и Guha et al. (2013) PLoS ONE 8(6): e67782.
Введение может осуществляться любым подходящим способом, в том числе, например, внутривенно, внутримышечно или подкожно. В некоторых воплощениях представленные здесь антитела против PD-L1 и/или их варианты или мутанты (либо фрагменты) и/или композиции вводятся в комбинации со вторым, третьим или четвертым средством (включая, напр., противоопухолевые средства, средства, ингибирующие рост, цитотоксические средства или химиотерапевтические средства) для лечения заболеваний или расстройств, связанных с аномальной активностью PD-L1. К таким средствам относятся, напр., доцетаксель, гефитиниб, FOLFIRI (иринотекан, 5-фторурацил и лейковорин), иринотекан, цисплатин, карбоплатин, паклитаксель, бевацизумаб (антитело против VEGF), FOLFOX-4, инфузионный фторурацил, лейковорин и оксалиплатин, афатиниб, гемцитабин, капецитабин, пеметрексед, тивантиниб, эверолимус, CpG-ODN, рапамицин, леналидомид, вемурафениб, эндостатин, лапатиниб, PX-866, Imprime PGG и ирлотиниб. В некоторых воплощениях антитела против PD-L1 и/или их варианты (либо фрагменты) конъюгированы с дополнительным средством.
В некоторых воплощениях представленные здесь антитела против PD-L1 и/или их варианты или мутанты (либо фрагменты) и/или композиции вводятся в сочетании с одной или несколькими дополнительными терапиями типа лучевой терапии, хирургии, химиотерапии и/или целевой терапии. В некоторых воплощениях представленные здесь антитела против PD-L1 и/или их варианты (либо фрагменты) и/или композиции вводятся в сочетании с лучевой терапией. В некоторых воплощениях комбинации из представленных здесь антител против PD-L1 и/или их вариантов (либо фрагментов) и/или композиций и лучевой терапии применяются для лечения рака, выбранного из группы, состоящей из меланомы, NSCLC, рака головы и шеи, рака уротелия, рака молочной железы (напр., тройного отрицательного рака молочной железы, TNBC), рака желудка, классической лимфомы Ходжкина (cHL), неходжкинской лимфомы - первичной B-клеточной лимфомы средостения (NHL PMBCL), мезотелиомы, рака яичников, рака легких (напр., мелкоклеточного рака легких), рака пищевода, карциномы носоглотки (NPC), рака желчных путей, колоректального рака, рака шейки матки и рака щитовидной железы.
В зависимости от показания, подлежащего лечению, и факторов, относящихся к дозировке, которые должны быть известны врачам - специалистам в данной области, представленные здесь антитела против PD-L1, их варианты, мутанты или фрагменты должны вводиться в дозировке, эффективной для лечения этого показания, сводя к минимуму токсичность и побочные эффекты. Для лечения рака (типа меланомы, NSCLC, рака головы и шеи, рака уротелия, рака молочной железы (напр., тройного отрицательного рака молочной железы, TNBC), рака желудка, классической лимфомы Ходжкина (cHL), неходжкинской лимфомы - первичной B-клеточной лимфомы средостения (NHL PMBCL), мезотелиомы, рака яичников, рака легких (напр., мелкоклеточного рака легких), рака пищевода, карциномы носоглотки (NPC), рака желчных путей, колоректального рака, рака шейки матки, рака щитовидной железы) типичная доза может составлять, к примеру, от 0,001 до 1000 мкг; однако дозы ниже или выше этого типичного диапазона входят в рамки изобретения. Суточная доза может составлять от 0,1 мкг/кг до 100 мг/кг общего веса тела (напр., около 5 мкг/кг, 10 мкг/кг, 100 мкг/кг, 500 мкг/кг, 1 мг/кг, 50 мг/кг или в пределах, заданных любыми двумя из вышеприведенных значений), предпочтительно от 0,3 мкг/кг до 10 мг/кг общего веса тела (напр., около 0,5 мкг/кг, 1 мкг/кг, 50 мкг/кг, 150 мкг/кг, 300 мкг/кг, 750 мкг/кг, 1,5 мг/кг, 5 мг/кг или в пределах, заданных любыми двумя из вышеприведенных значений), более предпочтительно от 1 мкг/кг до 1 мг/кг общего веса тела (напр., около 3 мкг/кг, 15 мкг/кг, 75 мкг/кг, 300 мкг/кг, 900 мкг/кг или в пределах, заданных любыми двумя из вышеприведенных значений) и еще более предпочтительно от 0,5 до 10 мг/кг общего веса тела в день (напр., около 2 мг/кг, 4 мг/кг, 7 мг/кг, 9 мг/кг или в пределах, заданных любыми двумя из вышеприведенных значений, включая любые интервалы между вышеприведенными значениями). Как отмечалось выше, терапевтическая или профилактическая эффективность может контролироваться путем периодической оценки проходящих лечение пациентов. Для повторных введений на протяжении нескольких дней или дольше, в зависимости от состояния, лечение повторяют до тех пор, пока не произойдет требуемое подавление симптомов заболевания. Однако могут применяться и входят в рамки изобретения и другие схемы дозировки. Нужные дозы можно вводить путем однократного введения композиции болюсом, многократного введения композиции болюсом или непрерывного введения композиции вливанием.
Фармацевтические композиции, содержащие антитела против PD-L1, их варианты, мутанты либо фрагменты, можно вводить один, два, три или четыре раза в день. Композиции также можно вводить реже, а не ежедневно, к примеру, шесть раз в неделю, пять раз в неделю, четыре раза в неделю, три раза в неделю, два раза в неделю, один раз в неделю, один раз в две недели, один раз в три недели, раз в месяц, раз в два месяца, раз в три месяца или один раз в шесть месяцев. Композиции также можно вводить в составах с замедленным высвобождением типа имплантатов, из которых постепенно высвобождается композиция на протяжении какого-то периода времени, что позволяет вводить композиции реже, как-то раз в месяц, один раз в 2-6 месяцев, раз в год или даже всего один раз. Препараты с замедленным высвобождением (типа гранул, наночастиц, микрочастиц, наносфер, микросфер и т.п.) могут вводиться посредством инъекции.
Антитела и/или их варианты или мутанты (либо фрагменты) можно вводить в виде одной суточной дозы или же общую суточную дозу можно вводить в дробных дозах по два, три или четыре раза в день. Композиции также можно вводить реже, а не ежедневно, к примеру, шесть раз в неделю, пять раз в неделю, четыре раза в неделю, три раза в неделю, два раза в неделю, один раз в неделю, один раз в две недели, один раз в три недели, раз в месяц, раз в два месяца, раз в три месяца или один раз в шесть месяцев. Антитела (либо их фрагменты) также можно вводить в составах с замедленным высвобождением типа имплантатов, из которых постепенно высвобождается композиция на протяжении какого-то периода времени, что позволяет вводить композиции реже, как-то раз в месяц, один раз в 2-6 месяцев, раз в год или даже всего один раз. Препараты с замедленным высвобождением (типа гранул, наночастиц, микрочастиц, наносфер, микросфер и т.п.) могут вводиться посредством инъекции или имплантироваться хирургическим путем в различных местах.
Лечение рака можно оценивать, напр., но без ограничения, по регрессии опухоли, уменьшению массы или размера опухоли, времени до прогрессирования, продолжительности выживания, выживаемости без прогрессирования, общей восприимчивости, продолжительности реакции, качеству жизни, экспрессии и/или активности белка. Для определения эффективности терапии можно использовать подходы, включающие, к примеру, измерение реакции на лечение посредством радиологической визуализации.
В некоторых воплощениях эффективность лечения измеряется в виде процента ингибирования роста опухолей (% TGI), который рассчитывают по уравнению 100 - (T/C ×100), где T - средний относительный объем опухоли после обработки, а C - средний относительный объем опухоли без обработки. В некоторых воплощениях % TGI составляет примерно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 91%, 92%, 93%, 94%, 95% или более 95%. В некоторых воплощениях % TGI у антител против PD-L1 и/или их вариантов или мутантов является таким же или больше, чем % TGI у контрольного антитела против PD-L1, как-то примерно в 1,1 раз, 1,2 раз, 1,3 раз, 1,4 раз, 1,5 раз, 1,6 раз, 1,7 раз, 1,8 раз, 1,9 раз, в 2 раза, 2,1 раза, 2,2 раза, 2,3 раза, 2,4 раза, 2,5 раза, 2,6 раза, 2,7 раза, включая любые интервалы между этими значениями, или более чем в 2,7 раза больше, чем % TGI у контрольного антитела против PD-L1.
Лекарственные формы
Антитела против PD-L1 и/или их варианты или мутанты (либо фрагменты) могут быть составлены с подходящими носителями или наполнителями с тем, чтобы они были пригодны для введения. Подходящие составы антител получают путем смешивания антитела (или его фрагмента), имеющего требуемую степень чистоты, с необязательными фармацевтически приемлемыми носителями, наполнителями или стабилизаторами (Remington’s Pharmaceutical Sciences, 16th edition, Osol A., Ed. (1980)), в виде лиофилизированных форм или водных растворов. Приемлемые носители, наполнители или стабилизаторы нетоксичны для реципиентов при используемых дозировках и концентрациях и включают буферы типа фосфатного, цитратного и других органических кислот; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (как-то октадецилдиметилбензиламмония хлорид; гексаметония хлорид; бензалкония хлорид, бензетония хлорид; фенол, бутиловый или бензиловый спирт; алкилпарабены типа метил- или пропилпарабена; катехол; резорцинол; циклогексанол; 3-пентанол; и м-крезол); низкомолекулярные (менее 10 остатков) полипептиды; такие белки, как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры типа оливинилпирролидона; такие аминокислоты, как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелаторы типа ЭДТА; такие сахара, как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы типа натрия; комплексы металлов (напр., комплексы Zn-белок); и/или неионные поверхностно-активные вещества типа Tween™, Pluronics™ или полиэтиленгликоля (PEG). Типичные лекарственные формы антител описаны в WO 98/56418, прямо включенном сюда путем ссылки. Лиофилизованные формы, адаптированные для подкожного введения, описаны в WO 97/04801. Такие лиофилизированные формы могут быть восстановлены с подходящим разбавителем до высокой концентрации белка, и эти восстановленные формы можно вводить подкожно млекопитающим, подлежащим такому лечению.
Приведенные здесь лекарственные формы также могут содержать более чем одно активное соединение, если это необходимо для лечения данного конкретного показания, предпочтительно из числа обладающих комплементарными активностями и не оказывающих вредного воздействия друг на друга. Например, может потребоваться дополнительно обеспечить противоопухолевое средство, средство, ингибирующее рост, цитотоксическое средство или химиотерапевтическое средство. Такие молекулы соответствующим образом присутствуют в комбинации в количествах, эффективных для их назначения. Эффективное количество таких других средств зависит от количества антитела, присутствующего в препарате, типа заболевания или расстройства или лечения и других факторов, обсуждаемых выше. Обычно они применяются в тех же дозировках и теми же способами введения, как описано здесь, или же от 1 до 99% от применявшихся до этого дозировок. Активные ингредиенты также могут быть заключены в микрокапсулы, полученные, к примеру, методами коацервации или межфазной полимеризации, к примеру, микрокапсулы из гидроксиметилцеллюлозы или желатина и микрокапсулы из поли(метилметакрилата), соответственно, в коллоидных системах доставки лекарственных средств (напр., в липосомах, альбуминовых микросферах, микроэмульсиях, наночастицах и нанокапсулах) либо в макроэмульсиях. Такие методы приведены в Remington’s Pharmaceutical Sciences, 16th edition, Osol A., Ed. (1980). Можно приготовить препараты с замедленным высвобождением. Подходящие примеры препаратов с замедленным высвобождением включают полупроницаемые матриксы из твердых гидрофобных полимеров, содержащие антагонисты, причем такие матриксы имеют вид формованных изделий, напр., пленок или микрокапсул. Примеры матрикса для замедленного высвобождения включают полиэфиры, гидрогели (к примеру, поли(2-гидроксиэтилметакрилат) или поли(виниловый спирт)), полилактиды (U.S. Pat. No. 3,773,919), сополимеры L-глутаминовой кислоты и этил-L-глутамата, неразлагаемые этилен-винилацетаты, разлагаемые сополимеры молочной и гликолевой кислоты типа Lupron Depot™ (микросферы для инъекций, состоящие из сополимера молочной и гликолевой кислоты и лейпролида ацетата) и поли-D-(-)-3-гидроксимасляную кислоту.
Для доставки полипептидов и антител (либо их фрагментов) или композиций по изобретению в клетки можно использовать липофектины или липосомы. При использовании фрагментов антител предпочтительными являются наименьшие ингибирующие фрагменты, специфически связывающиеся с доменом связывания у целевого белка. Например, на основе последовательностей вариабельной области антител можно разработать такие пептидные молекулы, которые сохраняют способность к связыванию с последовательностью целевого белка. Такие пептиды могут быть синтезированы химически и/или получены по технологии рекомбинантной ДНК. Напр., см. Marasco et al., PNAS USA, 90: 7889-7893 (1993).
Активные ингредиенты также могут быть заключены в микрокапсулы, полученные, к примеру, методами коацервации или межфазной полимеризации, к примеру, микрокапсулы из гидроксиметилцеллюлозы или желатина и микрокапсулы из поли(метилметакрилата), соответственно, в коллоидных системах доставки лекарственных средств (напр., в липосомах, альбуминовых микросферах, микроэмульсиях, наночастицах и нанокапсулах) либо в макроэмульсиях. Такие методы приведены в Remington’s Pharmaceutical Sciences, supra).
Можно приготовить препараты с замедленным высвобождением. Подходящие примеры препаратов с замедленным высвобождением включают полупроницаемые матриксы из твердых гидрофобных полимеров, содержащие антитела (либо их фрагменты), причем такие матриксы имеют вид формованных изделий, напр., пленок или микрокапсул. Примеры матрикса для замедленного высвобождения включают сложные полиэфиры, гидрогели (к примеру, поли(2-гидроксиэтилметакрилат) или поли(виниловый спирт)), полилактиды (U.S. Pat. No. 3,773,919), сополимеры L-глутаминовой кислоты и этил-L-глутамата, неразлагаемые этилен-винилацетаты, разлагаемые сополимеры молочной и гликолевой кислоты типа Lupron Depot™ (микросферы для инъекций, состоящие из сополимера молочной и гликолевой кислоты и лейпролида ацетата) и поли-D-(-)-3-гидроксимасляную кислоту. Хотя такие полимеры, как этилен-винилацетат и молочная кислота-гликолевая кислота, обеспечивают высвобождение молекул в течение более 100 дней, некоторые гидрогели высвобождают белки в течение более короткого периода времени. Если инкапсулированные антитела остаются в организме в течение длительного времени, они могут денатурировать или агрегировать в результате воздействия влаги при 37°C, что приводит к потере биологической активности и возможным изменениям в иммуногенности. Для стабилизации могут быть разработаны рациональные стратегии в зависимости от конкретного механизма. Например, если обнаружится, что механизм агрегации заключается в образовании межмолекулярной S-S-связи посредством тиодисульфидного обмена, то стабилизация может достигаться путем модификации сульфгидрильных остатков, лиофилизации из кислых растворов, контроля за содержанием влаги с помощью соответствующих добавок и разработки специальных композиций полимерного матрикса.
В некоторых воплощениях лекарственные формы содержат описанные здесь антитела против PD-L1 и/или их варианты или мутанты в концентрации более 0,5 мг/мл, более 1 мг/мл, более 2 мг/мл, более 3 мг/мл, более 4 мг/мл, более 5 мг/мл, более 6 мг/мл, более 7 мг/мл, более 8 мг/мл, более 9 мг/мл, более 10 мг/мл, более 11 мг/мл, более 12 мг/мл, более 13 мг/мл, более 14 мг/мл, более 15 мг/мл, более 16 мг/мл, более 17 мг/мл, более 18 мг/мл, более 19 мг/мл, более 20 мг/мл, более 21 мг/мл, более 22 мг/мл, более 23 мг/мл, более 24 мг/мл, более 25 мг/мл, более 26 мг/мл, более 27 мг/мл, более 28 мг/мл, более 29 мг/мл или более 30 мг/мл, включая любые интервалы между этими значениями.
В некоторых воплощениях антитела против PD-L1 и/или их варианты или мутанты составляют (напр., в концентрации более 0,5 мг/мл, более 1 мг/мл, более 5 мг/мл, более 10 мг/мл, более 15 мг/мл, более 20 мг/мл или более 25 мг/мл, включая любые интервалы между этими значениями) в буфере, содержащем цитрат, NaCl, ацетат, сукцинат, глицин, полисорбат 80 (Tween 80) или любые комбинации из вышеперечисленных. В некоторых воплощениях антитела против PD-L1 и/или их варианты или мутанты составляют (напр., в концентрации более 0,5 мг/мл, более 1 мг/мл, более 5 мг/мл, более 10 мг/мл, более 15 мг/мл, более 20 мг/мл или более 25 мг/мл, включая любые интервалы между этими значениями) в буфере, содержащем от 100 мМ до 150 мМ глицина. В некоторых воплощениях антитела против PD-L1 и/или их варианты или мутанты составляют в буфере, содержащем от 50 мМ до 100 мМ NaCl. В некоторых воплощениях антитела против PD-L1 и/или их варианты или мутанты составляют (напр., в концентрации более 0,5 мг/мл, более 1 мг/мл, более 5 мг/мл, более 10 мг/мл, более 15 мг/мл, более 20 мг/мл или более 25 мг/мл, включая любые интервалы между этими значениями) в буфере, содержащем от 10 мМ до 50 мМ ацетата. В некоторых воплощениях антитела против PD-L1 и/или их варианты или мутанты составляют в буфере, содержащем от 10 мМ до 50 мМ сукцината. В некоторых воплощениях антитела против PD-L1 и/или их варианты или мутанты составляют (напр., в концентрации более 0,5 мг/мл, более 1 мг/мл, более 5 мг/мл, более 10 мг/мл, более 15 мг/мл, более 20 мг/мл или более 25 мг/мл, включая любые интервалы между этими значениями) в буфере, содержащем от 0,005% до 0,02% полисорбата 80. В некоторых воплощениях антитела против PD-L1 и/или их варианты или мутанты составляют в буфере со значением рН от 5,1 до 5,6. В некоторых воплощениях антитела против PD-L1 и/или их варианты или мутанты составляют в буфере, содержащем 10 мМ цитрата, 100 мМ NaCl, 100 мМ глицина и 0,01% полисорбата 80, причем данные формы имеют рН = 5,5.
В некоторых воплощениях лекарственные формы (типа форм, включающих буфер, содержащий 10 мМ цитрата, 100 мМ NaCl, 100 мМ глицина и 0,01% полисорбата 80, причем данные формы имеют рН = 5,5), содержащие описанные здесь антитела против PD-L1 и/или их варианты или мутанты (напр., в концентрации более 0,5 мг/мл, более 1 мг/мл, более 5 мг/мл, более 10 мг/мл, более 15 мг/мл, более 20 мг/мл или более 25 мг/мл, включая любые интервалы между этими значениями), являются стабильными при комнатной температуре (как-то при 20-25°C) в течение 0,5 недели, 1,0 недели, 1,5 недель, 2,0 недель, 2,5 недель, 3,5 недель, 4,0 недель, 4,5 недель или 5,0 недель, включая любые интервалы между этими значениями. В некоторых воплощениях лекарственные формы (типа форм, включающих буфер, содержащий 10 мМ цитрата, 100 мМ NaCl, 100 мМ глицина и 0,01% полисорбата 80, причем данные формы имеют рН = 5,5), содержащие описанные здесь антитела против PD-L1 и/или их варианты или мутанты (напр., в концентрации более 0,5 мг/мл, более 1 мг/мл, более 5 мг/мл, более 10 мг/мл, более 15 мг/мл, более 20 мг/мл или более 25 мг/мл, включая любые интервалы между этими значениями), являются стабильными в ускоренных условиях (типа хранения при 37°C) в течение 0,5 недели, 1,0 недели, 1,5 недель, 2,0 недель, 2,5 недель, 3,5 недель, 4,0 недель, 4,5 недель или 5,0 недель, включая любые интервалы между этими значениями.
Эксклюзионная хроматография по размерам (SEC) является хорошо известным и распространенным методом, который применяется при исследовании стабильности белков для выявления потенциальной фрагментации и агрегации, соответствующей физической и химической нестабильности. В некоторых воплощениях лекарственные формы, содержащие 5 мг/мл, 10 мг/мл, 15 мг/мл, 20 мг/мл или 25 мг/мл описанных здесь антител против PD-L1 и/или их вариантов или мутантов, проявляют возрастание высокомолекулярных частиц (HMWS) менее чем на 1,6%, 1,4%, 1,2%, 1,0%, 0,8%, 0,6%, 0,4%, 0,2% или 0,1% через 1 неделю при 37°C по сравнению с исходным % высокомолекулярных частиц при измерении методом SEC, включая любые интервалы между этими значениями. В некоторых воплощениях лекарственные формы, содержащие 5 мг/мл, 10 мг/мл, 15 мг/мл, 20 мг/мл или 25 мг/мл описанных здесь антител против PD-L1 и/или их вариантов или мутантов, проявляют возрастание высокомолекулярных частиц (HMWS) менее чем на 2,0%, 1,8%, 1,6%, 1,4%, 1,2%, 1,0%, 0,8%, 0,6%, 0,4%, 0,2% или 0,1% через 2 недели при 37°C по сравнению с исходным % высокомолекулярных частиц при измерении методом SEC, включая любые интервалы между этими значениями. В некоторых воплощениях лекарственные формы, содержащие 5 мг/мл, 10 мг/мл, 15 мг/мл, 20 мг/мл или 25 мг/мл описанных здесь антител против PD-L1 и/или их вариантов или мутантов, проявляют возрастание высокомолекулярных частиц (HMWS) менее чем на 3,3%, 3,2%, 3,1%, 3,0%, 2,9%, 2,8%, 2,7%, 2,6%, 2,5%, 2,4%, 2,2%, 2,0%, 1,8%, 1,6%, 1,4%, 1,2%, 1,0%, 0,8%, 0,6%, 0,4%, 0,2% или 0,1% через 4 недели при 37°C по сравнению с исходным % высокомолекулярных частиц при измерении методом SEC, включая любые интервалы между этими значениями.
В некоторых воплощениях лекарственные формы, содержащие 5 мг/мл, 10 мг/мл, 15 мг/мл, 20 мг/мл или 25 мг/мл описанных здесь антител против PD-L1 и/или их вариантов или мутантов, проявляют возрастание низкомолекулярных частиц (LMWS) менее чем на 1,6%, 1,4%, 1,2%, 1,0%, 0,8%, 0,6%, 0,4%, 0,2% или 0,1% через 1 неделю при 37°C по сравнению с исходным % низкомолекулярных частиц при измерении методом SEC, включая любые интервалы между этими значениями. В некоторых воплощениях лекарственные формы, содержащие 5 мг/мл, 10 мг/мл, 15 мг/мл, 20 мг/мл или 25 мг/мл описанных здесь антител против PD-L1 и/или их вариантов или мутантов, проявляют возрастание низкомолекулярных частиц (LMWS) менее чем на 2,0%, 1,8%, 1,6%, 1,4%, 1,2%, 1,0%, 0,8%, 0,6%, 0,4%, 0,2% или 0,1% через 2 недели при 37°C по сравнению с исходным % низкомолекулярных частиц при измерении методом SEC, включая любые интервалы между этими значениями. В некоторых воплощениях лекарственные формы, содержащие 5 мг/мл, 10 мг/мл, 15 мг/мл, 20 мг/мл или 25 мг/мл описанных здесь антител против PD-L1 и/или их вариантов или мутантов, проявляют возрастание низкомолекулярных частиц (LMWS) менее чем на 2,4%, 2,2%, 2,0%, 1,8%, 1,6%, 1,4%, 1,2%, 1,0%, 0,8%, 0,6%, 0,4%, 0,2% или 0,1% через 4 недели при 37°C по сравнению с исходным % низкомолекулярных частиц при измерении методом SEC, включая любые интервалы между этими значениями.
