ОБЛАСТЬ ТЕХНИКИ
Изобретение относится к новым биспецифическим антигенсвязывающим молекулам, содержащим по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с белком активации фибробластов (FAP), по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с CD40, и Fc-домен, состоящий из первой и второй субъединиц, способных к стабильной ассоциации. Изобретение дополнительно относится к анти-FAP клону 212. Дополнительные аспекты изобретения представляют способы получения этих молекул и способы их применения.
УРОВЕНЬ ТЕХНИКИ
Во время генерации сильного адаптивного иммунного ответа необходимо множество молекулярных сигналов. Сигнал один включает связывание Т-клеточного антигенного рецептора (ТКР) со своим когнатным антигеном, презентируемым на поверхности антигенпрезентирующих клеток (АПК). Сигнал два состоит в привлечении костимулирующих рецепторов с соответствующими лигандами между Т-клетками и АПК. Одним из наиболее хорошо исследованных и наиболее важных костимулирующих эффекторов является представитель семейства факторов некроза опухолей (TNFR) CD40 и его лиганд CD40L (Elgueta R. et at., Immunol Rev. 2009; 229(1): 152-72). Несколько представителей семейства TNFR, включая CD40, функционируют после начальной активации Т-клеток для поддержания АПК и Т-клеточных ответов и, таким образом, имеют ключевую роль в организации и функционировании иммунной системы (Watts Т.Н. (2005) Annu. Rev. Immunol. 23, 23-68). Комбинация разных костимулирующих представителей семейства TNFR позволяет осуществлять последовательную и временную регуляцию активации и выживаемости АПК и Т-клеток, что приводит к повышению иммунных ответов при сохранении жесткого контроля функции АПК и Т-клеток. В зависимости от патологического состояния стимуляция посредством костимулирующих членов семейства TNF может усугублять или облегчать заболевания. Активация или блокирование костимуляторов семейства TNFR демонстрирует перспективы в отношении нескольких терапевтических применений в ряде областей, включая рак, инфекционные заболевания, трансплантацию и аутоиммунность.
Среди нескольких костимулирующих молекул представитель семейства TNFR CD40 играет ключевую роль в инициации иммунных ответов путем индукции созревания, выживания, антигенной презентации, выработки цитокинов и экспрессии костимулирующих молекул АПК, которые затем обуславливают антиген-специфические Т-клеточные ответы и активацию NK-клеток провоспалительными цитокинами. CD40 регулирует иммунные ответы против инфекций, опухолей и собственных антигенов, а его экспрессия была продемонстрирована на поверхности АПК, таких как В-клетки, дендритные клетки (ДК), моноциты и макрофаги, а также тромбоциты, и клеток негемопоэтического происхождения, таких как миофибробласты, фибробласты, эпителиальные и эндотелиальные клетки (Elgueta R. et al., Immunol Rev. 2009; 229(1): 152-72). Лиганд CD40 CD40L экспрессируется на активированных CD4+ хелперных Т-клетках, тромбоцитах, моноцитарных клетках, естественных клетках-киллерах, тучных клетках и базофилах (Carbone Е. et. al., J Exp Med. 1997; 185(12): 2053-2060, или Elgueta R. et al., Immunol Rev. 2009; 229(1): 152-72). Экспрессия CD40 и CD40L сильно повышается в ответ на разные иммунные стимулирующие сигналы, а взаимодействие CD40 CD40L между АПК и CD4+ Т-клетками способствует повышенной активации АПК и антиген-специфическим ответам CD8+ Т-клеток (Bevan MJ., Nat Rev Immunol. 2014; 4(8):595-602). Аналогичные иммунные стимулирующие результаты наблюдали при применении агонистических антител к CD40 (Vonderheide RH and Glennie MJ., Clin Cancer Res. 2013; 19(5): 1035-43).
Связывание трансмембранного рецептора типа I CD40 его естественным лигандом CD40L, трансмембранным белком типа II, или агонистическими антителами способствует кластеризации CD40 и индуцирует рекрутирование адаптерных белков в цитоплазматический рецепторный домен. Рекрутирование этих адаптерных белков, известных как ассоциированные с рецептором TNF факторы (TRAF), приводит к синергетической активации сигнальных путей митоген-активируемых протеинкиназ (МАРК), фосфоинозитид 3-киназы (PI3K), а также канонического и неканонического ядерного фактора κВ (NFκB) (Elgueta R. et al., Immunol Rev. 2009; 229(1):152-72). В свою очередь, это приводит к созреванию и активации АПК, что затем максимизирует антиген-специфические Т-клеточные ответы. Недавние исследования показали два разных способа действия агонистических антител к CD40 в стимуляции противоопухолевого иммунитета. Помимо непрямого способа действия путем опосредования уничтожения опухолевых клеток за счет активации адаптивной иммунной системы, агонистические антитела к CD40 могут индуцировать прямое уничтожение опухолевых клеток за счет индукции апоптоза CD40-экспрессирующих клеток солидных опухолей (Eliopoulos AG. et al., Mol Cell Biol. 2000; 20(15):5503-15). Прямое опосредованное антителами к CD40 уничтожение опухолевых клеток может обеспечить источник опухолевых антигенов, которые могут процессироваться и презентироваться АПК, активируемыми параллельно за счет связывания CD40 анти-CD40 антителами, которые затем могут индуцировать антиген-специфические Т-клетки, что представляет собой постулированный механизм, известный как эндогенная вакцинация. С учетом того, что связывание CD40 может генерировать эффективный противораковый иммунный ответ, агонистические антитела к CD40 успешно применяли в ряде доклинических опухолевых моделей, как в виде отдельного агента, так и в комбинации с химиотерапией (Vonderheide RH and Glennie MJ., Clin Cancer Res. 2013; 19(5): 1035-43).
На данный момент в клинических исследованиях проводят изучение шести CD40 mAb: Chi Lob 7/4 (агонистическое химерное IgG1 mAb к CD40; Cancer Research UK; Chowdhury F. et al, Cancer Immunol Res. 2013; 2:229-40), ADC1013 (полностью человеческое агонистическое антитело IgG1 к CD40; Alligator Bioscience and Johnson & Johnson; Mangsbo SM. et al., Clin Cancer Res. 2015 Mar 1; 21(5): 1115-26), APX-005 (полностью гуманизированное агонистическое IgG1 mAb к CD40; Apexigen; Bjorck P. et al. J Immunother Cancer. 2015; 3(Suppl 2): P198), SEA-CD40 (агонистическое химерное IgG1 mAb к CD40; Seattle Genetics; Gardai SJ. et al. AACR 106th Annual Meeting 2015; April 18-22, abstract 2472), a также RO7009789 (полностью человеческое суперагонистическое IgG2 mAb к CD40) проходят клинические исследования фазы I, а дацетузумаб (частично агонистическое химерное IgG1 mAb к CD40; Seattle Genetics; Khubchandani S. et al., Curr Opin Investig Drugs. 2009; 10,579-87) проходит клиническое исследование фазы II. Подлежащие включению в эти исследования пациенты имеют солидные опухоли, классическую лимфому Ходжкина (ЛХ), диффузную крупноклеточную В-клеточную лимфому (ДКВЛ) или индолентную лимфому (включая фолликулярную лимфому). Для этих агонистических антител к CD40 была продемонстрирована разнообразная активность, в диапазоне от Fc-зависимой цитотоксичности CD40+ опухолевых клеток посредством комплементопосредованной цитотоксичности (КОЦ) или антителозависимой клеточной цитотоксичности (АЗКЦ) до активации АПК с индукцией противоопухолевых Т-клеточных ответов, а также активации макрофагов с истощением опухоли и опухолевой стромы. Однако до сих пор нет окончательного пояснения для этой наблюдаемой гетерогенности. При этом недавние исследования показали, что это разнообразие способов действия может быть объяснено, по меньшей мере частично, разницей в специфичности эпитопов, изотипе или взаимодействии Fc:FcγR анти-CD40 антител. Например, оказалось, что агонистические антитела к CD40 in vivo требуют перекрестного связывания CD40, связанного своим Fab-фрагментом на целевой клетке, с Fcγ-рецептором, связанным своим Fc-фрагментом на клетке, отличной от целевой клетки, как было описано для агонистических антител, специфических к другим индуцирующим апоптоз или иммуномодулирующим членам суперсемейства TNFR (DahanR., Cancer Cell. 2016 Jun 13; 29(6):820-31; Li F. andRavetch J.V. Science, 2011; 333, 1030-1034; Teng M.W. et al., J. Immunol. 2009; 183, 1911-1920). Предложенный механизм включает опосредованную Fcγ-рецептором кластеризацию трансмембранных молекул CD40 на целевых клетках и последующее повышение сигнализации CD40 с достижением высокой in vivo эффективности.
Клиническая разработка агонистических антител к CD40 показала многообещающие начальные результаты. В первом клиническом исследовании СР-870,893 показал клиническую эффективность у пациентов с распространенным раком. Четверо из 29 пациентов с распространенным раком демонстрировали частичные ответы после получения одной внутривенной инфузии СР-870,893 (Vonderheide RH., J Clin Oncol. 2007Mar 1; 25(7):876-83). Один из этих четырех пациентов, прошедший лечение 9 последовательными дозами СР-870,893 в течение полутора лет, оставался в состоянии полной ремиссии более 5 лет. Однако наиболее распространенными побочными эффектами СР-870,893 являются синдром высвобождения цитокинов и тромбоэмболические события, поэтому при применяемых схемах введения доз и путях введения объединенные данные по клиническим исследованиями фазы 1 с более чем 140 пациентами показали только ограниченную клиническую эффективность, и было предложено локальное введение антитела (Vonderheide RH, Glennie М, Clin Cancer Res. 2013, 19(5), 1035-1043). Отсутствие ответов на один агент частично связано с серьезными целевыми/внеопухолевыми эффектами, вызываемыми масштабной экспрессией CD40, что приводит к дозолимитирующей токсичности (например, синдрому высвобождения цитокинов). Разработка агонистического антитела к CD40, которое специфически активирует АПК, когда CD40 перекрестно связан опухолеспецифической мишенью, может уменьшить побочные эффекты и повысить ограничения доз, предоставляя новые терапевтические варианты с потенциальной возможностью генерации эффективного длительного противоракового иммунитета.
Доступные доклинические и клинические данные ясно демонстрируют, что существует высокая клиническая потребность в эффективных агонистах CD40, которые способны индуцировать и усиливать эффективные эндогенные иммунные ответы на рак. Однако эти эффекты почти никогда не ограничены одним типом клеток или действием посредством одного механизма, а исследования, направленные на то, чтобы прояснить меж- и внутриклеточные механизмы сигнализации, выявили возрастающие уровни сложности. Известные антитела к CD40 можно вводить только в относительно низких дозах вследствие дозолимитирующей токсичности, такой как синдром высвобождения цитокинов, активация тромбоцитов/эндотелиальных клеток, что приводит к неэффективной активации пути на целевых АПК и узкому терапевтическому индексу. Таким образом, существует потребность «нацеленных» агонистов, которые предпочтительно действуют на один тип клеток.
Это изобретение относится к новым биспецифическим антигенсвязывающим молекулам, способным к специфическому связыванию с CD40 и белком активации фибробластов (FAP), и, таким образом, объединяющим фрагмент, способный к связыванию с FAP, с фрагментом, способным к агонистическому связыванию с CD40, причем активация АПК посредством CD40 обеспечивается за счет перекрестного связывания посредством FAP, экспрессируемого на клетках опухолевой стромы, и потенциально также посредством FAP, промежуточно экспрессируемого во вторичных лимфоидных тканях. В отличие от биспецифических антигенсвязывающих молекул, способных к специфическому связыванию с CD40, и рецепторов иммунных контрольных точек на активированных Т-клетках, таких как CTLA-4 или PD-1, нацеливание на опухолевую мишень, такую как FAP, делает возможной CD40-опосредованную активацию АПК преимущественно в опухолевой строме и дренирующих опухоль лимфатических узлах, где фибробласты экспрессируют повышенные уровни FAP по сравнению с другими тканями. Таким образом, антигенсвязывающие молекулы по данному изобретению могут быть способны приводить в действие рецептор CD40 не только эффективно, но также и очень избирательно в необходимом участке, в то же время устраняя необходимость в перекрестном связывании FcγR, тем самым снижая побочные эффекты. Новые биспецифические антигенсвязывающие молекулы дополнительно характеризуются тем, что они содержат новый антигенсвязывающий домен FAP, который не утрачивает своих превосходных связывающих свойств, когда он слит с С-концом Fc-домена.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к биспецифическим антигенсвязывающий молекулам, в которых скомбинированы по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с костимулирующим представителем семейства рецепторов TNF CD40, с по меньшей мере одним антигенсвязывающим доменом, нацеленным на белок активации фибробластов (FAP), содержащим новый мышиный клон 212 против FAP человека и его гуманизированные варианты. Эти биспецифические антигенсвязывающие молекулы обладают преимуществом, поскольку они будут предпочтительно активировать костимулирующие рецепторы CD40 в опухолеассоциированном участке, в котором экспрессируется FAP, так как они способны связывать FAP с высокой аффинностью.
В одном аспекте в изобретении предложена биспецифическая антигенсвязывающая молекула, содержащая
(а) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с CD40, и
(б) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с белком активации фибробластов (FAP), содержащий вариабельную область тяжелой цепи (VHFAP), содержащую (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 3, (ii) CDR-H2, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4, SEQ ID NO: 11 и SEQ ID NO: 12, и (iii) CDR-Н3, содержащую аминокислотную последовательность SEQ ID NO: 5, и вариабельную область легкой цепи (VLFAP), содержащую (iv) CDR-L1, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 6, SEQ ID NO: 13 и SEQ ID NO: 14, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 7, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 8.
В конкретном аспекте биспецифическая антигенсвязывающая молекула содержит (а) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с CD40, (б) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с FAP, содержащий вариабельную область тяжелой цепи (VHFAP), содержащую (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 3, (ii) CDR-H2, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4, SEQ ID NO: 11 и SEQ ID NO: 12, и (iii) CDR-Н3, содержащую аминокислотную последовательность SEQ ID NO:5, и вариабельную область легкой цепи (VLFAP), содержащую (iv) CDR-L1, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 6, SEQ ID NO: 13 и SEQ ID NO: 14, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 7, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 8, и (в) Fc-домен, состоящий из первой и второй субъединиц, способных к стабильной ассоциации. Конкретнее, Fc-домен, состоящий из первой и второй субъединиц, способных к стабильной ассоциации, содержит мутации, которые снижают эффекторную функцию.
В одном аспекте по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с FAP, содержит вариабельную область тяжелой цепи (VHFAP) содержащую (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 3, (ii) CDR-H2, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4, и (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 5, и вариабельную область легкой цепи (VLFAP), содержащую (iv) CDR-L1, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 6, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 7, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 8. В одном аспекте антигенсвязывающий домен, способный к специфическому связыванию с FAP, содержит вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность, которая по меньшей мере на около 90% идентична аминокислотной последовательности SEQ ID NO: 9, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность, которая по меньшей мере на около 90% идентична аминокислотной последовательности SEQ ID NO: 10. В конкретном аспекте антигенсвязывающий домен, способный к специфическому связыванию с FAP, содержит вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 9, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 10.
В другом аспекте антигенсвязывающий домен, способный к специфическому связыванию с FAP, содержит вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19 и SEQ ID NO: 20, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 21, SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 25 и SEQ ID NO: 26. В частности, антигенсвязывающий домен, способный к специфическому связыванию с FAP, содержит (а) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 15, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 21, (б) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 16, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 21, (в) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 16, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 22, или (г) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 19, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 25. В частности, антигенсвязывающий домен, способный к специфическому связыванию с FAP, содержит (а) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 15, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 21.
В одном аспекте антигенсвязывающий домен, способный к специфическому связыванию с CD40, связывается с полипептидом, содержащим или состоящим из аминокислотной последовательности SEQ ID NO: 1.
В дополнительном аспекте предложена биспецифическая антигенсвязывающая молекула, причем по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержит вариабельную область тяжелой цепи (VHCD40), содержащую (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 27, (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 28, и (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 29, и вариабельную область легкой цепи (VLCD40), содержащую (iv) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 30, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 31, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 32.
В одном аспекте предложена биспецифическая антигенсвязывающая молекула по определению выше в данном документе, причем по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержит
(i) вариабельную область тяжелой цепи (VHCD40), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 37, SEQ ID NO: 38, SEQ ID NO: 39 и SEQ ID NO: 40, и
(ii) вариабельную область легкой цепи (VLCD40), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 41, SEQ ID NO: 42, SEQ ID NO: 43 и SEQ ID NO: 44.
В другом аспекте предложена биспецифическая антигенсвязывающая молекула по определению выше в данном документе, причем по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержит (i) вариабельную область тяжелой цепи (VHCD40), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 45, SEQ ID NO: 46, SEQ ID NO: 47, SEQ ID NO: 48, SEQ ID NO: 49 и SEQ ID NO: 50, и
(ii) вариабельную область легкой цепи (VLCD40), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 51, SEQ ID NO: 52, SEQ ID NO: 53 и SEQ ID NO: 54.
Кроме того, предложена биспецифическая антигенсвязывающая молекула по определению выше в данном документе, причем по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержит
(а) VH, содержащую аминокислотную последовательность SEQ ID NO: 37, и VL, содержащую аминокислотную последовательность SEQ ID NO: 41, или
(б) VH, содержащую аминокислотную последовательность SEQ ID NO: 37, и VL, содержащую аминокислотную последовательность SEQ ID NO: 42, или
(в) VH, содержащую аминокислотную последовательность SEQ ID NO: 37, и VL, содержащую аминокислотную последовательность SEQ ID NO: 43, или
(г) VH, содержащую аминокислотную последовательность SEQ ID NO: 37, и VL, содержащую аминокислотную последовательность SEQ ID NO: 44, или
(д) VH, содержащую аминокислотную последовательность SEQ ID NO: 38, и VL, содержащую аминокислотную последовательность SEQ ID NO: 41, или
(е) VH, содержащую аминокислотную последовательность SEQ ID NO: 38, и VL, содержащую аминокислотную последовательность SEQ ID NO: 42, или
(ж) VH, содержащую аминокислотную последовательность SEQ ID NO: 38, и VL, содержащую аминокислотную последовательность SEQ ID NO: 43, или
(з) VH, содержащую аминокислотную последовательность SEQ ID NO: 38, и VL, содержащую аминокислотную последовательность SEQ ID NO: 44, или
(и) VH, содержащую аминокислотную последовательность SEQ ID NO: 39, и VL, содержащую аминокислотную последовательность SEQ ID NO: 41, или
(к) VH, содержащую аминокислотную последовательность SEQ ID NO: 39, и VL, содержащую аминокислотную последовательность SEQ ID NO: 42, или
(л) VH, содержащую аминокислотную последовательность SEQ ID NO: 39, и VL, содержащую аминокислотную последовательность SEQ ID NO: 43, или
(м) VH, содержащую аминокислотную последовательность SEQ ID NO: 39, и VL, содержащую аминокислотную последовательность SEQ ID NO: 44, или
(н) VH, содержащую аминокислотную последовательность SEQ ID NO: 40, и VL, содержащую аминокислотную последовательность SEQ ID NO: 41, или
(о) VH, содержащую аминокислотную последовательность SEQ ID NO: 40, и VL, содержащую аминокислотную последовательность SEQ ID NO: 42, или
(п) VH, содержащую аминокислотную последовательность SEQ ID NO: 40, и VL, содержащую аминокислотную последовательность SEQ ID NO: 43, или
(р) VH, содержащую аминокислотную последовательность SEQ ID NO: 40, и VL, содержащую аминокислотную последовательность SEQ ID NO: 44.
В конкретном аспекте предложена биспецифическая антигенсвязывающая молекула, причем по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержит VH, содержащую аминокислотную последовательность SEQ ID NO: 37, и VL, содержащую аминокислотную последовательность SEQ ID NO: 41.
В дополнительном аспекте предложена биспецифическая антигенсвязывающая молекула по определению выше в данном документе, причем по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержит
(а) VH, содержащую аминокислотную последовательность SEQ ID NO: 45, и VL, содержащую аминокислотную последовательность SEQ ID NO: 51, или
(б) VH, содержащую аминокислотную последовательность SEQ ID NO: 46, и VL, содержащую аминокислотную последовательность SEQ ID NO: 51, или
(в) VH, содержащую аминокислотную последовательность SEQ ID NO: 47, и VL, содержащую аминокислотную последовательность SEQ ID NO: 51, или
(г) VH, содержащую аминокислотную последовательность SEQ ID NO: 48, и VL, содержащую аминокислотную последовательность SEQ ID NO: 51, или
(д) VH, содержащую аминокислотную последовательность SEQ ID NO: 45, и VL, содержащую аминокислотную последовательность SEQ ID NO: 52, или
(е) VH, содержащую аминокислотную последовательность SEQ ID NO: 46, и VL, содержащую аминокислотную последовательность SEQ ID NO: 52, или
(ж) VH, содержащую аминокислотную последовательность SEQ ID NO: 47, и VL, содержащую аминокислотную последовательность SEQ ID NO: 52, или
(з) VH, содержащую аминокислотную последовательность SEQ ID NO: 48, и VL, содержащую аминокислотную последовательность SEQ ID NO: 52, или
(и) VH, содержащую аминокислотную последовательность SEQ ID NO: 49, и VL, содержащую аминокислотную последовательность SEQ ID NO: 53, или
(к) VH, содержащую аминокислотную последовательность SEQ ID NO: 50, и VL, содержащую аминокислотную последовательность SEQ ID NO: 53, или
(л) VH, содержащую аминокислотную последовательность SEQ ID NO: 49, и VL, содержащую аминокислотную последовательность SEQ ID NO: 54, или
(м) VH, содержащую аминокислотную последовательность SEQ ID NO: 50, и VL, содержащую аминокислотную последовательность SEQ ID NO: 54.
В дополнительном конкретном аспекте предложена биспецифическая антигенсвязывающая молекула, причем по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержит VH, содержащую аминокислотную последовательность SEQ ID NO: 45, и VL, содержащую аминокислотную последовательность SEQ ID NO: 51, или причем антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержит VH, содержащую аминокислотную последовательность SEQ ID NO: 48, и VL, содержащую аминокислотную последовательность SEQ ID NO: 51.
Конкретнее, предложена биспецифическая антигенсвязывающая молекула, содержащая
(i) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержащий вариабельную область тяжелой цепи (VHCD40), содержащую аминокислотную последовательность SEQ ID NO: 37, и вариабельную область легкой цепи (VLCD40), содержащую аминокислотную последовательность SEQ ID NO: 41, и
(ii) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с FAP, содержащий вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 15, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 21.
В одном аспекте биспецифическая антигенсвязывающая молекула представляет собой гуманизированное или химерное антитело. В дополнительном аспекте биспецифическая антигенсвязывающая молекула содержит Fc-область IgG, в частности, Fc-область IgG1 или Fc-область IgG4. В частности, Fc-область содержит одну или более аминокислотных замен, которые снижают аффинность связывания антитела с Fc-рецептором и/или эффекторную функцию. В конкретном аспекте предложена биспецифическая антигенсвязывающая молекула, причем Fc-область принадлежит подклассу IgG1 человека с аминокислотными мутациями L234A, L235A и P329G (нумерация в соответствии с индексом EU по Kabat).
В другом аспекте предложена биспецифическая антигенсвязывающая молекула по определению выше в данном документе, причем первая субъединица Fc-области содержит выступы, а вторая субъединица Fc-области содержит впадины в соответствии со способом «выступы-во-впадины». В частности, предложена биспецифическая антигенсвязывающая молекула, причем (i) первая субъединица Fc-области содержит аминокислотные замены S354C и T366W (нумерация в соответствии с индексом EU по Kabat), а вторая субъединица Fc-области содержит аминокислотные замены Y349C, T366S и Y407V (нумерация в соответствии с индексом EU по Kabat), или (ii) первая субъединица Fc-области содержит аминокислотные замены K392D и K409D (нумерация в соответствии с индексом EU по Kabat), а вторая субъединица Fc-области содержит аминокислотные замены E356K и D399K (нумерация в соответствии с индексом EU по Kabat). Конкретнее, предложена биспецифическая антигенсвязывающая молекула, причем первая субъединица Fc-области содержит аминокислотные замены S354C и T366W (нумерация в соответствии с индексом EU по Kabat), а вторая субъединица Fc-области содержит аминокислотные замены Y349C, T366S и Y407V (нумерация в соответствии с индексом EU по Kabat).
В дополнительном аспекте предложена биспецифическая антигенсвязывающая молекула, причем биспецифическая антигенсвязывающая молекула содержит
(а) по меньшей мере два Fab-фрагмента, способных к специфическому связыванию с CD40, соединенных с Fc-областью, и
(б) один антигенсвязывающий домен, способный к специфическому связыванию с FAP, соединенный с С-концом Fc-области.
Таким образом, предложена биспецифическая антигенсвязывающая молекула, которая обеспечивает двухвалентное связывание в отношении CD40 и одновалентное связывание в отношении FAP.
В другом аспекте предложена биспецифическая антигенсвязывающая молекула, причем биспецифическая антигенсвязывающая молекула содержит
(а) по меньшей мере два Fab-фрагмента, способных к специфическому связыванию с CD40, слитых с Fc-областью, и
(б) один антигенсвязывающий домен, способный к специфическому связыванию с FAP, слитый с С-концом Fc-области.
В конкретном аспекте антигенсвязывающий домен, способный к специфическому связыванию с FAP, соединенный с С-концом Fc-области, представляет собой Kpocc-fab-фрагмент. Таким образом, предложена биспецифическая антигенсвязывающая молекула, причем биспецифическая антигенсвязывающая молекула содержит
(а) по меньшей мере два Fab-фрагмента, способных к специфическому связыванию с CD40, слитых с Fc-областью, и
(б) Kpocc-fab-фрагмент, способный к специфическому связыванию с FAP, слитый с С-концом Fc-области.
В дополнительном аспекте биспецифическая антигенсвязывающая молекула содержит четыре Fab-фрагмента, способных к специфическому связыванию с CD40. Таким образом, предложена биспецифическая антигенсвязывающая молекула, которая обеспечивает тетравалентное связывание в отношении CD40 и одновалентное связывание в отношении FAP.
В одном аспекте предложена биспецифическая антигенсвязывающая молекула, содержащая
(а) две тяжелые цепи и две легкие цепи антитела, содержащего два Fab-фрагмента, способных к специфическому связыванию с CD40, и Fc-область, и
(б) VH и VL антигенсвязывающего домена, способного к специфическому связыванию с FAP, причем VH слита с С-концом одной из двух тяжелых цепей (а), и причем VL слита с С-концом другой из двух тяжелых цепей (а).
В другом аспекте предложена биспецифическая антигенсвязывающая молекула, содержащая
(а) две тяжелые цепи и две легкие цепи антитела, содержащего два Fab-фрагмента, способных к специфическому связыванию с CD40, и Fc-область, и
(б) Kpocc-fab-фрагмент, способный к специфическому связыванию с FAP, причем цепь VH-C-каппа слита с С-концом одной из двух тяжелых цепей (а).
В другом аспекте предложена биспецифическая антигенсвязывающая молекула, содержащая
(а) две тяжелые цепи и две легкие цепи антитела, содержащего два Fab-фрагмента, способных к специфическому связыванию с CD40, и Fc-область, и
(б) Kpocc-fab-фрагмент, способный к специфическому связыванию с FAP, причем цепь VL-CH1 слита с С-концом одной из двух тяжелых цепей (а).
Кроме того, предложена биспецифическая антигенсвязывающая молекула, содержащая
(а) две тяжелые цепи и две легкие цепи антитела, содержащего два Fab-фрагмента, способных к специфическому связыванию с CD40, и Fc-область, и
(б) два Fab-фрагмента, способных к специфическому связыванию с FAP, причем один из Fab-фрагментов соединен с С-концом одной из двух тяжелых цепей (а), а другой из Fab-фрагментов соединен с С-концом другой из двух тяжелых цепей (а).
В другом аспекте предложена биспецифическая антигенсвязывающая молекула, причем биспецифическая антигенсвязывающая молекула содержит
(а) две тяжелые цепи, причем каждая тяжелая цепь содержит VH и СН1-домен Fab-фрагмента, способного к специфическому связыванию с CD40, и субъединицу Fc-области,
(б) две легкие цепи, причем каждая легкая цепь содержит VL и CL-домен Fab-фрагмента, способного к специфическому связыванию с CD40, и
(в) один Fab-фрагмент, способный к специфическому связыванию с FAP, причем Fab-фрагменты соединены с С-концом одной из двух тяжелых цепей (а).
В другом аспекте Fab-фрагмент, способный к специфическому связыванию с FAP, представляет собой Kpocc-Fab-фрагмент, содержащий цепь VL-CH1 и цепь VH-C-каппа, и при этом цепь VH-C-каппа или цепь VL-CH1 соединена с С-концом одной из двух тяжелых цепей (а).
В одном аспекте предложена биспецифическая антигенсвязывающая молекула, причем биспецифическая антигенсвязывающая молекула содержит четыре Fab-фрагмента, способных к специфическому связыванию с CD40. В конкретном аспекте предложена биспецифическая антигенсвязывающая молекула, причем каждая из двух тяжелых цепей (а) по определению выше в данном документе содержит цепи VH-CH1 Fab-фрагмента, способного к специфическому связыванию с CD40, которые соединены друг с другом, необязательно, пептидным линкером.
Таким образом, в одном аспекте в изобретении предложена биспецифическая антигенсвязывающая молекула, содержащая
(а) две тяжелые цепи, причем каждая тяжелая цепь содержит цепи VH-CH1 Fab-фрагмента, способного к специфическому связыванию с CD40, которые соединены друг с другом, необязательно, пептидным линкером, и субъединицу Fc-области,
(б) четыре легкие цепи, причем каждая легкая цепь содержит VL и домен С-каппа Fab-фрагмента, способного к специфическому связыванию с CD40, и
(в) Kpocc-Fab-фрагмент, способный к специфическому связыванию с FAP, содержащий цепь VL-CH1 и цепь VH-C-каппа, причем цепь VH-C-каппа или цепь VL-CH1 соединена с С-концом одной из двух тяжелых цепей (а), необязательно, пептидным линкером.
В другом аспекте предложена биспецифическая антигенсвязывающая молекула, содержащая
(а) две тяжелые цепи, причем каждая тяжелая цепь содержит цепи VH-CH1 Fab-фрагмента, способного к специфическому связыванию с CD40, которые соединены друг с другом, необязательно, пептидным линкером, и субъединицу Fc-области,
(б) четыре легкие цепи, причем каждая легкая цепь содержит VL и CL-домен Fab-фрагмента, способного к специфическому связыванию с CD40, и
(в) Kpocc-fab-фрагмент, способный к специфическому связыванию с FAP, причем цепь VH-CL указанного Kpocc-fab-фрагмента соединена с С-концом одной из двух тяжелых цепей (а).
В другом аспекте предложена биспецифическая антигенсвязывающая молекула, содержащая
(а) две тяжелые цепи, причем каждая тяжелая цепь содержит цепи VH-CH1 Fab-фрагмента, способного к специфическому связыванию с CD40, которые соединены друг с другом, необязательно, пептидным линкером, и субъединицу Fc-области,
(б) четыре легкие цепи, причем каждая легкая цепь содержит VL и CL-домен Fab-фрагмента, способного к специфическому связыванию с CD40, и
(в) Kpocc-fab-фрагмент, способный к специфическому связыванию с FAP, причем цепь VL-CH1 указанного Kpocc-fab-фрагмента соединена с С-концом одной из двух тяжелых цепей (а).
В другом аспекте предложено антитело, которое специфически связывается с FAP, причем указанное антитело содержит вариабельную область тяжелой цепи (VHFAP), содержащую (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 3, (ii) CDR-H2, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4, SEQ ID NO: 11 и SEQ ID NO: 12, и (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 5, и вариабельную область легкой цепи (VLFAP), содержащую (iv) CDR-L1, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 6, SEQ ID NO: 13 и SEQ ID NO: 14, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 7, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 8. В другом аспекте предложено антитело, которое специфически связывается с FAP, причем указанное антитело содержит вариабельную область тяжелой цепи (VHFAP), содержащую (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 3, (ii) CDR-H2, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4, и (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 5, и вариабельную область легкой цепи (VLFAP), содержащую (iv) CDR-L1, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 6, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 7, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 8.
В дополнительном аспекте предложено антитело, содержащее (а) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 15, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 21, (б) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 16, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 21, (в) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 16, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 22, или (г) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 19, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 25.
В соответствии с другим аспектом изобретения предложена выделенная нуклеиновая кислота, кодирующая биспецифическую антигенсвязывающую молекулу, описанную выше в данном документе. Также предложена выделенная нуклеиновая кислота, кодирующая антитело, описанное выше в данном документе. В изобретении дополнительно предложен вектор, в частности, экспрессионный вектор, содержащий выделенную нуклеиновую кислоту по изобретению, и клетка-хозяин, содержащая выделенную нуклеиновую кислоту или экспрессионный вектор по изобретению. В некоторых аспектах клетка-хозяин представляет собой эукариотическую клетку, в частности, клетку млекопитающего. В другом аспекте предложен способ получения биспецифической антигенсвязывающей молекулы или антитела, описанных выше в данном документе, включающий культивирование вышеописанной клетки-хозяина в условиях, подходящих для экспрессии биспецифической антигенсвязывающей молекулы или антитела, и выделение биспецифической антигенсвязывающей молекулы или антитела. Также изобретение включает биспецифическую антигенсвязывающую молекулу, которая специфически связывается с CD40 и с FAP, или антитело, которое специфически связывается с FAP, полученным способом по изобретению.
В изобретении дополнительно предложена фармацевтическая композиция, содержащая биспецифическую антигенсвязывающую молекулу, описанную выше в данном документе, или антитело, описанное выше в данном документе, и фармацевтически приемлемый носитель. В одном аспекте фармацевтическая композиция содержит дополнительный терапевтический агент.
Также в изобретение включены биспецифическая антигенсвязывающая молекула или антитело, описанные выше в данном документе, или фармацевтическая композиция, содержащая биспецифическую антигенсвязывающую молекулу, для применения в качестве лекарственного средства.
В одном аспекте предложена биспецифическая антигенсвязывающая молекула, описанная выше в данном документе, или фармацевтическая композиция по изобретению для применения
(i) в индукции иммунной стимуляции CD40-экспрессирующими антигенпрезентирующими клетками (АПК),
(ii) в стимуляции опухолеспецифического Т-клеточного ответа,
(iii) в инициации апоптоза опухолевых клеток,
(iv) в лечении рака,
(v) в задержке прогрессирования рака,
(vi) в продлении выживаемости пациента, страдающего от рака,
(vii) в лечении инфекций.
В конкретном аспекте предложена биспецифическая антигенсвязывающая молекула, описанная выше в данном документе, или фармацевтическая композиция по изобретению для применения в лечении рака. В другом конкретном аспекте в изобретении предложена биспецифическая антигенсвязывающая молекула, описанная выше в данном документе, для применения в лечении рака, причем биспецифическую антигенсвязывающую молекулу вводят в комбинации с химиотерапевтическим агентом, лучевой терапией и/или другими агентами для применения в иммунотерапии рака. В одном аспекте биспецифическая антигенсвязывающая молекула, описанная в данном документе, предназначена для применения в лечении рака, причем биспецифическая антигенсвязывающая молекула предназначена для введения в комбинации с агентом, блокирующим взаимодействие PD-L1/PD-1. В другом аспекте предложена биспецифическая антигенсвязывающая молекула, описанная выше в данном документе, или фармацевтическая композиция по изобретению для применения в повышении регуляции или продлении активности цитотоксических Т-клеток. В дополнительном аспекте предложено антитело, описанное выше в данном документе, для применения в лечении рака.
В дополнительном аспекте в изобретении предложен способ ингибирования роста опухолевых клеток у индивида, включающий введение индивиду эффективного количества биспецифической антигенсвязывающей молекулы, описанной выше в данном документе, или фармацевтической композиции по изобретению для ингибирования роста опухолевых клеток. В другом аспекте в изобретении предложен способ задержки развития рака у индивида, включающий введение индивиду эффективного количества биспецифической антигенсвязывающей молекулы, описанной выше в данном документе, или фармацевтической композиции по изобретению.
Также предложено применение биспецифической антигенсвязывающей молекулы, описанной выше в данном документе, для производства лекарственного средства для лечения заболевания у нуждающегося в этом индивида, в частности, для производства лекарственного средства для лечения рака, а также способ лечения заболевания у индивида, включающий введение указанному индивиду терапевтически эффективного количества композиции, содержащей биспецифическую антигенсвязывающую молекулу по изобретению в фармацевтически приемлемой форме. В конкретном аспекте заболевание представляет собой рак. В любом из вышеприведенных аспектов индивид представляет собой млекопитающее, в частности, человека.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На Фиг. 1A-F показаны схематические изображения биспецифических антигенсвязывающих молекул, которые специфически связываются с CD40 и с FAP человека. На Фиг. 1А показано схематическое изображение биспецифического антитела к FAP-CD40 в формате 2+1, состоящего их двух CD40-связывающих фрагментов, объединенных с одним FAP (212)-связывающим фрагментом в виде кроссоверного fab-фрагмента, в котором цепь VL-CH1 слита в С-конце цепи Fc-выступа (двухвалентного в отношении CD40 и одновалентного в отношении FAP). На Фиг. 1Б показано схематическое изображение биспецифического антитела к FAP-CD40 в формате 2+1, состоящего их двух CD40-связывающих фрагментов, объединенных с одним FAP (4В9)-связывающим фрагментом в виде кроссоверного fab-фрагмента, в котором цепь VH-C-каппа слита в С-конце цепи Fc-выступа (двухвалентного в отношении CD40 и одновалентного в отношении FAP). На Фиг. 1В показано схематическое изображение биспецифического антитела к FAP-CD40 в формате 2+1, состоящего их двух CD40-связывающих фрагментов, объединенных с одним FAP (28Н1)-связывающим фрагментом в виде кроссоверного fab-фрагмента, в котором цепь VL-CH1 слита в С-конце цепи Fc-выступа (двухвалентного в отношении CD40 и одновалентного в отношении FAP). На Фиг. 1Г показано схематическое изображение биспецифического антитела к FAP-CD40 в формате 4+1, состоящего их четырех CD40-связывающих фрагментов, объединенных с одним FAP (212)-связывающим фрагментом в виде кроссоверного fab-фрагмента, в котором цепь VL-CH1 слита в С-конце цепи Fc-выступа (четырехвалентного в отношении CD40 и одновалентного в отношении FAP). На Фиг. 1Д показано схематическое изображение биспецифического антитела к FAP-CD40 в формате 4+1, состоящего их четырех CD40-связывающих фрагментов, объединенных с одним FAP (4В9)-связывающим фрагментом в виде кроссоверного fab-фрагмента, в котором цепь VL-CH1 слита в С-конце цепи Fc-выступа (четырехвалентного в отношении CD40 и одновалентного в отношении FAP). На Фиг. 1Е показано схематическое изображение биспецифического антитела к FAP-CD40 в формате 4+1, состоящего их четырех CD40-связывающих фрагментов, объединенных с одним FAP (28Н1)-связывающим фрагментом в виде кроссоверного fab-фрагмента, в котором цепь VL-CH1 слита в С-конце цепи Fc-выступа (четырехвалентного в отношении CD40 и одновалентного в отношении FAP). Черная точка обозначает мутации типа выступ-во-впадину. В альтернативном варианте во всех молекулах цепь VH-C-каппа кросс-fab может быть слита в С-конце цепи Fc-выступа.
На Фиг. 2А и 2Б показано клеточное связывание полученных вследствие иммунизации клонов FAP с человеческим FAP, экспрессируемым на трансфицированных клетках НЕК, в конкуренции с клонами FAP 4В9 и 28Н1. На Фиг. 2А показано, что все тестируемые полученные из гибридомы мышиные клоны (называемые 209, 210, 211, 212, 213, 214, 215, 216, 217 и 218) не конкурировали за связывание с анти-FAP антителом 4В9, а на Фиг. 2Б показано, что те же клоны не конкурировали за связывание с анти-FAP антителом 28Н1. МИФ измеряли методом проточной цитометрии. По оси х показана концентрация антитела к FAP.
На Фиг. 3А и 3Б показано схематическое представление конструкций антител, которые были созданы, чтобы определить, не утрачиваются ли связывающие свойства анти-FAP клонов, когда они связаны с Fc-доменом в С-конце. На Фиг. 3А показана конструкция, содержащая цепь Fc-выступа и цепь Fc-впадины, при этом домен VH слит с С-концом цепи Fc-выступа, а домен VL слит с С-концом цепи Fc-впадины (С-конц. слияние VH/VL). На Фиг. 3Б показана конструкция, содержащая цепь Fc-выступа и цепь Fc-впадины, при этом весь Fab слит в своем VH-домене с С-концом цепи Fc-выступа (С-конц. слияние Fab). На Фиг. 3В показана схема сортировки эпитопов, которую проводили, используя анализ на основе метода поверхностного плазмонного резонанса (ППР) на приборе Biacore Т200 (смотрите пример 1.9).
На Фиг. 4 показано связывание человеческих четырехвалентных или двухвалентных анти-CD40 антител в нацеленном на FAP (212), FAP (4В9) или FAP (28Н1) одновалентном формате с человеческими FAP-положительными клетками NIH/3T3. Трансгенная модифицированная клеточная линия мышиных эмбриональных фибробластов NIH/3T3-hFAP экспрессирует высокие уровни человеческого белка активации фибробластов (huFAP). Все изображенные анти-CD40 антигенсвязывающие молекулы с FAP-связывающим фрагментом эффективно связываются с клетками NIH/3T3-hFAP, но отличаются по силе связывания (значениям ЕС50, а также силе сигнала) с клетками NIH/3T3-hFAP. Конструкции FAP-CD40 с С-концевым FAP (212)- или FAP (4В9)-связывающим доменом связываются сильнее, чем конструкции FAP-CD40 с С-концевым FAP (28Н1)-связывающим доменом. Связывание показано в виде медианного значения интенсивности флуоресценции (МИФ) меченного фикоэритрином (РЕ) античеловеческого IgG Fcγ-специфического козьего Е(ab')2-фрагмента IgG, который использовали как вторичное антитело для обнаружения. МИФ измеряли методом проточной цитометрии и корректировали базовую линию путем вычитания МИФ холостого контроля. По оси х показана концентрация конструкций антител.
На Фиг. 5 показано связывание человеческих четырехвалентных или двухвалентных анти-CD40 антител в нацеленном на FAP (212) или FAP (4В9) одновалентном формате с первичными человеческими В-клетками с высокими уровнями поверхностной экспрессии человеческого CD40. Все изображенные конструкции связываются с CD40, но отличаются по силе связывания (значениям ЕС50, а также силе сигнала) с CD40-положительными В-клетками. Двухвалентные анти-CD40 антитела демонстрируют более высокие уровни ЕС50 и достигают более высокого плато связывания по сравнению с четырехвалентными анти-CD40 антителами, независимо от их FAP-связывающего фрагмента. Выявление связывания анти-CD40 антител с белками клеточной поверхности проводили с помощью античеловеческого IgG Fcγ-специфического козьего IE(ab')2-фрагмента IgG, конъюгированного с фикоэритрином (РЕ), используя анализ FACS. МИФ измеряли методом проточной цитометрии и корректировали базовую линию путем вычитания МИФ холостого контроля. По оси х показана концентрация конструкций антител.
На Фиг. 6А и 6Б показана in vitro активация человеческих В-клеток Дауди одновалентными нацеленными на FAP (212), FAP (4 В9) или FAP (28Н1) человеческими анти-CD40 конструкциями в присутствии покрытых FAP (Фиг. 6А) или непокрытых гранул Dynabeads (Фиг. 6Б) после 2 суток инкубации. В случае покрытых FAP гранул все изображенные биспецифические антитела, одновалентные в отношении FAP, индуцировали повышение экспрессии маркера активации В-клеток CD70. Повышение регуляции маркера активации В-клеток биспецифическими антителами к FAP-CD40 в формате 2+1 со связывающим доменом FAP (212) или FAP (4В9) было выше по сравнению с повышением регуляции, индуцированным биспецифическим антителом к FAP-CD40 в формате 2+1 со связывающим доменом FAP (28Н1), биспецифическими антителами к FAP-CD40 в формате 4+1 со связывающим доменом FAP (212), FAP (4В9) или FAP (28Н1) или FAP-независимым положительным контрольным антителом. В отсутствие FAP (непокрытые гранулы) не наблюдалось повышение CD70 в случае изображенных нацеленных на FAP биспецифических антител, двухвалентных в отношении CD40, тогда как четырехвалентные CD40-связывающие молекулы индуцировали повышение регуляции CD70, но в меньшей степени, чем в присутствии FAP. Показана процентная доля CD70-положительных живых клеток Дауди после 2 суток инкубации с указанными титрованными антителами. По оси х показана концентрация конструкций антител.
На Фиг. 7А и 7Б показана in vitro активация человеческих В-клеток одновалентными нацеленными на FAP (212), FAP (4В9) или FAP (28Н1) человеческими анти-CD40 конструкциями в присутствии покрытых FAP (Фиг. 7А) или непокрытых гранул Dynabeads® (Фиг. 7Б) после 2 суток инкубации. В случае покрытых FAP гранул все изображенные биспецифические антитела, одновалентные в отношении FAP, индуцировали повышение экспрессии маркера активации В-клеток CD86. По сравнению с FAP-независимым повышением регуляции CD86, индуцированным перекрестно-связанным анти-CD40 антителом (P1AD4470), повышение регуляции CD86, индуцированное FAP-зависимыми биспецифическими антигенсвязывающими молекулами, было сходным или немного меньшим. При меньших концентрациях антител повышение регуляции маркера активации В-клеток биспецифическими антителами к FAP-CD40 в формате 2+1 со связывающим доменом FAP (212) или FAP (4B9) было ниже по сравнению с повышением регуляции, индуцированным биспецифическим антителом к FAP-CD40 в формате 2+1 со связывающим доменом FAP (28Н1), биспецифическими антителами к FAP-CD40 в формате 4+1 со связывающим доменом FAP (212), FAP (4В9) или FAP (28Н1) или FAP-независимым положительным контрольным антителом. В отсутствие FAP (непокрытые гранулы) не наблюдалось повышение экспрессии CD86 в случае биспецифических антигенсвязывающих молекул, тогда как положительное контрольное антитело индуцировало повышение регуляции CD86. Показана процентная доля CD86-положительных живых В-клеток после 2 суток инкубации с указанными титрованными антителами. По оси х показана концентрация конструкций антител.
На Фиг. 8А и 8Б показано Т-клеточное примирование OVA-сенсибилизированных ДК, активируемых нацеленными на FAP анти-CD40 связывающими молекулами в присутствии (Фиг. 8А) или отсутствие (Фиг. 8Б) FAP. ДК, выделенные у трансгенных в отношении huCD40 мышей, обработанные конъюгатом DEC205-OVA и стимулированные FAP-зависимыми биспецифическими анти-CD40 антителами, а также покрытые FAP гранулы индуцировали сильную пролиферацию антиген-специфических Т-клеток. В отличие от этого, в отсутствие FAP (непокрытые гранулы) пролиферацию Т-клеток индуцировали ДК, стимулированные нацеленными на FAP анти-CD40 антителами. Пролиферация Т-клеток, которую индуцировали ДК, стимулированные человеческими биспецифическими антигенсвязывающими молекулами с двумя или четырьмя CD40- и одним FAP (212)- или FAP (4В9)-связывающими фрагментами, была сравнимой. ДК, сенсибилизированные высокими количествами SIINFEKL вместо конъюгата DEC205-OVA, индуцировали сильную пролиферацию Т-клеток. Показана процентная доля пролиферации (CFSE-низк.) живых CFSE-меченных мышиных CD3+CD8+ ОТ-1 Т-клеток, которые культивировали совместно с huCD40 tg ДК, которые предварительно инкубировали с указанными титрованными антителами в присутствии OVA (Фиг. 8А и 8Б). По оси х показана концентрация конструкций антител.
На Фиг. 9 показан эффект одновременного связывания биспецифических антител к FAP х CD40 в мостиковом анализе с двумя линиями клеток, описанном в примере 5.3. Показана оптическая плотность (ОП) в зависимости от концентрации биспецифических антител в разных форматах.
На Фиг. 10 показан потенциал антител к FAP х CD40 в разных форматах в отношении активации рецептора CD40 (независимо от FAP). В анализе с репортерной линией клеток передача сигнала приводит к индукции  зависимой выработки секретируемой эмбриональной щелочной фосфатазы (SEAP), и при этом измеряют активность SEAP. Построен график среднего значение оптической плотности (ОП) для n=6 и 3х СТДОТКЛ в зависимости от молекул.
зависимой выработки секретируемой эмбриональной щелочной фосфатазы (SEAP), и при этом измеряют активность SEAP. Построен график среднего значение оптической плотности (ОП) для n=6 и 3х СТДОТКЛ в зависимости от молекул.
На Фиг. 11 показан дизайн in vivo исследования на мышах для оценки профиля безопасности, фармакокинетики и фармакодинамики нацеленных на FAP анти-CD40 связывающих молекул в сравнении с ненацеленным анти-CD40 антителом в подкожной опухолевой модели MC38-FAP.
На Фиг. 12 показан рост опухоли после 1-ой и 2-ой (повторная стимуляция) инъекции опухолевых клеток MC38-FAP мышам, обработанным отдельной или комбинированной терапией на основе антител. Стрелки указывают на сутки проведения инъекции терапевтического средства и повторной стимуляции. По оси у представлен объем опухолей в мм3, а по оси х - сутки после 1-ой инъекции опухолевых клеток MC38-FAP. Наблюдали полную регрессию опухолей MC38-FAP после обработки антителом к FAP-CD40, независимо от формата и совместной инъекции анти-PD-L1 антитела. В отличие от этого, у мышей, обработанных одним анти-PD-L1 антителом, одним анти-CD40 антителом или анти-CD40 в комбинации с анти-PD-L1 антителом, рост опухолей был сравним с мышами, обработанные контрольным носителем. Кроме того, после повторной стимуляции в группах, предварительно обработанных антителом к FAP-CD40, опухоль MC38-FAP не росла, тогда как у ранее не обработанных мышей наблюдался рост 100% опухолей.
На Фиг. 13А приведена статистическая таблица со сравнением всех групп обработки на 28 сутки (6 сутки после терапии), а на Фиг. 13Б приведена статистическая таблица со сравнением всех групп обработки, начиная с 31 суток (9 сутки после терапии).
На Фиг. 14 показан фармакокинетический профиль 4+1 и 2+1 нацеленных на FAP анти-CD40 связывающих молекул, а также ненацеленного родительского анти-CD40 антитела у мышей, которым инъецировали FAP-экспрессирующую линию клеток мышиной аденокарциномы толстой кишки (MC38-FAP). По оси у представлена нормализованная к дозе концентрация в сыворотке, а по оси х - время после инъекции антитела. Наибольшую скорость клиренса наблюдали для ненацеленного анти-CD40 антитела. Скорость клиренса нацеленной на FAP анти-CD40 связывающей молекулы в формате 4+1 была ниже по сравнению с ненацеленной анти-CD40 молекулой, а наименьшую скорость клиренса наблюдали для нацеленной на FAP анти-CD40 связывающей молекулы в формате 2+1.
На Фиг. 15, 16 и 17 показана активация ДК, Т-клеток и В-клеток в дренирующих лимфатических узлах (ЛУ, Фиг. 15А, Фиг. 16А и Фиг. 17А), недренирующих лимфатических узлах (Фиг. 15Б, Фиг. 16Б и Фиг. 17Б), селезенке (Фиг. 15В, Фиг. 16В и Фиг. 17В) и опухоли (Фиг. 15Г, Фиг. 16Г и Фиг. 17Г) мышей, которым инъецировали FAP-экспрессирующую линию клеток мышиной аденокарциномы толстой кишки (MC38-FAP) и которых обрабатывали ненацеленным анти-CD40 (Р1АЕ2301), анти-CD40-FAP 4+1 (Р1АЕ2024), анти-CD40-FAP 2+1 (Р1АЕ2302) или одним носителем. Активация ДК и Т-клеток в опухоль через четверо суток после инъекции терапии (Фиг. 15Г и 16Г) была существенно повышена у животных, обработанных анти-CD40-FAP 2+1 (Р1АЕ2302), по сравнению с животными, обработанными носителем. Во всех остальных анализируемых тканях (дренирующие ЛУ, недренирующие ЛУ и селезенка) ненацеленные анти-CD40, анти-CD40-FAP 4+1 и анти-CD40-FAP 2+1 индуцировали значительную активацию ДК и Т-клеток по сравнению с группой носителя. В отличие от этого, только ненацеленное анти-CD40 опосредовало значительную активацию В-клеток во всех анализируемых тканях по сравнению с группой контрольного носителя (Фиг. 17А-17Г, *р<0,05, **р<0,01, ***р<0,001, ****р<0,0001, непарный двусторонний критерий Стьюдента).
На Фиг. 18 показана масса тела мышей, которым инъецировали FAP-экспрессирующую линию клеток мышиной аденокарциномы толстой кишки (MC38-FAP) и которых обрабатывали ненацеленным анти-CD40 (Р1АЕ2301), анти-CD40-FAP 4+1 (Р1АЕ2024), анти-CD40-FAP 2+1 (Р1АЕ2302) или носителем с или без совместной инъекции анти-PD-L1 антитела (Р1АЕ0099). По оси у показана масса тела как процент от массы тела до обработки, а по оси х показаны сутки после инъекции терапии. Только у мышей, обработанных ненацеленным анти-CD40 антителом, одним или в комбинации с анти-PD-L1, наблюдали явное снижение массы тела.
ПОДРОБНОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Определения
Если не указано иное, все технические и научные термины, используемые в данном документе, имеют значение, обычно используемое в области, к которой принадлежит это изобретение. В целях интерпретации этого описания применимы нижеприведенные определения, и в тех случаях, когда это целесообразно, термины, используемые в единственном числе, также включают множественное число и наоборот.
В контексте данного документа термин «антигенсвязывающая молекула» относится в своем самом широком смысле к молекуле, которая специфически связывает антигенную детерминанту. Примерами антигенсвязывающих молекул являются антитела, фрагменты антител и каркасные антигенсвязывающие белки.
В контексте данного документа термин «антигенсвязывающий домен, способный к специфическому связыванию с антигеном целевой клетки» относится к полипептидной молекуле, которая специфически связывается с антигенной детерминантой. В одном аспекте антигенсвязывающий домен способен активировать сигнализацию через антиген целевой клетки. В конкретном аспекте антигенсвязывающий домен способен направлять соединение, к которому он присоединен (например, агониста CD40) к целевому сайту, например, к конкретному типу опухолевой клетки или опухолевой стромы, несущей антигенную детерминанту. Антигенсвязывающие домены, способные к специфическому связыванию с антигеном целевой клетки, включают антитела и их фрагменты, которые определены далее в данном документе. Кроме того, антигенсвязывающие домены, способные к специфическому связыванию с антигеном целевой клетки, включают каркасные антигенсвязывающие белки, которые определены далее в данном документе, в основе которых лежат сконструированные повторяющиеся белки или сконструированные повторяющиеся домены (смотрите, например, WO 2002/020565).
В отношении антитела или его фрагмента термин «антигенсвязывающий домен, способный к специфическому связыванию с антигеном целевой клетки» относится к молекуле, которая содержит участок, который специфически связывается и является комплементарной с частью или со всем антигеном. Антигенсвязывающий домен, способный к специфическому связыванию антигена, может быть обеспечен, например, одним или более вариабельными доменами антитела (также называемыми вариабельными областями антитела). В частности, антигенсвязывающий домен, способный к специфическому связыванию антигена, содержит вариабельную область легкой цепи антитела (VL) и вариабельную область тяжелой цепи антитела (VH). В другом аспекте «антигенсвязывающий домен, способный к специфическому связыванию с антигеном целевой клетки» также может представлять собой Fab-фрагмент или Kpocc-Fab-фрагмент.
Термин «антитело» в данном документе используется в самом широком смысле и охватывает различные структуры антител, включая, но не ограничиваясь этим, моноклональные антитела, поликлональные антитела, моноспецифические и мультиспецифические антитела (например, биспецифические антитела) и фрагменты антител, при условии, что они демонстрируют необходимую антигенсвязывающую активность.
В контексте данного документа термин «моноклональное антитело» относится к антителу, полученному из популяции практически гомогенных антител, т.е. отдельные антитела, составляющие популяцию, идентичны и/или связываются с одним и тем же эпитопом, за исключением возможных вариантных антител, например, содержащих мутации природного происхождения или возникающие во время получения препарата моноклональных антител, при этом такие варианты в целом присутствуют в незначительных количествах. В отличие от препаратов поликлональных антител, которые обычно содержат разные антитела, направленные против разных детерминант (эпитопов), каждое моноклональное антитело препарата моноклональных антител направлено против одной детерминанты на антигене.
В контексте данного документа термин «моноспецифическое антитело» обозначает антитело, которое имеет один или более сайтов связывания, каждый из которых связывается с одним и тем же эпитопом одного и того же антигена. Термин «биспецифический» означает, что антигенсвязывающая молекула способна к специфическому связыванию с по меньшей мере двумя разными антигенными детерминантами. Как правило, биспецифическая антигенсвязывающая молекула содержит два антигенсвязывающих сайта, каждый из которых является специфическим в отношении отличной антигенной детерминанты. В определенных вариантах осуществления биспецифическая антигенсвязывающая молекула способна одновременно связывать две антигенные детерминанты, в частности, две антигенные детерминанты, экспрессируемые на двух разных клетках. Описанная в данном документе биспецифическая антигенсвязывающая молекула также может образовывать часть мультиспецифического антитела.
Термин «валентный» в контексте представленной заявки обозначает наличие конкретного числа сайтов связывания в антигенсвязывающей молекуле, специфических в отношении одной отличной антигенной детерминанты. Таким образом, термины «двухвалентный», «четырехвалентный» и «шестивалентный» обозначают наличие в антигенсвязывающей молекуле двух сайтов связывания, четырех сайтов связывания и шести сайтов связывания, специфических в отношении определенной антигенной детерминанты, соответственно. В конкретных аспектах изобретения биспецифические антигенсвязывающие молекулы в соответствии с изобретением могут быть одновалентными в отношении определенной антигенной детерминанты, что означает, что они имеют только один сайт связывания для указанной антигенной детерминанты, или же они могут быть двухвалентными или четырехвалентными в отношении определенной антигенной детерминанты, что означает, что они имеют два сайта связывания или четыре сайта связывания, соответственно, для указанной антигенной детерминанты.
В контексте данного документа термины «полноразмерное антитело», «интактное антитело» и «цельное антитело» используются взаимозаменяемо для обозначения антитела, имеющего структуру, практически аналогичную структуре нативного антитела. «Нативные антитела» относятся к молекулам иммуноглобулинов природного происхождения с различной структурой. Например, нативные антитела класса IgG представляют собой гетеротетрамерные гликопротеины массой около 150000 дальтон, состоящие из двух легких цепей и двух тяжелых цепей, связанных дисульфидными связями. В направлении от N- к С-концу каждая тяжелая цепь содержит вариабельную область (VH), также называемую вариабельным тяжелым доменом или вариабельным доменом тяжелой цепи, за которой следуют три константных домена (CH1, СН2 и СН3), также называемые константной областью тяжелой цепи. Аналогично, в направлении от N- к С-концу каждая легкая цепь содержит вариабельную область (VL), также называемую вариабельным легким доменом или вариабельным доменом легкой цепи, за которой следует константный легкий домен (CL), также называемый константной областью легкой цепи. Тяжелая цепь антитела может быть отнесена к одному из пяти типов, называемых α (IgA), δ (IgD), ε (IgE), γ (IgG) или μ (IgM), некоторые из которых могут быть дополнительно разделены на подтипы, например, γ1 (IgG1), γ2 (IgG2), γ3 (IgG3), γ4 (IgG4), α1 (IgA1) и α2 (IgA2). Легкая цепь антитела может быть отнесена к одному из двух типов, называемых каппа (κ) и лямбда (λ), на основании аминокислотной последовательности своего константного домена.
«Фрагмент антитела» относится к молекуле, отличной от интактного антитела, которая содержит часть интактного антитела, которая связывает антиген, с которым связывается интактное антитело. Примеры фрагментов антител включают, но не ограничиваются этим, Fv, Fab, Fab', Fab'-SH, F(ab')2; диатела, триатела, тетратела, кросс-Fab-фрагменты; линейные антитела; молекулы одноцепочечных антител (например, scFv); и однодоменные антитела. Обзор некоторых фрагментов антител смотрите в Hudson et al., Nat Med 9, 129-134 (2003). Обзор фрагментов scFv смотрите, например, в  в The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds., Springer-Verlag, New York, pp. 269-315 (1994); также смотрите WO 93/16185; и патенты США №№5571894 и 5587458. Обсуждение фрагментов Fab и F(ab')2, содержащих остатки эпитопа связывания рецептора реутилизации и имеющих увеличенное время полужизни in vivo, смотрите в патенте США №5869046. Диатела представляют собой фрагменты антител с двумя антигенсвязывающими сайтами, которые могут быть двухвалентными или биспецифическими, смотрите, например, ЕР 404097; WO 1993/01161; Hudson et al., Nat Med 9, 129-134 (2003); и Hollinger et al., Proc Natl Acad Sci USA 90, 6444-6448 (1993). Триатела и тетратела также описаны в Hudson et al., Nat Med 9, 129-134 (2003). Однодоменные антитела представляют собой фрагменты антител, содержащие весь или часть вариабельного домена тяжелой цепи или весь или часть вариабельного домена легкой цепи антитела. В определенных вариантах осуществления однодоменное антитело представляет собой человеческое однодоменное антитело (Domantis, Inc., Waltham, MA; смотрите, например, патент США №6248516 В1). Фрагменты антител можно получать различными способами, включая, но не ограничиваясь этим, протеолитическое расщепление интактного антитела, а также выработку рекомбинантными клетками-хозяевами (например, Е. coli или фага), как описано в данном документе.
в The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds., Springer-Verlag, New York, pp. 269-315 (1994); также смотрите WO 93/16185; и патенты США №№5571894 и 5587458. Обсуждение фрагментов Fab и F(ab')2, содержащих остатки эпитопа связывания рецептора реутилизации и имеющих увеличенное время полужизни in vivo, смотрите в патенте США №5869046. Диатела представляют собой фрагменты антител с двумя антигенсвязывающими сайтами, которые могут быть двухвалентными или биспецифическими, смотрите, например, ЕР 404097; WO 1993/01161; Hudson et al., Nat Med 9, 129-134 (2003); и Hollinger et al., Proc Natl Acad Sci USA 90, 6444-6448 (1993). Триатела и тетратела также описаны в Hudson et al., Nat Med 9, 129-134 (2003). Однодоменные антитела представляют собой фрагменты антител, содержащие весь или часть вариабельного домена тяжелой цепи или весь или часть вариабельного домена легкой цепи антитела. В определенных вариантах осуществления однодоменное антитело представляет собой человеческое однодоменное антитело (Domantis, Inc., Waltham, MA; смотрите, например, патент США №6248516 В1). Фрагменты антител можно получать различными способами, включая, но не ограничиваясь этим, протеолитическое расщепление интактного антитела, а также выработку рекомбинантными клетками-хозяевами (например, Е. coli или фага), как описано в данном документе.
Расщепление интактных антител папаином позволяет получить два идентичных антигенсвязывающих фрагмента, называемых «Fab»-фрагментами, каждый из которых содержит вариабельные домены тяжелой и легкой цепи и также константный домен легкой цепи и первый константный домен (СН1) тяжелой цепи. Таким образом, в контексте данного документа термин «Fab-фрагмент» относится к фрагменту антитела, содержащему фрагмент легкой цепи, содержащий VL-домен и константный домен легкой цепи (CL), и VH-домен и первый константный домен (СН1) тяжелой цепи. Fab'-фрагменты отличаются от Fab-фрагментов добавлением нескольких остатков в карбокси-конце СН1-домена тяжелой цепи, включая один или более остатков цистеина из шарнирной области антитела. Fab'-SH представляют собой Fab'-фрагменты, в которых остаток(ки) цистеина константных доменов несет(ут) свободную тиольную группу. Обработка пепсином приводит к получению Р(ab')2-фрагмента, который имеет два антигенсвязывающих сайта (два Fab-фрагмента) и часть Fc-области. В соответствии с настоящим изобретением термин «Fab-фрагмент» также включает «Kpocc-Fab-фрагменты» или «кроссоверные Fab-фрагменты», которые определены ниже.
Термин «Kpocc-Fab-фрагмент», или «xFab-фрагмент», или «кроссоверный Fab-фрагмент» относится к Fab-фрагменту, в котором вариабельные области или константные области тяжелой и легкой цепи поменяны. В биспецифических антителах по изобретению возможны и содержатся два разных варианта композиции цепи кроссоверной Fab-молекулы: С одной стороны поменяны вариабельные области тяжелой и легкой цепи Fab, т.е. кроссоверная Fab-молекула содержит пептидную цепь, состоящую из вариабельной области легкой цепи (VL) и константной области тяжелой цепи (СН1), и пептидную цепь, состоящую из вариабельной области тяжелой цепи (VH) и константной области легкой цепи (CL). Эта кроссоверная Fab-молекула также называется кросс-Fab (VLVH). С другой стороны, когда поменяны константные области тяжелой и легкой цепи Fab, кроссоверная Fab-молекула содержит пептидную цепь, состоящую из вариабельной области тяжелой цепи (VH) и константной области легкой цепи (CL), и пептидную цепь, состоящую из вариабельной области легкой цепи (VL) и константной области тяжелой цепи (СН1). Эта кроссоверная Fab-молекула также называется кросс-Fab (CLCH1).
«Одноцепочечный Fab-фрагмент» или «scFab» представляет собой полипептид, состоящий из вариабельного домена тяжелой цепи антитела (VH), константного домена антитела 1 (СН1), вариабельного домена легкой цепи антитела (VL), константного домена легкой цепи антитела (CL) и линкера, причем указанные домены антитела и указанный линкер имеют один из следующих вариантов упорядочения в направлении от N-конца к С-концу: а) VH-СН1-линкер-VE-CL б) VL-CL-линкер-VH-CH1, в) VH-CL-линкер-VL-CH1 или г) VL-CH1-линкер-VH-CL; и причем указанный линкер представляет собой полипептид из по меньшей мере 30 аминокислот, предпочтительно от 32 до 50 аминокислот. Указанные одноцепочечные Fab-фрагменты стабилизированы посредством естественной дисульфидной связи между С1-доменом и CH1-доменом. Кроме того, эти одноцепочечные Fab-молекулы можно дополнительно стабилизировать путем создания межцепочечных дисульфидных связей посредством вставки остатков цистеина (например, в позиции 44 в вариабельной области тяжелой цепи и позиции 100 в вариабельной области легкой цепи в соответствии с нумерацией Kabat).
«Кроссоверный одноцепочечный Fab-фрагмент» или «x-scFab» представляет собой полипептид, состоящий из вариабельного домена тяжелой цепи антитела (VH), константного домена антитела 1 (СН1), вариабельного домена легкой цепи антитела (VL), константного домена легкой цепи антитела (CL) и линкера, причем указанные домены антитела и указанный линкер имеют один из следующих вариантов упорядочения в направлении от N-конца к С-концу: a) VH-CL-линкер-VL-CH1 и б) VL-CH1-линкер-VH-CL; где VH и VL вместе образуют антигенсвязывающий сайт, который специфически связывается с антигеном, и при этом указанный линкер представляет собой полипептид из по меньшей мере 30 аминокислот. Кроме того, эти x-scFab-молекулы можно дополнительно стабилизировать путем создания межцепочечных дисульфидных связей посредством вставки остатков цистеина (например, в позиции 44 в вариабельной области тяжелой цепи и позиции 100 в вариабельной области легкой цепи в соответствии с нумерацией Kabat).
«Одноцепочечный вариабельный фрагмент (scFv)» представляет собой слитый белок из вариабельных областей тяжелой (VH) и легкой цепей (VL) антитела, соединенный коротким линкерным пептидом из от десяти до около 25 аминокислот. Линкер обычно обогащен глицином для гибкости, а также серином или треонином для растворимости, и может соединять N-конец VH с С-концом VL или наоборот. Этот белок сохраняет специфичность исходного антитела несмотря на удаление константных областей и внесение линкера. scFv-антитела описаны, например, в Houston, J.S., Methods in Enzymol. 203 (1991) 46-96). Кроме того, фрагменты антител содержат одноцепочечные полипептиды, имеющие характеристики VH-домена, а именно способность к сборке с VL-доменом, или VL-домена, а именно способность к сборке с VH-доменом в функциональный антигенсвязывающий сайт и, таким образом, обеспечивающие антигенсвязывающие свойства полноразмерного полипептида.
«Каркасные антигенсвязывающие белки» известны в данной области техники, например, фибронектин и сконструированные белки с анкириновыми повторами (дарпины или DARPin) использовали в качестве альтернативных каркасов для антигенсвязывающих доменов, смотрите, например, Gebauer and Skerra, Engineered protein scaffolds as next-generation antibody therapeutics. Curr Opin Chem Biol 13:245-255 (2009) и Stumpp et al., Darpins: A new generation of protein therapeutics. Drug Discovery Today 13: 695-701 (2008). В одном аспекте изобретения каркасный антигенсвязывающий белок выбран из группы, состоящей из CTLA-4 (эвитело), липокалинов (антикалин), полученных из протеина А молекул, таких как Z-домен протеина А (аффитело), А-домена (авимер/макситело), сывороточного трансферрина (транс-тело); сконструированного белка с анкириновыми повторами (дарпин), вариабельного домена легкой цепи или тяжелой цепи антитела (однодоменное антитело, sdAb), вариабельного домена тяжелой цепи антитела (нанотело, aVH), фрагментов VNAR, фибронектина (аднектин), лектинового домена С-типа (тетранектин); вариабельного домена бета-лактамазы нового антигенного рецептора (фрагменты VNAR), человеческого гамма-кристаллина или убиквитина (молекулы аффилина); домена типа домена Куница ингибиторов протеазы человека, микротел, таких как белки семейства ноттина, пептидные аптамеры и фибронектин (аднектин). CTLA-4 (ассоциированный с цитотоксическими Т-лимфоцитами антиген 4) представляет собой рецептор семейства CD28, экспрессируемый преимущественно на CD4+ Т-клетках. Его внеклеточный домен имеет укладку, подобную вариабельному домену Ig. Петли, соответствующие CDR антител могут быть замещены гетерологичной последовательностью для придания отличных связывающих свойств. Молекулы CTLA-4, сконструированные так, чтобы иметь разные варианты специфичности связывания, также известны как эвитела (например, US7166697B1). Эвитела имеют приблизительно такой же размер, что и выделенная вариабельная область антитела (например, доменного антитела). Дополнительные подробности смотрите в Journal of Immunological Methods 248 (1-2), 31-45 (2001). Липокалины представляют семейство внеклеточных белков, которые переносят небольшие гидрофобные молекулы, такие как стероиды, билины, ретиноиды и липиды. Они имеют жесткую бета-складчатую вторичную структуру с рядом петель в открытом конце конической структуры, которая может быть сконструирована так, чтобы связываться с разными целевыми антигенами. Размер антикалинов составляет 160-180 аминокислот, и они получены из липокалинов. Дополнительные подробности смотрите в Biochim Biophys Acta 1482: 337-350 (2000), US7250297B1 и US20070224633. Аффитело представляет собой каркас, полученный из протеина A Staphylococcus aureus, который может быть сконструирован так, чтобы связывать антиген. Домен состоит из трехспирального пучка приблизительно из 58 аминокислот. Путем рандомизации поверхностных остатков были созданы библиотеки. Дополнительные подробности смотрите в Protein Eng. Des. Sel. 2004, 17, 455-462 и ЕР 1641818А1. Авимеры представляют собой многодоменные белки, полученные из семейства А-доменных каркасов. Нативные домены приблизительно из 35 аминокислот принимают определенную дисульфидно связанную структуру. Разнообразие создают путем перетасовки природной вариации, демонстрируемой семейством А-доменов. Дополнительные подробности смотрите в Nature Biotechnology 23(12), 1556-1561 (2005) and Expert Opinion on Investigational Drugs 16(6), 909-917 (June 2007). Трансферрин представляет собой мономерный сывороточный транспортный гликопротеин. Трансферрины могут быть сконструированы так, чтобы связывать разные целевые антигены, путем вставки пептидных последовательностей в пермиссивную поверхностную петлю. Примеры сконструированных трансферриновых каркасов включают транс-тело. Дополнительные подробности смотрите в J. Biol. Chem 274, 24066-24073 (1999). Сконструированные белки с анкириновыми повторами (дарпины или DARPin) получены из анкирина, который принадлежит семейству белков, которые опосредуют присоединение интегральных мембранных белков к цитоскелету. Один анкириновый повтор представляет собой мотив из 33 остатков, состоящий из двух альфа-спиралей и бета-изгиба. Их можно сконструировать так, чтобы они связывали разные целевые антигены, путем рандомизации остатков в первой альфа-спирали и бета-изгибе каждого повтора. Их поверхность связывания можно увеличить за счет увеличения числа модулей (способ созревания аффинности). Дополнительные подробности смотрите в J. Mol. Biol. 332, 489-503 (2003), PNAS 100(4), 1700-1705 (2003) и J. Mol. Biol. 369, 1015-1028 (2007) и US20040132028A1. Однодоменное антитело представляет собой фрагмент антитела, состоящий из одного мономерного вариабельного домена антитела. Первые однодоменные антитела были получены из вариабельного домена тяжелой цепи антитела верблюжьих (нанотела или фрагменты VHН). Кроме того, термин однодоменное антитело включает автономный человеческий вариабельный домен тяжелой цепи (aVH) или фрагменты VNAR, полученные от акул. Фибронектин представляет собой каркас, который может быть сконструирован так, чтобы связывать антиген. Аднектины состоят из остова природной аминокислотной последовательности 10-го домена из 15 повторяющихся единиц человеческого фибронектина типа III (FN3). Три петли в одном конце бета-сандвича могут быть сконструированы так, чтобы аднектин мог специфически распознавать представляющую интерес терапевтическую мишень. Дополнительные подробности смотрите в Protein Eng. Des. Sel. 18, 435-444 (2005), US20080139791, WO2005056764 и US6818418B1. Пептидные аптамеры представляют собой комбинаторные молекулы распознавания, которые состоят из константного каркасного белка, как правило, тиоредоксина (TrxA), которые содержит пространственно-ограниченную вариабельную пептидную петлю, вставленную в активном сайте. Дополнительные подробности смотрите в Expert Opin. Biol. Ther. 5, 783-797 (2005). Микротела получены из микропротеинов природного происхождения длиной 25-50 аминокислот, которые содержат 3-4 цистеиновых мостика; примеры микропротеинов включают KalataBI, конотоксин и ноттины. Микропротеины имеют петлю, которая может быть сконструирована так, чтобы содержать до 25 аминокислот, не влияя на общую укладку белка. Дополнительные подробности в отношении сконструированных доменов ноттинов смотрите в WO2008098796.
«Антигенсвязывающая молекула, которая связывается с тем же эпитопом», что и референсная молекула, относится к антигенсвязывающей молекуле, которая в конкурентном анализе блокирует связывание референсной молекулы с ее антигеном на 50% или более, и наоборот, референсная молекула в конкурентном анализе блокирует связывание антигенсвязывающей молекулы с ее антигеном на 50% и более. «Антигенсвязывающая молекула, которая не связывается с тем же эпитопом», что и референсная молекула, относится к антигенсвязывающей молекуле, которая в конкурентном анализе не блокирует связывание референсной молекулы с ее антигеном на 50% или более, и наоборот, референсная молекула в конкурентном анализе не блокирует связывание антигенсвязывающей молекулы с ее антигеном на 50% и более.
Термин «антигенсвязывающий домен» или «антигенсвязывающий сайт» относится к части антигенсвязывающей молекулы, которая содержит участок, который специфически связывается и является комплементарной с частью или со всем антигеном. Если антиген является крупным, антигенсвязывающая молекула может связываться только с конкретной частью антигена, и эта часть называется эпитопом. Антигенсвязывающий домен можно обеспечить, например, одним или более вариабельными доменами (также называемыми вариабельными областями). Предпочтительно антигенсвязывающий домен содержит вариабельную область легкой цепи антитела (VL) и вариабельную область тяжелой цепи антитела (VH).
В контексте данного документа «антигенная детерминанта» является синонимом терминам «антиген» и «эпитоп» и относится к сайту (например, непрерывному участку аминокислот или конформационной конфигурации, состоящей из разных областей не являющихся непрерывными аминокислот) на полипептидной макромолекуле, с которым связывается антигенсвязывающий фрагмент с образованием комплекса антигенсвязывающий фрагмент - антиген. Применимые антигенные детерминанты можно обнаружить, например, на поверхностях опухолевых клеток, на поверхностях инфицированных вирусом клеток, на поверхностях пораженных заболеванием клеток, на поверхности иммунных клеток, в свободном виде в сыворотке крови и/или во внеклеточном матриксе (ВКМ). Белки, применимые в контексте данного документа, могут представлять собой любую нативную форму белков из любого источника, относящегося к позвоночным, включая млекопитающих, таких как приматы (например, люди) и грызуны (например, мыши и крысы), если не указано иное. В конкретном варианте осуществления антиген представляет собой человеческий белок. Когда в данном документе приводится ссылка на конкретный белок, этот термин включает «полноразмерный» непроцессированный белок, а также любую форму белка, которая является результатом процессинга в клетке. Этот термин также охватывает варианты белка природного происхождения, например, сплайс-варианты или аллельные варианты.
Под выражением «специфическое связывание» подразумевается, что связывание является избирательным в отношении антигена и может быть отделено от нежелательных или неспецифических взаимодействий. Способность антигенсвязывающей антигенсвязывающей молекулы связываться с конкретным антигеном можно определить с помощью ферментного иммуносорбентного анализа (ELISA) или других методик, известных специалисту в данной области техники, например, метода поверхностного плазмонного резонанса (ППР) (с анализом на приборе BIAcore) (Liljeblad et al., Glyco J 17, 323-329 (2000)), и традиционных анализов связывания (Heeley, Endocr Res 28, 217-229 (2002)). В одном варианте осуществления степень связывания антигенсвязывающей молекулы с неродственным белком составляет менее чем около 10% связывания антигенсвязывающей молекулы с антигеном при измерении, например, методом ППР. В определенных вариантах осуществления молекула, котораясвязывается с антигеном, имеет константу диссоциации (Kд) ≤1 мкМ, ≤100 нМ, ≤10 нМ, ≤1 нМ, ≤0,1 нМ, ≤0,01 нМ или ≤0,001 нМ (например, 10-8 М или менее, например, от 10-8 М до 10-13 М, например, от 10-9 М до 10-13 М).
«Аффинность» или «аффинность связывания» относится к силе суммарных нековалентных взаимодействий между одним сайтом связывания молекулы (например, антитела) и ее партнером по связыванию (например, антигеном). Если не указано иное, в контексте данного документа «аффинность связывания» относится к характерной аффинности связывания, которая отображает 1:1 взаимодействие между членами связывающейся пары (например, антителом и антигеном). Аффинность молекулы X в отношении ее партнера Y в общем случае можно выразить константой диссоциации (Kд), которая представляет собой отношение констант диссоциации и ассоциации (kд и kа, соответственно). Таким образом, эквивалентные аффинности могут включать разные константы скорости при условии, что отношение между константами скорости остается одинаковым. Аффинность можно измерять с помощью общепринятых методов, известных в данной области техники, включая описанные в данном документе. Конкретным методом для измерения аффинности является поверхностный плазмонный резонанс (ППР).
Антитело с «созревшей аффинностью» относится к антителу с одним или более изменениями в одной или более гипервариабельных областях (HVR) по сравнению с родительским антителом, которое не содержит таких изменений, при этом такие изменения приводят к улучшению аффинности антитела в отношении антигена.
В контексте данного документа «антиген целевой клетки» относится к антигенной детерминанте, представленной на поверхности целевой клетки, в частности, целевой клетки в опухоли, такой как раковая клетка или клетка опухолевой стромы. Таким образом, антиген целевой клетки является опухолеассоциированным антигеном. В частности, антиген опухолевой целевой клетки представляет собой белок активации фибробластов (FAP).
Термин «белок активации фибробластов (FAP)», также известный как FAP пролилэндопептидаза или сепраза (ЕС 3.4.21), относится к любому нативному FAP из любого источника, относящегося к позвоночным, включая млекопитающих, таких как приматы (например, люди), отличные от человека приматы (например, яванские макаки) и грызуны (например, мыши и крысы), если не указано иное. Этот термин охватывает «полноразмерный» непроцессированный FAP, а также любую форму FAP, полученную в результате процессинга в клетке. Этот термин также охватывает варианты FAP природного происхождения, например, сплайс-варианты или аллельные варианты. В одном варианте осуществления антигенсвязывающая молекула по изобретению способна к специфическому связыванию с FAP человека, мыши и/или яванского макака. Аминокислотная последовательность FAP человека приведена под номером доступа UniProt (www.uniprot.org) Q12884 (версия 149, SEQ ID NO: 2) или NCBI (www.ncbi.nlm.nih.gov/) RefSeq NP_004451.2. Внеклеточный домен (ВКД) FAP человека простирается от аминокислотной позиции 26 до 760. Аминокислотная последовательность ВКД His-меченного FAP человека приведена в SEQ ID NO: 78. Аминокислотная последовательность FAP мыши приведена под номером доступа Р97321 (версия 126, SEQ ID NO: 79) или NCBI RefSeq NP_032012.1. Внеклеточный домен (ВКД) FAP мыши простирается от аминокислотной позиции 26 до 761. SEQ ID NO: 80 иллюстрирует аминокислоты ВКД His-меченного FAP мыши. SEQ ID NO: 81 иллюстрирует аминокислоты ВКД His-меченного FAP яванского макака. Предпочтительно анти-FAP связывающая молекула по изобретению связывается с внеклеточным доменом FAP.
Термин «вариабельная область» или «вариабельный домен» относится к домену тяжелой или легкой цепи антитела, который участвует в связывании антигенсвязывающей молекулы с антигеном. Вариабельные домены тяжелой цепи и легкой цепи (VH и VL, соответственно) нативного антитела обычно имеют сходные структуры, причем каждый домен содержит четыре консервативные каркасные области (FR) и три гипервариабельные области (HVR). Смотрите, например, Kindt et al., Kuby Immunology, 6th ed., W.H. Freeman and Co., страница 91 (2007). Одного домена VH или VL может быть достаточно для придания антигенсвязывающей специфичности.
В контексте данного документа термин «гипервариабельная область» или «HVR» относится к каждой из областей вариабельного домена антитела, которые являются гипервариабельными по последовательности и которые определяют антигенсвязывающую специфичность, например, как «определяющие комплементарность области» («CDR»).
В общем случае антитела содержат шесть CDR: три в VH (CDR-H1, CDR-Н2, CDR-H3) и три в VL (CDR-L1, CDR-L2, CDR-L3). Типовые CDR по данному документу включают:
(а) гипервариабельные петли, находящиеся в аминокислотных остатках 26-32 (L1), 50-52 (L2), 91-96 (L3), 26-32 (H1), 53-55 (Н2) и 96-101 (Н3) (Chothia and Lesk, J. Mol. Biol. 196:901-917 (1987));
(б) CDR, находящиеся в аминокислотных остатках 24-34 (LI), 50-56 (L2), 89-97 (L3), 31-35b (H1), 50-65 (H2) и 95-102 (Н3) (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD (1991)); и
(в) антигенные контакты, находящиеся в аминокислотных остатках 27 с 36 (L1), 46-55 (L2), 89-96 (L3), 30-35b (H1), 47-58 (Н2) и 93-101 (Н3) (MacCallum et al. J. Mol. Biol. 262: 732-745 (1996)).
Если не указано иное, CDR определены в соответствии с Kabat et al., выше. Специалисту в данной области техники понятно, что обозначения CDR также могут быть определены в соответствии с Chothia, выше, McCallum, выше, или любой другой научно приемлемой системой номенклатуры.
«Каркасная область» или «FR» относится к остаткам вариабельного домена, отличным от определяющих комплементарность областей (CDR). FR вариабельного домена обычно состоит из четырех FR-доменов: FR1, FR2, FR3 и FR4. Соответственно, последовательности CDR и FR в общем случае расположены в VH (или VL) в следующем порядке: FR1-CDR-H1(CDR-L1)-FR2-CDR-H2(CDR-L2)-FR3-CDR-H3(CDR-L3)-FR4.
Термин «химерное» антитело относится к антителу, в котором часть тяжелой и/или легкой цепи получена из определенного источника или вида, тогда как остальная часть тяжелой и/или легкой цепи получена из другого источника или вида.
«Класс» антитела относится к типу константного домена или константной области, которые содержит его тяжелая цепь. Существует пять основных классов антител: IgA, IgD, IgE, IgG и IgM, а некоторые из них можно дополнительно разделить на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелой цепи, соответствующие разным классам иммуноглобулинов, называются α, δ, ε, γ и μ, соответственно.
«Гуманизированное» антитело относится к химерному антителу, содержащему аминокислотные остатки из нечеловеческих HVR и аминокислотные остатки из человеческих FR. В определенных вариантах осуществления гуманизированное антитело содержит практически все из по меньшей мере одного и, как правило, двух вариабельных доменов, в которых все или практически все HVR (например, CDR) соответствуют таковым из нечеловеческого антитела, а все или практически все FR соответствуют таковым из человеческого антитела. Гуманизированное антитело, необязательно, может содержать по меньшей мере часть константной области антитела, полученную из человеческого антитела. «Гуманизированная форма» антитела, например, нечеловеческого антитела, относится к антителу, подвергшемуся гуманизации. Другие формы «гуманизированных антител», охватываемые настоящим изобретением, представляют собой те, в которых константная область была дополнительно модифицирована или изменена относительно исходного антитела для обеспечения свойств в соответствии с изобретением, в особенности в отношении связывания Clq и/или связывания Fc-рецептора (FcR).
В данном документе термин «Fc-домен» или «Fc-область» используется для определения С-концевой области тяжелой цепи антитела, которая содержит по меньшей мере часть константной области. Этот термин включает Fc-области с нативной последовательностью и вариантные Fc-области. Fc-область IgG содержит СН2-домен IgG и СН3-домен IgG. «СН2-домен» Fc-области IgG человека обычно простирается от аминокислотного остатка приблизительно в позиции 231 до аминокислотного остатка приблизительно в позиции 340. В одном варианте осуществления к СН2-домену присоединена углеводная цепь. В данном документе СН2-домен может представлять собой нативный СН2-домен или вариантный СН2-домен. «СН3-домен» содержит участок из остатков, С-терминальных к СН2-домену в Fc-области (т.е. от аминокислотного остатка приблизительно в позиции 341 до аминокислотного остатка приблизительно в позиции 447 IgG). В данном документе СН3-область может представлять собой СН3-домен с нативной последовательностью или вариантный СН3-домен (например, СН3-домен с внесенным в одну цепь «выступом» («узлом») и соответствующей внесенной в другую цепь «полостью» («впадиной»); смотрите патент США 5821333, явным образом включенный в данный документ посредством ссылки). Такие вариантные СН3-домены можно использовать, чтобы стимулировать гетеродимеризацию двух неидентичных тяжелых цепей антитела, как описано в данном документе. В одном варианте осуществления Fc-область тяжелой цепи человеческого IgG простирается от Cys226 или от Pro230 до карбокси-конца тяжелой цепи. При этом С-концевой лизин (Lys447) Fc-области может присутствовать или нет. Если в данном документе не указано иное, нумерация аминокислотных остатков в Fc-области или константной области соответствует системе нумерации EU, также называемой индексом EU, как описано в Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD, 1991.
Технология «выступ-во-впадину» описана, например, в US 5731168; US 7695936; Ridgway et al., Prot Eng 9, 617-621 (1996) и Carter, J Immunol Meth 248, 7-15 (2001). В общем случае этот способ включает внесение выступа («узла») в контактную поверхность первого полипептида и соответствующей впадины («полости») в контактную поверхность второго полипептида, так что выступ может располагаться во впадине так, чтобы стимулировать образование гетеродимера и затруднять образование гомодимера. Выступы конструируют путем замены небольших аминокислотных боковых цепей в контактной поверхности первого полипептида на более крупные боковые цепи (например тирозин или триптофан). Компенсаторные «полости» идентичного или сходного размера с выступами создают в контактной поверхности второго полипептида путем замещения крупных аминокислотных боковых цепей меньшими (например, аланином или треонином). Выступ и полость можно создавать путем изменения нуклеиновой кислоты, кодирующей полипептиды, например, с помощью сайт-специфического мутагенеза, или с помощью пептидного синтеза. В конкретном варианте осуществления модификация узла включает аминокислотную замену T366W в одной из двух субъединиц Fc-домена, а модификация впадины включает аминокислотные замены T366S, L368A и Y407V в другой из двух субъединиц Fc-домена. В дополнительном конкретном варианте осуществления субъединица Fc-домена, содержащая модификацию выступа, дополнительно содержит аминокислотную замену S354C, а субъединица Fc-домена, содержащая модификацию впадины, дополнительно содержит аминокислотную замену Y349C. Внесение этих двух остатков цистеина приводит к образованию дисульфидного мостика между двумя субъединицами Fc-области, таким образом, дополнительно стабилизируя димер (Carter, J Immunol Methods 248, 7-15 (2001)).
Предполагается, что «область, эквивалентная Fc-области иммуноглобулина» включает встречающиеся в природе аллельные варианты Fc-области иммуноглобулина, а также варианты, имеющие изменения, которые соответствуют заменам, добавлениям или делециям, но которые практически не снижают способность иммуноглобулина опосредовать эффекторные функции (такие как антителозависимая клеточная цитотоксичность). Например, из N-конца или С-конца Fc-области иммуноглобулина могут быть удалены одна или более аминокислот практически без утраты биологической функции. Такие варианты можно отбирать в соответствии с общими правилами, известными в данной области техники, как имеющие минимальный эффект на активность (смотрите, например, Bowie, J. U. et al., Science 247:1306-10 (1990)).
Термин «эффекторная функция» относится видам биологической активности, характерным для Fc-области антитела, которые варьируются в зависимости от изотипа антитела. Примеры эффекторных функций антител включают: связывание Clq и комплементзависимую цитотоксичность (КЗЦ), связывание Fc-рецептора, антителозависимую клеточноопосредованную цитотоксичность (АЗКЦ), антителозависимый клеточный фагоцитоз (АЗКФ), секрецию цитокинов, опосредованное иммунных комплексом поглощение антигена антигенпрезентирующими клетками, понижающую регуляцию рецепторов клеточной поверхности (например, В-клеточного рецептора) и активацию В-клеток.
Эффекторные функции связывания Fc-рецептора могут быть опосредованы взаимодействием Fc-области антитела с Fc-рецепторами (FcR), которые представляют собой специализированный рецепторы клеточной поверхности на гемопоэтических клетках. Fc-рецепторы принадлежат к суперсемейству иммуноглобулинов, и было показано, что они опосредуют как удаление покрытых антителами патогенов посредством фагоцитоза иммунных комплексов, так и лизис эритроцитов и различных других клеточных мишеней (например, опухолевых клеток), покрытых соответствующими антителами, посредством антителозависимой клеточноопосредованной цитотоксичности (АЗКЦ) (смотрите, например, Van de Winkel, J.G. and Anderson, C.L., J. Leukoc. Biol. 49 (1991) 511-524). FcR определяют по их специфичности в отношении изотипов иммуноглобулина: Fc-рецепторы для антител IgG называются FcγR. Связывание Fc-рецепторов описано, например, в Ravetch, J.V. and Kinet, J.P., Annu. Rev. Immunol. 9 (1991) 457-492, Capel, P.J., et al., Immunomethods 4 (1994) 25-34; de Haas, M., et al., J. Lab. Clin. Med. 126 (1995) 330-341; и Gessner, J.E., et al., Ann. Hematol. 76 (1998) 231-248.
Перекрестное связывание рецепторов к Fc-области антител IgG (FcγR) стимулирует широкий ряд эффекторных функций, включая фагоцитоз, антителозависимую клеточную цитотоксичность и высвобождение воспалительных медиаторов, а также выведение иммунных комплексов и регуляцию выработки антител. У людей были описаны три класса FcγR, которые представляют собой:
- FcγRI (CD64) связывает мономерный IgG с высокой аффинностью и экспрессируется на макрофагах, моноцитах, нейтрофилах и эозинофилах. Модификация Fc-области IgG по меньшей мере в одном из аминокислотных остатков E233-G236, Р238, D265, N297, А327 и Р329 (нумерация в соответствии с индексом EU по Kabat) снижает связывание с FcγRI. Остатки IgG2 в позициях 233-236, замещенные на IgG1 и IgG4, снижали связывание с FcγRI в 103 раз и устраняли ответ моноцитов человека на сенсибилизированные антителом красные кровяные клетки (Armour, K.L., et al., Eur. J. Immunol. 29 (1999) 2613 2624).
-FcγyRII (CD32) связывает комплексированный IgG со средней или низкой аффинностью и экспрессируется повсеместно. Этот рецептор можно разделить на два подтипа - FcγRIIA и FcγRIIB. FcγRIIA встречается на многих клетках, участвующих в уничтожении (например, макрофагах, моноцитах, нейтрофилах) и, похоже, способен активировать процесс уничтожения. FcγRIIB, похоже, играет роль в процессе ингибирования и встречается на В-клетках, макрофагах и на тучных клетках и эозинофилах. В случае В-клеток его функция, по-видимому, состоит в подавлении дальнейшей выработки иммуноглобулина и переключении изотипа, например, на класс IgE. В случае макрофагов действие FcγRIIB состоит в ингибировании фагоцитоза, опосредованного FcγRIIA. В случае эозинофилов и тучных клеток В-форма может способствовать подавлению активации этих клеток посредством связывания IgE с отдельным рецептором. Сниженное связывание с FcγRIIA встречается, например, у антител, содержащих Fc-область IgG с мутациями по меньшей мере в одном из аминокислотных остатков Е233-G236, Р238, D265, N297, А327, Р329, D270, Q295, А327, R292 и K414 (нумерация в соответствии с индексом EU по Kabat).
- FcγRIII (CD16) связывает IgG со средней или низкой аффинностью и существует в виде двух типов. FcγRIIIA встречается на NK-клетках, макрофагах, эозинофилах и некоторых моноцитах и Т-клетках и опосредует АЗКЦ. FcγRIIIB на высоком уровне экспрессируется на нейтрофилах. Сниженное связывание с FcγRIIIA встречается, например, у антител, содержащих Fc-область IgG с мутациями по меньшей мере в одном из аминокислотных остатков E233-G236, Р238, D265, N297, А327, Р329, D270, Q295, А327, S239, Е269, Е293, Y296, V303, А327, К338 и D376 (нумерация в соответствии с индексом EU по Kabat).
Картирование сайтов связывания с Fc-рецепторами на человеческом IgGl, вышеуказанные сайты мутации и способы измерения связывания с FcγRI и FcγRIIA описаны в Shields, R.L., et al. J. Biol. Chem. 276 (2001) 6591-6604.
Термин «АЗКЦ» или «антителозависимая клеточная цитотоксичность» обозначает функцию, опосредованную связыванием Fc-рецептора, и относится к лизису целевых клеток антителом, описанным в данном документе, в присутствии эффекторных клеток. Способность антитела индуцировать начальные этапы опосредования АЗКЦ исследуют путем измерения их связывания с экспрессирующими Fcγ-рецепторы клетками, такими как клетки, рекомбинантно экспрессирующие FcγRI и/или FcγRIIA, или NK-клетки (экспрессирующие преимущественно FcγRIIIA). В частности, измеряют связывание с FcγR на NK-клетках.
«Активирующий Fc-рецептор» представляет собой Fc-рецептор, который после связывания Fc-областью антитела вызывает сигнальные события, которые стимулируют несущие рецепторы клетки осуществлять эффекторные функции. Активирующие Fc-рецепторы включают FcγRIIIa (CD16a), FcγRI (CD64), FcγRIIa (CD32) и FcαRI (CD89). Конкретным активирующим Fc-рецептором является человеческий FcγRIIIa (смотрите номер доступа UniProt Р08637, версия 141).
В контексте данного документа термин «CD40» относится к любому нативному CD40 из любого источника, относящегося к позвоночным, включая млекопитающих, таких как приматы (например, люди) и грызуны (например, мыши и крысы), если не указано иное. Этот термин охватывает «полноразмерный» непроцессированный белок CD40, а также любую форму CD40, полученную в результате процессинга в клетке. Этот термин также охватывает варианты CD40 природного происхождения, например, сплайс-варианты или аллельные варианты. Аминокислотная последовательность типового человеческого CD40 приведена в SEQ ID NO: l (Uniprot Р25942, версия 200), а аминокислотная последовательность типового мышиного CD40 приведена в SEQ ID NO: 146 (Uniprot Р27512, версия 160). Антиген CD40 представляет собой 50 кДа гликопротеин клеточной поверхности, который принадлежит семейству рецепторов факторов некроза опухолей (TNF-R). (Stamenkovic et al. (1989), EMBO J. 8: 1403-10). CD40 экспрессируется во многих типах нормальных и опухолевых клеток, включая В-лимфоциты, дендритные клетки, моноциты, макрофаги, эпителий вилочковой железы, эндотелиальные клетки, фибробласты и гладкомышечные клетки. CD40 экспрессируется во всех В-лимфомах и в 70% всех солидных опухолей, а в антигенпрезентирующих клетках (АПК) его регуляция повышена за счет сигналов созревания, таких как IFN-гамма и GM-CSF. Активация CD40 также индуцирует дифференцировку моноцитов в функциональные дендритные клетки (ДК) и усиливает цитолитическую активность NK-клеток посредством индуцированных АПК-CD40 цитокинов. Таким образом, CD40 играет важную роль в инициации и усилении иммунных ответов путем индукции созревания АПК, секреции хелперных цитокинов, повышенной регуляции костимулирующих молекул и усиления эффекторных функций.
В контексте данного документа термин «агонист CD40» включает любой фрагмент, который агонизирует взаимодействие CD40/CD40L. В этом контексте CD40 относится преимущественно к CD40 человека, таким образом, агонист CD40 предпочтительно является агонистом CD40 человека. Как правило, фрагмент представляет собой агонистическое антитело к CD40 или фрагмент антитела.
Термины «анти-CD40 антитело», «анти-CD40», «антитело к CD40» и «антитело, которое специфически связывается с CD40» относятся к антителу, которое способно к связыванию CD40 с достаточной аффинностью, чтобы антитело можно было применять в качестве диагностического и/или терапевтического агента для нацеливания на CD40. В одном аспекте степень связывания анти-СD40 антитела с неродственным отличным от CD40 белком составляет менее чем около 10% от связывания антитела с CD40 согласно данным измерений, например, методом радиоиммуноанализа (РИА) или проточной цитометрии (FACS). В определенных вариантах осуществления антитело, которое связывается с CD40, имеет константу диссоциации (KД) ≤1 мкМ, ≤100 нМ, ≤10 нМ, ≤1 нМ, ≤0,1 нМ, ≤0,01 нМ, или ≤0,001 нМ (например, 10-6 М или менее, например, от 10-68 М до 10-13 М, например, от 10-8 М до 10-10 М).
Термин «пептидный линкер» относится к пептиду, содержащему одну или более аминокислот, как правило, приблизительно 2-20 аминокислот. Пептидные линкеры известны в данной области техники или описаны в данном документе. Подходящими неиммуногенными линкерными пептидами являются, например, пептидные линкеры (G4S)n, (SG4)n или G4(SG4)n, «n» в общем случае представляет собой число от 1 и 10, как правило, от 2 до 4, в частности 2, т.е. пептиды, выбранные из группы, состоящей из GGGGS (SEQ ID NO: 82), GGGGSGGGGS (SEQ ID NO: 83), SGGGGSGGGG (SEQ ID NO: 84) и GGGGSGGGGSGGGG (SEQ ID NO: 85), но также включают последовательности GSPGSSSSGS (SEQ ID NO: 86), (G4S)3 (SEQ ID NO: 87), (G4S)4 (SEQ ID NO: 88), GSGSGSGS (SEQ ID NO: 89), GSGSGNGS (SEQ ID NO: 90), GGSGSGSG (SEQ ID NO: 91), GGSGSG (SEQ ID NO: 92), GGSG (SEQ ID NO: 93), GGSGNGSG (SEQ ID NO: 94), GGNGSGSG (SEQ ID NO: 95) и GGNGSG (SEQ ID NO: 96). Особенный интерес представляют пептидные линкеры (G4S) (SEQ ID NO: 82), (G4S)2 или GGGGSGGGGS (SEQ ID NO: 83), (G4S)3 (SEQ ID NO: 87) и (G4S)4 (SEQ ID NO: 88).
В контексте данной заявки термин «аминокислота» обозначает группу α-аминокислот природного происхождения, включая аланин (трехбуквенный код: ala, однобуквенный код: А), аргинин (arg, R), аспарагин (asn, N), аспарагиновая кислота (asp, D), цистеине (cys, С), глутамин (gin, Q), глутаминовая кислота (glu, Е), глицин (gly, G), гистидин (his, Н), изолейцин (ile, I), лейцин (leu, L), лизин (lys, K), метионин (met, М), фенилаланин (phe, F), пролин (pro, Р), серии (ser, S), треонин (thr, Т), триптофан (trp, W), тирозин (tyr, Y) и валин (val, V).
Под «слитым» или «соединенным» подразумевается, что компоненты (например, тяжелая цепь антитела и Fab-фрагмент) связаны пептидными связями, как напрямую, так и посредством одного или более пептидных линкеров.
«Процент (%) идентичности аминокислотной последовательности» относительно референсной полипептидной (белковой) последовательности определяется как процентная доля аминокислотных остатков в кандидатной последовательности, которые являются идентичными с аминокислотными остатками в референсной полипептидной последовательности, после выравнивания последовательностей и внесения, в случае необходимости, гэпов для достижения максимального процента идентичности последовательностей, и без учета каких-либо консервативных замен в качестве части идентичности последовательностей. Выравнивание в целях определения процента идентичности аминокислотной последовательности можно осуществлять различными способами, которые известны в данной области техники, например, используя общедоступное компьютерное программное обеспечение, такое как программное обеспечение BLAST, BLAST-2, ALIGN, SAWI или Megalign (DNASTAR). Специалисты в данной области техники могут определить подходящие параметры для выравнивания последовательностей, включая любые алгоритмы, необходимые для достижения максимального выравнивания по всей длине сравниваемых последовательностей. При этом в целях данного документа значения процента (%) идентичности аминокислотных последовательностей получают, используя компьютерную программу для сравнения последовательностей ALIGN-2. Компьютерная программа для сравнения последовательностей ALIGN-2 была разработана Genentech, Inc., а исходный код был подан вместе с пользовательской документацией в Бюро регистрации авторских прав США, Вашингтон, округ Колумбия, 20559, где он зарегистрирован под номером регистрации авторского права США TXU510087. Программа ALIGN-2 находится в свободном доступе от Genentech, Inc., Южный Сан-Франциско, Калифорния, или может быть скомпилирована из исходного кода. Программу ALIGN-2 следует компилировать для использования в операционной системе UNIX, включая Digital UNIX V4.0D. Все параметры сравнения последовательностей установлены программой ALIGN-2 и не варьируются. В тех случаях, когда для сравнения аминокислотных последовательностей используют ALIGN-2, % идентичности аминокислотной последовательности заданной аминокислотной последовательности А с или по сравнению с заданной аминокислотной последовательностью В (что в альтернативном варианте можно сформулировать как заданная аминокислотная последовательность А, которая имеет или содержит определенный % идентичности аминокислотной последовательности с или по сравнению с заданной аминокислотной последовательностью В) рассчитывают следующим образом:
100 умножить на соотношение X/Y,
где X представляет число аминокислотных остатков, оцененных программой выравнивания последовательностей ALIGN-2 как идентичные совпадения в осуществленном программой выравнивании А и В, a Y представляет общее число аминокислотных остатков в В. Следует понимать, что если длина аминокислотной последовательности А не равна длине аминокислотной последовательности В, % идентичности аминокислотной последовательности А с В не будет равен % идентичности аминокислотной последовательности В с А. Если специально не указано иное, все значения % идентичности аминокислотных последовательностей, используемые в данном документе, получены, как описано в предыдущем параграфе, с использованием компьютерной программы ALIGN-2.
В определенных вариантах осуществления предусмотрены варианты аминокислотной последовательности биспецифических антигенсвязывающих молекул, предложенных в данном документе. Например, может существовать необходимость в улучшении аффинности связывания и/или других биологических свойств антигенсвязывающих молекул, содержащих триммер лиганда TNF. Варианты аминокислотной последовательности антигенсвязывающих молекул, содержащих тример лиганда TNF, можно получать путем внесения соответствующих модификаций в нуклеотидную последовательность, кодирующую молекулы, или с помощью пептидного синтеза. Такие модификации включают, например, делеции, и/или вставки, и/или замены остатков в аминокислотных последовательностях антитела. Для получения конечной конструкции можно осуществлять любую комбинацию делеций, вставок и замен при условии, что конечная конструкция обладает необходимыми характеристиками, например, связывания антигена. Сайты, представляющие интерес для заместительного мутагенеза, включают HVR и каркасные области (FR). Консервативные замены приведены в таблице Б под заголовком «Предпочтительные замены» и дополнительно описаны ниже в отношении классов боковых цепей аминокислот от (1) до (6). Аминокислотные замены можно вносить в представляющую интерес молекулу и проводить скрининг продуктов в отношении необходимой активности, например, сохранения/улучшения связывания антигена, снижения иммуногенности или улучшения АЗКЦ или КЗЦ.
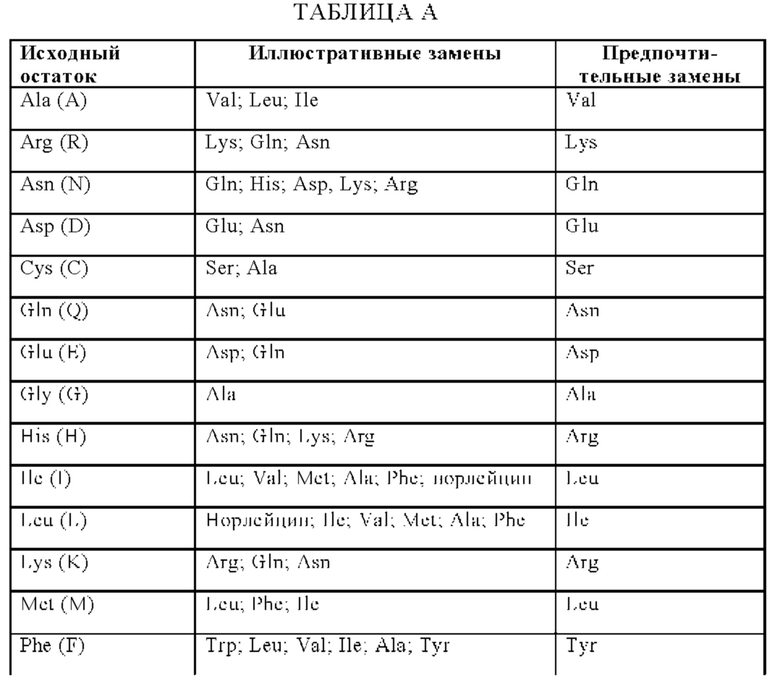

