Область техники
Изобретение относится к области физиологии и медицины, и в частности, к онкологии, иммунологии, неврологии и геронтологии. Изобретение применяется для лечения рака и других злокачественных новообразований (ЗНО), нейродегенеративных, аутоиммунных и геронтологических заболеваний, а также для продления жизни млекопитающих и человека путем терапии аутологичным биомедицинским клеточным продуктом (БМКП).
Уровень техники
На основе хорошо известных знаний и существующих биомедицинских технологий иммунологии, онкологии, неврологии, трансплантологии, клеточной биологии, молекулярной биологии и клеточной инженерии настоящее изобретение, исходя из нового теоретического понимания неразрешимой научной проблемы формирования канцерогенеза, нейродегенерации и аутоиммунного процесса, предлагает принципиально новый и достаточно универсальный методологический и научно-практический подход в лечении основных фатальных болезней цивилизации (рак и другие злокачественные заболевания, болезнь Альцгеймера, деменция, болезнь Паркинсона, сахарный диабет 1-го типа, миокардиодистрофия, системная красная волчанка, ревматизм, рассеянный склероз и другие), а также обеспечивает получение принципиально нового и выдающегося результата в персонализированной медицине и науках о жизни, основываясь на имеющихся и новых постгеномных знаниях о гемопоэтической стволовой клетке человека.
Известно, что иммунная система человека в норме является «дирижером» и «аранжировщиком» всех процессов регуляции, системной и локальной регенерации и саногенеза органов и тканей человека (Селедцов В.И., 2019). Она располагает всем необходимым спектром гуморальных факторов и клеточных инструментов для регуляции этих процессов. Это концептуальное положение в полной мере можно отнести к процессам регенерации и восстановления нервной ткани головного и спинного мозга человека при болезни поврежденного мозга, нейродегенеративных и аутоиммунных процессах и онкологических заболеваниях (Брюховецкий А.С., 2016). Роль иммунной системы в биологии рака и механизмах канцерогенеза при других злокачественных опухолях, а также при нейродегенеративных процессах является центральной и определяющей (Брюховецкий А.С., Хотимченко Ю.С., 2019).
Собственные иммунокомпетентные клетки (ИКК) иммунной системы человека абсолютно по-разному повреждаются в своей генетической и постгеномной эпигенетической структуре, а также по-разному функционально страдают при различных заболеваниях, и это по-разному проявляется нарушениями их эффекторных функций и многообразными клиническими проявлениями. Во всех случаях аутоиммунных, нейродегенеративных и онкологических заболеваний всегда выявляются различные генетические поломки, нарушения транскриптомного и белкового обмена и тяжелейшие метаболомные нарушения. Но они каждый раз разные и зависят в каждом конкретном случает от особенностей патогенеза болезни, топической локализации болезни, состояния больного в момент действия этиопатогенетических факторов и еще от целого ряда причин, которые мы не знаем или не учитываем. Например, в случаях онкологического процесса киллерные ИКК врожденного иммунитета, такие как NK-клетки, NKT-клетки, ϒδТ-клетки, цитотоксические лимфоциты крови перестают уничтожать опухолевые клетки (ОК), становятся абсолютно толерантны к опухолевым антигенам и трансформированным ОК и вместо выполнения генетически и эпигенетически детерминированных функций элиминации ОК, перестают их распознавать и уничтожать. Ведущие онкологи и онкоиммунологи страны считают, что ОК легко уклоняются от уничтожения клеточными системами врожденного иммунитета (Кадагидзе З.Г., 2017; Гривцова Л.Ю., Тупицын Н.Н., 2017), но на самом деле это именно ИКК перестают реагировать на ОК на молекулярно-биологическом уровне, так как получили транскриптомную и протеомную модификацию генома и эпигенома и всех своих внутриклеточных сигнальных путей под воздействием горизонтального и вертикального обмена опухолеспецифическими белками (ОСБ) из ОК, которые привели к информационному перепрограммированию их генетически детерминированных транскриптомных алгоритмов киллерных функций (Брюховецкий А.С., 2014; Bryukhovetskiy A.S., 2019). Подобные трансформации в результате канцерогенеза происходят и с ИКК приобретенного иммунитета. Так, макрофаги и дендритные клетки, которые должны уничтожать ОК, презентировать на своей поверхности опухолевые антигены и инструктировать цитоксические Т-лимфоциты, как уничтожать ОК, также перестают выполнять свои эффекторные функции, что приводит к их иммунотолерантности к ОК и беспрепятственному росту опухоли. Именно недостаточность иммунной системы, проявляющаяся гипореактивностью или ареактивностью ИКК врожденного и приобретенного иммунитета становится основной причиной беспрепятственного роста опухоли.
При аутоиммунных и нейродегенеративных процессах идут абсолютно противоположные молекулярно-биологические процессы, в результате которых собственные лимфоциты становятся, наоборот, иммунологически гиперактивными, аутоагрессивными и гиперреактивными к определенным специализированным высокодифференцированным клеткам и тканям организма человека и начинают уничтожать собственные клетки и ткани организма, что в исходе проявляется дегенеративным, дистрофическим или атрофическим процессом. При системной красной волчанке и полимиозите мишенью аутореактивных ИКК становится соединительная ткань, при рассеянном склерозе и рассеянном энцефаломиелите мишенью этих ИКК становится миелиновая оболочка аксонов нейронов нервной ткани. При сахарном диабете 1-го типа мишенью аутоагрессивных собственных Т-лимфоидных клеток становятся β-клетки поджелудочной железы. При болезни Альцгеймера и синильной корковой атрофии мишенью аутоагрессиивных собственных ИКК становятся корковые нейроны. При болезни Паркинсона эти агрессивные ИКК повреждают нейроны стриопаллидарной зоны, а при боковом амиотрофическом склерозе (БАС) они повреждают двигательные нейроны, поэтому в современных классификациях БАС даже стали называть болезнью моторного нейрона.
Главное, что объединяет все эти болезни это то, что они возникают в результате агрессивной экспансии собственных лимфоцитов в специализированные клетки, ткани и органы организма, в которые они проникают в результате нарушения гематотканевых барьеров или гематоэнцефалического барьера вследствие воздействия различных этипатогенетических факторов заболеваний и возраста. Это приводит к системному и/или локальному асептическому воспалению в определенных органах, тканях и системах организма, мощнейшему выбросу лейкоцитами провоспалительных цитокинов (цитокиновому шторму) и возникновению генетических мутаций и постгеномных (транскриптомных, протеомных, метаболомных, секретомных) нарушений в ядре специализированных дифференцированных или тканеспецифических стволовых клеток (СК), то есть эти специализированные и высокодифференцированные клетки разных тканей становятся клетками-мишенями для лимфоцитов, в которых запускаются патологические пролиферативные или дегенеративные внутритканевые процессы. Системное или локальное асептическое воспаление в специализированной (нервной, эндокринной, соединительной и т.п.) ткани в обоих случаях становится причиной эксайтотоксичности, нарушений метаболизма в клетках и развития оксидативного стресса во всех элементах тканевого континуума, вовлеченного в патологический процесс органа, что увеличивает спектр генетических и постгеномных нарушений в специализированных клетках. Это ведет к накоплению в клетках-мишенях различных патологических протеинов, от которых, как правило, безуспешно пытается освободиться клетка-мишень. В ряде клеток идет накопление опухолеспецифических белков и формируется ОК и опухолевая стволовая клетка (ОСК), а в других клетках идет накопление tau-белков (болезнь Альцгеймера), FUS и SOD1 белков (боковой амиотрофический склероз, синильная корковая атрофия, БАС-деменция), телец Леви (болезнь Паркинсона) и формируются глобальные атрофические и нейродегенеративные изменения в этих клетках. Эти патологические специфические белки - могильщики клетки, возникшие в результате мутаций генов, на которые в организме нет регуляторных инструментов. Это белки не нашего эволюционного вида (Homo sapiense), называемые также опухолеспецифическими белками (ОСБ), белками внутриклеточного накопления, таупатии и т.д.). Они накапливаются в клетке, не могут быть ею выведены наружу из-за их равномерной диффузии и глобального распределения по всем компартментам клетки и становятся главной причиной гибели клетки или гибели всего организма. Так, ОСБ при глиобластоме, раке молочной железы и раке легкого вовлечены во все пути сигналинга внутриклеточной трансдукции и только 10 путей лишь частично повреждены ими и способны к реальному функционированию (Брюховецкий А.С., 2014).
Другими словами, одним из фундаментальных механизмов большинства болезней цивилизации (рака и других ЗНО, болезней накопления и аутоиммунных заболеваний), старения и смерти человека в результате экстремального или хронического воздействия неблагоприятных факторов окружающей среды и эндогенных факторов является появление в специализированной клетке организма человека мутаций генома и нарушений транскриптома, что приводит к накоплению патоспецифических протеомных тел в клетке и к невозможности самостоятельного освобождения от этих вневидовых патологических белковых тел. Клетка всегда пытается освободиться от них достаточно стандартным путем в виде удаления их в форме микровезикул или деления и тем самым спасти себя. Примитивное, но обычно эффективное решение возникшей проблемы путем микровезикулярного транспорта, митозов и репродуктивного клеточного поведения, выбранное специализированной и высокодифференцированной клеткой организма при накоплении в ней патологических белковых тел генетически детерминировано и эволюционно отработано. Деление клетки, с одной стороны, позволяет освободиться от патологических белков путем выведения их наружу или в дочернюю клетку. Однако, при неэффективности данного инструмента саногенеза клетки путем многократного и безуспешного деления, она начинает и проводит пластическую перестройку всего внутриклеточного аппарата сигналинга клетки, что ведет к формированию неопластического процесса в одном из случаев и образованию опухолей. С другой стороны, когда деление пострадавшей клетки сразу невозможно из-за ее многообразных межклеточных связей и контактов, а также системной интеграции клеток в органе и ткани, то для выведения наружу этих патологических белков, специализированная клетка начинает готовиться к делению и перестраивать всю свою геномно-протемную структуру для процесса очищения от патологических белков, разрушает межклеточные связи, межклеточные взаимодействия и мембранные рецепторы, что приводит к ее дегенерации и атрофии, информационной блокаде и смерти. В этом заключается универсальный фундаментальный клеточный механизм развития первичных этапов как канцерогенеза, так и нейродегенерации и дегенеративно-атрофических процессов.
Эти болезни не возникают в один день или один-два месяца, они «текут» годами на фоне хронической функциональной патоспецифической недостаточности иммунной системы, а манифестируют только тогда, когда полностью исчерпывается ресурс регуляции и саногенеза иммунной системы или происходит сбой в витальных системах жизнеобеспечения и регуляции иммунной системы, вызванный экзогенными (травма, облучение и т.д.) или эндогенными (нарушение реологии крови, лимфообразования и т.д) факторами. Например, БАС и болезнь Альцгеймера проявляются и могут быть диагностированны только тогда, когда у пациента уже погибло 70-80% специализированных нейронов в головном и спинном мозге, что происходит в течение нескольких предшествующих лет, за которые болезнь формируется.
Однако сам по себе механизм появления и накопления патологических белков в специализированной клетке организма в результате мутаций закономерен, очевиден и имеет право на возникновение в связи с возможными погрешностями в процессах митозов клеток, количество которых в организме человека составляет несколько миллиардов в день. Процесс становится патологическим при нарушении механизмов регуляции и контроля за ним иммунной системой человека. Первая линия надзора и контроля это ИКК периферической крови (ПК), функцией которых является уничтожение патологических клеток. Вторая линия регуляторной и саногенетической защиты организма выполняется региональными СК, а третьей линией клеточной и гуморальной иммунной защиты является вовлечение в этот процесс гемопоэтических стволовых клеток (ГСК).
Центральным системным механизмом генерализации и хронизации канцерогенного, аутоиммунного и нейродегенеративного процесса в организме человека является вовлечение региональных и системообразующих СК в патологический процесс. В этих процессах участвуют как тканеспецифические СК, так и главные регуляторные клетки организма - ГСК. Считается, что ГСК это самая медленная клетка в организме человека и млекопитающего, ее клеточный цикл у человека составляет 360 дней и поэтому все клетки организма сверяются с ней и регулируются ей в соответствии с системным законом природы - управляющей в любом системном процессе является самая медленная фаза (Н.Винер, 1960).
Несмотря на то, что ГСК это основная кроветворная клетка и родоначальница всех кроветворных и иммунных клеток организма, она почти всегда участвует в процессах тканеспецифической регуляции всех органов и систем организма путем взаимообмена регуляторными белками. Показано (Милькина Е.В. с соавт., 2016; Брюховецкий И.С., 2017), что ГСК при межклеточном взаимодействии с ОК и ОСК различных типов рака (рака легкого, рака груди, мультиформной глиобластоме) секретируют огромное количество регуляторных белков, которые путем экзосомального транспорта попадают в поврежденные клетки и радикально меняют их судьбу, но при этом и сами зачастую получают внутрь цитоплазмы большое количество ОСБ, поэтому и сами аутологичные ГСК могут стать ОСК и быть основной причиной возникновения гемабластоза. Количество ОСБ в ГСК при разных типах межклеточного взаимодействия с ОК может колебаться от 15 до 67% протеомной структуры клетки (Брюховецкий А.С., Шевченко В.Е, 2016; Bryukhovetskiy A.S., 2019).
Этот универсальный механизм межклеточного белкового обмена между ГСК и патологическими клетками приводит к ее геномно-эпигеномным и глобальным протеомным изменениям и перепрограммированию ее функций. Этот научный факт подтвержден при протеомном картировании и профилировании нормальных клеток, здоровых СК и ОСК и изложен в ряде научных статей и монографий в России и США (Брюховецкий А.С., 2014; Bryukhovetskiy A.S., 2019). Потомки протеом-модифицированной ОСБ собственной ГСК в виде целого пула ИКК с патологическими свойствами наследуют эти изменения в своем геноме и эпигеноме и имеют патологические эффекторные функции, переданные им по наследству.
Таким образом, фундаментальным научным фактом и основной причиной системных и болезнеобразующих нарушений процессов регуляции и саногенеза иммунной системы человека является геномно-эпигеномная трансформация белковой, ДНК и РНК структуры ГСК патоспецифическими белковыми телами, полученными ей при горизонтальном и вертикальном информационном обмене из поврежденных ОК или дегенеративных клеток. Очевидно, что патоспецифические белковые тела (ОСБ, тельца Леви, tau-белки и т.д.) появляются в цитоплазме, митохондриях, геноме и эпигеноме высокоспециализированных клеток в результате накопления мутаций в них при различных онкологических, аутоиммунных, эндокринных и нейродегенеративных процессах в тканях организма. Этот феномен протеомного перерождения ГСК в результате патогенеза этих заболеваний характерен не только для рака и других злокачественных онкологических заболеваний, но и для формирования целого ряда аутоиммунных, нейродегенеративных, дегенеративно-атрофических и эндокринных заболеваний. Поэтому применение любых молекулярно нацеленных и таргетных онкоиммунных препаратов для лечения этих заболеваний будет только временно эффективно, но все равно приведет к рецидиву болезни и ее прогрессированию, так как не будет устранена главная причина болезни - геномно-эпигеномная трансформированная ГСК, продуцирующая или гипореактивные (толерантные), или гиперреактивные (агрессивные цитотоксические) ИКК.
Если в результате этих протеомных межклеточных взаимодействий между патологической клеткой и СК или ГСК произошла минимальная трансформация эпигенома, а геном ГСК практически не пострадал, то основные изменения эпигенома произошли преимущественно в периферических мононуклеарных клетках, и через 100-120 дней болезнь сама прекратится без лечения или на фоне проводимой терапии, так как это максимальное время жизни пострадавших аутореактивных или иммунотолерантных лимфоцитов ПК. Если же при выраженном эпигенетическом повреждении ГСК их геном не поврежден или пострадал частично и только у небольшой части этих клеток, то болезнь можно остановить путем трансплантации аутологичных ГСК после высокодозной химиотерапии. Это с большим успехом сегодня показано при трансплантации аутологичных ГСК у части больных с рассеянным склерозом, полимиозитом и системной красной волчанкой. Высокодозная химиотерапия перед трансплантацией позволяет «опустошить» костный мозг больного от патологических ГСК и убить все неспособные к миграции и мобилизации ГСК и восстановить нормальный гемопоэз и «перезапустить» заново всю работу иммунной системы от здоровых ГСК.
Если же при межклеточном взаимодействии с патологической клеткой ГСК получила необратимые изменения в своем геномном и постгеномном статусе, и глобальными протеомными нарушениями поражены все ГСК пациента, то трансплантация аутологичных ГСК будет неэффективной, а любое лечение будут эффективным лишь временно. Как показали исследования, если в результате межклеточного взаимодействия с патологическими клетками протеомная структура ГСК пострадала более чем на 30-35%, то процесс практически необратим (Bryukhovetskiy A.S., 2019).
Таким образом, установлен и доказан принципиально важный теоретический и практический научный факт, заключающийся в том, что большинство онкологических, нейродегенеративных, аутоиммунных, геронтологических и эндокринных заболеваний человека не являются уникальными болезнями определенных высокодифференцированных специализированных клеток и тканей (моторных, корковых или мозжечковых нейронов, нейронов стриопаллидарной зоны, фибробластов, нейроглии и микроглии, β-клеток поджелудочной железы, кардиомиоцитов или эндотелиоцитов, фибробластов и т.д.), как было общепризнано в мире, а являются геномно-протеомным заболеванием собственной кроветворной СК пациента, спровоцированы и активно поддерживаются ее потомками в виде аутологичных ИКК с соответствующими геномно-протеомными модификациями белковой структуры (Брюховецкий А.С., Хотимченко Ю.С., 2018). Канцерогенное, аутоиммунное и нейродегенеративное перерождение специализированных клеток в организме человека в результате патогенеза этих болезней является закономерным исходом болезни, а не ее причиной и инструментом для прогредиентности и рецедивирования болезни. Причиной системообразования и фактором хронизации патологического процесса являются геномно-эпигеномные нарушения ГСК и их потомков, а также подобные изменения в региональных СК, вовлеченных в патологический процесс.
С этих новых теоретических и методологических позиций и инволюционные дегенеративно-атрофические, и старческие изменения органов и тканей нужно рассматривать не как закономерный возрастной процесс дряхлости и изношенности клеток и тканей организма, а как накопительное приобретенное геномно-эпигеномное заболевание ГСК и ее дочерних клеточных систем, приводящее к дегенеративно-дистрофическим изменениям или пролиферативным процессам в специализированных клетках органов и тканей в онтогенезе человека. Старение это не возрастная норма, а реальная транскриптомно-протеомная патология ГСК и региональных СК человека, обусловленная снижением их качественных контролирующих, надзорных и регуляторных характеристик и свойств, дефицитом их количества с возрастом и болезнями, а также нарушением регуляторных, саногенетических и регенеративных способностей этих кроветворных клеточных систем и всего иммунитета. Снижение иммунного надзора и контроля ГСК и ее потомков за клетками специализированных тканей приводит к их несвоевременной замене или реставрации. Универсальная система контроля и надзора за эффективной работой всех специализированных клеток организма и их регуляция и замена генетически заложены в программе ГСК и тканеспецифичных СК, и именно поэтому в процессе эволюции для борьбы с раком и другими болезнями цивилизации не был заложен дополнительный противоопухолевый или нейродегенеративный механизм защиты. Он существует в гемопоэзе и иммунопоэзе человека в самой структуре ГСК и блестяще реализуется при здоровой ГСК, особенно в молодом возрасте. Повреждение геномной и постгеномной структуры ГСК у человека со временем обусловлено в первую очередь неблагоприятными условиями окружающей среды из-за вмешательства человека в природу, и только на последнем месте стоят индивидуальные повреждения ГСК этиологическими факторами болезни (травмы, инфекции и т.д). Этот альтернативный научный взгляд на основные болезни цивилизации, на происходящие возрастные изменения в клетках и тканях с возрастом и на сам процесс старения человека открывает новые пути к раннему предотвращению ряда болезней цивилизации и формированию путей долголетия всего человечества.
Манифестация клинических и параклинических проявлений большинства фатальных заболеваний человеческой цивилизации выявляет не причину болезни в форме поражения специализированных клеток органов и систем, а закономерный исход болезни, обусловленный нарушениями иммунологического надзора и контроля за клетками-мишенями в тканях организма, снижением саногенетического потенциала ИКК, являющихся потомками геном- и эпигеном-трансформированных ГСК. Клинические проявления онкологической, аутоиммунной или нейродегенеративной болезни - это манифестация глубокого дефекта ткани за счет повреждения специализированных клеток и нарушения их функции. Именно поэтому авторы настоящего изобретения убеждены, что БАС, на сегодняшний день абсолютно неизлечимое заболевание, является не болезнью моторного нейрона, как общепризнанно в мировых классификациях (МКБ 11 и МКБ 12) и современных учебниках по неврологии, а именно геномно-эпигеномной болезнью ГСК пациента, и в этом представлении у этого заболевания на ранних стадиях возможно полное излечение с использованием БМКП по настоящему изобретению. В частности, было показано, что что нейроспецифический профиль ГСК при семейном БАС позволяет выявить наличие компенсированного заболевания и у здоровых членов семьи, у которых еще нет клиники моторного повреждения (Брюховецкий А.С., Гривцова Л.Ю., 2018).
Таким образом, можно константировать важнейший биологический научный факт, заключающийся в том, что реальным главным дирижером и основным руководителем в самой иммунной системе человека является аутологичные ГСК человека. И если в процессе продолжительной жизни человека и/или в результате информационного регуляторного обмена патологическими белками с поврежденными соматическими специализированными клетками организма его собственные ГСК накапливают значительное количество патоспецифических белковых тел, то эти патоспецифические белки начинают негативно влиять на геном и эпигеном ГСК, повреждают их, вследствие чего формируется патоспецифичное (онкоспецифичное, тау-белок-специфичное, FUS-белок специфичное, SOD-1 белок-специфичное и т.д.) протеомно-транскриптомное перерождение всей атомарно-молекулярной структуры собственной ГСК.
В различных случаях мощного разового или многократного незначительного комбинированного воздействия на человека этиологических факторов окружающей среды, прессинга макро- и микроэкономических факторов общества и негативного влияния социально-психологического микроокружения эти разновекторные воздействия становятся этиопатогенетическими факторами и причинами основных заболеваний цивилизации, и в одних случаях развивается рак и другие злокачественные опухоли, а в других случаях эти же факторы приводят к аутоиммунным и дегенеративным процессам в клетках организма, исходом которого становятся инволюционные, дегенеративные или вообще дегенеративно-атрофические и фиброзные повреждения в специализированных клетках организма. Очевидно, что разнонаправленность повреждений высокодифференцированных клеток вследствие локального или системного асептического воспаления зависит от функционального состояния специализированной клетки, фазы клеточного цикла, в которую воздействует патологический фактор, и от степени иммунной недостаточности, обусловленной уровнем геномно-эпигеномного повреждения ГСК и состоянием ИКК организма, что приводит не только к различным геномным и постгеномным изменениям в специализированных клетках в тканях-мишенях пострадавших органов, но и к информационным пертурбациям в геномной и постгеномной структуре ИКК и ГСК.
