Изобретение относится к области биотехнологии, биоинженерии, биофармакологии, может быть использовано для конструирования и препаративной (непромышленной) наработки рекомбинантных полностью человеческих антител, специфичных в отношении антигена интереса.
Антитела способны специфично связываться с чужеродными молекулами и инфекционными агентами и нейтрализовать их. Поэтому медицинские препараты с доказанной клинической эффективностью, включающие мишень-специфичные моноклональные антитела как действующее вещество или как компонент терапевтической композиции, уже активно выпускаются, а данное направление успешно развивается последние два десятилетия. Наиболее перспективными для терапевтического применения являются полностью человеческие моноклональные антитела. Они характеризуются меньшей иммуногенностью по сравнению с гуманизированными и химерными моноклональными антителами. К первой половине 2020 года Управлением по санитарному надзору за качеством пищевых продуктов и медикаментов США (FDA) в совокупности с Европейским агентством лекарственных средств (EMA) одобрено около 40 полностью человеческих терапевтических антител в качестве терапевтических препаратов. Большинство из них сертифицировано за последние 5 лет.
Наиболее безопасными с точки зрения иммуногенности считаются нативно спаренные антитела, сформированные в живом организме (in vivo). Их фрагменты закодированы в геноме клеток зародышевой линии человека, поэтому будут минимально восприняты организмом как чужеродные молекулы. Существует несколько основных способов получения человеческих антител с нативно спаренными вариабельными домена тяжелой и легкой цепи (VH и VL). Один из способов предполагает работу с единичными иммуноглобулин-продуцирующими клетками. Как правило, это клетки периферической крови: плазмобласты или В-лимфоциты памяти с переключенным классом иммуноглобулина. Однако также можно использовать плазматические клетки (лимфоцитарные плазмоциты) из вторичных лимфоидных органов глоточного лимфатического кольца. Из изолированных по одиночке с помощью микроманипулятора или флюоресцетного сортинга (FACS) клеток амплифицируют кДНК, кодирующую вариабельные домены цепей иммуноглобулина, а затем, при необходимости добавляя недостающие (не амплифицированные) константные участки с помощью генной инженерии, получают гены полноразмерного рекомбинантного антитела в плазмидных векторах для эукариотической экспрессии.
Единичные первичные клетки периферической крови содержат крайне малое количество РНК: 10-30 пкг фракции тотальной РНК, среди которой доля мРНК не превышает 5%. Различные транскрипты легкой и тяжелой цепи иммуноглобулинов не имеют постоянного нуклеотидного состава в 5′-области, поскольку вариабельный домен кодируется набором генных сегментов V, D (только для тяжелой цепи), J, которые в процессе формирования домена подвергаются V(D)J рекомбинации. Каждый V-сегмент содержит три функциональные части: промотор и два экзона, которые кодируют сигнальную последовательность и N-концевую часть вариабельного домена. На данный момент известны, согласно базе IMGT (International ImMunoGeneTics information system), 55 функциональных генных сегментов IGHV для формирования полинуклеотида VH домена, 41 функциональный сегмент IGKV и 33 функциональный сегмент IGLV для формирования полинуклеотида VL домена (не считая аллельных вариантов сегментов). Среди V-сегментов одной группы встречается нуклеотидная гомология различной степени. За счет этого, а также использования по нескольку вырожденных нуклеотидов в процессе дизайна 5′ праймеров, можно существенно сократить количество праймеров для гибридизации с началом кодирующих последовательностей вариабельного домена или сигнальной последовательности. Нуклеотидное разнообразие терминальных участков J-сегментов, которые соответствуют концу вариабельных доменов VH и VL, представлено в меньшей степени, поэтому может быть охвачено суммарно 9-13 3′ праймерами для все возможных вариантов кДНК тяжелой цепи и легкой цепи обоих типов: κ и λ. Поскольку за вариабельным доменом следует константный, который кодируется относительно консервативной нуклеотидной последовательностью в рамках каждого типа тяжелой цепи (α, γ, ε, μ) и каждого типа легкой цепи (κ и λ), то количество 3′ праймеров можно сократить до количества этих типов, например, ограничится четырьмя для рутинных исследований: к тяжелой цепи γ и μ, легкой цепи κ и λ. В настоящее время для амплификации кДНК (комплементарной ДНК) вариабельной части иммуноглобулина от единичных клеток проведение двухэтапной (вложенной) ПЦР считается неотъемлемой операцией в протоколах по созданию рекомбинантных нативно спаренных человеческих антител. Однако способы исполнения вложенной ПЦР, описанные в литературе, отличаются друг от друга в той или иной степени, в частности, особенностями отбора единичных клеток, протоколом обратной транскрипции, праймерами, режимами и реактивами, а также векторами для встраивания полученных ПЦР-продуктов для дальнейшей экспрессии антител. Почти все известные способы исключают специальный этап выделения РНК, заменяя его прямым лизисом клетки в буфере, который затем в полном объеме задействуют в реакции обратной транскрипции и амплификации кДНК. Благодаря этому сокращается время и трудозатраты, повышается воспроизводимость результатов, РНК в меньшей степени подвергается риску деградации и контаминации. Существующие способы амплификации кДНК человеческого иммуноглобулина с нативно спаренными цепями рассмотрены ниже. Они либо напрямую нацелены на получение рекомбинантных антител, либо антитела можно создать post factum при помощи генной инженерии после получения результатов секвенирования нуклеотидных последовательностей VH и VL. Ни один из способов не является полноценным прототипом предлагаемого способа. Основные способы амплификации представлены ниже.
Первый рациональный протокол амплификации Fab-фрагмента иммуноглобулина G с κ и λ изотипом легкой цепи из единичных иммуноглобулин-производящих B-лимфоцтов и плазматических клеток представлен у (Coronella J. A. et al. Amplification of IgG VH and VL (Fab) from single human plasma cells and B cells //Nucleic acids research. – 2000. – Т. 28. – №. 20. – С. e85-e85.). Описан сортинг клеток непосредственно в реакционный буфер, обратная транскрипция с затравкой синтеза первой цепи при помощи олиго(dT)16, затем полувложенная ПЦР, где и в первом, и во втором этапе участвуют одни и те же 5′ праймеры по (Sblattero D., Bradbury A. A definitive set of oligonucleotide primers for amplifying human V regions //Immunotechnology. – 1998. – Т. 3. – №. 4. – С. 271-278.), соответствующие началу FR1 в V-сегментах. Праймеры 3′ для первой и второй ПЦР комплементарны последовательностям в константных регионах и заимствованы у (Burton D. R., Barbas III C. F. Human antibodies from combinatorial libraries //Advances in immunology. – Academic Press, 1994. – Т. 57. – С. 191-280.). Для амплификации VH и VL доменов IgG используют суммарно 29 олигонуклеотидов. В работе не стоит задача получения рекомбинантных полинуклетидов полноразмерных цепей с целью дальнейшей экспрессии антител. Несмотря на минимальное количество олигонуклеотидов и относительно высокий заявленный процент успеха одновременной амплификации VH и VL от единичной клетки (68% и 50% для плазматических и В-лимфоцитов, соответственно), публикаций с воспроизведением данного протокола немного.
В настоящее время для получения человеческих рекомбинантных антител из единичных клеток часто используют протокол, описанный в (Smith K. et al. Rapid generation of fully human monoclonal antibodies specific to a vaccinating antigen //Nature protocols. – 2009. – Т. 4. – №. 3. – С. 372.). Кратко, в данной работе человеческие плазмобласты сортируют в бессолевой 10 мМ Трис-буфер (рН 8,0), свободный от нуклеаз, и содержащий ингибитор РНКаз RNasin. Лизис клеток и выход мРНК в реакционную смесь происходит вследствие гипотонического свойства буфера для сортировки клеток с последующей однократной заморозкой-оттаиванием клеток в данном буфере. Далее с лизатом единичной клетки в качестве источника мРНК проводят реакцию обратной транскрипции с последующей первой мультиплексной ПЦР в той же пробирке с использованием набора OneStep RT-PCR Kit (Qiagen). В реакцию добавляют смесь из генспецифичных праймеров к константным областям тяжелой цепи типа γ и μ и легкой цепи κ и λ изотипов для затравки синтеза первой цепи кДНК, а также смесь внешних 5′ и 3′ праймеров для проведения первой ПЦР. Праймеры 5′ предназначены для спаривания с участком сигнальной последовательности (справедливо для группы 5′ L Vκ), либо с местом сочленения сигнальной последовательности и FR1 (справедливо для группы праймеров 5′ L-VH и 5′ L Vλ). Праймеры группы 3′ комплементарны кДНК соответствующих константных доменов. По завершении первой ПЦР выполняют вторую ПЦР, которую, в свою очередь, осуществляют в отдельных пробирках для каждого типа тяжелой цепи и легкой цепи. В ней задействуют праймеры, отжигающиеся внутри полученных ПЦР-продуктов и содержащие некомплементарные свесы (оверхенги) с сайтами рестрикции. Полученные продукты амплификации клонируют в вектора с открытой рамкой считывания и недостающими полинуклеотидами фрагментов легкой и тяжелой цепи для экспрессии антитела класса IgG1 в эукариотической системе. Суммарно, в процессе превращения мРНК единичной клетки в рекомбинантные ДНК, кодирующие легкую и тяжелую цепь человеческого иммуноглобулина, может быть задействовано от 50 до 52 олигонуклеотидов (до 1660 нуклеотидов).
Более поздняя публикация (Rudkin F. M. et al. Single human B cell-derived monoclonal anti-Candida antibodies enhance phagocytosis and protect against disseminated candidiasis //Nature communications. – 2018. – Т. 9. – №. 1. – С. 1-16.) описывает иной протокол получения рекомбинантных антител от единичных клеток. В частности, для синтеза первой цепи кДНК используют олиго(dT)20. Далее следует первый этап вложенной ПЦР, где используют 5′ праймеры, аналогичные (Smith K. et al. Rapid generation … С. 372.), а 3′ праймеры заменяют на авторские: два олигонуклеотида к кодирующей области CH1 домена тяжелой цепи γ и по три праймера к терминальным областям кДНК легких цепей каждого типа: участку нескольких последних кодонов константного домена и двум участкам, синтезированным на матрице 3’-нетранслируемого региона мРНК. Использование нескольких 3′ праймеров, вероятно, применено для усиления амплификации. Перечень праймеров для второго этапа ПЦР существенно расширен и доработан по сравнению с (Smith K. et al. Rapid generation … С. 372.) для охвата большего количества аллелей V-сегментов. В результате легкие цепи амплифицируют полноразмерными, а внутри тяжелой цепи амплифицируют только домен VH. Дополнительной особенностью данного протокола является отказ от использования рестриктаз в пользу In-Fusion клонирования, что влечет за собой дополнение каждого праймера большим оверхенгом по 14 нуклеотидов. Суммарно, в методе задействуют 104 праймера (4119 нуклеотидов).
Альтернативным способом можно считать работу (Ozawa T., Kishi H., Muraguchi A. Amplification and analysis of cDNA generated from a single cell by 5′-RACE: application to isolation of antibody heavy and light chain variable gene sequences from single B cells //Biotechniques. – 2006. – Т. 40. – №. 4. – С. 469-478.). Авторами разработана стратегия амплификации кДНК иммуноглобулинов с тяжелой цепью γ и μ и легкой цепью κ и λ от единичных В-клеток с помощью 5′-RACE метода. Она включает затравку синтеза первых цепей кДНК четырьмя генспецифичными праймерами в реакции обратной транскрипции, и последующее добавление адапторной последовательности на 5′-концы синтезированных кДНК через наращивание гомополимерной нуклеотидной последовательности у 3′-OH конца первой цепи кДНК ферментом терминальной дезоксинуклеотилитрансферазой. С этой гомополимерной нуклеоитдной последовательностью, в свою очередь, гибридизуется праймер с комплементарной гомополимерной последовательностью и адапторной последовательностью нуклеотидов. Эта адапторная последовательность в дальнейшем служит местом последовательного отжига двух 5′ праймеров: внутреннего и внешнего, – задействованных в первой и второй стадии вложенной ПЦР. Также задействуют по два 3′ праймера (аналогично, – один внешний, второй внутренний) к каждому константному участку легкой цепи κ и λ и тяжелой цепи γ и μ. Таким образом, при полном отказе от праймеров, комплементарных V и J генным сегментам, удается амплифицировать пару VH/VL от более 50% В-клеток.