В некоторых воплощениях лекарственные формы, содержащие 5 мг/мл, 10 мг/мл, 15 мг/мл, 20 мг/мл или 25 мг/мл описанных здесь антител против PD-L1 и/или их вариантов или мутантов, проявляют снижение содержания мономеров не более чем на 0,2%, 0,4%, 0,6%, 0,8%, 0,9%, 1,0%, 1,1%, 1,2%, 1,3%, 1,4%, 1,6%, 1,7%, 1,8%, 1,9%, 2%, 2,1%, 2,2%, 2,3%, 2,4%, 2,5%, 2,6%, 2,7%, 2,8%, 2,9%, 3,0%, 3,1%, 3,2%, 3,3%, 3,4% или 3,5% через 1 неделю при 37°C по сравнению с исходным % мономеров при измерении методом SEC, включая любые интервалы между этими значениями. В некоторых воплощениях лекарственные формы, содержащие 5 мг/мл, 10 мг/мл, 15 мг/мл, 20 мг/мл или 25 мг/мл описанных здесь антител против PD-L1 и/или их вариантов или мутантов, проявляют снижение содержания мономеров не более чем на 0,2%, 0,4%, 0,6%, 0,8%, 0,9%, 1,0%, 1,1%, 1,2%, 1,3%, 1,4%, 1,6%, 1,7%, 1,8%, 1,9%, 2%, 2,1%, 2,2%, 2,3%, 2,4%, 2,5%, 2,6%, 2,7%, 2,8%, 2,9%, 3,0%, 3,1%, 3,2%, 3,3%, 3,4% или 3,5% через 2 недели при 37°C по сравнению с исходным % мономеров при измерении методом SEC, включая любые интервалы между этими значениями. В некоторых воплощениях лекарственные формы, содержащие 5 мг/мл, 10 мг/мл, 15 мг/мл, 20 мг/мл или 25 мг/мл описанных здесь антител против PD-L1 и/или их вариантов или мутантов, проявляют снижение содержания мономеров не более чем на 0,2%, 0,4%, 0,6%, 0,8%, 0,9%, 1,0%, 1,1%, 1,2%, 1,3%, 1,4%, 1,6%, 1,7%, 1,8%, 1,9%, 2%, 2,1%, 2,2%, 2,3%, 2,4%, 2,5%, 2,6%, 2,7%, 2,8%, 2,9%, 3,0%, 3,1%, 3,2%, 3,3%, 3,4% или 3,5% через 2 недели при 37°C по сравнению с исходным % мономеров при измерении методом SEC, включая любые интервалы между этими значениями.
Катионообменная хроматография (CEX) является хорошо известным и распространенным методом для выявления таких явлений разложения белка, как деамидирование или окисление (Moorhouse et al. (1997). J. Pharm. Biomed. Anal. 16, 593-603). Продукты деградации обычно называют кислыми или основнымипо сравнению с gnts с более высоким кажущимся pI. Кислые продукты - это варианты, которые элюируются раньше, чем главный пик при CEX, а основные - это варианты, которые элюируются позже, чем главный пик при CEX. В некоторых воплощениях фракция кислого пика у лекарственных форм, содержащих 5 мг/мл, 10 мг/мл, 15 мг/мл, 20 мг/мл или 25 мг/мл описанных здесь антител против PD-L1 и/или их вариантов или мутантов, составляет не более 7%, 8%, 9%, 10%, 11%, 12%, 13%, 14% или 15% от общего белка через 1 неделю при 37°C при измерении методом CEX, включая любые интервалы между этими значениями. В некоторых воплощениях фракция кислого пика у лекарственных форм, содержащих 5 мг/мл, 10 мг/мл, 15 мг/мл, 20 мг/мл или 25 мг/мл описанных здесь антител против PD-L1 и/или их вариантов или мутантов, составляет не более 8%, 9%, 10%, 11%, 12%, 13%, 14%, 15%, 16%, 17% или 18% от общего белка через 2 недели при 37°C при измерении методом CEX, включая любые интервалы между этими значениями. В некоторых воплощениях фракция кислого пика у лекарственных форм, содержащих 5 мг/мл, 10 мг/мл, 15 мг/мл, 20 мг/мл или 25 мг/мл описанных здесь антител против PD-L1 и/или их вариантов или мутантов (либо фрагментов), составляет не более 8%, 9%, 10%, 11%, 12%, 13%, 14%, 15%, 16%, 17%, 18%, 19%, 20 %, 21%, 22%, 23%, 24%, 25%, 26% или 27% от общего белка через 2 недели при 37°C при измерении методом CEX, включая любые интервалы между этими значениями.
В некоторых воплощениях фракция основного пика у лекарственных форм, содержащих 5 мг/мл, 10 мг/мл, 15 мг/мл, 20 мг/мл или 25 мг/мл описанных здесь антител против PD-L1 и/или их вариантов или мутантов (либо фрагментов), составляет не более 39%, 40%, 41%, 42%, 43%, 44%, 45% или 46% от общего белка через 1 неделю при 37°C при измерении методом CEX, включая любые интервалы между этими значениями. В некоторых воплощениях фракция основного пика у лекарственных форм, содержащих 5 мг/мл, 10 мг/мл, 15 мг/мл, 20 мг/мл или 25 мг/мл описанных здесь антител против PD-L1 и/или их вариантов или мутантов (либо фрагментов), составляет не более 39%, 40%, 41%, 42%, 43%, 44%, 45% или 46% от общего белка через 2 недели при 37°C при измерении методом CEX, включая любые интервалы между этими значениями. В некоторых воплощениях фракция основного пика у лекарственных форм, содержащих 5 мг/мл, 10 мг/мл, 15 мг/мл, 20 мг/мл или 25 мг/мл описанных здесь антител против PD-L1 и/или их вариантов или мутантов (либо фрагментов), составляет не более 39%, 40%, 41%, 42%, 43%, 44%, 45% или 46% от общего белка через 4 недели при 37°C при измерении методом CEX, включая любые интервалы между этими значениями.
В некоторых воплощениях фракция главного пика у лекарственных форм, содержащих 5 мг/мл, 10 мг/мл, 15 мг/мл, 20 мг/мл или 25 мг/мл описанных здесь антител против PD-L1 и/или их вариантов или мутантов (либо фрагментов), составляет не менее 32%, 33%, 34%, 35%, 36%, 37%, 38%, 39%, 40%, 41%, 42%, 43%, 44%, 45% или 46% от общего белка через 1 неделю при 37°C при измерении методом CEX, включая любые интервалы между этими значениями. В некоторых воплощениях фракция основного пика у лекарственных форм, содержащих 5 мг/мл, 10 мг/мл, 15 мг/мл, 20 мг/мл или 25 мг/мл описанных здесь антител против PD-L1 и/или их вариантов или мутантов (либо фрагментов), составляет не менее 32%, 33%, 34%, 35%, 36%, 37%, 38%, 39%, 40%, 41%, 42%, 43%, 44%, 45% или 46% от общего белка через 2 недели при 37°C при измерении методом CEX, включая любые интервалы между этими значениями. В некоторых воплощениях фракция основного пика у лекарственных форм, содержащих 5 мг/мл, 10 мг/мл, 15 мг/мл, 20 мг/мл или 25 мг/мл описанных здесь антител против PD-L1 и/или их вариантов или мутантов (либо фрагментов), составляет не менее 32%, 33%, 34%, 35%, 36%, 37%, 38%, 39%, 40%, 41%, 42%, 43%, 44%, 45% или 46% от общего белка через 4 недели при 37°C при измерении методом CEX, включая любые интервалы между этими значениями.
Лекарственные формы, предназначенные для введения in vivo, должны быть стерильными. Обычно это осуществляется, например, фильтрованием через мембраны для стерилизации фильтрованием.
Способы диагностики и визуализации с помощью антител против PD-L1 и их вариантов/мутантов
Меченые антитела против PD-L1, их варианты, мутанты, фрагменты и их производные и аналоги, которые специфически связываются с полипептидом PD-L1, можно использовать в диагностических целях для выявления, диагностики или мониторинга заболеваний и/или нарушений, связанных с экспрессией, аномальной экспрессией и/или активностью PD-L1. Например, представленные здесь антитела против PD-L1 и/или их варианты или мутанты (либо фрагменты) можно использовать при диагностических анализах или визуализации in situ, in vivo, ex vivo и in vitro. Способы выявления экспрессии полипептида PD-L1 включают (a) анализ экспрессии полипептида в клетках (напр., ткани) или жидких средах организма с помощью одного или нескольких антител по изобретению и (b) сравнение уровня экспрессии гена со стандартным уровнем экспрессии гена, при этом повышение или снижение уровня экспрессии анализируемого гена по сравнению со стандартным уровнем экспрессии указывает на аномальную экспрессию.
Другие приведенные здесь воплощения включают способы диагностики заболеваний или расстройств, связанных с экспрессией или аномальной экспрессией PD-L1 у животных (напр., млекопитающих типа человека). Способы включают обнаружение молекул PD-L1 у млекопитающих. В некоторых воплощениях диагностика включает: (a) введение эффективного количества меченого антитела против PD-L1 и/или его варианта или мутанта (либо фрагмента) млекопитающим; (b) выжидание в течение какого-то времени после введения для того, чтобы меченое антитело против PD-L1 и/или его вариант или мутант (либо фрагмент) предпочтительно сконцентрировались в тех местах у субъекта, где экспрессируются молекулы PD-L1 (а не связавшиеся меченые молекулы рассеялись до уровня фона); (c) определение уровня фона; и (d) детектирование меченых молекул у субъекта с тем, что обнаружение меченых молекул выше фонового уровня означает то, что у субъекта есть конкретное заболевание или расстройство, связанное с экспрессией или аномальной экспрессией PD-L1. Уровень фона может быть определен различными методами, включая сравнение количества обнаруженных меченых молекул со стандартным значением, предварительно определенным для данной системы.
Представленные здесь антитела против PD-L1 и/или их варианты или мутанты (либо фрагменты) могут применяться для анализа уровня белка в биологических образцах с помощью классических иммуногистологических методов, известных специалистам в данной области (напр., см. Jalkanen et al., J. Cell. Biol. 101: 976-985 (1985); Jalkanen et al., J. Cell. Biol. 105: 3087-3096 (1987)). Другие методы на основе антител, применимые для выявления экспрессии белковых генов, включают иммуноанализы типа иммуноферментного анализа (ELISA) и радиоиммуноанализа (RIA). Подходящие метки для анализа антител известны в данной области и включают ферментные метки типа глюкозооксидазы; такие радиоизотопы, как йод (131I, 125I, 123I, 121I), углерод (14C), сера 35S), тритий (3H), индий (115mIn, 113mIn, 112In, 111In) и технеций (99Tc, 99mTc), таллий (201Ti), галлий (68Ga, 67Ga), палладий (103Pd), молибден (99Mo), ксенон (133Xe), фтор (18F), 153Sm, 177Lu, 159Gd, 149Pm, 140La, 175Yb, 166Ho, 90Y, 47Sc, 186Re, 186Re, 188Re, 142Pr, 105Rh, Ru; люминол; и такие флуоресцентные метки, как флуоресцеин и родамин; а также биотин.
К представленным здесь меченым антителам (либо их фрагментам) могут применяться методы, известные в данной области. Такие методы включают, без ограничения, применение бифункциональных конъюгирующих реагентов (напр., см. U.S. Pat. Nos. 5,756,065; 5,714,631; 5,696,239; 5,652,361; 5,505,931; 5,489,425; 5,435,990; 5,428,139; 5,342,604; 5,274,119; 4,994,560; и 5,808,003).
С другой стороны, можно измерять уровни кодирующей полипептид PD-L1 нуклеиновой кислоты или мРНК в клетках, напр., методом флуоресцентной гибридизации in situ с помощью зонда на основе нуклеиновой кислоты, соответствующего нуклеиновой кислоте, кодирующей PD-L1, или комплементарной к ней (FISH; см. WO 98/45479, опубликованный в октябре 1998 г.), методы southern-блоттинга, northern-блоттинга или полимеразной цепной реакции (ПЦР) типа количественной ПЦР в реальном времени (ОТ-ПЦР). Также можно изучать гиперэкспрессию PD-L1 по измерению выделения антигена в биологической жидкости типа сыворотки, напр., методами на основе антител (также см., напр., U.S. Patent No. 4,933,294 от 12 июня 1990 г.; WO 91/05264 от 18 апреля 1991 г.; U.S. Patent 5,401,638 от 28 марта 1995 г.; и Sias et al., J. Immunol. Methods 132:73-80 (1990)). Помимо вышеуказанных методов, квалифицированным специалистам доступны различные методы in vivo и ex vivo. Например, можно подвергнуть клетки в организме млекопитающих воздействию антител, необязательно помеченных детектируемой меткой, напр., радиоактивным изотопом, и определять связывание антител с клетками, например, путем наружного сканирования на радиоактивность или путем анализа образцов (напр., биоптатов или других биологических образцов), взятых у млекопитающих, подвергнутых до этого воздействию этих антител.
Промышленные изделия и наборы
Другое воплощение, представленное здесь, составляют готовые изделия, содержащие материалы, применимые для лечения рака типа меланомы, NSCLC, рака головы и шеи, рака уротелия, рака молочной железы (например, тройного негативного рака молочной железы, TNBC), рака желудка, классической лимфомы Ходжкина (cHL), неходжкинской лимфомы - первичной B-клеточной лимфомы средостения (NHL PMBCL), мезотелиомы, рака яичников, рака легких (напр., мелкоклеточного рака легких), рака пищевода, карциномы носоглотки (NPC), рака желчных путей, колоректального рака, рака шейки матки, рака щитовидной железы и рака слюнных желез. Изделия могут включать контейнер и этикетку или упаковочный вкладыш на контейнере или связанный с ним. Подходящие контейнеры включают, к примеру, бутылки, флаконы, шприцы и т.п. Контейнеры могут быть изготовлены из различных материалов типа стекла или пластика. Как правило, контейнер содержит композицию, которая эффективна для лечения заболевания и может иметь стерильный порт доступа (к примеру, контейнер может представлять собой мешок для внутривенного раствора или флакон с пробкой, прокалываемой иглой для подкожных инъекций). Как минимум одно активное средство в композиции составляет представленное здесь антитело против PD-L1 и/или его вариант или мутант (либо фрагмент). Этикетка или упаковочный вкладыш указывает, что композиция применяется для лечения определенного заболевания. Этикетка или упаковочный вкладыш также должны содержать инструкции по введению композиции антител пациентам. Также предусмотрены изделия и наборы, содержащие описанные здесь средства комбинированной терапии.
Упаковочный вкладыш означает инструкции, которые обычно вставляют в коммерческие упаковки терапевтических продуктов и содержат информацию о показаниях, применении, дозировке, способе введения, противопоказаниях и/или предупреждения относительно применения таких терапевтических продуктов. В одном воплощении в упаковочном вкладыше указывается, что композиция применяется для лечения рака (типа рака головы и шеи, рака легких или колоректального рака).
Кроме того, изделия могут дополнительно содержать второй контейнер, содержащий фармацевтически приемлемый буфер типа бактериостатической воды для инъекций (BWFI), солевого раствора с фосфатным буфером, раствора Рингера и раствора декстрозы. Он также может содержать и другие материалы, желательные с коммерческой и пользовательской точки зрения, включая другие буферы, разбавители, фильтры, иглы и шприцы.
Также предусмотрены наборы, применимые для различных целей, напр., для выделения или выявления PD-L1 у пациентов, необязательно в сочетании с промышленными изделиями. Для выделения и очистки PD-L1 набор может содержать представленное здесь антитело против PD-L1 и/или его вариант или мутант (либо фрагмент), конъюгированные с шариками (напр., шариками Sepharose™). Предусмотрены наборы, которые содержат антитела (или их фрагменты) для обнаружения и определения PD-L1 in vitro, например, методом ELISA или Вестерн-блот. Как и в случае с изделиями, набор содержит контейнер и этикетку или упаковочный вкладыш на контейнере или связанный с ним. К примеру, контейнер содержит композицию, содержащую по меньшей мере одно представленное здесь антитело против PD-L1 и/или его вариант или мутант (либо фрагмент). Могут быть и дополнительные контейнеры, содержащие, например, разбавители и буферы, контрольные антитела. Этикетка или упаковочный вкладыш может содержать описание композиции, а также инструкции для применения по назначению in vitro или для диагностики.
ПРИМЕРЫ
Пример 1. Разработка антител против PD-L1 человека и их вариантов
При разработке антител против PD-L1 вся стратегия сводится к следующему. Идентифицировали 7 положительных лидеров (т.е. PL1, PL2, PL3, PL6, PL8, PL12, PL15) путем скрининга библиотеки фагового дисплея человека с помощью PD-L1_ECD-His человека. В общем, после трех раундов пэннинга с помощью биотинилированного PD-L1_ECD-His человека, конъюгированного с покрытыми стрептавидином магнитными шариками Dynabeads® M-280 (Thermo Fisher Scientific #11205D), проводили скрининг Fab этих лидеров и измеряли их связывание с hPD-L1_ECD-Fc и hPD-L1_ECD-His методом ELISA, а последовательности Fab из семи отобранных лидеров затем клонировали в Fc-остов IgG1 человека с мутацией N297A, получая полноразмерные антитела, которые использовали для проведения дальнейших экспериментов по определению кинетических характеристик, активности связывания PD-L1 на целых клетках, активности блокирования PD-1 и функций in vitro и in vivo. На основании всех этих данных оказалось, что PL2 и PL3 обладают наилучшей противоопухолевой активностью по сравнению с другими отобранными лидерами, поэтому их сродство дополнительно оптимизировали путем создания библиотек фагового дисплея PL2 и PL3 и проведения по меньшей мере трех раундов пэннинга для созревания сродства in vitro. Затем идентифицировали происходящие из PL2 и PL3 варианты с повышенным сродством (то есть PL2#3 и PL3#7) и проверяли их активности и функции по такой же схеме, как это делалось при отборе лидирующих антител. Также отметим, что на этой стадии создавали варианты с различными изотипами IgG (т.е. IgG1, mtIgG1 (N297A), IgG2, IgG4) и в то же время сравнивали сродство и эффективность in vitro и in vivo у комплекта PL2#3 и PL3#7, конъюгированных с различными Fc-остовами. Наряду с моделями опухоле/антигенспецифичных ксенотрансплантатов Т-клеток, для проверки эффективности PL2#3 и PL3#7 использовали модель со вставкой hPD-1, и эти данные показали, что они оба сравнимы с контрольным антителом против PD-L1. Для дальнейшего улучшения аффинности PL3#7 создавали еще одну библиотеку фагового дисплея на основе PL3#7 для созревания сродства, как и раньше. При этом были идентифицированы три новых варианта (то есть PL3#7-19, -43, -54), которых использовали для сравнения их эффективности с наилучшим вариантом PL2, PL2#3. Данные по сравнению подробно описаны отдельно в нижеследующих примерах.
Так, в табл. 4 представлено сродство и кинетика связывания (ka, kd и KD) у 7 отобранных лидеров при измерении методом поверхностного плазмонного резонанса (SPR). Сначала на сенсорном чипе иммобилизировали Fc против IgG человека, а затем захватное контрольное антитело против PD-L1 и лидирующие антитела против PD-L1: PL1, PL2, PL3, PL6, PL8, PL12 и PL15 при Rmax ~ 250 RU. Эксперименты ставили при 25°C, а измерения проводили при серийных разведениях hPD-L1_ECD-His от 55,4 нМ до 11,1 нМ, пропуская их через захватные антитела в буфере HBS-P+ с добавлением 0,1% (масс./об.) BSA со скоростью потока 25 мкл/мин. Все данные анализировали с помощью вычислительной программы, а кривые подгоняли по модели связывания Ленгмюра 1:1. Представлены данные из двух независимых экспериментов, проводившихся в двух повторах.
Таблица 4
Совмещение аминокислотных последовательностей легкой цепи (LC) и тяжелой цепи (HC) отобранных лидирующих антител PL1, PL2, PL3, PL6, PL8, PL12, PL15 против PD-L1 представлено на фиг. 1A-1B. Определяющие комплементарность участки (CDR) LC и HC у этих лидеров отмечены жирным шрифтом и подчеркиванием.
Пример 2. Сродство связывания и кинетика у вариантов антитела PL2 против PD-L1
Для получения дополнительных клонов с улучшенными показателями связывания использовали лидирующее антитело PL2 в экспериментах по созреванию сродства на основе фагового дисплея in vitro. Сначала создавали библиотеки нуклеиновых кислот CDR-L1/CDR-L3/CDR-H3 (с упором на 3 CDR) методом ПЦР, клонировали в вектор для фагового дисплея и трансформировали ими клетки E. coli TG1 или SS320 для получения фаговых библиотек. После трех раундов пэннинга с помощью биотинилированного hPD-L1-His, конъюгированного с покрытыми стрептавидином магнитными шариками Dynabeads® M-280 (Thermo Fisher Scientific #11205D), проводили скрининг 40 клонов Fab методом ELISA, и четыре Fab (то есть #3, #4, #5 и #39) проявляли лучшие показатели связывания, чем исходное PL2. Затем измеряли кинетические характеристики методом поверхностного плазмонного резонанса (SPR), используя полноразмерные IgG PL2#3, PL2#4, PL2#5 и PL2#39, и обнаружили, что они имеют показатели связывания, которые эквивалентны или лучше, чем у контрольных антител против PD-L1.
Ниже в табл. 5 представлены аминокислотные последовательности вариантов PL2 с мутациями в районе L1 L3 и Н3.
Таблица 5
Измеряли сродство и кинетику связывания методом поверхностного плазмонного резонанса (SPR). Сначала на сенсорном чипе иммобилизовали Fc против IgG человека, а затем захватные контрольные антитела против PD-L1 и варианты PL2 при Rmax ~ 250 RU. Эксперименты ставили при 25°C, а измерения проводили при серийных разведениях hPD-L1-His от 19,1 нМ до 0,71 нМ, пропуская их через захватные антитела в буфере HBS-P+ с добавлением 0,1% (масс./об.) BSA со скоростью потока 25 мкл/мин. Все данные анализировали с помощью вычислительной программы, а кривые подгоняли по модели связывания Ленгмюра 1:1. Представлены данные из двух независимых экспериментов, проводившихся в двух повторах.
В табл. 6 представлена кинетика ассоциации и диссоциации, а также расчетное сродство (KD) у вариантов PL2 по сравнению с контрольными антителами против PD-L1 при измерении методом поверхностного плазмонного резонанса (SPR). В этом исследовании все варианты, исходное PL2 и контрольные антитела были клонированы в остов IgG2 человека. Также в табл. 6 представлено улучшение сродства у вариантов PL2 в сравнении с контрольным антителом против PD-L1. Представленные в табл. 6 данные показывают, что среди вариантов PL2 наилучшим сродством связывания с PD-L1-His человека обладает PL2#3. В табл. 7 представлены результаты по сравнению кинетики между различными изотипами IgG человека у PL2#3 в последующем исследовании. При этом различные изотипы PL2#3 проявляли близкое сродство связывания с PD-L1-His человека.
Таблица 6
Таблица 7
Пример 3. Сродство связывания и кинетика у вариантов антитела PL3 против PD-L1
Для получения дополнительных клонов с улучшенными показателями связывания использовали лидирующее антитело PL3 в экспериментах по созреванию сродства на основе фагового дисплея in vitro. Сначала создавали библиотеки нуклеиновых кислот CDR-L1/CDR-L2/CDR-L3/CDR-H1/CDR-H2/CDR-H3 (с упором на 6 CDR) методом ПЦР, клонировали в вектор для фагового дисплея и трансформировали ими клетки E. coli TG1 или SS320 для получения фаговых библиотек. После трех раундов пэннинга с помощью биотинилированного hPD-L1-His, конъюгированного с покрытыми стрептавидином магнитными шариками Dynabeads® M-280 (Thermo Fisher Scientific #11205D), проводили скрининг 20 клонов Fab методом ELISA, и два Fab (то есть #1 и #7) проявляли лучшие показатели связывания, чем исходное PL3. Затем измеряли кинетические характеристики методом поверхностного плазмонного резонанса (SPR), используя полноразмерные IgG PL3#1 и PL3#7, и обнаружили, что они имеют показатели связывания, которые эквивалентны или лучше, чем у контрольных антител против PD-L1.
Ниже в табл. 8 представлены аминокислотные последовательности вариантов PL3 с мутациями в районе L2, L3 и H1, H3. В табл. 9 представлена кинетика ассоциации и диссоциации, а также расчетное сродство (KD) у вариантов PL3 по сравнению с контрольными антителами против PD-1 при измерении методом поверхностного плазмонного резонанса (SPR). Также представлено сравнение кинетики между различными изотипами IgG человека у PL3#1 и PL3#7. В табл. 9 также представлено улучшение сродства у вариантов PL3 в сравнении с контрольным антителом против PD-L1. Представлены данные из двух независимых экспериментов, проводившихся в двух повторах.
Эти данные показывают, что среди вариантов PL3 немного лучшим сродством связывания с PD-L1-His человека обладает PL3#7. Различные изотипы PL3#7 проявляли близкое сродство связывания с PD-L1-His человека.