Поэтому транскриптомная и протеомная перестройка в ИКК и ГСК может сформировать как пролиферативный, так и дегенеративный вектор изменений молекулярной структуры ИКК и ГСК, что в клинике может манифестировать в виде злокачественных онкологических процессов и сопровождаться феноменами «плюс-ткань» (возникновение неконтролируемого количества ОСК, патологическое деление ОСК, избыточная репродукция ОК, миграция ОК, недостаточная иммунная противоопухолевая функциональность ИКК), так и пойти в противоположную сторону, с формированием дегенерации и атрофии и появлением тканевого феномена "минус-ткань" (дегенерация, атрофия, фиброзирование и склерозирование ткани, нарушение и выпадение функции и избыточная иммунная гиперреактивная функциональность).
Как отмечено выше, срок жизни ИКК ПК, даже патологических, ограничен 100-120 днями жизни лимфоцитов и гранулоцитов в организме человека, поэтому хронизация процесса, системность и выраженность хронического воспаления в тканях, наличие оксидативного стресса и эксайтотоксичности большинства фатальных заболеваний цивилизации может поддерживаться только фундаментальными причинами патогенеза болезни, приводящими к специфической иммунной недостаточности, а это возможно только на уровне формирования патологии генома и эпигенома ГСК. Поэтому основной фундаментальной причиной этих болезней цивилизации и формирование возрастной инволюции органов и тканей является не локальная патология специализированных клеток органов и тканей, а геномно-эпигеномное перерождение и трансформация молекулярной протеомной структуры собственной ГСК. Этот научный факт доказан протеомным картированием и профилированием ГСК и других СК при раке и целом ряде нейродегенеративных заболеваний (заявка на изобретение РФ № 2018145635, дата подачи 21.12.2018).
Обобщая все вышеизложенное, можно сделать следующее логичное заключение из всех имеющихся в нашем распоряжении современных доказанных фактов и научных знаний о геномно-эпигеномных изменениях ГСК. Решением проблемы излечения от большинства болезней человеческой цивилизации может быть, с одной стороны, только восстановление генетически и эпигенетически детерминированных функций ГСК, обусловленных ее атомарно-молекулярной структурой, и реставрация их функциональных гемопоэтических и иммунопоэтических свойств и характеристик. С другой стороны, необходимо восстановление достаточного количества здоровых ГСК. Качественная и количественная реставрация ГСК пожилого или больного человека и восстановление их регуляторного потенциала позволит полностью блокировать любые процессы канцерогенеза, дегенеративно-дистрофической инволюции и фиброза в тканях организма, остановить онкологические, аутоиммунные и нейродегенеративные заболевания и реально продлить активную жизнь органов и систем человека на 30-40 лет. Именно ГСК, а точнее реставрация или реконструкция поврежденной ГСК у любого человека, есть «золотой ключ» к его долгой активной жизни без старости и основных болезней цивилизации, которые он «насобирал» в геноме и эпигеноме ГСК за время жизни до 40 лет и более.
В качестве аналогов (уровня техники) настоящего изобретения можно указать на существующие в настоящее время стандартные технологии трансплантации клеточных ядер, применяемые при экстракорпоральном оплодотворении (ЭКО), в которых реализован принцип замены ядра (генома) донорской яйцеклетки на ядро (геном) соматической клетки реципиента (патентная заявка RU 2004117092 А, 20.11.2005; WO 0052145, 08.09.2000; патент RU 2280462 C2, 27.07.2006; патент RU 2252252 C1, 20.05.2005, Stojkovic M., et.al., 2005) и отечественные технологии создания гибридных СК человека (патент RU 2352637 C2 /WO/2009/002223) и применения их у человека (Тепляшин А.С. с соавт., 2009). Также аналогом настоящего изобретения является патент RU 2535972, 20/12/2014 (Противоопухолевый индивидуальный протеом-основанный таргетный клеточный препарат, способ его получения и применение этого препарата для терапии рака и других злокачественных новообразований), а также работы группы китайского профессора Hongkui Deng (Zhao Y., Zhao T., Guan J., Zhang X., et al., 2015, Xiang Li et al., Zhen Chai, Hongkui Deng, 2017). Все перечисленные известные технические решения не обеспечивают восстановление собственных поврежденных ГСК, которое привело бы к излечению млекопитающего, включая человека, от рака и других ЗНО, нейродегенеративных и аутоиммунных заболеваний.
Соответственно, целью настоящего изобретения является получение радикального решения проблемы излечения человека от рака и других ЗНО, нейродегенеративных и аутоиммунных заболеваний путем обеспечения технического результата, заключающегося в восстановлении саногенетического регуляторного и регенеративного потенциала поврежденных собственных ГСК, родоначальниц всех клеточных систем гемопоэза и иммунопоэза, и обеспечение их необходимого и достаточного количества в организме. Именно на биореставрацию молекулярно-биологической структуры ГСК, пострадавшей в результате негативного влияния факторов окружающей среды и этиологических факторов болезни ГСК, как основную системообразующую причину всех основных заболеваний современной цивилизации и инволюционных возрастных процессов в организме, направлено настоящее изобретение.
Сущность изобретения
Вышеуказанная цель настоящего изобретения с получением указанного технического результата достигается тем, что предложен способ получения биомедицинского клеточного продукта (БМКП) для лечения онкологических, нейродегенеративных, аутоимунных и эндокринных заболеваний и продления жизни млекопитающего и человека, включающий:
получение аутологичного лейкоконцентрата мононуклеаров (МНК) из эксфузата костного мозга или из лейкоферезного продукта мобилизованных мононуклеаров (ЛК МНК) периферической крови;
получение линии аутологичных гемопоэтических стволовых клеток (ГСК) с маркерами CD34+, CD45+, HLA DR+ и/или гемопоэтических клеток-предшественников (ГКП) с маркерами CD34+, CD45-, HLA DR- путем их выделения из ЛК МНК иммуносепарацией и культивирования в среде, препятствующей дифференцировке;
получение линии неповрежденных заболеванием аутологичных региональных мультипотентных стволовых клеток или соматических клеток и наращивание их клеточной массы;
перепрограммирование указанных аутологичных региональных мультипотентных стволовых клеток или соматических клеток в XEN-подобные плюрипотентные клетки посредством химической индукции;
проведение клеточно-инженерной гомотопной замены ядер ГСК и/или ГКП на ядра XEN-подобных клеток или на целые XEN-подобные клетки путем получения цитопластов из нативных ГСК и/или ГКП посредством обработки этих клеток цитохолазином В и трансплантации кариопластов, полученных из XEN-подобных клеток посредством их обработки цитохолазином В, или целых XEN-подобных клеток в цитопласты ГСК и/или ГКП с получением за счет этого восстановленных ГСК и/или ГКП, содержащих мультипотентные цитопласты ГСК и плюрипотентные кариопласты XEN-подобных клеток;
соединение восстановленных ГСК и/или ГКП с ЛК МНК, из которого были выделены первоначальные ГСК и/или ГКП, для получения целевого биомедицинского клеточного продукта, содержащего линию постгеномно восстановленных мультипотентных генетически однородных аутологичных ГСК с маркерами
CD34+Lin-, CD38", HLA-DR+, СD45+.
Кроме того, согласно настоящему изобретению восстановленные ГСК и/или ГКП, соединенные с ЛК МНК, преимущественно культивируют в среде, препятствующей дифференцировке, для увеличения их клеточной массы, формирования и созревания их свойств мультипотентности в гемопоэтическом направлении и контролируют эти свойства проточной цитофлориметрией.
При этом в случае онкологии или нейродегенеративных болезней головного и спинного мозга, а также в случае аутоимунных болезней в качестве неповрежденных заболеванием клеток используют мезенхимальные стромальные стволовые клетки (МССК) с маркерами CD10+, CD13+,CD44+, CD90+ (Thy-1), CD105+, CD34-, CD45- и CD117- и/или мезенхимальные стромальные клетки предшественники (МСКП) с маркерами CD10+,CD13+, CD44-, CD90+ (Thy-1), CD105+, CD34+, CD45+ и CD117+, а в случае онкологии солидных органов в качестве неповрежденных заболеванием клеток используют нейральные стволовые клетки (НСК) с маркером СD 133+ и/или нейральные клетки-предшественники (НКП) с маркером СD 133+.
Согласно настоящему изобретению перепрограммирование посредством химической индукции преимущественно проводят коктейлем, содержащим низкомолекулярные вещества, выбранные из группы, состоящей из VPA, TD114-2, CHIR, 616452, транилципромина, формсколина, AM580, EPZ004777, CH55 и витамина C (VC), а трансплантацию полученных из XEN-подобных клеток кариопластов или целых XEN-подобных клеток в цитопласты ГСК и/или ГКП проводят in vitro путем электропорации или лазерно-оптическим методом.
Кроме того, согласно настоящему изобретению предложен биомедицинский клеточный продукт (БМКП), полученный способом по настоящему изобретению и содержащий аутологичный ЛК МНК, обедненный собственными ГСК и/или ГКП в результате их иммуносепарации, с линией генетически однородных аутологичных гемопоэтических клеток с маркерами CD34+Lin-, CD38", HLA-DR+СD45+.
Перечень графических изображений
На фиг. 1 показаны зависимости выживаемости животных от их возраста, полученные в результате проведения сравнительных экспериментов по трансплантации костного мозга и БМКП по настоящему изобретению (см. Пример 11).
На фиг. 2А-2В представлены результаты GFP-флуоресценции ГСК у доноров, контрольной и экспериментальных групп животных, описанные в Примере 11.
Подробное раскрытие изобретения
При создании настоящего изобретения возник вопрос - что может быть предложено для восстановления молекулярно-структурных и функциональных возможностей ГСК с геномно-эпигеномными нарушениями? Любая попытка крайне сложного геномного редактирования патологических ГСК с помощью современных инструментов генной инженерии (цинковые пальцы, CRISP/Cas9 технологии и т.д.) представляется малоэффективной, так как в каждом конкретном случае будет иметь место как персонифицированный тип молекулярного повреждения ДНК-структуры генома ГСК, так и специфические эпигенетические изменения ядра ГСК у каждого пациента, и выяснение локации этих структурных нарушений в этих клетках даже путем самых ультрасовременных диагностических технологий секвенирования генома, полнотранскриптомного анализа с микрочипированием экспрессии 34000 генов или развернутого протемного картирования 8000 белков позволит только очертить и обозначить основные нарушения геномно-транскриптомно-протеомной внутриклеточной конструкции, но реального лечения этот подход не обеспечит и будет очень дорогостоящим. Поэтому очевидно, что поиск конкретных генов, нарушений метилирования и поврежденных гистонов, а также обнаружение транскриптомных, протеомных и метаболомных нарушений в ГСК для возникновения той или иной болезни не конструктивен и не даст желаемого результате на практике. В каждом конкретном случае повреждение ДНК в ядре ГСК будет индивидуальным и специфичным и будет зависеть от характера и силы этиологического воздействия патоспецифических белков, попавших внутрь ГСК и воздействующих на ее геном и эпигенетические структуры, а также от особенностей трансформации протеомного профиля патологической специализированной высокодифференцированной клетки-мишени в органе и ткани человека.
Поэтому автор настоящего изобретения отказался от самых современных технологий редактирования генома и решил подойти к процессу достаточно радикально, воспользовавшись более «старыми» и хорошо изученными и отработанными технологиями клеточной инженерии, применяемыми для трансплантации клеточных ядер, так как этот подход позволяет технологически убрать патологический геном и сделать невозможным реверсию аберрантных эпигенетических модификаций из ГСК и заменить его на здоровый геном другой соматической клетки организма (заявка на изобретение RU 2004117092 А, МПК C12N 5/00, 20.11.2005; WO 00/52145, МПК C12N 5/06, 08.09.2000; патент RU 2280462 C2, МПК A61K 35/55, 27.07.2006; патент RU 2252252 C1, МПК C12N 5/08, 20.05.2005). Такое предположение связано с тем, что именно ГСК и ГКП, а также тканеспецифические клетки той линии ткани, где разворачивается патологический процесс, являются основным инструментом регуляции в саногенезе. Другие клеточные системы, невовлеченные в патологический процесс, практически не подвергаются тому количеству неблагоприятных воздействий, и их геном и эпигеном практически может не иметь подобных повреждений. Перепрограммирование соматических и СК, невовлеченных в патологический процесс, может быть реализован с использованием современных технологий химического индуцированного перепрограммирования соматических клеток для получения мультипотентных ГСК.
Применение клеточной инженерии, как научное биотехнологическое решение, уже очень хорошо зарекомендовало себя в клинических технологиях экстракорпорального оплодотворения (ЭКО) и создания гетерогенных СК для клеточной терапии. Технические приемы и методики замещения генома одной клетки на геном другой клетки решены в современных медицинских биотехнологиях путем замены их ядер из соматических клеток, описаны и запатентованы (Тепляшин А.С. с соавт., 2009). В современном ЭКО это достаточно рутинная процедура, когда из одной донорской яйцеклетки извлекают геном, который упакован в ядре этой клетки, а на его место ставят ядро (геном), извлеченный из соматической клетки реципиента (будущей матери), и растят эмбрион, который потом подсаживают в матку самой женщины для приживления. Однако, как только мы начнем получать СК из искусственно созданных клеточно-инженерных бластоцист или эмбрионов человека разных стадий гестации (бластоцисты, зиготы, эмбриона и т.д.), то мы вынуждены разрушать уже живой организм человека, что законодательно запрещено в Российской Федерации. Все подобные процедуры создания генетически однородных СК относятся к репродуктивному и терапевтическому клонированию, так как в основе этих технологий лежит создание живого гибридного пациента совместимого эмбрионального организма, клетки которого при его разрушении используются для создания клеточных линий СК. Поэтому необходимо было предложить такую биотехнологию, в которой бы не использовались оплодотворенные или клеточно-инженерно-сконструированные ооциты человека и животных и не применялся эмбриональный материал различных стадий гестации.
Одним из современных подходов к реализации цели замены ГСК является получение химически индуцированных плюрипотентных стволовых клеток (ИПСК или iPSC) путем перерограммирования их из любой соматической клетки (фибробласта, нейрона, СК жировой ткани, клеток крови) с использованием современных известных факторов транскрипции генома (Oct, Sox2, Klf4 и c-Myc). Эти ИПСК с использованием других факторов транскрипции могут быть перепрограммированы в специализированные мультипотентные СК. Так, в результате применения четырех факторов транскрипции (Gata2, Gfi1b, cFos и Etv6) уже удалось получить из фибробласта кожи мыши жизнеспособные ГСК (СD34+, СD45+, СD133+), обладающие всем спектром мультипотентности и способные давать все типы клеток гемопоэза и иммунопоэза (C.F. Pereira, B.Chang, J. Qiu et al., 2013). В другом исследовании из лимфоцитов крови человека после перевода в ИПСК с использованием шести транскрипционных факторов (Run1t1, Hlf, Lmo2, Prdm5, Pbx1 и Zfp37) удалось получить ГСК человека, обладающие полной мультипотентностью и способные восстановить все типы клеток крови (J. Riddell, R. Gazit, B.S. Garrison et al., 2014). Сегодня плюрипотентные СК могут дифференцироваться практически во все типы клеток организма. Этот уникальный потенциал дает многообещающие возможности клеточной терапии для восстановления тканей или органов, разрушенных в результате травм, дегенеративных заболеваний, старения или рака. Открытие технологии получения ИПСК предлагает возможную стратегию создания тканеспецифических для пациента мультипотентных СК. Однако из-за опасений относительно специфичности, эффективности, кинетики и безопасности перепрограммирования ИПСК перед их эффективным клиническим использованием требуются улучшения или фундаментальные изменения в этом процессе, так как введение транскрипционных факторов в геном осуществляется с использованием аденовирусов, лентивирусов или плазмид, что является генетически небезопасным, как для самого больного, так и для его потомства (T.Ma, M.Xie,T.Laurent, S.Ding, 2013).
Существует еще один серьезный недостаток, который связан с данным способом получения ИПСК. Любой тип клеток в организме характеризуется своим собственным индивидуальным эпигеномом - определенным спектром и паттерном посттрансляционных ковалентных модификаций гистонов, метилирования ДНК, наличием набора малых некодируемых РНК. Комбинация данных факторов формирует уникальную структуру хроматина, присущую клетке того или иного типа. Хроматин плюрипотентных клеток характеризуется «открытой» архитектурой и декомпактизованным состоянием. Хроматин в такой конфигурации в процессе дифференцировки способен быстро подвергаться различным посттрансляционным модификациям гистонов, процессам метилирования и деметилирования ДНК. Плюрипотентные клетки содержат также «бивалентные домены», то есть области, обогащенные одновременно метками и активного, и неактивного хроматина. Большинство бивалентных доменов ассоциированы с точками старта транскрипции генов, вовлеченных в развитие. В плюрипотентных клетках эти гены характеризуются низким уровнем транскрипции, тогда как в ходе дифференцировки бивалентные домены превращаются в моновалентные, содержащие метки либо активного, либо неактивного хроматина, и соответственно наблюдается активация либо репрессия генов, обуславливающая специализацию клеток конкретного типа (цит. по Васькова Е.А., Стекленева А.Е., Медведев С П., Закиян С.М., 2013). Поэтому при выборе источника для перепрограммирования во внимание следует принимать тот факт, что ИПСК сохраняют эпигенетическую память о тканях, из которых они произошли, что влияет на их способность к направленной дифференцировке (J.G. Crompton, M. Rao, N.P. Restifo, 2013; J.M. Polo, S. Liu, M.E. Figueroa, et al. & К. Hochedlinger, 2010; K. Kim, A. Doi, B. Wen et al., 2010; K. Kim, R. Zhao, A. Doi et al., 2011; O. Bar-Nur, H.A. Russ, S. Efrat, N. Benvenisty, 2011; Denker H.-W., 2012; Jong-Hee Lee, Jung Bok Lee, Zoya Shapovalova et al., 2014; Васькова, Е.А., Стекленева, А.Е., Медведев, С.П., & Закиян, С. М., 2013; Kyttälä, R. Moraghebi, C. Valensisi et al., 2015). Остаточная эпигенетическая память не обязательно проявляется на стадии плюрипотентности. ИПСК, полученные из разных тканей, имеют надлежащую морфологию, в них активны гены, характерные для плюрипотентности, и они способны дифференцироваться в ткани трех эмбриональных слоев как in vitro, так и in vivo (Васькова, Е.А., Стекленева, А.Е., Медведев, С.П., & Закиян, С. М., 2013; A.Kyttälä, R. Moraghebi, C. Valensisi et al., 2015). Однако, эта эпигенетическая память может проявиться позже, во время повторной дифференцировки в специфические типы клеток, которая требует активации локусов, сохранивших элементы остаточной эпигенетической памяти (J.G. Crompton, M. Rao, N.P. Restifo, 2013; J.M. Polo, S. Liu, M.E. Figueroa, et al. & К. Hochedlinger, 2010; K. Kim, A. Doi, B. Wen et al., 2010; K. Kim, R. Zhao, A. Doi et al., 2011; O. Bar-Nur, H.A. Russ, S. Efrat, N. Benvenisty, 2011; Denker H.-W., 2012). Необходимо найти решение, как обойти механизм реверсии (возврата) эпигенетической памяти у этих клеток для широкого использования их в клинике.
В области биомедицины ИПСК являются наиболее стратегически выгодными, поскольку они могут быть получены из соматических клеток и являются аутологичными по отношению к донору соматических клеток. На сегодняшний день известно, что по своим свойствам ИПСК очень схожи с эмбриональными СК, то есть экспрессируют сходный спектр генов, формируют тератомы, содержащие производные всех трех зародышевых листков. В то же время много работ показывают, что в результате перепрограммирования линии ИСПК приобретают целый ряд генетических и эпигенетических аббераций, включая нарушения в работе импринтированных генов, изменения в количестве копий генов, точечные мутации, аберрантный паттерн метилирования ДНК и ряд других. При этом отличия в эпигеномах и транскриптомах эмбриональных СК и ИСПК обусловлены не только аберрациями, приобретенными в процессе перепрограммирования, но сохранением ряда эпигенетических меток исходных соматических клеток. Подобное наследование свойств эпигеномов и транскриптомов ИПСК получило название «эпигенетической памяти» (Васькова Е.А., Стекленева А.Е., Медведев, С.П., & Закиян, С.М., 2013).
Таким образом, несмотря на большие успехи современных биотехнологий по созданию и потенциальным возможностям применения ИПСК при создании ГСК из соматических клеток путем их перепрограммирования факторами транскрипции, недостатки этих технологий перекрывают все их достоинства, и нужен принципиально новый подход к этой проблеме, чтобы исключить вирусную интервенцию в геном человека, предотвратить образование тератом и найти механизм преодоления реверсии резидуальной эпигенетической памяти после трансдифференцировки одной клетки в другую.
Альтернативный подход к трансдифференцировке соматических клеток без использования факторов транскрипции и отказ от их вирусных носителей был предложен в 2013 году китайским профессором Хонкуи Денгом (Hongkui Deng ) и его научной группой ученых из Пекинского университета (Zhao Y., Zhao T., Guan J., Zhang X. et al., 2015). Они решили проблему перепрограммирования соматических клеток в требуемые фенотипы других клеточных линий путем создания химически индуцированных ИПСК, точнее путем создания инновационной биотехнологической платформы на основе XEN-подобного состояния (состояния, подобного эмбриональной эндодерме) перепрограммируемых клеточных систем. Полученное ими XEN-подобное состояние клеток, индуцированное определенными коктейлями из низкомолекулярных химических веществ стало мостом, который соединяет соматические клетки с плюрипотентностью при химическом перепрограммировании и как бы обходит стороной плюрипотентное эмбриональное состояние клетки. Внеэмбриональное XEN-подобное состояние опосредует процесс генерации ИПСК. XEN-подобные клетки транскрипционно и функционально сходны с аналогом in vivo. Клетки XEN готовы к экспрессии Oct4 и установлению плюрипотентности. Индукция ИПСК значительно усиливается путем высокоточного манипулирования маленькими молекулами. Процесс химического перепрограммирования требует более раннего образования клеток, подобных эмбриональной энтодерме (XEN), и позднего перехода от клеток, подобных XEN, к химически индуцированным ИПСК уникальным путем, который принципиально отличается от пути перепрограммирования с использованием транскрипционных факторов. Более того, точное манипулирование переходом клеточной судьбы поэтапно через XEN-подобное состояние позволяет идентифицировать низкомолекулярные ускорители и создать надежную систему химического перепрограммирования с выходом до 1000 раз больше, чем в ранее сообщенном протоколе (J. Riddell, R. Gazit, B.S. Garrison et al., 2014) с использованием шести транскрипционных факторов (Run1t1, Hlf, Lmo2, Prdm5, Pbx1 и Zfp37) и собственным протоколом авторов по химической индукции при получении ИПСК от 2013 года (Xiang Li et al., Zhen Chai, Hongkui Deng, 2017). Эти результаты показывают, что химическое перепрограммирование является многообещающим подходом к манипулированию судьбами клеток. Другими словами, в дополнение к возможности получения специфических для пациента аутологичных тканей и органов из ИПСК, другой привлекательной стратегией для регенеративной медицины китайскими учеными предложена стратегия химической трансдифференциации - прямого преобразования одного типа соматических клеток в другой через так называемый клеточный мостик в виде XEN-клеточных систем. Эта трансдифференцировка не только выявляет и использует развивающиеся пластические промежуточные продукты, генерируемые во время перепрограммирования ИПСК, но также продуцирует очень широкий спектр клеток, включая расширяемые тканеспецифические клетки-предшественники. Клетки в индуцированном XEN-подобном состоянии легко расширялись в течение по меньшей мере 20 пассажей и сохраняли стабильность генома и потенциал спецификации линии. Поэтому революционное исследование пекинских ученых устанавливает многофункциональный путь для химического перепрограммирования линии и может предоставить платформу для генерации разнообразных типов клеток посредством применения этого расширяемого XEN-подобного состояния (Xiang Li et al., Zhen Chai, Hongkui Deng, 2017).