Аминокислоты можно разделить на группы в соответствии с общими свойствами боковых цепей:
гидрофобные: норлейцин, Met, Ala, Val, Leu, Ile;
нейтральные гидрофильные: Cys, Ser, Thr, Asn, Gin;
кислые: Asp, Glu;
основные: His, Lys, Arg;
остатки, влияющие на ориентацию цепи: Gly, Pro;
ароматические: Trp, Tyr, Phe.
Неконсервативные замены повлекут за собой замену члена одного из этих классов другим классом.
Термин «варианты аминокислотной последовательности» включает заместительные варианты, в которых аминокислотные замены присутствуют в одном или более остатках гиперварибельной области родительской антигенсвязывающей молекулы (например, гуманизированное или человеческое антитело). В общем случае полученные в результате варианты, отобранные для дополнительного исследования, имеют модификации (например, улучшение) определенных биологических свойств (например, повышенную аффинность, сниженную иммуногенность) по сравнению с родительской антигенсвязывающей молекулой и/или в значительной степени сохраняют определенные биологические свойства родительской антигенсвязывающей молекулы. Типовой заместительный вариант представляет собой антитело с созревшей аффинностью, которое удобно получать, например, используя методики созревания аффинности на основе фагового дисплея, такие как описанные в данном документе. Вкратце, один или более остатков HVR подвергают мутации, а вариантные антигенсвязывающие молекулы отображают на поверхности фага и проводят их скрининг в отношении конкретного вида биологической активности (например, аффинности связывания). В определенных вариантах осуществления замены, вставки или делеции могут находится в одной или более HVR при условии, что такие изменения практически не снижают способность антигенсвязывающей молекулы связывать антиген. Например, в HVR можно проводить консервативные изменения (например, консервативные замены, предложенные в данном документе), которые существенно не снижают аффинность связывания. Удобный способ идентификации остатков или областей антитела, которые могут быть мишенями для мутагенеза, называется «аланин-сканирующим мутагенезом», описанным Cunningham and Wells (1989) Science, 244:1081-1085. В этом способе идентифицируют аминокислотный остаток или группу целевых аминокислотных остатков (например, заряженные аминокислотные остатки, такие как Arg, Asp, His, Lys и Glu) и заменяют нейтральной или отрицательно заряженной аминокислотой (например, аланином или полиаланином) для определения, повлияло ли это на взаимодействие антитела с антигеном. Дополнительные замены можно вносить в аминокислотные положения, демонстрирующие функциональную чувствительность к исходным заменам. В альтернативном или дополнительном варианте используют кристаллическую структуру комплекса антигенсвязывающая молекула антиген, чтобы идентифицировать точки контакта между антителом и антигеном. Такие контактные остатки и соседние остатки можно использовать для нацеливания или удалять из кандидатов для замены. Можно проводить скрининг вариантов, чтобы определить, имеют ли они необходимые свойства.
Вставки аминокислотных последовательностей включают амино- и/или карбокси-концевые слияния с диапазоном длины от одного остатка до полипептидов, содержащих сто или более остатков, а также вставки внутри последовательности одного или некоторого количества аминокислотных остатков. Примеры концевых вставок включают биспецифические антигенсвязывающие молекулы по изобретению с N-концевым остатком метионила. Другие инсерционные варианты молекулы включают слияние N- или С-конца антитела полипептидом, который увеличивает сывороточное время полужизни биспецифических антигенсвязывающих молекул.
В определенных вариантах осуществления предложенные в данном документе биспецифические антигенсвязывающие молекулы изменены с целью повышения или снижения степени гликозилирования антитела. Варианты молекул по гликозилированию удобно получать, изменяя аминокислотную последовательность так, чтобы создать или удалить один или более сайтов гликозилирования. Если антигенсвязывающая молекула, содержащая тример лиганда TNF, содержит Fc-область, можно изменять присоединенный к ней углевод. Нативные антитела, вырабатываемые клетками млекопитающих, обычно содержат разветвленный биантеннарный олигосахарид, который обычно присоединен посредством N-связи к Asn297 СН2-домена Fc-области. Смотрите, например, Wright et al. TIBTECH 15:26-32 (1997). Олигосахарид может содержать различные углеводы, например, маннозу, N-ацетилглюкозамин (GlcNAc), галактозу и сиаловую кислоту, а также фукозу, присоединенную к GlcNAc в «стебле» биантеннарной олигосахаридной структуры. В некоторых вариантах осуществления модификации олигосахарида в антигенсвязывающей молекуле, содержащей тример лиганда семейства TNF, можно осуществлять с целью создания вариантов с определенными улучшенными свойствами. В одном аспекте предложены варианты биспецифических антигенсвязывающих молекул или антител по изобретению, имеющие углеводную структуру, в которой отсутствует фукоза, присоединенная (прямо или непрямо) к Fc-области. Такие варианты по фукозилированию могут иметь улучшенную функцию АЗКЦ, смотрите, например, публикации патентов США №№US 2003/0157108 (Presta, L.) или US 2004/0093621 (Kyowa Hakko Kogyo Co., Ltd). В другом аспекте предложены варианты биспецифических антигенсвязывающих молекул или антител по изобретению с разделенными олигосахаридами, например, в которых биантеннарный олигосахарид, присоединенный к Fc-области, разделен надвое GlcNAc. Такие варианты имеют сниженное фукозилирование и/или улучшенную функцию АЗКЦ, смотрите, например, WO 2003/011878 (Jean-Mairet et al.); патент США №6602684 (Umana et al.); и US 2005/0123546 (Umana et al). Также предложены варианты с по меньшей мере одним остатком галактозы в олигосахариде, присоединенном к Fc-области. Такие варианты антител могут иметь улучшенную функцию КЗЦ и описаны, например, в WO 1997/30087 (Patel et al.); WO 1998/58964 (Raju, S.); и WO 1999/22764 (Raju, S.).
В определенных аспектах может быть желательно создавать цистеин-сконструированные варианты биспецифических антигенсвязывающих молекул по изобретению, например, «thioMAb,» в которых один или более остатков молекулы замещены остатками цистеина. В конкретных аспектах замещенные остатки находятся в доступных сайтах молекулы. За счет замены этих остатков цистеином реакционноспособные тиольные группы располагаются в доступных сайтах антитела и могут быть использованы для конъюгации антитела с другими фрагментами, такими как лекарственные фрагменты или фрагменты линкер -лекарственное средство, для создания иммуноконъюгата. В определенных аспектах цистеином может быть замещен любой из следующих остатков: V205 (нумерация Kabat) легкой цепи; А118 (нумерация EU) тяжелой цепи; и S400 (нумерация EU) Fc-области тяжелой цепи. Цистеин-сконструированные антигенсвязывающие молекулы можно создавать, как описано, например, в патенте США №7521541.
Термин «нуклеиновая кислота» или «полинуклеотид» включает любое соединение и/или вещество, которое содержит полимер из нуклеотидов. Каждый нуклеотид состоит из основания, в частности, пуринового или пиримидинового основания (т.е. цитозина (С), гуанина (G), аденина (А), тимина (Т) или урацила (U)), сахара (т.е. дезоксирибозы или рибозы) и фосфатной группы. Часто молекула нуклеиновой кислоты описана по последовательности оснований, при этом указанные основания представляют первичную структуру (линейную структуру) молекулы нуклеиновой кислоты. Последовательность оснований, как правило, представлена от 5' к 3'. В данном документе термин молекула нуклеиновой кислоты охватывает дезоксирибонуклеиновую кислоту (ДНК), включая, например, комплементарную ДНК (кДНК) и геномную ДНК, рибонуклеиновую кислоту (РНК), в частности, матричную РНК (мРНК), синтетические формы ДНК и РНК и смешанные полимеры, содержащие две или более из этих молекул. Молекула нуклеиновой кислоты может быть линейной или кольцевой. Кроме того, термин молекула нуклеиновой кислоты включает как смысловые, так и антисмысловые цепи, а также одноцепочечные и двухцепочечные формы. Помимо этого, описанная в данном документе молекула нуклеиновой кислоты может содержать нуклеотиды природного и неприродного происхождения. Примеры нуклеотидов неприродного происхождения включают модифицированные нуклеотидные основания с дериватизированными сахарами, или фосфатными остовными связями, или химически модифицированными остатками. Молекулы нуклеиновых кислот также включают молекулы ДНК и РНК, подходящие в качестве вектора для управления экспрессией антитела по изобретению in vitro и/или in vivo, например, в организме хозяина или пациента. Такие ДНК (например, кДНК) или РНК (например, мРНК) векторы могут быть немодифицированными или модифицированными. Например, мРНК может быть химически модифицирована для повышения стабильности РНК-вектора и/или экспрессии кодируемой молекулы так, чтобы мРНК можно было вводить субъекту для генерации антител in vivo (смотрите, например, Stadler ert al, Nature Medicine 2017, опубликованную онлайн 12 июня 2017 г., doi:10.1038/nm.4356 или ЕР 2 101 823 В1).
«Выделенная» нуклеиновая кислота относится к молекуле нуклеиновой кислоты, которая была отделена от компонента ее естественного окружения. Выделенная нуклеиновая кислота включает молекулу нуклеиновой кислоты, содержащуюся в клетках, которые обычно содержат эту молекулу нуклеиновой кислоты, но при этом молекула нуклеиновой кислоты присутствует внехромосомно или в хромосомном положении, которое отличается от ее природного хромосомного положения.
«Выделенная нуклеиновая кислота, кодирующая биспецифическую антигенсвязывающую молекулу или антитело» относится к одной или более молекулам нуклеиновых кислот, кодирующим тяжелые и легкие цепи (или их фрагменты) биспецифической антигенсвязывающей молекулы или антитела, включая такие молекулы нуклеиновых кислот в одном векторе или отдельных векторах и такие молекулы нуклеиновых кислот, присутствующие в одной или более локациях в клетке-хозяине.
Под нуклеиновой кислотой или полинуклеотидом, имеющими нуклеотидную последовательность, по меньшей мере, например, на 95% «идентичную» референсной последовательности по настоящему изобретению, подразумевается, что нуклеотидная последовательность полинуклеотида идентична референсной последовательности за исключением того, что полинуклеотидная последовательность может включать до пяти точечных мутаций на каждые 100 нуклеотидов референсной нуклеотидной последовательности. Другими словами, чтобы получить полинуклеотид, имеющий нуклеотидную последовательность, по меньшей мера на 95% идентичную референсной нуклеотидной последовательности, до 5% нуклеотидов в референсной последовательности могут быть удалены или замещены другими нуклеотидами или в референсную последовательность может быть вставлено число нуклеотидов, составляющее до 5% от общего числа нуклеотидов в референсной последовательности. Эти изменения референсной последовательности могут находиться в 5' или 3' концевых позициях референсной нуклеотидной последовательности или в любом месте между этими концевыми позициями, размещаясь по отдельности между остатками в референсной последовательности или одной или более непрерывными группами в референсной последовательности. На практике определение того, что любая конкретная полинуклеотидная последовательность является по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичной нуклеотидной последовательности по настоящему изобретению, можно осуществлять традиционным способом, используя известные компьютерные программы, такие как те, которые обсуждаются выше для полипептидов (например, ALIGN-2).
Термин «экспрессионная кассета» относится к полинуклеотиду, созданному рекомбинантным или синтетическим способом, с рядом определенных элементов нуклеиновой кислоты, которые обеспечивают возможность транскрипции конкретной нуклеиновой кислоты в целевой клетке. Рекомбинантная экспрессионная кассета может быть включена в плазмиду, хромосому, митохондриальную ДНК, пластидную ДНК, вирус или фрагмент нуклеиновой кислоты. Как правило, рекомбинантная экспрессионная кассета, как часть экспрессионного вектора, содержит, помимо других последовательностей, последовательность нуклеиновой кислоты, предназначенную для транскрипции, и промотор. В определенных вариантах осуществления экспрессионная кассета по изобретению содержит полинуклеотидные последовательности, которые кодируют биспецифические антигенсвязывающие молекулы по изобретению или их фрагменты.
Термины «вектор» или «экспрессионный вектор» являются синонимами с «экспрессионной конструкцией» и обозначают молекулу ДНК, которую используют для внесения и управления экспрессией конкретного гена, с которым она функционально связана, в целевой клетке. Этот термин включает вектор в виде самореплицирующейся структуры нуклеиновой кислоты, а также вектор, включенный в геном клетки-хозяина, в которую его внесли.
Экспрессионный вектор по настоящему изобретению содержит экспрессионную кассету. Экспрессионные векторы делают возможной транскрипцию больших количеств стабильной мРНК. После того, как экспрессионный вектор оказывается в целевой клетке, клеточная машинерия транскрипции и/или трансляции вырабатывает молекулу рибонуклеиновой кислоты или белок, кодируемые геном. В одном варианте осуществления экспрессионный вектор по изобретению содержит экспрессионную кассету, которая содержит полинуклеотидные последовательности, которые кодируют биспецифические антигенсвязывающие молекулы по изобретению или их фрагменты.
Термины «клетка-хозяин», «линия клеток-хозяев» и «культура клеток-хозяев» используются взаимозаменяемо и относятся к клеткам, в которые была внесена экзогенная нуклеиновая кислота, включая потомство таких клеток. Клетки-хозяева включают «трансформантов» и «трансформированные клетки», которые включают первично трансформированные клетки и полученное от них потомство вне зависимости от числа пассажей. Потомство может не быть полностью идентичным родительской клетке по содержанию нуклеиновой кислоты и может содержать мутации. Сюда включено мутантное потомство, которое имеет такую же функцию или биологическую активность, в отношении которых проводится скрининг или отбор изначально трансформированных клеток. Клетка-хозяин принадлежит любому типу клеточной системы, который можно использовать для создания биспецифических антигенсвязывающих молекул по настоящему изобретению. Клетки-хозяева включают культивируемые клетки, например, культивируемые клетки млекопитающих, такие как клетки СНО, клетки ВНК, клетки NS0, клетки SP2/0, клетки миеломы YO, клетки миеломы мышей Р3Х63, клетки PER, клетки PER.C6 или клетки гибридомы, клетки дрожжей, клетки насекомых и клетки растений, если называть только некоторые, но также клетки, принадлежащие трансгенному животному, трансгенному растению или культивируемой растительной или животной ткани.
«Эффективное количество» агента относится к количеству, необходимому для физиологического изменения в клетке или ткани, в которую его вводят.
«Терапевтически эффективное количество» агента, например, фармацевтической композиции, относится к количеству, эффективному в дозировках и в течение периодов времени, необходимых для достижения желательного терапевтического или профилактического результата. Терапевтически эффективное количество агента, например, устраняет, снижает, замедляет, минимизирует или предотвращает нежелательные явления заболевания.
«Индивид» или «субъект» представляет собой млекопитающее. Млекопитающие включают, но не ограничиваются этим, одомашненных животных (например, коров, овец, кошек, собак и лошадей), приматов (например, людей и отличных от человека приматов, таких, как обезьяны), кроликов и грызунов (например, мышей и крыс). В частности, индивид или субъект представляет собой человека.
Термин «фармацевтическая композиция» или «фармацевтический состав» относится к препарату, который находится в форме, обеспечивающей эффективность биологической активности содержащегося в нем активного ингредиента, и который не содержит дополнительные компоненты, которые являются неприемлемо токсичными для субъекта, которому предполагается вводить фармацевтическую композицию.
«Фармацевтически приемлемый носитель» относится к ингредиенту в фармацевтической композиции или фармацевтическом составе, отличному от активного ингредиента, который нетоксичен для субъекта. Фармацевтически приемлемый носитель включает, но не ограничивается этим, буфер, вспомогательное вещество, стабилизатор или консервант.
Термин «вкладыш в упаковку» используется для обозначения инструкций, обычно включаемых в коммерческие упаковки терапевтических продуктов, которые содержат информацию о показаниях, применении, дозировке, введении, комбинированной терапии, противопоказаниях и/или предостережениях относительно применения таких терапевтических продуктов.
В контексте данного документа термин «лечение» (и его грамматические варианты, такие как «лечить» или «лечащий») относится к клиническому вмешательству с целью изменения естественного течения заболевания индивида, подлежащего лечению, и может проводиться как для профилактики, так и в течении клинической патологии. Необходимые эффекты лечения включают, но не ограничиваются этим, предотвращение появления или повторного появления заболевания, смягчение симптомов, уменьшение каких-либо прямых или косвенных патологических последствий заболевания, предотвращение метастазирования, снижение скорости прогрессирования заболевания, уменьшение интенсивности или временное облегчение болезненного состояния и ремиссию или улучшение прогноза. В некоторых вариантах осуществления молекулы по изобретению применяют для задержки развития заболевания или замедления прогрессирования заболевания.
В контексте данного документа термин «рак» относится к пролиферативным заболеваниям, таким как лимфомы, лимфоцитарные лейкозы, рак легкого, немелкоклеточный рак легких (НМРЛ), рак легкого из бронхиолоальвеолярных клеток, рак кости, рак поджелудочной железы, рак кожи, рак головы и шеи, кожная или интраокулярная меланома, рак матки, рак яичника, ректальный рак, рак анальной области, рак желудка, рак толстой кишки, рак молочной железы, рак матки, карцинома фаллопиевых труб, карцинома эндометрия, карцинома шейки матки, карцинома вагины, карцинома вульвы, болезнь Ходжкина, рак пищевода, рак тонкого кишечника, рак эндокринной системы, рак щитовидной железы, рак паращитовидной железы, рак надпочечника, саркома мягкой ткани, рак уретры, рак полового члена, рак предстательной железы, рак мочевого пузыря, рак почки или мочеточника, почечно-клеточная карцинома, карцинома почечной лоханки, мезотелиома, печеночноклеточный рак, рак желчного пузыря, новообразования центральной нервной системы (ЦНС), опухоли оси позвоночника, глиома ствола головного мозга, мультиформная глиобластома, астроцитомы, шванномы, эпендимомы, медуллобластомы, менингиомы, плоскоклеточные карциномы, аденома гипофиза и саркома Юинга, включая рефрактерные версии любого из вышеперечисленных видов рака или комбинации одного или более из вышеперечисленных видов рака.
В контексте данного документа термин «химиотерапевтический агент» относится к химическому соединению, применимому для лечения рака. В одном аспекте химиотерапевтический агент представляет собой антиметаболит. В одном аспекте антиметаболит выбран из группы, состоящей из аминоптерина, метотрексата, пеметрекседа, ралтитрекседа, кладрибина, клофарабина, флударабина, меркаптопурина, пентостатина, тиогуанина, капецитабина, цитарабина, фтороурацила, флоксуридина и гемцитабина. В одном конкретном аспекте антиметаболит представляет собой капецитабин или гемцитабин. В другом аспекте антиметаболит представляет собой фторурацил. В одном аспекте химиотерапевтический агент представляет собой агент, который влияет на образование микротрубочек. В одном аспекте агент, который влияет на образование микротрубочек, выбран из группы, состоящей из паклитаксела, доцетаксела, винкристина, винбластина, виндезина, винорелбина, таксотера, этопозида и тенипозида. В другом аспекте химиотерапевтический агент представляет собой алкилирующий агент, такой как циклофосфамид. В одном аспекте химиотерапевтический агент представляет собой цитотоксический антибиотик, такой как ингибитор топоизомеразы II. В одном аспекте ингибитор топоизомеразы II представляет собой доксорубицин. Биспецифические антитела по изобретению
В изобретении предложены новые биспецифические антигенсвязывающие молекулы, содержащие новое анти-FAP антитело (клон 212). Биспецифические антигенсвязывающие молекулы, содержащие новое анти-FAP антитело, обладают исключительными преимущественными свойствами, такими как продуктивность, стабильность, аффинность связывания, биологическая активность, эффективность нацеливания, сниженный уровень интернализации, превосходные фармакокинетические (ФК) свойства, сниженная токсичность, расширенный диапазон дозировок, которые можно вводить пациенту, и, следовательно, возможно повышенная эффективность.
Типовые биспецифические антигенсвязывающие молекулы В одном аспекте в изобретении предложены биспецифические антигенсвязывающие молекулы, которые характеризуются нацеленным агонистическим связыванием с CD40. В частности, биспецифическая антигенсвязывающая молекула является агонистом CD40, который нацелен против FAP. В другом конкретном аспекте биспецифические антигенсвязывающие молекулы содержат Fc-область, состоящую из первой и второй субъединиц, способных к стабильной ассоциации, которая содержит мутации, снижающие эффекторную функцию. Применение Fc-области, содержащей мутации, которые снижают или устраняют эффекторную функцию, предотвратит неспецифическое агонистическое действие за счет перекрестного связывания Fc-рецепторов и предотвратит АЗКЦ CD40+ клеток. Описанные в данном документе биспецифические антигенсвязывающие молекулы обладают преимуществом по сравнению с традиционными антителами, способными к специфическому связыванию с CD40, в том, что они избирательно индуцируют иммунный ответ в целевых клетках, которые, как правило, являются раковыми клетками или опухолевой стромой.
Таким образом, биспецифические антигенсвязывающие молекулы характеризуются нацеленным на FAP связыванием с CD40. В присутствии FAP-экспрессирующих клеток биспецифические антигенсвязывающие молекулы способны активировать антигенпрезентирующие клетки (АПК), активировать человеческие В-клетки (пример 5.1.2), человеческие клетки Дауди (пример 5.1.1) и человеческие моноцитарные дендритные клетки (мДК).
В одном аспекте предложена биспецифическая антигенсвязывающая молекула, содержащая
(а) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с CD40, и
(б) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с белком активации фибробластов (FAP), содержащий вариабельную область тяжелой цепи (VHFAP), содержащую
(i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 3, (ii) CDR-H2, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4, SEQ ID NO: 11 и SEQ ID NO: 12, и (iii) CDR-Н3, содержащую аминокислотную последовательность SEQ ID NO: 5, и вариабельную область легкой цепи (VLFAP), содержащую (iv) CDR-L1, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 6, SEQ ID NO: 13 и SEQ ID NO: 14, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 7, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 8.
В конкретном аспекте предложены биспецифические антигенсвязывающие молекулы, содержащие а) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с CD40, и (б) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с белком активации фибробластов (FAP), содержащий вариабельную область тяжелой цепи (VHFAP), содержащую (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 3, (ii) CDR-H2, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4, и (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 5, и вариабельную область легкой цепи (VLFAP), содержащую (iv) CDR-L1, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 6, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 7, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 8.
В одном аспекте предложена биспецифическая антигенсвязывающая молекула, причем антигенсвязывающий домен, способный к специфическому связыванию с FAP, содержит вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность, которая по меньшей мере на около 90% идентична аминокислотной последовательности SEQ ID NO: 9, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность, которая по меньшей мере на около 90% идентична аминокислотной последовательности SEQ ID NO: 10.
В другом аспекте предложена биспецифическая антигенсвязывающая молекула, причем антигенсвязывающий домен, способный к специфическому связыванию с FAP, содержит вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19 и SEQ ID NO: 20, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 21, SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 25 и SEQ ID NO: 26.
В одном аспекте биспецифическая антигенсвязывающая молекула содержит
(а) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 15, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 21,
(б) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 16, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 21,
(в) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 16, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 22, или
(г) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 19, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 25.
В конкретном аспекте биспецифическая антигенсвязывающая молекула содержит вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 15, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 21.
В другом аспекте предложена биспецифическая антигенсвязывающая молекула, причем антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержит вариабельную область тяжелой цепи (VHCD40), содержащую (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 27, (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 28, и (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 29, и вариабельную область легкой цепи (VLCD40), содержащую (iv) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 30, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 31, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 32.
В дополнительном аспекте предложена биспецифическая антигенсвязывающая молекула, причем антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержит
(i) вариабельную область тяжелой цепи (VHCD40), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 37, SEQ ID NO: 38, SEQ ID NO: 39 и SEQ ID NO: 40, и
(ii) вариабельную область легкой цепи (VLCD40), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 41, SEQ ID NO: 42, SEQ ID NO: 43 и SEQ ID NO: 44.
В одном аспекте предложена биспецифическая антигенсвязывающая молекула, причем антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержит
(а) VH, содержащую аминокислотную последовательность SEQ ID NO: 37, и VL, содержащую аминокислотную последовательность SEQ ID NO: 41, или
(б) VH, содержащую аминокислотную последовательность SEQ ID NO: 37, и VL, содержащую аминокислотную последовательность SEQ ID NO: 42, или
(в) VH, содержащую аминокислотную последовательность SEQ ID NO: 37, и VL, содержащую аминокислотную последовательность SEQ ID NO: 43, или
(г) VH, содержащую аминокислотную последовательность SEQ ID NO: 37, и VL, содержащую аминокислотную последовательность SEQ ID NO: 44, или
(д) VH, содержащую аминокислотную последовательность SEQ ID NO: 38, и VL, содержащую аминокислотную последовательность SEQ ID NO: 41, или
(е) VH, содержащую аминокислотную последовательность SEQ ID NO: 38, и VL, содержащую аминокислотную последовательность SEQ ID NO: 42, или
(ж) VH, содержащую аминокислотную последовательность SEQ ID NO: 38, и VL, содержащую аминокислотную последовательность SEQ ID NO: 43, или
(з) VH, содержащую аминокислотную последовательность SEQ ID NO: 38, и VL, содержащую аминокислотную последовательность SEQ ID NO: 44, или
(и) VH, содержащую аминокислотную последовательность SEQ ID NO: 39, и VL, содержащую аминокислотную последовательность SEQ ID NO: 41, или
(к) VH, содержащую аминокислотную последовательность SEQ ID NO: 39, и VL, содержащую аминокислотную последовательность SEQ ID NO: 42, или
(л) VH, содержащую аминокислотную последовательность SEQ ID NO: 39, и VL, содержащую аминокислотную последовательность SEQ ID NO: 43, или
(м) VH, содержащую аминокислотную последовательность SEQ ID NO: 39, и VL, содержащую аминокислотную последовательность SEQ ID NO: 44, или
(н) VH, содержащую аминокислотную последовательность SEQ ID NO: 40, и VL, содержащую аминокислотную последовательность SEQ ID NO: 41, или
(о) VH, содержащую аминокислотную последовательность SEQ ID NO: 40, и VL, содержащую аминокислотную последовательность SEQ ID NO: 42, или
(п) VH, содержащую аминокислотную последовательность SEQ ID NO: 40, и VL, содержащую аминокислотную последовательность SEQ ID NO: 43, или
(р) VH, содержащую аминокислотную последовательность SEQ ID NO: 40, и VL, содержащую аминокислотную последовательность SEQ ID NO: 44.
В конкретном аспекте биспецифическая антигенсвязывающая молекула содержит антигенсвязывающий домен, способный к специфическому связыванию с CD40, который содержит VH, содержащую аминокислотную последовательность SEQ ID NO: 37, и VL, содержащую аминокислотную последовательность SEQ ID NO: 41.
В дополнительном аспекте предложена биспецифическая антигенсвязывающая молекула, причем антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержит
(i) вариабельную область тяжелой цепи (VHCD40), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 45, SEQ ID NO: 46, SEQ ID NO: 47, SEQ ID NO: 48, SEQ ID NO: 49 и SEQ ID NO: 50, и
(ii) вариабельную область легкой цепи (VLCD40), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 51, SEQ ID NO: 52, SEQ ID NO: 53 и SEQ ID NO: 54.
В одном аспекте предложена биспецифическая антигенсвязывающая молекула, причем антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержит
(а) VH, содержащую аминокислотную последовательность SEQ ID NO: 45, и VL, содержащую аминокислотную последовательность SEQ ID NO: 51, или
(б) VH, содержащую аминокислотную последовательность SEQ ID NO: 46, и VL, содержащую аминокислотную последовательность SEQ ID NO: 51, или
(в) VH, содержащую аминокислотную последовательность SEQ ID NO: 47, и VL, содержащую аминокислотную последовательность SEQ ID NO: 51, или
(г) VH, содержащую аминокислотную последовательность SEQ ID NO: 48, и VL, содержащую аминокислотную последовательность SEQ ID NO: 51, или
(д) VH, содержащую аминокислотную последовательность SEQ ID NO: 45, и VL, содержащую аминокислотную последовательность SEQ ID NO: 52, или
(е) VH, содержащую аминокислотную последовательность SEQ ID NO: 46, и VL, содержащую аминокислотную последовательность SEQ ID NO: 52, или
(ж) VH, содержащую аминокислотную последовательность SEQ ID NO: 47, и VL, содержащую аминокислотную последовательность SEQ ID NO: 52, или
(з) VH, содержащую аминокислотную последовательность SEQ ID NO: 48, и VL, содержащую аминокислотную последовательность SEQ ID NO: 52, или
(и) VH, содержащую аминокислотную последовательность SEQ ID NO: 49, и VL, содержащую аминокислотную последовательность SEQ ID NO: 53, или
(к) VH, содержащую аминокислотную последовательность SEQ ID NO: 50, и VL, содержащую аминокислотную последовательность SEQ ID NO: 53, или
(л) VH, содержащую аминокислотную последовательность SEQ ID NO: 49, и VL, содержащую аминокислотную последовательность SEQ ID NO: 54, или
(м) VH, содержащую аминокислотную последовательность SEQ ID NO: 50, и VL, содержащую аминокислотную последовательность SEQ ID NO: 54.
В конкретном аспекте предложена биспецифическая антигенсвязывающая молекула, которая содержит (i) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержащий вариабельную область тяжелой цепи (VHCD40), содержащую аминокислотную последовательность SEQ ID NO: 37, и вариабельную область легкой цепи (VLCD40), содержащую аминокислотную последовательность SEQ ID NO: 41, и (ii) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с FAP, содержащий вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 15, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 21.
Биспецифические антигенсвязывающие молекулы, связывающиеся с CD40 и FAP
В другом аспекте предложена биспецифическая антигенсвязывающая молекула, содержащая
(i) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержащий вариабельную область тяжелой цепи (VHCD40), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 37, SEQ ID NO: 38, SEQ ID NO: 39 и SEQ ID NO: 40, и вариабельную область легкой цепи (VLCD40), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 41, SEQ ID NO: 42, SEQ ID NO: 43 и SEQ ID NO: 44, и
(ii) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с FAP, содержащий вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19 и SEQ ID NO: 20, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 21, SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 25 и SEQ ID NO: 26.
В дополнительном аспекте предложена биспецифическая антигенсвязывающая молекула, содержащая
(i) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержащий вариабельную область тяжелой цепи (VHCD40), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 45, SEQ ID NO: 46, SEQ ID NO: 47, SEQ ID NO: 48, SEQ ID NO: 49 и SEQ ID NO: 50, и вариабельную область легкой цепи (VLCD40), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 51, SEQ ID NO: 52, SEQ ID NO: 53 и SEQ ID NO: 54, и
(ii) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с FAP, содержащий вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19 и SEQ ID NO: 20, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 21, SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 25 и SEQ ID NO: 26.
В конкретном аспекте предложена биспецифическая антигенсвязывающая молекула, содержащая
(i) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержащий вариабельную область тяжелой цепи (VHCD40), содержащую аминокислотную последовательность SEQ ID NO: 37, и вариабельную область легкой цепи (VLCD40), содержащую аминокислотную последовательность SEQ ID NO: 41, и
(ii) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с FAP, содержащий вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19 и SEQ ID NO: 20, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 21, SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 25 и SEQ ID NO: 26.
В другом конкретном аспекте предложена биспецифическая антигенсвязывающая молекула, содержащая
(i) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержащий вариабельную область тяжелой цепи (VHCD40), содержащую аминокислотную последовательность SEQ ID NO: 45 или SEQ ID NO: 48, и вариабельную область легкой цепи (VLCD40), содержащую аминокислотную последовательность SEQ ID NO: 51, и
(ii) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с FAP, содержащий вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19 и SEQ ID N0:20, и вариабельную область легкой цепи (VLFAP) , содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 21, SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 25 и SEQ ID NO: 26.
В дополнительном аспекте предложена биспецифическая антигенсвязывающая молекула, причем
(i) антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержит вариабельную область тяжелой цепи (VHCD40), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 37, SEQ ID NO: 38, SEQ ID NO: 39 и SEQ ID NO: 40, и вариабельную область легкой цепи (VLCD40), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 41, SEQ ID NO: 42, SEQ ID NO: 43 и SEQ ID NO: 44, и
(ii) антигенсвязывающий домен, способный к специфическому связыванию с FAP, содержит вариабельную область тяжелой цепи VH, содержащую аминокислотную последовательность SEQ ID NO: 15, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 21.
Кроме того, предложена биспецифическая антигенсвязывающая молекула, причем
(i) антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержит вариабельную область тяжелой цепи (VHCD40), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 45, SEQ ID NO: 46, SEQ ID NO: 47, SEQ ID NO: 48, SEQ ID NO: 49 и SEQ ID N0:50, и вариабельную область легкой цепи (VLCD40), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 51, SEQ ID NO: 52, SEQ ID NO: 53 и SEQ ID NO: 54, и
(ii) антигенсвязывающий домен, способный к специфическому связыванию с FAP, содержит вариабельную область тяжелой цепи VH, содержащую аминокислотную последовательность SEQ ID NO: 15, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 21. В конкретном аспекте предложена биспецифическая антигенсвязывающая молекула, которая содержит (i) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержащий вариабельную область тяжелой цепи (VHCD40), содержащую аминокислотную последовательность SEQ ID NO: 37, и вариабельную область легкой цепи (VLCD40), содержащую аминокислотную последовательность SEQ ID NO: 41, и (ii) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с FAP, содержащий вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 15, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 21.
Биспецифические одновалентные антигенсвязывающие молекулы (формат 1+1)
Один аспект изобретения относится к биспецифической антигенсвязывающей молекуле, содержащей
(а) один антигенсвязывающий домен, способный к специфическому связыванию с CD40,
(б) один антигенсвязывающий домен, способный к специфическому связыванию с белком активации фибробластов (FAP), содержащий вариабельную область тяжелой цепи (VHFAP), содержащую
(i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 3, (ii) CDR-H2, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4, SEQ ID NO: 11 и SEQ ID NO: 12, и (iii) CDR-Н3, содержащую аминокислотную последовательность SEQ ID NO: 5, и вариабельную область легкой цепи (VLFAP) , содержащую (iv) CDR-L1, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 6, SEQ ID NO: 13 и SEQ ID NO: 14, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 7, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 8, и (в) Fc-домен, состоящий из первой и второй субъединиц, способных к стабильной ассоциации.
В одном конкретном аспекте предложена биспецифическая антигенсвязывающая молекула, причем указанная молекула содержит (а) первый
Fab-фрагмент, способный к специфическому связыванию с CD40, (б) второй Fab-фрагмент, способный к специфическому связыванию с FAP, содержащий вариабельную область тяжелой цепи (VHFAP), содержащую (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 3, (ii) CDR-H2, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4, SEQ ID NO: 11 и SEQ ID NO: 12, и (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 5, и вариабельную область легкой цепи (VLFAP), содержащую (iv) CDR-L1, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 6, SEQ ID NO: 13 и SEQ ID NO: 14, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 7, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 8, и (в) Fc-домен, состоящий из первой и второй субъединиц, способных к стабильной ассоциации.
В одном конкретном аспекте в изобретении предложена биспецифическая антигенсвязывающая молекула, содержащая (а) первую тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 63, вторую тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 98, первую легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 61, вторую легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 62.
Биспецифические антигенсвязывающие молекулы, двухвалентные в отношении связывания с CD40 и одновалентные в отношении связывания с антигеном целевой клетки (формат 2+1)
В другом аспекте в изобретении предложена биспецифическая антигенсвязывающая молекула, содержащая
(а) два антигенсвязывающих домена, способных к специфическому связыванию с CD40,
(б) один антигенсвязывающий домен, способный к специфическому связыванию с FAP, содержащий вариабельную область тяжелой цепи (VHFAP), содержащую (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 3, (ii) CDR-H2, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4, SEQ ID NO: 11 и SEQ ID NO: 12, и (iii) CDR-Н3, содержащую аминокислотную последовательность SEQ ID NO: 5, и вариабельную область легкой цепи (VLFAP) , содержащую (iv) CDR-L1, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 6, SEQ ID NO: 13 и SEQ ID NO: 14, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 7, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 8, и (в) Fc-домен, состоящий из первой и второй субъединиц, способных к стабильной ассоциации.
Таким образом, предложена биспецифическая антигенсвязывающая молекула, причем биспецифическая антигенсвязывающая молекула связывается с CD40 двухвалентным образом и с FAP одновалентным образом.
В одном аспекте биспецифическая антигенсвязывающая молекула содержит (а) две тяжелые цепи и две легкие цепи антитела, содержащего два Fab-фрагмента, способных к специфическому связыванию с CD40, и Fc-домен, и (б) домены VH и VL, способные к специфическому связыванию с антигеном целевой клетки, причем каждый из домена VH и домена VL соединены посредством пептидного линкера с одним из С-концов двух тяжелых цепей.
В дополнительном аспекте предложена биспецифическая антигенсвязывающая молекула, содержащая
(а) две тяжелые цепи, причем каждая тяжелая цепь содержит VH и СН1-домен Fab-фрагмента, способного к специфическому связыванию с CD40, и субъединицу Fc-области,
(б) две легкие цепи, причем каждая легкая цепь содержит VL и CL-домен Fab-фрагмента, способного к специфическому связыванию с CD40, и
(в) Kpocc-fab-фрагмент, способный к специфическому связыванию с FAP, содержащий цепь VL-CH1 и цепь VH-CL, причем цепь VH-CL соединена с С-концом одной из двух тяжелых цепей (а).
В одном аспекте цепь VH-CL (VH-C-каппа) соединена с С-концом тяжелой цепи Fc-выступа.
В другом аспекте предложена биспецифическая антигенсвязывающая молекула, содержащая
(а) две тяжелые цепи, причем каждая тяжелая цепь содержит VH и СН1-домен Fab-фрагмента, способного к специфическому связыванию с CD40, и субъединицу Fc-области,
(б) две легкие цепи, причем каждая легкая цепь содержит VL и CL-домен Fab-фрагмента, способного к специфическому связыванию с CD40, и
(в) Kpocc-fab-фрагмент, способный к специфическому связыванию с FAP, содержащий цепь VL-CH1 и цепь VH-CL, причем цепь VL-CH1 соединена с С-концом одной из двух тяжелых цепей (а).
В одном аспекте цепь VL-CH1 соединена с С-концом тяжелой цепи Fc-выступа.
В одном конкретном аспекте в изобретении предложена биспецифическая антигенсвязывающая молекула, содержащая
(а) две легкие цепи, каждая из которых содержит аминокислотную последовательность SEQ ID NO: 62, одну легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 61, первую тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 63, и вторую тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 64, или
(б) две легкие цепи, каждая из которых содержит аминокислотную последовательность SEQ ID NO: 66, одну легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 65, первую тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 67, и вторую тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 68.
В одном конкретном аспекте в изобретении предложена биспецифическая антигенсвязывающая молекула, содержащая
(а) две легкие цепи, каждая из которых содержит аминокислотную последовательность SEQ ID NO: 62, одну легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 61, первую тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 63, и вторую тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 64.
Биспецифические антигенсвязывающие молекулы в формате «голова к хвосту» (2+1)
В другом аспекте предложена биспецифическая антигенсвязывающая молекула, содержащая
(а) тяжелую цепь, содержащую VH и СН1-домен Fab-фрагмента, способного к специфическому связыванию с CD40, и субъединицу Fc-области,
(б) тяжелую цепь, содержащую VH и СН1-домен Fab-фрагмента, способного к специфическому связыванию с CD40, VL и CH1-домен Fab фрагмента, способного к специфическому связыванию с FAP, и субъединицу Fc-области,
(в) две легкие цепи, причем каждая легкая цепь содержит VL и CL-домен Fab-фрагмента, способного к специфическому связыванию с CD40, и
(г) легкую цепь, содержащую VH и CL-домен Fab-фрагмента, способного к специфическому связыванию с FAP.
Биспецифические антигенсвязывающие молекулы, двухвалентные в отношении связывания с CD40 и двухвалентные в отношении связывания с антигеном целевой клетки (формат 2+2)
В другом аспекте в изобретении предложена биспецифическая антигенсвязывающая молекула, содержащая
(а) два антигенсвязывающих домена, способных к специфическому связыванию с CD40,
(б) два антигенсвязывающих домена, способных к специфическому связыванию с FAP, содержащих (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 3, (ii) CDR-H2, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4, SEQ ID NO: 11 и SEQ ID NO: 12, и (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 5, и вариабельную область легкой цепи (VLFAP) , содержащую (iv) CDR-L1, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 6, SEQ ID NO: 13 и SEQ ID NO: 14, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 7, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 8, и
(в) Fc-домен, состоящий из первой и второй субъединиц, способных к стабильной ассоциации.
Таким образом, предложена биспецифическая антигенсвязывающая молекула, причем биспецифическая антигенсвязывающая молекула связывается с CD40 двухвалентным образом и с FAP двухвалентным образом.
В одном аспекте предложена биспецифическая антигенсвязывающая молекула, причем биспецифическая антигенсвязывающая молекула содержит
(а) две тяжелые цепи, причем каждая тяжелая цепь содержит VH и СН1-домен Fab-фрагмента, способного к специфическому связыванию с CD40, и субъединицу Fc-области,
(б) две легкие цепи, причем каждая легкая цепь содержит VL и CL-домен Fab-фрагмента, способного к специфическому связыванию с CD40, и
(в) два Fab-фрагмента, способных к специфическому связыванию с FAP, причем один из Fab-фрагментов соединен с С-концом одной из двух тяжелых цепей (а), а другой из Fab-фрагментов соединен с С-концом другой из двух тяжелых цепей (а).
Биспецифические антигенсвязывающие молекулы, четырехвалентные в отношении связывания с CD40 и одновалентные в отношении связывания с антигеном целевой клетки (формат 4+1)
В другом аспекте в изобретении предложена биспецифическая антигенсвязывающая молекула, содержащая
(а) четыре антигенсвязывающих домена, способных к специфическому связыванию с CD40,
(б) один антигенсвязывающий домен, способный к специфическому связыванию с FAP, содержащий (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 3, (ii) CDR-H2, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4, SEQ ID NO: 11 и SEQ ID NO: 12, и (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 5, и вариабельную область легкой цепи (VLFAP) , содержащую (iv) CDR-L1, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 6, SEQ ID NO: 13 и SEQ ID NO: 14, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 7, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 8, и
(в) Fc-домен, состоящий из первой и второй субъединиц, способных к стабильной ассоциации.
Таким образом, предложена биспецифическая антигенсвязывающая молекула, причем биспецифическая антигенсвязывающая молекула связывается с CD40 четырехвалентным образом и с FAP одновалентным образом.
В одном аспекте предложена биспецифическая антигенсвязывающая молекула, причем четыре антигенсвязывающих домена, способных к специфическому связыванию с CD40, представляют собой Fab-фрагменты, и каждые два из них слиты друг с другом в тяжелой цепи, необязательно, посредством пептидного линкера. В конкретном аспекте пептидный линкер содержит аминокислотную последовательность SEQ ID NO: 83. В частности, антигенсвязывающая молекула содержит две тяжелые цепи, каждая из которых содержит фрагмент VHCH1-пептидный линкер-VHCH1. В конкретном аспекте пептидный линкер имеет аминокислотную последовательность SEQ ID NO: 83.
В конкретном аспекте биспецифическая антигенсвязывающая молекула содержит
(а) четыре легкие цепи, причем каждая легкая цепь содержит VL и CL-домен Fab-фрагмента, способного к специфическому связыванию с CD40,
(б) две тяжелые цепи, причем каждая тяжелая цепь содержит VH и СН1-домен Fab-фрагмента, способного к специфическому связыванию с CD40, слитые с VH и CHI-доменом второго Fab-фрагмента, способного к специфическому связыванию с CD40, и субъединицу Fc-области, и
(в) кросс-fab-фрагмент, способный к специфическому связыванию с FAP, причем VH-домен соединен с С-концом одной из двух тяжелых цепей пептидным линкером.
В другом аспекте биспецифическая антигенсвязывающая молекула содержит
(а) четыре легкие цепи, причем каждая легкая цепь содержит VL и CL-домен Fab-фрагмента, способного к специфическому связыванию с CD40,
(б) две тяжелые цепи, причем каждая тяжелая цепь содержит VH и СН1-домен Fab-фрагмента, способного к специфическому связыванию с CD40, слитые с VH и CHI-доменом второго Fab-фрагмента, способного к специфическому связыванию с CD40, и субъединицу Fc-области, и
(в) Kpocc-fab-фрагмент, способный к специфическому связыванию с FAP, причем VL-домен соединен с С-концом одной из двух тяжелых цепей пептидным линкером.
В конкретном аспекте пептидный линкер содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 82, SEQ ID NO: 83, SEQ ID NO: 87 и SEQ ID NO: 88. В частности, пептидный линкер содержит SEQ ID NO: 83.
В конкретном аспекте в изобретении предложена биспецифическая антигенсвязывающая молекула, содержащая
(а) четыре легкие цепи, каждая из которых содержит аминокислотную последовательность SEQ ID NO: 62, одну легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 61, первую тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 69, и вторую тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 70.
Биспецифические антигенсвязывающие молекулы, четырехвалентные в отношении связывания с CD40 и двухвалентные в отношении связывания с антигеном целевой клетки (формат 4+2)
В другом аспекте в изобретении предложена биспецифическая антигенсвязывающая молекула, содержащая
(а) четыре антигенсвязывающих домена, способных к специфическому связыванию с CD40,
(б) два антигенсвязывающих домена, способных к специфическому связыванию с FAP, содержащих
(i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 3, (ii) CDR-H2, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4, SEQ ID NO: 11 и SEQ ID NO: 12, и (iii) CDR-НЗ, содержащую аминокислотную последовательность SEQ ID NO: 5, и вариабельную область легкой цепи (VLFAP) , содержащую (iv) CDR-L1, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 6, SEQ ID NO: 13 и SEQ ID NO: 14, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 7, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 8, и (в) Fc-домен, состоящий из первой и второй субъединиц, способных к стабильной ассоциации.
Таким образом, предложена биспецифическая антигенсвязывающая молекула, причем биспецифическая антигенсвязывающая молекула связывается с CD40 четырехвалентным образом и с FAP двухвалентным образом.
В одном аспекте предложена биспецифическая антигенсвязывающая молекула, причем четыре антигенсвязывающих домена, способных к специфическому связыванию с CD40, представляют собой Fab-фрагменты, и каждые два из них слиты друг с другом, необязательно, посредством пептидного линкера. В конкретном аспекте пептидный линкер содержит аминокислотную последовательность SEQ ID NO: 83. В частности, антигенсвязывающая молекула содержит две тяжелые цепи, каждая из которых содержит фрагмент VHCH1-пептидный линкер-VHCH1. В конкретном аспекте пептидный линкер имеет аминокислотную последовательность SEQ ID NO: 83.
В другом аспекте предложена биспецифическая антигенсвязывающая молекула, причем антигенсвязывающие домены, способные к специфическому связыванию с антигеном целевой клетки, представляют собой Fab-фрагменты, и при этом первый Fab-фрагмент соединен посредством пептидного линкера с С-концом первой субъединицы Fc-домена, а второй Fab-фрагмент соединен посредством пептидного линкера с С-концом второй субъединицы Fc-домена. В одном аспекте два Fab-фрагмента, способные к специфическому связыванию с антигеном целевой клетки, представляют собой кроссоверные Fab-фрагменты, каждый из которых содержит цепь VL-CH1 и цепь VH-CL, и при этом цепь VL-СН1 соединена с С-концом одной из двух тяжелых цепей.
Модификации Fc-домена, снижающие связывание Fc-рецептора и/или эффекторную функцию
Биспецифические антигенсвязывающие молекулы по изобретению дополнительно содержат Fc-домен, состоящий из первой и второй субъединиц, способных к стабильной ассоциации.
В определенных аспектах одну или более аминокислотных модификаций можно вносить в Fc-область антитела, предложенного в данном документе, с получением, таким образом, варианта по Fc-области. Вариант по Fc-области может содержать последовательность человеческой Fc-области (например, Fc-области человеческого IgG1, IgG2, IgG3 или IgG4), содержащую аминокислотную модификацию (например, замену) в одной или более аминокислотных позициях.
Fc-домен придает благоприятные фармакокинетические свойства биспецифическим антителам по изобретению, включая длительное сывороточное время полужизни, которое способствует надлежащему накоплению в целевой ткани и благоприятному соотношению распределения в ткани и крови. Однако в то же время он может приводить к нежелательному нацеливанию биспецифических антител по изобретению на клетки, экспрессирующие Fc-рецепторы, вместо предпочтительных несущих антиген клеток. Соответственно, в конкретных вариантах осуществления Fc-домен биспецифических антител по изобретению демонстрирует сниженную аффинность связывания с Fc-рецептором и/или сниженную эффекторную функцию по сравнению с нативным Fc-доменом IgG, в частности, Fc-доменом IgG1 или Fc-доменом IgG4. В частности, Fc-домен представляет собой Fc-домен IgG1.
В одном таком аспекте Fc-домен (или биспецифическая антигенсвязывающая молекула по изобретению, содержащая указанный Fc-домен) демонстрирует менее 50%, предпочтительно менее 20%, более предпочтительно менее 10% и наиболее предпочтительно менее 5% аффинности связывания с Fc-рецептором по сравнению с нативным Fc-доменом IgG1 (или биспецифической антигенсвязывающей молекулой по изобретению, содержащей нативный Fc-домен IgG1), и/или менее 50%, предпочтительно менее 20%, более предпочтительно менее 10% и наиболее предпочтительно менее 5% эффекторной функции по сравнению с нативным Fc-доменом IgG1 (или биспецифической антигенсвязывающей молекулой по изобретению, содержащей нативный Fc-домен IgG1). В одном аспекте Fc-домен (или биспецифическая антигенсвязывающая молекула по изобретению, содержащая указанный Fc-домен) практически не связывается с Fc-рецептором и/или не индуцирует эффекторную функцию. В конкретном аспекте Fc-рецептор представляет собой Fcγ-рецептор. В одном аспекте Fc-рецептор представляет собой человеческий Fc-рецептор. В одном аспекте Fc-рецептор представляет собой активирующий Fc-рецептор. В конкретном аспекте Fc-рецептор представляет собой активирующий человеческий Fcγ-рецептор, в частности, человеческий FcγRIIIa, FcγRI или FcγRIIa, конкретно - человеческий FcγRIIIa. В одном аспекте Fc-рецептор представляет собой ингибирующий Fc-рецептор. В конкретном аспекте Fc-рецептор представляет собой ингибирующий человеческий Fcγ-рецептор, в частности, человеческий FcγRIIB. В одном аспекте эффекторная функция представляет собой одно или более из КЗЦ, АЗКЦ, АЗКФ и секрецию цитокинов. В конкретном аспекте эффекторная функция представляет собой АЗКЦ. В одном аспекте Fc-домен демонстрирует практически аналогичную аффинность связывания с неонатальным Fc-рецептором (FcRn) с нативным Fc-доменом IgG1. Практически аналогичное связывание с FcRn достигается, когда Fc-домен (или биспецифическая антигенсвязывающая молекула по изобретению, содержащая указанный Fc-домен) демонстрирует более чем около 70%, конкретнее, более чем около 80%, в частности, более чем около 90% аффинности связывания нативного Fc-домена IgG1 (или биспецифической антигенсвязывающей молекулы по изобретению, содержащей нативный Fc-домен IgG1) с FcRn.
В конкретном аспекте Fc-домен сконструирован так, чтобы иметь сниженную аффинность связывания с Fc-рецептором и/или сниженную эффекторную функцию по сравнению с не сконструированным Fc-доменом. В конкретном аспекте Fc-домен биспецифической антигенсвязывающей молекулы по изобретению содержит одну или более аминокислотных мутаций, которые снижают аффинность связывания Fc-домена с Fc-рецептором и/или эффекторную функцию. Как правило, в каждой из двух субъединиц Fc-домена присутствуют одни и те же одна или более аминокислотных мутаций. В одном аспекте аминокислотная мутация снижает аффинность связывания Fc-домена с Fc-рецептором. В другом аспекте аминокислотная мутация снижает аффинность связывания Fc-домена с Fc-рецептором по меньшей мере в 2 раза, по меньшей мере в 5 раз или по меньшей мере в 10 раз. В одном аспекте биспецифическая антигенсвязывающая молекула по изобретению, содержащая сконструированный Fc-домен, демонстрирует менее 20%, конкретнее, менее 10%, в частности, менее 5% аффинности связывания с Fc-рецептором по сравнению с биспецифическими антителами по изобретению, содержащими не сконструированный Fc-домен. В конкретном аспекте Fc-рецептор представляет собой Fcγ-рецептор. В других аспектах Fc-рецептор представляет собой человеческий Fc-рецептор. В одном аспекте Fc-рецептор представляет собой ингибирующий Fc-рецептор. В конкретном аспекте Fc-рецептор представляет собой ингибирующий человеческий Fcγ-рецептор, в частности, человеческий FcγRIIB. В некоторых аспектах Fc-рецептор представляет собой активирующий Fc-рецептор. В конкретном аспекте Fc-рецептор представляет собой активирующий человеческий Fcγ-рецептор, в частности, человеческий FcγRIIIa, FcγRI или FcγRIIa, конкретно - человеческий FcγRIIIa. Предпочтительно связывание с каждым из этих рецепторов снижено. В некоторых аспектах также снижена аффинность связывания с компонентом комплемента, в частности аффинность связывания с Clq. В одном аспекте аффинность связывания с неонатальным Fc-рецептором (FcRn) не снижена. Практически аналогичное связывание с FcRn, т.е. сохранение аффинности связывания Fc-домена с указанным рецептором, достигается, когда Fc-домен (или биспецифическая антигенсвязывающая молекула по изобретению, содержащая указанный Fc домен) демонстрирует более чем около 70% аффинности связывания не сконструированной формы Fc-домена (или биспецифической антигенсвязывающей молекулы по изобретению, содержащей указанную не сконструированную форму Fc-домена) с FcRn. Fc-домен или биспецифическая антигенсвязывающая молекула по изобретению, содержащая указанный Fc-домен, может демонстрировать более чем около 80% и даже более чем около 90% такой аффинности. В определенных вариантах осуществления Fc-домен биспецифической антигенсвязывающей молекулы по изобретению сконструирован так, чтобы иметь сниженную эффекторную функцию по сравнению с не сконструированным Fc-доменом. Сниженная эффекторная функция может включать, но не ограничивается этим, одно или более из следующего: сниженную комплементзависимую цитотоксичность (КЗЦ), сниженную антителозависимую клеточноопосредованную цитотоксичность (АЗКЦ), сниженный антителозависимый клеточный фагоцитоз (АЗКФ), сниженную секрецию цитокинов, сниженное опосредованное иммунным комплексом поглощение антигена антигенпрезентирующими клетками, сниженное связывание с NK-клетками, сниженное связывание с макрофагами, сниженное связывание с моноцитами, сниженное связывание с полиморфно-ядерными клетками, сниженную прямую сигнализацию, индуцирующую апоптоз, сниженное созревание дендритных клеток или сниженное примирование Т-клеток.
Антитела со сниженной эффекторной функцией включают антитела с заменой одного или более остатков Fc-области 238, 265, 269, 270, 297, 327 и 329 (патент США №6737056). Такие Fc-мутанты включают Fc-мутантов с заменами в двух или более аминокислотных позициях 265, 269, 270, 297 и 327, включая так называемый Fc-мутант «DANA» с заменой остатков 265 и 297 аланином (патент США №7332581). Описаны некоторые варианты антител с улучшенным или уменьшенным связыванием с FcR. (например, патент США №6737056; WO 2004/056312 и Shields, R.L. et al., J. Biol. Chem. 276 (2001) 6591-6604).
В одном аспекте изобретения Fc-домен содержит аминокислотную замену в позиции Е233, L234, L235, N297, Р331 и Р329. В некоторых аспектах Fc-домен содержит аминокислотные замены L234A и L235A («LALA»). В одном таком варианте осуществления Fc-домен представляет собой Fc-домен IgG1, в частности, Fc-домен человеческого IgG1. В одном аспекте Fc-домен содержит аминокислотную замену в позиции Р329. В более конкретном аспекте аминокислотная замена представляет собой Р329А или P329G, в частности, P329G. В одном варианте осуществления Fc-домен содержит аминокислотную замену в позиции Р329 и дополнительную аминокислотную замену, выбранную из группы, состоящей из Е233Р, L234A, L23 5A, L235E, N297A, N297D или P331S. В более конкретных вариантах осуществления Fc-домен содержит аминокислотные мутации L234A, L235A и P329G («P329G LALA»). Комбинация аминокислотных замен «P329G LALA» практически полностью устраняет связывание Fcγ-рецептора Fc-доменом человеческого IgG1, как описано в патентной заявке РСТ №WO 2012/130831 А1. В указанном документе также описаны способы получения таких мутантных Fc-доменов и способы определения их свойств, таких как связывание Fc-рецептора или эффекторные функции. Такое антитело представляет собой IgG1 с мутациями L234A и L235A или с мутациями L234A, L235A и P329G (нумерация в соответствии с индексом EU по Kabat et al, Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD, 1991).
В одном аспекте Fc-домен представляет собой Fc-домен IgG4. В более конкретном варианте осуществления Fc-домен представляет собой Fc-домен IgG4, содержащий аминокислотную замену в позиции S228 (нумерация Kabat), в частности, аминокислотную замену S228P. В более конкретном варианте осуществления Fc-домен представляет собой Fc-домен IgG4, содержащий аминокислотные замены L235E, S228P и P329G. Эта аминокислотная замена снижает in vivo обмен Fab-фрагментами антител IgG4 (смотрите Stubenrauch et al., Drug Metabolism and Disposition 38, 84-91 (2010)).
Антитела с повышенным временем полужизни и улучшенным связыванием с неонатальным Fc-рецептором (FcRn), который отвечает за перенос материнских IgG в плод (Guyer, R.L. et al., J. Immunol. 117 (1976) 587-593, и Kim, J.K. et al., J. Immunol. 24 (1994) 2429-2434), описаны в US 2005/0014934. Эти антитела содержат Fc-домен с одной или более заменами, которые улучшают связывание Fc-области с FcRn. Такие Fc-варианты включают варианты с заменами в одном или более остатках Fc-области: 238, 256, 265, 272, 286, 303, 305, 307, 311, 312, 317, 340, 356, 360, 362, 376, 378, 380, 382, 413, 424 или 434, например, замену аминокислотного остатка 434 в Fc-области (патент США №7371826). Также смотрите Duncan, A.R. and Winter, G., Nature 322 (1988) 738-740; US 5648260; US 5624821; и WO 94/29351 в отношении других примеров вариантов по Fc-области.
Связывание с Fc-рецепторами можно легко определить, например, методом ELISA или методом поверхностного плазмонного резонанса (ППР), используя стандартное оборудование, такое как инструмент BIAcore (GE Healthcare), и Fc-рецепторы, которые могут быть получены методом рекомбинантной экспрессии. Подобный подходящий анализ связывания описан в данном документе. В альтернативном варианте аффинность связывания Fc-доменов или активирующих клетки биспецифических антигенсвязывающих молекул, содержащих Fc-домен, с Fc-рецепторами можно оценить, используя линии клеток, которые экспрессируют конкретные Fc-рецепторы, такие как человеческие NK-клетки, экспрессирующие FcγIIIa-рецептор. Эффекторную функцию Fc-домена или биспецифических антигенсвязывающих молекул по изобретению, содержащих Fc-домен, можно измерить методами, известными в данной области техники. Подходящий анализ для измерения АЗКЦ описан в данном документе. Другие примеры in vitro анализов для оценки АЗКЦ-активности представляющей интерес молекулы описаны в патенте США №5500362; Hellstrom et al. Proc Natl Acad Sci USA 83, 7059-7063 (1986) и Hellstrom et al., Proc Natl Acad Sci USA 82, 1499-1502 (1985); патенте США №5821337; Bruggemann et al., J Exp Med 166, 1351-1361 (1987). В альтернативном варианте можно использовать нерадиоактивные методы анализа (смотрите, например, нерадиоактивный анализ цитотоксичности для проточной цитометрии ACTI™ (CellTechnology, Inc. Mountain View, CA); и нерадиоактивный анализ цитотоксичности CytoTox 96® (Promega, Madison, WI)). Эффекторные клетки, подходящие для такого анализа, включают мононуклеарные клетки периферической крови (МКПК) и естественные клетки-киллеры (NK). В альтернативном или дополнительном варианте АЗКЦ-активность представляющей интерес молекулы можно оценивать in vivo, например, в животной модели, такой как описана в Clynes et al., Proc Natl Acad Sci USA 95, 652-656 (1998).
В следующем разделе описаны предпочтительные аспекты биспецифических антигенсвязывающих молекул по изобретению, содержащих модификации Fc-домена, снижающие связывание Fc-рецептора и/или эффекторную функцию. Один аспект изобретения относится к биспецифической антигенсвязывающей молекуле, содержащей (а) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с CD40, (б) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с FAP, и (в) Fc-домен, состоящий из первой и второй субъединиц, способных к стабильной ассоциации, причем Fc-домен содержит одну или более аминокислотных замен, которые снижают аффинность связывания антитела с Fc-рецептором, в частности, с Fcγ-рецептором. Другой аспект изобретения относится к биспецифической антигенсвязывающей молекуле, содержащей (а) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с CD40, (б) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с антигеном целевой клетки, и (в) Fc-домен, состоящий из первой и второй субъединиц, способных к стабильной ассоциации, причем Fc-домен содержит одну или более аминокислотных замен, которые снижают эффекторную функцию. В конкретном аспекте Fc-домен принадлежит подклассу IgG1 человека с аминокислотными мутациями L234A, L235A и P329G (нумерация в соответствии с индексом EU по Kabat).
Модификации Fc-домена, способствующие гетеродимеризации
Биспецифические антигенсвязывающие молекулы по изобретению содержат разные антигенсвязывающие сайты, слитые с одной или другой из двух субъединиц Fc-домена, таким образом, две субъединицы Fc-домена могут содержаться в двух неидентичных полипептидных цепях. Рекомбинантная коэкспрессия этих полипептидов и последующая димеризация приводит к нескольким возможным комбинациям двух полипептидов. Таким образом, чтобы повысить выход и чистоту биспецифических антигенсвязывающих молекул по изобретению при рекомбинантном получении, целесообразно внести в Fc-домен биспецифических антигенсвязывающих молекул по изобретению модификацию, способствующую ассоциации необходимых полипептидов.
Соответственно, конкретные аспекты изобретения относятся к биспецифической антигенсвязывающей молекуле, содержащей (а) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с CD40, (б) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с FAP, и (в) Fc-домен, состоящий из первой и второй субъединиц, способных к стабильной ассоциации, причем Fc домен содержит модификацию, способствующую ассоциации первой и второй субъединиц Fc-домена. Сайтом наиболее активного белок-белкового взаимодействия между двумя субъединицами Fc-домена IgG человека является СН3-домен Fc-домена. Таким образом, в одном аспекте указанная модификация находится в СН3-домене Fc-домена.
В конкретном аспекте указанная модификация представляет собой так называемую модификацию типа «выступ во впадину», содержащую модификацию «выступа» в одной из двух субъединиц Fc-домена и модификацию «впадины» в другой из двух субъединиц Fc-домена. Таким образом, изобретение относится к биспецифической антигенсвязывающей молекуле, содержащей (а) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с CD40, (б) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с антигеном целевой клетки, и (в) Fc-домен, состоящий из первой и второй субъединиц, способных к стабильной ассоциации, причем первая субъединица Fc-домена содержит выступы, а вторая субъединица Fc-домена содержит впадины в соответствии с методом «выступ во впадину». В конкретном аспекте первая субъединица Fc-домена содержит аминокислотные замены S354C и T366W (нумерация EU), а вторая субъединица Fc-домена содержит аминокислотные замены Y349C, T366S и Y407V (нумерация в соответствии с индексом EU по Kabat).
Технология «выступ-во-впадину» описана, например, в US 5731168; US 7695936; Ridgway et al., Prot Eng 9, 617-621 (1996) и Carter, J Immunol Meth 248, 7-15 (2001). В общем случае этот способ включает внесение выступа («узла») в контактную поверхность первого полипептида и соответствующей впадины («полости») в контактную поверхность второго полипептида, так что выступ может располагаться во впадине так, чтобы стимулировать образование гетеродимера и затруднять образование гомодимера. Выступы конструируют путем замены небольших аминокислотных боковых цепей в контактной поверхности первого полипептида на более крупные боковые цепи (например тирозин или триптофан). Компенсаторные «полости» идентичного или сходного размера с выступами создают в контактной поверхности второго полипептида путем замещения крупных аминокислотных боковых цепей меньшими (например, аланином или треонином).
Соответственно, в одном аспекте в СН3-домене первой субъединицы Fc-домена биспецифических антигенсвязывающих молекул по изобретению аминокислотный остаток замещен аминокислотным остатком, имеющим больший объем боковой цепи, тем самым создавая выступ в СН3-домене первой субъединицы, который совместим с полостью в СН3-домене второй субъединицы, а в СН3-домене второй субъединицы Fc-домена аминокислотный остаток замещен аминокислотным остатком, имеющим меньший объем боковой цепи, тем самым создавая впадину в СН3-домене второй субъединицы, в которой может размещаться выступ в СН3-домене первой субъединицы. Выступ и полость можно создавать путем изменения нуклеиновой кислоты, кодирующей полипептиды, например, с помощью сайт-специфического мутагенеза, или с помощью пептидного синтеза. В конкретном аспекте в СН3-домене первой субъединицы Fc-домена остаток треонина в позиции 366 замещен остатком триптофана (T366W), а в СН3-домене второй субъединицы Fc-домена остаток тирозина в позиции 407 замещен остатком валина (Y407V). В одном аспекте во второй субъединицы Fc-домена дополнительно остаток треонина в позиции 366 замещен остатком серина (T366S), а остаток лейцина в позиции 368 замещен остатком аланина (L368A).
В дополнительном аспекте в первой субъединице Fc-домена дополнительно остаток серина в позиции 354 замещен остатком цистеина (S354C), а во второй субъединице Fc-домена дополнительно остаток тирозина в позиции 349 замещен остатком цистеина (Y349C). Внесение этих двух остатков цистеина приводит к образованию дисульфидного мостика между двумя субъединицами Fc-домена, дополнительно стабилизируя димер (Carter (2001), J Immunol Methods 248, 7-15). В конкретном аспекте первая субъединица Fc-домена содержит аминокислотные замены S354C и T366W (нумерация EU), а вторая субъединица Fc-домена содержит аминокислотные замены Y349C, T366S и Y407V (нумерация в соответствии с индексом EU по Kabat).
В альтернативном аспекте модификация, способствующая ассоциации первой и второй субъединиц Fc-домена включает модификацию, которая опосредует эффекты электростатического взаимодействия, например, как описано в публикации РСТ WO 2009/089004. В общем случае этот способ включает замену одного или более аминокислотных остатков на поверхности контакта двух субъединиц Fc-домена заряженными аминокислотными остатками так, чтобы образование гомодимера становилось электростатически невыгодным, но гетеродимеризация была бы электростатический выгодной.
С-конец тяжелой цепи представленного в данном документе антитела может представлять собой полный С-конец, оканчивающийся аминокислотными остатками PGK. С-конец тяжелой цепи может представлять собой укороченный С-конец, в котором были удалены один или два С-концевых аминокислотных остатка. В одном предпочтительном аспекте С-конец тяжелой цепи представляет собой укороченный С-конец, оканчивающийся PG. В одном аспекте из всех представленных в данном документе аспектов биспецифическое антитело, содержащее тяжелую цепь, содержащую С-концевой СН3-домен, описанный в данном документе, содержит С-концевой глицин-лизиновый дипептид (G446 и К447, нумерация в соответствии с индексом EU по Kabat). В одном варианте осуществления всех представленных в данном документе аспектов биспецифическое антитело, содержащее тяжелую цепь, содержащую С-концевой СН3-домен, описанный в данном документе, содержит С-концевой остаток глицина (G446, нумерация в соответствии с индексом EU по Kabat).
Модификации в Fab-доменах
В одном аспекте изобретение относится к биспецифической антигенсвязывающей молекуле, содержащей (а) первый Fab-фрагмент, способный к специфическому связыванию с CD40, (б) второй Fab-фрагмент, способный к специфическому связыванию с FAP, и (в) Fc-домен, состоящий из первой и второй субъединиц, способных к стабильной ассоциации, причем в одном из Fab-фрагментов проведен обмен любых из доменов VH и VL или константных доменов СН1 и CL. Биспецифические антитела получают в соответствии с технологией Crossmab.
Мультиспецифические антитела с замещением/обменом доменов в одном связывающем плече (CrossMabVH-VL или CrossMabCH-CL) подробно описаны в WO 2009/080252 и Schaefer, W. et al, PNAS, 108 (2011) 11187-1191. Они несомненно снижают количество побочных продуктов, возникающих в результате несовпадения легкой цепи против первого антигена с неправильной тяжелой цепью против второго антигена (по сравнению с подходами без такого обмена доменов).
В одном аспекте изобретение относится к биспецифической антигенсвязывающей молекуле, содержащей (а) первый Fab-фрагмент, способный к специфическому связыванию с CD40, (б) второй Fab-фрагмент, способный к специфическому связыванию с FAP, и (в) Fc-домен, состоящий из первой и второй субъединиц, способных к стабильной ассоциации, причем в одном из Fab-фрагментов константные домены CL и СН1 замещены друг другом так, что домен СН1 является частью легкой цепи, а домен CL является частью тяжелой цепи. В частности, во втором Fab-фрагменте, способном к специфическому связыванию с антигеном целевой клетки, константные домены CL и СН1 замещены друг другом так, что домен СН1 является частью легкой цепи, а домен CL является частью тяжелой цепи.
В конкретном аспекте изобретение относится к биспецифической антигенсвязывающей молекуле, содержащей (а) первый Fab-фрагмент, способный к специфическому связыванию с CD40, (б) второй Fab-фрагмент, способный к специфическому связыванию с FAP, причем константные домены CL и СН1 замещены друг другом так, что домен СН1 является частью легкой цепи, а домен CL является частью тяжелой цепи. Такая молекула связывается как с CD40, так и с FAP одновалентным образом.
В другом аспекте изобретение относится к биспецифической антигенсвязывающей молекуле, содержащей (а) две легкие цепи и две тяжелые цепи антитела, содержащего два Fab-фрагмента, способных к специфическому связыванию с CD40, и Fc-домен, и (б) два дополнительных Fab-фрагмента, способных к специфическому связыванию с FAP, причем каждый из указанных дополнительных Fab-фрагментов соединен посредством пептидного линкера с С-концом тяжелых цепей (а). В конкретном аспекте дополнительные Fab-фрагменты представляют собой Fab-фрагменты, в которых вариабельные домены VL и VH замещены друг другом так, что домен VH является частью легкой цепи, а домен VL является частью тяжелой цепи.
Таким образом, в конкретном аспекте изобретение относится к биспецифической антигенсвязывающей молекуле, содержащей (а) две легкие цепи и две тяжелые цепи антитела, содержащего два Fab-фрагмента, способных к специфическому связыванию с CD40, и Fc-домен, и (б) два дополнительных Fab-фрагмента, способных к специфическому связыванию с FAP, причем указанные дополнительные Fab-фрагменты, способные к специфическому связыванию с антигеном целевой клетки, представляют собой кроссоверные Fab-фрагменты, в которых вариабельные домены VL и VH замещены друг другом, а каждая из цепей VL-CH соединена посредством пептидного линкера с С-концом тяжелых цепей (а).
В другом аспекте и для дополнительной коррекции спаривания биспецифическая антигенсвязывающая молекула, содержащая (а) первый Fab-фрагмент, способный к специфическому связыванию с CD40, (б) второй Fab-фрагмент, способный к специфическому связыванию с FAP, и (в) Fc-домен, состоящий из первой и второй субъединиц, способных к стабильной ассоциации, может содержать разные замены заряженных аминокислот (так называемые «заряженные остатки»). Эти модификации вносят в скрещенные или не скрещенный домены СН1 и CL. В конкретном аспекте изобретение относится к биспецифической антигенсвязывающей молекуле, в которой в одном из доменов CL аминокислота в позиции 123 (нумерация EU) была замещена аргинином (R) и/или аминокислота в позиции 124 (нумерация EU) была замещена лизином (K), и в которой в одном из доменов СН1 аминокислоты в позиции 147 (нумерация EU) и/или в позиции 213 (нумерация EU) были замещены глутаминовой кислотой (Е).
Типовые антитела по изобретению
В одном аспекте в изобретении предложены новые антитела и фрагменты антител, которые специфически связываются с FAP. Эти антитела связываются с другим эпитопом, чем известные антитела к FAP 4 В) или 28Н1, что делает их в особенности подходящими для включения в биспецифические антигенсвязывающие молекулы, которые можно использовать в комбинации с другими нацеленными на FAP молекулами. Новые антитела дополнительно характеризуются тем, что их можно производить в больших количествах с высокими титрами, тем, что они демонстрируют высокую температурную стабильность (по данным измерения температуры агрегации Tar), что они предполагаемо обладают превосходными ФК-свойствами и что они связываются с высокой аффинностью с человеческим FAP по данным измерения в анализе Biacore.
В одном аспекте предложено антитело, которое специфически связывается с FAP (клон 212), причем указанное антитело содержит вариабельную область тяжелой цепи (VHFAP), содержащую (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 3, (ii) CDR-H2, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4, SEQ ID NO: 11 и SEQ ID NO: 12, и (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 5, и вариабельную область легкой цепи (VLFAP) , содержащую (iv) CDR-L1, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 6, SEQ ID NO: 13 и SEQ ID NO: 14, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 7, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 8.
В одном аспекте предложено гуманизированное антитело, которое специфически связывается с FAP, причем указанное антитело содержит вариабельную область тяжелой цепи (VHFAP), содержащую (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 3, (ii) CDR-H2, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4, SEQ ID NO: 11 и SEQ ID NO: 12, и (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 5, и вариабельную область легкой цепи (VLFAP), содержащую (iv) CDR-L1, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 6, SEQ ID NO: 13 и SEQ ID NO: 14, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 7, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 8.
В другом аспекте предложено антитело, которое конкурируют за связывание с антителом, которое специфически связывается с FAP, причем указанное антитело содержит любую из вариабельных областей тяжелой цепи (VHFAP), содержащих аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19 и SEQ ID NO: 20, и любую из вариабельных областей легкой цепи (VLFAP) , содержащих аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 21, SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 25 и SEQ ID NO: 26.
В одном аспекте предложено антитело, которое конкурируют за связывание с антителом, которое специфически связывается с FAP, причем указанное антитело содержит вариабельную область тяжелой цепи VH, содержащую аминокислотную последовательность SEQ ID NO: 15, и вариабельную область легкой цепи VL, содержащую аминокислотную последовательность SEQ ID NO: 21.
В дополнительном аспекте предложено антитело, которое специфически связывается с FAP, причем указанное антитело содержит
(а) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 15, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 21,
(б) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 16, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 21,
(в) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 16, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 22, или
(г) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 19, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 25.
В дополнительном аспекте предложено антитело, которое специфически связывается с FAP, содержащее вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 15, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 21.
Полинуклеотиды
В изобретении дополнительно предложена выделенная нуклеиновая кислота, кодирующая описанную в данном документе биспецифическую антигенсвязывающую молекулу или ее фрагмент, или выделенная нуклеиновая кислота, кодирующая описанное в данном документе антитело.
Выделенные полинуклеотиды, кодирующие биспецифические антигенсвязывающие молекулы по изобретению можно экспрессировать в виде одного полинуклеотида, который кодирует всю антигенсвязывающую молекулу, или в виде нескольких (например, двух или более) коэкспрессируемых полинуклеотидов. Полипептиды, кодируемые коэкспрессируемыми полинуклеотидами, могут ассоциироваться, например, посредством дисульфидных связей или иным образом, с образованием функциональной антигенсвязывающей молекулы. Например, часть легкой цепи иммуноглобулина может кодироваться другим полинуклеотидом, чем часть тяжелой цепи иммуноглобулина. При коэкспрессии полипептиды тяжелой цепи будут ассоциироваться с полипептидами легкой цепи с образованием иммуноглобулина.
В некоторых аспектах выделенный полинуклеотид кодирует полипептид, содержащийся в биспецифической молекуле в соответствии с изобретением, описанной в данном документе.
В одном аспекте настоящее изобретение относится к выделенному полинуклеотиду, кодирующему биспецифическую антигенсвязывающую молекулу, содержащую (а) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с CD40, (б) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с FAP, содержащий вариабельную область тяжелой цепи (VHFAP), содержащую (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 3, (ii) CDR-H2, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4, SEQ ID NO: 11 и SEQ ID NO: 12, и (iii) CDR-НЗ, содержащую аминокислотную последовательность SEQ ID NO: 5, и вариабельную область легкой цепи (VLFAP) , содержащую (iv) CDR-L1, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 6, SEQ ID NO: 13 и SEQ ID NO: 14, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 7, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 8, и (в) Fc-домен, состоящий из первой и второй субъединиц, способных к стабильной ассоциации.
В определенных вариантах осуществления полинуклеотид или нуклеиновая кислота представляет собой ДНК. В других вариантах осуществления полинуклеотид по настоящему изобретению представляет собой РНК, например, в форме матричной РНК (мРНК). РНК по настоящему изобретению может быть одноцепочечной или двухцепочечной.
Рекомбинантные методы
Биспецифические антигенсвязывающие молекулы по изобретению можно получать, например, путем рекомбинантной выработки. Для рекомбинантной выработки получают один или более полинуклеотидов, кодирующих биспецифическую антигенсвязывающую молекулу или ее полипептидные фрагменты. Один или более полинуклеотидов, кодирующих биспецифическую антигенсвязывающую молекулу, выделяют и вставляют в один или более векторов для дальнейшего клонирования и/или экспрессии в клетке-хозяине. Такой полинуклеотид можно легко выделить и секвенировать, используя традиционные процедуры. В одном аспекте изобретения получают вектор, предпочтительно экспрессионный вектор, содержащий один или более полинуклеотидов по изобретению. Для конструирования экспрессионных векторов, содержащих кодирующую последовательность биспецифической антигенсвязывающей молекулы (фрагмента) наряду с соответствующими сигналами управления транскрипцией/трансляцией, можно использовать методы, хорошо известные специалистам в данной области техники. Эти методы включают in vitro технологии рекомбинантных ДНК, технологии синтеза и in vivo рекомбинации/генетической рекомбинации. Смотрите, например, технологии, описанные в Maniatis et al., MOLECULAR CLONING: A LABORATORY MANUAL, Cold Spring Harbor Laboratory, N.Y. (1989); and Ausubel et al., CURRENT PROTOCOLS IN MOLECULAR BIOLOGY, Greene Publishing Associates and Wiley Interscience, N.Y. (1989). Экспрессионный вектор может быть частью плазмиды, вируса или может представлять собой фрагмент нуклеиновой кислоты. Экспрессионный вектор содержит экспрессионную кассету, в которую клонирован полинуклеотид, кодирующий биспецифическую антигенсвязывающую молекулу или ее полипептидные фрагменты (т.е. кодирующая область), в функциональной ассоциации с промотором и/или другими элементами управления транскрипцией/трансляцией. В контексте данного документа «кодирующая область» представляет собой часть нуклеиновой кислоты, которая состоит из кодонов, транслируемых в аминокислоты. Хотя «стоп-кодон» (TAG, TGA или ТАА) не транслируется в аминокислоту, он может считаться частью кодирующей области, в случае наличия, но любые фланкирующие последовательности, например, промоторы, сайты связывания рибосомы, терминаторы транскрипции, интроны, 5' и 3' нетранслируемые области и т.п., не являются частью кодирующей области. Две или более кодирующих областей могут присутствовать в одной полинуклеотидной конструкции, например, в одном векторе, или в разных полинуклеотидных конструкциях, например в отдельных (разных) векторах. Кроме того, любой вектор может содержать одну кодирующую область или может содержать две или более кодирующих областей, например, вектор по настоящему изобретению может кодировать один или более полипептидов, которые пост- или ко-трансляционно разделяются на конечные белки посредством протеолитического расщепления. Кроме того, вектор, полинуклеотид или нуклеиновая кислота по изобретению могут кодировать гетерологичные кодирующие области, слитые или не слитые с полинуклеотидом, кодирующим биспецифическую антигенсвязывающую молекулу по изобретению или ее полипептидные фрагменты, или ее варианты или производные. Гетерологичные кодирующие области включают, без ограничения, специальные элементы или мотивы, такие как секреторный сигнальный пептид или гетерологичный функциональный домен. Функциональная ассоциация имеет место, когда кодирующая область для генного продукта, например, полипептида, связана с одной или более регуляторными последовательностями таким образом, чтобы экспрессия генного продукта была под управлением или контролем регуляторной(ых) последовательности(ей). Два фрагмента ДНК (такие как кодирующая полипептид область и связанный с ней промотор) «функционально связаны», если индукция функции промотора приводит к транскрипции мРНК, кодирующей необходимый генный продукт, и если природа связи между двумя фрагментами ДНК не препятствует способности экспрессионных регуляторных последовательностей управлять экспрессией генного продукта или не препятствует способности матрицы ДНК к транскрипции. Таким образом, промоторная область функционально связана с нуклеиновой кислотой, кодирующей полипептид, если промотор способен обеспечивать транскрипцию этой нуклеиновой кислоты. Промотор может представлять собой клеточно-специфический промотор, который обеспечивает существенную транскрипцию ДНК только в определенных клетках. Для управления клеточно-специфической транскрипцией с полинуклеотидом могут быть функционально связаны другие элементы управления транскрипцией помимо промотора, например, энхансеры, операторы, репрессоры и сигналы терминации транскрипции.
Подходящие промоторы и другие области управления транскрипцией описаны в данном документе. Различные области управления транскрипцией хорошо известны специалистам в данной области техники. Они включают, без ограничения, области управления транскрипцией, функциональные в клетках позвоночных, такие как, без ограничения, сегменты промоторов и энхансеров из цитомегаловирусов (например, немедленно ранний промотор в сочетании с интроном-А), вируса обезьян 40 (например, ранний промотор) и ретровирусов (таких как, например, вирус саркомы Рауса). Другие области управления транскрипцией включают полученные из генов позвоночных, таких как гены актина, белка теплового шока, бычьего гормона роста и кроличьего а-глобина, а также другие последовательности, способные управлять экспрессией генов в эукариотических клетках. Дополнительные области управления транскрипцией включают тканеспецифические промоторы и энхансеры, а также индуцибельные промоторы (например, промоторы, индуцируемые тетрациклинами). Аналогично, различные элементы управления трансляцией хорошо известны специалистам в данной области техники. Они включают, но не ограничиваются этим, сайты связывания рибосом, кодоны инициации и терминации трансляции и элементы, полученные из вирусных систем (в частности, сайт внутренней посадки рибосомы или IRES, также называемый последовательностью CITE). Экспрессионная кассета также может содержать другие элементы, такие как точка начала репликации и/или элементы интеграции хромосомы, такие как ретровинусные длинные концевые повторы (ДКП) или инвертированные концевые повторы (ИКП) аденоассоциированного вируса (AAV).
Кодирующие области полинуклеотидов и нуклеиновых кислот по настоящему изобретению могут быть связаны с дополнительными кодирующими областями, которые кодируют секреторные или сигнальные пептиды, которые управляют секрецией полипептида, кодируемого полинуклеотидом по настоящему изобретению. Например, если необходима секреция биспецифической антигенсвязывающей молекулы или ее полипептидных фрагментов, ДНК, кодирующую сигнальную последовательность, можно помещать выше нуклеиновой кислоты, кодирующей биспецифическую антигенсвязывающую молекулу по изобретению или ее полипептидные фрагменты. В соответствии с сигнальной гипотезой, белки, секретируемые клетками млекопитающих, имеют сигнальный пептид или секреторную лидерную последовательность, которые отщепляются от зрелого белка после инициации экспорта растущей белковой цепи через шероховатый эндоплазматический ретикулум. Специалистам в данной области техники известно, что полипептиды, секретируемые клетками позвоночных, в общем случае имеют сигнальный пептид, слитый с N-концом полипептида, который отщепляется от транслируемого полипептида с получением секретируемой или «зрелой» формы полипептида. В определенных вариантах осуществления используют нативный сигнальный пептид, например, сигнальный пептид тяжелой цепи или легкой цепи иммуноглобулина, или функциональное производное этой последовательности, которое сохраняет способность управлять секрецией полипептида, который с ней функционально связан. В альтернативном варианте можно использовать гетерологичный сигнальный пептид млекопитающего или его функциональное производное. Например, лидерную последовательность дикого типа можно заменить лидерной последовательностью человеческого тканевого активатора плазминогена (TPA) или мышиной р-глюкуронидазы.
ДНК, кодирующая короткую белковую последовательность, которую можно использовать для облегчения очистки (например, гистидиновый тэг) или мечения слитого белка, может быть включена в середине или в концах полинуклеотида, кодирующего биспецифическую антигенсвязывающую молекулу по изобретению или ее полипептидные фрагменты.
В дополнительном аспекте изобретения предложена клетка-хозяин, содержащая один или более полинуклеотидов по изобретению. В определенных аспектах предложена клетка-хозяин, содержащая один или более векторов по изобретению. Полинуклеотиды и векторы могут содержать любые фрагменты, отдельно или в комбинации, описанные в данном документе в отношение полинуклеотидов и векторов, соответственно. В одном аспекте клетка-хозяин содержит вектор (например, была трансформирована или трансфицирована им), содержащий полинуклеотид, который кодирует биспецифическую антигенсвязывающую молекулу по изобретению (или ее часть). В контексте данного документа термин «клетка-хозяин» относится к любому типу клеточной системы, которую можно сконструировать для создания слитых белков по изобретению или их фрагментов. Клетки-хозяева, подходящие для репликации и поддержания экспрессии антигенсвязывающих молекул, хорошо известны в данной области техники. Такие клетки можно трансфицировать или трансдуцировать, сообразно ситуации, конкретным экспрессионным вектором, и можно выращивать большие количества содержащих вектор клеток для засевания крупномасштабных ферментеров для получения достаточных количеств антигенсвязывающей молекулы для клинических применений. Подходящие клетки включают прокариотические микроорганизмы, такие как Е. coli, или различные эукариотические клетки, такие как клетки яичника китайского хомяка (СНО), клетки насекомых и т.п. Например, полипептиды можно получать в бактериях, в частности, если не требуется гликозилирование. После экспрессии полипептид можно выделять из бактериальной клеточной пасты в растворимую фракцию и дополнительно очищать. Помимо прокариот, эукариотические микробы, такие как нитевидные грибы или дрожжи, являются подходящими хозяевами для клонирования или экспрессии кодирующих полипептиды векторов, включая штаммы грибов и дрожжей, пути гликозилирования которых были «гуманизированы», что приводит к получению полипептида с частично или полностью человеческим профилем гликозилирования. Смотрите Gerngross, Nat Biotech 22, 1409-1414 (2004), и Li et al., Nat Biotech 24, 210-215 (2006).
Клетки-хозяев, подходящие для экспрессии (гликозилированных) полипептидов, также получают из многоклеточных организмов (беспозвоночных и позвоночных). Примеры клеток беспозвоночных включают клетки растений и насекомых. Были идентифицированы многочисленные бакуловирусные штаммы, которые можно использовать в сочетании с клетками насекомых, в частности, для трансфекции клеток Spodoptera frugiperda. Культуры клеток растений также можно использовать в качестве хозяев. Смотрите, например, патенты США №№5959177, 6040498, 6420548, 7125978 и 6417429 (в которых описана технология PLANTIBODIES™ для получения антител в трансгенных растениях). Клетки позвоночных также можно использовать в качестве хозяев. Например, можно использовать линии клеток млекопитающих, которые адаптированы для роста в суспензии. Другими примерами применимых линий клеток-хозяев млекопитающих являются линия клеток почки обезьяны CV1, трансформированная SV40 (COS-7); линия клеток почки эмбриона человека (клетки 293 или 293Т, описанные, например, в in Graham et al., J Gen Virol 36, 59 (1977), клетки почки новорожденного хомяка (ВНК); клетки Сертоли мышей (клетки ТМ4, описанные, например, в Mather, Biol Reprod 23, 243-251 (1980)), клетки почки обезьяны (CV1), клетки почки африканской зеленой мартышки (VERO-76), клетки карциномы шейки матки человека (HELA), клетки почки собаки (MDCK), клетки печени серой крысы (BRL 3А), клетки легкого человека (W138), клетки печени человека (Hep G2), клетки опухоли молочной железы мышей (ММТ 060562), клетки TRI (описанные, например, в Mather et al., Annals N.Y. Acad Sci 383, 44-68 (1982)), клетки MRC 5 и клетки FS4. Другие применимые линии клеток-хозяев млекопитающих включают клетки яичника китайского хомяка (СНО), включая dhfr- клетки СНО (Urlaub et al., Proc Natl Acad Sci USA 77, 4216 (1980)); и линии клеток миеломы, такие как YO, NS0, РЗХ63 и Sp2/0. Обзор некоторых линий клеток-хозяев млекопитающих, подходящих для выработки белка, смотрите, например, в Yazaki and Wu, Methods in Molecular Biology, Vol.248 (B.K.C. Lo, ed., Humana Press, Totowa, NJ), pp. 255-268 (2003). Клетки-хозяева включают культивируемые клетки, например, культивируемые клетки млекопитающих, клетки дрожжей, клетки насекомых, клетки бактерий и клетки растений, если называть лишь некоторые, но также клетки, принадлежащие трансгенному животному, трансгенному растению или культивируемой ткани растения или животного. В одном варианте осуществления клетка-хозяин представляет собой эукариотическую клетку, предпочтительно клетку млекопитающего, такую как клетка яичника китайского хомяка (СНО), клетка почки эмбриона человека (HEK) или лимфоидная клетка (например, клетка YO, NSO, Sp20). Стандартные технологии для экспрессии чужеродных генов в этих системах известны в данной области техники. Клетки, экспрессирующие полипептид, содержащий тяжелую или легкую цепь иммуноглобулина, можно конструировать так, чтобы они также экспрессировали другую цепь иммуноглобулина, чтобы экспрессируемый продукт представлял собой иммуноглобулин, который имеет как тяжелую, так и легкую цепь.
В одном аспекте предложен способ получения биспецифической антигенсвязывающей молекулы по изобретению или ее полипептидных фрагментов, включающий культивирование клетки-хозяина, содержащей полинуклеотиды, кодирующие биспецифическую антигенсвязывающую молекулу по изобретению или ее полипептидные фрагменты, предложенные в данном документе, в условиях, подходящих для экспрессии биспецифической антигенсвязывающей молекулы по изобретению или ее полипептидных фрагментов, и выделение биспецифической антигенсвязывающей молекулы по изобретению или ее полипептидных фрагментов из клетки-хозяина (или культуральной среды клеток-хозяев).
Биспецифические молекулы по изобретению, полученные, как описано в данном документе, можно очищать известными в данной области техники методами, такими как высокоэффективная жидкостная хроматография, ионообменная хроматография, гель-электрофорез, аффинная хроматография, эксклюзионная хроматография и т.п. Фактические условия, применяемые для очистки конкретного белка, будут зависеть частично от факторов, таких как суммарный заряд, гидрофобность, гидрофильность и т.д., и будут очевидны для специалистов в данной области техники. Для очистки методом аффинной хроматографии можно использовать антитело, лиганд, рецептор или антиген, с которыми связывается биспецифическая антигенсвязывающая молекула. Например, очистки слитых белков по изобретению методом аффинной хроматографии можно использовать матрицу с протеином А или протеином G. Последовательные этапы аффинной хроматографии с протеином А или G и эксклюзионной хроматографии можно использовать для выделения антигенсвязывающей молекулы по существу так, как описано в данном документе. Чистоту биспецифической антигенсвязывающей молекулы по изобретению или ее фрагментов можно определять любым из ряда хорошо известных аналитических методов, включая электрофорез, жидкостную хроматографию высокого давления и т.п. Например, было показано, что биспецифические антигенсвязывающие молекулы, экспрессируемые, как описано в примерах, являются интактными и имеют надлежащую сборку, что было продемонстрировано восстанавливающим и невосстанавливающим ДСН-ПААГ.
Методы анализа
Предложенные в данном документе антигенсвязывающие молекулы можно охарактеризовать по их связывающим свойствам и/или биологической активности с помощью различных анализов, известных в данной области техники. В частности, их характеристики получают с помощью анализов, более подробно описанных в примерах.
1. Анализ связывания
Связывание предложенной в данном документе биспецифической антигенсвязывающей молекулы с соответствующими экспрессирующими мишень клетками можно оценить, например, используя линию клеток мышиных фибробластов, экспрессирующую человеческий белок активации фибробластов (FAP), и анализ методом проточной цитометрии (FACS). Связывание предложенных в данном документе биспецифических антигенсвязывающих молекул с CD40 можно определить, используя первичные В-клетки, как описано в примере 4.2.
2. Методы анализа активности
Биспецифические антигенсвязывающие молекулы по изобретению исследуют в отношении биологической активности. Биологическая активность может включать эффективность и специфичность биспецифических антигенсвязывающих молекул. Эффективность и специфичность демонстрируют с помощью анализов, показывающих наличие агонистической сигнализации посредством рецептора CD40 после связывания целевого антигена. Кроме того, проводят in vitro анализы примирования Т-клеток, используя дендритные клетки (ДК), которые инкубировали с биспецифическими антигенсвязывающими молекулами.
Фармацевтические композиции, составы и пути введения
В дополнительном аспекте в изобретении предложены фармацевтические композиции, содержащие любую из предложенных в данном документе биспецифических антигенсвязывающих молекул, например, для применения в любом из нижеприведенных терапевтических способов. В одном варианте осуществления фармацевтическая композиция содержит любую из предложенных в данном документе биспецифических антигенсвязывающих молекул и по меньшей мере одно фармацевтически приемлемое вспомогательное вещество. В другом варианте осуществления фармацевтическая композиция содержит любую из предложенных в данном документе биспецифических антигенсвязывающих молекул и по меньшей мере один дополнительный терапевтический агент, например, как описано ниже.
Фармацевтические композиции по данному изобретению содержат терапевтически эффективное количество одной или более биспецифических антигенсвязывающих молекул, растворенных или диспергированных в фармацевтически приемлемом составе. Выражения «фармацевтически или фармакологически приемлемый» относятся к молекулярным веществам и композициям, которые в общем случае являются нетоксичными для реципиентов в применяемых дозировках и концентрациях, т.е. не вызывают нежелательную, аллергическую или другую неблагоприятную реакцию при введении животному, такому как, например, человек, в зависимости от ситуации. Изготовление фармацевтической композиции, которая содержит по меньшей мере одну биспецифическую антигенсвязывающую молекулу по изобретению и, необязательно, дополнительный активный ингредиент, станет известно специалистам в данной области техники в свете настоящего изобретения, и как проиллюстрировано в Remington's Pharmaceutical Sciences, 18th Ed. Mack Printing Company, 1990, включенной в данный документ посредством ссылки. В частности, композиции представляют собой лиофилизированные составы или водные растворы. В контексте данного документа «фармацевтически приемлемое вспомогательное вещество» включает любые и все растворители, буферы, дисперсионные среды, покрытия, поверхностно-активные вещества, антиоксиданты, консерванты (например, антибактериальные агенты, противогрибковые агенты), изотонические агенты, соли, стабилизаторы и их комбинации, как известно специалисту в данной области техники.
Исходные композиции включают разработанные для введения путем инъекции, например, подкожной, интрадермальной, внутриочаговой, внутривенной, внутриартериальной, внутримышечной, интратекальной или внутрибрюшинной инъекции. Для инъекции биспецифические антигенсвязывающие молекулы по изобретению могут быть составлены в виде водных растворов, предпочтительно в физиологически совместимых буферах, таких как раствор Хэнкса, раствор Рингера или физиологический солевой буфер. Раствор может содержать вспомогательные агенты, такие как суспендирующие, стабилизирующие и/или диспергирующие агенты. В альтернативном варианте биспецифические антигенсвязывающие молекулы могут находиться в форме порошка для смешивания с подходящим носителем, например, стерильной апирогенной водой, перед применением. Стерильные инъекционные растворы готовят путем смешивания антигенсвязывающих молекул по изобретению в необходимом количестве в подходящем растворителе с различными перечисленными выше другими ингредиентами в случае необходимости. Стерильность можно легко обеспечить, например, путем фильтрования через мембраны для стерильной фильтрации. В общем случает дисперсии готовят путем включения различных стерилизованных активных ингредиентов в стерильный носитель, который содержит основную дисперсионную среду и/или другие ингредиенты. В случае стерильных порошков для изготовления стерильных инъекционных растворов, суспензий или эмульсий предпочтительными способами изготовления являются технологии вакуумной сушки или сублимационной сушки, которые позволяют получать порошок активного ингредиента плюс любой дополнительный необходимый активный ингредиент из стерильно профильтрованной перед этим жидкой среды. Жидкую среду, при необходимости, нужно должным образом забуферить, а жидкий разбавитель сначала сделать изотоничным перед инъекцией с помощью достаточного количества солевого раствора или глюкозы. Композиция должна быть стабильной в условиях производства и хранения, и защищенной от загрязняющего действия микроорганизмов, таких как бактерии и грибы. Следует понимать, что загрязнение эндотоксинами необходимо свести к минимуму и поддерживать на безопасном уровне, например, менее 0,5 нг/мг белка. Подходящие фармацевтически приемлемые вспомогательные вещества включают, но не ограничиваются этим: буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксид анты, включая аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензил аммония; хлорид гексаметония; хлорид бензалкония; хлорид бензетония; фенол; бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцин; циклогексанол; 3-пентанол; и м-крезол); низкомолекулярные (содержащие менее 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как ЭДТА; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; металлсодержащие комплексы (например, комплексы Zn-белок) и/или неионные поверхностно-активные вещества, такие как полиэтиленгликоль (ПЭГ). Водные инъекционные суспензии могут содержать соединения, которые увеличивают вязкость суспензии, такие как карбоксиметилцеллюлоза натрия, сорбит, декстран и т.п. Необязательно, суспензия может также содержать подходящие стабилизаторы или вещества, которые повышают растворимость соединений, для обеспечения получения высококонцентрированных растворов. Кроме того, суспензии активных соединений могут быть изготовлены в виде подходящих масляных инъекционных суспензий. Подходящие липофильные растворители или носители включают жирные масла, такие как кунжутное масло, или синтетические сложные эфиры жирных кислот, такие как этилолеаты или триглицериды, или липосомы.
Активные ингредиенты также могут быть заключены в микрокапсулах, полученных, например, с помощью методик коацервации или путем межфазной полимеризации, например, гидроксиметилцеллюлозные или желатиновые микрокапсулы и полиметилметакрилатные микрокапсулы, соответственно, в коллоидных системах для доставки лекарственных средств (например, липосомах, альбуминовых микросферах, микроэмульсиях, наночастицах и нанокапсулах) или в макроэмульсиях. Такие методики описаны в Remington's Pharmaceutical Sciences (18th Ed. Mack Printing Company, 1990). Можно получать препараты с замедленным высвобождением. Подходящие примеры препаратов с замедленным высвобождением включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащие полипептид, причем матрицы находятся в виде формованных изделий, например, пленок или микрокапсул. В конкретных вариантах осуществления длительное всасывание инъекционных композиций может быть достигнуто путем использования в композициях агентов, замедляющих всасывание, таких как, например, моностеарат алюминия, желатин или их комбинации.
Типовые фармацевтически приемлемые вспомогательные вещества по данному документу дополнительно включают агенты для интерстициального диспергирования лекарственных средств, такие как растворимые нейтрально-активные гликопротеины гиалуронидазы (sHASEGP), например, человеческие растворимые гликопротеины гиалуронидазы РН-20, такие как rHuPH20 (HYLENEX®, Baxter International, Inc.). Некоторые типовые sHASEGP и способы их применения, включая rHuPH20, описаны в патентных публикациях США №№2005/0260186 и 2006/0104968. В одном аспекте sHASEGP комбинируют с одной или более дополнительными гликозаминогликаназами, такими как хондроитиназы.
Типовые лиофилизированные составы антител описаны в патенте США №6267958. Водные составы антител включают описанные в патенте США №6171586 и WO2006/044908, причем последние составы содержат гистидин-ацетатный буфер.
Помимо ранее описанных композиций антигенсвязывающие молекулы также можно готовить в виде депо-препаратов. Такие составы пролонгированного действия можно вводить путем имплантации (например, подкожно или внутримышечно) или путем внутримышечной инъекции. Таким образом, например, слитые белки могут быть смешаны с подходящими полимерными или гидрофобными материалами (например, в виде эмульсии в приемлемом масле) или ионообменными смолами, или в виде умеренно растворимых производных, например, умеренно растворимой соли.
Фармацевтические композиции, содержащие биспецифические антигенсвязывающие молекулы по изобретению, можно готовить посредством традиционных процессов смешивания, растворения, эмульсификации, инкапсуляции, захвата или лиофилизации. Фармацевтические композиции можно готовить традиционным образом, используя один или более физиологически приемлемых носителей, разбавителей или вспомогательных агентов, которые облегчают процесс обработки белков в препараты, которые можно использовать фармацевтически. Надлежащий состав зависит от выбранного пути введения.
Биспецифические антигенсвязывающие молекулы можно вмешивать в композицию в форме свободной кислоты или свободного основания, в нейтральной или солевой форме. Фармацевтически приемлемые соли представляют собой соли, которые в значительной степени сохраняют биологическую активность свободной кислоты или свободного основания. Они включают соли присоединения кислот, например, образуемые со свободными аминогруппами белковой композиции или образуемые с неорганическими кислотами, такими как, например, хлористоводородная или фосфорная кислоты, или такими органическими кислотами, как уксусная, щавелевая, винная или миндальная кислота. Соли, образуемые со свободными карбоксильными группами, также можно получать из неорганических оснований, таких как, например, гидроксиды натрия, калия, аммония, кальция или железа; или таких органических оснований, как изопропиламин, триметиламин, гистидин или прокаин. Фармацевтические соли, как правило, более растворимы в водных и других протонных растворителях, чем соответствующие формы свободных оснований.
Композиция по данному документу также может содержать более одного активного ингредиента, если это необходимо для конкретного показания, подлежащего лечению, предпочтительно те, которые обладают дополняющими видами активности и не оказывают отрицательного влияния друг на друга. Такие активные ингредиенты приемлемо присутствуют в комбинации в количествах, которые эффективны для предполагаемой цели.
Составы, предназначенные для введения in vivo, в общем случае являются стерильными. Стерильность можно легко обеспечить, например, путем фильтрования через мембраны для стерильной фильтрации.
Терапевтические способы и композиции
Любые биспецифические антигенсвязывающие молекулы, предложенные в данном документе, можно применять в терапевтических способах. Для применения в терапевтических способах биспецифические антигенсвязывающие молекулы по изобретению можно изготавливать, дозировать и вводить способом, соответствующим требованиям надлежащей медицинской практики. Факторы, которые необходимо учитывать в этом контексте, включают конкретное нарушение, подлежащее лечению, конкретное млекопитающее, подлежащее лечению, клиническое состояние отдельного пациента, причину нарушения, место доставки агента, способ введения, график введения и другие факторы, известные практикующим врачам.
В одном аспекте предложены биспецифические антигенсвязывающие молекулы по изобретению для применения в качестве лекарственного средства.
В дополнительных аспектах предложены биспецифические антигенсвязывающие молекулы по изобретению для применения (i) в индукции иммунной стимуляции CD40+ антигенпрезентирующими клетками (АПК), (ii) в стимуляции опухолеспецифического Т-клеточного ответа, (iii) в инициации апоптоза опухолевых клеток, (iv) в лечении рака, (v) в задержке прогрессирования рака, (vi) в продлении выживаемости пациента, имеющего рак, (vii) в лечении инфекций. В конкретном аспекте предложены биспецифические антигенсвязывающие молекулы по изобретению для применения в лечении заболевания, в частности, для применения в лечении рака.
В определенных аспектах предложены биспецифические антигенсвязывающие молекулы по изобретению для применения в способе лечения. В одном аспекте в изобретении предложена биспецифическая антигенсвязывающая молекула, описанная в данном документе, для применения в лечении заболевания у нуждающегося в этом индивида. В определенных аспектах в изобретении предложена биспецифическая антигенсвязывающая молекула для применения в способе лечения индивида, имеющего заболевание, включающем введение индивиду терапевтически эффективного количества биспецифической антигенсвязывающей молекулы. В определенных аспектах подлежащее лечению заболевание представляет собой рак. Субъект, пациент или «индивид», нуждающийся в лечении, как правило, представляет собой млекопитающее, конкретнее, человека.
В одном аспекте предложен способ (i) индукции иммунной стимуляции CD40+ антигенпрезентирующими клетками (АПК), (ii) стимуляции опухолеспецифического Т-клеточного ответа, (iii) инициации апоптоза опухолевых клеток, (iv) лечения рака, (v) задержки прогрессирования рака, (vi) продления выживаемости пациента, имеющего рак, или (vii) лечения инфекций, причем способ включает введение нуждающемуся в этом индивиду терапевтически эффективного количества биспецифической антигенсвязывающей молекулы по изобретению.
В дополнительном аспекте в изобретении предложено применение биспецифической антигенсвязывающей молекулы по изобретению в производстве или изготовлении лекарственного средства для лечения заболевания у нуждающегося в этом индивида. В одном аспекте лекарственное средство предназначено для применения в способе лечения заболевания, включающем введение индивиду, имеющему заболевание, терапевтически эффективного количества лекарственного средства. В определенных аспектах подлежащее лечению заболевание представляет собой пролиферативное нарушение, в частности, рак. Примеры видов рака включают, но не ограничиваются этим, рак мочевого пузыря, рак головного мозга, рак головы и шеи, рак поджелудочной железы, рак легкого, рак молочной железы, рак яичника, рак матки, рак шейки матки, рак эндометрия, рак пищевода, рак толстой кишки, колоректальный рак, ректальный рак, рак желудка, рак предстательной железы, рак крови, рак кожи, плоскоклеточную карциному, рак костей и рак почки. Другие примеры рака включают карциному, лимфому (например, ходжкинскую и неходжкинскую лимфому), бластому, саркому и лейкоз. Другие клеточно-пролиферативные нарушения, которые можно лечить, используя биспецифическую антигенсвязывающую молекулу или антитело по изобретению, включают, но не ограничиваются этим, новообразования, находящиеся в: верхней части брюшной полости, костной ткани, молочной железе, пищеварительной системе, печени, поджелудочной железе, брюшной полости, эндокринных железах (надпочечнике, паращитовидной, гипофизе, семенниках, яичниках, вилочковой, щитовидной), глазе, голове и шее, нервной системе (центральной и периферической), лимфатической системе, тазовой области, коже, мягкой ткани, селезенке, области грудной клетки и урогенитальной системе. Также включены предраковые состояния или поражения и раковые метастазы. В определенных вариантах осуществления рак выбран из группы, состоящей из почечно-клеточного рака, рака кожи, рака легкого, колоректального рака, рака молочной железы, рака головного мозга, рака головы и шеи. Специалисту в данной области техники понятно, что во многих случаях биспецифическая антигенсвязывающая молекула или антитело по изобретению могут не обеспечивать излечение, но могут приносить пользу. В некоторых аспектах физиологическое изменение, приносящее некоторую пользу, также считается терапевтически благоприятным. Таким образом, в некоторых аспектах количество биспецифической антигенсвязывающей молекулы или антитела по изобретению, которое обеспечивает физиологическое изменение, считается «эффективным количеством» или «терапевтически эффективным количеством».
Для предотвращения или лечения заболевания соответствующая дозировка биспецифической антигенсвязывающей молекулы по изобретению (при применении отдельно или в комбинации с одним или более другими дополнительными терапевтическими агентами) будет зависеть от типа заболевания, подлежащего лечению, пути введения, массы тела пациента, конкретной молекулы, тяжести и течения заболевания, того, вводят ли биспецифическую антигенсвязывающую молекулу в превентивных или терапевтических целях, предыдущих или одновременных терапевтических вмешательств, анамнеза пациента и ответа на биспецифическую антигенсвязывающую молекулу, а также решения лечащего врача. Практикующий врач, ответственный за введение, в любом случае сможет определить концентрацию активного(ых) ингредиента(ов) в композиции и соответствующую(ие) дозу(ы) для каждого отдельного субъекта. В данном документе предусмотрены различные схемы дозирования, включая, но не ограничиваясь этим, однократное или многократное введение в различные моменты времени, болюсное введение и импульсную инфузию.
Биспецифическую антигенсвязывающую молекулу по изобретению удобно вводить пациенту за один раз или в течение серии процедур. В зависимости от типа и тяжести заболевания, начальная кандидатная дозировка биспецифической антигенсвязывающей молекулы для введения пациенту может составлять от около 1 мкг/кг до 15 мг/кг (например, 0,1 мг/кг 10 мг/кг), независимо от того, например, осуществляют ли одно или более отдельных введений или непрерывную инфузию. Одна типичная суточная доза может варьироваться от около 1 мкг/кг до 100 мг/кг или более, в зависимости от факторов, упомянутых выше. В случае повторных введений в течение нескольких суток или более, в зависимости от состояния, лечение в общем случае проводят до достижения необходимой степени подавления симптомов заболевания. Одна типовая дозировка биспецифической антигенсвязывающей молекулы по изобретению находится в диапазоне от около 0,005 мг/кг до около 10 мг/кг. В других примерах доза также может составлять от около 1 мкг/кг массы тела, около 5 мкг/кг массы тела, около 10 мкг/кг массы тела, около 50 мкг/кг массы тела, около 100 мкг/кг массы тела, около 200 мкг/кг массы тела, около 350 мкг/кг массы тела, около 500 мкг/кг массы тела, около 1 мг/кг массы тела, около 5 мг/кг массы тела, около 10 мг/кг массы тела, около 50 мг/кг массы тела, около 100 мг/кг массы тела, около 200 мг/кг массы тела, около 350 мг/кг массы тела, около 500 мг/кг массы тела до около 1000 мг/кг массы тела или более на введение, и любой диапазон, получаемый из этих значений. Из примеров диапазонов, получаемых из значений, перечисленных в данном документе, можно вводить диапазон, составляющий от около 0,1 мг/кг массы тела до около 20 мг/кг массы тела, от около 5 мкг/кг массы тела до около 1 мг/кг массы тела и т.д., с учетом описанных выше значений. Таким образом, пациенту можно вводить одну или более доз, составляющих около 0,5 мг/кг, 2,0 мг/кг, 5,0 мг/кг или 10 мг/кг (или любую их комбинацию). Такие дозы можно вводить периодически, например, каждую неделю или каждые три недели (например, так, чтобы пациент получал от около двух до около двадцати или, например, около шести доз слитого белка). В конкретном аспекте биспецифическую антигенсвязывающую молекулу будут вводить каждые три недели. Можно вводить начальную более высокую нагрузочную дозу, за которой следует одна или более меньших доз. При этом можно применять другие схемы дозирования. Эффективность такой терапии легко контролировать при помощи традиционных методик и анализов.
Биспецифическую антигенсвязывающую молекулу по изобретению в общем случае будут применять в количестве, эффективном для достижения предполагаемой цели. В случае применения для лечения или предотвращения болезненного состояния биспецифическую антигенсвязывающую молекулу по изобретению или ее фармацевтическую композицию вводят или применяют в терапевтически эффективном количестве. Определение терапевтически эффективного количества вполне соответствует возможностям специалистов в данной области техники, особенно в свете подробного описания изобретения, приведенного в данном документе. В случае системного введения терапевтически эффективную дозу можно оценить изначально по in vitro анализу, такому как анализ клеточной культуры. После этого можно составить дозу в животных моделях для достижения диапазона концентрации в циркуляции, который включает IC50, определенную в клеточной культуре. Такую информацию можно использовать для более точного определения доз, применимых для людей. Начальные дозировки также можно оценивать по in vivo данным, например, животных моделей, используя методики, хорошо известные в данной области техники. Специалист в данной области техники сможет легко оптимизировать введение людям на основании данных по животным.
Количество и интервал дозировок можно корректировать индивидуально, чтобы обеспечить плазменные уровни биспецифической антигенсвязывающей молекулы по изобретению, которых достаточно для поддержания терапевтического эффекта. Обычные дозировки для пациентов в случае введения путем инъекции находятся в диапазоне от около 0,1 до 50 мг/кг/сутки, как правило, от около 0,1 до 1 мг/кг/сутки. Терапевтически эффективные плазменные уровни можно обеспечить путем введения многократных доз каждые сутки. Уровни в плазме можно измерять, например, с помощью ВЭЖХ. В случаях местного введения или избирательного поглощения эффективная локальная концентрация биспецифической антигенсвязывающей молекулы или антитела по изобретению может не быть связана с плазменной концентрацией. Специалист в данной области техники сможет оптимизировать терапевтически эффективные местные дозировки без проведения необязательных экспериментов.
Терапевтически эффективная доза биспецифической антигенсвязывающей молекулы по изобретению, описанной в данном документе, в общем случае будет обеспечивать терапевтическую пользу, не вызывая существенной токсичности. Токсичность и терапевтическую эффективность слитого белка можно определить с помощью стандартных фармацевтических процедур с клеточной культурой или экспериментальными животными. Анализ клеточных культур и исследования на животных можно использовать, чтобы определить LD50 (дозу, летальную для 50% популяции) и ED50 (дозу, терапевтически эффективную в 50% популяции). Соотношение доз между токсическими и терапевтическими эффектами является терапевтическим индексом, который можно выразить как соотношение LD50/ED50. Предпочтительными являются биспецифические антигенсвязывающие молекулы, которые демонстрируют высокие терапевтические индексы. В одном аспекте биспецифическая антигенсвязывающая молекула или антитело по изобретению демонстрирует высокий терапевтический индекс. Данные, полученные в анализе клеточных культур и исследованиях на животных, можно использовать при составлении диапазона дозировок, подходящих для применения людьми. Дозировка предпочтительно находится в диапазоне циркулирующих концентраций, который включает ED50, с небольшой или отсутствующей токсичностью. Дозировка может варьироваться в пределах этого диапазона в зависимости от ряда факторов, например, применяемой дозированной формы, применяемого пути введения, состояния субъекта и т.п. Точные состав, путь введения и дозировка могут быть выбраны личным врачом с учетом состояния пациента (смотрите, например, Fingl et al., 1975, в: The Pharmacological Basis of Therapeutics, Ch. 1, p. 1, в полном объеме включенной в данный документ посредством ссылки).
Лечащий врач пациентов, которые проходят лечение слитыми белками по изобретению, будет знать, как и когда прекращать, прерывать или корректировать введение вследствие токсичности, нарушения функционирования органов и т.п. И наоборот, лечащий врач также будет знать, как скорректировать лечение до более высоких уровней, если клинический ответ не был достаточным (с предотвращением токсичности). Размер вводимой дозы при контроле представляющего интерес заболевания будет варьироваться в зависимости от тяжести подлежащего лечению состояния, пути введения и т.п. Тяжесть состояния можно, например, частично оценить с помощью стандартных прогностических методов оценки. Кроме того, доза и, возможно, частота введения дозы также будут варьироваться в соответствии с возрастом, массой тела и ответом индивидуального пациента.
Другие агенты и варианты лечения
Биспецифическую антигенсвязывающую молекулу по изобретению можно вводить в комбинации с одним или более агентами в терапии. Например, биспецифическую антигенсвязывающую молекулу или антитело по изобретению можно вводить совместно с по меньшей мере одним дополнительным терапевтическим агентом. Термин «терапевтический агент» охватывает любой агент, который можно вводить для лечения симптома или заболевания у индивида, нуждающегося в таком лечении. Такой дополнительный терапевтический агент может содержать любые активные ингредиенты, подходящие для конкретного показания, подлежащего лечению, предпочтительно те, которые обладают дополняющими видами активности и не оказывают отрицательного влияния друг на друга. В определенных вариантах осуществления дополнительный терапевтический агент представляет собой другой противораковый агент, например, разрушитель микротрубочек, антиметаболит, ингибитор топоизомеразы, интеркалятор ДНК, алкилирующий агент, гормональную терапию, ингибитор киназы, антагонист рецептора, активатор апоптоза опухолевых клеток или антиангиогенный агент. В определенных аспектах дополнительный терапевтический агент представляет собой иммуномодулирующий агент, цитостатический агент, ингибитор клеточной адгезии, цитотоксический или цитостатический агент, активатор клеточного апоптоза или агент, который повышает чувствительность клеток к индукторам апоптоза.
Таким образом, предложены биспецифические антигенсвязывающие молекулы по изобретению или содержащие их фармацевтические композиции для применения в лечении рака, причем биспецифическую антигенсвязывающую молекулу вводят в комбинации с химиотерапевтическим агентом, лучевой терапией и/или другими агентами для применения в иммунотерапии рака.
Такие другие агенты приемлемо присутствуют в комбинации в количествах, которые эффективны для предполагаемой цели. Эффективное количество таких других агентов зависит от количества используемого слитого белка, типа нарушения или лечения и других факторов, обсуждаемых выше. Биспецифическую антигенсвязывающую молекулу или антитело по изобретению в общем случае применяют в таких же дозировках и вводят такими же путями, как описано в данном документе, или в дозировках, составляющих от около 1 до 99% дозировок, описанных в данном документе, или в любой дозировке и любым путем, которые эмпирически/клинически определены как подходящие.
Такие виды комбинированной терапии, отмеченные выше, охватывают комбинированное введение (при котором два или более терапевтических агентов включены в одну или в отдельные композиции) и раздельное введение, в случае которого введение биспецифической антигенсвязывающей молекулы или антитела по изобретению может происходить до, одновременно и/или после введения дополнительного терапевтического агента и/или адъюванта.
В дополнительном аспекте предложена биспецифическая антигенсвязывающая молекула, описанная в данном документе выше, для применения в лечении рака, причем биспецифическую антигенсвязывающую молекулу вводят в комбинации с другим иммуномодулятором.
Термин «иммуномодулятор» относится к любому веществу, включая моноклональное антитело, которое оказывает воздействие на иммунную систему. Молекулы по изобретению можно считать иммуномодуляторами. Иммуномодуляторы можно использовать в качестве антинеопластических агентов для лечения рака. В одном аспекте иммуномодуляторы включают, но не ограничиваются этим, анти-CTLA4 антитела (например, ипилимумаб), анти-PD1 антитела (например, ниволумаб или пембролизумаб), антитела к PD-L1 (например, атезолизумаб, авелумаб или дурвалумаб), антитела к ОХ-40, антитела к 4-1ВВ и антитела к GITR. В дополнительном аспекте предложена биспецифическая антигенсвязывающая молекула, описанная в данном документе выше, для применения в лечении рака, причем биспецифическую антигенсвязывающую молекулу вводят в комбинации с агентом, блокирующим взаимодействие PD-L1/PD-1. В одном аспекте агент, блокирующий взаимодействие PD-L1/PD-1, представляет собой анти-PD-L1 антитело или анти-PD1 антитело. Конкретнее, агент, блокирующий взаимодействие PD-L1/PD-1, представляет собой анти-PD-L1 антитело, в частности, анти-PD-L1 антитело, выбранное из группы, состоящей из атезолизумаба, дурвалумаба, пембролизумаба и ниволумаба. В одном конкретном аспекте агент, блокирующий взаимодействие PD-L1/PD-1, представляет собой атезолизумаб (MPDL3280A, RG7446). В другом аспекте агент, блокирующий взаимодействие PD-L1/PD-1, представляет собой анти-PD-L1 антитело, содержащее вариабельный домен тяжелой цепи VH(PDL-1) с SEQ ID NO: 107 и вариабельный домен легкой цепи VL(PDL-1) с SEQ ID NO: 108. В другом аспекте агент, блокирующий взаимодействие PD-L1/PD-1, представляет собой анти-PD-L1 антитело, содержащее вариабельный домен тяжелой цепи VH(PDL-1) с SEQ ID NO: 109 и вариабельный домен легкой цепи VL(PDL-1) с SEQ ID NO: 110. В другом аспекте агент, блокирующий взаимодействие PD-L1/PD-1, представляет собой анти-PD1 антитело, в частности, анти-PD1 антитело, выбранное из пембролизумаба или ниволумаба. Такие другие агенты приемлемо присутствуют в комбинации в количествах, которые эффективны для предполагаемой цели. Эффективное количество таких других агентов зависит от количества используемой биспецифической антигенсвязывающей молекулы, типа нарушения или лечения и других факторов, обсуждаемых выше. Биспецифическую антигенсвязывающую молекулу, описанную в данном документе выше, в общем случае применяют в таких же дозировках и вводят такими же путями, как описано в данном документе, или в дозировках, составляющих от около 1 до 99% дозировок, описанных в данном документе, или в любой дозировке и любым путем, которые эмпирически/клинически определены как подходящие.
Такие виды комбинированной терапии, отмеченные выше, охватывают комбинированное введение (при котором два или более терапевтических агентов включены в одну или в отдельные композиции) и раздельное введение, в случае которого введение биспецифической антигенсвязывающей молекулы может происходить до, одновременно и/или после введения дополнительного терапевтического агента и/или адъюванта.
Готовые изделия
В другом аспекте данного изобретения предложено готовое изделие, содержащее материалы, применяемые при лечении, предотвращении и/или диагностике вышеописанных нарушений. Готовое изделие содержит контейнер и этикетку или вкладыш в упаковку на нем или связанные с ним. Подходящие контейнеры включают, например, бутылки, флаконы, шприцы, пакеты для в/в раствора и т.д. Контейнеры могут быть выполнены из ряда материалов, таких как стекло или пластик. Контейнер содержит композицию, которая сама по себе или в комбинации с другой композицией эффективна для лечения, предотвращения и/или диагностики патологического состояния, и может иметь стерильное входное отверстие (например, контейнер может представлять собой пакет для внутривенного раствора или флакон с пробкой, прокалываемой гиподермической иглой для инъекции). По меньшей мере один активный агент в композиции представляет собой биспецифическую антигенсвязывающую молекулу по изобретению.
На этикетке или на вкладыше в упаковку указано, что композицию применяют для лечения выбранного патологического состояния. Кроме того, готовое изделие может содержать (а) первый контейнер с содержащейся в нем композицией, причем указанная композиция содержит биспецифическую антигенсвязывающую молекулу по изобретению; и (б) второй контейнер с содержащейся в нем композицией, причем указанная композиция содержит дополнительный цитотоксический или иной терапевтический агент. Готовое изделие в этом варианте осуществления может дополнительно содержать вкладыш в упаковку, на котором указано, что композиции можно применять для лечения конкретного патологического состояния.
В альтернативном или дополнительном варианте готовое изделие может дополнительно содержать второй (или третий) контейнер, содержащий фармацевтически приемлемый буфер, такой как бактериостатическая вода для инъекций (БВДИ), фосфатно-солевой буферный раствор, раствор Рингера и раствор декстрозы. Оно также может содержать другие материалы, необходимые с коммерческой и пользовательской точки зрения, включая другие буферы, разбавители, фильтры, иглы и шприцы.
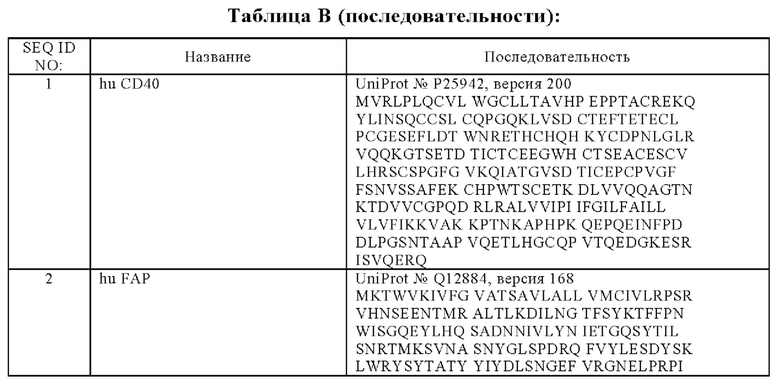
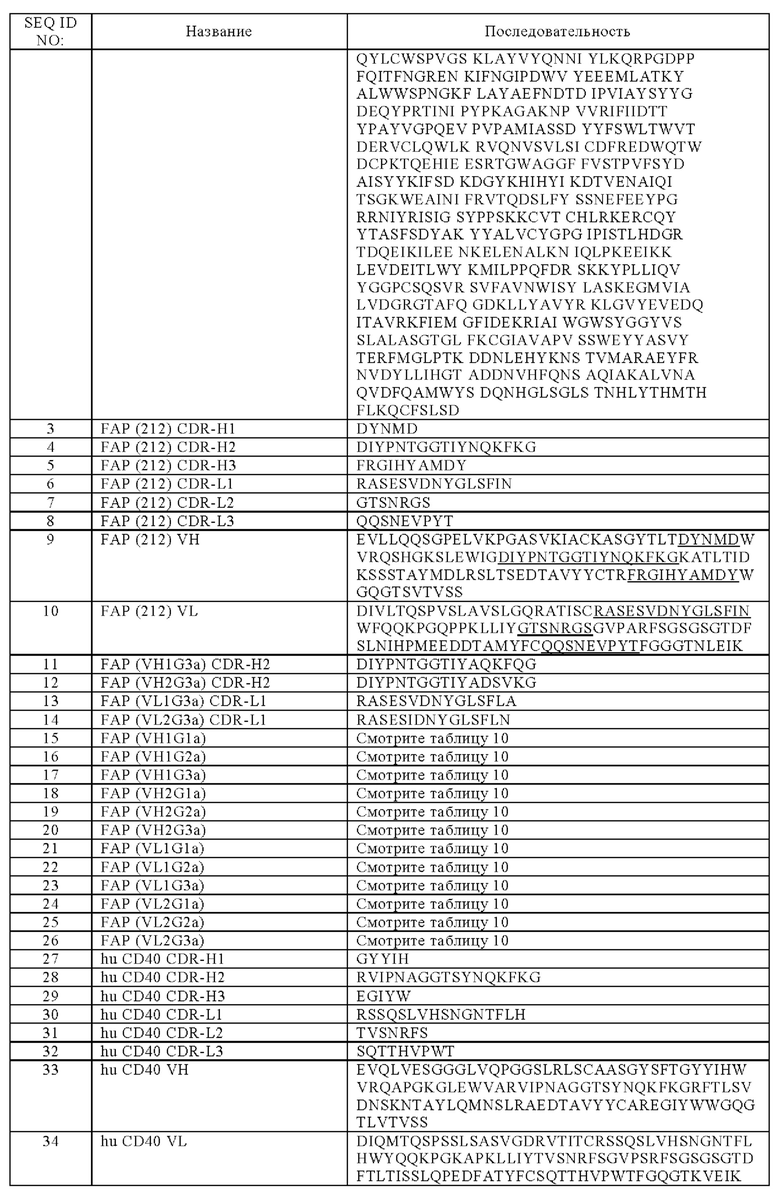