В работе (Rajan S. et al. Recombinant human B cell repertoires enable screening for rare, specific, and natively paired antibodies //Communications biology. – 2018. – Т. 1. – №. 1. – С. 1-8.) представлен способ генерации фаговых библиотек нативно спаренных scFv из человеческих В-лимфоцитов. Авторами сконструировано 92 праймера для гибридизации со всеми известными человеческими генными сегментами V и J: суммарно 542 аллели на момент публикации, полученные на основе консенсусных последовательностей из базы IMGT. Для способа, предлагаемого в настоящей заявке, разработан перечень праймеров к V-сегментам, сокращенный и рационализированный для удешевления синтеза на основе комплементарных частей внутренних 5′ праймеров групп VH_in_5, VK_in_5, VL_in_5 из данной работы.
Недостатками существующих способов амплификации полинуклеотидов пары нативно спаренных VH и VL можно считать:
А. большое количество синтетических праймеров и суммарную длину по нуклеотидам;
Б. тот факт, что некоторые способы не предназначены для последующей быстрой сборки рекомбинантных ДНК, кодирующих полноразмерные цепи человеческого иммуноглобулина;
В. тот факт, что для некоторых способов эффективность успешной одновременной амплификации кДНК и VH, и VL доменов составляет не более 70%.
Технический результат предполагаемого изобретения заключается в том, что предложен способ амплификации нуклеиновых кислот доменов VH и VL единичных иммуноглобулин-продуцирующих клеток человека, полученных из биологического материала доноров, позволяющий снизить вероятность ложноотрицательной ампилфикации, повышая вероятность успешной амплификации кДНК антител с редко встречающимися аллелями V генных сегментов. В дальнейшем рекомбинантное антитело, созданное в результете химеризации кодирующих последовательностей констатных доменов человеческого иммуноглобулина с амплифицированными полинуклеотидами, может быть протестировано и утверждено в качестве терапевтического агента.
Технический результат достигается использованием модифицированной технологии SMART (switching mechanism at the 5′ end of the RNA transcript) для синтеза кДНК и избирательного обогащения целевых низкокопийных матриц для следующей стадии ПЦР. Модификация заключается в замене заякоренного олиго(dT)30 праймера с адапторной нуклеотидной последовательностью на композицию из пяти генспецифичных праймеров к консервативным участкам цепей иммуноглобулина всех возможных типов. Адапторная последовательность остается идентичной таковой в олиго(dT). Результатом ОТ-ПЦР (ПЦР с обратной транскрипцией) является смесь из множества копий двух кДНК наряду с неспецифическими продуктами реакции. Одна кДНК кодирует вариабельный домен и часть константного домена тяжелой цепи типа α, γ, ε или μ; вторая кодирует вариабельный домен и часть константного домена тяжелой цепи типа κ или λ. При этом тип цепей после первой ПЦР неизвестен и не устанавливается, поскольку не имеет значения.
Также технический результат достигается тем, что предложен набор праймеров для составления трех композиций, участвующих в трех параллельных ПЦР второго этапа. Аликвота первой ПЦР, проведенной благодаря синтезу кДНК по технологии SMART, служит матрицей в трех последующих раздельных ПЦР с праймерами для амплификации регионов VH, Vκ или Vλ. Для каждой ПЦР второго этапа разработаны композиции 5′ и 3′ праймеров. Данные праймеры не имеют некомплементарных свесов (оверхэнгов) на 5′-концах для привнесения сайта рестрикции, однако каждый праймер группы 3′, гибридизующийся с консервативным участком, содержит сайт рестрикции с образованием липкого конца «внутри себя», без нарушения первичной аминокислотной последовательности цепи иммуноглобулина. В свою очередь, 5′ праймеры разработаны для гибридизации со всеми возможными аллельными вариантами V генных сегментов IGHV, IGKV, IGLV иммуноглобулина человека. После проведения второй ПЦР ампликон домена VH и результативный ампликон домена VL очищают и клонируют в векторы, которые при коэкспресии в клетках эукариот формируют полноценную молекулу IgG1.
Также технический результат достигается тем, что предложена изоляция единичных иммуноглобулин-продуцирующих клеток из биологического материала человека в процессе цитофлюориметрического сортинга (FACS) по признаку специфичного взаимодействия с антигеном. Таким образом, описанный способ направлен на получение специфичных антител к антигену интереса. Клетки сортируют непосредственно в растворы, способствующие скорому лизису in situ и не ингибирующие реакцию обратной транскрипции и амплификации.
Отичие предлагаемого способа в том, что для амплификации предложено использовать 32 или 34 синтетических праймера. Суммарная длина синтезируемых праймеров составляет 762 или 815 нуклеотидов – в зависимости от того, используются ли коммерческие наборы, где уже содержится 2 универсальных олигонуклеотида. Также предложены праймеры для амплификации VH от клеток, производящих антитела с тяжелой цепью типа α и ε.
Заявляемый способ предусматривает двухэтапную амплификацию кДНК вариабельных доменов VH и VL иммуноглобулина по принципу вложенной ПЦР, то есть в два этапа. Первую ПЦР выполняют для мультиплексной амплификации двух из шести потенциально возможных вариантов ПЦР-продуктов в одной пробирке: иммуноглобулин-продуцирующая клетка (плазматическая и В-клетка памяти с переключенным классом) может транскрибировать один из четырех типов тяжелой цепи и один из двух типов легкой цепи. Данную ПЦР удается осуществить благодаря обратной транскрипции по технологии SMART. Протокол технологии был разработан и зарегистрирован компанией Clontech Laboratories (Zhu Y. Y. et al. Construction of cDNA libraries from small quantities of total RNA using template switching catalyzed by M-MLV reverse transcriptase //Genetic Library Construction and Screening. – Springer, Berlin, Heidelberg, 2002. – С. 69-93. Zhu Y. Y. et al. Reverse transcriptase template switching: A SMART™ approach for full-length cDNA library construction //Biotechniques. – 2001. – Т. 30. – №. 4. – С. 892-897.). Технология предназначена для создания библиотек полноразмерных кДНК из клеток эукариот с целью дальнейшего NGS-секвенирования. В технологии SMART первая цепь кДНК синтезируется с использованием ревертаз семейства MMLV, которые по окончании синтеза проявляют активность терминальной трансферазы, добавляя нематрично на 3′-конец одноцепочечной кДНК несколько дезоксирибонуклеотидов, как правило, 3-4 нуклеотида dC. Стандартным праймером для затравки синтеза первой цепи является заякоренный олиго(dT)30VN с олигонуклеотидным оверхенгом из 20-30 нуклеотидов на 3′-конце – адапторной последовательностью. Второй олигонуклеотид, необходимый для SMART, представляет собой ту же адапторную последовательность с 3-4 рибонуклеотидными остатками rG на 3′-конце и имеет общепринятое название Template Switching Oligo (TSO). Нематрично достроенный олиго(dC) служит местом гибридизации олиго(rG)-участка олигонуклеотида TSO. Затем в присутствии ионов Mn2+ ревертаза воспринимает TSO как продолжение РНК-матрицы, синтезируя комплементарную цепь в продолжение 3′-OH конца кДНК. Таким образом, первая цепь кДНК получается фланкированной с обоих концов универсальной нуклеотидной последовательностью, комплементарной ПЦР-праймеру на 3′-конце, а на 5′-конце соответствующей ПЦР-праймеру в ориентации 3′-5′. ПЦР-праймер – третий необходимый олигонуклеотид для синтеза второй цепи и амплификации кДНК. Он выступает единственным, и прямым, и обратным, праймером по отношению к двуцепочечной кДНК.
Графические материалы, иллюстрирующие изобретение:
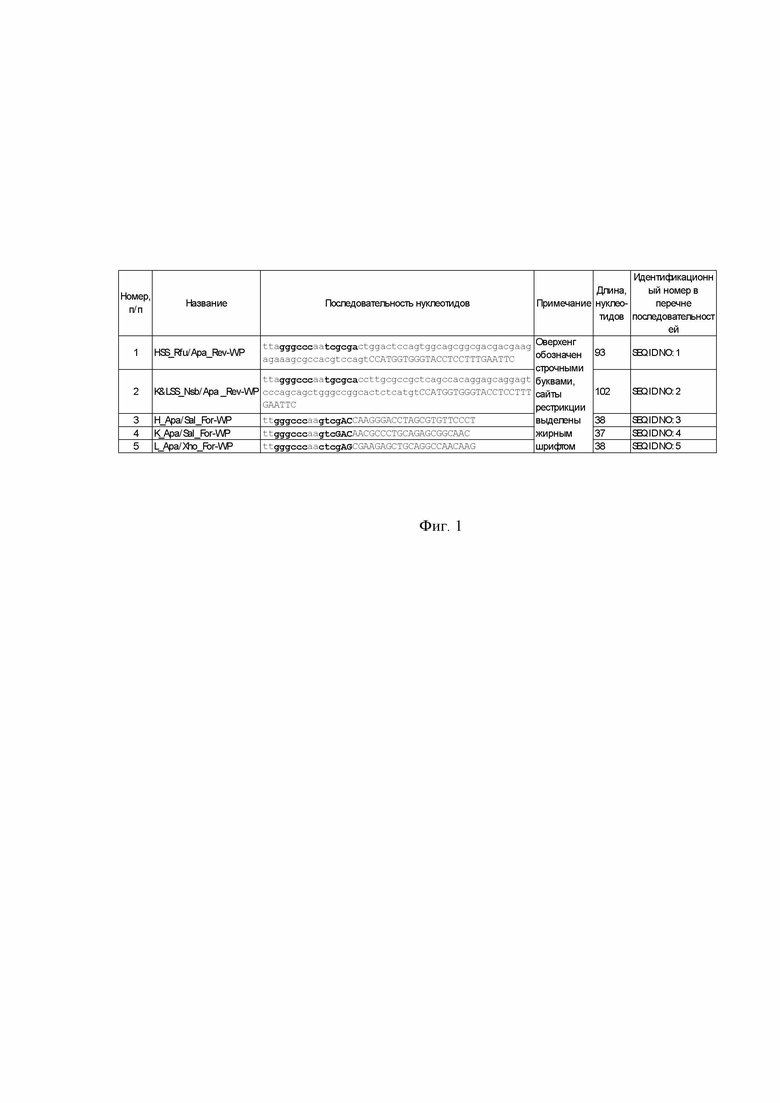
Фиг. 1. Перечень синтезируемых праймеров для внесения изменений в плазмиды из набора Human IgG Vector Set.
Фиг. 2. Перечень синтезируемых праймеров для амплификации кДНК, содержащих нуклеотидные последовательности вариабельных участков человеческого иммуноглобулина.