В способе по настоящему изобретению при получении генетически однородных ГСК используется не репродуктивное и терапевтическое клонирование и не трансдифференцировка соматических клеток с применением транскрипционных факторов. В предлагаемом способе для химической индукции с целью трансдифференцировки соматических и стволовых клеток в генетически однородные ГСК используют исключительно аутологичные клеточные линии взрослых соматических клеток (фибробластов, кариоцитов, жировых клеток) и линии мультипотентных негемопоэтических СК пациента (нейтральных СК (НСК), СК жировой ткани, мезенхимальные стромальные СК (МССК) и т.д.), выделенных от человека прижизненно. Способ по настоящему изобретению в принципе не предполагает получения зиготы, бластоцисты или эмбриона, как это принято во всех видах клонирования и последующего ее разрушения для получения мультипотентных стволовых клеток, а также не предполагает лишения жизни не родившегося эмбрионального гибридного существа или внедрения в структуру генома собственных клеток вирусных и векторных носителей и плазмид. В изобретении предлагается работать с одной из двух или трех линий взрослых соматических клеток или негемопоэтических стволовых и прогениторных клеток уже вторично дифференцированных организмом пациента в сторону определенной мультипотентности, извлеченных из одного человека и находящихся исключительно в постнатальном периоде онтогенеза. Все манипуляции с трансдифференцировкой клеток в ХЕN-подобное состояние следует производить путем геном-ориентированной химической индукции малыми молекулами по технологии группы профессора H. Deng (Zhao Y., Zhao T., Guan J., Zhang X., et al., 2015; Xiang Li et al., Zhen Chai, Hongkui Deng, 2017). Это позволит получить из соматических или СК полноценные ИПСК, минуя стадию эмбрионального развития, применяя постадийную алгоритмированную обработку клеток, находящихся в индукционной среде с низкомолекулярными химическими веществами (пертурбогенами) в заданной концентрации и экспозиции. Полученные ИПСК культивируют в специальной среде, препятствующей дифференцировке этих клеток и индуцирующей активацию экспрессии генов ядра клетки в сторону гемопоза и имммунопоэза путем применения каскада цитокинов, активирующих гемопоэтическую ориентацию полученных клеток также путем применения культивирования в среде с комплексом специфичных пертурбогенов - специально биоинформационно подобранных низкомолекулярных химических соединений, то есть из плюрипотентного XEN-состояния клетки ее нужно сделать мультипотентной ГСК. Для этого следующим этапом предложено сделать трансплантацию ядер (кариопластов) ХЕN-подобных клеток в цитопласты ГСК, полученных от пациента. Таким образом получают ГСК-подобные аутологичные клеточно-инженерные системы с генетически однородным XEN-подобным клеточным ядром, при этом целью является получение жизнеспособных мультипотентных ГСК из этого клеточно-инженерного микста собственных клеток пациента. На этом этапе авторы настоящего изобретения воспользовались методологическим подходом, предложенным ранее в патенте на изобретение RU 2535972, МПК A61K 48/00, 20.12.2014 на "Противоопухолевый индивидуальный протеом-основанный таргетный клеточный препарат, способ его получения и применение этого препарата для терапии рака и других злокачественных новообразований". В этом патенте раскрыт таргетный клеточный препарат, молекулярно нацеленный на путь интегринов и фокальной адгезии в ОСК путем подбора пертурбогенов (низкомолекулярных химических соединений) в базе клеточных транскриптомов Массачусетского технологического института (МИТ) США. Эта база данных транскриптомных профилей экспрессии генов клеток содержит результаты работ различных исследователей по картированию и профилированию экспрессии генов до и после манипуляций с клетками на аппаратах платформы Affymetrix. Поэтому в связи с настоящим изобретением было предложено снять полнотранскриптомный профиль экспрессии генов ГСК-подобных клеток с ядром, находящимся в ХЕN-подобном состоянии, и биоинформационным путем найти пертурбогены, которые бы путем химической индукции привели клеточно-инженерную систему к клеточным системам, имеющим транскриптомный профиль экспрессии генов здоровых ГСК. Проблему реверсии (возврата) эпигенетической резидуальной памяти в полученных клеточно-инженерных клетках предлагается решить путем добавления к культивируемой среде набора стандартных химических молекул, широко используемых для эпигенетического баланса клеток беременной женщины на ранних стадиях формирования эмбриона человека (фолиевая кислота, витамин В12, витамин С и т.д.). В результате клеточной инженерии в предлагаемомом способе осуществляют равноценную замену поврежденного генома и эпигенома (ядра) мультипотентной ГСК на геном и эпигеном (ядро) другой здоровой мультипотентной ГСК-ориентированной клетки в XEN-подобном состоянии, невовлеченной в патологические процессы в организме больного. Процесс слияния цитопластов ГСК пациента и кариопласта гемопоэтически ориентированной клеточно-инженерной (ГОКИ) ГСК может быть реализован путем их электропорации. «Дозревание» полученных ГОКИ ГСК осуществляют в среде, препятствующей дифференцировке ГСК, с иономицином и фолиевой кислотой, витамином С и витамином В12. Таким образом, в настоящем изобретении предложено новое решение по борьбе с резидуальной эпигенетической памятью ГСК, созданной из соматической клетки через химически индуцированное XEN-подобное состояние путем проведения клеточной инженерии аутологичных ГСК и изъятия из патологической ГСК пациента ядра этой клетки, в котором в компактном виде находится весь ее поврежденный геном, и замещение его на геном здоровой трансдифференципрованной соматической или СК негемопоэтического ряда этого же организма, но которая путем химической индукции стала ГОКИ ГСК.
При создании настоящего изобретения автор исходил из собственной научной гипотезы об измененной протеомной и траснкриптомной структуры ГСК у пациентов с онкологическими, аутоиммунными и нейродегенративными заболеваниями и, конечно же, патоспецифическом протеомном профиле мультипотентной ГСК пациента (Брюховецкий А.С., 2014; Bryukhovetskiy A.S., 2019). Поэтому возможным решением проблемы борьбы с этими болезнями является функциональное и молекулярное восстановление поврежденной ГСК за счет ее клеточно-инженерной реконструкции и восстановление их необходимого количественного баланса для гомеостаза в организме человека. Материалом для реконструкции ГСК был выбран геном и эпигеном (ядро) любой другой здоровой соматической или мультипотентной СК негемопоэтической линии в организме пациента. Очевидно, что найти здоровую соматическую или тканеспецифическую мультипотентную СК другой тканеспецифической линии в организме человека не представит труда. Каких-либо морально-этических, социально-правовых и юридических ограничений по использованию соматических клеток в качестве источника ядер для переноса их в другие соматические клетки нет. Это в полной мере касается и всех типов СК.
Очевидно, что эффективность способа значительно возрастает при использовании ядер собственных СК или прогениторных клеток. Более того, ткань, из которой как из источника могут выделяться клетки, должна быть доступной и универсальной. Наиболее доступной и универсальной является жировая ткань, костный мозг и обонятельная выстилка носа пациента. Но и аутологичные мультипотентные СК печени и СК, полученные из менструальной крови больной женщины или семенников мужчины, тоже могут использоваться в качестве региональных мультипотентных СК или соматических клеток для реализации поставленной цели. В связи с этим наиболее предпочтительными являются МССК, выделенные из костного мозга или жировой ткани. Данные клетки имеют в культуре уникальные характеристики: способность пролиферировать в течение длительного времени и при индукции к дифференцировке формировать клетки тканей, имеющих мезенхимное происхождение, их можно довольно легко получить от каждого человека, в том числе и от больного. При пересадке таких клеток в энуклеированную ГСК пациента можно получать клеточные объекты, генетически идентичные пациенту, т.е. обладающие генетически однородными характеристиками (Тепляшин А.С. с соавт., 2009).
При этом в случаях онкологического (например, мультиформная глиома), нейродегенеративного (например, БАС или БАС-деменция), аутоиммунного (например, рассеянный склероз) заболевания головного или спинного мозга страдают не только ГСК, но и НСК, поэтому лучшим источником для выявления здорового генома (ядра) клеток в организме в этом случае могут стать клетки мезенхимного ряда. Они выделяются из костного мозга пациента, жировой ткани или мобилизованного концентрата мононуклеаров (МНК) периферической крови (ПК). В случае же онкологического заболевания солидных органов (например, рак молочной железы, рак легких, рак почек и т.д.), как правило, страдают ГСК и МССК, но НСК, а также тканеспецифические мультипотентные СК, полученные из забарьерных органов (глаза, яичка и т.д.), СК печени и жировой ткани почти никогда не повреждаются. Таким образом, если у больного имеется процесс в нервной ткани, то для реконструкции ГСК целесообразно использовать МССК костного мозга и жировой ткани, которые не вовлечены в патологический процесс, а при локализации процесса в солидных органах лучшим источником аутологичного генома здоровых клеток станет НСК, полученные из обонятельной выстилки носа.
Сам процесс замены ядра ГСК на ядро другой тканеспецифичной мультипотентной СК в настоящее время технически реализован на соматических клетках в ЭКО и терапевтическом клонировании и не представляет каких-либо технических трудностей. Он хорошо отработан сегодня большинством молекулярных биологов, и эта технология сегодня стала рутинной. Сам биотехнологический процесс замещения патологического генома ГСК на геном другой здоровой мультипотентной СК равнозначен замещению ядра одной аутологичной ГСК на ядро другой аутологичной тканеспецифичной мультипотентной СК. Поэтому этот биотехнологический процесс в настоящем описании назвается не трансплантацией ядер (как при терапевтическом и репродуктивном клонировании), а заменой или субституцией генома или ядра ГСК. Субституция это хорошо известный в биологии термин (позднелат. substitutio, от лат. substituo - ставлю вместо, назначаю взамен), означающий замещение в процессе эволюции одного органа другим в том же самом организме, который занимает собой подобное положение в организме и выполняет биологически равноценную функцию. В случае с замещением генома целесообразно говорить о гомотопной замене или субституции (homotopic substitution) или равнозначной замене, что означает замещение в процессе эволюции одного органа другим, занимающим сходное положение в организме и выполняющим биологически равноценную функцию (замещаемый орган при этом редуцируется). Например, в целом ряде случаев происходит редукция замещаемого органа и прогрессивное развитие замещающего. Так, у хордовых осевой скелет хорда замещается сначала хрящевым, затем костным позвоночником; у кактусоподобных растений листья (фотосинтезирующие органы) замещены стеблями. Термин «субституция» введен Н. Клайненбергом (1886) (Гиляров М.С. и соавт., 1986) и этот термин очень точно отражает суть предлагаемого феномена искусственной замены генома (ядра), находящегося в одной аутологичной патологической клетке, на геном (ядро) другой здоровой аутологичной клетки, для выполнения той же функции.
Чтобы не путать предлагаемый в настоящем изобретении метод с разными формами репродуктивного и терапевтического клонирования, его можно назвать способом равнозначного замещения клеточного генома (ядра) или методом гомотопной субституции (замены) клеточного генома(ядра). Целью процедуры терапевтического клонирования является получение гибридных СК, генетически совместимых с донорским организмом, а целью метода гомотопной субституции клеточного генома в настоящем изобретении - реконструкция (восстановление) поврежденных собственных ГСК генетическим биоматериалом, полученным исключительно от самого пациента. Также будет неправильным использовать в настоящем изобретении терминологию терапевтического клонирования, называя результат клеточной инженерии «гибридом». Гибрид (от лат. hibrida, hybrida - помесь) это организм или клетка, полученные вследствие скрещивания генетически различающихся форм, а в способе по настоящему изобретению обрабатывают только однородный, аутологичный генетический материал.
В отношении манипуляций по блокированию эпигенетической памяти. Предлагаем обозначать факт возврата эпигенетической памяти в клеточно-инженерных ГСК можно назвать реверсией эпигенетической памяти. Ревéрсия (от лат. reversio - возвращение, возврат) - многозначный термин и может означать: Реверсия (биология) - склонность к возвращению частей со смешанными признаками к исходным родительским формам. Поэтому полученные в результате клеточно-инженерных манипуляций генетически однородные ГСК с гомотопной субституцией генома и заблокированным механизмом реверсии эпигенетической памяти можно обозначить как ГСК без реверсии эпигенетической памяти.
Итак, в качестве источника получения генома (ядра) в настоящем изобретении следует использовать только здоровые соматические или мультипотентные СК пациента, например НСК, МССК, печеночные СК, даже СК глаза. Однако применение СК и прогениторных клеток в реставрации ГСК наиболее предпочтительно для синхронизации клеточных циклов соединяемых клеточной инженерией клеток. ГСК как правило в 70-80% случаев находятся в G0 фазе клеточного цикла, и другие СК также находятся в клеточной культуре преимущественно в этой же фазе клеточного цикла. Поэтому их мембранные, цитоплазматические и ядерные белки и метаболические факторы практически сбалансированы по фазе клеточного цикла. Транскрипция факторов стволовости ГСК лучше обеспечивается в G0 фазе клеточного цикла, а также поддержанием этих клеток с проведенной субституцией генома культивированием в индукционной среде, препятствующей дифференцировке и благоприятствующей увеличению их количества с последующим созданием терапевтической линии здоровых ГСК пациента.
Настоящее изобретение базируется на равнозначном клеточно-инженерном замещении поврежденного генома аутологичных ГСК пациента с фатальным заболеванием на здоровый геном соматической или СК другой клеточной, но не гемопоэтической линии того же организма больного, переведенной в XEN-подобное состояние, путем химической индукции малыми молекулами и получения генетически однородных ГСК с гомотопной субституцией генома, и последующим формированием из них клеточной линии аутологичных клеточно-инженерных ГСК для изготовления БМКП. Полученные БМКП могут применяться для лечения целого ряда заболеваний с целью полной остановки прогрессирования и предотвращения рецидивов болезни у пациентов с онкологическим, нейродегенеративным, аутоиммунным или эндокринологическим заболеванием или обеспечивают антиэйджинговый (антивозрастной) клинический эффект и значительно продлевают активную жизнь человека. Технологически субституция генома ГСК (ядра) реализуется путем иммуносепарации ГСК на магнитных шариках с помощью иммуносортера из костного мозга или из ЛК мобилизованных МНК ПК, последующего замещения ядер ГСК на ядра соматически здоровых клеток (стволовой или прогенеторной) и дальнейшего культивирования и наращивания клеточной массы ГСК в среде, препятствующей дифференцировке.
Основным этапом способа по настоящему изобретению является получение двух аутологичных клеточных линий мультипотентных СК, одной из которых всегда будет линия собственных ГСК пациента. Источником получения линии ГСК будет или костный мозг пациента или лейкоконцентрат мобилизованных МНК ПК. Другим источником СК может быть жировая ткань, обонятельная выстилка слизистой носа, костный мозг или лейкоконцентрат МНК ПК и так далее. Клеточные линии выделенные от пациента культивируется стандартными путями.
Следующим и центральным этапом предлагаемого способа является проведение химической индукции соматической или мультипотентной региональной стволовой клетки по протоколам профессора Hongkui Deng (Zhao Y., Zhao T., Guan J., Zhang X., et al., 2015, Xiang Li et al., Zhen Chai, Hongkui Deng, 2017). Строгое выполнение данных протоколов позволяет соматическую клетку (например, фибробласт) или региональную СК (нейральную СК или МССК) перепрограммировать в ХЕN-подобное состояние путем геном-ориентированной химической индукции коктейлями, содержащими различные малые молекулы. Это позволит получить из соматических или стволовых клеток полноценные ХЕN-подобные клетки, минуя стадию эмбрионального развития, применяя постадийную химическую обработку клеток, находящихся в индукционной среде с малыми молекулами (пертурбогенами) в заданной концентрации и экспозиции. Полученные ХЕN-подобные клетки будут прокультивированы в специальной среде, препятствующей дифференцировке этих клеток и индуцирующей активацию экспрессии генов ядра клетки в сторону гемопоэза и иммунопоэза путем применения каскада цитокинов, активирующих гемопоэтическую ориентацию полученных клеток.
Следующим этапом способа по настоящему изобретению является трансплантация ядер (кариопластов) ХЕN-подобных клеток в цитопласты ГСК, полученных от пациента. Таким образом мы получаем ГСК-подобные аутологичные клеточно-инженерные системы с генетически однородным XEN-подобным клеточным ядром, то есть из имеющихся двух клеточных линий пациента (ГСК и мультипотентных СК или соматических клеток) предлагается несколько вариантов получения генетически однородных ГСК с гомотопной субституцией клеточного генома (ядра). Первый вариант получения подобной терапевтической линии ГСК осуществляется путем выделения из реципиентной клеточной линии ГСК пациента суспензии энуклеированных клеток (цитопластов ГСК) путем инкубации их с цитохалазином В, а из донорской линии ХЕN-подобных СК также путем 15 минутной обработки цитохалазином В проводится выделение ядра (кариопласта ХЕN-подобных МССК, ХЕN-подобных НСК и т.д.) и последующее проведение слияния их путем импульсной электростимуляции (электропорации). Другой вариант также предполагает возможность выделения из реципиентной клеточной линии ГСК пациента культуры энуклеированных клеток (цитопластов ГСК) как и в первом варианте, а в качестве кариопласта (генома) используется целые ХЕN-подобные мультипотентные клетки без выделения ядра. Последующее соединение цитопласта ГСК с донорской мультипотентной клеткой происходит путем слияния их мембран и цитоплазм с помощью электрической импульсной стимуляции (электропорации) с получением восстановленных ГСК.
Завершающим этапом способа по настоящему изобретению является соединение восстановленных ГСК с лейкоконцентратом МНК ПК пациента и совместное культивирование в среде, препятствующей дифференцировке, содержащей иономицин, факторы стимулирующие гемопоэтические свойства этих клеток и содержащей фолиевую кислоту, витамин С и витамин В12. Наличие микроокружения в виде всего спектра МНК ПК клеточно-инженерных ГСК с гомотопопной субституцией генома (ядра) с наличием факторов блокирующих эпигенетическую память (фолиевую кислоту, витамин С, и витамин В12) на самых ранних этапах их культивирования приводит к формированию гемопоэтического транскриптомного профиля экспрессии генов уже на 7-9 день культивирования в СБУ и появлению всех основных маркеров ГСК : CD34+CD45+CD133+HLADR+. Далее полученный БМКП криоконсервируется с ДМСО в жидком азоте при температуре -196°С.
Главной изобретательской находкой и принципиальным отличием способа получения БМКП по настоящему изобретению на основе линии генетически однородных ГСК с субституцией генома от технологий репродуктивного и терапевтического клонирования является то, что при любом виде клонирования всегда применяют в качестве донорской клетки ооциты, а реципиентом может быть любая соматическая клетка и в результате их слияния из него формируются зигота. В дальнейшем зигота разрушается и из нее получают стволовые клетки или так называемые пациент-специфические клетки. В результате реализации настоящего изобретения реципиентом и донором клеток будет исключительно сам пациент, что практически никогда не применялось ни в эксперименте, ни в клинике, ни при получении генетически однородных СК. Это снимает с повестки дня все морально-этические вопросы пересадки клеточных ядер, связанные с терапевтическим и репродуктивным клонированием и страхи создания клеточных и тканевых гибридов и мутантов, но именно это настоятельно ставит вопросы оценки гемопоэтической и иммунной функциональности полученных аутологичных ГСК и необходимости дотрансплантационного подтверждения факта способности их к полноценной кроветворной функции и возможности участия клеточно-инженерных ГСК пациента в полноценном гемопоэзе и иммунопоэзе.
Предложенное в настоящем изобретении получение аутологичных здоровых (восстановленных) ГСК объясняется уникальной биологией СК в одном организме, которую почему-то не учитывали ранее и тем, что 256 типов различных клеток человека формируются еще во время эмбриогенеза человека и возврата клеток на пройденные этапы уже произошедшего эмбриогенеза (рекапитуляции эмбриогенеза-повторение уже пройденных стадий эмбриогенеза предков) не существует, то есть, обход механизма рекапитуляции эмбриогенеза для соматических клеток в норме не запрограммирован в биологии человека в принципе. Однажды, получив во время эмбриогенеза определенный вектор дифференцировки, клеточные системы будут производить только себе подобные клеточные системы по «лекалам» этого фенотипа и его молекулярной структуре и никогда не переходят в другую клеточную линейность, то есть не трансформируются в другие типы клеток. Поэтому в эмбриогенезе сразу после формирования тотипотентных СК и преобразования их в плюрипотентные СК из них появляются мультипотентные СК, формирующие тканеспецифические СК (ГСК, МССК, НСК и т.д.), которые всю оставшуюся человеку жизнь будут абсолютно автономны в своей линейности, так как пути формирования и онтогенеза этих клеток практически не пересекаются. Так после рождения человека каждая из них отвечает исключительно за формирование клеток определенной линии ткани. Трансдифференцировка или переход одной СК в другую СК в обход рекапитуляции эмбриогенеза в природе также не возможен.
В искусственных лабораторных условиях было продемонстрировано, что в определенных специальных базовых условиях (СБУ) трансдифференцировка СК возможна и реальна. Было показано, что в особых условиях культивирования ГСК могут получиться из XEN-подобных НСК и XEN-подобных МССК. На экспериментальных животных, которым после высокодозной химиотерапии и опустошения костного мозга, была произведена трансплантация в кровь клеточно-инженерных ГСК, было показано, что донорские клеточно-инженерные ГСК, полученные из XEN-подобных клеточных систем, прижились в костном мозге, стали реципиентными ГСК и сформировали весь гемопоэз и иммунопоэз.
Придание генетически однородным ГСК с гомотопной субституцией генома пациентов гемопоэтических характеристик осуществляется в СБУ культивирования, в результате чего полученные ГСК приобретают отчетливые маркеры клеточной поверхности ГСК. Однако культивирование клеточно-инженерных ГСК в СБУ еще не может являться гарантией их доказанных гемопоэтических стволовых свойств. Поэтому предлагаемая клеточно-инженерная манипуляция, представляющая собой известный способ пересадки (трансплантации) ядер путем удаления ядра из патологичной ГСК больного и имплантации в нее ядра (генома) аутологичных XEN-подобных МССК или XEN-подобных НСК или других СК и создание здоровых клеточно-инженерных ГСК, не решает задачи гарантированного задания этим клеточным системам отчетливых кроветворных свойств, которым после трансплантации необходимо будет восстановить весь угнетенный гемопоэз и иммунопоэз в организме человека. Поэтому, чтобы придать дополнительные функции стволовости и функции кроветворных клеток полученным клеточно-модифицированным аутологичным ГСК. необходимо их активировать и прокультивировать в среде, препятствующей дифференцировке ГСК и повышающей (индуцирующей) кроветворные свойства полученных аутологичных ГСК. Такие среды известны и техника повышения гемопоэтической стволовости культуры ГСК уже реализована во многих клеточных лабораториях (Быковская С.Н., 2015).