Следующие пронумерованные параграфы (пар.) описывают аспекты настоящего изобретения:
1. Биспецифическая антигенсвязывающая молекула, содержащая
(а) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с CD40, и
(б) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с белком активации фибробластов (FAP), содержащий вариабельную область тяжелой цепи (VHFAP), содержащую
(i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 3, (ii) CDR-H2, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4, SEQ ID NO: 11 и SEQ ID NO: 12, и (iii) CDR-Н3, содержащую аминокислотную последовательность SEQ ID NO: 5, и вариабельную область легкой цепи (VLFAP), содержащую (iv) CDR-L1, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 6, SEQ ID NO: 13 и SEQ ID NO: 14, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 7, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 8.
2. Биспецифичная антигенсвязывающая молекула по пар. 1, дополнительно содержащая
(в) Fc-область, состоящую из первой и второй субъединиц, способных к стабильной ассоциации.
3. Биспецифическая антигенсвязывающая молекула по пар. 1 или 2, причем антигенсвязывающий домен, способный к специфическому связыванию с FAP, содержит вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность, которая по меньшей мере на около 90% идентична аминокислотной последовательности SEQ ID NO: 9, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность, которая по меньшей мере на около 90% идентична аминокислотной последовательности SEQ ID NO: 10.
4. Биспецифическая антигенсвязывающая молекула по любому одному из пар. 1-3, причем антигенсвязывающий домен, способный к специфическому связыванию с FAP, содержит
вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19 и SEQ ID NO: 20, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 21, SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 25 и SEQ ID NO: 26.
5. Биспецифическая антигенсвязывающая молекула по любому одному из пар. 14, причем антигенсвязывающий домен, способный к специфическому связыванию с FAP, содержит
(а) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 15, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 21,
(б) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 16, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 21,
(в) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 16, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 22, или
(г) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 19, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 25.
6. Биспецифическая антигенсвязывающая молекула по любому одному из пар. 1-5, причем антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержит вариабельную область тяжелой цепи (VHCD40), содержащую (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 27, (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 28, и (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 29, и вариабельную область легкой цепи (VLCD40), содержащую (iv) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 30, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 31, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 32.
7. Биспецифическая антигенсвязывающая молекула по любому одному из пар. 16, причем антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержит
(i) вариабельную область тяжелой цепи (VHCD40), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 37, SEQ ID NO: 38, SEQ ID NO: 39 и SEQ ID NO: 40, и
(ii) вариабельную область легкой цепи (VLCD40), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 41, SEQ ID NO: 42, SEQ ID NO: 43 и SEQ ID NO: 44.
8. Биспецифическая антигенсвязывающая молекула по любому одному из пар. 1-6, причем антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержит
(i) вариабельную область тяжелой цепи (VHCD40), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 45, SEQ ID NO: 46, SEQ ID NO: 47, SEQ ID NO: 48, SEQ ID NO: 49 и SEQ ID NO: 50, и
(ii) вариабельную область легкой цепи (VLCD40), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 51, SEQ ID NO: 52, SEQ ID NO: 53 и SEQ ID NO: 54.
9. Биспецифическая антигенсвязывающая молекула по любому одному из пар. 1-5 или 7, причем антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержит
(а) VH, содержащую аминокислотную последовательность SEQ ID NO: 37, и VL, содержащую аминокислотную последовательность SEQ ID NO: 41, или
(б) VH, содержащую аминокислотную последовательность SEQ ID NO: 37, и VL, содержащую аминокислотную последовательность SEQ ID NO: 42, или
(в) VH, содержащую аминокислотную последовательность SEQ ID NO: 37, и VL, содержащую аминокислотную последовательность SEQ ID NO: 43, или
(г) VH, содержащую аминокислотную последовательность SEQ ID NO: 37, и VL, содержащую аминокислотную последовательность SEQ ID NO: 44, или
(д) VH, содержащую аминокислотную последовательность SEQ ID NO: 38, и VL, содержащую аминокислотную последовательность SEQ ID NO: 41, или
(е) VH, содержащую аминокислотную последовательность SEQ ID NO: 38, и VL, содержащую аминокислотную последовательность SEQ ID NO: 42, или
(ж) VH, содержащую аминокислотную последовательность SEQ ID NO: 38, и VL, содержащую аминокислотную последовательность SEQ ID NO: 43, или
(з) VH, содержащую аминокислотную последовательность SEQ ID NO: 38, и VL, содержащую аминокислотную последовательность SEQ ID NO: 44, или
(и) VH, содержащую аминокислотную последовательность SEQ ID NO: 39, и VL, содержащую аминокислотную последовательность SEQ ID NO: 41, или
(к) VH, содержащую аминокислотную последовательность SEQ ID NO: 39, и VL, содержащую аминокислотную последовательность SEQ ID NO: 42, или
(л) VH, содержащую аминокислотную последовательность SEQ ID NO: 39, и VL, содержащую аминокислотную последовательность SEQ ID NO: 43, или
(м) VH, содержащую аминокислотную последовательность SEQ ID NO: 39, и VL, содержащую аминокислотную последовательность SEQ ID NO: 44, или
(н) VH, содержащую аминокислотную последовательность SEQ ID NO: 40, и VL, содержащую аминокислотную последовательность SEQ ID NO: 41, или
(о) VH, содержащую аминокислотную последовательность SEQ ID NO: 40, и VL, содержащую аминокислотную последовательность SEQ ID NO: 42, или
(п) VH, содержащую аминокислотную последовательность SEQ ID NO: 40, и VL, содержащую аминокислотную последовательность SEQ ID NO: 43, или
(р) VH, содержащую аминокислотную последовательность SEQ ID NO: 40, и VL, содержащую аминокислотную последовательность SEQ ID NO: 44.
10. Биспецифическая антигенсвязывающая молекула по любому одному из пар. 1-5, или 7, или 9, причем антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержит VH, содержащую аминокислотную последовательность SEQ ID NO: 37, и VL, содержащую аминокислотную последовательность SEQ ID NO: 41.
11. Биспецифическая антигенсвязывающая молекула по любому одному из пар. 1-5 или 8, причем антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержит
(а) VH, содержащую аминокислотную последовательность SEQ ID NO: 45, и VL, содержащую аминокислотную последовательность SEQ ID NO: 51, или
(б) VH, содержащую аминокислотную последовательность SEQ ID NO: 46, и VL, содержащую аминокислотную последовательность SEQ ID NO: 51, или
(в) VH, содержащую аминокислотную последовательность SEQ ID NO: 47, и VL, содержащую аминокислотную последовательность SEQ ID NO: 51, или
(г) VH, содержащую аминокислотную последовательность SEQ ID NO: 48, и VL, содержащую аминокислотную последовательность SEQ ID NO: 51, или
(д) VH, содержащую аминокислотную последовательность SEQ ID NO: 45, и VL, содержащую аминокислотную последовательность SEQ ID NO: 52, или
(е) VH, содержащую аминокислотную последовательность SEQ ID NO: 46, и VL, содержащую аминокислотную последовательность SEQ ID NO: 52, или
(ж) VH, содержащую аминокислотную последовательность SEQ ID NO: 47, и VL, содержащую аминокислотную последовательность SEQ ID NO: 52, или
(з) VH, содержащую аминокислотную последовательность SEQ ID NO: 48, и VL, содержащую аминокислотную последовательность SEQ ID NO: 52, или
(и) VH, содержащую аминокислотную последовательность SEQ ID NO: 49, и VL, содержащую аминокислотную последовательность SEQ ID NO: 53, или
(к) VH, содержащую аминокислотную последовательность SEQ ID NO: 50, и VL, содержащую аминокислотную последовательность SEQ ID NO: 53, или
(л) VH, содержащую аминокислотную последовательность SEQ ID NO: 49, и VL, содержащую аминокислотную последовательность SEQ ID NO: 54, или
(м) VH, содержащую аминокислотную последовательность SEQ ID NO: 50, и VL, содержащую аминокислотную последовательность SEQ ID NO: 54.
12. Биспецифическая антигенсвязывающая молекула по любому одному из пар. 15, или 8, или 11, причем антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержит VH, содержащую аминокислотную последовательность SEQ ID NO: 45, и VL, содержащую аминокислотную последовательность SEQ ID NO: 51, или причем антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержит VH, содержащую аминокислотную последовательность SEQ ID NO: 48, и VL, содержащую аминокислотную последовательность SEQ ID NO: 51.
13. Биспецифическая антигенсвязывающая молекула по любому одному из пар. 1-7, содержащая
(i) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержащий вариабельную область тяжелой цепи (VHCD40), содержащую аминокислотную последовательность SEQ ID NO: 37, и вариабельную область легкой цепи (VLCD40), содержащую аминокислотную последовательность SEQ ID NO: 41, и
(ii) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с FAP, содержащий вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 15, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 21.
14. Биспецифическая антигенсвязывающая молекула по любому одному из пар. 2 13, причем Fc-область представляет собой Fc-область IgG, в частности, IgG1, или Fc-область IgG4, и причем Fc-область содержит одну или более аминокислотных замен, которые снижают аффинность связывания антитела с Fc-рецептором и/или эффекторную функцию.
15. Биспецифическая антигенсвязывающая молекула по любому одному из пар. 2-14, причем Fc-область принадлежит подклассу IgG1 человека с аминокислотными мутациями L234A, L235A и P329G (нумерация в соответствии с индексом EU по Kabat).
16. Биспецифическая антигенсвязывающая молекула по любому одному из пар. 115, причем биспецифическая антигенсвязывающая молекула содержит
(а) по меньшей мере два Fab-фрагмента, способных к специфическому связыванию с CD40, соединенных с Fc-областью, и
(б) один антигенсвязывающий домен, способный к специфическому связыванию с FAP, соединенный с С-концом Fc-области.
17. Биспецифическая антигенсвязывающая молекула по любому одному из пар. 1-16, причем биспецифическая антигенсвязывающая молекула содержит
(а) по меньшей мере два Fab-фрагмента, способных к специфическому связыванию с CD40, слитых с Fc-областью, и
(б) кросс-fab-фрагмент, способный к специфическому связыванию с FAP, слитый с С-концом Fc-области.
18. Биспецифическая антигенсвязывающая молекула по пар. 17, причем VH-C-каппа цепь кросс-fab-фрагмента, способного к специфическому связыванию с FAP, слита с С-концом Fc-области.
19. Биспецифическая антигенсвязывающая молекула по любому одному из пар. 1-18, причем биспецифическая антигенсвязывающая молекула содержит четыре Fab-фрагмента, способных к специфическому связыванию с CD40.
20. Антитело, которое специфически связывается с FAP, причем указанное антитело содержит вариабельную область тяжелой цепи (VHFAP), содержащую (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 3, (ii) CDR-H2, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4, SEQ ID NO: 11 и SEQ ID NO: 12, и (iii) CDR-Н3, содержащую аминокислотную последовательность SEQ ID NO: 5, и вариабельную область легкой цепи (VLFAP), содержащую (iv) CDR-L1, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 6, SEQ ID NO: 13 и SEQ ID NO: 14, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 7, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 8.
21. Антитело no nap.20, причем указанное антитело содержит
(а) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 15, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 21,
(б) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 16, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 21,
(в) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 16, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 22, или
(г) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 19, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 25.
22. Выделенная нуклеиновая кислота, кодирующая биспецифическую антигенсвязывающую молекулу по любому одному из пар. 1-19 или антитело по пар. 20 или 21.
23. Экспрессионный вектор, содержащий выделенную нуклеиновую кислоту по пар. 22.
24. Клетка-хозяин, содержащая выделенную нуклеиновую кислоту по пар. 22 или экспрессионный вектор по пар. 23.
25. Способ получения биспецифической антигенсвязывающей молекулы, включающий культивирование клетки-хозяина по пар. 24 в условиях, подходящих для экспрессии биспецифической антигенсвязывающей молекулы, и выделение биспецифической антигенсвязывающей молекулы.
26. Фармацевтическая композиция, содержащая биспецифическую антигенсвязывающую молекулу по любому одному из пар. 1-19 или антитело по пар. 20 или 21 и фармацевтически приемлемый носитель.
25. Фармацевтическая композиция по пар. 24, дополнительно содержащая дополнительный терапевтический агент.
26. Биспецифическая антигенсвязывающая молекула по любому одному из пар. 1-19 или фармацевтическая композиция по пар. 24 для применения в качестве лекарственного средства.
27. Биспецифическая антигенсвязывающая молекула по любому одному из пар. 1-19 или фармацевтическая композиция по пар. 24 для применения
(i) в индукции иммунной стимуляции CD40-экспрессирующими антигенпрезентирующими клетками (АПК),
(ii) в стимуляции опухолеспецифического Т-клеточного ответа,
(iii) в инициации апоптоза опухолевых клеток,
(iv) в лечении рака,
(v) в задержке прогрессирования рака,
(vi) в продлении выживаемости пациента, страдающего от рака,
(vii) в лечении инфекций.
28. Биспецифическая антигенсвязывающая молекула по любому одному из пар. 1-19 или фармацевтическая композиция по пар. 24 для применения в лечении рака.
29. Применение биспецифической антигенсвязывающей молекулы по любому одному из пар. 1-19 или фармацевтической композиции по пар. 24 в производстве лекарственного средства для лечения рака.
30. Способ лечения индивида, имеющего рак, включающий введение индивиду эффективного количества биспецифической антигенсвязывающей молекулы по любому одному из пар. 1-19 или фармацевтической композиции по пар. 24.
31. Биспецифическая антигенсвязывающая молекула по любому одному из пар. 1-19 или фармацевтическая композиция по пар. 24 для применения в лечении рака, причем биспецифическую антигенсвязывающую молекулу вводят в комбинации с химиотерапевтическим агентом, облучением и/или другими агентами для применения в иммунотерапии рака.
ПРИМЕРЫ
Ниже приведены примеры способов и композиций по изобретению. Следует понимать, что на практике можно реализовать ряд других вариантов осуществления с учетом общего описания, приведенного выше.
Технологии рекомбинантных ДНК
Для манипуляций с ДНК применяли стандартные методы, описанные в Sambrook et al., Molecular cloning: A laboratory manual; Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1989. Молекулярные биологические реагенты использовали в соответствии с инструкциями производителя. Общая информация в отношении нуклеотидных последовательностей легких и тяжелых цепей человеческого иммуноглобулина приведена в: Kabat, Е.А. et al., (1991) Sequences of Proteins of Immunological Interest, Fifth Ed., NIH Publication No 91-3242.
Секвенирование ДНК
Последовательности ДНК определяли с помощью секвенирования двух цепей.
Синтез генов
Необходимые генные сегменты были созданы с помощью ПЦР с применением соответствующих матриц или были синтезированы Geneart AG (Regensburg, Germany) из синтетических олигонуклеотидов и продуктов ПЦР с помощью автоматизированного синтеза генов. В случаях, когда точная генная последовательность была недоступна, олигонуклеотидные праймеры конструировали на основании последовательностей самых близких гомологов и выделяли гены с помощью ОТ-ПЦР из РНК из соответствующей ткани. Генные сегменты, фланкируемые сингулярными сайтами расщепления рестрикционной эндонуклеазой, клонировали в стандартные векторы для клонирования/секвенирования. Плазмидную ДНК очищали из трансформированных бактерий, а концентрацию определяли с помощью УФ-спектроскопии. Последовательность ДНК субклонированных генных фрагментов подтверждали с помощью секвенирования ДНК. Генные сегменты конструировали с подходящими рестрикционными сайтами, чтобы обеспечить возможность субклонирования в соответствующие экспрессионные векторы. Все конструкции конструировали с 5'-концевой последовательностью ДНК, кодирующей лидерный пептид, который направляет белки для секреции в эукариотических клетках.
Очистка белка
Белки очищали из профильтрованных клеточных культуральных супернатантов согласно стандартным протоколам. Вкратце, антитела наносили на сефарозную колонку с протеином A (GE healthcare) и промывали ФСБ. Элюирование антител проводили при рН 2,8 с последующей незамедлительной нейтрализацией образца. Агрегированный белок отделяли от мономерных антител с помощью эксклюзионной хроматографии (Superdex 200, GE Healthcare) в ФСБ или в 20 мМ гистидине, 150 мМ NaCl, рН 6,0. Фракции мономерных антител объединяли, концентрировали (при необходимости), используя, например, центрифужный концентратор MILLIPORE Amicon Ultra (30 НОММ), замораживали и хранили при -20°C или -80°C. Часть образцов предоставляли для последующего белкового анализа и аналитического определения, например, с помощью ДСН-ПААГ, эксклюзионной хроматографии (ЭХ) или масс-спектрометрии.
ДСН-ПААГ
Гель-систему NuPAGE® Pre-Cast (Invitrogen) использовали в соответствии с инструкцией производителя. В частности, использовали 10% или 4-12% гели NuPAGE® Novex® Bis-TRIS Pre-Cast (рН 6,4) и NuPAGE® MES (восстановленные гели с добавление антиоксидантного подвижного буфера NuPAGE®) или подвижный буфер MOPS (невосстановленные гели).
КЭ-ДСН
Чистоту, целостность антител и молекулярную массу биспецифических и контрольных антител анализировали с помощью КЭ-ДСН, используя микрофлюидную технологию Labchip (Caliper Life Science, USA). 5 мкл раствора белка готовили для анализа КЭ-ДСН, используя набор реагентов НТ Protein Express в соответствии с инструкциями производителя и анализировали на системе LabChip GXII, используя чип НТ Protein Express. Данные анализировали, используя программное обеспечение LabChip GX версии 3.0.618.0.
Аналитическая эксклюзионная хроматография
Эксклюзионную хроматографию (ЭХ) для определения агрегации и олигомерного состояния антител проводили с помощью ВЭЖХ-хроматографии. Вкратце, очищенные с протеином А антитела наносили на колонку Tosoh TSKgel G3000SW в 300 мМ NaCl, 50 мМ KH2PO4/K2HPO4, рН 7,5, на системе Agilent HPLC 1100 или на колонку Superdex 200 (GE Healthcare) в 2 х ФСБ на ВЭЖХ-системе Dionex. Количественную оценку элюированного белка проводили по УФ-поглощению и интеграции площадей пиков. Стандартом служил стандарт гель-фильтрации BioRad 151-1901.
Масс-спектрометрия
В этом разделе описано определение характеристик мультиспецифических антител с обменом VH/VL или CH/CL (CrossMab) с акцентом на их правильную сборку. Ожидаемые первичные структуры анализировали с помощью масс-спектрометрии с ионизацией электрораспылением (ИЭР-МС) дегликозилированных интактных CrossMab и дегликозилированных/FabALACTICA или альтернативным образом дегликозилированных/GingisKHAN расщепленных CrossMab.
CrossMab дегликозилировали N-гликозидазой F в фосфатном или Трис-буфере при 37°C в течение до 17 ч при концентрации белка 1 мг/мл. Расщепление FabALACTICA или GingisKHAN (Genovis АВ; Sweden) проводили в буферах, предоставленных поставщиком, с 100 мкг дегликозилированных CrossMab. Перед масс-спектрометрией образцы обессоливали посредством ВЭЖХ на колонке Sephadex G25 (GE Healthcare). Общую массу определяли с помощью ИЭР-МС на системе maXis 4G UHR-QTOF MS (Bruker Daltonik), оснащенной источником TriVersa NanoMate (Advion).
Пример 1
Создание новых антител против белка активации фибробластов (FAP)
1.1 Иммунизация мышей
Для иммунизации использовали мышей линий Balb/c и NMRI. Животных содержали в соответствии с приложением А «Руководства по содержанию и уходу за животными» и аккредитованном AAALACi виварии. Все протоколы и эксперименты по иммунизации животных были одобрены органами правления Верхней Баварии (номер разрешения 55.2-1-54-2531-19-10) и проводились в соответствии с Законом и директивой о благополучии животных Германии 2010/63 Парламента и совета Европы. Мыши линий Balb/c и NMRI (n=5) возрастом 6 8 недель получали четыре раунда иммунизации рекомбинантно полученным внеклеточным доменом человеческого белка активации фибробластов альфа (аминокислоты 27-759; номер доступа NP_004451), ковалентно присоединенным к His-тэгу (SEQ ID NO: 78). Перед каждой иммунизацией мышей анестезировали газовой смесью кислорода и изофлурана. Для первой иммунизации 30 мкг белка, растворенного в ФСБ, рН 7,4, смешивали с эквивалентным объемом СБА (BD Difco, №263810) и вводили внутрибрюшинно (в/б). Другие 10 мкг белка, эмульгированного в адъюванте Abisco, вводили подкожно (п/к) на 6 неделе. Третью дозу в 5 мкг белка без адъюванта вводили в/б на 10 неделе. И наконец, за три дня до подготовки спленоцитов для получения антител с помощью технологии гибридомы, мышам проводили внутривенную (в/в) бустерную иммунизацию 50 мкг белка. Сыворотку тестировали в отношении выработки антиген-специфического общего антитела IgG с помощью ELISA. Через три дня после конечной иммунизации мышей умерщвляли и асептическим образом выделяли селезенку и готовили для генерации гибридомы. Выделяли мышиные лимфоциты и сливали с линией клеток миеломы мышей, используя стандартные протоколы на основе ПЭГ для генерации гибридом. Полученные в результате клетки гибридомы высевали приблизительно при 104 в плоскодонный 96-луночный микротитровальный планшет с последующей инкубацией приблизительно в течение двух недель в селективной среде, а потом проводили скрининг в отношении выработки антиген-специфических антител. После того, как начинался интенсивный рост гибридомы, секретирующие антитела гибридомы пересевали. Проводили скрининг гибридомных супернатантов в отношении специфического связывания с рекомбинантным человеческими белком активации фибробластов альфа (huFAP) методом ELISA с последующей оценкой кинетических параметров связывания с рекомбинантным huFAP с использованием измерений Biacore.
Культуры гибридом: Созданные гибридомы muMAb культивировали в RPMI 1640 (PAN - № в каталоге (кат. №) РО4-17500), дополненной 2 мМ L-глутамина (GIBCO - кат. №35050-038), 1 мМ Na-пирувата (GIBCO - кат. №11360-039), 1x NEAA (GIBCO - кат. №11140-035), 10% ФТС (РАА - кат. №А15-649), 1x Pen Strep (Roche - кат. №1074440), 1x Nutridoma CS (Roche - кат. №1363743), 50 мкМ меркаптоэтанола (GIBCO - кат. №31350-010) и 50 Е/мл IL 6 мыши (Roche - кат. №1 444 581) при 37°C и 5% CO2.
1.2 Конкурентное клеточное связывание анти-huFAP антител с клоном FAP 4В9 и 28Н1
Полученные в результате клоны тестировали в отношении их связывания в сравнении с клоном FAP 4В9. Создание и подготовка клонов FAP 4В9 и 28Н1 описаны в WO 2012/020006 А2, которая включена в данный документ посредством ссылки. Чтобы определить, распознают ли мышиные клоны FAP разные эпитопы, как клоны 4В9 и 28Н1, проводили анализ конкурентного связывания с человеческим FAP, экспрессируемым на трансфицированных клетках HEK.
Вкратце, целевые клетки собирали в буфер для диссоциации клеток, промывали буфером FACS (ФСБ+2% ФТС+5 мМ ЭДТА+0,25% кислого натрия) и высевали в 96-луночные планшеты с U-образным дном (1×105 клеток/лунка). Немеченные первичные антитела к человеческому FAP (mu IgG1) добавляли к клеткам (конечные концентрации от 60 мкг/мл до 0,2 мкг/мл; разведения 1:3) и инкубировали в течение 20 мин при 4°C перед добавлением меченного AlexaFluor647 анти-FAP антитела 4В9 или 28Н1 (конечная концентрация 20 мкг/мл). Через 30 мин инкубации при 4°C клетки промывали, фиксировали и измеряли интенсивность флуоресцентного сигнала AF647-меченных клонов 4В9 и 28Н1, используя Miltenyi MACSQuant.
Как видно на Фиг. 2А и 2Б, было идентифицировано 10 полученных из гибридомы антител (названия клонов 209, 210, 211, 212, 213, 214, 215, 216, 217 и 218), которые не конкурировали за связывание с анти-FAP антителами 4В9 или 28Н1.
1.3 Специфичность связывания мишени анти-huFAP мышиных антител
Белок активации фибробластов (FAP, FAP-α, сепраза) представляет собой трансмембранную сериновую протеазу типа II, принадлежащую семейству пролилолигопептидаз. Это семейство включает сериновые протеазы, которые расщепляют пептиды преимущественно после остатков пролина. Другими важными членами этого семейства, которые экспрессируются в человеческом протеоме, являются пролилолигопептидаза (PREP) и дипептидилпептидазы (DPP). DPP-IV является самым близким гомологом FAP. В отличие от FAP, DPP-IV экспрессируется повсеместно и играет роль в различных биологических процессах, таких как костимуляция Т-клеток, биология хемокинов, метаболизм глюкозы и онкогенез, и, следовательно, необходимые антитела против FAP человека не должны связываться с DPP-IV человека.
Связывание с FAP человека и DPP-IV человека определяли методом проточной цитометрии, используя трансфицированные FAP человека или DPPIV человека клетки НЕК. Вкратце, целевые клетки собирали в буфер для диссоциации клеток, промывали буфером FACS (ФСБ+2% ФТС+5 мМ ЭДТА+0,25% кислого натрия) и высевали в 96-луночные планшеты с U-образным дном (1×105 клеток/лунка). Немеченные первичные антитела добавляли к клеткам (конечная концентрация 10 мкг/мл) и инкубировали в течение 30 мин при 4°C. После промывания клетки инкубировали с козьим антимышиным IgG-РЕ F(ab')2 (Serotec) в течение 30 мин при 4°C в темноте. После этого клетки промывали, фиксировали и проводили измерения, используя BD FACS Canto™ II. Неспецифическое связывание с DPP-IV человека не было выявлено ни для одного из 10 полученных из гибридомы антител против FAP человека.
1.4 Создание анти-huFAP антител в формате huIgG1_LALA_PG
Последовательности ДНК новых анти-huFAP антител определяли стандартными методами секвенирования. На основании доменов VH и VL экспрессировали новые анти-FAP антитела в виде антител hulgGI с подавленной эффекторной активностью Fc (P329G; L234, L235A) для устранения связывания с Fcγ-рецепторами в соответствии со способом, описанным в WO 2012/130831 А1. А именно, экспрессировали антитела посредством временной трансфекции клеток HEK293-F, выращиваемых в суспензии, экспрессионными векторами, кодирующими разные пептидные цепи. Трансфекцию в клетки HEK293-F (Invitrogen, USA) проводили в соответствии с инструкциями поставщика клеток, используя препараты векторов антител Maxiprep (Qiagen, Germany), среду на основе F17 (Invitrogen, USA), PEIpro (Polyscience Europe GmbH) и начальную плотность клеток 1-2 миллиона жизнеспособных клеток/мл в бессывороточной экспрессионной среде FreeStyle 293 (Invitrogen). Клеточные культуральные супернатанты собирали через 7 суток культивирования во встряхиваемые флаконы или ферментеры с перемешиванием путем центрифугирования при 14000 g в течение 30 минут и фильтровали через 0,22 мкм фильтр.
Антитела очищали из клеточных культуральных супернатантов с помощью аффинной хроматографии, используя хроматографию MabSelectSure-Sepharose™ (GE Healthcare, Sweden). Вкратце, стерильно профильтрованные клеточные культуральные супернатанты иммобилизовали на смоле MabSelect SuRe, уравновешенной буфером ФСБ (10 мМ Na2HPO4, 1 мМ KH2PO4, 137 мМ NaCl и 2,7 мМ KCl, рН 7,4), промывали уравновешивающим буфером и элюировали 25 мМ цитратом, рН 3,0. После нейтрализации 1 М Трис, рН 9,0, агрегированный белок отделяли от мономерных молекул антител с помощью эксклюзионной хроматографии (Superdex 200, GE Healthcare) в 20 мМ гистидине, 140 мМ NaCl, рН 6,0. Фракции мономерных белков объединяли, концентрировали при необходимости, используя, например, центрифужный концентратор MILLIPORE Amicon Ultra (30 кДа НОММ), замораживали и хранили при 80°C. Аликвоты образцов использовали для последующего аналитического определения, например, с помощью КЭ-ДСН, эксклюзионной хроматографии, масс-спектрометрии и определения содержания эндотоксинов.
1.5 Клеточное связывание анти-huFAP антител
Связывание анти-FAP антител с IgG1 P329G LALA Fc человека с FAP человека определяли методом проточной цитометрии, используя трансфицированные FAP человека клетки НЕК. Вкратце, целевые клетки собирали в буфер для диссоциации клеток, промывали буфером FACS (ФСБ+2% ФТС+5 мМ ЭДТА+0,25% кислого натрия) и высевали в 96-луночные планшеты с U-образным дном (1×105 клеток/лунка). Немеченные первичные антитела добавляли к клеткам (конечные концентрации от 10 мкг/мл до 0,64 нг/мл: разведения 1:5) и инкубировали в течение 30 мин при 4°C. После промывания клетки инкубировали РЕ-конъюгированным фрагментом AffiPure F(ab)2 козьего античеловеческого IgG, специфическим в отношении Fcγ (Jackson Immune-research) в течение 30 мин при 4°C в темноте. После этого клетки промывали, фиксировали и проводили измерения, используя BD FACS LSR Fortessa™.
Все анти-FAP антитела демонстрировали сходное связывание с FAP человека, что наблюдали и ранее. Значения ЕС50 выбранных связывающих молекул приведены ниже в таблице 1.

