ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к новым антителам, имеющим сниженную способность к связыванию, посредством неспецифических и/или специфических взаимодействий, с технологическими примесями, такими как белки клетки-хозяина. Конкретнее, изобретение относится к новым иммуноглобулинам, имеющим сниженную способность к связыванию с такими технологическими примесями, благодаря модификациям аминокислотной последовательности антител.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Моноклональные антитела (mAb) являются биофармацевтическими средствами, используемыми для терапевтического лечения широкого спектра заболеваний. Для получения этих сложных рекомбинантных белков обычно необходимо использовать биологическую систему-хозяина, которая, при применении методов генной инженерии, способна экспрессировать продукт в подходящей активной форме. В связи с этим в качестве хозяина для промышленного получения множества mAb-продуктов повсеместно используются линии клеток млекопитающих, таких как клетки яичника китайского хомячка (Chinese Hamster Ovary, CHO), поскольку они способны обеспечивать сворачивание, сборку и необходимые посттрансляционные модификации этих белков, что, в свою очередь, обеспечивает их совместимость с человеческими системами.
Следствием этого, и ключевой проблемой при использовании клеточных систем для получения mAb, является необходимость отделения полученного белка от ряда других сложных примесей. Их называют технологическими примесями, и они включают широкий спектр белков, эндогенных для организма-хозяина или связанных с такими клетками (таких как вирусоподобные белки). Эти так называемые белки клетки- хозяина (HCP) представляют собой широкий класс молекул, которые могут отрицательно влиять как на стабильность, так и на безопасность биофармацевтического лекарственного средства. Некоторые HCP могут обладать протеолитической активностью, которая может отрицательно влиять на стабильность полученного белка.
Другие HCP, в случае их присутствия в готовой лекарственной форме, могут вызывать у пациентов иммуногенный ответ. Поэтому удаление HCP из биофармацевтических препаратов представляет серьезную проблему при их производстве.
Белки клетки-хозяина обычно удаляют с применением нескольких методик хроматографической очистки, в которых используется химия ортогонального разделения. Несмотря на то, что в большинстве случаев этакой метод эффективен, сложности возникают тогда, когда полученное mAb демонстрирует некоторое взаимодействие или аффинность в отношении одного или более чем одного белка, относящегося к HCP, в результате чего очищенный продукт содержит и mAb, и этот белок (белки). В подобных случаях стандартный подход состоит в разработке таких методик промывки хроматографической колонки, которые, в силу физико-химических свойств белков, позволят нарушить это взаимодействие. Поскольку склонность разных молекул mAb к взаимодействию с HCP варьирует, обычно принято считать, что это взаимодействие обусловлено в первую очередь гипервариабельными участками антител, определяющими комплементарность (complementarity determining regions, CDR). Тем не менее, конкретная последовательность и структурные мотивы этих участков, приводящие к усиленному взаимодействию, неизвестны.
Из пяти классов иммуноглобулинов (Ig), обозначаемых G, A, M, D и E, большинство биофармацевтических средств, представляющих собой mAb, относятся к классу иммуноглобулинов G (IgG), в котором есть четыре подкласса, нумеруемые от 1 до 4.
Недавно было показано, что такие HCP, как предполагаемый белок 2, подобный фосфолипазе В (phospholipase B-Like 2, PLBL2), также известный как белок 2, содержащий домен фосфолипазы В (phospholipase B domain containing protein 2, PLBD2), чаще других присутствуют в очищенных препаратах определенных вариантов иммуноглобулинов. Точная причина, по которой в очищенных препаратах одних изотипов IgG эти белки присутствуют чаще, чем в других, неизвестна; тем не менее, в предшествующих исследованиях утверждалось, то взаимодействие между IgG и PLBL2 происходит в первую очередь через CDR, но усиливается свойствами константной области IgG4, отличающими этот подкласс от подкласса IgG1 (Tran et al. (2016) J. Chromatogr. 1438:31-38). Однако точная природа этих различий остается неизвестной. Технологические примеси, такие как PLBL2, могут вызывать иммунный ответ у пациентов. Кроме того, установлено, что липазы, такие как PLBL2, могут быть причиной распада эксципиентов, присутствующих в композиции (Dixit et al., (2016) J. Pharm. Sci., 105(5):1657-66 и US 2016/0101181). Эти эксципиенты необходимы для стабилизации полученного белка в готовой композиции, и их распад будет приводить к уменьшению срока хранения и, вследствие этого, возможному снижению безопасности лекарственного средства. Поэтому присутствие технологических примесей и HCP, таких как PLBL2, в готовой лекарственной форме крайне нежелательно.
Повышенная склонность определенных иммуноглобулинов к взаимодействию с HCP, такими как PLBL2, привела к необходимости разработать специализированные схемы очистки для обеспечения достаточной безопасности готовых лекарственных форм. Несмотря на эффективность такого подхода, он не идеален, не только из-за роста себестоимости продукта, но и из-за необходимости проведения дополнительных работ по валидации методики и демонстрации того, разработанная методика очистки позволяет стабильно снижать содержание технологических примесей до приемлемых уровней. Кроме того, это приводит к повышению аналитических требований, поскольку было показано, что выявление PLBL2 традиционными методиками затруднено и часто требует применения специальных иммунологических анализов (WO 2015/038884).
Таким образом, сохраняется потребность в более эффективном, в том числе с экономической точки зрения, снижении уровней HCP в готовых лекарственных формах терапевтических иммуноглобулинов.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Согласно первому аспекту изобретения предложен вариант антитела IgG4, модифицированный в константной области тяжелой цепи по любой из или комбинации аминокислот в области между остатками 203 и 256 по нумерации Kabat, имеющий сниженный уровень связывания с белком клетки-хозяина (HCP) по сравнению с немодифицированным антителом IgG4.
Согласно другому аспекту изобретения предложена клеточная линия, кодирующая вариант антитела IgG4, как определено здесь.
Согласно другому аспекту изобретения предложен способ модификации антитела IgG для снижения связывания с технологической примесью, включающий стадии:
а) определения по меньшей мере одной аминокислоты, вовлеченной в связывание с технологической примесью; и
б) получения варианта антитела IgG заменой аминокислоты, определенной как вовлеченная в связывание с технологической примесью, на другую аминокислоту.
Согласно другому аспекту изобретения предложен способ получения антитела IgG со сниженным связыванием с белком клетки-хозяина (HCP), включающий модификацию последовательности антитела в константной области тяжелой цепи по любой из или комбинации аминокислот в области между остатками 203 и 256 по нумерации Kabat.
Согласно другому аспекту изобретения предложена композиция, содержащая вариант антитела IgG4, как определено здесь.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг. 1: Линейность разведения готовых нерасфасованных mAb при HCP-ELISA. Образцы готовых нерасфасованных гуманизированных IgG различных подтипов анализировали посредством HCP-ELISA для оценки нелинейности разведения в данном анализе. Для каждого разведения рассчитывали приведенные значения HCP, умножая измеренную концентрацию HCP на коэффициент разведения с последующим делением на концентрацию IgG. Приведенные значения HCP для каждого разведения образца наносили на график как функцию коэффициента разведения в двойном логарифмическом масштабе. Нелинейность отмечали в случае роста приведенных значений HCP при увеличении коэффициента разведения. В данном HCP-ELISA нелинейность продемонстрировало только mAb7.
Фиг. 2A-Б: Вестерн-блоттинг различных mAb с антителами против PLBL2. Образцы готовых нерасфасованных гуманизированных IgG различных подтипов анализировали вестерн-блоттингом для выявления присутствия PLBL2. (A) Окрашенный гель SyproRUBY, демонстрирующий равное количество внесенных образцов. (Б) Вестерн-блоттинг с антителами против PLBL2. PLBL2 выявлен только в образце mAb7 при приблизительно 60 кДа, на что указывает стрелка в правой части изображения.
Фиг. 3A-В: Связывание PLBL2 с различными mAb по результатам SPR. Сенсограммы поверхностного плазмонного резонанса (SPR) при связывании PLBL2 с различными mAb. (A) Сенсограмма демонстрирует сходные уровни захвата всех семи mAb иммобилизованным моноклональным антителом против Fc человеческого IgG, внесение mAb отмечено стрелкой при приблизительно -200 секундах, а внесение PLBL2 отмечено стрелкой при 0 секунд. Увеличенная сенсограмма, на которой показано связывание и диссоциация PLBL2 от mAb1, mAb3, mAb5 и mAb7 (Б) и mAb2, mAb4 и mAb6 (В), демонстрирующая, что PLBL2 селективно связывался с молекулами IgG4.
Фиг. 4A-Б: Аффинность связывания PLBL2 с mAb7. SPR-эксперимент связывания для расчета аффинности связывания PLBL2 с mAb7. (A) Сенсограммы, на которых показано, что возрастающие концентрации PLBL2 от 1,25 мкМ до 80 мкМ коррелируют с возрастающим связыванием с mAb7. (Б) График ответа связывания в равновесном состоянии как функция концентрации PLBL2. Для расчета KD была применена модель связывания 1:1.
Фиг. 5: Связывание PLBL2 с mAb3, mAb6A и mAb6B. Сенсограммы поверхностного плазмонного резонанса при связывании PLBL2 с mAb3 и mAb6, экспрессированными в двух системах экспрессии с разными клетками-хозяевами. mAb6A было экспрессировано с использованием системы с клетками-хозяевами CHO K1a, в то время как mAb6B было экспрессировано с использованием системы экспрессии с клетками-хозяевами HEK293. Результаты показывают, что обе молекулы IgG4 (mAb6A и mAb6B) взаимодействуют с PLBL2 сходным образом, несмотря на то, что они получены в разных клеточных линиях. В то же время, молекула IgG2 (mAb3) не взаимодействует.
Фиг. 6A-В: Аффинность связывания PLBL2 с mAb3, mAb6A и mAb6B. Результаты экспериментов биослойной интерферометрии (Bio Layer Interferometry, BLI), полученные для (A) mAb3, (Б) mAb6A и (В) mAb6B, на которых показаны приближенные кривые аффинности связывания этих молекул с PLBL2. Результаты по KD были рассчитаны с применением локального приближения к модели связывания 1:1. Следует отметить, что получить данные по аффинности mAb3 было невозможно из-за отсутствия взаимодействия.
Фиг. 7: Дифференциальный график HDX для образцов mAb7, самого по себе и связанного с PLBL2. Мечение дейтерием измеряли во временных точках 0,5 мин, 5 мин, 60 мин, 120 мин, 180 мин и 240 мин. Вертикальными линиями показаны суммарные различия HDX для каждого пептида в шести указанных временных точках. Область последовательности со значительным снижением взаимодействия с растворителем была определена как K218-F256 (тяжелая цепь).
Фиг. 8: Выравнивание последовательностей различных mAb.
Аминокислотные последовательности нижней части домена CH1, шарнирной области и верхней части домена CH2 mAb1-7 выравнивали с аминокислотами, расположенными в соответствующих положениях человеческих IgG1, IgG2 и IgG4 эмбрионального типа. Выравнивание было сосредоточено на шарнирной области IgG4, где, вероятно, расположен сайт связывания PLBL2, исходя из экспериментов по сравнению связывания PLBL2 различными молекулами mAb. Различия последовательностей выделены серым цветом. Интерес представляют консервативные аминокислотные остатки молекул IgG4, которые связывались с PLBL2, и неконсервативные (то есть, различающиеся) аминокислотные остатки других подтипов IgG, которые не связывались с PLBL2. Были определены 10 таких аминокислот, которые выделены черными рамками.
Фиг. 9: Выравнивание последовательностей различных mAb, полученных мутагенезом. Мутагенезом исходного IgG4 (mAb5) были получены в общей сложности 7 различных вариантов IgG4 с модифицированной шарнирной областью (mAb5-1B–7B). При данном выравнивании были сопоставлены аминокислотные последовательности вариантов IgG4 и исходного IgG4, а также двух других IgG4 с немодифицированной шарнирной областью (mAb6 и mAb7), IgG2 (mAb3) и IgG1 (mAb8). Суффикс «A» или «B» указывает на то, что антитело было экспрессировано с использованием клетки-хозяина CHO (A) или HEK (В), являющихся клетками млекопитающих. Различия аминокислотных последовательностей выделены серым цветом. Затем, серым цветом и черными рамками выделены аминокислотные остатки, предположительно влияющие на связывание с PLBL2. Мутации в последовательностях шарнирной области вариантов IgG4 выделены черным цветом.
Фиг. 10: Взаимодействие молекул IgG2, IgG4 и вариантов IgG4 с PLBL2. Результаты SPR-экспериментов (поверхностный плазмонный резонанс) по изучению связывания PLBL2 с вариантами IgG4 с модифицированной шарнирной областью (mAb5-1B – 7B) по сравнению с немодифицированным исходным IgG4 (mAb5B). В качестве дополнительного контроля в анализ были включены несколько других mAb, включая IgG1 (mAb8A), IgG2 (mAb3B) и IgG4 (mAb6B и mAb7A). Суффикс «A» или «B» указывает на то, что антитело было экспрессировано с использованием клетки-хозяина CHO (A) или HEK (В), являющихся клетками млекопитающих.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к антителам со сниженным взаимодействием с технологическими примесями, такими как HCP, включая PLBL2. Было обнаружено, что модификация (например, замена, вставка или делеция) аминокислот в определенных положениях приводит к снижению способности определенных антител к связыванию с HCP, такими как PLBL2. Это уменьшение склонности к взаимодействию приводит к снижению уровня технологической примеси, такой как PLBL2, в очищенном продукте, устраняя необходимость применения специальных методик очистки, направленных на удаление этой технологической примеси. В действительности, например, уровни связывания PLBL2 с молекулами IgG4 с изменениями этих аминокислотных остатков близки к уровням связывания PLBL2 с молекулами, в очищенных препаратах которых PLBL2 обычно отсутствует. Это, в свою очередь, повышает эффективность производственного процесса, уменьшает связанные с ним затраты, устраняет необходимость избыточного анализа продукта и, в конечном счете, способствует обеспечению безопасности пациентов. Кроме того, установлено, что липазы, такие как PLBL2, могут быть причиной распада эксципиентов, присутствующих в композиции, таких как полисорбат 80, которые необходимы для стабилизации полученного белка (рекомбинантного антитела) в готовой композиции. Снижение связывания с PLBL2 будет способствовать минимизации возможных уровней данного HCP в готовой лекарственной форме, обеспечивая посредством этого максимальный срок хранения полученного белка (рекомбинантного антитела), что дополнительно уменьшит расходы и обеспечит безопасность.
Эффекторная функция IgG4
В константной области тяжелой цепи последовательность IgG4 более чем на 95% гомологична изотипам IgG1, IgG2 и IgG3. По сравнению с IgG1, IgG4 имеет низкую аффинность связывания с активирующими Fc-гамма-рецепторами FcγRIIa и FcγRIIIa. По сравнению с IgG1, IgG4 обладает слабой или промежуточной аффинностью связывания с высокоаффинным FcγRI. IgG4 сохраняет сходную с IgG1 аффинность связывания с ингибирующим FcγRIIb. IgG4 также связывается с FcRn, аналогично IgG1. IgG4 демонстрирует отсутствие связывания или пренебрежительно малое связывание с белковым комплексом C1q и не способен активировать комплемент по классическому пути, в силу чего он имеет сниженную CDC-активность. ADCC-активность у IgG4 снижена или отсутствует. Это снижение связывания с эффекторами привело к выбору константного домена тяжелой цепи IgG4 для использования в рекомбинантных антителах, где эффекторная функция необязательна или нежелательна.
Ядро шарнирной области IgG4 содержит последовательность CPSC (SEQ ID NO:19), в то время как ядро шарнирной области IgG1 содержит CPPC (SEQ ID NO:20), которая менее подвержена восстановлению. S241 (по нумерации Kabat) IgG4 приводит к более гибкой шарнирной области, что делает возможным образование внутрицепочечной циклизованной дисульфидной связи и появлению «полуантител», содержащих нековалентно связанные тяжелые цепи разных антител, что обычно называют обменом Fab-фрагментами IgG4 (IgG4 Fab arm exchange). Ядро шарнирной области IgG4 может быть стабилизировано модификацией S241 с его заменой на пролин (то есть S241P), как в IgG1.
Ряд терапевтических антител IgG4 имеют конструированную шарнирную область, включающую замену S241P. Возможна одиночная замена IgG4 S241P без варианта L248E для сохранения нормального связывания с FcγRI. Вариант IgG4 L248E снижает аффинность связывания IgG4-PE с FcγRI по сравнению с IgG1, и его аффинность в отношении FcγRI приблизительно в 20 раз меньше, чем у IgG4 дикого типа.
Модификации последовательности IgG4
Авторы изобретения обнаружили неожиданную связь между консервативной областью молекулы IgG и способностью указанной молекулы к связыванию с технологическими примесями, такими как HCP, включая, без ограничения, PLBL2. Конкретно, авторы изобретения определили ключевые аминокислотные остатки высоко консервативной шарнирной области и прилегающих к ней последовательностей антител подкласса IgG4, которые приводят к связыванию этих антител с HCP, такими как PLBL2. При модификации этих аминокислот полученная модифицированная молекула IgG4 демонстрирует сниженную способность к связыванию с HCP, такими как PLBL2, по сравнению с исходной молекулой без указанной выше модификации (модификаций), сохраняя в тоже время те же эффекторные функции (например, свойства молекулы IgG4 по связыванию с FcRn), что и без указанной модификации (модификаций). Кроме того, модифицированная молекула IgG4 имеет те же (или улучшенные) биофизические свойства, такие как стабильность, свойства на границе раздела фаз, что и IgG4 без указанной модификации (модификаций).
Таким образом, согласно первому аспекту изобретения предложен вариант антитела IgG4, модифицированный в константной области тяжелой цепи по любой из или комбинации аминокислот в области между остатками 203 и 256 по нумерации Kabat, имеющий сниженный уровень связывания с белком клетки-хозяина (HCP) по сравнению с немодифицированным антителом IgG4.
