Далее в настоящем документе раскрывают активирующие антитела против BTN3A, которые специфически связываются с BTN3A и активируют цитолитическую функцию Vγ9/Vδ2 Т-клеток. Такие антитела полезны, в особенности, при лечении раковых заболеваний, таких как рак крови или солидные опухоли. Описание более конкретно относится к специфическим гуманизированным активирующим антителам против BTN3A с эквивалентными или улучшенными свойствами по сравнению с соответствующими родительскими мышиными антителами 7.2 или их химерными версиями с Fc-молчащими константными областями человеческого IgG1 или IgG4.
Уровень техники
Лейкоциты представляют собой клетки иммунной системы, участвующие в защите организма от патогенов. Среди этих клеток можно назвать лимфоциты, моноциты и дендритные клетки. Моноциты могут мигрировать из кровотока в другие ткани и дифференцироваться в тканевые макрофаги или дендритные клетки. Дендритные клетки играют роль антигенпрезентирующих клеток (APC), которые активируют лимфоциты. Среди лимфоцитов Т-клетки можно разделить на αβ Т-клетки и γδ Т-клетки. Vγ9-Vδ2 представляют собой подмножество γδ Т-клеток, которые служат важными эффекторами системы иммунной защиты. Они непосредственно лизируют инфицированные патогеном или аномальные клетки. Кроме того, они регулируют иммунные ответы, вызывая созревание дендритных клеток (DC), а также переключение изотипа и продукцию иммуноглобулинов. Эта важная клеточная подгруппа иммунной системы строго регулируется поверхностными рецепторами, хемокинами и цитокинами.
Примирование Т-клеток регулируется путем вовлечения специализированных клеток и секрецией хемотаксических цитокинов. Согласно гипотезе двух сигналов активация Т-клеток представляет собой результат двух синергических событий. Первое событие представляет собой взаимодействие между рецепторами Т-клеток (TCR) и основным комплексом гистосовместимости (MHC) в комплексе с процессированным антигеном на поверхности антигенпрезентирующих клеток (APC). Второе событие представляет собой ко-стимулирующий антиген-независимый сигнал с участием молекул CD28 и B7. Отсутствие ко-стимулирующих сигналов приводит к анергии и нечувствительности, в результате чего, не осуществляется пролиферация Т-клеток, секреция цитокинов и цитотоксическая активность. Изучение этих путей дает представление о запуске патологических событий, таких как аутоиммунные или лимфопролиферативные расстройства. Семейство B7 представляет собой расширенную группу ко-стимулирующих молекул (Coyle, Gutierrez-Ramos, 2001; Sharpe and Freeman, 2002). К семейству B7 принадлежат лиганды B7-1 (CD80) и B7-2 (CD86): их рецепторами служат CD28, что приводит к активации Т-клеток и CTLA-4 (CD152), который конкурирует с CD28 и передает ингибирующий сигнал (рассмотрено в Alegre et al., 2001). Критическую роль CD152 как негативного регулятора активации Т-клеток можно видеть по возникновению лимфопролиферативных нарушений у мышей с дефицитом CTLA-4. Важные данные об ингибирующей функции CD152 были получены в результате исследований пролиферации или продукции цитокинов наивными Т-лимфоцитами во время прайминга Т-клеток. В особенности, CD152 экспрессируется после активации Т-лимфоцитов, и он ингибирует цитолитические функции клонов CTL, полученных после стимуляции PHA или отбора Ag. Доказано, что B7-H1 (PD-L1, CD274) и B7-DC (PD-L2, CD273), рецептором которых представляет собой PD-1 (CD279), ингибируют пролиферацию Т-клеток и секрецию цитокинов (описано в Sharpe and Pauken, 2018). С другой стороны, различные исследования показали, что включение PD-L1 и PD-L2 увеличивает пролиферацию Т-клеток и продукцию IL-10 или IFN-γ. Другие молекулы, относящиеся к семейству B7, экспрессируемые на поверхности Т-клеток, включая B7-H2 (ICOS-L), недавно идентифицированные B7-H3, B7-H4, B7-H5, B7-H6 и B7-H7, также участвуют в качестве регуляторов контрольных точек иммунной функции (рассмотрено в Ni and Dong, 2017).
Henry et al. (1999) обнаружили, что область, кодирующая бутирофилин (BT), расположена в теломерной позиции от области MHC класса I на хромосоме 6 человека. В особенности, они описали два гена Bt2 и Bt3, кодирующие новую группу ко-стимулирующих молекул (BT2.1, BT2.2, BT2.3, BT3.1, BT3.2 и BT3.3), принадлежащих к суперсемейству Ig (IgSF) (Linsley et al., 1992; Williams and Barclay, 1988), и относящихся к семейству B7, согласно анализу сходства последовательностей: в особенности, они показывают сходство с Ig-V-подобными внеклеточными доменами CD80 и CD86.
Члены семейства BT3 встречаются в литературе под разными названиями: BT3.1 также называют BTF5 (Ruddy et al., 1997) или BTN3A1 (Rhodes et al., 2001), или, совсем недавно, CD277 (Bensussan and Olive, 2005); BT3.2 также называют BTF4 (Ruddy et al., 1997) или BTN3A2; и, наконец, BT3.3 встречается также как BTF3 (Ruddy et al., 1997) или BTN3A3 (Rhodes et al., 2001). BT3 имеет два Ig-подобных внеклеточных домена, которые характеризуют IgSF.
Было высказано предположение, что гены B7 и гены MHC классов I и II могут иметь общий предковый ген и кодировать белки, вовлеченные в осуществление сходной функции, такой как активация Т-клеток (Rhodes et al., 2001). Молекулы BT3 были обнаружены на иммунных клетках, таких как T, B и NK-клетки, моноциты и дендритные клетки, а также на гемопоэтических предшественниках и в некоторых линиях неопластических клеток. Что касается других ко-стимулирующих молекул, то их структура характеризуется тремя доменами: внеклеточным доменом для связывания лиганда, трансмембранным доменом и внутриклеточным доменом, называемым B30.2, который предположительно участвует в регуляции внутриклеточных концентраций супероксида. Пока что лиганд (лиганды) для CD277 все еще неизвестен (неизвестны) (смотри обзор Gu et al., 2015).
На сегодняшний день были предложены различные терапевтические и вакцинные стратегии, основанные на модуляции Т-клеток; несколько иммуномодулирующих антител к CTLA-4, PD-1 и PD-L1 уже одобрены для клинического применения множеством регулирующих органов по всему миру. Хотя эти препараты представляют собой значительный прогресс в лечении рака, все еще остаются неудовлетворенные медицинские потребности значительной части популяций больных раком, которые не реагируют на доступные в настоящее время способы лечения.
Патенты WO 2012080351 A1, EP2651441A1, EP2946791A1, US20140322235, WO2012080769A1 относятся к различным антителам против BTN3A, способным активировать или ингибировать цитолитическую функцию, продукцию цитокинов и пролиферацию Vγ9 Vδ2 Т-клеток. Однако эти мышиные антитела не подходили для терапевтического применения. Действительно, для введения пациентам-людям в настоящее время необходимо гуманизировать антитела, чтобы избежать иммуногенных реакций.
Гуманизация часто требует модификации аминокислот в каркасных областях, и нет уверенности в том, сохранятся ли активности гуманизированных антител на том же уровне, что и у исходных мышиных антител. Это особенно актуально при модификации аминокислот, непосредственно примыкающих к участкам CDR (смотри, например, Queen патент США №5,585,089).
Несмотря на сложность, авторы настоящего изобретения выбрали специальные гуманизированные антитела активирующих mAb 7.2, которые не только сочетают сохраненные функциональные свойства родительских антител mAb 7.2 с предсказанным снижением иммуногенности для человека, но также неожиданно демонстрируют превосходные свойства, обеспечивающие возможность разработки, такие как улучшенный выход при продуцировании клеточной линии, более высокая термическая стабильность по сравнению с исходным мышиным антителом и высокая устойчивость к кислотному и тепловому стрессу. Кроме того, гуманизированное антитело mAb1 по настоящему изобретению преимущественно связывается с BTN3A яванского макака и хорошо переносится приматом, яванским макаком, в дозах до 100 мг/кг/неделю, тем самым представляя собой отличного кандидата для применения в качестве лекарственного средства в терапии человека.
Раскрытие изобретения
Таким образом, настоящее раскрытие относится к изолированному антителу против BTN3A, включающему полипептид VH вариабельной области тяжелой цепи SEQ ID NO: 1 и полипептид VL вариабельной области легкой цепи SEQ ID NO: 2 или SEQ ID NO: 3. Такие антитела представляют собой гуманизированные антитела, в частности, с прогнозируемым снижением иммуногенности по отношению к их родительскому мышиному антителу. Такое изолированное антитело против BTN3A связывается с человеческим BTN3A. В частности, оно связывается с BTN3A человека с KD 10 нМ или менее, предпочтительно, с KD 5 нМ или менее, как измерено с помощью поверхностного плазмонного резонанса.
В конкретных воплощениях указанное антитело согласно настоящему раскрытию индуцирует активацию γδ Т-клеток, обычно T-клеток Vγ9Vδ2, в совместной культуре с клетками, экспрессирующими BTN3, с EC50 ниже 5 мкг/мл, предпочтительно, 1 мкг/мл или ниже, как измерено в анализе дегрануляции. Следовательно, указанное антитело индуцирует уничтожение опухолевых клеток-мишеней независимо от их тканевого происхождения.
В конкретных воплощениях, указанное изолированное антитело против BTN3A включает мутантную или химически модифицированную константную область IgG1, где указанная мутантная или химически модифицированная константная область IgG1 не обеспечивает связывания с рецепторами Fcγ или снижает связывание с ними по сравнению с соответствующим антителом с константной областью изотипа IgG1 дикого типа. Обычно указанный мутантный константный участок IgG1 представляет собой тройной мутант IgG1 L247F, L248E и P350S. Примеры указанного выделенного антитела против BTN3A представляют собой mAb1, включающее тяжелую цепь SEQ ID NO: 4 и легкую цепь SEQ ID NO: 6, или mAb2, включающее тяжелую цепь SEQ ID NO: 4 и легкую цепь SEQ ID. NO: 7.
В других воплощениях, указанное изолированное антитело против BTN3A по настоящему изобретению представляет собой антитело моновалентного формата, предпочтительно, выбранное из антител Fab или scFv.
Изолированное антитело против BTN3A по настоящему изобретению можно применять (i) в качестве терапевтического средства или (ii) в качестве диагностического средства. Например, они полезны при лечении рака, например, гематологического рака, а более конкретно, лимфомы или лейкемии. В других воплощениях они также могут быть применены при лечении солидных опухолей, в частности, рака простаты, яичников или эндометрия. В качестве альтернативы их можно применять при лечении инфекционных заболеваний.
Изобретение, кроме того, относится к фармацевтической композиции, включающей антитело против BTN3A, как описано выше, в комбинации с одним или несколькими фармацевтически приемлемыми вспомогательными веществами, разбавителями или носителями, необязательно включающими другие активные ингредиенты.
Другой аспект настоящего раскрытия касается состава лиофилизата, предварительно заполненного шприца или флакона, содержащего антитело против BTN3A, как описано выше.
Изобретение также относится к вектору экспрессии для рекомбинантного продуцирования антитела против BTN3A, как описано выше, в клетке-хозяине, обычно в клетке-хозяине млекопитающего, такой как клетка-хозяин СНО, включающему по меньшей мере одну нуклеиновую кислоту, кодирующую указанное антитело против BTN3A. Воплощение такого вектора экспрессии включает, по меньшей мере, нуклеиновые кислоты, кодирующие тяжелую и легкую цепи mAb1, как описано в настоящем документе. В настоящем документе также описаны клетки-хозяева, включающие такие векторы экспрессии.
В настоящем документе также раскрывают способ получения антитела против BTN3A по настоящему изобретению, включающий: (i) культивирование клетки-хозяина, как определено выше, для экспрессии указанного антитела клеткой-хозяином; необязательно, (ii) очистку указанного антитела; (iii) извлечение антитела.
Кроме того, раскрытие относится к мультиспецифическим антителам, таким как биспецифические антитела, включающим по меньшей мере одно плечо, включающее Fab или scFv, содержащим VH и VL антител против BTN3A, как определено выше.
Осуществление изобретения
Определения
Для облегчения понимания настоящего раскрытия сначала даны определения некоторых терминов. Дополнительные определения приведены в разделе «Осуществление изобретения».
Термин «BTN3A», как он используется в настоящем документа, имеет общее значение в данной области техники. В конкретных воплощениях он относится к полипептидам BTN3A человека, включая BTN3A1 по SEQ ID NO: 18, BTN3A2 по SEQ ID NO: 19 или BTN3A3 по SEQ ID NO: 20.
Термин «антитело», как он используется в настоящем документе, относится к молекулам иммуноглобулинов и иммунологически активным частям молекул иммуноглобулинов, то есть к молекулам, которые включают сайт связывания антигена, который иммуноспецифически связывает антиген. Таким образом, термин «антитело» включает не только целые молекулы антитела, но также фрагменты антител, а также варианты (включая производные) антител.
В природных антителах грызунов и приматов две тяжелые цепи связаны друг с другом дисульфидными связями, и каждая тяжелая цепь связана с легкой цепью дисульфидной связью. Есть два типа легких цепей: лямбда (λ) и каппа (κ). Существует пять основных классов тяжелых цепей (или изотипов), которые определяют функциональную активность молекулы антитела: IgM, IgD, IgG, IgA и IgE. Каждая цепочка включает отдельные домены последовательности. В типичных антителах IgG легкая цепь включает два домена, вариабельный домен (VL) и константный домен (CL). Тяжелая цепь включает четыре домена, вариабельный домен (VH) и три константных домена (CH1, CH2 и CH3, вместе называемые CH). Вариабельные области как легкой (VL), так и тяжелой (VH) цепей определяют распознавание связывания и специфичность по отношению к антигену. Домены константной области легкой (CL) и тяжелой (CH) цепей придают важные биологические свойства, такие как ассоциация цепей антител, секреция, трансплацентарная подвижность, связывание комплемента и связывание с рецепторами Fc (FcR).
Фрагмент Fv представляет собой N-концевую часть Fab-фрагмента иммуноглобулина и состоит из вариабельных частей одной легкой цепи и одной тяжелой цепи. Специфичность антитела заключается в структурной комплементарности между сайтом объединения антитела и антигенной детерминантой. Соединяющие сайты антител состоят из остатков, которые в основном относятся к гипервариабельным или определяющим комплементарность участкам (CDR). Иногда остатки из негипервариабельных или каркасных областей (FR) могут участвовать в сайте связывания антитела или влиять на общую структуру домена и, следовательно, на соединяющие сайты. Определяющие комплементарность участки или CDR относятся к аминокислотным последовательностям, которые вместе определяют аффинность связывания и специфичность природной области Fv сайта связывания природного иммуноглобулина. Каждая легкая и тяжелая цепи иммуноглобулина имеет три CDR, обозначаемые как L-CDR1, L-CDR2, L-CDR3 и H-CDR1, H-CDR2, H-CDR3, соответственно. Следовательно, антигенсвязывающий сайт обычно включает шесть CDR, содержащих CDR из каждой V-области тяжелой и легкой цепи. Каркасные области (FR) относятся к аминокислотным последовательностям, расположенным между CDR. Соответственно, вариабельные области легкой и тяжелой цепей обычно включают 4 каркасных области и 3 CDR со следующей последовательностью: FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4.
Остатки в вариабельных доменах антител условно нумеруют в соответствии с системой, разработанной Kabat et al. Эта система представлена в работе Kabat et al., 1987, в Sequences of Proteins of Immunological Interest, US Department of Health and Human Services, NIH, USA (Kabat et al., 1992, далее в настоящем документе «Kabat et al.»). Эту систему нумерации применяют в настоящем описании. Обозначения остатков по Кабату не всегда прямо соответствуют линейной нумерации аминокислотных остатков в последовательностях SEQ ID. Фактическая линейная аминокислотная последовательность может включать меньше или больше аминокислот, чем в строгой нумерации Кабата, соответствуя укорочению или встраиванию в структурный компонент, будь то каркас или определяющая комплементарность область (CDR), основной структуры вариабельного домена. Правильная нумерация остатков по Кабату может быть определена для данного антитела путем выравнивания остатков по гомологии в последовательности антитела со «стандартной» пронумерованной последовательностью по Кабату. CDR вариабельного домена тяжелой цепи расположены в остатках 31-35 (H-CDR1), остатках 50-65 (H-CDR2) и остатках 95-102 (H-CDR3) в соответствии с системой нумерации Кабата. CDR вариабельного домена легкой цепи расположены в остатках 24-34 (L-CDR1), остатках 50-56 (L-CDR2) и остатках 89-97 (L-CDR3) в соответствии с системой нумерации Кабата.
В конкретных воплощениях, обеспеченное в настоящем документе антитело, представляет собой фрагмент антитела и, более конкретно, любой белок, включающий антигенсвязывающий домен антитела, описанного в настоящем документе. Фрагменты антител включают, но не ограничиваются ими, Fv, Fab, F(ab')2, Fab', dsFv, scFv, sc(Fv)2 и диатела.
Термин «специфичность», как он используется в настоящем документе, относится к способности антитела детектируемо связывать эпитоп, представленный на антигене, таком как BTN3A. В некоторых воплощениях он относится к антителу, которое связывается с BTN3A человека, экспрессируемым на клетках костного мозга периферической крови (PBMC), предпочтительно с EC50 ниже 50 мкг/мл, и, более предпочтительно, ниже 10 мкг/мл, как определено в разделе «Примеры» (смотри таблицу 4). В других воплощениях оно связывается с рекомбинантным полипептидом антигена с KD 100 нМ или меньше, 10 нМ или меньше, 1 нМ или меньше, 100 пМ или меньше или 10 пМ или меньше, как установлено с помощью SPR-измерений, как определено в примерах (смотри таблицу 4).
Антитело, которое «перекрестно реагирует с антигеном, отличным от BTN3A», предназначено для обозначения антитела, которое связывает этот антиген, отличный от BTN3A, с KD 10 нМ или меньше, 1 нМ или меньше или 100 пМ или меньше. Антитело, которое «не реагирует перекрестно с конкретным антигеном», предназначено для обозначения антитела, которое связывается с этим антигеном, с KD 100 нМ или больше, или KD 1 мкМ или больше, или KD 10 мкм или больше. В некоторых воплощениях такие антитела, которые не реагируют перекрестно с антигеном, демонстрируют практически неопределяемое связывание с этими белками в стандартных анализах связывания. В конкретном воплощении, гуманизированное антитело согласно настоящему раскрытию, например, mAb1, перекрестно реагирует с BTN3A1, BTN3A2 и BTN3A3 яванского макака с последовательностями SEQ ID NO: 21, SEQ ID NO: 22 и SEQ ID NO: 23, соответственно, например, как измерено в анализе Biacore (см. таблицу 21).
Термин «изолированное антитело», как он используется в настоящем документе, относится к антителу, которое по существу не включает других антител, обладающих другой антигенной специфичностью (например, изолированное антитело, которое специфически связывается с BTN3A, по существу не включает антител, которые специфически связываются с другими антигенами, кроме BTN3A). Однако изолированное антитело, которое специфически связывается с BTN3A, может обладать перекрестной реактивностью с другими антигенами, такими как родственные молекулы BTN3A других видов. Более того, изолированное антитело может практически не включать другого клеточного материала и/или химических веществ.
Термин «моноклональное антитело» или «композиция моноклонального антитела», как он используется в настоящем документе, относится к препарату молекул антитела с одним молекулярным составом. Композиция моноклонального антитела демонстрирует единственную специфичность связывания и сродство к конкретному эпитопу.
Фразы «антитело, распознающее антиген» и «антитело, обладающее специфичностью к антигену» применяют в настоящем документе взаимозаменяемо с термином «антитело, которое специфически связывается с антигеном».
Термин «Kассоц» или «Ka», как он используется в настоящем документе, предназначен для обозначения скорости ассоциации конкретного взаимодействия антитело-антиген, тогда как термин «Kдис» или «Kd,» как он используется в настоящем документе, предназначен для обозначения скорости диссоциации конкретного взаимодействия антитело-антиген.
Термин «KD», как он используется в настоящем документе, предназначен для обозначения константы диссоциации, которую получают из отношения Kd к Ka (т.е. Kd/Ka) и выражают как молярная концентрация (M). Значения KD для антител могут быть определены с применением хорошо известных в данной области техники способов. Способ определения KD антитела заключается в применении поверхностного плазмонного резонанса или в применении биосенсорной системы, такой как система Biacore®.
Специфичность может дополнительно проявляться, например, соотношением аффинности/авидности связывания со специфическим антигеном, равным примерно 10:1, примерно 20:1, примерно 50:1, примерно 100:1, 10 000:1 или более, по сравнению с неспецифическим связыванием с другими нерелевантными молекулами (в этом случае специфический антиген представляет собой полипептид BTN3A). Термин «сродство», как он используется в настоящем документе, означает силу связывания антитела с эпитопом.
Термин «авидность», как он используется в настоящем документе, относится к информативному показателю общей стабильности или прочности комплекса антитело-антиген. Она контролируется тремя основными факторами: аффинностью эпитопа антител; валентностью, как антигена, так и антитела; и структурным расположением взаимодействующих частей. В конечном итоге, эти факторы определяют специфичность антитела, то есть вероятность того, что конкретное антитело связывается с конкретным эпитопом антигена.
Как он используется в настоящем документе, термин «активирующее антитело» относится к антителу, способному прямо или косвенно индуцировать иммунные функции эффекторных клеток. В частности, как понимается в соответствии с настоящим документом, активирующее антитело против BTN3A обладает, по меньшей мере, способностью индуцировать активацию γδ Т-клеток, обычно, T-клеток Vγ9Vδ2, при совместном культивировании с клетками, экспрессирующими BTN3, с ЕС50 ниже 5 мкг/мл, предпочтительно, 1 мкг/мл или ниже, как измерено в анализе дегрануляции, как описано ниже в разделе «Примеры».
Термин «субъект», как он используется в настоящем документе, включает любое животное, относящееся или не относящееся к человеку. Термин «не относящееся к человеку животное» включает всех позвоночных, например, млекопитающих и не относящихся к млекопитающим животных, таких как не относящиеся к человеку приматы, овцы, собаки, кошки, лошади, коровы, куры, амфибии, рептилии и т.д.
Термин «оптимизированный», как он используется в настоящем документе, означает, что нуклеотидная последовательность была изменена для кодирования аминокислотной последовательности с применением кодонов, которые предпочтительны в продуцирующей клетке или организме, обычно в эукариотической клетке, например, в клетке яичника китайского хомячка (СНО) или в человеческой клетка. Оптимизированная нуклеотидная последовательность была сконструирована так, чтобы сохранить полностью или в максимально возможной степени аминокислотную последовательность, изначально кодируемую исходной нуклеотидной последовательностью. Аминокислотные последовательности, кодируемые оптимизированными нуклеотидными последовательностями, также называются оптимизированными.
Процент идентичности между двумя последовательностями, как он используется в настоящем документе, представляет собой функцию количества идентичных позиций, разделяемых последовательностями (т.е. % идентичности = количество идентичных позиций/общее количество позиций x 100), с учетом количества пропусков и длины каждого пропуска, который необходимо ввести для оптимального выравнивания двух последовательностей. Сравнение последовательностей и определение процента идентичности между двумя последовательностями может быть выполнено с применением математического алгоритма, как описано ниже.
Процент идентичности между двумя аминокислотными последовательностями можно определить с помощью алгоритма E. Meyers и W. Miller (Comput. Appl. Biosci., 4: 11-17, 1988), который был включен в программу ALIGN (версия 2.0), с применением таблицы весов остатков PAM120, штрафа за длину пропуска 12 и штраф за пропуск, равный 4. Альтернативно, процент идентичности между двумя аминокислотными последовательностями можно определить с помощью алгоритма Needleman и Wunsch (J. Mol, Biol. 48: 444- 453, 1970), который был включен в программу GAP в пакете программного обеспечения GCG (доступен на http://www.gcg.com), с применением либо матрицы Blossom 62, либо матрицы PAM250, и штрафом за открытие порпуска, равным 16, 14, 12, 10, 8, 6 или 4, и штрафом за удлинение пропуска, равным 1, 2, 3, 4, 5 или 6.
Процент идентичности между двумя нуклеотидными аминокислотными последовательностями также может быть определен с помощью, например, таких алгоритмов, как программа BLASTN для последовательностей нуклеиновых кислот, с применением по умолчанию длины слова (W), равного 11, ожидания (E), равного 10, M = 5, N = 4, и сравнения обеих нитей.
Рекомбинантные гуманизированные активирующие антитела против BTN3A
Антитела по настоящему описанию включают выбранные гуманизированные рекомбинантные антитела mAb1, mAb2, mAb4 и mAb5, которые структурно характеризуются вариабельными аминокислотными последовательностями их тяжелой и легкой цепей и человеческими константными областями (изотипам), как описано ниже в таблице 1:
Таблица 1: Вариабельные аминокислотные последовательности тяжелых и легких цепей mAb1-mAb6
VL
(VH2 7.2)
(Vk1 7.2)
L247F/L248E/P350S
(VH2 7.2)
(Vk2 7.2)
L247F/L248E/P350S
L247F/L248E/P350S
(VH2 7.2)
(Vk1 7.2)
(VH2 7.2)
(Vk2 7.2)
mAb3 и mAb6 представляют собой гуманизированные антитела другого родительского мышиного антитела против BTN3A, называемого mAb 20.1 и описанного в WO2012/080351, для применения в качестве сравнительных примеров.
Соответствующие аминокислотные и нуклеотидные кодирующие последовательности константных изотипических областей IgG1, IgG4 и их мутантных версий IgG1 L247F/L248E/P350S и IgG4 S241P/L248E, применяемые для генерации антител от mAb1 до mAb6, хорошо известны в области данной техники (Oganesyan et al., 2008; Reddy et al., 2000). С-концевой лизин, обнаруженный в IgG, может отщепляться естественным путем, и эта модификация не влияет на свойства антитела; таким образом, этот остаток может быть дополнительно удален в конструкциях от mAb1 до mAb6.
Полноразмерные легкие и тяжелые цепи и соответствующие кодирующие последовательности mAb1, mAb2, mAb4 и mAb 5 приведены ниже в таблице 2.
Таблица 2: Кодирующие последовательности ДНК тяжелой и легкой цепи полной длины
Легкая цепь: SEQ ID NO:6
Легкая цепь: SEQ ID NO:10
Легкая цепь: SEQ ID NO:7
Легкая цепь: SEQ ID NO:11
Легкая цепь: SEQ ID NO:6
Легкая цепь: SEQ ID NO:10
Легкая цепь: SEQ ID NO:7
Легкая цепь: SEQ ID NO:11
Примеры аминокислотных последовательностей VH CDR1 (также называемых HCDR1), VH CDR2 (также называемых HCDR2), VH CDR3 (также называемых HCDR1), VL CDR1 (также называемых LCDR1), VL CDR2 (также называемых LCDR2), VL CDR3 (также называемые HCDR3) некоторых антител в соответствии с изобретением приведены в таблице 3.
В таблице 3 области CDR антител согласно настоящему раскрытию изображены с применением нумерации Кабата (Kabat et al., 1992, далее в настоящем документе «Kabat et al.»).
Для удобства чтения области CDR далее в настоящем документе называют HCDR1, HCDR2, HCDR3, LCDR1, LCDR2, LCDR3, соответственно.
Таблица 3: Области CDR в mAb1, mAb2, mAb4 и mAb5 и родительских мышиных антителах mAb 7.2 в соответствии с нумерацией Кабата
mAb1
mAb2
mAb4
mAb5
В конкретном воплощении указанные рекомбинантные антитела против BTN3A, как определено выше, обладают одним или несколькими из следующих свойств:
(i) оно связывается с BTN3A с KD 10 нМ или меньше, предпочтительно с KD 1 нМ или меньше, как измерено с помощью SPR, например, как описано в примерах ниже;
(ii) оно перекрестно реагирует с BTN3A яванского макака с KD 100 нМ или меньше, предпочтительно с KD 10 нМ или меньше, как измерено с помощью SPR, например, как описано в примерах ниже;
(iii) оно связывается с человеческими РВМС с ЕС50 50 мкг/мл или ниже, предпочтительно, 10 мкг/мл или ниже, как измерено в анализе проточной цитометрии, как описано в примерах ниже;
(iv) оно индуцирует активацию γδ Т-клеток, как правило, T-клеток Vγ9Vδ2, в совместной культуре с клетками, экспрессирующими BTN3, с ЕС50 ниже 5 мкг/мл, предпочтительно, 1 мкг/мл или ниже, как измерено в анализе дегрануляции, как описано ниже в разделе «Примеры».
В определенных воплощениях это воплощение может быть объединено с предыдущими воплощениями, причем антитело, как упоминается здесь, представляет собой фрагмент антитела из определенных выше антител.
Фрагменты антител включают, но не ограничиваются ими, Fab, Fab', Fab'-SH, F(ab')2, Fv, Unibody и фрагменты scFv, диатела, однодоменные или нанотела и другие фрагменты.
Предпочтительно, он представляет собой моновалентное антитело, такое как Fab из фрагментов scFv.
Термин «диатела» относятся к небольшим фрагментам антител с двумя антигенсвязывающими сайтами, которые включают вариабельный домен тяжелой цепи (VH), связанный с вариабельным доменом легкой цепи (VL) в той же полипептидной цепи (VH-VL). При применении линкера, который слишком короткий для образования пары между двумя доменами в одной цепи, домены вынуждены спариваться с комплементарными доменами другой цепи и создавать два антигенсвязывающих сайта.
Однодоменные антитела представляют собой фрагменты антител, включающие вариабельный домен тяжелой цепи полностью или частично или вариабельный домен легкой цепи антитела полностью или частично. В определенных воплощениях, однодоменное антитело представляет собой человеческое однодоменное антитело (Domantis, Inc., Waltham, MA; смотри, например, U.S. Patent No. 6,248,516 B1).
Фрагменты антител могут быть получены различными способами, включая, помимо прочего, протеолитическое расщепление интактного антитела, а также продукцию рекомбинантными клетками-хозяевами, как описано в настоящем документе.
Антитело по настоящему изобретению представляет собой гуманизированное антитело. Как правило, антитело, не относящееся к человеческому, гуманизируют для снижения иммуногенности по отношению к человеку, при этом оно имеет, по крайней мере, такое же сродство (или превосходящее сродство) по сравнению с родительским антителом, не относящимся к человеческому. В предпочтительных воплощениях антитела согласно настоящему раскрытию представляют собой гуманизированные антитела родительского антитела mAb 7.2, как описано в WO2012/080351. Сравнительные примеры включают гуманизированные антитела родительского антитела mAb 20.1, как описано в WO2012/080351.
Обычно гуманизированное антитело включает один или несколько вариабельных доменов, в которых CDR (или их части) происходят из антитела, не относящегося к человеческому, например, мышиного mAb 7.2, и FR (или их части) происходят из последовательностей мышиных антител с мутациями для снижения иммуногенности. Гуманизированное антитело необязательно также будет включать, по меньшей мере, часть константной области человека.
Предпочтительно, рекомбинантное антитело согласно настоящему описанию представляет собой гуманизированное молчащее антитело, как правило, гуманизированное молчащее антитело IgG1 или IgG4.
Термин «молчащее антитело», как он используется в настоящем документе, относится к антителу, которое не демонстрирует связывания или имеет низкое связывание с FcγR и/или с C1q, как измерено в анализах связывания, таких как описаны в разделе «Примеры».
В одном из воплощений термин «не имеет или имеет низкое связывание с FcγR и/или C1q» означает, что молчащее антитело демонстрирует связывание, которое составляет, по меньшей мере, менее 50%, например, менее 80% от связывания с FcγR и/или C1q, которое наблюдают с соответствующим антителом с изотипом человеческого IgG1 или IgG4 дикого типа.