Таблица 8
Таблица 9
Клон PL3#7, обладающий гораздо лучшим сродством, чем исходное PL3, в дальнейшем использовали в экспериментах по созреванию сродства на основе фагового дисплея in vitro для получения дополнительных клонов с улучшенными показателями связывания. Сначала создавали библиотеки нуклеиновых кислот CDR-L1/CDR-L2/CDR-L3/CDR-H2/CDR-H3 (с фиксированным CDR-H1 и с упором на 5 CDR) методом ПЦР, клонировали в вектор для фагового дисплея и трансформировали ими клетки E. coli TG1 или SS320 для получения фаговых библиотек. После трех раундов пэннинга с помощью биотинилированного hPD-L1-His, конъюгированного с покрытыми стрептавидином магнитными шариками Dynabeads® M-280 (Thermo Fisher Scientific #11205D), проводили скрининг 57 клонов Fab методом ELISA, и три Fab (то есть 19, 43 и 54) проявляли лучшие показатели связывания, чем исходное PL3#7. Затем измеряли кинетические характеристики методом интерферометрии в биослое (ForteBio Octet RED96), используя полноразмерные IgG PL3#7-19, PL3#7-43 и PL3#7-54, и обнаружили, что они имеют показатели связывания, которые эквивалентны или лучше, чем у контрольных антител против PD-L1.
Ниже в табл. 10 представлены аминокислотные последовательности вариантов PL3#7 с мутациями в районе L1, L2, L3 (SEQ ID NOS 125, 133, 134 и 125, соответственно, в порядке появления), H2 и H3.
Таблица 10
У вариантов PL3#7 измеряли сродство и кинетику методом интерферометрии в биослое на установке Octet RED96 (ForteBio) со стрептавидиновыми (SA) сенсорами при 25°C и скорости перемешивания 1000 об/мин. Вкратце, готовили черный 96-луночный планшет с колонками, содержащими по 200 мкл на лунку реагентов, необходимых для определения сродства. Обычно сенсоры Octet SA сначала помещали на 180 сек в лунки, содержащие 1×буфер для кинетики (PBS, 0,1% BSA, 0,02% Tween-20, pH 7,4), чтобы установить исходный уровень. Затем сенсоры переносили на 300 сек в лунки, содержащие по 10 мкг/мл биотинилированных вариантов PL3#7 (соотношение антитело:биотин = 1:1 с NHS-PEG4-биотином EZ-Link®, Thermo Fisher Scientific #21329), чтобы нагрузить покрытые SA наконечники. Затем сенсоры регенерировали путем инкубации 3 раза по 5 сек в буфере для регенерации (10 мМ глицина, рН 1,5) с последующей инкубацией по 5 сек в 1×буфере для кинетики. Регенерацию повторяли перед измерением каждого цикла ассоциации/диссоциации. Затем сенсоры помещали на 180 сек в 1×буфер для кинетики, чтобы установить исходный уровень. Затем измеряли ассоциацию путем инкубации сенсоров 5 мин в лунках, содержащих hPD-L1-His, после чего переносили на 10 минут в лунки, содержащие 1×буфер для кинетики, для измерения диссоциации. Кинетику (ka, kd и KD) измеряли путем глобальной аппроксимации четырех циклов ассоциации/диссоциации по данным, полученным при серийных разведениях hPD-L1-His 1:3 от 38,28 нМ до 1,42 нМ (один цикл ассоциации/диссоциации для одной концентрации) по модели связывания Ленгмюра 1:1. Все данные анализировали с помощью Octet Data Analysis Software v.9.0 и представляют два независимых эксперимента, проводившихся в двух повторах.
В табл. 11 представлена кинетика ассоциации и диссоциации, а также расчетное сродство (KD) у вариантов PL3#7 по сравнению с контрольными антителами против PD-1 при измерении методом интерферометрии в биослое на приборе ForteBio Octet RED96 (Menlo Park, CA, США). Также в табл. 11 представлено улучшение сродства у вариантов PL3#7 в сравнении с контрольным антителом против PD-L1. Представлены данные из двух независимых экспериментов, проводившихся в двух повторах. Представленные в табл. 11 данные показывают, что PL3#7-19, PL3#7-43 и PL3#7-54 обладают лучшим сродством связывания, чем исходное PL3#7.
Таблица 11
Таким образом, были получены наилучшие варианты PL2 (PL2#3), PL3 (PL3#7) и PL3#7 (PL3#7-19, -43, -54) с наиболее высоким сродством и превосходной функциональной активностью в экспериментах по созреванию сродства на основе фагового дисплея in vitro. В общем, проводили три раунда пэннинга с использованием биотинилированного hPD-L1-His, конъюгированного с покрытыми стрептавидином магнитными шариками Dynabeads® M-280. Затем проводили скрининг Fabs лучших вариантов методом ELISA и клонировали в Fc-остов IgG1 человека с мутацией N297A, получая полноразмерные антитела. Совмещение аминокислотных последовательностей легких цепей (LC) и тяжелых цепей (HC) этих самых лучших вариантов против PD-L1 представлено на фиг. 11А-11В. Определяющие комплементарность участки (CDR) в LC и HC этих наилучших вариантов антител против PD-L1 отмечены жирным шрифтом и подчеркнуты.
Пример 4. Связывание антител против PD-L1 с рекомбинантными слитыми с Fc белками PD-L1 человека и активированными T-клетками CD3+
Для оценки связывания выбранных антител с рекомбинантными слитыми с Fc белками PD-L1 человека проводили анализ методом ELISA. На 96-луночном микропланшете для EIA фиксировали белки PD-L1/Fc человека по 15 нг на лунку в течение ночи при 4°C. После блокирования 5% обезжиренным молоком добавляли серийные разведения антител и инкубировали при комнатной температуре в течение 1 часа. Не связавшиеся антитела удаляли и лунки дважды промывали PBST. В лунки добавляли конъюгированное с HRP вторичное антитело, а после инкубации смывали избыток вторичного антитела. В лунки добавляли TMB, а после инкубации останавливали реакцию и измеряли активность HRP, отслеживая повышение поглощения при 450 нм.
Выделяли Т-клетки человека из РВМС с помощью набора для обогащения Т-клеток человека MagniSort™ (eBioscience). Выделенные Т-клетки активировали фитогемагглютинином (PHA) при 5 мкг/мл в течение 3 дней для стимулирования экспрессии PD-L1. Активированные Т-клетки собирали и инкубировали в буфере FACS (PBS с 2% FBS) с блокатором Fc человека (eBioscience) в течение 20 мин при 4°C. Определяли связывание моноклональных антител против PD-L1 путем инкубации активированных Т-клеток с серийными разведениями антител в буфере FACS. Клетки промывали проточным буфером и детектировали связывание с помощью меченного FITC кроличьего Ab против Fcγ IgG человека. Клетки также окрашивали с помощью мышиного PE-Cy7 против CD3 человека (eBioscience) для стробирования CD3-позитивных Т-клеток. Проводили анализ методом проточной цитометрии на приборе Cytomics FC 500 (Beckman Coulter Inc.).
Таким образом, отобранные антитела против PD-L1 тестировали на связывание с рекомбинантными белками PD-L1 человека методом ELISA и с активированными T-клетками CD3+ методом проточной цитометрии. В качестве положительного и отрицательного контроля использовали контрольные антитела против PD-L1 и против PD-1, соответственно. Из фиг. 2А и 2В видно, что все выбранные антитела против PD-L1 способны связывать как рекомбинантные белки PD-L1 человека, так и Т-клетки, экспрессирующие PD-L1.
Пример 5. Блокирование связывания PD-1 с PD-L1 антителами против PD-L1
Экспрессирующие PD-L1 клетки CHO-S суспендировали в буфере FACS (PBS с 4% FBS). В суспензию клеток (3,5×105 клеток на лунку) добавляли различные концентрации исследуемых антител и инкубировали при 4°C в течение 30 мин. Отмывали не связавшиеся антитела, добавляли меченный биотином слитый белок PD-1-Fc и инкубировали при 4°C в течение 30 мин. Клетки промывали, а затем окрашивали стрептавидином-РЕ при 4°C в течение 30 мин. Проводили анализ методом проточной цитометрии на приборе Cytomics FC 500 (Beckman Coulter Inc.).
Отобранные антитела против PD-L1 тестировали на способность блокировать связывание PD-1 с экспрессирующими PD-L1 клетками CHO-S методом проточной цитометрии. В качестве положительного и отрицательного контроля использовали контрольное антитело против PD-L1 и авастин, соответственно. Представленные на фиг. 3 данные показывают, что все выбранные моноклональные антитела против PD-L1 блокируют связывание PD-1 с трансфецированными PD-L1 клетками CHO-S при измерении по средней интенсивности флуоресценции (MFI) при окрашивании.
Пример 6. Влияние антител против PD-L1 на продукцию цитокинов и пролиферацию Т-клеток при реакции смешанных лейкоцитов (MLR)
Для демонстрации эффекта блокирования пути PD-L1/PD-1 для эффекторных клеток лимфоцитов использовали реакцию смешанных лейкоцитов. При анализе тестировали Т-клетки на пролиферацию и секрецию IFN-γ или IL-2 в присутствии или в отсутствие антител против PD-L1.
Выделяли Т-клетки человека из РВМС с помощью набора Lympho-kwik T (One Lamda, Inc.). Выделенные Т-клетки суспендировали в PBS и метили с помощью 1 мкМ CFSE при комнатной температуре в течение 10 мин. После промывки клеток полной средой (RPMI-1640 с 10% FBS) помеченные CFSE T-клетки суспендировали в полной среде при концентрации 1×106 клеток/мл.
Аллогенные дендритные клетки получали из PBMC. Выделенные РВМС инкубировали с 200 ед./мл рекомбинантного IL-3 человека (eBioscience) в течение ночи, чтобы популяция моноцитов/макрофагов смогла прикрепиться к лункам. Не прикрепившиеся клетки удаляли и лунки дважды промывали полной средой. Затем клетки на планшетах культивировали в течение 6 дней в полной среде, содержащей 200 ед./мл IL-4 человека (eBioscience) и 200 ед./мл GM-CSF человека (eBioscience). Полученные из моноцитов дендритные клетки подвергали созреванию путем добавления в культуры TNF-α (100 ед./мл) на 6-й день и инкубации в течение ночи. Зрелые DC обрабатывали трипсином, собирали и суспендировали в полной среде при концентрации 1×105 клеток/мл.
Каждая реакция содержала 10E5 помеченных CFSE Т-клеток и 10E4 аллогенных дендритных клеток в общем объеме 200 мкл. В каждую культуру добавляли антитела при различных концентрациях. В качестве отрицательного контроля использовали либо без антител, либо антитело против VEGF (авастин). В качестве положительного контроля использовали контрольное (ref) антитело против PD-1 или против PD-L1. Клетки культивировали в течение 5 дней при 37°С. На 5-й день из каждой культуры отбирали 100 мкл среды для измерения цитокинов. Уровни цитокинов измеряли с помощью наборов для IFN-γ или IL-2 человека ELISA MAX™ Deluxe (BioLegend). Клетки собирали и анализировали на пролиферацию Т-клеток методом проточной цитометрии.
Представленные на фиг. 4A-4B данные показывают, что все выбранные антитела против PD-L1 усиливают секрецию IFN-γ и стимулируют пролиферацию Т-клеток при анализе по реакции смешанных лейкоцитов. В качестве положительных контролей использовали контрольные антитела против PD-L1 и против PD-1. В качестве отрицательного контроля использовали авастин (антитело против VEGF). На фиг. 4A представлены гистограммы секреции IFN-γ, а на фиг. 4B представлены гистограммы пролиферации T-клеток CD3+ при указанных концентрациях антител.
Кроме того, на фиг. 6A представлена секреция IL-2 при MLR, индуцированная исходным антителом PL2 против PD-L1 и его вариантами, а на фиг. 6B представлена секреция IL-2 при MLR, индуцированная при указанной концентрации исходного антитела PL3 против PD-L1 и его вариантов. Эти данные показывают, что выбранный вариант PL2 - PL2-3/3 (PL2#3) и вариант PL3 (PL3#7) проявляют большую эффективность, чем исходные антитела в стимулировании активации Т-клеток.
Кроме того, на фиг. 8A-8D и фиг. 12A-12B представлена эффективность различных IgG-форм антител PL2#3 и PL3#7 против PD-L1 и вариантов PL3#7 при реакции смешанных лейкоцитов (MLR). На фиг. 8A, 8B и 12A представлены гистограммы секреции IFN-γ и IL-2, индуцированной этими антителами против PD-L1. На фиг. 8C представлены гистограммы пролиферации T-клеток CD4+ при различных концентрациях антител L2#3, PL3#7 против PD-L1, а на фиг. 8D и 12B представлены гистограммы пролиферации T-клеток CD8+ при указанных концентрациях этих трех антител против PD-L1. В частности, из фиг. 12A и 12B видно, что варианты антитела PL3#7 против PD-L1, напр., PL3#7-19, -43 и -54, проявляют превосходную активность в усилении секреции IFN-γ и пролиферации T-клеток CD8+.
Пример 7. Активность ингибирования роста опухолей у антител PL2 и PL3 против PD-L1 на модели ксенотрансплантатов клеток A375/антиген-специфичных Т-клеток
Исследовали активность антител против PD-L1 человека in vivo на моделях ксенотрансплантатов у мышей, используя мышей с ослабленным иммунитетом NOD/SCID (с диабетом без ожирения/тяжелым комбинированным иммунодефицитом). Мышам подкожно имплантировали клетки раковых линий человека, экспрессирующих PD-L1 человека, и PBMC или антиген-специфичные Т-клетки человека. Мышам с инокулированными клетками линии A375 меланомы человека или клетками NSCLC линии NCI-H292 человека вводили дозы антител внутрибрюшинно. Отмечали влияние антител на рост опухолей до достижения объема в 2000 мм3 или макроскопического некроза опухолей.
Для получения антиген-специфичных Т-клеток выделяли Т-клетки из РВМС от здоровых доноров с помощью набора для обогащения Т-клеток человека MagniSort® (eBioscience). Выделенные Т-клетки культивировали вместе с обработанными митомицином С (50 Ед/мл) клетками A375 и rhIL-2 в течение 10-14 дней для обогащения антиген-специфичных Т-клеток. РВМС человека выделяли из цельной крови здоровых доноров с помощью Histopaque®-1077 (Sigma-Aldrich).
Клетки A375 и антигенспецифичные Т-клетки смешивали непосредственно перед подкожным введением при указанных соотношениях эффектор-мишень (E:T). Клетки NCI-H292 смешивали со свежими РВМС человека при указанном соотношении E:T и инокулировали подкожно мышам. Первую дозу исследуемых антител вводили внутрибрюшинно через 2 часа после прививки раковых/эффекторных клеток. Мышей обрабатывали антителами два раза в неделю в течение 3-5 недель. У каждого животного проверяли образование опухолей по два раза в неделю. Опухоли измеряли штангенциркулем, а объемы опухолей (V) рассчитывали по следующей формуле:
V (мм3) = 0,5×(длина (мм) × ширина (мм) × толщина (мм)/2).
Мышам (n = 4 на группу) прививали подкожно смесь клеток линии A375 меланомы человека и антиген-специфичных Т-клеток (Т-клетки: раковые клетки = 1:100). Исследуемые антитела вводили мышам внутрибрюшинно два раза в неделю, начиная со дня 0. Эффективность всех лидирующих антител против PD-L1 проверяли in vivo на модели ксенотрансплантатов A375/антигенспецифичных Т-клеток. Кривые роста опухолей у мышей, получавших PL2 и PL3, представлены на фиг. 5A и 5B. Данные показывают, что PL2 и PL3 могут значительно ингибировать рост опухолей A375 in vivo.
Кроме того, на фиг. 7A и 7B представлена активность ингибирования роста опухолей у лучших вариантов антител PL2 и PL3 против PD-L1 на модели ксенотрансплантатов A375/антиген-специфичных Т-клеток. Данные показывают, что PL2#3 IgG2 ингибирует рост опухолей A375 в дозах 30 мг/кг и 10 мг/кг. PL3#7 IgG2 ингибировал рост A375 только в дозе 30 мг/кг.
Кроме того, на фиг. 9A (PL2#3) и 9B (PL3#7) представлена активность ингибирования роста опухолей у различных IgG-изотипов PL2#3 и PL3#7 на модели ксенотрансплантатов A375/антиген-специфичных Т-клеток. Данные показывают, что мутантные формы IgG1 и IgG4 антитела PL2#3 против PD-L1 проявляли хороший противоопухолевый эффект в дозе 20 мг/кг. IgG1 дикого типа, мутантные формы IgG1 и IgG2 антитела PL3#7 против PD-L1 ингибировали рост опухолей A375 в дозе 30 мг/кг. На основании этих данных в качестве Fc-остова был выбран мутантный IgG (N297A).
На фиг. 15A-15B представлена активность ингибирования роста опухолей у мутантов антител PL2#3 и PL3#7 против PD-L1 на модели ксенотрансплантатов A375/антиген-специфичных Т-клеток. Мышам (n =4 на группу) прививали подкожно смесь клеток линии A375 меланомы человека и антиген-специфичных Т-клеток (Т-клетки (E):раковые клетки (T) = 1:100). Исследуемые мутанты антител PL2#3 и PL3#7 вводили мышам внутрибрюшинно два раза в неделю, начиная со дня 0. Кривые роста опухолей у мышей, получавших mAb, представлены на фиг. 15A. Объемы индивидуальных опухолей на 31-й день представлены на фиг. 15B. Данные показывают, что PL2#3 mtIgG1 проявляло противоопухолевую активность при 20 мг/кг и 1 мг/кг на модели A375 (меланома)/антиген-специфичные Т-клетки; а PL3#7-43 проявляло отличную противоопухолевую активность в дозе 20 мг/кг.
Пример 8. Активность ингибирования роста опухолей у мутантных антител против PD-L1 на мышах hPD1 KI
Исследовали активность антител против PD-1 человека in vivo на мышах C57BL/6 со вставленным PD-1 человека (мышах hPD1 KI). Мышам инокулировали подкожно транс фецированные PD-L1 человека раковые клетки MC38 мыши (5×105 клеток на мышь). Обработку антителами начинали при достижении объема опухолей примерно 86 мм3. Перед обработкой в каждую экспериментальную группу отбирали по 6 животных. Животные получали дозы антител против PD-L1 два раза в неделю в течение 3 недель. У каждого животного проверяли образование опухолей два раза в неделю. Опухоли измеряли штангенциркулем, а объемы опухолей (V) рассчитывали по следующей формуле:
V (мм3) = 0,5×(длина (мм) × ширина (мм) × толщина (мм)/2).
Мышам со вставленным PD-1 человека (hPD1 KI) (n = 6 на группу) подкожно прививали клетки MC38-huPD-L1 (MC38, трансфецированные PD-L1 человека). Обработку антителами начинали при достижении объема опухолей примерно 86 мм3. Исследуемые антитела: PL2#3-mtIgG1 и PL3#7-mtIgG1 вводили мышам внутрибрюшинно два раза в неделю в течение 3 недель. Представленные на фиг. 10 данные показывают, что противоопухолевая активность PL2#3-mtIgG1 и PL3#7-mtIgG1 сравнима с активностью аналога атезолизумаба, что эквивалентно контрольному антителу против PD-L1.
Пример 9. Связывание антител против PD-L1 с клеточной поверхностью экспрессирующих PD-L1 клеток
Выделяли Т-клетки человека из РВМС с помощью набора для обогащения Т-клеток человека MagniSort™ (eBioscience). Выделенные Т-клетки активировали фитогемагглютинином (PHA) при 5 мкг/мл в течение 3 дней для стимулирования экспрессии PD-L1. Активированные Т-клетки собирали и инкубировали в буфере FACS (PBS с 2% FBS) с блокатором Fc человека (eBioscience) в течение 20 мин при 4°C. После удаления блокирующего реагента Т-клетки суспендировали в буфере FACS для процесса окрашивания. Опухолевые клетки (A375 и NCI-H292) собирали путем обработки трипсином клеток из культуральных чашек и дважды промывали буфером FACS для окрашивания клеток.
Связывание моноклональных антител против PD-L1 определяли путем инкубации Т-клеток с серийными разведениями моноклональных антител против PD-1 в буфере FACS (PBS с 2% FBS). Клетки промывали проточным буфером и детектировали связывание с помощью меченного биотином кроличьего Ab против Fcγ IgG человека и стрептавидина-PE. Проводили анализ методом проточной цитометрии на приборе Cytomics FC 500 (Beckman Coulter Inc.).
На фиг. 13A-13C представлено связывание вариантов PL2#3 и PL3#7 с клеточной поверхностью экспрессирующих PD-L1 клеток: активированных Т-клеток (фиг. 13A), клеток линии A375 меланомы человека (фиг. 13B) и клеток NSCLC линии NCI-H292 человека (фиг. 13C). В качестве положительного и отрицательного контроля использовали контрольное антитело против PD-L1 и HLX01 (mAb против CD20), соответственно. Ранжирование по активности связывания для активированных T-клеток: PL2#3 mtIgG1 > PL3#7-54 mtIgG1 > PL3#7-43 mtIgG1 > PL3#7-19 mtIgG1. Ранжирование по активности связывания для клеток A375 меланомы человека: PL3#7-54 mtIgG1 = PL2#3 mtIgG1 > PL3#7-43 mtIgG1 > PL3#7-19 mtIgG1. Ранжирование по активности связывания для клеток NSCLC линии NCI-H292 человека: PL3#7-54 mtIgG1 ≥ PL2#3 mtIgG1 = PL3#7-43 mtIgG1 ≥ PL3#7-19 mtIgG1. Эти данные свидетельствуют, что все варианты PL3#7 и PL2#3 связываются с поверхностью экспрессирующих PD-L1 клеток.
Пример 10. Активность ингибирования роста опухолей у вариантов антител против PD-L1 на модели ксенотрансплантатов NCI-H292/PBMC
Исследовали активность антител против PD-L1 человека in vivo на моделях ксенотрансплантатов у мышей, используя мышей с ослабленным иммунитетом NOD/SCID (с диабетом без ожирения/тяжелым комбинированным иммунодефицитом). Раковые клетки и выделенные PBMC человека смешивали непосредственно перед подкожным введением при указанном соотношении эффектор-мишень (E:T). Каждой мыши инокулировали билатерально смеси раковых клеток и PBMC человека. Первую дозу исследуемых антител вводили внутрибрюшинно через 2 часа после прививки раковых/эффекторных клеток. Животные получали дозы исследуемых антител два раза в неделю в течение 3-4 недель. У каждого животного проверяли образование опухолей по два раза в неделю. Опухоли измеряли штангенциркулем, а объемы опухолей (V) рассчитывали по следующей формуле:
V (мм3) = 0,5×(длина (мм) × ширина (мм) × толщина (мм)/2).
Мышам (n = 4 на группу) подкожно прививали смесь клеток NSCLC линии NCI-H292 человека и свежевыделенных PBMC человека (раковые клетки (T): PBMC (E) = 3:1). Начиная со дня 0, мышам два раза в неделю вводили внутрибрюшинно антитела против PD-L1. На фиг. 14A представлены кривые роста опухолей. На фиг. 14B представлены объемы отдельных опухолей на 28-й день. Эти данные показывают, что все исследованные варианты антител против PD-L1 ингибируют рост опухолей при добавлении эффекторных клеток (hPBMC). Варианты PL2#3, PL3#7-19 и PL3#7-43 проявляли близкую эффективность ингибирования роста опухолей in vivo, которая превосходит вариант PL3#7-54 и контрольное антитело против PD-L1. Противораковый эффект mAb против PD-L1 проявлялся через иммунные клетки.
Пример 11. Перекрестное связывание моноклональных антител против PD-L1 человека с клетками меланомы мыши
Определяли связывание моноклональных антител против PD-L1 путем инкубации клеток меланомы A375 (1,5×105 клеток на пробу) с серийными разведениями антител в буфере FACS. Клетки промывали проточным буфером и детектировали связывание с помощью меченного FITC кроличьего Ab против Fcγ IgG человека. Проводили анализ методом проточной цитометрии на приборе Cytomics FC 500 (Beckman Coulter Inc.).
Варианты PL2#3 и PL3#7 тестировали на связывание с экспрессирующими PD-L1 мыши клетками меланомы (B16-F10) методом проточной цитометрии. В качестве положительного и отрицательного контроля, соответственно, использовали контрольные антитела против PD-L1. Представленные на фиг. 16 данные показывают, что варианты PL3#7-43 и PL3#7-54 перекрестно связываются с PD-L1 мыши, тогда как варианты PL2#3 и PL3#7-19 не обладают значительной перекрестной реактивностью с клетками меланомы мыши.
Пример 12. Перекрестное связывание вариантов антител против PD-L1 с PD-L1 макаки-крабоеда (cynomolgus monkey)
Рекомбинантные слитые c Fc белки PD-L1_ECD макаки-крабоеда приобретали у Sino Biological Inc. На 96-луночном планшете для анализа иммобилизировали PD-L1_ECD/Fc (9 нг на лунку) путем инкубации в течение ночи при 4°C. Сайты неспецифического связывания блокировали 5% обезжиренным молоком в PBS в течение 1 часа при комнатной температуре. После трехкратной промывки планшетов PBST инкубировали антитела против PD-L1 и HLX01 (отрицательный контроль) при указанных концентрациях с иммобилизованными белками в течение 1 часа при комнатной температуре. Планшеты трижды промывали PBST, а затем инкубировали в течение 1 часа при комнатной температуре с меченным пероксидазой козьим F(ab′)2 против IgG человека (Jackson ImmunoResearch Laboratories), разведенным 1/10000 в PBS. После промывки планшеты проявляли с помощью TMB (eBioscience). Считывали поглощение при длине волны 450 нм на считывающем устройстве для микропланшетов Varioskan LUX (Thermo Scientific).