В одном воплощении одна аминокислота или комбинация аминокислот расположены между остатками 203 и 243 по нумерации Kabat. В другом воплощении одна аминокислота или комбинация аминокислот расположены между остатками 222 и 243 по нумерации Kabat. В альтернативном воплощении одна аминокислота или комбинация аминокислот расположены между остатками 222 и 256 по нумерации Kabat.
В одном воплощении одна аминокислота или комбинация аминокислот выбраны из группы, состоящей из: (1) одной или более чем одной аминокислоты шарнирной области между остатками 226 и 243 по нумерации Kabat; и/или (2) остатка 203 по нумерации Kabat; и/или (3) остатка 222 по нумерации Kabat.
В одном воплощении немодифицированное антитело IgG4 имеет последовательность шарнирной области ESKYGPPCPSCP (SEQ ID NO:21) или ESKYGPPCPPCP (SEQ ID NO:22) (то есть положения 226-243 по нумерации Kabat).
В одном воплощении шарнирная область расположена между остатками 203 и 256 по нумерации Kabat. В одном воплощении шарнирная область расположена между остатками 226 и 243 по нумерации Kabat. В другом воплощении шарнирная область расположена между остатками 226 и 238 по нумерации Kabat. В альтернативном воплощении шарнирная область расположена между остатками 234 и 250 по нумерации Kabat.
В одном воплощении вариант антитела IgG4 содержит последовательность CPPC (SEQ ID NO:20) (остатки 239-242 по нумерации Kabat). Она известна как «ядро шарнирной области». В альтернативном воплощении вариант антитела IgG4 содержит последовательность CPSC (SEQ ID NO:19) (остатки 239-242 по нумерации Kabat).
В одном воплощении модификация включает делецию любой из или комбинации аминокислот в области между остатками 203 и 256 по нумерации Kabat.
В одном воплощении модификация включает вставку одной или более чем одной аминокислоты в области между остатками 203 и 256 по нумерации Kabat.
В одном воплощении модификация включает замену одной или более чем одной аминокислоты в области между остатками 203 и 256 по нумерации Kabat.
Ключевыми аминокислотными остатками IgG4, присутствующими в шарнирной области или рядом с ней, являются: серин в положении 197 (S197), лейцин в положении 198 (L198), лизин в положении 203 (K203), треонин в положении 207 (T207), аспартат в положении 211 (D211), аргинин в положении 222 (R222), глутамат в положении 226 (E226), серин в положении 227 (S227), тирозин в положении 229 (Y229), глицин в положении 230 (G230), пролин в положении 237 (P237), пролин в положении 238 (P238), глутамат в положении 246 (E246), фенилаланин в положении 247 (F247), глицин в положении 249 (G249), глицин в положении 250 (G250) и пролин в положении 251 (P251). Последовательности, представляющие интерес, показаны на Фиг. 8 и 9.
Ключевыми аминокислотными остатками IgG4 являются: лизин в положении 203 (K203), аргинин в положении 222 (R222), глутамат в положении 226 (E226), серин в положении 227 (S227), тирозин в положении 229 (Y229), глицин в положении 230 (G230), пролин в положении 237 (P237) и пролин в положении 238 (P238). Модификация шарнирной области IgG4 может включать проведение одной или более чем одной аминокислотной замены по этим положениям, что приводит к получению модифицированной молекулы IgG4 со сниженной способностью к связыванию с HCP, такими как PLBL2.
Кроме того, модификация шарнирной области IgG4 может включать удаление тирозина в положении 229 (Y229) и/или глицина в положении 230 (G230), и в определенных воплощениях эти делеции комбинируют с по меньшей мере одной аминокислотной заменой или дополнительной делецией по одному из указанных выше положений, что приводит к получению варианта антитела IgG4 со сниженной способностью к связыванию с HCP, такими как PLBL2, по сравнению с исходным антителом IgG4.
В другом воплощении модификация шарнирной области IgG4 может включать удаление тирозина в положении 229 (Y229) и/или глицина в положении 230 (G230), в комбинации с по меньшей мере одной аминокислотной заменой или дополнительной делецией пролиновых остатков в положениях 237 и 238 (P237 и P238), что приводит к получению варианта антитела IgG4 со сниженной способностью к связыванию с HCP, такими как PLBL2, по сравнению с исходным антителом IgG4. В одном воплощении модификация включает замену YGPP (SEQ ID NO:23) (остатки 229-238 по нумерации Kabat) на SCDKTHT (SEQ ID NO:24) или CCVE (SEQ ID NO:25).
В одном воплощении модификация включает замену на эквивалентную аминокислотную последовательность в последовательности антител IgG1, IgG2 и/или IgG3 эмбрионального типа. В другом воплощении последовательность антител IgG1, IgG2 и/или IgG3 эмбрионального типа является человеческой. Последовательности антител IgG эмбрионального типа хорошо известны в данной области. Например, последовательность IgG1 эмбрионального типа между положениями 203 и 256 включительно показана на Фиг. 8 и как SEQ ID NO:8. Последовательность IgG2 эмбрионального типа между положениями 203 и 256 включительно показана на Фиг. 8 и как SEQ ID NO:9.
В одном воплощении модификация шарнирной области IgG4 включает замену одного или более из следующих ключевых аминокислотных остатков: лизин в положении 203 (K203), аргинин в положении 222 (R222), глутамат в положении 226 (E226), серин в положении 227 (S227), тирозин в положении 229 (Y229), глицин в положении 230 (G230), пролин в положении 237 (P237) и пролин в положении 238 (P238); на аминокислотный остаток из соответствующего положения последовательности эмбрионального типа человеческого антитела альтернативного изотипа. Альтернативный изотип может представлять собой IgG1, IgG2 или IgG3. В таких воплощениях, где проводят более чем одну замену, им не обязательно быть выбранными на основе только одного альтернативного изотипа IgG, и они могут представлять собой комбинацию остатков более чем одного альтернативного изотипа человеческих антител. Таким образом, вариант IgG4 по такому воплощению может предсказуемо содержать в своих ключевых положениях аминокислотные остатки последовательности IgG1 эмбрионального типа и последовательности IgG2 эмбрионального типа, или последовательности IgG1 эмбрионального типа и последовательности IgG3 эмбрионального типа, или последовательности IgG2 эмбрионального типа и последовательности IgG3 эмбрионального типа, или последовательности IgG1, IgG2 и IgG3 эмбрионального типа.
В тех воплощениях, в которых последовательность эмбрионального типа альтернативного изотипа не содержит аминокислоту в положении соответствующем указанным выше ключевым остаткам последовательности IgG4 эмбрионального типа, аминокислотный остаток или остатки в указанном положении (положениях) будет удален из белковой последовательности варианта IgG4.
В одном воплощении модификация включает любое из следующего или их комбинацию:
(1) замена одной или более чем одной аминокислоты, включающая замену K203 на R, E или Q; R222 на T, K или Q; E226 на L или I; S227 на R, P, A, N или T; Y229 на S, C, F, W или H; G230 на C, A, N или S; P237 на H, E, D или V; и/или P238 на T, K или E;
и/или
(2) замена ESKYGPP (SEQ ID NO:26) (остатки 226-238 по нумерации Kabat) на EPKSCDKTHT (SEQ ID NO:27), или ERKYGPP (SEQ ID NO:28), или ERKCCVE (SEQ ID NO:29), или ELKTPLGDTTHT (SEQ ID NO:30); и/или
(3) замену YGPP (SEQ ID NO:23) (остатки 229-238 по нумерации Kabat) на SCDKTHT (SEQ ID NO:24) или CCVE (SEQ ID NO:25). Эти модификации представлены в Таблице 1.
В одном воплощении лизин в положении 203 последовательности эмбрионального типа варианта молекулы IgG4 заменен на глутамин (K203Q).
В одном воплощении аргинин в положении 222 последовательности эмбрионального типа варианта молекулы IgG4 заменен на остаток, выбранный из группы, состоящей из: лизина, треонина, аланина, аргинина, аспарагина, аспартата, цистеина, глутамата, глутамина, глицина, гистидина, изолейцина, лейцина, метионина, фенилаланина, пролина, серина, триптофана, тирозина и валина. В другом воплощении аргинин в положении 222 последовательности эмбрионального типа варианта молекулы IgG4 заменен на остаток, выбранный из группы, состоящей из: лизина и треонина. В еще одном воплощении аргинин в положении 222 последовательности эмбрионального типа варианта молекулы IgG4 заменен на треонин (R222T). В еще одном воплощении аргинин в положении 222 последовательности эмбрионального типа варианта молекулы IgG4 заменен на лизин (R222K). Как показано в Примерах, описанных здесь, модификации по этому положению продемонстрировали наибольшее снижение связывания с PLBL2.
В одном воплощении глутамат в положении 226 последовательности эмбрионального типа варианта молекулы IgG4 заменен на остаток, выбранный из группы, состоящей из: изолейцина и лейцина. Например, замена представляет собой E226L.
В одном воплощении серин в положении 227 варианта молекулы IgG4 заменен на остаток, выбранный из группы, состоящей из: пролина и аргинина. Например, замена представляет собой S227P.
В одном воплощении вариант молекулы IgG4 не содержит тирозин, расположенный в положении 229 последовательности человеческого IgG4 эмбрионального типа.
В одном воплощении вариант молекулы IgG4 не содержит глицин, расположенный в положении 230 последовательности человеческого IgG4 эмбрионального типа.
В одном воплощении вариант молекулы IgG4 не содержит пролин, расположенный в положении 237 последовательности человеческого IgG4 эмбрионального типа.
В одном воплощении вариант молекулы IgG4 не содержит пролин, расположенный в положении 238 последовательности человеческого IgG4 эмбрионального типа.
В одном воплощении тирозин в положении 229 варианта молекулы IgG4 заменен на остаток, выбранный из группы, состоящей из: лизина, аспарагина, треонина, аргинина, метионина, изолейцина, глутамина, гистидина, пролина, глутамата, аспартата, аланина, глицина, валина, тирозина, серина, триптофана, цистеина, лейцина и фенилаланина.
В одном воплощении глицин в положении 230 варианта молекулы IgG4 заменен на остаток, выбранный из группы, состоящей из: лизина, аспарагина, треонина, аргинина, метионина, изолейцина, глутамина, гистидина, пролина, глутамата, аспартата, аланина, глицина, валина, тирозина, серина, триптофана, цистеина, лейцина и фенилаланина.
В одном воплощении пролин в положении 237 варианта молекулы IgG4 заменен на остаток, выбранный из группы, состоящей из: лизина, аспарагина, треонина, аргинина, метионина, изолейцина, глутамина, гистидина, пролина, глутамата, аспартата, аланина, глицина, валина, тирозина, серина, триптофана, цистеина, лейцина и фенилаланина.
В одном воплощении пролин в положении 238 варианта молекулы IgG4 заменен на остаток, выбранный из группы, состоящей из: лизина, аспарагина, треонина, аргинина, метионина, изолейцина, глутамина, гистидина, пролина, глутамата, аспартата, аланина, глицина, валина, тирозина, серина, триптофана, цистеина, лейцина и фенилаланина.
В одном воплощении вариант молекулы IgG4 не содержит тирозин в положении 229, глицин в положении 230, пролин в положении 237 и пролин в положении 238 и вместо этого содержит следующую пептидную последовательность: SCDKTHT (SEQ ID NO:24). В альтернативном воплощении вариант молекулы IgG4 не содержит тирозин в положении 229, глицин в положении 230, пролин в положении 237 и пролин в положении 238 и вместо этого содержит следующую пептидную последовательность: CCVE (SEQ ID NO:25). Например, вариант IgG4 содержит делецию 229-238 по нумерации Kabat YGPP (SEQ ID NO:23) и вставку CCVE (SEQ ID NO:25). Например, вариант IgG4 содержит замену 229-238 по нумерации Kabat YGPP (SEQ ID NO:23) на CCVE (SEQ ID NO:25). Как показано в Примерах, описанных здесь, модификации по этим положениям продемонстрировали наибольшее снижение связывания с PLBL2.
В одном воплощении фенилаланин в положении 247 варианта молекулы IgG4 заменен на лейцин.
В одном воплощении модификация включает замену EFLGGP (SEQ ID NO:31) (остатки 246-251 по нумерации Kabat) на PAAAS (SEQ ID NO:32) или PAAAP (SEQ ID NO:33).
В Таблице 1 приведены примеры возможных аминокислотных модификаций (например, замены, и/или вставки, и/или делеции) определенных остатков по нумерации Kabat, которые могут быть проведены в антителе IgG4 для получения модифицированного антитела IgG4 со сниженной способностью к связыванию с HCP, такими как PLBL2. Эти модификации могут быть проведены применительно к любой одной аминокислоте или к более чем одной аминокислоте (то есть к комбинации аминокислот).
Таблица 1: Примеры аминокислотных модификаций тяжелой цепи IgG4
тяжелой цепи по нумерации Kabat
IgG4 эмбрионального типа
238
(SEQ ID NO:23)
CCVE (SEQ ID NO:25)
238
ERKCCVE или ELKTPLGDTTHT
(SEQ ID NO:27, 28, 29 и 30, соответственно)
(SEQ ID NO:31)
PAAAP (SEQ ID NO:33)
В одном воплощении вариант антитела IgG4 является человеческим или гуманизированным.
В другом воплощении замена на эквивалентную аминокислотную последовательность антител IgG1, IgG2 и/или IgG3 эмбрионального типа включает последовательность человеческих антител эмбрионального типа.
В одном воплощении проводят модификацию обеих тяжелых цепей варианта молекулы IgG4. В альтернативном воплощении проводят модификацию только одной из тяжелых цепей варианта молекулы IgG4.
В одном воплощении константная область тяжелой цепи не содержит дополнительных модификаций по сравнению с немодифицированным антителом IgG4.
В одном воплощении модификации не затрагивают аминокислотные остатки, необходимые для эффекторной функции IgG4. Например, F247 (по нумерации Kabat) IgG4 дикого типа (EU F234) необходим для ослабления ADCC- и CDC-активности.
Кроме того, модификация E248 (по нумерации Kabat) может ослаблять эффекторные функции, а именно связывание с FcγRI.
В одном воплощении вариант антитела IgG4 содержит дополнительную замену S241 на P и/или L248 на E. В одном воплощении вариант антитела IgG4 содержит дополнительную замену S241 на P. Эта модификация способствует повышению стабильности молекулы IgG4. В одном воплощении вариант антитела IgG4 содержит дополнительную замену последовательности EFLGGP (SEQ ID NO:31) (остатки 246-251 по нумерации Kabat) на последовательность PAAAP (SEQ ID NO:33).
Согласно другому аспекту изобретения предложена нуклеотидная конструкция, кодирующая вариант антитела IgG4, как определено здесь.
Эта нуклеотидная конструкция может быть использована для трансфекции линии клетки-хозяина. Таким образом, согласно другому аспекту изобретения предложена клеточная линия, кодирующая вариант антитела IgG4, как определено здесь.
Дополнительные модификации последовательности
Согласно другому аспекту изобретения предложен вариант антитела IgG, модифицированный в константной области тяжелой цепи по любой из или комбинации аминокислот в области между остатками 203 и 256 по нумерации Kabat, имеющий сниженный уровень связывания с белком клетки-хозяина (HCP) по сравнению с немодифицированным антителом IgG. Например, вариант антитела IgG, модифицированный по 1-25 аминокислотам константной области тяжелой цепи между остатками 203 и 256 по нумерации Kabat. Например, вариант антитела IgG, модифицированный по одной, двум, трем, четырем, пяти, шести, семи, восьми, девяти, десяти, одиннадцати, двенадцати, тринадцати, четырнадцати, пятнадцати, шестнадцати, семнадцати, восемнадцати, девятнадцати, двадцати, двадцати одной, двадцати двум или двадцати трем аминокислотам константной области тяжелой цепи между остатками 203 и 256 по нумерации Kabat. Например, вариант антитела IgG, модифицированный по любому одному из или комбинации положений 197, 198, 203, 207, 211, 222, 226, 227, 229, 230, 232, 233, 234, 235, 236, 237, 238, 246, 247, 248, 249, 250 или 251 константной области тяжелой цепи по нумерации Kabat. Например, вариант антитела IgG, модифицированный по любому одному из или комбинации положений 203, 222, 226, 227, 229, 230, 232, 233, 234, 235, 236, 237, 238, 247, 248 или 251 константной области тяжелой цепи по нумерации Kabat.
Белки клетки - хозяина
Белки клетки-хозяина или «HCP» относятся к белкам, не имеющим отношения к интересующему белку (то есть рекомбинантному белку / варианту IgG4), продуцируемому клеткой-хозяином во время ее культивирования или ферментации, включая внутриклеточные и/или секретируемые белки. Примером белка клетки-хозяина является протеаза, которая может повреждать интересующий белок, если она все еще присутствует во время и после очистки. Например, присутствие протеазы в образце, содержащем интересующий белок, может приводить к образованию родственных по отношению к нему веществ или примесей, которых исходно не было. Присутствие протеаз может приводить к распаду интересующего белка со временем в процессе очистки и/или в готовой композиции.
В одном воплощении белки клетки-хозяина образуются в / имеют происхождение от клетки млекопитающего или бактериальной клетки. В другом воплощении клетка млекопитающего выбрана из клетки человека или грызуна (такого как хомяк или мышь).