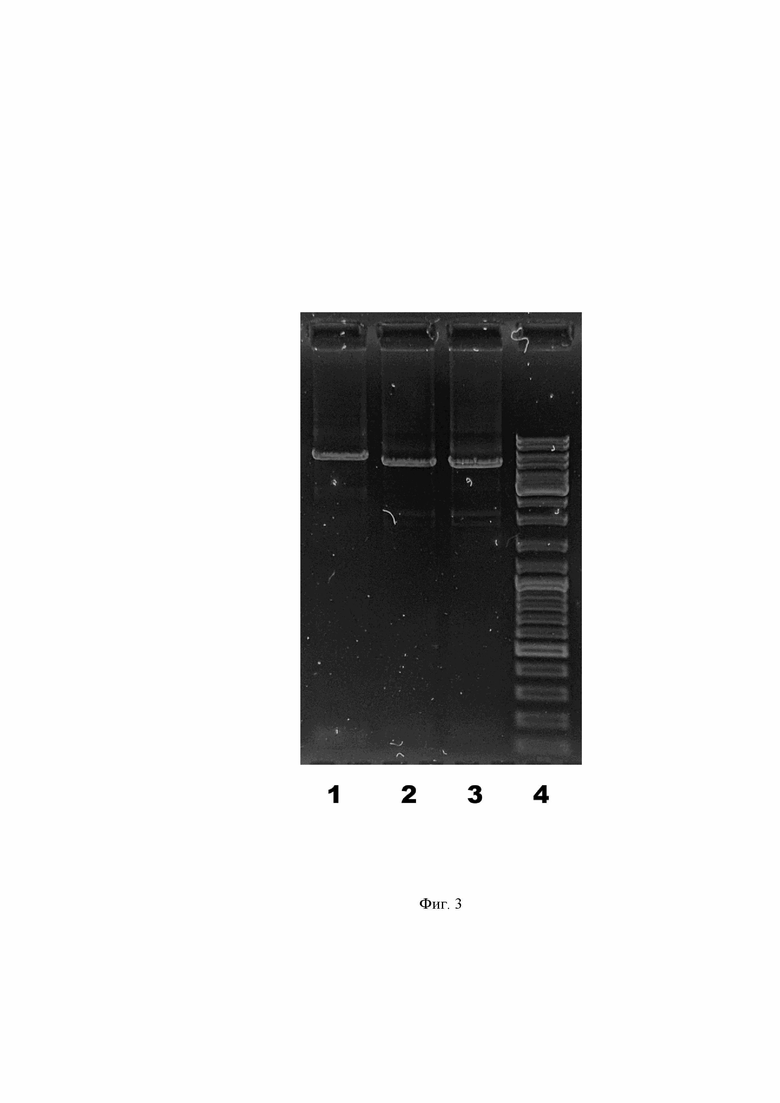
Фиг. 3. Графическое представление результата амплификации плазмид с целью внесения модификаций в виде электрофоретического разделения ПЦР-продуктов в 1% агарозном геле.
Дорожка 1 – pSF-CMV-HuIgG1_HCmod. Дорожка 2 – pSF-CMV-HuKappa_LCmod. Дорожка 3 – pSF-CMV-HuLambda_LCmod. Дорожка 4 – Маркер молекулярной массы ДНК GeneRuler DNA Ladder Mix (Thermo Fisher).
Фиг. 4. Горизонтальные генетические карты модифицированных плазмидных векторов pSF-CMV-HuIgG1_HCmod, pSF-CMV-HuKappa_LCmod и pSF-CMV-HuLambda_LCmod.
Фиг. 5. Гибридизация генспецифичного праймера 3' HuIgSMART-G&E с мРНК тяжелой цепи IgE (Homo sapiens immunoglobulin heavy chain V-D-J-Ce region mRNA, partial sequence, GenBank: DQ680069.1). Графическое представление, выполненное по алгоритму выравнивания нуклеотидных последовательностей BLAST.
Фиг. 6. Выравнивание праймеров 3′ Cλ_XhoI и 3' XhoI Cλ по (Smith K. et al., 2009).
Фиг. 7. Стратегия многоэтапного гейтирования субпопуляции В-клеток памяти с применением панели CD19 APC/CD38 PE-Cy7/CD27 BB515/IgG BB700/ антиген «А» РЕ.
Фиг. 8. Графическое представление результата амплификации кДНК цепей иммуноглобулинов от единичных В-клеток памяти после электрофоретического разделения ПЦР-продуктов в 1,5% агарозном геле.
Дорожка 1 – негативный результат амплификации тяжелой цепи из пустого слота A1. Дорожка 2 – негативный результат амплификации легкой цепи изотипа κ клетки из пустого слота A1. Дорожка 3 – негативный результат амплификации легкой цепи изотипа λ из пустого слота A1. Дорожка 4 – кДНК тяжелой цепи иммуноглобулина от клетки из слота A2. Дорожка 5 – кДНК легкой цепи иммуноглобулина изотипа κ от клетки из слота A2. Дорожка 6 – негативный результат амплификации легкой цепи изотипа λ от клетки из слота A2. Дорожка 7 – кДНК тяжелой цепи иммуноглобулина от клетки из слота A3. Дорожка 8 – негативный результат амплификации легкой цепи изотипа κ от клетки из слота A3. Дорожка 9 – кДНК легкой цепи иммуноглобулина изотипа λ от клетки из слота A3. Дорожка 13 – Маркер молекулярной массы ДНК GeneRuler DNA Ladder Mix (Thermo Fisher).
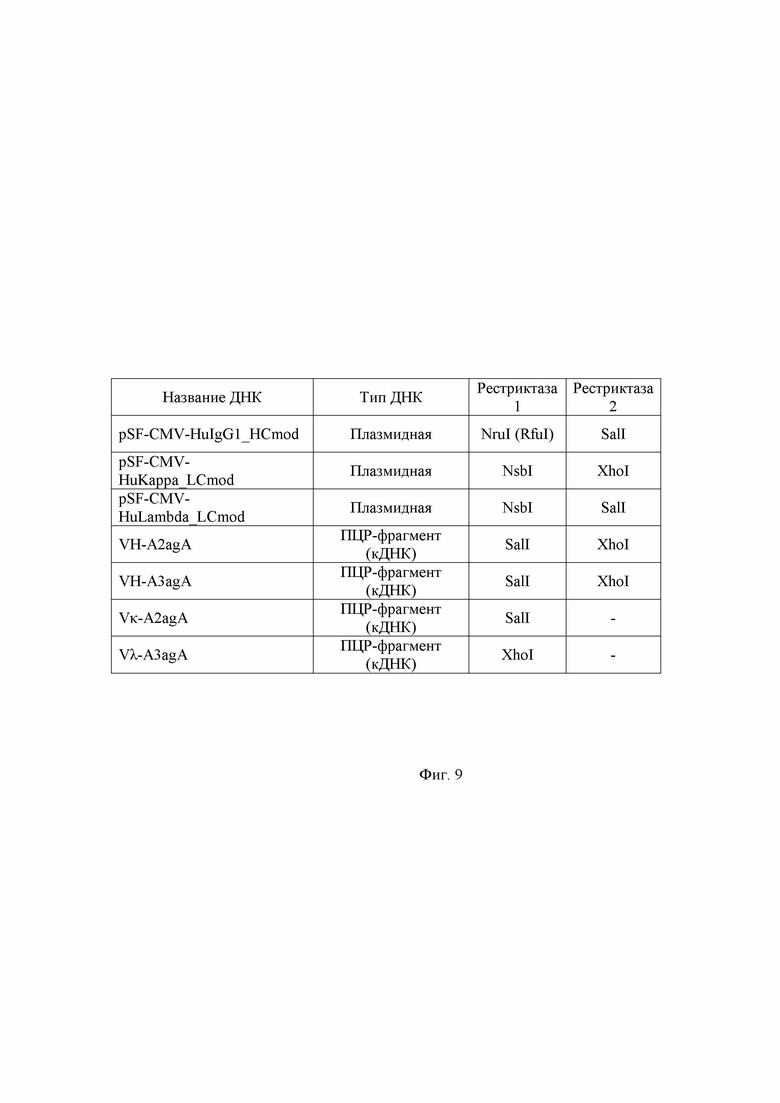
Фиг. 9. Обработка плазмид и вставок (кДНК) различными эндонуклеазами рестрикции перед лигированием: наглядное представление в виде таблицы.
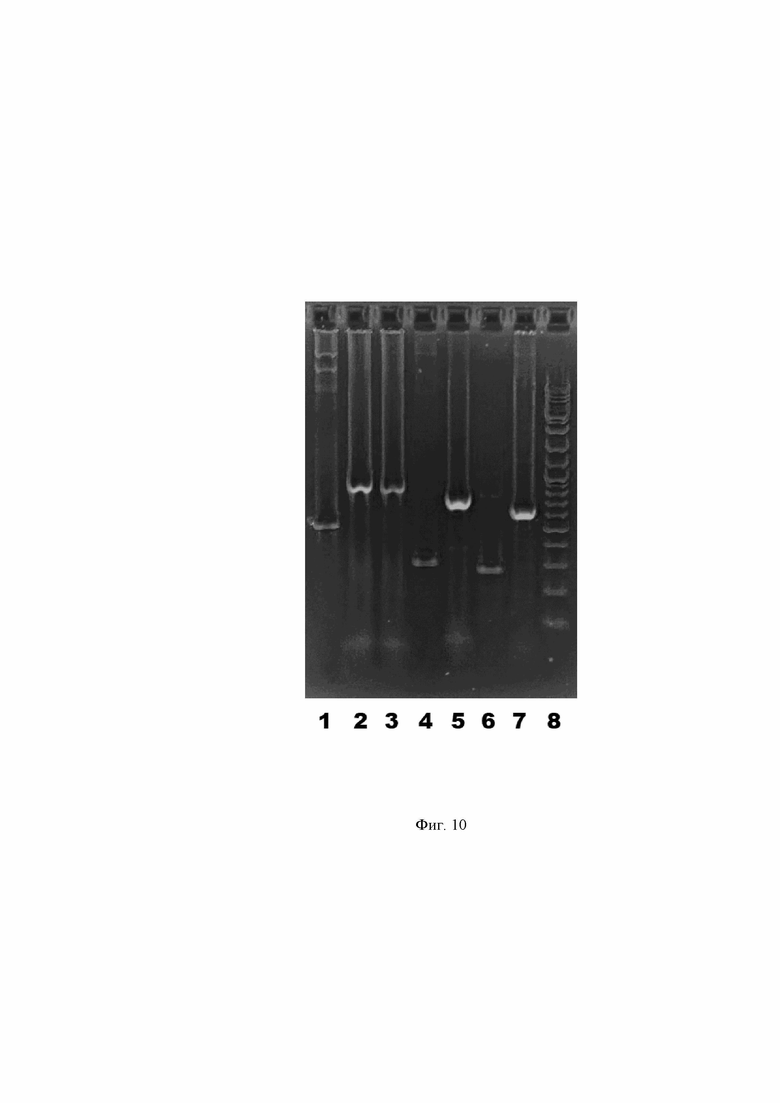
Фиг. 10. Графическое представление результата скрининга колоний E. coli на наличие вставки в плазмиду после электрофоретического разделения ПЦР-продуктов в 1,5% агарозном геле.
Дорожка 1 – продукт с плазмиды pSF-CMV-HuIgG1_HCmod без вставки (контроль, 519 п.о.). Дорожка 2 – продукт с колонии E. coli, трансформированной плазмидой pSF-CMV-HuIgG1_HCmod с клонированной кДНК VH-A2agA (883 п.о.). Дорожка 3 – продукт с колонии E. coli, трансформированной плазмидой pSF-CMV-HuIgG1_HCmod с клонированной кДНК VH-A3agA (853 п.о.). Дорожка 4 – продукт с плазмиды CMV-HuKappa_LCmod без вставки (контроль, 309 п.о.). Дорожка 5 – продукт с колонии E. coli, трансформированной плазмидой pSF-CMV-HuKappa_LCmod с клонированной кДНК Vκ-A2agA (746 п.о.). Дорожка 6 – продукт с плазмиды pSF-CMV-HuLambda_LCmod без вставки (контроль, 281 п.о.). Дорожка 7 – продукт с колонии E. coli, трансформированной плазмидой pSF-CMV-HuLambda_LCmod с клонированной кДНК Vλ-A3agA (639 п.о.). Дорожка 8 – Маркер молекулярной массы ДНК GeneRuler DNA Ladder Mix (Thermo Fisher).