Конструирование каркасного участка или Fc
Антитела согласно настоящему описанию включают модификации, внесенные в остатки каркаса в VH и VL, для снижения иммуногенности антитела по сравнению с соответствующими мышиными антителами mAb 7.2.
В одном из конкретных воплощений, антитело согласно настоящему описанию представляет собой гуманизированное моноклональное антитело исходного мышиного антитела mAb 7.2, включающее, по меньшей мере, следующие аминокислотные мутации в каркасных областях VH: V5Q; V11L; К12В; R66K; S74F; I75S; E81Q; S82AR; R82BS; R83T; D85E; T87S; L108S; и, по крайней мере, следующие аминокислотные мутации в каркасных областях Vκ: T5N; V15L; R18T; V19I; K42N; A43I; D70G; F73L; Q100G.
В другом конкретном воплощении, антитело по настоящему описанию представляет собой гуманизированное моноклональное антитело исходного мышиного антитела mAb 7.2, включающее, по меньшей мере, следующие аминокислотные мутации в каркасных областях VH по сравнению с mAb 7.2: V5Q; V11L; К12В; R66K; S74F; I75S; E81Q; S82AR; R82BS; R83T; D85E; T87S; L108S; и, по крайней мере, следующие аминокислотные мутации в каркасных областях Vκ: T5N; V15L; R18T; V19I; K42N; A43I; S63T; D70G; F73L; Q100G.
В дополнение к модификациям, сделанным в каркасных областях, антитела по изобретению могут быть сконструированы для включения модификаций в области Fc, обычно для изменения одного или нескольких функциональных свойств антитела, таких как период полужизни в сыворотке, фиксация комплемента, связывание с рецептором Fc и/или антиген-зависимая клеточная цитотоксичность.
Кроме того, антитело согласно настоящему описанию может быть химически модифицировано (например, к антителу может быть присоединено одно или несколько химических фрагментов) или модифицировано для изменения его гликозилирования, опять же, для изменения одного или нескольких функциональных свойств антитела. Каждое из этих воплощений будет более подробно описано ниже.
Термин «изотипная константная область» или «Fc-область», как он используется в настоящем документе, применяют взаимозаменяемо для определения С-концевой области тяжелой цепи иммуноглобулина, включая область Fc с нативной последовательностью и вариантные области Fc. Fc-область тяжелой цепи человеческого IgG обычно определяют как содержащую аминокислотные остатки от положения C226 или от положения P230 до карбоксильного конца антитела IgG, где нумерация соответствует системе нумерации EU. C-концевой лизин (остаток K447) области Fc может быть удален, например, во время продукции или очистки антитела, или его соответствующий кодон был удален в рекомбинантных конструкциях. Соответственно, композиция антител по настоящему описанию может включать популяции антител с удаленными остатками K447, популяции антител без удаленных остатков K447 и популяции антител, включающие смесь антител с остатком K447 и без него.
В одном из конкретных воплощений, шарнирную область CH1 модифицируют таким образом, что количество остатков цистеина в шарнирной области изменяется, например, увеличивается или уменьшается. Этот подход дополнительно описан в патенте США № 5,677,425, выданном Bodmer et al. Число остатков цистеина в шарнирной области CH1 изменяют, например, для облегчения сборки легкой и тяжелой цепей или для увеличения или уменьшения стабильности антитела.
В другом воплощении, шарнирную область Fc антитела мутируют для уменьшения биологического периода полужизни антитела. Более конкретно, одну или несколько аминокислотных мутаций вводят в область интерфейса домена CH2-CH3 Fc-шарнирного фрагмента, так чтобы антитело демонстрировало нарушение связывания протеина A Staphylococcyl (SpA) по сравнению со связыванием природного Fc-шарнирного домена SpA. Этот подход более подробно описан в патенте США № 6,165,745, выданном Ward et al.
В других вариантах осуществления область Fc изменяют путем замены по меньшей мере одного аминокислотного остатка другим аминокислотным остатком для изменения эффекторных функций антитела. Например, одна или несколько аминокислот могут быть заменены другим аминокислотным остатком, так что антитело имеет измененное сродство к эффекторному лиганду, но сохраняет антигенсвязывающую способность родительского антитела. Эффекторный лиганд, сродство к которому изменяется, может быть, например, рецептором Fc или компонентом С1 комплемента. Этот подход более подробно описан в патентах США №№ 5624821 и 5648260, оба выданы Winter et al.
В другом воплощении, одна или несколько аминокислот, выбранных из аминокислотных остатков, могут быть заменены другим аминокислотным остатком, так что антитело имеет измененное связывание с C1q и/или уменьшенную или полностью утраченную комплемент-зависимую цитотоксичность (CDC). Этот подход более подробно описан в патенте США № 6,194,551, выданном Idusogie et al.
В другом воплощении, один или несколько аминокислотных остатков изменяют, чтобы тем самым изменить способность антитела фиксировать комплемент. Этот подход дополнительно описан в публикации РСТ WO 94/29351, выданной Bodmer et al.
В других воплощениях, область Fc модифицируют для уменьшения способности антитела опосредовать зависимую от антител клеточную цитотоксичность (ADCC) и/или для уменьшения сродства антитела к рецептору Fcγ путем модификации одной или нескольких аминокислот. Такие антитела с пониженными эффекторными функциями и, в частности, с пониженной ADCC включают молчащие антитела.
В определенных воплощениях, применяют домен Fc изотипа IgG1. В некоторых конкретных воплощениях применяют мутантный вариант Fc-фрагмента IgG1, например, молчащий Fc IgG1, который снижает или устраняет способность гибридного полипептида опосредовать антителозависимую клеточную цитотоксичность (ADCC) и/или связываться с рецептором Fcγ.
В определенных воплощениях, применяют домен Fc изотипа IgG4. В некоторых конкретных воплощениях применяют мутантный вариант Fc-фрагмента IgG4, например, молчащий Fc IgG4, который снижает или устраняет способность гибридного полипептида опосредовать антителозависимую клеточную цитотоксичность (ADCC) и/или связываться с рецептором Fcγ.
Молчащие эффекторные функции могут быть получены путем мутации в константной части Fc антител, и они были описаны в данной области техники (Baudino et al., 2008; Strohl, 2009). Примеры «молчащих» антител IgG1 включают вариант тройного мутанта IgG1 L247F L248E P350S. Примеры «молчащих» антител IgG4 включают вариант двойного мутанта IgG4 S241P L248E.
В определенных воплощениях, домен Fc представляет собой молчащий мутант Fc, предотвращающий гликозилирование в положении 314 домена Fc. Например, домен Fc содержит аминокислотную замену аспарагина в положении 314. Примером такой аминокислотной замены представляет собой замена N314 на глицин или аланин.
В еще одном воплощении, гликозилирование антитела модифицируют. Например, может быть получено агликозилированное антитело (т.е. в антителе отсутствует гликозилирование). Гликозилирование можно изменить, например, для повышения сродства антитела к антигену. Такие углеводные модификации могут быть выполнены, например, путем изменения одного или нескольких сайтов гликозилирования в последовательности антитела. Например, можно произвести одну или несколько аминокислотных замен, которые приведут к устранению одного или нескольких сайтов гликозилирования каркасной области вариабельной области, чтобы тем самым устранить гликозилирование в этом сайте. Такое агликозилирование может повысить сродство антитела к антигену. Такой подход более подробно описан в патентах США №№,5,714,350 и 6,350,861, выданных Co et al.
Другая модификация антител, которая рассматривается в соответствии с настоящим изобретением в данном описании, представляет собой пегилирование или гезилирование или родственные технологии. Антитело можно пегилировать, например, для увеличения биологического периода полужизни антитела (например, в сыворотке). Для пегилирования антитела антителу или его фрагменту обычно позволяют реагировать с полиэтиленгликолем (PEG), таким как реакционноспособный сложный эфир или альдегидное производное PEG, в условиях, при которых одна или несколько групп PEG присоединяют к антителу или фрагменту антитела. Пегилирование можно проводить с помощью реакции ацилирования или реакции алкилирования с реакционноспособной молекулой PEG (или аналогичным реакционноспособным водорастворимым полимером). Термин «полиэтиленгликоль», как он используется в настоящем документе, предназначен для охвата любых форм ПЭГ, которые применяют для дериватизации других белков, таких как моно (С1-С10) алкокси- или арилоксиполиэтиленгликоль или полиэтиленгликольмалеимид. В определенных воплощениях антитело, подлежащее пегилированию, представляет собой агликозилированное антитело. Способы пегилирования белков известны в данной области техники и могут быть применены к антителам по изобретению. Смотри, например, EP 0 154 316, выданную Nishimura et al., и EP 0 401 384, выданную Ishikawa et al.
Другая возможность представляет собой слияние, по меньшей мере, антигенсвязывающей области антитела согласно настоящему описанию с белками, способными связываться с белками сыворотки, такими как человеческий сывороточный альбумин, для увеличения периода полужизни полученной молекулы. Такой подход, например, описан у Nygren et al., EP 0 486 525.
В определенных воплощениях С-концевой лизин, обычно присутствующий в константных доменах тяжелой цепи человеческого IgG, удаляют генно-инженерными способами для уменьшения гетерогенности, поскольку обычно наблюдают отщепление этого остатка во время производства или хранения. Такие модификации не вызывают заметных изменений в желаемых функциях этих антител, но придают этим молекулам преимущество в стабильности.
Молекулы нуклеиновой кислоты, кодирующие антитела по изобретению
В настоящем документе также раскрыты молекулы нуклеиновой кислоты, которые кодируют антитела против BTN3A по настоящему изобретению. Примеры нуклеотидных последовательностей вариабельной легкой цепи и тяжелой цепи представляют собой нуклеотидные последовательности, кодирующие аминокислотные последовательности вариабельной легкой цепи и тяжелой цепи любого из mAb1, mAb2, mAb4 и mAb5, причем последние последовательности легко получить из таблицы 1 и таблицы 2, и с применением генетического кода и, необязательно, с учетом предпочтения по кодонам в зависимости от вида клетки-хозяина.
Настоящее изобретение также относится к молекулам нуклеиновых кислот, которые происходят из упомянутых последними последовательностей, оптимизированных для экспрессии белка в клетках млекопитающих, например, клеточных линиях СНО.
Нуклеиновые кислоты могут присутствовать в целых клетках, в клеточном лизате или могут быть нуклеиновыми кислотами в частично очищенной или практически чистой форме. Нуклеиновую кислоту «выделяют» или она «становится по существу чистой» при очистке от других клеточных компонентов или других загрязняющих веществ, например, других клеточных нуклеиновых кислот или белков, стандартными способами, включая обработку щелочью/додецилсульфатом натрия (SDS), формирование полос в CsCl, колоночную хроматографию, электрофорез в агарозном геле и другие, хорошо известные в данной области техники (Ausubel et al., 1988). Нуклеиновая кислота по настоящему описанию может представлять собой, например, ДНК или РНК, и может включать или не включать интронные последовательности. В одном из воплощений нуклеиновая кислота может присутствовать в векторе, таком как вектор фагового дисплея, или в рекомбинантном плазмидном векторе.
Нуклеиновые кислоты настоящего описания могут быть получены с применением стандартных способов молекулярной биологии. После получения фрагментов ДНК, кодирующих, например, сегменты VH и VL, этими фрагментами ДНК можно дополнительно манипулировать с помощью стандартных способов рекомбинантной ДНК, например, для преобразования генов вариабельной области в гены полноразмерных цепей антитела, в гены фрагментов Fab или в гены ген scFv. В этих манипуляциях фрагмент ДНК, кодирующий VL или VH (например, VL и VH, как определено в таблице 1), функционально связывают с другой молекулой ДНК или с фрагментом, кодирующим другой белок, таким как константная область антитела или гибкий линкер. Термин «функционально связанный» в данном контексте означает, что два фрагмента ДНК соединены функциональным образом, например, так, что аминокислотные последовательности, кодируемые двумя фрагментами ДНК, остаются в рамке считывания или так, что белок экспрессируется под контролем желаемого промотора.
Выделенная ДНК, кодирующая область VH, может быть преобразована в ген полноразмерной тяжелой цепи путем функционального связывания ДНК, кодирующей VH, с другой молекулой ДНК, кодирующей константные области тяжелой цепи (CH1, CH2 и CH3). Последовательности генов константной области тяжелой цепи человека известны в данной области техники (Kabat et al., 1992), и фрагменты ДНК, охватывающие эти области, можно получить с помощью стандартной ПЦР-амплификации. Константная область тяжелой цепи может быть константной областью IgG1, IgG2, IgG3, IgG4, IgA, IgE, IgM или IgD. В некоторых воплощениях константную область тяжелой цепи выбирают из изотипов IgG1, например, изотипа человеческого IgG1. В других воплощениях константную область тяжелой цепи выбирают среди изотипов IgG4, например, изотипа человеческого IgG4. Для гена тяжелой цепи Fab-фрагмента ДНК, кодирующая VH, может быть функционально связана с другой молекулой ДНК, кодирующей только константную область СН1 тяжелой цепи.
Выделенная ДНК, кодирующая область VL, может быть преобразована в ген полноразмерной легкой цепи (а также в ген Fab легкой цепи) путем функционального связывания ДНК, кодирующей VL, с другой молекулой ДНК, кодирующей константную область легкой цепи, CL. Последовательности генов константных областей легкой цепи человека известны в данной области техники (Kabat et al., 1992), и фрагменты ДНК, охватывающие эти области, можно получить с помощью стандартной ПЦР-амплификации. Константная область легкой цепи может быть константной областью каппа или лямбда.
Чтобы создать ген scFv, фрагменты ДНК, кодирующие VH и VL, функционально связывают с другим фрагментом, кодирующим гибкий линкер, например, кодирующим аминокислотную последовательность (Gly4-Ser) 3, так что последовательности VH и VL могут быть экспрессированы как непрерывный одноцепочечный белок, с участками VL и VH, соединенными гибким линкером (Bird et al., 1988; Huston et al., 1988; McCafferty et al., 1990).
Создание трансфектом, продуцирующих моноклональные антитела
Антитела по настоящему изобретению могут быть продуцированы в трансфектоме клетки-хозяина с применением, например, комбинации способов рекомбинантной ДНК и способов трансфекции генов, хорошо известных в данной области техники (Morrison, 1985).
Например, для экспрессии антител или фрагментов антител ДНК, кодирующие частичные или полноразмерные легкие и тяжелые цепи, могут быть получены с помощью стандартных способов молекулярной биологии или биохимии (например, химическим синтезом ДНК, амплификацией ПЦР или клонированием кДНК с применением гибридомы, которая экспрессирует интересующее антитело), и ДНК могут быть вставлены в векторы экспрессии, так что гены будут функционально связаны с последовательностями, контролирующими транскрипцию и трансляцию. В этом контексте термин «функционально связанный» предназначен для обозначения того, что ген антитела лигируют в вектор таким образом, чтобы последовательности контроля транскрипции и трансляции в векторе выполняли предназначенную для них функцию регулирования транскрипции и трансляции гена антитела. Вектор экспрессии и последовательности контроля экспрессии выбирают так, чтобы они были совместимы с применяемой для экспрессии клеткой-хозяином. Ген легкой цепи антитела и ген тяжелой цепи антитела могут быть вставлены в отдельный вектор или, что более типично, оба гена могут быть вставлены в один и тот же вектор экспрессии. Гены антител вставляют в вектор экспрессии стандартными способами (например, лигированием комплементарных сайтов рестрикции фрагмента гена антитела и вектора или лигированием тупых концов, если сайты рестрикции отсутствуют). Вариабельные области легкой и тяжелой цепей описанных в настоящем документе антител можно применять для создания полноразмерных генов антител любого изотипа антитела путем вставки их в векторы экспрессии, уже кодирующие константные области тяжелой цепи и константные области легкой цепи желаемого изотипа, так что VH сегмент будет функционально связан с сегментом (сегментами) CH в векторе, а сегмент VL оперативно будет связан с сегментом CL в векторе. Дополнительно или альтернативно рекомбинантный вектор экспрессии может кодировать сигнальный пептид, который способствует секреции цепи антитела из клетки-хозяина. Ген цепи антитела можно клонировать в вектор таким образом, чтобы сигнальный пептид был связан в рамке считывания с N-концом гена цепи антитела. Сигнальный пептид может представлять собой сигнальный пептид иммуноглобулина или гетерологичный сигнальный пептид (то есть сигнальный пептид из белка, не представляющего собой иммуноглобулин).
Помимо генов цепей антитела, рекомбинантные экспрессирующие векторы, раскрытые в настоящем документе, несут регуляторные последовательности, которые контролируют экспрессию генов цепей антитела в клетке-хозяине. Термин «регуляторная последовательность» включает промоторы, энхансеры и другие элементы контроля экспрессии (например, сигналы полиаденилирования), которые контролируют транскрипцию или трансляцию генов цепей антитела. Такие регуляторные последовательности описаны, например, в публикации Goeddel (Goeddel, 1990). Специалистам в данной области техники будет понятно, что конструкция вектора экспрессии, включая выбор регуляторных последовательностей, может зависеть от таких факторов, как выбор клетки-хозяина для трансформации, желаемый уровень экспрессии белка и т.д. Регуляторные последовательности для экспрессии в клетках-хозяевах млекопитающих включают вирусные элементы, которые управляют высокими уровнями экспрессии белка в клетках млекопитающих, такие как промоторы и/или энхансеры, полученные из цитомегаловируса (CMV), обезьяньего вируса 40 (SV40), аденовируса (например, аденовируса большого позднего промотор (AdMLP)) и полиома. В качестве альтернативы можно применять невирусные регуляторные последовательности, такие как промотор убиквитина или промотор Р-глобина. Кроме того, регуляторные элементы состоят из последовательностей из разных источников, таких как промоторная система SRa, которая содержит последовательности из раннего промотора SV40 и длинного концевого повтора вируса Т-клеточного лейкоза типа 1 человека (Takebe et al., 1988).
В дополнение к генам цепей антитела и регуляторным последовательностям рекомбинантные экспрессирующие векторы по настоящему изобретению могут нести дополнительные последовательности, такие как последовательности, которые регулируют репликацию вектора в клетках-хозяевах (например, точки начала репликации) и гены селективных маркеров. Ген селективного маркера облегчает отбор клеток-хозяев, в которые был введен вектор (смотри, например, патенты США №№ 4,399,216, 4,634,665 и 5,179,017, все выданы Axel et al.). Например, обычно ген селективного маркера придает устойчивость к лекарственным средствам, таким как G418, гигромицин или метотрексат, клетке-хозяину, в которую был введен вектор. Гены селективных маркеров включают ген дигидрофолатредуктазы (DHFR) (для применения в dhfr-клетках-хозяевах с селекцией/амплификацией метотрексатом) и ген neo (для селекции G418).
Для экспрессии легкой и тяжелой цепей вектор (векторы) экспрессии, кодирующий (кодирующие) тяжелую и легкую цепи, трансфицируют в клетку-хозяина стандартными способами. Подразумевается, что различные формы термина «трансфекция» охватывают широкий спектр способов, обычно применяемых для введения экзогенной ДНК в прокариотическую или эукариотическую клетку-хозяина, например, электропорацию, осаждение фосфатом кальция, трансфекцию с использованием DEAE-декстрана и т.п. Теоретически возможно экспрессировать антитела по настоящему изобретению в прокариотических или эукариотических клетках-хозяевах. Экспрессия антител в эукариотических клетках, например, в клетках-хозяевах млекопитающих, дрожжевых или нитчатых грибах, предусматривают, потому что такие эукариотические клетки и, в частности, клетки млекопитающих, с большей вероятностью, чем прокариотические клетки, собирают и секретируют правильно свернутые и иммунологически активные антитела.
В одном из конкретных воплощений, вектор клонирования или экспрессии согласно настоящему описанию содержит одну из кодирующих последовательностей тяжелой и легкой цепей любого из mAb1, mAb2, mAb4 и mAb5, функционально связанных с подходящими промоторными последовательностями.
Клетки-хозяева млекопитающих для экспрессии рекомбинантных антител по настоящему описанию включают клетки яичника китайского хомячка (клетки СНО), включая клетки dhfr-CHO (описанные в работе Urlaub and Chasin, 1980), применяемые с селективным маркером DHFR (как описано у Kaufman and Sharp, 1982), линии клеток CHOK1 dhfr+, клетки миеломы NSO, клетки COS и клетки SP2, например, линии клеток GS CHO вместе с системой экспрессии генов GS XceedTM (Lonza). Когда рекомбинантные векторы экспрессии, кодирующие гены антител, вводят в клетки-хозяева млекопитающих, антитела получают путем культивирования клеток-хозяев в течение периода времени, достаточного для экспрессии антитела в клетках-хозяевах и, необязательно, секреции антитела в культуральную среду, в которой выращивают клетки-хозяева. Антитела могут быть выделены и очищены, например, из культуральной среды после их секреции с применением стандартных способов очистки белков (Shukla et al., 2007).
В одном из конкретных воплощений, клетка-хозяин согласно настоящему раскрытию представляет собой клетку-хозяина, трансфицированную вектором экспрессии, включающим кодирующие последовательности, подходящие для экспрессии mAb1, mAb2, mAb4 и mAb5, соответственно, функционально связанные с подходящими промоторными последовательностями.
Например, настоящее раскрытие относится к клетке-хозяину, включающей, по меньшей мере, нуклеиновые кислоты по SEQ ID NO: 8 и 10, кодирующие соответственно тяжелую и легкую цепи mAb1.
Последние клетки-хозяева затем можно дополнительно культивировать в подходящих условиях для экспрессии и продукции антитела согласно изобретению, выбранного из группы, состоящей из mAb1, mAb2, mAb4 и mAb5, соответственно.
Альтернативно, бесклеточные системы экспрессии можно применять для продукции любого из mAb1, mAb2, mAb4 и mAb5. Обычные способы бесклеточной экспрессии белков или антител уже описаны (Stech et al., 2017).
Иммуноконъюгаты
В другом аспекте настоящее раскрытие описывает антитело против BTN3A, как раскрыто в настоящем документе, или его фрагмент, конъюгированное с терапевтическим компонентом, таким как цитотоксин, лекарственное средство (например, иммунодепрессант) или радиотоксин. Такие конъюгаты называются в настоящем документе «иммуноконъюгатами». Иммуноконъюгаты, включающие один или несколько цитотоксинов, называются «иммунотоксинами». Цитотоксин или цитотоксический агент включает любой агент, который разрушает (например, убивает) клетки. Примеры включают таксон, цитохалазин B, грамицидин D, бромид этидия, эметин, митомицин, этопозид, тенопозид, винкристин, винбластин, колхицин, доксорубицин, даунорубицин, дигидроксиантрациндион, митоксантрон, митрамицин, актиномицин D, 1-дегидротестостерон, глюкокортикоиды, прокаин, тетракаин, лидокаин, пропранолол и пуромицин и их аналоги или гомологи. Терапевтические средства также включают, например, антиметаболиты (например, метотрексат, 6-меркаптопурин, 6-тиогуанин, цитарабин, 5-фторурацил декарбазин), абляционные средства (например, мехлорэтамин, тионэпа, хлорамбуцил, мейфалан, кармустин (BSNU) и ломустин (CCNU), циклофосфамид, бусульфан, диброманнитол, стрептозотоцин, митомицин C и цис-дихлордиамин платины (II) (DDP), цисплатин, антрациклины (например, даунорубицин (ранее дауномицин) и доксорубицин), антибиотики (например, дактиномицин (ранее актиномицин), блеомицин, митрамицин и антрамицин (AMC)), монометилауристатин E и антимитотические средства (например, винкристин и винбластин).
Цитотоксины могут быть конъюгированы с антителами по настоящему изобретению с применением линкерной технологии, доступной в данной области техники. Примеры типов линкеров, которые применяли для конъюгирования цитотоксина с антителом, включают, без ограничения, гидразоны, тиоэфиры, сложные эфиры, дисульфиды и линкеры, содержащие пептиды, такие как валин-цитрулиновый линкер. Может быть выбран линкер, который, например, чувствителен к расщеплению при низком pH в лизосомном компартменте или чувствителен к расщеплению протеазами, такими как протеазы, предпочтительно экспрессируемые в опухолевой ткани, такие как катепсины (например, катепсины B, C, D).
Для дальнейшего обсуждения типов цитотоксинов, линкеров и способов конъюгирования терапевтических агентов с антителами смотри также работу Panowski et al., 2013 в отношении обзора конъюгатов антитело-лекарственное средство.
Антитела по настоящему изобретению также могут быть конъюгированы с радиоактивным изотопом для создания цитотоксических радиофармацевтических препаратов, также называемых радиоиммуноконъюгатами. Примеры радиоактивных изотопов, которые могут быть конъюгированы с антителами для применения в диагностических или терапевтических целях, включают, но не ограничиваются ими, йод131, индий111, иттрий90 и лютеций177. Способ получения радиоиммунконъюгатов разработан в данной области техники.
Биспецифические или мультиспецифические молекулы
В другом аспекте, в настоящем документе дополнительно раскрыты биспецифические или мультиспецифические молекулы, включающие антитело против BTN3A по настоящему изобретению. Антитело может быть дериватизировано или связано с другой функциональной молекулой, например, с другим пептидом или белком (например, с другим антителом или лигандом для рецептора) для создания биспецифической молекулы, которая связывается по меньшей мере с двумя разными сайтами связывания или молекулами-мишенями. Фактически, антитело может быть дериватизировано или связано с более чем одной другой функциональной молекулой для создания мультиспецифических молекул, которые связываются более чем с двумя разными сайтами связывания и/или молекулами-мишенями; такие мультиспецифические молекулы также охватываются термином «биспецифическая молекула», как он используется в настоящем документе. Для создания биспецифической молекулы антитело по изобретению может быть функционально связано (например, путем химического связывания, генетического слияния, нековалентной ассоциации или иным образом) с одной или несколькими другими связывающими молекулами, такими как другое антитело, фрагмент антитела, пептид или миметик связывания, в результате чего получается биспецифическая молекула.
Соответственно, настоящее описание включает биспецифические молекулы, включающие, по меньшей мере, одну первую специфичность связывания для BTN3A, например, одну антигенсвязывающую часть любого из mAb1, mAb2, mAb4 и mAb5, и вторую специфичность связывания для второго целевого эпитопа.
Кроме того, для воплощения, в котором биспецифическая молекула является мультиспецифичной, молекула может дополнительно включать третью специфичность связывания в дополнение к первому и второму целевому эпитопу.
В одном из воплощений, описанные в настоящем документе биспецифические молекулы включают в качестве специфичного связующего по меньшей мере одно антитело или его фрагмент, включая, например, Fab, Fab', F(ab')2, Fv, Unibody или одноцепочечный Fv. Антитело также может представлять собой димер легкой цепи или тяжелой цепи или любой его минимальный фрагмент, такой как Fv или одноцепочечный конструкт, как описано в Ladner et al.в патенте США № 4,946,778.
Другие антитела, которые можно применять в описанных в настоящем документе биспецифических молекулах, представляют собой мышиные, химерные и гуманизированные моноклональные антитела.
Биспецифические молекулы по настоящему изобретению могут быть получены путем конъюгирования составляющих специфичных связующих с применением способов, известных в данной области техники. Например, каждое специфичное связующее биспецифической молекулы может быть получено отдельно, а затем они могут быть конъюгированы. Если спцифичные связующие представляют собой белки или пептиды, то для ковалентной конъюгации можно применять различные связывающие или перекрестно-сшивающие средства. Примеры сшивающих агентов включают протеин А, карбодиимид, N-сукцинимидил-S-ацетилтиоацетат (SATA), 5,5'-дитиобис(2-нитробензойную кислоту) (DTNB), о-фенилендималеимид (oPDM), N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP) и сульфосукцинимидил-4-(N-малеимидометил)-циклогаксан-1-карбоксилат (сульфо-SMCC) (Karpovsky et al., 1984; Liu et al., 1985). Другие способы включают описанные в работах Brennan et al., 1985; Glennie et al., 1987; Paulus, 1985.
Альтернативно, оба специфичных связующих можно закодировать в одном и том же векторе, экспрессировать и собрать в одной и той же клетке-хозяине. Этот способ особенно полезен, когда биспецифическая молекула представляет собой слитый белок mAb x mAb, mAb x Fab, Fab x F(ab')2 или лиганд x Fab. Биспецифическая молекула по настоящему описанию может представлять собой одноцепочечную молекулу, содержащую одно одноцепочечное антитело и детерминанту связывания, или одноцепочечную биспецифическую молекулу, содержащую две детерминанты связывания.
Связывание биспецифических молекул с их конкретными мишенями можно подтвердить, например, с помощью твердофазного иммуноферментного анализа (ELISA), радиоиммуноанализа (REA), анализа FACS, биоанализа (например, ингибирования роста и апоптоза) или анализа вестерн-блоттингом. Каждый из этих анализов обычно выявляет присутствие комплексов белок-антитело, представляющих особый интерес, с применением меченого реагента (например, антитела), специфичного для представляющего интерес комплекса.
Антитела по настоящему изобретению также можно применять для получения искусственного Т-клеточного рецептора (также известного как химерные Т-клеточные рецепторы или химерные антигенные рецепторы (CAR)). Например, вариабельные области антител можно применять для образования Fab или scFv, который связан через спейсер с трансмембранным доменом (обычно трансмембранным доменом CD8 альфа) и сигнальным эндодоменом TCR (например, CD3 зета), и необязательно костимулирующим сигнальным доменом (например, из 4-1BB или CD28), который может продуцироваться на поверхности Т-клеток. Такие CAR можно применять в терапии адаптивного переноса, например, для лечения пролиферативных расстройств.
Фармацевтические композиции
В другом аспекте, в настоящем описании представлена композиция, например, фармацевтическая композиция, содержащая одно антитело или комбинацию антител, описанных в настоящем документе, например, одно антитело, выбранное из группы, состоящей из mAb1, mAb2, mAb4 и mAb5 или их антигенсвязывающих частей, составленных вместе с фармацевтически приемлемым носителем. Такие композиции могут включать одно антител, или иммуноконъюгатов, или биспецифических молекул, как описано выше или комбинацию (например, двух или более разных) из указанного.
Описанные в настоящем документе фармацевтические композиции также можно вводить в комбинированной терапии, т.е. в сочетании с другими агентами. Например, комбинированная терапия может включать антитело против BTN3A согласно настоящему раскрытию, например, одно антитело, выбранное из группы, состоящей из mAb1, mAb2, mAb4 и mAb5 или их антигенсвязывающих частей, в сочетании по меньшей мере с одним противовирусным, противовоспалительное или другим антипролиферативным средством. Примеры терапевтических средств, которые можно применять в комбинированной терапии, описаны более подробно ниже в разделе, посвященном применениям антител по изобретению.
Термин «фармацевтически приемлемый носитель», как он используется в настоящем документе, включает любой из и все растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые средства, изотонические средства и средства, замедляющие абсорбцию, и тому подобное, которые представляют собой физиологически совместимые средства. Носитель должен подходить для внутривенного, внутримышечного, подкожного, парентерального, спинномозгового или эпидермального введения (например, путем инъекции или инфузии). В одном из воплощений носитель должен подходить для подкожного введения или внутриопухолевой инъекции. В зависимости от способа введения активное соединение, т.е. антитело, иммуноконъюгат или биспецифическая молекула, может быть покрыто материалом для защиты соединения от действия кислот и других естественных условий, которые могут инактивировать соединение.
Стерильный фосфатно-солевой буферный раствор представляет собой один из примеров фармацевтически приемлемого носителя. Другие подходящие носители хорошо известны специалистам в области данной техники (Remington and Gennaro, 1995). Композиции могут дополнительно включать одно или несколько вспомогательных веществ, консервантов, солюбилизаторов, буферных средств, альбумин для предотвращения потери белка на поверхности флаконов и т.д.