При этом, понимая все существующие риски трансплантации клеточно-инженерных генетически однородных ГСК с гомотопной субституцией генома (ядра), необходимо снизить риски получения отрицательного результата применения полученных ГСК при трансплантации костного мозга (ТКМ), а также быть уверенным, что эти вновь полученные здоровые аутологичные ГСК смогут сами восстановить заново весь гемопоэз и иммунопоэз организма человека после высокодозной миелоаблативной химиотерапии во время аутологичной трансплантации ГСК и ТКМ. Необходимо не только оценить иммунофенотипически их стволовость по маркерам их клеточной поверхности и их соответствие существующим стандартам ГСК, но сделать биологические пробы с данным биоматериалом на экспериментальных животных (иммуно-дефицитных мышах, крысах), на моделях нарушения гемопоэза путем смертельного облучения костного мозга у этих животных и оценить in vivo гемопоэтические функции этих клеточных систем ГСК. Однако даже при позитивных результатах биологических проб необходимо сделать дубликат трансплантационного биоматериала из неманипулированных ГСК и мобилизованного лейконцентрата МНК ПК или костного мозга, осуществив второй сбор ГСК на 6-й день стимуляции гранулоцитарным колониестимулирующим фактором (Г-КСФ) для возможности проведения повторной трансплантации пациенту, если гемопоэз не восстановится через 1-2 месяца после трансплантации клеточно-инженерных аутологичных ГСК.
Важнейшим научно-практическим вопросом настоящего изобретения был вопрос молекулярнонацеленного влияния цитоплазмы донорской ГСК на ядро XEN-подобных плюрипотентных СК и ее способность активировать в пересаженном ядре гены, отвечающие за гемопоэз и иммунопоэз в организме пациента. Положительные результаты экспериментов показали возможность индукции реципиентной цитоплазмы на ядро донорской клетки в XEN-подобном состоянии, когда было высказано предположение о том, что в клетках млекопитающих есть вещества, способные прямо или косвенно влиять на клеточное ядро, изменять его функциональное состояние с помощью положительной или отрицательной регуляции экспрессии определенной части генов. Однако комплекс объективных причин (гетерогенность генома зачастую со значительным дефицитом хромосом, а также с произошедшими хромосомными перестройками, которые происходят в окружении гетерогенной цитоплазмы), присущих методу гибридизации соматических клеток, не позволил идентифицировать вещества, участвующие в экспрессии генома, а также установить их химическое строение и механизм действия. В случае настоящего изобретения эти неблагоприятные факторы исключены, так как ядро будет перенесено в аутологичную цитоплазму в соответствующей фазе клеточного цикла клеток и будет окружено родными, то есть аутологичными компартментами клетки.
Оказалось, что в ядрах, предварительно изъятых из дифференцированной клетки и помещенных в цитоплазму яйцеклетки, начинались процессы клеточной дифференцировки, и во многих случаях развивался нормальный организм, то есть ядра специализированных клеток содержали полный объем информации, необходимой для развития полноценного организма. Эти эксперименты показали, что цитоплазма регулирует активность ядра в специализированных клетках, и многие функции ядра тормозятся компонентами цитоплазмы. Активирующее влияние на транскрипцию факторов стволовости и кроветворной дифференциации окажут факторы цитоплазмы ГСК. Было полностью подтверждено, что ингибирующее влияние, идущего к ядру со стороны цитоплазмы дифференцированной клетки, в пригодном по химическому составу цитоплазматической окружении устраняется, и в ядрах могут снова проходить процессы, определяющие клеточную дифференциацию (Gurdan JB, 1977). Долгое время стоял вопрос, будет ли преодолено факторами цитоплазмы ГСК ингибирующее влияние пересаженного ядра негемопоэтической мультипотентной СК. Сегодня мы получили ответ и знаем, что помещение ядра здоровой (без геномных и постгеномных изменений) мультипотентной СК, переведенной в XEN-подобное состояние, в цитоплазму ГСК окажет свой эффект включения генов ГСК в геноме трансплантированного ядра, что показали полнотранскриптомные исследования экспрессии генов этих клеток. Важнейшим моментом этих исследований будет синхронизация по клеточному циклу между донорским ядром и реципиентной цитоплазмой ГСК.
В настоящее время в разработке современных клеточно-инженерных методов пересадки клеточных ядер млекопитающих существенную роль сыграло использование цитохалазинов - веществ, синтезированных грибами. Главным действием цитохалазина В является то, что он резко увеличивает устойчивость клетки к механическим повреждениям в процессе энуклеации, вызывая деконденсацию микрофиламентов цитоскелета и резко повышая пластичность и адаптивность поверхностной мембраны к различным механическим воздействиям. При этом после обработки клетки цитохолазином В формируется цитоплазматический тяж, окончательное истончение которого ведет к образованию как бы двух "клеток" или, точнее, двух изолированных компартментов - цитопласта (лишенный ядра ГСК) и кариопласта (ядро мультипотентной ГСК), т.е. по существу происходит искусственное деление. Другими словами, цитохалазин В, разрушая структуру микрофиламентов, способствует уникальному расположению ядра, и оно остается подключенным к клетке тонким «стебельком» в виде цитоплазматического тяжа типа «мини-пуповины». При центрифугировании этот цитоплазматический тяж в большинстве таких клеток разрывается, и образуются энуклеированные (безъядерные) клетки (цитопласты) и ядра, отделившиеся при центрифугировании (кариопласты или мини-клетки), окруженные тонким слоем цитоплазмы и плазматической мембраной. Эффективность энуклеации можно контролировать по методу Гимза, окрашивая клеточный монослой, находящийся на поверхности одной из чашек или дисков.
Методы выделения кариопластов несколько сложнее и включают в себя ряд операций по центрифугированию, разделению в градиенте плотности и т.д. В некоторых случаях к смеси клеток и кариопластов добавляют частицы тантала диаметром 1-3 мкм. Они проникают в клетки и никогда не попадают в кариопласт, поэтому более тяжелые клетки осаждаются быстрее кариопластов. Цитопласты содержат все виды органелл, присущие нормальной клетке, сохраняют способность прикрепляться к субстрату, образовывать складчатую мембрану, передвигаться, осуществлять пиноцитоз. Кариопласты окружены тонким слоем цитоплазмы (около 10% от всей клеточной цитоплазмы), содержат компактный эндоплазматический ретикулум, несколько митохондрий и рибосом. У некоторых клеточных линий 1/10 кариопластов способна восстановить весь утраченный объем цитоплазмы и восстановиться в жизнеспособные клетки.
Для реконструкции клеток суспензию кариопластов в солевом буфере добавляют к монослою культуры цитопластов из пропорции 100 кариопластов на 1 цитопласт. Раньше слияние цитопластов с кариопластами осуществляли путем покрытия их инактивированными вирусными частицами и инкубировали их 45 мин при температуре 4°С, а затем еще 45 минут при температуре 37°С и отмывали раствором Эрла для удаления неслившихся кариопластов. Сегодня есть более щадящие процедуры слияния кариопластов и цитопластов разных клеток путем электростимуляции электрическим током в культуральном растворе.
Таким образом, используя известную технологию трансплантации ядер соматических клеток, модифицированную под цели получения здоровых аутологичных ГСК самого пациента путем применения способа разрушения структуры микрофиламентов в ГСК цитохалазином В, можно добиться энуклеации ядер патологических ГСК, выделенных из ЛК МНК ПК путем иммуносортировки с моноклональными антителами CD34+, CD45+. Применяя технологию клеточной индукции соматических и мультипотентных СК негемопоэтической линии, в заявленном способе получают ГСК с ядром, находящимся в XEN-подобном состоянии. В результате предобработки собственных ГСК пациента цитохолазином В, выполнения серии лабораторных этапов, выделения клеток на феколе и центрифугирования получают цитопласты (энуклеированные клетки), а кариопласты (выделенные ядра этих клеток) из ГСК удаляют и отправляют на протеомное картирование (при необходимости). Цитопласты аутологичных ГСК будут использованы далее в текущем биотехнологическом клеточно-инженерном процессе. Аналогичные действия осуществляют с донорской культурой здоровых XEN- подобных СК (НСК, МССК и т.д.) и разделяют их на цитопласты и кариопласты, но от них в технологическом процессе выделяют и оставляют аутологичные кариопласты здоровых клеток. При дальнейшем соединении цитопластов ГСК и XEN-подобных стволовых кариопластов тканеспецифичных СК добиваются их слияния в новые ядросодержащие аутологичные ГСК путем культивирования в СБУ. Создать СБУ можно путем их совместной инкубации, а также с использованием известной технологии электростимуляции электрическим током или путем применения неинвазивных оптико-лазерных приемов пересадки ядер у млекопитающих (Свиридова-Чайлохян Т.А., Кантор Г.М., 2010). По завершению процесса слияния кариопластов ГСК с цитопластами СК получают препарат здоровых клеточно-инженерных ГСК и очищают его путем удаления несоединившихся кариопластов, цитопластов и обломков из разрушенных клеток с использованием этапного центрифугирования. Полученные ГСК проверяют на жизнеспособность трипановым синим и передают в культуральную лабораторию для дальнейшего культивирования и получения клеточной линии в среде, препятствующей дифференцировке ГСК, и наращивания их клеточной массы до требуемого количества (1-2% от общего количества клеток МНК ПК). Цитофлуориметрическим методом на проточном цитофлуориметре оценивают их качественно-количественные характеристики через 4-5 дней культивирования и при их соответствии стандарту замораживают в жидком азоте с ДМСО.
При проведении ТКМ после высокодозной стандартной химиотерапии в миелоаблативном режиме пациенту проводим разморозку концентрата МНК ПК больного и культуры здоровых генетически однородных ГСК с гомотопной субституцией генома, отмываем их от криопротектора и ДМСО двухкратным центрифугированием с 0,9% раствором натрия хлорида, смешиваем их в пакете для трансфузии и проводим внутривенное введение при ТКМ в стандартном режиме или используем для интратекального введения. В других случаях, когда дозревание клеточно-инженерных ГСК происходило в концентрате МНК ПК, то для применения размораживаем готовый БМКП и применяем для трансплантации костного мозга или для интратекальных введений.
Технически изготовление БМКП по настоящему изобретению в форме клеточно-модифицированного ГСК с гомотопной субституцией генома состоит из создания и получения его основных клеточных компонентов для БМКП, их совместного культивирования и криоконсервации и последующего размораживания и смешивания (по необходимости) их непосредственно ex tempore перед клиническим применением или применением для трансплантации ГСК. Получение и применение БМКП для аутологичной ТКМ при лечении целого ряда фатальных и пока неизлечимых заболеваний человека начинается с диагностики наличия протеомной патологии ГСК у пациента. Для наиболее ранней диагностики патологии ГСК может быть использован способ ранней молекулярно-биологической диагностики злокачественных новообразований и бокового амиотрофического склерозаза, раскрытый в заявке РФ на изобретение № 2018145635, дата подачи 21.12.2018, решение о выдаче патента от 13.09.2019).
В зависимости от локализации патологического процесса необходимо определиться с тем, какие клетки будут использоваться для изготовления клеточно-модифицированных ГСК с гомотопной субституцией генома. В случае патологии центральной нервной системы, то используют аутологичную линию МССК пациента из жировой ткани. В случае заболевания солидных органов соединительной ткани или других заболеваний, когда ведущая роль в реставрации повреждения или пролиферации тканей принадлежит МССК, то донором ядер выступят НСК, полученные из обонятельной выстилки носа или ядро печеночных СК, СК глаза или других мультипотентных СК.
Способ получения БМКП по настоящему изобретению описывается далее более подробно.
Стадия 1. Получение лейкоконцентрата МНК из неманипулированного костного мозга, мобилизованной периферической крови
Получение лейкоконцентрата МНК неманипулированного костного мозга
Требования к донорам костного мозга. Доноры должны быть предварительно обследованы на инфекции ВИЧ 1 и ВИЧ 2, синдром иммунодефицита; сифилис, гепатиты В и С; острые инфекционные заболевания.
Получение костного мозга. Забор материала для последующего выделения клеток осуществляет врач-хирург, имеющий практический опыт проведения данной процедуры. Эксфузия костного мозга производится в операционной посредством пункции дистальной части гребня подвздошной кости под общей или местной анестезией. Пункцию проводят стернальной иглой; забор костного мозга производят шприцем типа "Луер", костномозговую суспензию переносят в мешок с изотоническим антикоагулянтом.
Мобилизация ГСК в ПК
С целью увеличения количества СК в ПК пациент получает 8 инъекций Г-КСФ подкожно с интервалом в 10-12 часов в течение 4 дней. В первые три дня доза препарата составляет 2,5 мкг/кг, в последний день доза удваивается. Ежедневно производят общий анализ крови и на 4-5 день делается УЗИ брюшной полости и фиброгастроскопия.
Сбор ГСК и ГКП
Осуществляется сбор № 1 на 5 день от начала стимуляции Г-КСФ, а сбор № 2 на 6-й день от начала стимуляции на сепараторе крови СОВЕ-Spectra с использованием одноразовой системы для сепарации и стандартных растворов. Длительность процедуры 3-4 часа в зависимости от скорости процедуры, веса донора и параметров анализа крови. Процедура проводится путем забора крови из одной вены, обработки ее внутри сепаратора, забора определенного объема СК и возврата остальных компонентов крови донору через другую вену. Венозный доступ осуществляется путем пункции двух периферических вен или через двухпросветный центральный катетер, установленный в подключичную вену на время проведения сеанса. Средний объем собранного материала 300-400 мл. Собранный материал оценивают по двум параметрам: по общему количеству ядерных клеток в сепарате и по количеству СD34+ клеток на килограмм веса больного. Ядерные клетки в сепарате определяются путем подсчета в камере Горяева до проведения каких-либо манипуляций. Процент СD34+ клеток в клеточной суспензии, полученной в ходе цитофереза, определяют методом проточной цитофлуориметрии. Весь полученный материал собирают в криомешок № 1. Весь материал, собранный на 6-й день стимуляции пациента собирают аналогичным способом в криомешок № 2. Дальнейшая процедура манипуляций не с биоматериалом в мешках не отличается друг от друга.
Определение периферических СК и ГКП
Установление субпопуляционного состава CD34+ клеток проводится цитофлуориметрически с применением метода тройной метки (одновременная окраска клеток антителами к трем разным антигенам, нагруженными различными флуорохромными красителями).
Cтандартизация препарата
Количество клеток-предшественниц определяют цитофлуориметрически в прямой реакции иммунофлуоресценции (РИФ).
Проводится метод двойной метки с одновременным окрашиванием клеточного субстрата моноклональными антителами к антигену CD34+ как основному маркеру клеток пула аутологичных мультипотентных СК гемопоэтического ряда и к молекуле CD45- - лейкоцитарному антигену, определяющему ГСК. Подобная методика позволяет сразу рассчитать соотношение количества CD34+ клеток и всех ГСК (CD45+) в материале. Для оценки уровней неспецифического связывания часть клеток окрашивают изотипическими контролями. В качестве изотипических контролей стандартно применяются мышиные иммуноглобулины IgG1 изотипа, меченные красителями, аналогичными метке используемых моноклональных антител (PE, FITC, PerCP).
Подготовка клеточных проб. Перед постановкой реакции клетки ПК и цитаферезного продукта освобождают от примеси эритроцитов путем стандартной процедуры лизиса и последующей отмывки в ЗФР-БСА центрифугированием при 1000g в течение 5 минут. ЗФР-БСА может быть заменен средой Хенкса или 199.
С выделенными клетками проводят прямую РИФ в 96-луночном планшете, при этом для определения количества клеток в любом материале используется панель из 3-х лунок: неокрашенные клетки; клетки, окрашенные изотипическими контролями с меткой, соответствующей метке использованных моноклональных антител (МКА); клетки, окрашенные одновременно МКА к антигенам CD34 и CD45.
Следует обратить особое внимание, что МКА к CD34+ предпочтительнее использовать с фикоэритриновой (РЕ) или перидининхлорофиловой (PerCP) метками. Данные флуорохромы имеют более высокий уровень специфического сигнала по сравнению с флуоресцеинизотиоционатом (FITC).
Для определения количества CD34+ клеток оптимальными являются антитела к антигену CD34 клона HPCA-2 (8G12), изотипа IgG1.
Таким образом, стандартная панель имеет вид
1. контроль IgG1 PE+ контроль IgG1FITC;
2. контроль IgG1 PE+ МКА к CD45 FITC;
3. МКА к CD34 PE (HPCA-2) + МКА к CD45 FITC.
Постановка РИФ
1. В лунки вносятся клетки в количестве не менее 500 000 на лунку.
2. Далее в каждую лунку вносят коктейль антител в соответствии с панелью и аккуратно ресуспендируют пипеткой. Каждое МКА берут в количестве 10 мкл на лунку, суммарный объем МКА в лунке - 20 мкл.
3. Клетки инкубируют с антителами в течение 30 минут при 4°С.
4. По окончании времени инкубации клетки дважды отмывают от несвязавшихся антител путем центрифугирования в течение 5-7 минут при 1000g.
5. Клетки переносят в специальные пластиковые пробирки для счета на проточном цитометре.
6. Объем клеточной суспензии в каждой пробирке доводят до 200-500 мкл путем добавления к клеткам ЗФР-БСА.
Счет должен быть осуществлен сразу после постановки реакции.
Счет и запись на проточном цитофлуориметре
Оценка реакции на проточном 5-параметровом цитометре. Данная схема применима для проточного цитофлуориметра любой конфигурации. CD34+ клетки в ПК представляют собой малую клеточную популяцию. Даже в условиях предварительной стимуляции кроветворения максимальный процент данных клеток составляет 1,0-3,0%. Поэтому при подсчете данных клеток в каждой анализируемой пробе следует накапливать не менее 20 000 клеточных событий.
Сбор и анализ клеток осуществляется в гейте CD45+ клеток, который включает все ГСК. В данном контексте гейт подразумевает область накопления событий, ограниченную определенными параметрами. В данном случае SSC/ FL-1 (CD45 FITC). То есть, по оси абсцисс (FL-1) на точечной цитограмме будут видны все CD45+ события, а по оси ординат (параметр SSC - боковое светорассеяние) клеточные события будут расположены в соответствии с их гранулярностью (цитометрический, а не морфологический термин). Подсчет абсолютного числа CD34+клеток в 1 мкл крови и цитоконцентрата проводили на основании числа лейкоцитов в гемограмме на день исследования.
Принцип определения CD34+ клеток в гемопоэтической ткани. Запись проб.
1. Выбор области анализа - гейта.
В приборе открыто меню записи клеточных образцов Acquisition. На первом этапе в режиме Setup (режим просмотра клеточного образца без записи) просматриваются пробы № 1 (IgG1PE+ IgG1 FITC) и № 2 (IgG1 PE +CD45 FITC) и выявляются CD45+ события. Данные события расположены правее значений 101 (среднее пороговое значение уровня специфического сигнала для флуоресцеина FITC) по оси абсцисс - FL-1 (CD45+клетки) и достаточно четко визуализируются по сравнению с контрольной пробой № 1.
По пробе № 1 ограничивается вся область, расположенная правее основной клеточной плотности - гейт, включающий только CD45+ события в пробе № 2 и содержащая минимальное количество клеточных событий в пробе № 1.
2. Прибор переводится в режим записи образцов (normal), и проба № 1 собирается и записывается на 20000 клеточных событий без гейта (вся клеточная популяция). Запись данной пробы в гейте не имеет смысла, так как в ней отсутствуют МКА к CD45 антигену. Как показано выше, данная проба необходима для правильного выбора гейта, содержащего только CD45+ события.
3. Пробы № 2 и № 3 собираются и записываются в гейте CD45+. Данный гейт идентичен (то есть в процессе сбора изменения параметров гейта не происходит) и для 2-й, и для 3-й пробы. Минимальное количество клеточных событий - 20 000 для каждой пробы.
Анализ записанных проб
1. Переходим в меню анализа записанных образцов Analysis. Открываем цитограмму (dot-plot) пробы № 2 в параметрах SSC/FL-2, в гейте R1 - CD45+ (гейт выбран нами ранее в момент записи сбора и автоматически перенесен в режим анализа). При этом по оси ординат будет отражен параметр SSC - гранулярность событий, попавших в анализируемый гейт, а по оси абсцисс FL-2 - детектор, отражающий уровни неспецифического связывания для фикоэритринового красителя, выявляемые анителами к иммуноглобулинам мыши IgG1PE.
2. По данной цитограмме устанавливается контрольный маркер таким образом, чтобы в правом нижнем квадранте практически не оказалось клеточных событий. В среднем, для вертикальной планки маркера значения будут около 130 (правее 102), а для горизонтальной планки маркера средние значения составят 60, что соответствует понятию SSC-low. То есть клетки с минимальным количеством клеточных включений. Таким образом, уровни неспецифического связывания будут равны или близки к нулю.
3. Автоматически переходим к пробе № 3. Все показания прибора, принятые для второй пробы, сохраняются. То есть цитограмма открыта в параметрах SSC/FL-2, сохранен гейт, выбранный ранее при сборе по CD45+ клеточным событиям, и сохранены значения контрольного маркера, установленного по пробе № 2. Только на данной цитограмме по оси абсцисс будут отражены не уровни неспецифического связывания, как в пробе № 2, а специфичные CD34+ события. Процент CD34+ клеток выводится прибором автоматически.
Выделение гемопоэтических CD34+ СD133+ Lin- клеток из ЛК МНК ПК
Данный этап культивирования ГСК в предлагаемом способе выполняется в случае выявления малого (менее 0,5-1%) количества ГСК в ЛК МНК ПК и необходимости увеличения их количества и активации ГСК. Если количество ГСК в ЛК МНК ПК более 1,0-1,5% от общего количества лимфоцитов, то можно сразу перейти к следующему этапу предлагаемого способа - иммуносепарации (см. пункт В данного раздела).
А. Культивирование в условиях ограниченной дифференцировки
как способ увеличения количества ГСК из ЛК МНК ПК в криомешке.
Цель этого биотехнологического этапа - увеличение количества собственных ГСК пациента для проведения дальнейших манипуляций по клеточной инженерии. Основной проблемой культивирования высокопластичных недифференцированных ГСК с маркерами CD34+, CD133+Lin- является их быстрый выход в дифференцировку, так как для решения описанных выше задач необходимо увеличение количества этих клеток без развития колоний дифференцированных клеток. При этом способность к последующей дифференцировке у таких культивируемых клеток должна быть сохранена, по возможности, в первоначальном виде. Задачей этого этапа является как увеличение количества недифференцированных ГСК пациента ex vivo, так и полное освобождение от эритроцитов и гранулоцитов и незначительное их увеличение путем их культивирования в ростовой среде, содержащей смесь цитокинов, состоящей из фактора роста СК (SCF), лиганда тирозин киназы 3 из фетальной печени (Flt3L) и тромбопоэтина (ТРО), отличием которой является то, что указанная выше смесь цитокинов дополнительно содержит иономицин при следующем соотношении компонентов на 1 мл ростовой среды: SCF - (50-150)⋅103 нг, ТРО - (50-150)⋅103 нг, Flt3L - (10-50)⋅103 нг, иономицин - (10-100)⋅103 нг.