Фиг. 11. Анализ отсеквенированного участка pSF-HC-A2agA инструментом IMGT/V-QUEST.
Фиг. 12. Анализ отсеквенированного участка pSF-HC-A3agA инструментом IMGT/V-QUEST.
Фиг. 13. Анализ отсеквенированного участка pSF-LC-A2agA инструментом IMGT/V-QUEST.
Фиг. 14. Анализ отсеквенированного участка pSF-LC-A2agA инструментом IMGT/V-QUEST.
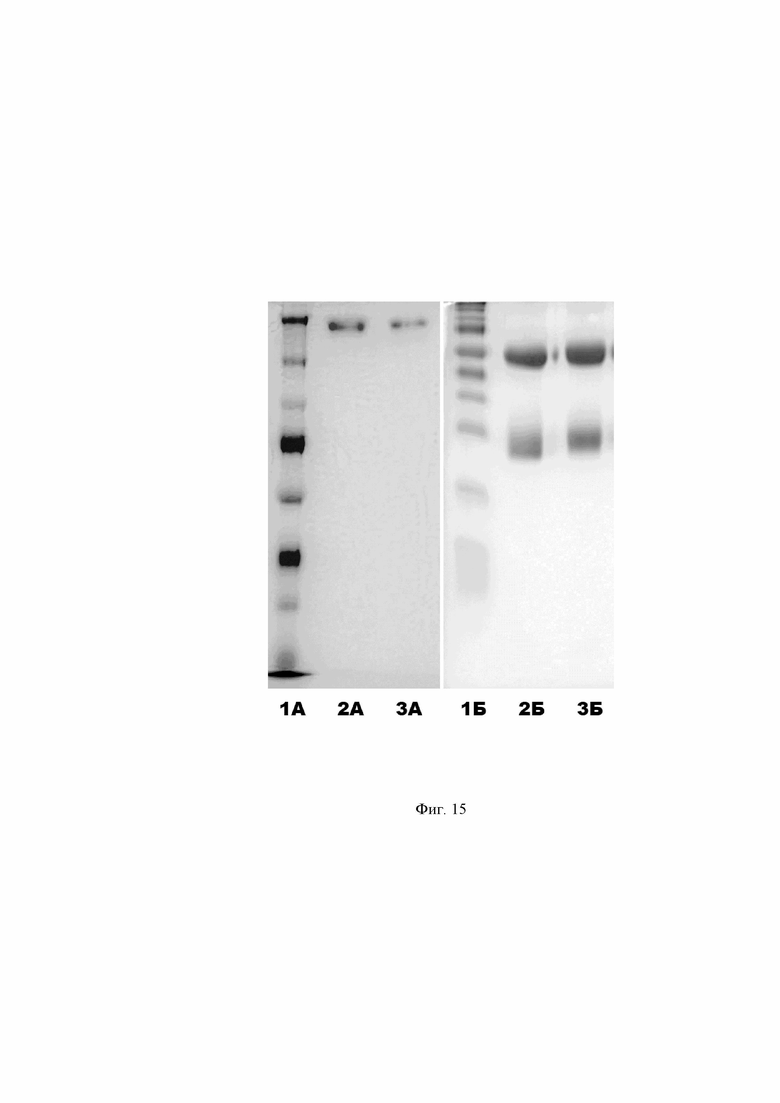
Фиг. 15. Графическое представление результата очистки рекомбинантных человеческих антител A2agA и A3agA из культуральной жидкости после электрофоретического разделения в ПААГ. Часть А – разделение в 7,5% ПААГ в нередуцирующих условиях. Часть Б – разделение в 12% ПААГ в редуцирующих условиях. Окраска гелей кумасси бриллиантовым синим R-250.
Дорожка 1А, 1Б – маркер молекулярной массы белков Spectra Multicolor Broad Range Protein Ladder (Thermo Fisher). Дорожка 2А – образец очищенного антитела A2agA. Дорожка 3А – образец очищенного антитела A3agA. Дорожка 2Б – образец очищенного антитела A2agA с добавлением 2-меркаптоэтанола. Дорожка 3Б – образец очищенного антитела A3agA с добавлением 2-меркаптоэтанола.
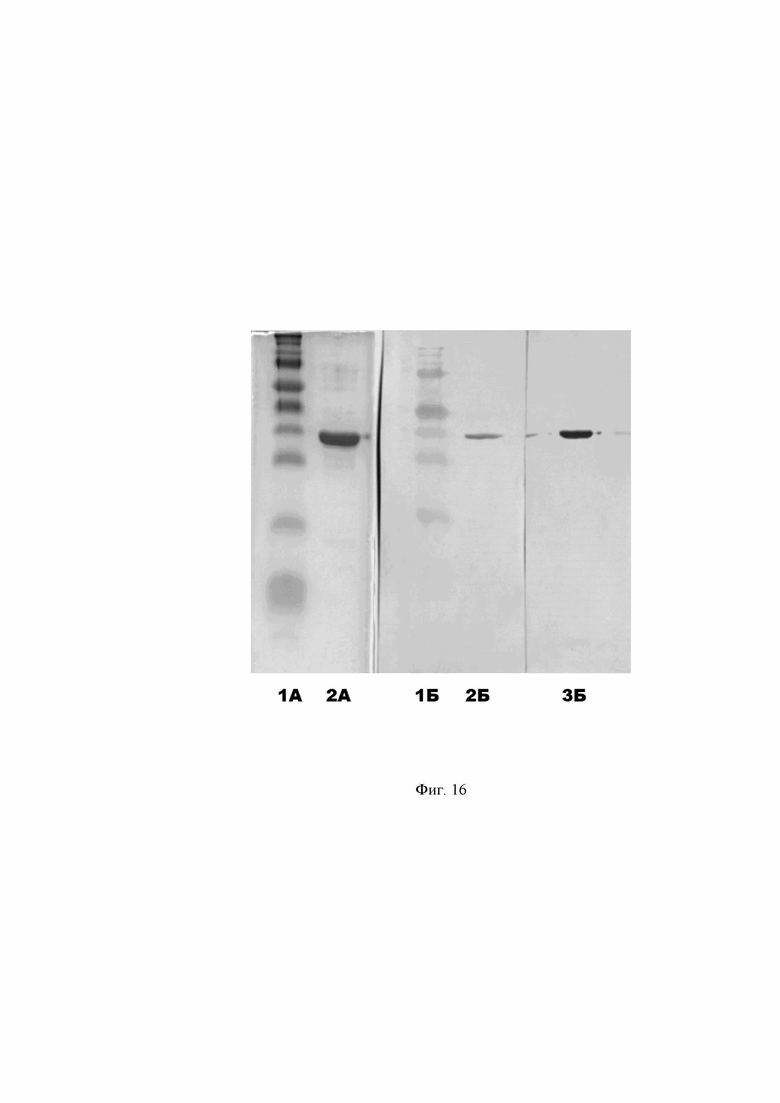
Фиг. 16. Электрофоретическое разделение антигена «А» в 12%-ом ПААГ. Часть А – окраска геля кумасси бриллиантовым синим R-250. Часть Б – результат иммуноблота по связыванию человеческих рекомбинантных человеческих антител A2agA и A3agA с антигеном «А».
Дорожки 1А, 1Б – маркер молекулярной массы белков Spectra Multicolor Broad Range Protein Ladder (Thermo Fisher). Дорожка 2А – антиген «А». Дорожка 2Б – связывание антитела A2agA антигеном «А». Дорожка 3Б – связывание антитела A3agA антигеном «А».
Сущность изобретения.
Для практического исполнения предложенного способа предлагается использование набора SMART-Seq HT Kit (TaKaRa Bio), который позволяет выполнить реакцию обратной транскрипции и ПЦР в одной пробирке (ОТ-ПЦР) с минимального стартового количества РНК 10 пкг, получаемого прямым лизисом клетки без этапа выделения. В данном наборе в качестве олиго(dT) с адапторной последовательностью содержится праймер 3′ SMART-Seq CDS Primer II A, а в качестве TSO содержится олигонуклеотид SMART-Seq HT Oligonucleotide; ПЦР-праймер включен в состав реакционного буфера One-Step Buffer. Настоящий способ предусматривает отказ от использования 3′ SMART-Seq CDS Primer II A в протоколе с заменой его на композицию праймеров 3′ HuIgSMART (группа SEQ ID NO: 6, 7, 8, 9, 10). Композиция представляет собой эквимолярную смесь генспецифичных праймеров для затравки синтеза первой цепи кДНК на матрице мРНК тяжелых цепей γ, ε, α, μ и легких цепей κ и λ. В 5′ области каждого праймера данной композиции находится адапторная последовательность (AAGCAGTGGTATCAACGCAGAGT), которая затем служит местом отжига ПЦР-праймера. В итоге, после проведения обратной транскрипции и первого этапа ПЦР в пробирке содержится обогащенная кДНК вариабельной части иммуноглобулина независимо от его класса и изотипа легкой цепи. Не исключено использование наборов или отдельных компонентов наборов для синтеза и амплификации кДНК юлагодаря применению технологии SMART с композицией генспецифичных праймеров 3′ HuIgSMART вместо олиго(dT)n: других наборов TaKaRa Bio линеек SMART-Seq и SMARTer, наборов Template Switching RT Enzyme Mix (NEB #M0466), SmartRT Reverse Transcriptase Kit (MCLAB, США), Mint (Евроген, Россия) и др. Наборы отличаются декларируемой чувствительностью к минимальному стартовому количеству РНК в реакции, разными названиями компонентов, наличием этапа выделения РНК из клетки и/или разделением этапов обратной транскрипции и амплификации кДНК, если это предусмотрено. В отсутствии специального лизирующего буфера для сортинга Ig+ клеток человека можно использовать 0,3% раствор детергента NP-40 в деионизованной воде, свободной от нуклеаз и нуклеиновых кислот, с добавлением 160 ед/мл ингибитора РНКаз. Также не исключено исполнение способа без использования готовых коммерческих наборов, с использованием подходящей MMLV-ревертазы, термостабильной ДНК-полимеразы, синтезированных TSO и ПЦР-праймера.