Варианты PL2#3 и PL3#7 тестировали на связывание с рекомбинантными слитыми с Fc белками PD-L1_ECD макаки-крабоеда методом ELISA. В качестве отрицательного контроля использовали HLX01 (mAb против CD20). Представленные на фиг. 17 данные свидетельствуют, что варианты PL2#3 и PL3#7 перекрестно реагируют с PD-L1 макаки-крабоеда.
Пример 13. Конструирование дегликозилированных версий варианта PL3#7-19 и варианта PL3#7-43
PL3#7-19 и PL3#7-43 - два лидирующих варианта, происходящие из PL3#7, но их нужно было подвергнуть дальнейшему мутагенезу, чтобы устранить нежелательные сайты N-гликозилирования на участке L-CDR2. N-гликозилирование происходит по сайтам N-X-S/T. Поскольку сайты N-гликозилирования в исходных PL3#7-19 и PL3#7-43 кодируются в виде N-S-T и N-R-S, то конструирование проводили либо путем мутации второго S/T от N-конца на идентичную или близкую аминокислоту в PL3#7, то есть мутации N-S-T и N-R-S на N-S-N/Q и N-R-P для PL3#7-19 и PL3#7-43, соответственно (т.е. это PL3#7-19 деглико1, деглико3 и PL3#7-43 деглико1), либо путем прямой мутации N на Q для сайта N-X-S/T, то есть мутации N-S-T и N-R-S на Q-S-T и Q-R-S для PL3#7-19 и PL3#7-43, соответственно (т.е. это PL3#7-19 деглико2 и PL3#7-43 деглико2). После этого проверяли устранение сайтов N-гликозилирования на L-CDR2 методом жидкостной хроматографии-масс-спектрометрии (LC-MS) и SDS-PAGE (данные не приводятся).
Совмещение аминокислотных последовательностей вариабельных областей легкой цепи у дегликозилированных версий варианта PL3#7-19 и варианта PL3#7-43 представлено на фиг. 18. Приводится совмещение последовательностей легких цепей этих дегликозилированных вариантов, а жирным шрифтом и подчеркиванием отмечены CDR (определяющие комплементарность участки). Тяжелые цепи у них были неизменными и идентичны своим исходным вариантам (совмещение последовательностей не приводится). У этих дегликозилированных вариантов определяли активность связывания с PD-L1 человека методами проточной цитометрии и Octet в последующих экспериментах.
Пример 14. Связывание дегликозилированных версий варианта PL3#7-19 и варианта PL3#7-43 на целых клетках
Определяли связывание исследуемых дегликозилированных вариантов путем инкубации экспрессирующих PD-L1 клеток CHO-S (2×105 клеток на лунку) с серийными разведениями антител в буфере FACS (PBS с 1% FBS) при 4°C в течение 30 мин. Затем клетки промывали проточным буфером, а затем окрашивали с помощью Fc-FITC против IgG человека (1:500) при 4°C в течение 30 мин для детектирования связывания антител на клеточной поверхности. Проводили анализ методом проточной цитометрии на приборе CytoFlex (Beckman Coulter Inc.).
Связывание дегликозилированных версий варианта PL3#7-19 и варианта PL3#7-43 на целых клетках представлено на фиг. 19A и 19B. PL3#7-19 и PL3#7-43 - два лидирующих варианта, но им потребовалась инженерия для удаления нежелательных сайтов N-гликозилирования на участке L-CDR2 (см. выше пример 13). У полученных трех дегликозилированных вариантов для PL3#7-19 (т.е. PL3#7-19 деглико1, деглико2, деглико3) и двух дегликозилированных вариантов для PL3#7-43 (т.е. PL3#7-43 деглико1, деглико2) определяли активность связывания на целых клетках с трансфецированными PD-L1 клетками CHO-S методом проточной цитометрии. Все исследованные варианты дегликозилированных антител были в Fc-остове IgG1 человека с мутацией N297A. В качестве отрицательного контроля использовали наше собственное антитело против PD-1 (т.е. HLX10).
Представленные на фиг. 19A и 19B данные свидетельствуют, что удаление сайтов N-гликозилирования на участке L-CDR2 не влияло на активность связывания с целыми клетками у дегликозилированных вариантов PL3#7-19 и PL3#7-43.
Пример 15. IgG-изотипы PL3#7-19 деглико1 и PL3#7-43 деглико2 в реакции смешанных лейкоцитов
Тестировали эффективность различных IgG-изотипов PL3#7-19 деглико1 и PL3#7-43 деглико2 по реакции смешанных лейкоцитов (MLR), а результаты представлены на фиг. 20A и 20B. В качестве антител отрицательного и положительного контроля использовали HLX04 и HLX20 (PL2#3), соответственно. Представлены гистограммы секреции IFN-γ (фиг. 20A) и IL-2 (фиг. 20B), индуцированный исследуемыми антителами.
Как и гликозилированные PL3#7-19 mtIgG1 и PL3#7-43 mtIgG1, изотипы mtIgG1 и IgG4 дегликозилированных PL3#7-19 и PL3#7-43 значительно усиливают секрецию цитокинов при MLR. PL3#7-19 деглико1 и PL3#7-43 деглико1 с IgG1 дикого типа проявляли меньшее усиление высвобождения цитокинов, что может быть обусловлено возможным эффектом типа ADCC на Т-клетки у IgG1 дикого типа.
Пример 16. Активность ингибирования роста опухолей у дегликозилированных вариантов PL3#7-19 и PL3#7-43 на модели ксенотрансплантатов клеток A375/антиген-специфичных Т-клеток
Мышам (n = 5 на группу) прививали подкожно смесь клеток линии A375 меланомы человека и антиген-специфичных Т-клеток (Т-клетки: раковые клетки = 1:100). Исследуемые антитела вводили мышам внутрибрюшинно два раза в неделю, начиная со дня 0. Кривые роста опухолей у мышей, получавших дегликозилированные варианты PL3#7-19 и PL3#7-43, представлены на фиг. 21А и 21С, соответственно. Объемы отдельных опухолей на 28-й день представлены на фиг. 21B и 21D, соответственно. Все точки данных представляют средние значения ± SEM.
Представленные на этих фигурах данные показывают, что противоопухолевая эффективность у вариантов PL3#7-19 деглико1 mtIgG1 и IgG4, а также PL3#7-43 деглико2 wtIgG1, была сравнима с таковой у HLX20 (PL2#3) на модели ксенотрансплантатов A375/антиген-специфичных Т-клеток.
Пример 17. Активность ингибирования роста опухолей у mAb против PD-L1 с mAb против VEGF на модели ксенотрансплантатов NSCLC у мышей
Исследовали активность антител против PD-L1 человека in vivo на моделях ксенотрансплантатов у мышей, используя мышей с ослабленным иммунитетом NOD/SCID (с диабетом без ожирения/тяжелым комбинированным иммунодефицитом). Раковые клетки и выделенные PBMC человека смешивали непосредственно перед подкожным введением при указанном соотношении эффектор-мишень (E:T). Каждой мыши инокулировали билатерально смеси раковых клеток и PBMC человека. В каждой экспериментальной группе состояло 4 животных. Первую дозу исследуемых антител вводили внутрибрюшинно через 1 день после прививки раковых/эффекторных клеток. Животные получали дозы исследуемых антител два раза в неделю в течение 3-4 недель. У каждого животного проверяли образование опухолей по два раза в неделю. Опухоли измеряли штангенциркулем, а объемы опухолей (V) рассчитывали по следующей формуле:
V (мм3) = 0,5×(длина (мм) × ширина (мм) × толщина (мм)/2).
Мышам (n = 5 на группу) подкожно прививали смесь клеток NSCLC линии NCI-H292 человека и свежевыделенных PBMC человека (раковые клетки:PBMC = 3:1). Начиная со дня 1, мышам два раза в неделю вводили внутрибрюшинно антитела против PD-L1 (PL2#3) и против VEGF (HLX04). На фиг. 22A представлены кривые роста опухолей. На фиг. 22B представлены объемы отдельных опухолей на 21-й день. Все точки данных представляют средние значения ± SEM.
Эти данные свидетельствуют, что mAb PL2#3 против PD-L1 в сочетании с mAb HLX04 против VEGF подавляет рост опухолей у ксенотрансплантатов NCI-H292 более эффективно, чем каждое из них по отдельности.
Пример 18. Определение равновесной константы диссоциации (KD) у дегликозилированных вариантов PL3#7-19 и PL3#7-43
N-гликозилирование происходит по сайтам N-X-S/T. Следовательно, сайты N-гликозилирования у исходных PL3#7-19 и PL3#7-43 располагаются в L-CDR2. Удаление сайтов N-гликозилирования проводили путем мутации S/T в этом сайте обратно на идентичную или сходную аминокислоту, как у PL3#7 (т.е. PL3#7-19 деглико1, деглико3 и PL3#7-43 деглико1), или же прямой мутации гликозилируемого N на Q (т.е. PL3#7-19 деглико2 и PL3#7-43 деглико2). За исключением L-CDR2, который подвергали инженерии для устранения N-гликозилирования, участки L-CDR1, L-CDR3 и H-CDR1, H-CDR2, H-CDR3 у дегликозилированных вариантов оставались неизменными. Все исследованные здесь дегликозилированные варианты были клонированы в Fc-остов IgG1 человека с мутацией N297A для параллельного сравнения с исходными вариантами PL3#7-19 и PL3#7-43.
У дегликозилированных вариантов против PD-L1 (т.е. PL3#7-19 деглико1, деглико2, деглико3 и PL3#7-43 деглико1, деглико2) измеряли сродство и кинетику методом интерферометрии в биослое на установке Octet RED96 (ForteBio) с захватными сенсорами AHC против Fc человека при 25°C и скорости перемешивания 1000 об/мин. Вкратце, готовили черный 96-луночный планшет с колонками, содержащими по 200 мкл на лунку реагентов, необходимых для определения сродства. Обычно сенсоры Octet против Fc человека сначала помещали на 180 сек в лунки, содержащие 1×буфер для кинетики (PBS, 0,1% BSA, 0,02% Tween-20, pH 7,4), чтобы установить исходный уровень. Затем сенсоры переносили на 600 сек в лунки, содержащие по 10 мкг/мл вариантов против PD-L1, чтобы нагрузить наконечники против Fc человека. Затем сенсоры помещали на 180 сек в 1×буфер для кинетики, чтобы установить исходный уровень. Затем измеряли ассоциацию путем инкубации сенсоров 3 мин в лунках, содержащих hPD-L1-His, после чего переносили на 10 минут в лунки, содержащие 1×буфер для кинетики, для измерения диссоциации. Затем сенсоры регенерировали путем инкубации 3 раза по 5 сек в буфере для регенерации (10 мМ глицина, рН 1,5) с последующей инкубацией по 5 сек в 1×буфере для кинетики. Регенерацию повторяли перед измерением каждого цикла ассоциации/диссоциации. Кинетику (ka, kd и KD) измеряли путем глобальной аппроксимации четырех циклов ассоциации/диссоциации по данным, полученным при серийных разведениях hPD-L1-His 1:3 от 21,3 нМ до 0,79 нМ (один цикл ассоциации/диссоциации для одной концентрации) по модели связывания Ленгмюра 1:1. Все данные анализировали с помощью Octet Data Analysis Software v.9.0 и представляют два независимых эксперимента, проводившихся в двух повторах.
В табл. 12 представлена кинетика ассоциации и диссоциации, а также расчетное сродство (KD) у дегликозилированных вариантов при измерении методом интерферометрии в биослое на приборе ForteBio Octet RED96 (Menlo Park, CA, США). Также в табл. 12 представлена степень отличия в сродстве у дегликозилированных вариантов в сравнении с исходными вариантами с сайтами N-гликозилирования в L-CDR2. Представлены данные из двух независимых экспериментов, проводившихся в двух повторах. Эти данные показывают, что дегликозилированные варианты имеют сродство связывания, очень близкое к исходным PL3#7-19 и PL3#7-43. Устранение N-гликозилирования на участке L-CDR2 не влияло на сродство.
Таблица 12
Предшествующие примеры приводятся только в иллюстративных целях и никоим образом не предназначены для ограничения объема настоящего изобретения. Различные модификации изобретения в дополнение к представленным и описанным здесь должны быть очевидными специалистам в данной области из предшествующего описания и должны входить в рамки прилагаемой формулы изобретения.
Список воплощений
Предусмотренные изобретением воплощения включают, без ограничения:
1. Антитело против PD-L1 (PL1), содержащее последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 35; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 41; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 44; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 51; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 59.
2. Антитело против PD-L1 (PL2), содержащее последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 36; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 42; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 45; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 52; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 56; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 60.
3. Антитело против PD-L1 (PL3), содержащее последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 37; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 41; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 46; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 53; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 61.
4. Антитело против PD-L1 (PL6), содержащее последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 37; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 41; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 47; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 54; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 57; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 62.
5. Антитело против PD-L1 (PL8), содержащее последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 38; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 43; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 48; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 51; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 58; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 63.
6. Антитело против PD-L1 (PL12), содержащее последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 39; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 41; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 49; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 51; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 64.
7. Антитело против PD-L1 (PL15), содержащее последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 40; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 41; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 50; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 53; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 61.
8. Вариант антитела против PD-L1 (PL2#3), содержащий последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 65; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 42; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 71; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 52; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 56; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 77.
9. Вариант антитела против PD-L1 (PL3#7), содержащий последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 37; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 41; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 72; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 75; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 78.
10. Вариант антитела против PD-L1 (PL3#7-19), содержащий последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 66; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 68; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 73; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 75; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 79.
11. Вариант антитела против PD-L1 (PL3#7-43), содержащий последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 35; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 69; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 74; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 75; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 80.
12. Вариант антитела против PD-L1 (PL3#7-54), содержащий последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 67; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 70; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 72; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 75; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 76; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 81.
13. Вариант антитела против PD-L1 (PL2#4), содержащий последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 94; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 42; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 95; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 52; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 56; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 96.
14. Вариант антитела против PD-L1 (PL2#5), содержащий последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 97; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 42; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 98; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 52; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 56; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 99.
15. Вариант антитела против PD-L1 (PL2#39), содержащий последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 100; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 42; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 95; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 52; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 56; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 101.
16. Вариант антитела против PD-L1 (PL3#1), содержащий последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 37; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 106; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 107; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 108; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 109.
17. Вариант антитела против PD-L1 из воплощения 10, при этом данный вариант антитела против PD-L1 включает CDR-L2, содержащий одну или несколько мутаций по сайтам N-гликозилирования.
18. Вариант антитела против PD-L1 из воплощения 11, при этом данный вариант антитела против PD-L1 включает CDR-L2, содержащий одну или несколько мутаций по сайтам N-гликозилирования.
19. Антигенсвязывающий фрагмент антитела против PD-L1 или его варианта по любому из воплощений 1-18, при этом антигенсвязывающий фрагмент выбран из группы, состоящей из Fab, Fab′, F(ab′)2, одноцепочечных Fv (scFv), Fv-фрагментов, диател и линейных антител.
20. Антитело против PD-L1 либо его вариант или антигенсвязывающий фрагмент по любому из воплощений 1-19, при этом антитело представляет собой полиспецифичное антитело.
21. Антитело против PD-L1 либо его вариант или антигенсвязывающий фрагмент по любому из воплощений 1-20, конъюгированное с терапевтическим средством.
22. Антитело против PD-L1 либо его вариант или антигенсвязывающий фрагмент по любому из воплощений 1-20, конъюгированное с меткой.
23. Антитело против PD-1 либо его вариант или антигенсвязывающий фрагмент согласно воплощению 22, при этом метка выбрана из группы, состоящей из радиоизотопов, флуоресцентных красителей и ферментов.
24. Выделенная молекула нуклеиновой кислоты, кодирующая антитело против PD-L1 либо его вариант или антигенсвязывающий фрагмент по любому из воплощений 1-19.
25. Экспрессирующий вектор, кодирующий молекулу нуклеиновой кислоты из воплощения 24.
26. Клетка, содержащая экспрессирующий вектор из воплощения 25.
27. Способ получения антитела либо его варианта или антигенсвязывающего фрагмента, включающий культивирование клеток из воплощения 26 и извлечение антител из клеточной культуры.
28. Композиция, содержащая антитело против PD-L1 либо его вариант или антигенсвязывающий фрагмент по любому из воплощений 1-23 и фармацевтически приемлемый носитель.
29. Способ выявления белка PD-L1 в образце от пациента путем контактирования антитела против PD-L1 либо его варианта или антигенсвязывающего фрагмента по любому из воплощений 1-20 с образцом и детектирования антител против PD-L1, связавшихся с белком PD-L1.
30. Способ согласно воплощению 29, при этом антитело против PD-1 либо его вариант или антигенсвязывающий фрагмент применяется при иммуногистохимическом анализе (IHC) или анализе методом ELISA.
31. Способ лечения рака у субъекта, включающий введение субъекту эффективного количества композиции из воплощения 28.
32. Способ из воплощения 31, при этом рак выбран из группы, состоящей из меланомы, NSCLC, рака головы и шеи, рака уротелия, тройного отрицательного рака молочной железы (TNBC), рака желудка, классической лимфомы Ходжкина (cHL), неходжкинской лимфомы - первичной B-клеточной лимфомы средостения (NHL PMBCL), мезотелиомы, рака яичников, рака легких, рака пищевода, карциномы носоглотки (NPC), рака желчных путей, колоректального рака, рака молочной железы, рака шейки матки, рака щитовидной железы и рака слюнных желез.
33. Способ из воплощения 32, при этом субъекту дополнительно вводится терапевтическое средство, выбранное из группы, состоящей из противоопухолевых средств, химиотерапевтических средств, средств, ингибирующих рост, и цитотоксических средств.
34. Способ из воплощения 32, при этом субъекту дополнительно проводится лучевая терапия.













--->
SEQUENCE LISTING
<110> HENLIX, INC.
HENLIX BIOTECH CO., LTD.