Весь ЛК делят на пробирки по 50 мл и работают с каждой пробиркой индивидуально. Иономицин был использован в качестве стимулирующего агента роста недифференцированных СК клеток в условиях ех vivo. Фактор роста СК (SCF) стимулирует пролиферацию СК, взаимодействуя со специфическим рецептором (протоонкогенный c-kit), обладающим тирозин-киназной активностью (Witte O.N., 1990), индуцирует активацию киназы и трансфосфорилирование цепей рецептора. Фосфотирозиновые остатки рецептора функционируют как места стыковки протеинов, связывающих c-kit с сигнальными путями, что индуцирует активацию пролиферации (Aronica SM et al., 1996). Flt3-лиганд (Flt3L) стимулирует пролиферацию ГСК в не меньшей степени, чем SCF. Flt3 представляет собой III класс рецептора тирозин киназы, экспрессированного на ранних ГКП. Взаимодействие Flt3 с лигандом (Flt3L) активирует сигнальные пути и последующую пролиферацию СК (Matthews Wea, 1991). Тромбопоэтин (ТРО) - это гликопротеиновый гормон, который регулирует продукцию тромбоцитов стволовыми клетками костного мозга. ТРО был сначала охарактеризован как лиганд, который связывается с поверхностным рецептором с-Мр1. ТРО относится к классу I гемопоэтических цитокинов. В суспензионной культуре ТРО в комбинации с SCF стимулирует пролиферацию мультипотентных СК и повышает их количество (Ku et al., 1996). Иономицин повышает концентрацию внутриклеточного Са2+, начиная с мобилизации внутриклеточных запасов, которые затем поддерживаются притоком внеклеточного Са2+, индуцирует гидролиз фосфоинозитидов и стимулирует активацию протеинкиназы С, стимулируя вход катиона регулируемыми запасами кальция, но не прямым действием на плазматическую мембрану. (А.J. Morgan and R. Jacob, 1994, Biochem. J. 1994, 300 (665-672)). Таким образом, иономицин является агентом первичной активации клеток (сферилизация клеток), см., например, патент RU 2108579. Иономицин использовался как агент роста дифференцированных клеток, таких как гибридом и дендридных клеток. Необходимый уровень стволовости ГСК обеспечивается тем, что из равноценного перечня известных цитокинов была выявлена такая их смесь, которая в определенных подобранных концентрациях оказалась способной вместе с новым фактором стимуляции пролиферации для СК - иономицином, индуцировать только клеточную пролиферацию и таким образом не потребовалось использовать дополнительные агенты для подавления дифференцировки клеток. Кроме того, неожиданным результатом для данного технического решения было достижение очень больших скоростей пролиферации клеток, что позволило сократить время культивирования клеток до 7-9 дней и избежать таким образом проблем длительного культивирования клеток. Более того культивирование в этой среде позволило не изменить базовые процентные соотношения фундаментальных клеточных элементов в ЛК МНК и при этом нарастить количество ГСК до 1,5-2%.
Таким образом, обобщая все вышеизложенное, ГСК при данном способе не выделяют из аутологичного костного мозга или аутологичного концентрата мобилизованной ПК, как это было принято в ряде исследований, а культивируют прямо в ЛК МНК ПК. Полученную суспензию культивируют в среде Dulbecco M (IMDM) с добавлением инсулина и трансферрина, а также фактора роста СК (SCF), лиганда тирозин киназы 3 из фетальной печени (Flt3L), тромбопоэтина (ТРО) и иономицина (Са2+ ionophore). Через 3-7 дней культивирования в выращенной клеточной суспензии содержится около 1,5-2% клеток CD34+. Методика позволяет за короткий период времени очистить ЛК от примесей крови (эритроцитов) и гранулоцитов, незначительно нарастить увеличенное количество низкодифференцированных ГСК, которые могут быть использованы в терапевтических целях как компонент БМКП по настоящему изобретению.
Проводят следующие процедуры:
1. Транспортировка мешков с клетками костного мозга, мобилизованной крови или ЛК МНК ПК осуществляется в термоконтейнерах ТермоКонт при комнатной температуре (18-25°С).
2. Забор клеток из мешка с мобилизованной кровью производится в ламинарном шкафу класса II, расположенном в стерильном блоке;
2.1. Клеточную взвесь забирают шприцем, прокалывая мембрану соединительной трубочки на мешке и делят на пробирки по 50 мл.
3. Получение фракции МНК: клеточную взвесь, взятую шприцем, разводят в соотношении 1:4 фосфатным буферным раствором (PBS) без ионов кальция и магния (PBS Ca2+Mg2+ free) и затем наслаивают на градиетный раствор (d=1,077 г/мл, MP Biomedicals) в 50 мл пробирках.
3.1. Пробирки центрифугируют при 400g в течение 30 мин при 20°С;
3.2. В результате градиентного центрифугирования эритроциты и гранулоциты опускаются на дно пробирки, а фракция мононуклеаров в виде плотного кольца концентрируется между двумя слоями на поверхности градиента;
3.3. фракцию МНК отбирают пипеткой, разводят в свежем PBS и три раза отмывают центрифугированием в течение 10 мин при 300g и температуре 10°С.
4. Подсчет клеток и фенотипический анализ мононуклеарной фракции:
4.1. Для определения количества живых МНК к 20 мкл клеточной суспензии добавляют 20 мкл трипанового синего (Gibco). Подсчет клеток производят в гемоцитометре, пропорция живых клеток не должна быть ниже 95%.
4.2. Для оценки количества клеток CD34+ и CD133+ мононуклеарной фракции 0,1 мл клеточной суспензии окрашивают соответствующими антителами и определяют пропорцию ГСК в исходной клеточной взвеси на приборе FACSCalibur (Beckton Dickinson).
Пример 1. Данные фенотипического анализа МНК в одном показательном эксперименте, проведенном на клетках ЛК донора Н. Пропорции клеток в ЛК МНК ПК: ГСК (СD34+,CD45+) донора составляла 0,5%, количество нейральных клеток - 5% , Т- лимфоциты 84%, В-клетки -1,2%.
Культивирование . Жидкая культуральная система состоит из среды Искова в модификации Дульбекко (Iscove's modified Dulbecco's Medium IMDM, Gibco, UK) с добавлением 0,5 г/л бычьего сывороточного альбумина (Sigma Aldrich), 0,39 мкг/мл человеческого инсулина и 60 мкг/мл трансферрина (Gibco). В культуральную среду добавляют фактор роста СК (SCF), другое название c-Kit-ligand (c-KitL, R&D Systems), в концентрации 100 нг/мл, а также Flt3-ligand (Flt3L) в концентрации 20 нг/мл, (R&D Systems), и иономицин (Sigma Aldrich) в концентрации 0,1 мкМ/мл.
Способ культивирования. Клетки культивируют при 37°С во влажной атмосфере с притоком 5% CO2. Свежую среду и цитокины добавляют каждый третий день. Время культивирования составляет 3-7 дней. Использованные факторы роста способствуют пролиферации СК, при этом не нужно добавлять агенты, подавляющие дифференцировку. Получаемые суспензии СК клеток любого индивидуума могут быть сохранены в замороженном виде либо могут быть введены в БМКП непосредственно после культивирования и применены больным.
Таблица 1
Оценка чистоты клеточной суспензии после культивирования
Заключение. Материал сохранный. На основании данных проточной цитометрии возможные для анализа лимфоциты составляют 84,7%. Среди них зрелые Т-клетки составляют 81,2% с преобладанием CD4+ субпопуляций. γδ-Тклетки составили 1,2%. Гетерогенные NK клетки представлены популяциями CD94+, CD16+, CD56- и СD94-, CD56+, CD16- и CD16+, CD56+, суммарно составляющими 14,0% от лимфоцитов. Популяции CD16+, CD56low не выявлено.
Эффективность получения предложенного компонента БМКП, позволяющего культивировать СК с цитокинами, которые способствуют пролиферации, но не дифференцировке СК, открывает перспективы для применения собственных СК больного для лечения таких заболеваний, как ишемическая болезнь сердца, ожоги, долго незаживающие раны и травмы конечностей, травмы спинного и головного мозга. Аутологичные здоровые ГСК, полученные в результате клеточной инженерии, могут быть выделены и храниться в замороженном виде, чтобы быть использованы в случае острой необходимости. При этом способность к последующей дифференцировке у таких культивируемых клеток сохранена на уровне первоначально выделяемых СК.
ЛК МНК ПК пациента, обогащенный ГСК, в условиях стерильности в ламинарном шкафу из пробирок собирают в мешок и передают на этап иммуносепарации.
Б. Иммуносепарация как способ выделения ГСК из ЛК МНК ПК
Этот этап осуществляется либо в условиях стерильной лицензированной операционной, либо в условиях культуральной лаборатории путем использования сертифицированного и разрешенного к клиническому применению в России иммуносепаратора CliniMACS® plus (Германия) с использованием антител CliniMACS TCR 34+/133+ Kit и специализированных расходных материалов к данному прибору в соответствии с инструкцией производителя. Для этого полученный ЛК помещают в расходный пакет для криоконсервации объемом 400 мл, в него добавляют стандартные антитела к маркерам CD34+ CD133+ на магнитных шариках, подключают к аппарату и проводят иммуносепарацию в обычном режиме. ГСК собирают в иммуносепараторе и передаютя в культуральную лабораторию для дальнейших манипуляций, а истощенный от ГСК CD34+ CD133+ лейкоконцентрат передают в банк костного мозга для криоконсервации в криостойком пакете объемом 400 мл с ДМСО.
В. Криоконсервация клеточного материала
К охлажденной суспензии клеток в аутологичной плазме добавляется стерильный раствор ДМСО в конечной концентрации 10%. Суспензия расфасовывается в криопакеты или необходимое количество криопробирок, маркируется, упаковывается и замораживается. После замораживания пакет или криопробирки с МНК ПК переносятся в сосуд Дьюара для хранения в парах азота.
Г. Криоконсервирование ЛК МНК без ГСК
Традиционный метод криоконсервирования заключается в добавлении ДМСО к клеточной суспензии в конечной концентрации 10% и стандартного замораживания на 1 градус в минуту до -80°С или -120°С с использованием стандартных программ электронного программного замораживания и хранением в жидком азоте или парах жидкого азота.
Д. Выделение клеточной фракции ГСК
Сепарированные ГСК концентрируются путем центрифугирования при 2000 об/мин в течение 10 мин при +18°С. С помощью ручного плазмоэкстрактора из контейнера максимально удаляется плазма, клетки остаются в объеме 40-60 мл.
Е. Добавление криопротектора
В качестве криопротектора кроветворных клеток используется высокоочищенный диметилсульфоксид (ДМСО). К полученным клеткам добавляют при постоянном перемешивании равный объем полиглюкина с ДМСО. Смешивание ДМСО с полиглюкином происходит по типу экзотермической реакции с выделением умеренного количества тепла. Концентрация ДМСО в полиглюкине - 10-12%. Таким образом, его конечная концентрация в замораживаемом материале составит 5-6%.
Применение полиглюкина позволило снизить количество ДМСО вдвое, до 5-6% конечной концентрации, так как полиглюкин обладает следующими свойствами: во-первых, способностью дезагрегировать клетки и тем самым улучшать проникновение криофилактика в клетки, а во-вторых, полиглюкин (6% декстран) сам по себе является криофилактиком.
Ж. Подсчет количества криоконсервируемых клеток
На следующем этапе обязательно производят подсчет ядросодержащих клеток, а также CD 34+ клеток, оставшихся в ЛК МНК, подлежащего заморозке.
Выбор оптимального объема замораживаемого материала
При криоконсервировании для создания наилучших условий важное значение имеют соотношение объема пластикового контейнера и объема замораживаемого в нем материала, а также концентрация замораживаемых клеток. Как было установлено, оптимальная концентрация замораживаемых клеток составляет (40-100)⋅106 клеток/мл.
Замораживание ЛК мобилизованных МНК
В зависимости от конечного объема замораживаемого материала выбирается количество пакетов или полимерных ампул (20-25) для глубокого программного замораживания. Далее клеточную взвесь переводят в эти термостойкие пластиковые пробирки.
Для замораживания ампул с клеточной взвесью их помещают в электронный программный замораживатель. Скорость охлаждения биоматериала при температуре от 0°С до -40°С составляет 1,1 градус в минуту, а затем 1 градус в минуту до -80°С или -120°С. После завершения цикла программного электронного замораживания пробирки с замороженным материалом переносят в хранилище для длительного хранения. Затем пробирки с замораживаемым материалом помещают в пары жидкого азота при температуре от -165° до -170°С, где они будут находиться в жидком азоте или его парах до использования.
В предлагаемой методике учтены и реализованы основные требования, предъявляемые к биоматериалу, подвергнутому криоконсервированию, а именно максимальное сохранение жизнеспособности СК; минимальный объем (100-120 мл) замораживаемого материала при максимальном количестве содержащихся клеток (до 100⋅106/мл), для ПК минимальный объем 50-60 мл; минимальный объем аутотрансплантата. Преимуществами криоконсервирования в парах жидкого азота являются хорошая безопасность хранения пакетов и пробирок с клеточным биоматериалом, а также значительно меньший расход азота.
Стадия 2. Получение линии (линий) аутологичных региональных СК
для клеточной инженерии
Линию МССК изготавливают из 50 мл эксфузата костного мозга или ЛК МНК ПК, их криомешка № 1, полученном при первом сборе ГСК.
А. Способ выделения и культивирования линии МССК
МССК выделяют и изолируют из разных источников по известным международным протоколам: из костного мозга (P.Penfornis, R. Pochampally et al., 2011), из пуповинной крови (Can A. and Balci D., 2011) или из жировой ткани (Zachar V., Rasmussen J. G., and Fink T., 2011) самого пациента. Для изоляции МССК из жировой ткани использовали отечественную модификацию (Тепляшин А.А., 2012), а для изоляции и выделения МССК из костного мозга использовали модификацию профессора Коноплянникова А.Г. (2014).
Для получения МССК использовали материал забора костного мозга. Осуществляли эксфузию 50 мл костного мозга из гребня подвздошной кости специальными биопсийными иглами.
Пример 2. Выделение МССК из костного мозга.
После отстаивания образца костного мозга в течение 1-2 часов при комнатой температуре выделенная верхняя фракция клеток отбирается пастеровской пипеткой и ресуспендируется в ростовой среде. В качестве ростовой среды служит среда RPMI-1640, содержащая пенициллин-стрептомицин (100 ед/мл), амфотерицин (100 нг/мл), L-глютамин 2 мМ, 20% эмбриональной телячьей сыворотки. Культивирование проводится в пластиковых флаконах c площадью дна 25 см2, в которые вносили (2-10)⋅105 клеток/см2 в 8 мл ростовой среды. Флаконы помещаются в CO2-инкубатор (5% CO2). Через 48 часов среда с неприкрепившимися клетками удаляется и заменяется на свежую. При достижении конфлюентного монослоя (90-100%) клетки пересевают с использованием 0,25% раствора трипсина-ЭДТА в новые флаконы, вначале с площадью дна 75 см2, а впоследствии при нарастании клеточной массы - в большие культуральные флаконы с площадью дна 175 см2 . Такой метод позволяет к концу 5-6 недели добиться получения популяции МССК в количестве (1-2) ⋅ 108 клеток, достаточном для трансплантации в организм реципиентов. Изучение методом проточной цитофлуориметрии специфических маркеров полученных культур МССК показывает, что они относятся к популяции CD10low CD34- CD45- CD44+ CD90+ CD105+ клеток, что характерно для клеток, происходящих из мультипотентных мезенхимальных СК взрослого организма.
Производится отмывание полученного клеточного биоматериала от среды культивирования 0,9% физиологичеcким расствором NaCl путем двухкратного центрифугирования клеток при 2000g, и материал может быть использован сразу для клеточной инженерии. Он отмывается от среды культивирования физиологическим раствором и переводится в 200 мл физиологического раствора с 10 ед/мл гепарина. Если материал планируется для интратекального введения, то он должен быть расфасован на аликвоты по 1⋅106 МССК в криопробирки по 2 мл и заморожен в программном замораживателе. Если материал будет использоваться для внутривенного или внутриартериального введения, то криоконсервируется весь объем (1-2)⋅108 МССК в криопробирки по 5-10 мл или специальные мешки для хранения костного мозга.
Криоконсервация клеточного материала
К охлажденной суспензии клеток в аутологичной плазме добавляется стерильный раствор ДМСО в конечной концентрации 10%. Суспензия расфасовывается в необходимое количество криопробирок, маркируется, упаковывается и замораживается. После замораживания криопробирки переносятся в сосуд Дьюара для хранения в парах азота.
Использование клеточного материала для трансплантации
1. Материал, предназначенный для трансплантации, отмывается от среды культивирования физиологическим раствором и переводится в 200 мл физиологического раствора с 10 ед/мл гепарина.
2. Транспортировка клеточного материала в лабораторию клеточной инженерии осуществляется на льду в термоизолированном контейнере.
3. Материал для клеточной инженерии передается в культуральную лабораторию и лабораторию клеточной инженерии.
Б. Выделение МССК из жировой ткани
Пример 3. Жировую ткань забирали биопсией в объеме 10 г. Полученные 10 г трижды промывают PBS (Gibco) при рН 7.2, без ионов Са2+ и Mg2+, содержащим однократный раствор антибиотиков и антимикотика (Gibco), конечная концентрация пенициллина составляет 100 ед/мл, стрептомицина 100 мкг/мл, амфотерицина В 0,25 мкг/мл, и удаляют примеси плотной соединительной ткани. Далее проводят механическую фрагментацию ткани медицинскими ножницами в культуральных чашках диаметром 10 см (Costar) до получения мелкодисперсной массы, которую переносят в две пробирки объемом 50 мл с коническим дном (Costar) и суспендируют каждый образец в 25 мл среды DMEM, содержащей антибиотики и антимикотик. Далее проводят стадию обработки ферментом: в полученные суспензии вводят по 1 мл 2% раствора коллагеназы I типа (Gibco) в буферном растворе PBS без ионов Са2+ и Mg2+ до конечной концентрации фермента 0,075%, инкубируют при температуре 37°С в течение 30 минут при медленном покачивании. Полученную смесь тщательно перемешивают, затем добавляют эквивалентный объем DMEM, содержащую 10% FBS, и антибиотики и антимикотик, и центрифугируют в течение 10 минут при 1000g. Надосадочную жидкость и жировые капли удаляют, осадки объединяют и суспендируют в 10 мл холодного лизирующего буфера (+4°С), содержащего 155 мM NH4Cl, 10 мМ КНСО3, 0,1 мМ Na2EDTA. Смесь тщательно перемешивают и инкубируют 3-5 минут при комнатной температуре. Далее в клеточную суспензию добавляют 10 мл среды DMEM, содержащей антибиотики и антимикотик. Клетки осаждают центрифугированием в течение 10 минут при 1000g. Надосадочную жидкость удаляют и проводят стадию промывания. Клеточный осадок ресуспендируют в 30 мл среды DMEM, содержащей антибиотики и антимикотик, и центрифугируют в режиме 1000g в течение 10 минут. Полученный осадок ресуспендируют в 25 мл среды DMEM, содержащей антибиотики и антимикотик. Полученную клеточную суспензию пропускают через фильтр с размером пор 100 мкм и центрифугируют в течение 10 минут при 300g. Осадок ресуспендируют в среде DMEM, дополненной антибиотиками и антимикотиком, 10% FBS и однократным раствором заменимых аминокислот (Non-Essential Amino Acids, Gibco) в объеме 25 мл. Суспензию пропускают через фильтр с размером пор 10 мкм. В результате получают гомогенную фракцию клеток, свободную от клеточного дебриса и клеток крови.
Полученную клеточную суспензию оценивают подсчетом в камере Горяева и высевают во флаконы площадью 75 см2 из расчета 106 клеток/см2. Доля прикрепившихся клеток составляет приблизительно 1-1,5%. Таким образом, количество МССК, выделенных из 1 г жировой ткани, составляет около (1,5-3)⋅104 клеток. По истечении 24 часов проводят смену среды на DMEM, содержащей антибиотики (100 ед/мл пеницилина, 100 мкг/мл стрептомицина, Gibco), 10% FBS и однократный раствор заменимых аминокислот (Nоn - Essential Amino Acids, Gibco). По достижении монослоя клетки субкультивируют, проводят визуальную оценку морфологии клеток посредством фазово-контрастного микроскопирования, оценивают митотический индекс и время цитогенерации. По результатам морфологического анализа в полученной после выделения фракции клеток выявлено две субпопуляции. Первый тип клеток представляет собой субпопуляцию малых веретеновидных клеток диаметром 10-15 мкм с четко выделенным ядром и гомогенной цитоплазмой. Второй тип представлен клетками округлой формы с вытянутым с одной стороны плоским выростом цитоплазмы, размер клеток достигает 40 мкм; отмечается темное ядро, смещенное к одному краю, гетерогенная цитоплазма и повышенная гранулярность в ядерной области. В фазе логарифмического роста клеток подсчитывают митотический индекс и время цитогенерации. Соотношение количества митотических клеток к общему количеству клеток составляет 34%. Время удвоения 54-62 час. Полученные клетки иммунофенотипируют методом непрямой иммунофлуоресценции. Посредством проточного цитофлуориметра (Beckman Coulter) выявлен высокий уровень экспрессии следующих антигенов CD10, (CALLA), CD13 (APN), CD44 (hyaluronic acid receptor), CD90 (Thy-1), CD105 (endoglin) (Becton Dickinson). Также показано отсутствие экспрессии маркеров ГСК CD34, CD45 и CD117 (Becton Dickinson). Результаты иммунофенотипирования показывают, что по экспрессии поверхностных антигенов популяция полученных клеток соответствует МССК.
В. Способ выделения и культивирования НСК CD133+
из обонятельной выстилки носа человека
Требования к донорам обонятельной выстилки носа. Доноры, то есть сами пациенты, должны быть предварительно обследованы на инфекции ВИЧ 1 и ВИЧ 2, синдром иммунодефицита; сифилис, гепатиты В и С; острые инфекционные заболевания. Методика и протокол получения НСК описан в работах Zhang X. et al., (2004-2006). Ткань обонятельной выстилки носа получают от пациентов и обрабатывают по стандартному культуральному протоколу C.T. Marshal et al., (2005-2006). В отечественной практике используется модификации метода, предложенная проф. И.В. Викторовым (2012).