На основе плазмидных векторов pSF-CMV-HuIgG1_HC (OG527), pSF-CMV-HuKappa_LC (OG528) и pSF-CMV-HuLambda LC (OG529) из коммерчески доступного набора для инженерии химерных или человеческих иммуноглобулинов Human IgG Vector Set (PP2409, Oxford Genetics) создают рабочие вектора для клонирования кДНК иммуноглобулина, которую получают по предлагаемому способу, в существующую рамку считывания. Изменения сводятся к добавлению старт-кода ATG в контекстном окружении последовательности Козак и полинуклеотида сигнальной последовательности с сайтом рестрикции на конце, а также отсеканию части ДНК, кодирующей константные домены CH1, Cκ, Cλ с введением в них «тихой» мутации с целью открытия сайта рестрикции. Обязательным условием является подбор такого сайта рестрикции на конце кодирующей последовательности сигнального пептида, чтобы после ферментативного разрезания образовывался тупой конец непосредственно после последнего кодона этого сигнального пептида. В качестве примера используют сигнальную последовательность аллели IGHV1-69*31 для вектора экспрессии тяжелой цепи и аллели IGKV1D-8*01 для векторов экспрессии легкой цепи κ и λ (международная заявка на патент WO2014058389A1. Haryadi R. et al. Optimization of heavy chain and light chain signal peptides for high level expression of therapeutic antibodies in CHO cells //PloS one. – 2015. – Т. 10. – №. 2. – С. e0116878). Для модификации плазмид используют метод WP-PCR (whole-plasmid PCR), в котором линеаризованная плазмидная ДНК используется в качестве матрицы. Для формирования тупых концов после полинуклеотидов сигнальных последовательностей тяжелой и легкой цепи используют, соответственно, рестриктазы RfuI (изошизомер NruI) и NsbI. В нуклеотидных последовательностях константных доменов выбирают участки, где возможно введение сайта рестрикции с образованием липкого конца без нарушения аминокислотной последовательности данной цепи иммуноглобулина. Учитывают, что желаемый сайт рестрикции в плазмидном векторе после модификации должен встречаться только один раз, и одновременно с этим этот же сайт рестрикции не должен встречаться в последовательности кДНК цепи иммуноглобулина, предназначенной для клонирования в соответствующий вектор. В результате в вектора pSF-CMV-HuIgG1_HC и pSF-CMV-HuKappa_LC внедряют сайт SalI, а в вектор pSF-CMV-HuLambda_LC сайт XhoI. Все новые генетические конструкции вносят с помощью прямых и обратных праймеров.. На 5′-конец каждого праймера добавляют сайт рестрикции ApaI для легкого замыкания модифицированных плазмид. Данные праймеры синтезируют согласно нуклеотидным последовательностям как указано в группе последовательностей SEQ ID NO: 1, 2, 3, 4, 5 и в таблице, отраженной в Фиг. 1. Получают новые плазмидные вектора pSF-CMV-HuIgG1_HCmod, pSF-CMV-HuKappa_LCmod и pSF-CMV-HuLambda_LCmod. Генетические карты плазмид представлены на Фиг. 4. Неполные нуклеотидные последовательности плазмид представлены в перечне последовательностей под номерами SEQ ID NO:38, SEQ ID NO:39 и SEQ ID NO:40, для перечисленных выше плазмид соответственно.
Осуществляют дизайн генспецифичных праймеров группы 3′ HuIgSMART: 3′ HuIgSMART-G&E (SEQ ID NO: 6), 3′ HuIgSMART-A (SEQ ID NO: 7), 3′ HuIgSMART-M (SEQ ID NO: 8), 3′ HuIgSMART-κ (SEQ ID NO: 9) и 3′ HuIgSMART-λ (SEQ ID NO: 10), которые необходимы для затравки синтеза первой цепи кДНК в реакции обратной транскрипции. Они являются генспецифичными, поскольку комплементарны только целевым мРНК. Принцип отжига генспецифичных праймеров на мРНК отражен в Фиг. 5, где в качестве примера представлен графический результат выравнивания последовательности мРНК цепи ε (GenBank: DQ680069.1) и праймера 3′ HuIgSMART-G&E по алгоритму BLAST. Каждый праймер конструируют с добавлением некомплементарной последовательности (оверхенга) в 5′ области: (5′ AAGCAGTGGTATCAACGCAGAGT), – которая впоследствии служит местом отжига ПЦР-праймера на первом этапе ПЦР с кДНК, синтезированной по технологии SMART. Данные праймеры синтезируют согласно нуклеотидным последовательностям из таблицы, отраженной в Фиг. 2, строки №№ 1 – 5.
Осуществляют дизайн 5′ и 3′ праймеров для ПЦР второго этапа, где происходит раздельная амплификация участков кДНК тяжелой и легкой цепи иммуноглобулина, содержащих кодоны вариабельного домена. К группе 5′ праймеров относят праймеры для гибридизации с кДНК в области IGHV, IGKV, IGLV генных сегментов, начиная с первого кодона регионов FR1. Данные праймеры синтезируют согласно нуклеотидным последовательностям, представленным в SEQ ID NO: 11 – 31 и таблице, отраженной в Фиг. 2, строки №№ 6 – 26. Поскольку 5′-концы ампликонов после проведения ПЦР не подвергают гидролизу рестриктазой, то полученный тупой конец должен иметь фосфатную группу у 5′-атома углерода. Для этого 5′ праймеры проектируют как фосфорилированные на 5′-конце. Праймеры группы 3′ создают гибридизующимися в консервативной части полинуклеотидов, кодирующих γ, ε, α, μ, κ и λ цепи иммуноглобулина. Внутри этих консервативных последовательностей находят участок, где сайт рестрикции возможно привнести с помощью праймера как «тихую» мутацию без изменения аминокислотной последовательности. Примерное место отжига праймера и желаемый сайт рестрикции известны заранее, поскольку это учитывают при инженерии модифицированных плазмидных векторов. Данные праймеры синтезируют согласно нуклеотидным последовательностям, представленным в группе SEQ ID NO: 32 – 37 и таблице, отраженной в Фиг. 2, строки №№ 27 – 32. Нижележащую от сайта рестрикции кДНК рассматривают как участок, подлежащий удалению, а затем воссозданию или подмене (в случае амплификации вариабельного домена тяжелой цепи иммуноглобулина, отличного от IgG1) с помощью соответствующего плазмидного вектора.
Предлагаемый способ в качестве примера описывает отбор IgG+ В-клеток памяти из периферической крови человека. Способ также применим к В-клеткам памяти, которые производят иммуноглобулины IgA и IgE, если таковые изолируют. Клетки памяти секретируют мембарнные (поверхностные) иммуноглобулины в составе В-клеточного рецептора (BCR). Поэтому популяция данных клеток может быть окрашена флюоресцирующим антигеном интереса в дополнение к иммунофенотипирующим красителям перед процедурой флуоресцентного сортинга. Забор крови предлагается производить у доноров, имеющих напряженность иммунитета в отношении антигена, к которому с большой долей вероятности могут вырабатываться антитела в результате вакцинации, инфекции, пищевой токсикоинфекции, инвазии и т.д. Способ не исключает получения моноклональных антител от единичных плазмобластов, которые производят специфичные иммуноглобулины IgG, IgM, IgA и IgE, и даже представляется более продуктивным при заборе крови на 5 – 14 день после заболевания или иммунизации донора, но не рассмотрен в примере предлагаемого изобретения. Плазмобласты производят секретируемую форму иммуноглобулина, поэтому прямое окрашивание жизнеспособных клеток антигеном не представляется возможным. Необходимо применять специальные техники окрашивания, например, предусматривающие создание «ловушки» молекул антител вокруг секретирующей клетки. Одна из таких методик, именуемая авторами «ICA» (immunoglobulin capture assay), описана в первоисточнике (Pinder C. L. et al. Isolation and characterization of antigen-specific plasmablasts using a novel flow cytometry–based Ig capture assay //The Journal of Immunology. – 2017. – Т. 199. – №. 12. – С. 4180-4188.). Для получения специфичных антител антиген интереса конъюгируют с флюорофором, спектральные характеристики которого позволяют использовать его в многоцветной панели с моноклональными антителами для фенотипирования популяции В-клеток памяти. Обогащенные В-лимфоциты донорской крови окрашивают флюорофор-меченными антителами и антигеном. На основании фенотипа сортируют единичные В-клетки памяти методом проточной цитофлюорометрии, совмещенной с флюоресцентным сортингом. Клетки сортируют по одной непосредственно в емкость, предназначенную для проведения реакции обратной транскрипции и первой ПЦР, куда заранее вносят лизирующий раствор, не ингибирующий данные реакции, смесь генспецифичных праймеров группы 3′ HuIgSMART и ингибитор РНКаз. Затем, после шоковой заморозки и оттаивания, ко всему объему лизата в ту же емкость добавляют следующие компоненты:
- вода, свободная от нуклеаз и нуклеиновых кислот;
- реакционный буфер;
- олигонуклеотид TSO с последовательностью нуклеотидов 5′ AAGCAGTGGTATCAACGCAGAGTACGC(rG)3 3′ или аналогичной, поскольку при использовании TSO из коммерчески доступных наборов участок олиго(rG) может отличаться и является коммерческой тайной производителя;
- ПЦР-праймер с последовательностью нуклеотидов 5′ AAGCAGTGGTATCAACGCAGAGT 3′;
- ингибитор РНКаз;
- термостабильная ДНК-полимераза;
- обратная транскриптаза (ревертаза) семейства MMLV.
Проводят реакцию обратной транскрипции по технологии SMART с последующей первой ПЦР, получая в одной пробирке нуклеиновые кислоты – кДНК и фрагмента тяжелой, и фрагмента легкой цепи, фланкированные с обоих концов адапторной последовательностью. Второй этап ПЦР осуществляют параллельно в трех пробирках с использованием продукта первой ПЦР в качестве ДНК-матрицы, что соответствует концепции вложенной ПЦР. Данная ПЦР предназначена для амплификации полинуклеотидов, которые включают домены VH и Vκ или Vλ цепей иммуноглобулина, притом полинуклеотиды предназначены для клонирования в соответствующие плазмидные вектора со сборкой полноразмерного гена тяжелой и легкой цепи человеческого IgG. Для каждой реакции используют свою смесь 5′ и 3′ праймеров и 0,1 - 1 мкл продукта реакции первой ПЦР без очистки. Для амплификации нуклеиновой кислоты домена VH в реакцию добавляют смесь праймеров с последовательностями как указано в Фиг. 2 в строках под №№ 6 – 14 и 27 – 30, а также в SEQ ID NO 11 – 19 и 32 – 35, для которых определяют температуру отжига 50 °C. Для амплификации нуклеиновой кислоты домена Vκ в реакцию добавляют смесь праймеров с последовательностями как указано в Фиг. 2 в строках под №№ 15 – 18, 31, а также в SEQ ID NO 20 – 23, 36, для которых определяют температуру отжига 55 °C. Для амплификации нуклеиновой кислоты домена Vλ в реакцию добавляют смесь праймеров с последовательностями как указано в Фиг. 2 в строках под №№ 19 – 26, 32, а также в SEQ ID NO 24 – 31, 37, для которых определяют температуру отжига 55 °C. Амплификацию проводят с использованием термостабильной ДНК полимеразы с 3′ - 5′ экзонуклеазной активностью. Выполняют 30 циклов: денатурация, отжиг праймеров в течение 15 с и элонгация ДНК в течение 30 с. Время и температуру денатурации, а также температуру элонгации ДНК, выбирают согласно оптимальным значениям для используемой ДНК-полимеразы. Результаты реакций оценивают на основании данных электрофореза в агарозном геле. Регистрируют успешно проведенные реакции, отвечающие требованию: один четко различимый фрагмент ДНК следующего размера (в парах оснований): VH 360-390 п.о., Vκ 470-490 п.о., Vλ 360-380 п.о. От одной единичной клетки может быть успешно амплифицирован только один вариант фрагмента легкой цепи, определяемый ее типом: κ или λ. Продукты позитивных реакций очищают и подвергают рестрикционному гидролизу. Нуклеиновую кислоту домена VH разрезают рестриктазами SalI и XhoI, домена Vκ рестриктазой SalI, домена Vλ рестриктазой XhoI. Параллельно обрабатывают рестриктазами сконструированные плазмидные вектора, предоставляющие Козак-последовательность, полинуклеотид сигнальной последовательности и недостающего участка соответсвующей цепи иммуноглобулина. Например, плазмиду pSF-CMV-HuIgG1_HCmod разрезают рестриктазами NruI (RfuI) и SalI, pSF-CMV-HuKappa_LCmod рестриктазами NsbI и SalI, pSF-CMV-HuLambda_LCmod рестриктазами NsbI и XhoI. Подготовленные плазмиды и ПЦР-продукты сшивают в реакции лигирования ферментом T4 ДНК лигазой. В результате от данной клетки получаю два плазмидных вектора с экспрессионными кассетами тяжелой и легкой цепи рекомбинантного человеческого специфичного иммуноглобулина.