Форма фармацевтических композиций, способ введения, дозировка и режим, естественно, зависят от состояния, которое необходимо лечить, тяжести заболевания, возраста, веса и пола пациента и т.д.
Фармацевтические композиции по настоящему описанию могут быть составлены для местного, перорального, парентерального, интраназального, внутривенного, внутримышечного, подкожного или внутриглазного введения и т.п.
Предпочтительно, фармацевтические композиции включают носители, фармацевтически приемлемые для препарата, который можно вводить путем инъекции. Это могут быть, в частности, изотонические, стерильные, солевые растворы (мононатрия или динатрия фосфат, хлорид натрия, калия, кальция или магния и тому подобное или смеси таких солей) или сухие, в частности, лиофилизированные композиции, которые при добавлении, в зависимости от конкретного заболевания, стерильной воды или физиологического раствора, позволяют приготовить растворы для инъекций.
Дозы, применяемые для введения, могут быть адаптированы в зависимости от различных параметров и, в частности, в зависимости от применяемого способа введения, соответствующей патологии или, альтернативно, от желаемой продолжительности лечения.
Для приготовления фармацевтических композиций эффективное количество антитела может быть растворено или диспергировано в фармацевтически приемлемом носителе или водной среде.
Фармацевтические формы, подходящие для инъекций, включают стерильные водные растворы или дисперсии; составы, включающие кунжутное масло, арахисовое масло или водный пропиленгликоль; и стерильные порошки или лиофилизаты для немедленного приготовления стерильных растворов или дисперсий для инъекций. Во всех случаях форма должна быть стерильной и жидкой до такой степени, чтобы ее можно было легко вводить шприцем. Она должна быть стабильной в условиях производства и хранения и должна быть защищена от загрязняющего действия микроорганизмов, таких как бактерии и грибки.
Растворы активных соединений в виде свободного основания или фармакологически приемлемых солей можно приготовить в воде, смешанной подходящим образом с поверхностно-активным веществом, таким как гидроксипропилцеллюлоза. Также можно приготовить дисперсии в глицерине, жидких полиэтиленгликолях и их смесях, а также в маслах. В обычных условиях хранения и применения эти препараты включают консервант, предотвращающий рост микроорганизмов.
Антитело по настоящему изобретению может быть включено в композицию в нейтральной или солевой форме. Фармацевтически приемлемые соли включают кислотно-аддитивные соли (образованные со свободными аминогруппами белка) и которые образованы с неорганическими кислотами, такими как, например, соляная или фосфорная кислоты, или органическими кислотами, такими как уксусная, щавелевая, винная, миндальная и тому подобное. Соли, образованные свободными карбоксильными группами, также могут быть производными неорганических оснований, таких как, например, гидроксиды натрия, калия, аммония, кальция или железа, и таких органических оснований, как изопропиламин, триметиламин, гистидин, прокаин и тому подобное.
Носитель также может представлять собой растворитель или дисперсионную среду, включающие, например, воду, этанол, полиол (например, глицерин, пропиленгликоль, жидкий полиэтиленгликоль и т.п.), их подходящие смеси и растительные масла. Подходящую текучесть можно поддерживать, например, путем применения покрытия, такого как лецитин, путем поддержания необходимого размера частиц в случае дисперсии и путем применения поверхностно-активных веществ. Предотвращение действия микроорганизмов может быть достигнуто с помощью различных антибактериальных и противогрибковых средств, например парабенов, хлорбутанола, фенола, сорбиновой кислоты, тимеросала и т.п. Во многих случаях будет предпочтительно включать изотонические средства, например сахара или хлорид натрия. Пролонгированное всасывание композиций для инъекций может быть достигнуто путем применения в композициях агентов, замедляющих абсорбцию, например моностеарата алюминия и желатина.
Стерильные растворы для инъекций готовят путем включения активных соединений в необходимом количестве в соответствующий растворитель с различными другими ингредиентами, перечисленными выше, при необходимости, с последующей стерилизацией фильтрованием. Обычно дисперсии готовят путем включения различных стерилизованных активных ингредиентов в стерильный носитель, который содержит основную дисперсионную среду и другие необходимые ингредиенты из перечисленных выше. В случае стерильных порошков для приготовления стерильных растворов для инъекций предпочтительные способы приготовления представляют собой способы вакуумной сушки и сублимационной сушки, которые дают порошок активного ингредиента плюс любой дополнительный желаемый ингредиент из его раствора, предварительно стерилизованного фильтрованием.
Также предполагается приготовление большего количества или высококонцентрированных растворов для прямой инъекции, если ожидают, что применение ДМСО в качестве растворителя приведет к чрезвычайно быстрому проникновению, доставляя высокие концентрации активных агентов к небольшой области опухоли.
После приготовления, растворы будут вводить способом, совместимым с лекарственной формой, и в таком количестве, которое представляет собой терапевтически эффективное количество. Композиции легко вводят в различных лекарственных формах, таких как описанные выше типы растворов для инъекций, но также можно применять капсулы для высвобождения лекарственного средства и т.п.
Для парентерального введения в водном растворе, например, раствор должен быть соответствующим образом забуферен, если это необходимо, и жидкий разбавитель сначала делают изотоническим с помощью достаточного количества физиологического раствора или глюкозы. Эти конкретные водные растворы в особенности подходят для внутривенного, внутримышечного, подкожного и внутрибрюшинного введения. В этой связи стерильные водные среды, которые можно применять, будут известны специалистам в данной области техники в свете настоящего описания. Например, одну дозу можно растворить в 1 мл изотонического раствора NaCl и либо добавить к 1000 мл жидкости для гиподермоклизиса, либо ввести в предполагаемое место инфузии (смотри, например, «Remington's Pharmaceutical Sciences» 15-е издание, страницы 1035-1038 и 1570-1580). Некоторое изменение дозировки обязательно произойдет в зависимости от состояния пациента, которого лечат. Лицо, ответственное за введение, в любом случае определяет подходящую дозу для отдельного субъекта.
Антитела по настоящему описанию могут быть включены в терапевтическую смесь, содержащую от примерно 0,0001 до 1,0 миллиграмма, или от примерно 0,001 до 0,1 миллиграмма, или от примерно 0,1 до 1,0, или даже от 1,0 до примерно 10 миллиграммов на дозу. Также можно вводить несколько доз.
Подходящий состав для раствора для инфузии или подкожной инъекции антител описан в данной области техники и, например, рассмотрен в работе Cui et al (Drug Dev Ind Pharm 2017, 43 (4): 519-530)
В дополнение к соединениям, составленным для парентерального введения, такого как внутривенная или внутримышечная инъекция, другие фармацевтически приемлемые формы включают, например, таблетки или другие твердые вещества для перорального приема; капсулы с замедленным высвобождением; и любую другую форму, применяемую в настоящее время.
В определенных воплощениях, предполагают применение липосом и/или наночастиц для введения антител в клетки-хозяева. Образование и применение липосом и/или наночастиц известно специалистам в данной области техники.
Нанокапсулы, как правило, могут удерживать соединения стабильным и воспроизводимым образом. Чтобы избежать побочных эффектов из-за внутриклеточной перегрузки полимерами, такие сверхмелкие частицы (размером около 0,1 мкм) обычно конструируют с применением полимеров, способных разлагаться in vivo. Биоразлагаемые наночастицы полиалкилцианоакрилата, которые удовлетворяют этим требованиям, рассматривают для применения в настоящем раскрытии, и такие частицы можно легко изготовить.
Липосомы образуются из фосфолипидов, которые диспергированы в водной среде и спонтанно образуют многослойные концентрические двухслойные везикулы (также называемые многослойными везикулами (MLV)). MLV обычно имеют диаметр от 25 нм до 4 мкм. Обработка MLV ультразвуком приводит к образованию небольших однослойных везикул (SUV) с диаметром в диапазоне от 200 до 500 Å, содержащих водный раствор в сердцевине. Физические характеристики липосом зависят от pH, ионной силы и присутствия двухвалентных катионов.
Применение и способы с антителами по изобретению
Антитела по настоящему изобретению имеют диагностическое и терапевтическое применение in vitro и in vivo. Например, эти молекулы можно вводить в клетки в культуре, например in vitro, ex vivo или in vivo, или в субъекта, например, in vivo, для лечения, предотвращения или диагностики различных заболеваний.
Термин «лечит», «лечить» или «лечение», как он используется в настоящем документе, относится к одному или нескольким из следующего: (1) подавление заболевания; например, ингибирование заболевания, состояния или расстройства у индивидуума, который испытывает или демонстрирует патологию или симптоматику заболевания, состояния или нарушения (т.е. прекращение дальнейшего развития патологии и/или симптоматики); и (2) облегчение болезни; например, облегчение заболевания, состояния или расстройства у человека, который испытывает или демонстрирует патологию или симптоматику заболевания, состояния или расстройства (т. е. обращение патологии и/или симптоматики вспять), такое как уменьшение тяжести заболевания или уменьшение или облегчение одного или нескольких симптомов болезни. В частности, применительно к лечению опухоли термин «лечение» может относиться к ингибированию роста опухоли или уменьшению размера опухоли.
Антитела согласно настоящему описанию представляют собой активирующие антитела против BTN3A, которые могут активировать цитолитическую функцию, продукцию цитокинов и/или пролиферацию Vγ9 Vδ2 Т-клеток и, таким образом, могут быть применены для преодоления иммуносупрессивных механизмов, наблюдаемых у онкологических пациентов и во время хронических инфекций.
Термины «рак», «гиперпролиферативный» и «неопластический», как они используются в настоящем документе, относятся к клеткам, обладающим способностью к автономному росту, то есть аномальному статусу или состоянию, характеризующемуся быстро пролиферирующим клеточным ростом. Гиперпролиферативные и неопластические болезненные состояния могут быть отнесены к категории патологических, то есть характеризующих или составляющих болезненное состояние, или могут быть отнесены к категории непатологических, то есть представляющих собой состояние, отклоняющееся от нормального, но не связанное с заболеванием. Термин предназначен для включения всех типов раковых образований или онкогенных процессов, метастатических тканей или злокачественно трансформированных клеток, тканей или органов, независимо от гистопатологического типа или стадии инвазивности.
Термины «рак» или «новообразования» включают злокачественные новообразования различных систем органов, такие как поражение легких, молочной железы, щитовидной железы, лимфоидных, желудочно-кишечных и мочеполовых путей, а также аденокарциномы, которые включают злокачественные новообразования, такие как большинство видов рака толстой кишки, почечно-клеточная карцинома, рак простаты и/или опухоли яичек, немелкоклеточная карцинома легкого, рак тонкой кишки и рак пищевода.
Примеры рака включают, но не ограничиваются ими, гематологические злокачественные новообразования, такие как B-клеточное лимфоидное новообразование, T-клеточное лимфоидное новообразование, неходжкинскую лимфому (NHL), B-NHL, T-NHL, хронический лимфолейкоз (CLL), малая лимфоцитарная лимфома (SLL), лимфома из мантийных клеток (MCL), лимфоидное новообразование NK-клеток и новообразования миелоидного происхождения, включая острый миелоидный лейкоз.
Примеры негематологических видов рака включают, но не ограничиваются ими, рак толстой кишки, рак груди, рак легких, рак мозга, рак простаты, рак головы и шеи, рак поджелудочной железы, рак мочевого пузыря, колоректальный рак, рак костей, рак шейки матки, рак яичников, рак печени, рак полости рта, рак пищевода, рак щитовидной железы, рак почки, рак желудка, рак яичек и рак кожи.
Примеры хронических инфекций включают, но не ограничиваются ими, вирусные, бактериальные, паразитарные или грибковые инфекции, такие как хронический гепатит, инфекции легких, инфекции нижних дыхательных путей, бронхит, грипп, пневмония и заболевания, передаваемые половым путем.
Примеры вирусных инфекций включают, но не ограничиваются ими, гепатит (HAV, HBV, HCV), простой герпес (HSV), опоясывающий герпес, HPV, грипп (грипп), СПИД и комплекс, связанный со СПИДом, ветряную оспу (ветрянку), простуду, цитомегаловирусную (CMV) инфекцию, оспу (натуральная оспа), колорадскую клещевую лихорадку, лихорадку денге, геморрагическую лихорадку Эбола, ящур, лихорадку ласса, корь, геморрагическая лихорадку Марбург, инфекционный мононуклеоз, эпидемический паротит, норовирус, полиомиелит, прогрессирующий мультифокальный лейкоцит (PML), бешенство, краснуху, ОРВИ, вирусный энцефалит, вирусный гастроэнтерит, вирусный менингит, вирусную пневмонию, болезнь Западного Нила и желтую лихорадку. Примеры бактериальных инфекций включают, помимо прочего, пневмонию, бактериальный менингит, холеру, дифтерию, туберкулез, сибирскую язву, ботулизм, бруцеллез, кампилобактериоз, сыпной тиф, гонорею, листериоз, болезнь Лайма, ревматическую лихорадку, коклюш (судорожный кашель), чуму, сальмонеллез, скарлатину, шигеллез, сифилис, столбняк, трахому, туляремию, брюшной тиф и инфекции мочевыводящих путей. Примеры также включают бактериальные инфекции, вызываемые Coxiella burnetii, Brucella abortus, Tropheryma whipplei, Mycobacterium tuberculosis и Mycobacterium canettii.
Примеры паразитарных инфекций включают, но не ограничиваются ими, малярию, лейшманиоз, трипаносомоз, болезнь Шагаса, криптоспоридиоз, фасциолез, филяриоз, амебные инфекции, лямблиоз, инфекцию острицами, шистосомоз, теноз, токсоплазмоз, трихинеллез и трипаносомоз. Примеры грибковых инфекций включают, но не ограничиваются ими, кандидоз, аспергиллез, кокцидиоидомикоз, криптококкоз, гистоплазмоз и микоз стопы.
Соответственно, данное изобретение относится к способу лечения одного из описанных выше расстройств у субъекта, нуждающегося в этом, указанный способ включает введение указанному субъекту терапевтически эффективного количества антител против BTN3A, как описано выше, обычно одного из mAb1, mAb2, mAb4 or mAb5.
В определенных воплощениях указанного субъекта выбирают из числа пациентов, имеющих опухоли, экспрессирующие BTN3A.
Антитела для применения, как раскрыто выше, можно вводить в качестве единственного активного ингредиента или в сочетании, например, в качестве адъюванта или в комбинации с другими лекарственными средствами, например, с цитокинами, противовирусными, противовоспалительными средствами или цитотоксическими, антипролиферативными, химиотерапевтическими или противоопухолевыми средствами, продуктом клеточной терапии (например, композиция γδ Т-клеток), например, для лечения или профилактики упомянутых выше заболеваний.
Например, антитела для применения, как описано выше, можно применять в комбинации с клеточной терапией, в особенности терапией γδ Т-клетками, химиотерапией, противоопухолевыми агентами или иммунотерапевтическими агентами.
Термин «клеточная терапия», как он используется в настоящем документе, относится к терапии, включающей введение in vivo, по меньшей мере, терапевтически эффективного количества клеточной композиции нуждающемуся в этом субъекту. Клетки, вводимые пациенту, могут быть аллогенными или аутологичными. Термин «терапия γδ T-клетками» относится к клеточной терапии, при которой клеточная композиция включает в качестве активного начала γδ T-клетки, в частности, Vγ9/Vδ2 T-клетки.
Продукт клеточной терапии относится к клеточной композиции, которую вводят указанному пациенту в терапевтических целях. Указанный продукт клеточной терапии включает терапевтически эффективную дозу клеток и, необязательно, дополнительные наполнители, адъюванты или другие фармацевтически приемлемые носители.
Подходящие противоопухолевые средства могут включать, без ограничения, алкилирующие средства (такие как циклофосфамид, мехлорэтамин, хлорамбуцил, мелфалан, нитрозомочевины, темозоломид), антрациклины (такие как даунорубицин, доксорубицин, эпирубицин, идарубицин, митоксантрон, валрубицин), таксаны (такие как паклитаксел, доцетаксел), эпотилоны, ингибиторы топоизомеразы I (такие как иринотекан или топотекан), ингибиторы топоизомеразы II (такие как этопозид, тенипозид или тафлупозид), аналоги нуклеотидов и аналоги предшественников (такие как азацитидин, азатиоприн, капецитабин, цитарабин, флуроурацил, гемцитабин, гидроксимочевина, меркаптопурин, метотрексатин или тиогуанин), пептидные антибиотики (такие как карбоплатин, цисплатин и оксалиплатин), ретиноиды (такие как третиноин, алитретиноин, бексаротен), алкалоиды барвинка и производные (такие как винбластин, винкристин, виндезин, винорелбин), таргетные терапевтические средства, например, ингибиторы киназ (такими как ибрутиниб, иделалисиб, эрлотиниб, гефитиниб, иматиниб, вемурафениб, висмодегиб), ингибиторы протеасом (такие как бортезомиб, карфилзомиб), ингибиторы гистондеацетилаз (такие как вориностат или ромидепсин).
Примеры иммунотерапевтических средств включают, без ограничения, фосфоантигены (например, золедроновую кислоту или другие бисфосфонаты), антитела против PD-1, антитела против PD-L1, антитела против BTLA, антитела против CTLA-4 и цитокины (такие как интерлейкин 2 (IL-2) (Choudhry H et al, 2018, Biomed Res Int., 2018, 6 мая), интерлейкин 15 (IL-15) (Patidar M et al., Cytokine Growth Factor Rev., 2016 Oct; 31: 49-59), интерлейкин 21 (IL-21) (Caccamo N. et al., PLoS One,2012; 7 (7): e41940) или интерлейкин 33 (IL-33) (Duault C et al., J Immunol., 2016, 1 января; 196 (1): 493-502)), или их рекомбинантные формы и их производные, или любые цитокины, способные индуцировать активность лимфоцитов (например, пролиферацию или продукцию цитокинов или метаболические изменения). Термин «производное» применяют для любых модификаций цитокинов, которые могут зависеть от ПЭГилирования (например, конъюгации с цепями полиэтиленгликоля (ПЭГ)), мутации, такой как делеция, замена или вставка аминокислоты, или ассоциации с потенцирующими агентами (например, комплексы IL15/IL15Ra, слитые с Fc IgG1, в котором IL-15 дополнительно мутирован (asn72asp), что дополнительно увеличивает биологическую активность, делая этот комплекс суперагонистом IL-2 и IL-15Rβγ (Rhode PR et al, Cancer Immunol Res, 2016; 4 (1): 49 -60)) (Barroso-Sousa R et al, Curr Oncol Rep., 2018; Nov 15;21(1):1).
Термин «IL-2» имеет общее значение и относится к человеческому интерлейкину-2. IL-2 представляет собой частью естественного иммунного ответа организма. IL-2 в основном регулирует активность лимфоцитов, связываясь с рецепторами IL-2.
Термин «IL-15» имеет общее значение и относится к интерлейкину-15 человека. Подобно IL-2, IL-15 связывается и передает сигнал через комплекс, состоящий из бета-цепи рецептора IL-2/IL-15 (CD122) и общей гамма-цепи (гамма-C, CD132). IL-15 регулирует активацию и пролиферацию Т-клеток и естественных клеток-киллеров (NK).
Термин «IL-21» имеет общее значение и относится к интерлейкину-21 человека. IL-21 приписывают плейотропные свойства, включая, без ограничения, усиление цитотоксичности NK-клеток и CD8+ T-клеток, модулирование дифференцировки плазматических клеток и ингибирование Treg-клеток.
Термин «IL-33» имеет обычное значение и относится к человеческому интерлейкину-33. IL-33, рассматриваемый как алармин, высвобождаемый при стрессе или повреждении ткани, представляет собой член семейства IL-1 и связывает рецептор ST2. IL-33 известен как эффективный стимулятор иммунных клеток TH1, естественных киллеров (NK), клеток iNKT и Т-лимфоцитов CD8.
Термин «PD-1» имеет общее значение в данной области техники и относится к рецептору программируемой смерти-1. Термин «PD-1» также относится к трансмембранному белку типа I, принадлежащему к семейству сигнальных рецепторов CD28-B7, которое включает CD28, цитотоксический Т-лимфоцит-ассоциированный антиген 4 (CTLA-4), индуцибельный костимулятор (ICOS) и аттенюатор В- и Т-лимфоцитов (BTLA) (Greenwald RJ et al., 2005, Riley JL et al., 2005).
Термин «BTLA» имеет общее значение в данной области техники и относится к аттенюатору В- и Т-лимфоцитов. Термин «BTLA» также относится к CD272, члену семейства сигнальных рецепторов CD28-B7, которое включает CD28, цитотоксический Т-лимфоцит-ассоциированный антиген 4 (CTLA-4), индуцибельный костимулятор (ICOS) и рецептор программируемой смерти-1 (PD-1) (Greenwald RJ et al., 2005, Riley JL et aL., 2005).
Термин «антитело анти-PD-1» или «анти-PD-L1» имеет свое общее значение в данной области техники и относится к антителу со сродством связывания к PD-1 или PD-L1, соответственно, и антагонистической активностью к PD-1, т.е. он ингибирует каскад передачи сигнала, связанный с PD-1, и ингибирует связывание лиганда PD-1 (PD-L1; PD-L2). Такое антитело против PD-1 или антитело против PD-L1 предпочтительно инактивирует PD-1 с большей аффинностью и активностью, соответственно, чем его взаимодействие с другими подтипами или изоформами сигнального семейства рецепторов CD28-B7 (CD28; CTLA-4; ICOS; BTLA). Тесты и анализы для определения того, представляет собой ли соединение антагонист PD-1, хорошо известны специалистам в данной области техники, как описано в работе Greenwald et al., 2005; Riley et al., 2005.
Примеры таких антител против PD1 или PDL1 включают, без ограничения, ниволумаб, пембролизумаб, авелумаб, дурвалумаб, цемиплимаб или атезолизумаб.
Примеры такого антитела против CTLA4 включают, без ограничения, ипилимумаб.
Термин «антитела против BTLA» имеет свое обычное значение в данной области техники и относится к антителам, которые обладают аффинностью связывания и антагонистической активностью по отношению к BTLA, т.е. оно может ингибировать каскад передачи сигнала, связанный с BTLA. Тесты и анализы для определения того, представляет собой ли соединение антагонист BTLA, хорошо известны специалистам в данной области техники, как описано в работах Greenwald et al., 2005; Riley et al., 2005.
В некоторых воплощениях, антитела против BTLA выбраны из тех, что описаны в международной заявке на патент WO2010/106051; WO2011/014438; WO2017/144668.
В некоторых воплощениях, антитело против BTLA представляет собой антитело против BTLA (BTLA8,2), которое можно получить из гибридомы, доступной под номером депозита CNCM I-4123, например, описанной в WO2010/106051.
В некоторых воплощениях, антитело против BTLA представляет собой mAb 4C7, описанное в WO2011/014438.
В некоторых воплощениях, антитело против BTLA представляет собой mAb 629,3 mAb, раскрытое в WO2017/144668, или его гуманизированную версию или вариант.
В соответствии с вышеизложенным настоящее раскрытие обеспечивает в еще одном дополнительном аспекте:
Способ, как определен выше, включающий совместное введение, например, одновременно или последовательно терапевтически эффективного количества антитела против BTN3A по настоящему описанию и по меньшей мере еще одного второго лекарственного вещества, причем указанное второе лекарственное вещество представляет собой противовирусное или антипролиферативное средство или иммунотерапевтическое средство (такое как антител против PD-1, антитело против PD-L1) или цитокины, например IL-2 или IL-15, или продукт клеточной терапии (такой как γδ T-клетки), например, как указано выше.
В одном из воплощений, антитела по настоящему описанию можно применять для определения уровней растворимого BTN3A или уровней клеток, экспрессирующих BTN3A. Это может быть достигнуто, например, путем контактирования образца (такого как образец in vitro) и контрольного образца с антителом против BTN3A в условиях, которые допускают образование комплекса между антителом и BTN3A (в виде экспрессированного на поверхности клеток или растворимого BTN3A, например, в образце крови). Любые комплексы, образованные между антителом и BTN3A, обнаруживают и сравнивают в образце и контроле. Например, стандартные способы обнаружения, хорошо известные в данной области техники, такие как ELISA и проточно-цитометрические анализы, могут быть выполнены с применением композиций по изобретению.
Соответственно, в одном аспекте изобретение дополнительно обеспечивает способы обнаружения присутствия BTN3A (например, человеческого антигена BTN3A) в образце или измерения количества BTN3A, включающие контактирование образца и контрольного образца с антителом или белком изобретения, или его антигенсвязывающую область, которая специфически связывается с BTN3A, в условиях, которые допускают образование комплекса между антителом или его частью и BTN3A. Затем детектируют образование комплекса, при этом разница в образовании комплекса в образце по сравнению с контрольным образцом указывает на присутствие BTN3A в образце.
Также в объем по настоящему изобретению входят наборы, состоящие из композиций (например, гуманизированных антител, конъюгированных антител и мультиспецифических молекул), раскрытых в настоящем документе, и инструкций по применению. Набор может дополнительно включать по меньшей мере один дополнительный реагент или одно или несколько дополнительных антител или белков. Наборы обычно включают этикетку, указывающую предполагаемое применение содержимого набора. Термин «этикетка» включает любые письменные или записанные на носитель материалы, поставляемые с набором или вместе с ним или иным образом сопровождающие набор. Набор может дополнительно включать инструменты для диагностики того, принадлежит ли пациент к группе, которая будет отвечать на лечение антителом против BTN3A, как определено выше.
Другая терапевтическая стратегия основана на применении гуманизированных антител, описанных в настоящем документе, в качестве средств, которые селективно увеличивают и/или активируют Vγ9 Vδ2 Т-клетки, выделенные из образца человеческого субъекта.
Таким образом, изобретение относится к способу лечения нуждающегося в этом субъекта, включающему:
(a) выделение клеток крови, содержащих Vγ9 Vδ2 Т-клетки, например, PBMC, из образца крови субъекта,
(b) размножение in vitro Vγ9 Vδ2 Т-клеток в присутствии любого из mAb 1, 2, 4 и 5 и, необязательно, других опухолевых или вспомогательных клеток,
(c) сбор размноженных Vγ9 Vδ2 Т-клеток,
(d) необязательно, приготовление размноженных Vγ9 Vδ2 Т-клеток и введение субъекту терапевтически эффективного количества указанных Vγ9 Vδ2 Т-клеток.
Изобретение также относится к применению описанных в настоящем документе гуманизированных антител (таких как mAb1, mAb2, mAb4 или mAb5) в качестве средств, которые избирательно размножают Vγ9 Vδ2 Т-клетки с химерным антигенным рецептором (CAR). CAR γδ Т-клетки и их применение в адоптивной Т-клеточной иммунотерапии рака описаны, например, в работе Mirzaei et al (Cancer Lett 2016, 380 (2): 413-423).
Изобретение также относится к антителам против BTN3A для применения in vivo в качестве потенцирующего агента в отношении опухолевых клеток при терапии γδ Т-клетками у субъекта, нуждающегося в этом, как правило, страдающего раком.
Термин «терапия γδ T-клетками», как он используется в настоящем документе, относится к терапии, которая включает введение субъекту, нуждающемуся в этом, по меньшей мере, эффективного количества γδ T-клеток. Такие γδ Т-клетки могут быть аллогенными или аутологичными. В конкретных воплощениях γδ T-клетки могут быть генетически сконструированы путем делеции, или нокаута, или вставки, или нокина определенных генов. В конкретных воплощениях указанные γδ T-клетки включают γδ T-клетки, экспрессирующие химерный антигенный рецептор. γδ Т-клетки могут быть размножены и/или очищены ex vivo. Альтернативно, γδ T-клетки также могут быть включены в клеточную композицию, включающую другие клетки крови и, например, другие клетки иммунной системы. Для ссылки относительно терапии γδ Т-клетками, пожалуйста, смотрите работы Pauza CD. et al, Front Immunol. 2018 Jun 8;9:1305. doi: 10.3389, и Saudemont A. et al., Front Immunol. 2018 Feb 5;9:153. doi: 10.3389.
В самом деле, не ограничиваясь какой-либо конкретной теорией, предлагаемый способ действия антител против BTN3A по настоящему изобретению заключается в том, что их связывание с BTN3A, экспрессируемым на поверхности опухолевой клетки, запускает конформационное изменение, которые позволяют им передавать свои сигналы на контррецептор на Vγ9Vδ2 Т-клетках.
Таким образом, изобретение относится к способу лечения субъекта, страдающего раком, например, гематологическими злокачественными новообразованиями, в частности, лейкозами, такими как острый миелоидный лейкоз, и имеющего опухолевые клетки, например, опухолевые клетки крови, причем указанный способ включает:
(i) введение указанному субъекту эффективного количества антител против BTN3A, как описано в настоящем документе, обычно mAb1, mAb2, mAb4 или mAb5, и,
(ii) введение эффективного количества композиции γδ T-клетки указанному субъекту,
в котором указанное эффективное количество антител против BTN3A обладает способностью усиливать противоопухолевый цитолиз, опосредованный указанной композицией γδ T-клеток, против указанных опухолевых клеток.
Изобретение дополнительно относится к способу лечения субъекта, страдающего раком с солидными опухолевыми клетками, например, раковые клетки яичников, причем указанный способ включает:
(i) введение указанному субъекту эффективного количества антител против BTN3A, как описано в настоящем документе, обычно mAb1, mAb2, mAb4 или mAb5, и,
(ii) введение эффективного количества композиции γδ T-клеток указанному субъекту,
в котором указанное эффективное количество антител против BTN3A обладает способностью усиливать противоопухолевый цитолиз, опосредованный указанной композицией γδ T-клеток, против указанных опухолевых клеток.
Изобретение также относится к способу лечения нуждающегося в этом субъекта, указанный способ включает комбинированное (одновременное или последовательное) введение CAR T-клеток, например, CAR γδ T-клеток, и гуманизированного антитела, как описано в настоящем документе (такого как mAb1, mAb2, mAb4 или mAb5).
Полностью описанное изобретение теперь дополнительно будет проиллюстрировано следующими примерами, которые представляют собой только иллюстративные примеры и не предназначены для дальнейшего ограничения.
Описание чертежей
Фигура 1: A. Человеческие Т-клетки Vγ9Vδ2, размноженные из мононуклеарных клеток периферической крови (PBMC), культивировали совместно с линией клеток Дауди (лимфома Беркитта) при соотношении E:T 1:1 с указанными концентрациями mAb1 (или соответствующего изотипического контроля) в течение 4-х часов. Клетки окрашивали антителами к CD107a и CD107b, и пороги для детекции положительной экспрессии определяли по нестимулированным контролям. Эксперимент проведен с 3-мя здоровыми донорами. B. Клетки Дауди предварительно инкубировали в течение 1-го часа при 37°C с указанными концентрациями mAb1 (или соответствующим изотипическим контролем). После интенсивных промывок клетки Дауди, подвергнутые импульсному воздействию mAb, культивировали совместно в течение 4-х часов с размноженными человеческими Т-клетками Vγ9Vδ2 при 37°C перед измерением активности каспазы 3/7 на клетках Дауди. Для A и B, аппроксимация кривых была получена с применением сигмоидального уравнения 4PL из программного обеспечения GraphPad Prism. C. Тот же протокол был применен, как описано ранее в (A) и (B), для оценки эффективности mAb1 (и соответствующего контроля изотипа), применяемого при 10 мкг/мл на других линиях опухолевых клеток (L-IPC: протоковая аденокарцинома поджелудочной железы, HT29: колоректальный аденокарцинома, A549: карцинома легких) по сравнению с линией клеток Дауди.
Фигура 2: mAb1 опосредует экспрессию BTN3A, убивая клетки-мишени с помощью T-клеток Vγ9Vδ2.