1.6 Клеточная интернализация анти-huFAP антител
Интернализацию FAP-связывающих молекул определяли, используя трансфицированные FAP человека клетки НЕК в качестве мишеней. Вкратце, целевые клетки собирали в буфер для диссоциации клеток, промывали холодным буфером FACS (ФСБ+2% ФТС+5 мМ ЭДТА+0,25% кислого натрия) и ресуспендировали при 1,5×106 клеток/мл в холодном буфере FACS. Клетки распределяли по 15 мл пробиркам (каждая пробирка содержит 3×106 клеток в 2 мл). По 2 мл раствором антитела против FAP человека добавляли к клеткам (конечная концентрация 20 мкг/мл) и инкубировали в течение 45 мин при 4°C. После этого клетки промывали, ресуспендировали в холодном буфере FACS и незамедлительно высевали клетки для момента времени «0» в 96-луночные планшеты с U-образным дном (1,5×105 клеток/лунка) и держали при 4°C, тогда как остальные клетки центрифугировали, ресуспендировали в теплой среде RPMI1640, содержащей 10% FCS и 1% Glutamax (1,5×106 клеток/мл) и переносили в увлажненный инкубатор (5% CO2) с температурой 37°C. После каждого указанного момента времени по 100 мкл/пробирка клеточной суспензии переносили в планшеты, незамедлительно охлаждали холодным буфером FACS и хранили в холодильнике до тех пор, пока не были собраны образцы для всех моментов времени. После сбора образцов для всех моментов времени клетки промывали холодным буфером FACS и инкубировали с РЕ-меченным вторичным антителом в течение 30 мин при 4°С. После этого клетки промывали, фиксировали и проводили измерения, используя BD FACS Canto™ II.
Сигналы, генерируемые меченным вторичным антителом, оставались приблизительно постоянными с течением времени, что означает, что с течением времени не наблюдалось потери антител, ни одно из тестируемых анти-huFAP антител не было интернализовано.
1.7 Кинетика связывания анти-huFAP антител
Чтобы оценить кинетику связывания FAP человека, биотинилированный FAP человека иммобилизовали на чипе Biacore CAPture серии S (GE Healthcare 28-9202-34) в соответствии с инструкциями производителя с получением в результате поверхностной плотности приблизительно 20 резонансных единиц (РЕ). В качестве подвижного буфера и буфера для разведения использовали HBS-P+ (10 мМ ГЭПЭС, 150 мМ NaCl, рН 7,4, 0,05% сурфактанта Р20). Последовательно вводили серийные разведения анти-huFAP Fab (3,7-300 нМ, разведение 1:3) в течение 120 с каждое, диссоциацию отслеживали в течение 1800 с при скорости потока 30 мкл/мин (кинетика одного цикла). Поверхность восстанавливали путем ввода 6 М гуанидина-HCl, 0,25 М NaOH в течение 120 с. Объемную разницу показателя преломления корректировали путем вычитания контрольных введений и путем вычитания ответа, полученного от контрольной проточной ячейки без иммобилизованного FAP человека. Аппроксимацию кривых проводили, используя 1:1 модель связывания Ленгмюра в оценочном программном обеспечении Biacore. Данные по аффинности приведены ниже в таблице 2.
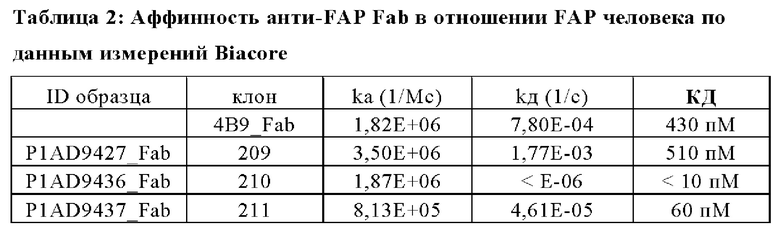