Полученные плазмиды доставляют в линейные клети млекопитающих, например, CHO, HEK293, COS-7 или др., при помощи совместной липосомальной трансфекции. Для этого используют смесь плазмидных векторов, несущих гены нативно спаренных легкой цепи и тяжелой цепи, в соотношении 2:1 по молярной концентрации ДНК. Клетки культивируют 7 – 14 дней в бессывороточной питательной среде, после чего осветленную культуральную жидкость пропускают через хроматографический сорбент Белок А для связывания иммуноглобулинов. Затем очищенные антитела элюируют с колонки при помощи буфера с pH 2,4. Очищенные антитела тестируют на специфичность в отношении антигена интереса в ИФА и/или иммуноблоте.
Для лучшего понимания сущности изобретения ниже следуют примеры его конкретного выполнения.
Пример 1. Подготовка векторов.
Плазмидные вектора pSF-CMV-HuIgG1_HC, pSF-CMV-HuKappa_LC и pSF-CMV-HuLambda_LC в количестве 1 мкг линеаризуют (релаксируют) по сайту рестрикции NcoI. Для этого готовят соответствующие отдельные смеси плазмид в 20 мкл 1X буфера Tango и инкубируют с 10 ед. рестриктазы NcoI (Thermo Fisher) 1 час при 37 °C. Затем фермент инактивируют нагреванием при 65 °C в течение 20 мин. Далее линеаризованные плазмиды амплифицируют со следующими парами праймеров: pSF-CMV-HuIgG1 HC с парой H_Apa/Sal_For-WP и HSS_Rfu/Apa_Rev-WP; pSF-CMV-HuKappa LC с парой K_Apa/Sal_For-WP и K&LSS_Nsb/Apa_Rev-WP; pSF-CMV-HuLambda LC с парой K&LSS_Nsb/Apa_Rev-WP и L_Apa/Xho_For-WP. Нуклеотидные последовательности праймеров указаны в таблице, отраженной в Фиг. 1. Для приготовления ПЦР смеси на одну реакцию объемом 25 мкл используют воду, свободную от нуклеиновых кислот и нуклеаз, по 5 пкМ прямого (For) и обратного (Rev) праймера, 0,1 мкл линейной плазмидной ДНК и 12,5 мкл смеси 2X Phusion Hot Start II High-Fidelity PCR Master Mix (Thermo Fisher). Амплификацию проводят, используя следующие параметры: предварительная денатурация 98°C в течение 1 мин, далее 20 циклов: 98°C 15 с, 60°C 15 с, 72°C 3 мин, затем финальная элонгация при 72°C в течение 5 минут. Качественный результат реакций оценивают электрофоретически (Фиг. 3). Регистрируют три фрагмента ДНК размером более 4000 пар оснований. После проведения ПЦР ДНК подвергают фенол-хлороформной экстракции и спиртовой преципитации. Далее по 50-100 нг очищенных ПЦР-продуктов гидролизуют эндонуклеазой рестрикции ApaI FastDigest в буфере FD (Thermo Fisher). Спустя час инкубации при 37 °C фермент в реакциях инактивируют температурой 65 °C в течение 5 минут, затем добавляют АТФ до 0,5 мМ, дитиотреитол до 10 мМ и 5 ед. Вейсса T4 ДНК-лигазы (все реактивы Thermo Fisher). Выдерживают 2-3 часа при комнатной температуре, после чего подвергают фенол-хлороформной экстракции и спиртовой преципитации. Осадки ДНК растворяют в 1 мкл деионизованной воды и используют для электротрансформации компетентных клеток E.coli DH12S. Из позитивных трансформантов, выросших на агаризованной среде LB с канамицином, выделяют плазмидную ДНК, которую впоследствии секвенируют по Сэнгеру с праймера CMV Forward (5′ CGCAAATGGGCGGTAGGCGTG 3′). Новые векторы именуют pSF-CMV-HuIgG1_HCmod, pSF-CMV-HuKappa_LCmod и pSF-CMV-HuLambda_LCmod.
Пример 2. Дизайн праймеров для амплификации целевых кДНК.
Дизайн праймеров для затравки синтеза первой цепи кДНК. Комплементарные части праймеров 3′ HuIgSMART-κ и 3′ HuIgSMART-λ оставляют идентичными праймерам 3′ Cκ 494-516 и 3′ Сλ по (Smith K. et al. Rapid generation … С. 372.), соответственно. Отжигающиеся участки праймеров 3′ HuIgSMART-G&E, 3′ HuIgSMART-A и 3′ HuIgSMART-M конструируют как гибридизующиеся с участкам мРНК тяжелой цепи в областях константных доменов CH1 человеческого иммуноглобулина G, A и M. Праймер 3′ HuIgSMART-G&E также способен к отжигу на мРНК домена CH1 иммуноглобулина E с тремя малозначительными несоответствиями нуклеотидов. Праймеры группы 3′ синтезируют согласно нуклеотидным последовательностям в таблице, отраженной в Фиг. 2, строки №№ 1 – 5.
Дизайн 5′ и 3′ праймеров для ПЦР с целью клонирования полученных продуктов реакции. Праймеры группы 5′ синтезируют согласно нуклеотидным последовательностям в таблице, отраженной в Фиг. 2, строки №№ 6 - 26. Праймерам в рамках одной подгруппы (5' VH, 5' Vκ, 5' Vλ) назначают сквозную нумерацию, не привязанную к подгруппе V сегмента или конкретному сегменту. Данные праймеры фосфорилируют путем внесения 5′-Pi модификации при синтезе.
Праймеры группы 3′ синтезируют согласно нуклеотидным последовательностям в таблице, отраженной в Фиг. 2, строки №№ 27 - 32. Праймеры 3′ JCHG_XhoI, 3′ JCHG&E_XhoI, 3′ JCHA_SalI и 3' JCHM_SalI конструируют как гибридизующиеся на стыке J генных сегментов и CH1 домена, а именно с последними 9 нуклеотидами региона FR4 и несколькими нуклеотидами константой области так, чтобы температура плавления составляла не менее 55 °С с учетом частичной некомплементарности. Праймеры 3′ Cκ_SalI и 3′ Cλ_XhoI конструируют для гибридизации с кодирующими последовательностями константных доменов Cκ и Cλ. Праймер 3′ Cλ_XhoI конструируют по аналогии с праймером 3′ XhoI Cλ (Smith K. et al. Rapid generation … С. 372.), с внутренним сайтом узнавания рестриктазой XhoI (Фиг. 6).
Пример 3. Клеточный флюоресцентный сортинг и пробоподготовка для него
Антиген «А», полипептид с молекулярной массой 33 кДа и семью аминокислотными остатками лизина в составе молекулы, метят R-фикоэритрином (PE) из набора R-Phycoerythrin Conjugation Kit (Abcam).
Все работы проводят в «чистой зоне» в строгих асептических условиях. Используют только одноразовые расходные материалы со степенью чистоты «PCR clean» (Eppendorf) и наконечники с аэрозольным барьером. Подготовку буфера «CDS Sorting Solution» для сортинга и лизиса клеток проводят с использованием компонентов набора SMART-Seq HT Kit (TaKaRa), но заменяя праймер 3′ SMART-Seq CDS Primer II A на смесь генспецифичных праймеров для синтеза первой цепи кДНК. Для приготовления данной смеси праймеры 3′ HuIgSMART-G&E, 3′ HuIgSMART-A, 3′ HuIgSMART-M, 3′ HuIgSMART-κ и 3′ HuIgSMART-λ объединяют в общий стоковый раствор, где концентрация каждого праймера составляет 10 мкМ. Далее подготавливают буфер на 12 реакций для сортировки 12 единичных В-клеток: смешивают 126 мкл воды (свободной от нуклеаз), 12 мкл раствора смеси праймеров, 11,4 мкл десятикратного лизис-буфера, 0,6 мкл ингибитора РНКаз. Буфер помещают в ПЦР-пробирки по 12,5 мкл (можно объединенные в стрип) или в первый горизонтальный ряд ПЦР-планшета и немедленно используют для сортинга.
Из донорской крови объемом 40-50 мл получают обогащенную В-лимфоцитарную фракцию с использованием набора RosetteSep Human B Cell Enrichment Cocktail (STEMCELL). Для выявления субпопуляции В-клеток памяти используют многоцветную панель: CD19 APC / CD27 BB515 / CD38 PE-Cy7 / IgG BB700 / антиген «А» PE. В-лимфоциты окрашивают комбинацией моноклональных антител, конъюгированных с флюорохромами: anti-CD19 APC (клон HIB19), anti-CD38 PE-cy7 (клон HIT2), anti-CD27 BB515 (клон M-T271), anti-IgG BB700, – и антигеном «А»-PE в фосфатно-солевом буфере (137 мМ NaCl, 2,7 мМ KCl, 10 мМ Na2HPO4, 1,76 мМ KH2PO4, рН 7.4, далее: ФСБ) с добавлением 2% эмбриональной телячьей сыворотки. Не связавшиеся молекулы удаляют трехкратной отмывкой клеток в ФСБ. Иммунофенотипирование и сортинг целевых популяций клеток проводят в день взятия крови при помощи проточного цитофлюориметра FACS Aria III (BD Biosciences), оснащенного тремя лазерами с длинами волн излучения 405, 488 и 633 нм. Сначала проводят гейтирование по прямому (FSC) и боковому (SSC) светорассеянию, что позволяет определить размер и гранулярность, соответственно (Фиг. 7А). После исключения дуплетов (Фиг. 7B) и гейтирования CD19+ В-лимфоцитов (Фиг. 7С), на основе коэкспрессии CD27/CD38 молекул выделяют В-клетки памяти с фенотипом CD19+CD27+CD38−/+ (Фиг. 7D). Таким образом, в анализ отбирают и «активированные», и «покоящиеся» В-клетки памяти. Далее в этой субпопуляции дополнительно выделяют только IgG+ клетки (Фиг. 7Е). Таким образом, данная панель позволяет выделить IgG+ В-клетки памяти с фенотипом CD19+CD27+CD38−/+IgG+. На последнем этапе в субпопуляции IgG-позитивных В-клеток памяти проводят дополнительное гейтирование по каналу PE, выделяя специфические клетки памяти к целевому антигену с фенотипом CD19+CD27+CD38−/+IgG+антиген «А» PE+ (Фиг. 7F). Клетки данного фенотипа сортируют по одной в слоты начиная с A2 и далее. Слот А1 пропускают для контрольно-демонстрационной реакции.
По завершении сортинга пробирки или планшеты с клетками подвергают шоковой заморозке выдерживанием в жидком азоте или на сухом льду. Затем образцы хранят при -80 °С.
Пример 4. Синтез и амплификация целевых кДНК.
Все работы проводят в «чистой зоне» в строгих асептических условиях. Используют только одноразовые расходные материалы со степенью чистоты «PCR clean» (Eppendorf) и наконечники с аэрозольным барьером. Амплификатор располагают в непосредственной близости от рабочего места, а все режимы инкубации и амплификации объединяют в одну непрерывную программу согласно дальнейшим указаниям.
ПЦР-пробирки, стрип или планшет с отсортированными клетками по примеру 3 извлекают из низкотемпературного холодильника и немедленно помещают в предварительно нагретый до 72°C амплификатор с разогретой до 105°C крышкой на 3 минуты. Затем температуру в амплификаторе понижают до 4°C и инкубируют еще 2 минуты. Во время инкубации подготавливают реакционную смесь для ОТ-ПЦР, не превышая суммарного времени инкубации 5 минут на эту операцию, поэтому замороженные компоненты смеси размораживают заранее. Для осуществления 12 реакций в микропробирку последовательно вносят следующие компоненты из набора SMART-Seq HT Kit:
Вода – 8,4 мкл;
One-Step Buffer – 96 мкл;
SMART-Seq HT Oligonucleotide – 12 мкл;
RNase Inhibitor – 6 мкл;
SeqAmp DNA Polymerase – 3,6 мкл (строго контролируя количество фермента при наборе и внесении);
SMARTScribe Reverse Transcriptase (100 U/µl) – 24 мкл.