Пример 4. Для получения здоровых НСК берут ткань обонятельной выстилки носа путем эндоскопической резекции куска слизистой (10мм х 10 мм) верхних хоан носа донора. Полученную ткань доставляют в лабораторию в охлажденном растворе Хенкса без Ca2+ и Mg2+ (HBSS), содержащем антибиотики и антимикотики (1:100; Gibco). Время доставки не превышает 2 ч. После повторной промывки в том же растворе из ткани опухоли удаляют кровеносные сосуды, после чего ткань измельчают и инкубируют в течение 40 мин при 36,5°С в растворе трипсина/ЭДТА 025%, приготовленном на 0,01 М фосфатном буфере (PBS, рН 7,4). Действие ферментов блокируют средой DMEM (Gibco), содержащей 3% сыворотки, ткань промывают в трех сменах сбалансированного солевого раствора Хенкса (HBSS, Hanks’ balanced salt solution, Sigma) и диссоциируют повторным пипетированием в питательной среде. Состав среды: минимальная среда Игла (MEM, Sigma) 90%, эмбриональная сыворотка телят (Fetal bovine serum, FBS, Gibco, Ivitrogen), глюкоза 0,8%, глутамин 2мМ (Gibco), добавок В27 (Sigma), HEPES 20 мM, ростовые факторы (только для первичных культур) - фактор роста фибробластов (FGF2, 1ng/ml, Sigma), нейроростовой фактор (NGF, 2 ng/ml, Sigma).
Полученную суспензию клеток центрифугируют (3 мин при 1200 об/мин), осадок ресуспендируют в питательной среде того же состава. Количество и жизнеспособность диссоциированных клеток определяют в камере Горяева после окраски 0,1% раствором трипанового синего. Для последующего культивирования используют клеточные суспензии только с 85-95% жизнеспособных клеток. Диссоциированные клетки (5⋅105 кл/мл) культивируют в 12-луночных планшетах на полилизин-ламининовом субстрате в течение 14 суток (36,5°C, 5% СО2). Частичную смену 1/3 питательной среды производят два раза в неделю. Первичную культуру после формирования сливного монослоя снимают с помощью раствора трипсина/ЭДТА. После промывки в HBSS и центрифугирования клетки ресуспендируют в питательной среде. Клеточную суспензию (10000-12000 кл/см2) переносят в 12-луночные планшеты или флаконы (площадь 25 см2). Таким способом культуры пассируют 4 раза до формирования сливного монослоя. Формирующиеся в клеточном монослое прикрепленные к субстрату и свободноплавающие нейросферы отбирают с помощью пастеровских пипеток и диссоциируют описанным выше методом ферментной обработки. Выделение нейросфер позволяет отделить их от прикрепленных к субстрату обкладочных глиальных клеток, фибробластов и стромальных (опорных) клеток. Клеточную суспензию клеток нейросфер после промывки и центрифугирования ресуспендируют в питательной среде и культивируют в 12-луночных планшетах (10000-12000 кл/см2) и на покровных стеклах (18х18 мм) в чашках Петри до формирования сливного монослоя. Полученные культуры используют для цитологических и иммуноцитохимических исследований. Часть клеток последних пассажей замораживют в среде для криоконсервирования (90% сыворотки, 10% диметилсульфоксида) и хранят в жидком азоте.
Клеточный монослой фиксируют в 4% растворе параформальдегида, приготовленном на 0,01 М фосфатном буфере (рН 7,4) в течение 30 мин. После промывки в PBS (30 мин) клетки инкубируют 24 ч при 4°С с первичными антителами к β-тубулину (1:300; Chemicon), нестину (1:100, Chemicon ) и нейрональной специфической енолазе (1:100). После промывки в PBS клетки последовательно обрабатывают биотинилированными антителами с авидин-биотиновым комплексом (ABC, Vector Laboratories, Inc), раствором диаминобензидина, приготовленном на фосфатном буфере (DBA 0,5 мг/мл, перекись водорода 0,03%,). Препараты обезвоживают и заключают под покровные стекла в синтетическую смолу (Entellan, Merk).
Материал отмывается от среды культивирования физиологическим раствором и переводится в пробирку с физиологическим раствором. Поскольку материал планируется для клеточной инженерии, то он должен быть расфасован на аликвоты по (0,5-0,6⋅106 НСК в криопробирки по 2 мл и заморожен в программном замораживателе.
Стадия 3. Химически индуцированная трансдифференцировка (перепрограммирование) аутологичных соматических и мультипотентных МССК и НСК в XEN-подобные клеточные системы
Подробный протокол проведения химической индукции соматических или региональных СК приведен в работах Xiang Li et al., Zhen Chai, Hongkui Deng (2017) Direct Reprogramming of Fibroblasts via a Chemically Induced XEN-like State. Cell Stem Cell DOI:10.1016/j.stem.2017.05.019
(https://www.cell.com/cell-stem-cell/fulltext/S1934-5909(17)30185-6?_returnURL=https%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS1934590917301856%3Fshowall%3Dtrue).
В используемом методе химической индукции получения клеток в XEN-состоянии из соматических и СК есть три существенных этапа процесса химического перепрограммирования. На этих стадиях коктейль из пяти небольших молекул «VC6TF» (VPA, CHIR99021, 616452, транилципромин и форсколин) использовался на стадии 1 в течение 16-20 дней; другая небольшая молекула, DZNep, была добавлена в начале стадии 2 в течение следующих 20-24 дней; и 2i-среду (с двойным ингибированием ERK и GSK3 с PD0325901 и CHIR99021 соответственно) использовали на стадии 3 в течение последних 12-16 дней. В целом, процесс химического перепрограммирования может занять до 48-60 дней. Чтобы проанализировать этот процесс, необходимо тщательно следить за изменением морфологии клеток во время химического перепрограммирования на каждой стадии. Примечательно, что в конце стадии 1 образовалось несколько эпителиальных колоний, и эти эпителиальные клетки быстро размножались во время стадии 2. Что еще более важно, нужно отслеживать динамические изменения судьбы клеток во время химического перепрограммирования, так как iPSC преимущественно появляются изнутри этих колоний эпителиальных клеток. В некоторых экспериментах 100% iPSC было получено из этих колоний, даже когда клетки были повторно посажены с более низкой плотностью; колонии эпителиальных клеток выросли до слияния <20%. Эти данные указывают на то, что iPSC могут генерироваться из этих эпителиальных клеток.
XEN-подобное состояние есть основная технологическая платформа для промышленного получения химически индуцированных ГСК. Этому предшествует тонкая настройка химического рецепта для получения XEN-подобных клеток в надежной манере. Чтобы индуцировать iPSC из фибробластов или региональных СК, XEN-подобные первичные колонии можно индуцировать на ранней стадии перепрограммирования, и с этой целью предложена модификация метода трасндифференцировки клеток с помощью семикомпонентного коктейля (VPA, CHIR99021, 616452, транилципромин, форсколин, AM580 и EPZ004777; VC6TFAE) (Zhao Y., Zhao T., Guan J., Zhang X., Fu Y., Ye J., Zhu J., Meng G., Ge J., Yang S., Cheng L., Du Y., Zhao C., Wang T., Su L., Yang W., Deng H. A XEN-like State Bridges Somatic Cells to Pluripotency during Chemical Reprogramming. (англ.) // Cell. - 2015. - Vol. 163, no. 7. - P. 1678-1691. - DOI:10.1016/j.cell.2015.11.017. - PMID 26686652) (см.Таблицу 2 ниже). Модификация XEN-подобной индукции позволила оптимизировать химический рецепт, заменив каждую из семи маленьких молекул функциональными аналогами. TD114-2 был идентифицирован как более предпочтительный ингибитор GSK3-бета, чем CHIR99021 (Таблица 2). При замене CHIR99021 на TD114-2 выход первичных колоний был увеличен в 3 раза без изменения установления XEN-подобного состояния. Таким образом, было идентифицировано, что низкомолекулярный коктейль индуцирует XEN-подобные клетки из фибробластов значительно улучшенным образом. Низкомолекулярные коктейли, которые используются в химическом перепрограммировании, были идентифицированы фенотипическим скринингом и не включают прямую активацию классических факторов перепрограммирования, такие как Oct4, Sox2, Klf4 и c-Myc. Кроме того, основным преимуществом химического подхода является то, что малые молекулы могут быть точно настроены с точки зрения их концентраций, длительности, структур и комбинаций, предоставляя возможность более точно управлять процессом генерирования iPSC на каждом этапе перепрограммирования. Следовательно, идентификация ключевых молекулярных событий и промежуточных состояний клеток поможет значительно повысить эффективность перепрограммирования, позволяя проводить тонкую настройку малых молекул и условий культивирования на основе маркеров каждой стадии перепрограммирования.
Длительность обработки низкомолекулярных клеток и плотность перезаряжаемых клеток были крайне важны на более поздних стадиях перепрограммирования. Хотя старый протокол поддерживает повторную посевную плотность 300 000 клеток на лунку, новый протокол поддерживает плотность 50 000-100 000 клеток на лунку 6-луночного планшета. Кроме того, модифицированный протокол химической индукции имел оптимальную продолжительность 2-й стадии 12 дней. Таким образом, был значительно улучшен переход клеток от XEN к плюрипотентным СК с добавлением новых малых молекул, изменением структуру малых молекул и оптимизацией плотности перезарядки и время обработки малых молекул.
Таблица 2
Малые молекулы, используемые для проведения трансдифференцировки
соматических и региональных СК в XEN-подобные клеточные состояния
Надежный протокол индукции iPSC был создан путем модуляции клеточных переходов через XEN-подобное состояние, так как были объединены оптимизированные условия перепрограммирования для этапов №№ 1-3. С использованием этого нового протокола была индуцирована лунка из 50 000 исходных фибробластов, и клетки были размножены до > 1 000 000 или более повторно посаженных клеток (пересаженных в 10-15 скважин). В конце периода репрограммирования было получено в общей сложности 1 000-9 000 колоний iPSC с общим временем индукции 40 дней (16, 12 и 12 дней для этапов №1, №2 и №3 соответственно). Кроме того, этот новый протокол был воспроизведен независимо более 30 раз, и iPSC также можно было генерировать из дермальных фибробластов и фибробластов легких взрослых со значительно повышенной эффективностью. Используя условия культивирования на основе TD114-2, XEN-подобные колонии могут быть расширены в течение длительного периода времени (по меньшей мере, 20 протестированных пассажей) даже при выборе одной колонии и отдельных клеток в среднем соотношении 1:20-30 каждые 3-4 дня. Несмотря на их высоко расширяемые свойства, онкогенез не был обнаружен после инъекции XEN-подобных клеток мышам с тяжелым комбинированным иммунодефицитом (XEN-подобные клетки при тестировании пассажа 6 и пассажа 19). Кроме того, как показали результаты анализа кариотипа и сравнительной геномной гибридизации, долгосрочные расширенные XEN-подобные интермедиаты сохранили генетическую целостность и стабильность генома. Что наиболее важно, помимо сохранения XEN-подобных паттернов экспрессии, эти расширенные XEN-подобные клетки сохраняли свой функциональный специфический потенциал без снижения их эффективности индукции ГСК даже после 20 пассажей.
Протокол для химически индуцирующих ХЕN-подобных клеток
из фибробластов и региональных СК
Подготовка малых молекул.
Малые молекулы, использованные в этом протоколе, в том числе: VPA, TD114-2, 616452, Tranylcypromine, форсколин, AM580, EPZ004777 и другие, перечислены выше в Таблице 2. Низкомолекулярные соединения были получены в соответствии с инструкцией производителя. TD114-2 был синтезирован в соответствии с химической структурой (Kuo et al.,2003, Storm et al., 2007).
Приготовление питательной среды.
Этап № 1 приготовления питательной среды для всех типов соматических и региональных стволовых клеток.
KnockOut DMEM (GIBCO) с добавлением 10% KSR (GIBCO), 10% FBS (Pan-Biotech), 1% GlutaMAX ™ -I (GIBCO), 1% NEAA (GIBCO), 0,055 мМ β-ME (Sigma), 1% пенициллин-стрептомицин (GIBCO), 100 нг / мл bFGF (Origene), содержащий низкомолекулярный коктейль для химической индукции: 0,5 мМ VPA, 2-4 мкМ TD114-2 или 20 мкМ CHIR, 10 мкМ 616452, 10 мкМ Tranylcypromine, 10-50 мкМ форсколин, 0,05 мкМ AM580 и 5 мкМ EPZ004777. Для значительного повышения эффективности индукции фибробластов рекомендовано 1 мкМ CH55 и 50 мкг мл витамина C (VC).
Этап № 2 приготовления питательной среды.
А. для соматических клеток и клеток мезенхимного происхождения.
Среда Dulbecco M (IMDM) с добавлением инсулина и трансферрина должна содержать смесь цитокинов, состоящую из фактора роста стволовых клеток (SCF), лиганда тирозин киназы 3 из фетальной печени (Flt3L) и тромбопоэтина (ТРО), иономицин при следующем соотношении компонентов: SCF(50-150)⋅103 нг , ТРО(50-150)⋅103 нг , Flt3L (10-50)⋅103 нг, иономицина (10-100)⋅103 нг на 1 мл ростовой среды, 1% пенициллин-стрептомицин (GIBCO), 25 нг/мл bFGF, а также содержать низкомолекулярный коктейль для химической индукции: 10-50 мкМ форсколина, 3 мкМ CHIR, 1 мкМ дорсоморфина.
Б. Для нейральных СК.
Нейробазал (GIBCO) с 0,5% N-2 (GIBCO), 1% B-27 (GIBCO), 1% GlutaMAX ™ -I (GIBCO), 1% пенициллин-стрептомицин (GIBCO), 25 нг / мл bFGF, 20 нг / мл BDNF, 20 нг / мл GDNF, содержащий низкомолекулярный коктейль: 10-50 мкМ форсколина, 3 мкМ CHIR, 1 мкМ дорсоморфина.
Химическая индукция.
1. Соматические клетки (фибробласты) или региональные СК высевали с плотностью 30 000-50 000 клеток на лунку 6-луночного планшета или 300 000 клеток на 100 мм чашки (день 1). Культуральные чашки или чашки предварительно покрывали 0,1% желатином.
2. Изменили среду на культуральную среду стадии 1 (день 0). Культуральная среда обновлялась каждые 4 дня. XEN-подобные колонии будут сформированы на 6-8 день.
3. На 12-й день индуцированные клетки должны быть переведены в культуральную среду гемопоэтической индукционной стадии № 2. Обновляли среду каждые 4 дня.
Необязательно: XEN-подобные колонии в конце стадии 1 были компактными. Операция повторного посева на 12-й день может расширить XEN-подобные колонии и увеличить выход клеток. Трипсинизировали и повторно засевали первичные XEN-подобные клетки в соотношении 1:10-20 в культуральной среде стадии 1. Ксеноподобные колонии реформировали через 4-6 дней после повторного посева и затем переключали в среду гемопоэтической индукции стадии 2. Учитывая, что FSK, используемый в высокой концентрации 50 мкМ, был токсичным для пассированных XEN-подобных клеток на стадии 2, была рекомендована концентрация 10-20 мкМ в гемопоэтической индукционной культуральной среде на пассированных XEN-подобных клетках и 50 мкМ на стадии 2 для непроходной индукции.
4. Через 8-12 дней в среде стадии №2 XEN-подобные колонии становятся гемопоэтически-позитивными. Увеличение периода индукции может привести к появлению большего количества гемопоэтически-позитивных колоний и повышению зрелости, что проявляется в появлении маркеров гемопоэза. Эти гемопоэтически-позитивные клетки могут быть подвергнуты характеристике или пересажены для совместного культивирования с первичными МНК ПК в условиях низкой дифференцировки для дальнейшего созревания.
XEN-подобные клетки: Пассирование и расширение.
На 12-й день индуцированные XEN-подобные колонии трипсинизировали из всей лунки и повторно высевали на покрытые желатином культуральные планшеты в соотношении 1:10-20 в среде стадии 1 для первого пассажа. После того, как XEN-подобные клетки в пассаже 1 станут 100% слияниями, пассивировали клетки в соотношении 1:20-30 каждые 3-4 дня в среде стадии 1. Клетки, подобные XEN, могут быть длительно размножены на среде стадии 1 для более чем 20 пассажей с потенциалами, определяющими клон.
Для одного первичного XEN-подобного пассажа колоний необходимы питающие слои MEF, обработанных митомицином С. Брали одну первичную XEN-подобную колонию с помощью 1 мл шприца, трипсинизировали в течение 5 минут и нейтрализовали культуральной средой стадии 1, затем пипетировали вверх и вниз, чтобы получить суспензию отдельных клеток. Сеяли клеточную суспензию на питающие слои с питательной средой стадии 1 для дальнейшего расширения. Последующее пассирование XEN-подобных клеток может быть без фидера.
Для гемопоэтической индукции от пассированных XEN-подобных клеток клетки высевали в количестве 50000-100000 клеток на лунку 6-луночного планшета (с желатиновым покрытием) в среде стадии 1 (день 0). До тех пор пока пассированные клетки не сойдутся на 70-80% в день 2 или 3, переключали XEN-подобные клетки в среду гемопоэтической индукции стадии 2. Через 12-20 дней появятся гемопоэтически-позитивные колонии.
Стадия 4. Клеточная инженерия и получение генетически однородных гемопоэтических стволовых клеток с гомотопной субституцией генома
А. Получение клеточно-инженерного препарата ГСК с гомотопной субституцией генома для трансплантации
Вариант 1. Клеточно-инженерный способ реконструкции поврежденной ГСК путем слияния цитопласта ГСК с кариопластом химически индуцированных клеток в ХEN-подобном состоянии (Сi XEN-like SC).
Для энуклеации ядер использовали технологию обработки культуры клеток цитохалазином В, когда концентрация цитохалазина В рассчитывалась из такого расчета, чтобы на 1 мл обычной питательной среды, в которой культивировались клетки, количество цитохалазина В достигало 10 мкг. При центрифугировании в большинстве таких предобработанных цитохолазином В клеток разрывается цитоплазматический тяж и образуются энуклеированные (безъядерные) клетки (цитопласты) и ядра, отделившиеся при центрифугировании (кариопласты), окруженные тонким слоем цитоплазмы и плазматической мембраной, то есть, следующая клеточно-инженерная операция должна была быть направлена на получение популяции цитопластов или получение безъядерных ГСК клеток с маркерами клеточной поверхности CD34+, CD45+,CD133+, и она производилась с использованием отделения карипластов от целых клеток. В некоторых случаях эффективность энуклеации не превышает 50%. С целью повышения процента энуклеации энуклеированные (цитопласты) и целые клетки, содержащиеся на поверхности пластиковых чашек и стеклянных дисков, снимали и обрабатывали трипсином, в результате чего в 1 мл солевого раствора с фосфатным буфером содержалось 106 целых клеток и цитопластов. Клеточно-цитопластическую смесь помещали на 14 мл линейного градиента ренографина-76, который находился в центрифужных пробирках, объемная концентрация которого изменялась от 15 до 30%, и центрифугировали при 1000g и температуре 25°С в течение 5 мин. Четко разграниченные слои целых клеток и цитопластов удаляли из центрифужных пробирок и использовали по назначению. Полученные в градиенте плотности ренографина-76 цитопласты не окрашивались трипановым синим, что послужило контролем эффективности разделения. Они сохраняли способность снова прикрепляться к поверхности культуральной чашки и могли использоваться в дальнейшем для реконструкции жизнеспособных клеток путем слияния с аутологичными кариопластами.
Б. Способы получения биоматериала ядра из СiXEN-like SC (кариопластов ) варьировались.
Вариант 1. Методы энуклеации из СiXEN-like SC были аналогичны тем, что были применены для ГСК и в той же дозировке цитохолазина В (10 мкг/мл) для получения кариопластов и цитопластов. Раствор цитохалазина В готовят добавлением стокового раствора на основе DMSO к среде ТСМ 199, дополненной 4% BSA и антибиотиками в конечной концентрации 5-7,5 мкг/мл. Клетки СiXEN-like SC, предназначенные для выделения кариопластов, за 2 дня до энуклеации высевали на вырезанные из культуральных флаконов пластиковые пластинки, перед помещением в 50-миллилитровые центрифужные пробирки, заполненные обычной питательной средой, составляли так, чтобы поверхности этих пластинок с прикрепленными клетками находились снаружи. Линии СiXEN-like SC пациента центрифугировали при 9500g и 35°С в течение 15 мин. Назначение этой процедуры сводилось к тому, чтобы в осадке кариопластов, который должен быть получен, уменьшить количество целых клеток. После удаления слабо прикрепленных к субстрату клеток пластиковой пластинки с клетками монослоя, оставшиеся переносили в пробирки, где концентрация цитохалазина В из расчета на 1 мл обычной питательной среды достигает 10 мкг. Содержимое пробирок инкубировали при 37°С в течение 15 мин, продолжительность центрифугирования при 35°С и 7000 об./мин составила 45 мин. Образовавшуюся надосадочную жидкость сливали, а осадок, представленный в основном кариопластами СiXEN-like SC, ресуспендировали в обычной питательной среде.
В изготовленных по описанной методике препаратах карипластов находились фрагменты цитоплазмы, погибшие кариопласты и целые клетки. Отделение фрагментов цитоплазмы проводится осаждением в градиенте фиколла 1-6% концентрации при 1g и 37°С в течение 90 мин в увлажненной атмосфере, содержащей 5% СО2. Фрагменты цитоплазмы, находящиеся в верхнем слое, отсасывали и удаляли, а осадок кариопластов СiXEN-like SC, образовавшийся после удаления и разведения питательной средой, снова осаждали центрифугированием и ресуспендировали в свежей питательной среде. Целые клетки (их 0,4-4%) удаляли путем двукратного 90-минутного инкубирования суспензии кариопластов в культуральных чашках. Инкубирование в течение 3 часов позволяет резко уменьшить загрязнение кариопластов целыми клетками, и оно обязательно для всех экспериментов с трансплантацией ядер.
Следующая операция - отделение жизнеспособных карипластов от тех, которые погибли. Она включала ресуспендирование осадка в питательной среде до концентрации 107 мини-клеток в 1 мл, наслоение суспензии на Ficoll-paque (раствор содержит 5% фиколла и 9% диатризоата натрия), находящийся в центрифужных пробирках, и осторожное добавление свыше 1 мл среды так, чтобы добавленная среда и суспензия кариопластов образовали два самостоятельных слоя. Следующее центрифугирование при 800 об/мин (130 g) в течение 75 мин при комнатной температуре позволяет получить осадок, который на 99% состоял из погибших кариопластов. На границе питательной среды и Ficoll-paque находится слой, состоящий на 98% из жизнеспособных кариопластов, которые и использовались далее в работе.
Для очистки кариопластов можно использовать и другие методы, в частности частицы тантала размером 1-3 мкм. Ко времени энуклеации практически все клетки содержат более 12 частиц тантала. В связи с тем, что плотность тантала более чем в 15 раз превосходит плотность клетки, компоненты, содержащие частицы тантала, осаждаются значительно быстрее кариопластов, в которых тантал отсутствует.
Вариант 2. В качестве второго варианта получения биоматериала для клеточной инженерии могут использоваться нативные тканеспецифические СiXEN-like SC пациента, полученные путем культивирования по описанной выше технологии.
В . Клеточно-инженерная биосборка аутологичных реконструированных ГСК.