1.8 Формат-зависимое связывание анти-huFAP клонов
Чтобы определить, не утрачиваются ли связывающие свойства анти-FAP клонов, когда они слиты в С-конце с Fc-доменом, получали конструкции, содержащие цепь Fc-выступа и цепь Fc-впадины, где VH-домен слит с С-концом цепи Fc-выступа, а VL-домен слит с С-концом цепи Fc-впадины (Фиг. 3А, С-концевое слияние VH/VL), конструкции, содержащие цепь Fc-выступа и цепь Fc-впадины, где целый Fab слит посредством VH-домена с С-концом цепи Fc-выступа (Фиг. 3Б, С-концевое слияние Fab). Цепь Fc-выступа имеет аминокислотную последовательность SEQ ID NO: 97, а цепь Fc-впадины имеет аминокислотную последовательность SEQ ID NO: 98.
Аффинность конструкций в отношении биотинилированного рекомбинантного FAP человека и биотинилированного рекомбинантного FAP яванского макака в сравнении с антителами приведена ниже в таблице 3.

Также определяли клеточное связывание конструкций с FAP-трансфицированными клетками HEK, как описано выше в данном документе. Значения ЕС50 приведены в таблице 4. Конструкции с С-концевым слиянием всех анти-FAP антител были способны связываться с FAP человека и яванского макака, при этом конструкции, в которых весь Fab слит посредством VH-домена с С-концом цепи Fc-выступа, превосходили те, в которых VH-домен слит с С-концом цепи Fc-выступа, а VL-домен слит с С-концом цепи Fc-впадины.
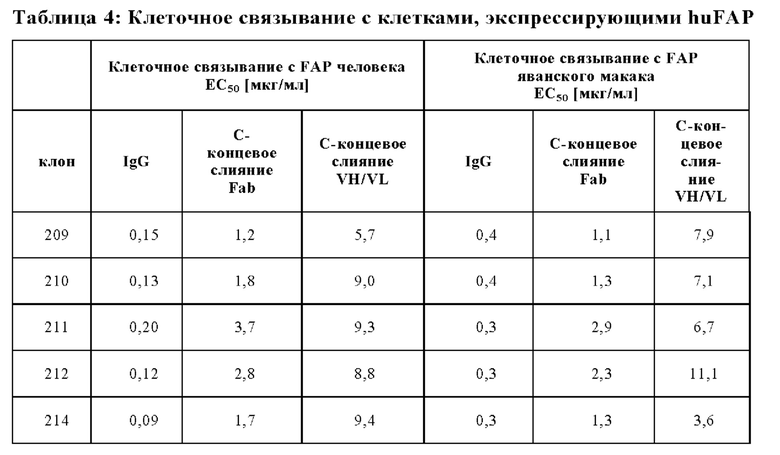
1.9 Конкурентное связывание клонов против FAP человека по определению методом Biacore
Сортировку эпитопов проводили, используя анализ на основании метода поверхностного плазмонного резонанса (ППР) на инструменте Biacore Т200. Проводили захват антигена FAP иммобилизованным анти-His антителом. На первом этапе связывающую FAP молекулу вводили до насыщения. После этого вводили вторую связывающую FAP молекулу. Дизайн анализа схематически показан на Фиг. 3В. Повышение сигнала связывания после добавления второго антитела указывает на его связывание с отличным эпитопом, чем в случае первого антитела. Отсутствие дополнительного связывания указывало, что первое и второе антитело распознают одну эпитопную область.
Анти-His антитело (набор GE Healthcare 28-9950-56) в концентрации 20 мкг/мл иммобилизовали посредством аминного сопряжения (набор GE Healthcare BR-1000-50) на поверхности сенсорного чипа СМ5 (GE Healthcare BR-1005-30). Время ввода составляло 600 секунд при скорости потока 10 мкл/мин с получением 12000 единиц ответа (ЕО) на двух проточных ячейках, одну из которых использовали как референс, а другую использовали как активную проточную ячейку. Подвижным буфером был HBS-N (GE Healthcare BR-1006-70). Для измерений в качества подвижного буфера и буфера для разведения использовали PBS-P+ (GE Healthcare 28-9950-84). Температура проточной ячейки была установлена на 25°С, отделения для образца - на 12°С. Скорость потока составляла 10 мкл/мин в течение всего эксперимента.
Захват His-меченного антигена FAP на активной проточной ячейке проводили при концентрации 20 мкг/мл в течение 180 секунд. Первое и второе антитело (FAP-связывающие молекулы) проводили последовательно, каждое в течение 120 секунд в концентрации 10 мкг/мл через обе проточные ячейки. После каждого цикла поверхность восстанавливали 10 мМ глицином, рН 1,5, в течение 60 секунд (GE Healthcare BR-1003-54).
Результаты приведены ниже в таблице 5:
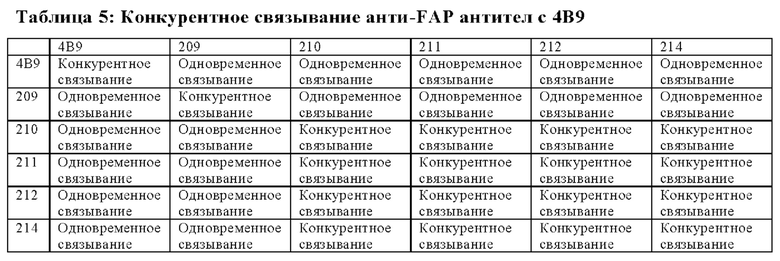
Таким образом, были идентифицированы группы эпитопов. Согласно требованиям, ни одно из анти-FAP антител не конкурировало за связывание с антителом 4В9 (группа эпитопов 1). Антитела 210, 211, 212 и 214 конкурировали друг с другом за связывание и, таким образом, формировали одну группу (группа эпитопов 3), тогда как антитело 209 не конкурировало за связывание ни с одним другим антителом (группа эпитопов 2).
1.9 Оценка термической стабильности анти-FAP антител
Образцы готовят в концентрации 1 мг/мл в 20 мМ смеси гистидина/хлорида гистидина, 140 мМ NaCl, рН 6,0, переносят в оптический 384-луночный планшет путем центрифугирования через 0,4 мкм фильтровальную пластину и покрывают парафинным маслом. Гидродинамический радиус повторно измеряют методом динамического рассеяния света на планшет-ридере DynaPro (Wyatt) при нагревании образцов со скоростью 0,05°С/мин от 25°С до 80°С. В альтернативном варианте образцы переносили в 10 мкл микрокюветную матрицу и записывали данные по статическому рассеянию света, а также данные по флуоресценции после возбуждения лазером с 266 нм на инструменте Optim1000 (Avacta Inc.) при нагревании со скоростью 0,1°С/мин от 25°С до 90°С. Температуру начала агрегации (Таг) определяют как температуру, при которой гидродинамический радиус (ДРС) или интенсивность рассеянного света (Optim1000) начинают увеличиваться. Температуру плавления определяют как точку перегиба на графике зависимости интенсивности флуоресценции от длины волны. Температуры начала агрегации выбранных анти-FAP антител приведены в таблице 6.

Анти-FAP клон 212 был выбран для гуманизации, поскольку он связывается со сравнимой с антителом 4В9 высокой аффинностью с FAP человека и демонстрирует благоприятные свойства для разработки. In silico анализ его последовательностей показал только одну предсказанную горячую точку деградации (Trp в позиции 401). Последовательности мышиного клона 212 приведены в таблице 7.

1.10 Гуманизация анти-FAP клона 212
1.10.1 Методология
Подходящие человеческие акцепторные каркасные области идентифицировали, проводя запрос по базе данных BLASTp относительно человеческих последовательностей V- и J-области для мышиных вставочных последовательностей (обрезанных до вариабельной части). Критериями отбора для выбора человеческой акцепторной области были гомология последовательности, одинаковая или сходная длина CDR и оценочная частота зародышевой линии человека, но также консервативность определенных аминокислот на поверхности контакта доменов VH-VL. После этапа идентификации зародышевой линии CDR мышиных вставочных последовательностей прививали в человеческие акцепторные каркасные области. Каждое аминокислотное отличие между этими исходными привитыми CDR и родительскими антителами оценивали в отношении возможного влияния на целостность структуры соответствующей вариабельной области и в случае необходимости вносили «обратные мутации» в направлении родительской последовательности. Структурная оценка основывалась на моделях гомологии Fv-области родительского антитела и гуманизированных вариантов, созданных по внутрилабораторному протоколу моделирования гомологии структуры антител, реализуемому с применением Biovia Discovery Studio Environment версии 17R2. В некоторые гуманизированные варианты были внесены «прямые мутации», т.е. аминокислотные замены, которые изменяют оригинальную аминокислоту, находящуюся в заданной позиции CDR родительской связывающей молекулы, на аминокислоту, находящуюся в эквивалентной позиции акцепторной зародышевой линии человека. Это призвано придать больший общий «человеческий» характер гуманизированным вариантам (помимо каркасных областей), чтобы дополнительно снизить риск иммуногенности.
In silico инструмент, разработанный в лаборатории, использовали для предсказания ориентации домена VH-VL гуманизированных вариантов со спаренными доменами VH и VL (смотрите WO 2016/062734). Результаты сравнивали с предсказанной ориентацией доменов VH-VL родительских связывающих молекул, чтобы выбрать комбинации каркасных областей, которые близки по геометрии к оригинальным антителам. Обоснование является выявление возможных аминокислотных замен в области контакта VH-VL, которые могут приводить к разрушительным изменениям в спаривании двух доменов, которые в свою очередь могут оказывать негативное действие на свойства связывания.
1.10.2 Выбор и адаптация акцепторной каркасной области
Были выбраны следующие каркасные области:
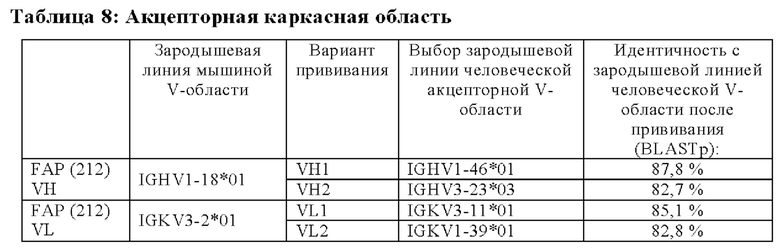
Пост-CDR3 каркасные области адаптировали из IGHJ6*01/02 (YYYYYGMDVWGQGTTVTVSS) человеческой зародышевой линии IGHJ и IGKJ4*01/02 (LTFGGGTKVEIK) человеческой зародышевой линии IGKJ. Часть, относящаяся к акцепторной каркасной области, выделена жирным шрифтом.
На основании структурных соображений были внесены обратные мутации с человеческой акцепторной каркасной области на аминокислоту из родительской связывающей молекулы в позициях Н43 (Q>K), Н44 (G>S), Н48 (М>1), Н71 (R>I), Н73 (Т>K), Н93 (А>Т) [VH1], Н49 (S>G), Н71 (R>I), Н73 (N>K), Н78 (L>A), H93 (A>T), H94 (K>R) [VH2], L36 (Y>F), L43 (A>P), L87 (Y>F) [VL1] и L36 (Y>F), L42 (K>Q), L43 (A>P), L85 (T>M), L87 (Y>F) [VL2].
Кроме того, позиции H60 (N>A), H64 (K>Q) [VH1], H60 (N>A), H61 (Q>D), H62 (K>S), H63 (F>V) [VH2], L33 (I>L), L34 (N>A) [VL1] и L27b (V>I), L33 (I>L) [VL2] были идентифицированы как перспективные кандидаты для прямых мутаций. Все позиции приведены по схеме нумерации EU по Kabat.

Примечание: Обратные мутации обозначены префиксом b, прямые мутации - f, например, bM481 относится к обратной мутации (аминокислоты человеческой зародышевой линии на аминокислоту родительского антитела) с метионина на изолейцин в позиции 48 (Kabat).
Полученные в результате домены VH и VL гуманизированных антител к FAP на основе акцепторной каркасной области приведены ниже в таблице 10.

1.10.3 Новые гуманизированные анти-FAP Fab
На основе новых гуманизированных вариантов VH и VL экспресиировали новые анти-FAP Fab.
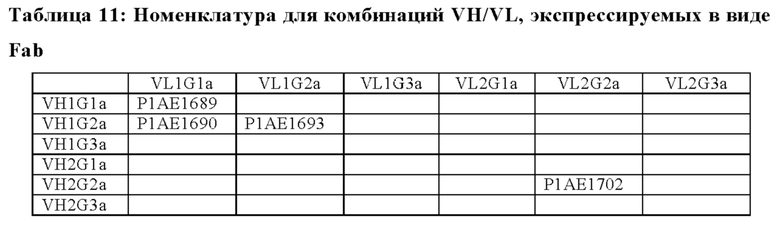
Аффинность новых гуманизированных анти-FAP вариантов на основе клона 212 анализировали в сравнении с анти-FAP антителом 4В9. Кроме того, рассчитывали степень «человечности» гуманизированных вариантов и измеряли температуру начала агрегации.

1.11 Связывание FcRn/гепарина и in silico распределение заряда
Распределение заряда антител 4В9 и Р1АЕ1689 в ФСБ, рН 7,4, рассчитывали в in-silico модели. В соответствии с моделью, 4В9 имеет большой положительно заряженный участок, что иногда коррелирует с повышенным связыванием гепарина. С другой стороны, Р1АЕ1689 имеет большой отрицательно заряженный участок, который может указывать на слабое взаимодействие с гепарином.
Эти прогнозы были подтверждены хроматографически для обоих антител с использованием FcRn-аффинной колонки и градиента рН, а также гепарин-аффинной колонки и градиента рН. В WO 2015/140126 описан способ предсказания in vivo времени полужизни антитела на основании времени удержания, определенного для FcRn-аффинной хроматографической колонки, тогда как связывание гепарина коррелирует с неспецифическими взаимодействиями со структурами клеточной поверхности.
Пример 2
Создание и получение гуманизированных вариантов анти-CD40 антитела S2C6
2.1 Создание гуманизированных вариантов анти-CD40 антитела S2C6
2.2.1 Методология
Для идентификации подходящей человеческой акцепторной каркасной области во время гуманизации анти-CD40 связывающей молекулы S2C6 использовали комбинацию двух методологий. С одной стороны использовали классический подход поиска акцепторной каркасной области с высокой гомологией последовательности, прививания CDR в эту каркасную область и оценки того, какие обратные мутации предположительно можно провести. Точнее, каждое аминокислотное отличие идентифицированных каркасных областей и родительского антитела оценивали в отношении влияния на целостность структуры связывающей молекулы и в случае необходимости вносили обратные мутации в направлении родительской последовательности. Структурная оценка основывалась на моделях гомологии Fv-области как родительского антитела, так и его гуманизированных версий, созданных с помощью внутрилабораторного инструмента моделирования гомологии структуры антител, реализуемого с применением Biovia Discovery Studio Environment версии 4.5.
С другой стороны, использовали разработанный в лаборатории in silico инструмент для предсказания ориентации доменов VH и VL гуманизированных версий в отношении друг друга (смотрите WO 2016062734, включенную в данный документ посредством ссылки). Результаты сравнивали с предсказанной ориентацией доменов VH-VL родительской связывающей молекулы, чтобы выбрать комбинации каркасных областей, которые близки по геометрии с исходным антителом. Обоснование является выявление возможной аминокислотной замены в области контакта VH-VL, которая может приводить к разрушительным изменениям в спаривании двух доменов.
2.2.2 Выбор и адаптация акцепторной каркасной области
Были выбраны две разные акцепторные каркасные области, описанные ниже в таблице 16 и таблице 18.

Пост-CDR3 каркасные области адаптировали из IGHJ6*01/02 (YYYYYGMDVWGQGTTVTVSS) человеческой зародышевой линии IGHJ и IGKJ4*01/02 (LTFGGGTKVEIK) человеческой зародышевой линии IGKJ. Часть, относящаяся к акцепторной каркасной области, выделена жирным шрифтом.
На основании структурных соображений были внесены обратные мутации с человеческой акцепторной каркасной области на аминокислоту из родительской связывающей молекулы в позициях Н43 (Q>K), Н44 (G>S), Н69 (M>L), Н71 (R>V), H73 (Т>K), H88 (V>A) и H105 (Q>H) VH-области и позициях L2 (I>V), L4 (M>V), L87 (Y>F) и L104 (V>L) VL-области. В одном варианте была внесена мутация T70S (VH) для исследования эффекта немного более гидрофильного остатка в этой позиции.
Все варианты содержали мутацию N54A (VH) для разрешения предполагаемой возможности образования горячей точки (дезамидирование аспарагина). Все позиции приведены по схеме нумерации EU по Kabat.
В нижеприведенной таблице 14 проиллюстрирована матрица спаривания VH-VL гуманизированных вариантов:
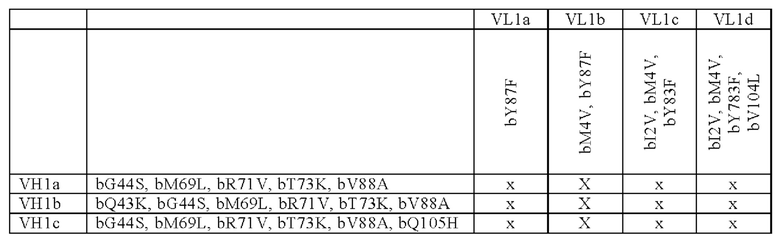
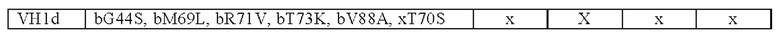
Мутация N54A применима ко всем вариантам VH и явным образом не указана. Обратные мутации обозначены префиксом b, прямые мутации обозначены префиксом f, а другие мутации обозначены префиксом х

Пост-CDR3 каркасные области адаптировали из IGHJ6*01/02 (YYYYYGMDVWGQGTTVTVSS) человеческой зародышевой линии IGHJ и IGKJ4*01/02 (LTFGGGTKVEIK) человеческой зародышевой линии IGKJ. Часть, относящаяся к акцепторной каркасной области, выделена жирным шрифтом.
На основании структурных соображений были внесены обратные мутации с человеческой акцепторной каркасной области на аминокислоту из родительской связывающей молекулы в позициях Н44 (G>S), Н49 (S>G), Н71 (R>V), Н78 (L>A), H94 (K>R) и H105 (Q>H) VH-области и позициях L42 (K>Q), L43 (A>S) и L87 (Y>F) VL-области. Кроме того, четыре позиции в CDR-H2 были идентифицированы как перспективные кандидаты для прямых мутаций, т.е. аминокислотных замен с родительской связывающей молекулы на человеческую зародышевую линию с целью повышения общего «человеческого» характера, а именно Н60 (N>G), Н61 (Q>D), Н62 (K>S) и Н63 (F>V).
Все варианты содержали мутацию N54A (VH) для разрешения предполагаемой возможности образования горячей точки (дезамидирование аспарагина). Все позиции приведены по схеме нумерации EU по Kabat.
В нижеприведенной таблице 16 проиллюстрирована матрица спаривания VH-VL гуманизированных вариантов:

Обратные мутации обозначены префиксом b, прямые мутации обозначены префиксом f.
2.2.3 Домены VH и VL полученных в результате гуманизированных антител к CD40
Полученные в результате домены VH и VL гуманизированных антител к CD40 на основе акцепторной каркасной области 1 приведены ниже в таблице 17, а полученные в результате домены VH и VL гуманизированных антител к CD40 на основе акцепторной каркасной области 2 приведены ниже в таблице 18.


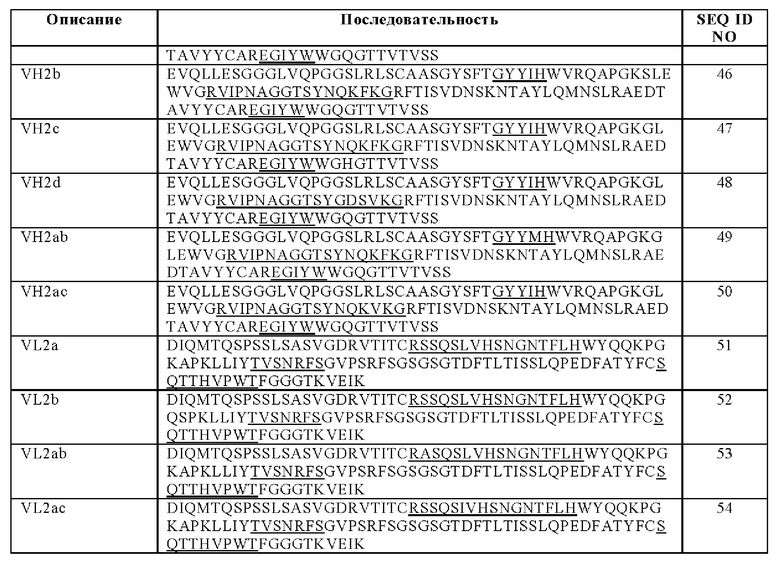
2.2.4 Новые гуманизированные антитела к CD40 в формате huIgG1_LALA_PG
На основании новых гуманизированных вариантов VH и VL экспрессировали новые антитела к CD40 в виде антител huIgG1 с подавленной эффекторной активностью Fc (P329G; L234, L235A) для устранения связывания с Fcγ-рецепторами в соответствии со способом, описанным в WO 2012/130831 А1.
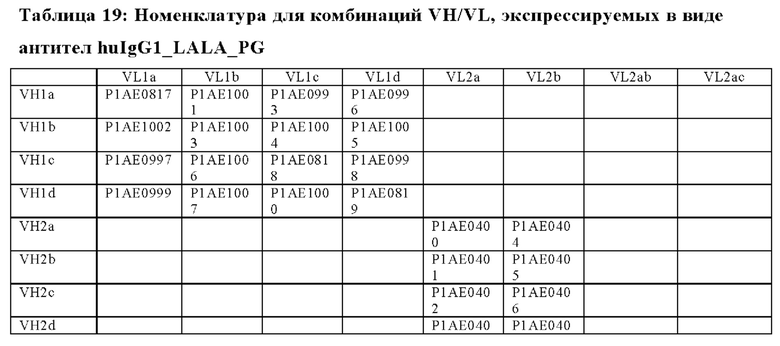
Типовые полноразмерные последовательности гуманизированных антител к CD40 в виде человеческих антител IgG1_LALAPG приведены в таблице 20.
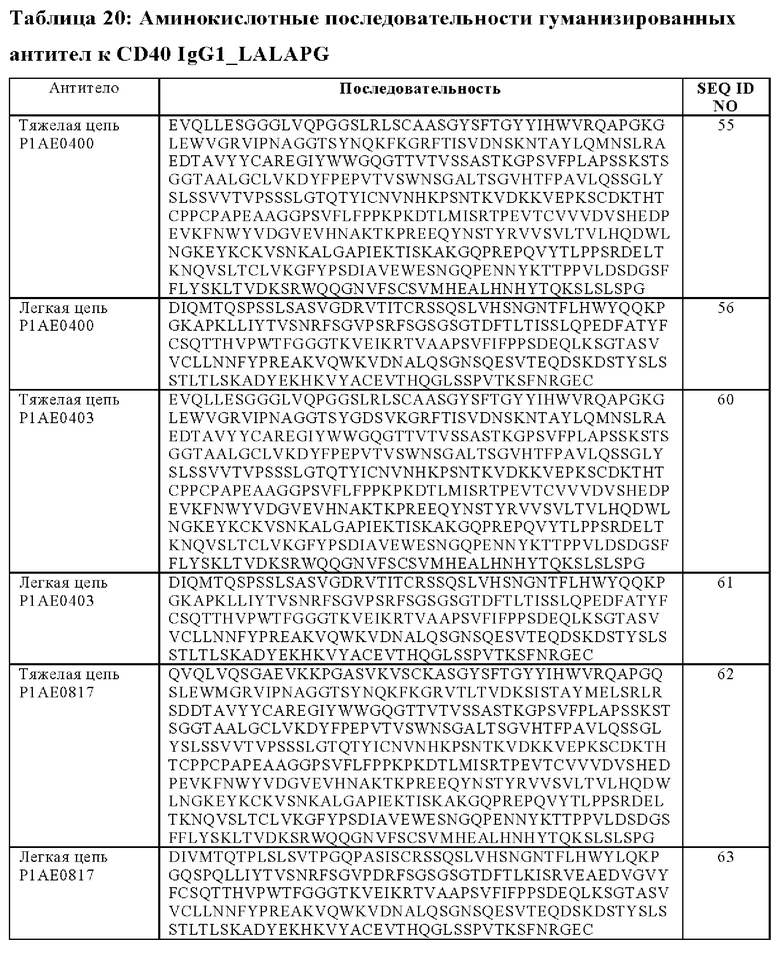
2.2.5 Получение новых гуманизированных антител к CD40 в формате huIgG1_LALA_PG
Антитела экспрессировали посредством временной трансфекции клеток HEK293-F, выращиваемых в суспензии, экспрессионными векторами, кодирующими разные пептидные цепи. Трансфекцию в клетки HEK293-F (Invitrogen, USA) проводили в соответствии с инструкциями поставщика клеток, используя препараты векторов антител Maxiprep (Qiagen, Germany), среду на основе F17 (Invitrogen, USA), PEIpro (Polyscience Europe GmbH) и начальную плотность клеток 1-2 миллиона жизнеспособных клеток/мл в бессывороточной экспрессионной среде FreeStyle 293 (Invitrogen). Клеточные культуральные супернатанты собирали через 7 суток культивирования во встряхиваемые флаконы или ферментеры с перемешиванием путем центрифугирования при 14000 g в течение 30 минут и фильтровали через 0,22 мкм фильтр.
Антитела очищали из клеточных культуральных супернатантов с помощью аффинной хроматографии, используя хроматографию MabSelectSure-Sepharose™ (GE Healthcare, Sweden). Вкратце, стерильно профильтрованные клеточные культуральные супернатанты иммобилизовали на смоле MabSelect SuRe, уравновешенной буфером ФСБ (10 мМ Na2HPO4, 1 мМ KH2PO4, 137 мМ NaCl и 2,7 мМ KCl, рН 7,4), промывали уравновешивающим буфером и элюировали 25 мМ цитратом, рН 3,0, с последующей нейтрализацией 1 М Трис, рН 9,0. В зависимости от качества продукта, получаемого после очистки с протеином А, включали этап хроматографии с гидрофобным взаимодействием (ХГВ), используя смолу Butyl-Sepharose 4FF (GE Healthcare, Sweden). Перед ХГВ-очисткой проводили диализ белка против уравновешивающего буфера ХГВ. ХГВ-очистку проводили, используя 40 мМ ацетат, 1,5 М сульфат аммония, рН 5,5, в качестве уравновешивающего/промывочного буфера и 40 мМ ацетат, рН 5,5, в качестве элюирующего буфера, и применяли для очистки линейный градиент. После этого агрегированный белок отделяли от мономерных молекул антител с помощью эксклюзионной хроматографии (Superdex 200, GE Healthcare) в 20 мМ гистидине, 140 мМ NaCl, рН 6,0. Фракции мономерных белков объединяли, концентрировали при необходимости, используя, например, центрифужный концентратор MILLIPORE Amicon Ultra (30 кДа НОММ), замораживали и хранили при 80°С. Аликвоты образцов использовали для последующего аналитического определения, например, с помощью КЭ-ДСН, эксклюзионной хроматографии, масс-спектрометрии и определения содержания эндотоксинов.
Выход при получении разных гуманизированных антител к CD40 приведен в таблице 21 в виде значений титров, рассчитанных по выходу после препаративной аффинной хроматографии с применением хроматографии MabSelectSure-Sepharose™.
Чистоту и молекулярную массу молекулы после финального этапа очистки анализировали с помощью анализа КЭ-ДСН в присутствии или отсутствие восстанавливающего агента. Систему Caliper LabChip GXII (Caliper Lifescience) использовали в соответствии с инструкцией производителя.
Содержание агрегатов молекул анализировали, используя аналитическую эксклюзионную колонку TSKgel G3000 SW XL (Tosoh) в 25 мМ фосфате калия, 125 мМ хлориде натрия, 200 мМ L-аргинина моногидрохлориде, 0,02% (масс./об.) NaN3, рН 6,7, подвижный буфер при 25°С.
Для прямого сравнения всех антител отслеживали термическую стабильность методом статического рассеяния света (СРС) и путем измерения характерной белковой флуоресценции в ответ на применяемый термический стресс. 30 мкг профильтрованного образца белка с концентрацией белка 1 мг/мл вносили в двух повторностях в инструмент Optim 2 (Avacta Analytical Ltd). Температуру пошагово повышали от 25 до 85°С при 0,1°С/мин с получением данных по радиусу и общей интенсивности рассеяния. Для определения характерной белковой флуоресценции образец возбуждали на 266 нм и измеряли испускание между 275 нм и 460 нм. Для всех антител температура агрегации (Таг) составляла от 64°С до 69°С и приведена ниже в таблице 21 или таблице 22.
Выход при получении гуманизированных антител к CD40 с разными каркасными областями приведен ниже в таблице 21 или таблице 22.
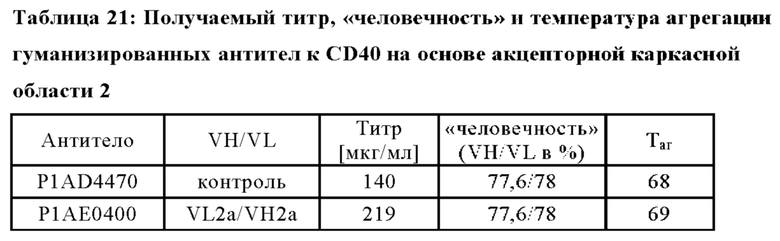
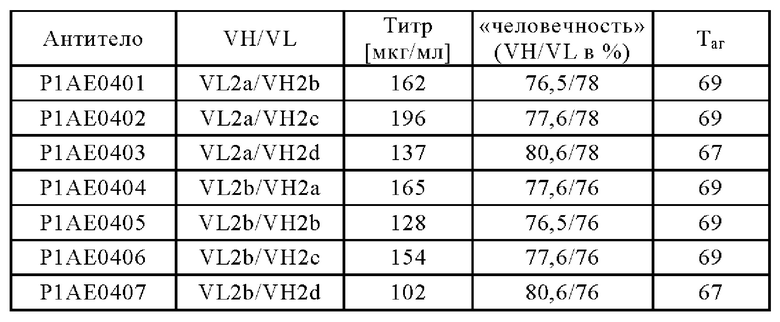
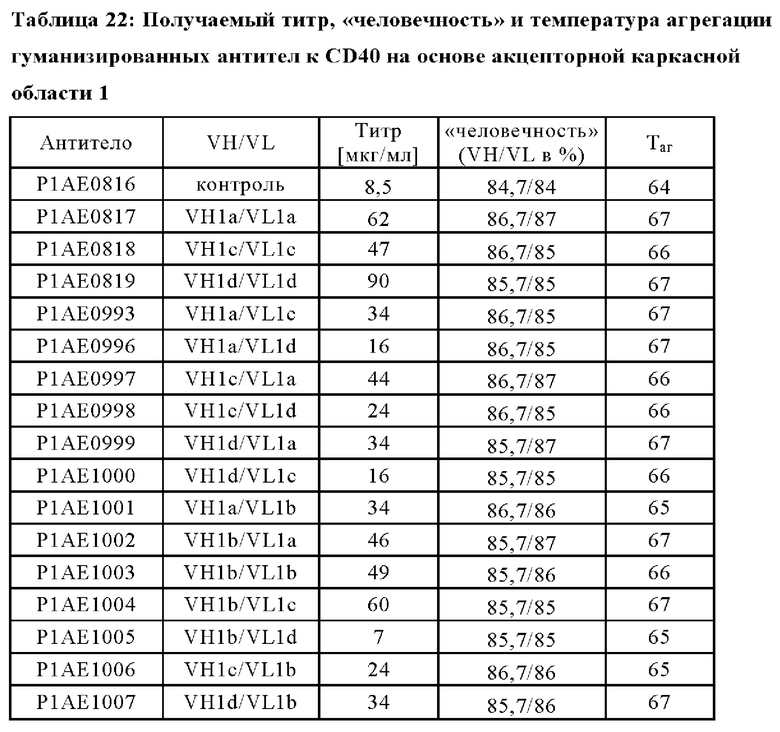
2.2.6 Создание рекомбинантного белка внеклеточного домена CD40 человека и яванского макака
Следующие конструкции клонировали и экспрессировали посредством временной экспрессии в клетках HEK293:
1) Внеклеточный домен CD40 человека (аминокислоты 21-193 SEQ ID NO: 1, номер доступа NCBI NP_001241) с С-концевым тэгом His-AviTag™ (SEQ ID NO: 266)
2) Внеклеточный домен CD40 яванского макака (macaca fascicularis) CD40 (аминокислоты 21-193, последовательность внеклеточного домена CD40 была взята из базы данных кДНК яванского макака Roche, неопубликованные данные) с С-концевым тэгом His-AviTag™ (SEQ ID NO: 267)
Антигены внеклеточного домена CD40 для анализа связывания создавали путем генного синтеза (сервис Eurofins Genomics GmbH, Germany), клонировали с помощью уникальных рестрикционных сайтов во внутрилабораторный экспрессионный вектор Roche's, используя стандартные процедуры клонирования. Клонирование всех конструкций подтверждали с помощью секвенирования. Все антигены экспрессировали под управлением CMV-промотора. Для временной экспрессии конструкций внеклеточного домена CD40 адаптированные для суспензии клетки HEK293-F (Life Technologies, USA) трансфицировали соответствующими плазмидами: В целом, 1 л клеток HEK293-F приблизительно при 2×106 клеток/лунка трансфицировали в целом 500 мкг плазмидной ДНК в комплексе с трансфекционным реагентом PEIpro (Polysciences Europe GmbH, Germany) в соответствии с инструкциями производителя. После трансфекции клетки HEK293-F инкубировали в течение 6 суток. После этого клетки собирали путем центрифугирования и фильтровали содержащий белок супернатант, используя 0,22 мкм вакуумную фильтрационную систему (Millipore). Меченные His-AviTag™ белки очищали с помощью аффнной хроматографии IMAC, используя смолу complete-His-Tag (Roche Diagnostics). После промывки 50 мМ Na2PO4, 300 мМ NaCl, рН 8,0, содержащие His-AviTag™ слитые белки элюировали, используя промывочный буфер, дополненный 500 мМ имидазолом при рН 7,0. Агрегированный белок отделяли от мономерных слитых белков с помощью эксклюзионной хроматографии (Superdex 75, GE Healthcare) в 20 мМ Трис, 150 мМ NaCl, рН 7,4. Фракции мономерных белков объединяли, концентрировали при необходимости, используя, например, центрифужный концентратор MILLIPORE Amicon Ultra (10 кДа НОММ), замораживали и хранили при -80°С. Аликвоты образцов использовали для последующего аналитического определения, например, с помощью КЭ-ДСН, эксклюзионной хроматографии и масс-спектрометрии.
Биотинилирование внеклеточного домена CD40:
Специфическое в отношении ферментативного сайта биотинилирование конструкций внеклеточного домена CD40 человека или яванского макака, содержащих С-концевой AviTag™, проводили, используя набор с биотин-протеинлигазой BirA (Avidity LLC, USA), в соответствии с инструкцией производителя. Вкратце, 1/10 объема BiomixA (10Х концентрация: 0,5 М бициновый буфер, рН 8,3) и BiomixB (10Х концентрация: 100 мМ АТФ, 100 мМ MgOAc, 500 мкМ d-биотин) добавляли к содержащему AviTag™ белку с последующим добавлением 2,5 мкг лигазы BirA на 10 нмоль белка. Реакционную смесь инкубировали при 30°С в течение 1 ч и очищали с помощью эксклюзионной хроматографии на предварительно упакованной колонке Superdex75 HiLoad (GE Healthcare, Sweden).
2.2.7 Анализ методом поверхностного плазмонного резонанса связывания CD40 человека/яванского макака
Около 12000 резонансных единиц (РЕ) системы для захвата (10 мкг/мл козьего античеловеческого F(ab)'2; код заказа: 28958325; GE Healthcare Bio-Sciences АВ, Sweden) связывали на чипе СМ5 (GE Healthcare BR-1005-30) при рН 5,0, используя набор для аминного сопряжения, поставляемый GE Healthcare. Буфером для образца и системы быт ФСБ-Т (10 мМ фосфатно-солевой буферный раствор, содержащий 0,05% Твин 20), рН 7,4. Температуру проточной ячейки устанавливали на 25°С, а температуру блока для образцов устанавливали на 12°С и дважды примировали подвижным буфером. Захват антитела проводили посредством ввода 50 нМ раствора в течение 30 с при скорости потока 5 мкл/мин. Ассоциацию измеряли путем ввода внеклеточного домена CD40 человека или внеклеточного домена CD40 яванского макака в различных концентрациях в растворе в течение 300 с при скорости потока 30 мкл/мин, начиная с 300 нМ с разведениями 1:3. Фазу диссоциации отслеживали в течение до 1200 с и инициировали путем переключения с раствора образца на подвижный буфер. Поверхность восстанавливали посредством 60 с промывки раствором глицина, рН 2,1, при скорости потока 30 мкл/мин. Объемный показатель преломления корректировали путем вычитания ответа, полученного от покрытой козьим античеловеческим F(ab')2 поверхности. Также вычитали контрольные вводы (=двойное сопоставление). Для расчета кажущейся KД и других кинетических параметров использовали 1:1 модель Ленгмюра.
Кажущуюся Kд рассчитывали, используя оценочное программное обеспечение BiacoreTM В4000 (версия 1.1).
2.2.8 Клеточный анализ связывания для получения характеристик CD40-специфических гуманизированных антител
CD40-положительные клетки (клетки Raji) отсоединяли от культуральной колбы, используя трипсин, и пересчитывали на счетчике клеток Casy. После осаждения при 4°С клетки ресуспендировали в буфере FACS Buffer (2,5% ФТС в ФСБ), доводили до 2,0Е+06 клеток/мл и распределяли в 96-луночный РР планшет с V-образным дном (25 мкл/лунка=5,0Е+04 клеток/лунка).
CD40-специфические антитела доводили до 20 мкг/мл в буфере FACS с получением конечной концентрации 10 мкг/мл. 20 мкл добавляли к 25 мкл клеточной суспензии и инкубировали в течение 1 ч при 4°С. Потом клетки дважды промывали в буфере FACS. После промывки клетки ресуспендировали в 50 мкл буфера FACS, содержащего вторичное антитело (<huIgG>-Alexa488, с=10 мкг/мл) и инкубировали в течение 1 ч при 4°С. Потом клетки дважды промывали в буфере FACS и ресуспендировали в 70 мкл/лунка буфера FACS для измерений с применением FACS Canto (BD, Pharmingen).
В таблице 23 приведены аффинность гуманизированных антител к CD40 (измеренная с помощью Biacore) и данные клеточного связывания с экспрессирующими CD40 клетками (клетками Raji).
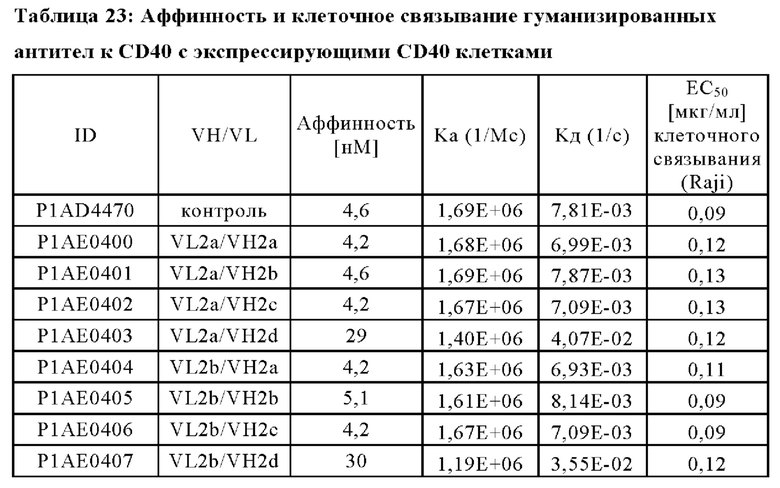
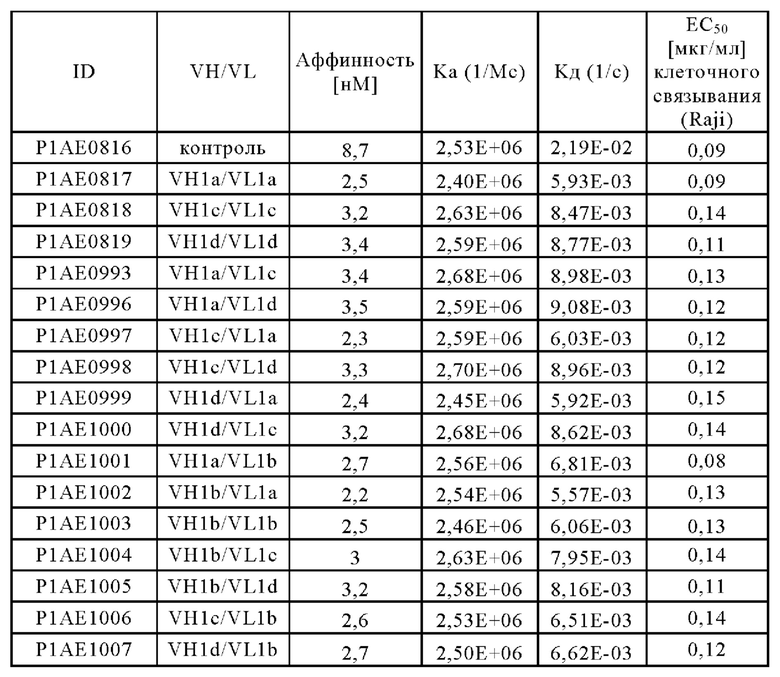
2.2.9 Получение характеристик антител методом масс-спектрометрии UHR-ESI-QTOF
Образцы обессоливали с помощью ВЭЖХ на колонке Sephadex G25 5x250 мм (Amersham Biosciences, Freiburg, Germany), используя 40% ацетонитрил с 2% муравьиной кислотой (об./об.). Общую массу определяли с помощью UHR-ESI-QTOF MS на системе maXis 4G UHR-QTOF MS (Bruker Daltonik, Bremen, Germany), оснащенной источником TriVersa NanoMate (Advion, Ithaca, NY). Получение данных проводили при 900 4000 m/z (ISCID: 0,0 эВ). Оценивали исходные масс-спектры и трансформировали в индивидуальные относительные молярные массы, используя разработанный внутри лаборатории программный инструмент.
2.2.10 Оценка термической стабильности антител
Образцы готовят в концентрации 1 мг/мл в 20 мМ смеси гистидина/хлорида гистидина, 140 мМ NaCl, рН 6,0, переносят в оптический 384-луночный планшет путем центрифугирования через 0,4 мкм фильтровальную пластину и покрывают парафинным маслом. Гидродинамический радиус повторно измеряют методом динамического рассеяния света на планшет-ридере DynaPro (Wyatt) при нагревании образцов со скоростью 0,05°С/мин от 25°С до 80°С. В альтернативном варианте образцы переносили в 10 мкл микрокюветную матрицу и записывали данные по статическому рассеянию света, а также данные по флуоресценции после возбуждения лазером с 266 нм на инструменте Optim1000 (Avacta Inc.) при нагревании со скоростью 0,1°С/мин от 25°С до 90°С. Температуру начала агрегации определяют как температуру, при которой гидродинамический радиус (ДРС) или интенсивность рассеянного света (Optim1000) начинают увеличиваться. Температуру плавления определяют как точку перегиба на графике зависимости интенсивности флуоресценции от длины волны.
Пример 3
Создание и получение биспецифических конструкций с новым анти-FAP клоном 212 и его гуманизированными вариантами
3.1 Создание биспецифических антигенсвязывающих молекул, нацеленных на белок активации фибробластов (FAP) и CD40
Биспецифические антитела к CD40-FAP получали в формате 2+1, состоящем из двух связывающих CD40 фрагментов в комбинации с одним связывающим FAP фрагментом в С-конце Fc (Фиг. 1А-1В), или в формате 4+1, состоящем из четырех связывающих CD40 фрагментов в комбинации с одним связывающим FAP фрагментом в С-конце Fc (Фиг. 1Г-1Е). Биспецифические антитела к CD40-FAP содержали новый анти-FAP клон 212 (Фиг. 1А и 1Г), однако для сравнения получали соответствующие молекулы с клонами FAP 4В9 (Фиг. 1Б и 1Д) и 28Н1 (Фиг. 1В и 1Е). Создание и подготовка связывающих FAP молекул 28Н1 и 4В9 описаны в WO 2012/020006 А2, которая включена в данный документ посредством ссылки. Для создания молекул 4+1 и 2+1 применяли технологию «выступ во впадину» для обеспечения гетеродимеризации. Мутации S354C/T366W вносили в первую тяжелую цепь HCl (тяжелая цепь Fc-выступа), а мутации Y349C/T366S/L368A/Y407V вносили во вторую тяжелую цепь НС2 (тяжелая цепь Fc-впадины). Независимо от биспецифического формата, во всех трех случаях использовали подавление эффекторной активности Fc (P329G; L234, 234А) для устранения связывания с Fcy-рецепторами в соответствии со способом, описанным в WO 2012/130831 А1. Последовательности биспецифических молекул приведены в таблице 24.
Все гены временно экспрессируются под управлением химерного промотора MPSV, состоящего из корового промотора MPSV в комбинации с энхансерным фрагментом промотора CMV. Экспрессионная кассета также содержит синтетический сигнал поли-А в 3' конце кДНК. Экспрессионный вектор также содержит область oriP для эписомальной репликации в клетках, содержащих EBNA (ядерный антиген вируса Эпштейна - Барр).