В определенных воплощениях клетка-хозяин, используемая для экспрессии антитела, выбрана из группы, состоящей из клеток CHO, клеток NS0, клеток Sp2/0, клеток COS, клеток K562, клеток BHK, клеток PER.C6 и клеток HEK (то есть, белки клетки-хозяина имеют происхождение от этих клетки-хозяина). Альтернативно, клетка-хозяин может представлять собой бактериальную клетку, выбранную из группы, состоящей из E. coli (например, W3110, BL21), B. subtilis и/или других подходящих бактерий; эукариотические клетки, такие как клетки грибов или дрожжей (например, Pichia pastoris, Aspergillus sp., Saccharomyces cerevisiae, Schizosaccharomyces pombe, Neurospora crassa). Например, клетка-хозяин представляет собой клетку CHO. Альтернативно, клетка-хозяин представляет собой клетку HEK.
В одном воплощении белок клетки-хозяина представляет собой предполагаемый белок 2, подобный фосфолипазе В (PLBL2). PLBL2 также известен как белок 2, содержащий домен фосфолипазы В (PLBD2).
Снижение связывания молекул IgG4 с HCP, такими как PLBL2, приводит к сопутствующему снижению уровня этих технологических примесей в очищенном IgG4. Это, в свою очередь, устраняет необходимость применения специальных методик очистки, таких как жесткие условия промывки во время хроматографии на колонках, для удаления HCP из готовой лекарственной формы. Это повышает эффективность производственного процесса, уменьшает связанные с ним затраты и устраняет необходимость избыточного анализа продукта с применением специальных иммунологических анализов. В конечном счете, эти модификации способствуют снижению риска присутствия HCP, таких как PLBL2, в готовой лекарственной форме и, в результате, минимизируют вероятность того, что эти белки вызовут иммуногенный ответ у пациентов.
Снижение способности этих модифицированных молекул IgG4 к связыванию с HCP, такими как PLBL2, может быть оценено количественно методами, представленными в Примерах.
Существует несколько способов, позволяющих измерить снижение уровня связывания с белком клетки-хозяина. Например, снижение уровня связывания можно оценить простым измерением количества белка клетки-хозяина, присутствующего в растворе с антителом после его очистки, например, аффинной хроматографией (снижение этого количества по сравнению с немодифицированным IgG4 является показателем снижения связывания с белком клетки-хозяина). В одном воплощении антитело IgG4 имеет сниженную аффинность и/или активность связывания с белком клетки-хозяина по сравнению с немодифицированным антителом IgG4.
В одном воплощении активность связывания у варианта антитела IgG4 по меньшей мере на 10% ниже, чем у немодифицированного антитела IgG4, например, по меньшей мере на 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 98%, 99% ниже.
В одном воплощении аффинность связывания у варианта антитела IgG4 по меньшей мере в 10 раз ниже, чем у немодифицированного антитела IgG4, например, по меньшей мере на 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 98%, 99% ниже.
Композиции , содержащие варианты антител
Согласно другому аспекту изобретения предложена композиция, содержащая вариант антитела IgG4, как определено здесь.
Композиции по настоящему изобретению могут быть изготовлены с использованием фармацевтически приемлемых носителей или разбавителей, а также любых других известных адъювантов и эксципиентов в соответствии с обычными методиками. Фармацевтически приемлемые носители или разбавители, а также любые другие известные адъюванты и эксципиенты должны быть подходящими для выбранного соединения по настоящему изобретению и выбранного способа введения.
В одном воплощении концентрация варианта IgG4 составляет по меньшей мере 10 мг/мл, например, по меньшей мере 20 мг/мл, 25 мг/мл, 30 мг/мл, 35 мг/мл, 40 мг/мл, 45 мг/мл, 50 мг/мл, 55 мг/мл, 60 мг/мл, 65 мг/мл, 70 мг/мл, 75 мг/мл, 80 мг/мл, 85 мг/мл, 90 мг/мл, 95 мг/мл или 100 мг/мл.
Фактические дозы активных ингредиентов в фармацевтических композициях по настоящему изобретению можно варьировать для получения такого количества активного ингредиента, которое позволит эффективно достигать желаемого терапевтического ответа для конкретного пациента, композиции и способа введения без токсичности для пациента. Выбранная доза будет зависеть от множества фармакокинетических факторов, включая активность конкретных используемых композиций по настоящему изобретению, путь введения, время введения, продолжительность лечения, другие лекарственные средства, соединения и/или вещества, используемые в комбинации с конкретными используемыми композициями, возраст, пол, массу тела, состояние, общее состояние здоровья и анамнез пациента, проходящего лечение, и аналогичные факторы, хорошо известные в области медицины. Квалифицированный врач или ветеринар может легко определить и назначить эффективное количество необходимой фармацевтической композиции.
В одном воплощении концентрация предполагаемого белка 2, подобного фосфолипазе В (PLBL2), в композиции составляет менее 500 млн-1, например, менее 400 млн-1, 300 млн-1, 200 млн-1, 100 млн-1, 90 млн-1, 80 млн-1, 70 млн-1, 60 млн-1, 50 млн-1, 40 млн-1, 30 млн-1, 20 млн-1 или 10 млн-1.
В одном воплощении концентрация предполагаемого белка 2, подобного фосфолипазе В (PLBL2), в композиции составляет менее чем приблизительно 200 нг PLBL2 на мг продукта (то есть нг/мг); менее чем приблизительно 150 нг/мг; менее чем приблизительно 100 нг/мг; или менее чем приблизительно 50 нг/мг.
В одном воплощении композиция дополнительно содержит буфер и/или сложный эфир жирной кислоты. В другом воплощении сложный эфир жирной кислоты представляет собой полисорбат 20 или полисорбат 80.
Как указано выше, установлено, что PLBL2 может быть причиной распада эксципиентов, присутствующих в композиции, таких как полисорбаты, которые необходимы для стабилизации полученного белка в готовой композиции. Поэтому снижение связывания с PLBL2 будет, в свою очередь, минимизировать уровень PLBL2 в готовой лекарственная форме, обеспечивая посредством этого максимальный срок хранения. Таким образом, согласно другому аспекту изобретения предложена композиция с увеличенным/улучшенным сроком хранения, содержащая вариант антитела IgG4, как определено здесь.
Кроме того, согласно другому аспекту изобретения предложен способ увеличения срока хранения антителосодержащей композиции, включающий получение антитела IgG со сниженным связыванием с PLBL2 посредством модификации последовательности антитела в константной области тяжелой цепи по любой из или комбинации аминокислот в области между остатками 203 и 256 по нумерации Kabat.
Применение вариантов антител
Согласно другому аспекту изобретения предложен вариант антитела IgG4, как определено здесь, для применения в терапии. Специалисту в данной области будет ясно, что настоящее изобретение может быть применено к множеству антител IgG4, направленных на разные мишени, поэтому указанные варианты IgG4 могут быть использованы для лечения множества заболеваний.
Антитела, описанные здесь, могут также быть использованы в способах лечения. Лечение может быть терапевтическим, профилактическим или превентивным. Лечение включает облегчение, уменьшение или предотвращение по меньшей мере одного аспекта или симптома заболевания и включает предотвращение или излечение заболеваний, описанных здесь.
Антитела, описанные здесь, могут быть использованы в эффективном количестве для терапевтического, профилактического или превентивного лечения. Терапевтически эффективное количество антитела, описанного здесь, представляет собой количество, эффективно снижающее степень выраженности или уменьшающее один или более чем один симптом заболевания или приводящее к предотвращению или излечению заболевания.
Способы получения вариантов антител
Здесь предложены варианты антител и способы получения указанных вариантов антител, которые были модифицированы посредством изменений аминокислотной последовательности с целью снижения уровня связывании с технологической примесью. Эти модификации оказывают полезный эффект, уменьшая количество технологической примеси, присутствующее в изготавливаемом лекарственном средстве.
Таким образом, согласно другому аспекту изобретения предложен способ модификации антитела IgG для снижения связывания с технологической примесью, включающий стадии:
а) определения по меньшей мере одной аминокислоты, вовлеченной в связывание с технологической примесью; и
б) получения варианта антитела IgG путем замены аминокислоты, определенной как вовлеченная в связывание с технологической примесью, на другую аминокислоту.
Согласно другому аспекту изобретения предложен способ получения варианта антитела IgG со сниженным связыванием с технологической примесью, по сравнению с немодифицированным антителом IgG, включающий стадии:
а) определения по меньшей мере одной аминокислоты, вовлеченной в связывание с технологической примесью; и
б) получения варианта антитела IgG путем замены аминокислоты, определенной как вовлеченная в связывание с технологической примесью, на другую аминокислоту.
В одном воплощении аминокислоту, вовлеченную в связывание с технологической примесью, определяют с использованием метода или комбинации методов, которые могут быть применены для изучения белок-белковых взаимодействий. В одном воплощении метод изучения белок-белковых взаимодействий выбран из группы, состоящей из масс-спектрометрии водородно-дейтериевого обмена (hydrogen deuterium exchange mass spectrometry, HDX-MS), кристаллографии, двугибридного (Yeast 2-Hybrid) скрининга, компьютерного моделирования 3D-структур или любой их комбинации. Для таких воплощений может быть необходимо сочетание результатов, полученных несколькими из этих методов, и при их соответствующем научном анализе может быть определена аминокислота или аминокислоты, вовлеченные в связывание с технологической примесью.
В одном воплощении аминокислота, определенная как вовлеченная в связывание с технологической примесью, расположена в константной области. В другом воплощении аминокислота, определенная как вовлеченная в связывание с технологической примесью, расположена в константной области тяжелой цепи.
Выравнивание последовательностей изотипов IgG, взаимодействующих с технологической примесью, с изотипами, которые с ней не взаимодействуют, в области, определенной методами изучения белок-белковых взаимодействий (например, HDX-MS), позволяет определить аминокислоту (аминокислоты), вовлеченную в связывание с технологической примесью. Затем консервативные изменения аминокислот одного иммуноглобулина эмбрионального типа на аминокислоты другого иммуноглобулина эмбрионального типа изотипа IgG, не связывающегося с технологической примесью, проводимые в IgG, который с ней связывается, позволят получить вариант, не связывающийся с технологической примесью. Таким образом, в одном воплощении аминокислоту, определенную как вовлеченная в связывание с технологической примесью, заменяют на эквивалентную аминокислоту в последовательности альтернативного антитела IgG эмбрионального типа.
В одном воплощении аминокислоту, определенную как вовлеченная в связывание с технологической примесью, модифицируют посредством консервативных изменений аминокислот одного иммуноглобулина эмбрионального типа на аминокислоты другого иммуноглобулина эмбрионального типа того же вида, проводимых в консервативной области последовательности антитела.
Таким образом, здесь предложен способ получения антитела IgG со сниженным связыванием с белком клетки-хозяина (HCP), включающий модификацию последовательности антитела в константной области тяжелой цепи по любой из или комбинации аминокислот в области между остатками 203 и 256 по нумерации Kabat. В одном примере белок клетки-хозяина представляет собой предполагаемый белок 2, подобный фосфолипазе В (PLBL2). Способ может дополнительно включать очистку антитела IgG4 с использованием метода аффинной хроматографии. Способ может дополнительно включать дополнительную очистку антитела IgG4 с использованием по меньшей мере одного другого метода ортогональной хроматографии. В одном примере метод ортогональной хроматографии представляет собой ионообменную хроматографию.
Таким образом, согласно настоящему изобретению предложены варианты антител, полученные сайт-направленным мутагенезом, а не очищенные более традиционными хроматографическими методами, для снижения уровней связанной с ними технологической примеси. Согласно другому аспекту изобретения предложено антитело IgG, полученное способами, определенными здесь.
В одном воплощении антитело IgG представляет собой антитело IgG4. Таким образом, в данном воплощении аминокислоту, определенную как вовлеченная в связывание с технологической примесью, заменяют на эквивалентную аминокислоту в последовательности альтернативного антитела IgG1, IgG2 и/или IgG3 эмбрионального типа.
Способы получения вариантов антител IgG4
Согласно другому аспекту изобретения предложен способ получения антитела IgG со сниженным связыванием с белком клетки-хозяина (HCP), включающий модификацию последовательности антитела в константной области тяжелой цепи по любой из или комбинации аминокислот в области между остатками 203 и 256 по нумерации Kabat.
Способы получения антител хорошо известны специалисту в данной области. Например, способ может включать получение суспензионной культуры рекомбинантных клетки-хозяина, трансформированных или трансфицированных рекомбинантным полинуклеотидом, кодирующим указанное антитело IgG4; и культивирование указанной культуры клетки-хозяина в условиях, позволяющих проводить экспрессию указанного антитела IgG4. Способ может дополнительно включать очистку антитела IgG4 с использованием метода аффинной хроматографии. Способ может дополнительно включать дополнительную очистку антитела IgG4 с использованием по меньшей мере одного другого метода ортогональной хроматографии. В одном примере метод ортогональной хроматографии представляет собой ионообменную хроматографию.
В одном воплощении способ дополнительно включает очистку антитела IgG4 (например, после культивирования), например, с использованием метода аффинной хроматографии. В другом воплощении метод аффинной хроматографии представляет собой аффинную хроматографию с суперантигеном. В одном воплощении суперантиген выбран из белка A, белка G и белка L. Таким образом, в другом воплощении аффинная хроматография с суперантигеном выбрана из аффинной хроматографии с белком A, аффинной хроматографии с белком G и аффинной хроматографии с белком L.
В одном воплощении способ дополнительно включает дополнительную очистку антитела IgG4 с использованием по меньшей мере одного другого хроматографического метода, такого как ионообменная хроматография. В одном воплощении одна или более чем одна стадия хроматографии выбраны из группы, состоящей из: анионообменной хроматографии, катионообменной хроматографии и смешанной (mixed-mode) хроматографии, в особенности анионообменной хроматографии. В одном воплощении по меньшей мере один другой хроматографический метод не включает хроматографию гидрофобного взаимодействия.
Способ может также включать стадии фильтрации, такой как глубинная фильтрация (для удаления клеток и клеточного детрита) и нанофильтрация (для удаления вирусов). Для веществ, полученных в системах экспрессии с клетками млекопитающих, стадия очистки может также включать любые стадии инактивации вирусов.
Согласно другому аспекту изобретения предложен способ получения варианта антитела IgG4, как определено здесь, включающий экспрессию нуклеотидной конструкции, кодирующей указанное антитело, в клетке-хозяине и, возможно, очистку указанного антитела.
Согласно другому аспекту изобретения предложено антитело IgG4, полученное способами, определенными здесь.
ОПРЕДЕЛЕНИЯ
Если не указано иное, все технические и научные термины, использованные здесь, имеют то же значение, в котором их обычно понимает специалист в данной области (например, в области клеточных культур, молекулярной генетики, химии нуклеиновых кислот, гибридизационных методик, очистки белков и биохимии). Для применения молекулярных, генетических, биохимических и химических методов используют стандартные методики.
При использовании здесь термин «антитело» относится ко всем иммуноглобулинам или антителам IgG (таким как IgG1, IgG2, IgG3 или IgG4), IgM, IgA, IgD или IgE, которые имеют происхождение от любого вида, вырабатывающего антитело естественным образом, или получены технологией рекомбинантных ДНК; выделенным из сыворотки, B-клеток, гибридом, трансфектом, дрожжей или бактерий. Антитело может представлять собой моноклональное, рекомбинантное, поликлональное, химерное (например, из разных источников (например, человеческое/мышиное химерное антитело) или антител разных типов (например, антитело IgG2/4)), человеческое, гуманизированное, мультиспецифичное (включая биспецифичные) или гетероконъюгатное антитело. Данный термин также включает отдельный вариабельный домен (например, VH, VHH, VL), однодоменное антитело (dAb®), антигенсвязывающие фрагменты, иммунологически эффективные фрагменты, Fab, F(ab')2, Fv, дисульфидно связанный Fv, одноцепочечный Fv, мультиспецифичное антитело с закрытой конформацией, дисульфидно связанные scFv, диатела, TANDABS™ и так далее. В одном воплощении антитело представляет собой IgG4. В другом воплощении антитело представляет собой рекомбинантный IgG4. В другом воплощении антитело представляет собой вариант IgG4. В другом воплощении антитело представляет собой рекомбинантный вариант IgG4.
При использовании здесь термин «CDR» определен как аминокислотные последовательности областей, определяющих комплементарность антитела, представляющих собой гипервариабельные участки тяжелых и легких цепей иммуноглобулина. В вариабельной части иммуноглобулина присутствуют три CDR (или CDR-участка) легкой цепи и три CDR (или CDR-участка) тяжелой цепи. Таким образом, при использовании здесь «CDR» относится ко всем трем CDR тяжелой цепи или всем трем CDR легкой цепи (или, по необходимости, ко всем CDR тяжелой цепи и всем CDR легкой цепи). В силу структуры и конформации антитела, существуют и другие остатки, признанные частью антигенсвязывающей области, которые будут рассматриваться специалистом в данной области как часть антигенсвязывающей области.
При использовании здесь термин «домен» относится к свернутой белковой структуре, третичная структура которой не зависит от остальной части белка. Обычно домены обеспечивают дискретные функциональные свойства белков и во многих случаях могут быть добавлены, удалены или перемещены на другие белки без потери функции остальной части белка и/или домена.
Антитело представляет собой получаемый белок, то есть интересующий белок. Например, получаемый белок представляет собой вариант IgG4. Например, получаемый белок представляет собой рекомбинантный IgG4. Например, получаемый белок представляет собой рекомбинантный вариант IgG4.
При использовании здесь термины «шарнирная область» и/или «шарнирная последовательность» относятся к тому, что обычно понимают под шарнирной областью антитела, то есть к домену молекулы IgG, охватывающему аминокислоты в положениях 226-243 включительно белковой последовательности данной молекулы при нумерации аминокислот по нумерации Kabat или в положениях 216-230 включительно по нумерации EU. Эти области могут также быть названы «генетическая шарнирная область». В дополнение к этому, при использовании здесь термины «шарнирная область» и/или «шарнирная последовательность» могут также включать аминокислоты в положениях 203-223 включительно белковой последовательности данной молекулы при нумерации аминокислот по нумерации Kabat или в положениях 196-215 включительно по нумерации EU. Кроме того, при использовании здесь термины «шарнирная область» и/или «шарнирная последовательность» могут включать аминокислоты в положениях 244-256 включительно белковой последовательности данной молекулы при нумерации аминокислот по нумерации Kabat или в положениях 231-243 включительно по нумерации EU.