A. 10000 клеток HL60-WT или BTN3AKO (острый миелоидный лейкоз) в течение 24-х часов совместно культивировали с размноженными in vitro Т-клетками Vγ9Vδ2 (соотношение E:T 1:1) в присутствии возрастающей концентрации mAb1 (или соответствующего контроля изотипа hIgG1 ) +/- rHuIL-2 (20 МЕ/мл). Жизнеспособность клеток измеряли с помощью биолюминесцентного анализа для определения уровней ATP.
B. 10000 клеток HL60-WT совместно культивировали в течение 4-х дней с размноженными in vitro Т-клетками Vγ9Vδ2 (соотношение E:T 1:1) в присутствии увеличивающейся концентрации mAb1 (или соответствующего изотипического контроля hIgG1) +/- rHuIL-2 (20 МЕ/мл). Жизнеспособность клеток измеряли каждый день.
C. 10000 клеток HL60-WT совместно культивировали в течение 4-х дней со свежими Т-клетками Vγ9Vδ2, выделенными из PBMC человека (соотношение E:T 1:1 и 1:5) в присутствии возрастающей концентрации mAb1 + rHuIL-2 (20 МЕ/мл). Жизнеспособность клеток измеряли каждый день. * отметка сигнала.
Фигура 3: 10000 опухолевых клеток из различных тканей в течение 24-х часов совместно культивировали с размноженными in vitro Т-клетками Vγ9Vδ2 (соотношение E:T 1:1) в присутствии различных концентраций mAb1. Жизнеспособность клеток измеряли с помощью биолюминесцентного анализа для определения уровней ATP. Указаны значения биолюминесценции. 4 столбца слева направо относятся к: (1) Нет mAb, (2) mAb1 0,1 мкг/мл, (3) mAb1 1 мкг/мл, (4) mAb1 10 мкг/мл.
Фигура 4: mAb1 способствует размножению и активации T-клеток Vγ9Vδ2 яванского макака
A. Цельную кровь яванского макака от 3-х животных обрабатывали буфером для лизиса эритроцитов. После интенсивных промывок клетки высевали при концентрации 1,5 М/мл в среде, включающей 200 МЕ/мл rHuIL-2 и mAb1 (10 мкг/мл). Процент Vγ9+ Т-клеток оценивали с помощью проточной цитометрии на 0, 3, 6, 8 и 10 день с применением специфического антитела. График показывает кинетику процента Vγ9+ Т-клеток среди живых клеток. Каждая кривая представляет отдельное животное.
B. После 10 дней размножения клетки от каждого животного совместно культивировали в течение 4-х часов с Дауди, K562 или Раджи, применяемые в качестве клеток-мишеней (соотношение E:T 1:1) в присутствии культуральной среды, mAb1 или контроля изотипа (10 мкг/мл), и анализировали на дегрануляцию (CD107a/b) проточной цитометрией.
Фигура 5: Образцы крови яванского макака, собранные в моменты времени после дозирования ICT01, окрашивали специфическим коктейлем антител для количественных определений субпопуляций Т-клеток (CD4, CD8, Vγ9 Т-клетки, регуляторные Т-клетки), В-клеток, моноцитов, NK-клеток, клеток mDC, клеток pDC и гранулоцитов и анализировали проточной цитометрией. На верхней панели показано процентное содержание Vγ9δ2 Т-клеток среди CD3+ Т-клеток для животных с однократной дозой. На нижней панели показано процентное содержание Vγ9δ2 Т-клеток CD3+ Т-клеток для животных с повторной дозой. Данные представлены в виде средних значений ± стандартное отклонение (SD) для каждого случая отбора проб и каждой группы. Вертикальные пунктирные линии указывают время дозирования ICT01.
ПРИМЕРЫ
Отбор гуманизированных вариантов
1. Описание стратегий гуманизации
a. Дизайн последовательностей вариабельных областей составных человеческих антител (Composite Human Antibody)™
Структурные модели V-областей мышиного антитела 7.2 и 20.1 получали с применением Swiss PDB и анализировали для выявления важных «сдерживающих» аминокислот в V-областях, которые, вероятно, важны для связывающих свойств антител. Большинство остатков, содержащихся в CDR (с применением определений Кабата и Чотиа) вместе с рядом остатков каркаса, считались важными. Из приведенного выше анализа были созданы составные человеческие последовательности антител 7.2 и 20.1.
b. Предотвращение образования CD4+ Т-клеточного эпитопа
На основе структурного анализа отобрали большой предварительный набор сегментов последовательности, который можно было применять для создания гуманизированных вариантов 7.2 и 20.1, и проанализировали с применением технологии iTope™ для анализа in silico связывания пептидов с аллелями человеческого МНС класса II (Perry et al., 2008), а также с применением TCED™ из известных эпитопов Т-клеток, связанных с последовательностью антител (Bryson et al., 2010). Отбрасывали сегменты последовательности, которые были идентифицированы как существенные связывающие элементы зародышевой линии, не относящейся к клеткам человека, с человеческим MHC класса II, или те, которые имели значительный успех против TCED™. Это привело к уменьшенному набору сегментов, и их комбинации снова проанализировали, как указано выше, чтобы гарантировать, что соединения между сегментами не содержат потенциальных эпитопов для Т-клеток. Отобранные сегменты последовательности собирали в полные последовательности V-области, которые, как было предсказано, были лишены значимых для Т-клеток эпитопов. Затем выбирали несколько последовательностей тяжелых и легких цепей для синтеза и экспрессии генов mAb 7.2 и 20.1 в клетках млекопитающих.
2. Создание гуманизированных вариантов и предварительная характеристика
a. Конструирование плазмид гуманизированных вариантов
Гуманизированные варианты 7.2 и 20.1 синтезировали с фланкирующими сайтами рестрикционных ферментов для клонирования в систему экспрессионных векторов для экспрессии тяжелой и легкой цепи каппа человеческого IgG4 (S241P, L248E). Все конструкции были подтверждены секвенированием.
b. Экспрессия антител
Химерные 7.2 и 20.1 (VH0/Vκ0), две контрольные комбинации (VH0/Vκ1, VH1/Vκ0) и комбинации гуманизированных тяжелых и легких цепей были временно трансфицированы в клетки FreeStyle™ CHO-S (««ThermoFisher»», Лафборо, Великобритания) с применением системы для электропорации MaxCyte STX® (MaxCyte Inc., Гейтерсбург, США) из соответствующей свободной от эндотоксина ДНК. Трансфекцию проводили для каждого антитела с применением наборов для обработки OC-400. После выделения клеток клетки разводили до концентрации 3×106 клеток/мл в среде CD Opti-CHO («ThermoFisher», Лафборо, Великобритания), содержавшей 8 мМ L-глутамина («ThermoFisher», Лафборо, Великобритания) и 1 х гипоксантина-тимидина («ThermoFisher», Лафборо, Великобритания). Через 24 часа после трансфекции температуру культивирования снижали до 32°C и добавляли 1 мМ бутират натрия («Sigma», Дорсет, Великобритания). Культуры ежедневно подкармливали добавлением 3,6% (от начального объема) подпитки (2,5% CHO CD Efficient Feed A («ThermoFisher», Лафборо, Великобритания), 0,5% дрожжевого экстракта (««BD Bioscience»s», Оксфорд, Великобритания), 0,25 мМ глутамакса («ThermoFisher», Лафборо, Великобритания) и 2 г/л глюкозы («Sigma», Дорсет, Великобритания)). Титры IgG в супернатантах контролировали с помощью IgG ELISA, и трансфектанты культивировали вплоть до 14 дней перед сбором супернатантов.
c. Предварительное измерение аффинности: одноцикловый кинетический анализ связывания гуманизированных вариантов с BTN3A
Для оценки связывание всех вариантов составных антител человека (Composite Human Antibody™) 7.2 и 20.1 и выбора антител с наивысшим сродством к BTN3A, проводили одноцикловый кинетический анализ супернатантов от трансфицированной клеточной культуры с применением Biacore T200 (серийный номер 1909913) с помощью программного обеспечения для оценки Biacore T200 V2.0.1 (Упсала, Швеция).
Антитела разводили в 2%-ном БСА/ФСБ до конечной концентрации 2 мкг/мл на основе концентраций, полученных из супернатанта, титрованного с помощью ELISA. В начале каждого цикла антитела загружали на Fc2, Fc3 и Fc4 чипа с протеином A (GE Healthcare, Little Chalfont, Великобритания). IgG улавливали при скорости потока 10 мкл/мин, чтобы получить уровень иммобилизации (RL) ~ 146,5 RU, теоретическое значение для получения RMax ~ 50 RU. Затем поверхности давали стабилизироваться. Кинетические данные за один цикл получали с применением BTN3A1-His в качестве аналита («Sino Biological», № по каталогу 15973-H08H) при скорости потока 60 мкл/мин для минимизации любых потенциальных эффектов массопереноса, а также с применением HBS-P+ (GE Healthcare, Little Chalfont, Великобритания) в качестве рабочего буфера. Множественные повторы с химерным антителом были выполнены для проверки стабильности поверхности и аналита в течение кинетических циклов. Сигнал от контрольного канала Fc1 (без антител) вычитали из сигнала от Fc2, Fc3 и Fc4, чтобы скорректировать различия в неспецифическом связывании с контрольной поверхностью. Применяли трехточечный, четырехкратный диапазон разбавления от 1,56 нМ до 25 нМ BTN3A1 без регенерации между каждой концентрацией. Фазу ассоциации для трех инъекций увеличивающихся концентраций BTN3A1 отслеживали каждый раз в течение 240 секунд, а одну фазу диссоциации измеряли в течение 2000 секунд после последней инъекции BTN3A1. Регенерацию поверхности протеина A проводили с применением двух инъекций 10 мМ глицин-HCl, pH 1,5, с последующим периодом стабилизации в 240 секунд.
Сигнал от каждой холостой пробы опыта с антителами (без CD277) вычитали для корректировки различий в поверхностной стабильности. Одноцикловая кинетика продемонстрировала, что все гуманизированные варианты связываются с BTN3A.
d. Очистка антител
На основании аффинностей, рассчитанных с помощью Biacore, а также оценки с помощью iTope™ и процента гуманизации каждого гуманизированного варианта, для дальнейшего анализа были отобраны гуманизированные варианты 7.2 и 20.1 с лучшими аффинностями и лучшими оценками по iTope™.
Отобранные гуманизированные варианты вместе с их химерной версией и наиболее консервативно гуманизированным вариантом (VH1/Vκ1) подвергали очистке для дальнейшего анализа. Антитела очищали из супернатантов клеточных культур на колонках с протеин A-сефарозой с последующей гель-фильтрационной хроматографией (SEC) (GE Healthcare, Little Chalfont, Великобритания) с применением 10 мМ ацетата натрия, 100 мМ NaCl, pH 5,5, в качестве подвижной фазы и буфера для конечной композиции. Образцы количественно определяли по OD при 280 нм с применением коэффициента экстинкции (Ec (0,1%)) на основе предсказанной аминокислотной последовательности.
Антитела анализировали с помощью SDS-PAGE, нанося по 2 мкг каждого антитела на гель, и наблюдали полосы, соответствующие профилю типичного антитела.
e. Подтверждение свойств связывания: конкурентный анализ ELISA между гуманизированными и химерными 7.2 и 20.1 mAb
Очищенные варианты тестировали на их связывание с рекомбинантным BTN3A1-His («Sino Biological», № по каталогу 15973-H08H) в ходе конкуренции с соответствующим мышиным антителом. Химерный (VH0/Vκ0) и нерелевантный человеческий IgG4 (S241P, L248E) тестировали на каждом планшете для сравнения.
BTN3A1 разводили в 1x PBS до 0,5 мкг/мл и наносили по 100 мкл/лунку на 96-луночный планшет для ELISA с выдерживанием в течение ночи при 4°C. На следующий день планшет промывали 3 раза 1x ФСБ/0,05%-ным твином (ФСБ-T) и блокировали 200 мкл 2% молока/ФСБ в течение одного часа при комнатной температуре. В 96-луночный планшет для разведения добавляли в равном объеме фиксированную концентрацию мышиных антител 7.2 или 20.1 (конечная концентрация 0,15 мкг/мл) к серии четырехкратного титрования тестируемых антител (начиная с 80 мкг/мл (конечная концентрация 40 мкг/мл), разбавленных блокирующим буфером). После трехкратной промывки планшета Nunc для ELISA с помощью ФСБ-Т на планшет для ELISA добавляли 100 мкл смеси мышиное/тестируемое антитело. После одного часа инкубации при комнатной температуре планшет промывали 3 раза ФСБ-Т и для обнаружения связанного мышиного антитела наносили на один час при комнатной температуре 100 мкл вторичного антитела против мышиного Fc HRP-меченного антитела («Sigma», Дорсет, Великобритания), разведенного 1:1000 в блокирующем буфере. Для проявления цвета планшет промывали 3 раза ФСБ-Т, после чего добавляли 100 мкл субстрата ТМБ и инкубировали в течение пяти минут при комнатной температуре. Реакцию останавливали с помощью 100 мкл 3,0 М соляной кислоты и сразу же считывали оптическую плотность при 450 нм, применяя планшет-ридер Dynex.
Значения IC50 рассчитывали для каждого варианта, и относительные значения IC50 рассчитывали путем деления IC50 гуманизированного варианта на IC50 химерного антитела, анализируемого на том же планшете.
3. Отбор гуманизованных кандидатов
a. Многоцикловый кинетический анализ
На основании данных, полученных в результате конкурентного анализа ELISA и оценки термостабильности, был проведен многоцикловый кинетический анализ большинства гуманизированных вариантов 7.2 и 20.1 вместе с химерным антителом VH0/Vκ0 с применением прибора Biacore T200 (серийный номер 1909913) с помощью программного обеспечения для оценки Biacore T200 V2.0.1 (Упсала, Sweden).
Очищенные антитела разводили до концентрации 2 мкг/мл в 2%-ном БСА/ФСБ. В начале каждого цикла каждое антитело было захвачено белком А с плотностью (RL) ~ 146,5 относительных единиц (RU) (теоретическое значение для получения RMax ~ 50 RU). После захвата, поверхности давали возможность стабилизироваться перед инъекцией антигена BTN3A1 («Sino Biological», № по каталогу 15973-H08H). BTN3A1 титровали в 0,1% BSA/HBS-P+ (электродный буфер) в диапазоне двукратных разведений от 25 до 0,78 нМ. Фазу ассоциации наблюдали в течение 400 секунд, а фазу диссоциации - в течение 35 минут (2100 секунд). Кинетические данные получали при скорости потока 50 мкл/мин, чтобы минимизировать любые возможные эффекты массопереноса. Регенерацию покрытой протеином А поверхности проводили с применением двух инъекций 10 мМ глицин-HCl, pH 1,5, в конце каждого цикла. Два образца с холостыми пробами (без BTN3A1) и повторение одной концентрации аналита выполняли для каждого тестируемого антитела, чтобы проверить стабильность поверхности и аналита в течение кинетических циклов. Сигнал от эталонного канала Fc1 вычитали из сигнала Fc2, Fc3 и Fc4 для корректировки различий в неспецифическом связывании с эталонной поверхностью. Кроме того, для каждого Fc вычитали пустые серии, чтобы скорректировать любые антиген-независимые изменения сигнала, такие как дрейф. Сенсограммы были подогнаны с применением математической модели связывания «один-к-одному» с глобальным параметром RMax и без общего сигнала (константа RI = 0 RU). b. Анализ связывания с помощью проточной цитометрии на PBMC человека
Гуманизированные варианты 7.2 и 20.1 были охарактеризованы на предмет их связывания с человеческими PBMC, выделенными из крови здоровых доноров. PBMC выделяли из лейкоцитарных плёнок с помощью центрифугирования в градиенте плотности Lymphoprep («Axis-shield», Данди, Великобритания). Затем PBMC замораживали и хранили при -80°C или в жидком азоте до тех пор, пока они не потребовались.
100 мкл клеток при концентрации 1×106 клеток/мл переносили в каждую лунку с U-образным дном свежего 96-луночного планшета, затем планшет центрифугировали и супернатант отбрасывали.
Серийное разведение антител от 0,001 мкг/мл до 150 мкг/мл готовили в ФСБ, 2 мМ ЭДТК. Человеческие РВМС ресуспендировали в 50 мкл приготовленных серий для титрования разведенных тестируемых антител.
После инкубации в течение 30-ти минут при 4°C в темноте планшет центрифугировали и дважды промывали 150 мкл/лунку ФСБ, 2 мМ ЭДТК, после чего содержимое лунок ресуспендировали в 50 мкл смеси, состоящей из козьих антител против иммуноглобулинов человека (маркированных как PE), разведенных 1/100, и красителя Live/dead для ближней ИК области спектра, разведенного 1/500 в ФСБ, 2 мМ ЭДТК.
После инкубации в течение 15-ти минут при 4°C в темноте планшет центрифугировали и один раз промывали ФСБ 150 мкл/лунку, 2 мМ ЭДТК, после чего содержимое лунок ресуспендировали в 200 мкл ФСБ, 2 мМ ЭДТК. Клетки анализировали на цитометре BD LSR Fortessa Cytometer. Данные анализировали с помощью программного обеспечения FlowJo (версия 10, FlowJo, LLC, Ashland, США) (данные не приведены).
Тот же протокол выполняли для PBMC яванского макака и на линии клеток Дауди лимфомы Беркитта.
c. Функциональная эффективность in vitro: анализ дегрануляции γδ Т-клеток
Анализ состоит из измерения активирующего или ингибирующего эффекта гуманизированных вариантов 7.2 и 20.1 и их химерных версий на дегрануляцию γδ Т-клеток, направленную против линии Дауди лимфомы Беркитта (Harly et al., 2012). γδ Т-клетки размножали из PBMC здоровых доноров путем культивирования с золедроновой кислотой (1 мкМ) и IL2 (200 Ui/мл) в течение 11-13 дней. IL2 добавляли в день 5, день 8 и каждые 2 дня каждые 2 дня после этого. Процент γδ Т-клеток определяли в начале культивирования и оценивали в течение времени культивирования с помощью проточной цитометрии, пока он не достигал, по меньшей мере, 80%. Замороженные или свежие γδ Т-клетки затем применяли в анализах дегрануляции против линии клеток Дауди (соотношение E:T было равно 1:1), посредством чего клетки совместно культивировали в течение 4-х часов при 37°C в присутствии 10 мкг/мл гуманизированных вариантов 7.2 и 20.1 и их химерных версий. Активация с помощью PMA (20 нг/мл) плюс иономицин (1 мкг/мл) служила положительным контролем для дегрануляции γδ Т-клеток, а только среда – отрицательным контролем. В конце 4-часовой совместной инкубации клетки анализировали с помощью проточной цитометрии для оценки процента γδ Т-клеток, положительных по CD107a (LAMP-1, лизосомно-ассоциированный мембранный белок-1) + CD107b (LAMP-2). CD107 мобилизовался на поверхность клетки после вызванного активацией экзоцитоза гранул, таким образом, измерение поверхностного CD107 представляет собой чувствительный маркер для идентификации недавно дегранулированных цитолитических Т-клеток. Результаты не показали каких-либо значительных различий между тестируемыми кандидатами, которые продемонстрировали такой же активирующий эффект в анализе дегрануляции, что и химерное антитело 7.2 или 20.1.
Тот же протокол выполняли с применением бластов ОМЛ, полученных от пациентов, в качестве клеток-мишеней вместо клеток Дауди.
d. Анализ термостабильности
Чтобы оценить термостабильность выбранных вариантов сложных антител человека (Composite Human Antibody™) 7.2 и 20.1, температуры плавления (температура, при которой разворачивается 50% белкового домена) определяли с применением анализа теплового сдвига на основе флуоресценции.
Все очищенные гуманизированные антитела и химерные (VH0/Vκ0) антитела разводили 1:1000 до конечной концентрации 0,1 мг/мл в буфере (10 мМ ацетат натрия, 100 мМ NaCl, pH 5,5), содержавшем SYPRO® Orange («ThermoFisher», Лафборо, Великобритания), для получения состава и подвергали воздействию температурного градиента от 25°C до 99°C в системе ПЦР в реальном времени StepOnePlus («ThermoFisher», Лафборо, Великобритания) в течение 56 минут. 10 мМ ацетат натрия, 100 мМ NaCl, pH 5,5, применяли в качестве отрицательного контроля. Кривые плавления анализировали с применением программы для оценки термостабильности белков (версия 1,2).
e. Отбор гуманизированных кандидатов
На основании всех результатов, полученных для описанных выше экспериментов, 3 варианта из 35 (15 гуманизированных вариантов, созданных для 7.2; 20 гуманизированных вариантов, созданных для 20.1) были отобраны для дальнейшей характеристики: 7.2 (VH2/Vk1), 7.2 (VH2/Vk2) и 20.1 (VH3/Vk1).
Результаты различных экспериментов, описанных выше для mAb 7.2 и 20.1, представлены в таблице 4 и таблице 5 для 3-х вариантов и их химерных версий.
Таблица 4: Отобранные гуманизированные кандидаты обладают той же эффективностью, что и мышиные родительские антитела.
VH0/Vk0
VH3/Vk1
VH0/Vk0
(x 10-10)
(KD, M)
(EC50, мкг/мл)
яванского макака
(EC50, мкг/мл)
(EC50, мкг/мл)
(на основе γδT-клеток, EC50, мкг/мл)
(на основе γδ T-клеток, EC50, мкг/мл)
(на основе γδ T-клеток, EC50, мкг/мл)
Таблица 5: Гуманизированные отобранные кандидаты mAb7.2 как с VH2, так и с Vk1 или Vk2 обладают более высокой термостабильностью по сравнению с мышиными кандидатами.
(°C)
Процесс гуманизации привел к созданию множества вариантов 7.2 и 20.1 с предсказанной пониженной иммуногенностью.
Выбранный набор из трех вариантов (7.2 VH2/VK1, 7.2 VH2/VK2 и 20.1 VH3/VK1) продемонстрировал свойства, эквивалентные их химерной версии с точки зрения аффинности, связывания и эффективности в функциональном анализе: модификации, сделанные в последовательностях вариантов для снижения иммуногенности, не влияют на функции антител.
Удивительно, но термостабильность выбранных гуманизированных вариантов 7.2 VH2/VK1, 7.2 VH2/VK2 была улучшена по сравнению с химерными антителами, и такая улучшенная термостабильность была неожиданной в этом процессе гуманизации.
Константная область антитела: сравнение молчащих фрагментов Fc
Несколько частей Fc были протестированы для подавления или снижения эффекторной функции антител. Связывание этих фрагментов Fc с различными рецепторами Fcγ оценивали с помощью анализа Biacore; их связывание с комплексом C1q оценивали с помощью анализа ELISA.
1. Связывание сконструированной части Fc с различными рецепторами Fcγ с применением анализа Biacore
Способность различных изотипов (IgG1, IgG1 [N314A], IgG1 [L247F, L248E P350S], IgG2, IgG4 [S241P] и IgG4 [S241P L248E]) химерного антитела 20.1 связываться с разными рецепторами Fcγ определяли с применением очищенных антител и одноциклового анализа Biacore. Рецепторы Fc человека, FcγRI, FcγRIIA (полиморфизм Arg167) и IIB, и FcγRIIIA (полиморфизм Phe176) и IIIB были получены от «Sino Biological».
Рецепторы Fcγ разводили в HBS-P+ (GE Healthcare, Little Chalfont, Великобритания) до конечной концентрации 0,5 или 1,0 мкг/мл. В начале каждого цикла рецепторы Fcγ загружали на Fc2, Fc3 и Fc4 чипа анти-His CM5 (GE Healthcare, Little Chalfont, Великобритания). Рецепторы Fcγ захватывали при скорости потока 5 мкл/мин для получения уровня иммобилизации от 30 до 180 RU в зависимости от молекулярной массы рецептора Fcγ. Затем поверхности давали стабилизироваться. Данные одноцикловой кинетики получали с применением химерных антител в качестве аналита при скорости потока 30 мкл/мин, чтобы минимизировать любые потенциальные эффекты массопереноса. Сигнал от контрольного канала Fc1 (без антител) вычитали из сигнала от Fc2, Fc3 и Fc4, чтобы скорректировать различия в неспецифическом связывании с контрольной поверхностью. Для каждого химерного антитела применяли пятиточечный трехкратный диапазон разведения, причем этот диапазон концентраций варьировали для каждого индивидуального рецептора Fcγ из-за ожидаемых различий в аффинности. Сигнал от каждого холостого опыта (без антител) вычитали для корректировки различий в стабильности поверхности. Фазу ассоциации для каждой из пяти инъекций возрастающих концентраций химерного антитела отслеживали в течение от 25 до 180 секунд (в зависимости от лиганда рецептора Fcγ), а отдельную фазу диссоциации измеряли в течение 25-300 секунд после последней инъекции антитела. Регенерацию анти-His поверхности проводили с применением двух инъекций 10 мМ глицин-HCl, pH 1,5, в течение 15 секунд каждая при 30 мкл/мин с последующим периодом стабилизации 180 секунд.
Кинетические константы одного цикла определяли, где это было возможно, с применением стандартной модели анализа 1:1. Для сильных взаимодействий обычно более подходящим было определение сродства с помощью кинетических экспериментов. Однако для нескольких рецепторов Fcγ взаимодействие было очень слабым, и в этом сценарии данные были проанализированы с применением анализа сродства в стационарном состоянии (который особенно подходит для измерения взаимодействий от слабых до умеренных). Были получены сенсограммы взаимодействий рецепторов Fcγ с химерными антителами (данные не приведены).
Как и ожидалось, рецептор FcγRI с высоким сродством связывается с немодифицированным IgG1 и IgG4 (S241P) с хорошим сродством. Модифицированные изотипы IgG1 вместе с IgG2 и IgG4 (S241P, L248E) не смогли связаться с FcγRI. Остальные рецепторы Fcγ показали гораздо более низкие аффинные взаимодействия с различными химерными антителами по сравнению с FcγRI. Как и ожидалось, немодифицированный IgG1 показал наиболее сильное связывание со всеми четырьмя рецепторами с более низким сродством, тогда как модифицированные версии IgG1 продемонстрировали значительно сниженное связывание с этими рецепторами. IgG2 и IgG4 (S241P) продемонстрировали некоторое связывание с FcγRIIA и B, но только незначительное связывание с FcγRIIIA и B.
2. Связывание сконструированной части Fc с комплексом C1q по анализу ELISA
Химерное антитело 20.1 тестировали в форме разных изотипов IgG на связывание с комплексом C1q для определения их способности активировать систему комплемента.
В 96-луночном планшете с лунками с U-образным дном готовили серию 2,5-кратных разведений (от 10 мкг/мл до 0,04 мкг/мл) в 2% BSA/DФСБ очищенного химерного 20.1 в различных изотипах. 96-луночные планшеты для микротитрования с плоским дном Nunc Immuno MaxiSorp («ThermoFisher Scientific», Лафборо, Великобритания) предварительно покрывали 100 мкл/лунку из этой серии титрования и инкубировали в течение ночи при 4°C. На следующий день планшет дважды промывали ФСБT и блокировали в течение одного часа при комнатной температуре 2% БСА/ФСБ Дюльбекко перед пятикратной промывкой ФСБ-T. Очищенный белок комплемента C1q («Pathway Diagnostics Ltd», Доркинг, Великобритания), разбавленный до 5 мкг/мл в 2% БСА/ФСБ, добавляли в планшет (100 мкл/лунку) и инкубировали в течение одного часа при комнатной температуре. После пятикратной промывки ФСБ-T связывание комплекса C1q обнаруживали с помощью анти-C1q-HRP («Abcam», Кембридж, Великобритания) (100 мкл/лунка, разведение 1:100 в 2% BSA/DФСБ) в течение одного часа в комнатной температуре. После пятикратной промывки ФСБ-T было обнаружено связывание с субстратом ТМБ («ThermoFisher» Scientific, Лафборо, Великобритания), после чего реакцию останавливали 3 М HCl, оптическую плотность считывали при 450 нм на планшет-ридере Dynex Technologies MRX TC II и строили кривые связывания.
Как и ожидалось, только немодифицированный изотип IgG1 показал хорошее связывание с C1q, тогда как другие изотипы продемонстрировали минимальное связывание или его отсутствие (данные не приведены).
3. Выбор сконструированных фрагментов Fc
Относительное связывание всех протестированных изотипов с рецепторами Fcγ и комплексом C1q описано в таблице 6.
Таблица 6: Относительное связывание всех изотипов с рецепторами Fcγ и комплексом C1q
Два сконструированных Fc-фрагмента IgG1 L247F, L248E, P350S и IgG4 S241P, L248E были единственными фрагментами, которые продемонстрировали полную потерю связывания с рецептором FcγI и комплексом C1q.
На основании полученных результатов были отобраны два сконструированных изотипа IgG1 L247F, L248E, P350S и IgG4 S241P, L248E для дальнейшей характеристики.
Создание 6 гуманизированных антител
Три выбранных гуманизированных варианта были клонированы для слияния с двумя выбранными сконструированными фрагментами Fc, что привело к созданию 6-ти различных кандидатов: от mAb1 до mAb6.
Примеры mAb1-mAb6, как описано в таблице 1, можно получить с применением обычных способов получения и очистки рекомбинантных антител.
Например, кодирующие последовательности клонируют в вектор-продуцент для рекомбинантной экспрессии в линии продуцирующих клеток млекопитающих.
В следующих ниже таблицах 7 и 8 представлены подробные аминокислотные и нуклеотидные последовательности, полезные для осуществления изобретения и, в частности, для получения нуклеиновых кислот, экспрессионных векторов и гуманизированных антител, полученных из мышиного 7.2 по настоящему изобретению.
Таблица 7: Краткое описание полезных аминокислотных и нуклеотидных последовательностей для осуществления изобретения
Таблица 8: Краткое описание полезных аминокислотных и нуклеотидных последовательностей для практического применения изобретения
Свойства, обеспечивающие возможность разработки 6 гуманизированных вариантов
1. Создание клеточных линий
a. Конструирование экспрессионного вектора
Последовательности гена, кодирующие тяжелые и легкие цепи, клонировали в вектор. Систему генов применяли для экспрессии антител в клетках СНО. Экспрессия антител находилась под контролем промотора EF1 alpha. Экспрессионные векторы несли уникальные генетические элементы, которые защищали трансген от эффектов подавления экспрессии окружающим хроматином (Girod et al., 2007). Транскрипцию поддерживали на максимальном уровне, и она не зависела от сайта интеграции трансгена, что приводило к стабильной и высокоуровневой экспрессии белка.
b. Создание клеточных линий
Линию клеток-хозяев CHO получали из клеток CHO-K1 CCL-61 из Американской коллекции типовых культур 124 (ATCC) и адаптировали для роста в суспензии в химически определенной культуральной среде BalanCD Growth A (Irvine Scientific). Клетки трансфицировали электропорацией с применением системы трансфекции Neon (Invitrogen).
c. Клонирование одной клетки с применением устройства ClonePix FL.
Эту же среду (BalanCD Growth A, Irvine Scientific) применяли в качестве базовой среды для трансфекции, клонирования отдельных клеток и продукции, чтобы сохранить среду клеток неизменной на протяжении всей процедуры. После трансфекции каждого вектора применяли отбор под давлением пуромицина для создания стабильных пулов. Разбавленные клетки помещали в полутвердую среду (CloneMedia ©; Molecular Devices) и планшеты инкубировали при 37°C с 5%-ным CO2 в увлажняемом инкубаторе. Разросшиеся колонии отбирали с помощью Imager ClonePix™ FL от Molecular Devices и переносили в 96-луночные планшеты, затем размножали сначала в 24-луночных, а затем в 6-луночных планшетах TC.
d. Оценка производительности культивирования с подпиткой
Рост и продуктивность индивидуальных клонов оценивали во встряхиваемых колбах объемом 125 мл для отбора лучших клонов по критериям производительности и показателям роста клеток в 10-дневном периодическом процессе с подпиткой на основе Cell Boost7 A+7B (GE Healthcare, США). Культивирование с подпиткой инициировали при концентрациях клеток 0,3 × 106 клеток/мл.