3.2 Получение биспецифических антигенсвязывающих молекул, нацеленных на FAP и CD40
Биспецифические антигенсвязывающие молекулы, нацеленные на белок активации фибробластов (FAP) и CD40, экспрессировали посредством временной трансфекции клеток HEK, выращиваемых в суспензии, экспрессионными векторами, кодирующими 4 разные пептидные цепи. Трансфекцию в клетки HEK293-F (Invitrogen) проводили в соответствии с инструкциями поставщика клеток, используя препараты векторов антител Maxiprep (Qiagen), среду F17 (Invitrogen, USA), Peipro (Polyscience Europe GmbH) и начальную плотность клеток 1-2 миллиона жизнеспособных клеток/мл в бессывороточной экспрессионной среде FreeStyle 293 (Invitrogen). Клеточные культуральные супернатанты собирали через 7 суток культивирования во встряхиваемые флаконы или ферментеры с перемешиванием путем центрифугирования при 14000 g в течение 30 минут и фильтровали через 0,22 мкм фильтр.
Биспецифические антитела очищали из клеточных культуральных супернатантов с помощью аффинной хроматографии с протеином А, используя хроматографию MabSelectSure-Sepharose™ (GE Healthcare, Sweden). Вкратце, стерильно профильтрованные клеточные культуральные супернатанты иммобилизовали на смоле MabSelect SuRe, уравновешенной буфером ФСБ (10 мМ Na2HPO4, 1 мМ KH2PO4, 137 мМ NaCl и 2,7 мМ KCl, рН 7,4), промывали уравновешивающим буфером и элюировали 25 мМ цитратом, рН 3,0. После нейтрализации 1 М Трис, рН 9,0, агрегированный белок отделяли от мономерных молекул антител с помощью эксклюзионной хроматографии (Superdex 200, GE Healthcare) в 20 мМ гистидине, 140 мМ NaCl, рН 6,0. Фракции мономерных белков объединяли, концентрировали при необходимости, используя, например, центрифужный концентратор MILLIPORE Amicon Ultra (30 кДа НОММ), замораживали и хранили при 80°С. Аликвоты образцов использовали для последующего аналитического определения, например, с помощью КЭ-ДСН, эксклюзионной хроматографии, масс-спектрометрии и определения содержания эндотоксинов.
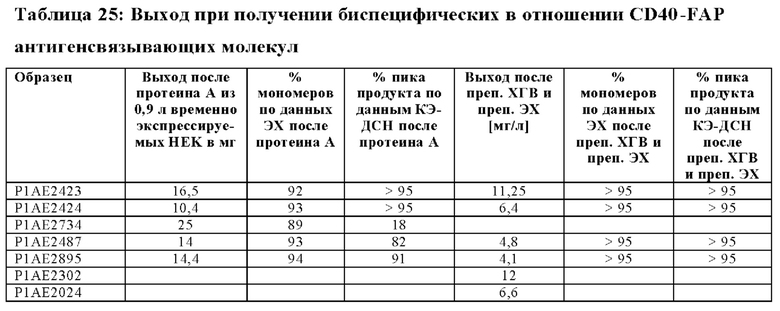
P1AE2734 представляет собой биспецифическое антитело, в котором цепь VH-C-каппа кросс-fab слита с С-концом одной из тяжелых цепей Fc-домена. Это биспецифическое антитело было намного труднее получить и очистить, чем биспецифическое антитело Р1АЕ2423, в котором цепь VL-CH1 кросс-fab слита с С-концом одной из тяжелых цепей Fc-домена. Необходимо было провести дополнительные этапы очистки, т.е. ионообменная хроматография, а также дополнительный КЭ-ДСН привели к получению 1 мг вещества.
3.3 Кинетика связывания биспецифических анти-huFAP антител
Чтобы оценить кинетику связывания FAP человека биспецифических нацеленных на FAP антител, кинетику связывания измеряли, как описано в примере 1.7, при этом использовали HIS-меченный эктодомен FAP человека в качестве лиганда. Данные по аффинности биспецифических 2+1 антител к CD40 х FAP с разным слиянием кросс-fab приведены ниже в таблице 25А.

Аффинность связывания FAP биспецифических антител в виде С-концевого Fab-слияния находится в сходном диапазоне для двух вариантов перекрестного связывания (по данным измерений ППР).
Пример 4
Получение характеристик биспецифических конструкций, нацеленных на CD40 и FAP
4.1 Связывание с мышиными клетками фибробластов, экспрессирующими FAP человека
Связывание с FAP клеточной поверхности тестировали, используя человеческий белок активации фибробластов (huFAP), экспрессирующий NIH/3T3-hFAP клон 19. NIH/3T3-huFAP клон 19 был создан путем трансфекции линии клеток мышиных эмбриональных фибробластов NIH/3T3 (АТСС CRL-1658) экспрессионным вектором pETR4921 для экспрессии в селективных условиях с 1,5 мкг/мл пуромицина huFAP.
Клетки NIH/3T3-huFAP культивировали с 1× средой Игла в модификации Дульбекко (DMEM) (gibco, кат. №42430-025), дополненной 10% фетальной бычьей сыворотки (ФБС) (life technologies, кат. №16140, партия №1797306А). 1,5 мкг/мл пуромицина (gibco, кат. № A11138-03) добавляли в среду для селекции FAP-экспрессирующих клеток. Клетки NIH/3T3-huFAP удаляли из культуральных флаконов, используя не содержащий ферментов буфер для диссоциации клеток (gibco, кат. №13151014). В каждую лунку круглодонного 96-луночного планшета (greiner bio-one, cellstar, кат. №650185) добавляли 0,3×105 клеток NIH/3T3-hFAP клона 19 в 200 мкл 1× DMEM с 10% ФБС. Планшеты центрифугировали 5 минут при 1700 об/мин и сливали супернатанты. Клетки один раз промывали 200 мкл охлажденного до 4°С буфера FACS (eBioscience, кат. №00-4222-26). Все образцы ресуспендировали в 50 мкл/лунка охлажденного до 4°С буфера FACS, содержащего биспецифические антигенсвязывающие молекулы (первичное антитело) или изотипическое контрольное антитело DP47 в указанном диапазоне концентраций (в двух повторностях) и инкубировали в течение 120 минут при 4°С. После этого клетки три раза промывали 200 мкл охлажденного до 4°С буфера FACS. Клетки дополнительно окрашивали 25 мкл/лунка охлажденного до 4°С раствора вторичного антитела (1:50 разведение вторичного антитела), содержащего конъюгированный с R-фикоэритрином (РЕ) фрагмент AffiniPure F(ab')2 козьего античеловеческого IgG, специфический к Fcγ-фрагменту (Jackson ImmunoResearch, кат. №109-116-098) как вторичное антитело, и инкубировали в течение 60 минут при 4°С в темноте. Клетки промывали 200 мкл буфера FACS и ресуспендировали в 85 мкл/лунка буфера FACS, содержащего 0,2 мкг/мл DAPI (Roche, кат. №10236276001), а данные получали в тот же день, используя 5-лазерный LSR-Fortessa (BD Bioscience с программным обеспечением DIVA). Анализ данных проводили, используя программное обеспечение FlowJo версии 10 (FlowJo LLC).
Как показано на Фиг. 4, биспецифические антитела, одновалентные в отношении FAP, связываются с экспрессирующими FAP человека целевыми клетками. Следовательно, нацеленные на FAP анти-CD40 антигенсвязывающие молекулы демонстрируют свойства прямого нацеливания на опухоль. Конструкции FAP с С-концевым связывающим доменом FAP (212 (Р1АЕ1689)) или FAP (4В9) демонстрируют более сильное связывание, чем конструкции FAP с С-концевым связывающим доменом FAP (28Н1), что объясняется более высокой аффинностью связывания связывающих FAP (212) и FAP (4В9) молекул в отношении FAP человека по сравнению со связывающей FAP (28Н1) молекулой. Самое сильное связывание FAP наблюдали для 2+1 конструкции со связывающим FAP (4В9) фрагментом (Р1АЕ2487). Связывание изотипического контрольного антитела DP47 (контроль зародышевой линии человека) с клетками NIH/3T3-hFAP выявлено не было. Значения ЕС50, измеренные для разных биспецифических антител, приведены ниже в таблице 26.

4.2 Связывание с экспрессирующими CD40 человека первичными В-клетками
Связывание с CD40 клеточной поверхности тестировали, используя человеческие первичные В-клетки, выделенные из мононуклеарных клеток периферической крови (МКПК). С целью выделения МКПК получали лейкоцитарную пленку от Stiftung Zürcher Blutspendedienst SRK. 50 мл лейкоцитарной пленки разводили в таком же объеме ФСБ (gibco, кат. №10010023). 50 мл полипропиленовые центрифужные пробирки (ТРР, кат. №91050)наполняли 15 мл Lymphoprep™ (STEMCELL Technologies, кат. №07851) и аккуратно наносили слой 25 мл раствора лейкоцитарной пленки/ФСБ на пробирку поверх Lymphoprep™. Пробирки центрифугировали при 2000 об/мин в течение 24 минут при комнатной температуре с низким ускорением и без перерыва. После этого с поверхности раздела собирали МКПК, три раза промывали ФСБ, ресуспендировали в 10 мл ФСБ и анализировали клетки в отношении числа и типа клеток с помощью счетчика клеток Beckman Coulter Ас⋅Т™ 5diff OV (Beckman Coulter, кат. №6605580). Перед выделением В-клеток из МКПК CD14-положительную фракцию удаляли путем магнитного мечения CD14-положительных клеток гранулами CD14 (Miltenyi, кат. №130-050-201) и последующего выделения с помощью сепаратора autoMACS® Pro (Miltenyi, кат. №130-092-545). CD14-отрицательную фракцию использовали для последующего выделения В-клеток с помощью набора для выделения В-клеток Miltenyi II (кат. №130-091-151) и сепарации autoMACS®. В каждую лунку круглодонного 96-луночного планшета (greiner bio-one, cellstar, кат. №650185) добавляли 0,3×105 В-клеток в 200 мкл среды R10, состоящей из среды Мемориального института Розуэлл-Парк (RPMI) 1640 (gibco, кат. №31870-025), дополненной 10% (об./об.) ФБС, 1% (об./об.) пенициллина-стрептомицина (gibco, кат. №15070-063), 1% (об./об.) L-глутамина (gibco, кат. №25030-024), 1% (об./об.) пирувата натрия (gibco, кат. №11360-039), 1% (об./об.) MEM с заменимыми аминокислотами (gibco, кат. №11140-035) и 50 мкМ β-меркаптоэтанола (gibco, кат. №31350-010). Планшеты центрифугировали 5 минут при 1700 об/мин и сливали супернатанты. Клетки один раз промывали 200 мкл охлажденного до 4°С буфера FACS (eBioscience, кат. №00-4222-26). Все образцы ресуспендировали в 50 мкл/лунка охлажденного до 4°С буфера FACS, содержащего биспецифические антигенсвязывающие молекулы (первичное антитело) или изотипическое контрольное антитело DP47 в указанном диапазоне концентраций (в двух повторностях) и инкубировали в течение 120 минут при 4°С. После этого клетки три раза промывали 200 мкл охлажденного до 4°С буфера FACS. Клетки дополнительно окрашивали 25 мкл/лунка охлажденного до 4°С раствора вторичного антитела (1:50 разведение вторичного антитела), содержащего конъюгированный с R-фикоэритрином (РЕ) фрагмент AffiniPure F(ab')2 козьего античеловеческого IgG, специфический к Fcγ-фрагменту (Jackson ImmunoResearch, кат. №109-116-098) как вторичное антитело, и инкубировали в течение 60 минут при 4°С в темноте. Клетки промывали 200 мкл буфера FACS и ресуспендировали в 85 мкл/лунка буфера FACS, содержащего 0,2 мкг/мл DAPI (Roche, кат. №10236276001), а данные получали в тот же день, используя 5-лазерный LSR-Fortessa (BD Bioscience с программным обеспечением DIVA). Анализ данных проводили, используя программное обеспечение FlowJo версии 10 (FlowJo LLC).
Как показано на Фиг. 5, все изображенные клоны связываются с CD40, но различаются по силе связывания (значениям ЕС50, а также силе сигнала) с CD40-положительными В-клетками. Вне зависимости от FAP-связывающего фрагмента двухвалентные анти-CD40 антитела демонстрировали более высокие уровни ЕС50 и достигали более высоких плато связывания по сравнению с четырехвалентными анти-CD40 антителами, что объясняется более занятыми сайтами связывания CD40 на антитело и приростом авидности в четырехвалентном формате CD40 относительно двухвалентного. Связывание отрицательного контрольного антитела с В-клетками выявлено не было. Значения ЕС50, измеренные для разных биспецифических антител, приведены ниже в таблице 27.

Пример 5
Функциональные свойства нацеленных на FAP связывающих молекул против CD40 человека
5.1 CD40-опосредованная активация человеческих В-клеток нацеленными на FAP связывающими молекулами против CD40 человека
Лигирование CD40 индуцирует созревание, а также активацию В-клеток и дендритных клеток (ДК) и способствует выживаемости этих типов клеток. После сигнализации CD40 повышается выработка цитокинов и экспрессия костимулирующей молекулы на поверхности В-клеток и ДК (S. Quezada et al., Annu RevImmunol. 2004, 22, 307-328; S. Danese et al., Gut. 2004, 53, 1035-1043; G. Bishop et al., Adv Exp Med Biol. 2007, 597, 131-151).
Для тестирования агонистических свойств и FAP-специфичности разных FAP-зависимых анти-CD40 антител клетки Дауди или первичные В-клетки, полученные из человеческих лейкоцитарных пленок, инкубировали с FAP-зависимыми агонистическими антителами к CD40 человека в присутствии покрытых FAP гранул и измеряли активацию В-клеток с помощью FACS. 5.1.1. Активация человеческих клеток Дауди нацеленными на FAP связывающими молекулами против CD40 человека с применением покрытых FAP гранул Dynabeads в качестве источника антигена
Добавляли 1×105 клеток Дауди, линии клеток человеческих В-лимфобластов с высокими уровнями экспрессии CD40 человека (АТСС CCL-213) в 100 мкл 1х среды Игла в модификации Дульбекко (DMEM) (gibco, кат. №42430-025), дополненной 10% фетальной бычьей сыворотки (ФБС) (life technologies, кат. №16140, партия №1797306А), на лунку 96-луночного плоскодонного планшета. Стрептавидиновые гранулы Dynabeads® (ThermoFisher Scientific, кат.№: 11205D) покрывали биотинилированным FAP человека (полученным в лаборатории) (связывающая способность 6,5×104 гранул: 0,01 мкг белка) в соответствии с инструкциями производителя и добавляли к клеткам Дауди в соотношении гранул и клеток 2:1 в 50 мкл lx DMEM с 10% ФБС. В качестве контроля к клеткам Дауди добавляли непокрытые гранулы. Нацеленные на FAP антитела к CD40 человека добавляли в 50 мкл среды 1x DMEM с 10% ФБС к клеткам Дауди в концентрациях в диапазоне от 6,7 нМ до 0,003 нМ (3х серийные разведения). В качестве положительного контроля использовали FAP-независимое агонистическое антитело к CD40 человека SGN-40 (IgG1, INN: дацетузумаб). Это антитело является двухвалентным в отношении CD40. Поскольку в литературе описано, что для биологической активности антитела SGN-40 необходимо перекрестное связывание Fc-рецептора (С.Law et al., Cancer Res 2005, 65, 8331-8338), антитело инкубировали с перекрестно-связывающим козьим специфическим к Fcγ-фрагменту IgG человека фрагментом F(ab')2 (Jackson ImmunoResearch, кат. №109-006-008), в течение 30 минут перед добавлением антитела к клеткам Дауди. Через 48 часов клетки переносили в 96-луночный круглодонный планшет, один раз промывали ФСБ и инкубировали с 50 мкл 3 мкг/мл блокирующего Fc-рецептор мышиного изотипического контроля IgG (ThermoFisher Scientific, кат. №10400С) в ФСБ. Через 15 минут инкубации при 4°С клетки промывали ФСБ и добавляли к клеткам 50 мкл смеси флуоресцентно меченных антител в ФСБ. Использовали следующие флуоресцентно меченные антитела: против человеческого CD83 BV421 (Biolegend, клон НВ15е, кат.№305324), против человеческого CD80 BV605 (BD Biosciences, клон L307.4, кат. №563315), анти-HLA-ABC FITC (BD Biosciences, клон G46-2.6, кат. №555552), против человеческого CD14 РегСР-Су5.5 (Biolegend, клон HCD14, кат. №325622), против человеческого CD3 PerCP-Су5.5 (Biolegend, клон UCHT1, кат. №300430), против человеческого CD70 РЕ (Biolegend, клон 113-16, кат. №355104), против человеческого CD86 PE-CF594 (BD Biosciences, клон FUN-1, кат. №562390), анти-HLA-DR АРС (BD Biosciences, клон G46-6, кат. №559866) и против человеческого CD19 АРС-Н7 (BD Biosciences, клон SJ25C1, кат. №560177). Чтобы отличить живые клетки от мертвых, в смесь антител добавляли краситель для определения жизнеспособности Zombie Aqua™ (Biolegend, кат. №423102). Через 30 минут инкубации при 4°С клетки дважды промывали ФСБ и ресуспендировали в 200 мкл ФСБ. Клетки анализировали в тот же день, используя 5-лазерный LSR-Fortessa (BD Bioscience с программным обеспечением DIVA). Анализ данных проводили, используя программное обеспечение FlowJo версии 10 (FlowJo LLC). Живые (отрицательные по aqua) клетки, отрицательные в отношении CD14 и CD3 и положительные в отношении CD19, анализировали в отношении экспрессии CD70, CD80, CD83 и CD86.
Клетки Дауди, которые анализировали после 2 суток инкубации с агонистическими анти-СО40 антителами, продемонстрировали повышение экспрессии CD70 для всех проиллюстрированных антител (смотрите Фиг. 6А и 6Б). Повышенная регуляция этого маркера активации зависела от FAP в случае разных нацеленных на FAP антител. При более низких концентрациях антител повышение экспрессии, индуцированное этими FAP-зависимыми антителами, было выше по сравнению с повышением, индуцированным перекрестно-связанным антителом к CD40 (P1AD4470). Кроме того, повышение регуляции CD70 биспецифическими антителами к FAP-CD40 в формате 2+1 со связывающим доменом FAP (212) или FAP (4 В9) было выше по сравнению с повышением регуляции, индуцированным биспецифическим антителом к FAP-CD40 в формате 2+1 со связывающим доменом FAP (28Н1), биспецифическими антителами к FAP-CD40 в формате 4+1 со связывающим доменом FAP (212), FAP (4 В9) или FAP (28Н1) или FAP-независимым положительным контрольным антителом. В отсутствие FAP (непокрытые гранулы) не наблюдалось повышение CD70 в случае изображенных биспецифических антител, двухвалентных в отношении CD40, тогда как четырехвалентные CD40-связывающие молекулы индуцировали повышение регуляции CD70, но в меньшей степени, чем в присутствии FAP, что указывает на низкую, но выявляемую FAP-независимую активацию CD40 четырехвалентных связывающих CD40 молекул в клетках Дауди.
5.1.2 Активация человеческих В-клеток нацеленными на FAP связывающими молекулами против CD40 человека с применением покрытых FAP гранул Dynabeads® в качестве источника антигена
В-клетки выделяли из лейкоцитарных пленок, как описано в примере 4.2 и добавляли 1×105 В-клеток в 100 мкл среды R10 на лунку 96-луночного плоскодонного планшета. Стрептавидиновые гранулы Dynabeads® (ThermoFisher Scientific, кат. №: 11205D) покрывали биотинилированным FAP человека (полученным в лаборатории) (связывающая способность 6,5×104 гранул: 0,01 мкг белка) в соответствии с инструкциями производителя и добавляли к В-клеткам в соотношении гранул и клеток 2:1 в 50 мкл среды R10. В качестве контроля к В-клеткам добавляли непокрытые гранулы. Нацеленные на FAP антитела к CD40 человека или положительные контрольные антитела (описанные в примере 5.1.1) добавляли к В-клеткам в 50 мкл среды R10. Через 2 суток В-клетки анализировали с помощью FACS после процедур окрашивания и анализа, описанные в примере 5.1.1.
В-клетки, которые анализировали после 2 суток инкубации с агонистическими анти-CD40 антителами, продемонстрировали повышение экспрессии CD86 для всех проиллюстрированных антител (смотрите Фиг. 7А и 7Б). Повышенная регуляция CD86 зависела от FAP в случае разных нацеленных на FAP антител, а повышение, индуцированное этими FAP-зависимыми антителами, было сравнимым или немного меньше повышения, индуцированного перекрестно-связанным антителом к CD40 P1AD4470. Биспецифические антитела, одновалентные в отношении FAP с одним связывающим FAP (212) доменом индуцировали сходное повышение экспрессии маркера активации с молекулами с одним связывающим FAP (4 В9) фрагментом. При более низких концентрациях антител форматы 4+1, независимо от FAP-связывающего фрагмента, индуцировали большую активацию В-клеток по сравнению с форматами 2+1 со связывающим фрагментом FAP (212) или FAP (4В9).
5.2 CD40-опосредованная активация ДК нацеленными на FAP связывающими молекулами против CD40 и последующее примирование Т-клеток
Чтобы продемонстрировать способность ДК, активированных FAP-завивимыми антителами к CD40 человека к эффективному примированию Т-клеток, разрабатывали in vitro анализы примирования Т-клеток. Для этих анализов выделяли ДК из селезенок трансгенных мышей, экспрессирующих человеческий рецептор CD40 (мыши huCD40tg; мыши со сходным профилем экспрессии человеческого и мышиного рецептора CD40; линия C57BL/6; полученные Taconic), импульсно обрабатывали пептидом SIFNFEKL или овальбумином (OVA; DEC-205 рецептор-опосредованное поглощение антигена) и инкубировали с разными агонистическими антителами к CD40 человека. FAP обеспечивали посредством покрытых FAP гранул Dynabeads®, чтобы показать FAP-зависимость биспецифических антигенсвязывающих молекул. Через 24 часа выделяли CDS-положительные Т-клетки из селезенок мышей ОТ1 (все CD8-положительные Т-клетки обладают трансгенным ТКР, распознающим SIFNFEKL в контексте H2-Kb; C57BL/6-Tg(TcraTcrb)1100Mjb/Crl, Charles River), метили сукцинимидиловым сложным эфиром карбоксифлуоресцеина (CFSE) и добавляли к импульсно обработанным ДК. На четвертые сутки эксперимента анализировали пролиферацию Т-клеток с помощью FACS.
5.2.1 Примирование Т-клеток посредством OVA-обработанных ДК, активированных нацеленными на FAP связывающими молекулами против CD40
ДК выделяли из селезенок мышей huCD40tg. Для выделения селезеночных ДК селезенку мыши huCD40tg помещали в одну лунку 6-луночного планшета, содержащего 2,25 мл сбалансированного солевого раствора Хэнкса (HBSS) с кальцием2+ (gibco, кат, №14025-05), 250 мкл 10 мг/мл раствора коллагеназы D (конечная концентрация 1 мг/мл) (Sigma-Aldrich, кат, №11088866001) и 12,5 мкл 10 мг/мл раствора ДНКазы (конечная концентрация 0,05 мг/мл) (Sigma-Aldrich, D5025-150KU, партия № SLBR0535V). Селезенку надували, используя 3 мл шприц (BD, кат. №309658) с иглой 21G (Braun, кат. №4657527) и после этого разрывали ножницами на маленькие кусочки. Через 25 минут инкубации при 37°С добавляли 50 мкл 0,5 М этилендиаминтетрауксусной кислоты (ЭДТА) (Applichem, кат. №А4892.1000) с последующим вторым этапом инкубации при 37°С в течение пяти минут. Раствор, содержащий спленоциты и маленькие кусочки ткани селезенки, фильтровали через 40 мкм фильтр (Corning, кат. №352340) в 50 мл полипропиленовую центрифужную пробирку. Оставшиеся кусочки ткани селезенки продавливали через фильтр концом колпачка 3 мл шприца. На следующем этапе 50 мл пробирку центрифугировали при 1500 об/мин в течение 5 минут при комнатной температуре, сливали супернатант и добавляли к спленоцитам 1 мл 1х буфера для лизиса клеток (разведенного 1:10 дистиллированной водой) (BD, кат. №555899), чтобы лизировать красные кровяные клетки. Через четыре минуты инкубации при комнатной температуре добавляли 20 мл R10 с последующим этапом центрифугирования при 1500 об/мин в течение 5 минут при комнатной температуре. Супернатант удаляли, спленоциты ресуспендировали в 30 мл R10 и определяли число и жизнеспособность клеток с помощью автоматизированного счетчика клеток EVE (VWR, кат. №734-2675). Микрогранулы с мышиным CDllc UltraPure (Miltenyi, кат. №130-108-338) использовали в соответствии с инструкцией производителя для выделения ДК посредством сепарации autoMACS®. После этого 0,25×105 ДК высевали в 50 мкл R10 на лунку 96-луночного плоскодонного планшета.
Потом ДК импульсно обрабатывали 1 нг/мл SIINFEKL (остатки овальбумина 257-264, Eurogentec, кат. № AS-60193-5, партия №1360618), для которого не требуется поглощения и процессинг ДК, в качестве положительного контроля, или загружали белком OVA в качестве антигена. Чтобы стимулировать поглощение OVA путем, независимым от стимуляции толл-подобным рецептором (TLR) (дополнительные стимулы TLR могут приводить к общей высокой активации ДК, делая невозможным выявление других состояний активации вследствие стимуляции агонистическими антителами к CD40), использовали реагент для доставки антигена OVA (Miltenyi, кат. №130-094-663) в комбинации с биотинилированным антителом к DEC205 мыши (Miltenyi, клон NLDC-145, кат. №130-101-854) в соответствии с протоколом производителя. Вкратце, ДК инкубировали с биотинилированным антителом, которое связывается с рецептором DEC205, который характеризуется высокой экспрессией на СВ8-положительных перекрестно-презентирующих ДК (М. Lahoud et al., Int Immunol. 2000, 12(5), 731-735). После этого к клеткам добавляли реагент для доставки OVA, антитело к биотину, сопряженное с FITC и OVA, что приводило к опосредованному рецептором DEC205 поглощению OVA. Чтобы обеспечить отрицательный контроль, ДК метили только анти-DEC205 антителом без добавления OVA. Кроме того, к ДК добавляли покрытые FAP человека или непокрытые гранулы Dynabeads в 50 мкл R10 в соотношении гранул и клеток 2:1, как описано в примере 5.1.1. На следующем этапе добавляли разные агонистические анти-CD40 антитела в 50 мкл R10 в концентрациях в диапазоне от 6,7 нМ до 0,01 нМ (10х серийные разведения). На этом этапе эксперимента биспецифические 2+1 и 4+1 антитела к CD40 человека, содержащие FAP-связывающий сайт FAP (212) или FAP (4В9), сравнивали с перекрестно-связанным антителом к SGN-40.
На следующий день выделяли селезеночные CD8-положительные клетки мышей ОТ1. Чтобы это сделать, селезенку мыши ОТ1 продавливали через 40 мкм фильтр концом колпачка 3 мл шприца в 50 мл пробирку. Фильтр промывали R10 и центрифугировали спленоциты при 1500 об/мин в течение 5 минут при комнатной температуре. К клеткам добавляли 1 мл 1х буфера для лизиса клеток (разведенного 1:10 дистиллированной водой) и через четыре минуты инкубации при комнатной температуре добавляли 20 мл R10. Пробирку центрифугировали при 1500 об/мин в течение 5 минут при комнатной температуре и сливали супернатант. Спленоциты ресуспендировали в 30 мл R10 и определяли число и жизнеспособность клеток с помощью автоматизированного счетчика клеток EVE. CDS-положительные клетки выделяли в процессе отрицательной селекции, используя набор для выделения мышиных CD8a+Т-клеток (Miltenyi, кат. №130-104-075) и сепарацию autoMACS® в соответствии с инструкциями производителя. Потом CD8-положительные, которые находились в отрицательной фракции после сепарации, промывали предварительно нагретым ФСБ, подсчитывали с помощью счетчика клеток EVE и доводили число клеток до 2×107 клеток/мл в предварительно нагретом ФСБ. 10 мМ раствор CFSE (набор для клеточной пролиферации CellTrace™ CFSE, ThermoFisher, кат. №С34554) разводили в 5000 в предварительно нагретом ФСБ и добавляли к клеткам, ресуспендированным в ФСБ в соотношении 1:1 (конечная концентрация CFSE 1 мкМ). После краткого перемешивания на вортексе клетки инкубировали в течение пяти минут при комнатной температуре. Реакцию мечения останавливали добавлением к клеткам 40 мл предварительно нагретой среды R10. После двух этапов промывки ФСБ CDS-положительные клетки ресуспендировали в R10 и добавляли 0,5×105 клеток в 100 мкл R10 к импульсно обработанным ДК. На четвертые сутки эксперимента анализировали пролиферацию Т-клеток с помощью проточной цитометрии. Следовательно, клетки переносили из 96-луночных плоскодонных планшетов в 96-луночные круглодонные планшеты, один раз промывали ФСБ и инкубировали с 50 мкл 3 мкг/мл блокирующего Fc-рецептор мышиного изотипического контроля IgG в ФСБ. Через 15 минут инкубации при 4°С клетки промывали ФСБ и добавляли к клеткам 50 мкл смеси флуоресцентно меченных антител в ФСБ. Использовали следующие антитела: против мышиного CD4 BV421 (Biolegend, клон GK1.5, кат. №100438), против мышиного CD86 BV785 (Biolegend, клон GL-1, кат. №105043), анти-I-A/I-E PerCp-Cy5.5 (Biolegend, клон М5/114.15.2, кат. №107626), против мышиного CD70 РЕ (eBioscience, клон FR70, кат. №12-0701-82), против мышиного CD3 PE-CF594 (BD Biosciences, клон 145-2С11, кат. №562286), против мышиного CD25 РЕ-Су7 (eBioscience, клон РС61.5, кат. №25-0251-82), против мышиного CD11c АРС (BD Biosciences, клон HL3, кат. №561119), против мышиного CD44 Alexa Fluor 700 (BD Biosciences, клон IM7, кат. №560567) и против мышиного CD8 АРС-Су7 (Biolegend, клон 53-6.7, кат. №100714). Чтобы отличить живые клетки от мертвых, в смесь антител добавляли краситель для определения жизнеспособности Zombie Aqua™. Клетки инкубировали в течение 30 минут при 4°C с 50 мкл окрашивающей смеси антител. После этого клетки два раза промывали ФСБ, ресуспендировали в 200 мкл ФСБ и анализировали, используя 5-лазерный LSR-Fortessa. Анализ данных проводили, используя программное обеспечение FlowJo версии 10. Жизнеспособные CD3- и CDS-положительные клетки анализировали в отношении сигнала CFSE, а также экспрессии CD25 и CD44.
На Фиг. 8А и 8Б показано, что ДН, которые инкубировали с реагентов для доставки OVA и стимулировали биспецифическими антигенсвязывающими молекулами, нацеленными на человеческий CD40 и FAP, сильно повышают пролиферацию CDS-позитивных Т-клеток ОТ1. Эти эффекты были FAP-зависимыми. Повышение пролиферации Т-клеток, индуцированное проиллюстрированными FAP-зависимыми антителами, было немного меньшим по сравнению с повышением, индуцированным перекрестно-связанным антителом к CD40 (P1AD4470). Уровни пролиферации, индуцированные ДК, которые стимулировали 2+1 или 4+1 биспецифическими анти-CD40 антителами с одним связывающим фрагментом FAP (212) или FAP (4 В9), были сравнимыми. 5.3 Мостиковый анализ двух линий клеток в отношении связывания FAP и передачи сигнала CD40
Способ действия биспецифических антител отображен в мостиковом анализе двух линий клеток: антитела связываются с FAP в экспрессирующей FAP линии клеток и с рецептором CD40 в линии клеток с репортерным геном. В случае кластеризации рецептора CD40 передача сигнала приводит к индукции NF_В-зависимой выработки секретируемой эмбриональной щелочной фосфатазы.
Вкратце: Репортерную линию клеток HEK-Blue™ CD40L (Invivogen, трансфицированных плазмидами, кодирующими рецептор человеческого CD40 и NFkB-индуцибельную секретируемую эмбриональную щелочную фосфатазу (SEAP)) отсоединяли с помощью аккутазы (Life Technologies) и переносили 6,8×106 клеток в стерильную 50 мл центрифужную пробирку (Greiner) и помещали на роллерный миксер (Heidolph) до применения.
Нативные целевые клетки (WM-266-4, АТСС), экспрессирующие FAP на поверхности, использовали как готовые к применению замороженные клетки. Каждый флакон содержал 2×107 клеток/2 мл DMEM, 10% ФБС и 5% ДМСО. Поддержание культуры клеток следует осуществлять в DMEM с высоким содержанием глюкозы и ГЭПЭС (Life Technologies). Целевые клетки размораживали в течение 2 минут в предварительно нагретой до 37°С водяной бане. 2 мл клеточной суспензии незамедлительно переносили в 50 мл центрифужную пробирку с 30 мл DMEM, 10% ФБС (Life Technologies). Аликвоту в 13,6×106 клеток переносили в 50 мл центрифужную пробирку и помещали на роллерный миксер до применения. Размораживание клеток проводили после получения серийных разведений антител.
Среду для выявления HEK-Blue™ (Invivogen) использовали для получения клеточных суспензий и разведений антител. Три пакета растворяли в 150 мл дистиллированной воды (Life Technologies) и фильтровали через 0,22 мкм мембрану (Millipore) в стерильную стеклянную колбу (Schott). Добавляли 150 мкл гентамицина (50 мг/мл, Life Technologies).
Разведения антител концентрировали в 2 раза в среде для выявления HEK-Blue™. Восемь концентрация в ряде серийных разведений (1:4) давали конечные аналитические концентрации от 518 пМ до 0,0316 пМ. По 100 мкл антитела каждой концентрации переносили в прозрачные 96-луночные микротитровальные планшеты для тканевого культивирования (Greiner). Оценивали по две повторности в 3 планшетах на молекулу.
Обе линии клеток (репортерная и целевая линия клеток) центрифугировали 4 минуты при 180 × g при комнатной температуре. Супернатанты удаляли, а каждую линию клеток ресуспендировали в 17 мл среды для выявления HEK-Blue™. После объединения обеих линий клеток конечная смесь содержала 2×105 клеток HEK-Blue™ CD40L на мл и 4×105 клеток WM-266 4 на мл. В каждую лунку аналитических планшетов добавляли по 100 мкл клеточной смеси. В результате в каждой лунке находилось 20000 клеток HEK-Blue™ CD40L и 40000 клеток WM-266-4. Планшеты предварительно инкубировали в течение 15 минут при комнатной температуре, чтобы обеспечить распределение и оседание клеток, а потом инкубировали в течение 20 часов при 37°С и 5% CO2 (клеточный инкубатор). Активность SEAP измеряли с помощью среды для выявления HEK-Blue™ на 650 нм (Victor Х4, Perkin Elmer). Строили график зависимости оптической плотности (ОП) от концентрации антител (смотрите Фиг. 9). Значения ЕС50 приведены в ниже таблице 28.

Значения ЕС50 очень схожи для молекул 2+1 Р1АЕ2423 и Р1АЕ2302, содержащих два CD40-связывающих Fab-фрагмента и FAP-связывающий кросс-fab-фрагмент. ЕС50 существенно выше для 1+1 кросс-fab конструкции P1AF0873 и 2+2 конструкции Р1АА9663. Кроме того, 1+1 конструкция P1AF0873 демонстрирует существенно менее выраженную большую асимптоту, что означает меньшую эффективность.
5.4 FAP-независимый CD40-специфический анализ
оценивали потенциальную активацию пути CD40 без кластеризации посредством FAP. В случае активации рецептора CD40 передача сигнала приводит к индукции NFIB-зависимой выработки секретируемой эмбриональной щелочной фосфатазы.
Вкратце, репортерную линию клеток HEK-Blue™ CD40L (Invivogen, трансфицированных плазмидами, кодирующими рецептор человеческого CD40 и NF ZB-индуцибельную секретируемую эмбриональную щелочную фосфатазу (SEAP)) отсоединяли с помощью аккутазы (Life Technologies) и переносили 3,4×106 клеток в стерильную 50 мл центрифужную пробирку (Greiner) и помещали на роллерный миксер (Heidolph) до применения. Использовали среду для выявления HEK-Blue™ (Invivogen) из «мостикового анализа двух линий клеток в отношении связывания FAP и передачи сигнала CD40». Разведение антител концентрировали в 2 раза в среде для выявления HEK-Blue™. Конечная аналитическая концентрация составляла 518 пМ. Из трех независимых серийных разведений «мостикового анализа двух линий клеток в отношении связывания FAP и передачи сигнала CD40» 100 мкл антитела в наибольшей концентрации переносили в двух повторностях в прозрачный 96-луночный микротитровальный планшет для тканевого культивирования (Greiner). Это приводило к оценке 6 значений на молекулу в 1 планшете.
Репортерную линию клеток центрифугировали 4 минуты при 180×g при комнатной температуре. Супернатант удаляли, а клетки ресуспендировали в 17 мл среды для выявления HEK-Blue™. В лунках находилось по 100 мкл, содержащих 20000 клеток HEK-Blue™ CD40L. Планшеты предварительно инкубировали в течение 15 минут при комнатной температуре, чтобы обеспечить распределение и оседание клеток, а потом инкубировали в течение 20 часов при 37°С и 5% CO2 (клеточный инкубатор). Активность SEAP измеряли с помощью среды для выявления HEK-Blue™ на 650 нм (Victor Х4, Perkin Elmer). Построен график среднего значение оптической плотности (ОП) для n=6 и 3х СТОТКЛ в зависимости от молекул (смотрите Фиг. 10).
Наибольшую концентрацию, используемую в FAP-зависимом анализе, использовали для инкубации CD40-специфических репортерных клеток с молекулами. Без кластеризации посредством связывания FAP активация рецептора CD40 и последующая передача сигнала была очень низкой. Она была даже ниже для P1AF0873 (1+1 кросс-mab) и Р1АА9663 (2+2 кросс-mab), чем для 2+1 молекул Р1АЕ2423 и Р1АЕ2302 (перекрывающиеся планки погрешностей).
Пример 6
Оценка противоопухолевой эффективности нацеленных на FAP связывающих молекул против CD40 человека в комбинации с PD-L1
Было разработано in vivo исследование (Фиг. 11), чтобы определить эффект агонистических анти-FAP × CD40 антител на FAP-экспрессирующие опухолевые клетки у иммунокомпетентных мышей. В частности, оценивали разницу между биспецифическим антителом к FAP-CD40 в формате 4+1 с FAP-связывающим доменом 28Н1, связывающимся с мышиным FAP, биспецифическим антителом к FAP-CD40 в формате 2+1 с FAP-связывающим доменом 28Н1 и FAP-независимым контрольным антителом к CD40 с или без обработки aPD-L1.
В целом 97+9 (необходимо для экспериментов по повторной стимуляции) самок мышей huCD40 tg НО возрастом 6-9 на момент начала эксперимента (приобретенных у Charles Rivers, France) и изначально полученных Taconic, держали в специальных непатогенных условиях с суточными циклами 12 ч день/12 ч ночь в соответствии с принятыми руководствами (GV-Solas; Felasa; TierschG). Протокол экспериментального исследования был рассмотрен и одобрен местными органами управления (ZH225-17). После поступления животных держали в течение одной недели для привыкания к новой среде и для наблюдения. После этого мышам подкожно имплантировали передатчик с левой стороны спины для идентификации и держали еще в течение одной недели для восстановления. Постоянное наблюдение за состоянием здоровья осуществляли регулярно. Животных ежесуточно осматривали в отношении клинических симптомов и выявления нежелательных явлений.
Клетки MC38-muFAP invipa (CRC) были получены из in-vivo-пассирования, проведенного в Roche Glycart AG, и после размножения депонированы во внутренний банк клеток Glycart. Клетки MC38-muFAP invipa культивировали в DMEM, содержащей 10% ФТС (РАА Laboratories, Austria), 1 мМ пирувата, 1х NEAA и 6 мкг/мл пуромицина. Клетки культивировали при 37°С в насыщенной влагой атмосфере при 5% СО2. Клетки вводили при 10 in vitro пассаже и жизнеспособности 94,5%.
Для инокуляции опухолевых клеток мышей анестезировали и иглой 29G (BD) подкожно инъецировали в левый бок 100 мкл раствора, содержащего 2×106 суспензии из одиночных клеток MC38-muFAP в среде из 50% RPMI+50% матригеля. Для повторной стимуляции той же самой опухолевой линией клеток 45 мышей инъецировали в правый бок, придерживаясь той же процедуры, через 93 суток после 1-ой терапии.
На 22 сутки, когда опухоль достигала в среднем 200 мм3, мышей рандомизировали на 8 разных групп по 9 мышей в каждой со схожим средним размером опухолей (таблица 29). После рандомизации мышам иглой 29G внутрибрюшинно инъецировали разные соединений (таблица 30) в соответствии с группами исследования (таблица 31). Через 7 суток и 14 суток после 1-ой терапии группам В, F, G и Н внутрибрюшинно инъецировали вторую и третью дозу анти-PD-L1.
После рандомизации и инъекции терапевтического средства размер опухоли проверяли 3 раза в неделю с помощью измерений штангенциркулем. Ежедневное наблюдение проводили, чтобы гарантировать соблюдение благополучия животных. Согласно лицензионным критериям (ZH225-17) мышей умерщвляли в случае изъязвленных опухолей, опухолей, достигших объема более 3000 мм3, или мышей, которые имели потерю массы более 20% от исходной массы тела (на 0 сутки). При необходимости умерщвления мышей анестезировали изофлураном и проводили механическую цервикальную дислокацию.

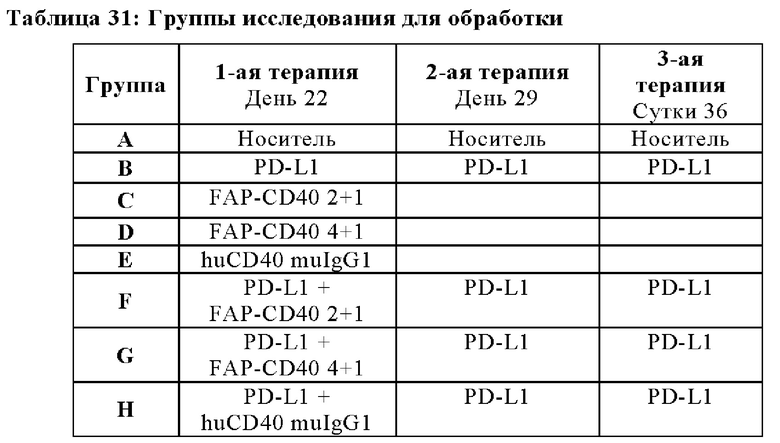
Из исключенных животных 12 животных отобрали и лечили терапевтическими средствами из группы А, С, D и Е (3 животных на терапию), чтобы умертвить для предварительного исследования для анализа иммунофармакодинамических эффектов на 4 сутки после инъекции 1-го терапевтического средства.
6.1 Нацеленные на FAP связывающие молекулы против CD40 индуцируют полную ремиссию опухоли и эффективный противоопухолевый иммунный ответ памяти
Размер опухоли проверяли 3 раза в неделю с помощью измерений штангенциркулем. Программное обеспечение JMP12 использовали для статистического анализа данных по росту опухолей. Чтобы исследовать значимые разницы в средних по группам значениях для множественных сравнений применяли стандартный дисперсионный анализ (ANOVA), используя метод Тьюки - Крамера. Метод Тьюки - Крамера обеспечивает критерий для всех попарных разниц среди средних значений; он является точным критерием уровня значимости в случае одинаковых размеров выборок.
Через 93 суток после 1-ой терапии мышам, которые демонстрировали полную регрессию опухолей, в противоположный бок подкожно инъецировали опухолевые клетки MC38-FAP, чтобы определить формирование памяти иммунных клеток против этой опухолевой линии клеток. В качестве контроля 9 не проходившим обработку трансгенным мышам C57b1/6-huCD40 инъецировали такие же опухолевые клетки MC38-FAP и отслеживали рост опухоли с помощью измерений штангенциркулем 2 3 раза в неделю в соответствии с ветеринарной лицензией.
Как показано на Фиг. 12, обе биспецифические молекулы FAP-CD40 (2+1 и 4+1) индуцировали полную ремиссию опухоли с и без обработки PD-L1. У мышей, которых обрабатывали нецелевым CD40, PD-L1 или ими обоими в комбинации, рост опухоли не был замедлен по сравнению с обработанной носителем группой. Кроме того, после повторной стимуляции в группах, предварительно обработанных антителом к FAP-CD40, опухоль MC38-FAP не росла, тогда как у наивных мышей наблюдался рост 100% опухолей. Эти результаты указывают на сильную противоопухолевую MC38-FAP эффективность и регрессию роста опухоли при нацеленной на CD40 терапии в отличие от не нацеленной на CD40 терапии. Поскольку все мыши, обработанные анти-FAP-CD40 антителом, демонстрировали 100% ремиссию опухоли, очевидный добавочный эффект анти-PD-L1 у этих мышей отсутствовал. Кроме того, не было продемонстрировано разницы между форматами 2+1 и 4-1 в терминах ингибирования роста опухоли. Данные по повторной стимуляции указывают на формирование эффективного иммунного ответа памяти против опухолевых клеток MC38-FAP у всех мышей, обработанных анти-FAP-CD40 антителом.
Статистический анализ показал, что данные становятся статистически отличными по сравнению с носителем на 28 сутки (6 суток после терапии) для группы, обработанной 2+1 антителом к FAP-CD40 (Фиг. 13А, статистический анализ, сравнивающий все группы на 28 сутки) и на 31 сутки (9 суток после терапии) для группы, обработанной 4+1 антителом к FAP-CD40 (Фиг. 13Б, статистический анализ, сравнивающий все группы на 31 сутки).
6.2 4+1 нацеленная на FAP связывающая молекула против CD40 имеет более высокий сывороточный клиренс по сравнению с 2+1 нацеленной на FAP связывающей молекулой против CD40
Чтобы отслеживать фармакокинетический профиль лекарственных средств после внутрибрюшинной инъекции, брали кровь у 3-4 мышей на группу. Образцы сыворотки получали чрез 6 ч, 24 ч, 48 ч, 72 ч, 96 ч, 8 суток и 15 суток после первой терапии антителом для анализа уровней анти-CD40 в крови. В один момент времени брали максимум 100 мкл крови. Температуру тела мышей немного повышали с помощью нагретого до 39°С бокса, чтобы гарантировать хорошее расширение кровеносных сосудов. Пункцию хвостовой вены проводили иглой 22G. Кровь собирали в 1,1 мл микропробирки Z-Gel (SARSTEDT), а сыворотку экстрагировали, собирая супернатант после центрифугирования. Образцы хранили при -20°С и позже анализировали в отношении уровней анти-CD40, используя соответствующее антитело для калибровки.
Концентрации анти-CD40 в сыворотке исследовали, используя ELISA для химерного CD40/Fc (R&D, 1493-CD-050), для количественного определения анти-CD40 huIgG в соответствии с протоколом (Roche-Glycart, Switzerland). Использовали реагенты для захвата белка: человеческий IgG-Fc биотин (Abcam, ab98561) и химерный CD40/Fc (R&D, 1493-CD-050), антитело для выявления: антитело против мышиного IgG (HRP) (Abeam, ab97040) или против huCH1-DIG (Roche-Penzberg, Germany) и против DIG-POD (Roche, 11633716001). Образцы сыворотки анализировали в серийных разведениях в зависимости от вводимой дозировки (мг/кг) и времени сбора после инъекции анти-CD40 (6 ч, 24 ч, 48 ч, 96 ч). Поглощение измеряли, используя длину волны изменения 405 нм и референсную длину волны 490 нм (микропланшет-ридер SpectraMax i3, Molecular Devices).
Для ФК-анализа использовали программное обеспечение Phoenix 64, WinNonlin 6.4, чтобы рассчитать площадь под кривой, а также Смаке. Поскольку фармакокинетику отслеживали только до 96 часов, такие ФК-параметры, как клиренс, не приведены.
Как показано на Фиг. 14 и в таблице 32, больший клиренс наблюдали для формата 4+1 по сравнению с форматом 2+1. Этот больший клиренс можно объяснить CD40-специфическим мишень-опосредованным распределением препарата (МОРП), которое более выражено для четырехвалентной связывающей CD40 молекулы (формат 4+1), чем для двухвалентной связывающей CD40 молекулы (формат 2+1).
Также не нацеленные на CD40 молекулы продемонстрировали больший клиренс по сравнению с обеими молекулами FAP-CD40. Этот клиренс нельзя объяснить гипотезой МОРП, но он может быть связан с образованием противолекарственных антител (ПЛА) после терапии CD40.
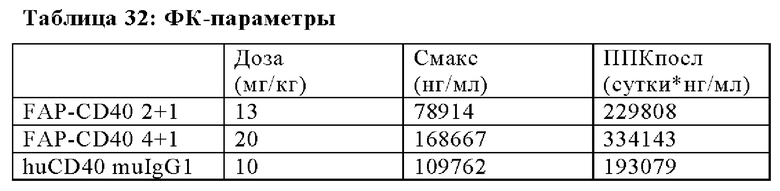
6.3 CD40-опосредованная активация ДК, В-клеток и Т-клеток нацеленными на FAP связывающими молекулами против CD40 через 96 часов после обработки
Через 96 часов после 1-ой терапии умерщвляли по 3 мыши на группу и вырезали следующие органы в ФСБ для анализа методом проточной цитометрии: опухоль, селезенка, паховые дренирующие лимфатические узлы и паховые аксиальные и брахиальные недренирующие лимфатические узлы.
Для анализа методом проточной цитометрии готовили суспензии из одиночных клеток всех вырезанных органов, как описано в примере 5.2.1, и окрашивали флуоресцентно меченными антителами. С этой целью подготовленные суспензии из одиночных клеток переносили в 96-луночные плоскодонные планшеты, промывали ФСБ и инкубировали с 50 мкл 3 мкг/мл блокирующего Fc-рецептор мышиного изотипического контроля IgG (ThermoFisher Scientific, кат. №10400С) в ФСБ. Через 15 минут инкубации при 4°С клетки промывали ФСБ и добавляли к клеткам 50 мкл смеси флуоресцентно меченных антител в ФСБ. Использовали следующие антитела: против мышиного CD3 Pacific Blue™ (BD Bioscience, клон 500A2, кат. №558214), против мышиного CD86 BV605 (Biolegend, клон GL-1, кат. №105037), CD45 Alexa Fluor 700 (eBioscience, клон 30-F11, кат. №56-0451-82), против мышиного CD19 BUV395 (BD Biosciences, клон 1D3, кат. №563557), против мышиного CD11c BV785 (Biolegend, клон N418, кат. №117336), против мышиного В220 АРС-Су7 (Biolegend, клон RA3-6B2, кат. №103224), против мышиного CD69 BUV737 (BD Biosciences, клон H1.2F3, кат. №612793), анти-I-A/I-E PerCp-Cy5.5 (Biolegend, клон М5/114.15.2, кат. №107626). Чтобы отличить живые клетки от мертвых, в смесь антител добавляли краситель для определения жизнеспособности Zombie AquaTM или Fixable blue (оба от Life technologies). Клетки инкубировали в течение 30 минут при 4°C с раствором антител для внеклеточного окрашивания. После этого клетки два раза промывали ФСБ, пермеабилизировали и внутриклеточно окрашивали в отношении Ki-67, используя РЕ-Су7 против мышиного Ki-67 (eBioscience, клон SolA15, кат. №25-5698-82), с окрашивающим буфером Foxp3/транскрипционный фактор (eBioscience, кат. №00-5523-00) в соответствии с протоколом производителя. Клетки ресуспендировали в 200 мкл ФСБ и анализировали в тот же день, используя 5-лазерный LSR-Fortessa. Анализ данных проводили, используя программное обеспечение FlowJo версии 10. Жизнеспособные CD45+, CD3+ и CD8+ Т-клетки анализировали в отношении экспрессии Ki-67. Жизнеспособные дендритные клетки (CD45+, MHCII+, CD11c+) анализировали в отношении экспрессии маркера активации ДК CD86, а жизнеспособные В-клетки (CD45+, CD19+, В220+) анализировали в отношении экспрессии маркера активации В-клеток CD69.
Как показано на Фиг. 15, оба нацеленных на FAP формата анти-CD40 антител (2+1 и 4+1) индуцировали существенное повышение активации (экспрессии CD86) в дренирующих опухоль лимфатических узлах, не дренирующих опухоль лимфатических узлах и селезенке через четверо суток после обработки, хотя в меньшей степени, чем ненацеленное анти-CD40 антитело. В противоположность этому, только нацеленное на FAP анти-CD40 антитело в формате 2+1 индуцировало существенную активацию ДК в опухолях по сравнению с группой носителя. Как показано на Фиг. 16, такой же профиль наблюдали для активации Т-клеток (экспрессии Ki-67). Существенную активацию В-клеток (экспрессию CD69) во всех анализируемых тканевых образцах по сравнению с группой носителя наблюдали только у мышей, обработанных ненацеленным анти-CD40 антителом (Фиг. 17).
6.4 Нацеленные на FAP связывающие молекулы против CD40 индуцируют меньше побочных эффектов по сравнению с ненацеленным антителом к CD40 у мышей
6.4.1 Мыши, обработанные нацеленным на FAP анти-CD40 антителом, не демонстрируют снижение массы в отличие от мышей, обработанных ненацеленным анти-CD40 антителом
После инъекции терапевтического средства массу каждой мыши измеряли ежесуточной в течение 15 суток. Мышей со снижением массы более 20% от их исходной массы тела (на 0 сутки) умерщвляли (в данном случае не было необходимости в умерщвлении мышей).
Как проиллюстрировано на Фиг. 18, мыши, обработанные ненацеленным анти-CD40 антителом, демонстрировали явное снижение массы после инъекции антитела, и это снижение массы длилось дольше в случае одновременной инъекции анти-CD40 с анти-PD-L1 антителом. Снижение массы не наблюдали в группах с инъекцией отдельных агентов 4+1 или 2+1 FAP-CD40, одного агента PD-L1 и в комбинированных группах FAP-CD40 (4+1 или 2+1)+PD-L1. Этот результат указывает на безопасный профиль нацеленных на FAP связывающих молекул против CD40 по сравнению с ненацеленным анти-CD40 антителом. 6.4.1 Мыши, обработанные нацеленным на FAP анти-CD40 антителом, демонстрируют меньше опосредованных анти-PD-L1 нежелательных явлений по сравнению с мышами, обработанными ненацеленным анти-CD40 антителом
Некоторые нежелательные явления наблюдали после 3-ей инъекции анти-PD-L1 антитела. Необходимости в умерщвлении мышей не было, но были описаны симптомы снижения активности мышей в течение 5 минут после инъекции терапевтического средства, сгорбленной спины и взъерошенного меха. В таблице 33 указан процент мышей, демонстрирующих эти симптомы после 3-ей терапии.