Альтернативно, «структурная шарнирная область» определена как положения 234-250 по нумерации Kabat (или положения 221-237 по нумерации EU), исходя из того, что началом шарнирной области является остаток, расположенный после дисульфидной связи, соединяющей тяжелую цепь и легкую цепь, а концом – остаток, расположенный перед Fc-доменом.
Шарнирная область может быть структурно разделена на верхнюю часть шарнирной области (положения 234-238 по нумерации Kabat или положения 221-225 по нумерации EU), представляющую собой конец Fab-домена до первой внутрицепочечной дисульфидной связи, образованной в результате одного оборота спирали; и нижнюю часть шарнирной области (положения 243-250 по нумерации Kabat или положения 230-237 по нумерации EU), от последней дисульфидной связи до начала Fc-домена. Серединой или ядром шарнирной области человеческого IgG1 является мотив «CPPC» (SEQ ID NO:20) (положения 239-242 по нумерации Kabat или положения 226-229 по нумерации EU), содержащий две параллельные полипролиновые двойные спирали, связанные дисульфидными связями.
Следует отметить, что в большинстве случаев нумерация, используемая в данной области, основана на человеческом IgG1 и будет варьировать в последовательностях конкретных шарнирных областей из-за вставок и делеций. Например, мотив «YGPP» (SEQ ID NO:23) рядом с ядром шарнирной области IgG4 эквивалентен мотиву «SCDKTHT» (SEQ ID NO:24) IgG1, поскольку «YGPP» (SEQ ID NO:23) IgG4 представляет собой положения 229, 230, 237 и 238, соответственно, а не просто положения 229-232.
При использовании здесь термин «с модифицированной шарнирной областью» в контексте описания молекулы антитела относится к любому антителу, в белковой последовательности которого аминокислотные остатки между положениями 203-256 включительно изменены относительно соответствующих аминокислотных остатков последовательности немодифицированного и/или исходного антитела. Эти изменения могут включать замену остатка или остатков на альтернативную аминокислоту или замену целых пептидных последовательностей на альтернативные последовательности. Изменения могут также включать делецию аминокислоты или пептидной последовательности из белка.
При использовании здесь термин «немодифицированный» относится к антителу до модификации, снижающей связывание с технологическими примесями. Он может включать антитело в его естественном формате эмбрионального типа. Он может также включать антитела, в которых уже проведены модификации для улучшения свойств, отличных от снижения связывания с белками клетки-хозяина, такие как замена S241P (то есть приводящая к последовательности CPPC (SEQ ID NO:20) в положениях 239-242 по нумерации Kabat), которую используют во многих антителах IgG4 для повышения их стабильности. Таким образом, в одном воплощении немодифицированное антитело содержит замену S241P. Он может также включать антитела, которые уже модифицированы путем замены L248E, которую используют во многих антителах IgG4 для улучшения их эффекторной функции. Таким образом, в одном воплощении немодифицированное антитело содержит замену L248E.
Термин «модифицированный» использован здесь для описания любого из или комбинации замены, и/или делеции, и/или вставки одной аминокислоты или комбинации аминокислот в области между остатками 203 и 256 константной области тяжелой цепи по нумерации Kabat. Например, модификация может включать одну или более чем одну аминокислотную замену, и/или одну или более чем одну аминокислотную вставку, и/или одну или более чем одну аминокислотную делецию. Термины «делеция», «удаление», «замена», «исключение» использованы здесь взаимозаменяемо.
Например, вариант антитела IgG, модифицирован по 1-25 аминокислотам константной области тяжелой цепи между остатками 203 и 256 по нумерации Kabat. Например, вариант антитела IgG, модифицирован по одной, двум, трем, четырем, пяти, шести, семи, восьми, девяти, десяти, одиннадцати, двенадцати, тринадцати, четырнадцати, пятнадцати, шестнадцати, семнадцати, восемнадцати, девятнадцати, двадцати, двадцати одной, двадцати двум или двадцати трем аминокислотам константной области тяжелой цепи между остатками 203 и 256 по нумерации Kabat. Например, вариант антитела IgG, модифицирован по любому из или комбинации положений 197, 198, 203, 207, 211, 222, 226, 227, 229, 230, 232, 233, 234, 235, 236, 237, 238, 246, 247, 248, 249, 250 или 251 константной области тяжелой цепи по нумерации Kabat. Например, вариант антитела IgG, модифицирован по любому из или комбинации положений 203, 222, 226, 227, 229, 230, 232, 233, 234, 235, 236, 237, 238, 247, 248 или 251 константной области тяжелой цепи по нумерации Kabat.
При использовании здесь термин «клетка-хозяин» относится к любому организму, как прокариотическому, так и эукариотическому, который может быть подвергнут генно-инженерным манипуляциям для экспрессии полипептида, не экспрессируемого этим организмом без проведения генно-инженерных манипуляций. В определенных воплощениях клетка-хозяин выбрана из группы, состоящей из клеток CHO, клеток NS0, клеток Sp2/0, клеток COS, клеток K562, клеток BHK, клеток PER.C6 и клеток HEK. Клетка-хозяин может представлять собой бактериальную клетку, выбранную из группы, состоящей из E. coli (например, W3110, BL21), B. subtilis, и/или другие подходящие эукариотические клетки, такие как клетки грибов или дрожжей (например, Pichia pastoris, Aspergillus sp., Saccharomyces cerevisiae, Schizosaccharomyces pombe, Neurospora crassa).
При использовании здесь термин «белок клетки-хозяина» и аббревиатура «HCP» использованы взаимозаменяемо для обозначения любых полипептидов, кроме иммуноглобулина (то есть интересующего белка), для экспрессии которого клетка- хозяин была подвергнута генно-инженерным манипуляциям, экспрессируемого клеткой-хозяином.
При использовании здесь термин «технологические примеси» может быть определен в соответствии с руководствами Международного совета по гармонизации технических требований по регистрации лекарственных средств (International Council for Harmonisation of Technical Requirements for Registration of Pharmaceuticals for Human Use, ICH) (например, ICH Q6B), и относится к примесям, присутствие которых является результатом технологического процесса получения интересующего белка. Таким образом, это определение включает примеси, образующиеся при получении белка, то есть имеющие происхождение от клеточных субстратов (например, белки клетки-хозяина, ДНК/РНК клетки-хозяина), клеточной культуры (например, индукторы, антибиотики или компоненты среды), или при его дальнейшей обработке.
Термин «технологические примеси» не включает примеси, присутствие которых обусловлено самим интересующим белком (например, агрегаты и/или фрагменты антител).
Все «аминокислотные» остатки, указанные здесь, могут быть в естественной L-конфигурации. Сокращения для обозначения аминокислотных остатков, соответствующие стандартной номенклатуре полипептидов, показаны в Таблице 2.
Таблица 2: Сокращения для обозначения аминокислот
сокращение
сокращение
Все аминокислотные последовательности указаны здесь формулами, ориентация которых слева направо соответствует традиционному направлению от N-конца к C-концу.
При использовании здесь термин «эмбрионального типа», используемый в контексте описания аминокислотной последовательности антитела, описывает аминокислотную последовательность любого антитела, полученного из системы, в которой использованы последовательности человеческих иммуноглобулинов, например, иммунизацией трансгенной мыши, несущей гены человеческих иммуноглобулинов, или скринингом библиотеки генов человеческих иммуноглобулинов, где аминокислотная последовательность выбранного человеческого антитела по меньшей мере на 90% идентична аминокислотной последовательности, кодируемой геном иммуноглобулина эмбрионального типа.
При использовании здесь «консервативные изменения аминокислот одного иммуноглобулина эмбрионального типа на аминокислоты другого иммуноглобулина эмбрионального типа» определены здесь как консервативное изменение антитела, где по меньшей мере одна аминокислота исходной последовательности антитела эмбрионального типа изменена на другую аминокислотную последовательность, определенную выравниванием и эквивалентным расположением аминокислот последовательности другого антитела эмбрионального типа того же вида.
При использовании здесь термин «исходный» в контексте описания антитела относится к любому иммуноглобулину, содержащему аминокислотную последовательность природного человеческого иммуноглобулина эмбрионального типа. Термин «исходный» в контексте описания антитела подкласса IgG4 относится к любому иммуноглобулину, содержащему природную человеческую аминокислотную последовательность эмбрионального типа консервативных константных областей тяжелой цепи и легкой цепи молекулы человеческого IgG4 вместе с любой аминокислотной последовательностью в вариабельном домене, включая гипервариабельный участок, где шарнирная область иммуноглобулина содержит аминокислотную последовательность цистеин-пролин-пролин-цистеин или цистеин-пролин-серин-цистеин.
При использовании здесь термины «модифицированный IgG4», или «вариант IgG4», или «рекомбинантный IgG4» использованы взаимозаменяемо и относятся к любому антителу IgG4, содержащему ту же аминокислотную последовательность, что и «исходное» антитело IgG4, но отличающемуся от него одной или более чем одной аминокислотой. В определенных воплощениях эти различия могут составлять по меньшей мере одну из модификаций, указанных в Таблице 1 или описанных здесь.
При использовании здесь термин «связывание» в контексте связывания антитела с белком клетки-хозяина (HCP), таким как PLBL2, относится к специфическим и/или неспецифическим, обратимым и необратимым взаимодействиям между антителом и указанным белком клетки-хозяина (HCP). «Уровень связывания», «способность к связыванию», «склонность к взаимодействию», «взаимодействие» использованы здесь взаимозаменяемо. Специалист в данной области может провести количественное определение таких связывающих взаимодействий между антителом и HCP. Например, может быть проведен вестерн-блоттинг с антителами против PLBL2, как в Примере 2.
Такие связывающие взаимодействия могут также быть количественно определены, например, с применением технологии поверхностного плазмонного резонанса (SPR) в приборе BIAcore™ 3000 или BIAcore™ T200 с использованием антитела в качестве лиганда и белка клетки-хозяина в качестве аналита.
При использовании здесь термин «аффинность» относится к силе связывания молекулы, например, антитела по изобретению, с HCP, таким как PLBL2. Аффинность связывания антитела с его мишенью или с примесью (такой как HCP) может быть определена равновесными методами. Методы количественного определения аффинности связывания включают биослойную интерферометрию (Bio Layer Interferometry, BLI), например, в сочетании с прибором Octet® RED 384 (см. Пример 6). Другие методы включают твердофазный иммуноферментный анализ (ELISA), радиоиммунный анализ (RIA) или кинетический анализ (например, анализ BIACORE™).
При использовании здесь термин «активность» в контексте связывания антитела с белком клетки-хозяина, таким как PLBL2, относится к количеству HCP, связанному с антителом. Например, к числу антител, связанных с PLBL2. Методы количественного определения активности связывания включают: твердофазный иммуноферментный анализ (ELISA) для определения количества HCP, связанного с антителом (например, PLBL2, нг/мг), см. Пример 3; технологию поверхностного плазмонного резонанса (SPR) в приборе BIAcore™ 3000 или BIAcore™ T200 для определения степени связывания HCP с антителом (например, связывания PLBL2 (RU)), см. Пример 4.
При использовании здесь термин «сниженный», используемый в контексте описания изменения связывания одного антитела с белком клетки-хозяина по сравнению со связыванием другого антитела с тем же белком клетки-хозяина, относится к относительной разности связывания этих двух антител с указанным белком клетки-хозяина. Разность аффинности или активности может быть количественно определена, например, с применением технологии поверхностного плазмонного резонанса (SPR) в приборе BIAcore™ 3000 или BIAcore™ T200 с использованием антитела в качестве лиганда и белка клетки-хозяина в качестве аналита, в случае чего одно антитело определяют как имеющее «сниженный» уровень связывания по сравнению с другим, при условии снижения активности связывания более чем на 10% и/или снижения аффинности связывания более чем в 10 раз. Таким образом, в одном воплощении вариант антитела имеет активность связывания по меньшей мере на 10% ниже, чем у немодифицированного антитела, и/или аффинность связывания по меньшей мере в 10 раз ниже, чем у немодифицированного антитела.
При использовании здесь термин «миллионная доля» или «млн-1» в контексте описания концентрации или количества компонента, такого как PLBL2, относится к концентрации указанного компонента относительно концентрации продукта, такого как антитело IgG4. Значение «млн-1» по существу представляет собой молярное соотношение компонента и продукта/антитела и может быть рассчитано, например, делением концентрации компонента (измеренной в нг/мл) на концентрацию антитела (измеренную мг/мл). Таким образом, результат этого расчета представляет собой число частей компонента на миллион частей антитела/продукта. Альтернативно, количество обнаруженного HCP может быть измерено в «млрд.д.» («миллиарная доля»), что эквивалентно пг/мг.
При использовании здесь «аффинная хроматография» представляет собой хроматографический метод, в котором для хроматографического разделения используются не общие свойства биомолекул, такие как изоэлектрическая точка, гидрофобность или размер, а специфические обратимые взаимодействия между биомолекулами.
«Суперантиген» относится к универсальным лигандам, взаимодействующим с представителями суперсемейства иммуноглобулинов в сайте, отличном от сайтов связывания лигандов, являющихся мишенями этих белков. Стафилококковые энтеротоксины являются примерами суперантигенов, взаимодействующих с T-клеточными рецепторами. Суперантигены, связывающиеся с антителами, включают, без ограничения: белок G, связывающийся с константной областью IgG; белок A, связывающийся с константной областью IgG и доменами VH; и белок L, связывающийся с доменами VL. Таким образом, в одном воплощении суперантиген выбран из группы, состоящей из белка A, белка G и белка L.
При использовании здесь «белок A» включает белок A, выделенный из его естественного источника (например, клеточной стенки Staphylococcus aureus), белок A, полученный синтетическим путем (например, пептидным синтезом или рекомбинантными методиками), и его варианты, сохраняющие способность связываться с белками, имеющими область CH2/CH3.
«Аффинная хроматография с белком A» или «хроматография с белком A» относится к конкретному методу аффинной хроматографии, в котором используется аффинность IgG-связывающих доменов белка A в отношении Fc-части молекулы иммуноглобулина. Эта Fc-часть содержит константные домены CH2 и CH3 иммуноглобулинов человека или животных или домены иммуноглобулинов, в значительной степени сходные с ними. На практике хроматография с белком A включает использование белка A, иммобилизованного на твердой подложке. Белок G и белок L могут также быть использованы для аффинной хроматографии. Твердая подложка представляет собой неводную матрицу, к которой прикреплен белок A (например, колонка, смола, матрица, гранула, гель и так далее). Такие подложки включают агарозу, сефарозу, стекло, диоксид кремния, полистирол, коллодированный уголь, песок, полиметакрилат, поперечно сшитый поли(стирол-дивинилбензол) и агарозу с декстраном, увеличивающим площадь поверхности, и любое другое подходящее вещество. Такие вещества хорошо известны в данной области. Для прикрепления суперантигена к твердой подложке может быть применен любой подходящий метод. Методы прикрепления белков к твердым подложкам хорошо известны в данной области. Такие твердые подложки с и без иммобилизованного белка A или белка L могут легко быть получены из множества коммерческих источников, включая такие, как Vector Laboratory (Burlingame, Calif.), Santa Cruz Biotechnology (Santa Cruz, Calif.), BioRad (Hercules, Calif.), Amersham Biosciences (часть GE Healthcare, Uppsala, Sweden) и Millipore (Billerica, Mass.).
Термин «буфер» означает буферный раствор или буферный агент, стабилизирующий pH раствора. Буфер обычно содержит слабую кислоту и сопряженное с ней основание или слабое основание и сопряженную с ним кислоту. Буферизация раствора при pH, близком или равном оптимальному, способствует обеспечению правильного сворачивания и функционирования белка.
Термин «сложный эфир жирной кислоты» означает любое органическое соединение, содержащее цепь жирной кислоты, связанную с головной группой через сложноэфирную связь. Сложноэфирная связь образуется при замене гидроксильной группы (например, спирта или карбоновой кислоты) на алкоксигруппу. Примеры сложных эфиров жирных кислот обычно включают фосфолипиды, липиды (например, головная группа представляет собой глицерин, включая моноглицериды, диглицериды и триглицериды), а также поверхностно-активные вещества и эмульгаторы, включая, например, полисорбаты, такие как полисорбат 20, полисорбат 60 и полисорбат 80, являющиеся неионными детергентами. Поверхностно-активные вещества и эмульгаторы полезны в качестве сорастворителей и стабилизаторов и могут быть добавлены в белковые композиции для повышения стабильности белков в условиях механического стресса, таких как на границе раздела газообразной/жидкой фаз и границе раздела твердой/жидкой фаз.
Теперь изобретение будет описано со ссылкой на следующие неограничивающие примеры.
ПРИМЕРЫ
Пример 1 – Линейность разведения готовых нерасфасованных mAb при HCP-ELISA
В предшествующих исследованиях было показано, что в образцах антител, полученных с использованием клетки-хозяина, являющихся клетками млекопитающих, может присутствовать определенный белок клетки-хозяина (HCP) – белок 2, подобный фосфолипазе В (PLBL2). Кроме того, присутствие PLBL2 может приводить к нелинейности разведения при количественном определении уровней общего HCP в образце посредством ELISA. В попытке определить mAb-продукты, которые могут содержать PLBL2 в готовой нерасфасованной субстанции, был проведен HCP-ELISA для оценки линейности разведения образцов. Для определения общего количества иммуногенного HCP в образцах продуктов, имеющих происхождение от CHO, был применен ELISA собственной разработки. В данном HCP-ELISA используются специальные козьи поликлональные антитела против HCP CHO и собственный стандарт HCP, предназначенные для анализа ряда продуктов, имеющих происхождение от CHO, получаемых в GSK, и этот анализ был применен как базовый метод мониторинга удаления HCP при очистке образцов ряда биофармацевтических mAb-продуктов. Чувствительность данного анализа составляет 2,0 нг/мл.