<120> ANTI-PD-L1 ANTIBODIES AND VARIANTS
<130> 000020.0006PCTLF
<140> PCT/US2017/056689
<141> 2017-10-14
<150> 62/414,785
<151> 2016-10-30
<160> 145
<170> PatentIn version 3.5
<210> 1
<211> 216
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 1
Gln Ala Val Leu Thr Gln Pro Ser Ser Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Val Gly Ala Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Tyr Asp Ser Ser
85 90 95
Leu Ser Gly Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu
115 120 125
Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe
130 135 140
Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val
145 150 155 160
Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys
165 170 175
Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser
180 185 190
His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu
195 200 205
Lys Thr Val Ala Leu Thr Glu Cys
210 215
<210> 2
<211> 214
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 2
Gln Ser Val Val Thr Gln Pro Pro Ser Met Ser Ala Ala Pro Gly Gln
1 5 10 15
Asn Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Tyr Ile Glu Ser Ser
20 25 30
Tyr Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Asp Asp Asp Met Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Thr Leu Ala Ile Thr Gly Leu Gln
65 70 75 80
Thr Gly Asp Glu Ala Asp Tyr Tyr Cys Glu Ile Trp Asp Ser Gly Leu
85 90 95
Gly Gly Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Ser Gln Pro
100 105 110
Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu
115 120 125
Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro
130 135 140
Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala
145 150 155 160
Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala
165 170 175
Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Lys
180 185 190
Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Arg Thr
195 200 205
Val Ala Leu Thr Glu Cys
210
<210> 3
<211> 217
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 3
Gln Ser Val Val Thr Gln Pro Pro Pro Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Tyr Asp Ser Ser
85 90 95
Leu Ser Ala Pro Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
115 120 125
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
130 135 140
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
145 150 155 160
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
165 170 175
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
180 185 190
Ser His Lys Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
195 200 205
Glu Lys Thr Val Ala Leu Thr Glu Cys
210 215
<210> 4
<211> 216
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 4
Gln Ser Val Val Thr Gln Pro Pro Ser Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Tyr Asp Ser Ser
85 90 95
Leu Ser Gly Gly Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu
115 120 125
Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe
130 135 140
Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val
145 150 155 160
Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys
165 170 175
Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser
180 185 190
His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu
195 200 205
Lys Thr Val Ala Leu Thr Glu Cys
210 215
<210> 5
<211> 215
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 5
Leu Ser Ser Glu Leu Thr Gln Asp Pro Ala Val Ser Val Ala Leu Gly
1 5 10 15
Gln Thr Val Arg Ile Thr Cys Gln Gly Asp Ser Leu Arg Ser Tyr Tyr
20 25 30
Val Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Leu
35 40 45
Tyr Gly Lys Asn Asn Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser Gly
50 55 60
Ser Ser Ser Gly Ser Thr Ala Ser Leu Thr Ile Thr Gly Ala Gln Ala
65 70 75 80
Glu Asp Glu Ala Asp Tyr Tyr Cys Asn Ser Arg Asp Ser Thr Gly Asn
85 90 95
Leu Leu Arg Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Ile Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Leu Thr Glu Cys
210 215
<210> 6
<211> 216
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 6
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Arg Ser Ser Ser Asn Ile Gly Ala Gly
20 25 30
His Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu
65 70 75 80
Gln Ala Glu Asp Gly Ala Asp Tyr Tyr Cys Gln Ser Tyr Asp Ser Ser
85 90 95
Leu Thr Gly Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu
115 120 125
Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe
130 135 140
Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val
145 150 155 160
Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys
165 170 175
Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser
180 185 190
His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu
195 200 205
Lys Thr Val Ala Leu Thr Glu Cys
210 215
<210> 7
<211> 216
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 7
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Tyr Ser Ser Asn Ile Gly Ala Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln His Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Tyr Asp Asn Ser
85 90 95
Leu Ser Val Ser Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu
115 120 125
Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe
130 135 140
Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val
145 150 155 160
Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys
165 170 175
Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser
180 185 190
His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu
195 200 205
Lys Thr Val Ala Leu Thr Glu Cys
210 215
<210> 8
<211> 451
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 8
Gln Ile Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Ile Pro Ile Leu Gly Ile Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Gly Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ser Ser Gly Trp Leu Gly Val Leu Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Ala Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 9
<211> 453
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 9
Glu Val Gln Leu Val Gln Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Gly Ser Asp Tyr Leu Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Glu Leu Arg Trp Tyr Pro Gln Ala Gly Ala Phe Asp Ile
100 105 110
Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Ala Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly Lys
450
<210> 10
<211> 451
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 10
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Thr Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Ile Pro Ile Leu Gly Ile Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Arg Asp Gly Tyr Ser Phe Gly Ala Phe Asp Ile Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Ala Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 11
<211> 451
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 11
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Thr Ile Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Val
35 40 45
Gly Lys Ile Ile Pro Ile Leu Gly Ile Ala Asp Tyr Ala Gln Met Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Lys Phe Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Asn Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Tyr Val Gly Tyr Leu Asn Ala Phe Asp Ile Trp Gly
100 105 110
Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Ala Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 12
<211> 448
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 12
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Val Leu Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr
100 105 110
Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 13
<211> 451
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 13
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Ile Pro Ile Leu Gly Ile Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ile Gly Ser Tyr Ser Phe Gly Ala Phe Asp Ile Trp Gly
100 105 110
Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Ala Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 14
<211> 451
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 14
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Thr Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Ile Pro Ile Leu Gly Ile Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Arg Asp Gly Tyr Ser Phe Gly Ala Phe Asp Ile Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Ala Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 15
<211> 327
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 15
cagtctgtcg tgacgcagcc gccctcaatg tcagcggccc caggacagag agtcaccatc 60
tcctgctctg gaagcagctc ctacattgaa agttcttacg tcgggtggta ccagcaactc 120
ccaggaacag cccccagact cctcatttat gacgatgata tgcgaccctc agggatccct 180
gaccgattct ctggctccaa gtctggcacg tcagccaccc tggccatcac cggactccag 240
actggggacg aggccgatta ttactgcgag atatggcgga gcggcctggg aggcgtcttc 300
ggcggaggga ccaagctgac cgtccta 327
<210> 16
<211> 109
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 16
Gln Ser Val Val Thr Gln Pro Pro Ser Met Ser Ala Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Tyr Ile Glu Ser Ser
20 25 30
Tyr Val Gly Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Asp Asp Asp Met Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Thr Leu Ala Ile Thr Gly Leu Gln
65 70 75 80
Thr Gly Asp Glu Ala Asp Tyr Tyr Cys Glu Ile Trp Arg Ser Gly Leu
85 90 95
Gly Gly Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 17
<211> 369
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 17
gaggttcagc tggtacaatc tgggggaggc ctggtcaagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt caccttcagt agttatacta tgaactgggt ccgccaggct 120
ccagggaagg gcctggagtg ggtctcatcc attagtagtg gtagtgatta cttatactac 180
gcagactctg tgaagggccg attcaccatc tccagggaca acgccaagaa ctcactgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaaacgaa 300
ctacggtggt atccacaagc aggtgctttt gatcgatggg gccaagggac aatggtcacc 360
gtctcaagc 369
<210> 18
<211> 123
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 18
Glu Val Gln Leu Val Gln Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Gly Ser Asp Tyr Leu Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Glu Leu Arg Trp Tyr Pro Gln Ala Gly Ala Phe Asp Arg
100 105 110
Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 19
<211> 336
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 19
cagtctgtcg tgacgcagcc gcccccagtg tctggggccc cagggcagag ggtcaccatc 60
tcctgcactg ggagcagctc caacatcggg gcaggttatg atgtacactg gtaccagcag 120
cttccaggaa cagcccccaa actcctcatc tatggtaaca gcaatcggcc ctcaggggtc 180
cctgaccgat tctctggctc caagtctggc acctcagcct ccctggccat cactgggctc 240
caggctgagg atgaggctga ttattactgc cagacttatg acagcagcct gagtgcccgg 300
gtggtattcg gcggagggac caagctgacc gtccta 336
<210> 20
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 20
Gln Ser Val Val Thr Gln Pro Pro Pro Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Thr Tyr Asp Ser Ser
85 90 95
Leu Ser Ala Arg Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 21
<211> 363
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 21
caggtccagc tggtgcaatc tggggctgag gtgaagaagc ctgggtcctc ggtgaaggtc 60
tcctgcaagg cttctggagg caccttcagc agctatccga tcagctgggt gcgacaggcc 120
cctggacaag ggcttgagtg gataggaagg atcatcccta tccttgggat agcaaactac 180
gcacagaagt tccagggcag agtcacgatt accgcggaca aatccacgag cacagcctac 240
atggagctga gcagcctgag atctgaggac acggccgtgt attactgtgc gaggtctaga 300
gatggctacg cttttggtgc ttttgatatc tggggccaag gaaccctggt caccgtctca 360
agc 363
<210> 22
<211> 121
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 22
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Pro Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Arg Ile Ile Pro Ile Leu Gly Ile Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Arg Asp Gly Tyr Ala Phe Gly Ala Phe Asp Ile Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 23
<211> 336
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 23
cagtctgtcg tgacgcagcc gcccccagtg tctggggccc cagggcagag ggtcaccatc 60
tcctgcactg ggagcagctc caacatcggg gggggttatg atgtacactg gtaccagcag 120
cttccaggaa cagcccccaa actcctcatc tatggtaaca gcacgcggcc ctcaggggtc 180
cctgaccgat tctctggctc caagtctggc acctcagcct ccctggccat cactgggctc 240
caggctgagg atgaggctga ttattactgc cagacttatg acagcagcct gagtgccacg 300
gtggtattcg gcggagggac caagctgacc gtccta 336
<210> 24
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 24
Gln Ser Val Val Thr Gln Pro Pro Pro Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Gly Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Asn Ser Thr Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Thr Tyr Asp Ser Ser
85 90 95
Leu Ser Ala Thr Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 25
<211> 363
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 25
caggtccagc tggtgcaatc tggggctgag gtgaagaagc ctgggtcctc ggtgaaggtc 60
tcctgcaagg cttctggagg caccttcagc agctatccga tcagctgggt gcgacaggcc 120
cctggacaag ggcttgagtg gatgggaagg atcatcccta tccttggtat agcaaactac 180
gcacagaagt tccagggcag agtcacgatt accgcggaca aatccacgag cacagcctac 240
atggagctga gcagcctgag atctgaggac acggccgtgt attactgtgc gaggtctaga 300
gatggctacg cttttggtgc ttttgatgtg tggggccaag gaaccctggt caccgtctca 360
agc 363
<210> 26
<211> 121
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 26
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Pro Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Ile Pro Ile Leu Gly Ile Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Arg Asp Gly Tyr Ala Phe Gly Ala Phe Asp Val Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 27
<211> 336
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 27
cagtctgtcg tgacgcagcc gcccccagtg tctggggccc cagggcagag ggtcaccatc 60
tcctgcactg ggagcagctc caacgtgggg gcaggttatg atgtacactg gtaccagcag 120
cttccaggaa cagcccccaa actcctcatc tatggtaaca gcaatcggtc ttcaggggtc 180
cctgaccgat tctctggctc caagtctggc acctcagcct ccctggccat cactgggctc 240
caggctgagg atgaggctga ttattactgc cagacttatg acagcagcgg gagtgcccgg 300
gtggtattcg gcggagggac caagctgacc gtccta 336
<210> 28
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 28
Gln Ser Val Val Thr Gln Pro Pro Pro Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Val Gly Ala Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Asn Ser Asn Arg Ser Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Thr Tyr Asp Ser Ser
85 90 95
Gly Ser Ala Arg Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 29
<211> 363
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 29
caggtccagc tggtgcaatc tggggctgag gtgaagaagc ctgggtcctc ggtgaaggtc 60
tcctgcaagg cttctggagg caccttcagc agctatccga tcagctgggt gcgacaggcc 120
cctggacaag ggcttgagtg gatgggaagg atcatcccta tccttggtat agcaaactac 180
gcacagaagt tccagggcag agtcacgatt accgcggaca aatccacgag cacagcctac 240
atggagctga gcagcctgag atctgaggac acggccgtgt attactgtgc gaggtctaga 300
ccgggctacg cttttggtgc ttttgatatc tggggccaag gaaccctggt caccgtctca 360
agc 363
<210> 30
<211> 121
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 30
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Pro Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Ile Pro Ile Leu Gly Ile Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Arg Pro Gly Tyr Ala Phe Gly Ala Phe Asp Ile Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 31
<211> 336
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 31
cagtctgtcg tgacgcagcc gcccccagtg tctggggccc cagggcagag ggtcaccatc 60
tcctgcactg ggagcagctc caacatcggg cagggttatg atgtacactg gtaccagcag 120
cttccaggaa cagcccccaa actcctcatc tatgctaaca gcaatcggcc ctcaggggtc 180
cctgaccgat tctctggctc caagtctggc acctcagcct ccctggccat cactgggctc 240
caggctgagg atgaggctga ttattactgc cagacttatg acagcagcct gagtgcccgg 300
gtggtattcg gcggagggac caagctgacc gtccta 336
<210> 32
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 32
Gln Ser Val Val Thr Gln Pro Pro Pro Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Gln Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Ala Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Thr Tyr Asp Ser Ser
85 90 95
Leu Ser Ala Arg Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 33
<211> 363
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 33
caggtccagc tggtgcaatc tggggctgag gtgaagaagc ctgggtcctc ggtgaaggtc 60
tcctgcaagg cttctggagg caccttcagc agctatccga tcagctgggt gcgacaggcc 120
cctggacaag ggcttgagtg gatgggaagg atcatcccta tccttggtat agcagattac 180
gcacagaagt tccagggcag agtcacgatt accgcggaca aatccacgag cacagcctac 240
atggagctga gcagcctgag atctgaggac acggccgtgt attactgtgc gaggtctaga 300
ccgggctacg cttttggtgc ttttgatatc tggggccaag gaaccctggt caccgtctca 360
agc 363
<210> 34
<211> 121
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 34
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Pro Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Ile Pro Ile Leu Gly Ile Ala Asp Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Arg Pro Gly Tyr Ala Phe Gly Ala Phe Asp Ile Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 35
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 35
Thr Gly Ser Ser Ser Asn Val Gly Ala Gly Tyr Asp Val His
1 5 10
<210> 36
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 36
Ser Gly Ser Ser Ser Tyr Ile Glu Ser Ser Tyr Val Ser
1 5 10
<210> 37
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 37
Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly Tyr Asp Val His
1 5 10
<210> 38
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 38
Gln Gly Asp Ser Leu Arg Ser Tyr Tyr Val Ser
1 5 10
<210> 39
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 39
Thr Arg Ser Ser Ser Asn Ile Gly Ala Gly His Asp Val His
1 5 10
<210> 40
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 40
Thr Gly Tyr Ser Ser Asn Ile Gly Ala Gly Tyr Asp Val His
1 5 10
<210> 41
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 41
Gly Asn Ser Asn Arg Pro Ser
1 5
<210> 42
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 42
Asp Asp Asp Met Arg Pro Ser
1 5
<210> 43
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 43
Gly Lys Asn Asn Arg Pro Ser
1 5
<210> 44
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 44
Gln Ser Tyr Asp Ser Ser Leu Ser Gly Trp Val
1 5 10
<210> 45
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 45
Glu Ile Trp Asp Ser Gly Leu Gly Gly Val
1 5 10
<210> 46
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 46
Gln Ser Tyr Asp Ser Ser Leu Ser Ala Pro Val Val
1 5 10
<210> 47
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 47
Gln Ser Tyr Asp Ser Ser Leu Ser Gly Gly Val
1 5 10
<210> 48
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 48
Asn Ser Arg Asp Ser Thr Gly Asn Leu Leu Arg Val
1 5 10
<210> 49
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 49
Gln Ser Tyr Asp Ser Ser Leu Thr Gly Val Val
1 5 10
<210> 50
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 50
Gln Ser Tyr Asp Asn Ser Leu Ser Val Ser Val
1 5 10
<210> 51
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 51
Ser Tyr Ala Ile Ser
1 5
<210> 52
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 52
Ser Tyr Thr Met Asn
1 5
<210> 53
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 53
Ser Tyr Thr Ile Ser
1 5
<210> 54
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 54
Ser Tyr Thr Ile Asn
1 5
<210> 55
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 55
Arg Ile Ile Pro Ile Leu Gly Ile Ala Asn Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 56
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 56
Ser Ile Ser Ser Gly Ser Asp Tyr Leu Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 57
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 57
Lys Ile Ile Pro Ile Leu Gly Ile Ala Asp Tyr Ala Gln Met Phe Lys
1 5 10 15
Gly
<210> 58
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 58
Arg Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 59
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 59
Glu Gly Ser Ser Gly Trp Leu Gly Val Leu Asp Tyr
1 5 10
<210> 60
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 60
Asn Glu Leu Arg Trp Tyr Pro Gln Ala Gly Ala Phe Asp Ile
1 5 10
<210> 61
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 61
Ser Arg Asp Gly Tyr Ser Phe Gly Ala Phe Asp Ile
1 5 10
<210> 62
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 62
Gly Gly Tyr Val Gly Tyr Leu Asn Ala Phe Asp Ile
1 5 10
<210> 63
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 63
Glu Gly Val Leu Asp Ala Phe Asp Ile
1 5
<210> 64
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 64
Gly Ile Gly Ser Tyr Ser Phe Gly Ala Phe Asp Ile
1 5 10
<210> 65
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 65
Ser Gly Ser Ser Ser Tyr Ile Glu Ser Ser Tyr Val Gly
1 5 10
<210> 66
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 66
Thr Gly Ser Ser Ser Asn Ile Gly Gly Gly Tyr Asp Val His
1 5 10
<210> 67
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 67
Thr Gly Ser Ser Ser Asn Ile Gly Gln Gly Tyr Asp Val His
1 5 10
<210> 68
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 68
Gly Asn Ser Thr Arg Pro Ser
1 5
<210> 69
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 69
Gly Asn Ser Asn Arg Ser Ser
1 5
<210> 70
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 70
Ala Asn Ser Asn Arg Pro Ser
1 5
<210> 71
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 71
Glu Ile Trp Arg Ser Gly Leu Gly Gly Val
1 5 10
<210> 72
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 72
Gln Thr Tyr Asp Ser Ser Leu Ser Ala Arg Val Val
1 5 10
<210> 73
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 73
Gln Thr Tyr Asp Ser Ser Leu Ser Ala Thr Val Val
1 5 10
<210> 74
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 74
Gln Thr Tyr Asp Ser Ser Gly Ser Ala Arg Val Val
1 5 10
<210> 75
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 75
Ser Tyr Pro Ile Ser
1 5
<210> 76
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 76
Arg Ile Ile Pro Ile Leu Gly Ile Ala Asp Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 77
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 77
Asn Glu Leu Arg Trp Tyr Pro Gln Ala Gly Ala Phe Asp Arg
1 5 10
<210> 78
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 78
Ser Arg Asp Gly Tyr Ala Phe Gly Ala Phe Asp Ile
1 5 10
<210> 79
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 79
Ser Arg Asp Gly Tyr Ala Phe Gly Ala Phe Asp Val
1 5 10
<210> 80
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 80
Ser Arg Pro Gly Tyr Ala Phe Gly Ala Phe Asp Ile
1 5 10
<210> 81
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 81
Ser Arg Pro Gly Tyr Ala Phe Gly Ala Phe Asp Ile
1 5 10
<210> 82
<211> 327
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 82
cagtctgtcg tgacgcagcc gccctcaatg tcagcggccc caggacagag ggtcaccatc 60
tcctgctctg gagttagctc ctacattgaa agttcttatg tctcctggta ccagcaactc 120
ccaggaacag cccccagact cctcatttat gacgatgata tgcgaccctc agggatccct 180
gaccgattct ctggctccaa gtctggcacg tcagccaccc tggccatcac cggactccag 240
actggggacg aggccgatta ttactgcaag atatgggata gcggcctggg aggcgtcttc 300
ggcggaggga ccaagctgac cgtccta 327
<210> 83
<211> 109
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 83
Gln Ser Val Val Thr Gln Pro Pro Ser Met Ser Ala Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Val Ser Ser Tyr Ile Glu Ser Ser
20 25 30
Tyr Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Asp Asp Asp Met Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Thr Leu Ala Ile Thr Gly Leu Gln
65 70 75 80
Thr Gly Asp Glu Ala Asp Tyr Tyr Cys Lys Ile Trp Asp Ser Gly Leu
85 90 95
Gly Gly Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 84
<211> 369
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 84
gaggttcagc tggtacaatc tgggggaggc ctggtcaagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt caccttcagt agttatacta tgaactgggt ccgccaggct 120
ccagggaagg gcctggagtg ggtctcatcc attagtagtg gtagtgatta cttatactac 180
gcagactctg tgaagggccg attcaccatc tccagggaca acgccaagaa ctcactgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaaacgaa 300
ctacggtggt atccacttgc aggtgctttt gatatctggg gccaagggac aatggtcacc 360
gtctcaagc 369
<210> 85
<211> 123
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 85
Glu Val Gln Leu Val Gln Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Gly Ser Asp Tyr Leu Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Glu Leu Arg Trp Tyr Pro Leu Ala Gly Ala Phe Asp Ile
100 105 110
Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 86
<211> 327
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 86
cagtctgtcg tgacgcagcc gccctcaatg tcagcggccc caggacagag ggtcaccatc 60
tcctgcagtg gaagcagctc ctacattgaa agttcttatg tctcatggta ccagcaactc 120
ccaggaacag cccccagact cctcatttat gacgatgata tgcgaccctc agggatccct 180
gaccgattct ctggctccaa gtctggcacg tcagccaccc tggccatcac cggactccag 240
actggggacg aggccgatta ttactgcgag atatgggata gccggctggg aggcgtcttc 300
ggcggaggga ccaagctgac cgtccta 327
<210> 87
<211> 109
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 87
Gln Ser Val Val Thr Gln Pro Pro Ser Met Ser Ala Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Tyr Ile Glu Ser Ser
20 25 30
Tyr Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Asp Asp Asp Met Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Thr Leu Ala Ile Thr Gly Leu Gln
65 70 75 80
Thr Gly Asp Glu Ala Asp Tyr Tyr Cys Glu Ile Trp Asp Ser Arg Leu
85 90 95
Gly Gly Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 88
<211> 369
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 88
gaggttcagc tggtacaatc tgggggaggc ctggtcaagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt caccttcagt agttatacta tgaactgggt ccgccaggct 120
ccagggaagg gcctggagtg ggtctcatcc attagtagtg gtagtgatta cttatactac 180
gcagactctg tgaagggccg attcaccatc tccagggaca acgccaagaa ctcactgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaaacgaa 300
ctacggtggt atccatttgc aggtgctttt gatatttggg gccaagggac aatggtcacc 360
gtctcaagc 369
<210> 89
<211> 123
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 89
Glu Val Gln Leu Val Gln Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Gly Ser Asp Tyr Leu Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Glu Leu Arg Trp Tyr Pro Phe Ala Gly Ala Phe Asp Ile
100 105 110
Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 90
<211> 327
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 90
cagtctgtcg tgacgcagcc gccctcaatg tcagcggccc caggacagag ggtcaccatc 60
tcctgctctg gaagcagctc ctacattacg agttcttatg tctcctggta ccagcaactc 120
ccaggaacag cccccagact cctcatttat gacgatgata tgcgaccctc agggatccct 180
gaccgattct ctggctccaa gtctggcacg tcagccaccc tggccatcac cggactccag 240
actggggacg aggccgatta ttactgcaag atatgggata gcggcctggg aggcgtcttc 300
ggcggaggga ccaagctgac cgtccta 327
<210> 91
<211> 109
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 91
Gln Ser Val Val Thr Gln Pro Pro Ser Met Ser Ala Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Tyr Ile Thr Ser Ser
20 25 30
Tyr Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Asp Asp Asp Met Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Thr Leu Ala Ile Thr Gly Leu Gln
65 70 75 80
Thr Gly Asp Glu Ala Asp Tyr Tyr Cys Lys Ile Trp Asp Ser Gly Leu
85 90 95
Gly Gly Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 92
<211> 369
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 92
gaggttcagc tggtacaatc tgggggaggc ctggtcaagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt caccttcagt agttatacta tgaactgggt ccgccaggct 120
ccagggaagg gcctggagtg ggtctcatcc attagtagtg gtagtgatta cttatactac 180
gcagactctg tgaagggccg attcaccatc tccagggaca acgccaagaa ctcactgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaaacgaa 300
ctacggtggt atccaaaggc aggtgctttt gatatatggg gccaagggac aatggtcacc 360
gtctcaagc 369
<210> 93
<211> 123
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 93
Glu Val Gln Leu Val Gln Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Gly Ser Asp Tyr Leu Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Glu Leu Arg Trp Tyr Pro Lys Ala Gly Ala Phe Asp Ile
100 105 110
Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 94
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 94
Ser Gly Val Ser Ser Tyr Ile Glu Ser Ser Tyr Val Ser
1 5 10
<210> 95
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 95
Lys Ile Trp Asp Ser Gly Leu Gly Gly Val
1 5 10
<210> 96
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 96
Asn Glu Leu Arg Trp Tyr Pro Leu Ala Gly Ala Phe Asp Ile
1 5 10
<210> 97
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 97
Ser Gly Ser Ser Ser Tyr Ile Glu Ser Ser Tyr Val Ser
1 5 10
<210> 98
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 98
Glu Ile Trp Asp Ser Arg Leu Gly Gly Val
1 5 10
<210> 99
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 99
Asn Glu Leu Arg Trp Tyr Pro Phe Ala Gly Ala Phe Asp Ile
1 5 10
<210> 100
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 100
Ser Gly Ser Ser Ser Tyr Ile Thr Ser Ser Tyr Val Ser
1 5 10
<210> 101
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 101
Asn Glu Leu Arg Trp Tyr Pro Lys Ala Gly Ala Phe Asp Ile
1 5 10
<210> 102
<211> 336
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 102
cagtctgtcg tgacgcagcc gcccccagtg tctggggccc cagggcagag ggtcaccatc 60
tcctgcactg ggagcagctc caacatcggg gcaggttatg atgtacactg gtaccagcag 120
cttccaggaa cagcccccaa actcctcatc tatggtaaca gcaggcggcc ctcaggggtc 180
cctgaccgat tctctggctc caagtctggc acctcagcct ccctggccat cactgggctc 240
caggctgagg atgaggctga ttattactgc cagacctatg acagcagcct gagtcgtccc 300
gtggtattcg gcggagggac caagctgacc gtccta 336
<210> 103
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 103
Gln Ser Val Val Thr Gln Pro Pro Pro Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Asn Ser Arg Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Thr Tyr Asp Ser Ser
85 90 95
Leu Ser Arg Pro Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 104
<211> 363
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 104
caggtccagc tggtgcaatc tggggctgag gtgaagaagc ctgggtcctc ggtgaaggtc 60
tcctgcaagg cttctggagg caccttcagc agctatagga tcagctgggt gcgacaggcc 120
cctggacaag ggcttgagtg gatgggaagg atcatcccta tccttggtat agcaaactac 180
gcacagaagt tccagggcag agtcacgatt accgcggaca aatccacgag cacagcctac 240
atggagctga gcagcctgag atctgaggac acggccgtgt attactgtgc gaggtctaga 300
gatggctaca gtgtgggtgc ttttgattcg tggggccaag gaaccctggt caccgtctca 360
agc 363
<210> 105
<211> 121
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 105
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Arg Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Ile Pro Ile Leu Gly Ile Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Arg Asp Gly Tyr Ser Val Gly Ala Phe Asp Ser Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 106
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 106
Gly Asn Ser Arg Arg Pro Ser
1 5
<210> 107
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 107
Gln Thr Tyr Asp Ser Ser Leu Ser Arg Pro Val Val
1 5 10
<210> 108
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 108
Ser Tyr Arg Ile Ser
1 5
<210> 109
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 109
Ser Arg Asp Gly Tyr Ser Val Gly Ala Phe Asp Ser
1 5 10
<210> 110
<211> 336
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 110
cagtctgtcg tgacgcagcc gcccccagtg tctggggccc cagggcagag ggtcaccatc 60
tcctgcactg ggagcagctc caacatcggg gggggttatg atgtacactg gtaccagcag 120
cttccaggaa cagcccccaa actcctcatc tatggtaaca gcaatcggcc ctcaggggtc 180
cctgaccgat tctctggctc caagtctggc acctcagcct ccctggccat cactgggctc 240
caggctgagg atgaggctga ttattactgc cagacttatg acagcagcct gagtgccacg 300
gtggtattcg gcggagggac caagctgacc gtccta 336
<210> 111
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 111
Gln Ser Val Val Thr Gln Pro Pro Pro Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Gly Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Thr Tyr Asp Ser Ser
85 90 95
Leu Ser Ala Thr Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 112
<211> 336
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 112
cagtctgtcg tgacgcagcc gcccccagtg tctggggccc cagggcagag ggtcaccatc 60
tcctgcactg ggagcagctc caacatcggg gggggttatg atgtacactg gtaccagcag 120
cttccaggaa cagcccccaa actcctcatc tatggtcaga gcacgcggcc ctcaggggtc 180
cctgaccgat tctctggctc caagtctggc acctcagcct ccctggccat cactgggctc 240
caggctgagg atgaggctga ttattactgc cagacttatg acagcagcct gagtgccacg 300
gtggtattcg gcggagggac caagctgacc gtccta 336
<210> 113
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 113