Следующей операцией была биосборка и реконструкция клеточно-инженерных ГСК. В первом варианте для реконструкции (восстановления) ГСК суспензию кариопластов СiXEN-like SC в солевом буфере добавляли к монослою культуры цитопластов из ГСК в пропорции 100 кариопластов СiXEN-like SC на 1 цитопласт ГСК. Цитопласты ГСК должны быть уже покрыты инактивированными частицами вируса Синдай (по методу McGras и Solter, 2005). Инкубируют 45 минут при температуре 4°С, а затем еще 45 минут при температуре 37°С. Отмывают раствором Эрла для удаления неслившихся кариопластов, так как 10% кариопластов некоторых клеточных линий способны восстанавливать весь объем потерянной при энуклеации цитоплазмы и снова превращаться в жизнеспособные клетки. Способность кариопластов регенерировать утраченную в процессе энуклеации цитоплазму и формировать жизнеспособные колонии клеток зависит от количества цитоплазмы, окружающей ядро. Во фракции кариопластов СiXEN-like SC, у ядер которых сосредоточено 2-4% количества цитоплазмы, содержащейся в интактной клетке (мелкие кариопласты), только один из 106 очищенных элементов может сформировать жизнеспособную колонию клеток. Для слияния цитопластов и кариопластов СiXEN-like SC применяли электропорацию (электрослияние) с использованием мультипоратора - прибора, способного воздействовать импульсами заданного напряжения. Во время воздействия импульса вследствие образования пор в цитоплазматических мембранах объединяемых клеток их цитоплазмы объединяются. Реконструированные клеточно-инженерные XEN-подобные ГСК пациента помещают в камеру для электрослияния, которую предварительно заполняют слабым гипотоническим буфером (200-240 мОсмоль/кг), содержащим сорбитол. Слияние проводят после предварительного элигмента в 0,3 кВ/см продолжительностью 10 секунд посредством двух последовательных пульсов продолжительностью 30 секунд напряжением 1-1,4 кВ/см. После этого полученные аутологичные ГСК с гомотопной субституцией генома тщательно отмывают от буфера в промывочной среде и переносят в постактивирующий раствор. Использование слабого гипотонического буфера в сочетании с низким напряжением, с одной стороны, позволяет повысить процент слияния кариотипов СiXEN-like SC и цитопластов ГСК пациента, а с другой стороны, обеспечить наилучшую выживаемость полученных реконструированных клеток. Можно для повышения эффективности слияния цитопластов ГСК и кариопластов СiXEN-like SC применять технологию оптико-механического лазерного слияния, предложенную российскими учеными Л.М Чайлохяном, Т. А. Свиридовой-Чайлохян (2010).
Вариант 2. Клеточно-инженерный способ реконструкции ГСК путем слияния цитопласта ГСК с цельноклеточными СiXEN-like SC.
Данный вариант клеточной инженерии предполагает удаление ядра из ГСК пациента и пересадка ядер соматических клеток не путем выделения кариопласта из СiXEN-like SC, а путем слияния цитопласта с целой СiXEN-like SC. Процедура получения цитопластов из ГСК не отличается от первого варианта. Полученные в результате энуклеации цитопласты ГСК переносят из среды, препятствующей дифференцировке и созреванию, в среду ТСМ 199, содержащую 0,1% гиалуронидазы, и инкубируют при 37°С в течение 1 минуты. Цитопласты ГСК удаляют осторожным пипетированием в промывающей среде и переносят в раствор цитохалазина В на 5 минут. Раствор цитохалазина В также готовят добавлением стокового раствора на основе DMSO к среде ТСМ 199, дополненной 4% BSA и антибиотиками в конечной концентрации 5-7,5 мкг/мл. Следующий этап предлагаемого варианта клеточной инженерии заключается в слиянии цельноклеточной СiXEN-like SC с цитопластом энуклеированной ГСК с получением клеточно-инженерных генетически однородных ГСК пациента.
Для создания клеточно-инженерных ГСК пациента, способных к развитию, цитоплазму перенесенной СiXEN-like SC следует объединить с цитоплазмой энуклеированной ГСК с последующей активацией полученных клеточно-инженерных ГСК пациента. Для этого обычно используют электрослияние, описанное выше. Активацию проводят одновременно с электрослиянием. Процедуру слияния и активации проводят при температуре 25-28°С. Соблюдение данного температурного режима обеспечивает оптимальные условия для воздействия буфера и, как следствие, увеличивает проницаемость цитоплазматических мембран, обеспечивая лучшее воздействие создаваемого напряжения без вреда для жизнеспособности ооцитов и клеток.
Г. Постактивирование ГСК с гомотопной субституцией генома.
Известно, что для предотвращения процессов метилирования полученные реконструированные ГСК постактивируют и только затем культивируют до нужной стадии. Активированные клетки культивируют сначала в течение 30 минут в растворе цитохалазина В вышеуказанного состава, а затем в среде для культивирования, содержащей DMAP (среду получают добавлением 100 × стокового раствора DMAP на основе DMSO в конечной концентрации в среде 2 мМ). Через 2-4 часа постактивации реконструированные ГСК анализируют и отбирают те, у которых произошло объединение цитопластов и кариопластов, после чего реконструированные ГСК, в которых визуализировано слияние, переносят в среду подходящего состава для дальнейшего культивирования. Состав среды для культивирования в ростовой среде, содержащей смесь цитокинов, состоял из фактора роста стволовых клеток (SCF), лиганда тирозин киназы 3 из фетальной печени (Flt3L) и тромбопоэтина (ТРО), отличием которой является то, что указанная выше смесь цитокинов, также как и при культивировании ЛК МНК ПК, дополнительно содержит иономицин при следующем соотношении компонентов на 1 мл ростовой среды: SCF (50-150)⋅103 нг , ТРО (50-150)⋅103 нг, Flt3L (10-50)⋅103 нг, иономицин (10-100)⋅103 нг. Постактивацию и культивирование проводят в средах, не содержащих феноловый красный. Данное вещество добавляют в качестве индикатора уровня рН, но оно обладает токсическими свойствами, что снижает жизнеспособность клеток.
Эффективность получения предложенного компонента БМКП, позволяющего культивировать СК с цитокинами, которые способствуют пролиферации, но не дифференцировке СК, открывает перспективы для применения собственных СК больного для лечения таких заболеваний, как ишемическая болезнь сердца, ожоги, долго незаживающие раны и травмы конечностей, травмы спинного и головного мозга. Аутологичные СК могут быть выделены и храниться в замороженном виде, чтобы быть использованными в случае острой необходимости. При этом способность к последующей дифференцировке у таких культивированных генно-инженерных клеток сохранена на уровне первоначально выделяемых СК.
Стадия 5. Получение линии генетически однородных ГСК с гомотопной субституцией генома (ядра) без реверсии эпигенетической памяти
Цель этой биотехнологической стадии является "дозревание" полученных клеточных систем до полной гемопоэтической дифференцировки и мультипотентности, позволяющей этим клеткам дифференцироваться во все типы клеток крови, увеличение количества полученных клеточно-инженерных ГСК пациента и полная блокировка в этих клетках реверсии эпигенетической памяти соматических и СК негемопоэтической направленности, из которых они были получены. Задачи этого этапа состоят в осуществлении «дозревания» полученных клеточно-инженерных клеток до мультипотентного гемопоэтического состояния и появления у них высокопластичных недифференцированных ГСК с маркерами CD34+, CD133+Lin-, но без развития колоний дифференцированных клеток. При этом способность к последующей дифференцировке у таких культивируемых клеток должна быть сохранена, по возможности, в первоначальном виде. Эту задачу способно обеспечить влияние собственной цитоплазмы ГСК на траснплантированное ядро клетки, находящейся в ХЕN-подобном состоянии, соответствующее гемопоэтическое микроокружение клеточно-инженерной клетки клетками крови при культивировании и наличие в культуральных условиях факторов, стимулирующих созревание клетки в гемопоэтическом направлении. Также очень важно на этом раннем этапе формирования гемопоэтической ориентации клетки применить факторы, блокирующие в ней эпигенетическую память о модификациях ДНК и эпигенетических изменениях гистонов, связаных с предыдущими этапами дифференцировки клетки в "прошлой" жизни, до клеточной инженерии.
Самой главной составляющей этого этапа является соединение клеточно-инженерных ГСК с ЛК собственных МНК ПК для совместного их культивирования. Весь криоконсервированный ЛК размораживают, отмывают от криопрофилактика и 10% ДМСО центрифугированием с физиологическим раствором, делят на пробирки по 50 мл и работают с каждой пробиркой индивидуально. Клеточную суспензию, состоящую на 98,5-99,5% из ЛК МНК ПК пациента и 0,5-1,0% клеточно-инженерных ГСК, культивируют в среде Dulbecco M (IMDM) с добавлением инсулина и трансферрина, а также фактора роста стволовых клеток (SCF), лиганда тирозин киназы 3 из фетальной печени (Flt3L), тромбопоэтина (ТРО) и иономицина (Са2+ ionophore). Через 3-7 дней культивирования в выращенной клеточной суспензии содержится около 1,5-2% клеток CD34+. Отличием этой среды является то, что указанная выше смесь цитокинов дополнительно содержит иономицин при следующем соотношении компонентов на 1 мл ростовой среды: SCF (50-150)⋅103 нг, ТРО (50-150)⋅103 нг, Flt3L (10-50)⋅103 нг, иономицин (10-100)⋅103 нг. Как и на стадии 1 способа по настоящему изобретению (при наращивании количества ГСК), иономицин добавляют в культуральную среду в качестве стимулирующего агента роста недифференцированных СК в условиях ех vivo. Свойства компонентов используемой культуральной среды описаны выше при описании стадии 1 способа по настоящему изобретению.
ИПСК (iPSC), полученные из нейральных предшественников и фибробластов, характеризуются остаточным метилированием локусов, ответственных за образование кроветворной линии, что вызывает снижение уровня дифференцировки этих ИПСК в соответствующем гемопоэтическом направлении. Обработка малыми молекулами вальпроевой кислоты (ингибитор гистондеацетилазы 1 (HDAC1)) клонов клеточно-инженерных ГСК, в которых репрессирован импринтированный локус Dlk1-Dio3, приводит в ряде линий к реактивации генов данного локуса. Такие ИПСК были способны давать вклад в развитие организма при тетраплоидной комплементации (Stadtfeld M., Apostolou E., Akutsu H., Fukuda A., Follett P., Natesan S., Kono T., Shioda T., Hochedlinger K. // Nature. 2010. V. 465. № 7295. P. 175-181).
Для блокирования эпигенетической памяти клетки, получившие клеточно-инженерную реконструкцию, с самых первых дней их культивирования предобрабатывают вальпроевой кислотой, витамином группы В12 и аскорбиновой кислотой. Известно, что аберрантное молчание импринтированного локуса Dlk1-Dio3 наблюдалось в определенных клонах iPSC, в том числе полученных из ГПК, которые также характеризуются низким уровнем транскрипции этого локуса. Предполагается, что этот эффект вызван "эпигенетической памятью". Из-за транскрипционных нарушений в Dlk1-Dio3 локусы ИПСК становятся неспособными к эффективному образованию химеры и не могут образовывать новый химерный организм посредством тетраплоидной комплементации. Лечение вальпроевой кислотой, ингибитором гистондеацетилазы, приводит к активации транскрипции в локусе Dlk1-Dio3 и восстанавливает способность ИПСК к тетраплоидной комплементации и эффективному образованию химерных животных. Эти низкомолекулярные вещества высоко себя зарекомендовали как индукторы правильной клеточной дифференциации клеток в эмбриогенезе плода при приеме этих веществ беременными женщинами в самом раннем периоде беременности. Сегодня показано, что аскорбиновая кислота (витамин С) влияет на паттерн метилирования ДНК. Так, в ИПСК, культивируемых с добавлением в среду аскорбиновой кислоты, в зависимости от ее дозы, наблюдалась реактивация импринтированного гена Meg3 локуса Dlk1-Dio3. Однако аскорбиновая кислота не приводит к масштабному деметилированию всего генома, она в состоянии только предотвратить аберрантное метилирование локуса Dlk1-Dio3 в процессе репрограммирования, но не способна привести к деметилированию ДНК в уже стабильных клонах ИПСК (Stadtfeld M., Apostolou E., Ferrari F., Choi J., Walsh R.M., Chen T., Ooi S.S., Kim S.Y., Bestor T.H., Shioda T., et al. // Nat. Genet. 2012. V. 44. № 4. P. 398-405). В то же время способность клеток к дифференцировке в определенном направлении и профиль метилирования можно восстановить путем повторных раундов репрограммирования. Показано, что, например, ИПСК, полученные из клеток-предшественников нейрального ряда, обладали очень низкой способностью формировать колонии ГСК. Но когда такие колонии подвергались репрограммированию, то формирование гемопоэтических колоний "вторичными" ИПСК значительно возрастало (Kim K., Doi A., Wen B., Ng K., Zhao R., Cahan P., Kim J., Aryee M.J., Ji H., Ehrlich L.I., et al. // Nature. 2010. V. 467. № 7313. P 285-290). Если ИПСК, полученные из нейрональных предшественников, дифференцируются в линии ГСК, и вторичные ИПСК в последующем получают из этих производных, эти вторичные ИПСК будут иметь более высокий потенциал дифференцировки в клетки крови.
Таким образом, дополнительной изобретательской находкой в настоящем изобретении стало культивирование клеточно-инженерных ГСК вместе с ЛК МНК ПК в условиях ограниченной дифференцировки и в присутствии факторов, стимулирующих гемопоэтические характеристики этих клеток и обеспечивающих формирование генетически однородных мультипотентных ГСК с гомотопной субституцией генома. При подсчете клеток и фенотипическом анализе мононуклеарной фракции для определения количества живых МНК к 20 мкл клеточной суспензии добавляют 20 мкл трипанового синего (Gibco). Подсчет клеток производят в гемоцитометре, пропорция живых клеток не должна быть ниже 95%. Для оценки количества клеток CD34+ и CD133+ мононуклеарной фракции 0,1 мл клеточной суспензии окрашивают соответствующими антителами и определяют пропорцию ГСК в исходной клеточной взвеси на приборе FACSCalibur (Beckton Dickinson).
Пример 5. Данные фенотипического анализа МНК в одном показательном эксперименте, проведенном на клетках ЛК пациента С. Пропорции клеток в ЛК МНК ПК: ГСК СD34+CD45+ донора составляла 0,7%, количество нейральных клеток - 15% , Т- лимфоциты 74%, В-клетки -10,2%.
Культивирование . Жидкая культуральная система состоит из среды Искова в модификации Дульбекко (Iscove's modified Dulbecco's Medium IMDM, Gibco, UK) с добавлением 0,5 г/л бычьего сывороточного альбумина (Sigma Aldrich), 0,39 мкг/мл человеческого инсулина и 60 мкг/мл трансферрина (Gibco). В культуральную среду добавляют фактор роста СК (SCF), другое название c-Kit-ligand (c-KitL, R&D Systems), в концентрации 100 нг/мл, а также Flt3-ligand (Flt3L) в концентрации 20 нг/мл, (R&D Systems), и иономицин (Sigma Aldrich) в концентрации 0,1 мкМ/мл. Для блокирования эпигенетической памяти добавлен коктейль из вальпроевой кислоты (Merck.) (VPA) в концентрации 1 μM (Sigma), Витамин В1 (Vb1) (Sigma) в концентрации 9 мг/л, и аскорбиновой кислоты (VC) (Sigma) в концентрации 25 или 50 мкг/мл. VC добавляли со второго дня и до конца культивирования на данной стадии. VPA добавляли с 3 по 7 день генерации ГСК человека соответственно.
Способ культивирования. Клетки культивируют при 37ºС во влажной атмосфере с притоком 5% CO2. Свежую среду и цитокины добавляют каждый третий день. Время культивирования составляет 3-7 дней. Использованные факторы роста способствуют пролиферации СК, при этом не нужно добавлять агенты, подавляющие дифференцировку. Получаемые суспензии СК любого индивидуума могут быть сохранены в замороженном виде либо могут быть введены в БМКП непосредственно после культивирования и применены больным.
Таблица 3
Оценка чистоты клеточной суспензии после культивирования
Заключение: Материал сохранный. На основании данных проточной цитометрии ГСК составляют 1,8%, а ГМК - 1,6% без нарушения процентных отношений субпопуляций всех лимфоцитов.
Стадия 6. Криоконсервация клеточного материала
Криоконсервацию на этой стадии по существу проводят также как на стадии 1 способа по настоящему изобретению.
Стадия 7. Использование клеточного материала для трансплантации
Материал, предназначенный для трансплантации, в виде концентрата МНК ПК с линией клеточно-инженерных ГСК пациента размораживается на водяной бане, отмывается двухкратным центрифугированием от ДМСО и среды культивирования физиологическим раствором и помещается в стерильный пакет с 200 мл физиологического раствора и 10 ед/мл гепарина. Количество клеточно-модифицированных ГСК должно быть в количестве не менее 1,5-2% от общего объема трансплантируемых клеток мобилизованных МНК ПК.
Транспортировка клеточного материала осуществляется на льду в термоизолированном контейнере.
Клеточный материал вводится внутривенно с использованием трансфузионной системы в течение 1 ч. За 10 мин до начала процедуры пациент получает 4 мг метипреда.
Данное изобретение иллюстрируется далее следующими примерами, которые не должны рассматриваться как ограничение объема данного изобретения.
Пример 6. Больной С-вой Л.В., 42 лет с диагнозом "боковой амиотрофический склероз", высокая форма с выраженными мышечными атрофиями конечностей, псевдобульбарным синдромом и вялым тетрапарезом. С информированного согласия начата стимуляция гемопоэза теваграмтином в дозе 300 мг внутримышечно в течение 4 дней, а на 5 день введена двойная доза Г-КСФ. На 6 день после катетеризации центральной вены проведен лейкоцитоферез на аппарате Spectra Coba. Получено 480 мл ЛК МНКПК, которые были переданы в Банк костного мозга, где после оседания клеточного ЛК на градиенте фиколла произведена очистка ЛК от эритроцитов и гранулоцитов. Очищенный ЛК передан в операционную клиники, где с использованием моноклональных антител CD133+ CD34+CD45+kit на магнитных шариках на клеточном сортере CliniMACS произведена иммуносорбция ГСК пациентки. Получены ГСК в объеме 5мл в физиологическом растворе с концентрацией клеток 5⋅109 в 10 мл Концентрация ГСК в ЛК МНК ПК составила 1,5%, поэтому ГСК направлены на исследование в лабораторию иммунологии гемопоэза для проведения проточной цитофлуориметрии.
Пример 7. Энуклеация ГСК из ЛК МНК ПК у С-вой Л.В., 42 лет.
ГСК, полученные в примере 6, обрабатывали в течение 1 минуты 0,1% раствором гиалуронидазы (Sigma), приготовленного добавлением 90 мкл 10 × стокового раствора к промывочной среде, препятствующей дифференцировке, с иономицином. Многократно отмывали в данной среде и переносили в свежую промывочную среду, содержащую цитохалазин В. Раствор готовили добавлением 100 × стокового (конечная концентрация 5-7,5 мкг/мл) раствора цитохалазина В. В данной среде аутологичные ГСК культивировали в условиях инкубатора при 38,5°С и 5% СО2 в течение 30 минут. Прокультивированные в растворе цитохалазина В аутологичные ГСК помещали в пробирку и центрифугировали двухкратно при 2000 об/мин в среде, в которой проводили процедуру энуклеации. В результате центрифугирования цитопласты ГСК плавали в надосадочной жидкости, а кариопласты оседали внизу пробирки. Пипеткой цитопласты переносили в культуральную среду. После этого цитопласты окрашивали Hoechst 33342 (5 кг/мл). Далее цитопласты тщательно отмывали в манипуляционной среде и переносили в каплю свежей среды под слоем минерального масла. На микроскопе Olympus в ультрафиолетовом свете проводили оценку качества удаления ДНК. Для слияния использовали только ГСК, несодержащие собственной ДНК. После подтверждения факта энуклеации цитопласты замораживали со средой ДМСО в программном замораживателе до температуры -196°С и помещали в дюар с жидким азотом.
Пример 8. Получение мультипотентных МССК у пациентки С-вой Л.В., 42 лет.
ММСК получали из жировой ткани пациентки, для чего жировую ткань забирали открытым способом во время хирургической операции из подкожно-жировой клетчатки передней стенки живота пациентки через разрез размером 3 см, сделанный под местной анестезией 1% раствора новокаина, и тщательно промывали в фосфатно-солевом буфере, измельчали маленькими острыми ножницами и подвергали ферментативной обработке 0,075% раствором коллагеназы типа I на среде ДМЕМ при 37°С в течение 30 минут. Коллагеназу отмывали эквивалентным объемом питательной среды (ДМЕМ, дополненная 10% эмбриональной сывороткой теленка) и центрифугировали при 1000g в течение 5 минут. Полученный осадок ресуспендировали и пропускали через фильтры с диаметром пор 100 мкм для удаления клеточных остатков. Концентрация клеток для посева составляла 5×106 клеток на культуральный флакон 75 см3. Культивировали МССК в среде ДМЕМ (фирмы Gibco) с низким содержанием глюкозы, дополненной эмбриональной сывороткой теленка, однократным раствором заменимых аминокислот и антибиотиками при 37°С и 5% СО2 в воздухе. Через двое суток культивирования были замечены колонии прикрепившихся клеток. Среду меняли каждые 4 суток и по достижению 90% прочности монослоя их субкультивировали и замораживали. В нужное время аликвоту замороженных клеток размораживали и отмывали от криопротектора двукратным центрифугированием при 1500 об/мин в течение 5 минут в питательной среде ДМЕМ, содержащей 10% эмбриональную сыворотку теленка. После этого клетки высевали в культуральные чашки Петри и культивировали в условиях, используемых перед замораживанием, до получения плотного монослоя. Последние двое суток клетки культивировали в бессывороточной среде. Непосредственно перед процедурой пересадки ядер клетки снимали с субстрата раствором трипсин-ЭДТА, отмывали и разводили в небольшом количестве манипуляционной среды ТСМ 199 с получением суспензии клеток.
Пример 9. Электрослияние и активация полученных реконструированных (восстановленных) аутологичных ГСК.
Аутологичные цитопласты (энуклеированная ГСК) и аутологичный кариопласт (МССК в XEN-подобном состоянии, полученная из жировой ткани С-вой Л.В.,42 лет, объединяли методом электрослияния в микрокамере с использованием мультипоратора фирмы Eppendorf. Реконструированные ГСК переносили в камеру для электрослияния с 50 мкл раствора сорбитола (200-240 мОсмоль/кг). Слияние и активацию проводили после предварительного элигмента в 0,3 кВ/см в течение 10 минут посредством двух последовательных импульсов в 1-1,4 кВ/см продолжительностью 30 секунд. Температура раствора сорбитола при этом составляла 25-28°С. После воздействия импульсов реставрированные ГСК тщательно отмывали от раствора сорбитола в среде для манипуляций.
Пример 10. Культивирование полученных восстановленных ГСК.
Подвергшиеся процедуре электрослияния восстановленные ГСК сначала культивировали в среде NCSU-23, содержащей 5 мкг/мл цитохалазина В в течение 30 минут, а затем 3 часа в среде NCSU-23, к которой добавлен DMAP (стоковый раствор DMAP, разведенный в DMSO, добавляли к среде в конечной концентрации 2 мМ). По истечению данного периода отбирали ГСК, в которых произошло полное объединение цитопласта и содержимого МССК. Отобранные клеточные комплексы культивировали в течение 5-6 дней в условиях инкубатора при 38,5°С, 5% CO2 в воздухе и максимальной влажности. В процессе культивирования проводили оценку количества полученных ГСК. В течение 5 дней вследствие оптимизации приемов энуклеации и пересадки МССК из жировой ткани были успешно реконструировано 54,7% ГСК. Режимы электрослияния и составы буферных растворов обеспечивают слияние цитоплазматических мембран ГСК и пересаженных ММСК в 75% случаев. Активированные клеточные комплексы обладают способностью к полноценному гемопоэзу, о чем свидетельствовали маркеры ГСК в гейте CD34+, что представлено в таблице 4 - результаты иммунофенотипического исследования культуры ГСК пациентки С-вой Л.В., 42 г., дата поступления материала: 10.01.2019 г.).