Содержимое несколько раз аккуратно перемешивают пипетированием, избегая образования пузырей. Приготовление смеси на количество реакций менее 12 не желательно. При большем количестве единично отсортированных клеток количество каждого компонента высчитывают пропорцией.
Пробирки или планшет извлекают из амплификатора после завершения инкубации при 4°C. Устанавливают температуру термоблока 42°C и время 90 минут. По достижении желаемой температуры программу ставят на паузу. В каждую ПЦР-пробирку/лунку добавляют по 12,5 мкл реакционной смеси и аккуратно перемешивают пипетированием 2 - 3 раза, избегая образования пузырей. После этого пробы возвращают в амплификатор и запускают следующую программу: инкубация 42°C 90 мин, затем нагрев до 95°C в течение 1 мин, затем 20 циклов: 98°C 10 с, 65°C 30 с, 68°C 1 мин, - далее финальная элонгация 72°C 5 мин и хранение проб до выемки 4°C ∞.
Продукт реакции ОТ-ПЦР используют в качестве матрицы ДНК в ПЦР второго этапа. Сначала смешивают праймеры для составления трех композиций – стоковых 50-кратных растворов с концентрацией каждого праймера 10 мМ.
1) Композиция «VH-JCH»: праймеры 5′ VH-1, 5′ VH-2, 5′ VH-3, 5′ VH-4, 5′ VH-5, 5′ VH-6, 5′ VH-7, 5′ VH-8, 5′ VH-9, 3′ JCHG_XhoI, 3′ JCHG&E_XhoI, 3′ JCHA_SalI, 3′ JCHM_SalI с последовательностями нуклеотидов как указанов в строках под №№ 6 – 14 и 27 – 30 Фиг. 2.
2) Композиция «Vκ-Cκ»: 5′ Vκ-1, 5′ Vκ-2, 5′ Vκ-3, 5′ Vκ-4, 3′ Cκ_SalI с последовательностями нуклеотидов как указанов в строках под №№ 15 – 18, 31 Фиг. 2.
3) Композиция «Vλ-Cλ»: 5′ Vλ-1, 5′ Vλ-2, 5′ Vλ-3, 5′ Vλ-4, 5′ Vλ-5, 5′ Vλ-6, 5′ Vλ-7, 5′ Vλ-8, 3′ Cλ_XhoI с последовательностями нуклеотидов как указанов в строках под №№ 19 – 26, 32 Фиг. 2.
Пподготавливают три варианта ПЦР смеси для каждого продукта реакции ОТ-ПЦР по технологии SMART. На одну реакцию объемом 50 мкл используют 1 мкл композиции праймеров VH-JCH, или Vκ-Cκ, или Vλ-Cλ, 0,5 мкл продукта ОТ-ПЦР без очистки, 25 мкл смеси 2X Phusion Hot Start II High-Fidelity PCR Master Mix (Thermo Fisher), а оставшийся объем 23,5 мкл заопляют водой, свободной от нуклеиновых кислот и нуклеаз. Амплификацию проводят, используя следующие параметры: предварительная денатурация 98°C в течение 1 мин, далее 30 циклов: 98°C 15 с, 50°C для амплификации VH или 55°C для амплификации VL (κ и λ) 15 с, 72°C 30 с, затем финальная элонгация при 72°C в течение 5 минут. Качественный результат реакции оценивают электрофоретически, что отражено в Фиг. 8. Истинные ампликоны идентифицируют, если они выражены в виде четкого монопродукта и имеют следующую длину в сравнении с маркером молекулярного веса: VH 360-390 пар (Фиг. 8, дорожки 4 и 7) оснований, Vκ 470-490 п.о. (Фиг. 8, дорожка 5), Vλ 360-380 п.о. (Фиг. 8, дорожка 9). Чистая дорожка или наличие 1 – 3 слабо различимых полос, образованных в результате неспецифического отжига праймеров с контаминирующей ДНК в отсутствии целевой матрицы, свидетельствует о непрошедшей реакции (Фиг. 8, дорожки 1, 2, 3, 6, 8). Данное явление должно иметь место при амплификации одного из изотипов легкой цепи (Фиг. 8, дорожки 6 и 8). В случае неуспешной амплификации пары VH и VL от единичной клетки клонирование кДНК не начинают, как и в случае одновременной амплификации и Vκ, и Vλ. При одновременном отсутствии продукта амплификации фрагмента тяжелой и легкой цепи, подобно электрофоретическому разделению ПЦР-продуктов серии реакций с содержимым лунки (пробирки) А1 по примеру 3 (Фиг. 8, дорожки 1, 2, 3), косвенно судят об отсутствии клетки в лизирующем буфере.
По результатам амплификации, устанавливают наличие следующих ПЦР-продуктов:
1. VH-A2agA – полинуклеотид вариабельного домена и части константного домена тяжелой цепи антитела A2agA к антигену «А», полученный в результате амплификации кДНК единичной человеческой В-клетки памяти, отсортированной в ячейку A2 по примеру 3 (Фиг. 8, дорожка 4)
2. VH-A3agA – полинуклеотид вариабельного домена и части константного домена тяжелой цепи антитела A3agA к антигену «А», полученный в результате амплификации кДНК единичной человеческой В-клетки памяти, отсортированной в ячейку A3 по примеру 3 (Фиг. 8, дорожка 7)
3. Vκ-A2agA – полинуклеотид вариабельного домена и части константного домена легкой цепи изотипа κ антитела A2agA к антигену «А», полученный в результате амплификации кДНК единичной человеческой В-клетки памяти, отсортированной в ячейку A2 по примеру 3 (Фиг. 8, дорожка 5)
4. Vλ-A3agA – полинуклеотид вариабельного домена и части константного домена легкой цепи изотипа λ антитела A3agA к антигену «А», полученный в результате амплификации кДНК единичной человеческой В-клетки памяти, отсортированной в ячейку A3 по примеру 3 (Фиг. 8, дорожка 9)
Пример 5. Клонирование ПЦР продуктов в вектора.
Плазмидные вектора, полученные по примеру 1, в количестве 100 нг гидролизуют эндонуклеазами рестрикции линейки FastDigest (Thermo Fisher) в буфере FD с добавлением щелочной фосфатазы FastAP (Thermo Fisher). Вектор pSF-CMV-HuIgG1_HCmod разрезают по сайтам RfuI и SalI. Вектор pSF-CMV-HuKappa_LCmod разрезают по сайтим NsbI и SalI. Вектор pSF-CMV-HuLambda_LCmod разрезают по сайтам NsbI и XhoI. Спустя час инкубации при 37 °C плазмидную ДНК подвергают фенол-хлороформной экстракции и спиртовой преципитации. Осадки растворяют в 10 мкл воды.
Продукты ПЦР по примеру 4, интерпретированные как удачно прошедшие, очищают на одноразовых колонках с мембраной из диоксида кремния, например с применением наборов QIAquick PCR Purification Kit или MinElute PCR Purification Kit (Qiagen). Для элюции ДНК используют воду (pH 7,0). Далее подготавливают реакционные смеси для рестрикционного гидролиза общим объемом 20 мкл каждая, 16 мкл которой занимает элюат ДНК. К пробам добавляют десятикратный FD буфер и ферменты рестрикции линейки FastDigest в соответствии с таблицей, отраженной в Фиг. 9. Спустя час инкубации при 37 °C ДНК подвергают фенол-хлороформной экстракции и спиртовой преципитации. Осадки растворяют в 10 мкл воды.
Сшивку подготовленных плазмид с соответствующими вставками производят ферментом T4 ДНК-лигазой (Thermo Fisher). В реакцию берут по 10-20 нг плазмиды и 5 мкл вставки. Реакции выдерживают при комнатной температуре в течение 12 - 16 часов. Продукты реакций лигирования подвергают фенол-хлороформной экстракции и спиртовой преципитации, после чего ДНК-содержащие осадки растворяют в 1 мкл воды и используют для электротрансформации клеток E.coli DH12S. После этого клетки высевают на агаризованную среду LB с ампициллином и растят в течение ночи. Затем проводят предварительный скрининг встройки ампликонов в плазмиды методом ПЦР с универсальным прямым праймером CMV Forward и обратными праймерами
HuIgG-const-anti (5′ TCTTGTCCACCTTGGTGTTGCT 3′),
3′ Ck 494-516 (5′ GTGCTGTCCTTGCTGTCCTGCT 3′) и
3′ Cλ (5′ CACCAGTGTGGCCTTGTTGGCTTG 3′)
для единичных колоний E.coli с плазмидами pSF-CMV-HuIgG1_HCmod, pSF-CMV-HuKappa_LCmod и pSF-CMV-HuLambda_LCmod, куда были клонированы ПЦР-продукты, соответственно. В качестве контроля параллельно осуществляют амплификацию данных плазмид без вставок с тех же пар праймеров. Результаты скрининга оценивают электрофоретически в агарозном геле, что отражено в Фиг. 10. Из позитивных трансформантов выделяют плазмидную ДНК с помощью набора PureLink HiPure Plasmid Midiprep Kit (Thermo Fisher). Получают следующие плазмидные вектора:
1. pSF-HC-A2agA как результат клонирования полинуклеотида VH-A2agA в плазмидный вектор pSF-CMV-HuIgG1_HCmod;
2. pSF-HC-A3agA как результат клонирования полинуклеотида VH-A3agA в плазмидный вектор pSF-CMV-HuIgG1_HCmod;
3. pSF-LC-A2agA как результат клонирования полинуклеотида Vκ-A2agA в плазмидный вектор pSF-CMV-HuKappa_LCmod;
4. pSF-LC-A3agA как результат клонирования полинуклеотида Vλ-A3agA в плазмидный вектор pSF-CMV-HuLambda_LCmod.
Корректность клонирования продуктов ПЦР подтверждают секвенированием плазмид по Сэнгеру с праймера CMV Forward и SV40pA-R (5′ GAAATTTGTGATGCTATTGC 3′). Результаты секвенирования отражены в последовательностях SEQ ID NO: 41, 43, 45, 47 для плазмидных векторов, указанных выше, соответственно. Прочтенные нуклеотидные последовательности также анализируют, проверяя на наличие открытой рамки считывания цепи иммуноглобулина (SEQ ID NO: 42, 44, 46, 48) и подтверждая принадлежность к полинуклеотидам, кодирующим вариабельный домен тяжелой/легкой цепи, с помощью автоматизированного компьютерного алгоритма IMGT/V-QUEST, общедоступно представленного на сайте: http://www.imgt.org/IMGT_vquest/input . Результаты представлены на Фиг. 11, 12, 13 и 14. В каждой нуклеотидной последовательности алгоритм определил регион V(D)J-рекомбинации и названия аллелелей генных сегментов, участвовавших в рекомбинации.
Пример 6. Препаративная экспрессия, очистка и валидация антител.