Эти результаты показывают явное повышение количества нежелательных явлений в комбинированной группе PD-L1+CD40, тогда как процент нежелательных явлений, наблюдаемых в группах анти-PD-L1+FAP-CD40 (4+1 и 2+1), был только немного повышен по сравнению с группой, обработанной одним анти-PD-L1 агентом.
Наблюдаемые нежелательные явления могут быть результатом сильной иммунной реакции (цитокинового шторма) вследствие образования ПЛА против анти-PD-L1, которая дополнительно возрастает в присутствии ненацеленного анти- CD40 антитела. Это наблюдение указывает на лучший профиль безопасности нацеленной молекулы FAP-CD40 по сравнению с ненацеленным анти-CD40 антителом, когда обе молекулы комбинируют с терапией анти -PD-L1 антителом.
В целом, нацеленные на FAP анти-CD40 молекулы с FAP-зависимой активацией CD40 в формате 4+1 и 2+1 индуцируют сильный противоопухолевый иммунный ответ у несущих опухли мышей со снижением системной токсичности по сравнению с ненацеленным анти-CD40 родительским антителом.
--->
SEQUENCE LISTING
<110> F. Hoffmann-La Roche AG
<120> Bispecific antigen binding molecules comprising anti-FAP clone
212
<130> P35043-WO
<150> EP18197866.9
<151> 2018-10-01
<160> 110
<170> PatentIn version 3.5
<210> 1
<211> 277
<212> PRT
<213> Homo sapiens
<400> 1
Met Val Arg Leu Pro Leu Gln Cys Val Leu Trp Gly Cys Leu Leu Thr
1 5 10 15
Ala Val His Pro Glu Pro Pro Thr Ala Cys Arg Glu Lys Gln Tyr Leu
20 25 30
Ile Asn Ser Gln Cys Cys Ser Leu Cys Gln Pro Gly Gln Lys Leu Val
35 40 45
Ser Asp Cys Thr Glu Phe Thr Glu Thr Glu Cys Leu Pro Cys Gly Glu
50 55 60
Ser Glu Phe Leu Asp Thr Trp Asn Arg Glu Thr His Cys His Gln His
65 70 75 80
Lys Tyr Cys Asp Pro Asn Leu Gly Leu Arg Val Gln Gln Lys Gly Thr
85 90 95
Ser Glu Thr Asp Thr Ile Cys Thr Cys Glu Glu Gly Trp His Cys Thr
100 105 110
Ser Glu Ala Cys Glu Ser Cys Val Leu His Arg Ser Cys Ser Pro Gly
115 120 125
Phe Gly Val Lys Gln Ile Ala Thr Gly Val Ser Asp Thr Ile Cys Glu
130 135 140
Pro Cys Pro Val Gly Phe Phe Ser Asn Val Ser Ser Ala Phe Glu Lys
145 150 155 160
Cys His Pro Trp Thr Ser Cys Glu Thr Lys Asp Leu Val Val Gln Gln
165 170 175
Ala Gly Thr Asn Lys Thr Asp Val Val Cys Gly Pro Gln Asp Arg Leu
180 185 190
Arg Ala Leu Val Val Ile Pro Ile Ile Phe Gly Ile Leu Phe Ala Ile
195 200 205
Leu Leu Val Leu Val Phe Ile Lys Lys Val Ala Lys Lys Pro Thr Asn
210 215 220
Lys Ala Pro His Pro Lys Gln Glu Pro Gln Glu Ile Asn Phe Pro Asp
225 230 235 240
Asp Leu Pro Gly Ser Asn Thr Ala Ala Pro Val Gln Glu Thr Leu His
245 250 255
Gly Cys Gln Pro Val Thr Gln Glu Asp Gly Lys Glu Ser Arg Ile Ser
260 265 270
Val Gln Glu Arg Gln
275
<210> 2
<211> 760
<212> PRT
<213> Homo sapiens
<400> 2
Met Lys Thr Trp Val Lys Ile Val Phe Gly Val Ala Thr Ser Ala Val
1 5 10 15
Leu Ala Leu Leu Val Met Cys Ile Val Leu Arg Pro Ser Arg Val His
20 25 30
Asn Ser Glu Glu Asn Thr Met Arg Ala Leu Thr Leu Lys Asp Ile Leu
35 40 45
Asn Gly Thr Phe Ser Tyr Lys Thr Phe Phe Pro Asn Trp Ile Ser Gly
50 55 60
Gln Glu Tyr Leu His Gln Ser Ala Asp Asn Asn Ile Val Leu Tyr Asn
65 70 75 80
Ile Glu Thr Gly Gln Ser Tyr Thr Ile Leu Ser Asn Arg Thr Met Lys
85 90 95
Ser Val Asn Ala Ser Asn Tyr Gly Leu Ser Pro Asp Arg Gln Phe Val
100 105 110
Tyr Leu Glu Ser Asp Tyr Ser Lys Leu Trp Arg Tyr Ser Tyr Thr Ala
115 120 125
Thr Tyr Tyr Ile Tyr Asp Leu Ser Asn Gly Glu Phe Val Arg Gly Asn
130 135 140
Glu Leu Pro Arg Pro Ile Gln Tyr Leu Cys Trp Ser Pro Val Gly Ser
145 150 155 160
Lys Leu Ala Tyr Val Tyr Gln Asn Asn Ile Tyr Leu Lys Gln Arg Pro
165 170 175
Gly Asp Pro Pro Phe Gln Ile Thr Phe Asn Gly Arg Glu Asn Lys Ile
180 185 190
Phe Asn Gly Ile Pro Asp Trp Val Tyr Glu Glu Glu Met Leu Ala Thr
195 200 205
Lys Tyr Ala Leu Trp Trp Ser Pro Asn Gly Lys Phe Leu Ala Tyr Ala
210 215 220
Glu Phe Asn Asp Thr Asp Ile Pro Val Ile Ala Tyr Ser Tyr Tyr Gly
225 230 235 240
Asp Glu Gln Tyr Pro Arg Thr Ile Asn Ile Pro Tyr Pro Lys Ala Gly
245 250 255
Ala Lys Asn Pro Val Val Arg Ile Phe Ile Ile Asp Thr Thr Tyr Pro
260 265 270
Ala Tyr Val Gly Pro Gln Glu Val Pro Val Pro Ala Met Ile Ala Ser
275 280 285
Ser Asp Tyr Tyr Phe Ser Trp Leu Thr Trp Val Thr Asp Glu Arg Val
290 295 300
Cys Leu Gln Trp Leu Lys Arg Val Gln Asn Val Ser Val Leu Ser Ile
305 310 315 320
Cys Asp Phe Arg Glu Asp Trp Gln Thr Trp Asp Cys Pro Lys Thr Gln
325 330 335
Glu His Ile Glu Glu Ser Arg Thr Gly Trp Ala Gly Gly Phe Phe Val
340 345 350
Ser Thr Pro Val Phe Ser Tyr Asp Ala Ile Ser Tyr Tyr Lys Ile Phe
355 360 365
Ser Asp Lys Asp Gly Tyr Lys His Ile His Tyr Ile Lys Asp Thr Val
370 375 380
Glu Asn Ala Ile Gln Ile Thr Ser Gly Lys Trp Glu Ala Ile Asn Ile
385 390 395 400
Phe Arg Val Thr Gln Asp Ser Leu Phe Tyr Ser Ser Asn Glu Phe Glu
405 410 415
Glu Tyr Pro Gly Arg Arg Asn Ile Tyr Arg Ile Ser Ile Gly Ser Tyr
420 425 430
Pro Pro Ser Lys Lys Cys Val Thr Cys His Leu Arg Lys Glu Arg Cys
435 440 445
Gln Tyr Tyr Thr Ala Ser Phe Ser Asp Tyr Ala Lys Tyr Tyr Ala Leu
450 455 460
Val Cys Tyr Gly Pro Gly Ile Pro Ile Ser Thr Leu His Asp Gly Arg
465 470 475 480
Thr Asp Gln Glu Ile Lys Ile Leu Glu Glu Asn Lys Glu Leu Glu Asn
485 490 495
Ala Leu Lys Asn Ile Gln Leu Pro Lys Glu Glu Ile Lys Lys Leu Glu
500 505 510
Val Asp Glu Ile Thr Leu Trp Tyr Lys Met Ile Leu Pro Pro Gln Phe
515 520 525
Asp Arg Ser Lys Lys Tyr Pro Leu Leu Ile Gln Val Tyr Gly Gly Pro
530 535 540
Cys Ser Gln Ser Val Arg Ser Val Phe Ala Val Asn Trp Ile Ser Tyr
545 550 555 560
Leu Ala Ser Lys Glu Gly Met Val Ile Ala Leu Val Asp Gly Arg Gly
565 570 575
Thr Ala Phe Gln Gly Asp Lys Leu Leu Tyr Ala Val Tyr Arg Lys Leu
580 585 590
Gly Val Tyr Glu Val Glu Asp Gln Ile Thr Ala Val Arg Lys Phe Ile
595 600 605
Glu Met Gly Phe Ile Asp Glu Lys Arg Ile Ala Ile Trp Gly Trp Ser
610 615 620
Tyr Gly Gly Tyr Val Ser Ser Leu Ala Leu Ala Ser Gly Thr Gly Leu
625 630 635 640
Phe Lys Cys Gly Ile Ala Val Ala Pro Val Ser Ser Trp Glu Tyr Tyr
645 650 655
Ala Ser Val Tyr Thr Glu Arg Phe Met Gly Leu Pro Thr Lys Asp Asp
660 665 670
Asn Leu Glu His Tyr Lys Asn Ser Thr Val Met Ala Arg Ala Glu Tyr
675 680 685
Phe Arg Asn Val Asp Tyr Leu Leu Ile His Gly Thr Ala Asp Asp Asn
690 695 700
Val His Phe Gln Asn Ser Ala Gln Ile Ala Lys Ala Leu Val Asn Ala
705 710 715 720
Gln Val Asp Phe Gln Ala Met Trp Tyr Ser Asp Gln Asn His Gly Leu
725 730 735
Ser Gly Leu Ser Thr Asn His Leu Tyr Thr His Met Thr His Phe Leu
740 745 750
Lys Gln Cys Phe Ser Leu Ser Asp
755 760
<210> 3
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP (212) CDR-H1
<400> 3
Asp Tyr Asn Met Asp
1 5
<210> 4
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP (212) CDR-H2
<400> 4
Asp Ile Tyr Pro Asn Thr Gly Gly Thr Ile Tyr Asn Gln Lys Phe Lys
1 5 10 15
Gly
<210> 5
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP (212) CDR-H3
<400> 5
Phe Arg Gly Ile His Tyr Ala Met Asp Tyr
1 5 10
<210> 6
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP (212) CDR-L1
<400> 6
Arg Ala Ser Glu Ser Val Asp Asn Tyr Gly Leu Ser Phe Ile Asn
1 5 10 15
<210> 7
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP (212) CDR-L2
<400> 7
Gly Thr Ser Asn Arg Gly Ser
1 5
<210> 8
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP (212) CDR-L3
<400> 8
Gln Gln Ser Asn Glu Val Pro Tyr Thr
1 5
<210> 9
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP (212) VH
<400> 9
Glu Val Leu Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ala Cys Lys Ala Ser Gly Tyr Thr Leu Thr Asp Tyr
20 25 30
Asn Met Asp Trp Val Arg Gln Ser His Gly Lys Ser Leu Glu Trp Ile
35 40 45
Gly Asp Ile Tyr Pro Asn Thr Gly Gly Thr Ile Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ile Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Asp Leu Arg Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Arg Gly Ile His Tyr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Ser Val Thr Val Ser Ser
115
<210> 10
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP (212) VL
<400> 10
Asp Ile Val Leu Thr Gln Ser Pro Val Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Leu Ser Phe Ile Asn Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Gly Thr Ser Asn Arg Gly Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Ser Leu Asn Ile His
65 70 75 80
Pro Met Glu Glu Asp Asp Thr Ala Met Tyr Phe Cys Gln Gln Ser Asn
85 90 95
Glu Val Pro Tyr Thr Phe Gly Gly Gly Thr Asn Leu Glu Ile Lys
100 105 110
<210> 11
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP (VH1G3a) CDR-H2
<400> 11
Asp Ile Tyr Pro Asn Thr Gly Gly Thr Ile Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 12
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP (VH2G3a) CDR-H2
<400> 12
Asp Ile Tyr Pro Asn Thr Gly Gly Thr Ile Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 13
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP (VL1G3a) CDR-L1
<400> 13
Arg Ala Ser Glu Ser Val Asp Asn Tyr Gly Leu Ser Phe Leu Ala
1 5 10 15
<210> 14
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP (VL2G3a) CDR-L1
<400> 14
Arg Ala Ser Glu Ser Ile Asp Asn Tyr Gly Leu Ser Phe Leu Asn
1 5 10 15
<210> 15
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP (VH1G1a)
<400> 15
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Leu Thr Asp Tyr
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Asp Ile Tyr Pro Asn Thr Gly Gly Thr Ile Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Met Thr Ile Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Arg Gly Ile His Tyr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser
115
<210> 16
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP (VH1G2a)
<400> 16
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Leu Thr Asp Tyr
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Lys Ser Leu Glu Trp Ile
35 40 45
Gly Asp Ile Tyr Pro Asn Thr Gly Gly Thr Ile Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Met Thr Ile Asp Lys Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Arg Gly Ile His Tyr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser
115
<210> 17
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP (VH1G3a)
<400> 17
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Leu Thr Asp Tyr
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Asp Ile Tyr Pro Asn Thr Gly Gly Thr Ile Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Ile Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Arg Gly Ile His Tyr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser
115
<210> 18
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP (VH2G1a)
<400> 18
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Leu Thr Asp Tyr
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Asp Ile Tyr Pro Asn Thr Gly Gly Thr Ile Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Arg Gly Ile His Tyr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser
115
<210> 19
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP (VH2G2a)
<400> 19
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Leu Thr Asp Tyr
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Asp Ile Tyr Pro Asn Thr Gly Gly Thr Ile Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ile Asp Lys Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Arg Gly Ile His Tyr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser
115
<210> 20
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP (VH2G3a)
<400> 20
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Leu Thr Asp Tyr
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Asp Ile Tyr Pro Asn Thr Gly Gly Thr Ile Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Arg Gly Ile His Tyr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser
115
<210> 21
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP (VL1G1a)
<400> 21
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Leu Ser Phe Ile Asn Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro
35 40 45
Arg Leu Leu Ile Tyr Gly Thr Ser Asn Arg Gly Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Phe Cys Gln Gln Ser Asn
85 90 95
Glu Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 22
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP (VL1G2a)
<400> 22
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Leu Ser Phe Ile Asn Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Arg Leu Leu Ile Tyr Gly Thr Ser Asn Arg Gly Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Phe Cys Gln Gln Ser Asn
85 90 95
Glu Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 23
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP (VL1G3a)
<400> 23
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Leu Ser Phe Leu Ala Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro
35 40 45
Arg Leu Leu Ile Tyr Gly Thr Ser Asn Arg Gly Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Phe Cys Gln Gln Ser Asn
85 90 95
Glu Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 24
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP (VL2G1a)
<400> 24
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Leu Ser Phe Ile Asn Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Gly Thr Ser Asn Arg Gly Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ser Asn
85 90 95
Glu Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 25
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP (VL2G2a)
<400> 25
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Leu Ser Phe Ile Asn Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Gly Thr Ser Asn Arg Gly Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Met Tyr Phe Cys Gln Gln Ser Asn
85 90 95
Glu Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 26
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP (VL2G3a)
<400> 26
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Ile Asp Asn Tyr
20 25 30
Gly Leu Ser Phe Leu Asn Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Gly Thr Ser Asn Arg Gly Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ser Asn
85 90 95
Glu Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 27
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> hu CD40 CDR-H1
<400> 27
Gly Tyr Tyr Ile His
1 5
<210> 28
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> hu CD40 CDR-H2
<400> 28
Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe Lys
1 5 10 15
Gly
<210> 29
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> hu CD40 CDR-H3
<400> 29
Glu Gly Ile Tyr Trp
1 5
<210> 30
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> hu CD40 CDR-L1
<400> 30
Arg Ser Ser Gln Ser Leu Val His Ser Asn Gly Asn Thr Phe Leu His
1 5 10 15
<210> 31
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> hu CD40 CDR-L2
<400> 31
Thr Val Ser Asn Arg Phe Ser
1 5
<210> 32
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> hu CD40 CDR-L3
<400> 32
Ser Gln Thr Thr His Val Pro Trp Thr
1 5
<210> 33
<211> 114
<212> PRT
<213> Artificial Sequence
<220>
<223> hu CD40 VH
<400> 33
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Leu Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Leu Val Thr Val
100 105 110
Ser Ser
<210> 34
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> hu CD40 VL
<400> 34
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
35 40 45
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 35
<211> 114
<212> PRT
<213> Artificial Sequence
<220>
<223> CD40 (S2C6) VH
<400> 35
Glu Val Gln Leu Gln Gln Ser Gly Pro Asp Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Lys Gln Ser His Gly Lys Ser Leu Glu Trp Ile
35 40 45
Gly Arg Val Ile Pro Asn Asn Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Ile Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly His Gly Thr Thr Leu Thr Val
100 105 110
Ser Ser
<210> 36
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> CD40 (S2C6) VL
<400> 36
Asp Val Val Val Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Ala Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Gln
100 105 110
<210> 37
<211> 114
<212> PRT
<213> Artificial Sequence
<220>
<223> VH1a (CD40)
<400> 37
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 38
<211> 114
<212> PRT
<213> Artificial Sequence
<220>
<223> VH1b (CD40)
<400> 38
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 39
<211> 114
<212> PRT
<213> Artificial Sequence
<220>
<223> VH1c (CD40)
<400> 39
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly His Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 40
<211> 114
<212> PRT
<213> Artificial Sequence
<220>
<223> VH1d (CD40)
<400> 40
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Ser Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 41
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> VL1a (CD40)
<400> 41
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 42
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> VL1b (CD40)
<400> 42
Asp Ile Val Val Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 43
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> VL1c (CD40)
<400> 43
Asp Val Val Val Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 44
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> VL1d (CD40)
<400> 44
Asp Val Val Val Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 45
<211> 114
<212> PRT
<213> Artificial Sequence
<220>
<223> VH2a (CD40)
<400> 45
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 46
<211> 114
<212> PRT
<213> Artificial Sequence
<220>
<223> VH2b (CD40)
<400> 46
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Ser Leu Glu Trp Val
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 47
<211> 114
<212> PRT
<213> Artificial Sequence
<220>
<223> VH2c (CD40)
<400> 47
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly His Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 48
<211> 114
<212> PRT
<213> Artificial Sequence
<220>
<223> VH2d (CD40)
<400> 48
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Gly Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 49
<211> 114
<212> PRT
<213> Artificial Sequence
<220>
<223> VH2ab (CD40)
<400> 49
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 50
<211> 114
<212> PRT
<213> Artificial Sequence
<220>
<223> VH2ac (CD40)
<400> 50
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 51
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> VL2a (CD40)
<400> 51
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
35 40 45
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 52
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> VL2b (CD40)
<400> 52
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 53
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> VL2ab (CD40)
<400> 53
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
35 40 45
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 54
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> VL2ac (CD40)
<400> 54
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Ser Ile Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
35 40 45
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 55
<211> 443
<212> PRT
<213> Artificial Sequence
<220>
<223> P1AE0400 Heavy chain
<400> 55
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440
<210> 56
<211> 219
<212> PRT
<213> Artificial Sequence
<220>
<223> P1AE0400 light chain
<400> 56
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
35 40 45
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 57
<211> 443
<212> PRT
<213> Artificial Sequence
<220>
<223> P1AE0403 heavy chain
<400> 57
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Gly Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440
<210> 58
<211> 219
<212> PRT
<213> Artificial Sequence
<220>
<223> P1AE0403 light chain
<400> 58
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
35 40 45
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 59
<211> 443
<212> PRT
<213> Artificial Sequence
<220>
<223> P1AE0817 heavy chain
<400> 59
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440
<210> 60
<211> 219
<212> PRT
<213> Artificial Sequence
<220>
<223> P1AE0817 light chain
<400> 60
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 61
<211> 226
<212> PRT
<213> Artificial Sequence
<220>
<223> (P1AE1689) light chain cross VH-Ckappa
<400> 61
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Leu Thr Asp Tyr
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Asp Ile Tyr Pro Asn Thr Gly Gly Thr Ile Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Met Thr Ile Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Arg Gly Ile His Tyr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe
115 120 125
Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val
130 135 140
Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp
145 150 155 160
Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr
165 170 175
Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr
180 185 190
Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val
195 200 205
Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly
210 215 220
Glu Cys
225
<210> 62
<211> 219
<212> PRT
<213> Artificial Sequence
<220>
<223> VL1a (CD40) light chain (charged)
<400> 62
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Arg
115 120 125
Lys Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 63
<211> 679
<212> PRT
<213> Artificial Sequence
<220>
<223> VH1a (CD40) (VHCH1 charged) Fc knob_PGLALA_(P1AE1689) (VL-CH1)
<400> 63
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Glu
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Glu Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Gly Gly Gly Gly Ser
435 440 445
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu
450 455 460
Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly Glu
465 470 475 480
Arg Ala Thr Leu Ser Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr Gly
485 490 495
Leu Ser Phe Ile Asn Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro Arg
500 505 510
Leu Leu Ile Tyr Gly Thr Ser Asn Arg Gly Ser Gly Ile Pro Ala Arg
515 520 525
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
530 535 540
Leu Glu Pro Glu Asp Phe Ala Val Tyr Phe Cys Gln Gln Ser Asn Glu
545 550 555 560
Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Ser Ser
565 570 575
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
580 585 590
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
595 600 605
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
610 615 620
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
625 630 635 640
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
645 650 655
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
660 665 670
Lys Val Glu Pro Lys Ser Cys
675
<210> 64
<211> 443
<212> Белок
<213> Искуственная последовательность
<220>
<223> VH1a (CD40) (VHCH1 заряженная) Fc-впадина_PGLALA
<400> 64
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Glu
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Glu Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440
<210> 65
<211> 216
<212> Белок
<213> Искуственная последовательность
<220>
<223> (P1AE1689), легкая цепь, перекрестно связанная с VL-CH1
<400> 65
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Leu Ser Phe Ile Asn Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro
35 40 45
Arg Leu Leu Ile Tyr Gly Thr Ser Asn Arg Gly Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Phe Cys Gln Gln Ser Asn
85 90 95
Glu Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Ser
100 105 110
Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser
115 120 125
Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp
130 135 140
Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr
145 150 155 160
Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr
165 170 175
Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln
180 185 190
Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp
195 200 205
Lys Lys Val Glu Pro Lys Ser Cys
210 215
<210> 66
<211> 219
<212> Белок
<213> Искуственная последовательность
<220>
<223> Легкая цепь VL1a (CD40)
<400> 66
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 67
<211> 689
<212> Белок
<213> Искуственная последовательность
<220>
<223> VH1a (CD40) (VHCH1) Fc-выступ_PGLALA_(P1AE1689) (VH-C-каппа)
<400> 67
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Gly Gly Gly Gly Ser
435 440 445
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln
450 455 460
Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala Ser
465 470 475 480
Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Leu Thr Asp Tyr Asn
485 490 495
Met Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile Gly
500 505 510
Asp Ile Tyr Pro Asn Thr Gly Gly Thr Ile Tyr Asn Gln Lys Phe Lys
515 520 525
Gly Arg Val Thr Met Thr Ile Asp Thr Ser Thr Ser Thr Val Tyr Met
530 535 540
Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys Thr
545 550 555 560
Arg Phe Arg Gly Ile His Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
565 570 575
Thr Val Thr Val Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile
580 585 590
Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val
595 600 605
Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys
610 615 620
Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu
625 630 635 640
Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu
645 650 655
Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr
660 665 670
His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu
675 680 685
Cys
<210> 68
<211> 443
<212> Белок
<213> Искуственная последовательность
<220>
<223> VH1a (CD40) (VHCH1) Fc-впадина_PGLALA
<400> 68
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440
<210> 69
<211> 907
<212> Белок
<213> Искуственная последовательность
<220>
<223> VH1a (CD40) (VHCH1 заряженная_VH1a (CD40) (VHCH1 заряженная)-Fc выступ_PGLALA_(P1AE1689) (VL-CH1)
<400> 69
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Glu
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Glu Lys Val Glu Pro Lys Ser Cys Asp Gly Gly Gly Gly Ser Gly
210 215 220
Gly Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys
225 230 235 240
Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser
245 250 255
Phe Thr Gly Tyr Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser
260 265 270
Leu Glu Trp Met Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr
275 280 285
Asn Gln Lys Phe Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile
290 295 300
Ser Thr Ala Tyr Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala
305 310 315 320
Val Tyr Tyr Cys Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr
325 330 335
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
340 345 350
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
355 360 365
Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
370 375 380
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
385 390 395 400
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
405 410 415
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
420 425 430
Asn Thr Lys Val Asp Glu Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
435 440 445
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
450 455 460
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
465 470 475 480
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
485 490 495
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
500 505 510
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
515 520 525
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
530 535 540
Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr
545 550 555 560
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
565 570 575
Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys
580 585 590
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
595 600 605
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
610 615 620
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
625 630 635 640
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
645 650 655
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Gly
660 665 670
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
675 680 685
Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu
690 695 700
Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Glu Ser Val
705 710 715 720
Asp Asn Tyr Gly Leu Ser Phe Ile Asn Trp Phe Gln Gln Lys Pro Gly
725 730 735
Gln Ala Pro Arg Leu Leu Ile Tyr Gly Thr Ser Asn Arg Gly Ser Gly
740 745 750
Ile Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
755 760 765
Thr Ile Ser Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Phe Cys Gln
770 775 780
Gln Ser Asn Glu Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu
785 790 795 800
Ile Lys Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
805 810 815
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
820 825 830
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
835 840 845
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
850 855 860
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
865 870 875 880
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
885 890 895
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
900 905
<210> 70
<211> 671
<212> Белок
<213> Искуственная последовательность
<220>
<223> VH1a (CD40) (VHCH1 заряженная)_VH1a (CD40) (VHCH1 заряженная)-Fc впадина_PGLALA
<400> 70
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Glu
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Glu Lys Val Glu Pro Lys Ser Cys Asp Gly Gly Gly Gly Ser Gly
210 215 220
Gly Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys
225 230 235 240
Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser
245 250 255
Phe Thr Gly Tyr Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser
260 265 270
Leu Glu Trp Met Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr
275 280 285
Asn Gln Lys Phe Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile
290 295 300
Ser Thr Ala Tyr Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala
305 310 315 320
Val Tyr Tyr Cys Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr
325 330 335
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
340 345 350
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
355 360 365
Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
370 375 380
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
385 390 395 400
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
405 410 415
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
420 425 430
Asn Thr Lys Val Asp Glu Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
435 440 445
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
450 455 460
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
465 470 475 480
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
485 490 495
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
500 505 510
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
515 520 525
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
530 535 540
Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr
545 550 555 560
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu
565 570 575
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Ser Cys
580 585 590
Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
595 600 605
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
610 615 620
Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser
625 630 635 640
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
645 650 655
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
660 665 670
<210> 71
<211> 213
<212> Белок
<213> Искуственная последовательность
<220>
<223> 4B9, легкая цепь, перекрестно связанная с VL-CH1
<400> 71
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Thr Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Asn Val Gly Ser Arg Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly Ile Met Leu Pro
85 90 95
Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Ser Ser Ala Ser
100 105 110
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
115 120 125
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
130 135 140
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
145 150 155 160
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
165 170 175
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
180 185 190
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
195 200 205
Glu Pro Lys Ser Cys
210
<210> 72
<211> 687
<212> Белок
<213> Искуственная последовательность
<220>
<223> VH1a (CD40) (VHCH1) Fc-выступ_PGLALA_4B9 (VH-C-каппа)
<400> 72
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Gly Gly Gly Gly Ser
435 440 445
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu
450 455 460
Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser
465 470 475 480
Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Ala
485 490 495
Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser
500 505 510
Ala Ile Ile Gly Ser Gly Ala Ser Thr Tyr Tyr Ala Asp Ser Val Lys
515 520 525
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
530 535 540
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
545 550 555 560
Lys Gly Trp Phe Gly Gly Phe Asn Tyr Trp Gly Gln Gly Thr Leu Val
565 570 575
Thr Val Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile Phe Pro
580 585 590
Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu
595 600 605
Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp
610 615 620
Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp
625 630 635 640
Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys
645 650 655
Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln
660 665 670
Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
675 680 685
<210> 73
<211> 915
<212> Белок
<213> Искуственная последовательность
<220>
<223> VH1a (CD40) (VHCH1) _VH1a (CD40) (VHCH1)-Fc-выступ_PGLALA_(4B9)
<400> 73
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Gly Gly Gly Gly Ser Gly
210 215 220
Gly Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys
225 230 235 240
Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser
245 250 255
Phe Thr Gly Tyr Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser
260 265 270
Leu Glu Trp Met Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr
275 280 285
Asn Gln Lys Phe Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile
290 295 300
Ser Thr Ala Tyr Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala
305 310 315 320
Val Tyr Tyr Cys Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr
325 330 335
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
340 345 350
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
355 360 365
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
370 375 380
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
385 390 395 400
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
405 410 415
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
420 425 430
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
435 440 445
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
450 455 460
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
465 470 475 480
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
485 490 495
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
500 505 510
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
515 520 525
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
530 535 540
Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr
545 550 555 560
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
565 570 575
Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys
580 585 590
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
595 600 605
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
610 615 620
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
625 630 635 640
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
645 650 655
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Gly
660 665 670
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
675 680 685
Gly Gly Ser Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln
690 695 700
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
705 710 715 720
Ser Ser Tyr Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
725 730 735
Glu Trp Val Ser Ala Ile Ile Gly Ser Gly Ala Ser Thr Tyr Tyr Ala
740 745 750
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn
755 760 765
Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
770 775 780
Tyr Tyr Cys Ala Lys Gly Trp Phe Gly Gly Phe Asn Tyr Trp Gly Gln
785 790 795 800
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Val Ala Ala Pro Ser Val
805 810 815
Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser
820 825 830
Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln
835 840 845
Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val
850 855 860
Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu
865 870 875 880
Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu
885 890 895
Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg
900 905 910
Gly Glu Cys
915
<210> 74
<211> 671
<212> Белок
<213> Искуственная последовательность
<220>
<223> VH1a (CD40) (VHCH1)_VH1a (CD40) (VHCH1)-Fc-впадина_PGLALA
<400> 74
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Gly Gly Gly Gly Ser Gly
210 215 220
Gly Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys
225 230 235 240
Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser
245 250 255
Phe Thr Gly Tyr Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser
260 265 270
Leu Glu Trp Met Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr
275 280 285
Asn Gln Lys Phe Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile
290 295 300
Ser Thr Ala Tyr Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala
305 310 315 320
Val Tyr Tyr Cys Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr
325 330 335
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
340 345 350
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
355 360 365
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
370 375 380
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
385 390 395 400
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
405 410 415
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
420 425 430
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
435 440 445
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
450 455 460
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
465 470 475 480
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
485 490 495
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
500 505 510
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
515 520 525
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
530 535 540
Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr
545 550 555 560
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu
565 570 575
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Ser Cys
580 585 590
Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
595 600 605
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
610 615 620
Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser
625 630 635 640
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
645 650 655
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
660 665 670
<210> 75
<211> 223
<212> Белок
<213> Искуственная последовательность
<220>
<223> 28H1, легкая цепь, перекрестно связанная с VH-C-каппа
<400> 75
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser His
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Trp Ala Ser Gly Glu Gln Tyr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Lys Gly Trp Leu Gly Asn Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile Phe Pro
115 120 125
Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu
130 135 140
Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp
145 150 155 160
Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp
165 170 175
Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys
180 185 190
Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln
195 200 205
Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215 220
<210> 76
<211> 676
<212> Белок
<213> Искуственная последовательность
<220>
<223> VH1a (CD40) (VHCH1 заряженная) Fc-выступ_PGLALA_28H1 (VL-CH1)
<400> 76
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Glu
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Glu Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Gly Gly Gly Gly Ser
435 440 445
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu
450 455 460
Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly Glu
465 470 475 480
Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Arg Ser Tyr
485 490 495
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
500 505 510
Ile Gly Ala Ser Thr Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser Gly
515 520 525
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu Pro
530 535 540
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly Gln Val Ile Pro Pro
545 550 555 560
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Ser Ser Ala Ser Thr
565 570 575
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
580 585 590
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
595 600 605
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
610 615 620
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
625 630 635 640
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
645 650 655
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
660 665 670
Pro Lys Ser Cys
675
<210> 77
<211> 904
<212> Белок
<213> Искуственная последовательность
<220>
<223> VH1a (CD40) (VHCH1 заряженная) _VH1a (CD40) (VHCH1 заряженная) Fc выступ_PGLALA_28H1 (VL-CH1)
<400> 77
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Glu
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Glu Lys Val Glu Pro Lys Ser Cys Asp Gly Gly Gly Gly Ser Gly
210 215 220
Gly Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys
225 230 235 240
Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser
245 250 255
Phe Thr Gly Tyr Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser
260 265 270
Leu Glu Trp Met Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr
275 280 285
Asn Gln Lys Phe Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile
290 295 300
Ser Thr Ala Tyr Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala
305 310 315 320
Val Tyr Tyr Cys Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr
325 330 335
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
340 345 350
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
355 360 365
Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
370 375 380
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
385 390 395 400
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
405 410 415
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
420 425 430
Asn Thr Lys Val Asp Glu Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
435 440 445
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
450 455 460
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
465 470 475 480
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
485 490 495
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
500 505 510
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
515 520 525
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
530 535 540
Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr
545 550 555 560
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
565 570 575
Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys
580 585 590
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
595 600 605
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
610 615 620
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
625 630 635 640
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
645 650 655
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Gly
660 665 670
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
675 680 685
Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu
690 695 700
Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val
705 710 715 720
Ser Arg Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
725 730 735
Arg Leu Leu Ile Ile Gly Ala Ser Thr Arg Ala Thr Gly Ile Pro Asp
740 745 750
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
755 760 765
Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly Gln
770 775 780
Val Ile Pro Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Ser
785 790 795 800
Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser
805 810 815
Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp
820 825 830
Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr
835 840 845
Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr
850 855 860
Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln
865 870 875 880
Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp
885 890 895
Lys Lys Val Glu Pro Lys Ser Cys
900
<210> 78
<211> 748
<212> Белок
<213> Искуственная последовательность
<220>
<223> Эктодомен hu FAP + поли-lys-тэг + his6-тэг
<400> 78
Arg Pro Ser Arg Val His Asn Ser Glu Glu Asn Thr Met Arg Ala Leu
1 5 10 15
Thr Leu Lys Asp Ile Leu Asn Gly Thr Phe Ser Tyr Lys Thr Phe Phe
20 25 30
Pro Asn Trp Ile Ser Gly Gln Glu Tyr Leu His Gln Ser Ala Asp Asn
35 40 45
Asn Ile Val Leu Tyr Asn Ile Glu Thr Gly Gln Ser Tyr Thr Ile Leu
50 55 60
Ser Asn Arg Thr Met Lys Ser Val Asn Ala Ser Asn Tyr Gly Leu Ser
65 70 75 80
Pro Asp Arg Gln Phe Val Tyr Leu Glu Ser Asp Tyr Ser Lys Leu Trp
85 90 95
Arg Tyr Ser Tyr Thr Ala Thr Tyr Tyr Ile Tyr Asp Leu Ser Asn Gly
100 105 110
Glu Phe Val Arg Gly Asn Glu Leu Pro Arg Pro Ile Gln Tyr Leu Cys
115 120 125
Trp Ser Pro Val Gly Ser Lys Leu Ala Tyr Val Tyr Gln Asn Asn Ile
130 135 140
Tyr Leu Lys Gln Arg Pro Gly Asp Pro Pro Phe Gln Ile Thr Phe Asn
145 150 155 160
Gly Arg Glu Asn Lys Ile Phe Asn Gly Ile Pro Asp Trp Val Tyr Glu
165 170 175
Glu Glu Met Leu Ala Thr Lys Tyr Ala Leu Trp Trp Ser Pro Asn Gly
180 185 190
Lys Phe Leu Ala Tyr Ala Glu Phe Asn Asp Thr Asp Ile Pro Val Ile
195 200 205
Ala Tyr Ser Tyr Tyr Gly Asp Glu Gln Tyr Pro Arg Thr Ile Asn Ile
210 215 220
Pro Tyr Pro Lys Ala Gly Ala Lys Asn Pro Val Val Arg Ile Phe Ile
225 230 235 240
Ile Asp Thr Thr Tyr Pro Ala Tyr Val Gly Pro Gln Glu Val Pro Val
245 250 255
Pro Ala Met Ile Ala Ser Ser Asp Tyr Tyr Phe Ser Trp Leu Thr Trp
260 265 270
Val Thr Asp Glu Arg Val Cys Leu Gln Trp Leu Lys Arg Val Gln Asn
275 280 285
Val Ser Val Leu Ser Ile Cys Asp Phe Arg Glu Asp Trp Gln Thr Trp
290 295 300
Asp Cys Pro Lys Thr Gln Glu His Ile Glu Glu Ser Arg Thr Gly Trp
305 310 315 320
Ala Gly Gly Phe Phe Val Ser Thr Pro Val Phe Ser Tyr Asp Ala Ile
325 330 335
Ser Tyr Tyr Lys Ile Phe Ser Asp Lys Asp Gly Tyr Lys His Ile His
340 345 350
Tyr Ile Lys Asp Thr Val Glu Asn Ala Ile Gln Ile Thr Ser Gly Lys
355 360 365
Trp Glu Ala Ile Asn Ile Phe Arg Val Thr Gln Asp Ser Leu Phe Tyr
370 375 380
Ser Ser Asn Glu Phe Glu Glu Tyr Pro Gly Arg Arg Asn Ile Tyr Arg
385 390 395 400
Ile Ser Ile Gly Ser Tyr Pro Pro Ser Lys Lys Cys Val Thr Cys His
405 410 415
Leu Arg Lys Glu Arg Cys Gln Tyr Tyr Thr Ala Ser Phe Ser Asp Tyr
420 425 430
Ala Lys Tyr Tyr Ala Leu Val Cys Tyr Gly Pro Gly Ile Pro Ile Ser
435 440 445
Thr Leu His Asp Gly Arg Thr Asp Gln Glu Ile Lys Ile Leu Glu Glu
450 455 460
Asn Lys Glu Leu Glu Asn Ala Leu Lys Asn Ile Gln Leu Pro Lys Glu
465 470 475 480
Glu Ile Lys Lys Leu Glu Val Asp Glu Ile Thr Leu Trp Tyr Lys Met
485 490 495
Ile Leu Pro Pro Gln Phe Asp Arg Ser Lys Lys Tyr Pro Leu Leu Ile
500 505 510
Gln Val Tyr Gly Gly Pro Cys Ser Gln Ser Val Arg Ser Val Phe Ala
515 520 525
Val Asn Trp Ile Ser Tyr Leu Ala Ser Lys Glu Gly Met Val Ile Ala
530 535 540
Leu Val Asp Gly Arg Gly Thr Ala Phe Gln Gly Asp Lys Leu Leu Tyr
545 550 555 560
Ala Val Tyr Arg Lys Leu Gly Val Tyr Glu Val Glu Asp Gln Ile Thr
565 570 575
Ala Val Arg Lys Phe Ile Glu Met Gly Phe Ile Asp Glu Lys Arg Ile
580 585 590
Ala Ile Trp Gly Trp Ser Tyr Gly Gly Tyr Val Ser Ser Leu Ala Leu
595 600 605
Ala Ser Gly Thr Gly Leu Phe Lys Cys Gly Ile Ala Val Ala Pro Val
610 615 620
Ser Ser Trp Glu Tyr Tyr Ala Ser Val Tyr Thr Glu Arg Phe Met Gly
625 630 635 640
Leu Pro Thr Lys Asp Asp Asn Leu Glu His Tyr Lys Asn Ser Thr Val
645 650 655
Met Ala Arg Ala Glu Tyr Phe Arg Asn Val Asp Tyr Leu Leu Ile His
660 665 670
Gly Thr Ala Asp Asp Asn Val His Phe Gln Asn Ser Ala Gln Ile Ala
675 680 685
Lys Ala Leu Val Asn Ala Gln Val Asp Phe Gln Ala Met Trp Tyr Ser
690 695 700
Asp Gln Asn His Gly Leu Ser Gly Leu Ser Thr Asn His Leu Tyr Thr
705 710 715 720
His Met Thr His Phe Leu Lys Gln Cys Phe Ser Leu Ser Asp Gly Lys
725 730 735
Lys Lys Lys Lys Lys Gly His His His His His His
740 745
<210> 79
<211> 761
<212> Белок
<213> мышиный
<400> 79
Met Lys Thr Trp Leu Lys Thr Val Phe Gly Val Thr Thr Leu Ala Ala
1 5 10 15
Leu Ala Leu Val Val Ile Cys Ile Val Leu Arg Pro Ser Arg Val Tyr
20 25 30
Lys Pro Glu Gly Asn Thr Lys Arg Ala Leu Thr Leu Lys Asp Ile Leu
35 40 45
Asn Gly Thr Phe Ser Tyr Lys Thr Tyr Phe Pro Asn Trp Ile Ser Glu
50 55 60
Gln Glu Tyr Leu His Gln Ser Glu Asp Asp Asn Ile Val Phe Tyr Asn
65 70 75 80
Ile Glu Thr Arg Glu Ser Tyr Ile Ile Leu Ser Asn Ser Thr Met Lys
85 90 95
Ser Val Asn Ala Thr Asp Tyr Gly Leu Ser Pro Asp Arg Gln Phe Val
100 105 110
Tyr Leu Glu Ser Asp Tyr Ser Lys Leu Trp Arg Tyr Ser Tyr Thr Ala
115 120 125
Thr Tyr Tyr Ile Tyr Asp Leu Gln Asn Gly Glu Phe Val Arg Gly Tyr
130 135 140
Glu Leu Pro Arg Pro Ile Gln Tyr Leu Cys Trp Ser Pro Val Gly Ser
145 150 155 160
Lys Leu Ala Tyr Val Tyr Gln Asn Asn Ile Tyr Leu Lys Gln Arg Pro
165 170 175
Gly Asp Pro Pro Phe Gln Ile Thr Tyr Thr Gly Arg Glu Asn Arg Ile
180 185 190
Phe Asn Gly Ile Pro Asp Trp Val Tyr Glu Glu Glu Met Leu Ala Thr
195 200 205
Lys Tyr Ala Leu Trp Trp Ser Pro Asp Gly Lys Phe Leu Ala Tyr Val
210 215 220
Glu Phe Asn Asp Ser Asp Ile Pro Ile Ile Ala Tyr Ser Tyr Tyr Gly
225 230 235 240
Asp Gly Gln Tyr Pro Arg Thr Ile Asn Ile Pro Tyr Pro Lys Ala Gly
245 250 255
Ala Lys Asn Pro Val Val Arg Val Phe Ile Val Asp Thr Thr Tyr Pro
260 265 270
His His Val Gly Pro Met Glu Val Pro Val Pro Glu Met Ile Ala Ser
275 280 285
Ser Asp Tyr Tyr Phe Ser Trp Leu Thr Trp Val Ser Ser Glu Arg Val
290 295 300
Cys Leu Gln Trp Leu Lys Arg Val Gln Asn Val Ser Val Leu Ser Ile
305 310 315 320
Cys Asp Phe Arg Glu Asp Trp His Ala Trp Glu Cys Pro Lys Asn Gln
325 330 335
Glu His Val Glu Glu Ser Arg Thr Gly Trp Ala Gly Gly Phe Phe Val
340 345 350
Ser Thr Pro Ala Phe Ser Gln Asp Ala Thr Ser Tyr Tyr Lys Ile Phe
355 360 365
Ser Asp Lys Asp Gly Tyr Lys His Ile His Tyr Ile Lys Asp Thr Val
370 375 380
Glu Asn Ala Ile Gln Ile Thr Ser Gly Lys Trp Glu Ala Ile Tyr Ile
385 390 395 400
Phe Arg Val Thr Gln Asp Ser Leu Phe Tyr Ser Ser Asn Glu Phe Glu
405 410 415
Gly Tyr Pro Gly Arg Arg Asn Ile Tyr Arg Ile Ser Ile Gly Asn Ser
420 425 430
Pro Pro Ser Lys Lys Cys Val Thr Cys His Leu Arg Lys Glu Arg Cys
435 440 445
Gln Tyr Tyr Thr Ala Ser Phe Ser Tyr Lys Ala Lys Tyr Tyr Ala Leu
450 455 460
Val Cys Tyr Gly Pro Gly Leu Pro Ile Ser Thr Leu His Asp Gly Arg
465 470 475 480
Thr Asp Gln Glu Ile Gln Val Leu Glu Glu Asn Lys Glu Leu Glu Asn
485 490 495
Ser Leu Arg Asn Ile Gln Leu Pro Lys Val Glu Ile Lys Lys Leu Lys
500 505 510
Asp Gly Gly Leu Thr Phe Trp Tyr Lys Met Ile Leu Pro Pro Gln Phe
515 520 525
Asp Arg Ser Lys Lys Tyr Pro Leu Leu Ile Gln Val Tyr Gly Gly Pro
530 535 540
Cys Ser Gln Ser Val Lys Ser Val Phe Ala Val Asn Trp Ile Thr Tyr
545 550 555 560
Leu Ala Ser Lys Glu Gly Ile Val Ile Ala Leu Val Asp Gly Arg Gly
565 570 575
Thr Ala Phe Gln Gly Asp Lys Phe Leu His Ala Val Tyr Arg Lys Leu
580 585 590
Gly Val Tyr Glu Val Glu Asp Gln Leu Thr Ala Val Arg Lys Phe Ile
595 600 605
Glu Met Gly Phe Ile Asp Glu Glu Arg Ile Ala Ile Trp Gly Trp Ser
610 615 620
Tyr Gly Gly Tyr Val Ser Ser Leu Ala Leu Ala Ser Gly Thr Gly Leu
625 630 635 640
Phe Lys Cys Gly Ile Ala Val Ala Pro Val Ser Ser Trp Glu Tyr Tyr
645 650 655
Ala Ser Ile Tyr Ser Glu Arg Phe Met Gly Leu Pro Thr Lys Asp Asp
660 665 670
Asn Leu Glu His Tyr Lys Asn Ser Thr Val Met Ala Arg Ala Glu Tyr
675 680 685
Phe Arg Asn Val Asp Tyr Leu Leu Ile His Gly Thr Ala Asp Asp Asn
690 695 700
Val His Phe Gln Asn Ser Ala Gln Ile Ala Lys Ala Leu Val Asn Ala
705 710 715 720
Gln Val Asp Phe Gln Ala Met Trp Tyr Ser Asp Gln Asn His Gly Ile
725 730 735
Ser Ser Gly Arg Ser Gln Asn His Leu Tyr Thr His Met Thr His Phe
740 745 750
Leu Lys Gln Cys Phe Ser Leu Ser Asp
755 760
<210> 80
<211> 749
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мышиный эктодомен FAP + поли-lys-тэг + his6-тэг
<400> 80
Arg Pro Ser Arg Val Tyr Lys Pro Glu Gly Asn Thr Lys Arg Ala Leu
1 5 10 15
Thr Leu Lys Asp Ile Leu Asn Gly Thr Phe Ser Tyr Lys Thr Tyr Phe
20 25 30
Pro Asn Trp Ile Ser Glu Gln Glu Tyr Leu His Gln Ser Glu Asp Asp
35 40 45
Asn Ile Val Phe Tyr Asn Ile Glu Thr Arg Glu Ser Tyr Ile Ile Leu
50 55 60
Ser Asn Ser Thr Met Lys Ser Val Asn Ala Thr Asp Tyr Gly Leu Ser
65 70 75 80
Pro Asp Arg Gln Phe Val Tyr Leu Glu Ser Asp Tyr Ser Lys Leu Trp
85 90 95
Arg Tyr Ser Tyr Thr Ala Thr Tyr Tyr Ile Tyr Asp Leu Gln Asn Gly
100 105 110
Glu Phe Val Arg Gly Tyr Glu Leu Pro Arg Pro Ile Gln Tyr Leu Cys
115 120 125
Trp Ser Pro Val Gly Ser Lys Leu Ala Tyr Val Tyr Gln Asn Asn Ile
130 135 140
Tyr Leu Lys Gln Arg Pro Gly Asp Pro Pro Phe Gln Ile Thr Tyr Thr
145 150 155 160
Gly Arg Glu Asn Arg Ile Phe Asn Gly Ile Pro Asp Trp Val Tyr Glu
165 170 175
Glu Glu Met Leu Ala Thr Lys Tyr Ala Leu Trp Trp Ser Pro Asp Gly
180 185 190
Lys Phe Leu Ala Tyr Val Glu Phe Asn Asp Ser Asp Ile Pro Ile Ile
195 200 205
Ala Tyr Ser Tyr Tyr Gly Asp Gly Gln Tyr Pro Arg Thr Ile Asn Ile
210 215 220
Pro Tyr Pro Lys Ala Gly Ala Lys Asn Pro Val Val Arg Val Phe Ile
225 230 235 240
Val Asp Thr Thr Tyr Pro His His Val Gly Pro Met Glu Val Pro Val
245 250 255
Pro Glu Met Ile Ala Ser Ser Asp Tyr Tyr Phe Ser Trp Leu Thr Trp
260 265 270
Val Ser Ser Glu Arg Val Cys Leu Gln Trp Leu Lys Arg Val Gln Asn
275 280 285
Val Ser Val Leu Ser Ile Cys Asp Phe Arg Glu Asp Trp His Ala Trp
290 295 300
Glu Cys Pro Lys Asn Gln Glu His Val Glu Glu Ser Arg Thr Gly Trp
305 310 315 320
Ala Gly Gly Phe Phe Val Ser Thr Pro Ala Phe Ser Gln Asp Ala Thr
325 330 335
Ser Tyr Tyr Lys Ile Phe Ser Asp Lys Asp Gly Tyr Lys His Ile His
340 345 350
Tyr Ile Lys Asp Thr Val Glu Asn Ala Ile Gln Ile Thr Ser Gly Lys
355 360 365
Trp Glu Ala Ile Tyr Ile Phe Arg Val Thr Gln Asp Ser Leu Phe Tyr
370 375 380
Ser Ser Asn Glu Phe Glu Gly Tyr Pro Gly Arg Arg Asn Ile Tyr Arg
385 390 395 400
Ile Ser Ile Gly Asn Ser Pro Pro Ser Lys Lys Cys Val Thr Cys His
405 410 415
Leu Arg Lys Glu Arg Cys Gln Tyr Tyr Thr Ala Ser Phe Ser Tyr Lys
420 425 430
Ala Lys Tyr Tyr Ala Leu Val Cys Tyr Gly Pro Gly Leu Pro Ile Ser
435 440 445
Thr Leu His Asp Gly Arg Thr Asp Gln Glu Ile Gln Val Leu Glu Glu
450 455 460
Asn Lys Glu Leu Glu Asn Ser Leu Arg Asn Ile Gln Leu Pro Lys Val
465 470 475 480
Glu Ile Lys Lys Leu Lys Asp Gly Gly Leu Thr Phe Trp Tyr Lys Met
485 490 495
Ile Leu Pro Pro Gln Phe Asp Arg Ser Lys Lys Tyr Pro Leu Leu Ile
500 505 510
Gln Val Tyr Gly Gly Pro Cys Ser Gln Ser Val Lys Ser Val Phe Ala
515 520 525
Val Asn Trp Ile Thr Tyr Leu Ala Ser Lys Glu Gly Ile Val Ile Ala
530 535 540
Leu Val Asp Gly Arg Gly Thr Ala Phe Gln Gly Asp Lys Phe Leu His
545 550 555 560
Ala Val Tyr Arg Lys Leu Gly Val Tyr Glu Val Glu Asp Gln Leu Thr
565 570 575
Ala Val Arg Lys Phe Ile Glu Met Gly Phe Ile Asp Glu Glu Arg Ile
580 585 590
Ala Ile Trp Gly Trp Ser Tyr Gly Gly Tyr Val Ser Ser Leu Ala Leu
595 600 605
Ala Ser Gly Thr Gly Leu Phe Lys Cys Gly Ile Ala Val Ala Pro Val
610 615 620
Ser Ser Trp Glu Tyr Tyr Ala Ser Ile Tyr Ser Glu Arg Phe Met Gly
625 630 635 640
Leu Pro Thr Lys Asp Asp Asn Leu Glu His Tyr Lys Asn Ser Thr Val
645 650 655
Met Ala Arg Ala Glu Tyr Phe Arg Asn Val Asp Tyr Leu Leu Ile His
660 665 670
Gly Thr Ala Asp Asp Asn Val His Phe Gln Asn Ser Ala Gln Ile Ala
675 680 685
Lys Ala Leu Val Asn Ala Gln Val Asp Phe Gln Ala Met Trp Tyr Ser
690 695 700
Asp Gln Asn His Gly Ile Leu Ser Gly Arg Ser Gln Asn His Leu Tyr
705 710 715 720
Thr His Met Thr His Phe Leu Lys Gln Cys Phe Ser Leu Ser Asp Gly
725 730 735
Lys Lys Lys Lys Lys Lys Gly His His His His His His
740 745
<210> 81
<211> 748
<212> Белок
<213> Искусственная последовательность
<220>
<223> Эктодомен FAP яванского макака + поли-lys-тэг + his6-тэг
<400> 81
Arg Pro Pro Arg Val His Asn Ser Glu Glu Asn Thr Met Arg Ala Leu
1 5 10 15
Thr Leu Lys Asp Ile Leu Asn Gly Thr Phe Ser Tyr Lys Thr Phe Phe
20 25 30
Pro Asn Trp Ile Ser Gly Gln Glu Tyr Leu His Gln Ser Ala Asp Asn
35 40 45
Asn Ile Val Leu Tyr Asn Ile Glu Thr Gly Gln Ser Tyr Thr Ile Leu
50 55 60
Ser Asn Arg Thr Met Lys Ser Val Asn Ala Ser Asn Tyr Gly Leu Ser
65 70 75 80
Pro Asp Arg Gln Phe Val Tyr Leu Glu Ser Asp Tyr Ser Lys Leu Trp
85 90 95
Arg Tyr Ser Tyr Thr Ala Thr Tyr Tyr Ile Tyr Asp Leu Ser Asn Gly
100 105 110
Glu Phe Val Arg Gly Asn Glu Leu Pro Arg Pro Ile Gln Tyr Leu Cys
115 120 125
Trp Ser Pro Val Gly Ser Lys Leu Ala Tyr Val Tyr Gln Asn Asn Ile
130 135 140
Tyr Leu Lys Gln Arg Pro Gly Asp Pro Pro Phe Gln Ile Thr Phe Asn
145 150 155 160
Gly Arg Glu Asn Lys Ile Phe Asn Gly Ile Pro Asp Trp Val Tyr Glu
165 170 175
Glu Glu Met Leu Ala Thr Lys Tyr Ala Leu Trp Trp Ser Pro Asn Gly
180 185 190
Lys Phe Leu Ala Tyr Ala Glu Phe Asn Asp Thr Asp Ile Pro Val Ile
195 200 205
Ala Tyr Ser Tyr Tyr Gly Asp Glu Gln Tyr Pro Arg Thr Ile Asn Ile
210 215 220
Pro Tyr Pro Lys Ala Gly Ala Lys Asn Pro Phe Val Arg Ile Phe Ile
225 230 235 240
Ile Asp Thr Thr Tyr Pro Ala Tyr Val Gly Pro Gln Glu Val Pro Val
245 250 255
Pro Ala Met Ile Ala Ser Ser Asp Tyr Tyr Phe Ser Trp Leu Thr Trp
260 265 270
Val Thr Asp Glu Arg Val Cys Leu Gln Trp Leu Lys Arg Val Gln Asn
275 280 285
Val Ser Val Leu Ser Ile Cys Asp Phe Arg Glu Asp Trp Gln Thr Trp
290 295 300
Asp Cys Pro Lys Thr Gln Glu His Ile Glu Glu Ser Arg Thr Gly Trp
305 310 315 320
Ala Gly Gly Phe Phe Val Ser Thr Pro Val Phe Ser Tyr Asp Ala Ile
325 330 335
Ser Tyr Tyr Lys Ile Phe Ser Asp Lys Asp Gly Tyr Lys His Ile His
340 345 350
Tyr Ile Lys Asp Thr Val Glu Asn Ala Ile Gln Ile Thr Ser Gly Lys
355 360 365
Trp Glu Ala Ile Asn Ile Phe Arg Val Thr Gln Asp Ser Leu Phe Tyr
370 375 380
Ser Ser Asn Glu Phe Glu Asp Tyr Pro Gly Arg Arg Asn Ile Tyr Arg
385 390 395 400
Ile Ser Ile Gly Ser Tyr Pro Pro Ser Lys Lys Cys Val Thr Cys His
405 410 415
Leu Arg Lys Glu Arg Cys Gln Tyr Tyr Thr Ala Ser Phe Ser Asp Tyr
420 425 430
Ala Lys Tyr Tyr Ala Leu Val Cys Tyr Gly Pro Gly Ile Pro Ile Ser
435 440 445
Thr Leu His Asp Gly Arg Thr Asp Gln Glu Ile Lys Ile Leu Glu Glu
450 455 460
Asn Lys Glu Leu Glu Asn Ala Leu Lys Asn Ile Gln Leu Pro Lys Glu
465 470 475 480
Glu Ile Lys Lys Leu Glu Val Asp Glu Ile Thr Leu Trp Tyr Lys Met
485 490 495
Ile Leu Pro Pro Gln Phe Asp Arg Ser Lys Lys Tyr Pro Leu Leu Ile
500 505 510
Gln Val Tyr Gly Gly Pro Cys Ser Gln Ser Val Arg Ser Val Phe Ala
515 520 525
Val Asn Trp Ile Ser Tyr Leu Ala Ser Lys Glu Gly Met Val Ile Ala
530 535 540
Leu Val Asp Gly Arg Gly Thr Ala Phe Gln Gly Asp Lys Leu Leu Tyr
545 550 555 560
Ala Val Tyr Arg Lys Leu Gly Val Tyr Glu Val Glu Asp Gln Ile Thr
565 570 575
Ala Val Arg Lys Phe Ile Glu Met Gly Phe Ile Asp Glu Lys Arg Ile
580 585 590
Ala Ile Trp Gly Trp Ser Tyr Gly Gly Tyr Val Ser Ser Leu Ala Leu
595 600 605
Ala Ser Gly Thr Gly Leu Phe Lys Cys Gly Ile Ala Val Ala Pro Val
610 615 620
Ser Ser Trp Glu Tyr Tyr Ala Ser Val Tyr Thr Glu Arg Phe Met Gly
625 630 635 640
Leu Pro Thr Lys Asp Asp Asn Leu Glu His Tyr Lys Asn Ser Thr Val
645 650 655
Met Ala Arg Ala Glu Tyr Phe Arg Asn Val Asp Tyr Leu Leu Ile His
660 665 670
Gly Thr Ala Asp Asp Asn Val His Phe Gln Asn Ser Ala Gln Ile Ala
675 680 685
Lys Ala Leu Val Asn Ala Gln Val Asp Phe Gln Ala Met Trp Tyr Ser
690 695 700
Asp Gln Asn His Gly Leu Ser Gly Leu Ser Thr Asn His Leu Tyr Thr
705 710 715 720
His Met Thr His Phe Leu Lys Gln Cys Phe Ser Leu Ser Asp Gly Lys
725 730 735
Lys Lys Lys Lys Lys Gly His His His His His His
740 745
<210> 82
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Пептидный линкер G4S
<400> 82
Gly Gly Gly Gly Ser
1 5
<210> 83
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Пептидный линкер(G4S)2
<400> 83
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210> 84
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Пептидный линкер(SG4)2
<400> 84
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
1 5 10
<210> 85
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> Пептидный линкер G4(SG4)2
<400> 85
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
1 5 10
<210> 86
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Пептидный линкер
<400> 86
Gly Ser Pro Gly Ser Ser Ser Ser Gly Ser
1 5 10
<210> 87
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> Пептидный линкер(G4S)3
<400> 87
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 88
<211> 20
<212> Белок
<213> Искусственная последовательность
<220>
<223> Пептидный линкер(G4S)4
<400> 88
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser
20
<210> 89
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Пептидный линкер
<400> 89
Gly Ser Gly Ser Gly Ser Gly Ser
1 5
<210> 90
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Пептидный линкер
<400> 90
Gly Ser Gly Ser Gly Asn Gly Ser
1 5
<210> 91
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Пептидный линкер
<400> 91
Gly Gly Ser Gly Ser Gly Ser Gly
1 5
<210> 92
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Пептидный линкер
<400> 92
Gly Gly Ser Gly Ser Gly
1 5
<210> 93
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> Пептидный линкер
<400> 93
Gly Gly Ser Gly
1
<210> 94
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Пептидный линкер
<400> 94
Gly Gly Ser Gly Asn Gly Ser Gly
1 5
<210> 95
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Пептидный линкер
<400> 95
Gly Gly Asn Gly Ser Gly Ser Gly
1 5
<210> 96
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Пептидный линкер
<400> 96
Gly Gly Asn Gly Ser Gly
1 5
<210> 97
<211> 227
<212> Белок
<213> Искусственная последовательность
<220>
<223> Цепь Fc-выступа
<400> 97
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys
225
<210> 98
<211> 227
<212> Белок
<213> Искусственная последовательность
<220>
<223> Цепь Fc впадины
<400> 98
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys
225
<210> 99
<211> 20
<212> Белок
<213> Homo sapiens
<400> 99
Tyr Tyr Tyr Tyr Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Thr Val
1 5 10 15
Thr Val Ser Ser
20
<210> 100
<211> 12
<212> Белок
<213> Homo sapiens
<400> 100
Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
1 5 10
<210> 101
<211> 20
<212> Белок
<213> Homo sapiens
<400> 101
Tyr Tyr Tyr Tyr Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Thr Val
1 5 10 15
Thr Val Ser Ser
20
<210> 102
<211> 12
<212> Белок
<213> Homo sapiens
<400> 102
Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
1 5 10
<210> 103
<211> 20
<212> Белок
<213> Homo sapiens
<400> 103
Tyr Tyr Tyr Tyr Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Thr Val
1 5 10 15
Thr Val Ser Ser
20
<210> 104
<211> 12
<212> Белок
<213> Homo sapiens
<400> 104
Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
1 5 10
<210> 105
<211> 219
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD40 light chain (charged)
<400> 105
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
35 40 45
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Arg
115 120 125
Lys Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 106
<211> 676
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD40 (VHCH1 charged) Fc PGLALA FAP (VL-CH1)
<400> 106
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Leu Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Leu Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Glu
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Glu Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Gly Gly Gly Gly Ser
435 440 445
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu
450 455 460
Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly Glu
465 470 475 480
Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Arg Ser Tyr
485 490 495
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
500 505 510
Ile Gly Ala Ser Thr Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser Gly
515 520 525
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu Pro
530 535 540
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly Gln Val Ile Pro Pro
545 550 555 560
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Ser Ser Ala Ser Thr
565 570 575
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
580 585 590
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
595 600 605
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
610 615 620
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
625 630 635 640
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
645 650 655
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
660 665 670
Pro Lys Ser Cys
675
<210> 107
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> VH (PD-L1)
<400> 107
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Ser
20 25 30
Trp Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Trp Ile Ser Pro Tyr Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg His Trp Pro Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 108
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> VL (PD-L1)
<400> 108
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Ser Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Leu Tyr His Pro Ala
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 109
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> VH (PD-L1)
<400> 109
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Tyr
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asn Ile Lys Gln Asp Gly Ser Glu Lys Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Gly Trp Phe Gly Glu Leu Ala Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 110
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> VL (PD-L1)
<400> 110
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Arg Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Asp Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Leu Pro
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| БИСПЕЦИФИЧЕСКИЕ 2+1 КОНТОРСТЕЛА | 2018 |
|
RU2797305C2 |
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2837821C1 |
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2808030C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, СПЕЦИФИЧЕСКИЕ В ОТНОШЕНИИ КОСТИМУЛЯТОРНОГО TNF-РЕЦЕПТОРА | 2016 |
|
RU2761115C1 |
| АНТИТЕЛО К CD40, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2779128C2 |
| СЛИТЫЙ БЕЛОК, ВКЛЮЧАЮЩИЙ IL-12 И АНТИТЕЛО ПРОТИВ FAP, И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2831612C1 |
| ТЕРАПЕВТИЧЕСКИЕ АНТИТЕЛА ПРОТИВ ЛИГАНДА CD40 | 2018 |
|
RU2770209C2 |
| АНТИТЕЛА ПРОТИВ CD40 И ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2796413C2 |
| АНТИТЕЛА, КОТОРЫЕ СВЯЗЫВАЮТСЯ С РАКОВЫМИ КЛЕТКАМИ И ОБЕСПЕЧИВАЮТ НАПРАВЛЕННЫЙ ПЕРЕНОС РАДИОНУКЛИДОВ К УКАЗАННЫМ КЛЕТКАМ | 2020 |
|
RU2833191C2 |
| УЛУЧШЕННЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ РЕЦЕПТОРЫ | 2018 |
|
RU2825816C2 |
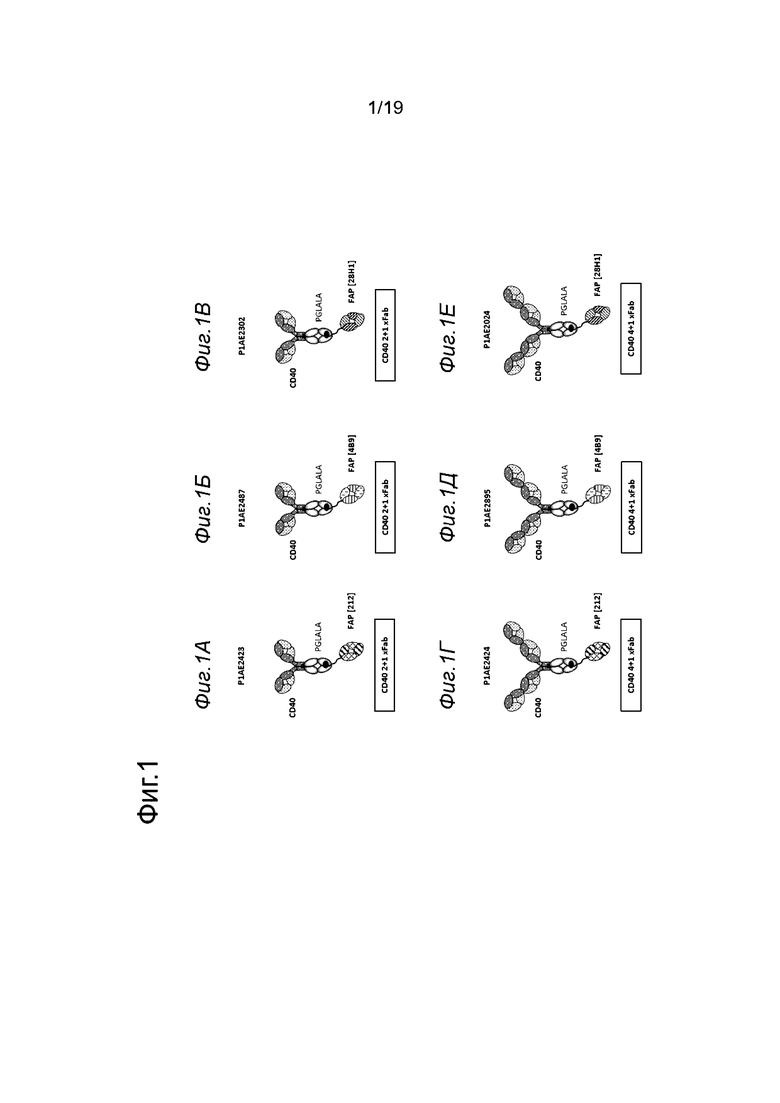
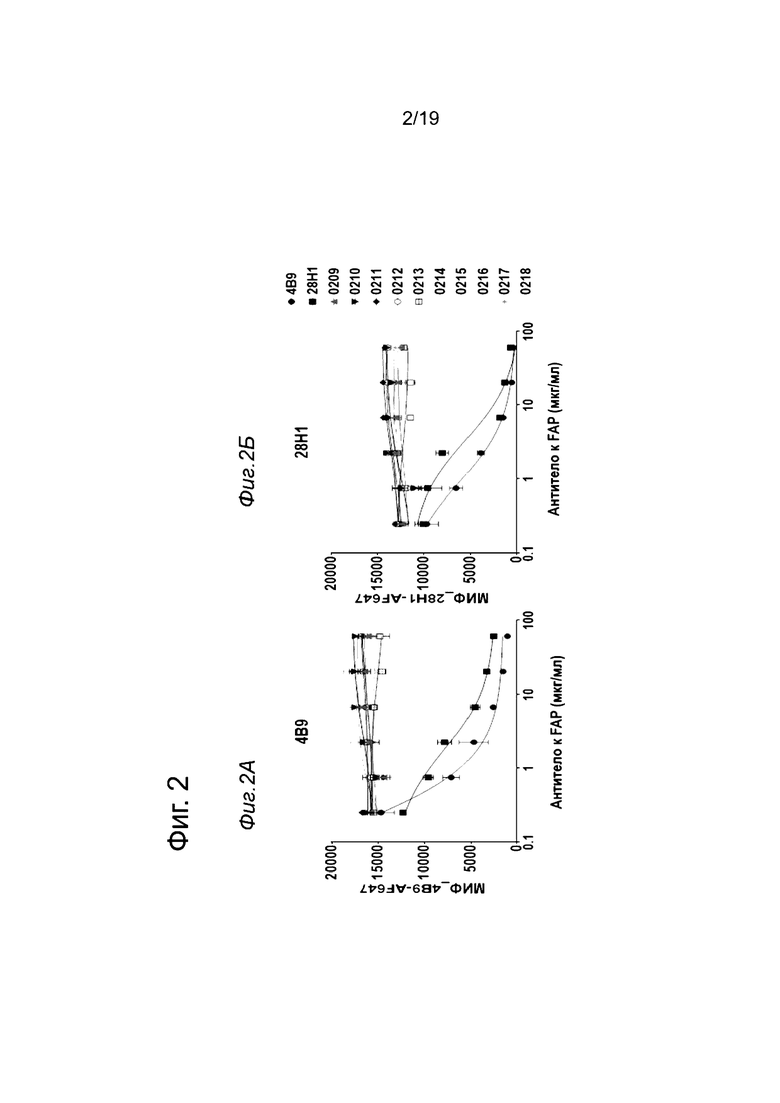
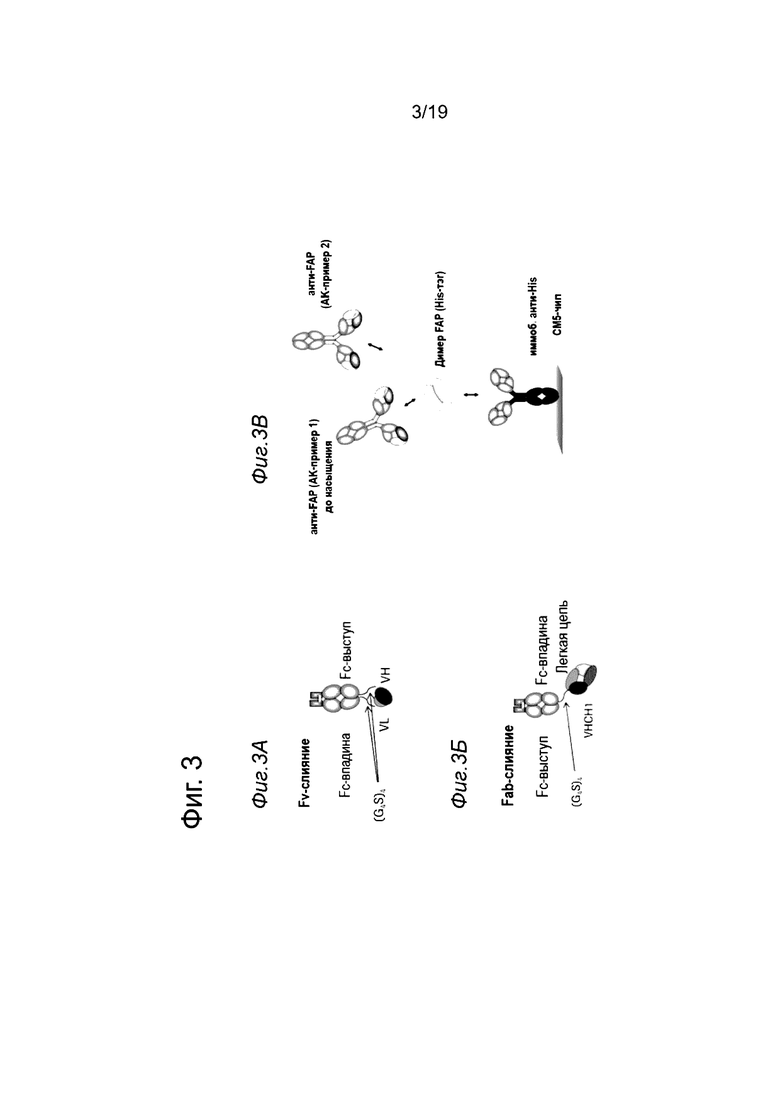
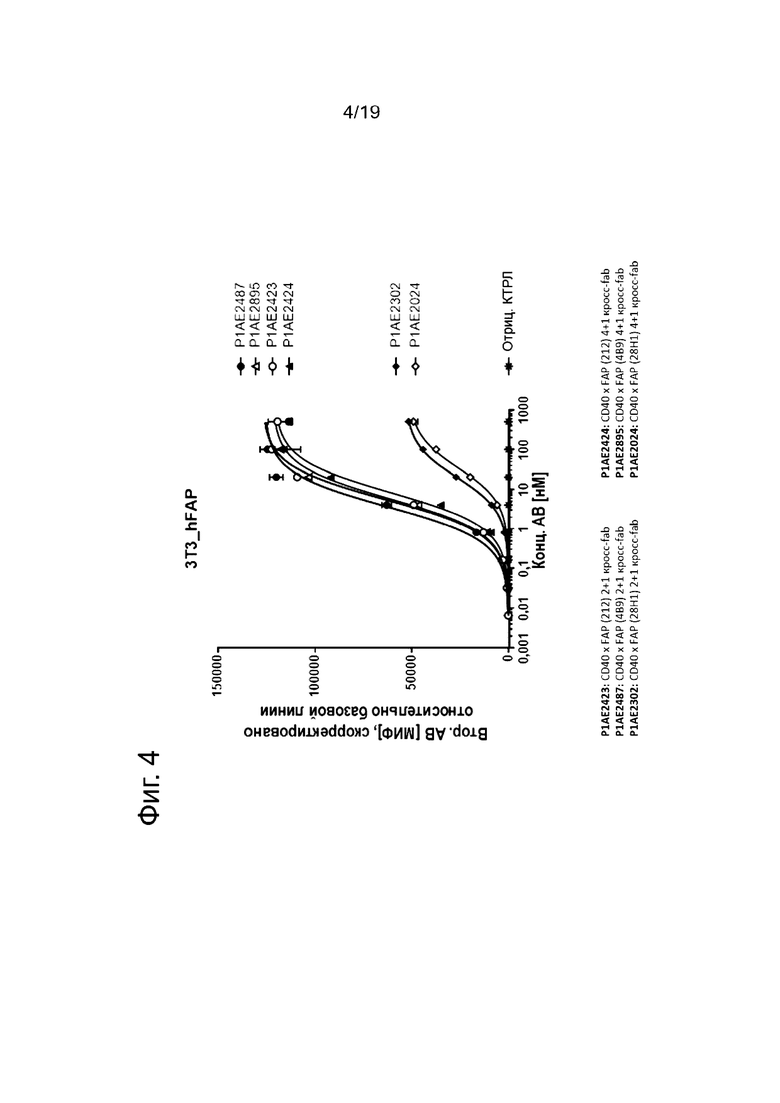
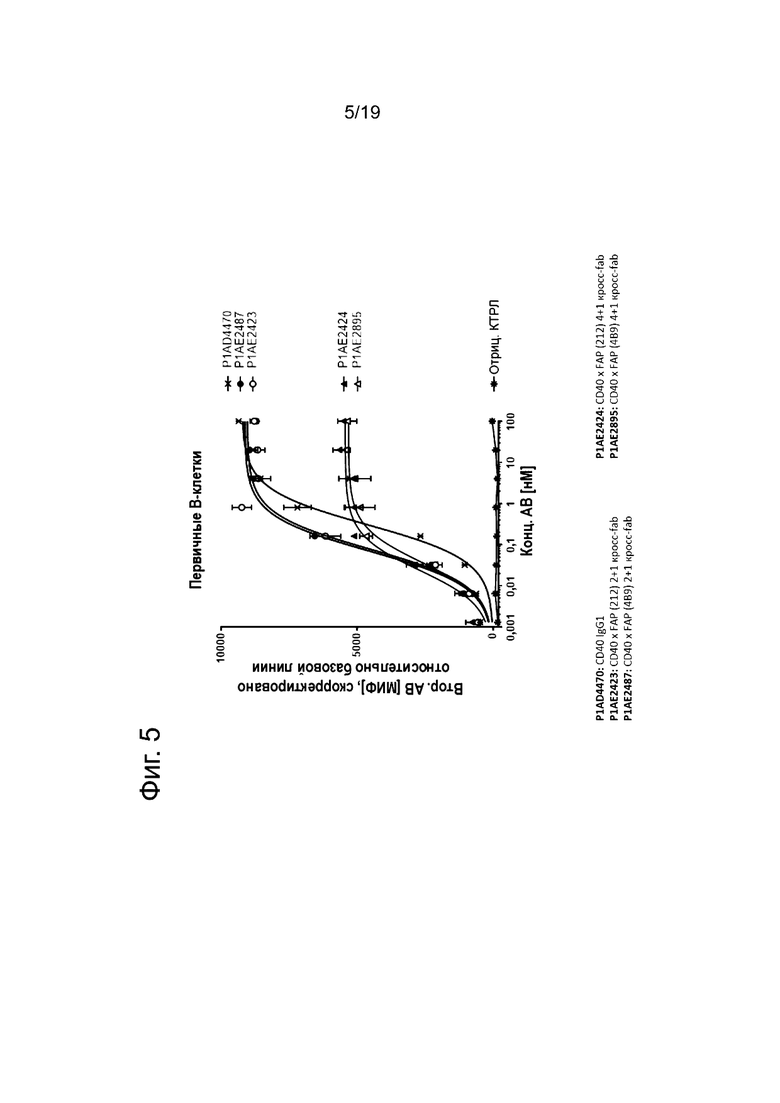