Таблица 4
Результат иммунофенотипического исследования восстановленных ГСК
CD38
CD13
CD117
CD56
CD61
CD10
CD62L
CD44
CD28
Полученные в результате клеточной инженерии или трансдифференцировки линии генетически однородных ГСК с гомотопной субституцией генома всегда должны трансплантироваться только в комбинации с ЛК мобилизованных МНК ГСК, так как изолированное применение клеток этой линии будет клинически неэффективно.
Так и при трансплантации генетически однородных ГСК с гомотопной субституцией генома принципиально важно, чтобы клетки полученной линии при трансплантации были в микроокружении мобилизованных лимфоцитов ПК, так как эта сбалансированная комбинация лимфоцитов работает как единый костно-мозговой кластер, механизм действия которого был описан японским исследователем K.Ono (2002).
При этом необходимо при трансплантации соблюдать количественные параметры содержания клеток полученной линии ГСК и не допускать снижения их количества менее 1% от общего клеточного содержания МНК ПК.
Пример 11 . Проверка эффективности БМКП.
Исследование на животных проводилось в соответствии с рекомендациями Московского частного медицинского университета "Реавиз" и протоколом доклинических исследований, который был утвержден локальным этическим комитетом этого университета (протокол 02-2018). Три группы реципиентов были использованы для экспериментов. Все реципиенты были GFP-отрицательными мышами линии B10-GFP, которая является гетерозиготной для трансгена GFP. В контрольной группе первоначально было 20 животных. В первой экспериментальной группе было 56 животных, пять из которых были использованы для контроля уровня химеризма и были исключены из статистики смертности, а во второй экспериментальной группе было 50 животных. Начиная с возраста 15 месяцев, когда около 50% мышей оставались живыми во всех группах, экспериментальные животные первой группы получали серию внутривенных инъекций костного мозга от молодых (от 3 до 15 недель) мышей-доноров, которые были GFP-положительными особями той же линии B10-GFP, а животные второй экспериментальной группы получали внутривенные инъекции БМКП, полученного на основе их (аутологичных) ГСК согласно настоящему изобретению. Мыши в возрасте 3-15 недель и гетерозиготные по зеленому флуоресцентному белку-трансгену (т.е. экспрессирующий GFP) той же B10-GFP линии были использованы в качестве доноров костного могзга. Все мыши были приобретены в том же самом питомнике, где эта линия мышей содержалась как маленькое стадо, тем самым была увеличена их инбредность и уменьшена их средняя продолжительность жизни.
Выделение и трансплантация костного могзга проводились в соответствии с процедурой, описанной Colvin et al. (2004) с несущественными модификациями. Животные были умерщвлены вывихом шеи и стерилизовались в течение 3-5 минут в 70% этаноле. Кости всего скелета (череп, позвоночник и бедра) были очищены от мышц и измельчены в стерильной ступке с пестиком (обработаны этанолом и ультрафиолетом) в 5 мл стерильного сбалансированного солевого раствора Хенкса (HBSS) (Life Technologies Gibco BRL) с 10 ед / мл гепарина. Полученную суспензию фильтровали через фильтр 70 мкм (BD Biosciences), затем фильтр промывали 2 мл буфера, и два фильтрата были объединены и центрифугированы в течение 5 минут при 340g. Осадок ресуспендировали в 8 мл HBSS/гепарина и центрифугировали в течение 5 минут при 340g. Шарик ресуспендировали в тот же буферный раствор (общий объем 2-4 мл), и клетки подсчитывали в камере Горяева или с использованием автоматического счетчика клеток. Обычно выход составлял (2-4)⋅108 ядросодержащих клеток ((5-6)⋅107 ядросодержащих клеток на мл) в зависимости от возраста донора. Непосредственно перед инъекцией (в течение 20 минут) клетки дополнительно фильтровали через фильтр 40 мкм (BD Biosciences). Всего (15-20)⋅106 клеток на животное-реципиента медленно, в течение примерно 1 мин, вводилось в объеме 200-300 мкл на животное инсулиновым шприцем (игла 0.33GX 12.7 mm) через хвостовую вену; хвост предварительно прогревался в ванне с водой в течение 2-3 минут при температуре 44°C. У 15 животных клетки костного мозга пересаживались в течение каждого дня трансплантации. Трансплантации повторяли 6 раз в течение 3 месяцев с 10-20-дневные интервалами у экспериментальных животных первой и второй групп. Для контроля уровня химеризма (степени приживления донорских клеток) бала проанализирована доля клеток GFP+ в клетках реципиента. Для этого 200 мкл клеточной суспензии, содержащей 10-20 миллионов клеток на 1 мл, было помещено на предметное стекло, 10 мкл раствора NucBlue (Thermo Scientific), и после инкубации в течение 5 минут клетки покрывали покровным стеклом. Изображения были получены с помощью флуоресцентного микроскопа NICON 2000, на котором было сделано 10-20 фотографий с возбуждением длины волн для NucBlue (Hoechst 33342) и GFP с использованием Cellinsight CX7 (Thermo Scientific), которые равномерно распределяются по всему препарату. Средний процент флуоресцентных клеток определяли автоматически с помощью программного обеспечения. Для анализа флуоресцентных структур в твердой ткани кусок ткани объемом 1×3 мм был раздавлен между предметным и покровным стеклами, и были сделаны фотографии на флуоресцентном микроскопе NICON 2000. Максимальная продолжительность жизни для опытных групп животных (MПЖ exp) была определена как средняя продолжительность жизни 10% большинства долгоживущих мышей. MПЖ для контрольных мышей (MПЖ К) был определен двумя способами. Первое значение MПЖ К1, была прямая оценка средней продолжительности жизни 10% наиболее долгоживущих животных из первой группы мышей (т.е. 2 из 20) и во-вторых, MПЖ К2 был рассчитан с использованием приближения выживания. Кривую выживаемости в экспериментальной группе мышей, умерших от естественных причин до начала трансплантации костного мозга см. на фиг. 1. Приближение было сделано с помощью нелинейной функции приближения из Nonlinear Model Fit программного пакета MATHEMATICA. Доверительный интервал для P = 0,05 рассчитывался стандартным способом с использованием критерия Стьюдента. Два значения (MПЖ C1 и MПЖ C2) полученные таким образом были сопоставлены и было рассчитано полученное значение MПЖ К. Этот метод позволил увеличить точность расчета MПЖ К, так как количество животных было ограничено.
Результаты. На фиг. 1 показано влияние сингенной трансплантации костного мозга молодых доноров (первая экспериментальная группа, незакрашенные круги) и сингенной трансплантации БМКП по настоящему изобретению (вторая экспериментальная группа, черные круги) на продолжительность жизни реципиентов. Контрольная группа (черные квадраты) была использована для непосредственного определения максимальной продолжительности жизни необработанных мышей (MПЖ К1). Возраст смерти последних 10% этой группы (две мыши) было 17,6 и 19,3 месяца, что дает прямая оценка для контрольной группы мышей: MПЖ C1 = 18,45 ± 2,6 месяца. Это значение MПЖ К1 имеет высокое стандартное отклонение из-за довольно небольшого размера контрольной группы. Для того, чтобы повысить точность MПЖ К мы использовали данные о смертности в экспериментальной группе до трансплантации (например, до возраста 15 месяцев), когда смертность была вызвана естественными причинами. Принимая стандартную форму кривой выживания в соответствии с законом Гомперца-Макхема (Makeham, 1860):
f (t) = N0 ⋅ Exp - Exp (μ0 + μ1t) μ1, где
N0 = 100% ⋅ Опыт - Exp (μ0),
μ1, μ1 и μ2 - параметры, позволяющие на их основе минимизировать квадратичное отклонение модельной кривой от естественной гибели животных.
Численные значения параметров были определены следующим образом:
μ0 = -11,4 и μ1 = 0,7.
Это позволяет рассчитать максимальный срок жизни, то есть среднюю продолжительность жизни животных, которые жили с t10 до 1, для мышей в первой экспериментальной группе до лечения (трансплантация), что таким образом служит дополнительным контроль (MПЖ К2):
MПЖ К2 знак равно ∞. R t10 t ⋅ f (t) d t∞ R t 10f (t) dt = 17,2 ± 0,6 месяца,
где t 10 - момент времени, когда 10% животных осталось в популяции.
MПЖ К2 значение находится в хорошем согласии с MПЖ К1, полученным с использованием первого метода оценки MПЖ в контрольной группе. Между двумя методами имеется небольшая разница. Оценка MПЖ нужна из-за меньшей надежности первого метода в условиях, когда случайные отклонения становятся значительными, то есть, если количество мышей мало.
Следовательно, чтобы компенсировать малое число в конце контрольного исследования (две мыши), данный факт можно исправить, усредняя значения, полученные из двух методов определения MПЖ:
MПЖ C = 12 ⋅ MПЖ К1 + 11 ⋅ MПЖ К2( 11 + 12) = 17, 4 ± 0,5 месяца,
где 11 и 12 - стандартные отклонения MПЖ К1 а также MПЖ К2 соответственно.
В возрасте 15 месяцев (начало инъекций донорского костного мозга) выжила 31 мышь первой экспериментальной группы и 26 мышей второй экспериментальной группы. В первой экспериментальной группе 9 мышей умерли во время инъекции на операционном столе (эмболия) при трансплантации КМ молодых мышей. Еще 5 мышей были забиты для отбора проб ткани, а остальные (42 мыши) умерли естественным путем. Во второй экспериментальной группе 5 мышей погибли от эмболии (костный мозг смешивался БМКП по настоящему изобретению) во время трансплантации, 2 мыши были забиты для отбора проб, а остальные мыши (19 мышей) умерли естественным путем. Как видно, кривые летальности животных первой и второй групп практически не отличаются. Немедленная смерть была вызвана у 9 эмболических мышей первой экспериментальной группы из-за неточности исследователя в устранении агрегатов из введенных материала, а также путем индуцированной активации тромбоцитов в кровяном русле. Дополнительный контроль проведен на 15 старых животных аналогичной породы, которые получили в общей сложности 75 инъекций по 300 мкл HBSS-гепарина, и не показал летальности. Таким образом, эмболический риск требует дальнейшего изучения. Возможно, что самые старые особи умерли после инъекций костного мозга. Приняв это соображение во внимание, было решено включить эмболических мышей в статистику анализа для расчета MПЖ. В статистическом анализе эмболические мыши учитывались так, как будто они умерли от естественных причин. Ступенчатые кривые на фиг. 1 отражают динамику смертности с поправкой на эмболических животных. Забитые для отбора проб мыши не учитывались в статистике и при построении графиков на фиг. 1.
Для расчета MПЖ после трансплантации костного мозга использовали максимальный возраст 10% наиболее долгоживущих мышей из экспериментальных групп, что соответствует возрасту 5 последних мышей. Максимальная продолжительность жизни после трансплантации костного мозга 21,0, 22,5, 22,7, 23,8 и 24,2 месяца, что дает MПЖ exp = 22,8 ± 0,8 мес.
Таким образом, неабляционная сингенная траснплантация костного мозга увеличила максимальную продолжительность жизни мышей в первой экспериментальной группе на 100% × (22,8-17,4) /17,4 = 31 ± 5%, а время выживания с начала трансплантация увеличилась в (22,8-15) / (17,4-15) = 3,25 ± 0,3 раза.
Время максимальной продолжительности жизни во второй экспериментальной группе увеличилась на 100% х ( 50 -26) (-11,4 + 0,7) = 25,6 ± 8%, а время выживания с начала трансплантация увеличилась в (22,8-15)/ (25,6-19) = 1,2 ± 0,5 раза.
В возрасте 19,3 месяца, когда последняя мышь контрольной группа умерла сидячей, почти неподвижной (сгорбленный вид, плохие волосы), а мыши экспертиментальных групп были активны, имели ровный позвоночник и блестящую ровную шерсть.
GFP-флуоресценция (зеленая) ГСК в костном мозге молодых мышей-доноров показана на фиг. 2А. GFP-флуоресценция ГСК у старых мышей контрольной группы показана на фиг. 2Б. GFP-флуоресценция реципиентных клеток костного мозга после траснплантации ГСК как нативных (первая экспериментальная группа), так и клеточно-инженерных в составе БМКП по настоящему изобретению практически не отличима (фиг. 2В). В обоих экспериментальных группах через шесть месяцев после трансплантации GFP-флуоресценция была обнаружена у 7% реципиентных клеток костного мозга. Учитывая, что только четверть (25%) клеток-предшественников флуоресцируют в костном мозге гетерозиготного донора (фиг. 2А), степень химерности пересаженного реципиента был (7/25) × 100% = 28%. Таким образом, более четверти реципиентных клеток костного мозга через 6 месяцев после неабляционной трансплантации были донорского происхождения. Следовательно, продление срока жизни должно быть следствием не только паракринного эффекта ГСК, но также и клеточной замены. Флуоресцентные структуры также визуализировались в других тканях реципиентов, таких как печень, селезенка, почки и мышцы через 6 месяцев после последней инъекции.
Перечень сокращений
БАС - боковой амиотрофический склероз
БМКП - биомедицинский клеточный продукт по настоящему изобретению
ГСК - гемопоэтические стволовые клетки
Г-КСФ - гранулоцитарный колониестимулирующий фактор
ГКП - гепомоэтические клетки-предшественники
ГОКИ - гемопоэтически ориентированная клеточно-инженерная (клетка)
ЗНО - злокачественные новообразования
ИКК - иммунокомпетентные клетки
ИПСК (iPSC) - индуцированные плюрипотентные стволовые клетки
ЛК - лейкоконцентрат
МКА - моноклональные антитела
МНК - мононуклеары
МССК - мезенхимальные стромальные стволовые клетки
НСК - нейральные стволовые клетки
ОСБ - опухолеспецифические белки
ОК - опухолевые клетки
ОСК - опухолевые стволовые клетки
ПК - периферическая кровь
РИФ - реакция иммунофлуоресценции
СБУ - специальные базовые условия
СК - стволовые клетки
ТКМ - трансплантация костного мозга
ЭКО - экстракорпоральное оплодотворение
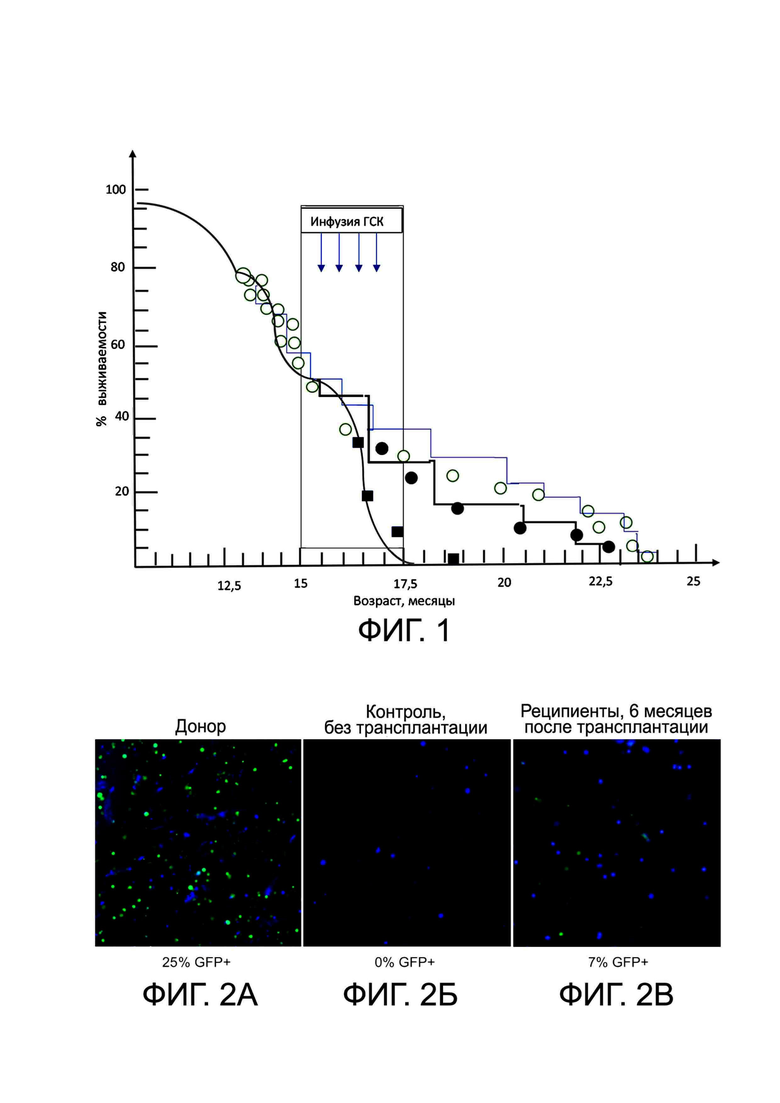
Настоящее изобретение относится к области клеточной биологии и медицины, в частности к способу получения биомедицинского клеточного продукта для лечения онкологических, нейродегенеративных, аутоимунных и эндокринных заболеваний и продукту, получаемому указанным способом. Для осуществления способа сначала получают аутологичный лейкоконцентрат мононуклеаров (ЛК МНК) из костного мозга или из периферической крови. После чего получают линию аутологичных гемопоэтических стволовых клеток (ГСК) или гемопоэтических клеток-предшественников (ГКП) с маркерами CD34+, CD45+, HLA DR+. Далее получают линию неповрежденных заболеванием аутологичных региональных мезенхимальных стволовых клеток (МСК) или соматических клеток и наращивание их клеточной массы. На следующей стадии осуществляют перепрограммирование указанных МСК или соматических клеток в XEN-подобные плюрипотентные клетки. Затем проводят клеточно-инженерную гомотопную замену ядер ГСК и/или ГКП на ядра XEN-подобных клеток или на целые XEN-подобные клетки. На последней стадии осуществляют соединение восстановленных ГСК и/или ГКП с ЛК МНК. Настоящее изобретение позволяет восстанавливать саногенетический регуляторный и регенеративный потенциал поврежденных собственных ГСК и обеспечивать их необходимое и достаточное количество в организме. 2 н. и 5 з.п. ф-лы, 4 ил., 4 табл.
1. Способ получения биомедицинского клеточного продукта для лечения онкологических, нейродегенеративных, аутоимунных и эндокринных заболеваний,
включающий получение аутологичного лейкоконцентрата мононуклеаров (МНК) из эксфузата костного мозга или из лейкоферезного продукта мобилизованных мононуклеаров (ЛК МНК) периферической крови;
получение линии аутологичных гемопоэтических стволовых клеток (ГСК) с маркерами CD34+, CD45+, HLA DR+ и/или гемопоэтических клеток-предшественников (ГКП) с маркерами CD34+, CD45-, HLA DR- путем их выделения из ЛК МНК иммуносепарацией и культивирования в среде, препятствующей дифференцировке;
получение линии неповрежденных заболеванием аутологичных региональных мультипотентных стволовых клеток или соматических клеток и наращивание их клеточной массы;
перепрограммирование указанных аутологичных региональных мультипотентных стволовых клеток или соматических клеток в XEN-подобные плюрипотентные клетки посредством химической индукции;
проведение клеточно-инженерной гомотопной замены ядер ГСК и/или ГКП на ядра XEN-подобных клеток или на целые XEN-подобные клетки путем получения цитопластов из нативных ГСК и/или ГКП посредством обработки этих клеток цитохолазином В и трансплантации кариопластов, полученных из XEN-подобных клеток посредством их обработки цитохолазином В, или целых XEN-подобных клеток в цитопласты ГСК и/или ГКП с получением за счет этого восстановленных ГСК и/или ГКП, содержащих мультипотентные цитопласты ГСК и плюрипотентные кариопласты XEN-подобных клеток;
соединение восстановленных ГСК и/или ГКП с ЛК МНК, из которого были выделены первоначальные ГСК и/или ГКП, для получения целевого биомедицинского клеточного продукта, содержащего линию постгеномно восстановленных мультипотентных генетически однородных аутологичных ГСК с маркерами CD34+Lin-, CD38", HLA-DR+, СD45+.
2. Способ по п. 1, отличающийся тем, что восстановленные ГСК и/или ГКП, соединенные с ЛК МНК, культивируют в среде, препятствующей дифференцировке, для увеличения их клеточной массы, формирования и созревания их свойств мультипотентности в гемопоэтическом направлении и контролируют эти свойства проточной цитофлориметрией.
3. Способ по п. 1, отличающийся тем, что в случае онкологии или нейродегенеративных болезней головного и спинного мозга, а также в случае аутоимунных болезней в качестве неповрежденных заболеванием клеток используют мезенхимальные стромальные стволовые клетки (МССК) с маркерами CD10+, CD13+, CD44+, CD90+ (Thy-1), CD105+, CD34-, CD45- и CD117- и/или мезенхимальные стромальные клетки-предшественники (МСКП) с маркерами CD10+, CD13+, CD44-, CD90+ (Thy-1), CD105+, CD34+, CD45+ и CD117+.
4. Способ по п. 1, отличающийся тем, что в случае онкологии солидных органов в качестве неповрежденных заболеванием клеток используют нейральные стволовые клетки (НСК) с маркером СD 133+ и/или нейральные клетки-предшественники (НКП) с маркером СD 133+.
5. Способ по п. 1, отличающийся тем, что перепрограммирование посредством химической индукции проводят коктейлем, содержащим низкомолекулярные вещества, выбранные из группы, состоящей из VPA, TD114-2, CHIR, 616452, транилципромина, формсколина, AM580, EPZ004777, CH55 и витамина C (VC).
6. Способ по п. 1, отличающийся тем, что трансплантацию полученных из XEN-подобных клеток кариопластов или целых XEN-подобных клеток в цитопласты ГСК и/или ГКП проводят in vitro путем электропорации или лазерно-оптическим методом.
7. Биомедицинский клеточный продукт для лечения онкологических, нейродегенеративных, аутоимунных и эндокринных заболеваний, полученный способом по одному из пп. 1-6 и содержащий аутологичный лейкоконцентрат мобилизованных мононуклеаров, обедненный собственными ГСК и/или ГКП в результате их иммуносепарации, с линией генетически однородных аутологичных гемопоэтических клеток с маркерами CD34+Lin–, CD38", HLA-DR+СD45+.
| Биомедицинский клеточный препарат | 2017 |
|
RU2647429C1 |
| CICUTTINI F.M., et al "Characterization of CD34+HLA-DR-CD38+ and CD34+HLA-DR-CD38- progenitor cells from human umbilical cord blood", Growth Factors, 1994, 10(2), 127-134; DOI: 10.3109/08977199409010986 | |||
| История клинической и фундаментальной нейрофизиологии в России: вклад в европейскую нейронауку (под ред | |||
| К.А | |||