Клеточную линию ExpiCHO-S, являющуюся частью набора ExpiCHO Expression System Kit (Gibco, Thermo Fisher), выращивают в среде ExpiCHO Expression Medium в одноразовых колбах Эрленмейера объемом 500 мл с вентилируемой крышкой (Corning) на шейкере Minitron (Infors HT), режим 125 мин-1 в атмосфере 8% CO2 при 37°C. По достижении плотности 6×106 кл/мл и количества жизнеспособных клеток не менее 95%, которую оценивают по окрашиванию трипановым синим, проводят совместную трансфекцию клеток плазмидами, кодирующими легкую и тяжелую цепь иммуноглобулина единичной клетки, которые были получены по примеру 5. В день трансфекции частично или полностью производят замену среды на аналогичную новую. Трансфекцию проводят с использованием реактивов и методики набора ExpiCHO Expression System Kit, а затем клетки выращивают согласно протоколу «Max Titer Protocol». Кратко: в каждую колбу объемом 500 мл, в которой находится 100 мл культуры ExpiCHO, добавляют смесь из 80 мкл смеси плазмидной ДНК, где содержатся плазмиды pSF-LC и pSF-HC в молярном соотношении 2:1 с общей концентрацией нуклеиновой кислоты 1 мг/мл, 7,7 мл среды OptiPRO SFM и 320 мкл ExpiFectamine CHO Reagent. Культуру растят на шейкере при 32°C в атмосфере 5% CO2. Через 24 часа после трансфекции в колбу добавляют 600 мкл энхансера ExpiCHO Enhancer и 16 мл подкормки ExpiCHO Feed. На 5 сутки после трансфекции добавляют еще 16 мл ExpiCHO Feed. На 14 день культуру центрифугируют 10000 x g в течение 10 минут. Супернатант, содержащий антитела, дополнительно фильтруют через мембрану с диаметром пор 0,2 мкм, затем концентрируют десятикратно в ячейке Amicon Stirred Cell (Merck Millipore). При необходимости замораживают -20 °C или сразу переходят к очистке.
Выделение рекомбинантных антител из фильтрованной и концентрированной культуральной жидкости проводят методом аффинной хроматографии на колонке HiTrap rProtein A FF (GE Healthcare) емкостью 1 мл с использованием хроматографической системы ÄKTA Pure 25 (GE Healthcare). Колонку уравновешивают буфером нанесения (50мМ Tris, 100мM NaCl, 0,02% NaN3, pH 8,0). Производят нанесение культуральной жидкости со скоростью 2 мл/мин на льду. Далее колонку отмывают от не связавшихся компонентов буфером нанесения, и проводят элюцию иммуноглобулинов буфером состава: 100 мМ глицина, 100 мМ NaCl, рН 2.4. Уровень pH элюата доводят до 7,5 – 8,0 добавлением 1 М раствора Tris основания. Впоследствии элюат иммуноглобулиновой фракции вносят в коммерчески доступную колонку HiTrap Desalting (GE Healthcare) емкостью 5 мл, уравновешенную фосфатно-солевым буфером ФСБ на скорости потока 5 мл/мин. Иммуноглобулиновую фракцию собирают через некоторое время, идентифицируя ее по изменению А280 (величине поглощения света с длиной волны 280 нм белком). Для расчета концентрации антител [мг/мл], замеряеют показатель А280 раствора белка на спектрофотометре, после чего полученное значение делят на коэффициент массовой экстинкции, который для иммуноглобулина равен 1,37. Как правило, выход составляет 15 - 20 мг белка с 1 литра культуры. Чистоту полученных иммуноглобулиновых фракций оценивают методом SDS-электрофореза по Лэммли (Laemmli U. K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4 //nature. – 1970. – Т. 227. – №. 5259. – С. 680-685.) в 7,5% полиакриламидном геле (ПААГ) в нередуцирующих условиях (электрофореграмма отражена в Фиг. 15, часть А) и 12% ПААГ в редуцирующих условиях с добавлением меркаптоэтанола (электрофореграмма отражена в Фиг. 15, часть Б). Согласно результатам данного анализа, очищенные человеческие антитела A2agA и A3agA обладают стандартным молекулярным весом для человеческого IgG1, который приблизительно равен 150 кДа, а также имеют характерную картину разделения в редуцирующих условиях, распадаясь на 2 идентичные легкие цепи массой 25 кДа и две идентичные тяжелые цепи массой 50 кДа.
Оценку иммунологической специфичности очищенных рекомбинантных антител проводят методом иммуноблота с антигеном «А». Антиген подвергают SDS-электрофорезу в 12% ПААГ в количестве 0,5 мкг на дорожку в двух повторах (по числу полученных антител) и в количестве 3 мкг на дорожку. После чего часть геля с дорожкой 3 мкг антигена «А» окрашивают кумасси бриллиантовым синим R-250 (Фиг. 16, часть А). С остальной части геля осуществляют горизонтальный перенос белков на нитроцеллюлозную мембрану Hybond-C Extra (GE Healthcare). После завершения переноса мембрану блокируют погружением в обезжиренное молоко с массовой долей жира не более 0,5 % и инкубируют в течение 1 ч на орбитальном термостатируемом шейкере при 300 об./мин и температуре 37 °С. Далее, после трехкратной промывки буфером ФСБ с добавление 0,5% детергента Tween® 20 (ФСБ-Тв), мембрану разрезают на две части для изоляции дорожек и каждую часть инкубируют с разными рекомбинантными антителами, разведенным до концентрации 0,5 мкг/мл в ФСБ в течение 1 ч на термостатируемом шейкере. После трехкратной промывки буфером ФСБ-Тв мембраны инкубируют с антителами козы к Fc фрагменту человеческого IgG (Sigma-Aldrich A0170) в разведении 1:10 000 в буфере ФСБ-Тв в течение 30 минут. Затем мембраны промывают 5 раз в ФСБ-Тв. Визуализацию реакции проводят раствором субстратной смеси на основе диаминобензидина (0,05 % диаминобензидина, 0,015 % Н2О2 в ФСБ рН 7,4). Реакцию останавливают после отчетливо различимого окрашивания образцов путем погружения мембран в дистиллированную воду. Как показывает проведенный анализ, результаты которого отражены на Фиг. 16 в дорожках 2Б и 3Б, полученные человеческие антитела A2agA и A3agA специфично взаимодействуют с антигеном «А» белковой природы с молекулярной массой 33 кДа.
| название | год | авторы | номер документа |
|---|---|---|---|
| Штамм гибридных культивируемых клеток животных Mus musculus 5B9 - продуцент мышиного моноклонального антитела 5B9, антитело моноклональное мышиное 5В9 и антитело рекомбинантное химерное (мышь-человек) xi5В9, нейтрализующие рицин Ricinus communis | 2022 |
|
RU2802436C1 |
| Универсальный интеграционный вектор SB7G_HIGH и рекомбинантная плазмида SB7G_HIGH_DEV_57, обеспечивающая синтез и секрецию рекомбинантного человеческого антитела к рецептор-связывающему домену (RBD) коронавируса SARS-CoV-2 в клетках млекопитающих и полученная с использованием универсального вектора SB7G_HIGH, и рекомбинантное моноклональное антитело DEV_K57, обладающее вируснейтрализующей активностью в отношении SARS-CoV-2 | 2023 |
|
RU2829359C1 |
| Способ получения димерной формы мутантного иммуноглобулина IgA2m1-изотипа в клетках млекопитающих | 2023 |
|
RU2822889C1 |
| ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА ЖИВОТНЫЕ СО СКОНСТРУИРОВАННЫМ ЛОКУСОМ ЛЕГКОЙ ЛЯМБДА-ЦЕПИ ИММУНОГЛОБУЛИНА | 2017 |
|
RU2757665C2 |
| Антитело моноклональное мышиное 1Е10 и антитело рекомбинантное химерное (мышь-человек) xi1E10, нейтрализующие летальный токсин Bacillus anthracis, и штамм гибридных культивируемых клеток животных Mus musculus 1E10 | 2020 |
|
RU2745116C1 |
| Способ получения димерной формы иммуноглобулина IgA1-изотипа в клетках млекопитающих | 2023 |
|
RU2822890C1 |
| МЫШИ С ГУМАНИЗИРОВАННОЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2664181C2 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ИММУНОГЛОБУЛИНА IGA2m1-ИЗОТИПА В КЛЕТКАХ МЛЕКОПИТАЮЩИХ | 2022 |
|
RU2801178C1 |
| ЖИВОТНЫЕ, НЕ ЯВЛЯЮЩИЕСЯ ЧЕЛОВЕКОМ, ИМЕЮЩИЕ СКОНСТРУИРОВАННУЮ ЛЕГКУЮ ЦЕПЬ ЛЯМБДА ИММУНОГЛОБУЛИНА, И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2778410C2 |
| МЫШИ, ЭКСПРЕССИРУЮЩИЕ ГИБРИДНУЮ ЛЕГКУЮ ЦЕПЬ ИММУНОГЛОБУЛИНА | 2011 |
|
RU2601297C2 |










Изобретение относится к области биотехнологии, в частности к способу амплификации нуклеиновых кислот тяжелой и легкой цепи человеческих иммуноглобулинов. Изобретение позволяет снижать вероятность ложноотрицательной амплификации, повышая вероятность успешной амплификации кДНК антител с редко встречающимися аллелями V генных сегментов. 16 ил., 6 пр.
Способ амплификации нуклеиновых кислот тяжелой и легкой цепи человеческих иммуноглобулинов, который включает
(а) изоляцию человеческих единичных иммуноглобулин-продуцирующих клеток (В-лимфоцитов), полученных из биологического материала доноров с помощью проточной цитофлюориметрии, совмещенной с флюоресцентным сортингом по признаку взаимодействия с флюорофор-меченным антигеном, притом клетки сортируют непосредственно в емкость с лизирующим буфером для выхода мРНК из клетки в раствор;
(б) реакцию обратной транскрипции на матрице мРНК единичной клетки по технологии SMART с использованием в реакции TSO-олигонуклеотида и MMLV ревертазы, а затем амплификацию полученной кДНК, которая при использовании данной технологии синтезируется фланкированной с обеих сторон универсальной адапторной последовательностью, причем реакция обратной транскрипции отличается тем, что затравка синтеза кДНК осуществляется в присутствии композиции из пяти генспецифичных праймеров как представлено в группе последовательностей SEQ ID NO: 6, 7, 8, 9, 10, которые в 3′-области комплементарны мРНК цепей иммуноглобулина типа γ, ε, α, μ, κ, λ соответственно, но в 5′-области имеют одинаковую адапторную последовательность для гибридизации с ПЦР-праймером, что позволяет произвести избирательную амплификацию двух полинуклеотидов легкой и тяжелой цепи иммуноглобулина данной клетки в одной реакционной смеси с помощью этого ПЦР-праймера;
(в) три последующие полимеразные цепные реакции с амплифицированным образцом кДНК в качестве матрицы, которые отличаются использованием для каждой реакции трех разных композиций олигонуклеотидных праймеров с последовательностями нуклеотидов как представлено в группах (SEQ ID NO: 11 – 19, 32 – 35), (SEQ ID NO: 20 – 23, 36) и (SEQ ID NO: 24 – 31, 37), предназначенных для осуществления амплификации нуклеиновых кислот, кодирующих вариабельные домены VH, Vκ и Vλ, начиная с первой аминокислоты региона FR1, при температурах отжига 50 °C, 55 °C и 55 °C соответственно для каждой группы, причем результатом трех реакций является два продукта: один полинуклеотид домена VH длиной 360-390 пар оснований с сайтом рестрикции XhoI или SalI на 3′-конце и один полинуклеотид либо домена Vκ длиной 470-490 пар оснований с сайтом рестрикции SalI на 3′-конце, либо домена Vλ длиной 360-380 пар оснований с сайтом рестрикции XhoI на 3′-конце, которые затем можно клонировать в подходящие плазмидные векторы по «тупому» и «липкому» концу между кодирующей последовательностью сигнального пептида и недостающей (неамплифицированной) частью константного домена цепи иммуноглобулина соответствующего типа для экспрессии рекомбинантного человеческого иммуноглобулина в клетках млекопитающих.
| SMITH K | |||
| et al, Rapid generation of fully human monoclonal antibodies specific to a vaccinating antigen, Nature protocols, 2009, т | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Электромагнитный счетчик электрических замыканий | 1921 |
|
SU372A1 |
| RUDKIN F | |||
| M | |||
| et al, Single human B cell-derived monoclonal anti-Candida antibodies enhance phagocytosis and protect against disseminated candidiasis, Nature communications, 2018, т | |||
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |

