Область техники, к которой относится изобретение
Настоящее изобретение относится к антителам к латентному (находящемуся в неактивной форме) TGF-бета 1 и способам их применения.
Предпосылки создания изобретения
Трансформирующий ростовой фактор-бета (трансформирующий фактор роста бета; TGF-бета) является представителем цитокинов суперсемейства TGF-бета, которое состоит из изоформ TGF-бета, активинов, ингибинов, Nodal, костных морфогенетических белков (белков, участвующих в остеогенезе) (BMP), анти-Мюллерова гормона (АМН), а также факторов роста и дифференцировки (GDF). Члены этого суперсемейства представляют собой димерные белки с консервативными структурами, и они обладают плейотропными функциями in vitro и in vivo (NPL 1, 2). Изоформы TGF-бета участвуют в многочисленных клеточных процессах, включая ингибирование роста, клеточную миграцию, инвазию, эпителиально-мезенхимальный переход (ЕМТ), ремоделирование внеклеточного матрикса (ЕСМ) и иммуносупрессию (NPL 3). Однако, хотя в норме изоформы TGF-бета динамически регулируются и участвуют в поддержании гомеостаза тканей, для них часто характерна постоянная сверхэкспрессия при болезненных состояниях, включая рак, фиброз и воспаление, и указанное избыточное производство TGF-бета инициирует прогрессирующее развитие заболевания путем модуляции роста, миграции или фенотипа клеток
У млекопитающих идентифицированы три различные изоформы TGF-бета (TGF-бета 1, TGF-бета 2 и TGF-бета 3) и для них характерна 70-82% гомология на аминокислотном уровне (NPL 4). Все три изоформы TGF-бета связываются с рецептором TGF-бета типа 2 (TGFR2) в виде гомодимеров (их активная форма); затем TGFR2 вовлекает и активирует рецептор TGF-бета типа 1 (TGFR1) в отношении стимуляции передачи сигналов рецептором (NPL 5). Однако уровни экспрессии трех изоформ варьируются в зависимости от ткани (NPL 6) и их функции являются различными, что продемонстрировано с использованием фенотипов мышей с нокаутом генов (NPL 7-11).
Аналогично другим членам суперсемейства TGF-бета TGF-бета синтезируется в виде белка-предшественника, образующего гомодимер, который взаимодействует с ассоциированным с латентностью пептидом (LAP) и связывающим латентный TGF-бета белком (LTBP) с образованием более крупного комплекса, который называют большим латентным комплексом (LLC). Ген TGF-бета кодирует последовательность препробелка, состоящую из сигнального пептида, пропептида, который заканчивается сайтом расщепления конвертазой (РРС), и последовательности зрелого TGF-бета. Гидролиз фурином сайта расщепления РРС приводит к образованию отдельных гомодимеров, полученных из TGF-бета и пропептида. Два гомодимера сохраняют нековалентную ассоциацию и секретируются. Указанный латентный комплекс сохраняет TGF-бета в неактивной форме, в которой он не может связываться со своими рецепторами (NPL 12, 13). Процесс активации TGF-бета включает высвобождение LLC из ЕСМ с последующим дополнительным протеолизом LAP для высвобождения активного в отношении его рецепторов TGF-бета (NPL 3). Латентный TGF-бета расщепляется с высвобождением активного TGF-бета с помощью широкого спектра протеаз, включая плазмин (PLN), плазматический калликреин (PLK), матриксную металлопротеиназу (ММР) 2 и ММР 9 (NPL 14), и с помощью тромбоспондина 1 (TSP-1) (NPL 15). Не рассматривая какую-либо теорию, ММР2, а также ММР9, протеолитически расщепляют латентный TGF-бета 1 и высвобождают зрелый TGF-бета 1 из латентной формы. И ММР2, и ММР9 синтезируются в виде неактивной про-ММР. Про-ММР2 активируется комплексом мембранно-связанной типа 1 ММР (МТ1-ММР/ММР14) и тканевого ингибитора металлопротеиназы 2 (TIMP-2). Про-ММР9 активируется посредством взаимодействия протеазного каскада, включающего плазмин и стромелизин 1 (ММР-3). Плазмин приводит к образованию активной ММР-3 из ее зимогена. Активная ММР-3 отщепляет пропептид от имеющей молекулярную массу 92 кДа про-ММР-9 с образованием обладающего ферментативной активностью фермента с молекулярной массой 82 кДа. Сайты расщепления ММР не являются специфически предопределенными; однако известны данные о том, что ММРЗ специфически расщепляет сайт между 79 Ala и 80 Leu латентного TGF-бета с активацией TGF-бета (WO 2005/023870). Альтернативно этому, при механическом растяжении интегрины могут активировать TGF-бета путем связывания в RGD-мотивом, присутствующим в LAP, индуцируя высвобождение зрелого TGF-бета из его латентного комплекса (NPL 16, 17).
После активации лиганд, представляющий собой димерный TGF-бета, связывается с внеклеточными доменами рецепторов типа I и типа II и индуцирует сильное сближение, придавая внеклеточным серин/треонинкиназным доменам рецепторов конформацию, которая облегчает фосфорилирование и последующую активацию рецептора типа I. Указанная активация рецептора типа I приводит к распространению сигнала посредством по меньшей мере двух по-видимому независимых путей: SMAD-зависимого канонического пути и SMAD-независимого или неканонического пути. В SMAD-зависимом пути активация TGFR1 (который обозначают также как ALK5) приводит к фосфорилированию белков SMAD. SMAD2 и SMAD3 являются субстратами для TGFR1. После фосфорилирования рецептором белки SMAD вместе с общим медиатором SMAD4 транслоцируются в ядро, где они взаимодействуют с другими факторами транскрипции, регулируя транскрипционные ответы (NPL 18). В неканоническом пути комплекс активированного рецептора TGF-бета передает сигнал через другие факторы, такие как ассоциированный с рецептором фактора некроза опухоли (TNF) 4 (TRAF4), TRAF6, активируемая TGF-бета киназа 1 (ТАК1, которую обозначают также как МАР3К7), активируемая р38-митогеном протеинкиназа (р38 МАРК), RHO, фосфоинозитид-3-киназа (PI3K), АКТ (которую обозначают также как протеинкиназа В), регулируемая внеклеточным сигналом киназа (ERK), JUN N-концевая киназа (JNK) или ядерный фактор-каппа В (NF-каппа В). Таким образом, клеточные ответы на сигналы TGF-бета являются результатом динамической комбинации канонических и неканонических каскадов сигналов.
Фиброз или накопление молекул ЕСМ, что приводит к образованию рубцовой ткани, является обычным последствием хронического повреждения ткани. Легочный фиброз, фиброз почек и цирроз печени относятся к наиболее часто встречающимся фиброзным болезням, для которых в совокупности существует огромная неудовлетворенная клиническая потребность. TGF-бета значительно усиливает создание внеклеточного матрикса из мезенхимальных клеток, при этом одновременно подавляет рост эпителиальных клеток, которые принимают участие в патогенезе склеротических заболеваний. Сверхэкспрессия активной формы GF-бета 1 в печени трансгенных мышей является достаточной для индукции мультиорганного фиброзного заболевания (NPL 19). С другой стороны, TGF-бета играет также важную роль в поддержании здоровья людей. Например, TGF-бета подавляет избыточное образование протеаз в легком и предупреждает деструкцию легочной ткани, приводящую к эмфиземе. Кроме того, у мышей с удаленным в результате делеции TGF-бета 1 обнаружена пренатальная смертность (примерно 50% через 10,5 дня после совокупления) или смерть их потомства вскоре после рождения, сопровождающаяся массивными воспалительными повреждениями, обнаруженными во многих органах, включая легкие (васкулит, преваскулярное образование скоплений лейкоцитов вокруг кровеносного сосуда и интерстициальная пневмония) и сердце (эндокардит и миокардит), это позволяет предположить, что TGF-бета 1 играет решающую роль в поддержании иммунного гомеостаза (NPL 7).
Результаты исследований с использованием нейтрализующего антитела к TGF-бета и животных моделей продемонстрировали, что склеротические заболевания можно предупреждать или излечивать путем подавления действия TGF-бета. Поскольку TGF-бета образуется в виде белка-предшественника, то существует несколько опубликованных подходов к предупреждению активации из латентной формы. Другим методом предупреждения активации из латентной формы является применение ингибитора или антитела, которой/которое связывается с латентным TGF-бета, блокируя расщепление протеазами, такими как PLK и PLN. Описано несколько антител, которые применяли в таком методе подавления активации TGF-бета для предупреждения или лечения фиброза/цирроза печени (PTL 1). Кроме того, известно несколько документов, в которых упоминаются антитела к LAP, предназначенные для лечения рака (PTL 2), и связывающие TGF-бета 1 иммуноглобулины для лечения связанных с TGF-бета 1 нарушений.
Перечень процитированных документов
Патентная литература
[PTL 1] WO 2011/102483
[PTL 2] WO 2016/115345
[PTL 3] WO 2017/156500
Непатентная литература
[NPL 1] McCartney-Francis N. L. и др., Int. Rev. Immunol. 16, 1998, cc. 553-580.
[NPL 2] Massague J. Annu. Rev. Biochem. 67, 1998, cc. 753-791.
[NPL 3] Derynck R. и Miyazono К. изд-во Cold Spring Harbor Press, 2008.
[NPL 4] Yu L. и др., Kidney Int. 64, 2003, cc. 844-856.
[NPL 5] Xu P., Liu J. и Derynck R. и др., FEBS Lett. 586, 2012, cc.1871-1884.
[NPL 6] Millan F. А. и др., Development 111, 1991, cc. 131-143.
[NPL 7] Kulkarni А. В. и др., Proc. Natl Acad. Sci. USA 90, 1993, cc. 770-774.
[NPL 8] Shull M. M. и др., Nature 359, 1992, cc. 693-699.
[NPL 9] Dickson M. С.и др., Development 121, 1995, cc. 1845-1854.
[NPL 10] Sanford L. P. и др., Development 124, 1997, cc. 2659-2670.
[NPL 11] Proetzel G. и др., Nature Genet. 11, 1995, cc. 409-414.
[NPL 12] Dubois С.M. и др., J. Biol. Chem. 270, 1995, cc. 10618-10624.
[NPL 13] Nunes I. и др., J. Am. Optom. Assoc. 69, 1998, cc. 643-648.
[NPL 14] Annes J. и др., J. Cell Sci. 116, 2003, cc. 217-224.
[NPL 15] Schultz-Cherry S. и др., J. Biol. Chem. 269, 1994, cc. 26775-26782.
[NPL 16] Munger J. S. и др., Cell 96, 1999, cc. 319-328.
[NPL 17] Shi M. и др., Nature 474, 2011, cc. 343-349.
[NPL 18] Shi Y. и Massague и др., Cell 113, 2003, cc. 685-700.
[NPL 19] Sanderson N. и др., Proc. Natl Acad. Sci.USA 92, 1995, cc. 2572-2576.
Краткое изложение сущности изобретения Задача, положенная в основу изобретения
Задачей изобретения является получение межвидовых гуманизированных и оптимизированных антител к латентному TGF-бета 1, которые ингибируют обусловленную протеазами активацию латентного TGF-бета 1 без ингибирования обусловленной интегрином активации латентного TGF-бета 1. В изобретении предложены также комбинированные терапии, включающие антитело к латентному TGF-бета 1 и один и несколько ингибиторов иммунных контрольных точек.
Решение задачи
Для создания настоящего изобретения проведены тщательные исследования в описанных выше ситуациях и в результате созданы межвидовые гуманизированные и оптимизированные антитела к латентному TGF-бета 1, которые ингибируют обусловленную протеазами активацию латентного TGF-бета 1 без ингибирования обусловленной интегрином активации латентного TGF-бета 1. Кроме того, у антител к латентному TGF-бета 1 обнаружено противоопухолевое действие при введении в комбинации с одним или несколькими ингибиторами иммунных контрольных точек.
В настоящем изобретении предложены:
А1. Антитело к латентному TGF-бета 1, содержащее:
(а) HVR-H1, HVR-H2 и HVR-H3, которые содержат аминокислотные последовательности SEQ ID NO: 20, 21 и 22 соответственно;
(б) HVR-H1, HVR-H2 и HVR-H3, которые содержат аминокислотные последовательности SEQ ID NO: 26, 27 и 28 соответственно;
(в) HVR-H1, HVR-H2 и HVR-H3, которые содержат аминокислотные последовательности SEQ ID NO: 32, 33 и 34 соответственно; или
(г) HVR-H1, HVR-H2 и HVR-H3, которые содержат аминокислотные последовательности SEQ ID NO: 38, 39 и 40 соответственно.
А2. Антитело к латентному TGF-бета 1 по п. А1, которое содержит также:
(а) HVR-L1, HVR-L2 и HVR-L3, содержащие аминокислотные последовательности SEQ ID NO: 23, 24 и 25 соответственно;
(б) HVR-L1, HVR-L2 и HVR-L3, содержащие аминокислотные последовательности SEQ ID NO: 29, 30 и 31 соответственно;
(в) HVR-L1, HVR-L2 и HVR-L3, содержащие аминокислотные последовательности SEQ ID NO: 35, 36 и 37 соответственно; и
(г) HVR-L1, HVR-L2 и HVR-L3, содержащие аминокислотные последовательности SEQ ID NO: 41, 42 и 43 соответственно.
A3. Антитело к латентному TGF-бета 1, содержащее: (a) HVR-H1, HVR-H2 и HVR-H3, которые содержат аминокислотные последовательности SEQ ID NO: 20, 21 и 22 соответственно, и HVR-L1, HVR-L2 и HVR-L3, которые содержат аминокислотные последовательности SEQ ID NO: 23, 24 и 25 соответственно;
(б) HVR-H1, HVR-H2 и HVR-H3, которые содержат аминокислотные последовательности SEQ ID NO: 26, 27 и 28 соответственно, и HVR-L1, HVR-L2 и HVR-L3, которые содержат аминокислотные последовательности SEQ ID NO: 29, 30 и 31 соответственно;
(в) HVR-H1, HVR-H2 и HVR-H3, которые содержат аминокислотные последовательности SEQ ID NO: 32, 33 и 34 соответственно, и HVR-L1, HVR-L2 и HVR-L3, которые содержат аминокислотные последовательности SEQ ID NO: 35, 36 и 37 соответственно; или
(г) HVR-H1, HVR-H2 и HVR-H3, которые содержат аминокислотные последовательности SEQ ID NO: 38, 39 и 40 соответственно, и HVR-L1, HVR-L2 и HVR-L3, которые содержат аминокислотные последовательности SEQ ID NO: 41, 42 и 43 соответственно.
А4. Антитело к латентному TGF-бета 1 по одному из п.п. А1-А3, содержащее:
(а) (I) последовательность VH, которая идентична по меньшей мере на 95% аминокислотной последовательности SEQ ID NO: 12, (II) последовательность VL, которая идентична по меньшей мере на 95% аминокислотной последовательности SEQ ID NO: 13, или (III) последовательность VH, указанную в подпункте (I), и последовательность VL, указанную в подпункте (II);
(б) (I) последовательность VH, которая идентична по меньшей мере на 95% аминокислотной последовательности SEQ ID NO: 14, (II) последовательность VL, которая идентична по меньшей мере на 95% аминокислотной последовательности SEQ ID NO: 15, или (III) последовательность VH, указанную в подпункте (I), и последовательность VL, указанную в подпункте (II);
(в) (I) последовательность VH, которая идентична по меньшей мере на 95% аминокислотной последовательности SEQ ID NO: 16, (II) последовательность VL, которая идентична по меньшей мере на 95% аминокислотной последовательности SEQ ID NO: 17, или (III) последовательность VH, указанную в подпункте (I), и последовательность VL, указанную в подпункте (II);
(г) (I) последовательность VH, которая идентична по меньшей мере на 95% аминокислотной последовательности SEQ ID NO: 18, (II) последовательность VL, которая идентична по меньшей мере на 95% аминокислотной последовательности SEQ ID NO: 19, или (III) последовательность VH, указанную в подпункте (I), и последовательность VL, указанную в подпункте (II).
А5. Антитело к латентному TGF-бета 1 по п. А4, содержащее последовательность VH SEQ ID NO: 12, 14, 16 или 18.
А6. Антитело к латентному TGF-бета 1 по п. А4 или п. А5, содержащее последовательность VL SEQ ID NO: 13, 15, 17 или 19.
А7. Антитело к латентному TGF-бета 1 по одному из п.п. А4-А6, содержащее:
(а) последовательность VH SEQ ID NO: 12 и последовательность VL SEQ ID NO: 13;
(б) последовательность VH SEQ ID NO: 14 и последовательность VL SEQ ID NO: 15;
(в) последовательность VH SEQ ID NO: 16 и последовательность VL SEQ ID NO: 17; или
(г) последовательность VH SEQ ID NO: 18 и последовательность VL SEQ ID NO: 19.
А8. Антитело к латентному TGF-бета 1, содержащее:
(а) последовательность VH SEQ ID NO: 12 и последовательность VL SEQ ID NO: 13;
(б) последовательность VH SEQ ID NO: 14 и последовательность VL SEQ ID NO: 15;
(в) последовательность VH SEQ ID NO: 16 и последовательность VL SEQ ID NO: 17; или
(г) последовательность VH SEQ ID NO: 18 и последовательность VL SEQ ID NO: 19.
А9. Антитело к латентному TGF-бета 1 по одному из п.п. А1-А8, которое представляет собой человеческое, гуманизированное или химерное антитело.
А10. Антитело к латентному TGF-бета 1 по одному из п.п. А1-А9, которое представляет собой полноразмерное антитело IgG-типа, предпочтительно полноразмерное антитело IgG1-подтипа.
A11. Антитело к латентному TGF-бета 1 по одному из п.п. А1-А9, которое представляет собой биспецифическое антитело.
А12. Антитело к латентному TGF-бета 1 по одному из п.п. А1-А11, где антитело к латентному TGF-бета 1 содержит модифицированную Fc-область IgG1, обладающую пониженной эффекторной функцией по сравнению с Fc-областью IgG1 дикого типа.
А13. Антитело к латентному TGF-бета 1 по п. А12, в котором модифицированная Fc-область IgG1 содержит аминокислотные замены в положениях EU235 и/или EU236 согласно EU-индексу.
А14. Антитело к латентному TGF-бета 1 по п. А12 или п. А13, в котором модифицированная Fc-область IgG1 содержит аминокислотные замены в положениях L235R и G236R согласно EU-индексу.
А15. Антитело к латентному TGF-бета 1 по одному из п.п. А12-А14, в котором модифицированная Fc-область IgG1 дополнительно обладает повышенной связывающей активностью с FcRn по сравнению с Fc-областью IgG1 дикого типа.
А16. Антитело к латентному TGF-бета 1 по п. А15, в котором модифицированная Fc-область IgG1 содержит одну или несколько аминокислотных замен в положениях, выбранных из группы, которая состоит из: EU428, EU434, EU438 и EU440 согласно EU-индексу.
А17. Антитело к латентному TGF-бета 1 по п. А 15 или п. А16, в котором модифицированная Fc-область IgG1 содержит аминокислотные замены M428L, N434A, Q438R и S440E.
А18. Антитело к латентному TGF-бета 1 по одному из п.п. А1-А11, где антитело к латентному TGF-бета 1 содержит модифицированную Fc-область IgG1, в котором модифицированная Fc-область IgG1 содержит аминокислотные замены K214R, L235R и G236R.
А19. Антитело к латентному TGF-бета 1 по одному из п.п. А1-А11, где антитело к латентному TGF-бета 1 содержит модифицированную Fc-область IgG1, в котором модифицированная Fc-область IgG1 содержит аминокислотные замены K214R, L235R, G236R, M428L, N434A, Q438R и S440E.
А20. Антитело к латентному TGF-бета 1 по одному из п.п. А1-А11, которое представляет собой фрагмент антитела.
А21. Антитело к латентному TGF-бета 1, содержащее:
(а) полноразмерную последовательность тяжелой цепи SEQ ID NO: 47 и полноразмерную последовательность легкой цепи SEQ ID NO: 60;
(б) полноразмерную последовательность тяжелой цепи SEQ ID NO: 48 и полноразмерную последовательность легкой цепи SEQ ID NO: 61;
(в) полноразмерную последовательность тяжелой цепи SEQ ID NO: 49 и полноразмерную последовательность легкой цепи SEQ ID NO: 62;
(г) полноразмерную последовательность тяжелой цепи SEQ ID NO: 50 и полноразмерную последовательность легкой цепи SEQ ID NO: 63;
(д) полноразмерную последовательность тяжелой цепи SEQ ID NO: 51 и полноразмерную последовательность легкой цепи SEQ ID NO: 64;
(е) полноразмерную последовательность тяжелой цепи SEQ ID NO: 52 и полноразмерную последовательность легкой цепи SEQ ID NO: 65;
(ж) полноразмерную последовательность тяжелой цепи SEQ ID NO: 53 и полноразмерную последовательность легкой цепи SEQ ID NO: 66; или
(з) полноразмерную последовательность тяжелой цепи SEQ ID NO: 54 и полноразмерную последовательность легкой цепи SEQ ID NO: 67.
А22. Антитело к латентному TGF-бета 1 по одному из п.п. А1-А21, где латентный TGF-бета 1 представляет собой человеческий латентный TGF-бета 1, мышиный латентный TGF-бета 1 или латентный TGF-бета 1 обезьян циномолгус.
А23. Антитело к латентному TGF-бета 1 по одному из п.п. А1-А22, где антитело к латентному TGF-бета 1 связывается с человеческим латентным TGF-бета 1, мышиным латентным TGF-бета 1 и латентным TGF-бета 1 обезьян циномолгус.
А24. Антитело к латентному TGF-бета 1 по одному из п.п. А1-А23, где антитело к латентному TGF-бета 1 связывается с областью ассоциированного с латентностью пептида (LAP) латентного TGF-бета 1.
А25. Иммуноконъюгат, содержащий антитело к латентному TGF-бета 1 по одному из п.п. А1-А24 и цитотоксический агент.
А26. Выделенная нуклеиновая кислота, кодирующая антитело к латентному TGF-бета 1 по одному из п.п. А1-А24.
А27. Вектор, содержащий нуклеиновую кислоту по п. А26.
А28. Клетка-хозяин, содержащая нуклеиновую кислоту по п. А26 или вектор по п. А27.
А29. Способ получения антитела к латентному TGF-бета 1, включающий культивирование клетки-хозяина по п. А28 таким образом, чтобы получать антитело.
А30. Способ по п. А29, дополнительно включающий выделение антитела из клетки-хозяина.
Б1. Антитело к латентному TGF-бета 1 по одному из п.п. А1-А24 или иммуноконъюгат по п. А25, предназначенное/предназначенный для применения в качестве лекарственного средства.
Б2. Антитело к латентному TGF-бета 1 по одному из п.п. А1-А24 или иммуноконъюгат по п. А25, предназначенное/предназначенный для применения для лечения фиброза или рака.
Б3. Применение антитела к латентному TGF-бета 1 по одному из п.п. А1-А24 или иммуноконъюгата по п. А25 для приготовления лекарственного средства, предназначенного для лечения фиброза или рака.
Б4. Антитело к латентному TGF-бета 1 по одному из п.п. А1-А24 или иммуноконъюгат по п. А25, предназначенное/предназначенный для применения в комбинации с дополнительным терапевтическим агентом, предпочтительно ингибитором иммунных контрольных точек, для лечения рака.
Б5. Антитело к латентному TGF-бета 1 или иммуноконъюгат по п. Б4, где ингибитор иммунных контрольных точек представляет собой антагонист аксиального связывания (антагонист по оси связывания) PD-1, предпочтительно антитело к PD-1 или антитело к PD-L1.
Б6. Антитело к латентному TGF-бета 1 или иммуноконъюгат по п. Б4, где ингибитор иммунных контрольных точек представляет собой антитело к PD-L1.
Б7. Антитело к латентному TGF-бета 1 или иммуноконъюгат по одному из п.п. Б4-Б6, где ингибитор иммунных контрольных точек вводят одновременно с антителом к латентному TGF-бета 1 или иммуноконъюгатом.
Б8. Антитело к латентному TGF-бета 1 или иммуноконъюгат по одному из п.п. Б4-Б6, где ингибитор иммунных контрольных точек вводят до или после введения антитела к латентному TGF-бета 1 или иммуноконъюгата.
В1. Фармацевтическая композиция, содержащая антитело к латентному TGF-бета 1 по одному из п.п. А1-А24 или иммуноконъюгат по п. А25 и фармацевтически приемлемый носитель.
В2. Фармацевтическая композиция по В1, содержащая также дополнительный терапевтический агент, предпочтительно ингибитор иммунных контрольных точек.
В3. Фармацевтическая композиция по В2, в которой ингибитор иммунных контрольных точек представляет собой антагонист аксиального связывания PD-1, предпочтительно антитело к PD-1 или антитело к PD-L1.
В4. Фармацевтическая композиция по В2, в которой ингибитор иммунных контрольных точек представляет собой антитело к PD-L1.
В5. Фармацевтическая композиция по одному из п.п. В1-В4, предназначенная для лечения фиброза или рака.
В6. Фармацевтическая композиция по одному из п.п. В1-В4, предназначенная для применения в комбинации с дополнительным терапевтическим агентом, предпочтительно ингибитором иммунных контрольных точек, для лечения рака.
В7. Фармацевтическая композиция по п. В6, в которой ингибитор иммунных контрольных точек представляет собой антагонист аксиального связывания PD-1, предпочтительно антитело к PD-1 или антитело к PD-L1.
В8. Фармацевтическая композиция по п. В6, в которой ингибитор иммунных контрольных точек представляет собой антитело к PD-L1.
В9. Фармацевтическая композиция по одному из п.п. В6-В8, где ингибитор иммунных контрольных точек вводят одновременно с фармацевтической композицией.
В10. Фармацевтическая композиция по одному из п.п. В6-В8, где ингибитор иммунных контрольных точек вводят до или после введения фармацевтической композиции.
Г1. Фармацевтическая композиция, содержащая ингибитор иммунных контрольных точек и фармацевтически приемлемый носитель, предназначенная для применения в комбинации с антителом к латентному TGF-бета 1 по одному из п.п. А1-А24 или иммуноконъюгатом по п. А25 для лечения рака.
Г2. Фармацевтическая композиция по п. Г1, в которой ингибитор иммунных контрольных точек представляет собой антагонист аксиального связывания PD-1, предпочтительно антитело к PD-1 или антитело к PD-L1.
Г3. Фармацевтическая композиция по п. Г1, в которой ингибитор иммунных контрольных точек представляет собой антитело к PD-L1.
Г4. Фармацевтическая композиция по одному из п.п. Г1-Г3, где антитело к латентному TGF-бета 1 или иммуноконъюгат вводят одновременно с фармацевтической композицией.
Г5. Фармацевтическая композиция по одному из п.п. Г1-Г3, где антитело к латентному TGF-бета 1 или иммуноконъюгат вводят до или после введения фармацевтической композиции.
Д1. Способ лечения индивидуума, имеющего фиброз или рак, включающий введение индивидууму в эффективном количестве антитела к латентному TGF-бета 1 по одному из п.п. А1-А24 или иммуноконъюгата по п. А25.
Д2. Способ по п. Д1, дополнительно включающий введение индивидууму дополнительного терапевтического агента, предпочтительно ингибитора иммунных контрольных точек.
Д3. Способ по п. Д1 или п. Д2, в котором ингибитор иммунных контрольных точек представляет собой антагонист аксиального связывания PD-1, предпочтительно антитело к PD-1 или антитело к PD-L1.
Д4. Способ по п. Д3, в котором ингибитор иммунных контрольных точек представляет собой антитело к PD-L1.
Д5. Способ по одному из п.п. Д1-Д4, в котором ингибитор иммунных контрольных точек вводят одновременно в антителом к латентному TGF-бета 1 или иммуноконъюгатом.
Д6. Способ по одному из п.п. Д1-Д4, в котором ингибитор иммунных контрольных точек вводят до или после введения антитела к латентному TGF-бета 1 или иммуноконъюгата.
Краткое описание чертежей
На чертежах показано:
на фиг. 1А - результаты оценки связывания антител с расположенным на поверхности клеток BaF3 латентным TGF-бета 1. IC17-hIgG1 обозначает применяемое в качестве отрицательного контроля антитело к KLH. hT0947AE04-SG191, hT0947AE07-SG191, hT0947AE08-SG191 и hT0947AE09-SG191 обозначают антитела к латентному TGF-бета 1;
на фиг. 1Б - результаты оценки связывания антител с расположенным на поверхности клеток FreeStyle™ 293-F латентным TGF-бета IC17-hIgG1 обозначает применяемое в качестве отрицательного контроля антитело к KLH. hT0947AE04-SG191, hT0947AE07-SG191, hT0947AE08-SG191 и hT0947AE09-SG191 обозначают антитела к латентному TGF-бета 1;
на фиг. 2А- результаты оценки связывания антител с мышиным зрелым TGF-бета 1. GC1008-F1332m обозначает применяемое в качестве положительного контроля антитело к зрелому TGF-бета. IC17-hIgG1 обозначает применяемое в качестве отрицательного контроля антитело к KLH. hT0947AE04 SG191, hT0947AE07-SG191, hT0947AE08-SG191 и hT0947AE09-SG191 обозначают антитела к латентному TGF-бета 1;
на фиг. 2Б - результаты оценки связывания антител с человеческим зрелым TGF-бета 1. GC1008-F1332m обозначает применяемое в качестве положительного контроля антитело к зрелому TGF-бета. IC17-hIgG1 обозначает применяемое в качестве отрицательного контроля антитело к KLH. hT0947AE04 SG191, hT0947AE07-SG191, hT0947AE08-SG191 и hT0947AE09-SG191 обозначают антитела к латентному TGF-бета 1;
на фиг. 2 В - результаты оценки связывания антител с мышиным LAP. GC1008-F1332m обозначает антитело к зрелому TGF-бета. IC17-hIgG1 обозначает применяемое в качестве отрицательного контроля антитело к KLH. hT0947AE04-SG191, hT0947AE07-SG191, hT0947AE08-SG191 и hT0947AE09-SG191 обозначают антитела к латентному TGF-бета 1;
на фиг. 3А результаты оценки активности антител в отношении спонтанной активации мышиного латентного TGF-бета 1. mSLC обозначает рекомбинантный мышиный латентный TGF-бета 1. IC17-hIgG1 обозначает применяемое в качестве отрицательного контроля антитело к KLH. hT0947AE04 SG191, hT0947AE07-SG191, hT0947AE08-SG191 и hT0947AE09-SG191 обозначают антитела к латентному TGF-бета 1;
на фиг. 3Б - результаты оценки активности антител в отношении спонтанной активации человеческого латентного TGF-бета 1. hSLC обозначает рекомбинантный человеческий латентный TGF-бета 1. IC17-hIgG1 обозначает применяемое в качестве отрицательного контроля антитело к KLH. hT0947AE04 SG191, hT0947AE07-SG191, hT0947AE08-SG191 и hT0947AE09-SG191 обозначают антитела к латентному TGF-бета 1;
на фиг. 4А - результаты оценки активности антител в отношении опосредуемой плазмином (PLN) активации мышиного латентного TGF-бета 1. mSLC обозначает рекомбинантный мышиный латентный TGF-бета 1. IC17-hIgG1 обозначает применяемое в качестве отрицательного контроля антитело к KLH. hT0947AE04-SG191, hT0947AE07-SG191, hT0947AE08-SG191 и hT0947AE09-SG191 обозначают антитела к латентному TGF-бета 1;
на фиг. 4Б - результаты оценки активности антител в отношении опосредуемой плазмином (PLN) активации человеческого латентного TGF-бета 1. hSLC обозначает рекомбинантный человеческий латентный TGF-бета 1. IC17-hIgG1 обозначает применяемое в качестве отрицательного контроля антитело к KLH. hT0947AE04-SG191, hT0947AE07-SG191, hT0947AE08-SG191 и hT0947AE09-SG191 обозначают антитела к латентному TGF-бета 1;
на фиг. 5А - результаты оценки активности антител в отношении опосредуемой калликреином (PLK) активации мышиного латентного TGF-бета 1. mSLC обозначает рекомбинантный мышиный латентный TGF-бета 1. IC17-hIgG1 обозначает применяемое в качестве отрицательного контроля антитело к KLH. hT0947AE04-SG191, hT0947AE07-SG191, hT0947AE08-SG191 и hT0947AE09-SG191 обозначают антитела к латентному TGF-бета 1;
на фиг. 5Б - результаты оценки активности антител в отношении опосредуемой калликреином (PLK) активации человеческого латентного TGF-бета 1. hSLC обозначает рекомбинантный человеческий латентный TGF-бета 1. IC17-hIgG1 обозначает применяемое в качестве отрицательного контроля антитело к KLH. hT0947AE04-SG191, hT0947AE07-SG191, hT0947AE08-SG191 и hT0947AE09-SG191 обозначают антитела к латентному TGF-бета 1;
на фиг. 6А - результаты оценки активности антител в отношении опосредуемой матриксной металлопротеиназой (ММР) 2 активации человеческого латентного TGF-бета 1. hSLC обозначает рекомбинантный человеческий латентный TGF-бета 1. IC17-hIgG1 обозначает применяемое в качестве отрицательного контроля антитело к KLH. hT0947AE04-SG191, hT0947AE07-SG191, hT0947AE08-SG191 и hT0947AE09-SG191 обозначают антитела к латентному TGF-бета 1;
на фиг. 6Б - результаты оценки активности антител в отношении опосредуемой матриксной металлопротеиназой (ММР) 9 активации человеческого латентного TGF-бета 1. hSLC обозначает рекомбинантный человеческий латентный TGF-бета 1. IC17-hIgG1 обозначает применяемое в качестве отрицательного контроля антитело к KLH. hT0947AE04-SG191, hT0947AE07-SG191, hT0947AE08-SG191 и hT0947AE09-SG191 обозначают антитела к латентному TGF-бета 1;
на фиг. 7А - результаты оценки активности антител в отношении опосредуемого плазмином (PLN) расщепления мышиного латентного TGF-бета 1. Cam обозначает применяемый в качестве контроля ингибитор протеазы камостат.АЕ04 (hT0947AE04-SG191), АЕ07 (hT0947AE07-SG191), АЕ08 (hT0947AE08-SG191) и АЕ09 (hT0947AE09-SG191) обозначают антитела к латентному TGF-бета 1;
на фиг. 7Б - результаты оценки активности антител в отношении опосредуемого плазмином (PLN) расщепления человеческого латентного TGF-бета 1. Cam обозначает применяемый в качестве контроля ингибитор протеазы камостат.АЕ04 (hT0947AE04-SG191), АЕ07 (hT0947AE07-SG191), АЕ08 (hT0947AE08-SG191) и АЕ09 (hT0947AE09-SG191) обозначают антитела к латентному TGF-бета 1;
на фиг. 8 - результаты оценки активности антител в отношении опосредуемой интегрином активации мышиного TGF-бета 1 в мышиных РВМС.IC17-hIgG1 обозначает применяемое в качестве отрицательного контроля антитело к KLH. GC1008-F1332m обозначает применяемое в качестве положительного контроля антитело к зрелому TGF-бета. RGE обозначает пептид RGE. RGD обозначает применяемый в качестве положительного контроля пептид RGD. hT0947AE04-SG191, hT0947AE07-SG191, hT0947AE08-SG191 и hT0947AE09-SG191 обозначают антитела к латентному TGF-бета 1;
на фиг. 9 - кривые роста опухолей в группе, обработанной контролем изотипа (закрашенный кружок), группе, обработанной антителом к PD-L1 (закрашенный квадрат), группе, обработанной hT0947AE04-mF18 (закрашенный треугольник), и группе, обработанной hT0947AE04-mF18 плюс антитело к PD-L1 (крестик). Каждая точка соответствует среднему объему опухоли в каждой группе. (N=10);
на фиг. 10 - кривые роста опухолей в группе, обработанной контролем изотипа (закрашенный кружок), группе, обработанной антителом к PD-L1 (закрашенный квадрат), группе, обработанной hT0947AE04-mF18 (закрашенный треугольник), и группе, обработанной hT0947AE04-mF18 плюс антитело к PD-L1 (крестик). Каждая точка соответствует среднему объему опухоли в каждой группе. (N=10);
на фиг. 11 - кривые роста опухолей в группе, обработанной наполнителем (закрашенный кружок), группе, обработанной антителом к PD-L1 (закрашенный квадрат), группе, обработанной hT0947AE04-mF18 плюс антитело к PD-L1 (X) hT0947AE07-SG181 плюс антитело к PD-L1 (закрашенный треугольник), и группе, обработанной hT0947AE08-SG181 плюс антитело PD-L1 (незакрашенный кружок). Каждая точка соответствует среднему объему опухоли в каждой группе. (N=10);
на фиг. 12 - результаты оценки содержания гидроксипролина в почке. Моноклональные антитела изучали с использованием модели индуцированного односторонней обструкцией мочеточника (Unilateral Ureteral Obstruction (UUO)) фиброза почечной ткани мышей. Sham-группа (группа, подвергнутая плацебо-операции) обозначает неиндуцированный болезнью контроль. IC17dk-SG181 обозначает применяемое в качестве отрицательного контроля антитело к KLH. hT0947AE04-SG191, hT0947AE07-SG191 и hT0947AE08-SG191 обозначают антитела к латентному TGF-бета 1;
на фиг. 13 - кривые роста опухолей в группе, обработанной наполнителем (закрашенный кружок), группе, обработанной антителом к мышиному PD-L1 (закрашенный квадрат), группе, обработанной hT0947AE07-SG191 (10 мг/кг) плюс антитело к мышиному PD-L1 (закрашенный треугольник), и группе, обработанной hT0947AE07-SG191 (30 мг/кг) плюс антитело к мышиному PD-L1 (незакрашенный кружок). Каждая точка соответствует среднему объему опухоли в соответствующей группе. (N=10 в каждой группе).
Описание вариантов осуществления изобретения
I. Определения
Для целей настоящего описания «акцепторный человеческий каркасный участок» означает каркасный участок, который содержит аминокислотную последовательность каркасного участка вариабельного домена легкой цепи (VL) или каркасного участка вариабельного домена тяжелой цепи (VH), имеющего происхождение из каркасного участка человеческого иммуноглобулина или человеческого консенсусного каркасного участка, указанного ниже. Акцепторный человеческий каркасный участок, «имеющий происхождение из» каркасного участка человеческого иммуноглобулина или человеческого консенсусного каркасного участка, может иметь такую же аминокислотную последовательность или может содержать изменения в аминокислотной последовательности. В некоторых вариантах осуществления изобретения количество аминокислотных изменений составляет 10 или менее, 9 или менее, 8 или менее, 7 или менее, 6 или менее, 5 или менее, 4 или менее, 3 или менее или 2 или менее. В некоторых вариантах осуществления изобретения последовательность человеческого акцепторного каркасного участка VL идентична последовательности каркасного участка VL человеческого иммуноглобулина или последовательности человеческого консенсусного каркасного участка.
Понятие «связывающая активность, активность связывания» относится к суммарной силе всех нековалентных взаимодействий между одним или несколькими сайтами связывания молекулы (например, антитела) и ее партнером по связыванию (например, антигеном). В контексте настоящего описания «связывающая активность» не ограничена строго взаимодействием по типу 1:1 между компонентами связывающейся пары (например, антителом и антигеном). Например, когда взаимодействие между компонентами связывающейся пары соответствует одновалентному взаимодействию по типу 1:1, то связывающую активность обозначают конкретно как присущую им аффинность связывания (аффинность). Когда компонент связывающейся пары обладает способностью и к одновалентному связыванию, и к многовалентному связыванию, то связывающая активность представляет собой сумму каждой силы связывания. Связывающую активность молекулы X к ее партнеру Y, как правило, можно характеризовать с помощью константы диссоциации (KD) или «количества аналита, связывающегося с единицей количества лиганда» (ниже в контексте настоящего описания можно обозначать как «связывающееся количество»). Специалистам в данной области должно быть очевидно, что, как правило, более низкая величина константы диссоциации (KD) означает более высокую связывающую активность, и более высокая величина «количества аналита, связывающегося с единицей количества лиганда» или «связывающееся количество» означает более высокую связывающую активность. Связывающую активность можно оценивать с помощью общепринятых методов, известных в данной области, включая те, которые представлены в настоящем описании. Ниже представлены конкретные иллюстративные и приведенные в качестве примера варианты измерения связывающей активности.
Понятия антигенсвязывающая молекула или антитело «с созревшей связывающей активностью», «с созревшей аффинностью», или антигенсвязывающая молекула или антитело «с повышенной (увеличенной) связывающей активностью» или «с повышенной (увеличенной) аффинностью» относятся к антителу с одним или несколькими изменениями (например, заменами) в одном или нескольких гипервариабельных участках (HVR) по сравнению с родительской антигенсвязывающей молекулой или родительским антителом, которая/которое не содержит указанных изменений, указанные изменения приводят к улучшению связывающей активности антигенсвязывающей молекулы или антитела в отношении антигена.
Понятия «антитело к латентному TGF-бета 1» и «антитело, которое может связываться с латентным TGF-бета 1» относятся к антителу, которое обладает способностью связываться с латентным TGF-бета 1 со связывающей активностью, достаточной для того, чтобы антитело можно было применять в качестве диагностического и/или терапевтического агента, мишенью которого является латентный TGF-бета 1. В одном из вариантов осуществления изобретения «антитело, которое может связываться с латентным TGF-бета 1» представляет собой антитело, которое специфически связывается с латентным TGF-бета 1. В одном из вариантов осуществления изобретения уровень связывающей активности антитела к латентному TGF-бета 1 с неродственным белком, не представляющим собой латентный TGF-бета 1, составляет менее чем примерно 10% от связывающей активности антитела с латентным TGF-бета 1 по данным измерений, полученным, например, с помощью радиоиммуноанализа (РИА). В некоторых вариантах осуществления изобретения антитело, которое может связываться с латентным TGF-бета 1, имеет константу диссоциации (KD) 1 мкМ или менее, 100нМ или менее, 10нМ или менее, 1нМ или менее, 0,1нМ или менее, 0,01нМ или менее или 0,001нМ или менее (например, 10-8 М или менее,
например, от 10-8 М до 10-13 М, например, от 10-9 М до 10-13 М). В некоторых вариантах осуществления изобретения антитело к латентному TGF-бета 1 связывается с эпитопом латентного TGF-бета 1, который является консервативным для латентного TGF-бета 1 из различных видов.
В контексте настоящего описания понятие «антитело» используется в его наиболее широком смысле и относится к различным структурам антител, включая (но, не ограничиваясь только ими) моноклональные антитела, поликлональные антитела, мультиспецифические антитела (например, биспецифические антитела) и фрагменты антител, при условии, что они обладают требуемой антигенсвязывающей активностью. Понятие «антитело» включает также любую антигенсвязывающую молекулу, которая содержит структуру(ы) вариабельной тяжелой цепи и/или вариабельной легкой цепи иммуноглобулина.
Понятие «фрагмент антитела» относится к молекуле, отличной от интактного антитела, которая содержит часть интактного антитела, связывающуюся с антигеном, с которым связывается интактное антитело. Примеры фрагментов антитела включают (но не ограничиваясь только ими) Fv, Fab, Fab', Fab'-SH, F(ab')2; димерные антитела (диабоди); линейные антитела; молекулы одноцепочечных антител (например, scFv) и мультиспецифические антитела, образованные из фрагментов антител.
Понятие «антитело, которое связывается с тем же эпитопом», что и референс-антитело, относится к антителу, которое блокирует на 50% или более связывание референс-антитела с его антигеном при оценке с помощью анализа в условиях конкуренции (конкурентного анализа) и, наоборот, референс-антитело блокирует на 50% или более связывание антитела с его антигеном при оценке с помощью конкурентного анализа. Пример конкурентного анализа представлен в настоящем описании.
Понятия «рак» и «злокачественное заболевание» относятся или описывают физиологическое состояние у млекопитающего, которое характеризуется нерегулируемыми клеточным ростом/пролиферацией. Примеры рака включают (но не ограничиваясь только ими) карциному, лимфому (например, лимфому Ходжкина и неходжкинскую лимфому), бластому, саркому и лейкоз. Более конкретные примеры указанных раков включают плоскоклеточный рак, мелкоклеточный рак легкого, немелкоклеточный рак легкого, аденокарцирному легкого, плоскоклеточный рак легкого, перитонеальный рак, печеночноклеточный рак, рак желудочно-кишечной системы, рак поджелудочной железы, глимому, рак шейки матки, рак яичника, рак печени, рак мочевого пузыря, гепатому, рак молочной железы, рак ободочной кишки, колоректальный рак, карциному эндометрия или матки, карциному слюнных желез, рак почки, рак печени, рак предстательной железы, рак вульвы, рак щитовидной железы, печеночную карциному, лейкоз и другие лимфопролиферативные нарушения, а также различные типы рака головы и шеи. В одном из примеров рак является устойчивым к ингибиторам иммунных контрольных точек и/или дает ограниченный ответ на ингибиторы иммунных контрольных точек.
Понятие «химерное» антитело относится к антителу, в котором часть тяжелой и/или легкой цепи имеет происхождение из конкретного источника или вида, а остальная часть тяжелой и/или легкой цепи имеет происхождение из другого источника или вида.
Понятие «класс» антитела относится к типу константного домена или константной области, который/которая входит в его тяжелую цепь. Известно пять основных классов антител: IgA, IgD, IgE, IgG и IgM и некоторые из них можно подразделять дополнительно на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелой цепи, которые соответствуют различным классам иммуноглобулинов, обозначают как альфа, дельта, эпсилон, гамма и мю соответственно.
Понятие «цитотоксический агент» в контексте настоящего описания относится к субстанции, которая ингибирует клеточную функцию или препятствует ей и/или вызывает клеточную гибель или деструкцию. Цитотоксические агенты включают (но не ограничиваясь только ими) радиоактивные изотопы (например, At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32, Pb212 и радиоактивные изотопы Lu); химиотерапевтические агенты или лекарственные средства (например, метотрексат, адриамицин, алкалоиды барвинка (винкристин, винбластин, этопозид), доксорубицин, мелфалан, митомицин С, хлорамбуцил, даунорубицин или другие интеркалирующие агенты); ингибирующие рост агенты; ферменты и их фрагменты, такие как нуклеолитические ферменты; антибиотики; токсины, такие как низкомолекулярные токсины или ферментативно активные токсины бактериального, грибного, растительного или животного происхождения, включая их фрагменты и/или варианты; и различные противоопухолевые или противораковые агенты, указанные ниже.
Понятие «эффекторные функции» относится к тем видам биологической активности, которые связаны с Fc-областью антитела, которые варьируются в зависимости от изотипа антитела. Примеры эффекторных функций антитела включают: связывание C1q и комплементзависимую цитотоксичность (CDC); связывание Fc-рецептора; антитело-обусловленную клеточнозависимую цитотоксичность (ADCC); фагоцитоз; понижающую регуляцию рецепторов клеточной поверхности (например, В-клеточного рецептора) и активацию В-клеток.
Понятие «эффективное количество» агента, например, фармацевтической композиции, означает количество, эффективное в дозах и в течение периода времени, необходимых для достижения требуемого терапевтического или профилактического результата.
Понятие «Fc-область» в контексте настоящего описания относится к С-концевой области тяжелой цепи иммуноглобулина, которая содержит по меньшей мере часть константной области. Понятие включает нативную последовательность Fc-областей и варианты Fc-областей. В одном из вариантов осуществления изобретения Fc-область тяжелой цепи человеческого IgG простирается от Cys226 или от Pro230 до карбоксильного конца тяжелой цепи. Однако С-концевой лизин (Lys447) или глицин-лизин (остатки 446-447) Fc-области может присутствовать или отсутствовать. Если специально не указано иное, то нумерация аминокислотных остатков в Fc-области или константной области соответствует системе нумерации EU, которую обозначают также как EU-индекс, описанной у Kabat Е.А. и др., Sequences of Proteins of Immunological Interest, 5-ое изд., изд-во Public Health Service, National Institutes of Health, Bethesda, MD, 1991.
«Каркасный участок» или «FR», означает остатки вариабельного домена, отличные от остатков гипервариабельного участка (HVR). FR вариабельного домена, как правило, состоит из четырех FR-доменов: FR1, FR2, FR3 и FR4. Таким образом, последовательности HVR и FR, как правило, имеют следующее расположение в VH (или VL): FR1-H1(L1)-FR2-H2(L2)-FR3-H3(L3)-FR4.
Понятия «полноразмерное антитело», «интактное антитело» и «цельное антитело» в контексте настоящего описания используют взаимозаменяемо для обозначения антитела, имеющего структуру, практически сходную с нативной структурой антитела, или имеющего тяжелые цепи, которые содержат представленную в настоящем описании Fc-область.
Понятия «клетка-хозяин», «линия клеток-хозяев» и «культура клеток-хозяев» используют взаимозаменяемо, и они относятся к клеткам, в которые интродуцирована экзогенная нуклеиновая кислота, включая потомство указанных клеток. К клеткам-хозяевам относятся «трансформанты» и «трансформированные клетки», которые включают первично трансформированную клетку и полученное из нее потомство безотносительно к количеству пересевов. Потомство может не быть полностью идентичным по составу нуклеиновых кислот родительской клетке, а может содержать мутации. Под объем изобретения подпадает мутантное потомство, которое обладает такой же функцией или биологической активностью, которая обнаружена в результате скрининга или отобрана у исходной трансформированной клетки.
«Человеческое антитело» представляет собой антитело, имеющее аминокислотную последовательность, которая соответствует последовательности антитела, продуцируемого в организме человека или человеческой клеткой, или имеющее происхождение из нечеловеческого источника, в котором присутствует спектр человеческих антител или другие кодирующие человеческое антитело последовательности. Из указанного определения человеческого антитела специально исключено гуманизированное антитело, содержащее нечеловеческие антигенсвязывающие остатки.
«Человеческий консенсусный каркасный участок» представляет собой каркасный участок, в который входят наиболее часто встречающиеся аминокислотные остатки в выбранных последовательностях каркасных участков VL или VH человеческого иммуноглобулина. Как правило, выбор последовательностей VL или VH человеческого иммуноглобулина осуществляют из подгруппы последовательностей вариабельных доменов. Как правило, подгруппа последовательностей представляет собой подгруппу, описанную у Kabat и др., Sequences of Proteins of Immunological Interest, 5-ое изд., публикация NIH 91-3242, Bethesda MD, т.т.1-3, 1991. В одном из вариантов осуществления изобретения VL-подгруппа представляет собой подгруппу каппа I согласно Kabat и др., выше. В одном из вариантов осуществления изобретения VH-подгруппа представляет собой подгруппу III согласно Kabat и др., выше.
Понятие «гуманизированное» антитело относится к химерному антителу, которое содержит аминокислотные остатки из нечеловеческих HVR и аминокислотные остатки из человеческих FR. В некоторых вариантах осуществления изобретения гуманизированное антитело может содержать практически все из по меньшей мере одного и, как правило, двух вариабельных доменов, в которых все или практически все HVR (например, CDR) соответствуют участкам нечеловеческого антитела, а все или практически все FR соответствуют участкам человеческого антитела. Гуманизированное антитело необязательно может содержать по меньшей мере часть константной области антитела, происходящей из человеческого антитела. Понятие «гуманизированная форма» антитела, например, нечеловеческого антитела, относится к антителу, которое подвергли гуманизации.
Понятие «гипервариабельный участок» или «HVR» в контексте настоящего описания относится к каждому из участков вариабельного домена антитела, последовательности которых являются гипервариабельными («определяющие комплементарность участки» или «CDR») и/или формируют петли определенной структуры («гипервариабельные петли»), и/или содержат контактирующие с антигеном остатки («контакты с антигеном»). Как правило, антитела содержат шесть HVR; три в VH (H1, Н2, Н3) и три в VL (L1, L2, L3). Примеры HVR включают
(а) гипервариабельные петли, включающие аминокислотные остатки 26-32 (L1), 50-52 (L2), 91-96 (L3), 26-32 (H1), 53-55 (Н2) и 96-101 (Н3) (Chothia и Lesk, J. Mol. Biol. 196, 1987, cc. 901-917);
(б) CDR, включающие аминокислотные остатки 24-34 (L1), 50-56 (L2), 89-97 (L3), 31-35b (H1), 50-65 (H2) и 95-102 (Н3) (Kabat и др., Sequences of Proteins of Immunological Interest, 5-ое изд., изд-во Public Health Service, National Institutes of Health, Bethesda, MD,1991);
(в) области контакта с антигеном, включающие аминокислотные остатки 27 с-36 (L1), 46-55 (L2), 89-96 (L3), 30-35b (H1), 47-58 (Н2) и 93-101 (Н3) (MacCallum и др., J. Mol. Biol. 262, 1996, cc. 732-745); и
(г) комбинации остатков, указанных в подпунктах (а), (б) и/или (в), включающие аминокислотные остатки HVR 46-56 (L2), 47-56 (L2), 48-56 (L2), 49-56 (L2), 26-35 (H1), 26-35b (H1), 49-65 (Н2), 93-102 (Н3) и 94-102 (Н3).
Если специально не указано иное, то остатки в HVR и другие остатки в вариабельном домене (например, остатки в FR) нумеруют согласно Kabat и др., выше.
«Иммуноконъюгат» представляет собой антитело, конъюгированное с одной или несколькими гетерологичной(ыми) молекулой(ами), включая (но не ограничиваясь только им) цитотоксический агент.
«Индивидуум» или «субъект» представляет собой млекопитающее. К млекопитающим относятся (но не ограничиваясь только ими) одомашненные животные (например, коровы, овцы, кошки, собаки и лошади), приматы (например, человек и приматы кроме человека, например, обезьяны), кролики и грызуны (например, мыши и крысы). В некоторых вариантах осуществления изобретения индивидуум или субъект представляет собой человека.
«Выделенное» антитело представляет собой антитело, которое отделено от компонента его естественного окружения. В некоторых вариантах осуществления изобретения антитело очищают до чистоты, превышающей 95% или 99% по данным, например, электрофоретических анализов (таких, например, как ДСН-ПААГ, изоэлектрическое фокусирование (ИЭФ), капиллярный электрофорез) или хроматографических анализов (таких, например, как ионообменная хроматография или ЖХВР с обращенной фазой). Обзор методов оценки чистоты антител см., например, у Flatman и др., J. Chrom. В 848, 2007, сс.79-87.
«Выделенная» нуклеиновая кислота представляет собой молекулу нуклеиновой кислоты, которая отделена от компонента ее естественного окружения. Выделенная нуклеиновая кислота включает молекулу нуклеиновой кислоты, содержащуюся в клетке, которая в норме включает молекулу нуклеиновой кислоты, но в которой молекула нуклеиновой кислоты присутствует вне хромосомы или в которой ее локализация на хромосоме отличается от ее встречающейся в естественных условиях локализации на хромосоме.
Понятие «выделенная нуклеиновая кислота, кодирующая антитело к латентному TGF-бета 1» или «нуклеиновая кислота, кодирующая антитело к латентному TGF-бета 1» относится к одной или нескольким молекулам нуклеиновых кислот, кодирующих тяжелые и легкие цепи антитела (или их фрагменты), включая указанную(ые) молекулу(ы) нуклеиновой кислоты в одном векторе или в различных векторах, и указанную(ые) молекулу(ы) нуклеиновой(ых) кислоты(т), присутствующую(ие) в одном или нескольких положениях в клетке-хозяине.
Понятие «моноклональное антитело» в контексте настоящего описания относится к антителу, полученному из популяции практически гомогенных антител, т.е. индивидуальные антитела, входящие в популяцию, идентичны и/или связываются с одним и тем же эпитопом, за исключением возможных вариантов антител, например, содержащих мутации, встречающиеся в естественных условиях или возникающие в процессе производства препарата моноклонального антитела, указанные варианты, как правило, присутствуют в минорных количествах. В отличие от препаратов поликлональных антител, которые, как правило, включают различные антитела к различным детерминантам (эпитопам), каждое моноклональное антитело из препарата моноклонального антитела направлено против одной детерминанты на антигене. Таким образом, прилагательное «моноклональный» определяет особенность антитела, характеризуя его как полученное из практически гомогенной популяции антител, а не сконструированное в соответствии с требованиями к получению антитела с помощью какого-либо конкретного метода. Например, моноклональные антитела, которые можно применять согласно настоящему изобретению, можно создавать с помощью различных технологий, включая (но не ограничиваясь только ими) метод гибридом, методы рекомбинантной ДНК, методы фагового дисплея и методы, основанные на использовании трансгенных животных, которые содержат все или часть локусов человеческого иммуноглобулина, указанные методы и другие, приведенные в качестве примера методы получения моноклональных антител, представлены в настоящем описании.
Понятие «голое антитело» относится к антителу, не конъюгированному с гетерологичным фрагментом (например, цитотоксическим фрагментом) или радиоактивной меткой. Голое антитело может присутствовать в фармацевтической композиции.
Понятие «нативные антитела» относится к встречающимся в естественных условиях молекулам иммуноглобулинов с различными структурами. Например, нативные антитела в виде IgG представляют собой гетеротетрамерные гликопротеины с молекулярной массой примерно 150000 Да, состоящие из двух идентичных легких цепей и двух идентичных тяжелых цепей, связанных дисульфидными мостиками. В направлении от N-конца к С-концу каждая тяжелая цепь имеет вариабельную область (VH), которую называют также вариабельным тяжелым доменом или вариабельным доменом тяжелой цепи, за которой следует три константных домена (CH1, СН2 и СН3). Аналогично этому, в направлении от N-конца к С-концу каждая легкая цепь имеет вариабельную область (VL), которую называют также вариабельным легким доменом или вариабельным доменом легкой цепи, за которой следует константный домен легкой цепи (CL). Легкая цепь антитела в зависимости от аминокислотной последовательности ее константного домена может принадлежать к одному из двух типов, которые обозначают каппа (κ) и лямбда (λ).
Понятие «листовка-вкладыш в упаковку» в контексте настоящего описания относится к инструкциям, которые обычно входят в поступающие в продажу упаковки терапевтических продуктов, содержащим информацию о показаниях, применении, дозе, пути введения, комбинированной терапии, противопоказаниях и/или мерах предосторожности, которые связаны с применением указанных терапевтических продуктов.
«Процент (%) идентичности аминокислотной последовательности» относительно полипептидной референс-последовательности определяют как процент аминокислотных остатков в последовательности-кандидате, которые идентичны аминокислотным остаткам в полипептидной референс-последовательности, после выравнивая последовательностей и при необходимости интродукции брешей для достижения максимального процента идентичности последовательностей, и не рассматривая какие-либо консервативные замены в качестве компонента при оценке идентичности последовательностей. Сравнительный анализ для целей определения процента идентичности аминокислотных последовательностей можно осуществлять различными путями, известными в данной области, используя, например, публично доступные компьютерные программы, такие как программа BLAST, BLAST-2, ALIGN, Megalign (DNASTAR) или GENETYX® (фирма Genetyx Co., Ltd.) Специалисты в данной области могут определять соответствующие параметры для выравнивания последовательностей, включая любые алгоритмы, необходимые для достижения максимального выравнивания по всей длине сравниваемых последовательностей.
Авторство компьютерной программы для сравнения последовательностей ALIGN-2 принадлежит фирме Genentech, Inc., и исходный код был представлен в комплекте с документацией для пользователей в U.S. Copyright Office, Washington D.C., 20559, где он зарегистрирован как U.S. Copyright Registration №TXU510087. Программа ALIGN-2 публично доступна от фирмы Genentech, Inc., Южный Сан-Франциско, шт. Калифорния, или ее можно компилировать на основе исходного кода. Программу ALIGN-2 можно компилировать для применения в операционной системе UNIX, включая цифровую систему UNIX V4.0D. Все параметры, требуемые для сравнения последовательностей, устанавливаются программой ALIGN-2 и не должны изменяться. В ситуациях, в которых ALIGN-2 применяют для сравнения аминокислотных последовательностей, % идентичности аминокислотных последовательностей данной аминокислотной последовательности А по сравнению с или относительно данной аминокислотной последовательности Б (что в альтернативном варианте можно обозначать как данная аминокислотная последовательность А, которая имеет или содержит определенный % идентичности аминокислотной последовательности по сравнению с или относительно данной аминокислотной последовательности Б), рассчитывают следующим образом:
100 × дробь X/Y,
где X обозначает количество аминокислотных остатков, определенных как идентичные совпадения при оценке с помощью программы для сравнительного анализа последовательностей ALIGN-2, где с помощью программы осуществляли сравнение А и Б, и где Y обозначает общее количество аминокислотных остатков в Б. Должно быть очевидно, что, если длина аминокислотной последовательности А не равна длине аминокислотной последовательности Б, то % идентичности аминокислотных последовательностей А относительно Б не может быть равен % идентичности аминокислотной последовательности Б относительно А. Если специально не указано иное, то все величины % идентичности аминокислотных последовательностей, указанные в настоящем описании, получали с использованием описанной в предыдущем параграфе компьютерной программы ALIGN-2.
Понятие «фармацевтическая композиция» относится к препарату, который находится в такой форме, которая обеспечивает биологическую активность входящего в ее состав действующего вещества, которое должно обладать эффективностью, и которая не содержит дополнительных компонентов, обладающих неприемлемой токсичностью для индивидуума, которому следует вводить композицию.
«Фармацевтически приемлемый носитель» относится к ингредиенту в фармацевтической композиции, отличному от действующего вещества, который является нетоксичным для индивидуума. Фармацевтически приемлемые носители включают (но не ограничиваясь только ими) буфер, эксципиент, стабилизатор или консервант.
Если не указано иное, то понятие «TGF-бета 1» в контексте настоящего описания относится к любому нативному TGF-бета 1 из любого позвоночного животного-источника, включая млекопитающих, такие как приматы (например, люди) и грызуны (например, мыши и крысы). Под понятие подпадает «полноразмерный» непроцессированный TGF-бета 1, а также любая форма TGF-бета 1, являющаяся результатом процессинга в клетке. Под понятие подпадают встречающиеся в естественных условиях варианты TGF-бета 1, например, сплайсинговые варианты или аллельные варианты. Аминокислотная последовательность приведенного в качестве примера препробелка человеческого TGF-бета 1 представлена в SEQ ID NO: 68 (NCBI RefSeq: NP_000651.3), а нуклеотидная последовательность приведенного в качестве примера препробелка человеческого TGF-бета 1 представлена в SEQ ID NO: 69 (NCBI RefSeq: NM_000660.6). Аминокислотная последовательность приведенного в качестве примера препробелка мышиного TGF-бета 1 представлена в SEQ ID NO: 70 (NCBI RefSeq: NP_035707.1), а нуклеотидная последовательность приведенного в качестве примера препробелка мышиного TGF-бета 1 представлена в SEQ ID NO: 71 (NCBI RefSeq: NM_011577.2). Аминокислотная последовательность приведенного в качестве примера препробелка TGF-бета 1 обезьян циномолгус представлена в SEQ ID NO: 72 (NCBI RefSeq: XP_005589396.1), а нуклеотидная последовательность приведенного в качестве примера препробелка TGF-бета 1 обезьян циномолгус представлена в SEQ ID NO: 73 (NCBI RefSeq: ХМ_005589339.2). Понятие «TGF-бета 1» относится как к латентному TGF-бета 1, так и к зрелому TGF-бета 1.
Понятие «латентный TGF-бета 1» в контексте настоящего описания относится к любому TGF-бета 1, который образует комплекс латентного TGF-бета 1 («находящийся на клеточной поверхности латентный TGF-бета 1», LLC или SLC (см. ниже)) и/или который не обладает способностью связываться с его рецепторами. Трансформирующий рост фактор-бета 1 (TGF-бета 1) представляет собой TGF-бета, являющийся членом суперсемейства TGF бета. Аналогично другим членам суперсемейства TGF-бета TGF-бета синтезируется в виде белка-предшественника, образующего гомодимер, который взаимодействует с ассоциированным с латентностью пептидом (LAP) и связывающим латентный TGF-бета белком (LTBP) с образованием более крупного комплекса, который называют большим латентным комплексом (LLC). Аминокислотная последовательность приведенного в качестве примера латентного человеческого TGF-бета 1 (гомодимер TGF-бета и его LAP) представлена аминокислотами 30-390 в SEQ ID NO: 68. Аминокислотная последовательность приведенного в качестве примера латентного мышиного TGF-бета 1 (гомодимер TGF-бета и его LAP) представлена аминокислотами 30-390 в SEQ ID NO: 70. Аминокислотная последовательность приведенного в качестве примера латентного TGF-бета 1 обезьян циномолгус (гомодимер TGF-бета и его LAP) представлена аминокислотами 30-390 в SEQ ID NO: 72.
Комплекс, образованный гомодимером TGF-бета и его LAP, называют малым латентным комплексом (SLC). В указанном латентном комплексе TGF-бета находится в неактивной форме, в которой он не может связываться с его рецепторами. SLC может быть дополнительно связан с другим белком, т.е. связывающим латентный TGF-бета 1 белком (LTBP), с образованием большого латентного комплекса (LLC). Известно четыре различные изоформы LTBP, LTBP-1, LTBP-2, LTBP-3 и LTBP-4. Опубликованы данные о том, что LTBP-1, LTBP-3 и LTBP-4 связываются с SLC (см., например, Rifkin и др., J Biol Chem. 280(9), 4 марта 2005, cc. 7409-7412). SLC также может быть ковалентно связан с другими дополнительными белками, такими как белок с преобладающими повторами гликопротеина A (GARP) или содержащий богатый лейцином повтор белок 33 (LRRC33). GARP и LRRC имеют трансмембранный домен и находятся в ассоциации с LAP на клеточной поверхности (см., например, Wang и др., Mol Biol Cell. 23(6), март 2012 г., cc. 1129-1139). Касательно LLC опубликованы данные о том, что LLC ковалентно ассоциирован с внеклеточным матриксом (ЕСМ) через N-концы LTBP (см., например, Saharinen и др., Cytokine Growth Factor Rev. 10(2), июнь 1999 г., сс.99-117.). В некоторых вариантах осуществления изобретения латентный TGF-бета 1, ассоциированный с ЕСМ на клеточной поверхности, обозначают как «находящийся на клеточной поверхности латентный TGF-бета 1»
Понятие «активный TGF-бета 1», «зрелый TGF-бета 1» или «активный зрелый TGF-бета 1» в контексте настоящего описания относится к любому гомодимеру TGF-бета 1, который не образует комплекс латентного TGF-бета 1 (LLC или SLC) и который обладает способностью связываться с его рецепторами. Процесс активации TGF-бета 1 включает высвобождение LLC из ЕСМ с последующим дополнительным протеолизом LAP для высвобождения активного в отношении его рецепторов TGF-бета. Известно, что широкий спектр протеаз, включающий плазмин (PLN), прекалликреин (PLK), матриксную металлопротеиназу (ММР) 2, ММР9, ММР13, ММР14, тромбин, триптазу и калпаин, расщепляют латентный TGF-бета и высвобождают активный TGF-бета. В контексте настоящего изобретения указанные протеазы можно в целом обозначать как «(латентный) TGF-бета-расщепляющие протеазы» или «(латентный) TGF-бета 1-расщепляющие протеазы». Помимо протеаз латентный TGF-бета активируют тромбоспондин (TSP-1), нейропилин-1 (Nrp1), ADAMSTS1 и F-спондин. Альтернативно этому, при механическом натяжении интегрины (предпочтительно интегрин альфа V бета 8 и/или интегрин альфа V бета 6) могут активировать TGF-бета путем связывания с RGD-мотивом, присутствующим в LAP, и индуцировать высвобождение зрелого TGF-бета из его формы в виде латентного комплекса.
В контексте настоящего описания понятие «лечение» (и его грамматические вариации, такие как «лечить» или «процесс лечения») относится к клиническому вмешательству с целью изменения естественного течения болезни у индивидуума, подлежащего лечению, и его можно осуществлять либо для профилактики, либо в процессе развития клинической патологии. Требуемыми действиями лечения являются (но не ограничиваясь только ими) предупреждение возникновения или рецидива болезни, облегчение симптомов, уменьшение любых прямых или косвенных патологических последствий болезни, предупреждение метастазов, снижение скорости развития болезни, облегчение или временное ослабление болезненного состояния и ремиссия или улучшение прогноза. В некоторых вариантах осуществления изобретения антитела, предлагаемые в изобретении, применяют для задержки развития болезни или замедления прогрессирования болезни.
Понятие «вариабельная область» или «вариабельный домен» относится к домену тяжелой или легкой цепи антитела, который участвует в связывании антитела с антигеном. Вариабельные домены тяжелой цепи и легкой цепи (VH и VL соответственно) нативного антитела, как правило, имеют сходные структуры, при этом каждый домен содержит четыре консервативных каркасных участка (FR) и три гипервариабельных участка (HVR) (см., например, Kindt и др., Kuby Immunology, 6-ое изд., изд-во W.H. Freeman and Co., 2007, с. 91). Одного VH- или VL-домена может быть достаточно для обеспечения специфичности связывания антигена. Кроме того, антитела, которые связываются с конкретным антигеном, можно выделять, используя VH- или VL-домен из антитела, которое связывается с антигеном, для скрининга библиотеки комплементарных VL- или VH-доменов соответственно (см., например, Portolano и др., J. Immunol. 150, 1993, cc. 880-887; Clarkson и др., Nature 352, 1991, cc. 624-628).
Понятие «вектор» в контексте настоящего описания относится к молекуле нуклеиновой кислоты, которая обладает способностью к размножению другой нуклеиновой кислоты, с которой она связана. Понятие включает вектор в виде самореплицирующейся структуры нуклеиновой кислоты, а также вектор, включенный в геном клетки-хозяина, в которую он интродуцирован. Некоторые векторы обладают способностью контролировать экспрессию нуклеиновых кислот, с которыми они функционально связаны. Указанные векторы в контексте настоящего описания обозначают как «экспрессионные векторы».
II. Композиции и способы
Одним из объектов изобретения являются, в частности, антитела к латентному TGF-бета 1 и варианты их применения. В некоторых вариантах осуществления изобретения предложены антитела, которые связываются с TGF-бета 1. Антитела, предлагаемые в изобретении, можно применять, например, для диагностирования или лечения фиброза, предпочтительно фиброза миокарда, фиброза легких, фиброза печени, фиброза почки, фиброза кожи, фиброза глаза и миелофиброза. Антитела, предлагаемые в изобретении, можно применять, например, для диагностирования или лечения рака. Примеры рака включают (но не ограничиваясь только ими) карциному, лимфому (например, лимфому Ходжкина и неходжкинскую лимфому), бластому, саркому и лейкоз. Более конкретные примеры указанных раков включают плоскоклеточный рак, мелкоклеточный рак легкого, немелкоклеточный рак легкого, аденокарцирному легкого, плоскоклеточный рак легкого, перитонеальный рак, печеночноклеточный рак, рак желудочно-кишечной системы, рак поджелудочной железы, глимому, рак шейки матки, рак яичника, рак печени, рак мочевого пузыря, гепатому, рак молочной железы, рак ободочной кишки, колоректальный рак, карциному эндометрия или матки, карциному слюнных желез, рак почки, рак печени, рак предстательной железы, рак вульвы, рак щитовидной железы, печеночную карциному, лейкоз и другие лимфопролиферативные нарушения, а также различные типы рака головы и шеи.
А. Примеры антител к латентному TGF-бета 1
Одним из объектов изобретения являются выделенные антитела, которые связываются с латентным TGF-бета 1. В других вариантах осуществления изобретения антитело к латентному TGF-бета 1 связывается с областью ассоциированного с латентностью белка (LAP) латентного TGF-бета 1. Приведенная в качестве примера LAP-область содержит аминокислоты 30-278 человеческого препробелка TGF-бета 1 (SEQ ID NO: 1). LAP является компонентом латентного TGF-бета 1, что описано выше. В некоторых вариантах осуществления изобретения связывание антитела к латентному TGF-бета 1 с латентным TGF-бета 1 характеризуется константой диссоциации (KD) 10-8 нМ или менее 10-9 нМ или менее или 10-10 нМ или менее.
Согласно одному из объектов изобретения антитело к латентному TGF-бета 1 связывается с латентным TGF-бета 1, формирующим LLC, и/или латентным TGF-бета 1, формирующим комплекс с GARP или LRRC33. В некоторых вариантах осуществления изобретения антитело к латентному TGF-бета 1 связывается с находящимся на клеточной поверхности латентным TGF-бета 1, который представляет собой латентный TGF-бета 1, ассоциированный с внеклеточным матриксом (ЕСМ) на клеточной поверхности. Согласно другому объекту изобретения антитело к латентному TGF-бета 1 связывается с латентным TGF-бета 1, когда LAP-область латентного TGF-бета 1 не связана с LTBP, формируя малый латентный комплекс (SLC). В некоторых вариантах осуществления изобретения SLC находятся в растворимой форме. В некоторых вариантах осуществления изобретения связывание антитела к латентному TGF-бета 1 с латентным TGF-бета 1 (находящийся на клеточной поверхности латентный TGF-бета 1, LLC или SLC) характеризуется константой диссоциации (KD) 10-8нМ или менее 10-9нМ или менее или 10-10нМ или менее.
Согласно одному из объектов изобретения антитело к латентному TGF-бета 1 ингибирует активацию латентного TGF-бета 1. В контексте настоящего описания понятие «активация» латентного TGF-бета 1 относится к любому процессу, в котором зрелый TGF-бета освобождается от LAP, являющегося компонентом латентного TGF-бета 1. Активацию латентного TGF-бета 1 можно оценивать, например, измеряя уровень зрелого TGF-бета 1 и/или измеряя активность зрелого TGF-бета 1, с помощью различных методик, известных в данной области или указанных в настоящем описании. В некоторых вариантах осуществления изобретения антитело к латентному TGF-бета 1 ингибирует высвобождение зрелого TGF-бета 1 из латентного TGF-бета 1. Как описано выше, известны данные о том, что зрелый TGF-бета 1 высвобождается из латентного TGF-бета 1 с помощью активаторов, таких как протеазы, интегрины и другие не относящиеся к протеазам активаторы. Примеры протеаз, которые активируют латентный TGF-бета 1, включают (но не ограничиваясь только ими) плазмин (PLN), прекалликреин (PLK), матриксную металлопротеиназу (ММР) 2 и ММР9. В некоторых вариантах осуществления изобретения антитело к латентному TGF-бета 1 ингибирует опосредуемое протеазой и/или опосредуемое интегрином высвобождение зрелого TGF-бета 1 из латентного TGF-бета 1. Как описано выше, протеазы расщепляют LAP-область латентного TGF-бета 1, что приводит к высвобождению зрелого TGF-бета 1. В некоторых вариантах осуществления изобретения сайты расщепления PLN и/или PLK локализованы внутри фрагмента, состоящего из аминокислот 56-59 полипептида LAP.
Согласно одному из объектов изобретения антитело к латентному TGF-бета 1 ингибирует опосредуемое протеазой высвобождение зрелого TGF-бета 1 из латентного TGF-бета 1 без ингибирования опосредуемого протеазой расщепления LAP-участка латентного TGF-бета 1. В некоторых вариантах осуществления изобретения антитело к латентному TGF-бета 1 ингибирует опосредуемое протеазой высвобождение зрелого TGF-бета 1 из латентного TGF-бета 1 и позволяет протеазе расщеплять LAP-область, при этом антитело к латентному TGF-бета 1 связывается с LAP-областью латентного TGF-бета 1. В некоторых вариантах осуществления изобретения антитело к латентному TGF-бета 1 не блокирует доступ протеазы к латентному TGF-бета 1, прежде всего к сайтам расщепления PLN и/или PLK. В других вариантах осуществления изобретения антитело к латентному TGF-бета 1 не связывается с сайтами расщепления протеазами LAP-участка латентного TGF-бета 1, прежде всего с сайтами расщепления PLN и/или PLK.
В некоторых вариантах осуществления изобретения антитело к латентному TGF-бета 1, которое ингибирует опосредуемое протеазой высвобождение зрелого TGF-бета 1 из латентного TGF-бета 1, представляет собой антитело, которое (I) ингибирует расщепление LAP-области, опосредуемое одной или несколькими протеазами, но (II) не ингибирует расщепление LAP-области, опосредуемое другими протеазами. Например, антитело к латентному TGF-бета 1 (1-I) ингибирует опосредуемое ММР2 и/или ММР9 высвобождение зрелого TGF-бета 1 путем ингибирования опосредуемого ММР2 и/или ММР9 расщепление LAP-участка латентного TGF-бета 1, и (1-II) ингибирует опосредуемое PLN и/или PLK высвобождение зрелого TGF-бета 1, не ингибируя опосредуемое PLN и/или PLK расщепление LAP-участка латентного TGF-бета 1. Альтернативно этому, антитело к латентному TGF-бета 1 (2-I) ингибирует опосредуемое PLN и/или PLK высвобождение зрелого TGF-бета 1 путем ингибирования опосредуемого PLN и/или PLK расщепления LAP-участка латентного TGF-бета 1, и (2-II) ингибирует опосредуемое ММР2 и/или ММР9 высвобождение зрелого TGF-бета 1, не ингибируя опосредуемое ММР2 и/или ММР9 расщепление LAP-участка латентного TGF-бета 1. Альтернативно этому, антитело к латентному TGF-бета 1 (3-I) ингибирует опосредуемое PLN и/или PLK высвобождение зрелого TGF-бета 1, не ингибируя опосредуемое PLN и/или PLK расщепление LAP-участка латентного TGF-бета 1, и (3-II) ингибирует опосредуемое ММР2 и/или ММР9 высвобождение зрелого TGF-бета 1, не ингибируя опосредуемое ММР2 и/или ММР9 расщепление LAP-участка латентного TGF-бета 1.
В некоторых вариантах осуществления изобретения антитела, «которые ингибируют активацию латентного TGF-бета 1», включают антитела, которые вызывают уменьшение активации TGF-бета 1 по меньшей мере на 5%, 10%, 15%, 20%, 25%, 30%, 35% или 40% или более. В других вариантах осуществления изобретения антитела, «которые ингибируют опосредуемое протеазой высвобождение зрелого TGF-бета 1 из латентного TGF-бета 1», включают антитела, которые вызывают уменьшение опосредуемого протеазой высвобождения зрелого TGF-бета 1 из латентного TGF-бета 1 по меньшей мере на 5%, 10%, 15%, 20%, 25%, 30%, 35% или 40% или более. В других вариантах осуществления изобретения антитела, которые ингибируют опосредуемое протеазой высвобождение зрелого TGF-бета 1 из латентного TGF-бета 1, «не ингибируя опосредуемое протеазой расщепление LAP-области латентного TGF-бета 1», вызывают уменьшение опосредуемого протеазой расщепления LAP-области латентного TGF-бета 1 на 50% или менее, на 45% или менее, на 40% или менее, на 35% или менее, на 30% или менее, на 25% или менее, на 20% или менее, на 15% или менее, на 10% или менее, на 5% или менее.
В некоторых вариантах осуществления изобретения антитело к латентному TGF-бета 1 стабилизирует структуру LAP-области латентного TGF-бета 1, не ингибируя опосредуемое протеазой расщепление LAP-области латентногоt TGF-бета 1. В контексте настоящего описания в ситуации, когда антитело к латентному TGF-бета 1 «стабилизирует» структуру LAP-области, LAP-область, связанная антителом к латентному TGF-бета 1, сохраняет определенную структуру, из которой зрелый TGF-бета 1 не может высвобождаться. В других вариантах осуществления изобретения латентный TGF-бета 1, стабилизированный антителом к латентному TGF-бета 1, может активироваться интегрином (предпочтительно интегрином альфа V бета 8 и/или интегрином альфа V бета 6). В некоторых вариантах осуществления изобретения LAP-область, стабилизированная антителом к латентному TGF-бета 1, либо расщеплена, либо не расщеплена протеазой. В некоторых вариантах осуществления изобретения антитело к латентному TGF-бета 1 стабилизирует структуру LAP-области латентного TGF-бета 1 и позволяет протеазе расщеплять LAP-область, а (при этом) антитело к латентному TGF-бета 1 связывается с LAP-областью латентного TGF-бета 1. В некоторых вариантах осуществления изобретения антитело к латентному TGF-бета 1 стабилизирует структуру LAP-области латентного TGF-бета 1, не блокируя доступ протеазы к латентному TGF-бета 1, прежде всего к сайтам расщепления PLN и/или PLK. В некоторых вариантах осуществления изобретения антитело к латентному TGF-бета 1 стабилизирует структуру LAP-области латентного TGF-бета 1, не блокируя доступ протеазы к латентному TGF-бета 1, прежде всего к сайтам расщепления ММР2 и/или ММР9.
Согласно одному из объектов изобретения антитело к латентному TGF-бета 1 не связывается со зрелым TGF-бета 1. В некоторых вариантах осуществления изобретения антитело к латентному TGF-бета 1 связывается с латентным TGF-бета 1 с более высокой связывающей активностью, чем со зрелым TGF-бета 1. В некоторых вариантах осуществления изобретения антитела, предлагаемые в настоящем изобретении, связываются с латентным TGF-бета 1 с более высокой по меньшей мере в 2, 3, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 200, 400, 1000, 10000 раз или более связывающей активностью, чем со зрелым TGF-бета 1.
Согласно одному из объектов изобретения антитело к латентному TGF-бета 1 не ингибирует или не ингибирует существенно опосредуемую интегрином активацию TGF-бета 1, т.е. опосредуемое интегрином высвобождение зрелого TGF-бета 1 из латентного TGF-бета 1. Предпочтительно интегрин в настоящем описании представляет собой интегрин альфа V бета 8 и/или интегрин альфа V бета 6. В некоторых вариантах осуществления изобретения антитела, «которые не ингибирует или не ингибирует существенно опосредуемую интегрином активацию TGF-бета 1» включают антитела, которые уменьшают на 50% или менее, на 45% или менее, на 40% или менее, на 35% или менее, на 30% или менее, на 25% или менее, на, 20% или менее, на 15% или менее, на 10% или менее, на 5% или менее опосредуемую интегрином активацию TGF-бета 1, т.е. опосредуемое интегрином высвобождение зрелого TGF-бета 1 из латентного TGF-бета 1.
Согласно одному из объектов изобретения антитело к латентному TGF-бета 1 вызывает пониженные или малочисленные токсические и/или побочные действия, ассоциированные с антагонистами к TGF-бета. В некоторых вариантах осуществления изобретения антитела к латентному TGF-бета 1, которые указаны в настоящем описании, имеют улучшенный профиль безопасности-эффективности по сравнению с агентами, которые обладают активностью в отношении зрелого TGF-бета 1, или агентом, который обладает активностью в отношении латентного TGF-бета 1, но ингибирует опосредуемую как протеазой, так и интегрином активацию латентного TGF-бета 1. В некоторых вариантах осуществления изобретения антитело к латентному TGF-бета 1, указанное в настоящем описании, обладает пониженной кардиотоксичностью, но при этом обладает эффективностью, более высокой или сопоставимой с эффективностью антитела к зрелому TGF-бета 1. Не вдаваясь в какую-либо теорию, антитело к латентному TGF-бета 1, указанное в настоящем описании, не ингибирует или не ингибирует существенно опосредуемую интегрином активацию TGF-бета 1, и поэтому вызывает пониженные или малочисленные токсические и/или побочные действия, которые связаны с: (I) опосредуемой интегрином активацией TGF-бета 1 или (II) ингибированием передачи сигналов TGF-бета 1 в сайтах, в которых TGF-бета 1 активируется интегрином. Таким образом, антитело к латентному TGF-бета 1, указанное в настоящем описании, можно вводить индивидуумам, которые нуждаются в этом, в терапевтически эффективной дозе, не вызывая побочных действий, прежде всего кардиотоксичности. Таким образом, указанный подход должен расширить диапазон доз, при использовании которого можно обеспечивать как эффективность, так и безопасность/переносимость для пациентов. Таким образом, в изобретении предложены способы лечения заболевания, ассоциированного с передачей сигналов TGF-бета 1, включающие введение индивидууму в эффективном количестве антитела к латентному TGF-бета 1, которое не ингибирует или не ингибирует существенно опосредуемую интегрином активацию TGF-бета 1. Под объем настоящего изобретения подпадает применение антитела к латентному TGF-бета 1 для снижения токсических и/или побочных действий, ассоциированных с ингибированием TGF-бета 1 у индивидуума. В некоторых вариантах осуществления изобретения токсические и/или побочные действия могут включать токсичность в отношении сердечно-сосудистой системы, токсичность в отношении желудочно-кишечного тракта, токсичность в отношении иммунной системы, токсичность в отношении костей/хрящей, токсичность в отношении репродуктивной системы и токсичность в отношении почек. В некоторых вариантах осуществления изобретения токсичность в отношении сердечно-сосудистой системы включает (но не ограничиваясь только ими): повреждение клапана сердца, например, кровоизлияние, воспаление, дегенерацию и пролиферацию интерстициальных клеток клапанов. В некоторых вариантах осуществления изобретения токсические и/или побочные действия могут включать кровотечение. В некоторых вариантах осуществления изобретения токсические и/или побочные действия могут включать повреждения или опухоли кожи. В некоторых вариантах осуществления изобретения токсические и/или побочные действия могут включать прогрессирование опухоли.
В некоторых вариантах осуществления изобретения антитело к латентному TGF-бета 1, предлагаемое в настоящем изобретении:
связывается с латентным TGF-бета 1;
связывается с латентным TGF-бета 1, формирующим SLC; связывается с латентным TGF-бета 1, формирующим LLC; связывается с латентным TGF-бета 1, формирующим комплекс с GARP или LRRC33;
связывается с находящемся на клеточной поверхности латентным TGF-бета 1;
связывается с LAP-областью латентного TGF-бета 1; связывается с LAP;
связывается с латентным TGF-бета 1 с константной диссоциации (KD) 10-8 нМ или менее, 10-9нМ или менее или 10-10нМ или менее;
ингибирует опосредуемое протеазой высвобождение зрелого TGF-бета 1 из латентного TGF-beta 1;
не ингибирует опосредуемое протеазой расщепление LAP-области латентного TGF-бета 1;
не ингибирует или не ингибирует существенно опосредуемое интегрином высвобождение зрелого TGF-бета 1 из латентного TGF-бета 1; и/или
вызывает пониженные или малочисленные токсические и/или побочные действия, ассоциированные с антагонистами TGF-бета 1, например, антителом к зрелому TGF-бета.
В следующих вариантах осуществления изобретения антитело к латентному TGF-бета 1, предлагаемое в настоящем изобретении, представляет собой: моноклональное антитело;
человеческое, гуманизированное или химерное антитело; полноразмерное антитело IgG-типа; и/или фрагмент антитела.
Одним из объектов изобретения является антитело к латентному TGF-бета 1, содержащее по меньшей мере один, два, три, четыре, пять или шесть HVR, выбранных из:
(а) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 20;
(б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 21;
(в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 22;
(г) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 23;
(д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 24; и
(е) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 25.
Одним из объектов изобретения является антитело к латентному TGF-бета 1, содержащее по меньшей мере один, два, три, четыре, пять или шесть HVR, выбранных из:
(а) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 26;
(б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 27;
(в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 28;
(г) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 29;
(д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 30; и
(е) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 31.
Одним из объектов изобретения является антитело к латентному TGF-бета 1, содержащее по меньшей мере один, два, три, четыре, пять или шесть HVR, выбранных из:
(а) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 32;
(б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 33;
(в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 34;
(г) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 35;
(д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 36; и
(е) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 37.
Одним из объектов изобретения является антитело к латентному TGF-бета 1, содержащее по меньшей мере один, два, три, четыре, пять или шесть HVR, выбранных из:
(a) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 38;
(б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 39;
(в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 40;
(г) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 41;
(д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 42; и
(е) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 43.
Одним из объектов изобретения является антитело к латентному TGF-бета 1, содержащее:
(а) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 20;
(б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 21;
(в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 22;
(г) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 23;
(д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 24; и
(е) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 25.
Одним из объектов изобретения является антитело к латентному TGF-бета 1, содержащее:
(а) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 26;
(б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 27;
(в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 28;
(г) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 29;
(д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 30; и
(е) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 31.
Одним из объектов изобретения является антитело к латентному TGF-бета 1, содержащее:
(а) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 32;
(б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 33;
(в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 34;
(г) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 35;
(д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 36; и
(е) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 37.
Одним из объектов изобретения является антитело к латентному TGF-бета 1, содержащее:
(а) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 38;
(б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 39;
(в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 40;
(г) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 41;
(д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 42; и
(e) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 43.
Одним из объектов изобретения является антитело к латентному TGF-бета 1, которое содержит HVR-H1, HVR-H2 и HVR-H3 VH-последовательности, представленной в SEQ ID NO: 12, и HVR-L1, HVR-L2 и HVR-L3 VL-последовательности, представленной в SEQ ID NO: 13, где HVR определяют согласно (a) Chothia; (б) Кэботу; (в) MacCallum; или (г) комбинациями (а), (б) и/или (в).
Одним из объектов изобретения является антитело к латентному TGF-бета 1, которое содержит HVR-H1, HVR-H2 и HVR-H3 VH-последовательности, представленной в SEQ ID NO: 14, и HVR-L1, HVR-L2 и HVR-L3 VL-последовательности, представленной в SEQ ID NO: 15, где HVR определяют согласно (a) Chothia; (б) Кэботу; (в) MacCallum; или (г) комбинациями (а), (б) и/или (в).
Одним из объектов изобретения является антитело к латентному TGF-бета 1, которое содержит HVR-H1, HVR-H2 и HVR-H3 VH-последовательности, представленной в SEQ ID NO: 16, и HVR-L1, HVR-L2 и HVR-L3 VL-последовательности, представленной в SEQ ID NO: 17, где HVR определяют согласно (a) Chothia; (б) Кэботу; (в) MacCallum; или (г) комбинациями (а), (б) и/или (в).
Одним из объектов изобретения является антитело к латентному TGF-бета 1, которое содержит HVR-H1, HVR-H2 и HVR-H3 VH-последовательности, представленной в SEQ ID NO: 18, и HVR-L1, HVR-L2 и HVR-L3 VL-последовательности, представленной в SEQ ID NO: 19, где HVR определяют согласно (a) Chothia; (б) Кэботу; (в) MacCallum; или (г) комбинациями (а), (б) и/или (в).
В одном из указанных выше вариантов осуществления изобретения антитело к латентному TGF-бета 1 является гуманизированным. В одном из указанных выше вариантов осуществления изобретения антитело к латентному TGF-бета 1 содержит HVR, указанные в любом из приведенных выше вариантов осуществления изобретения, и дополнительно содержит человеческий акцепторный каркасный участок, например, каркасный участок человеческого иммуноглобулина или человеческий консенсусный каркасный участок.
В другом объекте изобретения антитело к латентному TGF-бета 1 содержит последовательность вариабельного домена тяжелой цепи (VH), идентичную по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% аминокислотной последовательности SEQ ID NO: 12, 14, 16 или 18. В некоторых вариантах осуществления изобретения последовательность VH, идентичная по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%, содержит замены (например, консервативные замены), инсерции или делеции относительно референс-последовательности, но антитело к латентному TGF-бета 1, содержащее указанную последовательность, сохраняет способность связываться с латентным TGF-бета 1. В некоторых вариантах осуществления изобретения в целом от 1 до 10 аминокислот заменено, встроено и/или удалено путем делеции в SEQ ID NO: 12, 14, 16 или 18. В некоторых вариантах осуществления изобретения замены, инсерции или делеции затрагивают области вне HVR (т.е. FR). Необязательно антитело к латентному TGF-бета 1 содержит последовательность VH SEQ ID NO: 12, 14, 16 или 18, включая посттрансляционные модификации указанной последовательности. В конкретном варианте осуществления изобретения VH содержит один, два или три HVR, выбранных из: (a) HVR-H1, который содержит аминокислотную последовательность, представленную в SEQ ID NO: 20, 26, 32 или 38, (б) HVR-Н2, который содержит аминокислотную последовательность, представленную в SEQ ID NO: 21, 27, 33 или 39, и (в) HVR-H3, который содержит аминокислотную последовательность, представленную в SEQ ID NO: 22, 28, 34 или 40. Посттрансляционные модификации включают (но не ограничиваясь только ими) модификацию, полученную путем пироглутамилирования глутамина или глутамата в N-концевой области тяжелой цепи или легкой цепи в пироглутамовую кислоту.
В другом объекте изобретения антитело к латентному TGF-бета 1 содержит последовательность вариабельного домена легкой цепи (VL), идентичную по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% аминокислотной последовательности SEQ ID NO: 13, 15, 17 или 19. В некоторых вариантах осуществления изобретения последовательность VL, идентичная по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%, содержит замены (например, консервативные замены), инсерции или делеции относительно референс-последовательности, но антитело к латентному TGF-бета 1, содержащее указанную последовательность, сохраняет способность связываться с латентным TGF-бета 1. В некоторых вариантах осуществления изобретения в целом от 1 до 10 аминокислот заменено, встроено и/или удалено путем делеции в SEQ ID NO: 13, 15, 17 или 19. В некоторых вариантах осуществления изобретения замены, инсерции или делеции затрагивают области вне HVR (т.е. FR). Необязательно антитело к латентному TGF-бета 1 содержит последовательность VL SEQ ID NO: 13, 15, 17 или 19, включая посттрансляционные модификации указанной последовательности. В конкретном варианте осуществления изобретения VH содержит один, два или три HVR, выбранных из: (a) HVR-L1, который содержит аминокислотную последовательность, представленную в SEQ ID NO: 23, 29, 35 или 41, (б) HVR-L2, который содержит аминокислотную последовательность, представленную в SEQ ID NO: 24, 30, 36 или 42, и (в) HVR-L3, который содержит аминокислотную последовательность, представленную в SEQ ID NO: 25, 31, 37 или 43. Посттрансляционные модификации включают (но не ограничиваясь только ими) модификацию, полученную путем пироглутамилирования глутамина или глутамата в N-концевой области тяжелой цепи или легкой цепи в пироглутамовую кислоту.
Другим объектом изобретения является антитело к латентному TGF-бета 1, где антитело содержит VH, указанную в любом из представленных выше вариантов осуществления изобретения, и VL, указанную в любом из представленных выше вариантов осуществления изобретения. В одном из вариантов осуществления изобретения антитело содержит последовательности VH и VL, представленные в SEQ ID NO: 12 и SEQ ID NO: 13 соответственно, включая пост-трансляционные модификации указанных последовательностей. Пост-трансляционные модификации включают (но не ограничиваясь только ими) модификацию, полученную путем пироглутамилирования глутамина или глутамата в N-концевой области тяжелой цепи или легкой цепи в пироглутамовую кислоту.
В одном из вариантов осуществления изобретения антитело содержит последовательности VH и VL, представленные в SEQ ID NO: 14 и SEQ ID NO: 15 соответственно, включая пост-трансляционные модификации указанных последовательностей. Пост-трансляционные модификации включают (но не ограничиваясь только ими) модификацию, полученную путем пироглутамилирования глутамина или глутамата в N-концевой области тяжелой цепи или легкой цепи в пироглутамовую кислоту.
В одном из вариантов осуществления изобретения антитело содержит последовательности VH и VL, представленные в SEQ ID NO: 16 и SEQ ID NO: 17 соответственно, включая пост-трансляционные модификации указанных последовательностей. Пост-трансляционные модификации включают (но не ограничиваясь только ими) модификацию, полученную путем пироглутамилирования глутамина или глутамата в N-концевой области тяжелой цепи или легкой цепи в пироглутамовую кислоту.
В одном из вариантов осуществления изобретения антитело содержит последовательности VH и VL, представленные в SEQ ID NO: 18 и SEQ ID NO: 19 соответственно, включая пост-трансляционные модификации указанных последовательностей. Пост-трансляционные модификации включают (но не ограничиваясь только ими) модификацию, полученную путем пироглутамилирования глутамина или глутамата в N-концевой области тяжелой цепи или легкой цепи в пироглутамовую кислоту.
Следующим объектом изобретения является антитело, которое связывается с тем же эпитопом, что и антитело к латентному TGF-бета 1, представленное в настоящем описании. Например, некоторыми вариантами осуществления изобретения является антитело, которое связывается с тем же эпитопом, что и:
(1) антитело к латентному TGF-бета 1, которое содержит:
(а) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO:20,
(б) HVR-H2, содержащий аминокислотную последовательность SEQ ID NO:21,
(в) HVR-H3, содержащий аминокислотную последовательность SEQ ID NO:22,
(г) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO:23,
(д) HVR-L2, содержащий аминокислотную последовательность SEQ ID NO:24, и
(e) HVR-L3, содержащий аминокислотную последовательность SEQ ID NO:25;
(2) антитело к латентному TGF-бета 1, которое содержит:
(а) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO:26,
(б) HVR-H2, содержащий аминокислотную последовательность SEQ ID NO:27,
(в) HVR-H3, содержащий аминокислотную последовательность SEQ ID NO:28,
(г) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO:29,
(д) HVR-L2, содержащий аминокислотную последовательность SEQ ID NO:30, и
(е) HVR-L3, содержащий аминокислотную последовательность SEQ ID NO:31;
(3) антитело к латентному TGF-бета 1, которое содержит:
(а) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO:32,
(б) HVR-H2, содержащий аминокислотную последовательность SEQ ID NO:33,
(в) HVR-H3, содержащий аминокислотную последовательность SEQ ID NO:34,
(г) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO:35,
(д) HVR-L2, содержащий аминокислотную последовательность SEQ ID NO:36, и
(е) HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 37; или
(4) антитело к латентному TGF-бета 1, которое содержит:
(а) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO:38,
(б) HVR-H2, содержащий аминокислотную последовательность SEQ ID NO:39,
(в) HVR-H3, содержащий аминокислотную последовательность SEQ ID NO:40,
(г) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO:41,
(д) HVR-L2, содержащий аминокислотную последовательность SEQ ID NO:42, и
(е) HVR-L3, содержащий аминокислотную последовательность SEQ ID NO:43.
Следующим объектом изобретения является антитело, которое связывается с латентным TGF-бета 1 человека, обезьян, мышей и/или крыс. Некоторыми вариантами осуществления изобретения является антитело, которое связывается с латентным TGF-бета 1 человека, обезьян и мышей. Некоторыми вариантами осуществления изобретения является антитело, которое связывается с латентным человеческим, обезьяньим и мышиным TGF-бета 1, образующим SLC. Некоторыми вариантами осуществления изобретения является антитело, которое связывается с латентным человеческим, обезьяньим и мышиным TGF-бета 1, образующим LLC. Некоторыми вариантами осуществления изобретения является антитело, которое связывается с латентным человеческим, обезьяньим и мышиным TGF-бета 1, образующим LLC. Некоторыми вариантами осуществления изобретения является антитело, которое связывается с латентным человеческим, обезьяньим и мышиным TGF-бета 1, образующим комплекс с GARP или LRRC33. Некоторыми вариантами осуществления изобретения является антитело, которое связывается с находящимся на клеточной поверхности латентным человеческим, обезьяньим и мышиным TGF-бета 1.
Следующим объектом изобретения является антитело, которое связывается с тем же эпитопом, что и любое из антител к латентному TGF-бета 1, представленное в настоящем описании. Эпитоп может находиться на TGF-бета 1 человека, обезьян, мышей и/или крыс. Например, некоторыми вариантами осуществления изобретения является антитело, которое ввязывается с тем же эпитопом, что и референс-антитело, где референс-антитело представляет собой:
(1) антитело к латентному TGF-бета 1, которое содержит:
(a) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO:20,
(б) HVR-H2, содержащий аминокислотную последовательность SEQ ID NO:21,
(в) HVR-H3, содержащий аминокислотную последовательность SEQ ID NO:22,
(г) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO:23,
(д) HVR-L2, содержащий аминокислотную последовательность SEQ ID NO:24, и
(е) HVR-L3, содержащий аминокислотную последовательность SEQ ID NO:25;
(2) антитело к латентному TGF-бета 1, которое содержит:
(а) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO:26,
(б) HVR-H2, содержащий аминокислотную последовательность SEQ ID NO:27,
(в) HVR-H3, содержащий аминокислотную последовательность SEQ ID NO:28,
(г) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO:29,
(д) HVR-L2, содержащий аминокислотную последовательность SEQ ID NO:30, и
(е) HVR-L3, содержащий аминокислотную последовательность SEQ ID NO:31;
(3) антитело к латентному TGF-бета 1, которое содержит:
(а) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO:32,
(б) HVR-H2, содержащий аминокислотную последовательность SEQ ID NO:33,
(в) HVR-H3, содержащий аминокислотную последовательность SEQ ID NO:34,
(г) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO:35,
(д) HVR-L2, содержащий аминокислотную последовательность SEQ ID NO:36, и
(е) HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 37; или
(4) антитело к латентному TGF-бета 1, которое содержит:
(а) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO:38,
(б) HVR-H2, содержащий аминокислотную последовательность SEQ ID NO:39,
(в) HVR-H3, содержащий аминокислотную последовательность SEQ ID NO:40,
(г) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO:41,
(д) HVR-L2, содержащий аминокислотную последовательность SEQ ID NO:42, и
(е) HVR-L3, содержащий аминокислотную последовательность SEQ ID NO:43.
Следующим объектом изобретения является антитело, которое конкурирует с антителом к латентному TGF-бета 1, представленным в настоящем описании, за связывание с TGF-бета 1 человека, обезьян, мышей и/или крыс. Например, некоторыми вариантами осуществления изобретения является антитело, которое конкурирует за связывание с TGF-бета 1 человека, обезьян, мышей и/или крыс с:
(1) антителом к латентному TGF-бета 1, которое содержит:
(а) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO:20,
(б) HVR-H2, содержащий аминокислотную последовательность SEQ ID NO:21,
(в) HVR-H3, содержащий аминокислотную последовательность SEQ ID NO:22,
(г) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO:23,
(д) HVR-L2, содержащий аминокислотную последовательность SEQ ID NO:24, и
(e) HVR-L3, содержащий аминокислотную последовательность SEQ ID NO:25;
(2) антителом к латентному TGF-бета 1, которое содержит:
(а) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO:26,
(б) HVR-H2, содержащий аминокислотную последовательность SEQ ID NO:27,
(в) HVR-H3, содержащий аминокислотную последовательность SEQ ID NO:28,
(г) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO:29,
(д) HVR-L2, содержащий аминокислотную последовательность SEQ ID NO:30, и
(е) HVR-L3, содержащий аминокислотную последовательность SEQ ID NO:31;
(3) антителом к латентному TGF-бета 1, которое содержит:
(а) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO:32,
(б) HVR-H2, содержащий аминокислотную последовательность SEQ ID NO:33,
(в) HVR-H3, содержащий аминокислотную последовательность SEQ ID NO:34,
(г) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO:35,
(д) HVR-L2, содержащий аминокислотную последовательность SEQ ID NO:36, и
(е) HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 37; или
(4) антителом к латентному TGF-бета 1, которое содержит:
(а) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO:38,
(б) HVR-H2, содержащий аминокислотную последовательность SEQ ID NO:39,
(в) HVR-H3, содержащий аминокислотную последовательность SEQ ID NO:40,
(г) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO:41,
(д) HVR-L2, содержащий аминокислотную последовательность SEQ ID NO:42, и
(е) HVR-L3, содержащий аминокислотную последовательность SEQ ID NO:43.
Согласно следующему объекту изобретения антитело к латентному TGF-бета 1, указанное в любом из приведенных выше вариантов осуществления изобретения, представляет собой моноклональное антитело, включая химерное, гуманизированное или человеческое антитело. В одном из вариантов осуществления изобретения антитело к латентному TGF-бета 1 представляет собой фрагмент антитела, например, Fv, Fab, Fab', scFv, димерное антитело или F(ab')2-фрагмент антитела. В другом варианте осуществления изобретения антитело представляет собой полноразмерное антитело, например, интактное антитело IgG1-, IgG2-, IgG3- или IgG4-подкласса или антитело другого класса или изотипа, указанного в настоящем описании. В следующем объекте изобретения антитело к латентному TGF-бета 1 включает также любую антигенсвязывающую молекулу, которая содержит структуру вариабельной тяжелой цепи и/или вариабельной легкой цепи иммуноглобулина.
В следующем объекте изобретения антитело к латентному TGF-бета 1, указанное в любом из приведенных выше вариантов осуществления изобретения, может обладать любыми особенностями, индивидуально или в комбинации, описанными ниже в разделах 1-7:
1. Связывающая активность антител
В некоторых вариантах осуществления изобретения антитело, указанное в настоящем описании, имеет величину константы диссоциации (KD), составляющую 1 мкМ или менее, 100нМ или менее, 10нМ или менее, 1нМ или менее, 0,1нМ или менее, 0,01нМ или менее, или 0,001нМ или менее (например 10-8 М или менее, например, от 10-8 М до 10-13 М, например, от 10-9 М до 10-13 М).
В одном из вариантов осуществления связывающую активность антитела измеряют с помощью анализа связывания несущего радиоактивную метку антигена (РИА) и выражают в виде KD. В одном из вариантов осуществления изобретения РИА осуществляют с использованием Fab-версии представляющего интерес антитела и его антигена. Например, измеряют в растворе связывающую активность Fab с антигеном путем уравновешивания Fab минимальной концентрацией 125I-меченного антигена в присутствии полученных титрованием разведений немеченого антигена, осуществляя затем «захват» связанного антигена на сенсибилизированном антителом к Fab планшете (см., например, Chen и др., J. Mol Biol 293, 1999, cc. 865-881). Для обеспечения условий, необходимых для проведения анализа, многолуночные планшеты MICROTITER® (фирма Thermo Scientific) сенсибилизируют в течение ночи захватывающим антителом к Fab (фирма Cappel Labs) в концентрации 5 мкг/мл в 50 мМ карбонате натрия (рН 9,6) и затем блокируют с помощью 2% (мас./об.) бычьего сывороточного альбумина в ЗФР в течение 2-5 ч при комнатной температуре (примерно 23°С). В неадсорбирующем планшете (фирма Nunc, №269620) смешивают взятый в концентрации 100 или 26пМ меченный с помощью 125I антиген с серийными разведениями представляющего интерес Fab (например, пригодного для оценки антитела к VEGF, Fab-12, см. Presta и др., Cancer Res. 57, 1997, cc. 4593-4599). Затем представляющий интерес Fab инкубируют в течение ночи; однако инкубацию можно осуществлять в течение более продолжительного периода времени (например, в течение 65 ч) для того, чтобы гарантировать достижение равновесия. После этого смеси переносят в подготовленный для захвата планшет и инкубируют при комнатной температуре (например, в течение 1 ч). Затем раствор удаляют и планшет промывают восемь раз 0,1% полисорбатом 20 (TWEEN-20®) в ЗФР. После просушки планшетов в них добавляют по 150 мкл/лунку сцинтилляционной жидкости ((MICROSCINT-20™; фирма Packard) и осуществляют считывание планшетов с помощью гамма-счетчика TOPCOUNT™ (фирма Packard) в течение 10 мин. Для осуществления анализа связывания в условиях конкуренции выбирают концентрации каждого Fab, обеспечивающие уровень связывания, составляющий менее или равный 20% от максимального.
В одном из вариантов осуществления изобретения для оценки связывающей активности антитела используют метод на основе «захвата» лиганда, например, с помощью устройства BIACORE® Т200 или BIACORE® 4000 (фирма GE Healthcare, Уппсала, Швеция), при использовании которого принцип измерения основан на методах анализа поверхностного плазмонного резонанса. Для управления устройством применяют программное обеспечение BIACORE® Control. В одном из вариантов осуществления изобретения применяют набор для аминного сочетания (фирма GE Healthcare, Уппсала, Швеция) согласно инструкциям производителя для того, чтобы позволять молекуле, например, антителу к метке, антителу к IgG, белку А и т.д., фиксированной на сенсорном чипе (фирма GE Healthcare, Уппсала, Швеция), покрытом карбоксиметилдекстраном, захватывать лиганд. Захватывающую лиганд молекулу разводят 10 мМ раствором ацетата натрия при приемлемом значении рН и инъецируют, используя соответствующую скорость потока и соответствующую продолжительность инъекции. Измерения связывающей активности осуществляют с использованием содержащего 0,05% полисорбата 20 (другое название Tween®-20) буфера в качестве буфера для измерения при скорости потока 10-30 мкл/мин и температуре измерения предпочтительно 25°С или 37°С. Для измерения, осуществляемого с использованием антитела, захваченного с помощью захватывающей лиганд молекулы в качестве лиганда, антитело инъецируют так, чтобы обеспечивать захват требуемого количества антитела, а затем инъецируют серийное разведение антигена и/или Fc-рецептора (аналит), приготовленного с использованием буфера для измерения. Для измерения, осуществляемого с использованием антигена и/или Fc-рецептора, захваченного с помощью захватывающей лиганд молекулы в качестве лиганда, антиген и/или Fc-рецептор инъецируют так, чтобы обеспечивать захват их требуемого количества, а затем инъецируют серийное разведение антитела (аналита), приготовленного с использованием буфера для измерения.
В одном из вариантов осуществления результаты измерений анализируют с использованием программного обеспечения BIACORE® Evaluation. Кинетические параметры рассчитывают путем одновременной аппроксимации сенсограмм ассоциации и диссоциации с использованием модели связывания 1:1, что позволяет рассчитывать скорость реакции ассоциации (kon или ka), скорость реакции диссоциациие (koff или kd) и константу равновесия реакции диссоциации (KD). В случае слабой связывающей активности, например, в тех случаях, когда диссоциация является быстрой и кинетические параметры трудно рассчитывать, для определения константы равновесия реакции диссоциации (KD) можно применять модель стационарного состояния. Дополнительные параметры, касающиеся связывающей активности, т.е. «количество аналита, связывающееся с единицей количества лиганда», можно рассчитывать путем деления количества аналита (резонансная единица: RU) в конкретной концентрации на количество захваченного лиганда.
2. Фрагменты антител
В некоторых вариантах осуществления изобретения антитело, указанное в настоящем описании, представляет собой фрагмент антитела. Фрагменты антител включают (но не ограничиваясь только ими) такие фрагменты как Fab, Fab', Fab'-SH, F(ab')2, Fv и scFv и другие описанные ниже фрагменты. Обзор некоторых фрагментов антител см., у Hudson и др., Nat. Med. 9, 2003, cc. 129-134. Обзор scFv-фрагментов см., например, у Plueckthun в: The Pharmacology of Monoclonal Antibodies, под ред. Rosenburg и Moore, изд-во Springer, New York, т.113, 1994, cc. 269-315; см. также WO 93/16185; и US №№5571894 и 5587458. Обсуждение, касающееся Fab- и Е(ab')2-фрагментов, которые содержат остатки эпитопа, связывающегося с рецептором спасения, и обладают удлиненным временем полужизни in vivo, см. в US №5869046.
Димерные антитела представляют собой фрагменты антител с двумя антигенсвязывающими сайтами, которые могут быть двухвалентными или биспецифическими (см., например, ЕР 0404097; WO 1993/01161; Hudson и др., Nat. Med. 9, 2003, cc. 129-134; и Holliger и др., Proc. Natl. Acad. Sci. USA 90, 1993, cc. 6444-6448). Тримерные и тетрамерные антитела описаны также у Hudson и др., Nat. Med. 9, 2003, cc. 129-134).
Однодоменные антитела представляют собой фрагменты антител, содержащие весь вариабельный домен тяжелой цепи или его часть, или весь вариабельный домен легкой цепи антитела или его часть. В некоторых вариантах осуществления изобретения однодоменное антитело представляет собой человеческое однодоменное антитело (фирма Domantis, Inc., Уолтхэм, шт. Массачусетс; см., например, US №6248516 В1).
Фрагменты антител можно создавать различными методиками, включая (но не ограничиваясь только ими) протеолитическое расщепление интактного антитела, а также получать с помощью рекомбинантных клеток-хозяев (например, Е. coli или фаг), указанных в настоящем описании.
Настоящее изобретение относится также к антигенсвязывающим молекулам, связывающимся с TGF-бета 1, которые включают (но не ограничиваясь ими), например, минитела (низкомолекулярные антитела) и каркасные белки. В настоящем изобретении приемлемым является любой каркасный белок, если он представляет собой пептид, имеющий стабильную трехмерную структуру и обладающий способностью связываться по меньшей мере с антигеном. Указанные пептиды включают, например, фрагменты вариабельных областей антитела, фибронектин, домен белка А, А-домен LDL-рецептора, липокалин и другие молекулы, описанные у Nygren и др. (Current Opinion in Structural Biology, 7, 1997, cc. 463-469; Journal of Immunol Methods, 290, 2004, cc. 3-28), Binz и др. (Nature Biotech. 23, 2005, cc. 1257-1266), и Hosse и др. (Protein Science, 15, 2006, cc. 14-27). При ссылке на такое антитело, в контексте настоящего описания, например, понятие «антитело к латентному TGF-бета 1» можно заменять на понятие «антигенсвязывающая молекула к латентному TGF-бета 1».
3. Химерные и гуманизированные антитела
В некоторых вариантах осуществления изобретения антитело, указанное в настоящем описании, представляет собой химерное антитело. Некоторые химерные антитела описаны, например, в US №4816567; и у Morrison и др., Proc. Natl. Acad. Sci. USA 81, 1984, cc. 6851-6855. В одном из примеров химерное антитело содержит нечеловеческую вариабельную область (например, вариабельную область из антитела мышей, крыс, хомяков, кроликов или приматов кроме человека, таких как обезьяны) и человеческую константную область. В другом примере химерное антитело представляет собой антитело «переключенного класса», класс или подкласс которого был изменен относительно родительского антитела. Химерные антитела включают их антигенсвязывающие фрагменты.
В некоторых вариантах осуществления изобретения химерное антитело представляет собой гуманизированное антитело. Как правило, нечеловеческое антитело гуманизируют для снижения иммуногенности для людей, сохраняя при этом специфичность и связывающую активность родительского нечеловеческого антитела. Как правило, гуманизированное антитело содержит один или несколько вариабельных доменов, в которых HVR, например, CDR (или их участки), имеют происхождение из нечеловеческого антитела, a FR (или их участки) имеют происхождение из последовательностей человеческого антитела. Гуманизированное антитело необязательно может содержать также по меньшей мере часть человеческой константной области. В некоторых вариантах осуществления изобретения некоторые остатки FR в гуманизированном антителе заменены на соответствующие остатки из нечеловеческого антитела (например, антитела, из которого имеют происхождение остатки HVR), например, для сохранения или улучшения специфичности или связывающей активности антитела.
Сведения о гуманизированных антителах и методах их создания обобщены, например, у Almagro и Fransson, Front. Biosci. 13, 2008, cc. 1619-1633, и описаны также у Riechmann и др., Nature 332, 1988, cc. 323-329; Queen и др., Proc. Natl. Acad. Sci. USA 86, 1989, cc. 10029-10033; US №№5821337, 7527791, 6982321 и 7087409; Kashmiri и др., Methods 36, 2005, cc. 25-34 (описание трансплантации SDR (a-CDR)); Padlan, Mol. Immunol. 28, 1991, cc. 489-498 (описание «нанесения нового покрытия»); Dall'Acqua и др., Methods 36, 2005, cc. 43-60 (описание «перестановки FR»); и Osbourn и др., Methods 36, 2005, cc. 61-68 и Klimka и др., Br. J. Cancer 83, 2000, cc. 252-260 (описание подхода «направленной селекции» для перестановки FR).
Человеческие каркасные участки, которые можно применять для гуманизации, включают (но не ограничиваясь только ими): каркасные участки, выбранные с использованием методов «наилучшего подбора» (см., например, Sims и др., J. Immunol. 151, 1993, cc. 2296-2308; каркасные участки, имеющие происхождение из консенсусной последовательности конкретной подгруппы вариабельных областей легких и тяжелых цепей человеческих антител (см., например, Carter и др., Proc. Natl. Acad. Sci. USA 89, 1992, cc. 4285-4289 и Presta и др., J. Immunol. 151, 1993, cc. 2623-2632); человеческие зрелые (с соматическими мутациями) каркасные области или каркасные области человеческой зародышевой линии (см., например, Almagro и Fransson, Front. Biosci. 13, 2008, с. 1619-1633); и каркасные участки, полученные в результате скрининга FR-библиотек (см., например, Васа и др., J. Biol. Chem. 272, 1997, cc. 10678-10684 и Rosok и др., J. Biol. Chem. 271, 1996, cc. 22611-22618). 4. Человеческие антитела
В некоторых вариантах осуществления изобретения антитело, указанное в настоящем описании, представляет собой человеческое антитело. Человеческие антитела можно получать, используя различные методики, известные в данной области. Человеческие антитела описаны в целом у van Dijk и van de Winkel, Curr. Opin. Pharmacol. 5, 2001, cc. 368-374 и Lonberg, Curr. Opin. Immunol. 20, 2008, cc. 450-459.
Человеческие антитела можно получать путем введения иммуногена трансгенному животному, модифицированному таким образом, чтобы оно продуцировало интактные человеческие антитела или интактные антитела с человеческими вариабельными областями в ответ на контрольное заражение антигеном. Указанные животные, как правило, содержат все или часть локусов человеческого иммуноглобулина вместо эндогенных иммуноглобулиновых локусов, которые либо присутствуют вне хромосом, либо интегрированы произвольно в хромосомы животного. У таких трансгенных мышей эндогенные иммуноглобулиновые локусы, как правило, инактивированы. Обзор методов получения человеческих антител в трансгенных животных см. у Lonberg, Nat. Biotech. 23, 2005, cc. 1117-1125 (см., например, также в US №6075181 и US №6150584 описание технологии XENOMOUSE™; в US №5770429 описание технологии HUMAB®; в US №7041870 описание технологии К-М MOUSE® и в публикации заявки на патент США 2007/0061900 описание технологии VELOCIMOUSE®). Человеческие вариабельные области из интактных антител, полученных с помощью указанных животных, можно дополнительно модифицировать, например, путем объединения с другой человеческой константной областью.
Человеческие антитела можно создавать также с помощью методов на основе гибридом. Описаны клеточные линии человеческой миеломы и мышиной-человеческой гетеромиеломы, предназначенные для получения человеческих моноклональных антител (см., например, Kozbor, J. Immunol. 133, 1984, cc. 3001-3005; Brodeur и др., Monoclonal Antibody Production Techniques and Applications, изд-во Marcel Dekker, Inc., New York, 1987, cc. 51-63 и Boerner и др., J. Immunol. 147, 1991, cc. 86-95). Человеческие антитела, созданные с использованием технологии на основе человеческих В-клеточных гибридом, описаны также у Li и др., Proc. Natl. Acad. Sci. USA 103, 2006, cc. 3557-3562. Дополнительные методы включают методы, описанные, например, в US №7189826 (описание получения моноклональных человеческих антител IgM-типа из клеточных линий гибридом), и у Ni, Xiandai Mianyixue 26, 2006, cc. 265-268 (описание человеческих-человеческих гибридом). Технология человеческих гибридом (технология Trioma) описана также у Vollmers и Brandlein, Histology and Histopathology 20, 2005, cc. 927-937 и Vollmers и Brandlein, Methods and Findings in Experimental and Clinical Pharmacology 27, 2005, cc. 185-191.
Человеческие антитела можно создавать также путем выделения последовательностей вариабельных доменов Fv-клонов, отобранных из фаговых дисплейных библиотек, для создания которых использовали человеческие антитела. Указанные последовательности вариабельных доменов затем можно объединять с требуемым человеческим константным доменом. Методики отбора человеческих антител из библиотек антител описаны ниже.
5. Антитела, получаемые из библиотек
Антитела, предлагаемые в изобретении, можно выделять путем скрининга комбинаторных библиотек антител с требуемой активностью или видами активности. Например, в данной области известны разнообразные методы для создания фаговых дисплейных библиотек и скрининга указанных библиотек в отношении антител, обладающих требуемыми характеристиками связывания. Указанные методы обобщены, например, у Hoogenboom и др. в: Methods in Molecular Biology 178, 2001, cc. 1-37 (под ред. O'Brien и др., изд-во Human Press, Totowa, NJ, 2001), и описаны также, например, у McCafferty и др., Nature 348, 1990, cc. 552-554; Clackson и др., Nature 352, 1991, cc. 624-628; Marks и др., J. Mol. Biol. 222, 1992, cc. 581-597; Marks и Bradbury в: Methods in Molecular Biology 248, 2003, cc. 161-175 (под ред. Lo, изд-во Human Press, Totowa, NJ, 2003); Sidhu и др., J. Mol. Biol. 338, 2004, cc. 299-310; Lee и др., J. Mol. Biol. 340, 2004, cc. 1073-1093; Fellouse, Proc. Natl. Acad. Sci. USA 101, 2004, cc. 12467-12472 и Lee и др., J. Immunol. Methods 284, 2004, cc. 119-132.
При осуществлении некоторых методов фагового дисплея популяции VH- и VL-генов клонируют по отдельности с помощью полимеразной цепной реакции (ПЦР) и рекомбинируют произвольно в фаговых библиотеках, которые затем подвергают скринингу в отношении антигенсвязывающего фага согласно методу, описанному у Winter и др., Ann. Rev. Immunol. 12, 1994, cc. 433-455. Фаг, как правило, экспонирует фрагменты антител либо в виде одноцепочечных Fv-фрагментов (scFv), либо в виде Fab-фрагментов. Библиотеки, полученные из иммунизированных источников, включают антитела, обладающие высокой активностью в отношении иммуногена, при этом отсутствует необходимость в создании гибридом. Альтернативно этому, можно клонировать необработанную («наивную») популяцию (например, из организма человека), получая один источник антител к широкому спектру чужих, а также своих антигенов без какой-либо иммунизации, что описано у Griffiths и др., EMBO J, 12, 1993, сс.725-734. И, наконец, «наивные» библиотеки можно получать также методами синтеза путем клонирования неперегруппированных сегментов V-гена из стволовых клеток и с помощью ПЦР-праймеров, содержащих случайную последовательность, для кодирования гипервариабельных CDR3-участков и для осуществления перегруппировки in vitro согласно методу, описанному у Hoogenboom и Winter, J. Mol. Biol. 227, 1992, cc. 381-388. Фаговые библиотеки человеческих антител описаны, например, в следующих патентных публикациях US №5750373 и публикациях заявок на патент США 2005/0079574, 2005/0119455, 2005/0266000, 2007/0117126, 2007/0160598, 2007/0237764, 2007/0292936 и 2009/0002360.
Антитела или фрагменты антител, выделенные из библиотек человеческих антител, в контексте настоящего описания рассматриваются как человеческие антитела или фрагменты человеческих антител.
6. Мультиспецифические антитела
В некоторых вариантах осуществления изобретения антитело, указанное в настоящем описании, представляет собой мультиспецифическое антитело, например, биспецифическое антитело. Мультиспецифические антитела представляют собой моноклональные антитела, которые имеют специфичности, связывающиеся по меньше мере с двумя различными сайтами. В некоторых вариантах осуществления изобретения одна из связывающих специфичностей обеспечивает связывание с TGF-бета 1, а другая с любым другим антигеном. В некоторых вариантах осуществления изобретения биспецифические антитела могут связываться с двумя различными эпитопами TGF-бета 1. Биспецифические антитела можно применять также для локализации цитотоксических агентов в клетках, которые экспрессируют TGF-бета 1. Биспецифические антитела можно получать в виде полноразмерных антител или фрагментов антител.
Методики создания мультиспецифических антител включают (но не ограничиваясь только ими) рекомбинантную совместную экспрессию двух пар тяжелых цепей-легких цепей иммуноглобулинов, обладающих различными специфичностями (см. Milstein и Cuello, Nature 305, 1983, cc. 537-540, WO 93/08829 и Traunecker и др., EMBO J. 10,1991, cc. 3655-3659), и технологию «knob-in-hole» (обеспечение взаимодействия по типу «выступ-во впадину», см., например, US №5731168). Мультиспецифические антитела можно получать также путем создания регулируемых электростатическими силами воздействий для получения молекул антитела с гетеродимерными Fc (WO 2009/089004); перекрестного связывания двух или большего количества антител или фрагментов (см., например, US №4676980 и Brennan и др., Science 229, 1985, cc. 81-83); применения лейциновых молний для получения биспецифических антител (см., например, Kostelny и др., J. Immunol. 148, 1992, cc. 1547-1553); применения технологии «димерных антител» для создания фрагментов биспецифических антител (см., например, Holliger и др., Proc. Natl. Acad. Sci. USA 90, 1993, cc. 6444-6448); и применения димеров одноцепочечных Fv (sFv) (см., например, Gruber и др., J. Immunol. 152, 1994, cc. 5368-5374); и получения триспецифических антител согласно методу, описанному, например, у Tutt и др., J. Immunol. 147, 1991, cc. 60-69).
Под объем настоящего изобретения подпадают также сконструированные антитела с тремя или большим количеством функциональных антигенсвязывающих сайтов, включая «антитела-осьминоги» (см., например, US 2006/0025576А1).
Антитело или его фрагмент может включать также «Fab двойного действия» или «DAF», содержащий антигенсвязывающий сайт, который связывается с TGF-бета 1, а также с другим, отличным от него антигеном (см., например, US 2008/0069820).
7. Варианты антител
Некоторые варианты осуществления изобретения относятся к вариантам аминокислотных последовательностей антител, представленных в настоящем описании. Например, может требоваться повышать связывающую активность и/или другие биологические свойства антитела. Варианты аминокислотной последовательности антитела можно получать путем интродукции соответствующих модификаций в нуклеотидную последовательность, кодирующую антитело, или путем синтеза пептидов. Указанные модификации включают, например, делеции и/или инсерции, и/или замены остатков в аминокислотных последовательностях антитела. Для получения конечной конструкции можно использовать любую комбинацию делеций, инсерций и замен, при условии, что конечная конструкция обладает требуемыми характеристиками, например, способностью связываться с антигеном. а) Полученные путем замены, инсерции и делеции варианты Некоторыми вариантами осуществления изобретения являются варианты антител, которые имеют одну или несколько аминокислотных замен. Сайты, представляющие интерес для замещающего мутагенеза, включают HVR и FR. Консервативные замены представлены ниже в таблице 1 под заголовком «предпочтительные замены». Более значимые замены представлены ниже в таблице 1 под заголовком «приведенные в качестве примера замены», и они описаны ниже со ссылкой на классы боковых цепей аминокислот.Аминокислотные замены можно интродуцировать в представляющее интерес антитело и продукты подвергать скринингу в отношении требуемой активности, например, сохранения/повышения способности к связыванию антигена, пониженной иммуногенности или повышенной ADCC или CDC.
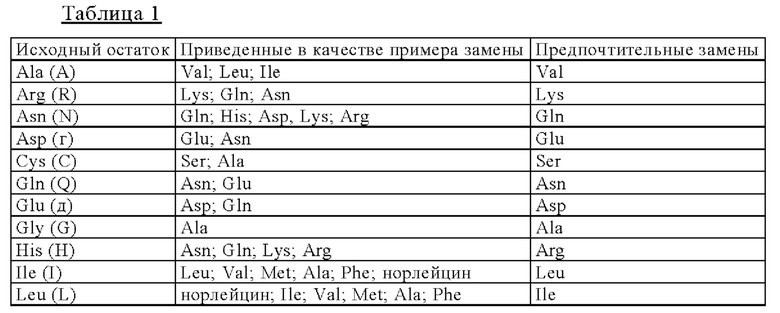
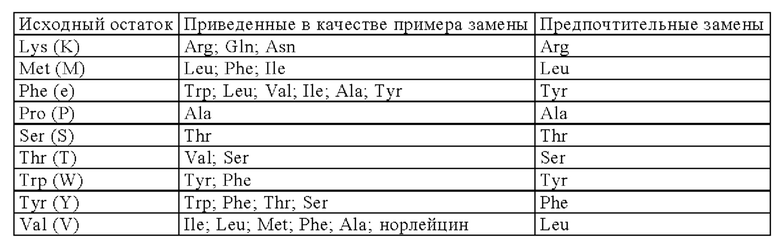
Аминокислоты можно группировать на основе общих свойств боковых цепей следующим образом:
(1) гидрофобные: норлейцин, Met, Ala, Val, Leu, Ile;
(2) нейтральные гидрофильные: Cys, Ser, Thr, Asn, Gln;
(3) кислотные: Asp, G1u;
(4) основные: His, Lys, Arg;
(5) остатки, которые влияют на ориентацию цепи: Gly, Pro;
(6) ароматические: Trp, Tyr, Phe.
Под неконсервативными заменами подразумевают замену представителя одного из указанных классов на представителя из другого класса.
Один из типов полученного в результате замены варианта включает замену одного или нескольких остатков гипервариабельного участка родительского антитела (например, гуманизированного или человеческого антитела). Как правило, полученный(ые) в результате вариант(ы), отобранный(ые) для дополнительного исследования, должен(ы) иметь модификации (например, улучшения) некоторых биологических свойств (например, повышенную связывающую активность, пониженную иммуногенность) относительно родительского антитела, и/или должен(ы) иметь практически сохраненные определенные биологические свойства родительского антитела. Примером полученного в результате замены варианта является антитело с созревшей связывающей активностью, которое можно создавать, например, с использованием технологий созревания связывающей активности на основе фагового дисплея, например, указанных в настоящем описании. В целом, метод состоит в следующем: один или несколько остатков HVR подвергают мутации и варианты антител экспонируют на фаге и подвергают скринингу в отношении конкретной биологической активности (например, связывающей активности).
Изменения (например, замены) можно осуществлять в HVR, например, для повышения связывающей активности антитела. Указанные изменения можно делать в «горячих (активных) точках» в HVR, т.е. остатках, кодируемых кодонами, которые с высокой частотой подвергаются мутации в процессе соматического созревания (см., например, Chowdhury, Methods Mol. Biol. 207, 2008, cc. 179-196), и/или в остатках, которые контактируют с антигеном, с последующим тестированием полученного варианта VH или VL в отношении связывающей активностиния. Созревание связывающей активности путем создания и повторной селекции из вторичных библиотек описано, например, у Hoogenboom и др. в: Methods in Molecular Biology 178, 2002, cc. 1-37 (под ред. O'Brien и др., изд-во Human Press, Totowa, NJ, 2001). В некоторых вариантах осуществления процесса созревания связывающей активности интродуцируют разнообразие в гены вариабельной области, выбранные для созревания, с помощью любого из широкого разнообразия методов (например, ПЦР пониженной точности, перестановка цепи или сайтнаправленный мутагенез с использованием олигонуклеотидов). Затем создают вторичную библиотеку. Далее библиотеку подвергают скринингу для идентификации любых вариантов антител с требуемой связывающей активностью. Другой метод, применяемый для интродукции разнообразия, включает подходы, мишенью которых является HVR, при которых рандомизируют несколько остатков HVR (например, одновременно 4-6 остатков). Остатки HVR, участвующие в связывании антигена, можно специфически идентифицировать, например, используя аланин-сканирующий мутагенез или моделирование. Часто мишенями являются, прежде всего CDR-H3 и CDR-L3.
В некоторых вариантах осуществления изобретения замены, инсерции или делеции могут затрагивать один или несколько HVR, если указанные изменения не снижают существенно способность антитела связываться с антигеном. Например, в HVR могут быть осуществлены консервативные изменения (например, консервативные замены, указанные в настоящем описании), которые не снижают существенно связывающую активность. Указанные изменения, например, могут находиться вне принимающих участие в контактировании с антигеном остатков HVR. В некоторых вариантах осуществления изобретения в последовательностях вариантов VH и VL, указанных выше, каждый HVR либо является неизмененным, либо содержит не более одной, двух или трех аминокислотных замен.
Ценным методом идентификации остатков или областей антитела, которые можно подвергать мутагенезу, является так называемый «аланин-сканирующий мутагенез», описанный у Cunningham и Wells, Science 244, 1989, cc. 1081-1085. При осуществлении этого метода остаток или группу остатков-мишеней (например, заряженные остатки, такие как arg, asp, his, lys и glu) идентифицируют и заменяют на нейтральную или отрицательно заряженную аминокислоту (например, на аланин или полиаланин) для решения вопроса о том, будет ли это влиять на взаимодействие антитела с антигеном. Дополнительные замены можно интродуцировать в те положения аминокислот, для которых продемонстрирована функциональная чувствительность к начальным заменам. В альтернативном или дополнительном варианте можно анализировать кристаллическую структуру комплекса антиген-антитело для идентификации точек контакта между антителом и антигеном. Указанные контактирующие остатки и соседние остатки можно рассматривать в качестве мишеней или исключать из рассмотрения в качестве кандидатов для замены. Варианты можно подвергать скринингу для решения вопроса о том, обладают ли они требуемыми свойствами.
Инсерции в аминокислотные последовательности включают амино- и/или карбоксиконцевые слияния, варьирующие по длине от одного остатка до полипептидов, содержащих сто или большее количество остатков, а также инсерции одного или нескольких аминокислотных остатков внутрь последовательности. Примером результата осуществления концевых инсерций является антитело с N-концевым метионильным остатком. Другие инсерционные варианты молекулы антитела включают слияние N- или С-концов антитела с ферментом (например, для ADEPT или полипептидом, который удлиняет время полужизни антитела в плазме.
б) Варианты гликозилирования
В некоторых вариантах осуществления изобретения антитело, представленное в настоящем описании, изменяют с целью повышения или понижения степени гликозилирования антитела. Добавление или делецию сайтов гликозилирования антитела можно легко осуществлять путем такого изменения аминокислотной последовательности, которое позволяет создавать или удалять один или несколько сайтов гликозилирования.
Если антитело содержит Fc-область, то можно изменять присоединенный к ней углевод. Нативные антитела, которые продуцируются клетками млекопитающих, как правило, содержат разветвленный биантенный олигосахарид, который, как правило, присоединен с помощью N-связи к Asn297 СН2-домена Fc-области (см., например, Wright и др., TIBTECH 15, 1997, сс.26-32). Олигосахарид может включать различные углеводы, например, маннозу, N-ацетилглюкозамин (GlcNAc), галактозу и сиаловую кислоту, а также фукозу, присоединенную к GlcNAc в «стебле» биантенной олигосахаридной структуры. В некоторых вариантах осуществления изобретения модификации олигосахарида в антителе, предлагаемом в изобретении, можно осуществлять для создания вариантов антитела с определенными улучшенными свойствами.
Одним из вариантов осуществления изобретения являются варианты антитела, имеющие углеводную структуру, в которой снижено содержание фукозы, присоединенной (прямо или косвенно) к Fc-области. Например, содержание фукозы в указанной Fc-области может составлять от 1% до 80%, от 1% до 65%, от 5% до 65% или от 20% до 40%. Содержание фукозы определяют путем расчета среднего содержания фукозы в сахарной цепи на Asn297 относительно суммы всех гликоструктур, присоединенных к Asn297 (например, комплексных, гибридных структур и структур с высоким содержанием маннозы), которое измеряют с помощью MALDI-TOF-масс-спектрометрии согласно методу, описанному, например, в WO 2008/077546. Asn297 обозначает остаток аспарагина, присутствующий примерно в положении 297 в Fc-области (EU-нумерация остатков Fc-области); однако вследствие минорных вариаций последовательности антитела Asn297 может располагаться также в положении, находящемся на расстоянии примерно ±3 аминокислоты в прямом или обратном направлении от положения 297, т.е. между положениями 294 и 300. Указанные фукозилированные варианты могут обладать улучшенной ADCC-функцией (см., например, US 2003/0157108 (Presta L.); US 2004/0093621 (фирма Kyowa Hakko Kogyo Co., Ltd). Примерами публикаций, касающихся «дефукозилированных» вариантов антител или вариантов антител «с дефицитом фукозы» являются: US 2003/0157108; WO 2000/61739; WO 2001/29246; US 2003/0115614; US 2002/0164328; US 2004/0093621; US 2004/0132140; US 2004/0110704; US 2004/0110282; US 2004/0109865; WO 2003/085119; WO 2003/084570; WO 2005/035586; WO 2005/035778; WO 2005/053742; WO 2002/031140; Okazaki и др., J. Mol. Biol. 336, 2004, cc. 1239-1249; Yamane-Ohnuki и др., Biotech. Bioeng. 87, 2004, c. 614). Примерами клеточных линий, которые обладают способностью продуцировать дефукозилированные антитела, являются Lec13 СНО-клетки с дефицитом белкового фукозилирования (Ripka и др., Arch. Biochem. Biophys. 249, 1986, cc. 533-545; US 2003/0157108 (на имя Presta L.) и WO 2004/056312 (на имя Adams и др., см., прежде всего, пример 11), и клеточные линии с «выключенным» геном, например, СНО клетки с «выключенным» геном альфа-1,6-фукозилтрансферазы, FUT8 (см., например, Yamane-Ohnuki и др., Biotech. Bioeng. 87, 2004, с. 614); Kanda и др., Biotechnol. Bioeng., 94(4), 2006, cc. 680-688; и WO 2003/085107).
Кроме того, описаны варианты антител с бисекционными олигосахаридами, например, в которых биантенный олигосахарид, присоединенный к Fc-области антитела, бисекционнируется с помощью G1cNAc. Указанные варианты антител могут отличаться пониженным фукозилированием и/или повышенной ADCC-функцией. Примеры указанных вариантов антител описаны, например, в WO 2003/011878 (на имя Jean-Mairet и др.); US №6602684 (на имя Umana и др.) и US 2005/0123546 (на имя Umana и др.). Предложены также варианты антител по меньшей мере с одним остатком галактозы в олигосахариде, присоединенном к Fc-области. Указанные варианты антител могут обладать повышенной CDC-функцией. Указанные варианты антител описаны, например, в WO 1997/30087 (на имя Patel и др.); WO 1998/58964 (на имя Raju S.); и WO 1999/22764 (на имя Raju S.).
в) Варианты Fc-области
Согласно конкретным вариантам осуществления изобретения можно интродуцировать в Fc-область антитела, представленного в настоящем описании, одну или несколько аминокислотных модификаций, создавая тем самым вариант Fc-области. Вариант Fc-области может содержать последовательность человеческой Fc-области (например, Fc-области человеческого IgG1, IgG2, IgG3 или IgG4), включающую аминокислотную модификацию (например, замену) в одном или нескольких аминокислотных положениях. В другом варианте осуществления изобретения вариант человеческой Fc-области может содержать последовательность химерной человеческой Fc-области (например, Fc-область человеческого IgG1/4 или человеческого IgG2/4) или последовательность химерной человеческой Fc-области, которая дополнительно содержит аминокислотную модификацию (например, замену) в одном или нескольких аминокислотных положениях.
Некоторые варианты осуществления изобретения относятся к варианту антитела, который обладает некоторыми, но не всеми эффекторными функциями, что делает его перспективным кандидатом для путей применения, для которых является важным время полужизни антитела in vivo, однако некоторые эффекторные функции (например, связанные с комплементом и ADCC) не являются необходимыми или являются вредными. Можно осуществлять анализы цитотоксичности in vitro и/или in vivo для подтверждения снижения/истощения CDC- и/или ADCC-активности. Например, можно осуществлять анализы связывания Fc-рецептора (FcR) для подтверждения того, что антитело не обладает способностью к связыванию с Fc-гамма R (и поэтому, вероятно, не обладает ADCC-активностью), но сохраняет способность связываться с FcRn. Первичные клетки, опосредующие ADCC, т.е. NK-клетки, экспрессируют только Fc-гамма RIII, в то время как моноциты экспрессируют Fc-гамма RI, Fc-гамма RII и Fc-гамма RIII. Данные об экспрессии FcR на гематопоэтических клетках обобщены в таблице 3 на с. 464 у Ravetch и Kinet, Annu. Rev. Immunol. 9, 1991, cc. 457-492. Примерами анализов in vitro ADCC-активности представляющей интерес молекулы являются (но не ограничиваясь только ими) анализы, описанные в US №5500362 (см., например, Hellstrom I. и др., Proc. Nat'l Acad. Sci. USA 83, 1986, cc. 7059-7063) и Hellstrom I и др., Proc. Nat'l Acad. Sci. USA 82, 1985, cc. 1499-1502 (1985); US 5821337 (см. Bruggemann M. и др., J. Exp.Med. 166, 1987, cc. 1351-1361). В альтернативном варианте можно применять нерадиоактивные методы анализа (см., например, нерадиоактивный анализ цитотоксичности АСТ1™ на основе проточной цитометрии (фирма CellTechnology, Inc. Маунтин-Вью, шт. Калифорния); и нерадиоактивный анализ цитотоксичности CytoTox 96® (фирма Promega, Мэдисон, шт. Висконсин). Пригодными для таких анализов эффекторными клетками являются мононуклеарные клетки периферической крови (РВМС) и естественные клетки-киллеры (NK). В альтернативном или дополнительном варианте ADCC-активность представляющей интерес молекулы можно оценивать in vivo, например, на животной модели, например, как описано у Clynes и др., Proc. Natl. Acad. Sci. USA 95, 1998, cc. 652-656. Можно осуществлять также анализы связывания Clq для подтверждения того, что антитело не может связываться с Clq и поэтому не обладает CDC-активностью (см., например, описание ELISA для оценки связывания Clq и С3 с в WO 2006/029879 и WO 2005/100402). Для оценки активации комплемента можно осуществлять анализ CDC (см., например, Gazzano-Santoro и др., J. Immunol. Methods 202, 1996, cc. 163-171; Cragg M.S. и др., Blood 101, 2003, cc.1045-1052 и Cragg M.S. и M.J. G1ennie, Blood 103, 2004, cc. 2738-2743). Определение FcRn-связывания и клиренса/времени полужизни in vivo можно осуществлять также с использованием методов, известных в данной области (см., например, Petkova S.B. и др., Int'l. Immunol. 18(12), 2006, cc. 1759-1769).
Антитела с пониженной эффекторной функцией включают антитела с заменой одного или нескольких остатков Fc-области, таких как 238, 265, 269, 270, 297, 327 и 329 (см., например, US №6737056). К указанным мутантам Fc относятся мутанты Fc с заменами в двух или большем количестве из следующих аминокислотных положений: 265, 269, 270, 297 и 327, включая так называемый мутант «DANA» Fc-области, несущий замену остатков 265 и 297 на аланин (US №7332581).
Описаны некоторые варианты антител с повышенной или пониженной способностью связываться с FcR (см., например, US №6737056; WO 2004/056312 и Shields и др., J. Biol. Chem. 276, 2001, cc. 6591-6604).
Согласно конкретным вариантам осуществления изобретения вариант антитела содержит Fc-область с одной или несколькими аминокислотными заменами, которые улучшают ADCC, например, замены в положениях 298, 333 и/или 334 Fc-области (EU-нумерация остатков).
В некоторых вариантах осуществления изобретения в Fc-области осуществляют изменения, которые приводят к измененной (т.е. либо повышенной, либо пониженной) способности связывать Clq и/или комплементзависимой цитотоксичности (CDC), что описано, например, в US №6194551, WO 1999/51642 и у Idusogie и др., J. Immunol. 164, 2000, cc. 4178-4184.
Антитела с удлиненным временем полужизни и повышенной способностью к связыванию с неонатальным Fc-рецептором (FcRn), который ответствен за перенос материнских IgG эмбриону (Guyer и др., J. Immunol. 117, 1976, cc. 587-593 и Kim и др., J. Immunol. 24, 1994, cc. 2429-2434), описаны в US 2005/0014934А1 (Hinton и др.). Эти антитела содержат Fc-область с одной или несколькими заменами, которые повышают связывание Fc-области с FcRn. Указанные варианты Fc-области включают варианты с заменами одного или нескольких следующих остатков Fc-области: 238, 256, 265, 272, 286, 303, 305, 307, 311, 312, 317, 340, 356, 360, 362, 376, 378, 380, 382, 413, 424 или 434, например, замену остатка 434 в Fc-области (US №7371826).
Дополнительные примеры, касающиеся других вариантов Fc-области, описаны также у Duncan и Winter, Nature 322, 1988, cc. 738-740; в US №5648260; US №5624821 и в WO 1994/29351.
г) Варианты антител, сконструированные с использованием цистеина
В некоторых вариантах осуществления изобретения может оказаться желательным создавать антитела, сконструированные с использованием цистеина, например, «тиоМАт», в которых один или несколько остатков в антителе заменены на остатки цистеина. В конкретных вариантах осуществления изобретения замененные остатки находятся в доступных сайтах антитела. Путем замены этих остатков на цистеин реакционноспособные тиольные группы помещаются в доступные сайты антитела и могут использоваться для конъюгации антитела с другими фрагментами, такими как фрагменты, представляющие собой лекарственное средство, или фрагменты, представляющие собой линкер, связанный с лекарственным средством, с созданием иммуноконъюгата, указанного ниже в настоящем описании. В некоторых вариантах осуществления изобретения любой(ые) один или несколько следующих остатков может(гут) быть заменен(ы) на цистеин: V205 (нумерация по Кэботу) легкой цепи; А118 (EU-нумерация) тяжелой цепи и S400 (EU-нумерация) Fc-области тяжелой цепи. Антитела, сконструированные с использованием цистеина, можно создавать согласно методу, например, описанному в US №7521541.
д) Производные антител
В некоторых вариантах осуществления изобретения антитело, представленное в настоящем описании, можно дополнительно модифицировать таким образом, чтобы оно содержало известные в данной области и легко доступные дополнительные небелковые фрагменты. Фрагменты, пригодные для дериватизации антитела, включают (но не ограничиваясь только ими) водорастворимые полимеры. Примерами водорастворимых полимеров являются (но не ограничиваясь только ими) полиэтиленгликоль (ПЭГ), сополимеры этиленгликоля/пропиленгликоля, карбоксиметилцеллюлоза, декстран, поливиниловый спирт, поливинилпирролидон, поли-1,3-диоксолан, поли-1,3,6-триоксан, сополимер этилена/малеинового ангидрида, полиаминокислоты (либо гомополимеры, либо статистические сополимеры) и декстран- или поли(н-винилпиролидон)полиэтиленгликоль, гомополимеры пропиленгликоля, сополимеры пропиленоксида/этиленоксида, полиоксиэтилированные полиолы (например, глицерин), поливиниловый спирт и их смеси. Полиэтиленгликольпропиональдегид может иметь преимущество при производстве благодаря его стабильности в воде. Полимер может иметь любую молекулярную массу и может быть разветвленным или неразветвленным. Количество полимеров, присоединенных к антителу, может варьироваться и, если присоединено более одного полимера, то они могут представлять собой одинаковые или различные молекулы. В целом, количество и/или тип полимеров, применяемых для дериватизации, можно определять с учетом таких особенностей (но не ограничиваясь только ими), как конкретные свойства или функции антитела, подлежащие усовершенствованию, предполагается ли применение производного антитела в терапии в определенных условиях, и т.д.
Другим вариантом осуществления изобретения являются конъюгаты антитела и небелкового фрагмента, которые можно избирательно нагревать, воздействуя на него излучением. В одном из вариантов осуществления изобретения небелковый фрагмент представляет собой углеродную нанотрубку (Kam и др., Proc. Natl. Acad. Sci. USA 102, 2005, cc. 11600-11605). Излучение может иметь любую длину волны и включает (но не ограничиваясь только ими) длины волн, которые не повреждают обычные клетки, но которые нагревают небелковый фрагмент до температуры, при которой уничтожаются клетки, ближайшие к конъюгату: антитело-небелковый фрагмент.
Б. Методы рекомбинации и композиции
Антитела можно получать, используя методы рекомбинации и композиции, например, описанные в US №4816567. Одним из вариантов осуществления изобретения является выделенная нуклеиновая кислота, кодирующая антитело к латентному TGF-бета 1, представленное в настоящем описании. Указанная нуклеиновая кислота может кодировать аминокислотную последовательность, содержащую VL, и/или аминокислотную последовательность, содержащую VH антитела (например, легкие и/или тяжелые цепи антитела). Следующим вариантом осуществления изобретения является(ются) один или несколько векторов (например, экспрессионных векторов), который(е) содержит(ат) указанную нуклеиновую кислоту. Еще одним вариантом осуществления изобретения является клетка-хозяин, которая содержит указанную нуклеиновую кислоту. В одном из указанных вариантов осуществления изобретения клетка-хозяин содержит (например, в результате трансформации указанными векторами): (1) вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VL антитела, и аминокислотную последовательность, содержащую VH антитела, или (2) первый вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VL антитела, и второй вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VH антитела. В одном из вариантов осуществления изобретения клетка-хозяин представляет собой эукариотическую клетку, например, клетку яичника китайского хомячка (СНО) или лимфоидную клетку (например, Y0-, NS0-, Sp2/0-клетку). Одним из вариантов осуществления изобретения является способ получения антитела к латентному TGF-бета 1, включающий культивирование клетки-хозяина, содержащей нуклеиновую кислоту, которая кодирует антитело, описанное выше, в условиях, пригодных для экспрессии антитела, и необязательно выделение антитела из клетки-хозяина (или среды для культивирования клетки-хозяина).
Для рекомбинантного получения антитела к латентному TGF-бета 1 нуклеиновую кислоту, кодирующую антитело, например, описанную выше, выделяют и встраивают в один или несколько векторов для дальнейшего клонирования и/или экспрессии в клетке-хозяине. Указанную нуклеиновую кислоту легко можно выделять и секвенировать с помощью общепринятых процедур (например, путем использования олигонуклеотидных зондов, которые обладают способностью специфически связываться с генами, кодирующими тяжелые и легкие цепи антитела).
Приемлемые клетки-хозяева для клонирования или экспрессии кодирующих антитело векторов включают прокариотические или эукариотические клетки, представленные в настоящем описании. Например, антитела можно получать в бактериях, в частности, когда не требуется гликозилирование и связанная с Fc эффекторная функция. Сведения об экспрессии фрагментов антитела и полипептидов в бактериях см. например, в US №№5648237, 5789199 и 5840523 (см. также у Charlton в: Methods in Molecular Biology, под ред. Lo В.К.С., изд-во Humana Press, Totowa, NJ, т.248, 2003, cc. 245-254 описание экспрессии фрагментов антител в Е. coli.) После экспрессии антитело можно выделять из пасты бактериальных клеток в виде растворимой фракции и можно дополнительно очищать.
Помимо прокариотических организмов в качестве хозяев, пригодных для клонирования или экспрессии векторов, которые кодируют антитела, можно использовать эукариотические микроорганизмы, такие как нитчатые грибы или дрожжи, включая штаммы грибов и дрожжей, пути гликозилирования которых были «гуманизированы», что позволяет получать антитело с частично или полностью человеческой схемой гликозилирования (см. Gerngross, Nat. Biotech. 22, 2004, cc. 1409-1414 и Li и др., Nat. Biotech. 24, 2006, cc. 210-215).
Клетки-хозяева, которые можно использовать для экспрессии гликозилированного антитела, получают также из многоклеточных организмов (беспозвоночных и позвоночных животных). Примерами клеток беспозвоночных являются клетки насекомых, а также можно применять клетки растений. Были выявлены многочисленные штаммы бакуловирусов и соответствующие пригодные для них в качестве хозяев клетки насекомых, прежде всего для трансфекции клеток Spodoptera frugiperda.
В качестве хозяев можно применять также культуры растительных клеток (см., например, US №№5959177, 6040498, 6420548, 7125978 и 6417429 (описание технологии PLANTIBODIES™ для получения антител в трансгенных растениях).
В качестве хозяев можно применять также клетки позвоночных. Например, можно использовать клеточные линии млекопитающих, которые адаптированы к росту в суспензии. Другими примерами приемлемых линий клеток-хозяев млекопитающих являются линия клеток почки обезьяны CV1, трансформированная с помощью ОВ40 (COS-7); линия клеток почки эмбриона человека (НЕК 293-клетки или клетки линии 293, описанные, например, у Graham и др., J. Gen. Virol., 36, 1977, с. 59); клетки почки детеныша хомяка (ВНК); клетки Сертоли мыши (ТМ4-клетки, описанные, например, у Mather, Biol. Reprod., 23, 1980, cc. 243-251); клетки почки обезьяны (CV1); клетки почки африканской зеленой мартышки (VERO-76,); клетки карциномы шейки матки человека (HELA); клетки почки собаки (MDCK); клетки печени бычьей крысы (BRL ЗА); клетки легкого человека (W138); клетки печени человека (Hep G2); клетки опухоли молочной железы мыши (ММТ 060562); клетки TRI, описанные, например, у Mather и др., Annals N.Y. Acad. Sci., 383, 1982, cc. 44-68; клетки MRC 5 и клетки FS4. Другими ценными линиями клеток-хозяев млекопитающих являются клетки яичника китайского хомячка (СНО), включая DHFR- -СНО-клетки (Urlaub и др., Proc. Natl. Acad. Sci. USA, 77, 1980, с. 4216); и клеточные линии миеломы, такие как Y0, NS0 и Sp2/0. Обзор конкретных линий клеток-хозяев млекопитающих, которые можно применять для производства антител, см., например, у Yazaki и Wu в: Methods in Molecular Biology под ред. В.К.С.Lo, изд-во Humana Press, Totowa, NJ, т.248, 2003, cc. 255-268.
В. Анализы
Антитела к латентному TGF-бета 1, представленные в настоящем описании, можно идентифицировать, осуществлять их скрининг или характеризовать их физические/химические свойства и/или виды биологической активности с помощью различных анализов, известных в данной области.
1. Анализы связывания и другие анализы
В одном из объектов изобретения антитело, предлагаемое в изобретении, тестируют в отношении антигенсвязывающей активности, например, с помощью известных методов, таких как ELISA, Вестерн-блоттинг, поверхностный плазмонный резонанс (например, BIACORE®) или аналогичная методика (например, KinExa или OCTET®), и т.д.
Согласно другому объекту изобретения можно использовать анализы в условиях конкуренции для идентификации антитела, которое конкурирует с любыми антителами к латентному TGF-бета 1, указанными в настоящем описании, предпочтительно hT0947AE04-SG191, hT0947AE07-SG191, hT0947AE08-SG191 или hT0947AE09-SG191, за связывание с латентном TGF-бета 1. В некоторых вариантах осуществления изобретения указанное конкурирующее антитело связывается с тем же самым эпитопом (например, линейным или конформационным эпитопом), с которым связывается любое из антител к латентному TGF-бета 1, указанных в настоящем описании, предпочтительно 1TT0947AE04-SG191, hT0947AE07-SG191, hT0947AE08-SG191 или hT0947AE09-SG191. Подробное описание приведенных в качестве примера методов картирования эпитопа, с которым связывается антитело, представлено у Morris, Epitope Mapping Protocols, в: Methods in Molecular Biology, т.66, изд-во Humana Press, Totowa, NJ, 1996. Методы картирования эпитопа включают (но не ограничиваясь только ими) рентгеновскую кристаллографию и методы аланин-сканирующего мутагенеза.
В некоторых вариантах осуществления изобретения, когда указанное конкурирующее антитело присутствует в избытке, оно блокирует (например, снижает) связывание референс-антитела с латентным TGF-бета 1 по меньшей мере на 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75% или более. В некоторых вариантах осуществления изобретения связывание ингибируется по меньшей мере на 80%, 85%, 90%, 95% или более. В некоторых вариантах осуществления изобретения указанное конкурирующее антитело связывается с тем же самым эпитопом (например, линейным или конформационным эпитопом), с которым связывается антитело к латентному TGF-бета 1, указанное в настоящем описании. В других объектах изобретения референс-антитело представляет собой hT0947AE04-SG191, hT0947AE07-SG191, hT0947AE08-SG191 или hT0947AE09-SG191.
В качестве примера анализа в условиях конкуренции иммобилизованный латентный TGF-бета 1 инкубируют в растворе, содержащем первое меченое антитело (референс-антитело), которое связывается с латентным TGF-бета 1(например, hT0947AE04-SG191, hT0947AE07-SG191, hT0947AE08-SG191 или hT0947AE09-SG191), и второе немеченое антитело, подлежащее тестированию с отношении способности конкурировать с первым антителом за связывание с латентным TGF-бета 1. Второе антитело может присутствовать в супернатанте гибридомы. В качестве контроля иммобилизованный латентный TGF-бета 1 инкубируют в растворе, содержащим первое меченое антитело, но не содержащим второе немеченое антитело. После инкубации в условиях, приемлемых для связывания первого антитела с латентным TGF-бета 1, удаляют избыток несвязанного антитела и подсчитывают количестве метки, ассоциированной с иммобилизованным латентным TGF-бета 1. Если количество метки, ассоциированной с иммобилизованным латентным TGF-бета 1, существенно снижается в тестируемом образце по сравнению с контрольным образцом, то это свидетельствует о том, что второе антитело конкурирует с первым антителом за связывание с латентным TGF-бета 1 (см., например, Harlow и Lane, Antibodies: A Laboratory Manual, глава 14, изд-во Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, 1988).
В некоторых вариантах осуществления изобретения связывание антитела к латентному TGF-бета 1 с находящимся на клеточной поверхности латентным TGF-бета 1 можно тестировать с помощью известных методов, таких как ELISA, Вестерн-блоттинг, BIAcore, проточная цитометрия и т.д. Например, клетки, экспрессирующие латентный TGF-бета 1, можно приводить в контакт либо с антителами к латентному TGF-бета 1, непосредственно конъюгированными с РЕ или АРС, либо с неконъюгированными антителами к латентному TGF-бета 1, а затем с конъюгированными с РЕ или АРС вторичными антителами, и таким образом можно выявлять окрашивание находящегося на клеточной поверхности латентного TGF-бета 1 (см., например, Oida и др., PLoS One. 5(11), 24 ноября 2010 г., е15523; Su и др., Hum Mol Genet. 24(14), 15 июля 2015 г., cc. 4024-4036).
2. Анализы активности
Одним из объектов изобретения являются анализы, предназначенные для идентификации антител к латентному TGF-бета 1, которые обладают биологической активностью. Биологическая активность может включать, например, ингибирование активации латентного TGF-бета 1, ингибирование высвобождения зрелого TGF-бета 1 из латентного TGF-бета 1, ингибирование опосредуемого протеазой высвобождения зрелого TGF-бета 1 из латентного TGF-бета 1, ингибирование опосредуемого протеазой высвобождения зрелого TGF-бета 1 из латентного TGF-бета 1 без опосредуемого протеазой расщепления LAP-области латентного TGF-бета 1, ингибирование опосредуемого протеазой высвобождения зрелого TGF-бета 1 из латентного TGF-бета 1 без блокады доступа протеазы к латентному TGF-бета 1, ингибирование опосредуемого протеазой высвобождения зрелого TGF-бета 1 из латентного TGF-бета 1, позволяя при этом протеазе расщеплять LAP-область латентного TGF-бета 1, ингибирование опосредуемого протеазой высвобождения зрелого TGF-бета 1 из латентного TGF-бета 1 без ингибирования или с частичным ингибированием опосредуемой интегрином активации TGF-бета 1 и т.д. Предложены также антитела, обладающие указанной биологической активностью in vivo и/или in vitro.
В конкретных вариантах осуществления изобретения антитело, предлагаемое в изобретении, тестируют в отношении указанной биологической активности.
В некоторых вариантах осуществления изобретения определяют, ингибирует ли тестируемое антитело активацию латентного TGF-бета 1, т.е. ингибирует ли высвобождение зрелого TGF-бета 1 из латентного TGF-бета 1, путем детекции зрелого TGF-бета 1, используя известные в данной области методы, такие как электрофорез, хроматография, анализ методом иммуноблоттинга, твердофазный иммуноферментный анализ (ELISA) или масс-спектрометрия, после контакта активатора латентного TGF-бета 1 (например, протеазы, интегрина, другого не относящегося к протеазам активатора и т.д.) с латентным TGF-бета 1 в присутствии тестируемого антитела или без него. В одном из примеров активатор может быть выделенным (например, выделенная протеаза или выделенный интегрин) и/или невыделенным (например, мышиные, обезьяньи или человеческие РВМС, содержащие интегрин). Известно также, что активация латентного TGF-бета 1, т.е. высвобождение зрелого TGF-бета 1 из латентного TGF-бета 1, происходит также в отсутствии активатора (спонтанная активация латентного TGF-бета 1). В некоторых вариантах осуществления изобретения, ингибирует ли тестируемое антитело спонтанную активацию латентного TGF-бета 1, определяют путем детекции зрелого TGF-бета 1 с использованием метода, описанного выше, после инкубации латентного TGF-бета 1 с тестируемым антителом или без него. В некоторых вариантах осуществления изобретения, если определяют пониженное количество зрелого TGF-бета 1 в присутствии тестируемого антитела (или после контакта с ним), по сравнению с количеством, определенным в отсутствии тестируемого антитела, то тестируемое антитело идентифицируют как антитело, которое может ингибировать активацию латентного TGF-бета 1. В одном из примеров либо пониженное, либо повышенное количество зрелого TGF-бета 1, можно оценивать по концентрации зрелого TGF-бета 1 (например, в г/мл, мг/мл, мкг/мл, нг/мл или пг/мл и т.д.). В другом примере либо пониженное, либо повышенное количество зрелого TGF-бета 1, можно оценивать по оптической плотности (ОП) (например, при определенной длине волны в мм или нм и т.д.) метки, прямо или косвенно ассоциированной со зрелым TGF-бета.
В некоторых вариантах осуществления изобретения ингибирование активации TGF-бета 1 включает снижение по меньшей мере на 5%, 10%, 15%, 20%, 25%, 30%, 35% или 40% или более количества зрелого TGF-бета 1 в анализе по сравнению с отрицательным контролем в сходных условиях. В некоторых вариантах осуществления изобретения это относится к ингибированию активации TGF-бета 1, т.е. ингибированию высвобождения зрелого TGF-бета 1 по меньшей мере на 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90% или 95% или более.
В некоторых вариантах осуществления изобретения, ингибирует ли тестируемое антитело активацию латентного TGF-бета 1, т.е. ингибирует ли высвобождение зрелого TGF-бета 1 из латентного TGF-бета 1, определяют также путем детекции активности зрелого TGF-бета 1, например, активности связывания с рецептором TGF-бета 1, или активности опосредуемой трансдукции сигналов в клетке, экспрессирующей рецептор TGF-бета 1, и т.д. В некоторых вариантах осуществления изобретения связывание зрелого TGF-бета 1 с рецептором TGF-бета 1 можно определять, используя анализ связывания рецептора. В некоторых вариантах осуществления изобретения активность опосредуемой TGF-бета 1 трансдукции сигналов можно определять, оценивая активацию пути TGF-бета 1/Smad. Пригодные для такого анализа клетки могут представлять собой клетки, которые экспрессируют эндогенный рецептор TGF-бета 1, или которые созданы путем трансфекции клеток геном рецептора TGF-бета 1. Например, можно применять клетки HEK-Blue™ TGF-beta, которые использовали в рабочих примерах, указанных в настоящем описании, или которые кратковременно или стабильно генетически модифицировали для экспрессии трансгена, кодирующего рецептор TGF-бета 1. Опосредуемую TGF-бета 1 трансдукцию сигналов можно оценивать на любом уровне пути трансдукции сигналов, например, изучая фосфорилирование полипептида Smad, изучая экспрессию регулируемого TGF-бета 1 гена, включая репортерный ген, или измеряя зависимую от TGF-бета 1 пролиферацию клетки.
В некоторых вариантах осуществления изобретения активность опосредуемой TGF-бета 1 трансдукции сигналов можно определять также путем детекции активации пути TGF-бета 1/Smad, путем оценки фосфорилирования пептида Smad (см., например, Fukasawa и др., Kidney International. 65(1), 2004, cc. 63-74 и Ganapathy и др., Molecular Cancer 26, 9, 2010, с. 122). В других вариантах осуществления изобретения активность опосредуемой TGF-бета 1 трансдукции сигналов можно определять, оценивая способность TGF-бета ингибировать клеточную миграцию в «поврежденные» монослойные культуры ВАЕ-клеток, оценивая способность TGF-бета ингибировать клеточный рост, оценивая способность TGF-бета подавлять активность активатора плазминогена (РА), оценивая способность TGF-бета осуществлять повышающую регуляцию ингибитора-1 активатора плазминогена (PAI-1), и т.д. (см. Mazzieri и др., Methods in Molecular Biology 142, 2000, cc. 13-27)
Ингибирование активации TGF-бета 1 можно также определять и/или измерять с помощью методов, изложенных и проиллюстрированных в рабочих примерах. С помощью указанных анализов или других анализов соответствующих типов тестируемые антитела можно подвергать скринингу в отношении их способности ингибировать или активировать TGF-бета 1. В некоторых вариантах осуществления изобретения ингибирование активации TGF-бета 1 включает снижение по меньшей мере на 5%, 10%, 15%, 20%, 25%, 30%, 35% или 40% или более активации TGF-бета 1 в анализе по сравнению с отрицательным контролем в сходных условиях. В некоторых вариантах осуществления изобретения это относится к ингибированию активации TGF-бета 1, т.е. ингибированию высвобождения зрелого TGF-бета 1 по меньшей мере на 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90% или 95% или более. В некоторых вариантах осуществления изобретения ингибирование активации TGF-бета 1 включает снижение по меньшей мере на 5%, 10%, 15%, 20%, 25%, 30%, 35% или 40% или более количества зрелого TGF-бета 1 в анализе по сравнению с отрицательным контролем в сходных условиях. В некоторых вариантах осуществления изобретения это относится к снижению количества зрелого TGF-бета 1 по меньшей мере на 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90% или 95% или более.
В некоторых вариантах осуществления изобретения определение того, ингибирует ли тестируемое антитело расщепление LAP-участка латентного TGF-бета 1 осуществляют путем детекции продукта расщепления латентного TGF-бета 1 и/или нерасщепленного латентного TGF-бета 1, используя различные известные в данной области методы, такие как электрофорез, хроматография, анализ методом иммуноблоттинга, твердофазный иммуноферментный анализ (ELISA) или масс-спектрометрия, после контакта протеазы с латентным TGF-бета 1 в присутствии тестируемого антитела или без него. Например, в случае, когда белковую метку (например, FLAG-метку и т.д..) добавляют к N-концу LAP-области латентного TGF-бета 1, то участок, к которому добавлена белковая метка, отщепляется, если имеет место опосредуемое протеазой расщепление. Таким образом, продукт расщепления латентного TGF-бета 1 можно выявлять путем детекции латентного TGF-бета 1 (или LAP-области латентного TGF-бета 1) без белковой метки и/или нерасщепленный латентный TGF-бета 1 можно выявлять путем детекции латентного TGF-бета 1 с белковой меткой.
В другом примере, если белковую метку (например, FLAG-метку и т.д.) добавляют к N-концу LAP-области латентного TGF-бета 1 и если локализация сайта расщепления протеазой не находится вблизи N-конца LAP-области латентного TGF-бета 1, то LAP-область с белковой меткой укорачивается, если имеет местно опосредуемое протеазой расщепление. Таким образом, продукт расщепления латентного TGF-бета 1 можно выявлять путем детекции латентного TGF-бета 1, имеющего укороченную LAP-область (или укороченную LAP-область TGF-бета 1) с белковой меткой.
В некоторых вариантах осуществления изобретения, если обнаружено пониженное количество продукта расщепления латентного TGF-бета 1 в присутствии тестируемого антитела (или после контакта с ним) по сравнению с количеством, обнаруженным в отсутствии тестируемого антитела, то антитело идентифицируют как антитело, которое может ингибировать расщепление латентного TGF-бета 1. И, наоборот, если количество продукта расщепления латентного TGF-бета 1 не снижается существенно в присутствии тестируемого антитела (или после контакта с ним) по сравнению с количеством, обнаруженным в отсутствии тестируемого антитела, то антитело идентифицируют как антитело, которое не ингибирует расщепление латентного TGF-бета 1. В некоторых вариантах осуществления изобретения, если обнаружено повышенное количество нерасщепленного латентного TGF-бета 1 в присутствии тестируемого антитела (или после контакта с ним) по сравнению с количеством, обнаруженным в отсутствии тестируемого антитела, то антитело идентифицируют как антитело, которое может ингибировать расщепление латентного TGF-бета 1. И, наоборот, если количество нерасщепленного латентного TGF-бета 1 не снижается существенно в присутствии тестируемого антитела (или после контакта с ним) по сравнению с количеством, обнаруженным в отсутствии тестируемого антитела, то антитело идентифицируют как антитело, которое не ингибирует расщепление латентного TGF-бета 1. В конкретных вариантах осуществления изобретения, блокирует ли тестируемое антитело доступ протеазы к латентному TGF-бета 1, определяют методами, предназначенными для детекции белковых взаимодействий между протеазой и латентным TGF-бета 1, например, с использованием ELISA или поверхностного плазмонного резонанса, например, BIACORE®) или аналогичных методик (например, KinExa или OCTET®). Если выявлено пониженное взаимодействие между протеазой и латентным TGF-бета 1 в присутствии тестируемого антитела (или после контакта с ним) по сравнению с взаимодействием, обнаруженным в отсутствии тестируемого антитела, то тестируемое антитело идентифицируют как антитело, которое может блокировать доступ протеазы к латентному TGF-бета 1.
В некоторых вариантах осуществления изобретения отсутствие ингибирования расщепления латентного TGF-бета 1 включает повышение количества продукта расщепления латентного TGF-бета 1 по меньшей мере на 5%, 10%, 15%, 20%, 25%, 30%, 35% или 40% или более в анализе по сравнению с отрицательным контролем в сходных условиях. В некоторых вариантах осуществления изобретения отсутствие ингибирование расщепления латентного TGF-бета 1 включает повышение количества нерасщепленного латентного TGF-бета 1 по меньшей мере на 50% или менее, на 45% или менее, на 40% или менее, на 35% или менее, на 30% или менее, на 25% или менее, на 20% или менее, на 15% или менее, на 10% или менее, на 5% или менее в анализе по сравнению с отрицательным контролем в сходных условиях.
В некоторых вариантах осуществления изобретения антитело к латентному TGF-бета 1 можно подвергать другим анализам биологической активности, например, для оценки эффективности в качестве терапевтического средства. Такие анализы известны в данной области и зависят от антигена-мишени и предполагаемого применения антитела. Например, биологические действия блокады TGF-бета 1 антителом к латентному TGF-бета 1 можно оценивать с использованием мышиной модели индуцированного односторонней обструкцией мочеточника (UUO) фиброза почечной ткани (например, описанной у Chevalier R.L. и др., Ureteral obstruction as a model of renal interstitial fibrosis and obstructive nephropathy. Kidney Int. 75(11), июнь 2009 г., cc. 1145-1152.), мышиной модели индуцированного диетой с дефицитом холина, наличием определенных L-аминокислот, высоким содержанием жира (CDAHFD), подтвержденного с помощью НЭШ-теста фиброза печени, мышиной модели индуцированного блеомицином (BLM) фиброза легких и/или сингенной модели опухоли (которые описаны, например, у Mariathasan S. и др., TGF-beta attenuates tumour response to PD-L1 blockade by contributing to exclusion of T cells. Nature. 554(7693), 22 февраля 2018 г., cc. 544-548.). В других вариантах осуществления изобретения антитело к латентному TGF-бета 1 можно подвергать указанным в настоящем описании анализам биологической активности.
3. Методы скрининга
В одном из объектов изобретения способ скрининга антитела, предлагаемого в изобретении, включает различные анализы, указанные в настоящем описании и известные в данной области. Например, способ скрининга антитела к латентному TGF-бета 1 включает:
(а) приведение в контакт биологического образца, содержащего латентный TGF-бета 1 и протеазу, с тестируемым антителом;
(б) определение: (I) ингибирует ли тестируемое антитело расщепление LAP-области латентного TGF-бета 1 и (II) ингибирует ли тестируемое антитело активацию латентного TGF-бета 1; и
(в) отбор тестируемого антитела, которое ингибирует активацию латентного TGF-бета 1 без ингибирования опосредуемого протеазой расщепления LAP-участка латентного TGF-бета 1.
Альтернативно этому, вместо указанных выше стадий (б) и (в) способ скрининга антитела к латентному TGF-бета 1 включает, например, указанные ниже стадии (б) и (в):
(б) измерение (I) количества нерасщепленного латентного TGF-бета 1 и (II) количества зрелого TGF-бета 1; и
(в) отбор тестируемого антитела, которое ингибирует опосредуемое протеазой высвобождение зрелого TGF-бета 1 из латентного TGF-бета 1 без ингибирования опосредуемого протеазой расщепления LAP-области латентного TGF-бета 1, если количество нерасщепленного латентного TGF-бета 1 не повышается существенно и количество зрелого TGF-бета 1 понижается по сравнению с вариантом, в котором отсутствует тестируемое антитело.
Альтернативно этому, вместо указанных выше стадий (б) и (в) способ скрининга антитела к латентному TGF-бета 1 включает, например, указанные ниже стадии (б) и (в):
(б) измерение (I) количества расщепленного латентного TGF-бета 1 и (II) уровня активности зрелого TGF-бета 1; и
(в) отбор тестируемого антитела, которое ингибирует опосредуемую протеазой активацию латентного TGF-бета 1 без ингибирования опосредуемого протеазой расщепления LAP-области латентного TGF-бета 1, если количество расщепленного продукта не понижается существенно и уровень активности зрелого TGF-бета 1 понижается по сравнению с вариантом, в котором отсутствует тестируемое антитело.
Кроме того, в настоящем изобретении предложен способ получения антитела к латентному TGF-бета 1, который включает, например, указанные ниже стадии (г) и (д) в дополнение к указанным выше стадиям (а)-(в):
(г) получение информации об аминокислотной последовательности антитела к латентному TGF-бета 1, отобранному на стадии (в); и
(д) интродукция гена, кодирующего антитело к латентному TGF-бета 1, в клетку- хозяина.
В этом контексте понятие «не повышается/не понижается существенно», например, фразы «количество нерасщепленного латентного TGF-бета 1 не повышается существенно» и «количество расщепленного продукта (латентного TGF-бета 1) не понижается существенно» означают, что уровень/степень повышения/понижения может быть нулевым, а может быть не нулевым, но близким к нулю, или может быть столь низким, чтобы специалисты в данной области могли технически им пренебрегать или фактически/практически рассматривать в качестве нулевого. Например, в анализах методом иммуноблоттинга, когда исследователь не может выявлять или обнаруживать какой-либо существенный сигнал/полосу (или относительно высокий или сильный сигнал), соответствующий/соответствующую нерасщепленному латентному TGF-бета 1, то он считает, что количество нерасщепленного латентного TGF-бета 1 «не повышается существенно» или количество расщепленного продукта (латентного TGF-бета 1) «не понижается существенно». Кроме того, понятие «не повышается/не понижается существенно» применяют взаимозаменяемо с понятием «не повышается/понижается значительно».
В некоторых вариантах осуществления изобретения определение того, ингибирует ли тестируемое антитело расщепление LAP-области латентного TGF-бета 1, и ингибирует ли тестируемое антитело активацию латентного TGF-бета 1, можно определять с помощью различных анализов, указанных в настоящем описании и известных в данной области.
Г. Иммуноконъюгаты
В изобретении предложены также иммуноконъюгаты, содержащие антитело к латентному TGF-бета 1, представленное в настоящем описании, конъюгированное с одним или несколькими цитотоксическим агентами, такими как химиотерапевтические агенты или лекарственные средства, ингибирующие рост агенты, токсины (например, белковые токсины, обладающие ферментативной активностью токсины бактериального, грибного, растительного или животного происхождения или их фрагменты) или радиоактивные изотопы.
В одном из вариантов осуществления изобретения иммуноконъюгат представляет собой конъюгат антитело-лекарственное средство (ADC), в котором антитело конъюгировано с одним или несколькими лекарственными средствами, включая (но не ограничиваясь только ими) майтансиноид (см. US №№5208020, 5416064 и ЕР 0425235 В1); ауристатин, например, фрагменты DE и DF монометилауристатина (ММАЕ и MMAF) (см. US Ms 5635483, 5780588 и 7498298); доластатин; калихеамицин или его производные (см. US №№5712374, 5714586, 5739116, 5767285, 5770701, 5770710, 5773001 и 5877296; Hinman и др., Cancer Res. 53, 1993, cc. 3336-3342; и Lode и др., Cancer Res. 58, 1998, cc. 2925-2928); антрациклин, такой как дауномицин или доксорубицин (см. Kratz и др., Curr. Med. Chem. 13, 2006, cc. 477-523; Jeffrey и др., Bioorg. Med. Chem. Lett. 16, 2006, cc. 358-362; Torgov и др., Bioconjug. Chem. 16, 2005, cc. 717-721; Nagy и др., Proc. Natl. Acad. Sci. USA 97, 2000, cc. 829-834; Dubowchik и др., Bioorg. & Med. Chem. Letters 12, 2002, cc. 1529-1532; King и др., J. Med. Chem. 45, 2002, cc. 4336-4343; и US №6630579); метотрексат; виндесин; таксан, такой как доцетаксел, паклитаксел, ларотаксел, тесетаксел и ортатаксел; трихотецен и СС1065.
В другом варианте осуществления изобретения иммуноконъюгат содержит антитело, представленное в настоящем описании, конъюгированное с обладающим ферментативной активностью токсином или его фрагментом, включая (но не ограничиваясь только ими) А-цепь дифтерийного токсина, несвязывающиеся активные фрагменты дифтерийного токсина, А-цепь экзотоксина (из Pseudomonas aeruginosa), А-цепь рицина, А-цепь абрина, А-цепь модецина, альфа-сарцин, белки Aleurites fordii, диантиновые белки, белки Phytolaca americana (PAPI, PAPII и PAP -5), ингибитор из Momordica charantia, курцин, кротин, ингибитор из Sapaonaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трихотецены.
В другом варианте осуществления изобретения иммуноконъюгат содержит антитело, представленное в настоящем описании, конъюгированное с радиоактивным атомом с образованием радиоконъюгата. Для получения радиоконъюгатов можно применять широкое разнообразие радиоактивных изотопов. Примеры включают 211At, 131I, 125I, 90Y, 186Re, 188Re, 153Sm, 212Bi, 32P, 212Pb и радиоактивные изотопы Lu. Когда радиоконъюгат применяют для детекции, то он может содержать радиоактивный атом для сцинтиграфических исследовании, например, TC99m или I123, или спиновую метку для визуализации методом ядерного магнитного резонанса (ЯМР) (известный также как магнитно-резонансная визуализация, МРВ), такую как йод-123, иод-131, индий-111, фтор-19, углерод-13, азот-15, кислород-17, гадолиний, марганец или железо.
Конъюгаты антитела и цитотоксического агента можно получать с использованием разнообразных бифункциональных связывающих белки агентов, таких как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан- 1-карбоксилат (SMCC), иминотиолан (IT), бифункциональные производные сложных имидоэфиров (такие как диметиладипимидат×HCL), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаровый альдегид), бисазидопроизводные (такие как бис(пара-азидобензоил)гександиамин), производные бисдиазония (такие как бис(пара-диазониумбензоил)этилендиамин), диизоцианаты (такие как толуол-2,6-диизоцианат), и обладающие двойной активностью фторсодержащие соединения (такие как 1,5-дифтор-2,4-динитробензол). Например, иммунотоксин рицин можно получать согласно методу, описанному у Vitetta и др., Science 238, 1987, cc. 1098-1104. Меченная с помощью С14 1-изотиоцианатбензил-3-метилдиэтилентриаминпентауксусная кислота (MX-DTPA) является примером хелатирующего агента, применяемого для конъюгации радионуклеотида с антителом (см. WO 94/11026). Линкер может представлять собой «расщепляемый линкер», облегчающий высвобождение цитотоксического лекарственного средства в клетке. Например, можно использовать неустойчивый в кислой среде линкер, чувствительный к действию пептидаз линкер, фотолабильный линкер, диметильный линкер или содержащий дисульфид линкер (Chari и др., Cancer Res. 52, 1992, cc. 127-131; US №5208020).
В контексте настоящего описания под иммуноконъюгатами или ADC подразумеваются (но не ограничиваясь только ими) указанные конъюгаты, полученные с использованием перекрестносшивающих реагентов, которые включают (но не ограничиваясь только ими) BMPS, EMCS, GMBS, HBVS, LC-SMCC, MBS, МРВН, SBAP, SIA, SIAB, SMCC, SMPB, SMPH, сульфо-EMCS, сульфо-GMBS, сульфо-KMUS, сульфо-MBS, сульфо-SIAB, сульфо-SMCC и сульфо-SMPB, а также SVSB (сукцинимидил(4-винилсульфон)бензоат), которые поступают в продажу (например, от фирмы Pierce Biotechnology, Inc., Рокфорд, шт. Иллинойс, США).
Д. Способы и композиции для диагностирования и обнаружения
В некоторых вариантах осуществления изобретения любое из антител к латентному TGF-бета 1, представленных в настоящем описании, можно применять для обнаружения присутствия TGF-бета 1, например, латентного TGF-бета 1, в биологическом образце. Понятие «обнаружение (детекция)» в контексте настоящего описания предусматривает количественное или качественное обнаружение/измерение. В некоторых вариантах осуществления изобретения биологический образец содержит клетку или ткань, например, сыворотку, цельную кровь, плазму, полученный с помощью биопсии образец, образец ткани, клеточную суспензию, слюну, мокроту, оральную жидкость, спинномозговую жидкость, амниотическую жидкость, асцитную жидкость, молоко, молозиво, секрет молочных желез, лимфу, мочу, пот, слезную жидкость, желудочный сок, синовиальную жидкость, перитонеальную жидкость, жидкость хрусталика глаза или слизь.
Одним из вариантов осуществления изобретения является антитело к латентному TGF-бета 1, представленное в настоящем описании, для применения в способе диагностирования или обнаружения. Другой объект изобретения относится к способу обнаружения присутствия TGF-бета 1, например, латентного TGF-бета 1, в биологическом образце. Например, способ обнаружения присутствия латентного TGF-бета 1 включает:
(а) приведение в контакт биологического образца с указанным в настоящем описании антителом к латентному TGF-бета 1, предлагаемом в настоящем изобретении, в условиях, обеспечивающих связывание антитела к латентному TGF-бета 1 с латентным TGF-бета 1; и
(б) определение, образуется ли комплекс между антителом к латентному TGF-бета 1 и латентным TGF-бета 1.
Указанный способ может представлять собой способ in vitro или in vivo. В одном из вариантов осуществления изобретения антитело к латентному TGF-бета 1 применяют для отбора индивидуумов, которых можно лечить с помощью антитела к латентному TGF-бета 1, например, когда TGF-бета 1, например, латентный TGF-бета 1, является биомаркером для отбора пациентов. Это означает, что антитело к латентному TGF-бета 1 можно применять в качестве диагностического агента, таргетирующего TGF-бета 1.
Более конкретно, антитело к латентному TGF-бета 1 можно применять для диагностирования фиброза, предпочтительно фиброза миокарда, пульмонарного фиброза/фиброза легких, фиброза печени, фиброза почки, фиброза кожи, фиброза глаза и миелофиброза. Антитела к латентному TGF-бета 1, предлагаемые в изобретении, можно применять также для диагностирования рака.
В некоторых вариантах осуществления настоящего изобретения предложен способ ингибирования в биологическом образце опосредуемого протеазой высвобождения зрелого TGF-бета 1 из латентного TGF-бета 1 без ингибирования расщепления LAP-области латентного TGF-бета 1, включающий приведение в контакт биологического образца, содержащего латентный TGF-бета 1, с антителом к латентному TGF-бета 1, предлагаемому в настоящем изобретении, в условиях, обеспечивающих связывание антитела с латентным TGF-бета 1.
В некоторых вариантах осуществления изобретения, например, для целей обнаружения/диагностирования, предложены меченые антитела к латентному TGF-бета 1. Метки включают (но не ограничиваясь только ими) метки или фрагменты, которые можно обнаруживать непосредственно (такие как флуоресцентные, хромофорные, электронноплотные, хемилюминесцентные и радиоактивные метки), а также фрагменты, такие как ферменты или лиганды, которые подлежат косвенному обнаружению, например, посредством ферментативной реакции или молекулярного взаимодействия. Примеры меток включают (но не ограничиваясь только ими) радиоизотопы 32Р, 14С, 125I, 3Н и 131I, флуорофоры, такие как хелаты редкоземельных металлов или флуоресцеин и его производные, родамин и его производные, дансил, умбеллиферон, люциферазы, например, люцифераза светлячка и бактериальная люцифераза (US №4737456), люциферин, 2,3-дигидрофталазиндионы, пероксидаза из хрена (HRP), щелочная фосфатаза, β-галактозидаза, глюкоамилаза, лизоцим, оксидазы сахаридов, например, глюкозооксидаза, галактозооксидаза и глюкозо-6-фосфатдегидрогеназа, гетероциклические оксидазы, такие как уриказа и ксантиноксидаза, сшитые с ферментом, который использует пероксид водорода для окисления предшественника красителя, такого как HRP, лактопероксидаза или микропероксидаза, биотин/авидин, спиновые метки, метки в виде бактериофагов, стабильные свободные радикалы и т.п.
Е. Фармацевтические композиции
Фармацевтические композиции антитела к латентному TGF-бета 1, представленного в настоящем описании, получают путем смешения указанного антитела, имеющего необходимую степень чистоты, с одним или несколькими необязательными фармацевтически приемлемыми носителями (Remington's Pharmaceutical Sciences, 16-е изд. под ред. Osol А., 1980) в форме лиофилизированных композиций или водных растворов.
Фармацевтически приемлемые носители, как правило, являются нетоксичными для реципиентов в используемых дозах и концентрациях, они включают (но не ограничиваясь только ими): буферы, такие как фосфатный, цитратный буферы, а также буферы на основе других органических кислот; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония; хлорид бензетония; фенол; бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцинол; циклогексанол; 3-пентанол и мета-крезол); полипептиды с низкой молекулярной массой (содержащие менее приблизительно 10 остатков); белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поли(винилпирролидон); аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как ЭДТА; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; содержащие металл комплексы (например, комплексы Zn-белок); и/или неионогенные поверхностно-активные вещества, такие как полиэтиленгликоль (ПЭГ). В контексте настоящего описания примеры фармацевтически приемлемых носителей включают также диспергирующие агенты интерстициальных лекарственных средств, такие как растворимые активные в нейтральной среде гликопротеины гиалуронидаз (sHASEGP), например, человеческие растворимые гликопротеины гиалуронидазы РН-20, такие как rHuPH20 (HYLENEX®, фирма Baxter International, Inc.). Некоторые примеры sHASEGP и методы их применения, в том числе rHuPH20, описаны в публикациях патентов США №№2005/0260186 и 2006/0104968. Согласно одному из объектов изобретения sHASEGP объединяют с одним или несколькими дополнительными гликозаминогликаназами, такими как хондроитиназы.
Примеры лиофилизированных композиций антител описаны в US №6267958. Водные композиции антител включают композиции, описанные в US №6171586 и WO 2006/044908, последние композиции включают гистидин-ацетатный буфер.
Одним из объектов изобретения являются фармацевтические композиции, содержащие антитело к латентному TGF-бета 1, предназначенные для лечения фиброза, предпочтительно фиброза миокарда, пульмонарного фиброза, фиброза печени, фиброза почки, фиброза кожи, фиброза глаза и миелофиброза. В изобретении предложены также фармацевтические композиции, содержащие антитело к латентному TGF-бета 1, предназначенные для лечения рака.
Указанная в настоящем описании композиция может содержать также более одного действующего вещества, если это необходимо для конкретного подлежащего лечению показания, предпочтительно вещества с дополнительными видами активности, которые не оказывают отрицательного воздействия друг на друга. Например, может требоваться применение дополнительно ингибиторов иммунных контрольных точек, которые описаны ниже в разделе «III. Комбинированные терапии».
Действующие вещества можно включать также в микрокапсулы, например, полученные с помощью методов коацервации или межфазной полимеризации, например в гидроксиметилцеллюлозные или желатиновые микрокапсулы и поли(метилметакрилатные) микрокапсулы соответственно, в коллоидные системы введения лекарственного средства (например в липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсии. Такие методы описаны в Remington's Pharmaceutical Sciences, 16-е изд., под ред. Osol А., 1980.
Можно приготавливать препараты с замедленным высвобождением. Приемлемыми примерами препаратов с замедленным высвобождением являются полупроницаемые матрицы из твердых гидрофобных полимеров, включающие антитело, такие матрицы представляют собой изделия определенной формы, например, пленки или микрокапсулы.
Композиции, предназначенные для применения in vivo, как правило, должны быть стерильными. Стерильность можно легко обеспечивать, например, путем фильтрации через стерильные фильтрующие мембраны.
Ж. Терапевтические способы и композиции
Любое из антител к латентному TGF-бета 1, представленных в настоящем описании, можно применять в терапевтических способах. Одним из объектов изобретения является антитело к латентному TGF-бета 1, представленное в настоящем описании, для применения в качестве лекарственного средства. Другими объектами изобретения является антитело к латентному TGF-бета 1, предназначенное для применения для лечения рака или фиброза (такого как фиброз печени, фиброз почки или фиброз легкого) и т.д. В некоторых вариантах осуществления изобретения предложено антитело к латентному TGF-бета 1, предназначенное для применения в способе лечения. В некоторых вариантах осуществления изобретения предложено антитело к латентному TGF-бета 1, предназначенное для применения в способе лечения индивидуума, который имеет рак или фиброз (такой как фиброз печени, фиброз почки или фиброз легкого) и т.д., включающем введение индивидууму в эффективном количестве антитела к латентному TGF-бета 1. В одном из указанных вариантов осуществления изобретения способ включает также введение индивидууму в эффективном количестве по меньшей мере одного дополнительного терапевтического агента, например, описанного ниже. В других вариантах осуществления изобретения предложено антитело к латентному TGF-бета 1, предназначенное для применения для ингибирования опосредуемой протеазой активации латентного TGF-бета 1. В некоторых вариантах осуществления изобретения предложено антитело к латентному TGF-бета 1, предназначенное для применения в способе ингибирования опосредуемой протеазой активации латентного TGF-бета 1 у индивидуума, включающем введение индивидууму в эффективном количестве антитела к латентному TGF-бета 1 для ингибирования опосредуемой протеазой активации латентного TGF-бета 1. «Индивидуум» в контексте любого из указанных выше вариантов осуществления изобретения предпочтительно представляет собой человека.
Другим объектом изобретения является применение антитела к латентному TGF-бета 1 для производства или приготовления лекарственного средства. В одном из вариантов осуществления изобретения лекарственное средство предназначено для лечения рака или фиброза (такого как фиброз печени, фиброз почки или фиброз легкого) и т.д. В другом варианте осуществления изобретения лекарственное средство предназначено для применения в способе лечения рака или фиброза (такого как фиброз печени, фиброз почки или фиброз легкого) и т.д., включающем введение индивидууму, который имеет рак или фиброз (такой как фиброз печени, фиброз почки или фиброз легкого) и т.д., в эффективном количестве лекарственного средства. В одном из указанных вариантов осуществления изобретения способ включает также введение индивидууму в эффективном количестве по меньшей мере одного дополнительного терапевтического агента, например, описанного ниже. В другом варианте осуществления изобретения лекарственное средство предназначено для ингибирования опосредуемой протеазой активации латентного TGF-бета 1. В другом варианте осуществления изобретения лекарственное средство предназначено для применения в способе ингибирования у индивидуума опосредуемой протеазой активации латентного TGF-бета 1, включающем введение индивидууму в эффективном количестве лекарственного средства для ингибирования опосредуемой протеазой активации латентного TGF-бета 1. «Индивидуум» в контексте любого из указанных выше вариантов осуществления изобретения может представлять собой человека.
Следующим объектом изобретения является способ лечения рака или фиброза (такого как фиброз печени, фиброз почки или фиброз легкого) и т.д. В одном из вариантов осуществления способ включает введение индивидууму, который имеет указанный рак или фиброз (такой как фиброз печени, фиброз почки или фиброз легкого) и т.д., в эффективном количестве антитела к латентному TGF-бета 1. В одном из указанных вариантов осуществления изобретения способ включает также введение индивидууму в эффективном количестве по меньшей мере одного дополнительного терапевтического агента, например, описанного ниже. В некоторых вариантах осуществления изобретения введение антитела и агента осуществляют одновременно. «Индивидуум» в контексте любого из указанных выше вариантов осуществления изобретения может представлять собой человека.
Следующим объектом изобретения является способ ингибирования опосредуемой протеазой активации латентного TGF-бета 1 у индивидуума. В одном из вариантов осуществления изобретения способ включает введение индивидууму в эффективном количестве антитела к латентному TGF-бета 1 для ингибирования опосредуемой протеазой активации латентного TGF-бета 1. В одном из вариантов осуществления изобретения «индивидуум» представляет собой человека.
Следующим объектом изобретения являются фармацевтические композиции, содержащие любое из антител к латентному TGF-бета 1, представленных в настоящем описании, предназначенные, например, для применения в любом из указанных выше терапевтических способов. В одном из вариантов осуществления изобретения фармацевтическая композиция содержит любое из антител к латентному TGF-бета 1, представленных в настоящем описании, и фармацевтически приемлемый носитель. В другом варианте осуществления изобретения фармацевтическая композиция содержит любое из антител к латентному TGF-бета 1, представленных в настоящем описании, и по меньшей мере один дополнительный терапевтический агент, например, описанный ниже.
Антитело, предлагаемое в изобретении (и любой дополнительный терапевтический агент), можно вводить любыми приемлемыми путями, включая парентеральный, внутрилегочный и интраназальный пути, и при необходимости применять для местной обработки, введения в повреждение. Парентеральные инфузии включают внутримышечное, внутривенное, внутриартериальное, внутрибрюшинное или подкожное введение. Дозирование можно осуществлять любым приемлемым путем, например, путем инъекций, таких как внутривенные или подкожные инъекции, в зависимости, в том числе, от того, является ли лечение кратковременным или хроническим. Можно применять различные схемы дозирования, включая (но не ограничиваясь только ими) однократное введение или несколько введений в различные моменты времени, болюсное введение и пульсирующую инфузию.
Антитела, предлагаемые в изобретении, можно включать в состав композиций, дозировать и вводить в соответствии с надлежащей клинической практикой. Рассматриваемые в этом контексте факторы включают конкретное нарушение, подлежащее лечению, конкретное млекопитающее, подлежащее лечению, клиническое состояние индивидуального пациента, причину нарушения, область введения агента, метод введения, схему введения и другие факторы, известные практикующим медикам. Антитело можно, но это не является обязательным, включать в композицию в сочетании с одним или несколькими агентами, которые в настоящее время применяют для профилактики и лечения рассматриваемого нарушения. Эффективное количество указанных других агентов зависит от количестве антитела, присутствующего в композиции, типа нарушения или лечения и других указанных выше факторов.
Для предупреждения или лечения заболевания приемлемая доза антитела, предлагаемого в изобретении (при его применении индивидуально или в сочетании с одним или несколькими другими дополнительными терапевтическими агентами), должна зависеть от типа заболевания, подлежащего лечению, типа антитела, серьезности и течения болезни, применяют ли антитело для превентивных или терапевтических целей, предшествующей терапии, истории болезни пациента и ответа на антитело, а также предписания лечащего врача. Антитело можно вводить пациенту однократно или в виде серии обработок.
Следует понимать, что любые изделия, указанные в настоящем описании, могут включать иммуноконъюгат, предлагаемый в изобретении, вместо или в дополнение к антителу к латентному TGF-бета 1.
III. Комбинированные терапии
В терапии антитела к латентному TGF бета-1, предлагаемые в изобретении, можно применять либо индивидуально, либо в комбинации с другими агентами, предпочтительно для лечения рака или фиброза, более предпочтительно для лечения рака. Например, антитело, предлагаемое в изобретении, можно вводить совместно по меньшей мере с одним дополнительным терапевтическим агентом. В некоторых вариантах осуществления изобретения антитело и агент вводят одновременно. В некоторых вариантах осуществления изобретения дополнительный терапевтический агент представляет собой один или несколько ингибиторов иммунных контрольных точек, например, ингибитор CTLA-4, PD-1, PD-L1, PD-L2, CD160, CD57, CD244, LAG-3, CD272, KLRG1, CD26, CD39, CD73, CD305, TIGIT, TIM-3 и/или VISTA. В некоторых вариантах осуществления изобретения ингибиторы иммунных контрольных точек представляют собой, например, антитело к CTLA-4, антитело к PD-1, антитело к PD-L1, антитело к PD-L2, антитело к CD 160, антитело к CD57, антитело к CD244, антитело к LAG-3, антитело к CD272, антитело к KLRG1, антитело к CD26, антитело к CD39, антитело к CD73, антитело к CD305, антитело к TIGIT, антитело к TIM-3 и/или антитело к VISTA. Предпочтительно ингибиторы иммунных контрольных точек являются антагонистами аксиального связывания PD-1. Более предпочтительно ингибиторы иммунных контрольных точек представляют собой антитело к PD-1 или антитело к PD-L1. В некоторых вариантах осуществления изобретения антитело к PD-1 представляет собой ниволумаб, пембролизумаб или цемиплимаб. В некоторых вариантах осуществления изобретения антитело к PD-L1 представляет собой атезолизумаб, авелумаб или дурвалумаб, предпочтительно атезолизумаб. Предпочтительно комбинированная терапия включает применение антитела к латентному TGF бета-1, предлагаемого в изобретении, и атезолизумаба. В некоторых вариантах осуществления изобретения комбинированная терапия, включающая антитело к латентному TGF бета-1, предлагаемое в изобретении, или один или несколько ингибиторов иммунных контрольных точек, обладает аддитивной или синергетической эффективностью, например, аддитивным, комбинаторным или синергетическим противоопухолевым действием, по сравнению с монотерапией на основе антитела к TGF- бета или монотерапией на основе ингибитора иммунных контрольных точек.
Согласно одному из объектов изобретения комбинированные терапии, предлагаемые в изобретении, предназначены для лечения рака или фиброза, предпочтительно рака. В одном из вариантов осуществления изобретения рак является устойчивым к ингибиторам иммунных контрольных точек и/или дает ограниченный ответ на ингибиторы иммунных контрольных точек. Не вдаваясь в какую-либо теорию, отсутствие ответа некоторых устойчивых к ингибиторам иммунных контрольных точек видов рака и/или ограниченный ответ на ингибиторы иммунных контрольных точек ассоциированы с профилем передачи сигналов TGF-бета в фибробласты, прежде всего у пациентов с CD8 Т-клетками, которые исключаются из паренхимы опухоли, а вместо этого находятся в богатой фибробластами и коллагеном перитуморальной строме. Таким образом, антитело к латентному TGF-бета 1 в комбинации с ингибиторами иммунных контрольных точек могут снижать передачу сигналов TGF-бета в стромальных клетках, усиливая проникновение в центр опухоли, и могут обладать повышенной противоопухолевой активностью.
Белок запрограммированной клеточной гибели 1 (PD-1; известный также как CD274 или В7-Н1) представляет собой мембранный белок типа I, который принадлежит к семейству CD28/CTLA-4 Т-клеточных регуляторов. PD-1 имеет два лиганда, т.е. PD-L1 и PD-L2, которые принадлежат к семейству В7. Считается, что PD-1 и его лиганды осуществляют отрицательную регуляцию иммунных ответов, таких как Т-клеточные ответы. Для PD-L1 и PD-1 характерен высокий уровень экспрессии при нескольких типов рака, и, вероятно, они играют роль в ускользании рака от иммунологического надзора. Ингибиторы, такие как «ингибиторы (иммунных) контрольных точек», которые ингибируют, например, взаимодействие между PD-1 и PD-L1, могут усиливать Т-клеточные ответы и повышать противоопухолевую активность.
Понятие «антагонист аксиального связывания PD-1 (антагонист по оси связывания PD-1)» относится к молекуле, которая ингибирует взаимодействие партнера на аксиальному связыванию PD-1 либо с одним, либо с несколькими его партнерами по связыванию, для того, чтобы устранять Т-клеточную дисфункцию, являющуюся результатом передачи сигналов на оси передачи сигналов PD-1, результатом этого является восстановление или повышение Т-клеточной функции (например, пролиферация, производство цитокинов, цитолиз клеток-мишеней). В контексте настоящего описания антагонист аксиального связывания PD-1 включает антагониста связывания PD-1, антагониста связывания PD-L1 и антагониста связывания PD-L2.
Понятие «антагонист связывания PD-1» относится к молекуле, которая снижает, блокирует, ингибирует, элиминирует или препятствует трансдукции сигналов, сформировавшихся в результате взаимодействия PD-1 с одним или несколькими его партнерами по связыванию, такими как PD-L1, PD-L2. В некоторых вариантах осуществления изобретения антагонист связывания PD-1 представляет собой молекулу, которая ингибирует связывание PD-1 с одним или несколькими его партнерами по связыванию. В конкретном объекте изобретения антагонист связывания PD-1 ингибирует связывание PD-1 с PD-L1 и/или PD-L2. Например, антагонисты связывания PD-1 включают антитела к PD-1, их антигенсвязывающие фрагменты, иммуноадгезины, слитые белки, олигопептиды и другие молекулы, которые снижают, блокируют, ингибируют, элиминируют или препятствуют трансдукции сигналов, сформировавшихся в результате взаимодействия PD-1 с PD-L1 и/или PD-L2. В одном из вариантов осуществления изобретения антагонист связывания PD-1 уменьшает негативный костимуляторный сигнал, опосредуемый PD-1, или опосредуемый PD-1 сигнал, проходящий через белки клеточной поверхности, экспрессируемые на Т-лимфоцитах, снижая дисфункцию Т-клеток с нарушенной функцией (например, усиливая эффекторные ответы на распознавание антигена). В конкретном объекте изобретения антагонист связывания PD-1 представляет собой MDX-1106 (ниволумаб), МК-3475 (ламбролизумаб), СТ-011 (пидилизумаб) или АМР-224, или АМР-514 (MEDI0680). В другом объекте изобретения антагонист PD-1 выбирают из группы, которая состоит из PDR001, REGN2810, BGB А317 и SHR-1210.
Понятие «антагонист связывания PD-L1» относится к молекуле, которая снижает, блокирует, ингибирует, элиминирует или препятствует трансдукции сигналов, сформировавшихся в результате взаимодействия PD- L1 с одним или несколькими его партнерами по связыванию, такими как PD-1, В7-1. В некоторых вариантах осуществления изобретения антагонист связывания PD- L1 представляет собой молекулу, которая ингибирует связывание PD- L1 с его партнерами по связыванию. В конкретном объекте изобретения антагонист связывания PD-L1 ингибирует связывание PD-L1 с PD-1 и/или В7-1. В некоторых вариантах осуществления изобретения антагонисты связывания PD-L1 включают антитела к PD- L1, их антигенсвязывающие фрагменты, иммуноадгезины, слитые белки, олигопептиды и другие молекулы, которые снижают, блокируют, ингибируют, элиминируют или препятствуют трансдукции сигналов, сформировавшихся в результате взаимодействия PD-L1 с одним или несколькими его партнерами по связыванию, такими как PD-1, В7-1. В одном из вариантов осуществления изобретения антагонист связывания PD- L1 уменьшает негативный костимуляторный сигнал, опосредуемый PD- L1, или опосредуемый PD- L1 сигнал, проходящий через белки клеточной поверхности, экспрессируемые на Т-лимфоцитах, снижая дисфункцию Т-клеток с нарушенной функцией (например, усиливая эффекторные ответы на распознавание антигена). В некоторых вариантах осуществления изобретения антагонист связывания PD-L1 представляет собой антитело к PD-L1. В конкретном объекте изобретения антитело к PD-L1 представляет собой YW243.55.S70 (атезолизумаб), MDX-1105, авелумаб, MPDL3280A или MEDI4736 (дурвалумаб).
Понятие «антагонист связывания PD-L2» относится к молекуле, которая снижает, блокирует, ингибирует, элиминирует или препятствует трансдукции сигналов, образовавшихся в результате взаимодействия PD-L2 с одним или несколькими его партнерами по связыванию, например, PD-1. В некоторых вариантах осуществления изобретения антагонист связывания PD- L2 представляет собой молекулу, которая ингибирует связывание PD- L2 с одним или несколькими его партнерами по связыванию. В конкретном объекте изобретения антагонист связывания PD-L2 ингибирует связывание PD-L2 с PD-1. В некоторых вариантах осуществления изобретения антагонисты связывания PD-L2 включают антитела к PD-L2, их антигенсвязывающие фрагменты, иммуноадгезины, слитые белки, олигопептиды и другие молекулы, которые снижают, блокируют, ингибируют, элиминируют или препятствуют трансдукции сигналов, сформировавшихся в результате взаимодействия PD-L2 с одним или несколькими его партнерами по связыванию, например, PD-1. В одном из вариантов осуществления изобретения антагонист связывания PD-L2 уменьшает негативный костимуляторный сигнал, опосредуемый PD-L2, или опосредуемый PD-L2 сигнал, проходящий через белки клеточной поверхности, экспрессируемые на Т-лимфоцитах, снижая дисфункцию Т-клеток с нарушенной функцией (например, усиливая эффекторные ответы на распознавание антигена). В некоторых вариантах осуществления изобретения антагонист связывания PD-L2 представляет собой иммуноадгезин.
Указанные выше комбинированные терапии предусматривают совместное введение (при котором два или большее количество терапевтических агентов включают в одну и ту же или в разные композиции) и раздельное введение, в этом случае введение антитела, предлагаемого в изобретении, можно осуществлять до, одновременно и/или после введения дополнительного(ых) терапевтического(их) агента или агентов. В одном из вариантов осуществления изобретения введение антитела к латентному TGF-бета 1 и введение дополнительного терапевтического агента осуществляют в пределах примерно одного месяца или в пределах примерно одной, двух или трех недель, или в пределах примерно одного, двух, трех, четырех, пяти или шести дней относительно друг друга. Антитела, предлагаемые в изобретении, можно использовать также в комбинации с лучевой терапией.
В одном из объектов изобретения, согласно которому указанные выше комбинированные терапии предусматривают совместное введение, и два или большее количество терапевтических агентов включают в одну и ту же фармацевтическое композицию, то представленная в настоящем описании фармацевтическая композиция содержит, например, антитело к латентному TGF-бета 1, предлагаемое в изобретении, и один или несколько ингибиторов иммунных контрольных точек, описанных выше. Предпочтительно представленная в настоящем описании фармацевтическая композиция содержит антитело к латентному TGF-бета 1, предлагаемое в изобретении, и антагонист аксиального связывания PD-1 (предпочтительно антитело к PD-L1, более предпочтительно атезолизумаб) и фармацевтически приемлемый носитель.
Одним из объектов изобретения является антитело к латентному TGF-бета 1, предназначенное для применения в комбинации с дополнительным терапевтическим агентом для лечения одного или нескольких заболеваний. Другим объектом изобретения является фармацевтическая композиция, содержащая антитело к латентному TGF-бета 1, для применения в комбинации с дополнительным терапевтическим агентом для лечения одного или нескольких заболеваний. В одном из вариантов осуществления изобретения одно или несколько заболеваний представляют собой рак и/или фиброз, предпочтительно рак. В одном из вариантов осуществления изобретения дополнительные терапевтические агенты представляют собой один или несколько ингибиторов иммунных контрольных точек, описанных выше. Предпочтительно в изобретении предложено антитело к латентному TGF-бета 1, предназначенное для применения в комбинации с антагонистом аксиального связывания PD-1 (предпочтительно антителом к PD-L1, более предпочтительно атезолизумабом), для лечения рака.
Одним из объектов изобретения является антагонист аксиального связывания PD-1 (предпочтительно антитело к PD-L1, более предпочтительно атезолизумаб), предназначенный для применения в комбинации с антителом к латентному TGF-бета 1 для лечения одного или нескольких заболеваний. Другим объектом изобретения является фармацевтическая композиция, содержащая антагонист аксиального связывания PD-1 (предпочтительно антитело к PD-L1, более предпочтительно атезолизумаб), предназначенная для применения в комбинации с антителом к латентному TGF-бета 1 для лечения одного или нескольких заболеваний. В одном из вариантов осуществления изобретения одно или несколько заболеваний представляют собой рак и/или фиброз, предпочтительно рак.
IV. Изделия, наборы
А. Изделия
Другим объектом изобретения является изделие, которое содержит продукты, применяемые для лечения, предупреждения и/или диагностирования указанных выше нарушений (например, фиброза и рака). Изделие представляет собой контейнер и этикетку или листовку-вкладыш в упаковку, которые прилагаются к нему. Приемлемыми контейнерами являются, например банки, пузырьки, шприцы, пакеты для внутривенного (IV) раствора и т.д. Контейнеры можно изготавливать из различных материалов, таких как стекло или пластмасса. Контейнер содержит композицию, которая сама или в сочетании с другой композицией является эффективной для лечения, предупреждения и/или диагностирования состояния (например, фиброза и рака), и может иметь стерильный порт доступа (например, контейнер может представлять собой пакет для внутривенного раствора или пузырек, снабженный пробкой, которую можно прокалывать с помощью иглы для подкожных инъекций). По меньшей мере одно действующее вещество в композиции представляет собой антитело или иммуноконъюгат, предлагаемое/предлагаемый в изобретении. На этикетке или листовке-вкладыше в упаковку указано, что композицию применяют для лечения выбранного состояния (например, фиброза и рака). Кроме того, изделие может включать (а) первый контейнер с находящейся в нем композицией, где композиция содержит антитело или иммуноконъюгат, предлагаемое/предлагаемый в изобретении; и (б) второй контейнер с находящейся в нем композицией, где композиция содержит дополнительное цитотоксический или иной терапевтический агент. Согласно этому варианту осуществления изобретения изделие может содержать также листовку-вкладыш в упаковку, которая содержит информацию о том, что композиции можно использовать для лечения конкретного состояния (например, фиброза и рака). В альтернативном или дополнительном варианте изделие может дополнительно включать второй (или третий) контейнер с фармацевтически приемлемым буфером, таким как бактериостатическая вода для инъекций (БСВИ), забуференный фосфатом физиологический раствор, раствор Рингера и раствор декстрозы. Кроме того, оно может включать другие продукты, необходимые с коммерческой точки зрения и с точки зрения потребителя, в частности, другие буферы, разбавители, фильтры, иглы и шприцы.
Как должно быть очевидно, вышеуказанное изделие может включать иммуноконъюгат, предлагаемый в изобретении, вместо или в дополнение к антителу к латентному TGF-бета 1.
Б. Наборы
В настоящем описании представлены наборы, предназначенные для применения в способе лечения, предупреждения и/или диагностирования нарушений, указанных в настоящем описании, в частности, для лечения индивидуума, имеющего фиброз или рак, которые содержат антитело к латентному TGF-бета 1, иммуноконъюгат, содержащий антитело к латентному TGF-бета 1, выделенную нуклеиновую кислоту, кодирующую антитело к латентному TGF-бета 1, или вектор, содержащий нуклеиновую кислоту, указанную в настоящем описании или полученную способом, указанным в настоящем описании. Наборы могут дополнительно содержать любые терапевтические агенты, приведенные в качестве примеров в настоящем описании в разделе «III. Комбинированные терапии», такие как ингибитор иммунных контрольных точек, включая антитело к PD-L1. Наборы могут быть упакованы вместе с дополнительным фармацевтически приемлемым носителем или средой, которые указаны в настоящем описании, или инструкцией по применению, в которой указано как применять наборы, и т.д. Аналогично указанному в настоящем описании изделию, набор может содержать продукты, которые можно применять для лечения фиброза или рака; контейнер и этикетку или листовку-вкладыш, прилагаемые к контейнеру; композицию, которая сама по себе или в сочетании с другой композицией является эффективной для лечения фиброза или рака; стерильный порт доступа и т.д. Наборы также могут содержать листовку-вкладыш в упаковку, которая содержит информацию о том, что композиции можно использовать для лечения фиброза или рака. В альтернативном или дополнительном варианте наборы могут дополнительно включать второй (или третий) контейнер с фармацевтически приемлемым буфером, таким как бактериостатическая вода для инъекций (БСВИ), забуференный фосфатом физиологический раствор, раствор Рингера и раствор декстрозы. Наборы могут включать также другие продукты, необходимые с коммерческой точки зрения и с точки зрения потребителя, в частности, другие буферы, разбавители, фильтры, иглы и шприцы.
Примеры
Ниже приведены примеры методов и композиций, предлагаемых в изобретении. Как должно быть очевидно, различные другие варианты осуществления изобретения можно применять на практике с учетом общего описания изобретения, представленного выше.
Пример 1: Экспрессия и очистка антигенов
(1-1) Экспрессия и очистка латентного TGF-бета 1
Для экспрессии и очистки применяли следующие последовательности: меченный с помощью Flag человеческий латентной TGF-бета 1 (SEQ ID NO: 1, 2) и меченный с помощью Flag мышиный латентный TGF-бета 1 (SEQ ID NO: 3, 4), и меченный с помощью Flag латентный TGF-бета 1 обезьян циномолгус (SEQ ID NO: 5, 6). Каждый из этих меченных с помощью Flag латентных TGF-бета 1 имел в направлении от N-конца к С-концу сигнальную последовательность, полученную из крысиного сывороточного альбумина (SEQ ID NO: 7), Flag-метку и последовательность латентного TGF-бета 1. Остаток Cys в тридцатом (30-ом) положении каждого из этих меченных с помощью Flag латентных TGF-бета 1 был заменен на Ser, что соответствовало «C33S-мутации» (см., например Yoshinaga К. и др., Perturbation of transforming growth factor (TGF)-betal association with latent TGF-beta binding protein yields inflammation and tumors. Proc Natl Acad Sci USA. 105(48), 2008, cc. 18758-18763).
Меченный с помощью Flag человеческий латентный TGF-бета 1 (ниже обозначенный как «человеческий латентный TGF-бета 1 (SLC)» или «человеческий латентный TGF-бета 1»), меченный с помощью Flag мышиный латентный TGF-бета 1 (ниже обозначенный как «мышиный латентный TGF-бета 1 (SLC)» или «мышиный латентный TGF-бета 1»), или меченный с помощью Flag латентный TGF-бета 1 обезьян циномолгус (ниже обозначенный как «обезьяний латентный TGF-бета 1 (SLC)» или «обезьяний латентный TGF-бета 1») экспрессировали кратковременно с использованием клеточной линии FreeStyle293-F или Expi293F (фирма Thermo Fisher Schientific). Кондиционированные среды, содержащий человеческий, мышиный или обезьяний латентный TGF-бета 1 (SLC), вносили на колонку, упакованную аффинной смолой с антителом к Flag М2 (фирма Sigma), и элюировали латентный TGF-бета 1 (SLC) с помощью пептида Flag (фирма Sigma). Фракции, содержащие человеческий, мышиный или обезьяний латентный TGF-бета 1 (SLC), собирали и затем внесли на колонку для гель-фильтрации Супердекс 200 (фирма GE healthcare), уравновешенную 1х ЗФР. Затем фракции, содержащие человеческий, мышиный или обезьяний латентный TGF-бета 1 (SLC), объединяли и хранили при -80°С.
(1-2) Экспрессия и очистка мышиного ассоциированного с латентностью пептида (LAP)
Для экспрессии и очистки применяли следующую последовательность: меченный с помощью flag мышиный LAP (SEQ ID NO: 8, 9), который имел в направлении от N-конца к С-концу сигнальную последовательность, полученную из крысиного сывороточного альбумина (SEQ ID NO: 7), Flag-метку и последовательность ассоциированного с латентностью белка (LAP). Остаток Cys в тридцатом (30-ом) положении меченного с помощью flag LAP был заменен на Ser, что соответствовало «C33S-мутации». Экспрессию и очистку меченного с помощью Flag мышиного LAP (SEQ ID NO: 8, 9) (ниже обозначен как «рекомбинантный мышиный ассоциированный с латентность белок (LAP)») осуществляли строго с соответствии с методом, описанным в примере (1-1).
Пример 2. Гуманизация и оптимизация антитела к TGF-бета 1
(2-1) Гуманизация
Родительское антитело к TGF-бета ТВА0947, представляющее собой химерное антитело, гуманизировали следующим образом. Сначала вариабельные области тяжелой и легкой цепей гуманизированных антител создавали, используя вариабельные области ТВА0947 и каркасные участки человеческой зародышевой линии. Затем полинуклеотиды каждой созданной вариабельной области тяжелой цепи и вариабельной области легкой цепи клонировали в экспрессионных векторах, содержащих последовательность константной области тяжелой цепи SG181 (SEQ ID NO: 10) и последовательность константной области легкой цепи SK1 (SEQ ID NO: 11) соответственно. Гуманизированные антитела экспрессировали кратковременно в клетках FreeStyle 293-F (фирм Thermo Fisher Scientific) и осуществляли Biacore-анализ. Отбирали гуманизированное антитело, связывающая активность которого по данным Biacore-анализа было по меньшей мере такой же как у родительского антитела.
(2-2) Оптимизация
Гуманизированное антитело, полученное согласно методу, описанному в примере (2-1), оптимизировали с получением hT0947AE04, hT0947AE07, hT0947AE08 и hT0947AE09, которые обладали улучшенной связывающей активностью в отношении латентного TGF-бета 1 (SLC). В целом, метод состоял в следующем: осуществляли комплексный мутагенез всех остатков в определяющих комплементарность участках (CDR) и тяжелой, и легкой цепей. Каждую аминокислоту заменяли другой из 18 встречающихся в естественных условиях аминокислот, исключая исходную аминокислоту и цистеин. Варианты экспрессировали кратковременно в клетках FreeStyle 293-F (фирма Thermo Fisher Scientific) и очищали из супернатантов культур для оценки с помощью Biacore. Отбирали представляющие интерес варианты с улучшенной связывающей активностью в отношении как человеческого, так и мышиного латентного TGF-бета 1 (SLC). Затем создавали антитела с комбинацией таких мутаций в CDR. (2-3) Аминокислотная последовательность оптимизированного антитела Аминокислотные последовательности вариабельных областей hT0947AE04, hT0947AE07, hT0947AE08 и hT0947AE09 идентифицировали, и они описаны ниже:
- Вариабельная область тяжелой цепи hT0947AE04 содержит аминокислотную последовательность SEQ ID NO: 12 (hT0947AE04H), а вариабельная область легкой цепи hT0947AE04 содержит аминокислотную последовательность SEQ ID NO: 13 (hT0947AE04L).
- Вариабельная область тяжелой цепи hT0947AE07 содержит аминокислотную последовательность SEQ ID NO: 14 (hT0947AE07H), а вариабельная область легкой цепи hT0947AE07 содержит аминокислотную последовательность SEQ ID NO: 15 (hT0947AE07L).
- Вариабельная область тяжелой цепи hT0947AE08 содержит аминокислотную последовательность SEQ ID NO: 16 (hT0947AE08H), а вариабельная область легкой цепи hT0947AE08 содержит аминокислотную последовательность SEQ ID NO: 17 (hT0947AE08L).
- Вариабельная область тяжелой цепи hT0947AE09 содержит аминокислотную последовательность SEQ ID NO: 18 (hT0947AE09H), а вариабельная область легкой цепи hT0947AE09 содержит аминокислотную последовательность SEQ ID NO: 19 (hT0947AE09L).
Аминокислотные последовательности CDR (HVR) hT0947AE04, hT0947AE07, hT0947AE08 и hT0947AE09 идентифицировали согласно Кэботу, и они описаны ниже:
- hT0947AE04 содержит CDR1, CDR2 и CDR3 тяжелой цепи, которые содержат аминокислотные последовательности SEQ ID NO: 20, 21 и 22 соответственно, и CDR1, CDR2 и CDR3 легкой цепи, которые содержат аминокислотные последовательности SEQ ID NO: 23, 24 и 25 соответственно.
- hT0947AE07 содержит CDR1, CDR2 и CDR3 тяжелой цепи, которые содержат аминокислотные последовательности SEQ ID NO: 26, 27 и 28 соответственно, и CDR1, CDR2 и CDR3 легкой цепи, которые содержат аминокислотные последовательности SEQ ID NO: 29, 30 и 31 соответственно.
- hT0947AE08 содержит CDR1, CDR2 и CDR3 тяжелой цепи, которые содержат аминокислотные последовательности SEQ ID NO: 32, 33 и 34 соответственно, и CDR1, CDR2 и CDR3 легкой цепи, которые содержат аминокислотные последовательности SEQ ID NO: 35, 36 и 37 соответственно.
- hT0947AE09 содержит CDR1, CDR2 и CDR3 тяжелой цепи, которые содержат аминокислотные последовательности SEQ ID NO: 38, 39 и 40 соответственно, и CDR1, CDR2 и CDR3 легкой цепи, которые содержат аминокислотные последовательности SEQ ID NO: 41, 42 и 43 соответственно.
(2-4) Создание полноразмерных тяжелой и легкой цепей
Множество аминокислотных замен интродуцировали в константную область тяжелой цепи SGI (SEQ ID NO: 44). SGI представляет собой константную область тяжелой цепи человеческого IgG1 дикого типа с делецией по меньшей мере двух С-концевых аминокислот т.е. G1y-Lys (GK). В результате создавали SG181 (SEQ ID NO: 10) и SG191 (SEQ ID NO: 45). SG181 включает аминокислотные замены L235R/G236R (аминокислотные замены для снижения эффекторной функции) и K214R согласно EU-индексу. SG191 включает аминокислотные замены L235R/G236R (аминокислотные замены для снижения эффекторной функции), M428L/N434A (аминокислотные замены для повышения связывающей активности в отношении FcRn), Q438R/S440E (аминокислотные замены для снижения связывания с ревматоидным фактором) и K214R согласно EU-индексу. Кроме того, создавали mF18 (SEQ ID NO: 46), константную область тяжелой цепи мышиного IgG, которая включает P235K/S239K (аминокислотные замены для снижения эффекторной функции).
Каждую из вариабельных областей тяжелой цепи объединяли с константной областью тяжелой цепи SG181 (SEQ ID NO: 10), SG191 (SEQ ID NO: 45) или mF18 (SEQ ID NO: 46). Таким образом создавали последовательности полноразмерных тяжелых цепей, которые имели следующие аминокислотные последовательности:
(a1) полноразмерная тяжелая цепь, содержащая аминокислотную последовательность SEQ ID NO: 47, которая содержит hT0947AE04H (вариабельная область тяжелой цепи) и SG181 (константная область тяжелой цепи),
(а2) полноразмерная тяжелая цепь, содержащая аминокислотную последовательность SEQ ID NO: 48, которая содержит hT0947AE07H (вариабельная область тяжелой цепи) и SG181 (константная область тяжелой цепи),
(а3) полноразмерная тяжелая цепь, содержащая аминокислотную последовательность SEQ ID NO: 49, которая содержит hT0947AE08H (вариабельная область тяжелой цепи) и SG181 (константная область тяжелой цепи),
(а4) полноразмерная тяжелая цепь, содержащая аминокислотную последовательность SEQ ID NO: 50, которая содержит hT0947AE09H (вариабельная область тяжелой цепи) и SG181 (константная область тяжелой цепи),
(б1) полноразмерная тяжелая цепь, содержащая аминокислотную последовательность SEQ ID NO: 51, которая содержит hT0947AE04H (вариабельная область тяжелой цепи) и SG191 (константная область тяжелой цепи),
(б2) полноразмерная тяжелая цепь, содержащая аминокислотную последовательность SEQ ID NO: 52, которая содержит hT0947AE07H (вариабельная область тяжелой цепи) и SG191 (константная область тяжелой цепи),
(б3) полноразмерная тяжелая цепь, содержащая аминокислотную последовательность SEQ ID NO: 53, которая содержит hT0947AE08H (вариабельная область тяжелой цепи) и SG191 (константная область тяжелой цепи),
(б4) полноразмерная тяжелая цепь, содержащая аминокислотную последовательность SEQ ID NO: 54, которая содержит hT0947AE09H (вариабельная область тяжелой цепи) и SG191 (константная область тяжелой цепи),
(в1) полноразмерная тяжелая цепь, содержащая аминокислотную последовательность SEQ ID NO: 55, которая содержит hT0947AE04H (вариабельная область тяжелой цепи) и mF18 (константная область тяжелой цепи),
(в2) полноразмерная тяжелая цепь, содержащая аминокислотную последовательность SEQ ID NO: 56, которая содержит hT0947AE07H (вариабельная область тяжелой цепи) и mF18 (константная область тяжелой цепи),
(в3) полноразмерная тяжелая цепь, содержащая аминокислотную последовательность SEQ ID NO: 57, которая содержит hT0947AE08H (вариабельная область тяжелой цепи) и mF18 (константная область тяжелой цепи),
(в4) полноразмерная тяжелая цепь, содержащая аминокислотную последовательность SEQ ID NO: 58, которая содержит hT0947AE09H (вариабельная область тяжелой цепи) и mF18 (константная область тяжелой цепи).
Каждую из вариабельных областей легкой цепи объединяли с константной областью легкой цепи (каппа) SKI (SEQ ID NO: 11) человеческого IgG или константной областью легкой цепи (каппа) mkl (SEQ ID NO: 59) мышиного IgG. Таким образом создавали последовательности полноразмерных легких цепей, которые имели следующие аминокислотные последовательности:
(г1) полноразмерная легкая цепь, содержащая аминокислотную последовательность SEQ ID NO: 60, которая содержит hT0947AE04L (вариабельная область легкой цепи) и SK1 (константная область легкой цепи),
(г2) полноразмерная легкая цепь, содержащая аминокислотную последовательность SEQ ID NO: 61, которая содержит hT0947AE07L (вариабельная область легкой цепи) и SK1 (константная область легкой цепи),
(г3) полноразмерная легкая цепь, содержащая аминокислотную последовательность SEQ ID NO: 62, которая содержит hT0947AE08L (вариабельная область легкой цепи) и SK1 (константная область легкой цепи),
(г4) полноразмерная легкая цепь, содержащая аминокислотную последовательность SEQ ID NO: 63, которая содержит hT0947AE09L (вариабельная область легкой цепи) и SK1 (константная область легкой цепи),
(д1) полноразмерная легкая цепь, содержащая аминокислотную последовательность SEQ ID NO: 64, которая содержит hT0947AE04L (вариабельная область легкой цепи) и mk1 (константная область легкой цепи),
(д2) полноразмерная легкая цепь, содержащая аминокислотную последовательность SEQ ID NO: 65, которая содержит hT0947AE07L (вариабельная область легкой цепи) и mk1 (константная область легкой цепи),
(д3) полноразмерная легкая цепь, содержащая аминокислотную последовательность SEQ ID NO: 66, которая содержит hT0947AE08L (вариабельная область легкой цепи) и mk1 (константная область легкой цепи),
(д4) полноразмерная легкая цепь, содержащая аминокислотную последовательность SEQ ID NO: 67, которая содержит hT0947AE09L (вариабельная область легкой цепи) и mk1 (константная область легкой цепи).
Затем объединяли соответствующие полноразмерные тяжелые цепи и легкие цепи и создавали антитела, представленные в таблице 2. Созданные антитела обозначали согласно указанному в таблице 2, и в настоящем описании применяли их соответствующие обозначения.
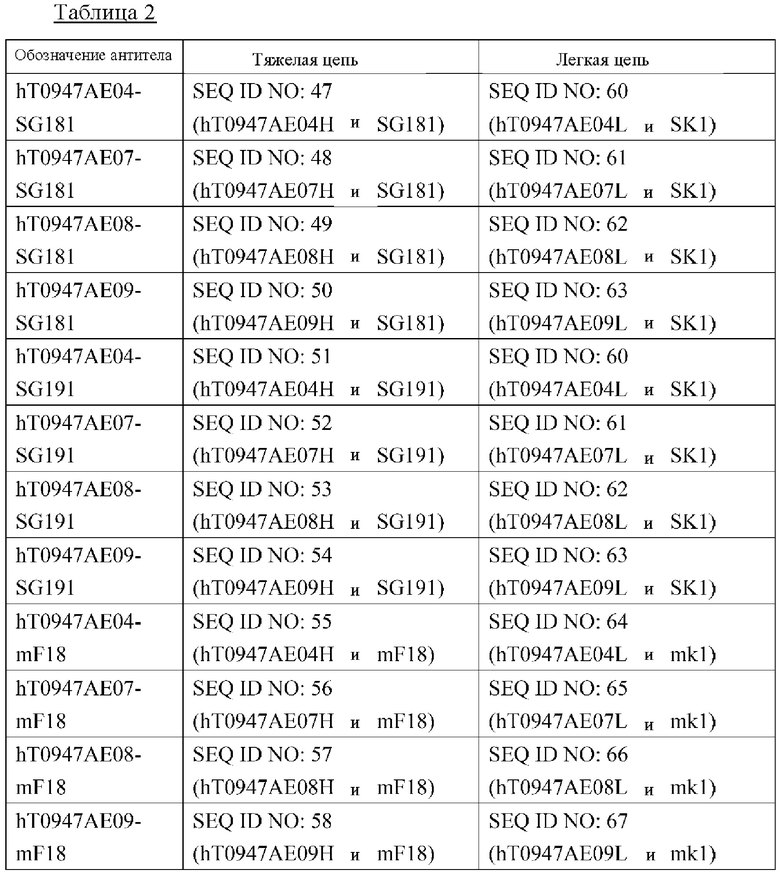
Пример 3. Biacore-анализ для оценки связывающей активности антител к латентному TGF-бета 1
Связывающую активность антител к латентному TGF-бета 1 (hT0947AE04-SG191, hT0947AE07-SG191, hT0947AE08-SG191 и hT0947AE09-SG191) в отношении латентного TGF-бета 1 (SLC) человека, обезьян циномолгус или мышей измеряли с помощью устройства Biacore 8k (фирма GE Healthcare). Мышиное антитело к легкой каппа-цепи человеческого Ig (фирма BD Pharmingen) иммобилизовали на всех проточных ячейках сенсорного чипа СМ5, используя набор для аминного сочетания (фирма GE Healthcare). Антитела захватывали на сенсорных поверхностях с антителами к каппа-цепи с уровнем захвата, соответствующем примерно 20 RU (резонансных единиц), после чего на проточную ячейку инъецировали латентный TGF-бета 1 (SLC) человека, обезьян циномолгус или мышей, который получали согласно методу, описанному в примере (1-1). Все антитела и аналиты приготавливали в ACES-буфере, рН 7,4, содержащем 20 мМ ACES, 150 мМ NaCl, 0,05% Твин 20, 0,005% NaN3. Температура анализа составляла 37°С. Поверхность сенсора регенерировали в каждом цикле с помощью 10 мМ глицин-HCl, рН 2,1. Связывающую активность определяли путем обработки и аппроксимации данных к модели связывания 1:1, используя программное обеспечение Biacore Insight, версия 1.1.1.7442 (фирма GE Healthcare).
Данные об активности связывания (ka, kd и KD) антител к латентному TGF-бета 1 в отношении латентного TGF-бета 1 человека, обезьян циномолгус или мышей представлены в таблице 3.
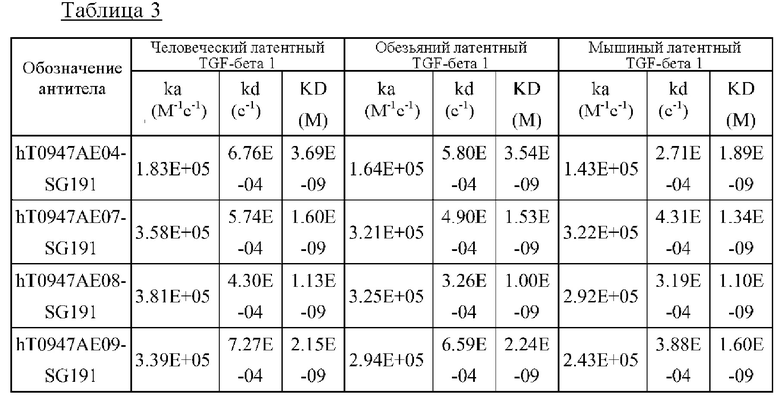
Пример 4.Характеризация антитела к латентному TGF-бета 1 (4-1) Антитело к латентному TGF-бета Е связывающееся с расположенным на клеточной поверхности латентным TGF-бета 1
Связывающую активность антител к латентному TGF-бета 1 (hT0947AE04-SG191, hT0947AE07-SG191, hT0947AE08-SG191 или hT0947AE09-SG191) с расположенным на клеточной поверхности латентным TGF-бета 1 тестировали с использованием FACS, применяя клетки Ba/F3, которые экспрессируют мышиный латентный TGF-бета 1, или клетки FreeStyle™ 293-F, которые экспрессируют человеческий латентный TGF-бета 1 (фирма ThermoFisher). Антитела к латентному TGF-бета 1 (каждое в концентрации 10 мкг/мл) инкубировали с каждой клеточной линией в течение 30 мин при 4°С и промывали FACS-буфером (2% FBS, 2 мМ ЭДТА в ЗФР). Антитело к KLH, имеющее Fc-область человеческого IgG1 (IC17-hIgG1), которое не связывается ни с мышиным латентным TGF-бета 1, ни с человеческим латентным TGF-бета 1, представляло собой применяемое в качестве отрицательного контроля антитела. Затем добавляли козий F(ab')2 к человеческому IgG, мышиный ads-PE (фирма Southern Biotech, каталожный №. 2043-09) и инкубировали в течение 30 мин при 4°С и промывали FACS-буфером. Сбор данных осуществляли с помощью устройства FACS Verse (фирма Becton Dickinson) с последующим анализом и расчетом средней интенсивности флуоресценции (MFI) с использованием программного обеспечения FlowJo (фирма Tree Star) и программного обеспечения GraphPad Prism (фирма GraphPad). Как продемонстрировано на фиг. 1, все антитела к латентному TGF-бета 1 связывались с мышиным расположенным на клеточной поверхности латентным TGF-бета 1, который экспрессировался на клетках Ba/F3, и человеческим расположенным на клеточной поверхности латентным TGF-бета 1, который экспрессировался на клетках FreeStyle™ 293-F.
(4-2) Антитело к латентному TGF-бета 1 не связывающееся со зрелым TGF-бета 1, но связывающееся с мышиным LAP
Связывающую активность антител к латентному TGF-бета 1 (hT0947AE04-SG191, hT0947AE07-SG191, hT0947AE08-SG191 или hT0947AE09-SG191) со зрелым TGF-бета 1 тестировали в помощью ELISA. 384-луночный планшет сенсибилизировали мышиным или человеческим зрелым TGF-бета 1 в течение ночи при 4°С и затем промывали 4 раза ЗФР-Т. После промывки планшет блокировали с помощью блокирующего буфера (1× TBS/Tвин-20+0,5% БСА+1x Block асе) по меньшей мере в течение 1 ч при комнатной температуре, а затем промывали 4 раза ЗФР-Т. После промывки в планшет добавляли раствор антитела и инкубировали в течение 2 ч при комнатной температуре и затем промывали 4 раза ЗФР-Т. После промывки в планшет добавляли разведенное вторичное антитело (козье антитело к человеческому IgG-HRP, фирма Abcam, каталожный №ab98624) и инкубировали в течение 1 ч при комнатной температуре, а затем промывали 4 раза ЗФР-Т. После промывки в планшет добавляли раствор ТМВ и инкубировали в течение 15 мин при комнатной температуре, после чего добавляли 1н. серную кислоту для прекращения реакции. Абсорбцию (оптическая плотность; ОП) измеряли при 450 нм/570 нм. Антитело к KLH (IC17-IgG1) представляло собой применяемое в качестве отрицательного контроля антитело, а антитело к зрелому TGF-бета GC1008 (описанное в U.S. №8383780), имеющее Fc-область человеческого IgG1 (GC1008-F1332m), представляло собой применяемое в качестве положительного контроля антитело. Как продемонстрировано на фиг. 2А и 2Б, антитела к латентному TGF-бета 1 не связывались ни с мышиным зрелым TGF-бета 1, ни с человеческим зрелым TGF-бета 1.
Кроме того, связывающуюся активность антител к латентному TGF-бета 1 (hT0947AE04-SG191, hT0947AE07-SG191, hT0947AE08-SG191 или hT0947A09-SG191) с мышиным ассоциированным с латентность белком (LAP) тестировали с помощью ELISA согласно описанному выше методу. Как продемонстрировано на фиг. 2 В, антитела к латентному TGF-бета 1 связывались с мышиным LAP.
(4-3) Антитело к латентному TGF-бета L ингибирующее спонтанную активацию латентного TGF-бета 1
Мышиный латентный TGF-бета 1 (mSLC) и человеческий латентный TGF-бета 1 (hSLC), полученные согласно методу, описанному в примере (1-1), каждый инкубировали при 37°С в течение 1 ч в присутствии антитела к латентному TGF-бета 1 (hT0947AE04-SG191, hT0947AE07-SG191, hT0947AE08-SG191 или hT0947AE09-SG191) или без антитела. Антитело к KLH (IC17-IgG1) применяли в качестве отрицательного контроля. Спонтанную активацию латентного TGF-бета 1 и опосредуемое антителом ингибирование спонтанной активации латентного TGF-бета 1, изучали с помощью ELISA, предназначенного для анализа зрелого TGF-бета 1 (набор Human TGF-beta 1 Quantikine ELISA Kit, фирма R&D systems) согласно предложенной производителем процедуре.
Как продемонстрировано на фиг. 3, спонтанная активация латентного TGF-бета 1 подавлялась антителами к латентному TGF-бета 1.
(4-4) Антитело к латентному TGF-бета 1, ингибирующее опосредуемую плазмином (PLN) активацию латентного TGF-бета 1
Мышиный латентный TGF-бета 1 (mSLC) и человеческий латентный TGF-бета 1 (hSLC), полученные согласно методу, описанному в примере (1-1), каждый инкубировали при 37°С в течение 1 ч с человеческим плазмином (фирма Calbiochem) в присутствии антител к латентному TGF-бета 1 (hT0947AE04-SG191, hT0947AE07-SG191, hT0947AE08-SG191 или hT0947AE09-SG191) или без антител. Антитела предварительно инкубировали с мышиным или человеческим латентным TGF-бета 1 (SLC) в течение 30 мин при комнатной температуре, после чего инкубировали с плазмином. Антитело к KLH (IC 17-hlgG1) применяли в качестве отрицательного контроля. Опосредуемую плазмином активацию латентного TGF-бета 1 и опосредуемое антителом ингибирование изучали с помощью ELISA, предназначенного для анализа зрелого TGF-бета 1 (набор Human TGF-beta 1 Quantikine ELISA Kit, фирма R&D systems) согласно предложенной производителем процедуре. Как продемонстрировано на фиг. 4, опосредуемая плазмином активация латентного TGF-бета 1 подавлялась антителами к латентному TGF-бета 1.
(4-5) Антитело к латентному TGF-бета 1, ингибирующее опосредуемую плазматическим калликреином (PLK) активацию латентного TGF-бета 1
Мышиный латентный TGF-бета 1 (mSLC) и человеческий латентный TGF-бета 1 (hSLC), полученные согласно методу, описанному в примере (1-1), каждый инкубировали при 37°С в течение 2 ч с человеческим калликреином (фирма Enzyme Research Laboratories) в присутствии антител к латентному TGF-бета 1 (hT0947AE04-SG191, hT0947AE07-SG191, hT0947AE08-SG191 или hT0947AE09-SG191) или без антител. Антитела предварительно инкубировали с мышиным или человеческим латентным TGF-бета 1 (SLC) в течение 30 мин при комнатной температуре, после чего инкубировали с калликреином. Антитело к KLH (IC17-hIgG1) применяли в качестве отрицательного контроля. Опосредуемую калликреином активацию латентного TGF-бета 1 и опосредуемое антителом ингибирование изучали с помощью ELISA, предназначенного для анализа зрелого TGF-бета 1 (набор Human TGF-beta 1 Quantikine ELISA Kit, фирма R&D systems) согласно предложенной производителем процедуре. Как продемонстрировано на фиг. 5, опосредуемая калликреином активация латентного TGF-бета 1 подавлялась антителами к латентному TGF-бета 1.
(4-6) Антитело к латентному TGF-бета L ингибирующее опосредуемую ММР2 и ММР9 активацию человеческого латентного TGF-бета 1
Человеческий латентный TGF-бета 1 (SLC), который получали согласно методу, описанному в примере (1-1), инкубировали при 37°С в течение 2 ч с активирующей металлопротеиназой 2 (ММР2) или ММР9 (фирма R&D systems) в присутствии антител к латентному TGF-бета 1 (hT0947AE04-SG191, hT0947AE07-SG191, hT0947AE08-SG191 или hT0947AE09-SG191) или без антител. Антитела предварительно инкубировали с человеческим латентным TGF-бета 1 (SLC) в течение 30 мин при комнатной температуре, после чего инкубировали с ММР2 или ММР9. Антитело к KLH (IC17-hIgG1) применяли в качестве отрицательного контроля. Опосредуемую ММР2 и ММР9 активацию человеческого латентного TGF- бета 1 и опосредуемое антителом ингибирование изучали с помощью ELISA, предназначенного для анализа зрелого TGF-бета 1 (набор Human TGF-beta 1 Quantikine ELISA Kit, фирма R&D systems) согласно предложенной производителем процедуре. Как продемонстрировано на фиг. 6, и опосредуемая ММР2 активация человеческого латентного TGF-бета 1, и опосредуемая ММР9 активация человеческого латентного TGF-бета 1 подавлялась антителами к латентному TGF-бета 1.
(4-7) Антитело к латентному TGF-бета 1, ингибирующее активацию латентного TGF-бета 1 без предотвращения расщепления пропептида латентного TGF-бета 1 плазмином (PLN)
Мышиный латентный TGF-бета 1 (mSLC) и человеческий латентный TGF-бета 1 (hSLC), полученные согласно методу, описанному в примере (1-1), каждый инкубировали при 37°С в течение 1 ч с человеческим плазмином (фирма Calbiochem) в присутствии антител к латентному TGF-бета 1 (hT0947AE04-SG191, hT0947AE07-SG191, hT0947AE08-SG191 или hT0947AE09-SG191) или без антител. Антитела предварительно инкубировали с мышиным или человеческим латентным TGF-бета 1 (SLC) в течение 30 мин при комнатной температуре перед инкубацией с плазмином. Камостат мезилат (фирма TOCRIS), представляющий собой один из ингибиторов сериновой протеазы, которого известна способность ингибировать активность плазмина, применяли в качестве контроля. Образцы смешивали с 4-кратным буфером для образа (фирма Wako), применяемым для ДСН-ПААГ, затем выдерживали при 95°С в течение 5 мин, после чего вносили в систему для гель-электрофореза в присутствии ДНС.Белки переносили на мембрану с помощью системы для переноса Trans-Blot® Turbo™ (фирма Bio-rad). Пропептид латентного TGF-бета 1 выявляли с использованием мышиного антитела к FLAG, M2-HRP (фирма Sigma-Aldrich). Мембрану инкубировали с ECL-субстратом и визуализацию осуществляли с помощью устройства ImageQuant LAS 4000 (фирма GE Healthcare).
Как продемонстрировано на фиг. 7, расщепление пропептида латентного TGF-бета 1 плазмином не ингибировалось антителами к латентному TGF-бета 1.
(4-8) Антитело к латентному TGF-бета L существенно не ингибирующее опосредуемую интегрином активацию латентного TGF-бета 1 в мышиных РВМС
Анализ с использованием совместного культивирования мышиных РВМС и клеток HEK-Blue™ TGF-beta осуществляли для обнаружения опосредуемой интегрином активации латентного TGF-бета 1. Мышиные РВМС выделяли из крови мышей с использованием среды с градиентом плотности Histopaque-1083 (фирма Sigma-Aldrich). Клетки HEK-Blue™ TGF-beta (фирма Invivogen), которые экспрессируют индуцируемые связывающими элементами Smad3/4 (SBE) репортерные гены SEAP, позволяют выявлять биоактивный TGF-бета 1 (как мышиный TGF-бета 1, так и человеческий TGF-бета 1) путем мониторинга активации Smad3/4. Активный TGF-бета 1 стимулирует производство белка SEAP и его секрецию в клеточный супернатант.Количество секретируемого SEAP оценивали с помощью реагента QUANTI-Blue™ (фирма Invivogen).
Клетки HEK-Blue™ TGF-beta поддерживали в среде DMEM (фирма Gibco), дополненной 10% фетальной бычьей сыворотки, 50 ед./мл стрептомицина, 50 мкг/мл пенициллина, 100 мкг/мл нормоцина, 30 мкг/мл бластицидина, 200 мкг/мл HygroGold и 100 мкг/мл зеоцина. В процессе осуществления функционального анализа среду для клеток заменяли на среду для анализа (RPMI1640 с 10% FBS) и вносили в 96-луночный планшет. Затем в лунки вносили антитела к латентному TGF-бета 1 (hT0947AE04-SG191, hT0947AE07-SG191, hT0947AE08-SG191 или hT0947AE09-SG191) и мышиные РВМС и инкубировали вместе с клетками HEK-Blue™ TGF-beta в течение ночи. Затем клеточный супернатант смешивали с QUANTI-Blue™ и измеряли оптическую плотность (ОП) при 620 нм с помощью колориметрического планшет-ридера. Известно, что пептид RGD (GRRGDLATIH, фирма GenScript) связывается с интегринами и служит в качестве «ловушки» для лиганда интегрина, что приводит к подавлению опосредуемой интегрином активации TGF-бета 1. По этой причине пептид RGD применяли в качестве положительного контроля. Кроме того, контрольный пептид RGE (GRRGELATIH, фирма GenScript), для которого известно, что он не служит в качестве «ловушки» для лиганда интегрина, применяли в качестве отрицательного контроля. Антитело к KLH (IC17-hIgG1) применяли в качестве отрицательного контроля. Антитело к зрелому TGF-бета 1 (GC1008-F 1332m) применяли в качестве положительного контроля. F1332m обозначает константную область тяжелой цепи человеческого IgG1, которая включает аминокислотные замены для снижения эффекторной функции.
Как продемонстрировано на фиг. 8, антитела к латентному TGF-бета 1 не ингибировали существенно опосредуемую интегрином активацию TGF-бета 1 в мышиных РВМС.
Пример 5. Противоопухолевая активность антитела к латентному TGF-бета 1(1)
Эффективность in vivo моноклонального антитела к латентному TGF-бета 1 hT0947AE04-mF18 индивидуально или в комбинации с антителом к PD-L1 оценивали на мышиной сингенной модели, созданной с использованием клеток карциномы молочной железы мышей ЕМТ6 и мышей линии Balb/c, на которой установлено, что лечение только ингибитором иммунных контрольных точек оказывало ограниченное действие на рост опухолей и выживаемость (см. Nature 554(7693), 22 февраля 2018 г., сс.544-548.).
(5-1) Создание мышиной сингенной модели
Линию клеток карциномы молочной железы мышей ЕМТ6 получали из Американской коллекции типовых культур (АТСС CRL-2755). Клетки культивировали в среде RPMI-1640 (фирма SIGMA), дополненной 2 мМ L-глутамином (фирма SIGMA) с 10% фетальной бычьей сыворотки (FBS; фирма SIGMA). Свободных от специфических патогенов самок мышей линии Balb/c возрастом 6 недель покупали у фирмы Japan Charles River Inc. и давали пройти акклиматизацию в течение 2 недель перед инокуляцией. Клетки линии ЕМТ6 на логарифмической фазе роста собирали и промывали сбалансированным солевым раствором Хэнкса (HBSS; фирма SIGMA), ресуспендировали в 50% HBSS и 50% Матригеля (фирма CORNING) в концентрации 1×106 клеток/мл. Мышам инокулировали в левую жировую подушку молочной железы из пятой пары (№5) 1×105 ЕМТ6-клеток в 100 мкл HBSS:Матригель (1:1).
Когда средний объем опухолей достигал примерно 100-300 мм3 (7 дней после инокуляции), мышей произвольно разделяли на группы на основе веса опухоли и массы тела. Объем опухоли измеряли с помощью кронциркуля и объем опухоли рассчитывали следующим образом:
Объем опухоли (мм3)=(1/2) × длина (мм) × ширина (мм)2
(5-2) Оценка противоопухолевой активности
После создания описанной в примере (5-1) мышиной модели мышей обрабатывали антителами, применяемыми в качестве контроля изотипа (мышиное антитело IgG1-изотипа в комбинации с крысиным антителом IgG2b-изотипа, которые покупали у фирмы Bio X Cell), антителом к мышиному PD-L1 (крысиный IgG2b, клон 10F.9G2, покупали у фирмы Bio X Cell), hT0947AE04-mF18 или комбинацией hT0947AE04-mF18 с антителом к мышиному PD-L1, как указано в таблице 4. Антитела вводили трижды в неделю в течение 3 недель. Первую дозу вводили внутривенно, а вторую и последующие дозы вводили внутрибрюшинно.
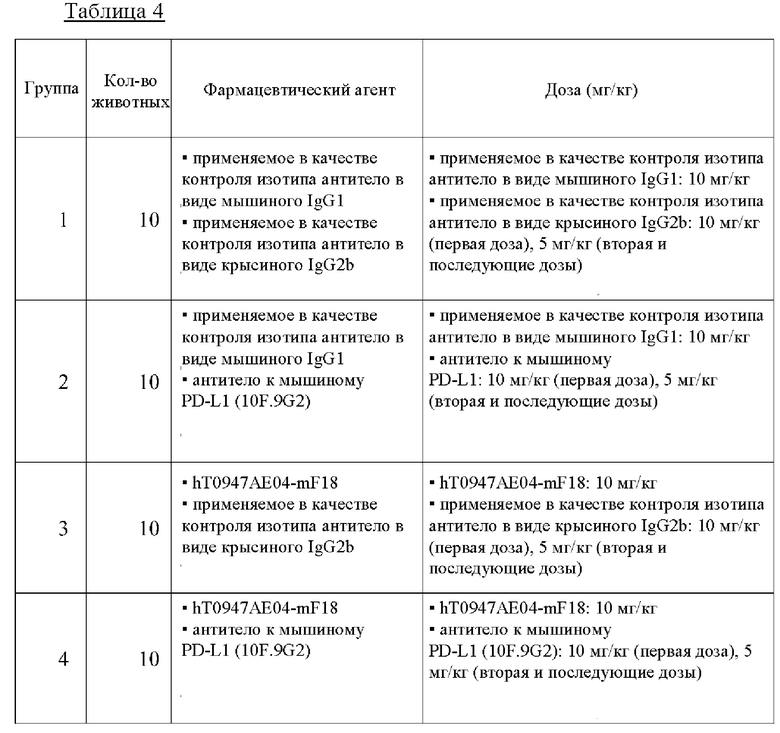
Объем опухолей измеряли дважды в неделю. Результаты представлены на фиг. 9.
Противоопухолевую активность оценивали также по ингибированию роста опухоли (TGI[%]). TGI[%] для конкретной группы в конкретный день рассчитывали следующим образом:
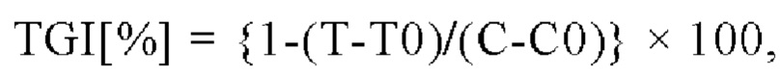
где Т' обозначает средний объем опухоли в группе в день измерения, 'Т0' обозначает средний объем опухоли в группе в день рандомизации, 'С' обозначает' средний объем опухоли в группе 1 (контроль изотипа) в день измерения и 'С0' обозначает средний объем опухоли в группе 1 (контроль изотипа) в день рандомизации.
В результате установлено, что величины TGI[%] для антитела к мышиному PD-L1 (группа 2), hT0947AE04-mF18 (группа 3) и комбинации hT0947AE04-mF18 и антитела к мышиному PD-L1 (группа 4) в день 14 после введения первой дозы составляли 51, 12 и 83 соответственно. Таким образом, обнаружено синергетическое противоопухолевое действие при применении антитела к латентному TGF-бета 1 (hT0947AE04-mF18) и антитела к PD-L1.
Строили также кривые выживаемости для оценки выживаемости в каждой группе. Мышь рассматривали как «выжившую», если объем опухоли у нее не превышал 1955 мм3. Как продемонстрировано на фиг. 10, комбинированная терапия hT0947AE04-mF18 с антителом к мышиному PD-L1 (группа 4) существенно повышала выживаемость мышей по сравнению с мышами, которых обрабатывали только антителом к мышиному PD-L1 (группа 2) и только hT0947AE04-mF18 (группа 3).
Пример 6. Противоопухолевая активность антитела к латентному TGF-бета 1(2)
Эффективность in vivo моноклональных антител к латентному TGF-бета 1 hT0947AE04-mF18, hT0947AE07-SG181 или hT0947AE08-SG181 в комбинации с антителом к PD-L1 оценивали на мышиной сингенной модели, созданной с использованием клеток карциномы молочной железы мышей ЕМТ6 и мышей линии Balb/c.
(6-1) Создание мышиной сингенной модели
Линию клеток карциномы молочной железы мышей ЕМТ6 получали из Американской коллекции типовых культур (АТСС CRL-2755). Клетки культивировали в среде RPMI-1640 (фирма SIGMA), дополненной 2 мМ L-глутамином (фирма SIGMA) с 10% фетальной бычьей сыворотки (FBS; фирма SIGMA). Свободных от специфических патогенов самок мышей линии Balb/c возрастом 7 недель покупали у фирмы Japan Charles River Inc. и давали пройти акклиматизацию в течение 1 недели перед инокуляцией. Клетки линии ЕМТ6 на логарифмической фазе роста собирали и промывали сбалансированным солевым раствором Хэнкса (HBSS; фирма SIGMA), ресуспендировали в 50% HBSS и 50% Матригеля (фирма CORNING) в концентрации 1×106 клеток/мл. Мышам инокулировали в левую жировую подушку молочной железы из пятой пары (№5) 1×105 ЕМТб-клеток в 100 мкл HBSS:Матригель (1:1).
Когда средний объем опухолей достигал примерно 100-300 мм3 (7 дней после инокуляции), мышей произвольно разделяли на группы на основе веса опухоли и массы тела. Объем опухоли измеряли с помощью кронциркуля и объем опухоли рассчитывали следующим образом:
Объем опухоли (мм3)=(1/2) × длина (мм) × ширина (мм)2
(6-2) Оценка противоопухолевой активности
После создания описанной в примере (6-1) мышиной модели мышей обрабатывали наполнителем (буфер 150 мМ NaCl/20MM His-HCl, рН 6,0), антителом к мышиному PD-L1 (крысиный IgG2b, клон 10F.9G2, покупали у фирмы Bio X Cell), комбинацией hT0947AE04-mF18 с антителом к мышиному PD-L1, комбинацией hT0947AE07-SG181 с антителом к мышиному PD-L1 или комбинацией hT0947AE08-SG181 с антителом к мышиному PD-L1, как указано в таблице 5. Антитела вводили трижды в неделю в течение 3 недель. Первую дозу вводили внутривенно, а вторую и последующие дозы вводили внутрибрюшинно.

Объем опухолей измеряли дважды в неделю. Результаты представлены на фиг. 11. Противоопухолевую активность оценивали также по ингибированию роста опухоли (TGI[%]). (TGI[%]) рассчитывали как {1-(Т-Т0)/(С-С0)} × 100, аналогично описанному в примере (5-2).
Величины TGI[%] для антитела к мышиному PD-L1 при его индивидуальном применении (группа 2), для комбинации hT0947AE04-mF18 с антителом к мышиному PD-L1 (группа 3), комбинации hT0947AE07-SG181 с антителом к мышиному PD-L1 (группа 4) и комбинации hT0947AE08-SG181 с антителом к мышиному PD-L1 (группа 5) в день 14 после введения первой дозы составляли 64, 89, 86 и 76 соответственно. Таким образом, для антител к латентному TGF-бета 1 установлено наличие комбинаторной эффективности с антителом к PD-L1.
Пример 7. In vivo эффективность антител к латентному TGF-бета 1 на мышиной модели индуцированного UUO почечного фиброза
Эффективность in vivo моноклональных антител hT0947AE04-SG191, hT0947AE07-SG191 и hT0947AE08-SG191 оценивали на мышиной модели, созданной с применением односторонней обструкции мочеточника (UUO), что, как известно, индуцирует прогрессирующий почечный фиброз.
(7-1) Создание мышиной модели индуцированного UUO почечного фиброза
Эффективность in vivo моноклональных антител hT0947AE04-SG191, hT0947AE07-SG191 и hT0947AE08-SG191 оценивали на мышиной модели, созданной с применением односторонней обструкции мочеточника (UUO), что, как известно, индуцирует прогрессирующий почечный фиброз.
Свободных от специфических патогенов самцов мышей линии C57BL/6NTac возрастом 6 недель покупали у фирмы Invivos Pte Ltd (Сингапур) и давали пройти акклиматизацию в течение 1 недели до начала обработок. Животных содержали при 20-26°С и световом цикле 12:12 часов света/темноты,и кормили поступающим в продажу стандартным кормом (5Р75; фирма PMI Nutrition INT'L (LabDiet), шт. Миссури, США), и животные имели свободный доступ к водопроводной воде.
Хирургическое вмешательство для осуществления UUO осуществляли в условиях анестезии изофлураном. На левой стороне брюшной области сбривали шерсть и на коже делали вертикальный надрез. Второй надрез осуществляли через брюшину и эту кожу также оттягивали для того, чтобы обнажить почку. С помощью пинцетов почку переносили на поверхность и левый мочеточник дважды перевязывали хирургическим шелком ниже почки. Почку с лигатурой осторожно возвращали назад в правильное анатомическое положение, после чего на брюшину и кожу накладывали швы. Добавляли обезболивающее средство для уменьшения страдания животного. У группы, подвергнутой плацебо-хирургии, брюшину и кожу только надрезали и зашивали.
(7-2) Оценка эффективности in vivo
Все моноклональные антитела вводили в дозе 15 мг/кг с помощью внутривенной инъекции три раза в неделю, начиная с одного дня до хирургической операции. В этом опыте антитело к KLH (IC17dk-SG181) применяли в качестве отрицательного контроля. Группе, подвергнутой плацебо-хирургии, вводили антитело к KLH (IC17dk-SG181). Животных взвешивали и затем через 7 дней после операции умерщвляли посредством обескровливания под анестезией изофлураном. Образцы крови собирали из полостей сердца или посткавальной вены и выдерживали при -80°С до анализа. Почку быстро изымали. Часть почечной ткани подвергали шоковой заморозке в жидком азоте или на сухом льду для молекулярных анализов.
Содержание гидроксипролина в почке, который представляет собой одну из аминокислот, входящих в состав коллагена, измеряли для оценки отложения внешнего матрикса в ткани. Влажную ткань почек сушили при температуре вплоть до 95°С в течение 3 ч и взвешивали. Затем к высушенной ткани добавляли 6н. HCl (100 мкл/ 1 мг сухой ткани) и кипятили в течение ночи. Образцы очищали, пропуская через фильтр, и 10 мкл каждого образца вносили в 96-луночный планшет. Планшет с образцами сушили при 60°С и содержание гидроксипролина измеряли с помощью набора для анализ гидроксипролина (фирма BioVision). Результаты этого эксперимента представлены на фиг. 12. Существенное увеличение содержания гидроксипролина обнаружено в почке с индуцированным заболеванием, и все антитела (hT0947AE04-SG191, hT0947AE07-SG191 и hT0947AE08-SG191) ингибировали почечный фиброз. Данные представлены в виде среднего значения ± стандартная ошибка среднего (SEM). Статистический анализ осуществляли с помощью t-критерия Стьюдента. При значении Р<0,05 различия рассматривались как значимые.
Пример 8. Оценка токсичности антител к латентному TGF-бета 1 Потенциальную токсичность антитела к латентному TGF-бета 1 оценивали на основе токсичности повторяющейся дозы на здоровых мышах и обезьянах циномолгус, осуществляя сравнение с антителом к зрелому TGF-бета GC1008-mF18 (антитело к зрелому TGF-бета GC1008 (описанное в US №8383780), имеющее Fc-область мышиного IgG mF18). Поскольку антитело к латентному TGF-бета 1 дает перекрестную реакцию на антиген мышей и обезьян циномолгус, то мыши и обезьяны циномолгус выбраны в качестве видов животных для опытов по оценке токсичности in vivo. Обобщение результатов всех токсикологических исследований представлено в таблице 6.
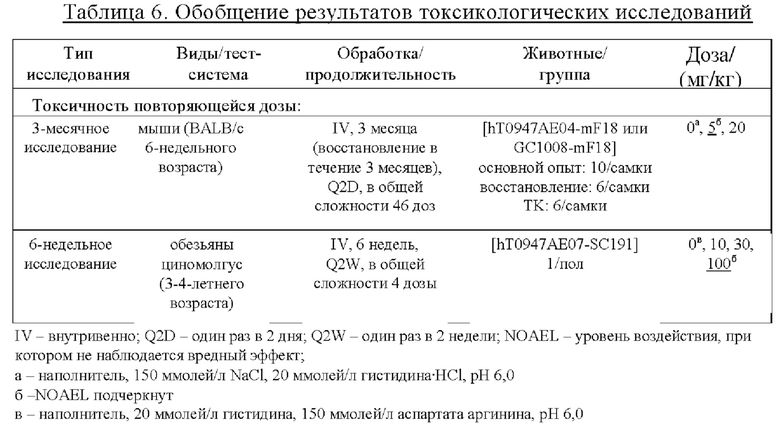
В 3-месячном опыте на мышах (IV; 5 или 20 мг/кг, Q2D, в целом 46 доз) обнаружены характерные для анемии изменения (в группе, обработанной hT0947AE04-mF18 в дозе 20 мг/кг; группах, обработанных GC1008-mF18 в дозах 5 и 20 мг/кг) и поражения сердца (в группах, обработанных GC1008-mF18 в дозах дозой 5 и 20 мг/кг; см. таблицу 7). Можно предположить, что полученные данные являются результатом нецелевой токсичности ингибирования TGF-бета. Принимая во внимание нецелевую токсичность, установленную в 3-месячном опыте на мышах, NOAEL (уровень воздействия, при котором не наблюдается вредный эффект) для hT0947AE04-mF18 составлял 5 мг/кг IV Q2D. Кроме того, с учетом неблагоприятных воздействий GC1008-mF18 в группе, обработанной дозой 5 мг/кг, NOAEL для GC1008-mF18 не был установлен в условиях 3-месячного опыта на мышах.
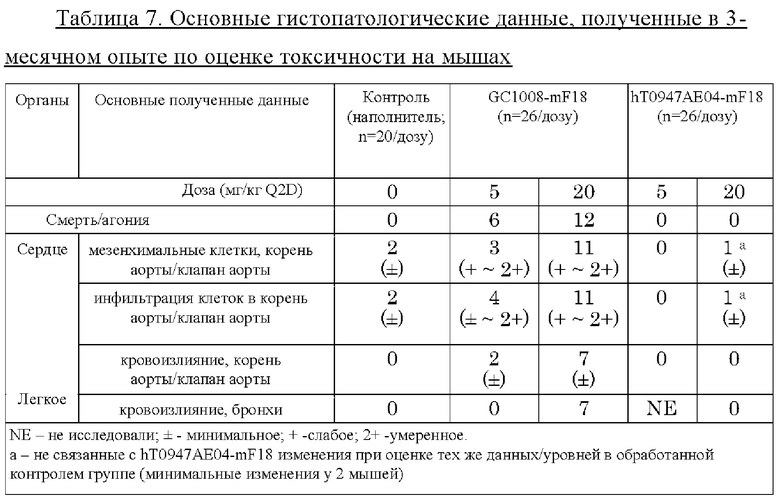
В 6-недельном опыте на обезьянах (IV; 10, 30 или 100 мг/кг, Q2W, в целом 4 дозы) не обнаружено никаких релевантных токсикологических изменений, связанных с IV-введением hT0947AE07-SG191, и NOAEL соответствовал наиболее высокой из протестированных доз 100 мг/кг Q2W.
Пример 9. Противоопухолевая активность антитела к латентному TGF-бета 1(3)
Эффективность in vivo моноклонального антитела к латентному TGF-бета 1 hT0947AE07-SG191 в комбинации с антителом к PD-L1 оценивали на мышиной сингенной модели, созданной с использованием клеток карциномы молочной железы мышей ЕМТ6 и мышей линии Balb/c.
Линию клеток карциномы молочной железы мышей ЕМТ6 получали из Американской коллекции типовых культур (АТСС CRL-2755). Клетки культивировали в среде RPMI-1640 (фирма SIGMA) с 10% фетальной бычьей сыворотки (FBS; фирма Nichirei Biosciences Inc.). Свободных от специфических патогенов самок мышей линии Balb/c возрастом 6 недель покупали у фирмы Japan Charles River Inc. и давали пройти акклиматизацию в течение 1 недели перед осуществлением инокуляции. Клетки линии ЕМТ6 на логарифмической фазе роста собирали и промывали сбалансированным солевым раствором Хэнкса (HBSS; фирма SIGMA), ресуспендировали в 50% HBSS и 50% Матригеля (фирма CORNING) в концентрации 1×106 клеток/мл. Мышам инокулировали в левую жировую подушку молочной железы из пятой пары (№5) 1×105 ЕМТ6-клеток в 100 мкл HBSS:Матригель (1:1).
Когда средний объем опухолей достигал примерно 100-300 мм3 (7 дней после инокуляции), мышей произвольно разделяли на группы на основе веса опухоли и массы тела. Объем опухоли измеряли с помощью кронциркуля и объем опухоли рассчитывали по формуле:
Объем опухоли рассчитывали как (1/2) × 1 × w2, где 1 обозначает длину, w обозначает ширину.
Мышей обрабатывали наполнителем (буфер 150 мМ NaCl/20 мМ His-HCl, рН 6,0), антителом к мышиному PD-L1 (крысиный IgG2b, клон 10F.9G2, покупали у фирмы Bio X Cell, доза 10 мг/кг для первой обработки, доза 5 мг/кг для последующих обработок), комбинацией hT0947AE07-SG191 (10 мг/кг) с антителом к мышиному PD-L1, или комбинацией hT0947AE07-SG191 (30 мг/кг) с антителом к мышиному PD-L1. Антитела вводили 3 раза в неделю в течение 2 недель, первую дозу внутривенно и последующие дозы внутрибрюшинно.
Объем опухоли измеряли дважды в неделю. Противоопухолевую активность оценивали по ингибированию роста опухоли (TGI[%]), указанную величину рассчитывали по формуле {1-(Т-Т0)/(С-С0)} × 100, где средние объемы опухолей в группах обозначали как Т в день измерения и Т0 в день рандомизации, что соответствует С и С0 для обработанной наполнителем группы. Результаты этого эксперимента представлены на фиг. 13.
Величины TGI[%] для антитела к мышиному PD-L1 при его индивидуальном применении, комбинации hT0947AE07-SG191 (10 мг/кг) с антителом к мышиному PD-L1 и комбинации hT0947AE07-SG191 (30 мг/кг) с антителом к мышиному PD-L1 в день 14 после введения первой дозой составляли 60, 77 и 80 соответственно. Для hT0947AE07-SG191 установлено наличие комбинаторной эффективности с антителом к мышиному PD-L1.
Хотя выше изобретение описано для лучшего понимания с указанием некоторых деталей с целью иллюстрации и примеров, описание и примеры не должны рассматриваться в качестве ограничивающих объем изобретения. Описание всей патентной и научной литературы, процитированной в настоящем описании, специально полностью включено в настоящее описание в качестве ссылки.
Промышленная применимость
В настоящем изобретении предложены межвидовые антитела к латентному TGF-бета 1, которые ингибируют опосредуемую протеазой активацию латентного TGF-бета 1 без ингибирования опосредуемой интегрином активации латентного TGF-бета 1. В изобретении предложены также комбинированные терапии, включающие антитело к латентному TGF-бета 1 и ингибитор контрольных точек. Ожидается, что антитела к латентному TGF-бета 1, предлагаемые в настоящем изобретении, которые можно применять в комбинации с ингибитором контрольных точек, могут оказаться ценными для лечения связанных с TGF-бета 1 заболеваний, таких как фиброз и рак.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ЧУГАИ СЕЙЯКУ КАБУСИКИ КАЙСЯ
<120> МЕЖВИДОВЫЕ АНТИТЕЛА К ЛАТЕНТНОМУ TGF-БЕТА
1 И ИХ ПРИМЕНЕНИЯ
<130> C1-A1921P
<150> JP 2019-155278
<151> 2019-08-28
<160> 73
<170> PatentIn version 3.5
<210> 1
<211> 387
<212> PRT
<213> Artificial Sequence
<220>
<223> flag-tagged human latent TGF-beta 1
<400> 1
Met Lys Trp Val Thr Phe Leu Leu Leu Leu Phe Ile Ser Gly Ser Ala
1 5 10 15
Phe Ser Asp Tyr Lys Asp Asp Asp Asp Lys Leu Ser Thr Ser Lys Thr
20 25 30
Ile Asp Met Glu Leu Val Lys Arg Lys Arg Ile Glu Ala Ile Arg Gly
35 40 45
Gln Ile Leu Ser Lys Leu Arg Leu Ala Ser Pro Pro Ser Gln Gly Glu
50 55 60
Val Pro Pro Gly Pro Leu Pro Glu Ala Val Leu Ala Leu Tyr Asn Ser
65 70 75 80
Thr Arg Asp Arg Val Ala Gly Glu Ser Ala Glu Pro Glu Pro Glu Pro
85 90 95
Glu Ala Asp Tyr Tyr Ala Lys Glu Val Thr Arg Val Leu Met Val Glu
100 105 110
Thr His Asn Glu Ile Tyr Asp Lys Phe Lys Gln Ser Thr His Ser Ile
115 120 125
Tyr Met Phe Phe Asn Thr Ser Glu Leu Arg Glu Ala Val Pro Glu Pro
130 135 140
Val Leu Leu Ser Arg Ala Glu Leu Arg Leu Leu Arg Leu Lys Leu Lys
145 150 155 160
Val Glu Gln His Val Glu Leu Tyr Gln Lys Tyr Ser Asn Asn Ser Trp
165 170 175
Arg Tyr Leu Ser Asn Arg Leu Leu Ala Pro Ser Asp Ser Pro Glu Trp
180 185 190
Leu Ser Phe Asp Val Thr Gly Val Val Arg Gln Trp Leu Ser Arg Gly
195 200 205
Gly Glu Ile Glu Gly Phe Arg Leu Ser Ala His Cys Ser Cys Asp Ser
210 215 220
Arg Asp Asn Thr Leu Gln Val Asp Ile Asn Gly Phe Thr Thr Gly Arg
225 230 235 240
Arg Gly Asp Leu Ala Thr Ile His Gly Met Asn Arg Pro Phe Leu Leu
245 250 255
Leu Met Ala Thr Pro Leu Glu Arg Ala Gln His Leu Gln Ser Ser Arg
260 265 270
His Arg Arg Ala Leu Asp Thr Asn Tyr Cys Phe Ser Ser Thr Glu Lys
275 280 285
Asn Cys Cys Val Arg Gln Leu Tyr Ile Asp Phe Arg Lys Asp Leu Gly
290 295 300
Trp Lys Trp Ile His Glu Pro Lys Gly Tyr His Ala Asn Phe Cys Leu
305 310 315 320
Gly Pro Cys Pro Tyr Ile Trp Ser Leu Asp Thr Gln Tyr Ser Lys Val
325 330 335
Leu Ala Leu Tyr Asn Gln His Asn Pro Gly Ala Ser Ala Ala Pro Cys
340 345 350
Cys Val Pro Gln Ala Leu Glu Pro Leu Pro Ile Val Tyr Tyr Val Gly
355 360 365
Arg Lys Pro Lys Val Glu Gln Leu Ser Asn Met Ile Val Arg Ser Cys
370 375 380
Lys Cys Ser
385
<210> 2
<211> 1167
<212> DNA
<213> Artificial Sequence
<220>
<223> flag-tagged human latent TGF-beta 1
<400> 2
atgaagtggg taacctttct cctcctcctc ttcatctccg gttctgcctt ttccgactac 60
aaggatgacg atgacaagct atccacctcc aagactatcg acatggagct ggtgaagcgg 120
aagcgcatcg aggccatccg cggccagatc ctgtccaagc tgcggctcgc cagccccccg 180
agccaggggg aggtgccgcc cggcccgctg cccgaggccg tgctcgccct gtacaacagc 240
acccgcgacc gggtggccgg ggagagtgca gaaccggagc ccgagcctga ggccgactac 300
tacgccaagg aggtcacccg cgtgctaatg gtggaaaccc acaacgaaat ctatgacaag 360
ttcaagcaga gtacacacag catatatatg ttcttcaaca catcagagct ccgagaagcg 420
gtacctgaac ccgtgttgct ctcccgggca gagctgcgtc tgctgaggct caagttaaaa 480
gtggagcagc acgtggagct gtaccagaaa tacagcaaca attcctggcg atacctcagc 540
aaccggctgc tggcacccag cgactcgcca gagtggttat cttttgatgt caccggagtt 600
gtgcggcagt ggttgagccg tggaggggaa attgagggct ttcgccttag cgcccactgc 660
tcctgtgaca gcagggataa cacactgcaa gtggacatca acgggttcac taccggccgc 720
cgaggtgacc tggccaccat tcatggcatg aaccggcctt tcctgcttct catggccacc 780
ccgctggaga gggcccagca tctgcaaagc tcccggcacc gccgagccct ggacaccaac 840
tattgcttca gctccacgga gaagaactgc tgcgtgcggc agctgtacat tgacttccgc 900
aaggacctcg gctggaagtg gatccacgag cccaagggct accatgccaa cttctgcctc 960
gggccctgcc cctacatttg gagcctggac acgcagtaca gcaaggtcct ggccctgtac 1020
aaccagcata acccgggcgc ctcggcggcg ccgtgctgcg tgccgcaggc gctggagccg 1080
ctgcccatcg tgtactacgt gggccgcaag cccaaggtgg agcagctgtc caacatgatc 1140
gtgcgctcct gcaagtgcag ctgatga 1167
<210> 3
<211> 387
<212> PRT
<213> Artificial Sequence
<220>
<223> flag-tagged mouse latent TGF-beta 1
<400> 3
Met Lys Trp Val Thr Phe Leu Leu Leu Leu Phe Ile Ser Gly Ser Ala
1 5 10 15
Phe Ser Asp Tyr Lys Asp Asp Asp Asp Lys Leu Ser Thr Ser Lys Thr
20 25 30
Ile Asp Met Glu Leu Val Lys Arg Lys Arg Ile Glu Ala Ile Arg Gly
35 40 45
Gln Ile Leu Ser Lys Leu Arg Leu Ala Ser Pro Pro Ser Gln Gly Glu
50 55 60
Val Pro Pro Gly Pro Leu Pro Glu Ala Val Leu Ala Leu Tyr Asn Ser
65 70 75 80
Thr Arg Asp Arg Val Ala Gly Glu Ser Ala Asp Pro Glu Pro Glu Pro
85 90 95
Glu Ala Asp Tyr Tyr Ala Lys Glu Val Thr Arg Val Leu Met Val Asp
100 105 110
Arg Asn Asn Ala Ile Tyr Glu Lys Thr Lys Asp Ile Ser His Ser Ile
115 120 125
Tyr Met Phe Phe Asn Thr Ser Asp Ile Arg Glu Ala Val Pro Glu Pro
130 135 140
Pro Leu Leu Ser Arg Ala Glu Leu Arg Leu Gln Arg Leu Lys Ser Ser
145 150 155 160
Val Glu Gln His Val Glu Leu Tyr Gln Lys Tyr Ser Asn Asn Ser Trp
165 170 175
Arg Tyr Leu Gly Asn Arg Leu Leu Thr Pro Thr Asp Thr Pro Glu Trp
180 185 190
Leu Ser Phe Asp Val Thr Gly Val Val Arg Gln Trp Leu Asn Gln Gly
195 200 205
Asp Gly Ile Gln Gly Phe Arg Phe Ser Ala His Cys Ser Cys Asp Ser
210 215 220
Lys Asp Asn Lys Leu His Val Glu Ile Asn Gly Ile Ser Pro Lys Arg
225 230 235 240
Arg Gly Asp Leu Gly Thr Ile His Asp Met Asn Arg Pro Phe Leu Leu
245 250 255
Leu Met Ala Thr Pro Leu Glu Arg Ala Gln His Leu His Ser Ser Arg
260 265 270
His Arg Arg Ala Leu Asp Thr Asn Tyr Cys Phe Ser Ser Thr Glu Lys
275 280 285
Asn Cys Cys Val Arg Gln Leu Tyr Ile Asp Phe Arg Lys Asp Leu Gly
290 295 300
Trp Lys Trp Ile His Glu Pro Lys Gly Tyr His Ala Asn Phe Cys Leu
305 310 315 320
Gly Pro Cys Pro Tyr Ile Trp Ser Leu Asp Thr Gln Tyr Ser Lys Val
325 330 335
Leu Ala Leu Tyr Asn Gln His Asn Pro Gly Ala Ser Ala Ser Pro Cys
340 345 350
Cys Val Pro Gln Ala Leu Glu Pro Leu Pro Ile Val Tyr Tyr Val Gly
355 360 365
Arg Lys Pro Lys Val Glu Gln Leu Ser Asn Met Ile Val Arg Ser Cys
370 375 380
Lys Cys Ser
385
<210> 4
<211> 1167
<212> DNA
<213> Artificial Sequence
<220>
<223> flag-tagged mouse latent TGF-beta 1
<400> 4
atgaagtggg taacctttct cctcctcctc ttcatctccg gttctgcctt ttccgactac 60
aaggatgacg atgacaagct ctccacctcc aagaccatcg acatggagct ggtgaaacgg 120
aagcgcatcg aagccatccg tggccagatc ctgtccaaac taaggctcgc cagtccccca 180
agccaggggg aggtaccgcc cggcccgctg cccgaggcgg tgctcgcttt gtacaacagc 240
acccgcgacc gggtggcagg cgagagcgcc gacccagagc cggagcccga agcggactac 300
tatgctaaag aggtcacccg cgtgctaatg gtggaccgca acaacgccat ctatgagaaa 360
accaaagaca tctcacacag tatatatatg ttcttcaata cgtcagacat tcgggaagca 420
gtgcccgaac ccccattgct gtcccgtgca gagctgcgct tgcagagatt aaaatcaagt 480
gtggagcaac atgtggaact ctaccagaaa tatagcaaca attcctggcg ttaccttggt 540
aaccggctgc tgacccccac tgatacgcct gagtggctgt cttttgacgt cactggagtt 600
gtacggcagt ggctgaacca aggagacgga atacagggct ttcgattcag cgctcactgc 660
tcttgtgaca gcaaagataa caaactccac gtggaaatca acgggatcag ccccaaacgt 720
cggggcgacc tgggcaccat ccatgacatg aaccggccct tcctgctcct catggccacc 780
cccctggaaa gggcccagca cctgcacagc tcacggcacc ggagagccct ggataccaac 840
tattgcttca gctccacaga gaagaactgc tgtgtgcggc agctgtacat tgactttagg 900
aaggacctgg gttggaagtg gatccacgag cccaagggct accatgccaa cttctgtctg 960
ggaccctgcc cctatatttg gagcctggac acacagtaca gcaaggtcct tgccctctac 1020
aaccaacaca acccgggcgc ttcggcgtca ccgtgctgcg tgccgcaggc tttggagcca 1080
ctgcccatcg tctactacgt gggtcgcaag cccaaggtgg agcagttgtc caacatgatt 1140
gtgcgctcct gcaagtgcag ctgatga 1167
<210> 5
<211> 387
<212> PRT
<213> Artificial Sequence
<220>
<223> flag-tagged cynomolgus monkey latent TGF-beta 1
<400> 5
Met Lys Trp Val Thr Phe Leu Leu Leu Leu Phe Ile Ser Gly Ser Ala
1 5 10 15
Phe Ser Asp Tyr Lys Asp Asp Asp Asp Lys Leu Ser Thr Ser Lys Thr
20 25 30
Ile Asp Met Glu Leu Val Lys Arg Lys Arg Ile Glu Ala Ile Arg Gly
35 40 45
Gln Ile Leu Ser Lys Leu Arg Leu Ala Ser Pro Pro Ser Gln Gly Glu
50 55 60
Val Pro Pro Gly Pro Leu Pro Glu Ala Val Leu Ala Leu Tyr Asn Ser
65 70 75 80
Thr Arg Asp Arg Val Ala Gly Glu Ser Ala Glu Pro Glu Pro Glu Pro
85 90 95
Glu Ala Asp Tyr Tyr Ala Lys Glu Val Thr Arg Val Leu Met Val Glu
100 105 110
Thr His Asn Glu Ile Tyr Asp Lys Phe Lys Gln Ser Thr His Ser Ile
115 120 125
Tyr Met Phe Phe Asn Thr Ser Glu Leu Arg Glu Ala Val Pro Glu Pro
130 135 140
Val Leu Leu Ser Arg Ala Glu Leu Arg Leu Leu Arg Leu Lys Leu Lys
145 150 155 160
Val Glu Gln His Val Glu Leu Tyr Gln Lys Tyr Ser Asn Asn Ser Trp
165 170 175
Arg Tyr Leu Ser Asn Arg Leu Leu Ala Pro Ser Asp Ser Pro Glu Trp
180 185 190
Leu Ser Phe Asp Val Thr Gly Val Val Arg Gln Trp Leu Ser Arg Gly
195 200 205
Gly Glu Ile Glu Gly Phe Arg Leu Ser Ala His Cys Ser Cys Asp Ser
210 215 220
Lys Asp Asn Thr Leu Gln Val Asp Ile Asn Gly Phe Thr Thr Gly Arg
225 230 235 240
Arg Gly Asp Leu Ala Thr Ile His Gly Met Asn Arg Pro Phe Leu Leu
245 250 255
Leu Met Ala Thr Pro Leu Glu Arg Ala Gln His Leu Gln Ser Ser Arg
260 265 270
His Arg Arg Ala Leu Asp Thr Asn Tyr Cys Phe Ser Ser Thr Glu Lys
275 280 285
Asn Cys Cys Val Arg Gln Leu Tyr Ile Asp Phe Arg Lys Asp Leu Gly
290 295 300
Trp Lys Trp Ile His Glu Pro Lys Gly Tyr His Ala Asn Phe Cys Leu
305 310 315 320
Gly Pro Cys Pro Tyr Ile Trp Ser Leu Asp Thr Gln Tyr Ser Lys Val
325 330 335
Leu Ala Leu Tyr Asn Gln His Asn Pro Gly Ala Ser Ala Ala Pro Cys
340 345 350
Cys Val Pro Gln Ala Leu Glu Pro Leu Pro Ile Val Tyr Tyr Val Gly
355 360 365
Arg Lys Pro Lys Val Glu Gln Leu Ser Asn Met Ile Val Arg Ser Cys
370 375 380
Lys Cys Ser
385
<210> 6
<211> 1167
<212> DNA
<213> Artificial Sequence
<220>
<223> flag-tagged cynomolgus monkey latent TGF-beta 1
<400> 6
atgaagtggg taacctttct cctcctcctc ttcatctccg gttctgcctt ttccgactac 60
aaggatgacg atgacaagct atccacctcc aagactatcg acatggagct ggtgaagcgg 120
aagcgcatcg aggccatccg cggccagatc ctgtccaagc tgcggctcgc cagccccccg 180
agccaggggg aggtgccgcc cggcccgctg cccgaggccg tgctcgccct gtacaacagc 240
acccgcgacc gggtggccgg ggagagtgca gaaccggagc ccgagcctga ggccgactac 300
tacgccaagg aggtcacccg cgtgctaatg gtggaaaccc acaacgaaat ctatgacaag 360
ttcaagcaga gtacacacag catatatatg ttcttcaaca catcagagct ccgagaagcg 420
gtacctgaac ccgtgttgct ctcccgggca gagctgcgtc tgctgaggct caagttaaaa 480
gtggagcagc acgtggagct gtaccagaaa tacagcaaca attcctggcg atacctcagc 540
aaccggctgc tggcacccag cgactcgcca gagtggttat cttttgatgt caccggagtt 600
gtgcggcagt ggttgagccg tggaggggaa attgagggct ttcgccttag cgcccactgc 660
tcctgtgaca gcaaggataa cacactgcaa gtggacatca acgggttcac taccggccgc 720
cgaggtgacc tggccaccat tcatggcatg aaccggcctt tcctgcttct catggccacc 780
ccgctggaga gggcccagca tctgcaaagc tcccggcacc gccgagccct ggacaccaac 840
tattgcttca gctccacgga gaagaactgc tgcgtgcggc agctgtacat tgacttccgc 900
aaggacctcg gctggaagtg gatccacgag cccaagggct accatgccaa cttctgcctc 960
gggccctgcc cctacatttg gagcctggac acgcagtaca gcaaggtcct ggccctgtac 1020
aaccagcata acccgggcgc ctcggcggcg ccgtgctgcg tgccgcaggc gctggagccg 1080
ctgcccatcg tgtactacgt gggccgcaag cccaaggtgg agcagctgtc caacatgatc 1140
gtgcgctcct gcaagtgcag ctgatga 1167
<210> 7
<211> 1824
<212> DNA
<213> Rattus norvegicus
<400> 7
atgaagtggg taacctttct cctcctcctc ttcatctccg gttctgcctt ttccaggggt 60
gtgtttcgcc gagaagcaca caagagtgag atcgcccatc ggtttaagga cttgggagaa 120
cagcatttca aaggcctagt cctgattgcc ttttcccagt atctccagaa atgcccatat 180
gaagagcata tcaaattggt gcaggaagta acagactttg caaaaacatg tgtcgctgat 240
gagaatgccg aaaactgtga caagtccatt cacactctct tcggagacaa gttatgcgcc 300
attccaaagc ttcgcgacaa ctacggtgaa ctggctgact gctgtgcaaa acaagagccc 360
gaaagaaacg agtgtttcct gcagcacaag gatgacaacc ccaacctgcc acccttccag 420
aggccggagg ctgaggccat gtgcacctcc ttccaggaga accctaccag ctttctggga 480
cactatttgc atgaagttgc caggagacat ccttatttct atgccccaga actcctttac 540
tatgctgaga aatacaatga ggttctgacc cagtgctgca cagagtctga caaagcagcc 600
tgcctgacac cgaagcttga tgccgtgaaa gagaaagcac tggtcgcagc tgtccgtcag 660
aggatgaagt gctccagtat gcagagattt ggagagagag ccttcaaagc ctgggcagta 720
gctcgtatga gccagcgatt ccccaatgct gagttcgcag aaatcaccaa attggcaaca 780
gacctcacca aaatcaacaa ggagtgctgt cacggcgacc tgttggaatg cgcggatgac 840
agggcggaac ttgccaagta catgtgtgag aaccaggcca ctatctccag caaactgcag 900
gcttgctgtg ataagccagt gctgcagaaa tcccagtgtc tcgctgagat agaacatgac 960
aacattcctg ccgatctgcc ctcaatagct gctgactttg ttgaggataa ggaagtgtgt 1020
aagaactatg ctgaggccaa ggatgtcttc ctgggcacgt ttttgtatga atattcaaga 1080
aggcaccccg attactccgt gtccctgctg ctgagacttg ctaagaaata tgaagccaca 1140
ctggagaagt gctgtgctga aggcgatcct cctgcctgct acggcacagt gcttgcagaa 1200
tttcagcctc ttgtagaaga acctaagaac ttggtcaaaa ctaactgtga gctttacgag 1260
aagcttggag agtatggatt ccaaaacgcc attctggttc gatacaccca gaaagcacct 1320
caggtgtcga ccccaactct cgtggaggca gcaagaaacc tgggaagagt gggcaccaag 1380
tgttgtaccc ttcctgaagc tcagagactg ccctgtgtgg aagactatct gtctgccatc 1440
ctgaaccgtc tgtgtgtgct gcatgagaag accccagtga gcgagaaggt caccaagtgc 1500
tgtagtgggt ccctggtgga aagacggcca tgtttctctg ctctgacagt tgacgagaca 1560
tatgtcccca aagagtttaa agctgagacc ttcaccttcc actctgatat ctgcacactc 1620
ccagacaagg agaagcagat aaagaagcaa acggctctcg ctgagctggt gaaacacaag 1680
cccaaggcca cagaagatca gctgaagacg gtgatgggtg acttcgcaca attcgtggac 1740
aagtgttgca aggctgccga caaggataac tgcttcgcca ctgaggggcc aaaccttgtt 1800
gctagaagca aagaagcctt agcc 1824
<210> 8
<211> 275
<212> PRT
<213> Artificial Sequence
<220>
<223> flag-tagged mouse LAP
<400> 8
Met Lys Trp Val Thr Phe Leu Leu Leu Leu Phe Ile Ser Gly Ser Ala
1 5 10 15
Phe Ser Asp Tyr Lys Asp Asp Asp Asp Lys Leu Ser Thr Ser Lys Thr
20 25 30
Ile Asp Met Glu Leu Val Lys Arg Lys Arg Ile Glu Ala Ile Arg Gly
35 40 45
Gln Ile Leu Ser Lys Leu Arg Leu Ala Ser Pro Pro Ser Gln Gly Glu
50 55 60
Val Pro Pro Gly Pro Leu Pro Glu Ala Val Leu Ala Leu Tyr Asn Ser
65 70 75 80
Thr Arg Asp Arg Val Ala Gly Glu Ser Ala Asp Pro Glu Pro Glu Pro
85 90 95
Glu Ala Asp Tyr Tyr Ala Lys Glu Val Thr Arg Val Leu Met Val Asp
100 105 110
Arg Asn Asn Ala Ile Tyr Glu Lys Thr Lys Asp Ile Ser His Ser Ile
115 120 125
Tyr Met Phe Phe Asn Thr Ser Asp Ile Arg Glu Ala Val Pro Glu Pro
130 135 140
Pro Leu Leu Ser Arg Ala Glu Leu Arg Leu Gln Arg Leu Lys Ser Ser
145 150 155 160
Val Glu Gln His Val Glu Leu Tyr Gln Lys Tyr Ser Asn Asn Ser Trp
165 170 175
Arg Tyr Leu Gly Asn Arg Leu Leu Thr Pro Thr Asp Thr Pro Glu Trp
180 185 190
Leu Ser Phe Asp Val Thr Gly Val Val Arg Gln Trp Leu Asn Gln Gly
195 200 205
Asp Gly Ile Gln Gly Phe Arg Phe Ser Ala His Cys Ser Cys Asp Ser
210 215 220
Lys Asp Asn Lys Leu His Val Glu Ile Asn Gly Ile Ser Pro Lys Arg
225 230 235 240
Arg Gly Asp Leu Gly Thr Ile His Asp Met Asn Arg Pro Phe Leu Leu
245 250 255
Leu Met Ala Thr Pro Leu Glu Arg Ala Gln His Leu His Ser Ser Arg
260 265 270
His Arg Arg
275
<210> 9
<211> 831
<212> DNA
<213> Artificial Sequence
<220>
<223> flag-tagged mouse LAP
<400> 9
atgaagtggg taacctttct cctcctcctc ttcatctccg gttctgcctt ttccgactac 60
aaggatgacg atgacaagct ctccacctcc aagaccatcg acatggagct ggtgaaacgg 120
aagcgcatcg aagccatccg tggccagatc ctgtccaaac taaggctcgc cagtccccca 180
agccaggggg aggtaccgcc cggcccgctg cccgaggcgg tgctcgcttt gtacaacagc 240
acccgcgacc gggtggcagg cgagagcgcc gacccagagc cggagcccga agcggactac 300
tatgctaaag aggtcacccg cgtgctaatg gtggaccgca acaacgccat ctatgagaaa 360
accaaagaca tctcacacag tatatatatg ttcttcaata cgtcagacat tcgggaagca 420
gtgcccgaac ccccattgct gtcccgtgca gagctgcgct tgcagagatt aaaatcaagt 480
gtggagcaac atgtggaact ctaccagaaa tatagcaaca attcctggcg ttaccttggt 540
aaccggctgc tgacccccac tgatacgcct gagtggctgt cttttgacgt cactggagtt 600
gtacggcagt ggctgaacca aggagacgga atacagggct ttcgattcag cgctcactgc 660
tcttgtgaca gcaaagataa caaactccac gtggaaatca acgggatcag ccccaaacgt 720
cggggcgacc tgggcaccat ccatgacatg aaccggccct tcctgctcct catggccacc 780
cccctggaaa gggcccagca cctgcacagc tcacggcacc ggagatgatg a 831
<210> 10
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> SG181
<400> 10
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Arg Arg Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 11
<211> 107
<212> PRT
<213> Homo sapiens
<400> 11
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 12
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE04H VH
<400> 12
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Asn Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Tyr Thr Ser Gly Thr Thr Tyr His Ala Asn Trp Ala Arg
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gly Thr Phe Ile Trp Asp Tyr Tyr Tyr Trp Val Met Asp Leu Trp
100 105 110
Gly Pro Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 13
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE04L VL
<400> 13
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Ser Ile Ser Thr Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Thr Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Ser Tyr Ser Asp Ala Asp Ser
85 90 95
Val Gly Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 14
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE07H VH
<400> 14
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Glu
20 25 30
Ala Met Asn Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Tyr Thr Ser Gly Thr Thr Tyr Arg Ala Asn Trp Ala Arg
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gly Thr Gly Ile Tyr Asp Tyr Tyr Tyr Trp Val Met Asp Leu Trp
100 105 110
Gly Pro Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 15
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE07L VL
<400> 15
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Ser Ile Ser Thr Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Ser Tyr Ser Asp Gly Asp Ser
85 90 95
Val Gly Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 16
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE08H VH
<400> 16
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Asn Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Tyr Thr Ser Gly Thr Arg Tyr Arg Ala Asn Trp Ala Arg
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gly Thr Asp Ile Tyr Asp Tyr Tyr Tyr Trp Val Met Asp Leu Trp
100 105 110
Gly Pro Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 17
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE08L VL
<400> 17
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Ser Ile Ser Thr Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Ser Tyr Ser Asp Gly Asp Ser
85 90 95
Val Gly Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 18
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE09H VH
<400> 18
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Asn Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Tyr Thr Ser Gly Thr Arg Tyr Arg Ala Asn Trp Ala Arg
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gly Thr Asp Ile Tyr Asp Tyr Tyr Tyr Trp Val Met Asp Leu Trp
100 105 110
Gly Pro Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 19
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE09L VL
<400> 19
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Ser Ile Ser Thr Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Ser Tyr Ser Asp Ala Asp Ser
85 90 95
Val Gly Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 20
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE04H HVR-H1
<400> 20
Ser Tyr Ala Met Asn
1 5
<210> 21
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE04H HVR-H2
<400> 21
Tyr Ile Tyr Thr Ser Gly Thr Thr Tyr His Ala Asn Trp Ala Arg Gly
1 5 10 15
<210> 22
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE04H HVR-H3
<400> 22
Gly Thr Phe Ile Trp Asp Tyr Tyr Tyr Trp Val Met Asp Leu
1 5 10
<210> 23
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE04L HVR-L1
<400> 23
Gln Ala Ser Gln Ser Ile Ser Thr Tyr Leu Ala
1 5 10
<210> 24
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE04L HVR-L2
<400> 24
Lys Ala Ser Thr Leu Glu Ser
1 5
<210> 25
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE04L HVR-L3
<400> 25
Gln Ser Tyr Ser Asp Ala Asp Ser Val Gly
1 5 10
<210> 26
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE07H HVR-H1
<400> 26
Ser Glu Ala Met Asn
1 5
<210> 27
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE07H HVR-H2
<400> 27
Tyr Ile Tyr Thr Ser Gly Thr Thr Tyr Arg Ala Asn Trp Ala Arg Gly
1 5 10 15
<210> 28
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE07H HVR-H3
<400> 28
Gly Thr Gly Ile Tyr Asp Tyr Tyr Tyr Trp Val Met Asp Leu
1 5 10
<210> 29
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE07L HVR-L1
<400> 29
Gln Ala Ser Gln Ser Ile Ser Thr Tyr Leu Ala
1 5 10
<210> 30
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE07L HVR-L2
<400> 30
Ala Ala Ser Thr Leu Glu Ser
1 5
<210> 31
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE07L HVR-L3
<400> 31
Gln Ser Tyr Ser Asp Gly Asp Ser Val Gly
1 5 10
<210> 32
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE08H HVR-H1
<400> 32
Ser Tyr Ala Met Asn
1 5
<210> 33
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE08H HVR-H2
<400> 33
Tyr Ile Tyr Thr Ser Gly Thr Arg Tyr Arg Ala Asn Trp Ala Arg Gly
1 5 10 15
<210> 34
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE08H HVR-H3
<400> 34
Gly Thr Asp Ile Tyr Asp Tyr Tyr Tyr Trp Val Met Asp Leu
1 5 10
<210> 35
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE08L HVR-L1
<400> 35
Gln Ala Ser Gln Ser Ile Ser Thr Tyr Leu Ala
1 5 10
<210> 36
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE08L HVR-L2
<400> 36
Ala Ala Ser Thr Leu Glu Ser
1 5
<210> 37
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE08L HVR-L3
<400> 37
Gln Ser Tyr Ser Asp Gly Asp Ser Val Gly
1 5 10
<210> 38
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE09H HVR-H1
<400> 38
Ser Tyr Ala Met Asn
1 5
<210> 39
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE09H HVR-H2
<400> 39
Tyr Ile Tyr Thr Ser Gly Thr Arg Tyr Arg Ala Asn Trp Ala Arg Gly
1 5 10 15
<210> 40
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE09H HVR-H3
<400> 40
Gly Thr Asp Ile Tyr Asp Tyr Tyr Tyr Trp Val Met Asp Leu
1 5 10
<210> 41
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE09L HVR-L1
<400> 41
Gln Ala Ser Gln Ser Ile Ser Thr Tyr Leu Ala
1 5 10
<210> 42
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE09L HVR-L2
<400> 42
Ala Ala Ser Thr Leu Glu Ser
1 5
<210> 43
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE09L HVR-L3
<400> 43
Gln Ser Tyr Ser Asp Ala Asp Ser Val Gly
1 5 10
<210> 44
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> SG1
<400> 44
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 45
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<223> SG191
<400> 45
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Arg Arg Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 46
<211> 324
<212> PRT
<213> Artificial Sequence
<220>
<223> mF18
<400> 46
Ala Lys Thr Thr Pro Pro Ser Val Tyr Pro Leu Ala Pro Gly Ser Ala
1 5 10 15
Ala Gln Thr Asn Ser Met Val Thr Leu Gly Cys Leu Val Lys Gly Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ser Leu Ser Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Asp Leu Tyr Thr Leu
50 55 60
Ser Ser Ser Val Thr Val Pro Ser Ser Thr Trp Pro Ser Glu Thr Val
65 70 75 80
Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys Lys
85 90 95
Ile Val Pro Arg Asp Cys Gly Cys Lys Pro Cys Ile Cys Thr Val Lys
100 105 110
Glu Val Ser Lys Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Val Leu
115 120 125
Thr Ile Thr Leu Thr Pro Lys Val Thr Cys Val Val Val Asp Ile Ser
130 135 140
Lys Asp Asp Pro Glu Val Gln Phe Ser Trp Phe Val Asp Asp Val Glu
145 150 155 160
Val His Thr Ala Gln Thr Gln Pro Arg Glu Glu Gln Phe Asn Ser Thr
165 170 175
Phe Arg Ser Val Ser Glu Leu Pro Ile Met His Gln Asp Trp Leu Asn
180 185 190
Gly Lys Glu Phe Lys Cys Arg Val Asn Ser Ala Ala Phe Pro Ala Pro
195 200 205
Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Arg Pro Lys Ala Pro Gln
210 215 220
Val Tyr Thr Ile Pro Pro Pro Lys Glu Gln Met Ala Lys Asp Lys Val
225 230 235 240
Ser Leu Thr Cys Met Ile Thr Asp Phe Phe Pro Glu Asp Ile Thr Val
245 250 255
Glu Trp Gln Trp Asn Gly Gln Pro Ala Glu Asn Tyr Lys Asn Thr Gln
260 265 270
Pro Ile Met Asp Thr Asp Gly Ser Tyr Phe Val Tyr Ser Lys Leu Asn
275 280 285
Val Gln Lys Ser Asn Trp Glu Ala Gly Asn Thr Phe Thr Cys Ser Val
290 295 300
Leu His Glu Gly Leu His Asn His His Thr Glu Lys Ser Leu Ser His
305 310 315 320
Ser Pro Gly Lys
<210> 47
<211> 450
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE04H-SG181
<400> 47
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Asn Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Tyr Thr Ser Gly Thr Thr Tyr His Ala Asn Trp Ala Arg
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gly Thr Phe Ile Trp Asp Tyr Tyr Tyr Trp Val Met Asp Leu Trp
100 105 110
Gly Pro Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Arg
225 230 235 240
Arg Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro
450
<210> 48
<211> 450
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE07H-SG181
<400> 48
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Glu
20 25 30
Ala Met Asn Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Tyr Thr Ser Gly Thr Thr Tyr Arg Ala Asn Trp Ala Arg
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gly Thr Gly Ile Tyr Asp Tyr Tyr Tyr Trp Val Met Asp Leu Trp
100 105 110
Gly Pro Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Arg
225 230 235 240
Arg Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro
450
<210> 49
<211> 450
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE08H-SG181
<400> 49
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Asn Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Tyr Thr Ser Gly Thr Arg Tyr Arg Ala Asn Trp Ala Arg
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gly Thr Asp Ile Tyr Asp Tyr Tyr Tyr Trp Val Met Asp Leu Trp
100 105 110
Gly Pro Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Arg
225 230 235 240
Arg Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro
450
<210> 50
<211> 450
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE09H-SG181
<400> 50
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Asn Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Tyr Thr Ser Gly Thr Arg Tyr Arg Ala Asn Trp Ala Arg
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gly Thr Asp Ile Tyr Asp Tyr Tyr Tyr Trp Val Met Asp Leu Trp
100 105 110
Gly Pro Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Arg
225 230 235 240
Arg Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro
450
<210> 51
<211> 450
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE04H-SG191
<400> 51
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Asn Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Tyr Thr Ser Gly Thr Thr Tyr His Ala Asn Trp Ala Arg
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gly Thr Phe Ile Trp Asp Tyr Tyr Tyr Trp Val Met Asp Leu Trp
100 105 110
Gly Pro Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Arg
225 230 235 240
Arg Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Leu His Glu Ala Leu His Ala His Tyr Thr Arg Lys Glu Leu Ser Leu
435 440 445
Ser Pro
450
<210> 52
<211> 450
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE07H-SG191
<400> 52
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Glu
20 25 30
Ala Met Asn Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Tyr Thr Ser Gly Thr Thr Tyr Arg Ala Asn Trp Ala Arg
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gly Thr Gly Ile Tyr Asp Tyr Tyr Tyr Trp Val Met Asp Leu Trp
100 105 110
Gly Pro Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Arg
225 230 235 240
Arg Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Leu His Glu Ala Leu His Ala His Tyr Thr Arg Lys Glu Leu Ser Leu
435 440 445
Ser Pro
450
<210> 53
<211> 450
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE08H-SG191
<400> 53
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Asn Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Tyr Thr Ser Gly Thr Arg Tyr Arg Ala Asn Trp Ala Arg
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gly Thr Asp Ile Tyr Asp Tyr Tyr Tyr Trp Val Met Asp Leu Trp
100 105 110
Gly Pro Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Arg
225 230 235 240
Arg Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Leu His Glu Ala Leu His Ala His Tyr Thr Arg Lys Glu Leu Ser Leu
435 440 445
Ser Pro
450
<210> 54
<211> 450
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE09H-SG191
<400> 54
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Asn Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Tyr Thr Ser Gly Thr Arg Tyr Arg Ala Asn Trp Ala Arg
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gly Thr Asp Ile Tyr Asp Tyr Tyr Tyr Trp Val Met Asp Leu Trp
100 105 110
Gly Pro Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Arg
225 230 235 240
Arg Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Leu His Glu Ala Leu His Ala His Tyr Thr Arg Lys Glu Leu Ser Leu
435 440 445
Ser Pro
450
<210> 55
<211> 446
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE04H-mF18
<400> 55
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Asn Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Tyr Thr Ser Gly Thr Thr Tyr His Ala Asn Trp Ala Arg
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gly Thr Phe Ile Trp Asp Tyr Tyr Tyr Trp Val Met Asp Leu Trp
100 105 110
Gly Pro Gly Thr Leu Val Thr Val Ser Ser Ala Lys Thr Thr Pro Pro
115 120 125
Ser Val Tyr Pro Leu Ala Pro Gly Ser Ala Ala Gln Thr Asn Ser Met
130 135 140
Val Thr Leu Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Thr Trp Asn Ser Gly Ser Leu Ser Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Asp Leu Tyr Thr Leu Ser Ser Ser Val Thr Val
180 185 190
Pro Ser Ser Thr Trp Pro Ser Glu Thr Val Thr Cys Asn Val Ala His
195 200 205
Pro Ala Ser Ser Thr Lys Val Asp Lys Lys Ile Val Pro Arg Asp Cys
210 215 220
Gly Cys Lys Pro Cys Ile Cys Thr Val Lys Glu Val Ser Lys Val Phe
225 230 235 240
Ile Phe Pro Pro Lys Pro Lys Asp Val Leu Thr Ile Thr Leu Thr Pro
245 250 255
Lys Val Thr Cys Val Val Val Asp Ile Ser Lys Asp Asp Pro Glu Val
260 265 270
Gln Phe Ser Trp Phe Val Asp Asp Val Glu Val His Thr Ala Gln Thr
275 280 285
Gln Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Ser Val Ser Glu
290 295 300
Leu Pro Ile Met His Gln Asp Trp Leu Asn Gly Lys Glu Phe Lys Cys
305 310 315 320
Arg Val Asn Ser Ala Ala Phe Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Thr Lys Gly Arg Pro Lys Ala Pro Gln Val Tyr Thr Ile Pro Pro
340 345 350
Pro Lys Glu Gln Met Ala Lys Asp Lys Val Ser Leu Thr Cys Met Ile
355 360 365
Thr Asp Phe Phe Pro Glu Asp Ile Thr Val Glu Trp Gln Trp Asn Gly
370 375 380
Gln Pro Ala Glu Asn Tyr Lys Asn Thr Gln Pro Ile Met Asp Thr Asp
385 390 395 400
Gly Ser Tyr Phe Val Tyr Ser Lys Leu Asn Val Gln Lys Ser Asn Trp
405 410 415
Glu Ala Gly Asn Thr Phe Thr Cys Ser Val Leu His Glu Gly Leu His
420 425 430
Asn His His Thr Glu Lys Ser Leu Ser His Ser Pro Gly Lys
435 440 445
<210> 56
<211> 446
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE07H-mF18
<400> 56
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Glu
20 25 30
Ala Met Asn Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Tyr Thr Ser Gly Thr Thr Tyr Arg Ala Asn Trp Ala Arg
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gly Thr Gly Ile Tyr Asp Tyr Tyr Tyr Trp Val Met Asp Leu Trp
100 105 110
Gly Pro Gly Thr Leu Val Thr Val Ser Ser Ala Lys Thr Thr Pro Pro
115 120 125
Ser Val Tyr Pro Leu Ala Pro Gly Ser Ala Ala Gln Thr Asn Ser Met
130 135 140
Val Thr Leu Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Thr Trp Asn Ser Gly Ser Leu Ser Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Asp Leu Tyr Thr Leu Ser Ser Ser Val Thr Val
180 185 190
Pro Ser Ser Thr Trp Pro Ser Glu Thr Val Thr Cys Asn Val Ala His
195 200 205
Pro Ala Ser Ser Thr Lys Val Asp Lys Lys Ile Val Pro Arg Asp Cys
210 215 220
Gly Cys Lys Pro Cys Ile Cys Thr Val Lys Glu Val Ser Lys Val Phe
225 230 235 240
Ile Phe Pro Pro Lys Pro Lys Asp Val Leu Thr Ile Thr Leu Thr Pro
245 250 255
Lys Val Thr Cys Val Val Val Asp Ile Ser Lys Asp Asp Pro Glu Val
260 265 270
Gln Phe Ser Trp Phe Val Asp Asp Val Glu Val His Thr Ala Gln Thr
275 280 285
Gln Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Ser Val Ser Glu
290 295 300
Leu Pro Ile Met His Gln Asp Trp Leu Asn Gly Lys Glu Phe Lys Cys
305 310 315 320
Arg Val Asn Ser Ala Ala Phe Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Thr Lys Gly Arg Pro Lys Ala Pro Gln Val Tyr Thr Ile Pro Pro
340 345 350
Pro Lys Glu Gln Met Ala Lys Asp Lys Val Ser Leu Thr Cys Met Ile
355 360 365
Thr Asp Phe Phe Pro Glu Asp Ile Thr Val Glu Trp Gln Trp Asn Gly
370 375 380
Gln Pro Ala Glu Asn Tyr Lys Asn Thr Gln Pro Ile Met Asp Thr Asp
385 390 395 400
Gly Ser Tyr Phe Val Tyr Ser Lys Leu Asn Val Gln Lys Ser Asn Trp
405 410 415
Glu Ala Gly Asn Thr Phe Thr Cys Ser Val Leu His Glu Gly Leu His
420 425 430
Asn His His Thr Glu Lys Ser Leu Ser His Ser Pro Gly Lys
435 440 445
<210> 57
<211> 446
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE08H-mF18
<400> 57
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Asn Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Tyr Thr Ser Gly Thr Arg Tyr Arg Ala Asn Trp Ala Arg
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gly Thr Asp Ile Tyr Asp Tyr Tyr Tyr Trp Val Met Asp Leu Trp
100 105 110
Gly Pro Gly Thr Leu Val Thr Val Ser Ser Ala Lys Thr Thr Pro Pro
115 120 125
Ser Val Tyr Pro Leu Ala Pro Gly Ser Ala Ala Gln Thr Asn Ser Met
130 135 140
Val Thr Leu Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Thr Trp Asn Ser Gly Ser Leu Ser Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Asp Leu Tyr Thr Leu Ser Ser Ser Val Thr Val
180 185 190
Pro Ser Ser Thr Trp Pro Ser Glu Thr Val Thr Cys Asn Val Ala His
195 200 205
Pro Ala Ser Ser Thr Lys Val Asp Lys Lys Ile Val Pro Arg Asp Cys
210 215 220
Gly Cys Lys Pro Cys Ile Cys Thr Val Lys Glu Val Ser Lys Val Phe
225 230 235 240
Ile Phe Pro Pro Lys Pro Lys Asp Val Leu Thr Ile Thr Leu Thr Pro
245 250 255
Lys Val Thr Cys Val Val Val Asp Ile Ser Lys Asp Asp Pro Glu Val
260 265 270
Gln Phe Ser Trp Phe Val Asp Asp Val Glu Val His Thr Ala Gln Thr
275 280 285
Gln Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Ser Val Ser Glu
290 295 300
Leu Pro Ile Met His Gln Asp Trp Leu Asn Gly Lys Glu Phe Lys Cys
305 310 315 320
Arg Val Asn Ser Ala Ala Phe Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Thr Lys Gly Arg Pro Lys Ala Pro Gln Val Tyr Thr Ile Pro Pro
340 345 350
Pro Lys Glu Gln Met Ala Lys Asp Lys Val Ser Leu Thr Cys Met Ile
355 360 365
Thr Asp Phe Phe Pro Glu Asp Ile Thr Val Glu Trp Gln Trp Asn Gly
370 375 380
Gln Pro Ala Glu Asn Tyr Lys Asn Thr Gln Pro Ile Met Asp Thr Asp
385 390 395 400
Gly Ser Tyr Phe Val Tyr Ser Lys Leu Asn Val Gln Lys Ser Asn Trp
405 410 415
Glu Ala Gly Asn Thr Phe Thr Cys Ser Val Leu His Glu Gly Leu His
420 425 430
Asn His His Thr Glu Lys Ser Leu Ser His Ser Pro Gly Lys
435 440 445
<210> 58
<211> 446
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE09H-mF18
<400> 58
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Asn Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Tyr Thr Ser Gly Thr Arg Tyr Arg Ala Asn Trp Ala Arg
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gly Thr Asp Ile Tyr Asp Tyr Tyr Tyr Trp Val Met Asp Leu Trp
100 105 110
Gly Pro Gly Thr Leu Val Thr Val Ser Ser Ala Lys Thr Thr Pro Pro
115 120 125
Ser Val Tyr Pro Leu Ala Pro Gly Ser Ala Ala Gln Thr Asn Ser Met
130 135 140
Val Thr Leu Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Thr Trp Asn Ser Gly Ser Leu Ser Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Asp Leu Tyr Thr Leu Ser Ser Ser Val Thr Val
180 185 190
Pro Ser Ser Thr Trp Pro Ser Glu Thr Val Thr Cys Asn Val Ala His
195 200 205
Pro Ala Ser Ser Thr Lys Val Asp Lys Lys Ile Val Pro Arg Asp Cys
210 215 220
Gly Cys Lys Pro Cys Ile Cys Thr Val Lys Glu Val Ser Lys Val Phe
225 230 235 240
Ile Phe Pro Pro Lys Pro Lys Asp Val Leu Thr Ile Thr Leu Thr Pro
245 250 255
Lys Val Thr Cys Val Val Val Asp Ile Ser Lys Asp Asp Pro Glu Val
260 265 270
Gln Phe Ser Trp Phe Val Asp Asp Val Glu Val His Thr Ala Gln Thr
275 280 285
Gln Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Ser Val Ser Glu
290 295 300
Leu Pro Ile Met His Gln Asp Trp Leu Asn Gly Lys Glu Phe Lys Cys
305 310 315 320
Arg Val Asn Ser Ala Ala Phe Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Thr Lys Gly Arg Pro Lys Ala Pro Gln Val Tyr Thr Ile Pro Pro
340 345 350
Pro Lys Glu Gln Met Ala Lys Asp Lys Val Ser Leu Thr Cys Met Ile
355 360 365
Thr Asp Phe Phe Pro Glu Asp Ile Thr Val Glu Trp Gln Trp Asn Gly
370 375 380
Gln Pro Ala Glu Asn Tyr Lys Asn Thr Gln Pro Ile Met Asp Thr Asp
385 390 395 400
Gly Ser Tyr Phe Val Tyr Ser Lys Leu Asn Val Gln Lys Ser Asn Trp
405 410 415
Glu Ala Gly Asn Thr Phe Thr Cys Ser Val Leu His Glu Gly Leu His
420 425 430
Asn His His Thr Glu Lys Ser Leu Ser His Ser Pro Gly Lys
435 440 445
<210> 59
<211> 107
<212> PRT
<213> Mus musculus
<400> 59
Arg Ala Asp Ala Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu
1 5 10 15
Gln Leu Thr Ser Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe
20 25 30
Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg
35 40 45
Gln Asn Gly Val Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Met Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu
65 70 75 80
Arg His Asn Ser Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser
85 90 95
Pro Ile Val Lys Ser Phe Asn Arg Asn Glu Cys
100 105
<210> 60
<211> 215
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE04L-SK1
<400> 60
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Ser Ile Ser Thr Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Thr Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Ser Tyr Ser Asp Ala Asp Ser
85 90 95
Val Gly Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 61
<211> 215
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE07L-SK1
<400> 61
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Ser Ile Ser Thr Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Ser Tyr Ser Asp Gly Asp Ser
85 90 95
Val Gly Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 62
<211> 215
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE08L-SK1
<400> 62
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Ser Ile Ser Thr Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Ser Tyr Ser Asp Gly Asp Ser
85 90 95
Val Gly Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 63
<211> 215
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE09L-SK1
<400> 63
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Ser Ile Ser Thr Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Ser Tyr Ser Asp Ala Asp Ser
85 90 95
Val Gly Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 64
<211> 215
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE04L-mk1
<400> 64
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Ser Ile Ser Thr Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Thr Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Ser Tyr Ser Asp Ala Asp Ser
85 90 95
Val Gly Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Ala Asp Ala
100 105 110
Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu Gln Leu Thr Ser
115 120 125
Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe Tyr Pro Lys Asp
130 135 140
Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg Gln Asn Gly Val
145 150 155 160
Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser Thr Tyr Ser Met
165 170 175
Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu Arg His Asn Ser
180 185 190
Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser Pro Ile Val Lys
195 200 205
Ser Phe Asn Arg Asn Glu Cys
210 215
<210> 65
<211> 215
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE07L-mk1
<400> 65
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Ser Ile Ser Thr Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Ser Tyr Ser Asp Gly Asp Ser
85 90 95
Val Gly Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Ala Asp Ala
100 105 110
Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu Gln Leu Thr Ser
115 120 125
Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe Tyr Pro Lys Asp
130 135 140
Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg Gln Asn Gly Val
145 150 155 160
Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser Thr Tyr Ser Met
165 170 175
Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu Arg His Asn Ser
180 185 190
Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser Pro Ile Val Lys
195 200 205
Ser Phe Asn Arg Asn Glu Cys
210 215
<210> 66
<211> 215
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE08L-mk1
<400> 66
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Ser Ile Ser Thr Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Ser Tyr Ser Asp Gly Asp Ser
85 90 95
Val Gly Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Ala Asp Ala
100 105 110
Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu Gln Leu Thr Ser
115 120 125
Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe Tyr Pro Lys Asp
130 135 140
Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg Gln Asn Gly Val
145 150 155 160
Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser Thr Tyr Ser Met
165 170 175
Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu Arg His Asn Ser
180 185 190
Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser Pro Ile Val Lys
195 200 205
Ser Phe Asn Arg Asn Glu Cys
210 215
<210> 67
<211> 215
<212> PRT
<213> Artificial Sequence
<220>
<223> hT0947AE09L-mk1
<400> 67
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Ser Ile Ser Thr Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Ser Tyr Ser Asp Ala Asp Ser
85 90 95
Val Gly Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Ala Asp Ala
100 105 110
Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu Gln Leu Thr Ser
115 120 125
Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe Tyr Pro Lys Asp
130 135 140
Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg Gln Asn Gly Val
145 150 155 160
Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser Thr Tyr Ser Met
165 170 175
Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu Arg His Asn Ser
180 185 190
Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser Pro Ile Val Lys
195 200 205
Ser Phe Asn Arg Asn Glu Cys
210 215
<210> 68
<211> 390
<212> PRT
<213> Homo sapiens
<400> 68
Met Pro Pro Ser Gly Leu Arg Leu Leu Pro Leu Leu Leu Pro Leu Leu
1 5 10 15
Trp Leu Leu Val Leu Thr Pro Gly Arg Pro Ala Ala Gly Leu Ser Thr
20 25 30
Cys Lys Thr Ile Asp Met Glu Leu Val Lys Arg Lys Arg Ile Glu Ala
35 40 45
Ile Arg Gly Gln Ile Leu Ser Lys Leu Arg Leu Ala Ser Pro Pro Ser
50 55 60
Gln Gly Glu Val Pro Pro Gly Pro Leu Pro Glu Ala Val Leu Ala Leu
65 70 75 80
Tyr Asn Ser Thr Arg Asp Arg Val Ala Gly Glu Ser Ala Glu Pro Glu
85 90 95
Pro Glu Pro Glu Ala Asp Tyr Tyr Ala Lys Glu Val Thr Arg Val Leu
100 105 110
Met Val Glu Thr His Asn Glu Ile Tyr Asp Lys Phe Lys Gln Ser Thr
115 120 125
His Ser Ile Tyr Met Phe Phe Asn Thr Ser Glu Leu Arg Glu Ala Val
130 135 140
Pro Glu Pro Val Leu Leu Ser Arg Ala Glu Leu Arg Leu Leu Arg Leu
145 150 155 160
Lys Leu Lys Val Glu Gln His Val Glu Leu Tyr Gln Lys Tyr Ser Asn
165 170 175
Asn Ser Trp Arg Tyr Leu Ser Asn Arg Leu Leu Ala Pro Ser Asp Ser
180 185 190
Pro Glu Trp Leu Ser Phe Asp Val Thr Gly Val Val Arg Gln Trp Leu
195 200 205
Ser Arg Gly Gly Glu Ile Glu Gly Phe Arg Leu Ser Ala His Cys Ser
210 215 220
Cys Asp Ser Arg Asp Asn Thr Leu Gln Val Asp Ile Asn Gly Phe Thr
225 230 235 240
Thr Gly Arg Arg Gly Asp Leu Ala Thr Ile His Gly Met Asn Arg Pro
245 250 255
Phe Leu Leu Leu Met Ala Thr Pro Leu Glu Arg Ala Gln His Leu Gln
260 265 270
Ser Ser Arg His Arg Arg Ala Leu Asp Thr Asn Tyr Cys Phe Ser Ser
275 280 285
Thr Glu Lys Asn Cys Cys Val Arg Gln Leu Tyr Ile Asp Phe Arg Lys
290 295 300
Asp Leu Gly Trp Lys Trp Ile His Glu Pro Lys Gly Tyr His Ala Asn
305 310 315 320
Phe Cys Leu Gly Pro Cys Pro Tyr Ile Trp Ser Leu Asp Thr Gln Tyr
325 330 335
Ser Lys Val Leu Ala Leu Tyr Asn Gln His Asn Pro Gly Ala Ser Ala
340 345 350
Ala Pro Cys Cys Val Pro Gln Ala Leu Glu Pro Leu Pro Ile Val Tyr
355 360 365
Tyr Val Gly Arg Lys Pro Lys Val Glu Gln Leu Ser Asn Met Ile Val
370 375 380
Arg Ser Cys Lys Cys Ser
385 390
<210> 69
<211> 1173
<212> DNA
<213> Homo sapiens
<400> 69
atgccgccct ccgggctgcg gctgctgccg ctgctgctac cgctgctgtg gctactggtg 60
ctgacgcctg gccggccggc cgcgggacta tccacctgca agactatcga catggagctg 120
gtgaagcgga agcgcatcga ggccatccgc ggccagatcc tgtccaagct gcggctcgcc 180
agccccccga gccaggggga ggtgccgccc ggcccgctgc ccgaggccgt gctcgccctg 240
tacaacagca cccgcgaccg ggtggccggg gagagtgcag aaccggagcc cgagcctgag 300
gccgactact acgccaagga ggtcacccgc gtgctaatgg tggaaaccca caacgaaatc 360
tatgacaagt tcaagcagag tacacacagc atatatatgt tcttcaacac atcagagctc 420
cgagaagcgg tacctgaacc cgtgttgctc tcccgggcag agctgcgtct gctgaggctc 480
aagttaaaag tggagcagca cgtggagctg taccagaaat acagcaacaa ttcctggcga 540
tacctcagca accggctgct ggcacccagc gactcgccag agtggttatc ttttgatgtc 600
accggagttg tgcggcagtg gttgagccgt ggaggggaaa ttgagggctt tcgccttagc 660
gcccactgct cctgtgacag cagggataac acactgcaag tggacatcaa cgggttcact 720
accggccgcc gaggtgacct ggccaccatt catggcatga accggccttt cctgcttctc 780
atggccaccc cgctggagag ggcccagcat ctgcaaagct cccggcaccg ccgagccctg 840
gacaccaact attgcttcag ctccacggag aagaactgct gcgtgcggca gctgtacatt 900
gacttccgca aggacctcgg ctggaagtgg atccacgagc ccaagggcta ccatgccaac 960
ttctgcctcg ggccctgccc ctacatttgg agcctggaca cgcagtacag caaggtcctg 1020
gccctgtaca accagcataa cccgggcgcc tcggcggcgc cgtgctgcgt gccgcaggcg 1080
ctggagccgc tgcccatcgt gtactacgtg ggccgcaagc ccaaggtgga gcagctgtcc 1140
aacatgatcg tgcgctcctg caagtgcagc tga 1173
<210> 70
<211> 390
<212> PRT
<213> Mus musculus
<400> 70
Met Pro Pro Ser Gly Leu Arg Leu Leu Pro Leu Leu Leu Pro Leu Pro
1 5 10 15
Trp Leu Leu Val Leu Thr Pro Gly Arg Pro Ala Ala Gly Leu Ser Thr
20 25 30
Cys Lys Thr Ile Asp Met Glu Leu Val Lys Arg Lys Arg Ile Glu Ala
35 40 45
Ile Arg Gly Gln Ile Leu Ser Lys Leu Arg Leu Ala Ser Pro Pro Ser
50 55 60
Gln Gly Glu Val Pro Pro Gly Pro Leu Pro Glu Ala Val Leu Ala Leu
65 70 75 80
Tyr Asn Ser Thr Arg Asp Arg Val Ala Gly Glu Ser Ala Asp Pro Glu
85 90 95
Pro Glu Pro Glu Ala Asp Tyr Tyr Ala Lys Glu Val Thr Arg Val Leu
100 105 110
Met Val Asp Arg Asn Asn Ala Ile Tyr Glu Lys Thr Lys Asp Ile Ser
115 120 125
His Ser Ile Tyr Met Phe Phe Asn Thr Ser Asp Ile Arg Glu Ala Val
130 135 140
Pro Glu Pro Pro Leu Leu Ser Arg Ala Glu Leu Arg Leu Gln Arg Leu
145 150 155 160
Lys Ser Ser Val Glu Gln His Val Glu Leu Tyr Gln Lys Tyr Ser Asn
165 170 175
Asn Ser Trp Arg Tyr Leu Gly Asn Arg Leu Leu Thr Pro Thr Asp Thr
180 185 190
Pro Glu Trp Leu Ser Phe Asp Val Thr Gly Val Val Arg Gln Trp Leu
195 200 205
Asn Gln Gly Asp Gly Ile Gln Gly Phe Arg Phe Ser Ala His Cys Ser
210 215 220
Cys Asp Ser Lys Asp Asn Lys Leu His Val Glu Ile Asn Gly Ile Ser
225 230 235 240
Pro Lys Arg Arg Gly Asp Leu Gly Thr Ile His Asp Met Asn Arg Pro
245 250 255
Phe Leu Leu Leu Met Ala Thr Pro Leu Glu Arg Ala Gln His Leu His
260 265 270
Ser Ser Arg His Arg Arg Ala Leu Asp Thr Asn Tyr Cys Phe Ser Ser
275 280 285
Thr Glu Lys Asn Cys Cys Val Arg Gln Leu Tyr Ile Asp Phe Arg Lys
290 295 300
Asp Leu Gly Trp Lys Trp Ile His Glu Pro Lys Gly Tyr His Ala Asn
305 310 315 320
Phe Cys Leu Gly Pro Cys Pro Tyr Ile Trp Ser Leu Asp Thr Gln Tyr
325 330 335
Ser Lys Val Leu Ala Leu Tyr Asn Gln His Asn Pro Gly Ala Ser Ala
340 345 350
Ser Pro Cys Cys Val Pro Gln Ala Leu Glu Pro Leu Pro Ile Val Tyr
355 360 365
Tyr Val Gly Arg Lys Pro Lys Val Glu Gln Leu Ser Asn Met Ile Val
370 375 380
Arg Ser Cys Lys Cys Ser
385 390
<210> 71
<211> 1173
<212> DNA
<213> Mus musculus
<400> 71
atgccgccct cggggctgcg gctactgccg cttctgctcc cactcccgtg gcttctagtg 60
ctgacgcccg ggaggccagc cgcgggactc tccacctgca agaccatcga catggagctg 120
gtgaaacgga agcgcatcga agccatccgt ggccagatcc tgtccaaact aaggctcgcc 180
agtcccccaa gccaggggga ggtaccgccc ggcccgctgc ccgaggcggt gctcgctttg 240
tacaacagca cccgcgaccg ggtggcaggc gagagcgccg acccagagcc ggagcccgaa 300
gcggactact atgctaaaga ggtcacccgc gtgctaatgg tggaccgcaa caacgccatc 360
tatgagaaaa ccaaagacat ctcacacagt atatatatgt tcttcaatac gtcagacatt 420
cgggaagcag tgcccgaacc cccattgctg tcccgtgcag agctgcgctt gcagagatta 480
aaatcaagtg tggagcaaca tgtggaactc taccagaaat atagcaacaa ttcctggcgt 540
taccttggta accggctgct gacccccact gatacgcctg agtggctgtc ttttgacgtc 600
actggagttg tacggcagtg gctgaaccaa ggagacggaa tacagggctt tcgattcagc 660
gctcactgct cttgtgacag caaagataac aaactccacg tggaaatcaa cgggatcagc 720
cccaaacgtc ggggcgacct gggcaccatc catgacatga accggccctt cctgctcctc 780
atggccaccc ccctggaaag ggcccagcac ctgcacagct cacggcaccg gagagccctg 840
gataccaact attgcttcag ctccacagag aagaactgct gtgtgcggca gctgtacatt 900
gactttagga aggacctggg ttggaagtgg atccacgagc ccaagggcta ccatgccaac 960
ttctgtctgg gaccctgccc ctatatttgg agcctggaca cacagtacag caaggtcctt 1020
gccctctaca accaacacaa cccgggcgct tcggcgtcac cgtgctgcgt gccgcaggct 1080
ttggagccac tgcccatcgt ctactacgtg ggtcgcaagc ccaaggtgga gcagttgtcc 1140
aacatgattg tgcgctcctg caagtgcagc tga 1173
<210> 72
<211> 390
<212> PRT
<213> Macaca fascicularis
<400> 72
Met Pro Pro Ser Gly Leu Arg Leu Leu Pro Leu Leu Leu Pro Leu Leu
1 5 10 15
Trp Leu Leu Val Leu Thr Pro Gly Arg Pro Ala Ala Gly Leu Ser Thr
20 25 30
Cys Lys Thr Ile Asp Met Glu Leu Val Lys Arg Lys Arg Ile Glu Ala
35 40 45
Ile Arg Gly Gln Ile Leu Ser Lys Leu Arg Leu Ala Ser Pro Pro Ser
50 55 60
Gln Gly Glu Val Pro Pro Gly Pro Leu Pro Glu Ala Val Leu Ala Leu
65 70 75 80
Tyr Asn Ser Thr Arg Asp Arg Val Ala Gly Glu Ser Ala Glu Pro Glu
85 90 95
Pro Glu Pro Glu Ala Asp Tyr Tyr Ala Lys Glu Val Thr Arg Val Leu
100 105 110
Met Val Glu Thr His Asn Glu Ile Tyr Asp Lys Phe Lys Gln Ser Thr
115 120 125
His Ser Ile Tyr Met Phe Phe Asn Thr Ser Glu Leu Arg Glu Ala Val
130 135 140
Pro Glu Pro Val Leu Leu Ser Arg Ala Glu Leu Arg Leu Leu Arg Leu
145 150 155 160
Lys Leu Lys Val Glu Gln His Val Glu Leu Tyr Gln Lys Tyr Ser Asn
165 170 175
Asn Ser Trp Arg Tyr Leu Ser Asn Arg Leu Leu Ala Pro Ser Asp Ser
180 185 190
Pro Glu Trp Leu Ser Phe Asp Val Thr Gly Val Val Arg Gln Trp Leu
195 200 205
Ser Arg Gly Gly Glu Ile Glu Gly Phe Arg Leu Ser Ala His Cys Ser
210 215 220
Cys Asp Ser Lys Asp Asn Thr Leu Gln Val Asp Ile Asn Gly Phe Thr
225 230 235 240
Thr Gly Arg Arg Gly Asp Leu Ala Thr Ile His Gly Met Asn Arg Pro
245 250 255
Phe Leu Leu Leu Met Ala Thr Pro Leu Glu Arg Ala Gln His Leu Gln
260 265 270
Ser Ser Arg His Arg Arg Ala Leu Asp Thr Asn Tyr Cys Phe Ser Ser
275 280 285
Thr Glu Lys Asn Cys Cys Val Arg Gln Leu Tyr Ile Asp Phe Arg Lys
290 295 300
Asp Leu Gly Trp Lys Trp Ile His Glu Pro Lys Gly Tyr His Ala Asn
305 310 315 320
Phe Cys Leu Gly Pro Cys Pro Tyr Ile Trp Ser Leu Asp Thr Gln Tyr
325 330 335
Ser Lys Val Leu Ala Leu Tyr Asn Gln His Asn Pro Gly Ala Ser Ala
340 345 350
Ala Pro Cys Cys Val Pro Gln Ala Leu Glu Pro Leu Pro Ile Val Tyr
355 360 365
Tyr Val Gly Arg Lys Pro Lys Val Glu Gln Leu Ser Asn Met Ile Val
370 375 380
Arg Ser Cys Lys Cys Ser
385 390
<210> 73
<211> 1173
<212> DNA
<213> Macaca fascicularis
<400> 73
atgccgccct ccgggctgcg gctgctgccg ctgctgctac cgctgctgtg gctactggtg 60
ctgacgcctg gccggccggc cgccggacta tccacctgca agactatcga catggagctg 120
gtgaagcgga agcgcatcga ggccatccgc ggccagatcc tgtccaagct gcggctcgcc 180
agccccccga gccaggggga ggtgccgccc ggcccgctgc ccgaggccgt gctcgccctg 240
tacaacagca cccgcgaccg ggtggccggg gagagtgcgg agccggaacc cgaaccggag 300
gccgactact acgccaagga ggtcacccgc gtgctaatgg tggaaaccca caacgaaatc 360
tatgacaagt tcaagcagag cacacacagc atatatatgt tcttcaacac atcagagctc 420
cgagaagcag tacctgaacc tgtgttgctc tcccgggcag agctgcgtct gctgaggctc 480
aagttaaaag tggagcagca tgtggagctg taccagaaat acagcaacaa ttcctggcga 540
tacctcagca accggctgct ggcgcccagc gactcgccgg agtggttgtc ttttgatgtc 600
accggagttg tgcggcagtg gttgagccgc ggaggggaaa ttgagggctt tcgccttagc 660
gcccactgct cctgtgacag caaagataac acactgcaag tggacatcaa cgggttcact 720
accggccgcc gaggtgacct ggccaccatt catggcatga accggccttt cctgcttctc 780
atggccaccc cgctggagag ggcccaacat ctgcaaagct cccggcaccg ccgagccctg 840
gacaccaact actgcttcag ctccacggag aagaactgct gcgtgcggca gctgtatatt 900
gacttccgca aggacctcgg ctggaagtgg atccacgagc ccaagggcta ccatgccaac 960
ttctgcctgg gaccctgccc ctacatttgg agcctggaca cgcagtacag caaggtcctg 1020
gccctgtaca accagcataa cccgggcgcc tcggcggcgc cgtgctgcgt gccgcaggcg 1080
ctggagccgc tgcccatcgt gtactacgtg ggccgcaagc ccaaggtgga gcagctgtcc 1140
aacatgatcg tgcgctcctg caaatgcagc tga 1173
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА К МИОСТАТИНУ, ПОЛИПЕПТИДЫ, СОДЕРЖАЩИЕ ВАРИАНТЫ FC-ОБЛАСТЕЙ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2789884C2 |
| АНТИТЕЛА К МИОСТАТИНУ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2778945C2 |
| АНТИТЕЛА К МИОСТАТИНУ, ПОЛИПЕПТИДЫ, СОДЕРЖАЩИЕ ВАРИАНТЫ FC-ОБЛАСТЕЙ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2799522C2 |
| АНТИТЕЛА К С5 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2746356C2 |
| АНТИ-CD137 АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2829536C2 |
| АНТИТЕЛА ПРОТИВ СИГНАЛ-РЕГУЛЯТОРНОГО БЕЛКА АЛЬФА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2771964C2 |
| АНТИТЕЛА К С5 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2789788C2 |
| МУЛЬТИСПЕЦИФИЧЕСКАЯ АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА, ОБЛАДАЮЩАЯ ЗАМЕЩАЮЩЕЙ ФУНКЦИОНАЛЬНОЙ АКТИВНОСТЬЮ КОФАКТОРА КОАГУЛИРУЮЩЕГО ФАКТОРА КРОВИ VIII, И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ УКАЗАННУЮ МОЛЕКУЛУ В КАЧЕСТВЕ АКТИВНОГО ИНГРЕДИЕНТА | 2018 |
|
RU2812909C2 |
| БИСПЕЦИФИЧНЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2800779C2 |
| АНТИТЕЛА ПРОТИВ ТАУ-БЕЛКА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2732122C2 |
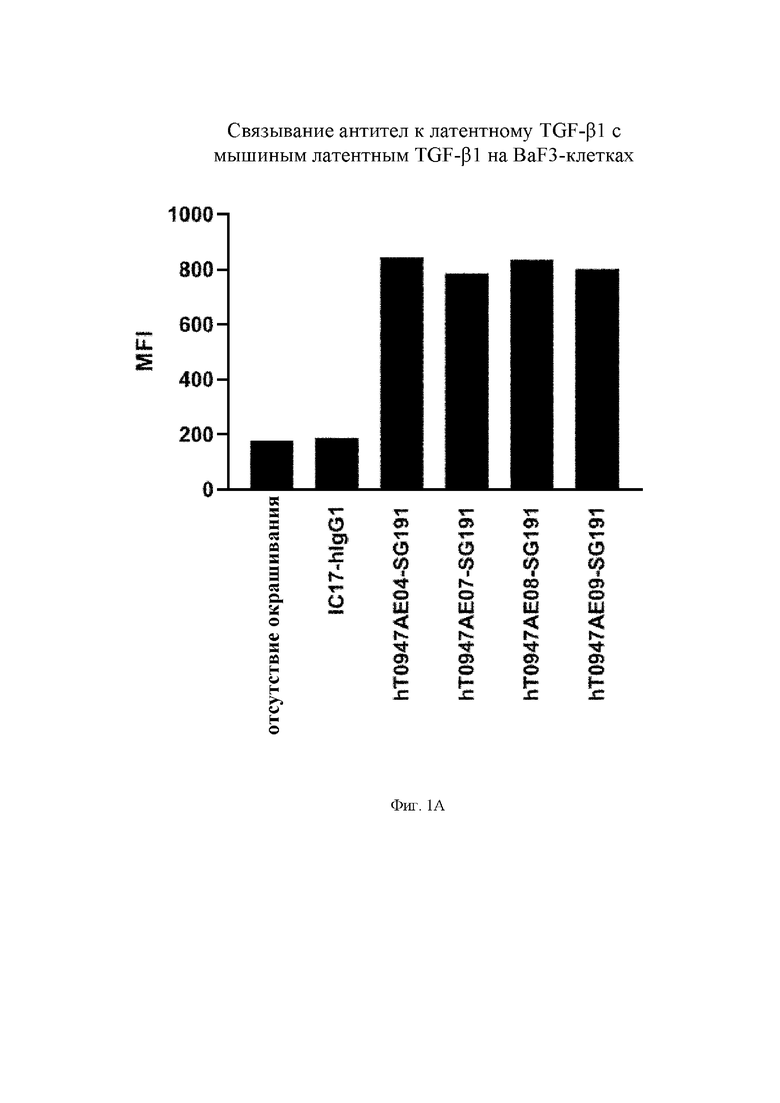
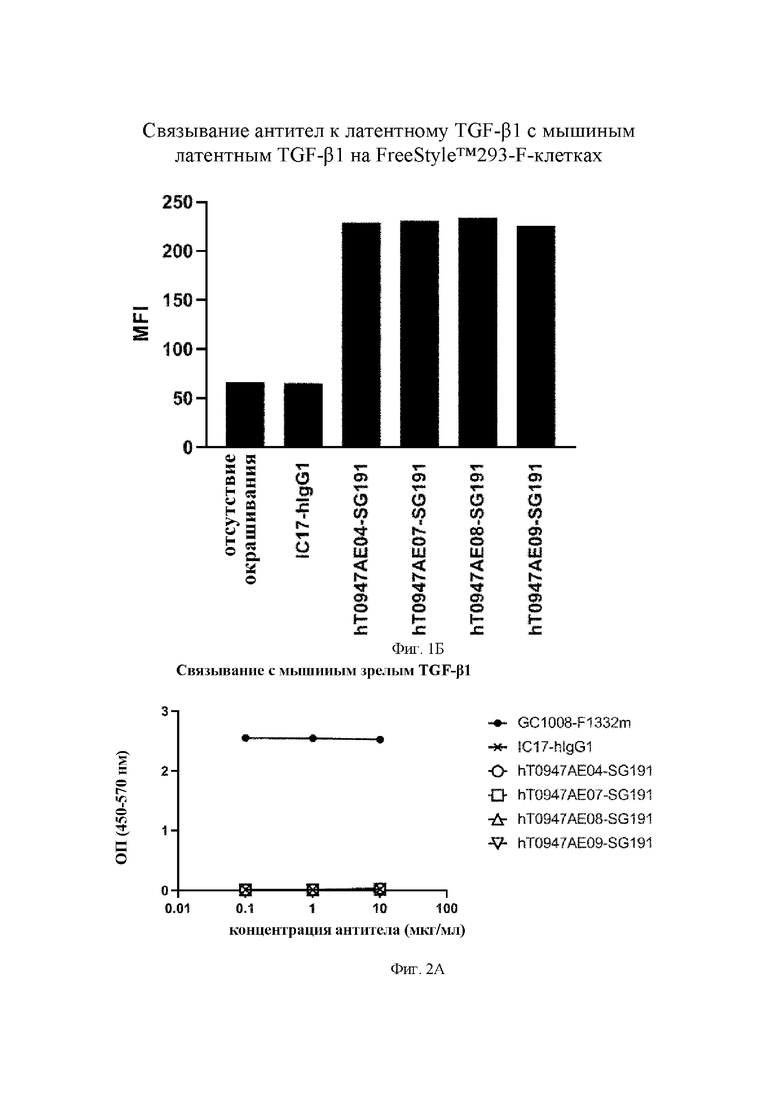
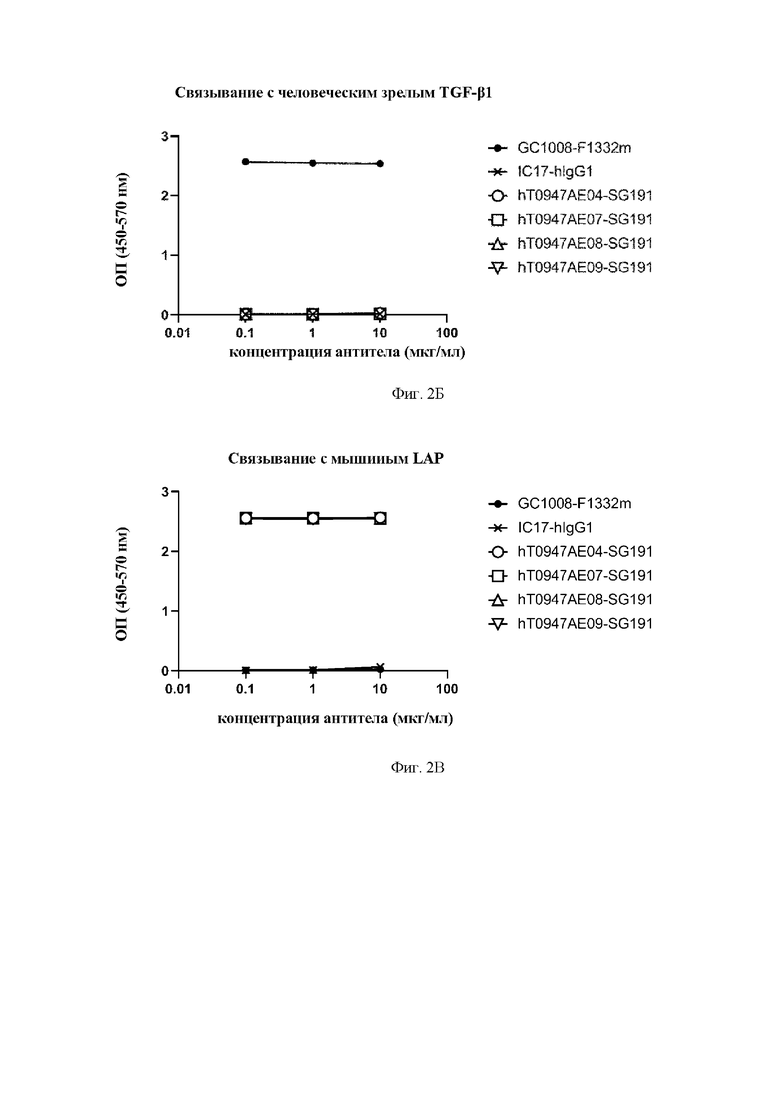
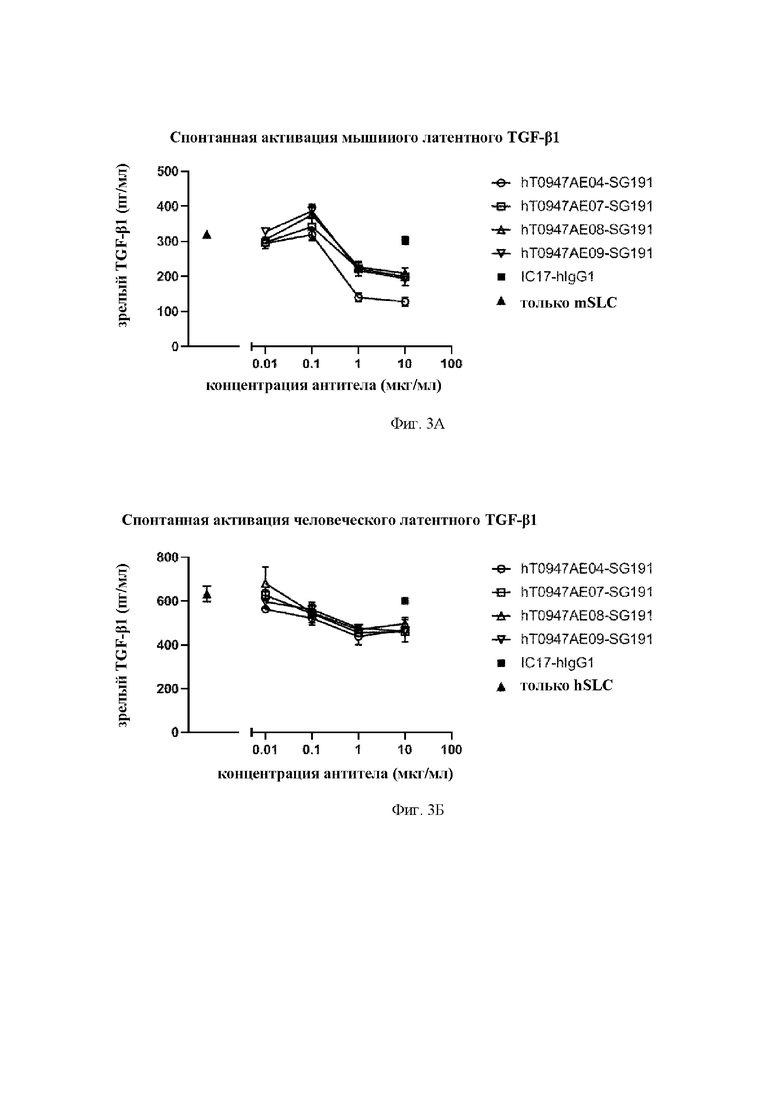
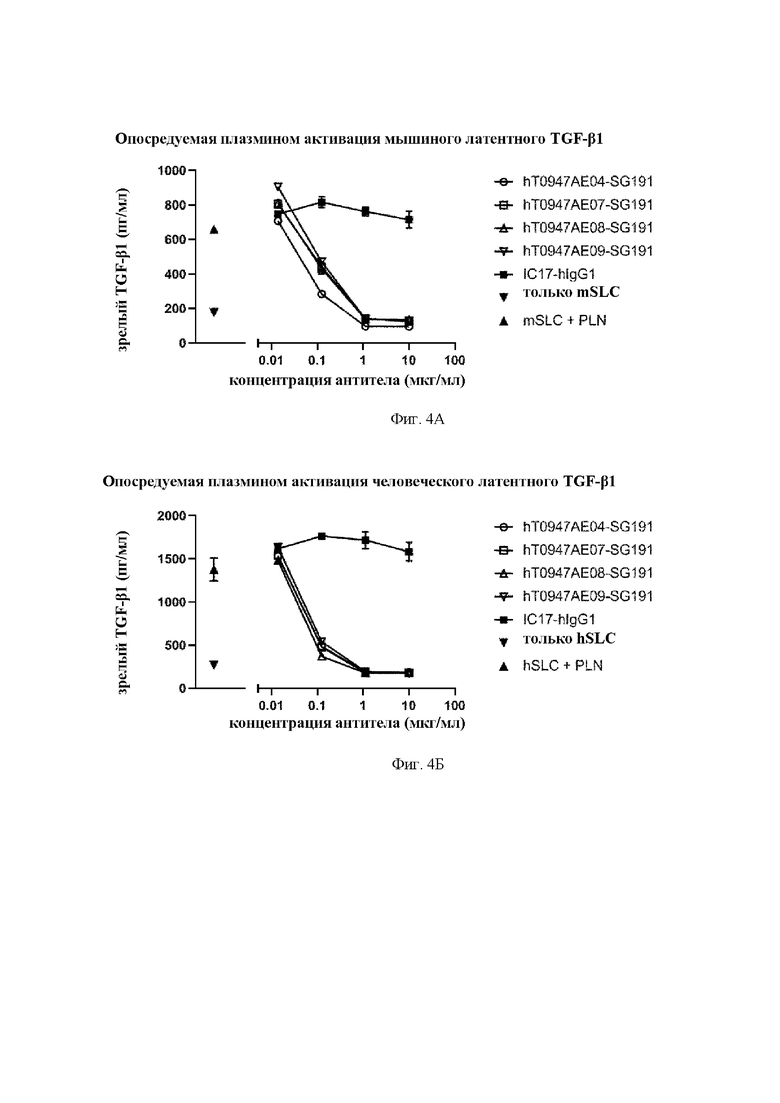
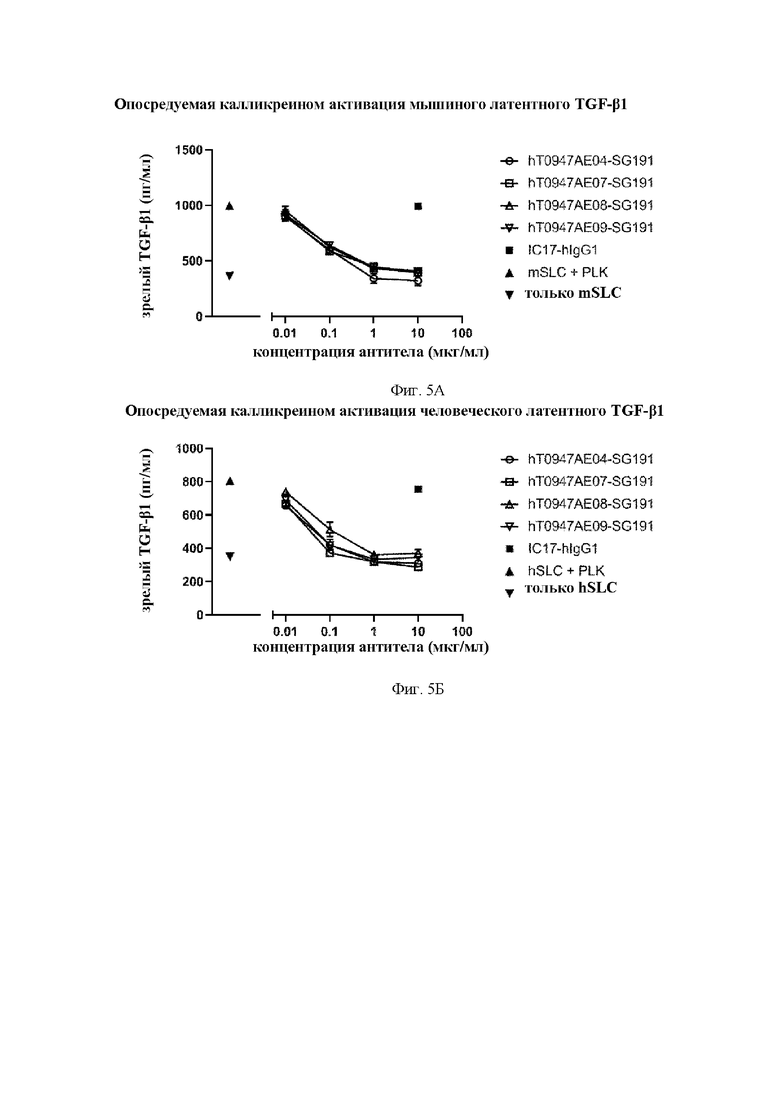
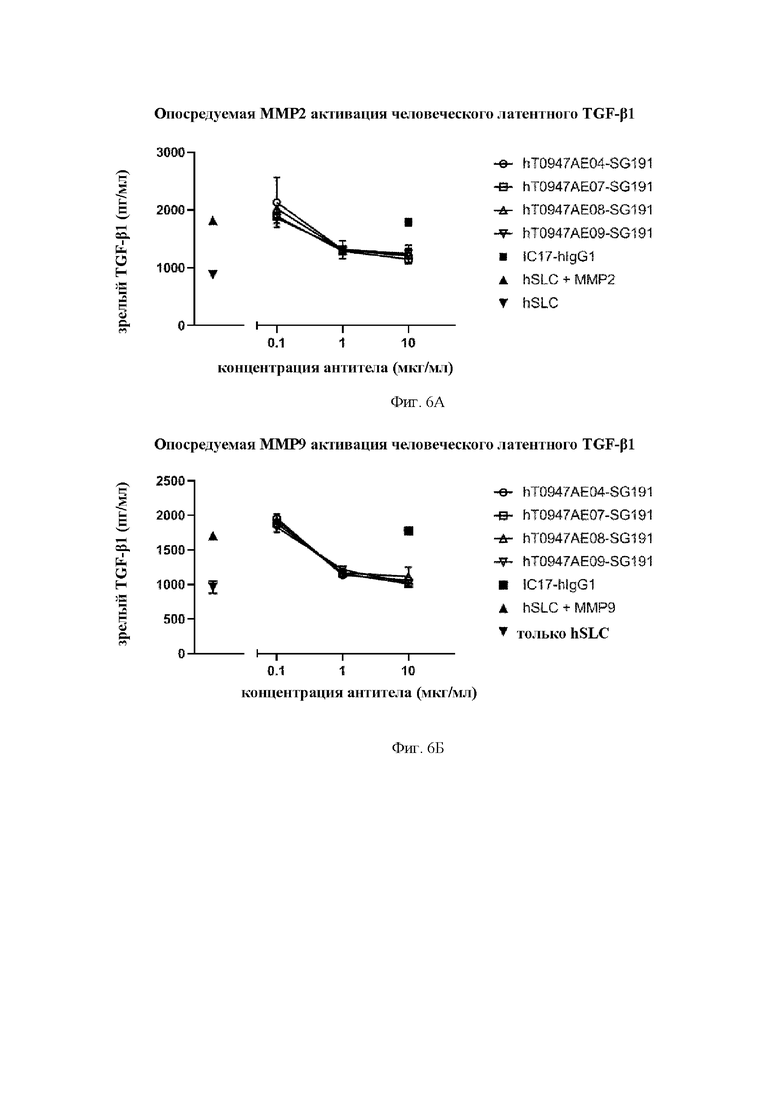
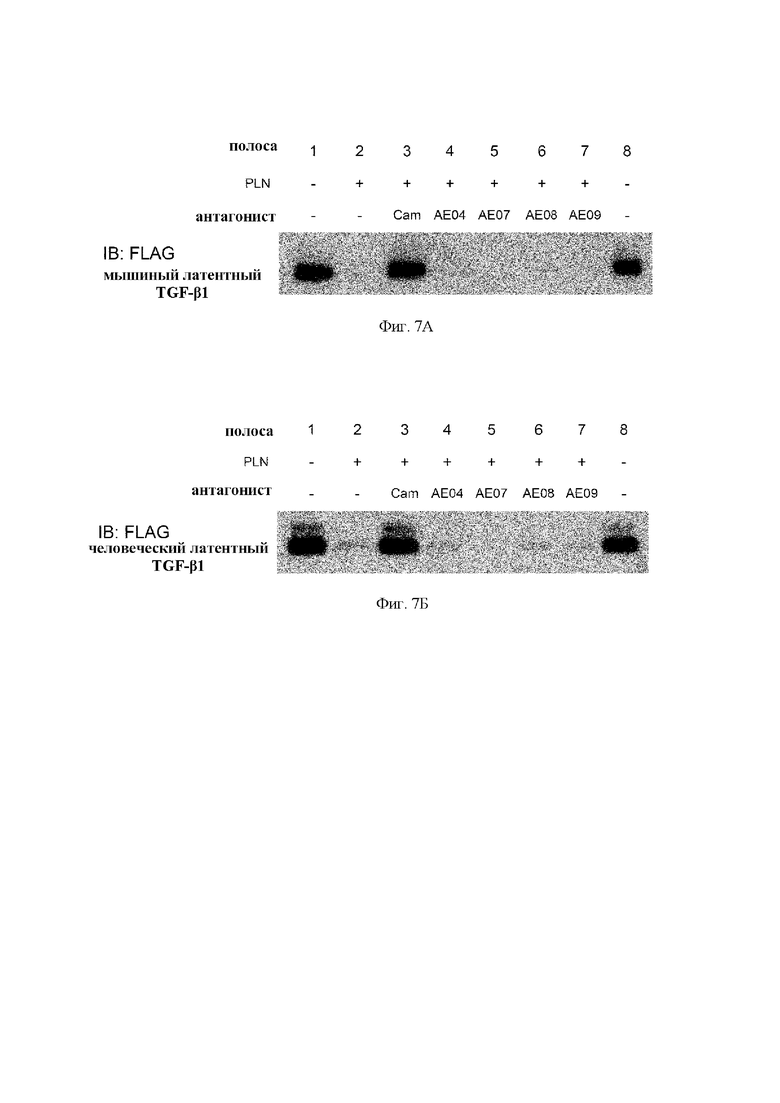
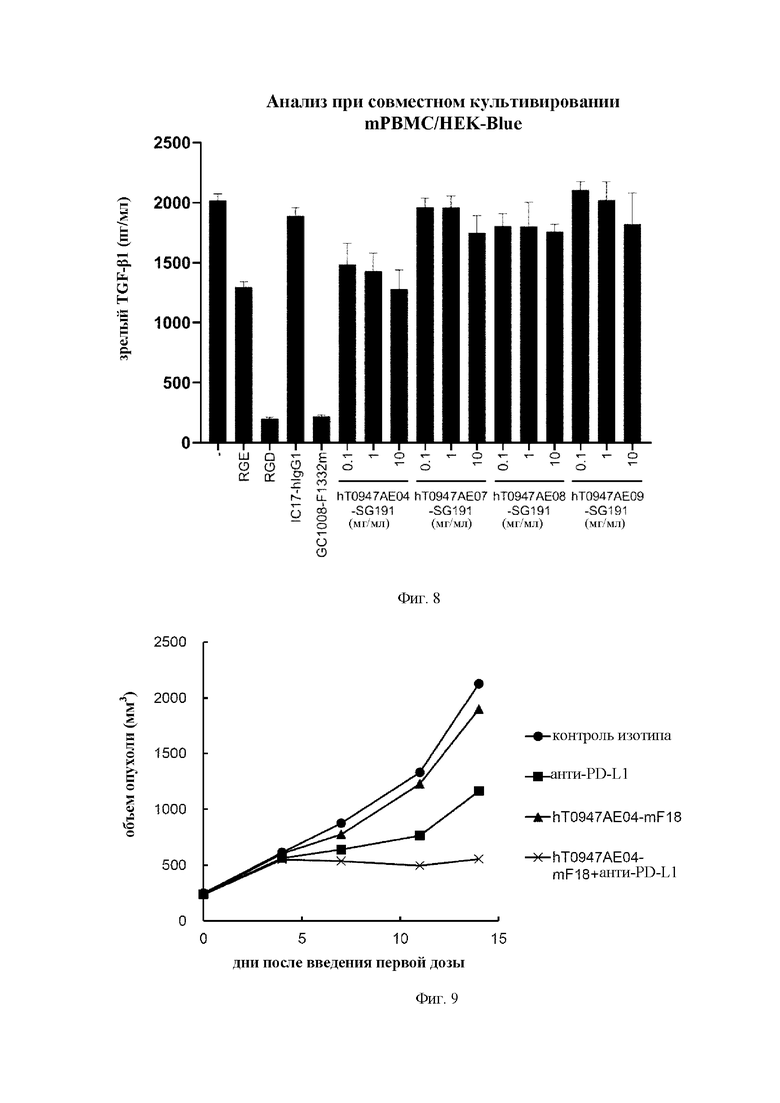
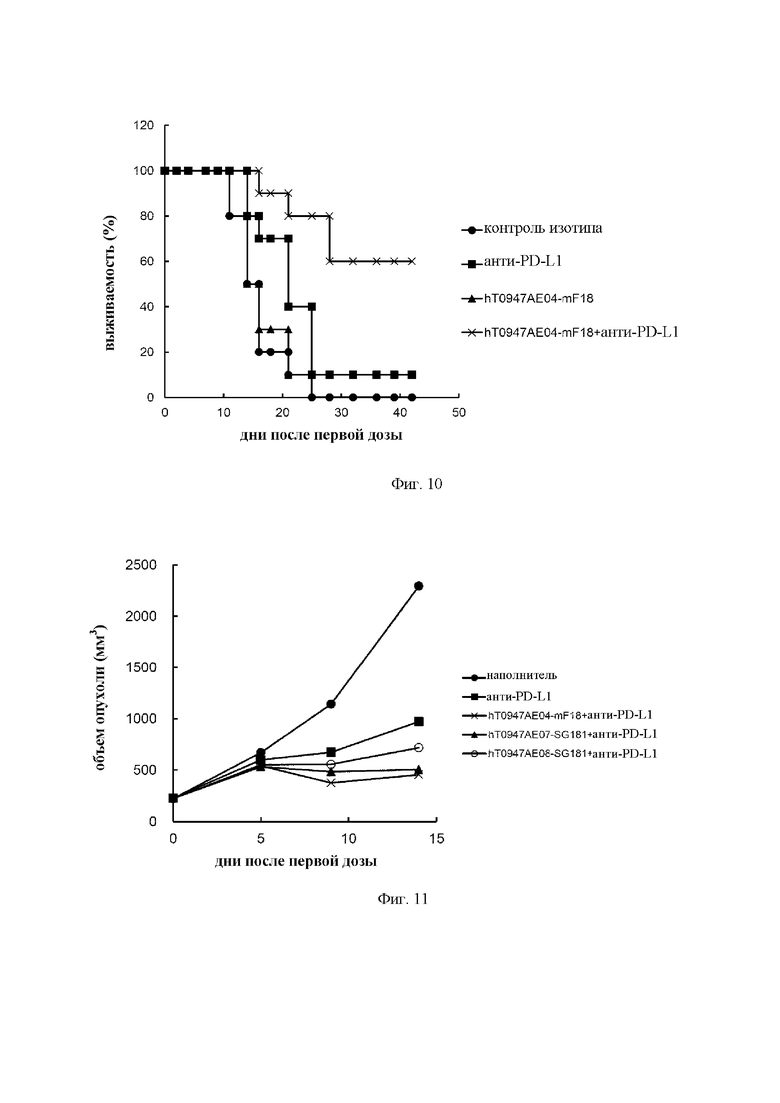
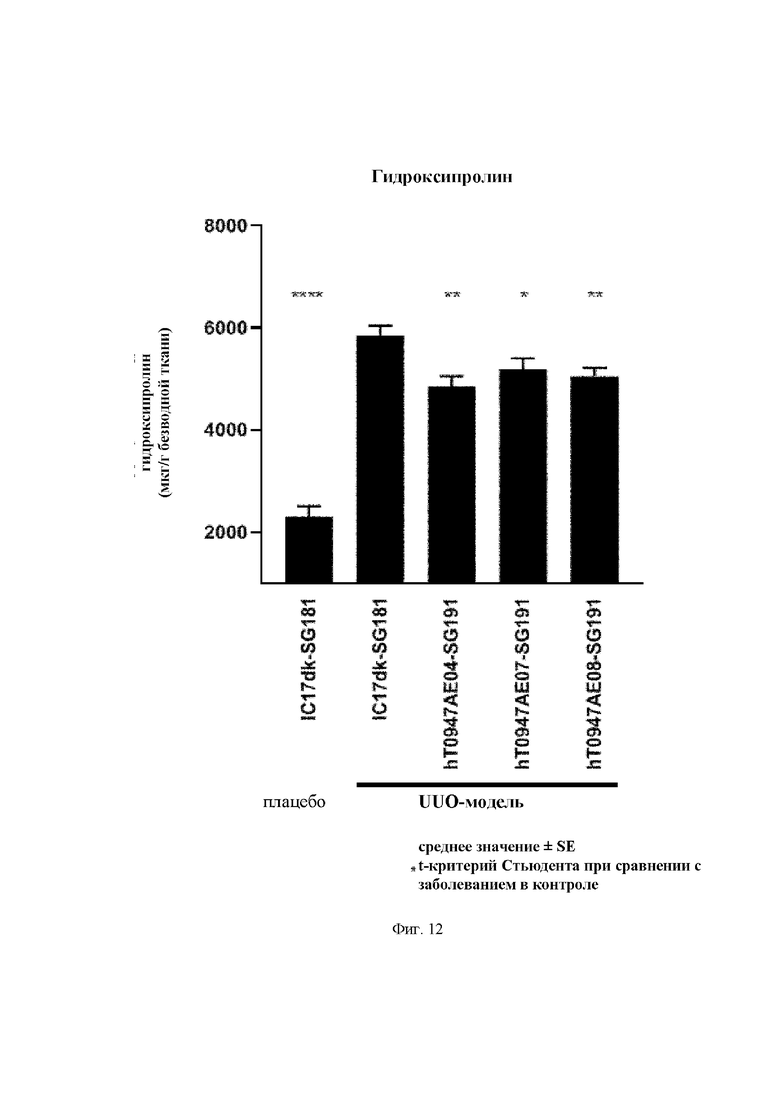
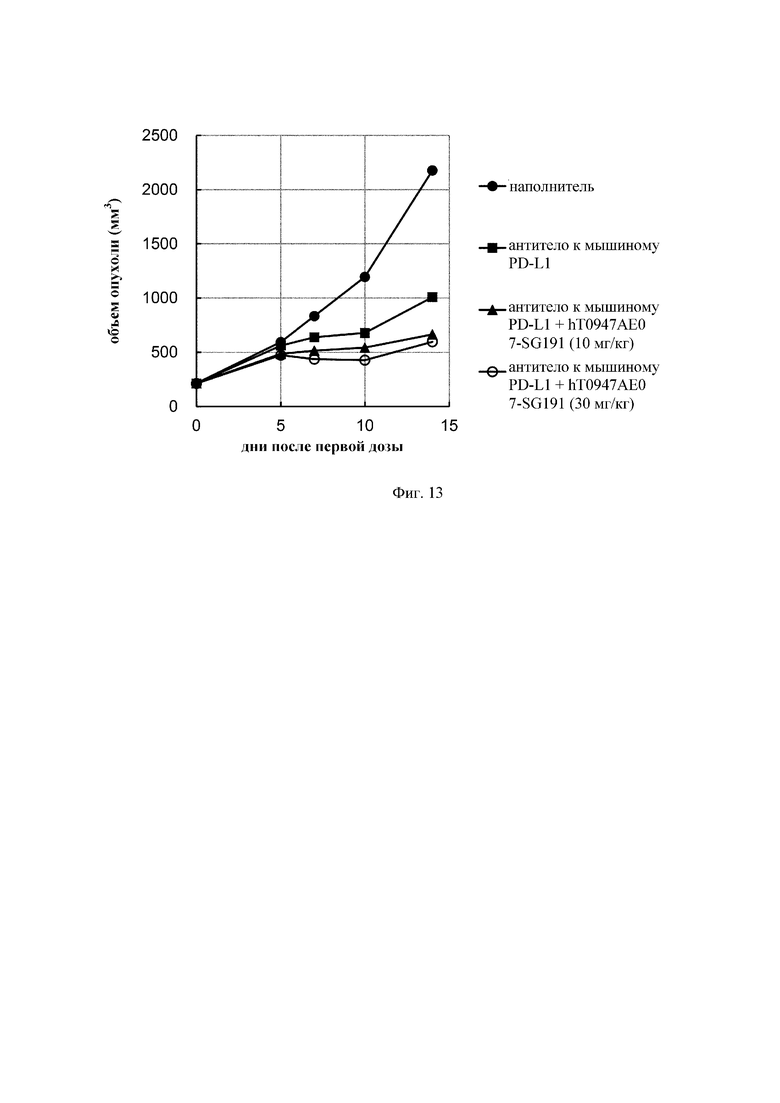
Изобретение относится к области биотехнологии, а именно к антителу к латентному TGF-бета 1, способу его получения, а также к иммуноконъюгату и композиции, его содержащим. Также раскрыты выделенная нуклеиновая кислота, кодирующая вышеуказанное антитело, а также вектор и клетка-хозяин, ее содержащие. Изобретение эффективно для лечения фиброза и рака. 9 н. и 5 з.п. ф-лы, 13 ил., 7 табл., 9 пр.
1. Антитело к латентному TGF-бета 1, содержащее:
(а) HVR-H1, HVR-H2 и HVR-H3, которые содержат аминокислотные последовательности SEQ ID NO: 20, 21 и 22 соответственно, и HVR-L1, HVR-L2 и HVR-L3, которые содержат аминокислотные последовательности SEQ ID NO: 23, 24 и 25 соответственно;
(б) HVR-H1, HVR-H2 и HVR-H3, которые содержат аминокислотные последовательности SEQ ID NO: 26, 27 и 28 соответственно, и HVR-L1, HVR-L2 и HVR-L3, которые содержат аминокислотные последовательности SEQ ID NO: 29, 30 и 31 соответственно;
(в) HVR-H1, HVR-H2 и HVR-H3, которые содержат аминокислотные последовательности SEQ ID NO: 32, 33 и 34 соответственно, и HVR-L1, HVR-L2 и HVR-L3, которые содержат аминокислотные последовательности SEQ ID NO: 35, 36 и 37 соответственно; или
(г) HVR-H1, HVR-H2 и HVR-H3, которые содержат аминокислотные последовательности SEQ ID NO: 38, 39 и 40 соответственно, и HVR-L1, HVR-L2 и HVR-L3, которые содержат аминокислотные последовательности SEQ ID NO: 41, 42 и 43 соответственно.
2. Антитело к латентному TGF-бета 1 по п. 1, содержащее:
(а) (I) последовательность VH, которая идентична по меньшей мере на 95% аминокислотной последовательности SEQ ID NO: 12, (II) последовательность VL, которая идентична по меньшей мере на 95% аминокислотной последовательности SEQ ID NO: 13, или (III) последовательность VH, указанную в подпункте (I), и последовательность VL, указанную в подпункте (II);
(б) (I) последовательность VH, которая идентична по меньшей мере на 95% аминокислотной последовательности SEQ ID NO: 14, (II) последовательность VL, которая идентична по меньшей мере на 95% аминокислотной последовательности SEQ ID NO: 15, или (III) последовательность VH, указанную в подпункте (I), и последовательность VL, указанную в подпункте (II);
(в) (I) последовательность VH, которая идентична по меньшей мере на 95% аминокислотной последовательности SEQ ID NO: 16, (II) последовательность VL, которая идентична по меньшей мере на 95% аминокислотной последовательности SEQ ID NO: 17, или (III) последовательность VH, указанную в подпункте (I), и последовательность VL, указанную в подпункте (II);
(г) (I) последовательность VH, которая идентична по меньшей мере на 95% аминокислотной последовательности SEQ ID NO: 18, (II) последовательность VL, которая идентична по меньшей мере на 95% аминокислотной последовательности SEQ ID NO: 19, или (III) последовательность VH, указанную в подпункте (I), и последовательность VL, указанную в подпункте (II).
3. Антитело к латентному TGF-бета 1, содержащее:
(а) последовательность VH SEQ ID NO: 12 и последовательность VL SEQ ID NO: 13;
(б) последовательность VH SEQ ID NO: 14 и последовательность VL SEQ ID NO: 15;
(в) последовательность VH SEQ ID NO: 16 и последовательность VL SEQ ID NO: 17; или
(г) последовательность VH SEQ ID NO: 18 и последовательность VL SEQ ID NO: 19.
4. Антитело к латентному TGF-бета 1 по одному из пп. 1-3, которое представляет собой человеческое, гуманизированное или химерное антитело.
5. Антитело к латентному TGF-бета 1 по одному из пп. 1-4, которое представляет собой полноразмерное антитело IgG-типа, предпочтительно полноразмерное антитело IgG1-подтипа.
6. Антитело к латентному TGF-бета 1 по одному из пп. 1-5, где антитело к латентному TGF-бета 1 содержит модифицированную Fc-область IgG1, обладающую пониженной эффекторной функцией по сравнению с Fc-областью IgG1 дикого типа.
7. Антитело к латентному TGF-бета 1, содержащее:
(а) полноразмерную последовательность тяжелой цепи SEQ ID NO: 47 и полноразмерную последовательность легкой цепи SEQ ID NO: 60;
(б) полноразмерную последовательность тяжелой цепи SEQ ID NO: 48 и полноразмерную последовательность легкой цепи SEQ ID NO: 61;
(в) полноразмерную последовательность тяжелой цепи SEQ ID NO: 49 и полноразмерную последовательность легкой цепи SEQ ID NO: 62;
(г) полноразмерную последовательность тяжелой цепи SEQ ID NO: 50 и полноразмерную последовательность легкой цепи SEQ ID NO: 63;
(д) полноразмерную последовательность тяжелой цепи SEQ ID NO: 51 и полноразмерную последовательность легкой цепи SEQ ID NO: 60;
(е) полноразмерную последовательность тяжелой цепи SEQ ID NO: 52 и полноразмерную последовательность легкой цепи SEQ ID NO: 61;
(ж) полноразмерную последовательность тяжелой цепи SEQ ID NO: 53 и полноразмерную последовательность легкой цепи SEQ ID NO: 62; или
(з) полноразмерную последовательность тяжелой цепи SEQ ID NO: 54 и полноразмерную последовательность легкой цепи SEQ ID NO: 63.
8. Иммуноконъюгат для применения при лечении фиброза или рака, содержащий антитело к латентному TGF-бета 1 по одному из пп. 1-7 и цитотоксический агент.
9. Выделенная нуклеиновая кислота, кодирующая антитело к латентному TGF-бета 1 по одному из пп. 1-7.
10. Вектор экспрессии, содержащий нуклеиновую кислоту по п. 9.
11. Клетка-хозяин для получения антитела к латентному TGF-бета 1 по одному из пп. 1-7, содержащая нуклеиновую кислоту по п. 9 или вектор по п. 10.
12. Способ получения антитела к латентному TGF-бета 1, включающий культивирование клетки-хозяина по п. 11 таким образом, чтобы получать антитело.
13. Фармацевтическая композиция для применения при лечении фиброза или рака, содержащая антитело к латентному TGF-бета 1 по одному из пп. 1-7 или иммуноконъюгат по п. 8 и фармацевтически приемлемый носитель.
14. Фармацевтическая композиция по п. 13 для применения в комбинации с дополнительным терапевтическим агентом, предпочтительно ингибитором иммунных контрольных точек, для лечения рака.
| WO 2017156500 A1, 14.09.2017 | |||
| WO 2018043734 A1, 08.03.2018 | |||
| MURPHY-ULLRICH, J | |||
| E., & SUTO, M | |||
| J., Thrombospondin-1 regulation of latent TGF-β activation: A therapeutic target for fibrotic disease, Matrix Biology, 2018, 68-69, 28-43, doi:10.1016/j.matbio.2017.12.009 | |||
| DAVID A | |||
| LAWRENCE, Latent-TGF-β: An overview, Molecular and Cellular |