Внутрилабораторная прецизионность для образцов промежуточной и готовой нерасфасованной субстанции варьирует в пределах 5,7-14,9% CV, а повторяемость – в пределах 3,5-8.8% CV.
С применением ELISA анализируют по меньшей мере четыре разведения каждого образца. Для каждого разведения рассчитывают приведенные значения HCP, умножая измеренную концентрацию HCP на коэффициент разведения с последующим делением на концентрацию IgG. Приведенные значения HCP для каждого разведения образца наносили на график как функцию коэффициента разведения в двойном логарифмическом масштабе. Угловой коэффициент разведения каждого образца рассчитывают с применением следующего логарифмического (log-log) уравнения:
Log (приведенное значение HCP) = A + B * Log (коэффициент разведения),
где A представляет собой отрезок, отсекаемый на оси Y, а B представляет собой угловой коэффициент. Увеличение приведенного значения HCP при большем разведении указывает на нелинейность разведения и возможное присутствие PLBL2.
Были проанализированы гуманизированные IgG разных подтипов, включая IgG1, IgG2, IgG4 и химерный IgG2/IgG4. Химерный IgG2/IgG4 представляет собой антитело с последовательностью человеческого IgG4 эмбрионального типа, шарнирная область которого была заменена на шарнирную область IgG2. Полученные результаты демонстрируют нелинейность разведения, определяемую как увеличение приведенного значения HCP при большем разведении, только в случае IgG4-молекулы mAb7 (Фиг. 1). В Таблице 3 ниже представлены значения углового коэффициента разведения, полученные для каждого из этих mAb в HCP-ELISA.
Таблица 3: Значения углового коэффициента разведения, полученные для различных mAb в HCP-ELISA
коэффициент
Эти результаты показывают, что в готовой нерасфасованной субстанции mAb7 есть HCP, присутствующий в избытке и насыщающий антитела против HCP CHO, используемые для выявления в ELISA. Для подтверждения того, что HCP представлял собой PLBL2, были необходимы дополнительные эксперименты.
Пример 2 – Вестерн-блотинг различных mAb с антителами против PLBL2
Пример 1 показал, что mAb7 демонстрирует нелинейность разведения при HCP-ELISA. Для определения того, связана ли эта нелинейность разведения с присутствием PLBL2, был проведен вестерн-блоттинг тех же гуманизированных IgG, что были ранее проанализированы посредством ELISA (Пример 1), с использованием антител против PLBL2.
Образцы, содержащие по 100 мкг mAb, разводили 1:1 2x буфером для образцов (Novex) и затем вносили в 4-20%-й градиентный гель (Novex). SDS-PAGE проводили в условиях постоянного тока при 24 мА на гель в течение 30 минут и затем при 36 мА на гель в течение 50 минут. После электрофореза гели фиксировали и окрашивали белки с использованием SYPRO® Ruby (Thermo Fisher Scientific). Гели, окрашенные Sypro® RUBY визуализировали с использованием анализатора FLA-3000 Fluorescent Image Analyzer (Fujifilm Corp.). Изображение геля, окрашенного Sypro® RUBY (Фиг. 2A), демонстрирует равное количество каждого из внесенных в гель mAb.
Вестерн-блоттинг проводили, перенося гели на PVDF-мембраны (Bio-Rad) с использованием модуля XCell II™ Blot Module (Novex) при постоянном напряжении 25 В в течение 105 минут. После переноса мембраны блокировали в течение ночи блокирующим буфером Fluorescence Blocking Buffer (Rockland Immunochemicals), разведенным 1:10 в TBST (Sigma). После блокировки мембраны промывали в TBST и инкубировали с поликлональным антителом против PLBD2 (Abcam, ab138334), 1 мкг/мл, в течение двух часов при комнатной температуре. После инкубации мембраны промывали в TBST три раза по 10 минут. Затем мембраны инкубировали с конъюгатом мышиного антитела против кроличьего cy3 (Jackson Immunoresearch), 1 мкг/мл, в течение одного часа при комнатной температуре. После инкубации мембраны промывали в TBST три раза по 10 минут. После промывки мембранам давали сохнуть в течение 30 минут. После сушки мембраны визуализировали с использованием анализатора FLA-3000 Fluorescent Image Analyzer. Нижний предел обнаружения при данном вестерн-блоттинге составлял 20 нг PLBL2. Изображение, полученное при вестерн-блоттинге (Фиг. 2Б), демонстрирует, что PLBL2 выявлен в готовой нерасфасованной субстанции mAb7 и не обнаружен ни в одной из других IgG субстанций. Этот результат свидетельствует о том, что наблюдаемая нелинейность разведения mAb7, описанная в Примере 1, обусловлена присутствием PLBL2, и необходимо провести дополнительный анализ для определения количества PLBL2, присутствующего в образце.
Пример 3 – Количественное определение PLBL2 в образцах mAb
С учетом выявления PLBL2 в mAb7, был разработан ELISA для точного количественного определения концентрации PLBL2 в образцах. Он представляет собой ELISA собственной разработки, в котором используются рекомбинантный хомячковый PLBL2 в качестве стандарта и специальные поликлональные антитела собственной разработки против PLBL2 для выявления. Чувствительность данного анализа составляет 2,0 нг/мл. Внутрилабораторная прецизионность для образцов составляет 3,23% CV.
Данный PLBL2-ELISA применяли для количественного определения концентраций PLBL2 в образцах, полученных в процессе очистки четырех различных mAb, проанализированных ранее. Концентрации PLBL2 определяли после сбора, после первой стадии хроматографии (стадия 1) и в готовой нерасфасованной субстанции (субстанция), и они показаны в Таблице 4 ниже.
Таблица 4: Концентрации PLBL2 в образцах, полученных при очистке mAb, определенные посредством PLBL2-ELISA
Результаты, показанные в Таблице 4, демонстрируют, что значительное количество PLBL2 остается в готовой нерасфасованной субстанции mAb7, но не других проанализированных mAb. Интересно, что в случае mAb7 первая стадия хроматографической очистки собранного материала приводит к удалению лишь 52% PLBL2, в то время как в случае mAb1 этот показатель составляет 99,92%. С учетом того, что для очистки обоих этих продуктов используется одна и та же смола, PLBL2, по всей вероятности, взаимодействует непосредственно с молекулой mAb, а не с хроматографической смолой.
Пример 4 – Связывание PLBL2 с различными mAb по результатам SPR
Вероятным объяснением представленных выше данных является то, что PLBL2 предпочтительно связывается с mAb7, но не с другими mAb. Как указано выше, mAb7 представляет собой молекулу IgG4, которая может обеспечивать сайт связывания с PLBL2, которого нет у других подтипов. Для дальнейшего изучения причин неудаления PLBL2 при очистке mAb7 был проведен SPR-эксперимент (поверхностный плазмонный резонанс) связывания. В данном эксперименте антитело против Fc человеческого IgG (GE Healthcare) иммобилизовали в двух проточных ячейках (FC4 и FC3) сенсорного чипа CM5 серии S (GE Healthcare) с использованием прибора Biacore™ T200 (GE Healthcare) до уровня приблизительно 7000 единиц ответа (response units, RU). Иммобилизованное антитело использовали для захвата mAb в 1x буфере HBS-EP+ (GE Healthcare) до уровня приблизительно 2500 RU на FC4 с использованием FC3 в качестве контрольной ячейки для вычитания фона. Рекомбинантный хомячковый PLBL2 разводили до 5 мкМ с использованием буфера HBS-EP+ и вносили его в течение 60 секунд, 30 мкл в минуту. Диссоциацию PLBL2 измеряли в течение 180 секунд, после чего поверхность восстанавливали, используя протокол, рекомендованный изготовителем.
Семь различных гуманизированных IgG разных подтипов были проанализированы на предмет связывания с PLBL2 посредством SPR с применением данного протокола, включая IgG1, IgG2, IgG4 и химерный IgG2/IgG4 (см. Таблицу 5 ниже). Полученные результаты демонстрируют, что PLBL2 связывается со всеми тремя проанализированными молекулами IgG4 и не связывается ни с одним из других подтипов (Фиг. 3Б, Фиг. 3В), несмотря на приблизительно равный захват mAb на сенсорном чипе (Фиг. 3A). Уровни связывания PLBL2 с каждым mAb представлены в Таблице 5 ниже.
Таблица 5: Уровни связывания PLBL2 с mAb согласно SPR
(RU)
Результаты, представленные в Таблице 5, показывают, что PLBL2 более склонен к связыванию с антителами подтипа IgG4. Кроме того, поскольку mAb2, 3, 5 и 6 содержат одинаковую вариабельную область, эти результаты показывают, что повышенную способность к связыванию с PLBL2 обеспечивает именно константная область молекулы IgG4.
Следует отметить, что PLBL2 не связывался с mAb4, представляющим собой химерный IgG2/IgG4, содержащий шарнирную область IgG2 эмбрионального типа, в то время как оставшаяся часть константной области представляет собой константную область IgG4. Эти результаты показывают, что неудаление PLBL2 при очистке mAb7 обусловлено непосредственным связыванием с mAb.
Эти результаты свидетельствуют о том, что связывание с PLBL2 преимущественно обеспечивают не просто константный домен молекулы IgG4, а, вероятно, именно шарнирная область этого константного домена.
Для расчета аффинности связывания рекомбинантного PLBL2 с mAb7 был проведен второй SPR-эксперимент. В данном эксперименте был использован тот же чип с иммобилизованным антителом против Fc человеческого IgG, только при уровне захвата mAb7 приблизительно 170 RU. После захвата PLBL2 вносили в течение 120 секунд, 30 мкл в минуту, раздельно для концентраций от 1,25 мкМ до 80 мкМ. Аффинность связывания (KD) определяли с применением аналитического программного обеспечения Biacore T200 и модели связывания 1:1 в равновесном состоянии.
Результаты демонстрируют усиление связывания PLBL2 с mAb7 при повышении концентрации (Фиг. 4A). Для расчета аффинности связывания использовали ответ связывания в равновесном состоянии (приблизительно через 110 секунд после начала внесения PLBL2) (Фиг. 4Б). Расчетная KD связывания PLBL2 с mAb7 составила 40,0 мкМ. Несмотря на то, что такое связывание является относительно слабым, полученный результат был полезен для проведения HDX-MS-эксперимента, в котором молекулу mAb7 насыщали PLBL2.
Пример 5 – Влияние системы экспрессии на связывание с PLBL2
В предшествующих примерах была продемонстрирована повышенная склонность антител подтипа IgG4 к связыванию с PLBL2, по сравнению с другими подтипами антител (например, IgG1 и IgG2). Было проведено исследование для определения того, влияют ли система экспрессии клетки-хозяина, в которых получены антитела, на их способность связываться с PLBL2. Исследования были проведены с использованием mAb6 (IgG4) и mAb3 (IgG2), имеющих одинаковый вариабельный домен, но относящиеся к разным подклассам IgG. Таким образом, различия в свойствах этих молекул могут быть непосредственно связаны с константными доменами антител.
Кратко, проводили раздельное культивирование клеток CHO K1 и HEK293, экспрессирующих IgG4 mAb6 (обозначенное mAb6A и mAb6B, соответственно), а также отдельное культивирование клеток CHO K1a, экспрессирующих mAb3 (IgG2). Эти культуры собирали в подходящее время до проведения стандартной предварительной и последующей очистки mAb. Очищенные образцы каждого из этих антител затем анализировали и сравнивали на предмет их способности связываться с PLBL2 с применением поверхностного плазмонного резонанса (SPR) на Biacore™ T200.
Образцы (Таблица 6) разводили до 20 мкг/мл в буфере HBS-EP и пропускали над активной проточной ячейкой с чипом с предварительно иммобилизованным белком A (GE Healthcare) в течение 60 секунд со скоростью 10 мкл/мин. Затем как над активной проточной ячейкой, так и над контрольной проточной ячейкой пропускали PLBL2, разведенный до 100 мкг/мл в HBS-EP, в течение 120 секунд со скоростью 5 мкл/мин. Обе проточные ячейки восстанавливали с использованием 10 мМ глицина при pH 1,5 в течение 10 секунд, 30 мкл/мин, и затем HBS-EP в течение 30 секунд, 30 мкл/мин.
Таблица 6: Молекулярный формат и системы экспрессии клетки-хозяина, использованные для получения антител
клетки-хозяина
Результаты (Фиг. 5) показывают, что обе молекулы IgG4 взаимодействуют с PLBL2 сходным образом, несмотря на то, что они получены в разных клеточных линиях. В то же время, молекула IgG2 не взаимодействует. В Таблице 7 показаны произвольные значения, полученные для каждого образца.
Таблица 7: Связывание PLBL2 с молекулами IgG4 и IgG2, экспрессированными в разных клетках-хозяевах
Пример 6 – Количественное определение аффинности связывания антител подтипов IgG2 и IgG4
В предшествующих экспериментах была продемонстрирована повышенная способность антител подкласса IgG4 к связыванию с PLBL2, по сравнению с антителами других подклассов (например, IgG1 и IgG2). Также были проведены исследования по количественному определению аффинности антитела IgG4 в отношении PLBL2.
Образцы получали, как описано в Примере 5. Затем оценивали аффинность связывания PLBL2 с антителом в этих образцах посредством биослойной интерферометрии (BLI) с использованием прибора Octet® RED 384. Образцы (те же, что в Таблице 6) разводили до 10 мкг/мл в PBS-T. Имеющиеся в продаже биосенсоры с белком A (Pall) погружали в каждый образец на 120 секунд при 1000 об/мин. После этого биосенсоры погружали в PLBL2 в различных концентрациях (1538 нМ с серийным 2-кратным разведением до 16 нМ) на 300 секунд при 1000 об/мин для оценки ассоциации перед погружением в PBS-T на 300 секунд при 1000 об/мин для оценки диссоциации. Биосенсоры восстанавливали с использованием 100 мМ глицина, pH 1,5, и PBS-T, следуя инструкциям изготовителя.
Результаты (Таблица 8) рассчитывали с применением локального приближения к модели связывания 1:1. Получить данные по аффинности mAb3 было невозможно из-за отсутствия взаимодействия (Фиг. 6A). Как mAb6A, так и mAb6B имели низкую аффинность; аффинность mAb6A была приблизительно в 14 раз выше аффинности mAb6B из-за более высокой скорости ассоциации. В то же время, скорости
диссоциации были очень сходны. Приближенные кривые для mAb6A и mAb6B
оказаны на Фиг. 6Б и 6В, соответственно.
Эти результаты показывают, что, несмотря на то, что система экспрессии не влияет на способность экспрессированного IgG4 к связыванию с PLBL2, она может потенциально влиять на аффинность связывания молекулы.
Таблица 8: Аффинность связывания молекул IgG2 и IgG4
Пример 7 – Взаимодействие между mAb7 и PLBL2 по данным HDX-MS
Для дополнительного понимания научных основ взаимодействий mAb7 и PLBL2, был проведен HDX-MS-анализ образцов mAb7 и mAb7, связанного с PLBL2, с использованием системы Waters HDX Manager System в сочетании с масс-спектрометром Synapt G2-S. Исходя из KD 40,0 мкМ по результатам SPR, после мечения дейтерием с PLBL2 связывается приблизительно 70% mAb. Мечение дейтерием измеряли во временных точках 0,5 мин, 5 мин, 60 мин, 120 мин, 180 мин и 240 мин. Данные анализировали с использованием программного обеспечения Waters DynamX и построением дифференциального графика HDX (Фиг. 7). Вертикальными линиями показаны суммарные различия HDX для каждого пептида в шести указанных временных точках. При сравнении HDX-профиля несвязанного и связанного с PLBL2 mAb7 область K218-F256 демонстрирует снижение мечения дейтерием при связывании с PLBL2. Это указывает на стабилизацию данной области, благодаря связыванию PLBL2 с mAb7 и коррелирует с результатами других экспериментов связывания.
Пример 8 – Выравнивание последовательностей шарнирной области различных mAb
Полученные ранее результаты демонстрируют, что PLBL2 присутствует в образцах mAb7 (молекула IgG4) из-за непосредственного связывания с антителом, приводящего к неэффективности очистки. Сходным образом, в предшествующих примерах было показано, что mAb5 и mAb6, также относящиеся к подтипу IgG4, тоже способны связываться с PLBL2. Анализ связывания PLBL2 с mAb различных изотипов показал, что, наиболее вероятно, это связывание происходит в шарнирной области
молекулы IgG4, на что также указывают данные HDX-MS. С учетом того, что разные изотипы IgG различаются небольшим числом аминокислот, высока вероятность того, что удастся определить те отдельные аминокислотные остатки, которые вовлечены в связывание с PLBL2. Для определения аминокислотных остатков IgG4, вовлеченных в связывание с PLBL2, было проведено выравнивание аминокислотных последовательностей шарнирной области mAb.
При анализе выравнивания последовательностей интерес представляют консервативные аминокислотные остатки молекул IgG4 ((mAb5, mAb6 и mAb7), которые связывались с PLBL2, и неконсервативные (то есть, различающиеся) аминокислотные остатки других подтипов IgG, которые не связывались с PLBL2. В этом анализе были определены 10 таких аминокислот, которые выделены черными рамками (Фиг. 8). Конкретно, остатками, представляющими интерес, являются: K203, R222, S227, Y229, G230, P237, P238, E246, F247 и E248.
Следовало ожидать, что мутации этих 10 остатков с заменой аминокислот, обнаруженных в IgG4, на аминокислоты, обнаруженные в IgG2 или IgG1, предотвратят связывание PLBL2 с молекулой IgG4.