Результаты, полученные для роста и продуктивности клеток, суммированы в таблице 9.
2. Подготовка образцов для оценки технологичности
Каждое антитело-кандидат очищали путем захвата протеином А из пулов очищенных супернатантов клеток СНО: для каждого варианта требовалось два пула, чтобы обеспечить достаточный материал для тестирования. Концентрацию белка определяли для всех образцов после захвата измерениями в УФ-диапазоне. Извлечение образцов и выходы для большинства образцов были равны более 80%, и все варианты показали аналогичные выходы. Затем для каждого антитела заменяли буфер на 25 мМ гистидин, 125 мМ NaCl, pH 6,0, центрифугированием с фильтрами с номинальным отсечением по молекулярной массе (MWCO) до 30 кДа до достижения pH в проходящем материале целевого значения. Во время стадии обмена не было никаких указаний на осаждение белка или замедление потока во время обмена для какого-либо из вариантов. После завершения замены буфера концентрацию каждого варианта образца доводили до 1,0 мг/мл с помощью буфера для композиции и добавляли 10% PS-80 до конечной концентрации 0,02% PS-80.
3. Оценка термической устойчивости
Дифференциальный сканирующий флуориметрический анализ проводили для оценки и сравнения термической стабильности тестируемого варианта антитела. Каждый вариант был проанализирован в трех повторах, и определяли среднее значение Tonset, Tagg и Tm для каждого наблюдаемого теплового перехода (данные не приведены).
Никаких существенных различий между значениями Tonset и Tm, полученными для всех тестируемых вариантов, не наблюдали. Определенное значение Tm1 для всех антител составляло 61°C, а значения, определенные для Tonset, составляли от 54 до 55°C для всех антител. Значения Tagg, определенные на основании графиков коллоидной стабильности, варьировались от 71 до 78°C.
В целом, все 4 отобранных варианта продемонстрировали сравнимую термостабильность: наблюдаемые вариации не приводят к значительным изменениям термостабильности между тестируемыми антителами.
4. Исследования по принудительной деградации
a. Перемешивание
Образцы для каждого варианта подвергали перемешиванию на орбитальном шейкере, установленном на 500 об/мин при комнатной температуре. Один образец для каждого варианта перемешивали в течение 24-х часов, а другой образец - в течение 48-ми часов. Один флакон каждого варианта хранили при комнатной температуре до 48-ми часов в качестве контроля. Для взбалтываемых образцов не наблюдали никаких изменений внешнего вида по сравнению с контрольными образцами: все образцы были прозрачными, бесцветными и не содержали видимых частиц (данные не приведены). Кроме того, не было значительных изменений в общем содержании белка, определенном измерениями в УФ-диапазоне (данные не приведены).
Влияние стресса от перемешивания на стабильность панели вариантов оценивали гель-проникающей хроматографией (SEC), капиллярным гель-электрофорезом (CGE) в восстанавливающих условиях (R-CGE), CGE в невосстанавливающих условиях (NR-CGE) и динамическим капиллярным изоэлектрофокусированием (icIEF) (данные не приведены). Не наблюдали никаких значительных изменений в стабильности тестируемых антител или заметных тенденций в накоплении продуктов разложения с течением времени между контрольными образцами для перемешивания, хранящимися при комнатной температуре, и образцами, подвергнутыми стрессу от перемешивания. Величина основного пика, определенного в анализе SEC, составляла ≥ 99,2% для всех контрольных и перемешиваемых образцов. В анализе R-CGE величина основного пика составляла ≥ 98,5% для всех контрольных и перемешиваемых образцов. Анализ NR-CGE не выявил значительных тенденций или изменений в % основного пика между контрольными и подвергнутыми стрессу образцами. Таким образом, значимых различий в стабильности между всеми вариантными кандидатами не наблюдали.
b. Стресс замораживанием-оттаиванием (F/T)
Три образца каждого кандидата помещали в пробирки типа Эппендорф и подвергали стрессу замораживанием-оттаиванием. Образцы хранили при -75±10°C, а затем оттаивали при комнатной температуре. Один образец для каждого кандидата был подвергнут 3-м циклам замораживания-оттаивания; другой 6-ти циклам замораживания-оттаивания; и третий образец 10-ти циклам. Обнаружили, что все подвергнутые стрессу образцы были прозрачными, бесцветными и свободными от видимых частиц (данные не приведены), и не было значительных изменений в общем содержании белка, как определено измерениями в УФ-диапазоне (данные не приведены).
Влияние стресса замораживанием-оттаиванием на стабильность панели вариантов оценивали способами SEC, CGE в восстанавливающих условиях, CGE в невосстанавливающих условиях и icIEF (данные не приведены). Стресс замораживанием-оттаиванием не влиял на стабильность антител на основании анализа SEC, R-CGE и NR-CGE.
Анализ icIEF выявил заметные изменения в гетерогенности тестируемых антител по заряду. Концентрация основных вариантов снижалась при последовательных циклах F/T, с самой низкой концентрацией основных вариантов при 10-кратных циклах F/T по сравнению с общими контролями. Единственным исключением был вариант IgG1 7.2 VH2/VK1, для которого снижение концентрации основных вариантов было на одном уровне для всех тестируемых циклов F/T (см. таблицу 9). В вариантах mAb1, mAb2 и mAb3 после 10X циклов F/T количество основных видов уменьшилось на 1,0, 1,3 и 3,4%, соответственно. Уменьшение концентрации основных видов связано с увеличением % основного пика. В вариантах mAb4, mAb5 и mAb6 уменьшение у основных видов составило 4,8, 2,8 и 4,7%, соответственно. Изменение в варианте mAb4 в основном связано с увеличением % основного пика, в то время как изменения в варианте mAb5 и mAb6 связаны с увеличением, как в % основного пика, так и в % кислого варианта.
В целом было определено, что самую высокую устойчивость к изменениям при стрессе замораживанием-оттаиванием наблюдали для вариантов mAb1 (IgG1 7.2 VH2/VK1) и mAb2 (IgG1 7.2 VH2/VK2).
c. Стресс кислым рН
Образцы для каждого кандидата подвергали стрессу кислым pH при комнатной температуре: каждый образец доводили с помощью HCl до pH 3,5 и выдерживали при комнатной температуре в течение 2-х часов, 4-х часов и 24-х часов, после чего образцы нейтрализовали 1 М трис, pH 7,0. Все образцы были прозрачными, бесцветными и свободными от видимых частиц (данные не приведены), и не было значительного изменения концентрации белка, как определено измерениями в УФ-диапазоне (данные не приведены).
Эффекты от стресса кислым pH на панели вариантов оценивали SEC, CGE в восстанавливающих условиях, CGE в невосстанавливающих условиях и icIEF (данные не приведены). Никаких значительных изменений в накоплении продуктов разложения с течением времени не было обнаружено для образцов, подвергнутых воздействию низкого pH, по сравнению с 48-часовым контролем RT с помощью анализа R-CGE или NR-CGE. При анализе гетерогенности заряда концентрация основных вариантов снижалась во всех образцах, подвергнутых стрессу кислым pH, однако четких тенденций с течением времени не наблюдали.
В целом наименьшее влияние кислого стресса на уменьшение основных вариантов наблюдали для вариантов mAb1 (IgG1 7.2 VH2/VK1) и mAb2 (IgG1 7.2 VH2/VK2). Этот результат хорошо согласуется с результатами, полученными с помощью icIEF при исследовании замораживания-оттаивания. В анализе SEC для всех вариантов наблюдали некоторое накопление агрегатов при воздействии кислого стресса, который был связан с уменьшением % основного пика, однако четких тенденций с течением времени не наблюдали. В целом, накопление агрегатов было примерно в 9 раз больше для вариантов IgG4 по сравнению с вариантами IgG1. Накопление агрегатов для вариантов IgG1 составляло от 0,3 до 1,0% для всех образцов кислотного стресса, а для вариантов IgG4 накопление составляло от 4,3 до 7,8% для образцов, подвергнутых стрессу, что значительно выше (смотри таблицу 9).
В заключении было установлено, что наибольшую устойчивость к кислотному стрессу наблюдали для вариантов mAb1 (IgG1 7.2 VH2/VK1) и mAb2 (IgG1 7.2 VH2/VK2).
d. Тепловой стресс
Образцы для каждого варианта подвергали тепловому стрессу в тепловом блоке при 50°C в течение 3-х дней, 1-ой недели и 2-х недель, а затем сравнивались с набором общих контрольных образцов, хранившихся при 2-8°C. Тестирование внешнего вида образцов проводили периодически, и не обнаружили признаков разделения фаз, изменения мутности или осаждения. Все образцы были прозрачными, бесцветными и не содержали видимых частиц во все моменты времени (данные не приведены). Кроме того, не было значительного изменения концентрации белка, определенного измерениями в УФ-диапазоне (данные не приведены).
Влияние теплового стресса на панель вариантов оценивали SEC, R-CGE, NR-CGE и icIEF. Результаты показали, что все варианты были подвержены тепловому стрессу.
В анализе SEC наблюдали четкие тенденции увеличения концентрации агрегатов в зависимости от времени для всех протестированных вариантов. Для вариантов IgG4 наблюдалось значительно большее накопление агрегатов по сравнению с IgG1. Через две недели увеличение % общих агрегатов колебалось от 33,7 до 44,7% для вариантов IgG4, но только от 13,0 до 18,2% для вариантов IgG1 (смотри таблицу 9). Кроме того, варианты mAb1 (IgG1 7.2 VH2/VK1) и mAb2 (IgG1 7.2 VH2/VK2) накапливали меньше агрегатов, чем контрольное mAb3 (IgG1 20.1 VH3/VK1).
В анализе R-CGE наблюдали четкие тенденции к снижению чистоты (определяемой как уменьшение % LC + HC) как функции времени для всех тестируемых вариантов, однако между ними не наблюдали значительных различий в стабильности. Уровень деградации образцов был сопоставим для всех протестированных антител (данные не приведены). Аналогичным образом, данные NR-CGE выявили четкие тенденции к снижению чистоты (определяемой как уменьшение основного пика) как функции времени для всех протестированных вариантов. Для вариантов IgG4 наблюдали значительно более низкую чистоту по сравнению с IgG1 (данные не приведены). Через две недели % чистоты снизился на 34,8-41,7% для вариантов IgG4, но только на 20,0-30,0% для вариантов IgG1 (данные не приведены). Кроме того, варианты mAb1 (IgG1 7.2 VH2/VK1) и mAb2 (IgG1 7.2 VH2/VK2) продемонстрировали более низкую деградацию, чем mAb3 IgG1 20.1 VH3/VK1. Для антител IgG1 уменьшение основного пика в первую очередь сопровождалось увеличением примесей (фрагментов) переднего основного пика. Для антител IgG4, однако, наблюдали накопление примесей, как переднего основного пика (фрагментов), так и примесей заднего основного пика (агрегатов) как функцию уменьшения основного пика (данные не приведены). Этот результат хорошо согласуется с данными SEC, где для антител IgG4 наблюдали значительно более высокую концентрацию агрегатов по сравнению с IgG1.
Однонедельные образцы для вариантов IgG4 были слишком деградированы для проведения анализа с помощью icIEF, и двухнедельные образцы для всех вариантов были слишком деградированы для анализа. Таким образом, для оценки изменений гетерогенности заряда между тестируемыми антителами применяли данные только для трехдневных. Разница в чистоте, определяемая по % основного пика, составляла от 12,4 до 14,4% для образцов IgG1 и от 16,0 до 16,6% для образцов IgG4 (данные не приведены).
В целом, было продемонстрировано, что деградация при тепловом стрессе была больше для вариантов IgG4 по сравнению с вариантами IgG1, и это наблюдали во все моменты времени. Анализ показывает, что варианты mAb1 (IgG1 7.2 VH2/VK1) и mAb2 (IgG1 7.2 VH2/VK2) наименее восприимчивы к тепловому стрессу, и этот результат согласуется с данными, полученными для стрессов замораживанием-оттаиванием и кислым pH.
Подбор лучшего кандидата с точки зрения свойств, обеспечивающих возможность дальнейшей разработки
Что касается продуктивности клеточных линий, наилучшие результаты были получены с гуманизированным вариантом mAb1 (7.2 VH2/VK1) с увеличением плотности жизнеспособных клеток до 54 x 106 жизнеспособных клеток/мл за 10 дней для mAb1.
При исследовании термостабильности образцы оценивали в стандартной матрице дифференциальной сканирующей флуориметрией (DSF) для определения Tonset, Tm и Tagg для каждого кандидата. Никаких изменений в Tonset и Tm1 не обнаружили, и Tagg была выше 70°C для всех вариантов. Результаты показывают, что mAb1, mAb2, mAb4 и mAb5 демонстрируют сравнимую термическую стабильность.
В исследовании принудительной деградации образцы подвергали перемешиванию, замораживанию-оттаиванию, кислому pH и тепловому стрессу. Результаты показывают, что панель вариантов кандидатов была подвержена деградации в различной степени в соответствующих стрессовых условиях:
- Не наблюдали значительной реакции на стресс при перемешивании ни одним из аналитических способов.
- Не наблюдали значимого ответа на стресс замораживанием-оттаиванием SEC или CGE, однако, анализ icIEF выявил различия в гетерогенности по заряду между тестируемыми вариантами и показал, что варианты mAb1 (IgG1 7.2 VH2/VK1) и mAb2 (IgG1 7.2 VH2/VK2) проявили наибольшую устойчивость к изменениям, вызванным стрессом замораживанием-оттаиванием.
- Реакцию на стресс кислым pH наблюдали с помощью icIEF и SEC, при этом все кандидаты накапливали некоторые примеси при воздействии. Варианты mAb1 (IgG1 7.2 VH2/VK1) и mAb2 (IgG1 7.2 VH2/VK2) проявляли самую высокую устойчивость к стрессу кислым pH.
- Наиболее значительную реакцию на стресс наблюдали при хранении образца при 50°C. Анализы SEC, NR-CGE и icIEF выявили значительную тенденцию к снижению чистоты образца с течением времени. Наблюдаемое снижение чистоты было стабильно больше для вариантов IgG4 по сравнению с вариантами IgG1. В группе вариантов IgG1 наибольшую устойчивость к кислотному тепловому стрессу проявили mAb1 (7.2 VH2/VK1) и mAb2 (IgG1 7.2 VH2/VK2).
Результаты анализа свойств, обеспечивающих возможность дальнейшей разработки для различных гуманизированных кандидатов, суммированы в приведенной ниже таблице 9:
Таблица 9: Гуманизированный кандидат mAb1 имеет более выраженные свойства, обеспечивающие возможность дальнейшей разработки, по сравнению со всеми другими протестированными гуманизированными вариантами.
(увеличение % агрегатов при pH 3,5 через 24 часа)
(разница в % агрегатов, измеренная с помощью анализа SEC через 2 недели при 50°C)
VK1 IgG1
550 мкг/мл
1,2
1,0
VK2 IgG1
330 мкг/мл
0,6
1,3
VK1 IgG1
2,0
3,4
VK1 IgG4
780 мкг/мл
1,9
3,0
475 мкг/мл
1,8
2,8
VK1 IgG4
430 мкг/мл
2,4
4,4
1 Изменение в % от основных вариантов было рассчитано на основе результатов без округления, вычитая % основных вариантов образцов, подвергнутых стрессу, из соответствующего контрольного образца.
В итоге вариант mAb1 показал лучшие результаты относительно возможности дальнейшей разработки, и он был выбран в качестве ведущего кандидата.
Функциональные свойства вариантов гуманизированных mAb 7.2 как одновалентных и двухвалентных молекул
1. Получение фрагментов Fab и Fab2
a. Расщепление пепсином для генерации F(ab')2
Иммобилизованный пепсин в виде 50% суспензии (набор «Thermo Scientific», № по каталогу 44988) подвергали замене буфера на буфер для расщепления (20 мМ ацетат натрия, pH 4,4) путем центрифугирования суспензии при 5000 g в течение 2-х мин. Супернатант удаляли и суспензию ресуспендировали в 1 мл буфера для расщепления с последующим повторным центрифугированием при 5000 g в течение 2-х мин. Этот этап повторяли еще четыре раза (всего пять промывок смолы). Затем смолу ресуспендировали в буфере для расщепления до исходного объема суспензии.
mAb 4, mAb 5 и mAb 6 (варианты 7.2 VH2/VK1, 7.2 VH2/VK2 или 20.1 VH3/VK1 IgG4) подвергали замене буфера на буфер для расщепления с применением колонки Zeba Spin 5 или 10 мл (в зависимости от масштаба) и концентрировали до 3 мг/мл с применением концентратора Vivaspin (10000 MWCO) в соответствии с протоколом производителя.
mAb 4, 5 и 6 в концентрации 3 мг/мл (буфер был предварительно заменен на буфер для расщепления) смешивали с пепсином, иммобилизованным на смоле, и инкубировали при 37°C при вращении в течение 1,5-2 часов.
Расщепленные смеси центрифугировали при 5000 g в течение 2-х мин. Супернатанты отбирали и отфильтровали в свежую пробирку с применением подходящего шприца и небольшого фильтра 0,22 мкм (Merck Millipore, Millex № по каталогу SLGV004SL). Смолы промывали 1 мл ФСБ, центрифугировали при 5000 g в течение 1-ой мин. Супернатанты отбирали, отфильтровали и объединяли с предыдущими супернатантами. Стадию промывки повторяли.
Прошедшие через ферментаивное расщепление и фильтрацию супернатанты очищали с применением гель-хроматографической колонки HiLoad 16/600 200 pg, применяя 10 мМ ацетат натрия, 100 мМ NaCl, pH 5,5, в качестве подвижной фазы. Фракции, соответствующие элюированному фрагменту F(ab')2, объединяли, концентрировали и стерильно фильтровали.
F(ab')2 фрагменты анализировали с помощью SDS-PAGE, аналитического SEC и считывания OD280nm.
b. Расщепление папаином для генерации Fab
Иммобилизованный папаин в виде 50% суспензии (набор «Thermo Scientific», № по каталогу 20341) подвергали замене буфера на буфер для расщепления (20 мМ фосфат натрия, 10 мМ ЭДТА, 150 мМ цистеин, pH 7,0) путем центрифугирования суспензии при 5000 g в течение 2-х мин. Супернатант отбрасывали и суспензию ресуспендировали в большем объеме буфера для расщепления с последующим повторным центрифугированием при 5000 g в течение 2-х мин. Эту стадию повторяли еще четыре раза (всего пять промывок смолы). Затем смолу ресуспендировали в буфере для расщепления до исходного объема суспензии.
mAb 4, mAb 5 и mAb 6 (варианты 7.2 VH2/VK1, 7.2 VH2/VK2 или 20.1 VH3/VK1 IgG4) подвергали заменебуфера на буфер для расщепления с применением колонки Zeba Spin 5 или 10 мл (в зависимости от масштаба) и концентрировали до 3 мг/мл с применением концентратора Vivaspin (10000 MWCO) в соответствии с протоколом производителя.
mAb 4, 5 и 6 в концентрации 3 мг/мл (буфер был предварительно заменен на буфер для расщепления) смешивали с папаином, иммобилизованным на смоле, и инкубировали при 37°C при вращении, 42 часа.
Смеси после расщепления осаждали при 5000 g в течение 2-х мин. Супернатанты извлекали и фильтровали в свежую пробирку с применением подходящего шприца и небольшого фильтра 0,22 мкм (Merck Millipore, Millex, номер по каталогу SLGV004SL). Смолы трижды промывали ФСБ, центрифугировали при 5000 g в течение 1-ой мин, собирая и объединяя в каждом цикле супернатант. Затем объединенные фракции фильтровали в свежую пробирку, применяя подходящий шприц и небольшой фильтр 0,22 мкм.
В прошедших через ферментативное расщепление и фильтрацию супернатантах сначала заменяли буфер на 1xФСБ Дюльбекко, pH 7,4, затем сначала очищали с применением колонки с протеином A для удаления Fc и непереваренных mAb, после чего следовала стадия очистки с применением гель-фильтрационной колонки (SEC) с 10 мМ ацетатом натрия, 100 мМ NaCl, pH 5,5 в качестве подвижной фазы. Фракции, соответствующие элюированным Fab-фрагментам, объединяли, концентрировали и подвергали стерильной фильтрации.
Fab-фрагменты анализировали с помощью ДНС-ПААГ, аналитической SEC и измерения OD280нм.
2. Измерение аффинности гуманизированных фрагментов 7.2 и 20.1 Fab, F(ab')2 и формата IgG с применением Biacore
Для оценки сродства ведущих гуманизированных антител 7.2 и 20.1 к BTN3A1 в их различных форматах (Fab, F (ab')2 и IgG), был проведен одноцикловый кинетический анализ с помощью прибора Biacore T200 (серийный номер 1909913) с применением программного обеспечения Biacore T200 Control V2.0.1 и программного обеспечения для оценки V3.0 (GE Healthcare, Упсала, Швеция). Все одноцикловые кинетические эксперименты проводили при 25°C с рабочим буфером HBS-P+ (pH 7,4), содержавшим 0,1% БСА (GE Healthcare, Little Chalfont, Великобритания).
His-меченный антиген BTN3A1 («Sino Biological», Пекин, Китай) разводили в рабочем буфере до конечной концентрации 0,4 мкг/мл. В начале каждого цикла BTN3A1-His был захвачен на Fc2, предварительно присоединенный к сенсорному чипу CM5 с применением набора для связывания His (GE Healthcare, Литтл Чалфонт, Великобритания) с помощью стандартной аминной химии при скорости потока 10 мкл/мин. Уровень иммобилизации (RL) ~ 34 RU, 16 RU или 11 RU, различные теоретические значения для получения RMax ~ 50 RU применяли для аналитов Fab, F(ab')2 и IgG, соответственно. Затем поверхности давали стабилизироваться. Данные одноциклового кинетического анализа получали с очищенными образцами (Fab, F(ab')2 и IgG) при скорости потока 40 мкл/мин, чтобы минимизировать любые потенциальные эффекты массопереноса. Сигнал от контрольного канала Fc1 (без связывания антигена) вычитали из сигнала Fc2, чтобы скорректировать различия в неспецифическом связывании с контрольной поверхностью. Сигнал для холостых прогонов BTN3A1-His (без аналита) вычитали для корректировки различий в стабильности поверхности. Фазу ассоциации для пяти инъекций возрастающих концентраций отслеживали в течение 240 секунд каждый раз, а одну фазу диссоциации измеряли в течение 1400 секунд после последней инъекции аналита. Регенерацию поверхности чипа проводили с применением двух инъекций 10 мМ глицин-HCl pH 1,5 с последующим периодом стабилизации в 240 секунд. Исходные сенсограммы были подогнаны к модели 1:1 для образцов Fab и к модели бивалентного аналита для образцов F(ab')2 и IgG в соответствии с различными валентностями аналитов. Кинетические константы рассчитывались для каждого варианта (смотри таблицы 10, 11 и 12).
Таблица 10: Одноцикловый кинетический анализ IgG
(1/сек)
Таблица 11: Одноцикловый кинетический анализ F(ab')2
(1/сек)
Таблица 12: Одноцикловый кинетический анализ Fab
3. Выбор лучшего кандидат по аффинным свойствам
Гуманизированные варианты 20.1 и 7.2 показали значительные различия с точки зрения аффинных свойств. Эти различия можно наблюдать с форматами IgG и F(ab')2, но разрыв еще выше с Fab-фрагментами: действительно, 20.1 показал средний KD, равный 20,30 нМ, тогда как 7.2 показал средний KD, равный 3,22 (VH2/VK1) или 3,01 (VH2/VK2).
4. Авидность связывания mAb1 с первичными Т-клетками и другими клеточными линиями.
Затем авидность связывания mAb1 была оценена с помощью проточной цитометрии на первичных Т-клетках человека, а также на различных клеточных линиях, включая WT и нокаут BTN3A, восстановленные с помощью изоформ BTN3A1, BTN3A2 или BTN3A3 по отдельности (данные не приведены). Значения EC50, полученные для каждого протестированного типа клеток, суммированы в таблице 13.
Данные, полученные на клетках BTN3A KO, ясно показали, что mAb1 специфически связывается со своей мишенью. Кроме того, восстановление отдельными изоформами подтвердило, что mAb1 распознает BTN3A1, BTN3A2 и BTN3A3 с сопоставимой авидностью. Все другие протестированные клетки оказались положительными по связыванию mAb1 с диапазоном значений EC50 от 9,6 нМ для линии клеток Дауди лимфомы Беркитта до 112,3 нМ для линии клеток колоректальной аденокарциномы HT29.
Таблица 13: Авидность связывания mAb1 на клетках человека.
мкг/мл
(+/-sem)
(x10-9 M)
(+/-sem)
(+/-0,57)
(+/-3,8)
(+/-0,16)
(+/-1,04)
(+/-4,4)
(+/-29,3)
sem - средняя квадратическая ошибка среднего
5. mAb1 не демонстрирует нецелевое связывание
Возможность нецелевого связывания mAb1 с другими молекулами, не относящимися к BTN3A, оценивалась с помощью технологии Retrogenix. Эта работа была направлена на демонстрацию отсутствия нецелевого связывания путем скрининга клеточного массива, экспрессирующего >5000 мембранных рецепторов человека или секретируемых белков, экспрессируемых на поверхности клеток HEK293 (платформа Retrogenix).
Исследование уровня связывания mAb1 с нетрансфицированными клетками HEK293 и с клетками, сверхэкспрессирующими BTN3A1, до или после фиксации клеток, показало, что 5 мкг/мл на фиксированных клетках представляет собой подходящее условие скрининга. В этих условиях mAb1 подвергали скринингу на связывание с человеческими клетками HEK293, индивидуально экспрессирующими 5528 белков человека, включая белки мембран клеточной поверхности и секретируемые белки, привязанные к клеточной поверхности. Анализ выявил десять основных попаданий.
В случае каждого основного попадания проводили повторную экспрессию вместе с двумя контрольными рецепторами (CD20 и EGFR) и повторно тестировали с 5 мкг/мл mAb1, 5 мкг/мл антитела изотипического контроля и другими обработанными образцами положительного и отрицательного контроля. После удаления пяти неспецифических попаданий осталось пять специфических взаимодействий для тестируемого антитела. Все 5 конкретных совпадений были связаны с BTN3A; две изоформы BTN3A1, две изоформы BTN3A2 и одна изоформа BTN3A3.
Выводы исследования заключаются в том, что для mAb1 не было выявлено нецелевых взаимодействий, что указывает на высокую специфичность mAb1 в отношении его эпитопа BTN3A.
6. mAb1 опосредует активацию Vγ9Vδ2 Т-клеток и уничтожение опухолевых клеток
Первоначально было показано, что мышиные mAb против BTN3A запускают распознавание BTN3A Vγ9Vδ2Т-клетками, опосредуя активацию, ведущую к (i) пролиферации этой специфической подгруппы в человеческих PBMC, (ii) продукции цитокинов (IFNγ и TNFα) и (iii) цитолиз инфицированных или трансформированных клеток-мишеней (например, перфорином, гранзимами, TRAIL) (Harly et al., Blood, 2012 & Benyamine, A. et al. 2016, Oncoimmunology 5, e1146843).
Не ограничиваясь какой-либо конкретной теорией, предлагаемый механизм действия mAb1 заключается в том, что его связывание с BTN3A, экспрессируемым на поверхности опухолевой клетки-мишени, запускает конформационное изменение, которое позволяет ему передавать сигнал его контррецептору на Vγ9Vδ2 Т-клетках. Активность антител против BTN3A обычно оценивают с помощью анализа in vitro, основанного на совместном культивировании линии опухолевых клеток (мишени) с первичными человеческими Т-клетками Vγ9Vδ2 (эффекторными), ранее размноженными из PBMC здоровых доноров в течение 10-14 дней, в присутствии rHuIL-2 (200 UI/мл) и аминобисфосфонатов («Zometa», 1 мкМ). В конце фазы размножения чистоту T-клеток Vγ9Vδ2 оценивали с помощью проточной цитометрии, и эти клетки затем замораживали для будущего применения. За день до эксперимента размноженные Т-клетки Vγ9Vδ2 оттаивали и культивировали в течение ночи с 200 МЕ/мл rHuIL-2 для поддержания выживания in vitro. После совместного культивирования активацию T-клеток Vγ9Vδ2 отслеживали либо с помощью проточной цитометрии, детектирующей экспрессию CD107a/b на γδ Т-клетках, либо путем количественной оценки активации Caspase3/7 в качестве меры уничтожения клеток-мишеней.
Исходно, человеческие Т-клетки Vγ9Vδ2, размноженные из 3 различных здоровых донорных PBMC, культивировали совместно с линией клеток Дауди (ATCC-CCL213; лимфома Беркитта) с возрастающими концентрациями mAb1. Через 4 часа клетки анализировали на экспрессию CD107a/b Т-клетками Vγ9Vδ2 с помощью проточной цитометрии. Результаты показали связанное с концентрацией увеличение процента Т-клеток Vγ9Vδ2, экспрессирующих CD107a/b, со средним значением EC50 0,89 нМ (+/- 0,39) (фигура 1A). Параллельно мы оценивали цитолитическую активность T-клеток Vγ9Vδ2 в отношении клеток Дауди, активированных mAb1. Как показано на фигуре 1B, mAb1 индуцировало апоптоз клеток-мишеней зависимым от концентрации образом с EC50 0,35 нМ. Кроме того, активация T-клеток Vγ9Vδ2 (CD107a/b; фигура 1C верхняя панель) и лизис опухолевых клеток (Casp3/7; нижняя панель фигура 1C) были протестированы с применением линий опухолевых клеток из различных тканей, и их сравнивали с клетками Дауди. Результаты показали, что активация T-клеток Vγ9Vδ2 и последующий лизис опухолевых клеток были вызваны связыванием mAb1 с клеточными линиями L-IPC (протоковая аденокарцинома поджелудочной железы), A549 (эпителиальные клетки карциномы легких) и HT29 (колоректальная аденокарцинома).
7. mAb1 усиливает уничтожение T-клетками Vγ9Vδ2 широкого ряда линий клеток человека, экспрессирующих BTN3A, независимо от того, из каких тканей они происходят
a. Материалы и методы
Культура опухолевых клеток
HL-60 представляют собой клеточную линию промиелобластов человека, полученную при остром промиелоцитарном лейкозе. Дауди представляет собой линию B-лимфобластных клеток человека, полученную из лимфомы Беркитта. Джуркат (Jurkat) представляет собой клеточную линию Т-клеток острой лейкемии.
HT-29 и HCT116 представляют собой эпителиальные клетки человека, полученные из колоректальной аденокарциномы.
PC3 и DU145 были получены из карциномы простаты (место метастазирования, кость и мозг, соответственно).
SUM159 и MDA-MB-231 представляют собой клеточные линии трижды негативного рака груди (TNBC).
Клетки HL60, Дауди, Джуркат, DU145 и MDA-MB-231 культивировали в RPMI Glutamax, 10% ФБС; 1 мМ пируват натрия при 37°C/5% CO2. PC3 и HT-29 культивировали в минимальная основная среда Игла, модифицированная по способу Дульбекко (DMEM) 10% ФБС, 1 мМ пирувата натрия при 37°C/5% CO2. HCT116 культивировали в среде McCoy 5a 10% SVF. SUM159 культивировали в среде F12 Nut Mix 1X + глутамакс, 5% SVF, гидрокортизон 2 мг/мл, 2 мг/мл инсулина гумалог и незаменимые аминокислоты.
In vitro размножение Vγ9Vδ2 T-клеток
Мононуклеарные клетки периферической крови (PBMC) выделяли из периферической крови, полученной из Etablissement 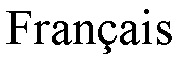 du Sang Provence Alpes Côte d'Azur (Франция), центрифугированием в градиенте плотности на основе фиколла. Для размножения T-клеток Vγ9Vδ2 50х106 PBMC ресуспендировали при 1,5х106 клеток/мл в RPMI1640 с добавлением 10% ФБС и 1% пирувата натрия в колбах площадью 75 см2 в течение 10-14 дней в присутствии rHuIL-2 (200UI/мл) и аминобисфосфонатов (золедронат, 1 мкМ). Начиная с 5-го дня, rHuIL-2 обновляли каждые 2 или 3 дня, а количество клеток сохраняли при 1×106/мл. В конце фазы размножения чистоту T-клеток Vγ9Vδ2 оценивали с помощью проточной цитометрии, и если количество T-клеток Vγ9Vδ2 достигало 80% от живых клеток, то эти клетки затем замораживали в ФБС 20% ДМСО для будущего применения.
du Sang Provence Alpes Côte d'Azur (Франция), центрифугированием в градиенте плотности на основе фиколла. Для размножения T-клеток Vγ9Vδ2 50х106 PBMC ресуспендировали при 1,5х106 клеток/мл в RPMI1640 с добавлением 10% ФБС и 1% пирувата натрия в колбах площадью 75 см2 в течение 10-14 дней в присутствии rHuIL-2 (200UI/мл) и аминобисфосфонатов (золедронат, 1 мкМ). Начиная с 5-го дня, rHuIL-2 обновляли каждые 2 или 3 дня, а количество клеток сохраняли при 1×106/мл. В конце фазы размножения чистоту T-клеток Vγ9Vδ2 оценивали с помощью проточной цитометрии, и если количество T-клеток Vγ9Vδ2 достигало 80% от живых клеток, то эти клетки затем замораживали в ФБС 20% ДМСО для будущего применения.
Очистка Vγ9Vδ2 Т-клеток из человеческих PBMC
Свежие человеческие РВМС ресуспендировали при концентрации 50×106 клеток/мл в ФСБ + 2% ФБС + 1 мМ ЭДТК. Vγ9Vδ2 выделяли с применением набора для выделения гамма/дельта-Т-клеток человека EasySep™ («Stemcell», # 19255) в соответствии с инструкциями производителя. В конце процедуры чистоту клеток измеряли проточной цитометрией. Живых CD3+ Vγ9+ клеток было более 80%.
Анализ уничтожения Vγ9Vδ2 Т-клеток
10000 размноженных или свежих T-клеток Vγ9Vδ2 совместно культивировали с линиями опухолевых клеток в 96-луночных планшетах при указанном соотношении (E:T 1:1 или 1:5) в RPMI 1640 + глутамакс, 10% ФБС + 1 мМ NaPy в присутствии mAb1 или соответствующего изотипического контроля. При добавлении к совместной культуре rHuIL-2 применяли 20 МЕ/мл. По истечении указанного времени ATP измеряли с применением реагента Glo («Promega» # G7572), который генерирует люминесцентный сигнал, пропорциональный количеству живых клеток.
b. Результаты
В дополнение к способу, основанному на окрашивании клеток-мишеней каспазой 3/7 для мониторинга уничтожения при совместном культивировании с Т-клетками Vγ9Vδ2, мы разработали дополнительный подход, основанный на оценке количества жизнеспособных клеток в культуре на основе количественного определения присутствующего АТФ, индикатора метаболически активных клеток.
С помощью этого анализа мы отслеживали выживаемость линии клеток острого миелоидного лейкоза HL60-WT, или -BTN3A-KO, через 24 часа после совместного культивирования с размноженными Т-клетками Vγ9Vδ2 (соотношение E:T 1:1) в присутствии mAb1 или соответствующего изотипического контроля +/-rHuIL-2 (20 МЕ/мл) (фигура A). Результаты показали зависимое от концентрации снижение выживаемости HL60-WT в присутствии mAb1, что свидетельствует об эффективном BTN3A-зависимом уничтожении опухолевых клеток-мишеней активированными Т-клетками Vγ9Vδ2. Подобные эксперименты проводили с размноженными in vitro или со свежевыделенными Т-клетками Vγ9Vδ2 (фигура B и C, соответственно), и жизнеспособность клеток измеряли в течение 4-дневного периода.
На фигуре B показано, что размноженные in vitro T-клетки Vγ9Vδ2 контролировали пролиферацию HL60-WT зависимым от концентрации образом. Добавление rHu-IL2 со временем улучшает способность эффекторные клетки к уничтожению, вероятно, обеспечивая сигналы выживания, необходимые для культивирования первичных Т-клеток in vitro. Аналогичные результаты были получены со свежими Т-клетками Vγ9Vδ2, выделенными из PBMC человека, совместно культивированными в течение 4-х дней с HL60-WT при соотношении E: T 1:1 и 1:5. Эти результаты указывают на способность отдельных T-клеток Vγ9Vδ2 многократно взаимодействовать и убивать множество опухолевых мишеней при опосредованной mAb1 активации (фигура 2C).
Затем мы применяли этот анализ для мониторинга опосредованной mAb1 активности по уничтожению Т-клетками Vγ9Vδ2 экспрессирующих BTN3A клеточных линий из различных тканей (толстой кишки, молочной железы, простаты, Т-лимфомы и лимфомы Беркитта). Клетки HT29, PC3, DU145, MDA-MB-231, HCT116, SUM159, Джуркат и Дауди совместно культивировали в течение ночи с размноженными in vitro Т-клетками Vγ9Vδ2 в присутствии возрастающей концентрации mAb1 и измеряли жизнеспособность клеток. Результаты подтвердили, что mAb1 усиливает опосредованное Т-клетками Vγ9Vδ2 уничтожение широкого ряда линий клеток человека, экспрессирующих BTN3A, независимо от того, из каких тканей они происходят.
Таблица 15: 10000 опухолевых клеток совместно культивировали в течение 24-х часов с размноженными in vitro Т-клетками Vγ9Vδ2 (соотношение E:T 1:1) в присутствии различных концентраций mAb1. Жизнеспособность клеток измеряли с помощью биолюминесцентного анализа для определения уровней ATP. Указаны значения биолюминесценции ×105.
0,1 мкг/мл
1 мкг/мл
10 мкг/мл
левых клеток
10. mAb1 улучшает терапию Vγ9Vδ2 Т-клетками на мышиной модели, трансплантированной клеточными линиями AML человека
Поскольку BTN3A, представляющие собой мишень для mAb1, не экспрессируются у грызунов, а субпопуляция Т-клеток VgγVδ2 специфична для приматов, то экспериментальное подтверждение концепции противоопухолевой активности T-клеток Vγ9Vδ2 обычно тестируют на мышах с ослабленным иммунитетом NSG, привитых линиями опухолевых клеток человека, которые экспрессируют BTN3A и в которые адоптивно перенесены Т-клетки Vγ9Vδ2 от здоровых доноров-людей (Т-клетки Vγ9Vδ2 в таких реконструкциях представляют собой аллогенные клетки по отношению к линии опухолевых клеток). Эти модели широко применяют для проверки противоопухолевого терапевтического потенциала T-клеток Vγ9Vδ2 в широком диапазоне солидных, а также гематологических злокачественных новообразований (Pauza, C. D. et al. 2018, Immunol. 9).
Кроме того, было описано, что мышиное антитело против BTN3A человека, m20.1, вводимое мышам, получившим человеческие Т-клетки Vγ9Vδ2, увеличивает выживаемость животных и снижает лейкемическую нагрузку в крови и костном мозге мышей, несущих NSG (Benyamine et al 2016, смотри выше). В этой модели m20.1 вводили в комбинации с RLI, слитым белком rHuIL-15/IL-15Rα, который, как было показано, увеличивал количество противоопухолевых лимфоцитов и улучшал их выживаемость у мышей, что приводило к лучшей противоопухолевой активности перенесенных человеческих T-клеток Vγ9Vδ2.
Для дальнейшей характеристики эффективности mAb1 в моделях in vivo на мышах мы исследовали потенциал переноса человеческих T-клеток Vγ9Vδ2 в сочетании с mAb1 для задержки роста опухоли и улучшения выживаемости животных у мышей NSG, несущих AML.
a. Модель U937
Здоровые самки мышей в возрасте 6-8 недель (n = 30) получали 0,2×106 трансдуцированных люциферазой клеток U937 в день 0, как описано в работе Gertner-Dardenne, J. et al. 2012. J. Immunol. 188, 4701-4708. В день 1 опухолевую нагрузку оценивали с помощью биолюминесцентного изображения, и мышей случайным образом распределяли по шести группам для внутривенных инъекций размноженных in vitro человеческих T-клеток Vγ9Vδ2 в день 1 (3×106 клеток) отдельно или в сочетании с mAb1 против BTN3A или с соответствующим изотипическим контролем (10 мг/кг, 200 мкг/мышь). Обработку повторяли на день 7. В группах 2 и 4 антитела также вводили на день 4 и 10. Поскольку комплексы rHuIL-15/rHuIL15-Rα позволяют пролиферацию T-клеток Vγ9Vδ2 (J. Immunol. (2006) 177, 6072-608), эти комплексы вводили вместе с Т-клетками Vγ9Vδ2.
Все группы описаны в 16.
Комплексы hIL-15/IL-15R-Fc предварительно формировали при комнатной температуре (RT) в течение 30 минут перед инъекцией (0,2 мкг hIL-15 + 1,2 мкг IL-15R-Fc на мышь) и перед инъекцией смешивали с T-клетками Vγ9Vδ2. Конечный объем для каждой инъекции составлял 100 мкл. Лечебные mAb вводили за 4 часа до приживления Т-клеток Vγ9Vδ2.
Наши результаты подтвердили, что инфузия T-клеток Vγ9Vδ2 плюс rHuIL-15/rHuIL15-Rα снижает опухолевую нагрузку. Этот эффект был еще более поразительным и сопровождался значительным увеличением выживаемости при добавлении mAb1 против BTN3A вместе с Т-клетками Vγ9Vδ2 (таблицы 16 и 17). Важно отметить, что цитарабин (Ara-C), один из наиболее эффективных препаратов для лечения острого миелоидного лейкоза, примененный в этой очень агрессивной модели на мышах (10 мг/кг у мышей NSG с опухолью U937), улучшил выживаемость мышей на 2-3 дня (~10%), как наблюдали для mAb1 в комбинации с переносом человеческих T-клеток Vγ9Vδ2.
Эти данные подчеркивают мощный антилейкемический эффект, оказываемый mAb против BTN3A в комбинации с иммунотерапией Т-клетками Vγ9Vδ2 in vivo.
Таблица 16: Мышинная модель U937 AML: описание групп
(3x106 клеток) +IL-15a
+hIgG1 (10 мг/кг)
(10 мг/кг)
(3x106 клеток) + IL-15a
+hIgG1 (10 мг/кг)
(10 мг/кг)
(3x106 клеток) +IL-15a
+mAb1 (10 мг/кг)
(3x106 клеток) +IL-15a
+mAb1 (10 мг/кг)
(3x106 клеток) +IL-15a
+mAb1 (10 мг/кг)
(10 мг/кг)
(3x106 клеток) + IL-15a
+mAb1 (10 мг/кг)
(10 мг/кг)
(3x106 клеток) + IL-15a
+ mIgG1 (10 мг/кг)
(3x106 клеток) + IL-15a
+ mIgG1 (10 мг/кг)
(3x106 клеток) + IL-15a
+ m20.1 (10 мг/кг)
(3x106 клеток) + IL-15a
+ m20.1 (10 мг/кг)
Таблица 17: Активирующее Mab, направленное против BTN3A, (mAb1) обладает антилейкемической активностью in vivo в моделях ксенотрансплантата AML. Данные биолюминесценции по модели U937
ФСБ
LTγδ+hIgG1 (День 1, 4, 7, 10)
LTγδ+mAb1 (День 1, 7)
LTγδ+
mAb1 (День 1, 4, 7, 10)
LTγδ+mIgG1 (День 1, 7)
LTγδ+m20.1 (День 1, 7)
Таблица 18: Активирующее MAb, направленное против BTN3A, (mAb1) обладает антилейкемической активностью in vivo в моделях ксенотрансплантата AML. Выживаемость после приживления клеточной линии U937.
смерти
смерти
ФСБ
LTγδ+mAb1 (День 1, 4, 7, 10)
LTγδ+hIgG1 (День 1, 4, 7, 10)
LTγδ+mIgG1 (День 1, 7)
LTγδ+mAb1 (День 1, 7)
LTγδ+m20.1 (День 1, 7)
b. Модель MOLM14
Эффективность mAb1 in vivo против MOLM14, линии клеток, полученных из устойчивого к AraC AML человека, оценивали на модели ксенотрансплантата с применением мышей NSG, которым трансплантировали линию опухолевых клеток человека и Т-клеток Vγ9Vδ2 человека. Целью этого исследования было подтвердить влияние повторных внутривенных инъекций человеческих T-клеток Vγ9Vδ2 в комбинации с mAb1 на рост опухоли и выживаемость мышей.
Самкам мышей NSG в возрасте от шести до восьми недель в день 0 вводили внутривенно (iv) через хвостовую вену клетки MOLM14 (CVCL_7916), экспрессирующие люциферазу (luc2), в количестве 0,2x106 в день на мышь в объеме 100 мкл. Биолюминесцентный анализ проводили в день 0 с применением PhotonIMAGER («Biospace Lab») после добавления не содержащего эндотоксина люциферина (30 мг/кг), и мышей рандомизировали в гомогенные группы по 7 мышей на основании силы сигнала биолюминесценции. 3×106 человеческих Т-клеток Vγ9Vδ2, размноженных in vitro, и комплексы hIL-15/IL-15R вводили внутривенно в дни 1, 8, 15, 22 и 29. mAb1 или hIgG1 вводили внутривенно в дни 1, 5, 8, 12, 15, 19, 22, 26 и 29.
Различные экспериментальные группы суммированы в таблице 19.
Комплексы hIL-15/IL-15R-Fc предварительно образовывали при комнатной температуре (RT) в течение 30 минут перед инъекцией (0,2 мкг hIL-15 + 1,2 мкг IL-15R-Fc на мышь) и смешивали с Т-клетками Vγ9Vδ2 перед инъекцией. Конечный объем для каждой инъекции составлял 100 мкл. Лечебные mAb вводили за 4 часа до приживления Т-клеток Vγ9Vδ2.
Сигнал биолюминесценции, испускаемый клетками MOLM14, измеряли на день 0, 7, 14, 21 и 28 после инъекции клеток, чтобы проследить рост опухоли. Забор крови проводили на день 19 для оценки количества циркулирующих клеток MOLM14 с помощью проточной цитометрии. Перед окрашиванием проводили лизис эритроцитов. Мышиные клетки mCD45+ исключали из анализа, опухолевые клетки MOLM14 детектировались по их экспрессии GFP. Сбор данных выполняли на цитометре LSRII SORP (Becton Dickinson), а анализ выполняли с применением программного обеспечения FlowJo. Ежедневный мониторинг мышей на предмет симптомов заболевания (значительная потеря веса, взъерошенная шерсть, сгорбленная спина, слабость и снижение подвижности) определял время умерщвления инъецированных животных с признаками дистресса.
Как показано в таблице 20, Т-клетки Vγ9Vδ2, инъецированные нерелевантными контрольными изотипами (hIgG1), существенно не контролируют рост опухоли. Напротив, как показывают сниженные сигналы биолюминесценции, рост опухоли сильно снижался, когда анти-BTN3A mAb1 вводили последовательно с человеческими Т-клетками Vγ9Vδ2. Результаты также показали значительное уменьшение количества циркулирующих бластных клеток (по оценке с помощью проточной цитометрии в периферической крови) на 19 день протокола (таблица 20). Важно отметить, что зависимое от mAb против BTN3A уменьшение роста опухоли приводит к значительному увеличению выживаемости мышей на 45% (по сравнению с соответствующим изотипическим контролем) (таблица 22).
Таблица 19: Модель мышей MOLM14: описание групп
Таблица 20: Модель мышей MOLM14: измерение биолюминесценции на день 0, 7, 14, 21 и 28 после приживления опухолевых клеток
ФСБ
LTγδ+hIgG1
LTγδ+mAb1
Таблица 21: Модель мышей MOLM14: количество циркулирующих бластных клеток на 19 день протокола. Данные представляют собой количество клеток/мкл крови.
Таблица 22: Модель мышей MOLM14: выживаемость животных. День смерти для каждого животного указан вместе с медианой выживаемости для каждой группы.
13. mAb1 улучшает терапию Т-клетками Vγ9Vδ2 на модели солидных опухолей, представляющей собой мышей, с трансплантированной клеточной линией рака яичников человека SKOV-3.
a. Материалы и методы
Размножение T-клеток Vγ9Vδ2 in vitro.
Аллогенные человеческие Т-лимфоциты Vγ9Vδ2 размножали из мононуклеарных клеток периферической крови (PBMC), полученных из образцов крови здоровых доноров, предоставленных Etablissment du Sang (EFS, Нант, Франция), и после центрифугирования в градиенте плотности на основе фиколла (Eurobio, Les Ulis, Франция). Исходно, для специфического увеличения периферических аллогенных Т-лимфоцитов Vγ9Vδ2 человека, PBMC инкубировали с 3 мкМ пирофосфата бромгидрина (BrHPP), любезно предоставленного компанией «Innate Pharma» (Марсель, Франция), в среде RPMI с добавлением 10%-ной инактивированной нагреванием фетальной сыворотки теленка, 2 мМ L-глутамина, 10 мг/мл стрептомицина, 100 МЕ/мл пенициллина (все от компании «Gibco») и 100 МЕ/мл рекомбинантного человеческого IL-2 (PROLEUKIN, «Novartis», Bale, Suisse). Через 4 дня в культуры добавляли 300 МЕ/мл IL-2. На 21 день чистоту измеряли проточной цитометрией (чистота > 90%). Чистые человеческие Т-лимфоциты Vγ9Vδ2 дополнительно размножали с применением питающих клеток (смешанные и 35 Gy облученные, трансформированные вирусом Эпштейна-Барра В-лимфоциты и PBMC) и PHA-L в среде RPMI с добавлением 10%-ной инактивированной нагреванием фетальной сыворотки теленка, 2 мМ L-глутамина, 10 мг/мл стрептомицина, 100 МЕ/мл пенициллина (все от компании «Gibco») и 300 МЕ/мл рекомбинантного человеческого ИЛ-2 («Novartis»). Через три недели покоящиеся, размноженные ex vivo Т-лимфоциты Vγ9Vδ2 применяли для экспериментов in vivo.
Мышиная модель
В день 0 мышам NSG в возрасте 6-8 недель вводили внутрибрюшинно (ip) 1 × 106 клеток SKOV-3 (линия клеток рака яичников, SKOV-3-luc-D3, Perkin Elmer, Waltham, MA), экспрессирующих люциферазу, на мышь в объеме 100 мкл стерильного ФСБ. Через 7 дней мышей рандомизировали на однородные группы по 5-6 мышей. На 7-й и 14-й дни внутрибрюшинно вводили mAb обработки в количестве 200 мкг mAb1 на мышь или соответствующий изотипический контроль (hIgG1) в 100 мкл стерильного ФСБ. Через 4 часа 5×106 человеческих T-клеток Vγ9Vδ2, размноженных in vitro, также вводили внутрибрюшинно в 100 мкл стерильного ФСБ на мышь.
Сигнал биолюминесценции, испускаемый клетками SKOV-3, измеряли на 16, 23 и 30 дни после имплантации опухолевых клеток для отслеживания роста опухоли. Биолюминесцентное изображение получали через 8 минут после внутрибрюшинной инъекции 1,5 мг D-люциферина («Interchim», Сан-Диего, Калифорния) на мышах, анестезированных 2%-ным изофлураном, с помощью Biospace Imager (Biospace Lab, Неслес-ла-Валле, Франция). Конечная точка эксперимента была достигнута, когда мыши потеряли 10% от своего первоначального веса.
b. Результаты
Эффективность mAb1 против рака яичников in vivo оценивали на модели ксенотрансплантата с применением мышей NSG, которым трансплантировали клеточную линию рака яичников человека SKOV-3 и человеческие Т-клетки Vγ9Vδ2. Целью этого исследования было оценить влияние двух внутрибрюшинных инъекций человеческих T-клеток Vγ9Vδ2 в комбинации с mAb1 на рост опухоли и выживаемость мышей.
Мышам NSG вводили внутрибрюшинно (ip) в день 0 SKOV3. Через 7 дней мышей рандомизировали в гомогенные группы в соответствии с лечением. Группы описаны в таблице 23.
Таблица 23: Мышиная модель рака яичника: описание групп
Результат показал, что человеческие Т-клетки Vγ9Vδ2, перенесенные вместе с mAb1, значительно замедляли рост опухоли (таблица 24), что приводило к значительному улучшению выживаемости животных (таблица 25). Следует отметить, что этот эффект наблюдался в отсутствие цитокинов выживания про-Vγ9Vδ2 T, таких как IL-2 или IL-15.
Таблица 24: Мышиная модель рака яичников: измерение биолюминесценции через 16, 23 и 30 дней после приживления опухолевых клеток
ФСБ
LTγδ+
hIgG1
LTγδ+
mAb1
Таблица 25: Мышиная модель рака яичника: выживаемость животных. День смерти для каждого животного указан вместе с медианой выживаемости для каждой группы.
ФСБ
LTγδ+
hIgG1
LTγδ+
mAb1
16. Влияние in vivo на Vγ9Vδ2 Т-клетки яванского макака
Из-за отсутствия BTN3A и подмножества T Vγ9Vδ2 у грызунов и на основании предыдущих данных, документирующих in vitro и in vivo PAg-опосредованную активацию T-клеток Vγ9Vδ2 у яванских макак, причем яванский макак (Macaca fascicularis) был выбран в качестве единственного подходящего вида для доклинической оценки безопасности mAb1.
a. Материал и методы
Biacore
mAb1 оценивали на предмет связывания с рекомбинантными белками BTN3A1, BTN3A2 и BTN3A3 человека или яванского макака (SEQ ID NO 21, 22 и 23 соответственно) с помощью анализа многоцикловой кинетики Biacore с применением прибора Biacore T200 (серийный номер 1909913). mAb1 разводили до концентрации 2 мкг/мл в 2% БСА/ФСБ. В начале каждого цикла антитело было захвачено на поверхности протеина А при плотности (RL) ~ 150 RU (теоретическое значение для получения RMax ~ 50 RU). После захвата поверхности давали стабилизироваться перед инъекцией антигена BTN3A. BTN3A титровали в 0,1% BSA/HBS-P + (рабочий буфер) в диапазоне двукратных разведений от 25 до 0,48 нМ. Фазу ассоциации наблюдали в течение 420 секунд, а фазу диссоциации - в течение 2000 секунд. Кинетические данные получали при скорости потока 50 мкл/мин, чтобы минимизировать любые возможные эффекты массопереноса. Полученные данные были подогнаны с применением модели связывания 1:1.
ELISA
Очевидное сродство mAb1 к BTN3A1 человека и яванского макака проверяли с помощью ELISA. Вкратце, связывание mAb1 с мишенью оценивали с применением рекомбинантного BTN3A1, иммобилизованного на планшете при концентрации 1 мкг/мл в фосфатном буфере (1× ФСБ), с последующей стадией насыщения блокирующим буфером (2% молоко/ФСБ)). mAb1 титровали в блок-буфере в диапазоне четырехкратных разведений от 0,00122 до 20 мкг/мл. Для обнаружения применяли вторичные антитела (козьи антитела против цепи Igκ человека, конъюгированные с HRP, Millipore AP502P, разведенные 1:4000 в блокирующем буфере) и раствор TMB. Кажущееся сродство выражали как ЕС50% (концентрация антитела, необходимая для получения 50% сигнального плато).
Авидность связывания mAb1 с клетками CD3+ человека и яванского макака
После лизиса эритроцитов периферическую кровь (PBL) человека или яванского макака инкубировали с возрастающей концентрацией mAb1 или изотипического контроля в течение 30-ти минут при 4°C, промывали два раза и окрашивали вторичным антителом, конъюгированным с фикоэритрином (PE) козьим антителом против человеческого IgG (eBioscience™, #12-4998-82). После двух промывок клетки окрашивали mAb против CD3-PC3 («BD Bioscience», #557749) и реагентом Live/Dead («Life Technology», #L10119). После промывок клетки ресуспендировали в 200 мкл буфера Flow. Затем клетки анализировали на цитометре Cytoflex («Beckman Coulter»). Данные анализировали с применением программного обеспечения FlowJo (версия 10, FlowJo, LLC, Ashland, USA), контролируя популяцию живых CD3+. Затем рассчитывали значения средней интенсивности флуоресценции (MFI) по каналу для PE и наносили на график в зависимости от концентрации. Аппроксимация кривых была получена с применением сигмоидального уравнения 4PL из программы GraphPad Prism.
Поверхностная экспрессия BTN3A на циркулирующих клетках человека и яванского макака
Для поверхностной экспрессии BTN3A на лейкоцитах 100 мкл цельной крови яванского макака и человека помещали в 96-луночные планшеты с коктейлем специфических антител (реагент Aqua live Dead, CD20-V450, CD8-BV605, CD4-BV650, Vg9 TCR-FITC, анти -BTN3A-PE (клон 20.1) (или mIgG1-PE для контроля изотипа), CD3-PeCy7, CD45-AF700 и CD14-APC-H7) и инкубировали 15 минут при комнатной температуре в защищенном от света месте. Затем эритроциты лизировали 900 мкл лизирующего реагента («BD Bioscience», # 349202) в соответствии с инструкциями производителя. Клетки промывали и анализировали с применением многопараметрической проточной цитометрии. Для поверхностной экспрессии BTN3A на эритроцитах и тромбоцитах 100 мкл цельной крови яванского макака или человека разводили 100 мкл ФСБ и инкубировали со специфическим коктейлем антител (Aqua live dead, CD41-APC, CD45-AF700 и анти-BTN3A (клон 20.1) (или mIgG1-PE для контроля изотипа)) в течение 15-ти минут при комнатной температуре в защищенном от света месте. После промываний клетки анализировали с помощью многопараметрической проточной цитометрии. Чтобы получить относительную количественную оценку поверхностной экспрессии BTN3A, калибровочные шарики (CellQuant Calibrator Biocytex, # 7208) и козьи PE-антитела против мышиного IgG (H + L) применяли параллельно в соответствии с инструкциями производителя. Для каждого подмножества клеток регистрировали MFI по PE-каналу для анти-BTN3A и изотипического контроля. Анализ выполняли путем вычитания MFI изотипического контроля из MFI окрашенных анти-BTN3A, и относительную поверхностную экспрессию рассчитывали на основе стандартной кривой, полученной с помощью калибровочных шариков.
Размножение и активация Т-клеток Vγ9Vδ2 яванского макака in vitro
Цельную кровь яванского макака от 3-х животных обрабатывали буфером для лизиса эритроцитов. После обильных промывок лейкоциты высевали при 1,5 М/мл RPMI 10% SVF в 6-луночные планшеты в присутствии rHuIL-2 (200МЕ/мл) и mAb1 (10 мкг/мл). rHuIL-2 добавляли на 6 и 8 день для имитации обычного протокола размножения клеток, применяемого для человеческих PBMC, и улучшения долгосрочной выживаемости T-клеток Vγ9Vδ2 in vitro, чтобы получить достаточное количество клеток для выполнения функциональных анализов. Процент Vγ9+ Т-клеток оценивался на 0, 6, 8 и 10 день с применением смеси специфических антител и анализа проточной цитометрией (CD3-PC7 «BD Bioscience» # 557749, Vg9 TCR-FITC клон 7A5 Invitrogen # TCR2720, Live Dead для ближнего ИК, «ThermoFisher» # L10119).
На 10-ый день подсчитывали размноженные Т-клетки Vγ9Vδ2 и совместно культивировали при соотношении Е:Т 1:1 с линиями опухолевых клеток человека. 100000 линий опухолевых клеток-мишеней (Раджи, ДАУДИ и K562) смешивали в 96-луночных планшетах со 100000 T-клеток Vγ9Vδ2 яванского макака в присутствии контроля изотипа mAb1 или hIgG1 (10 мкг/мл) или PMA (20 нг/мл)/иономицина (1 мкг/мл) в качестве положительного контроля. Дегрануляцию T-клеток Vγ9Vδ2 отслеживали через 4 часа с помощью окрашивания CD107a/b («BD Bioscience» # 555800) и анализа проточной цитометрии.
Исследование in vivo на яванском макаке
Прижизненное исследование
В этом исследовании применяли яванских макак (Macaca fascicularis) в возрасте от 4 до 6 лет, весом от 3 до 5 кг, вьетнамского происхождения. Всех животных содержали и применяли в соответствии с рекомендациями комитета по уходу и применению животных в учреждении GLP. В рамках процедуры по охране здоровья заводчиков все животные были проверены на туберкулез, а профилактическое лечение было зарегистрировано в записях заводчиков. После прибытия животные были акклиматизированы для процедур исследования за по меньшей мере 2 недели. Были проведены клиническое обследование на ухудшение состояния здоровья и тестирование. Оценку здоровья животных ветеринар проводил перед началом фазы до приема препарата, чтобы подтвердить пригодность каждого животного для исследования.
mAb1 вводили внутривенно (иммобилизация в кресле, инфузия в течение 15-ти минут) после дезинфекции кожи не голодавших животных. Животным групп 1, 4 и 5 вводили дозу в день 1, 8, 15 и 22. Животным групп 2 и 3 вводили дозу один раз в день 1.
Фармакокинетика
Квалифицированный фармакокинетический анализ был разработан для количественного определения mAb1 в сыворотке яванского макака. Вкратце, mAb1 определяли количественно спектрофотометрически способом ELISA. Планшет, предварительно покрытый стрептавидином, применяли для захвата конъюгата человеческого IgG-Fc биотина PK. Затем mAb1 захватывали на поверхности планшета, и связанный аналит обнаруживают с помощью козьего антитела против человеческого IgG-HRP (Fc-специфического), меченного пероксидазой антивидового антитела. Целевой диапазон количественного определения составляет от 90 нг/мл до 10000 нг/мл в чистой сыворотке.
Иммунофенотипирование
У всех животных брали образцы крови (1,0 мл) из vena cephalica antebrachii или vena saphena в пробирки с Li-гепарином. Иммунофенотипирование проводили с помощью коктейлей специфических моноклональных антител. Был проведен анализ относительного количества клеток (процентное содержание лимфоцитов/лейкоцитов). Общее количество гранулоцитов/лимфоцитов/лейкоцитов определяли в тот же день с помощью гемоанализатора и применяли для расчета абсолютных чисел. Абсолютное количество субпопуляций лимфоцитов вычисляли из относительных чисел.
Заполнение рецепторов
Образцы крови (400 мкл) брали у всех животных из vena cephalica antebrachii или vena saphena в пробирки с Li-гепарином. Меченое антитело, неконкурентно связывающееся с BTN3A (клон 103.2) на клетках, которые предварительно инкубировали с избытком ICT01, применяли для определения общей поверхностной экспрессии BTN3A на CD3+ T-клетках и CD19+ B-клетках. Для обнаружения свободных сайтов связывания BTN3A применяли конкурентное связывание немеченого mAb1 с BTN3A, которое ингибирует связывание меченного флуоресцентным красителем антитела против BTN3A (mAb1) на CD3+ T-клетках и CD19+ B-клетках. В зависимости от количества BTN3A, которое блокировалось mAb1, измеряли среднюю интенсивность флуоресценции (MFI) конъюгированного mAb1 по мере уменьшения среднего геометрического. Следовательно, интенсивность окрашивания Ab7.2, близкая к интенсивности окрашивания изотипического антитела, свидетельствовала о полном насыщении.
b. Результаты
mAb1 связывается с BTN3A яванского макака
Поскольку дифференциальная идентификация 3-х генов изоформ BTN3A была невозможна из общедоступных баз данных, ImCheck выполнила целевую ПЦР на кДНК, выделенной из PBMC яванского макака, с применением высококонсервативной последовательности кДНК, близкой к трансмембранному домену, для создания соответствующего праймера. Секвенирование продуктов ПЦР позволило идентифицировать последовательности эктодоменов BTN3A1, BTN3A2 и BTN3A3 яванского макака. Рекомбинантные эктодомены изоформ BTN3A1, BTN3A2 и BTN3A3 яванского макака, слитые с меткой 6xHis, были продуцированы в клетках СНО на основе этих последовательностей (SEQ ID NO 21, 22 и 23 соответственно).
Рекомбинантные белки тестировали на связывание mAb1 с помощью BIAcore и ELISA (таблица 26). Результаты BIAcore показали, что mAb1 преимущественно связывается с 3 рекомбинантными BTN3A1, BTN3A2 и BTN3A3 яванского макака, хотя и с более низким сродством к изоформе BTN3A1. ELISA проводили с изоформой BTN3A1. Интересно, что таблица 26 показывает сопоставимые EC50 для связывания mAb1 с рекомбинантным BTN3A1 человека или яванского макака.
Таблица 26: Связывание mAb1 с рекомбинантными белками BTN3A человека и яванского макака
BIAcore (MCK)
ELISA
(«Sino biological», # 15973-H08H)
Параллельно, связывание mAb1 с PBMC яванского макака оценивали с помощью проточной цитометрии. Среднее значение EC50 связывания mAb1 с CD3+ Т-клетками яванского макака было сравнимо со средним значением ЕС50 для CD3+ Т-клеток человека (таблица 27). Целевую экспрессию в различных субпопуляциях иммунных клеток яванского макака по сравнению с цельной кровью здорового донора-человека исследовали с помощью многопараметрической проточной цитометрии с применением PE-конъюгированных mAb против BTN3A (клон 20.1) вместе с панелью фенотипирующих антител с известной перекрестной реактивностью для маркеров поверхности клеток человека и яванского макака (данные не представлены). Эти результаты показывают, что BTN3A экспрессируется в широкой панели популяций клеток периферической крови у обоих видов, хотя очевидно, в целом более низкую экспрессию наблюдали у яванского макака.
Таблица 27: Связывание mAb1 на CD3+ Т-клетках человека и яванского макака
(+/-sem)
(x10-9 M)
(+/-sem)
mAb1 способствует размножению и активации Т-клеток Vγ9Vδ2 яванского макака in vitro
Затем функциональную активность mAb1 оценивали in vitro на Т-клетках Vγ9Vδ2 яванского макака. Во-первых, мы оценили, способствует ли mAb1 размножению T-клеток Vγ9Vδ2 яванского макака при инкубации в течение 10-ти дней с PBMC яванского макака. Как показано на фигуре 4A, mAb1 способствовало размножению T-клеток Vγ9Vδ2 у всех 3-х протестированных животных; эта популяция достигает 60% через 10 дней, что сопоставимо с уровнем, наблюдаемым для клеток человека. После 10-ти дней размножения клетки совместно культивировали в течение 4-х часов с клеточными линиями Дауди, K562 или Раджи, применяемые в качестве клеток-мишеней в присутствии mAb1, и анализировали на экспрессию CD107a/b с помощью проточной цитометрии. Этот эксперимент показал, что mAb1 индуцирует значительную активацию T-клеток Vγ9Vδ2 при совместном культивировании со всеми 3-мя линиями опухолевых клеток (фигура 4B).
В заключение, результаты показывают, что (i) mAb1 связывается с клетками яванского макака с такой же авидностью, что и с человеческими клетками, (ii) BTN3A экспрессируется на одних и тех же клетках крови у людей и яванских макак, хотя у последнего вида наблюдался более низкий уровень экспрессии, и (iii) mAb1 способствует размножению субпопуляции с Т-клеток в PBMC яванского макака, и размноженные клетки представляют собой реакционноспособные клетки против опухолевых клеток-мишеней, сенсибилизированных mAb1.
mAb1 влияет на компартмент Т-клеток Vγ9Vδ2 яванского макака in vivo
Исследование in vivo было проведено на здоровых самках яванских макак в возрасте от 4 до 6 лет, которые получали однократные или многократные внутривенные инфузии mAb1 (таблица 28).
Был выбран внутривенный путь введения, поскольку он предназначен для лечения человека. Животных лечили mAb1 в соответствии с планом ступенчато возрастающей дозы.
Таблица 28: Дизайн исследования ФК/ФД/переносимости для mAb1
(мг/кг)
в неделю
пол животных
Оценивали следующие конечные точки: клинические признаки, масса тела, клиническая патология (гематология, клиническая химия и коагуляция), иммунофенотипирование популяций лейкоцитов периферической крови, маркеры активации/пролиферации/дифференцировки, фармакокинетика и фармакодинамика (заполнение рецепторов BTN3A на циркулирующих Т- и В-клетках).
Все животные выжили до запланированного вскрытия трупа на 29-ый день исследования после однократной или 4-х повторных 15-минутных инфузий mAb1. Не было обнаружено влияний, связанных с исследуемым препаратом, на массу тела или потребление пищи. Клинические признаки соответствовали таковым, наблюдаемым на яванских макаках в лабораторных условиях содержания, и, следовательно, не относились к исследуемому препарату.
Фармакокинетика
mAb1 продемонстрировало приблизительно пропорциональное дозе фармакокинетическое поведение после внутривенного (IV) дозирования в диапазоне доз от 0,1 до 100 мг/кг (данные не представлены), а также длительный период полувыведения, типичный для mAb IgG в отсутствие опосредованного мишенью клиренса. После повторных доз по 10 или 100 мг/кг/неделю для 4 доз (данные не представлены), воздействие сохранялось у всех животных в течение периода лечения с минимальным накоплением в течение 4-недельного периода лечения. Фармакокинетические профили для контрольного животного после введения доз 1 и 10 мг/кг внутривенно в увеличивающемся еженедельном режиме дозирования продемонстрировали некоторые доказательства увеличения клиренса к концу недельного интервала дозирования, что может быть результатом образования ADA к mAb1; нет доказательств увеличения клиренса после последней внутривенной дозы 100 мг/кг.
Связывание рецепторов (RO)
Согласно профилю экспрессии BTN3A и представленности клеточной популяции в крови, RO mAb1 измеряли на CD3+ T-клетках и CD20+ B-клетках.
Результаты показывают, что BTN3A быстро связываются после инъекции mAb1 как на CD3+ T-клетках, так и на CD20+ B-клетках (Данные не представлены). Повторное введение 100 мг/кг mAb1 оказалось необходимым для полного связывания рецепторов в течение недельного интервала дозирования.
Иммунофенотипирование
В выбранные моменты времени после каждой дозы кровь животных, получавших mAb1, окрашивали коктейлем из конкретных mAb для количественного определения субпопуляций Т-клеток (CD4, CD8, Vγ9 Т-клетки, регуляторные Т-клетки), В-клеток, моноцитов, NK клеток, mDCs, pDCs и гранулоцитов, а также связанных маркеров активации (CD69, CD86, CD95, Granzyme B, Ki67), и анализировали с помощью проточной цитометрии. Анализ включал относительное количество клеток (процент лимфоцитов/лейкоцитов) для каждой популяции вместе с абсолютным количеством клеток, экстраполированным из общего количества лимфоцитов/лейкоцитов, определенного на основе образцов крови, собранных в одно и то же время и проанализированных с применением гематологического счетчика клеток.
Основные наблюдения, полученные с помощью этого широкого анализа:
Т-клетки Vδ9+ (% среди CD3+) значительно сокращаются после введения дозы у всех животных, получавших mAb1, и постепенно восстанавливаются у животных с однократной дозой. Этот эффект, по-видимому, специфичен, поскольку его не наблюдают в Т-клетках CD4 и CD8 αβ, и он свидетельствует об активации и маргинализации Т-клеток γδ на тканях, как это наблюдают для биспецифических антител, взаимодействующих с CD3, у обезьян и людей (Smith et al., 2015 Sci Rep. 2015 Dec 11; 5:17943. DOI: 10,1038/srep17943). Результаты показаны на фигуре 5.
Это исследование показывает, что mAb1 при внутривенном введении, по-видимому, хорошо переносится в дозах до 100 мг/кг/неделю. Более того, среди подмножеств Т-клеток, mAb1 специфически и в значительной степени влияют на Т-клетки Vγ9Vδ2.
Список литературы
Alegre, M.-L., Frauwirth, K.A., and Thompson, C.B. (2001). T-cell regulation by CD28 and CTLA-4. Nat. Rev. Immunol. 1, 220–228.
Ausubel, F.M., Brent, R., Kingston, R.E., Moore, D.D., Seidman, J.G., and Struhl, K. (1988). Current Protocols in Molecular Biology (John Wiley & Sons).
Baudino, L., Shinohara, Y., Nimmerjahn, F., Furukawa, J.-I., Nakata, M., Martínez-Soria, E., Petry, F., Ravetch, J.V., Nishimura, S.-I., and Izui, S. (2008). Crucial Role of Aspartic Acid at Position 265 in the CH2 Domain for Murine IgG2a and IgG2b Fc-Associated Effector Functions. J. Immunol. 181, 6664–6669.
Bensussan, A., and Olive, D. (2005). T-cell: Section report. Cell. Immunol. 236, 3–5.
Bird, R.E., Hardman, K.D., Jacobson, J.W., Johnson, S., Kaufman, B.M., Lee, S.M., Lee, T., Pope, S.H., Riordan, G.S., and Whitlow, M. (1988). Single-chain antigen-binding proteins. Science 242, 423–426.
Brennan, M., Davison, P.F., and Paulus, H. (1985). Preparation of bispecific antibodies by chemical recombination of monoclonal immunoglobulin G1 fragments. Science 229, 81–83.
Bryson, C.J., Jones, T.D., and Baker, M.P. (2010). Prediction of Immunogenicity of Therapeutic Proteins. BioDrugs 24, 1–8.
Chapoval, A.I., Ni, J., Lau, J.S., Wilcox, R.A., Flies, D.B., Liu, D., Dong, H., Sica, G.L., Zhu, G., Tamada, K., et al. (2001). B7-H3: A costimulatory molecule for T cell activation and IFN-γ production. Nat. Immunol. 2, 269–274.
Collins, M., Ling, V., and Carreno, B.M. (2005). The B7 family of immune-regulatory ligands. Genome Biol. 6, 223.
Coyle, A.J., and Gutierrez-Ramos, J.C. (2001). The expanding B7 superfamily: increasing complexity in costimulatory signals regulating T cell function. Nat. Immunol. 2, 203–209.
Dong, H., Zhu, G., Tamada, K., and Chen, L. (1999). B7-H1, a third member of the B7 family, co-stimulates T-cell proliferation and interleukin-10 secretion. Nat. Med. 5, 1365–1369.
Freeman, G.J., Long, A.J., Iwai, Y., Bourque, K., Chernova, T., Nishimura, H., Fitz, L.J., Malenkovich, N., Okazaki, T., Byrne, M.C., et al. (2000). Engagement of the Pd-1 Immunoinhibitory Receptor by a Novel B7 Family Member Leads to Negative Regulation of Lymphocyte Activation. J. Exp. Med. 192, 1027–1034.
Glennie, M.J., McBride, H.M., Worth, A.T., and Stevenson, G.T. (1987). Preparation and performance of bispecific F(ab' gamma)2 antibody containing thioether-linked Fab' gamma fragments. J. Immunol. 139, 2367–2375.
Goeddel, D.V. (1990). [1] Systems for heterologous gene expression. In Methods in Enzymology, (Academic Press), pp. 3–7.
Gu, S., Nawrocka, W., and Adams, E.J. (2015). Sensing of Pyrophosphate Metabolites by Vγ9Vδ2 T Cells. Front. Immunol. 5.
Harly, C., Guillaume, Y., Nedellec, S., Peigné, C.-M., Mönkkönen, H., Mönkkönen, J., Li, J., Kuball, J., Adams, E.J., Netzer, S., et al. (2012). Key implication of CD277/butyrophilin-3 (BTN3A) in cellular stress sensing by a major human γδ T-cell subset. Blood 120, 2269–2279.
Huston, J.S., Levinson, D., Mudgett-Hunter, M., Tai, M.S., Novotný, J., Margolies, M.N., Ridge, R.J., Bruccoleri, R.E., Haber, E., and Crea, R. (1988). Protein engineering of antibody binding sites: recovery of specific activity in an anti-digoxin single-chain Fv analogue produced in Escherichia coli. Proc. Natl. Acad. Sci. 85, 5879–5883.
Kabat, E.A., Wu, T.T., Foeller, C., Perry, H.M., and Gottesman, K.S. (1992). Sequences of Proteins of Immunological Interest (DIANE Publishing).
Karpovsky, B., Titus, J.A., Stephany, D.A., and Segal, D.M. (1984). Production of target-specific effector cells using hetero-cross-linked aggregates containing anti-target cell and anti-Fc gamma receptor antibodies. J. Exp. Med. 160, 1686–1701.
Kaufman, R.J., and Sharp, P.A. (1982). Construction of a modular dihydrofolate reductase cDNA gene: analysis of signals utilized for efficient expression. Mol. Cell. Biol. 2, 1304–1319.
Khattri, R., Auger, J.A., Griffin, M.D., Sharpe, A.H., and Bluestone, J.A. (1999). Lymphoproliferative Disorder in CTLA-4 Knockout Mice Is Characterized by CD28-Regulated Activation of Th2 Responses. J. Immunol. 162, 5784–5791.
Klocke, K., Sakaguchi, S., Holmdahl, R., and Wing, K. (2016). Induction of autoimmune disease by deletion of CTLA-4 in mice in adulthood. Proc. Natl. Acad. Sci. 113, E2383–E2392.
Latchman, Y., Wood, C.R., Chernova, T., Chaudhary, D., Borde, M., Chernova, I., Iwai, Y., Long, A.J., Brown, J.A., Nunes, R., et al. (2001). PD-L2 is a second ligand for PD-1 and inhibits T cell activation. Nat. Immunol. 2, 261–268.
Linsley, P.S., Greene, J.L., Tan, P., Bradshaw, J., Ledbetter, J.A., Anasetti, C., and Damle, N.K. (1992). Coexpression and functional cooperation of CTLA-4 and CD28 on activated T lymphocytes. J. Exp. Med. 176, 1595–1604.
Liu, M.A., Kranz, D.M., Kurnick, J.T., Boyle, L.A., Levy, R., and Eisen, H.N. (1985). Heteroantibody duplexes target cells for lysis by cytotoxic T lymphocytes. Proc. Natl. Acad. Sci. 82, 8648–8652.
McCafferty, J., Griffiths, A.D., Winter, G., and Chiswell, D.J. (1990). Phage antibodies: filamentous phage displaying antibody variable domains. Nature 348, 552–554.
Morrison, S.L. (1985). Transfectomas provide novel chimeric antibodies. Science 229, 1202–1207.
Ni, L., and Dong, C. (2017). New B7 Family Checkpoints in Human Cancers. Mol. Cancer Ther. 16, 1203–1211.
Oganesyan, V., Gao, C., Shirinian, L., Wu, H., and Dall'Acqua, W.F. (2008). Structural characterization of a human Fc fragment engineered for lack of effector functions. Acta Crystallogr. D Biol. Crystallogr. 64, 700–704.
Panowski, S., Bhakta, S., Raab, H., Polakis, P., and Junutula, J.R. (2013). Site-specific antibody drug conjugates for cancer therapy. MAbs 6, 34–45.
Paulus, H. (1985). Preparation and biomedical applications of bispecific antibodies. Behring Inst. Mitt. 118–132.
Perry, L.C.A., Jones, T.D., and Baker, M.P. (2008). New approaches to prediction of immune responses to therapeutic proteins during preclinical development. Drugs RD 9, 385–396.
Reddy, M.P., Kinney, C.A.S., Chaikin, M.A., Payne, A., Fishman-Lobell, J., Tsui, P., Monte, P.R.D., Doyle, M.L., Brigham-Burke, M.R., Anderson, D., et al. (2000). Elimination of Fc Receptor-Dependent Effector Functions of a Modified IgG4 Monoclonal Antibody to Human CD4. J. Immunol. 164, 1925–1933.
Remington, J.P., and Gennaro, A.R. (1995). Remington: the science and practice of pharmacy (Easton, PA: Mack Publishing).
Rhodes, D.A., Stammers, M., Malcherek, G., Beck, S., and Trowsdale, J. (2001). The Cluster of BTN Genes in the Extended Major Histocompatibility Complex. Genomics 71, 351–362.
Ruddy, D.A., Kronmal, G.S., Lee, V.K., Mintier, G.A., Quintana, L., Domingo, R., Meyer, N.C., Irrinki, A., McClelland, E.E., Fullan, A., et al. (1997). A 1.1-Mb Transcript Map of the Hereditary Hemochromatosis Locus. Genome Res. 7, 441–456.
Saverino, D., Tenca, C., Zarcone, D., Merlo, A., Megiovanni, A.M., Valle, M.T., Manca, F., Grossi, C.E., and Ciccone, E. (1999). CTLA-4 (CD152) Inhibits the Specific Lysis Mediated by Human Cytolytic T Lymphocytes in a Clonally Distributed Fashion. J. Immunol. 162, 651–658.
Sharpe, A.H., and Freeman, G.J. (2002). The B7–CD28 superfamily. Nat. Rev. Immunol. 2, 116–126.
Sharpe, A.H., and Pauken, K.E. (2018). The diverse functions of the PD1 inhibitory pathway. Nat. Rev. Immunol. 18, 153–167.
Shukla, A.A., Hubbard, B., Tressel, T., Guhan, S., and Low, D. (2007). Downstream processing of monoclonal antibodies—Application of platform approaches. J. Chromatogr. B 848, 28–39.
Stech, M., Nikolaeva, O., Thoring, L., Stöcklein, W.F.M., Wüstenhagen, D.A., Hust, M., Dübel, S., and Kubick, S. (2017). Cell-free synthesis of functional antibodies using a coupled in vitro transcription-translation system based on CHO cell lysates. Sci. Rep. 7, 12030.
Strohl, W.R. (2009). Optimization of Fc-mediated effector functions of monoclonal antibodies. Curr. Opin. Biotechnol. 20, 685–691.
Takebe, Y., Seiki, M., Fujisawa, J., Hoy, P., Yokota, K., Arai, K., Yoshida, M., and Arai, N. (1988). SR alpha promoter: an efficient and versatile mammalian cDNA expression system composed of the simian virus 40 early promoter and the R-U5 segment of human T-cell leukemia virus type 1 long terminal repeat. Mol. Cell. Biol. 8, 466–472.
Tseng, S.-Y., Otsuji, M., Gorski, K., Huang, X., Slansky, J.E., Pai, S.I., Shalabi, A., Shin, T., Pardoll, D.M., and Tsuchiya, H. (2001). B7-Dc, a New Dendritic Cell Molecule with Potent Costimulatory Properties for T Cells. J. Exp. Med. 193, 839–846.
Urlaub, G., and Chasin, L.A. (1980). Isolation of Chinese hamster cell mutants deficient in dihydrofolate reductase activity. Proc. Natl. Acad. Sci. U. S. A. 77, 4216–4220.
Williams, A.F., and Barclay, A.N. (1988). The immunoglobulin superfamily--domains for cell surface recognition. Annu. Rev. Immunol. 6, 381–405.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> IMCHECK THERAPEUTICS
<120> АНТИТЕЛА ПРОТИВ BTN3A И ИХ ПРИМЕНЕНИЕ ПРИ ЛЕЧЕНИИ РАКА ИЛИ ИНФЕКЦИОННЫХ ЗАБОЛЕВАНИЙ
<130> BEP180083 EP
<160> 23
<170> PatentIn version 3.5
<210> 1
<211> 122
<212> PRT
<213> Homo sapiens
<400> 1
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Ile Phe Thr Arg Tyr
20 25 30
Tyr Met Tyr Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn Pro Asn Asn Gly Gly Thr Lys Phe Asn Glu Lys Phe
50 55 60
Lys Asn Arg Ala Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Glu Asp Asp Tyr Asp Gly Thr Pro Phe Ala Met Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 2
<211> 107
<212> PRT
<213> Homo sapiens
<400> 2
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys His Ala Ser Gln Asn Ile Asn Val Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Asn Leu His Thr Gly Val Pro Ser Arg Phe Thr Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly Gln Thr Tyr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 3
<211> 107
<212> PRT
<213> Homo sapiens
<400> 3
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys His Ala Ser Gln Asn Ile Asn Val Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Asn Leu His Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly Gln Thr Tyr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 4
<211> 451
<212> PRT
<213> Homo sapiens
<400> 4
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Ile Phe Thr Arg Tyr
20 25 30
Tyr Met Tyr Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn Pro Asn Asn Gly Gly Thr Lys Phe Asn Glu Lys Phe
50 55 60
Lys Asn Arg Ala Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Glu Asp Asp Tyr Asp Gly Thr Pro Phe Ala Met Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Phe Glu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Ser
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly
450
<210> 5
<211> 448
<212> PRT
<213> Homo sapiens
<400> 5
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Ile Phe Thr Arg Tyr
20 25 30
Tyr Met Tyr Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn Pro Asn Asn Gly Gly Thr Lys Phe Asn Glu Lys Phe
50 55 60
Lys Asn Arg Ala Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Glu Asp Asp Tyr Asp Gly Thr Pro Phe Ala Met Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
210 215 220
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Glu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 6
<211> 214
<212> PRT
<213> Homo sapiens
<400> 6
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys His Ala Ser Gln Asn Ile Asn Val Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Asn Leu His Thr Gly Val Pro Ser Arg Phe Thr Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly Gln Thr Tyr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 7
<211> 214
<212> PRT
<213> Homo sapiens
<400> 7
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys His Ala Ser Gln Asn Ile Asn Val Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Asn Leu His Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly Gln Thr Tyr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 8
<211> 1356
<212> ДНК
<213> Homo sapiens
<400> 8
caggtccaac tggtgcagtc tggggctgaa gtgaagaagc ctggggcttc agtgaagttg 60
tcctgcaagg cttctggcta catcttcacc agatactata tgtattgggt gaagcagagg 120
cctggacaag gccttgagtg gattggagag attaatccta acaatggtgg tactaagttc 180
aatgagaagt tcaagaacag ggccacactg actgtagaca aatccatcag cacagcatac 240
atggagctca gcaggctgag atctgacgac acggcggtct attattgttc aagagaggat 300
gattacgacg ggaccccctt tgctatggac tactggggtc aaggaaccct ggtcaccgtc 360
tcctcagcct ccaccaaggg cccatcggtc ttccccctgg caccctcctc caagagcacc 420
tctgggggca cagcggccct gggctgcctg gtcaaggact acttccccga accggtgacg 480
gtgtcgtgga actcaggcgc cctgaccagc ggcgtgcaca ccttcccggc tgtcctacag 540
tcctcaggac tctactccct cagcagcgtg gtgaccgtgc cctccagcag cttgggcacc 600
cagacctaca tctgcaacgt gaatcacaag cccagcaaca ccaaggtgga caagagagtt 660
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gcccagcacc tgaattcgag 720