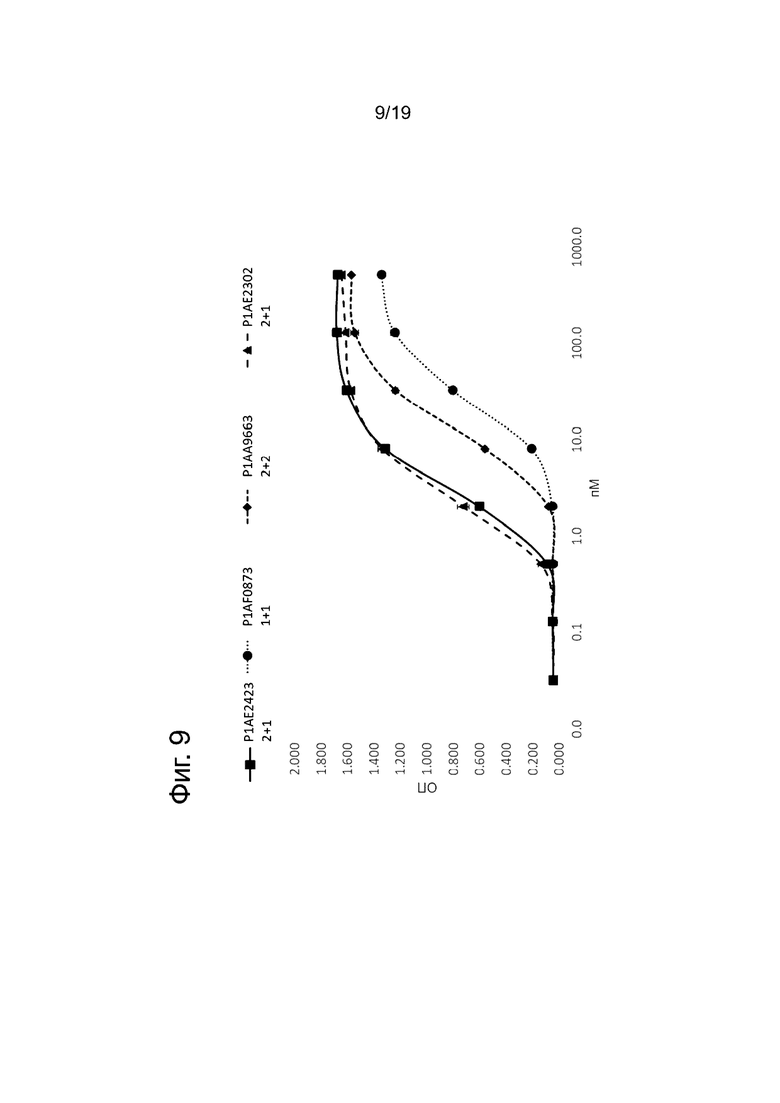
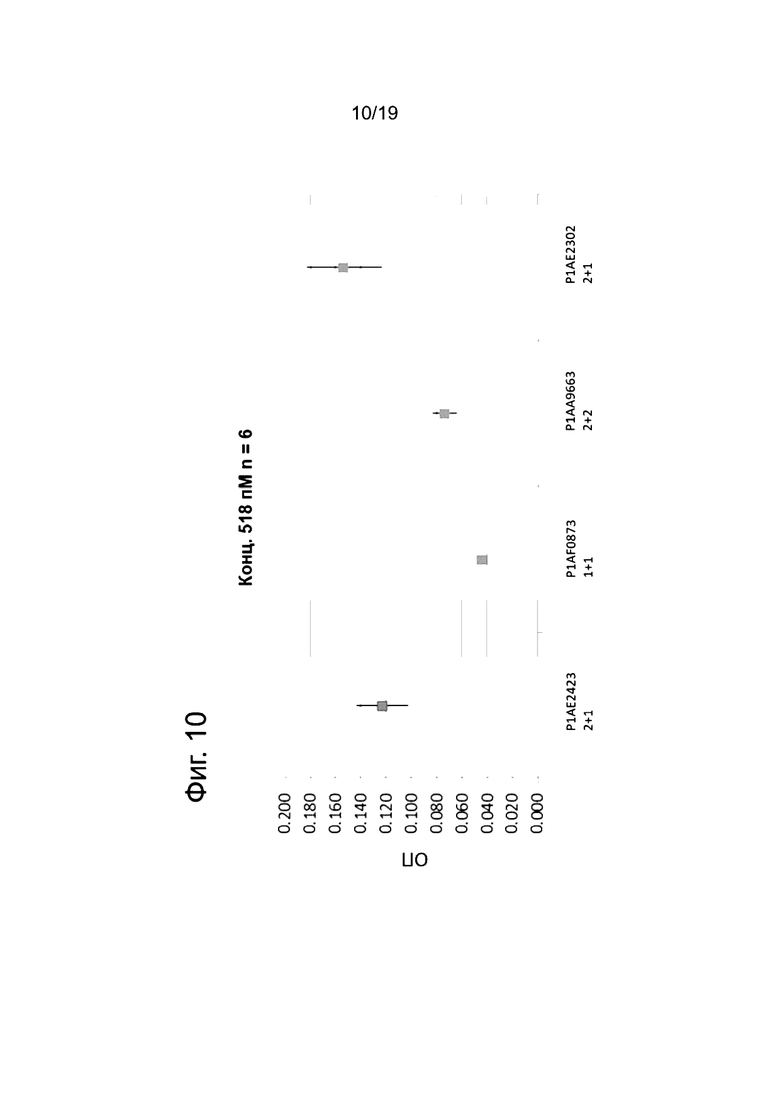

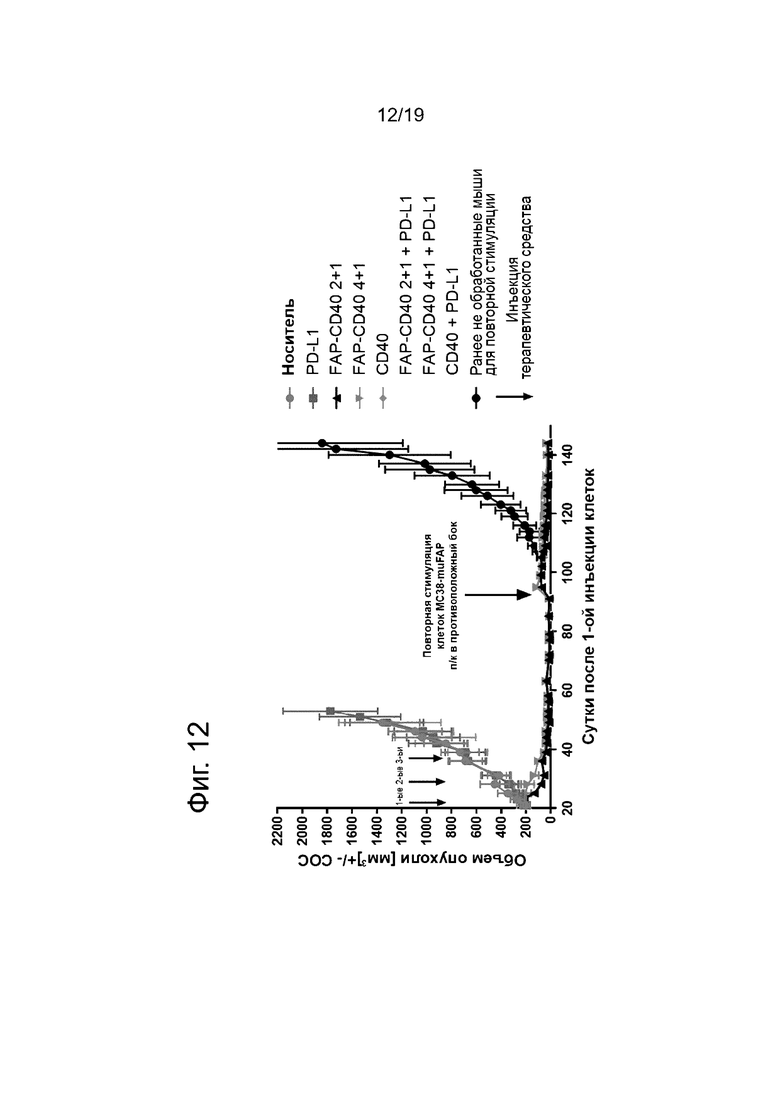
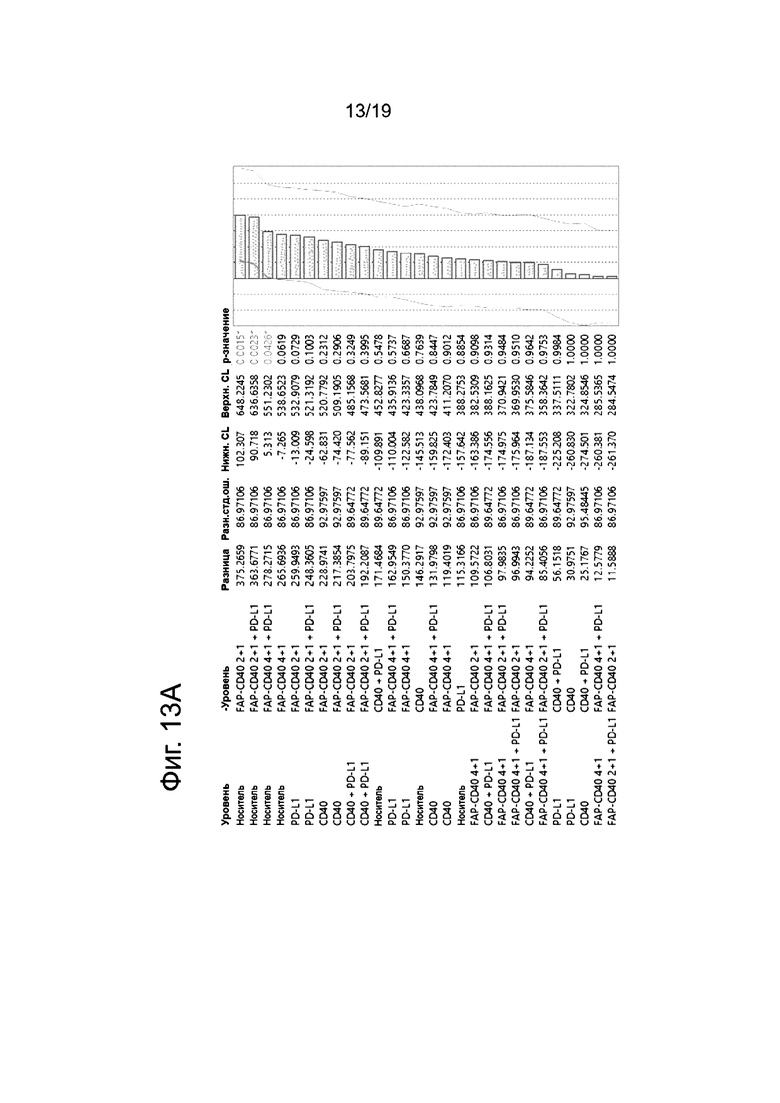
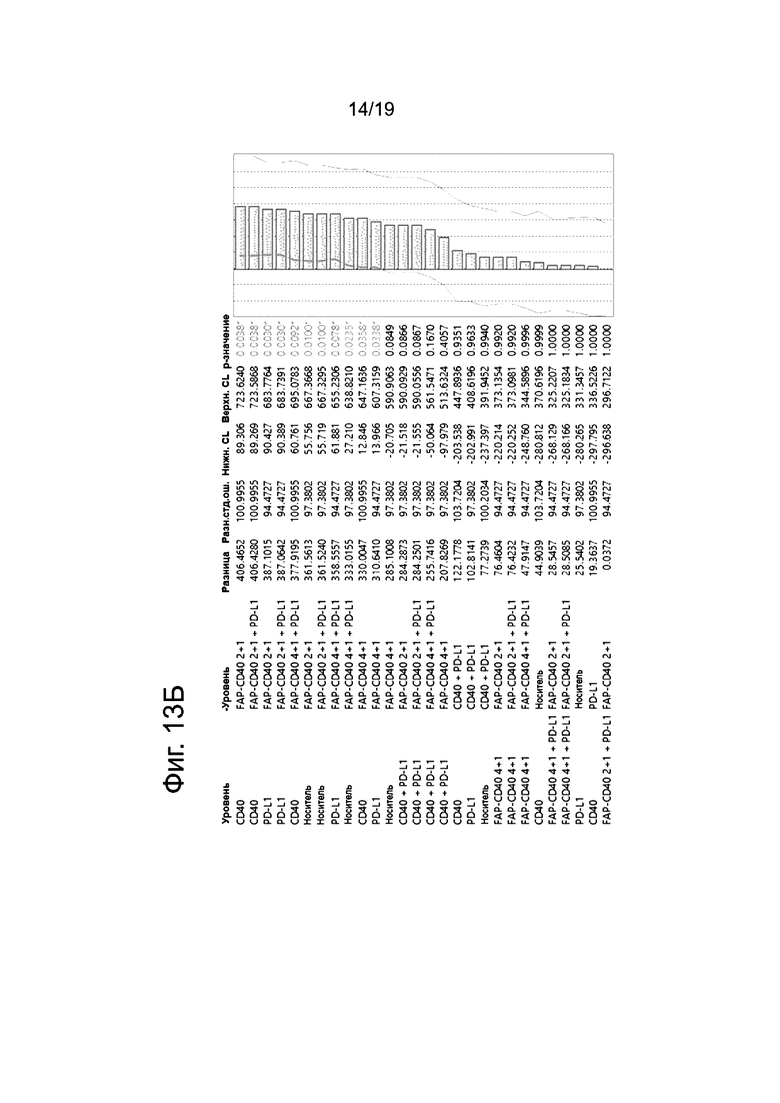
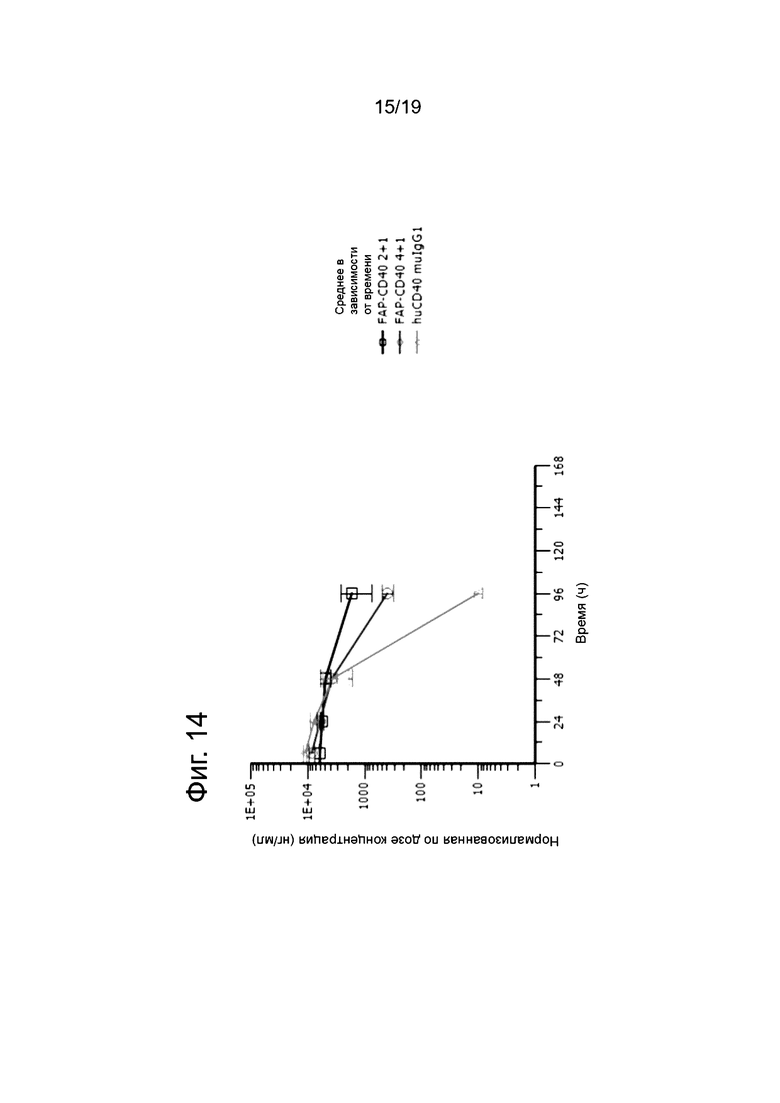
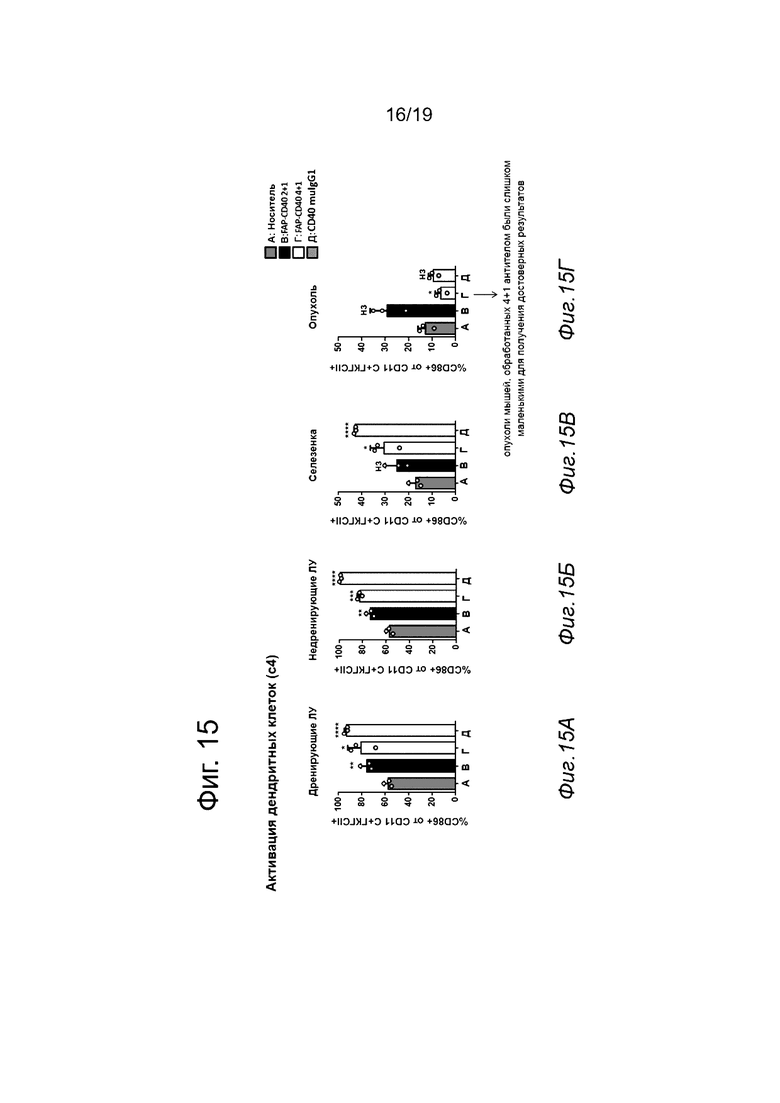


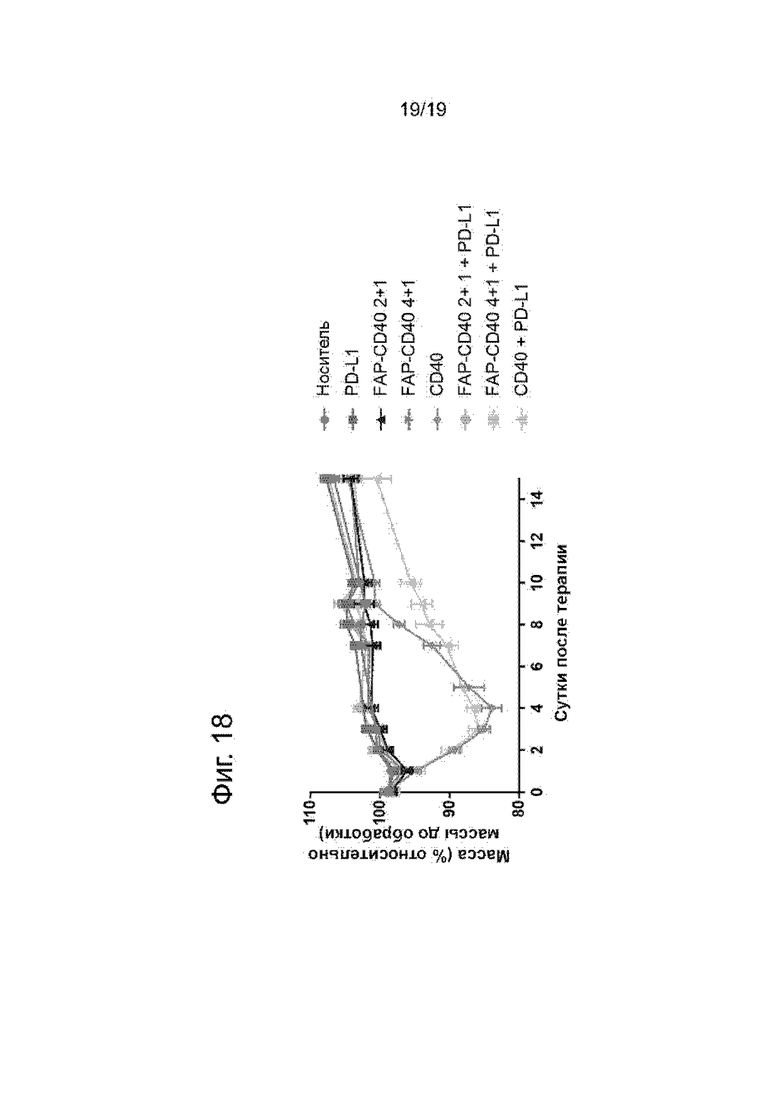
Изобретение относится к области биотехнологии. Описана группа изобретений, включающая биспецифическую связывающую молекулу, способную специфически связываться с представителем семейства рецепторов фактора некроза опухоли (TNFR) CD40 и с белком активации фибробластов (FAP), антитело, которое специфически связывается с FAP, выделенную нуклеиновую кислоту, кодирующую биспецифическое антитело, выделенную нуклеиновую кислоту, кодирующую антитело, которое специфически связывается с FAP, экспрессионный вектор, который обеспечивает экспрессию биспецифической антигенсвязывающей молекулы, клетку-хозяина для получения биспецифической антигенсвязывающей молекулы, способ получения биспецифической антигенсвязывающей молекулы, фармацевтическую композицию для лечения заболеваний, связанных с FAP и CD40, применение биспецифической антигенсвязывающей молекулы или фармацевтической композиции в лечении рака, ассоциированного с FAP и CD40, применение биспецифической антигенсвязывающей молекулы в лечении рака, ассоциированного с FAP и CD40 (варианты). Изобретение расширяет арсенал средств, специфически связывающихся с CD40 и FAP. 11 н. и 23 з.п. ф-лы, 18 ил., 35 табл., 6 пр.
1. Биспецифическая антигенсвязывающая молекула, способная специфически связываться с представителем семейства рецепторов фактора некроза опухоли (TNFR) CD40 и с белком активации фибробластов (FAP) и содержащая
(а) два антигенсвязывающих домена, способных к специфическому связыванию с CD40, и
(б) один антигенсвязывающий домен, способный к специфическому связыванию с FAP, содержащий вариабельную область тяжелой цепи (VHFAP), содержащую (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 3, (ii) CDR-H2, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4, SEQ ID NO: 11 и SEQ ID NO: 12, и (iii) CDR-НЗ, содержащую аминокислотную последовательность SEQ ID NO: 5, и вариабельную область легкой цепи (VLFAP), содержащую (iv) CDR-L1, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 6, SEQ ID NO: 13 и SEQ ID NO: 14, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 7, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 8, и (в) Fc-область, состоящую из первой и второй субъединиц, способных к стабильной ассоциации.
2. Биспецифическая антигенсвязывающая молекула по п. 1, причем антигенсвязывающий домен, способный к специфическому связыванию с FAP, содержит вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность, которая по меньшей мере на около 90% идентична аминокислотной последовательности SEQ ID NO: 9, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность, которая по меньшей мере на около 90% идентична аминокислотной последовательности SEQ ID NO: 10.
3. Биспецифическая антигенсвязывающая молекула по п. 1 или 2, причем антигенсвязывающий домен, способный к специфическому связыванию с FAP, содержит вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19 и SEQ ID NO: 20, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 21, SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 25 и SEQ ID NO: 26.
4. Биспецифическая антигенсвязывающая молекула по любому из пп. 1-3, причем антигенсвязывающий домен, способный к специфическому связыванию с FAP, содержит
(а) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 15, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 21,
(б) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 16, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 21,
(в) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 16, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 22, или
(г) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 19, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 25.
5. Биспецифическая антигенсвязывающая молекула по любому из пп. 1-4, причем антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержит вариабельную область тяжелой цепи (VHCD40), содержащую (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 27, (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 28, и (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 29, и вариабельную область легкой цепи (VLCD40), содержащую (iv) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 30, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 31, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 32.
6. Биспецифическая антигенсвязывающая молекула по любому из пп. 1-5, причем антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержит
(i) вариабельную область тяжелой цепи (VHCD40), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 37, SEQ ID NO: 38, SEQ ID NO: 39 и SEQ ID NO: 40, и
(ii) вариабельную область легкой цепи (VLCD40), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 41, SEQ ID NO: 42, SEQ ID NO: 43 и SEQ ID NO: 44.
7. Биспецифическая антигенсвязывающая молекула по любому из пп. 1-5, причем антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержит
(i) вариабельную область тяжелой цепи (VHCD40), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 45, SEQ ID NO: 46, SEQ ID NO: 47, SEQ ID NO: 48, SEQ ID NO: 49 и SEQ ID NO: 50, и
(ii) вариабельную область легкой цепи (VLCD40), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 51, SEQ ID NO: 52, SEQ ID NO: 53 и SEQ ID NO: 54.
8. Биспецифическая антигенсвязывающая молекула по любому из пп. 1-4 или 6, причем антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержит
(а) VH, содержащую аминокислотную последовательность SEQ ID NO: 37, и VL, содержащую аминокислотную последовательность SEQ ID NO: 41, или
(б) VH, содержащую аминокислотную последовательность SEQ ID NO: 37, и VL, содержащую аминокислотную последовательность SEQ ID NO: 42, или
(в) VH, содержащую аминокислотную последовательность SEQ ID NO: 37, и VL, содержащую аминокислотную последовательность SEQ ID NO: 43, или
(г) VH, содержащую аминокислотную последовательность SEQ ID NO: 37, и VL, содержащую аминокислотную последовательность SEQ ID NO: 44, или
(д) VH, содержащую аминокислотную последовательность SEQ ID NO: 38, и VL, содержащую аминокислотную последовательность SEQ ID NO: 41, или
(е) VH, содержащую аминокислотную последовательность SEQ ID NO: 38, и VL, содержащую аминокислотную последовательность SEQ ID NO: 42, или
(ж) VH, содержащую аминокислотную последовательность SEQ ID NO: 38, и VL, содержащую аминокислотную последовательность SEQ ID NO: 43, или
(з) VH, содержащую аминокислотную последовательность SEQ ID NO: 38, и VL, содержащую аминокислотную последовательность SEQ ID NO: 44, или
(и) VH, содержащую аминокислотную последовательность SEQ ID NO: 39, и VL, содержащую аминокислотную последовательность SEQ ID NO: 41, или
(к) VH, содержащую аминокислотную последовательность SEQ ID NO: 39, и VL, содержащую аминокислотную последовательность SEQ ID NO: 42, или
(л) VH, содержащую аминокислотную последовательность SEQ ID NO: 39, и VL, содержащую аминокислотную последовательность SEQ ID NO: 43, или
(м) VH, содержащую аминокислотную последовательность SEQ ID NO: 39, и VL, содержащую аминокислотную последовательность SEQ ID NO: 44, или
(н) VH, содержащую аминокислотную последовательность SEQ ID NO: 40, и VL, содержащую аминокислотную последовательность SEQ ID NO: 41, или
(о) VH, содержащую аминокислотную последовательность SEQ ID NO: 40, и VL, содержащую аминокислотную последовательность SEQ ID NO: 42, или
(п) VH, содержащую аминокислотную последовательность SEQ ID NO: 40, и VL, содержащую аминокислотную последовательность SEQ ID NO: 43, или
(р) VH, содержащую аминокислотную последовательность SEQ ID NO: 40, и VL, содержащую аминокислотную последовательность SEQ ID NO: 44.
9. Биспецифическая антигенсвязывающая молекула по любому из пп. 1-4, или 6, или 8, причем антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержит VH, содержащую аминокислотную последовательность SEQ ID NO: 37, и VL, содержащую аминокислотную последовательность SEQ ID NO: 41.
10. Биспецифическая антигенсвязывающая молекула по любому из пп. 1-4 или 7, причем антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержит
(a) VH, содержащую аминокислотную последовательность SEQ ID NO: 45, и VL, содержащую аминокислотную последовательность SEQ ID NO: 51, или
(б) VH, содержащую аминокислотную последовательность SEQ ID NO: 46, и VL, содержащую аминокислотную последовательность SEQ ID NO: 51, или
(в) VH, содержащую аминокислотную последовательность SEQ ID NO: 47, и VL, содержащую аминокислотную последовательность SEQ ID NO: 51, или
(г) VH, содержащую аминокислотную последовательность SEQ ID NO: 48, и VL, содержащую аминокислотную последовательность SEQ ID NO: 51, или
(д) VH, содержащую аминокислотную последовательность SEQ ID NO: 45, и VL, содержащую аминокислотную последовательность SEQ ID NO: 52, или
(е) VH, содержащую аминокислотную последовательность SEQ ID NO: 46, и VL, содержащую аминокислотную последовательность SEQ ID NO: 52, или
(ж) VH, содержащую аминокислотную последовательность SEQ ID NO: 47, и VL, содержащую аминокислотную последовательность SEQ ID NO: 52, или
(з) VH, содержащую аминокислотную последовательность SEQ ID NO: 48, и VL, содержащую аминокислотную последовательность SEQ ID NO: 52, или
(и) VH, содержащую аминокислотную последовательность SEQ ID NO: 49, и VL, содержащую аминокислотную последовательность SEQ ID NO: 53, или
(к) VH, содержащую аминокислотную последовательность SEQ ID NO: 50, и VL, содержащую аминокислотную последовательность SEQ ID NO: 53, или
(л) VH, содержащую аминокислотную последовательность SEQ ID NO: 49, и VL, содержащую аминокислотную последовательность SEQ ID NO: 54, или
(м) VH, содержащую аминокислотную последовательность SEQ ID NO: 50, и VL, содержащую аминокислотную последовательность SEQ ID NO: 54.
11. Биспецифическая антигенсвязывающая молекула по любому из пп. 1-4, или 7, или 10, причем антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержит VH, содержащую аминокислотную последовательность SEQ ID NO: 45, и VL, содержащую аминокислотную последовательность SEQ ID NO: 51, или причем антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержит VH, содержащую аминокислотную последовательность SEQ ID NO: 48, и VL, содержащую аминокислотную последовательность SEQ ID NO: 51.
12. Биспецифическая антигенсвязывающая молекула по любому из пп. 1-6, содержащая
(i) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с CD40, содержащий вариабельную область тяжелой цепи (VHCD40), содержащую аминокислотную последовательность SEQ ID NO: 37, и вариабельную область легкой цепи (VLCD40), содержащую аминокислотную последовательность SEQ ID NO: 41, и
(ii) по меньшей мере один антигенсвязывающий домен, способный к специфическому связыванию с FAP, содержащий вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 15, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 21.
13. Биспецифическая антигенсвязывающая молекула по любому из пп. 1-12, причем Fc-область представляет собой Fc-область IgG, в частности, IgG1, или Fc-область IgG4, и причем Fc-область содержит одну или более аминокислотных замен, которые снижают аффинность связывания антитела с Fc-рецептором и/или эффекторную функцию.
14. Биспецифическая антигенсвязывающая молекула по любому из пп. 1-13, причем Fc-область принадлежит подклассу IgG1 человека с аминокислотными мутациями L234A, L235A и P329G (нумерация в соответствии с индексом EU по Kabat).
15. Биспецифическая антигенсвязывающая молекула по любому из пп. 1-14, причем биспецифическая антигенсвязывающая молекула содержит
(а) два Fab-фрагмента, способных к специфическому связыванию с CD40, соединенных с Fc-областью, и
(б) один антигенсвязывающий домен, способный к специфическому связыванию с FAP, соединенный с С-концом Fc-области.
16. Биспецифическая антигенсвязывающая молекула по любому из пп. 1-15, причем биспецифическая антигенсвязывающая молекула содержит
(а) два Fab-фрагмента, способных к специфическому связыванию с CD40, слитых с Fc-областью, и
(б) кросс-fab-фрагмент, способный к специфическому связыванию с FAP, слитый с С-концом Fc-области.
17. Биспецифическая антигенсвязывающая молекула по п. 16, причем VH-C-каппа цепь кросс-fab-фрагмента, способного к специфическому связыванию с FAP, слита с С-концом Fc-области.
18. Биспецифическая антигенсвязывающая молекула по любому из пп. 1-17, отличающаяся тем, что биспецифическая антигенсвязывающая молекула содержит
(а) две легкие цепи и две тяжелые цепи антитела, содержащие два Fab-фрагмента, способных специфически связываться с CD40, и Fc-область, и
(б) кросс-fab фрагмент, способный специфически связываться с FAP, где цепь VL-CH1 слита с С-концом одной из двух тяжелых цепей (а).
19. Биспецифическая антигенсвязывающая молекула по п. 1, содержащая
(а) две легкие цепи, каждая из которых содержит аминокислотную последовательность SEQ ID NO: 62, одну легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 61, первую тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 63, и вторую тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 64, или
(б) две легкие цепи, каждая из которых содержит аминокислотную последовательность SEQ ID NO: 66, одну легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 65, первую тяжелую цепь содержащую аминокислотную последовательность SEQ ID NO: 67, и вторую тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 68.
20. Биспецифическая антигенсвязывающая молекула по п. 1, содержащая две легкие цепи, каждая из которых содержит аминокислотную последовательность SEQ ID NO: 62, одну легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 61, первую тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 63 и вторую тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 64.
21. Антитело, которое специфически связывается с FAP, причем указанное антитело содержит вариабельную область тяжелой цепи (VHFAP), содержащую (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 3, (ii) CDR-H2, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4, SEQ ID NO: 11 и SEQ ID NO: 12, и (iii) CDR-Н3, содержащую аминокислотную последовательность SEQ ID NO: 5, и вариабельную область легкой цепи (VLFAP), содержащую (iv) CDR-L1, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 6, SEQ ID NO: 13 и SEQ ID NO: 14, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 7, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 8.
22. Антитело по п. 21, причем указанное антитело содержит
(а) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 15, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 21,
(б) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 16, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 21,
(в) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 16, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 22, или
(г) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO: 19, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO: 25.
23. Выделенная нуклеиновая кислота, кодирующая антитело по п. 21 или 22.
24. Выделенная нуклеиновая кислота, кодирующая биспецифическую антигенсвязывающую молекулу по любому из пп. 1-20.
25. Экспрессионный вектор, который обеспечивает экспрессию биспецифической антигенсвязывающей молекулы по любому из пп. 1-20, содержащий выделенную нуклеиновую кислоту по п. 24.
26. Клетка-хозяин для получения биспецифической антигенсвязывающей молекулы по любому из пп. 1-20, содержащая выделенную нуклеиновую кислоту по п. 24 или экспрессионный вектор по п. 25.
27. Способ получения биспецифической антигенсвязывающей молекулы, включающий культивирование клетки-хозяина по п. 26 в условиях, подходящих для экспрессии биспецифической антигенсвязывающей молекулы, и выделение биспецифической антигенсвязывающей молекулы.
28. Фармацевтическая композиция для лечения заболеваний связанных с FAP и CD40, содержащая эффективное количество биспецифической антигенсвязывающей молекулы по любому из пп. 1 20 и фармацевтически приемлемый носитель.
29. Фармацевтическая композиция по п. 28, дополнительно содержащая дополнительный терапевтический агент.
30. Биспецифическая антигенсвязывающая молекула из пп. 1-20 для применения в качестве лекарственного средства для лечения заболеваний, ассоциированных с FAP и CD40.
31. Биспецифическая антигенсвязывающая молекула по любому из пп. 1-20 или фармацевтическая композиция по п. 28 для лечения заболеваний, ассоциированных с FAP и CD40, для применения
(i) в индукции иммунной стимуляции CD40-экспрессирующими антигенпрезентирующими клетками (АПК),
(ii) для стимуляции опухолеспецифического Т-клеточного ответа,
(iii) для инициации апоптоза опухолевых клеток,
(iv) для лечения рака,
(v) для задержки прогрессирования рака,
(vi) для продления выживаемости пациента, страдающего от рака,
(vii) для лечения инфекций.
32. Применение биспецифической антигенсвязывающей молекулы по любому из пп. 1-20 или фармацевтической композиции по п. 28 в лечении рака, ассоциированного с FAP и CD40.
33. Применение биспецифической антигенсвязывающей молекулы по любому из пп. 1-20 в лечении рака, ассоциированного с FAP и CD40, причем биспецифическая антигенсвязывающая молекула или фармацевтическая композиция предназначены для введения в комбинации с химиотерапевтическим агентом, облучением и/или другими агентами в иммунотерапии рака.
34. Применение биспецифической антигенсвязывающей молекулы по любому из пп. 1-20 в лечении рака, ассоциированного с FAP и CD40, причем биспецифическая антигенсвязывающая молекула предназначена для введения в комбинации с агентом, блокирующим взаимодействие PD-L1/PD-1.
| WO 2018127473 A1, 12.07.2018 | |||
| WO 2017060144 A1, 13.04.2017 | |||
| WO 2016110598 A1, 14.07.2016 | |||
| WO 2012020006 A2, 16.02.2012 | |||
| RU 2017116020 A, 12.11.2018. |