Пример 9 – Мутагенез остатков шарнирной области IgG4 для минимизации связывания с PLBL2
Исследования, подробно описанные в Примерах 1-8, показали, что способность молекул IgG4 к связыванию с PLBL2 обеспечена главным образом константной областью этих иммуноглобулинов. Кроме того, сравнение IgG4 и химерного IgG2/IgG4 позволило локализовать эти взаимодействия с PLBL2 в шарнирной области молекулы или рядом с ней. Выравнивание аминокислотных последовательностей различных изученных молекул позволило, посредством удаления аминокислот, определить конкретные аминокислоты шарнирной области, которые играют роль в связывании с PLBL2. Завершающая стадия состоит в подтверждающих мутагенетических экспериментах, чтобы показать, как модификация шарнирной области молекулы IgG4 может минимизировать связывание иммуноглобулинов этого подкласса с PLBL2.
Были проведены исследования, в которых 7 из 10 аминокислотных остатков, определенных как приводящие к связыванию PLBL2 с антителами подкласса IgG4, были заменены на альтернативные аминокислоты, такие как аминокислоты, обнаруженные в последовательности шарнирной области человеческих антител IgG1 и/или IgG2 эмбрионального типа. С использованием mAb5 в качестве модельной молекулы были проведены мутагенетические эксперименты, в которых систему экспрессии с клетками-хозяевами HEK293 транзиторно трансфицировали векторной ДНК, кодирующей ряд вариантов mAb5 с модифицированной шарнирной областью.
Мутации в шарнирной области mAb5 были проведены в тех положениях аминокислотной последовательности, которые были ранее определены как играющие некоторую роль, влияя на связывание с PLBL2 (Пример 8). Мутации проводили как единичные точковые мутации, как описано в Таблице 1, а также как их различные комбинации, что привело к получению 7 разных вариантов молекул IgG4 с модифицированной шарнирной областью (mAb5-1B, 2B, 3B, 4B, 6B и 7B). Параллельно проводили культивирование для транзиторной экспрессии немодифицированного исходного IgG4 (mAb5B). В дополнение к этому, были также проведены мутации в шарнирной области mAb7, также представляющего собой IgG4, но направленного против другого антигена-мишени, как единичные точковые мутации, а также как их различные комбинации, что привело к получению 3 разных вариантов с модифицированной шарнирной областью (mAb7-2B, 4B и 5B).
Суффиксы этих вариантов отражают проведенные мутации, и они одинаковы в обеих группах вариантов IgG4 с модифицированной шарнирной областью. Поэтому шарнирная область mAb5-2B идентична шарнирной области mAb7-2B.
Кроме того, проводили культивирование для экспрессии IgG2 (mAb3B) и IgG4 (mAb6B) в качестве дополнительных контролей, а также варианта IgG1 с модифицированной шарнирной областью (mAb2-αB). На Фиг. 9 показана модифицированная шарнирная область этих вариантов IgG4 и их выравнивание с последовательностями шарнирной области других молекул mAb, включенных в исследование. Подробные данные о различных молекулах, включенных в этот анализ, приведены в Таблице 9. Суффиксы «A» и «B» в названиях молекул, указывают на то, в каких клетках, CHO K1 или HEK293, соответственно, было экспрессировано антитело.
Таблица 9: Подробные данные о IgG1, IgG2, IgG4 и вариантах IgG4, включенных в анализ
модифицированной шарнирной областью,
HEK293
HEK293
HEK293
модифицированной шарнирной областью, экспрессированный в HEK293
модифицированной шарнирной областью, экспрессированный в HEK293
модифицированной шарнирной областью, экспрессированный в HEK293
модифицированной шарнирной областью, экспрессированный в HEK293
модифицированной шарнирной областью, экспрессированный в HEK293
Вставка SCDKTHT (SEQ ID NO:24)
модифицированной
экспрессированный в
HEK293
модифицированной шарнирной областью, экспрессированный в HEK293
модифицированной шарнирной областью, экспрессированный в HEK293
HEK293
CHO K1
модифицированной шарнирной областью, экспрессированный в HEK293
модифицированной шарнирной областью, экспрессированный в HEK293
модифицированной шарнирной областью, экспрессированный в HEK293
Материал, полученный из клеток HEK293, очищали с применением стандартной методики очистки mAb. Кратко, после культивирования, достаточно продолжительного для транзиторной экспрессии mAb, удаляли клетки и проводили очистку осветленного супернатанта клеточной культуры с применением аффинной хроматографии с белком A. Элюат, полученный после хроматографии с белком A, титровали до нейтрального pH c использованием раствора трис. Затем нейтрализованные элюаты, полученные после хроматографии с белком A, содержащие различные исследуемые молекулы mAb, подвергали серии анализов связывания, как описано в Примерах 10-12.
Пример 10 – Анализ взаимодействия IgG2, IgG4 и вариантов молекул IgG4 с PLBL2
Взаимодействие различных mAb, полученных и описанных в Примере 9, с рекомбинантным человеческим PLBL2 оценивали поверхностным плазмонным резонансом (SPR) с применением метода связывания Biacore™ T200 (GE Healthcare).
20 мкг/мл mAb пропускали над проточной ячейкой 4 имеющегося в продаже чипа с предварительно иммобилизованным белком A (GE Healthcare) в течение 60 секунд со скоростью 10 мкл/мин и давали ему стабилизироваться в рабочем буфере HBS-EP+ в течение 10 секунд. Вариация уровня захваченного антитела составляла менее 10%. Затем над проточными ячейками 4 и 3 пропускали 100 мкг/мл (1538 нМ) рекомбинантного человеческого PLBL2 в течение 120 секунд со скоростью 5 мкл/мин. Проточная ячейка 3 действует как контрольная, сниженная скорость потока 5 мкл/мин была использована для стимуляции низкоаффинных взаимодействий. Сенсор затем восстанавливали 10 мМ глицином, pH 1,5, 10 секунд, 30 мкл/мин.
По результатам предшествующих экспериментов было известно, что mAb8A (являющееся IgG1) не взаимодействовало с рекомбинантным человеческим PLBL2. Оно было использовано само по себе в качестве отрицательного контроля для нормализации результатов.
Результаты SPR-анализа (Фиг. 10) указывают на приблизительно 90%-е уменьшение числа RU PLBL2 для mAb5-2B и mAb5-4B по сравнению с их исходной молекулой (mAb5B), и приблизительно 80%-е уменьшение числа RU PLBL2 для mAb7-2B и mAb7-4B по сравнению с их исходной молекулой (mAb7A). mAb5-3B не продемонстрировало изменений связывания, указывая на то, что сама по себе эта
мутация не позволяла эффективно снизить взаимодействие. mAb5-2B содержит единичную аминокислотную замену в положении 222 с аргинина на лизин. mAb5-3B содержит единичную аминокислотную замену в положении 227 с серина на пролин. mAb5-4B содержит как мутации, присутствующие в mAb5-2B, так и мутации, присутствующие в mAb5-3B; указывая на то, что, несмотря на отсутствие снижения взаимодействия с PLBL2 при замена на пролин в положении 227, эта замена не оказывает никакого нежелательного эффекта, и что наблюдаемое снижение связывания с PLBL2 обусловлено заменой на лизин в положении 222. Действительно, замена лизина в положении 222 mAb2 (Фиг. 9) на аргинин, соответствующая переходу с IgG1 на IgG4 эмбрионального типа по этому положению, привела к мутанту (mAb2-αB) с повышенным связыванием с PLBL2 по результатам SPR с сравнении с исходной молекулой (mAb2), не демонстрировавшей связывания с PLBL2. Замена аргинина в положении 222 mAb5 (Фиг. 9) на треонин, соответствующая переходу с IgG4 на IgG2 эмбрионального типа по этому положению, привела к мутанту (mAb5-8B) с 30%-м снижением связывания с PLBL2 по результатам SPR с сравнении с исходной молекулой (mAb5).
Несмотря на то, что, согласно этим данным, аргинин в положении 222 является существенным фактором, способствующим связыванию с PLBL2, другие остатки шарнирной области могут также влиять на связывание с этим HCP, на что указывает сниженное связывание с PLBL2, которое демонстрирует mAb5-5B (Фиг. 10). В то же время, mAb8A не продемонстрировало взаимодействия с PLBL2; mAb6B продемонстрировало наибольшую степень связывания.
Пример 11 – Анализ аффинности IgG2, IgG4 и вариантов молекул IgG 4 в отношении PLBL2
Аффинность взаимодействия PLBL2 с различными mAb, полученных и описанных в Примере 9, анализировали так же, как описано в Примере 6, посредством биослойной интерферометрии (BLI) с использованием прибора Octet® RED 384 (ForteBio).
Имеющиеся в продаже биосенсоры с белком A погружали в образцы, 10 мкг/мл, на 120 секунд при 1000 об/мин. Для диссоциации слабо связанных белков биосенсоры погружали в аналитический буфер (HBS-EP+) на 30 секунд при 1000 об/мин. После этого биосенсоры погружали в рекомбинантный человеческий PLBL2 в различных концентрациях (1538, 769,2, 384,6, 192,3, 96,15, 48,15, 24, 0 нМ) и, в завершение, в HBS-EP+ на 5 минут.
Скорости ассоциации и диссоциации измеряли с применением модели связывания 1:1 с общим приближением; тем не менее, из-за высокой скорости диссоциации были проанализированы лишь первые 50 секунд кривой диссоциации. Результаты представлены в Таблице 10.
Таблица 10: Аффинность связывания IgG1, IgG2, IgG4 и вариантов молекул IgG4
(по сравнению с исходной молекулой)
(1/Мс)
mAb5B
*кратность изменения рассчитана, исходя из KD mAb5B
Кинетические параметры mAb5-2B, mAb5-4B, mAb7-2B, mAb7-4B и mAb8A рассчитать не удалось из-за невозможности построения приближенных кривых, указывающей на отсутствие взаимодействия PLBL2 с этими молекулами. Эти результаты согласуются с данными, полученными в Примере 10, которые также демонстрируют слабое связывание PLBL2 в этих образцах. Для mAb5-6B было отмечено значительное снижение KD (более чем в 10 раз) по сравнению с исходной молекулой (mAb5B). mAb5-1B и mAb5-7B демонстрировали несколько более высокую аффинность в отношении PLBL2 из-за несколько большей скорости ассоциации (kon), что нежелательно. Все остальные мутанты демонстрировали снижение связывания с PLBL2 менее чем в 10 раз по сравнению с исходной молекулой, и само по себе это изменение аффинности связывания было признано незначимым.
Пример 12 – Связывание антигена молекулами IgG2, IgG4 и вариантами молекул IgG4
Молекулы антител mAb3, mAb5 (включая все варианты с модифицированной шарнирной областью) и mAb6 содержат одну и ту же вариабельную область, поскольку эти антитела направлены против одного и того же антигена. Для демонстрации того, что проведенный мутагенез не оказывает нежелательного эффекта на антигенсвязывающую активность каждой молекулы, была проведена оценка связывания антигена с применением поверхностного плазмонного резонанса (SPR) и Biacore™ T200 (GE Healthcare). Активность оценивали, используя сенсор с предварительно иммобилизованным белком A (GE Healthcare) для захвата лиганда. Для восстановления поверхности сенсора перед его использованием во всех проточных ячейках проводили 5 начальных циклов (с использованием 10 мМ глицина, pH 1,5, в течение 60 секунд со скоростью 30 мкл/мин). Каждый образец mAb разводили до 10 мкг/мл в PBS-T и пропускали над проточной ячейкой 4 в течение 60 секунд со скоростью 10 мкл/мин с периодом стабилизации 10 секунд. Затем над проточными ячейками 4 и 3 пропускали антиген-мишень, 10 мкг/мл, в течение 60 секунд со скоростью 10 мкл/мин, после чего внесение антигена прекращали и пропускали аналитический буфер со скоростью потока 10 мкл/мин в течение 100 секунд для диссоциации. Затем поверхность сенсора восстанавливали с использованием 10 мМ глицина, pH 1,5, в течение 60 секунд со скоростью 30 мкл/мин, подготавливая к следующему циклу.
Для расчета активности каждой молекулы ответ связывания антигена (RU) делили на ответ захвата антитела (RU) и полученный результат выражали в процентах. Рассчитанный процент отражает количество антитела, присутствующего в образце, которое все еще способно связываться со своим антигеном. Погрешность анализа в данном эксперименте составляет 10%. В Таблице 11 показано, что активность всех mAb в отношении антигена-мишени составляла 100 плюс/минус 10%, указывая на отсутствие снижения или изменения связывания. Активность всех мутантов была сопоставимой. В частности, различий в связывании mAb5B или любых его вариантов с модифицированной шарнирной областью (mAb5-1B – 7B) не было.
Таблица 11: Связывание каждого mAb подклассов IgG2 и IgG4 с антигеном-мишенью
Пример 13 – Снижение связывания IgG4 с белками клетки-хозяина (HCP)
Проблемные белки клетки-хозяина обычно не удаляются из продукта в процессе очистки, и, несмотря на то, что они обычно присутствуют лишь в следовых количествах, бывают случаи, когда уровни этих белков в полностью очищенной фармацевтической субстанции могут представлять проблему с точки зрения безопасности и эффективности. В то время как связывание с PLBL2 является важным примером таких случаев, есть данные о других проблемных белках клетки-хозяина, взаимодействующих с интересующими антителами.
В результате, могут быть проведены исследования, направленные на демонстрацию эффекта модификаций шарнирной области IgG4 по снижению связывания с другими HCP. Первым этапом будет определение HCP, склонных к взаимодействию с IgG4, например, с использованием mAb6 как модельной молекулы. Эти HCP могут быть определены с применением метода обогащения HCP, при котором Fc-область mAb будет связываться с имеющимися в продаже гранулами с белком A и проходить многократные 10 минутные периоды инкубации с осветленным необработанным неразделенным (clarified unprocessed bulk, CUB) материалом нетрансфицированных CHO K1. Затем из гранул элюируют готовый связанный комплекс и анализируют его с применением LC-MS/MS. Для определения количества присутствующих белков клетки-хозяина может быть применен метод мониторинга множественных реакций (multiple reaction monitoring, MRM).
Когда эти HCP будут успешно определены, способы, подробно описанные в предшествующих Примерах, могут быть применены для оценки связывания этих HCP как с исходной молекулой mAb5, так и с модифицированными мутантами mAb5, получение которых описано в Примере 9. Эти исследования позволят продемонстрировать возможность не только снижения связывания PLBL2 с IgG4 с использованием модификаций шарнирной области, являющихся объектом данного изобретения, но также более общего уменьшения склонности к связыванию с широким спектром HCP.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> GlaxoSmithKline Intellectual Property Development Limited
<120> АНТИТЕЛА СО СНИЖЕННЫМ СВЯЗЫВАНИЕМ С ТЕХНОЛОГИЧЕСКИМИ ПРИМЕСЯМИ
<130> PB66137
<150> US62/404849
<151> 2016-10-06
<160> 38
<170> PatentIn version 3.5
<210> 1
<211> 61
<212> PRT
<213> Human
<400> 1
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr
1 5 10 15
Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys
20 25 30
Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
35 40 45
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe
50 55 60
<210> 2
<211> 61
<212> PRT
<213> Human
<400> 2
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr
1 5 10 15
Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys
20 25 30
Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
35 40 45
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe
50 55 60
<210> 3
<211> 57
<212> PRT
<213> Artificial
<220>
<223> mAb3 (IgG2)
<400> 3
Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr Tyr
1 5 10 15
Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Thr
20 25 30
Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro Pro
35 40 45
Ala Ala Ala Ser Ser Val Phe Leu Phe
50 55
<210> 4
<211> 57
<212> PRT
<213> Artificial
<220>
<223> mAb4 (IgG2/4)
<400> 4
Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr Tyr
1 5 10 15
Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Thr
20 25 30
Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro Pro
35 40 45
Ala Ala Ala Ser Ser Val Phe Leu Phe
50 55
<210> 5
<211> 58
<212> PRT
<213> Artificial
<220>
<223> mAb5 (IgG4)
<400> 5
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr
1 5 10 15
Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg
20 25 30
Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu
35 40 45
Phe Glu Gly Gly Pro Ser Val Phe Leu Phe
50 55
<210> 6
<211> 57
<212> PRT
<213> Artificial
<220>
<223> mAb6 (IgG4)
<400> 6
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr
1 5 10 15
Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg
20 25 30
Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Pro
35 40 45
Ala Ala Ala Pro Ser Val Phe Leu Phe
50 55
<210> 7
<211> 58
<212> PRT
<213> Artificial
<220>
<223> mAb7 (IgG4)
<400> 7
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr
1 5 10 15
Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg
20 25 30
Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu
35 40 45
Phe Glu Gly Gly Pro Ser Val Phe Leu Phe
50 55
<210> 8
<211> 61
<212> PRT
<213> Human
<400> 8
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr
1 5 10 15
Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys
20 25 30
Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
35 40 45
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe
50 55 60
<210> 9
<211> 57
<212> PRT
<213> Human
<400> 9
Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr Tyr
1 5 10 15
Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Thr
20 25 30
Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro Pro
35 40 45
Val Ala Gly Pro Ser Val Phe Leu Phe
50 55
<210> 10
<211> 58
<212> PRT
<213> Human
<400> 10
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr
1 5 10 15
Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg
20 25 30
Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro Ala Pro Glu
35 40 45
Phe Leu Gly Gly Pro Ser Val Phe Leu Phe
50 55
<210> 11
<211> 58
<212> PRT
<213> Artificial
<220>
<223> mAb5-1B (IgG4)
<400> 11
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr
1 5 10 15
Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg
20 25 30
Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu
35 40 45
Phe Glu Gly Gly Pro Ser Val Phe Leu Phe
50 55
<210> 12
<211> 58
<212> PRT
<213> Artificial
<220>
<223> mAb5-2B (IgG4)
<400> 12
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr
1 5 10 15
Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys
20 25 30
Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu
35 40 45
Phe Glu Gly Gly Pro Ser Val Phe Leu Phe
50 55
<210> 13
<211> 58
<212> PRT
<213> Artificial
<220>
<223> mAb5-3B (IgG4)
<400> 13
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr
1 5 10 15
Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg
20 25 30
Val Glu Pro Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu
35 40 45
Phe Glu Gly Gly Pro Ser Val Phe Leu Phe
50 55
<210> 14
<211> 58
<212> PRT
<213> Artificial
<220>
<223> mAb5-4B (IgG4)
<400> 14
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr
1 5 10 15
Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys
20 25 30
Val Glu Pro Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu
35 40 45
Phe Glu Gly Gly Pro Ser Val Phe Leu Phe
50 55
<210> 15
<211> 58
<212> PRT
<213> Artificial
<220>
<223> mAb5-5B (IgG4)
<400> 15
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr
1 5 10 15
Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg
20 25 30
Val Glu Ser Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro Glu
35 40 45
Phe Glu Gly Gly Pro Ser Val Phe Leu Phe
50 55
<210> 16
<211> 61
<212> PRT
<213> Artificial
<220>
<223> mAb5-6B (IgG4)
<400> 16
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr
1 5 10 15
Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg
20 25 30
Val Glu Ser Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
35 40 45
Ala Pro Glu Phe Glu Gly Gly Pro Ser Val Phe Leu Phe
50 55 60
<210> 17
<211> 58
<212> PRT
<213> Artificial
<220>
<223> mAb5-7B (IgG4)
<400> 17
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr
1 5 10 15
Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg
20 25 30
Val Leu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu
35 40 45
Phe Glu Gly Gly Pro Ser Val Phe Leu Phe
50 55
<210> 18
<211> 61
<212> PRT
<213> Human
<400> 18
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr
1 5 10 15
Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys
20 25 30
Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
35 40 45
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe
50 55 60
<210> 19
<211> 4
<212> PRT
<213> Human
<400> 19
Cys Pro Ser Cys
1
<210> 20
<211> 4
<212> PRT
<213> Human
<400> 20
Cys Pro Pro Cys
1
<210> 21
<211> 12
<212> PRT
<213> Human
<400> 21
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro
1 5 10
<210> 22
<211> 12
<212> PRT
<213> Artificial
<220>
<223> Немодифицированная шарнирная область антитела IgG4 (S_P)
<400> 22
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro
1 5 10
<210> 23
<211> 4
<212> PRT
<213> Human
<400> 23
Tyr Gly Pro Pro
1
<210> 24
<211> 7
<212> PRT
<213> Human
<400> 24
Ser Cys Asp Lys Thr His Thr
1 5
<210> 25
<211> 4
<212> PRT
<213> Human
<400> 25
Cys Cys Val Glu
1
<210> 26
<211> 7
<212> PRT
<213> Human
<400> 26
Glu Ser Lys Tyr Gly Pro Pro
1 5
<210> 27
<211> 10
<212> PRT
<213> Human
<400> 27
Glu Pro Lys Ser Cys Asp Lys Thr His Thr
1 5 10
<210> 28
<211> 7
<212> PRT
<213> Artificial
<220>
<223> Модификация остатков 226-238 IgG4 по нумерации Kabat
<400> 28
Glu Arg Lys Tyr Gly Pro Pro
1 5
<210> 29
<211> 7
<212> PRT
<213> Human
<400> 29
Glu Arg Lys Cys Cys Val Glu
1 5
<210> 30
<211> 12
<212> PRT
<213> Artificial
<220>
<223> Модификация остатков 226-238 IgG4 по нумерации Kabat
<400> 30
Glu Leu Lys Thr Pro Leu Gly Asp Thr Thr His Thr
1 5 10
<210> 31
<211> 6
<212> PRT
<213> Human
<400> 31
Glu Phe Leu Gly Gly Pro
1 5
<210> 32
<211> 5
<212> PRT
<213> Artificial
<220>
<223> Модификация остатков 246-251 IgG4 по нумерации Kabat
<400> 32
Pro Ala Ala Ala Ser
1 5
<210> 33
<211> 5
<212> PRT
<213> Artificial
<220>
<223> Модификация остатков 246-251 IgG4 по нумерации Kabat
<400> 33
Pro Ala Ala Ala Pro
1 5
<210> 34
<211> 61
<212> PRT
<213> Artificial
<220>
<223> mAb2-alphaB(IgG1)
<400> 34
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr
1 5 10 15
Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg
20 25 30
Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
35 40 45
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe
50 55 60
<210> 35
<211> 58
<212> PRT
<213> Artificial
<220>
<223> mAb5-8B (IgG4)
<400> 35
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr
1 5 10 15
Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Thr
20 25 30
Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu
35 40 45
Phe Glu Gly Gly Pro Ser Val Phe Leu Phe
50 55
<210> 36
<211> 58
<212> PRT
<213> Artificial
<220>
<223> mAb7-2B (IgG4)
<400> 36
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr
1 5 10 15
Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys
20 25 30
Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu
35 40 45
Phe Glu Gly Gly Pro Ser Val Phe Leu Phe
50 55
<210> 37
<211> 58
<212> PRT
<213> Artificial
<220>
<223> mAb7-4B (IgG4)
<400> 37
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr
1 5 10 15
Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys
20 25 30
Val Glu Pro Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu
35 40 45
Phe Glu Gly Gly Pro Ser Val Phe Leu Phe
50 55
<210> 38
<211> 58
<212> PRT
<213> Artificial
<220>
<223> mAb7-5B (IgG4)
<400> 38
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr
1 5 10 15
Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg
20 25 30
Val Glu Ser Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro Glu
35 40 45
Phe Glu Gly Gly Pro Ser Val Phe Leu Phe
50 55
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ И КОМПОЗИЦИИ, ВКЛЮЧАЮЩИЕ ОЧИЩЕННЫЕ РЕКОМБИНАНТНЫЕ ПОЛИПЕПТИДЫ | 2014 |
|
RU2793783C1 |
| ВАРИАНТЫ Fc-ОБЛАСТИ С МОДИФИЦИРОВАННЫМ СВЯЗЫВАНИЕМ FcRn И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2714116C2 |
| МОДИФИЦИРОВАННАЯ КОНСТАНТНАЯ ОБЛАСТЬ АНТИТЕЛА | 2023 |
|
RU2837820C2 |
| МОДИФИЦИРОВАННАЯ КОНСТАНТНАЯ ОБЛАСТЬ АНТИТЕЛА | 2021 |
|
RU2797273C2 |
| ВАРИАНТЫ Fc-ОБЛАСТИ С МОДИФИЦИРОВАННЫМИ СВОЙСТВАМИ СВЯЗЫВАНИЯ FcRn И БЕЛКА А | 2015 |
|
RU2713131C1 |
| АНТИТЕЛА ПРОТИВ TFR И ИХ ПРИМЕНЕНИЕ ПРИ ЛЕЧЕНИИ ПРОЛИФЕРАТИВНЫХ И ВОСПАЛИТЕЛЬНЫХ РАССТРОЙСТВ | 2016 |
|
RU2737637C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА К CD19 ЧЕЛОВЕКА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2731156C1 |
| АНТИТЕЛА К CD38 И КОМБИНАЦИИ С АНТИТЕЛАМИ К CD3 И CD28 | 2018 |
|
RU2812910C2 |
| АНТИТЕЛА ПРОТИВ ПРЕДСТАВИТЕЛЯ А5 СЕМЕЙСТВА 19 СО СХОДСТВОМ ПОСЛЕДОВАТЕЛЬНОСТЕЙ И СПОСОБ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2785436C2 |
| АНТИТЕЛА ПРОТИВ BTN3A И ИХ ПРИМЕНЕНИЕ ПРИ ЛЕЧЕНИИ РАКА ИЛИ ИНФЕКЦИОННЫХ ЗАБОЛЕВАНИЙ | 2019 |
|
RU2800726C2 |
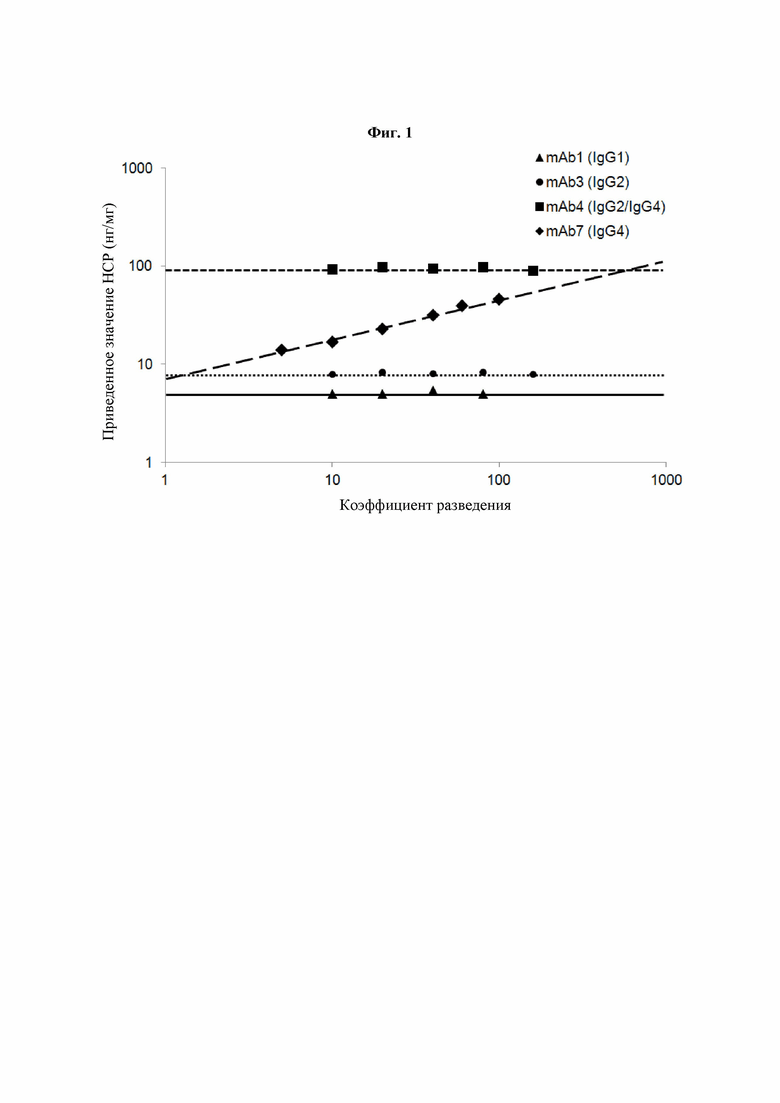

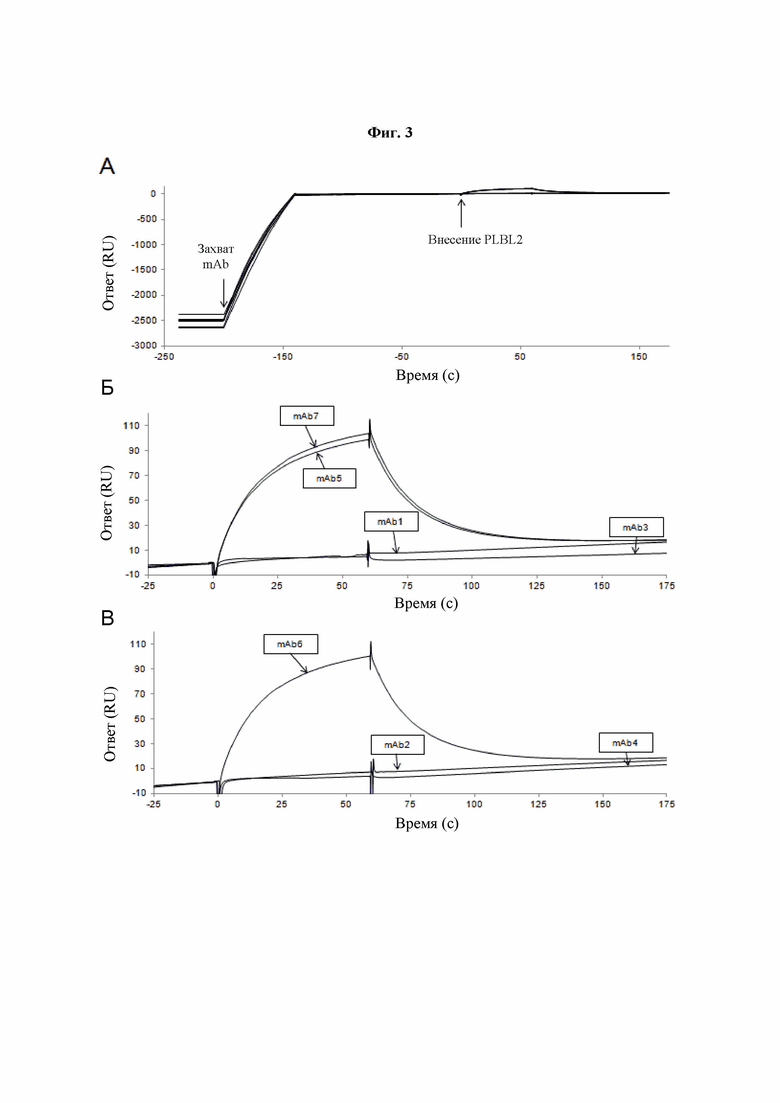
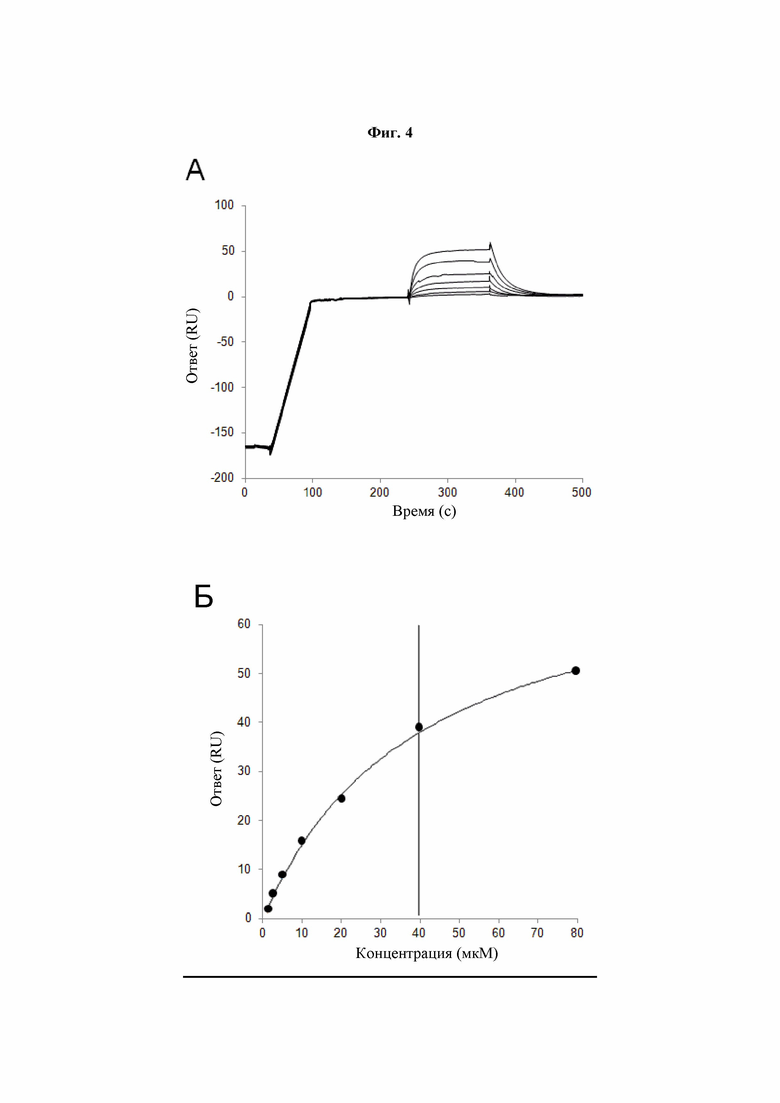
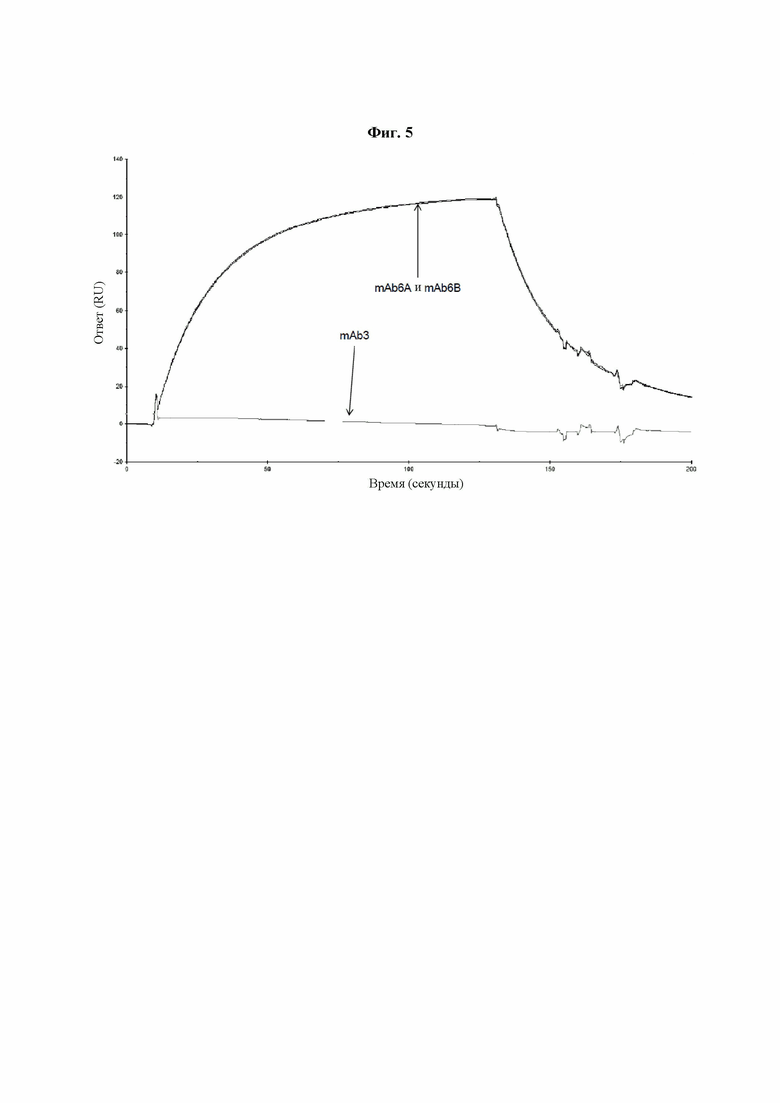
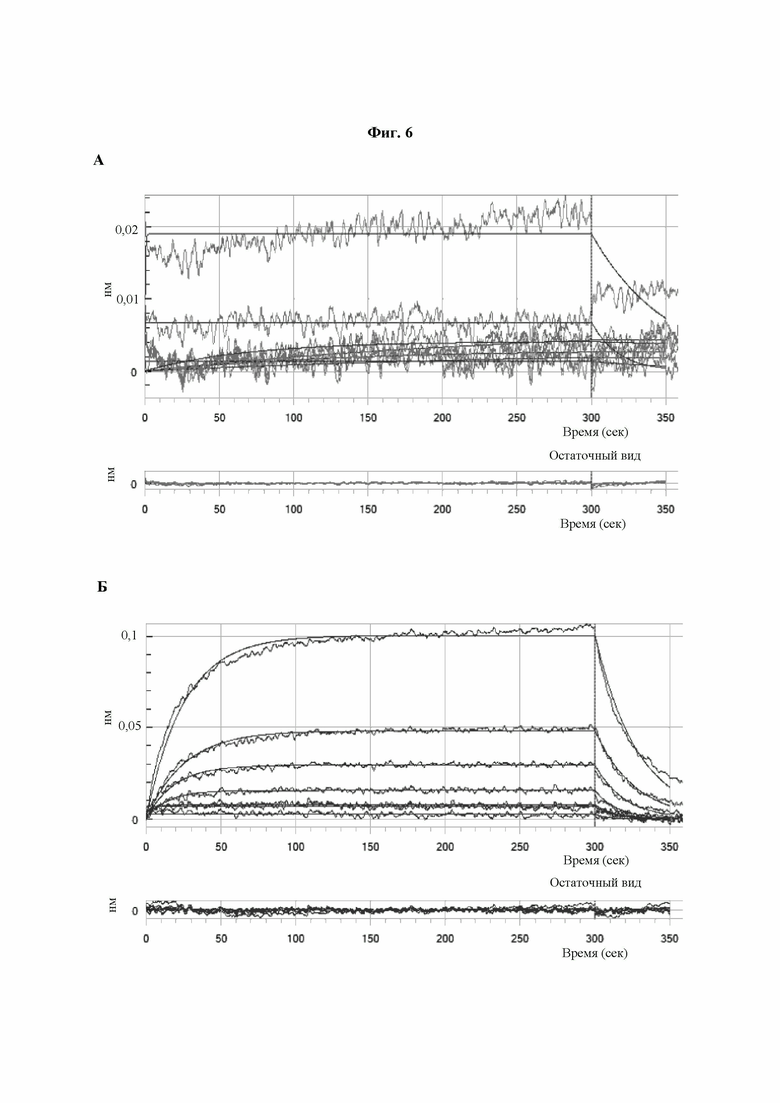

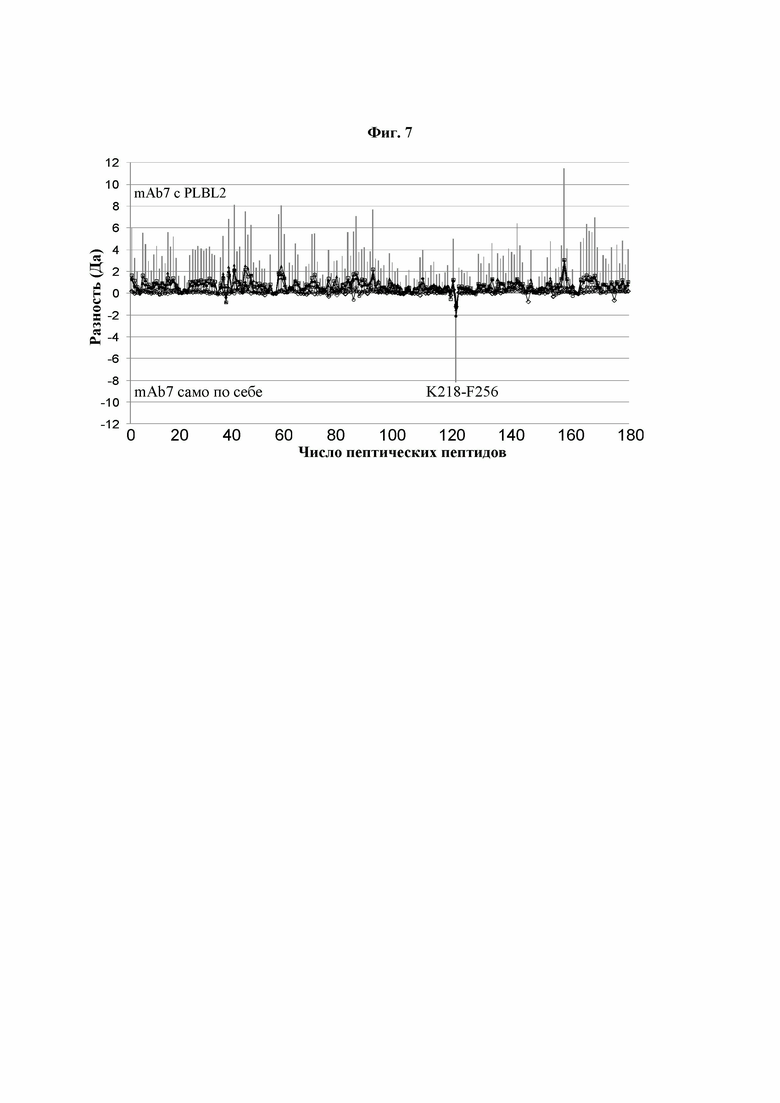


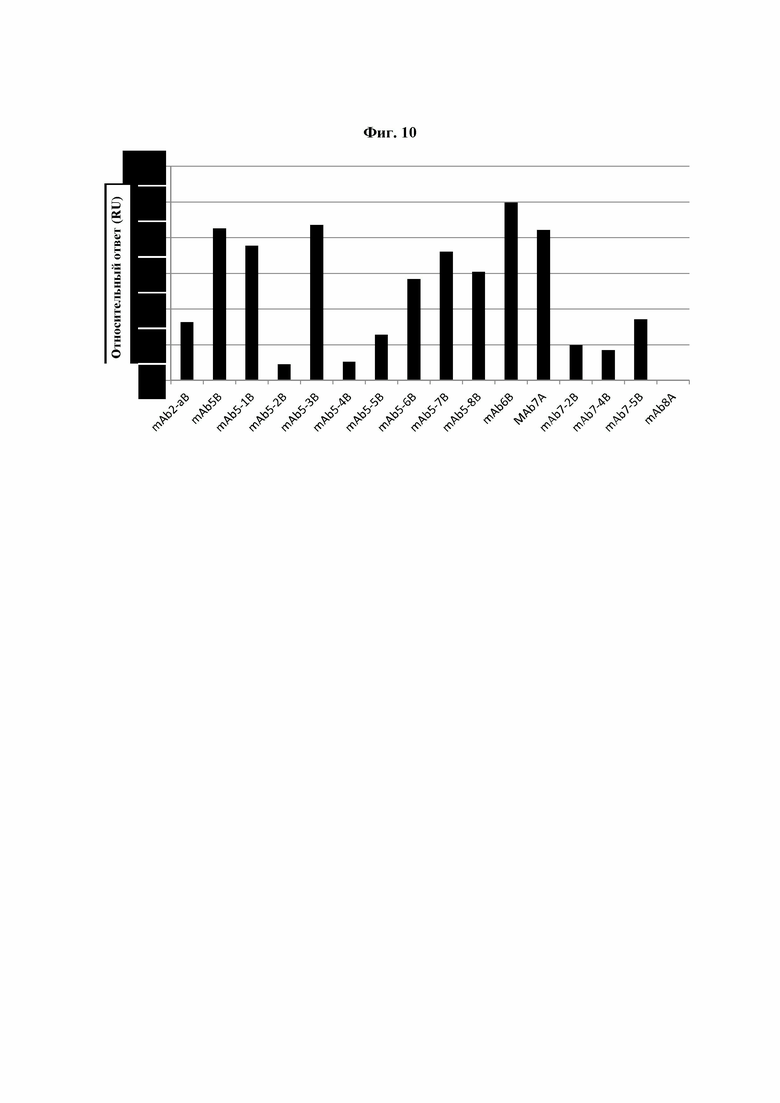
Изобретение относится к области биотехнологии, конкретно к вариантам антител со сниженным уровнем связывания с технологическими примесями. Полученные варианты антител IgG4, модифицированные в константной области тяжелой цепи, включают замену R222 на T или K и/или замену YGPP, как определено в SEQ ID NO: 23, по остаткам 229, 230, 237 и 238 по нумерации Kabat на CCVE, как определено в SEQ ID NO: 25, имеют сниженный уровень связывания с PLBL2 и могут быть использованы для терапевтического, профилактического или превентивного лечения. 5 н. и 16 з.п. ф-лы, 10 ил., 11 табл., 13 пр.
1. Вариант антитела IgG4 для терапевтического, профилактического или превентивного лечения, модифицированный в константной области тяжелой цепи, имеющий сниженный уровень связывания с белком клетки-хозяина (HCP) по сравнению с немодифицированным антителом IgG4, где белок клетки-хозяина представляет собой предполагаемый белок 2, подобный фосфолипазе В (PLBL2), и где модификация включает замену R222 на T или K и/или замену YGPP, как определено в SEQ ID NO: 23, по остаткам 229, 230, 237 и 238 по нумерации Kabat на CCVE, как определено в SEQ ID NO: 25.
2. Вариант антитела IgG4 по п. 1, где модификация дополнительно включает:
(а) модификацию остатка 203 по нумерации Kabat; и/или
(б) замену одной или более чем одной аминокислоты, включающую замену K203 на R, E или Q; E226 на L или I; S227 на R, P, A, N или T; Y229 на S, F, W или H; G230 на C, A, N или S; P237 на H, E, D или V; и/или P238 на T, K или E; и/или
(в) замену ESKYGPP, как определено в SEQ ID NO: 26, по остаткам 226-230, 237 и 238 по нумерации Kabat на EPKSCDKTHT, как определено в SEQ ID NO: 27, или ERKYGPP, как определено в SEQ ID NO: 28, или ERKCCVE, как определено в SEQ ID NO: 29, или ELKTPLGDTTHT, как определено в SEQ ID NO: 30; и/или
г) замену YGPP, как определено в SEQ ID NO: 23, по остаткам 229, 230, 237 и 238 по нумерации Kabat на SCDKTHT, как определено в SEQ ID NO: 24.
3. Вариант антитела IgG4 по п. 1 или 2, где модификация включает замену R222 на K.
4. Вариант антитела IgG4 по любому из пп. 1-3, где модификация включает замену YGPP, как определено в SEQ ID NO: 23, по остаткам 229, 230, 237 и 238 по нумерации Kabat на CCVE, как определено в SEQ ID NO: 25.
5. Вариант антитела IgG4 по любому из пп. 1-4, имеющий сниженную аффинность связывания и/или активность связывания с HCP по сравнению с немодифицированным антителом IgG4.
6. Вариант антитела IgG4 по любому из пп. 1-5, содержащий дополнительную замену S241 на P, и/или L248 на E, и/или замену EFLGGP, как определено в SEQ ID NO: 31, по остаткам 246-251 по нумерации Kabat на PAAAP, как определено в SEQ ID NO: 33, или PAAAS, как определено в SEQ ID NO: 32.
7. Вариант антитела IgG4 по любому из пп. 1-6, содержащий последовательность CPPC, как определено в SEQ ID NO: 20, по остаткам 239-242 по нумерации Kabat.
8. Вариант антитела IgG4 по любому из пп. 1-7, где константная область тяжелой цепи не содержит дополнительных модификаций по сравнению с немодифицированным антителом IgG4.
9. Экспрессирующая клетка-хозяин, содержащая нуклеотидную конструкцию, кодирующую вариант антитела IgG4 по любому из пп. 1-8.
10. Способ модификации антитела IgG4 для снижения связывания с технологической примесью, где технологическая примесь представляет собой белок клетки-хозяина, включающий стадии:
а) определения по меньшей мере одной аминокислоты, вовлеченной в связывание с технологической примесью, где аминокислота, определенная как вовлеченная в связывание с технологической примесью, присутствует в константной области тяжелой цепи; и
б) получения варианта антитела IgG4 путем замены указанной по меньшей мере одной аминокислоты, определенной как вовлеченная в связывание с технологической примесью, на другую аминокислоту.
11. Способ по п. 10, где аминокислоту, вовлеченную в связывание с технологической примесью, определяют с использованием метода или комбинации методов изучения белок-белковых взаимодействий.
12. Способ по п. 11, где метод изучения белок-белковых взаимодействий представляет собой масс-спектрометрию водородно-дейтериевого обмена (HDX-MS).
13. Способ по любому из пп. 10-12, где по меньшей мере одна аминокислота, определенная как вовлеченная в связывание с технологической примесью, заменена на аминокислотный остаток, находящийся в соответствующем положении в последовательности антител IgG1, IgG2 и/или IgG3 эмбрионального типа.
14. Способ по любому из пп. 10-13, где по меньшей мере одна аминокислота, определенная как вовлеченная в связывание с технологической примесью, представляет собой одну или более из следующих аминокислотных остатков: лизин в положении 203 (K203), аргинин в положении 222 (R222), глутамат в положении 226 (E226), серин в положении 227 (S227), тирозин в положении 229 (Y229), глицин в положении 230 (G230), пролин в положении (P237), пролин в положении 238 (P238), ESKYGPP, как определено в SEQ ID NO: 26, по остаткам 226-230, 237 и 238 по нумерации Kabat и YGPP, как определено в SEQ ID NO: 23, по остаткам 229, 230, 237 и 238 по нумерации Kabat.
15. Способ по любому из пп. 10-14, где белок клетки-хозяина представляет собой PLBL2.
16. Способ получения антитела IgG4 со сниженным связыванием с белком клетки-хозяина (HCP), включающий модификацию последовательности антитела в константной области тяжелой цепи, где белок клетки-хозяина представляет собой PLBL2, и где модификация включает замену R222 на T или K и/или замену YGPP, как определено в SEQ ID NO: 23, по остаткам 229, 230, 237 и 238 по нумерации Kabat на SCDKTHT, как определено в SEQ ID NO: 24, или CCVE, как определено в SEQ ID NO: 25.
17. Способ по п. 16, где модификация дополнительно включает:
(а) модификацию остатка 203 по нумерации Kabat; и/или
(б) замену одной или более чем одной аминокислоты, включающую замену K203 на R, E или Q; E226 на L или I; S227 на R, P, A, N или T; Y229 на S, F, W или H; G230 на C, A, N или S; P237 на H, E, D или V; и/или P238 на T, K или E; и/или
(в) замену ESKYGPP, как определено в SEQ ID NO: 26, по остаткам 226-230, 237 и 238 по нумерации Kabat на EPKSCDKTHT, как определено в SEQ ID NO:27, или ERKYGPP, как определено в SEQ ID NO: 28, или ERKCCVE, как определено в SEQ ID NO: 29, или ELKTPLGDTTHT, как определено в SEQ ID NO: 30.
18. Фармацевтическая композиция, содержащая эффективное количество варианта антитела IgG4 по любому из пп. 1-8 и фармацевтически приемлемый носитель или разбавитель.
19. Композиция по п. 18, где концентрация варианта антитела IgG4 составляет по меньшей мере 10 мг/мл, по меньшей мере 20 мг/мл, по меньшей мере 50 мг/мл, по меньшей мере 75 мг/мл или по меньшей мере 100 мг/мл.
20. Композиция по п. 18 или 19, где концентрация PLBL2 в композиции составляет менее 500 млн-1, менее 100 млн-1 или менее 10 млн-1.
21. Композиция по любому из пп. 18-20, дополнительно содержащая буфер и/или сложный эфир жирной кислоты; где сложный эфир жирной кислоты возможно представляет собой полисорбат 20 или полисорбат 80.
| PETERS SJ et al., Engineering an Improved IgG4 Molecule with Reduced Disulfide Bond Heterogeneity and Increased Fab Domain Thermal Stability, THE JOURNAL OF BIOLOGICAL CHEMISTRY, 2012, V | |||
| СПОСОБ ИЗГОТОВЛЕНИЯ ЧЕРТЕЖЕЙ ДЛЯ ОДНООБРАЗНОЙ РАСКРОЙКИ ПРЕДМЕТОВ ОДЕЖДЫ | 1919 |
|
SU287A1 |
| Солесос | 1922 |
|
SU29A1 |
| WO 2011066501 A1, 03.06.2011 | |||
| SCAPIN G et al., Structure of full-length human anti-PD1 therapeutic IgG4 antibody pembrolizumab, | |||