Gln Ser Val Val Thr Gln Pro Pro Pro Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Gly Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Gln Ser Thr Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Thr Tyr Asp Ser Ser
85 90 95
Leu Ser Ala Thr Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 114
<211> 336
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 114
cagtctgtcg tgacgcagcc gcccccagtg tctggggccc cagggcagag ggtcaccatc 60
tcctgcactg ggagcagctc caacatcggg gggggttatg atgtacactg gtaccagcag 120
cttccaggaa cagcccccaa actcctcatc tatggtaaca gccaacggcc ctcaggggtc 180
cctgaccgat tctctggctc caagtctggc acctcagcct ccctggccat cactgggctc 240
caggctgagg atgaggctga ttattactgc cagacttatg acagcagcct gagtgccacg 300
gtggtattcg gcggagggac caagctgacc gtccta 336
<210> 115
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 115
Gln Ser Val Val Thr Gln Pro Pro Pro Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Gly Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Asn Ser Gln Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Thr Tyr Asp Ser Ser
85 90 95
Leu Ser Ala Thr Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 116
<211> 336
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 116
cagtctgtcg tgacgcagcc gcccccagtg tctggggccc cagggcagag ggtcaccatc 60
tcctgcactg ggagcagctc caacgtgggg gcaggttatg atgtacactg gtaccagcag 120
cttccaggaa cagcccccaa actcctcatc tatggtaaca gcaatcggcc ctcaggggtc 180
cctgaccgat tctctggctc caagtctggc acctcagcct ccctggccat cactgggctc 240
caggctgagg atgaggctga ttattactgc cagacttatg acagcagcgg gagtgcccgg 300
gtggtattcg gcggagggac caagctgacc gtccta 336
<210> 117
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 117
Gln Ser Val Val Thr Gln Pro Pro Pro Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Val Gly Ala Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Thr Tyr Asp Ser Ser
85 90 95
Gly Ser Ala Arg Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 118
<211> 336
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 118
cagtctgtcg tgacgcagcc gcccccagtg tctggggccc cagggcagag ggtcaccatc 60
tcctgcactg ggagcagctc caacgtgggg gcaggttatg atgtacactg gtaccagcag 120
cttccaggaa cagcccccaa actcctcatc tatggtaaca gccaacggtc ttcaggggtc 180
cctgaccgat tctctggctc caagtctggc acctcagcct ccctggccat cactgggctc 240
caggctgagg atgaggctga ttattactgc cagacttatg acagcagcgg gagtgcccgg 300
gtggtattcg gcggagggac caagctgacc gtccta 336
<210> 119
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 119
Gln Ser Val Val Thr Gln Pro Pro Pro Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Val Gly Ala Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Asn Ser Gln Arg Ser Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Thr Tyr Asp Ser Ser
85 90 95
Gly Ser Ala Arg Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 120
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 120
Ile Gly Ala Gly Tyr Asp
1 5
<210> 121
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 121
Gly Asn Ser Asn Arg Pro
1 5
<210> 122
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 122
Gly Asn Ser Arg Arg Pro
1 5
<210> 123
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 123
Ser Tyr Asp Ser Ser Leu Ser Ala Pro Val Val
1 5 10
<210> 124
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 124
Thr Tyr Asp Ser Ser Leu Ser Arg Pro Val Val
1 5 10
<210> 125
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 125
Thr Tyr Asp Ser Ser Leu Ser Ala Arg Val Val
1 5 10
<210> 126
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 126
Ile Pro Ile Leu Gly Ile Ala Asn
1 5
<210> 127
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 127
Ile Gly Gly Gly Tyr Asp
1 5
<210> 128
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 128
Val Gly Ala Gly Tyr Asp
1 5
<210> 129
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 129
Ile Gly Gln Gly Tyr Asp
1 5
<210> 130
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 130
Gly Asn Ser Thr Arg Pro
1 5
<210> 131
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 131
Gly Asn Ser Asn Arg Ser
1 5
<210> 132
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 132
Ala Asn Ser Asn Arg Pro
1 5
<210> 133
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 133
Thr Tyr Asp Ser Ser Leu Ser Ala Thr Val Val
1 5 10
<210> 134
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 134
Thr Tyr Asp Ser Ser Gly Ser Ala Arg Val Val
1 5 10
<210> 135
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 135
Ile Pro Ile Leu Gly Ile Ala Asp
1 5
<210> 136
<211> 214
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 136
Gln Ser Val Val Thr Gln Pro Pro Ser Met Ser Ala Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Tyr Ile Glu Ser Ser
20 25 30
Tyr Val Gly Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Asp Asp Asp Met Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Thr Leu Ala Ile Thr Gly Leu Gln
65 70 75 80
Thr Gly Asp Glu Ala Asp Tyr Tyr Cys Glu Ile Trp Arg Ser Gly Leu
85 90 95
Gly Gly Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Ser Gln Pro
100 105 110
Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu
115 120 125
Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro
130 135 140
Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala
145 150 155 160
Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala
165 170 175
Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Lys
180 185 190
Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Arg Thr
195 200 205
Val Ala Leu Thr Glu Cys
210
<210> 137
<211> 217
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 137
Gln Ser Val Val Thr Gln Pro Pro Pro Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Thr Tyr Asp Ser Ser
85 90 95
Leu Ser Ala Arg Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
115 120 125
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
130 135 140
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
145 150 155 160
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
165 170 175
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
180 185 190
Ser His Lys Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
195 200 205
Glu Lys Thr Val Ala Leu Thr Glu Cys
210 215
<210> 138
<211> 217
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 138
Gln Ser Val Val Thr Gln Pro Pro Pro Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Gly Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Asn Ser Thr Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Thr Tyr Asp Ser Ser
85 90 95
Leu Ser Ala Thr Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
115 120 125
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
130 135 140
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
145 150 155 160
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
165 170 175
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
180 185 190
Ser His Lys Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
195 200 205
Glu Lys Thr Val Ala Leu Thr Glu Cys
210 215
<210> 139
<211> 217
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 139
Gln Ser Val Val Thr Gln Pro Pro Pro Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Val Gly Ala Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Asn Ser Asn Arg Ser Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Thr Tyr Asp Ser Ser
85 90 95
Gly Ser Ala Arg Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
115 120 125
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
130 135 140
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
145 150 155 160
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
165 170 175
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
180 185 190
Ser His Lys Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
195 200 205
Glu Lys Thr Val Ala Leu Thr Glu Cys
210 215
<210> 140
<211> 217
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 140
Gln Ser Val Val Thr Gln Pro Pro Pro Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Gln Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Ala Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Thr Tyr Asp Ser Ser
85 90 95
Leu Ser Ala Arg Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
115 120 125
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
130 135 140
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
145 150 155 160
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
165 170 175
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
180 185 190
Ser His Lys Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
195 200 205
Glu Lys Thr Val Ala Leu Thr Glu Cys
210 215
<210> 141
<211> 453
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 141
Glu Val Gln Leu Val Gln Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Gly Ser Asp Tyr Leu Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Glu Leu Arg Trp Tyr Pro Gln Ala Gly Ala Phe Asp Arg
100 105 110
Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Ala Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly Lys
450
<210> 142
<211> 451
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 142
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Pro Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Arg Ile Ile Pro Ile Leu Gly Ile Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Arg Asp Gly Tyr Ala Phe Gly Ala Phe Asp Ile Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Ala Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 143
<211> 451
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 143
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Pro Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Ile Pro Ile Leu Gly Ile Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Arg Asp Gly Tyr Ala Phe Gly Ala Phe Asp Val Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Ala Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 144
<211> 451
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 144
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Pro Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Ile Pro Ile Leu Gly Ile Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Arg Pro Gly Tyr Ala Phe Gly Ala Phe Asp Ile Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Ala Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 145
<211> 451
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polypeptide
<400> 145
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Pro Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Ile Pro Ile Leu Gly Ile Ala Asp Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Arg Pro Gly Tyr Ala Phe Gly Ala Phe Asp Ile Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Ala Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИ-TIGIT АНТИТЕЛА И СПОСОБЫ ПРИМЕНЕНИЯ | 2016 |
|
RU2732591C2 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ ИЗ АНТИТЕЛА К ПРОГАСТРИНУ И ИММУНОТЕРАПИИ ДЛЯ ЛЕЧЕНИЯ РАКА | 2018 |
|
RU2784604C2 |
| АНТИТЕЛА К ICOS | 2016 |
|
RU2742241C2 |
| Антитела против лиганда-1 запрограммированной смерти (PD-L1) и их применение | 2017 |
|
RU2721582C1 |
| БИСПЕЦИФИЧЕСКИЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СОДЕРЖАЩИЕ АНТИ-FAP КЛОН 212 | 2019 |
|
RU2799429C2 |
| КОМБИНАЦИЯ АНТАГОНИСТА PD-1 И ЭРИБУЛИНА ДЛЯ ЛЕЧЕНИЯ РАКА | 2016 |
|
RU2737216C2 |
| АНТИ-PD-1 АНТИТЕЛА | 2017 |
|
RU2752832C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА К CCL2 | 2020 |
|
RU2819613C1 |
| АНТИТЕЛА К TIGIT И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2827445C2 |
| МЕЖВИДОВЫЕ АНТИТЕЛА К ЛАТЕНТНОМУ TGF-БЕТА 1 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2020 |
|
RU2837609C2 |



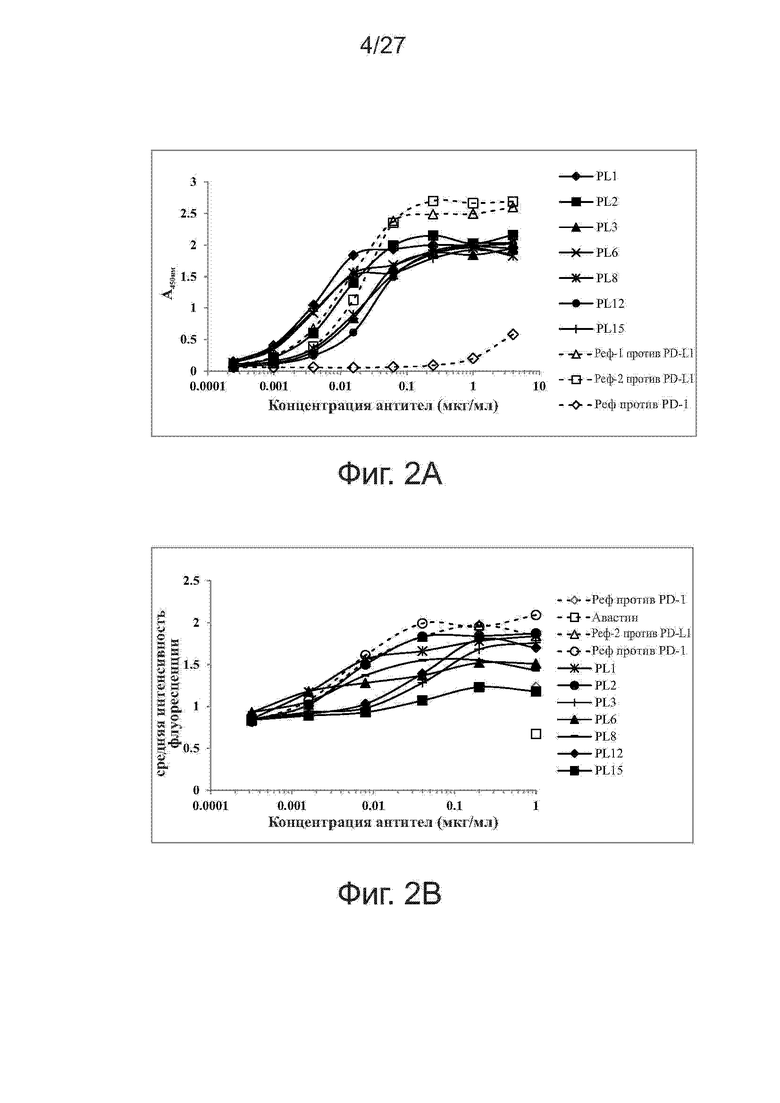
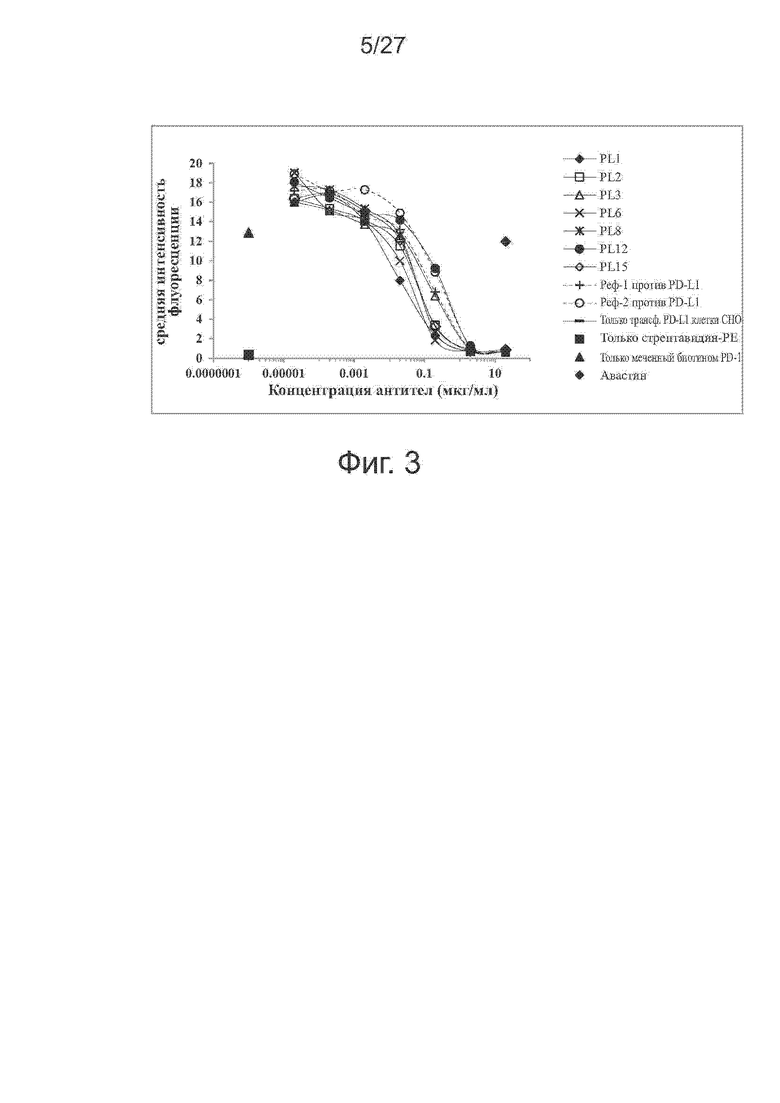
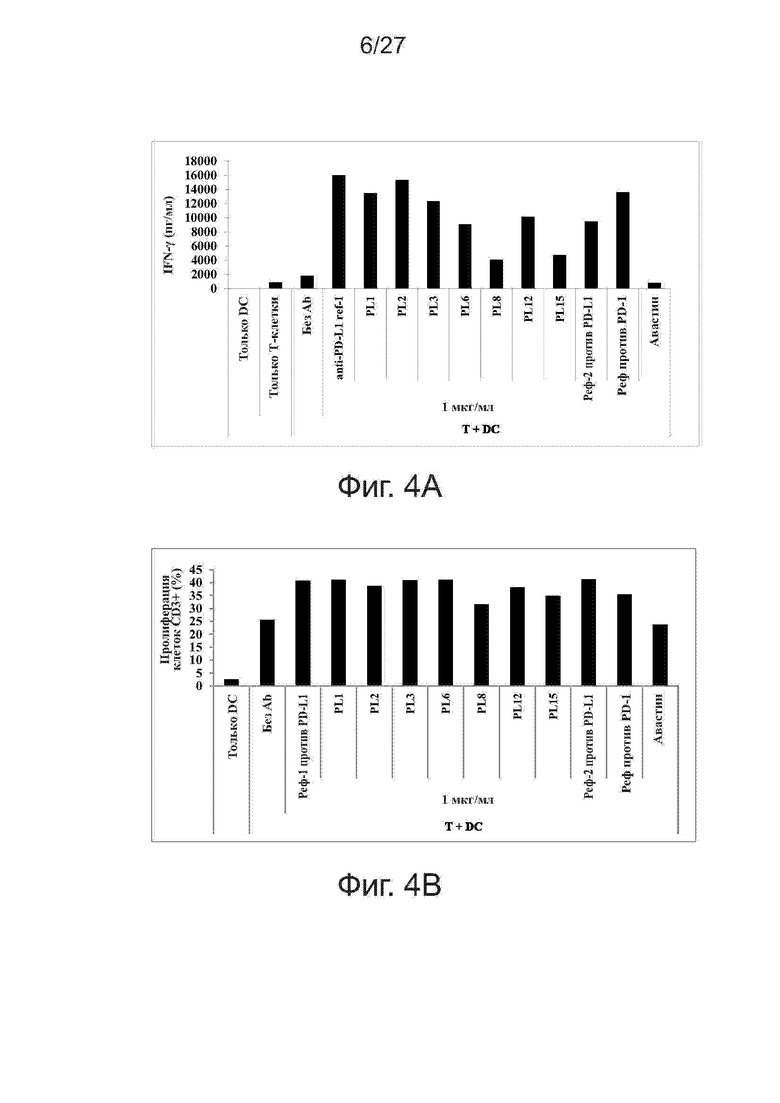
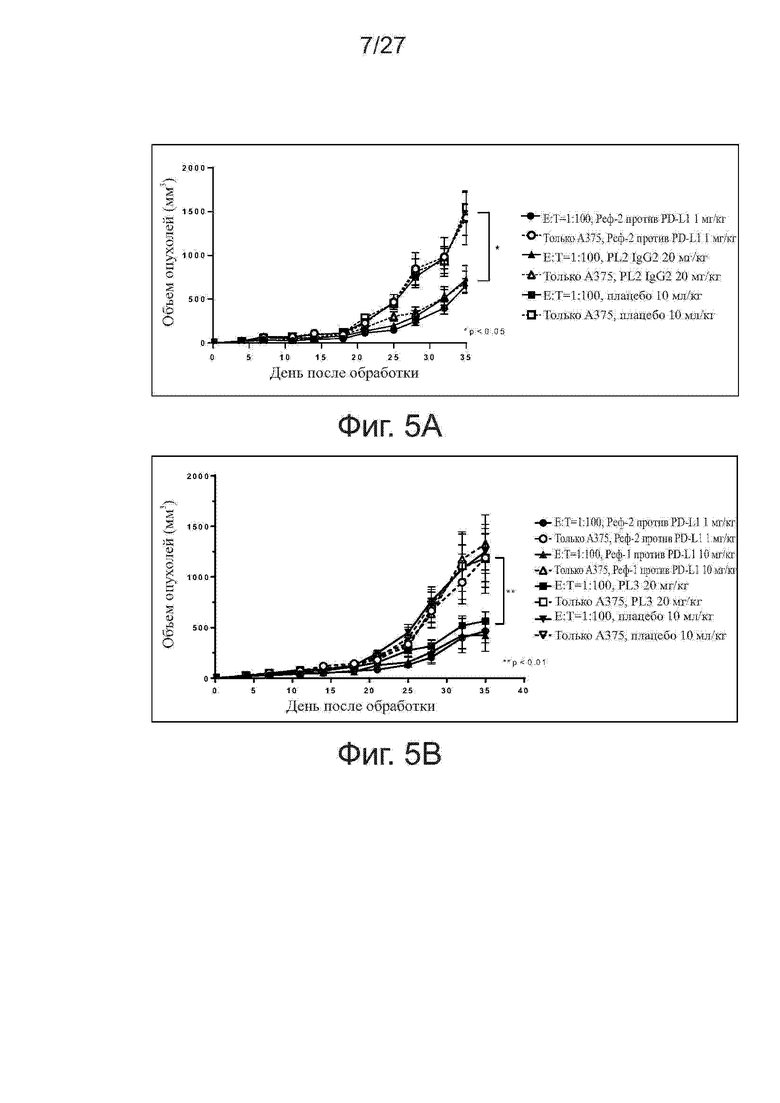
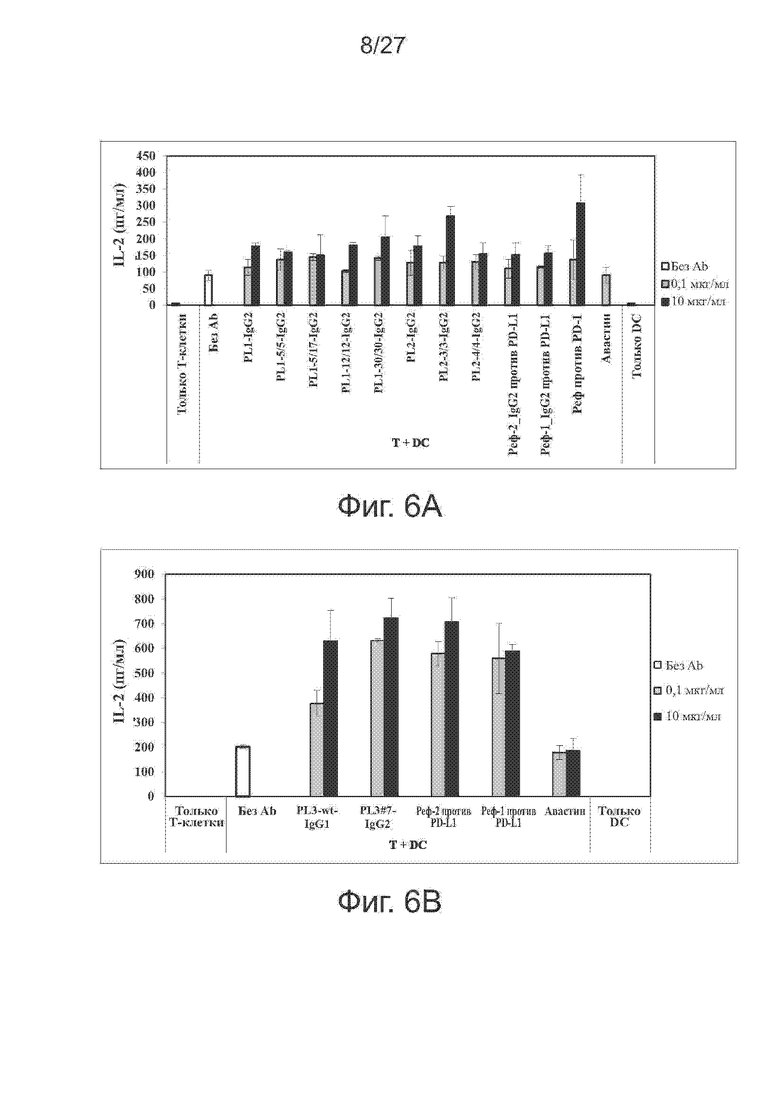
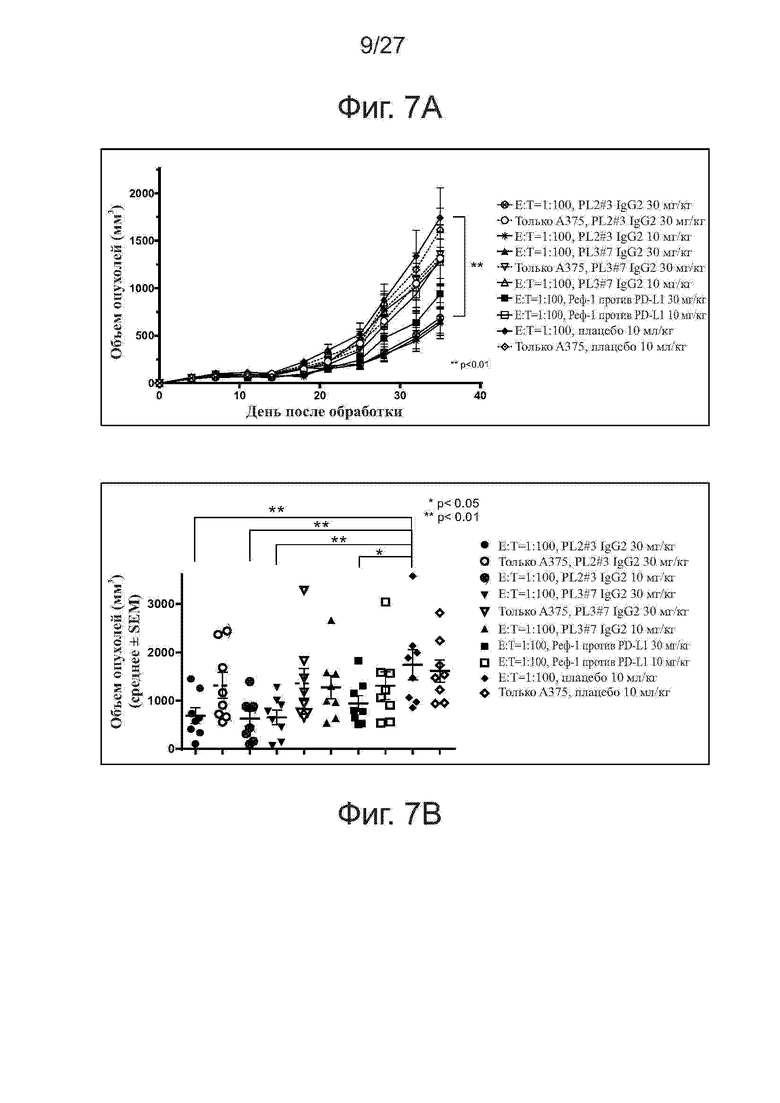
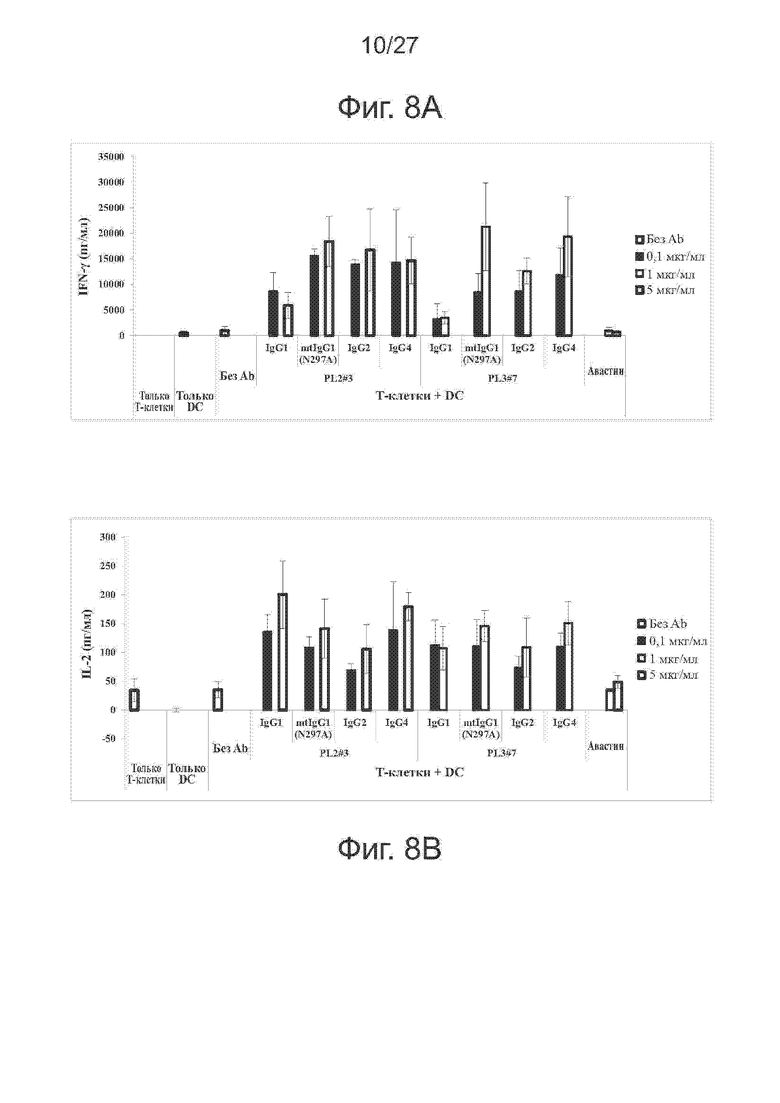
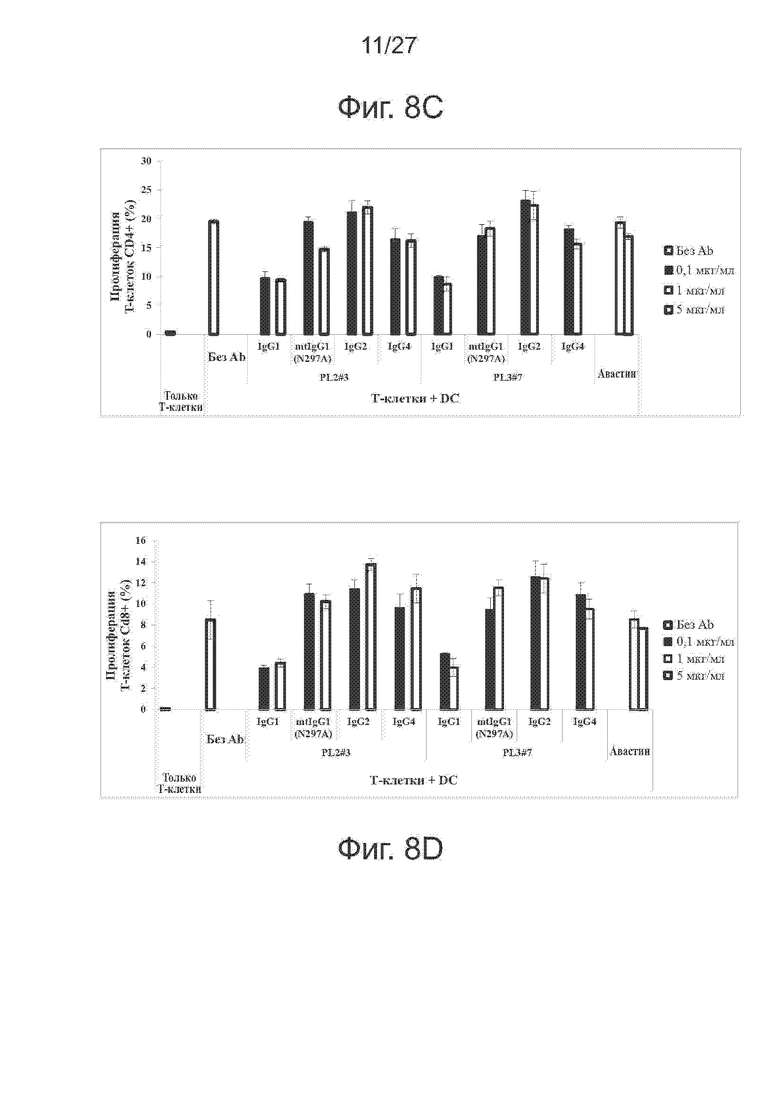
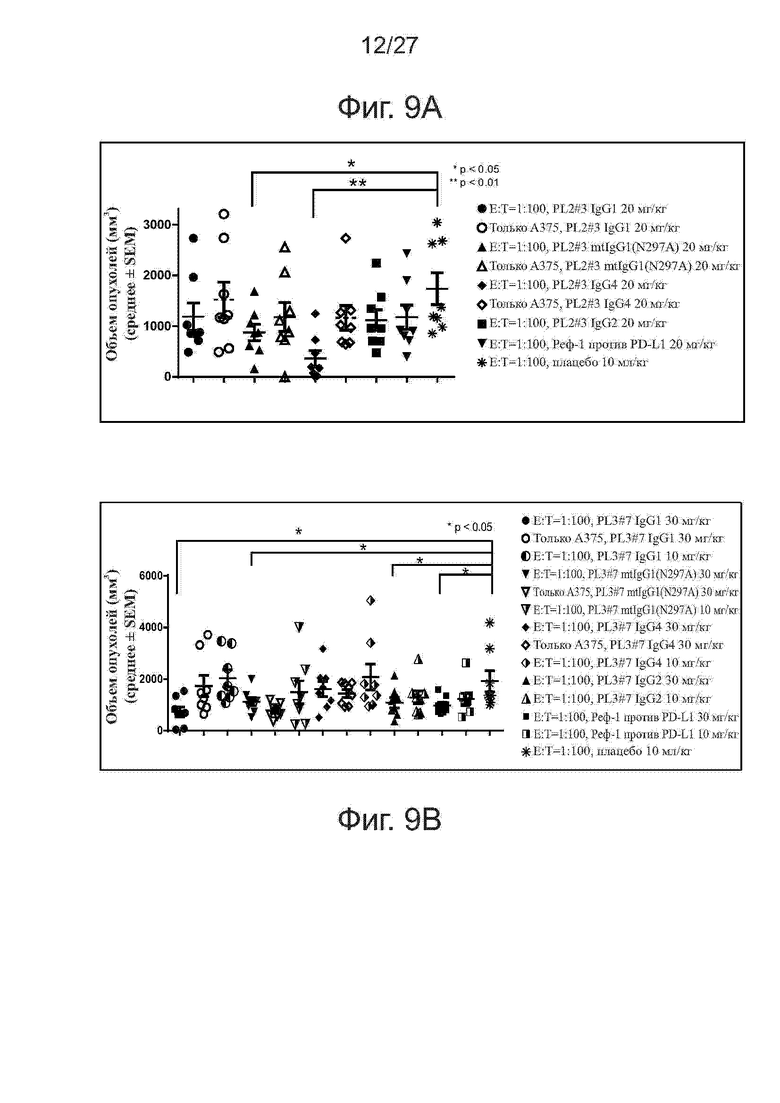
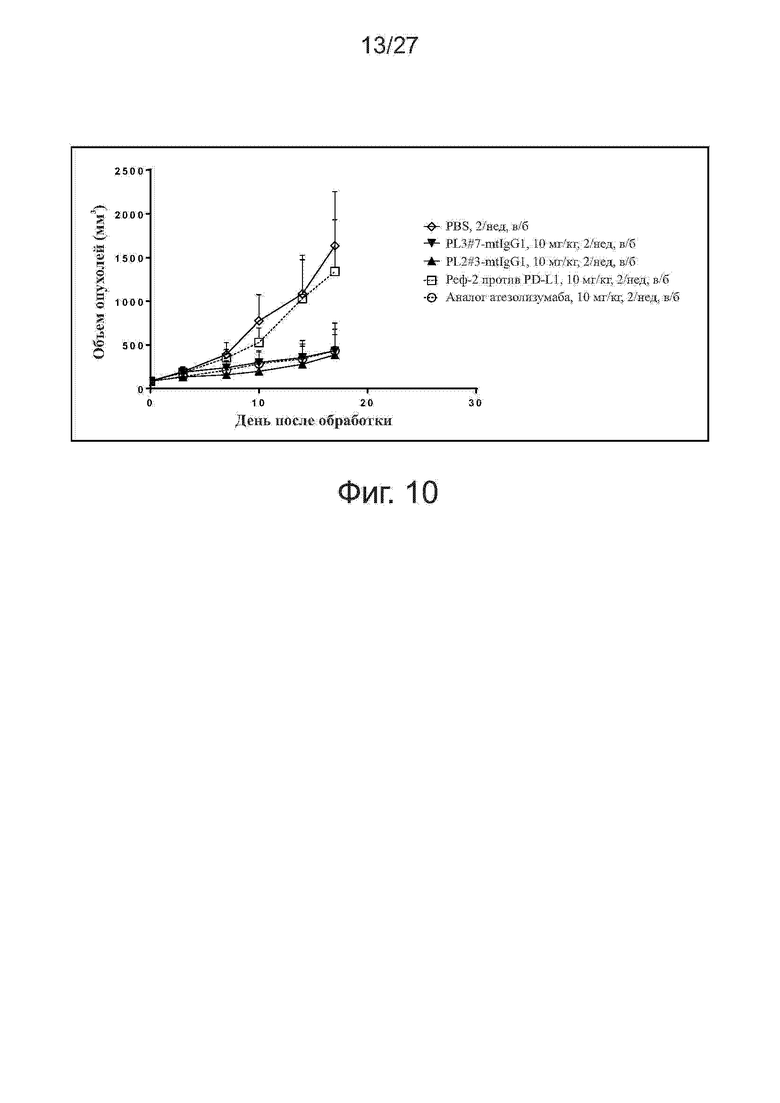



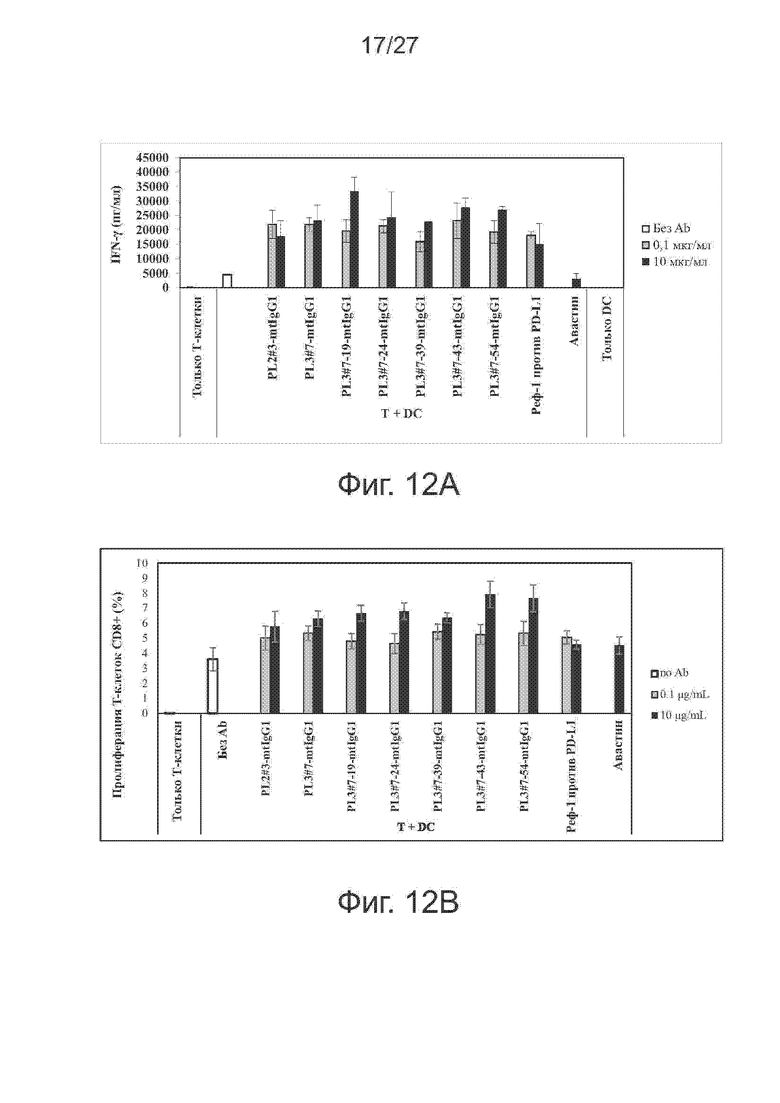
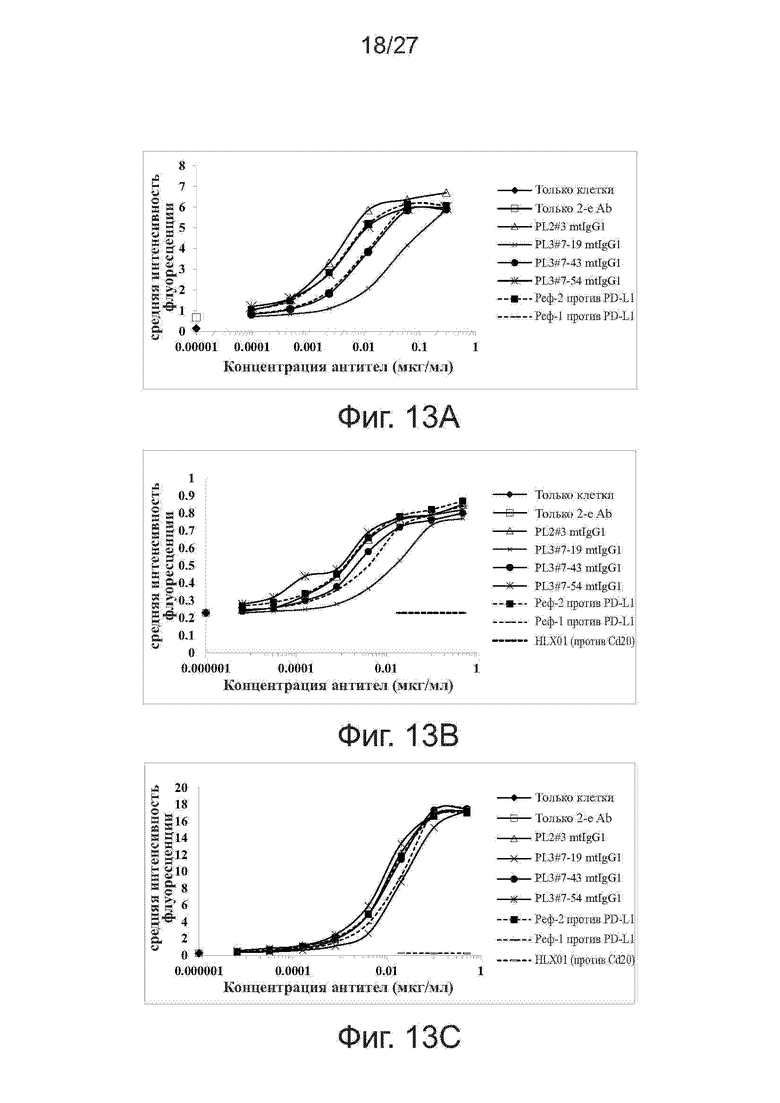
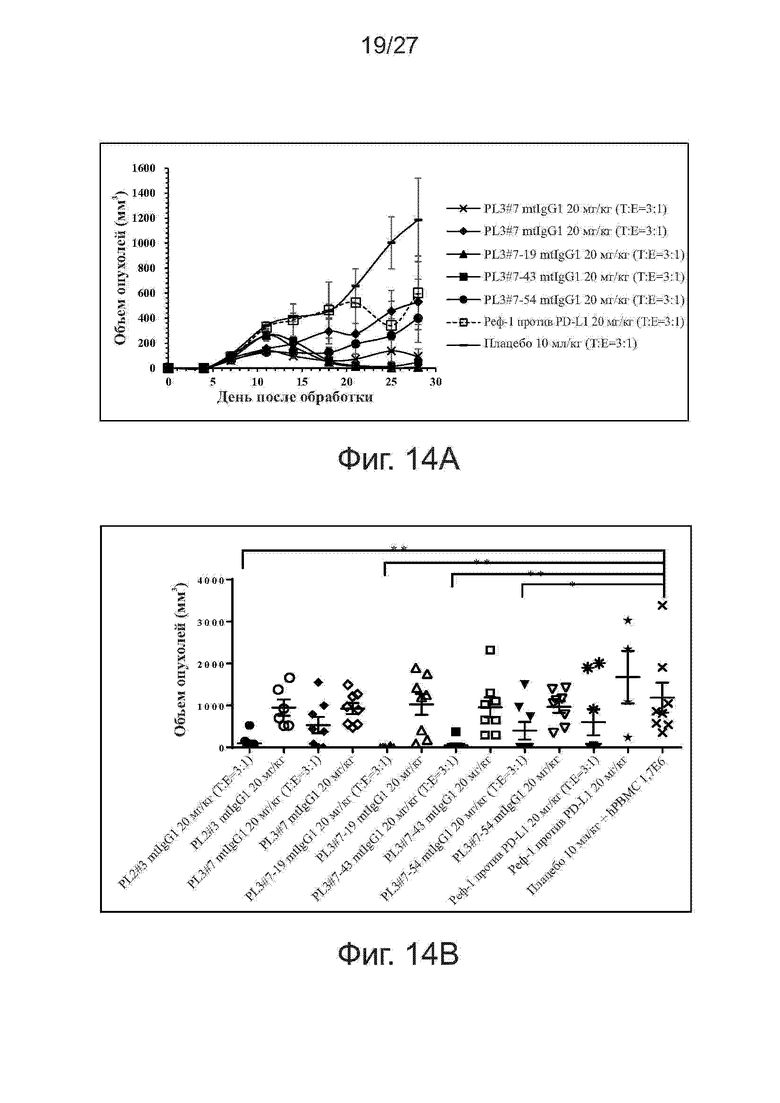
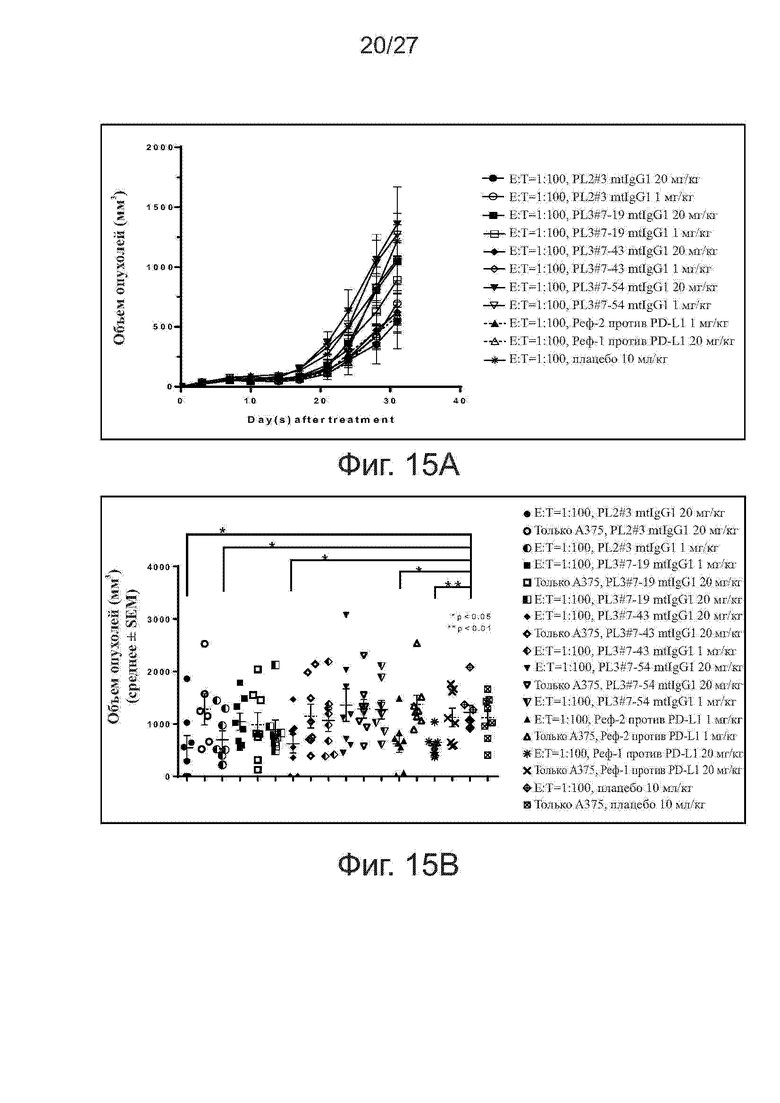
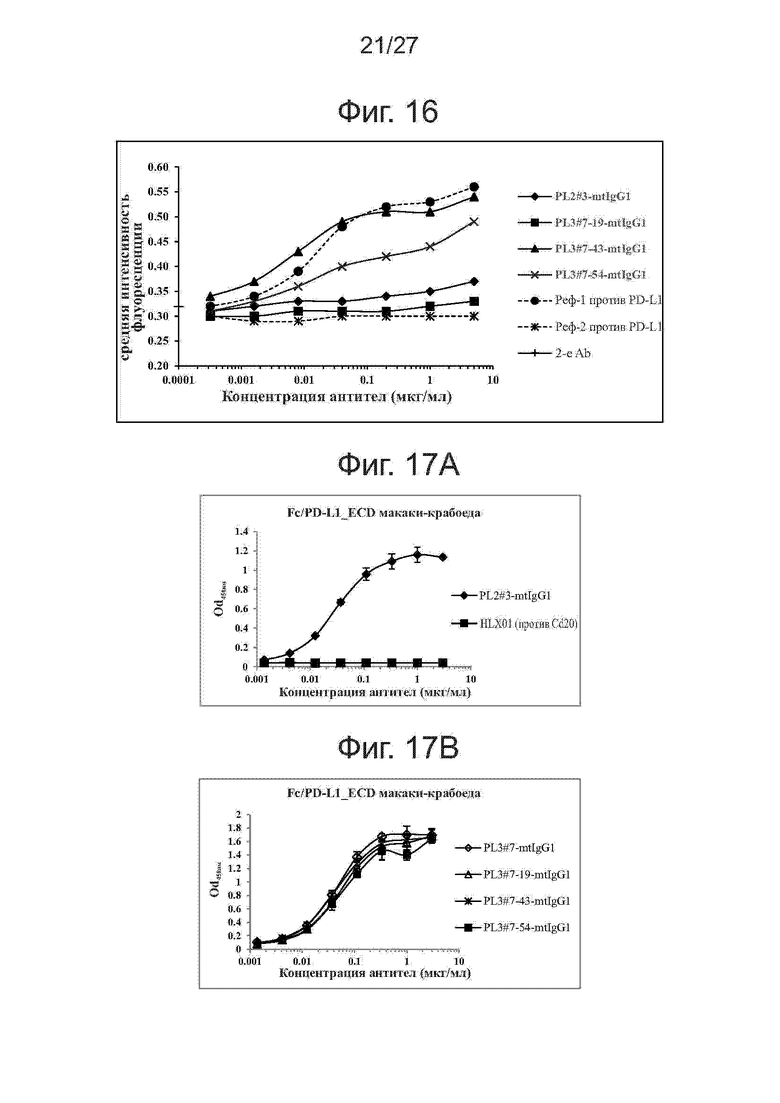

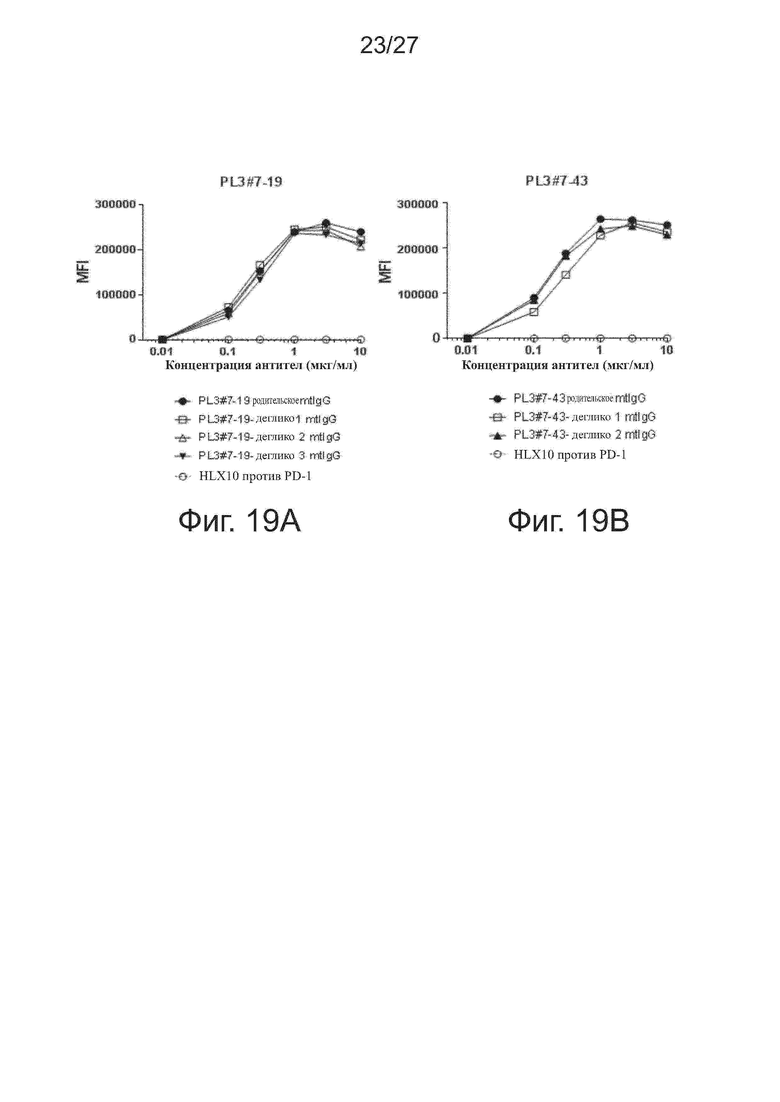
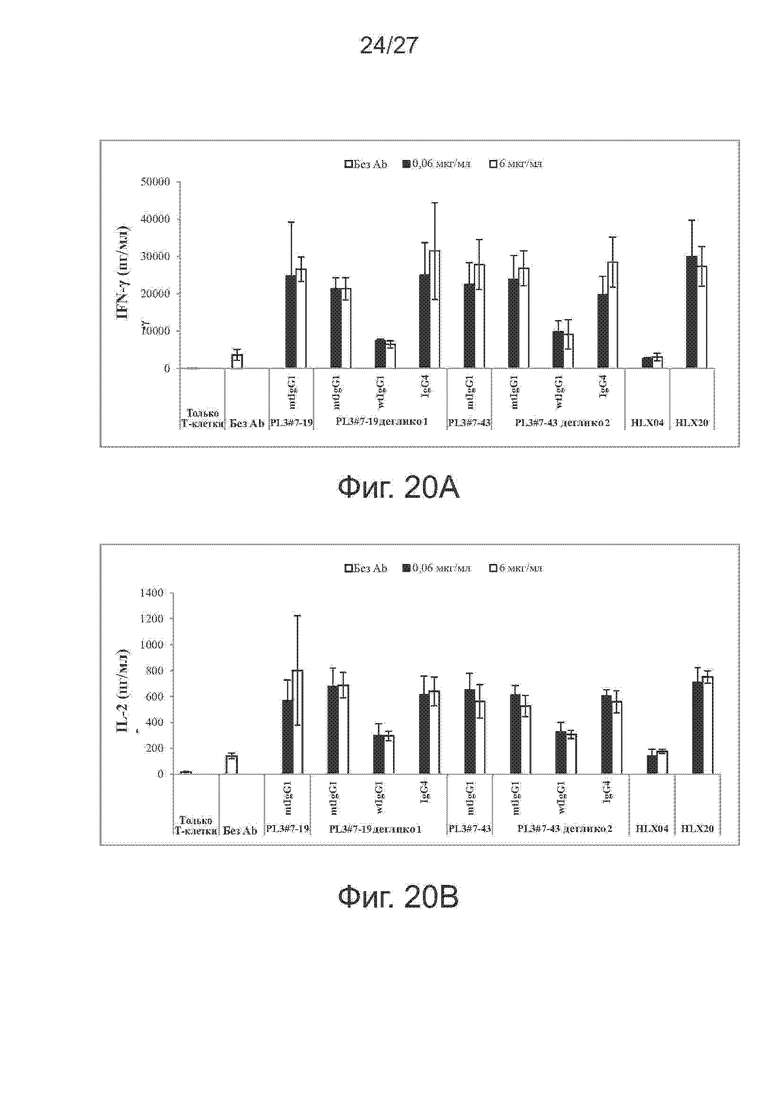
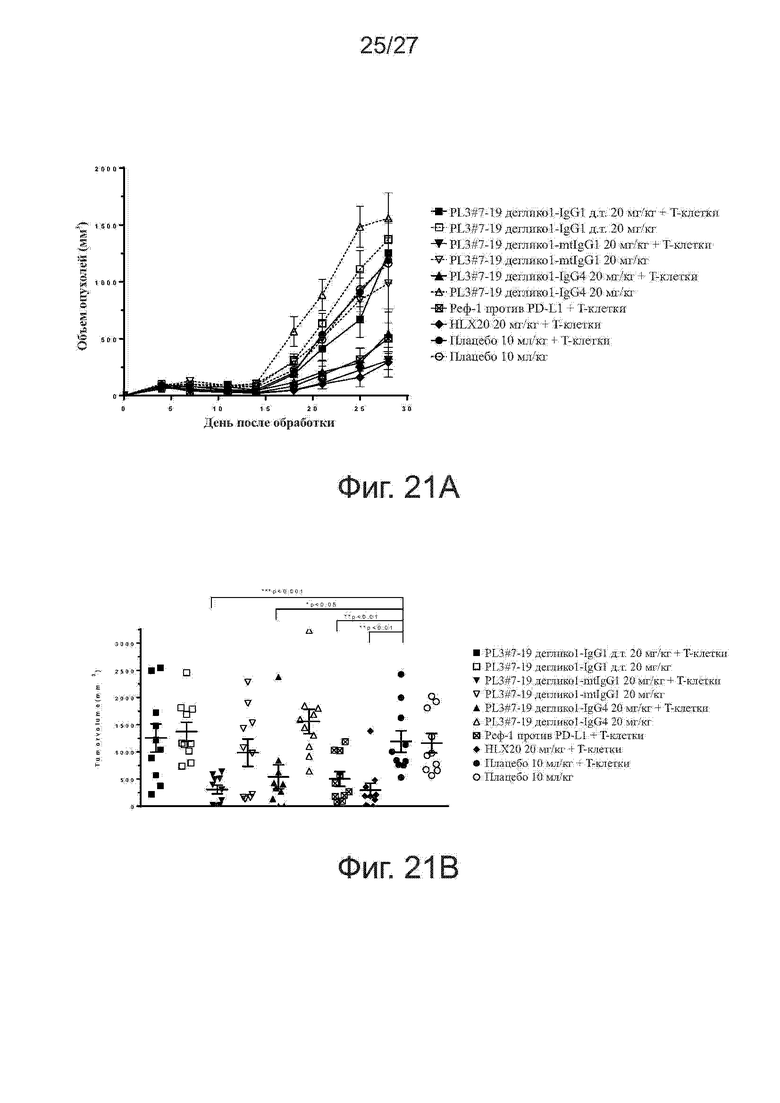
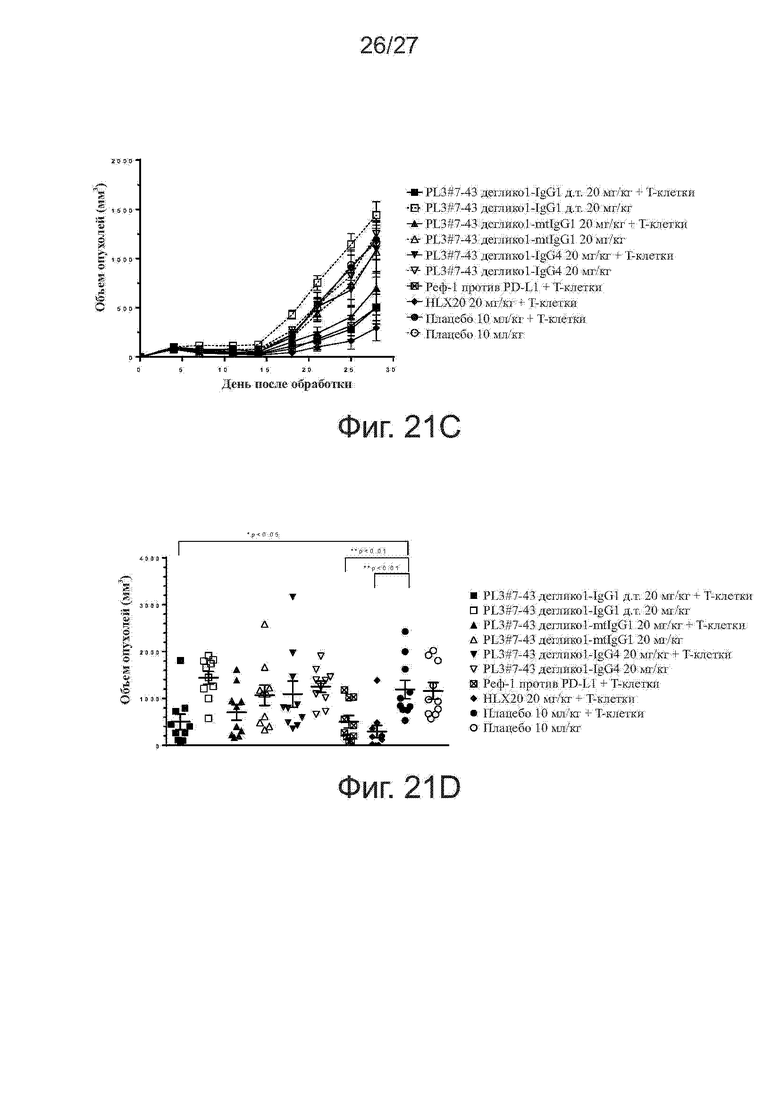
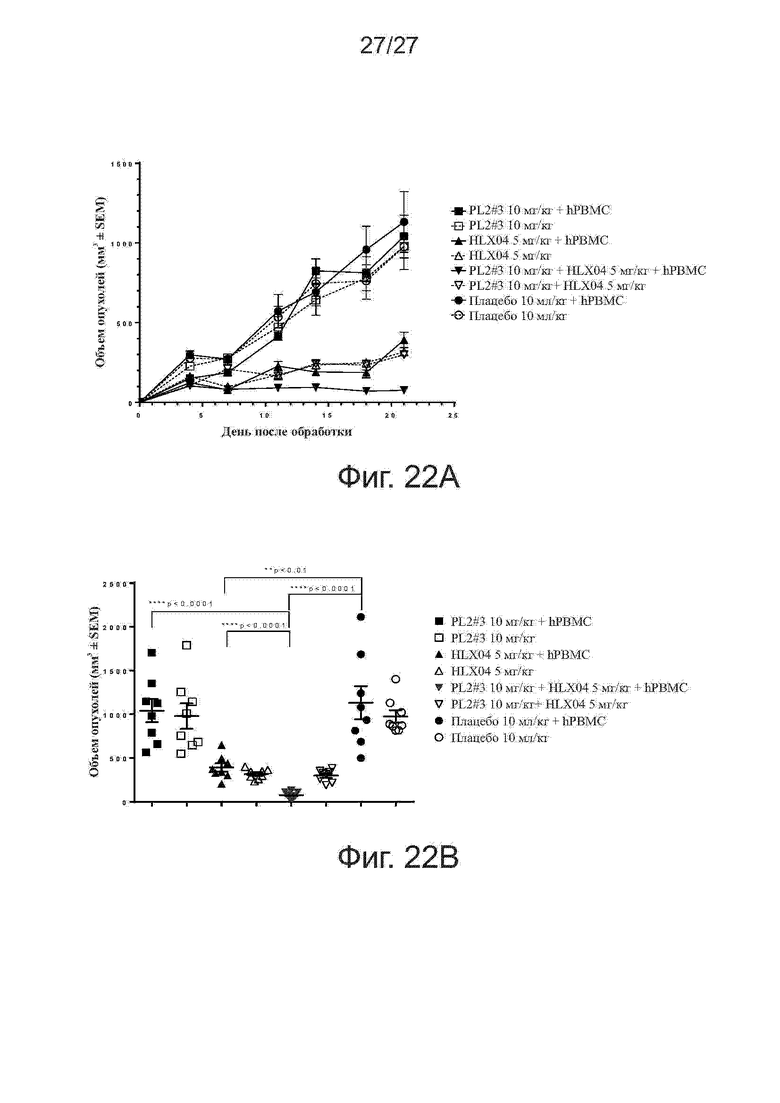
Изобретение относится к области биотехнологии, в частности к антителу против лиганда-1 запрограммированной смерти (PD-L1), которое специфически связывает PD-L1, а также к содержащему его иммуноконъюгату и композиции. Также раскрыт способ получения вышеуказанного антитела. Изобретение также относится к выделенной молекуле нуклеиновой кислоты, кодирующей антитело против PD-L1, а также к содержащему вышеуказанную молекулу вектору и клетке. Изобретение эффективно для выявления белка PD-L1 в образце от пациента, а также для лечения рака у субъекта. 10 н. и 26 з.п. ф-лы, 22 ил., 12 табл., 18 пр.
1. Антитело против лиганда-1 запрограммированной смерти (PD-L1), которое специфически связывает PD-L1, содержащее:
а) последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 35; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 41; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 44; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 51; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 59;
b) последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 36; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 42; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 45; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 52; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 56; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 60;
с) последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 37; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 41; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 46; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 53; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 61;
d) последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 37; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 41; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 47; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 54; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 57; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 62;
e) последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 38; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 43; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 48; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 51; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 58; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 63;
f) последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 39; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 41; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 49; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 51; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 64;
g) последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 40; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 41; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 50; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 53; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 61;
h) последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 65; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 42; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 71; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 52; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 56; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 77;
i) последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 37; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 41; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 72; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 75; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 78;
j) последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 66; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 68; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 73; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 75; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 79;
k) последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 35; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 69; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 74; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 75; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 80;
l) последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 67; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 70; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 72; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 75; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 76; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 81;
m) последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 94; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 42; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 95; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 52; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 56; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 96;
n) последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 97; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 42; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 98; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 52; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 56; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 99;
o) последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 100; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 42; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 95; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 52; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 56; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 101; или
p) последовательность вариабельного домена легкой цепи (LC), включающую (1) CDR-L1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 37; (2) CDR-L2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 106; и (3) CDR-L3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 107; и последовательность вариабельного домена тяжелой цепи (HC), включающую (1) CDR-H1, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 108; (2) CDR-H2, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 109.
2. Антитело против PD-L1 по п. 1, содержащее последовательность вариабельного домена легкой цепи (LC), содержащую (1) CDR-L1, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 36; (2) CDR-L2, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 42; и (3) CDR-L3, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 45, и последовательность вариабельного домена тяжелой цепи (НС), содержащую (1) CDR-H1, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 52; (2) CDR-H2, содержащую аминокислотную последовательность, указанную в SEQ ID NO: 56; и (3) CDR-H3, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 60.
3. Антитело против PD-L1 по п. 1, содержащее последовательность вариабельного домена легкой цепи (LC), содержащую (1) CDR-L1, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 37; (2) CDR-L2, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 41; и (3) CDR-L3, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 46, и последовательность вариабельного домена тяжелой цепи (НС), содержащую (1) CDR-H1, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 53; (2) CDR-H2, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 61.
4. Антитело против PD-L1 по п. 1, содержащее последовательность вариабельного домена легкой цепи (LC), содержащую (1) CDR-L1, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 65; (2) CDR-L2, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 42; и (3) CDR-L3, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 71, и последовательность вариабельного домена тяжелой цепи (НС), содержащую (1) CDR-H1, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 52; (2) CDR-H2, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 56; и (3) CDR-H3, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 77.
5. Антитело против PD-L1 по п. 1, содержащее последовательность вариабельного домена легкой цепи (LC), содержащую (1) CDR-L1, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 37; (2) CDR-L2, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 41; и (3) CDR-L3, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 72, и последовательность вариабельного домена тяжелой цепи (НС), содержащую (1) CDR-H1, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 75; (2) CDR-H2, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 78.
6. Антитело против PD-L1 по п. 1, содержащее последовательность вариабельного домена легкой цепи (LC), содержащую (1) CDR-L1, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 66; (2) CDR-L2, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 68; и (3) CDR-L3, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 73, и последовательность вариабельного домена тяжелой цепи (НС), содержащую (1) CDR-H1, содержащую аминокислотную последовательность, указанную в SEQ ID NO: 75; (2) CDR-H2, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 79.
7. Антитело против PD-L1 по п.1, содержащее последовательность вариабельного домена легкой цепи (LC), содержащую (1) CDR-L1, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 35; (2) CDR-L2, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 69; и (3) CDR-L3, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 74, и последовательность вариабельного домена тяжелой цепи (НС), содержащую (1) CDR-H1, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 75; (2) CDR-H2, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 55; и (3) CDR-H3, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 80.
8. Антитело против PD-L1 по п. 1, содержащее последовательность вариабельного домена легкой цепи (LC), содержащую (1) CDR-L1, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 67; (2) CDR-L2, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 70; и (3) CDR-L3, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 72, и последовательность вариабельного домена тяжелой цепи (НС), содержащую (1) CDR-H1, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 75; (2) CDR-H2, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 76; и (3) CDR-H3, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 81.
9. Антитело против PD-L1 по п. 6 или 7, при этом данное антитело против PD-L1 включает CDR-L2, содержащий одну или несколько мутаций в сайте N-гликозилирования, где указанный сайт N-гликозилирования находится внутри СDR-L2 в участке N-X-S/T, и где антитело против PD-L1 с одной или более мутациями в участке N-X-S/T устраняет сайт N-гликозилирования, но сохраняет ту же функцию.
10. Антитело против PD-L1, которое специфически связывает PD-L1, содержащее:
i) последовательность вариабельного домена легкой цепи (LC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 2, и последовательность вариабельной области тяжелой цепи (HC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 9;
ii) последовательность вариабельного домена легкой цепи (LC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 3, и последовательность вариабельного домена тяжелой цепи (HC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 10;
iii) последовательность вариабельного домена легкой цепи (LC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 16, и последовательность вариабельного домена тяжелой цепи (HC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 18;
iv) последовательность вариабельного домена легкой цепи (LC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 20, и последовательность вариабельной области тяжелой цепи (HC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 22;
v) последовательность вариабельного домена легкой цепи (LC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 24, и последовательность вариабельной области тяжелой цепи (HC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 26;
vi) последовательность вариабельного домена легкой цепи (LC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 28, и последовательность вариабельной области тяжелой цепи (HC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 30;
vii) последовательность вариабельного домена легкой цепи (LC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 32, и последовательность вариабельной области тяжелой цепи (HC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 34;
viii) последовательность вариабельного домена легкой цепи (LC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 117, и последовательность вариабельной области тяжелой цепи (HC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 30;
ix) последовательность вариабельного домена легкой цепи (LC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 119, и последовательность вариабельной области тяжелой цепи (HC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 30;
x) последовательность вариабельного домена легкой цепи (LC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 111, и последовательность вариабельной области тяжелой цепи (HC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 26;
xi) последовательность вариабельного домена легкой цепи (LC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 113, и последовательность вариабельной области тяжелой цепи (HC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 26; или
xii) последовательность вариабельного домена легкой цепи (LC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 115, и последовательность вариабельной области тяжелой цепи (HC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 26.
11. Антитело против PD-L1 по п. 10, где указанное антитело против PD-L1 содержит последовательность вариабельного домена легкой цепи (LC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 2, и последовательность вариабельного домена тяжелой цепи (НС), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 9.
12. Антитело против PD-L1 по п. 10, где указанное антитело против PD-L1 содержит последовательность вариабельного домена легкой цепи (LC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 3, и последовательность вариабельного домена тяжелой цепи (НС), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 10.
13. Антитело против PD-L1 по п. 10, где указанное антитело против PD-L1 содержит последовательность вариабельного домена легкой цепи (LC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 16, и последовательность вариабельного домена тяжелой цепи (НС), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 18.
14. Антитело против PD-L1 по п. 10, где указанное антитело против PD-L1 содержит последовательность вариабельного домена легкой цепи (LC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 20, и последовательность вариабельного домена тяжелой цепи (НС), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 22.
15. Антитело против PD-L1 по п. 10, где указанное антитело против PD-L1 содержит последовательность вариабельного домена легкой цепи (LC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 24, и последовательность вариабельного домена тяжелой цепи (НС), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 26.
16. Антитело против PD-L1 по п. 10, где указанное антитело против PD-L1 содержит последовательность вариабельного домена легкой цепи (LC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 28, и последовательность вариабельного домена тяжелой цепи (НС), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 30.
17. Антитело против PD-L1 по п. 10, где указанное антитело против PD-L1 содержит последовательность вариабельного домена легкой цепи (LC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 32, и последовательность вариабельного домена тяжелой цепи (НС), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 34.
18. Антитело против PD-L1 по п. 10, где указанное антитело против PD-L1 содержит последовательность вариабельного домена легкой цепи (LC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 117, и последовательность вариабельного домена тяжелой цепи (НС), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 30.
19. Антитело против PD-L1 по п. 10, где указанное антитело против PD-L1 содержит последовательность вариабельного домена легкой цепи (LC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 119, и последовательность вариабельного домена тяжелой цепи (НС), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 30.
20. Антитело против PD-L1 по п. 10, где указанное антитело против PD-L1 содержит последовательность вариабельного домена легкой цепи (LC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 111, и последовательность вариабельного домена тяжелой цепи (НС), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 26.
21. Антитело против PD-L1 по п. 10, где указанное антитело против PD-L1 содержит последовательность вариабельного домена легкой цепи (LC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 113, и последовательность вариабельного домена тяжелой цепи (НС), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 26.
22. Антитело против PD-L1 по п. 10, где указанное антитело против PD-L1 содержит последовательность вариабельного домена легкой цепи (LC), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 115, и последовательность вариабельного домена тяжелой цепи (НС), содержащую аминокислотную последовательность, указанную в SEQ ID NO: 26.
23. Антитело против PD-L1 по любому из пп. 1-22, включающее антигенсвязывающий фрагмент, который выбран из группы, состоящей из Fab, Fab′, F(ab′)2, одноцепочечных Fv (scFv), Fv-фрагментов, диател и линейных антител.
24. Иммуноконъюгат, который специфически связывает PD-L1, содержащий антитело против PD-L1 по любому из пп. 1-22, конъюгированное с терапевтическим средством или меткой.
25. Иммуноконъюгат по п. 24, при этом метка выбрана из группы, состоящей из радиоизотопа, флуоресцентного красителя и фермента.
26. Выделенная молекула нуклеиновой кислоты, кодирующая антитело против PD-L1 по любому из пп. 1-22.
27. Экспрессирующий вектор, содержащий молекулу нуклеиновой кислоты по п. 26.
28. Клетка-хозяин для продукции антител, содержащая экспрессирующий вектор по п. 27.
29. Способ получения антитела по любому из пп. 1-22, включающий культивирование клетки по п. 28 и извлечение антитела из клеточной культуры.
30. Композиция для лечения рака, содержащая эффективное количество антитела против PD-L1 по любому из пп. 1-22 и фармацевтически приемлемый носитель.
31. Способ выявления белка PD-L1 в образце от пациента путем контактирования антитела против PD-L1 по любому из пп. 1-22 с образцом и детектирования указанного антитела против PD-L1, связавшегося с белком PD-L1.
32. Способ по п. 31, при этом антитело против PD-L1 применяется в иммуногистохимическом анализе (IHC) или анализе методом ELISA.
33. Способ лечения рака у субъекта, включающий введение субъекту эффективного количества композиции по п. 30.
34. Способ по п. 33, при этом рак выбран из группы, состоящей из меланомы, НМРЛ (NSCLC), рака головы и шеи, рака уротелия, тройного негативного рака молочной железы (TNBC), рака желудка, классической лимфомы Ходжкина (cHL), неходжкинской лимфомы – первичной B-клеточной лимфомы средостения (NHL PMBCL), мезотелиомы, рака яичников, рака легких, рака пищевода, карциномы носоглотки (NPC), рака желчных путей, колоректального рака, рака молочной железы, рака шейки матки, рака щитовидной железы и рака слюнных желез.
35. Способ по п. 33, при этом субъекту дополнительно вводится терапевтическое средство, выбранное из группы, состоящей из противоопухолевого средства, химиотерапевтического средства, средства, ингибирующего рост, и цитотоксического средства.
36. Способ по п. 33, при этом субъекту дополнительно проводится лучевая терапия.
| WO 2015061668 A1, 30.04.2015 | |||
| US 2013130268 A1, 23.05.2013 | |||
| WO 2016073879 A2, 06.11.2015 | |||
| WO 2008130704 A2, 30.10.2008 | |||
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ЧЕЛОВЕКА К БЕЛКУ ПРОГРАММИРУЕМОЙ СМЕРТИ 1 (PD-1) И СПОСОБЫ ЛЕЧЕНИЯ РАКА С ИСПОЛЬЗОВАНИЕМ АНТИ-PD-1-АНТИТЕЛ САМОСТОЯТЕЛЬНО ИЛИ В КОМБИНАЦИИ С ДРУГИМИ ИММУНОТЕРАПЕВТИЧЕСКИМИ СРЕДСТВАМИ | 2006 |
|
RU2406760C2 |