gggggaccgt cagtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 780
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 840
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 900
tacaacagca cgtaccgtgt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 960
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcctccat cgagaaaacc 1020
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 1080
gaagagatga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 1140
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 1200
cccgtgctgg actccgacgg ctccttcttc ctctatagca agctcaccgt ggacaagagc 1260
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1320
tacacgcaga agagcctctc cctgtctccg ggttga 1356
<210> 9
<211> 1347
<212> ДНК
<213> Homo sapiens
<400> 9
caggtccaac tggtgcagtc tggggctgaa gtgaagaagc ctggggcttc agtgaagttg 60
tcctgcaagg cttctggcta catcttcacc agatactata tgtattgggt gaagcagagg 120
cctggacaag gccttgagtg gattggagag attaatccta acaatggtgg tactaagttc 180
aatgagaagt tcaagaacag ggccacactg actgtagaca aatccatcag cacagcatac 240
atggagctca gcaggctgag atctgacgac acggcggtct attattgttc aagagaggat 300
gattacgacg ggaccccctt tgctatggac tactggggtc aaggaaccct ggtcaccgtc 360
tcctcagctt ccaccaaggg cccatccgtc ttccccctgg cgccctgctc caggagcacc 420
tccgagagca cagccgccct gggctgcctg gtcaaggact acttccccga accggtgacg 480
gtgtcgtgga actcaggcgc cctgaccagc ggcgtgcaca ccttcccggc tgtcctacag 540
tcctcaggac tctactccct cagcagcgtg gtgaccgtgc cctccagcag cttgggcacg 600
aagacctaca cctgcaatgt agatcacaag cccagcaaca ccaaggtgga caagagagtt 660
gagtccaaat atggtccccc atgcccacca tgcccagcac ctgagttcga ggggggacca 720
tcagtcttcc tgttcccccc aaaacccaag gacactctca tgatctcccg gacccctgag 780
gtcacgtgcg tggtggtgga cgtgagccag gaagaccccg aggtccagtt caactggtac 840
gtggatggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gttcaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa cggcaaggag 960
tacaagtgca aggtctccaa caaaggcctc ccgtcctcca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga gccacaggtg tacaccctgc ccccatccca ggaggagatg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctaccccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aggctaaccg tggacaagag caggtggcag 1260
gaggggaatg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacacag 1320
aagagcctct ccctgtctct gggttga 1347
<210> 10
<211> 645
<212> ДНК
<213> Homo sapiens
<400> 10
gacatccaga tgacccagtc tccatccagt ctgtctgcat ccgtaggaga cagagtcacc 60
atcacttgcc atgccagtca gaacattaat gtttggttat cttggtacca gcagaaacca 120
ggaaaagccc ctaaactctt gatctataag gcttccaact tgcacacagg cgtcccatca 180
agatttactg gcagtggatc tggaacagat ttcacattca ccatcagcag cctgcagcct 240
gaagacattg ccacttacta ctgtcaacag ggtcaaactt atccatacac gttcggacag 300
gggaccaagc tggagatcaa acgaactgtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gttag 645
<210> 11
<211> 645
<212> ДНК
<213> Homo sapiens
<400> 11
gacatccaga tgacccagtc tccatccagt ctgtctgcat ccgtaggaga cagagtcacc 60
atcacttgcc atgccagtca gaacattaat gtttggttat cttggtacca gcagaaacca 120
ggaaaagccc ctaaactctt gatctataag gcttccaact tgcacacagg cgtcccatca 180
agatttagtg gcagtggatc tggaacagat ttcacattca ccatcagcag cctgcagcct 240
gaagacattg ccacttacta ctgtcaacag ggtcaaactt atccatacac gttcggacag 300
gggaccaagc tggagatcaa acgaactgtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gttag 645
<210> 12
<211> 5
<212> PRT
<213> Homo sapiens
<400> 12
Arg Tyr Tyr Met Tyr
1 5
<210> 13
<211> 17
<212> PRT
<213> Homo sapiens
<400> 13
Glu Ile Asn Pro Asn Asn Gly Gly Thr Lys Phe Asn Glu Lys Phe Lys
1 5 10 15
Asn
<210> 14
<211> 13
<212> PRT
<213> Homo sapiens
<400> 14
Glu Asp Asp Tyr Asp Gly Thr Pro Phe Ala Met Asp Tyr
1 5 10
<210> 15
<211> 11
<212> PRT
<213> Homo sapiens
<400> 15
His Ala Ser Gln Asn Ile Asn Val Trp Leu Ser
1 5 10
<210> 16
<211> 7
<212> PRT
<213> Homo sapiens
<400> 16
Lys Ala Ser Asn Leu His Thr
1 5
<210> 17
<211> 9
<212> PRT
<213> Homo sapiens
<400> 17
Gln Gln Gly Gln Thr Tyr Pro Tyr Thr
1 5
<210> 18
<211> 513
<212> PRT
<213> Homo sapiens
<400> 18
Met Lys Met Ala Ser Phe Leu Ala Phe Leu Leu Leu Asn Phe Arg Val
1 5 10 15
Cys Leu Leu Leu Leu Gln Leu Leu Met Pro His Ser Ala Gln Phe Ser
20 25 30
Val Leu Gly Pro Ser Gly Pro Ile Leu Ala Met Val Gly Glu Asp Ala
35 40 45
Asp Leu Pro Cys His Leu Phe Pro Thr Met Ser Ala Glu Thr Met Glu
50 55 60
Leu Lys Trp Val Ser Ser Ser Leu Arg Gln Val Val Asn Val Tyr Ala
65 70 75 80
Asp Gly Lys Glu Val Glu Asp Arg Gln Ser Ala Pro Tyr Arg Gly Arg
85 90 95
Thr Ser Ile Leu Arg Asp Gly Ile Thr Ala Gly Lys Ala Ala Leu Arg
100 105 110
Ile His Asn Val Thr Ala Ser Asp Ser Gly Lys Tyr Leu Cys Tyr Phe
115 120 125
Gln Asp Gly Asp Phe Tyr Glu Lys Ala Leu Val Glu Leu Lys Val Ala
130 135 140
Ala Leu Gly Ser Asp Leu His Val Asp Val Lys Gly Tyr Lys Asp Gly
145 150 155 160
Gly Ile His Leu Glu Cys Arg Ser Thr Gly Trp Tyr Pro Gln Pro Gln
165 170 175
Ile Gln Trp Ser Asn Asn Lys Gly Glu Asn Ile Pro Thr Val Glu Ala
180 185 190
Pro Val Val Ala Asp Gly Val Gly Leu Tyr Ala Val Ala Ala Ser Val
195 200 205
Ile Met Arg Gly Ser Ser Gly Glu Gly Val Ser Cys Thr Ile Arg Ser
210 215 220
Ser Leu Leu Gly Leu Glu Lys Thr Ala Ser Ile Ser Ile Ala Asp Pro
225 230 235 240
Phe Phe Arg Ser Ala Gln Arg Trp Ile Ala Ala Leu Ala Gly Thr Leu
245 250 255
Pro Val Leu Leu Leu Leu Leu Gly Gly Ala Gly Tyr Phe Leu Trp Gln
260 265 270
Gln Gln Glu Glu Lys Lys Thr Gln Phe Arg Lys Lys Lys Arg Glu Gln
275 280 285
Glu Leu Arg Glu Met Ala Trp Ser Thr Met Lys Gln Glu Gln Ser Thr
290 295 300
Arg Val Lys Leu Leu Glu Glu Leu Arg Trp Arg Ser Ile Gln Tyr Ala
305 310 315 320
Ser Arg Gly Glu Arg His Ser Ala Tyr Asn Glu Trp Lys Lys Ala Leu
325 330 335
Phe Lys Pro Ala Asp Val Ile Leu Asp Pro Lys Thr Ala Asn Pro Ile
340 345 350
Leu Leu Val Ser Glu Asp Gln Arg Ser Val Gln Arg Ala Lys Glu Pro
355 360 365
Gln Asp Leu Pro Asp Asn Pro Glu Arg Phe Asn Trp His Tyr Cys Val
370 375 380
Leu Gly Cys Glu Ser Phe Ile Ser Gly Arg His Tyr Trp Glu Val Glu
385 390 395 400
Val Gly Asp Arg Lys Glu Trp His Ile Gly Val Cys Ser Lys Asn Val
405 410 415
Gln Arg Lys Gly Trp Val Lys Met Thr Pro Glu Asn Gly Phe Trp Thr
420 425 430
Met Gly Leu Thr Asp Gly Asn Lys Tyr Arg Thr Leu Thr Glu Pro Arg
435 440 445
Thr Asn Leu Lys Leu Pro Lys Pro Pro Lys Lys Val Gly Val Phe Leu
450 455 460
Asp Tyr Glu Thr Gly Asp Ile Ser Phe Tyr Asn Ala Val Asp Gly Ser
465 470 475 480
His Ile His Thr Phe Leu Asp Val Ser Phe Ser Glu Ala Leu Tyr Pro
485 490 495
Val Phe Arg Ile Leu Thr Leu Glu Pro Thr Ala Leu Thr Ile Cys Pro
500 505 510
Ala
<210> 19
<211> 334
<212> PRT
<213> Homo sapiens
<400> 19
Met Lys Met Ala Ser Ser Leu Ala Phe Leu Leu Leu Asn Phe His Val
1 5 10 15
Ser Leu Leu Leu Val Gln Leu Leu Thr Pro Cys Ser Ala Gln Phe Ser
20 25 30
Val Leu Gly Pro Ser Gly Pro Ile Leu Ala Met Val Gly Glu Asp Ala
35 40 45
Asp Leu Pro Cys His Leu Phe Pro Thr Met Ser Ala Glu Thr Met Glu
50 55 60
Leu Lys Trp Val Ser Ser Ser Leu Arg Gln Val Val Asn Val Tyr Ala
65 70 75 80
Asp Gly Lys Glu Val Glu Asp Arg Gln Ser Ala Pro Tyr Arg Gly Arg
85 90 95
Thr Ser Ile Leu Arg Asp Gly Ile Thr Ala Gly Lys Ala Ala Leu Arg
100 105 110
Ile His Asn Val Thr Ala Ser Asp Ser Gly Lys Tyr Leu Cys Tyr Phe
115 120 125
Gln Asp Gly Asp Phe Tyr Glu Lys Ala Leu Val Glu Leu Lys Val Ala
130 135 140
Ala Leu Gly Ser Asn Leu His Val Glu Val Lys Gly Tyr Glu Asp Gly
145 150 155 160
Gly Ile His Leu Glu Cys Arg Ser Thr Gly Trp Tyr Pro Gln Pro Gln
165 170 175
Ile Gln Trp Ser Asn Ala Lys Gly Glu Asn Ile Pro Ala Val Glu Ala
180 185 190
Pro Val Val Ala Asp Gly Val Gly Leu Tyr Glu Val Ala Ala Ser Val
195 200 205
Ile Met Arg Gly Gly Ser Gly Glu Gly Val Ser Cys Ile Ile Arg Asn
210 215 220
Ser Leu Leu Gly Leu Glu Lys Thr Ala Ser Ile Ser Ile Ala Asp Pro
225 230 235 240
Phe Phe Arg Ser Ala Gln Pro Trp Ile Ala Ala Leu Ala Gly Thr Leu
245 250 255
Pro Ile Leu Leu Leu Leu Leu Ala Gly Ala Ser Tyr Phe Leu Trp Arg
260 265 270
Gln Gln Lys Glu Ile Thr Ala Leu Ser Ser Glu Ile Glu Ser Glu Gln
275 280 285
Glu Met Lys Glu Met Gly Tyr Ala Ala Thr Glu Arg Glu Ile Ser Leu
290 295 300
Arg Glu Ser Leu Gln Glu Glu Leu Lys Arg Lys Lys Ile Gln Tyr Leu
305 310 315 320
Thr Arg Gly Glu Glu Ser Ser Ser Asp Thr Asn Lys Ser Ala
325 330
<210> 20
<211> 584
<212> PRT
<213> Homo sapiens
<400> 20
Met Lys Met Ala Ser Ser Leu Ala Phe Leu Leu Leu Asn Phe His Val
1 5 10 15
Ser Leu Phe Leu Val Gln Leu Leu Thr Pro Cys Ser Ala Gln Phe Ser
20 25 30
Val Leu Gly Pro Ser Gly Pro Ile Leu Ala Met Val Gly Glu Asp Ala
35 40 45
Asp Leu Pro Cys His Leu Phe Pro Thr Met Ser Ala Glu Thr Met Glu
50 55 60
Leu Arg Trp Val Ser Ser Ser Leu Arg Gln Val Val Asn Val Tyr Ala
65 70 75 80
Asp Gly Lys Glu Val Glu Asp Arg Gln Ser Ala Pro Tyr Arg Gly Arg
85 90 95
Thr Ser Ile Leu Arg Asp Gly Ile Thr Ala Gly Lys Ala Ala Leu Arg
100 105 110
Ile His Asn Val Thr Ala Ser Asp Ser Gly Lys Tyr Leu Cys Tyr Phe
115 120 125
Gln Asp Gly Asp Phe Tyr Glu Lys Ala Leu Val Glu Leu Lys Val Ala
130 135 140
Ala Leu Gly Ser Asp Leu His Ile Glu Val Lys Gly Tyr Glu Asp Gly
145 150 155 160
Gly Ile His Leu Glu Cys Arg Ser Thr Gly Trp Tyr Pro Gln Pro Gln
165 170 175
Ile Lys Trp Ser Asp Thr Lys Gly Glu Asn Ile Pro Ala Val Glu Ala
180 185 190
Pro Val Val Ala Asp Gly Val Gly Leu Tyr Ala Val Ala Ala Ser Val
195 200 205
Ile Met Arg Gly Ser Ser Gly Gly Gly Val Ser Cys Ile Ile Arg Asn
210 215 220
Ser Leu Leu Gly Leu Glu Lys Thr Ala Ser Ile Ser Ile Ala Asp Pro
225 230 235 240
Phe Phe Arg Ser Ala Gln Pro Trp Ile Ala Ala Leu Ala Gly Thr Leu
245 250 255
Pro Ile Ser Leu Leu Leu Leu Ala Gly Ala Ser Tyr Phe Leu Trp Arg
260 265 270
Gln Gln Lys Glu Lys Ile Ala Leu Ser Arg Glu Thr Glu Arg Glu Arg
275 280 285
Glu Met Lys Glu Met Gly Tyr Ala Ala Thr Glu Gln Glu Ile Ser Leu
290 295 300
Arg Glu Lys Leu Gln Glu Glu Leu Lys Trp Arg Lys Ile Gln Tyr Met
305 310 315 320
Ala Arg Gly Glu Lys Ser Leu Ala Tyr His Glu Trp Lys Met Ala Leu
325 330 335
Phe Lys Pro Ala Asp Val Ile Leu Asp Pro Asp Thr Ala Asn Ala Ile
340 345 350
Leu Leu Val Ser Glu Asp Gln Arg Ser Val Gln Arg Ala Glu Glu Pro
355 360 365
Arg Asp Leu Pro Asp Asn Pro Glu Arg Phe Glu Trp Arg Tyr Cys Val
370 375 380
Leu Gly Cys Glu Asn Phe Thr Ser Gly Arg His Tyr Trp Glu Val Glu
385 390 395 400
Val Gly Asp Arg Lys Glu Trp His Ile Gly Val Cys Ser Lys Asn Val
405 410 415
Glu Arg Lys Lys Gly Trp Val Lys Met Thr Pro Glu Asn Gly Tyr Trp
420 425 430
Thr Met Gly Leu Thr Asp Gly Asn Lys Tyr Arg Ala Leu Thr Glu Pro
435 440 445
Arg Thr Asn Leu Lys Leu Pro Glu Pro Pro Arg Lys Val Gly Ile Phe
450 455 460
Leu Asp Tyr Glu Thr Gly Glu Ile Ser Phe Tyr Asn Ala Thr Asp Gly
465 470 475 480
Ser His Ile Tyr Thr Phe Pro His Ala Ser Phe Ser Glu Pro Leu Tyr
485 490 495
Pro Val Phe Arg Ile Leu Thr Leu Glu Pro Thr Ala Leu Thr Ile Cys
500 505 510
Pro Ile Pro Lys Glu Val Glu Ser Ser Pro Asp Pro Asp Leu Val Pro
515 520 525
Asp His Ser Leu Glu Thr Pro Leu Thr Pro Gly Leu Ala Asn Glu Ser
530 535 540
Gly Glu Pro Gln Ala Glu Val Thr Ser Leu Leu Leu Pro Ala His Pro
545 550 555 560
Gly Ala Glu Val Ser Pro Ser Ala Thr Thr Asn Gln Asn His Lys Leu
565 570 575
Gln Ala Arg Thr Glu Ala Leu Tyr
580
<210> 21
<211> 243
<212> PRT
<213> Macaca fascicularis
<400> 21
Met Gly Ser Ser Leu Ala Phe Leu Leu Leu Ser Phe His Val Cys Val
1 5 10 15
Leu Leu Leu Gln Leu Leu Met Pro His Ser Ala Gln Phe Ala Val Val
20 25 30
Gly Pro Pro Gly Pro Ile Leu Ala Met Val Gly Glu Asp Ala Asp Leu
35 40 45
Pro Cys His Leu Phe Pro Thr Met Ser Ala Glu Thr Met Glu Leu Arg
50 55 60
Trp Val Ser Ser Asn Leu Arg Gln Val Val Asn Val Tyr Ala Asp Gly
65 70 75 80
Lys Glu Val Glu Asp Arg Gln Ser Ala Ala Tyr Arg Gly Arg Thr Ser
85 90 95
Ile Leu Arg Asp Gly Ile Thr Ala Gly Lys Ala Ala Leu Arg Ile His
100 105 110
Asn Val Thr Ala Ser Asp Ser Gly Lys Tyr Leu Cys Tyr Phe Gln Asp
115 120 125
Gly Asp Phe Tyr Glu Lys Ala Leu Val Glu Leu Lys Val Ala Ala Leu
130 135 140
Gly Ser Asp Leu His Ile Asp Val Lys Gly Tyr Glu Asp Gly Gly Ile
145 150 155 160
His Leu Glu Cys Arg Ser Thr Gly Trp Tyr Pro Gln Pro Gln Ile Arg
165 170 175
Trp Ser Asn Asp Lys Gly Glu Asn Ile Pro Ala Val Glu Ala Pro Val
180 185 190
Phe Val Asp Gly Val Gly Leu Tyr Ala Val Ala Ala Ser Val Ile Leu
195 200 205
Arg Gly Ser Ser Gly Glu Gly Val Ser Cys Thr Ile Arg Ser Ser Leu
210 215 220
Leu Gly Leu Glu Lys Thr Thr Ser Ile Ser Ile Ala Gly His His His
225 230 235 240
His His His
<210> 22
<211> 243
<212> PRT
<213> Macaca fascicularis
<400> 22
Met Gly Ser Ser Leu Ala Phe Leu Leu Leu Asn Phe His Val Ser Phe
1 5 10 15
Phe Leu Val Gln Leu Leu Thr Pro Cys Ser Ala Gln Phe Ser Val Leu
20 25 30
Gly Pro Ser Gly Pro Ile Leu Ala Met Val Gly Glu Asp Ala Asp Leu
35 40 45
Pro Cys His Leu Phe Pro Thr Met Ser Ala Glu Thr Met Glu Leu Arg
50 55 60
Trp Val Ser Ser Ser Leu Arg Gln Val Val Asn Val Tyr Ala Asp Gly
65 70 75 80
Lys Glu Val Glu Asp Arg Gln Ser Ala Pro Tyr Arg Gly Arg Thr Ser
85 90 95
Ile Leu Arg Asp Asp Ile Ala Ala Gly Lys Ala Ala Leu Arg Ile His
100 105 110
Asn Val Thr Ala Ser Asp Ser Gly Lys Tyr Leu Cys Tyr Phe Gln Asp
115 120 125
Ala Asp Phe Tyr Glu Lys Ala Leu Val Glu Leu Lys Val Ala Ala Leu
130 135 140
Gly Ser Asn Leu His Val Glu Val Lys Gly Tyr Glu Asp Gly Gly Ile
145 150 155 160
His Leu Glu Cys Arg Ser Thr Gly Trp Tyr Pro Gln Pro Lys Ile Gln
165 170 175
Trp Ser Asn Ala Lys Gly Gln Asn Ile Pro Ala Val Glu Ala Pro Val
180 185 190
Val Ala Asp Gly Val Gly Leu Tyr Ala Val Ala Ala Ser Val Ile Met
195 200 205
Arg Gly Gly Ser Gly Glu Ser Val Ser Cys Ile Ile Arg Asn Ser Val
210 215 220
Leu Gly Leu Glu Lys Thr Ala Ser Ile Ser Ile Ala Asp His His His
225 230 235 240
His His His
<210> 23
<211> 243
<212> PRT
<213> Macaca fascicularis
<400> 23
Met Ala Asn Phe Leu Ala Phe Leu Leu Leu Asn Phe Arg Val Cys Leu
1 5 10 15
Leu Leu Val Gln Leu Leu Thr Pro Cys Ser Ala Gln Phe Ala Val Leu
20 25 30
Gly Pro His Gly Pro Ile Leu Ala Met Val Gly Glu Asp Val Asp Leu
35 40 45
Pro Cys His Leu Phe Pro Thr Met Ser Ala Glu Thr Met Glu Leu Arg
50 55 60
Trp Val Ser Ser Ser Leu Arg Gln Val Val Asn Val Tyr Ser Asp Gly
65 70 75 80
Lys Glu Val Glu Asp Arg Gln Ser Ala Pro Tyr Arg Gly Arg Thr Ser
85 90 95
Ile Leu Arg Asp Gly Ile Thr Ala Gly Lys Ala Ala Leu Arg Ile His
100 105 110
Asn Val Thr Ala Ser Asp Ser Gly Lys Tyr Leu Cys Tyr Phe Gln Asp
115 120 125
Gly Asp Phe Tyr Glu Lys Ala Leu Val Glu Leu Lys Val Ala Ala Leu
130 135 140
Gly Ser Asp Leu His Ile Glu Val Lys Gly Tyr Glu Asp Gly Gly Ile
145 150 155 160
His Leu Glu Cys Arg Ser Thr Gly Trp Tyr Pro Gln Pro Gln Ile Gln
165 170 175
Trp Ser Asn Thr Lys Gly Gln His Ile Pro Ala Val Lys Ala Pro Val
180 185 190
Val Ala Asp Gly Val Gly Leu Tyr Ala Val Ala Ala Ser Val Ile Met
195 200 205
Arg Gly Ser Ser Gly Glu Gly Val Ser Cys Ile Ile Arg Asn Ser Leu
210 215 220
Leu Gly Leu Glu Lys Thr Ala Ser Ile Ser Ile Thr Asp His His His
225 230 235 240
His His His
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА, СПЕЦИФИЧНЫЕ К BTN2, И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2821703C2 |
| РЕКОМБИНАНТНЫЕ КЛЕТКИ ДРОЖЖЕЙ, ПРОДУЦИРУЮЩИЕ ПОЛИМОЛОЧНУЮ КИСЛОТУ, И ИХ ИСПОЛЬЗОВАНИЯ | 2016 |
|
RU2758138C2 |
| ИНГИБИТОР ИММУНОСУПРЕССИИ, СВЯЗАННЫЙ С РАКОМ | 2018 |
|
RU2805232C2 |
| ИММУНОЦИТОКИНЫ НА ОСНОВЕ IL-15 И IL-15Rα ДОМЕНА SUSHI | 2012 |
|
RU2763298C2 |
| ВАРИАНТЫ КИСЛОЙ АЛЬФА-ГЛЮКОЗИДАЗЫ И ИХ ИСПОЛЬЗОВАНИЕ | 2017 |
|
RU2780410C2 |
| ЛЕЧЕНИЕ АНОМАЛЬНОГО ОТЛОЖЕНИЯ ВИСЦЕРАЛЬНОГО ЖИРА С ИСПОЛЬЗОВАНИЕМ РАСТВОРИМЫХ ПОЛИПЕПТИДОВ РЕЦЕПТОРА ФАКТОРА РОСТА ФИБРОБЛАСТОВ 3 (sFGFR3) | 2018 |
|
RU2794170C2 |
| ПОЛИПЕПТИДЫ, ЯВЛЯЮЩИЕСЯ РАСТВОРИМЫМИ РЕЦЕПТОРАМИ 3 ФАКТОРА РОСТА ФИБРОБЛАСТОВ (SFGFR3), И ПУТИ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2751483C2 |
| ПРИМЕНЕНИЕ АНТИТЕЛА ПРОТИВ О-АЦЕТИЛИРОВАННОГО ГАНГЛИОЗИДА GD2 ДЛЯ УЛУЧШЕНИЯ ТЕРАПЕВТИЧЕСКОГО ПОТЕНЦИАЛА ЛЕКАРСТВЕННЫХ СРЕДСТВ | 2017 |
|
RU2771173C2 |
| ГЕННАЯ ТЕРАПИЯ ГИПОФОСФАТЕМИЧЕСКИХ ЗАБОЛЕВАНИЙ, СВЯЗАННЫХ С ФАКТОРОМ РОСТА ФИБРОБЛАСТОВ 23 | 2020 |
|
RU2815545C2 |
| МУТАНТНЫЕ ВАРИАНТЫ Fc IgG1, ЛИШЕННЫЕ ЭФФЕКТОРНЫХ ФУНКЦИЙ | 2018 |
|
RU2764559C2 |
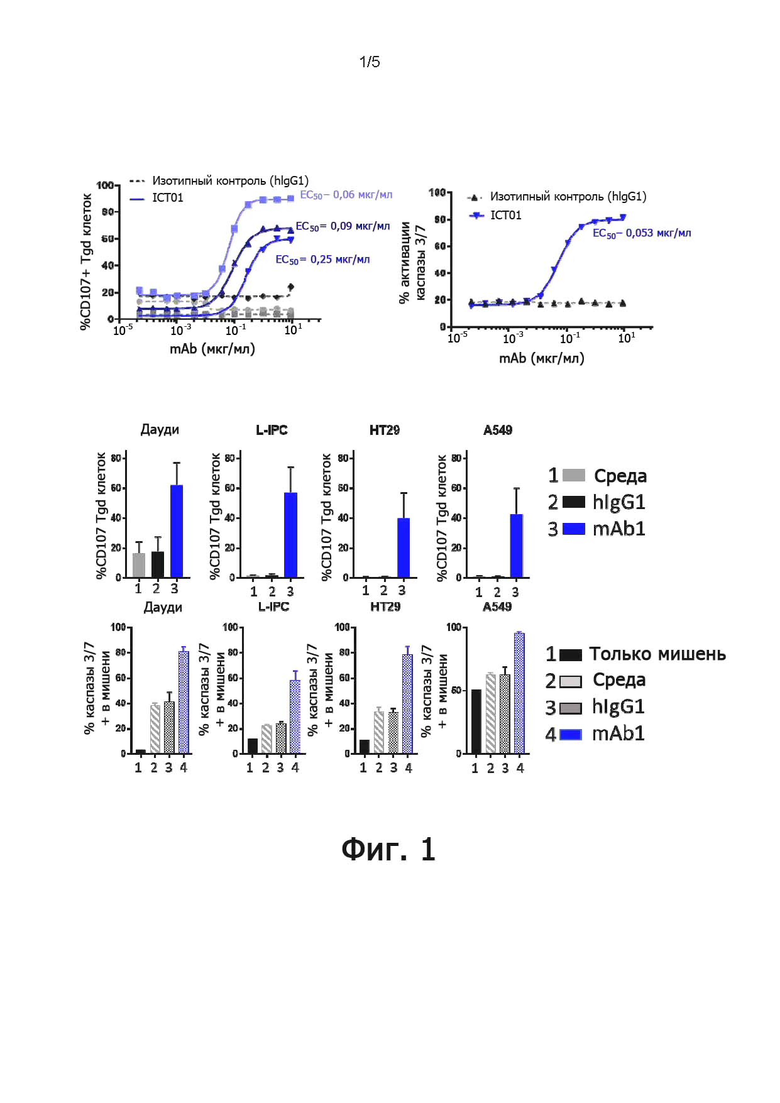
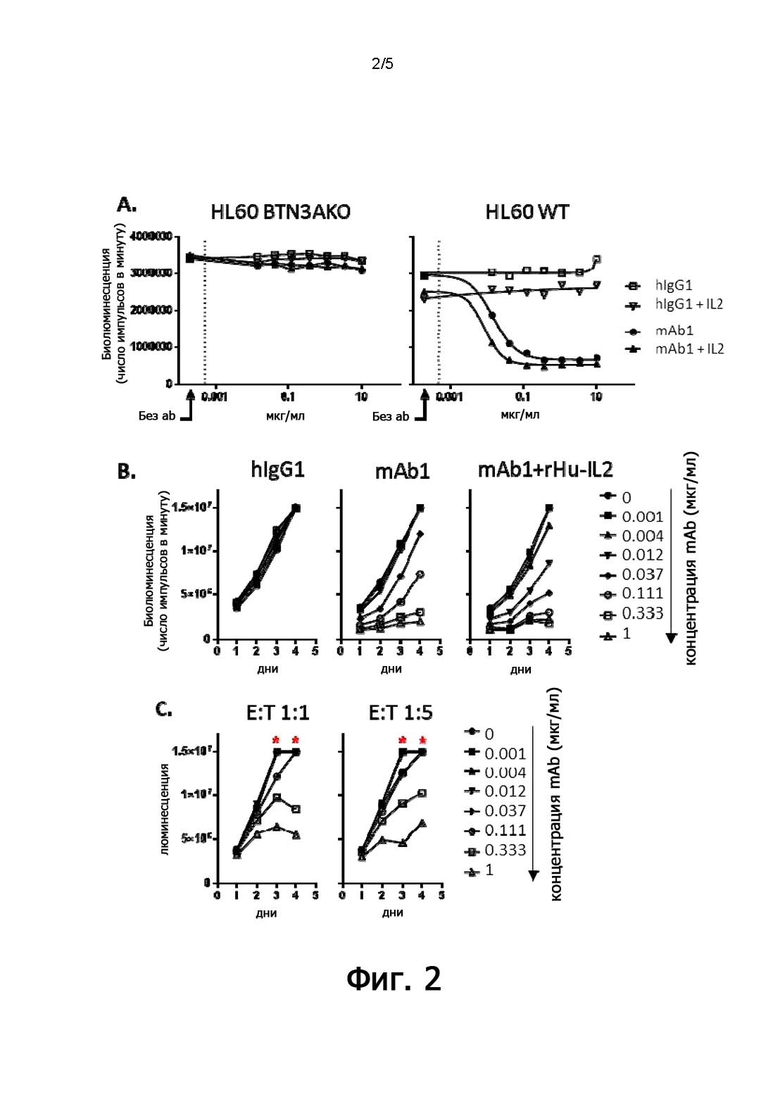
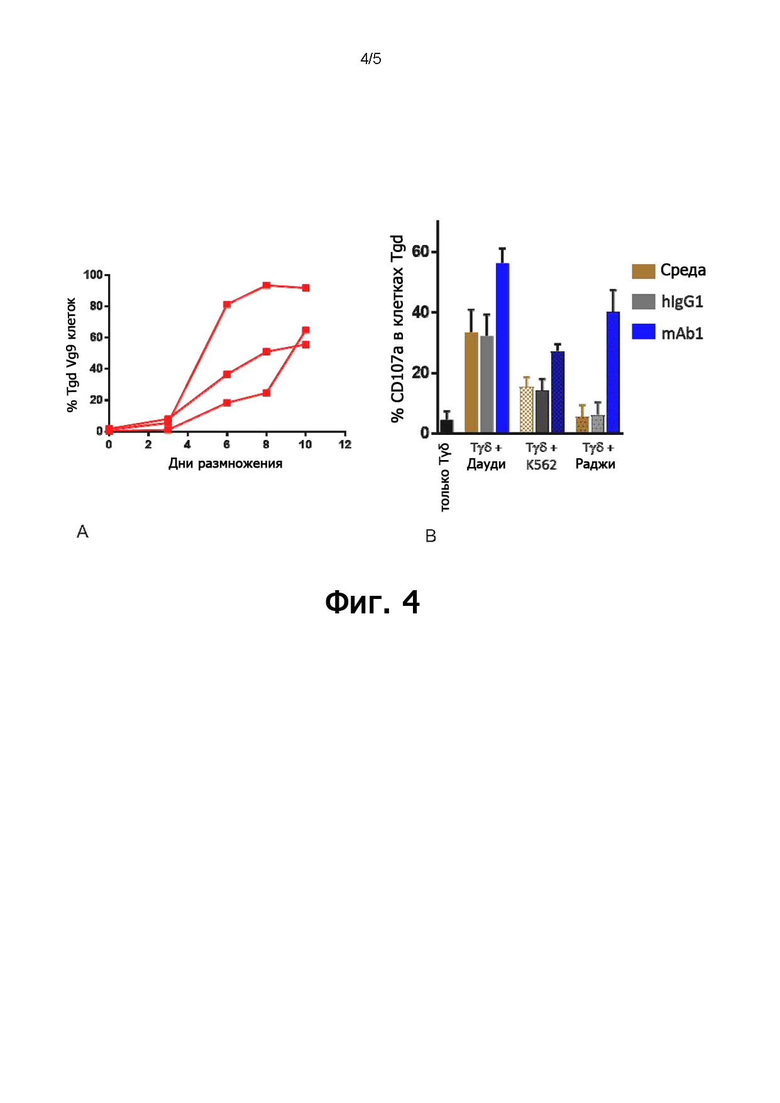
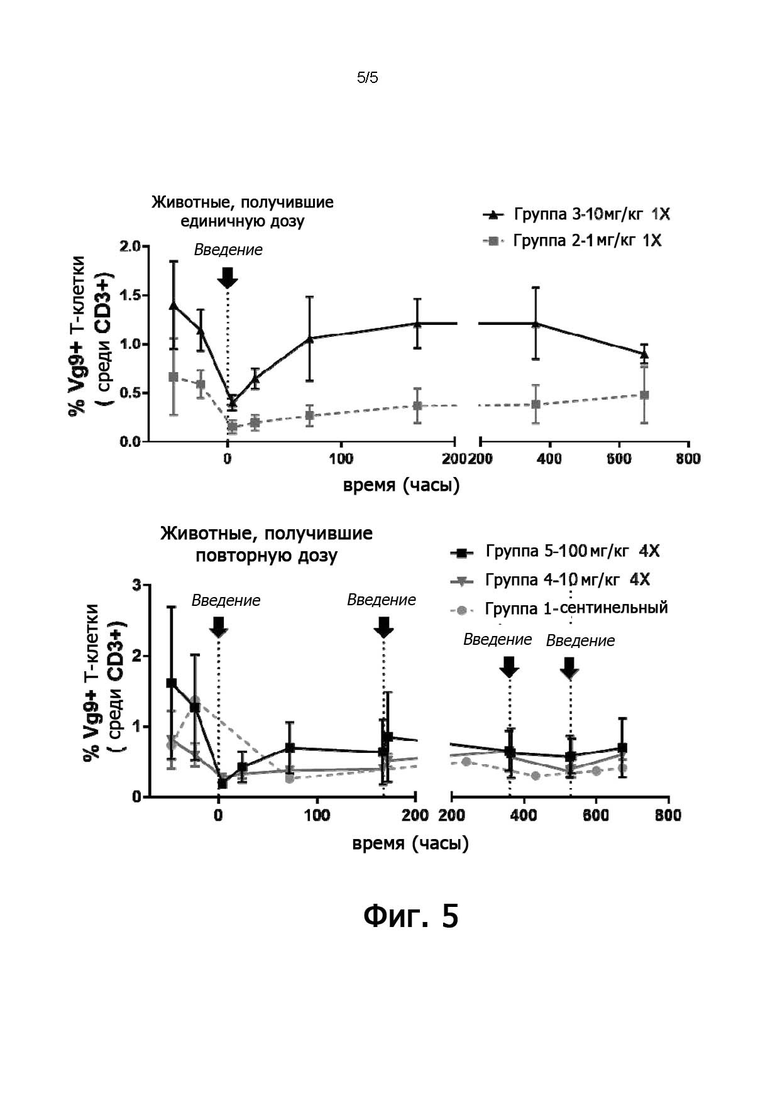
Изобретение относится к области биотехнологии. Описана группа изобретений, включающая выделенное антитело против BTN3A, фармацевтическую композицию, опосредующую активацию Т-клеток Vγ9Vδ2, которая включает антитело против BTN3A, экспрессионный вектор для рекомбинантного продуцирования антитела против BTN3A в клетке-хозяине, клетку-хозяин для рекомбинантной продукции антитела против BTN3A, включающую экспрессионный вектор и способ получения антитела против BTN3A, включающий культивирование клетки-хозяина для экспрессии указанного антитела клеткой-хозяином и выделение антитела. Выделенное антитело против BTN3A содержит в себе последовательность тяжелой цепи SEQ ID NO: 1 и последовательность легкой цепи, выбранной из последовательностей SEQ ID NO: 2 или SEQ ID NO: 3. Изобретение расширяет арсенал средств, связывающихся с BTN3A. 5 н. и 10 з.п. ф-лы, 5 ил., 28 табл., 1 пр.
1. Выделенное антитело против BTN3A, содержащее полипептид VH вариабельной области тяжелой цепи с SEQ ID NO: 1 и полипептид VL вариабельной области легкой цепи с SEQ ID NO: 2 или SEQ ID NO: 3.
2. Выделенное антитело против BTN3A по п. 1, которое связывается с полипептидом BTN3A человека.
3. Выделенное антитело против BTN3A по п. 1 или 2, которое связывается с полипептидом BTN3A человека с KD, равной 10 нМ или менее, предпочтительно, с KD, равной 5 нМ или менее, как измерено поверхностным плазмонным резонансом.
4. Выделенное антитело против BTN3A по любому из пп. 1-3, в котором указанное антитело вызывает активацию Т-клеток Vγ9Vδ2 при совместном культивировании с клетками, экспрессирующими BTN3, с ЕС50 ниже 5 мкг/мл, предпочтительно 1 мкг/мл или ниже, как измерено в анализе на дегрануляцию.
5. Выделенное антитело против BTN3A по любому из пп. 1-4, которое включает мутантную или химически модифицированную константную область IgG1, в которой указанная мутантная или химически модифицированная константная область IgG1 не обеспечивает связывания с рецепторами Fcγ или снижает их связывание по сравнению с соответствующим антителом с константной областью изотипа IgG1 дикого типа.
6. Выделенное антитело против BTN3A по п. 5, в котором указанный мутантный константный участок IgG1 представляет собой тройной мутант IgG1 с L247F, L248E и P350S.
7. Выделенное антитело против BTN3A по любому из пп. 1-6, которое представляет собой mAbl, включающее тяжелую цепь SEQ ID NO: 4 и легкую цепь SEQ ID NO: 6.
8. Фармацевтическая композиция, опосредующая активацию Т-клеток Vγ9Vδ2, которая включает антитело против BTN3A по любому из пп. 1-7 в комбинации с одним или несколькими фармацевтически приемлемыми вспомогательными веществами, разбавителями или носителями.
9. Фармацевтическая композиция по п. 8, отличающаяся тем, что находится в форме лиофилизированной композиции, включающей антитело против BTN3A по любому из пп. 1-7.
10. Выделенное антитело против BTN3A по п. 1, отличающееся тем, что находится в предварительно заполненном шприце.
11. Выделенное антитело против BTN3A по п. 1, отличающееся тем, что находится во флаконе.
12. Экспрессионный вектор для рекомбинантного продуцирования антитела против BTN3A по любому из пп. 1-7 в клетке-хозяине, включающий, по меньшей мере, одну нуклеиновую кислоту, кодирующую указанное антитело против BTN3A.
13. Экспрессионный вектор по п. 12, включающий, по меньшей мере, нуклеиновые кислоты, кодирующие тяжелую и легкую цепи mAb1, как определено в п. 7.
14. Клетка-хозяин для рекомбинантной продукции антитела против BTN3A по любому из пп. 1-7, включающая экспрессионный вектор по п. 12 или 13.
15. Способ получения антитела против BTN3A по любому из пп. 1-7, включающий:
(i) культивирование клетки-хозяина по п. 14 для экспрессии указанного антитела клеткой-хозяином и
(ii) выделение антитела.
| WO 2016191305 A1 01.12.2016 | |||
| US 20150353643 A1 10.12.2015 | |||
| WO 2012080351 A1 21.06.2012 | |||
| Benyamine A | |||
| et al | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Oncoimmunology, 2016, 5(10), p | |||
| Способ воспроизведения синдрома метрит-мастит-агалактия у свиней | 1983 |
|
SU1146843A1 |
| Olive D | |||
| et al | |||
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
